511-fear-stress
Rick Gilmore
2021-11-12 11:53:49
Fun
The brain bases of emotion
(Lindquist, Wager, Kober, Bliss-Moreau, & Barrett, 2012)
Locationist account
- Where in the brain is emotion processed?
![[[@Lindquist2012-jr]](http://dx.doi.org/10.1017/S0140525X11000446)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAgEAyADIAAD/4SBDRXhpZgAATU0AKgAAAAgABwESAAMAAAABAAEAAAEaAAUAAAABAAAAYgEbAAUAAAABAAAAagEoAAMAAAABAAIAAAExAAIAAAAbAAAAcgEyAAIAAAAUAAAAjYdpAAQAAAABAAAApAAAANAAAADIAAAAAQAAAMgAAAABQWRvYmUgUGhvdG9zaG9wIENTIFdpbmRvd3MAMjAxMjowNToxOCAxNzozODo1NQAAAAAAA6ABAAMAAAAB//8AAKACAAQAAAABAAAFeKADAAQAAAABAAADkgAAAAAAAAAGAQMAAwAAAAEABgAAARoABQAAAAEAAAEeARsABQAAAAEAAAEmASgAAwAAAAEAAgAAAgEABAAAAAEAAAEuAgIABAAAAAEAAB8NAAAAAAAAAEgAAAABAAAASAAAAAH/2P/gABBKRklGAAECAQBIAEgAAP/tAAxBZG9iZV9DTQAB/+4ADkFkb2JlAGSAAAAAAf/bAIQADAgICAkIDAkJDBELCgsRFQ8MDA8VGBMTFRMTGBEMDAwMDAwRDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAENCwsNDg0QDg4QFA4ODhQUDg4ODhQRDAwMDAwREQwMDAwMDBEMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwM/8AAEQgAaACgAwEiAAIRAQMRAf/dAAQACv/EAT8AAAEFAQEBAQEBAAAAAAAAAAMAAQIEBQYHCAkKCwEAAQUBAQEBAQEAAAAAAAAAAQACAwQFBgcICQoLEAABBAEDAgQCBQcGCAUDDDMBAAIRAwQhEjEFQVFhEyJxgTIGFJGhsUIjJBVSwWIzNHKC0UMHJZJT8OHxY3M1FqKygyZEk1RkRcKjdDYX0lXiZfKzhMPTdePzRieUpIW0lcTU5PSltcXV5fVWZnaGlqa2xtbm9jdHV2d3h5ent8fX5/cRAAICAQIEBAMEBQYHBwYFNQEAAhEDITESBEFRYXEiEwUygZEUobFCI8FS0fAzJGLhcoKSQ1MVY3M08SUGFqKygwcmNcLSRJNUoxdkRVU2dGXi8rOEw9N14/NGlKSFtJXE1OT0pbXF1eX1VmZ2hpamtsbW5vYnN0dXZ3eHl6e3x//aAAwDAQACEQMRAD8A9LwMJ+ILg7IsyPWtdd+lg7C/V1VUfRoa7+ar/wAH++j221UVPuue2uqsFz3uIDWtGrnOcfotVfqfVMXpeN9oyidpcGMa2NznHXa3cWt+i3d7nLhM/qnUur3OtsO+guPoYjTLGhp/RW+kdjMqx7h+idd/hK7bPT9P00lPRZf166TW15w67s01Dda5jDWxjP33W5Hp7t3+CZV6nqrPv+tn1qyGOf0/pTambNzH3tscCY3aE/ZP6rVisqoZ1J+PdRmEVODnBv6Sp5gOruyLLLPTZk4v6Rj2f4D1E2HgOyAzIvutt6j1BzjR06x27Hxqi8tdkHHdua306Pazd/2qf9BJSRn1hz+p+iy/q9leVmML6K8csx8WlxG59GVY39f9WhrX7K7LP0tv6NEtzM/F2HJ6nl31vd6Tb2ZDqXNsP8270rMb0ba/b+k/nvT+nYrWRg9M+wOzsmlrOnsO5lTPo1Vgw7Mv9Itfddp6r3N/mf8AAqOBgZ2ZnVdTZjPxun/Y/s1D7MguyyHOZfXlvLmWNbZ6Xs2Ps9b/AEqSlN+sH1lwnQ0nIbG6s5Ja1rx9Fzra4+0117j/AD3q17P5yzH9JdJ0f6z05+U7p2Zjv6b1Rg3fZLSDubzux7m7W27f8/8A63+kXPPbdkUYQsNV2RblX42RfbUHCxuN682em3bte/0mez+b9T1Fm5XTai2ge6u5jmtcwPLXV/pv1XJpuad/pO3s+z31fzX8ixJT6cq/UQD0/JB1Bps0P9VyxPqx9ZxnR03qLgzqle5odG1uQKy5r7KdGt9Zuz9YpZ/xlP6H6G31D+gZP/FP/wCpckpLVrWz+qPyKaA++rGwzkXO2VVV77Hcw1o3OOi5HK68epWVNybWV4OY4Nw8bcGi0nT08h4d6l9u76dFf6v/AIN/q/ziSnosj6xdLodsa9+Q/UBuPW60Ej6TfUrb6Pt/4xZeF9dLOoZVtGH05/6Mlv6a1lb5A3y6pvrba/5e9YmbVm5OZ1DFw3HGq+xsx6suwOrqocxz/W9N+wM2u3fTbsVT6ufbGYj342HjvuF3o2Zl+TpkvqYb3/Y7W1Pquooor9Sl9llVf+Ykp28X6wdc6vjZF4vq6S3FutptDGttLHUnaW3X5UVvbZ+b6eNWpi/6wZbRfX1ljq3EaYdNTqxMCN73W2t/7cesh2D01uJl5Y6dkWVZA+01jqFbWVUWn+fynZm+z1q73+lZXj003/pP5v8AQ2/owV4/UKrW7ejUU9cox7H1mt5ZXc+otHrfYsMfSf6lbmfavSZ+k9Ov1ElOti/WLr9OfmY7bm9TrxmVPANQa+bvV/0Dq/5v0Poem/etjpP1uwuoZjMF0NuuLhRZWS6uzYPUdt3spuZura61jnU+g/8AweRYuY6ZjU2nGxKraczE6i82dSu3xkWX+lZsoy8Ru37Nh+vX9m9H9H/N/Zfz0TLxGdXx8iumvEyKaGen9obSWsd6QGzGpyW5D9/2X6D/AE2+jjvSU+hKlXgWM6rbnnIe6uyv0xjGdgI2e/6ez8x3+D/wti5z6ufWoY2LTh9YurdXXSws6i1+9oEN21Z7j7q7mN+nmP8A0F3+E9Gxdgkp/9Dc+tvU6czqgwDLKcNr63OIPvuta0tbuY5vp1UV+9z9zPU/8+VsTG2MDsi2y972udS2129gpq7Pn3/4X1P0X6X/AIRVfrlXdhdbzA2QzK9LJY90uInbTc3HEja79H+k9L9P6Kr4uQ26oSXnCaCLMGoj0Xsd+j9RjqmerZUyxvo34+z/AI2tJTotwnWYjuj5Bc+rLp9CvOx9rdvev1q9z3Vfyrv0lV/9dH6Zmelh4Yy22ZDmNdhtqqaHfpGjZZ9ouljfWf6XpVN/7b+moW3W1+ji1W4eA8HgB2Q8AD6GMyttLadv+kfZ6ay+lY+Rdh5v2gWnpOM529oZ6eTlfZyNl1T9+xrvSDbMvKq9PJtyP9H+kSU9A/p5wvq8/Ae51lVOIKX2MINgp27Mws3fo/XqodY6lDxer5Fl+Pj9Gx39SZXiBmTlXF2JQ5g2/Y72NuY9zn/znqOpq9//AFtZlN/1dza8HLtvY19cud021+RZbYXa49LMNz3+tZU73/zV3rWK9m9f6nj0ZGRf07KxQ9jm4eQ8MOk+pVXl49LrrsZ/qOd/O/4D/hElJaxmY+X6d+HXX6A+2Tj2bWW5FjXUC2mm4D068f3/AGjbb6lttlN1io5GWHNxqaSyrLxchrrKXtdcK/f+krqrq919VrHfatns/RVersWkM/NyqGvbjfs+rY14syS1xcT9KnCrossY93t/pFtjKv8AgLVkZecMelzaQ2i3Ldtx6zb61ldlxb9quy7K/U3udWxn80+70a/U/wAH+jSUh6nW0vqvqva+sF7qbcRr9LWn1q2+973Vbbve5/qfoH/TXa9O6v8Atf6vX5Lm+ncyt9d7RqN4r37mO/ceyxljVx+/7S0Y/TcN9mNjMsoFNINh3P1s9Oxn6Oz1bffkZVr6/wCds/wv6NdZ0bpd3TPq7kNygG5WQ23IvraQ5rHPbpQxw+l6VbGM/wCMSU4n1v6zjnNowN25uMybNzHOoZedvpDJdUd3qMZ76/Z+gWBnZnp1W0vxMU5HUamusy6nPdDZ2k+jfXT6WQ/b7Nv/AB386ofWTB+yday7M2hjqDa65+Q21znlt8uxrfRq91foO/nK/wCf/wAJ76lnvc/ILsmRfU54m2oufWQ1rfYLXD2fo/39nvUvLxhLIBMit9f0v6qJE1o7fQqP2hivOQ2/qX6UDGote52NU5o/TZudbdvr9kbK6rfV/wCAx/U962sgWNtbi5ufVkuewsrxqcR3qFj4osqoNmQ2r07N2y19VaK3r/RsjDprw67m49c+nWGNa0Ro2v0nu9P2fv8A/XFUHVen3OFHUsVpw3P3b7C0hpPD7Aytu/3/ALj/AFEZ45ylKXBwjevl4QoEaC0mSR1D1sWs5L/oNuY3Fps0re17KN1NrNjHOr+h/o1a/ZvUGCzKbZ6lz9v6Xa6q2v0vfU11Y+0UWMZc912zZ6e+1Qey5uPXXlsc924BlFEVMAtdsq9OhltDGtfPsq/S2/6T1f0liyuqdL6xiXjqXR8t9jumvFn2Cx9lrxVJbeyqp/8AP1+jZc/0vU/4i3+bUKW9dRXkHZfQcd2K+j7NjgB3rl9jXZVt1tLf1mrOa57Lqv8AB7H5FzP0iuZ7cSmz9kUxUzMD7rq6BDnU7gy2vFrp9zrs76D7K/ZRT9ov/Rv2LHoyur9WZjdSwvRdjWF1VLbnuqLa2v8Adayin1/bZZ9L9bff/Nv/AEdKpdQyuodHuvOcS7I6m0g5rTva2uvZ+q7QDbj4mN6vrWs/nLN6SnXzHjLpOHk47KqrGl2M0lr6/RrLac3Z6PsxrcSl30Getjvq/m7rVpfVPrlm6voWa0NtorLMWwGS+ur9Gxto/wBJ6NfqMtb7L/0n83+fnN6v0bIpOXXayqyn3bd5bZWNvpNZiOrf6bqXV/omMZ+g/wBMhfU6rIy+psdXtdRi7MjIvJ95e6u/Hx8dohvs9Ozcza1lbKq/+GSU/wD/0fQPrB9XsLr2I2jJJrsqJdRc2NzCRtcNfp1P/wALV+euKb/i++sVNm2m2sNbJY6u97Wa/wAiyqyxn+f6a9KSQI+i4TIFUCPEPl+b9Vev9Dxn9UcxuSGEG4Yr32WtZ+/6fpU/o6/z/S3/AOk/m1Pp3UB9lezLyrdjZZZWyDvN7pPrPaH2urtfa5rfTXpqxMr6l/VrJe+x+EK7LN259L31fS+npS9jUgK7qlK6FAV2DzOJlgXYuPjBxwOmC71Mq+4kMc9orZj03PDn5b6Gb/3/ALNX6ddv6RRHVj0qiq7q1l2Ici219PqsEPYXOdVUNnu2Nq2/ztXqLax/qDh1WD1s3Iyqa9aa7dhLf7e3Z/m1MWf1r/F+/wBS7NwLX5e9pacW902BuvtxMon2+n+j9Ku1v/oQitcF/Vej2gPptZh1MsNpqosc611bxvNYxmVPwv0z/Z6Dt9lP/AqrTXn9Ssq9fJ2sqBLceh8FoI2uFr69zarbGex/p+klTj52Nl0dPyOmFubkPIY+4Mp9R4L9uy2z9Xa7/RM3fpP8H6q6Bv1e+uOS51ZooxanDa83WNcC0fmbMZtu5v8AWYxJSX/F6y3H6r1DEosccJlTLLKpcaxc57w19fqTtsfWyz1/9L+jXadQ/oGT/wAU/wD6lyyej9BHRq68WrIcb8jfblZDWtDnubtbVUxlov8ASx6W2fo6/wDp+pZb6l+3FttHoOzLiy1rw4batRAadp9H+WkpwPrX9XcvLfT1PptZfeKhVksY7Y9zW++mxnDbX1brWem/6e//AINcfOPu9Ztzqrzq+xjwx7v+CuosZ6Lmt/4Vnqr1NuNcGj9bugaSG1Rp/wBYUMahjsrIF8XvqLNltjGbwC0O27q2M/OTSDdgskZxqpRvsR6ZPl1L3epusyrbHEf4Msa0iZhwp33P9v5qJdfVZWKW0WdUpYAbfTDmi4k/oqWWVtcza6zb9rsY7+Y9lf6e1etAAcABYJ+qjMRlg6Nkvwg4ezHeBdjtP7zKbf0lf8jZf6Vf+hUkJyiJC/m0WS4TVAjzPF/3rxF2JY7NwK+oZhszMmysbiIFTyPTFTMcOZS1mPue3G9r7a7bP5xdRfTU4CizeWWNc1xeS5wDwWu2X/T+j+4qtn1D6lk27HX42HS+PtGTQ19mXZB3lxuv2trte79Jv/wdn+CXRv6U4WguJuZp73QXCP3/AN7/AK3/AJiah5PGfXjBnTI9CzCYyq0kHb6dZFdNptaz0t9tfurss9P1GWK7YDdd65G65zPQawEEGo7hZU3839PuZ6u3/Q0/zysdU6dYMht+LWzJhvo5NNglz6mF9lLqvzHX43qWekyz2W1fo6/TVOjKuyHNf0rAPUxa0ltocBQ0NI9I5Flmyr1G/T9Jr35H6P8AtpKcfD6Hh43XanXYGPfhZWYcf7PYZaxji8MfTjhz/Stqc3312+r/ANYXpGF0/B6fT6GDRXjVElxZW0NBceXu2/SdosPoP1YysayrK6tdXfdRrRTU2Gh8Fv2nJtPuy8ra9za7NjPS32/4T319Ikp//9L0WjLz8hrnUtoc1ri0ndYASP3C6keoz/hGfo0Xd1b/AEeP/nv/APSSNj4tGMHClu0PcXESSJPZu4nYz/g2fo0srKxsPHfk5VjaaKhL7HmGgfEpKQ7urf6PH/z3/wDpJMX9VAksxwByd7//AEmsPqH13rpMYmHa9oMPuyIoa2T+i/RWn7T+m2u276qlxebndc+sIY60ZDy58voYXFhhxr9SnA/RY1ONT/gft11mTbb/AIVJT6DmfWGrBcG5eVg1OJDdpudMu+hua2v27/zVWP1ywQYffRWN20vsbkV1z/x9uM2r+1vXM9P6Y/c+nApDc3Bd6HruyXbYc1rnb8elvrem3f6dlbb6KbL6k2Bi49+ezFv2vc6q11mSWh1rzWdNljm2+rc1/wD2lp/mv+tpKezweuN6jY6rBycHJsZq6uu8ucB+8WNZu2q7u6t/o8f/AD3/APpJcJ1jFrD2NtAo9wc6y6DaKyRX6lWPW132G31Hfo77LfVr/m/s130FodL+tvVsVwpzcK+/CYAG3PAGU1pHtF43elkWM+g/+j3bP51n2j9Gkp6cfbDnU/aG1NArt2+m5zjP6L99jES/1fXoDZaDvDyGgmI+g1zva3c73KvT1PFzvRzsAnMob6lTzUBua/8ARnZbXYa7K3e36Lmot2Y722HGvAqJeTtbwGvb/pP5SSmTbQHlrtYdtcTEB35o7fTb7kKs5ozMkY7anNHpz6jnNM7PBjHtQ2W3PcXlmQ+i2X7Cxsw4DaxrvU9lez/B7Faw/UdfkXOrfU15btD4BMNAP0XOSUrd1b/R4/8Anv8A/SSFk5mbiUuyMk4tNTPpPfY8D/z39JypZ/UupZBezprQKAdvrgjc6R7nV/uN/N3rNu6E3PFNfU6Bk1ncbBa71Nmnte3eXbbP6iSnTp+sPr61up2lrnh7vVa0tYN1jvdVu9s/up8b6wMyi5tN+JvYA5zHvsreAdA70rqa7P8AornsuizpdVeFTRZdQzFyMdtoO+6x9jfUb6bLfa5tfpe31bvp1VK10+unNstryaLm5Qo9J2c5rW7mz7B7HWN9diSnfacnJJsrbjWEaFzLX6Hn82tGYOqVsDGso2t0Hvdx4aVBcl1HEbg5Lsr1n19PpobU2mp5a+22f0fqR/hWv3u9X/hLFPoH1yZjXuxet5Laqrtgw3W2Cx7SS5r/ALRa1rP0T/0Xp3W/9cSU9Zu6r/o8f/Pf/wCkkGrOzbrrKKxjutp+m3dYP80mra/+VsWig1YmPTdZfWyLLtXukn/NBPs/lbElP//T9Te9rGl7yGtaCXOPAA5K80651t/Xc45ALm9PxyPsVLiWguAl2Y+o7P07nP21b/5ir+a2WXXL0Lq2Pk5XS8rGxS0X3VPrrL5DZcNvuLQ7b/mryB3UcHHdZXfkU0Prmr0WvDhW+v8AR7G+nv3MZs+nW59b0zITVBn5eMbMpEabA/m3jtDXEdhqSWy0fnPGvtV/pV5xw2jHdXi2lzrKcogFlpeQfQ6hId+hb9Ovbsf/AKOxlixGZmBcQxmYx5cB7Q6HEj6Ltn03P3JW5mEwkCwl5Guxr3aDx9Nj1HHiidiz5BCcaMh4G3uTl1MzDlfbG3P9NzLasUGwAe2yqmin9I536du+66z+c/wv+CXP5vU2Ydjb80UC1oDMbpYh9hL3+o5tgaLNt7nfzPo/8Van+r3T8LKwqH0XOc27IL73BtzGNDgyx+LstZj1/oY/pVbrvTstt/4Fb/1fq6d1Fv2v9ntpdiva3G3sBc0uYy517bXNY5+Tvftsu/qf8Ip2kd3HxOlPua27KfYcl73fZcYa+g57R6T8jb6rarKrq/Wrxmf0Wxa2IdmBUclrBYWtfb6pIh7vda17trmbvUL979n01q9Vx730ium94fY9oBbSLiyHB7i5te11bdrdvq/4N/8ApFSyMLrTCb6fQzKHzuqyWWVvE/u2hn6Nn/W0kIenB+J9ZqXV/o2ZjXVWtke/Yx9tfqNb+js9JzP1e5n+DttXVZf9Fu/4t35CuV6P067p+Zb1bOrbXjU1zj49Zc81vcNt8OfXS3bt3bGV/wCEttXV3/zFn9R35ElLYv8ARaf+Lb+QKn1S97wen47iy69pm1pj0x+9wfc6Fawv6Fj/APFM/wCpCxetW5OL1A2sEV2sa0Obr7hu3eoPzdzdmxJTnVMbh5bjZ064XsO4PxiRRZP5/pb9jbf9d66CsssxftDppe4Tsthpaf8AhB7tqzuodf6RgYjLMq0FxYHsrM7nD94t/Nas13VcvK6TXRkBnTuo5FjsjErraWuFbXD7P6wdu9O/I9/tSVetJOq9EFtOTn4l12bkZDHjGya/01dU/wCjx6Xe9zW+xrtn/Bq504XYdFI6tnB7bdtdP2hgqsss0a302Tvfv/d2qh0Hqp6h07MsyA7F6phEPssx9tdlwMtZ6lbv0V1z3M9H9K1/6T01V69lWZeXjdI6fYQbaWjPyt260h7Bd6VmR/oPSP2h/ou9Kz1UlOv1rpmFnMZXcXFjX79lT9smNo3/AJ7q2rlOq4Dsd12HVj0YXT7hFnUb373uaRtd6df0rbfzW0bVezOuV3dIzMDDzX47+mVtNWU2Nzw32trb9L+csb6P/bdn8hZlGU/6x0NOXNLsDc6+0tiWgj9LT/wvqM/ker6iSLF11q30D6v9Yf1Guyq4A3UNrd6rdG212Aupvaxwa6pztj/Vp/wb1rrkvqtZv6sXMYWY5wmNo3fSdtfNm+PzmOeutSS//9T07Nqvuw76sez0b7K3tqtH5ry0hj/7Ll5DRiV11MAqbW5gh9ZAEH9x0j/Bu9uz2L2Vct9ZfqpdmZTupdNLRkWNazIocdrbNs7Lmvh227b+jfu/na9n+i/SMnEkadGbBkjEkS2PXs8QK3s1axm3v7RWR/Z/rKYL3sA1jvE/2v3VaycHLx7hjuosNrQSWtqs0c0/zftr22V7W+p6lfqVqfRvsWRkPdc87cYV3bR7gGucQ2y2o+67H9v6XZ+Z+lUYgSaOjZOWIgZA2PBL0LIJptwqrYGNeHtsDodLh6tbBdey7Horqte/Z/p/Uvr/ANItLI6z17Gu3Cug0Nn7VDi655Ac9rsbb6VDL2f4Vj/53/g1YyMi9zbreoYg+y13vZRnP2tp9Bx3VvzGgm2rFqf+ha+7H2bP0/8AN/pFR6sLMbO9DBxfUuaxgDmlldRaXfzTnPLN9Fln+japw0SbJNVbojqPTer47H25Dm0stZZXksLmvbYyYbfU9tf2e1nu9Sq31Kf+EQfsdGSz13m7MaS4VU3WuMNH0dtIdW9n0fazeqIZazJ+2Ui2jMO1nU+n7APV2t3ZHoXR6b7XVv8AUx/Tu/TU/wCD/wBCfpXUcdzBjYbbn22bjXU1jocwE7bGV7mVsr2/6RJCPpeLl4vW8ZjHOcbMkOOMLHGptbmPde3Y92132f8AnWf8Kxd3f/MWf1XfkWT0Xo12Pk2dQzDORY3ZXXodjTG/e5vtdbZtZ9D9HXs/l2LWvE02DxafyJKYYX9Cx/8Aimf9SFmfWX1LsduJTWx9z3NePVe6ppaxzXPY2yr3b3f9tf6RaeCZwscjvUz/AKkLF6vW192QLXtbadrKnWHaNrvd+jf9Jr63bklPJZDWW/WvGsycB7clrdl28b6mHdGFfXa0Bt1VbW/zNra/1hS+sGNk39R+2UOc+wFrdkw6WaNfX++73fQ/66ukb0avIsws23JN/wBgYJDTNdlrRtrybtpbvfj+701LN6RiuD8h+9kjd69T3MMRHudU9u/9zY9RzycBHplLiNXAcXD/AHv0lRjrLUd9fJ5CgdRbkZGVnMONvrLA0/o3Oc5zGs2UNdutdu/O2/TROr4HWWZuQ/Aqeca4h/qsIEB219le4ncx2/2Pr+h6f8heh4fS8GmLxiV1ZL2BljwAXR+56jvfsVHq31b6bdQ6+uqxtlU2bKrHsD/znV2sre31GO/0akU8P0nozH2ubbZU92RW6l+OJ2yfe2qyxv8AN3+1r/o/8UqfS8TJqx8vC/R47GWv/a11ji5mhYMfGxmMf/Ot2P8AUv8A+F/wi7SvA6Vj1Mv2srY07mbBta1xPPpU7f0v8t6j1DGowm2OwqKH32PGTfW4wXT/ANqe/v3JkJ8Rl6ZR4Tw+ocPH/Wh/UUY6g30a31ZBo6+2umh9NdtTjuc5zzZWB/OWtf7cfbb6f0f5xduuH6DU/L6w1+N61NlUWX2W2B49KWl1VW33Wes9rd/qruE9T//V9VSXyqkkp+qlm9V+r3SOre7Mxwbg3a3IYTXaGifZ69W2z0/c79G79GvmdJJT7xlfUzqVDmuwct2ZQ1rqW4t9rqNlT2+m9rbcdlld/t9n6ej2MVLE6J9ZDfVRZhFlVFTaDdvqr31MY6qth9OzJ2v2na+xlX/C+n6i8USSU/QrOgdWzHtdmWU4NQIJx8YutJ2w2v8AS2Nx2foWN/RMfRfX7971sdM6RhdLre3FZ77Xb7rnRve7jdYWNY3+wxvpr5iSSU/VShb/ADT/AOqfyL5YSSU/UXT/AOgY3/FM/wCpahdT6czOrbo02VSaxY0Obu/NkfL6S+YkklP0T9m6uys4bMdjaHkANZta3YW/ptz2/Rs9T3sVzp/RhVU05jjc9plrD9EAfQ3hvtsevmpJJT9VJL5VSSU/SvU+hsy3CyktrfJL2OnY8nhzw391Zb/qnnX5L735DKfVBbaADZuBbscNrvTbt/cYvn5JJT9OdL6NgdKoFWJUGuiH2kDe/vL3f1leXyqkkp//2f/tJlpQaG90b3Nob3AgMy4wADhCSU0EBAAAAAAABxwCAAACAAIAOEJJTQQlAAAAAAAQRgzyiSa4VtqwnAGhsKeQdzhCSU0D7QAAAAAAEADIAAAAAQABAMgAAAABAAE4QklNBCYAAAAAAA4AAAAAAAAAAAAAP4AAADhCSU0EDQAAAAAABAAAAB44QklNBBkAAAAAAAQAAAAeOEJJTQPzAAAAAAAJAAAAAAAAAAABADhCSU0ECgAAAAAAAQAAOEJJTScQAAAAAAAKAAEAAAAAAAAAAjhCSU0D9QAAAAAASAAvZmYAAQBsZmYABgAAAAAAAQAvZmYAAQChmZoABgAAAAAAAQAyAAAAAQBaAAAABgAAAAAAAQA1AAAAAQAtAAAABgAAAAAAAThCSU0D+AAAAAAAcAAA/////////////////////////////wPoAAAAAP////////////////////////////8D6AAAAAD/////////////////////////////A+gAAAAA/////////////////////////////wPoAAA4QklNBAgAAAAAABAAAAABAAACQAAAAkAAAAAAOEJJTQQeAAAAAAAEAAAAADhCSU0EGgAAAAADYQAAAAYAAAAAAAAAAAAAA5IAAAV4AAAAFgBTADAAMQA0ADAANQAyADUAWAAxADEAMAAwADAANAA0ADYAXwBmAGkAZwAxAAAAAQAAAAAAAAAAAAAAAAAAAAAAAAABAAAAAAAAAAAAAAV4AAADkgAAAAAAAAAAAAAAAAAAAAABAAAAAAAAAAAAAAAAAAAAAAAAABAAAAABAAAAAAAAbnVsbAAAAAIAAAAGYm91bmRzT2JqYwAAAAEAAAAAAABSY3QxAAAABAAAAABUb3AgbG9uZwAAAAAAAAAATGVmdGxvbmcAAAAAAAAAAEJ0b21sb25nAAADkgAAAABSZ2h0bG9uZwAABXgAAAAGc2xpY2VzVmxMcwAAAAFPYmpjAAAAAQAAAAAABXNsaWNlAAAAEgAAAAdzbGljZUlEbG9uZwAAAAAAAAAHZ3JvdXBJRGxvbmcAAAAAAAAABm9yaWdpbmVudW0AAAAMRVNsaWNlT3JpZ2luAAAADWF1dG9HZW5lcmF0ZWQAAAAAVHlwZWVudW0AAAAKRVNsaWNlVHlwZQAAAABJbWcgAAAABmJvdW5kc09iamMAAAABAAAAAAAAUmN0MQAAAAQAAAAAVG9wIGxvbmcAAAAAAAAAAExlZnRsb25nAAAAAAAAAABCdG9tbG9uZwAAA5IAAAAAUmdodGxvbmcAAAV4AAAAA3VybFRFWFQAAAABAAAAAAAAbnVsbFRFWFQAAAABAAAAAAAATXNnZVRFWFQAAAABAAAAAAAGYWx0VGFnVEVYVAAAAAEAAAAAAA5jZWxsVGV4dElzSFRNTGJvb2wBAAAACGNlbGxUZXh0VEVYVAAAAAEAAAAAAAlob3J6QWxpZ25lbnVtAAAAD0VTbGljZUhvcnpBbGlnbgAAAAdkZWZhdWx0AAAACXZlcnRBbGlnbmVudW0AAAAPRVNsaWNlVmVydEFsaWduAAAAB2RlZmF1bHQAAAALYmdDb2xvclR5cGVlbnVtAAAAEUVTbGljZUJHQ29sb3JUeXBlAAAAAE5vbmUAAAAJdG9wT3V0c2V0bG9uZwAAAAAAAAAKbGVmdE91dHNldGxvbmcAAAAAAAAADGJvdHRvbU91dHNldGxvbmcAAAAAAAAAC3JpZ2h0T3V0c2V0bG9uZwAAAAAAOEJJTQQoAAAAAAAMAAAAAT/wAAAAAAAAOEJJTQQRAAAAAAABAQA4QklNBBQAAAAAAAQAAAAIOEJJTQQMAAAAAB8pAAAAAQAAAKAAAABoAAAB4AAAwwAAAB8NABgAAf/Y/+AAEEpGSUYAAQIBAEgASAAA/+0ADEFkb2JlX0NNAAH/7gAOQWRvYmUAZIAAAAAB/9sAhAAMCAgICQgMCQkMEQsKCxEVDwwMDxUYExMVExMYEQwMDAwMDBEMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMAQ0LCw0ODRAODhAUDg4OFBQODg4OFBEMDAwMDBERDAwMDAwMEQwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAz/wAARCABoAKADASIAAhEBAxEB/90ABAAK/8QBPwAAAQUBAQEBAQEAAAAAAAAAAwABAgQFBgcICQoLAQABBQEBAQEBAQAAAAAAAAABAAIDBAUGBwgJCgsQAAEEAQMCBAIFBwYIBQMMMwEAAhEDBCESMQVBUWETInGBMgYUkaGxQiMkFVLBYjM0coLRQwclklPw4fFjczUWorKDJkSTVGRFwqN0NhfSVeJl8rOEw9N14/NGJ5SkhbSVxNTk9KW1xdXl9VZmdoaWprbG1ub2N0dXZ3eHl6e3x9fn9xEAAgIBAgQEAwQFBgcHBgU1AQACEQMhMRIEQVFhcSITBTKBkRShsUIjwVLR8DMkYuFygpJDUxVjczTxJQYWorKDByY1wtJEk1SjF2RFVTZ0ZeLys4TD03Xj80aUpIW0lcTU5PSltcXV5fVWZnaGlqa2xtbm9ic3R1dnd4eXp7fH/9oADAMBAAIRAxEAPwD0vAwn4guDsizI9a1136WDsL9XVVR9Ghrv5qv/AAf76PbbVRU+657a6qwXPe4gNa0auc5x+i1V+p9Uxel432jKJ2lwYxrY3Ocddrdxa36Ld3ucuEz+qdS6vc62w76C4+hiNMsaGn9Fb6R2MyrHuH6J13+Erts9P0/TSU9Fl/XrpNbXnDruzTUN1rmMNbGM/fdbkenu3f4JlXqeqs+/62fWrIY5/T+lNqZs3Mfe2xwJjdoT9k/qtWKyqhnUn491GYRU4OcG/pKnmA6u7Isss9NmTi/pGPZ/gPUTYeA7IDMi+623qPUHONHTrHbsfGqLy12Qcd25rfTo9rN3/ap/0ElJGfWHP6n6LL+r2V5WYwvorxyzHxaXEbn0ZVjf1/1aGtfsrss/S2/o0S3Mz8XYcnqeXfW93pNvZkOpc2w/zbvSsxvRtr9v6T+e9P6ditZGD0z7A7OyaWs6ew7mVM+jVWDDsy/0i1912nqvc3+Z/wACo4GBnZmdV1NmM/G6f9j+zUPsyC7LIc5l9eW8uZY1tnpezY+z1v8ASpKU36wfWXCdDSchsbqzklrWvH0XOtrj7TXXuP8APerXs/nLMf0l0nR/rPTn5TunZmO/pvVGDd9ktIO5vO7Hubtbbt/z/wDrf6Rc89t2RRhCw1XZFuVfjZF9tQcLG43rzZ6bdu17/SZ7P5v1PUWbldNqLaB7q7mOa1zA8tdX+m/Vcmm5p3+k7ez7PfV/NfyLElPpyr9RAPT8kHUGmzQ/1XLE+rH1nGdHTeouDOqV7mh0bW5ArLmvsp0a31m7P1iln/GU/ofobfUP6Bk/8U//AKlySktWtbP6o/IpoD76sbDORc7ZVVXvsdzDWjc46Lkcrrx6lZU3JtZXg5jg3DxtwaLSdPTyHh3qX27vp0V/q/8Ag3+r/OJKeiyPrF0uh2xr35D9QG49brQSPpN9Stvo+3/jFl4X10s6hlW0YfTn/oyW/prWVvkDfLqm+ttr/l71iZtWbk5nUMXDccar7GzHqy7A6uqhzHP9b037Aza7d9NuxVPq59sZiPfjYeO+4XejZmX5OmS+phvf9jtbU+q6iiiv1KX2WVV/5iSnbxfrB1zq+NkXi+rpLcW62m0Ma20sdSdpbdflRW9tn5vp41amL/rBltF9fWWOrcRph01OrEwI3vdba3/tx6yHYPTW4mXljp2RZVkD7TWOoVtZVRaf5/Kdmb7PWrvf6VlePTTf+k/m/wBDb+jBXj9Qqtbt6NRT1yjHsfWa3lldz6i0et9iwx9J/qVuZ9q9Jn6T06/USU62L9Yuv05+Zjtub1OvGZU8A1Br5u9X/QOr/m/Q+h6b962Ok/W7C6hmMwXQ264uFFlZLq7Ng9R23eym5m6trrWOdT6D/wDB5Fi5jpmNTacbEqtpzMTqLzZ1K7fGRZf6VmyjLxG7fs2H69f2b0f0f839l/PRMvEZ1fHyK6a8TIpoZ6f2htJax3pAbManJbkP3/ZfoP8ATb6OO9JT6EqVeBYzqtuech7q7K/TGMZ2AjZ7/p7PzHf4P/C2LnPq59ahjYtOH1i6t1ddLCzqLX72gQ3bVnuPuruY36eY/wDQXf4T0bF2CSn/0Nz629TpzOqDAMspw2vrc4g++61rS1u5jm+nVRX73P3M9T/z5WxMbYwOyLbL3va51LbXb2Cmrs+ff/hfU/Rfpf8AhFV+uVd2F1vMDZDMr0slj3S4idtNzccSNrv0f6T0v0/oqvi5DbqhJecJoIswaiPRex36P1GOqZ6tlTLG+jfj7P8Aja0lOi3CdZiO6PkFz6sun0K87H2t296/Wr3PdV/Ku/SVX/10fpmZ6WHhjLbZkOY12G2qpod+kaNln2i6WN9Z/pelU3/tv6ahbdbX6OLVbh4DweAHZDwAPoYzK20tp2/6R9nprL6Vj5F2Hm/aBaek4znb2hnp5OV9nI2XVP37Gu9INsy8qr08m3I/0f6RJT0D+nnC+rz8B7nWVU4gpfYwg2CnbszCzd+j9eqh1jqUPF6vkWX4+P0bHf1JleIGZOVcXYlDmDb9jvY25j3Of/Oeo6mr3/8AW1mU3/V3Nrwcu29jX1y53TbX5Flthdrj0sw3Pf61lTvf/NXetYr2b1/qePRkZF/TsrFD2Obh5Dww6T6lVeXj0uuuxn+o5387/gP+ESUlrGZj5fp34ddfoD7ZOPZtZbkWNdQLaabgPTrx/f8AaNtvqW22U3WKjkZYc3GppLKsvFyGuspe11wr9/6Suqur3X1Wsd9q2ez9FV6uxaQz83Koa9uN+z6tjXizJLXFxP0qcKuiyxj3e3+kW2Mq/wCAtWRl5wx6XNpDaLct23HrNvrWV2XFv2q7Lsr9Te51bGfzT7vRr9T/AAf6NJSHqdbS+q+q9r6wXuptxGv0tafWrb73vdVtu97n+p+gf9Ndr07q/wC1/q9fkub6dzK313tGo3ivfuY79x7LGWNXH7/tLRj9Nw32Y2MyygU0g2Hc/Wz07Gfo7PVt9+RlWvr/AJ2z/C/o11nRul3dM+ruQ3KAblZDbci+tpDmsc9ulDHD6XpVsYz/AIxJTifW/rOOc2jA3bm4zJs3Mc6hl52+kMl1R3eoxnvr9n6BYGdmenVbS/ExTkdRqa6zLqc90NnaT6N9dPpZD9vs2/8AHfzqh9ZMH7J1rLszaGOoNrrn5DbXOeW3y7Gt9Gr3V+g7+cr/AJ//AAnvqWe9z8guyZF9Tnibai59ZDWt9gtcPZ+j/f2e9S8vGEsgEyK31/S/qokTWjt9Co/aGK85Db+pfpQMai17nY1Tmj9Nm51t2+v2Rsrqt9X/AIDH9T3rayBY21uLm59WS57CyvGpxHeoWPiiyqg2ZDavTs3bLX1Vorev9GyMOmvDrubj1z6dYY1rRGja/Se70/Z+/wD9cVQdV6fc4UdSxWnDc/dvsLSGk8PsDK27/f8AuP8AURnjnKUpcHCN6+XhCgRoLSZJHUPWxazkv+g25jcWmzSt7Xso3U2s2Mc6v6H+jVr9m9QYLMptnqXP2/pdrqra/S99TXVj7RRYxlz3XbNnp77VB7Lm49deWxz3bgGUURUwC12yr06GW0Ma18+yr9Lb/pPV/SWLK6p0vrGJeOpdHy32O6a8WfYLH2WvFUlt7Kqn/wA/X6Nlz/S9T/iLf5tQpb11FeQdl9Bx3Yr6Ps2OAHeuX2NdlW3W0t/Was5rnsuq/wAHsfkXM/SK5ntxKbP2RTFTMwPuuroEOdTuDLa8Wun3OuzvoPsr9lFP2i/9G/YsejK6v1ZmN1LC9F2NYXVUtue6otra/wB1rKKfX9tln0v1t9/82/8AR0ql1DK6h0e685xLsjqbSDmtO9ra69n6rtANuPiY3q+taz+cs3pKdfMeMuk4eTjsqqsaXYzSWvr9Gstpzdno+zGtxKXfQZ62O+r+butWl9U+uWbq+hZrQ22issxbAZL66v0bG2j/AEno1+oy1vsv/Sfzf5+c3q/Rsik5ddrKrKfdt3ltlY2+k1mI6t/pupdX+iYxn6D/AEyF9TqsjL6mx1e11GLsyMi8n3l7q78fHx2iG+z07NzNrWVsqr/4ZJT/AP/R9A+sH1ewuvYjaMkmuyol1FzY3MJG1w1+nU//AAtX564pv+L76xU2babaw1sljq73tZr/ACLKrLGf5/pr0pJAj6LhMgVQI8Q+X5v1V6/0PGf1RzG5IYQbhivfZa1n7/p+lT+jr/P9Lf8A6T+bU+ndQH2V7MvKt2NlllbIO83uk+s9ofa6u19rmt9NemrEyvqX9Wsl77H4Qrss3bn0vfV9L6elL2NSAruqUroUBXYPM4mWBdi4+MHHA6YLvUyr7iQxz2itmPTc8OflvoZv/f8As1fp12/pFEdWPSqKrurWXYhyLbX0+qwQ9hc51VQ2e7Y2rb/O1eotrH+oOHVYPWzcjKpr1prt2Et/t7dn+bUxZ/Wv8X7/AFLs3Atfl72lpxb3TYG6+3Eyifb6f6P0q7W/+hCK1wX9V6PaA+m1mHUyw2mqixzrXVvG81jGZU/C/TP9noO32U/8CqtNef1Kyr18nayoEtx6HwWgja4Wvr3NqtsZ7H+n6SVOPnY2XR0/I6YW5uQ8hj7gyn1Hgv27LbP1drv9Ezd+k/wfqroG/V7645LnVmijFqcNrzdY1wLR+Zsxm27m/wBZjElJf8XrLcfqvUMSixxwmVMssqlxrFznvDX1+pO2x9bLPX/0v6Ndp1D+gZP/ABT/APqXLJ6P0EdGrrxashxvyN9uVkNa0Oe5u1tVTGWi/wBLHpbZ+jr/AOn6llvqX7cW20eg7MuLLWvDhtq1EBp2n0f5aSnA+tf1dy8t9PU+m1l94qFWSxjtj3Nb76bGcNtfVutZ6b/p7/8Ag1x84+71m3OqvOr7GPDHu/4K6ixnoua3/hWeqvU241waP1u6BpIbVGn/AFhQxqGOysgXxe+os2W2MZvALQ7burYz85NIN2CyRnGqlG+xHpk+XUvd6m6zKtscR/gyxrSJmHCnfc/2/mol19VlYpbRZ1SlgBt9MOaLiT+ipZZW1zNrrNv2uxjv5j2V/p7V60ABwAFgn6qMxGWDo2S/CDh7Md4F2O0/vMpt/SV/yNl/pV/6FSQnKIkL+bRZLhNUCPM8X/evEXYljs3Ar6hmGzMybKxuIgVPI9MVMxw5lLWY+57cb2vtrts/nF1F9NTgKLN5ZY1zXF5LnAPBa7Zf9P6P7iq2fUPqWTbsdfjYdL4+0ZNDX2ZdkHeXG6/a2u17v0m//B2f4JdG/pThaC4m5mnvdBcI/f8A3v8Arf8AmJqHk8Z9eMGdMj0LMJjKrSQdvp1kV02m1rPS321+6uyz0/UZYrtgN13rkbrnM9BrAQQajuFlTfzf0+5nq7f9DT/PKx1Tp1gyG34tbMmG+jk02CXPqYX2Uuq/MdfjepZ6TLPZbV+jr9NU6Mq7Ic1/SsA9TFrSW2hwFDQ0j0jkWWbKvUb9P0mvfkfo/wC2kpx8PoeHjddqddgY9+FlZhx/s9hlrGOLwx9OOHP9K2pzffXb6v8A1hekYXT8Hp9PoYNFeNUSXFlbQ0Fx5e7b9J2iw+g/VjKxrKsrq11d91GtFNTYaHwW/acm0+7Lytr3Nrs2M9Lfb/hPfX0iSn//0vRaMvPyGudS2hzWuLSd1gBI/cLqR6jP+EZ+jRd3Vv8AR4/+e/8A9JI2Pi0YwcKW7Q9xcRJIk9m7idjP+DZ+jSysrGw8d+TlWNpoqEvseYaB8SkpDu6t/o8f/Pf/AOkkxf1UCSzHAHJ3v/8ASaw+ofXeukxiYdr2gw+7IihrZP6L9FaftP6ba7bvqqXF5ud1z6whjrRkPLny+hhcWGHGv1KcD9FjU41P+B+3XWZNtv8AhUlPoOZ9YasFwbl5WDU4kN2m50y76G5ra/bv/NVY/XLBBh99FY3bS+xuRXXP/H24zav7W9cz0/pj9z6cCkNzcF3oeu7JdthzWudvx6W+t6bd/p2VtvopsvqTYGLj357MW/a9zqrXWZJaHWvNZ02WObb6tzX/APaWn+a/62kp7PB643qNjqsHJwcmxmrq67y5wH7xY1m7aru7q3+jx/8APf8A+klwnWMWsPY20Cj3BzrLoNorJFfqVY9bXfYbfUd+jvst9Wv+b+zXfQWh0v629WxXCnNwr78JgAbc8AZTWke0Xjd6WRYz6D/6Pds/nWfaP0aSnpx9sOdT9obU0Cu3b6bnOM/ov32MRL/V9egNloO8PIaCYj6DXO9rdzvcq9PU8XO9HOwCcyhvqVPNQG5r/wBGdltdhrsrd7fouai3ZjvbYca8Col5O1vAa9v+k/lJKZNtAeWu1h21xMQHfmjt9NvuQqzmjMyRjtqc0enPqOc0zs8GMe1DZbc9xeWZD6LZfsLGzDgNrGu9T2V7P8HsVrD9R1+Rc6t9TXlu0PgEw0A/Rc5JSt3Vv9Hj/wCe/wD9JIWTmZuJS7IyTi01M+k99jwP/Pf0nKln9S6lkF7OmtAoB2+uCNzpHudX+4383es27oTc8U19ToGTWdxsFrvU2ae17d5dts/qJKdOn6w+vrW6naWueHu9VrS1g3WO91W72z+6nxvrAzKLm034m9gDnMe+yt4B0DvSuprs/wCiuey6LOl1V4VNFl1DMXIx22g77rH2N9Rvpst9rm1+l7fVu+nVUrXT66c2y2vJoublCj0nZzmtbubPsHsdY312JKd9pyckmytuNYRoXMtfoefza0Zg6pWwMayja3Qe93HhpUFyXUcRuDkuyvWfX0+mhtTaanlr7bZ/R+pH+Fa/e71f+EsU+gfXJmNe7F63ktqqu2DDdbYLHtJLmv8AtFrWs/RP/Rendb/1xJT1m7qv+jx/89//AKSQas7NuusorGO62n6bd1g/zSatr/5WxaKDViY9N1l9bIsu1e6Sf80E+z+VsSU//9P1N72saXvIa1oJc48ADkrzTrnW39dzjkAub0/HI+xUuJaC4CXZj6js/Tuc/bVv/mKv5rZZdcvQurY+TldLysbFLRfdU+usvkNlw2+4tDtv+avIHdRwcd1ld+RTQ+uavRa8OFb6/wBHsb6e/cxmz6dbn1vTMhNUGfl4xsykRpsD+beO0NcR2GpJbLR+c8a+1X+lXnHDaMd1eLaXOspyiAWWl5B9DqEh36Fv069ux/8Ao7GWLEZmYFxDGZjHlwHtDocSPou2fTc/clbmYTCQLCXka7GvdoPH02PUceKJ2LPkEJxoyHgbe5OXUzMOV9sbc/03MtqxQbAB7bKqaKf0jnfp277rrP5z/C/4Jc/m9TZh2NvzRQLWgMxuliH2Evf6jm2Bos23ud/M+j/xVqf6vdPwsrCofRc5zbsgvvcG3MY0ODLH4uy1mPX+hj+lVuu9Oy23/gVv/V+rp3UW/a/2e2l2K9rcbewFzS5jLnXttc1jn5O9+2y7+p/winaR3cfE6U+5rbsp9hyXvd9lxhr6DntHpPyNvqtqsqur9avGZ/RbFrYh2YFRyWsFha19vqkiHu91rXu2uZu9Qv3v2fTWr1XHvfSK6b3h9j2gFtIuLIcHuLm17XVt2t2+r/g3/wCkVLIwutMJvp9DMofO6rJZZW8T+7aGfo2f9bSQh6cH4n1mpdX+jZmNdVa2R79jH21+o1v6Oz0nM/V7mf4O21dVl/0W7/i3fkK5Xo/Trun5lvVs6tteNTXOPj1lzzW9w23w59dLdu3dsZX/AIS21dXf/MWf1HfkSUti/wBFp/4tv5AqfVL3vB6fjuLLr2mbWmPTH73B9zoVrC/oWP8A8Uz/AKkLF61bk4vUDawRXaxrQ5uvuG7d6g/N3N2bElOdUxuHluNnTrhew7g/GJFFk/n+lv2Nt/13roKyyzF+0Oml7hOy2Glp/wCEHu2rO6h1/pGBiMsyrQXFgeyszucP3i381qzXdVy8rpNdGQGdO6jkWOyMSutpa4VtcPs/rB27078j3+1JV60k6r0QW05OfiXXZuRkMeMbJr/TV1T/AKPHpd73Nb7Gu2f8GrnThdh0Ujq2cHtt210/aGCqyyzRrfTZO9+/93aqHQeqnqHTsyzIDsXqmEQ+yzH212XAy1nqVu/RXXPcz0f0rX/pPTVXr2VZl5eN0jp9hBtpaM/K3brSHsF3pWZH+g9I/aH+i70rPVSU6/WumYWcxldxcWNfv2VP2yY2jf8AnurauU6rgOx3XYdWPRhdPuEWdRvfve5pG13p1/Stt/NbRtV7M65Xd0jMwMPNfjv6ZW01ZTY3PDfa2tv0v5yxvo/9t2fyFmUZT/rHQ05c0uwNzr7S2JaCP0tP/C+oz+R6vqJIsXXWrfQPq/1h/Ua7KrgDdQ2t3qt0bbXYC6m9rHBrqnO2P9Wn/BvWuuS+q1m/qxcxhZjnCY2jd9J2182b4/OY5661JL//1PTs2q+7Dvqx7PRvsre2q0fmvLSGP/suXkNGJXXUwCptbmCH1kAQf3HSP8G727PYvZVy31l+ql2ZlO6l00tGRY1rMihx2ts2zsua+Hbbtv6N+7+dr2f6L9IycSRp0ZsGSMSRLY9ezxArezVrGbe/tFZH9n+spgvewDWO8T/a/dVrJwcvHuGO6iw2tBJa2qzRzT/N+2vbZXtb6nqV+pWp9G+xZGQ91zztxhXdtHuAa5xDbLaj7rsf2/pdn5n6VRiBJo6Nk5YiBkDY8EvQsgmm3CqtgY14e2wOh0uHq1sF17Lseiuq179n+n9S+v8A0i0sjrPXsa7cK6DQ2ftUOLrnkBz2uxtvpUMvZ/hWP/nf+DVjIyL3Nut6hiD7LXe9lGc/a2n0HHdW/MaCbasWp/6Fr7sfZs/T/wA3+kVHqwsxs70MHF9S5rGAOaWV1Fpd/NOc8s30WWf6NqnDRJsk1VuiOo9N6vjsfbkObSy1lleSwua9tjJht9T21/Z7We71KrfUp/4RB+x0ZLPXebsxpLhVTda4w0fR20h1b2fR9rN6ohlrMn7ZSLaMw7WdT6fsA9Xa3dkehdHpvtdW/wBTH9O79NT/AIP/AEJ+ldRx3MGNhtufbZuNdTWOhzATtsZXuZWyvb/pEkI+l4uXi9bxmMc5xsyQ44wscam1uY917dj3bXfZ/wCdZ/wrF3d/8xZ/Vd+RZPRejXY+TZ1DMM5Fjdldeh2NMb97m+11tm1n0P0dez+XYta8TTYPFp/Ikphhf0LH/wCKZ/1IWZ9ZfUux24lNbH3Pc149V7qmlrHNc9jbKvdvd/21/pFp4JnCxyO9TP8AqQsXq9bX3ZAte1tp2sqdYdo2u936N/0mvrduSU8lkNZb9a8azJwHtyWt2XbxvqYd0YV9drQG3VVtb/M2tr/WFL6wY2Tf1H7ZQ5z7AWt2TDpZo19f77vd9D/rq6RvRq8izCzbck3/AGBgkNM12WtG2vJu2lu9+P7vTUs3pGK4PyH72SN3r1PcwxEe51T27/3Nj1HPJwEemUuI1cBxcP8Ae/SVGOstR318nkKB1FuRkZWcw42+ssDT+jc5znMazZQ12612787b9NE6vgdZZm5D8Cp5xriH+qwgQHbX2V7idzHb/Y+v6Hp/yF6Hh9LwaYvGJXVkvYGWPABdH7nqO9+xUerfVvpt1Dr66rG2VTZsqsewP/OdXayt7fUY7/RqRTw/SejMfa5ttlT3ZFbqX44nbJ97arLG/wA3f7Wv+j/xSp9LxMmrHy8L9HjsZa/9rXWOLmaFgx8bGYx/863Y/wBS/wD4X/CLtK8DpWPUy/aytjTuZsG1rXE8+lTt/S/y3qPUMajCbY7CooffY8ZN9bjBdP8A2p7+/cmQnxGXplHhPD6hw8f9aH9RRjqDfRrfVkGjr7a6aH0121OO5znPNlYH85a1/tx9tvp/R/nF264foNT8vrDX43rU2VRZfZbYHj0paXVVbfdZ6z2t3+qu4T1P/9X1VJfKqSSn6qWb1X6vdI6t7szHBuDdrchhNdoaJ9nr1bbPT9zv0bv0a+Z0klPvGV9TOpUOa7By3ZlDWupbi32uo2VPb6b2ttx2WV3+32fp6PYxUsTon1kN9VFmEWVUVNoN2+qvfUxjqq2H07Mna/adr7GVf8L6fqLxRJJT9Cs6B1bMe12ZZTg1AgnHxi60nbDa/wBLY3HZ+hY39Ex9F9fv3vWx0zpGF0ut7cVnvtdvuudG97uN1hY1jf7DG+mvmJJJT9VKFv8ANP8A6p/IvlhJJT9RdP8A6Bjf8Uz/AKlqF1PpzM6tujTZVJrFjQ5u782R8vpL5iSSU/RP2bq7Kzhsx2NoeQA1m1rdhb+m3Pb9Gz1PexXOn9GFVTTmONz2mWsP0QB9DeG+2x6+akklP1UkvlVJJT9K9T6GzLcLKS2t8kvY6djyeHPDf3Vlv+qedfkvvfkMp9UFtoANm4Fuxw2u9Nu39xi+fkklP050vo2B0qgVYlQa6IfaQN7+8vd/WV5fKqSSn//ZADhCSU0EIQAAAAAAUwAAAAEBAAAADwBBAGQAbwBiAGUAIABQAGgAbwB0AG8AcwBoAG8AcAAAABIAQQBkAG8AYgBlACAAUABoAG8AdABvAHMAaABvAHAAIABDAFMAAAABADhCSU0PoAAAAAABDG1hbmlJUkZSAAABADhCSU1BbkRzAAAA4AAAABAAAAABAAAAAAAAbnVsbAAAAAMAAAAAQUZTdGxvbmcAAAAAAAAAAEZySW5WbExzAAAAAU9iamMAAAABAAAAAAAAbnVsbAAAAAIAAAAARnJJRGxvbmcjpi6sAAAAAEZyR0Fkb3ViQD4AAAAAAAAAAAAARlN0c1ZsTHMAAAABT2JqYwAAAAEAAAAAAABudWxsAAAABAAAAABGc0lEbG9uZwAAAAAAAAAAQUZybWxvbmcAAAAAAAAAAEZzRnJWbExzAAAAAWxvbmcjpi6sAAAAAExDbnRsb25nAAAAAAAAOEJJTVJvbGwAAAAIAAAAAAAAAAA4QklND6EAAAAAABxtZnJpAAAAAgAAABAAAAABAAAAAAAAAAEAAAAAOEJJTQQGAAAAAAAHAAgAAAABAQD/4R5yaHR0cDovL25zLmFkb2JlLmNvbS94YXAvMS4wLwA8P3hwYWNrZXQgYmVnaW49J++7vycgaWQ9J1c1TTBNcENlaGlIenJlU3pOVGN6a2M5ZCc/Pgo8eDp4bXBtZXRhIHhtbG5zOng9J2Fkb2JlOm5zOm1ldGEvJyB4OnhtcHRrPSdYTVAgdG9vbGtpdCAzLjAtMjgsIGZyYW1ld29yayAxLjYnPgo8cmRmOlJERiB4bWxuczpyZGY9J2h0dHA6Ly93d3cudzMub3JnLzE5OTkvMDIvMjItcmRmLXN5bnRheC1ucyMnIHhtbG5zOmlYPSdodHRwOi8vbnMuYWRvYmUuY29tL2lYLzEuMC8nPgoKIDxyZGY6RGVzY3JpcHRpb24gcmRmOmFib3V0PSd1dWlkOmVhZGU4MTdhLWEwZGQtMTFlMS1iZDI3LWNkYWEzZTA4YTgzNCcKICB4bWxuczpleGlmPSdodHRwOi8vbnMuYWRvYmUuY29tL2V4aWYvMS4wLyc+CiAgPGV4aWY6Q29sb3JTcGFjZT40Mjk0OTY3Mjk1PC9leGlmOkNvbG9yU3BhY2U+CiAgPGV4aWY6UGl4ZWxYRGltZW5zaW9uPjE0MDA8L2V4aWY6UGl4ZWxYRGltZW5zaW9uPgogIDxleGlmOlBpeGVsWURpbWVuc2lvbj45MTQ8L2V4aWY6UGl4ZWxZRGltZW5zaW9uPgogPC9yZGY6RGVzY3JpcHRpb24+CgogPHJkZjpEZXNjcmlwdGlvbiByZGY6YWJvdXQ9J3V1aWQ6ZWFkZTgxN2EtYTBkZC0xMWUxLWJkMjctY2RhYTNlMDhhODM0JwogIHhtbG5zOnBkZj0naHR0cDovL25zLmFkb2JlLmNvbS9wZGYvMS4zLyc+CiA8L3JkZjpEZXNjcmlwdGlvbj4KCiA8cmRmOkRlc2NyaXB0aW9uIHJkZjphYm91dD0ndXVpZDplYWRlODE3YS1hMGRkLTExZTEtYmQyNy1jZGFhM2UwOGE4MzQnCiAgeG1sbnM6cGhvdG9zaG9wPSdodHRwOi8vbnMuYWRvYmUuY29tL3Bob3Rvc2hvcC8xLjAvJz4KICA8cGhvdG9zaG9wOkNvbG9yTW9kZT40PC9waG90b3Nob3A6Q29sb3JNb2RlPgogIDxwaG90b3Nob3A6SUNDUHJvZmlsZT5VLlMuIFdlYiBDb2F0ZWQgKFNXT1ApIHYyPC9waG90b3Nob3A6SUNDUHJvZmlsZT4KICA8cGhvdG9zaG9wOkhpc3Rvcnk+PC9waG90b3Nob3A6SGlzdG9yeT4KIDwvcmRmOkRlc2NyaXB0aW9uPgoKIDxyZGY6RGVzY3JpcHRpb24gcmRmOmFib3V0PSd1dWlkOmVhZGU4MTdhLWEwZGQtMTFlMS1iZDI3LWNkYWEzZTA4YTgzNCcKICB4bWxuczp0aWZmPSdodHRwOi8vbnMuYWRvYmUuY29tL3RpZmYvMS4wLyc+CiAgPHRpZmY6T3JpZW50YXRpb24+MTwvdGlmZjpPcmllbnRhdGlvbj4KICA8dGlmZjpYUmVzb2x1dGlvbj4yMDAvMTwvdGlmZjpYUmVzb2x1dGlvbj4KICA8dGlmZjpZUmVzb2x1dGlvbj4yMDAvMTwvdGlmZjpZUmVzb2x1dGlvbj4KICA8dGlmZjpSZXNvbHV0aW9uVW5pdD4yPC90aWZmOlJlc29sdXRpb25Vbml0PgogPC9yZGY6RGVzY3JpcHRpb24+CgogPHJkZjpEZXNjcmlwdGlvbiByZGY6YWJvdXQ9J3V1aWQ6ZWFkZTgxN2EtYTBkZC0xMWUxLWJkMjctY2RhYTNlMDhhODM0JwogIHhtbG5zOnhhcD0naHR0cDovL25zLmFkb2JlLmNvbS94YXAvMS4wLyc+CiAgPHhhcDpDcmVhdGVEYXRlPjIwMTItMDUtMThUMTc6Mzg6NTUrMDU6MzA8L3hhcDpDcmVhdGVEYXRlPgogIDx4YXA6TW9kaWZ5RGF0ZT4yMDEyLTA1LTE4VDE3OjM4OjU1KzA1OjMwPC94YXA6TW9kaWZ5RGF0ZT4KICA8eGFwOk1ldGFkYXRhRGF0ZT4yMDEyLTA1LTE4VDE3OjM4OjU1KzA1OjMwPC94YXA6TWV0YWRhdGFEYXRlPgogIDx4YXA6Q3JlYXRvclRvb2w+QWRvYmUgUGhvdG9zaG9wIENTIFdpbmRvd3M8L3hhcDpDcmVhdG9yVG9vbD4KIDwvcmRmOkRlc2NyaXB0aW9uPgoKIDxyZGY6RGVzY3JpcHRpb24gcmRmOmFib3V0PSd1dWlkOmVhZGU4MTdhLWEwZGQtMTFlMS1iZDI3LWNkYWEzZTA4YTgzNCcKICB4bWxuczpzdEV2dD0naHR0cDovL25zLmFkb2JlLmNvbS94YXAvMS4wL3NUeXBlL1Jlc291cmNlRXZlbnQjJwogIHhtbG5zOnN0UmVmPSdodHRwOi8vbnMuYWRvYmUuY29tL3hhcC8xLjAvc1R5cGUvUmVzb3VyY2VSZWYjJwogIHhtbG5zOnhhcE1NPSdodHRwOi8vbnMuYWRvYmUuY29tL3hhcC8xLjAvbW0vJz4KICA8eGFwTU06SW5zdGFuY2VJRD54bXAuaWlkOjNDQzU3OTk1MkI1REUxMTE4Q0YyRjE3NTcxNzg2RTI3PC94YXBNTTpJbnN0YW5jZUlEPgogIDx4YXBNTTpPcmlnaW5hbERvY3VtZW50SUQ+eG1wLmRpZDoxMDQzMTFDMDVBNThFMTExQUM1RDlFNjVBMUJEOUE4RDwveGFwTU06T3JpZ2luYWxEb2N1bWVudElEPgogIDx4YXBNTTpIaXN0b3J5PgogICA8cmRmOlNlcT4KICAgIDxyZGY6bGkgcmRmOnBhcnNlVHlwZT0nUmVzb3VyY2UnPgogICAgIDxzdEV2dDphY3Rpb24+Y3JlYXRlZDwvc3RFdnQ6YWN0aW9uPgogICAgIDxzdEV2dDppbnN0YW5jZUlEPnhtcC5paWQ6MTA0MzExQzA1QTU4RTExMUFDNUQ5RTY1QTFCRDlBOEQ8L3N0RXZ0Omluc3RhbmNlSUQ+CiAgICAgPHN0RXZ0OndoZW4+MjAxMi0wMi0xNlQxMDo0OTozNCswNTozMDwvc3RFdnQ6d2hlbj4KICAgICA8c3RFdnQ6c29mdHdhcmVBZ2VudD5BZG9iZSBQaG90b3Nob3AgQ1M1LjEgV2luZG93czwvc3RFdnQ6c29mdHdhcmVBZ2VudD4KICAgIDwvcmRmOmxpPgogICAgPHJkZjpsaSByZGY6cGFyc2VUeXBlPSdSZXNvdXJjZSc+CiAgICAgPHN0RXZ0OmFjdGlvbj5zYXZlZDwvc3RFdnQ6YWN0aW9uPgogICAgIDxzdEV2dDppbnN0YW5jZUlEPnhtcC5paWQ6MTE0MzExQzA1QTU4RTExMUFDNUQ5RTY1QTFCRDlBOEQ8L3N0RXZ0Omluc3RhbmNlSUQ+CiAgICAgPHN0RXZ0OndoZW4+MjAxMi0wMi0xNlQxMTowMToyMyswNTozMDwvc3RFdnQ6d2hlbj4KICAgICA8c3RFdnQ6c29mdHdhcmVBZ2VudD5BZG9iZSBQaG90b3Nob3AgQ1M1LjEgV2luZG93czwvc3RFdnQ6c29mdHdhcmVBZ2VudD4KICAgICA8c3RFdnQ6Y2hhbmdlZD4vPC9zdEV2dDpjaGFuZ2VkPgogICAgPC9yZGY6bGk+CiAgICA8cmRmOmxpIHJkZjpwYXJzZVR5cGU9J1Jlc291cmNlJz4KICAgICA8c3RFdnQ6YWN0aW9uPnNhdmVkPC9zdEV2dDphY3Rpb24+CiAgICAgPHN0RXZ0Omluc3RhbmNlSUQ+eG1wLmlpZDozQ0M1Nzk5NTJCNURFMTExOENGMkYxNzU3MTc4NkUyNzwvc3RFdnQ6aW5zdGFuY2VJRD4KICAgICA8c3RFdnQ6d2hlbj4yMDEyLTAyLTIyVDEzOjQyOjMzKzA1OjMwPC9zdEV2dDp3aGVuPgogICAgIDxzdEV2dDpzb2Z0d2FyZUFnZW50PkFkb2JlIFBob3Rvc2hvcCBDUzUuMSBXaW5kb3dzPC9zdEV2dDpzb2Z0d2FyZUFnZW50PgogICAgIDxzdEV2dDpjaGFuZ2VkPi88L3N0RXZ0OmNoYW5nZWQ+CiAgICA8L3JkZjpsaT4KICAgPC9yZGY6U2VxPgogIDwveGFwTU06SGlzdG9yeT4KICA8eGFwTU06RGVyaXZlZEZyb20gcmRmOnBhcnNlVHlwZT0nUmVzb3VyY2UnPgogICA8c3RSZWY6ZG9jdW1lbnRJRD54bXAuZGlkOjEwNDMxMUMwNUE1OEUxMTFBQzVEOUU2NUExQkQ5QThEPC9zdFJlZjpkb2N1bWVudElEPgogIDwveGFwTU06RGVyaXZlZEZyb20+CiAgPHhhcE1NOkRvY3VtZW50SUQ+YWRvYmU6ZG9jaWQ6cGhvdG9zaG9wOmVhZGU4MTc5LWEwZGQtMTFlMS1iZDI3LWNkYWEzZTA4YTgzNDwveGFwTU06RG9jdW1lbnRJRD4KIDwvcmRmOkRlc2NyaXB0aW9uPgoKIDxyZGY6RGVzY3JpcHRpb24gcmRmOmFib3V0PSd1dWlkOmVhZGU4MTdhLWEwZGQtMTFlMS1iZDI3LWNkYWEzZTA4YTgzNCcKICB4bWxuczpkYz0naHR0cDovL3B1cmwub3JnL2RjL2VsZW1lbnRzLzEuMS8nPgogIDxkYzpmb3JtYXQ+aW1hZ2UvanBlZzwvZGM6Zm9ybWF0PgogPC9yZGY6RGVzY3JpcHRpb24+Cgo8L3JkZjpSREY+CjwveDp4bXBtZXRhPgogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKPD94cGFja2V0IGVuZD0ndyc/Pv/uAA5BZG9iZQBkQAAAAAH/2wCEAAEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQECAQEBAQEBAgICAgICAgICAgICAgIDAwMDAwMDAwMDAwMDAwMBAQEBAQEBAgEBAgMCAgIDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDA//AABEIA5IFeAMBEQACEQEDEQH/3QAEAK//xAGiAAAABgIDAQAAAAAAAAAAAAAHCAYFBAkDCgIBAAsBAAAGAwEBAQAAAAAAAAAAAAYFBAMHAggBCQAKCxAAAgEDBAEDAwIDAwMCBgl1AQIDBBEFEgYhBxMiAAgxFEEyIxUJUUIWYSQzF1JxgRhikSVDobHwJjRyChnB0TUn4VM2gvGSokRUc0VGN0djKFVWVxqywtLi8mSDdJOEZaOzw9PjKThm83UqOTpISUpYWVpnaGlqdnd4eXqFhoeIiYqUlZaXmJmapKWmp6ipqrS1tre4ubrExcbHyMnK1NXW19jZ2uTl5ufo6er09fb3+Pn6EQACAQMCBAQDBQQEBAYGBW0BAgMRBCESBTEGACITQVEHMmEUcQhCgSORFVKhYhYzCbEkwdFDcvAX4YI0JZJTGGNE8aKyJjUZVDZFZCcKc4OTRnTC0uLyVWV1VjeEhaOzw9Pj8ykalKS0xNTk9JWltcXV5fUoR1dmOHaGlqa2xtbm9md3h5ent8fX5/dIWGh4iJiouMjY6Pg5SVlpeYmZqbnJ2en5KjpKWmp6ipqqusra6vr/2gAMAwEAAhEDEQA/AN/j37r3RaPl98uOjvgx8e9/fJ75F7mqNqdTdcRYZs/kqDHVGay9RVbhz1NtnCYvCYSj/erKqprKuCOOCIXsWc2RGYe62BXoLPgB/Mb+LH8zLqPcXdXxP3fmN07M2nvrIddbji3JtzIbR3DiNzY/D0WeMNZgcradYJqWvp5aeotok9YU6o3A91ro9Xv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/dePDPXFdJF1+jc+9mvn1oAAY64xmQ6vIoWzWWxvcf19+PXhXz6ye9db6xCJY9bRr6nNyCTYn/AGPvfWqU4ddI0tz5EVVH0INz/r29+p6daqfPrN711vy6xK0gDGUIgBJvq+i/4n3vHWs9QpsriKYlqjJUEJHH7tZAhH/ILN79UnHWzStT1ihzOErjpgymPnZWFljrIS2r8WUNc+/cOvYPTtcWvfi17/Xj/Ye9dexx64q6uCVN7Gx+o5/pz73Qjj14EHh1zuPeut9cW1WOkAn8XNh73jz61ny64NNHEuqeSKK3LF5FVR/sWt70evDryTRSoHhlikU/R0dXQ/6zKfex8+vH5dcmLW9Gkn/E8f7x7914n065j6C/1/PvXW+ve/de697917roi4/x/H+v72OtEVHXFnWNdTkKOAT+Ln34Cpx14mgqeug5cak5X+p/NjY2597oPPrVT5dc1YMLj/kX+HvRBHHrYIPDrvn8/wC+HvXW8+fXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3RZfl9tKTe3QHYO3o2KtWYStW4vcWpX/pz7917rVx/wCExPUtV1V3h84KCp1/5ZvfKVSF9Vys2UBt6v8AW9+8+tV63J+b/wCH+x/3r3vy6913711vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917roi4IP0PH+w9+619vWNGiX9tWFx/Zub/AO8+90PXgR12THEpJOlb3P1+p/1vfsnrVAB1zBBAI+hAI/1vx711br1hq1fm1vqfp/re91NKdaoK167966311ce/dar148D37rxwOursASR9Bew5P+297NPLrQJ8+ukkSS+k3twR+Qf6H34gjrYIPDrkwDAqQCpFiP6+/A0z145x1xbWLBAv+Oq9gLcfT37HXjXy65c2H+wv79141x139feutkV6xPDHJbUDx9LMR73qPVdI67cskf7a6iLAAk/S9j79xOet8BjrkDwC3BP4v/vHv1PTr1cZ65e9dexx69791vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rogEWP09+rTrxAPHrv37r3Xvfuvde9+691737r3XvfuvdVn74/nK/wAq/rTem7Out/fPL43bS3zsXcWZ2jvDa2b7CxtFmNu7m29XyYrN4TKUknMdRS1MUkM0Z5V1IP096r1uh6gbc/nV/wApXdebxu3cJ/MP+KE2Wy1SlHj4a3t7a+Hp5qmU6YoWyOZmp6aMsbBfJMtzYDk+99ep1Znjsjj8xj6HLYmvo8pisnSU+QxuTx1VBW4/IUFZCKikraGspmaOWGWNleOSNirKQVJBB9+611M9+691737r3Xvfuvdf/9Df49+691pkf8Kb9/7j+Y/ys/lsfyUuqspMMt8ge3tudyd6Lj5ryYfYsOSqdr7Vqq9ILsYaHHQbw3BUwyfiipJQD6SPdbHQE/yX0p/5Tv8AwoG+eP8AKmnkrML0f8jaeo3/APHTH5TItUQrLtvEy9s9Y0NHU1FvPL/c/KZzE1k4AeapxkanUVHvXXvLret97611737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XXAH4A/2w9+69w64o4e5UHT+GPAP+t73TrQNeuQ/ItYD6f4+9de6See33tHbAkOfz+OxQjBL/AHk6xaQBcnn37rfVevyO/mj/AB76DlNO248duCdFbyrRzhwHVb2Uqef6e/de+3qpvf3/AAo42LQ5JMftvbeuNpAFlFPLK5s1uT79ny63imeiydwf8KDt55HGzUm1dtV0E08TLHUQ0c3Bceg6gffvLrX2dV0Zz+Zt8texKqor6HN5fG0s0nkCGKqTSD+LA+/YpQdaoa16hYr+Zb8sOrcjj87ktw5XK0sEwmlpglU5dAb2CEn36vl17PVzfxm/4UR7dyeMosN2dt+enqYIYYXqamnlgkaVRZ7OTz/sffut46OfkP57vRceNqK2gx4kaPUUUs73sL8qDz79nrQ+3ohHbP8AwozpaGqjg2/teUwtIUHho5iLarAk/wCt+ffs9bx0vtgfz+4avbz5bI4NvIICRFJTSfqA/Ifn34dexXolHdv8/XsHd2VqqHaeMrcfSpIFd46WcKQDYm9/z+Pfsnr2OHSf2h/woJ7D2Vjkx2SxNdlZl+lqKd2ufxwfe+HWq9C3tL/hRjuJJGkzO1qyNDchXoJ1BBN+bn/efes9e+3oR8d/wo4nymQFJT7Rq3u2kf5FLZrccG/v2R16g9OjD4X+e7mZKVKuTYdTPGyK2k0kt+R9fof9t79nreOlvt3+fFS1VasWY64njp/o7CnmjI/xvx7917ocNpfzuundwZilxVdiGxbTyiNnmZwIr/2mJPv3W8dWV7F+YfQW+cPS5CHsLbtJNUIjNR1dZHE6mQXAAb6/09+rTh1qlePRgdt7t2zu2lar2zmKHMUiGzTUMyyxqT+Lr79175dKFQQzekBeCDf68c8e9k16qBQ9d2N76j/rfj37rdDXj1y96631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdBr2+yp1xuwuNQ/g1fx/1Sv795daz5da5H8iZ0f5E/MUoukf3lrB/S/wDuS/w9+636162gffuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6911zc8cfjn3vFOtZr8uurKDfSLk/W3PPv3XqDrsgH6gH/XHvXW+uiBax4H044/2Hv3WiK9dgW9+68BTrwvzf+pt/rf4+9mnl14VzXrxNvwTz+Bfn/Ye/DPXiadeHP1FvfuvcePXfvXW+oVZXUOMhaoraiCkgH6pJWWNB/rk+/dap0ga3uDrKgl8NZvTB08oJBR6yNW4+vB9+qB14ivSby3yQ6QwlPJUZHsfbcMcS6mBroy5/wBZR79Udb8ukvgvl/8AHrcFQ9LRdkYFZkNrTVUaKx/2lgT79Udez0v6bvLqSrGqm39t2Yf826+Jvzb8e9V63Q8en+j7L2JkCFo90YmoLfQR1SMT/rW97610sKWrpq2JZqWeOeJhcPGwYWP09+691J9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcWvb08nj3sUrnrRr5dcveut9e9+691737r3Xvfuvde9+691857+VL/L7+IP8wb+d9/Ol2N8wOm8d3JtfYXbfdu6toYvIbl3ttlcLn675NZDE1eRhqNk5LGyyM9MzRFJ5HQA3ChrEa6sfPrYj77/4S3fyXt7dU71xG3vj9U9BZxdtZuoxPbWz+3O0/v8AY1bBj3nh3JVUW9s3ksTVU1IU8tRBX0bxNEHF0a0i7610R7/hGl3f3Lvv4f8Ayi6U3zujLb46n+OveOC290duLIVE9dj8fi924Cqye6tm7Zq6ksyY2mmpaPLU9GGKwNk3KhVlA9+68eHW4/791rr3v3Xuve/de6//0d+3IV9DiqCtymTrKbH43G0lTX5CvrZ46ajoaGjhaoq6yrqZiEjiijVnkdiAqgkkAe/de6+Xf8dv54vx12Z/O/8AmJ/NR+Q/VfePdOJzNNu3q/4o4fqrF7WyybN2dFPTbE2/nsrPu3IY1aaU7Rx/hWOk1F5snWs6qCC2ut9Bt/No/nZdH/Kz5+fB7+Y38Sul+9une3/i9WYCm38e18NtDGUW+cLsjfke8tk46ir9q5TI6/LDWbgxeRSqVNdNURIhIVgN9e6+pB1F2htHu7qrrXuTYGRjy+x+1tibT7F2hk4nV0rdubzwUG4cNOShIDGnqI9a/VWuDYg+/da6ET37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XV/959+69137917rq9iB/X6f097pivWqitOuiwBAP1P0/p/t/fqYr16uadcveut9dEgED8n+n+9n3unWq5p13711vr3v3Xuve/de66JAtf8AJAH+ufp7917ptyGbw+JAOTylBjweQayrhpgR/W8rD37rVemyLee0ZzaHc2ClN7WjytE5uf8ABX9+6307nIY56aSq+8pXpYk8ks6zRtEifXWzqSAPfutYPVIH8yP+cB1n8UsJkdp7IzNBkewbyxUvgqlkWOaNC2hUj/x+t/fs9eFOtd7/AKCHflzmUlgo6ugSGVWAbUElAuRp1KLjj3759b6Jh3b/ADM/lb3CJxPn5IzOJQwpMlOgtIb24I+n+Hv1OvdEblj7M7Pq3rN2ZzLTTSSayGyVS41E8gam+n9Le/dax0tsN01VI8REdfNNF6o5GZnu1r8lvr79nz690anavS+5q3F00rUcpVTEbSQK11B+vIPvfXvl0P1BtNMJi/sp6aKnqmTTrkp0Vb2uQWI496630Vbvje2Pw1AmMijppaxQUvCFa5vbm3+t791roomK2Xmt41LVsj1cFOz67QM0X1PHK2Pv3XsdCBLt+jwFOuPEuVeeQAc1EjAE8ci/19+PW/Lp9xPXFZlI0P29ZIJbCNnTykMfpbVz791qvl0Nu0eiN31bJQSR1ZpZrEBVsCrGwBA/p79Qder0PND8L9yTUz1FLAxaRbnXThm45/I/x9++XW+lFgPgrmK15GmxlTJV24YUgsH+tuR79jr1Ogq7C+Eu/qGodpKGq+3XUQkdKEIUD/Ae/DrXRWqnrjcHXmX8tdjq9Iqd7kyw29N/x791v7ejg9W99deUyU+L3FTBWjCI5l0AtYcnn34561XqyzpkfHHs0QRvLjqUyL+400sMY1H0kX49++XW8dCt2D8K+g6+OPIYHOY6OtcK4aCujRlZvUbaDf37r1Oihb8+Ju/4JSvXO4cwrwsTC9Lkp3hIT9Fgjf7Ye/deI6c9ifIj59/ELHTx4WurchjYgyv92081x+r+3cH36nn1qtMdDv0b/Pn71o90yR90Y6QYOhk11pWJow0EXD6bL9Tz9Pfqdb889XEfHz+ed8Oe/d4UvX+Jy9RiN0usKVCZCohipI55TpKq0gBsD/U+/dex1cRiN0bdz9NT1eHzeLyMFUgeBqOupqjWrC4sImJ/3j37r1acen737r3XibC/v3XiaddAgi497IpjrQNRXrv3rrfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdBf3Lc9abt03/AOLPXf4f8oz+/da4kHrXJ/kPFh8hPmMHN2O5qv6DiwyXv1Ot14jraG9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcGdVIBvdvpwT/t/e6V60SBx65+9db697917rwN/fuvddE2BJ/H9PfuvdeDA8c/15FveyKdaBB6796631737r3Xvfuvde9+6910SB9fe6daJp13711vr3v3Xuve/de6Zq3cWCx0Us9fmMbRxQ38r1FZBEIx9fVrI97PWvPoj/cv8x/419KS1EO49009a9OrkjG1cEupkNivF7f6/vXW/t6qH+RH8/LYtKk9P0+T9xCXRGllLNJZbIxAFuT/T3qlevV6qn3v/ADw/l/uSSSXCVkUEERZof3niAGo8cWv72OvdPvSP/CgH5Lbf3hQUPbNRSTbWWqp452SRmcU6sBN6zf8AF/fqderU9bZXw9+ePSfy82Em69l7kx9PVUUKJlaKvr6aCVJlFpZIxIVut/fvPr1R0Nna3yP6l6f25Vbl3Vu/CJS0sRmMEGTpHndAL3UKx9++XXs9Urds/wA/34+YKtrsNtCZZ6mkkqIPuGnDapY+F4tb/be/Z691XhvH/hQN2JWVtUNmz0kdMjnxmR7WUcDhh78B16vRYt6/zlvkl2k7Y2fNQQQS3jBiq2iIXVey6CAP9t78R1qvy6LjujvvvPeVd9/Duis8kylx48pUWYuOQQjf7b3vrxqcnoCc5P8AI3cVXIlfuPMyUsxsphyNUbi//BveqHr3UHEbE+Q9PXXw+c3DG7EHUK+ssCRxfn37rfl0Oe2h8sNsw/by53Nys/qjP3tW1ubm5v8A7f37rXn0rIe/PmfsqY1FBkMw0VMbsZKmpYWPAvf/AFvfqYp1sE16ETbX83/5q9ZziCpyC+KCwIqKmZmKx8EMH9+4da6M/wBcf8KB+5oahP7+1VG8ERQTaCAT6+TcD+nv3W+rKus/+FAXQLUcLb9ls02jXLBOQ8a6fUQrAj639+69ivVlvQX8y34w/IaohpdpbwoqCWZU0DLVtNApeQelNTaf9v7917o+WMzOJzUH3OIyVFkqe9vNQ1MVTH/yXESPfuvdOJIFv8fewK9aJp13711vrq/Nubn/AA97pivWq5p13711vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuvf63v3Xuve/de697917r3v3Xuve/de697917r5kPwk+KPzq+WH86H+cRtj4J/Nqr+Em8trd495Z3fG7KPG5fIvvbbVT8jslQUW3XXEEMggqmFTqY2NrAcm2ut+vVrnyf/AJCv/CgzubpnePX+c/nR/wCm7CZ3FVlLl+qNz7g7W6123vuganYy7bzWYwUNWk8NUQIWpchAaV9X7xCXPvfXq9Gv/wCEvfzV29Nszuf+U/vv4v7R+KXyX+DddlpN94DZD5F8T2rLRblTY2/t+bkOdq8hXPuany0dFDlqpsjVUtXDUUklA0FJGlLD7r3HrbT9+611737r3Xvfuvdf/9LYq/4UW/JvsD43fysu+KHp/bu89zdvfISnp/jnsel2LgM5n8xiqXsikqIuwNxvHgIJ5qdKTbNPmFhqiFCVUlMAwZh78etgE8OlP/IT+CWO+EX8rX4y9Ybt2lR0XaG+dtP3n27Dl8XSnLU+/e2wm5ThMpHPEHjqMNiWxeDlja5VqIgm9/fuvcMdDl/Nv+FOC+bX8uP5ZfHTFbZw8u8N1dWZfcPWhgxdFHUxdqdeld9ddClmRVaNqjKUFPRSOrA+GeVL6XYH3Xqnqsr/AISmfIHszsj+Wbj+ge59ob/2f2H8Rd95jrChg39tXO7arMr1ZuNn3l11V0TZyngaaOiNRksII1DeGKghBIDoB7rxBHHrZq9+611737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XRv+PfutGvl10zKqlibAfU/0/HvdOvVFOuwQwupBB/N/euvfZ10DbliAf6X/wB9/T3v5Dr3DJ66Dqx9JDW4NvwffqEcevAg8OufvXW+otTW0VGNVXV09MD9DPPHFf8A1g5Hv3Wvn1FlzeIgp3qpMlQrAilmkNVDpAAuf7Xv3W+is9h/N74/dYz1NPujd9JTSU3+cC1FM3N7WHr9+48Ovfb0TPfH86r4dbX8tPHvijaqQuqr9zSOxZeANGrj37rXQJyfz0Pj7OzHG52kqI7nSfLS3/T+bH37rePXohXfn/CjKt2TV1lP17tyHPiMN4DHFBKXIP1H9feqevXuqY/kp/OV+TXymhqqaOXcOwIXMipNjn8BRGGldIiI4H497p17PRNdkfNL5KbBrDWv29vXMfbknxz1r+oX1c6m9+611YNt3+fB8gsLsWv2Y8GYrHyNA2PWveSNpQxTxiTUTf37reeqidxbq35212fXb535lshmZsnXS10NJkz50h85/Quq/wBAffs9a6HXavUkeWlMlPQxKas3VUiIVL/2VW3+9e/de86dGj2B8Pq3ITCqlpCY5CoA8cxAv/QW/r7359bp0b3ZPwWw7yRvlUlpIjYmTxzKp59Nvp7116nn0anb/wAKcbSyUjUtNHNRAgJLJHKzMOObke/HrYp0O/8AoNwG3KL7NqWFfFDqNkKj0jk+/AU61jqrP5lbuxOzZTgsQ6pX1DCOIwFS/qW3+uPfh14n06rzx/VmY3VFDlswJJxIxcCZXYkE359+60OjC7S6vp6PFqIoE1KoXhX+oH+Hv3XqVz0qMF0cNw5NSaSORlb0hka99X0BsPfsdepXo4+zvj3LjqdJK3F0sdNTqJNX6SFA/wBU/v3VujB7Hw/XFFUolXU0YqoiqGMSRDSVNrHn37r3RpKV9jY+CCRZ6LQRwvmjseB/Un37rWeHQ89bPs+vqF8dPRyhyFuCjA3H+H/FfeqefW64oOhQ3F1Xs/OoGahomLKxVG025X6D378+vV9R0QTvf4dbf3Ej6MVSQLJwXSI8j/gyD3uvWqdVl7+/l6bbxwrK2KbwT2ldBCJFufqBcgfT37rxGOis0Xx23Jtyoq4MZunLYyGKQhXp5ZIyuk8AW/x/Pv3WqdQWp+3dvZSNKfe25MhFTuNKy1E5VubAHn37r3R1+nPlX2BsGWCn3JjZKvHxrGklTV/Uog9Tln/2/v3Vq+R6sY2J8mPj/wB3Ui7W3DJjY8pVgx+GVoAdbemw/P8AsfeuGet1BPUne3wj6p3LQSyY7GYn7SrQ/wCURxwvrWT1clfrwffq9apXh1Uz3T/LQy+3N1y53qGcYTJq0kyV2IjMFVcepSXiAPB/x92619vSM2B8mvm78FMmlRLmN1b7o4JA/wBtWzTTJEpa7KFlJFrD3rrWeHV5/wAGv59mG7h3JQbS7vooNp5KrqYqCITGnp5GmYBSSCRc3/I964db4jrZN2/uTE7uxOOz+3chTZDHV0CVMEsUqOrRSi4LeMmx97qKdaINelGL2u1gfza5Hv32de+3roMDYr6gb8gi3HvdOtauvKFsSpuCSb3uL/4e/Enz68oA4dcveurde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SP39iP47s3cmMBAepw2RWIm9vJ9o+kHT/X6e/fLrXzHWsH/ACE84s/yw+dm2mYfcYDeFbTyrxdSMnb6D37rfW1Z7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rib6hbgfUn+v+HveKdazXrl711vromwP++P+w97HHrR4dYYWmYt5FVVudP8AqiP8ffiAD1oEnrNf625I/H/Ee/U9et19OvckfSxP4vyP9j7959erjroLb8sf9c/7z79XrWnrsi4t9L/kfUf4j3rrfXgLf1PN+Tf37rwFOuMkkcSNJK6RoouzyMERR/Us1h791voK95919b7Doqmv3DuXHU0NIpaW1VTlgAL8AsL+/de6rr7r/m9/GLrjC1cmJ3ZS5HLwpIVphNTE+RCV0adR/P59+zw68PU9Uodwf8KIeyI6ytoOtdntk6YvJBFPTwxMWjYaVdWFz+ffqGvWuiL7i+e/yw+ULVUC1e4dsR5AMs60zyQiNfp9UIH5/Pv3Xq16CBegew8vUtU7q3Xms7JOxkeOvMtQLudRX1X9+63QnpVU3xBp5Ijl6qjjKwi5DQABvzzf37r1OuU/xu/iEQjpcXDFTKNOuOIj6fUG3v3Dr1OgL3v8ZfDM1HRY6CWcBm/zOog/48e/depnHQOwVHbvRdfFido763FtGmrWJlpMI8kETFuW1olhz7917pQ5bL/Ifsimkpsr2Xu/J0DxkNFVzzOmgD6MCT/tvfuJ611AwHx2E2Lknr2+8yRlZ5JZYLyPITdmLAe/U690s8P8c48pTzeakjgFOp5SmZC1vzdRz/j79Xrw6xN8d2mlWkxxaOaJgVdI5VNz+NQt/sffvPr3UTO7P7E61jElLSVNYiEEFo5Xuq/2be/de4dKTa/c2+KONDX7duY/UC1O3J/HFvfuvVPDox20vkHk0WKWfbkKWIViaUhvp9L2+nvx69XHQg13yxoMQFNVg4furaUX7fULn6Ai3v3W6+vXcXyJzu9aKqgxWyqWoEqaVdqT0mw+t9P+8+/YPXuir7+2TuzOvLX5TbcFGk8jDSkDXHkN7iwI97610XbfHS2Tw+MkrIMfG5lXyAGL+ovzce9de4dAbt+jgleuockII5IFcFGji/bsfqt/+I9+63XpT7f2H2lkXFT15v7cW2xTTCojkw7yQ6jEdSR/t8W9+61x6uB+Av8ANb+QfxJzFPsDuBs1uPassggqM5mbzBEaYASF5voQPfuPXq+vW490R80elu8Nl0W6sFuqhDy0sUtVTy1FOjRsYw7hBqtYfn36vkevU8+PSD7H/mR/GDrHKnEbg3pSrVq7ROi1FNpDobMLhj791vp865/mD/Gzs2RIdv7ypWmkNlSSemAYg24Jce/de86dG9we6tv7igSpxGVo6uOQKVEdTCzEMLqQqk/X/D37rXn0oSL/AJI/1vfuvEV6xNCpcOWcH/UhvSeb/T3up60VB6ykagOT9QeD/tvfgadbIr13711vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r5p/wDL0/mlfFX+Vn/Ot/nI9kfKqu39R7b7P7p7x2TtVtgbOfeVbLm8b8kslnapa6mSppvDEIEJWQsbtxb37rfr1ftvb/hYz/KbwG18xldn4j5O9gbmpaOd8JtGj6oxO3WzOS8RNJST5zP5mOnpYXk0rLORIyKSywykaD7r3QBf8JqvjP8AJ3uj5ifOv+dN8lurcl0hQfMKbdGD6X2FmKTI42uy23d8b/pN+7g3DjaHLRU9VJhsfDisPisXlKinQZJvuaiJRGgZ/dePW5v791rr3v3Xuve/de6//9Pf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691wJGoKVJB/NvSPzyfe+tedKdJ7O7v2ttdUO4c9i8Kr8ochVxUym/0sZCPej8+tjosvcfzY6J6iwFVma3euCy0lNFLKaWhro5mIiW5F0P1+vv3XqYqei69O/zPOtu7sfl6rZ+JkkbHipRGUyOHaFTZ7f0uPfuHXvPqiH5O/z4d+dOdybm2pTYPK1VLjazxrFDS1Mi2MpXkrx+OPfuvVp0VLev82D5Nd7+DJ7UGZwdE7qwDR1UakPyDYf09+p1qvp1Jq/ml8tMxgxil3Jl1aWLxvN/lICsU0s1/fuHW6nj0QztHJ9nZSoqa/fe92ysuTBBppaibVDqYkem9/z+fe+tGpz0VQ9O4fO5ZquuoGy8jMXLGaYgkm5P196690pk6hxkU8NLR4VqMEaQBLL/AFsPqffuvfLrivSFHHXySVELFhyAXLW44/UffuvV6dJOvcdj6aZTGEVInsbkDgcfX3vr3RIOzjNia6slopdUED2dUbi1yDyPdT1sV6SPTqZLeG5qhJVlenic+FLkgsOQAD72OvdWU9b9G57clZSt/DqgKoj/AHCDaxa1/e+tcerQemPj9TUBpxkWGpLhSxW4P+N/eutgevVq/UGztiYiaip8pWUWrXAjK/iP0AudJ96+zreK56sIptidT5GjpIY56CIPGrFiIV/1yD79WvXunrM7T2hQUNJRYupppLAKgQRm9/8AU29+4nrVcdFI+R82B672NkM/XSRxyfaT+MuADwvHvwx1atR1rE9h08nd+/q3M0dqmnx1YNBRxYBW44/wt731Tj0YbZfXUlfRRYg/5O6BVBuAQSOfe+HW+hrovj7lMZRJUrUNLA1pOGF7Hng/096J635V6lVEtH17C08kIWZQfqLlmte4v/re/Ur1rh0Dm9PkNu/J0VRjMPFPGssTRJIgdVN+Ab+/de6InurOdw4aefLRZipiLTNMELyKdH6vx79w61Xpup/ld2PQLT0mQzMrtFpBR/K3N7H37reePVr/AMRvlCGpaSbOZCxkMQbyavrp5Ylj791sdWk03f2CyFHDNSZ6mSyrxq5+vIIJv71jr3SyrfkZsKkxlMMxlqGV41GryBObi/59+8+vdEi7i+SOws5VT0GEmo5ml8igQ6Db/WA97GB1onz6Lpj9vVG9DJJSRsizEyXF9Vr82A/4j3oY69jh1gqusqLJGTHCE0lVRWElRZbtb6tzz9fe6nr3DHRZOx4cZteWow1fVx1OpJEWRv6fgf4f63v3z68aDogm4Fqdn7th3Ttrc8dBUQzLIlOkzB2s+sqEU/n37rXy6sz6N/mdV2MxEG2dzxPK9Eqxmrlik0SaECAhz79TrYJ6HOr/AJk22a9/sIII4p3U/wCUiMWGr8Bj798uvV6L9258ptlb/wARU4ybGxVOQq0KJUCJb6ipFwffuvV6pW35gq7Ab8ot1YqOWgmgyBqqSrDvGInMmtXspHvfWurofjB/Nu7o6KoMVisxl67ceIpKSCFY41qJkjjjsdH+296x1upHHq3Dpj/hQzsTIZ3Gbd3xg5Kf+IzaDPPFPCIbkKCXa3v329a/Lq/7qv5MdP8Acu2MZnNs74wSvkI4JGofvoBVRtMgcQmJjf8APPv1etkVGejDwOkkSPGQ0bAFGAsHX8Mv+B/HvZ60OHWX3rrfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691DyH/ACu/P+SVPH/TluPfutdakv8g0P/wAOFfzNPUNC9hVwVB/Z/wBytveut+vW3N7317r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuuhc/Xj3s08utCvn0ks/vLaOC/azm5sThm1C/3tZDTsD/AEvIRb37h16laHoqfdnzt6N6Wxj1NRurFZ6ohR3aGhq0mvo+oDRH3o8evKKDqvyu/nvfHWgqpKSaitJG2k/vSAk/63v2T1vHU/Efzz/j7lqlYFx7RBmsJGmktz9D7917HRl9ifzSOgN5mBTmaHH+Z41JlqSNIkNgSH596690bRflL0ScQuYPYm3xTtF5NBrYhLcLqK6SfezQdeFeiI94/wA3XobqemyBoKul3BU0sLtGKWcy3dSVvpQ/T37NevYp1r2fJ3/hQNvPez5Tb3XmNyGH1SVcMNRHS1KxhWGmNiw/p78c9aGOqXd3fPD5Hdi12Qj3N2LKlFXNZaOWaoRtN7FQpa3vfXuoOyNtp2BVNJuBzXyTEkyvLIVYuNRYEn37rfRxep/jPjKvKU5paXVAxQ6Fe/8Aatyfx78etefVovW3x4pcRSaqOAU72NtIUkm354/3n3Wnn1bHDo0+yOloghmyMIqiFFjZSRpH9Le/Vp14CvDqVuXpOTM0k1DQ1DUMDPyAFVQL8j+vv3W89Om3vjbN/AZ6ON3eVIioqAw9TW4I/wAfe69ap0AeW6ArNrVddX5OlauVkn0yELx+R+Obf4e9fZ14dEm3T8dpN7ZXI5KagYx01QTFJwoiAP05Hv1etUr0udl9BxYajcSUZmDKVFwHF7fUe99bp59KWh6npaF2aTGGzNcgqALA3Nlt7917oxvVPWe0M/T1NPV0NPSSrrWzhFZwOLgEfn/D3rr3y6WFb8ccFT5FGocWFid+JwF0883BI9+x1vh08ZX4n4TcMCQTUnlumouwjZbkfnj36tetUHSIb4P7WgraemlxiHzMFUFI+ST/AFt79UeXXqdYc/8ADXaVNK+Lp6GOGfRwVEa+oji5HvfWuHDp22H/AC/9mVkU0+4MXDXOWJjmk0EIt73uf959+69wz0KuF+H/AF7hJ5oMXBSQGEi8cbxWt/Wx/wBb3rrdB0Hu/wDpbbCpU4lKeKSpWNyltHJUcHj/AB+nvfHrRx1Xz2p0rUx4DMyyOY4oTOEBsBGixk2DH/evfhXr1Kda3u9dwVG2u58jhTWrLBJNInjvfV+4VsoH59+691fp8OevcRu3ZePZaRIZ5UpmaZkQatY9Rv8A7f34468KcOjV9tfCfa2/MVHTRIk9U8dyqlEJcDi+nn37r3HPRU8z8fO8ep8O2C6y3zVbdVEYNSJUzarfpCehh/tvfuPW+Ax0W+k6P7JzOdv2dPV7gyEkur7iaaZy7OfUwF/offutZPRgtq/FDs2WeCbr7cdXteUN+0UqJVsQbgAA3+vv1evUIz0JlP3v8yfiVmYKnLbry26cZSSpK9OjVExMUfp02B9+68T0fvrH/hQAMKKCi35tKuVYlhgqXqaeoXXITaQlm+n+uPfvl17Hl1cz8bP5l/QnyCoY50z+M21UyKSIK6q8YuB9CZTce/de6sBwu5cBuSBarAZmgy1PpDCWgqY6iMqwuDdD78CKdaPGnT7791vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917ouuZ+H/xK3HmMruHcPxc+Ouez+dyFXls3nMz0l1plMxmMrXzmpr8nlcnXYySaoqJ5GaSaaZ2d2JZiSSffut1PWTBfEX4obXytHndtfGH487dzmPkE2PzOC6W62xGVoZh9JaPI4/GxzRN/tSOD7916p6MKAAAAAABYAcAAfQAe/da679+691737r3Xvfuvdf/1N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XA+TVwF0W/N7397xTrWa46xwVMVRr8ZJ8baW4tz/h7916vl1n5/4r711vro8c3AA+t/6f6/v3XusD1lJGCz1VOgX9RaaNQP9e59+6903y7gwECs8+bxEKKCWaXJUcYUD6li7i3v3XqdVlfMf+aB0x8a8dkcRBmqLK7p8dRDSrSVcM0KTLGdBumofX8+/fZ177etUL5Q/wAwr5FfJDM1VVRZepoNuiQSUa0mTkgYRKfTwpFuPfutV9OiOz5/fe5I6ik3JuLOVMMqSh0ky08qesXI5PvfXq9XF/y6tybK6c6pzeay+VQHw1QIrKsBmdgSOT+fej1sdVn5aKg71+Ue8spUQwVGAqsi/ik0LLGU85PDm49+691aTUdZ9a7A69SejfDRvFRrKQRCsgKp6rfQ/wCx9+63QcR0Ryb5L9e4ioymGnihepjmmijZArLyCEuw9+8+tVHRBu1M9W7y3O9XiXdaWeQCEK7aU9Vx6fx791roX+q9s5THSxV2SaOSm0xuxdbgLa5vf37r3QtbqzW0o0D0vgNbFcWQKCrfUC5PvfXsDoPcSrZSpkqJVBjuSfoQB7117z6xbsxFFVYidaMHzMkgul9V7c/S/vfW+qxO08Q+EzDU1ZC/21dIPMzobcnj3rrXHpd/FdNqP2ImNHi0GrjReFIDH6rp/wCNe/db49bQ/T3WG3htCkycdLB5DTIVKwKCRf66j791vyqOhEyeO27iaaSSBfDUAC9v2wGt9ePfuHXsdEV7133vvb8Jy21KyUS07PKFjncG0J4uAffvPrVadBPsT+Y5urAL/D98V9XDNAUhXVWyA3BtcaiSPfuvVPVg3TPzopdxUtRkMlmEalpxrpDLXguABf6f4e9eXW/t6Az5C99b5+Rk39zNtVEk2MaUxSNTzGb9p20+rT/h738+vdLHoj4ny7eooWnhu9ZEHqmlhBcyn9Rs3v3Whjo3uC+MpFaK2FbJGynSsQUni5uR+D71jq1DXh0JOS2pSYXHfw+eECRUKLeIfUCw+vvfHrRr59F13J8fMjvqV5Y4neJmBVViPC/0IHvXXjTrLsz4hRCaopspjz44xqjL0gYBrci5HvfWumvdPwwpMtPPG+PV6fxSBQlFq+oOngD/AHn37y698uqjPk98Gd57cGYy+26CpjWm9cQSjb6C7WFh+ffsdaoRjqvfYm5+0toZn+BZVK2kaOoMSM3lhX0NpH9Pfvs695dH723mu2Hw6VNDXzsjxgi9Q35/33Nve+vVPThUY3uPcVRRQ11XkPtZyAximl5FwLCx964db+XRlut+hr11NPXvWSVLPCzNO7ObsPVce/V68B1ZltHY2A2Vtv7+qnpIXigZv3mVGA06iPUffuvV9OiA/Ib5HbIwByNJtiaN85ptIYZlLa7kEqF+nv2evY8+qWd7dm9gb33bVReKtLSPKEb9xlKufSbj3vz610yUnXW4q6oFXnRVtVk3p76/GOb8gk+/fLrfyHQz0HVtRVY0sn7dQqXfSoRgbfkj3qnXq9JGo2ccYWWaWp84ksCWtb/Y+/Uz1quehV2z1Nmd24iWtwMsklXSJ6QCXJdf8Pez14dAP3Ttje2Cw0MebpZGeEksY4DrsrWFrC/v2evdAntjfeWEBwOOoKtckWCg1EJ0kfk6mHvXXuJ6X1ftOtztF91PHXw5aFNQelDIVkAv6GT37rxoD0v+nu7fkT0dlYsngdxZ56eikE9NTVOXqFjBh/zY8bHj6e/fb1vrZP8A5c/88DcuUzeL2N8iq6mp6Y+GjjqZatWdfJIIkbW/+v79T069X162xdp7twO99v47c22shBksPlKeOppaqCRJFaORdQDeMmx/w9+68cdPKzSPU6AB4QpBNvUW/HJ/4ge7EUHVQanqX7r1br3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917qJX/8Aa3/AKhKn/rS3v3XutSX+Qb/ANvCf5mv+HYlffn/AKunHv3WhxPW3L791vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rxv8Aj/effutGvl1wdwi6jf8AA4F/r72BU068TQZ671WXU1h+T/h/sffseXXvLryuri6m496p14GvXElyRo02uQxa9+P6e90A49aqTw658/4W/P8Axr3rq3Xfv3XusErmP9xpoooVF3aWygD8ksSAPexSnVTqrg9NMm6NtQsyTbiwcTJwyyZagjZT/iGkHvXVuiRfJH+Y98bPjJBVyb43XSVMlLC8hixtXTz6nTgprQkDn8+/daz1QB3v/wAKKZsvka6l+PVPDVxxTSxRSVGh1KkFY2JK29+NTnrYwKdUs/Kf+Y/8tu0/DmczkaygXIMGmSgyUsCIGNxpCWA4/p7917iOiG7d7o7k37uo4nM7hzVTQysok+4y1RUC0h9Qsx/3j37rR9eh6yvTFHkKnH1D1+SapnVHkK1MpGoC5Bsfesdb6FTa/wAfppaikEdTlSjSKHKyyMdB5ve/vfXqHpcdl9Qbl2rioJtsZPOwVd0bTHVyobj9Bsp9+weHWsjHQg9TdXd77zxsMVdnty/bGMjSldVNcfQcA+/dbFelpuH4gbjjJmrqnO1U8ws61LzTAluSLNf3vr1Ogyznw0ytDj6jJ/w6SywyPqejsDZL3uR/vPvXXh1Ud3nsmbbO6XFRHPTmmmABA8aCzfm3H+8e/U690Kfx17Dil3NQ4aSbyKamKJlEmo6bAWsPfutYpXrZB6J2xQHFY2soYRrkhhkculydX1HP9feurY6su2TgsVLDRJ4wJX0ie6cDjm/vxJ68B59I35Kdp0PUu1o323CXyjen9oMOfoDZf9549769XqD8Zd9ZLs3bVXkNyoyzNZk1qbklvoQR718+tjo++2KSjhoZEMQMaqdLafrYe9H161gmh6AHtGSOpaShhp00MHU/tKTc8fW3uwPXqdANjdkSGoNJFTxrS1bAz/sgNe/JB9+x17J49CVgOpYqqWrgp6YFIELj9kWvb8H6e61p1sjqBX9Z0yU8zPRFpY2ZbLAC3HH0t73jr1KdNu0et4J2q6qGnmhmguQBGVBIP0IFvfq04dezTPS3L5SkEVLMIxDGwAMgGrjj3rrfQgYnO4/GRq9S0Lx6AHGoagb8297z1o9TJdz4StnWpjWMmncNH9CxA5Gr6e/dap5dFu3L2HHJvyKHUoSSoVCAVAVb2+nvfWxx6N9BX09NgKSSk8ZjnpQzsv1uy8nj3qmOqjJ6D3HY6kq6yqmpqj/KJbkhpPSCeeLfn3rq3QJ9ibZrcdWjJiRTI0mmRSQQY9XqPPvdc9aP8uixfIRMKu0JaamS33FNMawopILtASbMP98Pe+vV60v+8aXGUXyHzcimS8E0rxXB0/58/W/v3Wh1sW/Avc603WVHlatdcAp6eOMJ+okLZRce/eXXhjo1e4u0N54uaozGEcrTQPqRZm40H6kKfeut/LpB4f5E4GrztLNvCJDU1NSscrMbRWDc6vx7t1qvT13JvTbLz0eb21UYpFESEqrQqRcXNwp/25968+vdGC+Lu56DeVDPdqV8jTAlCjal1Cw/31vfjTj1sGvRoOweiju/b7V2SpaGQywmwanSRgPqGBt/th71XHXqdVgdkfCii3ZXVEMFIqQLJIXNPTKgDfQAaBxf3bj1rHRL+zPjJ2H1GPuNk1+4aUQAK60VRUQgg/W5Q+/da4cOlV0b/Mu+W3xeyVHRFq2u2/T1McdYMnVSzsaOEWfiS4/x9+69U9bNPww/nF9HfIKhxe39z5inw28po4xUmonihpzUsdDR+u1uefr711v7OrmMZlcbmaOHIYqupchRToskVTSTRzxOrDULPGSL2/Hv3Xup/N/8PfutZr13791vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r/1d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XRFwebf4j34Y60RUU6Qu7uwtmbBxdVmM/lqKhpKYM07eWLVdBqYEXHPv3XuJ6rg7g/mtfHzYWLzUu3dz0+YzVHTO8OPLwv+9HcaVF/yfx791unn1SFvv/hQj3FXZfI7f2z1vO1EJ6iGHI01Og1xLwj3/wB59+60ei3bh/nX/Jl6WvipdqZSVqm416BeHULA/Xi1/fqZ63WnRWNz/MX5M9vxzV9bv/c+0I5lJkgpql4wqsdXIVgPfqV61XPRT91Zvdm4axmzO4qze+UMtr5K88lz9Tdr8+/dez0IWx9gZzPUwp3gahnkBWOCMMiE/wBOPp791456HrCfHHJmknFYCk3i1DVq+rL/AMV9+69TojPyO3h2j0xhq7auNkrqfG1dQys8GoroP1LW9+69x6h/Hft6Xa2HGRlV6jLZKNn8pU+UyML3ueffut9C/Xdjdl9h10lBWZWvocWw0sgdgvib68e/da69T9Y7Ep4JaiszXmyjtrl8ly5k/wBc/wBPfut06g0e0sDRTVNTUVZREUtSfqtIfwLD8e99ax1yfeO50jbE4LHS1KN+0sio1itrKxNre9de6b8V1h2Jn64V9bSzxRuwdks9uW+vHHv329e6NxsbpzITUwhqVZCVGojWBwvN/fq062B6dCbSfHYOjzxuZXKk+L1/U34sffuvU6IH8svj/k6TE1+RXGKopF1eVUcsCFJ44/2Pv3Wvt6qb+P8AuD+6fc9MlZVNGxy4BVvStlbSTdveuHW/Lrcc+M3YlHuDD4XEpULJDJjYCw1La7Dg+99b+zowm99vbVq4KimqsstJUyDTGqyLdjawT/b+9fb16teq/uxevaiimrV8j1VEyy6fK4IIP6Stz9Pr731ryr1VJ3z1PFJU1VdTpHAyPJIdH1uov9B791r5dERpO8NzbPyVZtyGvq4khEkcYjJs3Om319+631sS/wAs/Dzblw1BufKwitqKoQHVMCSdY/Pv3XvLrYF2R1tDkHibQIQ4JA5AHPAA96NerYHQ6/6OaXEQpJ5UAOkt+6o4t+ST719vWqnosHalRtbGZHx11dFEA1ifMht/Xm/vY+XXsefSfwfbfWOyFgqJ8xBOum+hpYyBxf6X9+Na9exSnSH3188+o8NK6UlVTLKSVJjWO5I4/wCN+9/b1qg8uk1gP5hXUMEl8nV0yhweZEj/ALXv1D17HXeX+RXT/crTYjHGiqUrbx2VUswIsRx/vfvXWzw+fVQfza+O9NtykqN2bWxNOAnkq0eFebatfNvduq+Veii9C9tNklk25loVilp5o6dgzAWIaxsT791vq5LpXbuztyVeCx1e1OPOyorMyDk2/t3+vvVOt/Po0/f+xdkdF7KO9KWtgklWk80SCaNvUqXFgCbe/deweHVLm4/ktvXt/PTbewlRWUuMV2haaLUECN6Cbjj34da8+kEvxPy8maG4q6vqckal/IUqNbqQ3q/F/fuvU6HvbXxhxlTP98uLpvu0gBsqG7aFuL8f7f377Ot6adJ/L9ZwUVRUQVuOhhNNMVjsjC/P4Pv3WuHTNQ9XPXzmOgdzJMxXxpfTb+lh78TTr3HoM+1+kt24XHyVdJizP6XYsVYkLa5b6e99a6afjTv3EbE3Cu39xVGibIS6ZYZLgR6206QX9663wx0eztnoah7EwyZnD4ynr6aaKN1bRqBLDWOE9+4dbp6dEIz/AMbc7hsuaiPaNNTwRtY1KU7j6fTkD/effutUPE9ebAQbbpX/AIpi41IUlpDGQOB9QSPfuvdB3k9u7b3FT1EtHMv3DKw8H0ALX4tx9PfutfMdFxy/W2bwte9fCf4fHHUJUJXxg+RPEwdNJHP449++XXq4z1eX/K+/nE5/pXP47qDtuprMhtepq1oaOvyD6Y4oARGrxNIeCPr798+t+XW5V1h251525i6XcOytwUeVjqaSCoMUNRGzxCaMNp8ak8j6H3sE061QcehPmlVJYkaXxlzwtv1WP9fexwr1U11U6kc3P5HHH9P9j71inVs169bkm55/23+w9669TNeu/fut9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdRMh/wArf8AqEqf+tLe/de61Jf5Blv+HCP5mv8Aj2HX/wDu1/r7916mT1ty+/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rj6rgaQRblr83/wBb3vHVc+mOo0sJlYSpJqC3Hjv6CQef9j73w60c5HUXJZfG4SlarydVBRQgjU0kiIAT/rke9HrYx0Rnvb+Y18ceiRLTbl3dRrkE8irEJI2XyIOB6T/X3rz6t1Wv2f8AztsZRwyT9Z4dNwo/qpdESfuqRxpv79nrwpx6ITu3/hQR8lcFWmGj6XqKmMyEKRSow06rLxf8+99a6RW6f553ym7Awk2PPWGQ27DUxSRmqigVGjMy6dQ0/TT711sYz1W9uPs75X9rbmqdxDuDeW3IKqXymgpqmoSNeb6dIaw497x1rpG7m663pvdJqXf+8cluhhGxc5Xyylz9SCXJvc/X37rdD59BPiursJtWWWCmwtCg8lzJFAEJ0ng8D3rrXSb74moI9ngGJYzSU5ZSqgWsv14/3j37r3DPRavjlSvnM8aqEGQ/clA99R4Pv3Wz1bhsvr+GvpYaiskYTIF0amIPLWuB791rqzroP494XMY4ZGrdbRIH5Y3Nl+tz/j+PfqinW6Z6S/cPX+GpM9DjqdUmRZgoW4I44tx/X3r7Ot/M9HX+OXV8dHhKedcTAwMQLMUI4tcHn+g9+48evAADj0Pea2LjKyTx/wALpy0f9kqDbnm3v1T1ug4dBB2VtahXbVZQpiYRennUHQByUt9R78DWvWiKAdas/wDMS63mwUGUydNQxxtIdSMo9SkXPJ/H+t78OHWj69U+fF7dVZhuyIqnJO5vlSqiQixCPwBfj3vrxz1uI/FXfqZLA4mWWBVpxSQ/uXX68c2/4j37z695dWG0W8qmKAnB0wqpLXAUfT+p9+p14Hp2oepct2+pn3Hiovto/wBwedDYD63uR78SOt9CLs3ryh2a7YbCQRxKjeNkiW36W5+n+8e9cevdGfpJosXiBHOgWXRYhiL308e9dar6+fRet0V1PUZBrwobyH6WJ+vFz795dXHy6m0WDp50hqnIhcAFUVgNXH9P9b37HXq0x0Muxa7EUArUqwihoioZ2UEm1hc/8T78R1Xh0p8FicPXTSymOOWIs59RUglr8A+/da66qtsYjGU+QlpooovKSwWMiwuCTx791s9Fn31tueohM9K5RbsbqSCOeLn3v59ewRToIqTFVdVUCieqlL/lTxwPoLn3uvWqGtOniHb1fTGRPIxCtYkauQP9b3onrdKZ6L5ura9Sm646xXZHSUOeD9Qfe+vfI9Hd2KlXktlMkqlpIqchGINzpT6ge69ezWvn0Xeu3Rk9r56tM0siQpIfzYDn/ffX3YUp1rPSZy/YNJu6r+waubVwtywsWPA5B+vv2OtmvQGd91FNtXYeTrMhoe0M5iaS1rGA2PP+8e/de+XWnH3XU47OduZ/MwGO4M5BHANpSOLf8T791rq7r4Bbhkr+vMfiREXhaSliFgD9fTYW9+68PTq3PdvSE2X2lHU08XgMsAZwNaq3p+psPevs6seiNdjfH1JcTGlL+1Wo5IkVW1h9X1uOfr731oinRL9/dI9nYWjmyNPlshVUsBkZYtUrKFVSQLc/63v3WujAfy6O0cxjt05Db24Z5qaRaiSAeXUC3rsv6vfs9a6ut7f+RcXXO110zNPCKVNWp0IClf8Aavp71QdWBHRIcf8AP7rqkp62Ktr4f4i0xXQXgJ1E2IPPvYHWuhh65736d7liqaDM1lF5qm6osrQre3HGr/H34ggdeB6A/uf4obZ3U1U23KKkrKVo5pTIjCX0vz/Zv+D79149VId0/HDdXX33lf1hlKjb2ZpKhX82MR45leI+UqCLfW3Pv3HrRGerZ/5RP84HcWC3HR9Bd+5SrhkoKo46kr8q6KasKBGsgeQ3/H9ffhjPXjXh1uRbe3Did04ihzmErIa7HZCniqYJ4XDqUlQOvK/6/v3W/OnT37917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r/9bf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691Vf8+v5jmyfiriKvAUFsxvGeCVBRU95ZYHtpA0Rm/1+vv3ljr329a0HYny97y72ydVlanck2L2lXSGonxE8tRETHIp9Gkkfg+/cOvfPoim9MbtTEZWfLVLrlanIEl4o6mVzGXa5uCf6n37rWOmWapwtBinqqPG+ORgxVuCVB5Fyfe+vdBpS7nRpKgy1CRKzcK9jax+l/euvHrMmayeYm/hmMjaQSER+WIHS2ocm4+n497699vRmenPjxl6uZMpVeV5JCkzLw1hqvwPeuvdHj291zFQvDFDQs1ZGSAyouoMPozW96z1sDofcX15kKrF1c+Sqf4OtJTmZqirUKkgQXAB/HvfW/Pqhv50b0x2Y3kNgYyh/jNRJUtC2SpUEiRNex1N9B735da6x9EdASbixNNTSUk0FZAmmmuLPIwFzoHvXDrXHo3GB+MG96OpMctNWpE8Y0OY/wBSfgEkf09+63ToQMJ8MsvlpXkq1nhlZiwLMFHJ/Ib+vv1evBehowvwMyAhepyiSVNKg1RAkfgAix/1/fuvefQu7G+F8M02mHDTwhCNNQY0ZRbjg296Pz690L9P8V6zGTpElO8qD0nTElh/r2H497630tcT8ZsjTSzTvRTrEw4PisB/Q3It79177Onij6dbCO9VKDIi3Bj0qbEfQ2/w9+HXsjotfyD6bot54Kvoo2SmWdCrmyAH0H8H37rXWpL8lvjlmeqe2KrPULSiipKiap8wJCG0pNhp9+OOvdWY/BP5OQBqLF1E6/cU1PHTMzlyQ4a1tX+vyPfuvdWMb27zxW2ZZM3lMhBliz+SGlRy8kX5Asp9+69WvRROxvmHJuwNj8RjJaUHVEGMbAAE2DX4v78OvV6K1u/P53MY2pMlTrkqFdljHLG45Hq9+61Xqr7uXHtQ7hx1RS0zeYyg1jqCTbXc6rce/HrfW0t/LH3rtjaPUmIzWZyVIkKU9Gxp5HUFWVPVfke/V60OHVh29/5mmwthyDEYHGJlan1IJqRWk0n8Cy+9EdW+Z6J3uz5/9z9nV1RS7Tx+SxdM1wknjnVP1ekg/wCt9fe6DrQ9egYyeR763vXeXMZitDSEtdpJgBrPA+v49+69XpkqOou2KqrhFZnqmWnka5/cb0gnkE34t791rpfYT4ftm3FZlsiJ5uG8b1DamJ5IUE+/HrdOsuf+GUTkSCgqBDHe0nkYKdIvf6/T37r2OHS56o64w/WlcJXo3V6dr3dmYW/qLn/effutig6Tfy07ShqNmV9M1bFBAKKaNUdVAtbgKW549+615da39V2zldq7qyM+Ix09Y38RY+eBX0kBrggD3rr3RwtqfMXs7EQYeXF4nILUoFaOVVmBjb+pA9768MdHXx/ZXf8A8itu02K3HPkJcY4jUwnzcoy/oAbj6e/YPXujZ9IfFuTF4t6mPHyRVbWZ5XAvrBvzfn/Ye/cevYHR3cD1XLiaOJsl/lg0jTDZPSLfQj8/T37rdepsGxqybIySY8NQReJv29Cjj/Hj6e/der0A/YG2qPGPVvk1VnVzeZ1XTf8Arci3v3Xui97e3RiMFuGNqZErU84ForMEIP5+vvx68D59GkyO4sFuvGGCbGWElK0YDIvLMlhww9+49a6q07u+PdRh8rkOwMfRSg4+UVEdMh0l9J1jQq+99aIHVgPwM+RW3d8UVPsvO0iwV1M6UzwVCAyAxjQQQ/uuet+fVpu4uoNr7gxgakxaytURs4CxRWAP9CR79WnW6A8TTomHZPxDxOVo60GgZQ6my6YxpHPH09+rnrWT1VP2v8UN07Qqaus27BVaE8ziOK5sFPB49761T06IlvjO7i2xDVYncNBUTLdkJdCSCBY6uPx7914cei6VeY25kVlnjovJloATQtGzI9NKDdWuhB+vvfW+j+/A7+YX3x8VN6UWX3Ln5K7ZDVdNH/DfJVStHSRtyrqSR+ni/vXWut5/4Y/OLqj5obEps9tfJ0dLnIkUVWLapRKryKLM8MTeoi/1HvwJ63gdHpgSWNdErBwtgr/Qkf0t/h72aeXWhXh1I/P1/wBh711vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuomQ/4AVv/AFCVP/Wlvfuvdak38gwW/mEfzNuLf8ZDrv8A3a+/da8z1tye/db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917psyWSocTSS5HKV0ONoadC881VJHDEqqLsWkb/D+nu2AOtcT1V78mf5pfT3R81XhMLJDujMqzQqaGUTRpIyWDWivzf+vutTx69QDHVIfefzf+QPySmhoNgZWs23SVEoj5M8Vk13H+2Hv3VhXy6CXG/DrJ9n1cNX3JVyblrJAlQ7yVUpFzYv+o+/dap69OnYPx42J11RU1NtfCsGpEEaCM6yoUfQD/ivv3Xvt6LpL1pktw1kcn2k0QWTTpKrqIDXF7i/+29+J630Yranx/lymMRaumaBdKp5JkVV/pf36vXqeXQuY34w0eMp45RIkokAKpGiXFx9B/X/AFvfs+fWugm3x0xBj5qqR4DHaFyCQOTb/H37rYHRI83seVa+p1RnwCVl1MOLKfzx735dV6KB8ouvZ6jZGRfHyaCKJiQv+C34N/euvdFD+E2Xp8Bm5MflI/PIteYyT6j/AJyx/V7917h1eni6/BU+Ogq0qIIQUVwrsBbm4uPfut1HRnOvO/1w2OXGY8q4lHjDAG30sSCtr+/Ux17j0LWytvT9hblpspXS3haoifQ3B9TXsAfp798+tjq4/r3b2MwWyPtYKcfcmnsjAAknT9R7qc8evY6QrY+aOrllqKlIWLMdDkAkX/TY+/de8uklv3bj1O2auuRGYKk5LKoKsdP6h72MdawetaP+YpTR1OJykNWgjWIG5dFtbQbfX3v7OvdatTSVWI3/AEc+JqlaNMrJJIFc2AEp/C/T3rrfW1J8Cd3Rbk2ZiqCXJRx1HhpFkUvyPUAeT73WmetdbDXV22sF/DMbCRGssoAMx0kSEgeq/vXn1vgMcfXo22DxrYiCangrIhEadgGAUX9HAuPeuvGhz5dJjBfw/GLlK6vKNOJnZXIBv6uLX/r795U61XoEu4OxZKKGnakv45CNWgMOL8D3sDrfRaMt2tj6WL7+rrI42Ul2jdvV6fr9ffsdez59JKu+WO2I5aanTKUyNGAlmJUkjg/n/ePfqY63Xpyg+RlFkoi9DkEYkEFUJJPFwBbn36nWuhz6772MdJ+9UqjFhw5I+vN7H3rPl1ugOeh4p+1KHO0E0RrYY5Cpu1+Ax4P/ABv344618+nzCw0mYpPEclDVauBGrBiAeSD/AK39fe+HXqefUGu2DSxTfd0rBZfrcBfz+Cv4964dbr5dMs2JamjfzGxBN2bSL2/Pv3Dr1ei17hxNTV7nUwv5kEpJCc8av7VvewcdaI9M9HA2PBS0G1/XIqMIG1xufUCF5HvXXic9EV77zWPhfImnVfKRb0jkH88D6+7de+fRPNoVbvlY6xqrRoqlkdGa5Kq3K2P5t/h79WvWh0Evz27qpjs+Pb2OpWk10skUkqKbajAQWJH596HWz6nrUd3XnKgb5y0XiZnfz6FJJLXkN9I978+tdbRf8oDYUOe2zjanNU7IhejcJIACtze5J97OM9aAr1sT9jYnDYXbCRxTRQRRRlFUhPUNH9B7rTPVvPHDokWNq8BunI5DDLQLJLArqlTpQrqJtdSef8b+99azx6SGc6mR6Srjq5VlppfMFh0pYBwQAePfut/LqvTffUdX1NvGl3PhI3o4KicTSsLAWLayboB/vPv3Wuio/MD5YT122qbauMqPu81VEUTpDJJ5EJOi5sfz79w60Tnom2xvjhvfcsa7lyGQniatC1SUz1EitaT1aeT791uvS2zGz+1+p6mkzGArKsw07eSRVlmYcEEg6T7916nVlXxl+cdVjKCCg3jE00oEdPM8qy+rSoVlBf8Ar/j79TrVT0JXd3cGxtz4+XJYShipHmEks5Cx8lo7En37rfy6oa7ymxON3vjd3bYr4KHc1JNJUUzxymF5JvIGFhEQffutfLrfy/ks9i727C+I+3KzeqVUlZTQ0aQVs3kKyR+GwQPLyT+fz791vq4H37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/19/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcNfIX+2Rewvb/b+90HWqn8+o9TVLAUjHMsptGv4v9Bc+/AdaJpw65tI8aIGGqVyBZVbTc/Xkf09+69n8+ucok8bmLT5dPo1X03/AKG3v3WyPMdFQ+X3yBoOhuodybiXI0EO4IcbI1DTy1sUMom08FUJ1cn6ce6nrY60OeyvkDvLu/tnNdkbvqamqk/ieQpYKSaT7mlEQmIQqDxyPp731rz6eqCp3JueSGgoKYwwTr9IYyo0sdP0HvfXunfI/G/K4/wZeuaeUVDBijozgaubAH6f7b3rrfnjpWN1LRVOFNKaSQPotfwtct/Un377evU6Ix3R1Xl9q1DyUglEUjAhURg3J4AUe/Y69w6M18Veqq/PQU1RU0jazIhLSwargqLcsPfj14dW/bB6wqsVBClNQvLOYgi6ICyj8C4X/effqdbwDXpD961uX+PuFqtzZx6OOoq4zNQRMwifU6llBRv96Hv32dePz6qx3V85u2uzKGp29jIXp4ZPPS+WCTTdQdCt6APxyB791qvp0FXW/WWWyWbky254TWT1U/3Ek9TGZSrs1/1vz7359a6sG6xxH8IrIayKCKOPHOXhCoF8hXkDT70erDPVvfx6TBduoMTPS0MGVgiQEzxqgsq8f7H34nz690ZLFdMYGj3RDi9xrSiF3Oh6ZQUshsAbe9V8+vY8unDP43bOEyFVipfthi4LLTONPqX6DVb3rrYrTHT3gt49f42ibGxfaByDdwUD3+nBPv3DPWqV6CffvZeJ25ODjpKQrLYl2dbrc3Iv/T3sdePp0HOR+TsMePNMZaBG06HbyIvA/J/1vfh17oDsv8kNvwO8VZXUmhmZ201KHgnkcH3unp16vn0Xzs35AdeVtK9HR5CF6uoUgKlVGdJt+QOfeuveVOqhflrNsbcu2K9S1PJkXgnN1ZGcljxZh731XPHqmPrfdWf6z3xVLjRUfZvVlUIBX039NiPfut44dHbo917w3XUU8qHIVBqmuAdcsY4vY39++fWuhaw2yd8ZGoREx7hwEZpHpSF0sL8G3vfXh0KtH1PmKhEkq4NToCrosTW1fpJ0j8D3r5dbz0Ane3xor3wFdl8TQlqtqctf7UsVI54P+v7917ornVPdu/uv6dNhZeeupYhVmmRUlkiFr6LhR78D1ofLq3j4ybJp8/FFldwzSTvW3qEattJcE6hYv78et06tD2xitr4ZIIKGkoZZBpDBYEJF1t/ZHv3W64p0NuK27hsgqzvQxRsFuLQAAE88e/Y49a+XTNuHDJSSaaai1x3JOiK9iOQbj37rdek5XZjB7dioKqsE8cyyhjGNSr9edXvXXulHl+8NppjFinanSBYrEsy3vp+hPvfXqAcOiRd4/JbYO2sHkpqCrhWtEZMKiZfU2n8fn34V60adVH5jcvbHyJyTY/Hx1ZwDTyKJIw8imN3uLEf4e/de6Mn1d8NMeKeIZbGPJUOEeWSpoi3qB5Yu3v2OvU9ej5de/D7bVS9NSDD06/2VeWhUAD+oJHvfHr2OjvbP+P8ANsJIaejx9HJSjQRopVBsB9RYHke9db6MHi8T/DaZYYaKzMo1fskICxufoPejTrwqOlBVYjxUaVJVLmzOvJ+v+A/r79U9bNOge3Hvmgwc0wkMMcoR+NYQ3Uf097p1Un8uq7e8e6sXWZFsBKJGNc5VGgufqNP6h7916oHRSchjt1bUqI9w7fjnqaOZi8iyKzkKTe4HPveOvZ6Nj0l8g+u3MOK3rDPBkmEURaRSiCW9udVuL/ke9Y60OjRb8n6zyW3ZGqqrGPFWxMaJPPEWKFOPIpPvx6t1UxLkqPortT+/OKkIxE9Y0k6UjhYxGJLk+j/D3sDHWuHV2fx5+dvWe/6bG4fHZCAZgU0UTR1dUotIfSeGP196pXI68KcD0bXcW/dvxUjVOWlpGjqBctFKjAjTf6j3UjqwwK9E4392X1/F981qWeGSOZSXCuFv+Cf6+7da8+qju9YerN2jJLTJTitledkA0XF1NrAc+/Z60aeXVTu1+tp37JqaKNGajkqykWpP2wGk4/2Hv2OtdHc3L8d6mTAqaWKEaqa50xLxqW11H9ffuvU9Og/6B707m+Dva+J3DsLIZKejjqozPQTVTR4/1ShpNcR9P+wt7317/D1u8/y9f5kezvl5gxjctX4yg3vRxotdSJUxIrVCxgyIikj6n8e9db+zq1XUpXVcFSLhgeCD9Dcfj37rR6xySaXiS1xKSt+eLC/4+nvY4V8+tE5p5dcrOGFtIjAH1Jv/AI+94Izx69mvy65KwYXHIuR/sQbH3Xh1sGvXL37rfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3USv/wCANb/1CVP/AFpb37r3WpL/ACDP+3hH8zX63HYdcD/59feuveZ625fe+vde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691wkcouoKW/wAB72OPWiaCvXXkSwZjp/1z/wAU497oetah1y1rcC/J+lrm/wDtveqGlet6hWnXgwJKg8r9RYj6/T6+9deqK06xyzJDG0k0kMKi5DyyBIwBzdma34+vveOvZp0U7vj5p9C/HmkFTv7dtBHOV/Yp6Oqp52eQ8aCVbjn+o96r6dep69ayPzf/AJrfcPf26ajq3o6KeDYlXJLBJkqOUxzssjaVYyRC59P+PvwHW6+nRc+p/jxn87XUWb3XWZLJZKs0SVH3xlqEWUG51F7+/deoOrPeuOgafGmGraigCRWbUKdFHH4B496r1unr0sd51tHtB3qz4kjhpnUkMFA0r/rj3v59e8+iXw9mYzP5/IwVDRTL5yylnWQL6rf48e/ccde6EDamDpMhl2qIooTCDqUBARY+q4/Hv2eveVejCjEzV2L+xx8MQlvcFFH44+g+nv3WuuONpK/GzU8WWYCOBrHU1h/Sze/fZ1s8ei0977txorZKOleJSqMGCuCD+LkD6j37rXRLc8orKCoaEwCR2JPKXP8AiLfj34de4dFf7K24uT2fnoqhfI4pZAPSWF9PNve+tdUlbV3BNsDsyspVLRg5CVhb0gfuHm3vXn1vj1Ypiu2o8jioIJq2QFkjAAmIHJtawPv3Wq+nVinQFLR56ixBgleaR3AYt+5yRz9ffutjPVr238GNn4KkysIN41hkYjj9PJ+nv3XujQbN+RmCjgoKapqFvTRhJlMgte/OpT71TrZ9ehYg3t15vCoSUVkUdQxF18yrz9TbSQPfqHr1fLp33vm8RQbOqKSleKWGSKQAhgTbTx/sffs1691rifzJtp0+a2NuCvorBxTMzFOGDeMk2/p7359VPDrUGwNJHitx15q/I7PW1aoZFLC/lYC1/p791bq+3+XbR5moipZQ9THAHgZQzOFAElxx/T37qv2dbMPX2+IqahpKeoqDGaNQpdpNAJA5sffvn1Y9DRUd94WhpFimylMCg0m9Wt+P63PutM9bHDoBOy/lltXBUcmjKwFgGZlWrQAnTfm597p1qvy6IL2x/MF2XBj1p2yNOZUAF/voyb397p16ueqt+3fnkuQqamLD1jlWEoAjqtX14AFr+/UHVak9EKz/AMpN+zZf+IU9RkfAkgbSJnBIv/T6e/dep0YHqP5yZjGViDMTVQjVrnXKQRb6k3/3n37r3n0fTaHz823WyQRSV7RNZLgVQ5b6c29+p1utMdGW2p8zsP8AcRI+XYQT2sorFAsW/P8AsPfvPrxIp0fHp75VbOkqi8GU1SMoGl6u6klfwpPvx690aGi+Tm2HbRNWxtIbBAKgcgnn36nXsHHUrN93bfyeMmaCqivp9J8wDFv6i3vRHW+Geg76/wB50VTnZKypYSwTOVS51fU2v/xv3sgHrwr0cDHHGyYWauMrxx6JCFvp402HH0t715dezXqtruzKY/73NSB3ZYgWHN7AAt+fe/LPXiT0RnC7yp1ytW6ytHEvlGrVx/S59+p1qo6Jt8te1MJRYPINVzwyOol8YMgZxeMj6e/de+zqgjY8E/ZfcEAxtO89PNXyKzLEZFA8v0JH+v7917rdj+A/XMex+t8VMsQp6gUtGxIj8ZuqXIP09+J62M56Otvyprt2zU+JWWZVchSS50gEWvYe9DhnrxpWg6ZtldZ0u1KyeoqdJWQlncn1MD/j78et+eesu6ocIGacVCRU0YZpC8gjUaRc/qNvexwz1VuPVRHz1+RezNobcqaKhraFq6KNUXRUoW1aSAQBz7917h1T18UOpdz/ACS7XfN1SSVeNTIl4lnQzRFC+oEX9+611saYP4pY3D4ehaWj0tBRaWVKUhA0acA/7b36vVqUyOid9m7WhxdTmaKuoY0oYdS6poNKBAfqL/4e/Zp17oiG7cj13gKOSaKpiimjmkc+CRQQyv8AQW5/PvfVcVr0Dmf7Rk3DQvQbaWsmCoyahrcfTT9R7116p6KTVdcZzPb0xeZyqV3ixs7SSxur+MjXqIYe/dbPDPW3p8GP5w3x8+MHRGD643s8MFTi4aWGLRMkTeVIxG+pDa97e/de4jHRycR/Py+NEeWg/j+Uhgw1UyGKdaiG6xSvZSSLC4B9+60fl1b/ANEfJzqP5F7dp9x9a7lospSVKq0cP3NO1QdSBxaONifz/T34db6MFf8AH5/2Nve/n1qp4efXTA2JX9Vja5Nr24v79Xy69TNeuMblhZhaRba1/of6j/D34jz68D5HrJ711vr3v3XusE7TKq+FA5LqGubWX8n3sdVatMdZ/eurde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//0N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYGnQSeG9pSLqD+QfofexTqpr1GnqYaaCaqyIhpoaYazPI66QoFy1z9P9b375Dr1MZ6Kjv8A+b3x/wCu6qsxub3pQxZCmilYRGaEXkTjR+r+vvVQerAEY6qS+QX87Ruu8lVU2wNupuuiBljiqYKeOS11sjqV/obEe/fZ17rXm+Tfyt7++WO5arM5TM5jDYOpfyHCuJFi8ayF/FpBsAR7916pPTF1X09kc4IfLQ6YWeNibka5D+om/wDX349ayej8bE6tg2i1PVV+ORY404c8heNXF/8AW9+rXPVugp70+Te0uu6ymoskYVgldI49YjcLzoAB/wCJ9661UjpOUfyn6trcOzitpUqHh1KnoDXI+nv3DrwpxPVePf3ymwa7lpIxBHUY/wA0ep9AKaQwBJv/AIfX3vr3Hqxr4r/IzrTcmFxOOxr0VPkleJGji8aMXKcHjn/b+/deHp1dJtfu7q3qbrGp3puqeiM8VJM0K1JhY6lQsv1P9fp70cnrfWuR8yvkhu75p9hGg2ws9DtHAVqWlgUrBPBE9reji1h73w6r0luvetv4Q60lLjo52OkmcL6i7Dm5t/X37r329G02ptitgSLHT0HjaSzLJZjwf8fe+t+fQ74PbFRFPTTU0Bk+0IaaHU37oH4I/wCK+9dbx5dT9+fILMdM0B3FtqOXG5KGLVLBSqQzLCPzb+vPvXXj1K6a/mrDdNFVnckjJlaJmiWSdUEga5Fl1H3unWq9Pmf+d1JnKmqhgrUnnqiVUXjOkkcXN+Pr79nr3SRqflDBjqEyVeX+3qHOq7FNQDfhTf6e/Z690A2//mHiKSMrV7iLu/MYLIx9Q4/Pv1M569Xoq+c+TO9N0zpQbagnq1rGMEMyI3LE2BBX37HWun7E9d9tZCP+Mbjra2jgqI9aBhIFHkFwLH3vrdOkpN8beycjWVGWp8rWzQFi8ZLyD03vx/xHvR61TpAZL4tbxz+RaPL5Gr+3ZiriR5FXSD9Pe+t/Z0pMZ8DtupJDVyT+WrunBZuWvckD3rHn16h6Nv1t8ZqXaYghbDw1YlYBJCCSt/oBcce/cRjrVOjn0HTWNrcNHQUuIgiyaKAzpxIRb8n68+9efVqYp0Du/tjVvVkL5ufG+akiP7itcra1zfV73x615dISLsXrnee3a6KuajgnMDAwHxKVIHI0/X37r2OqJPk0mH2p2PDncVRQ1FJFkmk9KXXR5L3sOPfuvdGe218vUx+3cN/CqX7f7OhjSQRxMpBC/UD3759ez0N/Sv8AMewFHuePH7imRR59J86oOBx/b9+69nq9PpT5Ddd9q0FImMrKZXmghPpaIeqQW+gP9ffq4+fXv8HRpYcVhaClnlZUrvLqKFtLkAi/APvWet0HRX+3dqUWSpJ64KkHhjaREUEAEH+g97611Rf8te/cp14tXR0kbCngeZfOq8KEXkG/v3l141r1WLiO5m773Tjts1GSaCKplWKZ9PNlbSeVPH+PvwPXj1sb/Ejo3b209s4mnoKSHKzFad5pSfXpZAT/AI/X37h1oD16saqtv4Kgx0aw4ynhqbLdFADfWx9+x5dbPoOhDxMdHV4qCLF0yJkI0AvFpVg9vyfr70et48uhX2dkqrGwx0u4KQOzEeuWxOn6WBPv2K9eo1Ohirqbb0m36melihepKF1UAK/04AHvWevZ8uiwZivrsU1S9YjpTOCUUgm3J9NvdgK9eNR0Q/t/H5/cuVkmwsUhiJZCY42NyxsRx791qhNOgbpOmZ5K6lq9xY5ZJrqyyTghlP4tce/ZB63ToR4evIqMvDLjxUUky6IYz+j1Djj3rh17j0j9ydD7ZqYpKiejpsTVOrMsqqA6lvoQf8P6+9/Lr1KdEf7X6P7DxuSp2w+9MnV4tnFo1kkZKeM82X6/Qe99ap1ng6lyVTt2ODOyy5ISjQklQHbWxFrHXfj3rr3QG5frTMdT5M7xx2TkwUlM/kjgpw6qyIdY9I/4j37r2R0aLqD5e1u9fBtvdmWmgo4v2nrqjgWX06gW+n9ffqdeB8j0bKi2/wBMb089HH2ZE1Q8Oox+WwDyC7KOeffqenW+I6I33l0bhNrZaXKYHcz19Krl2ZJLrYC4JsffutEdEw3NuSh2DPT5inCzVIYSsyjU19VzqP8AX3v7OtdCftj5VZTL47RLj2eMReMXTgWFvp7117I6C7de+KTdctVDkqFKZp2LLOU0+M29Nh79Xr2OkT0x8i99/F/sFd17Ky1ZTUuOqTXTx0gf/LQpvoe31uBb37rZ+XW7R/LJ/m2defLPbeM2lu3JU+L3vBR0tOaWcxxTTSkaLSJcHn3vh14549XgUwOi/kWWJrNC62I0Eccj/ePfietAY65oW1urElfqpsLf4jj3s9VU56ze69X697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917qJX/wDAGt/6hKn/AK0t7917rUl/kGf9vCP5mptb/jIlf/7tffutVNTXrbl9+631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYvIvi8j+lQCTf8Ae9061Xz67ikWVA6cqbgf7Dj34inXga56Afub5G9VdJ4Oty+9d2UGJalheVI2miLu8fJjIJ96r1unWsp83P50O9t61lb1x0RjqqbEzzSUz7lxsBvCki+PyeRP+K+/der1Vnjuu+0+28nQZrtPfOUztLkpwTS5EyukKu2sWEh4tf37h17o8nWPSuB2tnaXG0uGppqDxRO+VVLHWovy9v+J96+3rw6sr6s2RjMjTtS0UUTlLhWX9SWPPP4/r78TTrYAGT0PObMOycG0EgUyGPgs1z9LauffuPXifXqrX5P9pwY/b2RJqSk96kBbrzdSP8AYj/D3vHDrXzPVcnRG4q/cmayk7uxjMzFX5Hp1/j/AJH79jr1anq2brbDKtDBL91eSQKLA2/s3sQOffh14jGejW7Sw0ePhavqJQiqjMLj8Wufr70et+XRde7OzsXt1KyZKoAx83Gj8g8e/fZ14+vVL3bfyBhyG4anw1wJ1uoA0/hrD8+99VOcdIDDb9yG4KyOCKtcI5BYAAfX6j/W/r731vz6VnZO99v7U2RlRkKyM1ElIw9bISXKfQA+9da6o2pcTT7+7JrMjRvdTVz203FyZCQePfqderjo1e2+nt2VtbRUlFTSywM0YLWccah9T791uuOrx/id1TX4Kkxa1txMjIfGxa4Ogfk8+/H5deA9erYaHatVlaGLFVaskDwhQSfTyLX596r1umc9IrL/ABwqRP5qKpZUlu2mN2sD+r6f7z79Xr1B13ieitz4quo56bL1YJmUGMSMQw1cAge9168R59GM3FtHM4XabNlWcxrTsSz6ixIX9Rv718+tk1HVA/z237NT7ezeJx9OKqJ4yk3HAuhXn3Y9VPWqvmJrb0p0kpFiSXKOCAtv1Sknn3Xr3Ww78K66Hbm2MfVUVEkhFLE9lAYNYaidI9769jo3fZ3yQ3XisbPTbfwjGp0WtFEdWu3HIHv3Xq+fRDs53D8j855p6XCV4hZ5NJWKWwuTYce/Z69notHZOf8AkPk6Wc1mPyEYsblkktfT/gPfutY6JhU7K7Y3VkxDkjUojSG6FZfrq/I9+6359G56s+EeZ3QsFVWrI7uYidbSAnV9R/T377evdG5i/lzpPQprpiPSL/5w/Qfn37A69npBZ/8Al8Pikd6SjUs4I51tY3+gsPfh14joOk+GmdwEjzpTMQl2DBHuCObXt79jr3QUbw687R2RU/dQUVTJSRuCp0yW0Dkf8i9+61XqNt35Jbz2LVU71QnhPlAcGI2sODfV/t/e+vY6Ovsr5ktW0dPV1FTeZQl4wi6yf8b+9Y63U9DbB8xBPLQ0gkaMVBAayi4HHLD37r3Dqzj4+7yh3FQY6sSTya5YZLMPrqW5/wBh+Pfut9WOVm4Yv4JT0QPgMlMVKrYXJT6G3vQHW+iN9i7NrM62WWFS4nBBA4JH9R72etdVm9t1tD1DicnLXSrDULDVSIG0F9Yu1jf+nvwz1o9a83eHcm4e0twZjFUpkamNcY4yA2loy2m1x/X6e/dap59WAfy8/jAqZOkzuRx6GSSZpgSD+pgG+re/cOt8eHW4H0D1tTSbLpYdP24hpYheNiOEWxvf3o9bp+3qLvCXE7byzQw1SvVwsVRQ12LXtyT79Tr3QfZPfORp0qarIh4qKFdckrgBRGOCeePe+teXVHfzt/mHQ7Iyc+zdmVr1VZUfc0/+Sp5WWRhpAYR/Tn37rx6o5zO2+8/kjmjlMq9f/DahlfxyLONKiS/I/wBb37r3WxH/AC3un6DrOkxMFWBBWNJTCQ83voAb68k39+88de8utkPH7RxNRteEiOOVpaYnV6SdJW9wT9PdRx62T1Ur879oUNDtPK0+BoomylXT+NXhF3LMpFyV5+vu3l1454da19P8Ue2N0Z41ORasShlykpETecARSSXCn/Dm3v3WqdWR9b/FGj2RQ49chiYZpJKdGkdwSWYrySG/4n36vXgPXqL2N0ti8PT11dFi4oUmjJQp+Dbj02+nvwPWvPqsnf8AsfFvVTNPQUk7+aQKkkVyGBsLX/xt79175dBlWbSxZjGLysUFOtVp8DvEP2wv6dH+t7917o5Pwu+bPcPwb7BxNamZyWb6/qMiHlpxGzU1HR6QCGvwFA+nv329e636vhd81+tPl515idy7XzFJJmHoqZ6+gWRPJ5nju5CKTY3+o9+630dsm1/6D6n+nF/e6dar11YD1gXJA5A5I/Hv3y698x1y96631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/0d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdRzUotStMVbW6Fw1vTYe90NK9V1Zp1nvf/Ye9dbrXr1/9j791uvRau/fk91Z8dtu1e4d6Zmjkq6ZJTHjoZ4xXSaE1iMKLn8W97Pp1oDz616/kL/NV3l3NNkMN1carAYichIZn88f7Yutyw0g8W596oetg9U8diVubzGXqMpvPKT5mumaWR5Iqyo9XkYuwNm9+61556REFNBX0j02Pp6hJGKhEd3kf62/tfn3vr2ehu696R3JueBJIS+iK/kTSjMQo+gH196r1sDo4e29hZHZWGiqKqkkp4qbRNI8kYUN4uW/H5/p791vovfyL+ZG3cfipNp4SgtnFX7dJogwbyaNJIIA9+60SOqt8l1xuzuuuiye546uampJjOiFnAYB9a8t791qvS8x/xhTKFJqWmqaZY1EZjMzAMVFjYA/T37r2eHWas+JFDnKiGjyWLmn5KrI2l7W+nLe99eHy6D/ePxn331BHJuPrgVsUmPH3kiRSOBpX1G6p/Qe/Yr1vosk3zV7C7K3OvSe8airjWOeOnmR5qmEt6tD/AJA+n196r1759WedWbcw+FwGPocFjpEmrINE8xXWHawuxcf488+/deGB0ZvAbaqsT9rKKV3kMiMdKj63uC/+Pv3W6efRhqD7ZI4ZJoSagRqLaVDAnggf71718+tefQgYKiqKZjOB40q+dLqLkniw/r/t/fq+XW/n0EvyD2thW2Nm62spSKpsXWFHP0DaDa1+fexjB68aHPWl/wBt9tdhdYds7no6Cvmp8ZJln8KNNMirGSSNNjYX9+6r9vSYpvmP2JiMxT1P8WeUPJdVE85Bt9TdSefwOPfuvVHRkX+ZG58/g4paw1LVZjCAiSe9yP6/n/be/db6VPVdN2H3TnKasqEr/sFkHpEkunSJBa4b/D37rXAY62Kfjd0HiP4LhlaivkKQxPeQKzFvGGNyw/r798utgYz1bBjOnqCp2SK/O06Gjp4RpR4Y+Ci/W1voLe/V8ut549FK3plqDFZJsRt4RLCGWPxIq3IJK6VX+n+w9+Py69TNOmzAbQzeZr1d8XPJCZBciEBbHn62t71Tr3l0ZXA9GQ5LwLT0+mq0LIyhEBXm5Avz7317oRV2gmCjFDVYhzUQDRFIYVILf7AX9+x17rBX5nCbRxs+VrWip6uGJpGWZbPZOGFvfuvHqpr5PfLejzlVV7Wosc9bDJI8LfboSt2GkMSAfz711qvRDNq9Pb/3plnr8LNVUtJVu1qYO2oBm1FbA29760OjWbX+B7boaM71x01UhCEGaONgSeSfX/vfv1PTrYHQl5T+WdgKvFSQ7fofDqja5TxgrYcGwv7914D06qD+T38tXe21qmryu2pK4VdGDMBBL4ybPexCH8+/U69gdAd8ce5+8+gt90+GzOOzD0FNVQU+t5Kh1KwtZvz/AIe/fb16uKjra4+LXydPZWGx38SxVTq8PrZ1diDwDe/9PfjkdeB8+js7t2njd1YGeWnqYaVpINXidl1gsL6fV70MdbJ8x1rQfzMOtjicBmZI6WSQq1WTPH9BZCbi34/p738utdUf/BqDFVfdmPx1aQJTWMCsh5Np7cav959++fXjXrd46b2//d3b1DWY6AsklLS20+oC8I5FvfvPr3AdCZnt3QYumknrqd5Zwj6bX4bT9CP9t79XrxHp1XrvH5hZrqzdzVdpnx7VAYoC7Ii6/oAvv3W+rDepvmn1v2DtSnyGXmo6Wu8cWrykqxbTz/xv3qnl16vn0P8AtPuPAb1gmgwE8EoS6r4WZr2Nr8/19+p1vielp9tRZxTBlEssYszHgG3+Le9Dj149JXObOwuPplqMXTLICxJ0Iri9+fwf9j73U+fXqdN2P27gslGr5WGIOo9PkVEIP+sR79XPWqV6TO9Nr0OMpDWQ0wWkhUyI4W4a3Isfe61wevEUz0CkFTtXeTS0eQq6bHtDqQNUlY/UosAC1vfuvVPUXCdVYrNVszPVQS4ulfS2rxlJUU39JJtb/Y+/de+fQh1nXXX+Ro1xFNQ06vENMcwSMIrj+0Lf7379w61joonfnxxpP7vV2RyM0VbRaZNCoIxoXT6VNr/T+vv1fTrZ6pH3D11LtzsLHUFLVrTYrJ1SoYlkKgK0lrcWtf8APvfVTx6sn2X0xtnb2Fiy1FBJVVs9PF+7FMZAGZBckgn6e6k9bA6QO7tu1jiogq5PJRvIQYy2ttJ5sbn3vrRHROO7tsbShp8dSyeKOSfSoVjpOonhffuvHoR+s/j3jcztKGWgkWFm0yFzpuQ3Ng3vfDr1PI9Bl271hRbMlWSZDP4zpax5Ygeo2/PvXXiKdA3uHaOM3Htd58VSGOaCAyThVCtYDkEjn/b+99eHr0iOh915fond1P2Ls+rqMVlqTIwvUN93MgkippNTgIh+vHvXXvs63wv5VX8xjb3yx6/p9v5vIwRbrw8S0q+ZnWWraL0MAZPr9OPfqdePVxuhy3jL+kerUANRufp9f95Pu+rz6rQcD1K906v1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3USv/wCANb/1CVP/AFpb37r3WpL/ACDLf8OEfzNbf8/Er/zf/l6/j37r2anrbl9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XRIAJP0AJP+sPfuvdY4ZknUsl7A2545HvZFOtA16y+9db66YhQWNyAL8Ak/wCwA9+69011+WxmOx0+Ry1TDjqCnRpKievkWnhjRBqJZ5CBawv72cda+3qmP5ffzZOuet487sDreRc1upValgr6CQzwRu10vGYbfQ/m/vXXqenWvjuvD/If5X52XK9g7mq32vUVU00eMNXNERTTPrClS39P8PfseXW6Y6MH1Z8PdtYoQUdDRtEWVVqHcrKXkU8sS9/979+z175dHlxPxcxlPj4XkkhdKdAUjVIgV4/Nufej6jrfA0PTodj02JjFHDEvhj9JkCgqdP4uP8Pfq+XXqGlehe68yNBtXFVop4CatlbS4sLHn34/PrfEU6C/uHfdXksYdblZKdQXIJuAD+R72DTj1rHEY611Pm13ZNT504mGcqj1LxSDW/6r2J0/4+/dePQufDqkp6vCQV8huaiLW17WGq3JP/FffutDq63pzA0tb9sotLGDH6RYj9PvRp1vJPQ19m7gw+0cNKroEC0kxcudC+iM/wBPehjrdetZP5r/ACnpky+Ww2JyUCTmTQAs0gZeSL2B92+fVT8uqY5ew925PcksklW1QgmeUqryMShbVwb/AE/wt79177Oh1wvdlVgkiq2V4GgTQ1y41EfUn37r3Qb7u7D3d3DkWxdHLN9tJdCyyOVC3sOPfvl17qwP4p/Famo6enymQx8lRNN45WcorkMwuWPBP1Pv3Dr3VuPW3x8p5GhqYKQQxxEK2uJABZvqAR/tvfs9eNOPR4dr9VVO1EocmReDUrH0gXUfUcfX37rfQtbg7KxuGx0bJS6ZogqNIBwNP+P9feqdbrXI6k7U7socnPTUTTU8ssossdwzC7f0+vv1OvdDDPviixE1JUTUQkAkRuEH5/w49+Hy60R0H/c/fOKy9CMEvjpjJTOhDXDG8f49+4de6oM+Y+GhrKCujoF+4lyS2VoyGAuOSb8+9ivWj8utcHtjrXNbcywyEtNP+zUyVGri4/cJt79177erPvgx21Q1NLSYCtqIY5ljhj0EkEc2J59+4jrXDq8XrbrLb29pWeeONj+oN6TrJ5spa/19+6v9vRp6D41YKSjWGmpIlbQDpWKC7G1rjjn/AFvesde6DndfxFWvZ5ZMW5o0JDI1NEARb8G1ve6ivWqenRSt6fDJaSrNXjcQYhckOsMQF7/0Ue/eeOtUHQudb9Uw7MxKz5WWCmCFTpkSEXEf1Jv79Q9boB0cLrjH7U3VhK1Y62gaan9KKqR3NuORb3o9b48ehb210XtzcStDNFTa7GxaGKx/xuR79WvWj6dJPffw8p5IZJaGCOVbSMEiijvY8g8g+/VHXhkU6I3298aqWixVXFXUAcRKVt9vCCpI4uLf7D3v59a6pc+SPxcpZMNLkcTTmnkXyyFVEaNdXNgLe/eXXuHRGdk9A9gZPImDHU+QEUcypqBIW17X49+610ebYXwl7Jz0lJXvPVn7Y+qMN6uDfTYn3v7Ot54Dq7D44db5PYuDx1Dko5jPA0IbVy5ESWPB96889epTo+smNkykNPXeT7WnpYR5FkuuvSOTY+9dWOePRLvkP8jtm9NUVVLkK2kZwBcu35sbAW97r1qgHHrVf+X/AMw5e4t91eG25MJqKV6mK1O8nLO1ha3v3y618+kR8dPjpuXdm4aTLVFLPNSTvDLIpGscyD66vfqdaz1tEfG7491O1cTgp0j+3imaMaCiKVBQcsQPfh1vA6ug2dDHtfaaY9GU1L0wu4tcBl+lx/vre9UHW69FL7ZNBjM7DX1jhmkcO73OlSBfm3vYz148eqlPmz8x6XbOFqtlbUlSozOSheiiSnZ/L5W9APpPv3WsdFE+IP8ALl3f3pln7M7Go66ubIVaV0KVH7oSOofWAolv79jrwB6vx2D/AC8NrbSw8ari4YY4k/tQU6sw03III96rnrYFOnah6Poto5yMYuiYGCQFVREXlG/Gj/eve8deoejFf3rzeKgpqA+SnhEKxgsGFtS6Tyf9f3odbNK9NE/TlHvmaLJZ4x5Cmdg5isrnSDcXBuPfuvYrjoR6b42dbVeORKPDU9PPEqtrNPCCHHBFgv8Ar+/da6Lz2t03S0k6RUcMZSACNdCKCABwOLe/eWetfZ0Ubf3QOXy+HrHeS0KxkohVb8XsPV/h7317qq3sXoOKlrKjyDXIJ5bCy2LK1z9Pfq9ePRUu0enpv4VLUSqaeeAf5PI5AsFHGm1vfuvUpnosdRX/AO4ibbecharhaKSmV0J41ej9Q/P+N/futfZ1Yr/Jy+VWQ+M3ydoNqTV8j7Rr5aKEYyWqqJf+BEmkjx3/AMf6e/de8+vofbU3FS7t29idx0ClKXLU0dVGrC5COPoT/wAT72OtHjjpSahfTcAkXA/r/re9dbqK0679+631737r3XQIIuPp+Dwf9iPfutceu/fut9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//9Lf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XBl51Kq6wLBmH9m9yLj3vrR64a/IWjAZeCGPIIvxcH3ugGetZPVe/za+eXW3xS2hkoa3JRT7plpJo6OkWdBLFOwKLqB5vf8+619Ot4Az1qP9nd0dofKfe9dvrdeVy1NtCKvlnpqM1Dmnkp3N01R3tb/AGHv3W6V4dB5vLs7E7Zo4sLiIYQoUI8yBEmtaxJa1/fh16vQb4ds3uSrjmiimqKKSS+tkaT0tyfV+ffs9e6Op1j1th6mOmr8jTuCqoG1wMosG9RIb8e/Z8uvZ6Phhtz9H9N7dmy2craaKTwCTQJUB9S8en3rr3Dj0Q3vL52be3NQ1eB2RH5aGQzUySIASCfSp1D3Y9aBrnojO1di5Hf+4TuDMQGVJJRN+5Fr41WFv8ffvt691Y91b1VjJ6CqiqaCNEp4tUbfbsoICcXNveurUHTpU7Ww2Fgkn8USBXYDSDc2P9B9Pfj1rh17HSYmrYWp0Ok/rMX5v/X+vv3W+om+qrCYzaW4ZFgo52lxcwKVAQhSU5Khv949+691rKrsKXcvyknzVPjmjp0yjeqmpmKEmXi5Qe/U60OHV/XV23BQYnGrHTA+GL1BkIa9uSQf+J96PXh0ZzGGniWMyU4MjMBYr+km2n6cce99e49CJRbUlrJI8iIQY1VfQqG1v6AD/eT7917p7ysksccEioYTRFWtbSDYcXBtf/W9+69g9EY+ZHfmF291xkIaqrjhrI6SpVx5EQMALfn8/wCHvw68etOzuiuyHce+cmNs0pq5pK0sJYo/MDfgsCnv3XqY6zbT+NO4KxqaTJ0MwkoDrmDUkn9Lm1xb377OtUH29GIw3SuWyktFjcTi5pSk0KOBRSEi3H0Ue/Z63Xq9/wCGvQkmB27TnL4ZUlaEC7UbI3H4BI9+z1r59W+bF2bV7TejyK02ihZ42sEb9JH5/A9+6t099/fKGh2bsqbDxzJAvieO4kRBrK2IN/fqdaqaU6r86g3XX793I+XfyzRmXUhb9xWQPccj/ePfuvD06uV65momxNJBJj6dZhHGCVhBa+iwDcf196z1uo6MfsvblLjqtc3VoiwopJVlKgD63t/rfT3ocOvfZ0ifkR251vsbCyZKpkpY65Iw0Kho0YnTwG/2Pv2Tg9ePr1r+dqfIzc/ce7a3b+yDO8/lngkWJ7xCItYABePp7t1XBPQrdFfGfG5EHK9gUgknltJK81OZGEmrlbkf6/v3W/n0ejZnx/2ZtqrbJ4SliaNW8iRtCVBsL259+4db6GNoqKYGnejhhREC60ite3HHHv3Wq56aqDdeBwuVTCMV11bWAlazeo29Nz9PfjTrw6f67pna+9ZKibJUdLNBUrpu8IkuCNQ596r1unp0BOe/l4dW5/LfxGHC44sX1FloASGPN7297B60QB0ZTqz4wbM6toDSwUtPF5FspWnCG39Bxx/sPeutjoQNydYUkdIK2ColFOBqMasQoUf8F9+r69ex1Q7/ADRo8LRbBzdLSxwyVQpqwWcc6hEeb/197pTrXWqz8JsaKn5NULzzGK1e40Kx0j/Kb24Pv3Xut+XpepoodoYegI8pNLRoHK35MI/J/wB79+690JW8utVr8Y1VDErK6MxVUvwV+o9++3rZFOq1O8vjXU5jH5Cvhoo2KISQae8gPJBQW+vv3WuPVNXZ1TvzrWappcVLl6aKmkmYrEssMdojbn8D37rXA06ND8EfnHT4Dc1Nt7duRnWpeSOOQVM4Fy0lrsHPv3W+HW0BsbO4bsvBR1+MlXTUxIVkiYEm6BgVZfeut/Z0/wAVPFgVOPrrvEWJDMC3F/rf3ry63x4dJTN4rH5CdFxsjRyMQQBwL/Xk+91zjrdMdBtvzPVDYWfAyrFqpIyrSahdrcX1H3v59VJ6pX7O3xlantOPY+Hr5qaolqS145wg/XaxIPB9+618j1ZJ0fiMyNvJRbor4I4njCCV6tUkYfkkk/09+8ut+dOjA/ddW4SlNFUZmFqtUAQrWRsxY/q9QN/fq9ewegR7ByGJ3Fi6zbmPqWqYahZHVjIHGnTwL+/cevdUgfI7Y2Q2/wBnbUpwjCkqKyIPIoIAUyAkh/e+tfb1dF0j0/h6rrejr0kWpmnpIFVXJkKO0QH1918+t8B0VnurqrcG18hVVVTCFw2uWRpBGVAQKW/V/gPe+vceHVUnZmDwXYe9cVjMbWyF6SqUVC+W9tMnJsPp791U8ej64/A4/ZnX1DSUUzmrWKBjocliFUXJ0+/efVugf3hgqHedK33h/dRCQZLE+leeD791o8Og+6g2DRZbcGV25KqPDMZIkV0JHJ08Afj37rWOkx3z8WX2pTTVFBTPFTypJUXEDhCzDURq9+630F3wt+R++viR35tvdK1NVFtPGVUX8QgjlEVKQ041eVPoePrf37rXz6+h/wDF35SdefJ/YeL3ZtHKUc1dJSU8lfj4qiOSWCZ4A0jBVN7XJuPx73XrdM9Gh96691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3USv8A+AFb/wBQlT/1pb37r3WpL/IM/wC3hH8zb/HsSvHP1Nsr9T791ocT1ty+/db697917r3v3Xuve/de697917r3v3Xuve/de697917rDNGZAoDaQDduLgj83HvY60RXrE+inAmUER/Qqg45/Nv9797zSh6rQAgjrDX5Cgw9HUZHIVUVJSRI0801RKkSIoGpjqkIH+w96+XVh69Vz9s/zWPiL1JlcltzPb8ppNwUcUwhoaZ4pRNUJdFj8gNh6v6+9db+3rXa+an83juLuaoyfW2yaWqx2w8vVSRRZbDvoqmo5vQjl4bH6H8e/dar6dFu6X66weNkx25dz1+Ry9XkSJJ5cqrzyksdYDGS/wBCffq9bHVquwsFtPLwRNi0SNRHFwqeO50/kD/eveut5pTobMdhI8HNGsA5a2kj888Xb3utRnr1KHpbmi3NS08lVJqalqFOgBm4B+nA/wB696NOPW6mvQdZHO1sBbGPExZixvoOu/1vc/4e9461VulTsSlr6mKeSSLUpLH1A3t/re9HjnrY9egC78q/4Ri8xKos605OlbC3H4Hv3l1Xz61H/l7vOoyHY5pWdipyzLcvfgy8c+91z1rq174Q0r1+0sfEpN2pha3JBsLFre/HrfyHV3/SNLPh5UQnWE8ZBNwCSv8Aj70QaUHVscOkj8r85USYashLaB9pVfo/A8Rv9Pz78KdaNa9aTXy9ro6LtSvlqq2sCGsBZTIQg/cNv9ce99a6DDYFfPXbhlkoIhPT6F1M41fQckr78OPXulBvrOl8hBiYYdHkdEl0RknUxseAP8fp798uvdWDfG7oijqsVRZlaVXmkQO5eJiblQdVvfutdXa/HXbtJj4oaGpp4k4iVdSW5UWH1/3r37Hn1sdWRbD2R9yAlPCvjk51CPSP1f1Hvx62PU9DJvPCV+PwdDRUkEZIABIABP0/A59669x4dIOLqw7lxUlLkorSPdj+0bDV+Rb6+/V69QDpA4foCmwW76GalnkEnkGkSMVQern9XvdevZp0b/O7R2zi8Kk+aniWeONGIDLc2H0B/wAbe6/Lr1STXqpjvCol3D2fBidteaSJnEK+MFlKsbDlfx73w690ICfDmp3bgEr81Ass/i1J5IWf6rxwf+I9+r17HVbnya+BCTYfJTw4pDLDTzOGjo5D9CQPoPe+tUp1r15Vd1/HLtSzxVdNj0r0jsI3hUKkl2ILW4964de+zrYc+FvySxm9/wC7kMVYDIWVaofcKx/A0m3+8e9463nh1sXbPxuNq8PQZSkOvVHDI5vq/sgkEj3oenXiT0NwgxOVwrww01OFVCJXZFuWC2b1e9de6BObb+CyEmQpJaenIp0I1aQxv+OffqDr1Tx6q4+YSHbG36+TDSNF40qXUREqPRcAce7Dh17qlzrj5u5zrnsRduZDJ1UcFTUqhDVQVLtJ/aDH6H/H37qvV8/SnyZG46OhnoqsvJUCO2mYPfVGCeRf37q1a4Oej9bK7Prqyrip65VeOWJeJHv+vj6+60xnr3HPUjsXZGI3Ti66do4AZQzBSt9Q0m4t73w698uqbu8Oj8vlK4Y3GUQmgaVx6YiUKlzxZf8AinvfWulx0v8AE+LApDkcxiKRaSyPKzUxBJ/tH1D36vXgK9WDdedWddJRVi0NLT/eIrBY/CQuoD8G3NvevLrfA9NuTwu39rTVVXkRDAIUZ4lsEQlfpf37PDreOI6Iv3l8s8bs/D5Wkasiplh+4iieOojVtGgn8H3ugpjrRPr1qv8AzQ+Uk/bOfkwFBlq5hJKqftVGu9nI+gPv3y61SueoHxu+Kbbgmpdw1lPLUtUTRSB54Gka8nPLfm9/fh17z62R/iB8V6bD4eKrr8ZAKdI9as1OytZWuAAR791sU6O9s/eWJpN4VO1JR4IaFxHSgqEQlbKLX9+pXHXhjPRm8puimxtFqEx8ppyUBeyAEcXB/wAPevl1vA6rA+XvfWL2TszcWRyFeiZCBS1FaoRXYmI2sCf9h78MY60c8eqgvhv0vub5R9vSb53fFNU7eTIvLRipRp4fGsmsaW5HvfDPWvOnW1b17vnqfobCYzbdbRxU6wUkEDSQ04RQ8Y5NwPx7r9nVjU9DpJ8k+oM7QpFRZKmRpFFlaeNCvptYg/j37T1rh030OW2Tkm+/paqhndm1j99Ga5N7Wvb3vr2aZ6bd2UuDy9BJMJaVXjuYykkYP+tx79jz69npr2fWpQxmGKqiew9Gqdbf0PBNvevt62esVV2VW4/NnHieNUdghtKoX62B4PvfXgPI9TN0ZLApQffZirpjJLE0qg1KBtWn8i/+9+9cetDHRIt49hzV2QfDYpqc007GMOZltp1WPqHvfDrfHor3Y3Te4a9hk8f9pKHbzOdYaysdRIt/vfvfDj1Xj0V7uHq6HL4daLTClUiaZ9H+rCWJNv8AH377evU8+qu+yesY9slUiiDy6iS3jY/Vr3JPv3WuHQe9M0lF1R3jg+292PLTbZpMhjfPLGpURrBMGkJY8fQH34de6+h/8G/mb8fvkf1jtyk6r3hQV9XicRFT1dDNVQCaKeBQkn55Gr34UHW+PDo+Th2CO7qCp1R+O5DnTcD+v+292AB4dUNRk9Z0aUsQ6BVH5vck/wCsPejTy62CeB65XV9S/UA2b/e7e/ZGevHux1yFhwPx+Pfj69bGMdd+9db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//T3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691xNxduW4/Tx/vHvYocdaOM9Aj8gu6cD0R1puDfubqIYlxdJLPBDIVvNIqFhZSRf6e9Z69QcB1o9/I3t6v+YXcmS31laozbXlrZ46SklVlhJjqTp0qPT791sjz6V1XQ4Xa+ymqITHS0cNN42pxpEblV+oHvXn17oh2ZjTeubnamXTSpJ6nW+lUVuDe/u1eq9H3+PVTsvAR01FmEpplV4gxk0Eiwsxu3vR6t9nRue1uyusNjdeVudxc1GKmGFysaiM3Jj9It791up4nHVQeSye//AJE5yJYTU0+3GlKKArLG8Sv9AR+Le/da6Gai6Y2ltWKLEVNLSmpIjcynTqMpHJGr8/19+49a4dGk6Y65x/8AEaSOR1ipm4tq40FuDb+v+Pv3E9W+3o4+5dw4Xr3bOYaGgiKU1CzmoCgekR8sSPfutHGeiGbN7awnZOTrMZDWRl1rJVaIBQP120/X/iPfut9GCoIMTiqGenaCIySa9MtgCBb6m315/Hv3WuPRb+xNnZPc0dZS0VcwSoieMRo5UPe/Fvp/sPfuvHpBdN/GvA7Wrpc3mKCGSuefUZXsXJLXLBj/AL2ffvs69w6PXtnYGGqiIhMlFHIf2+bX/wBb37rfQn4/YFNhahEWAV6SMoErguNR+mm3v3XsV6MftzZlJRUC1ddCsKePV42/RpIuOD71XyHWxnJ6Kv37l6XCUtVWY5UWOnGpglgCLEm/+9D3vrQyOtRr+Zt31Ubj17YwlWYq2uqp6JqeIMGu7aCfSf8AiPfuqn16Uf8ALm+H9TUYyl3buak+6mqoop/3izFgxvcBxx7916nl1dPB8TMBk/K1JioF86gOALEDR9baR+ffuvU6Mt0b8MNqYCrSvrsJTzi6Pd1BYEG/Fx791unVhe2+o9r46AR0uKgp1SwGlF0/XkgW96J6sB8+hHq9vYqlwVTHVRRxw09OTCTbTcLxa/v2a9eoB1rn/wAwHe2Njr329BVRwSy5R4wBYlgZLfUf7xx73w6rxx0Zb4S7YFPt3BTvSfdCoplZpDcXuLhiffvl14evV3fWOApZCkjUgbQkZIIFx6L396PHrYwvSr7U35jNh7Sra2qZadYYpQWbTpUIhIXT78OvZpXrXE+THf8Aku695QbN2y5qqeaoWCaaFHtAmvTqZlPFve6da+3oynxp+J+N2q8GdyEq/dy+KoeoZg0krONRTU3+PvfHrwwerMdu4DF0sEcZpoWSMaQhH67f2jxyT71w63SvS4ihpJY5IFiWmiXkEcLa1jf/AA9149W8uotJiqNZJBAiVbMrAAWNm/pc+/Enz6129F07P61qZcim7Ef+HnHyKTGbrrCm5BA928+tdGL6ky8dbg6bzSiQhOWNhcqtrC/vxoOvA9Gb21k8ZFH/AJWwjAWw/T9R/Qe69e8+mvc2ToYoZ5UmEh9RjHp+lvexw69mvRZ90dq1OGpKtasMaSNHbnTpsGsDyffqdeFOtZT+an35jczi8vj6F0jlZaxLoACSYyOCp974de617vhLOKf5CYPIVFa0fnqjIQ7EBiZ7+nn37rR9T19Bj48S4vI7Hw8o0Sy/aUV3YAEt4QQQfeifTrYxx6ORhhFU0TRtH5wqFRHwRb6ce9DrZ6DbdOxaWtaapcaYFGqSlLcOByQQfx735U68Mnoh/f3xh2B2Zt/JUqUlJja37WoPmXSjO5/GoD8n3vrX29aovyt6tzXxr7DTIYpWWjTKUo+8AeQrGJxdgeBb341HWutr3+Wd3THv7qPakSz/AOURU0ZqJgtmYLTi9/fuvA+R6turY8ZXY8MxjnlKfrKgNcjn3o9bFeHQN7miOJilnohdh+lo/wCxxyLn3oHr1ei5dp0Qr9rz18NaYa4RF59Bs/8AyEL/AO3928+vcetS75i/KKp6R+QcJpGkrMiZ5EQRr5JS6G4Ci9/futceHQx9NfKj5LfISahwWAxeYxVDV+lMj4541VTxqDD6f63vw68a+XR3Zure49qU8NXlt1ZHKZABJZKaWSZmuy3tzz79jz68a9GM6Z7Fr/JFtfdsX8Pq3AjXITC8gLEADU39R791scc9ZPlL1TWVOBTcuPpDk1o4klp60WJuo1Bhb/iPfs168fl0bH4Q5rJZHrijpMpOxkV0jKSXsmlbC3+t78fXrw6EP5Ebclyu2sjhgGm+7jnK1Iv+3qQ2AI59+63/AIOtfzI/HLNbO39W5yHIzSLNO8ioGbSB5NX09++fVD0bbadHLTUcVRnX10k0SU6NMbJe2njX71XrfSS7G67rccPvsRUOYaseZVVjYK4vYW49762T0AvV284+u9/x12dbQn3epy/+pD/Qk+/da889WO9kdudadw7NG36OSjjyf2OkSoEEhdo7D1D37068OqXex+nKyl3JNi6hWFDkJz4pWa6umq68+/de4cOj3/yqflTub4g9/wD92t17hqJtm7hyUVFR00zMaenik/aYes2sAePfutdb6ezd3YTfO3MXuXb9dBX47J0kFVFNA6uB5Yw+ltBNiP6e/cc9b+XSo9+6914f7b37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691Er/APgBW/8AUJU/9aW9+691qS/yDf8At4T/ADNf6DsSvtzew/ivv3WhxPW3L791vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6DntLs/avUOy8vvfeNfFjcPiaWeeSWZgqs8UZkWIE2+tvfj14CooOtOz5y/wA1/uX5ObyyHVPTFLldnbPoKk0/95aKOdKetg1GN5vOreq/1+vv3WvPqvbAfGvLZfJyZreFWd6ZacvOMnWapJfJJ6ypLXP1Pv2OvcR0dXon424VKpMhuPHQIYWUQ0stgAqtxYMPfut06P8AUnUW2Y8fK8VBC8KofHHdbRWX+zb8+/dbrXp52Zsus+8ipMMHQRyrdI73sG4Pp/p/X37r3Rgs7V47atNTUuZqFirnjXSJityfwR7159b4Dp327uajrIoo6qv8sD2EaMRYD8Wv79SnXscR06y4LEV+W1WjIdD6+L+r8n/Ae9Dr3HPSlosKmJpah6BRMWDaQp4+nP6f6e/HrwIPRKe/cfPW4bOtNFpYUrag/Fhz7txHWjSvWn18ucQIOypHBtpy8ht/ZsJueffutdW3fALctJRbdx8dWwRFpxd3IAW9h+ffuvcMdXs9e12PngWtoqtZdSK1kZCeB/T348KdbHQE/IqQ5CnqNbsY/BUqbkXN0tcWNveqY68aHrU++e3XdBT5DI5mBUM4ZZACbn63/HvZ4da4dEG+PPYL4/PZSiqcf5TGkqhiusegEC1/fuvHhQ9Gm68nxW991WrMZGs4qV0IwUC3ktcAj37r32dXqfHjbtHQ0VHStEsKBQoHptyALj+g9+8qdeHy6O9TUowUlHV0Yspkj1aSD9PqQB791vPVivU/YmEottLJUurVaw/Q6NRf/WJ96PXqY65bg7hDyNO0Jlgpxr50/g/0J9+pTj1vHTU/y62XgcRLNWmCnlhBQ6vHf0fg3P49+p1rone/Pnpt2uzqHB1kXmhkGkoYlJI4NgD73QcD16vp0/UnyTrOwKaipKnIPeokSMqT9Abf429+4Y69jo0fU/T2FzO4KTcFRHDUTGOKX6gnV9fp70TTh17NOjuwQ09I0dEsYjp6a6adQ0AWt+f9591699vTbu7r/bG4cTVCopYZFnpnjudNiW+vFvr7359eFOtY3+Zf/L9TctHltzbcxah4Xqp1MRI5CFlPpH+2t7t8utHIx1Tv/L+w/YXXfd1LtLOUs0FGmSkSN2ErLpNRpW5YW9+61xPW9n0JhJshi6ChMjPE1DSm31AZoR9PdTWtOrmnQ37l2xW7Zw1ZDSAsZ9Z1I1tFxex978+q9FYxm4o8XLmIq2XTO6uCxsTfn6gn34evVjnh1VP82d64qi2zlJKrIAIYawlSRa314APvfWs9ah/f+96QbznzmGlR56OrRkCck2e5J0n3r5daGOri/wCXT8p8fXUVFj89MsdTCdAMosbqlgtmPvfl1rh1sNdZ9s4rKPTVMU6snjjK30j6/T6H/Y+/dbFeI6MnkN+CfEyiOpKKyG2k8WAt71Qnh1sU6D3b2a2xJXNNlJYpJtZIWQoPV/hq9+z5deFOhA3N2pt7EbZmpI6SJUs1nAjvpAuSbf7378AevdF8oflPsPa0Fc6VlMatEOqCyF7jm1r+/U9OvcRnqub5Q/zDdviiyFNT1iQ1CwzaY1CamYH08Kbj3sevWiaig61zPkT8s9x9gTZKOGWWOI1DqvDWdXGm/wBT79w4de+fQW/Gn495PuLc8OVqtTkyvqe7r+l7k/Q+/de62gfjh0Hg9j7do4a6ZNVLT08ml7EnQn6CCL/X37PW8dH52F8jNo7EppsHk6anpaeP/J1kljTTb6Br/wCt70cmvXgcU6Su9Ny9cV+cxG78HmaWOWerjer8TRW0atRDgHkD3vr1PTrj3D33tCiwMlRjczDO8GOclozGQXjT6XB96+XXuP5dao3zh+TOc7X7JpdkYitlajmroI6gLfQyeUIw1A+9/LrVPPrZ0/lidbUO2+o9qWpVWeqSBZJAVudcIJPPvR62Orat79L7Xz+DaKWgpaqpmgYiqexaMutv1f4f196r17qqfvr4g7rwNPX57Z+56hJFHkioYZ5AE4LAAL9T7sOPXiAcnoguK7v+SXVdbLQVmFyuUpKd2UTeOZwwibSLav6+/da6n7g+fXdeLxM1TJtXIrDFe5+2ci4B4I9748etCvDoLcD/ADLe5sjk6elx+1MkzNJYqtLJ/W3It711vy6We6vnZ3LSeTLVWzshC0NO07Oad7EKL349+61XHRQs1/NB7e7JzrbbxGMyLT08ggaBYZkXhtJAt+ffvs63noWMf3r3mlDFXvtfIGZVEjftyhibA+/Y690l8989u9NtpJSVe0srMvqi0mGWwCi3F/x7916vl0GNP84d31ssn8ewVRTNUtqPkjZSmo6SrX9+69WvDoYZ9x7V31tWXNVc0MdUtO0pQldWu1yBf37r2COiMdhdkbfyWIrtpU9PDXP55oliZQbgenVx/vHv3HrXn0mehfkP3B8RNx0u6uuMzW0WFicS1+HoBOwl1zK7qVQ2/rxb37r3X0A/5ZH8wzafyz6pwUebysFLvSmoqdaqnq3SGVmWAFg4c3vcfn37h1vB6tljmjmjEsLpNGwOlo2DK1v6MOPe+tfZ1k+lzbk8n+v+t798uvcBXrwNxf3o468DUV679+631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//1N/j37r3Xvfuvde9+691737r3XvfuvdcWYKCTew/oLn/AG3vwFetE066V1ZdYPp5JP8AS31v7916uK9cgQQCPoQCP9Y+/db6bslmMXh4JKnKZCjoIYoZJ3kq6iKBFjjGp3LSEcD37r3WoD/OF+dVV2L2NQ9NbDrxVbfSuix2cbH1UjwyRA6JGvHwR731rh1Wtsxtr7Rggwz6EghlE6rI1n1yHW2on+vvXz631C7X3YcmkeOxlRGcO62kiR9X1FgvHv3zHXumLrbZctcAtBDIkU1xKxW9wfoxJ9+690IW+cdFsXB1FRGkwqkpXkEiMV9Sjg3Hv3WuiibPze6u3slWYDJZCX+EmrCmJ6hyCmvm4Y8cfj3vr3Hq1nqPa+3tn7dp8ZRiETU8didEZcnSP7R9662OgW31WLJvMCViI/PZV1H8H+nvwHXj8ujVdfVXi/hjQhjH4F/FtP4uSPfuvfLoyuVxOH3RtzIYnJxl/wCI0328bMDYalt+ffut9a6XyGlzfxB7t/vLA8r7Nnr3eos7iFLy3Juv+v791r7OrOuh+99j977axtdhaqmaeSmDyqsmo3K2Nyf8ffqde6Mgdv0NO0H2wWokUhn0BXstrn37rdOhRwu3MNm1SH7YRkKuq8aqdQHJ4+nv3Xv5dL6LZ2OpKikGkWgYAf4A8XNv949+68ehqxEOHpIfJUw+YRoGiVVBKsouDxz71k469gcekPvHdVXIkpirIqehiVx4WfxsFA4FuPfuvcfPqrX5O9kVP8Nr6ShdinjIkkDtoC2N21D3vrx61Xd77apewflDSUldKauCHKrM8QcyrzPdro3Hvx615dbZfxO6+2zRbL2/j6ShWP8A3HQqxEKhtQA4uB79XrwpTqyTF7CxOPpKeSngTWygy/tqxP5Pv3W8noetmbYoqymjhp6cayy3OhAQCOfdTUnrfDoe4tpbYwGMabLvDE2gkCXSg/Sb/wCPv3Ws+fRMu5uy9o4XC7hvWUkMFNQzPCzShAzICQB72AevH0HWqT2G3+nz5LtRJN5MPR5CRwwkLRMVk/oePe+q9Xq/F3EUuHxlNhIadg9AhhjcIACEsnpPv3VuA6tMjjO09oLnxkKaFhAkkqM4DaAOQR71nr1R1R98zfktmd+7hl672pkEf7mpMEwp6htRL+h+F/3j3sDHWq+XS8+MHxWwmHxlJuXN0EtRlKmLyyyyxq95G9XLPf8A3v37PXurCsfgKajSlhWACljaNURUC6QOABb349eFD0Z/aW0cNmKGJo4ikwjvyovcfk+9dbzXoPN7x0GENXTiaNfCpD/RWFjyCB79mvW646BLYfaW3Kveb7dWupUqFcLZ5bc/7H6/4+/HrQ+fRkd24DFZbCvFUVFPOlQNQC6Sfp9bfX3rgcdeGcHoMdqYmjxRnpaMhEpVawBPA+v1HvZz1YYHSU3t2F/d2nlqaivhgSHXcNIUGlf9j79w61U9EK7b+eu2NnrJSSZmiWVfSNdUeWAI+nvdAOtVxQ9VFfIP+ZNl8rT11Bt/NUw1RyqpWrbTbVYaSD/T37h1o8OqR+5O2N0dlVNXUZiokyJmmkZYIZmlZi/H096r14Y6Drp3Zu6KHeGO3JjcTkKIUkgZZJI/Gtg+om4/Hv3XutrT4l/Mn+6G2MZjNxUNXWzJFTRtpaRrBI9J4H9Peznr1R1Y1iPnPt6lhhmhw9aFkKgiz3Go2tY/096p1uvRrtmd14bsbHwVFJQSRmdf3VYHUeBxz+ffqenW+FOnzN7ewuTpZWSnankaNizSjTqvz9PfgevEDqj/APmTfHrC7r6+ztdDRrLWwuZIJYoULqY01Cz/AF/HvfHqpGOkR/Ji3tSUn3+x66qRZ8RJNTCOWQiQFfTaxP8Ah78eHXhx62ecEtBBRwtNE04kjUgrZhZv6H3rh1456ibh25T1sbPTkQxuCzIwFyAL2968utg+Z6J12zjqfG4PNAEWWlkN73Gnn6/j/H3YV63iuOtMv5G9a4/s35r0dJWI01NBWPIFsGUkyWII/wBb37j1WvWwz8Rth7Y67iwVPBiI4VjspaSljAY2FyDbn37r1MU8+rCN2bLxW8RVTY2nRapae4IgW2oLcW496Hz62KcB1WH39tDcG0aapySI4yFLV3hliUpaOM3/AFL/AIfj3scKdeNOhX6j7xxW+eltxbe3HW0iZKhxskMbVLqzq6xlQSX/AKe/fLr2ePQ1fA6ZZstUYmqq4quh+5cwCNgVuSdNrf7fj34568K06sD7Z29T01I0MlKNM6yMrMmoBbH+170ONOvfb1WF2Zt/AtVtFGYFnBI02S4Zm+o9769wz0COewD/AMKkp56hY6WnDSwk6YzrQXWxv9PevPrRyOmXr7LLnKavpMzUwOlGWipwxDftqdNiTf8AHvZ62PXojPf+3KeTeVHFRkLDNVKv7ZCAoZPwV/3j3v59V6MZs7o8xbegy+GeeOvWmjleVpCVICBjcXt9B711v7eiofIrfGVw1NNFJEZMpjbpDKqHUWA034/1ve+vdEw3Hld45va1NuCmWph3BSJJNjqiIvCy1S8xMZFsfrb3rPWutgH+R/8AzQ+xdt5ai6U+QGZSGnDU1LT1FdWylRE0gSIq03HA+vv3W/l1uqbf3Lgd1Y6HK7dytFlqCdA8dRRVEc6EMLgMYybH37r3Ty0ix6dbBdRCgfkk/wBLe99ar69cm1W9P1uOP6j8j34U8+vGtMdcveut9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdRK//AIA1v/UJU/8AWlvfuvdakv8AIMt/w4R/M15P/MxK/j/yK/ke/daHW3L791vr3v3Xuve/de697917r3v3Xuve/de6xySCIanuE/JAvY/4+9gV6qSQesTVKEftMHcjgD8X+hPvek9eLenTbn85S7awOTz2SOmkxNDNXVTXsBHCmpzf3o9bHCnWoz89Pm3vH5f9xS9Bdez1A64pqqakyklHNKE8kbeNg7Jxf3ofPrfQYYnpHbXX+x6nGQ0YOTpIDC0rxq8jsFuW8h5B/Pv3W6UyegO+NvZNJN3HldkbhRPtKKZvEJhwQGNjZv6e/daB49WE52SA7txr4zQMayLrEIAT68X0/n37rY6Xuc3ZBtyfEUzTRLBXSIrqXCkq3H+xPv1fTrXQnbXzBxeRfJ4wqIzAJFbhlJtc2Pv3W/PPQLdl53Jb23VTySeV1p3sTGx0jnj0j6D/AF/fseXXvt6VW2MLnMhaKlkmpo6S1i7mxI549+68c48+hvwEGWhKieqV5EVVtquTxb6n3o9bHz6W1BnchjMtS08sMj081/J6WKtf/W9+Hy68egC+RGUov4Zm2CACalsq2AIupvf3sde+3rVE+T2xzmN6VlbHCzxJWVMlwgYEhi1/fuqYB6cOgezxtWnnxGptcIMShZSlyp4AA9768ej84H5EdkY6ghh2398pkAA0tIwJI/r715dbzXqRn93d/bvxsskhq7TKxt629LDm/v2OvdEd7k6A7H3pjK2bMU9XVOULeNIC5vb8X+vvx699vVQu8+ld+dWZevy9Pi8hBC5mJLQaBYH8+9de6DTr7uLK7W3rDPVk0xSojEnkk0KX8l2uB/T37r1Or5un/kvj6jBYqRa+n+5KJyJj/qQT/T3brX2dDdkPl8Me8lFU5OmZI4WZNUxH0XhePesV6959Fd3T/Ml3TtnKyUeKyBmhDlVSGqm0qBwv6ffuvcekPW/zIe8s/IKLDUuQmiqbRMVlqHsCf9t79Xrwx0Fm/O9fkRm6WUSDIRQSo8pUzzx8uLn6H37r3RRpe7u2MDk5aqplrllhkGry1EuknVf/AB9+r59b+fVovxG+V1XlK7Gx7qylPCyzqoEtWwsVAB+v5v7914VHW078Ze4cblsTQ5ClydJKgpYVZY5tYtb+o96PW8Ho9mB3RjtxVJiRwxdgHdSSo/rce9fZ17pT1w8cqUccwWAkFrE2sbW+vv3Xuk1vbr3C7sxD4uujp6mGpQq94o5DpddJ+v1+vvwxnr2eij4b+Xt1vT70oNz0WLhhqEqBO8qUkKEkvq+qD3uvXhx6s82ztTb+xMfTrRxL5IYIkJKrcCNNIPHvw690F/bHYVPTRverp4E0MHLWVeE4vf8A3n37BNOvdUbfLX5YbM6poctV/wB4cU1fPE+mNKtUkVzcfQHjke99e61cPkB8t+zO5twV+LxS1s23pZalBUwSyyR6HYgG/wBD798utfPom9R1JurMznKPS1jwXDzkKXufyWJ9+69XPXHbO59xdWbgSXGtU07eU3TW8Wk/S50+/de49XlfFL5q4nF4Wjot2ZCI1wjgGqSpc6fwLkn37rw6sgovl/gMlSq8WboI6XSboaokBbcc39669jouPbHzc2zstJ8jDm6SZUDNGIq0mzA/4H3vr1R5dVy9hfzIu3d/1b4bryCuyGuQ06JRyTyFkk9CMFX37r3T3sDrv5U9gY+oz9emboHrFMhEv3AC+nUQb+/da6K93b1N2Jt6SqrN3/xKd0R2MmqQDg2+rH37r2R1X5UtW5Tc8OIXXHF93BGwYi51SAEEH3rrfW05/LU6Gwo2/jaySmRpJIy8haJCfpf88+99eHV7uI6XwMsUMa012ZEUlIwODxf0+9V6tToqHyw+PFHQYqWXEM0VVJG7KI20EMYrqbpzf3sdV61yu6aj5H7Crq+mwE2SmxserwASTt9GIsD/AK359761w6JluH5E9847F1WPz1PlQGEiSSSSz6FQiz2J/wB696J638ui6bIy8OX7DpMtlD5K6WeKR/I5cq5nB4DfS3+t795169jreY/l/ZwVPVu2IIGTWiQW5sAfCOb+9HrYAOOrfI8i1NtsGWNpZSlxpGr02ubn6+9fb17hw6DCabF53XRVEKpJOCLTIHsRwLX+h9769kdBPv8A6FxFXjxO1PTyLLckLSw3CsdR+g/3v37j17GPn0V3sH41bXm2hXhaOAK3kZm+3i+uk34A9+HHPXjSnRfei/jLsvLZKvip8Yn3tKz+NzTxn1KeCL+9nhTrQ9ehnz3xOhr6mpx+SoDLT1FO0Kg0cQGhxaxsLfn3okdbI6K7WfAXr7qKsr94PioVqpaj7ga6eL6l9X1H0/23vYPp1qnn0LuD2Bh8viIpKaliaJ19ISCEiw4uTb/D37rfQV70+OmKybSzy45Wjs5slLEfr+TcfX37Pn16g6rv+RPQuDxUV6LFyeVRYmOEJciO/Okf1976rjqqjem4d17UyMGAo5pKemqpfAY3kdbLq0kaR9T711vo7vx6+MGI3ZQxbiydOZquoiWdpGAYa2Fybv8A4+/dapmnQibl+N+Po6+opY4U+0ZyJFMcf0J5Nz791ulOg/od09r/ABfybbg6cys2Mn1M88Yq5okb+zxGht9Pe8da4dWj/C7+ev2XtDd2M2t3bkoZcIWpKWeSpqJCC7yaXbU4/wAfeqU4dexXrbz6C+SfWvyH2xTbh2LnsdXGeISNSU9VHLLGCoJIW9zYnn34HrZ6H1ZAhETai4W5axsbC5IPu3HPVAaYPWSORZV1L9Pp9PeiKdWBr136rm9rfi17/wCx9+xTHXs1zw65e9db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/1d/j37r3Xvfuvde9+691737r3XvfuvdYahpVjLQoHkuAFJsCCeT72Pn1og+XSb3VvDbmxcHV7g3PkaTFYqhiaarqJpY4wgC6m0oxF+B/X349aHp1Rn8t/wCdf111vN/AOimp95bggZo5kKxSxhlk8Z9I1f7Dn3ric9b4DHVW3a3z1+TnyV29Ux1MWQ2zBVo0YOPlaAiCddOkiO3Fvfvn17j0Qyl6a3VU5R8lUJNmcxVvrlqsgjSyq45v5Xvz/sfe+t0p0id+7VjwksiZypelyQWxjjfSAw4/PvXXvLpK7L2vks7kEohrqIGeMrJJqYhNX4I/w9+611Zl071BX1Io8Th6JJJ6o+N3MTAKQvJ1f6/vR6t0B/y6pIOvnfaeXVFyU8Zj8aWZrP6eR9ffhSuOtHotHVvVX8KikzVL6JKsLUAAFWAJve3+Pu3Ws9DxQ7gyuAqA1QZfCrckMSun6k3P+8+9db6D/fW5aGpycWXSRyVkDMArW1A3IB/p73w690MOw+9cXFS08EbM80MSpc21Ag8Cw5496+zr1RWnRk8H20+Zp1hBlDMBo03DA/4kfQ+/db6Jl8xejch3RsTKUc1GkrNTVk8dS0fklDePUnPJ9+Fa9ePy612+n/khv74UdnZHZW6ZauLAiuEFLNJqhjETS2UDyfgD/H37rVfTrZN6H+bfVm7sDRZmTOwTVNTHF5Uesg9OqIFibnk/6/v3Xq0OejFy/MfpvCAz/wAcp0ex1KlZACeLkjn36nWwekDW/wAw/q4ViQx52JgZBYfcwsR6vxYn8e/UPXqgdPsX8zHprCApks4i+UaI71ENyxFrWv8A7179SnWq+Y6Qmc+Xu2uwpnfA5Colo5yXQ051HSw/on+Hv1OvV8h0VXvvtrGY3bVWtSz6amHSZZl0MFYG5u/5vz79TrxNOqZegNrUO+/lDkMrjKl6tEq1k0vd0VvITYAe/V60a9baXx1pMjhsTj4Wpf8AMQaAdHIS/H15v718+rdWEbIhyGYrqemijLpOwWbyAhYwRz78etDo9OF2bhtoYL+LVDxiZaYSlCRwwW/A9+4jrfzPRA/kb3IasVUFJXSU9PTF1LJMFUBVP1/p73Tr3Wul8m/kLu/d2Yk2NtiurJiWenneGQvrUvY30n+nvfVT0JXxX+KdRXtBuerWQZeoaCpmlkV1eznnk+9db+3q6Xr3ZVDs2jhWNVaWMEVMmizarc3b36nW8+XSE+S/ZdXt/Y2SWlrZ4jPQSRwIHIUuB9APx79SnWq16qs+JXRme7U7Vq97blDyUCV5mUzByoQSXBF/fuvDPV+20MPQ4qB8NHCiRUo8cVkNrBbcH3rz6sOHQr4zZi1KJNIqCIes+nm31v7117UB0r6fJQYIiCkvqVSnpHp1Eccj37y6qc9F77JwNRVR5SsqZZFNevoIYjTfkD3sEcOtkenVEXa25M11N3EmTpchWLFJXspbWUQfucEEn8fn3uvWjjq33pXf8+9Nj0WSra8vMKQNZpQxuV+vBPvVOtg16hZvs6h2dLkqiuq0hRUudcyrdfweT72a8OtD16oM+evz4yOJqazB7OqJqmpd6iGOOmnVzq5VbCM/7379xGevcOHVT21+vfkL8nqwZWo/iVJRswcao5xdQeOR/X37r3n0O9D/AC4OwMhJTU1WJ5HmkVXLRyFjrHLXt9Pfvt69no6PUf8AKJlaekrsrQiVv2GdZIZWH6rkMre9deoerB6/+WbhcdteP+E4WjWrigs5SlbUSF+nAv73XPXtPTz0h8SsPisk+IymORKqKTSFelY+peAASPz79w69QefRs/8AZUKulq4P9xFL9gWD6hSm+gG9/fsdboSMdGm6/wBo7Q2BSwmtUQtCup0WL0k/Tke9cevEU6FSoyPX+70ako6v7aYoIwQREGc8W9+PoevDV0T35S9E19b1buKpgSCrpDDVyKwPkbQsJJ+nvda9ap5da038uHIVmJ+X++NpQVEtMIMzPG0KsI1N5yLab+/de4dbtu26GlxuAx6VPrlbHwNf9ZJKX+v491p1s8ekfuFMvU6xSqRTc2+q8fn3s8KdeBFeibd6wti9k7prK2TR4MZPISW4FlJ97HXiOtWbrHbH+k/5Y1mYpLTLSZV4yxBawWe3BB9+6r1szdddb0cuKxywxJ9zSxhXKpckhQP+I9+wOrD5dDtR4StwY1aCyyAA3BBHH0JPvRPXgKdAF3l13/e3C1UIpULyJK2oprPKHkEf73795dernPWv18kNqdjdODLrtk10VNkgfIIElVFDNzqtwPe+tcMdW5/yj6Kgq8JQZXd+USLISyQErNUorFyLsrKxv70etjq8HtvER5rHxw4iNJoDHIBMAHsum3BX8e9DHXhnPVSHcvS+cx1XNnU8hjg/dZQHNiCSRY/j3vr2eq8e391bikoRjMZA6zibxMVuTovpbkWsPr731o/LrBsja2Tjxy1YncSPEHqBdh6yt2+n197wOvGvQJ9n4erqty4OJAHlaqRebG3r+t/r71nrXyPVr/UnUubq+tY6kRIS2PQgcg6dHAB5+vv3W/tz1V/8oOpcumTrJWoo9Ec3r/aLcA/QkfX/AB9+HXiDx666+6Q23u/Y0SMlEK6hiMskV1DFkW5AUc/j6e99e8uiqdjdV53BZR8ztRpcPX4+dmSpxwMUx+3N1tIvP4/Pv3Hr329WL/y5f5u/aPx93bjutu1qmsr9v1lR4GqcpOHRYmkCK4M35A+nvVOvE9bs/Unam1O6dmYffW0shTVsFfSwyCOOojlWN5IxIyuiE2+pt79XrRHQuBgbj8j6j3sinXga9d2F9XP0/qbf1+nvXW+vA3vb6D8+9kU60DXh13711vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuolf/wArf8AqEqf+tLe/de61Jf5Bv8A28J/ma/+JEr7fn/l6+/daHn1ty+/db697917r3v3Xuve/de697917r3v3Xuo7liXWVVWDTbXq5/2I92oKdVqa9QWejxcM9fU1ccFFFG0jyzusaRqo1MS7WHvxOKHrwGa+XWvZ/Mg/mvUO0HzvR/UBgy+dzNLPh6yoheGRk8zGF2Ui5H+w96xTHW8g56rO+LPV1ds7bGX3xuKjhn3NuCufLtNMgNQDVP5WCsebC/vXVvl0YrM5PD1uKyBrnEVdOr+OMkWaTTwLH3759ep5dEF3J1fPsvNz9kYyJ0ra1pGLRqdVr8HV9PfuvfMdGD6Z3/lM7pXMq4MNgruLuvN+GPv3Xqjh0sd+Vw3Turb9HSVLCOmqY1mIkHCo4vcD6H8+9der5eXR1KWowWC25BSxVIlqzSxg+pSxOj8+9569jqFs/BPkUqsk0EbfullLKDcXve9vfq9epU9LumyNJReSmUKkzixEdgQfp+Peqdbr69LLa+EyFRKlc2vwGRSSwYixa5BPvx68Mfb0LW7arDYbGQ1SeA1UcdrnQCCF5/x9+88dap1Wp33u5ctR10byxgmNl0q68c2F7e99ax1R13ctDA1c5eFpS1Rp9aliTf37rX2dFP6Z2vBld3TVFUQifcagtwVI13N/fuvcer4+kertt1tDRSyU1PKqiE+pA1zpBJUj37rYp0d+j2FtqHHpDDjaIgIVsYwSOLc8e/cOvVPTHL1Tt+oEiy42maOS/p8RKm/BuPfq9e+fRVO9vhfs3e+DqtGKo1eeJ1ASmNxqNzYgf7H349e+3rXf+V38tiHaMGUzeEpnhnjmkmjMNNJ6Si6wLAfT36np1queqrYOwd3dV18WDlavVqRtB1hkWwbRwH4/F/fvLrdOhGl7cqtxxwyVOQljqZwIQFlXyMX4A0g+9de6P8A/EX4lt3JW09fnIZKmCoKSapYHcaWe19RHveOvdbCfSv8tzrLGRwPU4uhklBQ+uk1E+m/1YGxv7914UPQ47x/l3bCyELR0+LoY1MTLcUx5LD+gHvVadbPRIuyf5TG1sr55IMfAGf1AxQPybWuRbn36vDrVKdEa7b/AJUm/diYT+P7CjqYaikVp08MM4NxyLhfx73x69SnHoGusPlH8gPi/XHbu7KHK/a0cvh8xSVU0wGx/WPpYe9+fWuGerDeov5yWLxWYoqDP1Rp5KxgG8kiJYs9v6+60Hl1bNOrqOpPm51t2bQQVLZ+mSaojj0h62BPU6A2Bv79wyOtceh9j7xwUMfhpcxSTCQhkJyMH6T9De49+zXPW8DpRUvyFxGGjSWoydGATcn+I09gSP7PPv3Xugc7W/mBbF2Xia6omzVM00NPKQqVtO3K/wBnlvfgOvHrXz+U/wDNx3ZuSTK4LYUVZXyvUywRmkPmaz/tqQYvx78eq1I6JP1J8Ue8fmhmJ9w9hVmao8bM7SJSTfcRoULeRR6ve+t9Wh0v8vLp7qHrejG51ooqoyxJJLKiidjcKzMX5N/fuvdM3Yvxe6w2/tSKq2RHBXR1dIZpnEd9JaLVay/X37rRFOPVCXyg2ditnVsk1PAFq52KJGkdyrmQj/X/AN496630XDae2d/Vc4qKSnysNJdWV4aWoPp+oKsB7316o6HbEUXbmRo6jD4ifMLJJeJG8UwZTa1z+ffuvcOPQa4TrrfuU3zHtXsbIZJaSSo0ReZZgkjMeLs/+v8A09+699nWz5/L4/l5df4vDUHYGZoKCqo44KedjWRK1yDqBLOPr/j79gDreknJ6tS7FretNp4psNtfD4gT+MRqsEcNi2nTb0D/AGPvVD17Ax1VL8iepl39gq558TSQ+WOX1LCedTXBBH/Ee99e+3rXQ7g6cpeuu2qCOrRYYaivhchF0LZZARf37rXy62i/5YowOS2t44KoNJDAwRda+kqnAH/Gvfj6deWlOrOq7sx9k5Q0rapA48aC2r1f2bf096p6dbNOB6YN0U+Q7DopMxXxv9uqs6I4Yrpt+B738utfZ0Vio6Kw++q2roZMTRzKAUJem1Hgm5a49+4db446KR8g/gftqp29XUlLhKLzvBUsJEojcOy8WKj/AG3v1etU61uu4/jlWdGdjx5Svinp8atXEQRTyJGqmUG+puPfsde624v5Zdbhs71pt2fH1TS6o4PSpDm6wD+nvR62McOrz4hhsXs+Ssr9ZlWE2BiYm9v8f969+69XFOiX7k3NkajJ1NbhopFgpmuNK2uNV7m3v2K9eFQOhH2JvqLdMH8JyhIlQANcggC1iOffuHWsHI667WwWOotk1r0j3NpWtf8AGgk/7f34ceveVeqxML8jdl9E1+TyWaqmgdC72kZUDG/C3aw97I8ut4p0oKX+ad01nA4lydKk4/bRVqICxZeLLz/vHvVOvVHEdNWS+Ru2u8qWWjx1RM0M7AxGM+RSt9PFve+tY6MB1htvHUmEaBRIzRxNp8it9WF+Lj3r7Ot06dqrC1Ukc3lpgKZS5VyDcj6D/ePdsda6KD2ts3CZX7mnqoozKxfQmi7AspHpt/vPvXn16lRXqgD5p9Qy7TzeHzFJCEjkrFayxn8yf6o+/cevcOrdfgJ1vJvPYNJJ4i1qGAfpLWJj55Hv3XhU9Ch2f0/NicpWxSQmxl0qRHybH+n+Hv3W/PHRRuw+mqeejlqamK91JF4mPJ+n/I/futU6rS7X6nw0+SkoahmxkqyNJFPTx6JhInKFWHP1/wAffutfLoyP8uX5ldxfFH5HdfddzZfMZLY26Mj4pqmuqXWGGneZVKMXNhx799nXh19F3Zm88DvLb2JzeJyVDVR19DSzlYquCZ45JoFkaNtB+ov/ALH37rfn0q7xwj1EKGNxxx/t/djU9VGOsc0ywFG0lvM4Um/+2I9+48evcDUefUr6e69W697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/1t/j37r3Xvfuvde9+691737r3XvfuvdMWcylFt6gyWeyddHSUNDQzTytUOEiQQRmUnn8m30HPvdcdapnrT4/mIfPXszv3sXOdS9WV1VTbfoMm2OrpqQSrBMgvCXLIf6e9fb1vhgdF66R+Jm2KKnjzW7aeKfLuwlqKyZv3HZ/Wb+T/H36vXgOh1bE7cxGdXEU0sdHjIYyTOLeMhPooPHv2K9e6x1O/diUUVdSfxOkpqqAusE7FNR9J5uD9f8AYe/dbOcdVSd0bn/vLvuejjrfNTxysVqvqH9fK3HvwHVTx6Nr8eNr4ysq6BpKlNJjhDSXBAuwUm/v3W+rb8Fn+uukNpZDN1+dpGyRiaekEjoXVzHwEA9662MDqmzsPLy99dvV+461/v6DySx07kehT5LxAX4/p73gda49CZT9e5WkpY0pZGjiVFARW+q/4afx7917z6EDH9WrkcXKawAyCMkyMwBN1+vv1evdBtuPqDCjGPHJVQmokdlWMuoFybfUfn+nv1evUr0l9n/Hx6esE0cZeKdldXDm1i4sLf8AFPfuvU8uj47S6YoMPiYamdAJ9AYDUL3t9D791vp+qdjyZOjnoHQrEY5IwwIA0MLEXP8Ah79x68etVr+dR8a8TisriM/gQs1VFUwy1kaafVx6gdH1+vPv3y6oQD1Ur0BszufNTLT47KVWBxSyFYWQziNgptcXNuf8Pfut0NKHo9tP8Zu3NwCDyb3rSZFFx5pATq4vcH36nW+hy66+DG8hPHVZjcNTUXYkNJM5BP8AsT7917obcp/LtrNxSQVkmbkKwlZdIncLdRcm1/8AD37r3Vnfw5+NewNl0iJu3IUj/ZwaNM7qdZjF7Xf/AFvfvl16nRHP5qO+MJiaKqwPX23UrzZYVqaVQfGC2gvqTj37HXvOnRS/5W3UmYfsSs3JmYJAaupSRdZHp1m+k6vfh17rbn6a23TUdHD92oCMOSx/Grgm/vXW/l0cvFUON25S/wASoqtfJIgZFUglTptzb34563T06S+7+2shSYSoNdkCKNEl1a+FCKLEc/09+pnPWuqKPmX8n8XiKfJYvCZFJK+scRqiKS2pwVv6fdvLrRr0Wz4sdM/3xzdBvfPVAllyFX5ZY3YDSjNqI0t70T1oZ6vEwW26bbkNBS7aiEoEEIkMVmOoAAkBffut9GK2xShxBR5CEo1YpEksnBjYj6n3o9WHRLPnPsWgp9sxfY5uMuqs7wqwubnlQB+P6+9g16rjpq+E+Pan2bMiJ4WSZVM5sNQJ5IP+PvRPW6CnVpmJoMNLT0ZEiNU2Bka9jIRxyB/xX3qnl16vn0s6H777o0kMTGnFlBsbaTxx731rj0qZdn0kNK1dO6h7EkNbhrXtf3odbNR59F73lRvm6sUvl8dLA3LBuNIP9f8AivvfXuPHqqT5vfHCDPYSXP4Os8tfTeWpsjC6uov+PfutEdE1+OPyNquvJZ9o7oya089GDSw08+q8gHpBsfe88evA46AP51fMKbH4bJjC13jlemIVo1b6i4Fip9+69Wop1Up8R9obp+SPcJrtxU02SoPv0kjD6wpV5Of1X9++3rR9B1tsdRdBbe2FtKjho8EtJMtPpYoEDarD6gD34nqw6GraGz5KrKxLJi38YlUCUr+L/Um3v3W6ngOjybX2GlJRwVaO7KkS3hXQALfRb+9HPWqjh0uKSvn838PlwbtAx0vI6XDAcfqt79nrxpx6c8Z1/tlcq+WhoIlq2tJ4wUDBvr9P9f3o9e/ydPdWKtq5KaXHvFTqoUEr6dJt9eP8ffutdQsntHa08Es9fHC0jrfxMR+B/T36vn1vPr0Uff216tKmWXa9MaEIXKSwggagbqSV497r69epXh0FG6987q231nn6bdlRNUUS01WreYMUI8LD6t/vfvePLr1DXrVj+JklS/z537uLbs5NLWZd2WKI39QqDchR78Oq5PW9Z03RVOb2viqzMq2pcdSW8p06rIPqD70et+VemztrKNiIHp8VCzSPdRoA44+g0j/efevt631XL8g5MhU9Tb6rMpK0Crg6xwsn50xn6A+7DGOtGvE9a2fwWz9PP3PuOphkV2XctTAShub/AHJUC49+PWgOtszppIMbjYqzIx2WoQujkA/UXB5+vvx49WoaUHS+3XlY69VFBTsyrY3QfWx+pUe9dbAoM9JCSFaqldqin08Mp1fjjng+9dbFPPolXyF6WwHYG3cxSwwpPkZYyKdVKBlextb/ABv7vXqmeqImT5GfGHsbHSUdXkKbakeZ8sqapiiUom/Omw+nvXXutqH4pfL/AGn2V15i8PX1EE2cONjWZiAJPMwCkNq5v71Qjj1vHHpV75wtVno8nHJHI1JKjCM3Gmx/PPv2a9b8uqsO3OpRi6meeGj8gd5TG6kcsW+huOPe/s6rWmT0AscO4NtUE5qKOaOI8orC402+tgP+I9+68D0Ufdm456re2HLHS61wsoBBFpPpb/H/AFvfuvefV4/RGbzVbsiko2EsVM2PhBkJsDdAL/0+nvx69U9Fl+R+3KGSnyME1YpqKpnI9a6wxW359+635dUzZfcfYnSu46zKQ5GqqsJUPIy0o1snjL6dNgf6e/dV4GnS7wHyGw+/ojja/Gfw6WRD5Kh00lnf6m5Hv3y630WvvfBY+Kqp67B08ddWFkMdQnLQnVquSnPv3z691Yv/AC6P5mnZvxkz+GwW8cpVVe0566npFopUmaGmgACEtrJAFvfuHWqZ63dvjz8oes/kLtbFZfam4KCqydXRR1FTjopFEsbOOQiXv/sPfuvHoxtPHLG0vkcuCw0Fjc2/1vx72TXrwBHWcAgn+h5/wH+FvfjkdaAoeuXvXVuve/de697917r3v3Xuve/de697917r3v3Xuve/de6iV/8AwArf+oSp/wCtLe/de61Jf5Bp/wCxhH8zX/xIlda/1/4uvv3XhxI625ffuvde9+691737r3Xvfuvde9+691737r3XA3LFSoKFeSf63+lve/Lqp49Ud/zfvmhluj9m0nXWxqv/AHO7nhanlmowXekeZSqhniPBsfp+PevPPW/LrVx602TNu3ddHvHetaa7c0+QeaaeRj5f3JPILlzxb349bqePVrmMzM1AKDG/cl6RKKJQpF1sqgAHTxe3vVOtjpP5WGbdeeoqLFx2CS6HsCCfUNRt7317j0MHa2y6TGdd42nqoA0yrH5AwUkXHN/+R+9dep69BVsXbmLjoJhRwrFKVNwhAN7WDf4e9/z690F+Y2nurD56ozFMZnRJDMI1LWABuLj/AIp78evDpW4LP7yqquOqroqlKdWVdLKwUhRe3PvXy6359Hj6735QLhPs5SsdQ6FWBADaiLe/EdaqehI2HtiHL5arrayVZEa5jDG9h9RYn3snrQxnof68fwTbjvTREeOMlbWIFv7R96NOtjj1WX8jfkKu146qnqK8U0iEhQx5/T/QH3vhw6159VDdn/K6augqaeil+5lkBUuoa5INr39++fXq+nVbHYm+945+pnqEjlkjeSRtJD30sbjj37j1rpk6g3NuNN30NCYXiaeVAw9Qv6x9QPfuvEdbNnxzxmSXatDVSK4/biLAEk/5oXPPv1PXrY+XRyNrpUVsy07MTdgD9b8n/H37y6359GGxGyqNIUmrZVjVhcAstzxxbV7rxz1utOn/AC2w8VWYrVDOtwnHK/W35A9+r1r7eHRAO/8AYO1pqGspcpHFUKyzKQ/jsbxkfn/ifdq+fXiKY61ZvnV8UNqVkuTze3pI6epuZFjiKAg6idIKe/fZ1rh1r4TvuHBdqYHaKAlp9yU1Fou+t1M2kAEf7f37rXn1v9fyzOhzguo9v57I45jNNi6aa7gEnUAxNmHv1adWAr1e3sDrqnixlRm5/wDJ4o4fKqsVsLJcH3X59ez+zoAt6957exmcO3FqoEqfuPABZST69IPP9fewPIdeJpx6HfauGpM/iabJEmoSWIOAQtiP8B70cderXpbV3XeOqcNOa3GiameIkREIVta35B9+x14V6p5+YXwp2D2Jt7NZanwsFHVxiucNaJWJEZN7gXHu1etU9OtQfuvoz+5vYVdt7EYdcjWCpWOlkQNqjOuwsV9+690t9p9H/MOmpYptnz5TGUwKvGkT1IULpBQWH+H49+z1rHn0JNVtz+YHhoFmTOZiRoVsAxqzwvveet9MVZuj54SQ/a5HKZZHK+jUKoXI4HJ96z1rHSs2x8XPl93NRKc9lMwY5yRIqyTKCrqSfqffvLrfn1b18AP5US09HJnOwsc+UrIHDypVOHaRkk1Enyc39+qOvUPVzOd6xwPT+Lx8O2cYuIhp/GZxEE4UDSQ4Uf4e9Cnn1v7Oqd/ln3Pkux9/0vVmHqpIVSoQvLGDcWb1XI/3r3vh1o5PRk9jbCrcfsfG4uWNs7NUUIRrAs0bePTY3/23v32db8uq6O6vgtubd2+odwVuBnfEio8jQOQI41MmoGxFvfuq+eerAukvjB1BR7Up8VlduUBr1pUhaRhCSJSum1yPfh1unTu/wgwO2slVZzG4FJ6OqkaWO2iyo/NwALe9V63Sp6rm+d/x5xO0cNgt6YqlSgraasjqKhk069Ecnq1BQOPe+tGlK9Cx1D87aDafRK7Sw+SWpy6R01GIUW7hyBHY254Pv3z68Cejh9RUGY7KxGN3NlqhojUx+UoxILEjVax9+698+hr3T15UZPC1FDHE3FPpSTgc/QWP+9e/UPW69a4H8xDpzNbJyU+5aqOSoSmqfKsjniJUGr+nv3lXqvHh0J38rD5PU2DyVPjKmvQeaaSNovyRqCkC59+62Pn1sWPvTBbsz1CPtY5/JHTuGKjnWA1wR791via9HFoMF99tRKWhxrRRSwryqg34/UDb3XreAanqLtbrOoxRqJ1pXM8wOnSLsC35NvezU9awMjpUV/XeOrMJLBmqdRUsCLyFCSGPIF/8PeuvH16oa/m0/GPbdT1hm85iaWMVdJAkoeMrw0ceu/pF/ex1o9B3/I67cpqenpdnZRUqpKCqkpzEy3/zfoU83/p73nr3r1tqNmtvZjFR4SbDRAz0t1YqtzqH+291rjHWzx6LfldtYmhTNUsOPVWmZhGABweeV97HXiegF2jhanAbtlnmpWSnM2prgkBSfqR795U69xJPWf5EdrbV2rtGqeorYPTTztJA2n9axk2+v+w9+Az14mn59aq/ze+ReA3lFWYDauJhrcpUKsaiFDqaS5Xgr+R738+tdE7+N3xK7Y7GzVHUV2GrKClnyKyBy8yjxTHUPr/r+/fb1rj1s+/Gb4oUXU+Ow65qIVE0lLHcy+oobgn9X+9+/V6tpqOrOTtrB0eFEmOx0Ycw8sguWOj6DT71x60CRjpHV+HkyO25Y4qVo59T+q1vT+Dcf7H3rq/nQ9Ey3ts625KSnmFmkblTxqsLE2926pw6qq/mTbOoqTb+EjiQLIssZAuuprt9Lge/da48erBv5YtTS7e6hhnmpFkmEEKKpUFh+2PqD70etjhnozW86Bt25asm/hRMImLcoAP8dJ97xTHXvOvQXb86wx1Rt1kWhC1LJZI7LdiT9LH639++zreTx6qh7h+O+YrdwPUHEzIoaVlOn+z/AGQByffutceijbu6VrJcjST018bmsSwNBX6tL0kgN1ZSLH688e/daz5dDf1v8xfl/wDE2txe4qremY3ptOmrIJKjGQ/dSpDRQAAgpciwX68D37r2adbY/wDL5/mmdXfMDa9BS19fSYjdcNLDDU0c/wC08lSpETBkfkH8Hj36vW6dW1xT0lWAYJYKkIQR4nSQLf6H0n3vrRPUv3rrfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//9ff49+691737r3Xvfuvde9+691xOq62ta/q/ra3FvfutGvl1r3/AM375lbj24n+y+9aVop905RH+5npKhg3glQLIsjx/Qj+nv1Aet5p1TT1lgdvdXYJs5u+ohrt15uEVVVJMySStWONbWL3PB/N/fuvfZ0GO/fkfmqipnx23dcJsUUozhbg2uSvv3Xq+XRfKjsDtWvr2jqK0mklP1WR/Iqn6gE/09+61Wp6R26KHdM6NXT1dS35cRzPzcc3sfe89e6TW1dhbjz+UhnlhqpKd5bBtBY3v+WPv3XurV+lOsnxG3xVPHJDJFTF9TgKAUXWLt+Pp71XrfRQu4Mtuze++m21FkaiSgpagRPHTVMn6A2mxUH6W+vv3l1o9G+6e6dpMPjKRiEExMLu0n6gWW/qJ5PvR630Yar2fVQJGYJqYxrYEKFYgDkXAHv3z6304QY9mpftliJOjS7BDzz9fx9Pe8de+XSMn6qlzeRiKEiJZvIysPpzc8H6/wBPevs610ZHauzsRRiioBRaZVjCGV4hbUP7QI/x97pjr1ehrTreo8BklmiWmsCmoAKtx/U/09+r5db05+fVf3yY7q/0TGqxOOnhqa6WOpijFK95FfTZfSvv3XuqZNydIdrfJjcE+d3h5H2804mhp5oy5EV7gWf/AA59+61Q8ehq2R8N8HtdIoHxsS0cBBS1JGnIXnkD/evfvs69TodsX0Lih4zjqeBZI9IA8a8Wbj/X9+48OvEefQvbe6hqHEcBijurFW0xrpP4vf37j17ofqDo+YYtAnhVgn7noXhbcge9U630WrtPZybTYypUmGLzpHIUk8ajW1v7P0978+tEU6eO0fjF1rVfH/Kb4zVNFV5GqxJqaaWdFmbyNESul2H9fz79w68AK56Kn8DtnYug1Vi0yKq5WRFtGoPjSUqASP8AW9+68Orz8BFTPj40p9EIVSDf0i4PB49+69xx0wb031VbTpFE9TrprgH9y2lbfU/4W9+62OqjPlv84TiUqdn4Oq/yycTQKkM51amXSCLe98etdEf6N6O3j3xuhd0bpqJJ6GSUTCORvIAPJdR6z/T3r59e+XV6nTHRWH2zSfw+KJYvFGhT0hRdUsfp7959e8sdHK6+2TUYyoEuuMxA29ZDC2r68+9E9bHHoVNyY0U+OmqaeaEToCQU4INuLW968+t16qb+ZFfXtgYXWtCz6pCwaRubNwbX+nu3n1XPQs/BKolyOxZqWsYPIZF0stif1Xtf3o/Lqy08+rOttbfSkkarq3vFG1wuq1go/wBtx70OvE0PQ7bazO36krFH40qFsuo2+qiwuf8AH377etfLqFvk1ZpmjpaiNYWvazAKFIt+Pfhx69Q9IDCbYpqmmqDkpYWMqFVOoEW/rf37ga9a6Lz27sDHHE18NOvmEsE6hdGu2r+gN/ex1vBz1qJfzJduZ7qfeqbrwQqaSGCq8k+kmnVxbUSAlvdutdVAdgduZXs3AkNVk1WiW952YXS40nUfeuvcOr8v5MnT00uPoty5KmEqutK/kMC2+twQ3/G/fuvefWzpLhIIqKSPxpZTZAE+g/HHvVerUHS22NFjIHggqKM65GsrLELXJtf36vr141Ax0arC7cXwRsllidFcKVsQf8Le/daBFOnWf7eikSBoNQb6v4lI5NuTb3o+nXh1Il2tI0ZyOLKpKFvpbm5PN/fq0FOtedek/LV5YBosiqs0epQwUG34NyP6e9Gh4dWyM9BhkaWXI1UsXl+jWCqxv9fyB731oEdM9fFHiad4Z0EnpJF1BP8AvPv1Ovceq3fnfuhqTprdNPj5EgkeCe2kBG9UR5BH9fex149a7v8AL22jUUHdGT3ZIwlnbIzzzE8sR5CeSf8Ab+99aHHPW7n8fMhkN9bXo6ehdUWDHwALfltKW91pnrdR5dJvsSOuxmZaDIRPKsMjJqtcf7c+/Hrw6r0+YsOXz/U+9KPBKUJwlWrIg5IKH8j3sdePy61Z/gHQZLFd/wC4dqVYkhyUm55Z2EhKmxqixIDfW/vY619nW5xsamSo21jYJgQ0EAjf+zdlUC/+P+Pupr1ao65bgyf8Bhk8EbMulif7QBH4Hv1M9e+fQZU3YUVRI9HUjxh2KXe4uW4+p97p1qvWPIUlDiV/issZqfMNcQ/ULE3tb3brVOkvu/pzanb2253zOLgV3idYtVKgOojg3te/vVfLrdPPqonuDrTtr415qpz/AFrLWR4+nmMghjeRU8SevgL/AK39Pe/LrVM9Lv48fzKd2bhy0W1OwRJFUpL4ZDOzrcg6AdR+vvWOPXgc56spq9w7b33jYKqIwTB0Sf8AD2LjV9Pfvs630CfZ9HhRhaio8cRjpo5FZVRdXCfT377evHhjqnjcX8PyPZWPWiYBhX8KACbiTgAfT37rVc9bAfx8p1o9gUS5BBoaihA4F+UFjz9Pej6dbB9eikfLTHQ4zIx1EKvIlQ2oLFchbi4uR731r5dU9/IOvrFwfklhtCqOyBkUN9fp9OffutdNvxn60ouysXO8uKrY6hSStT4GVSb8EP8AS3v3XsdTu0Ota7r/ACzrX07vQI45kXUoW/4v/T37j1sj06Rm7sZs/cW0IEw0XgywsPNE2gI54LBl+nv3Xqjpw+NXyz79+HXY2KyuFzVbVbYjqKVJBJXTSRRwGUGQFL2ta/Hvx9etVPW+R8FPnZ138u+v8fXY3L0a7spKWNctRGpj1zTqgMjxKbEm97+/der69H6kmdXVUTWrC+pbtb+hIH492AB60xI4dZl1W9Vr/wCH096NPLrYrTPXK/8AvHvXW+ve/de697917r3v3Xuve/de697917r3v3XuomQ/4AVv/UJU/wDWlvfuvdak38gz/t4R/M2/8SHX/wDu19+61TJ625Pfut9e9+691737r3XEtp5JAX8f1v73SvVSSOPXGRgq6m/T+bAk/wCFgPfh8uvN8+u0dXAYHg/T+vvxFOtg1Fek7vDcFHtfbeYzdbKsMNDQVUwdjYeRIGZBf/X9668etD/5r9n717q+RW4pYMmtZjcNm5PtoRMZ9EaylQFXn6D37y63546KXujdm4dg5WHKziWGGArNIW1Rx6V5P+8e/darQ9HY6Q+RO2OysZBSRV9MuTjSKJrzAsCPSRYe/dbxxPRzdhV2MxNd96As1TqujfqW4+p1f4+/efWwfLoUN919fubEr90Q1MUCogP1uOD/AK/v3HrxHQXYZ6basc0tbKlNFpJvOxRQoF/qffuvdEV+R/zXwHVuQVTlaCWl8iCVFnQsEv6h9ffuvV6Ilu/+bDiISI8ZLEuP1EFjIFHp5LXHv3Ws9Hw+HnyUl79xy7lxmUiejiKiVY6kk6tfICg+9HrYz1eJ05uyhpgP4udUekCM6r2On8/T34jrXlQdCF2tvmaPbNZJhFkaAUkx9ALEkLwQPfhx69THWtz8gKHsTtnsU0MQr4setWFlNnClQ+kXv7t14nobNs/DPBUW1DX5vHzy1UtPqWV4gQWKX9J9669T16fOtvhVt7O+easoUNNrYKGp0awvwDqH+3t79Xr1B0J2F+Amx8Vm0ysFBFFLBIHRjDGB+q9xb37rdOrA+v8AYNLgMbFi4WhCxoqgrpAI0aeT70evUHS+THQ4MFo2j8oNxYg2tze/vXHreAOpS79pqemnTKzpGIUvEWcqAebXb+nvZHmOvVB6CDeHyU21tSgnMu4sZHHGjExPWKHAvb6X9+pjrVc9U3/LD52YCdKuixGYpHqvJIimOrBHItckHke90HWqjj1V3uTvzD7mwlacrG2QrZYG8fifzXNrC3J/3r3vrXHoI/h58Nx358iaXdGU2xWpjqXLU1XTPUUV4QfJqVxrFv8AYj3r7et9b4fQfUR2vtjbW2MfFDFR0+PhgkhWNVb9sAEED3XB4db4DozPbNS2ytj/AG1IpV3gVJNA0mxFuQPfhnrXDrUG+Z3eO99j/IKhkxNTOaObKqJKVZn1kmX8Rg+7dePWyj/Lv3/P2p17hZ82/wBq60AZ/uTYsQBcqH96PWxw6P8Adj752hsfFNFXSQNEI1VnVhYj6W4/HvXy631Q78s/ldhBuR9mbTqYL5CSWBkWT1EzjRwv+x976qOgZ6i+DGI31maLsHc+OjrpKxhVAmmSX+1rtcg8+91x177erP8Abnx82nhYKalp8NRRU8axp6qCIEgLp/A9+NfLrePMdD5gPj51VWIjZrBY6eExkuBQQnkixsSPr7rXz68TinSFzHxP6Zz+dCUuAo44IpLAfYQi/P0tb37JPXuA6VU/SmxevBGuMxVBHTuqoCaKIKSfT+B9f6+/Vr149LLE4uLbOPeoxNPBDHMjOUihVSxtccKPfuvU8+q4fnZ2xkdpbHyuRpJBFVR0Rcg3WxAN+B+fexnrxNB1rMdN9m5HfPc0u5MpPrqTlzT65JLrpMtreq9v8Pe/s60OPW1J8esRgabDYLJ5OFaqKamjkkAUSISbHgH+n59+J68K0r0djL4Prjd2MWjocXTQyzKIyz0iAk2sxUkfX3oVA63556ra722Q/VGfXIUdPKcW82seFWVA31W4X6e99e6GrpzsvF1m0Kqn3TNSymphf+HrdWeNWWyhg309669XNeqX/wCbJvzAYjrOop6OenFTLBPFGisoYl24IA/PvfWjXj1Tt/L26R3T2FvAVuVir58ZUZGmqFDM00Hjaa4Fjxa3v3l17ietxXYHV2G29tPD46GFYZYIVQgqFJYIPx/X3on06tSvHoWV2V5cdFEkaDWwUMyX9Nxbn37r2B0Wr5N/CzbXc+xsjisnj4p6+pp5mjk+3V+fFxc+916rjrUO79+L/Znwq7Bkz2EpsjHhoqoyoIo2iiUPKWvce/dez0eD4u/PRK2txi7iyEMFZDLSU7ipqfp41sQQ5+vv3Hr1fXq/3YHzq2RFhseKrO4nx/bKXU1kfHHP1b/be9dbr59Kyf8AmHdY42pVf49hl9Vx/lif14NgffqenWq4z0IeM+ZvWe/YVEOZxbTyJoRY6pDd3HHAPvwB69jotHyn2nX9t9ZbjhxkZrKSogcqYwZV0eI8/nj37h16lc9a8PwF3Hmuh/lrl9p18j09K+ZlEUbN4wuuYj9A/wB497+3rVKnreJ2fVtmduYbPRuDI+LgZbHUfUmoHj3U/Pq2BjoPtybiiwlXLkMvUwQU8bXkNQ2i4F2PvdOtfb1WB8o/5gPW/Wa5T7Cvomqkp5VXw1KCzoxXgjn34cOvZ61eflX/ADIOxO0twVGB2lkZXgqq0xIqVUpGidvGAoB/F/e+vdG4/l//ABgrux8lid09kU01e9RI0p86eVSL6xbVf37rwzjrar6b6Y6s2tT01NTYaji8FPA4IpIkOpEt9bf4e9VP7Ot/Z0pOzMntujtS4+nCVMZCRGJQBYfTj37rYOMdPuyosjkMRBq9KEfRwT9f6X9+4dayT0skoAiPBIYl9Njewvfn/Y+/cOtdV9fJaHL4reOJyGJjaSmhcecRLdQdP1Yj+vv3W/n1Tb/MS33jMnjtk0SOn38uRp4atC9yF8oB1D3vqpPRpvj53js7p3rKllyFZSNF9hTu1OkyrLrWL8D6/X3oivW69Ad23/Mpnp62op9h0s51PZRGjPqv+bj8g/n3vr1fToBcV/MR7MfIxSblxeSaiEuouUm0BAb3IP49+69WvRuNl/Njr7siopaDIxU8dc0UcbmcqpDP6SSH59+8uvVHDoRs51dtXelLPncLW0DEr5XjjaJpDb1EWHvXz639nQd7h6hkyW1paF6AGjliaGR2pwS0R9LHUw/p731r5Dqtzeux+yPjPmaneHx9q6/GZRJjUTIamaGnMgPlPpQgC5v73XrRxjownxU/nmfJTqjeONxPduXhno5p3FdetllXxBgtrte3HvVB17rdy+Jnye2f8oursLvzbVdSySVdHTSVFPHMjyK8kIcsVBv9b397690aNWbSxK+pdQsP7RH9PfqCuOvA465KSRyun/Y39+Ip14GvXL3rrfXvfuvde9+691737r3Xvfuvde9+691//9Df49+691737r3WGdpVS8S631LxcDi/J597FPPrRrTHXbu6lCQNFiZGv+n+n+w97ABx1okjPQa9wbzx+xuvty5uryKY+aLFVX2MnkCyNVeI+LxXP1v+fdfLq2OtJ7vPe+U3N2tnewc9UyZKsGQyUFPNVkPKEMpEelv6e/Ux17NadBDidnbq7IrXzmTnqKfGUjs0MepwhjXn/bW/w9+4depXpP1u2sTHk8hTY4LUVNMjFta8sV+tif6n3vrXTXtvAZLK5LxzUoQK+lbj834A9+690N1P1Q+TgaCSGMFgVA5uQPwPeut06M31N0dSUUNOJ6KDShDairXNx+L+/cet449HZzPTs1H03mM7h6VFkgpasegNe6QmxFv9496r16madUL9CtXZ/uff0WViaV8TkJlUSXOlVkNvr/rfn3uvWhk9WLy1tdArihVlmSDVHGn6WcCyKB/jb37rw6K9u3tf5GYXOlMTtmSqxSSW8tiQYwbsSP8AAfn37HXulltD5eTY+ohxu+aQ4+qZtEqtFaxPFgQPp7916vRzdhdiba3VLHX4urVg0aMELqtyRfkH37J62eHQrP2PBQJI80MEUUB0+bWgIC/2gx9+69THSD7P+UEeJ2w9Fh6oy1VVBojMcisVc8enSfx711s9V+YDY2f7O3bLuHcokq4nnadErFdlIYlwo1/1976qPl0bLbm2ZIqVqKmxdPS09LeNmjBjAA+pAFvx711vpn3HaMLjYYhqvpUr6tXNvr/h73177Ope19n5T7kXhbxS6Dfn6NYgC3096+3r3l0P9BshsfTrVONFlJ4Dfn+tve6enXq5HTTurcOV29iKiqjVvtkhf1l9PKiwNj+PfuvGleqhfkz3ismKrketdKmPIR2VJEDALINQ0jn37rXQwZ35HYfePSOyNgQZFmqsljoqSWNXQl2MemzKpv8An37rQoehz+K/TH90sfFIE/bkmWqsyuGvL6wf9v8AX37j1s/z6sPptuV09JakBAI5KqR/sLj36vXiD5dMG5emxuvC1lNkJ5FqJIiI9Wu4b6gC3v1R1vNOqj+2v5cstRuer3bW65445Jpl1rKwYg6gAPfutU6GH4w9Zvtf7nFvF4IaZmRSoZWAVuLX/wBhc+/de6PHhXy9LlzSpG/2oYWkAJJW1rAj6+/CnXujV4KKH+Cq/ndZwoIT8X+v096PVgOgS7V7Drdm0Uskzn7cAhiWAAut7+r3sdePp1Qd8tPkZUZzLw4ylqHeKSpaOySIVFntpAHvdOqE1z1aR/L8lqF66hyATU00cLAm5bUT9b8f7D3U9WHy6tawU9TUwvFknaJKgftqPz/Tn3rz68eHUaoxuVwlUKuiZ2ic3uOPz/Qe/fLrYofl0rpv4hl8YGqZZEKj9QIsCRYCw/r79jqvSTy0WQoKSJaGpkaQrwAx+v5B9+8+rCnSRVsxVIY8jGZI5DoYuD9D+r6+/f4Ot9U0/wA1j4yYzenXuU3DDEgaCB3corsdSwFh9P8AWtb3bB6qajrSP3Dt2s2tuyjwEBkC1FXUQkeNwGvPp4P5496611vSfyiesaXD/G7CZGaFRVyU9CxazGQalBub+/HrYHVysuzrUJrGJMYXUeG44+hHvXljrdc9LLZ+Ix00UDqsfkjN7ngkg/T/AA/2HvfWs1p0OuPqqWCELNL4lRQoAI+g/wBf3qvXqdOM8tPXxqIUVgg4Y2JYfge/da4569QZCaF/tiupTwFtwPxf6e/fb17rFnKcQ0rNDErySKzAKPzY/wDE+/Hhnr1Oiy1EWbxeXqKypiZIHcst7AFSeLf63v3n1vFOmfcGWirYKiQ6QyQyHn8sq/gHjn3vz68Mda/n8yXtndeL2znMTiqOSeJzKo0IzA3jIP0HvdOtA+R6q4/lsdn4Q9hVNBu2pahrKuumQxTHxgEsbW1W/wB59+6959bm/wAfN7ja1JQPt91qsfUUlOpkR0Is4BNtPvR49boKdGB31pztMKoU4kmqQZWOkMQSPq3vXHrXDJ6ITv7HXoNz4vJw3hraSanjVhwTJcDj3sU62ePWs9hNp0fU3zwjqiq0tFkKwOV/QhZ5uL6rDn/X9760cdbUmylqKnbGMyVKFannhEgdCDcMgJI03+nuppXqw6EZsBjsxirzANKym5N7sf8AEe99ar0RT5CUVTs6glqcTEVaOUvqiQggKNX1PvdetkDpIdG9z43sXIYzbeQqFatpW8MkMroblSFAK/6/9PfutAg9H13Vt3NUGLgiw9GiwhElYw2sVYXvcfT3Xzr1vjjoCd97Fxm9NtVVPm6aE1ul0CyqWJYrpPA+t/e/Og69+XVUfcPw9oMEX3PtiihgyUX7wFNGQ3kDFiTb3anVcfn0HvVXd+9dp5xdrbkhlhp1lWmWVxZdC+m9z+Lf4+9fZ17z6Pw2Dpt4bLrKymqxO1VFLK6s4NjoJt9ePfh8ut1HVKG58ZmsT3xj6Slid4Isn6wqlvT5vyB/vfvfVeHDrYY6kk3FU7bx1HHRaYBiqexZSlz4web+9fb1sg+XQHd1Y6pFXLFuCIFnZ/thINQtbiwP59++fW6eXVLXyfjzWZz+1tsYahV6epyscFVojJHhabSxb6/j3vqp6tl6V2Btrq7rbGwyY+kgqKnHU0rTCP1mR0BPJ/x966t5dFz+WW067JbRyGUp6CIwGASrOqjyW0Xv/tvfutHPDqsnr7aOez2LqKejjLxQtMSx9JUKebH+o9+618h1lpdoSV1VUbVyMH3KTGTXVS3Z4mH4F/8AiPe+PXunboHvPtv4ddkTZTaGVzH8KpKozLQ05KQShpBdSDwR70fTr3l1vO/y8/nttr5W7BxlPlqyGj35T0lMtZjnkhEkjeIX9IIN/wDYe/cOtnqzF5NDICLK17sfxYce9gV60TTrpkbWHVuP7SkcEW/HvdcUPWiDWo6y+69W697917r3v3Xuve/de697917r3v3Xuolf/wAAa3/qEqf+tLe/de61Jf5Bn/bwj+Zt/wCJEr7cf9XX8e/daHE9bcvv3W+ve/de697917qPND57Ix0orBrqfUxH4/w97HVT1mH0AH0+lz/Qf6/v329e+Q4dAp2n3h1T0Xjpc52BuijwdKVLsJqiIsik3OmIsCPej1sVp1ryfzDf51uyf7r5PYfRzpn1yTT42bJ07RySMXHiDWBOkc/j349bB6pX6U2hk9w56bfmUklqcjuhzXy009yEeRtekA/6/v3XgBWh6ErvHpybeOKrKLIUEdH5qYpA8MZ1MWFh+n377evHqpXP9f8AbHxjrKndGEirqjGrVmUqsc3EKNq4BH9PfutdH9+KPzgod5rTYvdM7UWSe6aZwkbEhrDiSx9+PW+rbqPsfb9ZgGnkqk8FNTpOJPJHYjTcD+vHv3W/Pqov5efMTI1FRWbM2JJJPXtLJTBqZkdgCNANkv79TrVeq+MR8Xdw94g5fsTJ5KOeQGSGnlEojLX1Dj8+/deqeuW9P5dFOm2akiICnWmlaGdYXD6wvHIHv3Wq+fSZ+GO5t1fEfeNRs/OSVabUqa1AKph40jR576SW/wAD78OHXvn1tf8AUvaG1d17XxWWx+TidKqNDGVljJvoH4B97HW+Ir0efaOS27k9uSUeTMMiPFoDTFT6WFvdeHVugdyPTHUzVFflj9olU8pkBDcg6tVgR79Xy61TPUDsDMbKotoUeGxMkElRFH4rBlLXHp/3j/W9+FetmnTX1VjZpMc6RJZWkDa724J+l/e69VpnoVdxbeqKOi8pcqCoPpY8fm5596HVjwr0w09XicXiZK6uyJi8SNI37qhwF+tyT72R6daFOPRP+z/mz01sCGuTI7n8ddTCoVYnnhsZI1JAGo/1/wAPfsdaJPn1RF8iv5xeRpclmcVthaiqxqMRDVUwje6m63XR9ffut9VQbq+Xnc/f+YfG7Ry+5VrqyWSJIUgqDEHka4W6i1vx791ro2/x2/l5/JnuXI0eR7ESt/g9SIpTM6VCOEdrEn/Yfn37r2a9Xk9Xfyfev8JHiaitqvuJTpNVFOZXsSAWuhuBz7916nVwXR3xP2D1NT06bexWPSrjjiHlSPS50LYc+9HPW6gdHc2ZT5THZKnipYfJKv8Am1tf83sPfvn16ueo/fmfkxm08xX7hSKAU1J5Y0eVLXVeAA3v3Drfy60ue9c/Vdk/Lemraal+8xlNmBCUZfLDxNa4CAj3bqmetp74s7YraHrzAVmLj/hpgx4LJSKY1aygkMv/ABX3U9W4YHQP/Nvtqp2Xsysqpa+oNYKViVLAadN/UAfeh1vrXw6u3nhu1+4f4pujLVKrTZGMqG5tpl+vHu2OqDjTrZ/+NO8duTYCkxODc1sNJGItUitp9Isbav8Ae/ej1bI6NHncrHS00UhiEKekllQg2P8Avvx7117oLd3/ACA25sqmSklriZX0ppZlHJ+ttXvdK9b+3pe7U7U2Zm8HFmMblY3rmjLyxmePVe1yCL/j37qvRee0/kLUVdbR4OiDPJ96qAiz3Hktfi/v1M0PW89GR2xk83k8Xip3pQ0L0Ad1ZTo/R+feqeZ69w6oW/mmdlZLEYXctBPCEp2pJUQekAfVeAfdh1o46of+LuEgyyQZo1JilfcUcjFeH0+e5v73w611t8fGTPbKpdlYGmyOQLMKBFkMhXhgoDED3X/J1b5dDpv/ALX2ftb+EDA1uuSSeMPawB5Fvp79x69w6f8Af2IwPaPXByVVHFJK1PqDMAzAsllP9b+/ZHXqdVf7twS9c47M5eLKT+OiLmKnaTQqLoLAKHPvZ9OtcRXrX3+RGV3L8pOxabZiTzy0FJkGWQK3k1IstiCY/wDevfuGOvdX0/AT42bb6swWHp66kiineko7u8Tq2oWux1f4+9HrfDq4nM9cUU9JRZKgnCU1OBJJpcBNNrn88e9U69XGOg5o+zdrzbkj2ZS1SyVsEiowDKxBHp/33HvYHWj0YiupqSmxsMTnXPUQFkBFzYrfkn/D3rPHrflTqsP5yfH7Z/bewsuMpjqOOpo6UNFIy+t3CEi9/wDH/H3v5deoafPrRo7K6j3bhe+5dl7TnqqRDl51vSpIVCebSvK/n3vrVfXq2vq74IdpblwOKqZtz5uEzUkbnTLPYMw/2n/evfj16les3av8tLuWgxa5LDbmzjzBQ366gjk/X8/X37r1K9E9xdJ8jfj/ALvo6bLVmbqcfTV0JkMiT6TDE9nJ1D8i/v3Xvs62lPg/8o+tOwusDtHdmSiOeraKSAxVckauXMejT6rH6n3qnXvz6IfvD4UZPHfKKTs/CwL/AAqbICeKSIu6sHm8g5HH0974nr3V8O2O4MX13sugTO1ywRUGJSNw00a2MMVytmN/9v71SvVjjPVGPzz/AJjedzEmT2v1aJ66p1+HVShWIFihuY7+9gUx1U/Lqlyu607Z7gxGR3FvKavjfwVFSY5FluFcmQCwHv3l1ryz0QbZu24YO6IsFXRgtT5KGK0sWkMUqNIa5HvXDrfl1uk/APZ1LJt3CRU1ND+1GAxU3sojF7W97PDrYoM9XK0e2MbTaVE2iTwICB9f02I91z1qgOegs3vt2BGNSq+Uo4AYrqJ5uPp/X34jz6sPTpYbafJRY2hFNS6A2kEhSODx9D/X37y6qTnpRbjw2XXHNkIk0sVuQpseRe9j/tve6+nXuiidsyGlwNfkshDE7UscpZ5NOpbRf1b35TnrZ4dah3zM7MXN9pS42CqeR4Khvs4FtJGJFmsAoXn/AI3731qvRmfi78b+w+7JKCt3BU5CnwDw01oWE3i8bACxHAtb37rVPXq0zBfy6tiUuSxipFS1cpIEwmQ31X5JNub+9V6sB8ujQbn/AJeXX0W0g0mExgM0ATWlPqZeODwL/wC8+/AnrxA6JnF/K32gMtU56gyRoZRJJJGlPrQ6gLoukW/P0Hv3XqdF/wCxure6fjvXz1+DauyuEhOuztKyNGTcg6uPoLe99ayMdKXanzDyFdt04/euKTG0YHi84iUPdBpYg2/r7914HoEOyu7+vczishj6KpSqep1nyMo9JZSAB7917qobuzYNDJkYs1TLEYatgWk0qzRq7XLKffutZ4Hq4T+U/wDP/L/HzeuG2RlMvWttKerx1BEZZFSmDMRGR6uPfuvdb6Ow95Ynfu1MLujDVcNXSZahgq1aJ0bSZEBZWC/T37rfSv5/p791rNeu/fut9e9+691737r3Xvfuvde9+691737r3X//0d/j37r3XvfuvddH6cce/daPXrXFjzxY/wCPv3XqYoeqo/5r2SzOP6boP4PWPRvLM6SFGZfIpkA0+n37z635danST1ueykuIrZzNKcmWPJ5Bk+tz/X37r3l0dbCSUG1NsxUlZGIYZKdhK7KLD0fk/wC9+9DreadVo90fI3ZnU256qpKU0qVbMjSCxFi5AuB738utde6x+W3W+4c5TRJkaCnecxWBZT65CLrb/X+nv3Xs9WN7My1FuI0tVjq5ZaeY6gUAKkGwJ1e/cOt9GewOYfDGNJpWeIAFTputgLWt7917y6OthOyqU9J52jahNXE9BWsdKAj/ADJ4968+t08+tZzqTs/A0HyI7SxktOtBPV5SZYY3UBn/AHTYge9/MdU9a9Wv7SocdkYY5XsZZEjYjSL2dbrYH37q3ljpbVm1qSaldGRhccgKnOrjk+/Y8uvZ6D2p+M+xNyrLXVmHiqa1lJ1WW4N9Vj/j/j735da+3oDc58fN5bNyVRmdt5GWhxkSu649JCQUU3AsP96968uvcT1Wv8sfl1vXrumn2D46mizddLHTUtQVkBkMhCgqQffvs690aP4e9a7u7B2vt/cW9cjNXiYiZlnkYqUZfJpKtf6e/deA6tFiwOK21FFJR04VUhVCoC8sgt9Pr79nreK06SG6M3LT4HJzUKijl0lgRyGuL/j37r3Qb9e00uWnjqsvOHDyMFMh/wAfTpJ9+HXvs6M9SYR41ikppiFUJfkElUNwBb34da6GDG19JHjRHWQiXQpEmoD62sSR9ffjx62MDPVaXzD7+wOyMTk6UV1PBGlNOoiLDlvpYe9ZPWj6DrWo7A7Mruydz1yUtYVoXq3lZ1LaDa5BP497r1rz6Fv4z72xuT7KwG26vLQVc9BWeKCAyFm4ccICf6+/db+3rai6urFpabH0jxEJ9tRkSMLXBjHPPvdOtmnRzMXRy1NFGaEMxdAbKL3vwPp7r9nW6+fQw7W2BUzU6VtasnoszhrEDi97H8e9U61WvDoPe4KKhGLqohAGSGCcmygfpT1XPvfE9eHDqsXae/MHFveqw1NFHFaqMc1tIIJe1/e+vdH32pt+izLIYoePTaUKoHqW97+9GtadeFKV6Fij2fTU66GrY0GlmK3Fxb6j/b8e9Dr1eqs/5ku5qvZPXeZr8ejFKOmLNLGGsP2ybll/H+v72D69ePqOtRPJd7PufJ46q/4FsMrUiQKSxHjlIJBv73XqvW2t/LO3Eu7ek6P7anZJPBANVrFdLWJufejnqw6tow9RT4bwrlp1YqP2xJ+GtYHn34cetnpSZrceOrKLxU00YcKdOllvf6jn/X96xw6186dBo25aujDU8lXoRm+hHI5+pPvf2dbx59KTEbhpMm0VEdLzFgA5Iv8ATi/+8e9fLr1PMdK3J7PnnohNFVhFH7hUDSf68f6/v2OvV9eicfLL+EVHTG6cVkaWOqn8MyeUqrWPhIvqP+9e99aPr1pA977Q2uvaG3oMdSwzVceXfypEqk81NyCPe/t6qOtxX+XFVQUnS+DxEdGyWo8ddQljcIPx70erAYr1bpQ0wr6QUbQtEHUqAwtcW/H+396PHr3lnpk3tmMb1rtyoyBgW8NO0pksOLc39+yevcOHVR27v5mO0Nv9gptPJZakovJXrThJHAJZpNIsL/n3sdaqOrZ+q+1cRuzbeEyeNqI6pcpAJI3Tm4YBiR/h7959ePz6H6mx0lSoq0WxNmAFxx9ffut0x1hyGTipEKVUf6Ra55N/z9feutdBNud0yKFkQtF9SQv1H+J/Hvw4562R6dFx3daCOSCnsZXDppX63Y2A9+63nol/aPxkpuxqeqny9C9UKhJG0voZSGQgHn3uo49aoeqD/kR8Ct09X7oq97dc/cQVcDmoWmpmYWYvq/TF7tWnHrVOjpfA75wZfbOVoOte0ZzQZOnmp6QffFwW0ERgjye9Gh69U9bSXXm7dsbj2SmVhqoa5pacyQrGVZrEfTj+vuv29ePReOy8RSZieH7eju8swWRVABA1/wC++vvY49bxSnWpL/NnzdT098gtu5bb1M/8SbI0wEMY/cI1i49PPvfWvPq/3+XT3Tk+wur9qQ7poZadpcbGF86sAPQB/a59+48Ot/Z1ZxmKDH4uNKiDSYSqvYABefwPda9ePoOiffIzb0W9NuVtLj08TNBMvnWwAcx2HP1597r16lOqA0wXYfRXbMe5cfJV1ePhrhLUGNnZUUy3a4H497x5dayDXrYA6A+VG3u0cBQ4irlhTKmCnp5VIHkDaNLXB5964dbp6dDtnNsUdErZElJ4Zgz6SoP44K/j3rrdSePRaN5bclzMkppaZmpWNpUC3BX8gAe948+vZrjqr75E9P2rqnIYynehqYY5pkk4T1jkAaP9697NOtEVz0VjY3yNz/V9DksNuauaOGN2hj8uohlYlRbV/vXv1K9VHRg/iNsvHd5dk1+7Mjjlmo4ZzPTzsqhXAOsWZvfvLrfE9XY1cNFgcHT0uJpPC8MCRB41uBoFrAD3ric9W4DHRMe8aSXNeOsnJElMD9eCxC/n3vrXVQO+a2HF9kYiXKUvjgOSQRTyWKhvLYMCeP8Abe/HrXVqlFSY7cWxcdUfeRzqlDCy6WX0ftiw0j/Ye/dbr+zou3Zcslbg6vCTTCppmi8XhFiANJXTx9PfuvHqt+obKdfZqojxsMgoKiRleNRcKrty3H49+6qccekxPuanod30VXEBN9w8ZmX8gyn1cn3vr3z6FXuXYNHJs+l3bjoDWSyxLNUU8YFwNOshtP09662ei3fEz5fb5+O3eGM3HjqybE4qqzdNSS0peYDxJKFfUpsLEf4e/da6+iT8S+/cb8hupcDvGkkWWoeip1rWW5DTFPrc/n37rZHRoL24HNrXH9B73TzPVa0wPLrsG4v711sGor13791vr3v3Xuve/de697917r3v3Xuolf8A8Aa3/qEqf+tLe/de61Jf5Bn/AG8J/maj/v4df/7tffutDz625ffut9e9+691737r3WJ08hC6rKpBYD6n+g/w92GBXqpqTTqvf5zfPTYXxG2jWyVlXTVW6ZaeVaPGhw0wlaI+MLGDe97e6549bxSnl1pYd0fIP5A/NjsPK5fO7jrcbsesmkWhw88lRTgxrKSqhSQLEfT37rZzjpPY/wCNOOxVRTSy0Ach0mkJd3aSQG5kOv8AJPvfWuj2dZ4iLHrjaZQKI00aogYgAgGyge9cercOHR3dgbAxW6sgoz1dDNGGTxpKyAMbfTn+nv3XqDqd3l8ftk5fAz4yvxMFVTSU0oEhWLx3KcEkj/be9deOD1rYfK345zdT7qp9zbCpmp4YplkcUzkKihtRuE92z1roII/mpvGlw8uzTS1C1jwPSfdEShWKro5f3rrw6En4g9KZXf3Yyb13a71lPUTLUiKZyyANJf6P/vXv3Xur+8D1HtWaGOOkxkEKU9gGshubW/A/3r37gOvE0FT5dZp9sYHcldNsY00UWiMgTFYwCw40i490LDj1SOVJfh6rm+YfxOxibTy5oolTKt5paWZLK940MisGTn26oZhUcOnvDbj1UL0p84t9fHPd67E3qlXLisbUGOnnlErRFBJ419bcfT8e081xFAQjnJ60qliQOrkNq/zR9tZHF0qwVsELGKHUlyPxySb/AO9+1qWV7LH4sUZZfXrzDTxPShzn80LaeMgWI1FPVySLZkVSSSR9CAfaB5LpH8MwmvTElxFHQHoMupf5gu2exuzBiK2WKlp56wxRpJqUctYkBj7MJ7K8s7T6+5QqlK56ujpJTQa9Xy9d7lxFXjKCoxFXFJFNTQy3hsQdQH09lVrfRXyF4vI9KRA4Grj0p+yewqbam1MhnczLopaGKSQrLwsgWPUPT7W6SDnpPJII+1uPWvP3J/MGgyG6M1iqLMR0GLAmiXU5WM6ZCCFsbf4e/EUND17VUV6p67337k+4twHEbXxsmeqK2q0GejllIvUNoDXT/X9+63X16PZ8Jv5KuX7TpRufsOWqxkFeGmWlq5yU0t6wqmU/4+/EU69x4dXydK/yz/jz0b9qJcFisjkqcRyGp00zP5EFidRF7+9V638zjo9+Iotn7YpYaTEpTUFNCFhjiVIlsg/SLge/EE9b4dDVgIMZFSx5SpqofEw1KSVAKgf09660elWOwdqxFYaaWlM3+bLKAW445A/r/T37r2R0tcBnaNW/iMddHG4ikeKxu3CFrD/ife/l177etfb+YV8oO697bwzfVWwsbkawVCtQirpUm0Rgvo1sye9461U9Ovwk/l2Z+DaEXYG/4ajJ7iqquOuMM9jMrSHyEWf1cX9+J68B1e71tt07L2s1DNTGlEEDJHAwAb9H6QT718z17qiL+Z3vfGzU7Ydq2GCoqS8AiZtJ5ksB/sfex15uiw/BD4cy7k3BFuirp3qqOokpqiwOpWRn1G9/6+/cOvDrZz6p6u2ZsGighioYaCQCzlgoLm3It/X3XjjrfAdDPm6Pa+dpDRwyQiVVFiugm4HvfXvt6rO+RPTFZld0RQ46kmngYOFljJK3df6D/H36vr14Zz0TXBfH/u/ZG5J8jDuGtGCqJS60KzyFIkLfQre1rfX3sdeIPDo5PWnXNJkM3TVuenSSeMxMxdhdpEHNr/4+/V8uvdWQ4tKWk2yxh0xR0tLKiMLEBVjNhx7r17rU2/nGbsERraVp1Van0K/01XkIIHu3lXrR49EM+DXVOe3dSUcdIsr00uSgdSPodbX1f0t731ryx1s/9XdGf3Z2xj6jJ5ERCOmBZZJACbC5BHupPV/t6ndnYPbD4qiaGeFpoCGSQOLmReRp9+Hz61ivQ2dK55IdoVgzLh8dT0sltZ9GlFPI1ce/Uz17rXv/AJl3ybqWzGa2P15UaqypqTTLFTEhhcFT+g/197x1onol38vrAbn2Z2N/eTs7EzTQV+Q8wmqgWQLI2oH9z6Dn37r3n1ts7dqdmZLY1LunDNTxJS0SMVjKhVKrcXC/63vVM163Xy6LD2X80cXsrb24sbPnqamkigZYkZyvOnSukn/Ye/U61inQYfCNa3tXfdZv+orfuqaad5FlDEqQSXB5Nv6H34463k9XS4yip8mUeWqE4owUuCLoEP0v7r1v5dVb/wAzTuih6z2Y1Bh5k/ieRp1jWBP85LIwKgKF5ub+7DrRqOqn/jB8NqzthR3Dn8a8GSqK0zqJQvEbtruAwv731oZ6vW6Q6epqLFR0dxIaOMRBAsbH0/191Pp1umOhxrNo42eWKgqaRWSJ1VozHGdQDWIbUP6H3vh1qvRMfkP8cth7qyFRTnaSMZKWfTV+CMAMwsTrI9+B9evEenWv18mOn95fGvIZDf3XzTx0+JlEz0kMj20f50jTH/re99ap59O/R/8AOPw0m1v4XvfHMuYoo2jvVQMHMlONDEFuTyPfvn1uuKdMGa+Rfcfy93B9jsIZDF7XqJjG1VCJkiMUzaRpYcfTn37rXn0f344fy2zQ0kuf3bO2cydcGmYSMJZNRIYcPf3rrYB6OZB8P8RQ0WSo6ilFHBU0TU6wssY+osPqPfq1FerU9OqDvlv/AC/s9sDfFR2BtLHzTLFXtU6oLaSobyarKPex1WlOHVn/APKx3nvWlpauk3HjqqJaLy6VlVjYD0ggn/W49+Pz62PTq8Gg3o02Q89RAUpvEBf6LfSf6/4+6+VOvUr0tsXksLmqedayNNQOoXvZvVa1j+PevLr3Stp8ti6aCGnpKVQIfpwDf83B97oevdJbdO75JaaagjTxJ43OoDj6ce/dex1Rx/MP+Tn+h7Zubp55iDPG4GofUvCQv5/x978+vE9a8vxD6vzfys7lk3XkKKSpxa5OVoOWZXUOXAAN+Pe/n1oDNOt03449JYHrrYWPQ4sRmCipwYmSP1FByfp9P9696635dQO8q2rxSJW7apmp5afQ1oxaxtc/T34DrfDPS36U7jiyG3aik3sodmgK07VCg2fTpH6v8ffiOtU6QtVmppdzzRU86w46aZpE+ioqMbgA/wDEe/eXW/Oh6RPcVHjt07XyeFqqZZzLTmOGcqlidPBX34V60fLqqus+POHx0dYMzFHV00xn8MDlbKWYkWFv8fe+tdFb3p8dMEIMhR4+lSCaqMskE6NzELelR/rW9+69TGOq1e99jbm2RVUFBVTS1tDIyRkm5CozWB1f8V9+63wHShpdqYTFbL29uGgrYabJ0+TpahbSWYzR2ZR6Ofr791qvW8t/Jg7E3Bv742wyZ6ead8aYaWlaTVbwp6eC3vw9OvfPq4wMCARyD9CPe6dVr6dcveurde9+691737r3Xvfuvde9+691737r3X//0t/j37r3Xvfuvde9+6910SFBJ4AuT9T/AK/HvfHrWFFeqq/5rUcLdJLO88ET06ySxiV1UuQ9wAG968+tjIx1qBbb3FfPV2Ua/ljyE6Kb+khH4cf8Rb3vj17owcO49wb5SDFSN/kttB8ZPKngk2/NveuHHrx6SW7vhB172NDqztDU1E7ixKo0hFzqJt+Offuvceq1vkl/L2yHXjHP9a5CfGSU7NUKpkaNj4PUi/i30+nv3Xq9BV138hfkv1yKbb601bXfaKIRMnkkVrD9Wrn37rx+XQj0n8xTvXFZ6oxe6MXkaeipluJpElRCqjixbj37r1acOjl9Q/zP9zbhwlRsmjEzPVQSxOHdiCZlKFeRx/h78OvVPRQxtnf1d3DXb6oRULV5CtM6iMMIyxfXZiPr79w61Xq0LY2/e0sdSU1Rk5DqSOHUqteyxrxb/H34+nW6noftv/I+daiLG5iCoEjWHlIYR3B+rsePfs9b+3oyey+1cVV1UfhyFGWmewiFSjOCwHBQf8T70evCnRl4KakzVC8lZCksbwXB0Bh6hcW97rnrXVE3z/8AjlS7w7Fw+51pY4afFVUUnMKBDoty3v3y618+jUfGXdFLhsFgtvx+IRQNHBJoFvQF0/UW4Hv2f29b4cOjrbjRKjw/YOrROilgGHDfU3t79jrYPSKzeJ1rBT1iN9nMq+YgAf4/ge/de+R6gviMbTRU0GJFlRwbqPVqP1Bt+P6+/cTTr1ehm2vTVzRxxzAhLIrXv9P6i/v1c9e4jHQ2QYOgOKm1AktG3Km3qC8j3Wpr1sgdavv82+jGLp6hsZJVpIZJteh2RWDy+rkfW3vZ6r1R1g8nU4/CzLRSEVkkbEsz3Yl47fX3uvWj0EnV+Z3d1t35tXdc9XIkNRkfNJrnZIyplDWKn/ivv2Ot9bsXxv7bbe+1cJlJZBKHpsfE3jbV/uoC4Pv3Xhnq1nrbPNDT08hIWPxXIkP1/og/I96PHq3lQ9GFp+xBDSzUwZP3U0pYi4J4A96p5jrx+fl1XH8ue959g4GsEc6LNVGohsZNJHkuPr/Xn3v59awMdEp+N21k3bVZDemRaU+WU1Nw1ybtr4PvfDHWuOerX9h5QUdKkdGpIKBQSOeFtb3r59WHCnQuY3D1dZeskcqrKfSWIHP1+vvVOvV6JD89OsI969Jb0xsdO0tVPQSpGwTW1zCRce9jrRoeHWgHvbGt07vuTbFUksdVFk699Et1IV6hudB59+695V624f5Qvb2Pi6oosXMQtU8ETJ5GFyS/HB+nvx60KU6u5TI4zcFWpr3PpYePQ9v9iR+fej6dW+zp7rMBEipLQLM62HNjYD8kj37z69X16T1RiRUVSLWLIp0/ji4+oN/fq9ex0qts4fHQVhdEk80TXX/XA97HWvl0OgkT+GNJUkIDFo5JC/Tg/wDFfejx61gdUy/zLO2aLrnrjcFLDULHPVRzNGpkC3LQEi35+vvY49eJqOtGrP8AcuRk7Xx+cqUqGievncsxLJpWo/BPv3Hr3Drd6/lldh025emcHkqMp90aShuA1zyAP9b/AFx78T1sCvV9/W1NJn8W09Y0fniU+AAgG/8Ajb3oceveVOom9thUO6KGtx+4vVQtA8ehBdtH+N/fvs61w49UCfOD+Wz1huWgrN7bOp8lDuairJK9JIldTqgPkGkp+OPz7tXr1OlN/L87dyONSPr3ctW3322lNHTrPOTJdD411Kfzx78R175Hq/TY+6XqykFUVKtHHzcWsV4Iv7qPXrZPkenLesNLNTll5v6gU4I4+nH+9e/Y630gabHyTY+Twxs3oPLJfn86SfeuvY4HoLa7bWMkqpZJ0/eXUVX8ar8X/wAL+7cetdILKS5BZWpI40FOAUHoAAB4vf3r5HrY9egK3jszETrUS5Gkp5zMDq8kKyD/AHkH3up4deIHHqvrsr4LbU3fnIt+bdppKbORVQlR6SHw2s1wD4/r738uq5PVgXxxxm69h4ekwucq5HSCEQxLLKWsoFrEH3o0p1sdDB2Pvej2htndO6clLFDDh8bJXRNK4QfsoXOknge/AZ68eHWtj13i9vfzBPl/NuHc8SZDaeByclJHwtVD9xDNpHPI4I974DquOtj3avSu3ersRSYra0UUEFCvipxFEsRCIBYEL/xA96PVhQ9K3KZHKrj1WsZmiUWBu1yB9OR798h1vHHoB9x70ppnO3Y1KzT39bjg6uD9ffgOtE9BznumMJndv5Kqq6anmkljYmQQI7hrXFifzf3vievZp0R3Ze06vr3f8s+CeqQiqfSmplj1B/SbCwt/Tj36vWurHdvdr5ST7LE7llbXLCoT1sbA/jn3ojrdTx6ETJ5mGCkEmMVXaZQG9N7j/Ye/fI9WJp0S/uUR1MNRU1sQuyTA3Tjk3sF/1ve+qfb1Rf8ALTahzUFTNhXETrNH/mjoNwfrx/T37PXurhv5R+2DR7Vp6fcLQsJI9KseJWvHYXY/X34nHWxnq3Xe2BoaE/bUEetHjdrKtyP9Sw91+3rfRSt17Yhyks33kLGNWYG6EDgW5H/E+7VrTrXVaPyW6Wpc0IqnGU7JJQsahJEjHpKNq+o/4n3rh14j06eOpspV4/YE+PrmlkqIF8C3bkeNbX9761XoI9y5uujyFQ8kc3hZjw19Om9re/dePHoFtz5HBzpVGohUzPE4HoF1Zhbg/wCv7917y6JtuymbFx1eTpGvJHMXj1XuADce/daJ6WOzO4MzXbarcblx5aYQtHZryLoC2HB/HvfXugtoOpZOwsrVZHFroSl81VTiKMBhOAWW3BP1968+vVPDrYU/kl/MPcPXu84vjrv+uMRyFQIaEVs5VfGZdETIkn+29+PXuJoOtvgaSNSkEOAQym4II4IPvdevU67AAAA+g9663137917r3v3Xuve/de697917r3v3Xuolf/wArf8AqEqf+tLe/de61Jf5Bn/bwj+Zt9OOw67j+n+5X36tadaHE9bcvv3W+ve/de697917oO+0t647r3Y+490V9VDSGhxVZLA88oiV544CYl1E8c+/eVevcTTrRs7t3dnPlp3turMb0r66eh2/nplxcSVTy0rxJKQl0JsRb37gM9awT0tsX13gsRUUcK0yRpCwMXihVbkfQMB/xPvXViKcOjb7Q6koNzQxtUpAD40IBRdWnj6+/da8umfsnYOO2fV0hgSzLYKUBTgcgen37rYr0xbXzuWiyCtSzuiU7CRAHKnUPoPx731odGIl3HuPc+CeOvRmhjiK6gCdQC8nUffut8OPRO+wessbvmWbFVFL5jIWjfyU/ke304J/3v3oHr3RXZPgRtKTOapcapPkL/8AAUfU8/Ui/PveevUB6Op1H8Z8ZtI0dPjaVacKiKCkAB4PAJH59+61w6Pltnq6aH7eCSIlZSB/m+bWsCT70fhNOvMNSlT59LPM/G/HYhH3YiRrO8RYAKFOojgt9OPaeOCWR9Z4dXtbJI8qKHonPc+zcIMLkcru+elipaOOcKkzKNSrEf8AVH+ns98W0S38FPjPRhJpSMg9akny02ttHfHZGRodqU6GOKcgS0saljaU3sy+zj2u9tt1595gk+oUrBCeJHb0Dd+3aSxjAtviPkOmHZHQuOoaJGr6rIxlgpCCRlIBW3JP+9e8+dm5D5Y2OyWz+mjnYABiwFK/LoKtvd7NHWQkHpdDpHbJkExrMoQpPDTav8bLce1B5L5PMvjNtsBb/S9Fk+4XEuGdq/b0zVPUce28gm4ds1FSmQp2MtPqdoz5Abi7r7L9+9u+VOY7M2E0CwKcdg4dGO3btPbkd1QPI8ejodWfPPsHqTbK02555negZIgDUO58UX5/23PvEj3C9rdt9v7zTZGsEgLA0oOh7Zb74qaWp1k7k/mdZzvrbNRtHaFW887xeCqhp6lnmDMvjYBF59wmLkSysPIcOl0kBuaSnoBekvg52L3hWpXZiDJrSSztNKJYZAzRzHyMutv9f27Utk9eAIweri+if5f+w+thSzjEF81EI2ElRRCT92I3UkuP6+99eoKdHK3B2T2N1I2Kws2PRMO+iOkNFTmLTHewLeP/AIn36vn17gOjHbByx3PSQ5vJ1GjzKt45ZfoxW54P+x9++zr3Hj0La7e21XoJZauPQCCQZeTbmw596z1ug6ETHRYbOQ0m36Gcra0ayCQgEWt9feurUx0FPaHW+c2JMuWpqvXTm0pCSHlb3tb6e7DqmM9AllPkhRbUqaPGVcdaamURoXRm0AyHSSQfeqDrdT0aPrrqnYOVpKXsqrx1HV5HO6XvPTJNJdhqDMX+lr+/fLrZ9esfeHa25+ndsx5HbUECwJMiCGBPGPEDYWVf+Ke/cT1r59LDrLvU9mddzZCs/ZzQpWcJ9LSCPTax/P8Are9cOvdU9d7/ABj3N3j2bR1Wb+4bHw5AycBnjZfNqB0/0/w92+zrR456uF+PXT2G6k2jj8fQR0qSRUNOjXjVGLIo41fj/Ye61PXseXQi7sqa+qlCUs4R9XCxMf1f64+nv3AdeGegHzG6N0bRrjVzzz6CT/bYj62sB73Trdfl1npe6/PWQy5FIrALczLzz+piW9+p5jrxPl0K9LvTbe6Io6eKmgvMNLMYla1x9Qbe9dePDHSe3BtzGYRYq2hYpK0iswU6eb/Xj+vv3WxUnpdnddQdoT0InEUxppdDM4HPj/J97611qQfzYKufdm46fDqz1c1NUr5hTEyMAs1hqC+9jrR49HF/lj7WocXtnGGopUSQGiZnnhCkWtqPP+8+/H068PXq4/5CPXN1zNUbIrKeOrgpyrCKUAhlT1EgHj/D3rzp1vyz0T344bkh7RyabW3ZUO1dhqhjXFpdQMatZgAfezjh17odfmd2zs3pbo6ui2fOsNa9NPTAJJ45mmdNANhzcn3759a6oh+MfQuQ7d3zWb47Fhnq3y1Q1ZjPukMqKhk1IBrH+9e/H168B69XXx/HjrCPB09BNQU8desUSxNT0yxusmmwJI968urfb0zfwLdvXdM2DikmqdrTOAyq7XSnP6uBxwPfh1r5dF6+Ufx/637i2/jKHYJrf701kUaVyQMzgSgXJYJ/j+D731rqL8Ood9/HncH+jbOM8EcjCOCWpRoeHGlCWb37rw6tM3nvXN9U7FrMrUZzHS1WQR56dErlMgEq6lH1v+feuPW/n0Q2HqTJfKXNUmZ3/IaiippvLQA/uwnxvqXXq4t/j79XrRHVu/UXSu1dl7FpcJTx0S6IoY9EUceqygAiy+9Hj1b7emPctFU7J3JR0G1opGWuKtMqRkjU3LcD377OvUrnp1zVWuAoZs5nJEpZETyP5mEVrLf+1a3v3VaE9UpfI/5+Gj7RXr/bE9NUySyPTEwT+SxLaR9Pdqderig6Su/sFlexuvayfM0RnTL0ztKz03l58JC/UfT3759b4461MvkD1ZVbS72psdOZ8dtuXJSa/rSRsrym6ki3HvXWvKh63Af5YXSXXlT05jKnDRY+oc0dG7y+OOSRnKgk6hze/vdevY6uAwm31wM8UGOiUGElWCoAvNj9Bx71Qcetk1x1l3BgcnuKp8aCMMti6oLMyj+oHvRznrwNOstV1D19uXbVTjd2Y2CScxPqM1Mj3KqQeT798uvGvRTdkbR2Z1/vCvxG3KWmp4J5mQGOJIwQWt+Le7GtKnrw6Ou+08dLgopLR6yiNqUANzyRx7r59eHy6ZKTbwSdWpJ1Cp6WjDjmxsdQH9fe+tVr0qKiheip1kU2Nhr1G5ufwCf949649eIx0Xvt3fNHtLb2UytRUwwpTUFXKWllCC8S3tqP+PvYz1sY600Plz3Fvj5pfIAdUbPFTVYqnykVPWS0hNRFoScRspK8e/AdaOT1sk/Bn4Vbf+OXWOIyNdj4o8x4UmkZ6NI5dckAJLG1739+NR14enVn21t3UlTt2SjkaOOeHVGqmynSv6fT/re9Y49bNR0C27snDX0mTik8UkgJVC3I4v8AQf4e/VzXrdMU6BKlqsZLAlJIxjqI3OgRPo9d+AQP9593+XVaDz6VMODzElOtXI6mG66SCC+i/wDX3qo63QnPSqzOJgkw0IaIvMYgrekm5I5JPvX2dbpinRc939fNlKJxDGS6am4QXB+o497610Tnfu1avFRVVT9s96dmUnRf6DkE+9dep6dV/fIHbWP3FQ46KSGN6iVo1NkVn5NuB9ePe/LrXVdXbmyt27QpKSojNT/AqWq+7MQ1hgkY18p791rPW4x/wnw+Qm0N79DVm0hX0tPmKWRdMEsyiaZo5dBXQefz+fevPrfl1sd+pGVQB4+F/Oocce74I6rkGnWX3Xq3Xvfuvde9+691737r3Xvfuvde9+691//T3+Pfuvde9+6910SALn3sCvWiaCp6xTTR08EtRMwSKKN5ZGY2CoilmPP+A9+68MjPWp5/Nz+VmU7B3HN1jtWulenw06RVsUDKUEazaWDaT/sT71mtet8Bjqj5VqqBVhWP/PTq7sPqXf8AV/vPvfWuj49E7OlmoIq6VbsY1kuRfSuq5B/p71xPWx0duhy/Xmzdu5rL7rykdFUw0by0CSuqeSVU9KqGIvz711anVNfZ2+N7d47rrqDBUsxwMWRlgEsbERPTh9IYAf1HvfVOjOdL9G9eQU1JS7ix9DNltOlvuVXWzgc3Lj+vv1erU6L98qOksFUVU1Bjtq4uGJg6vWQwrrKarA60H9PfutEenRYevumNr7PyEZxtFTSZCVo/QsY1Bi17Dj36nWqevVkXXPTdPNDSZGvo0haRfIgKn8j8H36teHVqCnRocd0pQ1CQ1GkCEaT4+dNrc3A9++fXqZp07bn6JpsvhpIcXiaYOEcGrEZWVTpsCCBf37PXuitdffGLdGA7ATJzZ/ImMVLSLRF5jCq6rgafp79WvWqUOerkNvLQ4fasMGQaNZ46SNLsRqJVfzf8+9efVq0HVWXz+z/8L603Ln6GNPvKBZZINDDWdENx9Pz731U+vVFHxE/mQ0tHuxtq78gOMT7yanhqqmNV9KSmPUrn/W9+695dbAnXnd20ty01HWYnNLkaOeOGXV5FcBnFwpKn6e/Hj1sGnQ653cSZikposZGssbquuQW9I/PI9+691G2jSSHIGOoQ2Ulhr/TYj8H37zz14joxNH44aREgRfLpX/b/AI4/Pv1OvVqOhT21h6vI46ZpgVXQfybcj+g968+t9VBfzF/iNWdqbVyE+HgFTWpTzyBVV2Ie9wbL72KHrRHWpl2H1XvPqDK5Ch3LRS01PFWNDG/imK6VNv7Q/p791rptzHUGa3picbu3b8DztioxUDQHYjT6jwv+259+691sD/ywOwqzcGKx2y8uhir6B6VXhdApBjXRyDzb37rfWwtiKeuhx6pRhiUU/p+vBt70aVz14V49NW5+yMfszE1NTlqsRVVPF5FjaRdTOvIHJ97x59e8q9VMdt7ky3yN3ccLAJPs46shSFazJq+vp/qPfq0HWuPDo8HRfVGS2hhKXEhGSFoFjfknURbk+/dbHDHVg21Ns0cOPpKdADUDSD/W4H1PvXn17oXKnFVeOw4kVLCwsQfoP9UPfh17iegc37RUec2/X0FaofzwurRuLg3Q3vf37reBjrRp+fnxpWu+VFM9BThYa3JvBoAbS5lqLXt/X8e9/b1X16va+HXxnr+qtn4GqgTwF8bSy6VDfmzfQ2/1/fj8uvCg49Wibaq6iKSATagy/qLXHPBvz/vPvxz17z6NFtrcNJNDHTSiMmyrq9LD8ACx9662RmvTvm8etTIk1NTL9AfTb6Hgcj62Pv3Xq+fTjjqCmxcKVRRXlkGqQMP7X4sR78MdeJ8+p2TrospjKiJX8Jp4XkbSQBZfqbe9eXXvPrVr/mr7iznY3ZWI6028JKuOtmSmm0K0gAtoe5jvb3bPWic9Vidk/wAvKTb2w4s8aANlKWmkqCDFKSHceQkcf4+/da6Ot/LH+VmO6tlh6r3LUGjqKSogo1jkTxkiNtA067ce/AVFOt+eOtsjpftDzihqaGoeopK9DIhVhp0m3+PvWeHWz6jowuQzuTrMlEJoStFUsq6+dOk8m1uPeqjrx6Zt/wCzsdW46qphCssdRjqjgjjXJEbEn/Y+/E+XXhjrWp3Jj8j0B8nBUTaqWgz2YDRqPTHoknuL3t7v8uq8M16vz693pNlcVjsjSBXjlgpfWPqf27/j3Xy6sfUdGDpMl/E4Y0qygNgfW17f4Ae9de+fSxOVwFHjjTxyxiUJpcArcX/tAH37r3y6LzuTxGseoScLFd2uHAWxP05/2/vdMda86dBxnc7jIYSAyMyg+ryRi5t9fr711sA9A1LkJ9xV6USRhkd9Aa4554uRx73w638uh62n1bLTwGWojHgaLyD1ELci9v8Ab+9Vpx60fl0hsrtPJY/NmpVGWKOQmNfUVYDki/09+62K9VSfzd+/arrnqLHbY27OxzG8Yf4RLBTurSA1J8H6V5tz72OqtXoJ/wCUX8VM/wBf7VfdeQhcVGZqUzMk7eRWJqm8zXv/AEv78evD59X9IxrQ8RcyGnYq97ggj/E+9db6R+7cqaOiEbxqYlBBYC4/2Pv1PTrYPr0WXeWL/icD5THRKtXEw0soAcAG559261X06fNh5g1mIqsdXzEyqhR1axs1rc+9GvHrw9OgYzW1qNN0iojVdXlD/jklrj6e99e4DPSh3Ni1imoK1x4mijTSV4PAH1t+OffuOB1odDTtarxjYJZZ5RNIkf0e1/p+L/0968+rfb0XHt3x5ejqooIQU0SgEaR/yFx73w60Pl1RB8rcpjNkPWCpriHkmDtFrUqOb/S49+6rTz6F34XfOPbXXMGMp67LCnhMoB1SKAVAseSffuPWwSOr1tlfNbqXe0dNPHuGnnrJII08LTQ/2+Bfnn3qh8uvCnQtybl2ru2ELQzU1qoXLRyx/wBr83U/71711boIuwtibOhw+Q+7yEXmlpZAmqVL6m/Tp974nr3AU6Klsnquf7SvlgVXoGqZSr6+CNRN+fexnqvn0ie0uuGgoZ3gpY2kKekqQWJsf6e/dbp1XNvilr8L921bSKkY12Okj6G1+fex1roqudq6fKQVULSkM0hHjufr+Db6e9da+fSp6y2xNksXl6ZqJFiWJgJCnBWx5VgPe+t9Cj0TE2E3fV4oIWhuSU5sVv8AQj3o9a4Hp57F3bVdLdx7b7mwBagmwtZRxz+AEM4M66i30Hv3l14db13wv7xxXfXQWxN6UmRjrMjVYemGUjMitPFUiMai6gk2P49+63506Nf7917r3v3Xuve/de697917r3v3Xuve/de6iV//AABrf+oSp/60t7917rUm/kG/9vCP5mv/AIkOvv8A+fXi3v3Wh59bcnv3W+ve/de697917qgb+dR8oKrYfXUew9o1sv8AHa12p6qmgYFykjaHLKOf+Ne/da6pA6f23R43b1Pm6prZnN0sVbVXUhzNINbar+9H0630K+Lr6M5LTm5vtolb9t3sA1jx9Pfut+XRgtnbpo6fJxiir3eIIlgsoFgv4+vvfW8U6i9z7ppq2KGdJVmNMLudQNjbkE+/dV4DoEtobiXIVccVEsbTCWzoXUkr/Qm/+29+62ejk4XcyUWDFFPSRKWSzMNJvccg+9Z69w6w4PF4epr2r7RK7Nqtx9b34v8AT3sdeOOlrT4XHVWSeR9IHFiGFuP8PfqjgOPWq+XShD0mCqIp4dLcD08fS/19+JAwevEgDodtrbypKtaQtEoVLam4Bt+T70Bnqy8anqV2LvxYcPUSz1HjxlNTvLfyBVPj5tz/AK3szF/aQwmBqVI6XiVFWtetWX+Y78xclndxv19svJVCrNUtTzmkYMF1DSbhD7O+Qvbve+a97WWFSYqgmtaUr0HN33ZYEJJ6rq2HsmopETL5ljU5GoDM7S31qGJN2B/PvoFy9y1tnKlgLLbECswHiMPxH/N0BJb8Xb6pvy6FtqRWiCopA/oBYj/Wv7OA7A561ohYeXWWCj0Kwbg/0P5v9femkY460IbYGo6bp0araaBF4g5Fj+oj8e30YKAT59IZIGWUyJw6Qtds/E5qiyLZlYkWSKdEWSxJYoQOPcP+93L777yz4lsKvCD+w9L7GUxXALHpefy1vgRWb47J3Du+ppFnw1PkpZIkeKRYjAs1xxa3094Ffu2a0k8JsUwepP22XxbevW3d0z0njcCtNhcTgqWLTHDE8kagWsgQ829vsCraevH4qnpV9tbZ/wBFSJkWoA+oBiNJe9+fe618+vY9OkZRLg+zcXDNksTSySLHqiMsa3Ww/wBqH9fejjr3HotHceQpuq6N6yOvNFTKHZYImGi6twAB7317Pn0TT/Zrd3Vte1PhDUVFIhALkHTYH63Pvxzx61w6f6j55bv2NEJhQa56Qa2dfGWuPoQb+/Uznr3Dp72p/M0xPaGQpsFvCtNJqmjgdZ3jUcmx4J49+63Xo5ssPTm9sNSZ1K6gndYUlaQvDrGjm1wf9j7917pU4X5R9dbBoocEc9E8NJZIoWljKJbj03NuPfqderXj0E3bXy42dn6GSmMlNX0RBK+VoSlv6Wv71Tr2D0Cmyvmp13tGp+zrchBjqYsF8URiCEH6cAi/vf29aGOhupfnF0YDUZE5+ENHF5AV031f1Ue9U8ut1pw6C/K/zRtjwVcuPwWamqzGzRxKIne/4HKj/D3vT16vn0l8X/MfzP8AHIH+xq6ihmku8rUsllX+t2Hv1Mdar1x7O/mLzSPAsWFknpgya5TRu9luNV+P9v7916o8+gW35/ME23NiVbHq0WSCKWhjpWuW+rD6e/fPr1QB0peuP5oOzMJiBHmapYMjDGfErw6DI9uF9Y/r+fevt69Xz6GbEfzK9m7op4J8hkVjJlASM6FUi/A/F+Pe6Z63Xpz7K/mBbOo9tPLDlVpn+2kEelohq/a4+h9+pnr1cZ6oy/024DtntLceTz1V95RrNKYGqCCLByRYm/v1Oq4Jz1Yv0x8hNgbGxAhhyMFCFiUL42jFnUW49+63Xy6i73+fKYqSowePyc+SpclJbghlCudIseePz791uteh6+NW4cVT5GLekWSENVnpVWaEOEZRKb2PPvWeHXuGegR/mI70kqqzE4WLItPS1FfSl4TIrKdcoBBAPu3VT8ujR9ZT021cX0zQ4+ihhTMY2lNVNGQBqcC7X/F/eurU6sYfBY5KuOaSv0iVIio8gIUso/x+vvVevUFellRYjZ+UmXa9RWR1OUqoSYkkkDfrUgHn36p62egqw/TQ6p3Hld1Uq/xOaKdp6aha7R8nVoAP+Pvx9PXrWPLol3yC7Ers7uHI5ysxS7dykFLKtKIo7OZojZGUpbn6WPv3y691U12P2X8xdzvWzY3GZbK7coKsRwNN53iNPGeCB9ACPds9VqfPqzH4Gd39ybwoodqbp2pNhf4erQipETIXsLA6iPejnrYJ6ut2dWS7WeKrz2XnRJY1PgkLmzMPpz7qeGOtkVPQ17dr4KqsTcUNHHkqem1yB51v6VFxa/H0/wAffh6nr3VOv81L5g0uydoZekw9ScdkWoWCwU5RTrBICkA/W/vdK9e4dUifAPo7c3yU39J2duTyVMz5NJIRUhzaJpNQ9P8Aj731rraPxHQMf9yGwaY2J5Yqdoo7ofp49P1+l/eut44DrWZ/nG/A04PYUm98YXoMzTRzVZamModXViwI0f097619nQ//AMg3sDI5LYEe09wZaplkovBSM80lnPiOhgFJ9+4Dr32dbHu6azduAqZpNs4Y5WmdjqmKFyAP8be9DPHrfDI6ibK3NWSZOWrzkbUlZx5Ka2mwtyLH62PHvXXsDpT7p3EMtSSRY6NY20unosGuRbkj37rXp0UbceyMpt/JJn5tQaSTyX9RuSb3497FPPrfHoQ8B26skAxdXK6WQQhgLEFVte/vxHWhTz6EfYUEmTr3lSulljmbXYvdfrf+t/evLqx6VO9qw46lnZnKLShWbmwFueSffvLrR61pv5n/AMzK/FY+TrfaVVJNmspUz4pI6UiRy1Q3iuVTm3Puwx1riOld/KX+AlXhKcdyb3xUc2YzAOWElUGErNI/m/tjj6+9H0631sDZ7yzlaGGmEdMixgR3shKAKAB79XrQHr0AG8qzI7crxPAnjjb0GOPlfVxcD3vHXiT024/G1uS8UtVeMVw1gMSS2ofn3rA4dWz59QsZ1tiq7cFSs+RMEsIMgQNpuQL/AEv/AF9+qetEdQt476g2TAcUZQ7h/Gg/WHVT9Sp59+HViaY6fNndnYTNUBTJOIgqcalC2/pYN78R1Wvr0+JLha1qh6CSOdHjbjUtvULg8f096oR1vooXeNTgcThcjBVPBHVzGR0UFCf0ni31v7txx1rAPVVtHtmq3xvSkTQ746mqwSxDlAge5Jtxb37h1rj1L+TXUmMi21UvjYY6tzj3XxfqUSGPSQVH19+630Av8rjuWH4qd2q+8s/Ubd2/PXK0kCMYoP3KoOwNyBa3J9+4jrXn1vydLfJrp7vfF0tTsPd+Py0/jiMlOlRGJhIIwTwD+Tfj3uvn16nl0Yf3rr3Xvfuvde9+691737r3Xvfuvde9+691/9Tf3JA+v+t79SvWiQMnrv37rfXVwTb8/wC++nvdDx61UHB6Lj8pO2sZ011BubdWSqIlaCiqFgjewMl4W4AHPH0uPej1sD160HO5OwM5uPuLdfZXmeoxG4ZpWhp7syRqZzc2Pv3Wjxx0Ac3Z0uY3fQ4egALGrgVoVvq5fklR7317j1fh8XdnT7kxWGokhaNamjUSy2sqPbkMT+f8fdeHVgeivfzOsRUbJxODxGLyKtO0scVQkUrBnDS6THZLXv731o4x0Wzo6jymFwsU6wvG0ixSMGALMSou3P19+x59azSg6ELdu6M/jJVydFDNFJDpIcBlJP1uLfX37HDreePQdZDuGfMU8qZucTTiNlCN9b8/XVz/AIe/dex1m6qxVPuLPJkzRlVEqWc2C/r+o/1/e69e8urXtg4SCsxsERjvHCCFFlGkj6gX/HvXW+jLbK2m08oDxs1NdSABcccXv/xX3qpHz69x6E7IYvHY6H7WHREzrzfSSOObi3+x9+HW+PQa7ifF7dxGSyxo1qammgaVSiJqva4Nhz73XrVOiAZ35VVrV1Zj6nHSpHG00MYKv6dLFVP/ABX37rVOiN95b1z/AG4tRsSOGZocySPKAT443Gmx1cX59+61x6KRW/ynqTN4KtylNUvBmkgklgmSREkEz3YFdP8AQn37rdD0y9JY7sP485Mdfbqp66qgiqhHFlahpTGIYn9Prbj6e99a4dXP9I9gU9dRRU00yTy1Ysr8t4h/ZBH+9e9dbB6NWmSoqBIC5RXJX1mwD3+huP6+/fLrfHoUsPuDHRwRu2gsNBIPJ/qLe/de4dGS2Vn6Orx7wiLx61ILlbDkcEn3UCnXiSc9JPdeMWQzCeJZ6aUMr+lWRkY/S3+PvY63w6qF+e3xD2p2JsfK5zFYRY6tPLKzxxRKdQQk2IH9fe+PWuqhfjNids7bqMz11naSL7i0lNGsyre+ooANY59+61xx0arq7YR+PHZz7/jfRhci8TLC3EaajwQE9+FOvcOPVpVP8vMPjcNFJRolVJLCSVjJLAunBt79TreOiwbmy+9O8c7rpjPS0LuQ0d3UFQb2A9+PXvLo5nTfQ9Ftijhy81Of4g6xlmIViXAvc39+61k56OjtPCVMrx0320iav0tp9IB/xHvXE9WrQdDntvb1VRVzyVPMcXrUkCxA/oB71T061WvQqVNTDmce1BGo1qtgQOCQLX/w/wBb341HXseXReN64WelSZir+NAwY8nnQRce/dbB61hfk9tOq3b8ncRFQwvKaXOJJLoW9gtQPr+fdj6jqnV9OxNt/wAP2vttZSDHFg6VJIyFF3CAm/8Are9V6tTzPWDcdX/D5TLTAlEIYhPx+fwPdvLrRGcdNu1ezqNs0lFLKiMJAoDtySPrYD3rPWwaZ6PBtTN0mVSmQhTeIHjnn8NYe6mvXqevS+OJT7esqZUBigVmRLj1f4AAe99e6Kh3F2JR7K2tmMsKiKiY0dZGI3Nm1BCBwLfX349e6qd+NPXWM767NznY24Kf79MPk5DC7KrLYTGxXyA2F/6e/V68M9WG796U23m6apoDhlmoqtFjRUijISyWt9D78D69bIPHqnbtT+Vtkf8ASlB2Lszz4+OOs+6amp2WNTpk12ZUt72Oq0PVzPxmx2X2ngcTiszSytUY+nWIs2oksoAvc+9E+nW6HgerJtp7jxe4XioKilELUehjJIp50jkXP+PvX29ep1m39uzDUatQQxr5fAyhzaxAW31/N/6e/de61jv5tWRfA7k2nvGhcxHGT088gWyawHDEEL9fduq+XRwvhL8qtvbm6wwqVtXAKoiCIpISXusYW/qP9fej1sfPqyfBb0pckgqYchEkbKWVATc/0I96pXj1ulPh6T+4+zcFtZaitzOSp4KePU5aZyoK2v8Ak+/fMdePz6px+Y/82Hrvqz7jG4rKUFTLCJxqic3BQ2/sH6+7fb1r7Oqytv8A82//AEj5yLGUWSjpo55Iv3TI6KA7BTcE+9Y69Ujq8P40/IzYmRxOHrM1uXFvVSFS7PKha5QG/J97PXhWn29WNDv7Z1fQQUeIzNFOulLvCwb/AKFPvVD17pAdw96bP2ls2r3FWZmjU0UEjSIxUaiIiRx/X36lePWxg161gslk9x/NT5M08UkU2V2dgc2ZqVY9UkI8dRqBAe4sLe9jrRz1t0/HnqDHYzrzF4TGBKGWHEUyyIERDeKED8D/AG/vXn1vyr1yyyUW2p8lRvTl5ldlaQA+o/QMQffuvZPRdt6Z+kronpGUUwXVeQ3APP8Aj78PUdbwB0CFTvDH4vy45ZFqnkV+UF7cfQ+946r0Fe392LDuk0UcoLZKYqqC4KktYAqPfurHOOhgqtl5QZWKsfyaJCjKD+AeQRf36o6rSg6XOR2n95SRU9Spd2QESH8cWFr/AI96r1ann0gslhazCokEMxaI8PYkgL/U2976qfn0BnaWZjweDrKhZVkdKeZyB6j6Rc+/fLrdcV61rPmBka3tXLZNqAMsVDUlJipIBI4JP/Ivfvl1rz6KX1l05uPf0r4TF1M1JNRFgZUkZOQeSCLe/darXo4m1PiZ8jNuQJlds7jyUzQlZEVKidlYJyqAA+/eXXs16FLGfIv5O9L11PQ5zHZisSmISSb/ACohje2on3v7Ot9LTL/NnujcYhq5sLlZqaNgZY2WosQp9Qsf+K+9de+zp1X+Yv2BjqVMJQ7Vy1OAoWRhFMFLKOTwPfqde4cOnnC/PrPVziPcuErFQfqE6SFbX44I9+x16vWbP9+bK7NQ47xQUM0lgWkAXUW5YE8e/DB69k9Fl37t+CnykBwMq1XmkRtFO2okE2Av791qtMDo9nSfXVV/ceoqKmilhlnpmYF7XBKX9+63SnQG7Zao2P25UzZZD/D5Ziqu99P67fj3vy61U9O/y9xmJ3Ts05LDTxlUCTyQRlVLlbOBx/j719vW8dXCf8J2fkJlMtT7l663DVt9rStLSY2KRpD42h4QgN/rAH3vr3W2f7117rgXAP8ArfU/ge7aT1UsAeuEs6QqrN9GIA/2PvVOt18+soNwCPyL+9dbBrnrv37r3XvfuvdRK/8A4A1v/UJU/wDWlvfuvdakv8gz/t4R/M1/w7Er7f1P+5X37rQ4nrbl9+631737r3XCRtEcj/6lGb/bC/49+HWiaCvWlP8AzOtx5DdPzIk27XSP/DkeZ1QuxUFZfyp4Hvw62fTp4wG3qVdq0ckMQk+1pQomQAogVbDUfwPfuvdF27I3E9PKKanpHneBv85GpN+frcf4/n3ode8qdFmz/wAnM31zX6XxdS8C+kyaZSFCm36hz72evdJ3O/zBNvTQR4uupVWqrUsHfV6CRbm/59+pTrx9B0W/HfPKLbG9ZY4alUp/JcWMhBBN/qD79WnWs+XVhuwP5guzsnioJclWU2opGG8jMFB/1XJ9+8+tgn06Mjtr5ddfZSjmraHPY2NkUs0ImB1H+lr+/Z8urVqehk2P8kdrbkJjp8zRrOvBCve5H5UX9tXsn0kCy8Sekn1AExSnQsUXYVHlUaRp1mCt6SpJJANvp/j+Pa2GOKWyN4WpTy6MY7cyivDpc0vaGH2xRmur6hIaeIXcPcA3F/ZfaXMV2kmkjUnAdbCIJBGTx6qv+bv8xSmiwtZs7ZrAVU0U9N54deksSUAuvsZ+3/tdunO25IzP4a6vM+XSDfZ49thLE1+zqlraGHrtwZqr3ruk/e11ZL9xAXYyeEO1yfXfn30H5Y5Vs+S9nTabM6nA73pk/IfLqMbm+a8lqwoPLoaDrQ61U+OS2hQLEf8AFPZyHU4Pl0kazk+MHj1KhDKdTNwRx+f9h7qzKcAdeW3lXOrrhOJSQ6kAC5+vJv78GXh04YpKca9RKmoWCINGoSRxZ2Nhcfm9vflTU3yHSpZtMYjI6RKUOX3PuXEYDHQSzipyVIkniBI8ckoVrn2n3mfbrXaJ2v3UKyMKEjjT069H3yAKPMdbcXw36CoOnetdtyw04hqNxY2OWqvHGCHkQE8jm9/fNPnLdYbbfJY7VdauxoR1IG1yFItPCvVgW3MFPh6gZGnqNChlfUQObC9ufZQral1Hz6MiScHpT7gxOF7Hpnoc3TrOkaMWlKiw0r9fV7159aBrnqvfvjtHYvSGPyeIwxp6jKKnio4IdJkV2FraV/x93yc9bqB1WlU4Lcnb0dRu/f8AuNMHtqBpKhaXIymGN4QdWi0hA+nv3WuJz0UPuP5P9QbLWbZPXO3afPZ2LXRmsxYE5ae2hZNcd+Sf8ffuvH5dElG0/kTvOprMy+z88+KyWpoYRHKVRGJIXkcC3v3y610X/efRHduMyv8AFKHbmZx00T+YlzMg1Kb/AEX37reOHSv233l8kNlUf8BqIsh4UTxaTJUcgcGwJ97615Y6nxb17d7NmejpKavoshCbM5lnBlYctp9669jz6MR1T8Wfln21UxYSmoM/T0Uskf8Al6GcxlG/Oo/4e/eXW/PqyrrT+Sj2bvOfHVG7921lI6AGaGoqfHYhudQb/D/H36uetU6sF2d/Jv2ftNUpc7XxZdWRUY+dGNitje/1v71XOOt0FOjKdf8A8t34ubBrkOe2PTZeYlXZikJYlTqbn6+9V630Zao+NfxHoKeNY+rKWGOIKGvBCQwtxdrf7z7359e8umqb44/EXMRGjj6spJUf06lhhNvxe9veyada49BZuv8Al5/FTIhq+DYlFjDIG0iSOAcuLC4I496Bp17j0Szsr+TJ1lvioFXtyngoYqhg0UkbRIF54KlbDj37j14gVr0Cu4/5CeUqcSIcLvNqSqj/AHY9NUqaf9SOD7316nRQu1/5KXc1HSvRrurJZZYgY0ihnMl1UfjT/vfv3WvLoFOtP5MfatRkKyklymSwkgLqZ5nC6r8csfyfx791vy6M5hv5HXYLw6Kvst49IElnrlUm3On6/wCwt78COvDqYf5P+ZxWtqzcLVlRRErFJ9yGLlTwRf6+/Gg68B0zbu+Lvd/VOLpk2xS5XJQ0pV4xTpI9yptxoH+8+/de4dVo/ITanyQ3Fn6KtyW2c0Y8dWQymMpP6hA4J/UP8Pp79jrXRqtn/LrMYOk2TtjcO066myWGpYaaKrmiktAy2Aux4+vPv32dbr1ZRgPkNL/d6LM1+RgqHljQpEZGLIxS4BAP49+69067T7rqMnlBuqGtVa+mkVUAdwRErX/Tf37rfy6Ozg+/MPvbGxYuaphpskU0yzuQGZ7Wub+9EdeGOg77E6uwu46FquotkKggv5IiCGUm4H+397BxQ9eIp0mtozbS29tys2dkMKj1NXKsaVDU8ZspbT9SPx7959e4jo6fW3Qe28Tgsbuvb1ZBBNpE9VCgiVm4120gXv71wHXvPoU6jK7TzbR4zKxJC8BWPzyEKrtHwefeqde4Co6ididlYHq/ZVY9JVQLTCnm9SkabeEkWI978+tdaiPzj3vP8jN9tt7bcL1r/cFKh6Zy3jUTG5cf7373Trxznq6X+VV0xkOvNtY2mqaCWSyUcsjFA39kFrkj8f4+/fZ17y6vWy+6MVRxxUNJR+GrIKu+j6N/qmt/j70evdEL+Yvx8fvHq/dtJWRNVMuEqhCgVGKu0ZsQG/Pvwr14+letW74H4rP/ABa+UuT2bmMsKbEVG4g0FFUSaW8f3XKKqkD/AGHvfXsjrea2Tv7ZO7Np4Ojx9HCKqviUS1KopDMyABr2/r71gY69mtege7G2RBh6yrkph5JpY3tJEF9Os8Hj6e/cOvDordM2fxW6IsSqzVKVUivdbkKhYcEf4/n37z6seHQ/bgxlLnaDHUFZGqkaI5FZQpFhze/vQp1XhnpOQdN7fyM1RRUdMEq/ESk1xcOf0n3r7Ot8OgrpcFvPq3crCpM89CZSFsGZdGq4/T9Pe8cevcT0DvzQ7vi616k3Huiasjpp5MS08EchKMXEZIC/4g+9gda4da1fwe6W3F82vkLVdk7wpp59u0mfLUglJMJEdQSJAstxzYc+91x16nW3ziKGh6rxe39qY1Ejo6SlWl8QREXSgC6Tot/vPvR+fW8g46hboz6VjFKArDLHpLNfm/8Arf7b3o9bA6LxuvF5zcdctP8AcOqBg2s/7Sbkce91x14jNOp6fdYuClnrMgsUGKUCQubKQv55/wCJ9+BHWqHieg4qt80b5mvr6KrSQTIyRtHbmRRyFI+vvf29eoePTvsXprLdlVj7vzMkkuOp5OI3J0kMfrY/4e9HGOvccnpUdgdIwUtCYsBJ4HdbXiIFrj62X36p8+vceHl0RLeu7+wemxkBJSVlbBDDLIs9pWQhPpYj3uvkOvfPqtTI/IrJ9s9kric7WDCUq1IRxVu8YJ16beogf7b37rXVnnSOxdqQ4XJVNNNT5KYxMYqiIxuLlL3uPfuPXhjos/c6PC1fTSMXjUVKhTySLm3H49+68eqx59jYLcW6aj+8CLHH5WZHlbxgWN1IIt7959aPHpw67+SfbHxS7k2zkevtzu20aTLxTZTHw1FTL56VWAMQVGP1HFre/dezXrbY6E/nU9RdgzYHDbnp4cBUPR42lqqird4GkrJIljkkDSm31ubW9+z9vW8cOrntk7+2r2Hg6fcG0stS5jHVESyLJSyLJoLC+h7fn3unXj0sQbgGxFwDY/UX/B9660MivXfv3W+sckqx6b3JdtIAte9r/n3sCvWiadZPeut9f//V397XHI/3n/ife+HDrXHj1x1rq0X5/A/r79Q0r1quadB52b2btHqna+S3Xu/K0uOpKCkmqEWaZEknaJCypEjkXuffq0GOvAeZ49acvzx/mN9g/Ifdec2XtqfxbEochJRMIZgqPTqNFwi8G4596p1avkOq/wDDY/GZPGnGznyEISHYBje1z9Prz7317Hn0JfUPxl2U25I93VqiSqaVCAVFgSfqfeuGOvdXs/HbZseIwscuONNDRQQuxllOjTYG3rP9Pej1utOqk/k1jJe0u+chgMzUisxeJqS0fjlE0do5eOBf3vrRz0O2A6twlBi6OGjiiVVp4hbSP7A/xH+8e9cetjh0m+y8JgKTCTRskJlVABpVT6tNrC3vY60eOeq6abrqXcu8DHErrTmof0JyunXewt73w611Zb1L05iMRhqdWSNJvHEb6CpJBvfj8+9cerU6OftTbAoqaKCjQlXGlvzyRz798+tE+XQ547OwbSoUFWqRnjWzDSNP55PvXVicdMmU39sbcGUgxeOyEcmYmsfGkyOAfzdQffuHWsDj1n37LtbDbWnp6x0bIVNOFUalYFyvAIH+9e/A+XWzToldD0Vhd2xy5CqiiWeSaVwEjU/tu1xc+99a6bk+O+38Hloa54IW8LcM0Q1gfX6t/vHv3HrWPLoaMPtjbsMbBol02svB0kgfpIHv3W/l0C/bPx/27vilqplx9PFULHO8ckcA1FwNSktbn3vr2OqkN5bi7B+OW6n8sNTJt6Go0KwDAJCWv9bf0966r8ujPdX/ACy252ikNCmTp4KmBlDJNVRo4dVAI0tz+OffurVPVhvXW4MDl8ZFJU5Gn8oCaP8AKIzcg8abe99az0Yzam5BDVwUazRfbsQgYSA3XVa/HvXWwadGbjpcRmMU0McsDTJCWvrUnVovc3PuuOA69kHop3akUa7fzGGro45YpY6tV9OrgxGxHPvY9R145wetTnveufrfvv72nEtNSz145QeJGBm/r731rqxHclUm++nsFW0xZpZDTiSQXZrFQSNY9+8+vfPoX+o+t8LPjKE1lR5W8UQZZGD2N9NmU/7H37hx699nVmOzepMFjMNjK7EQQmZwpmITkXUf09+rTr3l0ZPbmMxUFLBFVhFAKjSOP08Ef4e9E9Wp6dDTT/wyCGEY004kUfhgCBb8+/U6qa9RazfFLiSYMnV0sKtcFjIiHkf6o+9eeOrUBGescPY23KKB5qStimnYfSGYSH/E6R72T1qh4dBPvDtCmnwOayFU8UdJSqzSNI4U2AI+re/AHr2OPVafWnV23exe3MvvdYlqYvNJNTuU8gL6yws30497r1qnVpWy9hSZ3DGHSEFMhhiBU/ReB9PdfmerVxTpKbl6xqcYlXHMiuH4U21Fb3tz/sfe6569TopFZ1xNQ7oWrpvMky1IeQWtHpvzz731rh9vR7usoqePGQOJL1UUaKRqubr9ePeuvHA6H6fJRw0P7i6UZVMxPAA08nn+n496p16vVIn8yas3lUbaqf7l+cwj7ky+AMV0AHVYAf6/uw60T6dBl/LH3Np2hm6GolijyJlKVazMBMZdfq9J5+p9+PDrwGM9Xt7aw2LrsVFLUeEyeIH6qWdiL2v/AFv711vpslxmPgqyaihjeIORfxXBH0/1vfvKvWvOnT/SbfwmpaimghgY8gBAhJtfkH3qp63WvQUbi3xWbTzbrAGSN2CllBUH1WN297p5dewB1g3DvTET4iozNfWxxvFSyyjyzqLFYy3Go+/fIde4GvWot/Nd+WEO6s/UbWx9QlQKCqihXxTBrAOVB978utHjjqs/qL5hdk9cJTUeJrJRDBUeRYvuvGBY8XX8e/V8utHqwja382fs3DU8MGQq2UJBp4rFLXX/AB/p/T37rfn0Bfdv8yf5Hd4LJtXZFLk/8rHhSojd2X6aSwZB9fyffuvHpIdX/wAt/wCQHesMG5ewcjW1IrpxUSRVAkdQlQdbKFb+l/fqevXvs6s26j/kzYPF4oV9VZMjGAbiMK9weQCDce/Y63noyO4vgNufYW1J6zbORyK1FNTaqdIZJOG06QAoPv1QOHWuqZ+7fl38m/i5mKnEMchLTxVMsSNPNKnoiNlYs3v3Xhx6A/JfNr5VfInCTbfhkrFpskouaeqeZgp9LDShPNvfuPXvkerB/wCXx31/ss2YiXtbGVhNRUXkqZqRhIdY9RMhH9ffutA+fW0J1z85NlbqwNFW7Ar4o5ZI4Y5IZ6lEYCTggLe/vQ9OrU6HvJbxotx4+nrfuIGr6yPXPaVWBYrc2964deGc9Vb/ADR7J3xsTb9TU7as0vhlZfGSGdtXCgj/AIj3vrx416prx/yc+RdOtXuCro6iSghnKk2keyHg8kfj3uletV6OF8V+7DvTddBkdy1aQVonLiOomCEEH6Wb/H8e/HJ68Knq9PBZODckMZElN4kp4WRywsRpFrE8e69W6aczkq37z7VEIjiGlSoOk8W/UPe+OT144x0CW4MvlkqqqCYEQsxCsfz+L3/HvY+XWuPVaHyO7Y/guRk2tTPJUVlY0lN4oX8jfunT+ke/fZ1rgcdFpn+PdKNiZjcGVopjV5WOSsQy07B7lS5Av791r5nonHxi27uBe381hcfTqKb7mWPQ8djbWQLKeffutj062e+i+qoqHadH/FcXDLWvDEzh6YspDckWYe9dbAFM9Kjf3x0663ZTua/A0X3jE200KGzW+pYj34HrfQZYT4kdX0aSQ5HE0ixnVbVRov1/H09+r6depnpHbq+GXVshaqxeLogulibUUZsf9f34HPWiOiwdkfE/ragpnaSgjUheTFSANcjjke98eteXVcfZ3xuoaGqWp2q08DeY+kDRxrtzp/Hv3WqU6DXH4XJbL35g4szeoo/JSpICS6i7gG9+Pe+tV9etkPqPrrZe5OnoshQxU71MtAGCwoC4bw3PpH091z05x49Vjd49TNBW1c0UDRSJLUMp8RSQAOSpu3veeq/b0RrcNBmbTYrJNPLQ3K6HJYWtewB9+60a9P3xz+Qu+viVuyq3F18NMjztUeIMImLMblbfm/v2OvDrYk+FX87qp7Q3nj9ldspS42pmlpKISSSpFqklITUS1r+/cOt8etlPD5aiz2LoMxjZo6ihyVPFVQTRuro8Uq6gQy8H3vHVc+nU+WESeMXsI21AfUG3HPv1c169TFB1m966t1737r3XvfuvdRMh/wAAK3/qEqf+tLe/de61Jf5Bn/bwn+Zr/h2HX2/x/wByvPv3WgMmnDrbl9+631xHF2Nv6k/gAe9k9aApk9R4KmGq8ix+pV9JJ4DA8Hg8+/EU69XrVC/nMdQY3rPsGDuRZ6OM1k5Vh5ESVFkk5BBN/wA+9cMdbJrnosnXPYmLrNkY2jgnglTJ4+KaRvIHIMifQEf71798uvcR0xDblJkMjUBRDIhuTrGq6/1B/H+HvXXvn0jN8dJ7QzeLljrKONppA5/zIaQFv6f6/vdevUqcdEE7H+E2JyDS5DF08wkjJZG8P6ePovHv2OtfLqvDsf4RdkNkhUbcjrNTSm9qdySNVjYgHj37rYPTnTfC3uv+7ckUTZOKqEYtohlU6rcf09+69XoHR8WPmLtSrkkxFbm5KMljpkSbRa91At70cDr3DPn1Gotx/MPqTLNV1FPl6pY7F0iWcAaeDc8j3q+h+ptAi8R0wsGqTxDx6MPtD+ZD3BtRIaHckM9JWqUiC1Emg6ifyG/4p7QWlhe3SfQxVJPRi03hx46Gvc/y/wC/+xcPRnHOIsXVIvndJyJDGfrp/wBv7lLkP2e3W83u2a4VhEx7qg0p8+gve7oIgzA5HQbQbey24pVrNwTPPI7eZ/N+4wdvUw1f6/vO/ZOUdj5cKmxTuUAV4Co8+gVf7nc3g/UYkdCRjqSLHxpTQIAqi1rWtYf09iCVy51N0XRVZs9KNXJjUNb8Wtb+n449oqdx6NmC+GKdM1bUyRE6PSb82NuPahFBwek/29cKbIO7DXyOL3H+HJ97ZKdeB9OpuPeiyOexVBkm8VHVVsMMknCjxu4Dc+09wzwWsk0fFVJ/YOlluglYBuju9WbC2pR/IXbWCwtPDVY96fH1LtoEihwwbUW985+f/cfmq73e4s0kbQJGHHFK9Cnb9sDnh6dbSmxpsCuC25janSi0lOkQC/pSyix49hnlyza+tprrcMuK8ePQoS0SBcdL/cu58Pj6Y0tDINKoNXr+v9fp7ZcaXIHDpogHHRCu+vk/ketaKopsJVQiWUSoS04TSGXT9Rz/AK3vw611VFv/ALd2RJT5Pe3YuXarz06NPjKOGrWfXISWQeMf0Nves9aOOge6g2h8mvm7uX+5GKx1fhOs/OYI6tKeanklpHb0sJAALaf8fe+tZ4dW99W/ybeneoaaky+ZhOS3LNHFNO9XC1V/lCkM93kJ/P8Ah71XPVqefR/NudPbG27iExR27iWjgjCKzY+O3CheLj/iffvOvXvt6Y6v44dPburfHl8BQKsp0HRQpazHmxt71XHWqk4PTjUfyxfirnKU1tTh6VKhhrBFGtibXPvYr1vHSew38uP4obPyH3xxULTRlmQJSp9f8R79WuT16nkOjU7c2ztDrTCxRbGweHWliCRBpKKITaV9Iu55vb3rP7etYPUncu5KKkoBXVMSUs7rrb7cGMXI/AH+8+/db+fQVQbxqq6Xy007mLVcFpCW4b6WJ/3j37r3Tscx5qlKmeR1ZLAlibf7VyffqDr3E9Kb7nG5tYaZqmnETkCXTKmsAj6WHvQr16o6gtR0GLrGpsLV0zSx2Yo8ylvr6vT735U69XzHTbnBVZqM0lbkYKY2BANQsRIT6W59+8sda889RaPfH8DEGNetgkjpBp1mYN6bkGxB/wAPfut0J6e4+0I62q8FFOGlsLgyNp+n09++zr2fPrPk+wmx0JqnXHvOgItOwNz/AI39+OevDPHpGJuqbcQlqhT4+EoCZWpkRSLcnlffuvHpKZRqjKKWxs4WWK+oCVh9OSOD9Pe+PXhQddbep1ycmnIuklTD6VKuxF14Gv8A1/8AH3rrZrw6U5bLUlTLTnF4ypoLAXlgWU2ta3qHvYp59V+Q49NFT1j1Zu1nbcmAxQlkJ16KJOS/DfQe9db6CvffwQ+Me50SujxcMdSihmMNIikNYkAFR/vHvdT16g6Kj2B8JNq0NBU0+z1qFaOJ/ErI1r/g6Pfq9eFOiGZL49d1bOramqx8EsmOSSS6pGzFlH04H04+vvdetcOkrQ5Df23MosmVp8jCqvf0Qyx/n6h7c/63v3W+PRr8L37k6TBLTCKpllEYH7+ote1ub+9cOvdANvfvrcCTSiCmmWs8paBhHpX+o5P+39768T0IXVvys7q2/SOuWaZ8Y6lYwJWsUI9JUf63HHv3WuPQlZH5Z09ZRPNkapqaeJWlc+fxEMOWP9fx79wz16vRHu6vmpnOwag9d7drnqHrGECtHUmSyP8At/Ucg8+/der5DoxvxG+IO3aael3XulFq8llJNVV9wnkZTKNRPq/pf37rwFOrdsLkX6hmp8bsqmhEBgVf83pHA9Nj711vFOHS7w+6d1ZDJQV+YjcrUNrIVSVAJ+v/ABr37rxoMjow8uQjqtrZpTCripx7RhJF45S359660RTj1p9fI3bOPpfnTjqeMSU1dPljIgplZVs1Ra5C+7dePHrZ/wCpMFuDB9XYLIY1ppK6GhVodWq5bxgr/j7r59bPHpe4ne+Ukp5Jt6xESsrICwsARwL39+68enzaGMxddWtnlSnl8bMUOlXZVP0sfe+tZPWPdKeXKRywqNLPqsg41f4gcX9+r17oUdtDbmIpDmMi4WcQgnUQo1AfUknn3odeJz0Sn5X/ACu612BgcjUV9Xj4ayDzCHXPGrsVjLD6/wCPvfHrZx1qWfJH5V9t/M3tDb/UWz6eum2vWZA0FTUUoeaDwebSVOjgi3vfVc062iPg58Qtr/GvpfE/fUMMGcqKKjrXf7cQSGWWIO9z9fqfej6db8+hx3DQZDM5VK6RwKSna6DVclfwRe/vRx1uvSPmj0108kkipEhuWkOhdP0Iu3v3W/l0Vz5CfJPr/prA1OUyeTofuYFkDJFVxiUFB9NN7+9gde1fy6o++Qn80iryuLraPYq18qSqFP2rMwZQPqSv+34926rX16K91P8AzH89HWR026fvqUJM5Zqh2QfW/wBD71XrXWxB8Ivn/sHsLBQ7MlyFIK+qihOkVUesF2020nm/vVOt1rw6sjys+O/htRWwuJwVJhYHVxpvZbce/Vp1voq3YUuA3PhnxeUx8cjzq8Oo04Lcta4P+9+/D59ePWvd85uhH21vTHVO0lmo5qurglDU8ZgIV2BIuv8Ar+99ap1bJ8RetK7bvTWOyWRkqaisqqEGRpdRYkxC9r/X34kdb6LF3LTz1GcrKeNGv5Z1OpWtYPY8m3v3Drx4dVefJWddsVdJHTSSQ1NSV5iOhizcEAj37j1Xz6UHSPUFHubHw5/M/vN/ng0o8pH5v6v979++fXulnvbpSDNVsVVhqrJUMlI4liaiLQDyQ/o9Sfj37rXR2/hb/Mi7O+I3Y+1ut9919RV7Arnjgraivqi5SKSYKBd/obe/H16sD1uidXdn7U7d2fit67OyEOQxGUp4po3ikSQxmWMSaH0f6/B/Pv3XuhE9+698uuJVSQSASL2v+L+/V61QceuXv3W+v//W39kLFRrADc3A/wBf3vrQrTPSf3HnKLbuCy24qmzQYilnqpm/GmFLuCR/h79XyPWgvmOtMT+Yb84d999dwZbYG381XY3aVE9XTWgDLTv4pTGYwVPN7W9663x4dViZrE1V/tsZCWrZQplaMEPPJ9Gka319763Tpd7cw1fjKaleQMapbGqjH+6xb+1f3rr2D0cnqEGqnhKTMVUw/skqOR9fSfp78evV8uh67s+RVZ1N13X7foqyWgyNfAUppISC48iEenSb+/dbNQcdEu6Kocrlq6XfGWrpspW5Z3P+UMxca21Dhr+/da+fRwf7y5LGw3eJljMZUC11Uf4D6f7H37rfy6CbJPU7xmngaXQpYgL9RYf4j8e9VPXqeXWfZnV9NQZYT67uJFf6/X+0fe+vY6OXtnBPPFTpTu5ZFUKg5PpP+P8AvXvXXujG7XhkxqJHNDrkN1UfQ3I+pv78etinn0U/5T/JGl2hQPtSghSTNVKSUqoFUujyegNdfz9PfqdarXovXx32bv7+9EW+MnNVVEE+ipjjlL2jSRrhAP6e919OvcePR7Ny4HI7imWpnd3WRgTE5OmIWsbD36mOtfb1Iw+NmwcgR5mWABR+osAQfp799nXjw6at4tXVcf8AkKtKSbggjm/B4H19+68OsuLw+SgxtHLUwMGcDUSOf9fn37r1fPp/q6gmj+zWILI6lARw5J+gNve+vA9FR7q+OtNv/B5EZvGRH7hGaKWUXYjQbHn6e9fb1v59a53d3w17e6n3lLvLYuSr6PDwTyVJhp2nMckYlJKhF/w/r791XoQ+sflb2FgJqTDZOWsX7RoYKh2jdSzIbSMSbe/deqRw6s564+aGDq58PjarNGKtlASQHRq1EhbC59+63Xq0TrzsgZOGmqqTLSSpUeEGzC51KD9FP0596638+hzzuyJd2Yt60MXR4HZ254LLbm3vwPXiPMda5/8AMi+NtXRzLuDFUwappik2tblmOst+Ofe+tedeln8eRHk/j1RY3NH7fIUwC3kGllKLpBF+ffuHXvLow/QWz66aiyVetfLUR01TaONmLAKrXAAPv3nTrwp59WkdZVVctDTUtehijkUJGzEjkqALfj3o8OrY6Xu72h2/jRVtVmP6tbUDYWvz79TPWiaZ6KX2r8rMb1dQNXPk7iBNTamS4OnV9L8+9469U+fVUG7/AOY3n+7d2vs7r2pmq8pHOafw06lry69Om8Z9+610YHrntz5Ddd5aCq37tus/gs1OkgqKhJShQjk+sW9+69x6UXZPyiTfdHVbGxFQaXIZ20Yhh9LjUdNvT79wFOvY49H8+GnWE+19sUaZiR3rKgiSV3JvplTVzf8A1/ej1v16tB2rt8Y2i1UhJ1qWsC355HPvX29er0gN3LMs0yVIYhyTcn6A/j34de8/l0FeU2LWZmieegogzBXYzqPWwtwNQ9+yD1v7emzYUNZt2teOrLGVH0iGSxB5/offsV695U6MVly+5MUaejQpMyAMF+oJFvx79w49V+zopHcvV0Fds7K47IUiVVTLSVQAk5I1Keef6e/DHDrdePWs/unvOs+F/eKYevmmx+CzuTVpCNRhQyTXCkngD3bquethv48/JfB9nbUxmWwuZFUHjjcopSx/aBIAU+9UPVq46N7SdtY2SkEFTSRGXj1sqauOeT715169w6QG7u56ShkSaJ0p4Ih67BRwB9b+7dexxPRRu2Plfsymp3WaopzUxg8nxk6gfpcn+vvXXuqj/lN8+Wx23clQ4fJvAhhqY7xaSeUIt6fe+tE5x1qz9z93Ve6tx5zMZB2rJ5pPJAZgWLHXcEDn349e6Ff4v/FztH5F18Fdj8bUJj5ZFKuvnj9DC97Afn3qmevdXS7A/kwVufx9NkM1V1MFQojZ42ml0llYXTSfe+t9WKdTfy1tndbwUhr8RRtPAoIqDcvIQOOSPz791Wnn0fTavUNTt6jWDExSRUsUaqgjdtI0iwtp/wAPfvz63T5dPX8J3ninaaKWq8CGxj1PpIve/v3l1759dZXe258ZQE5HGy1NIVu3lDFdP0NyfeqZ6tnrXt/mgdZUPaeCrc1jMVHTVWqo9cV1bVY3NwPe8daPEHoFP5WHxlGRrkq8vSisWgPjaKYkg6ZPqQf6+/dex1sH7z+FnW+5KGOpqMLjo55ooxcoo8Z0cnge9A+nWqevRfa34fZrrqVsrs/JzJBCGkSip2cIxT1IoVePe616369JvH/JPuXr3KJjtw4KqShoZBHDPNC/7yj83P8AW3v2OtVPQd9zfJrcvZyU+Li239yrMI2YwtpW555tf34CnXq1x0tevdr0WT2PUUWZ21TRyVMbMS8RB1Mn05H0/wBf37Pl14fLqubuPr7fHWG/KXeu0JKmlxGMqBNU0kBfxNGG1EFQLfT37rx6te+Kfy0oOzsVjsHU5U0GSpDTRSoSEkZo1CuDY3P+x9+69nq0/H7hxWQo4Ka8T1Xh9MnpLSC3DX/PvR9erVz0WrvLeUG28Lk5VCpPBAzB7Am9v6+9jhnqp446rz+PHULd9dyVO5M8PucfSzmSJJrhCVe4Fvpz78aAdep5dH3+QNDtjaGxK7H1WPpqODHQyU6MFUXAjIWx/wBh79xHW8Vz1S/8YMGMz3rmsvt+D7qnirpXYRnj0yE/Ue/HrQrWnWz11RummpcFR0uUxkUJSkhQyOEJBCgWufevt63QEVHSsyWbw0dR51hikW9wo02AP059+62K9JTckD5ikNXTUwhgRdTOgtwv5PHPv1fTrw9D0E/38T+SnjnaRkuhW9hdRb375da6ALs/79qOoU49Xi+hfQSFB/I+vveOvY6rp3aTVZCpo0h06Q9zfSUF/wBXH5Hv3AV6159FM7GxlJi1kro5Pv61JhIgf1smk3sAf9497611Yb8DPkhQ4yhqcduvIeGlpQy/azFVjVQtrHWR715Y63U+fQV/Lj5T9epvudcdXU4p2jcNCmjxj1cnSp9+HHPW6jy6IHu/unZeSo5Mnj6qOeoUXEFgdRItYgc/X3v5da6Y+rtr1/be4KOcUjJSfcAshGlGXVfTpH4PvXXupfyE2tVdXbnw2R2XSJQ5zF1VPkJWpVIkZKQ+RgzD63t791rgetvv+S980K/5OdLx4TMSSSZDadMKJ2mAVw1MREy/1/B9++XW8dXcf4Dj37r3Xfv3Xuve/de697917qJkP+AFb/1CVP8A1pb37r3WpL/IMsP5hP8AM1Fv+aiV9v8AW/it/r798+veZHW3IRcfn/Yce9jrR66ZQ6sh+jAg2+tjwfeuvHOOi+fITvvafx+2TW7lztTFTtBA8kEbGMBiqm2oMeT+fex1vrQR/mufOjePyj3VWY/CV9W23aWsq0UICKYeOY6Aujj3r7OvcOHRQugvlfLszH0e29z1cgmgijpoGmsCAvpurN+PfuvVPVlHXnyB21nkMtPlUeYqCyBo+Da5A559+690Z3Z24qPc8gKyCa4FlLLbn6e/fb17HQlU+34cpKaRadGVrjiwuwNr8e/Y69nrDidk4SLOtR1uOgZlawD6eDf+z+P9c+/D5db6HdeutpwwRSHH03qCft3U6r/XkfX37PXsZx0I9N0htvcWG0UG3aSZ3jPq0i4Nr/QD37HXgDWvRPt+/HPZuHnzC7g25QE+CYp59Nh/S2oe/W048YxHIHS1I49Netan5sdK4X/SAH23j6WGnFazOIFGkBWvb0+xj7fSQXfNsVm4qCV/w9FO6yGKIkdP3Xm3UoNuYiHnXHEA6XsosLWt76MQ2tvYwrBboFCqOHHh69RrPOS9ONehQKiKyoLfQWB4/pf3vX5npP8ATMT8usvjEQ18E/XkDi/+t71q1YHWmj8Lj1Dqq96ZAzCwNrEn+p493EYOB1WKRmYhsgdJuqzMLMdZUH/fW9qFhPl09qoM9SaGohqFurAX/qRwB/h7o6sp69iv29d5tKuqXH0mKT/LmqI1hdR6tZa91I/PtDeUSwuJG4CNv8HT0M2mQL1dZ8Efj3nGrMfvjcySTZBYKYxSTatQjtqXg/i3vmLuN/bTcwXKTAV8Vv8AD1Kezf2Aanl1cjDm8PtmQUOVr1pXlbTCWdQVP4tf259eIo2ihwD1a8nCSBAePRau/PkttnqvEV1WcqlUywM4vJG5BBPJF/aFakVPVCTx61u/kl85M92LuWbE7djmqWqKxqWLxwsxvM2iM+i/9fe+tVrjo43wU/lwdk/IHNYbf/buQraXb8cn3NJQ1kkqUskKsHQGNuPfj14DGetsDrbrzZHSm0sXtjZm3qDHy08cEcmVo1VZZGjUIXLJz7qet9DBFOtTGDNO9drCnXIdfjJ4Iuffutdc6jC4yWOMhY2kk5ZCR6f9j7917pjrqfF4PTUtFEGvwbr+Dfi/vXW/s65PvKaSmC0qnxaGAZQAAqjn3v5db6AzKd17NwdRlk3BlYo5KSNn0SFPQRx+T+PfqVz1qhp0Hm1/k7sTdNTLjaLLRMkMjmyaLEq3/FPeyMY68egK+TvzFxvX+Ik/g9PHkniXSQERudBJ+v09+p69a8+iC9dfzKa3K5F6b+CDypI4EXhHNj9B+Pe6enXq9CTv/wDmE71gw7RY/aDLqRrSrT3ezDggqP8AY+9ADz699nQC7V+a3dMuUSviwta9GXLuvhYKijmxPvfXvs6YaP5/dnT9rVNHJR1kYUgCDxuqsSfpc/7x7916tMdRPkB86O4qKuoXo6Kux2pFKmCNzruOP9v+ffsdaqQKdI7D/OX5DGgoKyq2VW1GPZQzV7wPeRfyzMRx/X3v5DrdehX2F/MSzUueNDXYtqeqVVWRPDpdWA9Vuf8Ab+9U69Xy6MLV/Js73omE2UmxzyWa49JUE/j/AIn3rzoOt+WehH2V8odobBwORiyu4zVTvTsqeXSDqZbD9R+o9+869exToAcj8+ocfV1CYeQ1SySSoulb/U+gC3vdPMder69Dp0P8nJ87S19bn5TRVNS7SUytdSVL/wBhW9+p1qtR0dLC91YGSGnjqq5WlqyIxfSeT/W/vVKdbwejDbUxNJXUi5dWjqIJFDJ6r8v9PSPfj17pXth6mlljCUatDObov4AP+HuvW69B7vHGZOl1TRUFkkU3tcKAT/h72PTrXDoPaTG494ZZa6ijqHJYGmksQSf6Kfr7317Pl0kdwdPbC3UhmrMTRUsoAOkhB/vFvr/re/da49A3VfHDZsFQ8kcFOI2JAUFSP9t791by6QO/fijtDPYieLHx06ZIgvC8bAOWVfSAPxz/AE9+r14D06JLunYXbGx5qfD0uxHr8XRsEFU0TSeVR6Q1wPpb3vqv2dE5+Se2t8NtmtymLwrY7IimqC1FCjAr6eXsB72PTr1c9Vw/GbI1kG/ayfdYK5uGsUQwSqwe4msANfvXHrWK9bKvQHYTwwUkdTK0anRoViCFGgDi59+6tn7erAo6n+KY2DJwx/cH9oa76rEf776e9U63XoynW1LR5GlpmyUKxuiWRWsQQP6e9H061TzPTl2FnY9v4zKLQwq8S01xpt9f629++XWvPrV17KrI9yfOvFV01IjyxVSixC6recEkEfT3br3n1tYfHeCmzewYklW5o6YhYTpsGVBwB9feqV63XPTtvXYmH3HiZ6YtHR1SRt49LBSXPI4/1/eqefXq06AHZ9LmtiV0+IrTLPBNKfGZPoU1cHn/AA9++Z69WpoOhj2/DS5vKRRVgESySkavoBY8ab/n3r7evfZ0GXyT3PRbFwVbDS1KkQ0UspYAAjT/AIe7fLr3l1pp/O/svfffPakHXu0shU2qshFHKsAexjLhGBMf+B974daJJ6u8/lyfBLZnWu1dt7q3RiqWXccCpUtPOF8/lMYcm7i/196J9OtjHVu/Z++KKmwdNQ07iHxKlNGgF+Esq/7Ae9deHz6B2q3Ljdu4tMnnMj9rTND5DrkQAro139R97631TZ8yf5l2E2gK3aHX0gyeakiemiNNEJZTMH0jSYr+906rXquDrL41/ID5p7i/j2+6nJ0O3K2oE60chmjhMFQf7Yb/AA9+8+vD5dWqde/yfOt8Ph1pq9aSqqpIyv773fVb/ar/AE96rXr2Oi2fJv8AlDYKm27WVOz6CAZOGnd1FOx1awf6r+ffq+Z69T06pa2Xsztn4ddvJuLNmrpMPSV8FOEKzFPHDLctduLe9+fWs8OtsD4d/LDCdyYfCUVVURzNUxjyBwgBHAY8n3o0r1vy6PPvfaO2II5MhF9usUcH3Cm6nn9R+nvWetjjnqiH5zbux1RuCnroIlkhxdVFCzhbgaXA4K+7Drx49WZfCitHY3UNLCsF46OhZkBIJIWO9zb3o9ez0Un5H4WkwuerHSFFlLSqyC451fgf7372etV6p6+RPXr7oz2MrqqVoYY3Q21EfQav9hf3uvWult0/nKDHUM+Co60vNRRtphQ2JOnTbn6+9dbHRy+sKnBbgoajG5tYqCqEUhRuFZja4Nz7955630SzujrfHbl35Ft6WRI5KqqVaCrYetVRwUKn/inv3Wj6dXk/yhvkXu/Y2+a/o7dlZVVO3cc60uOlmJETftBE8Rc/X6e/Y695dbQ6OsiJKnKuiup/qrC4t738uqmtK9ZPeurde9+691//19/QqxcNq9ABBQgG9/zf3uuOqkEnol/zp7Pi606H3eY1CzZXF1FMrLcEeRSpChf6+9HrY60TaPISZPeGWzFSk16jL15OvU1w9Qx5LH36g69XPQj51oKPESZOiaOGujCsj+m44uPe+tkjy6i4LKVNRipKyocSTyofIwFtV+bj34de49JCT5IUfVT1MtVUpC4WXxI8nqDoLqQPrx70fn17oJ9r9obn+Rm7WyWXrZKzB01ReJFkdkEYcAfXj6e/daoT1Z717TUmExKU1EqxrAi+MAAaTpHvXn1byr04ZPPZGq1U/l1eoqEH1ANx+PfuvfZ0/bTw1QQdJ8cs1yWP5P40/wDE+/dePy6G/buFqKeeN50Mx1LrKKPVxa3+sfr738+vcBnoyGy6ULlaZIoGiBiW+oHSCW96PW+NOhG3zkl2tjHyTWkkiUMqDg3C3twPfh8+vHqjTtXNjf8A3L95WK8Phqyyo0h03WSw9JPvfVSPTq0fpXclPisHjqKph88cdPEgKJf0345tf6e9GtKdbAzU9DtvvORTbeebAwfb1QhBHA9Tgce/DrfHon9N2lm6Gc0WapZpVNQV8pVtIGsj6kc29761jj0M+A3dQVpgQKG8iarlLgD625/p799vXqeY6X5qKytsqTgwIAyRj62twLD37hw61Q9c5JIDpWSIpMmkCRgeCP7RPvw6907zYbJZ2gZZqxJIUW0cVlNxb6G/+8e9V62KnPQW53q3H7ho6rEZbGLPTSRsgJpoza9/oWHvdfXr3HgOqyu7fgPRNkKrLbeo3ooZGlmYhI1UkryCBb375dap1XHvf4vbp25n6SowZqTVwOtnikf0HUDfj34fLr3Dq3j4g0G66XGUtDuMzGamSPmQtqusdvz/ALD37rw4Z6uY6xzkQ2/NQVA8kpRtBI1WH0sb+/HHXuI6Jx8petcdvDam5zMqtVrTOKS8aEo9jYqfeh8urGvHqkjaE+X2mmU2pVrLUvTyVjpGoIsisdJIHHA926rT06PT8OcichszczVdXGlStZII4mYeQASGygG/vR630eDBZ7KxQJrjmZKQHQwDaTYcEFRz7917zp59IPt3e2487tuopaKKoM4ilVSge4sPSSR9PeuHWzxr1r6fLzb3duR25uKtlfImjpy3iUNKoVNBFjY+9+WOq/b0nP5NHUNLH2pkt17toZhOmVM7y1ZaVSAbsbyXA9+691sVfOPvvYOV61g632hh4p93S0wo4q6jgVXjJXxi7xj63/x9+p148Oil/Db4a5YVsG899tLW18/+U4/zEMYDq1qBrv8AT36vr1oDz6uv2rturwUtPEi61jEa6YwAAoS3496PHq2KU6NPhaiWXFClhJhnK8XPqH+vf3rHWs9I7cuFqDLHJPKJwbGRRyDz9L+99b+fWXbu4qSmkqcUIlRfCVRStzqta496610W/sL+LYrcP8QjidYPLqUAELpve3HvY63mnQr9fb2pa6nCaDFVAAM7EixC8sf8ffs9aPqPPrnunVXSymoT7qJ4yCQupWU/UG3vX29bHVBn80P4fYLurAVmaxGHkTL0X7sUsMaxyrJHGSjahY/X/H3YHr1PTqkDoHunuf4kZNcBn5snJhaOaRVMsk7gRK+hFP4+nv1OtAmvVuO2v5pnXMW14qnN19HDXiJUk807I3ktZgdRv9fe/t610F3ZP8zjZuRxNQMXCK8OpKvSySFSNPB49663WvDqlzur5Tdg9n54Q7Okq6FWmcNEaiYFl13Asbfjj377OvY6bsf0P3t3VTRxNTZgU8kStJUKJWTlfVy34/x9+698uhX6D/lq5LcvYmMotziTIU8VSY66mlVWvpcW1K3vwHXs9bdHxv8AiNsbp7aWOotr4CGjrIqam1slLBe4iAJNh71jrdPPo1OOwuSxlfHTy0zGHUqgGnjAsT+bAD3o9bHRjsH11jdy0airijL6bIPGgK/8a/1/fqevWvs6g1ux49vymiFKJYj9CIx/sOP6e/VHXj8+muo2CtWtjEAkg1mMKuqx/Fj/ALx72acOvAnpE5/rSKSmmpavHO9O4KgNEANJ+n+B9+HXq1x1VL8yvjFPmNoZZ9v0rKft6xxFHErMHKEqQPe/n1r5Dqrn4CZHOdLdmZPae86pKCOqyDLAlZ6CUabiwPv3Ede4mnW0VtOhxW5KCOpWSKemaOHxutin+bBuPej8+t1wCOn7KbKxMGPeXVCy6T6SqNx/rn3rHHrwPkeirb/6u2dusyDIUcMhjP1WOMW/FrkfX3vrZp0GWL+PvWeJkSq+wpxqYnTKsbeq/wDQj6+/Y8utcOhbpNhbGpKHWIaV4Fjt41VAbj+vv1fLr1BSp6JZ8muv9s1e2cwtDFTxxTwkMNEX0KG4uwJ9769nqnjorBR7K7Srf4fVinD1chQeVtIbWbHT/h731odXede7mqsTiYs5mMvG0dPAWTXISoUc8n3rHXvLoiXy0+W21JZJ8BQVkFXV1K+FkjlJcktp+g9+68ejqfAXs7rvBbChkrxSQZ6qaP1SlBIxk/Sbnn8+/Hr3l02/zDey9t4PqfcmSqJopZpYpJ6aNWs7aoiQVA96HWyMV61+vh98g90bZ3Lk85isNWrS1c9Q+vRI6keQ8qbHj3vqvDq1DDfzGMlS5KkwmTxFZCmpEaaRZVQ3NjyeB798ut46sQ64+UWyd1YqnlrsnR0tQ8YYRyz3b/bE+/dbHDj0avbPa2Fy1C2Op6iGeCePxrLH6lBdeG/239feqdernpKbkx2J27QTZpcvSh5HaQReTkD6/S/+8e9169Tz6QUG9Np7mx9Rj6itpPO6soLMtybW4v8Aj37PWuiT9vbVxO2VyOWjkidZopWVltZrnhQf+KH36nW+HDqtfN5GlyVXXSPSsY45JLE3KtYfRQfe+q+fRC+zt778ot10mL2HNUYqOepEdVJrkijZS9iF0Efj3r7OvCnQjbZ+OG8N3143Ju7JT10MsSOdUzOhLLqZRqP+29+AzXr3S9q/jzjqWjmempJo0gJvKz+j0/m3/G/fut58+jB/Diop9qb5jw+RmirKaauCRKdJEYvpAt/h78M9a/n0YH5v7Bokml3DilRZp8a66VVbhZIv8P8AX9+Bp1s8OuP8k/v+fpHv7GdXmr8FLu2vaWqjklexNRPydP0+p9+Pr1rrfGidZUSZWDJLGjoR9CrrqUg/43978qde869Zfeut9e9+691737r3USv/AOAFb/1CVP8A1pb37r3WpN/IM/7eEfzNf/EiV/8A7tffvTr3metuQ8An+n9Pr/sPfuvdNFdX0mNgmy2Sq46CipYJXnNU6xIqxKXZy7ED6D3s8KDqo4knrSa/nBfOTOdrdqZXqnZuUNRhcRWLT1L0c8ojlp0l0OQFsPevn1Y46pTyGxlq6SZIY5ESSNpnEjl2aZxdmJb/AB9+p1oZPRKux+sspJWVDUUFQ1ZG7eCaNioW30Ho/N/fut/Z0Hm1sp3BsCrlaM15i9dmZ5tK/wCN/wDevfs8Ot46P78bPl1ubb2Xhp9zVMrwqY1YySPZR+fr/T8j37qvVum0Pl9s6qginiyFIlRoudUo1A/1t/X36let19OsG5/mLtbb0kmXmWKscjUviZiSRwLEf7179Tr1elV1D8wh2flIoYaKemooZIyWkEgRgr2Nif8AeffuHXgc9Wvdc/LXZGIoYcAuGaXIsviNWiMwDldN72P596yeHWyOiRfKTsDM52sztVSVXhp0oZpVNioAN2W/+P49sXqBLcNb4c9WDNTj1rd7wzuc3b2pVPVV6y4qmq3hljJ1KXD6Tyfct+wvJW7XPOC79dIWt0WvyqOHQc3OYlSrmvQoU1KtGwWBl8P4UW4FvwPefGvUKNx6Bc6/qah646ekVSQTa9v99b20eHVvENaDrjKrf1Hpvx9R/sPe1x03INeW6Z6+NZggluVBvp/B/r7fRiOHTRVUGpeg53Bi5Wk8tMSqA86b2/x9r4JBSh62soB7sjrrFySRIF1C4Fif8fwR726gnPXp21LVMdDx0rtmr33vrBU1EWkXF18U1eUGsLGrAnyD/ivsMc0XcW37NcPNgMhAr9nSrb4RNIGfy62M8H3r191TtjH46fI46nngxkMUjOUj/cijAZSR9OffLnfzbnmKUxCgLk1/PqW9sMccIHDHVenyd+c+EJlO26mHO5ND/kkWPqGeSNtJAusZP59mNxAvhI0fSTcVVpgU416qIrp/lf8AJzdzY9Nubjp8BVzvFHIPuCnjdro9iLWt/j7otQoHWge3PVyHwc/lf0u3dw4rc3bGKnr43WnqpIq6NHkEyMGHpf8AN/e69bp59bG20NvR7YpqTC7Zgix+BowEhhESRhYrBbXA/p7qeHW89CkrxrKkckySpYehTci/B4HvXl1s8elBPl8XhKRqmaSJVRWJUuPwt/p731rooXYnytodvVc1PjcZNWSwsdAgEjlypI4AB97p17j0SDsX5Q9l7tytJTUVBkMDQNWIJaqqWaKBYncXOogCwH+Pv1OvdGpft/Abf63iaq7DwjZqWiu0JrU8qSMnqB5vcH37rWB1W9maDK9k7inrV3dTz0tXIwnSOrNpIy/0tex/23vdK9W6fa2TaXUlIwosXLWZRoGUzxySMHe3JGn8+/fZ1ry6J32vLuvsLb+drKXE5GGIBmWZ/KUQfi5Pv3z618ugz+N/ROWyFcKuqgqDU+aRixNwfzpJPv3Xvn1aPhupMeMVFSZbBz1c2hRrEetV45vx+B79Q0z1vHXPcXTix08FNtmm+yM6aJUEK+m4sbkD377OtCp49Em7q64xXXQpckKiH+89RKA5GkSB2b0/m/v3XvLoWulehZ+w5KHcu/KuOuxEFOkwpXCOdCgMOGvx798uvcc9Mvyw7j606621X9fbQoKSqzdTTCkxVNSIpqEnK+MelBe4Pv3Xieie/GTp7c2Nk/v32Tt3JViZGuJjhlia6QzPqS2oX+h49+zTr329H23T0p/Gpsflds4iqxeP8SSSQupUlSLm6/19+690XTuPpvI5mHF4nBY+tkrmkSOqMDPcm9muB79Xr3ljoQ9ifBrOrgYsnXUlXFMNEyxyj1FgL2AY+/GnXgPl0NWx/j5u+DduJx5oa1aCNlSR7EJYNyLjg+/fPr1aY6N/n/i7vCpCVGEmqY5IEDxIjsBrtccA39+qOt1r0KvVe4OyOs1jwe76evradGRQhRyulPqQfeiK9e6PnsjtHAblihpaiNKSflY/LZfGf6Wb34Y638j0udxYynqaBngnhyF0uI4bO1jyAbfT3o+vXvLPRW85QVUNc0ccDwOWYWZdP1+lh9ffuq44dJuswGVVTO7MVI1c3Cge99brjoK91Pk6SllmpmLNGrsVU3Hp+nP+PvwGadb6Dnr/ADu48tuJZJ4ZYqaB9LCRWIYg8mx97PDr328Oh9z1TJXVENNLCkkEjBSvhQki1jzb3rjw68OHRTe2dgUC5XyTbZnyFBUgxyJHTK66HI1lxa1ve+PWuHRDe+P5fmK3JTr2p1VCuJrMZGamsx9KkUc80yDyMpROb/649+rnrVMVPRXOt+z89sPcP8G3mXxr0LsG+8Zo/L420aRqPP0/HvxHn1sHHVxHU/dOO3Pt2kgoKqGGE+Au5c2a9rG9x71xz14Ho9Ozu1ttYfGxS5CeBmhjIY3C6uPqDzx/re/fPrx406Lt8jfk/sTbe1twZU7jxUZbHSvBSPUr5iyKSFAv7959eFOPWu50F2bku4/lbJu+mw9XJjqXI+COqClopF8/DISPpb3vy61xNetvT431+Rg25WmKnmiXQ55RrWIH5+nvXnXrdR0LRllrKoO0oR3bSNRsAfpcqfeq9ayekbv3FVMapW6jLUKDoKqA2m3v3DrdCeg7o90tQIGKPFPFyzkEWYcg3HHv3nXrVD1X58+t45p+qs7lcdkDHXfwyuUPqI/ShK+9j59b8uten+Wvt+o3923uTd+/KqOrqqDOSJTvPpcKiVHAHlv731oY620cNlsNjsXQeCsgpaWEJrkA0RqNAFzp/H+w96+XXiCei+d8/IrrLY2Lqa+pzWMzlXBBOY8dS1CvUGePkDxqSR/tvfqenW/Lqm/dfdPfvy4zNVtPrvCZ3AYY1HhgrTHOkPgLeMlWIH45v73w60a9G++M38rfbG2Kr+9Xc9N/eLN6vuDLVlZ2DuuuxWS/5+vvx68AOrRNuYDZPXmMOM25R02Ko6SAopaCJVCwjj1KPx/r+9cOt5I6Jd3L879udab9w+1KB4M3WVc8cTJRnW0Tu2jlUv791rt6PBt3cEu49lUG9clS6aXLrEPtJogXRZFufSw/x9++3rw6q7/mSfHjbfZHTmcye38RozA+4njlgp4llDCMsCpQXFve69ePz6oR+Kff1V8dMhU7ZzzzQZKhLxUrVEkgYMXsCL+/fZ17rYI6D7h318g8ZDQQSVEMLRxKazVMEeJ1+is3H09+4cevcRTpbd1fF3b2X68y9JWQrXbgnZ5knUK7h0Ute/Pv32db4DPSc+B/YNL1bV5TYGYqYqYRF6ZEqGKlgSUCqp+v+PvR69xx0NHyN2BiNyCbN0uhneN5o1UKAfJ6wbn/AHj36nXvPPVPXbfXOTqckmoH7WNyCSANIA5Ug/4e9jr2ePQN7e2Fj8LlTV0inzu5aRgQSxJ5Nhx791ro0+3sBTtB96AVqUpy3kuVXSovz/vfvRPW+HRJd+71ppu+Nq4/76FpKaqRH9VyLPyLe9+XVc9Gu3J2Tluhtz7e7Kw8reOKtgrK8wsyK8KC7D9vk8f19+62ettL4F/LPbvyq6fxO5cZNCtdQ0lLTVUGs+VyiaC5Dc/Ue/Dr3R6Oefp/h79jrWfPrv37rfX/0N/f6jn/AHj377OvdVa/zTvKelZ1QMV8Umuw4vza/v3Xh1peUtZLJm6+khRSFyNUSFXn/On6n37rQr0/bjocpXYaaKlSYkfUAE/644597690qOosdVVctFicwjRRPL4mMy6RpP8AUn8e9de+3qZ8vfhdt/K9fSbvxEzfeimlqCtNqb1BSzX0H37r3Dol3w8EO1KfNYavBE9JM0CGRSjNZ/1EN791uvVnONzU1LRJLDMojdRq/c/s2vcj/W97Ir1qtOHXeO3vjf4okUk8TMWAIZ14Oqx5966sDnowWJyAqftGoZImBAJ0OpAN7EG3vw61Ty6NvsmCnloIWKq9TpS+ocX03+vv3W+hKxktdQZBJWjjUCwVha9v8PfutVA6ft10b7io1SUF49A1BuQRbjj34YHWz69U+fIPZKbN3adwxRskfmeQlUKX9d+ffh6dap0av46b8x+ZwUKu2qaFEUBrcEtb37r1Oj7YjEQZvHvJZPGiXKmx/SL296NQetih4dAd2XtrESYqvFFFAlZSU9RUawiqQY1LfUj3v59a+XRDete5DPk87hp6uD7jH1b08cZnTyel7elPr+Px791uvRydo7/povA1a7XchXGokG44J/p78cdeyeHQw1GYxOWpllon/dKDSb2t/hf34V60ekjLvLKbZYswZ4FJP5K/1PI+h9+63jNOmSf5EIlTS0y0xHklWOaRozYC+km5H+8+/Y69U9CVm947WzW2DJNLTO80RDhZFYhmXkafehjr3HHRXqnrzEZ2qerxVPDO0huBLHqGr/aSf6H3vr2elPtinbatf4J6aOGVgqXjUAfS1+P9t7916lR0dfrR4koDVluDHqN+ALjn6+9de8ug17TpxkaLMF2ITQfTc6Wup49+HDrxB6oMq930OF713hicgqLSJjK5gJNIXUL+oFvexnrRwemf40d84WDe2fwFNVy/uZaXRAkto9PnNuB+PfuvcD1c5sHfdRmiMfFTrocKmtgQDdLj37z695dGBh2Wq4/72rpqd4WQsbop4P49+r1unQG9m9O7X7B2jm8NHi6RpqtZRzTDm0Z5+nvfWuqVti56H469m7l2jUU4x9O7zx000Sfb6mZiq+qw5/x96p1oUHR0OoqPH7t3XSZ7Lyfe08ssMimo/eVS7XFtV/eqdWHHq6bYG3cfQYCPIIkUFNBHeJUsmldNxYe/Z8+vegHSoxG/cPRVzGRg41WUkg+pRb8+9U69516XP9+xKRPSOqxBLixC/T+h9+63npPxdi66xoKl7mRtK3YG1z+R/wAU9+r59a09disWDLffqwEZsxPGm31/23veOt5PULfG9tsTwLDVBGkCkErpPrIt9T71QnHWuGegww+Tigd56BisbklLEC1zexHv2Rx63SvDpX0vYVHQu1PkG1sU/Lc/0/P09+FevEdAr2XmcBX4jJT1iwGkImaQVBULbxn6k+99ar1qZfzCu09s4zM5bFbegoZZryKi0gjlcnyfRQlz731Xoh3S/wAOOwPkc0ORilzNHQz1cUnjEVREnjkbgcWFveut16v5+PP8rvYm39t01HvpZ6iraABTUQPKCbcct738utefS03f/K/2DgTU7k2xjYHkp4mnQ/am5ccqtv8AjXvfXuPSI2h3btfory7Iz+JoIMhrahiMtOkRJPpGksL+9cTXrfDHVg/w66wx268vWb4lpYwcnM1TQkxEIAW1qV/B9+Pp1sU4nq5zYW3aaintk4k8KxIF49JKi3pHuvyPXvOvQg1m1KXKa5MbBH6NRuUsOB/X/D6e/V690hKLN1G18oYas6UhexUHSLfS4v795da8+lZUbuo8zOHhMbm1r+lmuOfoPfut9YabPQw5aAVSxrHaxuQBYD+nv3Xjjpa1mRxubglpaaGFn06R6Q3P5t/xPv3n17iKjotW/NnrIKmKqpYZoZYpQVaPWpDC1h72a9eBHVDnzQ+OeLwWYPY2Bhlo67HTLPahhaMkqfJpOn639760ccOjDfDX5JpndvUe26+asFZSgQSGcsrXRfHbn345z1sEcOrGYMvTZGlFOKh28oLWkk/J/Fv6e6nrdPMdIHcmCnpgQjHRN+bk3P6vewfXrxyOggzuGyFVGlLSSyBka3DXHJsSR731Xz6b4dv7kpLRyyyGnCclmJJ4/Pv3HI62fn0BHaWJiymGzEFbLIBDGdHJH0Bv9ffuveXVLO/MM/X2crN0wSTLTxS1E4cauNDX+v8AT37j1qtD0DGe+afZG5KCbbeyBU1FOjijlaNy7jX+23oXn37rWehM6j+JW6Ox3p98bokq5KuUiokhqg5VSf3LDXx/sPfut58ujZttGq6upxk4qypp4aJA5hgdlS9ML30/7D37z69wHRZt+9gby+V+98X1vjnqqnGI0cFU3qliZVYIVfT791qpPVnnSfw82dsDb9Nh67GUSZBI1Dk0oDMzpe/I9+63TpK9pfHHb8dZJD9jTxatbpOlKEYX/SQ1gR7917okvYvWOc2jHLk8Bk8hCtCNQjimdFYL6jwD79jy60QT0B/X/wDMX3vtDdEnX8D1U+Qjk8CF5btqT9sH1c+/A5z17PRy8P2t8jOyq+kglirf4VXGGQlTIy+OU3/HHv3HPW88B0eLC9A9l1WOxmRo3nWoZA8uoulnte1j711vqJ2tsPM0214qDdMxFRGPXpfUxA+t/wA+/de8uiS5DbmBpWliZAF/cBsg/wBR+fdutdFFXYcO8uxI6OjoWeGCqKlkgZyAJPywB96618ujvbo2/H1/synXxlGSBL+krbSov9eePfvPrfDoJaffGCy20spijZq6XWE0WaTkG9tPv3WsHHRaNi7zoes+ysRLlJpooqrIL4i50C7SXGon6f6/v3Hr3VlXdEg3ttCHcFJMKuilxyIoDCWzMnpFx78K9W49Vt9O5ql6c+UWzt1V7z0rRVNMYyhMQs04N9R9++zqvDr6LvQW70331JsrdCSeVMnh6WZXLaiR4l+rD6n34V62RXj0Mfv3Xuve/de697917qJX/wDACt/6hKn/AK0t7917rUl/kGf9vCf5mv8A4kOvB+nP+5X68e/da8z1tuVlXDQ0tTW1DKlPSQS1ErsQAscSF2JJ+nA9+69nrUz/AJr/APNszOByWX6j6rrHM33dRjahqCsUSq2kxNcxm4tz798z1v7OtdTG1OUzeVpt27jeSsy+clMldLVMZZFMj6vUzc/n6n37rR6HD+733rotKkT0zoLuvqPI549+690xZ3rzHUsRlEFO0j8t5F/P1/PvfXqdIDJ7DpK7F1iS0VFqMDCMiJNWq3p5P19669w6LO/WNLQ/cPUBKeUySadChCoN7EH/AHr37h1vqZsHrquqclIy5Gs8AfgCRiAt/wCn/Gvfq9a+zo1uK6xxuVakpaqaWaKB0aYvcgqo9QN/8PfuvdHE2PtrC7fpYKfbUCrxEryKgD6h9eQPp798+t06OLtumOLwj5SFVNWkRdrgawwFyeffh6depnHRJPmX23mdl9f0+ZX1vl3NFLoYCURu2gsy/wBPavYrT987ym3Lkk9buRoty1cjqrqg26lTFNmqMmSXJqlWWY3ZWkGthf8A1/fRnkPZoeXuXbewCBXpViBQkn1+zqO7+6qxNa9LfDlRCsc7N9wgAub2J/1/Ysm41Xh0UijjJz041NUYf0nm1xb8e2lXV1vIHWOnrzJcSXv/AL1/xPvbJTh1aop1zlemmSTzOEspK2Nvofxf35dSnHTcilhjpH5Gfx0zhdLLc83vwPxz7XxLVqnpDIdPaOgkr9wVNNk0paWNpJJCPT9Bz/T2sla1gj13LhB8z0rtI2mOjz6F3p/e3auxs7KnXmErcpntySGnf/I5pY4Gk9KkSAEC1/x7xE96PcFJrt9rsX7EXTgihPQx2/bWjALinVmvWvwc+VPdogruxGraHH5AJNpiWaPRHUclT/SwP9PeGk8U024GeTgTXoXQtoSlcdWS9Dfydus9mZKkzW755shVErLMlcsk4D6g1rPfj8D2KoiphCjrTKshqeretmfGvrHYdHA22duYJzCka62oIg50iw5t/sb+0bChoOveeOhCyO3cPJQXajgpKmFgkYpotAAH6SNP496HXq16kUf2ePxMz5CSnihijOl2kWJ/p9SWt79jreRw6Ax+0Nq0GakiSrknIZl9EnkUFSRwB73+XVRw65Z3duG3Ggi+4qIonFze4+vHI96+zqwB6D2t2dtPHz0+STxVczsD/lCBwTe9zfj36p60B1k3Hs7Adm4iq23LQUFGftXRZqeFI5dTLZWVlAI97yOvGnA9a8vzh+E3yE21mqjL9W7hzUmOSoeU0y1U7xrGi6yEQH6W/HvfWuid/G35H5vZG/KTYfa1bkaTJ0laIqpqmRoEYK2n9Ulvz79x61w6u/yfZHTFTi6GvmyVLU6qeCUhp4pCXYXPNyL+/dWHr0/5Xtrp2TqXcqUX8P8AIKcqlvEJWYr9QPz791roQPhvhtsbuwkFVT04AeV9EpiNiDyLsB71nrdKdWGV+O2nsvFyVGYFGI1ici5j8llBJADe/Dr1Qfy6IZ2J3RQVM+Tpet4RV5aKTTDGV1IX+lltx/r+98evEdVyV20N59ldgyVHYztRPBUGXwSvogVA+oFQ1h/re/Yr17Pn0JvavfGB6Q2suA2/US1OWNM1HDFQyeXU7x+NbBL/AJ9+61ivRPegvi13d3B2rTdxb0p6mbb8lZ99Q0tdDKUMPkEigq4/p7917q/E7cD4LHYxMHRQxUsVPGgjpFF5YkA4AH+HvXXvt6l1u1d71VPDFR4mFIljCRj7cqNP0uRb3vHn1v5dOGwuoKyjybZnO42neaOTyhJIA3I5sA3v1fTr2K1PQ25PIvKop0xUcUEK6QEpio9HH0A91695/b02R7/w+30MdZigtQCNEiUx1A/jm3vfz8+vU9OhIxveO0tu0EOUyEEhMlm/ciNrj8WI+nvQHXjjj0mdxd67W3XURy46hgJciLUIFv6za/Pv1OtEHpa4DYdFm/ta2kmenlqlDlUJi035H+t/h79w630ItP8AxXryZqisaSpo9Nx5SXAH5uT79Xr3y6UuHyG1t+VAqlkolqUHqjMkY9V+eP6/6/v1AevcOse6MHRyRmigWM2GkeNQeR/rfj37r3QDZXYc6PLeANDJq9TR3Xk/m/v3E9bzTpIybbG2aeWrgpYUIOp2EQU3P1sffutccdMi5NapvOixmWOxvb+v9B791vh0J+3MjjMnTyUmSoqeZpYmh1yxoxW621AEf7z79nrXQMbw2lXbDqZ67Ewmrwla7y1MTKzwori7LoHHveK9byeq4vkX8R8L3tX4jM4CJKDIGqR6haZPBcCS7atFv9t73w60c569QdEbo6pgo8Qkt6KJIT5VOlwsajVqJ9+68cY6MXtzYmR3xjY4cdkbNBGyzKKj6tpIP0P+8e/eXXvn1U78nviD2Hu/fv2L5zIx4lagmWGGpcxSRM/KlAeffutU8+rOPhd8MsHsjB0M9LjqU1iQUzy1ElOqytKi3Zi5H1/x9+PHrwx1aJRb/j6xpnwzRRq0ylGCKtvULDge6162KnpuouwTkcnTyyl0hlnUW5UEE3/3w9+8+vcOPQ555YchR0stGwfVArkOQQbp9Wv/AF9++zy690VPsF8ljp1hZI0SUqoaMC1ifpx79w68MmnVIv8AMf7Rze2dl12MhmTxT09TEVeVV5fg+k/6/uwxjrR+fVUXw63bQbDoM7uDJVkdI1TUGrkEc6oxZnLjTzz7316vRx8381e0Ow5U2L1glbVpP/kr1ERdygA0E61/24596x5der5Do5nxu/l7bp7AnpN9dp5PJyyTyQ1jUdVJMYQJfWyeN+P9h79XrVOrdcVsXZfR2Bgxu18HhTVJFoM6UkSTMbWuWte/vXHqw9B07RbkpnxUuTzssdIHQ2UShFtf/H37r2OqsflV8oxg5anaG0Z5Z8jXSTUkZpJPIymb0IAE5vz79ilD17pM/CP4F5XfW7Yu3+4TNUQTSPW0q10bvoUSeREHk+nHvfXuFT1clvuq23hKWk2ziFQYmmWKKBYQNOqJQt7L/tvdePHrw6Lv3JBRHr/IgxwSRy0tUPHKFZRrhIF1Pvw69X160xvmzsfIbZ7L/vLHC1JRTVUcmqFGiRh5rm5+nu3VeHWxB/LA7Q2zk+scZi4PDJlmhp4g6lDJ5PAFAFufr70fXqykAU6sU3VtbfcMNVkGpiaSXyvEGBIMbX/B9+FPPr2T59VAfIPbu8uu954zfNGtTTQLVLV1ZpUZVZNeohiv+8397610cfqjujb3bu34KKasJr44I4HSeZblwtiNN/6j37A63Q8ekx211+opZiIY3WQEh0W50hTaxHv1OvV6IHLtqfHZp0iVnu50ghrcngke/daz0qN07wodlbUrKrIyeGQ0c6Eq+j6JYEn+vvXlnrfVNmAiye/+9TuWgqJHp6fIF4y0jN6fL+D+Pe6dVPVx+/djPuHqiQ1iiUpixqZl1sv7FvSxv/r+/edet+VOjMfyMO0czhe8KjqOGtmkwpDOYWm9CurE8J/vFvfuvD59biPv3Xuve/de6//R39ywAuSAP6+/daqB1Wt/M2rcTRdF5aTKSLH/AJHN9uWH9vn9P+P+t78etjrSr65lpsnvHLMEWSIZCs0km9/3SQLe99a6Ntt2kphIcfVYtNFR+iQgfRjbUD/re9HrdehF3n0o1PthdyYOQ0320QmlaN+Yltc8D36h8+vdV/br+aWf22cj1xNiajclPGlTB5vE8gVo1KWLH37r3ljoAOht04zfO8cpFk6dNtT1ta7JEV8bPdzYc+/dex0ffPda7ijpIo8DXT1dM6qutGJABF7+jj34de6CHeHXu4dp4Wp3Aayc1NMsk7KdWoGNdf6R7917NOiw9afPKpwG85Ns5eZgtFP4XaSKT/V6bX+nv2OHXq06vk6S742vvHAUNTS5KIVMqwlkXShXVGDYg/W359+r1vo3GL3HTZERJBKJy2geUaTYcDk+/da+Z6MDgsXQz0UUrVKyTab+Ei5PpuAfejXq3yHRJvk31RPvuOSkx9FqqlWUGNODYn62H+397FPLrXHB6C7oXqiv2SJaPKQGE8MA7H9QP0ANvfuteXR1aLPVeHjNFSBpPL6PTyPUv097x16vRL/lr8g8D0/sbN1mUroqKvkx1YtnK6jdT6OTf3qvXutSfr75p7oHylqJEnlG0K/KiWaq0SmK0kw/tfT8+/dep1tFdQ9l43cuIgyUxjNJPFGaeoIBWQNEGOkX9662OjJbYy5rJ0iopQ8bWtoBsBfj/W9+630PmNxNDXUmnKRISb6hKQRyPrz731r16D/dewsDXwyUmLpIFnKFRPEBdGJJ1D/W9+61Xoqe5dhdgbJrWqaB6nMY/wAvkaAF2VUHqb0/4D8e/UA63U9CpsfsqkyKU2MqkXE10YCSRBQG1Wtb+vv3XuhqpMbHlauKJ18hurCc2LDVzyffh8uvVHQ7Y4rgMUKaKS8rJwp+trW964nHWzjPQX9l5psXtyuyji6xQ65ENvUbHg3+vvdOq561j+5t0tu7urckmAULkWo66BqeIHUbseG0+/da456Kh0jndydV95xz7ow5ioKquDieVXKnXPe5v7914dbQfV/Y+CzGAx9dhoITPUIhvEFBv4xYgi/Hv3VsUz0e3YtQ2a24i1tUyylVP27NyA3+v70fl1vAHT3V0uPx9DVKFjSQhgrcX5W3vXnjr3l1SD85OmTlBWbuxtCIaulR6p6pR6pWRtVjb/W4Hu3y6r8+i5fEX5M7bocyNoboroaWqop4oR5vSdcLWABf37jx68CRw6u+2r3nLuHHLjcMzzUbroSRLaSgFuWHvQ62a9KmDO46m0/f1qw1DG+hiuq5/Fib/n3vHWs9KNd6NBCsccoKN6QwtyDxwR70R1utenqjkMDwZWSQujes6uQt+dVvfqeXXq06c8x2FC0CrBKrKBZjYWFvpwPp78AevVHl0HM+68BNXIMpWxRq/wCrUy/72x97+zrWTx6xHf8Ag8fVOaSsSSkRrag6FQv9fr78RjPVhWuOgv3L3psHH5V6rK52mpaaJSZHZ47ALy9gT798h1XzqeqpPnj/ADDNpYqhOxOsMzBmMvkgII46IrI6yTJ4xfxk/n349az0RL4dfAjfHyV3rP2B21PUwUMlRLPT09S0qRtE58qiz8e/deOcjrZb6b+OG0OtMNRbe29hqcCCKBDWQhLkxi2oED34nz63TNOHRzMV17jIaARVUgeolH7eoglLjgD/AB96r1vHl08HYtXTUIp46Bq6nqP25bC6xRk/q/x96zXreK9UFfzIPjFTRbyxu5sHSoZBlKeWVYxezeQFw2n3YevVT1bN8TMRLtrq7Y0Qo/FLLjYzJIv0UtCtyfz/ALc+/fZ1vFOrKds0ymlgqJiZbqtwwBuLfQ/j3Xz69w6EKoydHQ0YWnQRSMpACjn6fUD37h17PQLb024mTppa7y+OVhf+hJHPNvfuvVp0XygzdXgq+WLSZQGYEC/ABtce99ePp06PuSbJ1K1AfxRREa5D/Yt9Rz711ugp0O2x8nSSpGYZhUSNe5Xnm31t791XFOlNmsfBXF/O2k6OE4DfT62Pvx63x6Kj2t1lgt0YutxtfTx1CVF7rJpsLgj62v72OveeeijbF+M239kZqqyWOpooA7swC8gXJ5/HPvZPn1uhpnoa62gqKKKOSgfVJGQugf4G4591qOvcB0tsVDWbgxp+/QwvAnpZhYekf48j3v7Ovfb0iP4B9tkKh0kMkg1FF5Oo/heePfvl16nn1GqVrapXpKymemQ8CQix0/TUD/T34UB695dFd74xFBhNuV9RBXK8rRXZSw54+gF/futHh69Uk987s26uzq+lrKiL7ysFVTQwPbU0puoCr9fe+q9JL+X58T6/O1+Q3NnMPqx1VWmqhaRW0NG0upWs3H0+nvx63xHV9tB1VjsLizSUDtRwqgCrHYKAFtp+n+39+690Q7v9aLasNbT1tQKmGriqKaJXKsvklBUf6x59+z5deNOlL/Lm+McdFLmd+ZChT7moq5KuhZrE+NpdQ06h+B79jrw+fVru7NsY3Fq2YapUzoiu0F1uLDgH/jXvQ9OvHhUdFQ7S39tGfEVCZGohoquMOqO2gMSOBzf3vzx1sU6q17i7DxmOpK0rVrUUThtTi2mxU+q59+p1o9UlQZPZknew3BPPTQwGvLtPYAkCS5P9D791rrcF/l/4jr/sHaOKq6MU2QEVFT/uLpNmVQARf/ivvR6sMDo9vYddR7NRqNG+zjjFoyCALW/qB70Ot1NKnqrPuvdVZX5KpkWolq6ddRC8leG/Pu3VeijbkqaTIUc9cESAxAo1uPxzYL/tvfvLr3Qv/CzYFJu3M5/I1WFjMVG8jrVFVNwpvqB/1vfj17qF8tarGRzVuAPjpqaCmnKTCwuyXAHFvfvPrxzw6r06MGJqN2zQZKSOSmEzqGf9JF7KRf37Pl1rz66786kos7lp8prGKpaSz4+r5UOwOpSv+PvfW+hY+L+66rImbr7N1719DTwhYqiU3QlBZQC1/rb3r59a+XQJfJHbmIoN90eRrJkx1RRZCk+wUnS0yipAW39ffvPHW+t9D+X7V/cfEbp6pYlw+3YAG/rwFBN/x79148K9HR9+691737r3XvfuvdRK/wD4A1v/AFCVP/Wlvfuvdakv8gz/ALeE/wAzUG1/9Idcfr/1dffuveZ62Jvmx25SdT9Ibsr3yKY+sr8RW01M5I1/uRFLr+frxx791qhrjr5xu+4a/cPdG9dzZ2b+JrW7jqKqlmk1EpHI5YaSfp79x69WnQixS4mSjkArkjdFIjXVbxMB/Z9+69w644HtAYCX7Caq+4iD2E1xb68Ae/de6l7x7EiySxTR13gQKuoKw/H9QD+fe+vdB9X9q02No40NYJmYWJNjx/sPeut9A/X7/ps7mGhFUIoj+bgfUfkD37r2OhX69zNHSTFIqrzM5/B+lzYe/da6Pv17gKOspErkkD1EqgmK4P1W/wBB78SBx69xOOjJ7Mx1LRT/AHFVCkCopGg2AJX6NY/7178v6hpHk9O+E9KgdLafc9LFBWZWeq+2w2KLPXyagI0jUclvx7TfVQ/UC3r3Hy6bYqnxHqqTtLszAd77/wA7s+asiGz8VDO+NriQYJqqNjZUYcXuPc1+z/t9zCOZv37eWxFsCCG9ei6/3Sy8E2+sB6evQKY2mbA+Sih1T08EhihA9QMamykH/W953jRIin4cDqM70yNKQmVr0/EQBBO6CJm5N+P9cce2TUnSD1eEFU7hTpqqrSHyBjobi9yRYC3txcY6sa1x1gZ4IIyzPp+pv+T+R7sAWNB141HDPSfr/uspojxz6tNvIV/AH159vKBHlx07DJHHUzGnp1gqaSSmpljm1NIw5QAsT/X6e9+PHGDLIQqqKknAA9ek80XinXEK9DP0d8W9zd77ioE29jnmXXEJpYwwZP3QGB494o+9fuvZQN+7tkuRJImKL69Crl3bXVvGmWlfXrbF+Gf8vvr7qvbdDkd7bao6zMpCkgerCSSJJ4gdV2F/r7xIj3Dcd2uWudwwzevQ3dYlFV6sMpcLQ0EQosVRfbxx+iJY9ICRrwtrD8D2aPBB4dQRXpMy6DUZ6zLjqiFwtTM7mU2RHN72/A9sxOVQjp5CCtTg9KrGQzTMaUBolQi5uBcW4BP/ABHunn1WnQOdtd17I6gpZkzVbTtVtHIVhfSxMgW6qAfzf6Ae/cR17HVX+9u6Oxe68rNTbbSrxGFZ9NPVQrIkcqK31JH+HvfXuPQu9b7Bn23SQ12eqBk6piHbz6mLMRc8v78Tnr2T0uN05JYYfNBRLSxxi9gpAIAv9Rb3oZ635U6A/cPd8onocUlEbQuEknVeLfTn3unWiT1you3c7j8xE9FRyGjlCKapEIA1fUAm/v1Ot1PQh5PMZbcbU01TBNkYJ4m8lO4uDrTTbkf7f3vqp9B1T38wf5d+X7SzFXvTaOAbDZEHztVUt45B69TXKC/+Pv3Xvt6riqfjj8pNnzS0tJQ5PP42kumpvKyBYeLc+9/Z16n8ugf3R212Z1xXJht44+oxNCWjjq4ZUlCG7jUGB496611sj/BL5idI0nUuNx9JkcZ/GaeniacK0QljcRjXquf6/X36nVgeg/72+WE+9N9R7e27lWq6KaYo8URuPHI2m90P096pQdaqPLpXUvYfVnTmKxm5MzkqJs1PGsktHIQXeRluVINz9f6+90HXq+XRNt89h9y/JzsBsZ1Rseso8VUSfbNmqOKRB42ayyCRR/Q39+69X06sK+Pf8rjKE0O6u06yXO1zrDVGkyEjOUcMHICv9LHi3vVRw63Tq2DBdXYPauOpMFR0CY2kokWKOOMIFZbW4A/1ufeqnr3HI6XmG2ftKunFJPNTxTU9pNBKBiR6rkH/AIp7917p5zM+06SWFPLFClOvi9LxBSB6ebn37yx1r5dYMc20k11jVsMyOCTGzxECw/1/fuHW+kxuHsTqjDxGTI1NBT6C2osYedP6vfvs69mnScxva3x83KWgefEzzqNIY/bn/Y3J9+8+vcB0odx7T6j3ph6ekxk+PZgAAI3gWxI5+h/Hv1fPrZx1F2z8a9vGlM1EkchVhKmllI9Jvxb3vJ61wz1P3PuPH9YwR/xKVcfHQgRpqYAtb/Bj71T068Oo1P2thuxNt5NYGSpCUcogclCGfT6QpHv3Xvn1Sj2b8mexOheyKivWGpXCLkWQRaWMbRh7G3P0t7txx149WR/GL5c7b7hjoHyeQhgr6sFlgbQpLcWW1/6+9EenXvLPVh9fi6OsovPEVZWgDgjSeGF+APfqDr2COi6b3oTHQ1cbx+gl7E+ofpuLg+60690V2hUR5GSJZQFLMAl7ck8/T/H3sHreOPQ4bZjpY2EdkafQGt+bkfgn3vqpJr0LGMxceboJ8TkaX7qmqNf7ji4QEWt/sPeuvAdFp7I6sz2yqXNZ3a1PNVJj6d6qCng1MdQBYKFUf63vf29W6qbX5NZ7sLsCp693PBLhMok8tKiSrodgGKBvVb3unWuhQq6neXTKGqp8jUTQZM60IZiAH5BNvp/Xj37rwx097Gp8t2BmIs7n8hNBTmW+p20qyjk3DH37rw9OrGdq7txe0Nvxpi51qQkSo8i2ZtSi1vdfPr2emL+M0m8stFNkFRCJLrrABJBv+fr798+tn59CVlMXQU1FSyUkSellZWQC/A+nH0976104Ve6hQ4cLUVf27qlgLi+lR6efpf3rr2eHRZO3u5dv4DCmvrshHMaRPITIVuNIuR72KdeqTw61B/5oHymqd95iqxe260zATzosMRvciThRp/1vez1r7eir/FfYHcHdGaxW3o6KspcPWLTeeZfKgN5ArcD/AF/fs9e49bg/wg/l8bc6mxlFuLP0cVZVTgSvNKQzqzIGY+sf196J691Z1m89iNqY77bCVKgQQmMUiEHSIxYf7H3r59b8qDoq+e3PW7nr5JWncRwsQ63AClBcr73T169UcOq5/mJ8s5NkYiPaG1p2rs9LH9ulJENTiW+kJdb+/Ux14mnXP4OfFnO9w1MXbfbdK9GDLFWQ01YzIuljrBAkH+8+9mnWurqH3FQbJwMu3sMAlBSK0EboRoWO3pItx9Pdet0JPQLTZ7FRx1GUrKqOpADSHUR+3zcmxPvx638h0T7sPtrH71zv9y9u5FaypmkMbUsPqZQzabWF/e+A61SuOqcv5t3W1PgNi7fcU602Tngpg1rhy7XNvx+fp72OHWiBXpT/AMn/AAGfwP8AC8nWCWqppK6AiN9WlV0jmxH+39+ORTrw49bWu6MljMxt2ho6qSPHNJSKrKbKeV+h491z5dep0TLuHpfG7p21WU1REK2KSAJA72IuR/j73w62cinVC2/Nu9g/HnsBsnhaSoGEXItK6wiTSsSyG+n/AA/PvfHrXVm/UnZ9B3PseOUgTV9LSWnSwDeXTYj37r2ePRed44T+D5itkqqfxorllY25sTbjj37rZHVWXy+7aBopNo4xBNkKiV4RGoBbS3pB9PvwHWjTrP8ADP4j7t321PuRGqaYOvmlRWkCiz3IIt7917PVvPY3Xr7D6zrMblqsoxxxijLXuW8egAk2+vv3WzSnREP5bvY0vS/zSir5ZSIauugiVzyNM1Rp/Pvxrx60Pn1vwbfy8Wew2Oy8NhFkKWKpQD6ASLe3v32dez59PHv3Xuv/0t/ZlDKVP0Nv94N/fq9aIqKdVCfzgKimh6FnFRJ4y0E4i9Wm7e/efW+HDrSu6m3E2CzeTrKu8lMMjVlTyfpKeeffutV6F7P/ACo2/tbNUiV7JpRlVYr6WI+oFv8AW9+62fToVMD8rc52lSS7b2xSz0eNqEMFQ0obxvHa10J/r798+vV6n4HoralXkmq8hiHnr6k+WWfwh1JlPrFz+Dz78fTrw+fSj3F8X9oYujl3DhMeaHJ06+VZ9CxKrL6r3X37z690BsfyhXrmep23lSKioplZA99dyvp49++fXukRXdq7n7enbG4qCWChqyyPdWAKy+k2v/r+/fZ1rpfdYfCjrOKebcW88OchX1bGV3WNf1X1A6vr791unQ7P1rDt7xQdcY+soI0AF7SBCVNrWH/Ee99e4dGB2DvnPbEgjTcrSO5CKC7Nb63uT/vfvX2db+Z6Pf1X3TtWtgSSvyNLTyNzGJ5tNj+LXPvR68McehKy279qPUS5SPMYyVpIyOJEe3FwP9h791s4NekBDubaVbVyVNXm8XTqpYhpJEiH+xJ92+XVaZ6Lt3l8qOs+qMdW1Q3Bhamop4hJGsVWhYnkA6Qef9h7114+vVJfZce7vm9uCrqIxV/3Qd50jZGk8EgcmxB+lve/Lrx49PPVf8tnamIoKuqr8WayaFj6kp0eUFGuoDDn3rh17JFT0fjrzrGt2TS0OLqqOpTCRFFpYmQhkQDSoPv3XujfYN8diY4f4fTTCQqgLEcFvp9f+I9++zreehswByNYYmq1YU5AuOQbX/r9PfhjrXQ1rtnFpj456KAmocAubAnkckD3WvW6VFOknNhKl2eGpgC08mpT5IQbBja/I97r6da09BNvPofF0QbceLp5DVACYiBBYsTqt6fe6iuet0Plw6w7abJ0dNFNLFLGyNZkdbEaOLG/v2T17paVu7YBSkv/AJ6NHsB9RZfrz791rNeqnPm98xKTrPZu5KKpyEMEzUbCmjlmIZnIK2W31PvfXjjooX8tDYdB3ruLI9i5+glqXr5naKSRPIpSViwI1/g+9A9ep0aX5qfHTa2K25kspicQaXMUryvSyiBInIjXX6Sovb37PXjTj0UT4W/JlafNT7FzSy09biJZItdTIwEpQ6F06vr/AIW9+z16nV83Wm+KqdIcgKjyU7QxnShaxVh6SAPfvLrdTWvQzJvSlyjtE6MpFwTyfpwAffvPrxqR0XLuTHvnsdXY6aE1FDPAY/GkYdgG4IuP8PfuveXWvb8quiodh1tXuraay47KLPPUm7tCSy+ocpb/AHn3v59V4cM9AB1L/MR7C6dqYsRuKreaOI6EZppW/UdI+vvXW8+XR2tvfzHqbdlZBXVpkkCsjFjM4X6Xt+Pfqde8+jebV+dm3MvRU8JlijfSoBMrennj6n/D+nv3Dr1anoyW2fmttiXHPSV1XSSIIyiaZySpt+eePfuvE5x0Evbfzm2LtPA1NTT19KZ/DKRGtT6yw+lhf6/T37r1cY6pB7k/mW9jZPcko2pRZOXHRzsPLTmdo9N+TqHv3y69noNx/M37MixtVSy1klLMkRUxzVEqSXt9CG9+69nope6fld8je7a/+AbVp8vUNXVBpvNSy1LaVnNg4K+/ZPXurWfgF/Lc3ruPI0HYnbC5StyThKoLkS8yoRJqFlm4H+29+691ta9RbG27sXCUuIjpIkjjiRLJBHG1xGFtqQc+9H5dbA6MJR4kfaCTGoI7tcG3qAvfj37z62ehX2PiYJGEuXYEp9CxAHB/P+296yOPWuPDoxdLLt2HG1UdPLTFzDYAgFrkfge/Yp175dU5fM/bhrC9THTGeEZFZA/jDAXbk/4D3sVp177OjAfH4Ct2Vt6mR40WjoUQx6RwAtuQPp798+vfLo4WI3DS0KxUcsbMqhbFeRwPes9e+XUPe244Y6US0Y0SkD1EtxxYH+nvdOvfLpEU27ZIaeH+LzCSGcfS91t/Zv71THXhWvTdlKLC16PkKSNF1qQbAcEjk8e/de88dIuOkx9LSVMFgxlZmtfUwP19+62K9d7LzdXhshII4pUgDEqW1W/wIPvX2deNeJ6GKTcb1empeSxYAXZvxbge9gefVeOOsVSMVV0U00+hprEg3uL/AFHHv3W/LoGcupDypEBpN9JC/Tn37h1uvTVQUUKFmmZNViRq/wCKe/fPrfTdPW5CCSdYn0U5PqvYLb6fX+nv3n1r7evPLQ/bwzx1MLVWrUQGDNq/x/p/tvfvLrfQdb83L9rSyfcV0FKY42IllYRpZVv+r3sU49aJxTqj35lfK+i2sKrD086ZCZyI/wDJp2ZW5Ivx731U+nVa3WmDrO/N648ZeY09EuTSRYZpDzrfVYq319+619vW1f8AGrqLD7P66oMdDFTozUUY1rFGpIRRY+n3o9WoOPSr7afF4XbNa9NJHBUU0BZnY6RqVeeB/X34cetmlOqKu1Tnu1dzY7B43y1kkGbWSUUxMhMIn5XSPex1Xq+DouPC9ddVYinqKdKKqhxapMZkWNmkEY1XNh718ut06KV3V3JWV9bVY/ByF2cmPVCzNYg2/Hv2evfPolGU2nntxZOeq3NWtHj9Ek2h3ZR9L3IJ5/x9761THVUvzHytfBVNtTZNR9xNUyLBoppCz+v0ngH37Pn1ry6B7qf+XN252DQ4vP1MGXUVMxkciJ72k9Ys/v3W+tuz+Wl8cMj0X1fDDm/KKiCCL01B9Z8f1BB/1vfj8uvAU6F35D56jzFRUxAaTESCwPAt/j715db+XVWHYYrI1nMVSoUCQkE3ZlB+n+vb37Nc9az0WfPVseYwdThcdKkOYnqI0jGrS5LNptY/192889ar1aj8S9l1fVmwoqrO6FlzdMNUjgLqEkd+W/r7qaEdWHqeq8/l5UUmY3vXYumfyo3msI3B5Zrkce99aJznoofWHV2Tm7JxdHTViQ0tXodqdms7EN/Qfj37rVKdGw+SG10wW2oKHJ07TxxxIqMiBdB08EsOT791s9Ex6wr4tubg/iNNUIsXlVWQPY2DXtx7914dT/lNJg+w4cTkcfpbK0M1CwMTc6hOv1Cn/e/e+vHref8A5bsFVT/DXpiKsJeUbehBVgbqthYN78OtH7Oj1e9db697917r3v3XuomQ/wCAFb/1CVP/AFpb37r3WpF/IOB/4cO/maNzb/SBXD/YjK8e9daHn0dP+e/vXI7a6lpEpZXRJQsRVJHXh2AIZVP597889e8sdag+YUTUYqlW1TVU6z6zfUSUv9Tz79nr1OiZby3Zujb2QqiqzywG9lUva2q3FvfsdeHp0G82+N25xWGOoaxJVBINnuxAvb37j17h0m4s921k3kpJaXIQwB9HlkWQKB9A1/fut9KRtu78raMq1TLKVF7RszNcf6/v3WgOgyyc+7dtVgEtNXTvrtqUMb3P9feut0r0Zvpjd74+ro33AklN9wA9qlmXgtxy3197z1o56uA6mzEUtNDWUlVGaZ1jMYD3BunNh706a162poQa9GgjysU2MMsrAOo9RB+oHJN/bdk5juKeVOjBJB4ZPVdXzs+S+P612a+xdv10SZLeEK0RhSdlqGknGlggTm/PtVyzy++982RIta6q/sPRNeMrGh6Jr0XhKuLZtLJXiR55Z2mMkjN5SX5JLnkgn+vvp3s9odr2a3smoCqCop1Gt9Ej37sOFcdD1pETKT9ALXP4t7VcetCgwOsOQmhZANS82Fr292jB6rT+Lz6T1XXpHDoVgRewANzx9be1CIS1T1TJx1FRo62PSQTfi34Hu5BQ1HW60+LpQ4bFz08iU2PppamerbQqxIZCGY25A9p5p1Cl5GAA9evGBrltPR7ej/h/uffNZjqmux9XUCqqKcvaDWqRTclf9hf3iX7ye8kdi7bDtp7TVWIOSf8AN0Jtm219WlxmuPTrZ5+JHxV2B8d9pUs8uJgfOV1OJld6eJnikb1AHi494vWu0xbjOdzcks2cmvQ7gttCZ6OaK4VpLy6BGoBCxgJpA4AsP8PayYaH016YcAN1EjyEH3dqeIlhZdRUgcfgj2mr5dap69KOSGlWD72sZVeMXjBP044JHu1fLrWeicfInvmTrnDSVeBlWetkja0VMxMmsEj9K88e9UqM9b+zonfXuyqz5NmTdW/kqhBBUahBJI6gjVe6q314/wAPdsAde49Geye0Np7JxNLisFjvF9nGI1YRLe1vrqH/ABPvXn14eo6Rv3FfX+GGOJ9KSAkgNa1+L+/cOt1J6fM/HD/CxDV07SsyFLAcg296B69T16AuTrOlrKn7wUzhGJOnRybm9jf3uo61noftodW4asx0cM1K0CDTZpIxYkfkE+/Ek561wx0IkO08Vg54aaFoWGkALZWIt9Pevn1vpzr4phjsjShYUhkp2VS8KWPp/F/fvl17zr0V3J1eM2vjqz76ChkSV5Q4NND9GPPLD3vJ69gDPVdvfXQ3TnceOyTVmFhnyVUzmLxwQqdbKQLEC/197PWvkeqaMz8Du9uu9zVtd1XHn6bE1jy/sRGfw6Gc2CovH09+68ej4fFv4F/IPdFdBk62TK0eWZYgZK6NrEsfUwaX+nv1aDr1M9Wq9c/yndz7gyf3/cORkzkML64KaNi5ADagAl+ePfq+fXuPVvHTXQHUfx/w0P2eMpMT4Fj1PXUkCltC6SS7j6ce9cMnrda4HS53d3T1xSUrrj8liXkiEi2hkjW1l4Ho+nvdR6daAxx6ru+QfzB2/sWCKqhiFcdSsUpmJdieeNPvQoePWz69VV9h/wAyHdbZOqqNp7YzqTP5IlmjiqNNv0jkD36nr1ropGe+aPye3FXSfbRZajjnkLDymdAAf0/X3unXj8uoVV3/APMyWlX+F5OtjWUcXlnAAIsbm/19++Z69ny6Q1Xub5Z7u1x57L1siPywE9QLKeTcX/23vxx17oOc32L2/wBbTLTTZapgq5eEkmq5UXX+D9f6+/fb1rgekJD81flvs7N01ZQ7pWuxCSapIIa6d3KDmxAP9Pfvl14Hq8j4efzfkosDDj+xaaomyMVEFkeQykeVU9Vif8ffqeY63WuOiM/zA/5p+e7JzNRiNh0ddRQedUEgM8UbKLgkOPr79nrxPp0VTqv+Zh2V19h4cdkso19V5BJUTMdLfi3v1OvVI6f99fOfDdlUE2U3VV01TTiGbVEJB5PLpsD/AF+vveKdar0Qbqz+ZBkOi+9sdNW1sj7Tlr9VBEtTPaKJpwqqyr+LH8+9cevZGR19CP4kdwUnfvx3212bjHKwZOmolIuWNmpwdVzz715Hq3njpQdkmGPFTqGV5WDX0kE3K2H096610USjoVSpmqCDqUlrk6b35sL/AOt739nVuHTxSZ+Snqo5aYnz61RuSeF4Ci3vX29eoOjx9RQPndvzCWMvWyIShAFxze4/N/dh6DqnDJ4de7lzeK6x6/3BktxVdJSoMcz2qgqm2gkC7e9EHh1YEHh1oudw9zZjcXzVG5dnVKVGJizLwTCje8dhUfUiP3vqvnXq+GesXsHrbCSVdXTrklx6OIiy+byKgFrHm/8AsPfut8eip5re289kyw4t5paei8hjWRgY0sfoQ3v3z630dHq3srFLsFmy2UpJKp9DWM4L3P4AJ/3j3rJ62KDh0pcb2HR/xGnkiyFOsQdbIJvUR+Lc/wCP09+8utVI6HfKdlzx4SGaB/JGqqSU5N/9b37J69gZ6L72F31QUmLqZMhULTvFHKn7khQAhCbH+n+v79Tz69U9UKfNT5lUtPiM1QY3KxyOYtKrBVsbkgjSNJ97+fWq+nVNvR2zs5372JDV18dTNSvl9VnLS6kke5A1+/Z69TrcE+FPxiwO1MNhqqlx0NLURUceuSWliX9DXJ1Ef4e/V63QcerYj2VjcdQja6hPPGghLoNIBtpuCv8Are6/Z16h6ArfmXXHUzvEzS1lQz+pWZgokPF/exnrZOnoqPdHZlH1L13msjV5OnizNbCZKOGSTTLqmQ2CA8/ke919OtcOPRZPhd8ZKz5E73j7N3/Q1dRQLWvUwyVMZkg0BtakeW4ta3vXl1oCvV0HYVHHtDHUW1dkzU9NS09OkD01OqJIfGNNgIv8Pevt6t0E+Rps3W4OaOSsFIzRESvK1uQv5v78OPXvKh6rd+R/yHx/XOMpNoUNdFkM9lpWx5WlmLyh5G0fpQ/i/u3VcDI6Md8Mfj7Sw4cdo7rglfN1ka1lMZwXKxy+sABv9f3o4630Ur+bL05ubtPbgyGAxlXVLi0ilVaWAvfxLqtx+ePx731rj0RD4C9/TdTVVHtTdmKqMVPRV6RNLXI0S6YjpLAtYDn36nXgaYHWyzsj5D9Q7ypcYmTy2MmqGpUIjWqUtf8AqVvf3rrfy6U27987SroFpsLlceIVICJ5ULBQbEDn3rPXuHRCPkdtnF7h2xVTaIZ20TepI1Y2YfW/vYqOt46KJ8e8vhutafMxfcR0IeSQFamTQLgm5APH+I976rTy6AH5E/ImhiqMp9vW01U6wloxTy3YsGP0A9++3rxPp1Wn1xh8l3Z3NT1lbQVRp1roBd0Z49DSfWx/Hv3Wutq/487S2v1XsSip46KKKolpLuTDGjayP8P+J96zw6uKAY6SnyHxNJvnaGQMNpft6YyBIQNQMZvyB78B1qvVA2xa2LbfyhxayxPCVztBCHYFBxVAfix978utfZ19CLpacVXVexqgG4l2/Qve976or/X34de6FD37r3X/09/ZdVvV9fezSuOtLWmeqTP52uMzlV8eZq/DwzzDHwTyzrDG8np5Nm0/1968+vHrQi637vbMZXJbZgjlGYTMVtO0MsZVgyTFbFT/AI+/VHXqenQ15LpWbP19JujeEjwLHokijX9Dc35Hv3W+rG/jrtbAz0dPDRUlMIKcXMwiCyMEXm5Hv3l14cKdH8osHh4sbHUUaRvLGELtY8KOeT/h791v7eih/Lb5GYHrjbkmIoJ1/iEsWjxI6hvI0ditvrb37r3l1UPt6WPsLItuLIFTLI8juj2PBYtz/X37qvVhPSNLtmSgijp4YUrECrwgU+n6Efn37PWwOje7ZyaU8FXBVgeOIlY+eAt/ze/vXW+PSopuycDRKYI46cyxi58ll5X/AH3Hv3y68Pn0GW7N8Y3dVT4Q8ELqSBpkQDg+9io61x6CncU1TSUwbH5menli5RYalQP6/g+9069XotPYfeHZOx8PV1sOYqJVhileJWrLm6Di6g+9cD17qvfCfMb5Edq79k2hi2zcFI8whNTEZTGRI+i4df8Ab+/depTh0c+n+DfYvaDY/Mbx3FnpIJmUzQSy1Bja412ZSfpzx791qlercOguo8J1fsuj2dDSRnxJEn3YitOWVbX1Dk+9dWFOjn9fbGpsZSzyLCskdRdmEikcX1Ac/n3vr3HpS5rE4asX7WppIYxH/mSii5P4FwP9h791rPSc/hNJQKq+NeGB5UEaQfp/h731uvQo4nKUDUkcRRY7KPoOfSPyPeqefWq0x0KGBylPEkIi/dRXGvVyAAOTY/717r59X4jpS7jy23qnG6gUhlQEkqACSBc/7z78Pl1r7fPoOYN74amR6CplSWM+n9xk4Frfk+90rnr1ekJuXcm36NWkieBEmDKArLa7fnj8+9/b1Xj0UTtbtPbuzKeoyUtdGHSGZlj8qgH9slbgn37rdf29aev80rufc3Ym5PuMX9+uCoJo5atoEdo3iSa7K5W4PHv3VSadXb/yJPkJ1xvDb1PsnF1lK2cx8NKs8UkkcUgmRLaWQ83v795dbB6uJ+W+Px9Zs7L5PNpBFNT/AHK0yqAyyJoIube/fZ17z61tNmbafKdxSVOBjaCT72Qt4YtIcmf6s6j6e/dazx6vc6yrN07ZxVD/ABKC1EKaBS1rekKLm3/GveurUx0ZXDbqwVXAXgmX7q1yGKiz2ufe6Z69Wop0Xb5H/KLY/TG2pq+vrqVq5oi6xeaFz5b8AoTcn8e/efXvLrXm7M+Tm/8A5KdgSYLa2Eq58bLUvEZRTSeBkmOkNcC3v3WuPRkeuP5Z2S7OXH5DcWLCvMoaQmmk1C3P1/2Pv3XurBtkfykdj0FDFFUQrE+hT6YWDX088f8AFfeq+nXqdChQfysdj0PMNXJGWtbSrKeTwLHj6+/VFOt0pjoRaD+Vxg6fHTT09fMfKuotdiRxbgg+9168F6KT2X/KvxOUrHMmQrpdLO7REyvFa/6Sp/Hv2OtU6jbY/ly9f7cwtRTZfFUcgWKb96Wk1Pwh5uR/tve6de8uqHfmX8Sqbb3Y8VDs+nRaeuqljWOKNgGUzWN1H+HvR691dh/Lf+C2zMbgcZntx4agkygjo53+4pgDq0hrksCPr9PfuHXh69X/AO09l7bwdAlFTUNLRxxKY0WnAQFVPF/9796r6dWp69CrgNk0uSnjKm0YYE/X8n6j/D3oUPWiacOhLyGFi29SqYNMiKv9eLj6Aj34Hr1PXpMHckOlqczpBNJYKomVCW+g/wBv798+vUp06Yel3GkjVK1DyUr2/t6uP6Bhx73SnXsdBp8hcNDU9c1lfURxtLEztdgC/pW9wffhx61mnQC/Fren31FkaKCZZHoUddBkUlRfhbD+nvx62fMdHw21L9yNdSBr0KwJ/s3HH196PDPWhg9Ivt3JzYra9fV0h1VEAlaMD62RCwsB/j78KHrZ6qv65+W2e3nvrJ7Ky5WFcdUPTwDUA3D25B/p7tQdaqerB8RuU/w+Okkkcs6RsQPr6l5Jt7rkdWA6ELbOGpq2J6uaUEahdWNzY/4e99aOD0K2YwGDjw9NJSKgqWQamUDUxt9Db3Xz690ikxs89IaeNryAk2tawHI+vu3DrXTDPFVY9xBO7BX54uR/re9dbxXphrJ49fj4JJIv9R/j/sffuPW/l021uHqZITVRsQqqX4JAt+bke/dernpB1GaasgqsaTZlvGXItz9L3PvYHXjTy6BXdVadn08uSeudrBnEfmU3seLKT72KEdVp1Xx3Lv7s/tivO0Now1avUSaFkiRzaJjpuXUfW3P19+x1vNek9iP5dQ3hjoave+uozDLeRasSOTJa/Bb/AB9+60fXpMbw+Ff+hWmpNxYCMw1NLN59NOHBYxG63Hv3WqHq0n4i70n3dsOVs9N4Mhi6cU8URkCeTRxfQfeurDoFPm9vV9s7fkp8dUyCqr4QrKji7av6D34dePQRfATqZsll5t7bmp1qI5JRNH9whaxJ1f2vez1rjgdHv7n+/wArBUYnb6iCFWeMCH0qEtbi3096Hr17NadEgzONxOxB91uGZWnc3cyMrMSxuDc+9062OiefIrt2hG3KmHb05hqnWaFPE6hvWtlC25P9PfutfZ0Wj4jfFbd/dG/ItzbggetpTUCRPu0dwoEt9Qvf37r1K562iOtOrcX13tuPES4THiOkpotLGIKSyxAEg+9V8+t/Z0pqXesGOpKigp/FTLeRdKMqgA/kDi/v3W6Z6Kv2/Xw1dHVz00mqUi7fRj/ri3vwrSnWuPVcfZFJlKLE1GVn1CmiWaZyxPKJySp+lvfutdBd8edl4ruDekefppp2o8fUIlQLN4tccnq9Q4/1/e+vU6tZ7b3bt7aPXf8AD4qtIajD0gWBY5VDalSy2sf6/X3rzp1vqrDYO3avtje2WzuX87Y2PzmKeaNtB0kkWb6e9/LrQr0DOclqti98YfJYx3bGUFSsM1mJTiQD/W97+zrXn0eXutsZvDr+LJ1iIHr6VfGdIJDGPix/4p711viOqjqXbNViNw1sUkoFJ5ZSjF7em9wAPx731rh0IfTezcf2N27hNkyTTVEmRqafxxEF1IE4t6Rz71nj17zoevoFfGnZg696S2HtQR+IYrD08Kx6WTTdBxpb34DrxNOh59+631737r3XvfuvdRMh/wAAK3/qEqf+tLe/de61JP5B3/bwv+Zpb9P+kKv5vzf+K8i3v3WhTj0PX/CgDLSrsOgoio8SzRsNX0IEoH/G/fvPr1cdat2aDvTYhqddSmip76VNrmMD37j17rPhOkF3yTNUUUbBz/ai1Xufqffut9CXtT41Y+kybUaYqB1jXXIft+NK/qH09+4dez0gu98dsfY2Nmx0FFRw5BxoISNUYORa3Ivx7317oIurduQ5KhnqaimWRZ0cQl0J1G1xb8W9+49a86dCzB0Tj8rhnrq7FwGUuWXVTEki/psbe9de/l0AnbXSGVqRTDBUT0/24AVoYXW4Tn8e/fPr3y6WXx53ZurCZZdqbhSqipqGQKkrq41gC3LH3R30fn1otnq0DF1sVZj2SNyUailP9LHRxY/6/ty3iYS+J0+r4oOtez5hbYzWe7/29WVlTUy4/G5EFYZCzxgCS1x+B7kb2hSK456gD+v+XonvpNJ4dHa2tS09FgqGKnULGlNABYW5CAnj30FlZmk7ugNd08c06mVzcG1wSD/r8njj3tB69JTU8Ok1PHK4vc2HNjc8+1KkDqtRSnn00inapn8f0AP5+hH+t7e1aRXqma/PpYY7E+BRew1D6j+ntFJLq6uB69Gf+LSYmr7EiweQpo6uoyTCCgR4/JaVyFH1+nPuDPfvmK95c5QW6sSVaSRVJHoejzYoRNMQRw62qOgtgUWwNvUcVZiqb+LPTU9RGpjsApTUrKSPeEU1qN9c3101W456kizhRUyOjG0W4Kyt8v3EbpJG5WGM30gA2UAf096huTa/4tEajq11Msa1Pl0JG3MNnK+nmq/GfGULCym3pHFx721WNTx6Qq/iDV1IoqfI0zMHpyWLkAmPg2P1ufx/h70Pl1s06RnZtXuOnw85o1YPp9GgEfVf6e/VJ69jqtutwO7d0Z+rFVAKoxtIxhqV1x2vewDe7dez0N3WmW/u6RhspTx41CyoVhAijJvb1Dge/N1qlcdGKy+Lx9bjRUkwyIVvG+tWNrce6/LrYwcdBxS05pgwpoImHOluBb83HvfW69NOSfIKjyyQROqm4B0/Q/ix/wAPej1vqNFUhBBNNFAqXBdQVUEHn6e/daHS1y26UOCp6bEmFZWkVGCOA9/6ce/de889PNJhfFR0+TzFTol8KyKpcHm1/wA/n3v5dar6dJHObhqK+GqpKMKY40IVlPL/AI5I9+68BQ9Fd3Xt/IZ2GWkfyjXKVDBGI9R97wevZBz1H2f0KafK0OQyzOKJWV2VtQUrq/x96rnr1Py6sy616n2zWUkBjwWMrKVVUiSohDkqF5+o9+48OvfPoyeD2Ht+jmRMNiKCmkWLUfBCsYBX63Kj/Y+/U60aAdBb2x3r170JQ1m4d95Wlx4xYMkcBq0QyWUkehzb6+95rTrfl1SB8hf5iW5vkpU1OA6WkkmolMtN56NxqXQ1rAx/4c+/U9eqjpI9NbG7DyFBPWb4y2VSSVw5WWWSwueRz/vXv3Dq1Oh5i6Q2tuaqi/jE7VccJsFqbOp54uG9+J69SvHpbVPS/VWHpRCmDw8zItyWpYWYt9bXt9feq9bHHPQdZLqDrrKVK+LEY6FQQCI6dR/rW0jj37y6959LCn6X2RHRRx02No2sOSYwSLC3Pv1cdep0m6/qPAQM/ix1GBY+pUA/1re/de6LZ2T8HNs9xU9SzKIcuoc0UcaMovp9Fzx73wPWqE8OqaPkf8Re6/jNLNnFwJyWBGpxqp5JyImc2a1iPp79jrVP2dFc2r39jyxo6qiWgyMEgSaP7ZIbOrWdfUAffutdGDoNx9Y9iU8MGdWhhqF4MipEjsRxqZhzf37r3n0Em7ukNr5Gvqzh5YzQhGaOQaT9ebi3vfXuiD9p7BzeAy70eJq6h6Al9agnR6RqIsPeuvdEf7Hx2VSSXKYmA1OQxSh/3YtQDpIG+hH9Qfp711vrfc/kDfO/aG/eitv9JbnytPSZ3GQ0dPJSS1MMXjlSnC6ArEfn3v5jr3yPV9+9ttV9QJZKBoqikqWeSJlcSqYzwvIv71wz17jg9AzHsavPmWoh0MeBZW45PN/dePW69Y9n9V1L7k81SVFNqBHkOlRfkHn3vr3Ho2uR35158d9hZLd+4cpRRDHU8zNGtVCSGSMsAUve/u3Dhx61Qk561Hv5mv8ANW3H31kZeuOqqqqalyTGgjkpHAA/dMV3Ke/U8+tVPRb/AIdfF6rxuQi3JvgrUZrIyxZB5KkFyDP62JZuL8+/deHr1ZLvGqj2NWYyXGV0kkNOF8lPHITF9eRpHH09+x1bp5TKbE7qpFwuTkjo8lCoETIFVmkH05+t7+/Y610I+xPipST2jkzFWIW0vHGJmCFSfSbE+/HrYGM9Krdnx2l2+0NVia+aWanIIGssLgX0kD3rB68cdJyv7DrdqYlqDKlfuIY9IDEfRfSCf8P8fe+vY6pt+cPyNylHRV9LRVLU7yfcBVgkAYjxmzWB9+611r+Llt19n7pkpqupqqmOplZdMwLrbXxwb+/cevdXy/Az4+0+BmocnUUCa9dNNcQsPqoJN/fvs615162ItlbshweEioqNDA8UPjGn0G44PvxB49W1dR6TcbZLLS3qYxUM/wDbkUfni1/949++3rXTN2vvFdo7br8zVypNLTUc8yKGRwWhXULD3rh1s5GetZ/tj5Ubs+SvyYwnWy1FXBiKOup4XgVmEE6JMEsyj8H3vrXW4Z8ZsUmxek8HgMTQU8NVFRwJJNGoWUs1OLlj/sfevPrfSbeirKbecuUytRJJH63KtL6Lj6jSfeut0x0Cnem9auTCZaDb61AmUMF+2RrXZCL6k+nuw615U6qA6K+Ou8e1vkK+Y3elZWY+DKCWkjq45ZYk/c1jTq49+r1XrZPqdhV+wtm4qmpaZIaaCgpo/HECE0Iv1IH+Huteredeo9didt7j20MfmcfR1T1sOlhNGH+osBY8+/fZ1qnRBe5v5cezt64ebLbUpaXH5mQSSKaWIxSB5H1g3X8+7deIHHquuq+GXdvV+8IqiDIZWShp7iJRJKUKqLqLX/qPfuvAGvQQdibi+SWwctU1FEmTqKSnfWQ3mJ03sf8AYf09+68T0E2Q+c3a1HTS4ncmNqEgRGRnmRgPSNLcsPfuvV9eiy7q7+3VvmplgwMssDzSXcQSaTdvwdP+v791rpe9R9IZjfNVPWboqKiRjdgtRqYEkarG/wBffuHW+jy9XdY0HW+dpshHQUiKjw3YRgX0G4Ym3vXWx1ZfQb0pstgIzHLp8MQX9tgQDa3HvfHrXDqPuComw20JsgrCdcpB4tMrjgPxwP8AY/j37reOqMexKOTFfInBVMaxxyVGfx8t1IBsapSTwPfqY61XNet+747TGo6T63mNiX2xjrm/9IrE+/Dr3Q0+/de6/9Tf3P8AS/J9+60fToMu3urNudx7C3DsPc1LDPQ52gmozLJGrPTtIpCyxsQSLE349+6318+T5lfAjbfxA+WedxdBNFJR1UtblI6jyKwDTzGVLDgfQj3odePTCrQV+J+1rqkGNltEZD/YY8FT/tvduvcejP8Axs+zxcs9LLIJKaTWsb6hYBh9b+/deHRmt99v7K652/VJVZSmjf7aWzNpFn025ufeutk0GOqBPkP2Dju294yyY6vhraeOq5aMkqF1EE8H37h1rj0mcHXptZKCGhdZ2Eg+4hW/Cnhg1vfutcePVhXU+cwxx8GSSRKOpeOONqcEBtR/qB+fz7316uOjdbcZJoRJLMHjqLt6m+gIuur3U18utinSX31sV8pBJ/Cq9qORlJMiPZiSbm+nn/W97HXiOijbm6g7Fp5ZJsRn6p1DMzOHe4FvobH36nXh0Xnd8Hc+2o5QstfXMB6TeU3v/Qf8b9+z1rqsPtzsPult64vGZ2qrqPD1uRWkeCQShGR5NLA3/wB79+62OthX4JfHrr2PaOL3blaakGWlpaeo1yGPW0uoMSSefr791qg6uGw23cfW0sVPRTBYU4jVCpDWW2kce/efVvLoTsfsWmpKNJ3p9c3pLE21W/B/2Hv3Xqnj0JOKxtOlGYYpwjMvCg3AFufp70a168Bio6Z62Oho5BHVKpYMLO9jf86re91OOtEdSP7n0mZAqo6xVSwOgMORa/0Hv1adbofI9eo6agw84pZkSZSbFzYn68/X349exwPS3lOIoqN6uKqjVzGNMN+S34HHvXn1vHRad+9mUGF+5fIVqUkKiQ+r0qQq31XPvfl1X59V676+Xex8fmXpW3RR0mhwpLOAfrY2F/fs+fW69Avvj5rbfxVG0+PzkOXUofEkbCT1/UcX9+60T0SbcXdGW703AlPU5NsRjS/MbakV1Y6bg3t79w698+m/tnpPY+a2PVYeDF0mara2mML1ICubstixNiffvs61TqqPpOu3p8Be75N74qKekwWRyaNMYhKUSJZfyF4sB79w62KU62Mj86Nu/IjruiQZSI1D0K+aMcSNI8dm1gn+vvw69Xy6QfX8e1tsZBM4IIvIjmWWUaPUGOog29749eHRyU+SGwWwKPV5qki8EWk0sjLx4xYj68+9cOt16KN3B/MF2LsbGVrYCppKiuhuiR04BkZ2SwsoP5J9+61XqvLrnrXuP5zdlR7k3ZU1+M2Ka3VS0szTRRVEJk1qSH4sR7917rYy+N3w56r2IuMxkO1qKfIpTUgatRYmdpE+rM1vyffs8evUHVk8OzcdsWGA0mMVYrXChUsv+8fge9Z63jic9PFRnqMURqgyw+NdcosPSoHJP+Pvw+XWyOi+5zu/DTbij25isnFUVYcDQltSm/K8fn34DrRJpjoftnb0ra2KGiaocubxlODz+ePfjx69UeXSjy22Jq/7hpkaP9ssZDpFy4uByPfuvYI6LLvOrxu1KLIjKqk0Ijqf3JArAERki1/969761nrWP7n7C2zvv5OY7bjVENLCmWRIFuoD2n+g/wBf8e/Dr3HrYa6S2mcHgcYlBPopzQ0UmtSCpHjBC8f19++fXjXo2m2IGrnEcsxBDWNz9bH/AB+nvR630M2DyEOJc08lQIWewRybC30PP+PvQ9evfZ1w3HXZKSKSOkWSuV42KlBf6/Q/0Hvfz68OHVNvzA7x3x092VtjGNLUY/G5aeDyzHXoiDtzqPvfz60cZ6s06R7Yr8/sPAyU00mVWv8ABHLVJ6lTyINTX5+nvR63g5PSz7f27Xbh2xUbcpq4u1bTTy6gx/beSOwW4/p79gdeGeq8fjP1f2L1v2JnYa5auoxVdVyaJHLsvjMhtyeLe/Y68MdWuyZCHF0yapRFIY4wQDYqdPJ/2/vwp59VofLoPN0TpnqCspJKoSLLBU2BYD6xkXsfevt6v1robooK/qD5J1WTqanw4/JZUm5LaQpmuOPp+fd+qU6vI6y3ht7cmMp6mCqiqXejpxb08P4gPx7qerDh0NWArnoDIZp2jhL3EVwNSg3uD71w62c9CHT7ho51URVazkciDVyCP+J978+tUPWasrZdAq4gYATp02I/H6jb37FKdayTjptnqaVoDJNMs1Qb6IyQXN/6f7f34/Pr3SYrKSlVBUzSrDITq8d+Rb6WB966t8ukrm96UuHoZB5RLpVlKWHP9b+9gnrRpxPRG+8PkttDYOCydY1bT0uQEbFIyVVmcg8Ae9jrxI6Jr1LvjdnyUzkUdPlJv4atUQI/WUK6voD/AEtz791oGvVznVXxzwe29sR56fGRtmI4VtU3UyFk4vyPdePW69SdyU/jjeohk8VRSi6QCwaUj8WFufe6Dreei47lkn3TFWUW4YWpoGjeOPzKLf0Fr+9gdVPRJUy+Y6c7IjnoK9/7vO7Gopw37LA+q7AG3v32db6B75CbxyncO8Nu0mDR66lnqoY6hIr6Yk1i68ce/cOvHPDq2P489dT7a2dRY4KaMmlp5GN7As0YJUnj3o+nXh0w907wqthR1AjxzVDOWP3Cp+nj9V7fX34evXjjqn3vjsifMSLWT1thruYGvxZzwR/X/Ye99aNOiG1EmZ7M7Co9uYmF5IRPE0hj1Cw1WJv+PfutY49bHXwn69otkbdolqXWmqBCBMx0HS1hf1e9E+nVh8+j0bvykUtBXPHWW8NMWityJyF4UW/J9+wevL69Vk7y3vuqHL1VYYJqalhqJItJWQB1VjZgL/0+nvfWvPpR7fyuP3/QeGor0pZ1UiQagDcDkNf37rZPp0Rz5N5eoiam2Htp3yM9TMaarMN3McUp0MxC/T37z69mnRgOidv4XpLqaqxtPRIcrl6f7qWvUKWglddbKW/wN/fq9a+fRPe199ZXe+78ZtXD5RslNXzrBWU6MxKgvpINj9ffuvHqyWh2dtTrboKnx7QwUG6ZaRZZTZTUHyx35J5/Pv2et/Lqn3e4nj3o2lTWyT1kcgXlrgyXYm3/ABHvfVT0dvdS46fp/Gtk6lMdLTUKtEkg0XYR34ufderH59UybqzmQq92VlHBXMtIs04inUnTJYkKo/r/AIe9+fVft6uc/k4/EjN9vduYTuDNUM0GP2nUnUSSEqUhmuHYN/Ufj37rXAV63WURYUip4o9MUaLGmkAIiINKgAfT8W97GM9bNTjrP711vr3v3Xuve/de6iV//AGt/wCoSp/60t7917rUm/kG2/4cJ/ma2+n+kSut/wCfT37rXEnow/8AwoLw1G3V+IrnmEUjVMQLXH6WluR7959b+XWuNFt2iG2MPWMgcrQ0un6eq8YsxHv3XqefRhuoRRUSU4nhVEn9NyAQLi3++59+PDrw6OJHX7F2liavLSGmqKt6JvSBGWDaeRz/AI+/Z63jrX/+Ye96Pc/ZaRUMKw071qlgq2WzN/Qe/da6Mf0nSYx8NhqeSBUUFRJI1gCSOST791odWZbX2ntKXb8TGSF41jRnBKEDjkAW96r1unSG3vjtoQSQQUVBBPYgMUCH1W/PH197Hz695dIao6z2NVU8eRp6Wlo8kx1P6UVy31sR70YPGb0p0y6a+GOsUFPSUcj41ZhCWhkUNqW4BWw93a9NrJ4Krq+fSqGMaM8equ/kRsOm/wBKGFL1HmjratfNMSLR+rn2fchbnJs/M8W5W41sDXT+fRPuERLdx6VMNNHQSnFwzCSCnRAsqm4Yaf6/T30hsblr6wivXXQZFDFfSorToDXWLgjrlLGGjY6gxB4tzyOPp7VA56SgHy8um2vb7amd/GSxUkD6c2ueB7cj7moOvAVFek7hopqmQzEFSGNkPBPtTMQo09VGR0sZa5KRB5QAVB4Nz+PaMIWOOr0GmterDf5TOw8V2z3dnKvJokg243npWkt6TGdQI1e4M+8daq/KUMDLWrq3Qi5Uo87uTwJHW2H/AA+OeSkooKe7U1NFEKofUpGNIBI/FveD0ks0QMUIoPTqQPFK4HU6+OpJI6YIhnFlY8Bi1/px7rbJKr+JIvRZco0xK149D115uujoCaSrVWjkUKRIPoLWJHtWX1MTw6vHHojCH9vQ21OJ2zk8c1TQtH9wyGQhNBP0uVAHvxAPVwadFe3hprKqXHeG8aNoL3sV/FyTxf3RjRa+nVvLoqu8di1O1Kts9jpmlMzF3pxybE3ICj2mjuC7aSOt8c9ARv7JZDOYx6ekxrY+uHrFbpIf088MP959mkcKMta06qT59ATW9+9h7O+02/R4Sr3IBaGWWNHkEd+C30NvbMgWPieqyvpGBnp7zHyO3/hMNFUU2wqyqq5P1wCCQkMR+LA+2AxJoOm0mLYIpTrjt3u3sLcWKqa3LbOrMbYMywtFINSgcWuPev1hgLXp8GpoOi670+R/Y8VZV0GL2vVz/b3CKkUpJb6WsB7eTwgKysFPp1Yr69JjbXevfdTVU0v9w8pKn3KME8E9jZuD9Le/aoKVDdVxXj1aP1bQ9s9zbcp6jLYLIbfWngUMro8asV4v6rfX6+/BlOAerUAHQgQdZ5Xb0zwV9U4Kn1F7nVf6gj2hvrqOzXUDXp1YgwrXpUx7RoxjfuPtBK6XYPb9RH9Pbu2zC/GcdUKgGpPTZTYjMbmmSghopKSmi/b1gECymx9rJ444mpqqetFNOePR2OrMOcBjIqOpqdUaBRLI1iFUix/1vbQ49U+Y6AP5VfP7p74qYOsgrs3j5s40VSkNOxjM3k8ZCKOb+948utGvE8etUr5Td79yfOLckq7cWvi2nkJlcvGJ44jT6+QCDb6e/deNT0aT4g/H6k6KxlLkso48lUyF9bhj5HX1H1f4+/HB68OrK4snj3oh9vKpjZb8WIF+f7PvWSa9WqPLqJHXyKS8BKKv0Kj9X+FvfvLPWqkZHUKXKT1LsrgyAjTex4/2HvdB17JPWWhmgxuqaazuxukZF7D8+/EV69WnTwueklB0L4Eb8344/Jt70F69qz10avyIFMwkcHUfzf8AwuffqdexwHTnQZ5sYBUx/stB6hN+RY3JJ9+p6dbJ6Ukub6573xdZsXdVLS5aoMX2oMqwmzHi/qB9+HWutef+YL/LXxOwa+v3Xs6FaUTSVNSsNPIqglvUosgHvfWvPqoPE9R9nUIqEgxdU3i4WoBkHH11A+/deBB49OWB3puzauRnw+W808oGho3DEqPof1e98evUHEdJbd+dNZWeCsojGk+pjUOtlXWPpqPv3HrQ6ATeHX9FkaGtrKSMQqqandTYTD63uPx/X3rrfSV+PG8ewuoewKbcPXe5Jdv1FHkUq6pIGkvVpF/ur0Ecm39Pfh1o9bZXxn/ne73wW0sbt3eGwMjn5KSlippMvNRSurFSA0usg8fn6+9dbr69WA4j+c31LkMaDlcVRY6vK+qnmiRZAwH0IP0/23v1PLrdcdFS7a/ncYLE1VTQ7bxKgVCSxRVUEBKxs/CvqX+n497p69arU9EcrPkd3h8nshUR0uars1tzKSvJLiAagRhJfQY7A2+h/p79Xr3SZ2/8G8ziM7kN15fb02PSQNNFNJfTRM51eUah/Xm/v3Xsj7Ohtx+6pdk4z+7FDkjlstF+2syHU6BeBGCPwBxb37j17gM9KTa2TzGflddy08kcRJtLMCoGn/gx9649eHy6RNZuqj2BvKWrxdVHKFlDDxEEqCb297zTr3n0abbfyrqJKKIxV/iniRY7KOb2/pf/AHn375dbJ6Eyg+VsdNSSHNSGdSp/eYKQvpt/a9649br0Qj5E/KnbsIrq+nqoZBJFIEVdJK3N7ce9/b1Xh1QV3v2rkeyNzvJAzS0jVDLpQPwrcce/GtetdCF8fOn6jL5GnysFAx8DGRipFyb3Nz791vrY8+JJoqeCmxc+PCTQwwgkqtzoQA/7H8H37j17y6N7v3e2E2vh6pZqWOkqtMojewvwpIa/v3y69THVV+5e7t75HeP2O3KipRWm0iWLWAQXsCCp978uvefRx8PtndG8dh1n98K+odTipZP32Khg0NyBq96631r/ANaMH1p8wsZX0NEqeHJoruoBLk1A/wBvz79jquetyz4/70fdvXFPVRg0fkooTFICFOr7cD6e9cOHWznPSJ3D/FaeaeWeveX/ACl/3GJJMYaxWx9+HHrZBpXoaet6LYWe2tksfmaKmmr6mNwKhwgkDEaeNXvX29ePr0+7C2vsHYWSergp6cT6yyyWiupv6TdR738+vdCduTsjF5dVxUlQkiFCqiwsBayi4Pv3n16nQKZLNHFZXHtApqaYOupbXCi/0449+8+t+XS1/vg1ZUn7eu+whRQ3jHpU2W4AHvQ698umHK5iiytQKmuQ1cFMjq7tp0sQvJ1H6j36vWgD0TnfuT2Pnc1V4QYemqXq2EJUiM6fVbm3+PvYr59eJ6rx+W/xu2pBsqvq6DEwUtU8FTOHTSHLNdha3+v73knrXDB6pZ6R6zzH+kGqhWGWeCOr0BOWuvk+oHv1OvD06uLwuzp9nYVstJC0BWESMrjhQE59669T16S79nYLK07UP8UijyKzFPB6dZ08KDzfk+9063XPRgur9y1FNip6OogMyVKftzW9IB5B59++3r3Dpb9l56Sm2hRXrSUhCyGDV9LEH6e/fZ17HVNW9chUb2+TO14qOpMaxZfGBo76QWFQouP8Pfuq+eOvoJfHWjOK6R64pJpdbRbboFLt+WaPVpHvwz16tOhsU6gCRa/497OOvA1Fev/V39+Dz/vP+t791rBz11dtQFvTb6+94p1qp1U60qf56vlT5PMyxkK9HpWRRYlriwv/AMV91HVm+XVYFLt6q/u7TZKul+3plpY2IlvGSAt7A/4+9/I9a+fSCr/kNRbThlw+1mMmSVSqeGQsS49JsV+p9+62eg8rutu8vkHSzzVWXqaCkkWSRYpJnQhJBdQASP6+/dar0WPJdO5zpbLy0GZq3q55ptJdnL2N735/3v34db6XWApI6OVspXI80dTbR6bqht/U/T375daNOhY2xm5Y8lTinrkipw8b+ITc21/S1/fuvZ6sR2R2JjJaKhpZZF8gjVWJYk3HBNx/U+/eXWxx6F77+mqY/JDURmIgGxcauRc/n3rrfSmxNViTjyJY4mJazagpuWbj6+/Hr3QiQdU7e3NiGqfs6Z30aktArkkC1rEe9/Z17oh/ya+HG3cphqXPfwTVWUk5qw0NGodfG+u/Av791qnp0tfjkI328MDRzfYT49IqZYJWMch8RsdK/wCw9+PXhkU6sT633ZR4JGp8tOokpb6GdjYkD+re9cD1v5dGZ252JgcnC0s+ToFUIVEbTIGFhYWF/wDbe9/IdeA6DPdncGJ23XOkWQpwmvgicKpB/wCKe/HrwHqekSezotyyrImdxwQEej7tC9geQOf6e/E+XWwBSo6E/B76ioqVGjyMTwKArDy31cckG/09+p1qmajpcjcuDzVC0lLW06VVjZmkHDW+tz79nrVegp3Bu4bcgqp8zkIaiEIxgWKc8G/F7H37h17qnz5fdy5qvjr4cTkhGh+4WNVqSGAKWVbXH9ffuvHGOtfff8G+9zbllOnN1gkkFzSTyup9V+Cp/Hvw690OmwukOz89j6aKlxm4LXQq0yzyDkX+p9+611YB0r8Puya9YanLw1kCr47+SAowGq9j791ulR1Zr138W4BRPQ1cLT1QGlXeMSAHR9G1fT377OvdVi/zH/iZX4fZtVWAwU8KpUSB1hSOwHIs9vqPfuvdVm/HPq3eGN21kMlgslUTR0jaWRZ3dbjngA2/2Hv3z690MbdndpYs1dDU43Kz00SlZZY45NIQem4YD37r1Ois9k9u7nQyRUM+TilqHeBYVqZVcSPxYLf+v+Hv3XuPQz/Fb4Ydo925uk3Rn3y8mHkZZ5Keo8kiOgkDchv8OPfqde+wdbYXxt6SxWwtsY3Ax4yOBqFIIw32yKTaMAlja/Hv3z63w6Pnsraj4/Iw1lJJGjhU9JsBwdVre/Vx14VOehdzWUYxpS5GRXY3sxHAuOACfdet06Kl39u+g2Zs7Iz0uRghqKujljijMwVxI17WAPu329aOR9nVYXxxoNxZ/s6o3VkqqWaj+/JTXKzppaT6gk+/daAzTq6vY+1at6qLLU0yCGJjI8eqwsOfeiaY6sPUdGZyeZwf8BElTLDE6RASFmVSdIsQT79Qjj1Xyx1Rv/MK7OzRxFfgeuYpqnKTrPHGaK8nqaMqosn1+vvY62etXtfjf8ka3tjDdm5PH5yJcRWNWVJNNMqsvl1jUSPfutcBTraz+Ju8Nx7j6/w6V5ZaqKGjp5Y5NRkUxoFN78+/fb1uuK9WI7cpnongknKjWAWZja5+vN/9f3o9bHQrV+0pMzSLWUM4DKoawYX/AK3B9+6rUdRMbmZ9ufs16meNSqkspK2B5/4r711aleHRCf5jPVOI7j69qt1YinAyeEpleN4YlDh4k1alI5+o+nvY9B1ogdE1+Dfyrx2zcbRdd7nyEcFdRTmnVauezhkugNn+nvfy60MY6ub21uzG5qODKvXU1RFKqyAI4caXP4v71nq2KZ6GWLI7Wp4IqiKGmSVxqZxCik8c+oD+vv3Vft6Qu5lizMcr0E6K2lrC4+o+hHv3n1sVp0BskmVw9RL91IXX1qNd2Fj9ef8AW+nv2CMde4HqiD+b9m32Di8Huvbit98fBPM8N0dWD3PrTn3vy60ehf8A5b3c/wDfPrfF5LKZaL+Iu0AkilqS8gIX9JQm/wDre/Ur14Hq6/bUo3JTrUeVQkSlbg/X8Ai/1964dW+fWGqxWUw1XJW00jlD6wAxN+f6e/da49OMW68jUw+CeQoFtw4P0/BN/euHHreOmbLZKognhr/v4oool9TM5CALzyffuvUNei7dy/KPY3XmMmrMvuDGTzpGSYoq5NYINrBQfr73TrRPp1R93/8AzL9x5PIVOH65xWXyU0sstNE1AJpo7ubIwZB+PexTj1U/Lotu0dnfIju7LQZnsmlzRwlW6yx0oWcaI9V9Mgb/AA978+vcerx/iXsnDdXpSCki+zZTEStUulyQnrDE+9Hq1OrvuvuwNu5HazU1XPTvN4baUIuDbj6e9Z69w6Lnv2rpVyVXW0zjwwsdKn6EDkgj8+/deGBnolfaXYDVReniZaYQFtRIKAqpt/tve8DrVPPqs75Kd5bewu3auB8hSjLC6xBp1WUto9Om5v8AX37r1emj+XP2DjNzbvnm3hJHPH92/wBo9S4kVRquLF/+I9++XXh8+tm/RtqTZkVRgaqk+6SiR9MLIz+lb2sv+9e6/M9b8+qo/lN2ZVUVPW0U8bs6al1aDYkLbgn3vrx6o87W3DPX0eSrVYgwxSMqsxABDE3HvfVep/wxyMEe5H3NkjFJJ9zHEY5BqbRrsbavfjnrY4562Geut67cTDaoZEWaZWYBW0hGt/QfT/W9662K9K6fdyzgCacJTR+o6pDZ1H0W597AAPXsnHl0g905TamfVqWNYWcxstwFtrIte59+690S/dW193bcqa+v25XCOEszBI3IOnkkWHvXXvl1WNuPt3dWwe2psnuaKpqochMaandw7KjltKtz7t1WpHRid49/5bAbKNfVVKtRVNOZEjWTlRJH6Rz/ALDj3rrda8Om/wCBux8t2J2HX9h1cvko4Kx6unjlPlsgfWB6vfuvdWO/I+qqakNU08ukLRiMxI5ABRdPCDj349ezTPVV+QNdR7uhyNVIpjSS41cgAG54Pv3Hr1OsvdHZ1duDbkOLirljp6SNRLolKhVUc+n/AFvfsda6Ll8W+r8/8i++Mf17t+CaqghylHJUTxx+VCjyDWC39D791sZ4dfQP+HXxqw3xx6uxm3qCnp4spU0kb17pTJCwlK3s7Lyffh8+teeOjbGcRrEJuJHIWy8jV/X/AFve6da1DqR711br3v3Xuve/de6iV/8AwBrf+oSp/wCtLe/de61Jv5Bv/bwj+Zr/AOJEr/8A3a+/HrQ4now3/ChWmap6qwgKOY46uBmsT9BKDe/4v7959exTrXehy+JO1cHBHNGDFjqRZF1gkMIx+u/vfXustV2bjNvYfyQVtMjUiFm/cANwLmx9+690Xje/yxx9RiKumgyCNUiOWMBKo2IH+APvXXuqkexO7qrI7/gnqDI6fcxlW1sQRrFuffuvdWn/AB87KxeaxONpjUwrNIFWzSW0tpHJv+ffvPrfy6sFxU+4KLFsaes8lD9t5VVJGYGwuouD7917oj3YXyF3Rtvcs1HHTVkkcM2lmUOy8Hk8+/Hhjr3UOl+WtLFpkynmjdNJYSSMtrfU8+3ocKSePVSKnoCOzPmlVSZxf7sSu7CN1PjkLC4HF9PHultD9S7F/KvSxKAdF7xHd25uwtyl84xSWGQ+BpGa/wCu1wW+nsUeykUW5+5ENhcDs1HBzw6Jtw+Krfy6Ojigxgju+t2ijYuTcsWUG5PvpHpVF0qKAYH2DqPrwk3bEdTBFUQv6ySpJa30+v596JVh0wK16bs7W3hKJa6jm3Lf4j27Amc9eYA5HTft6aXyF2WwJ+hv+PblwBSnXqAYPTvmqc1oXxXt/at+D7ZhbR8XXqUfHDq4P+RbR4yDtrsFKt41lkgdYw5sWLfge4D+8rdvByvDIorkDo95ZdYpG+Z62f58hT0NeMdTwOkjRkLKQVBDfQi/vCLb9zt5JtM3H59Dbxs9Nc+DeOVquWQPPIdUYU/Tm/s+3Bo1tfEgI69hjgU6EfbOLVqV56mOTXIjCNzwNX9AfZFC/iICenQPn0u9mZVcRkTRV8rLA4PqkYgG4tYA/X2+Mda49Mu+Ep6Orlq6ZkeOZ/Io4NwebqV9706jT1638ug2/iGDyRSHIRXswX1i9xfm+r228Hg1Y9aBHl1wzG0NmV8YUU0RVl/CKfqLNf6+0YnYtpHXv9L1DwfTfX9KJa2CgpmqJBdvJCj/AF5Om4PtZagM/wDjWB8+thQ47up9PsfYwqnjqsfSq9rBpaWMICPzYiw/w9ryLcH9LP2dXaNVWp4dMW5dibQpzrlyWDgoeS0J8MXpH40/63toi8VdUSE/l00skdegfyA+M+2IqqtyD4SoyCgs7Kac6j/T/YH2bWnI+8buonWNgD8ui6e6KyUrjoqm+PlV1JtaZht+jx7pDIxXxwRMP2zx9B7Elh7X7s7aZI2p9nTP1gpx6zbZ/me4TGY+THUlNDCoHjKpHo+vFxYD2JIfabcgmUPVhe1Fa8OltQfNvY27Kd6rISRJPpJ9T6ebXtz9fYN3v2z3Uy6FjbHy68u5VooPRm+mvkF1fu2n+xqqyjUtYAySob/4WPtJFyRuO2wGUoRQenSxLkMa1r0Y7F5Xa0ksi4ealkWRiFEelrC/11D2A5zNDuHhSA8elK3Abt6ZN7bvq8BTmKlmSP7wrEraioXUeTf2aEUbq2T1rLfzW/h32D2hVr2VSVuWqqKkqnrHgop5mVlSzsGVD9LD3sdaYenTR8Edw4Gh2tFtPJUS0eQxdL9tL/EoQJWkT0H9fP4vf34068PUdHQ7Y3BjTgaXF4OF3yIYENSgizfggJ791vPn019e02+3ol+985gJUqj6tRT+g9++3rWOjLQ43KyUFOkdFVNIyi9o2Y/Tn34EA169THXMYHP0S+aeiqkUgm7RMARb639+qOvU6TlbSZNpDUvHKIoyLix5ANvfseXXuljjsbPkKMaAVNuLAlvp+b+9Vz1bNOlrtvrjIVgE0syxqSCGYgCwP197r1Xh0O23el6LK0zU9bPDIsisHcabDj8H3UU62fTorPbXReR6Xq8hvnbNaSqg1PjjkJXgkn0qfr73Xr3VcG4e8N2dy7w/u3n8PW5DEwzfbyaIHlUqDpYm4P4+vv3lXr3DpT7w6/2Bh4MXTY6noI3qEArKd4YzOpK/pK/W9/fuB69THVLvy+2HQ4LcEuU25jnppWkLSyCEIhGq11P092x1XHHopP8AAZ9y0bLOQjpCWckLclRfi/vQ630UbfW+6vBT5LbcVmKHwrzbgm1+OPfj17PQRbJzNTj84amVhr8hdVf/AB5W3vXXqdW/fF7e2TzVAuPrsfNV0zxhSsFPqYqW0nTx/T3YdayPz6O5s74zbV7S3Msf8AzsFRNNpZykiICp/oLC3vXDrYHl0N24f5aVK+Up44hJHTeiT94XspAuHJ/p78DXr3Q8bd2lsP4f4H7h0pZK2OEm7IhKso1m3v3W6DoD9y/zK9t7nkr9tSfaRRyK1OdNkNrlb3/Hv1OtV8j1XdvvuVsLvJ9y7fycCU8jvKYWqQwuzXvpv79jr2TkdKjbfzP/AIlG1FkquHyLddaSW+n+I9669XpE5X5A4GTNTyzzmTyXHEpYWf68+99a67xnddGMzDLSSSNQ6QSBK1ifqeB79nr2PLpv7Y7r3lmIYKHaNNXmKZVWVofKfSRbVdffuHXq9ATRbG39uianXcMWSNLUVADeQytw5uffut9G/wBq/ErbFRjYZzQMap4Y5GvThjqtci5Hv3l1qhHQ8de9aQ7AWphigjhVi3DxhbX4AHvfW+HRluqewqXbOeLTSxxlAeNWkXX639668MdKDtPsSk7CyENJRZWmB5BjWcDnTYhlv/j791voM9hbbiod1UUrqsvhqgXkKhltqvfn37rVOrAt052nk2xU0dHLBEP4TImlSE+kdrWHv3W6DietUn5bncmI7zhzWAmE1RFlISIoDqawnDMOOffutHrZw/l/9+yZvrHD4LcVYlLkY4Ei0VEumRn8NiNLe9efXhw6P/HEmXrFpZiHgkOtSG4s3N+ePeq9WND0tIdlmig82KlsFF2Aci4tcm49+49ar0Gefx28xLJJTfcMigsCmtg3+BPvdRTrefLoF8lN2WmVWemhrRBFcOxEgF1+t/e/PrXl064rsjP0kjUudgn8hNk1atR+trE+9Y69XPTXme8qDBVJjrY6hWkOlRdhyR9SP6e9/LrZr0zVHd2Vq6VqXGQVMkdRwunUQquOCbf8U9661X16x4fb09N5d2VsUyVLDz2mvw1r8Kfe+tfLorHyo7Lq59h5CreWONI6epi0ycElQQpAPvw695dUvfHfsPMydnVjxyRPAa0/0a4Et/x79TrXVx2cz9TndtvTTzQASQqr/S+lowOB711bPVMPelPuzYW+zuLEvWHGLUtJIUd/GAj3/HFve6+fWujg9H/J8ZvB00LTKZaWHRMmsm7/AEsf9j799nXq+vQq9j9zxSbXr6mqqFZEo3aNTIboQL8AfT36nXifl0m/5W/SVZ8nPlNS5ivx1TLgaSrp6gVrxGWmBhqA5vKeB9PfvPrQHr1vsYLAUeBwWLwNNYU2Jp4aeAKNICQrZbD3sV49aNOlAPp711scOv/W39hpUhRYGxNvz/ife+OetcMdd3N7W4/rcf7179inXs161af56vRtYuQoO1kpEeKWVIml0MW0+Vb2b6e9DrZyK9UL96GZusKCHFM8DvjYNbQnS1zAP6f7H3vrVOiGdcbZosdlqPJ10v3dc0rMYpyJNZ1XsR/r/X37r3VpWysrA+HjcRxUEv28cYihYRg8WH0t711voj3ykxqw1n8TkkMj+QOhLlvqPx9f9v79w69516n/AB/2ph9/YeakzbJFqVo6dmPLNp4It/xHvfWuPSZ7D+Pe8dl5ObM4AzVOOTyTADWw0ISwHH4/Pv3DrZ6DvaPduVxVfNjMus8EtLJ42bxupBBsByPevl1qnmOjabK7lirXjUV8sjMVGiR9NrjlbH37rY+fRn9v7ySuijhEy6pND8Otxz+Pe+tfZ0bXYXYFVhWoYXHkiYBWW4bUOLn/AF/ejjq3Rpayv2tuPATHMR04Wel8ao9iFZx/qffut+X29Uzd9y5rpTtmizu245F21NUh6pYbiLSzar/09+8uq+eOje9c5zG9t4SHI0mSMNTMl5VSVQ3C/SwPvVOvV645vqntvHPPlcDJWS4eON5Y5F8ulgpuCfe/Lr1D0TfemU7Z3Bl5cC/3YqVlMYKlj9DYEEe/daz0wL1P8gMDbJ0NTlJGX1rAGmKtxq0/09+62AehEoN8/I3EYpVyOFqhFHaIuUluVXi54/2/v1PPr2eHQk7X7O7InwNdMy10VYgYiNUlBBt9Bb+v+Pv1OvfLoIMp2T8ic7WHGUOFq6umlJiEk0cjG17agCPfqde4dLLa/wAGu0O76yGv3FSVNOk+iWSPRMqjW3qBsLe/V9evU6PP1f8AyqMBiXp1mxdHU1YNiJ0fUzXudWv/AB9+r1umanqwXaHwtwWyqOKKXbGIXwhGY+IAhVAF1ve309668KdLeu6i2xS42WGix9HTyrqBEIAa4F7D8+9A9bx0D1LiqHbFRVJJpWRyRH6rer6C3u2Oq/I9UU/znOyYcD0bkhRz2yK0tcFCyc/pNgB/vfHv2OvH06pY/lu9x4mo2Dm13XWfuvVBdE8i/wBt7fRvqD+D78OtZ6ux3Bi9j4HrCbcdTicQ0GZog1FOViaVi0d78c839+8+tkY6qJ238eNx9n9vxZeLDRnZ4zCyrIIJCpUzX4sLWtz798+vCvDz62tfiZ07gdrbdwdDQ4qmFMlGiVcqxFSriwJP+++vvVfMdexw6sGn63jxUByUcUSUcqAxupGqxH9Pfq+vW6CtOi0dg9lVOxMj/kzsUVmDaXB0qluSPe/Lr3Sdn+SG3KigFXmsjHS+JNTu08aFeODYn37h1o16po+VfyNG8N5RYXbmcqqqgeqMdkmEkbfuWK2U/S3v1KdaOejt/GPHTxbeoagwqryw08he3Nz+ffs162OFT1Z913XZ+ERxTq60cl/Vc/pP5YH/AA96p59WHp0JO9evtxbvxS0m2ZZWklW0gQkEaxYkW9+GOtY6C3A/EYYyrTK7noospkdQYQ1imUar3IAP59+rjrwp0KW5vj3tvJbZq6KLaGJgqauAxho6XSb2/Fh9feuveo6Ld1/0Jn+tsnMFx6RY5pneNUjYIoLfi492+zr2eh2npMmVRGj0uFOgfS4HPuox1brhTb1zeELUxLeNDzd7DT+R9be90qMda889QNx9z7NxWEmqc/X09LIiyFi0sQbgXJsxvx79TrVc9VO/J/8AmCbE2xgMzgtuZNMmk4kjnQTROOYyv6Qfe6de1U61ld6d/wC7p+ypN67amnpaaKqlqHipiFUgSFuSP8Pfvs615dWGdKfzbNw4IUOIyNRUt9vHDC4ldSGKcP8Ar9+r16nn1YHi/wCbRDkoaeHz2ZgvAMdgfoR9ffuHDrdejLde/Omt3QYagOxglaO5LqFsw5AA96/LrX29GIr/AJGYivxjS1coSXxM1gyXLaePr78Ot1PHql355b/xnZNE2Krz9xEQEp0kIlWwP4A/x92x1U46qr6U+Qm4Pjj2BDRzVdZTbalyCeFVulMqhrWIPH+t711vraT+MPy4ot/bcoKykq43ikp0dyZo15aw9Qvf3rrdfTo7tH2qk5Rqp4jTPazPNFYhh+CT+PfqeXXiafPpBdld37J2rjKrI1eUpoPDBLIdFVCpDIt7fX34Dr1eqDvlf/NhyeFyFXsXrx6jKz1cv2/koyszxa/2w14r/T3s061XosHRvQHyK+W+64M1u3IZyn29VVWsU8n3Cp4nPl5Dcfn36uOvdbFfxl/l0dZ7JnoFzeIx2UrftYJJXrqbyMZEAuWLj6396J69T5dH/quiti7fp3hoMFjKeKMnSEhAVRb8ce9V6350PRSO6Njx7Vx1Vn8cwpYIoXltD6FCp9bW92BNOvU6Jbt/5t7f6+SrhymeeNKWZoCHqYwRpPIOo+/U/Pr3XHdv8xbrmfFT5CDOCVYk1uBNEx4HN1BNx791qvRG92fNTKdzy1GE6yo5slkLyxArTPyxPA1KPp791o9BbjPhF3B8gMgme30tZjSsqyR0yCdEaMHnUAP6e99bp0ffrL4l4TqHHRRUNdLTZekALhNStrtyTf6+9dbp0IM3yL7L6irDFMausxKWRjKxYCFOCef8Pehnr3DI6DHtP5K7I7Kx7a6iFcjIh80bGPUJCv00jn3vrQPVW3dmWyESpS4WETnKMYY1VR6hI1gLD6+/da6aep9s9v7AnGeqMTKmGlZJLpDNbk6uBb3vrQ6sS6v74q6GaGDMzy00s9ikUh0CxPFtfvWOrV6OPN2BXbqxdPDiX9EQVp5YnW/j4J129+r6db49caOuqmPno5WlgRW8shYemT+0L3/r79xx1qlOgt3f2a+3mnlr3Jgju0mtwQFsR9P+I9+z5dePDohvZW4NudkZJqijp6aWSFy6EKjHyKb8e/Y4deOekDJs/cfY1Odn46CaomZP2o2jkKAILWuB791rJNOjrfBOn3X1Pueo2huSkFHTPUNBrKsilG4+rW+nvfHrfRgvmJvSn2hM9VSSrLTPTalCOrEs340+9edevcB1ULujs3L5Ku+7SLxQWJvwCVI+vv3Ws0p0WPtXf24jQVcVAKh3qYHCiIMT9OL6b3/23v3XsdbWX/Cfj4j46DZdJ3znqJJM1UpA+uoidZg7C44kA+nvx62KcPXravtcg3It+Ba3+x9+61g9YZTCJEMpAYi0d/63/r/X3vy61+LPUj3rq3Xvfuvde9+691EyH/ACt/6hKn/rS3v3XutSX+QZ/wBvCP5mtv8An4lf/wC7Xj3rrXmerTf5xvx6yncvRdVPhqY1E+Lp5qh1VJJGBiOu9k/2/vfXvs60TdyZfcG0zn8TXNVRthKuSgIKuukxDTYBuffuvDonW6d/bhzNTUY2nratBWaoo1Usb3NuPfq9b6ZcT03n6lBLWVFQ5mYN69ZJ8nNjb37r1fLqPur4918KR160iySxgFWaJixCi5sSPfuvdJnYu88ts3dWPxM0lTTAVQjIVTGi2a17+/dePCvV23WHaFacFjYKpvuKOojp4mld1LEvx/r/AJ9+69wHRrqj4yYPfe3l3HT0NDNPURGYswXVqK39Q9+r5da6r/7L+IsdTX11OIRC4VxpgicqLk2+nvTNTtrx6q66uPQCUHw7gwYmrZIlqJtR0+ZCfqeeD7XW9YjpbzHSqHUV0t0ls18fV27uLD1MkKUlPVOC7RIUBDG/PtFypuzco84xbwpI0tX+fRLuGvXTh0Yas2lW7ex1LVU7eal0R2kvq4C2H/FPfTXYN8g3/bYdwjwJVDft6AV523Br5Y6ieaapQWUXC2IH1+n+Ps2oqcemaFsjpM/bSzTyl1uF5IPN/wDD2p1qoFOvUPDpwp18YChFDD/C3+Bv7oxqa9eAp06RMqIwf+19D/seb39tGpOOtj59D98Ue8cv8Z+1Mfu3HNKMbkqpf4oIyLLCXBJYH8ewfz9ytbc6bGdtuKaloVr8ujPaCVkI9etzHpnurr75BbBxm58JkKZ8s1FTeRBPCHDtECQbH+vvnR7gcmT8rbqY0BFD5A9CmFxipz0rKyugoHMdZUKJFYKqiRSCQbi59s2Ecl1aUPp59HcRoKHNel7FvYvi6Gmo4oWdWUMwKk/i9yPz/j7qIfBOg9WZaHt6SW6dzVpqEEWlKhVBOhl1D/An34EE16bZqDpEVfY8hKUtbLqlUaVV5EsbCw5v/X3ZT3hutgVHSOlz+TrKyQwBFUXIs6AD/W5963GXsGmtevGnn0qsTueKkpWky9YkSoGLapo/oguSOfdNstfqpRUH9h68BU06D/K/KfrvbD1sT5y89OSqxCaOwP0AAv7kO75DvN9tUG3qQQM8em528Knp0SLtv+YLlBPPR7ag80elkSdNII5sDcf4exHyx7Q7nEwF9SgPE9E818dZArTog2+vk32ZuyWY/wAby9MJWJEcczhVvxpFj7nXaeRdmsolE0auw9eit76Sppw6BM5TeGZeSXI7oy5Lk3jkmcgG9+efYsjtbK1QR28SgD0HSRpzXUx49Qo0rlmMEtXLVLzdp/Ve/wDifbxVSKgUPVluEGSenSlxjpIDEsd25I0A+2mzx6ub2MYB65VH3VMWUVctPqH6YzpAvwL+7eFGy1ZQ329FqXSyTmh6V2096ZbazR1FNnK1HEikhJLGwNx9D7Kdx2+G7iaJkFCD/g6EEBAjrXqyr41/IjNVlVS0s1dPUclWaaQE/qsb+8PeddnSy3N3QYB6ObZtVKdWg1sKdiYWAwzMaqJI3spuS2kH6j/fD2Dwaivl0YcOgo3ENWOqdl7jx8VXDLHLGpqlLpYrp51X/HvYOM9ayeqbfkr1anVlbXbt2ggoXVvMtPRqVVyTqswX6+7eXXqZzjpr+NvdFJuSpWXf0awgS+NPMlySnAvcfTj6+9V69xz1ZZt7N4CpWHIY2OKTHJb9KAiwa4/1xb3vr2PPoye2d97TpYoaiempdMK6iHVW/H9P+I96r5dep04bo7i6/wAvTGgSmpYnVdJZFQf7H8e/da+fQNZLfPV9FSSnM1kdNStfXIXjTT+fqfr72OrHpGU/bXU8az/wDOR1Soht+9GTb+vB/wB796oetV66XvPAGlWKDIiNWmCApMh4J/Fjx71TrZpXHQt7d7LlSOmjxtbNNDUpqklElyt/6G/veevYOekz2bl81ufC5OlgeWtijo5AUlYuCNJsSvvXXqdJz4q9f7HpsXmctujE42Ks8NYqPLEol87IdBFx/X3up8utH16IlJ8dezt3dub33FPDOm2qTLTSYVLyiGSmEhKFV/pb377et+fSF7X+I27+2mmwlJgEkmgjKibwSAuw+nNve+q8cdEk3H/LA7/wj1FVT7eKYwJNd4oZyVAW/wCB798+tU8uqUflJ8Zd19XZ3IVuboJI5I31TE08wtdjySw9+I62OiV7cwmRzubgTHJrdKkKwAP44t71Tr1cdbAvwRGJ2eMcm6MfSeUxRWWULyNYv+r3vr3V7O1u8entmKMlVw4mg0kM8kZgSRbi5N/eqenW69Fo+TH8wLrzC4upye1cxHNNFBMqrHUQn1R/TgH3scOtE9Uv7k+Xe+/kpn3xOFjqKykSYQlv85GdR8RJ0X+nvdOtHo2XVn8r7sXsSGk3TW4809JVssk0iRSKVje0hckD/Y/X3qvW/LotHy5/l9di7CyTz7PWsq6KCnmacaKhl1RjmwAPv3Hr3Dh1WCmEzu1K+bF5xa6nro2KOftplAc/X1Ef7z7917j1JNRJT1XklmqpdQIuInkcW/GlR7917o7fQ3UW8d+UEOUx2NmnxOqIvPNSzKwAbm1x9P8AD3v59e4jqxDD7J2zsOgp/wC8GPhMmm0jSUzcMRbnUPeut06wbozsBxf3O1sDDPDChcMlKT+n8hkHv3Wvs6BWH5SZ3bFQ0E2GqESDVEx+1mC3Xi3K+99arjpHbo+XeZyyTfY419QAPohZmFudXpHvVOt1r0AVT8l96NWVPigqI5Z1dR6Cliw/N7e/da6Ydi9pdkjdaV1fUVoill1hGcldJe55/p798uvfZ1YBtL5CSYaaAVZ/ymfSAWYEgkAXF/z791sEnoXt2/IOfG7YrcpWVhjibHVBF5lXkpxa5/3r37z69imeqz/jRhqT5M/Iaavr5Hq8ZTZKQN5wWiOia1+PfuvccdWV/IygynxXzGEze3ZZabEmpgkdKQMY9JIB+n9OffuvE9WC/G35YUPZWKx6vVL5xBSxtI0qB9ZUA3vzz79TrdfTqx3a+6WkoZI4pzUPODp1SKefqbX914deFDnoTcVuWipo4qfI0sRlchbsFYH8XI97695k9ChDFtg44mrx1CGkRnDaEJIYXFvevOnXuH5dFq3fsfCZzKpV46lp1SlcM2lQOQbi/vYxw69x6KpvLqyPdm7ZqSOCJBAh0hVGkkcrf3v5de8+lTtfaWM2Iqz5+jjWkgOlnkhuTbkG7D34nGOvUz0rN7ZfbeX2/NPhHhEKwaiokS1gLkWv+fehjrZPVBXyj7Ch3fm5+scdUMtVU1E0BijIIUSSWPK/4+99VPHPUTp74iQ9arS7mzLhGrIhUepjqN/WDz79Xr1M9D/lnFSwgxdSzRqVVwGAUKvp+l/fqDz62ei392U+CqsDVYirSKStMMzayutrlfqPrxz791oY6rr6uq5tr7xrKAuyUk9S1lHpAF/qAbD6e99a6Gbeu56rcuax+xsU08tTuOT+FUkcaNIPNMuhQRHf8n3rrx49bnn8mn4ef7Lv8fcdlt1YWni3hmVgqo5zE61CU8imZXDSWNzx7917q5+QKyr5CYwT/UA88AX97zwHWjQ5PWUCwA/px711br//19/j37r3XvfuvdU4fztsW9f8S2kp6cS1UWejAcAa1iKoWUE/j8+/db8j1qG94xVdJsbG1tOjGnpcZT/coObOKcCx976r0QDr/KPVbmpp5wVhWdiikGwAe/APvXl1s9WITbKz+a27Fntv5CSIJTq328TEE2F/0j8+99a6Ix3zm9wCCTH5WmnaaD9vySA3JVbXF/z791roMeiO7ExmYXEPVrQyUEpJ1sQZCONIv/vXvXHrfVuu1+4sJu7aMeIrKMSVEqJEZfGLssnpJBP9ffvPr3l1Hl+NvX256eWuWiggrasFyxEYcn/af8ffuvU9Oi7dj/FbLbVx9VmcLXSUkdLG88ZU2vp/Fr/T377Ot9FS2b27unCbhfF1dRJOlJUCmNQS+kaH5J9+611Zf1f2rDO+PkqagVj3F4gSbXtxyffqY69XqwTZGVi3YYoHIpaYqiqrkj1Fbhgfz79XrfyHQe/IbpobkxktJMpnSSN2jkuOAE4Iv71w63Soz1WJtHP71+PXYNDS1Us0e2pKohzIzvD4ne1ueLfj3vj1oY62AuuPkL1vuLrhpZMxQ+vEhPFqiuZWh9Vwefr+Pfut1x0SzY+ATdvYOYy2PjNfTfxFnhlhQFUXycEj6D37rXVoG2etKeXFQPVY01LOi6kKKdJKgElSP959662M9SavrbalWzYisw8X7lxcpELauPrb6e/E463Qjp1298duu8Os1VLi6aZJFJeG0RD3/wAD78T1qnXp+pOuauvgajoabECjkDuRHCjSAHlb/wCPv1a9boAOhywPYexdirHQUVBBO6RCIyCKMm6D/Ae/UJ6r506DveHyEGPyS1mO/wAjUPr0hQoPNxwPfqde+fUim+RNTnKV2krg7yKU+n0v9Pr79TrfAY6Bvc/atfj6t5EqNSuzEgXA+nAI97AHn16p49F3352WrwPlJasQGH9wqx5NuTz791rrWO/mkdqZLuXHZPY+GppJ5IYqtpKmJCbqpt9V/qPfuvZP2da++wexNw9TbiXHPTyQY2neOKsjPkVXeIghzb/Ee/dVBpjq4/4vfIDsP5MZrHbDhmmr9u4+cUywReRlp4kAWzX449++zq2Dnra7+Lvxr2xhdsY2lq4oWq5IaScwFYzIZCLsOeeT795dbHVp3X3X9HhKI0EdAaKCVG0zaAqgW+tx71x49eJpw6RnZ2fzm2sfkKVJ5K2hhgZqfTcqlv02t/T34U68fXqlTu/5JDbdTkYszjjWMWqI43ZHPjNuLe99a8uqV+9u299Z18pkMBuZsdRO3kjx6SyC68nRpB44/wAPfutdE86U70qMX2dSU+95BUxz5IRxPMWb82/t/wCPv3Wzwx1tN9BdiY2bA4etxkiTUr0kBKILgBiLAge/fLrfVomxt2LLQU0kQ+5SVASAA3huObgfQD3qnXq049Go2rvinxNJTzR1McUzMoZXtf8Apxf3rr1K9DHBv2h8aV1XEK82BBChrf1NgPfqHr3XKo7TwLwSEUEcTKt7FQdJ/JII9++3rx6Ru4N14rJYj7tFjLav0Kqgjn3snPXqdFb7F39R0LpVf8BKeFf3JSoUAKv59+oT1uuKdVN/Ir+Ynsrpr+KGpqabJNoawHqdLHSfSp974cOqk5618fkZ/Mwz/bNfXU+zcg9HTSSVEQiRpQDrFvoh9+rTr2eJ6I7tuXtnsHITmDH1m4WqpCZfDJUOFJNufr79nr3Dh0LX+y0d71cYem2NloaeRR5AqTAFDyb8e/deHTFL8Y93YuqWfM4eoxDAgvJKZEs1/UDe3v3XqjoddpbFGIpo3goXy0tMvPjYsbj+tvfutdHV6u7So6CmgxVTjTiJodAJluP0jTqF/wAe99e6MXkOz6JsKYYspFNUPfSFctpB/B596HHrdaCnQKbxo8HXUMebzWWp5RColMMjg6ber9LH3vrXHquXvfd+y9ywthsNhYa+vilfRNCvqWRTYMCvvXWxjpI9U9z9x9T1tNFjaqrp8SPHajDT8Ipvaw9+z17A6sGpfnF3LlsNHSUeJyLO0YWKoC1FrgW1X/4n37r2eimdv9i/K3fgqII3ya4ydJrgNUD0yKeCw/w/Hv3Xuht/ljfDut3/ANgDcXZtA+YqVqy0kVQWk5E9xYS3HH59+69jrdr6Q6H2ZsfF0ww+OhxMUccB8YjhA9MQHNh9PdSfXrfljof8tX7d27Q/eqsUlQg03GkHg2FgPz79Q+fWh8ugtzG5GzUBmiHggC3cEepha4NvfsdbHRCvl9vDIU3XGRoMZTPVVFVRTQKkSm6uwIB497A68T1qubw+NndXZWUzdKtPkqKkrsjLPHUB5VCJIbgKfduq59OhW+Pn8tvsap3NjqfdOQravDtLoq4JppGilj1XsdR+lr+9db4dbD3RXwv6c6ojpZ6TbtElf4oGkmVIixlCeokkX+vvRPp1sL0a2sxuC20q1FFNFFGiMBThY1NrfTjn34V4dex0VnszP4dZ3qp1ioi51CV9Khv9qHvfHr1eiN9q7iw6UtVU1Bjy8TQyoI1GoAEG1yPfscOtdVF9iVMeP3tHl8Uv22NWUvPTKz2t9dJH4t79mvWuI6ibC3rDvTs7BUc1N9xR0ORQtFYsNKsOOffvkOvUx1sT4FuusttDFYio23Boko6WMs0MenyFApN7e/Z6tSvHot3yY+IX8QwS7w2RKaEU0QmEcBC2Crr02X3vrXnnohOzvkzkeuKiXY+aopUqkZ6WfIurm6qTGWLn8e9Z8+tcOPQ5UHyCwuLxUjU246WplqQ0wpUkDuC/JXTf/H36met1x0WzsTu2fdsNZRU9G8ksvCrZ9Un4sLe9461XqX8cfjV2R2JuEZEU9bjcYswlN9YSRG9RHq9+68MGnVxe1Nj7H6fw0dXmaCmjzdPCUNXKkeu6/wBq5559663xPRJ/kR8gMHtfM0OWwVOi1DzJI80aBA1m5uV+vv1OvdFH3d25kOw5pszuGqM+JWEmKnkZtAIFwDf37rVeihbl31j66vkTGzRGSGYQx41D65udKaVHJ9+699vVknwI/l+79+Ve9MNlcrtyvotvUtarzs6kQSwAhiz6xa1vfuvdb1/x16N238futMLsHbdNHTQUNNAKgRqihp0jCtyn1/PPv3W8+fQ7+/depTh1xZFewYA2Nxf8H37r3XL37r3Xvfuvde9+691Er/8AgBW/9QlT/wBaW9+691qS/wAg3/t4T/M2/wDEiV9//Prxb37rQ4nrbJ3JgqDcuDymCydPHU0eToqikmikRXBE0ZQEagbEH8+/db+3rTV/mVfyzdy9cZvcm+9sbYq8rtnJ5CsyFVLToGp4hIpcK/jHv1evfZ1rJbg2CdubpE9XivF5JiBGf+UUiQi5BHFve+tDo3HX23IayGmdofuIj4QGVVIBNuOfeuvU6M7X9QU+V2+8keLa3ia7iMWtp+h9+62OGOq/O3vjhRqtXkqOg8NdTK0sUgUB/IDcfQe/U610B/WXaOZ2TmFwW55W8EFQkcCzMQLRtZSA39PfuvU6te6s+R2Who6RYs0RjyoAp9VwY/oFtf3v7OvfLo2u1e4dlZ5K9MnRQy1MkJCzMiahIyfW7f4+6lQevEV4+XQe1+2qTPGpqaOdRC0skghXRbSDcAD3cO1a16tqYUp0Vjv+mjg2zX0saBchTwaaSot6o3CmxU/6/tNcW8c/c4qeqlVc9+eiT9f90V9bAmxN0sYvtJmRclPqCyKGNgHbj/efeQntx7nT7FBFtt036KGmR5fb0FNx26OWclFzx6MIuDaop1qcXN95TFA7VEB1Io/xI95VbRzTtG8xiS0mVic6aiv7OiWW1emkDSfPpKyxmKd0hbytyJmTkJY2NyPYoV1ZRq/LoplgnU0XpxkpY/tFZDeQ29Q/rfm/ugar9Vq656wy0ZKoS/4ubn82v78G6dVgTx688UbxaKiRQtrICbDn6gH3pQa9o6s1x4FDWnRhug/lR2T0FlFiweSnlwiWYUSySsF0/wCoANvYL5t5B2Tm2Gt6lJP4hToxs786gWbqxbYv81GXL+aDdNG8FSGNqidHAHPBBP8AT3jhzX933d7X9Xl+Yuv8NaHoVWW5Rhe9q9Gz68/mD0uQhqzSU/3pWNmi8IJAOnj2DovZ7mKKKt4pVvQnoyO5Qnzr0ic1/ML3NBkpw+1a6oa8io4ilPov6fZbd+3O62x0BCetfvG34HoA9y/ObfmWyOqk2vX0+qQaXEMw08/Un/D2WQ8g79JeJ+k2gcem5dyg8IlOPSuoPlF2PXUETxtVU08tgxbyC1+bn3OXLvIO3JEG3GEEgefREL24d61x0w53tbt3Orpj3BUQwSAhxrkW4JsfY2tuX+X7Y0FumPQDpT48oSoOekTFt7N1z/d5Svarlk9UhMj3Zvrc39mqGzgXRboEHyHSBrm4J7mJHWefbtLGhc0wZyvNjf1f7H3ZZ29eveIzdvSLrMCfMSkbR2PHNj/sPa1Jxpz0wUJbqHPQmnUAygNp49Q/2/u6yljgdUaJDx6aFqY4JTqYM35I9vULcOqlFApTHTtTVhkuY+Db+zb/AG3tpgRx62IoqcOoOUZ5YytiXK8Ej1X92QkceHW44oQagCvr0naCJ6eUmrPkjJ4BuP8AWsfbrgSCg8+npZGRO006FnqbIZfF7zxjY+reKmaVWaJWNreQXHH9fcGe6OywQ2El0Ez69O7VPM9wAWx1sWfH3dmPkoKGGrnSGedYYyzckloxz6veMFvMhQA8anobsFPDpS/I/MbW2hgP4gtRBUZZ7lNOkyAEfUW59q5Y2MepR09Eq6c9VtdjYpOzcVSxyr6KmNW8dgNd/wAC/tNalwCJOmJQA1ekJsD4qLDkFqHxTmkEmsEWtY8/UD2902fQdHBx+xaHbVAlEioIQnMIIFmH9R79U9b016D7ckLUEwkSoEcINzBcgML/AKdI97HVSKGnSLzudx82PMVNQeCqQXaezeq39r37NevEU6Jn3PX5bNUE2Oxuou4cagzcEIRxb37rf2dEU2LXbyw1XuunqsjIDSQTtCpd/SLG3v3Veh7+PlZvPsGpGJmWpcpV/wDAwl/HpD3+vvx9et1HVv3W2wM1DiF81a/+Srpb1EG4+vvVetivQr0lDUqUondwtQwilkv+pTwefesdW6U6bHq8TIi0JkfHSBZprCy6vqV49+60CKdLOhr2NVR0sERgp4yFqkVFAex51Ee9/Idap0abbdXsSKgiSloKeDLekyTaItbMRyWNvfsde+XTvVZXFyqcdW06SUs6lGYxx6dD8Hm3596rXj177Oqlf5k38v8A2t371zmc1szGxRZOkpBJJHSpEJKlo4yx+g/r+Pewc9a60GN67Qz3x+7gzOy89j5sZVUtTWSQtOW9SxTFVYA+/de6Hvqj5NSbezcFLlLy+krFI97AX4597611L7l+TW4stPNDicxamlKqIEklFltbTZT7914dEo3TvXcdetshlHWmqpGjWF5ZQrySXAUFj9T711unV4n8p74/5vJbdqN7PtGqytLE6ztMIjIroJ9Ra5H097691uA/HTs7BLiY9o5HGrhSEWnVJo0QraMJzcfj3oimevfb0quzuoto7ipZ4TQw5IVEcsrsIomGiTlh9P6H34de6q27Z+BvS/ZC5H+G4Kgo8zESrFI4FkeQC3Nhf6n36uOt06IP1j/K+xXXnbDZ3e9IcttisrLxUdSqPBDEz8AI3+Hvda9ap1d5/su3UXX3W1NmNkbeoko0ihWWjpoYLFyBqJCC/wBfeuJz1sYFOgs358eNk792ZJWzYOLHTzU+tHZUT1Wvxf8Arz7916np09dIfGrrNdqLg8ht2mqpYA2udo428i/TSCw9+8+vf4OnPc/wV6G3xhcjh02xQYfKzCoENVLHCjamUqHHH9ffqnz69Xqr2p/lvDpjObqrf7sS70x1e7mjEaCaOmEhJABAPHvdevDj1VX8otn4DYxqBUbYG3KqNpba18bakfgWt+PfuPVadEmxG4HkgmrhKVEN/GwPBA+lre/H5db6zv21BjpI5KimFVMhDRnn0lePe69a6Tm+N9b23tijHA09PiyCGjPkCiIg3596638j0ef+WfHtvA5ORvuaeiyPmYyyuy6i5muSCeffqenWvt6uW+QuN2f2BsvIUGWnp8xUR49VprlGKSBeNJ/1/evn1anl1THsPL796N3hVZCKhrF20mRYpLaTwxwq91H9OBz739nVT8+rdOj/AJj4/Mx0qS5SLyhbmENdlOrkfX3v7Ovfb1Y5tjteg3BSR1Uci1NSEWRYwLkG11/5F7rSvDqxPr0MG0ty7h3JWKtRBPBAi6VV9YXQOAQPfqU69XHU7c254NuQ1kCyKlVLwE+hY/i1/fqdbx0l+rdr5zeW4JMhVJJjKOOTzvWvcRyoDqsGNr3/AKe/H060B59Cl21Bs+ooDinqqaZoo3ilYFPW6pYtx7159eA9eqPvlPu7J9UwV0O3NyCZckgWlx8ErE3b6KEU+7cc9aNeiO9Z9XVGXr37Y3bXGHJSzmSOKoa5Uk6gwV/9v798uvDhXocMzvjLZTx0i59auko1EMUCNwqD02NvfuveXSbgzqSSFErRSTRG7IW0l7fXi9zz7317pC59VyeRnqqqkNRGIJVJILXAHLc/X+vv2T1r7eiF7qwWZyvYUGD2Lipsvla6qQJTUinyLrk0afR/S/vXn1vrZh/lkfyhslJX4TuPuGkmZ1qoMjSY/IaWakk8YlTxrLz+R9PfuvdbW+HxtLg8Xj8PS6Up6ClipIEAC/twppWyj/Ae90rw60SBx6cJFLAAAfUEk/4G/vwx1ognrJ711br/0N/j37r3XvfuvdFT+ZHTcPdnSW5dsyIkj0tFV5GGN4xJrkhg1AKp/PHHvR9etjjTrRw7P2juPI4XtDZf20sddh8pVY6hieL9wrCpUeNP969249a6qXxmN3J15udKXdlLUwQR1DeqWIxA3ewvcf7H3rPn1rq3npncFLkNq08uOljeJoI4yjMHvdfyv9ePex17pMdpdW7f3NSVeUyUcCtGHcgxry2k/n/be/db+XVLW/8Ao/O0G8spm9qJUQwwGWaIqjiN3WQkAEcH3rr1en/rfuvfu08jFS7ghqI4YHSLU4dU0xty3v1evdWJbA+U2JrY4BUVccckf011Gkar2vyfex1o9GqxnbOB39jZMRW5GglhqIvB4/uEaSzJY/65I9+FOPWyekHlvi1tPJ4PI5HblOn8RqDNUKyxqzXIupBP+PPvXXqYx0RjbJ3n1x2ScRmzPHQ/dmOEylkQjycab+/fLrWOPVwfXWbyVXT0c9JMEjVaeUMJP9o5A/1/eurDh0Zh85V7koYsdIheqRdCSEagLLYgk/7373Xr3y6Lv3Z01jt87dq6SohjirqeFVWZYgHL2JDB/wAEf6/v1aDrVAeqRqSDv3Z3dK9eYitr5tryZAx+SOaVokjMmkLxxa3v1etY62c/iF19JtDZVJPltE2Tr6RZmZwJHDu17knk8+/Yr1bh0dej3JuHbDPNV+M4+QWiFrabjSL396NDw69Ujj0mc5vanqi09LLEtR6v7aj1Hk+9469noKcz21uHEyiN5bw/mzmx4tYe/da4dI3O9w/eUR01BhnIILCQqbn68/n+nv3W+Br0B2a7rkwUk1VPkV9KsQrT6r2XVzc8e/efXvl0WDIfKp9w5upo0kmmSKUp+07ugsdNx/tvr791qtehZwXZebkp6aehFUI5ZEFgzkjVze3/ABX377evA+Q6HNdxz5ShjlyBZZDDe7H1C4OoXb/evfutk9Ep+Uu+223tyqFNMVklgAjCyaWZjexAH19+6qeqkjgJ90Y6uz9TSyS1VQahZHmiLjxtfnUffvPrwB4dU1fKHr2tn3/Bt3aOKqKzIZSqjaRaKnEpDM3OpFFxz7914jHV7f8AJk+Ju7+s6587u/DVkP8AE6l5R9/QlLCQahpZh+L+/de4Y62zMPt6kx1PS1uGAjroqeKyCyqdK/lT791bPQ07I3pnKuKSiz1hChCKb6WsOLA+9fZ17oa6ram3dz7fq4ZYDI707CJtAYlmXhdX+v71w691S78k/iXSbgyGUDQkazVvGoiAsSPTcG1v9h7sD1ojrVr+d3XG8ekdwRmnWZcc88a2MZUFWax4H1HPv3XuiKSY6n3JT4rM0x8eUp6lZS0LhH1qb249669U9WifGv5dydafw3AbnqZI6aOKCINPOVX0tpH6vx73xHXuryOlfmftWqokekzFB46gXKyVq+lbW/rx/re98ePWq9Hp2Z8gtlZyOHy52g8x0FF++jtqIvf6+69W4DoaaXuWhpYw65zHNTAWFq2Oyg8XP19+pnrVcdQarvHa8KTST5vH6JQQB97HcH/Dn36nVqnoGuwvml1X11t2pqspuHGhYY5W0/xGNW1LyTa/vVOvV6oT+WH812feuRqtn9UQ1uVaokMRkx0ks6rcaSS8fH1t7t1TqnvdHXXyN75zjCpxW6kjyErIzGmqJIwrvcEG3v2a9bHViXxL/k877ylXSZjdEFRJRz/bzSx1lHZvW13UlvfsefXs9bIXxm/lydNdbUUXnwEJr7HzO+OjKl/qfWR71nrdKH16PofjV1+lKKemxOM8WkAxigg1EAW/p9be/V9etVHRe+1Pgz1Ju3HVn3WBQSNDMQ0OPQXkKkBgQPfgfXrfHqh7u74l7k6O3k+Sw2Omm2oZw00C0ZZhCW/sgfm3vfVcg9IfdO0Ovt17bNRhaKTH52OI+YyReA61/Ubcf4+99bp1W92buLN7EyctJSVjtKJLIBPdfSOL8/8AEe9cOvY4dB9Tbs3j2N48NTz1OuceKXxSO1jf+g9+8utV8+rQfid8DcFnYVzG64TNUSrFMfuYg5JddRILf4/X375dep59Ga3P8DMBkc5TxYnHRLBFIoJSkUgqrAD6X9+86dbpXPR09gfAzb8eBoIv4XAssS2Y/YRnV6bfUj8+/deHy6Eib4WYeLGz0smCik/akVWXGIQBaw5t79X0699vTV0V0rj+kt8qWoI6WkqZyxP2ohCkyauPx/r+/cePW6eY4dWzw7joaugL46aIIsKEKJAD6UsRx711qmeg5rc7j6kTU+RqBYFyoaUD1A8WF7+9fZ1bhnoL6/c9bTyyQRSr9iDZLOLlfxe3vdB1rUekDm1wm4Keqgyfhf8AacRLKVYayOAAb+9j0HWifXoCKLb1Lh5Kn7mlx5oDOWU/bop0g/QH37reOhx2RW7UCFoFx8TrcBtMcZFvpc+/Hh14canp7zu5sfj1klgnSRghN45AQFA/2n37j1rzqOgWyHYeNyQnMtQfNGxVUaU2P4tb36ppjrdOi59rZmm3JSrSPKsYjFgwcAjn+v8AT37gevfM56I92Tlcbt/Dz0/lMrMJFvq1H+lzf6e/eXVeHVdu+MxQvFWK7LqlJdHJGpbj6X9+690FXTO7cVtvsGlmldTJJWWLlgOGbi3+PvfW+tiLqbtLZNViMbDma+kRFpqdlLTqDqA1C5J+t/evt695dGJyHa+AyGHkxNNWUk9BLGU9UquAmnT9OQfeut8Oq8u6+p+qczSZDIRLTpl6mGS0kWkWkck3DD/Ye99aOc9VF7q63ym18/McfX1LyNVnwJ53ZFjJuvpHv3XurGPhz0pSbryOOrt8GAwJJqYzhQugEHUzN79jrQ41PV2027Op+pduS0eAqsHDNTUgYBZII3Lon1Nj70erY6qV79+TUG5KjIxKZamNaiVFXHyCTg/gCP3vrRIpTohO5cRvvtypoaDam2NyzsWCCY0NRKi+qwINre/Vr1ro6HV38tr5Cb623TUlRjclTRShGImx8iuVkXgG4+o9+r5deANKnqxn4xfyIqifN0O5d/QKGhlilP3NKoU6JAzBg/8Avfv3Hh1umnraK6M6K2P0XtGg2xtPFUNJJTU8cdXVU8EcckzhNJuyi/v1PPrXE9Dd+T/vv9sPfuvA9d+/db697917r3v3Xuve/de697917qJkP+AFb/1CVP8A1pb37r3WpN/IM/7eEfzNSL89iV/4/plfp71Xr3metuB/IbaLAXFzzew/oPdhTrRr5dJjeezdv7/27kNr7loochiMlEYaumljjkVlYWPDg2PP1Hv329eBrw6pJ+bX8s34h0nXW4dxYjF4Tbu4qWllfxzTUokmbSXuqmzA396yDU9b49agFbXVWwt/1G0cTDJNi6XLTxxzImqIxRTFVAYcWt791o+vVnWyNy4htiAV0KLO1KwIdVsSV+nP+8+9HjTqw6Jp2dV0NTXSpAYRTM37gbSQAT6gT735dV6qj+UO1aanqXzWFaLREXdvBbl/rYFfp78et46Bbq3uHcFBNBSVs0ohgUKLykcA2Hvw69jo6FH39S4+Cl8VWRUy6UktUEEkj+gPvfWj0crqPt37+iF6pGDRhvVLq4Yf4/8AFffut/I9ITtPcEu6czFioJov8pk0n13BubWH+9e/Ur1rhnpM4zpbZ1XK9JnqR5KvSWjlp49P7jC4IYf09sXEszx+BCSD5EdOfToV18enWv6O7twmArK3YVTGNtRiVvBOuqYxqCQBf/D2v5Y3Lfth3BbvxmKgj8R/z9F01lHKagZ6rwn7R7G2VvHI4fKYnI1WuYrWMKeRoVu1ro4HHvMLlr332R0gsN+oGONYOR9vr0VXO1EoxUVI6MHt/sSpyUUSpia8K2lmvG5AJ+vHuX7fnPlC9I+mvoyT5V6DJsJhllNOhHjy/wBxTa5UeAji0ilSLfgX9nsM1vcDVbOsgPmDXpk2bVrSn8uuMzLWxBnlXTEQw0t9OL/UH2qiOhsefSO/tGZQpGPTqLTZNNZRJEuDpN2F7D8e3XSoqR0jRGiPZ1iyE5YqsbH1nnR9Qf8AG3vUcY4npQbiRPM9GF+OO9qzE580Uk96dpvGUeTnSePofYe5isY5YddM9GtneSstM9Wh00tHUrTzCKB2kije5jQk3F7X9xw0YyreXSiV5ga0wepFVSUjgMIKJf8AVXijB/17+2wNINOrwTd4Qjj8usRijMAjpxAGBtdVW/8AvHtsM2qrV6OCiqtRw6yww1D07BlAGq3AFv6fX3aig9I3mP4TXrKWeKOy/QDmwH+w497oK9NKxY/PqOlXCSRPYcfSw5/2B97KmmOrVFemnKS0rKxj0gkWH0LDj27GGHHreaimOkcNvzZGRmBNj9Pz/tvarxxGOqEVPDppyey6mBPKgZrXJsCTx9bD27Hdq2D16noa9JAyVWOl0tG4sbfpI4H+PtWNEgr1RlOnHHp1+5jmjEr6Q1rWNr/42591KEYHSeJwGo3TXNB96SIeNPPHH5t9PewfD49K8MO7qXgdyvtXcGOqapgtPTSL5G+gA1fUsf8AefYN9wLJb/lqbSO4D8+HSjboVS5BU9HNqu5uxaSgxua2jkFNCjQyAxz2BC8m7D3gzJEbWXQwzU9DAHNF6V2C7F7F7l3LQR5+v89HCsaOiTPIuoWBBW559iiJkNtU+nTqyGuno82y9hVNTUUMc1LKtPSkKWeEhWQc3B9lb0LdvVXNTXofd1bnoNn4aKioKEtM8YUSJCb306b2A90Hy6rX16CjHVkuSWWurlZUYlhG1wTcfgH3rFet1xjh0Hu4MR/GclCYjogSQEqxtdQeQB+ffvn1o8aDpkrNoxS5JaZ4WWll0xayllOr8A/T3vyr1qndTod9r/FramRx4q6mnimlmgaWxhV29SXsPr9feq5x1Y9UVfzB9mno/dtFQ7PiEdTuqqWjMMcelw00ug3X6/n+nvfVT1ZX/L76RhxWw6LK7ppAldUxU1QTNTgEiSPXwx+o596Jp14UPHqwzIYDF4xZ0xgSGJyxYN6dRteyD3odb6KT8ud/V+yNhY2o6+p5ZtxIoNR4EZjq+l+Pex8+tZHDz6Dr45fIXfeW2bJN2AhgdJQn+U3Rwg+ttX09++zrfnQ9G3292JtTNEfaVNOJjy37qmzWsD71nr3l1xq9wZSgyqVNDUGSMyAsIpdS6L8cD37rYHkejL4HNw57bYE8qx13jW2s2a4/p798utcOPTTDu6fE1Ax1aUqcdIxWojf1BktZvSbg/X3v59a4/Z1rN/zdv5ftP2llcp2x13hpRk2pp5r0GPBcEyamHoH597/w9eoQetXCo6q3RtvdD4PdVBW42ppZVpxJWQNTAsvFwWtyffqZ69XpQdk9ZptrEUeXp6lah3CuQsvkNyLm/wBffuvcek5g+nMx2ZS4CSmjeMtl6ZVtHyzGSye9U699vW69/K92tkeh+oKHbu5MdDJS5TGwku9Kt9JAv9Rz/t/fj6nrdPTj1YvvHrmHOLT53Zn7NUxSRlhGizE3I9Pv1fI9eOeHQV4LvKv2l2FH1/u+pWCWWMQ6qyXQhQjTwW+v+Pv3Wuh7/untannOcxtZTVc+QZp/2J1miXV6jcgn3ry63mtOi0fJ/K5qXaLw7bpj/FKamkfyU8R1B1HB1j3YevXqUx1D+JG/a6u6vr8R2R5HrfvAI/OzMNIawIV/eqdeHD59HBqMPidz4R46AIlHTx2Fhp4C/wBPfvLrw416CCpyr4B6DDbXH+WCqC1JXnUuqzDj8/19+Hz63SvQp7uzeB2hiKTcG5lkgUUyNNIxMUfksCxLcD6+/eeOtcePTT1n2jjN4QZWopmx9Xh41d1VhHUN4wTYEte/+w9++fWwB5dVt/PT4Y7X+Q2zszn9r0ccOVpqarqZfFThfUh1mwW39Pfgc9VPp1px9m4fNdbbqzWx6jUklBXvRLdfHcpe5sf9b3utMeXWvt6Se3MXXZLIQrUMLawHD8jk3Pv3W+h+z2YxWA2+2KZIjKadgpVQDfT9ffvn1riegb6W7Czeyt3VFXRTyU8EtSzC0jRLpL3sT/T375depXq0favyTStmpkzNerpKQGDVRIsF+gv/AI/j3vA63Xo4eD3J1r2Xtx9qVMuME9XTMQ7vFr8ki2Ulj7159b8qdEK7a6t3J0nm5s5tmvknommDU8dLMXVUY6+NHHvf2dV6Fvon5n7iwFbRrlKhltKqOs9QR/mxpswb3rrea0HV6/QnzB27urHU8TV9DFkWp41BadByw4HP+8+9dex5dCzlc1jM7lRk8pXQzQCUMqwTK6lfqQQD/T3vy68OlnX964+gwZwOAnpIFhis8nlWOSy/Qlvr70R1vFeiG96fITEba2vmMpLlVetiaUWSt9RdkINuf6+9/Lrx49Vdvl8l2llqHdWZqJGw1POJ41mmLoyBtVrt/h731qvn0GHdHekVNkY9t7dk0Uyp4G+3kK2IGmwVPeutV9OmfZ24quDHvVf5RPUToWVFLOxNrk6fe+HXq9YNs5vN7g3cIJ4qqlQVBUNMropBNh9Rb3r7OvefQ89m7mo9j7WqJnMbZA0UoUj8jxn8D8+99b6NF/IW6m2p3j2bnd/bxxq5CsxmYqWpPuqYVESrHVEoPX9Pp718utcOt4mjpIceEoqGkp6WhhhjWKKnjWJECjSAEQAfQAe94p1rNepYiQOZbXc/Uk34/oPfq9bpTPXauWJsPSpsSeCSPqAPfqU49aqTw6ye9dW6/9Hf49+691737r3UatpkrKOqpJAGSqp5qdwfoUmjMbD/AGx9+69SvWpN/Mx6Qzfxt7toN8bawArth56t/iG4KsRu0UTzAvIZBb8X59+HXuq1u2+quue/MHT7gohS0MsirIDDZH1A8kheffuvfPqv7JdkZH487nG1qYtV45ZCqs3ItGeLX9+YgLXrwVjgZ6ELcffNJujZ1XWVVYaKqaPXHApCiS63AsP6+6+Knr1fwJSvDpPbZ3Dis/teJquli1wqWeYldctvVZv9f2+kMso1IKjppgYRSTHQC70xuG3LUS01JQU0IDupdFAbg2JDL7ZY6Tnra5FegC3f1ZlKcI216yX7lrFo42b62+nH9PfuPDrea0I6SHW8fcW1N70Jqq+vemesUPE3l8apew92owFSMdWMbDiKdbGXQtdl6rZ1FXvG9ZU/aQpJG5P1P6jY+9HqoPQH/MjZGLWjx26cfTpTZSmEc1THGvq1k6iCB79THWj69cOhuxZMntakjV7V8Mio8ZNiQgsLj/Ye/HrYPR8uvt4yOgjqaZIZBwJLC/J5N/eut1HT/wBm1tRNtuuGMTVVTU5AZRZ76fqLe99e+zonGw9gvFXPnczi45skaqR/uXQ+VRqJB1H34n9vWh1YZ1Xv+TFUYgqn0pCuiNCR6Av0Xn34dbp0I2T72nyZmxlVSqlLDdROWT9NuG5/r719nXv8nRd929oY3Hyu9DlDJVAuftw6W/Vcg8/7C/vfXq1z0Ee4vkbDSUrQ5JY1a1lb9vVyOD739vXvLotO8vkYk1OVxMpkmQagqhV9Wq9r+6s6Lx63HG8h0jPRUd3925rKiZcjJPTxszr6RquCLfVfdfFjrx6s0MqjI6VXx9zuPauq6jKwLLHK8hSaWIjUpNwST7sGHTdD59Wb7bz2Bhx9NNiIKeqd1QGFQpCG36rf191LqME56oZUBoePSlzOcq6hIzHTmB2UWhRfT9P6D3sHV8Oera1I+XQB949IZjsXFY2VFeVkCXQBwGA5IsPbhUjj17j0V/tTb+O6R6qnbI0cMNbUQTU8Kugu0xGlbavzf3r7Ot9JD+Xf8H6/uPctb3Vvjb0E9NTVv3FAKhDpNKJdaML8fS3vXXs9bPWx+sdn0+PgxuJw1Hj5KNU0mBFUsVUKfoOPeq+nXhTz6dNwYnI7YY1lPH5UA0iP9ShV4HHv3HrZBHTFQborpCTPTGBGN2cLz/sD79Qdez0aDZfZdNHjYaKsjWOKNRolIAMpAtz+ffqdb+zoD+4KiDLV8uTHogEUn6BYMCvH+w9+XrR61Y/5ucTZquxePpKCMvNNTrExFjJ+L297610RHpb4a1GewEWZq9aTteXwAvoVm9QFvfgOvdCNmPgTujeVLI2PpZIZoiUilQSazY2XkD36nXukXL8Kvkx12I3xlTk1o1IC6BUN6Tyfp791456YchP8p+u5XqGnzP20I1ArFU+oxcW9+z1rqPF81vkZjaeTESU2YmlF0UtBPY3/ACSR79Xr32dJKt+Y/wAhq5Z4q1crTxopuwWQEC/49+qaU63Tot82I+U3ys7Botn4TL5o4yethStS0+kU8zev1A/09+618utpT4Ifyotj9c7LxmY31SUuS3DJSxy1Mlcnlkae4ZheX8396r1anV1mw+hOr8FDRwwbMwn7JW0wgjDAKNNywH5+v196rTrdKY6NFTbI2pRY5Dj6WnoAqDStOQoGn+gX/ePfutddUW8cVho5sbNFEZDdFkuC9vpx/X37y691moMwhmasNQRTi7qpcC4P5v7959eyOHS2p98YTI0bUckUWvSY9fo1Eji9j/X377OtU9egH371ntvd61C5egpp6Oc2LTBWCi1rge/ZHDr2Cc9Vndu/CfbGXraj+6k6UwlMivFT3iHqb8afx/j7tWo63Sh6r27d/lQDMxTZmCpnqK8K0qp5JGGrRf8A3v37j1rokWO+J+8+i8vUZHI4PVR0rFxK0cjHRe5Y8f4e/de6Oj0x8l9oUdVDiK/JvjZYdERjJES6oxoK82+hHv3nXrXVifX/AHFtfJ1FMYquGpRrMswliJIvxc3/ANv79jrfHHR9thdk4aqNLTqYgFYAEOlmuv8Agf6e9dWJqMdGppN8bbixopGo6WWZ47EsUZ/ULXPvXlXrVDxPRfux9sYXcbtV0/jp6gkunjtxbn+z73gdbANOisZzsKu61qEpMjLMKGRtKS8tdQdN/e/t60OPTDlOwcTuGm++pMoVWxYnWEOom4WxPv3XvPj0jYN5ZStroqOMmWmBAaUsvIPpvcH8e/de6dspBTVAdv4m0c0C+TSsygMwFwBz/t/evs630CGZ32fvzhqipKNqKqdYOoDi/v3Wvs6wU9QyQVLtlZaUBSY2WYICR/Tkc+9jrXTPke2MBtPFTfxTNNM/je95Fk5Xiyi5/wB59+HW/n0V+TvmmzG4Hgws0syPIbIIiVa5/wAPx791X0HQfds9tZXFUymOOXzN+kJG/B/x49+63U9EJ7D+QNXDFIc1GY43Zk/cUDgm1yG97615dFvymfG8f8kwM3nq61lMSLybO1tKhf8AX9663w6Np1v8LMlW4Ci3RmfJTVkqiaJ1LX1ldagj3vr3QxRdH7sio/t/47W0ghIERSSQFgn6B9fp791rpPbi3R231pSNAgqq2hiWy1TlidAH1Jb3rrwqOgBz3ybzghkbL1M0ICuAWb6kfWxJ97PW6dI/bO9s12DkEqsbFUZGpMyeOMRSS6iWsoBS/wBfes9ax1ZL1fsT5WZrGUuP2bsyqQ1KaaeSOCZWYkWuDb6fn37rfDh0errL+Vb8mu1BTV/YlbmcMKxlE0JkqI1EcnNiOP8Ab+9VzjrdPXqyDqP+RZsXCmlrd1ZVq128cs61Mkkmpg1z6W5PveePVa+nVtHUPwf6H6ko6aHHbOxFdVUyqqVNRSRtewHLAjnn377et9Gno9r7cx6olFhcbSqgARYaWJAoX6WAHvwxw68c9PDQjwtFFaAFSFMahdF/yoFvfvt69w4dY4oGgU6XMjtbU0n5/wAbj3ao4HqtDT59SR9Ofr+bf1/PuvWx13791vr3v3Xuve/de697917r3v3Xuolf/wAAa3/qEqf+tLe/de61Jf5Bv/bwn+Zt/wCJErrf+fX37rQ4nrbglBaNlVgrMLA3/wB497HXj1GaVaKgnqJjoSnhnnkYm9kiQuxP+wHvZ49aUY60Cf5oXzC7Z3r8rt39b0G8s3hNq0WRaKGKmmkSnmiWoMeghTaxHunDp1kYjt6BzbGEwJxtJJVGLIZN2imlq5yHmdmAMhZjzz+fdj6+XTdNOD0aXE4XDZHa0wWqWExREBVNgDp+nvVfTr1QPPPRG+4trZmNKxMB5aqSRGAYX45/s2/p72cdV8RCaDj0QjdHV+/stiqqlraOaeRpJGCPrIAPP096x1c+nRZMt1jndqrK+QoGp2JOgrG1x/vHv3XugZyOdrsVVSFWnnaG50NE7aAPrfj37rY6Hzqf5ExY2mFJUVskMq2TQV0ng2sb+/daoOhopu1/4lnqHIrVP4EkV5JdQGkBuWP+Pv2D1riOrKetc7hc/iaGujkSpmfQNbMpJNuR/sD7st66fohPz6eElE0+XR59hUWPyGGeiq6rxxSIdUIeyEH8Ee/LbicananXqg8ePQLbu6S2JUZDIP8AwPHzvWEiSqeJGdQfze3tltvi0GjVbyPWwELAE8ekTQ9P7N27HIIaCjdTcA+MXFzwfp71YT7hZPqWZscOnJrW0YY49BRv/rHEvR1MlLpgkuzRxopA0kcEAex/tnuXzHtCAQTPjomubCN8oM9Fen2xuKMTQxRtDTwhg06qSCo45B9yJyn94LdNrlZN8ja5RzUVwV/Popl2zXjT0jIsbiI6pqf+OTS5Ym32yhh6r20295CbD71cs7xGDcMtvWnxHosn2aUGiL0sqDbO6I6Oab+G1E4B/YbQ13Um119i9eeuUJWFL+IV/pDosn2e8AOlK9HI+Pnx8WsNLubPV0lFJNJ5ZaXUy6F+oBH49w3zf77bLa7q20WK+PGuNYPE/L5dGVpts8MYZkz0fyeHCYZIKbHVf3DRRCMAyKxunFr/AF912fmO234a4agnND0YeERQOuOkjlMtVU8o85MaSHhhe1v9b2Jol18OrSQW6rUcepWNymhnkWTWtiBe/wBbce9vGo+3pE/iMNPl0rMdkpJoWTSbn+p+vP19syIoNT0nIc4TPT9R0bBXlk+lr2PP1/p7YlkXgOrwxynJHScylNAzs6MAwB9K8e3YmI49PsjcAOkDX1T0+sAlubc8sAT9T7XIA3VGDDB6fMHkVZEKkh73IJFgbfT2zNHnPXqk4/b0rTXI0Y88ani1/wAn8W/1v9h7TaDXB69U8B0l8tQUlUrFYVOrm4A9Jtf6e1EUjrivWtPr0DGah+yqdOorGD/sLH2bwsXXqjxrTUOssc4ip/LTkN/X+v0/r71SrUbqjHHb0hO05Gh6u3Rl4SUyVLTu8BX9WpE1Aj2XboFa0likFUKGvSyzXuDE+fRT/ip85Mruh6zq7JRtJWUMk1KTLp1XViiMOb+8Ieb1tE3IpbEE1OPToSqxLUXPVz/xfyM2DzlNX5dytJUywyqXNhd5NVrE/wBOPZU7y/R/pZPSlSwNWx1fxs7cWJ3BgaH7Kjp0ZIxaVQl3Ojkkj2V7cZe8zcerVBOOhEoevaDP080tVRU80kcZbTKAdNrnj/e/a8V49e6In3PuPG7Ezz44lKWISSL41Kqh8fNufe/n1vyp0VHL96Y/L7pwWLwUyykVASuMRX0jVb1aPe6gY698z0anee9toRbZwtLS1cQzhmpRIisplJcgHgc+9AD9vWh0AvyQ/mRbR+GK7eiz9bGKysooxBSVDRhZmeIFQBJ9fr9PfqA5PXuquqD5I9f/ADe7bwu9d6V8OPxGPyS1dPG7IsYWOXWukDi3u3WuJ62RuuMh1/lNmY2m68r6erSnoqWO0Lo3+bjAFtPuufPq3UrJ4XI5uuglmZ6aOjOiVASA5HBuB/W39Peqjrw6SuV69xdVPPJlaWGvp9N445lDrq/Gm/vZ690WDs7aW2Eo6jHUDRYyWQSJHTwDQpcj0gWt78CevGgz0S+TbHZ/XM9Vk6UVklBMxelYanDRk6hx/re90690ZbobuYZXJNR7sJikiYDTOLXsLH9X+PvXl17Pn0b+q39iy8SYetADaQVjdNPP9Of9j70evDzr0OOydqY/edMldFWLPUQKWnjLi6kckabn3vj17oQ6nr/bhoamHLY+myEMkBjkgmIZApPqJBuL+9A9ePWpP/Os+PGzMXlJ89sqmo8XWLLLMy0RWNg/i1kNotfn3brRA61m5a7LPQ02NyFfPWSQAq0MzFgObc/7Ae/de6Nh1BvWXb1Lg1ix+oUWTpKiXSlyscbXY8D+n59761x626/h13tt3s/rHFvHFAEw2Oip6l20Bo3QaeR9b8c+6nqwxx6PDs/tPE4TJU8dPU/dxSTi0TWKLz/ZB96p17h0Uz5vdK57sGsp+yNjPLS5tSLRUrEOVY3uFQ8Wv7tXrXHpr6Il3917t6Gn3hV1tXHLAoqJ6xnLU2r6hS/09+pXrYNOjw4uo2nuPa9bOVp64faMS7aGIIS55F/da9e8+gL2NRbayOWrcBFLDRSNVSMEUqiizHTb6e9nr1QM9Gh2jjzT4nNY5pmQQiSOB1a5kAX9XvRHXqgGvSExGHxGMzUVbUVazVi1WsRuylidX0IPPPv3Hj149Lb5C4/bvaHVdVtbLTDDVE0eiCeEaZPUmlQpH+Pv3XscB0U3pvqibofAVddnstO+3nikdamWQgPEwvqOs/097zw69jy6FnFdh7B3Bgc+2Fyf3VNVYuppYlU+kyspW3H+x9+p14daT/8AMr2X/c7u7IZmpUUtHkc3LNGy3u6OSVNj72etdF12D1vv/fdRi5djY2XIJNLyyo/0B/qBb/Y+/DrXRwNxfBH5JV23f47HtBJ1SkM7l0lLCMJdjcD6+/efW89V5bswGc2PmKnG5+ifHVlFMYaoeN10Sq3IW4Hv3WuoWRzmRlpKF8TXSs2tS2m4I+nPHv3W8dLzb3YG78BUU9VTZyuhkQpfxO5ZQp5AN/futdGjo/klJU4qGg3HVPk43j0zTVbBzFwORqv/ALx791vqPkcVt7MYptxbcrVNRIrSCGMqBrIvcD37rWeHTZsHt/fmyahpY6mrhWCcRxqmrlI24Pv3XgPPqyLpT5jVdTR1EW68pPALPoadwDbT/ifrf37rfQg7h+Q9Bk6eZtuZd56iWFkZUYEi/wDQg3v78Pn14nPr0STfeVzO7a+XH5rLzrQ1UpmdXcheDc+/da6ad1dw0W0dr0+1sTUAeGL7cSppu1xpube/db6L7RCTJTrlC/3dVLMXOuzEKx1AXP8AT37rXr0czpf7eWspKiSJahYVtJDIBo1A8g39768D0JG9chC+YSWkxlPjIaeUO88GkEi9zfSPfutkiteif9+9kVNRWUmGxztXLVyx0R9BmOqX9sAKL/196x17HW1p/wAJ7vj22xuqc1vLKUTU8+SeSeEOjx6mqJ9YOlrfS59+rnrXHHWyoAslpFc2tYafobN/j73kYPWqA5HWX3rq3WORWZCEOkki5H1t72D69VIxjrko0qATcgWufz/j711scOv/0t/j37r3XvfuvddMSASOSATb+th9PfuvHorPyS6EwPyX61zO08xRCGqaKeKnaWNCzt4iECuw45+nvx68OtMr5A9Lb0+JfYeS23nKeqo9rx1RjoKipuIJEecqNDfSw97XJHWmIUVPVNfyLxeW3h21T1WNZp8XcFpIf82Ltqtx7C2+bmbO58EGnQz5Y5en3YCaNSyk9Kis6onymDo4KWYpMIF1Krm9wORb2QpvqiQAmvU5WHtpE1n4kq5p0gZxn9hU8tHVNL9vErX13F1uQfYrsOY1Wker4sdQxzzypPt8n+LrUL0y4jsrAzmSm1xQ1ZL+pjzqJ59iHwfGXWCM56jhWkjOlhnqNTbmmxuU+7etjanZy1r3Gm97WPupKoaenT5k00Pn0MOI3bg8rUUk8dOk9R5FvIiK3+ub+/NKD1bx9QoerEup+16HD4qChWrhpyY418LEBjcgEj/Ye2+tjobN7Ve2uwdpVtK8CVFXLCUVxYkEr/Zv7917Hn1WDgt0VXVnYdRiZA6URqHYMb6VXUfx9Le/da6sX677UwM8EFQ9fTytIisw1AFWPOkj8+/HrYPkOjBS7xx24KIwUEyiTx6bra97cm/vR9et/Z02Y6eGggY1cyy8lvoL2vf8+/Hr3HpI7j7BpsZBO1KwgZWOk8m4sef+J9761XHRfcr3ZWQmYmrLix1KpI1c8An3vrXQQZzsOrz/AJJKN/tKoa/3CzANcXAP+PvXXvs6B5s5l6nLRLnqiSajRmDOWbTYfgn6e/dez59A13j2Ht+gWhxe262no8lUSLCXD+vUTYfQ/wBfbbRhznpiW6lt/wCy49Gt+NfUmB3DgIMpvvK01X5VimEc7Lzq5sdX+9e3UtVJFOtLuM03xY6O3neves8Zt9k21T0gmWPSGgC8m1rjT7VrbCmnq4nDY6U/x964rquvaoqKsikVmZYmN1Cr/gfdX24/F0ZJbWrxhiRq6OtS4TahqoKadYpZUAUv6SNSG3IPtp4fCz/LqrRQpgU6FWPZ1DULRz0sammpiGeNVVgw/UQPbZmVu3pLjUeqg/5qFXtWs2fgsLQRQ09emXiElgqMf3QCNK/63vVKdaPVgvwFyMWB6BoMXSKnlmxUBKoi6n/bHNx79TrfDh0eDHZl6SjjnhiNPOi6pTzc8e60z1uvTxTZWXcsZhqJC6qOFYEH/Wsfe6U63WvWbHYmhWqEFVDeMk31WH1PvX2deI9ehSPWMtTRrW0LMYlCuqKf6C/++49+x1qvy6TGWwFOaCelyUBCpFJ6m/qB/j/T37rfHrVM/nFRTY3em2qrCBhS0VXSmpMZuoXXdrj3bqvTr8S984fI7WxUc80Mx1RpJEqgsBoAbVf37PXhx6uN6gj2pWrBAlFFqkjVlbxR2Jv+r6f7x79XrfAdGzxu29oRXps7h4cjFUH9r9iI6AfxyPfj8uvClOnjJ/E/qLsbG+EbagieUWu0EFrvzxqHvQJ60RQ9Ep7d/ludXbeqpK8YeliTSzEMkAF9F/yPe+vUHVO3dfxG2/t2fcGQhMUdGI5HgjCxhFsSQbj37rwFOjB/y8fjpiaHLjd9GkRLTxhnCxE6lJH5H+2v7316nWw7tDbjPQeJ7SaQQpVV9P8AsB/xT3XBPVqkCnTtloJ9vwhwCykX0j8e9Up14EE9JGu7DkpaCRWcKblV+osBxz73Q9eqKdBc+cbMVDTSVqRsG9F2K/7Y+99VPShocnlj+xHX+SG1hpJYEf0N/eutivS0x4aBDUyVioVszAsRz/Tn6+6/Pq2OlNQZ+WvZqWap104svFyD/sfe8+fWseXUPJ0FLRvJVUzI2pdVmtck/Xn375dbB8+ktTZISyPHU8g3A9C2/wAQR79w610Ee/uqMLvqGppsjHFJFUKVK6UBKcixPvYI61T06ph+V38ufNPDXZ3rSOppayBZ6vVTNpLMDrW3j/3n3vjx61pz1U5h+zu7+h93rtrcVNkagU0yQ6pPM1wrjVYD375da+XVqfUPzjWtpqCiqKc0WQiKrJJL5EYkKLWv791avVo3VfyU29n8fBBk89RQZFljY+WWzBWH5596x16vkOjCR9x7VgxdRH/GaOtqmB8TxOJCht/S/vdBXrdcdF33Luva+95JqHLyRSgf5pyLBTfg3P8Aj791oep6K3vXorfu56tqfYm5nx9IZPOgjk9GkchBb+vv3Xj0nK7bvbXX+PXFTpXZPJFCiVCB3Lyabahp596610BW5cp39FFKIaHKrO6sBqSW5P4t7t9nWiT0EGDzvc9NndG4dtZerqZGIinKS2Xnj6+9cet8elHW9dfI3s7JJRYKuyO36aZyp8hkXTzb829+69noyezf5eHaUlPTVm+d3TZWBzHI8bzH1Kw1MCL2/PvXn16nRzthfFrrPY8cD12FiqauNArS2RiSDcsSR7314U6gdr9S9SV2OlnOAhVoUvcJGeQLc8e/de8uqAvln1TtbM19VR4toaCOBp30XVf0MTzb37rXRZPjb1PV5vtnC43Ga5qWBo0leM6l9EnL+/Ade6v+qqnGbO2//C8zXU9OMZTobShVJIj4JHvXE06t5dFzy3b+1qqhnanyVE7xSSXCBCx0/RQB7359aB6Kl233dh89i6nFfcwwFEMfqAHkOnTfj37h1rqontrL1NRNLT0sTyxEyCNl1aWZ29Nv9796JzTrfl1Z/wDyw/nF8ZPjvl8LtrvTrRM/kaisoqcVtXSiRCZZQBJ5GB4F/wA+9jr3Dr6EHxez3xs7b2Vgt7dR47ACGspUqloaaOnM9AxUMUdFHFuPr/tvfuvcOjjKIUKxqqLYelQoUAf4Ae90PHrVRWnWX3rrfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdRMh/wArf8AqEqf+tLe/de61Jf5Bv8A28J/mbf+JEr/APWH+5T37r3metuQgH6gH/XAPvdSOHWqA8esFXTpV0lTSyANHU080Dg/lZYyjD/efeut1pnr5u/80jrzO7U+Z/ZWUMMsONiyMr0khJEbA1JYAW/w91yTnowRgqD59AhsHsdolpxWzKBeKO7Xte1vbqyIBpbpG7oJKNjo7G1N504xyUsciSirQNrXlVv+D7uoU8OqBFY0rx6UyQ4+tN5DGpHJJ02N+eP6+3GiCrq62bdEz1Ertl0OSl8lNGrLo58aLcm/tLxNR1SvQY7o6FxG5KmOGqpRK7kXXQn0It9Pe+t8B0Fuc+CGBqfJLHgXBqgUVgiA3YfXge9Y8+vZ6J/23/LWy+Egkz+KoquliDPKNJCqbc/2bX9+690BmE+Mm9IqWox8b1LOw0LZiGUgcc+/UPXiR5dKPbme390hXQ43NmqGPp5gxmm16NI+gv71QV68erLum/krgtyYuGFMjTJUrEgIDerX9D+fdgT17zr0abZu8cXm6qSnqJ0maViqgkWv/X3o560AK/PoTc3t7GJRI3iBLEEG44/IHv3n1bypw6DnObJjy0FqekeW62BQX4Atx71jrQqB0GUvV9VG7QzY6bxzejmP6j/be/FUYUbrYp5jppg+Ke3hXHcUlAEnCmX1BeD9R9OfddAHwk/tPWvPHSiwWwfvq7+GxuIoID47kLZQp/3n3s+JxBP7T17HQg1+Bym3KeeGnyYWCJLjT6QbD6ceyZ0dLrxD/hPSoQCSOo6A18puSXMeejyepEms6q729JuVtzyfclcvc5vtGkLxFOiy5si3Hob6feMdZjo4aul89RElmk030uOOf9j7lG294bRUpKAX+Xn0WPayp2IOPQs7D2zT7ohIhqkhmIv4QQD9OBz+fZnY+7VlcSBblQor01Jt0yrWv5dKuv2BnsGC/gqBECSJSp0kA3+p9yFt++7buya7WQNXy6RM3hnSwp11DXNBAY3kEjrw/q5Btze/teY9TcOtCSndXpO1eQppNSADU1wf+K+3khcceHVjOpBIwek3UYpaqCSUHSRc/wCt7Wo2k9FdzcyDy6T2GMlNXtHq1KCRb/Y+3pxVK9OQS+ItehIZRIqk3AstuPrxf6D2XdPAkcOuMipHGyfTUPqf97/r72Kk9bNfiOegv3vjI/t1lHBKk3H+vx9PZjZSnVTqrqSMdISkdUoygubMLkD6fjn8+1zglq9Ng0x0wdkBT1luVXTVqp5OP9eI+y6/i8aCSP1Q9KLdvT161zupspVdefJ6HJNMYqTNZ14tJZlA1VFrBR/r+8HeaNs+n3uV/t/w9Cy0rwPWyzF39R7Wz+1MSVEkFXRY5iRewaQDnj2S2cxZjHXHSqfA62Afjr2ZRJs3HZeokVYWpFkSJjww03AF+ffmUBz0xEAc9Dxgu+56nK1oppDSUCxtdzq06V/1/wDD36lB08B1WH8490y7ipa7L7aysdRXwfdHRAxLq2i36Qf6+98OtGnVZvwV7EpY9+bsi7Jr0jq46ucUgq3IKkP6Cvk9+49aGD0dnB7kp675ARV2UzUX90oJaeVTI58AjWQG/Jt718urceqWv+FGm69k7/7V6gOyspSZDH4ySlTIRUdQwAMcIVg4iP8AX+vv3l1ropnw9yuSnxsGOw2OqmEYdFKM7aRptwwN/ex14162efgd2Nldl1VJTbhzC0dPIaZVpqiRieeCvqNuffiOvAnq5jcPZeFxOKjzEs8UdLKolMzABHRuQ1/9j7qBmnW/s6aaTemI3Ri46/F1kNbG/IjgsStvqPT78MZ69Q9BduXrabek5yVJC8BjbXciw9P59769jgekRnxPj2o8TlqCWrpKMCKTVGClrWJJI9++Z60fQ8Ogu7F6pxOcxIyOxrY7KOLsYSFYt/qfT799vVj0UfJVXZnXVXpyclSy+VY0lkZytr/UE+/da6N91t2zvTaeNocrS5KSaOuQSVEMbMQdQBIIHv329e+zo3fV3yj23u7I1uBzmUpaGripwrLO+ly/5HqPvVDx691qzfz2+2MhQdl4uh23lIv4VVVMqzJGSwkXx2JFj+fx7t5da+XVAO2YqrcNmp6KWepl1n0C5cg8/wBffh17q7j4AfDDJdt1MqZ/Gz0NNJSIYGqFAUtJHZbFh/j+PfutAVz1YZs/pjfPxK3o+zYKOuqsDnqtmUoWaPxs+q408WHvdfLq1KDqwDb/AGPsPGxrBkGpqbK6VWKKVrTCUrawBN73491NevAAcehi2j2ez1SU1fTvU0TlfHGylwYm/Sw1f09+pTr1ep/ce0ZuxNk5Ok2jXria+oj9FtKPGQtiPTa3P9Pfq9bNRg9AB8fcnlOo2l2r2Hkfvlnd4Y553/UrHTxq97PXjwp0z/Iva25Ns+PsbrfJNLG06u1NSsWZlkYEcR/8T71nrx6OL8ZN35jObKpZd10s0VdVUxaWSoUgq7L+dX+8e/EeXVfn0D2W21vNu6Fq6TKtU4N6xAtIjnSoL8H+nvdet06PJu7YNNkRhJamVSiUMbywlvrIsYPA/PvVfPr3AdEk+TGS3tn6Wj2ZhMTXT4eMJTTNAjBDH+kksB9Le/VHHrQGKdZ9iYrYfXPVeRpsnivtcjS0Eta8soNzKqlmuT/j7959b8utKf8Amt97L2L2dW4zBY1q6PF5+GmWSBWAWMzeO1197PWur2f5WfU236LpzB5rcOEFLX19CHo56lI2/daINqBYfXn37j14dXVbRyOLxOHrMHl4IqyOopZYI42iha8ci6VAuP8AH36nn1sVpQ9Up/OH+X9id74vdXZuLxzYylppZquayxxo+lC44A9+60R58OtYfNRQbY3NWYSnh1w0Usmtv1cJIUNx/sPfj17A6fqeTH5ZdMMixEISzfQA6fpf37r3SPqokoVrUllWqQNYRKxJYfgC3v3Dj1roRdpbtq8XQ0rR6oaVXXy0xLMRGODwffuvdDQ3Yu1s3R/bLBFDUBAjvYKTJaxJB/Pv3XuktKajy66atvTt9ArkcE3+gPvfHrfQhYnfdPtejSVAzzggu2pm4H5A596690lt2d0tl18lLIEdAQRc3+liffuvfMdIOlzEe4SXqNUkyeoDUSzcXIsT/sffh175dC7tbC1H2yVoyMdPyqtTOx1FD9LD3v59a+Z6G3Cb/i2pTmgjlWOqmFvubsLN9QffuHXs9MeY7frooa2M1AylVPC3hgjYl5GA/SFHvXXuHR7/AOWZ/L17E+Wva9BureW3slidm09XRZFaiqU/aNHG/kckyC30H09+oet/b1vrdRdS7T6a2XiNmbRx8FFRY2khgkeKNVeplRLPIxFvqeffutfPoTY4xHfSbKTcLb6Em5sfe69eA8+snvXW+ve/de697917r//T3+Pfuvde9+691737r3XG1iCDZQDdQAAb/n3vrR9eq4f5h3wnwXyu6yq6eCkpodx4qmeejnjgAqJ5I/Wg1L9be9rhwfn03NmFgPTrSQ706XrPjxmMltXd9KwzMdfWCCSeFklEMLlUN25+nuN+co5ZLwsvWWHsjDt7csh7gDWCOPRc8Xu+SjqFm4MIOoc8aT/h7j2ZZo2FK9SneXZVzHAMdct4VWE3bj5FlRBLKhVWCgG5+vszs5rhHV6nHHoPXGwW+6xyCdRqI8+iUbq6UysdXLk8PMyWd5F0ORYfgWH+8+5CtuaIhGIg1SB69QpuntrLFI0iJQVPl0HWUwm94ozTESuY1ClwGIBH+Ps8s9zW6ofPqNt35daxrQUp0w0namc2BKlPNFUvOHtJdSQAv+t7O9BKaz0BdeifQejBbC+RmOnyNPNmKipgkVUPj1lEv+Lg+2wa9GRx0bbbfyspqarhpaeqP27Nb1yCxB4tY/8AEe/dV6XWcfaPYEcuYhlh/issJYFHW+thew96Jp14D16BSmyOa2LPUTVdZUGiRyUCuf0Kb2/2HvfXvs6EzZXy3x+MqGpmnnFmIJkkP0HAuT799vWx0PEXykwFXTLIarUzqLAzg8kcgD/iPfuvefSC3D25Vbge1HLeNzcAtwBb6/4+/da8sdNNNW0jQ/cZSRrWLtY2/wBf6+/de+3pH5/fe2qKNzTzBGW4sGAOoXt/vPv2OvU6Lvu/uF/FPRQO2qb0xOPwTwLn8e9jJx1pqKK9BLtbovdm/s2Ny5apleigl+5o1ZywJHrHPsm3TclsQC3n1IPJXKD8ylxGKkfKvR1NmY7fdPNTYmConix9OkcRCO6CyGwHtFFzCg8+hBe+2E6E6Epx8uhnr97bq2vLRUiPVVCNZZNTO4sfyT7fHMceoZ4fPoKXPIV8pPhqaj5dGr6u+SmE21TeLKieOoMahirFBynqP09nEXMMEkYGrPQbn5W3uCQgKdP2dD/s3vzYedyHmNayuzXtJOqAc3AuT/t/ezeic6hw6Lpttv7f4wR0Y3B/ITEU7PSUtdTPBcrqNSpsLcgj3vwie5ekgZ43Ct+fVMXz53JTby3hjTFWCWIZFXMSTBwP3Ab6R7c8+ljU8urP/g/uhMbtDC0SSakGOgjZHYsLAAXsfp79jr3HqzuN0yUJMbRfupbSrC4JXn0+/VHXuPSQn3PFsqsV66QRUzPYlmCfT6/X3759e6FLD742fl6WOs+5jLAajaRfoBdbG/utOrVr1mo/kPTbUqaiCqqoP4YFYRa5xx+ACT73TrXy6Qu/vkhtHJYSsamraZKoxzBQs6A6nSwsB/j79Qde6oK+Wm0T25QbiqKyWCZ9RejaR1kcFQSNBP8AsPp73nrXVQmzew810JmajHV1Xpp4J5Wj1SrGFsxC8H375da6t6+G3zP23ubNUWHyGTiFW4hXSalNVmkt6Rf37rfWw9sFsTuSgpa5ZEkjZNanWGuLXtf3o1PW1HRvdn4qKGlieFW0IEtYcfS45HvRPCnXqV49Ar8mftpdrV1ZKwU00NQdd9JGiEnkjj8e9jr3njrUj+QHyNp812DVdZUs7TVdXIaONI5i8hvKV/QOfz731o9XP/BvYNPtHrzHrXwvFWSinn1Sq0bfuLquQ3+vf37rY49WVY3MT4tB9u6Mhv6QQTY88ke68Ot06a8zu5MiPBKQSOCAfz/h79Tz69gdAdvWSAwuFkMQXW2pjpFgLktf8e7DhQ9aPHHQL4beW2J8vHi6vKU4k16Aq1C3LX5Fgffhnr3l0YHH1mPpIpWopBIoj1agdX9m97+/ccdaPy65wZuGtopA8jKPIeEa7cf4e9CnW+sNRuAYmFfDIylgCCX0n/E397p16ppQdRqbfzMQlVKXGogevV/rA29+oOvZHShkz+NkiDlisjKB9QLk+9Co68W8x0JWwdux52ZZqprUrG5Jbk6fzc+/fLr1DSvn0KO4Nq4HHUM8dPSU1Sk0BR/MgkAVhZhY+/Zp1queqavl/wDFbZOXFZvOjxNItbA7Ss0dKttQUkm4HvYPXiOiM9J9Q9Kby3CmFqqympc9DM0ckZkjj0yA2IZf9f37r3Rx9zfAHN0jDcGzM4kYkp/2EjqdK6WX9u4B9+r14g8eiVdodRfJ/qFayrgrJ66m1M0YikeWygarXH+Hv3Hrxx0QPd3zJ7P64rFbdlNkoBFIfO6iROFbSST/AI+/HHWqdWBfFX+YvsHdFLTxVeQQVwWJWSpqlDLJ9DqBNz/re/dbqR1YQPlFtLM6KqA4yeojP7Gp4pAbfi/Pv3XhTpxpe7MTl5jJW47FgjlbRRkWA4HA9+wOPWxU9Zqfs7ZFdm4VyGLxwlUfrFNGFCj6tew9+rnr1Ol4O3uusf5Dj4cf91DyoSOMWa30BH5/HvWOvZ6D3cPyt+0SWm8lPFGAwC+dQQo4DaL/ANPe+tccdAxW/LOmkWeP76LyEnQTOAPofpc+/de+wdF1378m8pLDVxtXRvDIll0Tre349IPv3XuqhO+ey6jLZeplSpkLTtMpIk4u5IH+t799nXh6noQ/iV2NguraKs3RmpYPvomkkjaR11/1+puffqY69WnQTfJf5u5beeYyVJt+oqBFMDGDTzkKADYfp9+r1r7eis7d7J3RHRPOayvlqZ5dQj8zsxLG/wBB78D1v5dcdzzb2zSRVUsk8esKwUOwJJ+gb37r3UzC4isrKRVyUTSvT2cFk1tcG/Nx/h79nrXy6Zsrs85avjr0jlgeA3jeCMKyOnKMD/UH8+/dbFerFfgt/M97z+DO8Mbja3M19RsKpnBrpK/IEpBDJIF0rG5ta39PfuOD1rzqOvoh/Cv5YbK+WfUWA37t3J01VV1VHTSVMKVEcswd4RIzDT/sb+9569QdHM96631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3USv8A+ANb/wBQlT/1pb37r3WpL/IN/wC3hH8zXj/momQuf/Irb6e/da8yetuNtVvTa/8Aj72KefWm1eXXbcKf9b8e9dWArjrWv/nN/BSm3Ztiu7T2rj45MjIDUZSaKmLSkpdmZ2X345x0rjcHt8+tNDNY7J4xmxsSSR1VJlXhlVQQwEMmk8f7D2GLzdFguvAJ6ENvyVue4W4volJU/Lo2ezd84nbeNx6ZNaln+2USXBJ1Bfay23UHAPRbd8sblbA9px6jpxXuJMlk5o6Bpo6aM+ksdPpHAPs8ivVZKMa16IjaX0TfqA0HQlYrudcXSAtIGPHLSD6/Tn3r59WqD0oMX3SZq+CptrX62Davze49769x6Mvt3v7CVf2lPW04AgdCxItqAAutz/xHvXz6t8ulp2J25tfdG3DjqeGk8ZQBzpH002+v49+p1rh0WNq/rXatFPlcoKIsiF2AKXHB50/4f63vfXvt6JD3fisf3TSVCbUoh9vIkkUckUAZi1+OVH1/I96619nRHIus969KfcZaY5L7dZCwQLIq2X1ED/Ye/efW+hp6Q+VNFTbhp0y9ZLTsJbMs02kixtezH8+/VHWurX9td87V3itPTxZKFmEcVl+4UsLrawsf9v799nWyehKPa2M2/UwQh4GWRVAJdWFj+efoffuvdCXiN54vckMswNOpRbqWKgH+hv711sHOOktm95R0kc8HoZPG4Jja4Itb8+99er5dA5it7Y6KSuEblZndgrA8rc/g+918+tUr16Xe9JlBPjZnZnKshJfVzbkj24LZZBqbz6uLkINI6RK1mE21LJM5Z5JSxUNbkseLA+22sAe2vVzIsncvTBPu6toZ3qomX7Gdtbi4uFP4Htn92hHEwPw9WjRNQY06EvavcuO29GmSoqwrUx2dleYItx+CPepY3ujpQlT8unrkROKqBnoSR/MG2SKb+C7zqaSFQPCH80YbkafqT7FfLu63nL7iTxDQfP06DV5YNITo6csD2rsXdwnrcFk6UQVIvEZaqPlTyLc+555Y9zdq3oG3u3EUqniaUPRSdvnhYGYEr0rKKmomD1s2Qo/Dp1XE6EEHkW9yHHutrcALbyK9fQjrZhj1Yx0w5TcNCEkhoaymc6itkmVj/gOPZrbqSQzDHSWe0LigOT1CwFOZ5mlkYMW54P1J5/Ht25cUx03HEYe1+hAEghUIf7I/Jvz/AIe0NK56uWJNOmjJ1Z4a5Cg/63FrX/417cjUHqw1KOklueV6nGoACRoI1H6+1VsNMnVWzw8+g0p4HWIjljr/AKWuPZizCtR00B5N0nez8vjML11nTlXCxtBJ9SPp4zYc+ybeLpLKxmu3NFRD09BRW08KnrXxwuycv2F3KuVwMTmgxeTlqqd1VjrdZdVr2594VbvuEO8X8skJrx/w9C+2FVB62F/jH8ddy90bhwbZqlnhWgpKQCaogdUHhtzrI4+nsIW1Yrwh+lM1dFW49WxZ6kznXr4Xa+Nq4xjsbop6sxyWGhAFOr2ZuQzV6TwkEEDpcZvu/rzA7OyVAuVplzf8Olv/AJUiOHKW+pP9fdM9P8B1UbTfJPaWL3BuBN4ZVKmlkqqzxLJVrJcNfSLEn3vquKdVc793tuvcfZW4s/1Y1TDjoqkyRGmLIjqHLG+nj3vh17j0Em8vmR3dtySXb1alZDOYmh+7WR0ksOLmQ+9cOvdE23Dk949zblgTL1tflstVVVNJTiomerMCSSgXQG9hb3rj1vh1sufFnqzp7449M4rdXZUEUWVytCr04ZFSR5pIb2s3P192+XXui5dl9zdn5jedNlOo8flRtqiyAnZqeGZVaCKTUNLKP6D3r5Hr32dWudU/LKLuHqY7E3G1fR7rp6IUaCoYxP50XRzqsb39+69x6MD8cN07m6+kr6Xcld5cdTrI8fkn1EAi4srf4e/cetjpR7t+cWRxe5F2rtamqJzJIIyyqxTUTYW0g8e9DrVeh72fvTPb0wzzbionSWsQvGTEddyPpe3vxx1brHBQbiwNZ97TxvJQhjIUlU6Qv49P059+rXqtKZ6VmWw20ez8G9Fk4IIsyCApKItm+n1/r79Trfn0l5evqTZGJMVTEzwxx/tEISliLcMbj/X9+rXrxA49Eo7b63rsJ/xkvAVVcsbSF6iCjkZWEYku3CfQce98OvccnrXx/mbZyPsbJY3LYx55f4Kojrmqm1OJUiswu3+x9+zTrWOI6IR8Yt/7cj3/ALYwWSQSeaqaOVSFZWtKF0+/A9e8ut8b41bG2vjuiMHujZtPT0+WFLRTsUjCu1ow1jb36vVuHDoS6/I43eaLk900FKarDqY43+3DOWAsSpI/PvX2daHVcXyH6eyG5cxT7j63lqUq6KYVVRBHfRZW1EFFPvfWia9M21t59x00tNSmH/KafwUxBTUxCek3U+/cevD59HP2xvLN4HFxV+6RMrtGWl4dUBb+oPvRHr1uvl0hd9nDb+h/iVPI0RgDTQNExUs6+q5b6+/Z690t+p685Pbk+KyJ+6MCuY1qj5bpECRwePx78fXrYpXT1O6772jqcjuXaEkSU9Rj5pKaiWICMHQ2hdIUe/EHy68D5nozvV2BrzWPncrGsgKmSEyctcjUCCR78acOtGvn0YXA1H95p2jk1tLTalVQGb9tPzb+n+t70evDpLbu7N632tjs5j8k2Miy1LGVX7wwpIH0n9Jfn6/4+/Dr3WqF/MI+d/YtFu2p666zl+7qstUVOPWPHTBwkM0hRS6x82t+PdgAOtEk8esnxD/lrN3DtSfe/b9FHVZjNaMuWq6bU6uHEzWL8mw9+4deA9era+s95dB7ahi6VwhFHndlAQGMFYQzxDRYLxf6cD3o+nW/n0IEmWyA3BBLFr+0R0IBBAMKn9XP+H597I60D69ZvlpuPIbr+O+5cFsVKda2SiaKoR7eZpXhKMVX6nk+/Dr3nXrX42P/ACouzN4bRq955SnifIZiKpMJamfX5JJGZeT9fqPeq0635Z6rs7l+JHYXx9qqzHbppKq5qZjHLDTPHGIgb2JI/p73jrVeiy5HCrRPTzoH8slmtKODcagWH+8e/da86dNuOrZkyFXDWqUjeOygcICeBp/3j34db6hS1WLxVQ3mncPIxcBJBexP6SPeutdLvDbqiEI0Y3K1sRUgfbwSTk/T6aQfe+vfZ0u9o7T3p3Vk5dp9e7Y3HT5hR45JKvGVHi1EXGm6+/fZ1vj0fDoH+Tn8q+xq+Jp6CZIJxr/yrHTKL67HQSP979+690Zbcn8kL5PbRyELUNFqYyjUFoJWX+vAUcD36vXgD1zl/k7fLwqtZBT6SFH7S0M4Xgf6m3v3W6DpY7T/AJMnys3c6xV9MRMhCqXo5EIsdP0Ye/efVfLqwb40/wAg7d+M3biM12ulHNj6asSWogdBdobAkEMffuvcB1tM9PdM7D6P2fjtn7HwtFjKKgp4opJIIIopqh0TTqkdRf8ArYe9geXWiQM9Ck80bEKp1Sf2QL/qI4BI97FR1456yRLIFvIwLHmwHC/4e9E9bAp1l96631737r3Xvfuvdf/U3+Pfuvde9+691737r3XTDUCv9Rb37h1oiooevW9Ok8i1jfkH/Xv73XNevUFKda4X853+X5lOx8DX909b4v8AiW4qSmqjPjIhJbV49byFYx9Db2G95sPHPi9SRyZzfPsiiyXCEivWnHLj63CZGtwG5onx+fxjrTzUHhmQCbkMnI5sfcb7jbyRvRRjrKTlrd7Tc4Fkk4n16UNPjoauIQpLpqFFjGAVN7fXn2hS5cK0TLTVivRpuA0OrW5wONOnTF7AzNfMschlFK7C7knToPB9l8Nk8E/is5Nc9Xur2Oe28IxitKdLin6UxAKjUJ5Zr+RWfVYkcfX2MbDfHtFBpw6ireuS494LUOmvQT7v+O215qxnqaaBnJuS1iQfpf2K7LnE3R8GRQAOo7vPZSOEm5WQknyr0C2d+L+DNUMhQyJG0SkiJDwQBcLb2fJudvJSh49Bu99v5rJTpqegY3h13uTFyxjE07u1MRpKBuef6j2vhktXyz9Ay82a+tKiNSeouB7L3ZseaNsy88CqwGizcKvHAP8AX2oMUB7kavRZGko7bgaSOhbp+7cVu2pp8fWyg+YANrXi7mx968ND59VLgE44dDEOqdj5PFpW09fDDVzrdFQqGLEXt/xX3cRQkdzdUEpDZGOkBF1fvTF5BpKOkkrMUhLrIblSF/Tb3TRGRhunw9uy0LdPEe5twYZxFLi7NBZLaeP6+9+FEBhunP8AFOAfpym33uvKwmnixLrGV0+QKR6bc+6yRxJHqVqn060Y4KUVqnoMtz4rKvSSV8juko1sYQGtxzaw97tYUuRqY06QTTzIdAXpA7awuZ3nkEpJaARxxNpMxRwTpbg3t7S7hKm3AuprTo22zbLndTo0kV6PrtOlfbOGpscruWhCa73sbrb6+4q5k3RtypGvbTrKD2k2huXneSUVDDpd0m52iURxQBGJUF14J/2J9h6OSUDJ4dS3LPbS1BUcehNpc/hchRxpX08LTBP844XVf6Dn26JpB8z0WPZWjVIQZ6WNBszbGeogSKeN341ekm7D+o9qob+aOlc06QTbPt8yGqAE9JfI9O1FPKZMNknhLEkGFiOPoBwfZ3a8zNDxHDoDbp7c2m4Fih016Q9fiN+bS80keQrJwbHUS5H0Nj7PrTnNp2EBQZ6AO6ezcMFtJeJJQoK0r0QXema7DyvaEH3sVTVUX3aAiQvoADWPHsUwz+LQDz6hG71Wl0bRvI06va+JGWy+KwuNdqRmtSJ6bEhbG/HtcYVUDOelMsCxRiQGtej3YfuX+7+XmbM1LQRuw0RsQFUk8AA+0ytVio6TKzHiOgz7875wFbhZKiPIJEI0aTyao761Gr6D3f5dWJ8+q28j/MCTZ80uOpayaoSJ/GSiFrE8cW9+61q6fovkr2r2lR0suAwtTNSzWCTeF7ODyPx9ffuvfLpT4vAd37nq0jqcbVUsDorldMoNmPPv2OHWqZz0+5X4y9tbrlikjWvip4v8+gaVVfi1z+D798+t9FL7z/lY723vA+VphVfczLqt5JgSxa9uOffuvcOqzqr4x/If4r9xYnJ0GKrZ8UtTSQyemoc6FkBL8j37Pl1rrb5+Hm/NzV3X+3p9wxzU0tRRq7CQEEMUFhz9PfurA06uA6/3pFHhIoo0EzFEuzWJvptfn/H3X5de+fQIfIi24dl5qg1mFp6WubWLDSfA1gLf7f3v7OvAdaYHSHTkPafz43LHkKwvDs3NOwidjpmAqb2Zv+I97618utx3A9WYqTbmNp8dItC1PjaRdMTWJKRAfj6/7H3rNet4I6DfO1m4tpV4o/DJU0asVM5DHSB/a59+wevZHSag3fiTLVVeQqxC0Clwp4uQOeD731rNeipfIDuhYds5Crwjs9RHFUxJ4rgtpUhG4/2Hv3y69w61sexfmZ2b1h3LQNXy1woa2uj0JpcrZ5fob/19+61ny62DvjJ8kpN+7PpK6ZjNU18MKmN7h1Zor8D/AHv37rYzno721qepmoTU1KmFJSZOf8Tc/wCt79148c9S81iBm4jDTzEMlh6WH9CASffqdbr0HNdSQbST7jIVZax1KHYHkf4X9+p16vp1Hxm813BVx0tGAyhk9VrA2PGq3vdetY6OHtfPy4zFU8JJiAUAspvxb6g+6+fWxkdO8m8pal/tI5WqCR+f6/QD3rPWxStOkJvfas+68FXYqriCQ1sU2qQ/2dcZAPP+v79w68c/l1rw70+E/aG0O6srvLYmeyWl6t5lo6eSUxEmUsANPu/VerBdjRfJP+BU8GQ/i/7EccRu0wBEYt+f6/4e6n5deHz6HPZ6bhyVJU4rf9BLVzTlxF94C1g3pCgt/t7+9/b1v7OinfJL4K7S7FxOQqoMHRGWogb6BRpL8kkAfj349aoKda33dHxA3Z8f9z1ub23NUUVOtTNJ4oTIY9MZ1cD/AA9+p16uemPq75I7uxGZp6CrraupEbqsxZXOglvyD/vPv3XvLq0brP5Iw5HTDVSMqIq6pDGRYW5JJ9++R619nSr3v8m8bhqcrh2Ssrn/AG1ZUVpFZuALD37HXq9Mewt89t7oqBW0GClq4as6kujAWY3/ANj7317pW7o617fz06VUuLqaJZAAVXWB6uT/ALb3rPW/s6DvP9J53HWky+WqKCbQX8flPq9Nz9T731rI6Jn3BuHHdf3QbnkraoizQPIG0H6Wtf3rrfRC9wdn1OfzclKqMYrMRJosOT+oH37r3TZVbnENFJHNnp6Yt6RSDVZyTptYf196690KPUPVGQ3zKaysxamgclkq2VgWU+olife+vE9Ggxfx/pcXXxVeJiGQJ0I9NYsFa/4FvfutAjh0Iuc6PyIooqlqApIVVxEFYBR/Tn37rx6xUnUaYTHw5Cvi0GtIQow9IvwPr7316p6j5DY+3sTTOzSRJM8busfHPF/r+Pesder0RHuvbOPzc70dbP4aZiiq5vpK6r8MvHHv3W+r+v5DHzfk6O7IpOst27nmTac1bTY7Exytpp5Q4EaxqJCL/X37rR6+ghg8xR7gxGOzWPlWaiyVJDV08iMGVo5l1CxHv3Wx8+nX37r3XHWuopcagAbfmx+h97pivWq5p1y96631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3UTIf8AK3/AKhKn/rS3v3XutSX+QYf+xhH8zUf9/Drh/61b+/da8z1ty+/db66tc8m/wDh+P8AePfuvA06CruPYFF2N15unauRiWojy+Onp4g/+6S0ZAZbf7f3seY6tr0spHr189f5c9L0XRnydy+yauIDGT1uQqxM4JXWZydILC3uI+YnkTd+OOs1+QJtufkYBlBcUHl0m/4RhquaiiSgp5qYp/nCFta3HI9o4t6kjYKBWnSS52CDdCxCheoVV1xQJUTy0kMcCVAIGgBQL/09iix30ll1dA3evbq3kt30YNPXpLZvrWtgxzfZM0r6i9gWIsObD2MButqxBLeXUF7pyjebUjMoLCp6SVFk9z7YdfLhDOkVgCyXuB/ifa+Ka1lyG6CPh32vSI8dCRQ9n12SphGu346aeNR6ljsWsPz/ALH3ueS3hXVG1T0sNluIXWIyR9nUOu3vu6qoZ6SmxbqzK5R0U3HHsva/WvTXgX2ukiEDonmZy/Z2S3tSbayMNWuMys6xPI4kKxqz2/P+HtbbMLjjjpStstf1TpHVv3SvVOCwm1qSB0imqkijlY2BZmZNRHPtWYFU0PVXiiTuU19On3f/AExgt+YSsoqyhgiVkkQEgC3o03Bt/j7TtRMjy6Y0mvy6pO76+E2a23kazMbRRwYW8i+IvpI1FuLe2dZr616eSJWUscU6LJtzevZHVuXV8m9UggmC6SJCLR+ngn27w6TgV49HJ2f8nY9yT0gzEpp/CiKzuLG/5Pq9++XW6eY6N3g+/dqRUix0u4xE1v3EUr/T/X49768ellF3Xs+Shapq84rhwyet4/oeBwT7917pLP2LsyjqBU02TilWoJkbmM2vzY+/daz1Fj7P2TBWvWx5eJp2IJh1JccXsB7cWUgaemZIKnXXpvzvbGzKhDUVmThQoDpuyG1vdWlavVRGy5B6AnenyF23TU7nGZCOq+2F/Gukg2H+pHuuqQnjjpQKjNei71nfG5txypT4iil0VEhiISNgNJPHI97dqDsFD1sNQ1PShqOh919h49MnVTT0jNol1etSCefqPaRUmY/qMSOnhKp8uhU2V03vzEpTUlBubIxCj9OhHkAYf48+9TwuoDWx0H1HTNxpkUDo00m3uwqba/hn3BkItMIHlDyahpFvZntm9bvt7gidjTpCbJXwOPQDw7s3hsqSonmzdXkEWpRSZCxtqfkc+x/a8+b34YjW4YdeTa1dsnqzXo3OSbp23R5Ce/kmj1X+ljpvc+5c5Q58O+FbC6UmQY1+tPXohvYzaylRnoaJFXycMSAbXPI4/PuSA1eqGPw01Hj0x5aQkBLAD/VC3+xI9qI8Z6YAqft6jSUMdZQaOC6qdP8AQk+9xuRJ1u6Xwo6rnpAtTmKRqNk0SqSQLW4vyfZiwAGtTXpFb3LTdjClOiO/MrfkmI2/Js+FQ2SzEaQ0sQXU0jSDQOBz+fcT+5/MsVhssm2IO6UUJ+306N7Sx8aYNWnRif5Y/wDLp3f2ScHm9xYU0VNV1iyifTKiyrMQ41Ej8394h7TbtZamLatRPQqijEBp1sd99db9e/DDpJskstLi8/DQOA8bRiZjFFck3sfbzw1mMnn1d21inWuzv/55XXL1lJWPlWnDO0pKs0V786vx7eUUrXplF0dVO9pfKndueydbX0Gcq40qvLF9vHq0qSxFr3926c6CXr7G7v7JzqyZDJVRimmSRi5cBgz8/X+vv3XuHV1nxy6AwRwVRA0UMtZLG13J9RYpzc+/Hrwzx6LL8nfilVh8nUU+FjISmqJUqAD5AACQdXv2OPWhxz0Sv4ZdSSS93xUWcgErQVKxxwzAm4Sb06b/AO8D34Drderd/mvHn8bj+vMZR0BbDY2qpDUwnmMQBh9V/wAQPfutfZ0fj4+dlfF7E9Y0v96o8HRZZqWCKWF/AHMhjAe6n8/1964cOtilOvf6ONhdgbri3D1A9K1MsolnahUKpu30Yx/4/n37r1fM+XQs7m6m7B21ijlYKWerWSIF1bW4It9CB+B738ut9MPSk2OfeaU+6dr0ayo6n7qaD9wOreolnHv2OvVPVomB3NttYI4KbF00YgBEdkSzADi3uvDj178+mXOb3p6yeWgio4raStgoIA/r/sR731vgegW3NXT4mNqukkalAk8jSINJABuQSPexw6qehZ697JwHaO2KzbmSkiFZSRPDHNJp8juotck+9cOt8egwjwwx2Ryu18xAKrC1UMsUPlGqMNIbKQD/AL172eFevedOqFfnr8RWm3TU4/DLpxebmmqpZYtQVA6khbjj/Ye/daOOqA9xdSbp+OXauCzdRhWmw1LWa1rGjka6tKCG1EW9+69x49bvHwS7Qy29PjphcxR0/wDkJo6OIKgsvEQFm/4p70etigHRoMZuCCWplp8zSx0Bkdgq2AEwYaQSD/X349eB8j0ncXi6fAbiyWQqkVsRkTpXUbrGjGxI/HA9+r17h0UjtPflZ1/2fT1u3MGmU27JIvmqlTVGJWb6cXHv3z68TTo1WNzuF7Q2pA+QghxjSU+oqQqkNp4W319+4cOvYI6ADsDH1+z8W8eETz0saOfKBquh+vP+8e99e6Y+nOxKOpeehq5vtK864xAbBmJ4I9+Hz68R0KvW/WkNXvat3FAg0CoeaZgbljruxb3rh17zx1YVFmMVjsNO1FUCSopqQMaYMv6o0tYr9ebc+9DrWePRfdl/KHdGB3zLBRbaNakbS0wT7csja/SCTb3Yca9b6AjvH44b0+Q0u8N6ZHO12zFkDVEVNSu8C2VS1gBa3vVevda63WnxyzuD+Z2VotxNJurE0FQ08VZWs02kxyEWu17Xt739vWsdbMfVGTp4cHSwYxPs6ehopKc08Y0oLJpsVH496pTqwzjqofsuCmw3ygTKwVP8PkyWYQVHj1IakGYcSEW/x974dV6t2pspiTHjopo4wpx9LaQgepjHckn/AB96pXq1aHot/bOWyP8ApE25t+C9NhMm0P3MKgiKpXULl7ce99ar0bPLb0h2Nt3b9BCnioEkh9KcRsukf0+nvVPPrfRf/md0rsXuvpas3e1JTR1qU0l5OPIGKckG3vwNOq5OetUjufrbau2cqmNhrk+5j0RpFcXB+gsPdutV6AHc+wHXFwz0UeuaU2LDUWuTwLj3rr3ljoE6PYVRL2BisfnZWhopp6YyuwYKkTSASE/6wv7959b62+P5aHwk+BvYuMwsWZ7AwmS3XMlpcPUTUxYVGkXjKufwffutVBz1sidUfy8fjT1RlEz+B2bh2qCInSo+3h0yFU0o+sCxBH9PfvPrfDo6mI2vtvBRxjDYjH49IlKxmlgjjsv+DL78B1o0pU9Oc2PoKsh56WCc8WMkasePp9fe8jrQocjrgMVjQLCipgLWsIlt/tvfgacOt06zQUNHTMWp6aCFiLFo41Ukf0uPeut9SvfuvdRVpytQ02q4ZbaSPof6+91x1Wn7Os4jQHUFXVe97C/1vx78ST1sCnXP3rrfXvfuvde9+691737r3X//1d/j37r3Xvfuvde9+691737r3XvfuvdNuUoaLJ0lRQZOCCpx9XBJBUQTokiOsi6GGl7j6H+ntiRPFPhtwPVlLKdS+XVSvyV/lL9AdyV1XlNrbdoNt7kyUj1FXkkgpwgnZSQ9woNyefaC62WKZeGD0LNn52vdsYIpJA8qmnWtD86P5bPYPw3f+88dLW7qxVUzOKmhjEkCI0hUOSg4sP8AbewTumx+AG0Dh1NvL3uDFuAVZyBU06rmx3Y0dPE1IxRKqP0PCeXR7XKlRyCPp7AreP45jYUp1Ldpd2l0goeI6cIt11rs5jkEEkhvGWuOSPrz7eUtXT0ZCzQjUmemDI56rqXaKqmBmUH13vfnj29P4kMQeHj02wjDeHJQjoPa/JZNahY4ptSsTYLcg82v7Sw7luKmhr0ln2iwuxkDp9otsZPKxCUwM8jrx+1f6/Qj2Zw7zfAEEnoguOTtulb4RTplzvRdNuCnUZHFSSSlrF/Gov8A1vx7EFpv86rRz0Ct39s7eeUvCuD8umSi+K+KhljrIaZoXj0kXW305Fvaz+sMij4ug8/tWC2qmOlfP0znn+3gx1eYBT20er6Ef4e9DmN/4q9aPtamgrTjw6k5HD9i7fpDQfxlZofHYIFGr6W/Pux5kbyPRV/rI3c51xuQOgu/upuqtqXnrah3QNqYafqT9fr70OZG4McnrZ9i74Gokz0usfW0uFiFNUxLJIfTyoJBI+nuv9YXc0B6snspf2X+MM5YenSlx+Ax24SGqIgsRIOlkFyD/X2bWe/eGNZP5dWb27q2pkOPl0Lm1MH17tZgJMfCZje7FEHJF7i3sv3XezMhqePQ55d5LhtmBdadKTP0228qnlxcccBK8WAAt+PYHDmSUtx6k1rCKzhAjwfl0E2SpRSuVhFyPqwF/wDY+60PHpoL+XTBI+QFtMrqLE8XF7G/492Vj1vNehi2nuz+H0ccc0g1LbUC5PIHPveSKg9eoePQp4zsSJHGohwFt/qgLn68+6lVGePW6kYHSQ7A7JT+G1cghLGKMlTp+tvySPa3blX6tSvr0mvF8ezeI+Y6KH1Rlj2B2XU009CzrBIWU+MGxDcWv7l7bidQBHkOsP8AnvbUtNyLKKGp/wAPVrHWe+afYlbS0NbD4KYLpIkGlQAfqLezuUCnb0EpmLooX06gdz9l0G5hV023JVkyc6aKYQSEMJSLA2X/AB9l6fEersoWICnRZdr/AB27v7DmZM3maqLHSysywvK4vFIeFsx/3j2503To6HTn8vnYtMRLvnFjKzS+pncBwxBuDZh7916nViuyul+utlQQY/DbdSKmpQnjVYIyPSthbj37Hl16vQnrSYWmqozBQRwqoACmCNf0/kH3vy69U9Kin3JSYunlhjpEZZf7SwobC30PHvWOvZ8uhB2plsNXwj7+KnZRY6GhjJ+n+PvRB49WFDjoL+5+jNjdo0xqY8ZTGrjfVHMlLADqB4sbf19+r69e+VOm7ZXW0excJHj65AI4I/HSEoECj6Dgce91Pl1qnmehn2nXVWNjDpKzQ/SwJK2/Fv8AW91Pr1uhp0g++t5JjdgZnKNMkTRUtYtmJ5JgP9f6+99e+XWrf8LsvHF8wezdxiZG/iOUmc2IYH9/62/4n3anp1Xh1tR7N7BmMNGwk/aNNB6eeQBY291pjrfDB6W2c3Vg8lAEq6Hyu6/hASSRb6+/Ur1utB0EWU2Rt7OK3hodHlBV1FwRq/Fx+fe69a6Jd8gNobd2tiquA0TafFO17A2IW5PPvw68R1q//JzbQ353PgqPE42athiroS32yEhLSWOsj36nVR1sB/D/AKQr9vbNwtWaeaAItPJZ9QKgwgm9/r/h78erDhXqyejrJBRrjzKIyiBeSAfSPqffutcOg73VuXK4IGLHajJIOHGokfj8e/Hrf29FS3Sd/wC4srA9bkpEommsyFmWyM1wefx799vXq9Gm2HQ4LbeFgmleKWvZEBlLKSGvx9Pfh1qnRgcVUPW0ccvkHjcA6VNwLDj/AGPv1et9K3F/w6mHnYA1AvZr6uf9b3o8OvD5dPc2YqKimlVyAig8kcWt/W3vXnTrfSG21WbZpstWS5F6R5AW4aONibG9vV/vfuxHp1oH16EGr3dizSCOijhSIWBYRIBa/wBeB71Q9b1DoJ93ZjCvJHVNUQCaP+0uhdJ/AIHvY6qc9Fq3723JjoXp6asikUhlIFioAP8Avre/dbpnHVavf2OpuwcfWy1bxm8c/HiQfqW55P8AX37NM9ax59V+dF/GPA747Cmp4cWft46vRUyrGArESWv79TrWT1cJSfB3Zv8ACWocTilgrEpl1ShUDFinN/p+ffuHXqevVefcvT+C6S3C0mfxU1XTxzmRpDdkXR6wCf8AYe/deoOlbsz5u9XbYxMVBh8bDSTUcelmKAEELYm9vfqdbB9OuG6/n/Rz4meSiyVKpWJyigLqBAuB/Xj3vr3zGOqw+2vnNu/ctfUrSVXmCSNEjRl7aWGkfT3rr3RVP4LvLsivmzGWkmmatJaFNbH6tqsoPvXHr3Qtbf6WfE06VORx8zzVAEYDJqYFvp9fewOvcejB9d/E3ASo+6d2QrJSqTMlFOFGpf1KAre9UHXs8OhDhaoTP0O29k4uXGYuKdYZWWLTG8a2Ui6i3PvfWqdWabA2HgMNt6jyVZQJUVjRwlwIlLazYuT791senQjbqbYG38KcrnfsoFji8gjm8aGxTVyre9Z69THVcvbvZeE3oBj9mpEEo3ZvJT6dHpfk+n+vvZ69inSH2xsKs39TiCWtWOs4W17WvwRY+/fb1rPDpz3J8WsTDjJ59wVEEjRA6DIFBva4sW9+p1vohG7sZW9Xdl9a5Ha1RJTJSbupJWkgmeJZAko9JMX19+6919Mj+WV2fluyvjVs6ry7SSVFHh6CMSSF21DxW/U//Ee/de6sW9+691wMaFxJpGsf2vzb+nv3y61Tz65+/db697917r3v3Xuve/de697917r3v3Xuve/de697917qJX/8AK3/AKhKn/rS3v3XutSb+Qb/ANvCf5mv5/4yHX/+7X37rQ4nrbk9+6311b37rVAOuyLix5B4I9+631qkfz1vijpVu4Nu45zXWlkeaGNRou2ptRUX+nsCb5tZkuDMRinUt8j83yQyR7S5wSK9a+OyqyefGUkLPqngpo1nty0Uun1KT/X2Bp7bw5KDHWWe2wxS2STp5jowGyqVMrURwVciuImFwQPp9Lc+9mYxRkJx6QyQiS40vwPSk7Dho8PQr9kixkL9CoJP/I/ZfFuF/qzXos3nlqzu49NBQ/Z0BKMctdJlBuB9VW973uPZnDu1+ma46DVr7e7cZO5B+wdKODbePoY4pVjVmkIuVAPJ+vH+v7VjfLle6U9CWPkHaI4wHUU/LpV0+JpY40ljhBvwQEX1X/rx7tHvrO4FeizdfbnbZLdnhjyB6Dp3wXWOI3JuXE1dVSRwiGQFpXSMDhvqSR7G217srAZ6xz5w5JvrZyLcGnoOjR5N8RsspDTwfcQiJB5Yh6bhbWJHHsULdBxUmp6iDwby1uDazg1HQhbCy+3N1070Uvipp5FNvLp+pPF7+7a1JznoyiqfiPQW9q7LoaF54i8FVBIbFUSNwwYfT3Xw21VXh1cvpwOiG7v+LmC3rWyzjGDSxZriGO3qNz9B7e6bz69F/wB4/CCVmMeEppYCLgeJQrHj/afeutcT0XnNfEDfu1/uapJ6+QKC2jyPe3+pNj79Tr1ei9bl2D2hDqoIocikMTsNWqUAlDxyPfut4A6iUu3uxqWmEUstW7qulbvJdbDgc+/fb1rp0wmzd91k7n/LDMfqdcjEf639Pfvn1s+nSlh6L7O3NVLTlq4RORwrychja319+618+hs2r8MtwRxrLk4arkevy3u/N+L/APFffuvcejXdTfFWloMnEaqgkKJIpUGNWBa30HB974Dr3n0e6l6lp8VjY6ZKVIofHp9USj8fX36vWyKY66xWwMfjzUVDyQhlUlRpj1cf2fam1iEtQeqldXSjr9qUuX25VKDEoigdraVAYAf19k+4ubZ9K9Wp4Y1nqmT5LdgUGwqusw64552evSLVGL3u9r+n2hhubtv7L160koDZNOrMfh7XPmeu8ZXtC8CSUYKq97j9sNzf3kv7XbWyr9bIO4V/n0GL5xJc09D0Z/1Fio/JN/8AEX9zio6pcPjHHrFVUgn9BH1N7n6cc+3FanSQ1Hd1hnrYaaOCkp4CXDWeQDj/AF296CNqLsentazRBadI/cGSwWHmmra+rgiYU8japGAsQuq3+v7UK0rpTgBx6chtY488SeivdOfHbK/MH5M7XydOsuR2tt+uRap4FEtMBHMOZByvFvz7xj91dziudyaBTWh6NrJCpHW7VsLYvX/x56liiokocW+2cbTVrytDBGH8EAL+pbfW3uI8AU6Nq5z1p3/zcP5jX+mDeGS2diqwVWNop6/HSx000gT9u6fpX3vrX8uqCMLjtz76M9JgJJ4I5BokjWSRmYfkAH37PW+jH7H+MmRgoY6jOY6plP6yJE18sNRNz79jr3Rl+vOpxhchTtS0EkMUbR3LRAAKrX/H9Pe89az1a11bk9sYLb05hnpocjDFdyWQMCE5uD/xPvXW/PpG9s9kbfrtl1yVNRSPViCdTIwT1LY8An8+/eeevY6r3+LuIgznf0GRxFIwgWpJklRBo1eS5Nx+B79xPXhx6Vn8zbtrKdfVWLpGrk+2NTTRvGTYrGWsS1/6e/deOTTorvRHXO9vkNuHEU2BiyUmNmq6Lyz0084h0zEajYG1vfuvfZ1tz/Gf414v48dSxQ1Vq3MV9CHDEh5YX0arsTc3B96r1uhrToWtl7oq8rXzYrO0bVFAC0caNHqXT9OCR/S3vzDrYzx6Wdf0btnN1IyGMpFoKhx5NRjSMm/NiRzfn37Pl1rj0DPZcFZ1TT/u08sgIGmX1aVB/tEj3sZ49aI6S2xd8YbPxyVMzxJVKDqZm+tvxc+/EV63WnXPeU9NnMPUxUkik+tNKn68aRYe/U61Wp6CrYuIqtlZSLIRysqyv5JV1EjSTyCPr799vW/PHR4cvSYvc+yIM3SU4++iRWZlNmOkA86f6+60Iz16tcHqvD5G0j7g2JlKmHHuc1j5vFATGDJ41HqI/P0Hu3XvLorG5vjDsj5PdHS46gpaYbwwWNf74xwxfdCZEJN+L3uB79XrVK9Cj/Lq3DN05jX6M3ezUcWOqCIGrDpV1iJVdINv8PfuvCpFOrAuzMRJuWkqarA1Swzo5alqY7KoW/Fiv496HVj8+m3FUWafab0GYZ6ieCmK+XmzEKfo3149+qBk9azTpAbRo9uB6rH7roRUUheTxiVRJpcmytqa/wBPe/l17Hn0A/ZFVuHbHYuEo9rVbT7crJULUtK5KU8Rf1Kwj9+68Pn0NW4c1j6vDpjqiSKN5444ir8stwLnn6e/fPr2cV6LHDjaTCb5hoqCmkmlnZXFXFcoFZvqWHA9+69jo+fXklZhcFkKokhtDlvpqYEX4v70eI62OBPSLp81u3dOe+zw8smPQTmOpmYsEliLWIv/AK3veOPWhno2W3MbszamKjqcjFDU5wqrPPZNTSjljf8Ar/j7rx63muOmHvbvGh2h1VuKSCFJqqbHn7WFEtI7mM2AC8k+99a6oc+E3Z67w+TG5arfW1auko5aiqip6ispyqOGdgGDOOR/T3vrWa46u6x2Lw+OqZpsVSeCinaZ7cBdLnjgfX3Xq9Oq8/kr1vhpewtp7joaBjKmShlqplAICiXkk2926p9nRg8zu3A0OMpvEiVNXBjolSCJryh44h9VHv3l1vpH9W7cyncWdlzmQZqb+DzMtHHKOQkbcD1+9dbp0N/Z+AeXG02KqtUn2ekh1P0025/1vex17h027rmx2I6EzJzOSp4aGKiqz4p5FALLGdN7+/efWsEY60w+9M1LvTvzI0WEqFekp8kgiMLEqyiS3HNvfvPqvR3+tuq2yFAzZbHvNHTwJOoaO6taO9hq97630nd3/HqLc9RNurGULUcePWWL7Xxqpfxi2pR/j711roDNpQ9n9I7grN4ddHJYHJYx2qRJJU1KxTSa9ZKoDb8ce99er1tI/wAr/wDnnbX39Fjuke75lg3djpqSgqM1kpJY43KxiM6JJuDz9Pesjh1vjx62iNrby2zvPGUmW2xmaHKY6opxNGaSSOXUjC6/pP4+nvYz9vWmxx4dKg3bT42A0sNQI+o/I974ceq/Fw8usvuvV+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r/9bf49+691737r3Xvfuvde9+691737r3TPkopXZCG9AsbL9QQb8j/ifd0FT08JYUiKtxPXU1P54wyMykKPULqePr/sffpLlIwVJz0XtbtIarjoM+2uoNo90bIyWzd6YujyENXSSU9NNVRLK8TOnpdWb2kkiS8QqRj16MLK7nsHDqaEdaZP8AML/lJdj9I5DN9ldcY77jbX3VbXulNSSSkRAGQLpQG3H09hi55XhYFwB1LnLnPsplS3kJrgceqP6fdsv3kmKzFPV0GWxhEc8dTC1OPITaw8luPYQvtvFmSFXh8usiNk3Zbu3B1ju+fT3NOtSnmE0bSsBfTKrfUcCwPsPRXitOYXBoPl0ZPGzvpDrX7R1zxdBUzTLKwDqCttXIt+Df+ns1Z4FTh/LptoJYxUMPyPRgNr5+PGxxxSxRE2sSVHFrDn2nM9quQM9a1SotehCk3lQyRiERQa7CxCr9f6e07frd0Zx07FLOy8MdMku4KaRiJQqIbcBrfni3vYhfzPWnac/COo82SoU/cpZPV9W9R+p5v78IZDxPSaT6hc0z0kcjWY6acS1L/nkFrjg2+h91aFjhj0qhvpolpTPXOpmwxhjWJUIK3Yi1+RyfezAacenDfyMcjpIZDBbacGoZx9wfUo4AJP0N/wDX968MoK160dxcYkHaOklV1tfi7faW8YawIYABb2HPuxumRdQPTXjWk7HQP5dP1Kz5OGOSWoAl0huJACD/AE/1v8PbXjyTZkx1osi5QcOnJ5cjSQKIplKiwN5P7P1v7tGjRHWfPpsTtIdL8OpMFUJWtOyk2BPIN/d9QJoercc9cKsKVPiQm4Okhbjn62/x91ZgDQV6tor5gfn1Ap4JVIZmZeTcG4/wHuhemaED7OreF5h1/aOlJj6uOBQXlBHP0YEixudVvboYU4HqpRWwWFft6j7jyuNGKqpKpl8QiJYswJsPrx7Vbc6/WIQDUHpFuZW12+SYsO0evSg+Je3sLX78mzcMSfZSyohcqFF78sCePcvWJZpBXFQOsMObt3/eG5leNCRX8+ji/JLE43B4lKzDSoKgpcCG2sFhb8f4+zuRTTolnieOISDhTpBfGDqiqyeWptzbxaWSlmnMlOsoLC31W9/aJQNRp1VZfFjHl1bO2NwVJRUj4imgRYo4hqjQAkoPqbe7U63X06E/aCPmHp1hRQiDS3FrEcH3vqp6MhR4nEUtGFkEfmAH4ubkfS/196qSerACnQUbwphGTJTWCaiBp4uCfpf3vrXDpNtXtRpTR1IVkcC5AB4I/wAf9597B60eon8RraSpappWZac3JF9K6SPqfeut0PS5wPYCtB4UfXKnBDNwSDz78BU9erQV647q3jPuClFPEAk9KoUaGsCQLfj34daPp0FsPbeQ2oxo8sGFPf8AVfSNI+hB/wB79+4463U8eiw/Lf5A7fHV2ZWOtEZamqLgyqjXaE24P+PvXXj1rbfCff8AMvyL3PkPugtLXZKYrrmVRbzcWJPvfy6r1tkbC3XHU4fHTwVEDD7emB0zIb+i7A8+9cM9W49DlicoMlUwM7RuqgA2II+vN/e+tDjnpe5XN7b2/RNWT1EavpDadaqAbc2B/p70BXHVq9U0/Pzv1MHtbJZDGanQRVcSN/Z5UgHV7t59VPDqsr4CYNu4M3nt57jpoJzTVUklOZ08hVFluCpPvXz611sQ9ebiosVRQYt4Y4qVViSPSoUWVdI4A/PvXDq9OhRqqLHVOmrpmYXAcgXBueSfe+q5HSY3NQU1ZDGtFF5ahQCS0ZNj/gf9h7959eHXLC9M1266dDPFHG1/wulgCLixP+t71jrfHpWTdHU+IojqldpUvYEnkqOP969+rnr1Om/FTVmM10D3VIrquokcfn3vrWOPUz+8CU0wDzgBTcjUOf6+9+fXvKvWfI9m4WOkfHyVKJK0ZF1kFwdPB96oPLreeB6rk7a7azW0MzX5CGskSgQFzabTqXVfge/A560fTqFtn527Pn25JjqrKD+IpeMg1KBjIBpsAffqHr2B0Ee4PkfmM+1T/CK95YJXLo33INlPAAAPvdOvdJxN7170grM9PL4yCW1OSAPr9feh17oqfd3elJR0px2EmkmeSTxMI2u3qOmxA59+690dv4B5fr2l21kcjuKKSPcNaryUpeMhzIfUBcj/AG3v3Xhjq1HbG+do0kM8uRmCl4wELNZr6bL9f8Pfut9Vf/OjJ7R3Dt3KmnMLVTGfwsSpflOCD79nrXl1robkxtJgKXNzzVbrLMjmEI9mvc2sPz79177ei+bWwe9N3ZCaCjnrvtfI62mDhSpJAsx96690K1F1HVYeeOTIxtIjMvlAh8hZtV7AW97690djr7riRaCiya00a01GqyKpj0llAvYqf6e/da+zoQ5K5arJSNV00UOOpoyYi6aQZI+R9f8AEe99e6U+0a7M71+48qNDiqGQwqIWNnjDaQdK/X3rHHrw6N50/wBTw7gro2oqWBI6Vw009VGE9FuTqYe99bA6F/sfem0+rwKCWTy1Kx+LxwWkj8ii3AW/59663XHRDe5Y91duxqmPfIwYycKxMAlVdF7G9vfutHoROrPi3iNpbOOayMplklhaRzUn1rqW7Eavz79w61xx0k8PtyWDO1S7O1SVVO0pCAsARGSwIHv3Hh1sjy6DLcu+87uWuym38801JU46RoSgcxhmXjj+v+Pv3Wuip9iY3HVm8eqcPMWeabeVBGzldWoPMBYn37r3X0mv5dGzKbZvxp2NSU6gCow9BKWCaLgw3At791vo+JZRYEj1GwH9T798+tfLrsC3Hv3W/l137917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuomQ/4AVv/UJU/wDWlvfuvdakv8gz/t4R/M0/8SJX8Dkf8XX37rQ4k9bcvv3W+ve/de66/P8Ah/xPv3Wuio/MPpbGd2dObpwFTSRVNXBia2el1xCVtSxE2Vf6gj2nuYFmjKt0Y7XM1repcDFCOvnwbuxGY6i7W7C2bmKWppQuemp8cJYWhjEYkIsmv/iPcY7vbCORhTPWW3KHM7XVnFCWGB69CRgNzU+MWnnEpEykNJZxZrc/X2G0c6qN1IDapF1jien7dG7k3FEoQluNIUEEn8D/AG39PZiv0wWhoD0lQXJx1CxWLkSATSAcrqFvwP6+0zPAKjFOttPLEag/z6U0f2qxossjWB9IJ/2H596ECX6+HF+Hpo30850qeHShgk0Rq6gmG4BuDe1/wfZXNC9o1fToxh3WRU0SCv29C5Twfe7ekmw7KtdFCStjZwwX8W9rLPdTENVeHRPex7detWZQa/Loqu7e8907Sm/hm6aaU0ayEeWzMdAawNz7kra74XFkslePr1j9zlyahv3vYFolPIdOO1O/duV1bB/CKmtSoZV1BAyKCR9L/T2IrYk/F1B26MthOYiCKdD2m9JM9GjVFSWUAXE0g5/I+vs2VCFJx1u2K3Eetehr2bW4CTHEM8C1Gi3qdTc2P0B9p+PWxnrHBJGMk58MMkRY2ayv/ttPv3HrVRx6Te7cbjauOpdaSEgLeUGMfT8gD3sZ63SvDope6sPtJjMj0VLcs4b9pA2q/J97pXA61jpJba602ZuVahKaCD7kNwrRDkg3549+I61jj0IOxOodpx5yooqqjhM0V/rCAt/8Lj34468cdGGxHXG3cVkY/FQ0hUKrf5gc2+hufeurUHTjuw4CgVEEECOn0WMKLgngG3v2fPrWPLp02zVUH2kVRSQx60Opjp5sv+P/ABHvxB8+vde3pvB0xMrwrZoww4PPI+nvQPn1uleixVe7sxJN5C8qRn+lwv15vf8Aw9qIJjASR59PIg4EjoQNm7ortwVUOChWZkqZEgkb1abObHU39PZZc0urgKqlq+gr0xOvkcAdL/sj4K9c7ijpM5uyOneoqUSrIMYf9z9Y4PuWuTeSEv4hLOukccjomlZy9FNenXaGzML1/jI8Jt1FWigTxqANICgabW/1veQ2z7XBtdmLaEY6LH1eLqfj0p4oLsGHPPBH0v8Ang+zgGg6TTNVqHj1mqiES7LwB9eL3Hu0YLNTpNMQi1HQXb435g9i4bIZLLVEEYMBeDVKqtqtx9ffr11tLVruXCxiv29btZwxCLxPVFvc/wAjeweyd+nZ+x3rJqSaukiZ6Z2k0xO2kf5v3j7vPuvS7e2hY0FRg9C20su3W3W0F/I+xGA6l2dWUnYMlNDuTPrNLDPk2EEvldtQAeU3FvcNbvu0m6X5mbzPn0qEQjbHRg/5snzr2v1x15WbL2bnxLuOsojR1MVDXISTLdB+g8ge2KHz6fOeHWlxNiM5ubdWSy+blmqXy+Rlrg1ReVh9w2oi5/17e/dV+XRzfj/1rT4DcmOqqqGJqaeTXImn66mvyp97HXq5z1dvjekl3VtcZHCY6BoIqZXb9i3Cpc6SPeh1anp0UHeWQp9mZCpxbUjfcRSvERHAbgrwxP8AxHv3WjjoFa/N5eOmyVRgY8v5qhSzp4ZhGLi1k/2P097r1o5z0T/eOP8AkHvAvjMPQZNqSR5UVvt5mJ18LcAf63vXXurIPgN0P2X15iJ949g0cVP9ukkjTVMTQsAATYmQA3t78c9bHGvRavlF1o3zH7pXZG1ZDWvTV6RVSp/lCLolseF9++zr1etib4MfDLa3xh67x7blx9J/F1oKdkZ4AsvkWIWI1c/X6e9V62Pl0ahNwV89TXwTszUjTMKNCbosRbiw9+PWxxp0JO16PHLCah44hNGNRYLYi3P496+fWjwp0q1z8LL5SxSFH0XB0j0/m4Pvfz61Tph7Ex+E39gXp8hHG0ccWnWVDORb8H639+4dbHRWMR0xg6N6hMdJJENLlNRKA3N7c/7x79UHr1D5ddJ1vm6ESzx+qBS4AJuGAva/vfl14/LpKS7f3ApqpamNRTx3IKr+B+P+Ne/fLrXDoXesN2mKL+BTveAssTLIQFt9Dw3vVM16sOlfvXrTFy46pzzx082I8M8lTZVe7tGeNI/3v375da48ePVE8Pd9R8Uu+8pNlZJl2hvbINHAkrtHTJHNLpAAbgfX3viOvcDXo4nZG29sdh4lO0etKmOPJmjWsb+HONZldPJpJjP9ffvs60c8emT48fIrIv8AxHbXY/3MUtFKaaB5AVLBG0hiT/X37r3RhN9dq53Fx0ZwNK02Hqyup1jZm8bD9Nx9PeutnqfJENx7TOQxumPISRkyp+mQFhdiPz731qnRbm3LTbJ3bQ0GVJqGq3Bmkn9bRMx+iFvp/h791vpj7GyGUrsyKzDSOtC7LzrKrp+o0kce/V60OhL6ogpqyvhWsCz1GmMl30vICP8Aam+nv1PPreKdG0q6uLEYmckpHAqEMhsGtp+tv9b37zr1ryp0i8VvOCSIHBQj7suyE6dLFibcn3759b4dBJ218ltpdOxrP2DkXpapmVYohULa7/pBDH3rz61xz047S3Ge/wA7ayiK0u0ap1kuwLrJCDcax9Dx79wyOt8cdGF3x8fOrti0lHvLA0tJR1DiKSSWlgWOQsAC4JUD36vWuhL2jU4PP7Um+wkLTwRBQ72BuV+tz/xHv3W/MdF93zg6OSpSHKqjlnHgYjUVN+Le99ePGnQJ5HZNZiMy2XgaSanKkeOYEra3HDfj37rwz0tuu9w12FqpaamSGCpqXd1ETCNSSbkt79x49e4dLKDdGW3jmsngZY2NXSKVV7ageL8MPr78MY69x6r7+cO3e9h1dlcJhBUxUM71H7kWtR4WHI9P14+vv32daPVF/X/Sbbf33gqvdCt/E6p0apaZG1NIXv6i31v/AI+/de86Dq2PBpQqsWLxC0euCOL7vUyreIJzcf8AEe/der5dOWQ3R1vimNBV1FIjkESosiaTN9G4v/X3oVrnrfQW7w2DX9l4mum2TTY00FIpapkbxBmjRbk3H+Hu3WuJ6p07x2HuTr7NHOYaubEZfHTzVcU2KkFPPJNA3pRnjsSL/wBfeutCg62Gv5CH8wftjdvYuB6i3/X19TTGKGljeqrGqNSebxh7En8fX3r59ezw63dhHomaYMxEoUFfwLf2vdhnrRNDXqT711br3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//X3+Pfuvde9+691737r3Xvfuvde9+6910QD9QD/ri/v3XuutK2tYW/p7oyK/xDrYJHDri0asoX6WIKkfUEfke7oAmF6qw1ZPHpN7t2hg97YCt25uKjiyONroJYZoZ1DKfIhTUR/rH3smtR5Hq8bvEwdTkda9Hy1/kHded15fLbh2ZlF25VV03mENJIaZyQSbXUD2imsLef+0WvR9BzTvFmoEEhA6qA7I/kL9t9U0dZW7ZrcvuSaCOWSKnNRNMGKkhQo5+tuPaFuXtsyyoAerze4HMVsBJ4zN0QfcXxV+UHWVPWnP8AWddT4+klkj++kinv44hqZv0/QDn2VXmzQEFVQY6kPlH3KuLqgvHJOOPRd233PhKmsoc1Tfa1cJZZEeCUGM/T8j2DNw2YrhBSvU6WfOm2NCPEIr+XTniOxsBO6hcirVLEKI/FL9T9Ba3ssh2K9JoladebnXaENNYHQj4zA723aVnwWHmrKc8q6Qy2Yf7b2+eX9xpRa9U/rxs1KiUdcK3bXZeLqRSz7bnVy4Uaopv9h+Pz72OXNzYUJIp023O+yuamQdOlR1D23X0oyEG1qmSIgN/mpRbm4/HtRHy/fgZBPW/67bEOMg6aIeq+8pJBFSbGqqhgDZfHNyL8/j3v+rl+fUdaHPGxH8Yr0oqP4xfKjc00cGL6urpXc2QCKfm/54HvS8tbhwyR1RueuXytGcdLyT+Xt80a6kWOXqbKIxAYHxVPIP0IOn3p+VdwJ4dJ1545YUVWYDrNi/5cfzOhVvL1vl144tFVcG30Pp92HKm4YovDp4c+8tecw6my/wAvL5oQ6ynWuWl0i4Roqr/b8r7QXuw7nGQoFc9JLnn7YY6PHICOk1mfhF8vtnYWo3PuDrLJUtBRLJJKzxzhNMY1OeVt9Px7M7LYrkEGRa9GH+uDsD25VXGqnRU5u2JMNJlcdl8f9tk8KWSopZEsyuPwdX+PsXWmxwAd616hjm33DdXZLJyPSnQaVXyCq65agU2OYNpcKqoRp4tf2uXYrU5ZQB1FkfPnMclyNMzaa/PpQdbdmRZiOWnzE4pZ3ksgk9JALfXn20+xQLUBepU2XnC+ltw0shqB59KbsOodY6Sjx033SZG0bEG4VW4J4v7rb7PbpMpCjB6IOZOdb6RWthIaNjo0fUk0fXeyKevKiKdQJiwsD+m/59ioIsdNI6i4xiVy8nEmvXou1qjs/dFNjJagzUkMqLIh5Q+vTz7trY/LpxpXbtbh1Zh19RpHgqOjo4dIpFsQtmtdfxb/AHn3WlDWvVaUoRw6Gqm3K2KxqCeNQhdV5PJsbXB/x9+6tX06MF1xuehrYolgAhcr6ihUFb254/29/e6da/ydDDNl4YmYCo8j/Q83P04P+w9+p5dbJxXpEbgzn3NqZRZiL/jj/E/429+69k8ekTlMpBjsdNV1UgaSJLxI31JP0FvfvPr3n0k5OyqesxDRpCIpNOkkD62OkWt9ffutdIrD7wmpqpzosrsdTAghbngkj3rh17oUcRmoft6msnlVGa7qCwuwA/x97+zrfSB7FfDVe35ctNUxxtEjvdmXgKb2596J695da1H8w75LxYKrqdq46v1w1ErwmNOQSyFfqp9761nqsDoan7VTdNNuHaeOrJlqqhpFMazapSz6vwP8feut+Wetj74+9hfIOTC4+kr9r17MI6Yaikt9Ki17Ee946qK0x1YhtHtXtfA/b0lfs+oEU6EtUNFISpJ5NyOP8ffut/b0u65N/wDYMsZNLUQUvp1rZ1Cqeef68e/db+zqtf8AmToNodN1tHVYxZagecSTsAWsV5b/AFve+q9AB/Kyq8RJsbMzUdcrSSo7SQq4spZrlbX96631ctgdw0VNJFHMyuwZdNyCAw/oPp715db6HbG7sozDHdkEZCg3IFr2BFz/AL37314joS6HLYT7T7jTFcKS1yv1H9T79TrVenyj7GpKSFUpJlhN9PpYf6wt79T163Xzp1nq95PPYmoaUsOVZgQbj+nv1B17oLq/KS1dbJqh8KM3DmwB/wAR7914DoBOx83JhappVn/Z0/UNbT+Cbe9169Xz6KF2D3NSbdm+/WuWaSJWYwXuWYLc8e9Yr16vVXPdny0qN3ZSp2/TwMHmvEsYQ3YBrW49+x1U1Jz0nOpPi/2N2zl4KujiqqShqpopPJH5E9MzXLD/AHv37reOrgOq/wCWFur7ajklzdaUkjDykzOQh+pBuffq069TrP3T8WslsLEyYilnkyNR4TGLs7sGHA9768cY6JbsX4WZXcO5krt1RSQUhq45QJSyrzJexDfj3rrXVsOA+PW3tg4HHT4+ip41poAGmUKSVC/UW/PvXW+g/wB/VkFDj3qqKa5iXmNWBA0n6e9/LrxPn1S38wu6UokqaZaoLUKZIxTi12JU/T+vv329er5jqrTGUmc7U3Fjko3kmUSAS0i6vVZ/0n8e/cevUp1aBs746VNNtWnqJ8YcOywxyNVKQjN6bsSbf7f37rXy6dMPsjr7D1ElZmNxQVk9M2k0U8gILobi6n6+/cOvDrLW7qxlK9Q+MYJQQEskCL6JVHJAtxb37r3lnpp29t7c3e2ch23t7CyUcHnRWqYVZAyMeSWH19+6959WP7R+JS9Y7aimzlS1I7whpklkuJGA1EkH377OrAdInf3ZVTi6CDbHW0JOTAMNTUUinWbelmZo/wDivv3DrVeI6y9fdW1uahizHYmqqqZ5EYR1huF8h5/X78TTHWgKjo4mT6x2FtHYk+TYUsTGAyRRgpdRouoUf4e/U630RrdG5Nxbmpv4RgfMmLpzIJZ0uAYkaxPH+Hv3WuHSY2JR1uB3EcjSU5raYxtHPKObSEesN79Trf2dAx3lsXGVWUm3JTVi4yoeTzTxIwVna9zf+vvfWjUcOieU8se/O3eotuY+nWeqx++Mc886j1ssc6j1Ee9deOevp1/FjCDBdEdbUpBV/wC6+M1LzxaAfUH8+91x14Vr0YQqCQSAbG44HvVevUr13791vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6iZD/gBW/9QlT/ANaW9+691qS/yDAB/MJ/ma/1HYlf/hf/AHKfX37rXmetuX37rfXvfuvde9+69021bowlpZYxJFURtE44HpkXSwb/AA9sCYeL4fVWZkoR1ra/zZ/5ZK73grO1+tcW338RkrataW6v5lQyMWWMfk+0e47bBeREkUPQu5b3y6sLhVD061SMvPmdm5SXam6UkoMnj3dJFkjkDMUcoLkj3EW47dcQ3gUCgr1k/wAtcyJcxKsjVr0u9o5tKmsVXII0j9Qte30YA+0u4wSIo0enQymk1qBEcnoZ8fmIiHike4Y6V5/r+AfZLEs5fS/Dovaxu5vhr0sqXEx1aQOXBUuGH5PP0v8A6/s/gLW66o8Hq8FlLZMXk8+loMGY6ZVufGFvYfQf8g+9SSCY0cdOumrPUrbeQnxtetOZP2GYBkB9J+otf2lNtGDgdNiFVNT1L7B2BtreECNV0cDyMBcuF+p+pPtZHdzQUjQ0XpDebGb5SD8J6btpfHzZ9JSvUU9NTRTaSUZdIOoC/HsS7fusqsCzGvUZb17Ww3rkqta9BlvDrzcNHlV/g88/gjf/ADUbMVdQeR6fYri3tYxqLV6CMntPeopEKlQOktlcj2ZtlGqKfHVUkMaHgByGsOPaqPmG2mOlcHoMXfIN/bVJU465bS+SuewJkOfw8iCI6S0kbWvf2ZQzpcYjPQK3Db7iwqrL0N21PkbtXf0k1BUtDj25BYqEL2FiL8e1f07AVr0H7e7llcxgcOkxvDa+EyVS9bR5NRAVZ+GFiG55t7bNo7CoPSvxmTJHQDTdgYLrfctGkNfG41BZU+tyTzf3R9tunIKVI61+8I0StOhmw/yD2jVZEVkP261BZfIAF1Gw5v8A19vpC8K6HyejFLmCS3EmM9COPkPtKpq4qearggBUL5BpFr8H6+9MaCvRZI/iNpU06ecluXpvKUv32Y7AoaGaxZYXmQXa1/oT7Y11bT0riiIUyE1p0CcnyJ2Tgc3Dgdv5enzCTz+BXiIe92sLke1TxBE1Mcdajma5bwlGejHPQ0GdwcWSnnESVMSy+MkAHWt/aCTc4IDqfy6MF5Z3S9H6QOfTpGrsCHcAKU7iOKPjWpAuq8An23a8y7aJtMtD0th5F3yzkDzVo3rXocNrbAwW0cJDko5I2yUTI4bjXq+vHuXfaiXaN35huE0K5QAgGhA6LuaNourC0DvUYPSyyG6MzuOGOGonkMMKeNLkgBQLW95KmxgtV/SUCvkOo52y9PjFXNaHpItBOk4VVuoI1Eni3+w9uJ8GejOeWORu3j11JUaakRoL2PKji39eR7uCAlekwgaVqnoNuyO1dv7EjEuTq4oisbEo5XkhL2sfaW63Gx2+Iy3cgU+Q8+mvCcNo016or+WPybyfYWZl2ztieSSCYmKMRK2lbPY2KX9wDzZztuF5PJBE58E4A8uhNa7dbRRLIV7v59G0/l+fEzIzTRdk7gxbVqMsNU7y3sun1kkOPcFXlsr3DSn4mPRkCQtRw6MH80fktV9X1mLx+wMk23K7HOsP+S8NqK6SB4yPb8Fm0dJG6pxNa9VcZbfm/O1K2fc298xU51hqljNT5LkA6hfWfaw5PW+njrnccO4d1RYianWMpojBYWtY6RY+9efXq9Wn9KdB5fcFfS1tNUTTKjKY6cM1iuoHj37z63Tz6vu6jw9RtHY8OFyWLaLywJA0jLbho9N72PvXW606J93l09hNvZSbeTY+LIRNO87wOFkBP6itrf7b37rVOkdi989cjBH73Y2PpCIyTMYEUvYf63vfDrfl0gMZ8j+pdjZaSVNpY6ubU2iAxxMA97Lxb37rXRdPlr8+sodl1+C2ntd8HS1aOEmpYgiIsqFdXo/p79w68T0h/wCSFg5uwe39z7tz9YayqGTqZtTm7JpfVa7e/eXXutpTs3IzTZ6kx8Gp6SOkij/BRdKgE3+nuufPq1R0hcxLBiqWOo0AsFF78iwH0/w9+4462cdSNub0FVAYoIwWb0mw/px9B78ajj1WtfLpc07zVlK0Zi8K8sbg8/m49+63TrhQ18Ll45n0RUvpdSfTJp/Fj79TFOveeOkp2G9VTY2CvwURVSys/iuNXNyDb34deJ6S+B3FkMjSs1XrijRSjIT9WA/of8fe6deBqc9JrI7ivJUUxj/bY/U8/U2tf6e/Up1ok56kY7bcFdSR1VFUrBUyNcFTZgT6vx73X169THQ5bN2/k3xVRt3KVU1TTVqG2tiVAbi1z71xz1446qc/mZfCis3ns+ozuHpGFRh4VqKSWIn9uRQZARp/1vx78OtEDqsz4cfKPMdQoeu97ySVssVcce8VQCQIY2MTE+T/AA9769WuDw6uU2ZsrrbseGPdmC+z+6qV+6lhiMZYOfWVKj37y630sk3aMV/FcJPtZa+HGxExO8asPSDaxI/Hv3WgR0X/AG/3Yw3vNQV8AwuMLPH4GW0f1t6b8cfn34evW+OOuXYu0qDfOQizm3ZhXT07AlI/qCPz6fx7959e6Tm48Huun27FGmLmEscZYehl5C2+tvevl17oUfjTtXOVdW+TzVPLDFGFALhgvBJudX+929+4de4joVuys1SyZqDHR1wSnjkCTRhrBlv6gQPwfp738uvDj0xYjde28dXz46jaEVOiyKv18v4sf6+/D161x49FL+QXw/3H8gJ1zuVeX7GGqjnjjd20tGjgr/vHv1R149H5+PuxafrXYmE274kjTFU/iuLcAR6b8+9HPHrYr1k7V7WqKo0m06WnNVT+cIxAuqJqAJJ9+A69ivQlYM02F2vGtA4jnqYEd0jA9JK/pNvfuJr1scKdJOamestV5A+QwnWoktzY3/3j3senVTU5PQbb13XHkKR8TRw2qFDINFr2A4sffutj9nQE0MW4oNz4+ggjmb7tlHnAY+G45sffutV6Of1dhRh820tZSeSokkUyTso1PdbEkn3o9bGekz8xc/lcPsKqmp9smtxqxShqnQCicXJvb34enXq461s+5N5UzZamztNT/bT0Do5iABZCBexA+vu3VfOvRYYvkPvh9yVceDjqZJ64GmhjhSX1v+lVH+PPv3W89DFsrpbs7fcn8a3xW1m3hMwqYllaRNasdQPNvr79TrXDoY6td5dXYupxe381VVUNUpjJR3cyBxZrge/fPr3RBuy+vt97jrsnms3VVaUhgqHCSs+nm5JH+v7117PR9P5EuNNH8vtvKV86wTwIX5JJWs/Pv1Ot9fR0HKjj8A2/3n3vrVOuXvXXhkV69791vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/Q3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691wJC82JJ+tuf8AY+7AE9VJCnrxcfQFQ39G4/w+nvVOvaq8OsFVAs8TL44nJH+7I1kBH+sffq0z1WRS66R0GG5+qthb9xtRh904HHZCnnWQSI1HTi4lGlrPp/ofp7SmNZfiwet2yPbt4kdQeiZ5f+VR8K9x11XX1/WWPqKqoa8xURLZiL/pC+6tYwnLD+XRm+43lKayPz6bKX+UN8GKORZYupaATK2rXqjuW/Bto9vxwxRDtUfs6Tm7uC1Wck/aehn2v8A/jHs+FYMJ19jYIlFlVoYWsPwAdPu7eETwA68LqccGP7T0613wd+N2QmFRV7AxTyg3DGCEXP8AU+n3XTHXgOtG5m82P7elBTfEToamplo4dlYsQKAAgp4foOOTbn3ukY8h1sXc3kx/aepFN8TujqKUT0+zsbEy8j/J4LDn/FfdqxcCAeti6mXzP7T0JGD6k69wEkUuL27i4njYMrfaUzG4/wCQfdQY/IDrTXU5xqP7ehAaioFXmjpAFFhenhsAPoANPvZoOmfEfiSf29cEpceQf8mo7XsP8nhH/Ee66lpXh17xG8if29cjR44KXNJR6R9T9vCf9hwvvxSOTiAfy6sHkbAJ/b0jt97H2/vvaOa21XY+inpchQ1UKqaaFlSWSEqrKun68+7eCgoCoA+zqvjzKdSMaj59fO6/mtfC/cvQndW4Nx0WGrl29msojsUgEcIheUkP6ePofbptVShXh0z9PJeGr1JPVYGP27Qsk1fToFi0sCh+qlf1XA9q1ii0AMOnY4PDbwuLDj0gcnBo8tXj38csUp/S2m+k3H0/x9sSxqMjpc1+9olFNBw49GR+P+Ly+/ayOSvV54cY17EFwwTgjn2l8NK1Az0HpbmW5uAzVI6PNuqjpotuvjUo3VVpWQcEW9Ngf8Pz7vUHozODToCerqah2nkqzIVUJLmp1JfnSNV1Hv3Vc1z1ZJsrsQ/w2mlxz6fOB5FJ/JXkXH/FfeuPVh8+HQvJuJsrQIkjX/Om/J/r/vPvX29e6zUnYdZtSB5aVnLR2A0OeD+Rx/h7914mvQu7c7+xc+NWbI2jqNN5Gd2sbDn6/wC8e/cOvVFOghzXySoaneSUFHlaZVaQJp817AG30B/2/vfWq9GPwucw+86WnNRXwyWT1gOLMQL8f7H37HW/Lpsz2NoqeTxY5ToXkFR9f68f8T78OvfPpGVarSxMVQrKAeTwAeCPfutdIuv33UYNHFb5DAASD6tK3HOo/jj3r5de6q7+bXzkptjbTqsRgalZchKlRBFDDMxdpSbBQoP1J97698uquvj98Y+z/mnupN07ixuVTGyVkNRF5UZ0KGQXHr4sR7917gKDrab+Nf8AL72N1hhcGmUwqNNTBPIXpYWYegfW4v8AX8e9163Tq2/YXU/WuIp6Z0x9HAIlhXS9NChKr/sPda9eI8uhJ3XP01BRpiWw9PLXOmlJ4aeIhGA/JUfT3rPXugAzme2lg6GsTGRQQNDGdbsiiwvwbe9ivXqitOteP+Zp2ZtzdW1sltOKppKnLVUtVFHDE4MpZ/Qtlv8A7b3sda+XVYXxExXavSNLOtHicvLj8reWPSkjIUka/wDZ/wBfj37y611avt/sLe0tHHkqvE5JbKrEMsn4W5FiP6+/db+3pU0fyQraE/b5GmqKcIQt5S6opBt+f6e/fZ1rpZUPyoptP24yMKJJ6WDVBP8At7n37rYoD08x/JbEwhScnTXUhv8AgRe3+8+/U69XpcYj5U4icqRVwuFW1/IWHAsD9ffut16b9z/KaiVCIchBqC+kByNJP6eL+/der0XHe/yQpqjH1MlfWRyqI5bWkJNze9j79TrVeq68ru/dXcG+Y9u7VirJfup1jWSAyMiq76De3097PXvLqwXZ/wDLFyOIwtB2Hu2Y1MpVauSGVVLKGUPZr8+9DrfRw+oN97J62qIsLGKKmalMdOEmSNWBhFvSDz70ONevfLo/uyu/1qKZp6HJ0UVJDqDobcqeTY+9/PrwwKdMu7uw9j5kz1uUnpJZQpcagGuBzxf37HXjU56KDvDsikrcmKTbyxLBHJ9I0A4B4Oofj37r3Dpy3d8g8btnZMuPzNXCtSaXSrOxBS6/QsfeutY6p77t+XC4KirTQ18MqSrMqqkuoi7ECwB97611T7urK7s7o33HkJaphjjVKTHrYald7G4b37rfVoPTvVG0ev8AblNnzjRJkIoRK8gVCC2gMST/AK/v3Wj0LuIxveXdmT/gey4KqDCahT3ihY2iJsOQP6e/deyeHQ94b+XRm6MR57eWSfkJLU005KvJIDqcaTa/vdOtjo1G1PiT1FNiNNfHTU0dOn7ktUI0UgD1Ek/Ue6169QdBxunsbo/40yldrHGVuW1GOJaLxSS+VDZbaOfr7359eJ8umr+/XafyGxxylLHU0OIUakjKOgeL6G17fUe/Y8uvA1x0O3XXQOD2/i2zmVQJlGjMnkqEBV5GXm2r/H348OvcOgl7h3zFgsecZR0j1ddHMRGaVTf0f8E9+HXj1I6syGZ7PwkmN3IkwhCBIqdy2vSOLEH/AA9+8+vZp1M7Sx+1uvsDSYfCwx09fU/tVLMq+SzmzBj78B8+vdF83Lv3avXexK6njamfMVKySoylWfWy6uDyf9j79nr2Oqxdx783FunK1dRkcikGNdyUilkK+nVb+1791r59Zvh0KXcHyt2rQY6ikyH2u5qF5ZadfMg/eBNz79jr3X1AeoKf7XrHZMBRo/Ht/Hr42Gkr+yOCPeh1voSPe+vde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdRK/8A4A1v/UJU/wDWlvfuvdak38gw/wDYwj+Zt/4kOu4t/wBXX+vv3XutuT37r3Xvfuvde9+691EePXIdJIZeSSLjn8D2jSNvH1nHW2ytOoGRp6HK0lTisnTR1NHUxNT1EcqB43VxpN1YEezIw1XUM/LphLkxyDyp1Ub8rv5RHSveZqMztPF0W39zVl2mrPDAIixbVr1W/N/ZTc7dDcVLAV6Fe38zXdiVKkmnoetfv5Ufyhe7Oh0myu2JKzckAMkkQxMPmXxKPSAIrn/W9ha62Z2JAUj06lLZfck6l8c0+09Vx53qrufYFG9duzaG4cdBEAWnrKSaKMD63uwH49kNxtHgZIPUwbNz3Z3ihVYfOnSk2fu+appaS06SNqVTGGF0I4IP+t7KZ1VBpr0Kp79L6MBPLoxVLkvNTR6yvqj9RP8AiP8AD+vtCSymnSTUQeo9IitXxtcadX+3597FPTPWuJ6fq+SaSfxxyFUVV4va3HHt/SujPHp4S3CiicOskeZrMdT6A9gOBz/sOT7TPMyglOPVhfyRmjDrBT7kkM6zTFX0NqNwDfn+zf2jG4XJ/Trx6eXdVJ0sop9nQx4/c2z6/FacvRwyegKdUaXFzyfay1a6hfWSekO4w2N5ESiip+XQKb92z1nnsXXNSUMaTF3IARAeQRb0+xhYb19NQsc9RbvfIi7gxZFweisYnqHGUlZJXUMciBWZwASD9b8qPZtc83KqAKaHoJwe1ngyMwU5+XQv4bGRmIUlQkhFhES1yBxbm/vdpzitQrnpPee2ropGk9NO5/jLgN4wPkIaiKGrHrViwU3+hv7FFrzfbigJHQRu/babNFI/LrFtr4WqtFPUJl4VqXjOg+Zbg/i1z7pNvkU8hdTg9ENx7fbsnZCG0j5dFv7C+H3YNBkXqaTcgNKHcgJUWst+BYH6+0D7qnGvVLfkHdxL3Bv2dBnW/GjceVmpY8nW1s5hAVjDUSBeOLnn2m/fEK94pjobWHt7fsgR1ND8ujM9V/G3Zu1ZqbJZSCWeqR0kRpHMjBh9b6vaOfmdZFKA9CjbPbFoJQ7r/Loyu5MtHHFDjqD9ilhjCqGJXhR/T2E7/cZro0RqDqWNi5Zs7QqJV4U4gdJCTe2VwkIjx0oYEWc3J+n+I9hW4+qSdWBP7ehzuGz7ZuESJEoqvHHQyddbsqNyRGnr59bLYlNR/wB6b/e/eTP3cUnbmO6nY/Co6xx939otrawKIPI9DPHUJT2hVLhxa9vpb+h95vsdSBj1hlFbut02nhXrDUsY0d/KsaWu2riw+ouT7a1pTPRmIZEbWa9Fg7R722v15TVMk9ZTSVSpL9JBquOLm3sP7vzVs+0xkyyBnHBRw6NrUSSUAFK+fVMnyG79rOza2f7GpkWnEwRAsrLdXOkW94z8583XW73LNatgnyJ6ENtZRUzx6H74H/CCr7g3JSbhzTM2PiqvNI1SokQo3qtqf8ewWl9NIgSTj1do9DHPV6PfW9thfFnpabbe2pcemXjx7w6ogiv5Ej02vH7Ww2qudbdaBx1qxdt7s3H2tvGtzuUnc07VPniBdiltX19XA9vytFoKJx60eHU6gqsnFjftKKQFWj8ZC83uLXJ/HsuFadeHQ8fHnZkbbqp66vp3lkaeElgQeddz7917J62Gelamn25BjKyjhCCNAza1Fj9Of9f37PVurPNnZkb9w0sEUkcc1PTa1JspuqXFz718+vcRTot+60qclXV2C3ApqaBZZACFLAqn9bf19769/h6ArcGI6/fH5PGzUojECFImsFAFjfn/AA/Pvw+fXq9VX9kbc29PuSWkwlQkMkNQ0rEyWDDXewb/AGHv3VaDoFO7DiZdoyUdQYpZY42RiCDcqtrg/wCw9+63jy6M7/Iw3HiMfv3fVBCUjkSrq1RTx6vp795dbHz62YK2sBzSSVS6wyqy3Org/wBPdfs6txx045bb8WfSFIU0wMt3FrXJ/HPvw60fn1FxOz4NuzrK0Z8ZvcW/B59+qT16lOHQkSVlDJSGnpIby+FtWkciwvf3vrXRUd07qqaXOtSU06xRiW06s9iLmxv7317z6W9bu6L+7aQRulQ6JqIU6j/i3vVKHPW/5nosM3YGXbcJxlJE60jyMHIDABv999Pe+tUp0I1ITVaRUROjuLXN73J4I/r791Xz6EWgoKnG0UNQk4CBgygkfQc/T3o5PV6UFehFwu/DShBLKhlUBQ5bgWNz9PfqEdaqCehXoqzFdvbdzG3MglPK3i8IDIH1+gj0lvr71w68R1qN/Njpyn6k+TbKlI9Nj6+tmjRlURRiWaWwYhbfS/uw6qaV6HXE1vavRmExG7MJkpsht+ppI6malgkeRli0+Qoypfm3v3W/Op6EPan80jqg1H8C3HgJoc1U/sVU88Tp+5bQSS4+l/fuvVPRpdq5zoPsqhj3HUbjwGJlqCsojqqyCGRTLyFGog+/de6MN15uroXY0v27br27XGc+lUroJmNz+Bq9+Py62D0O6b26e3GRSx12JRJAFQl4SOR+Bf3rrw6dqbA4in1Q7dr6I0k0bECFktZh+Cv19++3rfQNbj6kNRkpcgyvVMWZj4/V9eb3HvdR1qlBjoJafr+HAbmbLV2Lrng8geI6GMYcH63Pv3WvLoZKuu3VnMelHgZ46SkUBWV09Vhx9OOR79wyOtgeR6fKvDbgxmFpoJa6MzzR6WZTpFyPyB79UcevdJKhwmKxkxrs20VTPcstrOdQP1H+x9++zrR49KGHcuMRdQQiFPSFN/p+Bb37rfyHDqNXZc18ZNFGUiYWuPr/AI2H+Pv3Dh1o549BrNS0iVrSmLVUkkXIuyk/7T/tvfut9M2QyVRgtwY6h+yklrq0qaWdUHoDf1Pv3Xq1GemA/JJdo7yG386VSqkqFgiZ2KFnY6QRf/evfuvZ4dIj5gfLHJ0ewm6+g2xksl/FYSkGQp4JJYUaoWyHyAcfX+vv1KdaJ6r86K/lv9w92SVG981m/ssFXf5VHjasqrLCDqCBX+vHHvxIrTrw9ejdbE/l0bS2/vCmevp6ZTi6mKoaeSOLxsyEagD/AIn377Ot06MN3ZtXr3FwUODosfBW1VNSrTj7GNCx0KASQnvXXuit7iwWz8bhpBU7OyFTVsgEEqUrMsbEcFiR78a8OvYoeqvO+8NuuspslDjKWakpHp6lUUw6SqsSFFrfj3vqpr0r/wCSpuml6t+ZG38Du1Feuq54Jone6tpessot/sfr7917r6OFJVRV1LBVU7hop40lRv8AaWGqx9+Hz68fl1J9+631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/R3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6910SR9Bf3sU8+tEkcOsMsCTW16vTewDEDn82/r79WnWitTXrqa8cNkuTwv5/PF/z72O456vHRSOo8MEusa1Aj0/X6Ne9gLe9eXSmSVRhOuUNFFTySSIXvISTdjxf/jfuksrKB59M6yeszRknVybKbAtzf3pSZF7sdUIU58+uKxu7FpBpAuBpY/X6cD+n592McdQR1WgIrXrC5qRKFWNWjv9Cfx/r+7eFGRU8er/AKeFrnqQUYi+gCw/BIPP1t7oY1J60wUDtPXBYddzIGB/pqP5FyR780aDh1QAHNevCFl/QBYC3J5PHvRjHlw6cVYwMk16yhLkarnj634v/re/CMAZ6qaV+XXD7WM3+v04sbD/AG3vxReqlQesckRjS0d31HkMTb/X9sy64gDF1dDoNeuqcTJq1JZbj0ji9xyR/re9xySMR4nVjppjj0QH56/C/AfKjYFZjxi6CbOpCVp5ZYgXdwpCm4/2HtWGJFOq+PLCh8PrRL+YXwU7o+LuQy8NRgqg4aOWvlVqeinfTEZiFIIH9Px70JXU6Tw6raySM3iy8T1VHRZQVtfLh5o6inrmqQrpPE0PrZrEaXt7ZZ2Y+vTd3H4rVXq1L4w7ZG1qOKeSGJkrlZpjYX5AN/bKNKXIbA6X21tbi3Ln4h0bTe1PhHwk01MqNKtI5NwP1W5F7f717Vmnl0jFTk9EJgyjz5ysoliVYfO1yoBHDfg+9db6F3Z3Z0uNyKYiJgYIm0tqsP6A2Pv3XvKnR/Otsjjc/Sxq8yBmiUhda3sRz78et9DdQ7L2tVUlS+Rm9I1En6iyj+n/ABPv3z638uifd8SjAYXKptB38kNLK0RXjkf6359+z1rqpbrXcPYec7UqVylTUKiVDaB5GsQGsLD/AB9+z17HVpXVG+N04LKQU+SqalKUyFQXZtJS9vzx78RXrwJ6PpiOwcQyh6mdXDIgVi68Arcnn37rdacOk9ufe+HpaaWreeL7ddbEl1FlXkcj34enWsdVZfLX5p4TZ+HrcJt1xWZirQRQQ0zLM/lYaeFW59+p175dVzfHz4x9z/KzsSk3DvDFznb/APE0qEgqqeUr9tK+sE6h/T37z691t0/GTpjrno3a+LwMWMx9PVpRxRyBYliYSxgf4Xvf377OvDo3lTuzEBQkGgJEPohB4A/A96016tWgp0l8hvuglXwU1dNHIPqiMRxax/3n3sAdaJJHSNzO7lo6SSr+58syIxQyyLq9I+vPveOtdVnd1fKGahyFft/E1b1GXyV6aKCKQOQ5bSPSP8feuvVp0U7bvwG7V7v3bR9g7nSokoJK2KtEEyytH4JJNR4+n09+68Aer7upfhr15TbTxGMmwlA2QpKRIZGemu2tFAF7j6+9VNet0p0Nz/Cnb1RQuEw+NEYT6LEBwfpwPeq9ep6dFS7K+CPWdfBVUdR4qWtfysDFHY6gtrAj3uvr16nVSHfPwkrOvhXZDE1zmmUFoSZSgC8n+0fe+tU6ph7H3luPY+4aignyNR4YnlVdEurlGKgC3vZ618umXFfJPIYejf7iqqAL8Fn5Y/i1/eq9eoeu1+QuSyUnnkrqhYXOpP3OSPze/wCPfuvY64ZjvGmqaJaSatl1NwQXB5YW59+690cL4gd39cdc5GPcG4JKWSsibyqajxg3DagLt731vPVim+f5q3X09FPhkyccdOyCJYkmUoLDSFC/T/be9deGOHVVvaPyjhz+75c/trK1CwszyBI5Ain829J97+09a6ybe/mCbrwNM2IiqalnPoBMlgSeCST9f9f37r1PToUNo/NLLZudjuTJVVPSA3kKz+nSRc2JP096630M2L+ZfWuNjeaiyj1c6xtr1srASAWIv79jr1c9FQ7a+SmV7WyE2K29U1FqiQrHoIKj8C5X6e/da+3oiG7Nu75qdwU2HneaqnqalUCHU6AyNxce9de6sB6g+KuXXF0OQylPFT1ckcU6f2AQPUeT/wAT7tw691YXsPY+Gpca2L3BPTpTRLplLSrwAPpYn3ry630YXZXys6b+NcE1Jg/4bV5UxERh2hYiRRbSAfz79Ty69XoCN7/ObevYGfkzdREMftCJ2aSSnmVQVHP6R78PTrVegC7H+RXcPalLJtfpmR5IJl8FRVCfQyqeGa6n6j37rfE9OXRXxJqKOvh3l3hn6yrm+4jrJKepnaeIMTrcBSSPr7914ca9WTnv/pbYVJR7T2wIzKIxD+3Gim4X+g9+62KefUzE9yY7svI0e2/JJSUsc6RF19F1uBz799vXvn0q9+7O6q2oy1dVVGrrXhDlZf3F1Fb/AJ9+68fn0DFJv3beNqDNtMAzQ6tUcSgLr/obe/dax0Qju7fvYW4N11xqKaSGks325VW0s+r0tf34de6LrnZkTGvW71r5llUlUi1hhb8WX377OtV6AXeO36DetBL/AHbqJY1jVf3F1IRb3s9e6s9/kedJ7RqPkNKdwRityNNk4Jg8yeVtam4sxv7r1unr19C/DUsNDisdSU66YKejgiiUfQIsYsPeyKY60OHTl791vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917qJkP+AFb/wBQlT/1pb37r3WpN/IMP/Ywn+ZsPqP9Ilfz+Ocr+PevTrQ4nrbk97631737r3XvfuvdcXJANuD+OL8/63vY6qxp1DRahtSzqgH4KgHUPwbj2mkkljfsOOrFEYU64eOpDkAsE+inj6EW+g+nu8ckjtRx9vTTIVwvUWrpaGWIR5Ghp61W/s1UUU4tfgWlBHtQIkPn1UytDjj1WD/NF6ffdvx93HPtPaWImr4KGciopqGJam4hJuPCoPH49kW7Wxcdoz1IHJm4xwXFJHP+brRW2L99tXdFdgc4KmDMUlbWGppqhWijRROw9Kye4y3iB7Sj3ApXh1lXy867lABbEMQOjXU+5Fkp446eTV6R+Rwfrx+fZYs0ZGpuPRzJaTI3eB0qcJlS88JqGACkcq3JP9T/AK3vYKPUr0mbSnSyeU1U7mFyQACeb3t/j7qZMUbpdBf2ix6Dx6b6urjJ8TsA4PPN/r+B70ihmr1V5EkOKZ6ZiGMg8ROnUAwBvbn+vtxLaEN4jcekLaC2nrhkclJFDJCkhQaL3BGn/Ece1GtG7Tg9Oi1VVrXpIUDVVW8wNQ5QtfTq4Nj9Pr7TSsW7U6cSWOLtPSjinWgQqeXNgVvYcfS3thbdrjsY8OvC7ikNNOB1mp8k0koQqqg/RltcA/S3uxs5Iz2nrTSQvgjp+FQsTIBWzIhHqUOQP9b3RvqQpVSa9J5bSykzQdPzZOpoqYSRZCoEOi5/eb8/j2ZW97cxxBCakdKVg20QhXQE9IHI7imrqrwLNNMpNmvIxBP59+e8nYUJ6K3W1WTtQV+zp7pEpqShkqZwhspuWPqBtxyfaU3lwAUHn0vtbq2iBDKK9NMWXopZFkMzIhf0i/F72tb2h/WB1GtOnTfwse0dNG7Jqitkjan9KBbagfxb6+1EcvhilcdeE/iGq9JZ5oaSOJal0YyWDXdSw/qOfdppkmkTwxw49aE0ts2tjjpb7B3Rg8HlZpKqrWFNIJ/cVQbD6nn3kT7B7zYbXv8Ac/XsI0kWgJ4dY/8AvFeNdWn6HcQDjoWMj8huusNRyzzZOHXCHWxljvqUcfX3lfuHOuwxRE284kIHl1intdncyXDCZaAnoh/Y/wA2ZaurymP21I80R1JG0bAix4/H/Ee4W3/3Nv3c29vhR5joXy2EGkBePRGNzpv7s+pkr6mSu8Eju4QhmBRzqtb3Gm47pf7mxkkc56qkCxDSB0nIen6loW8sEpqFKNYxONTI1xf/AGPsth8RAQ2elCzaVK9Hv6Q707l6fwq4bb2BkNJJEI0nijZXCadNyQPr7dt4f1KtgHpvJz0xdp1XcPb1q3NpXtBLLdopVkddMjXYWPs2mkCR6Y+vce7oGd5dGZWjwcSUlBL97LCAdFPJfV/S6j2RCW4aWjig62teJ6VfQ3xm3rmp0jzlCVpJJGCNJE4ax+gN/a3r3y6tE6x+JEGBaCqhpT5x43YeNuSvII/4rf37r1KdHf2ptA496airl+3giYKxsVup/HP+t711vzp0PW3d1DaFXWrSTN9oYCFKsLkldNre9/b17Plx6Azf3czU9LkWo1V60yS6CeWsb296r17h0RjcXcNfXtkKXLM1N5SQTH6bm1gR7tTrVT0Rve2TipsxLX09ZUOsjuSdZvct/sffuHHrR6A3sbLGuoTFDNLJ5EYm5LfUfkf4e/Z68OhM/lRdkYzrrvbKUubrPtEyWYkVR5FQOGfTcgn3ode8+tzRnxudgx+XxMizQvQ08msfQkoCb291+zq9a8enrbucpYpTHk9KRRkgtcADm3v3z68R0qdwwkUSV9KC9FMAYnALc/W1/e/PrQ406SGIzUVDWHyqS8qNEwK6h6hpv/sPfuvHPDoGOy+u6KKWfLRzSeTIN5fSD6CwuNI/2PvfHHXuGegs2xTZDFvUpUEz0bKyqJbtpFrWsf8Ab+/EenWq9ZaagxjTy1MMUbVJkZtWgEjn8X9+69XrNlt64SlnpKGNilUmlZgFAsTwSD79w69SvQu0uKqsxt+Kpp2cwlQR+fx9feq9WHy6SkOFq5qxaRXlE2oDm4Fwf6e7HqoHkejG9eYlcDV0bQzOJ2lU1CAmzW+tx7pxx1vgOquP5p/xp3Bu2mXsXD44OaPXV+dIXLjxtrLhlHBNvdhTy60anquvo/u3G5zAy9e51fPkaCIULQVoH6lXxmyyfj37r3Hj0Fnc3QmxKg12YpVpaTIqhlfwxohVidXD/wDFPfvLr3VRu89s7rym+YNu4zfm8MTjGyEdOseLmnEABk0qA0Z+n+HveevV6uN+OXwNylVisHufJ9gbqrVFOJGWsrKh2IBBJcFveuvAVHVgu7Om8F1vtqhzNdujLINKGNo5X1EgAXte/v3W+HQ8dIdhbSG2HZNxVNTX04uEqaizlF+llY39++3r2eA6EfC/KDb1LPkaSORqiWjJSQPZ1JDWAW/v1BXr2ehzwnaG2+wsEkcVLTGrCjVdV1C/Ivb8+9cOvCjY6QeVqMnj5WpsMkRkdy4VWAIAP0sPewMZ6959Juv3Pna+SlxbxuKq6rcggAj66f8AY+/cOvVH7Ok3mxVYmdXzkrrGpDNZr6UPPI9+610pNvVm09zyxUOJqfJUaQXD8AEfUm/+x9+6907bw3z1/wBYJS4vOVXjyGQIjpxa6u5Fhyf8ffs9e6UmG69yGdxK7ux0UctJLpmjJPJRxqBsP8Pfh8+vH06dcVt7GVmWpMhnqZI/4bdBeP1A/T0/639Pej1bj0G1f8Rtq9v9jtuOrm+0oqCoWop2toaUodQ0gW9+qOtZrToeu9Ov+oNndO6spjqCqrqBVjSoqKVGmtTj0nyOL/j6e/DrXRY+tO+9hJifsKaqegp6GIRCOkYomkcAEJb3454dbB9ehCyuQpt8Y2N9p1jGSQ/vTCUCUIxvc8349+Ap1456g4zrnZmJpmyObyAq84t9KVcoa7j8DUf6/j37PXuuNe2Ogw2TarxWIlphGxikZYmdU08EX9761XHVK/yTyeaq6vKwYPFUrUcENXLrRFH6WLWuB795deOOHQb/AMqTqSq7e+bm29xV8a0kmNkp4XWK4A8FUGY3H+t7914U4dfQwx9HFiqGjoIiSkEccCk/VtK2Le9jP5da4cOnH6e9db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv//S3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfutUHXvfut9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+690APd/x160762/kMHvLb9JWSVFPJAlZJTx6wZBf9ZBJ5/PvZGM9aBHl1q4fM3/hOvj8vkMp2F1plJqSWKeoyEdDQ1HjYCNDKqlEA4496ApnrdTXqlDAU27Ok951/Ve6sdP9xgpGplnmVi0umTxKST/W3Pv1M169qb16HHsLcFDHsuyOlLWvASyLYMFIvY/n37r3RIaDNQwyVbwwCpkeQlqhRcKx5Nz791rpOxVkmRrqt42NHJBz5FLDUTyWB9+698+hR6/7gz+y65WatmqYVKjTcm2hvp79Xr32dWQdJ9+0G9qd8fXgRvKGBLXWxvYcn36np1vjx6676pIY9v174imWpmajeyw8s5P9f+K+/Y68eqZNqV25cH2y9dlcfNj6H73SHdSEI183v/h+fe+tdW+bThxe98PRRUWiKeWM2qQArAhf1D/kfvVetjrFuHae4No00k0uQllpoUL+QuxUpe/BHv2D1rPRCO4e9t1s1TtDa6T5KrnZoAIdZN5F02Nr/k+/db+zpj+LPwP3b2hvqn3l2fTTS00tQ00dNVM+mNQ+oWWTjj37gevZ62lOkeqOsercDTYzE0FHDWpSwRkxrDq1JGBa4HvWRw63iufLoM+6tz/3XzEc8UX7V2fWoPpX/WHvx690FuC702/TMzVmWgklnPqpmIun4sFJ97z14U6n13cm2izyiWKntGxEw03vbgk3/wBh795der0U3s3u3I5utOE21WtPLOzKPAW4uNIDaf8AX+nv3Wj0H3S/xR3Fujsaj3ju2omeFaxagLM50qurXb18e/dep1sAdd1mL2nHjsRBCslFBSU8TABCt410k8C1/evOvW6Doy1DvfblFSP9tFFHUuGKkBdd7XsLe9HPWxXoPZu/avFV1ZS1cjRUzBkWRwLBefV/T3uh61jh0V/tT5HbG2s9VlMpnqXziOZhE7ISHKlgvJ9+A69WmOtd/wCa3z8fJTZTH4iuaSkDWiaNfQENx9R73w61k/LrX/3b2Vkd8bjkqpNdTqqZLqQSTqk1N7117qflcfDl6NImo/tXKDjT9WAsOf8AeffutcOmWl2llJ/HT02vTGQFIuRe/wDX37reOnut6/q6SAVVU/7ygOUJNiB/RT7917pNT1tbCy0pcwAFU4ZgOTYEj/e/fuvU6E3F9Xy5bHRZKSLzXXWJC7WYWuSAb+99ez0g6sS4bNCgeJo4QwUtc88/X3o9e+fS5j2zBmmghxkfmrJgCHU3ZT73Tr3QrbY6f3PUmHHPSTsak+Ek67gN+ffvl1rHn0Ju5+rMH0/tZ6nNU0T10yM2iTltTDg/4+/cOvefSJ6kqMZhYclnJ6SMy1BeehW9zpJ9Ngeb+/DhjrfQobYWu/vJ/fPK4t5KPyCSnDrxqU3Xi3v3DPWvPoxeY+SO4qyhixeHxc9C6IKeN0RhpU8KRb375jrfSFzOG7/zuLnyWCq8lN9wmsRoZgBcX/H+9e99a6DbaXQvbm48hJVbwpqzzlydUpmBAJPI1e9dbHQ2UfSu8ado9u1M08OPqAFeIu2lg35t79Tr3Qzbb+Nu8doQmt2Nl6g10w1SQUzsCrnk3HvfWhXo5/WXXHac+Dp4d4/d1PkkSOQT6rJEbAudX+396x1unRpMT8L9kVgptyrU01TW6BJLGJFZo3tcoR/r+/Z49exXPSvpvjlhcLMuRxcyU1REfI2kjlh79x698+mbtzYmJGyJcrVZNKjJRK0YiZhrVUX+yPfut+XRLthVFNi5a2ZMX96qFy2lbjj63H/E+/dar0C/ffZuJp6WpDYZaGdIiY38YU6l+n4/PvwwOvHooeweup+5cpNlcrlzHQRs2iieQhCAbAab+/daHSe7van6gweUgx9B4Y6dAoqY0A1AnSDq9+PWwPLrYQ/4T8fHKTcVWvc1fqkirmjqixP0UprBsf8Ab+/fb17rcbRFjRUUWVFCqP6ACw9+691y9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691EyH/AAArf+oSp/60t7917rUl/kGcfzCP5moH57Er/wCv4ynv3XqZNOtuX37r3Xvfuvde9+691737r3XvfuvdeP0/p/j7917pvli+7BP0Kmw5/H14/wAfdg2NPVZ4Mgg9M+XwdBubCV+38vAJ6WsikhlikUEFHUqLBv6e2Zhrrq9cdOWUzWzh1wR1ra/NL+SjF2NuvJ756+rHw9VMZJvFRyiJpXeQyaLJb6+yHdtlTdYhE44dS9y17ivy6vjBvixnqkbuH4ed6fHiokGT21kanD0bvG+TmErLoi/U4Nv6ewhecsfTKdIrTqR9r9zYtzkCyNlvl0D+H3Hj1iP3WSihrQPXSkkMrX4Xn/H2EbyKa2JVB1Jm3XdpuSip6U9PvBcb/lL1YYS8CMX4/F+fe7dGaLuGevS7cr3ZER7euNRummmIqWm03NzzYfX68e3dElMDo4h240z1yj3jTU8bnyq/mB0vb9JJ9+Ky8D0kl28mYAHqJVblpWoS3lDStccC59X0v70Y3+KnSk2EgxXHTVQbliohJLIwjViCpP0sf+J9+8NgdXn0mk21hx8+su5dzxR0dNWUk8coltqUXJUf4n3RxNH3x+fTMVmImJfHScpOzcXSAirq4kcW/WGvcfQX9sCS71VoadOfTxt0q6TfKZtFTGhKo/jSH/5KuPdy1x5L14W6efThkNy5ekpPFVQFI1Ti6yfT8fj3qtzWuk9a+mQ8emXC71xqVB87RCS9rMG/V+TyPfg84NSvV2tYePn095PdVVWaYaCETxvayIHsw/2A59+AnZvhPVI7SAsNYx69JqrG8axqSmxmBllnMw0RIkg1sTcA2HN/8PZjFZl8Hq15HtttHqDjHQt0vTfyf3XFSU+F6iys9LOIUNakU9lSU213C/7H2ZRbBLPgDoBbnzXYWFQHFR0eDqv+TZ3J2zTYyv3HWZLbctYA0lPJPJF4bC5uOCLezS25UYNVlNegjN7jQPGwdxQdVV/O/oDd/wAO+zch1/VZWoq0gpWYVUsjSFiOANY/r7P7XZprZtcJ0nqLOZN/ivgaHUD1W49NundMjmWukWjlIZn1OqWYWPsU2njwj9Rq9ACHQ0upMdGV6L+NUe6a7TN+6ZHssms2cH6cH27KEdtRFerS1DVrnqynBfFFdsY1HqKQfbmNdLPbT+m/5HuoCjhjpulTTpU7f+LNNuGY/Z49X5BJUrqHq+nA9+wevaQOPRudq/DbEwYuNKmgVptA9AIJDW+tre91I+XW/l0J0HxHx1Ng2DYteG1AkKQADe5Nveq1PHr1KceHSTzPRO0sX4Keqw8FTJZVCkIdLf15HvxA8xnrw6fdkdF4iqysK0SChVZR+wuj1f1tYf7x799vXs9GwTqtMK1Or04SEQC8p02ta9ybe/V69Q16A3uTHY3A/avjq9ZJG4lVCPQSOdVvfuvdFBzfZkFDUNSS1Fy76CPwQDbm596z1r5dITc+SxtTRPVxshZ1LXAFyCv05Pvw68eif78rcPaaZqhImXmQXXg/W9z739vXvToqe6s9t4UsssdZFM41hY9QBv791odA9S5N83HU01NQ/cVJ1rCqrdiG4Gn37rfQA0u3uwere6dl7rTH1NBj6rKI1W/rCpG0oNyBx/t/fvPr2Ot7H4kbnXevR2GycTiokOOoY2lFi2oRAMD791scehXym1K+toqp0kanfWCguQx5v9R7qMZ63506MptbbD1vX9BRSs09VTJdx9bWXkkn8+91zU9ayOkpRbBiqqqXUvrBYgcXGk/Q39+4deqT0mtwYlap5cXPF5PGdCG36bCwAP8AsPevs68K8egcfZL09fVwgl00MfGLWS4vf6e91HXqefSJfC4/FVEjVUyQNqb9trA8fQW/p739nWuHQJ9iT4qmyuMio6SOSSrkXyTgfpAPHI/2/vx62OjZdX1VUmMhoijVFOVjFv7Nilzb+lvejTr3T7uaKlxFfFkItKWuxAt/S5v79WuOtj1672fvBXzUDyNrV5RpuLjk+/cOvceHRn+1cXQ7/wCsKnBNilqmqaJoAQoI/dj+v0/3r3rzr1qvl1qn/Kv4t53499gzdlUWNkosR929TVBS2jQf3DdQPdvs60fl0Q7sfvOr7YijxW1ZHgkb9utngBQqqelmZv6D377OtEnz6sb+JHxL693HsGn3FuSWlrMuHhnaqlKFxMRqIBa/vfl1sDqyXae2qLbOPfGY2ZZKOlDRxxx2K6bcc/8AEe9de6SOeek341ZtbKUiNBSIwh1rYKQeCL+/fZjrfDogG89hR9cb/iydVvRdvYJ5GV6N5tKG7WKnkD37j1qtOjFYHF7WkxJye256fcDVqGWaphIfUWW5JIv+Pz711voZuj9xSLmchiDQ/aaYnMbEcj02vf8A3r3vjjrxpTHSkytfuba24m3BLPNVYlJSGU6mTTquLj8e/Vz16lehpod2bd3lR0lTjoo6euis0joFVwwHNyOb+/Z61jy49JHceOmqZpZsizVFMUNjJ9D/AEvf36np14H16DHb1PLS7ojkwkfgiD2d4j6RZub2+v8Are/dbPr0LHbPT1F2bjMNWTxh8njWSWKRjZpHUagB+fqPp7159e8ug6X5N5foqPHbM3fTSUWIeqp6FauVSYyC4jDa2/wPvfWhjqwfA0GA3tBtjeGEmirMRWUkdVWJGVMba0BGoD3qnW9WOk/3BUbg2RW4LN7YopUxE1VEa77cHxxwFvUzafoP9f36lcder0E3y87O2Vuro+op8XVUtTllo2aaBLa/OsV3Q2N739+zXr2AD6dUsfHDvrY8G5MnsretHS42pqpTDTTVQA1XfSrKze99aHVjWD2zNtisfP7f3H5cFXIrLFEwMcSONWrg/j37j17ypTpu3TtfJbvlDYbePklb1MsUhBVj/ZFj798ut16Dbc/XfdGExEs1Ochl6SOMBgGlZGUKblgP8Pr79nr3DPVUvyS7q3TsTE19JNtCplrXhqoZrRMzBeVZjce/Dh1onoev5E3YR3D8osNDJSnH1tZNFM0ZSzfuT8qf9v78fl14db9RANiRyPp/h+Pfuvdd+/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/09/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691xIBNz/rD/AGPvdcdVIBOeo87xEilljEkc6Orqw1IUIsysD+D79So69XTgda2383f4L4p6vE9vdZbceKuSpSt3GuPolYyKspZ2Z4x+fqb+9dWPqetbTvLI46owJihoqjF5CJpKSb7q8C2j9Lkhrfm/vfl1o9F32FTY1pHonraUioIaVjKpsT9Rcmw96690rt49erR0E9fg6ymZimuQxMpJuPobe/dez0W6nzkOMmeHJnVKJGW97W0mx59+69SvQ1bO7JpdtGOoopXRrBlIkIIBN7cce/der0cjbPduO3Vj6Snkq4i8RX7tZZA2pAPULH37rfQLfInFY7cuJ/iGzRDFVQnyyPDGisdHL+oWP+t79jrXQX9R/Iifa9djsLU1BSShVYasySlRrBsTY+/deyOjM7p78rd9NHt/GEulQqwtKhLCzjmzLx7916o6MN8ffj/17Q1MO7N3w09VVyATt5VWQrJfV9D731vo8cG7No4LVHhPsaamh/QAqRFVtzbSPdft62cdJur7S+2qRkKTJIFD+oCoNvT+NJ9769xGem7enaW394Yh4KuWJq1YHQPq51lbAX9+HWqjqsPsDGZSiyOSyOOrZYVjXXEzSsqA3vdeefe+vYp0GG36rsTc1R4H3VRw0asVZZK0K2m9rG7f7x711ro4fVabL2JRPldzVlLlcvELoY51mYkG4te497+XXqAdPO9fl5lMU8MO0sdUQx6gLojILD6nUB/T3rrxJHDo/Pxo7gze7trxZbNmXyMI2YMXJs3JGo+/cOrcejJ5bsrFChqKimrUpKml1czTFBqA/offuHW+J6rk7++XEOJpqnG0+QibJsrojwzm/kDWUm3v3Va04dUr929u763FNU1GZyVW1NNJIYliqpVBVx6eL+9+fWqnj0UFut9y78m8stRPLQzNwGYyen+hv70evdLvDfDncFZabB00omKhtRg13J+pDe90HXuPWbL/ABh37tf/ACvPQ1Apo1Y6ngKJYC/1A96690iNtVOFxm4VxdYI3by6WFg1hew9+690a/H9QYve7KKKESfsq5VYgTdhcWH+Pvf2de6Kb8h+la/aeWphT4+ohiBUu3iKISnJPA/HvXW+o23t9U1LgqbCU8YeqhjEcgVQWFhY3X37rXWTNddSZfBSbilpZELK7I/jAtcXHq9+x1scOk31BtTd0eRqczRRSzUlBKQF0eS4VuRx7916p6sO2XvOKGOn+6xrDIqyaLw2vIo5+g/r/j791o9Fr+UOL7G3jWUlfNQVQwSTp5NEbrH4tQ/IFvp78et+XQtbI6nw+f2BSV+LRHqcZRqKiKIBpNSILhgPfuvdGv6I2DjOwca22qmlWCTGKS/ljVWIQXIOr37rQFejTbU+PGwaqp+zqaGIVsbELIYVP6DpBA96r6dW6M7srpwYaUUtO+PagjOkxvHEbL+fSf8AD3vPXj0ud3dU480JqsdR08kkahn8NKguQPyAPoffvn17orW5Nz9f7aMuO3DhnjzSsEp53i0KPx6SR79THWq9DH0hU7azSvPhDSxVHLMagq4LDm4De9efW69GngeurEONjigeUKuqSKFSmg8GxX8+9jr2emaryy7UqRjacmOasDeQyMQgcj6/7f37PXjTz6Ih8j9+d77ZFRV7OmnqIJBcR0+txpLcfp/w9+xw61ny6rpyvePyNTyVe7KLKthPIyvHpm+n9v8AHv3XuhN6z+QNCtHPSVFLNBW1AYfvqwY6+AOffuPXukN2bsjdm/K6iy7uxwPnElQirf8AyfVqYEH/AA9761nj0Jj4naG0thHIbTeGmrKemtN6grmdE9VwP8feq+XW/LogW5590d675xfXsNPNkUyssUEoplMzkeYBuF9++fWuvoX/AMq348UHQfxc2XiTj/tcrV0NPJOZIBDMiCAWRh9fz798+vdWY+/db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917qJX/8AACt/6hKn/rS3v3XutSX+QYf+xhH8zX6gHsSvNj/2tPfutDz625ffut9e9+691737r3Xvfuvde9+6910wurAfUgj/AG49+62DQg9Q6YPEWjYfU3DD6fT3s56UTFZAHU8OskkJZw6mzAi/JHH091arLp6RFSG1DrjfzN43Rf22ubrqUgccX9+V/DNG/LrxHiGh8ug67K6e2D2xg58BvLA0GRoZw6vrpoTJaRdLesi/09syRRzijdLra6ltJA8Z4dVqdjfyePjTnzJXbX2/HisnKxJPjiKf1Buf8fZRNsNtKa0B/LoY2XPO4Wq6QSKehPVf3Zn8jfd2bmk/ujuLHUMWpjCJAiBAT6f+Iv7JpuXGWT9IY+XQvs/de5ijCyVJHz6K/uv+Rt8i8RFrg3HT18AYf8A1SSwH9be08mxSJihz8uhHb+8RaPTSn59PGB/kY995qip5Zdx0lBZTf7rShNvqNJ/3j3uPYZHwAf2dJrj3dIaoFaeh6EOj/kXdvUUaCq3PQVhut9BjOk/S5/w97fYJUwVr86dXX3kdl4U+VelxjP5Gu9nqqd8tuHHSUSmIzQKI9Rs13Fvzx7t/V6Urq05+zpI/vBMzZHDzr1Y3sb+Ub8ccdtnH43cuCirsnDEgqphBGQ7AWa1+Pr7WWGxxkkTrSnkR0GN89z95vCBZOU+delnH/KO+F+kfe9eQVj3uXfxx3P8ATSFPs2GxWXko/Z0Hf9cDmcChuW6VOF/lffDvAn/cb1vTQm1hdlI/6F9+PL9n6fyHXh7h8y/8pDHp9rv5cHxOr0MdT17SOCun+zwP6Eaffjy/aHy/kOvH3C5l4fUN0lj/ACrfhoJTM3W9MWJ/1ajkf6y+6jl+y8h/Idb/ANcPmYZNy3T/AEv8tT4jYzTLD11Sr4voSQ1iPza3vR5fsyKAU/Lrze4fMuk1uGp+fSioPgj8YaCqhq6PZNFHPTyCSNjDGwuPoLFfdY9itlbtOR8ukrc973IKSTEj5k9Gm21sfZe2MfDjcLg8ZTU0KrGgFDT6iq8KCSv49nMFskK6UHDoiut3u759c0hP5mnSuEUFOU8UEMS8reONE03/AOCgce3wKg/LpCzkcSc9asH/AAoM+N8NZtEdxUdEaipnQQPNDEpYaJOdTj/bn2qiMWijcekxMrvQfD1qdYFI4/t8YAojMUPmHF1NgSG/x9sshYmnSmEGE6vLo8nQ+9sTtrJUsRaJBSyDUWYeoC3PtqlMN1Z38Q6h1bhg+yts9g4KPEU70pq0pwbrpJB0fkH/AHv3r7etedB0rupM1FgdyDG1CIySyBEJQH+1bge/de+zqwTacMeQrGm8SGJSGPoGkLb8+9cOPVq+fQnbhz2AosA0Rhjkcgr6EW9xwfx791oCnRdK/DYXPVKyiNVkYhkZhZVP0N7+/cBkde4mvSvxXW0WKEWXpqyk9LiQxhlL8fggH6+/eXWxjpSZzsXArSjEV9GxnEXh81rc2sGuPfutefRCvknW7b25trJZszxhxH5I1aX1L6CwFr+7fb1r59a53YHfeWym654sTNK8a1EqJ45WI4lIX/eveutdMOf707Ex6xRSR1QpDC3qOu3097r16nRXNxdob83VlmoKVqgrOxXSjsxFjaxX3rr3Q+dV9KZzOywTZ6Gp+3laNj5r2JcAn6/j37r1c9H42N8f9vYqspJ6ehL1AQEu0QKA/jV79w62M9AF819lZjFbdSox8EaTRQ/5KY4FUxy/VSCB/re/VJ690eT+T18ushllj6H3BkYY83RvT3WaoJax4UaT78adeFetjyOaooayKnyjJLDUOfG4tpZVP1H+w91635Y6G7r3cNHDWzY+dbU037URsSlzwSnvfWupu55abbWZEvljp6SVDIzykIj6voRf6e9Ur17PDoFd0bkxs8/mxTRzSMxLtF6lv+bEe/fLqwz0DWTymYpZamujRmEqMBZSbgc8k+9/Lrxrx6Bhtubm3rlmqHeSGnUsSpuAb/X3v7OtZ6iQ7AmnybxZOnkf7OS0UjKSBY8EH37r3yHRitpS0+Do/CqEsFA0/wBqwH1Fvda+fWwPLpr3LULldWpHTU1gWv8A6x+vvYPXqDpvxeOgxhpqoXupJH1/1+b+/Enr3Do7nV27sXkcfHRT6ZJURDo/Vew41A+9daPHoL/mD0bg+7+kt2Qy0dP9zFT1n2/+TL5TopiLhrce/A+vWqenXzyu2f7y/HHu7P7SipqqmxsstSiVFUCkQ11BX0m3u3A9awR1s2/y+9oZDcXx+pN0VFUtZDMkEj09O5eQal1Em3+9+/dWFadGUzkNfh66Ovx0vgxdIw++gluGPPPB9+691C7YyuGxXWuR3dtdEnzrY6WVI6cgz+dI9X0W5+v9ffuvY6p225szsX5MUeZn389Ti46XMSrTR1DNTyNEsthpLEX4A9+Jp1r7ej+dVYLGdI7Y/grU02UtTmKMqfuDygF1YX+nv1et+XRmessFLkim4aSjnoUqnCnzxFLqR/U2/wBh79w699nS87S27mchtuXE4bT5ahfUyoDzb+v497rXh1WlOizUWI3fsCWho/uStVkjpBdjYO/F/wDD3rh1voZ6TeUOJWnxm7546uWQx30N6rMv149+GMdbp+XQs02Nw1XjP4jtyBYSyeS7JyeOOT799vWh1y23vGtp8nBT5ClmkippASdBKtpP4/Hvx63w6DD51/HbI/IXp7+IbOV8fk8ar5ESRR+N9VPaUWYc/j3oGnWugP8A5dvywbbeHyfRO/qSsO58TfE0VTVNIQzRSeIOpb+tr+/EVHXga46utwVbh6zAZDGb1ymLloKqmEdPFI8ZljEi+m9+Ra/v3Dh1vqjr5Oy7Z292NV7L2hWjIpkJp5DT01QZnHkb8Rgnjn3vPXq9FtHxSxG445ty1GPq4s9CfLRyxRmIxunqGsjm9/futY6h12+e2usYIMLlKfIVWG1eLWolYCFRoUk8j6e/U8uvV8ujSdL5ttx0iVmCdo68pHLLFJIWfWPUV0e/cOvdGtp/kjD13jqnF77oo2i0lS80PBBUgNcj8+9U691S/wDPrtXrHc2ArsltWOiTJVkNVEiARC8r3tpH9T+L+7cOvY6NP/wns+KW58vvZO9crTSw0lDJri1w2S3nuhVv6e9HPDrw9T1uw3HP5I5IH197p1qo68DcX96PXhkdd+/db697917r3v3Xuve/de697917r3v3Xuve/de697917r//1N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6910QD9f99bn3utOtEA8euK3NyygEEgfQ3H9f8Pfj8uvD59Mu4tt4fdWIrcJnaGDIY+ugeCeGojSQFXW1xqH1HvRz14CnVAPzr/kzba7Yw9XXdXwJTZCqeodqWPVERJM2u4WOwHvwHXj1r8dsfyVvlF1eWTbWMqZ45WT91UqJWDBb31qD/vfv3y69w+fQS1vwD+bG2MIEO1pqyKVdMhlpao8fTULj36vW6U6KZvn4id94JpW3JtuakZJHeQxUNTwVPr5I9+69w6B/M7byOz6fTmqbJRNEtmAx9U1ja/I0+/dar0j8Nv1cBXfc0s+YWOqJRlahqERQeDbUPx791voxOM7e21DhzTVlXVu88ZBEsTqQXH5uPfutdAhV7MG7MlV5XbbkrLIXZ1ujCzXJv791voW9pbzi6zjUZ4BpY9JEjFS4K8A6mPv3Hr2ehPoPmFXLKlHiqiZ4fpYSLpC/ji/vfXvs6eZPk9k6gMklXPHJL+FIP1+n/IvfuvZPUaX5A5pIUWOpqpFZhrAVmsL8nj3rrX29OtP3RU1yBoaitMvp1BY3/Ve2nge99er6dMm/cz2/uqHGYrE4aoNJktMS1QglDWPALkD3rr329GX6z/lydnbj2nDuisyOQo5aoJIUiknjs0g1EAD+nv3WyPPoxfVHwB3Fg8slRu7K1UuOiYGX7qZ3QrruT6/8Pfvs695dG93d8XdgUW26ipw0eNrKqlp9QVmiLFgtz9fr795dexXol/+zGRdGipwGSghpKWGSZY/GY1ULGf8PfuvdE37m+ddXuFpo9pVkpYsdSwyqoOrgkhffuHWs9Ejy++tz7ilOey1TUPKpM3jc6gRq1CxPvf29b6gVOWyO/KG7wBRAbarckJxcgf19661jpNN2lVdbaaL7czvbSixRtJzf+g/4j37rdOjUfHj5qRV+ep9uT4lfuBNHGBUQ6dRPH9oe/de6PB33Xb73r1/LPg9rQiOelkk88cDXCmP8Mo/2Pv3XvmOqG6vbefwu7K+syaSR1dPKzzRlGASz/i/v3Xvs6OZ0h3FkNu10FQYhLDrhUliCrBRYi5/Hv3WurAdxbcwPevX1Vm6uio4JaWml/dUpq1BOPp79XrfnjqqLZ/TE+N7HzFPBEtZTCZ/CHBcC8n1ueP9t79TrR6sA3P1PS0vUkTVlNHTyShUXSoX1MPSPe/l1759K34yfHiaHbuRhbHRVEmRDTU4eMsxU86rH6/7D3qtOt8T0oYOrUod4VWNfGwrPjn1yRNGVAAP9CPe+tZ6MFlNh7f7C2RU7Niw9H/HzSy+L9oXLFCFP+3+nvWa9bxTojnUnXO5ujN9ZTb/AGLDNT7dy1Yy07lJBEInk+gLcDj37r3Voe1ulcVQCDd3X7wzU+SEX3Ijdbqri7EqvP5Pv3XuhZotubToq2GKoyEkOcZbmBbgF2+oJPvwHVsHpxyW09xYeCbK0tVI1OAXULN6iv6lBHvxznqtMY6i7Z7er8KWXOU6vRrwzy8jSv1Nv6e/V62fl0m+wtsdZd2UM2Yjlgpa6nJskQRWLAX/ABzf37r3l0A9FsnJ9VEVNBUziil9UThj61Y/Qe/eVOtVp0PfWHfcmNrlpcjGpiOiN5nsz6Po1mP/ABPv3DPXsHoUuwtw7S3RTx1WEr1ORKa9GsEq1uV4+nPv3XuHHoGpez8JtKnNHuqiparyDxxrPplJY+kelr/nn34de8s9AT2J2pgcpRvhotq4xaKpcgTilQkB/ob6ffgOtnoq+79j7Sokiy0QhpKqYCSGKJAin82AHvw610nMr23ktqbZlo8hRxDHTwNBDVGwYBhpDC/v3Wuii7h7RoIcDW0NBlKqprK2okCU66pAHn4C6V/xPv32de+XVxX8jH4Qbw7Y7ETtbfO34nxGHr5KmGWtp5EBp1lDx8yC1yPp798uvU63rMbj6XE0dNjqCnipqKkgjhgiiAVEWNQgVVH+A9+xTrea9TibAn629+61Xrv37rfXvfuvde9+691737r3XRIH1NvfgK8OtEgceu/fut9e9+691737r3XRIUFmIAAuSfoB/U+/de64pIkih42DqfoR9OOD79177Oo+Q/4AVv8A1CVP/Wlvfuvdakn8g0gfzCv5mij89hV5/wAQRlrcn37rQ8+tuW4va4va9vzb+vv3W+u/fuvde9+691737r3XvfuvddG9jY2NjY/0P4PvYpXPWjWmOsUayKh1nW9z+f6n8H3ZypbtwOqxhlHd1yjV1BDG/PB/w/p7p085UmqinXOw/wBv7q66xQ9UGOvAW96jTQKdbOeu/d+tdRnhdpC3nkRTb0qbDgc+94pnpsoS1a4PXNywUaFEg+hvyT+D9PdSo4Eft6fQKfiNOuTLqUAMyEC4CkD8fTn3sY6bYAmleulQlRrZtX5sbDg8Hj3ogHj1tlFcHrsx3/tv/tx/X3vHVStfPryxhCSCxLfW5vf/AGHupGajrYAHXP3o669bx17/AGA9+q/XqDr1v8Bz72S/l16g69b/AAHvRL9ex10wDAggEH6j+vvY1Vz16gOOsCwhb2ijAIsRcm493Bpw694cI6yLEgA9AU/4e/aj1Uovl13KpdCn0DCxP9B/Ue2pA7CidWoDg9FM+Y/xyxfyR6Xzew66GKeUUs8tCJVZiZ9HpAA/r78FKgZ4dbXSOPDrQX+Tfwb7m+OvYefXM4OaHbDZSb7WpipZzppVYlWVgPpb2rilVemn1NhOHQFYqlo4lBoampFV/bDh4yCPr+q3590kKs+pfPryAqtD0L3WnbWX633IKquqahqKQrGNTekAfj/W9t06c6sM6/8AkLt3IZKgyclSsboFkv5UBuTYg39+IJ69Xz6sZ2H8otqQxx0S1sYqK8CGI+SO2plt9T70adb6OvtTbtNuba7ZzIzoIZVEqeoNcMNS8e9deJOOm3B9exbir56OknCw69IdRpa17GxHv2etmlOn7JdUybbk1NlJ3hj0uyNIQhBN+Rf3s5HXuB6Ld3purb+2sFVVjS00csEM37nli8n7a/jke/D59aPWvt3737me1MzPsXblbV1Zq5DTsFJkRfVo/s/T3vrVfLpG9a/DjJfeQVmZpXaUTLUSl4JWuknrN2sffvLr329GX3j8ZMblsItPh8XTz1UcXjbVCQQdOkfj36uOvEdMfSv8udKnKSbhzVDCiws0sgeNgqgHULL799nXqevRk+x+ndsbFwEZxqwRywALaOwIKf1vY+/Vr17pHbLz2EptvVpqJY2q4Syx6ioe4+nvR699nRPPlLvLG5rDU2PlEbONKC7hr8/m35978utHqrLrLfuS+N3yCx/amMqZoaasylFDWpASESBZBdyf8B/j791v+fW7H8fPkHtL5G9ebazW1MtDX5aDGxNWqaqEus5Ua7rf+vvRHWwfPo27Zqtw2Kop2iUVtDplkVNJJ02Njp/r7916mcdF87S7F7Q7cq029tTHkTRHxmVAVfxj6sLfj36lOtdLrrTaGawlHFQbh1y5Jo9E6y3OmX821e9Hj1bgK9L/ADlDRU0awrHE1Qwt4zYj6fSw/wAffuGetccdQIKBaagad6eOmk/HjS2of6w9+oR17Hn1jyc1DLTQWp40Nh55Qo1G4/J9+62K9JjEU0f8XL0/7sEhP6h6Af6C/v1PXr2el1W7QgylTTxxKoZ1BKqp/pYf7z72etVPTVuTbJ2/RMKoIp0lUDcc/iwPvVPTrYJ4dJrr7ceUxGcWSMK0LSqtgwsVvYD6+9mmKda9ejnrmnz+I/hNcFigrYZFkVB6HEiFfVb/AB96+3r32davX84T4C0kWMquyKDGQRU6wtVy1kUTecJrLMSwH192615Y6DT+V/8AJLG7U2C+yMDWSZmqp5BjvsZ21Is8QMYGhv8AWt79x68MdWInZveW8s7V1FZghS7bys5meRVKosLH+yR/gffvt63ToyfVvxwwuPrCdzVhq8dMVEtJVOTCqHlxpPFvfq+nXuA6z91fFTZVZEs2wHp8dGLtIMcQgMgH9pU/x96qCM9eHQJ4fq/L7Aijq8xQw12IplBqqiqjLlVHqJGv/Dn3v7OvZA6eNzd44BKPDYnatPEEmrYaaQ08QFgWCsdKj8e9U8+vV6MPNTwTYnHiiAnq6ykimcSHSys6Xb6+9fb1bhw6DjeWxcRkqWHIZdzBX0A1wKhJ9SjgXHvf2daP8+iFbk/iy77aaq8v8Ojm0wu4bSxVrAG/Fj7917HDo2eAz+4qHbCzwUIemSJLEKTqUc349+6159Lfa29ocw9HFU42CJlNpJPGFLAH+178R17o2VBXjN4hdvQ06LjqiEQu6AABZVswsv4/B96FePWz6noM4/gZsWHccW/dtxwRbmZmqUiiis08pufWQOefe+GOtefRLPlZ1r8m9hUW5dyxfd02MTHyS0axzvGFKA6AoJ4+nvfXq8T1Td8Id17x3t8s6rI9r1U8z01Q1NFS1rmWE6XIuNXv32dVGOtgKeTDY3MPipKeCFMtOVoAqrbSxsLW9+GMdbI8+k52RtLGUtBSUm6MJj48LXyJBDkGhQt+4bXDEf43v79WvXqdAlnNj4boiph3nteb7rBsBNVrf0ILBmUA+/DPW6eXRWflB8jetuydmVK4etp4cxRUxaoWJow4cKRYW5+vv3DrXHqqb4w/HDffzO7ppdk45q6sxGMzkclUrxymJqcyaidSi1re/da6+gh8Kvi5gfi31FiNl4qmggqno6f75oVIYSKnqUk/4+9/PreeHRxEXTfkm/5P1t/r+/E1HVQKE9c/r711bj1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf//V3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XBw5A0MFNwSSL3H5H+x9+60esNTR0tbGYqunhqIz/YmjWQf7DUPfut9M1VtPblZEsFTh6CWJforU0RFr3sePfutU6CbePxo6c3vE8eZ2fjGMmrU8dNCCS3BNivvXW+i6Zz+Wh8WdxO5ymw8dUBibh6WnOq4tz6fdhQdezTotnY38mP41bliKYDa2Mxx40/sU66AWubXA96+zr3l0Rfs7/hO9srdSTNgM5BjCRKyRxyJG2oj0KpFh791r7eqifk1/KI7e+LeLyuV2zFlcviqAapqlGMkIS1yWYD+nv3y63w4dUibw2tmd41Qw2XqWx9TT1EkcxdmRv23KkMePyPfutY49PUPTNHtjDLlUyaTOoVWHkvf+v+v7914dDr1d15tbcEUbZKup0eVeGd1Fj+D6vfuvHo7Ox+qOpKULSZTI42TSgIkkaOzDT9ASfe+vCnRgtodYdE0c61J/hs0UZDMh8RV7G4v/jx711bocMl2l8f8fBRY6nocXSyYrTebRCNJX8k/jn3rz610oar+YR13tDDDBYfJY+rMa6I6aLQzakFlWyX5P497p69bB6Lnvz5j9xdlA0HXGy8lX/cFREaGnlAkB41DSPfutcc9CJsv40/NXtzGYlhR7i21BlnVKlilTGEVubtq/pf37A4deFfPp075/kc92y9eVG+avcOX3Nmlpp6uTERzs8zME1EFRc8n3v7etZ/Z1rS9t9Q9idK7wqtt7o2hW7faCp8MctUsqmclyAfUPevn14jp2x+4pqyipcNT4g1VSxWGaRFJYACxP8Are/Y630q2xMOBhSlfKLjKuqXWlG7KJGeQXWPSf8AH/D37rVehU6U6ayO4d3Y6q37s6obATSl1y1VCRBJHqusgLC1rH37rfHoce/Ohuvers7tbfOxqKmmDZOkepjo9HqGsawVj/Pv3WurS6T5Fdep05QbfO04v4k+GjiM5hUv5Wgsfx/W/v3Hr3VN3YnXa57LbkzSY94IshqaMFRYXJNgP9b3vr3RS8ph6zaUJenBYRysfGt7gKeSwHvVet9LjC/JbcOB2lV7bxUshM5EbRR6zzIdJ9+690NOxqrJYSmw+6cjTyyT5VhJIGUkXcarE+/DrXnnqxjeu0Zt89B4rLUNR9pKaund4V9Labj8f8R791s8Ohy6M3NHsnO7Ixdbj2qKBaKGKrqyl0VmAVjIfp/t/fj1sHpW/NDZmN68ix/bW3KpGps08clVHEyhVjchnJC/j36vXjw6BHr7vnZ9NQQ7qwtLBlstD4oJIIVDyFrWcFRf8+/Z69UcejF5KDZvyA24zZ7G0+CzkkV6P7hEildtNxouB+ffqdeweq+6jtzs/wCLnYMmHqFrc/tJKhiqIskscMAewvbi1vfuq+fRvqH5I7M7Dipc9QYyGmrxToJJUUqUmYf2x/gffqdb6hZzsndOEp5ssMi+Vxuky/w5C0gKkXKBP9b377evfPoKD8icPuxlxeQxK4QSyeE+dClmJsW59+69UeXQl42KkwlEMtgskmQWQeZsfA+tnuL/AKR795063imOp9Z2zi85BHis/EmPkUaEjmGgqQLAANz79TrVekTWYaLGF83jJ1yUZUuKKE3YC9x6V9+r6dePQOZ7uSqxNeI/A+KkjbQRJrS5ta9vfutcOg0o+3ZMxuUPnYWytDBPrDEFlUKeDz7316vQzVXZezcrk6eBMPHBSiJAZSoKeQC1+fp711uvn0keysfQ5yjjyNFULDBSIWSJWADKBcAe/daOeq9e496yPiK3HThzTY6CaZWN7HxXIsfp7914dH4/lLfy+KL5qVtNumuqFioIK2mqXBkCoEgl9ZKvweB79XHW/s63z/jt8fdnfHXY9Hs3atNDEIokWoljjSMzFFAu2gC/I+p97ANK9VrnPQ6V9WmOoa2vnf8Aao6aeqk1WsqQxmRuf9Ye91HWgG9eqbuzP5yXTewd6ZDZkFLDlq7HVstDUeCRyUnhYqwOk/4e656uOkfP/Ot69iMYXZ8zeTgC89/979+611nP86fr4AH+6Et/qbmf8i4H19+z1vrgP51PX3P+/Rl/5KnH+v8An37r1R14fzqevif+PQl45tqn+n9bk+/fLrVR17/h6nr3i+z5jYXP/Ag/6359+z1vrBD/ADsOvJZGT+6Mo0mxJM9x/QWB9+8+vUx1Jf8AnUdfDhNoSsfrwZzYf61/fvs69TrqL+dT149y20pltxz5wb/7f37r3z6jf8PXdf3cHZ0xUHgfvXZfp9CefejXr3l1yX+dX18oAbZ0sSn9ItOF/wBYc+9569gU6mUv86HrauespKjaFYYnxtaY3iMo/fEDeJWJvwTa4/p791rPWvf/ACjfm4fjz8xPnN2HvTYFdT7f7R3fkMpgpzHInniNUaiJ42t9NXBH5Hv2fPr1Rx62B1/nWdeNCJf7nyhiORefjn88/wC8e9eXW/PrtP51fXzC/wDdCX8/mfjjj6n3vPXuuz/Op6/W3+/Ql5/2qfgD/WPv3Xuu1/nU9fEc7RkvySbzj6fgc+9Z69jr3/D1PX1if7oy2ufzP/T/AF/fs9ex69eH86nr7n/foSmxA/VPe5/2PvfXjTr3/D1HX1j/AL9GW/FvVPf/AB49+6914/zqevvr/dCaxFhzPwf9v71nr2Ovf8PU9ff88hKPoP1T/wC3PPvfXqjr3/D1PXpv/v0ZR/rtPx/hyffs9ex11/w9T1+AbbPlc8WAaf8A29r+/de6Hr4r/wA0jqv5NdiVfXGJjgx2cpSqSUryN5lc/UMrH/eLe/Zr1o8OrUFAHINw3P8AsT+bj3smvWlAHDgeuXF/8R/xPvXVuu/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691x4W5sT/gOfoPoB73x6rShr0AXePx2677329V4ndmEpKmpkhkSCqkhj8iOy6LsxH+8+9UHWx8uteX5NfyPHBqM513lHgExYrS0jopDFixGlRx795dbxXqoLub+Xj3Z1Hgp6us2bm89BTtM/3DRyOuiE316gP6D37r1D1XNlZ93YeaoofspsNVUrBWhcyqyMp5Q+/dap0KPXHyAqsDV0dNneZ6KS6zyO4sR+QT791v7OryfjN846HI4ik25lsvA9IY4kDOxIVbaQDf+nv3WwfPo/eA78wdNUwnB5GCT7hrtJHpYKCL8kH3rrfHJ6Yu4/lFi8NtjLy1OXg+4p6NpV9ViSoJGr36nWj69avXenzF7F7Z37XbL23VVE1NPkZqXyQmXT45X8fpI976r1YT8F/g7NWyxb43bUu9dIxqbTuGbVq8npD+/cOrAdXj7P6y2JjhVU1dDTmUUqxhmWI+oJpHHvWetn5dAFuvZ+GwmanmoGiFN9w7FQEK2Bv9B731onpmyXZuM2jha2NXRWeMgelP7Sm30/w966969Vcd8d0U+TjqaWCrXUZJiLNb+19Pr72Pn1omvRFc/wBn5HE4msehdnmOqyJq1NxYHj8e/da+zopG7N65LM6ajMVpjMh/ajkLekk2H19+63w6BfeeTwlbi2xuUEbSuSIKl+FaYr6FBH59+61w6tP/AJUXS3yqyENVvbY8GZTZWLlZ5KelNS8M9OkuouR9LW9+yOtj162kuhd4V2/KzJ7b3TQyU2RxUVqpJlIkkEaWYHVzyR71w631C3Buyp2Vv6OTbmLkpx5BC5SMkldViwA9+4jrWBx6H3PmsnwFLuOnLffVEQnkjHD6mXUQwH+Pv3Wxg0PQC7cz2fze5KiOso5mWInkhrG39q/v1B14E+fQzwJLWzqkkbGNVsYiOQR/gfeuPW+A6g5AOsrUsePJUixOkcX/AKH3unWq+fn1Ap6GSnkHhpyrhrqB+T/Tj3r7Ot9DTsHHVE0q19bGUESkLrAsLnj6/wBP6e9/LrVOgU+R+H3HlzHHg5pUVSoBQmw/qeP9jx78Pt695dAvsvbefoY1jrq6T7mNxIdZJ4Bubj349bHRyet8qtYsdLWt+9CoRS5PNvyL+/dVIzjoGfnvgX330tuLb0tE9RH/AAeSOM6R/bQqDqt78OPW6V61Cfhg2K6W+S+V2PU0aGWoz8k6RXLEF5zYkf7H3vHWh1tbf3zq9sbHj3BlK40WISDyhZQFjRNPABPvR6twGePSF6s3vlO7K7NLjKv+H4mjhkalyAusVUyA3VD9Cffsda+XXPZtL2BB2BJTVVXU1OChd103YxsUY3JB4vb3s068OHS73lnxuSpn2mkJmgqX8FTpQWVbaCG/p719vXvl0DcHUm3drZen0YtK0x1KzCNQHKkm5/x9+x5de65dwdhf3TrcWtNEcaPFFGi2IICgDi3+9e9jA62QCelVTZZKzF4HJ14+9irwjPq51AgE+9Y61X9h6C7tvE47IyUBx2FMampj1SKlgoLWZibf7f3vr1ejAbUw2FpNrUtHVlH81KhMbafSStgOf8fejx63xFOkzP1Y1TkaepxM32sBa7FWCqwJuRb8e/A460Rno4O19mT7a2rTZaFTkZIzGzooDMVT6/T3rr3HPQ3bd7gwNPjBXZTGxYVsZGymadAoJC8t6rfn3senXqdUc/zXPlbncjtenxmw9yjJLMFinoKIhiV8mkiyf0/offs9a8sdUHfHze1Thu8qfMZtmxdXNNDJJHMNMjOzBnYA/wBfe+tD06vvl7M2vWbch3PkslTQzYuHywOxAK6LEBbn6+9dWPWLt/5RbBynU215KzNUEzw1cVizp/Zccf7x731rotHd/wAkNr5/pus/h9bBNAlNJEUj5H+asDcH34Hr3lXqirZWyN3fIntjHbG2C0oTcFSKatlo5JLpebQbgcfn37h1qnW9V/LH/lyYb4j7WotxZVfNu3I0VLNNNKEea8lOCxZrX+p5uffutjq4Ej1A/wBAf9597rinWiMg9cVkDO6flCAf9iL+/deBPWT3rrfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//W3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XVx791oEHh13791vPWNWfUwZQACNLA3vf3vHl1WprQ9ZPeurdcSRfTc3PHF/6X97zx60aHHXbMFUsfoBc+9cevHA6x6zIhaK1/oC4IHH+HveAc561UkdvSb3dszbm+8BkNt7pxdLlMVlKdoKynnijcMrLYlS4NiPwfeutitOtef5h/yNdtdi11Xl+j1oMBU1Pkl0TRxRsJJZNbaWFr+90p17jnqmLs/wDkx/JXr2WbHZL7jJY5ixjNLSmVHCrfUpW/HvXW6dEl3l8A/kr160tTTYXcElCDdVp8dUKwB5t6R72M9a8+i6ZPpr5MHILRQ7Y3qEaTxRstFVj1g6dTWH096p175dGe63+DHzi3jQRS4ZNw0UU4jEYqaWpU6XbSNV/fj1scerKOjP5FPy13dEMv2Hub7ejrPU8VXJ4HdSLjQrG/v3WqdWm9DfyGeqNrZOnyfZbNlZIvFLIsZ8oaROXAuT9fe+PXsDq3HrX4N/Hjqv7VtrbLoI5aRQsclTTwTXZTfUVZfr719vWwfTo19Li8bRRRQU1BRQJEqqiQUsMSKANI0qo497zx61UDB6mPDDIrJJFG6MpVldFZWUixUgjke9db6qr+bv8ALC6q+VFszT4bF43cYYyGZaSBEeUA2drD+vv2fLrw9Otdrtv+T5vv49ZOs3RUUIyOEjMlQPs6JZUESvdTqX/D37h14Dqir5f9V5CbeVNmNtRVNFVYicFqWxiZ5ac6gvi4vyPfvn1rHDqdgvmB2Lhtp0G1t5YWaixeNpxTrUGnELyRhdAZZLX/AN59+638ul3je7sF2nhqDb+3qXIVFfSTK5lkR5k8hNzdiPfq9a6FrHZrclJlcbia+CpfWsEcaIjMoViFIK+/dez0e7JdQUL9dfxfJxxQPU0nki8qLGfVHcXv798uvUxXqjrvWoqttZiupoIvuIPNUJ+yuoW1kAgj/effutjoFdh0sE+TE00YEsrBwsg5BJuPSffutdWQ9M46q3SzUWf8S4/HgmiVlABAAAA9+69Sp6PxjKtv7nttuAjwU8TSQqf06lX0mw/r79xz1utBTos/+krsbAVWSpq5PFTU9RoondSpMSt6SGPP09+61n7OjdbG7Hw3emx6rYm9KlZ3hpTT06vL5GVilrKD/j78etjOOiTSdeydAb8mammkmwdRVvULGza4gmvUAFPH09+61Xo8Gzd77e33QwZqmqUhqsQoaOKKUIWKAcaVt/tves+XVqg9CLRZfpvfRlxW76WI5CRPDJLPGliCLMQ7f0974de8ui+b+6owfW2eXP7OWSs2Q5MlXT0V5WLWv6Qn+9e/dapnpw292NsHOQtSpG1GERleGuOg8cepZLf7179k9bBHDpA9pbW2dm8Q0u20MmYUM3loxeznleY/futHPQBbX7D3H1UJazPwV8i05KQ+YSNEIwfyrcfT3vrVesOd7m2l2l5chiZjFlaL9yZY5fH+4v19I9668anpL0vybn21GcdTGSbI3MTeQlkCD0i9/futmvWDL9kbc3XQyZPcxVKvllEbhSG/H0+vv3Hr1egjhz+Qr6uSPbBtSFrXYephfkaj799nWuhr27uvb2Owxxm4vJHkgvk8zMQNQ5tqPvfXsdBFvzvFceZaDGz+SmFo7CW5YcCwHv3Hr1MdYOlPjf258tN64fCbcw2UkwldkEpsjLHRNKjU8pBcal4It9fejjrfy634v5bPwS2p8N+osTQQUIhzlTQ08tX+yIpYpCA7KVH5uffhU/n1qv5dWdal0+UAksvH1ueLgW92z8PWqimodIrsmZouut71K3V49qZ2QH6EFMbIwt70fQdbXOT1qs/ysPhF1X8mt8d49kdmxVmVlxm/smI4JJPIvprGCoGkvpH+t71ny69gDq96X+Wh8VpHDjZgQC2lQ8ZA/wBY6ffqdbr1zb+Wn8WGAU7N+n19cfq/xtp9+691j/4bO+K9/wDjzSP8NcVv+hffuveVOuv+Gzvivf8A484/8lxf9G+/dbr1xH8s74sBtS7RNrWK64iPp9bhffutHj00/wDDdHw8jkkjO2YBKrfuL5Y7g88WC+/Z698+m/8A4b5+IVM1Sf7tTFQLkLCCAP8AaDp5/wB49+690A3efwr+JezOvW3NTYCtpga+OnVzTCMm7hSL24/2/vR62POvSz2r8G/h7X7HwG5KzbtVL91QRVDulIrksUBJPH+8+/fn1rIPTlj/AII/DzfkiYvb+CrIatGIdnpFGkryL/4e9jPWsgdP+T/l5fFXZFEP4lgaqpnktpMFKGYr+QdP9ffsefWzWmOsuG+B3wvaF6mHaApGnUNUySUiUzSk8ks4Xn/Y+9Ur59e6fIf5e/w1rQop9vU0pb9KxzxsQSfyipx7317p5T+Wh8VHAaPaAYW4KyRH/el9+Ip14H065L/LN+LIJJ2gWv8AgtHx/wAm+/de65f8Nn/Ff6/3NP8Ah6ouP+TffuvdeH8s/wCLA+mzj/sXit/ttPv3Xjnro/yzviwTf+531/GqO1/620+/dbqeu/8Ahs74r/8APHH6Afri/H/IPv3WuvD+Wf8AFcc/3NP4/txfUf8AIPv3Xjnr3/DZ/wAV/r/c43551xfn/kH37r3Xv+Gz/ixzfZx5/wBriH+9L7917rNB/LT+K0Msch2UJNDFirPGAwItYkLx7917rXk+NnTOzuof52PaO1dlSVlPt+KETRURkKwxSX4UQjjj6Xtz795daoK0HW4nD/mov6eNP9f9PvZ49aXh1kKgkH8j6cn/AIj3qvW6Zr13791vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuuIBF7m/9PezTy6qARx65fT3rq3DrE8UUtvJHHJb6B1VgP8AYH37PWqg9J3cuztubtxFXhc5iKGtoauF4ZI5aWB9KuNJKFl4+vv3Xuqtuzv5R/QG9M1U5qkxNPBPXTNNOn2kIQOfqTYe/de6A6u/kd9CZFmeaggErcalgRRf+t/fqdbqOiDfK7+UjuHprBVe5+rEqnxlFHNMY6CJmmAhGoA+Pnn8e/fb16mOqc8T3L2T1tuGbbuVgylLJSy+GSSvMkYB1aTp12/p731qvS67F3nX732xWM2RkkqamjZWWOpux1Jc3UH6/wCHv3W+g/8Aid8czVZ2XdOTgBK1iyrLUxKRxLqB1v711rj1fz1hunbOxsDIMlWUEcdFGbrHIqAWX6ED8e/Ur1YVB9OkFunv7FVlTX1W3qxAiQykFagkXT/WP09+pTr3E9EmzXyOz1dlaqnNejQo8llab6WJvf8A1/e89Vr0V/tH5KvU/c0CVdniOl9MxKggWuPeqU62eiMZ/f02Zr55TJI1y7G8hK8m9/e+tdBjlN6SUqyyTSqI01akdhzx9efeuvU6LVuXO1m58gwoqeorihvBDQgSMWDfQIn9Pfut4GB1Yn8Jf5aXbvzA3jgIKjCZjH4Cky2MrZpK2gkjhkpA4eQM7j6EcHn3759a+Q6+hv8AFD4s7B+L3UuD662tiKGIR42CPNTCmhX7uq8QEqkAcre4IJ9+690X35B9O0nW+aPYGzaF42yc18pFSw6VERe8gPj/AOKe9EcKdb48ei/12S2jm5I8lFTAZKNVEiOgMnm+pJB/xv711sA9PeKzkpqKeirApopDYIQLgfgaT/vXv3WzWnS8o4Nt0FbI9LTRLJKBrbxqGN/qD79nrRoPn0+Jt2ORv4hSgaCeVsLgn8ke/VPGnXqDqVDg4mcmZFBf6sUFrk8f7z+ffj69ex05/wByYJo/uKZ4/IoLWOnUTb629+yOvUHU/ERVNNSzUkkPrD6VZFvx9OQPfuvY8+lNUbVoKqhikq4FaWVVIVlDWa9uL+9fZjr3Re83tKPH7keSRFSmJNvSAp5/r7sOvE06d4MZT42RK6kbTp0nggAj68+9da6FOfG4DsPauRw+VWFzU0/gGoBmJ02/Pv3n17rTe+f3RmS+MfyZHbmAxlYcVT1stZUSU1OyxFI5dbMZEFvp7vXrVPPrDub+ZUO96vY3V+DqjSYKSnp6bcpeoMTgrZHHP596Hp1s1PV1Xx97f6d2jsfFbRw+UoTWxrH5m+6TzlpE9Yb8m/8Aj71Trda9DhT9kiTcIxuCenCVVNIySmx9Tg6bP/sffqderU46A7akHZm0+1J63dciVO38tXM8DCxAiZ9Shf6cce98evHB6NLU7h2vW7rkp8ch88aRG7+pNWm9r/6/vXW6nz6KB8zNrbkye2v7yYlRGtJURrdVIH6hzr/2Hv329ePQ09K4OXLdS7YrsswnkpKFXm0ESMNKXOojke/H7evDoSHxeI3DjHgo0VKimUn9wD+x/S/v2Rk9apXh0G+Lirpcv/D5vK8EcnjHiDFQqt9PT72ePWh8+hgqqlqSgeGiIU0yHXrNmJtzpv7qM9XyOhG2r2pPRbPNLYGZZNDeT12UG3+w9++fWuPRfvkZ3PtvFdabgGRydJDkXhfww09Ssc5cxm1lXn6+9jj1o8OtdzduaqMuZsyVrqiKTW1K9Xrmi1hywI18e7dax0QTc0mfbtenzU7+IQuNPh/bAQG9vTYe9da+XRxRvXM7q68ylBDXTRtTQNH/AJ51LWQ2tz798+vD5dE8z27mq9urtrNZOr/3GPJKi/eMCSGuLAkf7f37r3n0H21n7l7Qr4Nj7Axu4K/DVFfSU08lPFNVR+CaYRubrcWseT791s9bsv8AKl/lf7E6Y2Zgezt4YQneNRAKsCspEWaOdwJGdvICRY/4e/fb1qvl1f3TsGiTQhjjVdCqQQQE9It/hYce9kUx59eBrny6z+9db646QLkAAn6m31P4v73XrVPTrygqoBOoj82tf34mpr14Cgp1y96631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//19/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XQFr/4m59+r1oCn59d2/Pv3W6Zr1737r3XvfuvU697917r3v3Xuve/de66tz9f9h7917qFWYzH5AAV1HT1YUWUTxLIAD+AG9+r16nTFVbF2bXIY6zbOFqUP1Wegp5FP+wYe/de6Sr9HdRPOtS3Xu1zMraw/8Mg/Ve97W9+8qdapmvSwo9m7Ux6JHQ7exFIkYARaehgiCgfS2gD36nW+lGiKiqiKFRQFVVFgoHAAA9+691y9+691737r3Xvfuvddf7H37rVD14kLySByBz/U/Qe/deJA49JrdW18DvDE1eDz9HBWUdbTvAyzKrWEg03Qt+fe6GnXq5p1r3/I/wDkx4nffbce7sCiNhamu+6lpkd0jVZJNTqY14/r9R70OvEjiekx8sv5HnXW4eqMY2yMdTyZ+hoEWujSMqxmWG7G4HPII9+z59eoOtanduypPhPuWq2/uPacKNDUzqaj7Jy6JFJ41cuy+/fPreehQ6+7m6t3PkqbPNLTPXRIsn2roNSG9wAp/wAfr78cjrw9epvyQ+Tm8MxtxsTtHGyfZUEQRnhX06Atm4H+Hv3z68T0TTpfaFL2vkMhVdj3oaEiV45prrdySTy1hx739vXvs6CvtDYeB2LvqCLa1UK2j89gEYFdANw119660OhKwPaU+DFNHIGolRgNcS2L8WF/6+9/Z14V6GKm+S+To441xNP95LHpLKy/rVByG/2HvVet19OuGc+RWI35LTY7M0UGHrANBCKq63AsSxX/AIn3v59a8ukZT9j1Owck1ftyqkqJC2tlQjgnn6D3rr3DPS7qe6/9IdBq3HphrAukayL/ANLm/v3W6+vXPZu6/wC62VinxmQZ6RmBqIlYeO17m4v78Oq/b0Le8ewsTmaSKehrhj6trB5ISiuOPU1wffurefQmdWfIXD7LxjYvOSpuSnkj0n79klUE8Xs9x79Tr1Rw6Bzt+v2nu2orM1hcjHglqD5GioWWNIxe4WyW9+60fl0w7E7ioevsYiQyDcMhfQzVBEhP4/tX9768PTp83p2rJv7bVZRja1HHDULJqn8cetNa2uOPfuvH5dV4zYDcmz83UVeJpm8FdIS6RqQoVmuT70Pn1vpffb0DUaZLKQxxVLAB7qL6iLk+/U61WnWN9uYTOUck4yjRFBcRIQBwPzf37r2ePTUu8afYNK5CGZIxdHjUyM4Atc6b+/deA6b8Z2DN3NUf3W27R1jbgd9CgUNQjEObIdenn8e/dbp6dXA/AP8Akyb3+SolzHZ9JPjKCGS6yS+aFWiSTUCS9vqPfvPr3lXrcI+HvwI6k+I234aDa+Npa3K+GESZCaBXaKRYwrGJnH1/x9+68OHR7QL8kEfix+n+Bt73wwOtDPHruwta3H0t7116gpTpCdpW/wBGu/b8D+6G4fxf/l1y/j37rY6oj/kWOZ8X8gkQlEPYGTvY8WFa/wDQ/n3uuetEH+fWwx/xHvXW+vEgC54A+pP4Hv3XumTI5umoY7oRNJz6VNwPxc+94pXrWeHSJq89mMkGSihcEghdFxYD6/6/v1fTHW9Prnp9wCZX7do8l5YxMpS55C6+Lgn8+/UxnqpNCKdQP9HtF981b97PqaTWUtZef8AfeqdXr59LKnxVDTxCJaeJ7D1NIiuzG3JJb3uvVaCteim/NfFQVXSFfTxU8Q0ZOjeNFRVVSX9RCqPeut/Z0JvRGGpJOndp0dbRQSK2KiUq8atceIKCLj8e/V698+HULr2jp8dv3OUsFHFDEqSCN0QKb3NyPehx69XHQ9T0dLVALU08NQAbgTRrJb/W1D3vr3Scz20cXnMbPjhElFrUqJYI1Rl49J9Nve/L7etVqfs6LnQ9b5LrrPS1i11RX0M5Ywq5JW7/AEsD9Peqdb6GbD0+deL7pTKI/wBagG3BF+P9b3vJ49aNAOHTxit6UU71FLVMVnoriZwPwDYHT/vfv2D14VH2dKfHZjH5VS9FUJMB/Tg/7AH3qh69UdORNv8AW/r/AE9+631hmkkQxiOPyams3+0j+t/ewOqk5p1n/wBh/vv6e9dW697917roXub/AE4t73inWs16796631737r3Wpp1xTgfzzO1HDkH7c8G/5f37rXA9bY8QtFGP6RoP+TR791vrJ7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de64lQWDXPH4vx73XFOqlamvXL3rq3XQvzf/Ye9460K+fTdkaDH5qkqsXkqSOso6iMx1FPUIGjkRwQQQfevLr1eqg/lb/KT6m7rOU3Bt6nhxGand6lYIU8QZwtxpZf8fwPfs9bOePWvX3J/Lm+QXTGUni2vt2ozVDC0jDyRzSro8hC8kH8e/A9ap6dBzT/AOzHbDwU2Jk6/koGRSpmgpZbAqOWJA9+wet56LZvPujtvbMNdBuODKU0VYNLkU1Q/j1fX8fT37HWiaHpA7O+TM1CtZSVk1fIjo6iWWkmBLHggXHv2a9bHDoEd0d6TwZOpqo3yC00k0jFko52sGP5Kj/evfutZ6CLMdjUGQM1bUT1g8vquaOouwHIJuPfuvU6aNvbnl3DX/w7b9LW11XM/iRDQVGnW/0BNvfuvECvRwepv5eHyU72zdBTxbRmTAZExSPUCGoUiOV9J/s+/db8sdbDvwq/4T2bI2pVR7r7Uus6uswoneSUuzKGCCNuAPfutcetlrpzovr3o/bVHtzZGBoMfHTQpE9VFTRJUzBF0rrkAv791voXpEEyFGJUH9Wk2PH4v73wPVfiGemzMYPGZ7Hy4zJ00dTTSIUKyAMQCLalv+fejnj14eg6Ip2L8V6PD5KXc+2GaYlnlbH8hAL3PoHHvx9erA+vHouG4MLNhq6H9S1Ksvkhs3ob8rc8e69W65y1OSi8E7QehmUO4F7A8fT3utOq9CttfcNOZ4sakvkqJEBMTW44seD71Q+XW+hJgoqfMvNTIxin1aNINgT/AEB/B/Hv3HrWfLqRS7JztDVBmeU02oXuWK6D/U+/U630KGGxWPigMccaVEzW8hYXKtfgc/6/vY68eFen3IbdWoghBAh0EadN7fT6+9kV49arnoBd9bUhnmY+TSUN9QYn6G4B96OD1voMJqOWeWKjUHxqvjBU/W35596r14jqBSSZKgq6mCnLgRXtp41c8D3unn1rzoOik/J7q/avee1MptLdWLphWyUtXpqpkBlYsp/tf7178Mcet8etUfu/4M5rrvP7jznX9Gy/wuqbxS0sbqy6fUSGUfT3vh1roq3WXZncmw9/US1+SyU0tRX+H7aTXYKr6QLN+OPfqY691su9G5XtPc+zcdn4cUHlWlpXaoLDy6QLkW+v+v7914cKdH761x9X2bFFT5f/ACfJYkWIc2JkTi1/euHW+n3d2y6nY9RHlKONZ6kspqPqSI/ofp/h79177OgA7S712NuR4OnMtXQ0tblSmq7RqUkf8A34N/e+HHr3Ho0fSPXp21tmLFUlV99jqqEwRF51dVVk4tzx7116tOi7939ixdT5eTD0k2movI8qRuptHqsQSPfvt699nQY7b+YGxNs1MMmbqYEmkAZ2kMdwz/klvfiOvA049CNgu+9kdhZGpqMVmENNK/8AlGmRAiR83+h/3n3ulOtgny6LP8kfnLt7ouNsVtuSLKy1GqEohjlKyOdAYAX/AD71TrVfTqujf+7832xRtvuu3FWQUEyrWT0IkIhVSL+Px3t731o56fdnb92FvLbcW2Kn7aGWhQpHMAivK6ji5/2Hv3nUder6dEV+RGJh2tWvlMYwlqDKwgjjOsvdvQNK+98etcD0ktlzd45PBFsdtKp/hlREC060k4Dq3Fybf4+9db+Y6Nr8X/5Xvbvyu3bE2ZxNVjsN5/3pws9PeOT1NqYgfT+nv3XqYqetvH4O/wArPpz4sYDH1FfhqHNbhWmg1mqiWYRTR+rW5kHJv79SvDrRx1azFFT0McFLTQx08APjjhhRUjUW+gRfx72BjrRrXqUOOPx+Peurdd+/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r/9Df49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcWVWFmF+bj/Aj8gj37r1OuLxI4AZQSvKk/UEfQ+916rpFMdejLEHUQbMQCBbgf19+bB68vD167Ea2dSAyuSSGAIIYWIIP49+OetgU6rm+X/8ALr6a+TOCzNRX7col3JU0dQsNSsMILTP6gVNv6+9cOvdae3dP8s3efxg7VrsjHg6+t2yK6VlW7NAkCkt/ZFh79WvWyKdAfu7emzcB5qCrp6WnNQNE1LIRr+ugix9761005reG39wbC/u3s3bElBkArM2WhiKobm5YOo/x9+69X06ry3PWZTa+74KbMVhyTTSrZQbmLnkEn3rr3Q1V9Vt2bC01ZPSrq8atwLW449+690hk3HigxXGBYqgvpYA3IH+AHv1Ot09en6li29V0M71USvmXu1PP9Cj29+4da6DHTkKKpyD1WTuqoxhQudVxfTYe/db6hYfM1klLK8tUVm8rKuosDpB+o5+nvfXuPTlU7qy9AqolaYY5LFnL2Fjxz7117pTQVmSy+PiWhzsVRUm37aT+oE/j6+/Y8+tdMleu7KBhSV+SFKjsHWeWZlUD6jkn/ePfuvdKWly2RylEMdT5eOV41CNomJMv4Nrnn37rf29OmKi+zT7Wsro6WaIl280ttV/wB791XpVYzsNsXXxRDIQS45LJJTiQESWP6uffut09elRnN/Y3cEQpcPQpUVRUKscVi+q1uAPfuvGvSMp9h9s5ypRaPYeXzEFRIEjjgR2Hr/SRYe/U690cXrX+WD8lO6sUtRRbY3DtOKoUPGWjljBU2A9RAB/r79w691Zz8Mf+E/8Avpty/wAS7kzdTkcJFN5HpskVZfGedOlvfut9bCnV/wDKZ+JPWtXQ5eg2Jj3zNJDArVa01MrPLEOWYheeffvt69X06sO2hs/beysYcTtnFQYmjhsoWOJIg+kcN6AOPfhQdaNTw6Vqm6g3BuP1D6H/ABHvfn17iMdcveut9e9+690g+0v+Zbb9B/O0Nwj/AG+LlHv3Xhx6om/kToIsd8g0F/8Aj/8AJ2va4P3zk+9/Pqvp1sJXbVaw0/k83/1vfsU69U16Qm4dwo1b/AqWUJVkAswazC/0A9+86Drf29TcXt11AfIuZtS6tGo/U82J9+pTr1a/Z0p6aipaW/giRCeSQLtz/j79U9eoK16le9db697917r3v3Xui2/KU07dV1pqAvj+9puGFxcP/j78wpjrymuehE6ddH642x49OgY+JVA+gULwPfhgDrx49ONHteah3BUZWEgCoLDULEAN+WH192pmvVa/hz0uxcfU3/2H/FPeuPXgCOJ66Zb+pTZh9D+Df+oH19+B8j14iuR1FqqCCujWOqUSBTqFhbn36vXgD5nrnqgo6a5Kx08KWBNrALxb/H3s/PrwPQP7dwC1e4M1Ut6qSuLWP4Ksbm3+8e9ADj1Y4x065XaVfi4Gm2/O0QjfysgJubc/QfT/AGHvVBwHWq+Z6d9t7rWaH7XMH7SphtH5JzpEpH5u39fx799vW+lorx1Ol4JlZFa7NGb6uOAHX3atBnqvE46kFrG1j/r/AI596pivXtWaddkXI5+nv3WyKmvXfvXW+ve/de697917rU764P8A2PJ7V/r4Px+Rq97z1o062wIv81H/AMs0/wChfeuvDh1k9+631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3WIyhXCMCAbWb+zc/QX97pjrVc06yf63H/ABPvXXj8uvC/5/4p72evCvn01ZPBYjMIUyVBTVakWPliRyR/Qswv711voLs58f8AqncFxkdqY9w2rUFhjAbV9bi3v3Xs9AXu/wDl9fGfeqhcxsahkIN7iCAj/baR7917PRY92/ygPjTliww+06GiDFjqWGnU8n88e99aB9epO1/5PvxcoMZJR53Z+Pr5G/S7U9O5Fz+Tp/4n3rrx6RmV/kufGetrnkh2pj0o2YBYDBTWVb/63/Ee/fZ1voZevv5TfxJ2NNTVsGwsfJWU7pIH+2p1/cT6erT9Pfj1ofPo+2yuptg9f0sFJtbbtBjY6eMRxmOCPUqj6WIHv3W+hEGoE3K6fwLcj/XPv3Ws9eIa9w3H5B5H0/Hvf29eIPl15XViQDe3B/1/6e/Up14EHrl711vriyq4KsoZSLEEAggjn6+/de49AF2N0pjt2Rzy41Eoq2UlvKAos1rlr/09+pXhjrVadFFzGwdx7CSZK/G1OcgjDFJEQuLXNuLfT34162KdB7jK/HwVZz1Q0eJqoZNApZmCSC7c3U+9Y62a16EbJ9m7e2lQploa6lrqioUStHE4LIx/BX/e/daZ+fW8dGE6r7Dp98YWKrFpvIQgiVdRXj3cenVTxr0ImSx4o4XloozG5uzAD625II/p79TrwPTbDlKiQQw1IKDgXYfkcA3Pv3HrfDoJuxKOtqZLUUwAJ/s35BPPvRxk9aPoMdIunoZKfwiaNjKU9UhF/VawI96p1voNctHkKHKzVCEmItc25W1/r73XHWqdB/uSmir5TkFpTPMyGF1C3J1cXAHvXWxTz6CfIfHjEbxxuQvRJqrRK00ehdRJjN2tb/b397rjr3A461r++egarC987jwNFt2dBRNK1HUIgXTMZPTo0j/W97HVaZ6e+t/lB2r8ft10PX+6DVR4qpaFIpJw2lYS3pU34+nv3y69kdXGdYd9YGTFQ5THbho6OtykZkkk8iJ43YAknn375Hq1cVHQyZXvKihwTRT10O5q7JR/axmmIkeJ5OASBe1vevn14VOOqgfkv8XO1M3uyPtjbeYqqKsiqfuaakjle4Rm1hQqn/iPe+HVcnh0ajoDLfInEbOJzu46mOSnp2amE7S/qC8aQfe89bHQVb72N2puvMVG7dzZCqrI5GdGLFmGjVci34v9feuvdVxfJ2gl29UxSiuNPpcCSIykOx+vAB+vvfWuuXV29s1hNnVNdjcq1E8tM7a/K5LXX88+/Y69XoqO6N0z7s3QJdzZAVY+9KiRpmCpd+HOon6fX3qtevA9ChvveKbCwmKoKPcNJkcdX06eSjjnDaFcCyNpJ/1uffuvdJ3am1+3d0vR1vXmxMzkxXy2imoBK6qzHUG9A/PvfXqjq0X4l/ylPkH3x2Fht1dopmMPt2GWjqpcXkQ6RaEcPICkw/p7117rba6++B/QWzdm4fa8m0MdUzUVGkFTOYYC8zgepr6f8Pfvt6359GW2H1ZsbrSkNHtDCUmJiYaWMMaKzAf1Kge/AHy60SPPpfsis6sxuUvpUG3+xP597r5dep59dJKXdl0EBfoT/wAUPvxFBjrQycjrN711br3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6/9Hf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddWv8A7D37rVK9eF/z7914V8+sDpN5VaNgE41KfpYf4e91x1oLQ16ke9dW64kHTZbA/j+nv329a4cOgj7U6V2R2zhavF7kw9FPJPDKi1TU6PIGkjKAk/7G/v3WxjPWm1/Mf+AW1usN5ZHLJQV7U8816NaGmk0sC5IsF49+Hz68eiY5bqHsDavSv949rbZyc1KUdCz4yXzCNRx6rf097615V6q8y216/cGQrqnKUNbS52OclVrIGiCSA8ga7H3rr3WSiw2fr2hw1WGKq3jIj5DaeARb37r3U3cfSW59v0Lbhxccjq0ZbQUYm9r3Pv3W69Fvr9zbp2/mIzk9aRKRdblf9fj37zz17pl3T2xtykilrauWZZ1Qu9m9PpH05/Hv3XujbfFf4t9q/MCkSp6rqadBIUZEZTLIyk+kLGOf9f36nWurHto/yS/lnvOqOEroAskTeFpDQSBWI4/Va3v3XuHVjHQf/CeTtPbFZS5DetXQSJ54nkiazNotcgoD/wAR78et06sJzP8AIj663JjY6bK09F5RAqsY4UB8mmwIN/fhw60fn0Xib/hPwMRlGqNutQpTRyaoFkAuVv8A2gT/ALx79XrdPTppyX8gTcWayL1NdLjkjKkKIgoB/pqAPv1fPr2K9NdH/wAJ7czFmIZ5psc1ChXWLLqNjyfr70T69ep0en47fyM+lNk10+S7CoIK90l1wwQRLdjbgaibW97yePWurN9nfBD467ImppsNsugBpTG0YnpoZBeL9JNx7955630azHbfwOLpUosViMXRU9OgijigoaeNFAHAsq+94HWqk9PEcUMYIijjj/1QjRUF/wDEKPfs9ax10kaIp0liCQfqTzf8e91NevUFK9R3iqJWdJSv25+mmwew+l/eseXXs+fUoLoRVjsAtgL3PHvwpXPXs6e3rJ711br3v3XukH2kbdbb9P8ATaG4f8f+XXL7917qiP8AkVO0uO+QJvpcdgZPkD6gVr8AH3scT1U1pjrYEzOSjxOOqK2QgeKMlQfy1uPevn1s+nQRbYxM25c0+5pWYRGQgX4Fgfx/U+/YJqevcAAOhvtwAPp9D/rAe99ePp16x+txf6fm3v1R1qjceok+Qo6VvHPOiPYHSTzz70eFetg+R6xHL45WVGqY1LC4ufx72RTrwNeHTgrq6h0YMjC6sDcEH8g+9db49FG+a2UjxPSmQqncKoyVEv1tc6/8Pfjjrw+XQmfHrKU2R6o2jLHImpsZAxF7EgoLHn37y610N3P9eLf7H/Ye99ezXroni1xqI4tyf9t79ivWjWlPPrx1BfSATb6E2/3n37BOevZAx1xTyWvJpH+A/H+x9+x5dbz59BlvPdEB1bdxxMlfVFV1RepVN/0gr/vPvxPzz16melDs7E1mLx6iuK+ZlHFvV/Ulj/xHv3lTz69xNeleCp4Fv9b/AI178a+fXgQeHSazW1sbm+ahWikUel4SFJP4LAe9YpTreeg9yP8AejZ+k0jGfGRn9VtZKj/VD/ivvZr6168KHoQttbip8/RJKCqVAFpIrgEn8so96Hy60R69Kb37rfXQvzf/AGH+t791oV8+u/fut9e9+691qedc2P8APJ7UP/Nj625/X795daPGnW1/F/mo/wDlmn/Qvv3Xhw6ye/db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rgQrqR+P9b6Ef0v73lTXqpowp1yUEAAm5Atc/n3rq3Xgbkix4/wB5/wBb3unWq5p13711vr3v3XuurC9+f9vx/tve6+XWqCteuI162vbRxpt9f8SffsU69mvXIEH6e9Up14EHh14X5va3497NPLrwr59cHQuLaivN/T+R+B79WnDr1K8evBPSEJJFuWvYn834/wAffia560BTHXJkVhpbkf659+r1unXlXSAoPAHF/r7914Aj7OvKoW4UW5uf9c/X34568Kdcveut9cVFgB/Tj3s5NeqqKCnXIi4sfz711bqLU0VJVxPDU08M0bqUZZI1YFSLW597r1qg8ugF3T8a+ut0VL1lRTT01Qxd/wBlh4y7fkoLC3vXWxUdIjHfELrv/Kkyi1VUL6IfUQqqfqwDf7179SnWtVT1Ax+w5unM+gw6McBJIBFqB0gHgAX4B96JIPW+OejDVNXS1WIiqYLMzwlpABrGsrf6j+n9PdjSmOtCtc/l0GuQkerZAiaWU25B4PvXVs16CTe+UqcM4LC4U6iCSR9Lmx91PHrWem/b+4qHccJXUonjBFrgsT9Affs+XXvLPSf3bjqyGnneFFJEdrWv9OQePfvPrfl0DNOtfQ0tRkKnQUBb0Mov/jx78Pn1o54dOWxN30kNVUisBQySPoBsFYNf6D37q1KY6Anefx6wW9d+SbtShoy0s6tLJJApLqG1kc/X/X976r0GPym/l+9adhdc1O6cRjYv76UFHM8LU9MNQ8Merhl+nI9+rnrwFeJ6oV2JQ0+2t0Z7YG4q7IUeQoZmpafzytCtxJotGHt7314U6th+N+G64xqrT5PIGry8SrLpqZ1lRW06ltq/3n3rrflno0eZr9t1pvVpSyUkIZY4wquCqr6fR78Mdb49Jzb1bidz1FTQ0sENNSUJKi8YjVlHFrf7z78cdaAFei5fJbtvA7B27V4inkoVnSCU3EkalSD/AGrf4+99e4DrWZ+TvdNNu3dLQVDvLqqPQtLKHBNrKRp9+HWuPHrvZVD2zvrGY3b+yMDm5aeoAi8sVDPINJFjfQD791ro9fTf8oX5SdxZGjWKmlpIKh4KiV6uili/blGpru4/p79jrdCeruOhf5Am34KnET9zzQ18MEUbVcQ/dbXGwYqFJI/Fhb37rXlXq8/pf4T9B9D46kx+y9oY0/aaftnr6SCYh0TTc3B9+p5nrZ9B0amiooqSMCGioaQgBVSlhjiVU+lgUA+n9PewB59V7vLrklLA07VCF/KDyCx0hvzx72TnrwGOs7eQMxQXYgXvylv8B73inVc1zmnXYijMgnI9enmxJF7WJt/vHvR9OrCnxdZuP96/417r1br1uSffuvU8+u/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691/9Lf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdF37k+Nuwe7KihqN2UUdR9oytodAwcqb8+/efWhw6covjh1GmyDsOTZ2Gkwxp3gKmji13ZbBw1r3Hv3W/l1UX8kf5H3T/Zn8Xz+0K5sLlZDLVRUcKGFGIQsIlZP8fp799nWsUz1Q5XfyyexusexqzF5nBynAUlSQmTkEz6ow5UNqIt9Le/dbp69LnsD42ba2/jJKeKsFWopNL07B2COF9Q5H49++3r1Bw6p97h+NO3MhlK6pWtMemctpQP6b/gf09+610Sjfnxxws1HUUsUvlZo3RWKPqueLEn37rf2dL34Z/N7sT+XJ2FFX0GAr89g4Wi00kMTugSBtRupH59+698+tuP8Al7/8KKdnfJzs/bvWea2hDtnKZyp8bmSCKGfUziMkkgH8+/de+fW2HTVkNXR09bTsJIKmGKeJl9QZJkDqeP8AA+9gdarjqQoIvdixJJ5/H+A/w9662Pn1y9+691737r3Xvfuvde9+691737r3XrAfQWv7917r3v3Xuve/de697917r3v3Xuve/de697917pCdo/8AMt9+X+n90dwf+6uX37rY49UT/wAjCK9N8gXF49O/cndRazXrXHP+39+6r8+rxN8TvW1VHh4iSkzeOoI5ADnkH+nvx4dbFOPS4weJhwuOgoYQAsYubfkn6+9nrQ9enJX8gdRdSLrf6EX/ACPfjQdaBJHUDJZCPGUjSMdcoS0aE3Z2/BPv3z638ukHS4Kq3KxyNZVS06eQgRrcG39F96OePW+GOpOX2K1Volpa+VZIEGlCSNZXmx/43791rh9nWGkq8vTwfw5NUjIvjva4P44Iv/t/e8/s69jzxXoqXzpx8rfHeRMjUNE8eYpDI5Y3Alk4DE/096PHPXl4Y6ETonbldB1P1/U4iqkni/g8BYKxNiIwb+/AYx14mmD0NlFm8xR1M8VXA0gCixbV+ofQ29+zw69ilQesIqNwmZq/xSEatQjs3jCE2Fgfdqnj1qgGOlPQ5ysqIXMlGPKl7quofj8jn3Wh62SB0D24+y8+uS/gsNB4knlNO0yqQQCbXDH8+9aqcOtkdCFtzZtFjmhzORl89dMiS6pf91u41AFj/T3ankOtE1GeHQikg6CCNJIP+BFuLe/Dz60x4dcrC5NuT79U8Ot0HHr3J/w5/wAORb37HXsn5dY54op4ninRXiZSHVwCLW5+vvXW+gKyOFzOFyz5LDRuaHyF/HFqCkA8njj34A8V61UGlel/g94R1ZSDIaaecgccgg/Szj8/7D3scOvHpdAggEG4IuCPoQfeut9e+o/I/wB7974da4jrv3rrfWpz1sSP54/ahP8Axxbj+o1/T+nv3Wsk9bYcX+bj/wCCJ/0L7914cOufv3W+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rocAn6/X/H/Ye98etUpnpMbpwMe5MX9qQA4YSRluGHHIB/23vxHl16v7OgtjM203/hk5aoRxZdV2Cf7SCfz71w49bqD1KqXgdFqFjWP+0TYcX5tf8APvxpTr3nQ9Adv3GvnUnEQt445T9bAkC31968+vdFjxGTyG2cjOhiaOKOYrwDZgDc+9eXW8cD0K+L3ku5XNIVPJ0E8EH/AA974de6ZdzYSGJhAX0K4LGEG6+oXvz9ffqeR61XpCx7Xiavp1iITWRqK/n83968ut9P1bT5KjkSlptZRGOplH4H+t72D16nQubTqZUopYcjS/cUstM8Dh+QTKhX9J/rf3r59axw61h/5w3ROS6k3/gOwtlQmkizlbT1Va9KCpCyNrcnT/T3YcOvGnHoBum+zVx+Ag3A+deXI6F80ZcF/QlmU/7H377OvD1PRndhfJCnzday5KuISIlfGWUBwD6iwJ9+z16o4dPnY/y321tqllx+1qgfxyoVUjhgjLSyyfTTaP3vreOq8949V/MP5L52EY7ZmQ/gGSqvCK7w1IbwTtqV7gf0P9feqjqueJ6sk+NP/CeqLeEWN3R2nWzUVVLHBVVEdTJMdMgcM0YQ/wCH+Hv1fTrfDj1sTfGz+Xh0P8dsVDR47buNzlbCoCVVbSRyCMldLEK45+nv3Hj16vp0ePGYHDYYaMVi6LHqFCD7WnjhGkCwX0D3vrXn07+9db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//T3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvp791rh11fi9j/rW54/w97pmnXq4r1jikEqk2K2JBUjkf6/vxFDTrwNRXpKbl2HtXdkDwZvD0lWJF0s7QxeQj/FiPeut/Z0Tftv4KdebzxNXBt+hgx9bUpMPJojBVpB6eT/j7914Y49VFdhfycM/Q1OSy4rv4pSTSGRYIvG9geSoA9+r14CvRSd8fy1KjHY6dIdkZGqq0Rv3I6QMdSn6Dj3oH8+tkZ4dVbdr/AMonuTsrNPHidtZ/H00krx61pChCudIFwPoPe8daFR0Yb4Sf8Jzu4+v+9NldzruXL4/+BOklRBNMY4tPmEjakuOeP6e/daNBx63sNgYGv2rtDD4XJ1JrazHUNPTyyg6y5hhVOCf9b34deqB0sYpBMuvSy8kAMLG39feyKY68DXPWX3rrfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690hO0f+Zb78/8ADR3B/wC6uX37r3VEH8i13EPyABuUO/ctze4BFa/597z1rHHq8Ta9THmN17gNQgc0E5EIPKix0gg/7z71Uk563imOhW/H9PfuveWOuBIXgfUgn/bD6n3v1PWsDt6APNvnMtuUU0Ur+FJluFJC21fpt/T3rJPW6imOhzx8BpqOnhb9SRqH4sdRHN/e+tDI6lKysWt9QbN/gffj14dYPDBHI09kR7G7cKLfkke/VJx1qnmeiE/zHK2CL435SUTx6WzGPCPr4YrL6gCv9Pejx6sMdDT8UctSVHR2wIxLGJP4JTMQXF2/aHNyffhw68Tn5dGLlNGf3pfCdKk620khfz73nh1U049YUyuMdhGlbTFrf5vyLqA+liv/ABHvXA9b8upcQgN3hEfPBaMKL2/Fx73nr2K9Irde1afIwNWUcCx18RMgdFGpyOb2/r/re9cet8Ps6Ztr58zK2P3AfHJARFDJOxj4U2Fxf8+/A4z14jz6EcGKojj+1ljZYmFtJ1AhfooYe7DHVePUoBtRYnggenjgj/Ee9Y63nrn7117rhJJHGt5XVF+l3IAP+HPv3W/l1HiqaWW8UJWVfoQihkAP1B/HveePVagY6QW7sFQUVDVZmF1pXplMrljZLDng/j3r/D1b7epuztzY/JYalf72KV+V1K1xwbC59+GeHWuHS28ivGXidXH4ZSGBseeR72oFc9aY4x1kBuAf6i/vXVutTzrn/t+T2n/TwXA+gJ1/7z795Z611tgQ8xRH/m2n/Qo9+631k9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYo31NIv5Vuf9Y/T3Y8AeqqckdZP8Pdetj06T+cwVJlImkeLVURoTGR9SwHF/fh1utOgOrMTmqWpkWRZTTKfSLGwB5v70ajrfHh0z1sUTgpEEWUpZwefrx+ke/eXXvkekBmNi0GRp5pFiHlcMddlA1Wvx791roC6fB1O2Mu7KG0a9Q5PIv+AOPeuPXh1E3BmKusycbs1kVQGQnk8ce98Ot8R13HVVGg1UUbXiA1W+tl/xP+v791rpcYqrNfj2qXQeZVJLW5Nl4A96Iz1sdPO1c4I/IuQGqESqqoQbnmwPPv3Xj0Rf+ap8fNw999I5PO7PWQz7cx5kSOCMNJ+1CWABtf8AHvY49aOetQbp/cG4cFPV9dZnB19VuiGtrKcwev7glJjGvp/x924da+fR6On/AOXv8ou981Tzbdps/s2krJYZo56iOSKNqeSSzHUwAPHJ96+XXuB62HfiL/Jt2z1+tFlu8ZE3bmafVKHleOZtZAZSdV/z/tvfut06up2h1L1/sXH0+O2/tzG0lNSrGsV6WBmXxrpVtWkc8e/AenWiQBnoQ9JQKIVjWNeNAXSLf7SF4HHvdPI9aJxUddxyLJqIUgqdJuLf7b34inXhk9c1DC+ptRJv9LWH9Peut58+uXv3W+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv//U3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6910RcWP0Pv3DrRFRTriqBBZR+ebm/+9+99epmvXM3/HvXWz8uuvfuvdeF/wA2/wBh/X8+/Y60K+fXfv3W+uLKrjS6qyn6hgGH+2Pv3XuobYzGuSXoKNyfrqpoWv8A69x7917h1hXCYZDqXFY9T/UUcAP+30+/de4dTEpoIEZKeGKFW4IjRYx/S9l97HoetGvl1jhpBT3KO7sTyXa/+v73UHj1qhAxx6l83/Frf7G/vWKdbzX5dd+9db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XukH2j/zLbfn/AIaO4P8A3Vy+/de6oP8A5JuTgwm2vkLWvckdg5OO4+oLVrgXt7359a6vi2Zhqqira7MuF+3y/wC4p/thTZkLD/X9+APXiQOhK9662Om+uqkoKOoq5G4iUm7fj+g92NPy6oAePn0jdq0ddLXVOVqI4/tanW0LW9RB+lr/AO3968q9XJFehBBva30/r/vHvxFOPVQfTh0m85uKjwahXGuaS7BF+vP0JA/r71jreaY8ukpFJm8yXqVulLIG8uo6RGh5sPx9P6e98eHXjx6KZ88No0Ge+OtTiIp5JJFycE6ENqdiHDTAqD9Bxb3ogggHHWxQ1I6XnxJxeFrepdq0cctQ1RiMZBTTLrKEFVC3t78OtHj0ZHLxUuJgJeacxyqyOPqVX+gtz7314ZNekritvbVqGfIw19UagszeKRyoEpJICj+nv2OvVNOnTTuKpOnGMqwQkWYsV5H4497zwHWqAZPWf+8OUwkgjzKCRWt+j1FR/UEf4+6/b1aleHWDKUm2t0KkkkrUlZp1IEshZr3W9vz73QE06rUgV6TdRuCTr2ejgzUhNHVSBaJlJLSauNL+9VpjrdNWR0p6ndNVX+FqFGWGwdnUEgra/Pvf2dapmp6mz7ySmpw7xHyInq4LamA5uF+nv2OrAHh0HVFvWs7BzL4bFhoEoJD95qBj0qDYsNXvVanGOtUIGc9DhjsdBjYBFDqJsNbuxYsR9Tc+/fLrfnXoN+wMnQZmgqtpQyOavI/s6ojcKTwQdP459+J8vPrQ9emLBdXy7a20tLjKiWTJKhZ42Yjl/wBIUH36mOt1z087Ezv2rybWy0x/ipd3RWcatJ4K6Tz/ALH36vr1U9C2vCheTpAW5+psLX9+6t1qddbMf+HyO1Ab/wCYP+w9f1F/8P6e/Y61TzPW2JF/mo/+CJ/0L7914cOufv3W+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917roAAkgAE/U/19+69137917r3v3XusM1PDUIySxo4cWN1F7f6/v3XugP3fsqugmkrMXcxaWkbSOQByQR/W3vxAOR14HyboMI8nKvkpWjbWh0vqBAJtyRx9fz7r1v59J2uw9PkpXLKhf8XW5/r798+vcOgvzmyqtsgJI4x4hcni3+wt795deyOHT1idvxpSSUkigyyXWzAG/wCffga9exx6hw4+uxdU9MyBYb24Xgj+gt/X34kVz16np1KGMkepFrpGSpPBVb6uT71xx17oZsBTUuSwVbtqtijqqHIq8VUkyhgUK6SpQ/1Huw4U61xNeqz9ufy+fjZgvk3Vdh7go6aKSqrTPR07UwWB52fXpKtx+o+/deoK8OrycHt7buFx1FS4TFY6ioqemhjpBTUkEQEIQeOxRQeRz72Pl1o8Knp9QuRdwAb8Ac2H+J97x5deFTk9dFBJbUOFa4H9SPobj37rVKmp6ye9dW66/wB69+69nrv37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/9Xf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SD7R/5ltv3/w0dwf+6uX37rY49a/v8k9KarwnyBpqyQJSt2JkWZmPpv8AfPzz+b+/UHVQa9bGdEkMVJBHTEPFHEqxkEWIC8cj6e9mvXhTh1nSQMSLadJsbkcn/C3vxBpXrQYVI4U6S+9pXi25XMiazZBbngFvrx711alepW1JPLt/HPp0nwAEf4g8n37r3T7LIIo3f6lEZgt+SQLgD3vJ61hRToM8FQz5jL1NZkofNBDK+hZAGUer0rz/AE9+Ark9eanAdONdkZK6sfC4yMwIh0S+Maeb2Den6AfW3v3nTrXlWnQTd09bz5bYFbAsstdNDqnNOQTH9L+9U6tX+fSD+J+VpMfhs7javRRSY6QxyByqqoib1f7178D140rjozs+9tlTloajK0ElwVZZLMB/UHj6+/V60RnphaPaGSZmxObghdnuUik9J1fhdPNv6D3vHXqnrNXCDb1LrXM3aVQVHk/P9bA+/dePGh6w42skaB8g9L/F1KH9YDj63DLf37NPXr2K9QKOgp8rWtlq6U4eGnJCxNZVJBtpt79xz17hjpk7BwFdmKEZOigbLw4tfNS39QYx8nQvvR4dbHoenLrTeEdfh1hzuMjxVVE3i0yIBqUcDUGHvwOOtEZ6FyLH4yVfLHT08qyeoNpVwb/0/HvZ68KcR0h59nw4fLHL4OIUstVJ++IRYsWN7MP6f4+/Urkder5HpTbhykmKxDyhb1UqCJU4ZtbizNYfW3vxOa9eANKdIjaW0pYppclkIzNNXAzxVDm7U1zq9N/9696GM9eOcdCZV1Ax9DNO5Z/t4S2prXYgcX974Z69Q0p0W6DD1NRnpOyVnYR0lR4DDqNm9X5W/uuCdXXqeXHox2JyUWVoYayLgSLcgfg+9nrefPrVM635/nkdqn6fs/Qf4N9R795da9etsKL/ADUf/BE/6F9+62Oufv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3+8+/de64SIsiNG36WFmt+QfqPfuvdInK7FxNbFMYYlimkBJcDm9uLW/PveOtZp0VLd2I3RtDIvNQUE9dAWvqUMyix90IzTrdfTpOpveiqpBHmyMZU/pZHFiT9DYNz79wz1vqZV5fb1NLBJT5OORnAIIIvci9x7917pvqcnTSTRTpKZizf4fT/H37r3z6yZHORUc8QjiuSoBFhpH9LH37PWsDPStweWICZOmufthrkhHIY2+hHv1evfb0FvamBk3BVYzd1HMaaekrEqZYEawIia78D37j1uvR2+q970G9Ns0phkX7mip4qSoQMNTGKMR61H1/Huwrx6qaHHQoKfqCLabDkgk/4m3vZHXgfLrvk/4WPH+I9+698uHXfvXW+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/1t/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdITtD/mW+/Lf88juH/3Vy+9HrY49a/f8legbJ4D5BYen9FZL2LkXWoJsFUVr3F/e6dax+Wer8N0rnsVhKCkxMkj1FNGPO6K1nIFvqP6e/EmnVQM56z7P3THLQxUmZl8OQVypMw06ri4vf8A3v3upOOvFQDXpaZWnjyOMqIRaRJY7jT6gwBvxb6+/U8j1up4jpP4/IDC00FJUxtHGt7M5AAB+n+w9+xTrWa16VCvBXQCaJ1kjZGsVPF7fQ/63vYJGOvEA56DvGbkXHZWfH1SGCOSW3kmBVSb/VG/4r719vW/mOu6nI0mEy0uRpytVHUtaR4zrVVf66ivv2B1vj0r5ctjMliK6SKqp3j+xqGl1OumP9gk6w39PevmOtH0PVPnxj7JoaTszuvB7z3LjIaeTNViYiOapWPTEZmEekEj3rq1cdGNlGKpKvIZGnhmztF4XkiFA5mU35FtF/dT17/D0Eu3+5p8duZoItlZ9KdZjGH8E/jA1fXkfj8+9/Pr1amnRpcYa/sUwVUeQixUER1SUlVKI3sv9jSxv/h78M8evE/LpSZbd+VxsNPtTb1HUtVxOoetijZ42N9JOv6W/p7tU0p1ogV1HpJbpn3pRiClqYampSoVZXWPV9f1EED6e9GvDr1R0idwfJHL9cZDa+AqcJWz02RqI4Z1EbsdJIUgi3vwJr14io6Nm9Jh964JMriEWhrXp1qmpyBFNGzJrs6C1vzz7tjrWa/Z1E6wzdRUHIYutnDy0szRxqTdjpNvfhwp14jPQvSOsUbyOQEjVmJP00qL8n37rwxjoIp8t/ejcaUNMjpBEdJkJJUhDyT/ALz71WpoOt0oM+fQmfxHGUIjpZa6mieNVj0vKqm6i1ufe8nrQxgdBH2XvOKNsXisRVxVklXVJFVRUriV1R20knR/h70fTqwHmehKh2/QHb64pYSsE8CyuP7XneMOWb/Y/j3vHVanoPOvM3UPmslgwD9tQySRjV/RPoR70DXHXiKZHWtJ1wFH88jtMKwN4De1v9X+PfuPXvXrbBi/zUf/ACzT/oUe/Hj14cOsnv3W+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de66N7cc+/dePy68pJAJBBIBsfxcXt791oE+fXub/wCFv9jf37rfXfv3Xuo1RSU1UhjqIY5UPJDKD+b/AF97r1qnp0FW4OlNlbhqjW1VHoqCSSUC2uf8OPeiAet56R+T+P233kh+xSyoeCSOLf2mU+9afMdaJ8j0w5PoauPjFBUKmhhch1HpH9R/re/U6tXFOgs3x8d+xK1RU4nKonjYAoHGu30BAHvwB8utcOodH1J2XtHHxPU1MlUsqjzIlyR6bm//ABPv1D59brXhjpCbibd1NTVlKuMrpvLDLHGqROxVm4uB9PeuvdIXoCp7W2p2AkNXR5OPC1VQHdJUkCqHk/IP+HvZNOHXh8+rd6aY1FPBPpKeVFdkIsRccjn3b7eq8cjqT711vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/19+776i/5XKX/wA6If8Aivv3XuvffUP/ACuUv/nRD/xX36vXuvffUX/K5S/+dEP/ABX37r3Xvv6H/lcpf/OiH/ivv3XuvffUX/K5S/8AnRF/xX37r3XvvqL/AJXKX/zoh/4r7917rr7+h/5XKT/zoh/4r7917rv76iP0rKU/9VEP/Fffuvde++of+Vyl/wDOiH/ivv3Xuuvv6H/lcpP/ADoh/wCK+/de67++of8Alcpf/OiH/ivv3XuvffUX/K5S/wDnRF/xX37r3y6999Rf8rlL/wCdEX/Fffuvde++oh9ayl/86If+K+/de6999Rf8rlL/AOdEP/Fffuvde++of+Vyl/8AOiH/AIr7917r331F/wArlL/50Q/8V9+691776h/5XKX/AM6If+K+/de66+/of+Vyk/8AOiH/AIr7917rv76h/wCVyl/86If+K+/de66+/of+Vyk/86If+K+/de699/Qj61lJ/wCdEP8AxX37r3Xvv6H/AJXKT/zoh/4r7917rv76i/5XKX/zoi/4r7917r331F/yuUv/AJ0Q/wDFffuvde++ov8Alcpf/OiH/ivv3XuvffUX/K5S/wDnRD/xX37r3XvvqL/lcpf/ADoh/wCK+/de66+/of8AlcpP/OiH/ivv3Xuvff0P/K5Sf+dEP/Fffuvdd/fUX/K5S/8AnRD/AMV9+691776h/wCVyl/r/wACIfp/X6+/de6999Rf8rlL/wCdEP8AxX37r3XvvqL/AJXKX/zoi/4r79Ude6999Q/8rlL/AOdEP/Fffuvde+/oT9Kyl/8AOiH/AIr7917rr7+h/wCV2k4/6aIf+K+/de699/Q/8rlJ/wCdEP8AxX37r3Xf31De33lLf+n3EN/979+691776i/5XKX/AM6If+K+/de66+/of+V2k/8AOiH/AIr7917rv76h/wCVyl/86If9b+vv3XuvffUX/K5S/wDnRF/xX37r3XvvqH6feUv/AJ0Q/wDFffuvde++ov8Alcpf/OiH/ivv3XuvffUX/K5S/wDnRD/xX37r3XvvqH/lcpf6/wDAiH6f1+vv3Xuuvv6H/ldpP/OiH/ivv3Xuvff0P/K5Sf8AnRD/AMV9+69177+h/wCV2k/86If+K+/de67++of+Vyl/86Ifz/sffuvde++of+Vyl/8AOiH/AIr79Xr3XvvqL/lcpf8Azoh/4r7917rr7+h/5XKT/wA6If8Aivv3Xuu/v6H/AJXKX/zoh/4r7917r331F/yuUv8A50Q/8V9+691776h/5XKX/wA6If8Aivv1evde++ov+Vyl/wDOiH/ivv3Xuvff0P8AyuUv/nRD/wAV9+691776h/5XKX/zoh/4r7917r331F/yuUv/AJ0Q/wDFffuvde++ov8Alcpf6/8AAiH/AIr7917r331F/wArlL/50Q/8V9+69Q9e++ov+Vyl/wDOiH/ivv3XuvffUP8AyuUv/nRD/wAV9+691776i/5XKX/zoh/4r7917r331D/yuUv/AJ0Q/wDFffuvde++oh9ayl/86Iv+K+/de66+/of+Vyk/86If+K+/de67++of+Vyl/r/wIh+n9fr7917r331F/wArlL/50Q/8V9+69119/Q/8rlJ/50Q/8V9+691399Q/8rlL/wCdEP8AxX37r3Xvv6H/AJXKX/zoh/4r7917pD9m1VLL1vv3RVUpC7Rz5dvPEUQfwyQXdr2A/wBf37r3VAf8mR6nF7T+QtdStHI47EyJ1xOrWvXP9Cnv3Wh6Hq/PEb4hlpIBXxHU6qGYi4+luR735+nXqGnr6dSsntnBZyEZZZXpfSzo8ZEa+T62b/XPv3E5690y4vcVZg1liyV3pY20Qsp1XT8NcfQ29+4DPXjXyx0rhJh94UhVJDZP1abCQA8Xv/S/v3l17zx1DSCr20BTUl5qM2cPIbtf6EW/r78OtH5+vSY3ZWYLItTpXl6d2Ua5IV5B/qGHP+396NKdbAOem5sPqxM9PtFzkJZ4tDtUghotX1K6v979+p6dbqfPovO48F2LtzF5mlnaVUloa6bUjsLKYiSOP8PehUdb49atNRt7uXsb5mrgtlVmWTakuc8e6Kqkae0ZFVaQOycC3N/e6061xPW4n0h1Dg+uNnYymDtk6magpWqKiuAlIcxAPYSX+p97A9OvE9DCu3sApZ1w2MDOSWYUNPdi31N9P596p5daqKV6DDPdOYStnqcpjp6qirjqlghp2EVOXHIjKLYc/T36met1FOgk2/2Fndi7rbCb3xdPDippPt6KvMaGYsW0x3kYe9VIx16nn0bOKDH10cNYKaCVZ4kkjd4kY+ORQy21Djj3uvXqUx0gN67F2plIxma7G0T1WHAqYS0CadaeoX4+p/w9+48evdR9i42mqXqd0JMYxPFJTGkjsIFjjW19I4HH+HvQGa9bPCnSR2vDS0266+txsrPTLUTPWXNgpBJa1/qLe9gZ6qaUoekJX97zbl33XbJ20kzmjlEVezIfEsStZyHPH+Pv1fIdbpTJ6Mrt/CUONp0rU0maaJWeUWOkuPUAR/j738h1U1rU9JvMdeYjLZDzVeSqY5ZWLpCjkcn6Ec/8j9+p1utOgsxuwMb1tvKXP5WslrqLIShKNags6JIfppDXAt+fdcdb+zozkMkdRFFPEQ0U0auhH0KOLj3b5dapmvQVyY6Ha28sa2PQaNwVLCqBA1KXJDWv/t/euH59b49azPV1J/2PF7W1VFNCRAx1VU0cC213tqkIv73SvHrRI62tEraMRx3rKS2hbMKiIqbCxIN+R7114cOvCvovzWUd/wAWqIv+K+/GnXhXz65ffUP/ACuUv/nRD/xX37rfXvvqH/lcpf8Azoh/4r7917r331F/yuUv/nRD/wAV9+691776h/5XKX+v/AiH6f1+vv3Xuuvv6H/lcpP/ADoh/wCK+/de67++ov8Alcpf/OiH/ivv3XuvffUP/K5S/wDnRD/xX37r3XvvqL/lcpf/ADoh/wCK+/de6999RXt95S3/AKfcQ3/3v37r3XX39D/yu0n/AJ0Q/wDFffuvdd/fUP8AyuUv/nRD/wAV9+69119/Q/8AK5Sf+dEP/Fffuvdd/f0P/K5S/wDnRD/xX37r3XvvqL/lcpf/ADoh/wCK+/deoevffUX/ACuUv/nRD/xX37r3XvvqL/lcpf8Azoi/4r7917rr7+h/5XKT/wA6If8Aivv3Xuu/vqL/AJXKX/zoh/4r7917r331F/yuUv8A50Q/8V9+691776h/5XKX/wA6If8Aivv3Xuuvv6H/AJXaT/zoh/4r7917r339D/yu0n/nRD/xX37r3Xvv6E/SspD/ANVEP/Fffuvde+/of+V2k/8AOiH/AIr7917r339D/wArtJ/50Q/8V9+69177+iP0rKQ/9VEP0/2/v3Xvs699/Q/8rtJ/50Q/8V9+691399Q/8rlL/wCdEP5+n59+69119/Q/8rlJ/wCdEP8AxX37r3Xf31F/yuUv/nRF/wAV9+691776i/5XKX/zoh/4r7917r331D/yuUv/AJ0Q/wDFffuvde++of8Alcpf/OiH/ivv3XuvffUP/K5S/wDnRD/xX37r3XvvqH/lcpf/ADoh/wCK+/de6999Q/8AK5S/+dEP/Fffuvde++of+Vyl/wDOiH/ivv3XuvffUP8AyuUv/nRD/wAV9+691776h/5XKX/zoh/4r7917r331F/yuUv/AJ0Q/wDFffuvde++ov8Alcpf/OiH/ivv3XuvffUP/K5S/wDnRD/xX37r3XvvqH/lcpf6f8CIfr/t/fuvdcPvKDVr+8pdQFv+BMVgP9bV7916nXL76hvf7ylv/wBREX/Fffq9epmvXRraAkXrKS4+l6iH8/4avfuvddNV49xZ6mjcf0aaBhz/AIE+/Vp16leofhwGoyFMUWPBY/ak/wC8+/de49cRT7eEglEeJWQfRx9qGA/1x791qg6cBXUIAArKQD8WqIQOP6c+/dbp1776h/FZS/8AnRD/AMV9+69Tz67++of+Vyl/86If+K+/de66+/of+Vyk5/6aIf8Aivv3Xuu/v6H/AJXKX/zoh/4r7917r331D/yuUv8A50Q/8V9+69119/Q/8rtJ/wCdEP8AxX37r3Xf31F/yuUv/nRD/wAV9+69177+h/5XKX/zoh/4r7917r331F/yuUv/AJ0Rf8V9+691776i/wCVyl/86If+K+/de6999Rf8rlL/AOdEX/Fffuvde++ov+Vyl/8AOiL/AIr7917r331F/wArlL/50Q/8V9+691776h/5XKX/AM6If+K+/de66+/of+Vyk/8AOiH/AIr7917rv76i/wCVyl/86If+K+/de6999Q/8rlL/AOdEP/FffuvfLr331D/yuUv/AJ0Q/wDFffuvde++of8Alcpf/OiH/ivv3Xuvff0P/K5S/wDnRD/xX37r3XvvqI/Sspf/ADoi/wCK+/de6999Rf8AK5S/+dEX/Fffuvddff0P/K5Sc/T/ACiH/ivv3Xuv/9A5I+Gn81UX/wCM3bw/wH3NcR/tg3v2OtZ69/smf81P893bxJ+htVVx5/5K9+x1vu69/smn81S//M7d4H+v+U139eLWPv3l1rPl11/smf8ANU+v+m3eFvoP8rrgD/yd79jreevf7Jp/NVv/AMzu3hxz/wACq7/b2v791qh69/smn81Tn/jNu8LfUWqq4m1/r+r/AFve8dbz5nrv/ZM/5qnP/Gbd4fUAf5XXfnn/AFXvWOvdeHwz/mq2v/pt3gBf6/dVw+n9PV79jj17PXX+yafzVAb/AOm3eH45+6r7cf09Xv2OvZ67/wBkz/mqD6927vtcj/gVXGx/H9r36o69ny69/smv81QXt3bvC5HJNVXfU/j9Xv2OtZ66/wBk0/mqcL/pt3hYD/larr88/XV79jr2ePXf+yafzVP+f27w5+n+V13+x/te9Y63nrx+Gf8ANU/Hd27+Px91Xn/YD1e9449az5de/wBk0/mqj1Hu3eH4HNVX8f7z79jrZr14/DT+arf/AJnbvA/n/gVXAWP/ACF79jr3d17/AGTT+arxfu3eF+Ofuq4fX6n9Xv2OtcOvD4a/zVOP+M27w/P/AClV1hzc8avfsdez14/DT+apb/md28L/AONXXfn8D1e/Y4deyevD4afzViT/AMZt3hzwf8qrv+jvfqinW8066/2TP+aqOB3du8/T61VcP8Bzq9+x17PHrv8A2TX+ar+e7d325P8AwKrv971e/UHXs9e/2TT+aoef9Nu8B/h91XWJP/IX+8+/Vp17PXv9k0/mrH693bwP/VVWi3+I9Xvxp17J8+uh8NP5qgFj3dvDgXB+6rhz/iNX19+x1rI67/2TT+aof+a3bvv/AE+7rz+L8HV79jreSaddf7Jp/NU5/wCM27vte/8AwKrh9PzfV79jr2eu/wDZNP5qv0Hd27/z9aquv/Ww9Xv2OtGvl14fDT+ap9D3bvDni/3Vd9bcc39+x17PXv8AZM/5qn0Hdu8f6D/Kq4fn/g3v2OvZ69/smv8ANU5/4zbvDgD/AJSq7/o737HXs9e/2TT+ap+O7t4gG3/KVXcf8ne/Vpw63nz69/smn81QW/4zbvH6fT7qv/P4vf37HWu7j14/DT+apx/xm3d/04H3Vfz/AIfq9+x17PXv9k0/mqrwe7t4D8j/ACmuP+x/V70KdeNevD4afzVf+f2bx/oP8qrhz/sT73jz69k9e/2TT+amf+a3bw+pHFVXW/qf7Xv2OvZ68Phr/NU/5/bvD+v/AAJruD/yV73jrxr14fDT+aobH/TdvD6/8rVdf6fX9XvWOvHV10Php/NUvf8A027wBP8AWrrzx/yV79jh17Pn17/ZM/5qoPHd28L3HP3VdYf141e/UHXqmnXf+yZ/zVfx3bvD6fT7qu/r9eW9+x17PXj8NP5qn1/027w/Kk/dV5BX8f2vfuvZ69/smf8ANTJB/wBN28LkW/4FV5/1v7Xv2PLrdD14fDT+ar+O7t38GwP3deL2/H6vfsefWsjr3+yafzVfqe7d4D/H7qu9XPJI1e/deNfLr3+yZ/zVP+f27w/N71Vdcf4X1e/VB4dezXrr/ZM/5qn1/wBNu8L3t/wKrrcD/Bv8fexTr2evf7Jp/NUFv+M3bwtz/wApNdx/yd71jh1vPXv9k0/mqf8AP7d4D8j/ACquvzyOdX59+x1rI67/ANk1/mqDj/TdvAD/AKiq763vYHV79jr2euv9k0/mqEWPdu7+Pp/lVdxf+p1e/Y63mvXf+yafzVPx3bvD68D7qu4/xPq9+r1rPXv9k1/mqA/8zu3f+f8AlLrSP9cDV79jreeuv9k0/mqmw/03bw/x/wAqrwAbfj1e/Y69kddj4afzVfoO7t4WB+pqq4f7D9Xv2OvZ68Phn/NU+v8Apu3j9LD/ACqu/wBgbavfsdez11/smn81Tn/jNu8LAfmqrv8AW5uffsdeqeux8NP5qv8Az+7d55tb7uuuCP8AHV7959ez17/ZM/5qlgf9Nu77H6H7uuHA+o/V79Udaz14/DT+aoLf8Zt3gCPr/lVdYW5/1Xv2OvZ4ddf7Jn/NU+o7u3ha/wDytVw+n1/tf4+/Y69nrw+Gf81Tn/jNu8LfU3qq7/W+ur37HW6ny67/ANk1/mpj6d27v/8AOmvP+v8A2vfsdaz17/ZNP5qlz/xm3d/P9aqu/pf6avfsdeyOuh8M/wCapbnu3eHJF2+7rv8AX5uffsde7uu/9k1/mq8X7t3ha/4qq4/UfT9Xv2OvddH4afzVfx3bvAngH/K67j/k63v2OvZ9eu/9k1/mqH/mt27/AMX/AMqrvwP+De/dez11/smn81S1v9Nu8PqLf5VW/wDEN79jr2euQ+Gn81Sxv3dvAfm33NcL/wC2b37h17PGvXH/AGTT+ap+e7d4/wBQPuq7/etX59+63nrsfDT+arzbu3eBFjf/ACmu4t/jq9+wMdaz17/ZNP5qvB/027v/AKf8C642/pb1e/Y63mnTPu74a/zU0697Dnl7u3eKSm2llpaxTU1pR4RSNqDAtyPfsdayTQ9GR/4TW7L7Aoeg+2Nudh5aoy+5KPdddHla2qLiSqqlr3vIWP5P9PfuPHrxPWzZhk2/JPNhZhCaunPjHIDAgWtY/m/veDXrxr0o6nBySULY6OcQU6KWVhxdz9Af6Ae/cetCoOOkLA74eVsfl6E1tLIxAqnHoXV+dX09+pTHW61FR1IXbVREzV+3q8GGY3aCEk+O/wBAw9+p6deqOBHTpLj82KXyyCaeQAeg+prfkAe/A4p1qgJz1zxv2dW60eTxEbysf84yA6bn6Nq/4j340p1vzx1PyOEkongmwhakLSBHji/TYn+n/FffvmOvedD0CHZuXy2Imy33ML10YweQstyQLUzW96Ioetg9VV/ynsRh95dkfJDL53ARfxCi3TVRUE06AtCj1jqWjJHB/pb37readX1UtJ9vTRU7NrWKwXi1lX9Cn/WHvdeqAYz1M966t1737r3RcfkDtxsniabIwUJqJMcfu5JAOEEJ1arj88e/GtK+nXhTh009Q94Y/dGKOIhkSpyOLC0ehQNSGP06WA5Nvegc068R6Y6GHcmepziJKWqURPUxiOVieFuP1EW4/wBj7seHXhk9BticXlVovsMBljPBNKfJHE5sEc+q5H491APl17Ap17KDH7SylBtpq1Yshm1V2NxrZn/WLn/H3sGmOvdL2DrvblJS1dRjMPT0eclhHnySRBaiplK31GQXvf36i9aDevSV2Vk8ti8/LhszO4p9TLEZj6b39I549+Fa062acT0OZjh1ecqGYC4e2ogW+q+9knh1Wg49JHe2DizmIuUEr0v78KkXBNrgi/vXVq0647Dkq3w3jq1ZWhlMaagf0gfQX978utDjTpK74aRd77ECSmNTXDUv+r9fvXW6daQHzj66777E/m9b8oOgdz5HbGeSgkjranHmVZJHaX0sfDzx731oU4jo1Y+GH81c0tKR3ju+zxAoPuq8AD88Bvesdez59cT8Nv5q8Ohf9Nu72uQCfuq82NrfS/vxpx63nrN/smn81Uj/AJnbvC4JPFVXCwP9Dq9+x16h69/smn81T6/6bt4XP4NVW88/8G9+x1rPXj8NP5qhH/M7d4ccj/K67gfnm/8AX37r2euz8M/5qlhbu3d5/qBV15+vPHq9+qOvZ66Hw0/mp2P/ABm7d9/yfuq7/WA/V79Xr2euR+GX81W/Pd28DdbG1XXcf8ne/Y69nrx+Gn81S2n/AE3bw/8AOqu/1rjn37HXs1z10fhp/NUP/Nbd4Wt/yt130/Fzq9+x1vPXX+yafzVbi3du7z9LD7qu+oH/AAb37HXhXrv/AGTT+ap9f9Nu8Dybf5VX/g88avfutZHXX+yafzU/p/pt3ifzxVV35HB+vv2Otmp68Php/NUH17t3h/51VxFrW+t/r79jrWade/2TT+arz/xm7d/0uP8AK678C1/1e/Y6917/AGTP+an9f9Nu8ARwR91XWP8ArXb37HXu7y68Php/NVsf+M27wv8Akmqrv9iP1e/YHW+7j17/AGTT+aqeR3bu/j/prrj/ALxq9+wOvZ69/smn81Swv3dvAfW/+VV1r/0PPvxI69nrv/ZNP5qt+e7d4D8/8C67/o737HWs+XXh8NP5qnAPd28efx91Xf6/+q/3n37HW8jr3+yZ/wA1W1x3dvAX+v8AldcLfj+vv2OtZ69/smn81XkHu7eBv/00134549Xv2OvZ64P8Mv5qxN/9N+8Bb6gVdd/r3Hq9+x17PXQ+GH81b/n+G8SD/Sqrif8Aob3sEdez1y/2S/8AmrmwPeG7/wCo/wAqrgSD9ONXvWOPW89Yx8Lv5rALX7w3gRe4tV1xPP8AT1e/VHWs9dj4Y/zVvr/pw3gD/wBRVf8Aj6j9Xv2OvCvXIfDL+asP+a37xt/1FV/F/wCvPv2Ovdd/7Jn/ADVPr/pu3j/gfu64j6/UjV79jr2adcv9k0/mqfT/AE3bwFh+Kqut/X/Ve/Y693de/wBk0/mqAi/du8LX4/yuu4H/ACV+feq9ez10fhn/ADU/+f3bxuSTY1dd+fyPV73jreevL8NP5qlj/wAZs3gb/W9VX/8AFffsdeqeuv8AZM/5qn47u3gOTf8Ayuu/2P1Pvxp17PXI/DP+apf/AJnbvC30B+6rv9a36vfsU61nr3+yafzVOD/pt3hxwP8AKq48Wtb9X+w9+qOHW/Pr3+ya/wA1S/8AzO7eH1/FVXkm3+Gr3vHXs9dH4Z/zVPoe7t4D6f8AKVXfQj8er+nvWB14168fhp/NUP8AzW3eBsPr93Wn82+ur37HXs9e/wBk0/mq2A/027wIB+hqq6/04/te/Y61nrv/AGTT+apyB3bvDnk/5XXf0+o9X+9e/Y69nrr/AGTT+apf/mdm8OBa/wB1Xcc3/wBV79jreeuX+yafzVLc93bw+gterrvr/wAle/VHWu7rr/ZNP5qhFv8ATdvGw5v91XX/AK/6r37HWxUnro/DT+apc37t3h/tQNVXf7EkX9+x14169/smn81Qcf6bd3/633Vd+OePV79gnr2euX+yY/zVD/zW7d/9eKqu+n1v+r/H37HWvl1y/wBkx/mqG/8Axm/eBvf/AJSq+wt/yF7914V66/2TH+ap9P8ATfu88c/5TXm/5Njq97x17Pn10fhp/NU5/wCM27wHH/K1XAfX/BvesdeyOuP+ya/zVLf8zt3hwP8AlbruP9hq9+FOt568fhp/NV/Pdu8L8/8AKXXf65P6vfsde7uvf7Jp/NUvf/TdvAf1tVVxuf8AX1e/Y61nrs/DT+ap+O7d3/8AnTXfj6Dhvfsdbz1x/wBkz/mqH/mt28Dx/wArddx/h9ffsdeFeuR+Gv8ANU/Pdu7/AKc/5VXHj8AHV79jrWfLr3+yafzVLk/6bd4fT/lZrh+f+De/Yp17PXa/DH+aobH/AE37vve/NVXE/wDQ3v3Ws+vXY+GP81X6/wCm/d/+H+VV3IPFh6v969+x1up69/smH81Qcnu/d/8AXmrrvoOT/a9+x17NOuP+ya/zVD9e7t4f65qq76fU/n37A63nj14fDT+an9P9N28CB6uaqu/3n1e/E9az14fDT+ap/wA/u3hwLC1TXG39edXv3W8nrr/ZNP5qtwP9N28LfUH7qv8Aqf8AY+/Y693eXXZ+Gn81UfXu7d/+wqq4cj/kL37B60K9df7Jp/NU/Hdu8OPreqrfrbkW1e/Y8+t5OOu/9k0/mqf8/u3hwf8AlZrRz/yV79Udar10fhp/NUvx3bu/nj/gVX2B/I/V791sVPHrv/ZNP5qg/wCa27w44/4FV35/w1e/Y6117/ZNP5qlv+Z27w5HNqqu/A/4N79jr2euj8NP5qhP/M7N4f0/4F1w5vfj1e/VHW89d/7Jr/NUB/5ndvDn+lVXen8WPq9+oOtVPXX+ya/zVASR3dvC9z/ylV/0/oPV79jr3ceu/wDZNP5ql7Du3eFifp91XXP+3b37HXs9f//R39fFH/xzT/khffuvde8Uf/HNP+SF9+6917xx/wDHNP8Akhfx9Pfuvde8cf08aW/4Kvv3XuveKP8A45p/yQvv3XuveOMfSNP+SV9+6917xR/8c0/r+hfr7917r3jj/wBQn/JK+/de6944/wDjmn/JK+/de694o/p40/5IX37r3XvHH/xzT/klfx9Pfuvde8Uf/HNP+SF9+6917xx/8c0/5JX37r3XvHH/AMc0/wCSF9+6917xx/6hP+SV9+6917xx/wDHNP8AklffuvUHXvHGfrGn/JC+/de6944/+Oaf8kr7917r3ij/AOOaf8kL7917r3ij/wCOaf8AJC+/de6944/+Oaf8kr+Pp7917r3jj/45p/yQvv3XuveOP/jmn/JK+/de6944/wDjmn/JK+/de6944/8Ajmn/ACSvv3XuveOP/UJ/ySvv3XuveKP/AI5p/wAkL7917r3ij/45p/yQvv3XuveOP/jmn/JC+/de6944x9I0H/IC+/de694o/wDjmn/JC+/de694o/p40t/wRffuvde8cZ/3Wn/JK+/de6944/8Ajmn/ACSvv3XuveKP/jmn/JC/j37r3XvHH/qE/wCSV9+6917xR/8AHNP+SF9+6917xx/TxpY/X0L7917r3jjH0jQH/gi+/de6944/+Oaf8kr7917r3jj/AOOaf8kr9Pfuvde0Rn+wh/5BU+/de6944/p40t/wVffuvde8cf8AxzT/AJJX37r1OveOP/jmn/JK+/de66McQ5MacD66F+g/2Hv3XuuP7H10x8m36Ryf9t7917rloit+hLf8FX3vPWqjj1344/8Ajmn/ACQvvXW+veOP/jmn/JK+/de694o/+Oaf8kL7917r3ij/AOOaf8kL7917r3jj/wCOaf8AJK+/de6944/+Oaf8kL7917r3jj/45p/X9K/X+vv3XuuvFF/xzj/5IX37r3XfjjH0jT/klffuvde8cf8AxzT/AJJX37r3XvFF/wAc0/5IX37r3XvHH/xzTj/aV9+6917xx8/tpz9fSvNvpf37r3XvFH9PGn/JC+/de694o/8Ajmn/ACQvv3XuveOP/jmn/JK+/deoOveOP/jmnP8AtC+/de694o/+Oaf8kL7917r3jj/45p/ySvv3XuveOP8A45p/ySvv3XuveKP6eNLf8EX37r3XvFF/xzT/AJIX37r3XvHH/wAc04+npX37r3XvHH/xzT/klffuvde8Uf8AxzT/AJIX36p6917xx/8AHNP+SV9+690hez1Vet9+WRP+PR3BcaVsR/C5eCDwf9b349eHVFH8iySZ6P5A2VFh/v7lAQihBxWvp4H+Pv3Wsjq8XcGyJpMg+XwbeCumcO51aVDg3Jt73QHr1SOo3968rtxFp9yU01YW0xq0Cahx9GZgOf8AY+/VIPXiAeHSwhrsTnqC1V4okmS6xyssciowspBb34kDrVCD0hKzb28KGpLbWrokoyRcMyuGXVf08829+IPW6g9cXx/aAqUf+Iw+C6a1Upew/V/vh70AeHWyRSvQsUULinp3qlRqtY7SOAL6z+o3HvZOeqgCnWZ6iBBKzyKogXXIW4CLa9+ffqHrdQeisdpbkp88+TpMX/nY8XXRNMwGkgQFTz7qTXrY6r6/lqZDCbK3N35LlKunikkz9VPM+pVa8dSzW/2PvfDj17j1Z1u3fmUq8JHmtsPqoiC6OnJe39WX/evfifIdeHGp6T20N77u3pTy0kEq0VYG02lYxyMbWuL/AI96ycdboOPSyx2I7Kp5JPua9HRL6Rq1K3+K3PveetUA6YN57yyMGFyu3q2PyV2RoJqKKRkuomkXQHH+tf34kjHWqDDDoifRPT3YnUfYGS3BuLIfc4XMVz1sI1HRFHLIWCE/63vVc9b456sH3A1NuSleCCKQvkUWOnnjH7aOVsSzfT/Y+7Gh61QgZ6DBd+4jopWx+41mrqqsbxwPT/uNHIx9Cva/HPPutSMEdb8sdBvv7aW8txY6XuJ8zBTR44CrxkMkunxUg9QD/wBDbj3759bwMdDX0F3hQ9k4tMbURyQ5XHx+Oerma0FY0fpLQO/6j/rH3uvr1oinDoTt/wC1K3M0qVOEZYMjG4ZmQBXkVeRY/wBffjkdaGOmBOw323j4KLL4+qqqiliEczxglnYcH8e91pnr1K/Z0p9tdg4jcwZFikoAoOoV2mIMB+BrsOffhnPXj6dLOjnx8gMdDNTtZiWSJ1J4+pt9f9j791qnkOgu3jH95vrZX27CY0lYGqFQ6hENRN3twPejxz1YdazPWtXUYv8AnldqyFIp45qewSVFkCkv/qX49+PWh8utr6IJJHE4jjCtGrW0AWuL2A97x16px1kEUX/HJBz/AKhf9v711vj1344/+Oaf8kr7917r3ij/AOOaf8kL7917r3jj/wBQn/JK+/de6944/wDjmn/JK+/de694o/8Ajmn/ACQvv3XuveOP/jmn/JK+/de6944z9Y0P/IK+/de6944/+Oaf8kL7917r3jj/AOOaf8kL7917r3jj/wCOaf8AJK+/de694o/+Oaf8kL7917r3jj/45p/yQvv3XuveKP8A45p/yQv5+vv3XuveKP8A45p/yQvv3XuveOP/AI5p/wAkr7917r3jj/45p/ySvv3XuveOP/jmn/JK+/dex1144vp44/8AW0L7917rvxx/8c0/5JX37r3XvHH/AMc0/wCSV9+6917xR/8AHNP+SF9+6902V2VwuNUyZCsoaVRwTO8aWt+LHn36vXugW3H8hOvtuzyQzV1JUeMMWaGRCCFNvoAffiQOGeveVeg6b5ldVQzGKrmihINtbOiqfze7D36vWyB11B80OnaioFOlfTa2NlPmhte9hc29+rnrRHn0LO2e9etdzSrDSZrGRSOFIWSohF9QuB+PfqjrR6F+jqaDIQioopKephb6SxaWU/7G3veevY6leKP/AI5p/wAkL711vr3jj/45p/ySvv3XuveOP/jmn/JK+/de6944/wDjmn/JK+/de6944/8AUJ/ySvv3XuveKP8A45p/yQvv3XuveKP6eNLf8EX37r3XvHH/AKhP6/pX6/W/v3XuveOP/jmn/JC+/de6944/9Qn/ACSvv3XuveKP/jmn/JC+/deoOveKP/jmnH+0L7917r3ij/45p/yQvv3XuveOP/UJ/wAkj37r3XvHH/xzT/khf9f37r3XvHHa2hLf00rb37r3XvHH/wAc0/5IX37r3XvHH/xzT/klffuvde8cf/HNP+SV9+6917xR/wDHNP8Akhffuvde8cf/ABzT/khffuvUHXvHH/xzT/klffuvde8UX/HNP+SF9+6917xx/wDHNP8Akhffuvde8cf/ABzT/klffuvde8cf+oT/AJJX37r3XvHH/wAc0/5IX37r3XvHH/xzT/khffuvde8cf/HNP+SV9+6917xR/wDHNP8Akhffuvde8cf/ABzT/klffuvde8cf/HNP+SV9+6917xR/8c0/5IX8fT37r3XvHH/xzT/khffuvde8cf8AxzT/AJJX37r3XvFH/wAc0/5IX8fT37r3XvFH/wAc0/5IX37r3XvFH/xzT/khffuvde8UX/HNP+SF9+6917xx/wDHNP8Aklffuvde8cf/ABzT+v6V+vv3XuveOP8A45p/ySvv3XuveKP/AI5p/wAkL+ffuvde8Uf/ABzTn6+heffuvde8Uf8AxzT/AJIX37r3XvHH/qE/5JX37r3X/9Lf49+691hqamno6earq6iGlpaaJ5qipqZUgp4IY11ySzTSkKqqASzMQAPr7917pN4DfWyN2T1FLtbeO1dy1NIpeqp8BuHEZmemQMELVEWOmkZBcgXYDnj37r3Sq9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XXN/px/X37r3XFYwrFrsSb8FiRzz9PfuvddHyX4CkXH5sQv597x1rOa9eXya2LfosNIvf/X9+NKdaGquesnvXVuPXXP4/wB5/wAfe/t619nXEagSXKgfixIsP8b/AOPv2PLrwrWp66aaFFLPLGij6szqqj/XJPvXW+kZkewdp4yWaGuzOPhMfDH72nPB4vYNx73869a6Rzd59U0D/bnduMV2a1pK6nvqP4uze/eXXvPpUQdpbEqKb7qLcmLaErq1CspyLf64b3rHXs9M2Q7t62xkLT1m5sbFEBy33dObX/5C9+xXrdKjqvb5O/zavjT8eqGqhrN2Uk+QiSYraogf1RqW4Fzxf3vrQ6oR7e/4Uv1C1NZT9bUj5SmibTG8ESuHW/6gy+60HWznh0Vas/4Uo/JVzGcbsaslh1+pylgyA8fX/D3uuOtdC/tH/hSL29UxQNn9tzUjB4vKrBTpXVZ7/wCt799h68R1Yv1P/wAKJel8lQU0O98rTUWRZbTJLJErrIR+mx/3j3vI690Yyn/n7/Es6Fqd10Mbst9P3NMpBIvaxPvVT16nRmOpv5uvxf7OhWSi3VR6pGVUtVUw/U2kG1/p79XFevedK9WDbD7r677Hhjm2xuHH1okFwgq6fX9L2sG+vv1R1s9CjNNZCYXhZ/qFeRVDf4X9+x1o18uuC1USIDUz08TW9V5o1F/r+T791vPUZM1h5JTAmUx7y/8AHNayAv8A7YN7917pwV47emRSDz+sH6/Tn34mvWgKDrtWBJsynn6Agke99ezXPXL8/wCw/wB9z7117zp105YC6rqP9L2/2PPv3W+ux9B/re/de679+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SD7S/5ltvz/AMNHcH/url9+691RR/Iq/wCLf8gv/D+yYt/1Wv8AT3vrXljrYT96631gnpqacf5RBFKLW/cjV/8Abah73k461gZ6RmV2Jj8nUCb7qqplBuIoWsv+sOfp/h711vy64Pjc1hacw4xnqlAOkuxLW+gFv6+9+X2dVNK+nTQZ98S0+l6YBjcryw4B/P8Are/Anj1sgcOuNNNveOORTT8nhb83P+H1+vv1T14genTVn4t71NA4SmZQVPl0ElmXm9z9eB70a9eFAcdFvjGWlzuXxdXBopDiMgz1BTSwk+3a9yOfeurfPohv8vbqmh37vj5FQZXJ1kVOm46unRYXYN6qllVhyOB738utdWe0WPyWw2o9mCH7zblLOix1c3LSIDYhi3vXwmh68MivS83JhcHh6mi3Rj6g0cSpH5YqY2Rn4Jvb3vhkdazwPQt4fKQ5HH0VTGzMJ41sWFixt9ePe6V4daqQc9I/sLAUFViqnJPGi1FNEzhtN2Okarrb88e9HI62DQ06ADC5JN0bCz0lS7rHj6kwLUMSssYRrHR715UPW/OvUbM9ty7P2fR0mHp0rlhgtNVWWSWNfyb/AF97qQKDrRUVr0msjQUu+Ov6Te1JoyuVlrFSSKoHmaJdXLAc2t+OPdc0r1s8egu7H2/3lvAYbr7aFHM3X+Xhh/vDkF1p9keBKiAfj34db4How2R6kqNgdV4ak21GIMltaD7tq2nAWornCBytQy8m54592IxXrVc0HQpdF7/ym+tpioz8aU2XpJmppILgO0cY0iS31/HvwPXuhekoqKcvLUUFO7ITYvFHIWAH15HveetVHSW3HsbFblp4UVmxxQ6vJRKImYD+ywS3096I8uvD5dBru/B5HrzBvndrVNVkchEwhEErFkZWGkm4J5964cOt5PHpy6fo8vW01RuHcEbff1p8gWS58LP9QgP0497Fevda13XLK/8APH7T1AOop2AB4/t/UD3vrXr1tfw2EUVhYeNOP6en3omuevDA6ye/db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rgqBSxBJLG5ub2/1veya9aAoa9esS+q402/xuT/AI/4e/VxTrVKmvXZsPUTa39TYe/Z4dbIFanoHe0e7tk9V42eu3DlKSLxxGQKKmHULfhlvx711vqpbtr+aviMRX1Mez6lKtYWljCLLHoa3CkAH36vp16nVee+f5kW5eya+Wnr8lXYpQ9h9rIRwDY8J719vW8AdA1XfJDctXULBh8rXZOZ3sRUBn1ajcA+99a+3oTtv4TsrtGlSWuE9DEyq2uMMODwSLf8V9+63mvU6t6jy2IrKSmps5WtKHAqiruSoP15v79T069057jxuZ2TFQ5bG7vy3mglikkhjqXvZOSGUH6fTj349aGR1dJ8EfkfhN+bZptqZLKtPno40jQTSq8khX0cqef9j78PTrx4V6sqHAA/oLc+/de6Ln3/APIDbXS+2avJ1tZAauKCaVYxNGGUxKTYgn/b+/HrwyeqWcx/OwwmAzddi67SPFOY6cs6AOpaykX96z17HS/27/N6xO4AksMiiO12tIgUX59RPvfHrZ9R0MuB/mq7CLQjM1lPCHdEOqSGxDGzHk/j37r1K9HT2D80Ond9U9E1FnaUS1aBgPuYALk8+knj/b+/dVqejTYvN4rNU0dVi66mq4ZFDBopo3Ivz6lU+99a+Q6dfp/rWJ/xv9ffuPW+HXgPr/j7114Drhr0ltZVVFiGLC3+IN/pb3v7OvCvn1hNZR/pNVAD/TyoD/vfvw68aHHXa1dK3C1MDEf83UJ4/wBj79Qnr1QOs4dGFwykHkEMCLf14966317m4typvfn6f0t791rrHLKkIDMCdTAcC5+nvfHr3DrmjFhcrpB5H9SP8R78aeXXgSePXP3rrfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//9Peg737r6/+N/S3anfvauXGC646d2FufsXeeUCpJNT4DamJly9fHRQOyeaplWLw0sAYGWZ0jX1MPfuvdaF3xv6E/mC/8Kquze0Pkj8kfkR2H8Vf5aWy9+ZXZnWHS3WtZN49x1WPEdUdvYLCSvFjMlkKCkmpnz28s7T1n+WTGkx9J4Vlgofdb6Pj29/wjw6M6/2nLvr+Xx8xflP0R8mto42TKbE3PvTfGCrtv5XdFBH56OmrM11xiNuZnCipkURCvoqqf7bV5DTVAXxn3Xujsf8ACcH+aX3r81us++/id803lb5p/BveMWwuw81koKWjz+/Np/xKt2xTZfc9NQIlO+cw+VxddiMzUwALUD7OpfVPUTO3uvHrZg9+611737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddG/449+60eu/fut9cWZUVmYhVUFmJNgABckn37r3QOdjd/dTdWYutye8t64PDCjidxBWVkcc0rhNSIiMR9fp79Udar1R93L/PI65GdzOxOuI4cjW00dUgr6W812iJVXBBNr/09+6359Vbb/8A5n3yM3pLX47Z2cyMc1RLJ9vBCsytpPCjj/effuvGp4dFI3F3h80q3H5zOZ/d+XxsMULyK05qVUfU3uSPfuOOtCo6q53x8svlhXZaaDFdtzJLT1EwP7tQSGRvzZvfut9CbsX5h/McbSrIa3tqpmkSypMstSNNj9Dz7317o2GzN7fJXsbbFDUZLu+WnZ0DTB6mZbgjm9296+zrXSPyvQdHvPPCXs/ccG9kYfutLUO4uf1XBJHPvfXuhG2j8QfjRS6o55cJhnkY2jeSIcfUCzH37rZ6x5j4UdZbiqKyj2puSh/ZibSlLLEDe3Gm3vX29eOOHUvYn8seozFFUTx4+ozCFn0srfUfX6+9VHl16h6Qu8P5ZmIpMmpqNmS00kUnrkYyC7L9ST738+vEdEu7T+HuO2hmWEe13nS5UfuzXBVrfj8+/da+XUHEdVdg7Uplq9rTVO24Ywr2SacKFTkWN/fuvV6MTsf5w9/9KUUh2/2RNU1+NS5pIpZmlLgfp0qTzx72Ot/Z0Le1f553yzrzUUlfl83FPTIwDSx1ADLCLAr/AK9veuvVPTdD/O0+V+6Er6X+P5iCWKZ4kd1qVFg1ri/v1B1rgOkhW/zY/l5iaiLKndWV0xkSuy/c8i97XHvfXujT7C/nqd7U+AikzG48hJNGEV2lWYklfqovzx71jy63no9vxX/4UA5KTdlDjN8wVGTx8zqlRUyxSLEuqTQfWfobf4+/da62hukfmN0l3fhMfktv7wxEVdXRxE4yWqjWaOSSMOUFzc83Hv329b+XRpYpRIIzD+7DInkWdWVoyrcixH1v/X3vjnqoBGB1I966t1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdIPtL/mW2/eL/wC/R3B/7q5ffuvDqin+RV/xb/kFxb/f/wCT+v4/y17+/efWuthL37rfXvfuvddc3Bvxb6f4/wBfe/LrXn13711uo697917r3v3Xuve/de6B7dWxHr8rkMnSIIY3xNZGwSyq0jwEWUD8n+nvxHn1oHqvr4A7OfYHZvdNNkKu02c3BVTQQSGxv9wXW3v3nTrZoBno6HdtbkBXY6hxsbvJNLErhOAAxF3uvvTeXXhjpfYfZk0+JoqHLy+aKWGKZlbkqxXUQB73TyPXtXoOhKoqGloKeCjplVEp0ARRbVYcXPvYx1UivnnrFlvtBj6pa8p9vJC8b6rc61tZb+9Y49bzw6IlvrOUW1sLmsHjnSno8jPLJJIt9IJvbke61zjq1OgQ64ycPhy2Bz1Ula24I3psUJG1aTJ6VCX/ANce99ePHHQ/fH7Cbj2tvCs2TmsdM+FEbVdPLMpaA6hrUC/H09+Fa9aPDPR56ekgo1EVJFHBDdmaONQFLEfi3vfWvPoundXfG1dm4zJYSiqqXMbiaJ4ZcRA4kqItQtZ4xzf3o06tQnoh+3O1Owdmx/37fCVtFiqjJLH9myyBTHI/6iv+t79wyOvChHVqWwd2Rbz2xjM6iGJ6ymjkljItZmW5sPe/KvVcVp0shb6Wtb8f6/vZ68Og33/kolxpxlLaaqlkVvEliRY/0H596Pp14ZyOlBtGGWHD0wkXxnSNUf5Bt+R735Z699h61YettP8Aw+P2oVtxCxvwedfPv3yPWj1thxf5qP8A5Zp/0L711scOsnv3W+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de66v9f8P95/1vfutV6gxxVAqGkLlIOToZgb35/wBhz73XrQArjqtb52fPfZXx22hmsTjcnTz7skhenhSNxI8ErIfoqn63t71kdWHr1qvbn+VfbfeeTyFfuDdVScRLJOI6KRpUvG0hKjk/T36gp17oF6+GSpmapjqtGokuA7HW354v7317oZehtt4TcGVaHK0aTgylWmc/pANtfPvRPXujfVW2dj7Uy0LUVPTzSMyfo0tza3+9+/Y8ut19ejwddZBIdsJJTJ4UMahSALIGHFifeqZ63UeXTZnMTPjMbmc1U1OuSaEyUxLWIJUkW/4173g9Vp0Sw7nqK2oy65io80KwVQiRr/VASNP9Pfut/b0P/wDK+w+6c930u4aBp/7u0FWyT6XYwjTMQQw+nv3n1rhXrYf+RnctF0zsmrztRLHFLLE6ws3LB7aRx73Ujh1qgOD1redt9p7w+RuSyQkrZf4V5alAgLqhRmubAcfT8e9fPreOHVfu8vjPtjK5BkqKeOavL28hkOvVfgn37rZHn1npeg5do0ANNEypYkDUxBFrAf6/v3WqHpC5XZNZOssX27l1LMGDPwy8gg+/Dr3SNwu7OytlV7JRZyfGxUzaYf3Jbab8Dg+/fPrx4UPQu9d/zWu6ujd+Yii3BuKtyOAlro4nDeVoxCDY35/25Pv1BXr1T1tZfH3+YL192t1VRb0qKuBalaGnaZFYKxmZbEst/fievAenXe8fmlTVCadvRAxm3rjViSt7nke/dex59ArlvlnuSpVile0Ttf0WYWv+AP8AD3rPWxpHSOi+Ru8KqvRDlHiVvqCW1Hi9x73w60KV6403yM3auRmibMSBY2NhdiCP9ifes9bx0sqH5NbojkRWyz2BFgC1z/tI9+oa9ex5dLuL5jZfBQLNkFlkiULqZwbFb8m5/wBv78C3l1XSoPRxepPkNtDszH05FZTw10ll8ZIF2t/aU/Q/63uwOacD17TjOejFKVZQykFSAVKkEEH6EEe9dbx5dd+/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r5wXb3w43Z/M2/wCFNXz7+IGY+U3eXQuysRhK3svH5TrfcGTq2pKvavXOx6SHD02Bq66npIoJ2ykssjIAQyiw9RPvXWwT1ZvuL/hJH2PtnE1mf6F/m/8Ay12n2ti6eaq2hmNxxZ2mxCZSOMtTU9bktm7lo8jRxyOFV6qmaV4xdhBKRoPqderXpZfyOv5qPzV6/wDm92X/ACUv5reWqN2/JDr5M/J0l3Vl6yOvzm+Yds4Mbum2rndyNHTvn6XKbd1bj21n6mEVstPHNBXmSdoVi317rcU9+611737r3Xvfuvde9+691//Uvi/4VM7s3DtT+ST8rzt41MR3HmOjNp5uqpjKDTbezXeW30ygmaIECOoCLRvrIUibTe5APuvdDd/wnZ2jt3Zv8l74FUO26SlpabMdU5bd2TalSNPu9xbv3/mNw7gq6ho/1SmqqJVdmN/SAbWsPdePV1Hv3Xumah27t/GV9dlcbgsNj8plCWyWSocZRUlfkWaTzM1dWU6LJMS5LEyMeefr7917p59+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XRNhf34Z60TTrpWDAEXFxexFjY/1Hv1KdeBr13fm3++/1vfuvVzTrv37rfXvfuvdB1vftfYXX2NrMlufc2HxqUcTSvDU19PFMwX6gIxv/ALx798uvfPqiH5Wfz7+g+qs3UbF2hXRVm4lkmijnWcSq0kfCsPGLWv8A4+/fLrxHVZ2/P+FAvbcESptbxVE9aA1I0V3VGtf1WB/2x9+68ePRVcz/AD9/ndRV8QhSj+1qZSlMWKJcsfQDdffutdGh2P8AzXfmtujbLbw3nmKCmw/27s1DFUqlQwYf6hbE/wC29+p59er1Vd8sO8vkR8uM20aZDclLjEmUk0NXUxmVFa1rIRcW/Pv3Xuk/0R01TdRVj7l3jS52qmrynmeuWaVgW4kOp7n/AB9++fXvt6Hfc3f3VPWOUgyOISJMp9UhnRQdbcj0MOf9t791vHTTkO/9/wDd2JyGCx9LGKXLRNT0zwRfpDD6+ke/daJz0EHX/wABN/5jMTy1FPUSVFU71PkNOxTTN6tI4sP9j79x63no1+zv5fO6qWhqcNUY+plercnWlNfTZr6TpHHvx+XXqZ6NjsH4GZnEY5aKqpayONFOrVC62BH+Fvfut0A6Fxf5e+QfBHI42rSBml0sZgQ3J5Hq596r59apXoIOx/5buQenp8o+4EWWNQ0kcFWUdiEvYKp+vvYPXjxr0SvdHUW9OhZchl6LJPKssbrTCWoZgHHALXP0H19+8uvcOHSM68/mi756CyMuE3fPRVcGt1VonEnoB+jDnm3v2OvZ49CxX/zato9g0tdUyUi647s/7ZS5H+pNuf8AYe/Y690n6r5QdT7ywNLl8lRBqiochQ6AtdzcXuPfuveWOi799d+bTxGx6yrxNOurS8cccarrJkQrGFVefz79w61x6KJ8UegN7dpbnr975Tz/AMHyc7VENNUIxQRl7qNLcf09+A63Xq0jC/EfBw5Z6mux0Xikp1jGmljChivJNh799nXuvTfDXD0T1DUWOjZqhnkUpSoVUEkjUQPr/T37rxHp0CO9OikoY6vFT4tyxUxh1pV0K17Ak2+nvf2deNei3UnQ38aysmzqONKatkkYpUMiJENRsFvwPeuvZ4dP25/hx2v11tXIZehSqraYKZFegpy5QW1X8kY/3n37z60eg46r7j+RXSlfSZna2V3B9zS1HlWjmrakKkkZsAYgePp79SnW/s6vX+NP8/jt3YMeLwHdZvRQtRUzeV2eVYkYLKQxBP0/Pvxz1ry62R+jf5r3xl7o21SZfHblpqKpECmuhq6uKPRMEBbRrsTc+/db6PbsPunrjsamSo2zunDVxfTaGLIU7yksL2Cg+/da6FRWVgGUhlPII+hH+B9+6313cXtcXP0F+ffuvddM6oLsbC9v9v73SvWiQOuXvXW+PXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SD7S/wCZbb8/8NHcH1+n/Frl9+62OPVFH8ir/gB8g/8Aw/8AJ/n/AKbX/Hvfr1XrYT96631737r3Xvp7917riVB+o/N/9j72CR1UqD1y966t11Yf7b37rVB13791vrFL6opgBc+NxY/klTx73TrQPVT23MtUbX+RtdRyK1OMpln0oLp5Ly3+n590rTq1ARXo9eIzNDubsrKYmrpy0mOiSZdY9FxGP0397zXrWAOh50KNPpHoFlsPoALWAHvfy61wz1h0ReYONQlK/wC1C4ve59261xp0hN95OgSljxs/kMs8iKAupbayNJv+fejQDPWxWuOi99m9YUuQxi4SEN9/lEFRTzEgqoK3HqP9f6e60p1sHNR0wdU9AUoyVBPuGOUVG3pFlppAoCSOjXX88+90rx8uvVp8Pn0do01JC5q/BEJooSvmEa+XxRrwuoc/QcD3vPWsV6JH21v/ALyzO7IMR1Cgp4EkEM71EQaO19LHkWv+b+9edOrCnT5178VsYmV/v52DNJkt7ZBvLk/3PJT6zyQoJIHP9B71T1610ZHcPXu2M/tt9u1OMpvskhP26xxRhklRPQ/A5NwPez17opOwd87g6m3TkNq7nBfDz1Jjw0RBV1g1WjsP9a3vWRg9ewc9GrG/KTK080GLhqVq5IbRNIulVd14IP5t72Dnr3WDam1K6OaXKbgYTVjuWiQnUAh5F1P09+p69er6dCUkaRghFCgnUQP6+/dap1qd9bKE/nkdqqALGnJ4/wAXvf37iOtnB62w4v8ANR/8s0/6F9+60OHWT37rfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6910GDfT8EjnjkGx9+pTrQIPXibC/v3W+HXfv3XuvXt/vXv3XuuDukYLOyqB9SxAHv3WqgdRJMlj4rCSsp0ubDVKouffut9YJ85h6ZQ0+ToolPIL1EY4/wBv7917rENx4E2tmMdyLj/K4bWAve9/fuvdFJ+VXzE2H8c9n1WcrclSVdSsBkiSCdXYH6AAJf34HrR61n+8/wCff2TBTZp9jywQY1GrKVdb6ZgtiAQxHvVK563w4da9Havzs3X8gt91e5N4ZWq8YqxPPG9ZIYmYN+Ax/wBh73X161ToQ9md+7brTHSUk6AEqCPKBYD9X09+68AR0LNRvWKsgX+G1kN3ZTo8lyV/P+3/AKe/de6OZ8dq0SYurkCSrOEYySWYaj9boT791YdDpIrS19POPNJPJMqICxtc8i49669Tqynpmgc7RSnziM2qFHS1r2tcfT349eFeHTT3NLUUOJWKJtNEij0g2JS30b34fLrxwfy6qx7e3DU47EZCrwmtZhBWGSx08LGSANPvfWiaDq6f+QuIdxdI7s3NkaJf4mMzJS+WaL9xf3NRKs3Pv1TXr3l0Yf8AmzZCSg63283maGneq01Gh9OtC/Ib3rz69XHVPOx92bfw+ypKyijEjyzeN3sWuzi19fvfDq2Ohn6n6wxPYFemUYLJLM3lUNyAbn6j/iPfuHWscehR7M6YlpqNqampwFjRSSsINhwPr79177OiS7g2JPR1ssSQ6XGrUWjFrL9Tb349e6JL8gKaLF0NXXwwmNqMXmZV0gkIbkEe/da+3qm/Lb4m7I3/AEO2oXVo46/wkobzW12/2HvwI61kdbF/xY23U7N6+oMPLU1y0NRSUzuDO6i9gfofevOg639vRzcXWZCWndMHI8sUPDln1m35/wCR+9/b1vpKbh3JW0a+tm+7DEEBiFDfS3H9ffvn16nXPAbyrJZ46rIE+OMFBYnV+m3N/wAe/da8uudZvmlxlbLV1cgWnmYmIM1mFx9bn+vv3n1ulem8do0MlQjxSOVLqFAc21X4v/tre/dax0vMjumqy1BHFVyD7WSNT6GBIFrer34dePz6YqLtDKdWVFDmsJk2ioqSQ1NXE9Qw/bB1fg/Tj36nr17hw6vs+JHyG2/3p19Q19FXRSZCmiiimi8mpywSzfX/ABHv3y69wPRsKmrpqOMzVU0cEQvd5GCrwLnk+/de6CXOd47DwVXHR1GUp5ZHfQfHMhCte1ja/vVR17PS2xW99sZijSspMxRNG6glTOgddX0BH/FPe+vdKaGogqUElPNHMh+jRurr/t19+691m9+6911cXt+ffvn1qorTrv37rfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdaNXwZ/7jJP5hv8A4hjfH/vDdae/de63lffuvdaJH86Skg69/wCFSf8AJ+35sWBI9571pPjbjt1pRLpnyFHP37uPYk8tclP62aXDVE1KztyYY1S+hABrq3p1vb+99V697917r3v3Xuve/de6/9XcA/mi/D6T57fAH5R/E+hqaOi3H2x1nXU2xa7IELj6Tsfa9fT7166myEpZdFMc3jqFKmS/piZ2ANrH3Wx1rS/8Jkf5r3VPUPRWQ/lS/ODeOJ+NnyX+LXYW+9o7Bw3dOVpNj027NsZLdFXncnsVMzuBoKODP4DMVGQpf4ZNUK9TRvSvRio8dT49de62aPmP/Ml+GXwV6c3J3P393t1/hMVh8HU5bb+0MXurA5jsLsauFO0uL2917s+jqGrMlVVsgWKJ4o/BECZqiaGnSSVN9a6oQ/4TJZb5qfLXdHzb/mefKLsHuRurfkt2TmsD8Y+nt5dgbyzfXG2dsjeNZujeuW672dmas46lxuOlOL2zi62ioItZoshGpADhvdb622ffutde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691hVUDs2ssxNrFrgf0AHu3EYHVR8zXrlJGJLXZgBz6Ta/+ufegadbIrwPTfk8xjMJSSVmSrKejpohdpJ5o4wB/ryEe9db4dFq3V8zfj/tCpmo8tvjGpVQFxJCtTT6gU/UOW9+8+vZpXqqD5yfzy+mOgNu1rdfZikzufjQrTUqS08jzSMll0qpJPP09+611qW/MH54/If5UY6k3vQbx3FtOLNSyebHUk7qghM2oIUjawuPfvl1unRa9g9G7s3wkO790RLmIi0fmzFauqo1N+ty8nP8Ar+99a4dHf2p1N1VtTFpJlKqiqJJELSGcxk05AvpS/vXHrdc9Fx7ErNq1+5aTE4mKmkpKSqD0siBLSOr3Eeof1t791r59Ho6i6TynYsOOy+RrZMbiaamhDY1Z9FNLChGp2S9jce/deHz6PZiG+LXWmBq8jl8zi4czt+ItPSeSH1yIt7Mv55HPv1cY630SHvH5z9SZvBVtPtLH42umijnijjhgjaxiJVXsg9+63x6pf3oN8dv52XfdDjZYcPjaoXiWF1jIVrW0gW9+610dfp3tvMdU0uFlzG3kShnKkVBh9SCw9QNuPfutDHV6Xxh+XnUuZSnpM5LQ0skkUERmkaFJVZlsQS3PB9+63UefR9K35UfGzquglrK/N0M81UrSxtJJAxS66gFI9+oevVxXqvbub+cFhcBV1GO66wEObWe8MTw06yM3Om4IH+9e/deB9Ogdpv5g3yB3rgJJMbtOppKeUtJGqAoQrn62PHv1OvVJ49Ez7Z+cffe0NwUT5yPKRwzMv+SrK7oQfoGC+/cOvGp6CTe/fXb/AGxj1/3BVbUWYX7dZXiktB5eNY4/3r3vrwNePQVbL+Gi7mzpfdcjVM9Y33ZFUjSeMyG5jGr3qnXvLHQ40/w1wkEdWKTE0cSY8lQEh0+UD/Vj8+94690J22fiDS5XF+XwJEkQ1LCqkKukXBUD6e9V61TrJUfAj+9VJNkKqPzUVI5P25VzGxj+l1/xt731vgOg8ze69/8AxcyWG29htpCXDVcqRLNHCwVEZtF7ke9de4dH22buPtLdu14NwUW12ejnijJmERcAulzwo/Hv3W+lPszu3C7NzibT7LU498jIB5ZoyGjLG1gW+lh791qp6MZX7c6X3XGkNBkaWcZgLDHUNa6Fx+pSf9e9/eqenW6novXZHwlSi15DZM4qKmNXr1q4pbTMVXWkesH6f4e91PWqCuOhA+Ne/cPTYfL9Zdv4ymjbVJRw1FaqnUOYw2qX6+/db+Z6w7i+COyM3m8pujacNFW4+VDNHEroYxrPkOlQffifLrVPPqs3uv4lww7vnqWwtMkMAmjCmnYxlkHB4Fve+vU8+idTdV74weTqqDHbsy20qOWUiKPHCaCJkB4tpt9feuHWuPRjfjR8hvkb8c951lbh957l3dQY+9UtLPLMyARLbR6jb6e/U68DTrZy+B3853EdsvRbN7YiGGz8RipAKnRDKzNJ49barcDj375nreT8ur+9v7hwG9KGnz238lHW0/pcGGVHUjTq0Mqk2vf6+9g4p1Uih6USw+SQVILAldJhf9II4Nve/ketDIx59S40Ea2F+SSbm/1/HvRNT1YCg65+9db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XukH2j/zLbfn/AIaO4P8A3WS+/de6op/kVH/cf8g/6nf+T/1v+Br/ANPfvPrWPLrYS9+631737r3Xvfuvde9+691737r3Xvfuvde9+691jsxc8jRptb/H8397xTqvE46qe+TFXS7M+VHUUNOixLnq+AsE4L6pRcH/ABHvR6sK9WF4LH0kPY+bqYYUSSShgZnVQCdUSsBq/wBj78fiJ60KUHQte/db6wJKWkdGTSV/Sx/tj/D3umK9VB6R+4sJQZCtgmrZgpQo8YNvqhAAA9+AB68SQeut4YmGpxiVWpI5aKNTHMXCERqL+ljb34gdeBI49NG0N8bXywTC0uRgGTpBomTzRCRpE41E3uffq+R69T06EYLKylX02bg2GrUlvz/j/X3uor1qhp1CjxOOo2kqqSigSpIJ1otmZj/U+9efW2pp6cUZnQB9IcraQKw9JI9+xx69mnUarqI8dQz1LG6U8Ukg1NfUQCQtz/U8e9HrY4dVadkdhbp3T3ztmkmwSRYakq44zP4rCZFktyfzx71g5635dWgY6ixklHSSwUkCARRMpWNVZWCC4JX8+7cOHWuOD08+9db69a3+9+/de61OuuLr/PI7U/J8BP8AiPX/ALx791rz49bYMX+aj/5Zp/0KPfuvDh1k9+631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcSt/yR/rce9g060V64l1QhT9SOP8bf4+/UJ61Wg64VFTT0qeSomjhT/VSMFH+8+9dW6K92/wDKXYvWkclO+ShfIqraY/JEQXBtpCk/6/vx68KnojGe+bu5cnkNVLDImNZ7rIGH+bb6MP8AD3rJ69gdI/OfKfcNa0BiqZlS4JYP+lTwT/xr36h63gcOgr3r35m8tC0dNuKrimtYxxzcA/0sD73Tr3QE1/du+KQlH3PlOQVULM/9P9f8e/U8+vH06J33zV747HoZYsnka7LUfjI8dY7SoVHNtJ/2Pv3DrXE9VF9n/Gfde5PuqTC4OLxySSEokLAOzfk6f8ffutU6r/7S+FfcuE11NNt80dOTqZoo5QWABNyAPzb377Ot8Oid5xu0unqhZchi6sU0Uh1zR0s5Is1ubj3rr2D097f+V2cgytHPK9UkEUtOJVZWT0hhqGlve+vUr1sH/D/5JbV35tKOGOeFKuKDTKFaJXZwLEkD37jnrwPVg2ypqfOZagSK0i/cxEBjYn8nn/efeurefVtnXGHgbHUVNoGn7VBptf1aR9ffvt60MCvQLfJCNaOmNKCVF1S1jbn8KPfh14kdVV9xUkGLwf3s6/5PJJOJm0kr42Hq4/p72B1ryPWwN/JVOA/2X7KnBCLxvkxJN4raQ7H6G359+8+vDh0pv5u22K7OdM0dXReT/IZHkm0qWAVTqvx7151691r5dO7jpcjtGfBNIJqmGtdPG+n6g2Nx731sfLo9Xx83W208lHBVTOsTsCoLgabnlRf6f19+pXHXhTqyinnxm7aJyhSUzxqouVJ5X6WHvXyPWvn0Sbu/reqw081dBCUjLOQYwbMLfQEe9jh1vHHqu3uPqyo3XtfJUxplRqmN7MBYveMi5/PvXXqYp1SD098X8tjPkdXNVxyCkp8g0o1BwhBl1An3v59aHWxRhIcfiMFjMNSsDVCkghsou1xZCB/vXv3W+jpfH/rIZfE5KaruJWWRo1IIBOm4F/ej6cOvfPosvb1NUbT3jVUlVCBSDX42ZbAnV+b+98R6da4dBDPuT7ZWqYlvTJfUoIspAvfjj/b+/dbHQRb13u+d009AxvGbsot/ZPJHv3HrX+HoSOo9sVm6JqellVncyRElgxtq/wAR719nW+PR4c30/DtbbInqql/JJAWUMxvwlxa//E+9+fXuq7u0VeaiylCtbLqaF4o0DG1idI49+61npP8Awz/mA5H4v9lHZOcqJRjXnXSHZBGyBrE+r8n34/s69X0HVu/a/wDMxk3vjaPEbamMUOVA1zwMg8IZNJI0f0v795Z69Top+W7Y3PDEtemUqcg05cl5WLePUb3DEn6X9+oB1uuOo+I+Umb2zVRwVm6MgkvDrTrI3j4Fwum/vWOvE9GW6+/mW7lwNRFjamOaopeEFQ7i5F9IPJtb3v8Awda6sY6t+dW2N0mkjy9RDH9w0aWZow2px6jrU/j37y69To9WA3lt7c0EFTichBULNGHUCRNQDC9vr735daNK16VXvXW+saoUuQzNf6hjcX+pI97wetZ8usnvXW+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rQg+N3c3UHRP8Awr8/mGb47t7T666f2Y3VW78Mu7ez967c2HtpsxW7A65qKLErnN01NLSmpmSCZoacS63CNpU6T791vrbL7B/m6fyvusNpZfe27vn38To8FhKSasq02/3fsLeedqFhiaX7bD7V2bW1+Ur6hwpEVLQ0cssh9KIxNvfuvUPWp/8Ay+q7dH8+P/hQXnv5oeJ2Xufbnwl+DuIxu1eoMpuqikoJ9y57bOHr6LrHB2QNEMnU5TL5PfOQpIpXNBAKSkqHLTQtLrr3W+p731rr3v3Xuve/de697917r//W3+PfuvdVB/zCf5GP8uX+Zfnl378iOnqvE9wJQU+LPdfU2em6+7Jr6Cjg+2oKbcdbSxz4/NCmjCR0z5rHVckMarFC8cY0+/dbr1Xx0d/wkU/lE9P7xx+8Nz4Pvjv0Yqtgr6PafcnaFBLst5qZ/LCmUw3XOI29JXRagC9NV1MkEg9EsToSp916vWzLtbau2dj7bwWzdl7dwe0do7XxVDgttbX2ziqHBbe2/hMXTrSY3D4XDYyOKnpaWniRY4YII1RFAVVAFvfutdP3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6Z8xmMRt6hqcvma2mxeOo4nnqayqdYoI0QXYs7f4e7VxTqlM1615v5jX8+3pH4rY7M7Y2dV0+6dygy0cc+MY1bRSlDo0JETY3/PvXVqYoetTHuP+cv8AN/5C1zVe0d0ZHA7fyJZaSgqvu4G0CUlSqEj6r711vosGX3v8v96P/eDO7gyEzzXeULUVQDB/1H6/n37rWPPosnb1VX01ZSV3YHmqK2PSVeepmI8n9nhz9ffuPHrYHSw6rz+eMElbU1jZLbyrroscpZxAoF/p9Pp7917o1OI7k3zXbXl2/gaKrx2NBYGTxtoNr83Hv3Whwx0yYzE9g7xiqlrdxvRRR6g8jyOosn+qHv3Xug437V1O3UxuOx2RirMrQVCzPVJIdUxDcqGH+9e/H5de49DRt35J971mHp9obRFZR5B4EgjcCUHxsNOoW+o9+z17h0tdlfGfu/e4yeU31uCsd8lreemknlXy6udKqT9Offsdez0YrqP4bYbE11THX4Nqy0ZZkLmQ6tPqJDe/fb16nl0bXafRO1cNtzI4dcUtFTTyOWidEAvf+lvx79X063T8um7cnVHW1JtquGdq6HxUlOzU0UjRjx2XixPv3WuqnNw7S3XuffK4bq+unx1HHXlVrqaZxFbyWWzIQPe+vdWidU/EDeW6cFjY9+ZWfcErQR8tUyMU+l7C/vVevfZ0eLA/AXrHa+EgzGYxsVIYUWRpZyrD+pN3/wAffuPViOq/PlJ3zs/p6pm2XsWopZq8CaCNabSzCRRpX9H09+Hz60c8OgI+PG0c73FWT7r7LlOSooJDLT0swvpQPwLN/hx78cZ69xHV2GxeitgvtSkqabAxtDOkaUy+KP8AbfQLN9LDn37y638umbKdZYnalaZWpVMrcRShFOkXuqGw/px791o+vUKj2aldWRSx6IqaRiZYig1S3/AA+p9+p1vyp024Le+0tv77rNu5OvpKKGKMhaecorErwRpPvxwMda8+jM7UymEyNPItB4DjHuWZEUxG/wDaJHv3W+ilfLrZm3tx7dq4oKaMV0dOfsJ9CKI5B+kqR/vHv3n1ry6HH+XnmKHEbIXae9o0r1gdbPKgdRGv09TD+nvx68K06V3yU6F607GyZ3BicLDNU0k11aAIttPAJC/09+690DuZ6GgpdtYUYbcMWFrIgvpklCtCR+W59++3rY6ALsztHtDqDGDG0eefcThwjTU15VaEGxjBF+bfT37HWj0XzdXbWf3VjI8lDsnJQZdYw5rTG6GSQLcuGH+P59+69mnSB2h80u8+q8jEMhicnWYJJC9VTiKZ1SnU/wBr/Ye/Z691br0T8kOivkrtZaHJ46gxu5WgBmWpjjWYTEWcnXz9ffs8et8RToKO6em+sEq5Z6TJ42SeBj46RHh8j2H0Qe9Vp16leq7Kqr2xtXdldDIIaaB9ayxtb1JexBH+Pu329Vx1jwEOx59/02e2nRiHIxLraenYqA68hm0fi/vXXqeXVnHxM/mB706P7Ow22t4br+62zkqwRy0sjSMlPCz6dLajxYe/cOHXvkets/YnYG3+0tq47c2z8pDV0dZDTzloWB8ZljDtGSP9c2/r78DnrxApToRFFgP62F7/AFvb37rwFOu/fut9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SD7R/5ltvz/AMNHcH/url9+691RT/Iq/wCLd8ghf/mP8l/sf8tf37z615ft62Evfut9e9+691737r3Xvfuvde9+691737r3XvfuvddfT37rQFBTqnb5yxzt8v8A4tCNGaF8jF9xp4BAq7er/Ye9dWyB1Yr2Fl6va248fkaGIxCt8EE84HpKi0dj/rD34mmetYPQzUlVHU0dPVxOs/kgjbVHY6yy3I4+nP8AX3YenVSPxenU1W1qGta4vY/Uf4e/HBp1sEEV6BHsufMU0ktZSRyyRUEf3BCg8onqIFvfjwx15eqafkt869+5Te1N1fsqmq6aqdvs5JolkWzOPES2n835918ut9MvXvxi+VMefwXZtTv+sw+GlmWsyCTTTLG0LHyEMGP0sfz73Xr3yHV8XX+T8uzsVVV2XgyRipooZsgrAxvJGoRyX/Nz78OvHHSnyRmrMZOcbMPK0bNE8bD1EA8Bve6Hh1qo49V47e7X35tjuWo2/uSsnbGVlcaelSTUy2eTSOD/AIe6g56tpqOjJfI/sWr622bTbkio5q6gMscc9PEhNg9jdtP4sfdia4HWgOgh29uPAdsZXZmcwmA8BWCGWuKR2IcfqLcfX3XJPXj0eakpxTJ4oxpiVU0KSSy+kXBv7uSKU6qK1J6l/T3Xq3XvfuvdanfXAt/PJ7UubjwG5/B9f0P+t791o+fW2DF/m4/+CJ/0L7914cOufv3W+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rEwl1gqRoFuLfX+tz72KefVWr5dIbd3Y219nQTTZevhhkijkYIWUMHRbhfV/xHv329byfLqrD5AfMSoyEtVS7dq/t4KU2SRNaqb8AAj6+9Zrnq1PTqpje2f3R2Ln3y+QqZainileQkNJ+jVqNyxt/j7917j0oG3HNVYyLH4tTJJAixMEJY3Xg6vfuveVep1KmXqKBqbyOlSwAVNRLD8C1/fuvcOk7htk7vjzDVNbDUmlJJDPqIIJ+oJ+vv3Wq5r1y3TtDLTVcf27tHyNR5tx9Affut9c8Rtarprw5mYLBL6VMpujG39m/v3XuupdlfwTKLkoKP7jGXUtJGgZBc3Yk2/F/fvl16g49KvPbd2luZKJaihp6iAKPuIRFGbG1iDx72adezw6Bjsz4e9Gdi4U00u3qFJpF0uzRQC1zcsbj/AB96yetccda8fz0/lOZPDRVu6+qZpDBA1TUGjx7IFZY11hSsY+n+t7914jqrL4xZzt/qPt7EbJmxGTSOWo8dcC8rC/lCnj377Ovfb1uF/GiqrKqTCzZCgmglmkpNRlve5iBJGr37z68OHV8WwtuSRYygr14Q0qtY/gaeSQP94916350HQE/JTAU2SxzVKOmunjV3YWJuBe9h+fe/Prw4V6rK7R27Qbx2FkMVA8bVcFHWsrAgOrLGRc/n/W9768eHVk38hjcFFtXqvePX2cyKLmRnJJqOKViZJIxKQFFz/T34nPWsAdXd9+dcY7tHq7du16yiWrqKzEVaUF0DulUYj4ygP+J9+615160hP7pZ74+fIvObF3e01DBUZSunpY6u4/bM50ek2/w9+49b4HozubqshHCMjiMnYuRJCYyQAt9Rtb3rrwx0dn419vmjp6Slz+XjaoVwumU+phaw5v78et9H4y0WD7EwhgRY5i0BcEC1iRxx71w69U9V99tbSr8VLU0lLjZfFC2jVp9JAH4Nvz7txx1r/B0SiHYFDhNxVG46mh8VRVsdLsifrJ5sbc+/depToaNl7Qyud3BRVaU0opEeE/cEkRhdXJA/p791vq3DrDCLgMPSWXhgTI1hyNPHNvx7r9nXqZ6In8xcNTVlRJXUTI0y6tarYNy3JPvY698uq7czuqhxeCqsNKqrXShhdj6h6PoD7314noENk47K5fcDwx65UldgLXI/V+PfutfMdXT/ABo6gnoqClytTCUKpDKSQPwt+B711v59Lv5F7jXE4mSnli0LCjRprBGq6WBFvx7959e8uqgt501VWtNktZSnv5Dybadd+P8AD34cetdUXfKXtHEbA7ko9z5FfPjkqVp2QM1g4bSWOnj3utOvcerH+gezMBvvbWMyeJdHLUqukMchZlBAIFr+98R16vRv/wC9cs2NSh+5FPKBZFkJUlrcAX/2A91690wUuMMsj1WWx0k8pJZKptTLY/Q3/wAffq9bp0mNxYCvqZBPi6r7WKMAlQxGn+oJ97B69TqZgOyc/scwaWmrnjkXT4nZ7MDa4sf959+619vR2upPnRvbblZSUjT1EFOyxBtXlIRQ3P1P4964ZPXhk9Ww9P8A8wrblYaekzmWp6xpTaYFyTD/AK5vxb37NcZ69TqybYfam0exaZJtu5anqZNCSPBGyyOoZb2JHuwI60Q3Qle9db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuqKflJ/wnH/AJWPzH7+7K+THevVHYmf7Z7Zy9Dm96ZfEdzdg7bxtbkMfg6XbtLJSYTEVcdNTqKWjgUrEgBILHkn37rdegg2x/wlK/ko7bzmPzU3xx3vudKCZZ/4LufvnuGswdY6HUi5Ckx+XpZJUB+sZlCt9GDDj37rXV8/TvS3Ufx767251J0b1vszqbrLaVKaPbmx9g7fxu2duYuJ28k8kGNxccaNNM5MtTUSBpZpGaSV3kZmPuvdCd7917r3v3Xuve/de697917r/9ff49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdeN/x791418uutQJI/I/1/e6GletVFaddLru2q1rnTb62/x9669mueuXv3Xuve/db6RG+uxNo9c4HIbj3ZmqHFY7GwtPUPU1MMT6VF7Krkc+/de60+/wCaz/N8z3dT5LoP4t5CSWomasx9XWUFWQzG/haR3i/24Hv1OvH06o361/l0b97DyLbt31W5rN57LGOtqVynkrIUqTZnEay3sP6e/deHHo/ey/5eT00cNVkMRDHBgP3mH2AjVgq6jpUDn/Ye99eoRnoxOX6w6jwexVqKzHQ089ORBN5KYRr+16ZCSw/w96+zrwp1Vz8v9i/F3cGDeVEVstHHytOgYeULxwv05/r791rAwOiF9R4HD4edaKnp52xM0rJF5Im0+MnStr3/AB7917j0NvYfbmxetsGNuw0ZWunRmV/BpFyLk6iPfuvdAnjewq3NYXIPjS0a1KEroY6zf66bfX37rfzPRpvjz1v1VuFaKu7BE0tWZw4Eo8hLHmxB/wAf6+9/Z17jnq2XYPS3S9DVUG58RiUnWBIYU0UYkOm4sDYe9V68B0c+s2VsWD+7zR4iVP4oYzH4KVgqhvp5OOP9j79jrea56EnF9P47BVtVloKaExVFLaNfGCdTr/T8nn37yx1rzr0TvtLZfYUW4ZKHEUj/AGdVLK6ssRCjWLryLfT3r7etmh4dEW7C+L3yA3bkZ6Z2rhjKy6IkCSE6GP8AtP8Ahf3YdV6Od8Xfghk9rY6nk3BRA1GqOWSSopyXDfVjqYX96r1amOrGpMLtHqDa1XkcnPQ0/wBlG3ErJGbKuofqPv2TkdePVInzh/mY4uiw9ftDZ1Y0lYYXp41o6m5LqxW4CH/bD37rVcdVrfH2k2X2Lm03r2w+RnyNbkEZFqS8sSCd7j9f+vz78OtHq5DK7I2xsrY0WU2PJTJQzU6S6I3VZGQqDYqv59++3rfljo33xf3pmNy7WFNVNFBQYxDM71LhAVQchWf6/wBffh14/PpK9wd27JTITUsVXB5qVpUf98WMkXDWsf8AD37r1PXoiu/fmVitu0ddJi1nlrMcD4BCxZGb6Ekr799vXj0p/jz0BvH5cVa9hxVk0dfUMJVijlIYB/Wquq8+/cOvcejK7o2V3D0nkINozwVclEy6PIsUgOlDe2ofk+/db+zoM90ZjdW4ozR5HHVxMQCx64JCDf8Arxz78B1o9Jzrzs3dnWm46sZrH18G3442cyRQul1UHVfj+n09+61joz20/lPs/eG3cvQ7JpshPnTLJFeZHdVkLWvY/wCPv3Wxk9RaHZXae+6Wqq6j7mARo0jj9xBY8+gf19663U9Ct178eKDP4gjckQlqIpNV6pBKdS/X9V/qffuHHr3QtR9LbQfG1EK4uitRDxLeljGqw0jgD34nh14A16CbPfHbZNXjcmazHUAFXC8aE0sdwW4upt731rjx6pT7I20vx37kXIYGvqaSmnyAWVUlMFMInk9XpU24B9+61wPRvuz9/wDTeP6zpd7ybgkm3MtAKt4UrteqUxayvjv/AFv79TrdaV6qOzHY1T29V1cu28ZnJ8pUmaCB6emmePWCRGSyj6H37y69w6E7o/43fMOlqH3BSwyjGyEvEtTTyK6xOeA2r37r1D1J7o6n7u2fXQ5vcctQlRMVcGl1q0bltXpI9+49a62OP5a381L459MdR4zrrsrPV8G76OOnhqjV1Vypii0P6Zf8ffqZ63jq3nZP80f4xb9r48fg9zRNPKV0+WohRdJNrkk+/Go690dLaXcnXu84lkw25cTMzC4jFdTs30vbg+/Y69noTI5opYxLFIksbAEPGwcEH6EFffuvY65HVcFbW/N/r/h72KefVTXy65EgAk/QC/vXVuuCsXIZSPHY3/1RPvfWs1+XWT3rrfXvfuvde9+691737r3SD7R/5ltvz/w0dwf+6yX37r3VFP8AIqv9h8g/rzv/ACn9D/ymv+ffvPrXl1sJe/db697917r3v3Xuve/de697917r3v3Xuve/de66PJtxb+nvfWvOnVSHzbn8fyw+L6BQS+TQX/N/u/esefW/KvVlPZOMgyGIiMyjVFJ6Ta7KDyQp9+IqOtcD04bChWnwaRK7OA3GoliOLfU+/fLrfS2H/Ix/T37rQ6DXtnLnCbIz1ZHGjTNj6iNCVBYnx3AHv3kevcWHVCu3NhVe6M1uXfUdHTjc9Hl3egNVGmnSst0N3/3j37h1biej+4Bfkn2Pt7B7YSLH0220WOnzMsYVH+2/SfG30+nvQ69gdIfu3fu+OoMPD1hs+dkp6e9XNUVEuhzOvMiIxP0v78B5daJ6GL4m/JSHcVB/drfNT/D81cLDJXyeISFfSPH5LXB/r73ngevY/PpHfM/I022d99W5rCKtQK/PUn8RbHlZGEZqBdpGivxbn/W96pXh14UrU9GF7NzCdlwYfYGKpWqaesoaKoqZpITJHqeJQVNx+PfuPDrdPXoXesurcX1xhI6WijjfIGEAuygrG5/Cm3uwHVSehYhMpjXzACQcNb6X/qPfj14GvWX3rrfXvfuvdannXN/+Hye1DYf5k8/XgN78B1o0z1tfxf5qP/lmn/Qo9+PHrw4dZPfut9e9+691737r3Xvfuvde9+691737r3XX5P8AT37rXn1xeRIlZ5HVEUamZyFVQOSST7917ovHYfyX6868lko8nXCSrBKIsUilS30B4596r1bolnYXzpzFMkzbSWPxWa0kjC6Lfgi/049+49ap+fREt9947u7RaeaurJVYl/RDUELc/wCC+9463novrmpyE7UNTIzqTpd5GLsLHn/be/de+XShiGFxlK2OV4tcgKH6arv9effut09Osu3Nv0GAqmrVBljqGLtqF1Go/Sx9++fXsdLiKjpKurFdQqiyIdbAjjj8f0t/re/DqpxjobtsM+YjWhq4ac2TTqRFB+n9fej1sUOektvDaceOqPuCi+BT6rLcAfk397B68RTI6B7c8OPy1IIkurU4LJ4zYgr9Pp7914dRdtZ+R6N8BWxq1IQV1ul3/p+o+/fPrVOlxtHYtFlquWjx2sy1bMEuC1j+Le/HA69UnrLvzqnL7Xop2mSe0kfOkEfqH4PvWDx63x6LFegjpa3D5WnSpglFRE/3aeQ+saSAr+/fPr3yPVdO9fjLsOi7Pot4YvEUy1E1YJncUaekl9Vla3+39761Tq2D42da0+fytGZ6PRS0xg8bJCAp0oLG49+8ut5HVsDYx8RiKeioQPHHCENv1WAtz71x60a16Kh39DHQbZyEks0fnqYCArOoOog8KPr/AK3vwwet+XVYlBj4oaHKVErOJXiqgwlv4tJBvy3+Hv3l17z6Fj+WRUZKg+TuPx9LJKcZU1DNULA7fbljLwWA4974Z6rk9bYe4M9jds4iszeWnWnoKCIzVErEDSi8k8+/db6+fr/O7+Q8m/flDFuLpamkqqjHSVENScWurUKd9LO/h/Jtc+/de+R6CboL5MZjcu3qDG7kpslT5SnplhqBUpILSrbUArD3vrQrXo3uzd+STZaOSOeoj8T6l0uyWP1BNveut9Xc/FveMuewFP5KgtVCKNG8z3JA4P1/PvR63mnRocltjF7laVMlTwmJf8+5jB1fgnUPeuvDoE9y/HLAblrEWjijFHC+u6xi5ufx73jz698uhC2/1Lgdv0aY6GGMhQl20AEOvHFuffh16vQ07Y27VrTyU7hRRxAqhPB024APvVevfLonPyV6sqKuirK7ExSSymKQmyl11ar+9jr2OqU+xNkZVtxrT1FFXfeGZVXxwSab303PHvY61556PN8YPjo1S9HkcnSBWfU4MsBup+tzf3onqwp+XVt+K2n/AHXwccNGsSpHChOlbcKOPp9ffutGg49ET+UddS5GkmhmNpk9NlsCxC8ED37rwx1UjvndJhpZcOtlDkQluA4BYgKB738+tGtetbT574+fL9mY7Z2O8j1NdkkdtN2ZVklFyPfj14dWnfEPqXJ9fbBwleGnYmgieYShtIYgc8/6/v3WuPVnXV2wP9IWRp5a1JfRItii3Ww5Frf4/X37zp1vjno6OS6yoKLFR4ySkUR+EKHEfruBYG9v6+/de8ui67i6pyFJUEUsMpopiQSUY+j6n/H36gHW616AzIdd7nizMseLo3aEBv8AOU7FV5/Fx+ffuPXumnKYTM4enc5CnaMC4Z0jKEW+tj79g9e6Dahyc+BmravH5GqWSe7OpqG4JNjpBPv3DrVCejh/CX5sbi677hx2DqcpUTUtfX0tCy1VWzQ2chLKGNiOfp7914dbhu3MqucwOJzCMjDJUFPWBk/QfNGH9P8Ah7917p6va3B5IHH4/wAT73TrVeu/eut9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691/9Df49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdRaqIyKv7zRBSCQOA3+BPvYJrjqrAHj1zSRil3XQoB9TMB6QOGa/vxwetcRSnQEdn/ACU6m6lpKqbde5qKknprjwGeEszW/rqPH+w96rmnV6Y6rd3D/Oc+OuGylZjV3BRPJTiUKNUdy8TEf1/w9+69gdBBuL+eN1PT4HJVeCqKSuykEVQaamHjJcpGTGQv5uffuvVHWo58/wD+c38gvklvfcPXeJnz2ztuzN9t99EsiUskYlZToK2H0Pv3Xukv8GdgefP0dfUMm4cvU10NVPlZyGqJZJ21urObnkn37h175dbSvV2EgxWOxkmaxMOPpIqW7z641GkC5uWI4sPfut1z0guzPl50xtmsyu33y9HA+PXTUMJoB5F/Swbnn3qh68eqUflt8/dp7hx9VsLr+GmqmmlnU1VKIiVLm2vUlzf/AGPvfWuPVbiz0lfhq3JZrI/dVk41iCX1FDa+n37rx6m4bsP7KhoqXGbahmNMwOtI1JNm4ube/de6C3tnMv2JVwYmXb0FPkGdFEgj1S+r08WH49+Pp17oa+rugqxafEyTwskJjHkXRJZiALmxHv3Wvl0c7C9K5iiTz7eofuqiOPVDTKj2aULdRY+/fZ175dHK+P8A272F1bSCHsvYEFNg4JBH93Ux3Uqrj1kuPwOffurDh1cF053H1D2rjVSlTGyVJVhCLwB6VgONF/8AH37rVSehsqNp0UiU6Y+qSscsp8WtSqAnixHB914dW6GHA9C4DcuEORyEVOlfGLqGmiDf4/X36pHVR1wreuOpeu4osnubJUkQgtI0ctRCQCORa5+nv32db6ID8i/nb0j1pUV8GIzVEiU9POkYjkpyPJEOBx7t5de4cetaX5afzFd990bxOzNivWPgq2YRz1tOG8Sxm8ZOpP8Ab+/cOq8c9F42D8YpN5ZoZncczV0plMjGoDn9R1/2veuvfPo9WG6NpqCipsXg8ZFIyeNtSLypT6G49262B59CNufF7h2Xto/f5Kqkp4oFBpHdvEq/6ix4/wBh799nWvLoc9j57eu8+u8Ri9h0s1FIdMdfUUWpXeM8MXKf4e9UNa9br0Vvs7qXd0G94KHIZCpSCpYGolLOCJHP7gY/4/n3vrXH7OhNqvi7jMPsesylVSw5H7qm8vllAdl9Gocn34depQV6Fr+V73hXbJ7ryexlaSLE0tQIo4QyiIBTpNlJ9649bFOB62PewMbs/eVHS5quxtJPVeEyK0qozEEX5PvX2dbHRbclgdnZWupaKh21QeSIhXZYkN7G2puPfq9bFOoG9ek9nbhxs2NG2qD7qWBkv4lLAyLb6W9+Br1ogdI/of4ibV2VWV1TLgaKOSpmeUXjVdXq1f0+vv3Xuhb3nQ0mErRj8RRRwRhlWYQhVDD6G/v3n1vy6DisrKyhq1psZERqA1Kh4Or6397p5nrVfLpaww68NLKQBUkFmiBUF3IvyfesHq3RaO19yUW2MBX5PMZEY2OmheYL5k4Ccj0k+99UGOPWpZ/MO+a2EzeZq9tbSqPv8ytdPAk0CmWVHDadV4rn375dexx6CT45pubtqu25Rbw3PXpj5I4Unop1mWEgMLq6v/h9ffh1rrcW/l4fFn46Y/GPJm6LCVEn20bwSzLCD5TDfUC/5v78et+VOj87t692TgcLW0W16KkeINMIjC8ZASx06dP49186dW6qV+Q2wxloa77qjWoliF4ImGrlb2A926qeHVG3ZnUX+/iqqymposfkfJMCqxkPbXa+oe/da+3rP1riMtgM/S42p3RUYV5dAWojd0ZdR9IDX/H19+61jh1ad0Xiu5dvmvz2xuzc3uiWm1SpQfdysENtWgLqt/h791YV6tx+N38z3O9fxUu1e+aOTErHNHTCsrSqF9J0GQNJY/jnn3qlOvdXe9Rd27B7twsO4Nk5iDIU5QPoiqI3Nm/2lDzb/W92rig61TNT0L0s0UQHlcKG45/Pv3z68acOuSBQo0fpPI/2Pvx49eHDHXP3rrfXvfuvde9+691737r3SE7R/wCZb78/8NHcH/usl9+691RR/Iq/4t/yD/8AD/yduOf+Bz+/da62Evfut9e9+691737r3Xvfuvde9+691737r3Xvfuvddfkf8V/4j37rXn1VN8ysQ9d8p/jdVaQyU2RjIJJFrVNzyPfuPVvLqz/P0LV+NmiRdThSyj6/2ebf1976rxyekbszIijkkxNQQrFzoJP5vxyfeh6dbPGvQjPxNE1+Guh/pwNSj3scOqniM9MG69uUm58RUY2tYrA6OWsAQfTzqHvXHHW816q17L64ONzNRi9qSeKlkqi9RPThlIdWvYlf8feuJoerg46GfqDuWp66posFvmnegoY7R01bKpDVv4U6z+f8ffhjHVD69Er+dm7qzd+Rw24NkI8oiyNO87Qo/wC7TLICyPo+oI+vv2ePW8DHTTn9ibw7yynWVd1/Rzba/g2Mo4s1U49XhepqEQLK8zJbkm5597r16nVh/WHxzqaiCBOxKufNSYwo9JJVapGSQKNJUv8Ake9AY68eOOjfYLa+JwccaU9NA1TCgj+6MQ82gfpXWfpYe7UoK9a4mnSlPINvrY2P+PvXXvLrpSD6b3ZbBr/X/XNvfj14cOuXv3W+ve/de61O+uf+35Pan1BMJNv6+r8fj37rR49bYMX+aj/4In/Qvv3Wxw65+/de697917r3v3Xuve/de697917r3v3XumzK5ajwtFUZDIypT0tMnkeR3VRYfX9Vve6Yr1qprTqrH5BfLivnylTt3ZM8ksCySwPPCw9Kjg2ZPej6V6vSnRDN57nk3A8FfXVbVlSpD1CyHUUYckc/S3v1Oq16C7Pbtxz496WFYxLoKkLpHP0Hv2et8D0CtJvI7frZGkb0SkkKfpzzwPfuvCnn05PmauMtkyjKlZ+4rDgaWPGkf8U9+69020cVfl8klWZZAge5PqsNJvYH37rVCehjn3BT0WMWHWJJljVdNgSSP9b37rfXKHcdbS4SqrcbCZ6hYGbxgcg25Fvfuveeehw+N2Uze65nXMUzUpDkKzgjgfgX9+69kDoZOzYY8TQVUcv7hZm03F/7PHvQ9OvVx0UXH0lRFVVFZVRn7aXV4iQSPr9LH3vrxPADpirMwlLk0iggQXcKWUXvc/196690aXqRoaWro6uWyu3qHIuCTfi3Pvx69TB6ObntqUG7MIr1KIVeJApbm/p96z14dVdd/wDV1XgchJU4el1oskjOI1ZgyjkcD3br32dARt3ZOS3pksdjWxhDyaULeJhY6gLknm59+69Q9W1dK9d0mwcNBSVFLGtYyRrrtZxdQL391PXulrv3ceP2DharK5OqC6UmZVd1Nhp1KAG97HWuPVSnavZGW7IzuqCaSLEwTBuHISWMN/hwPe+HW+im/IjufCbI2RkIYPFTz0+OqnMyyR63ZISSD/X6e/dePRvf+E/ktP3lS7m7TZEnfDZCWKOokZS6BKkpYAf19+wePVa4oOrtP5jHZc/X3RGeWlqDT1GWo56aNwQramUqLE+/fLrfnXrU/wDjh8fsVvjceb3xvKmiyr1WRq2U1Y12WWQtxq978uvUHE9Dzuf4ydd4WqqcljqKioZNbMkMKKNZvxwPz711oDz6R23Omss2U8+Mx3lp3ayWRrH8D349b6PX1vh+z9j0EMmKwbPGpiJCahZQeb+9HrY6OJtvtvKtSRYvL0X2tZUL+4SLMHP9f6+/fz68M44dD5tzO45KNIXqwKiWw/VY3Ye9UJ69WnQkYjbkM8JyE05aP9fq/IP5J9+Pp1rp6gAlY0VK5VbkKV4DA/QG3v3Hrx49NW5MFGtFJDW0yzq8ZB1DUTe/49+GB1vPQAv0VszKvPmqvFUhljZmDOguCOTbj3uvWjUdLTae3cbRSiix0EdOiekeJQLD6XuPr79XrwNelTnRPSU8tMspc+Fxa5vb6g/8a964db4jqq35I1k1LkG86kI0r3ufp6eL397HXifLqpTsp51yM9ckZ0RsHBsBazcm3vY4da8+qpt+dVV/YnyEoNztRCajpXjJfSSo0sD9R7917PDq3DYsEVHtqi22sOh2p0hC2+lrAWt798+veVD1aN8Z9lRYDEw1tZAt2UPdlPF1vcA/7z70f8PW1FeHRqZcXHn6tIIadWiUcsQBYX4sfeq+nWyPXpEdgNtfbVMqV7U8bRAXBePggX+nvfWvs6K3uPufrzB080sZpXqGRgF/a9TDj/H3uvmOvfLop2/O0cdubF1hipI0R2kKSAAEK35Fvfs9aqOioV+KbK6mp6oxg2JVGsGAP5I/4n37rfRR+wu1k6U39sKuuI5KveGPpjPqGoEyheT791Xr6DXwp3+vYnQGwc2Kl6kybfx5LMQdOqEG3Hv3l1vFejaE2BP9Bf37r3XSMHUMPofeyKHrQNRXrl711vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//R3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3TVm8vjsDia/NZaeOmx2Lppa2rqJCAkUMK6mdiffutHrVS/mW/wA5yelytf1f8d9zU1HmYKufFyZCGZiRInoYs8Z/J9+699nVevX1Z2d8hI8PuftztKiaip4xLlqSXIyJ9wU9RWztzf37rYzx6mb46M+OWYzIjxGOhqKqdhE9ZBUEiV2P7lmuRyeffs9e6Czd/wAUdmbWytNlMfg6w0RgMjkO7RlQtzqH09++fXqenRFu0Oquu98f3gw228VFDuCFXiSVUXyxy82N/r9ffq049aNeA6Kb8eu192/GPtqTbO7MkGggqlqIFmJBEUb+kWv+B72KdbNfTq2PtL+Yvnt6bNXb+zsvHTVktKYkqEd1EYZNPJH9Pdc8Ovfbx6pU7e3RvrLZS9buF6jJ5eTx1UoqpvXrc3sL8fX37PA9ar59c9ldRZimWKtEdVUV9QyP5vI0gbyc+nVf+vvY62T0ayi6E3RkNtVEElDVxZKriL0jEEMbi40+/cetE0x0i8j1/ubrPblTLlY5lqoKeR2aVTcEA6S1/e/s69XPSy+IG28d3RvGOiq41qstHWwhmVQxjTyWNwfpYe9fPrf29X/5LYvUHSuDxNZuSbHVckES+eibxCXUVB06f6+/fPreOA64H5H9HQYZp9r7XigrZEMcEoiQgSqtgRxf37rXy6KNvnf27ezq+qwMlNULg5hK6KqeOwZLKF0j6fT36mOvVz0R+u7E7L+MGdyOXp83JSYkSs8VPNLKBoZtQA59+619nVgXTf8ANnx2b2bFRUzLW7n8QpzLH5HdZlGnVx/j78etjHHpq3b/ADCPkrQ4vI1OBzs8ImlZ6WDyShhE30UAf09+p16vRctw/J/5Xdv43w5XOVKRGMhv3qhW4/PJ/offuGevcegLznSG7t7UUdZuatqa2aeceVfuZNVnPr+p/PvfWuHQg7Y+KlLtqOly1DjJkEao0upAxb8m5I9663x49HB2NtSkip4aeKnFLK+lSWC3v9OB79UcevdGK2rsaow9Qa1X84MeoqAGOn82H+9e/de6AH5T7gozsjN4+GmP8XdAtLGq2lke3ACj+vvwx16vp0P3wf39idu9W4Sjy2LNNl5NKVLToDIfwCA3Pv3n16mOmf5K5rHR7jjy0YXwOxk0qoB1HkEe/fZ16vr0FsXbGSWgosXkJVqMRVKFCWuEjKmw/wBe3v1PXrxNeq9fjlundFP89dx0m35GhwTTI0KKzKnlaTkADj375HrXW41sbE7gyfV0NdkPIlT9opV5NX9PwW/r70fTq46l9fYOCmrDNVwNLIDqMhBIJB4sPeq560RQdDLW4+igljr9CIX08OAD/wAi9+61ny6Se7c9JhJIchDIq0qJ+4LBRyLfT3vrYAPRHO6+wsgmTxmRxTtHBNMhrJFVmj06vUDb3vr3A9BT2B8kuutlYcZau3lgqOuhhXy0lRVRJOzIvqUIxv731Xzz0RDtH+cLsTauHrcLgdr1O4c0Q0dLUYuKWcSllIVkMVx9fegKdbLdVF9j9t/Mv5hZOoxm2cXn9qbdynlgBqI6iJ1idtSsA1rce/da6OT8WP5PmBxW2hvbt5hnNx1LrV1MuQMcsplc6pLCS59++zrf29HWl+JvS2wVV8RjKSKoj5iMaRKYz/T0f09+8+t+XSyodndkbdxb5PZ+43xtEYz4IvMyH0j0IAp/p799vWukBsD5h9k9ddp0+y+ya2WfG1J8a1tS0hp7s1h6m4+n59+8uvDBp0eGnymJ3tHktwPUxZDHSlpYhCFdI1C3+o9+qT1vyp1Wv37t7asuXrsrSZShoZUEzmOR40YaTexQ2978utfPqtree+sFj0q6xHirqyjkKJLAb8r/AEKfjj3qtcdaoOPWXqL5zbz6wrwuB88kNS5E0SB3YAmxBHP49+690MXYPy6xHb1HJT7gwdXLllgZ4pUV4yJ2W6tdbc39+4der0In8un+ZV2Z8a+9MRtbdBrDsGrq0UQ1ElS8YglnAKsSbABT7914ft634+pu8+te6dp4Xc22Nw4isGSp4pTQJWwvV008kYdonhB1f63HvY68fn0NYtYW+luP9b/Y+9de+zrv37rfXvfuvde9+691737r3SD7S/5ltvz8/wC/R3B/7q5ffuvDqir+RWCMd8gr6v8Aj/8AJ/Ucf8DX/PvfWjjrYR96631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3VdXykhhk+QXR0jFQ8NZGVvbk/cX4v711sdWKDlRf8AIF/9iPe+tdBVvagjxCjNU5CN5AWvwFN+Rx+Pfvn14enS9wVYMjiaKqOli8atcG4DDi497qetUHTjVgmkqgOCaeYD/qWfeut9Fj2PtekyuRy8eQVZZHrZCjNpOn1E25/4j3occ9eNfLoRc50ts7c8cMWeo0rFpwPAFVU8ZH592x1oavPqPD0F1pHAtPPgoauFfpHOqMot9LC3vXW+lxtrYu19nwSQbcxNNjVk+vjRSQfqLXHHvwpXrRGMdKxFZVs7a2/1WkL/ALwPe+vCvn1iKOshkUkg2un+wtyf+Ke94OOq5GepHuvV+uCqAzNblrXP9f6C3+HvZPWgM165+9db697917rU864DH+eR2rYX/wAnv9L/ANv8A+/V681a162v4uIo/wAftp/hb0j37rXl1k9+631737r3Xvfuvde9+691737r3XvfuvdU4fzFvlNU7IWPZWCr0pqmRlgqoxIyOwdrHlffut0p1Wrht9Un8AkqnBfL1f7vlYs12kF7gn/X9+69x6DufcGWpJpY5vLNJkCWXQbldRvYD6e/da6RORo8oKj7otLZnLMlyGZb3/SPx7917yr050+JgziJ9xSsrxi7M3Goj/Yf7H6+/db6e5IpKqFMbAvmFKLKF/FuPSD9Pp7917y6faKknpaJoQfHIVJXVdW5Uf77j37z695dcMPR1C1RkyMMkkJYkH6i1/r/AMT78fTrf29CRgaMpVyCNG+1mFhGebD6cr7917o2PWOCmoZVrIKhKSHSr+M6ULcfS3v1c06rTFfXpebtpE3BDJHLE7hL/uclWIHLe68Mjq3y6KtvBRSf5FANKQGzW4P1sOB7tXrWOgqqJaBXVvFrqNYGr6kH+pHv3WuPQ8daU+VqsziSjtJS+nyItyQNX04/p78aU63546tHp6COfbEMEDLFMkKEgkBuE91690EFbs/G5xpafLUfnbWwDFAf8L3Pv1adeHUPBdV7W27UvV0tJEtUpJgOlLhibj6e/VPW+lDuTcNFgsPUV2QaOOaCAtEXsl2T9IHv3HPXvn1Ud8hu4Mxu+asoBkQmNhncH1sAVAtYc/S3uxp1rqvLur5Mbc6m2HWxUDRZPL/bERpTv5JlkKkerT+Qfp711qvWtT2h3X8mO+N15iiosXlsbtmV66BZ5vOsTQyagpT/AGHvfl1r59bIX/Cbn5D5L4/7wovjpuebwSbqdZZ6qrYqjPUTkjU7cXuffuGet0Jx1sH/AM77PVOC6HwlXTyFI5Kx1eQE6dJNxcj8e/U68CR1Sz8R94xVfXzz/cRyvJVAFQLsGbi1xz791sU6MDutJVeOolLVKTEPpS7aBe/P09+x149CniN+4DCbex6Q0iQ1YtqLRgODbnn37rQpTp6pvkpJjJUo7BoOAToDAAG1jYe9fLrfz6e4u49uZSeLKS0/jkp/3HLKV1m/Jt/t/fvKnXs/s6HXZ3Zm1s1JHLJUwY/6ESStpBIF/wC1b36h49eBHRn9u7gqchDHLQ5BavFppVhAxZbX/Nvejw61muehio54FhhlhXRKRctwf8bn3vrfy6jz1k9dWmnmPkjIPqHNhaxHvXy69mlT1Broo4oHpolCRsCDb6Fm/wAPfuvedem7buNEFW5ELAOSfKRxe31/1vfs1z1qgA6h7kop/wCITOPXH4G0leVNvqB791sdVXfLLH1OmrkjBM2t/HGB6jdeLA+99e49VDb1lqjRTQTRMkxVgwcHVcEgcH8e7cOtVr0HWy8PSpJc0mqvkmFn0qWAY3Fh791odWI9G9PNuPI42uqKdwkektqRbNpa9mv/ALx7r8+t/Z1Z/jNsw4SCkoYYdUYMaMFA9NwB+Px71k9W4CnSD7k7WxHUmHntNFBWNTylA9hJqCX/AB/j79x60eHVEncXyX3jvPO18hzsVJQJJe7SuiFQxH0v/T3anr1qvQNZTtPZC4eKbL7xxBqomDyRvXKGBU+pSC1/e+vdRKH5HdY5bHyYukzuLWRV0mT7lbEji9yffutfLoMt8/JbZPXuKqKhM1jK6QRh0ihnVnLWP4B9+PW6nqqHsvvSXvzeOCjx+Lqvt8Ln0yCyqGCkwvrV7/04968+vZ638v5JPy22tuLpjDde7kzdFjspQ0FPT0tPVThZdcNo1Ql/8PfuHWs062FopYamISQyJLE49LowdGBH1DDg+/de49c0XQoUfgf6/vZ68BQdcveut9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf//S3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYZVlawjYKP7RP1+v497x1o14DrmzBALn+gH+Pv1KnrROkddsSALfUkC/9L/m3vwpXrZOOsM7yKmmAK011sp/SAT9W/oPfqefWqgY67kL6VLMka2/cLGwFx+Cf8ffhTz68a09Oi39zfKfqboLFVeW7D3NiKKjooTOwjyVMagov1vESeffievAevVS+7v5+/wAVpMjU4vZGb/iVVTTS0g0TxyBpkOkEqo/r71U9bxXouHYn85juOoo5q/rPF4+opmDPTtVFNRQLe+lveh8+t9U0/J7+fN82N3Y7PdVVe26fHUOapZ8Y9bi5lgkZZSYtbeLk8H3v7evdUUbdwXY1Ru7JZXL1uRyOZ3BlHzErVrtI0T1D6mjjZuQBf6e/CvXujn43+/jYb7P+LZWiJQoy0tRIgH9OFPv3Ws+fS8xmV7q2li8VW4/76vp4KpJVlkMkruFII1H68+/de8ujXj5tnDbVXH9nUc1LMlE0GrxkP/mrGxI59+6tXy6Lt8ee1uic9v7dmXrskVq8nVSvjkq5QkZOq4AD+6sCR1XWI+8jh1XT829uTZ/umry+0mCrHSTuZaU618RN1Opfz/j7bUNWoOOtLeCQ1Toou3+28ntusG19dZV5SR1gAALlZGa1uPapCCKefXiQ5r0e3pf48737TrqTO5uCVIlkD03mjdVHAYar8ce7SfCAOvAFcnh1cH118d6HE4jHDI09Oa2IwXYorJZT9fbNerUNOjdZfDbE2Tt2DcO6TjKd8VTgQoHiQOAnGpTa59++XWzTBPVXvb3YPX3bE24qPHtTCJaeaOIRMugNci1x9f8AD34YHVTg9Vy9Ld60XxT7krcssuqOqkkjjQvZAA3BAP596638+jQbs+RWe74zseUFdlkoaifyLDFJI8RQsDbT739nWs9Wn/HrpCm3XtjDVVSJpCXiYiVCOCob1D/iffq062Bjo9dD0HHQoaiOjg8MFHMxk+3H+649QF/9h/X37r2OHWq9/Mw7aqqvsLJdbUP7VZHWLTwx0x/ckPl8YGge9de49HM/lx/CHISbXg3ruKmq2NXHFUBailPp8sfk/tD6+9469k9WmR/GKlysU8kFFGz00xRI5Kfl1DWJCke/VPXiOjBbO+Ke2IcFJLkqCGKoEJLAU4QA6fpx7917y6ccT8X9nTxh0S2mcWQodPBt7917z6GX/ZfcCcSKeKljaJU0n9n/AAsOPz71UdbpjpJVPxgxVFSS5TR42gQyw6UCgsORe3vfHr1D0AmWzNJsmepGVBRYhLGpcixVOLe/daB6LD/deLvLsChyONiafG4+ovVIV1wMEb1agOP9h78a069x4dKrsFRsav8AsMHGKd6G0jLAuhQFFiBb34dePQVZ/JVPYFCJK7X5IrIGAJAt/rf7H37r3Qa7xzGF2ttiqepkCzUMPoLkCxAte/497+3rR6Un8tjoWs7G72bsL7JpqCoyUTioEBdTGHvxJ71jr1K5628d1Y+nw+1sXgMUkaaaFEkGgKQyJaxA918+rDoMtv6KQEOqrLGx1lhYf7D37rXTVvHcVLBTmWoqYIYVVm9UyoBoH05978sdWHz6Kt2T3Nt6PbOSmqshTyQUiSACGqRpf20vZVB/r798+vfLqgbvj5kdwdgbmfrfoDAZXJVwlNNNVVFHK0ERDlSUkA/3o+906qfToSOt/wCVB2B3fhqTe/yHzGdpMhVtHVGjpZaiKmQTgOyeJSPp7917jjo8fXn8rjpbr6poKyWjgrkolUK2Qp1lZgOQXMgPvRNRXrYGejZL1/1hsGk1UOFwdPHTRgho6aGNgEWwHA9+r16nlx6Kx2T25RNWNjMNMIKMO0Zjp5NI44sqr9PezWnWvPokPdfZ9ZtyuxIoaieVarxtPqY2UE8qL+/de4dHq6xp6ncXV2MzeQSbwMsUsRKmzkrcA3/3n377Ot/b1X381qfAjatdm6p6Wgy9BJJ9tJCyR1B8SXT1CxuffutdVcYn+Yh2z1lgH2tgYp8lStCYUmMpkexGnm3+88+/fb175dM2Br++PkGmUz9W2Rpaf7SaoEYMoU61L29+44690Ge0uvt8xYzcFdm0qGo6GvkgmepRv7L6SQW/H9PfuvA9Gp686D/iOBk3Ht+Kknn8RlnEqq2my3JW/vfWuOevbNxVJjt2Od448R0KymMPDS8albTyQPeqdeJ8uhn3dhuqpq2gqqWNIJlUNHNHCFlQWup1j6H3vr1Ol10z81c/8YM9UVuztzZvIy6meKgqK+R6WNraVVIiSAPejTrw9Or9/h//ADxNu7gWhoO9aqnxyziCjinM8ayrLI2lXN/+J9+Na9bqKY62Eesu19k9ubepty7KzFJlcbVIJI2gnilcIRcFlQ8e/da4dCJoN7ggL9QLfU/6/u1RT59VIJ4HHXk1nl/wSAfoWH9SPfjQY62tTk9crvrtZdH9eb/T3rFOt5r0jOyYjN19vaFRdpdrZ2NeLm742RR711vrVI/la/LzbXx07c7l6837KcdT5TfmSdZJpFp9avVvpYCT8c3v79muOvYpnrYfk+d3x7jOobroW12CsKymGvi9jz79Xr3UX/Z+vj0H8Z3TSBv6fd09v8effq9e4dZP9n2+Pv43RSH6fSspv9v791unXL/Z9Pj7/wA9TSf633lN9f6fX3qvXqddD56/H387ppB/r1lNb/H8/j3uo6117/Z9fj7z/v6aT6i3+WU/P9fz+PfuvUPXh89Pj6Tb+9NGP+qyn9+6915vnr8elcRHdlHrb8Csp/p9b3v799nXqdAxvr5DfGXsPem1941G5qOSq2pIJIn++hC3VtY4B/1vevn1sDoZl+evx7aRoY91UbMg5tWUwFl+tufx79XrXTJuP5u/HfOYmahl3PRuHYHSK2AFSv0YlT79xwet0p1lwvzo+N+JoaXHRbvxsbqqqI/4jTszP9P7Rv8A7D3vrWR05y/PH48L5KVt00moo2pBV09iHFiL3/Pv1evUxjpLbe+YvxzxtTV18W46RC8jOSa+Hhj6r2v78OvZ6UcXz/8AjzJK0R3TSJY2DGspyD+PfuHW6Y6ln56fHwGw3VREW+orKaw/oPr791rrofPX4+n67poxxf8A4GU59+62R17/AGfX4+3/AOPppLfk/eU9h791rr3+z6/H3/nqaQf9VlN/xB9++XXqHrx+evx85/39VH+f+Uyn+v8AT37r3Xv9n1+Pv/PU0lv6/eU35+nF/ega9ez14fPT4+nn+9NGBe1/vKfn+n59769Q9Z6f53/HyaWONt2UUQckFzV07BOL3IuPeq9e61xvjr2Nhey/52HaW59ptLVYWWPxx1a/uQMS3pIlXj/Hj3uhp1rrcBi/zUer6mNL/wCvpHvfnjr2KZ6ye9db697917r3v3XuuiQPqbX497FfLrRIHHrHNNFTxNLM6xxRi7ux4UfS5PvXHrfDou3a/fuI2NjKufGSQVlbTBrKWDBm0HSNH+v791rrWv8AlFhewu8ex/75TrIKSpq9SoOVCiS4sPp9OPfs9boPLp6xWwI6GlpIqt7TRwxRtH9QCBybD377Ot06EWHrGarpBXQUxc066o7xFuPr9be9dbHr0iJtsu1XKlXDplFwoMRUcG9gPx731rpoqcDlKfUq048TGy+NeACfyf8AW96+zr1OnDHbakxyGtjXXI4u4Zbkfkhf+Ke98OvdOUeLNcWnnupt6QBbkfQW9+wRnrxx16npsrUzfbxUuqIMFJWO/pHF7+/Z690PWzNiVs5gkNOSeDpMZP8AS4PvVevfb0Ymj2RWpFC9pIkBjN0BAsPqOPeqdb1dL+qxKR4swIoaRo9J49Ruv9fe8dVFa1HRMN/YsU9ZOkcbl2uGuOVu3++597HDPWzx6BuDa9c2QWUKTG0n00km/wDrH+nv3XifXo6vR21/tniqZIi2kG5dD9b3/wBj70a8evV6OJEz6yY3IXSLr9B9PwB7rSvHr3WKUAKbAavyCBe/1J1e9nr2OmaOjnNUszGyq1yGJN/9pP8Ah7114nPRavk49cduSimLoogb1Rm3N/px/h7sBTj175DqmTdWNyuaFbSQl9TzMjuSC2ljZh/X3vI695dJTGfEXqjORjI7mq5aitqB/lMNR640Y88I/Hv3WgBx6Ue8PjH01hNmPT7fxmPFSsbWdKSLyFhwCSo/P59+699nVbWQwuV6E39Tdm7XVqOoxNdRIklOpidYxUgk+Rfp78Pl1qma9X/fzI+2G7//AJa3X248BU/xPddLt6nrMuFf7mVJYqMFmkIuf7P1PvXn1vy61IPhb/MZwexK3KbM39kBSTUOfqKIo9SqDVDMUJAY+99a6u22p83emM/BT6srHU/cDUlqmOQgH6fni3v3Vq+vl1i3r8w+mMHHrnramygECI3FifwBf37rVei+5j+Yb0JTP4jNlGkU21JE9+D+r0j8+/Z8+vVp01VP8yPqDwpFST5FTawEkTqCPwTqFvfsder0o8D818J2DNRY3buRrYisukmKZoyVP0DH37r3nXq934Y9tzZTBUmGqapp3mjhv5pPI4ueCQfqfx71TrY4Y6tLxSieFi1gDzyPUBb6e9cMHrx+XTuIYqdCw5f/AFVrt+Ppb377OtdM8zmdDE367tpP0IUfW/v3Wx6nqfLViCkigCKHsAXU88D8n37yr17j0xyVcciNDKVDabA2+v5+p9668B0VjtzqX+99V/E9CFIS1l06g/p5J/r72DjrfVZvZnxoqK3JzSw0pCluEWE/XVewt7t16nWDYfxbpaOujr8jAqMDGdLxECwNzqU+9VPWv59Hv2Bsej21AiU8SKoQ2NgvH149+638+lfuTcMe2KKfI1DppjjEiliONA+oPv3E9a8uqP8A5e9z/wB+dwzU0NVIPG08QWOTULA2HH+9+9+dOtfb0QCm68m3xBV0VTLJCs2pfIGKuQAeSfxf34da4dFh7A/ly1Wdqp6+j3Jm0WQO4jjq5hHdzcjSDa3v3W/OnSS2T/Lny2OrAlTmsn4iRdjUS6rXIJPN/futV8uhepv5bm26qujnzeUyNTCH9SzySSKRf+hNvfut9Gl6/wDhH01t144KGkp2rQqIWMI4bTa7G319++fXqV6MtsnqqTpnM0ub2Nlq2kyMTq0VLSTPFC3jfy8hD+be9Vx16merpvjd/MuzW0ZMZtvtyRVpNaxJPJNcsoAQFiffut06vF617d2X2nh4MxtfLUlVFNFFII1qYmf91b2Cg3/3j3vqvS+kqJQzR6SJyw8Q+iFfz/r+708vLqurz8+pl/RH5SFkNvpwNen8e6jjjrZ+fXOMuUBkADfkD6f7z78eOOtjh1z96631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//T3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691x0gnURza3P4/r73XyHVaZqevK2osLcLbn+t/wCnvxFOtg14dIrfPYuy+tsTPnN6Z6gwONgheeSorZkiBSMXbSGIJPvVfLr3Wtd/ME/nz7J6szc2x+l6pd1VMkrUnmxipUuzPEyh7xk2seefp791vrUm7n7T+YHy87IzO4clvzPw7Yy5kVNqzLVBI1aUuqk34Fja1vfvs61gdC70b0Rn+vpo67JdaU2WqiElklkpneSWYG5kZiPqT7917PRkcjQdxbvyVPisHsKbBY4kqzU8UqKIz6bAKPfuvdKnG/y4d47jr6Xc+WpaqoeWRJJBK72RWOs21f09++XW6HpaV/wNr6PcdPXQ0sipTwCNrFzYqPr/AMV9+NOHXtNeHSfyXRrbMTJS115Hi9UaOTcf4EH37j17ofui5+mNx0QwG5s9RUeTpSB9hJJErFl+iFWPvx68DmnWfuD40dddjRTJQ42hnp7SiKZdF3QLYMCP8PfvLr3n1RF8w/j03TlRSZLZVV/C6undGVaUtdyXJPC/X23IWCVXHTcqNMpRPPot+y937qz1BWpuPHyzVawPEcjKreR0VSo5P/FfaGNyWqTTpmLabqPiDTrL8c+vNubh7fWbL0UFRIawtaZdTXWW/wDa9rVdTTpUiGL4uthXZO2q9MPW0OB2+KKmoYf8nqIEK67RWBBAAPt8tUca9XahFR0F25/khXbIoKnaFPC9fu6EylKEjyTAKbjgc+69UqeiDdq9pfIDuuGrxyUuSxlPCRElKUlUTKPTYgC1v6e/da6a9qdDdu4XCUVTJgqjy5A6ZZtMl3183+nv3Xq+XSswn8tTdXbG66PJZ6gniiMkMxDmUAFzdrX+g9+63X06tg6y/l0YbYkGFo2pIi+lEOok6vpyb+908+vfI9WcbB60xeysbTYqn0QmlCGQIRYKq2tf3qnXifLoxcc+Fl2dlVpahJ6yLG1oChlv6YG4Nvejx6sPTrRw7M6rz/en8yVcdSYt6rB4jPacqxvoFqwcMRx9L/j3vqpIHW6v1X1/sXrzYeB2vanxxNHjogNUaszmFQRf3r7Otinn0LEfTRizdDkMd+5iqiNZpHjb0EEXvx78T1r5dOO8tuVGIoZxT0p8bx+lxcg8fm3vwHr1s8Ogo2/UhVNHJGEm8pJUg/QG4N/ez69eFPPoxuzsdT5DHyRzRjQRYOeLH6X9169w6Ydz4U1UNTRU8p0U6my3NiPqB7359b8q9VKfMfbsdDtiSqSb7eveskjdQyg+MtYvb3vz61inSt+JWxcPQdbV2axTx1+QNI0k+hhrV9Oo/wBf9h7916vRb+/dwYfay5HN7lePHh45CrSW9Rvaxv799nXjXz6r0oPl7tzH1dThcdTRVaiZx5hGG9P+qv8AT37qpOOisfLP5PQZza9Vitu0qisqIkjcxRnUjFgGuV9+6359bQ/8jQbTpehtuZLIeGXcM8NNJKjMhk1PDe2k8+9GnXh1cf2HXS/e/flfBTRFyFA4KfUn/be9dWpXA6I/3l8oNgdYUifcZmCCulBHh9JZiQRYj/X97pXqvDqqDuP5L/Int5lwnT+ya7N0NXO1P9/SQTaVWZrLJqQf0597HHr1SOhl+PPwZ7R3UaXMd17hyO3qetCz1WLrZZUi1lrvGVc/Qj36tOvZ8/Pq2fq34xfH3r9ScJtDB1+Wi5OXjjhM8rleZC9rk3591+zrfSv3Xuyhwcj4GKEJFFGxSL06Y0ThQB7917B6Kxv3s6hpcdWKa0RVEZYRRXTUeOFAv/tvfqdePoOq3e0+5spIFgSeQQl28xtZQofm9v8AD3b7OtY6LlQVtTm93pXwsZ8UVVpZCCFD/kH36lOvA+fQO/Ivc1Bid0bcppKNGjlkhBUgEMdYAuPfuvHq3na/bvVm0Pi5Q1mVydHQZKHHLJBTO0QLuIdSxhfxf6e9efXvLPWsr8rN6dg/JLcGXxe1pqyhx6ZGVIYoPIiVEFyquCv9fr7317Pn0pPit8Jc/uLHyturENVPjlZWkmDsXKm4Ziwv7917q2ZupsL0b1amZbGxUtPNTtFKygWCBNLEhh7916lMjqs3tztLa1btHO7Y2hTw1FZkawvI8EYLLJIfyUH9T791qvQsfHXrzsGTYDVES1YK0rMKYah5iVvpII9+690LG2dt5FopKfdGzEid5XjSaeG5d72DXI+vvfW/LqBu3qCf+HVcs2LSkNQSaR7abIeV0L/re/de4dIvr/4WYvdVU+QymQAepNgHeyr/AGraveuHXgOkHvT4p0WO3++06TLmnSGnepppQ7XWdBeLSQfrf6e/daI6Mv8AED54fJj4MVGc2dW0OZ3rhZql4sTKY5Zo46UyekLqP4H5HvdevY6v++N387frneTx0HbktPtGtARJUrfFTtGQvqY6iPz/AF968+t9XHdPfI7qXvTHx5LrrdNFm4HjD2hmiYjVxb0MffhnrXQ6c3+gt/W//Ee9469mvULJ0MeTx1djpv8ANV1JUUkn/BKiIxN/vB96631Rb2t/I96w7D3zlt7Y7fNftzI5WvmyEq0qSAiWZix/QRxcn37h17BOegzk/kK415hIe+dzKitdUDVIH+A/V79nrVB59Sn/AJDWGc6/9OW49drar1P+sLer3rrfXX/DDWJ/5/ruU8D81A+n/IXvdT14D59e/wCGGcT/AM/23KD+Tep+hN/oG9+r16nz69/ww1if+f67lt9LXqP6fj1e/dap14fyGsTzfvXct/xzUW/6G97JPl1vyz14fyGsUOR3ruW4sb/5R/0d71Xr3XH/AIYYxRn8zd6bkvp0/Wp+lv8Ag3v1evUAz1wg/kNUNMtQkPfe50WovqGqqIF/+QvfuvY650v8hnEUyuB3ruRpHUguTUFrH6/2v6+/dep516yUH8hrC0v3DT977oZHhmIANQwEpU6C12+l/fuPHrXVEE38t3db/NGHqsfIbNfwePP/AGS06101tP3Pj/Te/vfXiK9Xu1P8hjCzVMVVF3tuf/gPCr6jUC8mj9y1mNxf6H3rr1Osi/yHsYqNGvfG5lV2JIBqRbi3+q9+r8ut0z1g/wCGFcJxbvHcYa5Ja9Tc3/r6vfuvdc/+GGsT/wA/13Le/PNRYj/W1e/fLr3Xv+GGsT/z/Xco/wAL1P8Atv1e916917/hhrEk8967m/2BqP8AetXv1etU69/ww1ieP+M67l4/oaj/AG36veut/Z17/hhrFXv/AKdty/n/AJWfp/yV791qnXv+GGsT+O9dyj+nNR/S3Hq97r1vrv8A4YbxXP8AxnXct+Pqak/65/V70T16nXBv5DWMK2Xvfcym9wb1PH9ber37r1OjR/EP+Uh138Vd/wBR2PQbqrdw7lq5Fkqa+qV/NMVFvUz3/wB79+889e4YHVv6jSqr9bAC/wBPoLX976rWmOufvXVuve/de697917r3v3Xuiyd59q0e3aOow9JWBatoJNSpa5e1rG/9Pfjjh14DNT1VPuPejZirro8lUl/M50pJex+oA596oePViRw6CHLVtTUkUVFSWjT+2ouPrc20+98D1WvUjbfXOb3DmoGfymAvGxBJC8G/wCfeq+XW6dHx2311S43GU1BVQrpljCu7Akjjnk+9Z8uvY6QO8+jYKxqiXEwq50Ell55P5BHvw+XXvt49ApT9Zvi5JKbIpqYsVAkJuDew+v/ABPvdevUqanpSYjpOKpkeXzB4qi9or3Cf0A9+DGnXuJp0253oDI09Sj0aO8TMBpRjyCfwB79Xr2OHQm7W6Np6Oijkro9Mps2m/N/rzf36vXhx6HTbGwqXFIsjRBohyeP9jc+9efWjU56WNbR0FXD9tSrZxwFQD62tz799nWx0zQbaeFtNRcI2r9XIAt+PfuvY6AHenVxyeRq56RfIfwp/Fzfi3vwr5dex59IbbfWWROajpqrH/teRbubqLA/0/4n3upPW8A46N7hNj0mEoUESKCygnTxpP1/3x96+R61nj59K2mpIljsQupR/Ucj/H37HXqdQaiGEFuBYEH/AB/qef8Abe/U61w6RG5snFjoHaObSyi9lPFvpY2/PveOtg9BnuvCw7/2rU02oGf7eQJyBdyOffuHXvOvVZ1f0xnMFuSqWWld6ZqiRl1BuV/Hv3Xjxz0hN0df7uoq0SU1DL9szXUoW0gfgkf76/vfl1rpA7vkymJxrQT0zvIEsIyPq/0t791vjg9V9d81+RqdqZSjrMR44p51PlZLWs3DG/8AvHv3Vc9Wtfy5eoMp8iPil3JtTOTvUQ4naNfFhaaQm3kWiYqkV/yfpx78et58+tO/E/yfO39wfJneWKyOHq6XBTbsy89PWXmV1hkrGAIb6cfX37rVPPq8b4//AMmLEbSgp6vPb4nEkSjVDUVbsIiv1SxPv2B1uh8j0byt/lvdUgRQVmYosjpChhIyvq54vc/7H37j16lOm3K/Av4/7WgWoqsDg6sHhpHiia/9SSw9+z16g6Crs74V9CzYiObD4XCpLJGD+zHEGFx9PSPfvn149Fz2Z8V8DtbKPW4qngjVZGZDGLAaTx9Px9Pfq9ep1bD8LcHlcXvvHozOKNBAughyltYA/wAPesHrfV8H3AhhSzeMNwB9A1ha1veq9e48enJalkpkmLF1/UX+v0/Fj7917rC7pWEVMLIPGLMAbXJ5uPfuvV6ivmaVbrLy6C1jY/TgH36tetefSXytV5v8ph+lyDawAH+A/Pv3W646SdTmJ5ZVihLzRkFZV5AB/wAR/rce9jHWq+XQc1NTSVGVlWtoFSGFxdmXiw/te9Z6tmnQdbmSkqM7G2Ol8NIrDUF4BI55A97611M3HWx4zFxTUlUzNHGNYU8/TV78MnrZwMdEK+Q/yEhxm3Z8aJf3whjkva9wbfW/vfWvLqmncW5BuHPT1qr5C00j/SwYsfqB791r59Y6LdNZjplighaMudLEfg3tf3vHXs9CxiN3VMkGmoyEgOjhLmwFuAP8feuvDqbR5qukk8izyImo2cMQSP6n377evGtcdLF87LFT6jUs50k2JuSPqOfe8daz1lot2/a0ss+kRtoLCYcE2HJv+P8AH3qnW656Y8R3E4rJoJWM5V2VJWsNK3+oJ/P49+690Hu8+6cAta6ZKphimjOlJH0AxsTe4N/fuHXjnoTfj1/NLqfjv2Hh6Z90S1+AqchQUzUpIaCOIvof0k2Fh79TyHXq9bo/xx+SPXfyR2Dhd27JztHla6XHxVFZTRzR+SCpK3dGVD9Ln34V8+tHoxrwvPFEZD45UKuQhuNQ/HPveK9aIJ6lKSQCRY/kXvb/AGPvXVuu/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/1N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddE6QSfx72BU060TQV68DcA/1F/eutjIr143twbHj/e+fex8+tGpGOsTy6ZEi0sS4J1AelVA5JP4/wAPfvn1UtQ6T0Rz5t/N3rf4fdc5Tcm5czQRZmOkaSjoJZlEpJFlOkH63/HtppQrdVd9FBSoPp18/n5s/wA175VfM/fdZt3r+ryMWwazK1uJ8VPPVQv4pZPHG0Sj8W+nu6HxBUY6UIgdalqdDL8PP5cGazM+P3lv+DLVWTyUUdZJUZF5agK7sLtqmJ/B/B9vCInPVGAU8er/ALrD4G9U7Rgps5mMpgqZWMbslT9sjW0i9wfr9Pd1gL5B6ZadYsEV677O3L8fuvK9cTRxYjKSQpoaSijgk/zXB/R79JbGMfEOvQytIaKp6CbLfLv4+7T2/WQxbYiOXZCIJBSpdWIJFiB7SPJHGaM3RtBtl3cfAhFeiT0X8ywR7+xm3EomixVdkVpVUwkKI2ew/FgPp7ZF3CWCqa16pfbfeWEYaVCQerksdXbV3TsOm3fBU0NLHUY6OpmWZYxyyamAJ9m0Ni8y6lNOkqAEZIHVQ/y17C2xLVPjNvS033rMscjwMPU5Fj+n2hu2WzrrNadKk2+4lp4I1V9OqisvsDdNBuOXdmMrqmOseV5DoqHRTdtS+lT7DkvNVnDIYmGeh3Ye1m9bjaLeoxUN5enQwUfyE7P2ztp8SKieTICPwxOTIx+mkG/uo5s28/h6f/1ot/JoH6BGqo979qVr1u/JTUUyteKKQFvobgaW9tTc12RUgL0usvabeo7lWkfAPWLeuwcBt7arS0NKkUskbIwEYBa4tquB7rYbrBevpTo53vkm5sIC54AenSn+F/RlFmt0fx+WidytQG1hP6y3/PsRRwjTUY6hXdJI7eYxHiOrvO2c5hOnepsjNT0whylVjP8AIy0YVvKIrLp4/Jt7diarafToujDU1nh5dU1dH7H3DvfsfJ9p7thnnknrJ4KcOt0EDudACtx7e6c6sx2P0vR1PlklxipNVyGSnvTx8qzXBBt79XrfDo82F6fx9ft3HUb0MCzY8JJIft4uQq/gWv799vXjjofto9d0VPhlrKOlhV4mROIYw1k4NrD/AA9+4Y69k9CpR47ESfbNV0gkmp1tq8ai1uB7rU16tTpP5Ta5nqMjVRwMIp4XWI20gEjSPp73XrVK8Oio9l7tg6N2PuLdecro46RIMgvjmltbVE1lAv8A6/497qOvdau/x47brN3fKDsveWzUVmrcvLLTvGBKXYTXJQ8n3vqvVlffvZvycwGBwu9aN8jNjMZLT19bBEZgxo6azuBb/afeut+fVw/8v/5vYv5JdThaepigzGEp46CrgqCfuDVRHxSKQ/NwQePej69eGR1YOKLL7pjipRG+hAFlYLww+vHvXl1YU49YYunoKGvWsqiCkhFhYfqPH0Hv1evHGf8AiuhBh2s2Ep1MUbCnZCTYfhl4At9fe6160fn0Cm5zV4Woqql7+GsBEKkfpN/zf3rjx69XHVQPztrjS7WqKyWQLLG88zEsAPGB9PdqjrXlXosPxd+ZW2NqdV7v2tj6KWu3DUwyw09RAzSCGRho/F/z9ffj17poqvj32r8koaTJbpyEv8BknL/aMSt4XbyAf7b3v7OvV9ekZvb4PbQ2RXx1NJi5fGlPpqJVUFTIqeskn/Y+9fb16lOHRR98dK9ZRCtx8lHFHk6u6wPJoJ1n6Gze/de6Nz8AO78/8ct8U+JzlS822BVQrTIJHEUcS8WKjgcW9+p1r7OtsHZHaOwu89mU8+KzOMSslpBel8ytUF5Ra+i9/wA/091pTB6t0VTt74Eba7Gy9FlczDNXwxuGcIxC6Q2oXHu1fXr2CejL9RdR7K6TxFNTbdxEKSRCMESU0Urq0a6b3YE+61r16mOlF2pmxksVLW1qQ+KD6rEixW4sOVt/vHvfXhx6DvrmvWKnlrYFdaaMFwWJI0/0ufevLrxpWvRafkVvJ8THV7phf9sRzRKFcgFv0/j/AB97HXuHVRO+e19wZGSuAWqeed2NO4MhVVIJuB/xQ+90610XHaGR3VvXd74XJVDRQeZo3ExPrjJtpGr37r1a8ejl5DZ1Ls/BRUNBCfLLEszOLEHi/B9+pTPXvl1Xr8t44qyPFmnnSmzaLH9rK7aSsn4Jv/j78OvGvHoDdl7S7m7CocbtjP5WrqsW06RU/jnkEfJ9Ppva3vw61x4dWt9D/wAvjdeEoafduRR6mjSnE7a4VkBVRqNyfevl1sceju/Huj2dSZ7M7Vmgpaaoad4JbxRi3Ok349++zrfyOegU+W0J7Kyo6I2hE08sF/up6ReBE5sSdP4sefe+PWvl0GOxPgf1j1Ntttw71xyZaqcJJPAdMsxlHJ9BueD791vod5tjw1e2qeTrbGtgqbxDSJoRHdQLC3v2evdctl9Dbi3T5RuLcOOgZLtEjvCrqwNwbEj8+9fPr2emXevQ24pGkoKnc+LaGJtNPeWJSEUWAHP9Pe+vY6L3WdYdgY3KU9Jgdy0cUVNNeo0yppZQ34549+p1rI6T/YvU2fr5kylHmqf+8McHNSHUnUFueb88+/dbp0TTfsPbG2shBNlJRkqemIuYIw4F/qeB731X7egQzm3No71TJ1uWp6ymzNRTvGjR1ElPaU35CqR+feuvdGE/lzfKHsv4j9y0WMyWfqZNiTV9LHHQ1NbNM/iaYC1nP0t798uvdb9nR3fex+7No4jcG3s3j6mrrqZJJqGKZTPHIVBIMf19+630On5v+LfT3vr3WFKeJJGlAOtvySTb/Wv78ST1oCnXNSzM9xZL6VFuT/U+9de4jrtWuWGkjSbXI4P+t738z14U4Drn711vr3v3Xuve/de697917r3v3Xuve/daIr1737rfXVhe9ubWv/h791qgrXrHNfxSn/m09h/yCffuvfPrUaWND/NDLG9xvQn6sBcV9zYe/fb1v5Drblh/zMX/ACyj/wChR7917rIARe5vybf634Hv3Wh8+sUaSK8jM11YgqtyQv8ArX97NKdaANfl1m966t1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XXNzzx/T/iffutefXvfuvU68f6/gXJ9+6913791vrq/wDvv959+691inlEUTObgfS9v03HBPvfn1o8OqVvlPuSbHdoTQ/eedGWX9uNm4uf9SPdaDPVhXFOiuZwGu8dfCHjdApbVcXI5P19+8utdLPZ+59t0aeLJQLPVTei9gSh+lz/ALH37Pl1s0HRgdp7lwNAgVWghla0i3sGUH9Km/vVR1sgk9Dfgtzx5RSjsCgFo3AJAA/x9660OljSZ2mxv7XpkE5KA2uNTfQE+/cB17jg9J3K7bxlfVCrqHgWST9wAlB9foP8fe+HXsHj1wmxMOPp1NCVdyLroN7n/WH+9e9Hj1vFOnnEzTU8PmySCVRcgFb2I+lwfe8dVpmnWVa1cpU+OFPEmq3PC/64B9+6tSnSx1pS0v250ysy244tcW1Aj37rVekt9v8Aw2c131jdiwBN783P0/p7159e48OnU1rZcoIB47A/UHn+nuw68RmnHplixNdQV8tTU3aCU35XhrHkW96690ooaWkmmFVDHGoUAMAov/yF7917j06VE0UkDIFOoDggWUccfT8+/de4cOmT1IhOrWf9Yni31Hv3XumSukmdWEYIa5HII/wuSfes9ewOktJg4q9xFWNq8rWBYgWP+N/x73wx16nn59JnIYubA1axUb/tE2sL2PPIt/j73XrXUDcMGHlovuK2CJqlUJZ/Gt72ve/vXDPW6jy6B+qyG26iirzWLDGKRSYw6KA4Ufj3sZ69kHqtntHOUO4dzz0OJi0xU7yFmABUqG5/2PHv3WvOg6Ib8r8tRLtCpxtBT6siWSIFUt6r2H+3Pvea9ezx6tk/k67myvVHS2/q7e8UlLR5TDVhxz1IKRyFqc20k/Uc+9GnXs06b6fcuPyubyu5sYtGksmUrAZRBGGKGY/2rf097+XW+nLK5aprVVKSMl5h+60a2Go8auPfutfPpHZrDZkUaSUsU7Tfkgtb+t+PfsjrfHpGTbazG54HxVTFOr2YhiWBBt/U+/efXq46Dyk6t3BPVVWIniqCNRSBrErYHSDc/j/D3qnXhnPQ/bK+KuXngpmnVmV3BYaAbBvzc+9inXvs6sI6x6MwXW+3BnqqnU1cQQ3CKG1D8kj3Xzr17J6GOXJJkaKGopxoREvyOeB+ffsV69wGOnChyn31IKMNZgLXsebi1/eqder1lSq/hClXGuNr8fU6rcG3vdevfb0gqhp1yLVMzFKWV7hWP4v+mx559+4Y615V6i5SvlQF6XUYiCOLkX/w/wCJ9+NaZ62M8emShrYqWmqJpZV8hu1yRcm/v2KdePHqDnoKfK4sy0Mka1DoVcqRc2/x97r1rzz0HWC2/HI0sFUdcwDuGJva3+v71nq+KV6AztTONtajrkldtF2AIb9GpCR9fp72KDh1U1rTqgTvLe+V3DvqspFeSSi8rXZS5XiQj/Y+7da6DIURoa6CeMHxMoZyPpe1rc+9der0q6eip65WqiyEQi7LwP8AG/8Ah7317qZSY/8AiMrfazrEYrllJtcDj8e/de8+n8zPFQPFHLeWM6fqLkoPpx9ffqGnXh0lod4EySUM8cisp0hzqPP+H/Ee9de4dIndfZ0OHgmo5qxaZEjYkyORxf8Ar791o16JDv8A+UmP27VyUGJrYp62V2RTHNcBmHB4Pv3Dr3Rfqrc+/N8zyZCaqmaObmNUla4tz7959e6DDPbA3pkq3zmpqwIm8yETOCrLyCDfj37rfVof8uH+aHvD4J7xxOH3nnZxtVpoIqqGqq55daz1QVVKsbEEG3v3y6959fR6+M/yC2r8lupNsdo7SqqeWjz1HBO0UcgLQu8QkYMn1F7+9j59aPovRhfeut9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//1d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691wbXddIBW/qv8AW3+HvYp59VNfLrjIYowZ5WWNY1JZ3YKiqOSzFuBb+vv1TSnXqCurogvyn/mC9H/G3DZT+8O46A5mKnqkggjr6YFJlgJXUAx/Nre7R6WNDw6vE0bHuNOvnzfPj5f9j/O/vyuoGzuVpOv6KuV6Hx1rPTVqQ1BYRMqmxW3tYlkkmRkdLPGslBFany6O18eOoejtnbUx2eysVE+dp46aqKeGNi1RGgJJt+Sfaecxwto4AdETx3k91+mDpPCg6MznPmF2RFjGwfWm36RsdjwKaKeNBG4iH5BHskut0EJoD0K9s5bvrympSPy6R1T3Vv3fOHNFu3P5jD1Yj0eOlq5FA1H+yEI/NvZDc8xOgJQ/z6HO2+3000qidcE+h6COgx9dS5d6qozmRynku2qsleU8nkktf6+w7LzXdkkKST1JFp7ZWMZUsor9nXPM4N8xUxu4Ure1igN+L8j2UXHMl4e41r0MbPkGwiAAUdAxuvr2kgytLkqWmi+8oqhZomWMag4PBFvb+2b9O81ZD05vXJe2fSFWUaiMY8+jD03fnZkG0I9peWrp6RYhArJIVQJp0ggD+g9j635pKJpY/wA+sdt89td1lva2qnTXyBp0DuU23NVyDL11ZUVFXMPJaZmezfW9ifYd3jmMzGin8upT5F5JSyVRua5HqOsb0Sz0wUoHkQcqQfwPYQaQyyeKcV6mDRBb/oQAaBw6aVxVA86ippYjptcsgtxzfn3Vlr3V6qaHIHTXnZaChkhEEcaxrYsUGnj8kD3R66ePT8WpmGOi6dtb1VaaOkW32+uxBIIIJtb/AGHsVcsRgyLnPUe8/wB6tvZODxoehk+PfyA211bjgWK/cMgbQoBZjfVcA+5UArEOsOt2c3V80p9em/5U/NjN9lUuFxmKo62ShonQVPigZ18UfBDaR9PaaEaZD0sZAkCn16PV8RDguwev8XU0sMUdbrp5HidAj6jbUSh9v09emhwx1ZvjsOcfV4gRU0QWmiVX0KAGPF+PfhjPW+jg7SwdHLjZ6hkXXNARp0ni6fqsPz711sfLp821HHS+TFFQUaRje1j+r6A+9db4DoRcJtimr3mREXzO+mMEfW30597PVT0IeQ2CtJhUWaGISyqBawB/qP8AH3Xz69njx6qW+dvxV3v2zsbJYPDJOtLWSSKwhV2XRKukiy+7CnWs9V4fGX+WCnx4zGI3BkKMebISearaSmYs5kfUddx9f6e9jrdM56uZ3J0xt/dfWu4sZXYagOPG3KiKF3plPrFMVJ+nvVevEUFOtfz4S0dZ0n8vs11tQTT0+DzG4Jaj7aD0Uw1VJNtH0t/sPe69awOtvLZ+QGiaCjpw7DSusR3vaP63t7r16nr1zz4zoMR8Vow4P0PH9ffv8PXvLp+rMtUrtl5GhDPHFb9IvcL72PTrdPPorG881LlYYImi0iJ7m68gBufp79x62BTqmn+YNSHJbYraaIunkgqE9IYjUUsLn3v7OtEY6LJ/K++KlHuLHbrmykYqmq553VqhC5S0l7AtyL/4e/fb1oVJ6tv3Dt6t63pGwmIoo1pqBAZCqEWRRYm4/wCI9+z1uvQFb03RgdwbWqKFfG2SaQpJqI1qz+lwpP8AvXvR69jj1U38tesqXb0mDzuMqZFqpPFK6ITpZz6ipA/23vfEdaP2dOHUeDxmaw1DNllKVlXJHAkmizByNINz739nWhTox1cnfHxzjg331/W5Gvx8YWZKJppDEYl/cIEYNveuPViCB0cv4w/zdMvVw1dD3RjqygaMMGLQuQgHpuGA9+IJ60OrKOv/AJ3/AB13ejVUecRWmjCrHPURR/uOPoofn6/4e6062Dip6n5/s7ZO+MNX0uCysEy1M2qECqibi/0Gk+/Up17HSpxVTQYXYU0MM9M9SaVgxEy6wzD8W97+fXvOh6r57YyQ3DgqjAzOXIqpHPquCoe5BP8AT3s069nj0Sjfce39uU0U/jgaOCIrM5VQysq3sb+/fZ17FOq5Z+56KLuXb1FgwVSvzEVPK6kIoHlCkk/T3vrRz1a32zujBbZwGOrq2pprvhIp2BmjB1+G5uD+f8Peut9a5/zO7yyufzIye11nlgwk0Rk8fqQqJADYrx9PfvLqvHj1sKfyvOp9sd/dQbezlStOdwwxQVDJIyl/IsIbSQOfr799nWxw6ugoNxU3WOFqdrbrpIKfGpDJSrKoBGm2gW/4r70M9WI6oa+b/acvx53Sd3dXTT1n94ahJ5Ep2LOhlk5sB/xHvdPPqp6HH4qbpq8zgafufKUUtXujcCR0jxV0J1xmVeCC4/BPv3Xh0c6fCUeRpJZd1SumUrGNRR0XqaBgeUVk+nPv3W+HE9K3YvXfYG72OOgxdFBgqU2WWyxHxD6G/wDre9de49Y91fHykx+UElTmZqS5GpaWqKJqv6gdJ4t79X069Qk9JLcvx/wdbLSz1G4ZiiJ+pa0ktccajq+vv1evfPoJtwfH2Klp6yo25llqJREx/cqwx/qeCb397+3rR+XQG4bp/c9bX1FPXSszxlm1tNrXSt7i9/fqdbyOoVX1ZR19PlsfJSw1dbG5jXzqGsSLXDN79x61SvRP+yfixXY7x5qopzTK76kWlhfSfXxq0f4e/de6It2j1/ncTu2nrcJBKuQpY18YMThCEH154v798+tdCT8eP5gvfnxV7Bw5grcvW4dKlDkIpqhxTQK8gBVVbiwHvf29ezXrfM+GHymwPyd6k2/vCOto0ztTSU/39ElRC0jStCGZljU/W978e9db4Y6OKSACSeB9T78M8OvHHHrpWDi6/Q/T37rQNeujr1CwGn8n8/7b3YaaZ49aOqvy65+69W697917r3v3Xuve/de697917r3v3Xuve/de697917rFP/mZv+WUn/Qh9+691qRQ/wDbz978/wC/zH+vb776j37y6959bb0X+aj/AOWaf9Cj34cOvdZPfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+60OuDMqjWzBVUEkk2AsOSSfexXh1o0ND0XHsru/H4HHZuix7wyZCGBlhtKpYNYg+n3onqwHz61VPlN8i+48D3LXbgqcVW1OBjlqHMiQySKIFctcBQffuvHpl68/mD9Sbnk/gm4K+poM0reF4KlftlWUnTY+S3v2fPr1ejWbW3Xtvc9THX4TJUksbt5EH3kRuCAwIKn/b+/de+3oZYn+4q4ppKwowRQFikDAgfQC39ffuveeehbxe+8vhEgo6dg0MgCl2IDWtzf3rj17z6Xx7MioqWKSul9S2ZrOOR9SR/j78K9boOlxtfe22t3yCQ5Jo3QW0GdQA1vTwT70R1r7ehBwVRLJXywrIZqaNiFYkOCp/II97+XXvLoShHTzAJYFfoB/S/9ffuvdTH29GsDyUoKy6dQIFjxzzb3rrYz0gItwz0mTFJkAwCtb6GwF7A+/eVevY4dL+to48jSI9MwZLBzY34I/oPeuvH/B1zooolgKQjTLGCDb0gEDkX9+r5daGOu6ivaujNEUUOi2uOSbC34/Pvfy69XHUJYZ6KAsR9WuxU2v8Ajn377Ot1640lc9QzRTLpVjpuBY/Sw+vv2OtHHXikqTmOK7oP03Fxf+lrf8T798+vcOuc9OvkAcAFhb8fn8+/deI6b8hg446Z61ZAGjAcWJ5P9PeurV6DqsyeFip6ipyVSiNDG7/uSqD6R9QD78K9ar0R3sPvnHRZmfE0s9442dLh1K2HF+P6+7eVevfZx6KbvrtzI11UtDjJXRJzpf12DWPJ497HXuGekRmUpsNh5M9NPAKiaMtKTIvGr6kk/n/Y+9Dr2Oq1e1d1vvLsXGbVoDHWCsraQyLGyzGzThTdU59760ethH5UQbb+O/wZ6bqsSI8XmM/jKOCuanKQvN5YAHLKef8Ab+9U630Rboeuiz21J6uSoZ/JKZwS41EuNXNve/l16v8APo3nUNNSZyeop6tBrjcxxiVf7OrSuk+9Hh17z6NvR7CxCpHHU0sLIdN/272B/JI9+61XpJ5nY22aLMqKdEUt/qVsbn/W96JPAdbx0oYut8KkIrEp4vMylkJj+rfgE+9569XobdhYil8PhkijWSIWQEcXA49+PWqmvT7m6SpngkxJW1NIS3BOg83+n0HvWetj59JGSFMZTmlF+V0qPrx/Qf8AFT78DXrWB1wwkhp6ktpYKTZr/wC1fkk+/HrYPU/cFU0GiSJdaH9f1Njfgn34dePp0i93DIVGKhqKKP16QbRgk8f1/p78OPXvLPSexdXkRhSauO82oqNQ9Vr/ANPe+vHjjpJ1dDkZXkDllV9TAD6A/X6f6/uvW69B5HuSsoMv/CTIxjMgjP4B9VuPdvLr3n0+Z7NV2BnR4I2IljUlgDcBxzyPfhnrxPREvmB2HFhtrzFWtWVcYcKGGvUyfXj3sdaPVODn+K00uVqFT7iQyNra3kYE3Xk+91610hJczIS9JJ4zIC4U3BNgfTa3v3XuuGDyD66iOtleOG5uSbcX54/PvXWgeuK55abJyQY6eVl+jPfgqRwefe+t164VW9WxtasKSakkBMnkcWB/IuffutVoadAV3z8h9vdf4tKuikWbJtGreGnZZJGYi9gEub/7D3o9e6r9yW6++vkEznbOJyNJS1MrwrPLSTITG59LCwH49++zrfy6Mz0H/LW7E3JImb3wTPIwWZvOragb34uffvt695dH0pviXtPYsC09asYMC2YKPobWN/8AiPfutfb0l36r2FU109DCEDGN0GqM+pgOPr7317qtr5a/FyoqlqMpglZftqiKeJkiZhancS8Wt9CPeutg9bTf/CV75W7t7VpOy+l8/VVk0HWfnx8cdVNqjV6VEjDRx/jj6e/dexTrc69+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//1t/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+69015jM4zAUE+Ty9XFQ0FMpaapnYLGigXJJPv3XgCTQda2v81L+d/wBb/Hnb2a2V1lnIM5uKrpa2g/3HFJp45/8ANa18ZJ4PPvYjaQUAx0ml1ltANOtJ7s/vzvf5H7uyGf3Tma3M4nNVgrYIJ2qAKSGf/dbXP4v7SyXKwGh6VWm0XN52x56EfZXRVeDjKimq2oDUOPLKpYeO41Ekn2mbmFI1IU8OhBDyNuczqY0JHn1YhsLa1HtfHxUmQzq5S0aA6pNX45Xk/T2Fb7mEPMTq6m/l32/iSwV7hO758ehCpMhi8cssFBMlN525EZABuefz7D9zubTVZepG2vlyxtKawBTqbHNi4laoqqiORxc3cgEH/H2SvK7mrY6GSxbbFH+kATTrLT5zFsSFZHa/BFjwfwT7aUwVz0iaRl7qU6kfxWjimRjIqqxJ/HHF+fbxitzk56o+4yQrqA6T+Q8OTrC8DoxVubW+g5HHvwtgxrEPn05aq+7GrmmnqTPi4quJSnpdQF0gG9x9TYe22kmTB6V+NaWq6GQMR59QqiieCnOo+Ro1tp5J/wBb2jlSWbI6acw3WYe3plMJSLz+HQWFvoP949q4lKxaG49JpOwlfPpuZKT7aWaUIhuwBItwB+L+/GvHqoNMdAnuGKfM1clJR3PqC6lvYAEj6j3Ygnp63dvEAPRVO3ttZLGGJJg765QNXIBBP159jXltQJFI6g73cu2giYVxQ9C30F0KN95rFrVp5opo4tUZuAbtz/sfcpIFMPCvWL8Evj3B6vB6u+AGwMvjHpK3A0ks06+NS4VjdkFjyPZfwc16PZ1Koq9QM/8AHTL/ABYy8WdxdM8G2nmUGFbiGNFPLAAW+nPvdR0m+fl0bPrvsbB56LGtCUyQniV6maMC1K5+ob+nv3XvOg8ujgY3O09LFQJTkeKodEQjSRyOfp7rQ8erYHQl0+DEVfDWBfTMiMWB49Q+pHvxOevV9ehVwWJqqCaKtjjJuwZVBvquPfq56rmlepu4c9uCskSH7GYRxkckED0m3vYr148K9SqeaprqIxV2Pc09h+4yjTcfXgj3rA63X16YdybZxdfQg1CrIUUeAen0G3Fh79k9e6Lt2l2htfqbr3cVRuSsix1GmJrvFNNoCOVQ2HqPvYHXj69a1/xO7N2j3R86WrcFV09WtLlyokgIsQ1RYcr+Pe+tAkmvW6vsDbmHwtPHNW+MGQobswuQYhYc+6j59e6R3YG+MBBmTi6fxal+qrpJABtyR+ffvPrx49R8Tkcbk6F6BwmudTZG0m4YWBt79w69WvQMb12cytUfbUoVVBOq3BBP+Hv3y6tXPVV/zA67jye0aqqdFDI8wKknnjkW92Hp1o8c9Qf5bO3qjB4zOCEECN5NIHOq5PJ9+PWvLo33cRgpZKqSpAL1sTRMLDjVwefeh1unVZXaezK7C0tTmaCUrG0zz6EJHpHqN7e99e6It2xnKjs2uwOCijaSelMcbqB+oqdIvb3sDrRyOjl9ZdESYfamJq8jjhEA8MkLSD/dhAKt/j71XrdBToX9/Y+txeFgXNVDSY0LHCKZz+2Uf6WDf4e/Z68eh06n+NnUG49kT12W2jjnqcvTFqaokRAxLKeQf9596J69Q9E77F/l01Fdk66s2hvBtrRp554KSnqHjAJa8aKARz731ojzPROd1/7MV8YaWuix0mV3g9K7CAIZnMiIdepCb3vb37rWR1E6z/mo7wNSNu7+oazb1SrGKeKsRl0caOS9vz731vo5W3PlD19vaBftM/R1WUnQMaVHRpSZPrwp/r7r1uvSM31hsnvvVT0NM5xs9zNOgIVVtY3I9+611W93psPA9M5vBbkepQzw18c5clbw3kuWv7t8+tH06Bzf3a/Y/wAgN843Z+0slVVOKSjhjeWmaTTp0BRGdP8AX6W9669XoesD8KajcXXW4KPL0jLuCWFlhicnzVUiC4Kqb39+63Q+fR/v5aG6ar4+5Jdg5nMNh6+KcwxUTtpd7XRFs5Hvx62KdW+9w9kYupxBh3WUplqqeSWKsnIGq6XUhv8AH3rhx62SPLqpmDqyHvnuPA01Y4ye18fXovjc3gaFJb29VwePe+q+eOrnR1d11sDEY7b6QUeExUNPSmmlARY/MqgEi3F/eq9bHDHQX9h7uoNoywxY+gG56tlBo3iTWyx2/bsR+Pe+HXqY6Z9t787zz9HULgNtZLDQMps6JIo+n9APeqdeqfPpMZHbPemWWVcnJkYpX1/uv5Pqx+v+w9+NOvDpp2/8de2s9DVy5LfFdSxhm0RvLIAi/gcn37jw69w6Q+7fi/3pjZVk27vbJ5COQjyRwzymy/04P+v79UV60Qehh6y6K3XTYk0u5NxTQ5ox3YzOwlJ/Iu319+6t0ld29R7uxstYMLFUVspaxqELFmI5J497615Z6SGP3LlqCL+7e6dryZB6a4JqIy1gD+olvz79inXj8+i9d1bS2bnqWStFJTYCtYONZVFYEj9PH0Pv1fLr3Vd/bfx8iz2Fk/hdEJnliBFYB+u3OsFffutEdKv4cfMHs/4c7uxO3ZK+trsR/Eoo5KZfIyU8CEKb8/QD3vrw/l1vBfE75IYD5F9d4zcOOqElrGoopKuMFdQZh6lYD8j3rh1oj16NaAALL+PoP+I9+695dcULFQWFib8fW3PHPvZ68OHXP3rrfXvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYp/8zN/yyk/6EPv3XutSGI3/mfv+P8Af6fQfXiu/Hv3l14DPW2/D/mov+Waf9Cj37r3WT37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XX1/r73w61x49FA+UHe9P1VQRUS1QiqayJrKCA12Ti5P459+qeA63QefVWM2+81uzJHMxvJURVTkut2I0s19PuvWwc9TMvB13m43otybQpcnLPD4pDNFGeX4b9Q/r9ffh1vHVYXyr/AJd3WvZ6zZDYUdFsjK1N5VqaNvBMjhbqQY7c397r6dVpnqpyv6P+YPxay3k27WZ3fWFpZWkjYPPIJE1aVVSL/Qe/H59a+zo0HWv8wTsLaoig7O2PXYZqZY0lqa6GQKSn629Q9+63g8ejF0/8zjqbNTQQPuDHUFTGADC2kNqB/wAT7916oPQ+dffJ3b3a1U9BjMlBVx2A8kZ4AIuTf8/X36vXh69L/cOezW2qZMhgcjIimaMlI78ENz9P+I9763xFerI/jpvCfcu16OWpvJVGDVM3JJe3Jb/Y+6Gla9e6NljShEaufqRwR6v9Y29++3r1PLpd+CZKTyqpKFCNQH0vxa3vdOvcOkFldqQ5hJpERI6lVvrv+4WPOoH3rr1AOPSAw+RzW3sqcfUrJUUxbTqNyoQfQj34j060KDoT5HWdUloCWeQatKfVf63Hv3HrdcdRIXWOWzRFalz6vrf/AFz/AE97619vWSZpfIIpQSh+gP0/2J96zTHW69SaoUwjVIkCSkWBt9PxcH/H+vvwqevH06faCljpaPzzqWYi9zzza4N/8ffqefXsjHSOyVW0k5kQEKtxe31P5t78R17j0ic5udKCmmNbVeGmiB1sxGkC3+J9+zWo68aDqqj5D9tVK5eSDb+YLU0kjRyeFrgDVb6A292pTr3RRN0Z/GY7HtmKnII9YV1yAkM5NtTX1H/b+/dar69Fd378oOudk4+XJZTNUcNRAoZY5Wj1A2v9b/6/v3Drdeqxu7f5mWS3VTT7X6+pZMxLKZqaD7ONyA5JVTdffvs61noTv5bfW/de++/MF2h2Rha2LbsZgmanqBKIlVZhICQ/H49++3rw+XV/H8yPftd3DsXa+zMdC9LidmxwNGiArEqQqAfpx+PfvPrxAIr0V34272xK4xNuRVaeeAxLJYqf82LNf36nW606PRtjtfaW1czRwz1kNLOXVSh0AynUCSB/j791r59WK7Q7B2znsXTVJqIirBCTdRcaQebe69bHTzPi8Dn8kk1NLExT1EKR/Tix/wCI9+69U+XT/akjVaVbEQ+gk6bG39Pej69eA8j074d2p6lZoACC4vxzb/kXveeJ69jI6VWSqjVMphjGvSVt9b3/AKW9+8qda8ukhV420gkrBpu31bgfTgAH37gOrUB6lT4yL7Dy01mk06gFPN/fqevVa+nSfFHVVGKqUmRjMHsgtyw/F/fvPrdPLrLj4KiKkFPVUxP+pDfkHjm/+8e/Y8uvCvn0kKxYlrWpAlk5YL9ACDyD731rB6D3d1bUUtVAIUZY9IDH6mzfXke9dbAB6YINix100OaurOXElibE/wBq1v8AH3759bxXpx3QKKChqKzJwLDT09HKVmkAteNf6n/ePfvPrVadUKfJ7saLc+663Gu16OkqWjgHGhkUW/21vdvPHWuqku7/AJEUHX7y4+FlQpqCRhb35tpFv6+/da6CvrHuiXeeWFbUq0NOQCAVK6geSbn37r1PXoWtxdo4VcjT42hqYmqJnCtGvLgnj6f4/wCPv3Xj8sdLn+KUlBiFqwqPWzJwun1EkX5/1/fuvfLoId0YXf26cdU/wnE1UEshYRVSBhpV1IFrf7x7914Y6Y+gvhZkNz7tGc7XyslXSQTs60lczNEUB1AFX9++Z63xPVvGLg6k63xcGB25s7HmaOGONayCJAwdF06ywHvwB69gHHXBd0bliV1wImWOQkrDCDZFY3C8D8e9468D5DoOqzFb3zlfJJk4J3jZ72k120/1/wBf37r1OmPM7Sp8dpqDCKepuupyOWv/AIH6e/V699nQc9i4emq9sVSzQrKSjXY2twlr/n37rx6OL/wlf2om2fk38opUTxrkq/ITKin0+soxP+8e69eHW9n7317r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/19/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XR+nujsVWo68OotTVJSxTzSjTHT08tQ8jcRqkKF21N/rD36IlxnrxpUdaUX863+dRm9p5/cfR3T+ThOYpJVx2SpqOrlSUlmaBntGQQPaqREjXuPTU4l06rfy61het+ht8d27obtDsCty1ZWZGrkn8FXVzVMKpUv5SFjlJA+vsmuN2EI8Oo6Odi2W9v5Fd1JqR1Y/t/qDA7VxEdGlGjSBAurxISCOfrb2Fdx3XxeB6yG5U5KXwwzpn7OhIxO2GWmESoFW2lPQAQLcWt7BV3fS1IXgePUoRbSlkoVV/l070W0pEY6/Le4NiTYf0t7SRrr7ulyPoTQAOudZt6pikR1dwVHBuSf8AAG/093XBp1RlDqQeJ6wyYPJVCHVIzDT/AKr6heTcD3Zx4iaRjqkUIjz59MZosjj2LAsADa3Jv/j7SC1zWtelLymTt6bMjk8nHE0zGS0f55AJtx7cMIwV6YaKoo3T31xuMZDISxSt6r6fUb83/p7VxSvAOPHp61Y2tSmK9D7T07LMshuUYXNgODfgC3+8+2mYSGrZPTTqJTU+fUvIUUMnjMYA1cG/5PvaHRw62EC/D5dI7IUZjM3ldRFHGzgWte3PvTHUdR63XNW4nouO4dzM2UbGwyqYzIVOk8WA/I9+BHDqpAIr0IWyaXFwpJLWCMyvcoWA+pFxf/ePfjwoOllmoMyjy6BDvjGQ1C0syhTEKgDhQQefr7HPK4VpAp6x398FKMwHCnR0Ph1t2CaqxFRTIvkjihuCguSGFvcqURYaL1j7s8cTDVwPWwr1Hh5KSvwjTKCk80Qc6RZQVF9Q9lJPfRvLoznk1tpHAdGR+S/Xe0d+9d1m3ZKWBqr+FTSRzJFGWExiuDqt/X3rh8+kvy61Qd6dp7g+J2985t7NpWS4mtyTJSysZNEcRe9gfxx7v14/Lo8/x/8Am1sPsmCjxhylFQTYxgS9XVhNTIL2Bc+9Z69jq1jZXb+29xYWBqbK4+qlQRBWgnWS9uB9Pr7rTNOtnhXozW0d009W1J5JYyiW445t/Ue9/Pr2eHQ8U1VhKqCaWaFWAh/bIReTbm3vQ63w49A3ltyeLJyY2GI/ZkueBbgC4/1v8ffgOqn59V1/L35fYTonB5ChqZS+ar00YhFch1kcWQafyb+90635dUN7yh+XHzIoa7GZCqyNHsmraqiptHmjLQ1DFk9X+t/X3v7OtdF+6w+JnY/wJ3sO40auqKSCoikrZZQ5QaZRJIzPyePqffh17y62pvif809v/IzZeK/hlfTyZaCJTXJFMWkVkjAYMv45HPvVBWvXvLpT9nrnI8nLlscZjP4ncEsxuPxb37HDq2T0DWxu79wYXeNLS7gldYkkVVDu4U/uWN7/AOHv32daxXqz2k3Bt/dm2kqKSemeaohUMiMrODo5vf3759a+XVZHyjx6HH1+NlUGIfcyq9rKbKSBz72PXrxyR0jP5f2LqKnD7yqYk+3joJplDONIZVY2IPvR68RXHQmduKa6sZqueIRRPfUXsrBW+gJ+vvw62eI6rx+Qm8hRY+ago5YTCEMb/Qk3X1WHv3Xjnqsat7DwOwuz9n5GvVWpKueP70sbR2LgnUPe8da6vD617G2x2fjqWHF1uOGMpYKaSGFJFaUOEBIIHv3Xvs6RHc+IyO/62j2tgopXYTxDVCh02V7E+n/efe6068B6dH9wO0m2b1btSmq54krcfRxpJGSFkusY/Uvup6sOgO39u6jxtIuUnkGtpDGBqt6ibAf63vdKdV456Jr2hviKoy1JU1f2b0QiN42jR7hktcgjn/D3rz631UH8m+qNo7wny+aw1IYslNGWVqZVis973AT3vrRp5dE5+E+18nhvkrPjdwVdfNReWJIoZ55DGilwLLGTb37rWK9bB3f3cGyOgOq6nUsJyNdSSPTWI8gZoiVFxz9be/efViadazvY3ZXZ3f26D93549qmomEWosC6CW6hdX+Hv3l1Xh1aT8SesMNs3A0W4qajK1xFP5J6iNXuw/VeRve/s63w49Wr7ZqEyO5dsVsdMjQQsjVISMFZOLtdBwf9j718h1unVcn81c5HpDIbR7l6/qEx2Sly9JLWJHIadTF511+RUt+Pfvs6rno3HXnyMwXyI+ONAu5KmOo3hDh4RDPBNyrLAOAfz/sffut9OXwIzGYqc3vSAwyVC7elnSKYoXICk2IY/T3vr3R+c/j96dq1mPoZczHRY+jqkkkjklKOY0a5W9x+Pdet/Loz+J2RshanCwmphnr6Wmggmndklj8iAAksb+/Z610aKkwmUw1BSviK3EeCUD0xpAdItf3qo49b6Qm8t3PhorVgpKyRLs4p0jJ0j62AH59+Br17oN8f29tzMRy0NJh6uKQHTK/iIVmvz9B72cdeAqenfO9oUu0KCimpcZJUGWxkQQh3AvyCLX964469Q9S5ewNkZTb395KnC1S5U2QQpEVa5+p02v8AX37zz17NMdBNX9kph5hkBDElC/qZJYwGVG/Da/z72R14npsgxe2eyI8nlsRXYta0wuzwI0TS6rXtoHP197rTrxpjomXbfUSbl25lMbFBKubjkqGhlVdNygOixt9PfvLrXn1XHtSp31sjOZHb+9D97j4pWgo49GplQkqL3/w9+Oetj59Ivujq6lzFENw7bjFPkJFZpFVFEnq5It9f9b37rR6Nb/LH+Z+T6A7UxPWW5KySnxdY1PHKtZPKmtpJQt1Vjb6n+nv3DPWhkU63UduZyj3Ng8bnscQ1JlKaKriYcho5FuCD+f8AD3sU8+vZ6feb/wCHv2OvZr13711vr3v3Xuve/de697917r3v3Xuve/de697917r3v3XusU/+Zm/5ZSf9CH37r3WpFEf+xn7gn/mNL/4gfe/19+8s9a+zrbeh/wA1F/yzT/oUe/db6ye/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XusMkyxsqsG9XNwLgC9iT72BXqpah6yi3LA/X/bcf09+zw63jj1rx/wA17tWj2r2ftjGVztBTOYlcySlEJ0hRxf3rq329Bz1jumlr9uYeqwro6VKqXYHWBcAnke/eXWgejDUdBg8iitUlGrXUC6kCxb82/qD7qK9Xp69dZfqypraGXIxyfcLD6oVisxAA/IX3vquOg3q6WtaODGZPGQy0sLAN5KONm0g86iw97+fWs8KY6BDtT44dS9mo9JmMHFeohdXeCmijs8iWIOkD37r2Pt6rk3Z/KP6pl3CmZxdNUwxNKHZVk03UNq/SD79x69QdDd1v8YcF0xVgYWnqLcLdiWvpGkW+vPv3Wx8ujGDamXzYhoqSCd4XkiLGxP1b6W+nv1OvV9OrMOiNmT7VwFIGiZJZI7PqFr+mxuB70T5daHqOjL0pChZGBDD62/w+o96HWz0rqTcX7K0so9NtIUi/H9efe/LrXE56yz1sMYjFOpVpOG45H5/PvQ62ccM9RqqjoKiCTVGv3BRgGAFwW/NwPfh17ga9BS+QyO3cqo9TQu9vofpf/ePfvl149CH54JqX+IG4mkALf1JI/H+9e/Vr17qdjTFXIXnXleAbXNh/h798uveeOuc6UPm4+qfVb/Sw5B/2PvZ+XXvl025PLBIyiuVULa5ICi39f9h795da+XQQZjdy/dmgpHu7E3INzYmx96698uic/LLtGn2dgI8WZglXk4hGD5DGdTLbgj/ivvfDj1vj1SfuXtWgw5rJsxUrKB5pEVpizMdR4BJv79SvXq06rW7u7n7F3ll3w+wKPIimlkaLWglKWk9III976rjoH/8AZCO8e5Fo6/dWRyQoKkBpYFaRQF+tm5/p7917PVlXxe/lo9cbB+0q85QJU1cTxSu9TFHMdajlrv8A4+/DrdPXq33ae28HsHEDHYHHQQ08aeMeGnjSQqOPSVHv3W6VGekj3ZPgavZNZGsQFbU0pSQOAZL6Txz791qlOqCMp2/D0r2DXJ98kAlnnZVkmMfJckce/Y68TnpBb0+SO89z7oxmVw+SPgiljktHPISw1i/APv3Wjw6ue+LHem4s3timgyORIkWIBleZ+GEf5N/z799nWxTo4OB77zG36sj7gm5sG8zEBfoeT9feutn5Hoftqd+RzyCfIVAeOQa3UubXJ4+v09+p6de8ujY7E7Q21nGiipqmnic6QyvKNd7XNiffvPrxA6H0ZDGLTK1MqPIygmRLMgvz9R/j71XrVek7lDJkQF9JI+mkX+nHPvXW+nPCUZ8S0z3B1C5IH1P14PvfWvPpVthNFSkSoLOL308f4E+99ex015nFNC6qy2UEAHSARf6e9eWet8PPoNsht6OWueeGNrhWvx9Cfqf9v78K8OvH59Bpncaksz09Qn6W44vax5sx/Hv2SOtDGeotJKlMsVMoURpcKB9fryLe99bNa9Br8i5S/VOTNGGSUUlU3p9LcJe1x73kdaAHWqb2buLJ12681SEss8NSyR62N7ngnk8+/dex0Uqv6FTsHck1TuSCSojEhdLDUnJvwD7917hnobcB8bcTioFbF0ckKKgiPoCcWsPp791rp/xnxi2+lf8Axmop5pKqNhIAebsDf6+/eXW+l7S9UJX1oX7KoMcNjGAnoGlbDi3+Hv3WqU49L6h21u6gqIaDHYyWSk0qpH2oJtew5t/sffh8+t9CvtPp3fW5KtYzTzUiSuVJaDw2ANxyPfj17o1G1fjItFNB/HdU5crrOkGwI5tf37PXscejKUHSWyNr0SVbUqFwmoh4lYsfra3vXXjjqJk9n7GqqV5IKelpnAF2ZES/Jva49763nz6Ir3Xg9uok1PQyQiZWlIZSL8H/AA/1uPfvPr329EG39nEo4JMTUuAsilVe/pA0/Un3vrVehY/kPfIfFdS/MnsHAz19FDHuDNPSfuzBQ4lGi/8Ajf8AI96+zrQ/w9fQfoKuGvoqStgkSWGqp4p45I21RssiBro35Hv3W+pd7/717917j10NXOq314sPx/j791oV8+u/fut9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//0N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XFyQOBckgf61+L+9EBhQ9WUAnPVb/APM0+Y23PiF8c93bxrMnDBm2xdfDR06zxpMA9IwLFHPN/wAD3tCE4+XSecsVonXzTOt8LkPl/wDITsDvbcHlraDcFY9VSR1cZZYoo52caGPH+29ku4biYW446GfKmySbiNJFdXVwG1tl0eBw1PS0VPDFTwxxkNGAgARLG3+HuMN53aR7g6MdZG8ucr2u3QDxVGrHl0+lsPDE3mlVnUmw1Buf6WP9PZct9JMRrx1I9jJbWi6FAB9OoVPuPEpL4w1tDWHFh/Qe31QOSTw63d7pGToA49P3948TawkGqw5/Iv8Aj20VCPQdIqahqXpmr9w4+VtKyci1/rY/g3HvZ69RvMdO+N8MiB0bUWt6Sb2v/Qe6/b1s0PHqJk8a8hLpGCObL9eD78SSccOtDUBTpNV2Op6mFqaZF54IC/Tj8e/But0PGvSATFybayMNTRi0by8sAf68m/vx69imePRjtt5umrYoVdg0njUNciwNrNwfdSQOrCtKdKTIKIkEicr+Ppx/rf8AEe7Hh1sdF77C3bNSxPTUxbyS6oiNV/zYc+9cMnqpyakdA5hdmZfJ5IV8yMVkYSXYG9zyPe1ABz16pr6DoS5MJX0kbaNQaP8Aoebr/wAa97pXpRbMIpQy9BhvLF5XO0i0jRMft38mrSeNJuLexXsEphkBPUK+7u2PfwtKo4jy6N78Qt10OAylHR18wiqICkapIwS51gXN/wAe5OhuxJHUZx1iskUllMYnwK9XuUHdO3dpY7Dy1FXCJa0xCDTMjlWZQLC3upDFiejuQKY1b16EfP8AdynboqmmMqTxFFcsGYhxx/sPfgOmcdVUfJPpHE925WJcrQ0yR5FlkSo0XkAYcMW+v+t7359eIxU9VI9qfBrd+Nr6yh6YzOTp8rjNcswpTOgNmuQdFgffutV6Qm0u9/mL8Y5KXGbow2SrsJTVsEUteyTkiKNh5JGZh/T+vv3Wvt62UvhD8rtvd87Oo8lJlY4c1j6c/dU71EcbeZALhkY3vf8AB96I9OrDPVqu1NzSVyQRLJE/kCp/nkI5Fh71148adP8Avaq2Zsnb1duXdlfS0aR0lTIHE8Vy6RFlHJ+vvfHrwOetUjuXf+E+S3yhpsNkKmoba+GzCrTvoLwyxRz2GogEHj3sDHWvl6dXHYbK7M2Xtyg2htLEUb/b0tKfPHCFdmEYuSQL3Pv1D59bxSg6R/Ymz07r2fletq7EQebKUtVJHI8V2UCEkWB9++3r3VJ38vjN5742/MHfnUearagYpszLT42CRzHEFkm0qqa/969+6qOPW1pmooqiGn8cCzrU46CTVp1XMi3NvesHqw+fl0VDsDq5q+qOXgh8DQsGukZU8cgX9++R69Tz6ctgZfcWynQ1U1RJSTnTEjFmAB4+h9+p16or0F3ym3rTVO27mVPv5Wa6alHob63I9+HXj8unP4gZmPFde5+CmCQyZCOUyMrKH1ML3/1z78evdE6+ZfyLpNjU+PxMNdprmqND6JV1szSWsbH3uletEnoiO9t3T7q23Jm6mocOafzKrPbUCmq/vfy690Q+lxOU7n7N2/tLHxTSU7VKRVFSImcRaX0kq6/T/b+9depUdH/r+ve1Pi1NS1+Hlr6vF1CxGQXkIRGQX0j/AIj37rXDq3j4Y752FubbS7p3dV08e5Ik5pKqeNXMhNxYSf7e3vX2db6HntHLVWSMtfRVDDHR+pI42BiVAP7IB+nvYr1s46rx7X3XVV9PLTpK3gpTLIACCSyG9z71149V09l9q5usz1NRUSyS08JSGUi5t/Z+gvz7t1rJ6adzY6sO2Zc7RoahkhE1YsgJEaAXa45/2Hv3XvLPRIfj1vLA1nyYyOQq5lpoqVGBKFRaWNvUbe9da6M98haTdXyJ3Zi8NtIz5LBUUyxVhdXlQKp0kC3Fvfut8ehX2x8HctWYCgoaLDQxT0QVpZI4ijM2nUdR9++fXqdHU6i6Ey8OLj2PV0Iglj0uZAjKbIbX/wAffvOvW+jz9WdQDDTrDXQh/sy0YkeM30qL/n36vp17qjD+dfgNwVI2Tt6lhllx2dz9FjURAzqonqRHqAW9iAf6e/da4HpU9f8ARNB8b+n8Ll8tkZojXYekqBG7FbGWAMRZv9f3uvXjTo3Hws33Bs+m3PVYihFZJuoSPAXiuZPJc/W3P1+vvXWx69WI7a65z+fpW3DmqifErUIXVKd2iC6xcAWP1/2HvVR17zx0L2wuu8clBUipzlU0yyC0jyNrBLcAEn36vXvn0Yek2/jsdjaeM56skVksdUr+jj8XPvWeHXjTy65y7Z2XSUoyWQyUlSZhoAmJYajwBZvfs+XXuuNDg9rYf1U1FSH7seRHMa3YHkXPvfXuuq/GUqxTVYoKWsVQXSOZEYAD6AA+/cD1oV4dI+N52glrYsRSMoYoaURoIgoPL6Peh1s+nQb7g2vt7dCS0ddL9o9XdZQqlVivxZbf0/w9+Hp1by6K3n+kN2dS5Ofc/WWUr8zNWlvJj3kkMA1H9IU8fT3uny6qaUx0kI+86/beaWj7goUwfkTxakTSrGTglmP5/wBj7317PSO3DsPZ+9Mm25drSwZCmnfzswZZiE/UbBb+/cevV9Oid9t4PcW2cxJXU9I8mKHLxlW8aRo1jx/vfvw+fXjToiPZ9ZGm8MH2BgGakqMZlMZFUvENJuKldQJHv3VTTrfo+EO+affXxq6vya1HmrE27RwVeo3kMqQhi5/17+/dbPRsVZi7KVso+h/r72RivVQTWnWT3rq3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdY5v8zL/yzf8A6FPv3XutSGLn+Z/Je3/H6fX+n+XfQ+/E9a8+tt6L/NRf8s0/6FHvw631k9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6911cfT+nv3WqitOu/fut9cfSw/BB97yOtYPXCSWOBGkldY40GosxACqPqT79x69w61fP52+2Rv/L47K7XnaWvxQVL06lyzQxXuCn+I9663ngOqvPiR8pM3tFYNlb+pqylp6Z2gjqZIJFBUei+thx7914YFD1bztzduDz1BDktu5U1TPGkunzqWBZb2KX9+68cCvQzdfdqthlmo8kxnidmVhK1wBf6AN711vjnh0N1HVbW3evC00Xl+pAQHke/fb1rI4dccj1Lh2i/yBxJM12W3Ju3Kj36vWwT0Htf01k3nQVMkkaGxWxb6fi/v1SOHXsHj09U3QVDUIHntJwCWkB/3gn3qp68MdCrsvp3beFcT1KQM6AMoYAgEfT6/j3utetdCrSrSQyPT08arEhCpb6H+gHvXnjr1OnOWqiiUIQARb6fQcf197r17qbjPtp5UZiQDawt9T/QH+nvXW6V6VNTj0CxPENT6geBcAH8D36lOHXscOuPiiBHkOmUgekX+l/r735deNSfs6b6zAU+QlQ+IN/qS688f6/v2evYr1KptrVDAQspEdgFBvb36nXulLjtrtTB0vpH1/P1P09+oevEjphyuEakma31e5vfn6f4+/UPWsVxw6DXPhIo3hkOl2Ggkn8/1t/vXvXW+ghXAS0uWNcR5DzYm5upNxYfn37rXRBfnZsebN0dDlS0o+zVZNEaE/i/Fvx72OHXqV49UjSdK5nfu6GeoNUuOhmOpSkgUxq17sPpb3bz69x6NPs/o/a2EhgGOw1LXVkKp5DLTXcOnJN7XPvXXvLo1W1ziWpkw1PjUSuRTG8aQMApHp9BH+8e/eXW+J6FXbHTu5KurjqVgnSmZw1wkgGlvUPfq0690P0nW0WEw75CuVAYI2LBxzq03tz79U8OvCh49Vt975KSCTIyx/8AAUL6FVrKRc8ge/fLrXWoP8/d5ZVu4EpqGrlhjFRMzCFvUbP/AEH49+OevU6Y+uu2abFU+NfLzTl4IowSyEgsOQCR79XrXHqzD41fJXP5rKz4/bkFVJTwHSCIn0my2/A9+62a9WMQdt7lWgiaupJRN6Sf2jqH45v/AI+/U69XoVttd21P2iJkklii0j1LHb0g+/U8+t18ujKdfdm06yQ5GgzFSzEhimogAgcrb/D37rw6tS6B7ao904VcRNN5ciUURtIwLMp/qP8Ae/eiM9e4joxmJ1rkjDPq0l2DX5F73968uvfLoTWoEpVSoiB0gK5Nj9P9j7916h6VuOrIqqDzAAyQgKSRe9vpz/X3bz69x6TuVM9XOdahVDDj6cD6WPutc9e6TmRgSFNcQuTYG3+P1t79XrXQZZjEiofWijX9SD9Rf6nj34GnWznh0HM2JMNZdrgg8rY3+v1W/vYoetHGOmneuATcuAq8HKoZZqaaMAi4JdbG/v3n14Y6ow7s+C+5P721mbxOOLwT1Al9MT2KhiTf+nvfHrZA69tD4a7lnWnknxqR+oXIjcNYf149+610aLGfELFRYlY8hH4pyq3UKblvz79UdbAPl1IxvxNwUMjiZmEQuL2NmF/7XvVevU6UQ+OWzMUVaBI3cMpIC34H1B9+Br17PSrxnXOy8OwlegpZXQWGuIGx/B59+8uvVz1myNFiKFvuYKWmpo4zqBiCx8fj3vrXn0isx2Lh8MjVclRCwiB4MqE3U/6/vf29ep0X7eXydwlQ0ixVoUU5MZVZU0kr9ePes9b+3osO9fkbVZSM0uJqZFv6f23sSP6m3v3Wui07h3nl616iStq29KSTXkkFioF7XJ/2Pv1PXr3Vd3e/c1NXR12Cwc/nzisYKdInDuXIKcBebX9+690G3xZ6Z7x6535R9zYtayKvnrXr2RfIrObakBb/AFvfuvfPrfK/lu/zCX3lsvD7K7cqTRbipKajo4hVSqjLYaNWp/r79wGOtfPq8LFZfHZqkjrsZVQ1dNMNSyRSK/8At9JPv3Xhjpyt7916gHXfv3W+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//0d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3TXm8tSYHEZHM17iOjxlJNW1LngLFAmtjc/wCt72Mnr1adfOq/n/fzBqLvvv3H9NbbyC1G1ocrJiMjRIzBdCDxMHKn8/4+2rrSI6VpXp63SOeURg+fQOfGzZe2urdgYgY2GFBXUY1xppYKHUNa/uOt8utCnPDrJX262MBkdlx69C5l921U2uiogyRaSBYH6Hj8ewZDCt0xdj1N242kUSAxdNWGx1bXSsaiU6XYsSWP+29vi0RMVz0RtbsHDV6Wj7Qh0a103PJN78gfVvd9dAFGOnV23xCWJ65Um0llbSZP8LEkfX+h9ssc1PV9Phjwh5dTW2KNQZZASDe2ri9/+J9145633eeelPj8HJTKAW+g/rYEf4W/p79qHA9ap05FDEpVxz/Tjm/5t79qA62FNM9NkuPgnJIWzGxH0HJ+tv8AX97rXqufXpuyGASppyvj1lFJ5AP0F+P9j78O3j1unr0l6HH12PlEkIYBWA08mwB/3j346Tjrfn0Jy10j4pzMfWsZIJ4ubfQg/wC9+61INOPWwx4EdFVr2k3Bu9aC/CVHI/FtX5v7tqFacevccHoy0WPp8dFR00KDWYFDEW54tew9+rqGOvEmmeolTQENqk/t3uOB/t/e1w2eqM5QdM0lLRCQwtADrNmNr3U/gn2b2kpRgwPTG4bPHu+3tqyaHHQUb5x2V2hkqbde3S6Q0tnlhiJIOkamuB7Gu13YqKtTrEvnTli5sb5ikRoDx6XPTPyQy3Y25aPF7gqGSLF1Foo5ARp0H+h/1vYvVlmX9LNOgU7hUEb9tOrmNn71wdRQR/xuaJMVFRRukkoAjLpHe1mPupBXj1QKz/CK9BxU5/J9mbiei2orx0VHMIUq4OESEPpDah9OPdRqbgOvMrrQdHD2b0vjsBQ0dfKqS5StMYrKg6XaUMo4Yn/H341ByOvUwOgy+SPX/UNVtPJY3duPoaOZqCrkSrqkgUFxDx+oe/ccnrxGOtRzdXyR7E+LPeWRxvUdHV5LbVVk9JjxoklSSN5rsVEfFre99VA9Orwejv5k/aW59rUkEe1cnDlkghBZqeUS+QRj8nnn3vj1uvl0J2T7C+RnyPoZMFlTlMTj6mWxSbzxKYnOngn/AA9669k9Gh6P+F2C2otBlcnjUq8wAWedrM8sh9QN25PPvVc9bC9Hci6yxtA8NTJjvBKfGl2UfQCyix96z1vtPXOSkpdr7woanxrDP9nMESw1MrRkXt73xHXs8etdv5HYmTanzk2bufHwGKTPbkj+4lA/V5KgGzAf72fdsdVODXrbv6xx2HzG2sJLWLG0xwmMJJte5p1ve/8Aj7p9vW8Hpg39gMbEstOiqI3J9Nh9bce/deJAz0WLckdGlLNDJEFFGt1bhbBf7XvfW8dVLfKzekCqU/iCQNFVNGTf8are99a8qdLLpDsqLbfXlROa5VV6Rz5eLcx3Jb375da4DqjH5J9wUvYPeaYrJZmEUNDkS6l2Ko+mb6A3sffuHXj69CVvzMnJ4jF4va1V9xG9LBC0cHIcsgW/o97611Yj8A/j7isFgctvHcePIyyrLUU1TMBqR2GsadXP196639nRzewKfFbsxApsqiV8YtEoIX06Tb6+/ZI69Sh6pc+UWZ7M6O3/AI7ObDrKqj2pHUQGupYC5h0agzD0cfS49+69mtOrRfj58ntv9qddYmjqs9A+SNKq1cQYGVHEdmD3PFjxz79Tz68M9AV2PuCCl3PkMdHOslLJHNoN/wBWsmx/ofr78Otknou2C2vS1mfk+4pvIlRMXVmUWIv+q59760Oufdu+9o9abD3HQTVFOlZV49kp6c6dcrFNIVf9f3oZ49ewOHVRXw/6F7I7b7uzO5o8bW4nAVVZMYZ7yJFJFI5Kn6W5Hv3Dr2TgdbLXxo+PtP1lHUQ5miNQ1TI5asmIYKWa4JJ96+fW8eXR0MXsWYZqGXG5RYqOqlRWRNOkjgf63vZoOvZPQyUW0sRtbd9NJWVUQ8tOhNUxW2th9Peq1FOvcOhHzFZgNoYrIZTI1EMK1AeSmkkItIpQkFT70Ot549a9vy47P2l3R2bt7bE9JBPTbczkNZ92yrpiFPUhvLc/0t7sOqn0PTJ80t7YjdmxNu7d2xkYq+OkpaCnlSAqzRrCFV1bR+OPfuvHh0bj4xdhdU7H6z2jG+Oo8lnqOgQVMShGnWTQLgj6/Xj36nXgadGZyPyn3FnhS4rb2xauSjR1BSGB7FAbX9I+lvz71SvW69CfifkBlMb9rSVHX1bEJI0MsrU72Vgebtb377OvHJ6fcx8wKDHItHNtB/2wFLmIhfp9ef8AH36mevcePXGD5kbJr6GGiyGDhhKyK1pI/wBFm5a/v3Xh8+hQTvDr7emNhko85j8aYIwpIdV8WkfQ+/UPXhSvWfF9k9flZKBezcVU1cwKJSpPG0jOeCoF73H09+8uvefS827tzL1CfxKlqpavGOdYK3MboTe/+x96GOtmh6nZvGYwoXiokWtQi6C2qRhyeB73QHrQr59ITcGZdqKCgEQxkyEDURp1c8fX3rrY49Ew+S/VFJ2HtGviWAVmcSnqZ6epQjyKyRkpYrz9fdh1o9VD/Ej5N1XSHZm5eqe42NBTVVfLR4aoyIOkxNJojMbS/wDEe/EdarTqyfOwbQ7LqslSUeSp58a9CXgmUp4pGlTUFDD8+/Vr1vyHVXfyB6ck2tt3I19JDrpo8zDMUjJZSsUwYmw/wF/fvLqp+XW1V/KJ7W2/vboXEYDH1EJr8DQwwVECW1oUjCtq/wBt798ut/Pq3TUdenSbBblvxf8AA978utVzTrn711vr3v3Xuve/de697917r3v3Xuve/de697917r3v3XusU/8AmZv+WUn/AEIffuvdakUXH8z97n/mNPxxwa78+/daHHrbei/zUX5/bT/oUe/db6ye/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de66uOb8f6/+tf37rVc9eJtySAoBLE+99er+zoLt6drba2hE3nrqd5kNnUMCE/H9fx+fesefXvs6KD2D8hpMrBKmIqiIGZ0Dw/oVSbfUe9Ek9bA8+iW732bi+xp2rMrLHXqzapUup5PHOq/+t79Xrf29BRuv4p9M7h2hmY6PAUVDuGOjf7aqAjSSSfT6Sumx+tvp7916nkBx6qr2ztbuPo3fde9TSZGu22tTMYUPmeFYEc6NJPFre9k+fn1r/B0Y7bPyK2zlcolBnKmnw1Y0hAp5yqSO2r6WP+P049+4Z69X06Mrtjf0mPrPvqPIiWhvriVDxz6lH19+yePW6DiOjjdfdqQZiOKapqBAF0KZGAANv1A349660M9GLTOYrK08clPWxVJVeSgBINub296631gqqplhP21TdgOEFiSb/QD3rrdemKkylVJN455DGt/1N9Bfi3uxGOtBqmvSlpMlFSn9wBwTbyC2kj3rIz1rp3hqKWuk0pIpZrAkW4/1wffj1bpUUkNNSBW8ih7D+g0r/rf7z731XoQsE0E7xLM6mO99TW+o54/r78DXh14gdOeewqSSpPR2dSAW02N/9puPfqdb/l1PxtJTOadQE8gCgj/G/wBSfex1416X+PxkLXvECRe5ABKEf63HuwHTZJ9eoNTR3qzFENK/65v/ALG3vVCeHVq6ePSVz2GMK+V21AXNmtbgHi/vXyHW69Fg32YjVR3nEAL+m/8AaKn6D/jfuvn17y6yUdA1XHGAhZdMZD8AkW+tz71TrfTPv/qjE73wzUFfAjkxhQzgC9ltfke9ggdeoein0Hw6xVJXVK0bRUyya7FSn1PP9PewfTr1R59KrEfEKjwavWCVZizEkAoSQebn36pp148elztX41bYosguUejiEytrc3W7MDc3HvWa9eHDHQ01ePweGVaSmpkTRGo9IT8CwNvfj14Z6LZ3VBWzbcrvsSVhbWbp9ANFube/Ade8+qeO58fV5XCVlJTRSNURRFWaO/rKkm1x7t149al3yA6S352R8sots/wipSgeZlNSNageSWyi5/w9+61kZ6sy2Z/LGgw+3KCrz1K8slXCkieVr8cN/a97p17o+nx++KOzepdWRbDRN5RqJ9J1HT/h70D1vo4lLsLa+ahHi2+jIlgzBQAbfhjb37r3StpuuNh5CgbGyYKnp6ixVbhVJP6bgn36nXuA6T8XTEuHlAxMP7Tk2WO406uQP9e3v32de+Vej2fGXrnM7er4M7UwOKdPHqvf+ov6T70evdWPYjEvkqwVUakan1kACxH+t/X3r59eGehRrMbJBQAyDSix31H+lrc+7fLrWfPprwEsfhn+3ZZQp9QHP55/5H70Ot9N+4s1RxL4wUSbmy3Gpj/gR70evZ6Y6WZqmjMkqazqItYgWI4t/vfv3Xum0UHklOpCC1yAf+N/j375Hr3p0gN0YmogmSWOFiGIu3+pvxYn/be9inWuknkcbUimE8RImFiRfm3+PvXXhXprrZaCpoTTZAq07Jp5ChgwH4B97+XXvn0l4YqbHRFUjCBgdDAKbm31vb3rrfSanpqqsqC5kOgc24H+29762OPUKtCRxGFisbkHTewv/j711759Q6XDaIXq6iAvEVurn9Jv9fe/l17A4dA/vXO4PDQ1NVXPFRpF5CWfStwgv+fexXqpIPDqtzvn5bYbC0lTjcNUQ1MujSvhszFvoLaT7916o6r1yHfG4NwQzB694PK7qIm1A2Y3AsffgKda6RUWad5XqMnUMsbkuwe9m/N+T79177egu3X8metdhTyQ1uToPuYyPRKyar8i3J97PW69E07Y+XGd3hUPj9h4uepWq1UyVFNExQeUaVYFfxb3rr3z6cPjf8d9353dNPvTelLO0dQ61LrOz2X1ahcN9PfqV68T5dXVYikmp8bTY7EY9vtaVEVmRQRZVA4IHv3XqdL7C7hXAJFW4eoXH7hhlRwwJEi+I3AsLe/UzXrwx1ab8Vv5kGQ65NFgN91xqIGXxGeoUsEu4W92496yDjrxoePWwV1D3XszuHAUmW2xmKWumljjknggdC0d11EEAnj6+7DrRr5evQye9db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//0t/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcS6jgnn+n5/17e9gHqpI8+k7uvdWF2Tt/I7l3FWRUOJxcDz1NTKwRURQWA1Mfqbe/YPW8jrU0/mH/wA2fsrddfvzr349VySYiPFZCkqKiGpYhkjZo3GuPjm39fdSdPcetSNpWg49acWwerdwdqd25vfPZM9TXV9RnJayYzO0ypUNJzpL/i/sovpwQSPLow5a2a5vNxVwDQkf4erW8fizQQ0GKoi/2dOFiUX4RLc3/wBt7jTepDJUV6zc5X21rHagSKGnS4WhWLQird7cn/X+pv7I7IsgqnDpf40rPk46WOHomCA6Ta1zqFuDxx7Ul2Jr1rURWvSgnqkp4iPpYG5vzwLXPutCxqeqiR088HqPhM3j56owMR5D/X8C/wBPdSTXPWxk1bpZtVU0V9Fje5+oP04H+8H345PXjxqOsE9UChdGCta/15/5F71Sp602TXpsetEnM0iDi31sADx7uQTjy61XVjrgchRRcGeK9vy4/wBYCx/3v3Sh63RR1LpsnBL6Flit/wAGX6fUfQ+7cct1sAHA6kqlPcapISDzYsv+83968qgda0qMjpK7gzlDSQzR+eEAcWDqL8fS1/ewBTrROKDordDuKGk3vLVqQUeayt9VP4+vu1evA0w3RvMFWRZj7aoRlv4hexH5H05900n8ut07qHy6fZqCV2lci8a3bV/UD3pyStOvSKGUrx6QGQqUWchVb03AAH5Btz73Azg9L9vcQpoY46aauWXJUklBJGGhkDatQuTdbD6/19n1tOyDtPQS5s2y33FSdIJ+wdFd2/hJNlb8raqRJEWsnP2uj0AFn4Hp9jzlfc44i4uDx9esbeaeTJkcSopA+zo2g7wrZo6TYldUvD/m/wBwysh0N/ZDfnj2fPKs76o+HQGnR9tOlxSnr1bJ8NafEzYdoaVomlnRi7TeqVyDYsCf8Pb8cnhEF+HScXcdwaDo4e+N1S4mhqYMWNVVi4fMVT8MguAQPbbOJHqOHVWoG0jqoXveh7h+Q+dbGQz1NPhxUNC6wGRf22bSwJFube9A9UPCnUnYX8u3bVTX4SryGLWqrKcKK2WemSZiy/qLM1/fq+fXqdWH7C+J+zNo1sUlDiKdJYwhKikjI9KW5AHvVfTrdD0dLZHVWMmeGOLH0sZRF4WmjQizfmw9+r69b6GFNlxUlRClNBGpgaxslhqBt9Peq561k5PSf7Kp5abHQS0wXzxMGZQLNZD/AGgPfhwz17z6A7LYybPyLuSeVE/h1LIrIWUE6I/qQOf9h79U+fW6Dqj7fiYvtT5YbWx0MQkqMHn4y7KFbQY59Fzf+vu3VBnPW01szYlbRbdxc8ZaOGLE0I5uossIFufdfn1boL9+ZKFK/wAUkqKIiyO7t6eAbfX3sdapXojO+N3RQ1uXppqmKODSyqfIACC30X3umOt5rTrXz+YPYMFZvqLa9JO0k8mQaQiOVraRL9ePfutVr0pd0b4g2j0v9pSVBOUnxqqkKTfvM7x2AVByTf37r3yHRSth/BDf3dtPQb3ENfHV5OoZoWaJg8ZlbUrMx5t7917j0YSX4Rd79YVVHEs8lUyiJ49UflUInIBJv/tvfuHWvPo9PSE3b2P+x2flkamjrrws/jKAahpDcW49+r1uvn6dGX3FjpNsxyYqZmergh8zyclWJFzpP+v799nW60z0SvuXBUu/Nr5anrIolVVqfVOgLk+MgMNX+Pv1anrRrSo61+8T2Zv/AKG7dzGOwlVXPgWq3D6J5PBFGZLMQBwB791r59W07H35j9/7exufqchBNVVZp42QTh5Vd1AJYHn6+/de6HbJ5rbHXeGGf3BW0d46WR0XyoGt49QFv6+/cet46pi3bNvP5kd90GA2GuRi2/isoErHXyGnmSOa5B/BHv3Wutmz4sdQ7b6uxWL2hXYiGLNrj6R2qTSoqs4jFzr/AK3/AMfeureXp0fGk2ia5JMMaRklrmPgkEVl5NhZv8ffq+fW6U6VWA6c3Li6yCKd9NLTyB1eQFRa4/4j36tetdAP8xtzHrjb9JkUE8k/3UMIlgLBdQIsNafj3v59e1U6z4Xb+6u5NhbVqMs8woqzHRvEELBtBQcuT/gfr799vXuiR/IL4o7S2lS1uUw9NO+frI3SWQKGdWkNrhvr78PUdeI9etfTvbce6+l+wKXb8stXLTZPJQprqXZ4Y0lksw9XAtf3vqtOtk7+XF0J1BuHYVHvPetbDXVtbSmrWFKhJdLNHqAEQJ/3r3U8aU638+rNM5tTZe3aCmqdhY+BZnZYbS06E2HAIJH596zWp62Pl0hM7js8DFLVUkH2jqGkKQKQL8uL2v79Xrf2dOkOK6Sr6akos7iJ58lONLulOCur6EXt9b+99azXpu3F8Y+oq+EV9PC8C1C6VUgRkCQXHH/FPewetZ8ug4rfiHs7F4isGJnrB91d9KzMrC4vdLH3quM9eHHoqW6fhhWUbT53Z+RylPmqYNNB5qiQRmW9wBz79XrdPXrvrjub5S9HVbSdlpNk9gUzeIQwK8s5jh4uCtz9Pfqde889HR6y+ZPU3ZmThj+zqcLWIx8j5UGmiuTpDXmsPfqderUdDrubDY7ecVRkMPW0ddFSw+cHHSJNq418NHe9vfqenXgQOOeixZ6l3DLTVdRiwEMHlgaKcfuFF9LWVhz79XrxGa9UYfOT4wV3bW4sdlMGJMdumimiljno/wDJy8ynUSxSx+vvfVSK9Bxt3BfIjo7b+Jk3DPXVtBHPH5nHlL/brxdm/pb6+9+fW/Locd/7/qd+9ZTS0kYlKw/5VGF1y+YJ6iV55/r7159e8urC/wCQ1vR8PurdGHrp54vuqioiSnnkcDliLLG39ffvPrWaEjrbMBuAbfUA/i/PP49+63137917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XusU3+Zl/5ZSf9Cn37r3WpDFz/ADP5f/D04/8AO762Pv3XqGvW29D/AJmL/llH/wBCj37r3WX37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XB0WQaWFxcH/bf63vY60RUU6CDtvsOg2XgakGZFqpI2UAvpKgqT78T+3rwFMeXVWe6tytu2pq6irrJGSouEAmJ4ueAPdK06vToJ5MxBj1bExuzlnZQCdTAn8+99ewMdZ4IMxQJrpY6hkns3Icgg88e9Y61kcM9S6yPKutLXolQDTESVIsVUqvPI+h9+x1s9JzdOUod1QtRvRUYXwmOUtToZLWs9v8AH3YA9aqPLqtn5BfFnD7sabcuxnnpM7jyZB4nMQ8qDUfSp5Fx79ivWqHooGx+7d97A3PDsnecVbopZ/FJUtr8PjDaQxY/7x798+vA9Wk9c9i0m4Xo6LFZOnNPJDC7qk4MpdgL6hf37r3zHRraDeU22khjpKskOB5dbtb+hsP8feqE5PW8cOpNJ3DXxZCQtIxQ/S7Np/1vfuvCvn1jznc1WyAI9mv/ALrLDgH8ke/U9evYHDqZiu/KZqJqaqkXyKoA1O2of4n36nXqdPu2e88dTV956oFQwJDSm9r/AE/1ve6Hh16vn0O2P7w2lkJIlkqE1vbVaW1jfm3vVMdeHRjdlbhxe5UijoKuEhrKoV+b24Fz+ffh8+vdC3M1RhqfxSB5Aw4ve3qFhz+Pe+HWuOT13tijqJazzSK1nIYEfS1/rb8f7D34V8uvGgGehtgg0RafozrYni/I5+nu5PTY419ekblUqaCTz2LKSbHTwpBuL2/PvWRnrY7sdNlTWw5GglEmkvoNwPre3HPHutfLq/Dqtvv3D9gVGcxf8AjqftBWAy6FP6fJ/at/h711unRrusMBX1WJoYsrGRUClhEpIuwfSL/X35RU460TTJ6GWbZIqIiIo7gfUgC4H4NuPdivWgege3DtLKUFUz06sUBLHSp5F/rce60pgdb48ekjldyDB0jLWOikAg62AJa3Aa/vXW6dB1B2B4HmlllUxzG8OhuOeePfqU61g56TUu50rKuaomqIxEVIGpze9rgEH3vrYHTdUVFPm8DkaaeWKSE+RdNwxAKkAD3o+vXuq3eztrDAZhpljC0dZLpKugswLW4H+t7t16n59Es3N0tsgdgR7xTH0pyKFZTKsEZa6nUbsLn/AGHv1adap0aPbqxb+THYr7eJI6KNIVWNBdregggf63vw63xPRncH0XjshRRUskCoIgAbxqOCP8f9796r177T0Pe0OlNkYzFNT1cUXntYnSPr9Rbj36p8uvY8+uoegdr5DKpPD41jie5Nltb6k+/AkZPWuPShqNk7Iw1atF4Y5plKqmkK3qA5496zw69xNeHQ5bXwy0dAkFLSiOkcIeIQoAPN9X9PfvPr3Q7bTx0NOUkRSVVbuAvBv+QPex69b6FHI4+DKY14kXxs8Wm4U8Eg/wBfdzkdUHHpCbd2TPhI6t5Tqind2ta45/qfx7rTq1fLpE7h2DPWZSKqjDmISaj/AKm17ge9EenWx05zYiOhjMfAEcerkWFwvI/2Hv1OvdI+BjU1UjXGmI6R+D/Q3968uvH0645zxvTldCMyLcGwuCOLH3456959BG9WYqiVp1BiUPZSAT9OPeqVFB16uc9AVnZ/usrLJEzxpHJYXawP+Fv6e99bHCnXHIZSBaOJZZUslifV6iBx/tvfuvD1PTXR5M1FR/k9hBp5J4Fz9CvveevYHWQYufMzSSXukNwLfT/W96+fXienNa/7ilnwqqD9nFrYqLkhTzf/AFvfvLrVBWp6pS+e/Z9fiqubA4yokhLGZSEcpzptfg8+7DrR9R1SjlMvWyVsMtZUzzPIwLtJKzi5a/59+6r9nQf9idi4rY9FJla130RRl1WN7Esp/AH+9e/db6Izu/5U9hb+rHxmxaHIR08bGHzlZVTS40gggc+/dbofPoHaT4t9udobggze6q3IfbtN5HQSyKuknVYj8+/de48erWeh+gMNtakx9C9ItTVp9upaaNZG1qLG5N/futV6uJ6b6Zqszi46NqOOFJYx4/HCqnTq45A9+49b+fVjuwOgdr4PAyU2RpkaqkiKrriUnWy/639ffj1sZ6KV3J8davbslVufE09RIoeVwkaEAKCSvp9+690SrL0dTXM8GVSrpXjNlZWaJgR+NQt731Xj0cL4f/L/AHH8aN7YiCrys8m2sjkIadxVVbsixfpa2on3rrfl1uC9Sdpba7i2XiN37aro6qnrKSCaXxMGEcskQLKbcWvf3sGh60QT0KYAH09+Jr14CnXhf8+9deFfPrv37rfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf//T3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690mN27jxWzsJktz5edKejxtJLNM8kiRpoiQyHl7D8f197rinWqZr1qX/wAyr+bxjex8pmOhercvKuHknXHbkyGPmjP8PDExSM7If9c8n3qnXuiPdXdIdbUvVe4c7RZ6PcGXqsXWVU1XUVCNUO88ZkYOxJPBPushCrXrxwV09VVbGwsNJu7cqRqqePN1QXTz6fKbWI9gjebvQCAesi/bLZIZ9ErKK0B6NlgKBfDPI4u6303/AK/k8+wBLceNIVPWQt1LFaW/gjj0rsbj3mRXK6mB+h5PHF+PfjH4f2dEKUrr6WsMKQxBFBvp/pbkj3ShrXq59fPpO5GklYObn18Dj3fPVhnoN5oMnja41EEV1u3I44Bvyffvn14s1fl08nPZl4xogc2HHBN7/T37PXqgdNdVurcdPGyiiYhl+uk/nj8e/KOvUqajpj/im5siPGIZEJY3IB+h497oevUHHp4pdsZuq0GeeVAwGosWuP6WPutDwPW8dKeg2xW0d9FXK5b63ZrAj6n3418uqUqa9Z3wuSLH/KprG4tqa4B4597BAyerhqChz0lcp1/VZHWTWzXP9nU3BH9feiBx69WvSd/0StEqyqS8qevyfU8c/X3rB49bBU4bpYbcqqvBTRwOzeNSqXJItz+b+7A0GeqhfOvDocaPMCSlBK3Ei+o/qJ4+vH+v78NPn1YGvE46aFooKuoYKin6G4H+P0v/AK/vwWjVbA6q7AGiHrI+DRD5EUXGrgC9/wAWt9Pp9fagzeGO3h09BGsxo4x0Hm4dp09XURVrRIZqdxIvHLEcj/evb0e5NEwKGlek2+7PaTQadIqei2dqUW42qocji6MxT0kgd5o1OvxR8/Uf4exztG8KCATWvWOHO/J8j1aJaceHR2vhx8tI8fmMThZ6hoaiiQU9ShcJqkDaTqDf19iZ5vGWqdQy+0zWDlX6umxmbirqbLbjqNNRBl6S0KOysLvHcA/X2/EKJnq2T1x6c2Q+ZyFS6Y6NPJUs4ZQAbMxIPu9aDq3HHl0dDZmwaXGSzwSQjzSkhSf7Jvfk+/fb1qvkOht2x15SrVvNVaClgbMRe34H+t/h71T069Unj0s4sHSYzID7QqFUG4FgCf6D37rXUaprqen+6dmBkT6Dj6g/kj36tTjrfRXt17ykqM1X09YzJTrHIYxcaSQOAPfutjOT0Bdtx5tK5McsooXeVCVuFs4K8hfdsdaz0HnSHwNGN7Um7Tywu09aaxS5+nq1jk/7f37iOvfZ1bBv7tjG7O2lT46CVBLDTJTmzoGKxJp4HvVfLr3VZXd3fOOpcfWPHWBKiX1i0qAltPPN/fgKde49U+fIH5UY2gx0MWOy0jZUm0kaOpkLauRZb/7D3unWj8uqtcruKfefYJ3RlDK8pJEKPE5ux/TYi/19+610eH46fGLsXvrduM3BnKKaLZWGkBqFlSVYHhjcNd1PH09+63SvV7WzcZtPq+jXb2JoqQwxRRpThNAAlCabqP8AX9+49b4Z6yZXMUdQXhzFLE0p1SRPJpayE8AX/Hv3DrXzPRZd8dm4nEbjxdDjKOnjqjIFjeNYw2q9uD9ffuvZ8+kp2D2RTPA0lcyx1cUfklZmXUUHJ5/4j37rfyHVanf3ftIlNUxYaYJGheGTxSJ9SpUgge/fPrVeq7J9s4DP4vcOcnMdRXZKGSR5JSC8JsW9N/pz+ffutdFH2h21uzp3dFYK/IVbbbEsjY8FmaISI3oRQP8AePfuvUHQ0UvYHcnyTzVHh6SPIR4OaWFEJSXS8DyBSbD8W9+HXuB62V/hV8OthdM7Qw27MvTUsGarEWaaXx6JpJmjDFn1Dnk+/fLq2PPq2rbewto7vo4chjpYhlaYIV0susqhuASOfes9e6V1RvGhwWWx+GrqSGnqacCKOUadb6OA1/8AH/X9669xx06737YxkGGkiao+2aCIFZlkVWYgfQm/vwqDjr3VO/zC+Xe0KXZs+3csUqp4a55o5pYvMxMZuFHBPu1OvV9Oho+CXzBxvauzVxNRAlGmEp/tqC4SMSxxjSDbg+9Uxjr1R59Gh3Li9p7lyE7ZytS2QAijjdr6PIeCt/fqnh17FetcH+b31h1x12tI2Kro6jdeQydIlBEPXKPuXATR/S1/e+tV8urQ/wCXf8c937H6Y2DvEZzIV0WcxUdTUUUksjxRs8QYpZibAX+nv3Xh1aPLnI6WNYqxRE8SKEUDSC6rbj/G/uvDq3TbkN/V8mJng/h6PFqKJKwBbT9PfuP2da4ZHUbC5CdhR18mFppIY21PKyKSFBvcH37HDrYNR0q8vuTbeVnhR8o9IQUVadDZBJewUe99a4Y6l5sZuigpqqEmWn0L9umu4lQ/k+9cevfLrp5qbIY9Jsifs5lF9KekObXtx739nW89KfFbT25uPAOldiaLKLyDBVIHRz+Tpb+vv1etcTTotXbnxA2D2VhMkKWph2FlYgftDhl+2ZyouvqhtwT78CPPr3RM9p7l73+G2Vgoayirt2bEqqjxy5is8s4FJq0FiXv9B73jr3yPR2oN8bZ3/hYt9bcnjkpmgVq+njYWSqkX1xsg/IPvXnTrfAVHROO2dtzVmdpM/Qw38brK0YSwNjf3vh1XjnrzpU752/k8Pndu0iUK0EkVPUyRqSHCaARf37rw6rf24+G2Z2xUdY5N1aiy1RNKkb2MSgk+kL795deyMdWEfy4drV+O+RtU23oCmJhysgmaAELpMn1IHHvxGOvVI624I7hIwR/utbn/ABCj3vrWa06ye9db697917r3v3Xuve/de697917r3v3Xuve/de697917rFP/AJmb/llJ/wBCH37r3WpDET/w5+/HA3pyT/1HW9+8uvDrbfi/zUf/ACzT/oUe/de6ye/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6a8xlqXC0FRX1jqkUEbPywUsVFyLn37rXnTqoT5HdtvufNVVDj6gyxJPIjKpGkgCwAt79Suet8Bnoq1Fla15o4ZWdVBseLf8ne9069q8ulLtvAGu3RBUSlpIDKmrULixPJF/dTwp1ulc9WG4PYmEq8NStHTQyusIDEgMwIH1/wCNe/U8+vdNUvX+Ok+6ppIEEcqlTZbab/7D3rj17h0Uvf3T+SwWVqMhiYWnoykjMFVrAW5+n9Pe6+XXuiwZKnSGpqYIJWarLMHpgCAXPDA+9+fWuic909a4vL0GWmmxVNS5mSmbwTpEBM8p+nr9768R6dUkZT5Dd1/FfsSVq6gyNZtsVrkTiKVxHTxvcWJB+g9+69U+XR6dg/zTuvN6xUJyOdemyEKq09O4CMrgWZWU/wCPvXy699vRjI/nHtjJwq+KqRPqA9YUG/HFrD/b+/dex05Uvy6xc1O7VLnWb21R/wCFuAffuOevfLoO6n5VQJXT+RnjSUkxN42AYavwR79nrw9OlBhu+6rKTK8ckywsyEyBSAAf6H6+/dbr0Pe2OxJKxonjykxlJU6A34v798+veXVgHQPZOUos1i1+8neKSUB1LEg8gcW96IoOtdXWUlbQ5nadJkGIkqTDCCrfXlLgkfW/vZPbnrQ+Lp12s9VPExWnUGNQNQW3pBsAPexw601Ol4ZmpKdZJxwosQPrzzz7329VowNR0w1lXHllaBBpFmA/1X+x96+zq1fXpHx4KohqiC7KjH9B/SwbkX9+p1bV091u1aPJQxqcfTzOLfuOBYMv5ufezSmeqVNajqRidr/Z1Cu6LFElvSremw/At71QeXVqnJPSxVYokcQkMwFz6gT9bWJ97rU06rQAY6T2QWinjlWaOMswJ/pYn6j/AHj8+/EilKdeVTxr0THuXqzK7kMs2HMgAMkiohNrAXtZfdCKdX456JpmKDcGDIoKqGQNREAsyPz715dX45HRaN7914/beYNDm62WgjJCK/qRPra4J/x97GOPWiT094LtSnelWuxGQesoJFDO3lBFv9qBPv3Hh17pOb8zyb8pYUpkjZ4ivIKnSV5sbe/UHXgeiZblyEmI3ScbWHQHbRZpFve1v9t79mtOtdD31d5cBW02TijDUrgSObg/U/k+9k9ep0dWh7Go5KYuknjZ0t6WC3sAD9Le60PW8dJqu7NbGygvXyIryhQBJcWJ44B/x97APWqjoT6HdWTrcfBNg5qioknQEhAxsW4Pv2etgL0YHpbqLK7syi5bcXkS7K37gYC31Js3vwFetHGB0d2fY+Oo6JMVBGioFA8oIGmw51fke908+q18us+MwFPjNCqyuAhF7klhfm/+PvfDHWq149O+QrKCjpGdnSLRGW/V9D+Pr72TjHXgM9B9Hvyg+wrIo5xJIH4BYH/bD3TUKHqxFTUdZKbdVNUwxrLZSR+r8X/Pu1etU6ac8qVFM0sDXBBNwbmx4sPeurfbnoKJ0WhlZQxBZtRANrX/AK+6/wCXr3UZab7qX9yU6ZCPoeBzwfe+tcekruXb8CSEwyFiUOoDkatP1PvVadb4mnr0Am6MEtPT1NQ10I5sBxe172+vvdRx68MdvRQ9wbqlmyRxlLPIZKdzqUA/gn6297HXuOehj2pWx1OF8J4qjGCATY/4EH6+9cOtivQi7QqUaCuoXa08gcR3sGYkf737115hTh0jVqMrt3PVxqKcmmq0eLyMvOlv8T7314ceqyPmV8ccrviSo3PiKYzm8srKqsSupb2On34daIPVL+/Oqq/bMEr1dOyVNKPp43Nj/sR7t1WnRba/qOg7CiP8bkKwo7EwuraSAfyD71177On7bvVWwtuoMdjMVRPVDSusU/JIbgkgX+vvx69nozmzOtchl4UoKDEqkso8aFYnuTbgiw/2Hv3XvPo/nQ/w7yEdZBlN0QilgIjkQyBl4bkfq9+8utinVjuzdgYLZVRADLGtHGLF9agaQeOT78B1up/LpSbg7K2rTZGKlx+RjlZSFKiSMjg/nnj34D168f5dCE1Xt3d232gqBTzu8PKF42J1C1wPevt63WvVfXf/AEtijSVlZjoI6UA3LQqQV9J/p738+tdVm9ibf+1wcIWQrNjmlkWW/rUx35B/H9fe/LqvWwR/JY+Tc2XwkfUtXVyZBxHE0ckzqdGm6+nm/Hvw9D1759bGExdNLICx1WIF7G4tyPexkdVbBqOsw5F7WJAuPej6dWGRXrv3rrfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf//U3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691rbfz6/nPlOn+tano7Y1Y2O3puamcQ1CF1kK1MOkKChHBv71rAPVxExXVXHWpz138Wdx5TaVZ2Pm6qoostn6Ra/NVs8rlaqcqXaRi5/PvWoUr6dNCpbSOgYr9wd3bMrajEbb3jIu36nyUpjSaRkMd9GkWNvp7R315HbQeI3n0dbTst5utysUCmoPQ19OYOvx6vV5iQ1dZVOKieVjq1yu2pnN/cW71efVMfBPWVnI21zbRZqk4o1OjZ0lG2hXiuqOBwPp/sR/T2GLdHWUaj0J7szTTfLpYY5DSqLqNP4b8fT8W9mk4BXtPT6UC6T0raanV01lfrzc/Tn8g+0+SOPTwBA6i1tELfpNv7JHPH++/PvZNKdaKkkEdJ2fCPUknQ2gngfi9vz71Xr1euJxAiQRxoDdefw39OB/vXu44ceqj5jrIuKphEEmi1sVa3Fre9eXXq1OOpuOwtLF/uoX1A24sOL2v78a0r1sEnj0/GKCwTgfg+kcf7Ae61YdbINeHTVkKmlx4Qu63f6f7A/Qn/AF/d6161ShwOuEU6TqCLA8WYDix/HuvD59a49dSjxi6Dkgm4sTx9QR7rUnrQqD02tO6ElnJ1C1uLfT8/193AHW+PSO3XRhKF6mEeKQHWxH1sBfi3+HvxHW64z03bG3QuQjlolkM0lMGD2H6bfhj78KVFeqmtK9C/jHRUL6QCw+ptzcfj26+lsHNOtxrUZ6UFNG0lNKG4LElWP0F/yBb2mkBIJHTxYoO3pP5Cj9CjUGJvYDixv+T7SsrsO3FOk5eaYgNkDpK5DDQZCkmxzRf5RJEwM5AIKt9Rc+zCwuJIZQWwOrXm32txbESAEkdE5z/TGd2dvml3Ftmpkhh8yzVKxvZWOrW1wvuSNq322jQeMQOsf+c+T55i30anj5dWb9G/LqihWg2Xu2vhpnhaOCM1DEayqhTy5sfYqiuIrkeLEag9QhPbGxnNpMe9eIPVp/T2/wB6bcVJX4qtWpw81NHJeLmMBh9Tb+vt9kYCvVAoNaHo8W0exMXkqyUyPGkwkYr+b/m1j/vHtOZVTDdVIZULkcOhJqOxIKAkyViKlhcH08W+o9+EoY46ojahUDpIZjuHGINcGRiaQXBVDe1+Px7dVdRx1Y1A6D7MdsQI8NSKpWThpx+D+Tf/AI37syFetKQ2D0CR3Eu+N4SJSSeKkVj5HH6LH8X+nuv+Xq1MgdCbNuvBbLrqXD/dxJBUIDPK2gDX9SD70M4HW8U69uL5XYDaFHJRS5ujipwmmNjoCj8Agg+/U61wz1Vl3l82cdXZuvocfn6WsURVBSKOdi2r8cAnn/D3agr1qvVZm9+3e0+0pK6n25ishMBIUjqIfMUC2KixH19+69UHpNdd/EPtff8AlUqtx4bKOJJCWmkEpVNR1ajqFvfq+Z611aH0/wDAnbOBjpMlunwVRTwM0E2gsHHJFj/vXv3DrYHVgFIlB1rjKfbW0aJaDGVceipeONRGdS8kso9+pXPVuB6K5Bvg0/YeUoa6cVUAYrTclhG5aysv+sffs9a+fTN2L2TTYHIiHJZaFZGjLRK5IPjPNtN/fuvcOiWb+3lBujdmJr8DXowoZlaqkia+mxudQU/X375dar0XD5W/Jnbu29vriqPJU0m4RCY6gI48pA4YkDn34fPr32dVTPvXJb8yLGKpeOjlkV5rlrEn6kXNvfutDpY5/fe3dgbNy1PkcvSpNLSssfkZQVJXi5J/x9+6959BV8R+nJ/l5vyo29V10ZwmOrvOlYJB4nBfVwxFiPfhnj1s9bQPxl+GexuvoKGip2pqyvpQsK+JYpHPjcWube/Hh16n5nq3QfHvI9gbMbDYupfGVFLTBaYj9shwnpK2/PvQPWz/AD6SuxurOyem6Csp6+WsyM0Qlk+5s7AQoDa59+699vQNf6VMdvDclbteWrRt2SVEkVGl7To/9ACb/X+nvfXugF7bpey8TWSU2aarpKGhXy1FRMZFimiBuOT9ePeuvf4OoPVPSXVPyQqsjjMth6fJzUuOqKiSsurKJ44ixB1cA8e91Pn17HQAfG3a9Js7tzfu1Nt4xqOi27l56RPHfx6Um0/gW/Hv3z61+XVn9Zs3J7iplq6MOZ6NI6g6Sbjxi5JA9+qB14VJ6rr+Znwfy3yA2u++yHqMxt+rjmp6dGBmP2ZD3CNf/U/S3v2OvHIr0YX4Z/Jej2jtPB9QbleGnye36cUC089hIHQePTY/63v3XgOj+ZWs25mAss9ZTUfmVWVpGChdfq1cn3UV6tw4dI/M7beGkY4fOwZkyAutLTSBmWwuoCj8+9+fVaEinQfbX3Zu6HJVWIyWDraWjiYprljcRul7XBI49+oD1v5dCvNQ4Kvp4RFRBay6uZQf0MeS7X/p71XrfnUdN26Mhn8Nj0amy4zIjRRHSwN5DAPoEIHPvfXvmOoNNv18Xiaas3HjZUVhcLMpUAccWPv3WuhO25vTG7lxittnKQU1XqC/w+Nh5WvwPSD71TrfnnoSaOkqVxU8Wahk++mUmCaQEeo/Qi/1976qacB0Em/cbDlNvVOA3jTirxlXBJT0jyooSMvxqDEfj/X96z1biK9EU2RtTJ9b79famLnao2XXzGqkVGZ4I9TarX/HHu3n1roeO3qfDHbNR/CBGlTHCeUtquF/w59+49e+zoribqqqPZrUzuVqQHXVexb+luL+9fb17qsfsfDzp2TDvyqWSOalnIR24urHk8249748OtevV0P8s7sjY+E3nj4aqSkOY3DUkRAlNTOxAH1N/qffuvfLrZxQ6kRh9GVT/txf37r3XL37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYp/wDMzf8ALKT/AKEPv3XutR4SLH/NAK3/AFb05/wvXfX37rw49bcMP+Zi/wCWUf8A0KPfuvdZffuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6911cH8j/b+/de67vf/evfuvV6Jv8AKzsykwG2p8NR1aLkJI5Na3Ibni3H0/1/fq17evU8+qnMfTS5CWqylRJ5C0pbk3/V/T6/7z79wNOvCp6m1FHD4ZWRR5GF0H5LH62A9+8+t06X/Ws3iqYoaynOsSLZm5a30DX96PHPW8lerDtj1kdNiEIi8l1AFx+OLge/A9ap0pMjBTTU4qEUwvL+TYXJ9+p17pD1lDHVwy0Uy60dHQvYNcN+Ofejx619vRReyekIaSufJYymGudjIxW3Btqtx731vHRJOzOs8plpVjSJ45IG1uxBGqx/PvfWvKnRct5fGPYPYuNlxW7MZT1NVKjxGSZI+C40sdTD37PXuHROpP5QnVmPrazOYyehpXqn8kcCzovBOocD6f7b3rrfDoduvPhDsjaMEcdbDBNoKnl1fhRa3PvfWgKcehMyPx065p3jkTDxMikagNFj+Rx791v7euWW+P8A1rlqam+329DTyU4XW1lGsr9Sffs9eA+XThi+ittRw+GiwoDWFmUagLcD6W9+61Xy6ccb0vV0lapoMe6t5F0hPrbVwP8AYe/D59bPVjHxp6XrvvqeszNFLEkUgZTILf8AJIPvWDw611apiKCKlaHGwSr4kijGm9lBVeeB73w690MGMpY6JIwhBV4tTEW5I/P0vb/X9249N1oa9NeYr0lkFMHAUm2n6O1/+Ke9A+XV6efXHHvR0cp89o+C+pjex/P+w/1vfuHWjXqHWbnoAJiHR/EzDVYG4+ouR/rfT3s9eA6SMvdmz8a7U2QyFJTSR3uGkCEWP1IJ91JHn1sCmOlXt7sTbm5Y2qaDJ08tKFsHjYEFtVuWUn/be9gjy60Q3SmhlpKl3NJUlwVJYLyo/ryP96974nPWjgHqFW4ySdHKH6KRqb6kDm5v+P8AiffiKdeU1wOg0qczJjcgMcYy5lJUPYN9fSR71wNOrYpXrjnuv9p5ekWebHxPW1f6+E/UP8Dzx79Qdeqa/Loi/ff8uPC9w4yepoa5MTXukksCghW1m7KL2/23vVKderXI6qn3P8Fe/erHrcXiJcpmMckzRxfbtIwaNDxYqP8Aeffut+XSMh62712pEhfbWZZrgunjlLH8XsR79Xy69T9nVWXyeqPk3gexYM1BsDPNhYZvJPU+GcII0PqLG30t79imetGvDo2nQPyNyO6MPQbbkw065bwpDPCyN5klPpIIP5v791sdWCUXX3Zmew1O2GweSEsgW/jWTi4uDwP9v79jrdSOPQh9WfFDtncu5qZ9z0WQjx4kia8/kVFGrn9X9PfuvdXH9X/GzCbRx1PHklEsqxj0eliDwQDce/U8z1onowSLicFAtPjoYonjAUBE1NwNJBI/3n3sU6qa/wCr/N0m8nkpFDVNRIaSMX1TMLIFUc3/ANt79wz175DoFd4997M2cp+9zVDJoF25U6SfqW5496r1alOgGz/yR2puimkGFztJNI6MggjZSVJH1Kqf9591yet09Omnrh8hnq2TTVGdZ5CwUMeAG/SQP979+8utfb0NuayEeMiWms0E0Ngxa4DH37rYGesmG3L5I2ilcMNBIuSf8T/r+/cetcPs6YqjL416wrPMhvIbazYj+tr+/cet58+n9qzF/bBo7AgAqw/tEfQj3ry68ekvUTw1ZdAwZzzfn8D/AB9769w6CjdtFHPJ9rKf25P1Dg82974ih69XNT0WDIdY0NJnK3I+MHzBtBFiST9Pp/r+/Vx1759ewmOfDVwmlU+AAgav0ix/B96OePW6U4dPE2bgx1Z95Ao9LB7C97k8ge/Z4dePr0+RZaDdIb7giIIv1cWH+HPvdOvD5dM2TWjagqMTNoqBKHUCytcEW4+vvX+HrdK56Jr2V8U8DvlamqWKKJ3ALL6B+bnj3atePVSPPojm6vhS+OqStAgKvIwZI9I9JJHAA/23v3D7OvUr9vSg2l8J8bj56fJZGjS6qrPr0m5vybEe956r0bHbnUux9mQR1z00UJpwH8hSMKhA+tyPeurefXt6fIDa+1cRL9vXUtWIY3jSCPQxVo1IW4HPvx699vVbnZny73Dk4MpTYuokgXWRCV1qFUi31B9+xXPWq46Kfie498Ctmr6rOFpZNZRdcmrVf0mx+nvfy61w6PF8WO9t0ZPeFHjc7ky1K0kCaJGYgqzckE+9HrfVv2/sFh9w7P1U4jZqinu5XSSSU/PvQ+fVjw6pc7121T42mzdMgF2pqtQeP1G4v72Oq/b0K/8AJLy1Tg/k9HiqjWIJdJVhqtZ5CFF/pz79w60BXrdw9+631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//V3+Pfuvde9+691737r3Xvfuvde9+691737r3XvfuvdRa6shx9HU11QdMFLC88pH4SNdTe/de60A/50vc23u1Pnnt9Gq74jCtJTuhlvATTARgMrccWPvUMJkYs3Vb278KLw4ugo352sdy9d0WztgtTNCtEtNVGmYFwAmhjZfaTcJ47dSfTpZyxZT390FcEgn06AvdnXlDtvrHCVcoD5mWbyTluZGMjXPJ59xfu29fUMYAeHWX3KHJv7stl3HQO4eY+XSh65xZqMajFSJNKWIF/8fp7IUoFr0MnYE1pQ/LowU0FLhcZFPXHQBH9Ppxp/r7tqFKjPVCT5+fSWg7X2XC4oqryFkcWC3PINiOPdK0NetgErXpb0/a2yXomIEoKrcXB1cfj3c6eHXjWn2dJ+p7l2UEdHE10F0tq4P8ARveqheHXskZ6S/8ApfoqlmFFDN4bkD0k2vyDf340JovVeODw65xb5rKtr00bBrm3kHFx9Lf7f3oqB1sGuD1Eq8zu6WQSQr6CfoAbFfzb3sU61gdSo90bjo7edfR+SRYgfkXHvwpx62KnJ69mO1cTgMe9blS40pdiGtYkfUe/ECtSOvVJOOkHtTs7H9s5KWj2+ZXGPbXOSWYBVNyB/r+98e4cOtZrnowVAQYFRQQ0ahX/AB6gbG3+291PGo49aOD8+nJVjKnUbkXFgf6f6r3YHGOvUJ49cUoKcJLV1MkccMKGW7ME1AC9ufevLqyjFeid909r1M0FXgtoI8uTUyQxlLsvIsDcce/U61Wo6Af4+5vtDbuYzFbu+OT7OV5XZSjAaS1xpJ/qPehTqx49WH7X3TRbhgjeiJjUaQRIdIDAer9X4v7uBTqvcBU9CYtfClNoeWIm39mRT9Pxb3XPlx63wFemlZ4dbGVjJqB0WBIBPH096AJNOnoH0n5HqBLUhJzH9GtfU3H1+ht9ffqevTdzpPDPUtaagnx9RJOYpJRcLqVWYC1vz7bdSwoxI/PosCAyVcDovfY/Q53T/D83tOpmpMxTSCdxExjBYNq50n3IWy36w2yRA8Pn1jPzpyHu829zbnCpCPWlB0aTozt/fHWFLT4PdFQTBDHFD53kOq0fBGo/n2PbO5SYUPUV3lruO1yUmU0B9OjZSfLvDYGvoZVq3Ms5DAxz2W559VvdZLVJJRQ9eiv/ABR4bKan5der/mtHnquvx4q2SQQFYj9wVs1vTb2qFmi8PLpUqpCe7h0WHcne3cDZCepwWTjaiZ3kVHreQg5/Tf3VVEZIPT4ktpRQ46R2J+ZG96eny9PuGokvRBhJ/lBIa1+Rf220pkJHCnVJ0hjp4ea9PWC/mGY7A42A401U2Sml8TrCzvIW1WN9H09t9J606lbo+Q/bHaSxy7Vpsss9QFaNyJhp8npHNv6+909evZ6U+z/jp8luz5KL+99VVRYuocKLNIH0WuCbm/09+r1r5no1m1v5cewMFXRZXdlTPNWuEabysWW5/X6mP596r1amanozOK6x6J6iposbQQYySrnUGNpo4tV25+rf4+91690+/wClLC4CnmpsdRYoCddERhjjB4FuCPp7117h01bf3K2WrH/iUvhhu0oUNpTSefe+vdBv3B3lhNr4LJxRVMGqlDICZ1Lgabelj7914/PquZ+26LGyT7xylSPBVl3oiswLiQsSAT+effvLrXn0RDv/AOQuT3DmWqEnqfuUDpCsUxsYbem4H5t79175dE0rflHl+uYawUxr5KuuLEmdnADN9dDH36tOtdE5z/Zu3t8bkr9wbhyNW2ZqEkVIJKpngVmJPCk2+v8Ah719vXqHpHp2NnNv01RDiXiXXIwgd5dI0k2X1f7H3vrfRqPir8Ju1PnVuOCleWtfELMYq5KfVPE4Vhc3H4t9ffuvfIdbcXx6/lDUPxy6zpztKgWLdkmOiJkSn0OZvD/bZefr71Xy630gcb0x8uepOwDlqqKafDipMkaiJz+0D5OQfrx73jrWT0OlT8ye4uvq0iux9UnjYAqkbqthZeR78OvHPz6UuP8Amx3N2bLNisRidLVlM9NqmhNz5V06rkf7z71TzPWz6DrN0J8X94rvmp7g3eZVytPVNVU8LKwgJ8mrm/Fj/gPe/LrXRo/k7s3cXaO0EosXFRR1dfTrSzNAieQACxJ08+9de49BB8UekK/o18qmXLyTVdLUvK1zYK0ZLWb34nrenpu6V2Vser7G7CqsT9uuRyOVnlqNejUXaXUSL/4+/Hhjrwxk9WM4faOPwWBlaihSXIT05SS0auPWtgbC9h791759As2cwe2Islh81RiatrXnVIhEHT95SBdf9j791s5z0Q/F/EbF5LuFN/pDU01JWV/3MiJC6RKpk1WPvf2da6NnufYOEzebGAopKqNIYIo2kVmUalWx+h9669jy6wQbPi6rePLw1hqViNiKibUqkfhlY+/Z69XpYrux9yUgrZKehWnb1O0CJrYf66+9fZ1sdI2q3BhopJIKEMlRKGjdpDpAJ4JUn3vNM9ermnSErIq3acNTm4KqGpR2aZ45JRIoB5IUE+944dVNR0CW8u8sfvihlws6eKopFKXjUKo08f2ffhTrZrx6K5Tbv7J693TBuXbMtRLSCoS8XkZoTGrXFx+OP8PeuHHrwBOerR+tPlHh914Khh3oVpcusAKl30KZLW/Nvz79TNevcR0Le8qzH7t2bVvSz00qQUjyQGFwz6wmofp9+69nh0W/AYyOh6qzGfqYia6CvkiSWYWk0ByBpduePfvPr3QObur2pMElVNMWNXBq0O55DD+n59761/k6BHeEmPXaNHVwSotXI5uoYAX1f6n37reK9Ea7cqZKuYU9ePFDIyAOoC8GwB9+p1o9GT+Duwax+8urMnQVcz0tPlopHWOUkFFlBJKg+/de63T4OIYR/wA2o/r9f0D6+/de6y+/de697917r3v3Xuve/de697917r3v3Xuve/de697917rFN/mZf+WUn/Qp9+691qM41VqP5oFSZrp496toJ+hK1/CgH36tR14cadbc8P8AmYv+Waf9Cj37r3WT37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcAiglgPUfz+feySetAAdMm5c5Dt3D1eVm06KdGb1my3tfk+9depTh1Sz2xnqrsbfmTqDOz0gMqRqZLx8NyBY296rTPVqdIbG4OSlqBSAAo9hxYi3+I9+63Sgp0t5NnCJY6oLcLZiCp/pf6e/Z69jpb7Z26ZqmKoSIg3QDTFYXHHHvXDrxJ4dHA2jRzUGLVXU30LcEG9vr738z1XB6VU0lPk6cRgMoguxIB5sLH/kXv3W+HHpPGGNnKopAHOrm/pP049669WvSezEqvemkgD/AIX06vqORz799nWjTh0G+U66xmRjkqJqaMGVbcxgHk24/wCJ9+r6db889ABuLoWmq8g0lOjBSxPoT/H8W97B8+vefSOynQbRvHLHJMRCQdN2Ooj63Hv1evcM9Y6fpUSuDIkgH51Ibf0ub+/V9OvUoa9TaroiGoCrEuo3ANkuTYfU29+rTrxIpXpQ43onDpCiVy6SByAliRb8+/Hr2Ol3hOpdq0DBIolZibHVGGuf8Peq469x6E7F9YbUx7pXvSwEoob/ADak/wCxHvZ9B14V8+hPoMzisbF4aZKeCNQACAIzxxc29+691Nxu7RHkVkEweG4sVfV+eBb6+/A5r14/4Ohrxm8Pu4h4jc6Lfr544ABP4/PvdetAdZqiqhF6md1unq/WL/4AWPvfXugP7G7EqaGnnnp5WWOJJBpWSxOgf1H/ABHupJ68AOI6L91T3pQbv3VJtSpllFZUzNFG0jkxltWkC/vxGMdb6bflL03kcbiMjnqGorC6UhndaRmN1YavTp/w9+889aHDqizI/P3sfoavrcDTYTdlfi4KyWN5YqWrlt4m9RBAPHvdPXrxPRkOkv5t1dumqhpilfiGU/vrm5JKNiynkATW/wBh78B17z6t86g+cuy91JS0eYydJJUTmKNtFYGX1rqtcH+vv1SOPXiPTo6lBPtfctEm48ZWUct49aqsiuRf6EC9xf8A1ve69ayDQnrLTTwCojaaQFA19IP1AN/r9Pfh8+vdKqqqqas8bRSHSovYEj1D+yQPfvketUpwHXCor4iqo9FSSOfprjSRdI/ABH5/HveOvZ6a6rC4DJxmWvxGMBSxt9jEAwHPBtz/AE96IHn16p+3/J0gdz9T9T9h4au27mNs4aWSopp4FlbGw6kMq6Qytp5I/p79jgOtivE9V19dfy1tkbA7Prd0ihiFBNXPUQxilAiVPJqFl91yerAgdWk4DDbI2/GKDF42jWWBAt2pI7syR6DYkWHvenqoPn0poq2nQnxUlNGPwUWNPxwOB7tTrQPr1liNTO3kEgVi2pVDHSRa/A/Nve6eZ61Xy6CjsLsHaeyaWrr87XwQT08bSv5ahYwNHJC391Yjqw+XVIXyu/mi7a25/EMDhMjAIENVDqStCsSselQpv+fdePHrfDrW6+Qv8wntHeAzibKw+88iyghJ6GCrqIGAJtpkQEH/AF/e+vdF1/lz/Kf5d9n/ACZqNj5vF7jocHFloItWR+4TXDK3Nkk/A+nv1Sevdb2m0slhultk4fcO5aynTIyY5JpkkmRHMhXUbqxv7159b6D/ABfeOM7RzNfU44zGnp2Z2dCdDBPqVP8AvVvesDr3HPSwostkKupWTGyEQmyEE/j9JPveKda8+nzI7dyNdGtVCZPPYm41ct/hb/H3odbPDoJd55PsbEULRY5JXVE9FgxN/oBb34da6ROy+1dy46Vk3TDULMGAGpWAtex5I+nvdOtih6W2V3iuZH3EAZSCDbn6fi//ABX37rdPPpkXJirBE31W+nj6m35PvXXuHUSopKmshKqBoP5A5sOLX9+69WnTa23qfwSfdNd7egD+zb+vvY68eFOm2joZI3eFDohUnSeeRf6m3vVevdQ2x1SuQVg+tRz9S3vfAdb88dPf2lqad72bSSR/W3I4PvXXuGeg/kxQnLTtErGMliSt/p+bn3vqtPPos/ene22+u8VUQzzQrVIGUKrhSCF/p/vPveetCnVUG7/mZnNyvXYrH1LJSyXRJBOy2sSLH/X/AMPfscevVr8uiyZ3tegxdPUVmfzY8j+R9ElaCoLEmxDH3vrXRcj2ymfzL02M/eppZeHB1hgTe9/6f4e9fPr1KDoTMRja6uraVwGW8oJvcL/WxA9+69x6Or1Jgnod2YyeMuj3pb6QU/tC41D34+nW+r7dh0cFdsON6p5JHSmIsSWa/j/3w96OD1sdVXfIrFRtm62GKJ9DtNGAVOmzG17f7373w615dAb8dewMh8ce58PvJAlPQGroYXmX9pRqnFwz/wCxPvfWut43pDsSl7R6223u+mlSX+JUMUkhRtXqZA17/n6/X3rqxFMdC1791rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//W3+Pfuvde9+691737r3Xvfuvde9+691737r3XvfuvdI3sSdKbY266iRtKQ4OvkZv9SqwEk+9jj17r5a/80ne38Q+SmcrtuVj1FXT57LU5lj9RVvKfTce2b26NnFqA8ulu27a25zCFRXI6nfCGbddYkxzSS1gqGdgZFeygngC/uN913wTkxqaV6yD5N5H+kKTOvDo3XcWMyMppqVlcUoKERC+lfzb/AG/sGG1czGU+fWQX7zgi25bFcMOlN13jhQY9HdFCiNDz9eBcf7z7UfLohoQ2o5r1N3Xm4M6wxazFAlo2/HN7H3WhHTgIIqfPpJ0PXuMhlaZglRI92u4vYnn3ugpw6qQa44dOTbfpaX9k00QD8gaR9PpYe/Up1sN8uosG0cc8zvLSw2fkXVf62Fx/j79jqtKnPSjx+GxFDeI0EOmwCtpAFxz9Pe/n1smhp0+Q7dpqiUNBCkKXvxwLf4W9+61091hxWKp1WaVBIq3Cagb/ANbj6+9A5p1umPXpOVFVR1cDSiNNJuvqH+HB5/3j3vy61U8Ogi31tmkz2Kq4bIA6lQD+LD3qop05RaddfGPY8GzMnm5lgjY1KSBSAP7QP9fekwD1quoCvl0Zyip9EdRJwC0r+k8fVvxb34549V8qnruOnEUU1XVSCKJDcksvFhcn3ulRjrYGKnol/Y3cGb3Fuh9hbTMsgSRoppYbXVdVmZiv+x9686Dy68ACKnoRNt9X47EUMWQyyRz5GeJZmMjBn8v1I5/r7vWg68ag4z1MyZpkAp5sVFTQk2VwoBkH41H8+61zXr3w5HXLF+AL9tSP9pxceE6efwfexw+fWwQe4dSimUoPJUJWS1IDE6GJsQPV+ffgKDPWuJqOPQb7i+RWS2TUrHXYoyxX4Ype4vbgn34aAevAMMdKjBdwZDfscddRY/wxuFW6qLcjkcfT3UnyHWtOoivQh0FbVxkGvmeGFwS9+AP63/2Hu2nVkHpw2qsO3j0pYd1fbG1DOXRAGBUgX/qD71bXrRT6K9ev5raW1+kdAWGOmPOZxNwwfZMRFUlgA5tr/oSCP+K+xlZ76YVGeoy3LkW03WQlkGemur2xBQY8V09YKmqhGqFWa5va4t/xT2rXmdfFC6ukUntHYJatJEBrAwOgA3juPeuJhmyGGwpnm0sQyoQTpNgSPZ3HvySY1ceog5g5HvLJWJBqOiK7l+X/AGfs7fdPj8/Q1lDjpJEgJOsREOwB+vszjuROKA8fPqKprC9t5yDWteHVg/U1Nhuz6OkyOQq0p6bMJ5Kl3dbaG+oJJ9rRHoyTXpdEkqqPE6Oj1N8bOn8NuN8jX1tDW0AUSwpK8ZXzadQALf4+/A46eoOrGuq6/prbci0tRR4uEw6hA37C6lU+k3P5/wAPfq+Q63SnQuV3f2BxBrxjgkFNRpekaOSPSx+gC29+x1uvRb+xPlnmK/Eypi1llq0MgCqVYlUPBP8Ar/197HVeiuPv7eXZIlyc9VUQ19G6rDTkkF9JtwDb37hx69TPRktnUdY2FpK3PzmGSnVZJQ3JuBcC1/z7914VA647s7QpcTRVE2PqVukLwjSRqLBLKPT/AI+/db6rB7B3D2XvXclRiqWiqJsVkZ7ST6HKqjH63t791qp6y7a6S3juZqrA5sVMGNx6NJQlll0SOfUEH+x/p791oDrBn/iJX/wOp3JVUxetp5THFTvHIRJEp4ax9+63nonHdHRm3aikpYs7TQY0BV804UxsnHNieffuvZ6qJ7A6lkpd71VHsd5MrTRM7+RIpXsQ36dQHPv3Xun3bHxy7n7czOO2ngcDMFlmpjLUinqY2VGlCNZgPfqdernreX/kt/EOo+KmzVm3ZR68jkInkLz6zKskiA+kyf6/096Py699vWxvgsnt9VfKZipEVJHT64o2ZSC6j0jn/effhQcetmp4dAzuvsbZ+4ayopjjaWdoZGiiVkU6/qoYG35+tvfuPXugHyXx82n2PUzVTYKk5uWBi4H5/p799nXvt6U+zvihg9s1CV1JhqaJVbUHWLTwv1F7D37j14cMdL3c+2crjKJsfj6RkpZFIcrcBbi5HvXXq/l0X3YWK+53VlsZm8tLDDHrFPG7HSrX406vp735de+XU3L1sOB3JV4KCQ15mo6kI73ewkQgAM3v3l17ieqz8Vg+xNj921+TxlNUnD5PJmSYoXCIjy6mv+Pe6HrRoT1c/wBWbgFThstWVoFRURY1njhkI1CRYfp/r39169jh0G2ztuLvqryu58xj0WWjrpUhhkUAFI3Ok29+OerdLDK5bHYygrWWhSnSiW5ZUA0gfkH34YOetHI6B7G7t2buaeemw2Qjl3GxdJI0KiRWJsOBz72fU9e6at5daS5HZ+Qhy+Zlp8hMzvTwtIQWW3Fgf969+4Hr3HoBNpTPs2I0NfWPPDDcapbkafpY6vez14cadKCWPbW75RHTZEUcxuHMYKlQTyQR71kdbwa9NmU6ro6qleixu6Z8mZVPli8rMENuVtfj36uetYpXoue+eoKnaWOq6zD04ra942LjSWbWf6Aj+vvfXjxx0BtJ2BJg9vvjc/iUGTjqWYwyREnQh+oJHvXXq149JXe3cu0c1g3ipqxMFl6SPxxRwqsUjuo4Itb3vr3Hp16p+Xtbs5cNt/K1suSpq+qWmnlnYeiBzoNy/vxr1oHqyje++tr1XTyfwSoialrEjqZ2jZCFklGoiy/4+9db+fREO7t3vQbewc1HM0iR0sTNpYEW08Arf8f09768eiz7j3pX1+1KSud5I6anl8p+ulghv+PfutHoN8lXxdqYiRKZRHPTL4wUFm1ILXuPz7917j1YF/Ko2TmMn2tTpXSSTRbfrWsWMhAUNxpP09+I68OtuqOyxxrf6IoH+Nl9+oetVHWT37rfXvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdY5ReKRf6xuP9upHv3Xvl1qL93xnrL+aF18ZLwDdu8oCtrqJDJXDm5+t7+/cR17zFetuanN6eA/1hiP8At0B9+691m9+691737r3Xvfuvde9+691737r3Xvfuvde9+6910fxyRyPp72OtHrv3rrfQE/IqWoh6xzslOzI608put7/o4tb37y68Pi6pZ21VVtXE2lW+4NRNrk5Lfrt78etjGOhHpUejmhmqGu4sSWte496+fVvLoTcfUtm2pkgF0Sxf1fW30uPfuqcOjK7UwdJT0kU4hUkBDa/BNub+9V630Ja5KKKiZNCofoPp9P7Kg/7x795Z63w6b0yGgXVAFP62FgoU/k+/Vp1rHUmnr6TV+0kckhuLAj8n+h/3j3vrWOu2pqWaTzSQp5L8Di1/6W9+pjrZPl1wlooaiNoyqwgAj03PA/PP09+p145x0l6jFxCYwxtqBudX9Df63PvVevUoekxnMZDSvC4YvH/bHBFz/r+/U8+vfI9Raqmw8lCfFLGspTgA2F/6e9db6TuJno6GSRqydQFvpJYfj6W1e9+fDrVMdJ+tz2MrK6dGrPEikhSGH5P0Hv1OtdQZd0YnDHymrEg+oY88j/C/v1Ot54jpPZvuL7aItB641BA0qbD8gWHvw68R0D2V7lzmWlFPj6VwrnTqRGufwCbe9gU69x+fRieqYstV0sVTkS93ZSQ9+b8/Q+9eXXiR69GnxFLPHEpVCvpP4sCD9Ofex149dZeiyCQ+USPpPA5YL/Xj+vvxPXqevROu28rk7z0IjZYQJGkcA2Cjg/7f3rrYxx6rW3J8nsB1F3NtTaWGiin3FmJ4lSNYwZDI8gvY/X8+7eXVfPq87Hbmqd29e46t3NQLJJV00IqYZgCBE8YIJB/Fveicdb6m7L6Q+PW8MbPBXdf7XyVXKX+4NVRU00ja+G0sRcH6+9gYx1o8eiH/ADA/k69Td0Y6tzPWzjYWfo4zUUNJhojTR1EkSlhGWhI+p/w9+60etczfPX/zS+G+6HxFbsvI5PadFUSNHnnhqJJGgifQjB7EHgX+vv3Hr3+Dqwn4v/zFNwRijwWdrqpKzxwiajkjddDk8oVf+nvVOtjq2brH5j4jc1WKTJ1Qp01lbsEHANub/wBffj8utgUx0fnaO9Nubgo45MdkY5pGVWCiSPksL/7f3YH06qelX5tbFw40hrEg6uAPfsde6fYa9MjAlJENRgB182LWPH/E8e/Y49aApjpxhFFTRSGOOMVKqSG0jUDb0m/+v/T3brWTg9B/NuTJvkGppqdygchWa50ggi/P490qerU6aqvMmiqpFSJWlYDUwI4v/r/4+/dbpjPXAZKq8X3MsrIuq9tSj0/1P+v791vHSC7T+Qm3Ortvvkq+vWGemjLIhdCX/bJ+l/8Aevfq+nWqDh1rMfL752bz763TVbI2HLWJUiSanj+1ikIncSEBS0X9ffqde6TXQv8AKQ7V+VdXR57tb7vbmKllgrDM5nh8kerU9wbXuPe+tHHWw101/LS+MfRnXM238ltHC7hkjo2Spy+SpotYHjsSHlF7396pmp631V7urpzoXoHt7N9g7AxOHNcnmm8FIKcFWjc/RR/S3vXl16lD0V3uL5T43tvcEVLlN0y4OlxrmL7BZ1SNhGLMhAI97699vQmdFfLTr2lqjs3A1NLW1cOqnmqY/EzyH9BLke9Ux1uuerPOtK9txxwNFaMOY5gwIFww1cAe9dbINc9HGxVEhpYqeWFRHosXI+v9SD731r5npmy2ExzVCqIkqF1LqVwGW34HvRxnr3yHQb7r2Rt/IkpLj6alFuZVUX+n1vYH37r1Ogfye0MViGAppkkjIA08Nbn6+9/b17hjpL1eFEhElMF9JuwFh/sLD3rq3WCSqnp4jTqn7lv03F7e9AgjrwocdJ4PVTVP+UEoCw0g/Rufz/xHvfXhw64VdTJTmRUXULelv6Ej8j37gc9e6Z8dVVFPK09QWKXPFhbSTxx791rj1KrK+WpgqWjLKun8XBFh+D791v5dJrMZ+PE7PyM0CrJUillYci4NrC9/9b3vzz1qoPWt/wDKbeu69xb0yNJURyrSmtlCqQTdCCLAji1ve8dUz0QfeUm68bj6s4HFeaqWMlWVDqJJP9P8fe+t8cHqsjeVJ3rvjdwxeVFdi6D7w30+TSYy/wBL/T3rrw6sg6E6sfA0GPGYYzVZhQK0l7lhyCCffutcejv4LDrS1KFYVdlYBRa4+n9fe6dbqOjGbJyU+Ny9FO9PystOAWBN/V9Le/de6uq6czFZXbRhaWnIikpzb/U2I4/H591PW1OadFS+SW2or/xGCBS8hPCr/a1/X/X9++fXqYp1Wr3JiKyu22n2sRjrIchSThkH0WGXUTpH5t73x6qfl1tP/wApXtmPePTGN2tNVGWtwFCIJELqSDGFB4H+x49+62fs6t4/1/fuvD59e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/19/j37r3Xvfuvde9+691737r3XvfuvddX5tb8Xv/AMR73TFetVzTrBJULGHJB9Jt+fr711uT9NQx8+gY+QW6sXtrpjsDI5Srjo0fa2UjgMjaTJJNTMiBPditDUeXWoqy4Hn18ynu/YdBmuy97bqr0+5ppd4ZR45m9aaXmNvU3+HsH8yXzeEQp4V6mX2y2FLm8V5PUHo5nxB2ziqeJGFOvgYsV4UCxNxY29xA7O0+onj1lBcxxbbbBFFcdC98kIsTiaWKqjMYIC3A0/W/N7exETSzHqOimGyeVvqS2D5dAztDdtPkMNJDELyAWAF/z/S3tCp1Cp6UuM46g00CTV050kTs/BJvY31E+/Ur14DzPSyoYqqJrM+s8ED8j+l/e+tavIDHTtPSSzWlZW9IBF/oL/77n3qvXvn0l66qmV1EZ8YjJL2P6iP8PewB1sf0s9dtuWjSACQxiQGxJve9re908+vaM5PTrD2DisbiqiSeSMTBTovzbj8H3RhQg+XXvhPQJLuDJ7wzLtDIwpke6gsbNY/j3b4sjrZOmhA6GOni+2oAsqlyEXUPze359+Oeqk1NF6D/AHFuGHFUdU0sJY29II/23H9fescerUPDh1C6j7gxNJkMjHX0dk0MIyQRf6j8296DBhTrx/T49CTXdqUtTFKKGBgTIxGkN9CbgC3vfA56qCa+o6DrOdl5XJ0k2MhWSAyjTqKsOLW/31/fiB+HrYNOOei2YTaec2ruur3ZFVeSaUvM4/Ve5vb/AHx9+B/CR15u7J6GHF90DI5CKhyUWiWIrGCwI5vp4/H+x9+BqaDrQ1EfLoa5nhzNClR/nI0XUth+mw5A49+pQ9e4YPSPklWjfVGhDXtdbjkfT3tj+IdeoK0HTxSZGV0tKQVZb/i3P+v7suRnq2kHhjpDbu2Xid101SKmEM4SyH0ix+otf3XQBnj1oq1a16YOvJqTabNgdKoyMSshsQQObE+9HPDqyvpGeHQq12WTJU8kS1MaXuAbj8C3Huyg0r06s2ajpOwY/KtohoqhjqazOGNjf839pfp/1PE8+mZUSR9Xn1yr8DuGhtVpUSSSj1Ei55UXtb2pzp0r1sMDhcdJ2n7HqMZK9NuBDJpNgzaiAPoCfaJ7aTxBKDw619RPGfFbIHTxS73pM3KkFMYzTFlBXSL/AF+hv+Pa+3upEkz0xLtttzIpj00Pr0Q/5wbNxddiBk6KER1aOkiyIqoRIouDcW9jzZr8kgHqAvcrlSLl5WnUZ49NPxx3DvFMNgdtxVbs9ZCIYGBf9sWtq1f19jwsHUEdQRFffWLULpp0dd5+8NrVkdIKPKVmNpxHUNVIshiaMeojUPdfLq3U6buzddbnqF3efGQ0oSKpWaSVNUi8Hi/+9+/dexXowuJ7G3LPSpLVtPNTSreKxlKyC3AX+vv1OvdS6PsCqgq5HnxFUY3QqAwN7t+ffut9Om3e1Kzb2RaQ4ipkeWS8cCIxZtR9NgPfvl16vQ84Xde9t5RVdRUNPgscsZ0pUq0augFzpPHvxHn17Jx0xbe2rm9+blXb1FO0tMJQ8k8bkh2Dci5/3j3vr1MV6sd6s+N9I1PTffYx3NONM1QyIUNv7TNb/ifeqnrY6N5138V8Zl8rUSyU6/bQqGjYJGFey/QG3vVetUoesXZ3WWxdk09Su4Y6aKkghmBSdIxqKLccH37Jx1ulOHVB3yP2Hi+8dwV21thbemZBOIVqaVPToV9JfUB/j72OtefQ8fGT+UXQUGNps1ujGu80rrPP9ykTM6uNRF2H096r16nr1Yxt34X9a7C8Eu19vwUmYpnS1T4KfVdGDk8D36vWwOj1YLGy1eKwOMo6b7aox4VKqRVCCUqLNcqB798+vfb0INDsHdm4cqKR6uSLEqE/a1MAePoD798h1oY49DZgOmNq4qeGbIQJPMqguTyWYf1H9f8AH37Pn1uoIp0vYJ9obXqGjhEESynQI2Vbqfp+PfscevZ6cslu/BCkNNBNDGbEg2Urb6/X/b+91FOtAVPQf1WXx+Vp5qWOoikkk4VxY2Fvz/re68B1s14dAqOraKPOLkjIjPPKHcqQLAt+Le/dePXfZ3U+PpqJdx45ga+KGxKWJKhb+q3v1KZ49e+XVe3aefkw+OrKvFxfd19Kt5Up0DSq45NwB72PXrxrw6bOhu7M3mafK0ktNPTVUcEyukgcNIApBvf6e/GvXhx6OX03lq/J7TylVJOKSb7uT9iT0u6ljc2PvWOvGta9Zd8brwGNoTiq9EkfJL45pjbShIs3vw49eHCvRFuvdv4zb/duSy2MzNPUw1DF46JJPIyMzXF0U2978q9e+R6NLuquyNTuzFz10hmoljX/ACS5GtT/ALT795dbHSc3htrEZypp1o6LweawkjsFIH+Fvfq069xx0rdudA0opWqaSJ0mljCBhyFLi2v36vl1Xj1goPjlubbmRkyEWaSSGV9ZiWQMU1G9it/fq1638+sG7Nn1OAp5KvI0ktesgGlVRWJP5AHvX2db49Fiq9p7D3Fm5xmdtNSzyK0STTRhQS/5IYe9/PrQpwPRSe2/h5goq6fcOPdWR3MtPSo66iv6gFUe/deIz0SHunprL47CUtZiaWejqqYmVTcowKH0m/197+XWiK56j9TfIHL0uHPWO6ahoXDIFr53kES6PoNZNv8Ab+/H5deHS63fvk5hKfCxRvkaeMCJaiAl0IHFww/Hvw+fWznHS0x214d2bQkwEcf8OMEDSapBYOdH6QT/AF9++zrQ6B3D42LYGLyMEbD+ItWiCKBiPK/lfQCv/Ee/deoeti7+Ux0DmNr4TJdj7io5IxnF+4ofOq/7tF1tx/seffvn17gKdXdm1x/X6j3vNOtYr13711vr3v3Xuve/de697917r3v3Xuve/de697917r3v3XuuLfQm30B/3r3sdaPWpL/MTXzfzN/i+YCYtG8aBpfxrJrkPNvesde9KdbalJ/wFpv+oeH/AK1j37rfUj37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcSdK3Nzb3sCpp1onSKnoLu5sLNn+udx0NOrNOaGSRFX9RKoQQL/wCv70cdbGc9UW4muG1cvW4uvkEVVHUT3jkHrA8hsbe/HPXhjPSqkzSVKeUyBueBc/Q/Ww91NerA+nQj7GzMdBqd5Fbyji3JAP8Agffh8+vGp4dGXxO8ko8YjkegW9IBN/8AC/vdPLqtfM9PEW7oMlSySIQrIGstz9APeuHW+PQZVPYtRDVVFC8lla6IeQL/AEFre/deHU7DbkyNHULUOzPG7BrjUwC/UW9+p16tePQ2YHKy5LRVE+hfUykfX/W9+48evfLp4r8ujt4oUdXPDWBH1/s2/Hvf8uvEinWBFUxiV7gkG17XsfyPevt69xNR0l8hDJXiSPmy6iPzcf049+PDPWvn0FOSxWVhnIiDiMsbC54F/wCnv1RXr1D0k87hszUw2WRk/wBpvYAH6k2/PvfHh17hx6bKPrnK5EQOhdWABdr/AF/1/fqjq1T0JdL0p/EqVYpywkWzXJsWNv8AH3ocetVPTjT9FY1IjT1dM0yH6/pP55Bv78OvfLp4i6g2fiTAsdABM5+ulSQfqP8Aeffjnh16h4dDDt3ZdHRxxaQsUK6dKkKAQP8Aeve6E9MsaNpp0KEVBE4SGGyKFFyQoFh+T72eFB0yZmU6SM9NGbUU0YikKSICNYABHH1I/wCKe/Z6VKarq6J93FiFrZppqMhEEb+RbD6Ac3B/4r7rjj1bz6qf3/1b09L3BtnfmerKCkzeAnR4xUOis0iOG+jH+o928s9eNK46PT/sxuOloTSUeTppcfHTIiiNrgrHHoFj+OB7qa9ePoenfqz5IYvAZuPIrVRGkM8cbQ6iblnsTYH3vPEdawOPVtmzt243eOEo8xjpo5UqIhI6RkkoSL2I97GevE0PTbvfrrZXZmJqMLvDA0eXo542jZaunjMqI35jdgffiD1qo6qn7O/lR9dy7nk3hsKnTHOHklahgCpx+ocC3+8e/Z8ut1HHoC838NdybeaaTFiohmiuGZXKfQ8Xtx719vWwPToJH7T7P6IyYp6mCvrKSncGRlMjeiM8g2v73QcetY6Mj1z/ADN9jSZbH7Z3IqUNfOsaSPVFls7HSfr79TzHWyfXqxXr75F9d7vDrhMtjpalgFYROCSQL2IHv1fM9a09ClW7zwlBQmrq8lSxK4LFna1x9eP9b37j1qnQb5DtvZq63hzNJUSLcsEb9I5sLr/j71XrYp0FG7vkDsjCUclTUZGiM+i6q0mk3v8A0/x9+62eik7w+W+Rr6een27TSVKu7xxNT6ySSbJptx799vXugepum+3PkzK1Lk/4hTUlYbwFmdVjUenkt/h7316tMk9G0+MH8rnrzqDc9Ru/eNLS7iyTsJ6aOcJJ4ZyLksSDfnk+/cetdWXbn3NtXqzbDVdU1Fisdj6crT0yhIlKxJcKqpb/AG/v3l17z6pt+V38xLGiiqdu7WnVUmTxvLGJNPNxyy/n3rjx69QDPr1rjd3/ACW3FJmsgaKpkeSrEwkIeU2WR7n6n+nvfWiR1Xd2FvHMPFUZ2ATtO0peUI8nLP8AUBR9PfuvdGn+DVVjKvOpmKqGSmr5ZWaYyO7MzFr/AEY3+vvx4dbHWz30H2VFHWUWJEa3WOmtMAbfpHp596IB63Ujj1ZVh82tRjA2sSOVJFh9OLWv7917h1JinRQHYEM/1/2/1N/6+9dep6dB9vyqM1CaemfRUNdtX04/r78RivVJm0Ra+ggFD5YP8onVrI2tjwFsv5P49010x0/bKJYi7eXRQe5vkvtPp8zRVdXTSuqlQBzpa+n6j3oOBk9JDeW4mETHokVT/Mh2NFmRG5huZAoHrtY/778+1KKr5HRxdQwiESQkN0YvZPy92JvcQNHLBCz3sx4Fx/T+p97dAvRbVqcM9GTp81jMzQJX0VRHMjqGATn9Q1Wv/T2kMmlunkWoOrrPG8EtFI8gRQt/qCPoLgj271V1Cmnr0k4crHUvV0cA0fVQ7W5/BsR731Wvl0A2/wDeMWESrw81pVaGVQRcjnjn3vrQ41PVSndRwFZmKmTwI0jTO2sgCzW54PPvY60fXonFbTOcjMKZ4zCWI0ABrLfmwPvWOtZ8+gj35gaETRVaLHBOsq6nZI1HBu3v2evfZ0usPlsPBQUlQa2mM1NEg0BlBb/EW9+69jow/W1fT55vNdQqH18XJsL8f8R791vo0+w8VR5bclJS6PSkkT82NyGH9ffuvdXIdcimodv0eLgj0HweMGw59PHvR9R1YU8+gs+QG1RFt16uRNelCw+hH14It798uvfPqqnc7CaOt80FxE86qCBb0qQGF/x72Pl1U/Pqyz+QpunJZXd/ZuPqJmempMhWxwRkv6VEmkCx44978+vVxTraU5v9eLfT/H+vvXWvPrv37rfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf//Q3+Pfuvde9+691737r3Xvfuvde9+6911fn3QuA2nr3USRPIPHa2sueeBdRwbe34yB3Hy6pcAtRAfLqin+cv3/AFGzet8TsfB1jRVuXZaGsWCcJIRM/jZWUckeyvcL4W+qnS7Z7Oe+uFiRcCnWo93Ftamxm13xdy2Uytclcx+shM5DEn8+483K9Muqma9ZPcg7ULORHfBFOjh/FzYTQbVx4jVvuXp+fS2q4Xkcewc8ReTPUsbmdQx0E3zHw2ZwtJQGQuiSzoj8sFCGS3N/ZqCywaRw6RwswjCHosq5RtoYWKrppFlkeOIsquCbsLtwPaQ0OR1f0B6WW1N8Y6oWKorXEc89mIZ7AMR/Vvfia060RTh0uf8ASLt6nrUgNRHJMdICpIp5/pb3s9uT1UVPHoUsbmKbK0qSKDodLggcEe/UHXqMDTphz2KDBXhJs5/AN/rxf37FerAmnQP7iwlY8g8WsOWIKhTwP6n37I4cOt6a5HDoDezTk8bU0NFHM6mcR6k1m17/AFK+9Hr1Kcc9Gf672eafbeLyIKeacKXPF76QTz/j72Kg06rWmTw6FmHBzVDAu0RQKeCRbgWBt79Q0z14CmR0FG/9v0TAq8kXAAIVhb6cm3v2k8evZPDoH0wuEoEeRCFluT+3a7f05HvxWpr14Eg8K9cqHL01NIYwL88agDz/AEN/e+C1HXiD1NrK5pgWSNRxe4UA/wBdVx78Kca562Cq5HWKmjHiEkiX8tkKnkcm1hf3vyq3WjTiekZuPYc5k/i9EtmVg1o0tbm/IHuhLeXDq1QB0L/XOWqWpRj665axQauLcW+p97HqOq/DnpYZXFabnT6bFwSv15uBx/T3snq1Kip49J8IynSFOkE/7EWt7159aJ046kKrqulRfyCx+v0PHP8AxHvdPTqw1jpHZbasa1BrELCRubgG+pvx/re9UqPn1rjk8Om2fB5OGmaSnLjj83BFub291AYHrRzw6xYTdeRxrtTVEZ1J9DICOL/1/H9PdgfXj1U5+3oTaDdVXVUhnkWJgQQb2+n+Huw4U6uAKdBD2NHjMjRT1CDRUquqyWuf9gPdSQKjrVNR0Dh1g6uxMS0oqKkSi5JuykfTn8+9wxqXqOjrbAtuTw6CX5QUFHkds1dPE4NUBIyK5F2KrccH/iPY42KOjjGeoK95JkmjIJrx6Ib0f2Vu7am4bVlKPBiGJpnC/UCSwAJ/3n3InwoPn1jIsMaRgr1vGfA3qfa3yA6Cxm4d1YigaWsx9MPK1KjSuZIQf84Re/8AsfbZNM9aFDjz6JH86vhltPY+0dw5nbVMtJkoq6VqXTF4WJUkqAD/ALD6e/cetkdFq+N3Wu+90bdw9RuHHH+DYZddTL4DqaOP8lv9Ye/dVFeHRw63cvxKwoiodw1tPT5ZQkLwySKjGoU2YaffhXreB10NrdX7oylFk9g0y1zqoemVIRLE6jkXIFv9v79x63Q9Lf8A0Qdk9lOMBj8fTYulp2CTskSwOykcfpt+Peuvcehv6y+NGd6uy0FTWQrNTq0c0riMmU6Tdzq/p/h73Udep69Wm9cV+wc1s+uxcEYp8mkZjcvH43L/AE9JPvX29e+zowuy8Jj9pbUqcxWSRx01BTNVySM1nMagsRr9++3rXHh1Qn8wO19x97drf3S68knlwcNa1JWS0zMw1B/GwZo/6f197HW/kOju/FX4nbe2ZiMfls/Qw1GSnjL1ElRBqdmIDC7MLnn3UnFevUp0dGvWOJ/4PhaVIoo0CftR6eALDj37r3UCg2oXnvVKPISCAffvs68Pn0JGBwFJFKbxgGMgg6ODf3759e49KTcG6Itr45ZQEWUCylSA5H4vb3vr32dBPV9tZNGasu326gm5a3DLbg+/eVetD0HHouWe7G3FltwNJTPU+F5RZtTFQA1+AOP9j719vVh6npWVGa3FLTK7SyC6DgsQRcfg+99aoK8euW3tyVmGnJyVTJZyWGpz9Dza3+PvXHj1vpVJ2hSLNU+OdiVU6LubCw/A9++3rxFeA6Suf7xqKTE1aTSaoDDMvqcnSCv10n36gPWsjqvvD9obZfdWYhqahZv4hVSF1ncFBdjcaW/HvdaY69xz0Dad+4LZPaeSp0SGKhYsto7LHYmx+n497690cDa3yE23kqNHwNQRK8d2hjeyaj9fSPeqder01bt3RV7yiajgkk++qfRBpudL24sR73gdarXHVYbbj7G6Y75qcllK2R6HzeSWOaoPEQkv+lj/AE9+68Serbtnd0bT7B23TbypJxLW42nWKZFYNeUcG4/4n3Xj1uuOlVjuxsDXuKmYgTAhgF4sbcgr73TreBxPQjbR7mzCZj7IRxnDMAiuSdX1tx70RTr3Q747ceLqp1PlkLT+sBmYgEn6KD9ePfqHrVa/n0x5XOwZvKriqeGKZqaQKwdQ4PP0I9+A695dAp3dsdoaWHLR0ohuwJNNDpY2/qR72OveXRWN4CqkyGAFGs0kSJH51lBKEfnWPe89aFOkdvjB7e3HjKygytNEk32xEQWMBQ1uOT/j738utdUmfJ7rqowNDXRYpjS1X3U1RDU040OIlOoDyJz/ALD3rrfWb4c76oc3NHtvc7LPPTkweaoOtmINr3f/AFvfs9eHRtu7+x9t9fw0tHh5glRdBLobQQp+tx798utnpr+PHV2b+S3ee1oMPFNUbeZqOorWjiaWAyLKCzOw49+4dayRXrdY6z2dS7C2Pt7alLDFAmIx0FOywoEUuqAMzAfnjn36nWul1/aP+tz/AMQfe/LrQ+Lrl711br3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuum/S3+sf969+60eHWpN/MHt/wAOb/Gb+v8AfGgt+efvU4974dbzUdbaVJ/wFpv+oeH/AK1j3rr3Uj37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XBywtpAPNiD/AE/1/ex1o9c/r711vrBNDFUxSQToskUoZHQ8hlIsQfe+tVrjrVZ+WuY3ttv5abgxFNTS022w0hp5FiZIWPk49Q4PvXAdWr0NmxIajJUkDZBmDvCsg1EgXt/Z96Jx17FcdKnFZl8dnftpWYUyy2U86bX5+vvdMV69XOehZrN8yeaKhpDeGy+pDx9Peut0FelZDuykoaJVMy/cSL+nV9S39feh8utac9NNQpy+irhK64/XIfqSL8/T/be9nj1sY6EHbOWjqPFQHT5VZASxBK8WuPeutZrQdGRwNE1PRh7EKRxpHBueT/sfeqdeHSlpMUJWMxW9jqBsSf8AAW92HWvPruSiaWfQ6nSOPoOB9DYf8QPfuvHrhWYgUUeuEIQ/6jb8D63/ANa/vR+XW+k3LQib1sgB/wAVF+OB79w69XqGu2fv2BZF0D9XpAFwffvs60B0pKHFijCxrFHZQADY/j8n3vPXqjqfPkYKJf3WRNIvZSAeOOLe/Vz16mMdN6byjlbxjxcGysxt/rc/19649bzx6iT1MFTNHMKiLUpLaderm/8Ah79Qn8+rK6jB6fRl6giGPVpQMouh4NjYn3s1p1ddBbPQsUel6GOZChcxi51XLcf1/wCK+7gVWo49ONaoRq6T2ZEa0zvKbuwIF2BIN/rpPupxk9JaEMVHDoj3c24XwaZKpY/tLR1JTV9NQQ2PPuo63nj1ppfOztbubN/ILGQ7TrclT4iPKoJvtp5AhjD2Nwp9249V+fRpunu4Nyx0P8LzldP9zHCocSzNqvoF/qfz79SuetjH29DDju7JcTVpTy18mpqiNlKzjSDrAB5/3n36np16p6s66O+au8evptv0T1Jmw1eIxUNJPq0I1ha3+Pv3DPXvTq/XqrsjF9kbZo85Q1ELvPFGXRXUsWZAx4/wv79x61w49Ccb/Vbf43/I/wAD72PQ9aPqOmKtwGGyscwlpYXaXh5NPquPoL/09+Pz68Pljot2+Pi9tfdy1YnoqJzPHIgZ4kLAyG/9oX496p6dbrnqgL58/wAuncW1Mfk+wtjrWQVOPkmliWihfVZE12Gj8fn3ofPrfHqs34id69oddb5XHbpq8y/jrGSSJ5ZS2kPosUP097x17q2Hvb5N7kyuwq1tt11TBkIcNUSojzmK8qxXX8g3J96A62RTrW06o/mI/NDJ/IfLdRQ7e3LXUlblDBQZKKKoqqLxvP4+ZRcAgfj3vz61nrZp6N+I/wAg+2KXH5nsw11PR1CxOsbLInEiBxqH+x9++zrwrXPVs3Unwu2ztOGnOYp4p/EkZtNEGYuvJsjfn/H36h69UV6Ont3aeC2tTrS4qip6e17MkUaPb+gt79T061XyPTfvLd2M2LiavPZepRII42bTLIEQaeeAfeyVp1oaq561yfnv81JdxV1ZiMLlXTFRNUxSGnrLCyrp0hQfeuOern06oU3f39HV/eQx1U1TPHwHll1PcH37z6rX06AfCZ0bqzdXPkwdDK2lnX6C/F/e+vY49JDO+CPL/wANSFHpZ5TrBAZfrb6fT/Ye/de49Gq+OmAp6HPUk1EHjV5TrSMFVAv/AIe9fb177etlHpXa+CptlUe4PPGMkkEF0uBJdUHJ/PvRr1bj0aTrTfolaWkqHGiN9IJa9xqsOT9PeqenXj0MGRzDgo1PyjHgqeCAOOR7917pKZqtpWh8s0gMhUgC/HI/offgCxoOmLo/o9Fo7j3PPsvrbdWfpnKy08MksJZtJt4ifQT780ekUbqokkjtiw609/k38kt07w3RXJVV1StOs0ynVUFgdMxta/stmlKZPDqCea+Zp9uumcNQivn1X72N2XuCipBlcRU1Mk6SgsFck8HkA+7wXdOhPyJzqu5W/wDjTZ4cejifFPuvN7j2/NLWZOSDI0kZEUbVOhiy82Ivf2rEurJ6lSG7jmzCajq2T46/K7c9NmV25nqhvtEkRFaWoYroA/BPt1dLcB08+Bq6tNxHY9Bn6WBKSpiPlRSdMoNiwv8AQe/AdVrUY6Us8v8ADIPOtm81tTJzYfUkW97x1rIHRa+5lKYapy6hg7xyXbSfzz9f8ffuvY6p67SrZpnrqqKUs8c5JswYqSbEWH+v78evdIfHJtui29Pk6idHyDRPIyNIC97Xtb6+99aHRS97V2Z3ZDWQ4aOYlGmCEISCV4Ug+9de6LjsTbna+S3omOr1rFoBUBLetVYB7XPv329e6tB2TQ120mpKd0kDSOgewI4Ki5Nv8fe+t9WKdB7PyGZ3RQV0cT/bAQSMxUn86iTx79x69w6tjwcYoaaGOMeuBSr8fQ/XUD/sPdT1b5noO/kFuSlGzCruglEZ0qSLlh+Le/Dr1eqssxVUGUwOVqH0JNG1SFFtLng8D/D37h1rj0dv+QL5/wDSN2oCGWH+KVxU2+o8nAv7t1U0p1th+9db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/9Hf49+691737r3Xvfuvde9+691737r3UWRyk6/0K2t/seT/AL17RyahNXr1adcMjWQ4+iqa+oIEVJDJOx4+iITYX/r7WVNOthdTAdaZP8yLtcd2fIat2vTzl4Nv13k8VwAoiqfVpA/1vYB5kuzGzKOJHU/e2HK0d4TO44dVYd1xRJ2Nhaapm00qQ0yFCbqSoC8/7b2ARPIzd/n1O1rtsVk9OFOjsdeb/wBuda4LG1tRURiLwBgCVIbUt/apY1Pd07dMG4eXRQvln37hu1KQ4vBRpNVxjTH4wNQkVrqfT+fdJWPAHpNCWLU6Kf1/sHd+aLSZ1JhRNpKCUtpCA/i/HtoHz6foBnp03/s6TDokWPqWhlsAir9Qf8ffqAHUOvAt5jB6Ydk9fZuTJQZOvqHlXzIfWWPpJ/F/exVst1pq8Oj8bbioqPGQUx0axEByAOf6/wDFOfeqdVIJNOlG9DG8JZrNcen/AAJ+tr/7D3aqjrdKdIHcSUmKop6+qChY0kZTf6EC4v71UgU62DU0HRHt0z1e/Nyp9ijPDSygalB4C/Tkf196OTk9eBIOc9GKwuYzGLwlLjS0wFMAL8/S1vd8DrdPXqHk94biCmKkqJr8/pJv9LfT3qtDXy6rXy6Qk1RuTKS2rKmZbtfktY/19686+XW+A6yNhKyFoXLSTeVgrC5Nr/4e9EngOvYI6WUG0RojkeEKxCm/1J/3r3vJ68CBx6f/AO7QmhCqlgosxC2uSOCfewoHDrzKDnqV/AEhpEjZF1KQb/Q/SwA9+Irhut1U4rw6y01CwvFUAtGbABrW497rjHXhpAqM9SoMJFHW09VToI0VwzhRYWBv+PegBx6qSDgdCOYYKqI6rkhf9e9+Le9DrZqcdI6rxgFQQgGm/BH1Fv8AiR79UenWx2jPHrL/AAlAqG+m4uSG+n9fe604dVqfPrnPQUj06l5gDqPJ/qPx71xyOvce0dR6ump4UjSMh1KLf6EEHj/fW97zXPXhjHSJzGAjrEK0dMpnJALIPVe/14+vvxyOtmnHphg2/mKaX7OSKRI2vb6gWI/r71Q049e1ACg67Oxo5pllqZiQttcTEaTY3sQfe1Whqetgjz6XNNUbfxFIlLGsKyXVSnpHqHH++t7WQaXfGOrLctDqPl0Sz5TbYy09A+coWkSgD2bQWCaW4/Hsa7MpeQBescvdncVclVboH+punH3PmNr4iKAO24AqPNZvqXAvqH09yFPHpiUnj1CUdXgU9b3n8vXrGXrPp7be0opDK9MmOeSItf0GMenn2iJqOvUNc9Fm/mpVq1m8tv7Jok+zmr/B5KSIAecyWUswX+vuw4der0rOruoqLrL41ZrcuWoEhiiwkk8zyEWC+AsSdXvXy631p8/IDdNf2J3pV1u1smwwlLm6uBliB8ZeOU3S6/737t1XrZ+/lbbNxWQ6iny2apI8ll6SnZKeOUL5CRYC2q596NOHWwerN6PpLL5KrGapWl2+khDaYmMZcWuB6f8AD3qvr1v7OllR7bzODrFOWpGyOLRLNVTetLqObk+9AdeJ/wBXr055PDY3K0z5vbUMeMjx4LVgpQFWQoLnVp9+68OgK7++SH93ut321RV7mrylJJQto0htTDxni97e9gDr2egZ+FnR0U/3W7MnTLXVFbWfeCaQMWAkk1jn34kjrWOrbYaSOigWmigWNYxZUAHoBW3H+9+9DreesFDiYlqZajxElgbMSb3/ACb+/cD1rj13Fg56is86sbA20i/9bfj+nv3n1vy6V+QeHD0BbSvm06rm17lfx73wFetU6B1sVkt6VT08oP26FmDEH1XPHvw4db+XUSs67nSFqRkHjvpAN7BQeeD711r5jqCerocbEk60ia251G1zfm//ABT36vW+PSV3PhslR48tS0ZYi/ABtpH5497z1rFc9A1kaLL5V46bwtGzaQWAYFbfm5/3nn3rq3XdNtA0CustVqmkUgoW9Vz/AK/v3W/s6QG8dt1UWGySyRkolHVzBrf2UjLX59++fWutXXvf51UPU3dWQ2tjia3I0+S8JpFXU2oylQLe7dUyeh86qy/ZnfuYfNQbKlENUE0zGNg2l01A/T37h1sZ6sh2F1pv/Ym3nyZ2y9VUxqSKdgzMQPV9CPfq5x1umOu8L8i5dqZwDemFTB+GS6NJEyng241D37jx61Wgx0UXtXYfYfeHbVZv3borKnac6NqkiMogWP6lmA4+nv3HrR+XRs+nclsTqLaFYlTuWCrzkMU2rbjOh8s6LfQUJvckW9+r1voiGV/mUdgbO7Vy+B3D1bUYraM1S0VHmnpb07wl7Bg1v6fn37r1fPowFF/M52fgq7FJTCnqkqqmmSVQinx+RhrUj/D34ivXq0x1bTsb5M7J31t/D7joKuCn1UaSVARowFZlvbg/g+9U63jz6FXYO79vV2ebKwZNJjUyD060Ite/BB9+pjr32dDtvbdmKlxIWopUqoRCLKxRl4HBA/HvXnU9eGcDoqWTmw+aqGENDDAyH0lQLqCOSPe+rCp49AD2zQnF45ft4BqqiIxOLEjUbajz/vPvdajPVSM46qD+UNLPQ0dTj6+Vlo5/JIcpe/iLLfQC3v3WiKHPRGOns9RbE3zjXVw1NUz8VRsfICf1k/09+68DnpUd3dkU26ey6PAUdSKuTKyJQU6G5Hklsq/p/wAT791qvn1tufyfPitJ1b1bR7w3LQBcvX08U9FNJq8gSX1qVLC9re/de+XV3lxzz9OT/h79Q9eqOuk+l/yfr/xHvbcetLw65e9dW697917r3v3Xuve/de697917r3v3Xuve/de697917rpv0t/rH/evfutHh1qT/wAwf/t5v8Zf8d4UH/uan09+636dbaNL/wABab/qHh/61j37r3Uj37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XRYKLsQB/U+/de679+691737r3Wv1/Mi3bsvF9h0lHHT00Oearj80oCecqJRe/5596+zrdekvt6Ja3DYivo5DEhoYZDawv+3c3t73WmOtDqVmpqSopdNPpWshFyyr6nb6/X37h1s9QtvZOop4HethPlAOnWPoB9LX59+PDrwPTZFuCsrcsVlZ4olcj8iwBsLD/AHv36lMdaJxU9CDQb0OHmNK8t1qPQt+eD/Qe9U62OHQp7WleOrjykcrMGZCQOeTyb+9efW646OrsXK/xLHqJvSVFh/tVuAbfT36metE16X0M4pnU2uv4Fz9APofdvkOtdOsjQ1EPnjBuRc6T9G/33596z1bhw6ZvMTKI5ufqApF7qfp79XyPWuPWCpxjSSgouj6fQg2H1/3n36nWvl1gyFYuIjA082Gqy3N/9f34kDh14DprmzBhpjUaR+6Lqv19TfXn3qvW816BDdO5J6Npa2pmaKJNRKkiwUH6c+/VBwOvGo6J12z8rsbtCmmhxpEtZEShVFQnVY8/X37Setr3dFZh+e+UwCz5PKNKkEYMhVwgAUHjm9ve9VOPQfvbw28tTw6Oj8ZPnLtfuuqTFrLCtTdI+PGW1ngfQ/6/vwIrXp633SOYhQ1SOrQNr1UlRSsVnLRyfpHP6SeALfT3vu6PY7ksmcde3LCaWmlqJpSVCXRSfoR9D/xT34j16bLajXqvHvPIR7gpa3HG6FknjL2AtqFv9v8An3ode6oK+QHSOEw24pc9VsjyCbyo7G5XjVe59761TzHRON3wriIXymDqDJNLcWiNzxxf0+/Vx149ITB5LL14eereT7mJtaBr39Hq4B/x97B618+lTR/M6t2Tmsft3LY16iGJlgWd4gwjUMBwx964db+fV8XxD+etTiKLEP8AcSDHTyQDQNIRFKcmxPHv3XuthzqDuTbfauApa3G18UlZLCGliBQNdv6AH6+9jrR4fLoZ1UKNKiw/w9+Jrk9eAAFB10wP1DEfS/05t/r+/Ajz60QeI6Se98JidxbYytFl6KGvo3o6hvDMAUJ8RH5/r79TyPW61AI61j+lfjltrf3zF7AxGZxlPjMLRZWYUKFAkRQVB06bj6W918qdbNOhy/mG/A+XZ2zP787EyUooKeLx1tNCZFQIFGoFV/BHIPvflnr3Sb/lZdf/ABzhrAm49p4DL75JLDI1UEMlXDUGS4JLjVcH/H37r2etkakpKTH08dPQ00dPTIAI4oFCxqumwso/wt73x60SRwz1L96631heJWkSViQYg1hfg3/Lf63vdadapXJ6pP8A5nPyqo9o7frNoYmuMdYsMiSRpoJHq0lyb39662K061KuxOw8vuXMVxlrZJIqmaWUljYWe5P59+61XoCIcJH9/JPJIGWVmJHJ+vvfXq9CvJgqSkwUdVRRqtRMFjDLfWWP0Pp9669Tppo+v8jFQy5Cvgdp5JFaIvckhmuCpPv3XurAfjv1nVY7H02aqYSQ4LDUDxcauL+/dbHRzdv/ACIr9mZ6Lb0hZqJhHCItLFdP6fr9Pfut1p0eXYe6Ja+KmyVOQkdWqTHTdbajxx9PeiPTrwI8+js4LM0zYdHqHBYIGvwTbTewJ/r71Tr3QR7wyVSlQuRWRloxIbof02B/1I49vRDv6ZmXVHQZz0B/yBqJ99dcZDbmKiDzV1K0bBeLkwlTyPe7mgUnh0Yxwa7MimadagXys6ZzuwM7Wff0Two00rayjnh5ibEkewvcu5qqjrFT3M22UyuRivp0TKOgx0lR9pWlJImT9Lj+2VsbX49pIZHQUI6irYd8l2O4EDuakjpBfabp643VSZrATTxYVpRJUwQkmMozajcDj6ezWCYP2k9ZOcmcxC8ALtTq0LrDfWP3JhaTKUlQsOSCR6yhAYME9RJBBv7PICtKDPUuAeNEHQ16Px1F3Q23Md91X5R5GhKJZ2BsA/8Aj/h7q4oemwKYPVkvS3cmL3/QypLKszhGEYIQ/QXA5P1968+veVelJ2ljmy20MlTw0n0op2Q2FhdfSeB7917gada/27aHIwbmzuNr5pY43rphGpY2C82Av791rHQFUu1c1/eWaOevmOMkkssbs2gITchfx78evedOh3pdv4fEpDHR0kNRJIFVuP7RHJNve+vZHQi47ZWOoqQZhcVFFUldasFGoH88+/dbx0OPWvXEe/qmNRGvmDW/Oq+m/F/es9a6tj6A6updq0EcFZF46gRKysTdgqjjn3o16sMnPQ7VuQx23krDMVe5Yp/X6Ei9/fqZ631WP8iOychlq2agoVf7cM8QRP0lNdyfewOtH16Ipu/cuRxFDKiwusLB9YA4NwQbk+/cePWq+nVu3/Cf6qgrc/2LVLGFkkra9ibAEev+v/Ee/Hrw4dbUXv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r/9Lf49+691737r3Xvfuvde9+691737r3WGZbre1yOOPqA3Btb34IrOCeqt8J6QHZctQvX+5HiVmnXHVGhUvquFI9P592kAUnTwp07ZkmRdeM9fP87B3tW0/zY7Op9wI1DDEagwSVj2WQiqYBUV/cU81w3ElzqjOKZ6yh9utzjtrYRJQVPQCfJDNQ1OXgydIS8kZGmVBxwbqVI9hgTwqoB49S4yzSjWvA9Brkc5uTcG2oHarb7alpwCqyvcgD8C/uyyGlUPHpMttOrUk8+pGxduYOOGHK1XNUz6j5XuQxPP1/x90Gtnq3Sl0VBSnRmcJnoVRaaMR+BY9ICqtuPz7dNOkpBB1Dh0AfY+Yjl3JQxf7r8wDAgEfXng+60A49bNaV6HTbFDTVFLEyhAgVGAAvb0gkj3upPDrwJC0OelukKs6rEAGWwHOn9JsAPfuHXgcV6UTiohpw086lAA1rgckfT3qgbh1ommBw6KX3Lu7IZRhtvEswld2jYoedJNje3u2OB6vRQOvddbMfb2ND1sTS1tWocSldVifrz7qAOvVr3DoQJ8PXyhlAYmT0iw/qOCB73TpvLGtesdHsfKo/kmY8kEcE2H59+oPy63XUMDPT42zJnMcnFk5YAAXv/re/DHXs049S4NthpYg8RsjKWAH4H59+JqKDq2Bk9KqpoqVRENccYCW9RUfp+t7+/efVQdQqfLpOVdZTRt9tTSxSS/RfGQ35/p78BXr2lvM46itR5F01yRsbA/QH+t/+N+/VHW6L5dMlStajlWVv8Bax/wBh/wAT72SaY63hc9Z4p6/xlFVlAHBIP5+th79gjrVAeA6esYuRaPmRgTcH/G3IJHuvWuGB04NTTopL6tTX5P0/2A/x9+rjHW6Hj0wGXISu8SFgqE/Qf42HP+t9PduAx1vt4nz6yQ07AkVc6hL3IdgDx/T+n+w9+FKVHWiQvw9OBr8XSqHnCSKn0CsG9I/r7rnrwoBUdcJN1YGkVZY6cLI9gCQPrbn/AG/v3AcetZoCek/mt2JJCZYlDOVbToAsAeB9Pe+PWqitB0Buc3tk4KtIAkgEp4chwB+Be39PdJGOnHTi9zaT0ism+ekraOrWsZkeYMY1Y/Qngkf1Hu9lHM8gI4V6a3e4htrF2ODTpf8Ade48HL0VNiK1oIMvLLBGlRUSIhGogamZvcpbDGFKsR9vWF/Pd5Nd7iy17a9WEfy7+i+v8ns7b26dxZfDGsx9H5YXmnp2CstmWxJ49ji+ZCigdEWlY7dAPTraCxnbHSXSXx8k3tPuLA1mUp8WXhip6uASB6eH03VTf6j8eyyn7emB1Sh1TvqX54fKGLe1ROsu29t5QQRpr1xPFDPYEBvxx79gdeAz0dv+bR2zi+l+gKbrXbEWmr3TjUxsQiXxs7TxeOwKfXk+/Dreada2/wAYfhzVNgqnJbjopjkcll5sijzoJDescyqNTX4ufp73w6qB5dXMdC7b310rjTjsRRVMtPMLokKnTpvq4Ce/efW/s6ORR/Omj2TTDE76xU8bwrpEs6MixgLp1ubDj3ojrfy6HPbnyn2L2Fsl6ihymP8AtXLFiHX03FtIf+v9ffutdJ7Cdp4GqocpQ4GvpZ6SXyJWRwushJCm5Fj79nrfHqj/AHP22d4fKaq68rqgyYuHJNDS05Z7qxmsBo+nvfWvPrZS6D2hBtHa+LpkSOL7nHQSoNIVtLLcX91PHPW8eXQuVU95KqKxLsxEbAHg+/Z8uvHpS46Jnx8MLg+e66rDn/WsOfe6enXq46c5qYUUkdyFNgSt+efqL+9fb14V6aK6D7yZPOA0Jayrx6R+PfuvU6EDA4ahoaUSwwqJHW5JRQRfgH3ag60fTqJDinqsqIahbxMxYFRwwP049+P8uvdZanFxy1stCYvRF/m2Zbf7A/1Hv1D16op6dQchtWkSklFREsgMchCGMGxt+R/ifeqdewei75nB0+Mp8jP9k7Taz4GCCwH+BHvVPLrZ6B1aGoq6tKqamlIDgmy2uob37r1eou7trZHcVJV02MieCOpx9RSAMtrNNHovb/X97PoevV61hvkp/Ji3u/beX7mnqKmvFTWiujpogriwl8oXSPe+tUx0en4u5iu6nxsO36va1alTShYfO1FYXiTx3Jtz9P6+/de6sk2r2YarCytkcVrDA2vTgEK345H9Pevs63X16h4/qTp/t+tlyW68CHgoGZ5k8aKZVBvb+vv3Xj8upHYm3tk7e2bktt9T0dPhI/sJ4oxIiBvJoKg6jz9ffut8OPWvN291/v7aGJ3buR1rsjun+IzzY56d30mMMW9KL73XyHVeg22bu2q3913m4+1dhZWetxdBKYal6Ir+4ikKwkIub/19+699vRBjunYclVk8bj9nZaoyVPPVGnEYkcpIjnRY/UW97618+hA6H7+7vwE+QxCbf3AmD8zpTRsKtrQBvoPwLD3rPXujc7e+fO8+vckY8rTZGgijcGP7pp0Un6WGrn6+/dbrXoz+x/5t+MyWViwm4mfwlI01u0mgluCAze/UHXq+XRxtqfO7rLIIkL1VNT1NcdVO0klvqODcn34ivHrwJHTX2r8ndvT0NGwytBUwxyLIY0mUs/quEU/n/W9+49Wr6noi/wAiNwRd27Xkgw0gxCGNlvN+0XbTpBVjb37I6rWuOq86ToHfFHNBVz72oJIaIAwwipS6r+NRBv7317oevgb8cH7Z+VeOj3bn6X7PB5qkqIXqJ0MLiOQMbFzb37PWuPX0RevKDBbZ2ZtrA4uqoTS4/E0lPEad4gknjiCs/o/x96FT140HSyNfQhrGqp9X0/ziE/7x73kY69jj1JjZHUPGVZWFwym4Pvxr59eAHl1z96631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XTfpb/WP+9e/dePDrUm/mEXP8zf4y/8Ah40I/wDV5D78Mde/2ettKk/4C03/AFDw/wDWse/de6ke/de697917r3v3Xuve/de697917r3v3Xuve/de64sFIs1iDxz/vXv3WjTz68F08CwH9P8feya8evAUwOsFXVQ0VNNVVDaIYUZ3b+gUX49668TTrTa/mvb1rZu/BncXMTTJWHWFdvSEn/I/pb34ZGOvcOhu+O/ZEu69lY6NKuJ5oaCFSocs/Ci49+x1voY5M7D5Zoo6eVauMm7lSVZv8Affut8eHTdU7iqKanU1KMrE2LFbXU/QX9+4Z611yopnykiTUsixkgGx+th9SLe/V69w6eaVESuRMidbK48BuBzfj37rQz0bvr3AVs1HBMyNLAxjN1F7C3+HvWOPVjwp0bXCRQ01LBBSWik0AHix/1z715468eOOlXSrVCUiqPpUcFvyL/1HvfXvs6ntXmF/FECVNrgWP1/wHvWa9eHWVpImHmkDBwLrcfSx+nu3z611Ijq/LC8pYAj+za/05BHvw9evdJTLVkNYT5HVSt/rYaiv1/1/dTnrdBx6DTOb1xeLpJxVSxotKDbWeCR/QH3qQ6cjz62s0cWJOJ6rX7q76NbWZPH46siEaQyhQjkAWJHIH59+jFWr06iGYVXqjX5D921m2q2uyVRJLOgllIUPJ+Be5UX9qUddVT5dIrmOSLgc9Vxt8ktw9kSZrGRtUQ0apLHEC0i8A2/4j2T3sr+NSPA6AHOG9WttYMiH9SnRz/5f28dw7f7Kx1PTVs37uTpNaeaQnS7jUDz7dhZgQH6jTkTctzvdz0zNVNXW8X1PV1FXt7bc07gPNQRyyEseTpHBb/efa9Dqz1kjcxAW6mPjTpj723ZVY6hFLjyXk02YIP8eeR7qem4q+GK8eq4t71GdqoJpmWXW7MVZgwsW5AHvQGenOAx0STtTpXdO+YnqJqgrSsup9XGlQv+2/w9+z1406rE7D29SbUyNXt0IJZqZ3BcEEX1WN/e+tH08+i4T11Rjs60CuvqjkITjldPPHvfWui3bgoV3DuWp+5p3DJUNpcekCzf1PvXy690O+xd/bk2NPj6NaiY4tZ47Isj2VbWsSP9597x16vV5Hw3+Z8/XW7dv1lXkgu3WWjiqqeWok0mWSUBuGP+PvRHVsdbYuwd54vsDamI3XiJEkostTJOhRtQUsoJUH/Y+/fZ1rpYf1F+f96v9PfutfLqO9MktPLTS+uOZXRwfyr/AF97PXgDTqsPvTqKDqreMPYW3IhHNkKnzVckahC4Ml2uy8+606tjo0eys3trvfYM+0tyRQVEc1EsEkUiq7n0aSdMn1P1974jqpqDx6qr3l8eKH47fK/Yx2bUfZ4fcFZHLNCreNGWSUMQUWw/24966ser7KFw9FSOGDaqaA6h9CfGLn3vrQ6le/de6CDvDsrG9Wde53dGQqI4VpKSYoHP6j4yfoOffuvedOtE75r995vtHsvL7hhr2qcRVzVUUMcc0jolpyANNzx79SnWvPoieTq5aXHvXzya2ax0Dl+eLW9761gdP21ofu6M5CqOqEJqVDwwH1sR711voUKdjl8dR0uLhJZZ1A0g8Eflv9b3v5de6NlgOu8nltjNUSoZqhDCsYUC4AP5Pv1etjqzf489Wz1GzcZT18Ijdov0yIAf02uSR/tveq9bHoOhhpviRgqzP/xjKIpijIlAJSwP6gQB7116g6dOyNy7U6iwEkKVVOr0iiKKPWFbSv6bBfr71x699nTv0z3/AITd+LWBqmNndWRELkurDgWH9fezXy69joRt41tZWbekippwoGqVRfm3+uf949+UkGq9aagFT0F+wN1U0uUXFVumWe7ICx1WP6RcG/8AtvfnZm+Lh0utZm0FK8egK+XnxIxfbeIq6mmo1aqaEENHCl7sdQubf7b22LeB27h0AuaOX4b4Ehak9ar/AMmOhMp03vR8fXRyxwionKNp0qFQ+m//ABT2VbnAsZJjGOsTfcDlO4s90jniqqrXAx0BePyMFbD/AA3IhJIp7JGdIIA+g+vsljaUmgwOkWyb7uFlKkUb0/PqNht0Tda7jgVZjHjK6pWKJC5CAubcD/Y+xNt0500brMDkfdmv7MLM1TQdH3w2TWtxNKIZw6VkKTkq5/tDUPp+fZi+TjoYOpD9Dd013DlNhbhpIVqdNIs1nVpH9Sk2tYn3TqgOereusu6sf2PNPhRLEGmoFgtcWLOv4B9+PWx69VufMDqzM7I3HLn4/XSz1LTDQAAVcah9PfuvY6LZjsfLk8ZTZEenUpdx6QyqP6+/dap0ImAwn3scUtJE00gZVsPUb/T3vrfRwus+n9xbuhijrYJIaWyhRIlgUB/o3+3968uvUp0f3qrp7CbCP3M0KPKlmL6QbnTc2H+v71wGevfl0OeMzFNW7jjp6V0jiRNDaDpBAH9Pfjw6359Bd35uem29RtD9ZJrDUG0nlffhTrZz1V1vbMyFpK2xkWTWUXmRgWYm/vY9etHI6KJ3xusYbravycpEE3m8aib0GzcWF/futY/b1eF/wnWomq8DunOr6jV/dTO19V9RH5/1/fqevXsgY62pFNwDYi4BsfqPfutD1679+631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//T3+Pfuvde9+691737r3Xvfuvde9+691HmbxOspvosUbk2Fz6TYe7AVFB14k6T1hqaOGspqiiqY0kp6qORJEYXBWRbHg+9sQRXptQVNB9vWtL/ADQ/5SeR32+T7b6No413YkElZkRBTESsFlMr+lOW/wAfYe3HbDdI32dSFy5zMu3siyEgA9ae3em6d+dUbml2L2pisjRVlLWvRLOaGeOMtCdLEO49xrcbHMrkEGlT5dZEbLzlaXUSpq8h59Cx1bIm4tvGanZZKGWIm+rU2gj8qPaRraSLBGB1IEF7b3MQZTnrjHRZB8ytDRl1pEk4H0IGq3+vb2nEgLlek8suqoA6Mlt/ASw0cb6mJ8JJN+DYXNz7vQjj0mDU6Ld2RFVvuekjgVi/mUKAv51W91ZRXPVxnPQxbZ3JUYiOGkrPIrDQG+o+o92HqetDTTu6GjFZ2hkVC0qh3v8AqYAj/jXvXHPWqUGOlNVQ/wASp9ML3BFuGuP9uffjWtetD1HQIZTYITN/xB11kPquwOr6/n3rHHrfxHu4dCJi0p9EcLhB41AJ/pxZeT9Pe+PXsDA6VGOpIBMWZV0g3Qk3sb/VQfeifTr3HHTlV1UcS6CihiAF0i34+t/fhUY61npqaoipoXqJyAPqoB+v/G/9h7sKnr3HA6Dut33FFPLHSguSSosCTf6X97oRx63TGekBnMlunKK60auNQJ/INj9LWt7rQnB49eAFKr0n9nUO5qbPQS5nyGBWu2q5Fib8k+91pgdbpivRvkkpZo0MaxaSiW4X+nJPv1BWvVKqRQ9NdZi6eQl0WPVx/qSv9OPdcDPVs+fUQYm8bDxxkf1A5/pb/jXvf2+fXu4cOmtftMfNaocIgJuFNiP9b/D36h8utVHAdSpsph2jAR72+nPLAjgn375dbpQ46Y/KskjmmUG/1P5P5AHv3WzjhnpL5bG1k0mpmZIvqACQQDzwT73So69UAZ49Yo8Opp21uSLfk8n8fj36nAdaFBw6ZK7E006xxpIQyHkA8Dm1uPfqUwM9W0ny6zU2AVlCg614+ov9Bf3ulT17BHf0m9zYSili8cccZmAAUqvItzx7o66fh49OQJWdcdAxTVUFDvTam361izZrMQ0MScksZHAAC/k+xHsloZpAZPM9R97h7obOBkB8utiPE/ycdnd8dT4qszxmp5sjjaXIrF4JVuHUMpHuUbKzWCPUBnrFLc5frrgyN69Pm1/5WM/XWBm2rtPL5aCmo4fARCZk0qBY2F/b7yFzQ9IzSmkHh0I+H/lo7q3FthsBmdz7glxhDI0M01Q0Wl/1XRjb3qvVaft6XXUvxRxHw5p6yswhkKFnmmlVGRjIt3uf6+9ceHW6AD16LB3zlc38v+zdmYUxCoxW2snC1V5h5QEgkteQfj/Y+/Dj14/Po4G6un8L1/isaKGmpokhpYGbxRhVLog1G3vf29eNPLo3/wAYMJt3emIkqqikop56NTGokRZGNuB9feiK9eqRw6DP5u/GzYOS653Zl6lKbH1/8KmMLwIsbhyhKsrj8/63vwqDTr2OtXXq3sDdG1sluDrLG5mslgiyNeIZTVtqEaORGAb+7Y6qPl0ZboburcXX024Vz1ZWTF6ip0GaVmUrpNit/r71nrfn1WblPkJX4z5u4ndDLVJjavPxGaQqVh0CcFgz8D37r3z63jOiuzKLsnaW1s7h6hJKWLBUKStFKrp5PEuoFl491I49b6Hp8lGk6s9h6hcDk3Bvc+/V69TpUUWaiUrKPoGBAP0H+1X+nvXDr3XLI5dq6rj8dwthcg/gGx597+3r2T0pse1HUtTqXGpTZtRBH9Te/vfzHWvPpTZHJwUsMUcZsptcJ/qbWtx791vp+ws9PV+KoRr6E/ULagw4P1/P9Pdh1UjpxaWhNUx9LOpvw3qJve5Pv2OvZ8+uNbFHMPJ6TEeG4sv+x+vPvx68PTpH5fEYrIaKTxxaZVBdrL9bW5NiPejnqwr0nKjaG2aKmlDeHUEYk2Q8/UDj3qgHXq16RsGPwVTqoqaSAz620rrUG34uPqPehTr1PTr2Q2bi8lF/D8rBTTiT0KpIkFjxc+/U69w6DHIdHdfpP4pMXQxSsxAZKZVY6uR6re99aHUl+mNo46m8KwQCFwWJEV7cfQf4+9dbB8+kVW9bY7HrNHtwMPMD5hGpA5+oFvfvt690GuS6Mrc56I2lWUFnkAR7Mp5Nz731rj020vw/23m50qcxQU80cX+eSam1o7Dhi2oc+/VPW616ibw+LPWVXt7K7bxW3cbDPWU7wFoaBV/UpU3sPfq+R69w6rtwX8tXrzYu4sruHMYqkZZvuJo0NEulmclh9R79UdepnomvalNhOp94SbPxW2MY1Vk6hloB9gg1RuTpP0v72M9awD0EUvRW0+2KrIUe/wDHRYjI6T/CRTUugTVH1jU6R/X36orjr1D0U3uj4at19j2yVZTy0xac/ZvT0zCUoT+0Qyj3vietEnoDX6W7HSqwtbWVORpaIRq1AYVlR3Q2K6rW59663no1XUPx633ujO0024JsxJi6WeKWlWWOd0crzpcnjn36tOvDPRh+8OhO06rFxU+2sbPBTRwiJWhpZELErYN6R7917y6BTr74Td0ZGjq6vOfxMFwTEBHUeoHkWHvfy610cX4qfy/e8cB2HSbqijraHHmsil+5WKeGR11XN3/1veqjrdD1s99a4fc21MdiaDM5Crd4KGON/LK7kkfnkn/Y+68OHW65p1z3Tu/PU+RMeKqJnUPZj5DYc8iw/wCJ9+HWj8ulftv5CLt3w4/ccuga0UmRl/rZuW5971U69SuejW7V3pgt40S1eJqklVlDFPIhYA/00nn3sH068elaL3P9Px/xT3vrWa9d+9db697917r3v3Xuve/de697917r3v3Xuum/Sf8AWP8AvXv3Xjw61J/5gx/7Gb/Gf/w8KAf1P/A1Pr791o8R1to0n/AWm/6h4f8ArWPfut9SPfuvde9+691737r3Xvfuvde9+691737r3XvfuvddH/iR+be9jrR64yBihCEBj9Cf9596HWzw6QXZ1U1JszLyRkiUQN4wP1E6D9Pfjw68B5da3/eXxgh7k3BmMtlUYysKloy8TN6y5IIv/tveq9boR0THamyN7dC7menEU/8AduKYxtJ430rGrXsp/p73Xr3RsMPvzH7mMc9A8AnSzPGZEDNx+R79U9ep1Mz2UNd44qiNU1EJ6ACLX/w/4n37j17rlHK+J8D07uVIUn/bfke/Z610ssXU0WWImllAlp/Xa4ubfUA+98evUpjqwbojNUtXhFpZgmldCBnYBrAWA59163546MHTx0yVPljIYKTYX/xtx70OGevGnSqjqo6gGMgDixb8/wCsD7317plmqI6Sf9ty121ckfW/0PvX2db4ceuqrJeTSCQpYjSAwIJHv3EdernppObnpndHKhBxe9iAeORz78cefWvPoKN67qpaISSrVBSqubeVQTYXPHvw9evfLqvDufuimrWlxdJWkT/oYRzAi/0s2k/7f3cJqPSS7R3UaOq6t6Zn7aasqZaty8oe5aQW5a/PPtqT9PA6W2d0tvF+rxHVZvf9VT1j1AqpI5IXZyburcFeeP8AH2ladkNOgrvnMdrHUas58+iDLNFhKmqTDUyM9SG/zcYJYluCAPaAyGR69Y5cw7vLuN80KGo8qdXLfy0+iNwbp3bjNzVtJIkX3dJOGaFwAODy3tbCNRq/Q/5F2eWKUSSClaHh1uK7ShkxmGw0EXpWhpFjfSeFKJ7MEArQcOp+oWhVegb7I3/t+Cuq4KyrgaWAEukkqN/sbH6e/Nxx0mpTHQGZDMYfc2FkONWCRkck6dLXA+nI9662OgV3pn8dj8PU46Uxx1LLoGlhruVsbAe9jhUdeY16qp3x0yuezWWzriQ6/K6lkLC5bUbn83/Hv3l177OqyO58NV7P3I1bCsn7cjxW0nhRx+RYe/dV6B/I5Wl+2Wt8SrUzKHNgFcsf629+619vXPbO5lyU01LVxIPt0LKXA4IX8f7H37rfQ99e11Vlo1poKuWIw1sEilWIOmOW9rj/AFv6e/de8utwf+VZ3fLvDr9tl5Ot8k2Ch8FOJ6hWJ8IC2Ab8m3Hvw62eFerfwRa/9Rf3vrXDPXGMuVu40m54/wAL8e/Hrwr59Fz+TMWFj6/r8jlWs1FBI8V/oNKlrgf6/vR4dezWg6or2V8v8ztXeWRhwMsgp6aomjVTPoUiJiLAXHv3DrfQS78+Y+Y7L+TXXlHlJWFZBUwJBGkwlv8AvBSLAk+/U691tEdd1kuQ2dg6ubUJJqKFmDAg8xqfoffvLr3S04HP/E/8R791rHVAf86r5FV3Xe38D11jKh1l3akdM6wzAHVKp4ZByPfqdex5daqGaxGQx+ORMjqlnlnklUy3Lapn8nN/zz7917oHty0lXjp0mql/yYgEx3PA/J0+/de6VG1ppMxSvT4oF9QKsAp4/BA9+69nj0cHo/r3KZCphpRS+WeSRF0sl9OpfqAffj1sdW0dX9ZjB4yGhzNMlp1R1Xx3UWPAPHPv1PXr32dGopamTARY6nx0Kx00ZA9I0DT9Pevz63WlMdL3N9h4HEYCrrspX/bstFJo/eVGMiqbXF/6+9Adb+zrX5+UHb269174qIsfVVDYWOrcBhOWVoh9P8Pduq0z0FfS/wAhctsjfePoKyrlWimrFF2n0IbkAjn3vrVR1sJba3NT7x2JS5yjlEsMlBESyyBzqdOb6b8+9CletTH9PoHdv1EeJ3gtSzG5mueTe+v3oj+fWrabSaHy6O/iM5SZWlCTRiVXWNfWL8gfkH/ePeqg4HRkRDORXqiD+a70xR1+Iq910lPTxyw/cSXjQBv03NyPz7SXcZK9Q97obFG9q1wgBoD1rdJRtAtOB/nYkFx/jf8Ar/sPYfmVlaoFOsRLUP8AVsjDK9JzsDHS5vF08ykibGuKtyDyFi9XBH+t+fa6zmCNTh1PHtzzGY71bUtwIHHocei+69u5jFfwuTIo2SodFGEeoThozoKkE+z5aMAesmHYOqv6ivRja+uaOSmrIJVPk0sDHIrD+v4976bFOjLdJduzbLzuPrJatl8lVAjXlCgKPqrX9+68CePVsW99q435Edew5PGtDVVEdGWKkrISwT6aR+fe+PW/l1VNubauf2LlJ9vVNFMkCtJGpWBwqgtY/Qe/de6PF8X+nUzcdNXVNN5IEeGQmWFh9TcgA+9ceveXVmUO38bjaeGkxtPBCIlKSMFEZuP1fT37rfz6BDsDunC7KnbHZGrCsPSSkyDgnSQT79Xr1PXrB192Rh8/k1rsZVCQta5EoPDfQe/de6g/IV/4rifuCdUsQuq31XOj6gj37rRz1VxnN1z41qn+IqfDSmV/X9NKt9FB97+zr1RToknceSyXesb7P2xHL4vulMulGZTpa34/x96r17PWxX/IlyE3TNNVbG3AviqKwy066rQ6zcAAlv8AH37r2etrBW1WIHpZAwa9x6uQB/sPfutdc/fut9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//9Tf49+691737r3Xvfuvde9+691737r3XEi7c8rb6H+t/r7317FOsRjbyBg503uVvx7bAcN8utEefWNoWbzLN+9TyKytCwDBlIsykH6gj8e3jpIAHHqoLLUkY6pc/mJ/yjuovlzt/NblxuIosJuuKhyFUggiWF56gQMyurIP1Ej6X/PtPNawvVCB+XR5s28XFpcK7EqK9aRmK633r8Uu89w9K76op6XBrlTQYiuqvIVkjLlB63Fv6ewHvW2SKzGJcdT9y7zzYNGqSSjV59LDdZ/u9vt/s2L4+QoUlWxQlvqPYF/dV7HOXdSF6kSx3e2vnrE9ejH7aq46miiHkF3gDWuByV5Fvb8iMh0kdHxUcegi3ZhGTctHVPB5Y451YtbiwN/r7TGRUbu6eSB5ML59CFV7Woc3TJUQWSbxCyrYG9hYAD3Q3S1pXpLK307+Gwz0GOYx+cwEwYiVYUIsSSRpHP193jl8Q9vTqRvKNajHQh7V37AkCRTVIMkfLXIBv9Prf2qEMj/B0muLiK2NJDTpVVe5MfkUbRMDLpJ0gi/9SR70YZFOkjrS3duy6g+Okmu5cfTuxepUFDYg2Iax5978Fyer+NbAfGM9K/D7hgqpIjHKGR2uACAfryfr700Mi5p01Ld2kQ1Ful8UhqhqNmOi6m45/A9tsSOA6U2v+PDVB3DpG5Ckra2p+yAcQyNYt+NP0sD7prPljp14DC1Gwem+bZkOHmglNOtUZioe9joB/PP+9+/aweJ6cS2kmFVr0rf7txJ45YogiiMMwFuCeefftanh1RoHTBFOoNZtyKQFi5jfTYcqLj/Afn3bgKjpOxVD3dTIcTUQUIInfSo1C7fi3HA91qa9bBDdwGOk9PPloi7xJJKita6jgW+pv/j73jz6cWGSQY6k0OZr2UJJE4t/t7/T6n3sgHh1drd0Wp8uoNViarKVQMhZFY/QkfS9/wA+9O2kd2OqoA47ess21UpgCs4kcjhbg2P1/wBh73H+thMjpt2EQ7sdQIaqDGTIkth6x5PodI/rz/xHtQ9tKgBIp0kS+tnYoz0p1JzmSxtVCvhqFVlA9AZARf8A31/dPCkJ4dPC5tBxcdJOSvpKenfVWWsP9UPz/Q/73734MhNKY62buzGdY69DPg6am+7rMjHGJBePUy/q/H1NvevCeumnDrYuLdjUNjqFFvPbND5NeTg0gG13X6/g/X3YW8rYI69LcWy5ZukLWb623VxZCojyELNAWKkMnBA5tb24lpcPIEVa9JRv+22Mn1E8gCr/AIOih4vtvaU/yJ6obIZKmFHh950lVkdbqBBTJKCzPc/T/X9j3YbCaCRfGUinUAe53Ndhu8j/AEEgIPp/sdfQ360+d/xayGyuv8Tgd94KOSk2ljKarihkpRomip1jkQlW+uq/J9j4lRGRXj/m6hW1OsVJz0JuO+Q/RG5qmb7DduMTx3+4ZJaf9wf1Y39lYDBySOPVNDCQk9CLi+3epK2E4+m3XQxIy2SdZqex1D6Cx59v46tXz6AbuHG4PNYmumx9XFm8e8FTIZF0uo0wk/VfeuvVI619en+4KfZXe/ZOOnx4gjp6qoFK2kWQ+UgFfez17zp0ejdPY7b92bIlDP8AdZGKAuadD+4qAEj/AB9+69xHUr4fd15zb+Wrdsfw+RnkndWk0OxW7m9z/X3rj1vqZ/My+Ru49mdV1eNp8NPM+Xoft/OqEmAyKVD/AOH197rXPWjXrVu6X2vuHd27cg9ZkJsdPV5SWtbLsdLJHNJrMOr/AAHHv3DrXljq0/a3SW28rT01P/G4691UpVTiRTrcCzsSPz/r+/Z6t0Vr5YfGfrrZG3a/eNDWUQzVJRT1VIyuhl+4RdQ0H6/X37jx61w6te/k49lZ7P8AxgrnyTy1GQpa1KWm1hi/gSbQti3NrD3o9eB8+ryNuUP3+PSerYpK66jqvcMR+fevt630q6jHoKKBIVu6uNRHH+tce/cevcOnBKD1xSoAdK2Pp4+nN/6+/fZ1rrHS0dSlW5SRgGY8BiNNz/T6e/fZ1v8ALpZx4tp4D5ZQSFuCxP45/HvdOvHPHqdt0SRJOisQ2ohNJPJH5Hvw68aefS0xmH9UrzSjXMdQZgfqR+k+7AevVSa8Os9VRNHSvB5fXzwGv/iL+9fLr1fPpBVtCYaeon+7InS2hbk/QXK/X/Yi3v2aV6t9nQF7h3VV4ylyM9TL+iNyFJJvbi/Puvnnr3ljqiz5BfzOJugO15cfWIxolee76CRpjuSbj+nu1AOPWuGOlb8cv50HV3aeZmocpuLH008czRaJNKyRkNaxv+P6ce9U68adWp7Q+U3W3YBYUeaopFjiDisBT6ldWq9+Pfqdb446XWP7567q2OBOfoqipZzpcshZebEkg/7D36uOvAHz6EzBZDaFcyNSZWB2vdgjIVP0tyT71inXvPoWaHFbeVRUx1SBmS7C6XYEcj/H3bHl1rNepSQ4tkeKERqp/XbT6jfk/wDG/e6dbrQ9MVdsrFzRS1VOqpMRrDCwNwb2v71Qda+3oOa/rWj3GstPWNfUrKodl5J+lveqDrwr0Qfuj+XdQb739id6o4jbFsrRrrUMQvP0/wCK+/de49Mea+C1HRTUWZgUTVdBIs6x6hdmT1WsPr79Xz6300bq+Kg7KT7bceFKRUkQ8ccyqAfGPSVv/W3v1a9aNOi9TfCiDcGfpoKum/huLwrLDGrBUjZI2sLH/W9763w6P9sL4/bJweFosXQYGCtlpPGZKhRGSdIAZr2/3v3Wp8ut9Yu09k4fG4dEocAk8qyi8aKpIUHkmw9+r69aA6Fno3ZGxtyU+MgkoKY1MahaqlIQMH4uhBH1/wBf37BPW89GnzmEwWBi/hmMoFojEgZfGIwOBb8C4t7sRTquT0HCDXkNNXVXPKJrP0BHA9163mtOgt3bTHEV0k7sFSYko17A3P4/r/h79x690WrsyCPJUUtUJ/EaeOScyEkEaBquT72Kde8qHpn+MfyNgi3am06bNCpMdQsDxobkMJNOm5P+39+8+vccdXgYyqNbQUtURzNCjn8clb+7H19eqA8R6dTh711sfPrv37rfXvfuvde9+691737r3XvfuvddN+lv9Y/7179148OtSb+YR/282+M3/h40Frf9Rye/DPWvMdbaVJ/wFpv+oeH/AK1j37rfUj37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde/31/fuvdB72BSzVmKqYWQtTtGeBa309Vz/wAR73TGOtD4s9EA3PtenjkmMDiAetnKkXax90+XVvXoqHa+0sPuLGVeIaiQzSJJ+/YE6yvDKfe69bFCOqVOzsXv/ozeK5KhiqqnCvPqmC+Ro1iMl+bcfT3bqor0Z7rjsyk7PoofHojniRDJYAMHQeq5+v8Ah71jrw6E+mrvuKv7VmLpFdC1/SPx+f6e/db6eoR9hVq0Mn7btZgP6WsSffuvfI9GS2X2Z/AIYKakXV6oyxW3pKjn341rjr2B0cvZ2+49w00BhIaVlGsC1g30INvdRjh177ehZXImmpi9S/g0qGDt/S1/z739nWvl0H1Zu6netMNPOszarFAQTz/X/e/evPrwp1n/AI/C7LUyTCIU9iUuASR9bi/vdD1uoAr0Du8e28fR1VV/lKxpGhJPp0kg2+t/dQc9UVx69Vqd8fJKVMhJS4mtMgPkjYob2uLcEH8e3ADwA6uvdleiO5Tehq55cpU1pLsTJIpbkN9eQfp7cQmHL+fS6CNI0Lz4HRTe8O8aWjx08dJUAzrE4OnTq1KbD6e0c9xFQ5z1GHOPNm37UpUSAUr1WtluwstvCvmWpncxGUgBib6W4Fr+yK5kqp0nj59Y7bvzU+5TFbZ9VTinz6MN0N0DlN/5+i8dG88Ekn19VrFgfr9PbtrFLIoJz0IeVuVNzvpVu5ojpOQettn4b9OYbq/ZGOhlgjpskIqWym1wSn0sBf2ZpEyGvWSWx7bHaW6o4oQPz6Pvl950+y9r5CrysqwDxSmB3t9PGTf1e1qaRSuej1204Xy611+9vkRuHJdm5inxGRlloZpJYrpfSB5Oblffjx6YJ6OJ0DuuGp2YqTZcS5OZUvT3UyDWt7cn3U149WGPz65bt2RXV2cgyNbkHWnd9SxM1gwJ/IF/e+PVa06x7lwuNw2Cnq3VHjEJZzdbAW+vPvx698uqaPkRTUe5chWzYjGrVU0L1JkkTkK6j6G3+PvfWuqqd2VmTTKz00UDolE+kooYA82tb8e9Y63inSk2jVlysrU4jc8OSDyCLXZiPfutdDltqvrcZWwz4+4hJVpDHc3N+eR7917Pn1cr/K+7lzmM7lweDgqJI6fK5ExVSAnSwMgBB/x/HvxOOrD59bl8JDRRH+scbc/XlQb+/dVxw65SOUAIUtzzb8f7D3sCvXiSOHQEfIfao3d1ln4lYFaagmmkW9v20TU9/wDW96PCnXgc161IOxNoZP8AvHm49n+WWpira0TJThywCyHUSV/HvQ6twPTN8Q+gMv2B8utg5bI1UlTBjaqIZAszN9u6ThmVr/n8cj3uvWhx63Y8XjqfE0FLjqUWgpII4UH+EaBL/wCxt791odcMrX02MxldkqyVKeCkpJ55ZXYKqJFGXJu3vZNMdaGRUjrTO+fXblN8iO/MjTyD7il2Jl/DTOw/ziwOQukfT8e9dX6rn7KqIqvItSRQaRT6GI0i914Fj/sPe+q16L1n4P4xkEjmb9pFC6Dc6vxyPeuvYr07bUrYNs5OGlWkWL7l9KMF/WGbjj+vv3Xga9XPfGXr2WooqPctShpoZjEUlYWuSt+B791vqxfFotGkaSU33cQSwmK30j6ce9dbFOoudy9HFTzFnEJQDSLgBf8AYe/Lx62T5HqpL5M945CjzEu3afISLE0skZUE2A12J4P0976qOiV71y9JJTRFK9ZaqriEjMv9lmHII97690UTeeTqqCeOtp9T1FITKJR9Q173uPevs6r5dX/fy1+4KbfvX8O1c3kdFSsUSaJLC3psACT9T70zaRXrZAYUr0aXftLFtTeFOCnlpp5gFewtpLcG/wBPdlYOKjpPMQGAXo3u05qBtsJk0RAUSN7qQPov091eiZ8ulUSXAo5GOqKv5mffFBNBW7PSoXW5qk8ZKg306fr/AF9o5biKvceo19yOYLC2sGtZ5AshBwfs614XljJmmNvpdRe/AJ49kdw+pu04PWKVksTTyTSYDcPs6QW9Mq2M21k60D1VFFNGEH1HBUGw+nvUIYEV6EfJ1he/v/x4VqhPVW2K3lnto5vJ5PG18kOvINKVUt9der6fT2JoiREB59ZnWJrZxq2W0jqwnor5C7y3c9LS1tDUVEVM9vLpfSQBa5J9ufb05KREe/HR78HmMhm8ggk10axIJU+oBfTfkf4f19+bBr5dOQxtMniJkdWu/BrvnJYrPUuy8hVtURzARrE9yShfTcAn37iOqedOrYNzdU7O3xUmqq8VTrUykHysqBrlb3vb37r1ehQ6/wBoYnYdEuPx9LGS5AGkA2vxqv8A63vRPVh8+kL8g83/AHG29UZOiyHhrJoyyQi1yWS5sPfh1o9Ug733pl915OqrstUOw1Oy6y1mIc8G/u1OtdKro/tDJYXdtNTSBoqEyxJqKtob1WufejTrw49WY79Emf2nBXUitMtRCJL3JtdL+/cet8OHVYPZu06mrd6aQeFaiWWNmOr9JaxPvw6qelJ0x1XtjZkyVoFPVV0xSQoLNJcm9+fwPfutgDj1Zb0JPT7P3ZjdwU7pSAzeeQqApW7gkf7b37y631s5dU7pXeGyMNmllEwmpYl8n5bRGBz795da6Ef37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf//V3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6900F5IzJLIjSxEGORbFvST6vT/vfuwHSm4MYhoOqRP5p/wl6F3zsjM9uV1Rt/AbqwcX30ZqWpaaeedIzIpiLWY39orlGIxToujdLSUSajn59aAHZ3y72dgOx83sWeVKubC1FYoqady8Z+2nMaepf9b2TSWU9x26cdSZsnOC7coYNT8+l51f81tpVckdPU1McWkog8spUquri9/ZfPy8/wAIHHPQ2tvdBHNCwx8+jcr8jerMli3qaisoWnjjuoMouTa4tz7JpeV3Y0pnoX2fuZbhK6hXoEMn87urdn1bwVU0bFGYizmxCnj6f7D2WPyo5fgejaP3A5fn77xgG+3oPN0/zE+sc1A8SKrJ6hcarWI459mVnyzIhGlf5dM33uPtcduRYuv7egNqPmbsmOR56LWDIfSNUgtzf2Jrfl51pUdQRzVz7vE8tbVseVD0kH+ZVUldLU0VQwp2UqFMziw+g4/1/ZuvK0Lprfj0El505tjXDY+09YML8nMvnK1mmrlRHYkK07C4JtYAn3ZeVbcivXl545uP4qH7T0MmM+XdPgCKeWpSSWPgaZmJB+g/Pti45WiEJoPLr0fO/M6yqbuTsrnJ4dDNtf55bcp0EWWlUTkcapGvpvxx7C/9W5iaBep35U9yIYLMdw4CtT59Sqz597ap9y4unikUU07gu5clACRy3th+WXNe09P7h7owNPQuOhfyXzo67nNJRxTQPU1DJEjB2I1sBb2mHLbqaU49LbL3TihTLAjpYUPd1dmJo4sdlqONKmISohlJIVh6R9fakcs4yOq3PuhDIK6h02ZLLdo5wVEmIy0Ui05LKsRZ+F+guPamLls0oR0RXHuQjGmoft6ZKfe/bkcZx8q1MtSLhmWKQ/Tj2nPLzeKQQelCe5cKQ0Y56cEzfc5x0kEdFXa5WJ1fbvf1H6jj2/FysZO6nSSX3dWE1XNOoObo+9sJj6fMPUvFAEEskckbKwUfUEHn2YJyhHWvn0Uye9TyN4NOPUyl3f2RFgIs7NWxtDI4j03Nwx9LDjn2nvOUQwpx6Uwe65hw3+HqMvZ+5aKVJ8hXRBWUkIZD9CL839v7fymIMgft6Q3/ALs+ICB/l6Ll2R3juPFwZ3IjMUipDCz08ZqdBuLkD6/7b2IH2CCVQrgY6BN7z3ut9nbG0sONT0QKg+cW8hkJ4qupV0SokUWqmJsrWBAJ/P8AQe2v6uWVdIPReOcuaVwXH7enjMfNHdNbQztTGYyRgaT5XAsD/W/uw5YtiKEjq39b+Zz8TjPz6Drcny17Oz2LoqOiFVHJqUeRJJDc3sbAf7x7aXlu0LeGCKjrcPPPNavoDYHz49JGv7r7qSnEkwyjJIByPNaxFje3tQvLsAGDX9nS9+duZ5I6SkD8+m/C989i0nkpKqoqY2qydSTSyq/JsLAn2pXYreH9dMFf59B2737eb0m2nkqr8aNnqFDtffeey0+4aZ6uOoqPWagPIpXVzdG+o/1/aln1CuB0Vx7esLUqT9pr0u9tdl99db5GGPD7gzpAdE1TZaseBV12A1auB/h7aKqc16UBAmV4dHNrvk98uNq7Sj3Ft3eI0/biashfLVCufTdrXb3sE+fVpG1AU6FT4x/KX+YZ8iNw4Pb2xslXQKmWp4a2sFbVTpJAWswjN/z7959N563z/ihtnsTrb4e5DO971jVm4hiZCTUsxkLmI3K6/wDYe9fZ1sYx1UHg9q4vM9lbw3O9BOYcvLM9E8aWHMhIAPvf29bxxHQ97MwWZ2jWVOcrIqiPGzwNCqyg6bFTZbfT/Yj3vr3Hqyr4I9P7f3HiNx75r4QZIKionUmJWBHkJWx/p9feqVqB1okDj0Ev8zDE7OyPUm8ZZ/to58ZhZ5qEOFBLxoSLFvzf3oDPXiR1qS/FPG9jZxtx7lylSI9t0+4q6ngVTf8AyeKchQr/AOsPe8daz5dGu3l2xltq0s39woKqYQ6vvpIXkdVOn1cqeOfr7317qr7u/vjuDsPcm39oVVNkBTZvI/w6NmMpR/K+m1j/AK/vXy631uH/AMqHper6v6Ax1JuKklWqr6Skq/3Iwtw/7l7Afi/vRNOHW6Hq3LHyIXWNF0RL/sLW/wBpHvXn17z6Wi0/jg8g+gAFvre44NvfuvV6aY8nIKgQIOCxDD+ov/Q/n37h17Hn09RqyzxledZsbah+f639+p17PQk0tKrUQLxkWQElgRwRcgH3brXXLBCgHmWMASBjybG1zwPfhx683DPSvkeNVhsy/Ww4BNvryfe6dVr0l89lY6epZFdNbILDWL8C1yOb/wCt711YfPpHmaCRZZ6l1IF2Kg/Ugcj6/wC296qOt06L12dQQ5Ckn8J0xzKVW3A5/BPvXn1759Us9p/yzaj5E9tPmcoGbBSLIGGiMqTKLEaj/X/H3brxHp1G2X/Ia632pU5PJYylqabIyTNJHNHpRQfrckf4+/V61Tz6NrsH+XruLZ2Nq8Zh8lPHM0DRRSGYixCaEub/AF9+r14Y6QtB8AO8MZlKnKRbllMhqHeK9QSQuosB9ffvs69w49Kuq6U+TWyYzJRZqeYx3/zbsbk/iwPvVcZ630JHXNZ8io6yJM89a8SuFN/Lp0j6mxP59+oOvCnR6ds1O7XggNas4YKASVYc35v7114VPSxzlRumPGv9iZfI0fJ9V+RcH37y69Tz6Kvlsn3clfUNQNP44/I4IDkWXkW9+698umPF9k95VEdRHU09Wy0zFCTC/Ok8n3vr2OkXU9tdvLl44ZqarMUMvrXwyWtf6EW9+49eoK9c9z9rdo5OZKPFUVTFVNFp8iwuFBIte9vfsde4ZHQE7l3F37iQ89dBUVUEramEET3t9fVp59+49e889Tdhd/dibZeQVuNyDia6CMxSORfg39+p17pTf6bd3ZDNK1ZhshJQzfriMEh4Y/TSQffvPr35Y6G3r7cuRxuXhyeBx9dRCaTyVKzJIq3PJ4HvXyPWzSlel3uXuPc2T3HBjqairBO0kaSTBX0FSbf77n37ic9ePDHQz4htdTRHLB/uJKdXLMbWY82J9+61wPQfdu5ihiqqCiZdSzOqKENwoJ4JI+nv3n14eg6K929kaDC7ZyIkddMuOqR+r1amjI4I597pnr1cdV4/CnblVk/kEZ6ZKloZsl5VsxZArVFwQPfq9a62y8VTmkx1FTH6w00KEf0IQX92PWgCOp4+nJv/AI+9deHXfv3W+ve/de697917r3v3Xuve/de66b9Lf6x/3r37rR4dak/8wf8A7eb/ABmvb/j8KG3I/wCV1fx78Ot5620aT/gLTf8AUPD/ANax7917qR7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6YtyRtLiKqNRfUhuLX4tbj3uleteY6Izu3GKs1R5CVK+Q6TYfnkH/AA90+zq3HotO5cdTtLJUIhupYE24Nv6e/Z68D/LotG/+ucV2JjchiqmCIyzI0cbGJCwbT9QWHvdetkdVs1O0qvordkmPhjlWCaZ7MRZdLG1re/VPE9ap5dGhwAo6jBfxONdUs0fldhz625PB9+4dePSemqskkys0jCGUnTfjn6Dge99e6dItz1WJeJWDMWZbMAbEH83/AN79+69wPR5vjnn3nljl1F1ILMpYsPrfj3o8a9e8ujiZytGSjki1+BFjGstdVU2/r/h9feqivXjw6KZv/tvZPW0k7VdVTS1yrKbCY6rgc2A5v7354698uiB76+cEVLV1cVJKwikdvGS8mkg3H4I4978qevXiewjolfYPyrztfNLItdppp9SlRNILgtf6e2dDaq9UtoRI2TjosO4+8ca0jz1ExlqWu1zKzc2uefbolCmgwen7gpZrqJ6Lrvnvp4KWoFJLpMoOmzsPqOb29s3kpdQD5dRtzdzzBt8BTUP29E0yG6MvuzKTNVSySROz2XyMfSx+oB/3j2TMAT8+sYOY7v8ArNMfDLVNeB4f5Ohs6d6ZzW+87R0eOx9WVllhJdIwykF9JPHtp4S69vRzyhyTMzg6TxBqetnf4ifFuLrnC47LZ+mWN1QSAzQRhuVBH+v7O7IeHFpbrLjYLdrCwWAgYHVlG3IIK7Kh6NSlHSxxtdLpHeIXLce1NejM16Iz8+/krTbbxn93IqtRIqtBpimZXuY7XIHvYHXuA6oypd6CqrJMtI5JnZ2Bka7MWa5PPv3Wh0N3X3cOT21WxVsJnEMTox9cgiKqeffuHXq0x0cnE/KPG70rcRh2H+VEBCbseb6f1fj/AF/fqEder0LfZG5aXMbKrMFjQ38SejZCFJZ7svHI/P8AT37yr1unVcuC6v3bR0WY/ikczU081VKrTx6mKNdhZmH09+rjrVDw6r+3t141PufMqIDIZZzeycKCxv8AQe99a+zoOosE+MrHg8Tgf4gfQ8ce9de6fqXLPiplpSjBZP6DgXPH19+68K8R0bH4tdrN1X3LsbcJmWnpVyCz1TSOVTQJQfqv049+4deHW+t0h2jhu2thYXdGHmimjmoaMStC+tQ5pwT6v6/X37yr17oWZJUp4nlnkVUW5LEWCj8D3v7OvcBnosHyK7p2tsLZGZo6qpglqM9jpqSlVXsA1RGYzcf1HvzGuOtqPPqhvaGzVwOT3BvSumgajrzXyJDNpNo5yzDTq/1/det58s9cPhN2p1xsz5N0+1q+KGlzG4so70U9SBGp8s11KMf8T739nWqivW0UCGAZSGVgCGUgggi4YH8+/da6p9/nHfLum+Mnxg3LVYyu+33JWwTpCPK8bLCsdjZ0sbm/vx49b8utOv4/d7U/Zq5DdWTkY5POg1cjtIzmSRze+puT791rp63bX/7maqd0YiVbLx9Sfp79149B+McEY5erVlpQ1rHgcn08+/de8+he6j61qN/bio66WlmegpptURRdSlFYEXJ/Hv3l17j1ft1XicfRbRoMB4fGtLHD/YAOpUF+Rb349b+3pd5XeFHt9BjpNJDBtJ54ULe1/p71TrZJ4dEx7Z73weJFbTySLrAGnS7K3+0jj/b+99ax1Tb25u87m3fPlVlbwmSSzFib3bj6+/da6L7vTcctHLTyQTBtK8jyMR/tvfuvVxjoMclvOCto6lJVJkaFhqKnTq/2P+Pv2fPrfl0an4D9xVm0eyqalqKs0+PerplVGmdVOpubKOPdZQPD6tGA76W62Qu1MhR5zaeIz9Oyyn7dJxJH6rkgNcn3q0rkjpLLGUlqOk9k+3a7aHUVTkmlAhShd0UlgV0L/Xj/AF/drqmmrYB6rue8NZWlCaU61Wvkj27V9tdkZKsaZ5IqTI1icSOVuWNuD7Dd2tCesO/cW7j5h3dX1ElGOAcdFwqVkVZF5uD6Rf6fjm3tLHSoXoH3UJitgx/D+XWWTCUm4MNU4urW8skLxx3tyzDj/fW9rQp1CnDoX8k8zraXIjbBB6rU7a6Y3BtjK1ESE+KrrRLEdIHoaS6gW9iCIjwgB1ljy7uK30CtXNBnqw740bAGB2vjppqcSVFTESXVFNnK3Nz73gmnR7uiBl+3o3lHSiOo8QUxTaAb2A4tf6j3ap4L0r2qQpbFB0Ovx3yE+F7fwVbofRF4b2PDfuj3s9JnNXJ+fWzLsF5s/jqbIaSFaNHAYWt6Bfn3UnGevU6ETNg4/EzZOOVQ8Ubk6ja2kX+p/wBb3odbNQK9Vg9+dpNuiqlxksjOYZGj06mt6RYD/X97+Y6r5dEezFFR1s0UCIC+v1Ffzc/n37r1OsT484yopo6dgs4midACAfSb21f4e/de6sY6a3lNncVjsDVhnjEDRMGux/1P549+8+tivS+7G6Bgy+PjrKOEiRh5FCxjkub6hYe/cMDr3HJ6LM3UO4NrZuHITCVaKFgGXnSNPP0Hvx61w6GTa+UmrUqFjkMYpUJNzpGkED0ke/dbHWyR8KK6oquncWkxLrEsPjctqP8Am7Ee/DPWujhe/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//W3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690C3fPc22OiOuc/2Fu2qiosNicfWytPJIkQFTFA0kS6pLDki3vRanHrR7sdfNk/mlfzPu+vmD2hvDbe1dwZ7bXWu3MikcVdiq3RBX0KsV06YiARp+vuugOajpBLZuzaq9UOV3TuYylVPn8LNJlp66SV6irrATPI0jF3LyPySTf28kYTuOB1VY5q6Tw6i0nUGco5DUTVLUsi3JWO4sw5PN7e3vFiJ0np/wAOVDVK16YK6TL4bIJFJncn4YJAZApcoAPxa9re9rHA51dNNLuMZqhJ+XSK3XTTbhnklpp5ahHXSWdTcE/Ug/4+2JIYg1RjrSfvCVy8tSPn0lItrVFJA765zb9RKsVv9LBfp70FQEdKY5pkrUkAdKLHYp5I1LLLbj1eImwH0/HtYskaLxqeliXqstCK06faehxqSrHNPIoLDVeMra/J49oTdTq1Bw6st2DjSOl5icJt6TI0sYyE0Zddd0uv5/HvTXM9KV49a8QmpC9Lqi2htSPP0n3ORneF508zSXJCk83v7tHdTu+hzgdJ71/Gh8MDPS43Xs7riLJwyUGRmKCBdWiNiNYH5I9rneFM46tt8F1BblRUdIDN7ZxMtBPUUM7t4VJWXQRIot/Z/PtLrhavRDdzXbXWgHoMqNctAY5qOWqqHp38kReJiwdTcWJ9p2WJznHR1I1zBbqak16WuE7f7QxeVhngGQZIVCIgimC+k8Akj2/GYSdOOkvi3jcK/s6MvsL5id1bOSp8e2qrIrIWKBo2N78ni1/9b2t0xIBpA6sq3LmrVx8ujDbF/mE7rXIQHP7KMFQZU8geibi36udPukscarrYDpTGd0kbwY46r5Ho7mJ+cO4s3RQf3e2NRVTmI8vSqLsRx9V9k1xukNvUdCCz5U369pSPB+XQZ7/+RfyM3pSVOLo+vqWGCVNAdIrC30NgB7KG5utlbwlGehDD7S7/ACUnZAAM8Ogfn3Z3jhttQ0mY24IkeUsqpGxAZjcWuPa6HmGBxqbP59N3Pt5u6NVUr+XSLqsN3Xu+vpZFxLwUTIpcrHIeDx9Rb21ccyQRrjHTcPt3u8zUKVr8ulO3wm3BvenaTctXV08VaAvjVpVUgi9rD2Grzm4ynRA3D59Drln2naVm/eC6PTpNx/ywtj4+p81VUSF2cuWKsSSTzY+0h5ouq1DnoTf6z+3E6v8AP0tKb4A7Ho4DTwpFJqsBrjY6v6fX/iPe/wCtFx/Ef29eb2d22tQehDwvwv60poIaVaalarp2DOGhtyPwD7oOaZwa6j1eL2djHdGAfn0J9P8AGHr6Kj+3q8fQtGF0aniUgH+vtXHzRKeJPWrn2fk8IsFFOivd/fDPZ/8AC5tx7Zp4P4hQIJYkijIDFRqC6V+vPtdb8xzz3KQVoG6AW6e1M9nBJfUxHnouXUmP3BlKltsZfDR0wgkMBmMRTUiEi+ogexmaAUJz1H0kAVKDiD0ZTKfHrB5DF/w2lSFs1WENAptfyv6Yxf8AHNre9efRXMNFadGz+Kf8nb5Jd3TtDuzDldk1hK00ixzBTTD6Ev8AT9P19+GOksbmp09bV/wl/lbdIfFHF42oNPizuGiFJUVEUulCJYkHkY6/qb+9VPT1B69G0+V3ZOEzOw5tjbfkSGEIIGhoWEkdraWuEv78M9e6Jv1Vs7YtHjsbHmGigqIgS7ShYzIT6jq1/wC29+OOvdTfkBkMUNswYva8MDjWE8lPpZiDxe49+690Nnxf7Y/0cde1mCnZ4qmvp2IRTYMxX6Nf37I61SvVe3zuyO+O1KWTb2PFUtFlkalmESyNqRz9Do/w9+Ax17j0WXrH4iV+z+rJdu0dG0VXWympkbxyBmM3LaR+Dz73Tr3Q99QfEHbmK2Duyl3JT00+byIlNEtTHeQM3I035Pv3XqYz0Ge+PgBtSGo6xzz4mjFdSbhp6moP25Q2E4I/H09++XW+tiHYGHwm1drbcxNHTwwwU+EoojoSylkhAsbf7z7r51698ulbQaaiqd4rKisbCxA+vFvfuvdCJJE/8N1kekqL8H8f63vfXukjQwLJWksbFWPN9Q9+8utDpf4umSarj1C6RsD/AK4I/offhx62cjoY4aWlq8e8KGzLCQGt+dNtNx9B7uKHHTZqOgxmonwzVVSXYqrnixP+PNv97906vjoNsh29Sw16Y0TOH16CAv8AjYm55/2/v1a9b6i1GSFbXfdNPIVZdQN7gXF/euOetHpO1+5plqFpI3b7c/51h/W/4v8A8R79XrfTFuR4snDT0sD6tVgCBzqP5NvfuvUPSci7Aw3XM0VFmqhKNHKkTySLC2o/jUx/r738+tU6GDH9s4DIRU9LRZFJRkFDROKhHDBh9QVPvVfLrfDpa4+ei1SmSqKvIpsRNa1xxbn3vHDr1fXoOdw7jmweUWkpJXnMzcfuF/1cfj3qn8uvcehn2ZRDMUQqK6jgmYrys3rUkjj9XPuwz1o16UBgxVBVDXhqJWBtxGALj6/j37HWhw6c6utxIgUx0dLCACWWPQObX5ve3vfAVPXvPHSYn3DiYonDmJhpN0YoBce/HrfSFn3/AElHM4o8ZQToSwYsENogLN9fda+vWznqRjOwNmCCeNcfjfuZLmVf2r6/yPe6jrRPRYd/dy7Q29mJ0kxuNV3bhiIhyTwbH/iPdfmOtkDqG/cu2oduHOU+MxklW11RQIdXP04H+9+/DrXnjqFtbuLEVkNTHnsNjmNYS1OJRGws34FxwP8AYe/db8q9Ro8ttxsqZ0wWKaF2LIrJFYEm4tx7917h0NO1P7nV9ZHPWYTExPpUKrRRqPp/iPfh6Hr1M16Ej+E4pvO9FjMeouTAsSoCf6Wt79k9eFOmeXF4+kliqKjFUYqmlA1hFDg3/wBb34VHXj0y7tr5KSqpwkKqxjAjC8WFuPfuvUr5dAT2A4aOCqqTrm4dCbtoP19+p69b+XRAu8d0TV8DYqOSR5J2aHQp18MbWI/p731Xzr0bP+Xx0GKPKRb1qKZFaO0rM0ZBHquLE+99eJ6ut9+69Tr3v3Xuve/de697917r3v3Xuve/de697917rpv0t/rH/evfh1o8OtSb+YOB/wAObfGe3/PY0B/9XUueffh17zHW2lSf8Bab/qHh/wCtY9+631I9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdY5Y1mjeJxdXUqR/rj3sYPWiKjonvZG3ZxX1YjRtNnsAp/PIPup4162K9FuyGAadpaaZdJu9gQR9R9feqilevZr0BG4Ns1+3K9q1Vc04bUTpOkC/5v731ateiz9xdd47e1KcxHGv3MOpyQvPp5IJH+t73xx1rh0XvGZVsfSSYRCAY28On6WtxYD/AHv37r1elTVJHW4uNwNEtCNT24J0/k+/deNa9N2C8G46paR1H7bBQSp1arfTn3rr3ljo/Xx22nk6Crgk8B+0QEsdLaSoNx/xPvxPXvn03/Jrv+LYkeTxuNmVaxYSuhHRWDD0/T6+/D169+XVGXYvZWZ3VkKzMZfKzRxrNKwR5xp0k3IFz731ronHZXcW3qWopqVK6IyodLfuR3Zv9v7suWA69SvDor26e45q2rSgo3ZkL2NmUixPDce3JEAHTEtwlsms46g5KvpHpY62oqmWVoSwQEcEre3squJaA06jHmrmwpCyq9CK+fQGZLMtmKyWlIcRo3BsTcA/Xj6+yue5ZqBT1jRzFzDLvVx9OhJK/sx0Y/oTofM9ibgoKSio5JYpqmJWfwudQb8XUf4+/RESUHQ85J5YlvipkFc+nHraS+IfwgwWwsPjs7msfTrUx0kUo8sRU3U6+NX+9+zaG1VuHDrJPaeXYduhUsoGOjnbtmNfLTbfwMaRrEyx2gNgxHFvTf2qC6Dp8uj8AU7envcucoOoetq7J5WZYMiaGY+t1R9RU83a3vdK4HWq5r1q/wDyR7Ul7K33k5qiqklgirnMSs6upS50n/ivu3DrXHoMKDDLkVo1RtKow0qvp1D6+99ap0MsM+KxONNHWeFG8XBJXUbD+nvWOtj16TnXe7cbjOxsZD5V8LzrYkqPq/0H+Hv3Xq16tg2fjoMtlPvoyJKWUR2vYqbp+n/H3456t8z0J+8dq0c+3qiKCkhQNFKC8agm5X63Hv2P2dV8uqh+3NoU208rWVUkYb7mQt6xqIuSb8/T34deIp0VnMY+mqnaqSJRck2sb8n+o97690km22lfVpUBQI4/1H62I+vPvXWvt6QHcGffZm2K/N0DSpJiKNpxJECShWxuLe/de+XW49/Id7pftL4s4d6yukqatY4brNKrOoWHi6/W/Hv2Ot0PHq2X5A7/AKPYexMnX1VSKZ1hldGEoj4VCVsW+p9+PDrQ45616t0917h+R+7V25gJ56tMJUsGDEuhCSkclbj/AFveqU6t0/ZTr/eOSrabbNZ5KaJvFrijYgEE2IKjn37r3y6IB85Ng7j+PHYOwe2NqLUw1+B+0ll+0jcl7OrnUy/ni597B60a8eti3+Wv80qP5QdcUdJkKrzbnxNMiVyTTI1QPFEAwKXvxb3vr2fLrXn/AOFQPb0uLrcJ1stSy/xmmhiFNcFWMyqAxX68396+fXuI6pQ6N6xzmydl7Jy1PAwp67GQSSaAwX1qCQQffsdaNePRtKjFLnqeORolEsWku3HCgc2t73SnW/PpN7pwM+axCYPBJrq2qoQyop/TrAYkD/evfuvEdWl/GzY9Bs7ZON/iFNAmQems+tQH1lRf6+9deAxXo5lBmafE4laohI0IAU3CgD/D/H36nr17oId97qoqijqMgKjUIhKL6wADouLH/D37reRjqnzu3e0dXm6mMVJZTIBp1A255+nv3WjTj0TLsHdSY6ndom1zG/Fxe4/1vfutcePRZcvu/J1jmRkYqpuAxJt/hx791vqJQ5R8k3gaIRlwVYgcm/8Agfe+vdK/a+Ul2XuLEZJJpKc/xSiuyengyAnj/H23NTR1aI6ZNR4dbTPWm7KDdHxyTMTVLVE9FiAwJYE6liDfi9re3LUUXq5XxZKj9vRPt991Tbg67zG21DFKeiqU1HgBVBv+PdLkFkxw6B/OdvKtifLHVAGXeOnzedqCBqky1S2oAfVnt7DV1k6esML6SWPe3Vs9x6hpH5iJW0lH+vFwRf8Ap7TxgKQT0dX1p4toWpWo69Lqp6iCaE2tMpIU2Ui/NyvtbGTq+XQa260kS8WQYo38ugq+RYB23HuCaJFFHIgLA2JAIPs9gOqPHWU3I1+QqRk5wOjJfGbM0Oc2TjammWOZ4actYkEiy/T/AF/dwQ2OpjvE/RDevRjqnEGpigyKppnmlEQRBe9/px7vwx59MWTgLTqxD4sfHWfcNVQbiqqYDxJFIrGNi1tYa1/eqUPVyKMSer1NqYeDBYCGggVdSRJH6Rz+n8W96PWxw6L93Z2SNp4apx88xUyJMltYT0kWvY8/n3sAjrXHqobfu86ZqusrnnU+STWDqAPP09+x1rPn0HGDzIrqxKhH8iM34NwL/wBG97630LMG3kyVbBkZWKxoiAr/AGRY8/T3rr3RvOm/4ZiZoah5VjjiB1OSAPrckA/n8e9db4dGoru7sIIf4dTOszQoABrUi4H9L8e9469U8D0Du+O1aLMYuTGCKJMhKx0KpGsqTp1XHN/fj17pC7QEzTRUMlkkrgYlEfMjFiOLD/effuvevWz78N8K+E6YwdPIrh3SJyXUqx/a/IP+v791ro1vv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//X3+Pfuvde9+691737r3Xvfuvde9+691737r3XAuA4T8kf7b/X97pivVa5p13c6rWNv6/j36mK9bqa06oh/wCFC+4sntz4Db1qcZVNBI8NcGjXUPL/AJGxvdTe490ZGfI8uqmisDWh6+cBsXITT0uHNQVmXLUNPJW0pJZpi41HVf8A23PvSsyA0HThuBxI6WW5clkmY4LaeClpXRbOYkICX+rGw/p714rygqw0gda1vMf01yemim6M7N3XjGq6KorGeSVdaLqBUMdL3t9bew9um4ixHYa9C/YOVbrc2Alqtehgj+K2N2vtuOpy7pm8vWQBp6WRw8sUjDlSD/j7KbXmsyNoI6kBfbho0qa9Iyn+I0m4I0mpInxRmk0+AHRYFrA/717P494R11OadBTdORt1Eum1jLL8q9CpQfy+KdccKnJZohSnkZJJha6i/F/+J9vHdrfgGHRT/UDfCa+C38+mik+P+D23XHDR4dMopbxiceoAfQHV7YO5QV+PHVxyFvSnFu3U3L/Cahzh/iUEX2hZQ/hD6dNxf/ePald3toxxB6pJyBvpOoQMP29IaT4a1WOyMNdHI7rTWHj1k6h+ePfm3m1qK9VHIXMJ+KFh+3pXD41ReSmmnpxH61L8n6fn6+6/vq2B4jqy8hb6GzCxp9vS2yvUW3cJiUZcCmQnKrHqX1WJ4JJt70dzt5Dlx0aDlbfRF4X0zV4dQqD4xf3sgFdT038PpiA8lJfSH/wtb23+8ratAwHRI/t3vpl8YQt0o8P8XsRiqmNK2hjjikcLdrabHgnn2Q7nv/0lCncD1IXLHthdbzWPcAYgPXocqD4ubGekRKejp55pFBDLoJv+eB/xPsu/rY607KdCpfZqFTQSjpf7Z+Je2cbNFWVePSeP6+JtPAvcix9uDnSQYK16UD2htqaRKP29Clkejun4aWJpNoURqWsryCOMspAseQPbrc8a00aD16H2l8GYMJsDhnrHFsPYm3KY/wAIxVLTuRwEAuAObH2H7zeHuq0BFeh1tfLo22gY6qdP+EkxkBAkgUBvyAL/AOB+nsPtbys/iBuhom4RRReAVrX5dOlaNv18qU2RooahAQUDqvBP+HtbFNLH59FMkEUhNFx08Q0GCoYkanpooowvpQBfp9be255JbhdINOqxwwxNUL1m/i1zZGDRKbrHYHQAeALe0dtatAxZjq6XSzqUAjGk9cp6iGpiFRJpe5C2sLCw5v7VaaivTBeXhXqBOsbyR6EVL/U2HFz/AIe9BBTrfiyDplrYoUkYQRFZ2HqlAvcn+tvetArXpjVeq+tZaD06Zpqn7eJ4KhrtYtqb6A2vYE+3UIGOn2mvmGZTT06Qc1f/ABE1FLKhalJ0OCBYj6XH9R7diuzZOLrjp6K7+xudxtXsmkoJB0F28dnYWjoHyODpYqPIxqZHljChmI5J49jzZeaP3kwVkIrjqGN+9vhtcDy69VAT0X01u86qSny9JLPA9BlKRhUWbSY4ZgWWw/qB7Hyxgw6+JPWOW9Xr2l41uo4GnW3v8LPn9RbT6Gw+ObHyS5DC4sLV1X263eRYbEk2v7Rq9WIPS2CKkKy/xdAb2n89u2Ox83V0uzMFkIopjPEMlDDIiEMbLyg9uU6sTjqw/wCBnW1d2bsnI7h7MqzVZ1i0kGNqm/dY3NgEk5/3j37NOt/Z0nPmN03uPbu2Nxbk2jHNj3wdI9VHSxE3lZAWKAJ/X3oZx1418uiBfG7vKHe+25MbvNvt9w0VfLA9JUKROEgYqWs/Nj9fduPHrXz6s26i2rgu1qN4MVMlNPT3iEiMF5RrH6e6/Z1vowK/HzD070tJlYVrZ4HB87FT9fpyf9596yOt8ePUve3XmF2zhUnpKFJZ2KxrGgUt9ODYD37rXn0THsiLP7P7D2XMaKVcTXGGSePSSul+TqFre9jh1voRe5MhTmm2vXwgRQyVVOYoSPo3BsAPfhnPVeHRzdgQpk9qUNbN6ZFpYQqt9WATgf63vQp1vp+opjTVgUQaIwfWeSDz9R798uvdCjJVwy4pREFkdlACC3H+PvfW8dIiCWOkyBSY+PXZiW/qT9Pfh8+q9CFi6iKL1K+oSfUqAosPwSPfvLHW/l0vcRXiMn93UCtgDyP9e1+fewetEdT6w46SnlFU6N5LkR2ANyLfT/e/eya9aAI6ArMdZY+rqKjL09D64wZE0i+o3vZT+fderfLpG0+QpqasamqlSF4/2/AxsSF4Bt711ojrFm0oZqKolo4V8qAn0i7Mf+J96p1vz6DbC72xVBXR0uWRad0lAV5FHPqtbn3vFevZ4dVHfzjO55didST7n2hkhT5WllaZPCfUyx+oIFQj6297HXiKdFZ/l0fMjd/d+0MNJWRVZrsXRIhkeJwzyIdLPc/Xn3vFOtdXd4fsXd32qzVFJVGyIfIUcDgcX9663x8+uEPaNTLlEmq6V55o2HrIN10mxF/+I9+8qde05z0YzYvyExNNUQY6rqo6NpW0ASALa9hyT78K9ePQ5ZXeeFqac1dNXpUsIhKCmk3JGoi/v1fTrQwKdFxzPyFxNDWy4qskFJKGdE8ikFiBYHk/n37rfROO4PlhJtKqPl1UtA7emrZSI3U/29R9+A8z17otuY+bzU9FLX4epOTjsdRhGoKPyPT79jrw6Aen/mQph90QI+PklhLgVTaCVQv9Qw+nv1K9exw6w9mfIra3asRzGN3VSY+qhUSvSCRRIGHqI0g8e/cOvfIdJDr/AOTeKxsr02W3BDW08R0iiZhclTYMAT/vHvY60adKPOfNzaGOkkrZK+lhbHsft6Rrap1U3AHP+9e/f4OvHPDpJU380zZmNy2KhrPCkVVVxxSBkBEa6rFmueB79SvXq+fRv3+d2F3HWY2HZpWtSamileekQFUZwDp1Kfx79TrfRgOv/mfLS11HTZuVoQTpIlX6D+vPvRHWwB5dKbcXzr2nS7nNLXVkKQJp8bsFCkjn6n3qnWujA7I+R/V3Y0UaVG4KCGtMY0AtGrC4sACT78R17z6Q2++ydlTZX+ADL0stTUP4qOMMl5G/T6LH37NOvYrnoJm+O+5d37mpan+FVElC88Uy1R1BDG5upB/p73WvXvPq4TpTrqLrvaNLjVTxStBHr+hYELyT/wAb97p59aJ8hw6GRSfoSTYD1cC/H1497I8+qg1NOuQFhYm/+J966t137917r3v3Xuve/de697917r3v3Xuum/S3+sf969+HHrR4dak/8wbn+Zt8ZieSN4UP+B/4HJ9ffuvHiOttGk/4C03/AFDw/wDWse/db6ke/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6Se48BTZGGWYxa5CpDC4HFv1X/AB73x61kdFL3ftN6auNSg0qGJtwQw/2P+9+6EenV606CndWEXO4uopTCUbxlTKtgAALDn/D37rXRPN545No4ivWqmEkPhnsxIYf7f3vPHr3yHVYb7rxMG+p4VqkmEtSzFNQuLtcj3s9e6Fen3bRwVn2ZiDxVpEZuAdOrgE+9deJ6EnC4+jxtZT/aQCSSaWNrryQWPCgD3unWvLqybYWQi2b13VZ3Ij7crTSuocccxkj6+658ut1rjqgb5X9j5DcG8c7laeoaSkYP4kGqxAc8f8VHupcDFOvAdUN/J/vnd+2sdWvQyTwQxvODZW02A+ht7di7jU9W05z1WJF25vrsbMQHHNUT+NlM8g1gKwbn6f7f28YgFL1pTpm4uEtYTO34ejtbFxcmLxq5fOVH3FU8YYxOxJDab2APsrmvqdtK9Q1zPz/DCrRqP5dYs/uOryYK014lV9Cq17st7AcWP+t7LJpamo4enWPG88z3G6XZjhFAT0L3Q/U2c37uLHUK4+R0q5QrSDVZfX9Tx/T23aQfUOQejzkTkqTdbtp7g6RU+fHPW1/8Lvh3jOvMTjs/X0iTzaKabQ2liGKauLj6+zWLbxC1K9ZQ7Fs8GzFdGaU6sq3Pl5aDBNHRH7WOnjaLxgAcAWA49ro6R/Z0PvqhdIIgtOgp6TpK7N7tmq6zU0SVBOsgkH1XHpPvbNqPSZl0nT5jok38zjuM7ciqtuQ1WlFo5tcZ+psNPvQHVaj7etcxdxDK1lTK0RE00uoMAQri/Bv9efe+qjj0JVHuuoxtJToaV42BsJdP1I44v731v5dIHd278l9wK6SoZoQbCM3sV/2kX9+r1rps2zn5K3cuKzUBKJRsplQkjUQ9m4PvVet+derouk+5MblsPBjHZKGWCNP3WC/uFU5Fz/j79gdbycdGsxO9zmsRUYeCA1LOjlatQbC4stz/AK/v3Hr3RH+zth1m6MhX09TEy6HKq78EW49Nv+I9+630TDfGzf7qxy0LIWIDWkvcm5+gP497r1XPQdrgquh2xWZaPVJobUE+vp/1x711ry6QG4MbQb/693RhnxivUVWPkgaW3+bJXkk+/eXXurW/+E+HyQwvXXYGQ+PeXykcP8PcRxpJ9AZFKoNRt791v5dXF/zW+2qlaDH7F25kyK3MoscKQE3kV10EoQfeuvD16LX8Ueq8Z0X11X9jbknWTMV1JJVFZLNIHZNYuG/Pv3WwKceid7U/mCrvL5WVXX8NO4jpZnRX8dl0pJZfV/sPe6daHHqwrtzAbf7/ANi11DlKBKvICGVaVWVQ7FUIGkH3rgadb8qjqo/4W92V3wQ+XEW3dwTNR7c3buH7AQPqaOKCqqfE4u309J976rjoKv8AhRjksJ3Z8mus/wC5dWmYpDiMZVs8BB0/sITfTfkHj37r3QV9fUV+tNr4WsQx1FDjI4NBHqVlQKOPr79Xrfy6gUWPyOKyFVGQzRzhkUcsPWeD/wAT7914dKLZX3G2t0RTVlC1TDK6sZbalS7Xvc/09++XXurENmVsuTlx87yeGmv6Y29PpAuLe/Ur1sdCjv8A3djKPCrjklSMqn6zYAH63v8AT36tePXqU4dFJ3rutXwFfQ0dSs8syPpdALD08i/v3WuB6p67IymVptyVaTB5gJuDZiAxa4HvZr1rzr0EslNU5rLulbHaAxki+oAH+lj9T718+vDpJ7iwP8LgnlMdow3DEj6Xtxf/AG59+68OvUe36LH46jzQnRxMQ7xllLC/5sPfuvcOHXs7UUGXgo1p1UyU9TDOWFrgwnVyB7qwqKdeZggqer9vgtv6h3x0BuTBvGurGUUlMVK+pmRdDHn6+9owh7RnpTaAv+oD0zbs69ojtXLjHUgasqaWdGCfVS1+Tp9+dS4Hz6DnN0jXFqYgK46oP7P2vktn7ryeNrkcCpyFTLdibWLXAv8A8R7INyi8E+p6xJ5j2FrO/a9Y0qSadIubLRrTCCCxkVdIsPqfZdGxYgnh0QS73HEotyOPU+hqk+xaacqrQoZbkXJK82A9rFy1BjoWcu7TBuLpJqpqx+3omnyA7gTI4yr2gKezSTFbEHUQPo3sTwRKsPGnWQnLnK8dgqyq9eHQ4fAzfgrqar2+w8f2nkhHHBvxck+9OQgxnqSgBPGErXq2XbHh/jGOp5x5YPuYXEY9Vifzc+2hKa5GOkvgeC2lc9bAnxemwsm1KOjpRHTuaZORa9/rYn8e3Pn17jx6NzHL/C4qiVz5RALj6sDx+P8AjXuvE9XHDqnX5j9hz5jcU9BSo8CL5UuFsps2m5t7sOqHqqbs7IZ+OelipzNJTOsfmlUEqh41X/4n37rWejCdLpjKrFRmeojeenAaReNQbTc3/wAT7314dDxRblxjyHHU0yGROTEttXpP0P8AxX37zr1vh1Ipe18fhKlsPJMsTykrci5F/T9fx7116uelLJm8Zh4hnZM5E/mUOYWcfQm/0B96pjrYI6Y4N+4vMZAV8FTDO8ClVRGszEc/S/vdOtV6MV8Sd2Sb77w2zt/K45hRfxEQ+R1srIJQOCffvKvW/PPW5HtbCUG38FjcZjolip4KSAAD+0fELk+/da6UHv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//0N/j37r3Xvfuvde9+691737r3Xvfuvddf4fm3v3Wuo9OkgMjygB2f02N/SBx/h72T6daA8z0EPdfdO0+i9m5TeW8cjT01HSU0k8Mc0iw+RowfQtzzz78SApb068K6gnmetPv+YJ88B85qbN9PbcMk21WrKull0s0sJDgxkhSNNreyw8w2UZMNRUdCC35P3K8o6qc54dUN9pfGnB9T0eNyEXhVsfDEulFRSth9CPbEvMlhTyr0JbPkLcWoGBP5dOfRfWq7nr63OzRwGlqUMaeWNG/FhZj7QXfMdpJCBHSv29DDafbyS2l8adcfZ0K24NyYrqRZcPTUkVRWTu4j8cIblhwbD2Adz3Jrgmh6lvY9jtLahYAU+XQd4yDLV9RNuLJ08rR1TGWGFozpVTzwh9k1tOyS1OOhq0Vi8WkAdLjEyVdVVwzJHHDFrRdLIFIsb3tb2/f390SFgOOmYrixs8BAftHQsZuKdsYiSONJRb24uCLG1vaQ3l8eNenDu1nWhjX9nTDjtv4WihirZRB5idQLhSf9bn3r6u+B4n9vVju9nSgjH7OlCvknQTRLGYV4UWFiB+ePe/q9wB4463+9rIDVoWv2dOFLhaes5eKPU3FigN/8Qffhe3x+InrR3WyfJjH7B1Kn69hr9CLHGObelQL/k39++pvD5nrx3W0A7Y1/Z08x9Z4WOnWnqYomICtZkUkn88f4+9fVXyjFeq/vayp/Zr+zqXTbfx2KDQQxIqDgekfT/Ye/G8vuKk9b/etkM+GP2dYMjhcVkYvFMkYdR6PSOT+Dfj27FNcyf7kVp0zNdx3VPp1C/YKdJSHGV+Dn8lGSUvdfSTYKbjj29Uk9NZOBx6n13aL4GK+WRlRB9QGU8jj37H4etFadOO2c7BvMy1VM+mmdW0LqNr24J97qOPn14n16cl26PuSlQ5Kknhjf9P09+B6tqNMdZp8NR04LgMNPNwOAPp9f8fewT1ok8OmCSKN6oaQbB7cEH/efr7r8utgmnTvKpEahg1v9ibcfT+h9+zTqtRWg654yk1+QliAb/WwPvYqOtEjz6zSQOo8cbWuSfr9efqPeuvVby6Y8i+TS3ib/Nj6g8n/ABNvfqV63U8OkRX7hyELBPFK0ityyKxN/wAm/vZGMde0kmj9JPK5zI1Eqs9PUCOx/sN/sRz9fehkVHXgp8ukxX7nONmhg+2mBnt6gpAF/wCvusgDJp49bBoeoG6N34uh2/XyyRuahaWQ+of2tB/r7Odjk8KQIMUPQZ5lgEtm3zHRetgb4l3NtzLUELQRzCuk8XkIuArcH3LNldeJEB506ww5tsGTdGBqO7o4/wAc96dkQx5DCJlKdsQweOeFZxqKXsQgv+R7fMbBqt1tBpt1A8utiL4ZYnaWWwEVLlMUkskaRSM89KrSNIUu3rIuefe/n1Wg6Pem+cH0rSVG8KeM0dHQy6DEP2EKBr/oH/FPfhnA69wyehroO1dkfI/Zk8mLqKCVqmnIyMBdHdlC2a6H/evevOvWvl1r6fKH46b66i7HynZ2zoalNrOtXM8FLEY4+CSSFX/Y8+7de+fRlv5a3yW25WT5ChzNatJk46iRJIayfxyeUyW4Rj78flnr3EZ6vVXPUuTliySzRPSP43DAgjTpB/V/xPuvWzx6TNTuPA7q3ZFtmCWN5otEpRjqBsRe4+nv2a9exx6R/wAhdu4zLZDbiUkULvjViifxRqWBVRe9vfvPr2B0CfaeyBXbNpqrQxkxSfcQIo5Dxi6k/n6+9jrwHl1P+M3dWOyzPtHccyQZSncUsEUraCUQ6VIVv979+I690cnO0zUsL1MSq0N/QyA6SLX49+OOtfPppwW4aSZlpPIElVgGDkX4NiLH3r7erdRN9U1WAtbQtdQt2MZvaw+p9+xXquekFtzs1YqhqCrdlaI6W1MQbjj8n37rdOl/U9jRYzxTiX9tyovr4N/wffqnr3HpQtvOPMCF4JlDGO9lk+v5vx73Xr3DHTPR95wbfyy4fOLogmkEUMklwhubDk+/DHXuPDrFvTBwbgqUzuDmhvMFfTFICNLjVY6foffvXrXSB3FV5PbtPBwxUIPK31AJHOo+9DHW+PDoFt37eqt3RUdbiWZKjzIW8Ztclrn6e98M9a+R61n/AOczVdgYmm/gD/cT45pDG0YLMv0sxIHHvfl1onos/wAKe8U+POzaDI1NNIwlpEldIwS1zYkaVB/2Pvf2deFOr+OhP5guzN94OKKto5GlqIlgVLPrRin1YEce9U63WvHpF9z/AC1xm0ap6LCUNVJX1UvkikRHMaxv9RdR/T+nvwA62SSekPL8i6bceyq/K0j1UO46WmZ4nErrpmC35H45968qdaPQbdY/OXsDDQ1dLubJtVpB5dEKVbSOscd9IZPr739vXuHST3F8wf737h/jtZklpoKOdA8RrDGxVG5Gk2N/9h79jr1a56XvavyW6Y7J6+psDMzPl/tRErRSkuZNOn8/Xn37z68eA6IK2Vy+MxtRhNpUtfJPVyypCZdbi0rWj4/3r3vrX5dN20vjx2/WpWS5anrpTlnM8LrCx8Yc3A1AcWv/AF96690xdgfCnu/a1LHm8DV5USZEG0IZ7ENzYKDx798+t5HTj1h8I++ZCu5twVeQSMlFMD6hcH82Y+/deA8+hw3F8C9y7lno612raeaJdRDsVSQheSVJAPv3XgPTqvz5GfGWs2NV+F5qo1qPZ2ikay2a11I+h9++Y61wGegl607Z7E6V3RTUlHVVNRigkRdaqd5JNKG503v+Pfvt690eGD5lUe6azHOn3MU9Gf8ALFSRlJYAXA/rf3vrx6dNyblzPacgqNv1VXRyJpcySzyIDb8C5HvXXuPDoOMVvPuLZXZWLjhz1U1JCqI0cddKAQP9pDW/2Pv3Xs0p1ZB8ed07m7Q776xocjkKiT/ctBHVK9Q0ge8g4ZWPv3W/PrdOwGCx2Jw2NoIaSnC01FSxlvEl2eOEAuTb+vv2T1rAz09yI0iBUbQv54+o/oB7sKDj1o1bh1lVQqhR9AAP9t/re69W68Lfj/ivPvx+fWhTy679+631737r3Xvfuvde9+691737r3XFv0t/wU/7179148OtSf8AmDX/AOHN/jL/AF/vjQf4/wDKanPPvfWuttKk/wCAtN/1Dw/9ax711vqR7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917riSCdBFwV5/ob8W97HCo60SK0Pn0i9zbSpMtTStGirKFZrWXmy/2Tb/ePfuPXh28Oih72p5sCtTEYW8X0dtHFuTcW/wAPdOHW/n1Tf81ewMpgdsVrYvylNNRqMVwwW/0AHvY62eHWu1F2vvXI9nwRY2KvOuqTVdXIAZxq1X924dVJJPVx/Ve3stnsZjsnk0lEsQSSTyIQQdN+D/T+nvXVqYr0ffqrYa53JUtQihlhaEtcFv0Hn/jfvVfXrXHPQzfIzcS4vYz7cpX+3ZqVoiFsl/27caffh17qi3eGzKzKGqWYNIp1sGPqvdySST70VzXrYoTT06pa+avRu9c3SVVJgEK08ssusiEHg8GxHverSadUuJRFGXby6Kx011HSdT4KoO5Y4pclUIWR3iGoSN/r/m/tO90NDAnh1F/M3M6WtlKmrJHT1mP4pkG8tISKa5UBOL88cD/D2UAgtU+fl1itum9T7pMY7cmoJB6G/prpzPb5yFMGoah4TNTAuYta6S4+p/4372sIkz0JeUuV7i9nVpV408utjz4hfFPFYSkxle1BEtVCAx10yg6xYkkn2ttIPDNT59ZPbJyou124fTmnkM9XW7VkocDh0xreGN44VQD0qNQWw49mNQR0Jo4SMkZ6BHsTctctW9CjD7eSW3pPpIJsSfdaVPRpEdCg06GfrafGbb24+VaNfNIgYsPSSbX4I92K0+HqznVnrW4/mW7+O5uyaukE7CPVUIP3TYjX+bf097HTR4dV0YfEQJHDVghgiBmt+bC59PvfXuHTjmd5UGWihxNFGYp6MhXZlI/w4H59+68fl0UzevZcse+I9oGOWYshYFLlRxb8e9deHQx9a/dpWwiY3p3P6XvcC/05/Pvw69no3OD3VJSV9LQ4uZ42MiqdMmkG6/quPfut9WgdMb2iw+AjXITJPWSxJZvIDYm1iST791s5HQh1lBLXUOVyc/ieapUyUei1hcXB1W9+69wHRWt97Biy+IqKut8ZqKdHmkuBqMam/wBffutceiMZLfmCiau2vAoRUkeCXyDTyPT6R/T37r3SWweRoMXJU42mCf7lG8eogFBq/Vcj37r1OqzN59pdnfDz5Q03aeyZK2alyNdTtWpQyyRIsSSgk/t/0Hvx61x6vp6q+SO/vmZ2J1pv+saskwWDx1H/ABNJ5HnVp0sXvq/1vfvt639nVmvafYxyWGm2dRO8McsSU0axkra6ab2H+PvVOt8R1TLtDqbNbG+V825ZGYR1B8nkZTYAvqN2HJ/2Hu2OtZHV83UO6Yp8riYXfUxUhhq1Rm4sNSng/wC2918ut9VB/wA77Ys/X2Axvbm1qecZ+glgyVO2OQpJ5Y5PJe8XPvfWj1UR8UOzt+/IXc9N2V2/T1suOxMMeOpBkC8pRYh4/wBUv+tf6e/Dr3+Xqx45LGU+SvTzRrQK5MKqw0pH+Lj3vr1cU6n5PIRZGSAYl4QyuNYFjf8Ar6v9b3o9eHQm7cmxVa0GPqokNbpjOsKL3X63b37Pl14ca9Cnu/sTD7D275y6xyUUIIswQqdOq5/2Pvw+XXuq2Nx/Mmq3ZuKrwtLUuUiLoLTMdIDaQT7914k9Ttr9s11bkUpKioJhlUh9chI54tz791r7ekT2o+N++SqijVpHkR2KAG5/p7914/LpBJQRVFI9aqiNkQv+m36fwL+99ar0HNYBuqOrx8ELaomKaiurleDp96630lazDrQUEmLmuZY4yFBYgi/HAPvXz690Fcyz7djDTLIzT1HijB5UmQ6V/wB797wfi6pMjNH29XB/BubPdddZbhra9nSkykck8YYkHxytq/2PvVKU6U2AZBpPVl/U+Hg3fhZMj6HjqlIII1gXF7296LEHp+929bhKHIPVVPzn+P2QXOzZnFU37SNUOfFCoJ/JII9ll9H4j6z5dQjz9y0pakY9eqkqjF/wueSGrpp1njJuXU2DXNuD7QeEAaL1jTvexXcMhYKSFGDTqXDNRvFoq7+OwJUCx0W+lh+PbbM0T06X7Duk9joEh0gHoE+y+mdvb4p5cvhI2jykCyFbrzdQTyPzz7O4LtWj0nrIHYOZxLEsatmgxXor3VG5c1012ElFKJLz1QjqY4yVLF5Lago/r/T2oVgTnqYNquDPECp48er3uocrXbu/huUj1QoBTu0UjASABASSL/T24Y6nt6WykA06uP6G7HXAmgpUl0KkUaSXlYBmuL35/wBt73011ZJTdm7bq8DL5J4vuJogBeTm5X6e9U8+t+VOqkfkJSfxXc1TPAA6eWdgF5BBbUCB72MZ60TnoivYjJFQVFFHGPuHX06lHkNl/sg+99ar0TPb3dGc2duKswMNDlZ6mYvHE0KSlNTHSOAPx71w62OjbdYbb7WyNUu7qn7mnoJ1UiKYMptKb8qeffutdCzuj49do7y25mdzberSmQo0eWAKbMSqlh+b39+qOvZ6ri6+3/3LjO0MhsftWSr/AIbRzSJFK8jLE0aNpt6vfs9ex1bD1GvWKzUbPkKY1EkcTyRtWIxDs3IZb+99b6sk61yvX21M/gc5tg04ytNIrmaJk/zlwbkjn3rHW+PHrY4+Mnf+P7OxUOBqZ/Jm6GliLnWW1IsQsOR79nz618x0bhVC3tYAkn/Yk3PvZNetAU65e9db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6/9Hf49+691737r3Xvfuvde9+691737r3XRNhf34daJoK9euLgX5NyB+ePr791uvWtn/woR33nsL1Xt/D4rJVuMppmRalqfUqzLJLzrI496lH+LyU/h6tbEHcbdWFQzgda7vTNRitqbDqtwimgmqJImlasKr5nmMer1N/r8n+vuDb2O4F05U+Z6zZ2TZbUbZFLpHwjPRLuyt07i7T3DksZM1QtG09oAdTIy6rem3stlF0TQE9CKKzsoAO0Z6W2Mr6rq3bmKx1CXasq5Y4TGv6gz+m5/I+vthfqIz3E06VSw21wngx0qOhn251Z/eOqpdxbrjVnkiSojWoUkeoah+r2tiBkFWyeieSxeNtOR0t8zj9vYilkjkhhSCIWiCqCpFvr7UfSUGo9NaZAdJPRVM9vEUWXcUMdqKNyVZFsCVP0PuykLhhXpSltpGpuuVX2dX5OOKmpIXkYKosASOePd/GjfFOreADxA6UOGpc/n3p46jyQozWCLqAAv8AS3u2uHgRSnVfBBOAOjD4rbL0+PSja5kKqQbk24ub396M0QNAOrmFEGFHTjFjVx12eSwva17AEfTn3vxI+JHVGiDZoB090ecoINIaZdVxYaxwfwOfetcNKjj1YQn+HHTXns6Vl88UzFeLAMLf0t794sdKEDrwhUmlAOmY51XRX16m4LXYH6+964ga062YFGKdSYalK3RIzFLH6g2+n+9e25WVsr1oRhDwp0+DU5VlQShQOCfrc/4+2vs69wweoG5dl4rdNEIKhIopCoVvSL3t/Ue9/Prfn0lMRtZNhx6YKg+EsALMbEW+hv8A7x71xyet1FKHpRVG4ovCJVcl7f05uf6/8V92FDkdepT7OmiTc080UqKhdyp0A8n+g9+NOtlFIrXqJhpspUVNp4LevUCRcgH/AA91J8h1qnnXoQXitGPMhHA/FiLf4e98etdo67Iooo4yJyrMbsv0/wBcH37rQJ9MdN81RSJUBRJxp/x+h+hPvx62FNa9YYKjHzLKJZ7EEgA/7cE396xx63UjqVTR4PWxmiiZU51MFufydR9+qfLrxrTr2SbbNTSExQU/kS4sNOof4/7b3rJ+XXs8R0HGSxmAyCk+GAzQ/p4X6Hgf7b3sgk449aDevSPz3VWM3Hinjcxo9TGY7A8ENx9PZlZsEfvx0l3OASWbE+h6J3ujpufryrmpMTLIjVTyOqx60GphcDUP8fchbPIWAA6xH52tSl4zEYrx6M18Hend75ndVRktyTVEOOSeRoUZ20SJq4LD6Hj2LZsKD0DtSeGAD1tcfFmhxMdTR4GlggEkEVMGb0oWYR/Vj9efaY9VHw9Gj+R/VuI3H17XU1XItFO0UgihX6SNoIUkL9effuHHrRJ4dUr7O3/vT4q75gp8xWVEW3M3U6KbVL44GjeSyj1mwHvf2deB9erVdz9q9X9hdRTy5yWinjnxbBvNNA9nkiueL/7f3qnW+tQP5R9/T/Gn5JY2t60qq5tvVuVjOQTGRSPAiPKHJk8YsB739vWur8Olv5neAz/X+06GryVQK/ILBC6sumTV4wpJ1cjn36lT14cOh+298iZsLvo7vgqZZYZaPyJdx9XFwD71w62ejRdD9yZTufcFfkK2Mtj6SpfW8nqAUPbgj37HWujV7up6eSnqEcL/AA+SEJEotpcsORx719nW+Ar1UJ3U2f6u7Ip994FJKWiiqlE0cClVdVfVqNuOf6+9+XXvPqzXoj5O4HtXbuJxtRUxGuWHx1KySRA69IUhrnj/AB9+I69XpX7rp6jC5GfI451aARmVQkit+Lm+n/ivvwPWjx6i7U7cosgZcbmHUEaovWVHP0/P59649b4dJncO2YZ5q3MYORHveT0uF/xvYH/Ye/fb1uvSTydXlX27B500OJdJZWDMAGtc29760KVoOpuKzVVhaeOuaeUiNVBX/D/D/inv3XiOlhn48P2TiKXWVo69APBLGAHkcD63X6H3r59eHSVhqew+uoIRDHUZChaZF1vqe0Ia17n/AA9+PW6jz6HSTN4nd2LosdLoORrYFLoQLpIRa39fe6GvVajh1nh2pR7QommqJFLBQ6IW4Df2fSf9796PXuqhfm98cE+QsdSkeMinmV5tDOrNcH6WYe9jrf8ALqtvB/CP+49II9z0FOuMo49Da0ZlEYFyLMLe99ap0q/jHjupsH2/mMFlayClxLM0VJeLREspOkFC/vx4deAz1ZFvDrD4/DGpUtkaDIZapH+TrJ43YLJwulhf/YD3rPVqdFg7n6PxnVnSe/uwcSqyMMZNW0kCKW+kZcBQPe6561Tqq/4UdZdk9oUe7e1t4Y1qbbTx5NaATIwGmHVosrcfj8e/daHTbSfGjJdsUu781tnJVibixeZqIKLERCZIpUEpCkxrwR/j795de6MV118JN4bbxmIzHY0TY6S4kVWV1LN9RcP79UdapXHR1uuOocQmapk/hlNPRQeL994tXkdeP1e/cT1ulOrFtobYw3nxuIpsLRTRvGqu+hRoH0+vvXn1bhx6G2Xp/Zz1ELZhabxxOsggkIMaj6ng8ce9ZpjrX29RN5db7Slx+vbq04aBS4p4XGhyi/Sw4+vvYrx69w6KH2buOmoMBWy1UCY6fERmNfFYNKyrbULe/Z68aV+zqnvuHBPvgVOYrV8lKTIfK9wRZr3Bb3vrR9equ91YPEv2KtJTMtRGupGUgsoKixF/p731o8Olhsbr/Ff3pp/NBEkc093CxgqfV7916nVhVf1VHBtFq3CqKZ1pRJrpwQzWW4/T711v7OiJbeg3fW9xR0lbBK9JC7p5ZA3IQm7Anj37rXnno/fwrzMZ+UO3YGqGibFZyMhQwW1phe4/p795dbqK9b1uDqkrcPjKpGDrNQ0r6rg3JhW9yPz7917p19+691j1AMwJtxfnj/XI92oSOqVAJ65ixHH091PVhwx13791vr3v3Xuve/de697917r3v3XuuLfpb/gp/wB697HHrR4dalH8wf8A7ebfGX/w8aCx/wCq1ffhw696dbaNJ/wFpv8AqHh/61j3rrfUj37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XRFxa5H+I/HvYx14568BxY8/6/PvR60OGeg73psOg3Hj62MRqJ54z9QLCw5sR79SuB1sYz6dVCfIr44UNZjsnS1yRSUojqXJkU2AJJ4J96HXq9VLbU+MuzY961FRjqOkqpoaoh2MV+Ve/wBf+K+918x16mOjmU+Ihw9L/CIaaKAhNC+NSLkrpA49+FOvedOjQ9JY6XbVGa6tsI2sym9j/rc+9H5db+3oHfk/uBM/N9tjnJkZtPosfqObFfeqGnWxSor0RerwEtJQymsUK9iRf6/X6m/tQB+ngdJ0q05A4dVx/JLckW2sfVtLRJL6pgrmMWBtxdj7L7hwikg1PSffWWK1J+XVSe6cpW7hqJKqZGgpwdUar+ki/At7Jml0nPn1iNz5u8iTNCp+I46VnVe0TvPOUGIiGoGoVGXk2J/J9sRv+rjos5R5fW/mV5FyTnrZH+K3xbxGE23j6qeggMzwwTAlCCSCDcX/AD7PLdA1K9ZW8tcpwWcSyKBgdWe7Owq7XgVIokjhjPqKnTYWt/re17wqi1HQ5kkjakKcR0n8r2BQS7k/hcNcTPq0tGsit9D/AIe6fLphk08PLp/z+MpKqGnmqJTZ1VtVuebEG4/Pvxr+HpO5OnT0I2DqKGbbUuOSoDWj0LcgXNvqAPehWlOrLWnWsT/MfxrYXsGrqI2f0tUEMqswF3+pYe79bNfPohuyd5xpT+KeUu+i2hyOeLcX9+611Dy9dJDWVFelOkSScl0HPHP0H09+68OHSAxe26bMboG4Wo455wCvlaMFwCf6j37r3Q/4DBpTxT1ZHjkju0cQsOb8Wv7914Z6kx1GViroaoo0SLICHH1Nvz/r+/da4dGd2x2fX4unpmad2hQxB3ZuPT9b3+tv8ffurY8+jwbM7hxudwkFL98tjEFlBkRdJtyL/j/W9+4db6C7s3tbb2Cx9fCcmNT0sqovlVgzsbaf+I9+61jqozPbmbN7yqagqaOOSokZCllDi9wxI9749a6m/wB48nQrV2iJaMMaWQH1Fh+QT711vj0JnWHUGwO5phJ2BPC1aSf26n1aQR+oav6e/de+XVwnxQ6h2P1NhZ8btn7WamcEo0YHpGq91t/tvfvn1vy6Ufc2Vo8PnMVLGRG0tRFexsSS1rMD/vHv3z61506K/u3c9NJ2bBKg0zmGNQVIvcrYgn37yx17zz0ev4/z10mSo6qqBFKb3mJsUDNewv795dbxXp1+d2xdub82PSUmeCVuNljSJzVLrRUI5Nj+Pehw681K9VN5DrTqPrbqyvodqz0kVW0zuFpgq6Zbah6ePe/t615V6KtSZWnUtHVZCZQ5GhtfJF+LW9+618+s+O3dX4fJXpJHqYWcAlmvxfgD377evcOhy292TDRVkFU+la7xq3i4Nj/gD731v59I3vDsaTKbaybSlgzQngG9jptZfeqU68eqndj1dQN7VtXKpMUk8lnK2NiSSOffuvdGl/i8WOpRkYpf3YyOFYXFj9Rb3vrWOPSmizb5eKgqqn1K5vdzybEfX37r3nXpxyFdT1Yagpn0to5VPQORxa3v3XumPbrUW3quRp0QJKzF2Kjgn639+690xbgxNHlsnLkoZ7Qk67A2Q83t71Tr1egf3VGK/L4uiREaH+J0a3AHJ8o9+PDHTsKB30dXJ0aptbqXAUMcKxrX4yG+i35UX1W9+45PT0imDA6Op8XshVU+3Yk9RiccX5Uen8e9UINT0wL9hhuA6THfMgzGXfCvSLUR1JdWLRk2Mh0+m/vbRh+49IL60j3bBFeiK9m/D7Cy4Ws3DHFDHUMhkCgaeQpYix9lc6AHHQK3vkSKWAtpHVSm99r1G28zUUhjHjieUW/FkfSB7KpzQgdYvc67aNnkYpQEV6ZMXGEqfM48MSwOTGLBCSv1uOPdo5gjBekHJe9XM1yIqnFPy6Lb0NsTDdzfP3qHqvL6P4Vu3cr01XfldEcyKtyP66j7O7cllq3DrNHlNw9ip8+B627PmB/LKl+JdXh9ybJVqnblRiKCokanhmeNTJECwsot9bj2aIKrpHQkmKg06ArZ9IIYaZ45HScIjPcGMhj6tNnt+fbBpUjpsZz0POPymU+30fcN9ALar244UW/p7rnrZ9Ogq3bPVpVyy1io0YjYrI31B/Juffh8+vEZ6JNvaGozO76aHGxLPG0oDhfWLE6TfT731Xo0nXfx32WscW5MpiKCbIemXRJAC2rRq+rD37rfRiIdu0ddjxTUGPipY4iI1gij0oVU8H0D8f1PvXW69LTa+08xjqiChpIZXpKy3nSz6QG4YFf9b3v7etfMdQ91fy7+v+3MnLk66WDEZGpA8s0QaKRfIbkahbn34469SvDoLs5/Kh2jsaUZjb2/shUV6WZKQVc2klRqA5J9+8+vEDqHR9Yb06rkpzTtPkZUN4hI7Pq1G359+rXr1KHqwf4G979m7b7hMG78S+MwFTUU9PT1ZXQsl1021G3HvVadb8s9bQWKr48pjaLIREGOrp451sQf1rf8e99a6nj8/wCv791odd+/db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv//S3+Pfuvde9+691737r3Xvfuvde9+691737r3WNXDMy6SChtcj/D6j3ulB1UGppTqlj+db0eO3Pj/DNTUqy12GMs/lFg4iQ67X/PuslPp5PsPTkBC30DUrRh1pMbizWZ25s/JbRjqnpJoMq9GI2axIQ6CL+4GvFukvnauNRx1nFyhfLNtccb/wjoUOteuE/hdDuasBJigEsgaxErW1Ei/Pt9W4aujLdWCnsx0zzY2l3HvjzTWNJRzK8EJAGlkb66Tx+PdGCu1OrWRfwdQNG9ejHZHIVU+PpaSll8IihSNbi3A4UX90wPh68ZJG+I9RqfbJqKRnyh+7QqSAb/Qj8f4/7D3fxJKUJ6qzDh59BRk9j4WbITBaJGQ6rxkKSL/n3UgMOrAMwoT1Hx3XONgrBMlL4wObELa39n3plAyBnq1dOa9LSFKfETIRF+j8i1kN7Ef63vwCnB60WLDBp07V2+qSkhb6CXRYFufoP969+CgdUOrgTXoIdxdiSTlxFMpGr6D88f0HvWF8urEt69BvNuXLTyK0MrXduACxsTyAB70FWuqnXvEelK9S6nK7qbHvqinc6SVk9RAv9PfjSvDr1WPA9RNhV2ZqskTk6xhEjG9O7HkXJtY/0/1vdgnmerayMH9vRgjmadG0xAKI1UkW+o/PvZX06przQ9Nku9JZKxYqcGGNQF/Iv/jx78ccOtChHdx6UMO6JCUDzaWuLMTYf4/7b34EHj14D+XTvNlIKuMJVMJlFyB/X/Y+9Z63pByeoLTYu1vCCovdfxb/AG/veKde7jg8OsCVWORwYoVX1cGwPIPvdADjrWhqcenGky0L1iQp+259Or0gm5+nvxABwetUpnj0vlqqaKnBqlEhte31PH9Le9Z4jr1fl0l6jNY2ScRiDQA5sfweffqUGetjV+XTqBjZ01iAFtB+luOOPfq+vXqsO0dMctNSM5EUQjYfU882Fvp/re9U8+vE+XSVqFaKaePWLBSFF7gX/wBb3YcOthTXPTdj6SWSpZCwKNx9eDz9bH3qoUU69hTU9ZMptSt8byU0hjL8g8j/AHr3sGhqOvKRqqeu6Ta25IKSnqVrWl8FpnhBNyAfx7cWcaxXpTcBJISvy6BfvreLrgGSnwcxy1OdIrBGQfQPUSfr/j7Hew3sSuEPWP8A7gbC08TvCvr/AIOhB+EvyIpjnsTtjNzR0tVUO0WmUBWZtWn9J+vuR0uLeZRp8usaDtu52c7CcmnkOtjfYmAzFFNR7q23nDC0cdPVSxRnmVUQP4wFN/8AW9o7lkJ/THS+1LGSkladGer+78fuvb8jbnq4sZPioZIjDVkxvUNEn6wrHm9vaYMwOejiVY1WoHHrXS+f/dGZ7v3DTdf9a0UtZk8TKsNLU0V7xyJLYEMv+wPt4U6LiQTjPQq/GL4SfKvemyoafde98hjKSaON1pKueVCIZEuIyGP0t/h7917o6vX38qXbNTBWJ2TjE3Vk5nYpWzeObTY3B1SA+9Ejz62BXoYJv5Z+wcdSQRYPAiikoiHheMIBTso4a4H49+qOveXQRdodBbn6qw0+SgNRnoqWOUCkgu7JFCnNwt/p79nrfDpR/BL5L7Mxs2c2znXpcDkpJ5E+0qW0Ss7PYAhzf6+/Eda6s3i3/TZCXIGSp+4xscBamA5jUlbi3v1Ot549E/7j3HtnLYHKPlpYmgT7g6XAIuoNrBv6e/AevXq+Q6o07S+WtZ0jm6yXZuVXHwRSA+VXdVQFifVpP+9+90pjrRPS76U/n07Oz+Tpur90VUM2WgmjpqjKTI+iW3obTKxtx78c9e8+jtbr+eHXFbDQ/wB28njxka9I5UWJ9LFpCAouD+T79w69Xpdbm+UnaOytjUO4cfgcjlqOtpRIop1kkWRWXVYWv+Pfvl1v59NPTv8AMQXcEn8L3dtqXGlpvEsdYjoVcmzNZueD79QdePVmHWW68B2bh2WGWCOKoGpZAwsqk8Lf/D3Xh1759GQ2ZtChwj6XQ1yqbxMgDBT9Q359+pXr2a9CfXUVHXUyQSqJiBc0ZUalUfgj+h9+4de+fQT7lyODwGUovsMY0FdH6QLG+r8WHvZqevA9JjcVFujJSUuQrMi32tY4CURYgi59KBffuGT14Zx01b8MeyNqfxCpxrLKwt53S4YkXFiR7159ewOibbkoKntGSnwsMZjTKAKukBb6uAeB7tSnHr1ajHSazn8uqhG38nW4mL7TcX2U0kFYrqJVqXTUr3HPB/p71Ude6CzYfxE37guqM9HujMSZPdKZRzi6h5TJLDTRyHQoDf4e/V69kceh6x/Vub3F1FldmbtZ8rFUY5qVoGGpuY7Befejx635Z6KfsjaMmzqL/RBhymKxsVXNJNTMFQyQyPZo/pzf6e7dV4dHF696q2b15jjm8VtlTWOplqBHGhapmB1eQgD8nm/vXVgMY6Q3ZB3F3QZcVBTT4WPH+mItGIrBOBxxcf4+/fLrXHpw2Bsaq2niBjquRshW6uZUsSCR/h73WvDr1DxPS+pcTuimdqrGVMsMisWVkLDxr9SvH+8+9Y68cdJ7fveuZ2li1oJ6WfMZOQeI+IMz6r2v/t/e+tefTXsjtbdn8Olrsjh6yD7lHjVJUfSolHDG/wBLX9+oOt9BDuvb9TGmY3RuPMrksbM0lR/CCwJVbFgmn6/4fT37h14dVO/MLuvC0m3KfGbNhjxkkhaKWJAUdBr0sxsffuq16K/11sqg3VtOXK+mbNTHyDJqQQhb6qW/4g+/de8uhD2ftlNs56h/if8Al4aUXlFm8d2/w974de+fViOyclSZqnnxccAanSksI7DSTo+lz9fdfl1bj0Wfe2NxmyM/Vbskolilp2mVaHQPJNwfVb8+9+XWj0WHpbsOLbvyMxO7lq1oIa7Imc0jHS1jPewH49749arnrf8A/jdvCPe/Ue1M9E/kSoxtKoYf4QL+T/xX36tR16lDX16Hj3rrfXRVT9QDxbkA8f09+6912AB9Bb/W9+691737r3Xvfuvde9+691737r3XvfuvddN+lv8AWP8AvXv3Xjw61J/5g/H8zb4yi/P98aD/ANzk+lvfh1rz/b1tpUv/AAFpv+WEP/Wse/db6z+/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuuJJ1AaSQb3a4sLfTj3vrRPlTqqr5xdu7b6xTMy5uvhpKVcfVMI5gqoxVCSD70f59bFPy6p++L3dm2u3Jd1ZHCiKI0uRlEU0a8SASkBhf8e/db6NbkZ48dVQVVXCagu4IfTxzzb37BHWvPpXS7sq6yi+3pWamgCAKoGlTYXt711uvQL5iqarnmWogMsqsdEvJW35Jv8AT3s8OvDoAuxaXLztj8ZhYHytXlZPtkhpzd4nLcXt7u5ZYqDHXrWhuSCOgE+d/wAMNwbE+H2V7b3FNNT5MGSojoag2m0lNdgCL/T2V3HwknoL8+O8dm7xmgCn/B1rh7eqFyeIo5JlPqp0dgwINz+Df2RSMWenWG+9ObmRpZwSRw+XRlfi9SLH2hiij2hOUiLIAD6SRcG/19q4AuqnQ09vJma5FWzXrbI2nuPH4zCbfx1PEIDJjqZmkCgDlBf2d25VTTrMDa1l+hD18ulN2H2dQ7R2tURLPHV11ZDaC1iysycEaf6fn2ZS0KdFkRYXTM3r0WPpfYO5ty7rn3nkayUwTzM6xM5sEY6gdJ/FvaOvRkSWOej1sIJ4Fx8sfl8cbL5v7KlR+fe+q06B6k3U+IzVfQPLpiRyqG/BF+CB79TGOtVoa9ED+bHWOO3tturzi40VlUYagmZVBs1ib6v6+99aPCnWvlL11lod1SwQzNTQRTFWQsfSF+oA+nv3WgOl4MV9qZ6Kv1TrpCKWNjcjg+99a65QYyPB0RqopQqFz6RyVH1HPvXW+uJ3DOqAiSy2vp/J497691Ox+cq62VYiGkjVraVBBsfoffuvdCFA0hp9JktHoLGM8cgfm3vXXug4y/aOV27PLj8VX/au503DMbA/1AP+39+698+kLmt55XI0xky2SFazAlVLvwTzxc/X/X9+690m8eXrbzEnXcWe5/SBwBb8/wCv7917pXU2QXXHDVSq2mwBP9TwPz7317oVdrZpMTVLLRsI5+P3VuB+ni9veuvVr1a18MtyVWZydPTZCfzpIGGi/wBVvfgE+/dW6Fn5KihkzkCJSlVoZI3LfpF1P6v+K+9Z690UWnxUW4d8xVEBAkTSoNxc2/ABPvfWgQerIuk8FX0f28EkjSRMv6QQeQefoPfh1vpefMHGmDq6nQU7uTTrYkH0Fl/Vf3qnXvs615dyrVJU1+LqJyyvUTSKjSNwL30/X3vqtacOgUqIUeuMRJAhbT9bf73791rz6j5atfHBWQatFj9T+D73x68OPTND2lRY2rWWpozLLGNOog/2R/T3rreem3N9jUu5g9PJAUhlBFnDAWI5tc/7373177ekuvX1HUQnIYu0bsSxIABuebce/de4YHTTBQ1VIWirw80CPpKG9iQfr/xr3U9a6XFKHeGH7cNHClrKQdK/4WA97HW+sS5FKWvMa8SH0lzYm549+699vTvULTrTutTKkrz+pFPJ9RuB/wAV9+69ny6bEVMlHHQ0UiRyoQJAD+u3Frf8V9++zr3QK53GbhXfmOxNPQTeBKyCoarXUUAVwfz/AL173w62CQdS8erVcLmq/P4raeCkL1Ap6OKFwgJIKgLYe9cOrF2Yd5r0ffrLNybOoqOm+2ZY0Kl1N+VC3Nx79Xy6bMSHiK9GD/uhjuw5o9xQIsSwRa3B0j1KLksD7bkZgMHpVbUhjLDohny27gx+wsfJgqapi8iWicAeprqVN7ce0E71rQ8egRzPzStrG0YYdUL7q3dPunctbMys8TPKV44Opz+D7JZSA+esU+cJJd6m7chjw+3oIt/77x+ysLUV1VKnmCyRrTGwdyV4AHtUsGoalHDoU8ke380em4INTToUv5Jnxq7E+Vnzy637l27jK0bZ683RPPXVECExs0VWDJqNuFUA3/r7OIo5EUJx6yo2Pbxt1kErkZ6+rPvbrTZfZW3htjeOHpsvSNQxw6ZkRmgHjC601fT6f09rhqC14dKDIC+nz49aev8AOaodgfBavOY2xlqWaKtl838LjCeWlBj8ggaOI3DD6e6Yr1biM9UydPfzQOv9zYunmyddR42SXVqjqv22upKn9fv3XukX8hf5n2wcDjftsS1NlJanXCklJdvXJZQt1+nvw+XXqitehp+A++f9N1NUbxykHhgkLTwJID+gPe1n9+62MdWw0VZFka8YugjBWEaW0KQNNuL6fevs68OjX7C2DSyUMczxL5GRG0C31vzcfX3vy61mtOhVhpsHthWqa0RK0dyqsEsfyBc+/dbp0wZ/tPb+OpDWY6up0qDYGFLF/wChuB799vXuJ6LXuXv+prst/D4akRzE6UlCn6H8Hn3759a+R6DjL7xy1fNqqqk1DgkRMPov9Lge9db6Mj1TmJY4sHVVb/cz/wASg0WAutjcX088e99erjrZQ6EylTk+v8XNUXJSmgCg/XSE4Fj78M9aOMnoazc6SGKgG5FhyP6G/vfy61xz1y96631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf//T3+Pfuvde9+691737r3Xvfuvde9+6917/AHr37r3XQ/P9b8+9nrQHQGfI7ZeP3r0/vjH1tOJ5odv5Coo/QHInjhLKAp+t7e6tlCnketodEiyea9fNG+TFHn8P21uXH5CkqKOKLd1ckSTRNADEk5AATjj2AN02pIi0tM5PWQvIXMrOVgrUAAcejLUW5xj+uKMQcFKFdek/UhOSLewDcOInNeponb6hA46ROw6Cor55My+oLKzNa1ybc8H25GSw1jpdB224B6FVMk8kywAC6tp5JBax+oH/ABHvZ49U0049L7C5I1MUlHOAGIKJcgWuOD/t/fgBx6rSvHove9c7mdsbhcCGZ6XUzakRitr8/T/e/ftRr3dbBFaDh0oNv7+ossqQsNMpUelyAQf8QfezUmvXguqoHTtlHcxtP/nFI4CC9h+b296x17tpQ9BrVYvIZWVhCJRGWItpP5/qPejU8et0HAdP2C6fqK5hLV6wn6iLHgXt/wAR9fexXrXA46E/H9S4KlZWmUEoQQGUH1A/kj3vBx1rhWvSyrNtYCnxctOscBfwvpui3vpuPr71k9eWgHRD91NWYXdEn2yP4DObeBTaxb/D3qrg463VehDpMos9NAypIsjFddwQSLc8n/eve6k9a1D06h5dqhHWaiRh6TrsDcED34k+XWxTy6Tcu4KoVFNBMxR/odTEE82v78at8J68PXpfUGXnAjj1MwNjctq+o5APvygg0brRAORx6VlPK8qqzH/XP9efoLe9UHEdbBNKHrsssGuadtMa8/Wwv7314E1oem+nykdTXCWjJ4YAsPoCPr7t9nWmYA46EGPIVc0CKSSAv1ubHj6ED35qV6sAOoNT4o4jNJcMo1W/SSefeut5PavDqPh90xPOaY/glVt6iD/j/h73Tz6qVoaKenitnle7U6sL8jgg8/4f6/vWPPrwHkePSbrYK1jG4RgWa78Hn6c+/etOvE+R6c6ahqYikwB4sTx/sTf34CnXqqRnpcU7vIYIpFDK4BIP9LWAHvx7sDj1Q1Ax0occkyVM0YjUU5HGoXvx/Q+00sbpkdaUlumPdGzMFncdOKumidn1Kx8Sta4/1varb7x4ZME9Ir7bIL5CpH+XonW/uk/7jFuxtpCanrNvr92oiBjBK+sghP8AW9jzbd2c9pPUR87cnwW9v4ka/wAujO9cfzHdy4PrXESxVQm3C80dA9ItTrmvEfFp0Hn2OrI+Mg6xwug9veGP5/5ejh9d5Lu35KfZZeX7jGY+dIndSXhRo5TpfV9L8e/XYKHHDowumKW2oHqy7p34ndK9bT0G7dwilq9y+RZ616xI5AXIDNy/+P0HukdNII6QQnUoYefR26j5BdHbahSjqzQ04ghWNVp3jiH7QsBZP9b3b7Onegpzvzv6lwGUho6GaDwNqUEzC4sOLt73Tr3QEbx/mJYSjlySYx42gmRliYPc2P0sR71T069jhx6LfH868Nk5KmmykKTQVbyRMkxEhtN6SSG92z5deHQDbx6g2Vuapm7Z2NkTQZGB/vZ6eGoFPGWH7hDIhHF/x799vXqU4dBhuv5s726uxdJSV00s1PIywSPHIzNpQ6ByD/T37rVa9EV7+/mEZmv14PBDIu9bE7uwaQrrkS1rjj37rVa9Us989o9vbqir4o8XmqmHIixlpoZZmUG/+p5H19+638x0SgYDPbfpf4vU0u4KDLRPJUpLLFLBIZB67mWwI59+698+r/P5G/XUfyP70w0Pc2WrTgqeKn8UVdWFk0xVAsdLm3049+69kjre93nU/E/rLFU+ysvU4GTGYpI4II5ZKdpDGIwttJv7117PXPYfxA+GHd9L/eXb9PTiqmJZPtUjQBpBcMAthYn3unl16p49LGToTrzo+irIaOuiipIxMYIzMEmMcY1ghfeiCOvDI6qW73/mX43p3e6bawNPk6lIqnxyNGHdLB9PBAPvwHXqgU6Fbqv+YFS7qyqZuodqVp4EUw1MpRfWt7hW9+I9evCp4dH16d3Xju2UrN7ZZoxjsU7s8zkCAaTyGkPH496+XW8HoSMduja+890/YYmaCopsVKsoankEkfo+o9N/ofr78evD1HTp36cTurbFPho0iKBo4ykNi4I9J4HvZ+XWvPpD7Y6lw+Hk2vkqWBtUEMZI8fJI5u3v3l1vPRxcVtilq6KtqqtYVj+0LKrL9Qqiwsfz72FBHWic9FozGIx8NTWSLDGYfK6+IAFSb2uAP8fdT1sdBHuVW2jszd+fqFgiMFPLNQhtKBjpJAC+/Ur1rjk9Vp9WZ7Ddhb0qs1kIpUya1UsYdYyIQqyHSS39f6+9162BXqwzEOkFLCJKzGLoj0os08KjT9BqVj9ffsDh17j01pUbbpJ6+XIfZSPLexoyhAv9ApT3rFOt1PQevmMdSZJzjUfxudADi51HgX9+698j0p2bN0eBra6HwJG4ZlMgCNY3/J/3j37yz16orjoCcfPttq2tye4VjqaqItIigCUBgSRwL+7U8utefSPrOxMrXZadKeCkp9vRRSICyKhBX6ce9U68T0RzuPtWfHVlbRtO7QyyuFVZyIlDCwNv9b3vietcMdE5626GTvjeG55MpHNPRRQTtToEMsfIL3U/Qe/cOvcekfXbLy/TOam2xBSzjAmplVv22BUKeOffuPXuhd2Jg8TuWGpq1kTyR3ZY2N5b3/p79wPXhnoye0sXJtI42vkRxDVVUMRDmwKs1vz+Pfsnr1acOkz8h9rU+S3zhpljvjJaAzTRqh0sTFcnT9Pfut9Un9js+A7gnrqV56amxdXrp1uYwAJb/i3v3Vet6D+Ub3BkN+dF4fE1bGaOioqYxyM5Yi0PJBPv1M9bPCvVvvv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de64t+lv+Cn/evexx68eHWpR/MH/7eb/GX/w8KD/Yf5anvQ616dbaNJ/wFpv+oeH/AK1j37rfUj37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XX5v/X37rXnXpry+Zx2EpZKzI1UVPHGpb9x1UtYX4B97+3r3njj0Q35BfL1dm4PKf3QK1GThjJpPEQ2pwpsbr9efx719nW6daSP80P5M/L7vqpyO3MRQZaItPVhpaZJ/wDgO0h/1P4t+PfuvV8+q1eifl3398UKykwmbxGbkoajw/xCqME0YEgfVLrJ/wBj7917rYD+PH80bqDtiDB4HO1gizcjpFMtTNos4UBgddrc+/V691YFu/vvrzC4WKtpczjTC0KzECui1BCL2IBv7917HRcpfnD0lU7vxGx0zeNiyeVaFLy18agtI4j5JPvxx15T3dX4fFT4w7LyWMw/ZWT+xy8VT463HCPRURMzqJB6zcfkcj241NA6rEWWVmXoBv54O3mynxIy1BQwpFElPUosUcYEaqsRsNC8cD6ey+4XyHQQ51q+2PqzUHrQNx5bH00uPLATUsYjtwPoxHPsOSMA56xD3YAl0Xy6E7qLflLsreGCyNexjDZSIs19ChQR/a93hc6qdGvJF4LC9VpCBn162JcH3zis7tTC1uHradp4qKnjt9yGb1IALf4+zu2euX6y92LfYprYJUGoHn0ZXrTbX+kSClyO4Kn7hFuyLq12A/2m/s0Mh0U/Z0c+AdRl9ejf43D4/auLjTExqC4Cfptbi2qw/HtkCnHpzj1Poaeq0OrNq8+uS/4UsL/n3vPW/l0TvuVMpiZchVURcSIrEMptyDe4P5926pkV6TGxN04nee0Ztv7o8Rq5RJAolZb+v0g+r62/1vfhnr1fTqv75BfGRtr19TnMNTO0dU7zxvDBdbEauSB799vW/KvVdW9MdXYk2qInWZmtyh1X1Wuf6e/dU49I2rjrWxHkncaQ4a1/oPfutinTYpgkiQ/UKAGt9b35Fve+vdPmMY0h+4hUWI4BIv8A7Y+9de4celVQ5KpmZvLZUKsOfzc2I59+610E+4cDRVeSnq6h9OhuLfQWN7k+/U63nz6Q+UXECalhZnMfmVbqeNN7X9+pnr3S7zFJjqHEQvhyusxKW5ueRyePz7916h6StEVrGiY6/MnMgNxc/wCHv3Hr3Qt7VoZ6mpBlFogBp1cXsOB/vPv3Xj69Wd/CSky6b6xqhXagVfWwB0gBvyw9+PWx0a75GTUNTnMjSxXMwVf6n1W5BH19+699vRZ9gYf7PPx1U3+cMy/qv9CffuvZ6tB6feD77GQQ6R5bFuAeSeRz795Y6351PRj+89k026+vcxBLGHfH4wyqAgY3Cahpt/re9D1618utWjf+LSi3vl4qkNGsdfVRLrUofQxsLH3vrVM9Fx3YHociJaYcNISCv5/oT7317hjpN5f7megWVWBZhyDzYjk8D/e/futdBnlMO9RRS1Eer7kBiBb/AG1r8+9Z630BGerNz0Eq+FSFv6jYgkA+99bzWvQ89e7rrjhtFa2qQIBYm3IH+P8AtveutH16UGOycVfPPS1aKDI7Mv8AxA5/Pv3XunDFvVGrqqaMjwxG0er6af8AX9+z14dNdRjK7+KiZimgstuRY/43/wCI9+IqevUHSuq9sz1v2r0SyyygIumMFr6msfp/T37r2OjwdO/Eerz2Jg3BPC4lljMsitDbT6b+/de6FrO/Gqho8V94uNWSvjuolFKCxK/kOB799vW6dMeydjV+3Ny44T08mhnUKJIwAATb6H345HXh0dKhxM1dM6PCwQIArKn0AWxv/T37HDrxqR0ZDaOHyGP2RkJKBWutJVMvB1elCQBb23JQGp61MxS0anGh61gfltunP5ntvO4rKSTrTwZAKA7sBbWfoPx7L5qA9Yr+4FzeR3T6SaY6KnlYIcZI7UgLNp1Fvqfpcg/7H2TTYkr0CtoYTzI9x5MOPQb5HrHGdlMKjOSMIop42MYJFwGFwAf8PZrbSxqor1kls+/bfY2SjtBAA62Nv5KXevS/wo3Nj9q08OMjfdtZMtRJOkcbo07DW4a31ufr7OInQrjoUWfNNpd0jjYft62tvm/849jfHP4vZ3vChy+OkFZt6d8ZJ92ilJ5KfXCC6nggmw9vKpYaehPbQC4Pip6dfKG+Z38wrvb5o9wbyye88q9RtiHcNXLi6Y1cs8cVIHKQgsxs2pfVx9PbOmh6q5OrT6dF36a6NyPc28qTBYiXMwq9S0btRGUQgluQAnA+v49++XVadbCPW/8AJ3w9dsugqcxFUVlckVPVlquMytfTqIJck+/HrYFRXo+XR3Q8PSOAq8DjaeOFadTCAsYiNgedFvz7914V8ujzfHLa9dl9zSSVkT+AyeppUJDj/gx/3v349eFejzbgqsZsmoQRSxALTkn1qF1hfyo9+63TOeiC9y9s56rywEFQI6JJbECRlUrbm9vfvPrRwOiTdj9+bd2cJazLZqFJGDftffhR/iACf6/j37HXj0zdbdmUXZM4y2InebQyaWR/Kpu3Fm9+69no3uJBESNkAwawNgPyfyR/X3XrfR+vjBtjE7ny+MgmdRGK6DSrkXuP6qfp7t17y62SeucBBt/aWMoYNJU00bXQAC2jSov78MdVPn0u7gWBP14H9Sfe6E9eqBg9d+9db4de9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//U3+Pfuvde9+691737r3Xvfuvde9+6910b8WF+eebWHvYp59aNfLrv+nvXXvPqLW0cGQpKmhq0EtNVRNDNGeNcbizqfe+HXiKinWjF/Pp+NUPV3cFHvHa2JFJturkkr6yphjcKskoEkgcji4P+PtBu1v8AV2zeGO6hx0MeSdyj2+/WOdqAkZ6r+2BhKDdnVS1cVQHkjoFOi4Jb9v8AoPcIbraTWk5Mq06yw2y7ivYFFudXXDYl6SFMaiHUkjK6/Sw1WNyfx7tC4eEEdHzjw4gjdLfL416aZKunXU3LELe4P5uB7tjh59MZ4169Q1k0E0T2sb3Yf776+7DAr1oqDxNOhGlxeI3JQrFW0UTyyKUaVwC9z/j7oag6uvUpw6Bfd/TT4tXye25DLPfWYoiRawuANPvykkdbBIz0lMPubM4kGl3Hj/DFGLNI6n6fQH1e7Ejz69k9L/G7q2tNoalniMhKgxjT+on9AJ96BBOetZGehSx+4aVIFjUpHrUWIK83/wAB73X069lc8a9Y62uqf85c+OQ2Rgf6/T6e/YHXqVHQY7ozlbBKaeF3Zn9IUE/U/wCHv2ocOvArXPXHbPX0WZf+I5ynQeQh1Mn1I/rb3qtMHrYw3y6fdwbOwkKLHR+NGT9VhYG3HPveOtageIx0FWer8XhI3WRVNgw1H1c2917a0HXjqHw8Oip79zOQrMrSS4KMyBHUuI42F7G/On34AAV6sMCg6HHaM8lTiaaWsGitJQNGwsQSP8fd64q3VQADjoSo2MEQ1NbgHTe3+x9168ccOoOQirK6nkp40a0wsptyOPr79g462ADnp52rtH7Kl8k73mLA6STf+vA97qa9e7UPr0s6YmEugiEiDi5sP9jx70c9bPr0nMxFW1rtFTx6vMbDgm3Nvx73xxw60en/AGrsE0rCqyACMbPax5/PvxwOqhi3DpcjE0iTKIysiBuefwT791vz69NjqV3dbC68gX4PHPvX2nrdTx67paBJkaNEAIB5BA/TyOPfsDrRbqNj56OKqkMzKDTsfTqX8H3eFlWUM+FHV0QyOI1yW6eYMl5almCaac8K62swvwL/AOt7W3CwyjVEa16rdD6HE409OBmOrxxRmRX5YWuP8QfZcbSSPuIp1q0DTtqiyOgw7uz9Ht/qXeeRrEVYqPHSySqSoBVYiT6T7OtoimmasPdTj0Q+4DWllYgXzaSwx1XF/LB6Tznyn7erM00DzbPx2baemjdHSnZUmL3B+h+l+Pct7LWKKtxj5dYW73A8m4mWAVTUetw+n2XgNgbMpKHbqxY+qx1MKaRKUhS0kQtc6efqParcCJf7Lh1e6dWs/DHxdAZvDN72y1NTww1NSiKtpHAfmx/Ube0sQZU0MOHRfbqQgr0AW8dh7trYEqzVVTwu2kysJCCSbMP9b8+3fPpQfl0G+U6R3FUGCrV5J7KCWKOeT/j/AMT7917z6VWP+P8AUyxU0lWmozMAyMjAA/1sfeutU6k5D4vyHI/cx06Rw+EsCEt67XBFh731v7OgnzmxN9beosnQUmRrYaFmZfDG0ioU5tx+ffhkdaoa9BHuDrwZfC0cedolmEbajJOuu9jq1Eke/de49ZMf8Rtk7vwpzr46jSSMhQ5WxJH4BH+297r1r7enXAfGfFbarKejTZGOzWOrGtJVTxLJ4FPH1IP096630CXbnw46x3tmK/GZigo9vUpgdaQwR+ITTt9IwQAPr731r5dFo636w7D+GO/l3jtKOpptswt40rKYTMBEGurgDj/H3rrx6Ezu3ujvDsDMYDe2G3Flq3EeZKnKxuX0iJSCdQ/1v6+99e6vA+FXzJ3FgNmYlMK81bWv9nTTQKVFpAulywvfg+60Hn1sVPVj2UpOzO7q3HbsymQrsZjIULS03kcRSahZkZVNj/tvfuHW+PHoMt4/DXZWdr6bN5Tb+PrpZZFkeolRWd7G51cf7z79XrQA49F479+HM+DwUW6+tIlXJUp1nE02pVKxC9gq/wDFPfqnr1AOHHoxvQ/Z74j4Rdt7bz9tv9nw080GDo0Kw1FXLpK3VWsTc2+g97Pp17PHpVfy48bvXCbMzG5uzmngqaqGrkglqSxaxuUOp/ejTh1vgK9G765qzvnfM4mqDUYSKocXZgEBD/j/AIj377OvUp0dOmpdv080dMZYglMdK3kSwCjhbH3sUr141p0G++uzI8bWHE4yYiJmCStEwsE1aebf4e9V8utfZ0EeVknrsnE2LLVdPIBNOL+kH6txewP19669Xz6Ix8+O6H2xtah2rtRxW5rJQrTz4+IpIVlYWKMEJP597Hz630VX4/VXZWO2ujSbBSPJVkza5fFplRJeRL6he59+698x0ZnG/GndG/UbL5/euU2y8hM32kdRLGq83C6QR/tvfqjh17HSx2z05XbCqnhbOVO5KVm9VRUuZbKfqbv9PevLrYPUHfWe2rsZJMpV1Uauq3hjsG8lQP0pYf1PvfXvKvRcMv2/3L2HIcPjNrzY/bsjKIsiiMokp72D+kD8c+/efWh59KaDDY/Y+EauyWQavydTHeWml9el2HqAv9Ofe+vcOi+7pyO4s5S1bUlC2Px8YmczRekMoHB4tx/h7917ohWUxuc7C3rHtLGRS1yzziGqqCpfwsHtbUB791rqy3479OUvSEsv8UOubLKkLam/SZU4/V/r+/HPXhSvSL+RXXOMqM+aCmx0ExrYpajz2DNrkGogH+vvXWzjHVXOTwu/+sezsRSYajnrMRkKtVqYwJNEatJyCB/h7tx61w6syWDbm4MDiEeuSPN0j0089DqF1KqGb03HF/eqdb+R6i9mRYyajg3CZEeXH0TQCA6WB8cdrke/efWuHWvn8nKts1uHOVO2KZZ6+HU0kESEnX5LaSq8+/de88dbj38iPGZdfjpjslm6NqWtelgQqwKkfsj8Ee/U8+tGvV9fv3W+ve/de697917r3v3Xuve/de697917r3v3Xuve/de64t+lv+Cn/evfuvHh1qUfzCD/ANjN/jL+f9/hQ2v/ANRqX9+HXj1to0v/AAFpv+oeH/rWPfuvdSPfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYamdaaCWduViRnI/wUe/de6q4+THZmZzeabAY2vmpabymNzGzWVfpwQfevPrYGOi7T4TEQUmNlrZUyjnSavzjUVH9rVe/v3W/t6CHdvTPWW4cpNlBtzGyySxsrAooN2+v0HvVfXrRHl0Sru74S9Xbyw9dBBtfEmtm8jI3iVnUsv0Vre7A9aIx1V1nf5WkOEqZc5tq+GyMJaWnajDRSeS+oH0W49+69joE6/wCHPyEqsxPHk92Z1cRGjxxI8tR4rLcJe/49+68RmnRI+7Pgh3bRPXbzxe5sxj87hqiObGVML1HkIp3Ewt/UG3I9+49eGD1ex/LS/ntbs6C2ptrorvejqEp8AyUVTnMoigOkSCHzLLMQRe1z7cZGKButwxky92AerTPnf/M2+IXyV+NOQ2ptbs2gqt31tFOwxUVTSa/LLCQqBY3J/Nvp7LbmrfLoJ86xE2DCLODgdadFVhkXM5aWCTyQs7eFidXk9RN+PYdlB1kt5dYfb2phldHwT0wZGheoRY9KpNSEuptyCOQR7oJafLoosUvIGEkYrTo2Xx63/lVaDGZDKzR0yPFGFJst1NkXn2b2sytQdTdyRzE8ciJdNQ4x/k6v/wDh5uPcuSmWCvEi4xbikdrhZUvYG5/w9nYUlAxHWT8bxT2SSxGtR1ZBnMt/CjADEGjkEYNxcrfjUAPfuk9fPqf9xTU+O+5M/MiawC1tGofpF/euvHoAeyaCDLY+d0Al1r6hf/Yg3/4j3YV4darn16ITu7EZ/bjR5HDRuPFViVlj1cor3tYe/de6MntbcVL2TtRcfuCjjWop6cxuZbayxjtf1fT37j1rqtH5FdKS407gy8FAPsqSneWndQWB5LD6e/daqOHVUG2d9Q7i3NW7PqFaKSGeWIKYJAPSxXgke/V69Tpc5LFNgWeKRL+ZiI/8QBdbE29769034M19fkBBJEyUkbXeQcWUH6k/7f3rrw49KncOVose8cGGYVT6P3RwQGH6gLe/da+Y6DbP10FdSO3maOrUeqJTYkn+z795db6TWNwS1dE1RVECQFinBvf8C359+68OpWKkq6fyQ1qn7RWKh2A+l7DSffq9e6UWNp1FUJI4x4HNw2m91/HPv3XiehpokiWmoxQ3NRUSJCoCn1MwsBwPe89a+Xn1dL8PNq02yuuavcW4ESnzXhMlDG/65EI1Cwf+vvXDqwz0y9k5KTMVlfmTCGklBOixvYH6D+n+29+PW/lXoGuuK7J5neD0rUloYx6WIGm/45+v+v78evV6Or1huXM4Le2NpKmApTeVVVyLqLOACL+/fLrw6tWmp48ztWraVB4MhSrEz/Thk5596r17NetYj547CGz+w6hsEl6aepqJpGhViAzG5v8A6/vfHrVKY6r33Iwajp3lB82hTyLG9vzfn34da6CGo3isNR/D5gFVWILXsOTa/v3Xvn1zpsitXXokIEtK6kyMP0gkXtce/cevV6R+46CjrKtoU0l9VgvI+v5Yj3vr2OsdJjZMRGjkEK7AKFW4JP1sT7117pQNSViSQ1UEJHAN1QnVyDf6f096690rpJGlpqYY1DJkJOJo1UlgSf0kD3vr1ehs2l0lu/fWOooMZQSS5OWojDRgOGCOf1fT6e99e6tD6X+EC4Olx1XvOMQSSQI7rIxLFwdVzq9663Smeh13NWS9U3oqemEeERAsUyghWS2i9/fvPrfljoRurd0bf7Dg/hEsFPJA5ANSWUG7fUX/AAf8ffuHHrXHh0p99dC4+mqaXLYqKORYirXU6wBf+g/w96Hp1ulOlts/r/F1dA2vxxVGgq6g88C309769SvQsbTwOOoMfXYN2Ekk9NUqsZN7s6aVFv8Aintt11HHTc66oig8+tbL53/GXduG3/ndz0+HIpKus88MoRyWQkkEWHPtFMp4DqE+buXZ7pmdU6qxlxtXS188WRhZGj8i6Wjcfp+o9Q+vsqlgZ644dQ1c7de2Mp0rSnUCLH1U0rTUpanp1cqQgK3t+SPaXVJG2o4A6KZ963QEpFXGOlFtTK5bAbz2/XxVLo1PUaknCnXESwPF/wDe/ayO6JOD0f8ALW/bhHdp9Vjo238zz5kbqznwfpOu5MxWVkbQx05ZvVZNIBWwN/wPZrbyzMRnrLblndGm28MMmnWt10j0Nv8A7hq6ai2FiHycuQFNLOxhnVdTELIRZeT7Xk9GlMknz62kfgx8BaHp2gxmc3PRRxbgkZJaxJSweKWRQSLPzx791vy6uFhz0OIdcLAzJCIUhVlJ0FtNlUf4e/fLrdT5dNFB1jn9z5tJamkamxc8hZpyWAK3uW44968+vDhXoxTjEdQ4hP4cqy1MiAeRSCxbTYsb/j3vgOtcei4dmb4zGZxU+WSSRpkL6YtRsFJuPfvl1vh0QHuzsTJx7Yrf4bG02Z8P+TwKjBmk8ZAAt/j79T060SeqOe4+kvk32uIMvSUdbDCs80k9OPuAogL3U2X37r1erOPgThJtk4Cnwm6G/wBzkMcMMkEw0sZkNmJ8nP8AsffuvY6tYoUeteV6qFaemUnTILWI/qpHvXHrfQzdTbpzm2914l9vtLLTrXwGYxm/oB5YW/2PvfXhnj1tIdEbvXd2wMPVGXyVENJCk9yCyuV5U2/offhwr1XgadDP/wAR7917j14G4B+n+H1/3r3s4PXgaivXfvXW+ve/de697917r3v3Xuve/de697917r3v3Xuve/de6//V3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691Rn/Pf6Xp98fDXeW6aCiNRuDDxTmPxIvlKNTMyaGtfjTz71DrbuHn1eAKk6ux8x1pp/BfM5HOU0u0Mo5RqKIUtTTSk6o2UlNLavcWc4ozTNXrK324ulNuoGR0OO+8DX7O33MtJTSrQEhvJY+M6j+COOPYStFKxZz1KF2DJJXy6FHCNR5Sjjd9JcRc83Ooiw9vnGT0kOTpHQf5xDj615FNo0a54uLXt9PdxTz6sDXB6cMbu2IlYYmVrEKebfXi3vVfIda8q9Cdi8odAc6dBXkEXvqH9D78R14EnB6w5PbOA3L5RW06sGABsACQOeSf6+9Z+3r3Hgegc3l0oklJfat6Oa7OrIbG9+LD37rZqcdFe36nZ/XFTTSzmrr6VNAJXUVsSCSLe/EU+3rdDWnRm+sd+U+7MBSDJD7WqVbaJrq19PJ9X592FGHVSBXpU5LB009QtUsgmbhgBp5ANwP8AYe9HrdR6Z6e43ytU1LTU8bpGqheBYEf7D37PVc0p1K3BsbLVlKHp6lopZOGJNiT9R718PHj1sCoz0jKTpCoyE4nzNSJ6fUC8bEXt9WP+v+PfqDz6tw4dCVjuq+usTGgfGQyygHU2lGY8/nj/AIn3vHDrwOa16SG8utsY6fdYKIQC9lQAAg3sAAPfiBXqvE56L7ubFbqwVbGJaSpnpwV9ShioAHN7D/b+/EAdbA8yelJRbooYf4dFUIsEjFFYScNc/wBQ3PvwpTrQ8x0Jprqf0ywsrhlAstvq3P8AZ96yet48+uVOqVTlV/aLcsLWvzc/7H37zp1o44dKynioaYRAU+uRWB1KAf8AYEj8+/Uocde+Z6eazIQpBqkZUQIeCNOkA8KR72AK0PWiTw6DvJ7npaPWI5ogxvYBrXBFvz79UjrdB5jpPrvaGAqzsJTK1uDc3vxpt70T14ZxXpPZ7fGZppgMVR1BVlI1Rhrc/kkD345WnVvnx6kbNgz2UqWfIQzJ9wbnWCotq/r78QroV6tFIYpRKvEcOjP0+11jxlPZC/0JsORxfk/8R7TCQwfDw6Lt2NxuMg19Tv4THSU5nIKBVJLNa3Aub+zGC8+pGhul1qku22Xiny6qq+f/AGvTtWbY6hwdZFDJv6SLGVmhm9CynTIWP+t7HHKFq1vJIW4GvUFe6W/jdwkUh+CnVp3wU2ntX4ldN4akwklMmcq4oHnqAqszGojux1Jzfn2Oaas9QncSALpXo7e3OxqrP7jgpcpmoqalyLeZnmbTGPI1/wC0ffhTonnBqejhbWxW0fMlLV53H1oqW0R6HjcC44tY/j34mvDrycBXobH6pweco4sVjfHVldM9okRr3F7ED/be9V6v1Bz/AFti8Rg5qA4wx1emyMY1BJA4sCPfgT1vHl0G1BtWAwCOqjWOaL6FwosR9CP9j731rPTRlNt1zt+zOvhDEaVsePp/j+PfuvdB5vPZcEZpVqKbyxzKPIulSGuOebe/U699vRb997DpPt5oaamIjlRkhjAHoZuA1/fuvdJzblA22tpT7dnISqnm1xSyEelWP4/2/vx+fWs8OhG25iMzSUK5GoqBLQU6apY1VSGUfUX9+4jrYx0/dqdWbV3513R5nB0Sx5anHmZ41Gsyr6jqtz791roCur8FtXfdZUdU9h0UdRLLFLHTtOiagVUiOzv7917NMdV49+de5X4874zuFMDzbNyk5hooyA0VPA7f4cWt+PfutdAXtLtfNdV7lps7t/JgYCOoSpekiZ2VdB1OSF9+69nq+L4yfzMNs9gbfpNkRZOljybQpE9/1iS4Ui/v1Bx6tw6sk2j8jduLSPjc5U09a8K6YhYXJK8Wv9fdTx68KEdSn7bxmZlNNj4ooomY+QMofVGzepbH/D3uh69ivRDvmRtnL74ahx/U9advZqoaES1EZMfkluA50IbfX34dePVkPxm6131i+h9t7W3dkZMxmquBKWqroxzT+WPR5JGX8D6+9HJqOvYBoeil9pdx1vxQ7cg2D/HYsxPlZfKkULl5YWma9mEZJ9N/e+PWhx6NLhsl2Vu7AxbpodwGnirYzUxwF2V1DDVYj3444eXW/l69ShUZKhpaWszWUSWseUCo1Mf03sSfesde6bu2vkJhutuv8hPiVSXKJR1GmaEFmDCEsOV/PvYznrRqOPVAHx07a3n8nPkFujIbilm/hu3crL4RMZNAWOc82fi/vfy611fBt3eONxNXDjKaWnYrBCFZVThljtz/AI396oePVsHh0OmCz8lZJH93H91C3IRV4IP9Lf8AEe9H1696dJftHYu693UbJtDLDbo0kNrPjLcXNi1v9b34fPrVfToDaTp+LHY2SHsWX+80qB3hKOH/AH1BK/X/AB97J62Ope2dsblfCZWKgp5sfRxuy0XlhsEgB4Aa39Pfj69eBNPTpGZvGYrEUEsm5Y1rJwh/dYj/AHhT/Q+/D5de4ZPRFu3N1blyEYwWy8bVQUdTUNTGtjQtHplOkAMvH+w9+p177ejffFP434vYW06rdW76L7ncGUiaqgqZlUlGYagQxF/fq9eFa56fO2sPkqoY3I0c+mOlqIpW08Bo42vb/be/DhnrxpUdF4qd4028u3KDbT6YXhoNLSNexKR2b9XvY+fXs8OPQV5zbdPnOx/sAmkY6vaMyuigMEbll/1/euJ695dAT2ztrdXXW/Wz8NVIMbX6Y4VBYpdjYCwNv94978+tH16lZPN5dMWBk4ZJKespWLHmx1x8fXjm9vevOnW/KvVcuH60yO4+8YMFi8ZUVke6a7wo0I1+ANNYXB4sPduq463wPgl0o/SfRW3MBVRGOuno6aaYMFDKpiDBSAP6n3ry6359HT9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XFv0t/wU/7179148OtSf8AmD/9vNvjNbj/AH+FBxb6f5cg9+HXuttOl/4C03/LCH/rWPfuvdZ/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdBD3RvIbO2fXVY/XLDIqm5BsBawt/iffjw6159UyVu4a7cmcr8hUatBq5GGokixb+vuvl1fPDy6WeNxX8WpKkRuCyrfR9dXH0UH3v7OvV8uuMe0K2KgldYJTJ6zp/tc/S3vXHPXqgefQanbWdNbIZaacwq5tcEj/Xt7tUda7jnppy218nWVEMCU7LdlX1Kt+fwLj3qo49eoR0jNz7NlxjiKup1dCmr1Rxgi/1vce9jPXqdAfvjrnG7jxk8v28ZhiSQSKI42BtGfqbW96PCvXq+vWrf86+sMFh+w443xTyUlfViKXwN4hEjS21MYrf6/tQj9lfXrQYg6egt7H+OHXPXHSD9r7NzMEe6Yo0l+yXIySzsdIYr4S9/8Pp7L5lDVr0Xbva/UWxRfQ9Nvx8zWS7A2lLk5BJJPSU+qcAMxV1Hq1E+yie38Q1I6xz5u5PmfVehahc46Ek0oLySSR6ZHDKb/U/g2B9lk9u/DgOooWeWGoApTpR9eQTRbvx9FEGEEldRMwA/WGlGr6e1VmFDAHj0s2C/lk3NUJxqX+fW1/0XQ4vbuw9k1MSRiWaliMllAPKD6n/ivsVBiUAr1m9sZLbTEV4UHRyo6rD52nXzgHxorD6E8fgX/wAfdOlla8egz3TkfJKaPHqwiQlDpvaw+v097HWj6DoKs5uSHHUNTTVso8vjKIj3D3ItYD8/4e9461mvRfTmJJ5H/iEX+QvI/jDL+GPBuffqU4db+3phyu4odsUNRLjh4w5JZ1NuDyPpx/t/fievGnl0k9o7qpt41s2P3kY6nDVRaIxy2t47/S7ce/fb1454dQd7/DrpWcS7w2ZiKalyBiaWSaJIgWk/Vquv+Pv3WqeR6rL7v6uyFJVyNQyedqJxohiK3BAJ+g9+690Xj7fM42hdnhko55FKusg0s63tcE+/da6StPJBS+aoqpUhl9RJeQHn6kc/197690HkuXiy+XNHRQ2LylWmVwwI+l73496638+lJl6qbAwQU5/fLkWKtexPv3XunrHCSuhjSaAtHLp9Ab6k/wBT+PfutdDzs/r2szEUEFNQSMJPTHotdSB+f8Pfut56sO+Pnxbjnrqav3FCzU9KY6oJMFPK+qwB9+49eA6P9VUdBVpTYrGQKtPj4/tikdgAIxYcD36nXuHRZO35jtDDZmtrlNNBBTSSo83AvYkAMffuPHrfA9Fr+L3Z1Hu/dk3208LlawQFlItp12P+x9+PWgerFc/VrhsrjsivrKshP4t6/obe/db8urNNmZN831th6qKUHUiGWNfyNIJBB96+fXuGOiIfJvpvb++4auqWgVq0RTkSaVL6iODcj+vvw4dePVD/AG18fdyYfKVIpqKpenSX0AA20j/ePe+tEdEf3b1rlIckY3oJ45WZgdVlv6rce/deHHpWdf8AWueXJx0kuNqPt5I76+CtmHJuffs9a6n736jzOIq5KxKCYIWurfQfXg34+vvxz1vqZhetMlmcXDLNAzEMNKcfUfT3vrWehZ250/lPsHmrqKW0Y0xpb8fUfj/bc+9de6GT47/HqPPbyaTJUUlPSmqNmmAKab/W5/3ge/dbAz1YlHtKg6r3hSDCmMAiBQ6opRWI03sL+/edet8ejPVdVuOqXH1VfWrPTyxLIsaCwRCOFNv969+z17Hl0FPe1Iu49ovRwQu0kEBNwvI/Leo+/daA6IP1/vbPdfZNqeJ5IFWruS7G1ka4tf8Ap7917qznpTuyn3nQ/Y5Kuhd2VkKkc3va1z+ffuJ630ZXDpR01XqgAdHt9LgMrcn3ry69w4dLmHbyPVplYAU0oDZTYfW+n/W92VSwx1sUJp0EfcXX+M7DhWjymM+7SNbO3jQgcWHqI90Zagk9MXG3m6Q1Feqdu7vhztutyGSXD40Q1KxyvdUTVctwLD2kdD5DqO925QafU5Xqovs7qjKdc52XHTtIsDStZXH0A/It7JLhDrp5dQzvfLg24tKPU8egyaigq6yj8Bs0LXkYfgg3PPtPAv6lOo9tb5juQi9D1J7X6mk7w21htkU0jSs1fTpPHE3qKFhcEG9vYotlogpnrMjkKMybcrEeQ/wdXU/CT4jdc/HvZuJrqzDww5QY5Cs00UJcuBrPJH1/PHtTwNehS2WYH16NLXS5Dde5vBhqJ1haUKksS2QAcE8C1re9efW69GSx3UuLx2Kgr8xEJqlVjmZ2I9JAvbn8e9V69nyx03bw7Ix+2cRJRUDxGdIysSqB6Rp4BI97+fWqV6Kjk+wZdwpN99WAmMOUUsfSb3AHvXW8dBx/Ecjllko9bCnaQjSCeVY25/2Hv3zPXsdRX652/PV09TkYEdgb3dQw/H1B/wAfeya8evU9OlnSUOAxgFGlDAaWQrG14YyAh4IHH09+889e8qdFL73wmO2TmoNy7UojEQyy1DQgAEA6mJVPe8dVNOPS02l3zjt14CjxlJVwDIrCUqUBIkD2tz/sfesdb6PH0tS1dLQ46udj5p541GogsQ63Fz/vXvQ631sffDSnqafYf76OvkjRxqvb1Nf6n3sda6OX7917roAKAB9B9Peya560BQU6796631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//1t/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdBH3j1jt3tzrTdOytzUwqsdkcTkB42UOomFKwjYowN+T7tD2Y6q3ENXhnr5ylZtDIfG/5m9qbVrKeTHYGq3K8ODM0ZpIXiaoItEHsDx7jTnFKyFgMHrIj2wvQYkUn16sB37isXubbXnpYYpsgtKjGaNRKf0A21D8j2DoAVhHU9y0dAw6KJgslPiMlPjaglW8jKqMdNrHgAHn3Yd3SXFMdKTKwQ5BJI5CCX4FiCbkXHv2a9eqBjpkx2zDROJy7W16uRcn8/T37yx1btX4el/BIKeEJYnSoB+pvb/ifeh1Uiueu/wCMxUrXkcrfkC5FrG97e99eFeHUqk3LLJUKqsjQgqfWwP8Aj9D+PfqHz61+Lp4zqYTdVB9tkqSmclAOY0fm39feu79nW6+vQR53rCkpMZLPtljHVRIzIkYKgMPUNIHurY61Vq9Bfsmq3/DnRRZyJjSpNYMym3jU8m9v6e9ioHWzQZ6NmNw4zF0kLKimcLZvSAdX+B97z16q06xz7pNXBHMfSvFgDYW/r78KnBHWjjI6hVO5omit5Co5vZjzwOffhQih613nj00ploCxYTMSbFbubXYf0vzb3sEDHXsnh5dT4co8jArIjBLWBYN+nn8f19+ApnrdSOs9Tl4MgjU9fS0zKAwDFVJ+n6gfeqdb1Vx0F+U6823mah6ppfFLEdcXjIAJHNja3vfy6qDXHQDb83VVbCnjjp9csHk0ggsxsGAB964Hu6sBXpebI3r/AHkelKxVCu8QZhpb6n68ke98cr16igd3QnT5LLQNULRQszRLdgyH8f0H49669SnHh0jhU7n3BWGl8ciRlrWIOmxPN/dq4x17Cn59Luj6uhqVV8vK6k/6nj6fUH3XAHd1o+p4dPdV1ZtSnWjeGRi4cEhvUt/yOffuA62SfI9KYbYxFJGgjhilYAWLRrzxxp/23vZx14gcD1LpqdAreKCCNUHJVFVvrewI91UJqqOnLZA06huHShXOT46jVmeFYRyxkcBgv0Ngfa1LI3BqB03u+42G2tWUjoKex+79p4Db9cKiY/crBUACFwRr8ZK/T/ePZ1t+wvrBp/LoKbvztYNYNGjiv2/LrXG31uDeHcPfC7pjSrOG2nkfLStOraQkcuolGbgcfT2PtstxarpAz59Yw81but7cfpGtMnq3XaO9uystt/B1NDLNJjkakQKJGkYCMAcgH2bcOHDoJ/UM2Dno/G3dt9hb5xlBV0j1UEsdPErPHrRg3A/UPfgOvUZ2qBnoWa7ZffGxKbE1kE+Qk1lGgJaRuWI06iPr71itT14gg0PQqdf/ACo7/wCrdyRVO4KWaWiHiRvIzn0L9Tz/AFHvxoeq58+ra/j98l+u+8J6THbrnipc/ULdYZnSJTITpAs319669x4dGm3f0njfHLXULwvAyB0ams9wwuBx71XrY+fRec5saakgb7VWJWQryCACDxx731utekrldmVFTRx1uRWIQ0yXJI+luPfgevEZx0Du6dkYmJaaslETx1jiOAAahqZrAf8AGve8eXWjgdBF2F1U0SR1CBhKUEsaxodGk8re3v3Hh14fPHSewcWVp8NXY2tiIi8ZRfT6rW/3319+PXsjp6xeap8PtyspxJ/mopGMdQxVARe9g3v3XqZp0VjZVNLuLtxNxwMyRUkxVmph+2Src+pffutedelX8otj4TfW2cu2chiDUsH7ExTVIG8dwxf8e/dePWrr2KOzcRvyt2jtSlqa3CTT1MCSiJ5guuQoLN9APfutcRXqHtfE9wfH7d2O3FDUZAVlcUqFg1PFEA7B24P0t/T37r3y6td6H+YmX3ju3bG38rXzRZOrro4KwNUHSv0Ugk/1Pv3W69XsTbywO0aCir62sHkkoIZtUcq2JKA/X/e/fq5690mNo5TO9q7/AMTmNvK0mEx8ifdmT9J0uLm44P8Ah79nrYp1Y33h8iaz429YYmXHw+atzkMVPB4lLCOR00XBHAt/X3UZPW+I6pM3f1x2z2l2QO/spKainjkNRBTzO01lPrAW97ce7dapnPRmevvlluHA1NLtbNj7dYyIIgZBHHY+kCx+nv3XujIYXtLbm88o9BV5SFXS2tRVICQeSAAfr71Tr1egd77w9ZmIKik2yKipxjUdSZmdC8YIhN/dj16nRBv5dW3Xqt/9xU1TDHBPQ5CrUuIxG19Tcj3r59aHmOHVrOI2/EKqNoJZWq42DNcsRYH66h78T1vjw6MrtvK5DGY0ZOJUeKkURPqOoqwP+P191xTrZqTnoRqKHdG/II6mleGnplAZwJPGSB+b/wCw9++XWvPrDlcPh8RSSNlayF5KVTM4kmVgxjF2Auf9697r17os2a+Ta5TOjYO1qMFnfwNJFENPB0n1KLe9Up175efSsoOr63fEsdHn0aGFiBKxW3pbm9z72D1vpI752jtXaNZS7EwFFFV1qzRzNO0CuRZr28gH9ffq460adCzlMzVw4bCYSaKOC9IkTLCAALLYX0/7z7159bHGvQHZvOQ1ctfhHXyGiiZjxeyj88fT3sdaPp0Snc2zsqm4p947TUnIU7OOQVIX6NY/X6e95690EmP3nmD2HicfUIwyNbUKtYQGGpw1m/4N711rp/8AmvuPHbbwGx4ajT91PlaJZlTT5NLuoOsD/X59768eFOnXs6TaVF1XiMsojAfDRM7RgatbQBrFvx79TrZz0x/yr9tbb7A7vfJZ+gWqSiyUhxry0/mACy3W7/j34nHVfl1uTUsMVNS09PAixwwwRRRIosqxogVVA/1h7914cOpHv3W+ve/de697917r3v3Xuve/de697917r3v3Xuve/de66b9Lf6x/3r37rR4dak38wj/t5t8Zuef740H0/wCo5Lge9jh1vzHW2lSf8Bab/qHh/wCtY96691I9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690CfeW1zujaslGEZtIckKLj6cX9+IxXrw+LqtWt2BT4qSpo3TxyM72BjKkkf4+6ceraqY6D+kra3b2V8YBEAez3+ltVibn3umevZp0uF38Pvo6aOOPxMUDWN+D9effq9ap59DXiava9dRx+VaUTOqkk+PVc8G/vfWulNVbJ2zLRR10Ap2lUBwFUEm3IBt711v5HoGt1dfY7csNVrjVXWOXTZeSQOB/X3sV60OiL7pwVVtmpyGJmjcU8zvGCUI9DAj6/T3sjUOvfLqt/uf4d7c7SkzFdUwRzz+Nmp2MOt1YksCpH9PbZcjtHWxHQ6z59a33y1+OfY/VW4aimSpy9RtVquaM0f7z02i+lRoPFvdtOvI6uwV00nj0geparfPVlbicZh6JZMHuPS9eZF0tEsp5Fj+PfjbA5botv9sgvNvktgBVh6V6PVV7NkzOONdjKdnqBEJGEcZdQzLqNtP8Ajf2XXMIyp6x23/kGSzDyKvr5dIjr6qqsZ2HQ4/I07xTCupBGHUqSfKACAfZagEb08+ox2ra5LXeAGHBh/h62l+p6OvGwtqTTCURtSxmOwP5QHj+vs9geooes29kjcbPF6U6NVg41ajFi2sqA3NuLfi3t89LAfPr0kFLRrNK92mCSsAwB9Wni1vr79Wo69Tqsbd+4N+53taoxqRzJh4qwxqWUrGIvJb6n/D37HWuOPLocdyYGpqKCjx9MsYkEcRdgPoeNVyLc+9nr3nTpD7l2e5o46edlaJkXyEEajYeoW9+69SnSHq9s7dfEzQ0FS8NbSRl20ELYgf7D/ePevs699vQZYLs3NYuafbKzTyR+uNWkkb9P0/2x978utV6Y6XZ0OVzctfkQksUshd1f1g3b6Ef8V9++zrfS6p/ipt7squQQRQwxK6mQhAPT+o+n34de6TvYH8tfF5qtp6bCIrBtJlKIQvNg3Av791qg6kV/8sXB7YwlM9FTxNlHiBkbw8rIR9dX9L+/fZ16h6CJ/gNV1FY8GRpVZomJF4tS8Hix/wCJ9+r69eoeHS+2x8H6ehq4mrKRRTxMg4gJFlNuffuvUzTo3+A6G2NsahinNMhqlF1DQEC5Fx+Pfvt68OOB0IuFeGFkpqYRRrIwhAiFn0kWHA/w/Hv1Ot8elJuaHDdf445mcsjSqZHaUFQW06rXaw9+qOtUPVVvzl7hTdmwqqg2zIj1E1GUmNO4J5upB0/n37rx9B0UP+XPt/MU25ZzlPOTJWeQeTURctccf09+PWh69XuR7ZbMxk1OrTExC6yeLH8D377erdDP1p2VlMFJNtybV/D6QFYyTZOFtfn36metcRXpc5jN4rLQM50vM+rg+pfUeVJ/3r3qhHWzQ8eg1rOuNq7rDmroqZmAN2aEE3taw9+NOHXs9Fu3h8Qtl5DNRVU9JEsJn1XFOguNVz+Pex8uvHoY8R8R+sKShp6uGjpg6QpdlhGskD8Ae9V69QHqPnvin1nuaikpzTU6yopCsYwDf8fq/wAffut0r0VPcPxtwezq/wC1pKeJ6eGXWFEfBGvn6ccf097qeqkevl0k980uEwePQU9JF+0ojZUiAuyjgm3vXnTrfQHVfZ+48HSs21KMR1AB/REY21f1DW497699nT3huyd05fGQV2fB/i4mU2ZtRAB4+vNvfutfM9G46/7AzmcpqOKulp4Y4YQqeeVUFlFhqLe/fb1vFestVujIVeWrsXM9LUUrlktGyygC/BHvXn1vHSJ3R03is1jpquj0LkZA76QgWzH6/T3utOq0HRalxm+up8sKuFpUpVcNwzKpBN+Pfut1PVh3Q/dSbmWngyVQoqSUQlpQeQvNr/63vRFcdeBpnqw/GO02HWpgdXUqrEjm4I44/Pt+ADi3TTvpbj0/YunparE5EzwqZgraG0XN9PFr/n3uRBxPRlbTilCeq+974uWh3LuGuqogKaOjqJEDrpWym5Nzx7TFNVT1u6mt/Dc4wOtcf5k78jy+/ailx/i/Yq5orRMGtc2IJ9ktwhBJ6xd553WCS5a2jOakdEozu4IdoYWprJ5P8sngYw3YKWY88D2kgTvr0A+XeV5by/E/HPp0b/4Dbb3DvbcS7kzFMTQpPHNGZkPj0KNV9R49im1UGKp6zG5atBt23LHwNB/g6vLqIqrdMuPweLurQKkAWHhbK2m5t/h7rShp0qPxE9Hi2DsLb20tqx1FfDCcnHDqdnUatRW4Oo+9Z8ut0A49AhvvuDHQLWYYTIsirOi/uAKNPAvf/D+nvfWqnqtnsfsetizbJS1LTI8hFo5hKP8ADhb+/Adez0iaCHeGYyFNWQI4o5JLuFuTY8nV/vf09+60ano0+2cPHLJTQiImUxxrJ6Bcv+ePfuvZ8uhsxXVMmbkiLXS/1uGJFv8AbD37j1vh0w9p9drtbDCan1edkIU6Ler8cj36nXuOB0UPL4CfN7fyFJk0DuyVAjMo1NbRYEE+/V61T16rl682nmNndzvHJLUPjKyvI8XPjCmW9lUcf7f3vHWutjT467Srd35PDY3HUlTJBFJROzeFmUERgsCfdaDq3l1sr9N7bXbWy8bR+MJJ4Ig3p0n0Lax/2Pvflnqvn0LHv3W+ugbgH3s4PWgaivXfvXW+ve/de697917r3v3Xuve/de697917r3v3Xuve/de6/9ff49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3UPyRTmSCS3rjZCt+CjizC3uwr5dXkhoNVeOOtZH+cX/ACqsp3bnMb3D1NSSUeawdTDkamOhaVHqpof3GWVIhze3PsLb5tK34o2OhlyhzRLsdyF01H2dU+dZbryPX8snX3Z9CKDORM1JMtSjq2mI+HVqkH5I9gy92o2MGpO7rJTZOcYd5UQuNAx0E/d2xFw+bj3rgqhp6FmM0iK11VWF/wBI/PsKmaaNj29DKFbeRcPXpHYTLfxWGnqUPqT1Sr/qbckH2+kjMRXz6dmhjiUlDU9CNj65axgtwUIABA4ufbxA4dIkJOWx0o4aOFELOdVxf8f6978+6/Z1ah8umPK4eOrClCEtf8829+zSnWxQdBjkaPJYmr8sDs8QJsOSLA8WB9++fXsHA6V23s9HUx+CscQufqWtc/jj+nvYytevEenS3FW6RlYbyx/Q2Nrj6/X/AFvdcDrQHr1wpo6UnyPSoZLEaza4v9eR+ffmB4jrxPUGrxlPJqaRrAngcem/0HvYr1viOoMcUN1p1PoFhxybfgW97PWvl1JfD0rD1Tix5sQLi/8AX3oDy6tT+fTRW7R8iF4axlsOLN/sTYe9A+XXs8OHSDya5jAkOpkmUE2Nyb2P5+nvYPXqDyFR1mxWbyeccQilaK5ALAc/W17j8+/Go60aE14dLQbYr9Cv9xJGL3cBja315v739nVsHPQP7j2nR5TOx0lYi1aiYalkNxYsL/X+vupDHz60WzjPQ+7d2xgduvjvt8dT8wJqZQLLx+L8e7aqDHVfn0tamnxsrvItPHH5AdfP6uPobe6VPWxSnUGKLH0RLxU8Yf6lwBqv+Pr78tevV9esM+SaVxaSyi/pJ4uPoB73kCp63Wgp1ieokqwFUH9v1Fj+P6/63tb9MiQCWuT0/HbCQalOep0c0cFJ93NLeNGAkLcBAD6jY+0cKyzTeFTHr0nvZYLC3M87UIrjpr3Bu7blBTINt5CPK5OZR5KKMFrOTYqdN/zx7HFjycl1F4jPTqCt795pNl3AW9tD4i1404dB5Vdfdt9pRRwY3HVVAoY+mJZVLKTwfR7O7Tl2O2Iq1egVzJ7j3W/glF0V6dqL4Wb5zqLjNx4mR6eVCZKudJeD9H1E+xHFFFGummeo6lvr6WuqQ56Suf8A5c0dHjsmNuUKipkQ+fwawWb86rD3sJpPRYsDli5avSI2jsXenR0C0GWwElZj6d3YeSORgix8auR72a16UIhAr1Z/8X+xttbr2/JFKsdDWxuAKcWUqwbgWIB974Y6fV9PDierL8ViKDduOx9DW06LHTALDI9vUQOLH6+/E+fVSSxNegz7U6ew1PTiaeiikiJsZXsRY/6/496rXr3RKO2tsDrqWj31svMtjarEpG5pqNgGYqNZHoP5t731rh0bn4kfzOtv5mjn2x2RmvtqigQoZa7Spk8S6fq3+t71T063x6FPtP589YUkVQ2AytNWTMsjxxxCNjI/9kcf19+AoKdewT0UtPnJ2LvKjydFh9nTT4860SpETWKg2Dah79SmOvV6DDJfLTflA9BR5jbc60tHP5WdomKoFOotcj6e99a6Mntr5ddd7vxNPS5avpqXJCNIvFJ4w6sTYrz79w68TinS9WbB1iQZOlnR6CcFy900uLX/ABz79nr2eiKd79vJJm6naWygKvJzBqZ4IELMpN1Jsn09+63joUfjZsHObf2vWVm4KWRctW3qolkD6rOdY0huf8Le/de4dQfknuFcZ17uilnjEGQloW+0Uj1NIVICge/V68cjotnxO+MO0919eTb53dHTfxeSpmnC1P8AnBdi621Di3vx60Oi6fKPpeiydRVzY6AA0EkkdKVLE+MekWIH09+61w4dVm53Zu6epquk3xg8fJUV2Kf70qgcuWU3sAPr7917yx1ZF0J8rsx3ztelweVhqINwUQihkopQFk0RjSxF+be/V6txx1ar8ce5cXtaqp9oZeKPDQVQKVOTIVRGwPJY+/Hh1oZPRreye39p9rR0nXFHSwbohxgRIcg6K4ja3+cD82I9+p1vpR7N6gpcfhjRT7jk8E6A/wAMaW6RhxbQqfSwHv1ade+fn0CPa3w9jz9QuV2pWCSdDrmaFmJicc2On+h9669TqrvtDZvcvQW+cRnzVZGbEPk4zXSapWQU6yC5a/Fre7D59VyOHVp2D+U3Vtd1cktXV0Qrlw7RVMhMYkFSYLMDfn+v196pmvVq4oOqvfjh8hTsrujf8W28atfRbnyMz+eJNSkNKfqfp+fx791qvp1b7j+2aqkxK1tPhvNX1EKkxLH6rut+f+Ke/dbx1OwXcO/qmgqaGXas8VJNIW16Bptf68+/fLr3l0IS9ndi/a4/H4WgqaJJLLKYVdfSePqPeqDz63WvSgynWe/d4Y+mmnylZBLVSorqzuCVc+pSPfscB1roVtofGXF7Bmx+5MjSwyVoj8rVDC7s/wCosSffq+R698+lvuDfOMpRJRYqRBWyjx/tqAVb6X4/3j3qmevcTXrFt7amNqaafcFZDFX7gMTyJ5LNMFYXWwPvx698uglqKlgmdlya+PIU8rihgYcgG9go9+4dbHy6DjH7bmxyZDceZpzD/E4pEiMl7OzCy2H+292+XWvt6LjuHMybeq6mkSFw1RI5EY+hjc/qHv3XuI6BLNUWBxm9tt7qqhHAYJknnJtceq/q/wBj/X377OvdF17xlPb3Y9SySGrw2NjE9KhF4o3i5V0/xBHvw61jpN7h3dX12w5dqJCa4JJHQiRhfxXPiVQBzx7914+nV838oH4lR9WbTruwc7RJPV5+P7jHGZS/25ns5Khha9jx78cnr3DHV5ispHpIIHHH04/1ve6Hz61jh1y96631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XTfpb/WP+9e/daPDrUn/mDgf8ObfGX/w8aD+nFq1Le/db4dbaNJ/wFpv+oeH/AK1j37r3Uj37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddA3FyLf6/vZ60Dip64I+osNJAU2BP0b/Ee/EU68DXpo3Dk8bicTW1mUlijpYYJHcSsoDWXgDV711vrXo+T/wA3cbtLtgbdxFEk1AapopalFuqENpN2UH3rr1KGg6TFb8gMJk8fT18UiSy1qhmVV5ivybD36nW6+nT5tLf2By+phXJ91YnxnhgSLgAce/EenXh8+lTSbwqZaot969PHE2mMXsHAP1Fjz79Ty68TnoV9v9wV9DJHSSStVQFgt2/p9CP6e/U699nRitvZig3JEJ1nSBmRboNPLH6jj3Wn7OvVp0G3bvXtNm6YyUVJG0iglph+olV5Nx/vPu3Xjw6INkcb/dzI1FAJDNNI5WSP+nNrc+9GEt3A06MXCxWniHOOiX/KvqPZ24Nn1dVmaamFRIszr5dNzJo1Lpvz7cX9PB8uiSGVriTSOqxekfgj2p8g93/w3Z+0pa7btLOIzkoRKBBEsliyMosLD/H3ZLpdYUjHSshrWZdfA9bB/Vf8nTrjrXZ0WV7G3kuMqaini+4pauqA8OqP13Eh/B96ukjcaq06b320tNwtWU4xx6pN+dXSHSnQvceHrth7upM6qZOnEixTpIuoShgAAT7IfDAfUT1jDvVjZ7Tu6yI1TrHn8+r0/jxPR9gdObNnpKZQaTHRsxUAatMY5svs4s4VkyT1kTy7un1u2xxU0gD9vQpwUNXQVnMWmJjpCgXH15t7enj8LAzTo5A0mnTpkqIQ6asDy2X1REcfp5v7RGd1OkDr329ALvnZdDUpNmaPHR0tfGvlBRdLSMOR9PfjNJjHWh/Lor9Tv+KmrZaDNVn2E8RdU8jKpunAtfn3cStXI68Om+m3BBuGZvHkWnCEqqhwQV+lyPboYsK9epnpJZDbDVlXWBci1LcEBVb/ADn4tcfX3vjw69wHQcPsCvhrmdKUyEsQtSQbkf1J9+z1rHQhbZ2NXUMMrEtVvJ6iGYnTf8C/9Pfs9b4dGF2riKva9PDkVrZIp6rSGiVjwG4+h/PvfXvs6Fqk3plsTolgSavlfTYjkj/Ej6+/ccda4dLlNw53PUn7lJItTKP2kNw1/wCgB9+p16ppnpt2/tXc02VqJcnjpY4dJPkcHhT9Dd/x71Xz638ukB2X3h0x0wk0O/d4UmDqFErrDUTQIdare3rPvfWvt6qe79/m77FwGQhwPX+Np93QSSLFHXUkImABbSWZlB9+69WvSf6V/mBYTObolym65lwsFIgrYqOYKqyFV8hisf8AevfuvVpx6EPt7577h+U+Frti9cbMKtSs9FBkqelKeRFHjEoaMeq/1v78B16vQRdU/HPfGQq8ft/ueKXA43NS/bQZCqWZFdZjwyvIP8fr731qlDTqxrZvwsj+PmWo8/tZhuDbVWsEz14d5kV3AOkcEcX96Br1vI6ObQbQhyFLTzwuI45UDzBSV8ZPPI9+4dboSM9R8nsaTHgyY2H7qR19cqkMSPqQffgR59eIPTLPispQUIkjpmabWLpe4AJt9Pfq9aofPpW0VPXRU9KaSFnqZEVpY1tqUn6i3HvR4Z62K+XQhVu06qswEdRUqYasi6ozDUWtcDT9fevs63x6Q2Jrcri6t8dk4njQBvEHP1Ci4K+7H160D69BhunemcXMPR42leODyBXnT+yDwSbe/cOvVBz0kN0Geox5qGkapqZBZ0J5QH6lve+vU6AXNbUoaqhqKitKSt6iUcj6n6C5tz9feuvdA9lMRt6mQU9JTwTTSkK6Lp1KT9Dz9Le9deHy6KL2j29srqHJI2cycNLPNJ46WilKETTlrRxAf1J4A97618uiTfKf51dudd4ikrsRtKtxFBWpE+Mq4IGP3VPIoCSKE5sffuvdGH/lq9zdy96VkuY3viq+mxrzSGKpqElQSRgXDWb/AA9+68OPVzW8o6nH01PNjJ3eMKnk0E31Dki6+7JQvRsD16txx0CPaeUpsrtWdpkX7uCIc2uWcLb280cSHD9V48egi6YrJMfWUdU1YYGM5ZUUgfQ+kAH22FQnj1YAcOrferuy66qo6XHVEJNGYYlaoK3VgB+PbrFIcKa9IpkZpKDo22GyEYo7U0AnhmJLyW4W45Jt/t/bZmqKkdGdtY1i1M3Dok3y+mi2zsfO5qiRVknxtTyvNrg3Fh7oMqWHkOgxu91Jaxyac4PWnDu/Kz7j7Gz9TWyswTKVJUNYWuxPI9he6vC0pQDz6xK3it7vj6m/H/l6BXfGEruwt37f27T3io0qY45WUGzqHAsdPtRaxlyDWnWQftzy9bsolZqkdbA/xz2HiOuurcbRUZSKuWnpyZY7a3bw8rz7E0SaYq16lm7pajQOj6fHTD12cybNUwFdLeioNw4Ba9wfaeuT0wKkV9ejGdx7wXZ2KejMhJZVQyDljxpBBb+nvWOvf5Oqde9N5bkeKoq8FSy1FRNJNEZY1LOvkP6/T+P6e99aNegh6f2TnJauTI7nqJauSscSiKcsxhBNwBf3upGevDqwHZeyJKpo4MbSioSY6bIG9JK/U/096z59byOHRxNndJfwrHx5WtiBnIUhL8C/Jvfn34kcOvU6FCmw1PiYzKhCtpJ0ggH+ot79X061556AntirjytAaaVeI2Nyfr9fr791v59Ei3Mharejx6eUGOUPo+oJX6X/AKn+vvQ9evdFLwlBQ0fdWBw2QiV6nMV0aQI/6lJlAFr+99V63OPhz0Fgdp7QxW4qmggNVU0lJLEWF2JNOGBP+359+49bNej8LGsaqkYVI1FgqgAAf0H9Pe+tEHy6ye9de4de9+631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/0N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XjyCPfuvdN8MaiRlYgTWYg35H492yB0/OwdQB03S1GqRqOtgSelk1LKs0QdCrDkFWBBHtOc9rCo6L1neOTPVRPze/llbQ7hpdw9hbMggx+54MfV16pTRxRtNJGDMyRgC5PHFvZbcbWso0nI+fQx2jmOWwIav8Ah60w+4u3dxdPdlZXqLsSgqaOkgyVTj4Ja/UolEJKhkL/AI49k9zy5GELgdD7bfcFxIFZ/TpKbH7HxdXlqikxriqpauRhGIWLLGt7eo/j2Edw282p4Up1NfLG+xbnpDNk9GVx/kpxGySAxMUbQCSbsLn6fT2R6qmgHQzu4kQig49CNRymSJdRtdRz+P8AWI97bpAccOs8kLScKbH6c/U8fT3oMKZ61XpqraSCKJzUqGGg8fm/1PP+v73g5HVgxPnToCNyR1n3wGN1Ld+PHYcf0Fve8r9nW89LLBy7jSkCyrISFIJa54txwfeiAc161TOelFjxnZLlmkW5tbT+G/Nj79WvDrzUwB0/viMtMgvJa9vqbE3+t/fqkinWqdNM+2s3GC8M5uCWsCbWvyOffgDSp6sajj0m62trsWwatd2RTYjkfQ/g+9mhFetfNelbh8/FlYAITpIWxB/Txx9ffqCnVRWtD1H3NLE9DHDZTMTb6C1vryR71TPTgFeHHqZs/FrFB9wYxqtcKALj/Ee9kitOqmlcdKDI1rwpOBccG5IFyLcW/wAfegetVHp0FOLo2yW5ndrEI2rS3+H+v/sPe8062aV4Y6GONFsoa5MY0i4/A4Nx7pgZ6qaE1HXCpkX0jVYi4/w+nH++PuwqTXy63w6jyONBUOoP45uBf6/T6e9A1PXq+XUSmg1y3c3Bv+eD/j72SevHIx1LqauDFQyzSFRHouxJt6R+fdYJJZ5vC8h03dbhHtMBuZzQU6y9a4ufsbc38Oo5NeJIIljjsxIbhrj8+5D2TZdTiZh6dY1e4XuC98zW1i2akYPR/Oqvjn1Bg8VncvksMZcvSs7rJIqkhv6gH8X9j+OBbeLSvUP28jXAMl1lvn1Z10H8fMPmtsUedxNAkfnKXH20R9IXgk2/p7TkjgOnAKdGaHx+oJKZqeakj1203FPELn+1cge9Z8ut19egu3f8eMdga3HrQUila11E4EEZvc2N1t+feyTxPW/s6L73N8TcRuTHTU4xSiSSF19NNEGu4+ouPfgetfPquyH4kZnrvPynCQTRGSbyBUGkG3qNwvvfEde0gdGR2V2JF19T1FLvSqTz03EKytpdWIsL39+690V3v/5Eb330w25sGkqZy8xiSelR2XSzWViy/wBPfqjrWek7gPij3z2Fs6etzOfMa1URb7aR/wBxLrYgh/z71XrfEdA+n8r7fiVn8VGWqBLM5aYRVBTVc350297qD1ogjHQrYD4WU20ykG5KesrKlQoEjyvJZ1H6vV79nrdAOh22BtGPrjPUOHqYo/4ZkXIVGRCURj/aLf4W9+A69UcOhi7Q6Y27m8XPW09KksU0H+6Yk/K8jj3qvXjwz1VJ2z0njsVHV1eBZ6LJRzu8dpWT9xWuvCn+vvfl1o/LotdT333Z13uHb2z8tPV/wfIukEdXI8hijRmCet/pa3v3Ws8T1e/8avi/s3M7Zx/bOSiTKZaujgqGmAWYCSRBI1yf8fei3Vj8+hT3zjpcTnKeroovt8dSqsTxaNF1HBsv0+nvY68eiJ/LmkpNz5zYlDhHWf76aBcjDBy2kyAFXA/3n37rXRxdp7B2/tTrKioEpPtmNLExUoFOtoxcgC3J9+62MdFL7c65SbDZHKxUchiR2YEID6bXPPv3XjQdFp251Ltvsvbufx7UiRVlNSSjU6IbkA2A1D8+/Z61x6pD7c2t2f8AGHtCt3jtI1Ix5qp/NGoIj8EbksdK/wCHv3Xvt6Pl8Ru8an5ITU2HeCoXLvdZ5FLqRMHs3r4t7917zp1c313tGm61yUVMyyQ5OYx+WpnZirF1/LP/AE9+z16mMdHHwG5cdDWwU1VItZVyqgDRuWADcfT6e9Ur1sH8ujX7QhpsNjJ5yqvHXI0gUoGIBF7Wb36nWyfToiXzPwWNy23aNauBFgyMop0BhCldb2Njb3759aI9Oqw+5PjS22Oq6vK4aZkhqbVDlJiBpK6iLg8f63vfn1rpH/EU9b0VTFR1GIaXNxlklqiA2qUGxZmI5/1z799nXh69WmQZvC4Z6eWenOmoEaQEpZbtwo5/1/fuvZ4dDDgGr6memiWojENSokjjEYvpNrDge9HI62K1x0ZXrjbFZVVjmopmKQvdJHiuoA/xI96+fW2rw6MKmToMKuiogEzRgGMLDezj6cAcD3qhPXukTuLPbk3TKsC1IosehAWN10HRa2nn/D3vyr1oenSazO1NoYugjyElTTnIxqJHcyL62PP0J9+69noKIOwqqXJtjMDc1g1R+WO7AxjgAW497OR1sU6mQ4x6xqjIZp7VYZmKvwzOOeFP196p16ueg23fuOoykUmNnbTR41GkiBAQao+b3H197FetGnRPd75qKZqjcFWhjpaGOZDKwKx/trfk/n3s9b6I7h9z5T5B9ix7f2frfG4ir8GSnpyxjHjeza2H0t791U9C/vnZ9J1LNlElIlkmoWXWAGId0sxLc/T37rfQL/D/AG3lO1fkFRbCekmrcRkK+GpaREDwr++G0m/HHvxx1qvW7hsDatHsTZ+B2xRxLHDjaOCmCxIB6wgBLBQOfwfe6daJ6Wqqq30gC/Jt+T/j71WvXgAOHXL37rfXvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcW/S3/BT/AL1791o8OtSf+YOb/wAzf4zHkf7/ABoOP+q1Pr78Pl16uettKk/4C03/AFDw/wDWse/db6ke/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuk/ltx43DgiplVXHGgkLY6bjn3vHEnrX2DqsH5098Zii2hJj9sxz63jkRzTl7EX5uy2+vutanrdD1Wh0301tbtbE1+6d70EkmQErSa5xdidV73k/3v37HXj0I0/UGxKF5KOmoQIkISI3Fvfq46t0mqvq6jwglyOLDo6ozIFJ+oHBsP8AD3uoPWsjoL8VuTKRblXE5NJNDygITewFxa/+B96690YU1dFSRRxRAmeqCpEwDHS7D8n+nvfWgOlvtnMZ3aMsFbX1wahaVD4y5A0k/Qj+vvWPLrw+fR6dl5Ki3pgQ9M8WpoWP1B/Fvz+f9f3ryx141p0Tbvnrmm2lUS514jLLWXZNPGl2PAsPatMKOnJ5P8W0eXRAu2NgZHfe06gvI14nkkWMkavGqm1gfbMvxV69ssSljXj05fEP5sbG+G20N37czYoRm6hJko3mjj1qxB0EP9frb2iY6O7rfMl3DaWzy1FV4dVvfLn+ar2R2Zlspj8TmdONlWdIUgnliAUudJAQ2/2PsovNw0rpTgese969wJoYHjVuFfOnVKe898bi3hnKbL7jq5quqky9LNCHqZJLBpR6v3D7Qwzs7mp6x7veZbvc95Qkk6pBSh+fW21/LW3nU1HVuPpKtHZEodEAb1f7rFrX+nsQ2czRjPA9Zj8iCU2CmX06s9/uw+Qx7V4XjxmUJpW9yL8f4+zeIePSueh6xoaDh0GJc/dSU80DFUcoSwP0vYXH+HtQ0EMa92D1biM9JPfgipsLVNDHql0ExKoBa+n6Ae7pDBItR1Ug9UG726l7s7K+QFf9vkajCbYj8zI9SWhhYajblrD/AG3tLNAi5HWqEefQh0ex93dbVypW7ip6uGF9M0qzqYwFPJdwfZc5Kmg6sOPRtdkz9erjly+6d97bheRNYgnr6eNl4vyGYc39vAgjqvQj7U3h0/ncs2Lo947ZqTpKqsVVTOzN9AAFY+99ez59KqPacdJk2qcc4yGPZyf8nTXGVJ+oIv8Aj37Pl1uvl1Oq9v11W89T9tPHRUqGQK0TDRpF/rb/AA9+p17oO4+6dlbfyJosg9OZIHMbB3CsGjNj6T79WvXuB6HLq/5EdX5XLRU89XQR1SuRSwvLGHfmw0Kfr/tveqHrxNR1D+afyLrtgdbtW7MopZcrk6doKR6aPlZnH7TXT/Ye/AdePoOqBYPhX3T8m6+u7c77y9ZLtaKp+6gxElRJAoppDq06SRf0/wCHvdcdaA6VW+Pit0DsjaxTaWzZpsrFB+zJGqzO7WtcNz+ffvl17ocvij/LjxncmLpMvldn5Sn89UE8jwMo8Tjg3I+nvx62AOtjr4k/yvun+m6SlydXt+B5REpMM8MTO8gOr8j+v19+6rXoefkx8T9mdg4OBsZhIaRsPGj0iU9PHEY5EHpswAPP9PfiKdbB8uijdUbmxu288ent+Yh5MZHKsUM1QgKAKdIu7Dgf7H3rjw62Pn0vvk31/i9r9f1+Y2BX0UErwO9PTwvGXUCLUBpX/ePfsA0691Rjt75B9zbb3DVUmZo8hX0AlkRNMcjKV1kDTYe/de4cOjHYD5FVFadOSxNTEfCXIljZjcL9Df3vr1cU6Dpvl1UUW45aalxFUsdPOy62ikCMFP5BHv3Xuk12X8uN5k01bjMgIIIWEklMGcMFX8Mg/wCJ9+HqOvVxTpYdffzCeqauOLGb8FKudSEQ+aok0DVazMdXv1B17V69K/G9/wDW27chWtic5iNMzkwxLOjOp/I4N/8AD36letdBn2Z3LtnYtNPlMjuHHrTPGSkbzKjNc2FrHn8j3rrfy6IHvj5h4yumlo9vp9x5GJEkDEq1xYW/Fv8AH3vrVcdEl7y+Q3a9Tgax+udv5aXORws0csKzMrSgcG4H+8e/fPr1cUPQS9M/Hntvt7DQdq/IBKzNTCqE+P25GzGelnRtUTeJuSQbfUe/Drx6PTsb4d9ifI3cOHpd60NTS7Nwgjgx9FV0yqFpIWtHqZx/qR7917J6tv2H8ecV0tiqfb+04oYIYI1jl8cMa8FNLANGL+/cet8BTpx33LDgNvSR/wCdlZXdrC7XAP0/1vdX1FKLx69x6JfXbipsq1XTVasqO5XS4NiLfSx/x9sJBMx7uthTXPDprTES+THjCo4KzgsUuAwJubafbjwTIOw9eC1IJ6tN6Ux0s+xIYpyfvQkZBb9QC/Tn6+9QiRcSdNyKwcY6NDs/PSYilkoKu7GUEKTe4vx7eOePRikpWOh8+kb8mtn0e7+n8vBFplqTiqkhF0lyxUkcD+v492FFU/Z0FN3gM6SU9D1pH9p7Pz2yOwdz/wARo6mKCTN1KxSSgKujWef9b6ewxNa0lJHWIm92d1bb87aaqX6Ej4+dcvvPc9NkkK/5PO7E+lraTc8n2ZWsdACesgPbu6miiUDHVteF27nKaqxmMpmM0CvTqYomP6QBcWX2ciQeHQ9SffTO8gLmvDq37o/bsGFwdBPLTGmmakUuzoqMXIvfj6+2Dxz0r/CPs6Dn5S4WXK4pZqOmmlYRghowf1n/AFh72OHVT69VoQ4rIrP9lV4+aQNUaV1DWQSbfQ/7f37Hl1rPn0bzqL4/ZDOJT1VVSyxQyKrLqjVboW54t9Le9Z49b4cOrAcP17tDqvBtk8iKcSpCZArBLg6fyffuPXh0X/cnyNxDVFTQ0HqhiZ410Cw9PAAA/wCI97oPPr1T5dF23B8gKx6qSKAMser86v0/m496+3r3Qd5jtiLJwyRzudcilVvcEE/g+/Z63xOOgipM48eQmB1yTSByjhSwCm4t73TrQp1Xv8nNw7n2l2rsLc23aWoqaunrIXApg5Knyg86fp78PTrRPn1uwfy+PlFi92/H7ace8alaHcFLRUkUsdQ/jdh9uL6w/N+PfsDr2T1YXhOydrZ2XwUmRgaUsAoWRWDXNha3v2Dw61mtOl4LNZgbi3FuQfzce91xTr1KmvXL3rrfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//0d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdNbktUXOpSPyDYcfQG3tsMwOeqa6nT5dZKmx0q0d7fVh/h9Tcj/Y+1C6RUnrZgMgAFOuLxJHEY3Anp5x45EflfG40tcf4j6+2JpRGA3+Hq0MLaitfLrWk/nVfyottd37MzPc3XOLp6feGMhrq4R0VIzS+VaYs0n7XJ+ntVC6TJpGR0V3rT2cglXHWlH8V1zOxezN09c70NQc3hKqSnqFqlMZVjKVBVJOR7BnMtqoB8qcOpn9t+YpS6Fznq1OApFVIkTh4gqNe/H0/wPuMmWjn16ykS6+qgWQ+g6EnGTq0K3YfoBbkcG9re90zjj1Xh05+ZWPBuAb2Jtyf6f7b3XPWyfTpnyEVRkLwLcqf8eSP6+9109eI8z1yxu1aSF1lnUSSnjkBgTe354Hv1PxHrdQccOlUlMYivjgiCD6jSBcD+n9PetIOevEHyPXp6yCmI0xRjgXFrAG/Jv78M5HXs/iz1GbKwv6QUu319dyP6W9+4nr2OPUl62JKfhizaR/seb+/UJOOtHh8ugp31DUVtG700fJJ+g+pP1+n+PvfWwScDoPcHNlMPTTSSI4Krcekrx+P9f3sEAYz1rSaV6d8FmptwV6wS2JRgGUkfUce9UqKjrWo9DnQqaBY0tYEW0kED+nHv1R59eUj8+mXcdS6QysoPrAPAFxccj37hx63xOek3tmkWOpkrrG7HTbTb6/kn34V4de1VpTpbNOIg5JuWJNuL3/B5/2PvzCvWq06a6id2UuLkWNv6n3vPl1s9J8V87zCMknm31uOBYAW96rTrRrTh0qqGKrdQxDabGxF25HvTEqOH8unEVGNKj8z0BnfG76nBf3fxOObXPmaqOjqQsguiSHSfSPY22fYkmhW5Ycfl1APP/NUk88u0RNlAeB6tA+J/TuNwG0KHcNC/wBxmKuGmlkRgXIM3J9yHZoltEEHWN0yXK3ZluakE9Wddd9FS5mrphk1MdBlW11x0tYajfgf8R728+pqDoQQWfjQGaPgvVlO1qTGdS7Z/hOCCy00UCmEn/joVtx/r/091Hz6Sg+fRBO2vnbP1x2BDtDIqkUtVVKiq0qxi0jWUAH6+99bx0evrfdtN2HicHnMkYylSiyRHWrfgEc+/dePS73htmqqC09DSxSwCMkEoD9Rext798x16nl0Rrt8Ha4mzH2UclRBHMNIi1C6odN/x791sHqlKvxe5+9+2KvGzLV0WPesKSLFG6BVWW34sDx72MdVycDo/wBtDqXavUvhoVoqevrWhifXVQLI/kK/UFh78M163w6EnHZ7I0mSjo3p1hxtR6mCHSgUm/AHvRGOvA9C7h6vGTSukWiRItJYOTY/4Dn6e9ef29bp0z5jCY/cOQ0QxQmQA2FgSSPxx79X069T16DHe/UVDXw/fzaY6yhX9jQDq4F+Le91z1ryx0EMW6MthKiiwFdGZaatqEpEkk5WzNpW4b3vga9e+XRYflV1zJgMhQ19KpK1rxTMq30fuEMb249+HWvLrhvT4s4fs7qrFZSCipf4vHj0dJUgUyh1iDXR/re/5J9+x1v7egK+L3zN3r0v2BV9Cb2jli29iJDHj6qd9CSCMmNFVX/pb37rw9D0ebt/5I7Rj23PlMhM8JkileN1AAsUOkgr/wAR78BTr2OixfChcf8AIrtPJZCrmeqxmHrZXpDVN5I0EcmoFb+/daGeHVrHfu26TbGLpTRFFplhiBCfoGlfwPp70M9b6Ru3OvqTsDrHLVPihkgigcSMF8hDBf8AVL79nr3n1V1vTF57qbcdTHgadhQVUhFWSrC0TP6jc+99aPRV+2tp0PZkldDXUEElE9FUEyyQhyJ2j5APveadeHHHRHvjFvDHfFTvMJuOOKk21U17EzFRAFDTXsNQ/p7117Hn1doe78R3bnJqzZtXTtiokiYTrUpqCqoJ0n/evfut+XQq4jeMO24o8nT1Yq6iN442VqhZD6WtYf8AFPfsjr3R4+tu2ZM9QUklRJGKAIfuvJMo8elORcn3o+vWx1WX8hPkRuLtLu6i6ewNPNJh8fko0WqjVpIQBNpJ1x+906r59Gc7i2BV4zrug2xlVvja3GwyTzNqJRmj5vbke/cePW8Hoi23uvNvdc5SlqcJMrzVjki+m92PHB+v+t7916nRyszi8/n9lUcuJhD5WBElQFRcui3Xg/19+FevE+nRZ85278i9iZSKrnw0pioWSGHxxMylVPF+OePfutdGs2N8z++Z8NTxUW2iauoUciBixa1jfj3o063jpd1HdfywaLHZT+7CyU9RWQiTXHciF2Gokf0sffqefWvl0bPI5rtfc8G3Y48clLNV0cT1gjQJplZRqvb/AB9+NOt54dSMt1Xu2enhmzdVNEp0mTTI9gtubgf09+HHrxGKjrjSYjB7IphlMVIlXmeEdZiD9Pqefz71XrwGekXu7e0U1dSNUyeCpnAJhhI0lj9bhf8AX9763kCnSHyMf8XFTJUukNHDGzSSawpZLXN72/Hv3Ada456IP352HjszjqvrbZZE2QnMsUki8/qGlrMLe99a8+pPwU2psbobZnZGT3NNAdyZIVMqPVMFkSZwW/bMnN7/AEt71x68MCnRTuzu7Y947sy+HeaST7taumorP5WMhJESAD/evduvDq8f+Tx8W4cLs2p7V3djlTcr1WrGPPTMsvgaQsjgtawt7159aPDq/kG4Bt/rg/j+v09+68DXh12Dfke/db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuum/S3+sf969+60eHWpP/ADB7/wDDm/xmv+d40Fvpc/5anv3ljrxrUV620aT/AIC03/UPD/1rHv3W+pHv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6ZM9lo8JiaqvmYWhjYi7BebfUE/wBPezTj1XNOiFb37MXMVNX46lwkTcssl7D/AA91Oc9XHRWt77ixe5aOfHVccdQsaPpadRIxP54I/wBt78OvD0PQH4nJxYPH1uOplFPDLIQBEAosT+LW9768PTpebRwdHufTGkjtUsPSpvckf4+9dbr59d7ywr7bVaSrDeSRwn04I+liB/xT3odePQR5vZlA1ZBlBHGswiaRAqgXKrqAJHuwNRnrRGOqZfmx85uwvjFuKjmmxVVNgRUIFmp4XYLH5LAllB/H9ffq9a+zo3/xj+Yu3/krsXH18lWwrWSBvF5vUpZAxV4xyD79w691bX8c95oJUxaTuUN0AMgH6T9CP6e9deJIFfPoWO7caNw4ypDKJBQReYkqWFgLkAj/AHv28uVp09LF/i4bzPl1XigavkySMoWlijqoiv0BspX9PujcdJ6Yt7j6cFh1ri/zAIptu79ZsbW1EMdTWgPEjWFieRb2jmxGy8D1DPuLza8EZhJ4449Ed8UVRRCqqHLzAa2ZiNRsL29he4Q16xtvLi4vyVc1BPQv9DfHzcvee7qJMZQzPQRVNKxb7WR47JKCTqUW9qLS11MCR0e8pcpm5vlnkFaEUqvW3B8RenJOutu4jb8kAjniiEUy+IqQdIHII9iOe38GNSvWaG0bZ9Dt0YpTHVkc1BHhcWnm0qpjFwTpC3HP+39mdiaQ93DpYcevQI5+v2/rlUhfKdTF415v+OR9T7DO/wC8Laqf1FBH9IdaJYUoD+zoDMzmsfTTN/FBJ9urfs6kLBjfi/so23mVXSpkX/ex1pXc4of2dAlvrHYbcaI0VOlDBK4T7yCLxTEMbcuvsQW+6LdNh1NfRget9wXgf2dILcfxcwu4tvSUONyFRNUZKJmEkhdmQunBBH059nUdssy+IePWxwz5dVM/Ij+VJ2nn5VbDbz3RjqSV+VoK2rRQhe/6UYfj2ncaTTr1M1HREsp/Lj+W/SucOe6r3ZuzM5GlH3C09fU1k0LNCNSr6ifr70CCPl17H4ulNtr5q/zS+jYK2l3f1jHlMFidUEtYkcslQ6Kb69LL/QH6e6kjr2K46E7aX8+x3Wfae/8ABZPBZupikp6qOoo5IFRkXRIyF1F+foR73ilT1shh1dX8EOofj380thTdiZncNamdr3E8EBkewaqb0BUFvz/h731rz6GjtP8AlLb229vTb27usK6tqKCjlSZYkM2mWMEOTIFP+29+Jp14AEdG5ovihuTce3sdQ9g4qGV6DwtGk1MZVaSNQACDe3I96PHHXq4p0uMh8Xp8/tCfa82OjoKEqIkFNTtHqjtZbqoHvWfPqw6RHXf8vHBSZiBM3SiopIZNMZnp2ZWW4Oltf192Hy6oT59W29bdR7Q6zwNJhcFh6CFadEBeKmjT1KLXHH+8+9jHXuhSuqlUFhcHSoFhYfgAe/fPr1QMdcJoIqiNopkWSNv1KwuD/rj3rrfRb99fGzaW7ci+ZihhpMhd3WSOILIWP0/cH59+oOtVYY49Avuf4tZGsoJYhW1FXGgKxwTSPIrC3HouRz/j79Smet16Kxu74oZ6iSD+Gbbx9S7SfuMaFXbk88ke9V8+vfLh0XTsr4tdoxp5MHtykjkKFf2qK3J+q8D3uvr1vy6DXZPw07Kyck0u5NvU8BYsdS0Rvcj6l7ck+/VA69TohXyP+NXaGx9z5GekxkjYbxksRSvpEatzbj/iffhnrx6qz7Z6Kkyi1G6pqvJ49aTy+cUkc0beRR6gNPvfVfLoi9fvLdHT75HcGGz2fqI8dqqEimmmGpQeF9R96695dBhjtyfLr5t5aixmy485S4mGvWNpniqJI5I1NwRp459+zXreOtiv4R/ARqXbmOh7zXw51aaIfvU7IXKMNTEMPyPfuvU9erSk+L3x/wBswpSYjH4+uqrDy+ekRhcfX6+/Z8+vAgdSMP0L1lt7J/xpaalULZlxyxAUlx9D4h6f9496z5db+3h0KD1WMpqGSPDYyho44dQR6aNIibD68e/DJ63UcB0DeS3dTY81zVMmufxn0s1zzxxf3b7OtEdE8372Qk1VNSyLKyu0gUG7BVY8EAf19uRUR9TYHz4dXjBZuHQV43a9bvKpb+HUk93Nxop3Ib1fUWHsj3nmG22+tXT/AHodKjGagkH9nRk9l9UV2GprVdLI0yKWUPA/LafTz7Ktu5utrmUAOmT/ABDrTLmhH8ujndXbez1Pt9696ciKI6QqoyXUH6W9i9rhJ0BQg/Z1Rgp7T0L23VXL1TQ1CiOaNrKGFuQfoT7bAIwek8jlFoOlPkNs1NT5qGvYPQyRGNlZtS6W+tlH+Hv2riOqJEJEIandjqrL5Z/BPanYYqKvB0cArJGlmbx0xuZCLE6lH9efaYwa2qePQE33kO0kb6sKCTnh0STp74d7w6+y1RQ4/HuSzsvqppB+pvqCfdtOkV9Ol3LOypanTSlOrSekfjnW0WSgyO6IFDJ42AkjOmwA/J/PtwP69Ca7iCzCv5dHay23xSUsCY+MJBTgR+hLBgPybe7ZA6VYIHy6gS4DHZqh8OViiZCtiXUMLAfXn37rxzjpET/H3r5A+fZKa6fvadA4Zb/QH3vrXnTpy2/urDQs2Fw0MJ+21QqVAU+kW4t70R5nreOA6J18luw86XbARTVMd/SwWRgpBNrcH/be90oOtE+XVfGZ3jPtSpZa0vKJHD6zdj6vzce9061WnWOq7Dwc0MLyFw8oDg6fqT/j/t/eqGvXh0gdz7woKOJKxZXEPpLBeeL/ANT7317pjp+2cUUWbHNJJoXQ5Km4Yi1gSPp/X3vj1ryx1P2vT0W9tw0mWyVHDUyUc2um+4iWRLqdQHq96pivW+PVhG1d7ZbbkET09TLjqfxxKkNGxii9C6RZVPH+296+3rfRietfkruPbmZpJfv6qRRJEi65SAVaTj9X+9+98evU6vv+PfaNT2LtunnqmR5BDquGDMLLyOPfgetEefRjALAD6/4/8j97OT1oCgp13711vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/S3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6911YXvYXP149+48etUHXZAPBF/9f37rfDh1jkVWQqQLWt/re2LgViPVlYhtQ6YM5iqLKbey2MrYI6ymqcfXRvBIAwdJIGXSQf8Ae7e92IMaADGemrlBOh1iuOvmnfPrqsdN/NXsTckeKGHx2XzX7J0mzqapiCCBb2T8xJrQnoZ+3Hhwbiqy5FeHT/gq2XL08GRpZDKjpHZVBtwo4t7i6aNwxPWZ1qbeSxVowOA6EWlytVT+L7hPEgFiLcH/AF7+0Bk0MR0xJGRkdKOPNQTKCJlH0vY8G/Hszt3ikFKdUVTTOOnGky6h9PJ5Ua7j/WJHtPOAJKLw62MZ6VMFZE66y/8ARrhhc2FiOPbB7Tjq1Ac9dyZdUugGv8FiQfrxcke9kVyOvV6SubyDmKTxAuxRrEW+tr3/ANh738h1WlDXy6Rm1JK6uq6gTu+kPZAfxzwffjgV69RuPr0L0NCGVEJ1lbH+v1+vuyOqirdKoHQVD+fWeqpoNAgMClbi5P05+t7+0wOqrV/LqhtHdqpw6a8vtyhamjCqtpANSmxsD7fikWuR06i+BiXoEp8JUbczqVsIMdK8o5H6f1cgX92ZhXHDpmQoZNa5HQ2R1UeSFNLDJ/utSw/Nx/h7r5dNEhuHWaeOKpR6eRdXFvxcG31591rX5daFOosVPDj0Ed1HJIP5v9fe+P2deFT1AqSr3YOLD6i9/wDY+99bGB02TZSligemZlV5RpjJ/BPA9sTq5poPTTo7uCmaceoFPTfZIZqhg4uzazcmxF/Znt8DSsAwr0Y7nu+3WG2lnoHA6WOE3TJR46pXEYz+O13qEVPGrM3qXSukf6/sf7btNo4BmWvWLnN3Ot99QwspSucU6L7u7obtDceZj35l8PWUlJDJ93DQOJAtPY6l0rb/AG3sVQxxwpojFF8h1E813c3E5uZmJkPE+fVvHwq7BoFw9Jh8vOoyFMKam+0ewceM2vz7uRU06YkAmNXz1djs/ctMIKKOn/zbDlgOIRb8n/D3XSK56dSWSBCiGgP8+hBymbSMQh5vNEzoACLryfxb34Cg6brUY615f5plX/dbsXD79jg8NFSVlKJJfxqVhf8A3j3sdaPAAdWrfCruSm371XsmSkrFumPR7gcGyDi3+PvxGeveXVidJ2hFRKMbVPrLIAS1rg2sOB795db4YPSW3hhtvbyw1UlRHFJLOHZQdANipFwPeq569wwOihY3pXA7Zzc+SoKGGGoLszSoFUk31XuPe6nrwpw9Ovb1o6Z4452jWWqWwDCxYgHgH3rh1s5HTZjsRHlYYjJGEZEGmS1jxwP9h798uvAU6419BJjJ6eClcK0zhXK3AYH8E+9/Lrw9T1npY8hiNwCUBmjaENxcrcrfj/W9669x6Z83mKo/dQTzMGndjHfi1+Pr73x68cDoBu0aESYzEz0gvXUNQszSr+oFHBuB+fp738utHy6Q/bytvTq5JJlaXK0jxBZCbSL4uOPfuvdLDoGtyuS2fFiqtJIoqCAwlmuPSq2uL+/deHHqon5fYCj293RiMrTYxVlmy6Cev08lGnFy2n8e/dVrno329Nn7O3r1/hKOKenlqJcTCXS63MjQ8+/V6t0wfDWSg+PO68rROVgbNTyrSEWILSN6bXt79jrQwKdHq+WXd9BhuqKWsedZsjNGU0mxZA6/rAB449+APW8dCb/L13JFvPo/dVVUVhn8msqtx6A3NiL+9Hr3QQd+9WnJy1VRT0wewJufqyauSffh8uvdEvq+uTPj56CGkKSrr1OotYX+l/e/s69nqgb+ZFJ/oz3tt4UVD/Eq2WohUUCgu07Hi/p9+r1qteg96/8Akp2D1ZgcfnKigqNuUNcQZYvHIEEZFgNJ+nv3DrQyerMfjh8h6HszwZifcMc0QWIyY64F2BuSVJ/2/v3W+jnZ/vrcziLa2xKGaSjq2EdbkaZLR0gb0NqK/T34ep68ST0ebobpDDY3D47fBxkWe3VXvHJPWWDzxPINdz9Twfej177OjJdy7FafrqrzueyjU9TS0spio5X5ULGWUWb+nv1TwHW6jqhvHbmzWa7LqKV6pzR4qr000V+JQkn9i3vfl1rz6te2RmpqvAwedWoGigjs99JlKoPpf37r2OhVwdJQbkwNXNXbfGVkgk0xl0DagDYn37rfr0MezMft3EUcEibHikqUswiES3Bte2kD3rz695dCFLuvLVAjUbKmipFZQkQjARQp9LWI+vv3y69TFep2X7Dy2Pko3gwUsJRFFtH6bDgCw96GB17B8+kzmO294V9PLFPh51gZdKuynSBbg39768eg1AOagqZanJ/ZyBJH8bMAQQL2964HrfHoAMlunB7fgzWRyWTiyFXjJG+3gZwzmwJACn3b59VPRXt4987m3wtNgdqUcsE1fI1JIsKsGCk6QW0/717917pRJ03Tdd7Rl3zu8ikyciGo+8nazh2XUVu3+v711unr0QrtXsHP5urNPtwvHjZG/wAorYwQksZNi114P9b+99a6Ez4GfC7N/IzvCmmrqyQYrCZCGvedpHVJUVvIytfgj6+/HrXnnrdl632Fh+uNpYja+Hp44YMbRQU7sgH7jxJpLkgD375db6Xnv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuuLfpb/gp/3r37rx4dalH8wcD/AIc3+M35/wB/hQWF/p/lyH3vrVfTrbRpP+AtN/1Dw/8AWse9db6ke/de697917r3v3Xuve/de697917r3v3Xuve/de697917oAvkTlzith1TI5R5NdmvbgLa3HvxOOvADh1V5DNPW65FkY+blx9L/wBQR7p5dX88dJ7K4aSmUVDxFWlNvqR9Tyf6e9/LrWeg0zWOFLkI5GsYG5ZebXPI492FOqkdC91nWnHZWimhp/2dYBcD9I+t7e9H5dWpjoVO1cZDmqV8skIl8cOoGwBXSL6jb3rr3DoopzkeQWqpDJ+5C7RKlrabcW5/4j3alOHWq46Kh8legut+0ti5xd3Yakr6iKgfwSVCo3jaxKsNX5Hv3WuPHqqv4Sdc0fXHdGRwGMrIxh46yTw0KksihWOlABxx795deFeHWwR09uqHF7nhxP24hkqp9McvF7GS31Pvx49bORTqwjd1MsG06yVwJZKmiCs5sTZkvb/H20WYcOHWpCyx6SceXVZOXpHxsmUliBSNjVErzpJNz7TSO9aA9MqgaInzoetbr+YlHUVW+KFkjOn71S5AJXhv08e2GLaDqOesWvdm2lIYrxBGeiXbN2fnt97kx+3cNTSTrW1KUkwUMPEJPTr4/pf2WpG5ajdRxyTtVzfXQ8apQngets34E/GbA9F9b0dVlqFKvMVVPBOHkIaRWYarDUL+zq1jCDh1l5yzyzFbW6y6eA6tD2JTQCsbJy0ogiZy6aiFsB/qfx7OlCSKNfAdDZmklpEhwvl1g7JydfnKlMfjJmWJSFYITptf+o/HuBveL3t2v2/2eVAQH0mh+fQr2PbBOw8Ra9RsPs6mWkSWuRZZtABZyTdv68++QXPP3g+euYd9kn268ZINRxnh6cepGtuW7LBaMHpqzWysRlg0E2PjsOEYgD/Wt/xX27t3vLznLEsQvWUjic9Vm5Tty5dEx0HeW6soXpjRqixIpLIpIsCDxb/ivuZeS/vB79y7cxtfXBuBUYNT0VXnL9tFHXTnpLLTxbQHikrQzr6YgSLi3AsT9ffQz2y97oOcrFWkj8Ogyegfe7OTXwuHy6U1FPPmIllqakywnhUY/Tji1/cl3XPOwRP4TTLqHHh0GpNru0JAB6a6ukxNNORNArs/o1cNcE8jke72fMmz7g+mKcftHSdtqvTnI6TGY6y63ytPI2a29SZKmqyWnhmSMo+oW5Uj/H2MLa0tZY9aS6vz6TPt95GamtB0SXuj+Ul8cO6olzuK2dg8NkJtTGWNIEku7XJDKL+6XG2yMP0z07Erjj0Yv4efFWk+L01FtrAVa/w+KWmCxxNqQCN7WUD/AIn2XvbTwDuNelZIHb1sbbSlM23cXI0vmZqWPUbglSFto4/p7eQ1HTdenmooqWqULPAjj6i6gG/+w93rTrRFc9YVxlAP+UaK5ABBF+B9OPfiT1oAdSo6eCH/ADUUcdv9Sig/0+o9+qeHW6DrN711vr1h9ffuvde9+691737r3XvfuvdcGijcWeNGF72ZQRf+vPv1etEA9RZMdQS/5ykgf6/WNfz9fe65r16nXAYrGgWFDTAf0ES2/p711v5dFk786IwO+MBVGLHI8rQSLJ9B9Rcn6fQ/09+I60Ca06pE7K+KOMpDkcP/AHfSeOolqP2La1csLCwA9+r59WPVUvc38rLOb6yM322CehxlcVV40ZkjaMtyCLf7D37qtadWRfEb4c9f/FrYdAarAUgyAZNU37ZfUFseSL3/AK+/dbxToxu5aeLI1sVfiIdESXsygERqfxx+Peo5FW5CP0pgVWXUem6Wio2hLzMIKrSC0hIBZh9Tz/vPum8PSng46L7pipx0m8nkxQUgSNPvm1aQfwLHj2tsAEtPEn8+rW+tsnPTTAK7IyotOjRLMt3j5AH9Rb2nnu7C1BkmcAdX/d93I1UBp0hz01mtwZ6cOZfE7+lTqBP59xxzR7xcrctwkySrVR8ujq02eaUDUD0/f7JWuayEMtSDGupWkJe9/wAsNXvFznv74uyPYSbPtZAmyAwGehJabIyuCy9G22B8etl7Gx0ULUVPPURRkFjZmueefeEfNvv1zRf3DNDdtQ8OPQstdghYDWnQizbJ2xLC6rjqZG0mxsB+LXv7CvLfvdzfb7orT3b6a8Olc+wWiqOwdd7exNLQNJhqaASQy39IsRdvx76yewPPU3OXLiXMr62AHUb8020dnNpjWnUTJ7BEOVgnpSaLXIC9iByfeQOp9ZX06CiGq1PUjJbVyENTHIa154ja/N1C/m/tSMrTq2oqcY64ZHa0OQgVYl0kAB2BAvfhuffuryTNImlsjqFX7Nwe2qakrJqJGmkGp3OlmJPIJ/1veiB00iqh7MdcpKSPMUnlx8ghMZuwQhW0j8e/ADyHW37sNnqG9V56CXEKx+45XyHg3H1597qet8D0jcjlKfCQxUM0gaSQ6L3Ba54t6ufr+feuPXvmekPmKnIAvTpUSNBIh/bBNtLC4v72M468T0i9u46PFZNpWURyTux1D6m7fU397Oeq19ekB3h1PUbnjgyFFFqqGZSzqWLEfXmw96rih6tTzHRRN19E1EdEanKYxpVQ8yP6iP8AkXvYr1ojoJs31ZQ0uLkmXHoCkfpIFwLL+P8AD37r3QMT7Ehy0P2xhRo1JAjJ4Nvrfj6+99ap16m60xFNCaGDHxLVOw/aC/rJIANx+feq9epnoZth9P5ehkikioXQzE+NRqtdv9SSPfjXr2OjA5LaTYrH04ykphnXRqib66R+eP8AeePfvs690yuKf7ymFK62DQgsOb+u9wR/j7917Hl1fF/LzlrpMTVrUzmaNIX8akMAo02uAffuvdWge/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/09/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XCRgqkn6fT/b+6OmsaevEhcnqBMYo2MlgVZCji5N1YWJI9uIukBOnUAeMnqiv+ad/LOwHyE27V7y2himG44Y1qJDTU8LTSSIC/Bte/tDfQ+MoBHSzZro2N4HU061C5Nub66E3xVbB3lhcrBHS1c6R1NZF4o9EUmgH1fT6ewbfbU1SVHWRuxc6QxwLHK4Bx0YyggoM5RR1DyRSB4w2iMgsARe3HsEbhYvC1SOpGsN0ivF1qQR1jl2e7jVRSCMfhSwB+vtHCzg08ujJtRNSO3y6hNgsnSfV78n+0f7PI49qya9bUqR1xlgzZCiKRlUAE2v+f6W/w9+60VPFelPiaCqaL/KZAzW5tchSfyfdagcetHUOlBTYindWMouSPp9CAf6396Jp14CuD1EpMLTUVQ/24AMj6hwPr/Tj3sGuet9xPU5K2WkqmR9Wm+lSBcD+ptx7SzEBgB59VKniPLrFV5RFqFRp4gzWIDGxN+OR7MYLIsvAmvV495hgHhyEBuvfdvI6FzeMcH6m9zwQfpb2ju43iOBw6pLdrdAmM1+zpq3LTQ5KhkSFR+1GXUqAWBte+r3uIs0Yc9eTC0I6DXauebH17UFS/IbQuskm1/8AePboB4HrZrxHQv1MyIEnDC0nH5ufzxb3oEeXXqEdM2QaSaMsha2j+z/vBH+PveOt8fl0h6qtrIWaK7gN9L/UC1re/AefWwCesVHjJq2RaqoZikJ1EXNj/sPaiCMSPQDqxuobWFi2DToStkbWyXYG4IcLQq60upFey3BF7E3PsfbNtVQHK+nWMvuNzfMkrQQt5nh1aH078dNobDqcdXZWkE07rqcPGjXY/W6kexR9OLYdQlFeXN2+qSpr0dJOocPvSOpo3oII8ZVxiGJTToPqtgRxxx7fU1UdGFPM8eiab/8Ai3kent0Lu7btPVGhE5kaOOOyWU6iTo+v+Hu4OOtEdG16p7Cpq3DxUrymOuaH9xXkOtJP9SQDx719nWx0YuWtL4ugLThpNaM51Ekgm9+Pevt691Sd/OzpnX425rP4+J5K6km8qyxL6rRIHJBHI+nvYx1o06Cv+T18g13H1btmkSrR6rG43RUR+Z3dZEAUqw/1xbn3v7evY6vOi32tblUlqtSXVACWZVPFySD70eFT17HDoSKTfVMldTU4mIhZBwZCbjTbj8W96yevUPT5X5WnqY5GhkjLMPqDbm3HPHPvZ61TPQO5N/PU+KS5sb3/AFCxN7+9eXV/Pp3jUx0emldY5FUaRb+g5uB78eHz619nTDFmcStbDDkqqmaeOS9i/qBH+Df09+HXqinz6FxUw1XAlZaJ1aIKrgLYG3AJ978+q1Ix0Du8duTnJQ1EEbmnbSQQLqb8/j8e/cOPWxkdBHu/DTRwPKwZ1dCFUKGAb+g97rjrx6Kzu7dMmAppsdVcxF2fxvfkDk8fT37r3QmfHDsXF5ulzFDT6UljLrxYX/BuP8fx791sUPQKfLHqqg3JS09fHAXrFkMqyIinktqB1W4N/wA+916rjol9JVbh2zlKSkqapvt4Y0jVXdmCoo5UKD7117rn2F2BQ02V27koJAk2OmillKMU1kW9Pp/Hv2evGg4dCP8AJDeWOy3x9pt6ZCo8iy0xjig8hLhxHYFR799nXjw6MF/KV7Ay9T0vust51oWmkMYYtbxmWw5P4t78etgV6slzeRocxj5IQkc0roOANTcj3Xh1s9F5qdrLRyZSplp7A09Q63RV+iXABPu2KdV4HqnvOfFOk+R/yVxOQzuPesxWHyFpA0SyRjTJb1BgR9Pfhw63xPS2+anwd2Om3pMDjsWsUFJTIIhFTQoR6bcaR71nrxHVBNb1tv8A+Pu6mrds/fx4ZKu0tOXawhRrtpVT/T/D3vrR6uX+E/ZlBv4UdHTR6PKbZlqgF3Mmoa7O/wBP9h79nr329X/9cZugoIKPA7UCrWx+HyMTqS+kA8e9HrYp08fKbB7jm6XylTJIajIGGV0EGq4AiJPpH+Hv2OHXsg9UO9AU0GZ7TFNkqWRGpa21X5F+tpfqwPvfWvn1dZLsegrEpIsbp8BipwI4FC6WZffsjHW/sz1myON3jsWpphjI5ZMY66pIkS9za596wc9eoR1PxvauXxkpnqcdU3uSNUTBQB/hb36nXuPHoUl7zqpcRHMKZEckKVZOf8ffgp68adJPN9syVaLKwRW9JI0/4fg+9068CPLpG7q7+xuJwyw1SI7lQB41GokcAcfnn3qnXq9FeyvbOY3HMKTbmIyJqaiYxl4UkZQshsD/AEH19760Pn0oa74u74z1NRbglq2p6erjWeuglJU3/UwKE8/nk+/dbGD0rqPa/WHVeLOTqKaF83RJ53lfQSZIx6rA/wCP09649eFPz6Kl3P2llu9cdUYCkqFpcDSu4YoxSPRH/ZLcf0974dePRLqlF3DkaLqnYOPqMjnanTRPU0aioEcusIWLC9vfga9aIzjrbZ/l0fF+n6J6dwmQ3BRePeWSpo5a6eWFI5VRohYNxf8AP19+8+tHqyD37rfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddN+lv9Y/71791o8OtSf+YP8A9vN/jN/4eNB9P+o1Pfh17zHW2jS/8Bab/qHh/wCtY9+631I9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3QGd97cO4dnTxLyY1k4vbi1729+8utefVUGekqNrVf28al1hsGAFrljbj/AIn3Xz6v8z0PG3cRjd57WgY6PvAoJQhdeoC/N/e+tVpnoFN4dc5WTMpSRRSNEHHqUXsq/wCt7917DDoa+vtgR0dC0NTEfOU0xsy2ZTa99X/Ee9derXh0luyNxUu3qCpwMzKaqSKSP+h54F7+/DJx17h1X5kzUYHJ1FbIX01MxkUXOmzG6+ke7fZ1oDoOO2stVDalc8zl1rqdlSNSRb08D/H/AFvfq9b8qDqv74+7Ay9N27NuFYZkppam+tgQlmYm9/8AeLe90618z1dZ19tj7zOYvNIjOaMqZCn0LK/1I+vupPW+Ar0fWfKU2axAxzj6RLGQ1hyFsRp93EbMK9WnH6IbopHZWxjBBVLSRl2lWUjQFvdv9b8/09ppI6fZ0ntyJAVHWvX87+gN657N0f8ABcTXzTTTxOkkcAbljYm5HtP4dePDqI+etjN3KEIrqPRjvgP8G8vtaFd5b3x8slRMVnpxUUyFkk0XFgw/r7stuFzwPVeUOU1s5AXXBPoOrj9sY/JU88dHVRstNTkJCgXQDGpstgOPp7fRKNnh1PUEEMFsFHp0MWQylZSUKwUUTpZQuq3+H1v7S77f/Q7TLcL+EdFu26frGB4dZtqxl2aevYO55sSCQTz+ffE/7zPPl7ve/S7eQ2kFvM8K9SrsrwRAFePSxR5Kl2KOoiQkWBtwP6+8XrCeMQ/TxxnW37eh7b3MLLQHpiytfJ5Eipv1qbNYXv8A0/2/se8vch8x71MqWUTjX59Ib3mK2taxEivQd7orMrCv7d9TcCwI+v8Aj7zS9uPu7fTwLf8ANZogAPcOgRe7y1w5C8OkNSdfZTdlfDUVbSCO6tdjYfXm3sS84898o+2tq237C66xUYP+bqtm7SOFYHPQ6YrYdJjIEpSosgA1MB/T8294Vcwe+HNsm4SXkEp0NWncehemywyRCQj+XWPLdcUdXaaMesAt6bH/AGJHu/Lf3jObrS7HiSmlR+Juk7bHBxp/g6DjPbMrBTvDShvRYA3vaw/w/wAfeV3IH3q91a5jhvpaL51J9ei7ceXozCdI49I7F7jmxTfwecN51JTliAW+g9558g+7dtzGyiNwa06jncLD6QUIz0LWIQwUIyMzBasDyRkgXAHIF/c2Xc3jweIvmK9EHbr0jo1nx07aqq6oqsJm5wUDGOlFySvPBAPsntDrLAdedCmT59HdBBAI5BFx/rH2q6111pGoNzcC1vxyb+91xTrVM165e9db697917r3v3Xuve/de697917r3v3Xuve/de697917rgVYsCGsB9Rbn/WB97qKU61TNeuM0SzRPE4VldSpDDUOf6g+9dePQQ1/T+EyWX/iNXHDIhfWVMSk/wBeD79Qdeqegw71wWF29gIWxmOp0kiQBv2YySD9Px70fTrfVcG7qp9w0DU8oEYikchQAn0b8D34HOet0x0GGPyzUchxcSMWY+MaiW+n5v7RXaMbhXHDp1HCoenVtp1WSmV5n0xkg2ubn82Hu8beNKqEV6SyAuat01ZDB09JVR0Mcd3JC2Ive/HHunNe5Js20ySMdOhCf2Doys7djop5kdDVtDrqKP7SqqYrKUVhdVt/W5J/3r3yy9zfvJ38d9c7dZy5RitAx6l7b9lje1VtPEenQtU2ExdDUeSOKMPqF/QoLf1/H/E+8KuZvcTm7mG4YTSsFbhk9HsGzwRZA/l0oPMzOEptCKRp4Uf6/wDT2H02phAb+aQmU/0uln0cAHDPWBaazmWpa6m/A+vH0v7S2c8Bm8OfJ+3pfBCoXPAdeFNTyvI4UiJVFyfyPr9ferwxhybbjQ8Okl2FAFOg1Oeah3nBBTACE6Q/4AubG/vq/wDch+qPKtLgEDFKnqH+divj9vHPQqZqtgkjp6lipY2J5HBtf8e88ySJmPQFX4B1noTHkKb0i5sQQeeP6j28DUVHVq+vUWtpvs6SQwoVfVyQL3Ia1/fvPPXqDy6h5SmgzWOghqReRRY3sLEe7da4jpAJLDi6p8XTLpd7oxv/AF4uT7159b4ivSdzVTS7Yp5a2rYPLIXZTfi5/rb8e99er0VzsLf2MoZKXMVtRCsfkVwnkIawN/p731vjnoCs98m8JJVBcfZwhWMspYqQOLt/xr3rrVT0+Y3vvb9dJSPKQsujlgX0lvpYj3vFeveXRg9q9l4jdLwUUFVSgqyqwZgSBbn6/X3qnp1sdLPdFZtOvpk23JFBUV0ylw8aqw+nHBHv2fLrX29A9k+usHUxPQvToFlW3EYv9LW597+fWiOkRj/jxg4axZFpm8TNdrxqBa97A2/x/Hv1adbp59KLOfGrB05h3PRQKY6cLrREQi6jkaf9796r17pzanwO3MDUZSZKeM4uFpAjogZSF+gA/P8Aj731odEU7L37UbqYZKjlRIWmaJFUmxQPpuR70aHrY9esezKCtzOUx2PpwZZp5Kc2Q6jy41WP9P6e/Dr2OJ62avhp13UbR2XT1tVE8ctVByXAFyyjjj/D3b16rXPR1/eut9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/9Tf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691hmjMiaQfzyP6/7H3dCFNT03IpYUHUJoZhIq6boQOQL2P5v78xUmo6WW5VIaMcjri0Mys6vElTDJcNHIA6H8L6WuPekANQ56QMZEk1IKjquT5S/y1ekvknJlM3kMVSYvcctJUeF6WAJ/lDjVdyljyf6e9tFFSjCoPn079bfqQ0L0K8R59ak3yq6Oyfww7gh2LmBU/wAOyVU0eObxzGJqf+ydcnH09gre9rQ9yjB6nvkLmRdCxXb0NBxPSLlyVPURQV1LKdElmIVvqPryB9PYDls3RqDh1O9vuu2zxgBwfz6eKeX+LnwR6GKKGuXH9OBf3tduupMxLjpFd73tlmaSPQfaOg831u59og0zx6pDYL4hrfn62C+3xs+4t8KdE7c88to2kzZ+0dP2xs2+bxz1viquULEGB/6XAsR7udh3Rh8HDq6878ttlpf5jpRxVtSYJJ2p6gRpIVBFPJzY2/APug2LcitSv8uqnnnlgf6Nj8usTZU0kkLyxzhWN7tC403+nJHvf7i3PyT/AA9ePPPLQys1fzHQe7/7Wwu0qSasrJVjaSI+C5AJf+lm92XYNxZx+nUD5dLoOeuU5ImV5u4jAqOPQPdfbk3T2RnTkQskeLV/S9iF8d+CCP6+xFbi1skpc4PDqI993HeLq8Mm1gshOPs6MvEKimdaVrlVOlmN/wDb+yHc1tZyTAa16k3kdbuRFO4Ch869Z6hkpYpmZrgobliP6WNl9koXQNHp0NLwItyVTh0W3cuRjos6k1Kx1+UMQv8Ar6uD70Tmp6Tk0NOhbpN1w19DQokhLx2E1+SOLH3anp1oBielOterx3iAeIgA/wCIPvwOOtGlajqE1NBWE2HrJFgLcfm1z71Wmet58umDcFfXbeopWji1B4jay8fQ2v7N9oKCejefQN50mu7aw8a2BoB3HoxPxe3zQ4e2azMYjCOGLqnOleTf3NGy26pEHYUWg6xH3a8Teb9hWpBNR8+rN9hdsbb3/P8Ad4us8lLQP46ks6gIUb1AAn3fcvCkJEfTSwQ28fbxHR99rbmpMjS4alw7RSfvRCodWXVo4uWC/wCF/aCOoTT00O4VPRhc7j9nZegTE5ZYZmqKYKS5DhXdbcAj25XqvRT8j8av4FnWr9pM1QuRk8ixx6yqBjf6LwPe6169kZPUfM7Z7IwT+CWgkKR2P6X4UCw/HvVQetmvn0VH5k9NV/cPx53NiM1SpZ6asYrIhJJWA34PvYxgdePz61M/gH8kI/ij8m9y9L5qseGhly0sNCryIkYilqSuhVf6f7D3vPXutxKLeuE3LtbC5vC1MUs9VHRvIqSI5CyoGIOn/e/evt699nS6jE9dX416aVh+xH5PULarAm4/p/X37r2TkdLV8oMfLHTvMfNKygKXHLHi3PvXW6jy65PV0dFVTVWanigiWFpB+6q+kDV79x62adFF7U+U1HtqvmxO0PLkclraOGKNPKhJGkcre/PvdB1UnPRRNq575Odgb2q8o+EqoMb5maK0MyqBcsDa1ve+q/Pq2TqXG9kZbbdHi8lROK1DAZtaMp4IDfX/AHn3o06tw49Gzy+1KOk22hyAVaqOACb/AFSuEtcX/wCI9668OiPdlZhMQkSQjyRFiq/1vqtcn3sdbNadEC7dztHkNxPQT6Iqk0ksqxCwFjHe/HvfVa+XQP8AxG7JxA3zunBS1ojqIauWFEMqLcq9uR791vy6NP3n2Jh6Ohkx81Qhmii1sNYFl/wA/r7917yz1U92v27j6dqmox8uuqWQoioVY/Sw5B9+610A2S3lV56ChyNfIYaens87OQlx9T9ffvLrWa16Bz5H/LfHrsrbPXuOyBnpKnL09C8XmiZEWWQRsbA+/cOt1r1srfyzsbsXD/HyPHQVdOtRmsZHUsVmi1LLIgZgQf6+9EdbFRgdGg2+s9NuuWlkJlxsMwGp+QUv+T/re94p17z6FTNbdxudralKIR/aNQOJCDYBvHZgAfeut8ePQA9Q7Axm2N2ZeeigikqZK2Vg7KdQuxNgfeyetDpGfJnB1mXNRJ9vr8K6qhfGW9A+v++Pv2adernqoHuPrHA5XD1lXDRRyVKmRWRo2NjaxsLfn34daIFK9Fd6j3BVdKbwo8LiKZUizc6mcqqoYTI9rj+nv3Ws162EuicvQ4THUG5sjWqZsk0Cwh504leMWUXP9T79nqwI6OJvrdlBP17WDNEN5qaYRRuQ6lXjOlrH/D6+9UFevdU/9b7X23T753Zl2ljpZp6mR6LSUS3rJUf8i97698ujR4XsjdO06+klqIJJcXLNHHFMfX6f0g3/AN59++3r32dHLh7i2zHhaVsnHTyV00SFEkMdyXF/0N71Q9e8+hRwsOzd14qCor6SgpfuE/bAEYJJF+Peutmo6Zq3rzZcTuJKiGKA3KqJkA+tgQp/3n3senVTUdIPKdY7ZyMvhoatCpa361Fhe3AH19+qa9b4DpL5r497N0U9RmK1NIYMELhh/Xge9mvWq1x0qcbjut+uKH+J0VDjK5ok0kypH9V513P596rXrdKDoH+w/kVRT009DipNM0l1gp6Yho0uLaDo+g/w9+4dewOirZPDVuYw2f3DvutkoMY9FPJSsZApIKlgAre9+XWj6dEt6zwm/wDtTPVmwOuMZUVmHqcmYJcoaaYSLDLJ47rIg/oeT78c568MdbD/AMD/AOWFsrowwdgbxpkyu7Kpvu40qY2cwyyESXbzc2B/2/vXXvKnVyKIiIsaIqRooRUVQFVFFgqqOALfQe99a49c/fut9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691xb9Lf8FP8AvXv3Wj1qUfzA1Ev8zf40leSm8KC//ncv0Hvfl17hjrbRpP8AgLTf9Q8P/Wse9db6ke/de697917r3v3Xuve/de697917r3v3Xuve/de66P8Ah791o/LpJ70xcmVwFZBF6nWJ3A/JAXnge94oa9e4cOqjOy6WjoctkYZAWqI3Pof6Xv8Ai/uterDhTpLdc76fBZcpVzMtO76BGGAUL/re/deJFejj42rw2Xjhr4Ujldow1yQWuR+ffvOnWvmen2r8FDjKmvhQLNApkVRwCwF7e9db6rP7e3HVZ7PyVVQXidZnUKv0AV/8Pdh69eNOHQP7jqqTIxw/ccJEgUkgcEC17/09+61joIMrSvvGVcXpP8OoSAzkMbov1JPv3W8dKjbG1cPj54qfB06vVMyKzKhve+knj37rWOrCOmdszYyh82QgCpImollN1v8AX9X+396Oc9bI6HKDDrJPPNTX8ZHp/wBtwbD2+hFBXp6Yj6bSOPWFNtUGRkCVw1SXtZxcEn/D3SUKTVekNghViW6bst0rsbOTwT5XF0U08NvAJIg2u39QR7SgMHHSXdLBLu6QjgOPy6VtN13j6SjipIaGGioICCggXQCFH+pHta6oUr59GP0sVsgMQz1jfZtFVVUYoYYzbgkLa9vqD7TnAp0lkuLs9vl08rtLHVaGiZEWYKVYkfQ/0B9pLqwi3SM2dwaI+D16HxY219dU/WFJTB7SMrWLEfWwI+lveNXuL90nkvmQtuEdPFavlxr0KrDdHiALHr1PsQ+XxQsdJPLc354v7hjafuabNazmaRRgimDwr0IRzDooQ3l17P7Fx+26E5Cpb95l1oGudTHkcn6+8j7T2n5I9udgO6XCqHjWo/Z0Hbi6mvrrUpJqeggWjGUn8lTCogDkKSAQADxY+8Afef7zlzcXEvLuxDQo1LVSPLHUhbLy+twolm6V9PRwUUINOikxgW0i3+x/1x7wY3/cN33C5a/vp3JfOT0L02rbbcAscjrLLIsijWxWT9RBvqJtcf7f2SNuNxNGICoI9en1n/Cg7R59YzUyQKQrHn/VG5A/ofZptu1yO3ilcdUZ2bHTDU11OurzlQrX1ccc8fn2JIvqZ7hbXalJlwBQdMtIioWuMKOgwqtk0uUy75OkA/UGICfU3vcn/iPfSj7tG271a+Cu7hgQRWtR1GHM0lrMT9NnpPbmzNZiK6OgmDRwemMAA2sBYC3+PvpnHHafu1ApqdPUeqGEuo9KDZGbrsHnsZk6fWlN5FeVlFrjVySfZDHH4bGnA9KpWDKKHPVuGxNyQblwVLVxOGZYow3IJtp+pI9u8c9Jx6dLX37rfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737rXlnr3v3W+ujf8fX3v7etGvl0GvZe1v7yYWSNIhLIBbQRfi3B964jrXn1VB2HtyfbmbqqSqRYI18hXVdLjV/j7qc9ODHRdq7LUeP3FAItLKSLkgG1/pz/vXtXbpAyEzcetUPQ14R46oRVFSwjgchhY2K8f09p4FiS7DVwD1UV1cMdOsu0nrcrFkaKITUilW1sCfoeSbe4t95od43PaZ4dkXUfDb5+XR9ZTQxFNZ8x0M1PIq0kESKFaOPSQp+hA4uPfBnnbad72vmm7bdUIPiGuD69TXYXcb2qeHnHXaUyTMGZyHJ+n0HPPtJaXsNyPBiUFul6tK32dOAhEY4Pq+l/x/rce0klpcrdGOWoSletqZC9D1jmBZrX/AMfr9fYV3GEQ3Q8Mnj5dK6so7umnMZEU1N9vT3M0g02A/Nvpcfj3NftPyFLzXucUcikqzUNQegzut+sAJY46aMT17XVkJzNRGwqC4ZeDfST+R77Rezvt5acj8upDbAAlVPUR7/fpeynQa9Lf+5VZVUhNQWARb8n6D+oBPuYEDlixHQdGMdPmLw6YmlCo+oiwbUPwOL+364631LkfGSwtHKw1/kFr8/U8H36o63TpEVFLO1RdQVgUgBrWAW/+J97Hz61Ty6Ts2CgkyT1SMxk02W/01D83/wCK+99br5dBh2phahdnZLIVWpZIBKIhY2YLGWUgD34HPXuqJOzuxMxmMrXYSplqYKelZkjY3HGojgH3sde6LjlNw7rwjEYCmbIPdnYyIDZf8W9+49a8+om2+65ocrDjtyM9BUyuF0IuldRNgAWtb37j1oV6M3gN/wBZgKhMnQZGo8cxBQrKP02vpKg8e/D/AAdbJFKHo0HX3dayZCHJ5SokldUC/usCbD6k3968+t/b0MH+m7EVuTilM2mBZBrOoWsTe597wOHXj5dCG/eG3XiEEFVGSVVAfInF/rY391oK562OnNu2Kek27U/5XHJAVeRlaZSP03Nhf3YjHWhnqvjt3uuqylXNiMHNIYqwiKpRWBRQTySPp7916tD0DuCP3UseMmaRneUFUAYjU5v9B/r+9Ede6tc+FHxsrtzbvxOYydGf4eBDJeSKQLpWQMLEji/vY4060aU1dbGGDw1HgMZTYuhQR09LGFWwtcgWLH37r3Tqoso/2P8AvJv72ePWl4Z65e9db697917r3v3Xuve/de697917r3v3Xuve/de697917r//1d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3UaaEkrJG/iZWBdv9Ug+qn24r0GlsjpsrQ6l49VcfzBP5em2/mVSU+biqoaXcWFppI6apjZUlVo4rKAbXJuB+faSe28YaT59K7fcJbUa4jw61q97/AMtn5F9V1eXw+H21mt100SulJURmaQG7EAqVB+gt7JJtjU5UU6PLbnm9tyFBrTomee6y+R3W9TLjM7sDLYmQyOgqJ0qBYXt6tS/j21HaGDsA6Mr3mqS9s/EZ806FXpbquKbN0+d7WmhljB8poaxr3u19NpP8PZ1bKiUqOo8e5nnnLavPq1XY+yehalE/hdLjAkwVZKaMxHx+mxv7Nqx6KU6PkMn0/Hh0Yuj6k6Eq8GcdHQ4v711DaQILgn6njn2l7a4HRehkIycfb0GW6Pih1/uHB5GbF09NFLCr+Ioy6iVW9xb3vtHl14mahAbrXK+YPWE+C3am3ZYW+2hqnQMSdJ/eta596kmjhhbHEdK9ttZrm8jGrz6FPqbbUG39o0+OowI53ijcOLBrWBIv7iDf5HadiDivWW/Ju2RPaIJBqNB0I9SyRRpHoEk621kXuWUWJJH59h62uCho5x0PUhWzJ0rpp0h8quTqpGgSF1Qiy3B+lvoD7VFg7F/LpliZXMnr0mU66bIVPnqvSf1an/At9B71jh69UAFO4dSW2LLQ+YUktx+FU3ufp9feh8+tGnAGnUWmqavEN4KgMy6gChP0F/8AH3unl1bt4dLKgzdAijWUV2tpva9r3PPvdKYOetFRw6m5zK4lMbLNkI1lhMRIL6Qq/wCPtdtqg3IofPoi5qA/q/c1/gPRmfiJtTZfZ8lRhfuqcGSIosOpQVZrhbj3Om3gjbhX06wU2k/7tZj/AEm6M5gvhxuzYmSzFVhM/LT4usqZZft45W0MtyxFhx7K1Jacg+vRrMO4n59KfovvMbD7DzO0dzZFESkZkhkmOoKVOm41/wCt7VuB5dVU9WNUXZ+Ez1FD/DqxK+qdkKTQ6WKhjwLAnj23mmOvYrnz6NV1tuePFpRyZZPudagh2W4hH9OffvPr1fTpQbw3bhK+apbwJIvj9B0Kb/k2/p799nW+q1flT2GcVsbPr4TQ400tdGWK+j1QEC4Nv6+9j5daOOvnr9z9c5zN/KXeG/8AA1RnrEyCyY5opHRlaOoL+hU+lz739nVaDj1eN8IPnfktp1Y2P2hOtM1BT08cclZ5ApEUejVeTj36nWx1d91b8mdk7xoJK2i3NQBwzGPRIhIjB4uL8D37zp1vByOkr3r809mdV0UeQatp8lVQIJSqqrMGH04U8+/fZ1qvr0BexO7uwPl/URf3Qnq6CiMyo5iV4h4HPJLD8W9+4db49WwfHr4cbar4qbJbgghy2Yp0UTSuVaQzow+ob83+vuv2dbp1ZBt/pvae16bRHiIoX0WbUqfXTa3A97z59V8uudDhaPEZCWWliEa6H06QvFhwAB71XrfQL7wz8iSV9NO3l1u3iQi9734PvXXieq/u2Nx43DLWVOQKyBUZo6c2bxvf6gH6Ae7jAz14g9Vd7o31RZjfNVU3SUCnqYkfSfQWBUL9f9h795de869VKb17fyfxn+Q9FuKuqWpsHnsqJHL6zFpll1er6C1vfutU6E/t75sbc3nlajIUmepUpZaVI7Bhp1FfUCb/AI9+Hy69k9FWx/cW0s3lXnqM/RVEess0WsAlr/Qi/wBT791odM3ePc9RuXbybW63oFNc8PgMtIGuzEWJLIPfuvcOPRRI/hZ31ufb9Nu7I0+Rnmp6k5KFA8pCtq8kdh/h+PfurV9OrDvh980e0+gtwYvrzsGOrxOMpzBSCuq1mMKJ5AvqZ7D6f4+/da869baPSXb2weydq47J4PcVFlsnVxIZYYGjdy5iBt6T70a+fVh8+huqamvx1I0gElMSpJuLBo/qffvs691G2SaXN52GroWERp2/ynQR+41/USf6+/GvA9er59SewsJHWTZQSU3mWohZOfyCLAAj3utD1rFK9V97o6jglevpzQWWQzuik/TVc2Jt9ffsHHXuqo/kJ0znduZeXc9DTzImMfyKVLDSq+r6j37PWiOkl0t8vtw57de39i11fJTx4zKQKEYNY+FwLBvz9PfuvfLq/wCzPY+Jyew6BshVo2rFLGEP6ncxWF7n8+/Y8ut1J/LqlnvztHOdS9h4OtaWTHYLM1aFZ5EPiaN5ONJB+nv2OtefR8+v/lZsPdW2aLBTxU+QqKSKGT71Qpu4QEkn/X9+698x0K2MXF9l1cGcotwRUbY8qsWISQB5Qh4IS/5P+Hv3XuHQk5DfG+aBsfiaHE10VNTOi/dBG0lQLFgR/t/fsdbyB1Oz2Z3PLS01VU7iekBePWjtbgkek39+4cOvcTnrvN7nyGNXHVtJu1YhHCplRZBaQ6frb37HXs9MU3cG4YaepnrqiTI0+g+FgGZR+AR79TrdOgf3F23l87iJqKRJaGnaSRTVFSihX4vqPHv3WqDy6bdh1W0du4fKZ3K5yhz+SQs1NQNIHlv+AF5/P9PeuvYPQhdcdI9s/NLcNDhKWir9o7Nx1UDNUXeGmradTqKXsAQR79Xy618+tgP46/DnrLoXC0EWMxNLNmYIIVqawIja6iMetywHNz/j739vWjno4BU3TSdKqeQOARawFh735dePEU65fm9/9h715de8+u/fut9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddG/wCOPfutGvl1xe/ja3J0G1he5t/T37r3AZ61Kf5nb/3G/mY/EVshaE7h3jjlhJt6i9ahAHvfXvQ9baFGb0lKf600B4+nMQ96631J9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XFgJEZfqrKyn/WIsfe+Bz1riMdVZfMLaUe0chTZelAKZJ9bNc2vfm4H0P1968+reXREp6OpnFNkRMYI2lHOogWB/4n36vWs8ej19LZSlhxEKSyLVMI0Gm4J4/p7r8+rHj8uh7qpqPK0k9IyCnSVSDf8AoVtz791rHl0RPtLpytmyFRV0BaWO8jixuNV72At72D17ouNX11mmElLJRyk3IswI+h5IHvdRXr2elNgOk8qRGIaSSH7k6ZyPpY/6r3qtMdeOR0ZjZfx6x2ISGrn0PUHQ9iwB1e9V8+vY6MpiNt02Pp0pGQEFdP0BJ/1j79TrRIHTjT4w0tSyKlofr/hzzYH8e91PW/keuVbSLTsJVhH6gWNrEH63Fveq169w4dTqdfvo0mEYBp7njkm319+PqevZPUwPLVoYGHjUen8/Qf4H3vUfXr1STnqGy/wokxjVewP0B9X0+n09+rip619vUWVJQVq4XtJKbtbgrf6+9CgyOtjA4dS46+qUWke5PHN/rb6c+3BLL5nHVTXy6fcM01TIug3JYfjkG/vxkbyPVqkiles3be3a3IbfganV2eJLEA2NrD8+4+92tol3blGaKHJ00/l0YbdOI51r0WzFXpqdqKsUxyLJcsfrf+nPvgN7ncp7psvNM07RkAu3r69Tbsd0rRjOKDpyFRZwVI0Lf6/Rhb629h0Ml7GqTYYDo5uLcS56w1WRjUMxhUWA9X9T/r+19pswJ06emHZIYtI49JiryFXI3mhhYw3+o+hF+Pctck+2u97/AHapBESjED9vRNc7uluKcepNNRDPANGtjFbWPoSR+CPeePtj91uGCWPcbyPv40+fQa3PeUmhMYbJ8us2XzmO2pQCJ4rTkD8C/H5v/r+8zuXfbi02Nw6qFp0CJZdQKdAtUULb1yJrD6Y/IDc8Kq3sTf8A1vcilI7eHwic9EsuCfIdC7j9vUiUlPQQ6Xk0BCw+o5/w59lZPdUdNxqRkmvR/elseMZtxKdiusIlrH6i3I597HDHVjSo6Gf37rfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6911/vv8AX9+6159UV/zCtybg272BEmLp5Y6SZlWSSO+mxkF+B9L+/HFerA9ARQw0tbg6PKyWeqFMkj6vqGIBPJ/r73pJHVeGel7s/OS5RVolBPiBU2v/ALCxHtooeJx04GNKdG22BkqSGBsTVRqJSgCk2Iuw4tf+p9sgIA6OK6wR+3ppgxZSOFeu6rb+Wo66apHl+2Zy68krpJ+g94CfeB9jYLpZ91gjGqQlv5dSLsu8hIljrwx1nSokjIZ4yAQf63uRY++Ym48vbhy9uTp4Z7Tx/PqSLHco5UFePTqoP2v3RPpAuRwOAf8AH6+w9u+93DUjjHfw/b0s+pjVq16gioFZaOmGuQ3sF+v1+tx7lP2o9rNy55u0NxGaGh8/Xoo3Hekt1NT0qcRt6kQCqyttSDUBIeOP9e3vq77T/d4i5ZgjvClCKH+XUW7zvguSRq49KJM3jpXFDTVSIiegopA+n4A/p7yotEFtALYfhFOgiTqavUuumqIoNEDF1cWP9Ofxx7fPXh1DCSvR2As7qQT+VuLXOr37NOt46YqPb0kcrVM03kflhHqu39eR/X8W9++zrXnnrlNSS1DNGF8Sg/kcccWt79jreDw6aKuL7GymLWxPEgva/v3nTr2B0nM9i1zmNnoKoa6eZHVke2lSyWPB97BHXuqd/kl8bpo8pUZPbtGzuzBmjjP9XJbhfr7917y6I5mcDX7UUrkMY8ThmR2YG5A4axI/2HvYPVft6CvdGwcJuVocpBBHTViaWEosCDa/PvfW+kxS4PdGFnjlNRNWUED6vGCzDSD/AEHvXXgOhexu9qGeFafWtDUxqA6sdLFl5LH6c+/U691PHamHx0E1JLWRGdgwVyRfV+Pob+/U630HFd3C1PUyLDkwB6gCA1gfxzf/AHn3vzr1Wvp0rNp9y5PIE418q1ckgAEV2bhvSCTe/vRHW+l3j9vTZKresMRV5X1lzewN7W59++XXs9WYfFD4hN2HlKLM1CiZPPC+ksbKCl/UPfuvVpx62GOo+pcT1thaWkp4ESoiiCcBbLb83H59+GPt60c8ehm9+6314WP09+68KeXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//W3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcWGoW/BBB/wBY+6MpPDr3Ta0SUSMtLH6nNyXBYMSbn3dSB29ORQIxJPn1iJie2qjpnY/qvAjf617+7Gvr06LWGvl/LoLt/dD9YdjsJd2bax1U7Ly32VOH5+t2Cn/efbfhK51U6QyKAaE46Jd8gP5b/UO+dkZZNjYVMPuVaeRsdNCiAaxGQqjSB+QPfiirwHTKW6htSmvWpH3h1L8nfjD2XU4uQZZ8P906tUSLKlMsbTlA4f6WA91D1ag6MhcBE8M9CbtfP9xIKbdTbhjloFSLzxR1xJ49TqbH6+3SPXpgeo6PZ1b8kcWMSaLNV8UdYsZVzPVaQzEWI596889b8j5dVDfOfdX95N0GqwcRrn8zMn2i+Xny34K/X+ntHeRNIlF6X7fexWc4kYgdB31lu+tpqCCly9PUUtb44lQ1CtFZbf0b2B902osC1MdZDckc3QPpi1Dy8+h1jr8fOhnSSPyC5chrBrc839gm4sjC3DqaHuBeQho8168+ZokTyN472P1sfp9R/X35a6QD0mWoTT59Q3zUFRGxEsS2/GvTwB/h732jh1uh4npPTZ8Usy3KuHPFuf8AAe91PXgoIqOoNU1JkqnySaQdN/6X5/F/eqefXq/s6DfdCS006mi1KgNhpJsbf63vwzjrZFc9MGemzOTwLRNI/gjiPmt9QlrW49mG3gicDy6IOagBsVzTjoP+Dr3wm7QzO0++2xlDWSGlWWEmIzOTcN6rIPc6baD+7ARwp1gptNf3tPX+I/4etobZPZZ3FLi8fUJaOpQ+curAm/pJ59lqqDMejSU0ckdFz+UXxHr87T5beXXpkp8sKZ6mWSBSGkb9dgQeT7UNTV1oZFeq/vjZ8idyde9nt1bvaappszDVKiS5CaSOMxxPa/7nvXHr3V6+1e9sNW4+KiXIUs9TJdB4akFgQP6A+/eXXjTj05yb7qcpUR0NAZiYZFaeRixXx3ve/wDT3qh49WFK06qG/mud37mpNizbC2Lqrs9kmFOsWPJkqQ06iM3SPni59+Ap1pj5dFJ+D38s3de7oNrb47Fx08tRmIxVVa1NCNal21+suL/n3vrVK9Wm7w/k9dVbryElTRYV4Mi9LGvngpEj1SFOfUtvz70DnrxHRecl/KR7e2FVS0fXmRrKRJvI0IcuUCWJW5vb/X9+r69eI/b0Q/s3+WJ8q8rW59dzZauq4IlcQqqO6Dn0hBcj37iet08ugx+P3yG3t8DewaPYG98JkmpGrYqZsjNSyJTrGraJCzWtb/H3s9a4GnW4B8KO/Oue0dn024Nt5XHSVVbT/dT04qUkdZGNypQHg+9ZFOvVr0fpayDIGR5JYghX/VADkfgn3vj1sDoPNyzUNHHI8UkTcOGKtdrj/Ee69e8qnomO/wDMRQzS1rq9oSxuCbG4N7f19+PCnXh1Tf8AJvfdfNW5QRrP4HhmWJY2YXbVYen/AIp7t17h1Xls+trZstPJkRPGjzkAT3VgGb68+99a6BX5mfHGo7yyWzsXimUPLLANVMP3gRbkv9b+9de6ctsfygMqduY5sjJVOap4lcyjm0gBLHUffuvdDrhv5MuE25kaGolk+4p5o45p/EEdVuburEE8+/Dr3R3ev/gJ8ddnSUHlxEUuTiIE7S06PdwPrc/7H8e99a6Mbk+t9iUNE+2cJh6VacQeNAKGIX4sL8e9dW+XVdvyG+DO39+Y7IQHE+LLVXllpKilpEikQhbpZ0549+611VZt7c/fPwU3vTos+ZrdtU9WXCyzVDokWvSP6gC3v3Wj1fD8af5hWD74p6PC5arp48klNTx1CvU2cHT67g8j3r59WB8j0dyLubYOyJkelzeMpxKxao89bGl2J9X1PvdKnrwx0Le3u6tibySMU2fwk0kxVQFroXN7WIAJ96oetgjqfuDaaZJvvseaaeKRbloRrvr4BuP9f34Hr1K9E/7y6rfKYiqx0tKrtWIU4gH9pCLk8+91r1rqgPvP455rqHdeN3rg4ZQ8WTWpqhBFoKx+XUdXj/w+vv3l1Xoac18pWyO28Xg6KrMeSp46SKSNqkiUkEa0KD/W+nv2Ot9J35PblwndGC6ywjTwfd0iUyVzwsPIroBe7X4P9ffuHXjx6Z6zF4XrzbePxu0a3yZ6QxCUJVF3sw/tW/Hv3Ws/s6Ffrzdm/wDamUoMh9/IJnhR11TsIgfrYryPfiOtivRtD8pOzcZT+fMNDVY9OW8ahm0W/wBV9fp7317pz/2cnrfL4xYdwQziYONREjoNY+oFveqdbr1m/wBOPWe5oFraCoeGCBQHheqs7Bf0kK3v329a6Zsj3UMtRT4bauDylZI6GOmkpoJZlLkWBXQLW9+631M6e6T+SXbmc/u5WYnKUeCq5FaOSahkXSk7WUliBa39ffjSvXgOrtfi/wDyqcFtSNsn2xI2VlnPljpLoSqltSKykm3+xHv329a+zq33YvXmzuvMTFhtoYijxtJAFQeKKMSHQuka5FAPvf29aqDw6W4jRdVhbWSzfXkn6n36p49eoKU65+9db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6jwCQa9dtOo6OdRtf8Ar72eqqajrUf/AJ9Yel/mI/y0amnPjeq7Ax0U5X0tIqZSygkfWw/r7r8+t+nW27Qf8AaL/qEpv+tK+99b6l+/de697917r3v3Xuve/de697917r3v3Xuve/de697917ro8Di3vfE9aOBjoj3zIwn8axeHjCk+Mlj6dQIufeut9ViZ6hrKSlWjcAU0LFwotq4PI96FOPWz0OHSGTgKikjLJMAoVne44Nr88W96z17y6Mz/ABMQ1PhqqhOWHOsAX+lzb3oHr3DrFmd1YDHR2rZaaRCALXV73928sde/w9JqkzOxMhUI9qVZG9Q1aeSfpYH3rr1PIjpXUsuIFQFonpNNxbSIzYfkn/iPeuHXj/g6c8hSzoy1VNURGMBSQlvoObADj3v1I699vU7H5eKoZBLYPEAGP0JYfT34cOtcD06NkEMp8bAi/wDsVN/z/Ue/de65PkoKlhC45YheRx/jz78et56mQRTR2SCyJJbXqtfn68+/V6907yU8cdOTe76fovLH/Hj349eHSHqXbysJ/QoJALkj/Ee/efWuuElY8IRUA0NwrHn6/Vgffj6nr1OuUUqzWVpY73/qOb/63+9e99erXpXYh3o2R0a51A6iD9L/AEv70T1sdDbjZoMvQmKpVJVC2ZSA1xb8X9qJ4Ibu1MMoqrChHW1Yq4YcegI3z1zErSV9KNKlnIVVHBPP4/3j3iH7u/d/23fYnurWIFyCahR59DXad9FudDn+fQISYbJRu4aN9KNpLBLCwHvnhzZ933mDb9wJtIZKA+SdDKHmeLRk/wA+p2M2fkcylZZSqU8ZIOmwIHHuZvbX7ve57wIvr4mUA51J0UXO/pVip/n0FOGztTFv87MmgeSAtpLql0U6tJ599Dvb/wBo9i5RtUEsalgB+EeXQPv92ZyWU1/Powu6Nv0mxcR/EorDzp5HFhdbrfm3uXomgsRqtwAB5U6IDcyO9WPH+XRIt07lXdFbMkc4j0lgFL2uQ3+p9orvdpJjWMcPTpySRjQcep2ya2tgr4saKeacSOq64kZ1YMbA3A9lpuZ5G7q9NsAw6O/sXratr/BWNBKisPo8ZIB+v09qrcl6g+XVNKxnHRyNt4NcPQxQkWYKp0iwsbc6gPz7U8BQdVpXj0pfeurde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdV+fOPq+jz+0m3FFT+WtiDBisCu4KWIOr6/n348a9eFRjqrXa9E5xdXTVAZfAdARuCQot9P6e3ooyRXrxoTTz6UGwchS4jPaam0cRkYgs1hyfz79MlR14YJHDo6mHGFnaHI0c0TORGQUcGxtf6D8f6/tG8dSD5V63inQkT5SoqoYodKmMKoBAvcWte9vft/5etd/sRBMFNRTP2detJpYpQa46aqvHLLF6GUORa9hwf6+8YubPuybPuqPPHGtT6KOh1a7oYkDM1B9vSNrYcnTqYXkAgYnj8gfQ2t7xZH3THHOQZoiYQf4cdPy8zITpVs/b0qdoUFJj4JcjVMmmMMxDEcc8fX/b395n+33s3tfJqxyJGoI/ogdEW5bhNdita9AF2r3EFraiixlQojhDahHJouF4HA/HvI+LcYo7T6eMAUFOHQTkjfVqbou+P7cy02UEtFV3ZX9QWcsPT+D7JFDeKWbh06pHl0azrbugZKSHH5V0LsSoJck6voL39umla9X+zo09HJT1iK8BVlYAnSQbBufdfLr1DwPXOeiRmAgBSQ8kk2/2/wDsfex17PTRl5UxMHkqlDn6nQNVjb6cfn36vWqdB9UZ+groXCyxQsgY/usFYEfgBiPfvPq3SLpd00dS81CZo5JhIyLpYkEDji3vfE061QjPSHz1BTColkrIEnjkJ0hkD6fzY396+XWwaDoqHbXTm3t5UtS0FIkc5ilcFYlB1Ef0A97+R619nVXe9uqdw4LIVNLS08zRRyaIwFNioNuLfX3vyz1o8adJ2glGKgkostRuWtb9yH+vDfW3v3Hh177egy3FsuHNVTz4byQ1bt9L29LfSw9769noN9wdLZmYK/3LrVWBW7EXNvp791roPU6A7AaSaafI/wCTyIVRS3PN7Ae9dep1Exext77AqhWL56hQ6gkKz2AP5v8Aj3vrw49XO/y9+qs53rUznMwM9PSu6urQ6yqqQLc8e6n5dWHW0j0z07trq3b1JSYuhSKsaCPzTMiiQEJbgD6E+95HVRTy6GmxJNzx+Bb/AHs+91HWqEnPDrl711broC3A9+49e4dd+/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//X3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdY2j1EHUwsPoDx7sGp1Rl1GvXHwjnUzG9x/sP8fbZUE162AR59Y44GQtZ7Kf02/wCJB9teE4NQ3VgWrnqRpv8AUk8Wt+D/AIn28tQM8etg0FB15VC/S4H9Px7sTXj1rHRUPk38UNg/IjaOVxGWxdHFlqqkkihyIhAnDHlSHHN7/T3TSK1HHplogX1jrXC7S/lhd19P4/LUewI6zcNK9VWTwxyGeZFj1akXTzx9Pdq+vT+KY6qR7i6Z+XOxquVsntSqxVKrpeSnpKnVYm9xYf4e/daNfPqT00uC++pj2c8xrRMocVlJMbkGxHrH9fewB0XX0RlUBTQ9Dh2ztnrDP6JNvTLSzJSsY/toHDEheP0j+v8AX2mlt45VIPn0d8uT3O2Thg56Ig2T3VBn5sDgoK2tRJdGs0lSwKg6bagPYcueWluKt69T3tXudJZxCErqp0P23+gPkhuoU0mD2rLVQVRjSMtR1PHkHAvb2C7vaJoZzCBgdSLZ89bDPaLNcyaZDxFR05dwfFn5KdSbFqt67m2zNQ0NNE07tFS1BIRF1sRYe0/7ruRxHTy858ttnxf5jok+1O98Lk5YsZkamojygupimhkis6nSB6wPyD7bewuFUtTh0stuauX7uZbeKYEtilR0OeMzlPVzeSWUpF4wwK8sbjgWX/evaERXBJqvQhuHsYh2uCftHUmfL0MpLTazSqSrSNDJYAfTU30/3n37wrgfhP7Oi+TcrGIVLiv2joPO0OxsLsvauRqkkjdJ6U+CzIWJ0/0+vsTct7XLezkyY09Rn7ic5xbdtZtoaN4ooeijfCvsetPf827KxGFBNXRRIJV0oF8vBF/csW7+BB4Plw6xUs7cW9zJc+bkn9vW3BsDeeErdu4/cVFNEgpYFeYpJHdTe/tP4YWTUOlbHUSfXo720t0wZ3BY2WJ1rIMg0Uc6u4KlCPVqA4Hu1amvW/LHVJf8yvolNt7+pe1dswpi2SQiSooUIbU1ix1Jx72OqmvSt+JmFmzVNgslVbgqqiSSJZpVnmvyBc3BPv3XurH9y9i4zZ+LrqLBRR12cmoTEq6Q0iyaLA8c/wCx96p5nqwI4dF66M+L1X2p2K3ZfaVKs9FFV+eGlrI2aIfuXUhG497rT59apmvVyu2aXae06VKGhoKSjo6EWpBEEVNA+gC+6+fXsU6GXH7gxklFFV06QliyAyBgGt/Tj37Az17pRVVZRZWBKiNULRoFY6jfV9DyPe+PXq46DDdWAwr0xmalhnnlUlkY69Zb+t/ej17qqr5g/C/rfuHaeZrZdv4ym3B9jWNBUimH3CzMh0OHAve/092HWs8T1qsbL+VHb/8AKv7/AG2Xu6szNR19m8yI6Oql8op6KGaouIy0vAFvp791r5+nW3t8YPm1tv5G7QoMztvOLPU5GniKolTCVuYgTwD711box9bks1GjNWTOxc641Z9V0Jvew/HvXXukfuSCLM4WqeZEUxqbkHm9rEke/cevDB6rx7X6vx+SQVCxxzNqJa8d7jVfT/xT3bj1rquzuzbNNtBnraWnWnkjQ/txrp5C3v8AT3v5de49Ib4xQ5PsnsvDPXmSaChq/wBsNchdLjgfXj8e9daxXPV724sRSxYWbGQv4JKXHI0bBtLqyxWGkX/Hv3WyPTHVftd8g8511kK3EZKoqq+A1bQxyzPr0Iz2Wx9+HXujBbExWZ3jQRbypA8sVQv3DISTpH14A9+63SvQ6YTZ1RUpFm5qZS7MsWllt6h+ffutAdLyr68XJSwPV0ccUhQlQFte634P496zSnW8dVi/MD464/c9DlYpsDSzsYdKSPTlmBubEf197+3rRHp1UN8cOgpttd81uFatlxUNQzJEkOqEBmaygXsDa/Hv3Wh1YB25/Lu7S3sY6jF7qzVNj6smVZYamcaVPN108f4/X36vl1viK9FXzvwz+U3x8EGd2fuDcG4RTP50p5paqRZDfgAE/wBPfsdep6dHb+NXzw7J2UaTbnfGHnwlIslNQitnUqGk1aG9cg9+pXr1adXP4yj2x2vtqkz2CqIqyCvg80MgdX9BTUp9P0PvWet1HDqovvTBSJ2HX7PzmOSTG1ztSUss0bFdUj6FCkj3vr3VW3yc+JVb1Fuin3vSSyri6iP7kxDyCFda6gBfj/be/daPp0oviX8aY+/62qymRy8saUsjvCPMVEYT8qFP+8e/daxw6VXY/wARt/dd9jVWSwz1GZxIS8azNJLGmngFCbj34evW6dITc1V2NghHHNh3EMJVXl8bHRf+jD+nvfXjUdKnL9j53bO2sc2fxStTZKJQkjLfVqHGtT791qp6QqrBuCm81PQxolSxEWiCxMsn6VCgf196631YV8Jv5c+9/kFUfxHcH3OKwplVlKxzRJJEJPp9LHj36uadep1s69Cfy7en+nsdQw1eKoM3V06qXknpxIxbQARrkH9frb37r3R5cRsjaWBSNMRt7FUAiVVQ09HCjgL9LuBc+/de6VP+H49+691wVFW+kAXNz72TXrQUDrn711vrq3N/futUzXrxANv8Pp791vrv37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691qNfz8/wDt4R/LKP8ATsOg/wBj/uV9+61XI622sf8A8AKL/qEpv+tK+/db6l+/de697917r3v3Xuve/de697917r3v3Xuve/de697916leve/de6Lv3/g5snhY6iOMOYFOn0k8j6/T34jFetVz1VtuvFLGZzOn74L2jKmw9X1t718+rnIx027Nqjg/JWr+20dyFUaNX+Fvezx61THTdufsLPS1Vo2mSOVh61P6R9AePegB1o46DDMbsz7yLC088q3HqZyfqf8AH3vr1c56xVmVyUEUVdHXzRvGouiSfm30A9++XXs9LnavYeajppWeonYFbamk5H+IPvR+XXhxp0tKTvSsxNKVlqJZzqsUdgwBH9Affut48ulZtzvCnrleprnFOiEG4dVIP9LC3v1OvdCvs3fCbkr5Ps5TLF9QCwIP54t71w4dbpjPQtU0c01UmkeokckfS34Or+vv3Wvs6Eilo6iQQxiNtTAAG1yDxcm3v3XsU6c58VVUkfkddV+LEXv/AK3+HvdPXrR6SeWw8mQjZnTw2UkaAbmwtzb3rjx63T06Q1S04impQnEI0h2H1t/X37PXuke2Uo6UOKqreKSK7W1hRqHIA9+p16tOlTs7dn8WmNMj6o0biQsCWANr/wDG/fqnrxHQ0Yjc8uIqRAo1xyHmzDgt+T7oQ/FSerVFOhIrTQ5THrK8n1ALRFuAD9bf439qkZWXTMNQ+fTTK1eP59JmfDYaoo5YEWMSkH1gAsCOb/7H2nl2vYbhw1xboftHTiyzLgHoN9xVsu1cXUQ4+DW88TIWAF7n/VEf6/t+S32yGPRZQrH9mOtM8pNdVekH1ttvHCvn3RmqeL78l2R3UCQk8gXb2lj8Tg56bycHh0lO4d5I2NycEsreNCVhHBsCOAPd5KsdPl1UpXh1XHkWPlqMjDUPH4xLIArEBiCTYD26ipH5DraKR1Zn8KsDjN47YbP5WhhnkpmVA0iAs5U2BP8AsfdHRGxTp0nFOrFKejpaRQtNBHCoAFkULwOPx70qKgovWupPu3XusccZQG7FyTck/wC9Ae9k16qop1k966t1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XX19+61g8Okfvva9Hu3beQxNXGJA8EjRqwuPIENhx/X36lcde4Z6pW37sd9p7nyFCYzCj1UgiRY20kX/AK29uRyaVoOr+dfXoNK/aZqnLs7UzXvqQFTb+t/e2mJPWvXqfhtwZraVVDAks1RRh0Gti1rA83v/ALz7bY6jXrWRg9G823vakymLidXUzGJdQuLgnj8e2XExNQSB1dDoNKdK6Ef5mcy3SdlBUOLcn8+7RSzJ8RJHp1eSUsKA9M+7Zmpox4gDGEYl/wC1cCxv7UarVhqMY1+vRaLdxN4uroH8pu/JDHVFKmuKEqQ0gawAIsWB/wCI9sPU8OHRpBIIx3Z6Idv2fMLka2Sg11InWQOzXbSCeWBH9Pegg6vLKJF7RTpA7T1YuSSrqJ2NQX/zZNhqP1AB9uuoIxjpGg0inr0M+y8665VKqWQxhXuCPydQvcj6+65pTq3zPVrHXGUpKvbdPWLUa5GjSwLBgfRx9fdfl17oQIJYKimlqJJTHMhIWxsSP9Ye/Z690Gu4899hT1k2QCGnjBKs7IOLcEj36leveXVYPcve5xmZmpcTUMqa5FvG6kA67Xuvu1KdeB6TnX3eEVPX08lXVNI8mi+tweSef9v71QefWwfLo6GM3HSblpVq4irh1UspYWsebce9HrXQab2rPsRJNTt6tL+hSBx/S3vY62TTHRcqzx5eSaqr6CFVjY/50KCbi2q5+t/z735Z60Pl0Du6tnbCycjzVdVTU8g4sroPp+LD37r3HoLk642SK01VNlVVUVrJrt6h+fr70evdJTJ7IxVbkQkeRcqGAAEn4vyR79XGOtUHSX3ltCTG0Stj6p5Xp/V4xISGtz9L+99bNemPbogylDJHmaOH9gizutybHkm/vx68M9W+/wAq/sDa239x5fBhYUeeplWMKEUElgLC/vXz695U62KkYOiuvKuqsD/UMLj3vrXXdub/AOFvfutUzXrv37rfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/9Df49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691jkijlUrKiSIRYq6q6kf4hgffuvdBjvDpvrnfMbwbj21QVqyCzXhi+h/NiD73Tz61XNOiZ9lfy0ugN4qsuK2xRY6qUk6gkanUzXJ1qo4HutPn1pl1Gp6BCH+VDsGlzMVWFgko0AXxBkJ0/Qg6h79ngenkfTnowGxf5cHx62rK9XWbPoq2rdixlMUDNqve/K+7Bm9elYuFVaLxP29GnwXR/Wm2IYY8Tt+kpIoCmhBBDqDR/pvYe2Wt7d21ugJ6QmO5eSqyEV9D1w7Q6c2N21s6t2fuXDU1VjaqOWEo8MKrpdNFz+Lf4e7fTWxYEqP2dUnhuymlZCG+3j1r7/IX/hPbsXsfKT5fYGYh2nUStqiandImQlyxN0H+P493ktrORSpUdN7Z9dt14l4sxqhrTPRaesv+E+vZ+2N8S0uc7EqcjhUKyI8tWrxiMHgC4+n+t7QjaLIrqK9D+5593Sc115ApTo9/ZX8nrbO1vjPvTD4cfxvfn2dTPQ1MCq0w0wsU0Pa4sR9Afev3VZtjT0Wy82bk1XZ89aCnc+0eztm985rq3sSoqKDH7WqHP21bK2qqjFUyCOzW/AHtXFZ29kK2/nx6Ir7d7neCBc5C8Oh/6yxWNpNyQVGMpPsKRY4vUoCI0qgeoWt9fbnSXPVnvV2/d6Y3Gy40Vkz46X0iO7kFL3HI/wAB79SvXs06sM6W+VFDiYRgqvIQpNj9N4m5ZCq25uf6+/U9evVzjoce1O1uu+6usq3bOTqaH+LGOZoZHZA2oL6bX/r70K9ePCnVRW2uw929N77ZKeomk21QT6NaamiEWvnkcWt731qtD1Zb1D3psrsDO0tY5gnqamSKO4GohiLEEE+/dW+zqzXbO8FxklJi6dCtFUQKyqFCrdvobj3Xga9bwePSmirsi1c9PVSs1PVvpgB1ERqT/j/T37rwp0uafLviQmPbJoqrZiur1WYcc39++3rX2dLaj7To8ZAMQGWaWdT+4PXYn/VEe99e8uHXDD74/iGQf7vU0FM3q1qbFf688f7D3qnWzgdRN4ZrDZZ9NN4jGRaSwHKg+oEe/Dqv2dUd/wA1T4GbG+VOxavK47HQvlsdC0iNGsaS+eCEsHVlGq4I97HWyMdarHQXyf7c/lzdx4zYe6ZKyHaFHlJAk07VMqpAs+hFJvbgf7D3v7etDh1uIfGj5uYX5LJicnicvS/bS4qiVkRiuqSRB9VP5J96p6dbxXo+cSTPR1VPLN/wJLCI/S4/qOfeurdAruPERRCpFTIGjplaUkm4Cg/m9/e+qn16qV+UG5sNlMjV0MMkZVRPFcfhglv9b3vr3SK/lx4evp+zKysqg0+PjrZmjB+mhZL+/dVFK9WH/K7tZtq5sph6xIZK6BKdKVG9bORpCgD3ocOtn5dFr2d1jm9/VkdXujBVkdPUhKyKWdTZg3qD3b6j+nvefLrfHj0bzZfYg6WgqNt5HHvLjKhTDBMyakp0YBQfp9B71Tr3Rp9n75wm5MJCMXVQyrrjlAjsQGb6j3759bHp0JI3DRtGkb1KNUKulQVFww4P096+3rVM9Nm7OvaPe+HeoSMO1NGHqNISzWFyTf3vr3nnqhL5MbIr+uO16Hd+343poY8pGsjxgABFlBfheP8AG/vYp1o46ua6X3VW9lbO2lU0eWjijpMZEKuG66pHCAMGFje/vR639vQs7nxtPVRChyGnwKqhHdE0sPyBcfn3rh14Z6r8+TPxm2j2Bt6oSLGxx1SGSpjqFCpaWMakIZbc3/x9762eHTT8CcvuPYdfW7J3TmtdBSu9LjYqhnOmJW0Kq67/AEHvfWvt6MT8jetsRuLOYzL0yLroaqGrlq1C6bBg55H496zXrw4V6qx/mQ9gYd9gHGY+SOuqaWjNIsUdi3lWHQoNv8fe+tY6pJ6G+QHevxzqZs4mDyk+3KxmmVI1lKrEzavxx9Pfh8+vfPq1bqr+af1z2dANsbixMeMzWgU7zVcGl/MRpNmk/N/fuvfb0vN0b82Pldu10J+1qKyv8k9MQq6ypBZLH/ePp7959bPy6LlgY17czlFt2uov8gwcyookIVZAjfTn6j3vqvEdWY/Hj4LVncm+8RhcZj5sVhKCpoq6WrGlYXSOzuoNuQR71Xy631tbdTdV7c6j2fituYOihhagoYoKmeNED1EqqA7EqB+fex6dVOMnoT4ZvMpbQyAGw1f2h/Ue/HrY6ze9db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XutRr+fn/28I/llf1HYdDx/5FffuteY622sf/wAov8AqEpv+tK+/db6l+/de697917r3v3Xuve/de697917r3v3Xuve/de66Jtb/XHvY60cdd+9depmvTblMXTZalekqVBRwRewJFxY/X37r2fLohXY3T0tFuKrrWo5Hx7KxjuFKkE3uLcW96ofPq1fToqW/MCKR3pqOD7RdRvwRf8Aqbj3rrYqRjoCd6ZzE7Yx8D5CpiVNIM07myQ8fqZj73xz1o4weq4O/v5g3WPTFHVytNj82aaKZi8OmTmL8HTzf3vrVR0XPoL+aNsbvepq6WhSkp1iqDEEc6Ty9gBqt7917o/uI7zxkdOxp1hmWUDlGuvqH1FvyPfq9epXj0nX7vxMNe71UUZhu1g2oHUfobe/fZ1uvTrQ9pw5tjHRw+CGQ31WNrXt9feq9e6ON8V83mqrctTBNqkpI2YqbkqRa4P59+PXurUMHSpkJoDH+1JpT62+tvp79Q8OvDoXsNQmlZXmBfRf1WtcX59760elbNQw5GIunCqBqUgX45P0/wCKe7eXVekRkIYrSQBAoCtyAD9B/j/vfuvVwOih9q71p9r1JhQAa2AkYcaVJ+tx/j7r59eHr0Sbdu+6vK5ZVo6saGk9QRmvoLcg+9gUx14keXS5wu+ajbX2wppSzyKmsgkkEm/J9++zr3yPRsds7rTLYqGtlnQTMoYg8G4FyP8AkXuvl14j06XeD32JZPtnmGhTbT6je3+v/wAU97rTr3+TpX0u9MdDKRIYweeP97NvfsHrwB8um6tyePyMrySyLJEfogtwPwLe9UANevGo48ekJkc5TQ1DUcUiwxENciwUEfm3vfXvt6Kr3NmYpr0lKyymS2rT9DccEge9j068eHRcM/jgmAh8RCzOr6lBsTcci3u3HHWvn1a78E8c9J1S2pWheSZRci5IBJ1f8R7qOvHo8aggAM2o/k2Av/sB731odd+/db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuuDrrUrci/5HB97BoetEV65KNIAH4FvfiamvXgKCnXf+B/P+8/4e9db6LJ3D0jTbzqPv6KAJPZnLRgXD2+gAH+29+IrnrwNMdEE7Q69zG0oDTR0k1TIABqtc/wCB49662DnoCHiFLjJP4mtqj12iewYG9gNJ9+48Ot+Wek5gc5lqE1Bh1xxa28Q55UG/A92yetdDBtXsyukqKamyDkxRSAamBsPoBz7r14Zz0KW7d10lbRLJSSrKdHIS/wDTnj3UKOtgdATufdlMMPU0caKlTIAF451afdwPPrR6Lak05eWObWzzsY7/AFA1N9fpf8+/dex0V3f82Z2/vOKjp5pJUnbX4xewVuRb/evfuvDJ6GDbOXngipUqf25ahARrB4JIvb3vgc9e4ivVm3S2Umo9uwS1U4aBQrLc+kgpf6e6nrfy6Eus7FpIBJoISJFk1PyRdefejnrWOiB/Kb5DTLj5MVgKr950Ct4w2oEixY2974Y68K8eqqdw7yn+wkrctOHrJHflmYsNRvzf37z690qOr8tBmB9xNMl0YeOxI9IPF/r738utdWM9Vb1FPTrTj1ono1Akj6fm/vRr1sH16W244p84zTwyFYkiaRlU8Wt/T34fPrx446rn+QHctVtCpbEY93jsTHM9nHOmzXI/4r73177Oq494d7bopq2Gf7mR4ZJbMdUlguq4P19+60T0tKXvqjmwUbjIRw1jBVKGQ69RPLf7H36nXq46VtBuzNZLFfxChyhdyhcFGLc2+vHvfn17PTHRb33XU1LU9dUyzhjpP6vofp799nXuhi2YZK2oFFODaZdTFr21EfXn3ojrwOOrAPgXs6vTuCiqKCFzTxVrfcMjenmUGzAH3qvXvLHW1TTrpggW1tMMa2/pZALe99e6ze/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r/0d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xj/h7914/LrFEWOoP+oNb6WFvxb3sjHVVOadczYkqf6A/X37hkdb44PXL3rrfXDWgOm4BP4/qT73x60WFaHqP43eRhKt1A9JHH5vz7YbxQ1RkdWjkdTTrygya4XSyG/wBOLC/Fre9qJhx60HLMevGHxqqxABR+rkkkf7H35jMcdWdi4oeuv1cJGv0I1lSW/obH6+9UmpnprtOB1lSJVUxsAwZSGDepWBFmBDcc/n3ZfErRuHWwqjHWrB/OC/lAQdvZnI9zdZYyJdwVLLV5L7KgUytpmMrqojHP197RWUkt1ulMLjrWI3f0H3B1hJLR1u1dwFMfVOnkTEzLrFNwbEL/AIe79ePSx667oymLylBis3t3O0dPGviqHqaGSFb/AEJ1MB7317pYUW8tsUu9cxlTXyQx1YPjiM4QqWH0KX9669XoQ8b2Fg6muEdHlpRVhdaqazSpX6foB9763Xpj3z29tODAZTEVDrJWTq0ZeNg0gcqf0nk3v711roqHRncW/dr9lxHC/fPhaWv87AtISY9erhf9b37r3A9bJ/Q/ypwO92w1FkahKTIxUUMbmplEfrX631W9+x59Wr59HLy/aVG9Tjlo6+gZYnXUy1K6Tb83v71SvHrdafD0G+Z7lx9buOfHxV4NWqAa0n/b1f0HvdKDrXH5dFS7++bmC+P8UldurIxxmOOR4m+6CNpRdQHJ9++XXiT5dEh6X/nV7d7B3vV7doa+9LNO8Ku1SRrAfTdWJt/tveqDr1fPq2vYnyTxu6Ei+0rVeSphjcj7gOf3VuR/sPeyB1oE8B0KDb2oGpKhKxo5IqhJTKsr61N1I5B96A9Ot18utQ3+e1sjbtDjoNy4eOlpqrITBVlpUSOYM84uVkXn8+99VpTqy7+Rn0c1Z0Tg9zCvqKivaOgk/enLyWa1hYn348OtjrYS3S+awNXQU06sY47KCL/QD+0feqdW1efl0TDtHsbc712fxePgmANJJHGygm7WNyLe/YGOqn16pl7qx+98fTV+4cqlSinIMVWUMCVZvxf/AA92HWqdWAfy/avFYra9duDJUzI6QyTPMygWuLk6m+vvR+fW1NT0w/LLM43O7w2DmMRUhzVbko45k8uq6/cKDrW/09+p17q5vKY+ipNhbNajpaUTy7coZHeGJFYsYASTp/4n3qmet+R6KxvePC5Cgq6fcklPChUKrkojKL8WY+7E9az5dEb2F2XvbE9tVWyNiLVVmKkleOCaMvNDrZtMfC3AFves9er1Zh111l3Sc5jslu+QmgrNEvjGoBQzA/71+PeureeOrAH23UYHATGkkiZKuJVmBNyqsvqIHv3Vc+fVMvzFwuPyK0+JoI0nyUuUOtV/cmGp7GyfX3vreK9Gx+OGzYOu9i4pq3zLU1lAjqJSVsxUGwB+nvXXgPLpebx3aK8xR+RIo6Y3J1BRwf7Tf7D37j14jy6BDdnYmIalmoZchRFhG6lWqEvcL9LX+vvfDrfEdEF3jveLbG4VzuIrYlankV28FQFY+rUfp79muOteXS8m+VGT31g49u4WnqZ8tMn20kyK8xB024ZR78ePWvLqLtr4fydl0b5vsLyvBNN9z46lC31Nxw/9PeuvU9Ol/wBk/GjqbF7IbCLRYsxxUoijP28DMBo03Itce9jrfy61yvlf8VaXZ2UXdGxJZKWanrZakpQnx69EmoLZPx78c8OtdGe+A/Sna/yX3/t7AxY/MGnpqeCF55YJXpWbWI2ZpLW/2/v3WutqT4+fynKDZublyG+hSzRPJrHhij1gEaiD/wAb97+3rdT5dW+dddN7K6voo6Xa+NhhkRET7ho1WRgo0m5X+o/x9+60anoV/wDX96631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Wo3/AD8gT/MI/llcm3+kShvY82/inv3Wjx622aD/AIAUX/UJTf8AWlffut9S/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xub/4e/dez025TF0uWpXpapFdG4uQCV/rb37rWOq4Pk9teLECVcGU8+t+LANe1jb/invRpWnVhUZ6rOz3Vub37SVeMzszJj6hTHOVOlwn+0n37HXui11/8sr447llqKXc5qqqOo8hlMwEwLyXZwdf+PvxJHXqDrhsf+WF8Qeqa56jCUjwTzPr/AGYli9V9Q/T79Wox16nr0M9X0Z1hgft6XGJK1LOQigEsR+Ln/H/H3sUPHrRqMdMe6+iuuYqVGjSdpSQ91F2/231/2Hv3W+uGD6cpaujMG3qWq8oKhSYiWtf6k+/cB17jwHVl/wAa+rjtfEpVV1PpqynqdowGvp5PIHvRNePXjXo1eMlnpa0SRekA6eTYAA+n/Y/09686de6GXb9VLURu1TcAFit+CT+fr7sD1o1PHp9p8jDFUsgkUxgElQ30/rx73wx1ognI6S25K+Bi7UrJrMcp40i3p+h96Oetj5jqqL5FZ6qmyNfS0zHzsxUWYkBiSOPeh1v7Ois4KmrKCOWtyzuZTrZNVwb24t/xHu1PXrR9B1OwWTzNbkSagkRLJ+3difRf03B96r1unRptp5qthSKneWyj0hdf54FrD3Xrfl0JD5n+GAVLzqjML2Mmk/15B9+8+tUrw6ZKrfZ1C0r62PpJc2bn+vv3W6efn1En7PqsX4/NOGWQgWEtv9jf6+/U9OvUpnpObo7BqarHmooJD5SCOGNyf9f3senXj69BDR5evydUXyYeQmxAJLEf0v73TqvQi7S2BlN95mloaemqPt1nCkmJioUn6/091rTqxz1cl1BsWLYO0KPDxqI28cbuoTTZtP5Huw6qeOOhV9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XBGLF7iwVio/xt72etAmpr12wLCwJB/qPqPfh145HWMy6ZViIYll1Brccfg+/eVetA0wekpm9i7e3A0jZGjjlZ7c+NOCPzz711YY4dVt9/wDQtdicxPm6GnY4j9xlSKEkWvYHSvvXA9bGR0TbJ081E5jWIpGpAcMukixtyP8Ae/ewevdOjU9M2GkqYFK1CxMQ36TqAvcf4+9efW6U4dJnbu6ZIKadcg7u6tItmJPA+hA9+p59aqa9BJvHcNTPlRJSzKqB76S1geb8+99e6Deq3ZX09TVyNJFqjTVFYlgXUXUcf4/7H37gM9ePpToE6SbcW5t+R5jJxk0cJA9YIQKGuOT/AFHvWOPXs9DoaWozmSpExSFkp2VXEIuBpbk+n3s+nXurJ9lGixPX9JFWI4qEiTX/AGWA0j629+48evefRTO7u46XbVJVY7HykVMplC6ZeVuv+H096HCp62STgdVx1+758pUVWQzVWkik60DzliP7QBv/AL179x69gdABmslQ7rzz0KMyUyFrAEBfrxYe98Oq9K3BRvhKiFKBnWL0hzqKrYMLg397+3rWR0ePZ++Mbt7Cwz1Mo8zxm/7gvyP9V711bozmys1/evbc9VjJo3ZoZAo1gsbrwPejx68PTqr/AOSG0q2TIZV6yOXzmd9DBDzxe9z731romEO29u1FG9Pmk8k4UgWFzq+gP+v791roNqnqvGVtZIuONV5ELyogDcBeVsv/ABT37r3l04bKym5sPuWl2pHBUiCd/FeVDptq0j6+/deHp1YVtvpTMVsCV0lDI5KI5IgOm5XUbH36vVqevQtbT6jzhrop0xlXpV44yUp251GxPA9++Z61Tq8X4C9ExbX+6zeUopI5p2kmiMtOUa5IYHUfpz79xPWz1bILfQfiw9+6rXy679+631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/0t/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcSqkhiASPofe6+XWiATXrl711vr3v3Xuve/de66Kg2uPobj/X97BI60VB67+vvXW+o9TS01ZC1PVwRVMDizxTIskbD+hVuPfuvdARvr4x9L7/gkjzex8G0shYmeOiiViX/VqUAXv7917z6I523/ACmOgN/Uk7YjC4/E1pR/G8VP4zr0ELyv+Pv3XutSv52fyl/kD052DXZ/rzDVWT2wsrvK6iqeNYPKblQPpYe/eXXjx6rdyOzMnsbNx1OayVTQ5WJRTT0IhqQFkAs4uR79mvWj8uhM2j1vtvPV9PX5vNOY6lvK6ygqADY8lvfut+dOh46121s/a++a1UpaapxjFVFRJp0202Ju3+Hv3XuhX37ihO6ZLrOvNNXxDX46JgpuvJHo+n097691M6x7d3dNDW4/e2YqsdWYwFYmkJDTOv0N2/r9feuvVx0Iq9u7fw3mztVuJnqlUubsuqyfj6+/U611Rt/Mu77re09MeCyEtVBTVUVM0YUsCrsIyfT78eHWx1P+N3w1zM+wNt9l4LGqlXS0611dJEkge5XyHhR9f9f3759ex1Z38au2329uwxbjyM1PTUwSn0yI+lWg9BHqvyLe/deHr0ebcnybxdOVo8VWGsp5lk1yCO+i4t9bW9+PHrdeteD+afuTcfZ+MPhEtRQ0IMqBkfQojfVcBeD791U56ua/kU9iVGJ6axNNPUyaYY6SM0xYaLxKBYi/9ffutgAcetiiq3dBvOCpqpIEEtG5EYsCWsPof6+6kZ6suR0E24MdtqKBslPSU71hDGRSFJJv+f8Ainu2fPqp4VHVTHy8zFXuOpTbWE29AIXq4wxRAAV1DU3pHvwz1408ujD9Ix4LaXWdLg6mOGjnrqHx1P09LmOxFz795/LreKfPqubsnIZip7oxGHo5pKjC4/MpU07k+jWs2oabe/da8+thDZO4qvJ9YYiuyNjLR4aGJA5H6Y4gOP8AYe9YHVvLqqX5bduvTZOn25iK94shlXFPDFEdR8jGwsqc/wC297+3rR6sZ+AXQsO29r0vYG6MVDX5esFPJ56qM60Ei6rguOPx7qT17h1ZZW5qgWpSNSI4kuFW91iH10KP6D36nXugx3tv47axOVrKirZ6OKFpFVzdFFjzf36nW69VgdZ0tT3b8h6jKyK1XtymmZlEgPgDI97/ANPe+tdWCdjVOMw60VBTulNT0KmIhdOlQBb6/wBB79Sp62OGeqyflz8hsF1RtOsq4MoPvKylkSmRGS7SXIAHN/r/AE976r8+qn9h7u+Q/bdHXbgwuPrZ6GSrk8TFZVLU7tZSLfi3v3Xs9D/138cO4d8Z2kG5Yqumoi4+5LtIkajVc3DWv79nr3VuHTnSPWHS8SZPPrQzyhUZpJjHqVtNjbX/AI/n3quet0oOkB3n8yOs9r5f+6mEytPT643URwNFpU6bAcf4+/AdeJ6INle+q/c0O4QuSqJ6dg4pmb6f4FQP+IHvfWugO2D092P8id5YzbWKx9TlaKbKxxVbPHOFSnnk9TGwta3v3XuOOtx34OfDLYvxg66xNPQ4XHjctXRwzVlcItVRC7jU0YdxcEH+nv329a448uj6+/db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de61Gf5+f8A28J/llf+JEoP/drYW9+z1rzHW23Qf8AaL/qEpv8ArSvv3W+pfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuscgbRIUF3KEL/r2497HoeHVTwqOPVcPyApq6m3E09YX+21sbNqK/4k+6nq4pinROshFVyVTx0LuErCVjK8Dkm4Fveh1bHTX/AHH3D5RGDMzMwYfqNwx/r7t9vVD0oajpHM11ItbVO0ZCgga2tb/Y+9V63TrLjOklnC/dVSlo/UAX+jL9L+/VPXhTgOoud6igpp4mnqFMQZQT+AP9cfT37y61gnoznVfXe3MdhGqqaCGoqES4HHLfW/vWetn5dDFR1tFj0jSRFpy3oKIVAU/T9PveAOtfPqa9VFFULLEwKMVPDL9SPp71TrfDpcYvMzzU7RQKBrJAZTx9PyT7t17z6CzJ77bDZmejqpjGxDAEm1z+Bf3Wp6959SYcy+Sp5aoTMy+KU31D66OBb37rx6rb7NrPut35ASj0RVBsCLlgG/Huwp1rot3Zm7/4DEsxiKU4sSQLiwP1/wBj78etjrD13v3FbqQfayoJY1CsA6Dm/wBTY+/daPDoYKbcmRoshThLmKKQFiOVIHBv79SvWxjpVbvzlZkaGnqoZ3RVZAwU+g/1AHvQx149OOLqzl4qSlUevxAEgWJbTx9PfuvAg46TWZ25lZ8h4p3lWCM/X8Efg8e91x14g9CHjdrCXFiGnPnnKqNBDG5IsPes9boAOhl6l6Ez2465TkcX4qVmFpCHHp1ckkD37jw6qDjPVknXvUOB2PGskVNBJVBVIZVJCPpsxu3197AA4deqTg9C9cXtxf6f8a97zTrWK9d+9db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XumfN4WgztBNQ5CnSeJ43Ch7elmW1wT791rqrjvPpTKRZKsqcdjfDSeZjrj16Cg+p496ODTqy5HQBSbcWKhp6BS3n4WQAOPqdPqFves9W+XQM7xo8TtCsmqMvVLSUvikYsWCjXpvYBrc+7DPVDjh0QTe/ZtfVbjkptsxPX07zaUlVSVMdyo5Uf09++fW/QdKDC4TcWXEU9dBLEHZS6kMAQR/xT6e/UqOvfb0Jj4aNadMdSxeOokKgyIPVe/wBPp7917HRxvj303T0OOrMxmbMJA8kflNix/UNNz+PevkOvYp0MdflKOijy1NEqyQRUcghUAEBlUi4P9feuvfb1UZ3BPUZfNZZpFYlamZYxY8Kbqun/AGPvfzHWjU9FAqevd0ZXIk6poqIyXKhmCEauWP8Axv3v7evHpszu0ZNrMKiC7Sg6nZQSePrc2Pv3z6903U++I6eIRznTICBewuCOLC/vfHr3DoUaPc0uRw5vUHQsZ0DUF/HJAHupHXq9Gg+NfY1ZiK00FXK5x8llUsVCKW/JueffiK9bHQod17dwu6cTW5IskUza2Q6h6gE9P+H+t731rqoTN0SYrclVCG8yB2IBuVurcWJvx7317oVtmYb7ysTICkUqqC40kq3FrMB/X3r59e6OB8d/itP3pvygqqfExU9NSTjyzpHKhusg51Ae/Vx1vh1sI7Z+Ce3cXg6SimkiMywxB7u17hACpH/G/fqdarUdC9sr4o7L21Ir1NLTVChlbSAWJKnV/a4+vvw+fWs+XRncRhMVgqdaTF0kVLEoC2jWxNhxc+99e8+nb3rrfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691/9Pf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XRNvzb8D/X/Hv3XuvAH8m/8AsLe99aAPmevAWJt9D79WvWgKHHDpP7i2zgN14+oxGex9LkKWqjMcsU8UbvpI5szgke/dbxXquLuv+Vf8buz6WsqaTaNFRZupeaX7vxU62kkH1vpH5/HvXDh1vjx6pw+Qv8hfetTIuT643UaakgAb7SmljRlC3JQqtuPxf36vWjn5dVM9nfAb5K9IVOSEm2dx7hpoIpWNTCkkiaUJUnWBa3596qDw63QgdEtwm/d8debpkpNwUNVhzC7xS01azBgwN3Uq3+H1t731rh0s2+QmxKuethy1HG9bOSFmB4LWtcHjj3vr3lnoA8tuyp3rnqrb2CxtVLHKjxiWAylR5BdORx+ffuvU6EXYfwSqd44GrbNxStVVtSlZGZbOUCyagt3v9Peqdb6un+JOydvdcbJruus1TR16VFK1GjCKOTwBo9ALEDj37HWx0r8V8denE3BNjcji4IZJ5nnFa/ijiLuxYrqt9effutEU6W2U+HWxa+aFcIIkpXBYyIyFQLf1H19++fXqA9Vx/PH4f4/Edf5P+F0BrZ/sJVDxCN7FgQQbe/Urx68eHVev8tPvWn6Y3zVda7gkWgEGQ0LT1DlWKLJawBsPfuOOtfMcOtoHaXyC2XU48xUVTT+er1FSpuSzfQi319+639nSHzW+W3Vkaiix9eIDQqZak6mBZb35B/w9+OevU6AneOZwlfX2io/uKyK6NUhA/qUcsf8AX9663Tz6QGU3vQ46EY2Vx557InJXxM3Av/Qj37r1eit7sWpO5cbWUaN5IK9J5akDiRdeuwI/qPe+q9WFZT5DwbZ6VkrJMlBStR414Wgdyru6wkXF+bn37j1auOq7PhFh9x/Lr5GT5PP0NRLhMJl2+xkkkLQtGk9wwD+/HrXW4rBiMNtPaFBszDxRxVtLRUgdY1W50RAE3A968+vdBVmwKaCZ2BWRW5JPJP5vf3qg68DQ16LV3jS5HObIysFEJSWoHUlRa/pN+f6+91r1s8K9Jr4r7cx2xusKrLyxaM3JUy3dtJl5Y/U/X37rwHQbdy9hTpLVJPIY0nZz5GLADjk8+98B16grnqhP5SZVe1+xdmbOp6wVVPFm4o6uNJmN4vOLqy3/ANf6+/DrVerzOocZ130v15iMbIlDTMcPSTSJLHHrJWIM3Fr/AOx96699vQB9y/OfZGyVqWwE9DGaNrVCDSGcj6kAW97p16vmOiCdjfKftjvugpqLrzHZC08ph81OZlQ3b+zp9+Pr16vl0htofCT5Fb73jjt0Zva+5cjDJ4nZl87KNTc8H6f7H37r3VrXWX8r3s3e+V20kdPX7exqSIMnDUgIJVuCwbULf7f36o63TrYv+NXw766+P+GozRYulqdwLDB9xXNFGxEscelje3Jvzx79x60TjPRxgAAABYD6AcW9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Woz/AD87f8OE/wAsr/xIlBf/AGGV9+60fLrbbx//AAAov+oSm/60r791vqX7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuiyfJHYcu4tqVdfj4y1XErFvGAG4Tj/b+/HrQFD1WnQY6txqGGujeKqpgfGW+oIP15/4j3Xjw6v5dLXGbg+0pfua6RWlQXAJBbj6Cx/r72OtU9OkNn+1NwV0j0eNkaKBWZfSOABx9P9b34Dr3Qc1Ha2YwDOKmuWSRyQE+rK3+Hv3XsDj0qtq79qt2zRU1bNZXkUX/AANXAJ9+4cOveVOjibZyFFsvCvVVtUk0DRs2m9uCLjge9V8utD19eigdvd9NPXFcHKI44nJsmoH0tyeOfewOt9R9kd55iuWE1tRrhDxrdiwsAQOCf8Pr7917hgHqwTZm4UzGCTJ0cwbwxF5CnFiEuPeuHDr1Oq5u6+w8uN7VphqiggaRmjDEGyN/Qe9gde48Olb1L8gKWv04avKhmCQlnvyx9JF2PvR9evZ4dLTtrZmNraFdx4qEyFlEtSYwObDVe6+9gnr2D1X12fgl7FxdXtrHQfY13jkiFQTY6r8G/wBfe89ewOiG4z46939VZmozVLuWorcc05lSjjlLkJq120qffsdaznoym1+5psPC8W7v2J0T1PUEpcgAAEtb3rrfRievuxNu9lGPB4nKUsk4kTVAjgyXbgAke99apU9Gzx2warALTV0OqcxopYIQR/rk/wCHvRqetrQYPS1pdo5jdMifaUM3kdrNoUcWPJ9+69q9Ojo9OdCUmOo4chuCn1P6GWN1Gp7c/n6f6/vdPXrVa8OjYUGNocXCtNQ00dPEAdKxoBb+tyPe+tcMdTueOf8AX4+vvXW+vaRe9hf+v+v9fe6mlOtUFa9d+9db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6b8ji6DK070tfTRVEMgswdQT9LXB9+698x0F9X0lsepZ5EoFhlcg6wiEjm5sLD3ogdeBPWtv8AzHesOxT3xTbS2/kpaPb0rGU2YqjRMeALccj3sY4dbyRnqDtLqnD7Q2pRxV1J91mXpwDXGz2kK8m55HPvwx17p921gMukxSYtPDM2hEVf03PF7e/eXy69WnRjtmbI25jHXJ7heD0oH8cpGr0+rSb+9E+Q60fU9JXt35G4nbEYwe2XRFssdoRyRfn9Pv329bpnpT9Jbhg33j69q6QNO9M9gx51Fbgc+/de8qdFF7b2nVY7P17mkkEElVMwYjUvF+b+98RXr3y6BWtrRTwrToBe2m62uD/tv9v78etdIjcFDQVVBIahlMkkbDkXKsfofeq9er5dFP3JsKqSSoroi0iByyID9R9eP+Ke91690kMTm83Q1P2bh2gBKiHSWYLex97PXh0ZvrrPPT1ketjTxWRjE5IIYckke9Hr2a9Crv3s6WrxZoKGZZGVdBVSxtdNJNh7959b8uixY7a9Rl66asqonmkbU1uBYD1WsffvLrXy6P58TeiMt2RuijwsWMqPtWkgMk2kGMjV6l/1re9H5db+3rZk6D+O+1+msPCKGhiTJNEfORHETrbm+oD6/wBB731XoyKazdmPB+iWsV/1z72acOvCpz1k96631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/1N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdY3jWTTqv6WDDni4/r72DTrRFeuRDagb+m305+v9ffuvEGtR1y96631HWFkmaRWBVvqp/B/qPe6imeq0I4dZtA1a+b2t+o2/23v1TSnW6CteuyAQQQCCLEHkEH8Ee9db446Y8ptnb+apqiiyuFxldTVMTQzR1FHTy6kcWZfWp9+x1rPDqnz5Wfyfele6Z8huHbuKpMbnKl55xFHSxKvmdDa2iwtf37PVhTz61uvkh/JO7g2Jkq+vw9FK9DC+um+2oHe6E31XX6/wC39+r1qnp0TjZPxv7l6iz9RUZTamQqYIGN5ThpzdYfSfUw9++zrXn0NeP+RD7QyMeJymLqsdN/m2+5gemXUeL3cAce/dbr0Km1+86XDVc9fFXQFsu1owalCQW+hAJ9+68MdCHi+36HceYOKylZ43iUTLLFUAPxzYsP6+9UPXq9Drtn5BU+CjOFWrebkpBLJObhfxa559748etg9P25+zdl7627ksLlQtVVyUxUIx8p1W/ob/T37h1454dUF9lfDPd9f3NV9h7Gp6qmpUlqKj/J6d4lYAl1vp+v9B791r5dSdv9z9j9aZ+Og3I2SibGzeJfJ5I0azfW59+610bXYfyGzOXyNVW00zeSvRYptU34YWv/AI+/der0tH7TraOsOOhEk1fVXkBNyCzm55/H+397631Pp9sZvcVPVZiskKSxXkQFwCLC4t7917oIN2b6mwzQ4t1WStWTT6Dd7q2kEW9+61WnQ49Z/Hrs35IpSYwx1cGAnMDSKYnSIxswDXHF+D711sg9bBHxY+HvWfxS2xSZKSLH0+ZeLySuI4428jLcksefr+feq+nW6EdCdnu5ts0GelyrZCm8hVonDVcegRobelT/AIe9dbwOPQS7s+RvW82Qjx8uYoVmn9TD76MC5F7ab+90PWgQOPSIru6tnZBJcRS5GhmhmBjfXUxlRqFgBz7114kU6DPN9x7R2VQ1NCMnSJTrHLO0cdXGoJAJPoB4978+veVD59Vb98fK7Fbpos5QYSpWTIwSNFSCGZZGI0lb2X/ex731qtOiRdMbQy9Xuqp39uGOqqZ6SZ62NWDSMNLaxoHPvf29aHy6MLvzuTf3YFZT0O28duMpEi4z9ujqDGQP2lKi3vX2dezTHRjfjH/Ko7R+QuWgzW7Y8jHga9hNXLW0kqkLrDEWkt+P979+rmnW/t62bOgf5ZPx+6Y25iqGPb9PXZOlWCWWeSnhK+VEAZbEf1vf37r3R98DsPaG2qOKhxGAxdLDEoUFaKn1kA3F20+/cOvdKiOlpobmGngiPH+biSP6fTlR791qlOHWYXtyLH+n1/3n37rwr59d/T37rfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdajX8/K/8Aw4T/ACyh/XsOgI4H/O1t7917j1ttY/8A4AUX/UJTf9aV9+691L9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6910QCLH6e9g0z1oiooeotZSw1tPLSTorxzIylXGpTx9ffvn1r5dFP7C6SpKmomrKOnHJBOiIW55uB70R1sHPz6K3nOoqqOqkjaKbx6je0bWte3uv29WrToFN/wBHhOvYGNUBG7g3aS0fJF73a3vw69XolOVqH3DmpK6nZpKRnLIYzqWwa5I0+7deHCnQl7IkqhloaalEo0vESSpX6f4n/e/fj1rz6H7sbdlTS4ODHmaQu8egiNmI1Fbcr70OrcK0x0QTtXK1u1MFU5yYvoeF3TysVP1uLE/1/wAPeznrXSd6C7By29cHUVKqwihqtKvckCzcc+/Drxz1cB8ac5XS7Tzkc0hZII5Lhm1EWU8Ae9Eder69EO7Nr2rezMzEyyCMiW4YekDyH6X/AN69+rjr3QPZHOUG2Mkk9HUNFUa0ewk0gEN+L/n3vrw6Or1T2/FlsVHiNxyI0NXGUhLyC+kiwHPvXA162aEdOG8OtqGnhkzW3gsplVpLxqG+vq+o9+r1qnrjoFYVqxM8VbBG7hvHpkQEG/F7N735V6qR0Hm+vjfhOy6WeZSKWci8ghXx/i5tp964dXyc9BV8K/ipvDD/ACoq8ZG2Qm27GUeN5Vk+1IBt+s8e9nqvz62TMT0TDBClPViBkBYNcBuPwQfeqdeB6F3buwdubZRPtqOEzk2EjRi2o/0Uf8T7sOtE4oeluoKgKFVAPoFHAH9ABb37HXhXrn711vrrn/D6f7z73jrWevc/4e9dez59d+/db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de69791759ev+PfutVzTquX5k9Iw7orYt6Q0glq6eEgOset/SADZh9D79wz175dV+1WBkGPNNNE6vAmkCSOxBtxwffhx6uR0GsVdnNv1bmOmVoASwYqfwfwR9PfutDHSM3XuzPZmQ00ZqoBpZT4taKdQ/Fvfqde+fRe81tuZ6wTVTVEsjWY+TU9rc839+698x0az45GfGZgiaRo6Z3jBBYoujTz9fej1sdGP7gwm38lQ+SlFO1Q8bF9IDHWV4b37rWRx6rX35t5KB5XiIHFwEtcc8/p/r/h79jr32dAfU0GSrLoqOFU3YEH6f1/4178K9e+XSWrKOpkm+0ihZm+jB1ZlBH5N/e/t690iKjZkgyayCmPmeQAlIuOT+Pxx/h73jrXS2o9p1MdUrSRyKGVQPEtvrxzf3qvp14dL3FdW12QcfZ0VdUPJYj/ACV5V9XAIIHv3y68B0df49fDLdm9Mon32Llho5HIIlpHQEMtx9R711404nq+b48fGzb/AE1jIJhS0/8AFfEilkhVWjKj6sR+f8B73T160T0ad3CIXINh/sf9a9vewKmnWiaCp68pEiA24dfof6EfQ+/cD1viOvCNR9AQOBa5tx/h79qPWtK9cgAPp/vZ/wBb36vW6Add+9db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/9Xf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6903ZHEYzLxeHJ0NNXRWI0VEayLY/UWPv3y690HuV6V6nzEMsWS2Lt2ZJVZXZqGNWs36vX79TNevdUx/zB/5S2xu4sNX7o60xFHicrTxzSpTUH7beRYiVH7Y/r735da4/LrTm7x+MvyT+Pe8Go9w4quhwONmvHVSpVMujylQSSP6e9Y68cdBviO6clj9yBDVOaiPQsxEUqsGQAMNTD37r3Qyf7M1TUORoVqGAqLKNZiLM1z9bn37r2ejl9K9u7UyNV99k66DyVJt45AqsNQ4A1n348Ot/PqwbanaXWMNCKSWKlknkhF+YrlW4+ht/sffutinQCdx9A9Yd34zJZbDCiosjTFnRIWjR5GVeDZPfuvGnVZe5epd79V19CMVjJammSpLVEiksPEr2uxH+Hv3Vehhw+9MZFXUcuQoYhPHTqJZHVA6OANQJP9PfuvcOlPke26R6eelwlWZJmFjArcLx/aC+/db6Jds/sCfNd9x4zeMAosVR16SLNKpCSrr5DF+Le/Hr3WzL1n8m+revdgY2nwC0kNfHQRIXphCJGKj63Fj71Trdegp7t+d4yuFMUOekicRC4axK2FrALc8f63vdKcOtdVLdh/K3PV71K43c9Q00plVFBc+pjYAc+/daqQaV6KVU7s7q3BuKPJtuHJQUeoMswMttJNwBz7917oYaDtLsPAxaYdwVtdUSflllchgLFuPe+vVz11Tbb+UfdORWj2tTZLJCoPisI6q5jkNh9B/T3rr3VlPxF/lD9u76rYa/sqgqsQ058j+ZqhNZ1/Ulxz79Xr1Or1Ol/wCT9tHZFY9RnMgtVSMEDUryO6upSzJpsf8Aevfj14dWG7C+FHQWx1iaHYeDrJ4ghV56RHBkXnyNwLn36nW+jP4XbuD25TfZ4LF0eLpRb9ijiWKPj/Ae/de6effuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Wo1/Pz/AO3hP8sr6f8AMxKAc/j/AHKe/da8wOttrH/8AKL/AKhKb/rSvv3W+pfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rplVgVZQyn6hgCD/rg+/de6aqjCYqrUiahgJYW1eNQ4v/iPe+tAV+XVePzH+LtX2Ht6prNvO8Uqhm0RMwZQqXHC/wBfejxqOtg+vVY2wetMtsgPhszC081MzhmkDMfrb8+/Y69nh0IFJV02FyzSJSoDp/UB+mw4tYW9+pUdbqB1BymVkzE71cyCSKE8IeAQP8D79SnWjnomPzX3XSUOytv42NEifLSJTKthYs7abE/X8+/Uz1skeXTj8dcRFtbr9McYAlTWCKoDD6kSjUD/AF9+r69e9OrKPjpkZsDFWU1S5emyAbyX/SuoWF7/AO8+69e8q9AJ8gf4ditz1WQoI4yZyQzKRe5bk297631X/wBzRZSPGtnKESakkVrKDwDzb36nr1roTOpMvkd27fx7wyNT1eNh9WgNqBUcg+9549er0e7qTs1qXXhNwfvr4/EDLY/i3Af/AHv3rr1a9Q95wQrnVq6Ef5PNJrsv6QG+nvY68esiSV09TjKDDM8klYypMIg1wWP1JA96p177Orb/AI99VYba23qTOzY+P+PVUaNJVkfuWKXI9Qv+f6+9gdVP+Doy7g8HUVCn6C1mvxzf3sdaavHrlwbfQ25H5/2I9663x679+631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+69015fD0ObopaDIQpNBKrKQwvp1CxI9+60eiadk/GOmr5pKvEAKkhvojJBHH0PHv1PTrwJ6K3ub485agjIagZ1jJLu2oqR/hx70K9WqK0I6A/K9WSRTuP4egKFtRCm/p96qRnrdBx6Cqt65jklqWlgUNGSFBVjb8/097qOtUNeklT0dbhqt0o1MTpchlB454+nvY61gjrlWZ3N1D/bzzSsbEch/6W/Hv3Wz0FuV2lkMrXiaUuyFgAragCAfpa3vw69w6esb1lHVO0MNOrVDjSFUPq1NzYC1/fsefWvOnTzRfF7ec9R9zDtx5RKweNikvKH6fVfesDrf29LRfhhvet0VDbbkiY+o2En0+v0tbn36vXvLowPWn8vyu3JPTnPUzUCK0bMSXXgfUFj+PfhU8OvYHVkvWHxF692HSxRVmOpMlJEtgWQOC173Jdfe6evWujPYrbuDwaBMTi6SgUC3+TxBDa1vr9ffutU6evfut9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//W3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691HneMKElQyB/qApI+v5t7sPXqpPl1yihhjj0RxqIzclbagSfrfVf3WnWwajoAO3/AIxdP920r02+tq0GREg0s4pKcyfnk6lv73SvXq0HVLnyO/kPdZ7+erreqjQ7WragTFZNFPCytIfTwQPp70fTrY6KP17/AMJy8xjMq9VvLeVJlF9TROZKZxHb9A4v/tvfuvHogXzM/lMd39E7nXIbEiy+Vw1PPHJGmPjZojHrsbeL/D6e/deI9OiHZ3ZXfOx605DJbf3DS00ERhbzidBqjX1EhvfuvdBKe/8AuPZuajFJkHoqd5LyQ1FRJcgm36SffsHrQFel5D8tc1kI8hT7llhmklgaKJv1gOVsLX9+4db+3oGqLe+Rz9XPKK6KES1DFdUukiNm/HPv3Xs9DftbK7f220NRNF/E6ub1P4pNZLn6j37rXHj0tq7beI7BmSowO0a/+NsRaWCG0hf+yLqL8/j3759e+3oxPS/wU+TfaWYppMbQ7kxOGKqE8olWMIRxcEfT37rfVmfQ38ljtLcVbmj2DmqtqV3YQGqZNKAj02L8e/cevEUHS8zf/CfTOVWXmqsbuyjp4ijNFd4ABIfUp+thb37rWfLrntn+Qt2tQrLDkOxKKSlEo8cbPShvGDyAR791unVjPRH8oLqDZVHH/pGoaLcdZEp9Xjp5NTFfqWAP559+z17A6P8A9c/Evo7rKaOq2ls/HUU0WnRIaWnJun0YjT78QeB69WvRjIKSgoxHHBT00B5EYjhijP8AjbQB731rh1KUOCdTAg/QAWt7117PXP37rfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdajf8/P/ALeEfyyj/wB/Cobcc/8AF1/Hv3Xj5dbbOP8A+AFF/wBQlN/1pX37r3Uv37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3WMyASCOzXte4B0j/AFz73TrVc069JGs0TxSqrrIrKykXBVhY8H377Ovfb0QruzpucVmQyeNpSRUAsrRQqByb+mw91PVq9EGrOttw/wAWlNVTTrGHc+pbKLG3Fvfq4p1qmanqV/c1qTVRGBjJMDYWU8EfXj36vVgBToiHys6U3Hvuu2tTUFHVSR4eujncRoWtpkDfT37y6rQfl0OO19gVFJhMbDJC9NPS0VPE0ci6W1RqFNx73Udboehi2tkK7C6YnLaI7rcDkcfTj3o568MY6C3teenyz6k0tOx/Bufrxe/v3W/Ppho+t6XcuzJaatjUSyaVQOEJ1ONIIJ97J8utUqeld1b8W927Dp6jMyiWTF1yGSKPSpVUPNx/sPfvLrwzx6W9Z1tWmSPI0NNKX8liUGn6fX9Pv1evU9OhRwuwsvmaBI2xlS0w0oreM/61wbe69brjo5nRXxshxcsWdz8BPIliilUXB+oADc+9/b1Xo9kEENJBHBAixQwoFRVACqqj/D3vr3Drn6XH9f6H+ht73kHrXxDrmOAB711vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuuLhiLK2k/g2v8A7A+/daPy69yF9VmIHNhwf9h735469mmeoNXjKGvQrU00cgI0m6gG310n3o/Prw6DHP8AUO3soWelp4qZ31ahoXTcjgg29+62MCnQF5P4wrUvUtCYwZWJFhGP9sffqdeB9eglf4gZA5GaV9TI+oCyC1ibX/4r78BjrVa9RJPhDkJ6wSipSOO4PqMf9b8/096APW616V+G+F0dNVRNkKmCWFWuf80wA/JIH+9e90xTr1fQdD7tv4z9dYSSOplxkVRVR6bN40C3A+v0uffiBx60CfPoZqPaW3qERinxlMoiUKl4k9IH4+nveBw69Qnien0UtKFCrTwBR9AIo7f0+lveut8esyoiCyIqD+iqFH+2Hv3XuuXv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/X3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691xXUR6wL34tzx/U+9mnl1oVpnrl711vriQSVtbg3N/6W/HvY60R1y96631737r3TTlcFhs5CYMvi6HIwsLFKymin4/wLgkf7A+/de6LP2T8Nej+zaOajzW1MbFHNruaejiUgyCxJItf377OvAkdVPd+/yL+lt25WmymzKZIGZw08MkBAUi9wug2tf34V69UdEz3L/wAJ+llkkkxlPTAtqCao25b8Hg+/V9evCnEdATuD+QN2rSPqwVPTLGJVAKxOWKBv6A/T377evU9OjW/H3+RdUYurxlV2HHEyoWM+pCQORbgn/ePfuPDrxoOPVwfXP8sP469fz0lfTYWKproBEzs9PH4i6LZgAfrf+p9+690fbbWx9q7Sx8GNwODxtBTwRiNfDRwK5A/1T2v/ALz79177elQkccYtHGkY/oiKo/5N9+691z9+6910Abkn+vH197619vXfvXW/LrhGgjXSPpcn+n19+PHrw4U6xS00czxyNq1R30kH+v8AW/vYNOtU6kf4e9dbp5de+nv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de61Gv5+f/AG8I/llf+JEoP9h/uV9+60eI622qD/gDRf8AUJTf9aV9+631L9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdRqqkpq2JoaqGOaNhYq6hv8AYj37r3Qd5XqjamULs1IkTtf1KgJ9X1/3n37HXs+fQcVPx2wT5GOrj8ZRSvBB4/re59+oPLrVTTPUyb477SklaY0lHM5II8sV7W/IPvVOt9Fg7S6AyFHmDXYykVMcivcRQG3HNhp49+IpnrYNR0XmfYNdDUvB9tLpZm8jCFjx/S3vR+fW64qeg5PTVVX7jkmqoJxRo+sXhk06b/0/4j3vyp1roY8R0/8AxGso8bjopli8kBe9PIouGF7g/j3rr1cU6s0291tizsqgwWSpYxIlMIy4QFgNFlJB/wCK+7DAp1rz6Zcf0RtqjGl0idNZf/N3+v8AQH36g61U8OhAw2wNuYUWp6GF+APVEtjY3Fx79jrYr0tERUUIiqiKAFVQAAB+AB7917rxBJsQCtv9jf3vFPn1rNfl12ABwBYe9db679+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde5/2Hv3Xuve/de64soYEH6EEH/WP197Bp1ogHryoF/TccW+pP+9+/Vrx69T068uq3qAv/AIHg/wCPPvxp5deFfPrl711vrgqKpLC925Nyfr/re9160FANeufvXW+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//0N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+69176c+/de64qwZQw+h97OOtA1HXL3rrfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Wo3/Py/7eEfyyv/ABIlB/Sx/wBytvfutHiOttmg/wCANF/1CU3/AFpX37rfUv37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3WKaCGoQxzxJKjXBSRQykH/A+/de6SFTsHbNRIZDj4kZjdrKCDzz9ffsHj17PWOPrva0ZJ/h0TE/XUq2P9eLe949OtUPTtQ7VwGOlEtHjoIpVNwwAJH9Bz71SnW+lCBb/AIj/AAH9B7917rv37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/R3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XF/0t/wU/T6/T3scetHh1jp/80v1/P1/1/x78evDrN711vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rUb/n5f8Abwj+WX/4kSg+n/a1966159bbNB/wBov+oSm/60r731vqX7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuuj9D/rH6e9+fVfI9e/H+w/P0+n59+8+tjgOo8P+dm+n1H+v9P8Aevdm4dUXj1J906c697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/9k=)
Figure 1. Locationist Hypotheses of Brain–Emotion Correspondence. A: Lateral view. B: Sagital view at the midline. C: Ventral view. D: Coronal view. Brain regions hypothesized to be associated with emotion categories are depicted. Here we depict the most popular locationist hypotheses, although other locationist hypotheses of brain–emotion correspondence exist (e.g., Panksepp, Reference Panksepp1998). Fear: amygdala (yellow); Disgust: insula (green); Anger: OFC (rust); Sadness: ACC (blue). A color version of this image can be viewed in the online version of this target article at http://www.journals.cambridge.org/bbs.
Constructionist account
A psychological constructionist account of emotion assumes that emotions are psychological events that emerge out of more basic psychological operations that are not specific to emotion. In this view, mental categories such as anger, sadness, fear, et cetera, are not respected by the brain (nor are emotion, perception, or cognition, for that matter.
…emotions emerge when people make meaning out of sensory input from the body and from the world using knowledge of prior experiences. Emotions are “situated conceptualizations” (cf. Barsalou 2003) because the emerging meaning is tailored to the immediate environment and prepares the person to respond to sensory input in a way that is tailored to the situation
![[[@Lindquist2012-jr]](http://dx.doi.org/10.1017/S0140525X11000446)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAgEAyADIAAD/4RyIRXhpZgAATU0AKgAAAAgABwESAAMAAAABAAEAAAEaAAUAAAABAAAAYgEbAAUAAAABAAAAagEoAAMAAAABAAIAAAExAAIAAAAbAAAAcgEyAAIAAAAUAAAAjYdpAAQAAAABAAAApAAAANAAAADIAAAAAQAAAMgAAAABQWRvYmUgUGhvdG9zaG9wIENTIFdpbmRvd3MAMjAxMjowNToxOCAxNzozOTowMgAAAAAAA6ABAAMAAAAB//8AAKACAAQAAAABAAAFeKADAAQAAAABAAAEIAAAAAAAAAAGAQMAAwAAAAEABgAAARoABQAAAAEAAAEeARsABQAAAAEAAAEmASgAAwAAAAEAAgAAAgEABAAAAAEAAAEuAgIABAAAAAEAABtSAAAAAAAAAEgAAAABAAAASAAAAAH/2P/gABBKRklGAAECAQBIAEgAAP/tAAxBZG9iZV9DTQAB/+4ADkFkb2JlAGSAAAAAAf/bAIQADAgICAkIDAkJDBELCgsRFQ8MDA8VGBMTFRMTGBEMDAwMDAwRDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAENCwsNDg0QDg4QFA4ODhQUDg4ODhQRDAwMDAwREQwMDAwMDBEMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwM/8AAEQgAeQCgAwEiAAIRAQMRAf/dAAQACv/EAT8AAAEFAQEBAQEBAAAAAAAAAAMAAQIEBQYHCAkKCwEAAQUBAQEBAQEAAAAAAAAAAQACAwQFBgcICQoLEAABBAEDAgQCBQcGCAUDDDMBAAIRAwQhEjEFQVFhEyJxgTIGFJGhsUIjJBVSwWIzNHKC0UMHJZJT8OHxY3M1FqKygyZEk1RkRcKjdDYX0lXiZfKzhMPTdePzRieUpIW0lcTU5PSltcXV5fVWZnaGlqa2xtbm9jdHV2d3h5ent8fX5/cRAAICAQIEBAMEBQYHBwYFNQEAAhEDITESBEFRYXEiEwUygZEUobFCI8FS0fAzJGLhcoKSQ1MVY3M08SUGFqKygwcmNcLSRJNUoxdkRVU2dGXi8rOEw9N14/NGlKSFtJXE1OT0pbXF1eX1VmZ2hpamtsbW5vYnN0dXZ3eHl6e3x//aAAwDAQACEQMRAD8A8qSSSSUpXMLo3VM+p12JjuurY7aXNj6QG4t1P5rXLe+r/wBX66fs3Uc01XC4bqsd3uAnaa7LPzHu/wCA/wC//o10FVTam2uqEbotAbA3Onf+b+d6LfT/AOESU8BldI6nhmMjGsYAN26JbA5O9m5iqL0+51dtL6GvcW2dgNS0nXbu2t+j+8on6v8AROoYxrvoqoFbdrH1N22Atn9GH1j3v0/wrHqPNmhihxzvhutBxbsuDBPNIxhVxHFr/ivmSS6HrH1QzMbJLOnMtyaQwvcXAAtj83cDssd/U/Sf8GueIIMHQjkJ0JxnESibBAP2rZ45wJEgRRI/xVJLWd9WOo1jdc+imsMD3Pfa0BoO2N4G5/0n7Po/TVbP6TkYDGvseyxrnuYHV7nNlhLHfpdgq+k36O/enLGkkkkkpSSS6b6t9Couw7MzLxX3vc5n2Zn5pZudvt2NO+z31+n7v0aSmn0/6qdRzA82kYm1rXNbaDucHjdo1v0fb/pEPN+rHVMS4VhgurcCW3M+hprDy7bsd/qxdwH0MdkZDg02Nd+s2GQ/aBtYXT7XbGH6StEhobudFZ04ndP0W7klPlNtVlNjq7Wlj26OadCor0rqn1XweoYovaGMtILi1pDIjmz1He1m7+3/AMSuL6l9W83CeNazWRMm6rSePdvH0vzFHizQygmBvhJib09UWXPy88MuGdecflclJazOkdN9Ku23qlTd1fqWVtaXOboXem2He5/0W7P0ar5uFhU1GzFyhkgP2cNYfo7ifT9R1v0vz9no/wDDKRif/9Dypa31a6Y3Pz91umPjxZYTEEz7GO3fmu2rJXcfVLHFPRmWmCcu1ztezWTVu/sub7klOoyhuPWG1cMJe2lnsBLj7n11yzY9n+j3ogYWMY1wHptMwC4kuOtYYz27Hbnb/poNbLm7bbCbYhjKtCeZa2v+a9qi7IyBW8MLbfScHBzRLxDt7m7Zczd/o/5CBIAsroRMjwx1Jv8A5vqLbeBG1zHBwPAe2eO+rkq7TTaHMc4SImRvb+7tsHtt/wDPytMq6Y4WUmizLsFh23NeQ3Ux6jTX+jbz/Of98Va2t+PYcXKIdaw7bAYkNM7H7vou9v09v56qDmceYHHcQZaxjL1cUP64/Rb+Ll/ZnDIYyyAAmYiRGf8AehD9L+6zpsJvPrOL9pDpI1A+jv8Ab9Otc19cunHDstzum0tZVbpmW6PsD3ktndG2ll0+70P9J+ks966TdTXjB1kurd7qyARDj7fQFpbY13qN/wAD+Ysz6w2YP7Eyq7GXek3YB+kH0g5tja59F+32/vJnKxJzSmDwxiPb4K9E4x+Qw/qs/wAQMfYA4ZGzcTL5scvmnx/9B88S8lZ39MOgpuYSR7ja1wAn3ewY7N3t/lq/ZkfVgOIqxr9oOheZ3N3F/wBFttfou93p/Su/R1f6S31Vfcdx0lO81G6w0iKi4lgI26T7fZvt2/8Abtn/ABigkptdKwTn9QpxZhryS8jQhjQbLS2fzvSY/avRPQprcPsbPUx2Ha6hp2ubtDfTfjeps+j+fXu/Sfzta5L6kGsdUuJdtuOO8UNiSXSzds/lNq9R39RdofX9pdkPYQJcAGbBt0cXOsG73JKYsZYWyXusa4QKrWgH+qbHfRr/AJLmPUHY7JG7HLDt9M+jZHt/dFf6Pds/M9qLgiwZBb1DIruxtrv0lH6N4skBrfc9zPS2ep9D/Cqv1LB3frWNd+pwYsvIcG216tqbaw+11rFBl5iGOQiSATW//chsYeWlkjxa1tp3S05FzKHse42taSQ8N+m2fzq/8HkN/PYgdY6bX1DB9ep1bi5npvc+du0lr/a9u7btdt9yssoe+0Oo/SO2h4eD7XtGu3d/Kn2KYZ6N9rKf5kH1tnZpeZsr/d+n9D/MVQyvPAY5RjIn3ZSiPmjP/OR/ek65EBy8hwT4RDhljkeKuD5eGX/dvmGfhvwsl1LiHAaseCCHA9/bKrrpPrViPZX6llTazVd6bXNYatwcHu/m3E72s9JrWXf8YubWk4D/AP/R8qXRfVrq+cGt6TjMrdZY4mm2wwGAjc9rtP5v6Tvb+eudVnp2db0/Nqy6tXVOmOJH5wSCg+kYPTr8jdblBzA5rrGMaS2x7hAPtmz9C79z9Lf/ADanmUgMpx37nY7mj0C5rdzXH9J6f6D3W1e32P8A6RTd/wAYrfQr+l5FFebWYrsDyGyWts3bfV9WneWtrrf/ADm36d9X/GKxm+i6zHrqscbnXG1t2nqDT9K8H6TWfynITw1xZBkkKjrH5sfD+l6P3v8ADbHFHhNxHCB+jpLRr4ZysZttVFmzFyj6XpuA3kvBrYa69u57t21lnpbLP+C/wqz/AK2NZjXY2Vivbk04uODlVkBrg1rg39C5rf55rm++t/0/8Kue+sn1r0OB095c6l4acgaAem6f0X73ub9NZfXOvdWyMl7LMgtbYxjnisCsH1Kq/UDvT2+16oR5KRzDOCI3dx4Rc7HDc5fvf1lQ5gEAzBsAaD1RmYfzcpiX/obtU/XjAaXONN1bjI2ja5pHbe3dXv3f6N3sWf1j604nVMdmH9nsxqA4l5pcGB3Hpb8UD0f0f0/T3V+//CLm0lfEQNgxTyzyEmcjKyZfWW7qs6V051It/aNbnPG5lGjHke9rm2Osd6FNjX+j9O3+bfb/AKL9JSzsami4Mx7PXrIBFgjWfJpds3fT9Oz9Mz/CrY+p31dr61mWPyQ5+Ji7PUprkPsdZuFdTHta5rP5uy1/8ir0616l1WvCw+mYGL0oV24NBF1bWwCGndXjepU/b630bWfaLf8ACf0j9IgZxBIJFxHFIfpCJ/SpMMM5mIiPnPDEnSPF24nw1Je0fXb6udD6vi/aPUq+3nHa7FtoYWmDr6tvp7m241rm7K/V/mGWWej/AKNeLo2LIsWNwsMZACRBAldHvW71P1Y+rVhycLNzHvpryWOvwjUHOLnVP2WNf6fvbsY192yv3+n/AJi3+qenScZ1GVYDY42se87qnms7qtlvsZc31W+9j6Ktin9Qb8TqfQmdPDjVm9Lu9VjmuIcN7nPZbX/Js3WY17P/AEsugvwasPOdn2UuyK7JfDQ60ssn3bcXdtd9ob9N7PoWJssEzOGSGQgAi8Z+SuvytjGYiO24NkDi3/vfuuZ0vBbT1enqGA+u5lwsf9le7dWHu/nq2u1bi5Tn1fT+g/ZWr3XHUYVz/wBp014tOeWAX12Qw2Br97b3Wj06n2WbGfans9PZ6frKRFL7PVaWuZaHG4AtcN0mRur9nqU7fStZ/YXJfXP6y9ONDOjY7m59AcH3Ct011QPazEyIe37R79z7K/Ux6/5r9P8AplQz8mcvOccSOGIEJ/NoI+uHDLi+aOT9DgWjmZCRqxKGglHbX/uv7r0mT0/FxcOvKxMlt+N6f6O7fv3+4V7N9Z2/zizMrq3TsV4+2ZTK7AN3pkODTy0foqmvc/3t2rkcTL67RVVi9GyDkYtj3GlrWMLg8tL313VvD3V2bGvf9P0v8JQqGRgddybfVyaMm617Q4Psa9zi2Bs9zv5Kscvy+THYyTGXUkSMeGf9VklzcjiERPJxAcOp9HD6evzz+VN1nrTOoVVU1scG1uLt9jtx14ZSzX0af5HqXf8AGLKVu/pPUsdj7Lsaxldc73lp2iHCrduHt2eo709/76qK01H/0vLaabb7WU1N3WWHa1vGp8ytbF+rHVfVDsnEf6TRucxrmBzpabK6vp+z19uzd/0LLP0ax2uc1wc0lrmmQRoQQpetcDIsdJEEyeI2/wDUlJTu/sz611ZoyKqnY97gYDHtYxjayKW0u3P9Nja5YxjLEOj619Xw6s6iw78nKhjsh5O+vbuY5tW07G8rG9a6CPUdBiRJ7cKLWve7a0FzjwBqSgQCKOoUddE2DjNy8ltDrmY4cHEWWkNZua1z2Nc930PUe309ytdXx65GTTezIbIpfsAEGtra637fUtfsuYzeql+FmY1jar6LKrHwWMe0gmeNoKfJwM3Ej7TRZSD3e0gSe39b+SipAktDCq6I7Ha/NutbaC/fXXztAHpenNb2Oda/cx++2r0fp/pv5tV8xmGx7Psjy9jmAu3TIcfpNdNdW138lnrM/wCHsSU99/i86V/kK7qFW52VbkgbGlwcaaQ3ccfY3+frsufZ/hP0S6jqmPYxrMXKvaTXutx/a4viTtZdp+j9Rv8A27+fsXN/4sur4QwW4eS8NfhZPrem1m5767Q0erO5vsxrq/f6bX2++pdJ9ZemZzKWPfe63IsfsvuYA0E7j6X6Nx/R11sc2qyxv5igzY/cBGMiGX0xM9OKOO+L92Tb5bN7RjKdnH6pQAF/rB6f0vl/rMMqvLsGPnX2etXk2NraGh7Ghs7XM9zf0Hpu/wAGz+c/wfqLyPrvTj0zq+XhDVlNrhUToTWffRZt/wCEpcx69xzKLmdNsxi+K72M9Gona1voB12S/d79jLGurZs/0i8S+svUx1XreVmtO5ji2ut/G5lTGY9Vn9uupr0sPBGc8Y1kBCeSV/NPJx7/AOIt5iZyQxzNRFzhjgNOHHHh4flamB1DN6dksy8G5+PkM+jYwwdeWn95jv3HLY6l9e/rP1KoU2ZZx6gIc3GAp36bf0rqdr37p/m93pf8GufSU9lr2e7uVfWvKp+rh6FTSxm7c12TJ3+m9xssrDPot37tjn/6JYaS08T6v5+biNysba9jjBA3ae70xucG7N24fQ/Mr9Kz/CoCIF0NzZRVOdVdbS/fS91b4jcwkGDzq1Wq+sdUqnZk2S4uJJMmXAsc7c6Xb9rne/8AMVp31X6o230nBjLDSL2h7tksPnaGN3f1v/SiHf8AV/PpNG41ubk2elU9jw5pdp3H8r2/v/8AgW8qa1vU+o3VuptyrrKnAB1brHFpAO9oLHO2/T96rLXb9VupuGjqN52nYbWgw8+mx53exv6T2fS/4X+Y/SrKsY6ux1bvpMJafiDCSn//0/KkkkklJsPEuzclmNQNz3z4AAAbnuM/utC9CwOldM6bXFNbPVa0g3a+oR+frP8AJXL/AFLGN+0n+uXMc9hrY8M3gB389t1H6bZ/Ns/P/SLr8j7O47aBYaxJG8ATp7zt3fn/AEfTf/24op5uGYhwnuZ/5OI/vNjFy4nAy49aPDADikZbR/u8bh9eqYOq9JfZt2Pc8kRI3tLT/m7vTatyyuq8ux762vrJ9zHSRBPsLv8AX/Bqs6wi6pjmtNoALTbB2SLLN9TvzfYdu789WMPOcLxkVAOhwbYHOAMQHNO127876aZkzmImYgS0jwa+iX+EyYuVExDikAScnFC+HN+r4Y8EY/4v/hjy31p6TVg41V2HWWY+RbY+xvIYRtZU1n5+z+c/trml6Z9dH1ZvQ7vUa1mTWG2gMY9jedz3bvdTu1d+56vqLzNO5bMc2PjlA4z+6f8ApRl+lFgzYhjkIgnb1X+jIS4TFsdP6hldNy68zEfstqIInUGDu2vb+c1epN/xmYPU8YXvxL6sfGG7NY3ZaQXbmY7KnbmWZFe39LdbZXV/N/8AFLyVJSnw02YiSQBe38i+jdd+vber3VYOJ+r12UWerfS/3e2u306mOsb+ha539I/0n+D9/wCkXnKSSAjRJ6mrPkq5UAZEgDQdmVddltja6mmyx5hrGgkk+DWtW/0X6pPzqmZGVaaa3yRUxs2FoJZ7nO21U7nNd9L9J/wS1Pqx0izp9Dcs12HOyW7RAA9BhcWfpPUd9J+31X/n+l+j/wBItTMy6MTGLX3B+Q1pApqaQ8vgurNdLXF1Xu9+5G970XVqK9R7V/zWi3oeBjub6WC3FspeDVlZFvqNL49u6tu71tn85s2Us9RajxDNzdrMWtjn7gdjGuH57NG7a0JuOLXU42Qw2Naxlrr3Fr2OtaA1u1s7vpv9T6OyxVhU3KNT8pn2ix9h37rCxoNDnN9P7M79G/Hdt9/s/Sb0VrWd07B6i9+ZkV+o7J27BaXCGO+h9mcxzK6mu2b6fU9V657qfRLMPB+1Vsc+h17mtsP0mM2tdULmN9rH3ep/0F12S8ODhc6sM31WWgS42bnaM9v5u4ejWxVeoY+O9lWFk+3GbNj2M9s2XG1rnvcPe9tbm4r/AHP/ADElPCJImTj2Y176LPpVuLSRwYMS1DSU/wD/1PKkkkklPY/VBm3pj3Fn07nHc8w0gMDGuZ/Krc53vXR9Fy8erqLLssD0xrWx0EDcfTbuDy36LLPVcsH6nXD9lFrTLmWvBYCS73CvaWNP0F0tXRjbU+z05u9gaHEBhZHafdbZt/qM/wCMUeXD7uPJj1HuRMSR5cNWzYiTwiR/VwPyjf1eqRr/AAWdV+P0sWYXTMcZ2PSHNqNn6Ta8kwxlry3dW1jv0n85Uyz+Z/wiNk5HT34JzK201ZLaGMoaG7rGit2yzDsrcz9Ez1PbZb+esx9eZhvsq+0Bja63WNG2Nxn+aY11bd9isM6LlZDRa/a21wBu1ED2zXXtYHN9Vn+E3qpHkckskTxzjkxkSnk4tcn6cuP9CXuIxylxyBFxGwHp/ueppZFzMjHey5tI3h1ba9gI959Jw/RtZY7+R6a88flUkOa7DpDtRuBtBB4nb62z2/1F6WKLca0V3NbWN26txPBHucPUb+a3+cXl+S82ZFthd6he9zi/jdJndw36Sv7McgQdXRZ1XpX2aqq7pbLLK6xWbW2Gsn6W6xwrZ7rPd9Kzek7q3Sy66OlV+m/aaWGx0sc1hYf0jWtssY+x3rOZuWUkkh2D1fopeD+x2BjQQGi5/wDKLPdt92zd+f8AT/4r9GqbNmb1SsY1Ix222MDaWGQ36IO11s/1vch09Ozrgx1VD3tsIaxwGhLg9zQHf9Zt/wC20UdN6pjX17qXU3B7SzfDCHR6gd7y3bsb77H/AOC/wiSnv/Xs278cOy23lw32OFezbudxsDvd/bQ30uxqBQxpuzrWvFon3O2sc9oy7H/pPVZLdmH/AG/0Sz8P6w1tDK6stpyK2gvvdL3Ot27fTxGvcz8//Dv32/6D/SvJZ1XpeHTSJY3LxTZ9qtqsL7HudD3F7nyx3vc+j9H/AD3p7/8ARqHOSBGomevTvXp4mzypAMvUIGVR4v3Y/p7tmh1T7GNfb9putcWmA2ofov57WsH0sf8Amva33+p/XWfk0UGul2aRj2XWPd6dQfZZa57t1jZG3062N2M/RsRXX5OdU6vpuC2p7y112VY6bGGweq9jNfa57HfpK6/0iDiMa7feRa2uo+hfm2Pc620g+2jDZ/gXept3/uM/nP8ACJ+O+D5eCtBEni/6LHkEBk344neURw8X9yLrU4dVtzGNb6bYbsLy4O2s/Sf4Qt27r/Rr/wCEQetdO+w1g2GsXZFooc7c6wMdWGN9N/7z6t36b0fz0bFrvy62MxCW5LnFvph7HWWWV++1tt9/6JjGtH0f9Xy67R1d9uF9ox3Vspa5tRe5uhs/nLXN37LLtf5xz/01j1UyZssOYxwOSPDLU4xw+5Xr/R+b1y/6DZ9vl5g+0KoGJ9y+Pioesf3Xi+v0FjaHEHdWDS554eWEt3MH0vTb9H3LHWl1XIJHouebLJl24QW8ucNv5u57lmq80X//1fKkkkklPRfUvqleHmXYthYwZrWtZa/QB9bvVrr3fmMv/m/6/pr1LDyab6gwciJYeWx+btXha7Lov1updTVRn3OovrhovLdzXfmh9jmxa32hrX/9upwycOhGngy4jelgduLT/nPd5TsfIzQy1woGPFlb21h7nOcdnqFzvo0Uu2f8Z6iuYuTZZS11wDbI97RoAV5/m/Wf1esYGOx9ZGO51hyA7cC8seyqprnbW+nud71pdX+suHjUA2W1lzhuY2obnkHw3n/q0/jinjAJCvrz1mvHwXMpeG32EtqI+lLhtsc0j6Pp1H/PfUvNFc6t1O3qWW698trmKqy7dtb/AOSd+eqajJs2xzNlSSSSC1sVdRzqa21V3OaxgIYJ+ju+n6X+jc7d9OtRdmZdlRpfY59ftJB1jaG1t930mt2Mrb/1ur/RoKPi5uTh+t9nfs+0Vuot0Bmt8b2e8O27tv5qSkmGRitfmO0urAGMw873ztvj9yhvvY7/ALkeh/wil0awUZ7csiRiNdfEx7mCaQ7+S6/0mOTW9X6jkYowrbt2OBW1rCGwBWPTqg7fbtarGVV07Hxr29Py/tBdta8uG3fXuD/0bLGsex7Lq2bmf6P9IkkGjY6Oz/zp6W5oyXMubeystGJWBXS59n85Y57H73e7856u4XWMDMpxqacZ2S4OaTRIDax+e1rf8Eyv3fpFwqsdPzrsDMryqT7mHUcS0ja9n9pqaY6ADp/LVUiZHi63b6162P0+iu66x1+XS2Gtp/mQ50/zO1vvs9/9If8Apf8Ai1y3XOpfaXHex9Z26vsfuc1n0vdtLljZ31iqLjXiG1+OYcGuOyCeWv8Apblj5OffkDaYZX+43j+1+c9GMMQ9UMcYyIonhiJ8P7spMss8pfpHXXfr4rZ2ScrLsyCI9Qz93t1QEkkWF//W8qSXZJJKeNSXZJklPHJLsU6SnjUl2SZJTxyS7JJJTxqS7FOkp41Jdikkp45JdkmSU8ckuyTJKeOSXZJJKf/Z/+0inlBob3Rvc2hvcCAzLjAAOEJJTQQEAAAAAAAHHAIAAAIAAgA4QklNBCUAAAAAABBGDPKJJrhW2rCcAaGwp5B3OEJJTQPtAAAAAAAQAMgAAAABAAEAyAAAAAEAAThCSU0EJgAAAAAADgAAAAAAAAAAAAA/gAAAOEJJTQQNAAAAAAAEAAAAHjhCSU0EGQAAAAAABAAAAB44QklNA/MAAAAAAAkAAAAAAAAAAAEAOEJJTQQKAAAAAAABAAA4QklNJxAAAAAAAAoAAQAAAAAAAAACOEJJTQP1AAAAAABIAC9mZgABAGxmZgAGAAAAAAABAC9mZgABAKGZmgAGAAAAAAABADIAAAABAFoAAAAGAAAAAAABADUAAAABAC0AAAAGAAAAAAABOEJJTQP4AAAAAABwAAD/////////////////////////////A+gAAAAA/////////////////////////////wPoAAAAAP////////////////////////////8D6AAAAAD/////////////////////////////A+gAADhCSU0ECAAAAAAAEAAAAAEAAAJAAAACQAAAAAA4QklNBB4AAAAAAAQAAAAAOEJJTQQaAAAAAANhAAAABgAAAAAAAAAAAAAEIAAABXgAAAAWAFMAMAAxADQAMAA1ADIANQBYADEAMQAwADAAMAA0ADQANgBfAGYAaQBnADQAAAABAAAAAAAAAAAAAAAAAAAAAAAAAAEAAAAAAAAAAAAABXgAAAQgAAAAAAAAAAAAAAAAAAAAAAEAAAAAAAAAAAAAAAAAAAAAAAAAEAAAAAEAAAAAAABudWxsAAAAAgAAAAZib3VuZHNPYmpjAAAAAQAAAAAAAFJjdDEAAAAEAAAAAFRvcCBsb25nAAAAAAAAAABMZWZ0bG9uZwAAAAAAAAAAQnRvbWxvbmcAAAQgAAAAAFJnaHRsb25nAAAFeAAAAAZzbGljZXNWbExzAAAAAU9iamMAAAABAAAAAAAFc2xpY2UAAAASAAAAB3NsaWNlSURsb25nAAAAAAAAAAdncm91cElEbG9uZwAAAAAAAAAGb3JpZ2luZW51bQAAAAxFU2xpY2VPcmlnaW4AAAANYXV0b0dlbmVyYXRlZAAAAABUeXBlZW51bQAAAApFU2xpY2VUeXBlAAAAAEltZyAAAAAGYm91bmRzT2JqYwAAAAEAAAAAAABSY3QxAAAABAAAAABUb3AgbG9uZwAAAAAAAAAATGVmdGxvbmcAAAAAAAAAAEJ0b21sb25nAAAEIAAAAABSZ2h0bG9uZwAABXgAAAADdXJsVEVYVAAAAAEAAAAAAABudWxsVEVYVAAAAAEAAAAAAABNc2dlVEVYVAAAAAEAAAAAAAZhbHRUYWdURVhUAAAAAQAAAAAADmNlbGxUZXh0SXNIVE1MYm9vbAEAAAAIY2VsbFRleHRURVhUAAAAAQAAAAAACWhvcnpBbGlnbmVudW0AAAAPRVNsaWNlSG9yekFsaWduAAAAB2RlZmF1bHQAAAAJdmVydEFsaWduZW51bQAAAA9FU2xpY2VWZXJ0QWxpZ24AAAAHZGVmYXVsdAAAAAtiZ0NvbG9yVHlwZWVudW0AAAARRVNsaWNlQkdDb2xvclR5cGUAAAAATm9uZQAAAAl0b3BPdXRzZXRsb25nAAAAAAAAAApsZWZ0T3V0c2V0bG9uZwAAAAAAAAAMYm90dG9tT3V0c2V0bG9uZwAAAAAAAAALcmlnaHRPdXRzZXRsb25nAAAAAAA4QklNBCgAAAAAAAwAAAABP/AAAAAAAAA4QklNBBEAAAAAAAEBADhCSU0EFAAAAAAABAAAAAM4QklNBAwAAAAAG24AAAABAAAAoAAAAHkAAAHgAADi4AAAG1IAGAAB/9j/4AAQSkZJRgABAgEASABIAAD/7QAMQWRvYmVfQ00AAf/uAA5BZG9iZQBkgAAAAAH/2wCEAAwICAgJCAwJCQwRCwoLERUPDAwPFRgTExUTExgRDAwMDAwMEQwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwBDQsLDQ4NEA4OEBQODg4UFA4ODg4UEQwMDAwMEREMDAwMDAwRDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDP/AABEIAHkAoAMBIgACEQEDEQH/3QAEAAr/xAE/AAABBQEBAQEBAQAAAAAAAAADAAECBAUGBwgJCgsBAAEFAQEBAQEBAAAAAAAAAAEAAgMEBQYHCAkKCxAAAQQBAwIEAgUHBggFAwwzAQACEQMEIRIxBUFRYRMicYEyBhSRobFCIyQVUsFiMzRygtFDByWSU/Dh8WNzNRaisoMmRJNUZEXCo3Q2F9JV4mXys4TD03Xj80YnlKSFtJXE1OT0pbXF1eX1VmZ2hpamtsbW5vY3R1dnd4eXp7fH1+f3EQACAgECBAQDBAUGBwcGBTUBAAIRAyExEgRBUWFxIhMFMoGRFKGxQiPBUtHwMyRi4XKCkkNTFWNzNPElBhaisoMHJjXC0kSTVKMXZEVVNnRl4vKzhMPTdePzRpSkhbSVxNTk9KW1xdXl9VZmdoaWprbG1ub2JzdHV2d3h5ent8f/2gAMAwEAAhEDEQA/APKkkkklKVzC6N1TPqddiY7rq2O2lzY+kBuLdT+a1y3vq/8AV+un7N1HNNVwuG6rHd7gJ2muyz8x7v8AgP8Av/6NdBVU2ptrqhG6LQGwNzp3/m/nei30/wDhElPAZXSOp4ZjIxrGADduiWwOTvZuYqi9PudXbS+hr3FtnYDUtJ127trfo/vKJ+r/AETqGMa76KqBW3ax9TdtgLZ/Rh9Y979P8Kx6jzZoYocc74brQcW7LgwTzSMYVcRxa/4r5kkuh6x9UMzGySzpzLcmkML3FwALY/N3A7LHf1P0n/BrniCDB0I5CdCcZxEomwQD9q2eOcCRIEUSP8VSS1nfVjqNY3XPoprDA9z32tAaDtjeBuf9J+z6P01Wz+k5GAxr7Hssa57mB1e5zZYSx36XYKvpN+jv3pyxpJJJJKUkkum+rfQqLsOzMy8V973OZ9mZ+aWbnb7djTvs99fp+79Gkpp9P+qnUcwPNpGJta1zW2g7nB43aNb9H2/6RDzfqx1TEuFYYLq3AltzPoaaw8u27Hf6sXcB9DHZGQ4NNjXfrNhkP2gbWF0+12xh+krRIaG7nRWdOJ3T9Fu5JT5TbVZTY6u1pY9ujmnQqK9K6p9V8HqGKL2hjLSC4taQyI5s9R3tZu/t/wDEri+pfVvNwnjWs1kTJuq0nj3bx9L8xR4s0MoJgb4SYm9PVFlz8vPDLhnXnH5XJSWszpHTfSrtt6pU3dX6llbWlzm6F3pth3uf9Fuz9Gq+bhYVNRsxcoZID9nDWH6O4n0/Udb9L8/Z6P8AwykYn//Q8qWt9WumNz8/dbpj48WWExBM+xjt35rtqyV3H1SxxT0ZlpgnLtc7Xs1k1bv7Lm+5JTqMobj1htXDCXtpZ7AS4+59dcs2PZ/o96IGFjGNcB6bTMAuJLjrWGM9ux252/6aDWy5u22wm2IYyrQnmWtr/mvaouyMgVvDC230nBwc0S8Q7e5u2XM3f6P+QgSALK6ETI8MdSb/AOb6i23gRtcxwcDwHtnjvq5Ku002hzHOEiJkb2/u7bB7bf8Az8rTKumOFlJosy7BYdtzXkN1Meo01/o28/zn/fFWtrfj2HFyiHWsO2wGJDTOx+76Lvb9Pb+eqg5nHmBx3EGWsYy9XFD+uP0W/i5f2ZwyGMsgAJmIkRn/AHoQ/S/us6bCbz6zi/aQ6SNQPo7/AG/TrXNfXLpxw7Lc7ptLWVW6Zluj7A95LZ3RtpZdPu9D/SfpLPeuk3U14wdZLq3e6sgEQ4+30BaW2Nd6jf8AA/mLM+sNmD+xMquxl3pN2AfpB9IObY2ufRft9v7yZysSc0pg8MYj2+CvROMfkMP6rP8AEDH2AOGRs3Ey+bHL5p8f/QfPEvJWd/TDoKbmEke42tcAJ93sGOzd7f5av2ZH1YDiKsa/aDoXmdzdxf8ARbbX6Lvd6f0rv0dX+kt9VX3HcdJTvNRusNIiouJYCNuk+32b7dv/AG7Z/wAYoJKbXSsE5/UKcWYa8kvI0IY0Gy0tn870mP2r0T0Ka3D7Gz1Mdh2uoadrm7Q30343qbPo/n17v0n87WuS+pBrHVLiXbbjjvFDYkl0s3bP5TavUd/UXaH1/aXZD2ECXABmwbdHFzrBu9ySmLGWFsl7rGuECq1oB/qmx30a/wCS5j1B2OyRuxyw7fTPo2R7f3RX+j3bPzPai4IsGQW9QyK7sba79JR+jeLJAa33Pcz0tnqfQ/wqr9Swd361jXfqcGLLyHBtteram2sPtdaxQZeYhjkIkgE1v/3IbGHlpZI8Wtbad0tORcyh7HuNrWkkPDfptn86v/B5Dfz2IHWOm19QwfXqdW4uZ6b3PnbtJa/2vbu27XbfcrLKHvtDqP0jtoeHg+17Rrt3fyp9imGejfayn+ZB9bZ2aXmbK/3fp/Q/zFUMrzwGOUYyJ92Uoj5oz/zkf3pOuRAcvIcE+EQ4ZY5Hirg+Xhl/3b5hn4b8LJdS4hwGrHgghwPf2yq66T61Yj2V+pZU2s1Xem1zWGrcHB7v5txO9rPSa1l3/GLm1pOA/wD/0fKl0X1a6vnBrek4zK3WWOJptsMBgI3Pa7T+b+k72/nrnVZ6dnW9PzasurV1TpjiR+cEgoPpGD06/I3W5QcwOa6xjGktse4QD7Zs/Qu/c/S3/wA2p5lIDKcd+52O5o9Aua3c1x/Sen+g91tXt9j/AOkU3f8AGK30K/peRRXm1mK7A8hslrbN231fVp3lra63/wA5t+nfV/xisZvousx66rHG51xtbdp6g0/SvB+k1n8pyE8NcWQZJCo6x+bHw/pej97/AA2xxR4TcRwgfo6S0a+GcrGbbVRZsxco+l6bgN5Lwa2Guvbue7dtZZ6Wyz/gv8Ks/wCtjWY12NlYr25NOLjg5VZAa4Na4N/Qua3+ea5vvrf9P/CrnvrJ9a9DgdPeXOpeGnIGgHpun9F+97m/TWX1zr3VsjJeyzILW2MY54rArB9Sqv1A709vteqEeSkcwzgiN3ceEXOxw3OX739ZUOYBAMwbAGg9UZmH83KYl/6G7VP14wGlzjTdW4yNo2uaR23t3V793+jd7Fn9Y+tOJ1THZh/Z7MagOJeaXBgdx6W/FA9H9H9P091fv/wi5tJXxEDYMU8s8hJnIysmX1lu6rOldOdSLf2jW5zxuZRox5Hva5tjrHehTY1/o/Tt/m32/wCi/SUs7GpouDMez16yARYI1nyaXbN30/Ts/TM/wq2Pqd9Xa+tZlj8kOfiYuz1Ka5D7HWbhXUx7Wuaz+bstf/Iq9OtepdVrwsPpmBi9KFduDQRdW1sAhp3V43qVP2+t9G1n2i3/AAn9I/SIGcQSCRcRxSH6Qif0qTDDOZiIj5zwxJ0jxduJ8NSXtH12+rnQ+r4v2j1Kvt5x2uxbaGFpg6+rb6e5tuNa5uyv1f5hllno/wCjXi6NiyLFjcLDGQAkQQJXR71u9T9WPq1YcnCzcx76a8ljr8I1Bzi51T9ljX+n727GNfdsr9/p/wCYt/qnp0nGdRlWA2ONrHvO6p5rO6rZb7GXN9VvvY+irYp/UG/E6n0JnTw41ZvS7vVY5riHDe5z2W1/ybN1mNez/wBLLoL8GrDznZ9lLsiuyXw0OtLLJ923F3bXfaG/Tez6FibLBMzhkhkIAIvGfkrr8rYxmIjtuDZA4t/737rmdLwW09Xp6hgPruZcLH/ZXu3Vh7v56trtW4uU59X0/oP2Vq91x1GFc/8AadNeLTnlgF9dkMNga/e291o9Op9lmxn2p7PT2en6ykRS+z1WlrmWhxuALXDdJkbq/Z6lO30rWf2FyX1z+svTjQzo2O5ufQHB9wrdNdUD2sxMiHt+0e/c+yv1Mev+a/T/AKZUM/JnLznHEjhiBCfzaCPrhwy4vmjk/Q4Fo5mQkasShoJR21/7r+69Jk9PxcXDrysTJbfjen+ju379/uFezfWdv84szK6t07FePtmUyuwDd6ZDg08tH6Kpr3P97dq5HEy+u0VVYvRsg5GLY9xpa1jC4PLS99d1bw91dmxr3/T9L/CUKhkYHXcm31cmjJute0OD7Gvc4tgbPc7+SrHL8vkx2Mkxl1JEjHhn/VZJc3I4hETycQHDqfRw+nr88/lTdZ60zqFVVNbHBtbi7fY7cdeGUs19Gn+R6l3/ABiylbv6T1LHY+y7GsZXXO95adohwq3bh7dnqO9Pf++qitNR/9Ly2mm2+1lNTd1lh2tbxqfMrWxfqx1X1Q7JxH+k0bnMa5gc6Wmyur6fs9fbs3f9Cyz9GsdrnNcHNJa5pkEaEEKXrXAyLHSRBMniNv8A1JSU7v7M+tdWaMiqp2Pe4GAx7WMY2siltLtz/TY2uWMYyxDo+tfV8OrOosO/JyoY7IeTvr27mObVtOxvKxvWugj1HQYkSe3Ci1r3u2tBc48AakoEAijqFHXRNg4zcvJbQ65mOHBxFlpDWbmtc9jXPd9D1Ht9PcrXV8euRk03syGyKX7ABBra2ut+31LX7LmM3qpfhZmNY2q+iyqx8FjHtIJnjaCnycDNxI+00WUg93tIEnt/W/koqQJLQwquiOx2vzbrW2gv31187QB6XpzW9jnWv3Mfvtq9H6f6b+bVfMZhsez7I8vY5gLt0yHH6TXTXVtd/JZ6zP8Ah7ElPff4vOlf5Cu6hVudlW5IGxpcHGmkN3HH2N/n67Ln2f4T9Euo6pj2MazFyr2k17rcf2uL4k7WXafo/Ub/ANu/n7Fzf+LLq+EMFuHkvDX4WT63ptZue+u0NHqzub7Ma6v3+m19vvqXSfWXpmcylj33utyLH7L7mANBO4+l+jcf0ddbHNqssb+YoM2P3ARjIhl9MTPTijjvi/dk2+Wze0YynZx+qUABf6wen9L5f6zDKry7Bj519nrV5Nja2hoexobO1zPc39B6bv8ABs/nP8H6i8j67049M6vl4Q1ZTa4VE6E1n30Wbf8AhKXMevccyi5nTbMYviu9jPRqJ2tb6Addkv3e/Yyxrq2bP9IvEvrL1MdV63lZrTuY4trrfxuZUxmPVZ/brqa9LDwRnPGNZAQnklfzTyce/wDiLeYmckMczURc4Y4DThxx4eH5WpgdQzenZLMvBufj5DPo2MMHXlp/eY79xy2OpfXv6z9SqFNmWceoCHNxgKd+m39K6na9+6f5vd6X/Brn0lPZa9nu7lX1ryqfq4ehU0sZu3Ndkyd/pvcbLKwz6Ld+7Y5/+iWGktPE+r+fm4jcrG2vY4wQN2nu9MbnBuzduH0PzK/Ss/wqAiBdDc2UVTnVXW0v30vdW+I3MJBg86tVqvrHVKp2ZNkuLiSTJlwLHO3Ol2/a53v/ADFad9V+qNt9JwYyw0i9oe7ZLD52hjd39b/0oh3/AFfz6TRuNbm5NnpVPY8OaXadx/K9v7//AIFvKmtb1PqN1bqbcq6ypwAdW6xxaQDvaCxztv0/eqy12/Vbqbho6jedp2G1oMPPpsed3sb+k9n0v+F/mP0qyrGOrsdW76TCWn4gwkp//9PypJJJJSbDxLs3JZjUDc98+AAAG57jP7rQvQsDpXTOm1xTWz1WtIN2vqEfn6z/ACVy/wBSxjftJ/rlzHPYa2PDN4Ad/PbdR+m2fzbPz/0i6/I+zuO2gWGsSRvAE6e87d35/wBH03/9uKKebhmIcJ7mf+TiP7zYxcuJwMuPWjwwA4pGW0f7vG4fXqmDqvSX2bdj3PJESN7S0/5u702rcsrqvLse+tr6yfcx0kQT7C7/AF/warOsIuqY5rTaAC02wdkiyzfU7832Hbu/PVjDznC8ZFQDocG2BzgDEBzTtdu/O+mmZM5iJmIEtI8Gvol/hMmLlRMQ4pAEnJxQvhzfq+GPBGP+L/4Y8t9aek1YONVdh1lmPkW2PsbyGEbWVNZ+fs/nP7a5pemfXR9Wb0O71GtZk1htoDGPY3nc9273U7tXfuer6i8zTuWzHNj45QOM/un/AKUZfpRYM2IY5CIJ29V/oyEuExbHT+oZXTcuvMxH7LaiCJ1Bg7tr2/nNXqTf8ZmD1PGF78S+rHxhuzWN2WkF25mOyp25lmRXt/S3W2V1fzf/ABS8lSUp8NNmIkkAXt/Ivo3Xfr23q91WDifq9dlFnq30v93trt9OpjrG/oWud/SP9J/g/f8ApF5ykkgI0Sepqz5KuVAGRIA0HZlXXZbY2uppsseYaxoJJPg1rVv9F+qT86pmRlWmmt8kVMbNhaCWe5zttVO5zXfS/Sf8EtT6sdIs6fQ3LNdhzslu0QAPQYXFn6T1HfSft9V/5/pfo/8ASLUzMujExi19wfkNaQKamkPL4LqzXS1xdV7vfuRve9F1aivUe1f81ot6HgY7m+lgtxbKXg1ZWRb6jS+Pburbu9bZ/ObNlLPUWo8Qzc3azFrY5+4HYxrh+ezRu2tCbji11ONkMNjWsZa69xa9jrWgNbtbO76b/U+jssVYVNyjU/KZ9osfYd+6wsaDQ5zfT+zO/Rvx3bff7P0m9Fa1ndOweovfmZFfqOyduwWlwhjvofZnMcyuprtm+n1PVeue6n0SzDwftVbHPode5rbD9JjNrXVC5jfax93qf9BddkvDg4XOrDN9VloEuNm52jPb+buHo1sVXqGPjvZVhZPtxmzY9jPbNlxta573D3vbW5uK/wBz/wAxJTwiSJk49mNe+iz6Vbi0kcGDEtQ0lP8A/9TypJJJJT2P1QZt6Y9xZ9O5x3PMNIDAxrmfyq3Od710fRcvHq6iy7LA9Ma1sdBA3H027g8t+iyz1XLB+p1w/ZRa0y5lrwWAku9wr2ljT9BdLV0Y21Ps9ObvYGhxAYWR2n3W2bf6jP8AjFHlw+7jyY9R7kTEkeXDVs2Ik8Ikf1cD8o39Xqka/wAFnVfj9LFmF0zHGdj0hzajZ+k2vJMMZa8t3VtY79J/OVMs/mf8IjZOR09+CcyttNWS2hjKGhu6xordssw7K3M/RM9T22W/nrMfXmYb7KvtAY2ut1jRtjcZ/mmNdW3fYrDOi5WQ0Wv2ttcAbtRA9s117WBzfVZ/hN6qR5HJLJE8c45MZEp5OLXJ+nLj/Ql7iMcpccgRcRsB6f7nqaWRczIx3subSN4dW2vYCPefScP0bWWO/kemvPH5VJDmuw6Q7UbgbQQeJ2+ts9v9Relii3GtFdzW1jdurcTwR7nD1G/mt/nF5fkvNmRbYXeoXvc4v43SZ3cN+kr+zHIEHV0WdV6V9mqqu6WyyyusVm1thrJ+luscK2e6z3fSs3pO6t0suujpVfpv2mlhsdLHNYWH9I1rbLGPsd6zmbllJJIdg9X6KXg/sdgY0EBouf8Ayiz3bfds3fn/AE/+K/RqmzZm9UrGNSMdttjA2lhkN+iDtdbP9b3IdPTs64MdVQ97bCGscBoS4Pc0B3/Wbf8AttFHTeqY19e6l1Nwe0s3wwh0eoHe8t27G++x/wDgv8Ikp7/17Nu/HDstt5cN9jhXs27ncbA73f20N9LsagUMabs61rxaJ9ztrHPaMux/6T1WS3Zh/wBv9Es/D+sNbQyurLacitoL73S9zrdu308Rr3M/P/w799v+g/0ryWdV6Xh00iWNy8U2fararC+x7nQ9xe58sd73Po/R/wA96e//AEahzkgRqJnr0716eJs8qQDL1CBlUeL92P6e7ZodU+xjX2/abrXFpgNqH6L+e1rB9LH/AJr2t9/qf11n5NFBrpdmkY9l1j3enUH2WWue7dY2Rt9OtjdjP0bEV1+TnVOr6bgtqe8tddlWOmxhsHqvYzX2uex36Suv9Ig4jGu33kWtrqPoX5tj3OttIPtow2f4F3qbd/7jP5z/AAifjvg+XgrQRJ4v+ix5BAZN+OJ3lEcPF/ci61OHVbcxjW+m2G7C8uDtrP0n+ELdu6/0a/8AhEHrXTvsNYNhrF2RaKHO3OsDHVhjfTf+8+rd+m9H89Gxa78utjMQluS5xb6Yex1lllfvtbbff+iYxrR9H/V8uu0dXfbhfaMd1bKWubUXubobP5y1zd+yy7X+cc/9NY9VMmbLDmMcDkjwy1OMcPuV6/0fm9cv+g2fb5eYPtCqBifcvj4qHrH914vr9BY2hxB3Vg0ueeHlhLdzB9L02/R9yx1pdVyCR6LnmyyZduEFvLnDb+bue5ZqvNF//9XypJJJJT0X1L6pXh5l2LYWMGa1rWWv0AfW71a6935jL/5v+v6a9Sw8mm+oMHIiWHlsfm7V4Wuy6L9bqXU1UZ9zqL64aLy3c135ofY5sWt9oa1//bqcMnDoRp4MuI3pYHbi0/5z3eU7HyM0MtcKBjxZW9tYe5znHZ6hc76NFLtn/GeormLk2WUtdcA2yPe0aAFef5v1n9XrGBjsfWRjudYcgO3AvLHsqqa521vp7ne9aXV/rLh41ANltZc4bmNqG55B8N5/6tP44p4wCQr689Zrx8FzKXht9hLaiPpS4bbHNI+j6dR/z31LzRXOrdTt6lluvfLa5iqsu3bW/wDknfnqmoybNsczZUkkkgtbFXUc6mttVdzmsYCGCfo7vp+l/o3O3fTrUXZmXZUaX2OfX7SQdY2htbfd9JrdjK2/9bq/0aCj4ubk4frfZ37PtFbqLdAZrfG9nvDtu7b+akpJhkYrX5jtLqwBjMPO987b4/cob72O/wC5Hof8IpdGsFGe3LIkYjXXxMe5gmkO/kuv9Jjk1vV+o5GKMK27djgVtawhsAVj06oO327WqxlVdOx8a9vT8v7QXbWvLht317g/9GyxrHsey6tm5n+j/SJJBo2Ojs/86eluaMlzLm3srLRiVgV0ufZ/OWOex+93u/OeruF1jAzKcamnGdkuDmk0SA2sfnta3/BMr936RcKrHT867AzK8qk+5h1HEtI2vZ/aammOgA6fy1VImR4ut2+tetj9Poruusdfl0thraf5kOdP8ztb77Pf/SH/AKX/AItct1zqX2lx3sfWdur7H7nNZ9L3bS5Y2d9Yqi414htfjmHBrjsgnlr/AKW5Y+Tn35A2mGV/uN4/tfnPRjDEPVDHGMiKJ4YifD+7KTLLPKX6R1136+K2dknKy7MgiPUM/d7dUBJJFhf/1vKkl2SSSnjUl2SZJTxyS7FOkp41JdkmSU8ckuySSU8akuxTpKeNSXYpJKeOSXZJklPHJLskySnjkl2SSSn/2ThCSU0EIQAAAAAAUwAAAAEBAAAADwBBAGQAbwBiAGUAIABQAGgAbwB0AG8AcwBoAG8AcAAAABIAQQBkAG8AYgBlACAAUABoAG8AdABvAHMAaABvAHAAIABDAFMAAAABADhCSU0PoAAAAAABDG1hbmlJUkZSAAABADhCSU1BbkRzAAAA4AAAABAAAAABAAAAAAAAbnVsbAAAAAMAAAAAQUZTdGxvbmcAAAAAAAAAAEZySW5WbExzAAAAAU9iamMAAAABAAAAAAAAbnVsbAAAAAIAAAAARnJJRGxvbmdS78diAAAAAEZyR0Fkb3ViQD4AAAAAAAAAAAAARlN0c1ZsTHMAAAABT2JqYwAAAAEAAAAAAABudWxsAAAABAAAAABGc0lEbG9uZwAAAAAAAAAAQUZybWxvbmcAAAAAAAAAAEZzRnJWbExzAAAAAWxvbmdS78diAAAAAExDbnRsb25nAAAAAAAAOEJJTVJvbGwAAAAIAAAAAAAAAAA4QklND6EAAAAAABxtZnJpAAAAAgAAABAAAAABAAAAAAAAAAEAAAAAOEJJTQQGAAAAAAAHAAgAAAABAQD/4R5zaHR0cDovL25zLmFkb2JlLmNvbS94YXAvMS4wLwA8P3hwYWNrZXQgYmVnaW49J++7vycgaWQ9J1c1TTBNcENlaGlIenJlU3pOVGN6a2M5ZCc/Pgo8eDp4bXBtZXRhIHhtbG5zOng9J2Fkb2JlOm5zOm1ldGEvJyB4OnhtcHRrPSdYTVAgdG9vbGtpdCAzLjAtMjgsIGZyYW1ld29yayAxLjYnPgo8cmRmOlJERiB4bWxuczpyZGY9J2h0dHA6Ly93d3cudzMub3JnLzE5OTkvMDIvMjItcmRmLXN5bnRheC1ucyMnIHhtbG5zOmlYPSdodHRwOi8vbnMuYWRvYmUuY29tL2lYLzEuMC8nPgoKIDxyZGY6RGVzY3JpcHRpb24gcmRmOmFib3V0PSd1dWlkOjQwYTQwNWUzLWEwZTItMTFlMS1iZDI3LWNkYWEzZTA4YTgzNCcKICB4bWxuczpleGlmPSdodHRwOi8vbnMuYWRvYmUuY29tL2V4aWYvMS4wLyc+CiAgPGV4aWY6Q29sb3JTcGFjZT40Mjk0OTY3Mjk1PC9leGlmOkNvbG9yU3BhY2U+CiAgPGV4aWY6UGl4ZWxYRGltZW5zaW9uPjE0MDA8L2V4aWY6UGl4ZWxYRGltZW5zaW9uPgogIDxleGlmOlBpeGVsWURpbWVuc2lvbj4xMDU2PC9leGlmOlBpeGVsWURpbWVuc2lvbj4KIDwvcmRmOkRlc2NyaXB0aW9uPgoKIDxyZGY6RGVzY3JpcHRpb24gcmRmOmFib3V0PSd1dWlkOjQwYTQwNWUzLWEwZTItMTFlMS1iZDI3LWNkYWEzZTA4YTgzNCcKICB4bWxuczpwZGY9J2h0dHA6Ly9ucy5hZG9iZS5jb20vcGRmLzEuMy8nPgogPC9yZGY6RGVzY3JpcHRpb24+CgogPHJkZjpEZXNjcmlwdGlvbiByZGY6YWJvdXQ9J3V1aWQ6NDBhNDA1ZTMtYTBlMi0xMWUxLWJkMjctY2RhYTNlMDhhODM0JwogIHhtbG5zOnBob3Rvc2hvcD0naHR0cDovL25zLmFkb2JlLmNvbS9waG90b3Nob3AvMS4wLyc+CiAgPHBob3Rvc2hvcDpDb2xvck1vZGU+NDwvcGhvdG9zaG9wOkNvbG9yTW9kZT4KICA8cGhvdG9zaG9wOklDQ1Byb2ZpbGU+VS5TLiBXZWIgQ29hdGVkIChTV09QKSB2MjwvcGhvdG9zaG9wOklDQ1Byb2ZpbGU+CiAgPHBob3Rvc2hvcDpIaXN0b3J5PjwvcGhvdG9zaG9wOkhpc3Rvcnk+CiA8L3JkZjpEZXNjcmlwdGlvbj4KCiA8cmRmOkRlc2NyaXB0aW9uIHJkZjphYm91dD0ndXVpZDo0MGE0MDVlMy1hMGUyLTExZTEtYmQyNy1jZGFhM2UwOGE4MzQnCiAgeG1sbnM6dGlmZj0naHR0cDovL25zLmFkb2JlLmNvbS90aWZmLzEuMC8nPgogIDx0aWZmOk9yaWVudGF0aW9uPjE8L3RpZmY6T3JpZW50YXRpb24+CiAgPHRpZmY6WFJlc29sdXRpb24+MjAwLzE8L3RpZmY6WFJlc29sdXRpb24+CiAgPHRpZmY6WVJlc29sdXRpb24+MjAwLzE8L3RpZmY6WVJlc29sdXRpb24+CiAgPHRpZmY6UmVzb2x1dGlvblVuaXQ+MjwvdGlmZjpSZXNvbHV0aW9uVW5pdD4KIDwvcmRmOkRlc2NyaXB0aW9uPgoKIDxyZGY6RGVzY3JpcHRpb24gcmRmOmFib3V0PSd1dWlkOjQwYTQwNWUzLWEwZTItMTFlMS1iZDI3LWNkYWEzZTA4YTgzNCcKICB4bWxuczp4YXA9J2h0dHA6Ly9ucy5hZG9iZS5jb20veGFwLzEuMC8nPgogIDx4YXA6Q3JlYXRlRGF0ZT4yMDEyLTA1LTE4VDE3OjM5OjAyKzA1OjMwPC94YXA6Q3JlYXRlRGF0ZT4KICA8eGFwOk1vZGlmeURhdGU+MjAxMi0wNS0xOFQxNzozOTowMiswNTozMDwveGFwOk1vZGlmeURhdGU+CiAgPHhhcDpNZXRhZGF0YURhdGU+MjAxMi0wNS0xOFQxNzozOTowMiswNTozMDwveGFwOk1ldGFkYXRhRGF0ZT4KICA8eGFwOkNyZWF0b3JUb29sPkFkb2JlIFBob3Rvc2hvcCBDUyBXaW5kb3dzPC94YXA6Q3JlYXRvclRvb2w+CiA8L3JkZjpEZXNjcmlwdGlvbj4KCiA8cmRmOkRlc2NyaXB0aW9uIHJkZjphYm91dD0ndXVpZDo0MGE0MDVlMy1hMGUyLTExZTEtYmQyNy1jZGFhM2UwOGE4MzQnCiAgeG1sbnM6c3RFdnQ9J2h0dHA6Ly9ucy5hZG9iZS5jb20veGFwLzEuMC9zVHlwZS9SZXNvdXJjZUV2ZW50IycKICB4bWxuczpzdFJlZj0naHR0cDovL25zLmFkb2JlLmNvbS94YXAvMS4wL3NUeXBlL1Jlc291cmNlUmVmIycKICB4bWxuczp4YXBNTT0naHR0cDovL25zLmFkb2JlLmNvbS94YXAvMS4wL21tLyc+CiAgPHhhcE1NOkluc3RhbmNlSUQ+eG1wLmlpZDoyMEE1ODZDMzVGNThFMTExQUM1RDlFNjVBMUJEOUE4RDwveGFwTU06SW5zdGFuY2VJRD4KICA8eGFwTU06T3JpZ2luYWxEb2N1bWVudElEPnhtcC5kaWQ6MUVBNTg2QzM1RjU4RTExMUFDNUQ5RTY1QTFCRDlBOEQ8L3hhcE1NOk9yaWdpbmFsRG9jdW1lbnRJRD4KICA8eGFwTU06SGlzdG9yeT4KICAgPHJkZjpTZXE+CiAgICA8cmRmOmxpIHJkZjpwYXJzZVR5cGU9J1Jlc291cmNlJz4KICAgICA8c3RFdnQ6YWN0aW9uPmNyZWF0ZWQ8L3N0RXZ0OmFjdGlvbj4KICAgICA8c3RFdnQ6aW5zdGFuY2VJRD54bXAuaWlkOjFFQTU4NkMzNUY1OEUxMTFBQzVEOUU2NUExQkQ5QThEPC9zdEV2dDppbnN0YW5jZUlEPgogICAgIDxzdEV2dDp3aGVuPjIwMTItMDItMTZUMTA6NDk6MzQrMDU6MzA8L3N0RXZ0OndoZW4+CiAgICAgPHN0RXZ0OnNvZnR3YXJlQWdlbnQ+QWRvYmUgUGhvdG9zaG9wIENTNS4xIFdpbmRvd3M8L3N0RXZ0OnNvZnR3YXJlQWdlbnQ+CiAgICA8L3JkZjpsaT4KICAgIDxyZGY6bGkgcmRmOnBhcnNlVHlwZT0nUmVzb3VyY2UnPgogICAgIDxzdEV2dDphY3Rpb24+c2F2ZWQ8L3N0RXZ0OmFjdGlvbj4KICAgICA8c3RFdnQ6aW5zdGFuY2VJRD54bXAuaWlkOjFGQTU4NkMzNUY1OEUxMTFBQzVEOUU2NUExQkQ5QThEPC9zdEV2dDppbnN0YW5jZUlEPgogICAgIDxzdEV2dDp3aGVuPjIwMTItMDItMTZUMTE6MDU6MjErMDU6MzA8L3N0RXZ0OndoZW4+CiAgICAgPHN0RXZ0OnNvZnR3YXJlQWdlbnQ+QWRvYmUgUGhvdG9zaG9wIENTNS4xIFdpbmRvd3M8L3N0RXZ0OnNvZnR3YXJlQWdlbnQ+CiAgICAgPHN0RXZ0OmNoYW5nZWQ+Lzwvc3RFdnQ6Y2hhbmdlZD4KICAgIDwvcmRmOmxpPgogICAgPHJkZjpsaSByZGY6cGFyc2VUeXBlPSdSZXNvdXJjZSc+CiAgICAgPHN0RXZ0OmFjdGlvbj5zYXZlZDwvc3RFdnQ6YWN0aW9uPgogICAgIDxzdEV2dDppbnN0YW5jZUlEPnhtcC5paWQ6MjBBNTg2QzM1RjU4RTExMUFDNUQ5RTY1QTFCRDlBOEQ8L3N0RXZ0Omluc3RhbmNlSUQ+CiAgICAgPHN0RXZ0OndoZW4+MjAxMi0wMi0xNlQxMTowNToyNSswNTozMDwvc3RFdnQ6d2hlbj4KICAgICA8c3RFdnQ6c29mdHdhcmVBZ2VudD5BZG9iZSBQaG90b3Nob3AgQ1M1LjEgV2luZG93czwvc3RFdnQ6c29mdHdhcmVBZ2VudD4KICAgICA8c3RFdnQ6Y2hhbmdlZD4vPC9zdEV2dDpjaGFuZ2VkPgogICAgPC9yZGY6bGk+CiAgIDwvcmRmOlNlcT4KICA8L3hhcE1NOkhpc3Rvcnk+CiAgPHhhcE1NOkRlcml2ZWRGcm9tIHJkZjpwYXJzZVR5cGU9J1Jlc291cmNlJz4KICAgPHN0UmVmOmRvY3VtZW50SUQ+eG1wLmRpZDoxRUE1ODZDMzVGNThFMTExQUM1RDlFNjVBMUJEOUE4RDwvc3RSZWY6ZG9jdW1lbnRJRD4KICA8L3hhcE1NOkRlcml2ZWRGcm9tPgogIDx4YXBNTTpEb2N1bWVudElEPmFkb2JlOmRvY2lkOnBob3Rvc2hvcDo0MGE0MDVlMi1hMGUyLTExZTEtYmQyNy1jZGFhM2UwOGE4MzQ8L3hhcE1NOkRvY3VtZW50SUQ+CiA8L3JkZjpEZXNjcmlwdGlvbj4KCiA8cmRmOkRlc2NyaXB0aW9uIHJkZjphYm91dD0ndXVpZDo0MGE0MDVlMy1hMGUyLTExZTEtYmQyNy1jZGFhM2UwOGE4MzQnCiAgeG1sbnM6ZGM9J2h0dHA6Ly9wdXJsLm9yZy9kYy9lbGVtZW50cy8xLjEvJz4KICA8ZGM6Zm9ybWF0PmltYWdlL2pwZWc8L2RjOmZvcm1hdD4KIDwvcmRmOkRlc2NyaXB0aW9uPgoKPC9yZGY6UkRGPgo8L3g6eG1wbWV0YT4KICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCjw/eHBhY2tldCBlbmQ9J3cnPz7/7gAOQWRvYmUAZEAAAAAB/9sAhAABAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAgEBAQEBAQICAgICAgICAgICAgICAwMDAwMDAwMDAwMDAwMDAQEBAQEBAQIBAQIDAgICAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwP/wAARCAQgBXgDAREAAhEBAxEB/90ABACv/8QBogAAAAYCAwEAAAAAAAAAAAAABwgGBQQJAwoCAQALAQAABgMBAQEAAAAAAAAAAAAGBQQDBwIIAQkACgsQAAIBAwQBAwMCAwMDAgYJdQECAwQRBRIGIQcTIgAIMRRBMiMVCVFCFmEkMxdScYEYYpElQ6Gx8CY0cgoZwdE1J+FTNoLxkqJEVHNFRjdHYyhVVlcassLS4vJkg3SThGWjs8PT4yk4ZvN1Kjk6SElKWFlaZ2hpanZ3eHl6hYaHiImKlJWWl5iZmqSlpqeoqaq0tba3uLm6xMXGx8jJytTV1tfY2drk5ebn6Onq9PX29/j5+hEAAgEDAgQEAwUEBAQGBgVtAQIDEQQhEgUxBgAiE0FRBzJhFHEIQoEjkRVSoWIWMwmxJMHRQ3LwF+GCNCWSUxhjRPGisiY1GVQ2RWQnCnODk0Z0wtLi8lVldVY3hIWjs8PT4/MpGpSktMTU5PSVpbXF1eX1KEdXZjh2hpamtsbW5vZnd4eXp7fH1+f3SFhoeIiYqLjI2Oj4OUlZaXmJmam5ydnp+So6SlpqeoqaqrrK2ur6/9oADAMBAAIRAxEAPwDQABsb/wC9+/daIr16/BH9Tf37r1Ouvfut9dkW9+60DXroWvz9P8Pr791v7OvH68fT8e/de67sbX/F7e/darmnXXv3W+uViRf8Dj37rVQDTrj791vr3v3Xuve/de697917rs2vxe3+P1/3j37rQr59de/db67It/tr+/de669+6912bX4vb/H37rQr59esbX/F7e/dermnXaqW+nv3XiQOPXXF/wA/7x9ffuvZ68Lfm/8AsPfutmvl117917rkpJ9It+f959+6qaDJ66PBtz/sffutjrwNjf37rZz143vz+ef9v791odde/db67Nvx/vPv3Xvt69fkHjj37rVOuvfut9e9+6913fgj+vv3WqZr11791vr3v3XuuwCTYe/daJpk9eP+P44/23v3Xh11791vrssTa/49+60AB10Lfn6f4e/db+zr3v3Xuve/de65Ai1j9P8AD6+/daI8x15vx/rcf634v7914ddC3N7/AOFv+J9+63nrwt+b/wCw9+699nXX+t7917r3v3Xuve/de697917rvliB/sB791rhnr3Fje9/949+69ny669+631737r3XZt+L/7H37r329eAvx7917h1yCj6G4b/AHj37qpJ4+XXD37q3XvfuvddgXv/AIe/dar17i35v/vHv3Xs1669+6312Bc29+68cddlbDn/AHj37qoNTjrq3F/fut1zTrxBFr/kXHv3Xga9de/db67IIAP9fp791oGuOvAEgn+n19+68TQ068Bc29+62cdesBcH/ePfutfZ1ysW/px791rC9cQbH/Ee/db4jr3HN/r+PfuvZ8uvAX49+63w68QQbH37rQNRXrr37rfXvfuvdcgxAI/r/r+/daIqa9dcWP1v+P6f439+63117917r3v3Xuu7cE/0/wCJ9+69Xy65HgC39oc3/wCI9+6qMn7OuHv3VuuQUkXH49+60SB1x9+631737r3Xvfuvdd8W/N/94/x9+61mvXuLH63/AB/T/G/v3W89de/de697917r3v3XuvDjn37r3XZNzc/7x791rgOvG1+Pp791sV8+uvfuvde9+691737r3XIi1r/Q/wBP9b37rVa8Ouib2/w+n/G/fuvAU68CRe359+68RXru/Fj+P6f8T7916majrj791vr3v3Xuve/de674/N/8Pfuvdd6yOOPpb8+/dV0jrom5v791YY68Bfj37r3DrkBY2/tcW/pc/S/v3VTkfLri17m/19+62OGOu73sD9B/T37r1KZHXgbAg/n37rxFSOuPv3W+ve/de67BtyPfutEV69qJIP8AT37r1BSnXibm/v3WwKde4txe/wCffutZr12eRq/JPv3Whxp1x9+6t1yJBtb8e/daAp1x9+6312Rbj37r3Hrr37r3Xvfuvde9+691737r3XYt+b/7D37rRr5deHJH+v791s8OuyObD/fce/daBxXrj791vrmtvxe9v6ce/dVavXD37q3XfBH51X/2Hv3Ws/l14n8f0449+68PXrr37rfXv9f37r3XZt+L/wCx9+68K+fXXv3Xuve/de65MLG3P+x9+60DUdcffut9dk3N/fuvDHXLnR+Lf7z9ffuq41dcPfurddk3N/fuvAUx117917r3v3Xuve/de67sCB9dV7f4e/daz+XXuVP+I9+69xHXib/X37rwFOuvfut9e9+691737r3XZt+L2/x9+68K+fXXv3XuuVwDx/T8+/dapXj11b0k/wCNvfuvVzTrr37rfXvfuvde9+6913xY/wBfx791rNeuvfut9ciRaw5F78/X/W9+60AeJ64+/db67It/tr+/deBr17i35v8A7x791rPXXv3W+uQJAJFvxz791U0JoeuP59+6t12bfi/+x9+68K+fX//Q+f8A+/de697917rkLfn/AIn/AIj37rRr5dcffut9ciCBaw/17f7H6+/daFD1x9+631yIINvr+eL+/daqOPXH37rfXvfuvde9+691737r3XduL8f8T791quadetxfj/fce/dermnXXv3W+u/xa3+xtz791r59eHJsf98B7917yx121voLce/deFfPrj9ffut9cuWNvp/vXHv3WsDPXVyLj/effuvUBz119ffut9di1+fp791o/Lrx9+68Ou1t+Tb/ABF/fuvGvl1x9+6314cf4/6/v3Xuuzf8/kX/ANh791odde/db67vxaw/1/z791rz68LX5+n/ABr37rxrTHXbEG1v6e/daFfPrj791br3v3Xuve/de67AJ/w/1+L/AOt791okDr1iP9h+R9P9v7916oPXXv3W+uQtY/T/AGNv6e/dVNajrjY+/dW674t/j791rNeuvfut9d24v+PfutVzTrvjT/j7917Nfl1x9+6313fgjjn8259+61TNeuXouP6fn6/7D37rXdTroj6kfS9v8ffutg+R68SLi30/3319+68K0z17UOSQL/gW49+61Q8OuJ559+6t13/S39P959+619vXXv3W+u7H62NvfuvVHXXv3Xuve/de697917rkCfoLf6/5/wBv791ogefXH37rfXd+Le/de669+691y408/U/T6/T/AA9+6159dWNr/ge/deqOHXh9CfyLW/2Pv3Xjxp13c2FxwPpe9j7916g8uuib/wCH+A+nv3Xqde5/N/8AD37reOvA2v8A4i3v3XqdeIIAv+fp/h791oEHh17kf1Hv3Xsdde/db697917rsEj/AIp+PfutEV669+631737r3XJTY88f645Hv3WiKjHXfo/Nz/jz791ru8uuP1uR9P+K+/db+XXXv3W+uTW4sLfX+v/ABPv3VRXr1za1h9Prb/Y+/dbpnrofUfn/A/T37rZ4dcibW4Fxe/HH+Hv3VQOuJN/+Ne/dbAp11791vr3v3XuvXP9ffuvde9+6912TcAcce/dap11791vrlYqQeD+f6+/daqD1x9+631737r3XIX/AAAf9f37rRp1x9+6312Ra3+PPv3Wq1669+631yGn8/8AE/8AEe/daNfLroXsbC/+9j/W9+68addf4e/db69Yj6i3v3Xuu7n/AJH791qnXXv3W+ve/de67+h/HHv3WuI67Bu1zYf7Dj/be/depig68bar/i4/H+349+69mnXR+p9+62Ouvfuvdd2+n05/33Pv3Xuuvfuvde9+691y/Fze/wCDz/Tj37qvnQdcffurde9+6912Bf8Ap/sffuvE066PH/Gvfuvde9+691737r3Xvfuvde9+6912TcAWHH+39+61TrxN/wDin49+63Tr3Hp/3n6/19+61nPXja/H09+68K0z11791vr3v3Xuu+LD+vN/+I9+61mvXXv3W+uXAAPF/wCn/GvfutZrTrom5/4p7914CnXgbfgf7H37r3Hrwvzb+hP/ABv37rx68Db+h/1/fuvU67uAPoL/AOI9+69Qk9dAkfT37rxFevWP1/H9ffuvVHXQ+vv3W+u7XJtz/j791rrkStuP6j8f0H+Pv3WhWueuIv8Agf7f6e/dbNOvEWNv969+68DXPXXv3W+u7GwP9T791qo4deII+pv7914fLrtbc3/2Hv3XjXy64+/db697917r3v3XuvHk3+n+t9Pfuvde9+6912eeeB+LD37rQxjruxH4Bv8A7H37rVR1x9+6t1737r3Xvfuvdd3uLWHH5tz/AK1/futUzXr1ja/49+69XNOuvfut9dg2/AP+uL+/daIr14c3/HBP9PfuvcOuvfut9e5t/h/vHv3Xuu73FrD/AFwOffutUoa9de/db697917r3v3Xuve/de65gWNiPxf6En/ePfuqk4qOuHv3VuuXGm/N/wDY/wBffutefXH37rfXvfuvde9+691737r3XbWvx9Le/daFfPr/0dAAixt791oGvXfpt/j/AMb9+69mvXRFvyD7914GvXXv3W+uYu3p/wBuT/t/fuqmgz1xAuSL/S/+8e/dbrjr2o3v9T9OffuvUFKdde/db697917rkq6jb/Y+/daJp10Bc2/2Pv3XiaddX9+6312eLc/UX9+60M9de/db67Bt/tre/daIr11+Sffut9chptz9f949+61mvy66Bt7914ivXgOL3+nv3Xjxp14/7e4v/t/fuvDrwNv9t7914ivXXv3W+uzbi39Of9f37rQr59cgotc/4/7x731ony643+o/F7+9db+fXha/P09+68a0x15hY/Xgi4/wH4Hv3Xh14Wv6v6f7z791418uuekAc/7E8+99VqScdY/eur9dgX9+60TTr3Fj/W/+8e/dez11791vr3v3XuueoW54I+lv9b37qtD1xubW/wCR+/dbpmvXib2/1gPfuvDrr37rfWRuOLf8a/Pv3VFznrH791frnZTwPqf9f37qvcMnrjyLj8X9+638+u9Pp1f7x7916uadcffut9cgpIJ/p791omhp17T6dV/9h7916uadcbm1vx791vrmUtfn6e/dVDV64e/dW697917rsGxv79145669+691yLXFuPx/vHv3WqZr1x9+631yU/2bXuf99b37qpHn10Ryf8D791scOuypHN/9t9f9j791oGuOugtwT/T8e/dbrmnXXv3W+ve/de67Jvb/AA9+61TrwJAt+D7916g49d3XTYXubf7f8+/dazXrotcAf09+63TNeuvfut9cr3Fj9R9P+J9+61TPXH37rfXvfuvddk3t/gLe/daAp11791vrsC5t/vfv3WiaCvXiLG3v3Xhnr3Fj/Xi3v3Xuuvfut9cyQbk/U/6/9Pfuq0IwOuI+vPv3Wzw65em4+tvz9f8AYe/dazTrom/A+g+nv3WwPPrvluSfpx731rC9cSSbf4e9dbAp119Pfut9d2+hv9f+Rc+/dar/AC67sAeeRb37rVSRjr1gCCfob+/deqTjz67AXn+g+n1/2/v3XiW64m1+Pp791sVpnryi55/x/wB69+68cDrxHJt9L+/deHDrr37rfXvfuvdcrAi4/H1/2Pv3Va0ND10T9B/T37rfz67W5NgbX9+68aceuiLG3v3Xga9eH1F729+699nXja/H09+68Pn11791vrlytj/X37qvHHXRNzf/ABv791sCgp14te309+68BTrr37rfXIafzf8A41791o18uuja/H0/x9+68K0z10Tfk+/db697917rsC9zf6e/daJp14C9+bW9+68TTr1wbk/Xi3/E+/de4deNuLf7H/X9+68K+fXXv3W+uza/H09+60Pn16/Frf0/3j37r1M168LWN/r+PfuvGvl11791vrmoBH+9+/dVJI64m1+Pp791sVpnrxtfj6e/deHz669+631737r3XYNrj+vv3WqV68bWFvr+ffuvZr11791vr3v3Xuve/de697917rkq6vz791UmnXXFh/Xm/wDxHv3W81669+631737r3XLgAfXUCD/AIfX37quT9nXH37q3XY+v+vx79149eIsbXv791oGo669+631737r3XLUQLD/AG/59+61TNeuPv3W+uQNv9j791oivXRtfj6f4+/deFaZ69c2t/yP37r1BWvXIgWX+pt731oHj1w966t13c/8R791qg66vf37rfXvfuvde9+691737r3XvfuvdcuNPF7/AJ9+61mvXH37rfXv8PfuvdcgxH+I/p791ogHrj791vr3v3Xuu/xb/H/jXv3Wvn12Rb839+68DXrj791vrsC/v3Wiadde/db65AC1z/jYf4+/dVqa0HXSrq/w9+62TTrnYAc/T/iT791WpJx1wBsbj37q1KjPXZFwG/r+P9bj37rQOadcffurde9+691737r3XYJH+P8Ar+/daoOuvfut9d34t/vv6+/dapmvXXv3W+uzb8f0/wB59+60K+fXXv3W+uVuL3/2H+xt791quadcffut9f/S+f8A+/de65X9Nv8AffX37rVM164+/db67Fvze3+Hv3XjXy669+691737r3XLSb2/Pv3WtQpXrj791vrxt+P959+6912Dbke/daIr11791vr3v3Xuuxb83t/h791o18uuvfut9e9+691737r3XZt+L/7H37rQr59de/db697917r3v3Xuve/de697917ru/BH9ffutUzXrr37rfXd+Lf439+6951669+691737r3Xvfuvde9+691yJBA+twAPxb37rQrXroG3v3XiK9de/db697917r3v3Xuuzb8X/wBj791oV8+uySGvxf8A3j6e/depUU64+/db67sCQBfn+vv3Ws+fXiLG3v3Wwa5669+691737r3Xd+Lfg8+/dapmvXrm1vfuvUzXrr37rfXZBFvpzz791oGvXXv3W+uX0Fj9Dz/j/vPv3WuOR1x9+6312Be4/P4/p/sffutE0669+631737r3XZtxa/+N/8AiPfutCvn163F/fuvVzTrr37rfXZt+L/7H37r329eBIv/AI+/daIr12DY3P8Aj/vPv3XiKinXH37rfXvfuvddgXv/AIe/dar17i35v/vHv3Xs168OSL+/dePDruwuf6D/AG/v3Xq9dWuCR9P8ffuvV8uvA2N/futkVx17+v8AX8e/de669+691737r3Xvfuvde9+691yAtpb8E2/4j37qpNajrpvqf99+Pfutjh14m9h/T37r1KddD6j37rfXZFjb37rQyOvAkG49+68RUU68ST9ffuvAU68Bf+n+x9+68TTroi3Hv3W+PXduCf6e/de669+6912Rbj37r3Hrr37r3Xvfuvde9+691yDEC3Hv3WiK9dAXNvfutnHXIXAYf7f6+/dVNCQeuHv3VuuWk2vx/X37rVc066BsQf6e/dbOcdeJub/19+60BQU669+631737r3Xvfuvde9+6912Lfm/+w9+60a+XXXv3W+ve/de697917r3v3Xuu+Pxf/H37r3XXv3Xuu+L/m3+8+/daz163F/fuvVzTr1uAf6/8R791vrr37r3Xgbc+/de49ePPv3Xuve/de67vwB/T37rVM1669+6313bgH+t/futefXXv3W+u7cX/Hv3Wq5p11791vrv08/W34+n+8+/daz14W/N/wDYe/db+zr1uQP62/3n37rVfPrxFjb37rYz14C/v3Wiadde/db65KCeRbj+t/futEgY68SQT9Ofr791qgI64m34+n+Pv3Vvt697917r3v3Xuve/de67Btz791oivXidRuf949+68BQUHXXv3W+u+Lfm/wCf6e/daz11791vrsgj6+/daBr14ggA/wBffuvVzTrr37rfXvfuvde9+6913e5JP+8f19+61SgoOuvfut9d3uLe/dapmvXhb83/AMLe/deNfLrr/W9+631737r3XvfuvddgXPv3Wiaddkc2/wB79+69XFeuPv3W+uWo2t+P9j791qma9dcW/N/949+69mvXgLm3v3XiaddqAb3v/sPfuvE066Isbe/dbGeuvfuvddkW49+60DXr1+APwP8Ab+/depmvXRt+P959+631737r3XfFvzf/AGFvfutZr14G3+2t791vrr37r3Xfpv8Am3+wvf37rWeuvfut9e9+691737r3XfFvzf8AH9P8ffuvdde/de697917rkF5sf6X49+6qTivXXH05vfj+lvfut568Rbj37rfHr1uL/429+69XNOuvfuvddm1uL3/ADf6e/daz11791vr/9PQAII+vv3Xq1669+6912bfj/if+J9+60K+fXXv3W+u7G9v979+69Xz68QR9ffuvA14de5+vv3Xvl117917r3v3Xuve/de697917rwNjf37r3Hr3v3Xuvf4+/de69/h7917r3v3XuvWP9PfuvdciL8r9P8Aif8AY+/dVrTj1x9+6t12Te3+At7916lOvC1+fp/vv6e/daNfLrwv+Pr/AK1/fuvGnn11791vruxtf8e/daqK066sR9ePfut9esfrbj+vv3Xuve/de67AJ9+61WnXXv3W+uxyRf37rR4ddggBv6n6e/daINR1x9+6t12bA8X/ANj/AF/Pv3WuIz11791vr3v3Xuu7m1vfutUFa9de/db697917r3v3XuuX1sB+RyP8ffutfM9cffut9dgE/Qe/daqB14k/Q/8R/xHv3XgB5dde/db697917r3v3Xuve/de671H6f1/wAB791qg68AT9Px7914kDrr6e/db697917r3v3Xuve/de675+n9fxb37rWOPXiCPr7914EHh11791vr3v3Xuve/de699ffuvdcgDew+vv3WjSlT10b35+vv3Wx8uu9R/wB8B791qg66NrC31/Pv3XhXz68bWFvr+ffuvZrnrsWAv/a/Hv3XjWvy64+/db65cGwH1/Pv3Ws+fXrAAg/Xj37r1anHXJ/x/t/9b3vqq9Y/eur9d8WH9ffutZr11791vr3v3Xuve/de697917rlbSRf37qtajHXuNVz9CT/AIX9+635Y66I54+h+nv3Xh11791vrux4/wAfp791qo69+ef68+/db679Or/af+Ne/dazT59dG1+Pp/j7914Vpnrr37rfXZtYW+vN/wDiPfutZr11791vr3v3Xuve/de65Dj6/Q/X/kfv3Wj10bXNvp7914fPrr37rfXvfuvde9+691737r3XYtY3+v49+60a+XXXv3W+u7EH6cj/AGPv3WsEdde/db674sP6839+6112oBNj/sLe/deNRw665Fx7917jnrr37rfXf9L/AE/31/fuvdetc8f7D37rXAZ68Sfofx/xT37rwA4jrw/3j8+/db66Nr8fT37r3XYtzf8A2Hv3WjXy669+631737r3XYJH09+60RXrwBP09+63XrxFjY+/deGeuyAFB/r791UHNOvAixB/2H+v791s8cdcffut9ev9P8Pfuvde9+69176+/de67+h5/wAL+/da8sdde/db697917r3v3Xuve/de67sffuvdde/de675sP6c29+61ivQobD6g352QJn21gq6ppaeMs1b9pVGkYr9VSZEIJ/1vfuteeOlyvxi7MDf5RRR0yg2dpkqE0/1sGUX/2/v3W89CphfhDv/cdDFNiKyCSoccxyRtpv/T08/X3vr2epp/l895eQxJFjXkH9kiqX/elPvXWqkcR0yZP4Kd2Ym33dNj0/rzVi3NvzH791up9Ogf3d8fuwNnpJJX0K1IjuStIs0jG3PAK+/dbz59AxUUdXSOY6qmnp3HBSeJ42B/oQ4Hv3WqjrAbWFvr+ffuvCteurGwP4Pv3Xq5p11Y/X37rfXvfuvdcmCj6fX8+/dVFfPrj791bru5IA/p791qma9de/db697917r3v3XuuQt/a/4n37rRr5deJ/A+n49+68B5nrynm5v7914jFB1x9+63176e/de67Bt9Pfuvceuh9Rf37r3XZtfj/if+J9+60K+fXQ+vP09+6312R+QOP9j791ofPr31H9Tf8Ax+nv3XuB669+631737r3Xd+Lf439+69TNevaTa/44/3n37rVRWnXXv3W+ve/de675H+Hv3WsHrr/AB9+631737r3XvfuvdeFr8/T37r3XZtfj/if+J9+60K+fXXv3W+ve/de697917rs2vx9PfutD59de/db697917rskn6+/daAp11791vr/9T5/wDcn68+/de67Ckgn8D37rVQDTrr37rfXZtfj37rQr59eJub+/deApjrxv8Am/8Asffut48uuvfuvde9+691737r3Xvfuvde9+691yI/I+nH9fr791UHyPXH37q3XfP+Pv3XsdeNvxx791ofPrr37rfXLUQLfj37rVBWvXQJ/F/9h7917Hn11791vr3v3Xuve/de67Btz7917j117917rvn6c/63v3WsdeJv9f8AW9+68BTr1za3v3XqZr14XuP6/wCPv3XjSnXdjyf6Xv8AX37r1Rw64+/db697917r3v3XuuZIFiv+N/r791XJ49cPfurde9+691737r3XYNuffuvEV669+69176c+/de699effuvde9+691737r3XIX0m30v9ffutYr129uPpc8/j6e/daWvXD37q3XYtY3+v4+vv3WjWuOuwhP5HP+v+ffutagOuPv3VuvDjn37r3Xdz7916nXrG1z791qorTrvTcXH0AF/9f8n37rVc0PXmtxYEf1v791sV8+ujawt9fz7914V8+uwTcG30Fvp7916gp119SLn/AG/v3XuAx14WJ/oPfuvGtOvH6m3I9+62OHXja/H0/wCJ9+60K+fXd/02HIHPH5/2Hv3Wqca9dXN/6e/dW669+691737r3Xvfuvde9+691737r3XYFzb37rR4ddfn37rfXY5PP0/Pv3Wjwx13c2P5HPPv3XqDrrix/rxb/iffuvefXgLm3v3WzjrxFjb37rwz14fUX+nv3Wj8uvXte30P+9e/dep69de/db675IuTe39ffutYGOvE3sP6e/dep12b2U8W5t/t+b+/dexU9dE3Nz7914CmOvXP+PH+8e/dbx117917ru3AP9b/AO8e/dar11791vrsWsb/AF4t/wAT791rNeuvfut9cgpPv3WiQOuPv3W+ve/de67vxbj37rVM167bT+Pr791oV8+uPv3Vuve/de67Av8A0/2Pv3XuujwbfX/W+nv3Xuve/de65C3N/r+Pr791o166DEf7H+vv3XiAeuvfut9e9+6912Db37rRFeuvfut9cuQPx6h791rBP2ddX4A/p/xPv3W6efXX0+nHv3Xuve/de67BsCP6+/dap11791vrkbWFvrxf6/7H37rQrXPXRFgD/X37r1euvfut9e9+691yBIHA/wBjz791ogE9cffut9dm/wCb/wCx9+61jrwF+Pfut8Ou2Frf63Pv3Wh1x9+6312Bc2Hv3Wiadde/db67/wAf6/8AEe/de+XXXv3Xuve/de67Auf8Pz791o9dsbnj6D6e/deA66uef8RY+/dep11791vpfdXbTj312DtLaE5lEGfzNNjpjCSJFjmJ1MpH0tb6+/daPr1undEfG/YnSXUOB2PjcJjKyZ6GnqJ6+opYvuT9xT6iGe1za/8AX37z69jqRub4r7D3PQyF446aWRXZftkA9T8/gfj3759erT59F6f4zTbDqfPhXqJoVdiFLScre4JsfyPfvl17Pn1Nj2xmVnWamhk+4UgOJGccL/r+/dep05N17V7hn8eTiCeTg6Wb6/W4/wAR7917pg3X8WdtTULz1ESTM41fuqGvcXt6vevs63wHVfPc3w22zWUmSqYqaBSVLBUiVT9D9Lf4+99aGeqcu1OkM3sevqXoqaafHISbaWeRBqtx/Ue/V63SnQDMjKxRw0bL9UcFSD/Qg+/da66v9Afp/wAR7916nmOvNp4t/sfr/wAT791oV8+vKCxtpLE8AC9yTwLW9+639nXOSCaK3khliv8ATyIyX/1tQHv3Xq9Yvfut9e9+691y4sP6k+/dazXr2kg/jjn/AA4Pv3Wqgjrom/v3WwKdde/db697917rkbXGn/iffutCtM9dEEGx9+68OHXXv3W+uRA4I+n0/wBj791oV4Hrogj6+/deBr11791vru/Fv99/X37rVM168CV+nv3XiK8euzaw/r7917Neugbe/deIr11791vrvn6G/wDre/daxx669+6312Bf37rRNOvEk8n/AFvfuvAAY67BuCOOBf37rRwa9cffurddta/H++/2/v3WhWmeuvfut9e9+691737r3XJrX9P0/wB9/X37rQrTPXXHFv8AC/8Ar+/de699T/xTj37rfDrkAATq/p/j9b+/dVqSO3rhb37q3XvfuvddgE/T37rRNOv/1fn/AP8Avfv3Xuve/de697917r3v3XuuYAA1c8H/AHn8e/dVJzp64k3P+8D/AFvfutgU669+6312Bfge/daJp11791vr3v3Xuuai45+l/wDeffuqk064e/dW67Btf/EW9+69SvXYYjj37rRAPXRYt9ffuvAAcOuvfut9e9+6912DbkfX/iPfutEV49de/db697917r3v3Xuve/de674sfrf8f09+61mvXgSPp7914ivXXv3W+uwbG49+60RXHXbf4/Ui4/pY+/daHy66AJ+n9Pfutkgceuvfut9ZIoZZ5EhgjeaWRgqRxKzuzE2AVV5Pv3XujO7I+I3cu/KOCuw2DZYZ4xIoqI5ldVPJ1Kqn37qvHy6EOX4A96xel8WnktfSIagiw/xt7314auFOkhmvhh3bhL/cYJmAv9I5h+kX/K+9dbp8ukgPjJ2yW0/wKUG+k3jl4P8AyT79x49ep8ulXjPhv3NlGjEOKjUuB+oS3F/pxp9+61Q8KdCJQfy+O66xRLJSBIxbWY4JHI5/F7D37reesmR/l/dtUkZMcLkqCT5KZrEgX/BuPfuvfz6AHfHxw7R2JFNPlMJNPFC3rNJFLI4H+q0af94B9+r17hxHQG1NHVUbeOrpqilk/wBRUQyQt/yTIAffuvdRzb8G/HP+v7914V8+vEk/X37rwFOvX4t+PfuvUzXr3Fvzf/ePfuvZr14Gxv791s568QRa/wCeffutA1669+631737r3Xvfuvdd2ta/wCR+PfutVrw669+631yNyL/ANOP9t791rANOuiSfr7914CnXXv3W+uwQP8AX/3j37rRz11791vr3v3Xuve/de674t+b/wC8e/dez14Gxv7914iuOvcE8/7x791r7OvG34+n+Pv3WxXz669+691737r3XvfuvddkW/41791oGvXjb8X/ANj7914V8+vG34v/AI+/dez59de/db65G6i3HN/+Ke/dVwTXrj791brsC5t7914mnXiLce/de49eJvb/AA9+60BTrr37rfXYFwT/AE9+61XNOvAX49+68cZ669+631737r3XvfuvdcgQAQfzb6f4e/daIzXrjxf/AA/3n37rfXd7G4/H0v791qlRnr17nn/ePfuvUoMde45+v+H/ABv37rfXXv3Xuve/de697917rs24te/59+61nrr37rfXvfuvde9+6913f6D+nv3XuvC35v8A7D37r32ddf63v3Xuuyb2/wAPfutUp11791vr3v3Xuve/de67v9B/T37r3Xhb83/2Hv3Ws+XXYAsSb8f09+68a+XXE/4fT37rfXvfuvddm1+Pp7914fPrr37r3Xvfuvde9+691737r3Xvfuvde44/r+f+Ne/de67txf37rVc065avTYDm1if8Pfutac164e/dW65Fifr791oADh11yp/xH/Ej37r2COvE39+68BTrr37rfXIC/wBPr+f6W9+60TTj1x9+6312bfi/+x9+60K+fXXv3W+u+OPrf8/8a9+61nr17Xt+R+ffuvUrx6ywU89TKsNNDLUTP+mKGNpZG/wVEBJ/2A9+630YTZHxg7Y31jVymOwk1HSuToGQhmglZLXDhGW4B5tfn37rVDxA6FjCfCHseOqpqvckcMWL1g1CQCTysl7EXYC3v3Dr1DTPR7ejfjhsDY+79sZnEUbS5SGsjZ5atS5SQFSxR35AH+Hv3l1seh62OMVFVZPamMkWUPUJRwxqykGwSMaRwf8AD3sn160B03Ybc02Jrmx+WR9BYrG7fTk2tz/sPeiOt9CgIqKuiDmOCaNlU8hX+o/J9+OOPWxSvQU7qwtLBM88VPCijgaUC/4nge/cR1rgemPBYyGeqMri2k/TT/T6fT37Net+XU7c9Ilb4KSNjdD6lUfj+luPe6jy60QRk9Atu/q+PIRSMReN05Urcn8nj34/Prw6Id3J0NhP4dWVH8OWapEUh0+AMhN7qQCPfjTrQrXPVMnb3x3rquevyeDoZKaro0mnqEMRSKSNGJCgL+f8fdfn17zqOiMTwS000tPOjRSwyNHIjqVZXQ2YEHn3vrfQ/wDx++N3YXyG3KMHs/HSNTQ2krchJHJ4FjVv3Ehe1mYC9+ePfutdWs474A9Y7EoKOo3GJG3LAEaaOdnlgDKvqsrnSDf36vXh9vQc9l9Cdf1lMI2pIgiXUmGNVbSnA5Xn34/LrZHCvVeXafSeQ25U1GR21R1VRhAbLEEd5l/2oX5P+I9+60RQ9F3kjkhdo5Y3jkU2ZJFKMpH4Ktz791vj1w9+691yNxwf9f8A4j37rQoc9cffut9e9+691737r3XfFuL39+61nrr37rfXvfuvdd34t/jf37rXnXrxJIAP49+68AB14m5v791sCnXXv3Xuve/de697917r3v3Xuuxb83/2Hv3WjXy68Tf37rwFOvW4B/r7916uaddcW/x/3i3v3W+uzccH/X9+60KHPXXv3W+u7cA8c/7f37rVc0669+631yZtVv8AAW9+60BTrwuRp445/wBt79140GeuPFv8f94t791vrvi3+N/949+6916xuB/W3+8+/dar14ixt7914GuevX4t/jf37rfnXrxN/r/sLe/daAp11791vr3v3Xuv/9b5/wD/AL37917r3v3Xuve/de697917rkCNNjf/AGH+39+6qQa1HXH37q3Xvfuvdd34t/jf37r1M169bi/4/wCJ9+61XNOuWn8f2v8AeLf149+61q8/Lrjxb/G/+wt791bNeuvfuvde9+691zH19P1/x+lvfuqn59cbcX/xt791auadetYc/U8j+lvfutVz11791vrlcE+r+n4Hv3WqEDHXH37rfXO54bj08f4+/dVxw9euHv3VuuVvTq/N/futV7qdcffut9dm34v/ALH37rWfPrxN/fuvAU669+631737r3XJb8kW/I/4n37qppw642v/AL3791bqzr+X58Y8Z3BnF3HloWqIKKdY0jkQvCLVOgnT9CePr791rz+zraD656i2jsjCwUiYWgQ08JiQmnVSQAPqf8f8fe+PXiaU6WdPhtqGuU1GMxxN7WaNSOfp/re9Yp1uvp08ZDrfY+Zj/dwOLfWpIIp1N7jk3/p79Xy69k56D2t+P2zWLvTbfxgJDWtSrY8X+h/I97xw69SvHpDTdV4bD1Gg4ijS3ClacLxzp59+x14D59Lna21sZLBLE+MorqbC8I5H+J9+x16pr0ybk2liGnMS0FIAxa9oh9CORf3o+nXs06CHdvR2D3JQyQVGCx8qzL+p6UM1j+b+98OvdV/dv/y69m7pM9VHQmmqtDmP7WHwlX/2l0NwT798+tHJqeqoO4vhDv3YlbPLhaGrqcdG0gVHilkcqi6lKv8AU3/x9669wx0Auy/j12PvSrkpKTDVdA0M3ikaupZYxcGxKj8j37HXs+nRk6b4GbzFKr1MkrTMlyY0OkN+bBffuvdFu7c6I3Z1RURrkaeaoo5QzmdIW0wBef3WH0Hv3l1759AWbfgW/wAPfutjrr37r3XM3YX4sP8AkXv3VRQGnXEG3Pv3VuPXXv3Xuve/de697917r3v3XuuTX4vb/D37qop5ddE3t/h791ulOvE3/wBtb37rwFOuvfut9e9+6913xb83/wBhb37rWevE39+68BTrr37rfXIEDnkn/ePfuqkE9ea/BP8Aa9+62KcB5ddAEgn+nv3Xq5p14Wvz9P8AD37rZr5dde/de697917r3v3Xuve/de67Bsb+/deIrjrr37r3Xvfuvdd34A9+61TNevA259+63x69Ym5/p9f9j791rhjrr37rfXvfuvdcr3Iv9P8AD37rVKcOuib/AOw+nv3XgOuvfut9e9+6913b6n+nv3Xuuvfuvde9+691yLEi3v3WgAM9dEWA/wAffuvA9eFrc3v+P6e/dez11791vr3v3XuuxYc83H09+60a9eJvz791vh117917ru1rE/Q/09+61XyHXja/H09+62Pn117917rsGxv7914567FzqP8At/r+ffuqmmB1x9+6t12Rbj37r3Hry/UW/r791o8Ou2Bvdha5/Bv791oEUoOuJtfj6f4/X37q329e9+691768e/de67Isbe/deGeu1W/+9e/dVJp11x+L/wCPv3Vuuvfuvde9+6917/W9+691737r3XYtze/+FvfutGvl11791vr3v3XuuyLGx9+68DXPXXv3XuuQFmt/vvp791UnFevN9ffuvLw66AJvb8c+/dbJp17i35v/ALx791vrsW/N739+60a+XXRNzf37rYx117917rsC9/8AAX9+6904YnFV2ayFJisbTtU19fMlPSQp+qSZjYKPfutE9bFfwT/liYKDB4XtDuOlnWolvUUdLNCxQs6B44/E30U8C5HvxFR14V4n9nVm+7Ottj4ei+0w+Ex2PpaKEBfBTpEG0DSNVgOfe8darTooGfoKTJZQ46CNBToxU+Meng2t/sPfq9bqeI69UbLgx0UNVjrLPTESoQLesWNveuvHj1Yr0hmspX7IokyPLxkqGP8AaAXi59+rjPXvs6ELN46nytG7ehamNGMZUjWX+ilv9j739nXhg06Ydi56oiq5sPkiRJCXAMnpBA/pf6/63vxp1vh0pN1tEtPLJzwgYEni3/Ee9D069xz0ndrIKuL0MCQzE2uAPzb/AB97PXq0z1MNH4shK5FzpHBH4t9Rf+nvXy69g9R8pSySRDxq5JuGsLk/nkf63vw9evV6DbIbJoczrir6clTw4ZCeL/763v1fXrZzjoDN3/F3ae5Ish46MJ5YZVukNtRJuQf9f37rVPPy6qH7e/l7Yupz+RnoaSsp3lmkYSU0UkWonkEgcH/Y+/Up1oUrTo1/x7rdvfGTryl2pTYH/c9D5BJkmpCKyRXFv3JrEm/1P+PvfXvmOoW8N57n3/PLUeBoqSzGJtFmOo35v711uny6C07archMY65JSoP5BsSPr9Pe+tDqbW7AopMdNBLjop0kiIAeENYkW4HvWOt56ru7y+PFFO1XktuUskOVjVpZ40idYmCn9Ogcf63Hv3y68B0QOvx1diqiSmrqWamlRnjtNG0dyraSV1fX37rWDjqB791vrsgj6+/daBB669+6312Bfge/daJp1z5cWA+lr/j8e/dV+E564E8W/p/vfv3VqZr14C/v3Xiade4t+b3/ANhb37rfXuP8b/7x/j791rPXXv3W+uybm/v3WgKdeBI+lv8AY+/deIr17i35v/vFvfuvZr14G3v3XiK9de/db697917rvi35v/vFvfutZr11791vr3v3XuvDk2/r7917rsgg2P19+68DXI669+6912Lfm/8AsP6+/daNfLrr/evfut9crgfS/P8AX+n+w9+6rQnj1x9+6t1737r3Xvfuvde9+6912bfgn/G/v3WhXz69xb83/wBhb37r2a9de/db6//X0ACLcH37rQNevC359+68a+XXR/w+nv3W+uzb8e/daFfPrsKT/wAj9+68SB1x9+631737r3Xvfuvdd/4f19+698+u7WPJtx9Rf37rXEY64+/db674J54H+Hv3WsgY669+6313+Pzf/iPfutefXib/APGvfuvU68PqL/7z/T37rx4deNr8e/deFfPrr37rfXvfuvdd8W+pvf6fj37rWa9dp9f9h/xPv3Wm4dcffurddkWt/iL+/daBr13Yt9AOPfuvVA664t/jf/ePfut5r14j+nP+93/1vfutD59Lja/Xe6t3TePFYybSLDyzRTJGb/6khST/AK/v3Xq49elxnfj92DgMdJkqmjjkgjTWyw+Znta9gNNvfutZ8x0Ck9PPTSNDUQywSrw0cyNG4/11YA+/dW62Jv5QQp6jadVqCq0U5DWH6tOQdSTb/b+9+XVfPq/XI0jVFM6owA0lVsef8CCffjnrfQK5ugy9HO80bN4lIIufwPxf37rY6UW1t6zQjwZB/wBIVPqLjmwN/wDeh718uvVr0MNBloquJbNw1ipB+gP1JPvxHXq5z0lt20itE0wAvxpIv9Bzc/7H3759aFc9NGCVngfx2JW+qxsb3497qfLrdBSg6T+cp5WyUcb+nXyP6kE24tb3rHXvt6VEFLH9rGgAJC2uTyDb6ce99aPTHk8N96rII+Tf/goH1P059+PWxSnQfV3WGJzFxlaCnqAQ1hIpYWItwDf37Hl1qnSCm+OW2GlmloMbS0Rs7aqeHxXJF+SoBv7114jHRdN1da7z2zNWRUkSyU5Z2gJ1k6OSLE+/fb1o0r0TftLrnJbt27mMbuShh1vHVNHMUZmC6DYKW9+Ip1sEdULb225PtXcuWw8sTxx0tZNHTsylQ8QN1K3/AKXt7917pJ+/db67Bt9eR/T37rRFevHm54A/pcf717914Yx11791vr3v3XuuTLYX59+6qDXrj791brkRccG9hzf/AIj3vqtaceurG1/x711uorTruw0k/m/v3Xq5p10BcE/09+69Whp14g2HHH9f+K+/deFK9de/db65EC1weB/W/wBffutAngeuPv3W+ve/de652YgcfTgf74+/dVwOuPP+Nvfut4699fx9PyP+J9+6914fXgX9+68evH6/S3+t7914dde/db68P8fp7917rsj+l7fgn37rQ+fXYP4P0P1/r/sPfuvEefXH37rfXYNv+N+/deOeuib+/de67II/2P8AT37rVa9de/db67P4/r+f+I9+6111791vr3v3Xuu+OLX/AMf+Ne/daz14ixt7914ZHXXv3W+uyLW/x9+61Xrr37rfXvfuvde9+691737r3XvfuvdeJub+/de65WFgRzb63+nv3VamtD1xJv791br1j/T37r3Xvfuvdd2Nr+/dar5dev8A4A/649+69Trwt9D9D/t/fuvH5de/1r29+699vXX0/wAPfut9e+v+Pv3Xuve/de67v/sf9f37r3XjzY8c/wBP8PfutD0669+6313xxz/r/wCHv3Ws9eJ/H1t+fyf9f37rw669+631737r3Xvfuvde9+6913wCbWI/x9+61k8evC3Nzz+PfuvGvl178Dj+vP8AX37r3n11791vr3v3Xuve/de65BQfoefzf37qtSOPXr8m3P8AQn6/T37rflnrq55B/P8AX6+/dex14An6f7Hm3v3XiQOPXjccEf8AFffuvfPrr37rfXd/rwOf8Pp/re/dap14W/Pv3XjXy699T/tgf6W+nv3XqeXVk/8ALR+O47v7k/iNTBO9JssQ5ZLxuaWR0LNICwFiQF+l+L/S9vewOtYrnrb/AKejjhx1NSQxCChpaSmp46ePV4FaGMRlhGeATb+nv3Hh1vos3fNXTYLb9RUo7K80E36fwwP0sffiada48eq1cR2DRQZCR6qVWZ5GCksL31fU8+9dbpTHQm0+8o61DoYFGWy3tyTwOf8AH37h16nVgXV2WpqPrSmqGBEzBr2HPKcEE29762cdc9u7vqMhkxTo2uz+v/U6dX4Pv3WuPS+yeG0V9LkoF0yuyNJz9QSCb2+vHveOHXupG6p2nx8zJckRAEX44HIsfdeHXhTz6adhTrGqrITqZyLH+hAFuPez1vHSjrpGGX0hbo1tNr2Nz+R/T37rVa9KunxxZdboGJ/Fr8fU+9dbNOoFZj4zIWCIq3ANv6gW/Pvf29epmg6azFFGSiqCoNiQbXH+w96+fXvl0F+6tuwZKtulJEACCSb+q5tcge/Drxz0E+5Op6TL1ShsfS3OkEmEOTbgm9vfuvEU4dIXO9PU2JUrHToi2BKrGY1/3r37r3y6CPP7XoMMVkqIrqx9HjAJuD9WFvfutUznpHVtXhSn20alahrABuPqbCw9+69SuT0Fm79r42KmnqqqEapUIFgbG/0JsP8Ae/fut8TXqtrufrHCbkmqAsKwVMXlMEkQ0sH1G1jx71XrVKio6r+z+0M1gK+ainpJphGx0S08UkqlL+ktpHBt73177ekw6OjFZFZGU2KuCrA/0Knn37rfXH68e/de65EAD6+r+nPv3Vakn5ddWIF7/X37reDjr1uL8fX37r1c068CL/0/1vfuveXXvr/r/wBB7917h14gj6+/deBr14C/+2v791vr3Fv8ffutZr11791vr3v3Xuve/de697917rsgX9Nz791oV8+uvzb37rfXIgjg2+v1H/FffutAg5664t/jf/ePfut9dc/X/effuvfLr3v3Xuve/de697917rlbgH8k/wCw9+61XPXRBH19+63WvXgL+/daJp1yt6T9Pr/vR9+61+LrjYW/N/6e/dbzXrr37rfXZufx9P6D37rWB16x/wBube/deqOvEW9+68DXr//Q0AOSLk/4f4+/dawDTrxFiL/4H37rwNRjruwb6cW/r/vfv3Wq049cffurddg/g/T+nv3WiPPr31PHv3W+vEWNvfutA1HXXv3W+ux9R/rj37rR4deIItc8+/deBrw68LfkX/1vfuvGvl11791vr3v3Xuve/de697917rvj8f0/3n37r3XXv3XuuyCDY+/daBr14i3v3Xga9e4t9OffuvdeW9+Db8+/deNKZ669+631737r3XdyPoffutUHSp2zsvcm7q+lx+BxVZWzVU0cKSJC/hVpDZS0lrW9+698url/in/K0ze6aCPdHZsP295IJ6Wn5eJYw4YehvqePqR73Trw9OHVk8nxJ2jsCg8GOx9G608JjWTwRq50i4Y8cn37HXseXRbdzbUxqmqxslHTzRmRoyHjUj9Vv02/2B96GR1vy6Lnv74ZbZ39TTZCmpRT5DwTNA1Mnis5W6m6W/Pv3y61xzw6kfC+j3z8bt2zbVzaN/DKmt/antYMpqPINQH597qevUAz1sW7cyb5PBUmTlY+OeETK1zpVWUEe/Vz16nWSumoq6CWCLTKdLXsQ1iBb6e/deoegA3BFPi530kqrMW1c/g3t791qvl0p9kdjUSTJj66XQbhLsfzq4sT798ut4PDoac5UxVmNEsZ1xMjMpBvwBce9U6350PSe2RJHMaqJ3sRKbH6nTf6j3v5jrx4/LrHu/xUdbHMCRGlrt+P9v791ry6zYDIpXgAPqsOWBFv6fj8+/Y8+vVNKDpc0dAjN6r2Pq5H6h/jf3o+vWwKdZazHwSXsArKB9Bbj/Ee/Dh17ia9MU3ji4U/gk82Nx79w4daGc9B7u9aWZPJJGjaUuVIDcW+nP8Ah7316nRFu5aaiTE100UcakpOLgAfUEe/dexXHVOncnxzTsPCZbcmFi05iheR4SqhQ/Goq5H1BI59668eifdOfF/ffZ+7lwDUT0dJS1RirpWJDOEazrCf8f6+/U698+rfNp/yuOvcpiIxOszV0aaassha0oA/S597HWukhv7+UzDDQT1G1BUtOgdkXW6qbLdRa/8AX3rr2PM9Vm9j/DDuPr2rq4qjDvW08EjKjRJIJSoF/UoBH+8+/cOt8fLot2Q2VuzEyPFkNvZanaMgMWo5tHP51gW9+68TTpOSxywvonjkjYX9EilGH+uD791ofLrD791brsX+gP19+60aceu2uABe49+60KHPXV/6f7H/AF/fut9de/db67ubW/33Hv3WqefXXv3W+u+Lf4+/dazXrr37rfXamxva/Hv3WiK9dkkc3+vv3WhQ466BsCP6+/dbpmvXr8Ef1/4j37rfXXv3Xuve/de67AJv/h791omnXR4Pv3W+uyb24At/T37rQFOuvfut9dgX49+69w68RY29+68M9eItb37rQNevDkgX/wBv+PfuvHGevfQ/6x9+63x68Tc3P59+60MY669+631737r3XYtze9/x/h791o1669+631737r3Xvfuvde9+6913xz/X8e/de669+6912TcAf0v/ALz791qma9eFvyPfuvGvl11791vru5Fx/X6+/daoOuvfut9dliePfutAAdde/db67ubW/p/vXv3WqCteuwt+b29+68TTroW/P09+639nXdrWP9ebe/da4466Jvz791vh13xxp4P9T/re/dVz59dH6n/X9+6sOHXXv3XuuQBP5+n/ABPv3WiaddW+v+H/ABW3v3Xq/wA+uvfut9dgXv8A4e/daJp11791vru/BFvr+f8Ainv3WqZr14G1/wDH37rxFeuvfut9e9+691737r3Xv98Pfuvddi1+fp7914/Lrxtfj6e/deHz68Be/P0/3n37rRNOu7gWtcH8n37r1D59dA2N/futnPXiST/j791oCnXgSPfuvUB67I41X+vv3Xhxp1x9+631737r3XZtYWHP59+61mvT3tvAZPdWdxW3MPEJ8nm6yGgoYSbB6iZtKKT+PfqVPWj6+nW4B8F+kW+NXT2B/i2PpKbeeUo4ky7RxIkv286hrs1gTcH3vqwFBQdWI4rLwZWmRI2F2Fm/pc83H+t7114mvRUfklga/KbdykUCnxU9FOyH8kn/AB/1/eya9e8gOqAs/mMvt/cNVHUQ1jQRVE2khWYHTJbj3Xrw4dGO6UyuZ7E3Di8VjqSqRVmUTtKpVPGtgeT731rHVt+Sgq9q7VoMHbS3hj/TwNTLY+/deOBXp06zxM33K1Mim7OpY/7Sxvc/4+/Z63wFOjJ1sPkiVY+dChjzciw5A9761jy6SNbE00DJa9wQQbC3H9Pevs63mmemnGmPGsLi3rvxe/0uePfuHXjnh0qIWSsrYZnvYn0g2/2IJ/Hvda9aPHoQJJlhRAotddNjb6W4PvXW89NLxy1etUBFwefwD735/Lr2fLqGcdMltUfJ4JAubk8fX3qoBz1qnkepMe30YK7gFi1vUDcX/qT/AL379XqxUV6yNgIoHDsqsfrYfQc8X9+Br1qhp0hN74yGWgkm0KGWMhARySvFgD9fe8cOtcegJxHVeQ3EZ67KU4+0csIAyX9X0BsffqZ68fn0EXZvx2qKeCXK4WORq2IeSNEWy6xcj/Ye/fOvVdR8gft6I1vOo3rjvLjtwYiqCoCivHAx1KDpWzW/3n3ryr1atOi4V22INx1rROstJJqJYT+gm7XNifzf3rrfDPTbmep4aGnkngoYKxilnkeJJCeOeSPfjjrw+fRM+0ulRkhU5HDUjQ5SNS80aJogKpyV0rx79XrxHp0TWtoarHVMlJWQvBURMVZHBB4Nri/1HHB9769x6ij688/19+615Y669+6312DxY3/w/wBf37rVOuwL8D6+/deJpx66IsR/t/fuvVr12SCPob8c/wC9+/deAI64+/db697917rsm/8AvXv3WgKddrbm/wDqTb/X9+68euha/Pv3XjXy68QVNj7914GvXgbe/deIr165vf8AP1/4n37rflTr1yfr/r+/dap13dSfobW+n+P+w9+61Q067a4uvFvrx7314Zz1x4t/j/X3rrea9de/db697917r1z7917rsknk+/daAp14jT+fx+P969+68DXr3P0v9T791759e/Sf9b37r3Ede4t/jf8A3j37rfn14G3v3WiK9d6uLfn6/wC839+61TPXH37q3X//0dAAm/v3WgKdeJv/ALAW49+68BTrr37rfXvfuvddgXNh791omnXdyLrx/Q/7f37r1Ae7roixsffuvA168Rb37rwNeuvfut9cjc2Jt/Tj37rQoMddW4v/AI2/3j37r3nTrr37rfXvfuvde9+691737r3Xvfuvde9+6912bfi/+x9+69nz69c20/i9/futUzXr3IIP+x9+69x68Tf37rwFOuvfut9dm34v/sf6+/dez59Gt+Nfxi3V3nm6SSnoZ2wCVGmd0RtU4RrMAxtZf8R9ffutdbBPR/xN2j11W4LFpg6SorY0pJJBLSKdLxjkf6/vf29e+zq2HH4mHHYykpKOnSljihjTRAoRF0/iw9++XXq1z0jd47bXJY2sMRvKsEjAcEk6SePfuHHr1fXqpHctY1BvGtxGQBjf71wvkGkEajYj3oU60RinRh9kUVLNQLFpVtSgXsGPI4sffut0Jx0m+0OvIYKKLNU1JCk0VXCfuAgDgBgSNXv1evedT0dXZe5UHVdIPInkpMYiOVazBglweB79Xy6sQTRukr09uyTO5LJRzTeZYauSIFmuF555I9+6qcCh6EPsDER1MLvEB+l+VXjhTzx731scM9E8yE747OQw+VoyZ19QYr/atz70D1UDHR4sE7f3UpRIxdnpNSsxu36OL397630nNoZAU+QqkYnV5hb/AFNtXI/1/eutjHT12LMr4eSYcMFNuLDlLC359768MnpF9F5CXJCuSUl2ildVL3NgDbgn37r1OjQyFKaLWR+ker8f7yR71xPXq06SORzcK+QIV1/RvUDb8/W3+t73XrVM9I6fK6yS/Ngf0n+nvQ+XW/PoLt0506ZGJA0qVAvYHjj/AHr3vPHrxp5dEq7arJKvCVzKHN2k4FzckX4X3qvWuHQc9XbYpq7a2Wmq0Uwi+tZFstyDxcn8+/deB6Er4/8AS01buaprsZi0gozUOz1CwaCwL8kNzf8A1/e+tnq1jAbQxWCo4qXTEZnUtMbC+ofhr+2Zp44EMkmKdLdt2u83W5FrZDUzfn0+jG4yYFCsRW2lhYfQ/wCP/FPYbu+Z7ZD4cPHqaNi9jt8uoTc369tK46SmX6u2LnNS5HA42rLhtTS0iO1yLcliPZ7YXS3UAlcjqMOa9kfZtyNlbox0mhoK9F03X8NOrNzRVol29i4WkD6DHRICD9Ba3tUXSvEdB0W11Spjan2dVKfJf+WNATW5LauKkE+mZ4TTwlCAAWT9BGoX/B97DKxopr1VoZ4xqkQgH5dUkdodD9g9U5B6PcOFrTD5XjSpipZSvpGoeRVBtcX5+nH4961rXTXp42tyI/FKHT606BggqSCCCDYgixBH4IPu3Sfrr37r3XfFvzf/AHj37r2euvfuvde9+6912Bfj37r3Drtjc/7D37rQFOuPv3W+uXIH/Bh791rBP2ddA259+63x669+6912Ra3+Pv3Wq1669+631737r3Xd+CP6/wDEe/dep117917r3v3Xuve/de697917r3v3Xuve/de67Bsb+/deIr117917ru/AH9L/AO8+/dap59ev9R/X37rfXrA2A/2N/p/vHv3WuHHrv9Nx+fof+Ne/de4566Nvxf8A2Pv3XhXz669+631737r3Xdvof6+/de67sV549+6rUHHXH6+/dW67txc/n6W/4n37rVfTrwt+b/7D37r2fLr1+CP6/wDEe/db68Gt/T37rRFevE359+63w669+691737r3XIEAH/Ee/daIqevXFgDfi9rf4+/deoa1HXEW/P+8e/db67Jv/sPp791oDrr37rfXYNjf37rxFcde+tyfqf9t/j791rrr37rfXZubE/n/iOPfutD0669+631737r3XduAf6+/de669+6913fi3v3WqZr11x/sfz791vr3v3Xuve/de67Jvb/AA9+61TrxPAH9L/7z7916ma9de/db697917r3v3Xuve/de67Fvzf/C3v3Ws+XXQ4N/6e/db67JLG/wCT791oYHXiLG3v3WxnrkurhRyWtZQCWJPAAt791UgHo2fTvw17h7ipaTJYPCz0+NqDqaWpp3STwq3LKHtbUOQT/tvfut16MXuD+XDvvD0wb7WpWZUVpCwYpe3NuffvLr3yr0uvhR8Pq7G9842u3/SMYdvZajqsMTG/ieSKW7PICbav7P5+h+lyPfut08utkfs+rbE5BKySNYqV6SmghhiFoUEcIRWVF+nvZOetDh0ndmb2P3Ji8mlPoAzEXP0Fj7r1sEefS03z/v4cMlMipItShSfTZyUb63Xj3v7et8RXoAT8O+ut0061uQp4xOXMjkRDWNR1eq/9T73Tqv29CD1/8b9u7AyH3mBxtOscd7TeAB7ngkEfn36vW6AYHQk7t2fUZgwlY7hQAbr+AP6H3r7evY4dKvam0zjqdSyaSVAJK8Hi11+nv3DrxBPSqEIikdFIYMpBvY2sObe99ep0wVtOIxJIPpYmwB9++XWsdJKgp/va7Sf0K5b6c8m3Nv6D3rr3z6UOKrIpM3LjwtvtwDcfUEck2PvdOtEYBPS1eM1dRGi8KvH0/F7/APFfejw6sKdKFadEChQoIsGP9f6/7H3rqx49YpaimZ9AXkG2r6ggH6/09+6r5dY3r6WJiZHsAOLsBdvx791v59MWR3TjqaOR2lX0KeNV9Vhe1iBf34da6QVEtfvGu8rLoxFO+pCw06gDdgQfrfkW924Y68fXoRRVUtMiUdNFGIYAoIUAXYDkWX/efZbud89hF4iZPQx5I5Wh5s3H6W5YIq+pp1NH2NXHZ6aGS3BR0U3AH9D+PYZ2zfZ72+0TGg/Z1NXO/tbt3LXLS3e2r4rMOKjV/g6D3dXWm2N0aIqrA40ICpaT7VNdtVzdh7GviRGlGH7esaRZX4qZInwT+Hoi3evxFx1W0uV2vTNBJEruFpowgZibjhSP9b3fUhwCOmjHcjLRsB9nRGKjH5DZtdHhN5UckWOeoNOJ3iI4BtdmPtuSVYst0otbSe9JS3BJHQoVfTHX+cxIrMPLS1L1UQLqjLI5LDlXUfT/AGPuyFZF1IajpmeG4s5fCuVKn5inRGO6viXsqRKqvejkjrvHIY2gRo/UGut2jI/33+v731So49VQb/67zOyK+ZKiln/h7zyLBOY20KqtZFke55P9ffutZ8+g6UA3ubf09+62SRw64+/db697917r3v3XuuWkgX4/H+vz791quadcffut9e9+691737r3XIC1j+G9P+NvyR791Uny9OvMADYX9+62CTx64+/db671Hj6cW/3j37rVOvcm5/wv/tvfuvcOuvfut9dm34v/ALG3v3Ws+fXXv3W+uQ+vp/p+f+Ne/dVPDPXH37q3XZt+L/7H/jXv3WhXz69bi/v3Xq5p11791vr3v3Xuve/de67J4sPp/vPv3WqZr11791vr3v3XuvC35/3j37r3Xvfuvdf/0tAA2/Bv791oV8+uvfut9e9+691yAuOPrf8A3j37rRND8uuPPv3W+ve/de697917r3v3Xuu78W9+61TNevEk2/w4Hv3XqU68Rb6+/dbBr14m/v3WgKddg39J+n+H19+60R5jrzXvz791sUpjrj791vr3v3Xuve/de67Bt791oivXXv3W+u78Ef19+61TNeuvfut9dg259+68RXpTbQwM+59xYrDwoX+5q6dZQqs37BnVZSQv+Bt791qlOHW2h8OOm9s9a7MxWRhpI4mkwgmYunjHkMYNgDb8+/U69xwOjLdeZynyeer8pVxgGkrXhi440A2Fj/sPfvLrZxjo1tHkKOqpwYiCdJvb8X/Fve/Lr2Ogq3nnHwJkkCs8DBtYsbAH6j/iPfvPPW8Up0R3s7qeg7KWr3ft2RY8rSMZDFH9SY/U3pH1P+HvXn1rFK9Bz17uqrxFb/A8vS1AnpZBBqWFwp0HSeffutZrUdCv2HuKbMYKPDUNLMWlqEbV4nJsWCm9vp73XHXiSePQy4XA/wAJ6waUyOrHHHyKbhQ2j8g+/Hh14UHDoNvjOlVU5bPIt3VspLY/UAFyeD799nW6evRvt1I9HB4nVWDI9yR+SthY/wDE+9cePW6dEO7PjONzlLWAhddZFZb8AMwJA/xPv3Vccej27SWSr2bi53C3bGagB9D6AffqHj69bqAadJ3blCZcnUvpFhMTq/xDXt73w618j1i7RqxDiJKRCdRFr3v6tN+Le/UqM9eFeo/x7xElLSV80q2aVnZSfqNR+o96PDPWxXA6GLdGY+xjliVhrIYBSbkD8WHv3l1rzqOgthaqychEYf1sCxI+ljwL/wCPv3W+PT//AAOSOL1ltWgmxWwPHPvwr17Hn0D278BUzyNApPrBa4t/rX4/w9+8utGlcdF+3ltyFMDWpOUITyG7f7Sv15/r7316h4dA71Jj8ru7JzbPw0Ev8OnqV+4nWNiBpktww/H9fdTjh1vq2nZ20Mb19tumoqSGMVa04E8pWzmTi5JP1Puwoc9e6k09bLPO7O7FnY2BY25+oHtFewLPCRKaDoT8r7vPtF6slkmuSooKV6mzSy0wDr9ASTci30ubD3DXNF3ZbexNu1WHXRr2N2fmDmuLRvMZWKQDgDivSgxWQiqYLm3lC303Fjzz7Kdu54uPD8Gvyx0O+c/u0bLFem+K1DGpJB6y1NckcmmMamY3YXFh/rD2IY+Y75l1MaDqMH9leWFkMYQY+XWDK4+lr4UkraZZImjZTqWy2I5Nz7DcPu7tkW5naxMvij+kOlb/AHbdn3yHw4FpX5HoonbPxe657Op6lchjqQPIZCJDCS9nTRwwH+9+zi655n/tQRj0PRzs/wB1axSm3yISGxUqcfy6qG7u/lbbVp5q2o2lFVrWVBadRAJRGGIPGkXHP19rdh9x7jcLgWkiD0r0DPdf7m+08q7U292d0cipWhFDTh1VD218Qe0erpJ5qjFVddQxgupio6lpwt/ppVSDx/j7lu1uPqEB8z1gDv2xvtF00KnUq/t6KzVUFbQuY62kqaSQfVKiGSFh+LFZAPaqhHHoPAg8Oolv99ce/db697917rsWvz9PfuvH5dde/de697917rsgi3+P09+61Xrr37rfXZt+P6f7z791oV8+uvfut9e9+6912Bc29+68cdeva4/r/wAR791r59de/db697917rsgi1/z791oEHh11791vr3v3Xuu78W9+61TNeuvfut9e9+6912DYW/r791qnn17ix/r+PfuvZr11791vr3v3XuvfX37r3Xvfuvde9+691yA5sf99xf37rROKjrzAA8fT37rwNRnrj791vru30v9D7917rxtfj6fj37rQrTPXjbi3+x9+68K+fXXv3W+u7/Qf09+69117917rsG3Pv3XuPXib8+/de4devwB/T/iffuvU67X63P0H1/2P049+60eFOuuNX+F/wAce/de8uvED8fT8X/3n37rw+fXXv3W+ve/de697917r319+691y5sR+B9ffutYrXrj791vr3v3Xuu78Af09+61TNevX4t+D7916ma9ev8AQf09+63Trr37r3Xvfuvde9+691737r3XI6fxf37rQr59cffut9e9+6912QR9ffutAg8OvWNr/j37r1c0669+631737r3XIXIta/0sB9Sb/T37rXnXq2n4MfAap7Xaj7A7Bp6ukwCGCrw1G0E6ColFpE+7jkUX/wH09+69xx1sfbH6sxuysDBjsZiKShpaeBI1eCHxFgqabtwPra/vf2db49JffePpJqTxIgLldJJvcG/59+618+gv2ttnF4PM0OZEKI8VSsjyKCDYPc3Pv3WwDx6N7urB0u98JBk4yzoI4woX1D0rp+o96610Vqtoq3C5KWKK8ccLGzHhiAeQb+/deHS1xG8ZFko4JHLIJUEhbkaR9fe+tV6MBjakxyQV8B1UVUigqD6RxYkL71nq329DHRx0a0KSqNaFAVKkn6jm9v8ePevOnW6DTUdMkcdLU1ZVyUAJsLWAF+fe+tcM9P1TPQU1LoRluF0p/Um3196oerVHHpKRFmLHg8/UW/T/sfdqHz6qKdMVewmFVGv60Rj/T/eD7114ny6asBSNTwz1UgIYa7fj6n8j3vrRr5de23FGMtkcjL+oowBH0JH0v79x4deFelUmdSMkqtv8fyCP8ffiOvKa8OpEVbU1jBw2iG9x/X/ABHv1Ot5J6jVuQjo1d/JHcAj1OB/r3v7114fF0Gub3TCkNRK1VEgiUk6pVU+kXt79WvXiBx6Qe2J6vfOXNOruaOCQkyLcq6Kfyy8e9+fWh8+HRi2hixlJHQUAC2VdelbG4ABNh+b+9DBz1bPl1GiopUAkLE6731D6H639sXPgrEWnGoDPRpsh3GW/S32xzG7GlQeuP3hoKqMkllYkEEixBP9D7gHmjmH6Hci9n2566w+yPtG3MfJCw7/APrHRUasnpTfdJKmpPoSLA/61+SPaqz5ovriMMjdE2/+yHK22ztbyxitTwHUaWOCthMM8QZLNquDpsDe5/x9uXHPo2h0+ukCljQAkCp6CkvsLy7u8T2tqlHI9PXooveHQ2I3/j6gUsEUUp1PHKiHWr24YEeze6558WMM3CnHol2P7rMm33pjt1Ysx81NOqxc3sXsrpDMTyYlqjKY5JGaRJtbrHET6tI5+ntzZ+eGnk8OA1AP29F3uJ92KKwtXn3Psk01Hl0N21qrZfa22amnzEsdLuEU7I0UjLFpcjUdKvY/Ue5Js92gvB3Aqfn1hFzJybuHLtyYh3oD5ZPHoo/Zvx42/k8fWYbLQwyUxeZkmC6pGV24sf8AevZmuhvhYH8+gs0Vwgq8bD8j0QPePw7pYWmk269YIwzMA4ldQv4VeD9Pp7sVIz03qPAgj7R0W/PfHveWJqJUjjDQxrcNOksbE/QgEAg+6g16tnoGMthMnhKl6XJUc9O6MQGeKRY5APqY3YAEf4j37r3TWbfQfT6+/deFeJ64+/db67sbX/Hv3Wqjh14G3Pv3W+PXVuL/AOw9+698uve/de674t/jf/YW9+61mvy68Tf37rwFOu2Fjx9PfuvA164+/db697917r3v3Xuve/de697917r3v3XuvfX37r3XMmy2/P8AxB9+6rxNeuN+Lf7H37q1M168bfj37rQr59de/db697917rsC/wBPfuvcOuvfuvde9+6912Rb6+/de49f/9P5/wD7917rs24t/Qf1+v8AsffutCvn11fi3/I/fut9e+n09+691737r3Xvfuvde9+691737r3XdjYn8D37rVc066/x9+63165P159+691737r3Xh7917rtr35Fj791oUpjrr37rfXd+LWH+v8An37r3XL06efqB/yL37qua9cSb/0H+t791brwFzb37rRNM9eIIv8A4G3v3Xq9eIt+R/sPfuvA16cqXDZaut9ljK+pBFw0NNNIpH9dSrb/AHn37r1ehv6BpcphOzcRUVmIrVRCquZqaVEjLVCEEsy2/B9+69Wpx1tQbG3dHU7BxFPTuqVLUEUYVWHGpB6Sv/GvfvLHXiacehs2Jt2sj23W1qRDyNI05a3LfVixA/3v37r3Dj0ptob6SGskx8rN5Y20EOVtfV9QD735U690Ie4I6XP0klOEjeWaN1S63FyP9t79inW+BoOiU7lj3R1VmZJWhd8XVVF3VNWkJIbEW/pz+ffuq+eeh96/2rsreFCmaMFIK2YLJLZELh2FzqAH19+62MivQo1HXu3YIC7wU90BYMYlvYC5sSP+J9+rjr3E9FI7s7Sp8XC2xNukTVdRIsJETBkVCLMOPeurHGB0L3xp2X/AsO2Qq1P3FeVqGuDcO3q5P+HvfDh1oDofN5Ukk1OxKaksxDfVv034P+HvQ68R1Xj3cDFNTSOBoSthUNwP7Qt799nWscD0ezYDxy7Awj31FsWgFjyTp+gX37yp1sjPWfadGQ9e5QgeVjq4BuT7317oNN/SGszENGQrIXsbfW1/979+6150PQubRo0weD1IoV2QsLLY8/Q8f8U9+yOvYGT0w1sM+WyCvLdgZCLH9IH4v711vpd4/GUWOpQ8qIDpBv8A1t/T37rwznpize4KGmjJV1IGoD1D6W/2q/veOtVJwei8bx33jaQSSq7tKqEjngcfT37rx+XRR8/mtz9jZePbW3YJHiq5VE0gBCqrtZ+R7159eHVhvR/TWG6vwME7pHLlauBHqZHUCSKYHW2k/wCve/Pv2K9b8sdP++d3JTSGBJAST/Xkf159749VPUHatZUZN1nQsFQrrP05A/Hsh5gkeO0Og+XUq+0dlb33MEYuACNQ49K+tm+6kWnQ+oEhzf03H9QPePG9WNxdymRjjrr57fcybby7YxWyxKDQUoB1Jp3ix0Nyf3ANJAt9Tyfqf979h6NLfb31McjqWrht25vhMcaUjPn8ulLtfD1GfqlmYN4Q4F2H6gp1fq59w97ne+VrsNjJte0nVMRQnyFegBuPt+m1TVlarHPQuZTB0RoBQhVWTSwuAL/094WWvM+7Tbq27GRtRbVWp48ejvbF/dsiSKMDj6dBwdhVqLJKzqIRcr9foBe31/p7l6P303lY47XDFaDPE9SJHzBaNpCrRj0hINtpk8q1G8Jd1LLfm1g3AHvIrlr3W2vl3YBvm6sDORULjj1G3vdsW6c1crNY2BoHHrTy6dN0/HPbe5MVJFkMLj6yWSJkBniVyNQsDYj8ey1vvk3ImMcEAC1x1zmufuy7jcStLO5Yn7eqzO8f5Xmy9301XVT01LQCV5GVKeC3JH4K8i34sfchcj/eqff91j266jWkhpWox0G92+7DuEMBkgNCPLPVau+f5UtHio5XwstXMUusZE1UA5tcEjVb3mZtW6R7nAk0dKMK46x2515JuuTZTBd11dVj97/FzfvR9Y7ZmkaXHPZ4pI0dniiYXvIebj/H2a9AJWqKnosXv3V+ux9efp+ePfutHhjrk9uLf0/pb37rS18+uhz9TwP99wPfutnHDro2vx9OP969+68K0z14gj6/n37rwNeuvfut9cuLC3Lfke/dVz58Ouje/wDQ/wCHv3W/LrxFrH+vv3XgevX+v0/339Pfut9eH9LDn8n8e/daPr17+l/fuvfZ10f94/Hv3W+u7/63v3XuvE3/AAB/rC3v3WgKdetwT/T37rdeuvfuvde9+691737r3Xvfuvdd3+v05/33Hv3XuvAH6j37rRI4HrxN/wAD/ff19+68B176WNwfz/yP37r3HrxJJv7916mKdeFyf6/6/v3XsDrs834At/T/AB9+69w643+n+H4/Hv3W+slha7C1zYf0+n+Hv3VKngOsfv3V+ve/de67v/re/de66sf6e/de67H1Fhf/AA9+60eHXbf61j+bfT37rw64+/db67/B5/px/X37rXn11791vruxsP8AG9vfutYr11791vr3v3XuuyCAD/X6e/darXHXXv3W+u7n+p5+v+Pv3XqDrr37r3Xh7917rs2ubfT37rQ4dde/db697917r3v3Xuve/de67P8ArWt/vuffuvdde/de697917r3v3Xuuwbf0/339PfuvU67P0HJP+H9PfutDj10frx9Px7914fPr1rWv+f9f37r1evG1zb6e/deFaZ6Pd8DvjLmPkB2hjqmGATYbbGSpKrIRyRCSGdEDSSK4It6RYj/AB9+69xNOtwXYvX239q46gxeJpYaXH0VPTBIoYxHEXjjAayjj6/4e9+VPLr3S/y/iFMY0CgHSukH6g8W9663THRTd+Simrnj/sE8D/DV9Bb3sZ68R6dMlHHSz0ojZv8AOCwI/APF/wDYH3rrXHoeeu89FFRnb9Qx8EYtCzHhm+gHv3XhnpHdlbbMbNVCPxxGx1gEFg31vb+vvx68OHQBPKKKVCdQRZCBwRzf8e/de+zo0nWeRGcoJqI3IjhDLqN7NpsCLe99bxXoRqbcc2Chlx0/rjjVtDE8kk8fq/HvR9evccHoOq7samhq5A0jIym9gwA/2Fve80z1r5dZcdvcZWo8cchKsV+p/SD79nrdB0KFHVKFssiM5C+kcnn6e/deHUMwL97K7GxkOkA/T6Xv79nrxzw49SKyNaejmWMcyRkf1+v5H+Pv3Wvs6bsRSmnx8hYESNqvc/W/1Pv3lnrdc16xR0EshDWOjV6v9b82t+ffgOvGvXGtyz0ETQwD1aTb8AW4Nvz7114fPoH83kM7XMyQLKutiAbsByePr73Tr1c9BHk9j7v3JlYKFWmjp2lAqGUtbxk+rn/WPv3WqZpXHRvdl7TotiYCGhpdMtSU1POwJkZ2T1Ak/wCP9ffut8Ol1hqV6lnlkQsSf7Q4t9bg/wDFPdSadbGc9OtdClJCG08tyb/0/qPbc8YmhZD6dLdpvG27dYbpTwYV6DuvCzVQBUeltX1uQLfUe8d+bdrh+rd5cUz12C9hOe9yn2CC1sc+IAvHqdDkUDrTwBpHYaSPqFYD+n/FfYCuOZ7LYbV7qZtKRiv206yGvvb6fdQb2/NHPQvYHbavjJqmpFnlh0qCL+or/T8e8GPc/wB2b7mfmFHsWZIoWqtCRwPHoPWWww7NfgKagHJ6CvLbZydMtU1j4C8hBNwLX+nuYuV/eq1vrO32q/r4x0rUefl1MW33G1eIssajXQdBXW9S4/dbSrUU4mM40t5BrXn8AH/b+8qdg5n5P5UsU3DfpgpfNMVz1gb96jmHnXcd0fbthjJQAAHI6KN2v8Rf4FNLnNtVFTBXBWcQ0oeJCwbjVo/23sbWvvD7cbgBFHcha/Z1gPc7H7gJJ4l5b6/PNT0QLe+O7RxuYSlzVO6UcUrIHOskojaRe/8AUezO73S+a1O48uN40QFa1rj5f5ujXbdw2qR1sd9h8KTh8NM/b0OewdmjcmLjMn21wgLa7B9Q/V6fYFtveK92jcAm8xkRcKgHH7ehbvftltPMG0eNy+ytLStBSv8ALoLu3+pDPEVxcURniUtwraXe9uSB7nrl/mC05msF3Cx+FusZt82K+5euzZXqlWHr0ULIdYbe3MH21vehp6KW7CKaKEAu5NuXIv8AX/H2e/b0UHPRXu3PiFVbXo5MntaQ1EF2lUPJJIjRH1AAkm3H+29+OOvCvRIcjjK3E1UtHXwSQTROUYOrBWsbakJAuPfut9Qv66TYAX+p9+619vXuNP8Aj/sf6+/dezX5dcebf4f7xf37rfXvfuvde9+691737r3Xdz+ef9fn37rVB1yRGlcIis7uwVI0UszsxsFRVuSf6D37r3DqXWY+toGVaygrqFmAKrW080DMP6qJVW49+63nqGtvo30/4n/X9+60a+XXVr3/ANv7917rwtfn6f77+nv3XjXy6eqPb2eyKCTH4HM18bD0yUONrKmNrHkq0MbA/wC39+qOvAHy6lts7diAs21NzIB9WbC5JQBb8kw+/dbo3n0yTUVVBKYJ6aogqLqopp4pEqGLGwCxOob6/wCHv3Va+XT1FtHdEqh4tq7klRgCGjw2RdTfm6lYSLf09+qOt6WPXbbO3Yis77T3MqKpZmbC5JVUD6szGGwHv3W6N0wx008sogihmmnJZft4o3aYMpsVMYBN/wDC3v3Wq9KFNmbukiE0e0d0PDa5lTB5Jo7Hm4kWC3+8+/dbo3HpirKGtx8niraKsoZT/uusgmp5OPr6JVU+/da+3qL+r8Af63v3XuHXH6H/AFvfut9euT9effuvdf/U+f8A+/de670m1/futVzTrxFvr7914GvXjb8X/wBj791sV8+uvfuvddiwPP8Avj791o9eJubn37rwFOuzcccf19+68M566+lueDzx7917j13cHg3sLf6/HHv3WqenXQt+b/7D37q2fLrr37r3XdrW/wBa/v3XuPXXv3XuuwL+/daJp17gX/rf/Ye/de68Lfm/+w9+68a+XXXv3W+ve/de6yxRyTOkMMbyyyuEjjjUu7uTYKqjkk+/daPGvVh/xf8AhFuHs7LY7J7xoZ6fb0zRS6FWQNoPP7thz/rf737917hjq/TYnw0602tgaShp8DQVIWnjDzVFJGWGkfS5Hv3W6CladIHe/wAbtm0OQAxmFx8Uo1Mrw0yKQ1rgAge9mvn140r0kqPFbn2Y0ZaOY0UEgVVXUQI1+nA/2HvX2de6sX6I3rjNybTkoS0f3pTwNEzAObjn0/7f3v59ayRnpo39sebAzNmsara5JDJIEUgXt/h/sfeh17yx027Y3zJDVU6Vzm6lEsx54/rq97zTr3Q27ywOG3ttt5KinhmZ6Rir6VJDhbj1+9cePXqVHRJdpZTK9c72kxmuo/hcs2oI5YIFDcABrfT/AA9+69Q+fQpdv92T4jbL/wAGZvvajXEtjchZFt+Pe+tnPQEdQ9bVu5srHuXNq9RVVb/dDzAyKoc3NifesdVzWvVku28bDi6OGnhRURIghVVA+nFx73Xqw4U6dsnpmgZGW40MBcX/AB/X3rHW/Pqvv5D7fk/h8skaEutWsycfhSGsPfifTrRoTQ9DP8dN2LuDbNNjqiZRUUMS06RNIOQot+k8j37j15SejJ09IlIJ1VdOvVcC31tf6fX3vrx4dBJXbfqarPrVGPVGsgbkHgar/X37AHXhk16FZoQlBHEvA8YH1/29vr/tvfvLr1OJHUSLHmmVZtNifqSAbH/e/fscOtcOmXLZGqdTEhNgGAsD+Rbg/T/ePevn1ulMHpES7SyeWR5W8liTYchdJFhb/H34eh69xz0lK3p2myMLx1TsJi3pDX+h4Nz79mvXjSvS9656v2vsWN6jxUr1zlXWVwvlUg39LN9Pe+vY6W+6t00WKxs07VKalDKFWQEn0X4H+v71QdbrT7OimjM12787a7NTCbjTzwD+ffgc5618Xw8ejQYCCPDYtAlg7JY3AHNvz+fYU5nv7dIDFWpp1kD7Jcpbrc7qu4BCIww8upWKj+9qJZhwb8fT+vuILt6RsfWvXQjZEZrqGFvw0HT1TY5K/Kw087DxGQAqxtyD9SfeM3u7zHd7Ft8gs66m8x5VHWZOyyfQ7CrxCh08ejN4vGU2CwqPTxqoCE3AHP8Are8A76+ud53U/UMWYnJJ6jncr+a6ui0hrQ9Pm3Nr1+85Q2OiMkisoIUEmxNifZZvG+WnLMZF6wVfU9E28b3ZbPCsl2aKR0LknQm8qnFTMtIyRxIxfUulrBfoPcdL7t8rxbkkbTAs5xQ449A7/XS5atLhayVPlnouJwVNtTLz09VADXRTaTdb2bV6v9t7mr94Xe+bekkTnwiMZ8upKTmRd9skkhb9MjGelhHXR+MSnTY2uDYWvxx7DT20gfwxWvSJ4FPlj7OkHvLJY8UzrWFVjN+Ra17fS3sYcrxbhDcrJZ11gilOP5dKrTZBusoijUN5dATDj8dXtOs8ETwtrMLOF5X8Nc299UPaTmzdbbZLeLcmOvQBnj1Bnv8AexG073tpvY4wJEFTQeYHVb3zK6y23uWGbG1lHTTRy04XW0SMygxldKn/AGPvKawna6thOfPrkPzTtCbFvUm2pwQ061pPkJ0DV9bZF8jhaWaTCuXecAFlp7Nq1Lf6D+o9rOiA+h6Krf6/4m/v3XqdeJvb/D37r1Ovem35vb/C1/fuvZ669+631737r3XvfuvddkkgD+nv3WqefXXv3W+ve/de67IIAP8AX37rVeuvfut9e9+691yuWsP6fT37rVAM9dEWNvfutjPXha/P09+699nXrfW30H++Hv3Wuuvfut9e9+6913fgj+vv3XqefXiSbf4ce/daAp11791vrsEj37rRFevfU8+/de8sddkAAEG9/fuvAnz64+/db67F/oDa/wDU29+60acevf15/wCN+/db69xx/vPv3XuvEk/X37rQFOuvfut9dliQB/T37rQABr11791vrssT9ffutAAdeF/qPx791404Hz671Hn/AB+vv3XqD9nXH37rfXZtxa/+N/futCvn11791vrkovc3+n0/3v37qpNOuib8+/dW4dde/de697917r3v3Xuve/de697917rsW5vf/C3v3WjXy669+6313xb83/3j37r3XXv3Xuve/de67BINx791oiop10eeffut9e9+6912Rbj37r3Hr3H+N/futZ669+6313x/jf37rWevcW/N/wDePfut569c8f4fT37r1OvE35/Pv3WhjraH/kuHZMG1s2aKShGdegk+8BKCTWaZtZva+r6e9eYp1sHsz69XNxV9PSwqGdApZrXNrD82/Pu3XuB6Tuc3BTxWETqxYr9GFrn+v9PfuPXvPPn0VvfeTNXknOpSVIbSOBbVfj3rrx6TUGVMCgaipUXtzb+n19++zrVB0L+1q+N0oKpiEdHDEAEFze/J9+r1vTToe92x0+b2vBKQAscasSByxA+n+I9+869e4dEwzOOnq8oYY4nEKObHSbfq9+615+vRhuqaN8YshYadcai1ufp9D/yL3vr3Qg7hoqaopKyp1BZY49YW41Mb+9cOt1r0VDKYDK1tXNWBJVh1MQNJHpDWHA97p1o56ccbTZPGaZ4kks3L+k8WP+F/fvLHW816VW3d45CmzCu0hMY4kErkqFH5/wAPeuvY8+jFmpp8xjkraOSNpUUGTQ4JH+quB73nrWOnKii89CjyH13tZvrYL/j+Px71w62KHB6gyNIjmNEZkW3KqSv0ufp738x17FMdcky1OD9rbRIxK3f02a//ABv6e9dbpTrANvioqi899DANxypH4N/fuPWh2/Ppwq8JQ6FhSCLWLAEKNV7f4e91x16meuUNA9HHeKkXUPpIqAsLCw5/PvXWsjPr04YmknrKhUluU/N7Xt9PfifXq2Dw6EyCgigVUiVQFA5Ate1uSPesnh1qoPSc3KFjh9ZHANhe5/w+n+Hti6nFtbtIeNOjbYNqbe93isxwLDoKIXFTWShf0gMtyf8Ab+4H5kuPq5JHbz66u+yOzJy3FaWa8Rp6V+zqTHHKoKy12lsgPFz+feBfvluO8pIbO2DCFRxFafn1nHvE92duBh8lFejGyUp8dPS0i+qUosaAfU2twB/r+8SIpO955zhaknqKZrwIWklPDj1Oz3XO6shijBBiqhgqB2dadrEEXuG9p9m512Tb9zS6NwoZWwNQ4g9U2vmvZLa5EjzrnGW8+kdtzawwAMNfT6KgMQwkUAgf4A+xlzLztufM031AuGZKAABjpx6dK9/j2rmB/qPCR60zQHp9yODw9fBK01NTyMY2A1oLHi/B9kVjzBu1rKqpI3H1PQMn5P2Wc6ZYFz/RHRA+3Om8PuaWsijo6f7gNMQyIpIUn8ED31x+7DvEW48pRJcuXfFQx/l1hF95vkd+X7AX+zWwUOT3KtD1Xf2VszM9Yxzvt2aqNajMfAGZVW/9F/p7yI5g5K5Z5og8K+iVKZBUUJP29YV8t868y8n3ouLSRpDXIY4p9nRJt1d7di42oekyONqJGFxrSAt6Qb/X2/sPL9py5a/Q2ApGOHRhzhzjNzlMLu5UCSmaCnQdV27q3ddOZaullpaoXZX8eiTUeeG+v19npz0C+GOnPaO+vvJH2juhw9KV8dM87XZnIsqksL2/r7317oDu5uo8Fk6yqp56QQ6oTNRVFPGUvK3KfuL/AL4/n37h1oiv29Vybp2pl9qZCWiyVNLGoZvDPpbxSR6iEs9rXt9R79175HpMe/db675t/hf/AGP09+6159de/db675U/j/kfv3WuI67vY3X+n5/1/fuvUrx6V+xNh7n7J3Th9nbPxNZmM/m66moKKkpYJZj5qmURRtL4gSq3PJI9+PXhUmg6vh2N/wAJ8P5jOxs90n2ZubYG0TtXL772dW/aJuGnrsmMactTVc/3+DaNZRHJFdGDgfU839+6sKfF6dGi/wCFGXSOJ3d889ndS9Idc7X2TlKPrzYWEq9u7dx0WKoJs/Dg44q+V46VeZJJblnCer883PvQwT1t6kp5k16ItsD/AIT0/wAyLsXLw4HCbC2tRZOfGxZWCmz25FwpmpZY/JF4mroVDMw/SoNyePe/n1UCtfl0B2J/k1/OjK/Jqv8AieuwKKg7WxtMtTUJlK+Sg2+EZhwuZqIVj/N9Vrf4+9V/b17TXNMevSF+an8q/wCXHwKo6fI/ILa2IxNBU1a0UFfhcp/F6GSoc2CR1sMaxMf6gOT/AIe9060TQ062d/iz8ocP/L0/4T3de/Knrr49dJ9id0Znuuu2W26Oyto4/cbU2Iq4oHu8VSpLFGUhDfgM/BNiKcTTpRwQH5f5eiddNf8ACmPtvs3sTa/XfcfxG+J9bsne2dxO2ckdqdTYiizNNTZisShnlpZgoYOA91K3IP497p59U11IB/w9NP8APi+FHSPQ/wAn/j12/wBOR0VFt3snMdX1uU2k9OlG0a56ro8jPSnHfhUM7qCByCb/AE4tmhJ6ZwJFUeZz1at/N8/mwb8/lmdvdT9A/Gf4bfG3KbRl6N663RW7g3P1LQZ6pyeTzO34qqrvVxxsfSxJFxf63Yn3RRWo6UO2inz+f8uqVezf+FJny43vsLdOzNwfEX4p7fxW6sHW4Ory+O6ap8TkKOGvj8L1NDXmAeORf7LA8H3YAjpsyBv+L6Wv8lz4D/H7e/X+8/nj8tKmam6xwtdn5MZTQ06vQVe5qWWfIw4aONuAkjoA5Nzp+lhf3vyoOqAANV+Hl/s9Ou/v+FKG+9i78q9jdQfFz4wT9N7cysmD15jq7E12cy2Doas0qTJkGFxI0Sg+Q/qb/Cx96Kjq4fBH+XpG/wAzbd3wN+YXw4xHy06dp8Zsv5D1eWgpd27Aw+OixOGpYgwesqKGljA06SChW3vdPTh1TBHdxH7etX/37rfXK35P54/2P+Pv3Wq+XXRUjn37rwNev//V+f8A+/de697917r3HH9fz7917ru3F/8AG3v3Xvl17lT/AI/8V9+61hh16xI1f776+/deqOHXgL+/deJp11791vrsG309+60RXro8m/v3W+u78W/xv791759de/de697917rsG3I9+60RXrkV5Fj9Rfn37rQb164/mx/2Nvfut+WOvE8W/Avb+vPv3XqefXib8/n/AHi3v3XgKdTMdjqzK1lPQUFPLU1VTKkUUUSNIxaRgi8KDxc/X37rfVvfxJ+CUmTrcduHfFLM9U4jqaWnaFzAsbMCvoP5/wAffuPXq0OethjqXpXC7UoKKBKGGnip4kRUSHRqCrxcf4+/de6MFXYekp6F0iUIyxvYKLAC3A9+60eik7wUx5YxEerU1i3pBH59+J8x1vzz1HbbtFl8ZLHNDHJJIhAutzcjn3vyr171B6QnX+Cz/X2+op4hOuEkmRpBYrEoY8ix/Hv1T1ocej35AUe6cQrUrJUAxnyBGEml2FrG309+4db+VOiibzwUm3q9pNLreXUvBAAX1A3P09+698+jA9P5xNx4h4ZXEiRo0ZGoMeBz9P6e9V68BivQZb/2djqvelNFFojZiHJtpY8+o2H9ffq9a6QHanWtK1NTTw+WQIy61ZToBVeVv79177Oln1Xk6PG0tPRMEjESCMlrA2HHN/fhQjr3DozWLydHUpanmjaT+ge9/wDbf8T78evD59SMlI4FrEXU3HJF7fk+9gfn17otvbWJOYoJg6FgA3AFybJa1x70K0qettk46K11huefYO80pppZIqdqlRpY6YyC30Yf7x79x618J6s5weSXO42KujZZEkAYlCCPUL8ke/fLr2CeusgtNSBpCyq5v9Tb8XH19+B8utnBr0kJ94Y+llRJp4lA9P8AnB+Dzz/ife+tVPl070+7aGvKLDPTtGbAAShrk/W/vXWzXj0/HFUtU8c4a6OAxAsRY88W9+rnr1MY67y2Qp8JS6bR/S1r24A/N/8AevfsnreCegY3D2DFSs1ivlC2XSw4b6XsPe/IdVIPHoI8v2hkYlk0vJZjdfUbcf4D3quevYHQbZLeO4txyfaq0rK5Fl5P+AJHv3XuHQ49SbKnpYv4jl1KPZnRWWysL35Bt/vPtu4fw4WceXRhtFuLvc47f1I/w9C1nsnHEphhsLMEVb3J1fX3DG8Tvc3hzUDrpF7cbXb7Ny6gKAMw406Xm1aHRQCokBVpEW3/AEMfYau2BRkHUx8vQj6pHPE0/wAPXKjgkn3HTRQlheRWYpe9g30P/GveNXvGLWPle5mmoXBxX7OswrCRYuW1Z/4adGhyUojw1LSA+sxWb8sSPfPzb0Lbk9x5A9RLMuu4dvInoxnxTooRnlWrCmNnQ2cDSPV+SfcGfeRvZ15bka0JDAUqOP5U6ij3fcrsv6R7gDw6si3PLjYsVUwRGii1RMBZ41OgizEH3ixy27TbfafUxs0p0nVQk1Gcn/D1g6lxuLbugq7d3DP+DqtfsTZmw4qjJ5Nq0PlXmaRYxIpXVz/T3mTypzPzPLBBZCKkAUCtCD1mDyHuvNjiCCSMi3ApUjPl0UzNZZKMzxIbqGGizXNvpz7nTadse4KysOPHHU8Xd4qQD18+gz3VUPlcY7Lcsg+lvza1vci8qRQ7bvURlAIJHHh0ecl3pa/VT5/5+gwaslx1DJNJrVY4SCzAizfUc+85eX9z25JLYrIvAYDD/B0NOe7Bdx5euoY6FijEfs6IL3fug5nIy05OphpRbm/9R/vXvMDYby2k26PvWpGBUV6+fj3c2y4sOc7ldLN38aH59EE742HR53rvMQVMETTzRTaZGUa1Gg/pv7OtSnh1HnhyUrICPt6oEzOO/hOWyGNYlvsqqaAE8FhG1hf3vpuppjqFHSVUy64qWokT/VRwyOv/ACUoI9+63UddSU1TCLy088Q/rJFIg/27Ae/dbqOsHv3XuuwbAj+vv3WqZr10LX5+nv3W/s697917rlcrxx/X37quGz10Qfr/AF9+62D5deAubf19+62cZ69axsf9jb37rVaio669+6313aw5+ptb/jfv3Wq1PWanpqirlSClp5qqdzZIaeKSaVz/AEWOMEn/AGA9+630f/8Alt/EbE/Lf5sdEfG3tFN07S2f2pudcLlczS42vp6vG0skDP8AxCPyKvoUgBmvbn3o9bAHn5dQ/wCY38WdpfEz5edwfH/rOvy26dude7rrdvYzLTJPVVWQhpJpIRLIF1HU2kG3vw+Gp684o9PKgPRKF2rumVCybXz7pFfW6YbJsq/11MI7D3vqlPMdQ6PAZ2vkliocJlq6SA2nipMdW1Lwn/UypChKn/g3v3W+PDrg+KrqSsp6XJ0Nbj2mnijKVdNPSyaXkCMQs6g8A/09+691uB5X+Ul/Jh+KvxS+L3e3zt7/AO8ttbp+S+zU3bt3b3X9JJWxw0isYaiWYMGKqsiuvA/H490qenyij/Z6buov5bX8gf5i52Dqn4h/JTvfI91ZqcUu2dv9gAYmgytQeBErtoa7GwX8H3sV8+qkKR20r1U5tX+VjX4X+ap1/wDAHuOXLbb29vjsj+6MGYgeRatqFpCnmppSRqdeOb8gg+7GtKjptANelskdW7fIz4Af8Jsfib3FvToPt75QfJ6bsXrzKy4PdUeCxpqMfTZWnYpU0sUkavfxspUnVyfdanp4qg48eq/Plp1P/wAJ/NvdJbpynxN70+RG7+6oIids4PeePqKXCVEmjjzSKi86voCfexXz6owXy6SP8tb+TVuL5V7Q3P313nW1PX3x52Rj4Mxunc8tYMbVw4txqV6EyEeaV1/Si3t72TTHVEGrgaD/AA9Hdx3xC/4Tgbr3hT9PbW+S3yGXsmuqGxOPymQi8W0pM0EKCCXIkaAvk4/V9P8AH3rzp1uq6aih/M9VT/zSv5ZG4f5f259oZWlyVPm+rezqT+KdeZuDIR5JsniyGZJppo2bS1lIZT+ffutaTTV5H+XVSdwbk/7D3vrXDrr37rfXvfuvde9+6912CQCP6+/daIr14W/N/wDC3v3XjXy669+631737r3XvfuvdctPFz9LX4+vv3Wq+Q64+/db67twD/X/AIj37r3XXv3XuuQvZrf059+60aVHXV/oP6e/db669+691yDci/0H/IvfuqkYNOumt9Rfn6+/dbFfPr1uAf63/wB49+63Xy67UX/1h9ffutE066Jvb+gHF/r7914CnXXv3W+uyLAH+vv3Wq5p14Am9vx7914mnXdr/T8C5/417916vr1x9+6314W/P0/w9+699nXv9b37r3XvfuvddkEAH+vv3Wq1NOuvfut9e9+691yRHkYLGjOx+iopZv8AbD37r3VmX8tLtHfewu2o8Ht5Kk4/O11DT5BGVxFAsxaOViCPqwt/tveiadaHHranz0GRjMapqKfaQTC1/wBboC1/dutg+fQR5rcgo4pzUO3kiAIBv+oA+/VHWvmei61+7UrspqmkUKZT9XC3W9+b+/HrdOlRQTUVdLGIpAfoWtYqbWNuP9f3r5da8+hPw2UpPvaPGQsC5kUWU3tfj8fT36uOrccHoz1dIKTBQUkhteNSQSAPUv19++fVc16CiXHUBlMzGMMWNyWH0LfQn3v59eP8ultgp6enCrG8ZQWsFcG4H0HvVM9WqAOpWUWpFRFVAuaKRh9wqcqEvyfe+tV6Vse1cVm6WKbHxqItI1Lbktbnj3qo69SvDppqNn0SBqSSNUU3AYADi/1H+8+/fLr329FO7hq4NiT6aFm1zuyEn68j8H37rR49KTpTsGzRUuTqHaOscxKHkty39qx9+rTh1sZND0ZnKZKPEy0sZJNJU6bMPpZz/X3uuK9eA8uhXwtHjpqFHQRzBlBZjpe1xqIGn3XjnrdCB0mNybOp6lTV0RZamMa0RBYar3t72PTrR9ektic7NG7YvIjxzxEjU3p4tYfUe/dez5cOlXTPGmiWQ3Asx/IP9LH/AHn37r3z6UUVbSvHf0iO/JYryL83P1966sSKdcdOPZllpJVEg/WA4AF/pf8A2PvWetEDh1POTehiLGRWIBI9QaxH+HvfXqA9BbubPNUeR55NJ9eldVr/AOw9l291/d7EenQx9uGC81wqM1cf4eue0qVZsdLXuNJYMF/JH9ef9494/wC6uXV/n11s5IhEM9sxOKL1lwEJn3LThHdUjqVJAuB/U3t7xV97Z4Nv5eLaAZJARU8R1mBPOBsVSBUr0b7baJW7t2/TEHxrWUolH1Gm44PvADd5HtuXruccSj0+3PUEb6DFtNzKPi0tTq5vCbYwD4OhSPH0beaihjdjEjFiUsSx/r+ffMxDud/Lc7lJfulwJpAiF6AUPGn+oDrn9u/MO8RbtIpkcBXJwT69Ex7f+PDzZTI5ehampKRIjME1rGC1tX5/3r3kb7c+9YtIIOXd4DPeBtDEDUPtqOsjOQvdhYrGGxuA0jE0J49EC3TWSYH7ujLBpYBIoKtf9JsPeY/L9kN4McoGHp5evWSljPDdmGcYDkY6LgmZFVLPPOo8pmdSP6rqt76e+yVl/VvaoLaAkM2kmvz6D/3gOUdvvuTpJZ0BRELDGa06L12F1h/e7KTVoj1RTsQQV1La/wDT3mrZsXtUc5JA64Y8yxxxb7PFGCqBmABFPPoBd9fGzb0WLWf+EQzVLfqdqbU3+Iv7UmvRKBQY6KVuLoNYmK0eNaNmLIAlO39bA+9HOet1oaHoC9xfGPJQV8WWK1MM0Mglj0oynV/Q+/db8+pWX2XVZbDyYvI0h+9oISYJhEfI5QekM3+v79xHXsnor29OmIex9uZLCz0Yp81hYpq0TxxWn0w3KWP+8f63v3WsHqqTKY+pxOQrcZWRyRzUVTPTSK6lWDQOUPB/1r+/da86jpt/3v37q3Xvfuvde9+691737r3W0v8A8Ji+kdq5nvzvTvDsjbeH3JgOquhd8bz25j8lAlYDuXA42WtxdR4edNpQvJ/pf3U+XVowCpGRSg6Ld1R/Oj+VEP8AMRwXbOY3JNW4LI9lYvaX9yKqsml2zSYhd0pQ04ioJP2wQgQBQoA/B/rvyr59e4tpPw14dXH/AM3iTHZn+dj1PnIItNJuzD9X7jqoGX0pNmcdDVVKKPyoJIHvS8G68/8AaRdBb/woQ+f3eW1P5iHW/wAdeq81L19sLZ+H6lqIazbM8lDkc19zBEsq1U0BXjUGJvc2I+n535UPWidJDqfn9ueji/zrPnXuTZvwJ6U3rsDAYjbPd2Sptv4HcfZ+LjFPuzIY18ZHSRvV5GMeRyWOtrnkjn3sAChp14s2kpWnp8uiN7q3JmvmD/wnTxu4O48hPmt97I7wyFfi961rtV5qanNKHFFLUSksYQy6ghJtf3oZb8+tkkRmpqQPPo4PwM6W+KnyG/4T44HZPzb39nOtemtsd5VlXhNz7a8Yyj7hWnjMMSRycEEBmP8AS3vVO6nTmr9IMaAUFfl0XXrzoj/hN38LN043urId+9xdq5/blTBk9r7ZyFFT1OP/AIxQkVFEZtALFWkAD6Re3HvdSPLqpSMitRUZGf59U5/zRP5gGN/mI/zAOteyNm1U0ewqHcHXmB2zgo/LDj6BcXlKXG0bR0bgAP44o7m35Nvqffvn69UBqwFOH+fPW3D/ADYPkd/K+6n7D6M2h8ycdlqzt+i+P3W38ROOxprUjpH2/GaVCVVjwpHPvytj59WZatqY48utbD+Zf8mf5T/aXxsk2x8T6HNUfaQq/KZq7EyUURgSVWSOJmQAk255/r78SSR1XSoBPn5dHb+MFLlch/wmgzlNsdWfd0XyIy82Q8ALVIwf2lSslwvOjVpJ9+FdWPXqzaREfWn+XrTUyaVEWRyUVTbzpW1C1AA48qzsH/3m/vZ49NDgOlZTYrsafab1VDQ7sm2U00hkNJHkpsCJFez+Tw3iBuOSR79Xy63Tzp+fSEZSpsRY8grYgqf6EH8+/db64+/de697917r/9bQABt+Af8AX9+68c9eJv8AgD/W9+60BTrq3F/fut9e9+691788+/de67NvoCbf8T791ofPrr37rfXYBPA9+68TTj16xHHv3Xuuvfuvde+vv3Xuu7G9rc+/deqOPXrG9re/daqOuvfut9cjb8En37rQ+fXtJ/PA/r7916o8uuPv3W+u7W/4Nf6f4e/darXq0H4HdCUu46uPfWXp2mC1EUMEU8TmNI/uALgEWuSAb8+/de+Z62YerOuqHEY+kqvFGgWICNLudKX4A449+6351HRkEMVPCqqFBAAvY8gD/H3vj1ogAdNmRrkVDqbjS2r8i1/p79UcD1vJyOHRR+yqpWzKSwH6AqbH+o+p96+fWic9SdlZMSgQykf05Y+kfkk297+zrw4ZHQzLQ0FXh8hrWISLR1UqSNywKR3SxP8Aj791utT0Dvxn7Arpd1Zza1ZIaiE17CNmYMFUSkAD3qtePWhUdCx3rioUx9dWxKD4vKCbWIsv4v79XyHWycdBT8WMsZosok0hKLWyoD+RyeAFH+8e/V68DjpWdoyy0G9aHLRNalVkDWuBp1c3v79Tr3DPS63VLjM9tqmSh0PNJGpcsQbErzyP9f3vrw8z0D0e0qimZFgYhnFzoufUObcfT3qnWjx6cKPIZXa2TglnMrUqlTISrAWH1HI+vv2OtinRicHubFbooQ1O6NLHYSLcEhj9bgE+91Fcdepxr0h92Y/RT1AZLh9QHFvxxx791qvRD+xdvMlfJW06aJo5EcEAgixJ/PvXWjnHRhOl+1ZsNhGxWS1O50BCxBI0/gX/AN597p6dWHz6EHcG9p8spSAssTi+r6Nc8fj/AHn3rrxOc9Bfk8FX5FGY1Eio/q1K3quObf8AFPfutHoP8hkM9s9gaaSaZF9Z1XJ4+p9+630KfXPyAWtmWhyrFJUKINTKv50WIJ9+6r9nQw7qzRylIKqmfVF4ywBIP1HHvfz63ToEKHGzZmec1VwiSckixsOARb/iffutedR06TbKpaiRII1DB7Wv+Px/Z9+oet06UmK62ioZY6hol9CggFSD9bkn3rr3DpZ1GYjoKb7SO6FRYXNgLe2blTJbsq8T0abHOlru0VzJgKRX9vSNgyv8QzEULPrVmjBHHH9eB7iO/wBsnjd5SOug/KfO2031la2CMNdB5jo0dBMlDiIUIGkRfni3Fzb2B9wuI7dGZuPWS3K21XO4XMS24xjND1g2dKk24BM31DsV1X4F/eJfvxdPJtLInBiOso7+1ez2FbY8QM/b0YBjJVSxgC6g8WP+8+8MRot4yTg9RgsYDMT0J+0t35DaSFcWVWpZSwdiAOOf1Ej/AHv2C975asuZ3C7gNUYPD/Y6A/OG22t7aGW/NIxjqFV93b0y1bUUNRWTAa2T0yAryNJAt7OF9tuWtqsUlt4l4enQI5X9vNgursbiEBVTio6DPdG4Jqfyy1VVM80tm0s1xf8ApqJ9ibYNnS4KiFBpX5dTBM1nYRCGBAgUdAzDJkczUPckxM3DfX/WPHuT5FtNthHkQOg/G0905HkeHQl4XacU8XjqjwbcG/J/1h7Ae8cxOkga2OehTtRfbE8RR3evSc7O2JBBtmsGOS8si2AAPCheSRb2f8gc7bgm/wAUtxK1E8icdHR3ue8he3mOHBB6qT7/ANmV+2Vjy0cbyuyRvJGiFmU/U/Qf4+86/b3nncd63KMLKxCtQAcKdYTe6Hs/ay7nLudwgETdxYjojPaddmM1taTH0FLOa2ogcRr43sNS2tYC/wDvHvOHZxJ9GryZJ655+4BtYd6+isaeHGSKjqs7ZXw37E3tvOrq9zUjQYaWtklmeCOo80qvL6QbKNIC/WxN/wDD2ZO6oKseghBby3DeHANR+WerY+sfg/tikwlJioccksTIFZ5qeRnS9ubuCTf/AF/aV9xtI/iYdCS05H3+6oyxGjfI/wCbpY9l/wAvXY2ZwQpZqZYWMDC8VPIragvpuUFxz7Y/e9pWgbjwz0YN7c8wLQmMgfYeqO/kR8Ut1dP5KrqaGiqqvBRSOoIhneWNA3DqbHULHm5v7WRXEU4rGegzf7Pf7a5W6QinnQ/5uifkEEg8EEgg8EEfUEe3+i3rr37r3XdiBf8Ar791qo4de4tyTf8AHv3XvPrr37rfXvfuvddm3Fr/AON/futCvn11791vrsWJ5PFj/vXHv3Wvs62cf5D3w66L3F1R8lfn38gA8+0/ihHiq6HC1dK0uOzdRkJo4oaeRZh42BdrNwfpzxf3o/Pp1UJFR/xXV2/8q/8Am7fG35y/zBdoda706K6s6vr8blchhei8xsXbmOw+Qr0hpTBRNk5YkDSMxGsyAjkn8Ee/cDU9VB1DSuKevp8vToiPS3Su2O3v59PY3WfYmLos7jsh3pmMfk4clGssMsEWRnaUuXB40qfr/h78B2D7ettT6gg+S16OJuX+Z78Zul/5kdb8Hupvjf1NvPrLM77q9mdiZ7dO0sdUZHFhZjTVX8CrXjDRvEl2BW/IF7Hj3s+o/Z1QVHY1ft6avnz2J8Nf5G3ak2+vjvsHZ/d2U+R0UufrcD2LtyjzWL2tBUzGWWjx9LMpA0aeJFAsPT/r6AqcnqzPRaqvn5GnVYv8/PpbYu8+nvhN8zeodqYTbNH35scbs39gcBRpQ4/C56smjiNPQwxKF8Kk3AH45/HvYqcDrThV76dWV/zE/wCWr8o/5mH8u3+VznfiztnB7jo+sOnptsbpXN5mhw00GQmr5apXp6msYEx6ZR6FUgn8j3VR59Oy0JpXP+x0An8qv/hPh8lvid8ktjfKT5nZraXU/X3VObjzRhxm+sDNlcmKch1NMi1Ecj6gD6dHH1uTb3uoOOmqMDq+31r0XGn+VM3yW/4Ud9GZyjSnk2tsL5BQYvadXTyCpqcljnqV0VdVUAnUSmkcE/k/63iTTPW0C+JVfSnVnf8AMz/kE7T77+bvfnb3+kTbGJk37vSr3FJQVe6sXQVUDZOZ6loqikadWRhqs2pQeLH6e7LQYPVH1O1Rj8utaL+a5/LBwv8AL8h62mw+7MPuVt3IktSMVm6TMeFXFk8zUssgQkj6G3vxpWg68Aaaiertv5kG88l8c/5LfwVw2wayvwZ706lmrd3SYieeFK2SlqZaSKSvjUhWYafzfgD3oeh9K9XkXTHqX1A/KnWlJBV1FNPDWU881PWwzipirIpZI6iKdH1pLHKnqDBuQwN7+/dN08ujLdxfLbtvvbrvYHXHYmbrc9jOuqZKbA1eSyNTWzxRqhRwiTDSmq5JCm1yffsde7hxNeiwlStxa9/9v7916oPXHngW/wCK+/db699D/vj7917j11791vr3v3Xuve/de697917rsA/W3+PPv3Wqjh143PNrX/p9PfuvD068WJsD+PfuvAAdde/db67FrG5+n09+619nXvrxYf7D37r3DPXh9bHj+vv3XvKo69xf/D37r2adeuATaxH+Pv3XuPXja3B5/p7917Neuvfut9e9+6913fi3/I/futU69fi3H/E+/depmvXXv3W+ve/de67FrHkj+n+Pv3Wj11e3v3W+vHn/AA/1vfuvde9+691737r3Xvfuvddk3FvfutU8+uvfut9P22tt5Tdmcxm3cJAanKZaoSlpIfVZpW/1WgE2HJNh791qvV23VX8v7bmxNmY/c2+fM28JqVXnpZY5jCokQSKFibgA3/pf+vv3yHXhXj0Lvx760xu3O4KCfE4yBQ+TpwxWKwQK1i4NvfgPXr2B1sEVGJjkhiEiXLUsAVjz/uscD36vWxXotvZ3X0jRyvQofLOVJADEc8nm3+Pv1a4PWiOiB762dm8VVSaUmVgSx0I97X/Fh78K168B1g2hnc5Qv9m9NMzEhQzQvf8Ap9fp799nXqDo5/UXXeWrKxNyZFGEdlkRXDgcm/5Hv3nTr1K56XPZG6avHyzRoD4kjRUFheyj3vr3nXooua7er4Kw0rSNGoc/UhRYt/tR/wCI96r1voR9g9iTZSqWITl24uQwZefoLg297HWs16Ontqsp8lRrTShWYxgn+1/ZuR6ePfq049bA8+lHTebFvppSRBcEqOBf6mw96p1upXI6jZKrnrnBjWzLc3BC3v8Aj37gKDrQoTXoAOy+tYd5RCeucxvSXk+pufzz78OvELwHRaKYUe1s/DTLUcUklwFI+qtb6N/re/da+fp0fLbPh3rtMyakM0MAEBJUt5NF15W/+t/r+/E8B1vjw6cOu9wTY+omwWQkJaJiVLNY2BsP1cn/AGHv2evcOhmedZUDLqs3+Dfj88e9dWrkenQZbmwYq5lqYx45Vu4ZOCxBvYn3v5dazx6T1LumjhlbF1RkWdPSCxABt+Dq/wAf6e/deqOoeazGQhhcQ38K2IZRe6n1e9561gdIZd+1ePRmeRw3N7kD6Hge/fPrRAJp0tNu7xbMQqZZGYs3INiP6m/vw623y6wbtppZhSSwMbSS2I/Fj9ePaXcIxNYyKeAB6O+VLo2HM1pIPxSLX9vS7xFSlBt8RKRqsLjj+nqsPeNe+3cVrM0J41PXaX2z2C93u1trtB2aVNenHYn7uZaocfWW4H1sLcEfT3iB7/XDSbXEnz6yU3qMWm1JbV+Ff59Gi2rVDHZmnykwBWBllQfT1Dnm/wDre8Jd9gN1tr2MfFwQfz6h/dYDe2b2afiqD0djFd87pjxsVRT0gGPhjUI7HSSIxYWv7xP3H2P5Wkv3SWVxM7E0B/i6gC+9sdkkvWjlb9Zjw+3oEezu7M9u+CdY6uelhhU+cRy2DBOWBsR7ln2+9ptj5TmVliEkjnDMKtn5n/J1InKHt5tfLtJJI1cnIqOHQFUvSHa/Z+MbcO26NKrFVLGFJ3BYmQ/UHT/rH3nRyN7c7xdbbFuVhBWMOBWuMfl07de5HKnL/MQ2zdpNBjo1BQY6R83wu78pi8slFSsru5INPKLXNwb+8y+Wf3vC8NvBAdShQafLob8z+8ftPzJsj7ddyNpCU+IHNPTqQvxJ7tigucTGVQctpkAJAuCt/eTG2cwvtdmkF7iSgweuRXvFsOy7jzJNd8tf2BY06bl+LPcmRZqSfC07xAWBZG/H1I9nFtzP9XNoAx1EcnLM6qoiyxx03v8ABXtSolaQ4CAgi6kwy8N+PoPZvDeyzzaEHb69Ge4cq2e1bULy8ekx8ugw3t/L37orIpJaXCwAaf06ZQLX/wAR/r+zP0PQGHzP2dFryv8AL57rglYRYWkeodirghuRf3v5dWoeJI6ADeP8sv5P01Y+Y21t2mklrAIqtFuAYS/IIv8A0/HvXXgM5I6qe77/AJOnzOy2/q/K4nZmOagqgCA0stK0Ug5k4VH16m/JIt79XFevUzih6Bj/AIZk+bQIH9z8Xc/0yEp/3qP37rVPkOu/+GZPm0PrtDF/7DITHn/H9v3rr1Ps65R/yYfm7LIkabOxep2CrfISryxsL3j97r16maUHRCe/egOw/jbv6t617OoIcfuihhjnnp4JXkRY3+h1SKt7/gj37r3V3X/Cc75f9d/Gr5Q9jbS7irqhNg919Ubm6tW9RElNSZPd1G+JgqXFQdKopdWa1rfW496I8/IdWRqdowT69Hf2v/wm07jwfzW2lvXK7v67Pxprt8YTfmL3QN9bceSalq85FloKKZPutSaAwY6rc8cKOd/LrWQ1fn6fy6Mb/N1yO2K3+dj1fR7d3Ft6vwW3sX1rt2DJ0GVoKrHRxYXHQ0v7tZTyNGCun1ktx70ODdWemuP8+qxf+FB0mOk/m+7bkx2bw2Zom271CVyOGyNJkceDZA6/d0kjpqU/rGrj8+/dab/V+3o1n88GbFv8Eunkos5hMjPDV7XSaDH5Gjq6lNcMH64oHZgB9Dce94x1WnE/Prl0O2M/6ByM5G24MBHkm7byDrhXy1CmbMK0ZHlGMMgmKE/RhHb/AB96HxA/Pq7f2bfZ0z9gT0kH/CWvZJod44WlzP8As0VUZ9vUmVo/7wvRtRIpnlxok84hYBkMmiwv9bke/fj6t/oAqK4HWo7V5TI16olbWT1Sx/oEzl9HFvSW+nu1SemgqjgOhD6RVW7k6mDOkYPZOyw0kjBEjX+8dLd5GawCjkkk/T3rqy/EPy62Tv8AhVs2Fqvml1bXYjM4HcKSfH/quFslg62iyKBodspHLBLU0jyWKsvKM39CR9PdFxUdWlPw/n/h61YBbm4vxx/gf6+79U62qf5CXzj6uO2N0/BP5KTnG9M7toM3VYzIwzU8MtPufMU81LRVDvKRZFlkXWSeODf37Pl14aT8XHy+fqOmXtD/AITK/MDcvb9bmumsj1xnejNzZyozUW66nsTatLPhNu11WamKSSkmqQ7lY2uFIBH+1e98PLrwHqf5HHRz/m52D8Qf5W38s2D4Q9UbX2d2f8m8puD77e3YWSagysmOjr/2cjT4ytj8jFVc61CkfkcAi1eDYz69bJqmkmnkPXrSxyVc+Urq3ISpHHPXVc1XJHENMaNM5dlQH6AE8D3vqnDpu9+6t1737r3X/9f5/wD7917r3v3Xuu/wP9j/ALD37rXn163F/wDG1vfuvVzTrr37rfXvfuvde9+691y5U/j6e/da+Idd3BGo21A/7fm/09+61ShoOHXEkn6/j37rYFOu9QvcD8W/5Fb37rVMUPXRYk3+nv3WwKCnXiSefzb/AG/v3Xqdde/db697917rkbgWvfgH88f4e/dVFCa9cffurdZYXEc0UjDUEkjcg83CsGIt791qmOtgb4O7zxVT1tBLTfaR+NqcGG6iUukgDXQc+/DHXjkdW2bS7foIMfDHPOEKKECl7f2rcD34fPrdaY6F6DdrZejE1K7OGFxpJP8AieR/gPfq9e+Z6Ysplcj4Sh8gSxJfnhfz7916vl0XTeWchbJpAzFTZvU3A1fnn3vrVM16ctpVIjk8zOpjYBtQP4vaw/x9+pTrfWbe+/arF4+rjxs0n7lNNERrtfWluB/sfeutdJ/4l4mtizeTz9TdppqsyLr5b1Scn37HXvs6Hr5IbuXH7bqaRH/yuokYWU/UOtuffqderUU6DD4xp9pj6hg3qqqgzHjm7Xv79x49b6HnsTEVmSp0mMAaNVbkDk2HvflTrwz0E1Fk8nQRfZgPYWRb3AsLG1vfuqjHQr7MqzMyyVagsAeG5v8A7E/4e9fPq3lQdLbcVFistjailaBBI0MuhwouHI9J9+4deoSeik7cyud613VNTVnl+xqakMl9RTQZLj6/4e/Y69w49HNWpp914VZ4I42MkZkNrXDEfn3vrXnnoqfYW15VlmjeJA+oEcf2b39662aV+3pB7dxCwzlVS7Iyg3Fvzzc+/daGmnQuU9ENKlh+Obgni9+QPe+vCvAdKOkp42A1EqB9ADxa/wBeffutg9Qs3gcdVwSeWNHvGy8gEi/H0Pv3XuHRVd1bJOOq2rsWPHIspYBOAdJ1AD/X96690LXXWfzWdhhxNUAqRKsfqNrgcX9+619vRiZNntTY/wAtAoMzIXcf4qPz+PexXreOI6Dil3G2Gz8dJkU0AuoLMPSvNjc/T37r1Rw6MBR1cGRplaldJFdOdJBsLDg+/Y49eB8ugp7Axs1OTV0ykhbK4UEfUG/A/HvXDrR+XQMYDJyUebhnbUw8qhwfwb8/8V9lu528Mlm4IzQ9DLknd7+05kt3Eh0Ky+fz6OZjKmHLUVMfMojEd3XVex0/n/iPeNG92rPdOGNACeu13tjvsUOxW01sod2VSTSpHT1t1I1zMSQsugmzEfmx/wAPeLXvmFTbqAcOp2uLiW52oyzChI4dGRx8LLGlv6ar/QWHvCW7lDOa9RnOyoWJ6CPsTe1VhatKOgJ8r6RcNa1zzY+5L5L5bhvLc3NyBToBc1XEdwi2jHB6UW0JJHxZytYCzugkdz+Db6H/AG3so5lVWvxYW586AdGWxxCys1VcCnSG3VVVmarvHSJLNFrW7RqWFr2I49inl+1g2211TkKaefSDd9xgaQRs9CfU9CFtTb609LG8iaWVVdw/B1254PsF8ybu0s7RocHhT06EO1QxxQq65r59K2qqxSIHT0leABwD/S4HsMW1sbh9DcD0cNpCnVmvUKvykNXi5zPYgIV0tYi9rk8+1ltYy2t8gixU8em7S2W5u1VcDHRPt0bUoNzV9VFWwxTxuxUJIoZUUHSAAf8AD300+7psdhFYRXEgDO+ST1HH3o55bHkJksxpbRxHHh0H03QOzp6mCR6Kk1Rrpt41sBf8ce84qxRQ6lIAHXE3wb7ct2MMis0jt5/M9KuDpfaGMVJIKOlQrpuI0Qav9cD8+4o5u52htYXRWCgVz9g6z49g/u1y7lJFu26JVSA2RUcehIxGyMfjaJp44IYY1QsmpQhaw4AH+PvBrf8A7wHMP79ex2nvRW0jzrnNOsuL/wBsOX9kbwzEgC/IdNFVj6GtLpUaRAoPLAG39bf4+8ltg5gvZ9gi3S/JV3UGnpjoNWHItlvl8bO1hUitOHRau2esOvd0Yutoa2ipaxpVmBMsSOVZlsLah/rezzbuc5vFUIxp01zZ93Hbm26SSeFK0Pl8utc75ifC5tkS5XeWyqVzQiZ6mpp41IjZCNTCONeAfzf3Mux76L9VR+NOucPun7Vz8p3cs9mC0Yb0x1VoysjFXUqymzKwIZSPqCD9PYp6gzrj7917r3v3Xuuxb8/8U9+60a+XXj9Tb6X49+62OHXXv3Xuve/de65EDi31tzzf8e/daFfPrcg/lGJV/J3+S/8AzJ+gNovisX2XFS7Vq9vYSKRErNzR0s8fnK062aXSp1WsbEe9cW6cUBY/2/z6rj/kBfFXvHMfzYOhx/c3OYim6o37LVb3rq7H1dNFj4KBHiqY4pJUUOT+pSv4sfz72R1RG86Hh1cL8dJr/wDChftCehaZl/067lanlCuZWUVFWASLXv8AT3r/AEOvz622Lg/6Xqlvc8X/AGPQ3wni0s3e+ed0CFW1sSzll+uonk+/eXW2+LoxP/CjLzPvLpp53klaPFzRRNKGukfjc6EJ/Fxe3ux49UHRp/5pIki/lE/y4k0PHG/WWNI9DKhtVRcqTwfekGfy63NlD9o6FD+bb8ru7Piv/Lc/lN4vprdOd2ZSbm6dq8pnGw+QrcVHkK1shNaSoNKyF20IoW9+OfdVI889OS6iKqaGv+QY61Ve0/nT8o+3nphununsWSlp1YfZpu7NmmkLHhpIxKNVh9AePdqKOA6b1SH4jXo5X8imqqsx/Nx+HFXlJp8tVVfbGMlqZqt5KiepkMbEvK8l2Y8ck+9HPHq0dK/t6Ef+eV3D3Jtv+aH8tqDFdq79x+Ph7RziUuPotyZihpqOEV86xwwQQSqgVQLLYf7xb3pQCvW5GOug9B1TXufsrsDe4pU3lvfc+6oqWRWgTPZrIZQQaTe8QrXfT/sPduqEk8et2HHYXAfzTf5Oe3Ovumaqhy3ZPw/6thpshh8pOgrpGqjK0z4iHlnCll1hRcEe9g0NPP8AydeKs4+X+UdacWz/AIy91b47aoukMHsHcEvYVRlhinxUuOq40gYzeL7qaZksIfysn0b6Dnj3rrSgnIHV9/8AMy/lV/E7+Xx8Qfj3m+wex5av5Ydm7QbL5vZGBlWakw1YKbVAlc0BNrubOrgEEN+Le/Djk9bcALqjHn+3rWULNfkm/B59+61QHrokn37rwAHXX19+631yVb83A5/Pv3VSaddG1zb6e/dbHDPXXv3W+uwLm3v3Xjjrs8cf0/p+ffutDOeuwLj6nj6f0Hv3WiaHriBcE/0t/vJt791snIHXYtY3+v49+68a1FOuuOP6/n37rfXgbG/v3XiK467A1X+lz9L+/daJp10RY2/xt7914Gor14ixt7914Gor11791vrmFFrm3IuPfuq1zjrjb6H+vv3VuuwtwTcce/daJp11bi/v3Xq5p11791vrkovf/D37rRNOuPv3W+ve/de697917ru3F/8AG3v3Wq5p10ODf+nv3W+uySx/H/I/futAU656P8effuq6uuIHqAP9V/3n37q1aivVrP8AKc6uw/YfeNTVZWijq/7vijqoS8Ycx6i7NYm9r6Rz78RivXgRXPWx58gNq0VRnRT0UapRrFTJoUegCOMLYW49++fW6eh6LttTbGP2zvGgyQhGn7uN2ZQLgBuSbf7c+/fPrWOHVkFNuOmylNSmidJP2o0UfVwVW3Kj36nmevVPTiKKatBWogR+AyEryD+B79wGOrAV6RNd09R5KtesrYKdxKxNnAt/quB735daPp11/oY2tQKtWuOpGqANVvEvJUf2eP8Abe9A9e6f6OtjxsK0MVOIYVATREoUW+nvfWug03hs6DcMjvpCpzchTyALjke9db+zqubvTYVfSVdV/C6SY+NHKskZAPrA+o/2Pv3DrXy8+g16zy24Nuu33MMupzoGsEMBf+n49+63To9PWfZdbj51kyFzFOmldR9XPA4P+8e/darQ9Guw26KfJoHRlAZfo5HBPPFve6efW64zx6UkNVSEhQq/42H0P1+o+n+v71w68OHQKd1blG1sHNURayahSt4xcjj37y698+qlK3sM/wB7qmevqJY45pnCeQnjngC/v3XgOrDOg+xUanp6JKv9iWSMcvYcoSb/APFfe+PXhXPRitwYlqavgzVI5/cKvJp/x5uT+ePfq9aB6EnC7glqIIwycFFUMeebc3v70QBw6sKnHTxU1KtEwMalgDcGzAf4+/UNetA1x0D25MGMo7SwosMsZLRsg0tIwNwCfr72fXrw+fXe1MitS74TNokckIKl2tZh9ALn8+/V9OvUr0Gfam0K4eeoxMd4ETV+39ODf6j3rrxHl0AWyOwsthM8cVVppgaTxozflr6fz731XPR2qF/4niBO4DssKSrY3Ophf6+901o0f8Qp1aOY2t1FeD/QmDfsPXWFr6KXVS1MmiRFZtDmwJ/xv/re8buc9jNtuzzS10V67O/dj9z137ky12+2YG4AAp50Hy6FfYlElRkGaLQY1Ia6W54+nHvAP393dRuf7vj+FB1lRvs8osFFxhz0PQVoxE6KCIyDZvoePyPeLBOssrHj1Hyhcr616XH+kLJz4sYQ0cMVKqBQ6KobjjkD2Gf6pWMd8dz8QtITwJ6Dn9VLJL87l4hMh8q9BvuGYRY6oUegVKuhZeP1D/e/Y42OEy3yE58Oh/Z0bXM5iQCueHQudPfIPffW20ots4inimx0dQ06PJ+oMQfr/t/c1Qe8PMnLFiNl2soIgdWa1r+XUabv7Qcscz7i297izCdhTHp0u93/AC47RbEa8XS0zzfVrnj/AG/s55W+8Rzna7mWuGjUevd0X2nsVybrMczsQcDopHYnz87zwWJnp8Zj4ZcgqtwoDrrH+P8AxT3kPyjzl7g897gLi2Ksh8wDT+fRPzz7Acj7BsD7zcvoQAnOD1Xvkv5qfzIwm5EinwFKuLWoHkfxDWIi1jcge8ntk5c56SHxJfiI6wrg5i9ptl3zRuRrEjZpk0HGnQ/U3827v7IxQ0eLxUf3jIgYmDUnksL8kf1v7HvLh3bbWb9/MPl0j93F5O56toj7YpI+mmrUP8HSli+e/wAudyh3nx9NHTNFr9AEZ0/7D8+xM28WSnDinUOr7Z8xGIM8Jr+fQS7u+bPyToYKqrWjiNREjMvN7uOPz7t+97VjRGHSVvb/AH2IMZYTj7eiHbs/nC/NDrrNx+TB0MuEWo0VLSIjlYFexb1f4e18cyyLUMOgjd2FzaTeDLEaj5dCpkP5p3fPZm1jmtpY7Ez1q02uSJ1j8glZbvZRzwfbg4Vr0kOngR0QXe382X55bXq65qfZGFqKGkMrNMVhDrFH9CIx6j/rAe9UPr1uoA4dAM/8+v5ZRyPHLhdtJJGxSRGgdWR14IKlbg3+o9+/PrVfs69H/Pw+WEbxyLh9sakZXH7TfVDcf2ffuHn14laUx1V38p/k1vf5Zdp5DtrsCKkg3DkKaGlmjpC3i8cQsmkEC1hx7317ovVDXVmMqYK7HVlRQ1lO6yQ1NLM8E8Uim6vFJEQwIP5v791rNcjoaG+TnyIeghxDd4doNjKYqaekO8894YtIsoRPPcAfgXsPeqDq+pukFX9ldgZPLpn8jvjdFfm4yDHl6vM5CoyKEfQrWSyGQW/4N735dVOTXpuze790bnyqZ3cW5M1nc3EsKQ5fK5KrrchCkDaoglVUM0lkJ9Pq4/Hv3WmJPHqfnOxd+bmoI8VuLee5M9jIHjePHZXL19bRoYiNBSCodlBFhaw9+69jrFS9hb5o8Adq0m8Nx0u2mZnbb8GWro8QWb6uaBXEVzfk6b+/efWzwp1ik33vOXba7Ql3Xn5drJMJ023Jla18Ks19XlGOL+ENf86b+/dbzSnl0kx9efpf/W9+615dZ4JpaSaKqppZIKiCWOannhkZJYJ4nEkcsbryCpFwQeD791oE9KHc+9t37zqYavdu6M7ueqp4Up4KjO5Ksyc0UEa6Y4Y5Kx3YKo4ABsPfh1s5yeksDbke/db6csXl8phKuOvxGRrMXWxg6KuhqJaaoW5vZZYSCPoPz731ogHjnoZqb5QfI2jo/wCHUne3a9NQaBH9pBvrckdP41FgnhSoAsBxa3vVB1bUwwOgt3JvDdO7qs126NyZrclYx1NVZnIVeQnJP11S1bMT/t/fuq0qa06TQuTxx7914/Pr1jc/m1z/ALb37r1euvfut9f/0NAC3F/8be/dermnXXv3XuuzcWB/H/E+/daGc9de/db65EkDTx/vuffutUBNeur8W4/1/wA+/depmvXXv3W+u78W/wAb+/dapmvXXv3W+ve/de67Av791omnXQt+f949+639nXvfuvde9+691y5c/wCP+x9+61hR11bm3+Nvfut+VeuSq5cIql3awVVBYkn6AAck+/dVwR0IeI6p3zmqRqyiwVaYVjMwMsDxl0AuWXg/7C/v3W89LHqfuLfHTe4EgoWq1iFVHFPipy8QRzMFZliYcn+g+nv3Xur0uutw5ne21qPcNdLNSO0McpVCVvI4DWt+ffutmg6tT+O1JNlNoGorbu8DJGhcFtSlSAx9+60M9GCq9sUlRSyRPGt3DAaV5Abj6H3vj1smnRTuzum62slZ8dHKGOttaLyCBwLi3Bt79mvWsnot0H969q1EmLrKedgpZVco5awNhz70Ot+fT/jMHlt2OIpqefS0gAuh5DMRa5/4p731ro5XVHWU21aNap0MWqMNbgAn8XA/23vWK9e+Y6Av5JYHM1yvPBE8iq2vSq6rKFvfj34de6SXx33ZTUpGPrmWGWKTxMj+k6r2PB9749eGMjqwSlaiy1AJEMcqlLEXDra1geP969+63mvd0jKvZVBU1PkWNQLk+lAAeOfp799nXj04UW1IKRGZLr/gAPoOSSB791qhr03ZillxY++jJZAw1AgWtfm/v2Ot8egX7hqMXVYVMjRiE1cK3ckASAql/fqdaIx1746dmUWQSXF11Uomj/aWKR+RpNrC596p1sHoZ+wdvxZTVV08V2I9On1DT/r/AOPv3DrZNegap9vihlJCHWf1KVtyfqRb3vFetUr0pY8bI8YBVl1H6aLgf6x9+69Ty6mrjZEKqNTX4NlPI9+4dazw6dY8DNKBrDkEHVdTa/8Ar396690jN0bFdqaR1hup1NwLtfTccD3br1egJ8GU21kFmp4pIzE4IshW4Dcg+9efXuAr0aXrzsmhyUMVBkHC1JRkIkJHJ4A/1veqY63kdTN67FxO4qKrrKJgtUiu4eMgPcC62Yf1PvfWscD0EHW25avb2YmwmakmVPII4XmJ5Ae3DH/D37HWxWlOjMZnFwZjFt4QkizRlle/qPH4J/p79XPXui3NtPwVtRG0bKwlJU6f6Hjn3SWJZkKt0os7yaxmFxDxBB6fqWuyeJTwLJMqAC3JN7H+h9xZzTyYbqs1sD58Os7vYv7ycWwIm37yQVFBRvt6Ve0OyYKHKRNkHtZgLuNI4b8sf+J94ie83tvvd3trPDGxVeNBU8OujvLHu/yjz7t/gWU8aykcCwHRv8B2BRZylVaCaKRmRgCpB/wIJH+9e8Ftz5Qn2yfVdKRQ8D0jvraWJ2Jyp8/LoFd90tTXZ+mtqLMycgX/ALX0B9yhyxLDbbU3kAD1G+9wvLdqRkdGTx1AkWzoaVVvLJAgJ/2o+m3uFru7eTmhpmParHoShfB26o8hnodfjX1FBlZqh9yUcbxOJXiMqBzpN9Nr+wd7v89zbVt+vaH7xStD1hz7v89z2u7om2yEFCAQD132vtCj2luWSkoAoppJAVVRYBT9LAew5yTzHccxbOt5djvAzX1HWS/tVv02+cspNcGrqOPQSVuI+9VhcgcgEf4/4n2O7PcvpG1Ur1I8ra1oTTpFbhwlRR46c02tkAYNzcX034v7Fe07tBd3aiYAGuOrbcGW7T0x0VtshUxZOaNg2tXAvY2+p/P9f8PfRX2bvnttojMPDp/3d5ds+YuXRHdHBWmfs6ePNUTDyqrEqeByOb/m3vIO75pvYrJlFcjrCLl/2K5dvOZo5RpwwrT7enfCzCoq0/iD6YIwTJdrA2N7f717wz96+at68A2VlUCU5I49dGdk5dteWOXo7TbVGoKAKDps3fu6WrqxjsUf8khIi9APPHNz7IPaT27hVF5g3xamuoBvM9Rjuezb7vW5iBlOhzQnphq6qobHNTKQKl0sukeotb68+5t5l5ottttgrEJGvADGOpL2DlXbuUrX6yQ1cZJPQe4/bGczdTNDLTyFA9zdCSRe1z7j2X3M27aVE+oV6U3O8We9xNAxGk1/zdJPtzoM5/bVbQ5HGCamqIZQVaG/q8RC+xfyP76WO6bilgjgSMaAV456h/m/2f5a5r2y4WQKW0scjz0mn8+tSf5Y9F5rqHsbNGShkgw1fWGSlKxERw+Rb6ePoCBcX/JPvOTZ75b+xSYGpI64se5PKM/KPM9ztzKQqsaYx0VAiwv/AF9mvUfV8uvcW/N/949+63mvXr8Ef19+69Trr37r3XIHgjjm/wBb/wDEe/daI8+hE666q312nubE7R2btvLZfMZp2FClLj6ueN1RCzyFo0I0C3LX9+60SfLPTFvPaOb2DunN7O3HSmjz23618fk6Y3BgqFjV2jKsAQQGH19+63xGejT/AAt+cnc/wb7Io+x+pa9WngmjnrMFXSM2Ey4jIvBlaIq6TRsBpZHUgj37y68CQ3HHmPXq72L/AIVOfLLBbli3rsHoroDYm7pY5Tls3tzZWJx1VX1M8XglmmeGm9TEC5Ygc/j3rPVtQpT/AC9Vj9dfzafkF1r8t8z8ycLj8LUdrZrcdZumrkrAJaI5Wud3nlMJiKkFpCeV97zSnWq1fX6in5dFvrvmd2RkfltlPmNVUuPk7Iy+7avd9ZS2C0JytWlpmAVANN+baPehw60TU9LH5sfzAO1vnNX7XyHZ+Ow+OqNqwvDQ/wALXQjoyFCHUInPP19761w4dCF3/wDzS+9vkV8bul/jLvfHYUbG6LwdNgNmz0iiOsFHTMrJ926xqXYlRyWPHvwx1ZiGGn8+k/8AMH+Zb3j80Olvjn0b2hS4WHafxm2z/dXYEmNgjiq2xhUqRXyJGhkexADMxIUAfj3oCnV2fUKDqugC9/8AD3vpuvRlPiN8nd9fDj5C9a/JLrKOim3z1dm0zu3oskglozWxxGJTURsrBksxBUqR70RXrasVOeu/lx8n99fMbvvf/wAg+yKegpd5djZibN52LGoIqIVs7tJIaeJVUIt3NlCgAe9gUx15jU6+i1cC9+bfS30/3n37rWTw6PT8KPn93d8F90VW4+p68Sx18iyZHCVtTJ/BskigBqfIURV0ljOkEoy2/Pv2OHWs6q1x6evVyNT/AMKgPke1DUfw34ufF/C7oqaZoH3vjOv8RTbpQtGYzPDko6UOrm9yw5v79mv+x05rXTp/y9UUfKj5b9t/LrfU2++1c1WZGuLO1LRzVktVS0Ac2MVGkgUIgHAVRbj36nVM1446K4SSbnk+/db67LA/W/8AsP8Ajfv3VaEcOvaiLfTj/X9+69pHXH37q3XvfuvddgXNvfuvE0z117917rsC9/8AD37rVadde/db697917rw59+6912bjg/j/iffutD1669+631737r3Xvfuvddk35/P59+60Mdde/db697917r3v3XuuyQefz/vHv3Whjrr37rfXdvoT9D/AE/3n37rVeuvfut9e9+691737r3XvfuvdcgRe5vf/ePfuq0PDr1x9eb/AIt9OPp791unl10Tc39+62BTHXXv3XupVHSVddUw01DTzVVVK6rFDTxtLK7s2lQEQE/X37rXWy//ACjuhsr0hS7k7M3/AEy0S7txSw4lahPFKkpiPgj0OTyCxube/fPy62OGRnqyLe26qfJ5Bg80ckrStaz3Ng3H+8e/fLr1PPpvo8VTVASeRFJNivpub/4e/daNOlbi6zJbdqoqukLTJIyhka5VVBuNI9+OevZ6dd3957gokhixNMnnXQJSqj6cX5t798+vE+Q6SEHfu+ZHWKZAvP0HH+HPHv3Xqjow2w+xKzc8CQ1yRmdNOofmxH4Pv1Ot54jh0ssgaZ2K6Y9ZJBK2/P8AQj3odeOOHSdlGj0CxB/P5F/6+7de4YHUg7R2pnqGaHJ0VK87xkNI8YZyfx+Pz70SfLrQHn0WbKdAUAz0k8NNGlEjtIhRAB+q4Xjj3vrefLqHlOqKunnhqKSALTI44UW4B+gH+9+9dapnPSywu3suniFMH8SEX/s8AWN/rx731vHn0KcFDV0NHJUzkgpGGLfjgW5P/GvfutdBy2Kg7PNbh64BoqJWZSRdfTf6EHm/vxx16hOeqpflX1RPtfI1FTi4pYo6ZxLrjTSps5P49663XPQddGdzvRZSjxNXVPDJT1Uau0jstyo0m9+PfqkCg691dFsTsvBZzGQ01bWQkinhIZ5VK6iv+qPveOtUpnpcLvDa2ISSZ6gvGt2/ZPkQD6f2ffut16T6/IXrAVf2U2RaGUHTadynINje/wBB7117yx1mbtPZ2TrliocpREMfQFqQb6ueffvLrwoTXqVuKhpq2jjyeKni+7XTJqilUlx9R6hYn3anWq+Y6wYLNz5ahlx2ViBnZWgGsXuoFgbm9/esDHWyT0WXszY/8Br4snFEq6pC8bLx6tZIB9661w49Dp1Dm5qykFPXuqgRRjSzHUTp4Fm97HHrxoVoeB6c9z4Op/istXR6/G+o6UJAseeNNvZPvOxWm9weFPhvWnUme1vutzJ7Wb9Fum1MTGrZUnAH2dLXrHfsO2Mg1Nmn0wSsFBIPDfpAYn8e+cX3kPY/mBblt722JnipxAr59db+QPvGcp+7u1RWjzJHuCABgSACadHMoKylzFGtZRyK8LoGUobjkX/HvAG7tp9tuTbXK0YGhr1Ip1REA5r1ClqDHLoA5vpP+w/Nh/re1McQdNR6VhKpq6nnAvuKOGihBLrICQDybi9re9Wm6LtMz3EnCnQQ5o3EbdbrcNgV65VmGlwrNjJl8cirqC2+n+Ptpdxj3FvrYzUHo02O+j3GwW7iyG6SNTLWQUtVAQ2kxuFNr2J5BHs9ijtppEkUgGo6ObTwvrVZ+Fei0V+PpqmtrVrF1T+ViFcXFieAoPvp992Q7U20QAFagZ9a9Qn98C43n+oapswYrpzpz5dNSbB2vk3eSsxdC5sdXkiQ/wC3P/E+8ueYN+j2i2DxsMjyPXLr2m9qb/3F5g+iuEdjrzUft6m4TrXbcld4cdhKMOlv3I4Af+Tr/wCx94Se7/vjdbBIEs5QZCTUV4Drqvyz7AbHyby/GggQuQKkjPDPS2r9tRYiPwiKOOy86ALafoV9h72s93t+503NrS6B0AVqK06S7ryjs9mgZYlyeFPPoNd27Y23VY1xXSiAspvbgk2+hA/4j3kI/MRs5RHqJP29NRe0g3yya5jt1EZHHT1VR8lOucFLRVrY8RzFY5WAsCP1fXj2Otn3me4VXqePWLnuT7b7TtbSRBF1UPlmteq0Nj5zdXXu5HeiapfGGokWrpwf2o6cNzZP9bn3JlpdJNGKnPWFHMewz7XdsI1JWp6O/BtXZvbGEhrcVKJa2sjWKsgsLiRxaQeP6+1vQZBr1Tz8vOhK7qveUtbQUJTDVKK7vGhASUt+tlH455Pv3Xj/AIeiZHk/7b/evfuvDh11791vrsfUf6/v3Xjw6817m/19+60OGOvAkG49+68RXrr37rfXZtfi9v8AH6+/daz10bfj6f4+/db+3r3v3Xuuzp/F7/429+61nz68Db/evfuvHrxt+L/7H37rYr59eBtz791oivXr83/xv791vyp14m5v7914Y68xuf8AYe/daAp14W/N/wDYe/dbz5deABv9bWJ9+60a9eFvzf8A2Hv3W/s6/9HQANzz/sPfutCgx11791vrsj6W5B/4j37rQ68LkWH+v7917Az16xBt+ffut18+vEEfX37rwNeHXXv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XujJ/FvrWDs7tPGYiqiL0kLLMRpLKZdY0KQPr+T738+teWetlWi+NOO25sikq0wFEyLQKjN9swLjSFJPH59+PXuJr1W53f8StsZPcUG8KOmNNX0lSjmlp42SBzGwkUyoBY2Iv711759D/ALBfIYrE47HSUyw46GOGF9C2LEKADpPPvdOt16tr6B3JtzE7Wjx0lSgqZwkgV5EWxIvaxN/p+PevLrQpXHQ2VG88bDdw4ZRcAhhpt+Tq9+p1YVrQ8Okvk+zttxqyzPCW0MvqmjFuP9f3bh1UivDoFZt0bNyGQmkq4aaRmc6WurEAm/19162PTp7oN0bUw58tPTRKNWpT6R/iOf8AD37r2Dg9KmLufDRhYBKoT8/uppv+bX/x9749erTh1wr8vg916nqJad4ngYBWkQ8kaVt791qtB0Rnfm3q3ZG43zOEa9I1R5NEXqBLN/tP/FPevPr2TnoyPW/aWUosZBHko3VJ1jYagVPqHNiffutg+nRkMVvCjrYUIIGpdXNr+o/1Hv329b6Ua5VZACACo4Fvzb6Wt72R5jrQPGvSW3RkYDjqjVZbKzWPPAP1t/h799vWuJ6Jzk0qNz7kGChkcwVKkEfqALHT+kf6/wCPfjjh17FekbuLrLd3TmQh3ZQiVsYJgZhoa1mbWfr/AE96OM+nXvn0c7rDs3C74wVMk8yRVQiCy+WRFvJYWFmt78c9eHz6VNfhKATmdqmmC3Gj92MXP9QL+/UqOt1A4dTFosSkILVMBCrc2mQ/4m5J/wBj78AOvV6Z23Zs2gmemkmWWcHgRyI/qA4Fh73Tr3Hphr+wPCzy09M32asLyaCbKDpvcce/dewOnnD7421n1MImRZSAHWV0C6z6eLn3oHrx+XSa3bh8ZXuqwrA0j8L4irDj03On3vrXHj0j4NhvQyJWwExOPWNIC2P5Fz79jrY6EPBZGWFft55AwBIfU31A4IPv1evU6DbtCjx8csGXx1kkp31SabC7AXJv7117pedc9kY/NUlLjZ50WaCMRG8ijVb66h78OtZPQh1eFo6mUyR6NT+q/wBf9jcfk+9/YetnhnHTDX4OLS48VygsLr+q4ta/vRFR3DHXlLIwMR0t6joHN07YZ0eSiDRzAk2VdIFjzz7QXe02G4xmK5QEH1HQv5d5+5n5Uu0udvnftIxXqf1zubO7brIqUBpFkl8bKeSuo2/4pf3hR75+zNksMu7Wa0CjUQBjrqv93j3mtPcjaV2/epFEyAipYV6OpQ4WPNx0uUqCgkKKzDULg2/PvAa43C42tns41NK04dTDuX7qa/8ABjkDFTTBHQtxy0v8LSnSRCYVUCzi/oN/p7jdxOu5NcOp7j6evRktmZIdFDpII4dCNs/5Ax7JgSlnSNTArRI4dR9Rxf8A4p7Rb37aLzOhapIfNOsTuffZubdN9N5FXSzAnB6Se6OzP7/ZcZEyLpH6dLg2B/Bsfdtt5KHKtl9HEv8ALqfvb7YYOW9mFnXNM9c6aeGS1v6H/WsB/Ue0U8EsfEdDBjU9vQadg7shx9LJRxadcl1sT9W0/j3KHtlyLdcz7olKhAe4+g6F/LWyvdSid+A6LbFNFUTmoeNdTux4UH1D3099quWLS0jj21TiPHUQ/ek5t3Dk7lktt4NdPSjpKdZo30ILHkD/ABv/AE/I9zhvXLFuIKjA65+e2PvXui7p31aRyD60z03VtBKt1i9LN+BcWuORb3j9vHJe3312TcIHAPn1015c90zJs0TXQ7yo6hHAQ0VLNXVFjKBqN+Df6259pZ9rjsLbwrftRRw6HPLvMp3e8VdFK9NW16CXcWUOgMUhk0L6SR9bX4943e6m9pBMoLDtXyPRtzxeBLcWsR49GexW1qDD0n3c0KKyKC/Fmv8AW5v7xZ3Pf7ncZ/pomqCcdRxt0LoQiE1bpL7g3DiMpBVUSxRsER1A45OkqoHud/Zn253v9/Wu+XIIhXur1rmcbhstiWUmsox+Y6pK+dXxhTsXY+7szS0iNkoYJDRsIi7IUjYoy6f6H3005d3pLR47ct2/b1gJ7ye293v+2XG8LETKM10knHn1qrZvD1238rX4bIxmKsx9RJTTIwZfVG2nUob8H6j3K0brIgdeB65/3ED28zQSCjISD+XTT7v011yUG/4FgT6iB+P8ffutHh0u9h9a7z7K3HiNrbR2/lcrlc1UxwUgpKCtqkJlbSsjmnRrJci7e99aJ0ivHrY3+K//AAnh7BqNrL3F839x0HTnTEwpqiDI0edoo8/UUYTzTNT46oKSlilwV0ccc3968sdWC0IqcU4dG03j/NB/l5/yxdtZrr7+XVsTb/cm/MHi6jbD767ewNFW11NWSwmnrqzGBw3qV76SrXH1+vv2Tx61hQNGQT5eX29akncnaOd7s7Q3v2zueCgpNwb8z1Vn8tS4yIw0MNZVIolFLEf0p6eB+PfurE1PQZg2N/futEV675NyP9jz+ffutYGOuPv3W+uwbX/x9+61Trr37rfXvfuvddi/1/p791o04deJub+/dbGOuxqK2tx/sP8AX9+6rivXH37q3Xvfuvde9+6912PoeTfi3/E+/da669+631737r3Xhxz7917rl9Tc/T829+61wFB10bX4+nv3Wx8+vfUWP4+n+x9+615468CR7914ivXXv3W+u7/Qf09+69TrtfVYH6AH6fX37qpxkddEWJHv3WxkdeJvb/D37r1Kdde/db65kj6j6n6/0/2Hv3VQDwPXAC/A9+6tw675W4/2B9+61g5669+631737r3XY5IH9T79148OuyvJt9Bb/eRf37rQPr10CR9Pz7914gefXX09+6317/D37r3XZJ+n9P8AiPfutAefXXv3W+ve/de697917r3v3Xuu7G1/wPfutVHDrr37rfXZNxb37rVPPq7X+Xl8RKWTFf6YuxMetTi6uKCq27E9PI6SuieZIpCwtyT6v+J9+69jqxvsDsHcmJpfNiKEUGLoFAioYYylPCkXpXQn0+g9+69jz6QHX/bJ3fmaeKWY/dmYrNE0n0sb2C+95HXq16sEwbI9HSkkH0xn6cAEfXn/AF/fhTrdOhIo8RDURJICWVbEgcj/AFvfjSvXvLpIZ7bCPUSzmFdGoENpAFrfn3r7et4pXoFcljsnQ11RUeBfsW4gaxN7G17+9jqlfToQOvt2JhZZHqWCFgo+oFrf0P096HWyehSqOwoLiXyqQ7X/AM4hNh9P+Re/Z8+vVHTrQb8x9T6Hli1i2q7pxf8Apz72D16tTjp4h3bReZVgkQsxKgFlsb8c+/eWetjpQnILLEXZ1ZmT6Bhe1r/T/W9+8x17qMZmqYRC6fthho/2rm41e/dewB1Np8e0UY8KgaiSeTYD+lx791oj16atwVNSKCagi5knVowRf6/T6/7H37rfy6x7J26mBo2lLWqZz+61+SrHnkf6/vRoT1oeh6Dnt/qXb+/sXLTZAePyppaRUYsOf6+/db6JlSfy+NtVFaclja+ricStKpRXUlr6/r79Qcet8MdCtTfHfcm34afH01ZUCljKRmf1hiqi1yf6+9/b1XHQ/ba69osbiI6Oqmkq5ggBMwLEmxvq9748OtkU6Kn3T8eKjMT1eVx8s1ERGWjFMjAatVxcr71x61jj1WtuaHtLrHcCVD11dJjVnJLFnuI0fT+f6e9efXh6dWC9N/InbddiKCmzGUnWZUhSUSzKt3HDBgxB/wB49+6359GlHYu3ahaabFTxMrEWYSIXL/6prH3vj14gV6D3tTedNLj4JpXLFPXYm44Pvxp17HSC2F2X9xXwJRSBWWVRIisBZQfyPevs60Pn0eDF5ugy9LB5HQTPEnpYgEgjm1/8fewaDPWumjcW2hVR+akQLIPUrIv1b8+6XNpY7laPYblGHjkGkk5pX06W7TvG8ctbim8bHK0ckZ1aQaAkeR6XXW3ZmQ294tv5KM+HWqB2NmQn0nhv8ffMH7yP3bm2K6fmjYwWtZKnAPrX9nXWP7t/vbt/untv7l3uRY9ygUVBYAEAfzr0ZX01SxVSOpWRVkte4u3P1HvCABoC0DA1FRw6yYeZFJj1rjhQ9LvaeSgxeTp6ypdfD5V1WYcD/Y+w9utrLd2720I7iMdAnnfb5tz2Ro4B3KK9OO9Mlj8vuBqugcPH4lU2IKg8f8j9otlsbzbttFvdijVr0zyBDd23Lyw3QoQ3Seko4qgGMxqdQ0lj+B+be16XMkR1VOOhcJCh1dFd7P27/Dcoa6ilCvIwVkQ3ubc+kf6/vMb7t3ON/Dui2Y1aCPyGehBfbXa81csS7duUYZFQmrD5dINBXrGXFwGGl/8ACw/p7zm5q3DcpdlaVQSQpI+3qBvZ/lPlDZOeJls2VZAxwCOhu2fW4XFYg1Ezr97JYAErccfXn3zJ5m2zmfnDnM2MytlyDg0Ar1kjzOm5Ty6LYHQOHSY3BnY5ZZbDWCeSWH5+lr/j3mnyJyvtnIeyJDbAG40gs3nXz6CG38iz71cCbciVTyGei9bzqKrJzvFAJGiQeoKrFR/X6e3Zeatrl3bwblwHr6inUqLaQ7Xs77JY0LlaZ49Eq7owgTF1k2p7+CTh1I5/1PP+Pub+Xd5tJQkMNDX0NesFPdzkDebcTbnd1CmpFa+vVf3Xm1Ic/uPJ0dZTo4mMyAFSSFJIvx7kN7p7RQ6HHWIVty3Bv1w9tcjhXoc9q7Pn6nzn8WoTJLSN+5JSlSIlQ/VgD/xT2LNpuzd24d6V6gfn3l9OXd4a2hBK/YegU+R7YXsb7uqkpgwlpGhEc6iyy/S6g8ezMj06BIrSjDqkPsHYWT2blJ1niLUU08zQSojBI0Mh0Rsfp9LWPvXXhjHQegXHH6r/AE/w/r7916tD8uuja/BuPfuvCvn14G3+xFvfut9de/de697917ruxtf8e/darmnXhb88e/deNfLrsW+h+n14/r791414jrw4BI/1uf8AH37rRyadcfr791br3v3Xuu1tf1fT37rRrTHXrXJt/r/7D/Y+/derTj16wt/j+R/sffuvZr142/Hv3XhXz669+631737r3XZFvr7917j1/9L5/wD7917rsC/v3Wiade5+n9Pfut/PrwNvfutEV69yDzyf8ffuvYIx176n37r3AdcvqbEAfngf7x791rgKjrh791brsi3+2v7914GvXXv3Xuve/de697917rvi35v/AE9+61mvXr8WsP8AX/Pv3W/OvXYUt9PfutFgOPXaqWYIql3ZgiqouSxNlAA+pJ/Hv3XurcP5e3VORw27sbubMwfbSTzxyRRuCCYSQUJvbn34de62ctwzU/8AcijpyEaP+HqfqeCFt9PexTrZz0RrNbfx2amnimULH5WFwOPr9Rfn3rqvlXpK5Hr37KgaSmjBiQ6voQCB/vh791uvSm2xgq6tx4qsTVTrkqW1qYOwUqh9VgPx+Pfh8uvCvStG985BWxbeykUtO7IIzM62TUfTcsf+K+99eBB49B92JhNwUMsM0FS5gqDrLLIxADgC4N/eut9Cr1f1FV5qjhyk9W8hKqzqSX5P+Jv72OtefQ3N06ayL7Xy6SAyjjnSOfx70evdJ3IfHsaG0VzhrE3u1gR+Ljm3vY6159B7kevs9tfUYqqeSNTyQzH0jn+vvXW/s6aaLFTZethhySyTQiRdfm5H1ufr7916lOjGU2w8JlcfSpTtHEYolNgqgjQPp/X/AA9++XXqUz05Y7a7U8oiDlUhsoNyLgHn+n4/w97698+hKx9FBHEUMgbT9SxvyP6f4f7D37HXq9An2pk4sdC8cUpVpLJZSOSeOT78MHrxyOmfqfYk9RkotyzqLpoYBgf0k3HqPvRzx68MHHRj977cpN54g4itC/b2IIJJVbC3I9+HWzx6LBUdFvtuYyYOtkiQyeQrE7Ktzzfi1x731r7OpddszcORaniOSqUZFVDpmex/BI59+630uqLp6vlo41ky9T60GomaQ2JHJsT711qnUKm6Rp8PkVyEmRmqnBDaZNTAG/8AtX9ffh16mOhJTAUk1G1B9rBoZCrMUXUCwsfe+vDPQQZvppo5JqvGVksMjM0gWIlQD9VAtb37r3QYVlVvDZ87PLDNVwwMQrHW5Onm5P8Aj7116vScyfyDybH7dsfUoY7qdMTAf7A297p17UOmel7Zy+QqVampqlWNlIKsASeOb+/der0s6aty26IGp54ZQZARbS2kEiwN/wDD/D3rj14dBjWbe3VsHMjK0azyUZk1yABvp+rjj3vh1vPQp7d+RtNTvHTZETxzKwRvIAo/p+be9da4Y6NjtLeWD3jSR+CaPXIikhmHpJH5sfe/Lr2fw9PeZwNOaVlVFvpYalBvf/X96z5dbFPhbz6AOo1beyqzVEOlBKCp0/4/Uk+ybf8AZbfmCxewufhcUP2dCrlHnHduTLv6ra3Kj0Bpx6MtidxNNh4ZKaRjqiDehvyVuL294vc5/dz5XjtXniA1vXNOsuvYj3P3bmzmtP3q5CahWp+fUGLcOVgqVWSaX90sVUEkAX/r7x8m+7/aTahG3D5ddUDvvLMMEUBYaiB1nzeR+9xNQHsK4KTGQBqNh9T/AMj9x5ee3O6cv77HZ2wMkR4nr0nLVru0i3Fuexugeo+w83tl3iqI3WENfUP9Tf8AJ9iDd/bZpYVnlUZHQX5h5Q3Gw/W28ah6dCZhfkzgaYCKsMqyBNJP4v8A159xvd+yu5bnOEsxUk4HQAud8l2tD+8EKU49J/OdmYrd1YZ6SUBb3AYi9/ofeQPtT7Ub/wAp6lvUA1000r1IPLnufyxZ7Q9zNJTwxU/l1kw1VRyuIlmSQ6jcggBSTwLD3mHyVybfWMovp8A56wS+8795HlPm62l5e2sFnHbmnHh0JVCkUcfoAJtyD9P6+zvnDdZo2+nGB1F/3e+R9qvI33a4AZwagHrjUDxyrNIOOdXF7Ff8P6W+nuMp7tIkLSHPWa217NLf3KW9svaKfZ0HG7Ms0zGmp2IgZlDlT9Lmx+nuLuZb27ltJWsidYGB1lfyvy4u3bYrMg8QL6fLoVeqqTF0pQRXknnIJJFyWvfk/wDE+8FPcO73KeV/qTQqTXoG73ZX8szz3eFXh0Ou4ov9xE6C4cqdIHJ/R9bf09xLtMn+Pqx4D/P0S7Y4F4reQ6LFJgcs9TM6xFRqYr6eSt/7Vh7zY5U94dl5f2eKzb4lUA9SLuNvtO5wotya0HSQ3VtNc3jqrE5EFYqmNkkAP11IVsQf9f3LXJ3uLHzPdeJZGgU5r0HN35O2e/2eW1hSoYEftFOtYv8AmQfDKr6+zy742dSrVUVVHLU5cIGE3JL2H4LL/T3l5yfzPbbjB9LI36i449chvvFexu88l7m292UJe0epLKDgk1ofs6qt2R1xvDsPcuH2ltTb+VyObzVVHS0cUVHVSIWkOnW5ijJCg/VvY+Ar1iUSV4/s62Nvih/wnd7OrdoRd1fN/NYvpnpOZBVrkaXcdGdzPj4V8s01Ni3lSRyyfRTHce9E1GOPVtJVs8PT/Z6N7vL+Z/8Ay4/5Zu2Mr17/AC+tmYbvTfeIo/4ZNvLtzaGOqqikrzGIpZsNUvCwj0tc+hyVtzz73Qnjx69qVRReBPl1rkfLL+Yx8k/lxuZc/vffO4sFSK9RMu29v57KUOBjkmfUqQ0MEiIqKCVCW088j6W95daqxJzUHy9OiGTTSVEjzTO0s0rtJLK7FnkdzqZ3J+pJ5J9+68BTA6x/U8k/4n8+/de8sdeNr8fT37rf29d8kXHAH9OPfutYr1x9+6312QRY/wBffutAg9de/db697917r3v3XuuVlt9Te3+8+/dazXrq5/qffut0669+691yKgAHnn6/wCHv3VQa1669PP1/wAPfut5669+6312OOT/ALDjg+/daPXj9fpb/D37rY67vYWsP9iOffutceuhf8f7b+vv3XjTrr37rfXvfuvde9+6912Ra3+Pv3Wq166H1F/fut9d/Q8H37rX29de/db68P8AH6e/de65W+tvoOffutV9evLp51E/7D37rxr5dcffut9e9+691737r3XduCf6e/der117917rs34N/r/j/Tj37rWOHXXv3W+u7/8AI/z791qnXXv3W+uRUj6+/daBB4dcffut9d3/ANb37r3Xr/63v3Xuuvfuvdeufp+Pfuvde9+690p9m4R9ybrwODjQyNk8nS0uhASSJJADwP8Aeffs9a8ut3D4pdYU2A+OGysHk6WNHpzGwidDfQ0P10sOPex1sg+XUvtPqTEZHEV6RwRxxuvIROLW/A96+fXuqw26O3LsPdEm6sZEzYyGdpZdIKjQXuTz/sffutfZ0aPbvbFJUQ00Xl8Uy6I3WRgtiosbD37rVfLo2mxt00dZjYZJJ76rX5vYHkm359763Wox0vqs0+SpiiE+ORSFNyOTex/4n3759ez0mZtk02UpDQLNql9Vgx/J+hHvXW/KnQTZTqrM42rZW1eE8grf6k/m3v3XjTpirNoZanUmJmYQ2NmLXuPqBq+vvZB6qRTh0FWZrsxhal6kvINBuygkKFH1v7117z6aY+4ZKKWF5nlUs+kWP5B4I9++fVqeY6HHanbtLkTAZKk31LxrB/3gn6e/Y60D0ZzBbkxGZp0jhnXzKl+CL6gvI49762R0v6J3igNxqDAgfW/04N/9b3ompoOvAU6aM1DFT0ktVJbWELLc25+vHvfXhxPSGx+7aVUMc8kn6yAByLA8C4/HvR+fXs+XSlmyEGTpNNNqeTTclhYgfW9/fvn16vkenXbjukRjd2XSfoGIP04t738j14ZPTxWI1QCql2jNrG7E6jxf3XPW8dNKUohls2orYWJ/r9b8/wCw92619vUTKY9a+F6cLqGlgAxNgCOePevLHXqZr0T/ALT+O9TvBJ3jgRwut7Nf6E6jYf6x+nu1M9awOicV/wARtwQ1DTY9pacRyFrRBlAANyOPp7r17obOu+m91YpadJpZpkjfnWX+gH1sfe6Uz1unn0I2/wDYFdUYplWMuywW/QxAI4P19660a9Ej+63J1znpK4UxeEysGBBCgBudJPv3XvLoxmy+9pclJTL5UhlS1kZ1T1qbW5/x9+68Oj6ddb7p9x0UNJVvAtXazKXX1C1+Lc39+p1vV0pt04FQ1NkKELrjk8kjKBe19X1/p7Ld+2i05l2iTZ78akdSor5Vx0d8scy7nyZuv752dij1GrSaVA+zpb0O75lxCGN5TJFFGoS976BYD/YHn3gPz592Dadn13FnISXZjnyr10g+7p7hbv7lbwibmxCADz9OolL2BkpZmpZbqHNuD+lT/T3Bdz7C36RtcWo1FM/bTroRc7XszR+AjgsVpT8ulPjN+JiK2OOulLQSsoR2N2uRf/e+PcVb3yXcyB7dk0zR1qBw6AdxsV/t9fASqfLoZZ9wQviZKyllRv2dY9Q1ci4PHuLYtmkXcVtZlOWp/PpJt8RuLsQyYz59FVzWdq8vmSZWZgJpVCliQQBe+kcfn30K9meS9t2a0triFQZH01NM56H3Oaxcv+324XUR0sIGIPzoep9DSSTRyB1IF2FiPpccXv7za3/ZLR9mQNQdg65B+1HuZv0PuFNLEXc+MQfSlT12mPnW0YJKlvoT9Le8fpOUdvjumukjUPX4gM/t66i2XuYklnG06DWVH+Dpn3FA1KkKQtrkqJBEFFgxvxY29kHM2jZNsmupW/CaEnhjqReWeYY90t5JpV0hVJHp0Ju0uv6NMUarKIgknj48gHOpePrx7wE5k5wvJ90MdkxLaq1B+fQRl3i5fcmvErQHokfyZ2GsVPNFj4wySqx1ICQp16dPHvoL7DR3dxyzFf7mT4jevHh1CfvjzlJv23LtNsKFRnqs/A0X9x99QPkIzHHWVIij9NgXZrcg+8hbx9VoX/hz1h/y/arbb+ttN2+IwX8z0dKs2WMz9tUxwCWGeFB9NSnUv0YfT2R7Nzh4k30UPEGnUn+433e4o9rfmi/I0mPWPnjorvyI6ioMXt5snCVpanxuTDGCoLfUG3HP9fcz2pZrdWfiR1zV32OKHd5IIfhUkfs6q03ZtHHbuwGTxdTGrZGjSeVTICTZb2sfr/rc+3/OnRZSo6rMydFLjchWUUqMj01RNCQQQbRuQDb/AFvfuteWeoIBP09+68SB176E/T/e/fuvcR12APqfp9PfuvE+Q64/i3+8/n37rfXdz9Of9b37r2OvWFv8f6e/dazXr1uL/i9vfuvVzTrxN/8AD/AcD37r1OvX4tYf6/59+69TNeuvfut9dg2/AP8Ar+/de69yDf8A2PHv3WsEU68Tc3/3r37rYx142/Hv3WhXz69bi4/2Pv3Xq5p14D+tx/S39ffuvH5dev8A8j/Pv3W6df/T+f8A+/de697917r3v3XuuyLe/daBr12QSNXH++49+68KA064+/db67uf6n37r1Ouv979+691737r3XduL/g+/darmnXYF+Pzf/ePfuvHGeuPv3W+uyb2/wBa3v3Xuuvfuvddj/Xtbn37rXR4PiD8acn2zumnzGZx0y7eo5IpI3midVldJQ5mIYAWFuP63v8A09+69j8urxMLsaj27uDGwYGBIqXFwRwN4AApaKwu1h9ePe89b6H7cncc+OxCYuqMrPFEIhZWIKgWtf8A4r718utefSM29n4cz++rKHaVRoP6rMfyPfvt68ccOjByYI1mASLxh2kQNcAXPH1978+veVfLpKYCiqNt5ynBQKrzRho1/SyFjfVb37rQz0Mve21aDJ9cU+bxdDDBlI1En3NKgE5IGr1MOfejwx1siuD0VXBUu7N04ApNA80NIEhLst5AVFib/wDG/futA56HHrXdVRsqlbGV/kF7j1A2A/oL/T3vreadCjN2ZSqDJFLyeeGFwD9LW96+3rfDI6zJ2XRTIFeo0yFQCC6/X/Y+/CvWsdS4cxictJ4Z5YWLC/rYFfr+fe6463Spx1mqNsYye7UMcWsi/oAvrPABt711vyz0nf4RuPG1amHyCAc8XAKg82HvdQD1r7OlVT19QzRoUcSCwe4JOr83/wBf3rrwp0qhBVvE7hWQmJjz6fxxwPe+tYHRQO38nNRVw+6Z9C1EX1vpKh7fT375deJFajox3X26ce21qJImjQGCLUVPP0/NveuvZPS5ptzUbOF8wuLKRe+oAf1/w97x5db4dZaqeKuGqNyVIP8Atm9+8utDB6YhSyJUoYyWsbkGw+p/r79TrfnXoQMdUN4likYrb6t/j9Rx70c9eGOpMpuLH1rYj6g/X6D34Y68eoP2zxyBlF1a9/6/63v2OvZp11JEXYJY3IH9eAB79w63XrC+Bx1V6Kqjp6hWvq8qahz9b/8AI/fq9a4Y6SGT6s2nkZlc4egQm5OmNVBsBY3tx78Pn1o+nWD/AEUbXgUNTY+lQqR+mNeSOC1/e+t0PSqx2zsLQooho6cMV9TBBcfkj3rr3XPK7VxdfRyU0lHBN5OP3FDWBFjp96HHrZ4dFw3v8csNmo5J6GBIKkhiBClm1j6cj/efdh1Unz6K7l6HsjqGvU0KVT0kLDlfIV0IdVj/ALD3o0Ar1ZVdjpVST8h0a/p/vej3vRpR5GZYsnEgjkjqGCEyA6Wtf3oMrYB628U0eXUivy6EjeuBTLUxqERDaxDR2PqH0t/vftpry3Q6WYA9GUXL+8Tw+PBCzIfMCvTFsnKTYiT7Co1SR6gLSfQf0sT7Kt2jh3CLw9Q6Hnt9ue5coXv1QjZWqPI9DRIlPWpHNEilyAVt9QSOb2/p7B0vLVrFCzVA49ZG2Pvfv15uEUFHpUcQfXpO1UU0bSedSovwf6f69/cO79s1vFM04UMwPHroX7V+4d1f2MFre4LDz/y9JPJY+nro3EkMTi3F0BNj/gP979ggxTSPQrUenWSaXtl4YEzKQfU16CHM7CkmeSSOkVIzqYskdiLjg3v7FHLpS3uAWjGr7OoO92No2+82t3tgq8TUfZ0GFVhsxgZ5JKRpgqE3W5Fh+T9fcv2gMjLIV6wM5gla1s7i2hnoMilePThgN65LG1gaaRhZxr1XFrGxt7lbb2P0oHy6wB5vjH78dzQksM/n69Ga2j2HTVqqlRLZ3NkUkf7z7g73A3B7e7CjgeuoH3SOTo942B2bJoOhFrMjJko9NMGcm9ioB/P+HuFt65ssLIBb+QKD8+s5tj5StNjlMswFes2H2S2WBeoQLzyWuOQ1/wDb+8cOefd0Q3JttlOqOmT8+j/cOYY7AUQ9DLtHbeNwR4Yaw31PNjf3jlzTv99vjeJIKVHl5/b0Ad23y43U6VFB0IVRKlSVBAdf8eQbf0HsERI0OeB6Io0MQrwPTZOlNGh/ZjHB5sBcn8c+1kTTSHDHpSkjkirHoPqjYOW3HXiPF42uqqipcJD9tTvLHdjpQEqLAci/uXuS+at62thabKpeSQgUUEmp+wf4ehLHzLYbZamS7mREQZ1MAcfb0Kkf8pTJdqYT+O/Jyrx23OsZAJ/u4qiNchFTqPIQaY2YnT+PfSH272DdrLZU3zmadbMsAe5xqOKmi8esXPeP395N5uT+pnItmdwviCrKyVj1HHxcOPRZe8eyviv/AC2Nk5+H4FdVdf8AdHZlLj6mjG4uwdsUmRrMTUiG7S48TXCsGJAb8fX3MWw+5fLG7TDa9tdnaPBdxQMRg0/PrA7m/wC7R7nbZZS83b5ZrAkxLiOPgvnT9nWmR8w/5gny2+S2/wDI1/au9Nz7Ukp5JEm2Th8zkqHblFKxPppcbCyxqlrgAArb8+5GVldQ65r59YzXEM1vM8E4IKmhX06r7mnkneSWZjLPLI0ss7lmlkkc6mZmP1JPJ926aApgdY2YtyeT+T/X37rwFOuPv3W+uRFgD/X/AIp791oHriLfn6f4e/db+zrv/Wvb37r3Xr8Ef1t/vHv3WqZr11791vru30P9ffuvdeNvxf8A2Pv3WhXz669+631737r3XvfuvdcgQL8XP49+60RXrq9zz9P9v/tr+/de4Dro2/Hv3W/t67sbE/0/4n37rVc067DfTjkfT37rVOuibkn+vv3WxgU65G7C/wCB791oUBp11YgXvx/sffut1BNOuuOPr/j9P9jb37r2euwbXt9D/Xj/AHr37r1K8evK2m/v3XiK9e0k/n/e/futVA66Isbe/dWBrnrr37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvddj/G9vzb37r3XiLW/ofp791oGvXvwP8AH37rfXgLm3v3XjjrxFuPfuvceuvfuvdcw1h+SfpyeP8AYe/dVK1PXD37q3Xdvp/j791qvXiLGx9+68DXPXlFz7914mg66Nvx/vPv3W+nPDYiuz2UosPjIvPX5CdaeliBtrlYEhf949+60erwfh3/AC6c/RZXZvY28KKb7ynroq6Cnk1fb/pDKojtb/WLc+/Z61mtetmrCU/2OBoKRoEpjDDBH9vGAsaaI9N9I/r78eOOrjhTphzRWrZ4GUFSNLKeQV/oR731XPn0i81sWmz+KkxaUkPjkUI50gXH51f8V9+4de+fVe/dfx6ymEdq7bTTpUQa5TFFqCFg1wCF96+R699nQddb9xZjblYu3NxyGmqo2EYEp0D0HTYE39+62QerKevswM/jKeUSK10D6geCLcAH37rx6EWGB6ecTQE+QHkAi/8Aha/vf2deGOlNNE1VTpNMoZlW7Fhf/H/kXv3DHXsHpE5HFQVzvEq6JX9PAAH+8W+vv3Dh1oenQPbl67hrmlppkUEjlrDnk+9cevdF+zfSkE0k8EUbMQW06VNwP9pPvfXvs6AzcGzNxbPmmkpFqvHDdowqPYkfQe9de+3qbsjvOs2zULJlnqo3hJ1AqwB0/Qc+90630abEfMraLUsS1c7BgBdQBewHPvw6rxPSqx3dK9m1CUe3BUyRagJNaMqhGNwQ3+t/h70anq1OhfotqqsUInRgzgEsy2N2/UL+/de6EbEYiCngKWBfTe/+wsOB78T14Zz1gdTRza1uXLfQcC5/Hu2OtA06cKfKsCscotqax4I5P0vb3X7Otjjnp48MUyiQm5t9f6kjj6e9fLrfWL7ZlOskf1uBbj8e9/Z17yp1laSnYNGWFipDf6xFubD37PHrQI4dNwosdGpUxxEPe5IF7H6m9uffu4568acD1ypMbjkdjHFEAeOABc3459+8ut44dYsrh6SpganNPAwkUozFQbXPLe/fPrRqeiwdo9FY/OY6U0VNG0kXkdvQBfVyeR/xX37zp148M9Vg9h9Xbl2rk3q8aKmFaaYuVh1qNKNwLD340HE9a7mNI1J/Lp5677i3BhsvSirrZoZI5FDKXIPo4uRcfX3qo8urFXVqMKHq2PrXsL+92FR6iZXklijjUMw51Rjn/iffgyDurw634Ezjw9J1NwHQg4OVYK6bHVQFrFlJ/S7HlbH2DOadt/fMYMBDZ6yY9iueZfbe6LbmrRVXFRTqdVUENHM9Uq3JAKhQTyTawPuPbzlqW3tGXTSo6y95b98YN33mNlmYgsPPy6ZqxBXxhnJQobq1v9tb3CN/yvYQXMk1xECZKgkjrPLlXmfbt5tE8KjkgCh+zpvTcuax8ElLDLNLCqlVDsSOOL29xlce1HL9xuK3fBdVSPz6Vb7ynFMv7w24UmGaDpgx244o67/LRpleclSRb1MeST/vHvKf265S2wSRiJuxAKfl1g396P3j5t5d5ZfYViIVx4bUHlw6HShnielRomRxINV1INza/wBfYx5wurhJhbqaKop1B/3eNl2ebbW3h41eaRixJGakZ67M6U15Jio03JFjYj/D3H9zcpAmljx6yz2PZLreL1Ft1wDwp5dJvb9PHuPdg8p1Q08vkjQkFdQP9Pp/T3iL7683XcEDbVESI18/Wvl1kRfWk2ybGltboA5ADUHRgN7008O3ofstUTQxr/m/T+n6294jcmzwS8yRte9ytIK19K9Bvl/w2u2ilAOoHj6norebpo85Ey5FPKyE/rW5Nm+g99TOX992bb9ujhsZFC6VwCPT06C/MPtVcbpftcIo0HPVanyP2LkJMhDl8VQMIMVVGqLRRn9MTXP0/HuTdj3dL+F4ZCDqFB1iv7ochXvLl9Df2yEeC+pqegNehb6s7g2y/XvmyEsa5ShTQUcgPrgWzXQ8+0Ox7EbLmHVIBRjX9vRl7je6c3MPtcLSFyJIo9BFaHtHROO9e3f7+SzUdMXFIjNGCost7/m3vIBAqxKtRw65VXLTS3srsrGrHNK9EGzNNDjMwsqyKI6yQRSG49WtuVI97yemwfI8eiefJHrSTGZI7pxUH+QzqgmWJdVzJz5CF/p+ffuvfPopVyBa1jfn+vv3XqVNeuPv3W+u/wAX/F/p791rz68RY2/3r37rwNeuh9ffut9dk3N/futAUFOuvfut9d24v791quadde/db697917rkRYf7H37rQNT1x9+631737r3XZN/oLe/daGOu1vfi30/Pv3Xjwz14n6gjm5/3v37rwHn1x9+631//9T5/wD7917r3v3XuuVwx9V/p+PfutUpw648W/x/3i3v3W+ve/de67Fvze/+Hv3Ws+XXXv3W+ve/de67vwR/X37rVM1669+6312CR9P9b37rRFevWNr/ANPfuvVzTrr37rfXYI+h+n14/r791o+o6MT8Zem27p7Jxe3JUqPsEqKaSqaKMsjgyj9l2/xHJHv3XvOnW1f170Dt3q/Y8FNgcdDDPHjyssggEbl4lN/9697qet4OOo3Vez49xS5Ragn7n72UaxckDV+CPr/sPevLrQBB6Hys+Oe263HvJVM0k0kZ5ZdTam4Fv8fe+vdFD3x1pmettzU4w0U1Rj2lR3XQ5CqrBj9PevkePXq9Gz2rUU2XxNHHGyrOKeMyxMwVg+kFhY8+9+XXq06j5XDJ/FYmsQUZCbAkAg8G/wDT3rj17yp0MVDDFm9vTYeqRXi+1kVQw1KWKWF/+K+9jrZBGei47dqKXYmdrNv5yIx4ysqGKyaCALsdIDn8f4+9de86Hh0NR6221uWL7qmradBMmuIrMtwDyNRBv72B69eNK46Sdb0//D3aKGqE0LHUH1aiP9t7917geg83J1/WUUTPTSzGdWuoQGxH4N/fvt6qR0CmWye8tr1CyIlS8aH1HSxsLfj/AGHPvXW8fn0Iuyu+Pt5oIMoZRMtgfIHWMmxvq1e/fZ1s8aHo2W2N77c3XTKIKqn8pAWzToCNX1JHvfDrWeJ6WsWKxcTa/JSk3uSZF/HA9Xv3lnrw9eoGZ3JjMer00tRTRhUPq8yKLf0v79wPWwfMdEF783biqqcxUs8M8hkAAilWQk344Xm1/fq9eNPPpT9SVuTqMbDBVLMtPJHHourA2JsLe/cOtfb0ZbH7bdtMkkkiqbNylrge/Vx17oRMTjokZQCWAUfq/wAB798ut+fTnWUSGJtAs+oWP5t9efeq069pqek5MaqI6IVsw+nH+HIt/sD735deHxZ6lUVXUqAKhT9bACwuCefes+fWzQnt6VlPVRzIApOoWBvYD/be/fLrwPUjStiTx9PUQOLH6e/ZHXq9Nk9Y6Fo4LNfgNa/+NvfhTrRIAz02CasYtpDXvzcfQf0HvfXq4r1w01wN7Nb6f4m/4t/T36o60a068tTU07WIc6jcX54H9PfqdeJPl1PjyysRrVxawJANgTxb3ogeXXgT59OcTxyHUtzqH0PA9X49+Bp156AVHSW3Bs7D52knNbTRzykSAa0D2LLbi/sKb1cX5lCWw7ep99tNo5UW0N3vbAuaUBp0QDsTqTM7Byku7drLURwwzmV4IFYKV/WQbeyW33uW2JilPd8+pL3b2u27edG4bcn6RINAOhA6u7xk3MYMDknaKpQiN/O4Rtf0bh/r7jXf+ZNyW8MkZNAfLrMj2i9oeSJNiSz3CNTI6n4gMH8+jVU+AocgqtC8IlazXQqSRb8lfz7vb833Loqajq889e3v7u+yWs0lysS+HU0oPLp8oI6nGOIW1NGjWB/wB/B9mrbzdyR0LHPQFtfbblyxvfGSJaofTp5radciNCrYlTqNrf717J5dMinxc16kGzJsXU2XZoGKdBHlI6jC1jPL5REjXNwdIUm/1/1vx7bi2m1nYBQM9K7zn3ftujMjuSq/PpXYLcOCy0Yhj8UkyqVZCw4axGq3+HsY7NySrXCyycPs6x69zPvM3EO3PY2zVkFQQT+XTHu3YdJPTyVdOt3e7aFFx6hzb/W9ybBslrbgKKY6wi3f3O3/AHiVjVhrPAdFZ3NtSekmZ4opVZbMwEZCgjn/AHx9rvqbK1GlnUY9R0Fo9l5i31xJBbyMSRnQT59ZdsvWx1MMZSW6upJCNwAeTf3ix7w8yWVs5jtXBlINM1p11e+6hf3PKexSJuimNqClQV6N9tTMUmLpY5JFEkrKtg/44/p/r+8GOZxuW+3BM0jUFeBx1kXvPuNdXsxjTABwR0JVJucu6GnQLr5IQjTqt9PYCn2BEBEhrT16Jv37cXdBIa9LrC1jVcqs39s/T6ck8c+wpu1qsERp5dGVrc1Ib16XV1hS7X5ZUFxflj6fYNzI1B9v7OPRgO9sdHO67+GGazW38fv/ALdyFJtrritgWtSugrIo65qcHWCsd73t7yp5E+73ubWMHOPOMq2WyyJq1iQFyPkK14dY4c6feF2yw3KXlbkaJrzdojoKFSUB+Z4cehPm+Qfx++PlDJt/orEY3f8AXpG0dTX7igSaSKZxYvE0gP0Psc33uZ7cez8P0/tjapujSVrNcoCVJ4sKg8PLoE2Xth7ne61yNx9ybmTa4q1EduxAIHrT16Jh2T2h2B2tWVGSzm5snjKCUSP/AACjqZExsaN/utYVIFvx7xv5n96OcOY92e6nuGCvhURiqL9gBp1khyryXytyTZi02+zjlkUf2zKDISPOvVRPyxrNp7YxeTSWmpY6mcTiSf0iWUsn6j+Tf3l192Sa4ukkvb+Qs2NOo/t/n0H/AHN5gvt02dtsfMVDUfl5dahvy7x9DDvyTIUMHjSvkZ2k0FRJwxFmP199Ddol8S2BrXrkf7k7eu38wOirpBJ8uikccfX/AB/417Nuo7z11/re/db68Bc2Hv3XuHXvfuvde9+6912DY39+60cjrr37rfXfFuL3/Pv3Ws9d2JH04F/+Kn37r1QD9vXH37rfXhb8/T/D37r32de/1vfuvde9+6917/X9+691737r3Xvfuvdd2+p/p7917rwFzYfn37rxNM9dkW+v+wt/xPv3Wq14dcQLmw9+63w67Isbe/deBrnrr37r3XIKSLj/AH1vfutVAx10DY39+62c9de/de697917r3v3Xuu7/Uf19+69117917ru30P9ffuvdde/de68LX5+nv3Xvs68bX4+nv3Xvt67t9D/AF9+6914ggX/AK/8Tz791qoJp11791vr3v3Xuu7c2/Pv3Wq4r17SbE/0+vv3Xqjrr37rfXvfuvde9+691737r3XYF7/4e/dar11791vrmrAA3+v4Hv3VSCT0Yb4pYZM98ges6GeCSeB9wwGZUjMihPG0Z8oH0X1fX+tvfut/LreewO2cdg6LF4+lpoUgp8bj5IgFA0MaZb2Hvx+XWx0/VdWiApYl7c2+oNrD6/7370M9eNB0wNRTaBUyggElhcWvc/Qn/ifdzQHrVOnqgt4iVuCf1f64/HPunmet/IdJHdeCpK2kn88YmllifSCuqwP497xTreAaDqoL5N9SzUFdFncZ5aWtjmeRBAhUO2q4uR73T061Wg6V3x6+S0NNFT7Lzk602Uox4CZJPCZNChRYtb6+9Ag8evfPq0PaORjymPhqw4l1qp1qdV9a6gdXvfXh8ul7HUpHD6rG44UBSb/0I/1vegDXPXq+fTHkJPLT1EtJGPuI1LRqFtqI5t6frz9fex17j0i8W02cnkgql8VbETrVRyFH9b/196oD17p3G1qemf7iZVfUb6WAPP8ArH3vjjr3z6x5TYO3s5SqtTRwHXcFvGCf6WIH496GOt9Fk3l8bNqZKoqkp4RHquxKx2/1+R791rBz59Brivhrt/72GqmqagReTUYyraSCfp73mvWq06OVsDrXbOw6aKDG0UTSjRqmaMCRrLY3P5P+Hvxx1ulehSqmiBXWqRKLWPHBI/B96qOtefTbPnMbjB+5V0qMOSJJ0Xg8/U+/dbFOPUT+8uKrCrR1lC1ibf5THe4F+Rf6e/YHWsnrDNW0Mv7pqqYBf7Mc6MTp5FrfX3uuadexTp2xuUpfqsodTwCHDcfT3rB68B041WTp41QB19RH1fmx+o9+oerVHDqEKJ6h2kjdQGNwL/7Ajj3vHn1qvn1jlxkqldb/AFv/AMk+/deI9esMlNUweuFiwPP5P+vf37rfzPHqXTZBw+ioRhpHBC8G4t9f9h71TFOtDJ6nzQw5CIwktpkAUm3Nj7ZnkMMJkHEdLtqtF3HcI7SQ0VjQnovvZ3WWMraKqiipBNLKpAbRdiWubj2A7rfb/WxYUXrLPYfa3lIWcaROJJyASOOeql+0Os8htHPvkBHJDEshayoQBZ+PZvZb3GbAvKc9RvzJ7X3UXMixW8ZKkjABpTobOmuz6ihNLS+ZrrIiqpk03KjT/vh7Bu7c6TW8TxQ8adZG8h/d023eL+3uNx7QpU06sq2vLVZnH0uRlBFQyRtwf1D8G/sH7Jznfx3BExwT5nrIr3J+7Zyvf7Oi7euiRFFSq+g6EuikSePxVCjzCw0sLAL/AF5/PsTblzDJewBR+dOoS5P9nLPlfcmmlcmhxXpgzUb0gdo1upUtYC4/17+wPuFgL5Mcesl+V+Zjy1L4h+EUp+XSSpK2OeUK6ppJ08/1/JI9k45bkYU6klPeeAOC2kL9vUrP7FgylB99Rz6ZUHkVYjyW+oBI/wAfYy5QsNzsrsLCCQOse/vBc18kcwbBI+5eGZHBAIIOekjtbe2QwdbLiMuHVYHVImkuLqTYcn2JOfoL2OzS+Cmp446g77qm5cv3nM83KjzAISaVYAZGKdCzVZF8nEPtl1xyD9UaluD9L294971zRaWsbveSBCgrQsAeun2w8sWGwHWtCeI6XewNnT0tUuSlYxiTS35BP9AfeFfuhz+nMUj2kSAhSaN5npvmbfoJIvplAJFR0PWSpoa6jWkmt49JUk2v7gWzmktbj6mL4q16jy0lktp/Hj49BVntjY+lo5J6YXk9TEKPrzfkD3J3LfPu9RbjEJJm0BlqNRpToYbfzDczTiObh0WbsDC0dTt7JwT42FjPBPCWeP18i2oX595/cpc7WNzFCtlJqcBSc18s16Y5t5NseZ7OQSAElc+fVL/ZWFz3XmWr6zHrUfwmoqJbxLG/iVXPqt/S1/r7mm139pVFwn9oo4dYI+4vtje7PDLHaKxgckHBoB0GKbj23NQu1SwR5Ll2J0tqbn6f6/tVDzfvl9OFZdNDw6jjl7205Dttul/eJ/VevxDgei371hpnyEdTQVLSRLKXKltQA1XB/wBt7lPaN2lukC3AocdYye4nINlsly8+1NrjqTj7evbkxdPmtsB6yJquCaMRMhQyaRa2oW/p7EPlXqIAa1FOGOiH9k9O1uASXNYaGabFsS7RCNjIlzdiqj6Afkf7b37rfn0ARBBIIsRwQfqD/Q+/de67vxb8Xv791qma9de/db68OTb+vv3XuuyCDY+/daBrnr1+Lf43/wB49+695168Bf8A1/8AePfuvE0675tp49Nz/r+/da86+vXH37q3Xd+Lf439+61TNeuvfut9e9+691737r3XYFzb37rRNBXrsrbkfT6f7zb37rQPkeuPv3Vuv//V0AAbf0/2Pv3Xuuvfuvddi35NvfutGvl11791vr3v3Xuve/de697917r3v3Xuve/de65gcemxNuQfwPz9ffuqn59cSLfX37q3Hr3Fvrz/AE9+6159de/db65BGLBQPUxAA/qSbAe/daqCKjq+j+V71jhcPio99Z0JBVmrilUS/tuI3csAbn+nHvVevdXZdl9s7YoNsfa4U+WvmD04VWVxaRSt9K3P1PvfW/PHUj48bVqYMfPmMhGUarf7mNWDLdXN+L8/n+nv1fTrwxx6M9USLTxFSBbnSp/I/pf/AIr73xPXs9B9lMFQ56d5aynjd9LoocF7ArYH3oenXuGegWzXX2d25LNmsKXKRMzCIaxGVHNlHvfnjqvA9dbN3Wd01px2ZiFJko38Sg+hGCk83a3PvRPVhQjowmJxclGjaV/bKNZx/T/X/wAf6e9g068QDgdJTcewMXu5HgqwIZbkpOELOCBcC4uffvLrXn0E2U683htSlqP7uVs9SYkZoxK7gaV/Aufp7914ihx0VTcHyP7F2Bl3xu46U+BJink/V6QbfU/8V9+yOvcc9Gi6g7b232NTD+ITpHLIF/VJGDqci1he/vRz1sDoS8/tnD1BfRTw1EZJNyt1ta1/fqdeHQNZLpmkzdPUJTU8UEkrsyPFH6xz9LqL+9nh16mc9BtV9P8AYey5jUbflnlVfWFJe3p5APvXE9eoaY6ZKzdneNF+x9oxYAhSdVzbm5B9+HWiKcekXXHufd7mmqIZqcyEIzqrhgDxxb37rdSeHDpT7O+OGcqstR1+5KmeS7xyOkokdbhrkf09+69To9uD2XjMJFSLHEmmCJEC6Tzb6nj/AIn3vr2Rx6X7ssyITEkcKC2pRY2A4Nvfj6der59N9VnsbiYjLNMiKL/240IHFz6j/vPvXXhTj0iqzuXaFPIY5qwXHpYeWC2r6WBHv2K9eqOHTQ3dOxyWYVXquR/nIfpf8c+/V60aA9N8vdW0bhqWoDMCRZ3iPJPIHv3W6+fT1RdqbfnZHhqFUNdiPJHcN+bW49769wz041XZuKNOwiqfVz9ZEubi1r/T37h1onHTNF3Ht2gRhVSAte3Dobn/AFz719nXgRwPXTd/bNg5Zzqb+jJYkDj6+/ZPWz1wpe89pzytLNOVW/pUPHYqfoTz79T069XPTwO79hyL66g3J+oeK4FuP6H/AG3vfDrWK16dqTtjYNWoEU367Dl4/rbk/X3WoGT1dUZzpXielDQ7vwOQljix8uq4UA3Vr2/rb3Qyx+uelH7vvKaih0/YellD43Qeq+s8i55AP19tXKL4DNTy6XbRPcfvSOHWQKjFfn00bixFFk6CWCogRoXQoUPK3ZbXt7gDmu7e1mZkOa9dZPYvYbbe9tt4roAppFeiI716LnxuUn3DtjVFMrCXxQghTYk31Lx7AUe5+KdNxwPn1lJufICWpS72Y6WQcB8uktiu1OwNl10cWZhk+1iYevkmy/4t7VQxWzHxYDU9EEu57zbqbPdkolOPRu9g904LcMcJyTIh0jXrdAeTybH/AIp7E1tWWLByOoe3h4YL4sB2sfTobqfcG35V10coZTyLFW/21vd2jYYY9NRzwuP0x1gz+Hps9jZYY0TVKptJbm5HA592jkaFwwNOmL63hvbZoSOPRXc5h8j1xUGsi1SB3LG4JFi2o2vzbn3IOwczhSIZjgfPrEr3T9jnnQ7pYgk14UNM9C5tTfNNm6GBqjSZWiBaPWpIJH6SPZdzNzzLZho7c/n0NfZX7rtnzH4N9uo06RUih9fTpq3hjhmE1UaiP6B9Kizcfk+4dvuY9yvpSxkIwfPronyj7M8jcu2X0jWUTsAaEjP+DqNs/adPHTzGSJWmvbU63IF7WB+nvFHnPer797Olw5JBOa+XQV535bs9tmC7fCIk9FH+x0vYNrEjUDYLeyj+g/1vpb2Cpd604Pn0AIttQ0Y8eucitiCrMTpDAGx/BPAtf3pGG4Cg6u9LU/Z0M+zqpK6OCaMWC2B1H+n5B/3n3GnM0DWxaN/PoWbbMksAdehhSGN1W4uODa3AZf0+4xeRwSPy6XGRgajpZbw7C33urCY7b9VuLLRYbE0wp6fHJVymjeNF0hTDfTb/AGHsaPz/AMy3lpFtl9eSPbRKEWLWwQD/AEtc9BnaeUuWtnvJ9yhtIzcTtqaQqNdftpXoukh/h9czLGquHXU6qqs5+nNvZnFEt5aaScU4eQ6EklzIaBSaelehHpqgVWNaQ3uImBA55029gea3NvfiIeo/w9UdwI/Ep9vVMvzAxlZuXdDwTtKlDFPLq0qbaVWwB/r7z49oLhdt2WPwTRmA8+oY5huX3HeE2+lI2J1HrXl+fuAgxWS2vFj6Ziikh5I4mN707W1sosObfn3n17ezvc7KJpDUnj1gP95yxs9t5xjtbQUAQVPqaf4eq2yCPqLf6/sfdY0de9+691kIVR/iQSPfuqVJ64cc/wBfx/xPv3V+u1tyPyfp/tvfutGvHro/6wH+t7914dde/db67Fr8/T37rx+XXK4sRf6fT/G/1v791XNa9cQLm3+9e/dbJoOvH+nHFx/yP37rw9euvfut9dk3N/fuvAU669+6912bWFvr+ffutZ69xYc883/w9+69nr3Fj/Xj/W9+6959evb37rdOurk/U+/de697917rkbG1rk/n37rQ+fXH/D37rfXK5F1P+2P4/wBb37rVAc9cfr791vru31v9R/vj791r7OvWPH+P09+69Udesb2/P+uPfuvVHHroi3B9+63x697917r3JsP9t7917r309+691yN7AkC3+w5/1/futCladcffut9e9+6912SSAP6e/dapmvXXv3W+ve/de65XK825/qeT791XBx11fi3++P8Ar+/dbp11791vr3v3Xuve/de697917rv8cX/x/wCI9+6116/FuP8AX/Pv3XqZr12ARY/XkWH5P+t79145x1dr/K3+OVXPnqrs7edBFR44U/3W3DUo6ztNEpZWu4AGokED/AG/49760ONR5dbMe3M6ayJJahtTCGOFVvfhECIBb/iPehTq1fPpf43b0mRZayT0xgiysTyAb2Pv1c463givT5l8TGtE4Cp6dPAHHP8AT3qvWsV6CRc9HQVT0z8eoqBYWvfkj3vj16lOlLRwz5ZtfjHhtcXt9ALk29+wBTr1c16AXufrOm3VBFBBGDML6RpNrkfj/W97PCnXq56pp+RfTOc6/wAjHuLERmnraKcVDmFXUyXb+2Tb+nuvXqdHR+G3ygx27cLHtPcNQKXM40GJ1llijLPHHZb6yD9fex1ok8ejzU+9aU5DwPNEyS6dLCWMgavwAObn3vrXQnQimljWSKRG1ga9LA8H63K/T3rh1utOkXDDUYzKZSqpUD+aJ1hJBv5CL3N/fvPq2COsW3M1WZeqqKGuGmenubfgkDiw9760TXpQTVstJIsTEqur0/lSSbWPv1etY49OT0oqaUOIxrK8kC17/kn34YOevHhXrJRUoiTQV1MoJKk8WvxwfevPq1R59c5YmjDuwI0i/wCNNyPp791roG+xOxocZR/Y0t3yZOhESzHUOPUF9+69w6JJu+HtzcdRPNGKmCmbmMxiQHx/UDj37r2CKHoN46PtXES63myBjBJb/OXAtY/19+61Ty6VmG3P2DRyB6p6xobj9QlNiP8AEj3vrdR5dDft7t3M4qNRPFK5K860t/rmx9+60c9PGQ7vqJQHIYOpuAEA0/m30HvXWwOmmP5H7hpZNEUF4V4udN2t+Pe+tAnp4o/lFVSs0eUQRKnquGRTYf1/w96qD17gKenS4w3yi2p49NfI5vcXBQ25/wAOffq9b6EzB96dfZoeMSuoAGlj47kkfQ259+YgZPVoxI7UQVPQkYfceIyz/wC46RirDg8WFxcWH/GvdSY37Tnp4Jd2pMyAqR59KOSigmikMyhzpBBIJA/x/PsEc2QpDb6oxQdZM/d/3Kbct4EN25ZuAB9eidd7dUR7koqieKMeQxsbKrm/q9IAX/H3CN9zJJC4tIjgHrpdsntBZX9h++rhQZSKgZ6rortm5jZmRjqaaN9VNO0vj0OBZX+puPapLuG9i0zefn0HrnYd05euRc22qiHhny6OB1V8g64SUePycKQxwLHHYFRyo0/n/ifZFc7e1vJ4kRqK/wAupV2PnUbpZtZ3qgNpp0fjauUoN10qVVNMiSOqn9SXbi9iAefYktpCYAxHUTbxFH+9ZEjNPPpXVOGikpZaaXQ7upS5N7A8XH+w9uCVgajpA9tG6aG8x0VPfU9btetkhpFdgJJApAawN7gcj2J9l+nmnDT9Qn7kJuVht7x7XUsK56UXX+95JDHTZOX1MRZC62+twCL/ANfcr2FtZQoJ4gBjrA3mveOZNzujtd8z4bhnpV762tS7ggGQx2iGop0MzsoIL6fUASv19od/3LaTt0lveEGoIXhx6GHtHyN7hLzbb7ny6jalkUthhUDpbdGikqaSWiyIU1cbkaXHIT8H3yM9+1v7LmGWSzdvCJOAcceu0dnzHvNxtNvDvC+HOiKCPWgHQ5Vz/wAKkRFYiLUNABIAUnge4Msolv4yWHd0XXd2zvrkHSkoqo1UCvcm/wDX/jfshvLb6abQcdOxsjiq9cq0AwqHF1uL/Ugi/utoP1Tp49XDsh1Jx6B3e20/4wDFSRDwuCHABJBb6/T6e5n9tOdk5WvzcXzEilKY6G2xbskcBFycnHHoonaPQcGWxdZSSUaOrRSMGeN2sSv+I/23vJjYPeS03HcIobbJdgKdIN/stl3qzkt3A4Enqr7efxcdKitjptcYUO6okUg9VydIt7y82WRXt0nkQAsAeuf/ALjbHFFuElptpNBXh0S3OdR7qxWZei+1q5IFkcamp5yNIa1wdNvYyjvreJQ5IU9Y73PLG8Xc726o0goeIPQ49VbLo56mTCZ6ICDwegSoVtIfobN7Fm3Xa3UIYdQBzXsc2ybi8LrQVPSG7X6y/gVbVR01Mk+Lqo2j0sjeNVcWLAEEf7b2vNePQX+3qpPuTYD7O3BLLRwucZVlnEgR/Gk7yMzLqPHN7Af4e/db4HoGffuvde9+691ysLcXuOffutZrnrj791vrs2/Hv3WhXz669+6313xb6m/59+61mvXuLf43/wB49+69mvXZFlsRzf8Aw9+60DU464+/dW67vxaw/wBf8/19+61TNeuvfut9cvSBwTf+nv3Vc1yOurn+p9+6tTrr37r3X//W0AAbfgH/AF/futHPXZHF+PoPp791oHy64+/dW69+Lf7z7917rsWvz9PfutHhjrxP4sPr9fz791759de/db67Bt/T/Y+/de69f/Ae/da68Db37rxFeu2Fj9b8e/deGR1x9+631737r3Qh9Z7Gy3YG8cXgMTC08j1UM05ALBYI5Qz3t/X6e/cOqjy62MejunpMDt7EYs1k9CGgphNFGxQeSwBuPfqUz1c5x1Yf1/0NiTVUtbkKl61AiSBZyWUkNccn8+9jPWiKY6ODQY6loKaOClhWGGJAiqgsCqCw9188dbx59JvOZMLJp4VAArH9RBH+H+v/AE974dar6ddYc+X9xgNRI4/wt/X/AB97A68SfLpV1dJHVQmOT/NlbkX4t/sf9j7r59epjor/AGjsZ6eT+L7fX7eppnFRMYQVZkT1M3H597619vQrdS7wpN57baIyD7vFFaWdWYa3dBYkg8+/HJz1sYGOl7PNHS6mCqCpv/Q3Jtbn37r3SBzWQqZ1lRHZDJqA0tYBW/Fh/wAR7916g8uiYd0deLmKaokno45pZA7CVkvIDe/BP+x9++zrXnXoomIx+W2Llaeoo5qmGH7mGIol1AHkH1A/3j3v5de8+rRcTueii2lizLOZa+qpoGILanu4Gr/e/furfPoUMDH9vTQSSfWZRKD9LBubD36h8+tE+fShqqinYospJU2tyebf6/vVfXrwBOT0z1m28dUzx1JgTUBewFxz/vHNuffsda8+sAwVBHIGSmjiYEC6CxFjf1W9+63kdObvRUSD6agLDi5AH4H+x97+3rXHh1EhrPIGlI4Q3Ab6Ffr79XHXiDXHSX3LvzGYyjqDJJokVXConJ1Acce/ceveXRMty7j3RvjJvj8e1RHSySBAyFh6WNube/Hrw6UmL+NFZl4Y6rIZarjmdQ2nW9/pzq9++Q63Sg6fF+K0IH/F2qb/AJ9T/X/D3qnWgPPqXB8YIYLFchOQCC3qkN2vwefe/t68R0/03x9NMbffzAD6WZv+J9+69UdOZ6KlYafvpyDbi7j/ABF7e9cOvYPTfL8eBMfXWSlvoAxc/wC9e91pnqwHWH/ZaaU2JqG+l+QTyeeL+9DqvWCb4yQSnQK2VCebqWt/j9Pe+t8OPUdPi9DFJq/iM54vyX+v1A9668OmnJ9E5DEMXo6iaRUN/SzEmwufbdwR4LEenSza1J3CNT6jrPt2rzG0HdZo5X0MOXU20/4A+46uN1ktryjE0r1mTtHItnvXLFVjAkKmhpnh0LuI7comhLzSOs0ZACHi/P19jmzu4dwtMGhp1izzFy9uHKO/jxVNCwofsPS/w2/qLPQSRtMqStcqmofT829wNz/B4EpPz66pfdU3E7ltsQmwQoI/b1LkEUyOhdSjgAgsObC31/2N/cTYp1npkHI6R2e2Bgc/CUqoImFmGrSNVyOB/j7eikaHuVukO4WFpuSCO4jGPOnQAZrqGoxTzT4iZ1VHJWOLUBpUXHA9mMG9PG/cSB0BN49uLS7haSNRjPUvZW6a7b1WtHl3nIDFRqBItfi9/wCns4j3F53EhPaOgLccoW1pbtBGvf8AZ0cjbu5KWsoo6iB9fHKm17gcDn2bC7WeM+F5dAp9hls7lRcClSKdJDddHFuB2FYpdRr0A8rduLH/AFvYTl3S4guCYjQjqd7TkjZt22iOO6WoYdE83PWZ3rjdsFlk/g1RLdnQMVRDyOPp7VPP+9rctKe8dBZNrf2/3JUta/Tyenp0Yfae+8ZnqWIJN63P5Iv/ALx7D00U1pUUxTqXdr3a03VA8DZ9D0tmyFRTD/I0Zlbn0G4P+uPca7r7fWm93bXjOQx6VXuz2G4Z3AgUFM9ZsNuishqCtZqWLUVfXfj/AGJ9xnzfyQu06RbVboB77yVbIhksjWvCnU7cVSchTvLA11I1DTcE2Fwf+KewxtMYtn8OQZ6gnfre5tZWhYUI6k7D33NSTR42UBLOqKT/AEPpvf2xzPy1FdRG6XOM9J9l3oIxt5sHo3eFyUVVAgEitIVW41An6X4A948btYSW0xOkhfs6HyOsiBgR0/BieDYWtz/Xm3slp59epTpDbjxkcrmoQaWTSeP7TfX6f09i3Y9weNfAc1B6t4OtdQ6zYKr8sT0sgAAUqQTwb8e293tvDlFwnEnrTUKFQOHQDdw9O4vcdDXV8kcQYln1WAPK3PI/P9R7lDkP3Cutvnisqk8B/Poti5Utt2v0oNLHzHVaW/Ph5srfUdX/AByCKZ41bwyyoCselSOL++jfIXN29tt9vFaoSJBkdBX3U9gPbo7VJv8AzROFlRTQnjUA0HVH3yz+D8eyBlNxbKieWmoVLVCxqwjcoLkLGOL2/PvKHapJprNXuRR6ZHXG3nm023b+YprfZ21wg4Py6qqdXid45EKyIzI6uCHVlOllIP0II9mHQV+zrH791vrsC/Hv3XuHXM2UcWLf1v8AT/WHv3VBVjnh1xsSC1/p791atDTrj791vr3v3XuuWnk8jgX/AN4v791qvXQtY/14t/xPv3XjWvXZtxb6kc/6/v3XhXrj791vrsm/4A/1vfutAdeIItf8+/deBr14iwB/r/t/fuvVrjr30seD9ffuvccdeJv+AP8AW9+68Mdde/db697917rmLHk2H+xt+P6e/dVNRw64r9bfT/H/AI3791s9eP1/2Pv3W/Lrr37r3XM2ABU8/wCv791UVPHrhz+ffurddj8C9hf/AG1/z791rrsi3IJ5/Pv3XhnHXH6+/db67B+vA5/3j37rXXr/AE/w9+6316/N7D/W9+61TFOvEk/X6fgf09+68AB12ALG55tx/j79141qKddAX49+63w68RY2Pv3WgaivXXv3W+ve/de67JJ+pv791oADh11791vru/8Are/de669+6912B9L/T/be/daJ9Ou7D1f4fTke/daqcdcffurdchYAfT68/nj/W9+6qa16sF+APxbj+RW/qmoyiO+F2q0FZPHIv8Ak83jPkYMD+q39Pp/X37rdP2dbA42nTbQrcbt7DL/AAvb9B44qX7QeONhGApOlbAXtz795deJoejzbEw5THUNXSSGqUhSxc6h+nk397OBUde49GJpcikVBHqIR1X9I4H09162K06ROe3xQA/ZipRWdirKWC8jj34AcD16vkOg/wAjhY63x1tLIryKQ4AIa9zc8/6/u3Ws9LXAZQ01N4KlNEoFhYWv+CT70R59eBxTqBk2mlqYpo1Bs1xxxb+vvxz1vFPt6BDsro6PtAGCpQCOawlZjzYcnj37r1PTqvPsH4Yf6MMvU7l23kKqmqYwahY6VpE8rluQ5X6/7H37r3DPRecl2v3FszKLJVUWQloYntHLpkf0xsF1A/n/AG/vXl16vr0brqT5W5WohiGXMqrJGFYTekg2tYA/09760AOjc0Hd+GroIGknjImHBVl1fT/D3vFOvefTvsPcQy+7alscdcMjL5CbelSbnT/j718urD5dGF3FjonpzJEqa4ow97c3PNxb3qp4da6x4CdJ8ailgXTVfk34uoPvfW8cOvRzutSbqApYDj6Dn8+/UBOetfZ0y733DBhMVLIrD7h4m8K/XU31At79kdeNOHQNbH2Qu4axt0ZtdbVDOY4ZP0qT+llU+99a6HdMJQKCgx8LJYKBoA9P0B/23vXW6U6gy7Zw0hYPi6Vh9bNGCP8AeR7317I6b5th7dnUE4ylQ3vpEa2H4B968uvUzXpln6r25MzP9nTAt/WMcC3497FPPrxFek7UdFbeqGkYpCoa9rD/AGPvXWqcOk1U/HHASL+26g/Xhf6m/wBPfuA6359IjJ/F3HVDuY5QEb82a/8Ar+/fZ14ivHpB5f43Ph010kcc6Dlrgn/E8H3unVTjHE9I+DZsmLrggRoTAyu4UWBH6T/xHsKb3uvgfpIep59seRP3qwu7tew/LoyWzN34rFJHSCXTONCMW/wFjYn/AHv21sO4CaakpNejT3W5R/c+3h7BBo9fPocaXfuMiSKFp9bTsF/V/sLn/Dn2s5ngWaybV6dBz2L3ebbeaI/DWtXof5dOGVaCtjV0VZI5VUafr9eRcH3inusSx3z0Pr13Y5AuWuuX4ZJPNR9nDoE94da4/L09RKKaMysjlfSPq3qP49oILieF6k46EG47Ft25QuGXupjonGd6zyGFrpKingEaRysw0hhe/wDX2J4NxSSMK5qeoW3HlKezlZ4108aU6ETYG+dxYiqgpUkeMo4RhfSoU8G3Ps6h3KGOKhHUeXnKO5Xd7qQmtejxbQ3Hksj4xUvrDqlze9rrc2Pt+GdLhdQ6S7htk21HQ9S3U3sTBY+rxXkKxvVshZSQNWo88k/n3Q7ktlKKmmerJyZLzJZuVSuOiTVtZ/dnNtJWTmERykgarC1xxb2LY+aZDZ6Ij5dQFd+x9rJzJ9RuMYWjVpT59GI2bvpM1SiGOUtG6iMt9b3Xi/uK973e8muDrYnNadZ2e2PJnLe0bSklnCglWlSAK8OhI2vKuHzX3EZKpIya9PpFrW4945e8HLYutv8A3qlWZ+I49DbmPl+HcLYTwDTIvQ6ZrIxV9FFJTyIzKis3qu1x/X3ivtFm9pdssoIBOOop3KCeGKjrQjBx0pdnO1VQhXNypI4P1K/1P49kfNCeDd6l6bs5QYNXn0sZ6FJInBJPpNhzwbfn2GIbpkkB+fT4mbh69J/EyCOplpp41cqW0k2Jt+Pr7OtyQyW63ER61VwKBuPUfdGBSvpZpVjRYxGxewAJUDm3sRe3/NS8u71BcyjXpYcfIV6QTw3F0306uV14qOigZfadDUZWYR0qyJrKklOCb2Iv9PfROL3o2GPao7uSXTgYHHpbb+z+3z/4xfsDXNSc9NWZ6lw2QpmBwNCgZCDK9OgexN2Ie1/Yc/18Nk3C7WKGdizGgA9ehbt3t7yZYW7W8cKO7ChJAr+3oiHa3SL7fysuUx6Cnhhdpg0Pp1BeSvp95U+3nNK38SRzYJA65y/es9kpto171tqfpOSw+Q6ALfMa5zEx0bQj7mIeMlhY+njke5iB1DV69c8nRonaJvLHRIPkr09HXdYSVcFMv3UZaXyhfWGVrg397wD1rqmiRDE8kTAakdkP+BRrG3v3WuOeuHv3W+ve/de67Bt+Af8AX9+6917jT/jf/eLe/da8+u1A/P0/1/fuvGvl11a5Nrfn/be/derQdeAv7914mnXrk/U+/dboOu+NP+P+v/j791rNeuPv3W+ve/de697917rw+o9+691yYC/H09+60K0z1//X+f8A+/de69/h7917ru/0H9PfuvdeNvxf/Y+/deFfPrr37r3Xvfuvddm34v8A7H37r2fPrxN+T791oCnXXv3W+u78W9+61TNeuyBa4v8AW3NvfuvZrQ9cffut9dgEkAAkkgADkkn8Ae/de6ur/lo/H7z5CPfuaxkvmk1aEqYiLQ8+Kwb/AGB9+HHrR456t5q8e2EzSJHFojEyFF02UKG4/wBh738uvZr0dbr7J/cYqEyhBIqqB9ASL/09+p5dbrTpWbg3VRYailnqp4oY0jY+pgCXH459+p59a+XRa67s+hzWVEVNKTCkgQkEEXHJtyPfvPr3Qx4Hc1D4IvXclR9eOTwD78BTrfHj0qKjdUNPEWaRBYH+h4/AH09662ekRVbopsuamkh0zNNFJCQQOS4tcH34569Wg6B/a2Gy/VGaqs5Ul48HXVRkbk+Ms7/4n/effqdeJp0ZamkjzlGuRop454Z0WUiJ/JpMh1AED6e9461npvfGhCRISWN7fXn+nv1OvV8x00ZbaaZiJ9CCU6baWII4APAN/eutV6AbdXRlZlSHjxqBEkEl1jHBB4sffj17PSZXBZvbG48X/Fo5hhqeOOHUwJUEcAD3sdb6OLjqd66gpJ4F/aanVo+CfTYW49+69x6QW6MzPhKuKCcFVZlsxU/1/rf3rr2Bx6FDb9VFkcfFMHDMY1JAIb6mw49+Hy63SnThWqkETtYarXH0F/xx/vr+/A1PWj0iUoqiuqtbhhCvP0OkqD+Tx79nr1RxHSd3buKgwVO8aSqJBHIAAQbnSeP8D/j72fl175dFQr6zI7pyMviZmUyemPmzC9vx/vfvXn1r7ejC9Ydf01LAldW0yCcepbgE6lNzc2/33+Hv3Xh0Yamx6Ig8QAHFwPxf3416twx1OFOVXkD/AGJ/JAFjYe/de66VA17W9+69031ddRUcbSVMsUYQn9bhTa3P19+APVSQBq/wcek0++sFGHtIzlSRdRqvb62I97IVcs6j8+qxC5mNIYJSPkhI6Y6ztbb1JcM0uq1/TESRYf1Hth7uzj+KRf8Aeh0bQcv8wXf9jbSU/wCaZ6SFX8gdpUz6ZWrAV4a0DEXB/r70t3ZsaI4Nfn16bl/mG3Baa3cAf0D13S/InZMn0lqfpa7QlOSb/m/09v1VvhNf59FzRzR4lUr9op0q6PuTZVXGNFadTG9m9I5/A9+8s9VqCcdKmk3PtbK6GjrKdy31WSVeD9QL/n23MC8RVeJ6W7dIsN8k0mAueuGY2tgsyAyxU15E5MZUg6h/h7Bs/LTzOZG4nrI3aveqPabWOzjppAoegb3F0udZrKCM+MBvSgNjYEi/9fZTJcTbExjJ49SFZbVtnuzbi5VRrUjgOgQy+K3VtWvVqSKoECN6zocAAHnke4w5n3aO9nKOa6j1mn7M8iXvKm1+NEtDGKD59PeA3rlhOErpJQpYX1XH+uRf2CJ4QBVOsldt3qcuBeY9ehyxe7aGpjSM1EakL9CwUn8A8/n2VaXQmtT0MkntJ11RMK9KWKop6hNQMciuOeQ17/k2971Lqo3TxUlcGo6SWX2jiqurSqWnTUASzKP7X9ePfnnlTtjOD0nG3Wk7+JKor9nUfGiuw9RpQN9pqNybgAE/QD2YWu6G2j0eZ6Du88mQbjdJMnBKcOlxFVrVWZT9QBb82/4j2g8XxXZm8zXoSQ2n0dukIGFFB0mN47Wot0496eenjknVbRuVUuvFhY/717fileB9a/8AF9Itw2623K2MFwAfQniPs6K3l8Flutataqj8slMLOw5Kr/UD6/48ezaO4i3EGKQUIx1Hl3t93ytMtxbElD6dD505v/EbrWSKaoiFVC2l45m0kNb6Kp9qrCwNqrtIK+nRVzBzdNvFzBHYsVIoGHz6EbP45KmWZqWyEMNPjIt/Tn2GL2zhunYPGH48fs6mLZpQbCNbjNV4nqVgcfUVNJJFKWJUOOeRccWHvFjmGeOy3eRFAFGOPTqFvcLafDuzKFoGyOkZlMFk8fVyVVMhDRtqQgGxt/wX/ivs1tNwtLqARSGtePUGXVjdQS+LED0sNg9gZmlycMNXJIQjhdDE29JHAvcew/zPyvt93YuYlFSDkdGmz7xc+P4UpyPL16PFjKxMpRU9SCAzqCdJ+lx/T/ifeL+42jWF29v5A+fUkRs3hhm8+uGUhSWmeMkBwlgR+o8f19+2+Ro7gOOHT6E0+R6CeXMU+CqGWeXxMxOkngG3I+v+PuRk2+TdoQYhqHVVD6jQY+zoMN3dhz1qzUUMgeJm02DX1XH9Pcwe1/trY3l4Ly7B1RUIHkepT5a22zZFu5e0gdBC809SpjjViXJBsPqWuCPfQL2xsESZY2QUTA6xC++nv5t9jNrZTlRp/Cfl0DnaXVeNzu2shTV9HFKKyGRpFdAdV0Nrj/jfvJDSq/CKdcbxI8p1yEsT5nrVV+YXQlX1RvOoyFLRTx43LVU8jBIWMERLFlfUv6QQOfx711vomJt+L/7H37rwr59de/db65BiP9j791ogHrjc/T8e/db697917r3v3Xuu7n+p9+69Trr37r3XuP8AY/j37r3Xvfuvde9+691737r3XYvcf7x9ffutGlOuZLD6/n/X/Hv3VQAeHXEsSLce/dWoOuib2/w9+68BTrr37rfXvp7917ru5vf/AH39PfutUxTrvUbEf19+69QceurcA/1/4j37rdfLrr37r3XIkED63A/2Hv3WgCD1x9+631yAuOTwPfuqk064m34+n+Pv3Vvt697917r3v3Xuve/de67B/H4P1/r791o9eva9voeOfr7916lePXhb83/2Hv3Xs+XXXv3W+ve/de697917rkF1E2/3n37rRNOPXXFvzf8AH9Pfut568Da/+Pv3WqV669+631zvq4P1+o/2A/Pv3Vaac9dH62P4449+62PUdcTb8f7z791vr39f99+ffuvdbPf8mzF4w7VygFJDHNW4lBPOkYDvdB+px+fexx60Phr1Zl2RsbHKQzSFCspKspN/1Hjj/ivv3XiPXqPtbfs+2qGPEUcnmaM6Rr9R/oOfescet+VOhPbsCSmwrV2RcI+gsqKbk3jP449+NOt1z0Qvtns/KUmVgyNDUVCwSzXddRXSNV+FP/FPfh1UkHA6MF0P2nBuTxU9XVlp9BBSRweQAPqffuOOt8B0amprIA14lFyF4F/oVvwfz72OFOvH1HWWGvjdFU2LgWKkj8fTg8e/HOOtV6VOHrqUkq76JALgXsP9jxwPes9b6b81tzCbqm+3yLxvfglyCCv9L249++3rZp0H2++l9hVeENCuAx1TI0bBZjArEuwsLH34EHrVTTolm4fh9TTRPV4xDSy6mcRQ+kLduAoX6W/w9761Q9IfGdAbixNT4ZqistETpU6iv+vf3rr3lX16Mr1fh/7lS1E9aZDLZbBlJ/StgePfiT1ug8+jSYrKpnseGF/3hoJYEen6Dj/W96oaV62DmnSUlkfBZI05LfbztojJvZrm5sR731rBOOlhSU7yyrIxGlgGU/Uc825/I9769ToJOxz/ABDJ4/HJyqThXAtcgvYiw9+68acehcwWMio6KCFFVFSOM6AABcL/ALH3o8OvD59PxUKCT/S4/wBh+Peut/Z1AqKiOMXbgtwFHJ+n+HvfDh16vr1Fjm858UaG/wBbldI4/rf3r7evCvWCepWCXxyG0gP05sCeeP8Aej731XPUWfIlCtmF3a1uOP6Xv/xX2kvr62223N3eELGoqT8ujrYeX925pvl2zY4zLMxpQCuT00Vu7YsZKgqWUl7ABVJ/wP0+v+t7DNl7icn35P004IXj3DqT94+7n7ubLbi7u7FghFa6fLpxpt00VSAx1qCB9V0gccfX2ILHfNj3T/cK4Q/a46incdi33aHMd/byAj0Q9OPjx+RC65YyjKRpZwDYjn/bezOVNMLGN1bH4Wr0X2L676NZ0ZVBFarToPN0db4+sAlolRnb6sB+L8En3GW57dfXMxfSadZrclc58rbTtkdnrUPiuc8Oi27p2Lk8LXeSGKVYwSQ+hvwb/g+2tquBYXGmbB6X897U3N2069o/UBHlnj0hK3cOSxNVCj+ZirDUebLb8+03MvNcbM1srdG/sr7E3VvCm83ERVgScj59DDt/s6paCCCI+ckoG8nJHH4vf3Bu6BjctMeB4ddMeS7v6TZ4tup3LjobcXlv4hTiWcAEgcA8EkX/ADx7JyxrRupJWE+GHXz6bcrtqjyiTFolZ5FICkem5+lj9R78deoFDTqksUEsZSVQa+fRdc719WUuTc0cLq8btbQOGubgcezFLwKgWTj0B7rl24a5JtBUdCnsTK5DDSQ0uQ9D6lAJPNiun1f63szsb8GTRGcdBPmHlh4YGnuFrIQehvyNcmRgSMkOpUWP+pJP09s7zOkkvYc9HPt3t01vama4Sg9COiFfIjbGbGQSrxyzNA0g1tEpIKj82Ht7ZrnSpjc9EvuRs88si3dlFgnJA6SmyN9y7VSlpKlmWR5EUmQEC5H0P9Pb13Zicl+NOkHL+/ts8aQS1BJyD0cnbu70ydNHK0kXKIdSsCwJW4+nsIXtnHOTDdrqX0OR1OW23UG4QCRWBNBj8ulvDXZcRLU08s0sBN2CkldIPPJ9gnfPbDatwtGvbRAkgBIoOJ6LL9tgv2famoJqUH29Dn1hu2mqHFCZAJwSGViBZjww94he4XK17YFjOhDJ5dQ9uW1Nsl+1tLUKcjowYANl+qn6f4/kXPuFMj8ukta5HSQytMaOvjq4vpIwDD8AexNts63Vm1rJ+HqpZ8HrjueplGOvGSsWi7lP9Tpub+97BbxNe6Xy1cV6eErWp8cZpnqt/uXvrM7AySwYfFpU0ayap5mg1uBqs7E2/p7yx5N9vrLerIPeSHVTArj9nUZ81e4fNTXJg25WKD0HSu2T8ltqb4wdNRVFQlDXgp5mlQRDURdv1cce00vtvfcr77+9Y4jNGoJUDOfLoa8i8/bbBADzBI0cpp8QoK/PpQZ/Bbe3vjKyGmyVBKRGZELVEau7EfRQL395T+y24b3vN8txuEDQAUWlCBjzz0DPvNc6ct3Pt+8EdxFLKQdIVgTw/b0Rbf8A1FUUFbNMsI8Ubs7tD6l8Y5vce8zkp4agenXFW71NeSynFWOOg73J1zhN77Nr8IJac3p3iCGRTMJGW36Pr7v0xnrXD+Q/SGX6g3jkaQ0lW2Fmqp2p6mSM+OItLcQmQ/jn0k+9db4YPRdvfut9dk3+v9OLf8b9+60BTh11791vr3v3Xuve/de65EgkW4Fre/daAp11e36SQP8Aff09+699vXhb83t/h791418uuwLk/wCtf/effuvE064n6n37rfXvfuvde9+691yX8/8ABT791o9eB4sfpe/H1v7916majr//0Pn/APv3Xuve/de697917rsAn6e/daqB12CSLAf776+/dewM9cffut9e9+691737r3XYuOR+PfutGnDrxN+ffutgU669+6912Dbn37rxFehN6j2ZU753xiMRDG0iJUxVNQixtIGjjlHpIX+vv3Wvs62luja7E9dYDBYyHHxwBaCmimMcZGp9IuTwLf4+9/PrfRnjiF3rNBk8eimJVUsfq2oc2IH59+6qOlguQyO2IBEiWkRQAtuOOR791alegb3pkt0b1nFD+5DSmSzMmrSRfm/vXWs8B0KGwul6SLGw1VVKGnIR7m99RU3LavfvPrdPMdLeTa0uKcimBeNbgEg/U/4+9k9e49MtZg8vXyCJS6xswuSG/SeDf37jw60elJjtkfw0CoRmMo0uVIa5b8/X37rdPI9N/YsbZfa1RhZiVYLI8TcXDW4It/t/fs9axQnorXWndVT1Tn5dmbgqZaqhrZAyzO9yg1XULq/A/wAPeh8+tk06NtD2Jga+H7yGrj0S+tA08Q4IuOWPvfWvOnTXUdnUWMPkgqIpDYlVWWM3N7fQE+/dePTnhe3Z8i3ianjMRI5NjwTb8X96pXh14nA6dN11FHmsSklTTRAidHBt9Bf+g/4n3sV63ivQlYI0/wDCaFKdkKJTIpCMCOByCR7rWnWxgUHSE7JwS5ahLLGBPGpIcA/j/W97+3rRp59IXY24anEOuPqGPpISzcDhja/v3z690NFbI9csTqLq6g+n8E/63vdKDr1T1MqjFR42XgK/hYXX6/S4596qa163QUp0SzsKrqKjISQuxCEswII54tb+vveeqmgHUbr3HibIIAuo6gx/rcG4+vvXXhXo6OAphFTxRmMKwUcC3Nxcke/HrYNelZcKvH+w4/3nj3rrfTZVV0UAJZ1HGo6m0gAG/wCffs9aqBjoDN+9yYna6SssjSSrdQkRVyXI9IsnPtwKioZJGCqMmpA/w9UJmdhFEjMWNBRSf8A6DDbNdv7uSpet+2mpdvxyFDJGrROR/Tm1/eNnuj94rYOTZDtu2OJJOByDnz6zE+7592K95+uk3fmusFqGBrQio+zoVKDryqo1eneWYgcanTUwC8/U+4D3T7w/1kAmt3IkPlXGeun3L/sT7RcsWixyWkU4Wg1MBU/Ph0zVu2aSjqilXIXYkhVaJbfX+h9mew8/3vMcBnimYMMkV6k3bfbn2vlhrabXAR60H+bqXT7b2vIymqx9LMpCiTXGl7E/X6X9jCx5o3W2lDvKxA+fRLv/ALJch7zaPbLt0UZYGhC9O03WvX9fHb7alp3YXPjQav8AC3uYeVOd5J2CTmvXPP32+7RbbfG82zoEpWgAPl0lqnpbasrs1NkJYhzYILCwP4t7lVd3tZQCpyesD7v283zb2bxUOhK1ND02jo+ESasfn69GB1Kiu4UN9bmx49mSnUoYefQLlQxytEfwmnXCqi7T2eQMZTnI0MFi80wZ2ZR9eT7uD0w6AjpQYPvmZWXH7jo5YJAVR9EbAA3sbXH+N/YK5l2GXchri49ZK+y/urY8mkW16BQkenkehKqsrtDc9EvhMAaaMs3mkRW1W/Aax9xHvfKU1pbtcSZI+XXQT209/dv5l3aLaLMrR8UqPX7egsy/XFDNqqKNwGt6fGdYOr6fT/D3GbSOjFW4A9ZlDabS6iVxhiPLoOcvsPKY0iop5ZT+k2QsAD/Tn24t0GXQwHRZcbHcwDxrVySPLrhT7rye3I1jqTIQOCT9bgfS5+nvXgLPlB1eHeJ9tTTcitehA2hvqHK+SKodbyXZNbrwCdJH/GvbE8JhFWHR3tu8RbmcEKfLPQiTrHOll0lJOVI5sv4PtCaOcdHcTMhr6dQoKc0pLRMSD9Qx/wAfrz+PeiWBqPLp0kOCG6doqv06WAufz/aB/wAD7eSUnB6SvBnUPLpuyW2KfchWCaITAqyhdJe9+BceyveN9s+XbN7+7YKRwFePSG7S2kjP1Qqo9eikb6643L1duBN04VJocVFKXqUSKRE/VqOq9h7Y5V9zbPmJTZysAzcACK06x+3pbfbN+F7bf2QOfTj+zoyWwd60e7MXBKkiGoaNDMutCysT9Prf/b+xVLDJAWY/CQafs6nPZtytdzsUntiMLkA16NHtHC0lRi3kOgSWY/Uajb6394F8+bpcQ803AodOvoD84BricK4rTh007hwKojMEUrYj/eOL/wBP8fe9n3YyECvUX39gVWqjHn0AuRpWxuSSRVVfX+pfpbV7k+1mF3aFSa46AF3F9LdBo+PRrdh7iEWEWbIToipHaIawLqPpcX9wNzfszSblps1JLHOOpT5f8fcbdYlWp6cW7Bx7SygyAqp4OoEW+n1/x9o4+Sd18NXSJu7+iehx/Vm48MUHHoIex8lBnAi0jmNrfrQcgnm3Hub/AGr5F3SW413yERLxqD0INk5diSrXQ/4roI6Dbsks8bTVDsQwvfVySbAgH3lLsmx221XQjtR8dOjTfYbbbtrlmRwgVSa8Bw6FbHbcoKOn1k65DY82Pq95Qcl7PLbRrc6aV+XXIT7ynuDZb1ezbaJdRQ6eNfI9JzcG2Dk4jEzFYiLMbDi/4X/ifcnGnWDqrThw6rL+avxyxO4evM1US0cc88cEqxStEWdAU5K8e98B1YfLrU83Phm2/uDL4ViWONrp6a5FiVjewJB/w9661Xpit9T/AE9+63Xrvkj/AAH+t791rAPXH37rfXfAHH5+v+Hv3Ws1z14fX37rfXfFzq4+n09+6rmmOujbi3+x9+62K+fXXv3W+uxbm/8AsPfutZ68Dbn37rfHrr37r3XdiLf7x7917rwtY3+v49+61muOvC354Hv3Wz8uuvfuvdd2+l/offuvddsb/wCt791oCnXH37rfXvfuvde9+691737r3Xvfuvde9+691zXVbj6fU/T37qp01z1w9+6t1737r3Xvfuvddk3N/fuvcOuvfuvde9+691737r3Xdzx/h791qnXvqef9j791vy679PNyf8PfutZ69b0g/wCPv3Xq5p146bC31/Pv3XhWueuPv3W+uzpsLfX8+/daFa567A+hP0/417914n0699CD+L/7xf8Ap7917iOumsSbe/deHDrw/P8Arf8AE+/de62oP5POGq6DYZyksZFNU4uIRSFDoJ8Q+je/Dj1oUp1arv8Aw1ZladjCvpBLN/Wwvf6e/dXPRYqihrcdW/5PC0jo3r1qfwfx73TrRpWnQtbL2xUbolJy5daWEBhHY6HsLlfeq9eyOgo7T6hoczW1EFLEyxKxEfjiJsAP7Nh7917y6DnqvZFZtTc600AqNUcp0l0dEN/6lve/LqoNDU9WD0VHO1HFNVKok0C4/oNPFwPz/T37HViOosdFP5mZbkte1+B/hz78evCvTtTpJT6lBvKxtccixH0uffqDrVSOmGtzFVR1LRKzeVmtcWuOeB79Xr3Hh0JeBqnyeOYVgBfTdL/UG3A5/wBh7qBQjq1cVHSLyFRPR1xhl1CLXawPptfnj3emMdaJNa9PIwFJWotUqrJ5P6fW/wDjb3WtDTrwz0zV21YIWdpIEZbAj0mxH4PPveD1rNOoWMrEoav7XT4orgRqBYX/AKfgc+9HrdD5dO+4cetVBFMy3ens408/X6fT377OvEVPUjD14lpApPqjBB/wK8D6e9dbrk9BtPRGt3T5ZL6VkBH4/N+fduA6qKcOhqhAREUfRVUH/AWtx7rk9W4DqBW1RjbQjc82te4P1Auf9697Arx60zEZ6esPgjURrWVmrQbGzfg34B9pZr62gbQ7AH7ej7buV953OH6m0iLL60PXDI1VDTzPDRLGZAvrYWJ/xI/H19uxypOtUz0WXljcbdJouRQjHSAzAk0vU6zq+pJIJ5P149vcWA9ekZNFJ6Z8Kk+RlPOpFlFyfqFX6k2947/eN5zj5a5Y+jjaks6lQB6n5dZm/cw2m0uOb5dwu0DCM1FR6Dp4zD7Z8sSVCxF6dhIzEpcn8i5/HvnHt03Mio0sM7jX8z59dONz5/savY3UCSpSgBoR1HfMbYyB+zLwUqkBUlSSMMf+DEezi2v+c9rXxbW5l+zU3USbltnI+9yMbqyhUtXyHHoMN0ZCnwNdF/DciKzU6t4lqEAsfwAT9PeQftL7mc6W8xhumaRBUnXU06KIvure3fPMZmllW1D4Gkf5uljgd6Co0RVDxQ20el5kZmBH9k395c8t+5W3buRbX4pJwxih6xR96PuZc08hf7ueTZGuLYVYkhjgH/N0IddRYrcdAiaIGZQLsXUkm3qAHsm5raZZzLZZDVpT58OpF9hYrI7clnvx0vHQMGxw48ei2b86uiSplq4V1jkhQptY/QC3uIL2W8jmLXHE+vl1n/y/t+xXe2iDZtOkDyNft4dAk2Pq8RKzRQEGOzD0EfQ2sR7aZllFGPVxaXNg5aFcjpZYDsGtgnSCqUBVYC30JsLD6/T2jnsgO9D0INs5plQ/T3K8cdDpitxx1kQleRUAQOSGW5P+3/HtA1Ym6F8LLcr4ikUPTgVp8gxqIyrLe2oWPP0P09tmr5PSmN1jAAoek9kMDE8xqAWDqwcAH8g/i3492SVoPgx1S4tLa+NbgVHT7ia0SKYnLeRBYE/0HH+8e7B3ZtTGvVfpYoECQLpA9OsuaxFJlqcxVMMcy82Lrf8AFvr7uXdO5DTpM9vb3KmO4WoPRPu2NhQxB3oICksQ16lQ8HV/Ufj2fbVfO+HNR1FfO3LsEQraLkeg8+i7YrunJbKyJwlYzckxh2uLAcAC/s9m2ZL+Hxk8uox233GvOXL47dc+eM/s6sT6b7Hxue27FFLKJZpIV/U6H688+0NwEtLXwSOHQy2SS63XfhuUTVDZ6EnE+fFZv+JUrFQ0wbSpAQj6i3/G/ePPuvs1tcWQv3H9p2nqQudtq/eG1i4hH6sYqT546OPtbNtmcZHMxtKoCsOLiyfX3gxzFtS7ZfGNMoc/z6iCxuvqFKyYZcU6ccrAZqfyXLeP1A3vyOSPZft0vhz6B59GQIU0p0zp4cljZY5pFVEVlOpgNVri9z7NWL2N6rxgkn0HTfhNM+gZr0TDsnaOCy+WlpJaOnlg9QZyoe5v6if9f3mT7OWu6bhbrfTkheABx1Iey8qbFZ2Dbjuca0GSSPLoqXaPTeMTHIduzPjaliTejRkYm/5KW/23vN/k/liSak19Asi4+IV/w9YKfeN94OSbRZNm5fAilUMCQRWteib7ppu2usFbK4jN5Ws0OxSCVpRGVXlRZj7maHZ9rhQfTwrEafhFOuaF5zXvt1evLNcvNGSaKxJFPz64bJ+ZW45aqq2v2FQwpJLAaeOY6QxZxZSSfa9FEY0jh0UXNwbt/FYUJ406AndnYW99qbvn3Bt55KnFSBphTM2qEKW1C4Bt72ek3TH2Bk9qfJnYmQwueoaeg3PRU8s4eGwkkkUXAuvP197rU9eI6pB3ttSu2duXJYGtjMb01TMICQQJKYyHwuNVvwLH/W9+69XpJ2Nr/j37r1RWnXXv3W+uwbe/daIr11791vr1r/737917rsAn6e/derTrr37r3Xvfuvde+h59+6912bX9P0/x9+60K0z11791vrvi3+Pv3Ws1669+631//9H5/wD7917r3v3Xuve/de67vx9Te/8AsPfutefXrEc+/deqD13a6lub/n37rVaGnXQt+ffutmvl11791vr3P09+6912QR9ffuvA14ddix+pt+OP+J9+6rkcOuPv3VurXP5cvTgzuYbflRF5IVkkpT5AdIWKYqpH45+vv3Va562ItudY0FdHFTJBE0jQgqxXlQR6f+Ne9/Z1vNK9PmJosr15mFpSobHOwdmP0H+A9+49b4dLWurqXcFWkUKqzuA1zyLE2J49++zrwPr1hzmOx+ExrTFYVn03BW17j6lb/wBPfuvVqfToEpO5sptjIRowkfHmVY2k+tgW0+q/496NOvZ8ujbbP3HjNyYSLIRyJKzqsjqWBK6vxYE/7z7914cOneSenV/2lQMObKPwTe//ABv3vy61ivUn+J07qbkAqpLFraeOOCfeurYr0WntndS4/wC4FKwaQpKunUOARw1x731qvRSNv7Nh3tnmr8oymbX6HYXZQxsLf6w9+619vQ4zfH/NT02ugyc4UC8caM6qVt6QLe/U62fl0loeqtxYqvEWQqZZQCbK7OQVH5N/futGtc9DFtfFjGKkZUM4ZRci5tf+nvXW8cKdGAmwqZPbVSP82yU8koIP5RTpAHP197+zrXDj0D3Te7qw12VwmTmYSx18sNMrt/utWstr+9Drda9GaCxVKMswV9Sstzcg3HPvwyade8q+fQI57bEsOVE9MNMZdmY2sOGuOR7917jw6EnAVSiJIGYFlVQWJvYgW597+zr3A56lZseSF4hySpCi9h/QW9+60fXonnZdLJRV13AVyGN+QLWJ9+rU9bPDp76eVKiuV7am03Fhb6H37rQOMdHDx6qhIH1/x/HH5PvRx1sZ6mVUojiZ9RBF/wDWt78AT1otpz0U7urstsHCaKjd2nncoPHy/IsFFufz7K923W22i2a5umoig5+dOhjyhy1LzFcjQK8P8PSf6566ptwUUW593TqGkInipJwH8iXvco/vAj3W96+Yd2v5Nq2UskQOnUppj7R10X9oPZfkzZNpG5b8qyy8QrCv+Hoya7229t2hjxuHpoaOCNV1fbhEDWHLEL7xmblvcd1umvdycyOxJ7s0r1Pn9Ytv2uAWe1L4UY8lwP2dNlV2/g44Sjn9wWuRovf/ABHtVDyNd69S8Pz6ST88I6aXc1HQI757V24zLK07pJqve4FuPrb3LPIHLu42O4AfgPHqSOTee7KOApI51eXSJpe3dtso8lXIGuALaeSefp7m19suQxIXHUn23Pu1kUZ9J+Z6WeJ7IwNUqmOrcnj6lPzxx/xv2L+XrKeOvbTqB/d3mbbrkdkoK5rn/Z6XNDuujq/81NdbcG68+5G2aG7W8XXUjrDj3G3HY5uWpfptIfNeFa06X+PyULorxS/S1yT9B+fcvpXwlHy6553LA3spXNW6VkOUp2XRKfMpHKXJQ8W5Un3vps4GemrIYbaWV1NNiaKKUhh5FhUk8Wub+/HhTqhUHIx0Du6enHrtVbt7KTR1CH9qljdkQgC+n0249lu6bZDulq1s2K9DPkbnPceRt6i3i0JcoR+ytT0lKTcG7tlK1NuanJpYdCRsqu8hQcatX9f6e8c+Y+QN1spmktFqhNanrsP7Qfey5G5p2y32/fJfDugKH4aV6Enb28cBumnZYmIk/SUqdKNc/Swf2AZ7eezfwrhaN9mOss9v3Sw3eAXe1TpKh8gQT+yvUPcWzKPL01QsYj1tcxsoUm9vpx7bileNqr1u+22C+iKSChPr0WnK7dzG0sgTF5NOsEaQ1rfXi3s2WSG5TTLg9R9dbdebRPWI4+XQmYHe8yLTLVM48aqrK1uSRYk39lk9rQkKMHz6Gm379riSKbB8z0LNBl6XIKvhe+qxJ4Fv9c+0DI6Du6E8Ukcy64jWnTk6g208lQbc35I4v71gZHTgNOPSw2CzLlY/uAGUyfRgP6g+8fffB5yEhRiFK1p5dEXNFPoSYsY8uhW7T2vTbt2jW4mnoomkqIm0kgn1FP8AD639wByPvMmyb6l5cSnSp4dQTuVtJcW7IwyfPqrCLam/eld0NFV0sjYWuqCS6q5Eaa9QCfge88dg572bmSyXQ4qgp0Qcn81XXK+7jb9wr4chx6cejtbE3lX1FJT1EJkeN012H1P5swHuK/cDlTlneHNzC4SY/wAz1kdcQbXvFqtzEwOoV+zpXZ7dNfUU8kKIdVr3/wBTccewhy/7T3TSLclh4fr0Uf1TsrlKE06AHc+5KpUZJIzqFwrn6gr6j9fY7tuRL21lIQVToAb/AO3ojk8SPI+zpD4XtLKx10eNrJpY6W4VPV/tXJ5/HscbZ7Y7e7ruMiB5FyVPSrl7ddn5VlWDde2uK+nRoNtikyVJHIJ1cyAsWDC9v8QP9h7HVlytaMSHgVafLo35h5+sLbTJYsGVuBr0qJcJEVVxpdV4ufx/hz7N4eXkjbw7ZAtfToKN7oLFEZXNB51x1Fko6akfyKwBAuR/r8n6+xDsXLU6bgkkq1APUXe6fvFYzcpXFvbygOQcgivDpvkzLlzHFyFI+um3AsfeSdlGsNokaimPLrjLzPfSblv1xdO5fU1c9ONFU1FW6pptZSSG+h/qb/4f4+36U6JtVeq6PnV3lQ7B2jlKNwrAQ1AmLkBVOghv1e/Vx1Xzr1qEb43Am59257PRJ448nkKioRf9odrA2/x+vv3XgMdJP37rfXvfuvde9+691737r3XYt/Wx/FvfutHrvmx4v/j+ffuvYr1x9+631737r3XK/wCLD/Xtz/t/futU8+urXPHPv3XvLPXXv3W+vcn37r3Xvp7917rlwQbgA8WsPr/X37qua9cfr791brs3+h9+69117917rsW/P09+699nXiLc/g/T/H37rQPXvwOP9j/X37rfXXv3XuvW/wB49+6912Db8A/6/v3WiK9d6iP6f6349+69pHXH37rfXvfuvde9+6912bX4+nv3Xvt669+6913a/wDsPfuvdde/de67AuQPfutE0FeuyACBf/X/AMPfuvAkivXRtfj6e/deHz669+631737r3Xvfuvdd2P19+61Xrq59+6313zxf/YX9+61jro8/wDGvfut9KvY+067fO7cBtLGm1bnshBQQGxazStyQB/gD791omnW7h8MOkE6N6L2xt+sQLmzBTiqjIIZI2jGk2P+9e/H069gHHRrpIUkiMRCnVe/4+v9QPehg46uaEZ6Rk2yaColaR0RSxPIHFgebH3bqgr0rsBhoKCNYIo1I/tt9PSOAePeq9WAp06ybQpauo87xJ4ib6rfgHm1/fgSB1qmesVR17gB5KikiiWttrDAclv9e3vVTXrx4dYaLF1yB4ZItTKNKqRcab/Q/n3smuR1sYFOnBMDUSxt9wnhLXFxcFR+Dx7914fLrKmJhpIrN69POrnX9eTf3rPHrdaY6SeSwtPW1YNKoaQsA2tTcNf6A8e99V+fSjxWGqaUrq0hY1UuFJ+v5t79XrdDXrDuPBCrpnqEA1IAb86vob/8bHvVc9aoK9JbbeXNDNJR1TvZS2kNYAG9hb+nvdOvVA6Ws1XTTqqFlcki4ve4tex/334964ZPWz0hcjT00tYhiVRIjBm03AUX492Hr1o9SknmDNA4LROAoJH1FrG59661506aqdmoq140Hoe4t+P639+68SPLrr7WFMmJzpViVNjxyT/T/X9+qOtgUpXz6Xn9i4+pXg/6wv711vpM1MwSrV3PCuGIP0+v0F/fmbRGWPTltC1xeRRAV1Hpyyu946KjaCA2LRhQOLa7fS3uLL3Xe7kzlqKmT9nWdnLQtOXeSo4/ADzTjSopmpGOkliq+aqDVM1/UbsSTdgfYy5e3W0v2a0tG1aBn/i+sVfcnYd02e7+v3VPDaVjRfQE+nXeaqg1FUE+gCMkWuOf63H+8+xPCpMynyBz1GcrAQsOFR0G2I3LJjTPBTPG8tTqQBnUaS3H194D/eRvIt75tO3kkpbZP29Z8/dZsp9p2dtzoAZqgH8uoma2jn8lSTVsNVAJ5FZgq1AP6jcDg/8AEe4Cst5262mFu6nSPl1k/d7VuE8BnVhqPz6KlvqTfe2FZkWZ5dbKBGXa1jwRb3LfLNhYcwy+FarqxU9Bu25a5gkf6m4qsYPHy6Ais3zvmOoNTWx1YYG9nD2AH0+v9Pc17TyTHZx0toxnzp0KYvcWw5XjFpPPQr8xx6Xu0+zc5UTRmcOCGUan/wABx9fZ7ZcnyrNrVdB9R0m3/wC8NYttclqZfFUqRpLY/ZXo1Ozt+5aR1ZpXMZUWsQbm1/8Ab+5HsdknJWNxqUUyesNeZfc/aY4p7uybwZjqoqUyehqpdwy5KC0y3kAW4b1XN+fr9fYN9wuVbiJfq7RKpxNOskvuhe+Gz7nMdi5huDHcNULqIAr5ceolVt7H5Uu4iWN3QqVsLEj8ke4UJcGnAjrpAYrdkrh1ccQaj9vQN7g2aMXJPUogJAJVQt9P9CPapJ3oI34HoNXuzRqWuoxgdBW+5chRTNAWkjjGpbglf9gR7MhaRMmM9A/98XsEhjqQBjoXdibyjjKU80zSeQ3tcFRfi9j/AE/PssuoGjyBQdDTYd1juUEUzdx6G1ZYaldaMG9Or9XNravZaAfxdDCjJjrGYViIdFAN7krf+vJv70zUwOrA149TY6m50tY2H1P0/wAf9f25EzGiHph4gBUdRqzZD7ognEMKySFCCbE8n6XI9gLnH3Bs+VJktoWrITkenRJvFzZQ2rfUippjquD5GdI5DbcMmbnpFjN5mjcK1yQ5t7H/ALfe5FrzBKLYtxA4dYo877Ul3qv4VAYVPSf+PvZK437Wgq3KzJIsXjFhco1ub+x7v23uxLr8Jz0d+2PNaQqtpc/HWn8+rU9k1sedNG8mkJL47AkEANbk+8fvdmR7flUtGCxXPWSt8Q+2mQcHXy+fRlcCFwuTEKvaCVFsASUJIAv/ALe/vCndmbddu8QjvB/PqBGgSC9fFASehIqZoyjR6gda8i4Iuf6ewJBG6v4lPhPS9Ii41EY6BTe81ViYJTDIyRSXtpJ+jH6+5S5VW23G5iM6hirCtfkej7l+L/HgpXUPn0Coppq5zLr8jyEgl+fx+fefnKUVtHZQGyQKopgCnQ+5meBdlmtpOwFPLrDltuwy08ayopbWWPFjwf6n3mNyhcs+3AMPIf4OuDn3g9tjtub5Xjao1N/x49Ft7i2xja7Gz02lQyI9x9LEJ/h7FHzHUEgYp1UB2Hso1G+Y6OmhVZRUxqJVFyQWsLf8b9+OevfZ0qeyds5XZm0qeqqKZpklhVSzIxJ4tb349eHRRduZytxuakyFDTuhkciZVSwKE83sPeh1sj9vRaPlLgP4tWJviliEaEJBOEHP1sWYf4X9761TNeiXAn6fW/HP+P59+69TrwAvY8e/deJxUdde/db67IAPBv791oV8+vX4tx/r/k/4H37r1M168L34+v8AtvfuvGnn11791vr3v3Xuve/de674t/jf6f4e/de8+uwLg/W4/wBtb37rRND11xb/AB9+69mvXhb8+/deNfLr/9LQAIt791oGvXiLW/xF/fuvdde/db697917r1z/AF9+691737r3XIWHP1/Fv8PfutGpx1x9+631737r3XYBP/G/futcOvW4v/jb37rfy65xRPNLFDGNUkrrGij6lnbSo9+61XrZc/l8bNi2x1EI6qMRzytBIhIALarMWP5/1/8AH37rXDq3fruoEldALEgRqoI/IAt735dW49OHauKq6qAzUsZYqtiyi545H0968qjrwz0CGCzUmFnZJxJ94BpTUDxzcW/1vfuvefWeufNbqq0p283jeQcLwuhj/vPvx60RU0HS8zfTuDXZ9QZVWSranklLSEFlk0XPP9Qfe8de6Anp7JV+1s7U4L71pqZ6iypJJcIC3CgEn/W96p6db+R6O3RUSCD7ieoiGtdYGsfQ/wCP4978utdB1vTc1Di0elgqoPIRY+NwxJ+nPv329ex5dE87ay0sODmybyEyvIyIGPJ1Gwt/xT3rrwA6xdO0lW2MOSmL+VyjpfnhiD+f9f37z69wFB0azD7zyFJCsIdmCgAkm9h+R/re99ex17L5j+IMJXH7pHPHP9Cfx79XrxHXHA0MlXVKpW49LC315Nv98PfuvYp0PLxpj8JPGW0lqdvqwtyv0ufevs62OFG6rL3xv2p2Tvw1FE/jSStAYqRZmZ7W/wBf/E+906qOrBNg7iqsztiiysq3kngifi/9sfq49+PV8dKmsjjyFMWFhIFJPFje1hb/AGP+9+9dawekbQxTUVQ5YsV1cgfn+oPvfDrWRnpXeYVkIFgCgvcD+n1+vvXW89F+7nxhfFPWogJiLAsLFrKvNvfqU60T0HXRmbjXLLTkgkCxDGxvq/Hv3Xhwx0eOikVizLYgi9uBz/iR78erCvWHKnXBUAMFtTzNyfoQl/x7o8kcS65MDpRaWdzuEwtrVdRJ6rmq6psx2latQ1tNSTOnhYa4iysQDb3jD788wztH+7rJ9EbCpIP8uszfYzk8bQonv0q/Gn59GJjgqqpRLPN9hQwemOFH8aiMcABePeHbSRxHRGviSN58c9ZZFC8fiSN4cS+VaUHTVWT7fiR9NRPPIt1IVWZbn6c+z3bNi5i3Nv8AFocfs6Cm5838q7cCksxLD8+g5yVDG1SJqaKpkV7sdUT8X/wHsb23KPMhj0vDkfMdAq45+5bMmJDT7OkRuTZ5zRRkhqEA4IMbGwtybexXy3y/vljeKbuPTH5mvR7s3unyrYAmWQk+Qp0wr1dT+IErU69NrCNxYn829zZt77clPHpQevQP5w93ZrtH/cbNXy8uk/NtGvwVRqieq0W4BBAuP8D9PY728cuykeG4B6xs3v3D5+nSRbsMVPDjw6XO1M/PQuErJWVLgHUSG+vN/Yzhs7MUkiANOFOog3Lmne7wNBO7AE5B4dGTwGapp4InFQNLDgBuD+eQPz/W/tbXFOg5g9x4npcQ1rqLqSRbg3/Bv+f6n3rr2fPqUuVdQAb8Wv8A7wCLD37r2On3H5MkhopWDNbgEiwJ5+vvY9evNkU8uleabE5qIpkaCnqjpKmSdVZhdeSP9b8H35tEg0TKHX0PWoWurVxLYymBgeK1HQQbg6QoxUNlcDVzCoF5EooHMaAj1DgewnvvJu1b1EVVRESPLj1Pfth947nz22u1Mc73EdRXWSRTzx0GMu6d37EnlTc1G6Y6NtCMkZkm0Aggkm1/9v7gnfOQNx2ti1qhZQeJ66m+2H3sOUueIIbbeJVjunUV4ABjjoSqObBb9oIa+DxsDGGKS6RJcj8p9fYCZZI2aOSqsvHy6ygjk2/c7dLiB1mR+BBqOgv3JsSqSSSShhJEZNtH0Kg3tx9fbsVwRhsjouv9iLqZ7fFPLpMUGcrsLqhqUkREKqxA54NvdnhWVqx5PSGy3iXaf0pzT7T0J22d2x12katQBsTIRf8Apxf3SLbJ5XJ046NJOctsQhLlgpbhnofNiRpU5OBlIN2vcc8XuLf737xr9+HNvJHH/R6c3m6in24yxmqkY6NJBSosaekMQBxx+R9PeIEk7MxpjqKJJasR5dJjeWwcNu7Gyx5GjgeVI3MUjIpYWW4sR7P+W+bdx2C8U28h0Eior8+g5u21W24RkEAOBg+fRNdl19LsrcuX25mI9EclTIlBqU+NYwSUsT/sPeRe/wAFzzDtEG6be5qAC1DmuOgvypzdf7Vu37pvXOjVRa+nS2yE0U1ROYwDGbaNJ4F/999Pc68jSPJy7FHI9XUUNTnrLHbyJLZJQagivSFzO2Ya6J2sAzcg2v8AX8i/sd2FhcXbBUrToj5s5q2vYrFpr1sj/N0Bm4dntFK7ooARv1L+oED8e5G2zY7rbtMsgqp9esOebfcvl7nFJrOxk0zITQjHDqFhN4bl2zUJDGJZKZLDU+o21H+n+w9jFNlt7yLXFhz5DrHa79y965dvfp9wLNboeJzjof8AF9r1VXSxxsLvpGoaLWJHsy2PlpknLXC4HQT9zPeu1u9oWDZpSHYeWOn2Gvy+YIZtSRH1X5Clb+x3FtlrCwcKMdYpX3Ou/blA0E8zEN8z1MAjo5AJJPWzC4+vswxwHQUq2S2SePSuxtSJI5UjHr0Gxv8A4e/U69iletc/+bXuLcizGgoKcvj5GkjrnVSdCG6yMbf4E+9Gla9eJ8utf7/iPfuvdcrDjn6/X/Dj37rVT14r9bcgfn37r1fXrogD6G/v3XgT59evwR/W3v3XqdeJBAt+P8PfuvAdde/db697917rsC5t7914468RY29+68DXPXYawIt9ffuqkVPXRBFj/Xn37rYPXXv3W+uwCf6f7Egf737917rtmv8Ajkfm9/futAU66A+pva3/ABPv3Xj6deN/z/vYPv3XuuQbSLWvf/H/AIp791oipr1w9+6t12Qf6j/bg/Tj8e/da67J4Uf0ufqD9Tf8e/depx67NiC1+T+PfutCox1xBsCP6+/dbp14ngC30v8A7H37r1M9etwD/X37rdeuvfuvde9+691737r3XYtfn6e/daPDHXR+pt791vrv6Ac/X68+/da8+vAE/T8e/deJp11791vr3v3Xuve/de697917rtfqPfutHh15vqffuvDh12SCAP6f8U9+68BmvXiBYEfXj8/4f09+68Ca9cffut9eHv3XurNf5X/UWC7K7zxuVy/M21srj6uhjLekuyuzXX6H6W+nvx4daHxaT1uN5NAtdGkahEFNTxrEAAAViC3AHv3W6U4dRqilqQvlp/W/BcAji3HH+29+62fl01zZGajCGvBih/tPwLX+tx79TrVenGlz+M/a8NQWQkWNwCbDn6f7171mvW/Pp4rN2KqrDS20AAE2I+o/HvfqT14gDpqTdtJQuZKycJYarM1tQ/ob+9Urx699nSwxu4sRlIo6iGoRQ5ALFgLfg8jnn36h61516dZaykKspqEKsLKdf+H9ffvs62Rip6StVn8ZjpXWoqI2AItdhZr/AEAI/wAPfuvVp01y5XH1/wC9RuiMPUGjNjf/AAv73TPXgfPpzxuZ4MMza3BADHm9ha5I96OOvYPTrJkoGikDsgBU3XV9Dbi4Pv1KnrwxnoNslS43JTFaeZI6jV9EPPB5t9Pe/l1r59M7/d4RmepctD+G1gkqDYe/HrY9D0lKXclM24HDSnxSWVBe41f097z1oj14eXQsQpFP6w/FgfV/jz+feqdb6Y61F+5BUglT/ZII/p9fe6Yr1quemjMyzUjUkqjUGcayDyADaxt71SnXtVeliMjSLQrPNOka6AOXANyORz/vPvfaDViAB69VqzdiAkn0HQW5zsHaFDNIlXkNJj/KMptzb2CeZN5eKMw2x4+nWR3s7ybsd1eJf70wBBrRukRRdhbIzOUNOciQuoKjMygFj+Tz/vPvH/n3mjdtj2SV7BS0sqkCnr/m66I8kbJ7cGePcN6nj0QkFEJFKj5dK+l7Q6+lzkm2aLIp56GFWlcuuluOTqvz7G/3dLl5+Wjc7wdN45OoHFB1z5+9jup3v3All2xALFSQgTh8ukT2R3Bs/E0NZjqDIRy5F4iiIHGnU30PBP8Ah7GXOPuRHy+DZbWDJO2OFQPz6Bftr7PXvPd5Ebxljgb1NDTHHojVXuPfVfNNVQSyQxeR3jMUh1aSbqy2/wAPcNxcqW3NV6+67yaSzZb/AFHrNq45RX242GLatpmV9Ir2tXP5dKfbGU7eqpmegq66pQAEpJK2kccAD36+9quTQtHIB+wdRjfc4812DGVXb7K9LCv3n2Hi4G/jG3oq2SMcPNH5efqfr7Vcr8obBy1dtc2T6tWM9KLr3955k2UbI8YVBjVTP7egqy3b+In/AMl3ZioMaZHMYaKnAtzxcj3OOzbptMarGygnrF/nW/5p3q4a5E7rXNAepNHS4Kvp4q/CzI8LHVZDci9/qB/h7GSC0kXVEgp69RjLdb3G3h3Fw9ftPQtbOyIjlWGMhtJQtfgi3193AVfhFOkzPLKaysT9vRh8NkI5VAX0MAACoOq/5/31vfnWKVDFcKHU+uetwXF/t9yl9tc7W8qGoKEgn9nSshqvC4kZip+mtrhSPrcj3EHN3tql0zX2zr3UqVHXQ37vv30rrYIoeVvcJybfUAJGy3CnE9T2ejyWqnqEDuyGzaBo0g/TV7gW/wBuutvuDBdqUYddTeWOati5w2hN22CZZ7d6YqNQqK5FegE7C2GsEj1NPGCJHLWX8avpa3Ht6zn8MaHNeirf9kRv8Zg4HoNMPSTYio80uoLc6b3uG9q5mEy6eg7YQzbfN4rYHQv4Hcz+VQJ3YPpUg/63P19lLw0FKdD7bN4DvRzqJoKdDDS1EUqK2r1OASD+bj6c+0OA2ehQ6sRqAxTr0qF7CMkXupP0Iv8An21cS/T20k/8K1/Z1ZSB8fRg+sIPtaXXPGGRvoWAJNx9feCvuTuUu8bo9xr7qkfsPUU83Tl7nwE4DoCflRtDKbswVQtDj1elhilZiF/2q4uPY/8AZrerPablVnl/UJGK9Q5zT4sVqXUYAz1Rl/lOz9/uJxJSiOsddDAqupW/ofedtpvybhYrHPlSuCOsf9o5tt7LmcCVtADcPz6ti6Zz1dlcZjauGSQhDGvoP4Cg3YD/AG3uMeaL/YHhm268IIdSAG6z65M3uy3fZELuGBWg/Z0aTIb2q8dFFNOrkwaWZuS9lAHPvGxPa++uGdrFNcL10kH1r0Xbvygl8fqLTiOIHXKj7Rqc3KJKKU+kldIPII4Ooezba/YlXsyb3tkJ4Do0s+X7ZLEPOAKcepuXzmRzFIaaoQt+QSt72H4v7EvL3smmzbkt65JRTWnr16zi2uzuPERxUHpO4RZoKl1dWCq91uOLW+lveSHLO3Ga+S1t14UFOgp7wcy2GwcpSbnM4Haaf4elhXwJWQ2WwIX9X0bj63/1/eVOxbc23Wyxt5gf4OuFPujzdBzdvck8WaM3+HoBewdkGvoKqcSMJND6VBvcW4P+Ps94nqL/ACp0Qal6hrsh2GtTJTM1OtWpMjr/AKlvwfp70evDGOhc7s69hymFp8R9hFJDHAg1aQ3rt+ffvt63jom+K6DpoJ65pMfEkXikOrQBb/Yke/AdazTomvaHWCT5ncGEnow+Jho6qaJSoKGQJcWB/wB49+NK9bGR1TjnsdJiMxksfIhjNNWTogIt+2shEZH+FvfutD59NABJ/wCK+/deJp17i3+N/wDePfuvefXZ9IKkfXm/v3WhnI64+/dW67Jub/T37rQFBTrr37rfXvrx7917rsi3Hv3XuPXXv3Xuve/de65aTbV+PfutVFaddcWv+b/T/D37r3n1/9PQAII+vv3Xga8OvAX+nv3XuHXXv3Xuu+Lf43/3j37r3n117917r3v3Xuuybm5/3j37rQFOuvfut9e9+6912Bf6e/daJp11791voaOjdkf323rR0jRNJBSSQVDHTcCRZgUv/tiffvs619vWyN0bD/d/alJjo1ZCPFHoUWNuByB/sPfvLrxpwHVnHW2EkhpaWulDqzxRuB+BcXt79jrYqRToYqikjqoXSoiR1dSov6ravzb345x1unQWVvWtDVVEk/iUOz3BVb2F/oP+J97r1rp4xu08Rt1Tka9kjjgViWlKoo0/8G96B8+vUznoHOxO2462Cpxm3T5VWOWnJU3H10jke/U68SK46JFUVW4du5ebPM7iRnLhLnSLG/Pv3nXrRHUnLfJzfU1JJQUqEsisispINgPT/vfvfW/l0GO2+y87X5r7nc2QqViMhYq7NYc/QA+9Up16hp0pN57pq9/1dDt3DozY/wAkBknsSbgi+o/T3v7etDo4uy8VS4bb1JSIAXSkiSSwBbUo5uP639+zTr2mh6fUSN2JLaTqXSALE/19+8+vfLp+o8bNXTRwU41FrDlbi39Tf37r329DRt7bkWHRKipf+wGIPHqXkix964nq3SO7I3jFTU0sUE5RdJuVNr+n6X/r73jqpz1WF2F5dybrg4Ig+9hZnsb8Sckn/b+/cD14U4dWodURw0mycPSRuJCtJCpudXOnm/1+nv3W60wehKjoGX91f0m91H+9X/23vVfLr3z6TldA0JbQA2tiS3AYAn6f63vf2dePHPWSjR0S3HI/r+Cfp72BXqvn0GHZEbz4qel0oyv5SwNrXKG1re9Hj1atPn0TLC5uTZW4zKCqBp+dR0gAtyffuq1Hr0e7avZWAyWPp5Pu4llMQ8g8qqdWkf1/x97Abj1slSMnpN9jdv4TbuHqVoZhV5OdJIYYon8pPlXQpsv+Psr3p7azsHvL5wkaqfOhrToX8iXMq73HBaxmRyV8qileg86W6+lgoMtvzeEBiWqLVlIJ10MwkbUqgN/sPfNb3W57k3/fhte2NrbUVxnFeuonJPLsG17GN53RQg0as4zxx0/UeJzfZWeePFpNTYyGfxqIFbQ6A6bm39fch8kch2u1bem6b7mVhqVW8/t6g3nbn3cOYdwk2/ZyUhQ0JX5dHP63+MlEKaKXK0wqGcrfzpq1XAJuD/vHsX3O7wQdllGIx8ugnb7NJMtbti5PmejI0Hxh2jJGhbEQcgFh4QFH4t/h7K35ivAfi4dG0fLtrTKDqXP8W9vKL02DpmOk2vCDfi/APtscxXLgh3PW/wCr1rWhjHQf5j46Y+idjU4OmiuPSFiA9J/J49qId3d/xkjpLJsUMZqq06BbenxyoqmKSogxqLCFcEiL6Ej08W9m1pvEsTDS5B6QXWyQSoSyAinVfnbPSeQ2cJcpRLM0KuxZNDAKAuq1vcq8qc6yJOLa6NV9Seok5t5Hikt2ubRaMPIDoMdobpqQyU0jkeNtLXNtNja1j7mqOaK4QSwmobPUHTWtzZuYrhaEfLozu3cnDUwqGlHkKi124B0gkH27Q9Mg4oOlJNp+o5+l7fQG/wCL+9deoeoTVclIQVH1I/x+nPPv1D17y6VWG3GFdVqGCc3+o5B+h/3v37r2aVPQlUeQjlVWDmxGq4a4t/vfv1RXHXqYz1jzOGwm4qKSjytJBKsqkCWRQXBIKgj3V44pkKTqHHz6ftr/AHLbphc7bM0LqQRp+WeitZ3rzM9c5CbM7Yqqiuxpl8ssblmihBuTGEv9B7ivnH29hv4muttFGHkB1nR93b72+6cqXsWw83PrtmwS7VNCRkV6E3r3dWJ3ZBJHVaY6qmslTHKugBz/AKlW5I9wsOXbmCUxXQIIPp10sm919m3Pbk3PYZUeKQV+IE/n0+bk62xGZDSRRoosxLIBZza4NvYs2DlOe5m0lTQ+dOsfPdX3523YbDxFkUS+gYdFv3RtjI7QqlnxSySQiQ3FjYANc8fi3sx3qwg5erE9KnoN+2PNe7+65W9siSsdCaVPA9DN01veBqyGOtmWOqLBdDuASb/gH3hf77bNc7i63cakpp4gdZiibw9jFrKf1IxQg+vR9MdWJUwRShl0MAfqLEFfr7wovbV4JWQg46A5bVWvHp4dtaMBfkFbf1uLH6eyxBpcHzr0yVoCD0V/tXqyPIJV56mXxV0J8iMim5C83uPc78kc7+E0e1TGqEUOegBv3L2sncYcOmRT5dA1tfMU8NJWUGSlIr4XEcYlJEj24uobn3M1tuW4bVuUU9sT9K/x04DqVvbfnVNxtxtd84V4sCpoT0uaJf4jEacWLc6Txc3Fvx7yw5VvLaa1S8te4EDoAe6Lfve6ayRyEzX06SmfwBo1cyRhwxFwQb8/Qi/uY9ugut6hBK0VR6dYC8471sPtruckcU2u4mbAr69BtUbeSoct4Us97DRYf6/tJKr7Zeqpaig9CKxltueeWZJPCVpSpIxnh09YXbcFMw/bSw03FgbH629ydYzRz2waI9YQc0bfebXvEtvdBgKkAEEDj0IRrEpIhFCBqCBQP7P9PaofPoiwRUdNtLHLX1XlnYWXm1+ePx71506tQ06EDCT0lI7CZxYowFrA8iwH+w976rSvVTfzr6Qk3tis3WpStVRuZ2DeIyWXST/vPPvR8uvcetVjd+Ebbm5s1hCCP4dXz04BvqCo/AN/6D37rw6Thtxb/Y+/dez11791vr3v3Xuu78Ef19+61Trr37rfXYNjf37rRFRTrxN+bW9+68Mdde/db67FvyT791o18upENJVVAY09LPUBP1NBFLKq3F7MYwbf7H37r3HqR/Ccoq62xmQEdr6jR1IW39dRW3v3WzWmOoZVlIEikWJXRyHBtwCDz791r7OtlT+Sd/I22d/Mmo9/1PePYuY6fahxtLWdc+GONf70SShdauZyo0gG/pJNrWB+nv3A068pXTWnVJ/zT+Pg+LfyY7a6LhqK2tx3Xu68jgsfkqyJ0ORgpJDGlUjsLMGt9R78M562RpPz6Fz+Xz/L67l+fPauP2Z13gamp2jjsnTpvnciusVJt+hMInllqahuI7x+oOxsLe/Dqpxnq7ve/wDLp/kGdD5yv6275+c3adB2hg5Vo9xUO08QmZw2Or1ULNDT1sCssgV7gseOPda9PUUYPH7eil/zDv5NeB6Q6d2t8nvh3vqXuv475zHS5aq3TWZCkTKUeOddUEtTQggxsoB1oRc/j3sHptsGnRLf5cX8s3t/59b0qanb2Jq6LqHZtSKjsze5Jgo9u4aKMS1VU1VJZAVU3HPJFvfvl16mKk9XFbh/l6/8J8Ni7rqeq93fPDts9l0UiUdf/DcElRtykr2GgwR5JV0PZ+Ddvpz79ny6t2gVNP29ES/mb/yfMl8NNm7D7q6j3EOw+juxqObJbY3ScjTVFTW4r9UdTPBGbxuoA1If6/4e9jOOquCmW4dUVEc8D/Wv/h9ffutdeH10/wC3/wBh/T37rx4V66IsSPfuvA1Feuvfut9e9+691yuCOeLDj/H37rVCDjrj791vr3049+691737r3XdiLf4/T37r1evAEmw+vv3XiaZPXfIBH+sT/xHv3WsHPXRFgP8Rf37rwNeuvfut9e9+6912LX5+nv3Xj8uvG34+nv3Xvt669+6912bW4vf8/09+61nrr37rfXgbc+/de49XDfygRP/AKXspLGrFIqrHPKQCQF8UlySPpb37y6rWjDrbC3BVpPKlTTkBTBDpa4Iusf9B7sBjrZ6Qs26Z8O5kkbWosfUTb/D6e61rjqwx0Du+u7yZ46MwRCN7RlgwuOeD79TyHWsdYNkbrXKZNYRIWi1LZLkgXNxYe/VqevVz0PU63lXSBpsp/pz9be99a4nosfbu4shipJikmmFbcagPqbkj3rr2QKdAPH8jKnFU8WOp6lxLExDes3Bva3HvfWwPLpRbi+U1TjcHh1jq3aprZPG58h1AHi/Hv1Rw61w6Qe6Pk6II6CGSsledmUznWW9JFz7117Pl0t8J8ssBBSUsAq38nAbUxFzb37rQqeh/wBq/IDaGZpVK5CNK7gaZJ1QXI9P6v8Aefe/t62PXpTnsZpfKVq6dxIhC6ahGAFrA2X37j1v59JiXf0GNkeplrUEjA2/fUAE/Ww9+p16vl6dcKLs0ZlpKSSojlRhouzj0BuRYe/daBB6e8bisX9xBW/dgyu4KqXFuT/T349b4nobEZ3oogHjT0KSVkUsABYG/vX29a4cOkBkMnkMDXqZitRRVDqqSF9TWvz9Pfvs63jz49LBJaStpbOGllmW0KorNZ25B49l27btbbLaG4uSB6dGmybPdb7ei3gU6fM06g1fTe9N20gSJqyjpuWRoUYEg/Tj3A+7e4l5d3DCM6UHCh6yA2r22sbSFSRqanmOgyy3w/ztQzNWVuRDSf2ikh4/oT7JBzO0ranbUft6PRyzPAumBzGPljoO8p8Ls/Tsaigy2QjZLNqQODxyL39uHfbC6XTeRK4+eemX2jflH6d3IFHlU9JGb4nZzGmXKfxbJR1rqRJOA4d7f6pvau13mG1JWwAiHouOkVzy89yNd6TKw9c9Ajvr47bgWKXIjKZCWS1hIQ+oX+p5/wBb2tt7uxnm8S6QO3qePTXh7pt8YFhK0IXhpNOiyZLP7+6yqBFIlRkMfG58klQrGyA2Nr+xHCLWZP0qKT6dUj545n2+YNcSNMg46zXHQx7J+VNDSikpvDDFIXjWW3B4+t/9b2nl2PxgWcmvQtt+adu39w0raW+3HRuMF2TgN7qI5YqaTzxKFOpCdTDg+yG52qa0GqM4HHpff7XbyxeJCAQR5dBN271fhsjQVDpBCrPFrilQLrDNzcEe72N3IsnDoBbhtxUFNNfy6Kv1tW121dyVm366oeWlvpg8rWW59IC39yjy3uhJ8GU4+fUUcy7SGTXEvd8vt6M1imloa5ShP7gBFv0lW5sfY3qGyOgG8UsJ0yih6MdtKohdIzJKBI2kEXH1+vveOqdCsfGyKD6gwFjYe9qzIaqem5YY5h3DI4HzHzHUGWdqI3CmRQv9baf8b+wrzHyftnMUDHSI5s0IHE9Tx7N/eJ549nd0jEErXFkSA6uagKMHH2dO0KwZiitVsjggkeNQ1hay3940cwcsbhsF2yXCELXBPn12o9qPeTlL3d2OK82aZTMVDOhYVB86DoM90bNZUllhjJiXUy6VvyPzYeyKOYh89SLuG0CaItHinQSQ1MuHqtZQ2VjwwP4PPtY6GVe3oHwytts2p+Hz6EzCbxWtqI1QgMABpvcEW+gH+9e0p2+Z28NRk9Cn+t9naweJO4AA8z0NOHcV01MrW1SMtwBe1z+B7CvPMp2PYZnlxVSB5dGllvdpudo1xamtB0bTa2JFHjYrLYMAbfQ2t/T3zx3y/NxePnzP+HqMN4uxPdsfPp/yOEpcvQTUc9OkiSx6DqHLX/w9lu37tPtt4tzE5Gk16DV9BHdxtBIAQ3VEnzY6X/ge5jlsNSvEsda9RKYoza2oM30HvP32n5s/fGyp9QasVA49Ybe6/K8ljuP1VgClGqaenQ//AA/3RgZ8BFR10qtU06FNDNzrUfXSf6ewj7sbduLN41pUVzUdS37N853At4tuaUnOk1PRp8zLS5GSZFRTG6yJpsCLE2Hsae2Aul5YH1LamrxPHrOLb6taq4OGAr+zoJoYa/aWY+7hhZ6B3JYEHSFPLC/uWNnt3uJtMvQG573X9y2DG3kpX1NOjAbfzuKz9OJomjDJFZ01BTr/ALWlT7Gf7sErCELx6x0PPNxZxyXcs3alT8Xp1JqDSjW9OBf6ekXNxxyfYs5e5XuNuvluNPGnl1AHu5727XzZyu+0mfKBhTV/k6T8eRlFQEueTbT+LfT/AJF7msH9MBuua82k3UkkWQWP+HqZk0++ptEthFq9TA2JWwLKR711U/LovXYu6NtbNEaQfaRVuoF3eRFYEn8k+/de4dJ3G782vvCJIJ6qmlqCFVUjmRySRcCy/wCP09+691ly+1aaSCaOiekFZJGT4WljRtBHp9J97695dEi7I6zqkbPNJQf5R9lVaWWPVrLrcAH8j3rz68D1rad74mtxHYuXpa2m+1k1llj0FAV8jeoA/X34darnPQN+/db67txf/G3+8e/da86dde/db697917r3v3Xuve/de697917r3v3Xuve/de697917rsm/v3WgKdde/db6//U0ADf8/73f37rQp5deF7j6i9vp/Q+/dePDrxFj/vPPv3XhkdePPNh/sPfuvDGOuvfut9crjSP6j/D/H8+/dVoa9dE3/p/sPfurdde/de67Bt+Af8AX9+6916/5HH+t7917rr37r3VkfwW2dT5KtqMrILMJuW0t/Ybi5HHHv3Wq8R1f50Psk5mYSugMEEvIKsV9L2BJH+t735dep59WE/c0W38fElgPFEAE/A0WBvq/wB496p1uo8uk3J2LQQMxqHVYxq+rKBx/g3v3Hr1ep9B2Hga9GeOpU+PkhJIyfT/AID36nW+gU39urLb2eTC4YSRYyN2M1SNSvZT6gbEce/da+LHQMyY6mxcv2MJEsn1kc/qL/RgT/X+vvZ61wNOg73tS6qeS9yRcBRz9RY396690F209jTbgyYTxERM3qIRgdN7H37r3y6XnYfRcOJoYqzFl5piiPIoR7X+pHHveade6Qe3q2h23xUQeKqisGZVKksOCQLe9fLr3DI6Eal7nfHIyU8Lz/01oSL/AFH4t73U9e4Z64Uvbu5shXipTGslIr2b9tuR+bG3v2adeqOjNdfdmU1TJAKqB45AFBfQQFb6EXt/vfv3Dh17oZMrvfGzwiFKoqSpIBkQAG3Nr+9YrjrdfI9ApuL7TLeTyVaeE3Oryx6if6fX/ife+PWiB0U7sVcbineal1vJCTIrRLcEoeCbf6/vXn17yx1G2B8vsptyFsTPTeRIH8as2n6Rm1iG+nvx9B1vHn0OOK+bReaKnrqbxwMwBZfGQAx/2PvfXqdGF2x3Fit5LCVkVPOupPXHcC3INj/tvfutEAdCNLmIKenMkcyNpW4/dj/r/r+/DOB15u0VPRa+zu0IqBJYmkjvaQC0ifkf1HssvtwFk3dw6GfLPKjcxjsqD9h6IPvrsL76q10JWSoMgVFWzsXb/Bfrz7I7vm+ysIjNOaDoV3Xs7var4kKlvyPRmOiOpO196Y5MrVU8lJjZP82w8iHxnm9uD/j7gLn370e1cpzfQWWl2PqRx6kfkj7rm+czRfU7grRLX5jo6XWvxepYcxHmNyVMlc0TqwpKhXdAVa97OLX/AKD3ib7q/eb3vmWz+ktP01cH4Tg16y09q/u07FydcjcLs+M6EGjD0+3ped3tT0NPQ7PxcYpVqFUKiekaFFrBP8P8B7DHsDtzb/zB+9dyPiUJrXPR778b8Ns2VdnsqR6vIHypw6G748ddxUeNpWMKid4g19LAvJe9zf8APvL7fbs+J4Smqx4HyHWMGw2nhxiVhlsn5nqxTaOyWipvuaosqr6kBDEWtwB/xHsF3V1U6V6GltBq7vLoRqanjQrCguQAAfp+b+y8sTlulaooJ6fRTaQPrYfgH8H+p9t1PT40DphzOMp65bTqpsLAnm4HA/r7djkaP4emJIg/HoL8jtqBvJB6DC1wQxP+ta3tckzkV6RPCD2joonenVdFV00kEUaypNDK2kKzKGMf9PYi229eoZsU6Du6beugqfPqivsXb9Z1hv8Amx1bG0eOrqguJNLgRh2uBdh7n3lTmcxWgExqAPl1FN9yPFzFK1pEKSscdCjtuqk0081PPrimAdSrA8E/m17exfDzbaXDlUPA/LpFuP3feYdrtPqJkNCKjB6HvGVK1NOizABwo5HOq30Nx7EVtfRXYrGeof37li/5efTdKQPsPUypxyuCb/2ebXuP6e1gBrToNa1pWh6YZYHjOq9lS+n8/wCtc+96SOPXtRA4HpW4DKSIn7kh1ILKCeCv+t7rwPVsU6WS5N20kvbgkqDwLfge/Y69xz060OmvRlqFE1MbiWJ/0G4/tD8/7b3YPpwPPpto1c6vxDgfPoHOx9hVGCQ7r2iCskLK81HACqSAeptaqOeOfYc3bYba8HjRKA3p69TL7e+7O98st+7rmVmgatCTw+QHQhdfbri3NhoXlfx1lPEqVcJKgiQjSQoJv9fZhtVlHZwUIGoDoJ8/80X2/wC4a2ctGzinn59Yd1YuCeA+kOXdiQRbgi1v9j7gL3Qkc3xocddWfuQWcMHLjMFoSnp8+i2Zjb9Xt/KRZuiZo0hdH8aqwvaxN7f8V9xHuFrFu21vts9CzDBPWX++bE7NJcRmgOaDo03XvdNDlKelpZHMc9OEWVCUHqtpsAfz7w/5z9s7rbJ5HIqGqQeo7tduuJbh6cB0bbD18OSo0njIOpQSfqOR9f8AW948bjaS2VwY3xnpLOpSUoeI8uuOVip6mEwzm0cnotbglvSAR7c2ySeC4EkIqw6aaDxoijcOi19q/D7u/L5XD7t2Nt55sZJEJU8Uc2icAXJe3B95t8gbRvF1sC/vG3ZhOKqajh+zrHHmjnHYdm33xNtuNEkBo6/PrP1h8ffkU+QnjzO2UhCsAieOUMObHk/4e8lvbTlrcLAtA0TiJqU1eX2fLp7mb3h5Qv8All5YZ/8AG1BrQrxp0Mmd+L/ctZTyN/d7U1rglJL/AKf8PeV21W6WdoI0FKjPXLXnzeJuYeYXvJ216XqtfLPQVH4r97NK6DbWkAjQTFLcqTz9PYD5lsLqeesKk9ZV+zHN2x7Xtoi3CSmOGOnml+JfetgW28y3sWIjlF7fTg+1vLH18B8KdSAOgx73HlDdAL7a2Hiflx/LptyXxd73pw/g200jj6AxSXtf8X/x9j4kDrFFamo6ZKL45fINZHVtrFLmx/al9JvYj+vv1PPqw406W2K+MHe0zBp9uspHFjFL/X8H/H36tevEEdROxPiB3DktpZSKXa6Ss1NOAHjc2ZorKbD/AB+vvVVPXgD69ahHdH8q75l5vtbfGRxfXdPPQVecrJqSUVM1Mpp5HPjTxiKTkD6nVz9ePfqjrVKcegz/AOGlPm3Yn/RtTWH1/wByM/5+n/KP71XrfXv+Glfm3YH/AEbU3P8A1cZ/z/1T+99er14/ylPm2AT/AKNqaw+v+5Gf/wCp/fq9e68P5SvzbIJ/0bU3H1/3Iz8f+q/vVevZ67/4aV+blv8AmW9NbkW/iE/4H/UP79WnWsV66H8pX5t2v/o2phb+uRm/p/1D+/V68fTrx/lLfNsjUet6awsP+LjP/sPpT+/dbA8h1lg/lHfN+ofxxda0zNpdrfxGccIpdv8AlH/oD79Xr3RAN9bG3D1vu3M7I3ZRih3Dt+uegytKHZlhqIzpeMF1U8H+oHvfWj1trf8ACYP4/wDx17u69+atT8iduYet21s/adJl4Nx5GmimqMFMkOoGjaZSql7WJJ9+HHrZHbUevRqOh/n58L/kf8lW/lrby+NvUGy9k127JthbN7kx2EpBuutiqC1PTZCvyUVy7W0SFFAax4B596A82NOvFtTUUEin5H5dazf8yj4N7q+N/wA5d6dF7Xoxl8dl921NDsCelSRUy1GkwSORFYXN1s50/wBfpf3bJz1UhY+1eH+fq/z5r/Jbcf8AK5+Nf8urE9V46sx+/cl17Hk+3aqjSppBHWrMVNKlRIhUyeLQVjZhfnnj3X5HpxxpFV/1Y6K1/Pd6Tx+/vjd8L/mxsDCTS0vdmzKrcvZFe1DPBX02ZlVYr1hKAEAkkkn/AB59+BrjqrUGT9nRm/5WGzpfjD/Jv/mNdybQy1XTbz7G2HSxYXcFFO8ddg/tqUs0lNLEQEJQql1tcX/rz5q1x1aI1WrcanrTYzOay248lVZjO5GryuVrZGlq6+uleeqqZnYu8k8z8sSSbkn3vqhx0aPaXzT7v2p0luzoWLdOUr9jbno48emOq8jO1Fi6NeHipaEei1ibWt/jf34ADrdTSnl1tkdXrX/F3+R9uKu6xq59q5PufrKpky2XwkrU1ZI0SJKz1TREFtRuTc83P9ffjnrQqMHy/wAvWkXVVlXWVc1dWVU9VXT1LTz1lTK808szNraWSR7sWJ5uT7917hgdG/3R85O7N59Q4Xpnc+fyWa2pgMeMdi6auytTPBRxeMozU9Iy6I9V7kLYe9UHEY68dRGkmo6JqSRb/D/XH19768Ouh9fqR/j+ffuvHh14/U2JP+J+vv3Xhw669+631737r3Xvfuvde9+691737r3Xvfuvde9+6913yLH6X+h/3v37rWDjrthbnnn+v+8e/daB64+/dW67Bt+B+Pr+PfutEV68fqbe/deHDrr37rfXYtzc/wCt791o18uvC35NvfuvGvl14WvzwPfutn5deIt/rfj37rQPXrcE/wCt791vz62Hv5MvVj0rbo3lWRljXwK8LGJwESOI+IBjwT9eR/X37getAV7utgyiMtZR6NIYxxgFuP6cAD34Y62cmp6DHdcE8YKlbhja555Bv9PfqUx16teih72xUz5iO30Emq1vTYn8f8T731quehI6wpWiy8RFyPIl7f8ABfrb34deBI49GoytfDjqWSqma4ijVrFrXsPp6vezx610R7s3J5PeNe1LRU8hp3kKh9D24b63A+n+x9163Xz6DVugXmWOvn8qySepgI5CRfngH375dbJx0s8H8ZsZuZokrKmZTCw0Bon9J/JFxx/tvfuvCtaHpSZ34cbapUgkNa9TJcXDo5Cj/DUP8Pe+vdJas+Ke3aQxPFOdbsBZYjZbfn6e9AdernqfJ8Qqaejkr6TO1VE0QLr4vIpuBce9/Ida4Z6JPvDc3Z3U+563GRtVV+GpHZI6iXWxdVNl5b3qnXvKnQObo+Tuenq0t5lKmzx+rStjzbj36vXqdOm3fk5laSqjaXyCMkFioH4Frhre/V69TPRtNmfJ7CZBYhW5CaEoFIGtV9f55IHv3W/Og6Feq+V+GxqKsOTaVQF1EzxMwH+tf34060K9Qtu/KfGbw3JR4UVPl+4qY4IFd4z62P4sT9fe9QRGlbgoqetaTI6wji5oOrtPj/1XDkcRRZvJQrOahYWSKSN9ChogwI4/1ufeLXuRzTJue4NZ270jT5/PrLH225Uj2vbVu51/UcDy+XR7sTsWijgiUQxxxqB6FBF7fn3E7Xbg0HUsLaoAKYPTrP13i8hG0cqhbiwZQ1/9f6/8U91F5IhDDpz6NXx0EG6eqYcZJ5IJHlic8qQ5Fi1gOP8AeOPZjBftIMjpBPYrGa/y6DHNdYxZCmmjZVjRVJ1jVfkcmxt7XR3rIajovls9YwKDoru9uuaSnpa2llUtDGG0MUYAsCfpf/iPZ5a3ru4Pn0RXVmigoeqxe8usY5KLISJArReKQodD8nVccH2ONtviHWvQF3exGhmPDqtKXZUdBkKkSAQymR1hARls2qy2PsZi5LAaeHQKjiaGXVGdP2dCB1nvyq27uE4Oodg0NirM1h9bjk/8V9qLmFJbUSN59TFyrdyXqLZVq3RxM1n5shgvvJZ2JMFwoYEfS4Bt7C6G2Sbw149Dm55OvnjL+GSGHoeiH723NWY/P0uQVCix1IaZxYHQr6jz/vPPsQW728Z1I3d1He9ci3UC/UOnDPA9Hm2Jk6PeG3aHMUcqvL44lk9QZhZACfT7FVtzBbWkQW5bu6AVz7V71zTN4+3xdg9Af8nQ0YCGZHhaNiRcEi319r4eYLebK8OiDc/aDedtqJFIYfI9Dxiv3adAx1Hj/XF/oR7OYJ0uF1J1Gu57VdbRMYLkUI/1efWerohIGBPNrA/i1vb+VNRx6LDodSjioPTJC8+NkZVLGFv1KvHp59Nj7K982W05gsWt7kDXTDfPoee13uVzD7U8zw7ts0reAWGtAaKFrn5dCdtkUGTo5Yakq0j30oxvbj+p9407pyfPtV80DCorjHXabkr7wO0888tQ7vZsA+kBxqXyGeg43Z1gs001THGqRkM1lHCjVck8+xJtnJ0rWhnccB8+oX52+8TtcO9jaYGBZjTBB8+i25SOt23ltNHHdUkYOStuEP1ufZePobB2VyC/CnQoiTmTmu2huLcMIDQk58+jG9V5pMnXUcs7gGNkGgkDn/W/p7xt98bmefavAjGCfy6yW5aiXaeWhHWraaH16sBwtQk1JCR+nSARfjn6ce8BN0gaK4YHj0D5pGkkYnhXpScBPTcWHNj7I86s9Jc6s9FF756xTedDlGnhVg1PLoOlidRHBBHvIT215sTaBDAreY6jfnDl8brFKWHkeqjdsZat6S7Mq8VXK0eMnnZY3IsAWbSLFxb3lnfW0HNGxrLGatTrGvZ7+Xkvmfw5RSMNx/P59WdbTrKTL7ep80s8bmYarK6kgMtwbA/4+4ms+aL/AJb3UbHo/SLcc48uui3IPM0XMO0IYmHAeY9Onh6WLLQPBoDHSdJPJvb8+8tuUJ7dYkuGowYA9Q973QXO4hrG0kIcVpT16DSfH5LadeKqleTxM5LwBSqaT/h7lK0MFzMs0a9tRnrCbehf7Pt0+330p8cq2CfLoTcPnlyUSuy6CQqstgOSPqR+fco26RmBSB1gvvE94m5ypMxFScdZamWFKmOxAGv6L9Sf6f19qB0VcPz663Xl1xWIaqb0fttpUWHOjgkn37yz1aor1SB8wOxM9JmZ5MfPMkZ1KojcAXvbjTx71wHVaV6Lf0n3JnNm7tpMhnqqaTHSTwAa3Urfgt+r+hB9+6tgdD33x8jM/idwY/emza6qnhVYTLSCUCHQouQRcD/b+/da6M10l8jtt97YI4/MLFTboipAk0IaIawiHV9Pz/h79x601eJ6ps/mB9MVtLumffuMp2joAskUiLEyrIqyXZl4+o9+z17gK9Vce/db697917r3v3Xuu/rYWH+v+T791r59eII+vv3Wwa8Ouvfuvde9+691737r3XYt+eB791o18uuwLn37rxNOujwT/rn37rY4dde/de6//9XQBuNPPJvx/re/da8+uPv3W+uQBa9yeP8AY+/dVPb11fi3+N/fut0zXrs8C1hf+v8AvPv3XhnPXH37rfXZAB4N/wDH37rQNePXfBP4HHv3XuHz66It9Df37rwNevW4v7916uaddojO6oouzsqKP6sxsB791vq834LdcVtD1sa2opwlTI6z6udVpG1Wv/re/deFOHV4XxfpJIaKsjqYwCCSCbc2Pv3WgPToaNyCqr550PohVmVbXtZeLj37reK9Fn7Ww+QnpHpMXM8Q0kvMraSAfrcj3vj17hx6buitqRx5UJXZWapa12hd2dbg+q4N/evt610bSrw+PpoKtaWmigPjk5iUBnNvqbe99ez5dEr3DkIsRn6g1rEXmcLf6G7cfX6e/dePr13N/B6yi+/qp0AZjpRyLWP0PvXWqnz6dNoZLbmLyaHzwoNPAUr9Przf34evW6ivQ2NvHZEtDPHUVEU7yKygOyHk8WAPvdet48ui75zae08tmTUwTQKHk1aLqFsTex9+60PXpeU2w9pGGnRaWhZrJqJ0nUPz/vf596630J1Ds/bLQQ09LjaMJpQM6qoufyTYe9jrZ4U6E/FbA241H4YqClhYg3mRFVgbcm4966qB59JLcfS+MrIJZEyz0r2cgo5UgaTf6e98et0HHoqWe61qIM3FjKHP1k0bSqrMsrMLA2YWB96p1o56MRjehtrxYLVXTrkKmTHSu5n9RV2jB0m/vx+fW8dV37i6ho6XOZmOgpontVzMnp4VAx4X+nv3Xia5HSPpcHDSTmjq6GIaW062QXBVrAgn34nz68eHQpbbjyWIljlx8jiMMAFViFAvwLf8U9+4CvXlBdgnr0NX8a3NJi3VZZmuh5LH88EeyW/36y25GLnuAPUl8p+0/MXNlzGtulYmYZ6LFvWl3Dl8uMRG00tVKdQBLMFWTg+46j5hbfL3wY81NOsv7v2rtfbDlZdxv2CEjPrUCvWGq69j6yoqLduZSOrrBNCVo5bujDUGJKt/T2l5u5XuL7bXs4zplIr6eXQd5T9wIdwk8eEeJDGQOrdPj32zjN17VxC4yOKjjSmRJYYQq+u1tOlffMz3M5Ln23cppLursGOTmnWaXKPMcd/axRwdgIGB0fnrLa0m68xTUEQ1LO8Rck/pBIuePeKfPvMUXLu1y302PDB/kOhdzDuqbHtb3jnIBPRdvm/sig637f2bC06oJ6ON2QmwueSbe8r/ALh/Np555Pm3EJQl2FfPDEf5Ouf3O/Nl1zfzCQxJVDTozXSWUwK0mHZ6mIM9OrkawQfofeX+729x40qqDgnoy2mzmYKoHDo7X94MKmPTTWIAE4AdbWAv9fYUNrcmSmnoVG2kjTTp6ZaPdeGMrA1kZ559a3/2/tx7O4AFFPTKWswY1HShfduDjRf8tQn8eteT/r+2vpLkn4enzbSngD0it1dgYLC4uoyk1bGEiV2ChwxuQbWHtXa7bc3VwIAuT0x4NxNMLZBxp0SbJfIySty8tPQufEZ7ITwpUvbj2PoeVxHAPEHl0MU5YWO3DvStOhHrdz0OUxST5CdPLJT8XZeCVsb3/wBf2G5YTbTmM4APQM3HZr66mKWUZZR506rE+V2z8FmqWsqgYpKtB5YZRbWAFJADD2NdgczoYyaDols+RuYUu/3hbREFD0THoWtbMZSu21Xu0dWk5ho45foVUlRp9gHnbe975Mk+rhynxH7OsgNl5l/f22jl3dY9My9oJH+Xo4ePw1TRV32UyEPG2lTpNjbj6/09jjkj3y2GbbQ+5S+FLjHUTc8+zL8w3Y0oNK46WQoQt0cEH6C9vqRexJ9ylbe4tlukXjWUmpfUdAi0+7Q4rK8Y0j16S+TpWTWFSy8kgcfUc+zW35peSRVqaHog3v2JtbO0lnQDtB/wdI0ZH7WViSwtxcXFrcXHuRbWYXECyA56xA3rbZ9q3GS2lUgAmnSloMsZ0BEjahY3JPq5tY+3z69Ffy6FHbldG8PjYWdjzb6H/H34ivDrw+fSv0xOjQzIstO6srRN+ltQsPeq9bIB6LXujDVOxt4UuTx7tDi6lhLURRkiMknVYj6fn2VbxPJaWpnj4CvUge3W12fMO9R7Td8WYU/b0MLypmKWgnSwFUFkW9jqB4I494pe4nMRto5dzuvhjr/g67c+xHKlnyfywI7UDK5PS4XqFM3jFklVWWWI8E/QHn/jXvAzePee/wD300sLlFVqAD06kLcObGjmMHFfPotu7OrKnZ2YircchjgWQNJpuFNmvz7kPaOeF5nsWjvTqdhivHqPd65rlsiXt1op9OjTdV7lSXGrTTTHyCNOSeB+OR7g7n3ZGW58aJcV6J9n3Fb7VM5qSa9CllalYKN6g3IjKyKPrcqdfA9x/YRO10qDGehfZw+PKIxwbH7eld/s/wD2XtLb0GAodsrLR4aleKlleNPWoS17kf4e81+WOduYIdps9rtChMYCjPQPvvuvcmbzcz7rcXIEk9WYVPHoqe0v5nvc2S31VUT7XjSAVbgOI41BHl/3n3lXy3zPzLb2qfVFcgcB1At192fk+3u54JrjSKmmTnoyWW/mDdsLHdcHCqlR6dC35HFx7lGz50uzGBNSv2dY/wDM33bdiS+kNjJqAPGp6x7Z+d3cdfMZqjAQimQn1mNANIANybfTge024c2XLf2JGejblP2E2uMabtioHqT1m3B/MZ7Fo3ano8TTtOLqF0ofpwTwP9t7O+WtyvL5/wBamOo196eSeX+WYAtlIWanSDb+Yh2/I2qow0Cq1/qiA/Xjk+x9x6xMGOHUOt/mP9pUMsYTC0h12DHxxm5PFz79T59bBUeXSv2x/MK7frmZpcJTaCbqfGoFv8CB798utAitSMdBH8k/5o3cGxts1FPiMNTy188UnjjMaENaO9mPv2mmOrEg+XWu72T/ADwPldsfcmRizvXOLpKaapc0VXPEBHUoTcMJQpF/8Cb+/Gvn1Wvp0Ho/4UA/Ii1v7m7fPHP6ObD/AIL71+fWjT5ddf8AQQB8iLW/ubt//C2n/Yf2fe8+vXv2de/4f/8AkVa39ztv3J/2m3+20+/deqB17/h//wCRHA/ubt8Xt+E5/wCTffuvVHy67H8//wCRP52bgCOLfoFv+TfeutY+XXv+ggD5Ef8APG4Dn/gv+t/qfe88et4+XXH/AIf++RBv/vzdv/1t6OAP+Qffs+vW6jzp1Jpv+FA/yKppVlj2bt/Wqsq2Ki2pCt/0/i9/fq9ax506pN7m7PyvdHaG7ezs5BFTZbeGYlylbTxNdY552DPbgcX9+HXuPW1n/wAJu9i5vtb4e/zNus9rySw7k3N13jxjZ6ZmWoianiLN42Tn/Ye6kVPTgYKKn7P5dVGfy/8AoHsDK/zb+quro6DKvuHDdyU1DkMpW01Sn2M0TfbyZGepYWRQSdLk/S/+PuzUPVIwcD7ethDvbpql+Vn86PpzDYKenzuz/jRvxKDt/Pwv5qOngo6gR18s1QlwQAhsWPPv3EVXrYBWSj8Kf8V0q/n9/Px+NeG+RHZXQu4vjF0525sXqDc9VtvZldvLbdFXyxUmPK02tKllNy1i+n/G/vRGo1r1sOEOnSTTz6HKi+W3Wn87X+V78qutNmdf7T2Z2L0Bs+nk6q2Lt/FxYugho1lH3FNjaeFQf83ciw592OBQZ60tH7iCoPkf5Hquj+RpVT9v/Dv+Yj/LV7PqHxPbe69vU+3erMLk6hYZVz1LCzy0AaSxUFfRGV+oAP596Na9bjZQuTTJP7f9nrVS70+OHaXx/wC4Ny9K7y25lY95bby1RipqODH1TvUTwTND/k4VPX9Bcrxz/T37B4daII+Pj1f7s/8Akc7D2n/K37W+Z/yB34djdiDbFFnuo9oVMyx1W53QlqxBTy2aO1rKNJv+fx79xFV62oA7ZOJ4U8uj7/y9c1iPn7/KR71+OmwspS1nefV3X09DtHaeZnWnkrptCa4qBAdUjFAxsl7j6e/EjgOqLwq+OI/zdadm4ujO1tq9mVnT2b2Vm6TseiyxxVRt8UkslQtQW0EjQpun9ov9APr79x62cZPWxp35/I16e+Lv8ubaPyg757Q/ut3VvvZUGe2j13R1RkfIZAws0qVkbHUgJAIUqP6e/Z8utntBBHDz+fWre97KSbk3H+sFNh791oddG1lt9bc+/deFanrof0te/H+t79149cgt73Nrfj37rRb064kW49+6tx68T9OP+N+/da669+631737r3XZFrf4i/v3Wga9eA+vP/G/fuvdeAv9Tb37rxx14/0vce/deHXXv3W+ve/de67Av791omnXiLAH+vv3XgamnXrfQ/19+6314gAA35Pv3WqmvXXv3W+u73sPpbj37rVKdSKSknraqCipU8s9TKkUSAganbgC/v3Xvn1ukfy6+tIeu/i9s3Ly00aZPMU6xzv/AGvVSkEEn/H36oPXsgU6PXhovGrFhpPHp/BA+o9+PWxSnTHuLDnJyjQgCr6rA6QLDkcf6/vfWs8eiq9o4+DGOZCFDcEWH+NiOf6+9dePr111bUq0jzRgM6nj82tYah/j791scc9DtV0NXuDwRThkhH6xfgi9v9597HHr2KY6XFN13RxUNNHSYqlkY6T5yoDXI9XqA96x59eofLpTUfX1FoVaqKNQv6l/A/B+vvxbHXh69KDH7TwuNZpYEiv+NP1v/sfeqnq3n0m9zwUbahFZpEI1Ac/Tn3YdVOekQuMiqXj1qSFIb6XJt9b+9derTp3qaNTCKaJtKsukg24BX6j/AGPvfWuiodsdP0OYhq5p6SKU2ZyzKGZrj6Ej37j16np1Xduv42RZWaYUOKiSbWQulLAkH8ED3rr3THgPjTkmmNA+GiedWt6ov6/TkD34DPW6+Y6h9ifG3cu1saa2TGmjhdDpanRtf0/AX34068KnPRIs7j8vhKt1qpa/7fUVJkjk+gP51e9E4r175DoyPwv2InZPeG16WGuKpQ5ylkePUQHCsCysB7D/ADbfNtuxyXEfEqR0IuTrBNz3+KGXgrCnW+T1tsyh25s3GQRwRsY6GisAABYUwHHvB+9unuLlpGOSx/w9Z22NrHb2yLTgB/g6V1Owd9IsNJ/H0/p/vPtKRQV6UB+6lOlDEo08AADgm3B916eOBjpmycUUilJEVxY6dVuD7spIzw6bYBst0GGQxtIZ3QyRqrfqS4Fwfrx7MIi5Wo/LpFJCZDVRjoCu3tnUlTiInoVVp3IDeMA3H9Db2a7dPKknf0U7hZjwq0z1X1291e0222keHS2hzbSeTqvYn/H2L9vvwZqDoGbnt5MPd6dVA9t9YZakL1tJT6TTSSysQCL2e4+vuQNvvYnGlj1HO4be4BMQyOizLijmMnSyUi6MvFUKtSEFnKoQD7d33djs9iJZPhbh1I3tL4Em66Lo0Zc9HT2psvOZbERwRwzy3hUHWpKghefcKT8+Q2t2XuWABPkes++X49n3KxEbKMD8+i6dzdS5mkoqyRqUq4ilPpB4NzewHsQ7Xz7DdSgQnUteoo9xbWys45UgWtQelP8ACePMZDJV21p9TimjP7Ul/wDY6b+6c9c3223WIv1cgjh0a+we7bGqXG330YLaaivr1ZLFtHIYYxzyw2QnhRc+n/D2F+UPeKyu7tbK+bSCaA/M8K9Oc37DHeXMtykY0mtB0vsLTSOgkCWW1gCPz/sPeUG2807fbQqolBrnj69Yfc4e0G7b/uLXEUVFzTp8lo3IYMLDTckfXnj6+zNub7YnsOOglH93zejEZHSnSQyUUUSm3LngDmx/23tdacyQ3EgROPQW372e3PaLN7m4WigdQcTkHoquApI0Y8oLBTxYH829mG5bXBuYWRxUjPQS5N553fkmWa0tpGETArp8hXoeZJY8nj42RgwKgE3vc25B/wCI9pd6IsNkdYhQhT0f+3EMvN3uPDNfOWrIuD8z0W7ee30qPuVigV5dUul7C/14Fz7xMubqWTcmlc1yeu+e0bBb7dypb2dlGAfCj4f6XoLNo1VVtLclHLkXeGl+5XUAf0qWFjb8+w5z9tq7zsDRWa65FFT+zoJXMV5YROJsA8B1YxtzfuIq46WOjqg6FUt9OTpF7n3gpvXK17C0j3CUOeg7axzzsdIzXz6HKhmjqqdJVYkMAeDf6/T3FF1G8ExQihHXpVaN9LUr017gx4yFJJToPVICh/wW1j/t/Znsd99DdLM5wOkN1btcQlFHHqrD5UdA1FZG9fSUyvV6jIsyr6wb3HvMf2v59t7uIQu1FGKdY8e43JjSDXGvefPoEurtzbg2DBFhtxVFUMfFJGUZyx4UWK+r8ex1v+2WO8sbm1VTIRx6Rcg81bryVILOd20E9G22L2HjsxuCOGlkJgZUKk8eu3Jt/h7HXtlHdWyHbrxyzVxX06njeJ4bzYn5hDanC6jXoZN20EFbTpKqLpKhjc2PNz9PeXWz7ZHDsjNTIWoPXLD3H50udy9ykCyNoeUIV8iOg+oXipkkWBf0XBtc8/QA+1nLl8Z42tpD3A9EPvJyodrvot7t1CxSIv7TSvThjI/PVfcVDkgMOD/h9Tz7EnDqF61z1G7Yp/4vtgx4vU9THGysF+tgvJsPe+tYpjqoXvXrDLZiinaSmYVId9Bs1yfqo59+PXvn0C/Vvxe3HvKshpq6m0rHUL+rUQi3uCPeut9Kz5U9Kwda7Qo4qRknrPSksatrIAFiCPfjnrwyeiddHZHJbM7DoMvQ3Qz1kEU0KEgaGezi3+I9668erCPmVj8XvDpyCrNPFCBj5JWKgeqbxlmLED6397PHqoHWsXWRCGqqoh9Ip5Yx/rLIVHv3WweHUaxtf8fT/Y+/db+XXvfuvddj/euffuvddEk/X37r3XYF/futE068f9b/AI3/AI+/deHXXv3W+uzyL/T8W9+60PTrs2UWBvf6n/W9+60M8euPv3Vuve/de6//1tAE/X1f0/H/ABv37qo4Y64+/dW697917ru3F/8AG3v3Xvl119ffuvde9+691737r3XYNj9L/wCHv3WjnrkvJ9I/H0P/ABr37rR4Z64m/wBP8fp+Pfut9KPaGGqs7uPD46kjaR58hSBtIJ0xiYM7ED/Ae/dePW0z8YdmwY/YuNxyIFeWjp2ZfyDYA8/6/v1evU8urDOscJ/AoXPKKz2tyL88i3+Pv3n14V8uhcy9IkyeSEWDIbkC/qI449+63QdAD2DjJzSVCRBxMIZHOkWFlFxyffs9e+3oCuic7KN8TY2Z2aRXYFTyf85zb37rXR5slKsKTlzYeNvzYf1/PvfXq+vRBu48a2ZyUy0GvzCVm/bU3ve4PFvfutE56QuF2RufOUiYyU1cKXXTJpZeAebn3rrYPSjp+lsjHWotTkKmJNOkyaiOBxyfe6eXWs8OhKx3x2oq1BJJuaqRwNQQS31G1xyD7rTrZ65P0Q9IxljzNW4SSyHW5BUfQjnn3vy63Tpa4XqeuWAyHJVcgVSV1M3497p1rpG5Xc+69j5NoPtJJsfGxV5mQvZV4uPfuvY49LPCfI7CrB9tPULFVr+uOW0f+8e9Y68SCKcOmbevyOxBxLRUlZGtUzNFZXFxrGkW961gNQ9WVJHFV4dS+o8dJuaUZurnjlR/3VZpVJOo3/2Hu3VejQMESmmgEiBRTSopLKOAPpf/AB/HvfXuiVZTEoNz1tIGWSSolka8ba7BjwCw+nuvXvOnQf7r2IYpS7QNG12YMFN2/NwffqV60TTpgwlKaedIpyyhWCrrvYm/Bt7ST3sFtUSmmOhBtPLG67wFl29NXcOHRjNu4haqgb9pJEKGx+vJX+nuE+dN+tNbJGfXrpr92v2t39rSOW7jZcqegchxsNL2rTpUwqVaBrAgBQBcL6vb3tj4FxL4zgE6sft6KvvtfvDadqG3Rkqiqf206RvyGierQUqkNAjErGDcLx6bW/3v3I/NrNZq98PKo6w09kbxL1V2liNT5/4vp6+IG5MhgdwUWFqJG+3nqBZG5QKTcAj3gl7uWVvulnPdxjNCes8eU76Pb9ygs42rSgPWxj8bam+5MbKHAuafi45GocH3yi9+LWvLl5ERwV/8HUwe4x18sknJKn/B0An84Lr3OJl9sdjYoVL0tDRxRu8akxhjEHALL/rn3Ov929ztacucqHaSQ0pJfSePcxPD5dYE7JbQz81Pay4qa5+3oknUfYO4EwmInNTOJFpFsNR9J45599Xtvv8AauYvFmiKhwe4HFD1kZtu028IDMMD5dGcpe094V0SU8U85QrYm5H+H+x9gznXmPYOTbfxbl18U5VfXoWWXLce5tVFx59dz7u3nSxmoilqGa5uA7XFhf8AHuIrb3z2uS5EcsShfXoZ7R7bWV3MEnOkdNv+kXfdTdfJUroHNy5IF+SD9PYhf3g5aRAyaST8uq7/AMg2NhMLezo9cV6Rm4OwNzZON8dWVNQsXPkBZrf7G/19jLlnnrl/c7Y3zMqsDgcOq7X7USswuAuTnpD47IyTVumkXXJGQCVHOoH6k/19qeZvc3a9ltSVcM7AkDj0t3PkK4sZVS4btNP9npYVOW3pW6IoXnSOP0BByoBH09wXc+75eYyTIoB+fUm8u8s8oWVoqzKGZhk/PpJbn21nchjpqnLCWVAhUh9TWFv8ffrH3pmN4ttbKBU8a9CgbRymkZhgjXU2OA8+ikZPZGV2dnYN8YSmkRKKePyqkdg512Y8f4extuXONjzbafuq8ILOvE9Qh7hcubVsco3Lb1CyY4dWB7JmoN0YbG5Yoj1UlLHJV/h45nW5BA/PvFTmI3ezX0lqpKqGOn7OlWyiPcLSO5OTQV9a9Rt0YabHVazwX8EgJbk+kkf09zX7Dc5NPfnZL2Spk+GvQ13G/wBvttheEoA9OPQf5OO8LMCfIwuOPwB/T8fj3mgh8BqrkDrHLc4xepJGeDdA5mPJBO2r06r3Gng/kc+xxsW9NCtZmwPLrF73Q9srfdJAm2x1dhUkDqRia39+GMcXtcAc39jix3Nb4krw6xe5q5Kn5WQCeoPz6FvDSPCVbm7te304PPsz6A4OK+vQmUlQJvEgN2Kr9QRb6X/1vej8uvVA6BHu/cFItfjttUzpUZKtCmKONlkYN9Cp0/T2Euft3g2Plia+nYDRU5Py6lP2T2y53Hni3kt1OkECoHz6G7Z2ysrJiNvpPBIrLTxGQlSpTm/198u/cj3wtt4tLra4lUAkivXcnlKeHZeXkilbv08Py6NJjaY0VBDTn9SpoJP+PPvDK7mFxdNN8+g9dSC4uGk8j0EXbmNpRturnsPMDcf4X/w9yP7e39w+8JCfhp0H9+hRttZiOi2dc5iWDKR0okJVyFsDe9m+gHuaObLCOaxaYjI/zdA7l27Md0IgcHo5kEcNTj1NUy+Mx3NzbgD/AHw941ztJBeEQca9S9FJIhUxVrjovHa1fi4qGop8fHEzCNkZlsbEra1x/vfufvafZOYL3dIbyfWIlNSTWnUjcvwXZhaW6JoeFeiSbJxcNNuqSrlXQZKgnngG8nP+9++iOyxVse4V0gU6xa9xd4mXmEwQkqNRr0bikxC5WaLUy+AAF2ZrLpHPqPtWxABoOHQcjiN0RrPHiekrv3sDHbapzgsK8TVTApIY2u2ojTb082+nu9tavKfEfA6Qbru0Vmv0lqasf216Rm2tv5DIwtlcgrWl9Z8lxwTqHDe5A5XKrPoTh1it75WsrbcJ5uJHSa7CzNNh6J3ppAGiOnSr8qbD8H3JHy6wvU1rp6Qm3TXbono5yHMdxe4NiNQH59+p1vHRmMclHhKNPK6hwpLDVa1+R9feuvfLotncWIx29KCqqKyaNWplcQjUPX/tjz73jj14g9Us/Lfr/FvsTcNbJSwtUY+J/t5mQMwKqeUb8e9cTnrzfDjqkIC/Hv3W+HXZUj629+60CD1x9+631737r3Xvfuvddg2t/h+L/wBfr791oivXbNe3v3XgKddAXNvfutk0z1yXh1/4Mv8Avfv3WuI63Hf+EzXZWY6M+K38yzu/bi42q3BsPr7F1OOxmSnjiirnlX1IIpP85x9QPeq6TUdXChxpIrn/AADouu+f+FJO/cSm58TtD4dfHrafY1Y1RS1vcOG29T43sFapozG9ZT5SGJ2STm+sEE8+/ac168JKrQCn59Vz/Er+cV318T8r8lM9gtuYLeed+Sspl3JmtzStUZPBOZXmMuHrGid0kJblwVvb6e9+eMdVU0WjZPGvVXvZm/Ml2dvzdHYGaiRMruvKzZWvUSPJ/lM4Gt9Zte9rXt/sPfj1UV4E16Px/Lq/mcdsfy4tybg3F1htrA7mbcyCPL47OyMcdWwCLxiCrp/G4dQPwffuHXqV48OgtX52ds4v5iZv5l7OZNk74z+9Du+twm3aqShxhnYK0lEqQIq+LUCVBj9NyAOTfwwOtsamvV2uP/4Uz7kromynYfwV+MHYW+o0Vl3ruXatLkc7kqhY9AnylY8SFmuA2oox91I6cVqCmf29Vf8Az+/m0fIP59fwLF7pipOuNnYKKohptj7MrZqHbRp3bVFSpQwRxKscfICWII4PHHveB02TU1PRSfif8te2vh/2ngu0Oq85XUNZiq6CsrcPFWS0+OzSQE2pshGoZWRgdLAoQVJBHvdOt1IFPXq/9f8AhTdn63z5zcHwG+KuV7DaMCLf1RtGkk3DPIE0+aorfCnrv6tehjfn37zr1YMANOcfPqnv50fzLe+PnflMVP2HVNgMHhvL/DdrYrI1BwlFCzEx0lHTBYwkS3NksQeL/T3v7OmzUnJqPTquYsLgi/8AsT/t7e9dboTx64/U/wCv7917gOvHg+/db49dXP8AX37r3Xf55v8A8T79177OuwbAj8H/AHj37rVK564+/db697917r3v3Xuu7XHH4+v/ABr37rVaHrwIH1F/99/j7914ivXr8Ef1t7917rr37rfXvfuvde9+691659+6912DYEf1H+29+60RXrr37rfXvfuvdd8cWv8A4/6/+Hv3XulXsWeCm3ht2epKrBFlKdpSxAUJexJJ9+60et5X4vVtLXfGbreWjdXh/bK6bEf5g/n34cOtkitOjBUdYkDjWAEX9R/w/r/xr34j068PTplyu7qL7sUsXjFm0u3BJH1J/wCI97FevE0z0CPaWy6jc1OtZRa5IwAZCpJAF7nj3rh1rPSP2Tjht0xwLGJJGYK5sfoOOR/r+99bIoOjOYipoBFTecLGSQXa9rf7b3rPl17zx0LtPVwpSLNTuskKKCCrAkccj3rq9PXpjyef00dRMrgKiudQIuOOSffs1p1XotdZ3TLS51MXDOGEkgjOqx/taeL+9/b1rUAehE+/epC1JdXEgD6lbXe/qa9ve+OOvZrXqfSVySMGVAAODxY8e/deJqB1jyVQ/kjeHn6ah9SPr/T3qvXqefr0+UO3KfO07moNxIguHJte3AAHvRNOt0x0xxdTY+nqJJBSxFZC2k6F5N+fx73UUqetUFeo9P19QYmsmqxRQtI/CgqnHNgeR79UeXXqUPTbksZiquoFNnsXT1dFqA8cyBlt+R9Pez14enRTu+fjHtPfNFWVG3sLR00GhXJpoo9UYsdR0qP6+9EYx14E1z0Qz4y9U7u6F+QVHm4aSpOD/jMUxnlQhNAexs39PYe5ws2v9ieBMnSeht7ZyRDmuNJzp1MAOtv3Z3yIwGR2vQa6mASpSUyMFZeCIFBX/D3h/dcr3MU7YxU/4euhTcuXEdsjoKhlFP2dZoe9NvrVlTUIApJP0sSP8fev6s3DLgj9vTA5ZvKamU1+zpWwd77ZZOKuINp/TdR9P8PaZuWrquM9UfZLxPwN+zqRkuz8F/A6vKiqjNoHaEa7EuBewX2xHslybgQkY8+kabLeyziIqaV9OiJZruzP1ebnlpXf7d5XjjsxtpDWHuQ4OXIEtwrcepCt+WbdLYK4zjy6Mds3O0OU29FVbgqY11rqAkkUcWv9Dz7CG7Wz21wY4fLoIbjyve310YLKMmh9PLoPu05tm1WCkK1UBEaNZVYWPP5HtuxklScA/n0WT+2e/wA7CFYWNfl1T9348M9PVU+LpkaMmc60jHq9ZHPuUtnMCkPI38+t3nstudpa+I8Laj8uqz8PjKza/ZOEr8nTNFjMplViqWdCI1jeQAsT9Bxf2bc02VvzBy9NHC9HgUsKHzA4dRieTd55V3NtySJlWucGlOtmjbXRuxh0jtveez4qXLTZamhST7dVqHSVkGsNp+lj9ffG7mD3R3yD3Su+UN51W6QDWrGoVlqQKHz4dZM+1e8Pu0zichFUUoTTgOiHfIbqh8bijLV47RJUiQBHi0gAv/j7yP8AbjnKDdJtFvLqCU4H5dHHPWzJLb+IvdqBHr0Tn46beh2n2hW/tLT/AHDafwpIJ9y3zxcG+5dqM0Feor5Bsm23mRjWmrFOrRq3AQ5D7dX4TxIwW31Df094yW27yWhZ4+NTn7Osl5bJZSqyDFOsmQ2xHjccJ6OO5VLsLcAfQn2IOXfcLfI97iSS4cqWAoWJBHR/sXLm2XcwiuVVVB40HSDSaVwy6Bblb2sOOPwfefWxu9zZpcM1QwGfy6CvPke22FyLOwUY9OkLnGVHZfyCT/UX/wAP99f2MttuBayBzx6x3512eXerVrRB2nj9nQfTVIimZSzAm9mA5Un/ABPuV9q3FJ7cGUgdYE898lXG1bs8Vghfz9R0Kuws5I5koZpC6KoZST/tP9Pz9PYb523TwttaOLIYEdTT92PkOTcucYpryqOjqafZ0vKbaNRm/MYYS5Zna9rn1G4594Fc/e5tjyrcG0iIaevcD5Drtn+9bfbLSKKQ4VFX9g6A/fnVGV8ks7xSRiMF7kWA08ra1vaTYveC2vIQoClnwR0Ed93DbriIzu9KeXTF1/PmsdXmkkaTx076gzXAuB+CfZbza1huduJggUt6dApubdusoGWChb+fVg3Xmekq8UiSspkFlGq30tfge8TOc9oS2vdUYx0X2N0dyUzNxJ6Xcs9laZroqjUbnj+v09hCOKrCNck9GyR1IRck9A/u3MYDLSvSV3jkjTgtIo4I+vP49ydy3tO/WNsL2yU6T6dGUnJX70gEky1PVf3deJwstfIcdDGsKBnGi1idX04/w95L8kz7g9mDd11n1r1C3OHtldNchraMgL0mOnsfP/HVrShjpogApANifp+P+Ke525Th8C7W5cmo6NoOXL5OVpLGQmjLTJ6N1mM8TRwU68hwsYP1I/HHvJrbuYIztTRMaHTTrnPzt7RX8XPcd7CpKiYN/PrBR4vw0UhZT5JzqU2I+o/H09peUpw14zV8z0efeD2+WHlm2iCGqqK/lTpMVM1fjZLaSsbXtf8AIHHI9yQRnPWGKGgp0tsNGMjSfvKCXT1D6j/H3rq/y6AftfZkOQCwUlKuuSQK5A4UE2Jt/h7117h0gTuvZvTWGmWpqoBmTCGjUupfXYAD3v5dbx5dVQ9/d45TfGeqKWrBlpGmmMIYXGkt6Tz/AIe9derXpK9UbEfP56grERrtURFUVbqCzXvx78KdePQ7fO7d0PWHQ+PoR68nUL9uYJOL+a0KDm9uWHv3HrVcda3s0rTSyzPbVNI0jW+mp21G3+39+69TrHfi3+N/fut0zXrr37r3XY/P1+n4/wCJ9+6914W/N/8AYe/de+zrr6fT37r3Xvr9ffuvddg2N/r791oivXXv3W+ve/de67Btf/Wt791oivXXv3W+v//X0AOLf43+n+Hv3XvPrsAEH+vv3WiaddWPI/p9eR7916o669+6313xb68/09+61mvXXv3W+u2tf0/T37rQrTPXXv3W+ve/de67sbX/AB7916vl0YP41U8VV2ZjVljWQK0bAN9Lh/fjw61wIHW1N8Y8E1fQ0Ujho4Uhj0+kgDSfpz78M9bHHo80NElEFjWwVSePqT6rX9760RXpb0lOPtwzgMmkckgEC31PvR49W8ugz7AgojT1bU+lnNJMtgb3Pj/w9+z59apUdEl6pgOO7MnrZV0v5GABGlShfn6/kc+/HrXy6Obm6ubKQNHAGAdudI/Fvrx/xHvdc168K9J3GdcY+qq0yGSI9B1uGB5Cckktb/Y+9cevdStxZzaeDg8OPp4FkiDKZF0hiUFxx72ePW+i25vsLI5nLJQ0EAWDVYSL+q/0Iv7114fLoXdp43MVKwM88qXsSLN9CL/X3vrXHPQ00+N1JHFJdiEAufre1zb36nXulAKRaSi0EaDwPyOPxf37rf2dMFftjF5bG1Iq6SCYMG9b2JAI5Iv79muevGlOq3O7OuaZdyeHBzNSuWYv4P0/X6HT78ajh1Xjg9J/bnxxyu84U11tRF47EuqSG5v9SRx7oYwXD9OiUiPwxjodMF1l2b1tEkG23qMpCpCETF9PHJ4b3Y56b0kDpRbwz3ceM2xNNPizC8iWeREN1Rv1Wt/h78et9MPUDwVJlq89VySZOadS3nIV42ZuVXV/tvfqeXWq9GQzGz6bJUySWVlZDY2LG1vwR+ffuB62KHHDoDNzbDgxkyyxj1XUgWa4J+nsn3nbRd2zSg0IB6kb255zfl/eobSVNUbMBn5npz2mK+knMILMoKqVN7AN+APeK3M4b61kc8Ceu6fszLXlu2v7NQBKgJp5dZ9x9eVEua/vJEsitHS6QAGOkm5BuOPa7kzmaHl69QXLhYywySBn8+gL96D2sj595Lmu48zoGNKEmgX5dFV7YZ/3zI7sYW0uebBgLkc+8odzt4t62XWlCJFLCmcU+XXEvk/cpuUOcpLZ+3wX8POPPpl+PmbWr7AxiU19cMyRkC3LBuBx7wX5/sTa2V1DKKULfs6z75UvDdbva3Va6wG49bEvRz7ooK/DZKgp5Jo0ekMlo3K6FYGRtX0+nvmh7qwbLd2lzaXrhSwYDI4kYHWT24bls+7bE+33UgDBTxI406Wn8zH5KbCTqim62T7TJbvydPDIkQaKSSmaOIa0ZVub/i3+HsK/c09pOdIOaJuc92YwWUZMccYBVWQMdLHyqR5/Z1g2NsuJeaHbbl/TjY93mc8OqZOmd0wvDDQZjVR1JC+KM2UaQfpzb30b3ybdNpmN9sshHmaHj9vWR/LG528iLbXwowxnHR2dv1OPgSOWF0kDLzZ1NiB+PcF83bnvfMNzr3NiSvDqYds+jgTXCRQ9KwZehCkPLEdRuVMiC1/xb6+wU223JPapx8j0eJexxntcAn5jqO+XxqXtJTKbXsJEJsP9j/vPt1NtvG/i/YeqSX0DHUzhj6kjpDbgTF1qSmF4fLJx+3Kha5H9F59ijaG3GzIBLBfzp0stOaGtO0PgcMjpr2rgqejmLfqeRuSQfoPa3fN0nuUGs1oOk99vc28PpY4HQ4YXEtPaOnpHnd2HqEMjAc2FiB/r+4u3bcVi75nCgfMdK7eNLeKs0gA+0dLqr6vzWXxzQmglCSWJP28trD+vHsIwc8bdYXglMoqP6Q/z9a/rFtFtL/agkfMdIDcXVNHSY2fHVtHHYwSv42Qrd1S4YqR9b/T2MNn59muLtbm3c/EBUHyr0jvUtt/jaUnWv7eiebC7Koevd5Vu1twyfb0dRUslLqIjRFWWykFuPp7nnmnlafmjYI912zulC1PnXHQK23eYtmunt7oUjB8uHRqszltt7hoU/h1fTz+RQ6aJ43IUryLKb+wR7XbbzDtfONukkZV9XmCPPoW7hNbb1trS2rAqBmhHQRZSkWOFjYEL6SfyVAvx/r/n30pgZhaI8vxUz9vUSyWuubwl9adAZutwZ1IXhVP05t+OfZPHul3Ld+CmFB6kK55L2W35f/eVwwMmnAx6dJvDyn7+FhfgAEc/XVwbe5m5TmVEo5/aeudHv5tUlzcFbMVyaYr5fLoesY7yRx+jghQDcn6fU29jQ3tsGoT1jInLm8CMHwycfwnrHvrftLsHAzVyMk2UaFlp4g6twwst1HNx/re024bxZ7bam6dhjgKjoQ8qe3HMHNu9xbRbxNpcgE6W9ego6J2Zkd+7xh31ul5WKyrNR08xJQpJJewB+lveBP3mvdTcp9sO2WWEeoJH2ddYfaT7te0e3OzJul33XJANCDVTx8xx6tPiWkpqaJI0RfGgsPpYAce+Y8jXE8zO5JqepcIkL08uk1l90Y/G6jVTqmkXBLAD6fQD+vs823l+9vwPAWtfl1Sa4gtBWU0HRUO4O5KGelOHopFkaUW4dS3JsOBz7njkH2+ms5Pr7nBHy6AXM/MkU0f0Vl3V6DbYFbHjEbLVoBqW9cCfi5bg/wCw9jvfNtvN4lXbLBSQ2CQOl3tvsA3C+DXZofKv29DMnYddX0rUy3jDGwKkAgH+g/p7a2f2RjN2txuB4ZIp1k0vK9raMsgzTpI5CE5OOZZGvq4Ykc2H595DWO22e3WqWlkgQIOI8+jiPREugjBH+HoC87QphMgs8JACEnUSFBN7/wC39j3Yd4kUi1b7Osd/dT24iKtv0DUIqade3H3auBwYo6CQNWyx+PSjKz+oW+g9jyGx8VxI3DrGbcuZP3fbm1X+04dJrrnb2Q3Zk1z+WLv5JPIEkDW4N+fbt1OsSeFH0g2Oxnv7gXl2TnPRj905ODE4kxwftLDGVKoQLEL+QPZ5yeymYu56iz7wsMw25I7da4/ydEczeen3BuBqAOXhaoIcHn0l7fT3Jcu4W0I7j1hTt3Ke97nIVtoyT9h6N1sHbtFjMTT1CoocRXAItf8AqefblrdwXQLQmtOk+8bDuewyCLcUKE/I/wCXpk3xVgo5in0qFOohwNPH19qM06KBnoke+d0VdEtXElWzgmT06wbhbm9r/wCv7117qpH5od6wUeFfZ+NWOpmzCtDVP5ELQER+piBzb377et+VOqj/AH7r3XYtzc/63v3WjXy68bfg39+68K+fXgbG/v3Wznrr6+/de697917r3v3XuuR4At+Rz791UZPXasFv/X8ce/deIJ4dG16K+YnZ/wAfuse0+rtkVklJhe2KRaLcRhqHhE0CReNEkVP1AfUA+/Y49bBYfCaV6KdUVM9VPLU1ErzVE7tJNM7EvI7cksffuvU6xajawNv9bi/+v7916g49cffut9clvz9QPyR/vHv3Wj11e17fn8/n/b+/dep16/8AXk/g3+nv3W+vEk/Uk/6/v3XuvAke/dep12dXBN/8Ln/evfutY66Jvbkk8/X8e/de68Lfn6e/db+zrxt+PfutCvn11791vrkNNzc8fg/4+/daz1x9+6312Lc3+v49+61ny669+6312LX5+nv3WjWmOuj9Tb6fj37rfXYB5I/H19+61Udde/db697917rlqNrcfS309+61QceuPv3W+ve/de67Nvwb+/daFfPr1/oP6e/db68bWFjz+f8AD37rQrXPXa/W9uB9ffuvHhTrxFySBx/h9PfuvDAoevKxQh1JV1ZWVgbFSDcEe/db628v5YfyAxPYvRW3Ou/u6dsntOFPuNU8IkZ1p9LAAm97/j34DrwNc9WdVcE9RiqielALrGS3JuPT9f8AX97FK1PWvn0UbNbkqaLKzQF2acuQRc8EG1j79XOOtgjofNo7lhfbzQ5BVLzRFVuQTciynn36nWjXqBjtvBqx6kg+N2JTj/kIXPvXWxjrBubJT42SngsI0chSym3H+A9+p6dbr0KG1cjV/wAJk0uXjeMAaxx9OLX9+zgda9T1Bf7muSppQx/dV1seOTwffhjPW8dEf7RwOT2xno6p7oaiVijgNqU6uLcfX37qpA6FvqzdtZK8VDWO05kUJZxwnAta/wDj7314enQ/vRS0E/mbV4pFDAWNgG5HvXz62eFOn3E0pyDghFZeBz9QL+q3vdPPr3AU6E3H0cdEhWMWFgf6j/X490+Y63k9T5KoRJ6xdSbX/pcf097z1rh0HOczUgm8ECqQSQDwT9frc+/AdbYjj0nKlY6uMip4uOb39PFrg+7/AC6ocnqLS4Cb7eRqCVqhZAVkiJ9Ok/T0+9fLrfHh0gNz7FFcKVKPHrHXLKTrjjPqe9/x7o8X1ELwH8Qp0os7t9s3GDcozQwsGPzp0vtn43NYwU2MyUskD6kbSuoeki1tJ/p7w893Oc7fknx7Zl/V0tox59dVfaDfoueeWYbsMGddOoVBwB6DoylFsNaujWb71gXUN9G1XsD9R+ffPjc/frmyO+Kq5ABOAcdTtBtFqrLIyinp00jYGQiqmYVsxi1fjVyPxz/T2LbH7ze+21sI3jVyPMnoRX1hs13ZiNIgHHy49dZ3GbhpKRKWnnmlhC8p6yhB/BHsZ8l/eKjutxZt+VY08iCOiva+VNrnYm4AU9Bygmp6xIJ6fSzG36WBF+NQFvr7nBPebYLy0lurKUMI1rSoyehQ/t/tz2rTQvU06EsYLcVZjolo6qeKABWAu44I/p/re8bN6+8fey7g6iJVWpAzmgP+XpjY7PZ9lkK3EYd+FT0kMhsbc1WphmqZZIjfhtRBH+IPthfvCXRWojUH7R0N4N22FKOIVr9g6B7fHU6U9LGauO5kciwVjfV+Pp+T9fYs5V979w3W7aOWgRRU54dGkUm074/hCJaLx6LN3F8dv4ptGWWhpjDkPGzUskcTa45iAUZLc39jvY/ed7reTbK/6A454jzr1EPuVtWzCE28EKnqwL+UP2RUQVNd0D2ZKtWuJ+4egbIOqSPKw0xxokxuf9Ye8KPv68lTX/LX+uRycDFdLpZjEMlAasKj/CesI+ZrneuUtxF1ZgxQSGlRXFTjoxvzY64ojmxi0gjjhdTJEUVtIjeYFeR+PcWfdS54ub3lsX87lnU6Wqc1UUP+f8+sh+TXPM+wI8pLMfXPVPG8eqZdobzwuVx0sqvPkEEoUOAyaxYf63voLs/NUW87NNDKBhP8nQb3jleTat3huLckEvn9vR9MfGZaShLiz/a0yn+v6Lm4947XknhTyheGpv8AD1OdqivFGW40XpS5Gj0YqTWL64zpX6j6eyXarrxd3jC/hYZ/PozuAbaDXGaMRXoA6qkkhaWwIUFiQL/199QuRJDJyxAa1NFyfs6hjeXmnu2lmNTnj0HmbEYimkJuwU3/AMP8PYslvIrYiRzmvSK35fv95Rlt1ND50PQSGbySu4F/qDfn9J4vf2Z2u+S3DrHEaDoC737a2u0iS7v0DufUZ6XvXiyT5WZQDq03FgSL6SLWX2k9wuZ9q5d5fFzusmkNgZ6D3sjZTRe5DJarpRSK09Oj0daY+aGmaSogADKPqpBP/JXvkt7v75Z7zzHLdWD6lY4PWevM9wjsscbZA9enLeeChyNHXO0CqqwMwsL3IH09hfljd3sp4ow1SWp0C7mJ5oXDMaU6JFWY6aDIypTU7LaRvUqML82PI/w95Mw3KSWytK1cevUVT28/1J0VIr0OuwqqupligaI21JclWGkEfX3GfNdtazhpSw4HodbJLNGBCB6dDzPHJXUf249LypYkHnlfcPxultceL5Keh7A4gkEreWegN3ntKlxFPPVz1B8pBYgt/h/t/cz8n86Xdw0O2wxApqH59D/Zd8nu3WKJO0dEf33MtTM8S2cDWt+ORqIUG3+P195vcubbbSwR0iCk0JoPUdIud9yttts3uJKatJP59LPq7EJHRCZ00chrAWB/1P8AsPclxbRDbUYAA06xkl56u9xhe3QUFSOhtx22jlKxZ5ARChGgENa4+n14/wAPa0zNEmhDToNpYi/uPHuUBoa16F+DbEAohLOAGC2C2/AFgfa/YtyNjfAk4PQT90+RBzPy8wjFWRT5E9ALv6KOiYm7KFZ9Fub2JH+8n3OFncC7gEqny65icxbPJsG5PZTChBPl6HrvZNU4o3lkZli0fUkcD/An2q49EladAj3P2pjNpUFaRLGZ44nZGMiBgR9Pqb+9HHXs8Oqht378yPZ+fkmlnlWCOZrWOoEB7249+69wGOgU3nhvucvQ0yRHXJNHE0gRvoTa5Pv3yPXq1yerPfi50RElBis5IxkjQwzHUGvxzb/be/AdaPQAfzQer67eO2hDhojKKCnExCxu4RqZ/KrkR/Tlfz79144p1rQzwSU800EqlZIJHikUixV420MCD/iPfuvVHWH37rfXYNvfutEV669+631737r3Xvfuvde9+691737r3Xvfuvddi1+eB791o18uvG1+Pp791sfPrr37r3X/0Pn/APv3Xuu7/wCw/wBb37r1Ourn+vv3Xuve/de697917rsW/P8AT8f19+60a+XXuLf43/3j37rfXgL/AE9+69w669+6912Lfkm3/E+/daPy6MF8ZmZe08GB/aqIFIvbgy29+68eNetyr4/Yejo9k4mVEVJZaGBzpHJJseT79g9br0YIRIxJdblTYE/QC/F/e+tHh0CfZvcX90p/4RSp5asjQIU0t+rj6AX96pjrdeg72p2TUZ7LmhzkTU61S2jLqVFpRbTdvfjnHXgeuGS2TV4TdK5uGL/Im/cVwpNwf0/4e99aOM9D1tSvp8hEBMgQIoItwDYfT37reOkvvfeTUMrY2ht+4Gj1KVLAni3vxPWq9BzR7Jrs/HNUStw6GUhr2ta4/wBt71149JDaG2oP7+x4ep0RINTamFrlGve59+4DPXgfLo1oo48cYxCAFhOkFSAGA+lz79nreKfPp6grYnZJABYL9f6t+AB72OPWiB1GzWbWGkdzc6Q3qA/AHp9++zq329AHmu2Kikp6mhQWkYsqcAEC9r/X3rrRoOHQBlqjcGYFTI7PK8oIDE8r+bEe/dVoeHRpNqSNisbHDSRkTlV/SCCSOTf/AIj3vq2aV65ZTeuawyeWaAmz3ILNpA+tvfvt611CHamH3FAMXmikayKFYOVIH9Qb+9Yp1sGpp0F+7Nk42oyKZnbuSjhip4jUGGORUWQxDXYqCPez17/B0HlX8ppNvU01DPD5HoHNILBGZip0jke/da8/l097d7Tyu9Wiq8njmpqWYh4nZCODwpJtb23KR4DlvQ9KNvAbdoFHHWv+HoZtvU0FVVRtDpb1KSVa54N7HT7xM51lhbdWWPGT133+7XZ7hDyHBNd5TQKH0x0Y/b+36bN0NRTMQXEDLY8C+khVP9feJHuzzRe7XvFvbQsVjFGNPPPUkc6XEibdNCBUSKy/kQR1XH8gtpVmz3y75OnCU89RI0LgEkhk9IAP/Ee8z/YL3dtt+2yPYbxiz+HQN9nXFj319m7/AG3e35g21aK0mo8fPPQd/DnaOLyW94q7L1RoJ3yANMko0K0bE6Tqa3uOvvCzXe2ib6KIurKTgV/wdTH7GoP3d9VvD6ZIlOkH5dbU2wcztjqDojO7w3E1ARTUFSlC/lieWRvtz4TYkj625HvjFzty1vHuZ7rWHLu3tJHGrrJLQEDDV01Hr6dB/nbnbcYd/NrtBZqtpoPQmhPWuPhNxZrvruLdm59zFpqTF5mqXFRTfuLJTPL6NBPFrHi3vqdf7Pb+33J9ptlkugvGoNMcB59ZHclcjPb7Mm53S5mUNU18+ha3pszHRlajHstFVw6QixDRq0i4HH09hTZd4uJB4dwNaN69Lt32i2V/Eg7GHp01bb3RumnZMfBDJUFSY9TXN/7N7n2p3LatrmUzykL59Jdu3Lc4j9NGC3Qkx4HdeSdaiWaeDWLmNSwAv/gP9j7Dj7js9qDEAGp59CRLHd7hg7Er+3rDVbI3H6pRkassysFj1yXJv+QP9697i37ajRfDWnrjrUmy7ko8TWf59JmPYnb+NMmao6OrrqWnkBIkSbSUB5JH0+ns2/fPKd5Sxd1V2HkR1G26c0/1dv8ATPLqzwJ6M91LIm4a3H0OYBosgZYI5oZBps7sFIAb/H3EPPyTbNayXVmPEjAYg8eA6l7lLmrZ+Ytre7sZAZIwarUcer0+oejdn0GAo6t6emq5pYI3d3RWIJAb8398r+ePcjmPmXdbuziuHs1t3KinFsdY8c7e4u/rub2cbFFBIwfTob5dk7cgo5kjoIRaNyCEsBZeLAe4Y3W736yjG4jcJJCHUUqfPoBW3Ne73F4gaRskefVVPetEsO56yClQKiagEFwAGH+HvoD7RXj3XLdvc3RqzBST1m77fXLtsIZskjqnf5F9afe1k1VA329a8ksiVCD95QV1AA/X30U9peYluEjspT+nQDPDo6j2ay3WV7GRRrfIPRNsdvbenVFbFUTV9bX0sUyp45nYqFLXJ0t9PeQM+x2UN9FfWSKXGQQPP06j/e+X945PuG+jcyRscjNAP9jqxLa/YeK3rsynzELg1JptdUt1uspFiNI/p7kR7xIbaLxfiYZHS7YNlu9+Q3CChGT8ugxzv+USeS4If6C1gL/63szsbeB18VRnoK81bpuNo52t3JUY49NNDD9sVqXUKEb/AGFifyPZ7Fdy2y6IvPqMLrl603dvFu1rT1HQ14SZGw02T0grDG4A4+pHpIt/j/X2zLudyrULdGm38hbHcxgLGKnHDovn936vce4avJ7grb0EE16ekkYMkkYYlbqT/T8e445h5lurxzbqTQYPWXPtV7N8vcvwpvEkaeI3cp8x0O+yt2UuCrYaelRVhgCrGiWA0j/D8D8+8avdrajukaQDgRUnqUecNwtrbbzpNaf5uhgzHcE8cAEUYuwVB+nkvwpv7gXbfbq3M1XPWPFzzLMuoIvr0APbW7t57bShkzGIySjNQpPjWp6Oolhlie+l9aqRbn+vub+X/bp4RGqR6QwqCfxD5dBqC+uOafENpIumE0cFgCPyr0C2E21md01y5XIo9OUcFIZRoZgTflWF/c5bNyE/0/h3J0A9LrGz2na4nnuSJJB+fRicRtcJTIJCF0oByeB+bc/T2LU5U2varf8AxZA0p86Z6Wcqb3Ja72Lo9kQPTvLHSYyFnZ0Fuf7IHA/1Tew5dxy27FZOPWU+07um9Ra4TUAcT0BPYnc2M2XSySrL5agqTGiFXBP+Ki/t7b9vnvGpH59FHNXN228uxUY6npwFD+3opNb27vff1UaXEYmQw1Df5wqwIDGwI49jfaeX4oJg7nI6xu5191ty3Gye0CYatB8uhw6p6Nz+froclui6QrZ2WcNZebjk/wC9exjLdLGBbwCp4Y6x5tNilvbl933RtKLU0P7fPo4UFDidswtQ0TJphFiUHH0/F/b0+xXgtxdHgekG2e6GxDdm2WDJRtPl0X7s3eSQpJCJPRpbXci97Hj2zt3i25Ojj0Z82x2W7xg3lCtMdF56tw9Xu3eEssSn7dagkvzbT5L/AI9mO43kog/UND0DOTuXLX95/wCKxgqD1YQcHNjcStLEbssbD88krwB/T2Z8oXchfTWtegX94nl20S2W4RApUD/B0TbtnKbhwkVS6QSeGxLPYnj/AF/cmE1yesJkAC6R1W52P2bFisTl8/k52gTHLKWSXSFYWP01Wv7r5dX4DqiLs/eUu+d4ZjN+WV6Seqc0iyE2WIGwKg/S/wBffutdB7791vr3v3Xuve/de697917rkSLACx4545vb37qoBrnrj791br3v3Xuve/de697917r3v3XuuRAAFvz791oHrwtY3/px79141r1x9+6312LXsSQD9be/daPqOuvfut9e9+691737r3XK45PF+LC3Hv3Wvl10ST/xT8D/AFh7914CnXrfS/0Pv3W+vEAWsb+/daFfPrxN/wAD/Ye/db68foOP68/19+6159de/db674sf68W/4n37rWevG3Fv9j7917PXgbf0/wBj79149eNrm3v3Xhw68LWP+8e/de69fi3+9fn/AF/fuvU669+6313Y/W3v3Wqjrr37rfXd/rwOf94/1vfuvddgLzc2/px791o18uuha4v9PfutnhjrxHNhz791ocM9eAJvYfT6+/deqB148XsTb/e/fuvfb14GwI/r/r+/deIr11791vo2HxO+TOf+OO/KLLUUrHB11bSnLxoTqijDeNpAp4IsfWP6C49+61UjI63Lej+7Nu9xda4zdO36iOb+J08Szxq0RKaoh6iqfS/+t7317pN1fXceQ3BPVy2VPIXFx+rm9iR/tuffutkUyelZPttYJcdFEdKRSLdfw1l5/wBcf09+4dewePQpGmSCnhCrp0KNVgB+LE+9de49A9vujetyWNWBSwEw8lr2Av8Akj3vzz1ryx0NuDqKKmxUVDGq+VUGvgCxIF7n/b+9cT1vy6SldWNQZNGVT4mdblV4AZuTf37yp17zp0kewdjDd8UNapRkiOpTIxuSPr73T068Knj007D2FJjq/wArxrZDZWANibW+vvRpTr3RjKnFPJRNrjLgRqE9JuCP6e9D5cOrE0x0k8OKvHVrrIVjjb0lnOhBe35b/D3Y5OOq6qcR0KtIYXUeOqiqCApYQypKV/4MFvb3pUdjRR1YVk+AdRq2ajqlaCmq6VqlB6oDPGJFA/qpN/firKaMKdVNQwBHQWbhSPGFZ62ppYEJJu88akDV/iePewG9OtFX8uHTRFV0WRhDU1XDKhFgUlRuRx6iD73n7OvYHHp/w8rY28r1ETRMTZY5UbVcfQqDf6f19+pUdU8QA9oP7OlDkN009BTxVSUaSSXB1Mo1Cx4INvelqh1enW6iQFW4HoOst2I6ZKkq/EeZxrK2JC6rkW949feP5H5b3/lVdx3NxBOgYoRxZvQ9ZJfdi5q5o23mxtt2lTNASocHgq+v7OjW7U7D27XYyNp6yOB44lMilkUqbfgE++NW/clbzb3zeBGXVmNDnPXU+bcrRlDowBoCR6Gmekpm/kP1zhKyWjqK+byJwbKCuq/4v7ONv9pObL6ATqigH9vQfn5w2qCTw5CajpOy/KbqqlF6qukYN+GROB+Bb2ZD2V5wkP6IUft6SS8/7PFQlyOlJt/sHqPe0yVtDl4FmaxWOaSKMi5uP1Eey6/5S9wOX4TA0JK8CVqa9CDb/cW0ni8C2lwfU06G2kmo/tEONqaean0gKYZUcWvYf5s+4xuYbpbkreoyvXNQR/hHRkk63h8WuqucZ6lUOB3FmHCY6ieYaiNWmS30vyV9p7rdNp25NV3IFoPUdVut42jbx/jkoQ+mOs2X6R3Zl4opKrH2jjYONSyMP629s7f7n8u7fI8dvcDU2OI6ZsPcfYbN2SGXJx5dBNu/Zhoi9FVxhWphZo+SLp9Rz7kXlrmbxitxbnEnA/b0aSSW++2/1anUpz9vVXfZe49ydN9t4vf+yvNT1FHlKeoyIpyUd4kkBYMR+P8AX95kct7bt3OvJsvL+9AOksZUasjI6iDm/lyHf4msSgIXh0ZvtP8AmPbY3zjsdVZRJP49BTQQVEbLH+qJQHcEf1IH59whyV91McmXM8O2EJayOzgCvFj0X7BvsfIO1/R6CzLnh6dEc7D+Y8+XraSoosMklNRzCUyFVJsGuDwP6e8gOXfam22q2eFpjVxToF757s3F5cLIsPaprw6NF0H8pNkdp1lJgvvhS7hjQCWklaOMXRbWHIv7i/n32z3PYbWS/tV8SEZqK1z1KHI/uVtfMsq2bnRNTgaAY9OjR7tz1QsyYuBDoKLqlH0sfobj2GfaLkyw5g3cTbhJoVDWnqR5dZAS7dA20NeSN5Y6QtWY/BKXIJKngj8kc++jW2W8Nnt6QW3wqB/IdY9biPEu2CZoT/h6BzLUclV5I1LWZmAAub88gf7D2ivNulvJag46H3L3Ntnsdh4bx932dJV9vfbRySuAuoHSpBF2/wCN+zvbLVLBBJKfgyeo35s3Wfmi98C2X+07R9vRkOhthU0hkzNaAXYhQhBA/RfVf3gl95z3Pn3Tc/3FZV8GLz+dehTyZ7eNyc373nxcS8R6Do2kYgoYvBAqogPFvoSP6j/eveGz+Lcv4kpqepAYyTvrc1PUCaKTILJSxi4kUowtb0t9Tce1UUiWbrOx+E16dKxoh8TgekfH1rSCZneGNn1X1MPp+Rf+vsUNz1ceGFUkDora0sGOug/Z1IqsNj9vQmeVEBUEgD/AcfX2ni3W83mTwYyc9Gu3WUc0wSAdJs7+x8MMrKbyKCFBsDqHHsUW3t1vl5NGixGj09fPoWf1cuXkAbgegJ3luKvzf3Fy5RwyKp/SOOAP8feVvI3tVt3LtrHe3VGnwSD5dSDs+1QWMXbStP59FT3DQPR1XnnuS8pbSRf0kkcH3k3yyttpAagI4dY4+8ku7UKRglM5+Vehn2A1LVRQRQg6dKah/iPwbexPchly35dQds/huNCijefRptv42KOBW0ejSpA/r/sfZTM9ft6HtlbigrinSjyMyU8B/rpICn9IvwD7aTEgNel87qYGRuBFOiq9hVMNdUw0yraTzNqQXPo1XJsP6e5w5ReSS07zjrmH94GytbbmVzCKEk1/b07UGG82MTG0d1nkgAIW4HrH1IH+v7Fp456gEVp0U/vb4u5DO4mSvOSlMsoZnhGsldX1H+t711YjomG3/jhWYyqajponkk1nU+iQaT9Dz798h1rpTVfx6SKqeWsj/fpozMrFG+qi/wCfez1uvn69SetPkXurrjfEfXT4jy4J5EplrdAa2ptPAP8AT8e9VzjrRxno1XbGGxma2RubM5GOOaKXb1XPH5R+l2gZrAH3unXjkZ60yOwVhXfO7Ep1RIV3BlFjWP8ASAKtwbf7H3rrw4dI737rfXvfuvde9+6913xb/G/+8e/de69fi3H+vbn/AG/v3XvOvXhb8/0/3n37r3XuLf4+/dazXrxt+PfuvCvn14G34B/1/fuvEV669+6312Db+n+x9+691//R0ALcX/xtb/ib+/darmnXgbA8f7H37rxHXVvof6+/db65BSf6e/daJp1x9+631737r3XYNj9L+/daIr11791vr3v3XuuSgG9/97A/3v37rRJHDox3xYgWftnBK1rCogPP4Pk4Pv3XvxDrcf6TydPTbWw1G0isfsIOCfoOPz738+vHPDoc6/JQUlDWT+RbR08soa/N1BNgffuJ62cY6JNtOOm3v2XU5TJWqoaeokjWKXlDpe30b+n49++3rQOel12rDjMY1BV0VLFQNBURFngUBiiMC30/Hv2OvdDtT57B7q2bj4aEioqoqaFHYqNZawU3t/j/AF9++3rfUKhxk2LxlbMRZ0ppHCi97qpPI96oQOvefRV9uw7m3zvSoqfDL9lSVjxs4Uhf2yeOeP8AD3sde6ORtqnWGH7R4wjrGYiLAc/4i3vXy68eFfLosndeHymxKxd8UJdIfOIjID9NXJA0+/de+fSz2bu2fcm2osg85aQxK5Go31WvYj8+95HWqA9P+P3aIH+3mJDXAtpJ/PPvXHrfn1Ozmdp3o2jQ6jIpIvwASvFz78Mdeyei05jEVeYyDNEgUajqYfX37rQHS/2XtGCkqYJKu+oEct+bj+nv3HPWzjj0ZTF4+mpqdWSNWt+lmA1c8fTn37rx9Og17NgU0EhCopOq3AHJubj6e9+fXuI+fVfe84ainrGkTITQys9oxE5+hNh9D/sPejig60KV6FPrPp/tTfmLmrsZNkZKKON1LKZAGVl+t/8AW9mtvstzcx6wD0cW2y3VyniKDTpv2z8co4t3vHvASK6VN3p57lHYSXJYn/Dn2kubSa07XHSG6tZrR/CZej8v1xtPJbbiw+JpaGkNPTGNZqdkWTWi2Xleb+0o0spX1FOkqvJbzR3Cg1RgeHp0Fe3tn5bYste1RKZYmlaXVMWa0afpCD/W/wAfeP8AzxyLfybj9VYIXWh4evXW37r/AN6Lk/b+UTy9znP4EiUCeuPz6Uu3vkBt3EZBsVVVYgq2kEfqjCqfVblifeKHuN7L8x79ffXPGQFBx+XWWdt7me23N3bY3y0PkxA/y9Bf8hcjTbuzuDnqJIqnFyxxvIoKtGWb/VqP8PYk+7bsR2LcJoL1CsqEgVBrg+XWK/3iIrcX8MVqQ9uaZUgg9JDK7Apf4TR1+23GJqaaJJY5KI6HYpYqPTb3lfd29juDSQ7hAswao7hWgPUAS2em1VrJjHjgMdBT2T8hu8ZsEnWGRrMl/dc6Y2qneRgRbSTcGx49xIPYbk2w3U8ybZEv1FScAfbTh0TcqWW32HNsV7vvclVrX7fn0Knx/koaOGJqaRDL4IxUOzBXkcfV2v8AUn6+4g93kuHnWCVSAgoOs/J932mbYUXbWXRTABGB9nQ/5mnbNVQCEgarErc/649xFYuLGGjdRrdK13LpT5dCfszZVNSRxTSUwkmaxHAJNj7BvMHMLyFo0eijoXbNssUMYcqCT0YnF7ZjkWEGD1uqgRqoJ54sPcP7jzA6M2lsDzJ6G8NrCkYlegp0afq34t1m72gy+RpDBRKQ6qy6dYD3uQR/T3AXPXv/AG3LcrbRYP410R8IyeHr5dRLz57m7PsUb2FmQzmoJ406O3L0vtnHYEYdcJRlFp2jkZooyZGtb1EDk+4F2f3c5rk3E7puErxVfC14CvyPWB3OF7eb1ucl2JCQ2eqrPkV1WuyNyQbp27T/AGEVFKuqOmGhGdG1KWtxf30V9rubYudOXTtu4t4ski+foR0Scl857pyZvQYyt4B+IeWejCdcfLXcmztl0uV3HQKm3qCKKKavMZurED63+vHuCObfu08tb3zRI22uUvZSToDYP2jqQd83Sx3uX63b5dcsudP29GS2h80Oqd3Yp6gZkLNJC1oBGF1Ei1wb+8e/cL7sfuDZTCysIkMOsEsWNRT0x0t5b5P5svJluGtiVrg/KvRKO192024tyVOQxTCamkdtDgDkfgfX/be8h/b7lmbl/YYdtv8AteMAH8vPrPLkXbriw2JYbtdL06KRvXZ9RnFnydZEdMPk0jizC39D7yJ5d5kh20pZWj5NOhbtSCxuvrJBkcOqvfkThYKP7uNUBVJbi2m4IXUPp7zl9rtyl3batVz3FR59Pc7BbnbzdEcRX+XTV8b94wjAZimrKsRQU0kiWdrDSpvcXP8Ah7mCbZ2u2R6YHUFbXz6dltZ7OEhSTSvQg5/t/a6ZOLF0lajygc2IK8f4g+zgWLRweHFSvUf3vNWxrf8A1m93C6Cc9wr0x7l7owWMw7pHUpJVF0QIlmPP04X2SJBuUd3plNF6G/8AW72x3DZydsuUMwoAKgdGQ6n3XQ7g2DVSCdTJPSyMI3sreQLwVU/4/S3sou7513ARue37epb5e5TspeVDu1g8crsKgKwYj9nRasPNuyp3FnIJfuBQrVuIfqV0aiBb2Vbklkj60pU8ehVyXNv8qtDdagi4Azw+XQkYtMrSVQZQznj63Btfng+475m2gbrR4xwFOhPuGyXW5AwSE0J6V88WVySxCV3hEc8M4sbF/C4fSQOObW9h/b+Ukt28SX0I6rtfttbxyh5lDVBGfnjq3XpT56bSq9oYDqjtboLZeUw2Hov4Ym8snjYXycUHAM0NRa9wLkX95D8pcy2Mdlb7RuVpDMkI0h6fqAfb1it7ufdVuthnuOcOUt8njmlOs26OdFfQr0MNR8d/g53gRles+zpMdvJr/wC/YKrS0QqHF44kk1AW1G309yO+27HufdYTGM/wkUHWM9vzz7rcmMbbmGx8W2HGT4jTzNPs6K73F8I/kd1fTVG4DtujqtnIJHpMhRVkVa8lMq61fRDc8rz7KbnlrcIRqMZYZNRkdSBs/vhyZuIWA3Bjm4MpGmhr86dVFdnb/wBz1WSrtu0OMy0NVRSfb1LGgmjjV+QdMhHI4PPsAbjtaLcVl/Z1lbypzpNNsoWxkXS4wQwr+zpC7H6brd15H+I7mmadFcMaWpk9A5+mkngf6/tNDcqJhbQCnlUdGN9sUj2D7zubl6CoDVoejg7X642ltp46mKgo40hAJtpH6QDYXH+HHsUIWSPwxlvXqFp0hubs3EigKvAeVOn/ADu+8ZTRvTYmSKBYgUcIQFvb8AexVytaLJPW4Go/PqCPfTmCex2kx7XIY+AIXoF85vuKOGZmnHkYEah9b8kXtf8A3v2Id5vJIgbcDHAdRF7acu2t+V3q4asoya+Z6LjXjI78zK0GOWaWN3tO+kkKS1r39hdWEKmRsdTk8c+5TCCKpUdGu6V6+G1GkNRFokAW7kAm99X59k9/c+NgHHUhcqbSNuJLCjdGcliEgF1upF7sLk/4n2JeTRSbPUMfeMqdv/L/ACdB9vDZOM3RjJ6CekiIl4MpA1L+Pqfcs8esAADX8uqVf5g3xipaTYORymNqZKKOngn8kFM9o53KFgXH596+fVuOOtZ2ohenmlp5BZ4ZXjcccMjaT9Pfut/PrEOD9L/4e/dePXjyf6e/de68Tf8AFvfutAU67Av+bW9+68TTruwAN7cjj37rVa9cRx/j791br1tR4Hv3WuAz163Nvz7916uK9d6Tz9OP99x7916vXH37rfXO4Yc8W/1uffuq5HDrr6Egc34Bv791viK9esObnkWt/jxf37r1fTrocH+vv3Wz12pF/wAc/wC8e/dVIx101rm309+62OGeuvfut9dkWt/j791oGvXXv3W+u7Gw/wAfp791rrwIF7i/v3XiK8Ouvfut9e9+6912Bf37rRNOuiLce/db697917rsWsb/AOw9+60a9eIt7914GvXXv3W+ve/de65aT9P99z791qo66B/ryP6f7D37r1Ouj/tvfut9d/gf7H37rXXakA8i/v3XiCeHXRsDx7914cOvWIsf6/T/AGHv3Xq1x11c/wBffut9clP1Fr349+60eurc2/xt7916uK9eItx791vj14cm39ePfuvH16vZ/k9d9y4XdGb2FujMk42WngpcDSTyWiWR1CIg1H6g/Q29+wDU9a+XWyvHFDS1P+VaA2hXBB1DRJ6lN/8AW597I68D0111XTirFraFIKNwfofr719vW+lBTB69dMXquouPxz+Pfj175Hj0lspNj6OtSCqVTP5PRpAJuD9D72B1rFcdZodc9YIYlKNIQLrxwefr711uvr1m35X0O2tvszhZa6WAmMtyyt+Le95PHrVAMjPSI2dV5TcOEV6iWWBXZhwbcE8ce/U62M9CHis/Pgomp5aVZYacEmdl1OzG/wCR/wAV97VDI+lBU9bUO7BEFT0Hu5/lts/a80lBWsqzU4PoEQ5N9IHtZ+7J6VIz0u/d10B3Dj0S/u35bbl3di66g2FQyfcCGRaaamgYOXJsvK/X/b+1tvtwCfqcelsO2gJ+rx6MH8QU39tbrHPb+7Jqq1q3KY2YY+lrncqJ2UlNKv8A09nAex2fb2vLkD5V6GnLXLp3BvBCVr1XP2V3N3TtnsfPbxxdRkZMdJLI8dEHcU6RiQhSqnj3HMvM31dzriUaK9CDcOQobaryChHl07dd9oS/JjLxYDcO+q/b2bWZKd6NKlqeMyEXA5YD6j2MdrvNu3IrE5Cv/LoIz7HGhpSgHR/9v/H7sXZFA9LiMpW5+CqhAiqBO9QQCvDArf8A3v2X807XJBpltDUg1oOhJy3yRaby5F0Qin8ug3qtp917IzX8TqjkKmgp5RI8FQztFpPOmx/1vYNmfdSjXEqFAOpCi9u+WYWWyhYSN089c/JAdh9h5PYGcSmoJKArDFysQeRRY2P+v/h7UbFuEt5OsU/DqM/c7kyw5dszcWPxgVNOhuy+NXH5KaCQLMGVXpipDglgCun/AG/vG37z27vNb2u1o9PDYkgenU7/AHRttW2e63OSPMiYJHnTqVh9hZ6vlavkrKmkhkHEKllXT+Ba/vCm/wCZNttV+nCq7DzPWZllsF9cyG6lYgNXFfn0+VPVtHWsElolqquTgO6l5Hf2Xxc4SRDUsmhB86Dpfc8uWscZmuVFBxJ6Zz8Sd25+dqg7eaOgNjHII3VSD9DyPZknu1stsoiW5Bk8xUdQjzlv/LOzAxxupf0xjos3ZXxh7N2/X18uHrsniaSKOQx/bSSxgFB+ApH0/HuStg572LdIEEyrIxpWtD1E+6c4baLZXs7jQ7GlAf8AZ6LhsD5edsfHnfsO291VGQy+3Wqft6iordc3ij1ga7k/j2IOZvarlbnzaC8ESxzUqCtAa9CDk/3U3nlu+iF05ltpGAJbOOtvf4Jdw7A7o66o8thlxlZU+JHkL+MTg+MOxMdybjn8++I/3leSt/8Aa/nwy7w0hsJRpWlSgYmgJPkD0JvdK7k3Mx7ztNwxjlFaA/t6Pdk6DGfwytP2FMPHTSEaEAIIHFre4EEa3T6KlGWjAhv2ft6h7br+9N9EDI2WA6pq7ifVufMiNRxJOuj6aVuefeentslNktfEP4Vz69dB+UGMfLUNfNR/Pqtztjaa5iauZ6RJgyEsWFjYkn8+80OSN4+jWFQ9Mjo52nwWvfDKBmc0yOq9N69c4+meaRKGKOQM5+l/q1v+J95ibVdRXtkhkpkCvT3MfKFhMC00KkkdBJNS0VDTvTPSxO7XVUK/qtwAb+y67224N2RbklfI9Y9cwclWrzmK3iABrw6Bx9p7q21vHG752w9Tip6OuSqnioy6CWFWDaWAP0P59jC023bb/Zpdu3IBmZSBX16J4vbu92mRNwsVKOpBxXq97qTtWl3915jcnkwi50RpBOCo8l4o+WY/4294TXXIO8bN7iix2cssJcMaHAGrPWS+2S7xf8rB7gEBR/k6Ukwq67WI1JDEheLcfg/69veZNw1zZbdFbRVLUFf2dJOS9r2uW6ebcqUHr1yp9vmMKZISXk45W9iR+fZntAnCVnPRLzydsNwYtvUADhQdIXeuOMbwUd5I3V1LMqkggt/X2m33cHiiMUbDuwRXoU+0nJNruN2dyvgKJkA+vyHRiupq9EpY8ejhSqoBc/qOj8n3zj97dnmtd+ku3U6HyD5cepI5us9DCUDAx+zowdLi5XdXmI02BAJH1t9f+J94+T3sarSPj1HE12qrpTj06S1eMxULSVDwxyKPqSL8D/H2ijtr7cJRHCGYHpCkV7eNpjBIJ6CDcnY8gqPFjY9aKbF1F7m/B495C+3nsVu3Mkf1N8hjjPAnFehnt/L1vDDrvnAPoektls3UZmgP3DMrgG4PHB95Kck/d4sNkvPr7r9TTwB4dBqfe7fY91CwtVSegOq9YqZUQal1OTYcf7A+5fn2u0s2CLCBpwMdTftG52+4WaTBhUgdQvIshKFeQPTf6Bjx+R7YqSaHo60slGB6RuV2pJlZSZIwI9J034H9faywuJraUODivQf5m2ay3iyMBQMxBH7ehB2DteDDmIykC7Agcm/P5v7GsW8C4AVj1jjuPt6+0M0luuak9GQo4AKdWuqIEBtwAAOL29vNIGNVz0XLFJENLeXSF3VlYqaKcCUlo0LFSbfQcEf8iHtZFbOwDlcdEd3uVmsxty41jyr0XyikgzGbkqXN0j1EMwNhY/776+5h5OmRrXwj8XXOj7xe33Y5gbcCp8Ek0P59C7gmoxIGSRFlVQL3sSt+B/xPsYsesdVGM9Lr7Wly0DQ1MkTwupUh3Xn/AAW/5916uQKZ6SE+wdvUbvNDTwa2udVgTcf1Nvdq06rSp6CXsjE4/G4HJVcNHE1S1JUJGyrdi9uFBA9745PWuiSbN6soaihzXY25KKGF8caqqQTWBtCdaNpIv715depXiekBvnuej3X1VukUFTHHFT0GQoVUME4SNkGmx9+qT17161PtyyNLuHPSsbs+Wr3LfklqtyTf37r3p0ycE/0FvfuvZA64+/db67Fvybce/daPXXv3W+ve/de65fp+tjcf73791X4uuPv3Vuve/de697917r3v3XuuyLe/daBr1//S0AFOk3/w9+60RXHXdrnj8/X/AF/fuvVoM9cf8P6e/db69c/19+6912AD+ef6f8b9+60TTrr37rfXd+Lf43/x9+61TNevX4t/jf8A4j37r1M1669+631737r3Q3/H7cVBtnsjC1uRnEETVdNErE2BkMnpBb8e/da49bTvU/YDy4TCzQzgq1FC0ZSQMCpAIHp9+4jrx41HRgctuTPZzESUdDE7eVCheNSSVYEEkH/be/deNfLpNdX7CyuDyklW6yF5pDM+pSLFmuRz/T3vHWxWvQg9gbSnzUKCXyKuoXWx+t/6n34Up1454dLrrbZkGLold5HDKo/b+g4t795V69506EfIRxRUlXrYaGp5UubC6sD/AF/p78OtH0HSJ60w9BjcdlKpIY4S1XJIZLWuTfgE+9edOt+VT1iym46LG5MzCeBFH6/3kA/x9+pnrdPLoDvkN2Rt3J7CfGiqgml+6BEayB7Np/AHv32da8ug76NlnnwiwGUrCxDKGbRZDyAL/j34V60OHQ05VMfRyI3mgBVASTMgOofUm/vZx177OkXmd0Y/xeMTRhxe1pF+luL8+9E6etD7Ose1avH1Jkq6iqhQK/C+ZPVzcgj3alRUEU68G9elwu89tUDuHqEbxKWLBlYDTz+pfdTXAHXqgnHDoMcl8j8jTbgipsHSJV4uFgksgXWAQebkeze32xpYtbcejiDbvFiqeJ4dYt+dqTbship8EgeulQa4bEDyFbED/D3RNrnd9JHTC7ZcFipHSO2N0HvTfueopc3TTw0slTFIxSNtOgNex/w59nNlsYx4g6PbHZNNDKMdXbdZdd4zr7ZFLi4YUieSnRZHK6WZgtmJvz7GEEYihEailOhvFAIIgtKDpD706Pw+71mraGQ01e+u0kC+o6hp+q/4839tTWMNxiRa18+ktxYwXAppBPRHt59fdidNZFq6GfIV+KEyvK0iSaI0Y3Nj/QD3GvMewXO3sbyzqy16Pdr2PZ90iNjdgRsFJHkT0JOAz2K33gxWRPBNPAvhq4i6s8cn9oFf979llvKJ4e8AnzHUM7vZXG07pJBCxjAJ0n5dE0+S3StXW459wbVqJ6Kuh1zMKRTf9tdd7L71LZWU6lZIh05Zcy8z7XMstpeyACmASOHRL9t9p7nCf3f3FJPJPQssSy1OpXVY2/UNfuPty5O27bLz967dGFJ40HWS3Jnu5d83Wi7FzFJWRRRSxzj59G72V2dS5WjhojUKWijEekyfUiw4B9o2TWdQ/PqRbe/Cnwa4XA+zpTz4Oi3dOY3pY5LggMQGtf8AIA/PvSs0Wa46UG1juzkZ8j0nKvr7cOyaxchh5Kk049fgRW0aQdViB7CvMnKOz82QkT0RxWhHr0Y2e5b/ALGQI5GkT0NaAdC51n2jRZHKQ4bPhaLI30oJCI1e3AJLfn3iHz97d7xy4ryKhMdcGlcevUw8oc4WW6TJb3JCSfs6tV6I6wk7GVUpAk1reNoiHDKBwfT7wI93vcC35Fjae9YovnX16lre+Y4eVbMXE9CpAPHo9vXfximocvHNnIy1NCVYK4+hjcOoN/x7xF5t9/Bu+2tb8rgyTtg/IHFeok5t95lm28x2JClvT59HmxuLosXSRUdDClPDAippjUKCFHBIH5PuCIopnlN5eMZbhx3uTU1JrQfIeXWLm4bhc7lcNcXLFixrnrlWR+SJlC39JBP+v7MLYF5gGNR/l6KbhAYzQdV9fK7AwSbTrZzFHqV3Yiw/SOfr7zW+7bvM53pLbUSKAdRpvyARsfMHogO5N54yp+Me7sRJR0zVUWQpqWEjSZNNgCf6+8p7rZbmH3h2+9jlYKY3LDyr1IHtFy3uW6zHdmBaCGlcYwekN1Z13JTbPxWZpYJJBVwxlwis4VmI9Jt9Pabn7mn/AHdS2ErBSpNK4qOuhHt3v2y7lELN0RGhBB4CtPM9Hg2V8eNw7nxEeQ8Dxq6BkuhUn/YH3iTzb738uct7k1jPKC4OQM/tp0/zD7pbRsd6bOMqdPzGOg57q60l682/U/eyESvHLZG4/Sl/Y69q+fLbnjc0lsMoCM/n0Z8sc5Lza5ith+zqlXtDaVVv+mz1RAjJDTNMQ6gm+lDf31f9t4Tseyh7kgBwCOm/d/ny05H2X6e8IqVzX7OqvYslm8FX5vbmKqJ1Zqwwv4S3ku7lfUq/n3kLHuUv0aPEtFK11Hhw65U87feRs0uJrbaXEkzHSEU1NTgYHQp4HpDeJpP45X11XqqF8oZw2pVI1AC/sPvvRguESZTSQ4av+TqCPcO090LXl5+cLy5e3hdS6ocVFK+fSjoOnqzIATVWXl0ghirpcahc3ufZrcXloO1kJP8ApusJh79+40EpNhuLKq1xT0+fSm2jkNxbP3XTbYxlbWV8RjeX0uWgKhgNKqn0H+v7Dm+7RBFCu6wvo1EDT5566G/3eP3wvcbmb3Gh9td6jknjlejuasjKXC49OPGvr1YRgcDBJhaLITU6w1tZTrNUhRZhIwubj3H96ZVlIrUV6+lW2t7O3PhpGF9evS4go4kQC1vrYAD/AH1/afxzT06UfSxeJrXiOsazGnDEgE2t+OCPdipkIVTWvTj3It1LPgLxPWNsnKsMl1FvwB9DcH/eOfY22DZmUiZxjrHP3X9xLXwW2+0YaqU49JKHfW58DVs+1aqpxVYzXFRR60k1g/UFbe5OstsuJx+mCAPMdYTc188bRtv6O6FZXfgpIIz0bToP5XfJbY+Rin3Hlst2HgdSE4XcMslTRGItcxmGa4A08Hj6e1J3q42hzEjavKjZHRAvtjsnP1ut9FCtoGFdUYoftx1ZFRfIH4md60K4/uvqfaHUVVMBFW5zb9LDDVSykaRPLpAuT9T7sd12HcB/uzt1jY/jQZ/Z0ki9ufcPkO6E/Jt/LuAWhEUhOkedM9BpvP4B9Nb3pavK/DfsP+/mfqIyy4vJZCGmUyItwkcczAcfS3skj5Gt2uDf7Vc+OCahCNJHy+fUq3H3pubYdkHLXuXty2EcYoHRakj1qB1VL371R8u+ifuYew9gTUFPTGRXegpmqgyqfqGpwb3Fufa07bLakteoYwD59Be15723mRwnLM3i6uGo0/w9Ewj3Jv7Kg1BxGQphIb2moqiEm/FiHHHsRbIiPJSI/Z1EfundXtsn+7FQQOOajpb4DYW890zwx1UE0UMxBkbSeACL/wC9+za92iWb9R/LqPeW/cbb9tC2sJUA8fLo3+wOr8XtGm1NTxTVTkF5XADrbk/X2GrjYLyY0ANOps2f3Z5e21AzMpY8c9CzDHSQ+pDGDzq02F/9Yeyebl28U0CnqRNp93eWJozNPKqmmMjqQctFLaElQwXStrH6fS/sZ8r7FLZ/rSV6xt98Pcyx5i/xSwIYYFQesGTzOKweOmyuUr6Ojp6eN2cVFXFAzWXV6BIef9h7Hnn1iqoIFD1Q18+vm31rVbezuzcPlKeuybmeNaeOVJdciqyAJpv+R9T71x69wz1rW19U9dWVVbIqo9XPLOyr9FMj6rD/AG/v3W+oot+fp791v7OvG1+Pp79177euvr7917r3v3Xuu7n6X/3309+61QdeBsb+/dbOevXN7/T/AFvfutU8uuvfut9euf6+/de697917r3v3Xuve/de697917r3v3XuuySbf4e/daAp11791vrsEj37rRFeuvfut9e9+6912CBY/nm//Ee/da49e+p59+695Y669+631737r3XfIsf6/wCPv3WuOOvEk/X37rwFOuvfut9e9+69165/r7917rsW/P09+699nXrfW30H59+61X169c/1Pv3W6deNrcXv+ffutZ68PqLfW/v3Wzw653HN/qfqP9h791Snp1xW/wCAD/W/v3WzTz64+/dW675sP949+6111791vrl9SSf9j/tvfutcMdcf9b37rfXvfuvde9+690vetuwc71hvHB7z27PJBX4atgqmRZGRKpIpdbQygfggcf4+/daIrg9bjXw0+WO1vlBsLFPLlqOLe9NQxNlaCWsiRofFTqugKx5YW5Hv1fTr3z6OPW4GodbWJe3pKrdf8Dcce9/M9eFelTgEbH42X7iSnWSNeGkmRGuBYKQeR72V86/z60ZAG0gE0+RPQZLHQZTO1FTUZClLQszMhqogqWP0Fz72FBxUft6qDJSuhv8AeT0t8fmNn0VYslTl6RZQdKAVcVtQAFuT7qVAGSP29OKtwxqEb/eT0ht+x0W46+nY19F9oXslquGxUEG559+AJGKda7v4T+w9K3CU1Lg8Uohko3hZP22jnjIJAtxb/eLe7GKR+1OPWlLP2gGp4Y6zZnP4Tam358nXmmmlyEZSOGaRBp1EgML+xns22Jax+PcgamHn0NNnsI7SH6i6WrN69VjdmSbUy+XqMi/25lqJG1Ra1KC73Fvai48MselMzK7VGOjW/GfqXae8moI4MPQ1LyTRCZiqPpRh9SD/AE9q7SCJ+Ar0utYVmGRXo4HyX2nBsra+3duY2L7ajSRRJHAtoypte+n/AIn2D/cy3uItviFuOz8XUr+3n08N28UuD5dU7954enhpq0Qx3jMYuVAN/rcC3+8+4ZsHoo08OhvzLEKkEeXVQe+KvLYPP1NbtTKVW38pA7SJU0ZaOR5FcBdRX6+xPbmRe+M0PUKblRZipHV338qn57LW5k9X91VsdQI44qSgyeTmXzTSsAg/ck/r+PY/5Y3eKef6TcqEeVfXpRbz3MVv/i7FSPTqxf5q939T7JxNdR0OUotdVDGYXiqImctMpItpP059l/uHOIUFtZAUbjQ/5upI5HvLJC95uLnUPX161os1v/NR9uUe4dh1E0tZW5MFxTsWMn7lwGK/4e4+252sYjOTQpnok5mWDmDdBauaxytpz8z1ft8fqLcO9cTQ5XdkEq13iprxOrareK/0989vvC87TXXMMrF+FRx6zs9qPb/bOWOVreC0UazQsfkRw6sB2jsaqzdRS4rH0ck00pWLSsRYD8C9h7wa5l5tg2yGTcLyUIi1JJNOpkurvb9is2uLxlCqPMjo+PX/AMP8RSChzGfB+8QrK1KyegGwbSR7xK5q+8rv10JLTliAzQMSglrTIwf+L6xk5896Vull23bVULwBHH7ejAZPYOHipvsIcdBTw08a2ZY1F9P1v/sPYQ5f9wd5gkWfcZiZ5DlSeFesOd9+t3O8eeVyQanohHcezaSsqsjQRUCMumRdYjBawuOGt/vXvPH205jlG3w3U0mWpXPQDuo5Un1aj28B1Sr3v8PcdnKrMZWShZ5pg7wr4WYRsSfWvvNPlT3DCwRxK49Dno/2/maZ3js7he1fM+XQW/G3s7tv4fbmV8NVV9RiJKrS9BMZVp44SdH6SfbPuhyDyb7xbI+277EjkrhqAmvyPWR3Kl5HuVp4Dt4kYGB5D7Orx+rv5o/94ammw248XSRJUww0stTGt31OgEhJHvALmX7jGw7JaXG67Fcy+KwY6WYlfMgUPp0FY7HeNz5uXbtpiqisDw4Z6gb+z2N3TuSty+LcNQV6tIrD8eUX9s8o7Te8u7PFtt9mWAhftC9dC+WdtubDl2KzuxSQKK/b0XTfG31WhmnUX1qQxNvofc0ctb00tysdaaeHQh2cRbbcePIoZj6+X2dV5do4uKndkU21GQH+v6+L+82fbTdJ7228Kc1C8P2dCncJPq7MSPxPReP7q00ldTvKA15l403BBPP0/r7ln6jShoM9An9xJPOhYfi6F6p68x9RjwESmF4L2Z0HOm5+vspXc3Empq8ePUiS8oQyWgIKVI4EjoauhdvtQyTY5kCwRSFxo9Ua+n8Ece9FbB7sX8lBIfM4J/b0XrYyWtnJtiBdIBPHozdVvXZ22/NS1dQjVFMnksulwSBYjj2N4bc3MAmpUdQBu+8QbXuDWcchVs4Hn0Wvd3y0rcfXy0O3sDT1kUblUd4CTYG1/p/sQfe5Po4EI10b0r0ztz75fXgl+mLxV4kHPSeznyaTLYKihqcVTR5tjaZRGVkUH6n6e4Q3jl3mXc+ZjuEcjC1UgqAcfn0tu+ZPcbat7gXZLIi0DDUQp4efQudT72rpsfDlJCYZjOW0/pvGeVv/ALx7Q+4fI+3c12H0ZFJVXB8606yuU/1h2uO5mShZBqFODUz/AD6NBH3BNHEFIAIjAvcfUC1/eKH/AAPO+PcEFO3Uc/LoINyQjPqFePQd5neeQzFTf7t1jkb9IYkH/afeQft97UbHyzCr7hCssmK6h0I7TYrewtz2AsB1Ppp4DAAo1SgjU1r34+nvJbarSyhtVW2jCDyA6gHnDd9yg3J41Yqo8usUjS1OpQSo+gI44HBAA9mxIRdAPQDeWe5lE0nceoE2FneNyISfyXC3JJ49lO5Wcbwa1yeh7ydvV7DuSxXDlU9Ok8+NaF2WRWDKf9uP6g+wQylXIYU6yRjvVmiV0NQR1yMYiNy2lfxe3B0/19+K1694oK6vTj0/4Kpp0cOz6tJ1W44I/PPsSbXtLOomY06hznHniCF2sLcAnI6UdXukxCS0ioiR8AyKC3+Fj7FlraCWVYAK1NOoP37fG27b5tzLAaVLUr0B24NwzZyqamppAXZijqGBNjx+Px7Ht7Z2O17WsTZc9Yn8t8y8y8687vfQAi3B044cep6YL+C4p5EBNRLHqJt6gSL259u8nK7SlwcdIPvDNBHtqWk6jV6+dei37z7Q3BsqTyRJKY1lbUzKw9C/X6/0+vuTjUjrCZBSo456S1B8pquoykAkrUgp0dDLqmCW59fB+vupp1fI+zo3e1+0aLdtLFUY+qSoUqqPplDoGtdjcf8AE+/UqOt1I4dRd8biw9NjJf4rU08cJRtIaZNTP+VIPvdB5daBJOeiT9mbzrKzamUxOAfRj5oqiN/CTpZWFj9P8Pevl1rJx1Td3FmN4YHamUwe2vLokNRLUFdViHvrLEfX6+/dbHDqnaqeb7qpM/8An2mlM1/r5C5L/wC8+/daoCOo4BP0/wBf37rxNOvC35NvfuvGvl1631t+Of8AYe/der69eAJ+nv3XiadeNvxf/Y+/deFfPrr37rfXvfuvddkEfX37rQIPXXv3W+ve/de65A255Lf4/wC39+61Ty8uv//T0ADb8e/daFfPrr6e/db697917rs2/Hv3WhXz69fm9h7916mKddfm/v3W+uXBPPHH4/r791rI4ddcW/xv/vHv3W+vC35+nv3Xvs67uATaxH+I9+61kjPWWKd4J4aiI2kgkjmjPPEkTh0PP9CB791oA06vA+B3yEj3NQU+1M3UA5aklhgCySRf5tXAVkVjcKR7959bx1fx1/PRSUsRKIw0qAxsfxb37rZ4dDJS0NMJQwVY0PJZbA3+v0HveOtDJ6jZ2CkkKAEaRYg8kGx+hF/ehUdb4dTMfURU1MSgUAKCQSAFVP1G/vfWvn0Duc33FmtxDbONkLIpL1EqsrKoU+sAr+P9j711s0HDpt3H2RBRRptHbsfnyk1oplQArqawLllvyBz9f9v72OtV/l0AvcWwt3wbY/jGNyFS2TmJlkplMg0DTdvp7959WHCp6JvQ4ndmWkWLK/cyKkieVZEkK+QfW1x7bkljiFZDTp+2sry+fw7RCx+w/wCTo0e0sXvGKCmpcVRlIdCpqCso0kWJ5t7Tm/tQpctgfPo3j5S5gklWBYTqbyofP8unzcGwNyV0kf8AEsnUUcpAYrHKbWtcggE2490226febnwrMVQHJ6Hs3tlLsliLze28J2FQvqOg23ht+nwdJ4osxWT1Og86XJ1abfUD2dbpFbbfDpkbu/wdN7NyrDdkiMA14V6BGlh3o7yQ0VbVmNnutjICVJ/p7jibeZYJz4WUr59Cu19o2ubkSzYHyB6GXb+O3MMLJC9PJV1kysgMgYsAwt9W9qRzQIRqemPs6NV9i5bq6X6fUV/0rdTNsbW3lt+CqjGBirPupzKZZQQy3N7c+1qe5CxLpxQfMf5+hlF7A7iIRpRsfI9ONTi93UVZHk4cEsLxesrEjC5vf+z9R7Jbn332Wyl8IyLrrSlQejbZ/u2brus/BhQfPo7fxq+RuFx2ax+G35TR47xskN20qxsR6vXf3JHKvursW8lUmfLehHRRzL7Dcz8shpooyyLxJDHh1ZXubfm1M3FRSbbropqaWnUkeWG6sw4XSD7lgTRXEImtmBU+hB/wdQ9dPJaSG1ulIZcZBp097BLTyeVwJEDAgXBFtR59+hBGK9WiyusdZe99upn9hZakpMbBUTmlnYOy6nAERtx/h+PdrqNJrVoWFQQetuZI2E8WCP8AB59a7G0Kzs7rffW4gKaaTEnMSs0LmQRiEOdQAb3EP7iurad3StCSeifmCGz3tA9aSKPz6O7tffOB3zQNDL4zVFPHNTzMgAdvSwAax9t0Kt4b4PUa3dlPZtRxX8j0U35A/Hujq45Nw7bRIKxXu8UCPZ7r6iSv+9+096KWj41YOOrbRO0O4LLEdBGa+fRb9v7C3JtzGS5+DyymkfTNFpkJDA+oEf7D3Ddhu0YvJbO5NCWNOpd2f3Kt/wB7xWNy1KYJqM9Gn6g3XS1ao1QQk6PGJVlPiKsf1ABrH2b3EeKrkHrIzZL6KVFmU1DcMjo6cH8JytGsJiimLxkEllP14+o/4k+ywgoar0O08CaOjCpPRYuy+pXjq5crjh9nUpJ5IpoEbyC3KjUP6+/X0NrvFg1lfKHBHE9B9tqks70XNudJrXHQs/Fr5+7k+MW6qHDb2xiybcM8cMuRmA1+FjoY3f8A2/vAf7w/3ONp9zbN57XuZAWUZ06hwqBx6O+a943rfdnG3wrqdVoK54dbLfRvyx6l74w0OS2pn6R55o43emkqKeJkZxylnI/2HvjN7i+wPOftVeTtf2QW3UmjxI1SB6gA9QTLLPbEW+4Bll4cO2vRl09SLJG6OjgFWRwwIP8Aip9w2pYqHIKgivcCpp9hAPThUjHUWtlSCB2c2IWw/qT/ALD2stn8KUSEYOOmLkgRE9EF+VWUgG0MrTF1EkkFSYvUvqPjOnT7zJ+7Xt0p3xLuh01HkfXqMuYJRop6nqjyn3UkezN17e3BOaOeqyIloopGEflWM3DDy299A942eeXmqz3WxXWkakMR5V+zrNH7vf7pTkK9gYjxGXHDqy3+X5mtp7v2dX4jPzRH7CbxU4maLSAjcFC1veHv3stk5jtNytty2KqhsuVBJpTPCvUQX3N13yrzHPHBIQrsaUPVweETExUS0+HenNPTqqKsckZ4HAuqG/vndd7Ws9xLJuluZHoavIjCuONSB0UXG6zbpKbtpKlzXiPPqqf5z19XLkocf6lpZFqPI1zb9H1ube8w/uiWFrDs8lzHQuJMfZXHWavsjBBZ7M9+fjAHVO26M9t3Ye3M7DVTp5KlJ7EOhBaRDY/4fXnj31F5RuN93ncYLRCfCxg18vLqEPvl7rC/Ks25TyUZccR6dV19UbExW497ZfcU6o0VTkWlRmIsyq5ZPrxx7zBt1uL+0WxZtKQDI9SB1gf93f2P2TmrcH5wvGVnHfRvxacgdGX35uymxUEGOjSKKkpVaJ/C6sWjVbagouSfYW26CbdNzk+u7WhNI18uPHqGvvsc889W8sHL9lAyWCAo3h1bsAp8K1PRa9xdnNLB/Dtti0rkqVqKeRZCt7MUK24t+b+xZKtjtrar86pD8IUgivz49YYe13sj7he7O5Q2vLVuVtXID+JHKJD3UOgDT5cDXz6Hb427Vp8ojZ3K6jVUtS37MrBw4lJuVuSfr7Au/wBxc3t4J7htIA7VGOH+x19KP3G/ub8o+y/LkO8TxK1+tW0uCXRzkkls8fn0dSRRFGsUY0xxjSoANgv09h6Ylm1HrphHk6jxPUR4fuIxEpGo/S9voePbQhMraFGT1aW6js1M82FHHqJ/daUsJJGJU24AOn6/Q+zKwsbj6tIyPPoJ808ybbFsc93G47VPmPTqXPtWnVdLkXt/sBc8295J8vcsKdvV5eJFeuNvuv743H9ZprS2FVRiK466xe38LiahaquSKRQQdL2t9efr7HENhDbWxij4kdYzX/NN7vO8ruF6x0IeB+3pS1XYGGxyMtHpjCgqVUpb6Wt/sf8AW9gveeWUMTTk5Oesk/bn3ruxew7REn6QAXy6BDP7sl3JUSQR04qSzkLrQOBb6WNvYGW2RCQ2adZRS75eTqklqxTWK4/2OlBsqg7WxM6TbU3pm9nnyI4lxE81IUAa+nVAR73JdiMUiYgj0NOkicvQ7xN4u8QLcgn8Yr/h6tW6d+Y26Nl42kwXaGzqHuWmMEVNU1W72+/dlUANJqnYkm3+v7eteaL6OkVxGs0Y8nz0X8wexfLm725n2i7O2z8QIqr+WB0YqqwfwS+RMKy7s+w6h3BWJ5IsXg6aIUsc7DTHENI4BPsd7Fd7ZeuJLdfDf+EDHWK3ujy3zlyrZPYX5N3bDHivXVT1z0D+8f5du8oqObNdER0e8dr06tIuQ+6iWbxIt2fx6gf9hb6+x0fIHPWLyACrIadV9diYLdfW9e2H3ViquhrzqDItLUuupTY2bSR9f8fetKk1p1fxJxxY9BbBl6mqlITyJY20uNLHng2PP5+nvRSMmpUdOfVXqjSspHT8HamhmrJ2CLDDLKdRC8xRGQG/+w97AAwuB1RmdzWVqnrWf/mSfOHem4t4P13s/LVOGpsSpp66eilVDpU6Cvo+rN/Vh9PfuOetGhx1S/X5GtytVJWZKrmraqT/ADlTO5kkfkt6mPJ5J9+61wGOoRJP19+63SnXXv3XuuxexsAfyfpx791o068AbXH4/N/fuvEjgeuvfut9d2t/sffuvdete1uSb3Hv3Wq+vXtJvb8/64/3v37r1RSvXha/P0/Pv3XvLHXrfW3IH59+69X168Lfn6e/db+zrr37r3XYvzwDbk39+60evcnm31+gHv3XuHXiLfX37rfHr1rf7H37r3XXv3XuuwCfp/xHv3WiQOuvfut9e9+691737r3XYtfn6e/daNfLr1rmw/2Hv3W/LPXX09+6912DYWsD/rj/AHr37rVPPrv6/gAD6kfXn6X9+69w64n/AA+nv3W+ve/de697917rsDgn+lv959+61XPXXv3W+ve/de67Fvz9D/t/9h791o/Lr354/wBh791v7euvfuvdchbm5I/1vfutGvl11Y2v+Pfut18uvfgf7H37rXXXv3W+uQvyF5H/ABr37qp+fXH37q3Xvfuvde9+691y9PH5P5H+9+/daz0NPRnbPZXVO9cXl+t6ivfJfdBhiKQyaMixAURyiME2+gJPAH19+44HXutwL4t/Int7d/WP8e7c2jHgauXHL9ggjs7yrT+gsSObn8+zGDbZZV8VhQDo623Z572RVYUUnjnh0DO6u2t+5jP5GlhmqKWnlLKvikICam+oHuPt33Ke0u2iTy6yR5W9vNolswZFBJA4gnpNYzZ+9a+eepi3RkllqxyoeQD1ew7JzHJBlzx+Y6kmw9n7LcF028df9qf83WPLdJ9iVKGSn3Xky49S6Xm1X+txY+0j83wopkmeirxNR0cy+xrwQVWEH/an/N0g8lsTuGggMc+dy5jprlHD1ALC/HtHH7nbUr+Hayh29NQ/z9Vsfu03W7qZigRfPB/zdNEnc3cWzIKKmip6vJw4qQSSyTuzeRU+oYMb/wC39iTb/cQxsGNKnyx0R7t92kbaWuIwXKZoAegl7H+Zu+N8zR4iso5cYtOwCpGwVGZTY2/1/r7GD83319GGTy+zqH9x5cFhcNbXaaADQYOOguqt97hyUAm1yMVIYktck/0J59nG0b1Lefpy8egVv+x/SILi3NR1ZJ8HvkRlcBlI8Y0YMqvCpaR1W34/Psc7ddFO0dEW23bo9PMdXe7rxkvaWwMpnqyNZJ6PHiogBIfS5S4II/x/p7V8w2jblssoI4DHQ+5fujBukc5wCeqct77LqMglZDWhgRLKrAI9ggcgW94zrW1naA/hJ6nS6t/r7YTngw6rU786DqaKJ87iEMl1eSUKsnoAe5FvZ3aXqk06i3mDl54gbmLuHnjh0RaircvtrLR5HETzUOWo5S0csSsjtJG30ubW+n9fZpLr0+NEaEdA7bphb3OiXIbowOL3Vne1Ggj3tuavqKhVRRHUSmU+gWUDV7DO672YYibhiW6lzlnluz3jcI4Xl8NWP5dHS+I/xpXKb+qs5UypU4+hnFRTiQsf0Jq4X8f7AewZeczrLGLOI9zDPDo19xeVG5Z3za4rWjJNIg1DPEjrYz+IGyaPfO6Mnt9YhGKGGCKwU2BWIDgD/e/fJ775XM0nJMkm68anAB4kmgGes4eY7qTkPk2xvFyHVST+Xr1c11f0lh9hSS1bQRzVEqDTJIuoo3+0XHB98o+ded+Y+fSlnfK1pbIdRFf7QeSmhx1i3z17nXnMaCCFiFB8uh6IBH+PH4/w9htNMKCG3Xw0XgBw6hw9x1NknpMZ+DXAx02ZlYE/8j/2Hszt5A0sLadRQ1J8+iq9jorHhXotO5tgivknqWTUzE3OluR9bXHvI7ln3BSzhjtQ1AKefQOmszIxbotm9OtKZ1mFRRoy2IN0Y/k/Un3kPyrz40ujw5P5jonubYrUqKH16rw7t6Ep56HJ5daYRxQQSSR/tyfVWvz/AMR7yR5X56LSx2+qrMQDkdSp7ab08V3FtxyWah6A7427BpcruOqpZ8fFUzeTRArQk6ZFawNrH629mfuxzHNtuyCeOQoKVOfLrNflDbrDadzO43EYXtBrw8h1bbjvjnn1w1PWNTiGIpG2gLKulCOCAR757Xfvny2d2ewEwaQEitRkjyr0L7z3V2uPcDao1Qv2dFZ72oU2Pjp6SWUO4TnnkfUE2Y+8h/am5bma6W5jWgY46G2z7i3McaS2wwT1WrLgsh2DmJ6eiV5QspB4dhy17en30Y5R2Z+X9oW9uaDWMD5U6U81887VyjarBeP3j5joIOz6WHrHyU9UoqMgoIEF2UI1voS/I549jG0lutyMa2C11mhJHAevWLPun977YvbjZZNytmUyUNC2mlRw6JPufsftqtlnqMRh8kkEZPjiSQ+NkU3WxD/Qj+nsf2vK9pGUM8yVwSKHj+zrnNff3m3O24bk88cMqqrkDSYwtBwoNfA8fs6MB8WvltuWDcFRsXeeC+weoP2sFVJbzh5F0BtTE/k8ewDz17ay79f29xt1y0XhMGOk0VlBz6dZnewP3wNz9yXNvvDANINPFQQacDQkfz6sJHXc09T/ABl5vvo6tRNpkBcaJRr0XP8Ar8exxNKthtgs4zUqoFfsHWSmxbJ+8+Y1vLs6g5BznBz16TZWCd9Bw1GkouC/jGon88j3F9xLcSzE6ic9ZqbZtW12llGEiUDSKinSWzXX+3cUr5KfEUkjBSY2MS672uAG/p7eSe/0+GsjBfSuOnpRtUETMEGeC+VelHtSEHGpURwJTx6yFjVSFsPpYf4D2muFdZak59endrnV7QIE0Z4dK8Ndef1jgj8fX3ozy8KmnSwihxw6yQlywRQASbD+lz/j70CzGpNT03Kq6SzcOhZ2zi3kh/euSfoT9LH8XPsabO0vg9+B1jlz9BaS7iTCc9LODC068kAWsf8AC3+w/wAfZ9DFPO36Qqeo0v7vbtoiE144UfaB09iOjp4GFo2LKOOLCw59sXFtNEdEop0s2vdLC+iF5YOGA8wR/k6C3PtT+V5YwpU3AAAHN+LH6e0DbVHKxkboWpz1dW8a2y1Nccege3TuCDGwAyyqgUnUdSD0hrX5/PsO3Ft/jiqgxXqXNp3dF5flubpqPprxFeHQeN3DtvHI+qtGsXAAeLlvpz7kaysZPAVV9B1iDzBzPafvSWRm/E3+HoN8128tfO70dRI0bErGqyKdX9OB7FWz7ekLePOKUz1A/uRzXc7hbDbNqbV4h0tQg4P2dLfq2my2Vyv8brkZaRmUIrBrH8g/6/su3y9FxKV406GvtXy42zbchZKFiDX7ejS1ECVKr6boAAQfyBz/AE9ijkl1IIHUGfeZilDCQ1Iz/h6APtvrqLcFBNHTUyGySMWsb8rza3/FPci8esOFrpp1TN3z15uTC5oph554EklK2hWbjUbW9I96z1bgeh56k7PrentjJ/FKh6mvmdQqTGxBKACyy8+/Vx1rFeg0393TuDd+RgL1lRDBPUXWJHugV24sF49+869W8+h+xFOzdfh5W8s0tOSeSSdQ9+HHrXn0Tnf22lbDbgq6iFRF9jVEMwK3IUkWJ97x17jw61+88v8AuczATlVyVaB/wUVLAe9deqAOmf6e/db697917r3v3Xuu7/7D/W49+61Trr37rfXYtfk2Hv3WjXy66/1vfut9d3Nrf439+61QVr11791vr3v3Xuve/de6/9TQAJv9Bb37rQ68fx9PoPp/xPv3Xh11791vr3v3XuubEfQAfQc+/dVFeJ64e/dW67It/iP6/j/W9+60DXrr37rfXa2vybD/AH3Hv3WjWmOu+CfwPfuvZA64+/db6FjpfduX2hv/AAmQxEskUk1ZS083jYg+FqhSTx/S3v3Wuttz489kRZLbWH+/l/ekoqfWW/UWZRqP1/r72evDh0cn+OUYiiC1QsUuNJH5tbke9V63gZ6TOa3NSLE9qlbKrC5ZR9Pe/Prwx0X/AHz27JjMe+Lx1Szz1TGHyIblRKNH4P4v70cdaJHSBodxUfW2FapqKz7zcWZkVo3dg00aVPJAueLX9+69Sg6Ml1fSbdo8Kc5lft58xVgVUU8mlpUDC9lJ/p78PXrX29Yd37woLyGedftxqGhmUrpta2n/AB96kkjgiMshAp0qs7K53G7SytVJZiB+3oEMj2Ps/FlmioKVmBu7CNCC31J/4p7jHeLnd+YJ/pdnU6QcnrOv2r5O5O5A2j9987aWcrVVNCSfsPWPD9r7h3HJ9rgMVHT0V9BqEURFVJtqBHsY7N7fXbxIL+Qg8TnoI8ye7+yfvOSbl+0RlUkL2j8j0ItFjhIqz5bMStUMQzxvLqAueV9yptezWmyW/hQaR6nqGd/5l3Tmq9E9yWNcBPIdCBTYzravoJKauNPNXlCFkcKSDb8k/wCPsOcy838jbNA/77lGqh9DmnQr2PkbnSeJbqyjKpg1zw6KzufAVG285NLhKY1dEXd10i6ql/oLe8Rt290tnud1ePbSPp6mjdZJcpbZf20UZ3VaFaVr09bW3LuKuqVp6HDgzIwjAeFrEj6cgewNvvOJ1GSKWiH59T7a+5Xt/wArWQjvEUyAZwOPQn/a9qy2mTAEwrzZVcWH9Pp7A95zXHdwm3NwVJ8wc9BPdvvQ8oWymCyjXHyHSgoN07jomSkz23o4w48bMYtX6jb6kf09gK92SG4JuLe4Ytx4/wCz0WbB94jZpnLq+ip8qDpDdkbZwM6LkIIkoa+UCRZol0yRseQwIt7E3KHMW8bTJ4KMSFPHqbNo9zuU+Z9vKSsslRQhqdBNR773z1w61dFmK7L0ylXWGSRiqBOQACfeTHJPvLulhIIJ2Omo4mvUYc7+1XJXN8bz2DRxSEE0Wgz0bvov55y47K0+P3lB9jSyBEL2UnUX/BPvKrlr3Is910i5YAsOI6wv5u9ur/lSdhCC8YJ+fDqypO6Nt7zxcb4jIo8FZT6yNaC6uv0Iv/j7k0TxTKGhaob/AAdR2LgMPDaoPRKe4OsYqxqnOYOBZZSXeWnT9MgP6mIUf7f2hniPEdFtzb0q8eD6jomeV2VkHrVq9v1E2NzNKpd8fETEkzR+okj+vsL7rsi36Fou1/QdXsZ7ZZBFeIGU4JPz6fdodzSGom2h2FSrjp1hmhildb+aRRZWEh9xz+8ZLC4bbt2GkkEL/s9GG6e191fWL79ymPFjUEvnh6gdLbqGgxmWzWRx00EVXjaysfRHJpaNombg2948831g3J5IDSpqCOsY95+os9yOrsljOfWvSk7o+KNV+zufrhxDVJG0j46lZkjd1GvlU+vtvl/ndrdvpdxFRXieph9r/dC7tm+j3dzpBFGPp0X7bXYe6th5inwW8aWWlqxphS6sVYhvVct7kmKS03GLx7Vq1z1lly7zzt+6MPo319G8p9y4rcWOhWRonNRECLleCR+T7QyRvHUAdSra3cF2yrIeg93h0lt/dMLPW0lJVxSIShkAYq39kgD+ntAL24Rij1A9OhmmyQyRi4tG7gOgQx+yexOpczHlNg7pyePpoJEYY2hlliiPifWoABHH49gXm3252Pm61f6m3SVmBwwBrUdB/eLHY7uE297Eqtw1ederJOiP5l/amxpMfg98Y2avx0SpFLW1gLzW1adWpvqbf1986vdf7nfJm+Xjzxr9JcJUhUwD+zqH969v93sUfdNirNGPLJAHVqWP+cPWG5tvRZipyE1PO8GpqcRKAHZfpYf6/vCjc/umc62e8/S2Ko9sGwamtOoVu+Zysr216pSVMEUx1Xp8ivkfi95V8GOwtTPLRz1MaSOEF1R5AGAt+LfX3mp7Qe0c3KO2l7xFEqqaZ8wOgbeXoup88OgK+WXx52ZvjaWzt47H3W1HlqagpWyOMgl0GWpMY8usIebn6+xX7V87cz22+bhs/MtnSHxG8KTiClcdSRy3z43KtgbSzYnUM0/wdLj4+bFrdn7Uoqajy09NUz0oNTJC0mryEgEsVP1Hv3Pu62l/fMZ4FkVTgGnQF3Tdbrer17uYkFjUdGt2z2Hv7q3JQ5akytdn6TUrz0s7u8dvqw0t/Qe4Y5k5E5O9w9rl2e8t0tWdSodAFbOOI6bttwvtvkEsLFgDWnEY6BX5j/JCDf2DggwmOK7ikUxNH4goR5E0MAw/x9tfd++71/rcbjIrzmS0ZqrU1PHHWZPs77u7Pb7c9nushjwMdVQd29VxYPqKv3DvLPSQZzIRispaFZTIzI0esra/Hvpz7cbFJd7lGu0RBkplqfDTy6xz+8xzvD7jSS7RbOVhU6ccCKdUwQfMTaHXeQqsJLnqmkqKGXxGMppc3JsxB/1vr7yBbky/E7SiilhmnA9Y38gcwcy+3Nk237c7uvAZPA9G96smk77wtTuehzkk+MhMbzOJwstpDchFvyPcG84T3nKO6LaXIKtNWjccdZKe0/ttsnvlvQXm25WJ9Q/tKUyfn0bva/Wm1cXiklmMFdK8QXVPApmFxYKWPP8AsfZZ9TMWWckuTkGv8+uunt37F+2ftrtccWy2cL3CLTx1pX7RjqPgsPuDZWXnrdvlqmillaQwKx8cat/ZC/Sw9mMl1b30YW5wwHR9b7bu2y38lzt7GVJDUjjToye1N5pnYjTZFBS14uGQiykjji/59k9zAFB0ZXyPQ82reTcsEuRok9PLpQyJXwuZqZDJYWX8ji54t7W7FEGuwJR0Qe519Jb7ETbtQn06bKzduYpIvDLTvpvy1r2I/wAfcrWe32z3KuoFcdYQcw80bxb7XPG7sYyDWv2dI+q7Nr6VzDJEzA8Bj+oC30N/z/h7m3awVs1QeQ65jc9SJNzFNMOLNU/z6TdXvnIZU+ONXDclVsSGsf6e13zPQVPCnl1Pw+3MhmZ0kmLqh0kqbgNqPIPss3dJJrYxR5J6Hnt7eWe2byl5eGipx/b0PW39m43GCJ5IYmksGPFyT+Bz7jqTlq9di3r1mDZ+8/LlsqpUELjy6FehFNGAkSBRYAm1gLc8eyq+2CWzhMsuOh5yt7t7dzBfrYWXmc9Zp6+CD9bFbDnm9yOPYbjhllbQgqepjuby2sbc3Vw4AGek5kMrRCRaiFIxVxreOVP1gjnUCD9R7lDlTZJrYC4lFOsH/fz3Vst51bLYtroaHpw298lu5+vK2Kfbu/Nxw4+lbV/CYKyZaSZfqyPFcgggWPuQahh1iEoYdHQ2H/MhoN6Uq7R7f6R2lm8bP+1U7trMdC+Xp0kXxtLHMwv/ALV79SnVgamgqD/LoTW+Nfwp7yQ5nrntefEbynBmXbMsX29N9xKLrAkg4ADen36pA+fW6CvVfHzQ+EXy46uwNXlto7Qiy2z/ALGaSDL0NUKgzUxp2ZnZYrngD6e/Vpg9a0k5HWhR8o6fJx9w7ohy+LrqDNU1T4srHPTSRKageolNYBI/F/p791ocOi6FRpvqH+CkjVx/Ue/deqa9cPfut9d/14/417917ru5tpA/2359+61TNT14i3F/9ce/deBr11+Dz9Pp/j7917rr37rfXvp7917r1ze/59+691737r3Xr+/de65BQfzY+/daJp14gAHnkfjj37rQJPXQNv8AffX37q3Xif8AYf0H9PfutU67W1/VyPfuvGtMdev9bnhfoP63/A9+61/l66vze3+w9+635ddi5PFwDxxe3v3WsefXRABsDf8Ax9+62Mjrr37rfXvfuvde9+6914G3I9+69139bf7yffutcOu7C9geP6+/fb16pp10QRx/X/efz7914UOevAcHn/jfv3Xq9de/db697917rvn6D8/j37r3z69Y/wBD/tvfutVHXXv3W+ve/de67AFjc/63+J9+60T11791vr3v3XuuRa4sBb+tvz791oDz64+/db67H54v791o9de/db67twCOf9b8e/dar14/0tb37r3Xl+o4v/ha9/8AYe/dePDo9Px6+CnavdIo89VYqXE7QqFDmumk8M7I1mDKpHp45sef8B7W29lNcEUFAfM+XSmC2knNEB/Z1dj8c/hn0J0FW0+Tr56bdmbsS8GQVZ5Ked1AbTIw/BHFv6e39wv+XeWYDdbxMNK5NCD1JHK3tpv/ADDOPoYtY+dR1ZjmN47bfa5o6DGwoi0ojpqWOMaIWQWTTYf09xXzP943k+HbZbXYG1y0oooOPUx2Hs3zDZ3SLcpoCnOeq8tx7gyOK3FVVMGMnn8jsyjwuYwSxItYe4Ot/cO73J2uLzSNfz4dTLtUW0ctlI9xYAL/AJOhP2vvvOmjSY4vxvp4/bYMOP6EcewVvO9XdxdF4ZTprgV6mew+8B7d7DCtrBEpkAoTpH+HpeYTuCposhHFk6abxs4DjxelQePYM35Nx3W10JOVoOANK9GMvvjtu+0S0jVU+wDofpcngd0YWSVfGnkh4VwFYsRccEe4oji3bar8HWxoeOehry17g2V5ILeJwB5jHRTd4bSp6h6qGOOPQxbWeOQbnn/eveQXLvNcTWqR3xIYUz0NNwn2rdFMMTCpGeiIdldPwSVDVFDBGs/kL/t8E8/m3uaNj3t5VUxMSp8+scfcLkTa5VeUIoIzXoIJNv7jwYJFJ5Yx+q/Nlva/PuaOWh9QwccesIOfIjtjNCMqOh46Z3RiMXlKfVL9pkDLEH0ixJRiL/7f3Jts+ggH9vURwvGG1jietjXovseCq6nqsbDUirq6mgCAMQSxMfAP+v8A09ibU0m3SRLxYY6F+13YhmikfgDnoC5cJQV1ZXwZGCNKh2lZlfgcubafeK2+JNbb3KkmM9ZccvrbbjscbpQin+TornYnXyGevpXhWSgq0ZFVhdVBP1Fx7pDPpApxHRJuW1BpWip2tjqq3un49SYfITV1HTKtPM8jRvGDzc3/AB7Mf3mwSimtPLoGPyNCbn9UBQeB6Kq+0Nw4atElMsiBXuJV1XUDi3smv2tbyFmnoCOjluTdwtlDWrldPAg9G16S+Q+/OkquPJQQ1GTp53BmjlXWipo0Hg+4jvbKGe5L27kSDhToPbtcbqLiJNzcutuQyk5yPTq+D+V987tl7r7trsdn5YsLX5l6Ckp4mVIozVSIIyCTb6n3gn99T2S5h5y5CbcrJfFeCsjDNaKdWB60HU38we+Ww8z8h23K125FzbVyQMj7etsWOVZ4kmRxLHIkciOCGUqy6hYji1jf3xLS8W9UstaRkoQwoVZMEEeXUEFNHzBzUeh69Zfpq/HH0uPe6DqueI6hVVMKpVXSLfn/ABH4HtRZ3AhdlI49J7iHxh02zYaJ0ZNAH5JPI/23taJ42YBGof8AVn8ukbWIpWnQWbr2hBWpUJ409SkgkAXYH6+x9yzzdPtNwiSOWFaV+XQevLIitBjom3e2zafFbIrPulWMTRSJZfoxP0UX95Qe3XO/7z31IoG1EU+3ob+0fLdzunNaSRrVI2BY+QA+fRTPhvtj7Ts2okrMdEaJatWp5HX/ADl29zz94PYeY7n2rHMNvUQMpBPn1mp7ob3yZY8sR2e2XY+vK0dQR6dXRdl5+DbOzqutWFUH2hWONFAACx3UW/1/fJ3bOV4eZeaLJLVFj7x4pHE041+f+frGTkWwm3zf1hnck1rn0PWv13tX5Xf9bkXHk0PNIhBuFCq5tpt765e11nY8qWsAWlVAPXQLl9bflm1S1UVNBT9nTv8AHLqyhoocxlalY5XpqV5yCNTKyR31c/7D3Nc/uVvG87rb7BFq8NiKkcAK06xX9+drv90f6p3KCTAz8+q9PkLBh8vvjKmeOOpkEs4hp5bXsshGux/p/X3lVytFcxpHHESscYFXHmacOuQP3t+fOXtk5bHJkAFzuFT2sMkVyfsHQPJiqOlg/bpomVQWKjTbSByBf/evYwM0kr5Jqcdc5Rc+ApDNppmn5cB9nUPYHxnzvZG+Uz+KxRpKKmrIpzVCJ0dlj9bMCB9P6ew9ztz9tHK+0BLicCelBQ+fl1k192u/55i5kO5bYXFokgYAgjVQZ/L06tAp9sZrbGOhotPnpoYI0aWS5bXHGEPH+wt7ibavey1v4Db7kmlgcEeY675e133geUEs7deaJTDMoRSABXAA6iUuGMqCuyDRwwuzKjKTcn/WP+v9fYM5j92zBN4GxwmVq5qMU6zKt/ezkS4sUlsbuqkDiR0k964uL7anlLpLRmRSoVtTsSfof8PZpyj7mHd5habrEYXOK0xU9Guze6fIm9367ULoGcnAqKV6ak8C06/aqI4VFtKjSAbc/wCx9yhOjqRqNSc/l1LsUYQgUGeH2eXWNWsQdQBPFr/71b2mqCQOn2wDXgOneGmlUJVKgYAg2sb2/wAPa1LeYUYDj0SSbpYSM9qZBUDOehFxm40pIVDD1Mi+m1tJ+lvcx8o8sz7raeJJgdc8/fz3o2/kXfGtLVvEYHPn59cqrekgjKxG5JKsxABA/BHuVNm5bttuzIAT1gr7je8e783MI7OVo086fPptG6qeJC89YdRvqQsD9f6i/sl5i5ea51XESgAdSV7O+8EOxpDs17I0hcgZzk9BbvHsikhd4KRmkkvpCqOC5/1vcexWjAlH6y4u+YoTEtxbDVqAYdAHuDDbw7A1U1NDLEjE2caltqN7kj2U7pFDbFWjFTXocckXm5b6skd25SLT+XSAqfiju6VzOayRz9WRpGt6uTc+xxys31xVXHp1jL73xNyrFLc2rltRboQ9i/G2fEVcVVmqjXFEwcRSOxHBvY/19yNebUklv4UWD1h3y1z7cWm8G9viWUNwPRt8dRYzH0qUlOkcMcCAeiwBKjgkewDuHKt0gLxjUT1lryl78bDcutvet4aLTy6eo62JyYkYE6RYfTgfSwH+9+zrlLZ7qykMkwp8uou9+/cTYOZbVbPbSHIPHHTTm6qlhoZRUzxwRsGRpGYLZbWJufx7kCvWJgFOiW9nVnVWPWWrq6yCtromaQRehwZB9F59+rinXqdVxdoUlXv3LtUYmLwY+MEQxRXVfS919K/4W96+XXiPPpP7f6zzdTU0v3MTiOOWMq3qsNJ5+vv3W8dHZwG3MlPiqPDwIz6kjj0gHTyPyR+PfqdaHRJfn9v/AAnUnWibXoqmMbvqXKz08bBiI6j9tNRHNrkE+/Y8ut/5etd+WZ6ieaeVv3JpHlc/1Z2LH/eT791U8OsHv3VuvWI+vHv3Xuuxx+L8e/dePXX+P49+691737r3XIC/14H9T791omnXH37rfXvfuvde9+691737r3XfGn/G/wDX/iPfutefX//V0AGAB4549+60CTx669+631737r3XvfuvddkW49+68DXrofX37r3XJiPoPp9f9j791UA8T10oBNjx791smgr12ACSL2HPPv3XiSB13Yn0j8c39+61UDuPXD37q3Ttg8pJhctQZOMajR1UFQV/1axSB2Xn+oHv3WiOr3/jn8ldq5rD49YcxDDlaanhg+x1+OxAsdSsbjn6e/V690fag73jo4Y/uatJWdBo8T+YeocLdOPfvOnW8U6Tu4Oxs9ngTSu8NO2q0kYKto/qTf8Ap79XrXnXoMJ92YrF1AmyWQeprNWpIZCzDyD9J5/2HPv2evU6BTsvubF4rIQZ3eWYixtDSlBTx+UaeP8ANjSPz7oWJag6VLb0gMr8OuOP+f3XlFFSUK7t1oxio4Aglc6pDpQWH0/1z72dVO3poGFiNVehYbfud31BT1mKrKmemqwjxWDXYSmyjSf9f2HL/lrfN6lEeUjrWo9OpV5Xv9q2GL6yMB5aYB49DvtDqjJ1lAmSzhkSPQrlZTYMAf8Aavck7Jy9Y7HaBaVamWPGvWt45g3nmOWk0raK0CeXT3ks9SbXgeiwaRpJGCjeI2Or6E3Hst3jnOz28mOBwWHQg2D243PclWWSIqhPEDj0HM3YOXFWqzVssfkkEZYk2UMbH3C3OPuNubW7C2wSCBTrKXkH2g2Hb4BuG592mhofl0O+1NwYWlNIJciaquqjCuktrbU7f6m/1ube8QOYbzfN5d5Lx2ZRU5Jp1Kb73s1sptLeMIq8KD06s/6t+NKbs2xFuLJUyj7iETQLM6jyK63HDfg+8V+afdGz2Tdv3WstCDQ0HDqF+cOa9/mZototiVHmB0J2B6Ipdm+TJy7OoZqWKUD7oBHNwbA6lHsuu/cnbd1C2cN+RIw+HqANx27nLmK8MUsbBj+HI6Ep9nVFXjnqMZtaiaMoeAU+pFgSCOfZJDzTYR3YjuL5ga9Azc+XN121zDewENnyPQBZrqLc+ZmmnbasOlHJuqL+PyNIHuTbTnXYbNFQ3lSR5noliS8Tth1Rn7COgiz/AE9S5eujxu4YHxPqWPUsZGm/0JP9PYsteZEFsbvbSJcV49He180b7sJ8OKZqV9T1KyH8v+Hd2MM2Gzcs8TQOwHkAJfT6Rz/X2Gm987bZbvwNwj8Nq+nUlbD7ob/bMJZZWpUefVaffXxM3j0w0+UqIKlqOCodUqJCWW0Y1XDD3kf7de8VpvsiLYyipAx1PvK/Mm386kWe4EMzDz41OOk70/vvelBLR+HI1csETLqhLSaVhDeoFSfwPee/t5zTPuMK28pqSOPURe8nt0eVrj95WNfD444dXV9Kbu2lvXAQUlTWxVGRMaxTxTMofyMLMtj9fc1QNHIKE56hq0kjuUGc9Br331jSYWpTO7cjVKpQVaGnF9Yty1l/17W9tXEGlta46Yvbc5CDquPuzDJNjoK77VqTNRzwkTCMhzcjV6h/X3E3uftJn2ltygFJBio48Opd9l+eLDlaS4seYDW1eNxRuGog06HX41YjJY00k9crs9QY5Y3l+rKw5sfeJe7XCzQBXNXQUP29YU+4N5bX/N19dWVPCMpK09Knqy2jqvFTxaPUGjJZL8Xt7BEsettR6AMkksdDGaVI6KF3x1fgt+TGWKkjocgC7JVQrpkDFLAB1t9T7kbla9nsrYuG1AeXWfP3d+U7Le9vGqb9UgYr5k0/w9EiyCb16un8VbFUyYenkEYrGRnbx3+o+vsfWO8We4dkpCv6dZKcxcgcy8qUvERnt+OrjToxfX/aeIzOPQJkldwoVkkOhtRF7aT73d2nfqOR8s9ObJzBMYSgYgjBr0JAFBlFJujrIdRIN/1cc+9xVjFV69cBrqbU58+kvndlUOR0olMhkDK6EAc6fwbewVzTyzabxayXMvYQCdQ48OpG5V5lsNshO23iCRJBTI9cdDZ1Nsuqz5g249KsTzkxrY2uAdINuPeAfN/NUPLl3NM0hZIWIqfkeob95/ZSxML8y7OvZINZ05oT0InYHQldsAR11bjmmgmVmSVkLBQBcNf2k5U9zNu5uUxWUwLKaEA9YV3NjJaTFHFNPRbcrUZahzFLBBk6moppnW1ESSigtwqrcj3KtpHaz2rO8QVlHxdInWjV49WVdGdb5TK4zG1ElPJeopw6xkG1jY3J94l+6XPW3bNNMjSABDk/t6P7G1eWmMno3v8AoIkradUmpzZ15FlsR9CLe8aV97oIZi1u2qnmOj4bNMRUjoKt6/B/CZ2jqa2CARVoSWSMoOdYS62v/iPYz2D72U203kdruCkREqNR+Zp023Ll7oMtoxQ+g6ob+XHWG8djbvkwO6BV1eIVpIKaOqVjB4SlhpP5Fvp77R/dQ91eW+bthS1snRpZhVSDn9vQA3qwvHctQkrx+fWtN88+jMZh8jJvPAUAphHDGar7aMlZUYksJNPHH1B/HNvqfeZEqsrlXyR0Ey5qQ3Eca9FY6D+VO/ejJftcVVTVeGkmjaWhaQhEjDeuyfRgB9Af9v7CXMfKG0czBTuKBmQYNMj8+hDy/wAy7tyzdi82qUxsCDg04dXd9H/MbY/aNPSUn8egh3FURws+PaTxBXYlSND2tyCPp7iDd/be628tNaDVGlaD5ddGfY/70dvftFsXNE+l2AUE4zWmT1YnsrM09bTAGVZGtyoN1vx+n3F93azrcGIL3Dronse/bc1slz4itE4qCDUUx59CdNtqjqacVNMUopyQ2tLLe3PJB90slmW5EUw6Nd8l2+fbHvrMhdAJ1D5dLnbm6tvYil+0zdVTeRE0qzyAs9gQS1/cj2vLk8ai7iXHHOOsRt692ts3K4fZJ56shK/bTHXqqv2tuGd4MfJBIXvpIa/4tx7EG3W1y1ypXy9Oop533fY49jmLyKpocV+XSAr+uZJ6p2EIddVwDc+kc39zVYo8dsobj1zK5nuILve5pITVdX+fqRjNiwwSK09PFGY2IJPBP9Rp9quiLj0JlDjKajVWjWNAq/UEc2/oT79g8et1kX4DT7OnGPIQuWVJkst9TFwbW/x/1ve1oOI60zzH8R6YMt23szbavT1uSVJ04YK1yGA4+nsp3PbRuA0Hh0PuSucZOU3M6Grk1HQQ5X5GbPFQIjXMIiSDIIzYC9geD7KIeVreNw5xTqRdx99d2vrZrdnqG8us0XduxdCumSaRpLH1A/Qn8XJ/23sUwxiFBGvAdQNuN0243j3cpqzmvU6LtbZNa4SaeNUY/qK2Nx/X250nGMHpbYnP7VyLLFjqimlL29OsLe/+qHvwp1o9CJjcnUYJXqtv1zYmtjAKT0kjLKGXlWDKf9jx71x69Tz6RXbP8wnu/wCM2zshuqr3FWdhY3GR+X+7u5Kh6zGypp1+FqeUnUpAsR/Q+/eWOvA1Pf0SHZ/82b+Xl8+KXI4D5ldEdUfG+rEv2VV2F1/teKHO5WGYmNa6pSNtLSKDdiD9frz70MdXLKSF8vn0GHZX8h/4t99YfKbj/lu/I+f5BZfLwy5LD7YyD0O35aWWoBnWlc1UjAm50hb2/At78CTXqpUYCmn51HVGnyX/AJYHzW+I9Lk6zvXpzKbSo8TJoq6hXiyChTysqPRqwKkc3v72M9aYaT6/Z0QN6WeMB5IKiOMkeuSJ0Xn6ethb37rWesbAxlhwb8Ahgf8AevfutceuiQxFxb+p/P09+69QgY64e/dW67JuAP6e/daAp1njpKmbV4YJpgunUYInmC6hcaigNvfuvV9OsbxvGSHUowYqyMCrqR+GQ8j37r1fLrpEZyqoC7MbKqAs5P8AQKOT7916vU5cTlGUsmOyDIB62WiqGC/65C+/deoT5dRpKWogIE0M0BIJXzRPDe3J06wL/wCw9+699vWONHkdVjRpHY2CIpd2Nvwo5Pv3Xjwp1NGJyjgsuNyDKLlitFUEL/rkL7914A06iS088JAmhlhJBIE0bREgf0D29+69WnHrD791vrsGxv791o568Tc39+68MdeBt7914ivXr83t/sPfuvUxTrvV9ePr/j9PfutU4deC3A/3nn37rxNOuJ+p9+6t12QQAT+fp791oEHrr37rfXvfuvde9+691yAuCf6e/daJoadeAuDc/T6c/wDEe/dePHrtb8/Q8/m9/futN1xH5P5H++Pv3Wz12bgC5PI/r+Pp7917HXH37rfXYNjf37rRz11791vrsAEEn8f4+/daJ669+6313bi/v3Wq5p11791vr3v3Xuve/de69cj6e/de67+tyf8AjfPv3WurSfgX8LajtPKR9m9jUNbjtj4CSHI0HlhvDlRCDK5dCOVPGnn/AB/p7cZ7e0hN3euERc5NOj/lzl3cOZb9bOwSorQnyH59XE5zf1C5pNn9bUVRgsHQaYUNDSmCKXx+ltdgPcVc3+6kdnA0G2stPKhz/LrNL2/9mtj26IXW9yIpUAkFga9DX11tjG5GopIamdqzJTm8gqDps5AP594c83848w7h4sl1M7Rk8Kkj9nU5fvnlLlu2L2SxoqD4hSp6sk6/+MmNzdJSz5B440k03USCwUrfj3jNv/uBc7dIwQGo+XUDc2e+Ikne32oBwMaujCv8W+p8djf8tw2Nr6rx3MsyIXLAcEW9gS291OZ7m70wsypXHHqB955l3feZjPJKVr5dF63N03snGTVAoMJRonIQKgIuPwvHuYNp5u3W5iXx5DXz6BU4nMhkMhJ6CCq6XwUzGpbERDSxYWivze4HHsWLzJL8Bkz9vRjacw7vYpogkPRfey+vOwMaXn25iqpMfTqXQwxOFcXuQRb+g9iPa9y2WbtupFLnjUjqXORvcC7sWGqUlzxr0UfNdgbsxE09HlsVNG93RmeFkbjj6n2NrbarCUCS3ev5jrIPbPcPeUK3GSp6Cuo3fPWVEiSQ3Lnm6/oU/Uc+5i5Rmhii8F/wCteh5ebnfc17aPCrqp1yoKD+MyVCCl8kbghrj6H629z5yXerPP4acOsUfc/ly4sYmkuh+37egB3dgp9r7kFRCjwfualKggXEgNhf3KUiFHr1jc60ankCeroPhJTZ3O46hqKyqcULwpYNJYNZbAWv7EG3lsMx/Lo/sFdwAeBHR1O1OpstRyHc+HDyQoEmqwh4KAXY8ewTzzySNzQ7jYj9QZIHUxck84S7ROtlemkRNB0ClfDidzY1omSNK5EKMGsrBl/w/wBf3AcqT2kjQTAhl4jrIWGW13OJJrejK3A+vRa97dbTZSlkoJ6ETRqX8blb2BNgQSOPadLhVbjx6MLrZpGgWq5HDonm5ujp6CaUvQMYlcs5ZeAl/r9PZJv8dxPbGS0JqPTo62m3iltms7rD0oOkZP1JTyxKTSK1I5szaR6QOTx/xv3EX73aG4KSkh1Oeoj585SvRA72wLNQ4HTVT9TZTYeZxm/NgzVFDlMJVxZEGjLI9TJBJ5FjYob/AFFvZ4m62e62r7duNJI5VKkNwoRnrD/eU3Tb7krdIY2BOfXPW0n/AC/f5pWzt/7YxXXnc2WiwG76CGKljkniIapeJREBNK7cHj6++S33p/uZ7/ZbjLzl7T26yiZtUsRNFCk9xUAcRx6GXK3OUOgWO7Gijg3nX/N1dVitz7czdLBXYzLUFTSzqskUsdZAdSuNQuNX9PfOHfbPceWb07bvNpOsycdMLEcaGhAp1IttPbXieJBItPmwr1yrt0bcxt2rcvj6cfSz1kA4+hI59v7Tte576Ne22k7D5xMP8I6bnu7a2NJXFfkepFDmsPk4JKqiylDUUsK65Hjq4ZLIBybIT7an2/c7bcv3bcW00cv9KNgPyNKHpyKe2ljM0bggccivVcPzE+WWW6xxtS+wqKHKy0DFql/HrURj63tx9Qfecf3e/u67XzYyXPOBMYkHYK0Oeo35n3911Q2Y4dVy7Z/mD0PyNNDsDc4osPlqGrMVQsf7DS2PpViTzz7yv2j7pG0+3vNf9ZNpZ3t5itQSWUfYPL8upD5H949p5J5Mv1lp9fNEwQ0yCRinRxurctjNq7oxSRGIRT1EL+RQLMjEWOv/AFvc++9XLS737Xz7PYLSOCEtpXIqF/z9YT8u+4nNt5zZ9fvN28izSkBWJoAWNPP06sr7Ix0e8tiMKRo5YzTK91dSbGIXNh9ffAHaN6flPmlPrVaOl1KlCKcDx+zPXRj223iK03aG9U1DKufn1Rz2xRf3cyNXioYSap5ZERVTUS7Nxf31b9ntsvefHgFuf06AsfQevXRzla2s95s03e7akaqCSeFB0Yv4vde52HHVtZnKSQY7NR+BNaMLCRB/ZP8AsfeU28bDy/ylbxx2VJLpDVicnrC37zPuBtM96m27DIGMZyB8j0VP5Lfy8+1shuifc2xNvV2XpsnNKYmhiL+JZHvYccX9inln3O2zbAEvZMNTUvoR1yu+8F7Q2XuFt/7320A7jnNMiv8AkPTL1n/LG7NylVjq/srH1eAx9PL5ZCzGJWTjiSIWuBb6H2Uc/wD3j9j5T2+WWzIAZaAtQmvy9D1jLyR90vft43SK65tDBIX1AKSqkYpqApq+w1HVmOF6c2N1ntiLbeBx9NJNFCEfILGnmkfRpYs1vfN7f/c7mbnjmh94urhvp69qZApXj1n1y/yhsfKG1JtW1RKCgywAr0XTeuz3geYCDVGwcKtgfqTYX9zNy1zGlxGvdnFemrmwUnxKUI8+ix7jxc6K9PZokR+EA9K3P49y5tk9tIRKVBJ8+m1v97tl8OG7dVHAVPQVZGGZYngqZHeGPUYw3NiPwL+xVCtvIQYlCsaZHr0q2jmbmPl/d4d+t7p3eFtZBPGh4HpEQSVcFQ0SwN9kTckqWcuxvYf4e5U2PmOKSIbdujUk4K1eIpw67Lfd6+8VsvurtMe3bnKse5RhV0khcBfP1OOvVjSwN501aFswFri55tx7Ge12LXF4I5Bjj1kDzlva7NsbzgjUQRXqAu+Z6BTCVLpYqSwAKgn6i/09ynbbPFMEiC56wh3v3Av9suJr9pSEz59QJO0cfGwimZVsOWH1JB/r7mjlmwO3WQQinXNf3n5qTmvmWScN4n/F9M2R7CgqAUpZW9X0KD9X9Pp7EVa5PUQ00KaenTVRw7jzkhERnMbjhhqtZj9f9t7CPM26NBGYYck8eshfZLkWHdLxd13KioMgnGehe2p1lGVSoywMzghm8v6v959xXPJPqJAOes69ssNrEaq8ikKKUqPLobMfhcfjkC0tOkbAW1A88fT8eymdTI36o4dDmxm+ljEdmdIPmOpNWZVRrSEaVHAItYf14/2/sb8mSKLkKBx6xr+8bDI+xl2NTnz6S9XVaUZnkDWFyCwsbfW3uYMinXOzzYHjU9IOprpqiq8ULEC/9n6W/P0/Pvx0njnrw8VMoxXpwgNTTukjalVQCzWI9N+bn+nvQAGFFOtku2ZGLH59V6fLrujL0tb/AHZwde9MBL45JKdyHsRyCVP49763wHRL2rcuIqWpq6uoybVDqW82pyQ/H9ffuvdHo6W6Rk3zjoK77Z4YmMeoBSnLKGtYj6f196p1oZHRjqzoaLDBKemp43kAUXlZUW4NuWI9+Gcjr1KDoD+7u9+o/irszLVm6MxjxvR6GaKgxwmjqAlWYz4QiJzqv+AD73nrZIGetTTv/u/cne2/snvDO1EphlmkjoaQu4RKcSFlkMd7BmFv9YWHvXWvPoDPfut9dgA/U29+60a+XXr8EfX/AB9+69TNevE6iP8Abe/deAoOvEEce/deGc9etxf83/r/AMR7917zp14/4Hi3v3Xh8+uvfut9ctXN7fi3/E+/dVpinXH37q3XIm4At9PfutAUNeuPv3W+v//W0Afrwv0+v/Ee/da+Z66+h/1j791vj14m9j+fz/xHv3WgKdde/db69/r+/de65EL/AFP0/p+ffutZ66Bt7914ivXXv3W+ve/de697917ru/Fv8b+/de+fXXv3XunPF5nK4SY1OJyFTj5yLGSmkaNyB9OR791ro6HQ3y7zOyKinxW74lzlCZP+LlXSq0kSWv8AvGW3+sCPfuvcOjzn5udYSUHpytFBNInqhWogshb6gLf36nWxw6BneXy/2TRUFTXYU0eayLq4SASwuwLLYMov9QffuPXq9EMrMj2R3/uiWkokq69ppmlpscplakphf0AlQeQPpf8A2Hu0cbMaIOnkWabsQE9H66G+Dj4igpt19l0B/iEcqTU1DPHIYE/tRFUdQL/Tkj/W9mkG2yEh5MDo926wjjQzXK1I8j1cR0H1LRw0S5CppY4qWl8aQQFSEKI10PPHsWWcbBBQ0A6NoreNj4hFB0se4ez6PblOMBjmiWZtSKsUiFgv0sAvPuO/cDm2PY7RoAaE+fn1NHtryVFv94tzcYiXNeijyZutrdbI8slVO91UIxN2/BtyfePVtNf7xObiOrKx6zGW/wCUth25bagrGKeXl06U+0N21dM1Y9CWQqWDtFLe31DDj8f09iE8hbruNv4hQkH5HoJze8PKMEhs5HAXhSo6FroXYk2T3tRnNVMwMU8GiKUOkdxMPrqA/wB59wJ7r7TuXKG1yB4TQg5APp0N+U4uUea63e3TBm40JHWyRsbrLema2bjV2/VSCjpqGNEELMU0qnpHo/w98m+YOc+VrDmSRd6oJXc/Fjifn0Hub+bto5UZ7R7ZC4x5Z+fWVc7W7IxeR23vmWpjiYTSCWVJOCi/guP95v7PX5Wt+Zpot85X0sRTgR/k6gj/AF09sbmGO8eMR9wFBShz0UbJ/If+H5Krodu5WeenSZkVfKOLNYDSD7lrb/a+WW3W43GEB6ZIB6yWi2zlPnWyjupolUleOPPoWer/AJAVklekO4FAo5Pq8jKCTfi5P+8ewtzdyAEti+3k+IPkegPzX7KbJc2jSbTTXxoOjpYbrnZ/a+MfLpHSpM4LRurRs5v9DZef6+4Gv/cnmHkG/Xb5dTJwPGn8+sVuauSW2C4ME6mo+R6CPdPX27+vqxztmuqKqEBrUxLqoVP7Ok8e5b5d505b50sw28xKjGndgnPUfz201u2qIkj06JR8hpc52FtitwG5sd4JF8zhxG5uVi08Hn6/X3P3IG27Tsl0t3tEupTTz+fSvYuaty5b3RL6IkCoBH51r1T/ALapm2VuyuoaqL/I2neCPWrLZbkXu3vPr2158a03G2huKBaUJqOszt/37afc7kFUt2BnRBXI406G/C7iymwM7R7mw1XKca00bzRIzeO7nkG3HvNmyvoruBLu2NQwqesJr/brjY714HxpNOrFtn7xpexqGimrJEkaSKLUhdG5k4+ns7WYSLUZ9elcUvjAFjnpJ/JDqLDxbPXKUlLEztLExOgXBABAv7KuYdrW/wBllgHEgn+XSPebJrjbnEJ0sBx6S3VuIQYvHzeMRrSU6IbAgjTwLH3z335TabrcWhHByOsYrwSRXcscmTXowtBUO1lW5AFl5P0/2PsgcV7ukMqlUDU4HpPbpEBAYhVJLergEccW9inl4kijcOs7PuozymcBm4U/48egf3JisPnaF8Zk6eGqjm4DSWYqbWHs83Gzc27vZHTJTBHXTbad9juJf3fviiW3II7uHRHOz+mtzdeZNdz7NMtVhwPPVQRI7RxG9yFC/wBL+wVyb7lPt+5ycuc2HQzN2MxoSvlSvUK89ckrabk268u5hOSoGOs2xO7mZkpK+OWKSMqsxeJ00sDZgC4HueIFt7yPxbJwynIyP8/UYHcblZqTDTTj5Z6NRtjd9HnZYWimQxgC5LoTz/rH/efaW9tNVu1tLwbo2sLxJpwRkjocNt7qqtr5/E5ehYFIqmFCqsovdwSCfeJvvj7X8tScmX10qqspUkEca9ZU8jbV/WzlyXZ9xWqsrUJHy4dW17gfBdr9Rw12hJK6HH+ocOVk8XN9P+PvjvyPdb1yD7kybc7H6eRwR5Yr8+ucnu1yPLy1vlxA60TWxB+QOOqmsR1xWZXtako2hZqWGrCaNDlbLLY6vfRrcOZ4LHk97zVRihPEenUFJEzTgdX1dS7Io8Ht3GRrToskNMATp4HA4F/fIv3O5yuN85humEpKo/w1wePHqTdl24eGHccR0Ny08SqABwv0H0/P9fcSeNqYyL26vIdCoQoABxp1zkRfCyk2Fjx/Tj/H3eAeNcBZ/wBT0r5dWkAWI0x1Q/8AzQsdj8tkqCiggQVYFzUC2sDRYAH31e+4pvu7crbEd5LajGzaVPACvQZ2baLff+cU2c4SQZ618e8uo8Pktm18ObhjnvQyFfIhbVaMkcW5t7678he9C80XBsdyURy1oucHp/nD2Im2qWW7tiSmTwPWrZ2HjKfDb13NjKVPFT0mUqYoIwpQCMScaVt9P6e58U6lDDz6xvuIWgnaFvwMR+zpr2xufL7QzNHnMJVyUldSSKyuhtrQOGaNh/Q29+dFdSjZB69BPLbTLPAxV1NQR1sAfDL5nYve9HjcBnZoabcdPEacQTTxK02khQ8msjg/UH3FfMXJaPcG9tR8yOs7fZ77yVzFs6crb4+VGlGJAPl69WLb47WrqKgVaSogWQo3jWKoiZWt+kkofYCn5flN2rqmR8usrdv92Nv/AHJLBcXI0sCB3r5j7eil1nYu6KjIPXblr0x2MQs3kSqiuyD1HjV/T2r5kfmGa0TbdtjIqOIBr1ihv17yvYbrJvYuqsSSBUef59I0/PjrjqzctJTw7kjrfC/jqEqKqD0nVdiSx45/r7HPtxsm72G3n9/KNZ+H1/PrHP3L5/O9XSxbVKxj/Fn/ADdHy6w+YtD2/QxZDZn2VchDCW9TTmxJ03QKfpfj3KHDA4DqGaliXbiehLym6d+1UqTighgBszBJFsR9f6+/VPW/LrO+/cg1F9kZIUrWGgq08aWc8MOT/X3rr1fXpwwtJkXhdqrJAPVXJ0VEbBNQsbFSf6+9+XXukTuDp/CVlV/Ea7PGUsQzxNICL/UD6/7D3vI60VBoD1wj6j2NLGpaeCQ2P62TkgXPLe9V8utaF6ZK7rrZ+JlaYTwlYwTYvGQCeVF+ffqdWx+zoB9+by2Ts+CpbLV1LRi16cx1MGrT+CRf/e/fuHWqknHRZKf5ZbM2tk/vaXdDNHG91Q1cIVgG/wCDe9U699nQu0P8yvYFDRNNX5enVYIzdmq6azEDjUSePe+HVq+Z6rZ+cnz7buigTbWy6qJsVV04hq5aWojcIBGVLExE+o34966r8uqjARc6gT9fzbn37refLoYOvvkF3d1SqJ1r2nvTYoiZHj/uznKzEMrRtrQh6MqeDza/v3Hj14dvw9XifG3/AIUM92dVbdxe1u8up9lfKfGQ0KY3Kf6YfLnpK+A+iSRnqSTqC/Qk+9H5dXViPiz9mOrAZd5/yUP5kGIgoO1N1YP4db93DCrUWE67xcFZjKGvsPHC604JRRIeL/QfUW974daFHNT+VcHomnyB/wCE6vddTjf74/B+sq/kV15RCSTJ7giqaNZYqTSZI6o0sADIpWzG62/xHvVevacnPDqhOk+PnaNd3Q3QFLtupn7Rjzsu3Zdvxo80i5SI2aD9lSeOLm3HvfVSCDTqyrcf8g3+Z3tPYq9kZ749Zei2g9O1UuUaRnBgWMyl/AiF/wBIv+n3quetlaCtf59VHbs2lm9j7myW09xUT47O4esaiyFJKrqYZhYFWSQBh9fyPe+qjI63fv5B/wAPfib0psCqznzY6/2xvbHfIXDUdbsjLbwpqeBcKFjVmbEpWFdWq1gysebH8296wRUdWBbTT161kP5rnxqr/j78te0p8Zt+HCdZ7t3hmK3rj7eN46WbBFy0H2/pCFVTTYox+v49+HDPW2pqovRyP5Jv8uzqv5Qx9u/Kfv8A3VNtjpH4pSUO4N8rJSs9JkaGWj8tNGJGXSxaQGK2q39QSPe6561pLcOjl9j/AM/b4y9S7vzGwOi/5d3xk351phKpqPC7t3dhIq7PZ2hT9k1VZNB6dbKNQH4J91A9er6gMDpMfPvb/wDL4+b/AMRKH5Y9KT4PpPuXZ2BOT3h1FtlKRMKuRrBeWjpAg9SXUafqR/W/uw6o3meP+rz6DX+UJ/LM6k3P1Jv357fLTcI211p0rSf3owe0chTsYN7xIqinp5BIgAErtY3a2g/1+mjXrw00xk/6s9DduX/hQj8ctv7xrdj7K/lr/F7K9U0lYMfR7grcAsm6K+kLiCSvnqrBD6bsP8ffqVPl1sPRPPoK/wCbT1N8De1Ojeq/kh8Y87Q7X3tuPBTZve/XGLipRQ7cr51M5oKcw2DxkWCkfSwPvdfI4686gL4i0r/k61jibqCTz+Ba3A/1vfuqeeOuHv3Vuuxbm/8AsP8AX9+60a+XXXv3W+u7EW/x+nv3Xq9dC1+fp7917rs2/H09+699vXXv3Xuuzawsf9cf09+60K1z17k2Fv8AAf4+/dewOuiLce/db65Lzx/X/fce/daPr10RYkfX37rwyOuvfut9e9+691631/w9+6913a/+w9+6914Wvz9PfutGtMdetzYc/wBPfuveWeuzYW/qPqD/AF9+68M9dE/Q/n8j8C309+68B5ddmxF78/ke/deFeHXViP8AY+/dbr117917r3v3XuuwpYhVBZibAAEkk/QAD37r3ThHicnLHJKlDVska6nYU0xAH05IW3vVRw63pc5AJH2dGe+LPx2z/c+/cP5qBl2pjq9Zc9PURSAfawgM/pZbFeeTf6i1vaK63bb9rpLfOFX7R0e7Dyxu3M96m3bZGWaQ0qAf8nW658HOqesptuP19NRU3908bBFDTVSIgFUxgKSAi3Av75uffB99earO6hsuUHKRQHIU01/b1nrtXshJyXyOk0OL2VAXNKFSR8+jURfCLprI5nIxYGemi03bxqiKULf0Fh+feKEf3heaIrGKTdLY1PE1r1A3Muwc87JbrcSXkrI5PkegxyvwdfF5mWq2/kn1wOTHoWS9vqeVFv8AYe5B2/3z226slN/EF1jNSOo3uNx3+ZTbSzuy/OuenzKR9q9WYhRSU89cKSMNqdXOrRwAQfd7Sbk7nG6IdlXX6EdE5Se1FaZPQSYf5sZV82Nvbwx4oXEoiV9GnUS2jlm/F/r7El37I7bHa/X7RJqqK0r+fVF3KQnQ+OjFUOZTeUlNLRRLJHOVZNNnBD/4r/X2BriyOxownamnj5dKlpLlejBbU2CQqnIYqN4nCsS6fS/9CfcT7/zagYi0uCGFeB6XQW1RV16X+T2dslcfJSZChpY45Y2WxVeOOTz7B1nzHzRNdiW1kZtJ/b0ujEVqPETB6p/+bHXuxcFTJV4CjpmmmaVpDGEYr/TlefeX/tJvG/Xw/wB2ZIApg9Tn7ccz293Ku23VGOBk9VALjFqKqYiMKQ76QAR9D9B/xHvMzakSGyRxguOsztn2eC3tUeMBVpX9vRmutdjQtj/u5UBa2rlSSbr/AE95K+2e1qIBcN59Yg+/u8/42bOIYGP59A93VsXzO1ZDTD0ByfTf6Nf6e5SuUzUdYpyxgrqHr0ZL4i70r8NTU9JLUGCClX0oG0g6fxpP+HtRYyaR0tspmUY8urc6Xfcu49ux0auZIZotMilgSwtyCB7PUlLLTyPH59HpbxY6efr6dAVubqXL0wl3Jt9JJQmqWSmVXP512sPcZ858kJuKNe2Ao9CSB1JXI3O0+zSLaXp1IDgnpH0Wbp8pC1FkqMU1fACsno0AleCeR7x2vtuuLO5MEoKkfLrLTZuZrLdLAS4OPUdInce1oMuskXiAjfUC9r8fXkj2ut41iHfn16DN/fSPOfp8ehHQJZXruLGGSO7PTyXv6Wslze4HuAOfdpvY90k3Cyj/AEgKmnUjcv8AJ17v21tcu2p6YBPH5dCp170jjMzjZG+4MhkQ/tEXABX6ge4Tvec5bG5CkUoesNfeHlu+2/cHjlgpSuafPptyfxKw9PUtl8ZWTYXKAmQVVNFKkqve9wUHP+39n1t7lzyL9PMgljOKEih6x6NoACeB6FfZMfamz4I6Kl7L3BPBCNIRqqpAAHAAUt+B7DG9WnJ29OZrraoC7eZRT/OnTsM11CNKSEdPe7arsvcGPlB7Dz6VHjIiC1FQSWP+Cm/tJtO38o7dOPC2uELXNFUdXkluXWrSE9C78UMB3jt6syVZmN47gyOJqqZo0jqpaoxBSLXAlNr/AOt7Afu9ce3d9bRQRWMCzo1agLX+Wele3SX8daOwB6EXuKt+5w+SxGVg8zVkZglqHUhvq3LX/wADx7J+RbVYrmO7sm0quQo4eXW7o1FH49UJdp4OLqzsim3Ht2qlppUrxVTGO6hx5dViePebew3sm87P4FyKgrToNXVtby1WQVrjq4P489xU3Ze3aJ5qtIsnRR0wLNKiyeiMC63IvyPx7JlEdvr2a+GpJQRngQfXqJuYtmn2+5+rth2jIp69HAx3yj3jsTz4mvC1WIeLwQzSygsQBYEEm3vB33p+53sXNe6DetohowfXRQQBXPl1mn92b3E5W32aHlrmSURTrQA4BJ9Oktt3+Adn7yh3RnZxDjYqkVEh1KQVB1twPr7kr2l2e19qNoki3KTw5whGmtKACg66F8/b7uvLHKx2jl+ra0wR6EY6PXTfIn499fUSxZHJvDj8dCsugKqLqiUK3DWvf/W9qpNxvOZLsz2eqRnNAeP2dc2OYJ7mDc5Jt7b9Ykkg/M9StofzYvjjntw0GxNqVcVfkJ6pKGOOoeBdLlvGui5+t/ZfzZyzzNy1y/cb79MZGRC1CPQV6b2qCPemIsyDoyfWnRpdwDIbrkny71s0VHVQrLFRJfwhZE1qVA/1x75P717t71zjuV1LdrRIpWUxlsLoNMDh/s46P5dvaKJQpIFPLz6LxuXA1ETuEQsy3t6SCQD9T/h7kHlbmG1u4VYsAPt6DdxbtEx+fQI7jwT1EcvlgYyKsnGkkc/Q/T3NOw7xFCy6HGk08+iuWImvp0V/d210mBQwMkpZrkIfoP6/737nbYt6K0bUCtB59FMsVfLoANw7NqJGRIYCw1kMSjekDgmw/wAPcl7bvkQBZ2z5Z6RSRE46eKDr2nqcZJSrAoqNB8chRgxdlt9Lf19orzmR0uFnLfCa8eFD0a8s7zuvKG9w73s0rRMjBmCmlQPXoOanYk+0KeVM7Tt9pPI7RvJG/DM3BDEfT+nvK32s522Xm+2Fssg+tiAqtRwHn11F5a96I/c7leGCVwJ0VQw1DyGTx49AZvjEU2h5KNF8cjWV0Bbg/Tlfz7ya5TsHnmEknl1i77+8zxbJt7WFu3e2DQjz6Bw7OaqaxDlm5uUYn/AfT3KtAqhR5dYGM8k8rTyZJNel1t/rkRaXmTVGLG7o4/17XHvxpSnXhhgxzTodMPj6LGU6xxRgFV/V9CSPqDf2Vy7Tbztrkyehvt3P27bVCLeyJRR6dK6lrnZVVRZRa/F/959l93sNnHAz0BNMdC/l/wB1OYrrdIrVpCF1CufLrquz0dN6GbkDgj63/wCRn3FV1tVw95pRcE9Z4bPz1s9lsAknmBkVQeI49Jup3HM8bLHc6rrc88fm3uUdg2CHboBKfiIHWDXu17sbjzbuMm3wmkKlhUHoN8plco05ig1vrbTpFzYE2B9ifjg9QcKDjmvSq23iJEVJ6osZGcNZgdIX+1b37reenPsHdON2/tuskhVGqTTuq2K/q+gI/wBj791406os7JTP7y7BnkhimqHmrnARY5HUKzcWK3+nv3WqdGSoeusPsLY7b47FmpKHHUNG1XTI88KSTSxLqVGidr82t9PfuHHreTnoK+v/AObt19s+umwqwYihxdFO9OkgnpY9YhYxXuT/AIe/dax6HoDvkr/Og3HnBVYDrjE46WjqYJUXLU80OuIn0qRNCSQf9b37z68anh1R92j3Bvjt7P1Oe3pma3JPNO0sVPNO8kVPqv8Ao1fn/H37r3n8+gu+v0+guf8AYe/de4dde/db697917ru/FvfutUzXrr37rfXvfuvde9+691737r3Xvfuvddm44P+v791rBz11791vrskkAcce/dapQ1669+631//19AFiL+n+n44/Pv3VRwz1x9+6t1737r3Xvfuvde9+6912Lfk249+60a+XXXv3W+ve/de697917rk1uAOf6n/AI3791oV4nrom/4A/wBb37rwx14W+h/2/wCffuvH1HXvzx/X37rfl1y+p59PHA9+6r5Yz1x/H+N/fut+fS16/wBhbg7H3JR7a25SNVVtSys4AYrHD5VR3JUHn1cD8+/GtMdKrS2e7uFgTixHWzT8MPhbiuu9u0mRyuKp6jJLRpVyzSxt5Y5D65CXcf1912CK9uty1MpEYPHqdH5a2naNhWQENMR0LvZ2ew43BHt+JFRKZwjKliqiM2I9yBOyhhEvAdR7My6tA6a9yd9YzYu2/wCE4z11zRMqxx6eSF9It9fr9LeyrdeYbXZIC5bupw6Eewcu3/MVwttbrRcVOfXos+Dxub7Cy0m6M/JLAodnhjfVYhuR/gfeMXNF3PzRu4nlxGDw8us0eTuVLflzYG8R6MBw+fR2uhuoqLcGQgraykjemhkCetDYqDbV7lvkPlm3lRZdACL1BHPvMMkEr2kTVLE56s7wvRW25cfFGKGm0+NVtpuALfm/ucYrWOKIRRig9Oobe0WdvFkyT0g9xfGgNVCuwCrjpIW1g06lWZl5Xlf9h7It75T2LmK2a03OBZAwIqwrxx0c7bu+97HIJdsuXjA/Cppw6NR0d8kt99DYf+7e4MJJlsZC9jVz3dlRfyNX+t75ZfeN/uw+WfcXcG5k5cnP1SkuiKSqgnyNKY/l0Jb/AJ+3LdQG3BKsooSck9GmXuPof5T4mfAZKoh27uKqhko0keOGMrLKLCzm3F/z75s8wexf3gfup70Lma3a/sK6jGCx0qONOP7D0GbyTbt3YSECJ0IIoPPohu8Ph3j+mt7Lk6/Nx5LbORlM8dYrCULqOoKxW4/oPfST7l/ufyT72h9h32IW96CAYpBQjyNK8epa2f3GuNn2E2QrrHBh0lu6NrHamChzO1kFXQuFkE8PqKXXUNWj/ePeUHuz92LaEja92cKDWo0jH8sdDb2+9157q98LcXNK0z0nemPmhN1lJDDl5qzwQ+meEpdTb8+oH/be+Znul93GTebiS0mhCy5oaH+XU2808n7ZzvtRuoCodhg4r0J3Z38yHa2XkpZcTDVJImmOYCEH6mzcgfT2DuSvuv7lssDxzgupqRg46gm39mxFI8d24pmhr0qcL3f1l29hER2hhyUtNaQy+KNvJImi5vY/X2tXlDmbk++JTUYgeGeAPULc5cgXmzTvoXWlTQ8cdEk7l6Bw7VtVlsfWws7M86IjfXjUOR7nnlrnSeSGMTKUZaZ+Y6D3JG/bry7ughtQZIyaFfLPRe8MtIcbX7XyWlKmMyiESWBJT9JW/wDj76Yey3N9nzLy8IWakqUWnqadDP3GsGvok3mEUxV8cD0sfj7vafAbwbbmRqHVWqE+3DGy+JZObBvxb3M9sTFL4bdRRaSnWB5dHH737Kps/haHZWBY1WRqJIWKrY2CgK3K/wBPabm3fbbYtkkvJ2oQCKfl05v26xWNg0jGmKfy6c9qYF8RgcbBIdM0lKrTqAfTJYagT/sffO/dr395bvcX3k71H2dYy3c/1F1JMeBNR0uYHjplDBhdVN7/AFuByb+y1jXBx0gmMmAM1PQWb3y0shWGnBuxYMQL2stgePY02CKOOPU56z5+7RcbNtG1m+3CXQwpQY416DCKapEgEhZmFrXvb/bH2L0ow1L1m3BexX6C5s31oc9Cls4R1l6bMQLV46QlZaeYAxstubg/7z7w9+8rZ/TX0G47c5jnC0BXqReUx9TG9veLVGxnoyG1Pgj0t37g6yXFZCn21nB5GEFJEqvI7LYE6R7wl3f72/ub7R7rbbdc2ct5auQPEDYWh8x1jd71cqbhsO6jcNtp4DVJ9OHRAvkJ8Me3/ijk2zW3RUbk2hHKzVVf+9J4IyNZJQfT3nt7KffC5W91rSPbb2ltclcqSNVRjqOOS+bLS0u1Xe6oa/5ekZ1J2bBvfLUeM8sjVUc8aVEEoKaJ7jUqiw+ntb7ybrd3dhJDakmAjj69Z/cve6PI+x8ricSBW0HIpXq/jpOhhxvXNTFkJljElOzKpcX0mO/5Pvkrz3DdPzrDNbISVYVx8+ufnvdzxt/N28M23HUmc/b0DXX9HiG7GqapVjeKKta8h+qr5PSQfc7c53O4DkhYRUO0fD1x1jlaIouacRXq0/adXTPjovGy6BGujkH6ADn/AIj3zR3eznh3a4iuFpJI1eHHqUNtmX6cZwOlcrhr6eP6/j/X9kskMkLaJBTo1WRWyOoOTnMVHI63sEaxsbk6ebe9RM5mVY+JZR+RI6aun0wlj5A/4OqF/wCZdk5qJGzMEeuanZVUD9QAiBvYf4e+tH3SbEHloWbnB4/t6C/KO7R7Vz1Dfy4A6pSym9cPvHBPjqg6ql6VoZVdR6XddJt/rc/X3mjZ7bd7Rei5gNNJqCOPWYd3vtnvViQ2RIpx9vVbea/lu7f7I3jW5yXLPSJkZjM4hlnQEs1xdY+P9j7nO398rjZ9uWCaLxGUUqesdb/2ETft0a6t5hGrGpGfPpo7h/le7a6364y26aTJCqmpUdo2LzmQ2i1cFifbvLXv8+970lg8JUNx4etOkfNX3dBsGwSbr46uU8hWvCvVKlNlM3sLc9TPhK6XH5LF1U1OlRCfVZHsQb/g295RQSrc26zDg4B/b1inIslpcMiNRkJFR8ujMYL5m9jY2j+2yLyZaQKQJpqora4tfSVP09tmytWbUUFejSPmXe408Nbhqfaegh3V312VumqqpJ9x5Clo6hmK0MM37aK341Wv/tvbgtoAahR0Xz7he3P9vIWr8+gdnnmqZpJ6iR5ppWLySSMWd2Y3LMT7e6R9Dj0t8gN8dKZmnyG366oegikMsmME5hikckEkPY2vbkW9+691Y3SfzXt3vj1Wqw9UKiNQuhGjdSbf6sWv/jx79WnWvt6BTen8yDsXcwmSjxX8NLOWSojq1WW3+Kon+x+vvxz17PSt6p/mW78260WO3ElTUwM2n7xpxPoUn6sGAIt79177ejVZP+Yztytxn3C5iT7oJqMOqP8AVb6W+vv2Tx69UV6Ltuf+ZluWglC7fpZa/wD2ppEhiUf8Gs1/9t78Pn1454dJLK/zOt/5CgNKMAPJLGVlkarjXS5W2pCEJP8AW9h7916h4dEX373f2Bv/ACtZX5TPV6U9TJqjoVnPihXTbQCAL/n37r1OgrmrqyoBE1RLICbkM5Iv/Xn37r2kdRvqDdv9gb8+/de8+uPv3W+vck/1J9+69w67II+vv3WgQeHXtTWtc2/pc29+63TrLTzy0s0NTTu0U8EiSxSIbMkiNqR1I+hBsR791oiuDw6PN8fv5jHyz+O+68PuHZ/c+/IsVjQ8FVtiDcdZTYnJUjxGM01TSqShX6X9PNrHg+/edT14EgUXHVsf8jv5AdDbm/mGz9l/JKahg3v2TuqryOHr62OORcfW1KBYWSqqRYNqsxNxc3Nve8AdV7i1Serhvkb1V/O3+IHzC3T8sejafcvyb+MmX3LXZ3FbRyG6GzWyptty+SQ46qxSTsiIkJtoCHgcAfUUx+fT2pqZytOH+rz61Yeyqbef8wD+YJVYLcOxKLqDfnaG9p6XMbZxVGY6fGZSWoMslNS02kWT6rHYHjn3YYFD1RiXbUo49bTn83P4B/zFJupfgjsH4p9YZPLYnoHrmm27lqjbVfSpka7NRyGrSpy0Esq+kqxRg1mtpsRa/vXw9bP6i0+fQU/z8fgx3Pm/5fXwq797E2nTbT7N6o6uix/beIppRVXyUwQSS1zQlwsrAfVj9f8AD34V8+tvSmPXpJfCiHbMH8gj555TrWGItmdr447/ABRqvkjcxs1MlQkf9HL/AF/N/wCnvzUr+zrUVdFPt60sBf8AH5sCL2v+bH3vqp6VeMyO7qbb2YosZUZeHbNVLTPm4aX7gUExDWhFW8Y0lbjkMbHi/v3WiB59bp2/qWnh/kb4aLap81HkOspTu+CMftB1gTxifTx/t/fvIV69UUOn060idRA0nlPITpvYE2te/v3Wzx6WEm5N8NhYsVLXZv8AgawhIoGSoFH9uF0gK2nSVt/jb36tetUHl0jWFjx9LfT37ra8OuPv3W+ve/de697917rkbWFjf/C97e/daHXH37rfXvfuvde9+6916x5P9Pr7917rvnj/AHj37r3XjfgEc/1/Jv791ofLrr37rfXY5PP+39+695deNgeDce/daHDPXXv3W+ve/de697917rsAn6e/de4ddkWtzz/r/wCHv3Wga9dW+l/ofz7916vp142vxyPfuvD59eAJ+nv3XiQOPXr/AOx/1/fut0669+6913YkE/gWv/sffuvdXP8A8ub+X/W9wU9V2ZvmiFLg6SJK3BR1CuVyCKgk0FWAHq+p9qI9suNwGmI6R69CLYLKKafxbgVQdGS776RwWzp61qPaWPp6dv2kSKLiyem5H9PaDcNj3C1AWNi3z6kuC42JBR4VAHRWNq9r1+wKyLHYfELhqRnaOsmplEYqYtelkkHuO982CW9NLxiSOAPUp+3XMm07Hu0V1ZQrUH5dbQP8rjOYne+3ZqvKTR02LiYPPUOyKYYzAWZhf/Ec++Un3xotw5fvVXbIzLcSUCIK5NRQY6zh503JuYeS4N5240YIDpHmR0YbtTsqm25vrI0nX+UmrKeMWeaNhZyH+o0+4/5K5Tu9x5chn5pgEUrD4T5Y+fQX5f26x5r5ejh3mJVZfXocej+163Nt9tnHf7h1IjZyCzNblfYJ5+5Xj25PEsACg406h33F9otvtozcbZT506NzmesYNz4iOuqCHiqU9Ub3tYi/49wzs/ua2zbm1hEKMh49Ytbrsv0lwbdjw6rt70+I2285O1ViaanpslEZHMqJZ9V9akEc+8wvbr3pvhAq7gxMZoKE9A+725Qx8PoEuv8AfGX6NycWL3VjmkxlHJoiq2QszhTZbFh7kbmXl+z5+sDNtctJJBkA8P2dI4JWtm7xw6Ohj/mf1XVUiXrft28aqYyI002X6H3jpdfd75riuCyd4rxz0crvEBFDg9Bpvv5FbTztFUNjcogUowjZZYQVB5JAB9j/AJW9rt02mRRdR1IIrx6SXF6sxNMU6qs7p7ZxOTrKnH1Nc1WJtccJYqwDE6eD7yl5b5XlhtVdUCEcek+1b7Ps24LdwE4Nf59FXpdqhZKjIhrxyhmQG5Vbn6gfj2LDvFxbOlsxICHrO/kX3Lm3zZI7dSdYGfXoZut8hWySLRQqzwI2lyPpa1ri3vMD2w3ae6somhqUP+bqKvdCzSfVdXeCc56cuyMItTBO1ro0TFgCbDi5HudZFqgL4x1jVOgLkJw6ALYmQq9v58RxFhTM7DgEcE+rg+08MnhvTpLG2ltI6t76XyDZWgolF7aVBH0/sgm3s7g7hjo/tCa4NR1YfsvZr5ShjUx64JQA8ZDWYMv0YezNKIKvwP8APo1SHxBXoLu0PjPSVZlyOLjjo6iQu7rErKf9Ubkf737CHMPJe2b9GXipG48/XoW8vc2bly9L5yR+Y8h0RHcSy7OzUuCyaG8cnjWQXbm/BB943btYw7XuUm2B9TIc9TZsHPGwcxsIbWQfUL8S4x15cJS5svE7IyMoIP8Agefx7jj3AN3abBI23R+LKykaepk5S5jv9u3JF1kIDkVwR0YPofYWIrdyU2H/AIn49U0KmMnSpv8A2R75qe6278xcsbdJuW5WhSuojHn0c+9XLKb1y/8A1htoQwKkseNPXo83aPQNFi8TBWUCpKbKJdAIA4BJuPeP/tt71Sb7fvZXdUI4V65u7nt6idpEGCTjogG6doVuNyEsyQMlMGJIVWHpB4t/xPvLraN6t7y1CFqseg3KhVuHQq9N9NVvYeYppI4tVFBURGUvqFxa7A/7z7AfuR7j2PJO1PJctpdlNKU4+XSuysnu5AqCvVpGN6KoKLGQUdMPtgsUakRggXC2YE++fm5e9+53G5SXDwmVakgn0r0N4dhPhKtaHoBO6vjJDPgquvpJWmqQpbQ2ognSbNYe5j9pfvCNd7qlhfQmFCaAmmadFW7bE0Caga9UO97/ABeym4snULNG0BgZtT6JBwr/AINvfS/k33EsVs0ZGDBgPMenQFntWDcOHQUUnXG7Om8NT5zb1dUyiifz1UMRkAMUX4I/px7GNvzJZb1feDKArH4T0gu7Bbu3MUo4+vQsxd4nsXatPPd0ydMNFRBYegx+l2t/sPcgbexaQ2U+ARg/LoE8s28fJPN0O8NgB6g/n0M/x37LxGVzuP2jV5bxSU9YZK2HyJqKEjUjL/T/AGHvGT7xHtvvWybPJzlZVkSVdIGceh9Ou0ftj7j8q+4nJptGcPuHh6QppigwR59Hy7y+NlD2PtL+IbTMbRvRWkCKQG4DsSV/JF/eJfs975HZN0/c3MVY5VegqR9nn1gX7v8AJO5bVv0t3c5Vif2V6oWzPS+e+PfeG0N1SxOtEdzQvUMkbjRHFVAuSW4+nvoceYbD3B5Mu9vgYFmhIGeOpT1FHLm43HLm6GZKlZO2np1ug9QbxxPYXW22dw4mojqYanF0SEB42OuOnRXDBTwR7+dznzlq45A5/wBx5UudSyvNLMKghSrsTQE8fPqdEYXlqtxjgBQdKbJ7bgrSZCihj+pTfg/0HtFtu5yWNDC2mv4eim728TjGOknPsLH1AfXChc8EFQb/AOHI9iO3513i3oyOceXRd+6VoQei4dhbDoaKaqSOjQDQx1hORf6G/wDvPvIrkDna+v7aJ53OokVHQcvLJYpGj9Oif5emoMXkpacKsxYnWjH6D+vPvKLb5rq+sVmqV9Og+wVCQenbB4aHIVEb04ChyoVVF7EngA+y3dNyksoSJTWnn1eOJZOI/Lp+7n6+pc313Jj2jjiyaQu0NQf1hrXt7Jva7na/2nncbnZyEICOB8vToSbPv25cqSi625iteKjqrWjjxO36uo2tuKrDVNPUSktKQ3pDenSW/wBb32p9qfdXlvmPZ4/rG8G5CioFKGnn0S87Dm/nGX97mImJqca9TvHto1VqOdJgCPVdbj/Y+5gh3zZbk0hmB+0jqNptm3m2FJ4SPsB6dHko4QqxSqwI/wANI/w49+vN+2LbhqvJwo+RHVrXYd73DFnAWp8j/m6ltV4mnphLLUWkJF0uCPpa/tTYbht27ReLtcokH2jpNe2G4bU/h7nEY2+w9Yl3JiYIn0PzpIU2tcgccn2qeEsKPw6Sw3wtZPFiqG+XHpONUxZGX7h39H4+n+9/8U9pPobcPXSOj4c0bxJB4XitpPWVazHxExM1zYW/17X4/wBf2rFKU6I2rrLNknrEs1CjgvpLk2va555vce9dboOlStXCtDaJrvb0jjgHj/b/AI97HWiadA5vPb9XnY3plVnEwaNRZrFmNhyPp7914nNeiW917+61+H+1MjvLc6UeS3NUwzx0eMdY5nSpZf2joe7aiTYD36mOtV9etbv5AfMDtTvPcOSq6rN5DD7aqPJFS7dpqkilWBjYNIoH1K8WXge9de48eil+/db697917r3v3Xuve/de65WuODz/AE9+61Wh64+/db699ffuvdd2P09+61XrxBH19+63x669+6913fi1h/r/AJ9+61TNeuvfut9e+v19+6916xtf8e/de65WFr/n/X9+61mvXH37rfX/0NACxtf8Xt791quadde/db67IINj791oGvDrtrX4t9Px7914V8+urcXv/sPfuvVzTrr37rfXI2HAsfzf/iPfutD166/F783+n/E+/db8+u+NJ+l7/wCx9+61mvXQF73IHH5/3r37rxNOuvfut9e9+691737r3XYBb/bfn37rRIHWWGCSeaGnjGqSeSOKMf6p5HCKP9uffqZp16tetlb+Vv8AEClxtBT7y3NiaaXLz+GYSShQWiaXUp9QvZQbAf6/sW7RtsLJqlFa9DHYbNoj9R+LiOrt9/GLb1A+2toUaT5KthMLiBb+MOLEDT9Lez7wIbQFbdcnoU3d5PMugsW+XRQcl8ZqrH4zJbq3BM65Ko81SiSyfuIX9Wmx/p7RXFrILVpFy1D0htLKtwHnwK9EXrurqjLbvaSRHqaeGoA0nU66Q3uC+YNvvJ2aSRiVBOOsi+Q9x2rbAqxgazTo4e1evKSsgosPFTrBZY9WkAciwP4H19hKwtVlulhAxXqWb+6lNk85c0IJp0fXYGxn2jhIftKQahGGsBqN/qCwA+nvJzl20Ww21VUUqB1iHzTctebs7HNGPQl4ntOTGVKUFUixWIQ6yFJ/F/V7PvGIND0RLOQNLDoy20czSZiONiyuJAhuSCPWbXFvatGV16MIGVhXoZoOotv7ki8lfDDPG6H9uUi1mHPFvb2toKFGI+zpULdaVIrXoC9+/EyipayTJbQqv4FKkbTKaF2VzIoBB9JHN/p7D+/8rcv84Wcllvtqlx4ildTgGgP2jpHPYpq1x8eiQdy9q92dd4Kp2Pl6Gtz2HVpfFlq5WkmgsDoEcj8/7z7wI50+57tvIW+Lzz7eu0Fwjav0jpFOJGP8vQj5TsLHdNxFhub6NVBx+fQS/Hr5K5mpyEvX2/8AHNVYvJya0kqoWlMEZNlCM17cW4v7yZ9sfce55jsF2TmJayxAJkZJGKnqReefbG85Mii3SxP6bDVqBrjy6s52v8X+jt+UMeXapxkf3A8jwzBI7FhfSQT/AI+5NuuR+VrmUXE1kkx9Tx/wdA+05+5kS38C3unQLjH/ABfSzm+DnRUtBUtAmAeUQylW1xA69F1Iufdhyjysq+ENuQA4rT/Y6bfm7mZyWa+c/wCr7eqsu8/i7ujYWSrct1xVVOumnmkipaF2aJo41Miiym349w57hexOx72rTWcSockBaen2dCrl/nlZj9Lvg8QHGpvn0muiMX2r2jS11Pu+CroZ8Y0kLK5YFljbTc3/AK2984veKxtfaW6Flfx+G7mq18x6jof8o8h7JvXN8dxt+l4ny1OAPQYfJbYlf1yn976MyWoyI5wt/UNWlmIHsVfd89zzFv0EiylUrkeRr8upE92Pbq3sOX3FulFIPDovGyt0VG6d54eqwRD1jwQiUagjKXNmJP8AXn307vOa9rs9vXd2kGV1eXpXrndvd1/V64kE6mi1pivVnmytt7e2bSpuTdtfBU5FojKplmR5YW/XpAY/QfT3iVztzrvPO+4mztCRCDSnkw6hbfN+3LmG58C3R6emk0PTRuH5PbCxs8sVNXxyPG5VY/GGAJ4twf8AD2TJyLuscatOtAR69LNt9uuadyAMEJofXHUDE/IrBZ51hklWNJCoV1itfVwB/h78eSdykatuuojy6OL/ANpua9rtTeTQGgz/AKsdCTRmmzoWppyk8bKCL/Wx+l/ZLP8AWbbKbe5BRh5dBeLft72wfQwSNFkVA6ynaZq50lWMAIwBt9P8L+zay3oW8Wh2rXrMP2s97IeU9h+l3SfxZGGNWSOlhj9syUqA2KpYlzc3N/oAPYc5lt9q3q1eW8jWWQKQoI/Z1LVr96Xb4pUjhcDUwHD16M58Yt5Hbu+6bFwu1p/GklgSOZLENz9Pp75Ofee9v9yuOXru7aMxFS5Qg0pg0p1lduOnnb24O9XZWhFQaj06ta3hhcDvDb9TiNx0FJlaDI0jiWGrVJI7SxWudQNj7wJ5Z5mv+WZ7S72G4eO6j01Za6jQ5BzmvWCG62w+okRhhWND6U9Otbb5A9Ibd6K7jmzG0ooIKGqqxWmmp7FIy7ljHYW99l/an3HuPcPkmJN5UiQJpJbiSBx6XSclc27ly++47VK8qKD21NKD5dDLs35A5vL46PFwVLpEE8T6ByFtaxF/aTePbfZ4rn94aAzVqK9QlK1/ZytaX4McgwePHoQ9n71hw+W8Uc0kk9Y6ySF1YC5a59XsP8wcuNuNhqdQFjFB+zrUNwI30+fR5dldw1dNHTwO48QUKbm/P4sR7xO5y9pbDcJHnjFH41HHoRWe5SRihOOjV7Q7DosvEoeVAx51EqLf63PvGXmjkK/2WfQ2pojxPEj06FFjvCn4qDqbvPfGPxeIqz54gdEhGphe+j63v/xHtHyvydf7tvMVvDETDVSWoa8eA6dv90jNuQpz6da8nzL7Zp9072OBnEdRSvIyMhAKG66Sb8++wnsnyV/VjlFZY+19NegDtztecwxgcAR/h6rbbq+lmzzS0UawxTuHKxmwH59zaeYHS1pKakdZ07Py8kljbSIcFc9DfszaD4+ORWuJEICAkWZQf9b2Dd43ZZiCOB49SBs+2GJSvHpNfKyB4egN0a7l1WXmwJF4ACBb257eyq/N0WnhT/L0j9y4dHI9x58f8HWlpvf/AI+7cX/a1q/+tnvqRtP/ACTIP9Iv+Drk3uX/ACUJq/xt/h6S4Nvxf2YdIT148/4e/db669+691737r3XYIt9Lm31v/vPv3WqHr2k2v8AS39eD7916orTrr37rfXvfuvde9+691737r3Xdvr/AIe/dar11791vrsm/wCLe/daAp14f1+tv+J9+68fTrx/r/Xnj/b+/deHp12ASLk/T37rRNDTrj791br3v3XuuQX9NyLMQOPqOffutV6tJ/lefF/pP5Yd1YnrTsHu2t6b7Ay1ckewctDMKVHySIPtrVHp0tc/XV/sPe/LrQy1PLrZ6+A3xu/nbfDL5kwdc7sfe/bfw9od1VSZLdO690pk9iV+0BqkGShkqZJEQmBSHjAsCeD/AGfdcCoAz1ckmhY9vp/g+fQr9b9H/GTvz+Zlu/5k9W0+2Mdj/iLu58x2Zi8fPS0kDVNPPJ4h4SQJlaRSuoDke94IoOvZVtR88dUB/Nn+dl8uKr5f/IKu2P2ju/B7Oq9/5c4PHYfNT01LTUKVCikjp6ZToEYjsq2P+w59+oPPrRd61Q46uH/lO/K3f38074U/ND4afIXcbbg3fvrD069dZvP5RXrIK+jtVBKasrTcXtpe1h73TUaDHWgxGWNT/kP+bonP8nPt7rr4w9kfNb+U/wDJPdVLRbO773B/o8qMtVLDU0NDU42Jvt6qlq1OgN5WUyMosbkgWNhXBz1urL2MP83RWO9f+E3nz7j7s3Pj+g+o3310vPmZDtTfmMzNHUYyrwrzaxVmQn02jIYKTx9DYD3uueHWyoUUU/ZU9We/KTqX4M/yx/5WG9/jp2ht3Z24vmB3PtmiWGr0UuVyWyczjS16ahrqe9mLf5wj/ere9fEKnj14Hw8LkefrX/N0XT+Tr8selvk78Q+7P5bPyR3NS7TzfY+DXbnVW4fTExrJdJp45qhiDGGYeI24uQbi4tstU56qqlRQZOf2dE03H/wms/mOUfb1btfbvU9XmOoosxej7RGUxjYWpwgkGqq+6VlUP47mx/17e/edCOtlTp1KRnh1aJ/NCi/l/fBH+Xv1/wDFzZm0Nnbv+R+4dkRYbfG7Xpo63NbczdPC8Pho62K5LAkhmI+guT78B69eYkcM14D/AC9aTh+ii1iL3/rz9PfutdeC/S5Av7916vp10eD9b/4+/dbHXXv3Xuu+Of8AePfuvdP1VtrKUlFFXzRJ9vKAylXuwDC4uLD/AHv2ZXO039pax3lxGVjl+E+vTSzRsxRTUjj0wey3p3r3v3Xuve/de697917r3v3XuuXAseG/qPfutZPy68Ler8f0Hv3WjXHXVvpz9fr/AIe/db68eD/X37rfXiCAD/X37rQNeuvfut9dkW+hv/re/daGePXiLW5vf37rwPXXv3W+ve/de65EFeb8H+nv3VQQeuiLG3v3Vhnrwt9CPrxf+n+Pv3Wj0Ofxs2RS9jd49c7Nq6daqjze4aKmqYJACjxFrvrB4t/W/u8Y1OF9erxoZHCep63/APr/AKHwXX3Tm3dn7apYaBqCkVXNNoBYmLm5AFxx7kyxtEtrMKvHqTrKwS2tFA6Id310jV1kNW9bAZEDHSWFxzc8ce01zASKnPSS6tnCk+vVIXdPWQwlbN44GULKTa2n6SH6g+wTve3rLA0sYqw4dV2fcZNtvFBOK9G/+DnyG3NtTIUfWdDI9HQ7gqUx9TUL6Skcg8RcNfiw94Tc++3G3c283Wr7xGGdZVpXhx6y85R90twj2X92zgGBRj9nV6GW63xOx9qRbkqq81rVEYkSrZi7SMRqIJb/AIr7lv3H+7psU3LETWoELoAQU88dU5f90by43g2lov6daU9M9BjsPtYY7OQ19POvhjldtJIClVIF7E/4e+d3PXt3LaRy7dOp8wG9esmbTwt720w3OdQHVvOyflL19JsC+TzVNT5GnpP+A7Mlyyp9AL/n3gNvvsbzN/W4T2UBaFmy2fXrD3nrkXco+Y3isULxM3H0z0F+3+8dub6y9VTwy07epgral5BPFjzz7lq45B3Hl6xRs48ugJzByVd7TEJSp9T0H/aGz9u7q+5jq6SmlCAOknB/x5a3uQuTd83PaVQozCvEdRdcRKzlSMjqtbsb4+S1U9e+JnkoYlJ0NCWRbX4AP095N7Fz0giT6nuJ9eiiS2q2hBU9FF3V1Jv/AGzBUVsGXr5aSmV5CPNIVKr+CCfck2HMmyXzCMqoY9Nmxv69qGn2dFVzm4pJ6oPXyE1NHKWYsbXKNyOT7GkEcaJ+nwPTX0FzJ2Bc9CbjewKLIbfSlp50arjT1gEXFh9bH2Ftz2yl2JvInqc/Za9udu3lba6HaTToWeoN50FErR1LL9xM7LcgarkW495V+z1/aQWUdoAKj/N1MvvptyfRJdWx7XUcOhlzk0WShP6dMi2Km17N/T3kQaOoIyOsQXGklT0Gseyb10M8EQuX1WXm92/wH/E+2NFTQDqhhFa9Wf8AxywlNiqKkrc9JHRUA/VLM4jFgo59dv8AePZvaJTLYHRxYqAKNw6t961yOxazDU/8FyMNW6qo/aZXuwW5B0X+nu1wZmNU+HoQIV0BVPXux62CLb+VlKrD46aVopVAuSqkgnj2V7rcSWW0XN0pIMcbH86Y6T7tOLXappT5DH7Oqk9yYzG7pnr56sJJUSSTLHUMLyagxsQf8PfPG85h3G73+XdJJCx1EU+w9YrbbzTvew8wHd9vlbDElRwOeHHoBhT1uzcqyVDyyY+SQiKQ3ckE2sPchbfvlrulv+uBqHkeugvsv7ybdzu8e3XziO6wtDjNPPoT9l5urod24XL4ipkjSKqiabRcM+lr+pf94ufeLH3n4dn3HlcWMkS621f5euofL3Lxm5InsN2UOssdF8+I8ur2dg5RexdpxJV04kY0MQLMBy+ixY/W5Pvg/v8AFfck8369sYldZJAPBeuaXudyHa8uX8sUR4uaD8+ge7A6QpGo6ycUgdFRnFgDa/44t7yK5H95ZJb2KzmfS9aEdQNebVoQtTj0LHxz2Bjtu4jzxQokkhJZbD+ypF+fca+/HPF1ve8jbXOpFWuftHR5yvZKoLsOH+bozojUNwb2H4t+R7x38aUdurB6GWhQa06g5HFwZFDFOFkjYWZW5Ug/Xj2oXcbq3Ef0jeE0ZrqHHpPNZxz115r5dFI7i6RwlfTVdXSUUQZ4m1MqqDz/ALC/+PvJ32n94t3trpNu3GVmVSNLGtGx9vQH3rZY4u6Mceq496dN0oxeaxslOsiTwzRJqCn9X6v99b3nPy9z8bi4gukamQT0DZrNo1IPVOlXt4dadj1eJeLx0NVWyRBDcp6mP+8c+8w9u3v95bdDdo1GoMjosm5Yl3u0kaNNTRgt1Vr3x8odyfFH5aR7rxVRPVbdrquD7+lsZaekgdx5HKX54vwBfj3lDact7Z7h8inY93UNG8ekfbTB/Los9t+dd+5E5kXcbaQqYmyuaUrw63RP5anzC6h+VnVdHHg87j3y/wBvTR1VG8kMTtLLTAsI43JJF7jgce/nl++F93jnP2U9xRzUqytthDEPGrSU7u2oUcM5PyHWYfMvPO2e520x3qFRKaBlOKGlT/PoeO8vg/tbtmGtFTohcLM9JURkN4ZJDfyLpHHsp9uvvZbxyL4VvcwtIuA1R5D1zjqFr7lCSSpjI7c1B6JR0bvnfvxN7XfqbKVFblNhpWPS01XUq8sMfkI0shP0tx9D7ya91OQuTPvFe3K85wIsG4lBIGQhXxmhIzQ8KdEW07xuGzbkLZiXStM9XiYrMUObx9JkKGZJoqmCOYMpH9tAxDD8f7H3yw3Pa7zaL6Sxu00mJiueOMdS1HMkqArxI4dT/HrYW4/P/Fef9h7RI7qdQyR14qG6Dfem3Yq6GZnQHyKbNfnkc34Psacs79dbVfRylzpYjHz6D262gJLU6q67l2fVYXPzV1PI5idtJ54APvo77Y8zW+87IkMgGoDqONwiKS46EPpjANPAchUMWhptMgJtpBVQ3+39gz3T3tbYixg+OSo/ael1hGW724DpM/IjsakweErp0kWHTTyxQD+rqLBh/vV/a72n5TmaVBLUsSCT9vR9se2Pv+7x2qCqKRX7OqNd3VtbvLcFZlVqXjkmqJVDKbNa91tY/wCP9fecuzk7JapHFUFQMg9Zo7PyztlntUdkYVagHHp5wm2MhR0zFslUNNJ/tZJAP9Dfj25cc4bmsv6M7qB6MejyLkvYZodMtpGxb1A/zdCNgsTWRU/inqZJXLAgu5PI/Av7Dm682btdN+pcyMvoWPR/tPJvLm3qCtnHqr6DpP78r021TRVhqDJNLJ4/tyb3P1At7GHt1z3zNtG4eLZzO0ajKsSVPQM9z/bjlLmLbws0KQyHzAFekNld6ZCPArVHGn1D0OhGon+z9P6+8iNk97t+3Xc126VVAHp0G+RPuf8AK+7bZLuV3PpwdIJ49RNi94bDra5dq5nNR0O4VYMaWciJVuPSC7EX95J8v7od1shLL8Z/wdYQe7nI68ic0S7famsKEivzrToaMvktpY6nirMpnsbTo9jG0dbCzOh/TqAP59nnUYjhU9eTePWOPxFTns/uWhpcLR07TvVJUxGUhQSbJcX+n9ffq049epXoo9V/Md+KGM3JLhG3/TfZ00/28ruYVlDK2kixe9/e8da8s9Ze1f5oPxY2VtGtr9ibtos7uT7N2o6OoWJrShbroTWSWv8A4e9GvWvketWn5NfJfe3yT37kt1blq54sbLVyy47FLI4p4oyxEcxj/TqI+gtx/r+/dezx6LQST9fwLf7D37q3XXv3Xuve/de697917r3v3XuuQ5FgLH+t/fuqnGT1x9+6t12Bf829+60evH6/W/8Aj791vrsAnm/+39+61UDroC/5t7914467v/Zt+bX/ANj791r59dW5t/xr37rdcV68bHkcfi3/ABPv3Xh6dde/db697917rtTY3tf37rRFR1//0fn/APv3Xuve/de65XuOfr+D791qlDjroW/Pv3XjXy669+631737r3XvfuvdcmAFuf8AjXv3Wga9eAubL/T88e/derTj1x9+631737r3Xvfuvddi359+60a+XXvp9Pfut/b0p9mY6fLbp2/RQoztLl8ep0gkqv3KsWNvwLe7Ipdgo62qlmCjz629OnexE6u6vwjwSKtW2Ip0Cg2YsUH0t9OfY7tpPBtlI9Oh7bSGC3UVpjo33xw3mN4fcZ3PRiWsadlp2ltwGb6jV7MLWUyjUR0YWbiViz8ehl762zuefaM+Rx1P5KVopCSttKIRc2t/vHty8SVoSE4HpTOHXvHl0UjpzbG1qpqsbg8kNaJwSTAx5vyLsPp7DLbJBdRNHL59G20cxfR3IkpgdHo626W21uGYVuJkSZYWAJsFI0ngEH/e/aW25FsY38aLiDXocp7hXlxCYTleHR1Nt9S0CURSpWJyIzEAGVvxYGw/px7kC3t1ihWIjCinQLlUXEzTn8Rr0WLu/wCNT/5RuDDVskE0KyzimiYi5iGsKLfg+2J7UfGDjorurUirr0BvUnYOexWUOHy0TRvTTLDGr3BcI5Grn6/n2zA9G0nh0ltpGRvDPHqx3bnYFUYKZ14jKoCBe349mIYNhRXo6WZqV4joTJ95008cQd1DyBQbuv0b8c+7qIwcdOlwBUHoHO7eoMbvzbcsxgQzSRyOr2DFrx+k2H+PulyEuYWgkGpSCKfb59MSxujLcW7FZFINR8uq/uuOtNndbb1I7ExUYpJpCkFfLFzEC1kt/vHsDWHJGzbXuP18SBWJrjoW3/uXvu67cuyboS0ajSCfQdHZl6Hfc8EWV2DvGogxVQnlihppz6FIuqlUPHsalF4hsdA0WgyYm/Z0nqnoTtHGRTSU+7spOkUckjXlksQg1EAE+2nQIhfUTTP7OtraTySrAjGrED9vUXqXKrS5+rw+/A9YqrLDeqUkMRddd2/r7Kdn3uPcpmgHxKadCzfOTLzly3ju7htSOK1r8q9ZoMXs/D7oyM2DqqWiWoeof7OOaJBJqYn9F/r75Nf3kazNzzZW1tESBACxC4BqPMdTl93S78bcVGrBbFfToqHyb2jT7w2dmseQGMscpULY3stwbD/XHvFH2e3eTat1iZsKKdZg8/bOu67CYWNTpNOqe9j9UZrC7omix2RqqOuSXTAiEqdIPFr/ANPecF1z3dLZrJJIXiA4E4p1h6vtds2/XLbbuEYDscEjOTTowtX1p2RnaqGkrt0ZUQ3s4LOVsRZl+vsQclc9WO4OVjhWo8x1MmyfdE5bsdsO7OqtQVpQdCps74rpXzQmoaWvmNtRdHYl/wCrXv7mGLc7zcSI4AW+XRC3t/sOwyMZEWNVOOHRgKL41YzAQ/cV9LHSRRQu3kddBDRjUAL+xlyltG8HcQZ4zoJzUeXQS55l5YG0PArKe0jiPTpx60oqiHJVFBSF6vHpKYzIwNowGsAP8PYL989t2yyv4HtiFc1qB5465le5FvYWu7F7WgqScdGLTAyav2rgXFx9QD/r/T3Af1RABYdRz9RK1CzHp4p8XLGCCS4sRZjYA/1A9svKrjPHqiyzJKswY9rBqfZ1L2dkKXYufOeeNHmiOu7kfRTrsPcE+/3L1vzBy79KBhgRgeo6zg9rvevmPmyGDkSNzHDRQaH5gdCFu75kZGANFDMkaopQIsv9krx9PeDPLP3ctqgnMzRVLGtSOs7I/ZfbbzbISp1yOoJ8zXqv/tbs5ewMnNlcpOsjLE5F2v8ApQ6f98feV/J/Kg5btFsrNdIqOhlsvJcnL22vt8cdUINRTp5+E2y9s7+yubr8vnmp3o8lHDDRX1RyxtLpIsP6e3vePmjd+VNpRLK38QMhJbzBA6wh97OV6c0RQ2MOl3rUAV/wdXD7z+Hu3MjtA5jaFUz5uOAVESxLy9k1MOP9b3gVyb96+6n5uk2LfItNqjaHc8AxJAHUfbhyLLZ2gmWvjEVp0R6m3TkNt5OfbOcjakyGNmNOEkBiMhU2BOr63t+PeYs2zWm72SbxtxEkUy6qjIAPQDErROYpMMvl0I+H7iyGMJWG66T6QNXJB/J+n09gzc/bjb9yH64rXj0+L1osjpK777P37uqhnioo5VptMmuVS1tJGlrk/wCHs75X5D5T5euFd9OvFBjj0zLfXM4ooOfkeqru7gybtpapp/uqoN/lDagxjltyOL+8m+X7gHbGt4xpTy+zqSfbnlWS9nF/cggA1yPn0jMPmzQ1kbV6lFdgQWFuLcfX3W7sxPEVgNT1mrs9ytrbxwvwUUHQyYjOUMzo8Uqi4+lwD/r8+wfeWE6LRh0NLO/gdvIDpEfI6BNwdN7ixkFpJJoZm0j1MQtPcEW9mHIga05nimfAwP59FnPyC/5UngizUMf5daUfaWNlxPYW7aCVGRoMzVABgRdWbUCL/j+nvqlsjK+0W7Kagxr/AIOuR+9RtFu1xGwoQ7f4ekD7NOizr319+6912bfj37rQr59eB4I/rb/ePfuvUzXrr37rfXK5Y2/2FvfutUA64+/db697917rmCtrc8/X37qtDWvXD37q3Xvfuvde9+6912DYg+/daOR12T/Qnn6/j37rw+fXV/p/h7917rxNybXt7914Drr37rfXZBFv8fp7916vXvwLE3v9P6e/da8+n/bW6c/s7OY7cu2MrW4TcGJqEqsbl8fO9PWUcyCyvDInIP8Aj791vhw6N038xz5xPiXwL/JjtR8S8bQtRHc9aYfE66GjA1fQg2I9+63ranQHbU+RHd2x5d3zbS7L3XgJd+zmo3k+Oy1RTncUxJOvKFT+6eT+r36nWvKnr0E2SyddmK2pyeTqZqzIVszT1VXM5eWeRrXeRj9Tx9ffuqgAYHQidc919rdQVEtV1lvvcWyqie5lnwORloZXLLoYl4iDyPfvl14U1avPpgqew96Vu8pOw6rceVn3zNkf4rNueSrkbKSV4jEYqnqf1F7DliffutnPHo1WH/mRfOfb+KbB4j5Odr0OLKiNaSDdFaIY006NMXq9Itxx71QdWDkDot/ZXc3aPcWQp8r2hvfcG98jSCb7asz2Qlrp4jOQZCskpJ5sL+9/LqtPPpIbZ3TuDZubodx7Wy1bg85jpBNRZPHztBVU8wNxJFKliDex9+68MdHCpv5k/wA66TFSYKD5QdtJiZVMbUR3VXtAEK6SEUsbC39PeqdWDsBjore/Ozt/dm5Jcr2DurM7tyKeUpWZitkq5w0x1O3kk5JJ+t/dvl1Tj3dIMm+ot+o2sbf7f3rrfmKdesbDUG0j82/3r37r32dd8BTa5B4/w+nv3Wsk564e/dW697917ofN2GNdm0HpKs0EAuBb6Ri/uWObD/yDtsHy6LYSPqGp8/8AD0AdvcT9GXXJbC9/95Hv3Wj8uuPv3W+ve/de697917r3v3Xuve/de697917r3v3Xuu7/AEvew9+619nXjb8fT37rf29de/de697917r3v3Xuve/de67uffuvU671cH+p/Pv3WqZ66BsD/iPfuvEV6tN/lTdRxdj984zKhkSq2xkqOpgJZVblfIVXV9b2/Ht2NJHB8IVby6PuW7aO63qCOY0XVnre360xNflFbF5BGS0MQgkZSLkoENr/AO8e5J2MT/RBbr4vn1MW6wWq3Gi0NVp1D7c+P8+RxDmPySSSC5smrk/19mDwK6kDoluLMsuOqLvld8b58FRzVj0cjfUyOYySAXN7cfj2Q3lmFGRjz6DG4WpQBiKEefVVLUWV6y3DQ5/CNJ9wlWsiW1Ax6ZOSP6H3E/NfLkLut9Cneua08+hxyPzCsMosb34WNM9bG/xs7Dx3yV6gxWyqvKE7noaUB4GNpGl0abEt9bn8exVsG72fMOzfuO/f9dPX04dDjcdovdg3Ab7tq6rd/Ty8+usz8De5aWok/hFJk9Dsxj8KvYhiWBB/xHuPeYfYLa+YpfFml0EcD1KGze/M+12i24t1enEnos3Y3x1702IKtqvJZmmKIT9q4lCkqbW9grcfu27ft9v4oPiU4HSOjW397dt3WXRdWyKx8+gU2p3lvnqqsmp8o9R9wrWMr61I0tbi/vHbm32dlnnaAwVRSaGnHoykO283DwqgV/y9H/6s+XMO9KXGbdRkq8vkrQ6QfJNqb0/T6/X3jpzJ7avscsl5KPDihyfIY6iPf/Z65hu5LqAHwxn5dWG7l6G3JXdXR5ySklpauopRNq0ENZ11j8f0PvFi299eWLXnhuXI7hHZTTTqrwNOi/kb2yTe98EU2FBx8+qyt5bU3R9llMJUQTSRxrMjyspJA1WNz/vPvKjZeY9smeK9ikALUoK9ZE3fsdZwodKcB6dVAd17ByG3q+tngkk1F5S6c+k3v7yS2Dd1voFqcdQpzN7e2ezys8XHOKdFc25uPLYnLOHlk8bsystm+l/p7Gl9bwTWoZePQN2lv3ffiSJaMD0YvYe+qWDNJJXTGCJHV/UdIK25Nj7kr2oMa3JhkIFD0Mud7+7vthWaQlgBw406H3/TljqvL0mLpJleJnjjLg/W5sRx/h7ygS4jCqqEEY8+sX5LsNOVYGpPp0eHrjcXWlNFFV7oy4gEUaOE1Kf1Lcg+16z2ajVI4B+3o1t4WcVINPsPSX7j+QuK7LpK3rDYOdfBQwQino8pRv4pXkHBOoEc/T3WW8juR4URoPUdN3k5p4CnSfXpUfy+fmPu3p3smfp3syZsniRVR0uOz+RctLVGrOgNre/0v/X3SzupAxtJjgcD1ba9wktpRbTGoPAnq8/uHsvbNRt6qo8Xk4KmaagWQpHPG5/di1W4P49lnPE5sOV7lpCBrRgP2Y6Wc4XkcWxSgHLA/wCDquSnnqJQ8gXQTPI7KL8Atxx753QaWLyN5s3+E9YnIdQL+pPWOqx0WYcQVKK2n9LMASv+PtXHO1uwaLHSzbL/AHDZb+PdNokMMsbV7fPrngNnLjMq2RJcxw2KqSRGW/PA9x17p8sXnOW1LDZH9Ramg49dH/az7/e4bFy4vLPM8SsEAAkf4sCnHq6T44dg7FXZ+Nop8hDS5fwrDLAZFU3C8/X6H3xV99vZb3M5R5wn368tnbb2BppUs1fsA6T7p7s7T7rXz3G3sCa1oDgV6NNLTYrM0zRJJBNFMgDMHR9Stz+D7xv27cZ0uDLarJHLG2S6spBH2/4Oie5so2HhyU6hY7HU22X8cQAp2J0Bfpc/j/Yezi/upN0mFzfyapKUHRfDGu3safCelQjpKnkB9LWK259P9PZKyMjEOKdGqsHUMDg9djgkEXJ/xsBf6H3WnkfPqwNOkluqlkqKJ40j16oyCtr2vx9fZ/s10IbqBqkCNqn7OibdYmaNmHVdXdmLqtsU1VlZI2FOgld/R6dKsST/AK3vPL2s3yz3tEtYmBYU8/Og6BdvtE253f0icSadUhd0xYXeW6aXIY9wtSuS1OEAubNZveeHKtzd2FktvP8ADpHU08pchXWz7betepUeGSCR8j1rK/zQYmi7Vq0P9h6dSP8AWDD30I9qX18tQt8usKL9RHzDcxjyZv8AD0V74v8AzH7j+Ku56PPdebjyNNSU1QtU2JSslippJVa/ABsAebrax9iLnHkblnnza32jmW1S5icFe5QSAfQnoxtNxvLB9drIVp5Vx+zrbE+FP/Cm9sgF258hcJg8BRQQQU5y5fVUvpULJLqsNQP1+nvl77y/3W/IO/3Mu/cgSSQ3kgb9Nm/Tz8uGPI9DvavcbcLJfDu6MvVo+/P5o38vDsvrXIb/AMdv3F1G5qPHy5CCIoscwq1W6qHP+I948+333OvvRcgc2wctXrJLscjhCwPckfr6H06M5+Y9guGjvgCJFbUw8j0z/wAv7+bH0/3Vnjsqk3dQS1LZGShpo6mujiQAEiNUMhA/At7O/vUfcW3zYdpbm3lqMzrFHrkKrVq8TgZ6lF+ZuVd12iK72+TRcjDJwFB59bBtFW01fSw1VPMkkU0SSJJEyyIwkUMhVl+oP9R75CSwSWtxJBIGQxMyMGBUhgSMg5HDpRHKkyB1Na04dcMlTfcUsiyBbMhCm4HNv6H3WW/j2aP6u6yoI0jianrz2rXdEA/l1VD8otxzbey7UAjWRZXsn51E8C3vP37vd3HvWxR3yHSKVI9OpK2T2Si5k2ltxFdQFTjh0JfV71sHVUWSamMDyXaRmUoRGy/Xn+nsu5wltNw9wPpWk1BQKAGua9QNzPsv9XdybaYqniP59VNfLfsmpyNbPt+hk1JBNUGQo19QDcg6feZ/tZsEdvbLeTChIX/B1NvtXyebK2/es6ZYDiOiM4jM/bzrrH6ZCzA3A1XGo39zLdWniJjzHU2293pkpTA6Fyg3NDM0YFlAAA5F/pckn2FZ9sZQT0JLbdFLAdCJQZamipzNqLeNA5B+gX6k39hq4s5WkCUpU06EsF3Ho1E8M9FV7F3nT57dcNAswMEdVbRqIFwfx7lDYNpaw2wyU7iOou5j3hb3chCDUA9Yc5lKiszOL2/S80p+3UKvK3b6gsP+J9j/ANudnFxeG7Iq5JHU4bJzHHsXKLM9MLX+XVHH8xneWc2J2XDQ7Wy82Byar5J6nHVHiqZYww4WSPkWJH0/xHvNXlnbzY2SkniOuUPvdzavM3NEygCgZqkeeT1XfU/InuysjSKq7H3PPHGAFWTISsAFFh9fYlqeoU0ilOodX3x3BWUEmLq9/wC4pqGVDG9M9fIY2RuCCPeq9eoCMdBK8jSO8kjM8kjM7uzEszsbszE/Uk8n37rfXQBIuRf/ABv+B7914nPHru4NiR6QLWPNub29+61ngOuHv3Vuve/de697917r3v3Xuve/de697917r3v3Xuve/de67IH454v791r7evXP0uffut0HXgQDzz/vP+9+/daOevcf7z/th7917PXvzx79177euvfut9dm34N/futCvn14Dgn8f8T7916uadesbX/Hv3W65p1//9L5/wD7917rsi1v8QD791oGvXhb8n37rxr5deNvwb+/deFfPru3pv8A776+/dermnXQItY2+n1/N/fuvdeFvz791418uvfi/wDjb37r3nTrw+ot/vHv3Xjw67YAHi/+x9+68DUddX4tx9b3/P8At/fut+devC1+fp/rX9+60eGOvcXP9Obe/dezTrr37rfQ4fH6jes7FxSJH5Gjnp5NJ/2mYc+1VoKzjp+1zOvWx1te2RxuJp62/jjoolEbcrqtYcexWhLAKehhGAUCtjqyjobY9bPsWoyGIQmppXWRIkB9QTkCy/63s3tkPh9o6NbaEMlQeHXPszvbfWMx67bymKkhpIZkikfQ4LxodLah/iPr7rNcyr+nTHXpriX4aY6FXqntDqCvxcdLm8PQUNXJTDzVemNZDKy21HVY/Xn2phniZdDDp+Ce3ddNKdI7dfydw3R2ZmpttVCV1BXM6xOJIyA730Kv+8W9tNeLanSD01JeLaN2ZHQqdW/I/vWqpG3dHteSu27O4kWodJJUWKTnkrx9Pb8NzOV1lcHpTDfXVPGC1TpTdh/KmpamabIxSUtUKdxNSFdEQbT6gdQt7tJc0wR1uW/PxN0W7rjsCk7N3slTSRiB45jqWMCzHXpP6PaSJzJLUDpJCwmfV1aRsbBPUQ08DA2ZQPzx/Ui/s4jUhacOj6FarU+Xl1w7IwWX26qZagLyU8DI7KNX9gAkf09tyqyjWh6rOjJ3r0tdgdo4vceLioMg6R1EQWEo7IDqI0m+r+nt2OQPngerwzhxTpm7S6lxu78dNUxwpVNoZ45FRpXQhPSQIgT/AE92kjWWobqs1qJhTz4jon23ewuzPj/WVNM1NUV+AWp0mWZZAsMJNtKLJ+Lf4ey8+JACGFR0hSSa0JDZHRntu/ITJb6xhnx6i0gCTKHT0s4sylRz7UI6PHQDj0uivS7idOK/5Op0eLgyMc1S2PiFbKj/AOUAXcFwfz/gfae1260s5DLbrRjmo9ejm/37dN2thbXrlkAoAeq5u6di7929vRtyYPI1siU+qRaGMyiJ+bkEDj/D3Dvu77K7L7rWzSbnRZgMNSrY8gaHpbyPzpeci7qlzAutAf8AVXosOb+QvZOPrJqLdG3DDjl1RPUsGJZTxqs1x7wY3r7p93sEzy7SjOFNQaU4fl1mDtnv7tm8Wyw37BGIpSo8+gZm3zHV7upMtjIyHkdPIq/2QzW1HT7Zsfbne5Yv3RcxEVxUg9E95zDapdjdrYii0IyPt6tE6b2A3Y1NQyxMWqJIEkl4JYH6MODf3LPtv7BbntayXNzUFzUYPDqSLL7z1gmzttEtNSinEeXR9eu+oMftKKSpyYR5YS50uPrbkXJ95Ycm+31ttaia6AZh1jH7h+6M+9zPHaEqG6KD8ot8iXKptnCgwmRhfwkD0ltJBC+xVzPeWnLuzy38ShdIP+DrHbmbmG7t7FpriQnB672Bt6nwuIo2+tRWwpJIw+pe4LX/ANj7558zb9d8wbpLdXLEqrHTXyB6xU3W+l3K7aWc1zjoaKOmjCi4LE2JJtcewuTXPRZUD7OsVdqgDBVAAJIa66iB9ef9797UKT1sUJoOgf3XWyTLKIje4YC35Om3HtWuz2W6EJuQBQUwepQ9q99t+Xd9W+kbQVIz9h6LNujBV1Qz6GLGT1X9XHH9b/717XXXLXLqQiG3iQfOg66M8mfeo2jbQjXcgbwxQAlegJz+ArqanqbzMB4pL+oj6Kb3590teU7AutFH7Oh3d/fB5dlRnolSD5r0InwO3lR4LeGVxFTOqPNk0KiSSMcpNqNtZ947/el5ZuU2MNa1xGa0B8x8ure1/M2xe7txd7tJGjyxEha0PH062R9q/IHbeFxlPSSTRORCiMdcfHGlhxzzf3xek9rOZINwurm2jDeOxbuBr8v2dNb/AO1m5bhftLGpVamgoeB6It8s6PbO6a9ewtoNHHk6TTJUUMADCodVLM5WLk/4+8x/uz8zcz7FtB5N5zBdSWCSHyUnAz1EnNH3d92nc3u3qdXmKHqv2v8AlLhdr0ctFnMeIchTm2l4QrMf+ng/P+v7y9PIF9uk4n26WsbfP/N0Ucs+yU014Lbeax5+fTTuD5k4yv2jUY/C03iyVQpWMRKFZg6aben/AF/erD2qvIN2W5vX1Ivr/s9Cq59ndu2neI4EOuNuJpXoBtj7dzm9qyq3Ln0lWlLPP+6Gte9x+rj/AG/sY79udns6JtlkdUhoKDqd9i9sIrrwdu2he1wNRoRTpm3GuNGSnp6gJDHTyaIGNlDrbgi/+tb2dW+37nbWiTOhBkFepO3n2wk2+GGFCCQKGnr1ArKerpKWOvxrM8S24GohlH5BX2/bWU105imjNT8uk3+thvUUAnhUstK8D0IO34P7/wCEqcRJw8lPLHIl7kGRCn0X2E90f+rl6LpsAEfy6I5dmupo5NtuhQkEftHWtF/Ml+K2T6a37Vblp6cyUWWmFRUzASLpjZWbUNXBsSL2/HvOf2R9xLLmzZEti/6iYAweHXOr349t7vk/f3ukWsUma588/wAuqsfc99Y+de9+691737r3XvfuvdcuLf4/4D37rWa9dWP1/p+ffuvY4dev+fr/AK/v3W+uvfuvdd34tYf6/wCffuvdeJv+AP8AW9+60Mdde/db67sTz/T37rVR14An37r1R16xvb8+/dbr59de/de697917r3v3Xuuzf6H8f7x791759de/de67vxbj/iffuvfPrskcW4P5/239ffutCvn10Tf8D/X/J/1/fuvU69yLH8c2v8A8U9+691yHqPPH9B/yP37rXw9cT9T/r+/dWHDrr37r3Xvfuvddj/H/jf+w9+610NPV3Q+/e2mlG18bLOIm0XamqXDn+o8Y+n+N/fut9WX9Ifynd+b6KJuqnrKB5NJUrDWxJyL+mwv/tz78OvUI6sv69/4T10u4FpamszE8McoViJI697X/BVvfj8utA4z0PW4v+E8+AoNvSwQ5byyPCQHFJOrD0f6oC4/1/egK+fW6KeIz1Tl8qv5Q2+emEjl2qKvIxK378rxVcsajUQQdYv9B/X3anl1rPRJtp/CPsfM1go6+nkhlaTQixQzXNmsfqPeuvfaadWB7B/lETbjxtDPla+spamqRS6n7tCpuDa1h9Rf2zcMUhLKaEefW1GaHoXt4/yiNw7hxX8LwVTUVMmMjjSKKKKpuxCWubD/AHv3NHMQiXkHbw3cdPSCMu9ywFBToBaX+Sb35O7LHiMtKAWKt9vLyL8X0pb6Wt7htWqtSOjDQx8x0hOxP5PvfeycXUZA4XIM0MTyES01QyroW5uqKD7tWvWtJHn1WB2B1VvPrTKy4rcuLqIJY2ILrT1PiIX/AFZdAF/2/v3WuPQb3+vA5/3j/W9+69Trr37rfXvfuvdd2v8A7D37r3XXv3Xuve/de697917r3v3Xuve/de697917r3v3XuvEW9+691737r3Xjz/xr37r3XvfuvdH1/lydk5bYnyh60o6Svkocdndy0NPkdEixq66gArlrCxFxyfam0cx3CMPUdKrKVobuORTSh6+jvgK3BHAUGaxkyPLHDFIWV42DsFHAZOD7lyJozCCoGR1MNvIHhEta9CVQ7ipNxY0LNp8wXSVNj9BxYf8V97jUo1V6VFvFQevVfPyo2bQZfE5GCSBX8kRtdL2JJ/p7SXUatUN0V30FU7s9a2Pfew5tubkjMSF6R5yShVgqDy82vx7CF9bCSsR4HoKs7WdyssQ4Hqy74FbMi/ieFzWHrkx8kUsL1CxTKhlswIDWIPsH2vJ1ym7R3tm1AGBNPMdZG8sc/7ZJyzLtu4AO7LRa0waUx1sw4jcWVWio/I5ZlhiUyB9RPoA5IPuYJII2/l1GShFckCoqf8AD0EfbXXuK31R1M9ZMDVNEQA12uTzb3cxG5i+mbh034axv4qDPVS3evwKhyuKzG5YagpJDTzzwoiSEkj1D6e493/kdLiJpo2yKmnQ02Dm25sZ1VQQB59Fc+CvVW3Np96Us3YFXFQ0uOr2am+5KQKxWXjmoI/p75lfe6sOYdu5TubLl5C886suAcYOcdZGb/zvZ2vJqSznulUZ62ydx5HZ+6OvquDB5Gjmp1xgMHhqaZgCsAsNCMffzrScucx8l872LbjbySXBmZppSj92t/M0oKZp8uoy5B5jQ8wQ3Fq5Cs4FOqZN9YWCOTOxgDyWqbNxf9dhyPfSvlPdZ3+kNe2q4/LrocBHdbP4tO5kGfy6pc7+200tZkgyFmLS/wBljfkggW9548m3wNrGRwoOsTOd9v1TyKRXJ6JLtjqSoyuYqGaAWLN40Kk3J/H09yRdb0kEABPUTbdyw1xdFpBTJp0ImX+Mm5KpI6imhanEhAUokgJv9LEf4e7ct88Jb7isVm/eTkDqW9s9rdx5gsDEkdYwMmhpTpc7I+J+4YZY6upSXWGUhzHKSCD/AFt7yKg5xulhU1PDqP7z2Utbe9MbKKg+nRoMT8aslk40WSqneyhHi8M9uBwAbf8AE+3tp3Tcd53ERLIwBPSjfOSNs5e2gyvGpNOkhuH41VlLk4ocGktNk6eVGEkVPOrSP9bFgv8AvPvICw297e0VGNW8z1iDvUaz7m5hFADjqBlOj9y0VbRZavhmpczi5I6mKcrIssrxG6gN7Vm3aoPn0X/TPqDHBHDo0PQGe3nuvcmTodw1s+mipkASZyQwjXSOGN+QPcJ++e9XdpsEcCEirUP2dArny+njskirxwejm0uNjRZHtZSpUcccHm3vEVAFSh+3qHVOiijqRiMQs9UxC3N7X/w/FiPbkslFr15n0YHS1j285vGxsn1IPI55HtIt4yZjND0klgilH6ignr1RQV1Ckf2FfLjjGdXkp3Kuef6j2m3CDbN3tJYt4tluhTGsA9DPkPmuTke/a5tl1+JjT5dCvt75I7g67oIonr6nKzU/I803LccLZz75we5/s3sHMu/T29vbrbozcUWn+AddJvZbkiH3MtW3ncZfA1qaL5A06HjDfO/atXt5xn4YaXKRRlgjSxcvbj9R494/bh92Pd7XcFt9uYy2xPxEEkD5HqRL37vO+XeuPaKzU4YP+TofOjPkttTtEPi4KuCOuiGoQ+aE3BN0sb+449z/AGa3PkiBbpFaSE8TpNR61FOHURcycgc38jFRzHbmONjRTRs/t6NYLcEm4IBBBuCDzwR7gRkaNqSYPQbGc9YqiJZYyD/sT/gPpyfdork29XBx514U6bliEgoegO7c6xp94bVyNJVxskNRTyoknjcqCwPN7WP+39y5yFzTvnJd9bbyIHNtM6gNofT/AL0Rpp+fRdZILPdY7iLirVI/PrXF7E6qTZHZVZj9fkhjyM5S6kA3b021e+tXJnN39ZthtrmMdzhP59ZkNGs/t9dbvIgH6Pn9nWrD/Nbhjh7gyKqyg+eABBb6DVc2+vvp57TQywcswRyinaOuRt5MLnmO6nTgXb7OPVSY/wBcj+lv6+5U6fPXhYnk/wCx9+695Y6dKbN5ajo5sfS19RBRVAZZqZHtHIr8MGFvz70QGpXy69TpXdadn7x6n3Ri92bMzNZicji6+lrgtNO8K1Rp3v4pig+hFxf8fUe2L21t9ws5LC8QSRSqVZSKggih6cSWWM1jYqfl1vr/AMnX+dHT/ITAU/VnY8DU27sDR0kPleRJS8UFOI2YswBcEglW/P8AQG498QvvqfcMS2a59xPb4afEV/ETOkFjWtAaVHkaV+3qdvai+veZNzHL4FXJBHzoPXq27tL5HZqgr2rsNPK+PYaljSRdGkm30B94Hcgex+3T7Uu270ollU0JcE5/P+XXRPkv2o2qe1W13Ciz0HRYMvu8drZ7DV+VHjSCujabyWtpVwTe/ue9n5cHt/tVxZbd5oQoHljFOpUl5ZTlHaZbOzyXQgU6G/u35Ebf2X18Nubfp4WnSg8N43i4OgKeAfqSPYJ9ofajf+YecX3vdpGcySVVaGgHWMu2/d/3bm7mP94XgITXUkg5zXqjXObmqN2ZmurK+Jo5Jqie+oc6Xe/BHvpVHy9Ny/aRwaSKAfyHU/bh7Xts9om22A1BQK0Hp01xYCBjM0UesaS19B5P5t/j7cDXbqCFPSC39rNzmiLhM/YR0hkraigzTU8yGOENZSFax/Atfj2eT7bJ9CJiCCfXojv+Tb7aatOtCPkejLbX29V5nF1HiVyksQXyKGJ1Mv0A/wCKe4q3fc4LC6USHIPDq+27VcXMTqwpUdEq7Z683LtDcCZOCnkmRqxmJ0ygWLX/AKce5f5X3uw3q0ECMKkDzHUZbjyvuUO+KI1L92cHh0x7/wC5uv8Apnr/ACW+t75aOi3DLjXTD0DSwhnrRGRAoSQ3BJ+gH195Me2vKDQWyz0xqqT8ug172+59hyjtJ2GMnx2jA0inEj9vWr33h2/ne7d95HeObOl55JYqODWX0U5nZo3JIFmZdOofQW/2PvIKNBGgReA65vXt3JfXT3k3xOanoHrWNm4936S1xjrxtfgk/wCJ9+68OHXY+nIFvwf6n+h9+615466uf9b/AFuB791ug669+6313fi1h/xPv3WqdeFvz/T/AHn37rfXXv3XuuTAAW/P19+6qCTnrj791brxFvfuvceu/wAfT8/X8/63v3WvPr3Fv8ffuvZr14Hi30/xH1P+B9+69TNeuvfut9cj6j6f6f63v3VRgZ64+/dW68AT9PfuvcOuzb8G/v3WhXz669+6312Lni/H55/H59+60adeNvwTb/H37rwr59f/09AAm/4A/wBb37rQx13xYEm5FuD/AEv9PfutedB17hj+F9+69kD164+/dW65A8af6n6/n37rRGa9eYaTb/D37rwNeuPv3W+ve/de67vx/j/X8+/dap11e/19+631737r3Xvfuvde9+691yuL/pFv6c+/daoacehZ6T3PHtbf+FrZb6Z62lpSQLgeWYC7H2/buElDHp2F/DkDenWyts0xVuKw9TCVYS46KdipB5ZQfqP9f2LYWEkYI6GEREkYcenVrvw63xTQ4uegnC6Vcq6NYBlBt+fZ3YSdtD0fbc60oMgdDJ3YuwsvFNLNRU3neF7AIhJZhe/09qbjwSK9KLgxspp1W7unY/3dRNJgal6ZLuQEDKAbcAW9lLxmlV6JnQ+WOiednYvdVE1NFWRyVywV6P5XLHSkZDE8/wBfaGfVQahXpFMWGKV62L/gP3Z1PUdG0+B3eaVa2GKnpzS1AhCsyIV5D/63s4hdpYVWNqeo6F+0XFubQJJg9IH50ydWZLZ1RWbHxmLhyrLKP8mMYeRCOG9Ht66jVbcCvd69NboYHQ6AAeiSfETN7R23XSJuBlpMxLLH4otIPpL3Y3J+t/aWykVWIfj0U2Dopo3Hq93q+TF5LHxV9LNBJGygj1JqIPItz7PtQ8PGa9CiBagOD0/djRUxxM8crRmFoZLrdSPxfj35CShqM9WlAyGOOqvOxt0vszORSYNpwaiqSBIqdGbVNLIAgIX+pt7L5WZTq4dE1w7QPVOHV1nxdp8b071C3c/yAhhTKV9DH/dfaldpkOUpqtdUdU8EtuLEfj2SXN/LfTCxsyQPxMPKnRNd7ndblKNutDp9WHQC9h4nbPeiZqqoMPSY6hykk9RSQQqqoiyElAukcfXj2JkiVYFjkNTShJ6FFvaA2wjc6iBk+p6q/fE5npPuuk2K0rtj8pJHNpBJj0O3pXn/AF/aEL4M+gHB6LCGtroRrwPVk+Lwk9JQU1U4stTF5F5/skkg2P8Ar+zaMLpquejo6wBXgeg/3Ts5Mq0tRPEpTSwBPqJJFr+23Uklh03JCrjP7OiPdz9ZYSpw+Tilx1OziKdllIGoW5H4/wAfaO5USRmOTuU16KZbdoW8W3NCueqt9tYTGYve01DMU0rUehD9LK9xa/sAnlqy+u8ZlFa16OYOb9wji+mdyfLq4X4rZxMdkadKKQCJEKkqQL6m+nsfWXhRosQGB0X2z652mrxz0dnf2+KLD4KvqaqsSKYxyMi61vYi4/Pte5KDsHRjNIuirdVQVeSfsLsr+KQlpaGll8crsONStxb3j574czW1lsz7Srd8gNOoj9wt3hS0+hjbUzZ6M9halVljpuCsOmNeSBYE/j8+8LWQiIE+Y6g3TgE8T5dClTSLHTFmIJPIB+vH09o/xU6aYEtQdI7cOVjpIXczEuQQATew+vHtTBE0hHp06qUwBnoHK7JLIzSlr6udN+Bf82Ps1jjY49OlIJUVXHQf5rKRRwSsCNV/qbE2t+Pa1IgtK5PWwJW7UNB0VrtDdlPQY+qAkXW8Tj8f2lIufYg2+2dnVVrUkdL4UdiAGr5efRAMLvrP7I3ad1YfIMrLVp+xDKCXJk+ulT7k7mD222vm3lN7bcI1aQrgn7OslvYDnm95A5mhUuVt2YFvQ1Pn1eh0H2TUb42jR5jMVUkdQ1LC7h7glmP9L++UXu17YbhypvRs7a2OhmNCAaY6662vuRyrPYxXazLWRQTkYr0dHoapweb7WwmLysceTx85ZZqCYao5wSOGU3+vst9rOUbm55vtIry18RSTUMDT/B0DfcT3E2VOULk7bP4c1MMCKjoPf5k/SXVZ3NiH29tfGYKWWjV5Y6ZAonkADFjpA+vvJff7bmDa+eYNm2W3McEiUIXgD6not9hoLXnHlK7ud3uNUgJpI3EcadVaUNLtLbsqUmRwVGnhKxpOyqWJD/Xn+vs8v+QObrh6+KVDcaeXQ85S5c5esNxlbcpxOATp1HhTox9NuXb0e2z9sIaSiWnOsR6AGYL9dI/r7N+TvZiOz3Abrukn1EvEBvL7K9G26c6bdyzctPZIERSaFeq5u0+ycTXbxjgoJWNLBIkchVfQSCdRuOP9f3Nt5slrbW/07IC1MfLoKbR7j3vMe8i8mYi3jPdX0HQm0HcO08bgAlVU0QSKGzeVkVuF59J/PsOW3JW5zy1ihoG4dS9dfeM9uNjtDBf3FGjBFAAa0HSD6w+VHVO2NyVwTPxS1LzOftv29AYuTpsT+Pcfc/8AspzTu0Yt7aLUp8wcjrGG8+8f7f328SzwSmhYkVA6re/mz/IbafaFPTUeFrEdhSpAsceg63dbG5X+nuR/u+e1O88jpr3QaaMSOsV/vF+62yc6AWm1tqAAHAdUCm1lseebj+nPHvLXrD7z64+/db697917rsj6WN/9b37rQPr11791vr3+Hv3Xuve/de697917r3v3Xuuyb/QW9+60OvEWtze/+8e/deB669+6312Cfpe3v3WiB176Hg/7H37rfHrvgD+pI5/wPv3Ws/Z11f6cfS/+x9+69Trr37rfXIEEi4H5uT+Tb6n37qpBA66P14/3j37rY4Z669+631737r3Xvfuvde9+691yUXP1twffutE0HXVvr/r8D8n37r1euvfut9e9+690NXRHS25+8ewMHszblC9U+QrqeGdluo0PKqmJSvN2vb/AXPv329azwHW/F8Ef5W2K646925VZnAUUeaeBGq0I1MW0A2e6/Ue/V63Q+XVyG1fjpt7EQxRNgaSBoQioyxIGYqoUG9veq+nn1ulOjM7Q6o+1pkEEAEdhYKP088jke9Cvn14kUx1D7AwNFt/GyS1UyJ+2WILJ6bIT+fx9ffuPW/t/LqlX5a7g2rW4TJ0siwuWBGoiM3GogfX3apPVSPXqmLpTbsWd7a8Kwo9DHXtpFl0BDL+PfutdW+T9fskmKXEQAGJUtHFYFyfpcAe091U27f6vMdWUUah6Mn8YutsvnN0Z+CsxzOULC0i208+n6j6+5k5hYf1GsAvED/J0Wwj/ABt68OrDp+o6vb9DNVtj1UKim+gXN14/s+4gRuwDoy0iteiz712zQ7iknx1bjY5g4aKRZFW1mGm3K/7c+7g+fXjTy8uqrflR/LG667WxNbPQ7ZxsOTq0kvULGoe/iOn6L/X36vkOqkevWm58yv5d3aHx63PkajHYGoqturUTeqPW3jBlNmjLgDSB9Rf/AFvfuvE0ND1WlJG8UjwyoYpImZJEcWZXU2ZWB/IPv3XsjrH9Lj6/Tn37r3HPXh/S9r+/dbPXjx/j7917rwAP1NvfutE0669+631737r3Xvfuvddj68/7G/v3Wjwx14/U/wCv791vy699bf4/n37rXXiLG3v3Xhkdevxa3+x9+6959de/db6UW1NyZbaO4sRubCTmDLYatp6yhdQS3mif0hQOf8OPfgaZHWuHDHW+J8Mvk7m634l9Zbj3WtUMxX0UbVK1SkTtKyDTHKr/ANPofchbHdv9CPE8upC2S/kFoPFzTq0DpzctVurHUW4ghip5z+7GOFAH09HsQxNJIlV8+hVbOzrrOOlp2xtmgzeNeR6dAsqqATY6iOSf9593lj1DPV5lDqanqg75o9MVMVLXVuNhLMIppIgt/SddxYAew7uUNR2DoK39sRUrnolHxV7Xy2xt4tgszm63GKk6pCt2VSVlsRyR7L7G5kjfQxpTous3WN+4062YOjPkhgpaelx2ZzL1Q8KhJXIckmMW5J+vsVRypIlK/n0Mba+Vloxr0auo7E2zVwO1PVCYyR3iBAtqI4s3tzIQ6TnoxhniaZantBz014evTNwZSmzWj+FyxPHH5SCtihJA9s24uDq8cY6Mdzms3VRZmh86da0H8xvEbtpN5VFR1bkKvEtja13aSgZ0ZwHv9Y7e4L5+5Gst4kkmuIgw4ior0ILbmK23Szj2S6yExU9AZ8Wf5gfb3VGWiwvY+589UYnU9POap5JEMYAjBCyN9OPeCPuh7A7DzFBIkdhGJxwYKAePrTqQ+Tdtsts3SGSYDwkaoPVrmO+W/WvYtEJMdm/JV10fjcHQDqdbm4v7xTPsrzDy3d1eH9OM1HH16zk27mra7naFjgcUpT+XRZu38ht+sA+3nimmckubjkNyLn3MnKdvfwrSVSFGAOoi5snsZjWM6mJ6L5s2IVe6aSjx8RKioHmKL9FvzyP9f3It3t97LtrzpGzYxQHqO7W6ghvlWUhVBz1ZDt7YWSyopY6WlFTFBHG2gi5vpDE8D3r2w5F3+5vn3e6tyqk9tQa/b1l5yzz7ydYcrtaRTjxtOeHGnRyurujGyEC1GVpkgiYKRqFrfm9iPebHJvt3Nfx+NfLpXGD1iT7h+6FtYXbJYtqck5/Po6HXHSWyMfLLU1tHSVKhRdHVQP8AXFx+fcsbdyfYbK/iQICx8+oL373E3HeIPp5Klekt2vtPq7CzPXUGEoI6/wBJURxpd3HC/QextaofBrL5cOoxnSIsZKAHqu7s7H0FfmaCR6SKMVNV4/HYL6dYsth+PbcoAYMOimcIzAU49FRr6HLdfdtGqxiMcflZkgkWP9KxuwBJA49wl74bG247AksK1KHUfy6jrn/bWns1aP8ADno7TSwmNDFpKvTQsfzZmS7e8KoyXBB8iR+zqDBXz4g9PW2ol8hYDW1zY/4n8cf7f23cmoCjh15jXj0Ik0QSnBI0vYXP05/rz7RA5x01qNdIz0icrVhQYmNza+oEXBHNre1UaVFPI9ORgCVGPkeggzWBrMrK2nUIz9Dcgnm49mVnsvK6/wCMXdvG8h8yAes2Par3ytuRtoS08U6qUp0iMh1pJUF2lp9QA5Yl7/i1/ZZfbNss8/8Ai0KIPQAdZlcn/fY2Ll/bFjaUGX1oD11tnbe7Nk5qkzu1MjVY2SknSWaGnlkC1QQ38b/4eyPeeROWeY9sl2ndbeN1kUqGIFVr5jqnN33zuR+cLB9u36KOao7WZRVSfMdXDdA/LWLPQY/a++AKDJIi08VTIvNRKAACzH+v9ffKT7wH3M+YOUI7nmD2/T6i3Z/EcMxwp46RmnDA4cesTLn3J5Ru928CymBRyaUpivl0fdKilrKZZaeojeGdFKuHX6MNXvnful9YsZNonMkE9dDBkICsDwJPlX+XQzt4JZYxcwjWhFQR6dCTQ7869yOCj2DvSSlwvmXwUuU/b8ssko0qC3+vz76r/di93PZ/3k9to/YD3FsI9rNmFWK/0jVO9NI7qYNc0rWlD59Ba55R5qj3H+sWxI1xpNWjzQAZ61/fnH0FvvrrsX+8hxjV+xMplXXB56mvOKxZSGQMUBA4PvMzkr2D33kXco9rIaXb6qYJxlXj/Dn14dTh7je9vLNr7KwbZZSeHusgMc8FKFSBTh1on/zXsRuCm+SWdrqtJExDRxRwKzEKs7ux5jP+Fhf3085c2ttu2K3QrSigfy6517dazR2xuZxRpGJ+ec9Vbezrpb1737r3Xd7gC3/G/futdde/db62Hf5HeJ2Dj9/5jcElXTybmlxixTQzSKHhBh16FueBfkD/AB9xx7ucs3fOXJM3L1iSrScaYr1J/tLzfb8k8zDe7jOnA62b5s//ABLR5pyYlYhYT9CisbAD/Ee+bl792HnPZ45HtIA1KmoOes1Nu+8rtl9eRlH0yMQBTpE713HksWKR8FEYo0KvO0X1Cg8nj21yZ7B7tuc0y8yL4VKgV8+st+XOc7HcdlTcNxfWzjAPl0kMpkDvCnDSeSeRgEdpVYaWsCTz/j7yG5E9n9r5Vg8OFQ8pNa+fn1e59zIOX42eMhUPpToPqnaGKw04fIIuuoayBV1jn8j2Mt42eKCELNGHYevQg5E53h5lvfFZgyNw+3p+pMDjRZIkT1/TiykEA8+wrHaQO2hIgPl1N897HYweI4oo6TO6Np7NxUbZjc1dQYykWx8rzxo55tfQT7vfco7zvkS2+3rpp5+XUW85+4ft5ZWHjb7OsejicV6ZcJ82viR1mf4Rmt9U4qYAA0YEbqCnpIBvz7gXnL7sfu7vt54mzojRnzLEHrFnmD70HtJtU5t9ulZqV4KP8I6aN9/L34jdsYeqGF3lS+WmieWR3WKJkCDlgb+1nIH3ePejlXd4xukaGGoqQ5PTOw/eT9p76Zrq9mK6RX4fTrVK/mC9jYPefclVSbK3ZU7h2hRQg08RkPghqxIVJ0KdN7fT+nvpNynttxtezxW12KSACv29c/ffnnTZueOe7jd9gctasTpqKdEIueP8OB7E3UJ9dkEjUTf/AJHb6+/dVrQ066Jv+Le/dbHXhzcX/wAf9t7917r1ja9jb+v49+63UcOvEW/N/futA1669+631737r3Xvfuvde59+691y408fX/efr791XNeuhyeT/t/fut8Ouv8AD37rfXYHNjx/r+/dar6dde/db697917rla3IP+29+61xwR13xp/F/wDefr791rOrrh9Pfurde9+691yFjf6D/H37rR66+l7H/D/XHv3W+PXXv3Xuv//U+f8A+/de697917rsgC1jf37rQJPHrs8Hg/Ue/deGePXH37rfXKxIvf8A3u/19+61UVp1x9+631yAv9Pr/wAR791omnXQFzb37r1aCvXR49+631yItyPp791oGuOugLmw9+62TTPXdhb68i/H+sffutVNeuvxf83+nv3XvPrLTzPTVEFRGbSQSxzIRxZ4nDr/ALyPfut9bBvwi7ept8bGgp8lUKcnCq0kcEjAOojkETWAJP8ArH2JdruPEGh+hLtM5dSreXVmnWW6p9lbpioxL4qepVpbi+hmbm1/Z7GfDkwejyA+G/bgdGq2xlIt3btp4MjMGpZ9PpdvSbn6f737WJR2oelyN40o1cB0YfL9S7fqsYwoYoYZmDEun1sfzz9fal4V00HSt7dHXt6Kxu345T5R5fFC1Wx1sL8kf0A49opbRnyM9IpLJq9o6CXHfGDtxqtodtNlcXRiQtppvIicfkabe2ls5+EeOmPo7jVRMdOud6C7moIBHl63M1yKh9M3kcEf0IN/e2t7gDS1T1t7e5Aq9T0h8Fs7cmysqa3J4uddKkrPLG91tyObe6pEyZbj0yEaPiM9C31j8ud57F3nTbaheauop6j0hw8gQM1go593hv5IpdFKjpTDuMkEgic4PVu226/dPaGLx+RqI5oIaqGImNQyhjIt7AfX2eKTINfDo/TXOobgOjsdI/DvYNFQ1PcfcWPp5du4iJpKSiriFSqyMCmWmtG/qN2AF/ZBulz4rDb7LukPQf3q4Jf922J1SNxI6rT+ZfZ3avZPYEecgpavGbI23Kcbt/b1LG6458bAdFMyxJZbabc293stvO2x1Aqx4/5et2u3NtkYYCrHifn0nuo/kvlNqfbY/cVB9pRjTGZHha6Le31b2YrPrGmQY6MrfcGRqMOkr39uXG7z7Awm/sSI5xTR04aRQNQ8ZB/H+Hus2ksrJkDqt2RLMssflx6PV1zvmk3vgcY1OQXoqGGGYaSo12CN/vPsxgZdFV6NIphIgUGvSzyVCr0swsBpB1AC9/bjGq4HTpApXooHcGGjqMPlPEl5TDUAEAXNh+PaGYdtB0hmFFPVPVV1BJWZ/J7gkyMlJNTTTMkJfRrC+r9J9k5gJcvWtM9B4wBpCSel91b2juza71cGNpTOKWYwidSbnS1gSR7A9z7i7ZtF99BeOq1NM8eiSTm202+4+mlIFOhVzu99+b6ZYMjNPSUrkByGYC1v9f8AN7ew9zJ7x2Fpbsu3sJGINAP9Xl0Wbtz5bpGRasGJ8ulJtOGi23TmngZXlmIaWfgPr/PPvFnf92vuY75r69Ytk0B8h1D9/ez7lc/UTE/L5dC9g8jpmjnDav0seb35+tv6+w3LHjScdISMZGfLoQK7csMNOX8w1BP0A8D+o/2HtAltJq0EYPTNGY0Ip0C+4tyyVTuPLcarEBvwfrb2bQW4QU6UKlDTpB1mYEcLkvptf882H9PawA4IHVwg8+gh3Tuloo5rPbg+m5sbDg+zO0tQTrfpWiZpw6rN+Vvb/wDdHZucyslUqTwU7lY9RLCym1l+vuRuU9sF3dqSMDo6262PiA0x1Rjsz5Xb2wG7JstlaiXMYioyEUq0c54pYBPclQSdXp/BP1/PucUUJGIhwAp0LkZonDxmjLw62fviL8oNob82Zj/4LmKBJftKb7qHzpEscl7GMBj+Dx7DG58l8t73J4u5WySsPNgP83Qkj515kjjEa3DAKKAVP+fq3P4v7shqe7NoqtdRuZSCBHVRuxuw/C+0tr7c8obfMLu0tI0kXgQBUfy6sOcN+uW8G6uGaM8Qa0/w9Ln+Zfv7G4DduFevrqOFY8V5CJaqNHKhASdJPsm3Tkqyut0F+kQ1jANOskPbT3nTk3lybbVl0mQk9a4PaXyi2fntwNtrb+egfN+chYElW1g+m45+l/zb2bRcqxiLVNx6Ktz+8Lust8I7M0UniOhY2tuzclbtY0VRkI1SSDU7tUgAJb1NqJ/p/T2C7/brm3vNMCmlesiuWudtp3zl4T7vOurTU1Pp1Xx8ivlHs7p+WqxeIamz+dmBDqCssizufVb+gH5J9iPaOVTMwutwqfQHqGvcL32j2+J9k5WRUFKFxxPrnqrHsP5U9hb1LpR1c2ChlJ1rSym5U/ULfgf7b2PYLSC3XTGvDrFTc+Yt33aYzXcpNeOegJg3tu2nqmrodw5KOq5JlFS+oknk8/8AFPagqp4jonWWVGqrEE/PpvzO485uCRZc1k6vIyJ+lqmUuR/sPp78AAKDrTMznU5qemT3vqvXd+LW/wBj791rzr11791vr3v3Xuve/de67sLX/N7W9+61XNOvfg8f7H37rfXgLgn+nv3Wq9de/db67It9Pp791oGvXgOCf6W/3n37r1c068OD9L/4e/dbPXJh+Ra3049+6qp8j1w9+6t1737r3Xvfuvde9+6912Bc29+68cde5B/x/wAPfutcR12QAQf1ckHn37rQz8uuPv3Vuve/de697917rsn+gt/re/dap69e5/2Pv3W+nXD4PLZ+thoMRQ1FdVztpSOGNnuQLkswFgP9f37rRoB0Zrbvwn+R+6RSNh+vq+qStKCF1VuVkNg1gn+x97p1r9vWwT/Lv/lm9kdE1OJ7k3hjMlS5CjeLIzUtVH40QwkMqLGV/Ta/9SfyfeuPWwM1Pn1t+/Fb5L4HctTT4HMinpZ4WSPRIFQ3Nk9N/fjx63TGnh1bFFsyhzkNPXUSI0MiowMdiACuoEG3utK9b4DA6EOn27R7e25W1jxo7wwysqsoBuqFvwPeyKDrwcEgHqmT5PdmZ/M5atxGOEsUaaoyIlYLcoRbj34LjrxIrjqjP5Abe3tX09auiscG9wVb9Ie/197LKo1NgdbSOSaQQxirngPM9IL4b9YV1dvGrNfC0dVHKzIWWxJ13sur20lxBK2iJqnpddbPu1hGJ72EojZBI6vg6n6PrarcWNqci3+R6kLJK4CKF5vc/j3q4p4LA9F657hkdWGfHPZ+zMZv7ctLSmhaQS2ZUljv+rkAWv7kjcZJ25Xt0b4QuP2dJu3xajB8+j77i2PiMnj6ihWBCJUXSQVPGgHiw/x9x4g7K9Ka5z0QvfnR9LispLUxRAK0mrgAixN+ePfurcMdBTndpU8MWkwAabrew5sCDz71wGOteefLqsn5mdDbA7B2HmqbLYajlq/F+0zCLyFzf1A6b/4292rXHp1sinWi38mP5dXYFFvrdGU2Rhq2ejnq55qalhhLU/Eh9XpW4FrcD/X9+6pSg6rI311lvfrbI/wreOBrMTUkHT5IX8TFSAQshUc8jg2Pv3y61Xz6QJBH14/Fj9f9t791uvXXv3W+uS/k2vb8e/daPp12WBFrf8a/1vfutAGteuIBNyPxyffutkgdevwPwRfn+vv3Xqdde/db697917r3v3Xuve/de697917r3v3Xuhz+N2Ix2f7z62xOUpo6mgrdy0EVTTy2MckZbkNq/Hu6DU4HXkHcAfXrd7yOwsDtHYWwKDGU0dFgkjpj9tFbwxKVGthp49yFawLHarpwOpFt4UhtVZeGOrcOhcRhZOtqX+CPFKngFtDKXU+Pm6r9PZ7C4SMBeB6E9soMAMZr0rqQpmJajBVw0GEMtOWFtZBNhf2oLYI6eALCh49FL7m6Jm3IchFV05NPDFIbWB1KfoOR+faOa3DCo4dILi21qQcDrX1+Vvx8rNk5d9y7eglgmpZWmcxKR+l9XGn/AFvYcvbXwjqQdBO7tihLqOHQlfD/ALKq89W0eNzFTKs9OWSQyagw0qBzc/63u9nKW7Dgjp/b5vEIVsdXu7IxlPXUdIy1UjhkS1wxH6bjm/sQxgrQ8ehNCofzp0J2Xx09HiKhYa+RdcJAUM3B0/4/nn2pbA49KWQLHVfPqv8A371PRZupyVRkaUVclVqs8tjqbnn1X9lVxbxzoUYVB6K1EkE31CYbqtzuT4e7frIazJCNaaTSZAI9PFmJDWW3sDXvJdrPIZaAV6E39c76C38OpxwPRRNk9c5zbm6TisPNVMkM4CFVci/6f1D/AF/cdb3yLZzSshtw4+wdDnlX3FvIYlillIr5dHTxPSHYO5ZIg1PXT+RUFwJDfV+B7CI9nrOd/Ejh0j0A6GD88q8gMr16Oh1Z8Oc/hYoMtVUNUtROpIZoyxLEC3P+39y5yv7Z7bb2wivIFK0pQ06AXNfNM87UsnoT5jq07o3qaDb+PZ8zR65ZIwoEqc/otfkce5DtuWtk22AQWsCqPy6DVjv++W9SJ27vt/z9D2MOaXRBRxGKIWtpHFrkD2dQLBbx6IgAPTpNcT3F5IZbg6m+fSwp4ZKWhdYyyuEsTYgn8Fj7ZZg8meHTdBToofY8ksGRqK3KyH7WBvIBIfSV/AsfapqYJwB0guONa9EuytW+7d5QmmDfZRzhYCAbXDfge0bHU9Tw6LHOuUKBw6Ve+OiqtvFuc+aVY0WUIwOmPSt/aXedpTddqntphXUpp+zrW5bZ9ZYyRuKkjHSEx+bVC9FI9pYFZSCwuQvpsQf9b3zq5h2SXZN+n25wQoJI/aesXN0sms9we3cU0noTdm1qNKbOW9f6QwJ5H+8+w5dRNSq9F7YOehAyFaCLBiC3A5/IHHHtAiE/Z0yAx7uk6cJV1LCoeNmiLBiSOSt7/n2p8ZUGkdOVoKdOkeELH0IQBa4Atf20ZfXptvE9epowC1AKeMkEH6KRb+yOR7qJimeqHXwLf4evR7U8KNaEH6kAC9z9P1Hn3trlmI68BjubPTTUYOClXzQIKaviN4p1BDI4OpSCLf7D260qXMRs70eLC4oVNKU62plRxJbuVIyD0wVXefamzZoPttwV9WkMg/bkdivjQ2C8n3jpzz90/wBuecp5b1LOINLU0CLUE/l1mN7E/eE2vlimx85wK1ue3xX8hTjXpw3F8ld0brx+uqrTT1vjCxtGWV4ZAOXVgfrf6e4r5X+5zb8sboo2yEpCGB1/4D646zim9/fY3lrYzuu1XUc0pFdGCCfToy/xS+Ws2TqR1v3nhYd77EceH+LZtDM2ARrgV9LK9yGQcj/W99RfZq4u4bWLk++rfi1A0s+aU+fy65q87e4MPufz/c7nt1mtvCSdIQUU/P0z1rt/8KA/5WW9ctW5z5PfGGhqe0+ppZv4tlarB1ENXNg8fFG0tQZqKIlx4ybfT6A+8nL12iRbN10aaGny6T3s0hVbd00FRkdaYrRsjtG6PG8bMkqOCjo6MQysrcgi1iD+fZd0Wk9cDa/Hv3Xh8+vX4HFiPz7916meuvfut9D/APHXvndHQW/MfujAVkkFM9RAMpEhb9ynR7MwCkc24IP1HvYNOqsNXW1v0r/MT6K7B2th5cnvGiotyikpUrKBHVPGwS0jsmq4v7rMI5omjda1FOlO2T/QX8d3XCEHo003yv8AjpRbbmzWe3pSQ00MDyufKjXjUEm4Lf4ewceVIGlLgUB6yLt/f29stvWzhHwig6q+72/nGbC2C01H09TYzdMGt40fwI7s6yaA2oki1vVqv7Mrblqwt28SlT0Ct796eZ95iNrqKrnPQOdZ/wA7SbcGdpcZ2Ps3E4/FO6g5F4lPiS9mKtH9GA/qP9j7Q7xyft+6xn8LU49Cn22+8RzTyJcpqImiqK1FSB1c5sH5RfHffvX39+KLeeNp6M0X3VQ/kjDQeglwqlrm1uPcVXPt7fQ3OiFfPB6zt2n73XLO67MLncpgpC9ykcD5+eetYz+Yl8vc1vPt6txfU/ZeSqtj00TxslDPpiNQkmnTquwH5Nlsf6m3HuYOXNnG17esMyjxBxPn1zz95PcWXnXmyfcNtuH+kf4UBIUfl1Vxk9w5vNVDVWVytbWVDA6pZp3Zjc3N+R7Equyiimn2dQsyhjqYaifXPWTG7n3DiI5kxuXrqRKhTHMsVQ4DoRYqQb/7x78XZviNft6sp09q4+zpkkZ3dnldpJHYu7sxZmZjdmZjyTf6+69aFKY660m1/wDkfv3Xqjrjc/T8f09+631y02+v9ffutVr142H054I/1v8AH37rQqfl1x9+6t1y4J/oLf8AE+/dayB1x9+631ysAAfqb/Tj8e/dazWnXRN/xb37rwx12eOAbj6+/deHz64+/db697917rkBfk8D/iffutE+nXRHIuf+J9+695Y65FPypv791rV69cPfurddgFjYe/daJoK9e/Nv9h791vrxFuPfuvA1669+691yuCRx/sB+ffutU69+r6C1h/vj7917hx66/H+N/fuvefX/1fn/APv3Xuve/de67II+v59+60CDw68DYWsP9iPfuvU699D/AMV59+69x69cm4H05Jt/T37r2B11791vrvlT/Q+/dawevAkcj6+/deoD17V+LD/Yj37r1Ouv8Pfut9eBI5Hv3XuPXd/625Nyfz/t/futU67On8X9+68K+fXlAJ5/p7914mgx0Zv4v9wVHV2/aKSrrXixFXJHEY3ciFKkzqEsTwNVze/HA9qLaUwyBgadPQTNDIGBx59bJu1M/Sbu23jM9RSpLUvDDOGjdXKxsAT+j2MoJFmi1Dj0MoJvGiEqjowWA3PLA1Jk8XJeaiCLIEYXvGLsSP6e1qOunUPLpWknbrH7OjcbL73psrTRUFa6w1KBEJLoNX+uCfatLhThjnoyhu0oF8+jH7Zz+PrGilskg41WZTe5vfi/+x9qVcE46XK+rz6NZtjMbcgx8VqaFZivJHj9R/qf+N+1iMoyOPS5DHo4V6Vk1NtfM0rmopaZ3ZSBreIE3/qD7vRW+LrarHpI6Afsno/AblwNbNRU0azKspVUVf8AjmQLFP8AWHtiW3QpVePSaa3jmWlOiMdVfEikr+zFzeTXTBRVTfstG1joa+o3H1PsviswZdTdFNvtpM9XzThXrYM+MnSNBkKWmymVRcdsnAojVFcymNTLSrqEPrtfUR9fbW73v0qC3gzIeA6f3XcxYR/S2w1StgD5Hz6VndnZ0W+a1NtYVUo9p4cPRrQRMop68xjQtVIqcXNr+6bZYm1XxpMyNmvp1bZ9qFnH9Tcd0r5r5j5dE93djMDPTla+khmij9CodPAUWCjV/T2egBl/Uz0cOARqIr0V/efWe19zq0dLRxUSggCSNL2N73uvtiWJJBQY6QywLN5Up0icf8dqXIVUMSZiRaWN0vFZgtlP0s3tsWg4A9UWxFKk9G32jtTC7ExqUeOMZOhVZgQCzDm5H+39rERUFOHS2KFYk0celFV1iPTOC1rgn+lx9fd2Onq+QvDh0XnfX21QtRD6CHRwbW5LD8+08mmlek0o1ceqbvlDlMps/N6MGGAnkIdE+h18EkD/AF/Ye3K7jsIXuJDRaH/B0GNxn+kVpW4UPWHqPD1z4oVNTqMuQQVEgCkaWbnj3g9zffR7jvM0/EKxp1AW8zrd3zOPI8eh4/h9XTUzay4S1hcE2P4N/YRUoX1Dj0U/pk0HEdJapqKulmuAxS/1N7f7An2sVVfiadX/AA1p0utt7kkjZQ7XQgAk8WN7C3tLcW+o5x1RlBWo4npU1+UeaMsj3Yg8A/QHm9/9b2xGmnj5dUAotOkTK7h3ZrtckAX59qRnp0UVadJXL1EaI2t1BXVZfqRcf7H2/Epd+3h1dR2Z6LxvvcNNQ01UZGQyKjMNToNKKtzbURz7PbO3eeRYo/PpTBG8pAAyOtdn549yxZ3cEm1sJXCankUDIIsisI9P1HpP5PHPueuWNp+gtFaQdx6GVhAYo+7j1Wrf0WvyD9PYq6MM16XuzO0t+9fOW2luTIYhCwZ4KeUiB21a7lP9f+h9+69TNerkv5V/y77j3N8w+s8DmM7VVlHO0kZUzyt64ypDsHP0P9PeiMdWqejHf8KAe6uzKHvDr7G4zdOUxdJWbYeaojpahkMrII1AY/6n1G4t72ONeq8RQ+XWuIm7tzw5gZ9M1XjLxtcV/nbzGzarX/pf/D37j1oADAx0Zuk+ZnaFJtN9upWTfctB4BkBMwaxFixX639sNbQM2srno2h3vdIIjBFKQh8qnoq2cz2W3Jkp8tmayWur6ltUs8rEsebgL/Qe3gABQdFru0jF3NSfPpo976r1737r3XvfuvdcvTb/AB/43791rNeuPv3W+uyCPr7914GvXXv3Xuve/de697917r1uL+/de69f/effuvde+vv3Xuve/de697917rsWvz9PfuvH5dd3sTb6f4+/dapXj14k/kAf7D/Y+/deFPLrj791vruxH+x9+69Xrr37r3XYvfj6+/daNKZ68b35+vv3Xhwx11791vrsgj37r1a9eBtf/H37rVOuSkBSL8m/4/w9+60Qa9ceLHnn8D37rea9DF0r0tu3uvd+O2ztnH1NQtRVQw1dTFDJKIVZ1DKojBOog8f0Hv3XvOnW4j8PP5Ruz9q7RxWc3LiYKnLGmSRo5qRvIH8IY3Z15N/qb+/fb14jGOrU+pOvNobN3tgNsnZtJ9vS1EVMztTrb0tp4JHvVMY6uDQ56uV3t0ngcz13RyY7EQ0tPJQkNHFCSCvi+hKj+vvY8uq8K+XVOG9Ot8v1pv3H5rCCajpIqlJJ1jSRAbSajfj36ua9eNR1sI/D7fb732lSRVEup4KRNRZwWJWLTyDz+P8AeffgOvE56NDvdo4sFXRIw5hf0/66EEge/NSnWlBPxdU8dsUWDwNZlMzk4F8a3clkJvcH2zPMIIjMx4dGW17fNu16tjAO58dED7jzOxq3CCppEgMlVHdF/b1Frn+v+9eyTdd5tk20nVQv1KPIPttvN5zcgkj1R257sGnRPNjbuTYG44srQ0sarJUFybAektexPsC8rbksV9JNcydo4VPWU/vjyU19yza7dstmBKygEhSc0+Q6Pxtn5Q5bOQpR4colQIymqGRdasEIIYKbj2LE5v2zdJZLK0NWUgH9o6xk5j+7/wA58m8txcxbtCyRyLqFQRj8+mToX5B742l3DmJ8nUzSQVFYRMHdiAPJ9dRPvI3mKOJOR7Hw/i0iv7OoBgWUTs0oIDcMdbJ3TvY9N2BgYq1ZQ8hp0d7OCbiEXI03/P19w/F3KK9L2AXh0ie5Mt9vTzOqrqVTpH0uLWvf3tsGnW1FR0RXcG89ayxsV1i5sWH5FuB78DUdbp5jqtj5EbzqYZxQn1RVhIJ/VYaja/v1cdep0xdX9LbU3PjabIZLEQVbViE6ngDE6/qPp72DTJ69Q8B0HXyR/lRdM9w7crMxJt2gp8osEjwsKHyMX8LFT+k8g29+BFetMDSjZ60T/n78Pc/8Yuz8rT/YyxYCrrHSlH20saKSxUOnFrN+R/X37j1qgGB1XqTckj37rw4deBI+nv3W6V67uCObXAsPxf8A1/futZrjroWsb/7D37rxr5dde/db65G9hwB/j/X37rQpXrj791vrs2vxyPfuvD59de/de67NrC31/Pv3WhXz669+630vur92DY/YG1N1tfThcvSVjkC5CxuAzWH9Pe1bSdXp14Eg1HW/z8XcLS/Kn43bBzUdeIpZ6JdUkTCUqqrYNaO/B9yNtJF7YqDgjqStpK39mq+nRz+qMPm/j80mKyVZNX4erBiSWbWUhUi2pdXs2WHwqIxqB0fWwe0qvFT/AC6OxQbdwmew9PuXEVF5ZVEilWUlpLaiCB7ql3SQowx0vIxqHA9I3ISx5J5sRkkWGSa8TubB2H0F7/7f2uGkISpx03itG6Ir8wPjvjMhtON8bAKmadJGm0RFvqpPOkf7x7LriPx04dF25WsZjGnz6qY6x6gfYW9KH/JPF93VsDdCnDNY/W39f6eyuG3Eco9T0QRW4ilpw62C+vto0NFs3FTlEM8kEbcWNyY/wfYiiXGg56FkaKI1YdOuQwLVKWe4jX+x+LAfX3cpXj5dbBIJqKjoB96YSFhUCKFQKf6MF4Nvybe2GUhe3pNKAT0THeWJ/itRPSSp+zch+ABYMfaFwSc9FkiBia8OgqwWxto4rOmVqOB5mkUl7IpBv9ef979sLBCr6mFT00g8NtaGh6s46K2pt7ICKeSmh/aSMqF8djYA2/437MoIYVQsqjo7tXlcdzV6O9WV21cBilqaukhWmpIla2lQBx/iL/7x7YpOxopoOltFUVOeghX5NdaPWNQuY6VYpNGtAoLaTpPNvejEF/HU/PplbuCukmnQ0bO3jsfda/dUGRSWMAOUd47/AE1WAv7ZdZ9NVHT6uHHYa9d7w33tbExuKedWnC8RAjSfwV9Pu0MEwI8Xh1rWMnzHRBu1twV2+Kh6aGA01JCbMyKT5UBvYlfr7WuKjSei+d/Fxw6QfX+zqcbmx61K+CD7uMKWuqsSbE2a3+v7bEVAWPl03bxL4gJNKdHm7Co8VT7WqKCJIyEpE59J/wB1/wCHtRbO9S78OjWXSoqeqcdz47M43c+VroYHFG8soU6GCEFj9P8AYe8ZfejkYy13+xHcTU/Z59Qbz7y6Cf3nAOJz0q9h7qCVQhlYLJqK8kKeB+b/AO294u3EPix6weGOoikVWyOjM4OBco8cjDWDZr8EXt7IZNSCg6SM1K06EuGh0r4yvpUDi3FvqfaTUTk8emTqPceswoVvZUAH9B+Offg1OPXhX8R6lJShBa31A5H++/w91LA0615kcR105SNG+hsCLfnk/UX97GevACvQf7keJIZJVID2J5/FhZSAPamIHh06lWNB0WzMLLXVkgJLgMSwN/pf2e27+EgKmh6USRxTAJKuodMDYuGSW0FPrbgBQCbN/rD2+9/dJEUViSfhHqfTplreML4YyvkvQidlZzE/Hf48b27G3CsMFVmdtVa4KKomjhY1aQnQE8pHJNvp7yk9luUf3Ntb8z7gtJJlwCKZ+Veps5E2VdpsG3m5WmoYB/l1qbfE7+dV318evkxuHeG662bsno/emSqMJunqTc+Qlqtqf3bnqmhdqejkJjVhGxZj/tvY5uZjcXBnbiem7udrmZpmz6dWk/KX+Vp8YP5lOwsp8sv5cGax0HYM+LqMvvv477cjp0p9sukX3c2ToNC/vwsdWoIdQuTyLe03A08umjV11Clf9X7D1qU9jdcbv6p3dmth76wlXgtzYOskpK2iqomi5hcoZEMlrqSODb/A2PvfVBTj0g+SDwOPqRb37reAft64+/db67F+bf7H/W9+61jqdj8pkMTOKrGVlRRVAFvLTyGN7f0uPfuvEV49KSr7C3tX0T4+s3PlaiikQxyU0lQTG6H6qRb6f7H36vVdC14dI46bC1wfzzf37q2fPrq/9bn/AGPv3Xvs6WmL7B3tgsTJg8PubK4/DzCRXx9PUladllJZ1KEfm5J9++fXgxppzTpGMxdizElmJZiTclibkn37rfXH37r3XfP0/wAf959+698+vG9+fr791oU8uuyxP5/23v3XtI64g2/43791vrkx/ANx/j/X37rQ+fXQ/wB659+6310Tf37r3Xvfuvde9+691737r3Xvfuvde9+6913xb/H37rWa9de/db69/h7917ru5PJ59+61SnDrsnj68ngj8W9+61TPXVja/wCL29+6tXNOvC45H+t791o0OD11791vru5vf37rVPLrr37rfXvfuvdd3PJv9eD791qg669+631//9b5/wD7917rkLE8+mw/H5Pv3WjUfPro/X63/wAffuvDh11791vrvn68/wCv7917rvi178/0/wCK+/dazWnXR55tb/W+l/fuvD0669+631737r3Xvfuvdd/j/G//ABHv3WvPrr37rfXvfuvde9+69176e/de65glSpViGU6wwPKsObgj/W9+611dL/Lv+Ri1ssfWm4qySat8LLBNUMgDQr6EZS31/FwPZxtV14U2hz2no62m78OTwpDg9XLbcpvsMokKESUtZp5FtJEv5uPYrAAOpeB6E8YByOB6GyP4+b1zencGzUepAUzvBF5OAvPOj/ePbwtXI1LnpVHZzSd0QqOhw6izOXxuQXbe5InpcnCfEKdr3YobMWLc+1MJoaPx6egZlfS3EeXR9tu42atplaAamKelRe+r6iwHsxjA/b0bRrUVr0DPdlR2LsTGx7joUqVxq1SROUMgXSG5AH0+ntNcNJGajgOmbk3ESa/Lof8Apnfybx2MaqSVmqFp41muRcMw5Fh/sfamGXxE9OlVvLriFOPQ8fH7pPcHY/YCLFEaLa1NI1fkMs+uONI6c6pIyxsOefaLcdwj26Etxc4A6QbhfrtsRPF2wB9vRwe5O4cZBSJ1X1kBDt7HxilzNbS6VWavi4kZxH9eb8+0e12LyH6+9y54A+Q6R7Ltskkh3K+y5yAfToqNfPkYkK04ctpuXAJLn8n2IQE9K9CbNOg8yWHz+amGsyrFezLZrMb8sQOPdGHdk9MhWY06W+D2SkNIEqY1dyAbtxa/491LxqaDPTqIY+OeouS2pJSylqEaBp4C3Nv6ni3valWFeHWmXUag5HSXfEZY1AV3bxqwY31Af1PHuxBJ6r+p5jh1l3BM9FQnR67JoJ5Bv9be9PhadbbHRbdySVEzzOGfUQSAPpY/UAD+ntO/cdI6QyFmBI6rM+SlHr3JTJJCZS5jJLD6c+wbzpEbjYpI4+Of8HQT5lhebbmUYOf8HQ29WbSpzhqB3srPSRlUF73tcAH3gJukrR3s0Vcq1Osdbo6ZmHoehgn2mKpDCYVCWJFr2v8AQ/8AEeytbhVNQekwlrwHQN7y23LjC6eC8RB9Y1G3+sfZpbTK/cDXpTHJqx0GEE/8NlLuSBcWQ/U/m9j7MGBlWgHTpWrdvSgTNxsC7SaQfx6bD+gHtKIZOFOtFM9MGV3RSUuq01z9Ryv+xAJ9vxWrsanqyoW7RnoAt89q4zFRy/vFpjrCp6Df0/8AEezq0sGcgIOlUdu8jZHVUvyx+VKbNwdfSRzE5uuZo6GBHjZnWSOx+l7AXufct8rctBALm4WnQi26wAIkf9nVDmezVbuHL12ZyEjS1dfO88pZi2nUbhFJ/A9yUoCgKPLo/AoKdNRA/Buf6e99aBPn1x+n19+6t1Zv/KNsPnB1eb2AmqP+ifeutAdHW/4UF2/0/wDWv1/49Ob/AHuH6e9+fW+tfM/U/wBLn/e/fuvdeBI/3v37r1Ouvfuvde9+6913YWvfn+nv3Ws1669+6312QLCx+v1/w9+60K9e/H04v9f+I9+6959e5PPJ/wAfr7917A66+vv3W+vWP9Pfuvde9+691yNgBYmx+ov791ofProAWN/9h79141669+631737r3XY/ofz+f8Aih9+60fXrxH1te39ffuvA+vXh9f6+/dbPXjf/jX9Pfuvdde/de67N+L3t+PfutY669+6313zfj/E8fj37rXlnrvnm4v/AFv9R7917HXH37rfXd+LfX+n+H+t791qma9esfrbj37r1R11791vrnHG8siRRqzvIyoqqCzMzGwAA9+691trfyP/AInzbSo6jsLfuGppKfITmtoaidGuFGm2lZPoQoA4/p715060PXrbT2nvDY9XOMZTSQQJFpREXxqpCqF02Hvx9fPrY40p0MNF0xh8xlMdn8VHBK8UscrSKLseb3svvVT9nVuFT1ZNt3bgrti0uPmS7Q0rJpYGxbRpF/dgKCvVSQcdVzfITpKvya1P2tGLtYqyxvxzfiw/2/v3y68CR0ZT4Zdf5vZOJtXxuiNAQhIe/K3Is3+Hvymp68VxXo2G/dwYbEY6qlylYIY0RjIGZRwFv9GPP+x96btUt6dXjUySLEOLcOqgfkP3D1lk8Rn48TUx5Gtx6OJaZ/GVZtJAAA9k15uVr9JI6d5TiD1IXLfJm9Jv9nDckwLckaXFajPVLtVk5d5biSOW9LRySM0NOLaIh5DYC349xS053PcEic6VJ4eXWf8AJtUXIXJ824WoEs8airebGnn00bm221DVlInWWkjOksCdX059h7fuVt3sb4yRsViY8Rwp1LHtX74+3vMXLEdrewrPuUS0KsBhh0K3xp281RuOeKmiMkMtRIZZbFmiuDdhb6W9mHJ+yJa3MtwZNRNM/mOgF96j3OuN85ZtdsjgEaqmlU8ulzi6eFO4svtrBNLlqqsrBHUM0T/5IfJY2a39feXO/Xpg5XsIVbUHUV+WOsCL/lNt15VO73kQtWthVQD8eOth34d4fMYDGS0Fcjsq0wXU+r8wg2F/9h7A0Y0inUJq5kj1H/VmnQr907er8nQVLUkZLeOyrYk6rX/H5Pvb8fl04tAuOq1917WztE1TLPSyKq6udLm9lP8Ahz70MHHVjw6Izu/ZeT7A3PT46npWkMM2jWVbg6+fp+PeyKZPVASR1Y/1B0PHtPbOOqc0YqWnpYlch+PSVuSNf9fdSfLrYOeltvbfPXuOxMuNp54WmjikjuBF+oRkH3umc9e4cetTj+b/ANFYvuXA1lVtjHRZHMU6yGALGWbWQSDeMX974cetfIdaQW89lZ/YWerdu7ko3o8hRSOjqVcK2mQoCpcD62vb8e/dez0k7H6249+69Udde/db697917rs2sDxc/j37rQ9Ourn+vv3W+ve/de699Pfuvde9+691yAUjk2Pv3WiT5ddH68e/deHXgeCLXvb/ePfut9bd38iX51UUG3z0juCP7T+CUlJjMPJK8RWaR0SJSmrn1C/sVcubgYZTbv8LdCzlncDBL9Oc14dbRGclxe58NU4yrKvWLCrQs+klNY1DTf/AF/Y/kUMtAOPUh6hIpU+fDpUdMrX4mllxVTI/wBpTq5p7k244W1/aSdKIABnp+3XSulj0qN27c/iTpXU0xinjs3oJDO31+o93ifGk8OvOo4rk9QZ8G+5sC+NytOpaCP0yuLl7J9Cfz7cbQrD59VC1XvHVW/b3SW64t8Y+XAURmC1oaFUD2/zl1tb/D2guIWVwV/b0Sz20nihkFa9WE9Z4bdOO2rj6XdFI9JNBGlkcsTbx2HJ/wB4HtfbyqUoPi6NIY5EUB/9jpfSU/8Ak8ps36Tzb1E2tYD2/Wpp08eGOgNzuPln/iCCPhgy3sfTfj/Y+2HDcOmGUPUdFl3HsWs8FbWRQliqsT+rkaifaR4ycjy6QvA2SOiMbvfMYzLTSxRyII3a9g3Gl/oT7QyElqHHRXKdD9Cp0d8uqHYO9sLtrdMzwU+ZqoKKF38ekkyBDy3093ivfCOg+eB0/aXojmEbmgPV7mZxuE3PtFauleKox+Road4ZQQ4YyxB7XXj+1b2ohkfxfDfz/wAHQmOl1OjI6JBL0ZQVeZqvFRQ21uytpNiS1za3HtW1uobV5dFxtEZiadCHtnq2t286yU2RejiBUtFGzAAfTSL+3FT8x0/HGVWladCJFsilyM/lqalZj9G8hN/9ck/196clfiFenRGGwDw6c5OttsNrRaiEPCl5lBBCr9Sf9v8AT20ZGJ+HB60YEIyei8bzxsUu4cZR4CTxrjKkPPLH6TIFb8lfr7sw10C+XSd0q48Py6FTJzzZGlMTP5FaBYyTcg2Wx9vsO2g8un37hQ9Fs7F2lTUmLkdoYwZQx1f1N/YK9xVj/qrOx/g6CnNyhNjlY5oMdE9zG2azFBMvRBv25DI4AY3Ab6e+edlcCTXG3DU3+HrGGF1kBDDzPRk+ot1JkqZI5gFmXQukkAiy2J/2/ss3C2MbV8j0xcRU49GLglWQHkXt9RyP629k5BBp0mpTPWZQRc2J/wBj+fz703y6rX165uNKAAXNwef6H88e9DrflXpuqgfE5Itfi9rEE/4e7qe7rYr0Em5ImiEylyxYXAvYfW5v/j7XwFdQ6ejYYYcOgllo6iQuscR1+olufp/h7Xs6IaDPp0+XAyfPoVNg9eR0qHd+4mSl23hx99k6iUkaaVBrlJDWHA9y37be1+68zbjHu+5r4dpGQwJrn8uhxyjyhfbtcrd3C6IYzWp8+tXn+d//ADIabuLNr0D1bkqqn2ntPJSD+IUUyRiqgjtHJGzQHgErYfn6+8rN4vLdYo9tsBpihFAB508+pG3zcInA260xFHx9KjrWrDEXFzpa2oX/AFfnn2HOg2R0an4sfMnvb4hb8wu9unt75zb38OykGQrsPj6+WloszFE3roK8R8tFIPS68gjgj3oio62vaSR58fn1tPV9F8MP+FA/TMVXjJtp/G/53bSxscpwGNoqSCh7DyEcXiML1ThSZKlhcsb3J97PWhQNTyP8v9jrVg+VPw078+G+/wDL9e95bIyO1MvjK+WkAnRjHPGCftqhZLaQJFsy88/j37yr149raTx6KsVtb83F+ObA/wBffuvV66Bt/trf7f37rxFeuvfut9dg2/AP+v791oivXXv3W+ve/de69c/19+691737r3Xjz/xr37r3Xf4v+b+/da86ddkkjkf059+61ivXVhYf1/p791vz67A/1VwP629+68T6ddW/r/xT37rfXZAH6Tf+vv3VQT59cffurde9+691737r3Xvfuvde9+6912Tf+n+w9+69117917r3v3XuuwbfgH/X9+60RXrr37rfXvfuvde9+6917/D37r3Xvfuvdesfr+P6+/de67Bt+Af9f37rRFeuvfut9e9+691737r3X//X+f8A+/de65La/P8AT/iffutGtMddH6n/AF/x7914cOvAX+p/2J/3r37r3Dh14c8XsPfuvH167A5P54P/ACP37rxPDrq1hzxz9P8AiffuvVz11791vrsG34v791oivXR9+6312Rb839+60M9eHP8Ah791vrlYH8jji3HNub3/AMffuq564k3N7W9+62BQdeIt+b+/deBr16/Frf7H37r1M9KvZW7stsjcmM3Hhquajq6GojkZ4T6nhDAyxkXF7j3tTpOrqwJU1Hl1st/FX5L4XtbaGHqKiriGWoUp6ZorBJZHhGly63ve4sfYr2y+DqFkOR0LNtv1kQRtg9X3fGruOjo8Q1IRHq+28X6U51DSOD7Fdrc6RToV2l0FXT5dABvveUdN3lT1zxinpJWkZpUSwuWNueAPaeRqXFfLpJI5W61KOjvdW9n0FDVUkk8wanYqV1BTqFhzyfa6KYV6NLaeh7uHQq939gbd3jseTA0awTJMkjFQigCVlsGB/rf+nvcmnSTWtelV1KkkGivHrF8K+m89uhqna+KpZKqWqqoal9JYJHRiQeUlxwAF/wB6Ptrx4rK2aaY0Hl0W/VQ7faGSY09OrT+393YPqLYydPdbVEf8Sq40Oez9IB91S1Wj/KKIzLY2vcfX2WWNlLuU/wBfeDA+EHgR5Hor2yxn3S5/ed8KKvwqfP06JftV3pMgyTDyyVUhkqZWJJmlc3LOTfk/19iKRSE0jy6GAOkhKYHQxDGwOt9KjUL/AEB+v++/p7L/ABWBx05pFPl1i+2hpjoWNW5tq4FgT/re7a2bz63QAVHHqdDFrBtHpA+v0tx+fbTGh62BQd/WKeki0EHSSTYFrAgji/uyuQeq0BPSercZEEeUlUsLk3HNl/HtSkprTrRU1ycdATvCupV8kAYX1WIB4v8A649vyfDXz6TyOAaeXQLZJKVUmlcjR43a5I+g9t0OjV0lfjXquPv9aPIZaNqZleqSoCKg5cpqABsP979h/fZLePbZJbntWhGfs6Id1dRbsX+Gh/wdGJ6j2/UphcbUValf8kjKg34/xN/fOnma5jO93Rgypc56xi3Jka7lCGo1HobhQmOQsblCpI44/V9PYcLCmOkSt5HpD7soqaeCQSxI7aWC6gDYHg39rLdmBFD1dTRsefRQd7YQRu8sJC2ZrKOBpC39ieylNdLefRhGzMc9F9zG4KjFsUdyoUkAXv8A71/tvZ1FCJK9KVWpJ8ugQ3bvGukp6iSGRUgjR3lmdghVU/UwJI+ns6sNskuJAqjpbBb6x2jqsvvz5Y7U2TBW0uNycWczv70Ro/S8iOy2BAubWP5+nuVdj5YjhUS3C56ENpYaVBcZ6pr33vvO9gZ6qzebqpJnmldqenZtUdLG30iiH+9+xwiLGoRBQDo3AAGOkWT9OLW/3n3brw69z9ef9f37reOHXXv3XurNf5R3/Zb3WHNv3ai3+v6fejjry8Ojs/8ACgv/AJn91nyP+PTm+gtbmD3v1695da+Z5Y82uTz/ALH37rXAdcffut9d/wBffuvdde/de67sLXvz/T37rXn11791vr31+nv3Xuvc/T/ePfuvddk/04/w9+60B17kf1H9Pfut9dXP9ffuvde9+691737r3XvfuvdcuALcG/P+t791rz64+/db697917r3v3Xuuyf6cf63v3WgOu7Ackg/4f6/v3WsnHXH37q3XYP0vz/h791qnXVv8Pfut9e9+6913z/jz791rHXZFhe9/fuvA166I+nP/Gvfuvdde/db6ccTi6vM5KjxdDC89VWzxwRRxgsxLtYmw/oOffuvdbSfwP8A5J9F2Ts/bPaG4BO8lZRx1/21SzsGdLSafGRZef8AD3o+o60ONGFerlMRs/OfHakh2ZTUk0OFo0MMGmMmMRseCpsOBa3u1QcdeAPHoXNkZRMrkKeqo8nLHKzqXUalOv6sDY+9EEdWxx6uc+Ku4pamrx2JrpnniYKnq5ub25J9+IxXrVQTjq33C4WnioI1i0iKRf0gEf4/n3ZakY6qSAaN1AyO08NVy+Gpo4KksS51WIJ/1PI4/wBj70RTrYbHTjQYaloYzDQ0sdOkcbehdICgjgDT78B5daJ9eqk/mvujdVPX1GLx/wBxDSsxV2iD6SNJBJI9+ADHSc162WaMeMuGWlOqLu0xkcHPJI07K2Qb/KPUSZBybuL/AJ9xZz3MdnXxIcBhU9Z0fdV21PcG8+l3Ua3hNELClM+XQMYXIXyVLFTktPLqKWWxupuefcS7ZzTI+4x6UzXrPrnz2Ugi5PvWuJ9KKuc+VOlHlKiend46xn1TH6Ob8/jm/uVeZOZhNZJBedmB/g6wX9oPZOfbOarnc9jdrgFiSKkjj+zoYfjZ21iOn9xz5HL08FbSzSu7JOBIgRr3t9R7CW3cy7btUbySOCGp/h6m/wBz/ZnmznizSOO3KGNcEY/ydH7+Jm8dnd1d5birtu7Jx2iaqZ6muggCmACXmTUAfeWF5dQ3/Ju33cKAh14/l1ghz1ytvPJ0Mu07rdvVMCMnB62CdpbUpNuuY4FVBIsYIW1irRqefpa3sIqO3qA0OPT/AIvpUV1FR1BdZY0lQAggkH6Hj/Y+3DxAHWwDQk9Blu3rnbm5cdUUn2NPDJICocEAgldJNwPdSAeHXqkdAfs/4v7X2zlpctURwTO0jGNiwJPOo34J/Pv1D1avp0AHyxrN04+k/hW20qIKREWO8CkKUHBN091GT1bhjqq/N7d3jV3DS1s09Qz6iwkJFza3vY+XWqDoTtpfFtsvtury+58ZHW+VPITWOoMd1vYK1/8AevdwK8Omi4FVFT+XWnf/ADnPhjRbWzOV3ftjDpS6Kp6tnphqveRtYYIBcEX/AN796I04PXlcvkA/s61lXVkZka4ZGZGU/VSpsQR711frj/X37rfXXv3Xuve/de67sf6H/be/deqOvAfXmxH+8+/dar13Y2B5P/EW9+69UcOvAX5Jt/ifz7914mnDrj791vrsf617fX37r3XX5449+690aP4hd1VfRvd2yd0/dilxKZzHPlmJ4SCKe6u3IH+Fj/Ue3YXaOVXBpQ16cglMUyyrih6+jD0lu3Fd6dZ7a7Q2lko6uCvo6d5vC45UQgtqjUn/AHv3K9ldLdWizKa16l2xlS7tVkiNaDPRidrZ4U8z08wMbAlRcDkqdJF/azD4PRghPn0J6VzTQgBdWoCxH5N/pz7aMYVuniagdKrEQK1OWmbxnksvHPFrXNvaSdu7HVwdIz0FG6KVKfLLX00QnMMgcWAJjKm9xcf63tahLRDUK9MuDXHSqxlXUbjiTyubpYKCQLaR/T3R1WAVHn04rFiR1JqcTKqmFByOQRbn/b+9LMCdR60RTHTLJtPzRSfsrqcc3+t/rzx7cMyHj1QoQagdJqq61WXHV8bRKvkiKgarfUc/j3Uyoe1fPrXhk1r0Q7s7pcwJkSKUEtHKUcXN2ve/09pZreuR0U3NqCagdUtfJTrvMYiqhyeOhkGUxUrTUToWMiSrJqBUgfg/T2SXMJDal4joP3cLqajyPV9n8tXt3cPZnTWN2Rup5Jczh6SWR5aj0zaYowqKWY34A9q4paRrK/xDB6FW0XLTweG3Fej8/wABkxrmTQruxP6SGbni5+vs0+oEopXA6M6Ba0yeknmaipd/tY4gjMSrMWCgc8H2qjUBa16ayxx0DW8t9zbbWPF0Unkr5G0voGoqW/xHPv0rqvw56TyyeGQg49BDX9l5nayyzV88jTZdPEqNyy+Q8GxP4v7ZMmgAEcemXmaP7T0uNl0lRkKZclKt5JiWaQ/VtXqA9uICRXp2M+Z4noQV/ZjdiP24heRjwoUfUj26o1NV8Dp9VBweA6Kt23vFc3kaXA4mQyfb1C+coOAmr1fT/W94ze+fP1tBaHY9ufU9aMBnqHfcfmiJo/3XZmpHxfl03HBpLjvtJ11rLGFJYCwJFyb+8PEkMTVGM1/b1BmVOOgopsNX7RzhqqNn+3eThE9Sizf0H+Hs2MiXUOluPSotrWh8vXo1G1c/T1tJCS95tKhwfrqtzfn2QTwlGNekBU+eOhDhKMLgixF/x/sfaUg8OqEmunrI8kUaMGtezEX45I5HupqeHWwppx6R+RyDEsiKLDk/QXP0+vt9AFBJ6scmnDoO8uUq5DCC7TngJYnk8ix9rdvt7vcJhDt0Zd2NKUPHpTbxTXTiGBSSfl1n2hszOZbPUNEMczK9Qqytb0+NuPW30A/N/eSPtz7KX1zdJu/MieGiHUFJwR6dShyn7eX11cLc7mmlAQaHoj/85j5sYD4r9Gp1rs/JU53duCircTl6WjaKSWATgwoLqw5H9CR7yYvp4Npsvo7ELGrCgCjgKD/D1LPMN9a7VYLt23jQSKED9nWgPlsrXZjKV+YyE89VW5Crnq56md2eR5Khy7Fma/8AXgX49gokk1PUZcfPpq966t12CQQRwQbg/wBCPfuvdCF1l2nv3p7dmN3x1xufKbT3Li54poMniqqSmqD45RKIyY7Ercci/wDrEe/dVIr/AJOttD4pfzPPjB/Mv6+oviB/MWx22tt71rccuF2V8iszQpV7ryGYkg8OPoMxXhrvGSFAkY6lsT9OfeuB6v8AEmk8R+Vfs9D1SR/Mp/lXd2/AXe9RWzYTJ7o6F3GUruvu1aVIarEZ6gqGvGonpgFBUMLAX9PLW9786HqooRqHl/LqqS3+I/H1+vPv3XuvEAHg3/x9+68M9de/db697917r3v3Xuve/de65WsL/wBeLEfT/H37rVamnXH37rfXvfuvdcg1rg8j/X9+60R6dcffut9dg2+vP+Hv3WiOvH6/W/8Aj7914dde/db67uLWtz/X37rXn11791vr3v3Xuve/de697917r3v3Xuve/de67/F78/09+615069+LW/P1/4j37rfn12bAW+pP5/p791oVJr11fi1v9j7916ma9e/Frfn6+/db8+uvfuvdd24vfn+n5+vv3Wq5p11z9P949+6313bj/G/0/4n37rVeuvfut9e9+691yXg8/0/P/G/futHhjr/0Pn/APv3Xuve/de697917r3v3Xuve/de67A/P4H/ABHv3WifLrzG5/2Hv3XgKddqL8ngf19+68TTh11xfk8f19+69mnXj9ePfuvDrxFvr791vj14i319+69x669+6913fi3+N/8AD37rVM167FiLE25v/wAa9+68ag1HXRtfj8ce/deHXrG1/wDYe/de86dD98fO48x1HvfGV1PPI2LqKuCKqpC7GJWlfR5wo4/Pq/417ehlaJww6dhmeFw6dbTPxz7ppqzE42ujrkeGshgkldZQfG8hBMdx/vHsa2F2JUBHQ1sbsSRg9Ga7K3HjM5j6Salij+/aaMitRg0mk2sLj8ezGZloD59L5pFI6F7akdb/AALHyRyszLTx3YsQ19PPHt1alQR0pTXoHQoba2tv7fucxm09o0VXl8rX1dLCaeFHlKQTSaHm1Lx6Rz79LKkMZkkNAOvPL4EZkkwPXq6Sj3Xs74JdbUvXVBkKbOdubsoYauuyLOktXhDKmuagVl5FiSCPZHbiXerrxZe2FDw9eiS1Em8XIlmP6S8B5H59Fsk3rW52WfMV0rTVmQkNTMzFi3lkOpib+xopVVEaYAx0N42CoI0FABQdNq7rmgqEKgK4Km9wOPpb3XWa56sZAGrXHQm4fsNEEUdY6C45N/wR9PbbxI+eHTgn+WOlxBvHAVJDLWRlvyCwvf8AxB9pzC4NB06rr59Ycjv7BYqmklerjOkFgNXB/H1HvwgavdgdaaSMDUT0WTevyByf3T0uBpVn5YKVDEkj6Hj26CsZooqOkUt5pbSo6D5O1+x8mxE1DLHH/QIw1DTb3XXU4XpoTzMKkdZYpspk3NTkmeMn1Mpva45/PtzLEHqwLMKMOg33/uenx9FURPUJCiI3OsAkL/tPtqZyDQGg6ZmlUDPl1WllN2fxvuSgoIh9xjnePyORdb67fU8e459xFup+V5ls6lqNw+zoFc0+PJtjLBjjw6tS25jIqbG4toNIjFMosDe1h+Lfn3z0neQXUsctdYOa+vWOMmoTsr8R0/TyRpG4Nz+olh9bfTi/9fbQqc9aFfToHdz5GGMTqzXPqNyw4AHIHsztkJYMP2dPICSKDog/eHbeG2dS1EtTVQRohkLPJKqkWS5tf2Ntj2W53GUCNT0b2ltJcNoT9vVQnbXz72Lt2SrjoMjQ5OvjZgYHkRyHAN0AHuYdr5FWNQbon7OhPbbSqgCTqsDuT507/wCxfLS4CI7YpWDwmSmcCSWImxbQvAJH9fY5stlsrFaRrWnRpFaRRfCOiO19fWZOrnrq+okqqupcyTTzMWd3JuSSfZsMdKqU6h/Tke/db697917rkeALG4/pf/e/futDrj791vqzb+UcbfODq9v6TVB/1xZePfj1pejr/wDCgk37/wCtL/jacv8Atv2PfvPr3Wvq/Lm3NyT/AK3P09+68MDPXiCxuB/h791r4ePXG3Nvz791b59eHB59+60cjHXdxzxck3v/AE9+69Q9cffut9eBI+nv3Xuve/de697917r3v3Xuve/de65MQTxxx791oCnXQPBH9bf737916ma9de/db697917rtgB9Df37rQJPHrr37rfXvfuvde9+691737r3XvfuvddkEfUWv791qoPDr1za349+63TrwNgR/W3v3WqZ669+6312QQP8D/j791qoJ67sDpv6f8f9h791r1pnrsEAkfUHj/jfv3XjUivVm38rvorDdx9+beTMr56ekykFOaZtJjZHZJHYg/n8D37rdKmh6+mr8Y+t9m7C2dt7baR09HSUVBFCIl8Spo8Y4/pf37Fc9erRfPpH/LbbHSW36BcnXVNDU1FRCJBF5IW8Z0E8KPp/sfZLvW+2Oxw+NcOP2ivUl+3XtTzN7lXn0m2QvpJpXSR8uqlKnd+x8ZkEqsHNFTwl9V4rFT6/oLewL/rtbTq7StB8x1lTF9wXmySDU7yB6Vp1cj/L6FP2PXDI0lREVw6LLKWlVAyhv1Et7Huyb1BzBD49vSnHrFX3N9s7/wBr9zO13wYkHTUg+XV3kEscSJCj+lFAujKymw5N1/3j2d5GOoyNOJ65EqGLsbs1xq/2n3SuerUoMdctTJrcWKEWJ+hsB/xPv1T5deAHn0XLtvqHEb/pasvSiWrljk8UllJU6CP979+IPAcT1sEBxryoIr+3rWc+bPTm5+q90Wy0FX9jXyK9EPA7AI1yNIUf717hb3BhvDGyOK1GK+nXTT7oe48utfwtYlY2Rv1DUDz6JRhIZKCaKuKssqK2gsjI4DHkaW5B9xly7tJglF5cLlTUDrM73i58i3Kzk5a2tw0ctAzDqVnquor0WR2ZTqBF7g2H49u80z3VzMJpcJ5dN+w9py9tNs1pCQ87fETnPXhFj8jQLTGxmCWIuQSRwbkf63HuAodt5km5hkmnkP0uoUzimodTXut7NsyvOEqpB8vkerP/AOUV2htXanaO69oVGOikyOa1UlPKyaiknkC3Df199SrC4htfbva7cipKih/Lril95zYdx3zmm536NikSGpXh59bRs9NFT+OVmWxVGBPpt6R7IxgDrEHgSPTqBLVU8h0J6QSSWA/V/X/invdet0p516Ysg0KqywuNV7nSbEf6/wDxP+PvXAH59WySOmOWvghsKidRcWAdgukDixJ96690jNz0Gx9y0kkOSajaQAqztLDew/Nm96NK1638z0X6v2P1Bg55a7JT0KwQiR9RlhuNKF1P+8D3sFV7mOOtaJZ3EVuCWJAAp8x1R/8AIn5kbr232Jm9o7Oi17cSdoqV4SNDRq+gMSP6j3D/ADBznd2d+YbQgrWnXRf2m+7Ps3M3Jq73vA0OFr9pp1XX37T0Xe+wt0Qbuo4mmNMTAzhSUOlmNif8T7UbXzjLDMrX57W6Kec/u62G4bZLDywhaWMZ0j060bvkftak2Z3LvTb1DF4qaiyLiNAmgWe5LBf9549ypFPHdRLcR/C4qOsC9y2yfZr+ba7kESQOUavGoPQIFgSDYi1/bnSAKaU64k+q/wDj791umKdcmYG3Frf63Pv3WgCOuIufzwP8fpf37reB12Tew/pwT7914CmevE24BJA+n+x+vv3Xh6nrjc2t+Pfut9e9+6912Pzzbj/b/wCHv3Wj16xAB/B+nv3XqitOvC5IH+9+/de4dbRv8lf+ZquxpIegd/1tLRUU1KmP29JUSkipeQeJRGX/ALQ4BHsR7Ju5s5RDLlTw+R6EvL+8S2Un08h7G628qCjgraChzdHokgnSOcTIbq4lUSDkfX68H3IysJAHU9SYhSRBKpx0MGKolr6KnkjurD624tpH14/1/bEkuhiD08o1dPKUlQt4fKQNNjzb/Gw9sF1w1OreVOsLYeN0cOBIXB1E/Sw+v1938cg9aC6h03Y6gbD1MkqG8UpJROQAP8Pd5HWZAvp1tQU6fY6gySFn9LG1gBf/AG//ABHtgrQY68KHj0rKV4DCLIrMw5ewFj9PaRwdXHrYLA0PUSsaMKEFiWNiDwLH8e7IDWvW61PRfuycLHXRyIkd2dH+gFiL/T2ZJVo6HpiVNSkDqsrvLqmijp5qyoohUSPrKgpew+v4/p7STwinDolurfSNQz0G3UfZG4fjlQZfc2Ax33dVWUzxU9HJGQh1gjgD/AD2mXRCpJGodMW88lkpdRWvQx/GH+YzvXd3YOYxfbG3aXCYTySR0dW6eO31ClWa3+396S4+pbRKBGBwI6U2W7O0pSdaD16Mz3x3rQz7arszsbIwTzUVHJUJ4ZkZ3dRqKkKfa1pBHEdB1HpddXS6CYTw6rH64+du323XLSdiNTx5JZ3SJJrsxdX0j0n2ij3GMPSTojh3VTJ+rQno6VXkqPuCowmYwSGWkmmjaNUQqqjg39rtYmoVyOjIutzpZOjy7V20cXgqShClp9CDSRcg6OCPawEINPDozSLAPn0GHc+6Itj4BqeKxyOVDU6Rn9SFvTcAe499yOcoOVNgeYsA8oKr9vQW5y3yLYtrJBo8mBnz6KhtDA1U0n3+SBFZNI0ju99RV+Qefxb3z23LcrjdLuS/vGLSOTx9K46xcnuZr2Z7mU9zE9C69OkcWm5cKlhwOSB/tvZSWrk+vSUNXHn0lTTxVVWEliDq729Q+lx9P8P6+3w5UYNOngWAr0saDa7U8kc1MxQCx0Lex/J59sPNqw3TLSAmnSzjqjRi1S+jSABc8EgXNvafTqOOmypJzjqMcvHXyinoC09QxACBWa5J/wAP6+1u2bVuG7XH0u3RmR+FKGnSi3tLm7lENspZifTpxOw87XvF9xFJTCYg3UMCFv8A7x7nnlH2B3Lc3SbfS0K8ftHUlbB7Y7hfyCa+rGvQ0bT6hw2LgFdkWjqXQa3eqKoEBGosTJx7yb5f9v8AlvluJYreFZHXg5Gepq2fk7ZtkUN4YkYDJNK16r9+d/8AMw6G+GOytyY7CZTDV3Y9ViZo6ehaaOWWlq2/biEJjv67/j2Idy3JLSMRhhqXgBwp09u/MVvtsBit6FiML18/b5Q/KHsL5S9kZbfm+MhVSRVdXLLRYtqiRqeliklZ0bQxtrIPJH0+nsC3FxJcSF5Dx6iq5uZrqUzTGrH+XRZjxxe4/wBf8e2OmeOeuvfut9dm1+Pp791ocM9d2Gm9+f6f8T7916prTqRR1U9FUQ1dJLNTVtNNFUUdXBM0M1LUQyCSKeKRLEMpAKkHg8+/dePWzB/Lo/nQYEbFoviT8/NqYzu/pTJwvhcdube033uR2NSVA8QyuHqJ7lJ6fhxbhgPfqAinVlbQ1W4H/Vnpv/mQfyXsTg9jS/Lz4BbjqvkD8eMxLLkslkMPFFNLt9JkNS9NU0UfrR4RdSNNjYn6+/A6hXqpXSdI4cf+K61q5YZI5ZIZI5I5oneOaKRSkkcsbaZEdGsVKngg/Q+/derTj1h9+631yHFyebgj6+/daOcdcffut9ZLMVFgbDm/+tx9ffuqigPXD8Xv/vPPv3W/l11791vrsgD6G/v3WhXz65hhb6H6D/ePfuq6TXrgAefxYc8/j37q2Ouvfut9dgE/T37rVadde/db697917r319+691zKGwNjyAffuq1z1w9+6t1737r3Xek21WNj+fx7916vXXv3Xuve/de67U2Pv3WiK9de/db697917r3v3Xuve/de697917rsGxv791oiop11791vrkVH1Bv/AF5/r/re/dVB9euwSF+nHPN/fuvHJ64e/dW6/9H5/wD7917r3v3Xuuxb8+/daNfLrr37rfXZt+Pp7914V8+vC/Nv6c8/j8+/daNPPrr37rfXvfuvdchyRwPfutHA69xqN/p/h/vXHv3Ws0x15rnk/wCt7914U4Dro3/Pv3WxTy669+6312Tf+g/1uPfuvDHXXv3Xuve/de65LY/U2H/E+/daNRw66DMrBgSGUgqwPIINwQffut9Wl/CL5NQ4WSPr3dlW0MLvG1Jk55U1MPKFSMFrci4Fv6ezCxvDbsE8j0Y7feG3bQ3wnq6Gi3rMsNJqqBNTSlJKWYOkgaEn0+pSbexSlwHAbiD0KlnSQBvLo5PXW+8nkYsTh8TTvXV1dNT0cMCI0hJmOgEBL/4ezFJhGmp8AZ6WJcBF7sD162AOtJtn/ArpYdgdgLTTd8bvxn3W1ttTtT+elxldF+3WfuEuCAwI+nsklnfeLrwV7Y14noolnfdbgW6GkQ4n1PVZeS3zlN+burt7blyEmTyGTrJK6N6mTW9IlSxkMCsfwt7exJCqQRiKIUAx0I4EjgAjj4DofMHuFXo4irahptcsPTx9P+R+1iPinRrrFO3p2Mz1ZEgezXAXkAcfT3Y0Y460dJx0oqLF1NVHbylZCLWDL/rjke3FWhwa9OhSUp5evUGvw9bSHXHVshvyQ6i3HNj78QQ1SeqFSo49Jqrp6ur/AGJqsyKbC5fg34t7p3NXNeqshOTnp6xWAxONC1VR42cDkHSefwPdlQKM9XVEprp0s6J6KZDaBAg/Qf6gk8e7ChOOnaArUDpC76zFFgMXXVjFUWOGRgSygalW4F2/P+29tytpFR01KwjSrdVO7u3znew9yZFkklo8XSPLEWUkq2kn+h/NvZO0jSsT5dB2V2lkJB4dMeztpyVjVWchT96gqOJ7HUQsl73t+bfj219NHdRPDLlSpx+XVRCs8bRtkEHqxjpvc7Z/atSXPlnx2mGzHklTpPPvn17l7Mmx80uiYWZiescuZbMWG6Mi/iPSozeZWlp5DwH0sSAeP8fp/T2DI4tTgdEoJ6KF2vv2LDUVbM0tgkNRKx1KukRpqbnj2KNtsGnkVaUNQOjC3QlgAOtVH+YD8t8ju3c9RtPaWQaGnjlnSvnhmRnh0gRtGBH9C3+P495L8o7DHttkryCrnod7bZC3j1EZPVSsskk0jyyu0ssjF3kdizuzG5Zmbkk+xp0bdcdXFrD37rVM167tYc/U8gf4e/derU466It9ffut8evAE/T37rxNOvWNr/j37rVRWnXuLf4/Tj/ez7917NerNf5R3Pzf6w/5a1H/AET78evDh0df/hQVY9/9aD/s1Jgfr/WD+vv3Xq4619XsHaxPBP8Ar/X37rwyM9dE2PpJt/tvfuvcePXG5+v5/r791vrvi178+/dazXr1+Lcf8T791vrr37r3XYBP09+69WnXvpxYf8T791rjnrwNvwD/AK49+68RXruwJH15+v8Atvx791rIHXR+pt/X37qw4deHBvwf9f6e/de669+6913a/wDsPfuvdeFvz9PfuvfZ117917r3v3Xuu78WsP8AX/Pv3Wqdde/db65AA3ubf09+60a+XXRP+tx/vP8Ar+/deHXXv3W+ux9R79148Ou2t+PfutCvn1xHH/G/fut9crggkk3/AAPx791qhHDro24sSf8AX/Hv3Xh11/j791vq/v8AlDdXblwu5IN+UUEsyxzwZF2RJdKA6SigjjhVHPv3y6rWhr69blFT8lRgNgU5WaRMwcetluA6y+PTpF/zf2Eea+YY9p26QwZl6yG9hPZ+6595ws/3gNNoWFa1oRXz6JfTbp3X2zlquq3VkKmXHGUiOmnLsqrqtp5+vB/HvnV78+5e6RPHb2FwwZh3AH+Xy67V7XyByp7Z7bHBsECCYD41ArnoTqHoFc3TSNicak8S8oVhmPFrnke8Pbr3iu9rlVNwu2QnyLD/AAdJLn3L/dso+um0sccR0pNl9l9tfF6v/heExlVDTZZ0j1wGSP8AbBudYb3mV7T/AHqLGPlx/p3DyQKBxHd9nUP+4ftJyd7zJ++BMvix1Yg0NT+XVn+wf5pWF27trG43c6RyZqCJFq1eSHyCR2CkENzwL/n3mxyT7t2PMuwwbi+HkFSMU8usBebfuo8wW+7XE1ih+nBNDRuGT9nVqnS3d22O59qY/cGIq42nqY9bwo8J0WXVYrH9D/r+5Ysb6G/g8eM5PWLHNHK24cq7m9heKQFPHP8Al6eOzewqXrqh+8r3C0xUM2rQAOL8F/pce1ZJGCOg2KHI6DLrr5TdY78rv4TQ5qD+LRMYvAamiU+W+kCxI/23vYYAVPl15kr2+vHqR218eNk93S01du1KSpWjjIpvKYpCLQnTYpfn+vPtFebTa7qP8bAYDyPQp5a553rkvUNlYxFzkivrnrWF+UnTuU2F25X7dwmOIoJ6548aFilVJI/MVXQf9b8j3Du87abW98GFaKTjrod7bc4Qb5ywu5bhLqlVQXJIrWnRac3gMvj66HD5SmNLUykIgsQLj68Hn3FXP+8nZNqmvbhe2EVp1mD7Mbbtxhk3+1k14qR1Khw0LZrD7XpLLkslIkIcWJ1O4Vif+K+8b+Sd15h5w3V7pWK2rMKD5VHU43N2G2643i7XVDECSD6UPV238vz4u7X6VzWd7G3tU04qZAamjlmeNHicgSalaY8e+s21xLFyFYLdZMSjP5dcO/vGe539ceeLjZOW49KqaOq8MH5dWJ5/5kbAyu6KPaWEysdVV1M600ccc9M41KRHa0ZufZNFcRTL+lmn2dY0Xu1X22kfXpoLeoI/w9Gkx1TI2Jo6uQ6fLGHf1W0hl1L9fbvlU9IjSuOg13b2LhcCf8pq0iZif7cfN/wS/Pv1a8Ot4HRdN/8AYE+co2fA1pL2uTHLGST/AIaT/T34ZOem2kABYdVX9w/I3f8A1xlGjrZKo0rSsvkErC4LkC/P+x9hzf8AmBdgTxXA00qSfTqc/Z/2ak93J2tLWUpJwoK8R0WvdXye3XumSi/y+ojx0jgVTiT0BD6Tq9xvJ7ucubjcpaw3ADZDAEenWQ2z/dg3Hlma6tLlDJOn9nUGpNeizdhUMtZkf41QO9ctWysJTqI5PJ1f8b941b57u7N/W252qVgqxHDVGT1n97M8u3dtyyu1b6vgsgpT16DaUzzk46rXTFUkrIADZh+R6r/j2fQ8wfvYLLaPqUdTZt/KWybdDLJaxglxk0Hn1rA/zXusdu7O7Eo83iIoUq8rWSLVvGqhiDGXsx+tr+8n/brdZtx2rw5q/pgAdcYPvj8g7byV7gifb9Nb0GRqepqaGnVQ7WBIH0BNv9a/HuROsPuvWNr249+61UcOuvfut9e9+691737r3Xvfuvdd8W/x/Pv3Ws166Avx791vh1ysBa/1/I/p791qteuif9t+P+Re/deA65W/LcXta1v969+61XyHSz673tW9db0wG9MbEJ6zb9dHXU0TSGIO6XAGsA2/23uytpYN6dWBIII49fQ3/lr/ADOxvyY+Pu0oGdItxUeJiqslSvJA08SrTqQjhCbG31BPHuSOX9yF2miQUYCn7OpO2Dc1u7YRNgqP59WkbO3tRxhaKVlW5VbnSD/j+r2bzx+Ia+fQoRgBw6FIyRVKGWGRSjC91K2/r9B7S0ZDRh04BqGOs8GlY2Zjxb+0b3v+f9j70cnrfw0HUeZHlufGbfRSFvxb3ZSBw68a/i6xJSTvIFC835Ive39B7tqWlT02ailOlVRUMyRm4LD6fQ/gc29pHcV6sTmnWSTHCVixJH+A+g/23vwkoMda1Hh0lsxtlZlZiNQtYN/T86ufaqG5oKdeIFTToEN+9V4vcVCIagqrKWsSt78Xsbc/7C/tUHE3b0nlhLig6JL2Z08sVPFQwQ/sQuLyqjEFQ3F7ce2JYdPb0Xz2pICjonnZPUxp8TWw4zVT1bwFUqUVtaOeNQt7QywnTQY6Lri30LRePRFdtxdobF3DkKPL53IZHD1ZkQxzNN4kjJIJ0vcfT2XxiWF+41HRcomibJqD0+dY/G2Ltjt2lrYVWeBakTzEo7BVL3cnSLe7Q2qzXBJ4dWhtFnuQeFT1sodadO7Z6v25i6OlennmWnhCozKhRzGAeJD/AL17EEZC9ijSB59DGGCKBQvRjMBiI1gbJ1CqIoFDm5sNI/x/437R3lwFGkmg6VowT9R/hXJ6IL3JUxbt39V0/jElFQMz0xGkqrq39n3g37280ybtv/7jRtUcGQfLHWL/ALj7226by1rGeyMmnTNTUghRWtpewW3N9Kj63PP9PcL/AG9R+SxwOuVVUBIfSQSQdQP4/pYe/ACuevAitemrH+qpR1RSokB1W5BJtwf+Ne7setu2OhTgnENK8zEKI0NjwLW/1/aQirdNAHV0C+591T1NSaaBy2ssgN/oQbXFv949rBDqUIoy3Dp3SVFOJPRyumtl4ei2tSZudIqzJVoUssttUR08EFvebXtHyXt+37FFuUqBpphqJpwp1kv7ccs2dntKbrcKHkc0ofs6VO895bO68x9Tn97Zenw+MpIXmL/dUnoWIam9Lt7nKSZYotVRQevUgz3sVqpEraR6dazf8yT+e9g9p4vOdZ9EGmztVMZcScrTVsUFTS+hkaqZ4blbfjg82H+ICu472qhorTNfP/N0At45ndqwWn5nrTt7O7b352/uKr3Jv3cOQz1dU1U9RGa2by/bCZy3jiYi5ABtdrn2FHkeQ6nNT0CXd5G1uak+fQbEk/U34A/2A+g906r117917r3v3Xuu/pfj37rXHr1ja/49+69XNOuvfut9dkk2BJNhYc/T/W9+691bd/LO/msdu/AzfEGMr56nsTozca0mK3X1ZnK+WXbLY5pPt6idMYw8ZfxEqb/jnn34YOOqk9v2f6sdXOfK/wDlbfG3+ZH1/l/l3/LgzmKG+q2gqNx76+Ou3YacPtjVGaivyVGUP78OsEuFGpbEX/Hv1KHq57k1Dj/h/wAx61M+yOtt4dT7xzGwd+4aqwW6cFUGkrqCpQxeNl41AyKpIP8Aqrf7z791QH06QFz9Px791brkAeWH9mx/3m3v3Wj6dbMP8u7+QDSfOD4W7w+XOY7qTrmgweKzzYLFVlPFo3HuXFQeWDCUhljveQ2W63/ULce/ceHl1sUBAfFf9WetePtXq3dvT2+c3sLemMnxWbw9ZNA0NQrJ5YFmaOGoXWFOlwLgkD37rVfTy6DoAgni9r3/ADa3+I9+68euPv3W+ve/de675/x/x9+691z02XUbHgG1weD9OB791XJNOuF+bjj/AFuPfut09eu+Pxc/X62/2/v3Xs9cffut9T8VSx1uTx9FM7xw1ldSUssiLqeOOeoWJ3RQCSVBuABz7959aJoK+nVznyW/lU4Lob+Xh1V85cd2lJnajsfdK7cXZU1Isc2PIAL1LsYluPx9fr78ag062BVa/IH9vVKhNzf+vPv3Xuve/de6uuwP8p/E5j+VVD/Mek7VEVXUdg12xR12KdBOs1HGsxrhL47eMhuRqv8A0PvWajrekBCa5Ar1SrIuh3T/AFDsoPHIU25976qPXrH791vrsgi1/wAi/wDsPfuvVr14f0sOf9v791r59eII+vv3Wwa8Ouvfuvddnnmw/px791r5deBt+Af9f37rZ669+691737r3XJdNiGvzb6e/dVNfLrq/wCATb37rfXXv3W+v//S0APx9Pz9f+I9+6159de/db697917r3v3Xuu1tfn/AHx9+60a0x13e17f1PP+H9PfuvUrx64+/db697917r3v3Xuve/de67uf6+/deoOuvr7917rux/obf1t791qvXjxxwfzf/iPfuvDOevAAnk2/x9+68TTh16w55+h4/wAR7916p669+631737r3WenqJqSaKqppngqIJFkhljYpJG6HUrow/IPv3WvOnViPx7+beb2nBRbS3rFU5uGqnio6XKMgqJIDNKIqeNowdVrlQSo9rbe8khx5dLYLySLtJx1vPfAnrLr34ldDUfzA+TMWLO58viI8j171zlniafJ0eQi14/M/a1HIVTZl49r5L9rwC2BoD59GTXrXQFshp6n5dEW+RPfG7fkB2VW9j5fcNXW0fnkGBx8st4qDF6v8no6eNTpCItlAH9PZ/bQxW8IEGfU9H1vFDBFSPqFtDsOYpHBMzK6hRz9T6rWufx7MI5QePSxJTTo0u1t3yzQxqrgAqLC/Bv9Db2rVxSoPRikpYgHoQavdNZBT6onYWBOoHm9/wCvtzX4Y1Dp53auMdNdN3Hk8aCrPIWW449V7cc+/CcqfmeqC5dTpHDrDN2DvbdkgjxNPK0Ra5cEgr/rW91MruaAdeEkkxqvTpT0fZICyPRzkKQfq34/1/dv1RwHVwZx5dLTFf3mKo+YR4o4+Crki4HF+fd11sM9XQSglpMDp/ruwcNg6KT7qrSMRqedS8aBewufbgkXpxpVTzp0QnvLvVt1VrbdwlZIaZmXzSpwoUcH6H2XXM3iHQh6Krq6MvaOHRca3dWGxFGuHpJi1XVyI1TIi3Oomz/T/be05KRL3kD9nSOR1RaVAJ/wdGl6uoqTJbafHY1WkqazQZm06dJb0km/+v7LL7mXZNmtHmuZQCAcY9OqS7vt+325kkcA0PRoOv8AZsexMNWQGYM9cxmYKTdTe/5/23vAXn/mIc08wNcxCixsaH16x25g3IbpubTAdoOOkVv7OrSRSkNb0M3H+P8AX/b+yO0g10LYPRZGorUdUi/P/wCRNJsDZ+UanrGXKNTVUa0yW1yLIhQWF7839zJyHsf1c/jzDApT8uhPtFpqkDMKjrVRz2XqM/mclmar/gRkquWqlub2aQ8C/wDgLD3PaqEUKOA6GQFBTppB+vH/ABr3br3XVj791vrsm/8Atre/daAp11791vr17e/de67Bt/iP6fj37rRHXXv3W+rNv5Rv/ZcHV/Nv3qjm9rfp59+PWl6Ot/woJ47/AOtfz/v1Juf6/wCY9+631r6nljc/k3P/ACP37rXAY64+/db67ABPJt791o4699D/AF9+69xHXib/AEFvfuvDHXXv3W+ve/de65Cx4P5/P/I/futHGR1x9+6317ke/de697917r3v3Xuu/wCvv3XuvA8Hj6/7x791o8euvfut9dgcE3/2Hv3Wq569cWtYf6/v3XqZ68Rxe/8AsPfuvVzTrr37rfXvfuvde9+691737r3XrEfXj37r3Xvfuvddi4sbcf73791r5deP1+lv8PfuvDrr37rfXNR+m4+rj/Yj37rXn1vJ/wAizr7b+5uuZYq6GBi2HoQRIFDHVAvAv+ffq0NOtAalBHkerkO+viiKSkTO4OZZKCIJLPFG7AQDTqIPuCPeyw3TbeVLzftrBlaNSaD7Os+vun+72wbbfxbFuyiKcYjqB3Z49AbsbbeNjy+MwjBY0nlRZ5eBpJkH1P8Are+MvOe/brcQXO63eoyrXtPEYOKddHd+3m4k22Xc4TroCVHrg9XZddbH2rgtt42GjpaWokaCMu1kYu7IL3/qf6++YnNXPVzcbtdXNxK887tpSDSQEoTTrnRzjzDzHu29yy3LvEisfUCnRQfkd1nntwHJbgOEjpMbhPJNDInBdAOCAB/vHvIL2c5y2/l/6XZL6ci7v+MZ8ickD7OsgPaPnTa9vSLaluTLLOQCD5H7eqVN0UNNNuXJTSU5OiX1Ehh6w39f8Pfaf2i3a1teX7SHxe7SMV6zhu9uiveVWgQDW6mhx1dR/KEy2aq87ueiqMvU1NBA7Cno3LmKAB7WS5/2HvNjkK4eZmNTpI65Afe12a32mWGMRhZK5I8/t6tl+W+D/vRtPIUCSmGRYlVWuQVKxC3P+x9ybJh6Dh1g5DQoD1qZ9p7F736Q7Mqd57NzWWqqVMiapYKWaaxXy+QjQDb3Tj1ehBoOrg/hT/MQymfjotqdpCajqwWiMtUly3IBuzn/AF+ffqqncxoOnEimuG8GFdTNgDj1w+ZveXW2/wDtbYmO2FQUlbm8dIqT6I41NTJqF9TAG/J+p9x9zDuVjdblGlqAWXB+3rLz2l5H5m5f5UvL7fy0UEwqta4FOqvPkpVZqj3ZSVmRxa42qhqUZYYyCW1D6XX/AA9wf7m7b++4ZNpulCrKKE9dG/uznZm2B4kui4deB4DoMMdXR1GdwubgYJl6OVZo7mzhkOsLf/Ye8b+SuV+auWt9azsYy1oHFDXFKjqc+Yd1sNj2a4ttzcC1dTqPEUof8nS97M+bHaBoKjZVLLV07eMQRywE3f06ASUNx76pbltu7v7a2EZQoXUEn8uuPfMW5e0my+413vu3TpM+qpWg9evfy+9j9vb478w+5N2VeRTE0OUFX5ah5NEkflD6bMfz7Ceyba222oZ21FhkdY/e5fPEHOe5lbW3EMcZ7SBQH/UOtyPJTQLt+FacKI46GAKRYanWCzE2/wAfz7PTw6jRRTHVLnzH3luXbMktVE86wFS8TJrNwFJ+g9oL+7WxgMzenQo5S5cm5q3ZLGHhWhz8+q3di/MPcOBy0sGUlqGotTKJJv0/qtYXPuLh7jCzvWS7PbXHWd5+5tPv3LUV3s/dNQVAPyr0nN691f6atzybelxbGmq5Figq2isg1t9Q3uCPvA+8MdjypOsFCzqdJHEdTp7N/dr3P2gWLmiScxyJ3Ola/wAuh7qviVloOvjlY6NBRJRGqklDm5gWxZr/AOHvj5afeDS35rS2E7LLI+hfm+cf5epEHvJsd3zabOYhpy+mhA4+Q6Tm5cdsHB9Y46PESQVuYooLVUKqpZWCkWN/z7NtkvOceYPcCZ92qlvOw0PXjnj0f7Ndcy33NkovEMVs57D5dV/1OR/iUmSrftzDLRiZ1iA+mljY8f7D31N9teVI7CztrUS+KHAqftA6l73C3p+SeT5txiYMVSoNfl1qS/zJ+z8vvTuXIYKvpXgpsJWSmB2FhJa8YsP9j7za2DZbfZ7BFh4sBXrgN7u+4+8e4nOFzdbqxPguVSvpnh8uq3rXJ/H9f8P9f2edRbWg67+hsDx/r8e/de+fXRFjwb+/deGeuvfut9e9+6912RYA/wBb/wC8H37rVc069+Pp/sffuvefXd7AW+v9fyPfuvcTnrom/wDr/k/19+68BTrr37rfXIc/U/QcA/T37qp+XXVr3/H9B/xHv3W+HV5P8mP5g4zontR9nbqy8kFHumZcdjInXUgSRFAiDubD1XsP6ezjZ776K6DngcdG2z3wsbsFvhPHrdQxu4qTLRUeZxk6yU1XHBKkkbCxDpr4t9be5KjmWVBKmepShnSVBKhwehu21uquj8URdpE4BBPCj/kXt3SJKluPSsSnHQ84Mx5Mxs0hAPq0H6fS59op/wBKuOnganoTUoqOGFbqrAC4aw/pc8ey3W7HrVaN0n6ypigkZokB5PNhfj/W9vopYd3VgKdSqfLJKgS2k8BrAfX8c/4+6GKh60VoQD08wxpIurg8C1hb688+2iSD1Rq8OoddNT00b+VlsBwptc3H0HuyKzHt6uASB5dBLliayeQopWFLlbf0Bvb2bRDQuePWqgtg9BlvLG00+LndoVMiqPwP6+1FNYz1SRe3B6KNmdsiv8l6QMLuCGH4vx7Tsuacei94jTuHREe6Ng1M1fLHT0XiT1aSFI1HVxyB7QTpU0A6KbiFg1R0ab4LdFZCjy9TuDJD7WhWImad/SiIRcli1h9P6n25br9MpkpU+Q+fS/a7YAmSTCjz6rg/mj/zYn+NHee3NhbCrv4wMBnqaPPxU7xmOGiSRfLJIdVmsAePr/T2Q7pvckMypF5HPQe33fGtr1Y7Y1VeP+bq074xfzVug/kt1rLicRueKLfk2Ljp58VDHEqx1T0mrSGU/kj2Xb1ulq2zz3QfS4RjT5/Loxn5ptH2eap0sUx057dSoqZ6irrWLTzPI6F+W0Obob/m4987t4vJNw3iW8lySzD+fWKl1IZ7uS4c1LE/4elJUkRoPy3Nx+f8L29oQST00goM8OkZk6+QOsKHSddmt9T/AMa9qIoq95yerADh07YW0Lxh7soIYk/635H+v7blGrh1orUU6ybrzc1PTmGAlI5FILL/AFsTbj3e3QO1X68gyK9APnNyYjZ+Mrt27hq1psNgomr8jPIR6IEuzMA3B+n9fYg2iwfcN7tYEGC+fs6XWkJnu4lpXOeiLd1/z9+ruttnVGK6oko9zV1FHPTrGkcXkEsLePTcG97+88ds3CHatritLde5Fp8uHWQ0HMybftiWMC5A61s/lr/NM+QfyhriTuXObRxMrTrPjKGv0RS08h9ERMZsAB9faC63G5uz3tQeg4dBy93O7vmrMxp8uqy6qrqa2pnrKuaSoqqmRpZ6iVi8ksj/AKndm+pPtB0X06je/db697917rn6dP8Aj/r/AOPv3Vc6uuHv3Vuve/de67H1sTYe/daPqOu/ray3t9bfn/X9+69w4nrr8n8fXj/iPfuvddqfqL2B+v05F/6n37rx6Nf8U/md318O9/4ffPTW/c/to0FdFU5HE4+vlgoMxRq958dXQg6WilHpdTwQeffiK9bU6TX/AFHraYmg+Ff/AAoB6Xgqad9m/HH54bZxixJhsfS09MnZWVoYSkEEtSVA1VbDgm1yePfjUivmP59exqzw/mP9jrU1+TPxs7U+JvcG5uke5NvT7Z3vtiotW4yfmRaaaV1o5zwP84ihx/gR+D7914imekp0jtFN+dxdYbKen+8j3VvnbOBmptJcSxZPLRUkqMq3NtLG/wDh791oVbHW4L88/ljL/Lrk+L/8uXrioyGMOzt9bG3VvzBYWnYUk1BuP7SWpad0sZSyNbRY3AP9D79QcR1cHTRTw+fp1Xr/AMKMfjzQ0XySpfk/11SLB1N2VszZIo2WFoo1zzYuGOugjUAKCJGcsPehWmetNSop+fVk38uP4mfCXIfyZ4flX3vsbbg3tsbdlRUUWQrYIpKzfH2UJqI8DIXQ+gkBTzzf34UzXrxGQDw4nozXxe6z/lvfzpOlcnX7V+P/AFv8Ut3dA0GT3XvOt23RpHDuHa+0zorautTRfVIF1Pfi/wDr+/ZFPLrY0yVwSMfkfL8ugd+Pva/8sf5bdh9v/ALavxU63pKfa2y8/Q7Z+SNHj4Ys7W7mo4Xpldn0WZWkWwIPB9+oa/b1rUAtD5fzHROf5XnxP+OcXyn+UnwW7367wm4Mlt3qve+6tgbtyaQPONwtRzvt2jhmkUt6pCgX/AAe9n0HVUXJL+o/Z1TT8UPgruPsj57nord+LOPo9ib5bN7nxMyEx1W1MdmzMwDkH9poVAa4vbjj37y68D3U8vPq7Hdv8vT49/Nv+c7uXpHp7buG6q+PnSvVtJvzcctCka4yvpNnYz7jckk5RdJaRoi7KT/at7rkY6v8WacOPzPl0ZLbHzg/k69gfJXbvwv/ANlQ6owOxcRmcXtaDvSHHQfxjc2dSuTH1S1jMmkKxDEgc/n3sUrU8OvGrDQvxfy6Kp/OQ/lo9d4z+YX138fPjfsnFYPbmUrNrV9RDjVWOJsJno4KhaqYxqLBo5A5B+nvw4E9VZRVVONQyejt/IPsz+WD/K839sj4Z7h+LfVvyB7JyGJ2lTbi7EyNJFLW7fyuco0kqpaGoROHgdyl2sNQA/PvdPPz62Ci0Shp/qpXqX/woa2l1VtD+V98TMF0tkKRut94dkUWUq6mEXpdtS5YLJUUrBONNPqNwP6e9eY60C2lhThSnzHRFOvu7f5W38sv4s7JyK9S7I+bPe246yiqcnhN44lJcTh0q6P7yq81YVI9DMygXsLW+th78f8AVXqy9uSf9noyPyi6N+G38zP+WLP8t+i+gtofFTsnbWSmqcptza1EKehq4aRTIfGuhSY7ryAbEfQ/n3sZP2dackCvrw6L3s6WSo/4TMvjCQj0XyQ3PipJ0JKhloactKpNuBq4PupHcPt6tUmEkDiP8vQZ/CbfH8tD4RfBOj+Qvbmydr/Jb5C5Ddc2Pg6g3HiUqsWKJoS1PW1lYVIAaQhAt/8AXsL+91qadaARF1eZ+yv5dWP7D2v8Jv5yvww7J3r158Ruufif2j1ric1uCWTalCKWgzFFjYnm0I5jFxpUBSpsfqPfuHd1olX7MkH+R60VtzUKYvcm4MZHzHjs3lqGMj6FKSvkp1t/sF9761SmOmUn+gt/re/dap69ev8A154tz7916np16/Frfn6+/db8+u7kDTb839+61TNeuPv3W+ve/de697917r3v3XuuyLfm/v3Wh14C/wCbe/deOOv/09AC3pvf82t/sPr791quadeUEmw9+68SBx67tfjgEXuf6+/darTPXRFvfutg166+vHv3W+uyLce/deBr117917rsG34v791o9eIsf9cX/wBa/v3Xga9dhb83A/p/U+/daLU64+/dW67Jv+Le/daHXY54vb/evfuvHGeuPv3W+uQ/ra/4t791o+nXH37rfXvfuvde9+691mjQuURUZ3kJWJUGp2kPCKFHJubcD37qpBrXrZ+/lE/y0usdl7KyX8xP5+08W1/jz1193/BcHuRIoYtybnanNThqeOjkKvIzsEtbhb+/Y8+vDU/w/wCr5/5uq+f5if8ANi7m+YXepz23MrU7K6i2H49rde7Gw03gw7bVwlT4MY9TRqAoLxL9Afob3va2wxB1DB6sh8M1Q9KfoX+YpDNDQ7c3yYMVFTRxQmtqbMrqBZnWW/4/x9mtrukkHacjoztdzlioj8PXqzTYfde0t7RRVe1M/S16sFYlJkVeW4tz/rezqDcoHOTQ9HsG4wTjRWh6M5tvtjJYd4RKyyRKVJcHyD/Een2aR3FRVejWO4BXjw6NLtnuXD5qmhgqPEspUKRbSCT/AFv/AL17WJcAcc9LUudS6T0t4Uw2a9cbxLc/QEaRf8e3cHPVxUmv8+lBRbjTZKGWm0SAfRbg3P1+nuyPpzTh1dJFi4dOUvyrosfA1PNSxNMotpaMMTYfW493F7pwR1b940U1Gegn3Z8qquvjkjo6WKMFXC6YyODwB9fbD3dahOmGvyR0TXfnY25c/wCaaXImhp2dibyCPhhYfn2WzzeGpeRqL0XTXJoXkOOPTBsPauU3bWKqSyBXsGr1Zrnnk6j7h3mv3Rs9nDwWREjg0xx6A27c4w2hMcFGIx0dTrv4vbclqKevzNeKuW4a0jAkkC9jf6e4E3v3e5iuSy25KV/l0ALvm6/uGIQlejd4XZ+3tpwLBi6GCN4xp8qG7HT9CT/T3G9/zDzBuhP1tyzKfLoPT7hfXJpJISPTpr3FnRDFIWkOpQVHq/J/pb8e0ltCS2Bjz6YVa9E57V3UtDicpWTTKDDTzzKpYWCKL6uf959irbLfxJkj41I6XQx1ZQB1qIfzAO6ZOxuynxdDX+Sjxhnjq4Yj6fJr0KsnJ/oTb3k9yttabbtyrTuOepBsLcW8IHr1Xv7E/S/rsC4P+Hv3Wiadde/db65caT9L3/2Pv3WvxddEW9+68DXrxIsOLW+v+Pv3Xh11791vr3v3XurNv5Rv/Zb/AFh+f3ajj+v6ffj1ocOjsf8ACgoW7/6z/N9pzG3+F4Bb37z695da+bkFmsLcnj/Y+/deHDrj791vr3v3Xuve/de697917rsgj6+/daBr14i1ub3/AN49+68D12ORa3J/P9PfuvHjXroEWItyfz7914g1669+6312ASbD37rRNOvEEcH37rwNeuvfut9e9+691737r3XYP1Fgb/n+nv3Wj69dfT37rfXvfuvddk3PAt791oY69+Pr9PoPfuvdde/db67J+nFrf7z791rrq5P1N/fut9e9+691zHqX/gvP+w9+6rwP29cPfurde9+691yU8rc8Bgf9bn37rVPPrby/k1/JPGdc7chpKqrEUBoaRGYqWuqxixFiP9b3vjx62D2gdbLdX8o9v702pU4v75YoK2nt59BUi6Wvck8e0m4Q2V3t0tluABgcEMDwOOjnlmXcrXmO1vdnjMlwjLpABrxz0Slt50+N3HOKGo8ieRPtqkXJ5fSDf/be+VPvx7JfSbjNu2wp4tq5JkUcE/2Ou3/tpvK7ryXax79WO4K9ykcccOrU/i1nDmoo5clvCSpqUAMWOlltYab6QGPPvjh7+WMXL25RT2u0qsSt3zquVzSuB+fUHe9O3i2iIsLEKjcXA/w9HR3Rg6Pc+CyeCq5vHDkqdoJHBF1Vx9be4PtOa9kh3y23xpi0tq2pKhqAgelOsbeX72+2Dc4dyhSpiNadVAfKvojb3WNJDLh1WtqckA3qYNIGZtH6RyOT76C/di99N55v3cz39Y4LOQDFdLLx/wAAyOs9vaX3C3fnyOS2vWMAQUqMeXRmf5bezNy9UUWc3juCgehx2WDy0k0nCSgnXdPp/vHv6A/ae4j3TZY97t/7KVe0/l1gZ977mDarnfBs1tceNLAaN5n8+rNNzbspewIKmnp6lWapDE6XFxZbWAJ/4n3KrEFjTrCVE0xhT5dV29pbAjpsnURZGl+8hLtZXUEEMD+be9g46vQ8eiJ9q9cYzCUlZn9tQLiMhTwySCSHhg2gsDxa/P49h7nEzjl6drQkOPTj1L33f02qT3S25N+UPbGQageBGodEZxGa3NSZKHPz1zyZ+IyNHXjUHRhIdJAJ/HB941bdNu9vfap6lSfM9drvcLbvbzdeX2s9pCxxoFoijB7R1J3JuzcG5K77vctbLkpVZSrTAsV4+q6vyPaXeNzmF9rk/U+3y6NuQuSNpXlWOLaE+lYjLDievYVY6zKUkEB0TTnSjqLOpNxwfZ1tV/aXYHhAI1RUfmOol91uX+Y9p2i5S5kaa3AOSTSlD0ZDpboPBVu7q/LbpC5clg0MVQ1xGL3AX/D3n/uc5PtttiniF/ydcOeZYI/67XzIME/P16tg6g2fQYHM0R23jI6GJGQP4UUDSFA/AHuI1OpR1UADFOrO6PeWP/hVFiKipQ1bRBCt/WW0WI9uAmlD1XCnosfeOwsBvLB5Smr6aCrqnhkFKrOpIYREDSP9f2mu4I7q0eJxqPl+zo65a3a72DmW1v4HMcQYFzTFNQ61rewupclt/snKYnLUv2uOq5mSgVT9H8xvbTYci3vAf3l3e95ELXkqVZmOlT6fLr6Efafn/l/efb20ueX5RJLGgMtB/RHQibV2bj9o1EVZFGsktKySLIfUyafV7wN5v503PnFnS7YhHqAvkBw6c3nfbneomgc0D1FOja5D5s0MWyptiPTxLUfYtjpqkITaJho0kf7zf/W9472X3Rt7ueaYueFkleDxPGjjLdganEfl/l6hC2+71I/MK8zIxpr1gfOteitTPBkMNW1ETfs1KljINX9q7ce5wEdxt26R28uJIzw+zqZYxLa38cbjuTy6TlDszZEe0s9VU+Vjmys1PIzRSSLqRyCSoB99Xfu7wWu7bTb3G5H9VUqF9TTrDr75HuB7h22wNtkFqYLQ0HigUx69aY/8yvYE2G7TzuaEJCS18jawLhkaW17gf7H6+8sGGk6RgDrk2jNIPGkbWzE1b1PVXNz/ALf6+69O9e9+6913fgC30vz+T791qma9de/db697917rkAW+n4/4n37rRIHHro3HF/ofp+L+/deFOPXri1rc/wBf6e/depmvXrEAH+v0/wBh7916vl11791vr3v3Xuve/de6dcJmcjt7L4/OYqoamyWLqoqyiqFvqinhN0cWt9P9f37rRGKdbVv8s3+bLh85R4HqHuLIUmKydO9Pjsbkqs/uZF2TSpErsB+D/iPz7E+ybw0DCCY9p4dCbZN5e3kEE57fI9bROyMhSZakpsviaiKuxdUqNFURSLIjXF/7BP8AsPY+jpIA4ODw6kaORZAHTI6Mzt1HKRywyW9P0HF+LEc+27niQ3SxKVx0I8WRVY1hnYBjYLcgA+y0x51DqzL6dTxS00sOosCWvYA8ci/9fdNbA9apjpPpEVqtIFgG4+v0B+ov9PagntqerGhFBnpdp46bHtOxF0juSbcf4j+vtCas9B03XuoR0EdU+SzuRKRs4gWQLqBIVlvcfT2bJ4UEdTk9bOcDAHSibbki0piUFiUH+H1HJ9sfUgtU9eAFekpWbUkqQaZlL6mIKg34v/iPakXKgV62y44dQI+o6eQi0MIJHJd0Tg/1uP8AefbRv4x5daEOKnz6Lv3p1Ns/aWIqtz7pr8ficXQQTVc88lXTkJDAPI7lSQeALe/C5tpEMpwF416RXqQwprlI6op+Zf8AOu6i6X6lyXXfQOZxOf3LWCoxOQqaKJDXUmlTEZFKte9zweLfX2Gdy3+Omi1z5fZ0ENw5njjh+mtBU+vWmL2v2hunuHfOZ35vGtlyGYzFTJLLNKzNIIzIXjQsxJNtXJ9g+R2kYu5yegNJI8rl3NSerDf5TlVkYe+oY6Oukpo2rsMZY1ZtMmp5lIIB/I4P+w9h/mQsNomocaT/AIOi+/qbdh8j1vM0TDXCotGRQ0xbTwDaIfX3hPdD/GXI/iP+HqK2Wkh66r6uyWW5Y+kNwbfn/fH3WPLZHVgo6baLDyVrmV7kCzBm/tc8W9umUJ+fWmIpQeXT89OtBGCyrdA1zb635+vtmpY46rXyJ6CbcGRbJ5KGhRiA8mgn8KDx7XxxmJC58+nUDAVOeqLP5xHywXrvr2HqjZuVp2z2TSPH56GJy5WGYNrE+hgSAAfyPx7nH2u5aZmO7XanjVK9C7YLJmJuJBSvDrU0dzJI8j/WR2dvr9WbU1r+546F/ljribE8fT37rw4Z669+6312CB9Rf37rRFevW4vf/Ye/de8+uvfut9e9+691737r3Xrk/Xn37r3XrkfQ29+69176n/iffuvddkc2Bv8A4+/darivXiCp/wAf98PfuvVBHQpdP9wb+6O37gux+ttw5HbW5sBW01ZBX0E7xSyGCYS+N9BFwbcc3B5B9760R6cfLpf/ACo+S2/Pll21kO4uysjV5bdmUx+NoMhka6Yz1dUuPjMUTyynkkDi/vXWySePVxH/AAnW+L+C76+VO9N7bvwkNZtjoTYdb2zXZKtQClx67YDZMyNJIpBuI/0ggn3qvl69WVCSGHEdXLfLj+d7/Lf7F7/yPZG8PiZ1V2rv3aUUGJh7Cy0Uxrqqp24PtKUvDE1n0AKq2F7AD8e9aSerGVAaGuOhU/mCZvrP+cf/ACk9p9pdM7exGwt29K1ee3PmdsbXgUUhwuGpHqIZ6inGpxGgSxBN/wClj7sAPPy6qxBU+GMnhX19OiAbXz9TH/wmm2kaIyU8lP3ruLDVZKSRs4paQRsrA2/I91BIbqzAeEfsHTP/AMJt6mop+vvm2IJXQVXxq7S86prPkV6JtWq3+t783kPt61FwNPUdEN/kdxovzn3oFGnQc2FPqNlGUlsL+7HyPVMcPl0KvyY+Q2d+MX85an33hEWeDclfs7b24IWBVnxNfkxRz6SLfRb3B/Bvfj37hw69QlqHhgdbEO4/ihtn4u/OP5K/PlAKjqHsn4n5Si2PlpZYpMTHvHcu0WZIadraTLBOQQQbgg2tc39Ty6uKLny6qW/4Twd0ZLtv5MfNzYu5qunO/wDd3QHclPt/dtRPHT1uQkqY5YKTCLNK1y0pQqFX6gi1+ffjkAdejOnU1eJB617OovjF3pVfOvaPTdPsLNHsTDdy4PIZDDGnlWSmoKTd8VY+QeS3+aZBdG/JNv6+9EY60CNf59bs/wAys7h+sP5vex8VvbJwGryPW/W+2KWornQfbZ3+79HReDVJwDHL6b/4e9r59bfiqjiQetZD+ev1vu/YX80fJYHdEdW2brqrY9VBWkSzGqirqiGfHyU8ouHCrIpGi/HvwpXqjA6fy6tk/mr9eb32n/Jw+DnVm9mmXcm/+4cbR0NbkldmpaXNtHFSzsHsQqhh70xoerL8HzFB0s+zNj/Cv+Sx8atgR919G7O+XvcO7dt4DOUVDvGKoqcdRwZrDR1VPDNPTsqBQZOeQBbgfRffqY60rAN6D/Ceje7B+UWG+YH8ojtDtLY/xi2j8btlUC53DjD7JpJoMLPVCJljam8g1SKR9JPz78BTrchNO7NR1Vf15Cct/wAJy49qxs0Um6vl3ufDJUgEGneppKSMSBjwCNX1Pvx+IH59bGIDT+EdD1sPoP4efyqP5eezO+/kr15tz5Kbo3juEph9hbiheroRJU0vnp4J2pCqqrs4UC4A+pI+vvZBx1VNIqx4/wCU9WM/yxvmjtb5ofGz5Vbg6x+HGxvjF1zh+pt149MhsWgmgopKunopUSITSFzJqAsWDe9UwT1vUNYrxr+3r50u+rDe28QDcf3q3DY/1H8Wm59+HDrx49Je1jzx+f8AX976rXGOvEi/0+n9Pzz7914A069wx/1PHHv3XsgevXRN/fuvAU669+631737r3XZFuD791oGvXXv3W+uwL35tYE/8a9+60TTrr37rfX/1NAC5tb8fX37r1PPrwJH09+68RXj14WN7m3/ABX37rRqOHXiSfr791sCnDrwvfj6/X/be/daNPPru/BB+v0vYH37rVPTrj791br3v3Xuuzbixvx791ofPrwt9CePx/r+/dePqOuvfut9chptY/X+vv3WjWtR1x9+631yAuOPrf37rRNDnr3qUf0F/wDeffuvYPXH37rfXvfuvdd2I+v9L/j37rVR1fT/ACXf5Y8Xyy3vVd6d1zvs34xdZTNkNzb8rKaQY1MliT9+mNWadRG0k3j8aKGP1JP096J8hx6sqBj3cP8AD/sdOX86P+Zz/sz27KH48dJwU+x/jl1UibXoNn4KZRh85V7fcUUG4a5Kc+N6qYRiSRiCQTbj36g4nrbSVJVeA/n/ALHVApIIW31AsT/sfe+qAdd+m/JJ/wBvz731ru6Xm0u0N9bHZP7tbir8dFG4YU8Up8LWbVYr9bf7H34Ejh1ZSVNRjqyPoX+Yll8PVUGC3zQxVFJIgjnylROuhSv1YtIeP8PZhDuE0TDOOjCHcZYqBuHVxPVHfvXW+6Skrdt5qlnmm8bOqVMJVJHF/GCDz+fYhtdxiloK56EMG5RSqCDno3uA35JBEhSpitwQVqIj9D9RYn2bLMtcGo6NY7pSuDjpaf3xXIozVFRCQy/TzxW/wNr+3TKpFAf8HTjSKRk9B9uLMYCginqKh6QyAFixqIAR/sNXtqSWNFqx6akkjXuY9Eb7h+V3WfXMdQ1ZlqOKvjilaOlaqgVHZBdR9f68eyO53aNAVj49E91uscRomeqg+wfnfvbs3fOLwu30fF4qXMU9N5oJvTPEZAplVYybg/4+wtu9/PcWbgmlOg3e3880TCtOtmv41UssmydmTv6p6vDU8tQ5+ryMoLMf9c8+8Rt+KrfzV46uouvQvjOxznj1YntaBKanjZgA2n9Vuf8AAj6+wTdVLVHRQ5DGgx06ZWsljVijXHP5+nF7e240DCp49a0kZ6ATfG5FxlLPLM4ZyWCozAfi4IPs5tIdRp5dKYUFc9UwfOT5Lx7A2XnTFUxvlJKaZIaRJ4wzLLdQoF/qT9Pcv8k8u/VTrPMvYuftp0J9rsDI4kPw9as24c1U7jzeUzlYT9zlK2askBbVpMzltF/6AcD3OyIEQIOA6FyjSAo6ZfdurdcvUvH0B9+61g566sbX/H+uPfuvVFadeNvx/QXt/X/Y+/deFfPr17nk/wCx9+69wGOvWubDn37r1aDPXuLf4/8AG/fuvZr8uvWNr/j37r1Rw6s2/lGf9lwdX/18tRb/ABPpt78evDo63/Cgu57/AOtLn/mE5v8AYD9n+nv3W/IfZ1r6Ecsfxc/737916vXH37r3Xvfuvde9+6912LX5+nv3Xj8uvEk/X8e/daAA669+6312L/Ufj37rWOHXf0BBHJ+n0NvfutcTUdcffurde+nv3XuuySfr7916lOvG3Fv9j791oV8+uvfut9cjpA45Jt/sP9v791XNc9eJFwRx/vHPv3WwMUPXH37rfXvfuvddm349+60K+fXXv3W+uY0gfkm3+8/7H37qvceuJNz791scOuvfut9esef8Pr7917rkPpcf7H37rXnnrj791vr3v3Xuu7cAj/W/2Pv3Wq5z1tdfydehcbvnab1NWysUx1K6o12PriBAAHvwwetUqo+3q9ndmx9s7XxDYqkqxFVUkJjdFNiTpta3sF82SC7snht5NLD06yX9hLG42TmS13Dc7ITwswIYjyr0WwFqSvhYuXjR1BLfVeQRb/b+8XOd4rtdhurONy7upoPXHXZjk9No5isBPFZLHp4U+XRvdlbsyODpaTI4CtaGoRA7NGxDA6ebge+W3NfLtruVzNtu+QhlJIKsMcfn0R79sdnfSPZ7jGCnkCPLoVD372aTf+L1JP8AXXJwbWuCT/h7joezvt+MCyj/AN5H+boHn205PIp9Ov7B0ishuzcPaG5cXid25KVaSSaJBWSs1ogJQb3b3k393b2h5Fk3+KyjK2ys4JQUAbPn/l6D/N+j295cuLrli3BbST2jzofTq0Siy8OE2Bg9qYrIiso6GDR5wysZFKW50f8AE++2ewbPY8u7LDtG3AeFFwpw64189bze8w81XO67khSWQ5BBH+HqPtXPVuNrEkjqnRVPALkA6uPz7NaGnQU6FXsNcRV7WTKZGWKKaSEN5RJGzFivHCm/19tzSx2665TQdKtusbvdJfAskLt54PVeXcGClqNp18lETUfdwMadlF9XoI+n49ot3X6vaXEIrUY/Z0L/AG9YbJz1btuJ0CJhWuKdw8z1VTl8JncJOoqkVBIW0XV731n6j3ixvlrvlvdtqVlWuOu6ft5zD7Yb/sMQjkSWVQNdSD5DpoVJp9TVBUOLkfUXuLj6+w+bK4lDS3JIp1J6b9tdq0VjsgDqeIHl047N1LvHFKvrUyc3vYesXA9+2ud7e7Xwhq7hn8x0W+6W2QbvyLdC6fwqI3/HT1aH0fh5cnuGeFImYEKBpQtzx9Lf8U99K76QP7a7YzihK/5OvnC5tgFtz9uNrG2pUbj656tc27tam2jg2ylV+1IkCuLgKf8ANgi5a3uKk4Y6Rn5dE/7S7/qdhb1xE7vqx9XLcyl/TGn04P09l247kNvlSN+DdDvkvkiXnWxuZLckSQcAATWn2dFY7F+ZWawm56qqpas5CgrpP2EaQskKlSCEsfYM3HmtNqujcagV9Cw9OsoOQvYCTnfbI9rkiKSphjpNTkfLomknaFX2JveuyGVBkSWYyUmtriNi2o6CffPb7y1zu+9zpuRctCCaDyANeulfI3tNb+2HJ8VhZPVmUax+Xn0Jk/qhmRmsHWxa4H9n/H3hdHiRSBw6WR9sisPI9BhitqYFq/J1lS6VLFryBmB0WP09yluHOvMk222e0WQMQQUXTWrdCjc9/v7ayjUdiqOPU/ce4qTbWOgipohLRVA0EC5UC349oeWuU9x5r3eWK4YrcJk149JNis23+U3GvuGegDztTPk6esOLnagWpR2PiJ/pcg2+v9PfRD2WFzyTBHaX51kCgJ6h77yHtvec9cqS2EFwS0S/DSvDrVv/AJk++Mku7MhtiuxJfTNJTrkXRxqXV+tnP5/p7zJtphdWyXa8HHXDndttOxbzcbBJ8dsxBr9vVRvt3pL1yBW1iPfuqkHy66Nr8G49+62K+fXh9effuvHrx5JPv3Wx14Ej6H37rRAPHrom/J9+63w697917r1zx/h9Pfuvde9+6913Y3t+ffutVFK9etY2P+x9+695VHXja/H09+639vU3HV1bi6ymyOOqZ6KvpJ456Osp5DFLTzxtqSRGXkEEe/AkGo61Wh62P/5Y385bPdSTUPXPdE8OR2+iQwrnMrWgER6hFrTynhwOSPp7EW1b7LZnRL3L8+hHtW/zWVIpu5PLrb36Z+WvTHceLo8jsLddJXvWRoyQJVU6qCYw7ALqvwT7GttfW1+AY2GepCst1trmMOCK9GLq8lW1UaaWUaVBV45UkBB5BDIbezBEjB9ejHuNCDjpabdralqdBUSFmv8An62P1BB9obiNA1V6sDXB6WiwROwlH9ASfqSR/ifaLUeHWyCBjrlkhPWRR08N1Umzn+oHABHv0elSWPWh21J6kY+ghx8asyhbWLMdIF/yePdZJGkPHrZB8vPrBlNyUGOhkmnngigjUl5HnhQqLXFtZ9+SFmNB17tjUtKaDqpj5ffza+hPi593TR7kxVfuSnsj46rq6ZVE7XAj16rDkfk+0N5uNtbLoYhnHkOHRBuPMtrY9kfefTqgbt//AIUv79qqurotr7Fxk0B1xw1GOqxbSHsrlw9vpz7J5OYHI0pGoHQSn5tupW1RLTqnf5R/zZ/kl8kYnoX3LmNsYqoSeGqoabISvFLTTXvDoDEW/wAT7KrjcZ7gUJoPQcOiW83e+vcTNj0HVWlRUTVc01TUyPPU1EjzTTyNqeSRzqd3J+pJ59oOi3rGG/rz/T/D8e/dVI9OrUv5TXPfsAAveuwt/wDW1zeyHmSg2iYn+E/4OkW4Glu32HreTLqGTizrQ0zX/FjEP6e8KZ6/UPT+I/4eoteoc9RKZ/PVDWS0ev1XtcD8Ee/HtXIz16pAJ6EilolsI41AjCjT/j/sfactTLdMPU5HTduKOOlx83lIDGNtF/qTY3sPe4jqcMvDqyDUcnoinyM7Ywnx/wCrdzb+zVVTrkJ6KY4KKaWOJpKtVNgusg/09jflbZn37d44DXw1Pd0cbbbteTCP8I4/Z1ol/IbvHcXf3Zuf3/uGWS+RqpRS0vldkip1kJjYhjbUb3P9Pp7y4sLKHb7VLS3FFQU6keCFbeIRJwHQFe1nT/XZt+Pp79177euvfuvde9+6913Y2v8Aj37rVRWnXamx/wBiL+/deIqOujb8G/v3XhXz669+631737r3Xvfuvde9+6912P8AD8c+/de68ST9ffuvUp14Ej6G3v3WqA9eALEC/wBeLk8D/XPv3XuHV1Xws/nB5v4T/HbsPpPr3qHb9RnezdoZPZe6N/TNGuWqcRk4nhkgR1IYKA/096p1sEjh/qPVMeTq/wCIZCvyDIEbIVlVXFFYkRmqnafx3I5tcj3vrWT1bX/Lc/m39l/y9dt90bIx+zsT2ZsnuPrvcHX9btzcbs9FiafcVE9HU5GjjJAEqa9ScWP0PB96IJ6shCVNK9KLdn83rcO4PgVB8Dsf1Pt/DbJpt+5vf8W46bQMs2SzUrSS08hU28a6rDi/vVM168xqhX16R38ub+aru7+Xpt3uTBbX67we9P8ATD15uPrzJT5kg/w3H7ihMU9bSKCtpY73Qnj3Y568h01BzWh/Z0B/wp+cmX+G3dme7lxez6DdVZnfvX/hVZLphpzW1T1J9Rt+kvYe/dUOel72Bv3u7+Zv8waLsLq/rVxvauqMBG2CwtpYqEwVTLBU6UJchrWGkE6vrYc+/cMnrZBIoOPWxt/OR+SnZvSn8qD48/EHeeWq9s967f3NSVm8aJapBlDtytphHHSVQDGRSvAAb6D37zx1agC0NMin+z1r/wD8nzY3ce/vkzlsj0vuHIY3fm1dvvu2Giop2im3OaOeSolw82krqWYghl+hv78QD1oEginDrZE2D/MZ+aW9/kf15gYf5TXXO1u38fvPCYHeHcOMxM8O5sjtmjykcDZPJVDylVNl8rPYD+p/HvVaZ6tQHtzTqu//AIUwfISpxX8ybFV+ws8Wym1cVtbdVZWUNbHKYcwaWGaoxpnpmItDOkkf1v6f8PflrkevWnAbSR+EY6bMV/wom653Ntjbx+Q/wE6h7+7Z2tjcdQ4PuHeTGfddI+KpxT0MjSF7OEUWGrn8fT3oivVlc0o3RXf5jf8APT7F/mEdIdVdK53qbbOwcd1FvSm3dtfK4Nz91D9pKHgoD6iAiaRpsOP6+7ZJFfLqlFodOCfPoc9s/wDCgPZu4OpsB1d8lPhX1r8g325RY6gx+7N2v5c39pjaNaOGildnAMWhALf7x719nVgcd3Hp03X/AMKNstWdO7w6A6/+LeweuOn9y4GuwtFsnBsq4zDS1cJiGTgXUNUyXDAtfn34VGOtE1ap8uq+cD/Na3lgv5ddZ/L8puvcGMBL2ple16Lfgb/c9QZvIpHElNE5P+ajWNSPTe/v1M169Xs0/KnRpvjJ/PQoeu/jdR/Gv5PfFvYnyr2rjs0Mxj6/fMhevpQkfjjpIZGYaUIABFj+ffjXrYKiny6MVsX/AIUs5HpzHdj7S6S+I3WvXHWvYGx8psk9fYxmOCxMOTpWpWydPGrjVOga6lri/wCPfs0p16q6tRHDrV53DlDnM9m82YlgOZy+SypgU3WA5CterMIP+067D3vqtamvTVcsLfU/1NuP9b37qvA18uuHv3Vuu+Lf43/3j37r3XXv3Xuuxb8/04/3w9+60a+XXj9Tb+vHv3W/LPXiSfr7916lOvcWP9eLe/da8+vA2P8AvH+wPv3XiK9eNvx7914V8+v/1fn/APv3Xuve/de697917r3v3XuvfT/D37r3Xd/9j/r8+/dep117917r3v3Xuve/de697917rkRzZeffutA4z11yAbr/ALEj37r3E8euvfut9euf6+/de67uf6n/AG/v3XqDrr37r3XYF7/4C/v3XurSf5YP8s7s/wDmFdz4fb+P1bW6sw8s2W3pv7JoIsVQ4nDMtXkYUnmAQkxhlY34PH1+nuJp1rA6tK/m0fzKurOquvaL+XZ8AXXbXTW0KOPH9k7kwsopP70b0oSKLKVrfbW8jMyPZybAH377OvNqPxf8V8utXKaV5HkeSV5ZZXZ55HYu0sjNqZ2c8kk8kn6+/daH8vLrCATwOffurVpx66PP/Gvfuvdd2Fib8/09+61mvXXv3W+lttPsTeeyGZtr5+txN5BKVgk9BcfnQ1x/r+7KzLlTTrwJU1HRrtnfOztTblMkFfWVmUZQoMhqtAaxuTpI9vx3cyDjXp9LqdOB6ENP5j/YkcTRrSVl2BsfvUFifp9B7cN9P6np/wDeFx69AvvT5pdwbrSeODOVuNWa4NqgylFP1CggD6f19sNcTMcsek8lxNL8bdFh3FurcG66z7/cOVqspVhdIlqJC1geSAPoP9t7aJJyemak5PT11nf+/O3Lf87Olv8A63nW/tFuFPpHr6dMz/2Z63jvi/D5th7EsLkYOmBsR/qFt9f9494ocwqRuM5B/F1HV8f1HHz6sHoYIaemiBb6J+SPrYcewVKWdyOirFekpuPJLSQzMWAtrt9OQFuLe34YtZqOrjLdVw/J3uCh2TtXK5yvqhDBTJKiszoLSCK4Fj/j7H/K+0Pf3qwgY4no5sLcXMgQYp1qFfJLvfOdzb3yVZUVEy4emqZIaSDyllqBG9hNIBxb/Uj/AGP+tk1tu3w7farBEKUHQ/hiWGMImOi2+zDp3r3v3Xuve/de65f2f1f7C/8Aj/T37qvnw66vxbj/AF7c/wC39+63TNevEAfQ39+68K+fXr/S3Fv6e/dep69eAvyfp/X/AI37914mnDrx/IB4v/vjb37r3z6s1/lG/wDZcHV/5IlqCP8AbL78c8OvCnR1/wDhQVz3/wBa/X/j1Jhz/rw/T37r3l1r6NcMwN/qeP8AY+/deFOI64+/db6520gG17g/X/W9+6rWpp1xsfrbj37rfy669+631737r3Xvfuvde9+691ksv5b/AIn37qlT5DrGfr791frlp4uePr/vHv3Va5oOu+DybD/Ace/deyOuJP4sP9e3v3WwOvX4+g/1/fuvdde/db697917rx+vH09+6912T9OB791qnXib/gD/AFvfuvDHXXv3W+vW9+691737r3Xvfuvde9+691737r3XIhR9Df37rQJ8+uPv3W+nbC4PK7hyEGLw9HNXVtQ2mOGFGYk/1Nvp/r+/de628/5L+A7J64x32m6cdNRQ1VLBDAriUAhUWwBYc8W59+BArUdVoSykGgqOr19z/Hvc27qvJ7ogrCKeoJmEOp+AEuRYe47v+Wdxu7tpkeikk06zE5S98uVth5et9ruLcNJGAC1BWvRON67Xr8BkZMRUKVm1Aajq5P0uD+Pp7Ae4ckXN5dFqV01x69Zfchfek5d5Z26KS9/spPs8+lZtrcsu06AxVis6yKtmIZvxz9fx7wa91fZDmO6357uygojEnz49ZQ7HzVy57nRR3+xTAlhUrUV9ehGwO9cfmIncv4ggvzYcE8cH3jtzHyNvfLcyxXkRGrgc0PS3cNkms5PDQhz50PDrlu/KxQYaWSgqGgrWVTTzr+tSWFmBHtZ7cpuVtzbavbkqRIvCvr0kstpt7+Yw7nGHhzqB9KHow3S/YtXtehw9LurKyVi5OMGPzODov+QG+nvsnydu72uzwfXOSz+p/wA/XML7wPtnb7vzLeTct24jSA+QOf2dHJ/jGLz1G8WCrFermUeLQRqVtHFtP9PcjC4iuIysBqx6wxl2jcNkvFk3KOkanNft6C/ZM2+qrfL7U7CqKin29VVDQUs9QzmMIx9BXVxb2DW+s+tNveVCk4r1kfanlr+rCbry4qtcItXAwa9Cnv7rKv68wm4dy5SQV216aN5cWzkmNowhb0j6ez6EvtdtJPOdUa8Ooq3RLfnXdLXbtrHhXkho9ONeqYezt+43c+bQoq0kTznwIoADKsh+lv8AW9gvfNyg3zStrHUqOsn/AGq5N3r2w8aTf7xgJfhBJ/y9BtX+aPgJpje37vJNj9P949wZzH9TFOYWUoB5evXTv2jj2fcNmTcraUTyMATXiOpWzJI4N5YYyvphMo1m3BBf63/33Psp2q5itbhTIKjUPyyOhL7o7Heb/wAl3dvZOVfScevaer+/hXszDZzL1+TSaGeKkRXnDMnoFgSf9h76PTXiXXt5t5QdoUU+ynXzjc8bXe7N7gX1leqfE1HyOc9Gf747D2lQw1GGpMrFE8do5Y1ki9OkaCOD7jGO9shVS/w1rkdOjlPmd0SSO2JEvw4bPVGPya7Pod45BNqYH97I0TmNZktdAX/VdfePHvD7nbXtFu8qyU8EEeWT8uuo33Q/u83PLG0tzXzaoEdz3BGr+yh6CLEbGNThljzknnqgh0eS5ZWt9QTz75sc1+8/Mu77t4ttcOsII7QcU+fWXkUWz7LuzXWyQiNSc0HSOi2JkcJkWq4SY4QfS4YgAXvcA+xLuXu3a8w8vjZ7qHVIBTV86dD5+YrXcLbwJMt5jrLu/c+QpoKaOkqHeQWWUhueODe3tb7Te2m3czJdX25gIkeUr59Em1pancPAmjwxweg8pM5kllmAqHX7oEuAx/1XII/1vc97VyJy5Jf2jGFf8WOMccdHXOm3W45auWUUaNMH9nQhJmMdmMXDhqqMGdBaJ3HJJHP1/wCJ9xNzbyBzfs3N97zVsERW2U6iFrw9KdRfyvucXL+3Ws8ktTNxyMdJ2l2RkMjLU0lBqIUFUsTbn+gH+295Qe2m17pzvtsN2ylHUVYeeOo89zfvK8rcjJcW12od5BQEgeeOqf8A+ZX8dcbWbYy+UbEwNl6fHytJMyHyeQLcNcc3v7y42u3ay2uGzc1KDJ64r877pBzFzxuHMVqKR3TlgB8z1qzVdLPQ1M1JUxmOeCRo5UP4Ye1nQe6je/de69/h7917r3v3Xuux/SwN+P8AkXv3Wj176X4/2/v3Xuuvfut9e9+691737r3XZAsLHn8j37rQrXrsD6km3HB/r791onyHXV/r+b/19+63Trr37rfXfJ45Nube/dax12WuOSbgADk/T+nv3XqZ6MD1b8n+6uoa3E1Gzd9ZvG0eLlQrjqeseOCWAcNC1ubWHB/H+t7djmliIKMRTp2OaWE1jJFOtqD+Xp/Pc25WUdJs7vh1xL0gipI8rkJopnqj49IcSSHn/Yfn2J7DmF1UQ3NSPXz6Fu2c0yQsI7rPz62d+ofkp0j23tzH7g2rvCmqIa6JGQGamQDUvPF7jn2IEuhc90BBH8+h5b7jZTwh1cV/LoxVBWYisiE1Hl6OaIWbV93TC9+Rcavd31KaMOPSpZEZaocdQM32BsradPLWZ3PUtLBENRYVVK3H1/1X+v7bZJNOoU/b02bi3T+0anVanyP/AJuvxf6NapoZd70lTloUcihkamtI6kqI1IP59oJtwsIRpdquPIdE15zHZWDUU6+tV755fz3OyO4KjLbP6tjrdt4+N5o4s7jq0U/kVriJ1Ed9QAI+lh+PZDfb5PP+nD2qPTj0B9y5jur0kRdqnrXt3x2JvbsfKHNb43Hkdx5OQuzVmQnMsrNIdTk2sP8AePZEzMxqxqeg6TqYscnpEi9+Bc/77+nvXWjSmevEk/Uk/wCuffut9d6fTe/++vb37qtc066sfrbg+/dWqOrVP5TRYd/0wXm9fg7/AI41z+yHmQE7RN/pT/g6Q34Bt2r/AAnreOqrRSwsPp9lSggmw/zQH094VzCtw/8Apj/h6i0sNZA9euWPXyVN1UfWw0jjk2PumQuetHhXoUqWRaSC5UFrf70L8g+0jAuemaavl0G24c1DVTytPKFpaC8tWzMBHHGBcsxP4HtXDC9QqZLYp59PKOAAyetOr+cf8yKrs3sKo6Y21VTxbf2nkGeWamnCRTqv9kGI3IZl5H9L395We3fLUe0bYt1IP1JQDX06kLY9vW1gEjcWGeqKLXtpH0HP+v8Ak+5J6Pq049eIUfQ39+68CfPrj791vr3v3XuuS25vb6cX9+601fLrj791vru/FrD/AF/z791qma9de/db697917rkQQLEfT8255/x9+61ivHrj791vr3v3Xuve/de697917rs34uLcccfX37rQ669+6313c/1P+3Pv3XqDrwBP0F7e/daqB12CbEaiBa9geCffuvHrq5IsSbf0vx791ug68CQOCRz9Afx791rz67PpPpJ/wBcGx9+68M8ejxfBD5rbj+DnZuV7O2vhv4xlsjiocfT6ahKeSikp5nmiqUkcGzBmBFv6e/daIrjpDfLb5cdqfMLtHMdn9nZ7J5OuyU5lSnr616oxBmuASeGKiwXjge/fIdeyTnj5fLoLemu7uzeg95UW++qt25bZ+4qR4Q9fiKlqWeop45dbUzuv1VhcH/X9+6tnq5et/n1fIGTYtVhcZQV2L3zXYuTGVm/qfKIuVnR4fEWaVFDf4k/X37rXdWvn1SV2D2PvbtXc1dvDsLcmS3TuXInXUZXKVLVFQ2qQylCzfQAsbKPpf34enXjXpCkk2uSbfS5v791ulOuZN15ck/WxPF/fuqiteHXC5P1JP8AS5PHv3Vqddhn/DN/sCfxz791rHXVza1zb+l+P9t791vrsk20ni3Nuf6cce/daFOI68GYfRj/ALc+/deoOuPv3W+vXI9+6913fi1hz+fz791qnn119Pr791vr3v3XuuXFrk8/gf7H6e/da8+uPv3W+vfi3+8+/de697917rlxpA4vf/Y2/wBf37qua9dmy8gg/wCvz/vHv3XhU8cdf//W0AALm1wP8T791omnXXv3W+ve/de67It+b+/daGevE3N/fuvDHXXv3W+ve/de699Pr7917rkByP6fk/gf4e/daJ67uBzYf639Lfn37rVDw64e/dW67uT+T/t/fuvUHXubfm3+xt791rFeuvfut9e9+6917/ff7b37r3Rzfgt8N+zPm933tbprrfA12ZqMhVR1mWNMjKIMRTyA10rVBBVQsepmJPABPvxx1oBmNF/P5DrZH/mZfL3qn+Wp8aqb+XP8LMxik3dmMRQ1PaHbW14VjzUGckjRNybUgycRDCJCzRuwPqcXsRf3qpb5dXokYouT/g+f2nrT1q6upraiorKyolqqutmlqKuoncyTTzzSeWWWV25LMxJJJ5976pxPUX37rfXfI/qP949+69g9de/de697917rnbTyPV/sOPfuq1rjh1xJv+Le/dbGOuvfut9cubWHN+fx791r59dA2BFueef9hb37r1K9de/db6UW08s2D3Hh8oFVxS5Cld1b6GMTqXP+wHI9tyxiWMxt59Vcalp1uLfEfvfD5rYe0pKKtgIosRBDOupQVcAC3J945c3cvXNtfSuikqzVx0BdyspFkYgcc9WUxdwbdTFJJJkoFmKfTVHe/wBPyfccHar3xaCM06IvppNVadF+7Q7px8WPqZv4jBFTxxzyGXyIP0RFjzq/NvZ9tWw3VxMFWM1x/q4dLbaxkkatOtZr+YH8vzveeu662zXrVUTzH76eLjwhVCMCwJ9Tfgf7f3kLypy1HtMAmlH6hHQ12/b1t1DefVQnsa9G/Xvfuvddj+trj/ePfutH0669+6312SD9Bb37rQFOuvfut9cmXT/seffutA164+/db65f2fr/ALD/AI1791rz64+/db6s8/lDwPUfOPq+ONkVvJUtd3CKAoUn1N70a9apjo6//Cg+nkg7+6zLmM6tqT28ciSfpMF76Sbe/fLr3Eda9rG7H/XP+9+99b4dde/de67+v1Nh+L+/da+zrx/pf37rfXXv3Xuve/de697917r3v3Xuve/de697917rl9R+r/Ye/da8+HXH37rfXvfuvde9+6913bi/+8e/darmnXXv3W+uyLAH+vv3Wganrr37rfXZAB4N/futA149eAufrb/H37rxx14/0ve3v3W+uvfuvde9+691737r3XY4/wAffuvdeJub/T37rQwOuvfut9bVP8mr+XJt/sSjHYG7MfDkqeWVKiN6p04jFmVVUoeB9P8Ae/e6Zp1ry1dbPMvReC6wMD4HCQUtLR6AjQBdIVUH6QoHvWKcOrD59LvD9vUlDRviaioSnspjZX0qDxaxvx73ralB020ML/Eteivbyr9pbn3TGshgkmeVQCCCWYm45H+v7qFRTqAz0+1xdSRiCSQlF4D06HTE9Cbc3hi4o5RT06sgs5YKSCot+r639le77Nt+7RaJVCt68OpG9uvdjnj26v8A6raLiSSMcIxUgfz6Zm+HNbTbgxq0gl/hBc+aWOW0Wj6+qw5/1vcbb37T7HvEsS3UCSqvm1D+fDrKrlf75W+We3Xl1uwZrh/hVlyD8qnpL9qdU4rYePysWUlEbqt8dru3lAUlbewHZ+yfKvKF9PubQoWc1Sn4fsx/g6kHlX7xfMvPjwWu1ow1kCQgUIyAfPoleDbcu5c7TYKlr5qiZiBi4VJDQrqtZCPaHaZLm53aS2RyVDUUf5usqfcO02TYuQLfe7uBUcpWZzxY0rnHVh/W+XouoMHUT7kzT1G4ljBhoZS0jpNovpt/X3MVtuEXL0ZW7eshGBxp1zw3XlG993r9W2K1AtEPc4xUV6Ffq7u3bvdVTksRnaqnxOUx72oalgIpV8T39NyD9B7f2/mTb98uDbTuFfyPD+fSDm72R5s9tLBN22iB5belXUVIz8qdCF3h8nNhN1Bneppc/BkMpj6WSnQuis7t4jGQZCb/AI49ucwcybZb7TLYGTUy/wCr16KPan2Z503fne05nSzaKOVq+eM+lOqJJ9vx57MY6WnlYfavJogS9n/cLcge4Xf3C2jla1k3C7ZQoznrPnn72G5g5os08aRovDAzWnl9nS3rcTX5OpixVPSNrQKD9bnSPrf3C/N3vVy9fwfvJHXPDHU6+z/JSe23LiR3dz4ppTJr0mchjZsRkFgn/YqISAHB5TnVb6j+n9fbGx7/AAcw7V+8bFqjB+zI6m2G4S+s2aLuVgf8FOht6G+WW/el6/dGOxUlVk1y0ZgDIhcwg+kBeePeeO/c3nYPZvb7iNhhKsT5dvXN/ffu9bTzp7vSX25IIo1euTTVmvp0kt89i9p5zMT7jym462mp8xL5Fp2D/t631AaS3+t7wu5U9zV5n3K4it5iaE1zw6zIX2s9vxtUVhY2Eck1kMEfioPs+XS22nhaGkoRuavlWrr508stRMdTk2uD6r+8O/drmTdt75vuNkR2aNH0qPXqz315Nbx7NHGYI48BBwH7KdKbG7rpstO0SMf2zYWWwWx/P+v7jjdOV9w2aFZblNIfh8+qXezS2UQkYceu95LV1mMK0M9iiqX0sPeuUZ7Sw3ZJ7+PWgPA+fXthMEN1W4Xj69ALkcbIaAyxzNUTxgtOCSbG/wBLc/n3lxyVzxt0d09pcRi1jnxEKUDfZ0JfqJv3wlYKRng3y6futtg1O5qt62VQlNTBxMGa1rAm5B/1vr7ya5P5TuNzuorhB+nUGtcU6hj7wnvry5yZyxe7THMDdMhCilTWnU/GbSOT3LlcVQeqSmdlhZTY3vwQ3/FPc+3fJ1k1s0dAwIyvr1yj2b7yHM826RRXxcQAniDRRX7ej29U9JphMTJkcs4etlgLRo7qTr03+tv9b25yzy9HsVWs18IHio6Ifd33E27nFhbxxCTI76dVCfzB9kZqTEbylGOknpft6nS2ksukKfpx7FpNanqDUQIAi8B1pE9r0clDv/cdNLC0BSrFkKkceJbWv+P8fdaU6t0HXv3W+uwbG/19+60RUU68Tc39+68BQU68Da/H1/P9PfuvEV68TcAf0/3m/v3XqU68Bc29+68TivXiObDn/W9+69Xr3I/qP+J9+69g9dH37rfXd+LfX/iPfutU68PoeL/4/wBPfuvHrr37rfXvfuvde9+6913Y/W3+8e/dex14EgggkEEEEcEEcg+/de6MTsb5Y/ITrmmhodpdn7kxlBBoEVFHVn7eMJwNKppP/J3t1J5kFEYj8+nElli/s2I6Nttf+a18ocBjpqObsTc1TJJGI1kWqeyWFtQ1S39qk3G6UU1n9vS1d33FF0iToJd9/wAxH5U71R6eXtndNPSyXDqtZIJCNRIALO62/wANPtt765c5c/t6Ty3t3MT4jnooW6t7br3xX/xPdueyGer/AFf5TXzeSQajdrWAHP8Are0pJY1PSbJycnpM2AAYEfUccX966rk464k3/wAB/T37rY669+6312eD/X37rw65fRfre/4/p791Xz643/rz/h791unRwvhL3V/oW7t2tlpo1NDkMzjI6uZrEQRwSOdRUkfXUR9faDc7UXtjJb/xKR/Lpi4jEsTL8qdb8nXW+ML2rtnC7qwGQgrKWvoKMgxuhs3hGoMqk2t/j7ww37arnZtxkhnUgaiQePn1GF5bta3DK46HTB7akjbyykDUOOeBYcew89wGyOkBkFacOnfONBR0rRtNDGSoBZ5VXTxYmxPv0CyzkLEpJ+Q6qilmovVNX8yr5pbT+OHVmYxODzFPV7t3Vj6zGlKYrPPSNJeJHGhr6ifpzx7mP275Mub6+F9foVSMggHz8+hTsm2vPJ4k60ANR8+tIDc24stuvO5PcGcrKivyeTqpqmoqaltUreVy6qf8ADYD3kyiLGgRBQDoeqoVQq8OmH3fq3XZ5PAt/h791oYGeuwbfUf7f37rxFeuPv3W+ve/de67H54vwf8AYf4+/daPXh/rX9+6312GsSbA3/3j37rRHXH37rfXdyfyffuvY67IsBzyfx/T37rQNeuI+vv3W+uzx/j7917rr37r3XZ/17+/daHXh/xHv3W+uvfuvddi/wCDb37rR67J/HHH5459+68PXrj791vr3v3Xuve/de675H/Gx/xX37r3Hr1j/Q+/der117917rvkfUW/2Hv3Wsdde/db67Av+be/daOOvW4Jv/sPfuvefXXv3W+ve/de67JvyBb37rQ9D176/U/j/fD37r3Drr6e/db67Jub2t791oCnXXv3W+u7H/Hjn/W9+69jr34vfn6W9+6150669+631737r3Xf0PB/2Pv3WuPXvxe/5+n/ABPv3W/PrwNvxf37rR669+631737r3X/1/n/APv3Xuve/de697917rsAE8m3v3WjUcOvWNr/AI9+69Xy669+631737r3Xr3+vv3XuuxzwTYe/daPqOuj9ePfut9dkg/QW9+60Kjj12RfkCwH+9+/deGMHrsA2vfj+n+x+nv3WqitOvXueB/sPe+vcBk9cALkAfU8e9dW6FnpfpTsfv7sPBdXdWbYyW69356qigp8bjKc1M0dO06QzVjhfokWvU5/p9Pfutcetvnsbc3S/wDwn5+ILdT9f5fD7y+e/bm36HKZfeMMsYzGxMVuGjDZPbtKIbtEELlZHH+t73SnHrxaoovD/D8/83WnH2Bv3c3Zm787vnd+SqMrntxZKrylbUTyvIVnqqhpmjXX9FF+P6/U839660B/s9IqxNz/ALf37reOHXrcX/Hv3W/l14kn6m/v3XqU4deubW/H9PfuvU8+vEg2sLcf74+/daHXrn+vv3W6Drr37r3Xvfuvddi5PHv3Wjw669+631yvcc/X+vv3WqZ66+lr/wBL8fX/AA9+69x6HfrP5Fdl9YyQphM7UnHQi32DyMEI/Glh/wAT7TT2dtcqVmQN9o6o8Ub/ABCvRmov5ifZ6wiKWKV7Jpv90fqPoeT7Lv3Btta+GP2dMmztz+EdBD2F8yO3t7/tQ5yqxFGysssMUzSNIHXSQS3AH+t7WQbZY22YowD606tHbRR/CB0VSrq6mvqZqyrmeoqaiRpZ5pGLPJI5uzMT7XdKBjrAOPqPqOPfutceHXE/Xj37rfXa/UC1/wDD+vv3Wjw69a97e/db68QRb/H37rVa9de/db697917rIqlrKqsxIv6RqI/2A9+x1XJOOvGJwSCjrYXOsFTb/Y+/dbrTj1xVWa1lLXJACi5JA/AH+v791vpabH31vDrTcVHunY+XyO3tyUQb7XJURMdTCHFm8Ztf37rVM9KPs/uHtPubIUWV7P3Pm925DHQNTUNVky0ssELWvGrEfT0jj/D37HXj0F3glP0gn+nP7bfX+v09+69Q9YxFIb/ALch0mxsjGx/offuvZ66sVALDg3AB/wPNx7914/Lrh791vrsi3+xF/8Ab+/derXrr37r3XLjT+L/AO8/X37qudXXQ4PPPv3Wzw66H159+6314/X37r3Xek2vx+f949+61XNOuvfut9diwPIuP6e/daPDHXR+vv3W+u+SL/UDj37rWBjrwBP+t+ffuvE067tyBquOf9h791r5069wGH5H+v7917JHXuDc/T+g9+69kY64+/dW67JHFh/r+/daFeuvfut9e9+6912PoeL/AE5/p791rz669+631737r3XagFlBOkEgE/0BPJ9+6919BD+StU+PpfFU1LUatVIilRb0/tgC59+PWlBpTz62MMDsSDdNNJTV0ImDRkD6EXZL/n8e6+WercPt6rG+ZHSud2TDlMtt1Z4/HrlRY1Y8aCzAaf8AD3sZ68anqlTEd95HB75p6HOyOtRTVMausmtdREgH592+zrQNDU9W64ftfKbq2TSzbbq3papIUJ8OoHiK9yR/vXsp3iK4ktf8W4j06kD223HZ7DfQd8AMTevQ0dVfJzfGUy2A2Dk8OSilaWoyjxuXcK1g7Pb6n+vsNbVv+4G4Tb5o8VpXz6mrn72o5VbZbjnDbbr4hqWMEU4enQl/KJJsvBhtr0OAGXr8vThI6zRd4GaL+zb+n19v83RyXqDb4ly4w3mMdE/3fLqHl65fmm8uNEVs1TGeDUI6q0xlPkOmuxoMjkIpJarEuR9vIrEg67lf9v7hCytJuWt4L3GTqrU9dQ+bd+2z3z9tBabLpQFRUKeGOhLrO/MZk85W12Y2/FWyV8mqBJAdMDEADTf2s3fna2Bnu7+NWVa5PQP5V+7rvW1bLaHY75rdUA8RR+Ljg9JzD4DJyZbKbtwtZJhPvWMkUMTG0YP1AC/194Rc0+/V/Yb9LFtC0QMQCMdZHyfutNjt+XN3gW50CjlgO77euGD6sruztxyUNJVS1+dqpNEjqZPLK17G/wDvPsP73943deWLP988wALbAamLHFOt3W77ByLtqXwtkggjFQaDHRqdv/CDdO1cZUZeopaqWenXyaJLta63NveNHOf3sdv52vVt7YiO1bGoGin0z1FO4/eN2Her1bCORVVjTHQbTYdMNk5kqKMU9ZAzKxK6W1D0XAPt+O/bcLJWhk1xsPWo6GcV+1/Zq8MmqNsjoOM713Fn8i9bJVOjStrKi1gQp55/3j3J3K/ubfcrbW2228YZTiv5joVbdzO+22ot0QGg6DPaGx6TG7uy0kkjzRwSaijeq5DXt7ye92vejeN49q9o2e1XwhKgDH5U6ANnvaX293LNGA5OGHEdKzdBxG5K2LESTCkWCQAAegDTxYMf9694/cmT8z8qWMu92MBlRx3N8upR2cX21QG+RdZYfb17enk27hsZT0UzS0yIocKb+ZQL249qvb+1j513+7lvl0zzNVT5qetbD4e5300twNLGufTqFsvcdBXLNCKJaJ1JDTn0te319rvc/kPd+WTC89y10jcFIOPl0/v+2T2xWVpdYPAV6fdtVqZfczYGqr5Vo6qQoZ1VjpGqwIP+xv7k72d9k4+bpbfdt7jMEceQpGH+3qKfc/ne15F5fG620ay3CioSo6kdp7Lh66raGTE1MuWgyLqZUkuwYP8AgD/X/HvID3N9jNtvbeGHaGEciAFGQcP2dRh7S/eV/wBcATxcw2y2SwVGrhSmOgmym8dw7Uoq6rxokpUdCz0ygrckW02Fr/X3IntrFu/L9pa7PuTktFgufxdRl94DaeTOZOXrzdtrdby40kg8SDTpcdJbtpaQTbqztQsM84aSRHYKQRyBZv8AH3khUOA6cOuTDJJDI0M4KsK1FOjMn5UUUijH4kJWuo0rpB5NrHgf63vRqMjq6UpwPTfvrCxd07EzMWQweiWpopl1+MnUWiNibj3qhJqercBQY60ef5mPx6peo9/z5angeFq+qWMrpKjxsSRf/W9+6rgcOqrffut9e9+6912ODyL/AOHv3Wjw67JB+gt7914Ajj1x9+63165/r7917rko5B/APJ9+6q3CnXm5P++t791sYHXH6e/db67sRz791qo66ufp791vru1xcc/W/wDh791quc9de/db65EC178/09+61U1p11+Bz9b8f09+6959de/db67txf37r3XZ+i8fj/b+/daHE9dcWH9eb/4e/de8+vWFr35/p7917NevAE/T37r1QOuvfut9e9+6914gjg+/de49e9+6912QOLf7H37rQ6yRyvDIk0LvFLGyvFJGxR43Q3VlZfoQfpb37rwHVm/xN/mg9yfGSI0LiTeOJjCLS4+tqNMcCoLAEPwT/iPYZ3vlPad+H+OJn1HRdd7bBdmrjqxJP+FE/ZiQ+IdS4y4AAb7uLi359hE+03Lx4lui48tWR49FZ7n/AJ1HfHaMFbHiaA7TkqYDFE9FVgLAxPDgR+z/AG3kPYdtoY01EevSy32e0t+Cg9VSdi9v9i9r1iV+/tz1+4J1lklT7uQlUeVrkhf96v7F8NvDANMKhR8ujNURPgFOgzN78m/Ht7q4669+631y4ABB5v8A7b37rWa0PXgR+Rc3+p9+60QfLrogD6e/dbHz669+6314e/de65kgCw+v+qHv3VaEmp64e/dW65EgjgW9+60AfPrj791vr3v3Xuve/de697917rkQLgA8cc+/daBNK9ea3Frf429+60K+fXQ/P+t791s9de/db67tY2Pv3Wq+nXrcm34F/wDYD37r1cZ669+6312ACeTYe/daNQMdde/db697917ru5ta/Hv3WqDj12CBfjn8f4e/deI68wYAXP8AsD9ePfutClcdcffurddgX+nv3XiaddkC1788cce/daqa064+/db67II+vv3Xq169f8fj37r3XiQTcC3v3WgDTPXI/pB/JPv3Wh8VOuIIH159+62a+XXiSfqffuvUHXrn63/w/wCI9+69QdesLXvzf6e/de8+uvfut9dgA/Xj37rR+XXhYH+o5/417917JHXXv3W+uxb6n8Ecf1Hv3WjXy68bEj8D37r2QOvEAHg39+68Pn1//9DQAFr83t/h791o18uvC1+Tx7917NOuj/h9Pfut9dkEfX37r1a9eubW/Hv3WqefXXv3W+u+VPI/2B9+61x4de4t/jf/AHj37r2a9de/db697917r3v3Xuu7m1vx791qgrXr3qt+bf6//Ee/dexXrwJH09+62RXj0tuvevt1dn7rw2zNmYubK53OV9JjaWnj0281bUCniZifotyLn37rRPW5f1L1z1J/wn9+KJ7v7VxGI3N85uwcVKNn7LrUpUyGG29n8aUp87PJq1osflJhj/wub839Whx1bwyFq5/2fl/n60/u9u8uw/kN2duftbszP5HP7m3Nk63JeevqjUmhgq6tqmLH07OOIog2lB791WteHQNkkkk8k8n37rfXrm1vx791qgrXr1+Lfj37rfz669+691737r3XICwv+fwPqPfuq1qadcffurde9+6914C/Hv3XuudwAeBccDj+nHv3VaVPy64e/dW697917r3v3Xuve/de67uf6n/b+/daoOurk/X37rfXZtxb+nP+v7917PXrk/7D37r1Ouvfuvdd/SxH++Pv3WuPHrsWsb3/ANh9D/r+/deNevaj/h/rfj37r1Oujf8APH59+62OvC9+Bf37rR6vN/4T4dM9Vd8fzEupOue4dl47fezc/kpqWvwWTjD0tQDSO8Zm/OkMASARf6e9HpyMAsT6Do/P/Cob4O9RfH/5F7Vyvxb63xW1OrcXs9Y93xbeUmgxu4lkvUxSSC9rWtpY3B/w9+A9OqEigY8T1VN/Je+O+C7/APnV1Ht3sXbAzPU9VkpIN2zVsY/h8dLNMqRq00no1XUryfezUCvWhQmh8ur1v5/HW/xL/ls/zCPjvurrn457Ty3WuN2hja3cfWpGnBbukKCdqmqkJ1WlQWbS31+hF+KDiQOnW+FWOePQn/y7P5mf8uv54fK3qz4obh/lOdJbPxnY71NB/fHAVeTfNUFbR0TVLVEkU9Q0ZjOk3AX/AA4497pTPW9QIp01fOb+aT/Lm+G3eO++g9v/AMpXpXdFNtbLZnARbmz1Xk0yE/8ADpTSCth8FSqatZDWKW/r79pNOtCQA6M448Oi6/youm/jZ8l/iH88O8t+9J7ebcdFuSsymxqZAs1Ps+hqi0i0FC7KtwoIAuv497H+Dqp9QONa9arPc9FRY7tTfdDjqdKShpdwVkNLTRqFSGFbaUVR9B72em1+HoMgL8D37q1ade+h/wBb+vPv3Wuuvfut9d2Nr/j/AFx791qorTroC/09+631737r3Xvr7917ru5HH+8H6f7b37rVAeuvfut9e9+691737r3Xdza349+69Tz68CfoPz791rHHr30uP98Pfut9dAX4Hv3XuHXvp7917rscfgH/AF/fuvdesfrb/H8e/daqOHXYvbgA/wCJ9+60adcffurdd3PI/r9ffuvU674sD9T+Qfp791rNadcTz/h/re/db6WWwdj5vsXdWK2jt6Hz5PLVMcECAM1g8gRm0rybXvb8+/de6+iN/K2+K+9uoeicRVZMTx1stCrLAY5g2oRC49Y/B9+4jrVNPaPLq3TZvY25ttVywV1HMEUqhYhrH+z/AGv9596x1bPQ19i7doe1tkVU706vPLRyExmMEk+E3Atz78T14Y49ayHyK+LlJi+2aSsNOtNA9TG8n7bgACS5vce/MwRCzcB1eKJriZbeMVZuA6N5sKk2ptfC0NJFko4tKKJNNgNIQLb6fX2UNv8At8ZKM1KdSFD7T83XcSTRW9Q2Qc9Hy6kxuycrQJksXTQVWTplVjOFBZXXi/H9PbtqdtvGNxbAFh59It/g5y5chXat3kZYjgLXo1tLiMZXR4+vraKOStpo28Ezj1R+ggm3449mHhRM3iMKkcOgeu430EBtYHKxvTUPXPRFOzOpOpc/urMZDP7wgoK2afVLAYSxjctewIH9fceb3tG2X8rGeQBvL5dZi+1vuLzxyttcA2izaSADND8Q/wA3Tthv5fe1d6bYqdw7b3Ela0cJnpNKPeTjUNNrf09h1vayC/spJfHV1I+Ejj1NEP36932TeoNi3DbDGrEK5qMeXRRd39OdtdNtlHzWFqJdvQs329U0VQbwg2uL8fT+nvDb3L+7mxluN5tqwhCThe0/nTz6zK5T91vb33Ljt12q7UXkg7kquD0LXxTakqOxNu5WljWGWqnLSfpUg6r+oN75pfeHM9lyRuNjeVkSFCADXPAdEHvSk68qXdlISwUUHR6t/wDyebbW9ZdkQ4FshK8pgB8t0lvxcrcjn/invETkz2JvOauT4+aLvdHgiZdQjFQqgcBSvoMfLrG3lf2Zh3Ll5eY5bgRADUcZH59Fp371Bvrf+afO0W3TjoqpvOqIGsVYarG3F/c8cp+5vJPJm1Jst5uYneEaCSamox6dS7yzz5y7yxt4225uvFMYpXHlj7ei/wC4trZjZ+ROPzEDU86nSVZZALm5Auw9zJsu/bZzLYi+2yQSRtkEEH/B1KW1bxYb7Zi7sG1qegM2sgqN/wBZSP8A5uon0SMTwAz2B95Qc9p4nIWyRg6dSgV9K46DG3Fo769uEFSladHU7f8Ag3mcltDb269iMK6urYYqiZYVkLozIJG1+I+5y5V9qOZNi2DbZ7MtuFpuC/qDjoBHy4dALkD71+y2nNN/y5zZ/i8duxVSxFDQ0xXosL9C9kU+7NmbR3vi5cdT5ObwmqZKgKi39LnX+Lc+x1yV7ENy5zZ+9mDIkrCiFaAA8ehrzn94Hku35X3Dc+T7gXEwWtFK1B/LqVv7475jbXYGM2ztcSZCCrnjjqZY4pNK630knSP979zZvvt1t+4bokd1Gs6AigIrTPUKbN95S43TkmW53kmGRFOglhXA6Obsb4hYrZ8+PzWWqI62skCyzUjIbxs6hgLn+h49yrs/JtltCIIgFRfwgdYT8/feH3vm+CTbFLUWoV6/M9M/anSqZXNRZYRqlDQfuR02k6NK/QD/AB9nE2ywzXAuj8K+XUb7P7k7pte1SbLDUSz4MlfM+fVbnatTQf3irsPHTLDHSSFXAAs34sL+4k5z3+CCdvCXQyYHzp10P+7f7Vbnu+zQ3W5zG6huR31qaAjoD63b89cxjpsm9HTPwYlcgD/YC3v20e5sdvYLFcZYdIPcH7lVzuHMk17tDARMagAHo2Xw06jx2/d8LhZE+8NPMUaVuRIQBcm3sUcs853G/bj9OE7OIPUJe9v3cNu9rOU13Sa6BuWUHQONfTq9LM9C7b2Ztp6FKenjmNF6wAo+sJH+v/r+5Mkw5A8usI7ZjJCJD5k9aS/88jpSnSp/iMFOpRFle4jHoKRkglhx9fesnp08OtRWRPHI6H6o7If+QWt7917rjcD+hP5v791rrwtfn6e/dbPy68bfg39+60K+fXXv3W+uSgE2P+w9+60SR161gbnn+n9ffuvefXEfXj6+/db697917rlf+v4+gtx791Wnp1x9+6t12CR/sffutUr14G39P9j791vrwtY3vf8AHv3Ws165C2nkD/D+p9+60a1x1xFvz791s18uuxyOT9Pp7917zx11f6f4e/db669+691737r3XYJH09+61QHrr37rfXYNvwD/AK/v3XuvWJ5/4p791qoGOuvfut9dk/TgC39Pz/r+/dap11791vrlZl55H+x9+61UHr2pv9U3+3Pv3XqDr2t/9U3+3Pv3XqDriTf6+/db697917r3v3Xuuza/H09+68Pn117917ruxtf8e/der5dde/de65caR9L3/pza/v3Vc168foLDg/n839+68Pn1x9+6t1737r3XZFuD7916tevfWwsPx/r+/da4Z68fqffutjh117917rvi3+N/949+69117917rkORa30BPH1J/wAT791o4z10bjggf7xf/b+/de4567uD9eP9b37r1COHXV+LWH/En/A+/dep10ef+Ne/db65C1ufp/X83/p791o/Lrj791vr3v3Xuve/de67Nh9D791oV8+vG55P++/23v3XhTgOuvfut9dg25B5/wCI9+61x66H19+6312b3/of8Px791ocOvEk/X/b/n37rwFOuvfut9e9+6912bjg/wC2v791rHHrsWtzwL/UfW/9PfuvGvl1x9+631737r3Xvfuvde9+691737r3Xvfuvddi1+eB791o1pjrr37rfXvfuvddk3/p/sPfuvdf/9H5/wD7917ruwte/PHHv3WqmvXXv3W+uySfr7916lOuvfuvde9+6913fm55/wBf37rVMUHXrfT68/gDm39ffuvV65hbD9DH/Er791ognriV+h49XIA5P+PHv3W+uypPOlhx/qTb/X9+68K9daTYH63v9OTxx9Pfut9ctLWA0uBbmynk+/dVoePXEKT9Ax/1hf37q3St2bvfdXXmbpNy7Oy9Zgs5RSxzU2QpbJNDLC/kikjYg2KsAQffutUzXq83F/CPub5xfy++wf5l/e/yYy+8sl1zuyl62x+09x5GWvzAo6eEiA0ySX/bAXjTbgW/HvQxw4dOEagSTkCvVBtRGiTzohkeKOSSOJyliyI5VSwP04976bHD59YCvAb6XJAUcnj6n37rw9OvFT/qXAt/qT/T37rwr0db4D/FCD5m9/YbpSo3Iu1Y8pQ1tYMs4W0bU63CG4P1/pb37r3z6sc2B/Jgo9/fzK9u/AKk7Zo8cM7svNbpG9q1kjpoZcRSiq+1cabXcXVRb629+68ncc8Mfz6qO+UHSsHx97r3x1NT5cZ1doZvIYd8lHZkqmoKt6YzRlQLhtNx70MivXiCGI9Oi/kNpBJPJsBwSbf8U9761iuOuOn8EMD/AE08kf4D37reeu9B/wBS/wDyT7917PXgtj9Hv/gv+H9PfuvZ68wt+G5/1Q0+/dez1xsfpbn37rdeuvfuvde9+691737r3XvfuvdcjYC3BP1v/wAR791rzr10CB9Rf37rxB669+6313Y/0Pv3Xq9dn6X+nA4/r/j791rz64+/db67sLXvz/T37rVTWnXRJP19+631yub+kW4/Hv3WqevWxj/wl9plqP5pHUsrC/8ADaTc+YjX/VzYrbdTXRIP8SUHvR8vt6vHSrfZ1Z9232FmP5hvwS/mtT5Giat7g6n+RNfhdmUFUy/d1mCTPvT1q0mnU1kVb6QP9ce/Z62wWpr5dEa+BeLrviZ/Ku+Tnybz+Emwfau0Oz9l4HZlHOirl5aLKVJnrpY4TYjSQt7N9Dz72eNOqilNQPH/AGOhM/4U/btrOyMR/Lm7XygkTM9mfFPYu6MqswIl+9noWSRpAf7Rtz7ovxdXl+EelT/g6rh/4Tv3/wCHY/jR/X7/ADNuOeMd+Pdj02PP8+kv/PGP/OdHbwNv+P8Ad3G3F/8Ai4gXv72Mr1Vv7Y/YOri/5C1/+G0/nMb/AElBA/x8P196FKnpzyA61Oe8j/xl3sG4+u463/onn3vpsDA6Cq1uQeffut8ePXH37rfXvfuvdd8ngf7b37rWOPXuQf6Ee/dbwevE3/FvfutDrr6e/db697917r3v3Xuve/de697917rsi35v791oZ669+631737r3Xgbcj37r3Hr319+691737r3XLUbEfXiw/w9+61TNeugSP8AW/p7914ivXXv3W+u+LH/AHj37rXXhyeTYf19+68eHXjweOffutjo5fwDKL8pOszJbT/F4Abi4H+UR/Ue/VpnrR6+pj0nvzbWzeqtt1GQEEkQpfWjLHwDCPqPx7bL5p69bABP2dCfs/ffW/ZWSaGmpaWOQSHnSgvY8/j+vves1oet0FPt6NPitoY+kpRFSpEtNMhWNRbSQy/7H3qvW68Oqvvmn8f6ipin3JjqNJJIIxMdBBvZSxuAP8OPezpdCh4HpyKR4Jhcx/EvDqpj+8u3Y3Xb2XCUGT1FFJABLK2k3N/Yd3Dl61uEJhFCeph5M95d72ScLuDs8ajAIx1Z58bUx1Bi8dS00aaKiFBJLfmUEXuxPsy22yjsLdY0GfPoD86c23vN+8SXtwx8ImqA+XR0ayKanWExx2isF9P5VhYf7x7XF6Y6CZHmesWJ6C683R95ks7iqSoqqklzJKx1ah6gW9l8u1WM7a5Fyehjt/uJzLtVuLSzmYInADoaNkUGM2Yi4XCwx09DGvjSOPhQo4/PtbCkdugjiFAOg1uW43O83Bur0kyE1qesnalBh97bcqsRmaCnqqR6eRWMqx2UCMi5NvqPfr+3st1snstwTVGwIII+XSzlLmbfuS99i3TY52SUMDRSc5+3qh7f0UHSvZP+/Syvk+5qUEVLTsCKEM9iUAPFr8++WP3nfYraDHPf2iLJbuCZF4gY4Eddrfa3mmX3S5Fj/rXB4Uir8TDMlB0P8nX9FncGeyq7co/vFEi1McZlBkLFdX0v/X3y7HOlzs++pyFabc30T1UuF7AB+X5dFacyXNhuP9U4bP8AxQnSTTFOjv8AUfZu3q7Z+MizOYiWupqYJJ5yiltAsbHi5/1/eM/Mvtq9rzrfXE1oTaSEmMKCe5snA4cesbfcbkreIN8kk2qA+E7VFK9Fj+Tu/NiZiZcfiqelqskNSmsRVLXKHSdQ9zP7E8l807G9xdXTtHZs1Y4iTQAH58K9TL7Ncrcy7bCLrcJGWL+Ak/LqsTbSyRbwyZufOsxKObj+3fg++l3uC6nkHZ2GKIP8HUs7WQ28XQ/CTw6ss+OvzFfr3N0ezd6VLVOOr2+3pXqEWRaZLadSFvpa/vI37oXuluVsJdl3uYzwgUjDCukEUHH06xw+8N92i35v2Sfm/lxBFPANb6TQt9tOrCptz9U9wZbH1WMkostkMUUNO2iLVC2m9ho5HPvoLJdWG4spXuKgUx1zSFjzfyfHLb3QaKOUkNkkEfn1l3psLa+HemzseOpv4h6H8tl8mq1x+PqPe0t4Y28TTnpDPvu53Vv9EZD4fCnl0gamOormjrmS0YOpgbD0j/D2+XDGp6KI4xGNIHQH9m7jooaSopkKI4jZXPp9P4Yn3sPRSPI163Q+OjDiCP8AD1Sv3Dg6Q7qrshT1InlqpyXRG9KAsf6e8euedgF1feJC/E1/Prr391r3Zk2jlRdu3KDQirRTTjw6CaPE1mQqY8XjYzUZGpIWCEMwGo8clfx/r+4th2yaWeSBASyeXr1nFc88bdt1ha7xfEJbz/E2O3q374ZdJ1vScKdgbhnSCsrIvP8AZl9TI8iaraWt7yC9utguLCIXtyugkUoePXJf74/vHs/OO5Ns+zSeKiNSoxgH5Hoa+3PkvFksjVww1zNMUaNIAq2JUaQLA/7x7lUurNXrAmNfDTSete/+aF15uDtfrXNZmlpGklp6KZgv0Yk05Y2A5/PvdaGvViCTTrRR3Jhq/AZzJ4nJQNT1lJVzxyxtf8SmxBP1B9+Br1UdMfvfW+ve/de697917r1j/T37r3XgbfT37r3HrxJPJ9+69w67H1H45+vv3Xj14/U+/deHDrw5P9PfuvHrx/pxx+R+f8ffutD168AD9Tb6e/deJI4deIt/rfg/19+68DXrr37rfXf1H1+n0H+v791rgeuvfut9djj/AB9+6914m5+lv8PfutDHXVifoL+/db67P0taxH5/Pv3Wvn13cEWsB9OffuvUPHroj+nP+I9+69XrxIsBa1vz/X37r1M9euRx/vHv3XqDj11791vrscfUXv8A19+60c9de/db67uTwTf37rVAOuvfut9e9+6912OT/T37rx67I545/wBbn/evfutA4z1x9+6313cWtYf6/v3WqZr164ta3+x9+69TNevXNrfj37rdM1669+691737r3Xdz/tvfutUHXXv3W+ve/de67+p5P8Arn37rXAY669+631737r3Xdha9+f6e/dazXrr37rfXvfuvddg2N/futEV67/U3PF/fuvcB1x9+631737r3XYNvxf37rR66/w9+6312Bf8gf6/v3Wiadde/db67HB+l/8AD37rx67FiLGw5+v5/wBYe/daNfLrq/4/H4/4r7916nn11791vr3v3Xuux9fra3Pv3Wj14/Un8e/deHCnXXv3W+uwL/m3v3Wj176E/n+n/Fffuvceuvr791vrsm/+A9+60B14Hgi1/wDH+nv3XqZ68Pz/AKx+vv3Xj11791vr3v3XuuwL+/deOOu7C9r/AOx/4j37rVcV64/T6+/db697917r3v3XuuwQPqL+/daPX//S0AGFj9b8e/daGevXFxx9Bb8c+/dep11791vr3v3Xuve/de67/oLf8b9+6113wxP0HBNv9b37r3Dq7L+SR/Lnb54fJGPHbzaHBdTbTwuU3Xufc+RlWnx0VHt2Nq2upnkkBF3VCg/x97yOHn17Sj11fh/n1d5RfLD+TJvzufOfC1viz1zs7bmDxWewjfJaCSaXJTbhxyyUCTyq7+IgyBeFHBN/wR7rgZPDqwOqq+f8vsHVfPw5/k/7P7W/mKZLYcW7qPO/FrE43P74rOzf2zjKLC4wvkfsJWQEEmNdC/n8e91Pl1oKpPfWg4U8/l1YHR/Ob+SNubtqp+GdR8PNjYDCRQZDYVH8j4KavfMVW7o5DQU1bJGXMYLSra/jA/2Nx70BTj1auCBxr6Y+z16rn69/lz7I6J/m5YT4y9h0VNu/r/snbJ3ntkVaL46fAbgq0qMbUvqWxVI2BXi+m4Pvdem6eRHVvHz9pP5TH8vxT8ZJOkNjdk9qb5xqyUe/ZSy1G36zMU/ipKalWH9seOSVOXX6i3vQzUDq2pQAvGvp/l6L70h8WvhB/K++J1d8ofmJ1rh+891753PFmNm9S7gp5Gj/ALt5q89DO01MFYRhdCKAefyfe8k068AFGtv5f4B0kvkr1r8Cf5l/wb3z8gvhv0zgehO1dmZGpgq+r9vU9T45aKhBmeuhkqtbECxVwD/jf3oYND15irDWMHyrxr0cn/hP/sLqvsL+T78n9r971tLjtmbL7TbMVdFXsqRZHLYmGrqHxBZzbVL4mjH+v79SvVq9ob16T3x2n/lXfzXstvr4c9a/GnYPx07u2thtzSbc3xjamY5DeNVtmCTxmoasJj1TtHyYxyT79wz5/wCrj1olHJTNP2fsPVIn8vP+Ulu7ub5+7j6l7gw9Ts3qnYuX3dldwbqyKhMamF23PJXSJFIy2fXEoQcfX6e7Z/b1UKpOk17f5+nVtKfOD+SNV9w1fwsrfiBsjHYLH/xXY4+R8VNkTmavdkTPjaeoMJfRzKFOoJb/ABJ491oPy/n1cNWvrXhTH2Dz6JT0J8QF+DX85bYfWtFXHJbY7E23ld67PqSp/wCLBmEevx7K1h6GiZGQEfm30t79x6pXORTj0f3ockf8KNdhsvp09Tb0Yc/W2FBv/wAT723W4+J+wdFz+Bvxk6W+TH8w75JYbufZ1HvHGUP+k3KwU9ZpBjq8dTzz08oYg8hgCPeh8Kn7erUrK/qKU616OsNhbUy/zf3HsirxEMu1oOys5jYcSwQxR4+HdiUcUAJH9mI6QQPexwr1QjuA+fV//wApvhf8c9n/AM4v4wdD7b69xWM653h03trK7gwgCNR1OSytLG9TXyC31BYk+/L6HrzgihHz6PT8pcl/Kb/ld9y7W6K3/wDGnYXyBr+w8Ria6bPZGWWP+7km4lRRHTGjKhTTFwPXxf34itaeXVlZUCnPd6eXzPy6UnzP+IP8uX+XdgurPmpuDrvaXaW0e0MZQbgoeh6+RI6KjxWURZ1adobOUVZCAQQfT713FQOtkRpITn/V6dVt/wA1X4zfFDu/4OVHz/8Aib1jheqNmUmXxGIy+1MG5egxmVyqxSikiMhZjp12F2/x97rTqnxAH+fr9vWpEw0hCOLpc/6+oj37r3XVvTe/+w9+69XNOuPv3W+ve/de65EWtze/v3Wga9cffut9cgRYgjnnn37rRBr1x9+6313c/wBT7916g68BcgX/ANj791o4Feuvfut9dkg/QW9+60Kjj11791vr309+691sj/8ACWfH1OQ/mmdZQ06xk/wHerN5J4YBoG1KpW0tMyi/PAvz703Afb05Hxb7OjHfyuO05MF/N3+RPxI3HHSv1/3T8gN6xbgmnnhWCllp9xS+bXKxEemxurE2uLe/Amgp1oqNbVOP8vQi/wDClPLYP4s71wvw+6jNGdg9h11Hu/KVFLUQVMJNHWRRiBjSnSJAWUWN+Ln+nvfqD1QjVShx0G3/AApkwlRhesP5VdNIacxxfDnYlIngqIpgJIKENIriIm1ta/6/+w91X/J05LXSPtP+DqvH/hOnRz1/82z4yU1N4vM9fnCvmmjp4/TjLtqllIUcf1Pux6bXAP59Jf8Ano0c1F87u34JzGXXfm7QxjlSZCf4gG9Lxkg8H+vvw+HrR/tT9g6vB/4TodV9gdw/y9/m1svrLCJuTdmSqVjx+EWspqeqrHMVgsMc7DVc8Cw+vvwGSetsTQAdUwdofyK/5s2e7I3vlKf4YdoTw1WdramOopsfDLSSwOxMcsEuuzLpW5IHHvVR1sRvpo3+Hqsj5DfFfvb4rbkpdod9bAyvXu5a2Bp6fEZYRLVGNDZmdIybe/V6qcGnRem4NrWI4P8Arjj3vrw669+6312psb2v/h791oio68Tc+/deHDrr37rfXvfuvde9+6912SCeBb37rQBHHrw4/wAffut9de/de697917rkBcE3+n+8+/daJoaddW4J/pb/effuvefXXv3W+u78Wt/sffutefXXv3W+ve/de697917r3v3XuuxweRf37rR4deP19+62OlxtJMS6zDIPAr6rL5XC2X+tz/xHsQcvxbZLcEbk4RR69JbkyhaxCp6sB+CG06PO96bTqMIsE9ZDkoo0aJgzpapVrKBz9bex7DFyKkbrNMob8Pz6Rq16aVXrfJ21sHtOt2Piopo61KEwWTVq06fGLCx+ntFp5OBoZl6UVuBwXoQdgUW5OvphVIjrMNJ+pZmNyST7q0fKFaeMv29WBnp8PRztkd77vrWpqOSEsF0xjV/Z5t+fdSnJ4GJh/n68BcDFK1/l0PGbxO6+wcFJTzY5qhZoWQLxY3Qrfi/tIG5W1/2i09enKTU4dUk/KP4Q74otwLujD4yqpzC5ldIiTqYuW+ij2YIOUFHdKuekzrPKKMDjp86GyPYuNlxeDlxNVDNQhYCzxOGcq1gfe3TlAHUJVPVh9SBQClOrT6LAdhZDb9LMcZUl3jUkhSB9R9Tb/Y+09OUCaeIuer/AONcehQw23t9QYaEfw2oUlTraxNj+eD7TN/VVHoJFI6cHjnyoemj+G75gqPTQTa3uVYiwBB5vf8Ap7eVOVG/0Qfy6TyvcRivRNfkB8otxdR1bbdyOOWSavEkUbSrdwWUqtj/AK/sp3LeeR9urbXMygkHj9nUve3ftTvXOzpu9nqKwkEgcDQjqm/duQ33uzd2a3npnkpZHEtLDZtMZU/QDn6/X3hb7i87+1i7g/LV5fRyLdEhyTXQD12G9tYmflix2g2/gPbCjN/H0qds7w7JqYTDVZSoSnYWjozrsyjgCxP+w+nvB/3K5W9o7S9VOX2jkK8XWh4n16GO87UsUviQWhLDi/Stn3B2JQU0lTTVFVBGiEhV8nP9baT+fcb2PLfIO4XqWt3LGA5yxpjoiitjdTCKe21Enz6Te2927lze4ac19NNOI5NNQ8gYavyLlvY85x9u+QuXeXPqdq3JHZ1qqin+To+3Datw26wIhBUEYA+zoS5czjpZ8vBjqCMZUXF1A1qR/Qj/AB9m3Nm0cv8A+t9szXF1kKOJ+XWMfKtrz5HztdT3IY2xb8uPQT7nwu4qqkOSmgljraI66eouVMZJ/Fv9f2r9jt+5Y2fna2szOojnNCfQ/wCodT7ve7bxBsdzZ26lo5VAYeop0cT+W3Udg5TduZ+4kqK7xTt6bsQoVrAcn3195Yfk5zrhnVyVH+Drml95S1e3sYSsPhV49XPb623vzMULrHjagNEtkUAj+z+f+J9jCJ+WNZR3UdYcQifwQR0E+RwHYGN2xKXxlQsiIedPI0j88fnj2/p5UBp4i06t+v6dVDd+9i77wGSrMbHQzTz1bMlwjkw3crYe1KR8nsoDTKOmna5qHQZB/aeibbixe7IKaHN1/mafIHWIJNQMdzf6H3HHM1tyHBc1NwtW6zj9g925h3ba2t7i2KJEO00+LoV/j9QpFTbm3JmKZTkMSjS0CysAS49QCq31N+beyHZNo9vBPLe+OhZRXj59SZ7t89c7vBt3LFqjCGQhSQDjyr0ZrbnZvZXY0EuMp2qEjhUrDGisPSh0j9P+HuTdnvOSrlNCzrj7OsG/c3lTd9ivxcyoWDnJNTx651fWO7KEnL5WnlkcnW3kJ9N/Ubg+zfRyeMmVeo213BwF6Zd07Ir9/wC0sriJMb5ojTyxlNKkMpp2AH0/p7068nU0iVT1sG6U5FetGH57dFL1J3TuGry1IlHT5DJSQwCYhVVZKg2A4F7e10yciy26COVQ448K9Jm+uVqKKjokoh239DU0fI4tKv8AxB9pBaconPjL/Lr3iXlaBf8AD17wbaFz9xR83uBMvNzx+fevo+UP9/L/AC60Xvv4OHSX3YmHSgj+xmgefyrYQuGso+p49hrmSHZY1T91uGPnTpVamdq+OKenQeA2B/x9hPpYR1x9+631737r3XK4ta3Nvrx791qhr1x9+631yK2F7/8AG/8AW9+6qDXrj791bru/AFvpfn+vv3WvPrsC/F7W59+68TTPXH37rfXvfuvddkWAP9ffutVzTrwFzb6e/deOOvEWNvfuvA1699ADf6/8R7917j14C55Nv8T7914mg669+63139B9fr/xHv3WvPrr37rfXvfuvde9+69165/r7917r3v3XuuagHkn6H6fk+/dVJ8uuLG5v/h791sCnXXv3W+ve/de697917r3v3Xuve/de67txe/+w9+61XNOuvfut9crEC9/r/yP37rVamnXiL8/S5+luPfuvVpjrs2A08H/AGr/AGPv3WhUmvXD37q3Xvfuvdd3Gm1uf6+/daoa1669+631zNtI+n4+n/E+99VFa9cPeurdcgL/AJ9+60TTrj791vr3v3Xuve/de65E3Frci3P+Hv3WgKHrj791vr3v3Xuve/de697917rkADx9Dyb+/dVOM9dEWNvfutg1HXLhjxxb/ffj37rXAZ64gXv/AIC/v3W+uvfut9e9+691y08Xv/sPz791quadcffut9e9+691737r3Xhybe/de65Fbc3v791UGvXRIJ4Fh791sV8+uvfut9dkWNvr791oGvXvxb/H6+/db68oubXt791omg68Tzxx+OOP969+69Trr37rfXvfuvdd/g/4+/de67Wx4/3m1/z791U4z148cf7G/wDxHv3WxnPX/9P5/wD7917rkBbkji3+H+t791U5x1x/Pv3VuuROngH/AHo+/dVGePXEfX37q3XIkX9PHFj791oD166/r/rcf7f37rfW5n/wnKFPvnofvHrPZ9bHB2PJsTsLLCBJYlqqjE0uPllqIkS4c3Xmw9+PDrSAk48j1qw4zprsPtX5Fbx6x6+xtZU7un3nuqNaOmFWJ44afOyR1UhEKl7LrDcj/Yj3quB14ipz1vJ/yM9hZn4kf3x6Z7FxsPaXZp643Bu6fY2VkUyVuFpMa9TVYkxSOZShUEEaibe7Hh14KdQJ4dVdzfzm/hinc+d2ns/+Sh0bley6bsTIpi4lqspJkchuHHZR4Ya6spNatqaRfJa+kgjnn3ShIx074gVqmtfsHS2xPyz7s+Wn85bYO+/kd8eqP4479oeq8Vjtq7KpKOahWPalBAIMa0cNRdtGkjS2o3A5txfY6o5GKj16rI/ncu1X86dqzVDF5ZHwUUkhdjeNcrDpHqPFrnn3s+fVRin2dbYX8xr509CfGr4OfGvcHZfwf2J8qtkjrHYu3a7L7rlkSkGXlolh+zlqKMgKEuS2u5tf6/T3oAU6vqyaV49VG9Zfzos1uzpDsfBfCf8Ak+9Z7O2E+387hc9uPbENbWx4eorqBlqKgVssqx3hRtQsvFrfXj3upH2dVbSaE1r+VOpX8vLa29Oyf5EPzCTZFTNi97VfyEk3FnaWjWVZcairWtV0PjT1aVu9/wDgp96zXq4K6PtBp/I9Vo/8J5Ovt371/mj4vGYCaf8AiuGwm96/I5S0sf28VErNWTTyNbTqCtfWf6+94r02ldOPTrbK+DPf3XG8vk/2/tvZ+HxnYe69sYrftTldrNLSOM/DhKN5spStoIdhKikEXuR72c/n1ZQan5Ur+fVCm4P5y/wwoe8t3bO2x/JS6My/ZtN2PX09KY6vJy5TK7ixuVaOOtr6VHU+QyqHIvpseSPdNJI6sZVTJPy4DpY0fy27t+XH85HpbdvyL+OWP+NO68V1hSY3ZmzKKjnoYX2zQ440+PlhinZ/SY1FrPzyQoUr72OvOQTgeR6Hzon/ALiNNhm4H/GJd583AFzhQB9fez8uqR8TX0H+Hpt/lFhMp/M5+TO3qV0bMVOL7ekgpS6K0ipjalzYMb/Tnjn3U/AB1Yf2sn5da5nxi2NmuwP5lme2lhaaaozFX2rukw0scMrSMaPekbTjTbULAH3YZHVHBDA+h62RPnFSS43+fn8YMPUjx1uG6L2xQ1UZ4aKpp6CNHVv6EW96Apx6s51UI9D1TZ/Pcmar+bXXrzsZH+wwUTksSTGuRpwo5/wv78eJ6qowOj/f8KMcjXxdM/DDFQVjJif9l22fJ9krnQZDQRkuRe34H19+U0BA6tKB2n5npmwrCP8A4TG9zwahpHeGytCltRAEFPaxfn3r8RPVx/ZL9n+XrUWk+kf/ACzH/Q5926a64nk2HP8AtvfuvDhnrj791vr3v3XuvW4v+Pp79175de9+6913wCPyOCf+Ke/dazTrkpAv/j791ogmnXE/U/6/v3Vh14WvzwPfuvH5ddsQbW/H+Hv3WgKddXNrX49+69Qceuvfut9e9+690KPU3c/aHRm6Id6dT7zzOxd0wQzQ0+cwVU9HXwxzxmKZYqiL1DUpKmx+hPv1OvBiDVeuWC7o7N2z2FU9r4HeOZxnYlbkKvKVe6aapaPJTV1bJ5aipaoX1a3b1E3+vv3WjUmp6m9qd7ds93Zmh3D2tvrO73zONjdaCvzNY9XU0xd1kIWSbm11B/2Hv3Wq9ZO0/kD3H3XSbWoe0+wNw71pNlYuHC7WhzdbLWJhMVTp44KChEv6IlHAReB/T36gHDqxJbj0mutO0N+dPbuxu/utN0ZXZ28cQJRjc/hqhqXIUQmUJIaeeP1KSBYke/dayDjr3YfZ2+u18/Vbp7D3Lkd05+sklnqcpk5jPV1FRO+uaWWRuSzHlj7917zr69Cr0h8vPkl8bIK6m6M7e3l1lBkpfNWptfK1GNNS9/1StTkX/wBj79QefWwxHDo0sf8AOX/mfxRrDH81O8UiRCiIN45HSEIsUAv9CPeqDq2tv9VOia92/I3un5HZ6m3P3d2FuDsXP0cLQU+W3BVtWVqRsblWmk5PvfVDUmvQIHnk/U+/db697917rkDYcfX+v+x9+61Spz10Tf8A23P+J9+68BTrsEWIt7914g1r1x9+631737r3Xvfuvde9+691yA/rwD9Pp791qvp15vrYfQfT/fD37rw4dcffut9cvqDc/S1vfutcDjrskaQB9fz791qhrnrh791brncDTzew+nHFx791Wla9cD9Tb6e/dW67Bsb+/daOR11791vr3v3XuuVxYcfT/eR/T37rVD10Tc3/AB/xq3v3XhgdWi/yqNx0GJ+R+0aOojVpqjMUzhj9QhlRABf/AB9tSQxSZcVI6spIOOvpdQ1mnqnA1NM+jVSA3ULx+0OOPdTbW5/D14MwHRWTU5DK1VTDHUs4DlRbm3r/AN797FtCMaevaic16cI23DgFWeCaW5IOqxH0NweOfezbw0pp62GNcdGl6T+ReSxldTYnL1LEPJou5+l2AFtXvXgxAYXr1STWvR+6+mw3YWNgdvFUrMoMnETadQuPxb3U2sByV69rNMY6DKg+PW3sdlf4rTwojLIHKhIwbg3Frf8AEe/fTwcNPW9RA6MHSY77Siip0GmNEAVQFs1uLAf197+nhONPWqniepEkVTFCAkhEYuCotxqH5t/T3vwIgKkdaLdEY+aHcue6O23T57Ew/dOYdei4WwIJP9PZLzBdjZ7YXKKD9p6lf2b5Gg9yN8OyzNpJNBQV6oK7H7N3H8jslRbmqqSSJqabW63fko/I/PvCz3X97dv2uaSznQeMQQKHhj7euuftV7E7V7Y7U1rdTVMq4BHr0IGHpIqfFQ08kQX9s+VCo9R0/wBon3zo3/cpN13qXcGOXaozXz6kf6eK0k8O0wBwI6BV4MhV7u8dBUtFTwSeqIcIBq/oPeSNwOTtk9tka6t1e7uUw3E1p0L9sv5/3dKNzi/0rHz6GGDNU806YiZw0qAK6kj1EDmw940zbZNFEdwRaK2R0RyWUqRm+TAOR0kNy7lodv10UFBRqtQWHkZLeon88exryvyff8y7ZPfTSHw4RwPR1tO1XO5W5luZCVpivTHsyuSbcGRq3cRy1DXILA2N/pz7H3uVZG35O2q30GipxocinUW2cdxHzBeQR5RW8uhL3LUtNiaqOCUSzMFVUWzMT/rD3Gntjaa+eNvRkwZB0Yb4PA2qWR+0Acf+L6P7/Lo6m3ds2Wq3fkEqKegzLNJAXidEIZtWtS3199p+S9m+jtkuXSgKgfy65afeQ5127e5Rs1qwaSE0NCD1dPj6+oneUyzNIkg9IsDcex40UTGpXrEuIMkYU9QMzRnIUs9G7XjkFh6VAFh7p4MQxp6crjoiO7fh9j92btOTq2EkU0zOFaP0i51f0+nv3gQntK9eEmgiTjSmOq3fnn8f36bqMJUYysespK9BK8KRkrTLa5X0+4v5w2aK3uBIO8MK/Z1nZ93TnCPd9ufbmiEJixXA1dV70NVnmo53w88ix2/yhEVvULcg6Rx/sfYLig0KfAFK8aV6ySuIdvmuUXcVDMD2E0x1YX8PqPAV0zmTOq+WWNmmoiy3VwAWT1fm59yJyjb2bEUbv9K9YgfeJ/eMEZElqVhr2vTy9ehP+Se7pdvU5xlHU3qKiMKESxZSTawC+5Da3gJqV6w8hkqlV6h/HTbmYyuOqKvISySx1SOFVluf82eLf0/x90+ngArp6d1Hy61Jv+FEPXdNt7cO38lSUQWSXJ6ppEjA5aQpc6R9TqI92SGFGrGtD1QkkZ61a/8AXv7d6917j+h/2/v3Xs9e/wBb37r3Xjf6H8e/da+fXhweffuvHrvm+ofQW/pxxb37r3yPXH6+/db697917rsEc3H+t791o1669+6312CBe4v/AE9+60a9dg/15Hv3XiPTriPfut9ciQLj6i3H+F/r791Wh49cffurde9+691737r3XYBN7fj6+/daJA669+631737r3XLk8nkfX8f7H37rWBgddfUm3v3XuAz15vqeLf4f8i9+68OHXXv3W+ve/de675Bt+fp791rBz161jz/AL37917iMdeYgnj+n+++nv3XhUcevKATz9PfuvGtMdeNr8fT37rw4Z69c2t+PfuvUFa9d/QCx+v15B/H0t791ric9dgKQObH8+/deJbribXsPpx+ffutitM9cm0gWFjze/1/2F/futCpyeuHv3VuuS/ni/B/5H791o9dAE/T8e/deJHn14/Xj37rw4dcv7P1/wBhx791r8XXEW/PA9+6sfl13yR/gP8AePfutYB66BHP+I/p79149eAv9P8AX9+63WnXrG1/x9PfuvV8uuvfuvddi359+60a+XXZsW/oPp+B+PfutZA66BI+nv3WyAePXhbm/wDTj/X9+68a+XXXv3W+u/xf83+nv3WvPrtSB9f9hwD/AL37914gnh1x9+631737r3XZAB4N/futD59eJFgLc/19+69mvXXv3W+ve/de65WIF/x/X37rWDjroWN7/wBD/t/fuvHHDr1ha/5v/vH+t7917z669+631737r3XYF/p7917h117917r3v3Xuu2AH0N+P+J9+60Knj14An6fj37rxIHXXv3W+uyQbW/oB791oCnXha/Pv3Xj8uuzpIuOObW/4n37rQrwPXRBH19+62CDw68Lfn+nH+v791418uuvfut9f/9T5/wCAT9PfuvdcuNP6jf8Ap+Pr791Xz4dcffurddkAfTn37rQr59de/db67sLXvz/T/Y+/dazXrr37rfR8f5ffzq7B+AneFB3DsdqytC0kuPyWFhrDBT5LH1B0VlHUI3p0TR3jkH5B9+61Vh8P59Xp1P8AwoS+JWByOX7F6q/lqdYdc9153DZGgyu/cRUhq2oyeRhcS5qMSErHL5H8hCgAke6gdOMwbj1Ub8c/5rPffRnzAqvltW5vNbozddUZOObCz5LRCuIyExL4cA2URGJjE3HI+vuwA6bJatVx/m6t3qf+FBnw1p8xP2dtr+WJ1Vt7uqqxk0c+86WdDNJuGZbncgVjoWcP67BbX596p06WBGeqssV/Np7Jr/nY3zb7Iw02+s/TYGTbmHwORqIfHjMVcCnpIXAACRr6VA+nvfDh01k5P/FdFv8Anj8zqn5pdwU3bA2rHsmopKRYIqGCpWXTIswljljkXm6kDkm/v3z61wFOrOvhL/PUoekPj7X/ABm+WPx6wny96wqK2OppsTvquEkVCtOCtMKVxZlKAkAqfej1dWoQep/yh/nsbe3h8cs18a/hr8atu/EHZ+4q6oqctPsiqj+6qoqz0VUMtRJrf1J6b39+FQKdeYgmvHpOfy3v56mV/l//ABZ398X/APQhheycT2Nu87p3Fn8tVqaqZnd/NAdZvIJFdgxe97k/k+95Bx1rtZQCOHRl6r/hQd8feuNhblo/i18DNi9Edxbxoa6k3B3Ltmr8O5amPJxMmRgjLEhEl1HVoAvf3oD16tqIWi9Uu/Dz+YP2t8Q/lBUfJfa1bkKrLZXK5XIZfGxV7RNXU+TqGNTRTOTbTJExhk/1S297xSnVAWBqOPn8+rx6n/hQj8Nv46/aeJ/lh9VYfu2ahqVrN6006NNWZydCV3A6MdCzrIfLwv6vdQKcOnGbWM1r1VhS/wA2zsrMfOzF/NjsjDVG9svgcRNhMDtuvq4/FjMbZkpKKCQAKscSMI1A/AHu1B5dUJJJr+X+r16FrYn86TK7D/mOYH5/0HUdBV5HB7PzW149j1NXGaKZsxSilepaf8aBcjm9/fj8uvL2mpz0BPx9/mlb2+PXz2zvzZ2ntwrUZ/K5KsrdmJVKlG9HlT/ltC4/QUdWdG/qDb37yA9OvV7iwxq6tQ3H/wAKAfiXjtyp2h05/Lf626v7srMg+VznZWFqguUyFbPUGqq5FjkJWPyOzX0Ae9UzjHV3YMM1PRKe+v50+W71/mG7A+eWX6mo8dlNl7Oo9rS7Rp6xTBkno0CLWSVB/LWu3PvdTWvTRAoFAwOiWfOz5yzfNLuvbfcE+zIdovt2ChibFxVCSpUfaVcdT6WH5Pjtz/X37jnrYx0Pv8x7+a5k/wCYHtHp3bWU6zodjv1LsDD7CpaihqI5EydLiaZadK2ZV+jtpuR9PegKcetudVKClDXrDSfzV8pR/wAsXen8uKPrWibD7x3lhN4y78eoQ19JVYYRqtMkX10OE5P19+pnrwJ0gHyx/PqoKQn0AgehdFxyDyWvf/Y+99aFOsf09+631737r3XiCPr7917rv8fX8/T/AGH19+6159e4JF+P9bj37r3AY69e1xx/r+/de65BQbWP459+61UjrifqffurDh14XHNrgH8/T37rR9Ouvfut9e9+6914i319+69x697917r3v3Xuve/de697917r3v3Xuuyb/gD/AFvfutdde/db67Jub+/daAp11791vrmbFfwD/h/T37quQeuHv3Vuuzbixv8A19+60K+fXajUbe/deJoK9dEWJtyAbX9+68DXrr37rfXI2strfTn37rQrU9cffut9e9+6912LX54Hv3Wjwx14/wBL3A+nv3Xh11791vr3v3Xuuwbf4/6/v3Xj13YEE35549+61mtOuP09+6314cf8b9+69139TzwL+/da8sdeP149+68Ouvfut9e9+691yW1je304/r9Px791U1qKdcffurdWT/ywtn5LK/Ivae4aaIvSYzJU8U7D+wBPHM3H+wHv3Xhk06+kNg9101V1Ti8aslpYqTSfpx+0ABx79TPW/KnQKbbq2xuQrHmcyKzlrN9AAx+n+8+/VrkdbAxQ9CXLvvC1EH2M0SK7Lp1MFP1Fr8+/HP59aIp+XQXZaSGnrBkKGcqY2V0ZWItbkH3sgcOvD16Ox8Yu9J5a5cJXyvLY6FaSxC2Fvz/t/dfPr1QBXq0CCZayCGaMXSdQ6Afm6hrn/eve6A9aqfLqQ2UxePZFr5UUgAWJHpb/ABv70D1sgEY6nx1+KyN0p5I/Gys5IKgNpGo6re3oY1lkEZx0xcS+DEZD1Rj8/u26bfuVl64pk8X2cjUryG1mCtoPI/w94JfeB98t75a3M7MtqRENQD+R0/8AF9dSvufe0dry9YLzzeUlMoDqPSueq9Nl7WGz6JqFJBJfVc/WxY3987ObuZpebdxO4zAgk16zo3zdhvUyzadNOA+zpWD6FSOGBv8A6/0PsMfP06Jvn6dA1ncHV4fclNXUdWESsf8AdQHm17j3P3LW52fMPI15b38Gt7Nexj5Y8uhfb7/tkm3GxvsOB218+mnd+eNHW0k1HBoqkddcyj1yX4JPs89tfbKfmnapmvHpG4OgHyp6dP7K+3zRSwzzAqB8J8usG5NwYKsoqB6t0psjKnLEjWzWspPN/r7EHIXtfztY8yTbWqN9DqAJ/Cy1HD8uo55m91eXvbpnkvpw0WcAjA6Rmydn793DlcnPgEaqhjZm16nsAPz6ffRb3C9kdjn9utuZrJXZVHAedOoS5N+9F7bbhzXcm9kESSnB7f8AL0M/TNLlx25t7B7oDVFOa8R1dJIHaNh5RdWv/X6c+4Z5E9reVdp3GG5lsk8VWwaZB6t7++41tu/J91Pys3hxova4I7hTjjrZO2hUQ0uNxWJxNLHRY+lhhWKBAqKBo5It+L+8uNCRxrFGKIAKD8uuNE1zcXt7Ld3jF5XZiSft6MHivDFTpJMQCBybjjj3UY+zrVTw6nPJTVP/AAGYOVBN1Kn6fUG3u1B14mnWMVZi9CqS4Friwtf6iw/3j3WtD1sCvQId+dXbT7N2PlqDcFDTzV8lJIlDWzINVM3iJ1Kx/ofaO92633GIxSjJ4H06EvK3OO7cn7lHuFi5EakalH4s9a8Um2JOktx7o21X4Bc1RZSWSCgrmVmFPqcrrW/+v7x59zOatu9rdmuJ9xI8SVT4QxVvs66B8j7ofeK62/dNqnMZtCDNH608j0G2ztwZrqfdOR3DjsfUTw1cryAQXtGshvYj/D3APtx95Db4rl7jcqRsagA8OsrPcj2S2P3W5di2uS4W2kRRx8yB0L+15d3d/bxiqJaaYQeeMNG9yVXVYC309528pcwLzVtMe7xfBIKgjz64/wDvH7cR+1u/tssMnjKrEV8uPVyfWPVlLsjacQyTRwOI203sLERk8lvYk6irB61FP+FC2KwVZiIKkxRSPGztE7BT6tbG4/2PvwoTj0602F/PrSEf9Tf8GP8AvfvfW+uubXtx/X37rXn12trH6Xtxf/ex79148euPv3W+ve/de65C1vr/ALD8H/Ye/daPHrom/wDh/re/dbHXXv3Xuve/de69Yn/Ye/de697917r3v3Xuu7H624Pv3Xqjrr37r3XINb+yPp/T/effutU+fXH37rfXvfuvdd3/ANh/rce/dep14AE8m3v3WjXy669+6313c/S59+69Trr37r3XKwte5J/I/wAPfutedOuPv3W+ve/de69yT/j7917rmV4ub3/of9f37qoOaDrh791br1ifp7917rsfn/W9+6110Prz9Pfut9e9+691737r3Xvfuvde9+691737r3XL6C4PJ4I/w9+6159dqOCQTf8Ap/vXv3Wjxp1w9+6t139P+N+/de68OPxf/X9+6916/wDsP6gcA/6/v3Wqdev9eBz/AIfT/W9+69Trw/PNuD79149df4e/db69Yj/Y+/de67t9P6X+vv3XuvGwPBuPfutCtM9de/db67Bt+B/sR791qnXR5/41791vrvi315v9P8P6+/dazXr1j/T37rdR14fn/W9+6111791vrsC/v3Wj12wAtb/H37rQNeuPv3Vuve/de65FrgDj37rVM166sLA35/I9+69mvXXv3W+uaqD9T/vv9b37qrEjriTf8Af63v3Wx16/AA4/x/J9+69TNeuvfut9e9+691737r3Xvfuvde+nv3XuvWv9Pfuvde9+691737r3XJrfg39+60OuPv3W+ve/de6//9XQAU6Tf/jXv3WiKinXgeb29+69TFOuvfut9d8af8b/AO8W9+6159de/db697917r319+6914G3v3XuuQsebgH/AGB/2Pv3Vcjrr8+/dW68DY3tf37rR4devzf/AIge/dbp14/j/WH4A/2HHv3Wh11/h791vr3+Hv3Xuu7n/iP9h791qnXvx9R/rWHv3XvPrkSA1xY8D+n1H+Hv3WhWnXD/AB/4gf71791bru5P/Ih791qnXeo8fS4+hsP6Wt7916nXVueOf9bn/ePfuvVxnr17fQW/3n6f6/v3XqevXr/76w9+6314H/C/+8e/daI68Tf6/wC9Af71791ulOvfS1x791rrsXbi9h9bfj37rxxnr36T+D7917iOvGxF/p/h/X/H37r2QaddDn6n/b+/de4cOuj9ffut9e9+691737r3XYva4v8A04/p791rFaHrx4J5v/j7914cOvA2/wBb+n49+68RXrr37rfXdha9/wDYe/darmnXr8W/3n37r1M9dj6Hi/8Aj/T37r3nx66JuALWt/vPv3Xqdde/db67Jv8Ai3AH/G/futDrxN7cW/4n37rwFOuVtPPDf7D37rXHHDrixubgW9+62BQdet/Xj/X9+6317+o+v+P+t791rrr37rfXvfuvde9+6912L/i9/wDD37rxp59e/wACbf6/v3Wvn14AWJva34/r7916uevC31P4/H9ffuvH0669+631737r3XK/0NuB9ffutfn10OT+B9f9b/W9+69wHXjx/j791vrsHg8X/wAfrb37rRGeuPv3W+ve/de699ffuvde9+6912B9ebf8T791rrr37rfXZAH0N/futA167H0b/Ye/dePEddDk/wBPfutnrxFja9/8R791oZ6vB/k0bUrty9jSJRkgJmEDWsLkLEbEn37y68Pi63ocH1/uSnwEdHEJnZ47AgsQvpAPI/r70aefV1yMdRU6t3LAzmQVA8g/tauDb6n/AGPvwzjr1acemR+mNx12QhmiqZgqtdlDn6fnUPe/l1Wp9D+zoT6Lo3cU9KsbQSSBlIDhmHAHNif8fevPHWq0FG6FfpzoTcGI3JFWNBPHGsly5J/JvzYAe/fZ1bjmnVwG18O642ggkb92CAA3uSQkYvzf3viOqA0NK06Kb8g87/D8lHS01eaaUaQdLhSbC9rX/IHuvy6c4Gp49Fwy3yAyuAw38MxzyVFUsbK86glwSLHkH2otmCShzw6SXseu3Krxx/h6qY7o3BuJty1G6MhiJpWqKh5fP45CzanuWtb3zw+8B7Scz8z7+95bFmiLNQeQDHrsP903nXZ955Oi5furlYmiQChoMgfb0GuH7EXKVSx1cP2iOSrPImk2H+vb3iVzX7N7/wArWQumjLinl5dZZz7XZKtLGVZWHkDXp4r95UNNXU9LDIs0EzAPMBfR+L39hfauQN73LbJtyWIgQitOFemYtrbwyZzof8K+vTJvPcuOSmjemZKmdVtG4A9H9T7FPtlydzJvV623wq0du5/U4gEfPpu02O3a7S53RhEE4VNK9BLXuhpv4rkKmMRJGZJNTKfGSLj0kj3k9yhfbntHMCco2tqyRodOuhoR9vRlue58r7ftt1JFIi9pqwI/z9FD33XPX7ro63F7iNVBDIrCijc2sDcrpU395scuwxw2Wh4dLCmafMdch/vD3bX+4zzWl8ZYjXsBxT049G7+Lvdu4MBuurxTYp54KlhEVaMkaSbfUj3lZzIo/qFt4pxX/J1hLZK6XRkjJBHmCf8AP1arh9tYRa2HepwEUeRUpUh9PrDlQ4IAH159wJHs1iJfqQo11qPt6k9vcjmd9r/ckkzGEilDwp+3p/zHyt3vtqshoKDbs8tOjLGahadm0qDxcj6ezXiM+XQKoFJY8Tk9G16/+S0u5tuack4o6x4rFWjKsrhf9e/1+nvxFQAOtDiT5dP3SXaO48xvmuxk8ks1CsziJm5UoW+v1+h9+4dbGePR+ZoVhfWq6mkVb3/xFyR9ffsHHWvKvQH969mbT622dkstn66CGohppHpqOXSWmbxmwCkj2xdbjabXA11dGgUH/AehLypydu/O+6RbTtcZfxGAJHlkfLqg/fHY3+lLNS5eCFY6R5GNPpUKqjUTx/X3xn+8Vz7unNXO88N1MXt7Zisa+Q67E+zftVt/tjy7HEIglzIB4nmSaefz6AzJbso4KmrxVTGAV9Acp9SOByfcZWXKe43FtHuVqC4OaA9ZCRbFcXFstzC1NXTt0F30/UvZFPVVUa1GIkrYmcOgaMRtJYg3/wCKe+pPs/zY218l7dZXB0FFAYdYj+/f3eLnmm1l3G3iMkzAkEHNeNeB6uy3H2bR9tbYos5tCvCxeDVPR04CCMCPm6qf9v7yYsb+DcYRcW7VB65Ucz8rbpyjuT7XukZRkPn59afP89WnqqrbwNZVPH4C5CueHN2P0J/H9fawCjdBs5TrS+f9bf8ABm/3v37rfXXP05/1v+Ne/daxx66sf6e/db697917r3v3XuuybgC1rf7z791oDrr37rfXvfuvde9+6912P9e1+D791o9eIsfrf/H37rY669+691y1cWIv/T/D/W9+61TNeuPv3W+uyRYC1v6n+vv3WqZ69z+L/wCPv3W8dde/de697917r3v3Xuve/de697917r3v3Xuu78W/3n37rVM9de/db697917r3P4/HPv3Xuu7n+p/2/v3XqDrtf8AEXFxf/D37rR68RbkH8/j/jXv3Whnj1x9+6t1737r3Xvfuvddk3N/fuvDHXXv3Xuve/de697917r3v3Xuu7n8X9+69jrr37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XfJ/qbf7H37rWB11/h791vr3v3XuuyAPyD791oHr31FgPp+ffuvedeuvfut9e9+69165/r7917r3v3Xuve/de65WseG/F7/T/Ye/dVrjI64+/dW67vwBb/AGPv3WqZr11791vrkbEDkfT6f6/v3VRx64+/dW697917rw49+6912Tc3t/xv37rQFBTrxNz/AE9+68BTrr37rfXvfuvde9+6917/AB9+69176+/de697917ru3F78/0/P19+61XNOuvfut9d2P8AQ+/deqOvWP15t791rHXYseOB+b/8R79144z1/9bQAAv9PfuvcOuvfuvde9+691737r3XYPNzz/xPv3WiMUHXdxY8fU8f4D+nv3XqGvXH6e/db697917ruxtf8e/darmnXXv3W+ve/de67Av7917h11+be/de67It+Qf9b37rw669+69176+/de67A+vNv9f37rVeuvfut9e9+691737r3XduL3/P0/4n37rXn10Dbke/db67JBtb+nP+J/r791odde/db67Av+QP9f37rRx11791vr3v3XuvXt7917r17/X37r3Xf0FrcnkH/D37rXn12v5Frk/T/D/H37rx9eurcX/xt791uuadetxe4/1vz791quadde/db65cgDng82B+n+v791rieuPv3W+uQIH1F/futEHy64+/db67Av8AkD/X9+60TTrr37rfXMH0n6f8T9Pfuqnj1xAv9PfurcOuvfuvde9+6913YkX/AKe/dar12ASPrYc/U8e/deJAPXRFv6f7D37rfXX19+691737r3Xvfuvde9+691737r3XfI+h+v8AT37rXHrr37rfXvfuvddhS30BP+t7916vXZIuOPpe/Hv3VaGnXbI68spA/wCK+/dbqOvaH0FrHT/sbH8e/deqK9cVUsbD6/j/AB/w9+68TTrJ4n5XQb/63+x+vv3Wq5r14KwU+k88X/A9+68aE164hS5CqLn/AHv/AGPv3Xq049cgjAEFDfkfQ8H6e/dePGvWMgjgix9+6t117917rsC5twP9f6e/de699P8AjXv3XuPXQ549+691yJ/A4/Bt+be/daA8z1x9+6314C/v3XurcP5SPfJ6u76wG3fCGOcy8MvkcekBpIoSL/1BA9+rQY68ONevpW9S7ow1VtXEZKujhl+9jEjX02XUoP596OTUdbAIFT0Ke9d+7Awe067KUtFSVuRpoWMNIDGS7afyo+vPtPeSm2tzKo1EeXRry7t8W97vHYTP4SMct1UZJ8tMpia3edNVUCQVNVVyfwpGUr9qgc8R3/w9xseaLqJplk4k4+XWbEXsTst7BYXVodSIo1kU7vt6Ot8QPldt/fb/AN1t2rS02Q1+CCWUqjyNrCA3f8n2f8v8xC7Pg3BAPl8+ok93fZmbYFG5bQpZKVIHl+zq2/Fbep6OGKqhjQ082lo5VKFZFsH4I9jOnn1jXU10Nhhx8uhKwziCmqah1OmCCoINvwsV/exkV68wzinWs588vkxl9pdwVkFPkbUdNUhZIiSFRdRuP94HuuPLrdfXoLOmfkttXsjNwYJ8jC2Rq5liMSnW2ouBz/T377OtnPVxeX646io+sosrvDD0lTPBRB18/iBlcxBr+rn8+2Lm3tLuMi6XUAOjzYOaOZeXbtU2G5MBkIqRXH7OqIu8aBN37ylpOuNtrjaGiqmGqmUhZYw1uNH9fePXOe3C83LwTZl4OGmleuv33fOatks+S23DmneQ94yVAY5r+fSFi6+3jjKKoGUw0sbzKTFNNdCpt/Vv8PcHcw+2HMM++20+yK0Fmx/VjAww6mKH3m5CuY/qLi8TXZ/0h3dA/vSqq8PtjLSlx91QoSG1XOv6f7x7mHlXl6x2m9W1W18GvE049Rd7u8+2++cqT75yzegstDpU/D+zqprf/efYUVRXYtMtOaWeSRClpPSusgAH3P0HKnLsei8S3XxeOqgr1yf3X3h5/uLu526S8fQSRxanHqF07u7LpvHE1GWqWq4JKpDIkmsjS0gFvV7M7iCGK3bw1p/xY6AMu57rePW8nMleNer2+hKTblZlZc9TUUY+1Mc0ioAbfQ+q3uauYWY8h7efKn+TohhFLuSnCvV3nTFdsjfuKXFyLTx1fiCAakBuIwoHP9SPcPJ8I6Mixr0sd0/Gukq4Z5qOEtcM6aApDAC/1Hu1aderXHRDO0MjQ9QvUrlZjQJBcBnuL2Grk+9ilOtGoNOkN8VPmNiMr2kMJQzQVSvVNEZQTcBXsf8AeOffiK561U8OtkOmrkrcJiMuq3jq6eFwRxfVEDf36lM9aJ1LQdVqfzFehtwdlbCm3ti87U43HbZoGmrKSEvoqgi67Pp/J+nsD8+7bc3mzS3cBwgI0+uOsqvum89bTyvz7a7XuUKyNcSCjH8Ofn1R1tDc0VDiMdj6WPyzcpItiGiKsVJIPPvjd7g8t3I5pvZNwrGASRX8XXZfdNuS/uJL0NSMgFSOBx5dKjM4nCSUc1dXARVMqXFweWtcWJ9hPaN539LhNv25yyA0oOi2yv7+OUQQGqLx+zosNdRyTZ4Qql6VpU0SAN/ZkvcW99BuRbLcZtlsFuVKOQNXy6F++81bRtHLE93KyvNGjHQacadXGfG7OUm3NikGVVWSnZZVZrBD4z/qveaHLFgths0aA1Pr18+3vZzVPzP7iXszrpQHA61Xv5+vadFOlFQUNVHJ5KgRuqMGvrmKfQf70fZ+cHqKjXTQ461JDySf6kn/AHn37r3XQJHI9+63x675sfrb8+/dax11zwP9t/sffut9d8A/1A/3n37rWSOvHm5AsPfuvDGD11791vr3v3Xuve/de697917rs2sP683/ANvx791oVqeuxYWJsf8AD37rxqcddH68e/deHXXv3W+uyPz+D9PfutA+XXh/r29+6313wAQQCT9CPx791rJz1x9+6312b/U/n8+/daxw68frx7914de4sOORe5/r/T37r3n11791vr1r/wCw59+691737r3Xdz/X37r1B14H6/4i3+t791o9etza4/178e/derivXuR9D9Rzb37r3Hr1v8R7917rxFh9Qfp7916vXgRY8c+/deINeuvfut9e9+6914fUe/de65N9fpbj6f7H37rQ64+/db697917rsAn6e/daJpx669+6312Be/+Av7916tOuvfuvde9+691768n37r3Xvfuvde9+691yB03/P8ArfT37qpz11bi/v3W65p11791vr3v3Xuu7EC/9ffutVBNOvAE/T37rdaddggCxF+b+/dVIJOOuPv3VuuQIta1z+P9c+/dVINa9dW5seP9f37rdfPrr37rfXvfuvdePB/r7917rsC5+oH+v791omnXXv3W+ve/de67/H1/P0/4n37rXn11791vr3v3Xuve/de69e/19+691737r3XID83H+t+ffutV8uvMb/Tgf09+68BTj15f6kXH09+68fTr1vzxwf0/Q/X6W9+69Xy66Nr8e/deHz669+631yDf15HFx/re/dap6deJ/pcD+nv3XgPXrj791vr/19AAEj6e/depXrr37r3XYAP1NvfutGvl11791vrkCtrH/b29+60a1qOuPv3W+ve/de67JBPH/FPfutCvn17kC34PPv3XsVr14An6e/deJA49de/db68Db37r3Hrvkn/H6+/dawOuvfut9e9+691yBAH+P449+60RU9dMb8/4c/6/v3XgKdde/db67uLWtz/X37rWa9e4ta3N/fuvZrXrxBH1FvfuvAg8OvC1+eB79148MddH6m3v3W+ve/de67Nvxz791oV8+uvfut9e9+691737r3Xvfuvdd8mw+v8AT/W9+61gdeBt9Pfut8euv8Pfuvddi1+fp791418uuvfuvddm3Fv9j791rPXXv3W+u+Lf4+/dazXrxBH1Hv3XgQeHXdxptbn+th/X37r1DWvXH37rfXKwuLHj/H37rWade5XkfQ/T/Ee/dewePXH37rfXrW/2PPv3Xuuwbf635+n/ABPv3WiK9d8sSB+bm3Hv3XsDJ66II4Pv3XgQeHXbEXFrW4+g9+60K0z11ybgfn37rfDj14Dmx4/r79148Mdde/db697917rmAPxybfQ/T37qpJ8+HXEj6m30+vv3W/l16x+vv3Xq9Wgfyyf5c8/8xbtyi6po+wqbr+tr2yCw5OrhSWljGPofvGMq6HPqAtwP8ffvLrw408h0WXevxX33j/lRu34qdeUtV2HvTA78yuxsQuKp3lmzdVjak07VMMcK30m2r9P+w9+rjreklqDrYK21/wAJ5vjxsDYGC3X8yf5gWy/jRunI0NPNWdf7jxi1OYpK+VNb0EoiQMpUEarkWPF/eierqtQSfL8uga+QH8g3B0HSu7+8fhT8lsV8wdrbRpvuskNnUEUU1IFBaSKopUTy+kAjUOP8fflNa16q4pSnVTHwY+FO5/mP8yes/h/V5VuutydgbgnwNRkspTevBVMMBmc1dOVY8BbabH34mnXlWpp1f/2N/wAJ7/5dvTW7s11321/OL6m2dv3bVfNitw7eqNvtLJjMlTtomo5JfGoZ0PDgfQ3Huuonh05oXzqPzHRGvmh/Kx+AXxz6K3L2b05/M66++Q298QY1xnXO3sOtDX5RpONSSyJyAeCqm5/Hu1eqMAK06Lz/AC+/5SPb3zdpMnvbJ1FZ1l1Bgism4ux8lRePEYSiAGutrJKlCtrXIX6m3+PveBg9NrVvh4f6uHVmsH8hv+X7uLMS7F2F/Nm633Z2mqP4NjwYOOGpqpkTiGKoZLai/oAtyffunaLSv+bqj/52/Aft74Idix7N7IxtYmLy7TS7Vz1RAIoM/jEs0ORpGUBWSRTdSvv1eqEH7R0RH37r3XY+t/wOT9P+J9+60fTrxsSbfT/be/deHDrr37rfXvrz7917r3v3XuvA25Hv3XuPRj/idu2g2R3rsjceTqVpKShyULySsyooJnTSt245PvY6q3X0hPjt3FBv7pPD5Xb9T92Voi0ciSIWN4Rb9H+8e6PqMZ08en7fR9Uhl+Dz6SE26924iurMnuSqnGOjkZlppXkKSKSTpHNvp7IkuvoUkl3DK+VepSl2Feari0s+UhSX8TL5fs6AbtXcO0c88GRxcccNRKwabQL+s8Eej/ifcbb/AHO3XU3i2Y49Zn+0uzc27HZHbt8YsEFAT/s9JbYu1uxKrcOIrdmUdWk0k0TRzwrUBmIlHqAS3+39lVpDdGVTCCK+nUjb5uXLy2EsO7OraQcGnp1tc/Gih3vR9W4ePfxnbIGkTxmcyF9RgHF359zTs63AtFFzUn59czvcSTZ5d/lOy0CV8qU4/LoxNAWOLysY5H2VWbAkFf2bn6fj2aA1FKdAXTSnWi1/N27Bl293Nm6GBXSWWvMa6C121kqBx/r+9Chx1UjpYfylOgcxvjd535mI5DQ0M4qryLLpsH1jk/7x72TTrYFTXq9n5Ebkrcojbcx2v7Wl8cCxRlirKsYjJsn9Le/UBB6sGKEMpz5dMnTfUOJocPLncvjYhK0byXljBLWGq95PbZgszl46n8ulibzzLB2W14yp6VPRDfn12s+y9vVIwGO8AigFpYV0EEr9Lrb6+7rHCBQIKfYOm33bfzXTdMNXHJz1Ux1++V7YxFT91mJImrQzPG5YkC5JDX9obvarO5mWYKAV+XQh2PnzmTZbKbb5LhpY5vKpoOkjvD4s4aiiqK6qyUUsyXk8RUNqJ9RH09rlUIoUcB0F55WnmaduL9AfhdoYPbu4YIJdCu0qrBcaf7Yvb/euPbVz/uMw6opzU9XL/Djbq5iLc1KRcGm9BI+gA4K3+p/p7mTmI05B26np/k6L4h/jb/Poye0c1uXrvsGGGlln+3arCkKZFGnXYf4fT3DyCiAdGRIrnq8vrTeY3HtKjkdQakUpDsdJa5j/AK/j3sjrXrTqrj51bMoM5jcnNUhUfwsVvcXvGR+Pflr140Az1UN8KOvFg72X7F2k0ZBy6rrJX929+D7sT1Uccdbm+Chkp9nbeoZVN46SnupuSSYRx7qeGOr9JjtWTFp1vuGk3CI/4JNRyisWUKU0eAix1+ybmfebHl7l243fdMwQqSwP2GnQg5Nstzv+arW32Wq3TMNBHGuoenWtDvHbm26TduYq9spEce8zikWMABQJSeAvvil7nc8Q83c1XF7ZL4duWIX5jruzyHNvdpyhZ7ZvrEzxqNZPnj556FfO0excx1slJeBNwLTAafR5DIFtz+feOWwbhzZsnP67iAWslavnTj+zogsX5jtuY5HOr6dmNONKdFPwGwGSoNVVNcU8khRWH67HULX99qvYfftr9xdniktwFeADWMYIHWMv3jeet85QW5EWpknXFK46lbq75frbEZGklQwUccLgOH0C3hPP1t7ypjiWCMRJwHXMO9vZt0u33G4y7nNetNL+ZR35N2x2HU0CVbzU9LkHk8bS+QCNJHKWF+PVb3bpgmpHVX3v3W+u+Lf4+/dazXr1yP8AY+/dbp117917r3v3XuuR+gAP15I/x9+615564+/db67AHN+PfutGvl11791vr3v3Xuu+LD+v5/4j37r3XIaRYnk/04P+9+/dVNT1xH1Hv3Vj142J44Hv3Whwz165PHv3Xqdde/db69/r+/de67Nr8fT37rQ+fXr8W/Hv3W/n12SLWtz/AK3v3WqGvXH37rfXvfuvdeBI+nv3Xuve/de697917ruwueeLcf4+/daz14fUX5Hv3Wzw68SD9Bb37rQr59de/db67Fub/wBPfutHrr37rfXIDi7cA/T37rVfIddXFgLc+/deoa9de/db67JJ+vv3XqU669+691y+vpHIHN/fuq8MnroEj37rZFeuVhyTcD6j/H/D37rVTwHXEX/H54/2/v3VuvCwPI/2Hv3WjkY669+631737r3Xvfuvdd8W/wAb/wC8e/dazXrr37rfXdza349+69Tz68LX59+60fl11791vrvjSOTf+n49+61mvXgSPp791uleuvfuvddkEfX37rVa8OvGwPH4+nv3XhkZ66+vv3W+uyCPr791oGvXjpsLXv8An+nv3Xs1669+6312SCeP96t/vXv3WhXz669+6312bfj6e/de+3rr37r3XYBP09+61UDruwsf6j8f7H37r1c9dXFrW5/r7917Neuvfut9eH19+6912bH6e/daFfPrwFz7914mnXam3FyB/h7914jrifqffut9d2Nr/j37rVfLrr37rfXvfuvde9+6912SCeBb37rQBHHr/9DQAJv791oCnXgL/U29+68cdde/db67Av8Am3+P/Ee/daJp11Yn6c+/db697917r3v3Xuve/de65g/Uk3NrAH6/X37qpHkOur2Nwfrybf4/g+/dbpXj10Tf8W49+68BTr1uL3/2Hv3XvPrko4Jvb/Ye/daJzTrh791br3v3Xuve/de67AB/NvfutE9dg6T9P9gffuvEVHXRNze1vfuvAU669+6313y39Tb/AGPv3WsDrr37rfXvfuvde9+691yABHJtz/h791okjrj791vrsm9v8Bb37rXXVj791vr3v3XuvfT37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvddkk2vz/T37rVAOuvfut9dn8c34H/IvfutDrtvrxb/AGH/ABr37rS8Oujfi9/8L+/dbFPLrw5/w9+6314/697cD/W9+68OuWrgi3+8+916rpz1xBI+nvXVqV68ST9ffuvUp16xP0BPv3XqjrvUeLcWFvfuq0HXgQeDxfkn/jXv3XiKZHXH37q3XvfuvdctVrWFrD/b+/dV08evFib/AND+PfutgAdeDWBFvr7914ipr1szf8Jo7/7N3t8jgr/esg3/ACu2ZGB9+PAfb1aMdzfZ0fD+TzsPalJ82P5gXyozWBpMrvHpPuDdc+3MtWRpO+Kd8w6QvTQSgrcEgknnj35T1V61JHnjrWM/mE9+9rd8fKPuTOdk73zO6j/fzPVGPhr6mQ0lHBNVa4o6amuVXSpA/wBhxb34deqK1HVjn/Cdb5Ldk9XfzDOlOs4925YdO77zlTjewdiNMZsFuDGVtP4amGpopboSQeCRwebX96OOrKRQg8OP59WGdUdS0vxw/wCFNfTVZRJCm3ewu4Mrm8LSwhdNHS5VJIPAEW1gqshH+N/fiKAdbVtUhA9K9LP+Yl/wn07x7W+avyM7GwdfIMXvLs7c+4qQyViTA0+Tr2q4mQyXKgow4HH9PdlWigdUdy0hZeHVAX8wn+Wr2J/L9zuyMdviR5zurxVELSPExEbEMlljH9bjn3o0rTrQ1Uqetj/+Y52puL4Vfymvil1z1RmKnZ9N8nOloN27ujxYWM5SpEhpvJUyxAG5INhfj3rjg/b1d1KLVeBNPy8+tKfbW7tybV3Fj93bbzeQxW6sbX/xGkzNNVvFWRVKtrMomvcsTfVc8+7Z49UoBjy6Oz8xf5gvbHzT2d1Nt7tnI5HPZXqvBwYLG5jItA8slHDGVMauhLkXNxq596xx63UgUPDqvz37rfXY+tv68H37rR68Rb83/r7914GvXXv3W+ve/de67Av/AMV9+60TTruw1Wvx/X37r1TSvXlZkYMjMrKQyspKspU3Ugj6EHn37rfW5J/Jg+YC53ZsWwZ6iWrOLVaTxuhJDIAhFyb/APFfewSOHVdOpdLefWxDmdiJ2VLSU0jfa08ujVDYqG1R3UG3459k267Q264Z6L6dSNyJ7if1EkJtrcu7H46Vp0uelP5edNuHIZao3SYMdiFnElHNUykJLGrayUvxz7Au1cpSyX0sdyKIDgnh1lVz194OxtOWLG62aTxbqRB4qqMqaefVmWzNk9M9OY2lw9Jt/HZGvowEiyKpG7K1x6lYj+o9yFabXZWSCJUBK8D1h5v/AD1zJzFdNdyXDRq/FM9Gs23n/wCO4+nkB/yVV/ZjCqqogH4A/wAOPZgcmnp0E1GS9KseNelVg6qJMk8DqPDPFUxupHDB4iBz/re9jt68e7HWpb/Ns+GeT3t33jtw4ugMuNmq4J5yjOFYtNdlNh/T+nvQJ49bbyp1aH8IOkMf1L0zQx0tHFDX5GlYTCMAsA0fAJA+vvRqcdb6Hih6npa3J1OYy0avHG5kKSG+r8j68e6mvW8Vz0ld25+jWV9v4hViiQ+IhAABxp0i3vdK460DTHVaXzX60ps7s2aKenVvLATJIwP0MdgeR7316lOq4+sem8ZSYR0wuShp6qnjbyBJAHUgk/T3utetUoPt6WlL1KkkzS7gzqTodemKSS5b08Lb36tePXuHDok3Y3VEy9nY80xC0Qq4/BpPpKiS/H9fbFyP8XYnry5YenVsvw9xv8CyldAy2WWNUb/WAA4/4n3MXMJ/5Ae3r8v8nRfAP8bevViub6fgyc9PnKamjbTpldx9VuAx+g9xDHUJToyIFSa9Gj6gkbEUX2RNtKMpU/0C/ke98eteWOicfM6cVFBWQLbXJH6RzzdNI+n+v73WnXiKjoKP5aXxmkHYGT3nlaZZIHMk0PkJPOvyAkEe91xnrQ49bDFaqeYU8CBI6dY0jUDgBEC2t+PfqU63XoHfkjDj/wDQnvRKt0SZsZNpVmCsx8XBUe4i+8A0Q9od2RzQmI4rnh+3qU/Yr6g+6m1tEpIEgzThnrWlhCpJKqcKJHte9iNZHF/fEKWpUMeu4r6iqs3mB/g6iV9ZQ4l46ir9IkP6ieL/AOHtVY2N7utYLQaiPIdPW0FxeKYofLpkoBl8pnoamgqS2JfU7xD6WVDyP9695jfdU5jHLvMh2CVzHLO6gr6+vUOfeJ2rl4+1O4vuVsr3ccTaHIyppx6ry/mFdm7f2r19uKieRKTJ/ZOIWGlWZxAb2P1PPvqlcrpk09cNbAsYNTmuT1pQ7szlVuLcGTytZK00tTVzkMxJPjEzeMc/4e0/S3pOgc88e/dePDHXJv8AW4/B/HPPv3Wl64e/dW697917r3v3Xuve/de697917r1/z7917rsm5v791oCg669+631737r3XduL3/2Hv3WvPrwFyB/U29+62cCvXbcen+l+f9f37rQzn166ABvc2/p7914kjru/JNvqLf63+t7916nXH37rfXd+ALfS/Pv3WqZr11791vr1iPrx7917rsGwIt9f949+60RXrr37rfXYFzYm3v3Wjw68RY8c/wC+/wAPfuvA+vXXv3W+ve/de697917rsAH6m3v3WiSOHXiLf8V9+63117917rlfi3v3WqZr10b8X/2Hv3XhTy669+631737r3XJrEA35+lvfuqjBp14m4At9Pz791ulDXroX/H+9j/evfuvGnXXv3W+u7k/n6e/deoOvAfT8C/19+60euyOb/Uf1/5F7914dcffut9ch9bf14/2/v3Wj69dEWNvfuvDPXXv3W+ve/de67Jub2tf8D37rQwKdeHB5H+3/wB79+68cjrr37rfXK1rG4P049+61Xy69f1E2+otb37rVMU69YX/AFfi9+Pr7916ppw67PIBLXP9P9fn37rw40p1w9+6t1737r3XZJP19+69SnXXv3XuuwL/AF4H4P4v791onrr37rfXvfuvde9+6912Rb/Ef1/4j37rQPXgSPpx7916gPHrq/8AvPv3W+uwATybe/daPXXv3W+uQFuT+Pwfz/t/futccDrom5+lv8PfuvAUHXrkf4H37r2D11791vrkpsf9fj/effutEVHXiTyL8f74+/deA8+uPv3W+u78Ef1t/vHv3WqZ67v6bW/2P+xv7916ma9cffut9f/R+f8A+/de67Av791omnXiCP8AW5F/9b37r1evG44P+v7917Bz12B9COf8PfutE+R68eT9Lfg/69/futjA65aOPqPr9f8AiPe+q6s9Y/eur9e9+6912SLCwsfz791oA1669+631yCkjj/be/daJoevWNvr/sP9j791qor1x9+6t12QR9ffuvVr13p5AuORe45t791quK9dWPP9B+f9b37r1R12BqPJ/H+v7914mg64+/db697917r30+nv3Xuve/de67AubfT37rRwOu2IJ4Fv+J/23v3XgCOPXQNvxf37rx68OD/X37rZ65cFWP8AibfS/wDh791XNQOuIv8AQf73791s08+uvr791vr3v3Xuve/de67JBtYW9+60ARx669+6312Bcgf19+60cCvXXv3W+ve/de67BANyL/4e/daOR11791vr3v3XuvWI+v55/wBh7917r3v3XuuRB4sb/wDEe/dVB66H1+l/8Pfutnh14/U/6/v3Wxw69Ynn+nv3Xq9d6eL3/wBh/sffutVzTriBfge/db4ddjVYkXt+effutGlc9de/db697917r3v3XuuQF782t791omnXE8Ej37rfXvfuvde9+691s0/8JolLfLzbwBsWO6lFgSSW21IoHHvTcOtx/G32dHP/AJQ/Y20T/ME+c3xT3buim2vWd19ubuXGJWOtOtfImXd0hSeQqoN7C1/yOPewBQH8uqtqLlRw49a238xfoHsz4+/LLuTbXZG0MttYVO9twvt+eup2WlzONgrPHT1lBUn0yIyBWJH9ffvt68QPw8OrOf8AhOZ8UN3dlfN/rjv/AD1BWYvo3qPI1uY31vJw0GOxsNFTifTPUSAIQ9tCgNy3+t70eNOroCST5f5fTo5Pxy7oi+SH/CmbqOsMkddtbaHcmcw+1ZF5SfH46GWWlqeLixslrH6C/wCffiagdeVQJCV9OiTfzWf5knzq2F8//lBtrbXyM3vhMBju2940mJxmLrBFTUdDTZiSGCmWOdXsEUAKFsLfj3oZUV625o5A8qf4Oqdu6flj8hPknXYeo7z7R3F2LLiZYBQT7glgnlpI0cALHIiKQAPxf3bqpJPHrcC+ZuwF+fX8oLo3fnTVMexMl8UupYNobrhxTI9RgJpC1SIpoBqYgHVpIAFx78BTrzamX5VqP8vWlDtLYG7987sxuxNq7dyuZ3dksgMdT4KjpJXr3qTJ42jaG110n9Wr6e/cOPVQa5GR1ej/ADDP5MGK+Afxl6L7f7R7ZocH2j2/syk3RF1NPLBLl6I1FOJlhnRiWA5AJUCxuDyD78Knj1twqgaDX/V5da/h/rfkk3FrW9+69139RxwV/I+p9+6r5/b1x9+6t1737r3Xf0uLX/F/9b37rXHPXYHBN7W/3n37rxOeuPv3W+ux/wAQf969+68er/f5GO79q7c7KysG4qmCJqvI0/26SsBf0KDa597x59aAq2Ot/wD6kzXXsq09XUCmmVkheJrhrXiHPupBrjqwNOjR5nsGlpsE9TBPHRYeihctKNKqECXFz+fbhICFnIVQOqQWs9zcLbWql5JCAAKnieqn+3fmvBt/P1tLjqWHJQ00qIJlUt6S4X6KfceXvPUEFybe10vpOesweU/uq73vOyx7vuReFZASM0+fp1cD8Ke1P9LuwDk2pkpWipdWlbKbeMHlTzz9PYr2a+O6weORTrHz3G5R/qPuv7uVzIK0rn1p0bao/YkWVVuycf0/w49mvnToCeQPQQdudTbd7EoY8hXU0clVCi2J0ggr+kfT3on08uvVpnpGbY2wmCoKXFwxaaenXQBbgcWHvVa9aFTkjj0octTJFi6unSMF3jawH1N1+n+8+/fZ1sjj8uq/c4YsJuyrnrnCK1QWIY20gG/BPPvdOvefQOfIavw2f2bXRRyRTTCmcIi2/EJN7/6/vwB4nrRIrQdUY7P3Qu2d3Z7H5zM/wilqJZFp/ICAF8hHpv735Z618uhP3Fu7ZeHpf4hUb3p6hPFqjUsRckX0lgffq+dOt09D0Aey91v2r2bQUGLT7mhpK1I1niX0uBJ9dXti7P8Ai7EdbQ91OrbeutujbmRqjGPG8aoGBPJ+hN7+5h34kcjWAPDT/k6QRgfVOR1al0/msdm8C1BKUklEOkrcah6AB9f8fcQr8AYZ6MOOD0r6LGDHV8ui6rd+L8gH6Wt7scDqpoR0WDu7Zkm9czQUMMLSszorgc39YA5/r73jj1vNB1YN8eeucb1vs6nC06wVtRT6WHFyWj+n0/r73gjrWa9LndO/cVtSlqaytqIkqFjd4oHI/cYDhRz9PbkI1yBfPptzw9Cc/LqmX5G/IHtHtTdT7Mw1DVY3EyTClhWBWCZJGfx2uOPp75x/fb533PYkigubg2tjpbxADQMApNT8uulX3VOTeSLDY5OZr51nmjGqp4xnjivSN3X8WN143ZmPymHpqmuztTGGqcereqORl1kMAPfG/l/7wvLV/wAyz2W6MLeyUkRzsKLJQ07T1kFsvvVsd3zFLt986x26Giv656rd7Tr9w4Bzhd04uXE1tNIYwsivcuG0j6WPPvpP7Qco7BukMfM2zXq3ULjUVUgihFesr+TbfbN0X6/ZZhPG4rUenSh6Tpt3TJVZuo8h27j45pJZXUqqRiMkn1fj3l17c+3m0PzRHv1taaZAwbWPl+XWPH3v+Z+TNm9vr3aLmZUvZ4iFWoJJPWtJ/OC7b/je6a3Ebdyy+OCUxVSRMeU5EikKfyPeZ0ziR9Y64QWSPHDpf1P+HrXz9sdL+uYYW5Fz/Xj37qpBrjrjyeOT/h791vAz11791vr1v959+6914i3B9+69x699ffuvdd2PPH0+vv3Wqjrr37rfXvfuvdctJtf6cXH+Pv3Wq+XXH37rfXYIH1F/futGp4dde/db67Avc3+gvz+ffutE0669+6313Y2v+PfuvfLrxFrf4+/daBr11791vrkFv/gP6/1/wHv3Wiadcffut9e9+691737r3XYFzb37rRwOuzdeL/X37rwz1x9+631yP4stueD791UfM9euRwRc/wCP/G/fuvUrw64+/dW69c/T8e/de67I5sOffutVxU9etyAeL+/derio67va4PNuB791qlcjHXH37q3XvfuvdOGLxtfmMhR4vFUk9fk6+oipqCipovNNVVEraUhjjH1Yn6D37rR62Pfh7/IAftnp3E94fLTv7F/ELZ2RWSrjqd901Ii1VEiGRGgge0t3Fhxf68H3o8KjraZND/q+3oV8p/II+J/Z+E3PJ8PP5iexfkPv3b9DNLF1vt6hp6bL11Yikw09NJUli/kIAGn6kgD34VPW3AWnz61sO5el+yOg+ws91l2ptTK7P3ft2tqKSvw+YpmpqtPDIVDlWtcMADf/AB9+BrnrTArx6Cr3vr3XYBYhR+foCbD/AHn37rXz69z9P949+630uusdr43e/YOztnZjMx7dxO5dw4zD5LOyqrR4ijrKkQz5Bw4I0xKS5/1vfvLrwFTno3nzw+KvT3xT7Fg2V1R39iu+sbLh8Zkf7yYimipqX7ithWWqo1hQAgwFirE/Uj3oZHW3FG0johHvfWuu+bf4e/de67F+WFvTY/g8344Pv3XurBt7/A7c+zPgr1x846jJ1cm0Ox94VuzMTTPTRrStl8dJauhRymr9teTc25FvqPeqitOvaWpnh1XyfpyOSL34+h59761TPXH37rfXh9R/xH1/2Hv3Xuuz/r3/AMT7914de/Fv8fr7917z68Lfk2/2F/fuvdde/de697917rsgj6j37rVQeuvfut9d82F/ofp/t/fuvY669+691kf0sv54Bt+Oeffj1UZHXC/14+o/23v3W6dde/db6VeyNvnd+8dr7SWU0x3Nn8RgVmWMSPFLla9KKJ0Ug86pB+PfutUPlx6Nv8+fhdub4J9103SW7q2prM9NtPb+66gVkEcM0FLuGlWtoB6FUWZG1D/D6+/VB4deIIPd0R337rfXv9b37r3XIDWQouWJsL2A5/xPv3WuHXiNJIP6lYgj8cGx59+69k9ea/BItcf1v7914CmOuPv3W+ve/de697917r3v3Xuve/de67sQbHj37rVcVHXiLG17/wCPv3Xga9f/0tACxtf8fT/Y+/dermnXdhpvfn+lx/X+nv3WqmtOuPv3W+uySfr7914CnDrr6e/de697917r3v3XuuXA+o5/4j37quTw69xYm359+6359cffut9dkg2sLcf7z791oV8+ugSPp791vj1zAB4P6iLg/wCuf8PfuqkkfZ1w9+6t13yx/qffutYHXVje35+n+x9+638+ud7Cx/P+t+ffuq0qajrjxb/G/wDvHv3Vs1669+6913bgn+nv3Wq5p17i3+N/949+63117917rvi3+Pv3Ws167YAWsb/7G/v3XgSePXH37rfXdhpvfn+nv3WqmtOvG3Fv6f7z7914dclsAf6829+60a164gkG4+vv3ViK4PXj/vYuf9f8+/daHXXv3W+ve/de65AAj/H+l/futVIPy660k/717916o669+6313xb6HV791rNfl1malqUQSPBMsbC4donCkf8ABiLe/der1gsffut9d2Nr/j37rVfLrxv+TfgfT37rw669+6312Ljkfj/W9+60aHB69c3v+ffut08uuz/X63Fz/wAT791odclBchIlZnfgIq6mYniwAuf949+61Q+fT6Np7iMAqBiK3xEXDeFvpa97fX37rdemF0khcpIjRyISGV1KspHFirc+/db49dANYgfT8/T8/wCv791Ula9cffurdd2Nr/j37rVRWnXQF+Pfut9e9+6912Ra3+Pv3Wq9eFr8/T37rZ+XXXv3XurQf5ZH8xmo/l09uUPa9H19Tdg1mPfIPFi6yZIaVxkKD7JlkYuh4Bvx/re/Hh1pcMT6inRVux/kvvLc3yc3n8nNk1NX1/uvc2+8rvfG/wAJqGimwtRkao1L0kMsZPpF9P15H59+pjPW9Wajq/LAfz9Pjp2XsnY+3vmv8Cto/J3duzsXS0NPv7ceSjpsoamCPQ1XIY2LSA2H6lv+be/ZIz1tSBx8+g2+Sf8APmwO5ukt19DfDj41Yj4j7K3XCkOVptp10c5yLpcNUVdQH8pve+n6f4e/LjjxPXnOqlP83+Dqq/4A/NzOfCT5m9Y/MCt28nZuc67zFZmHwmVqCqZuqqKU03+VVAaNgOb6gwPvRr1pCFx5dBn8zvklJ8tvkf2n8gZtuQ7UqOzd3ZrddRgadleDHS5irNW9LCwZrqhJAN/fgKCnW2Opy3r0Vr3vrXVt/wDLf/mu9jfACLce2YcHLv7rTeEyndewqur04jPUdgWpK2OVwpFy5Bt+ffsZ9evKzDH4erUqL+f38D9o5et7A60/lS9c7L7grVcjsCmzMU9TSTuL+WnpZHsCH9YN+D71mvV6ihAJz9nVFHzw+eHa/wA8+zk7C7MyNfPFjPuoNuYusqnniweLnK+PHUia2RI0CgKF/wCJ9+pmvTdTShwBwp0RX3vrfXvfuvddkAWsb3+vv3WgT11791vr3v3Xuve/de697917rv8AB/2Hv3Wuhx+PvbOW6h7Fwm5KCskp6eOri+6jV3VH9Y0MSp4I/r/T37r3X0Ef5efyExncmzcBUvkIpKsUdKZ4vOGcN4FX6Fr/AI/PvYrw603CvDh0eT5m72z21NnYrGYCtaCnyFIjVMalrSBk5F09xj7lXG4W1p/iTlVpmn2dZzfcq2Pk/e99MXMtus0mrtJAqM4pXqqnbuDq92yMakPI8xBml9Z1FTcE6uPr7wy5393LLk8IUAmnPxCvD7eurW+cv2m2QvbRkeAPgSgwD9nR5fi98ld3/HbedJhK6aWbblU5iMUkkixCL9Nj+L2+nsf+zvvpBzNdJA58NyQNFeP2dYr+9nsXyxzDyXc7xbooulFQQBUHrY82R2XiOxMDi8/QSwkZeCOZYI3UgXQGwAPvNqNvGiE4wGAPXHHcLRtq3CXapTVoSQf29KyUyKrU7HUst9Kk30/778e/dM8OPSTylVS4aJnmVOQLfQAW55/4j3oA8etfb0m6PJY/PFhDKBIt1Ef+1N6Qf+R+99aI9OigfILpHcWfqlrMCZgzsGYRIx/4NcL791uuM9Artv44bgqYGj3BHNNHZgyyIbadJ4Ov/Y+/V8uvUoNQ6qA+evxqg2zV/wAbxVOaOOnkJlZAFBJYkklfex1ph1WBuvFDJ7aoqGKMyS6FR5QX/wBVb+vvfn1Ug9H4+DXTLYeegzksDMGcy6mUG5HItq549prsVt2A+X+HpxcHqw/Gz1mU3tl8VQxMT5QrFQbgA6Tx7mPmDHJNgPLT/hHRZDX6pyOrDOk9kZnClaqqWVI5FBsykAgqG4/w9xAh7AOHRkQAejBtjKioyYZRqVj+m3FgOR/r+7V9OtEUHThh9gR1GcXITRXMbhk1KOPVqFiffsmo63Qmg6MvQRrULBRxRgFLA2HICrY2A92Hy615VJx1VD8/dw5HAeSnx+YajrrAQQq7KzsW/AB9xr7u86xcg8rTbn/ooUlRWnU1+wPIj88c6Q2t3EXtddGNMU/PoPPh/Ed44aXK7liSty2IYmGeZQzRnWLMrOL+/m/++R7ic8+4+/xwXt+y2DdzRVOR/CD8/P8A1DrNr3W5e2/2zij2rlgeFHOKPpxX8uj7k8HkggcHkcAf6oe8KWsNuuYksp46wp8K+n2dY5a5QS4OT1Vd8gOqMR2R2dB91HGiw1avOCq2cIeCR76h/ce2vmfmiGLYdimaK2B00BJooNKZ6zE5a92Z/az2yfdJCSxjOnj59Ef+dHyF2B8a+rsztDb01LBlqjFTQSRJJDHI8jRGOx088/4e/oA5W5fh5W2GDaaapkUa28yadcu/cf3I3/3T5hk3vdZi0Oo+GpJxQ9aCHyd37mt99g5PP1NbI1JkXf8AyfWzKrhtTX1/1vxz7PegN50PRZffut9et/vPv3XuuQBGr+otzxxc+/dVwadcffurdcieF/wv/vfv3WvM9cSb8n37rfDr309+691zuSOPr9GP9f6e/dVoAc9cRYXuPfutmvl14WJF+B79141pjrkdJH1+nH1HP9PfutCoPXD37q3XvfuvdciAAObnj8/4e/daBNeurE3/AMPfut165WUAH6/4e/dVycdcbm1vx791ann17jj8f19+61nrx4PHPv3Xhw6750/UWv8AT8+/de8+ujb8fT37rf29dW+h/r7917rskWFhyPr/AI+/daFa9e0n/fEe/deqOuvfut9dkWNj7916vXjccX+nv3WsHPXjc8n/AG/v3XsDA669+631737r3Xgbcj37r3Hrs3PJ/wBv791oUGB13bgW5P55HHv3Wq+vXVhYH8+/db8+u7C1783+nv3XqmvVoX8oroSj70+avT1DkiXxe2t87czGRprIyT0kNdrmR1k4I0qwseOffqVHXlNHFeHVhH/CkL5P9jZT5+dqdD7Z3PksT0hsnEbcwm19kU8zR4ygp1x6+Z1hisgdmUkso/PvwJrXqzgaQvlxPz6pg+F3cfYPSPyQ6n3f1tuvJ7Ty0e89uCtqqKdokq6SHJrK9NUqn6o7X4/r799nVQc1by62af8AhQT1Rh+7p/i93fS4yipN/wDyb3HtvatVuj7aMeatZKWglq5Y4ApYlpLsLi/5I+vv2Ot0J/LH5dDBvnq7+Wv/ACXupurOq/lH8eNpfKzvHeEeJqt312YqIsXNtDFbgpkmgrJIwnkJRX12dRa3PHvRFeH59b1BasRjyxk/PqsP+aJ8RviduLEfHruv4h53AYvF/IveVDtKn2FikglXaNXkatEjdzS2MiAP6bkXsefz72eNB1qgajDj/I/Pqz7fvVH8tv8Aks9VdZdT/Kb487R+VveW8Ew0+7K3MVNPjJNnYzMwJVCsmVE1lwr3swAFvfjT7T5/Lr1dJqcDy86/P7Oq+fn98X/hz/eL4efKP4w5Hbq7K7t7hwWBzvU2GamK7ex1XV0zTxCSC2pGEmmNm/o349++zrdASD6/s+3oe/nd8H/j/tj+aP1109tTY1JRbN3BQdcwzYdliqdcmepaZ6sesEXZpDwfevwmnl140MiKfxDqwX+YFD/J+/lQ917U6h3N8Ttjd+bp3NDt7HbmwFbU0tBUbLiyNLEazKzeBSTIhcmx9P8At/e8eXWgdJOsY8jT/P1St/Oq/lx9P9Sdddf/ADv+N1ZQ4/ojvbIwUG2doY+GM0mErXiWapx8dTCAjGDyFSQSeOR9PeuH59bpj0/y/PrXd2flMXgN17dzWbxEefw+KytBkMnhZWCxZWhp6hZqigcngCRQUN/e+qV6+gL3983viJQ/8J3+mt95L4dbbyuyd6bvz+yNidezVEaUWzN7UdBDFUbxhlHDySO6sV+n+vYe6/L16UcFrWv+r/J1UF/LO+DvxW6h+K2Y/mk/NvG4TdPTG5MjmsLsXqavhSKDKZunneWLF09RMtrLYLrBHptxb3sEdMkFRU/8UOjY7UxP8rX+b90j3Jg/jb8ddm/E/u3qPrvc2+IcPj3FbPuehwEPki0mqRXBmU3BQfUEfUG3jju63VZKpQ/bSn7Pl0Vj/hPp/L/6F+UGe70x/wAj9qUE2M6qwO7dxVlTlqanK0tHgKeSbzztOCQgKfUH6e9nyHr1VQGViOIIHVjnxOoP5P8A84O4+wvgHtX4xbK23uGLGZalwXyWoauklkosxGz0tOsuPddGhZLC4NwRzY+60PD16uGBH+l444/Z0EFT1/8AyvP5anbsfwA7y6H2X8nOzty7tXC/6YHqqamlwUO4J1ShloqXT6vt7/7sIBJsPxfdCQT6dbJj1qgrQ/Kv7eq7vmN/Ly6/6O/nG7d6Dxe2KOr6X3NUbSqMZ9vTU64yel3M8c0VPAhAQsqyBbke91wT6dU0qHVf4j1cd/MHof5Qf8pvtfZfW26vijsTvnd+6MdtaLcO1a2qp8fU7Ygy9JBNVZSUQrcsglJJPAP1+o9648OrDtrr4eVB1TH/ADpf5dvS3X/UPXn8wT4xT0OD6G7ozFLgsLsegigaHBZeoj+4qqAVVONLeC5W+q5A5Hvwxwz1qh4kU/wHrWsf+yPwQp/2JFzz731QCleujYfTm3N/xb/H37rwqePW03/KO/l2fHrbPxk3B/Mr+XsWM3x0vjDVbdwfW+Sip4Yc3ualQzmg+5qBbU5AF1NwD9PyfYPHqxB4ev7AOjB5Jf5Vn8zXpXtN+m+mNj/ETs/rfbm4szRYTHVVNXzbiGDUimi1yqrq9QAStvyOOPeqEZ63VXag/bw/P7Oi8/yvP5cHQW0/iZvT+ZJ8v58Nk+q9u72zmzMB13moIlh3Nk8KrVEdFTVFSttUoUA2Nxf3uvWtJxX9vlTqwbpXr7+Vz/OW657L60+Ofx12P8W+8OvNm7o3FiYcdW0mRqN3rgaaWopY9MqhladYwFK8avpx71kUPn17sclc0xmlD/xXROf5F38s7qDtjvTuvrj5ZbMo5aLqzC7y3PkDlqaALHittCSV5pDUi6oUjJ497Pl8+vKPj9VIp0O3bXy8/k5dK9ldW7Q6V+HmwuyN70ncOG29Duajr6Yz4qshy0dMle1PGAbxuymxtz70Rio6sjgtoI/YOn//AIVRd2dH9jfKHqP4x4P4/wCEx/e2a2h1YmQ701xtlXxe8sbRRYfAfZryY6TzoAQbkJ9fevOvVmHaBxJP+Xpp3D1Z/LY/kj9O9d7P+XfRG1flz8hN+UmCz2T21l70Eu28BmqKPIGqPijMjEJJex/w/wAPe6dUFAc/7J/2Oih/zS/ir8RuyPiHsT+ZL8K9uYHauwcrvrH0Gc6kxSRyJgpImWesoqudFD+NRrWTVyLG4vx72TQ061QOusYP2f5OhV64+Dnxn/mg/wAsrbG4vjT17juv/mBtbeIxm4tu4Wlpq2uzuHok0tkkhp/3WiYi7Lwbe/efXtLFTppXy+3/ADdNPevxA+K/8r/+XTSJ3n19jOx/mBurdNTSfwTLU9HS5LbmFqgRFXzxzgSKod/QpuQoH1tz7FevFG092P8AP/sdamGSqI6uvrayKCOlirKqoqoaWI3jpo55jIkC/wCCg6R7959aFeHUIgA8G/v3Xhnj14W/Pv3XjXy669+631737r3XvfuvdcvqLk8/7D/evfuq8DQdcffurdf/09AC5tb8Xv791qma9ePP4t791vrr37r3XZAB4N/8ffutCp49dWJ+nPv3W+ve/de697917rskn6+/depTr1za349+61TNeurG1/x791vr3v3Xuu7G17cf19+61UcOvXPH+Atx7916nXX19+6312CQbj37rRFcdePBB+v5/wBj7917iOvE3N/fuvAU669+631737r3XIXIIAv791o0GT10Db8A/wCuPfuvEV669+6313cWtYf6/wCffutUz11791vr3v3Xuve/de65ACxN+fwP6+/daqa064+/db67vzew/wBb8e/dap16/wDgPfuvdcrAEW5/r+bc+99aqSM9cT9T/rn3rrY4ddre/pBY/gAE/wCH0Hv3Xjkdd2ZTzqVj+CCCb/6/v3XiBw6We3+u94boRZcLhqmrjc2VwulSb24v7916o6sG+LXwN3nvvJQ5rduKNLRwTReOjqFYoyluXYkWJ/3j3v7evZ6tYzP8u7beY27/AA2mwlHTVIiKeYwqLnRYG9v959++zrdPXqtPvP8Aljb82TTVuc26r1cCOW+2jRmiBILEW+oH+sfej8utUI86DqvPcnSHZO06Wrrcvt6eGjpCRNMt2UafqdP19+60aefl0En0PI/rcH37rdMU669+6313fi349+69TNepuOx1ZlauGhx8ElTVTsEjiQXJJNh791qvRndk/ETtTdtVRo+Kejp6kRvrVHkfQ7WP4tx78cdezxHVqfRP8vbb204KPIb0FLWTyqZZBUxh5A1wQvqHFv6e/Urw63jiejtVnxp6ejxoijw1AbQkX8MZ+o/1r+9nPXq8Oq6vkD8JsFuGeWp2jSwUjxtK6/bpoYnR6R6Bzz79THWvs6rS3H8W+zsJXT0tLiZK5IpGUOFkR7L+TYW96HXqdAzuDYW7dss4zWFq6PQfUzRsyi5/LL7916vSRJsNNufzccjn37r1M168D9BYD/H8/T+vv3XiPPrj791vr3v3Xuve/de697917rkrMpupIPP+88H37rVAeuiSSSeSfqffut9d6mta5t791qg67u2kgkgfgfggn37r2K468GtyLAgcEfW/9ffutUNeuJJPJ59+6tw65FVsfV/rf4+99VqfTriGI+hI9663QHrvUw/J9+69Qdeb6/Un88+/deHDrj791vr3v3XuvWI+vv3Xuve/de697917r3v3Xuve/de67/B/2Hv3WuvA2IPIsb3H1H+t791s1pjq2j+Wn84N29FdpYPB5fOVx29V1ESBmm9EUKMqmJg3+3B/p72MnqrHSvW5z3B2zB3X1nsXObdqxWHJ0lEylWViYyF8typP4v7hL3o39Ng297qYViVCT9tMddI/7v8A2bZ7me63XdaBoNRWvGo4fz6GraW39jpsfB4qiWnh3JVUwSYqqhxKVAuzDn6++EHOHMXNO4c63u9XGo2Iaqgk8Osxt43LmE79cXk+o2qN25xSp/ydF+7q2Lm9pS42pljNXrYFXRraST9br7yG+7lzhsm58wC61+EYj219ehPtG+7JzFs11a3FFAHA9HQ+CHY3aG6N5YDD0zVv8CxMiR1MJeQxpEraT/hb31+5T3a93SGNUJMagZ8iKdcvPvAcocn8tXlzeRFPGnJ00pWvWwlkKZkjp6i3rZU1L/qfQbn/AG/uSGGBTrDJWJJB8ugh7NpaibDmSK6sqDkfm4vYFfdT8+r4NCOg+6vo5BNqlJV2v6mueT+OffsV69ngejKyfbw06idUdFUl3KqSot9Tf3o/PrdBT59AJvvsnbOE89OKyFX5DaVjuLqfoPe6Hj1Xy6qS+VFHTdx4rIYjC6KmRl0llT6kMRf0/wBPe/n1YnqkLdOwptsZJttfZiSuhmMQXSSdXk/wHvfnnqopXq0D4qx4vEYTF0OZdKOoCurRsoVhcDghv9a/tNd1W2any/w9WU91B0bHpWu6/pezs993LBJJJP8AtkhW0kP+P9j7l/ftf9SrGvmv+ToviNLlurPRUUZo4XxqKICi6dKgWXxi1rf7D3EaU0dGB456l42K8kbhPUxHNvoD9Ofd6AZ60c8OhMpIDDGrKhaTSB9Prf8A1ve69ez59CDtanQVKSSC0pDNY2HIH1F/dky2Ombiqw0/b1QL/NBqar/Stt2CmqSkT1aLNGH4YeXm4HvEP73zsuwqCe3w+HXRb7lUNsYJpXSrAmh/LqX8W+wqHaFFkqKrhMnncNcXF7ngXH+PvgV72cu7hul1BfWLDswVPmOp497eUbnmOeGaFqBej4ZnfeLweBotxVi2oqxdf5Ghf8T7xnsYbi/3j91bfGHueJT0/wA3WL228mXu57rLtMJ74zT7eqg/lH3wmIylTura8jtDAxlkMVwVQMQSLf63vqp9yKHe/b7fIIJuwyMSVHkTmn8+szeVPZ605l5Mm5c5lUV8M6S3yFetI/8AmhfKzcHZnZmWx1FmauOeinDVERmJIiaXkFb2PvvdZ3TX9hFeyfFIBX9nXGvnjaI+W+fNz5ctR+haSsqkcOPl1TzkcvXZRg9bM0rAk3Y35P59vdB2ma9Nn+Pv3W+u+SABzb6ce/dawM9eJJv/AI/X37r1B11791vrsgX/AMP6/wC9+/darjrsgD6G/P8Avj791oEnj1x9+6t1zFjfnT9OBxf37qpx8+uJ+p/1/furDh16xHNuP8fpz791qoOOuj/tvfut9e9+6912Bc29+60cDrs8Ei3+3HPv3XhnPXVyOPp/xv37r2DnrwAJ549+68a+XXXv3W+uRIsOByOf6j37qvn11YWBvyb3H9Pfut5r16/FuP8Aiffut9de/de67P0HN/rx/Tn37rXn13wADwT+QffutZOOvXa3+Hv3W6Dr3NuAOPzb+nPv3XsV49cfr791vrux+pvz+ffutY4deubW/Hv3XqefXXv3W+ve/de65G30FuPz+T791oV4nrq5tb8e/depmvXgSPp7916leuvfut9e9+691Yd/LD+QNZ8e/mH0tuIkR4TKdgbYo9xyhwrw4h8gFqpADwQEZiQbce/HhTry0LgngOrWP+FKfxc3HifmPub5a7Hhmz3QfceI21ldq7tgRno5qk0KR1NIjRAoDE7HyC/Fveh1ZwRQ+XD8+qT/AIS9Jdi97fJHqjaPXG08puuu/vptt8rHQU7yQ43Hz5RYpquslX9CBbm5/p/S/v1T5dUx59bfP8zHcm0dv/KT4YfEdJ4d51PUHYGyKzIUyKJocPVzvSTVsQdbi8TqQ3+t78RgsOI6ujUkVWPHPVXP/CovrHtNP5mG/wDe9Xjcpkthbg29suLaWRgEstFLD/CI0ipoYo7jULgeke9ipx1qRkFD8ui4fFv4rdr9ES/DDvfv2bIRdP7w7u2xHtDa+TWRqSjqqfNRSVVRBHNwGIcFhawJPvzfDTz61Gp8VX8ujAf8Kk9ldiwfzMd9b7ysFbXbB3VhNqjZGUUM1DLAcRGUp4I4/QGsVC6Rza3vS1p1uQgkZ8v2U8uiXdX/AA5+RnWu2PhV33vyfLP1Hu7uzb8e29uVv3K01DUJl6eaapgifjUFdQ2oem50m177JFKdajUlwwGD/q/n1slfPrZO68j/ADkOnM1j8dL/AA+deo3p6wK+iJBTUf7t/p6be/D4T1uoEyfn1Sv/AMKYesOx8N/NE7FyWdw+RqKPdNFtyk2/ltE0sGXnFIB44XbgMS4AA4P1/r70oNOtyMBSvp0e35vYjMdS/wDCbL4YbJ7ZxpyG78h3JuGvxeKrpP8AKcNj6mGD7OobyXcDlSPx6ePeqmvyz+fWzQx0HGg/LPWoVUbVz9LhINxz4yeLC1baKeuZT4ZC36VVj/vHu9Dx6aqK062nPlEjr/wmH+Frknxn5Jb2BFydVqajNiR9P9c+6Dj08a+H/q+fVrHWHyL6S6W/4T1fH3sPenxGxfzH62l7OraSr66rKmroqfZNfRRWq89UNjiJhr0sLONP+x9+pU8PPHXmdVUZ8hXokPxm/nTfGvG5zsaP4CfyYNuQ9p7j2Hl6beeR27lMrmqmg2r9t48jUmKNmWFLFvqBexH0BHvfW61HmafZ0LX8oXsd+zq/5/77pdlP1Uc18eezI8nsOzQvQzVGJm+5iIWxFmJF/fmwB1SKp1E/xDqmr+RVek+cm/WpXaB6ejrPA6uwZPHm59B1fU2sPr72OI6oePSY/mNSTZH+bjgK2vmlqKyq7C2G08zyuS5gzcZisSfx/QfX8+/evW1yR+XV8n8ymRn/AJpXxrXWI5PsemwkxADofDTaXLHnj63J96Uko3Vm/tYwOqdP+FOew99Yb+Z5uzJ56hrKij3Ns3YkeAybeZ48oy4eJfHCW9IP7iKAPrz/AE9+HXpDQ93p/gPR4vlJh831F/wmO+NG0+2cc1fubOfIDJV2BxtdNrqsXQVkDGkqrynyBbFebfjj34cc+vXqgpg8QPyz1qJTbUz0eFXc74qoiwU8xSKs0EQ3ZyFVS31H4B92p59N6hXT59MERjWWMyJ5IkfU6g6S6D1Mmr/EC3vXn17NPn1uRds4HPdkf8JfOmF6YoZkottfIbM5HeWFx7COpWgNDGRVypCdZFlK+rjj3UfF+fTrMojof4RX55611/gP8Vu+PkjvfeMvT1blMLBsLa+R3LvPJUrTwo+38av3WUpahhZWUIp8gb6XH593qBk9NFC4ovWyL8uNt5Hs7/hN103/AKE4v47iNhfIzcVN2FQYvmSCso42etyE0NNy5ZQeX4/HutKtjpzUVjGo4oOq7f8AhNFsTe28vm7uubZ33NPLt/rDNZbK1iO8MUGLo4pqjILM/C28aG4Y8D/X978x1oV0mn+qvV9/xB3zit4fLP515zas6fYr8eO18NXyU2hFlmjxFRT1PMXHLX9X196auD1aM0DAeor1o69WIE+XeyIkvoj77xKIGa50pvdVW5P14H19+8utDL/n1su/z58tt/C/z1+l63dUaTYk7I+PUCtILxU1TV4Oggo6n1ekhZCB/gTf3ryP2dXOoOh8q/5+rWP54Hzx+IPxp+QXXW3fkr/LS238nYKnp/ZEFL3jmcvXY5czT1G3IGgoKY0rCC6xkICTq4/PvwGK068zr5Gta+XVXXcX83Db3Y3wIznXXQX8qCh6m+I1NXZGGm3jh3yVfj6bcsqBJqmTI1WuOUx/2iL6bHjj3v8AFjrTV0E54fl0/wD/AAmA673j1X3Hk/m/2BvVOuPjBHV5fb9S9XNKMGM9VFylEVmGjUoIU6FPHNr+9lhw6bVGANBStM/5egr/AOFOfSfYW9+76v5nbX3tJvr477ympsJt7NUzyfwv+JRFYzSxRwftKy/0IBtY/X37FOtspweNP5f8X1qXEWA/xGr/AGB9+6qOPXRN/wCn+w9+63117917rsgA8G/v3WhXz69+P8b+/de8+uRAX8XuPz+PfutDPXG/14HP+8f63v3W+uvfut9f/9T5/wD7917rkbAAcE2BuP8AH8e/dVHGvXHn8+/dW697917rsXHIv/rj/ivv3WscOuvfut9e9+6912DY/S/v3Wjnrr6n/iPfut9cgCeCbW55/wCKe/daJAyOuPv3W+u/x9f9h/X37rXn11791vr309+691737r3Xvfuvde9+691737r3Xvfuvdcg1gR/X8+/daIqa9cffut9e9+691737r3Xvfuvde9+6913yeef9f37r2OuvfuvddgX/NvfutHHXrcXv/sPfuvVzTrr37rfXJSQeBf/AA5/4j37rRFevWJIsCSxsAOSSTa1vfuvdWlfD34i03YGIO5dyUi6nMZjhqHsPFI/pKqw4459+8sdaHr0cHdP8vLA5WrpqzG4+nj8UqExo4BcKbkWCm4/2HvfXqAdHc6R+IO2tu0NG+UwtPRxU6oZC5ULIUA5AK/n3rq2K9HLo8xsfY4ixWMoqaOKFAGkjjT9Q/xHvY68aGhOOpdb3Ztyijv5IUCC1ylwCOfqffjSnWgadBhvDt2DdGGq8bTUsckEqswl8ShW4/1RB9661UdE2/urtPeuM3Lg9x4qmDSwV8kcsqIpuqFl/s+/dezxHWtb3XgaDbfZm6cTjdAo6fJTCJY7BEGsrZQP9b37qw6Cr37r3XvfuvdGc+JlHjazt3FJlIo5qWOLyFJACCyyLawPv3Wutpr4yYfam6aKraGnpUOOAjjXRGpA8mn6W9+63xyejTZPrShyayBdKBSFVgwGn8AD3vrxz0WvsHqnceMy1PBiqqWSlmVC/jc6RqHI49660KeXSh230/Uw+E1wEplVS92D2J5sQfe+vU6UeX+OmCnp5cgKGB5ApZk9F2YrduLe9dbr5dApX/GDqzd8dVRbk2vQ0sxDqtRMUTUbFQxutv8AePfqAcOvE+R6rL+Q38sWjmnrMxsXQkUYlkVKCaMXC3YI6aCD/tve+tfZ1S92Z1Vurq3NTYrcePmpUE7xU88gJEoTn1GwAP8Ah711vPQZ+/de67Av9Tb37rRx148H63/x9+62OuvfuvdcuDYcC35/r791rhnr2ni/9m9rj/b/AE9+69XroAm9ubC/+wHv3W6049dj+pN7fQH37rXyHXX1ufp/h7917rw4/wAffut9de/de65W4vcfjj/X9+61XNOuPv3W+uTcm/1Fvfuqr6dcffurddg2/wCIP9PfutEV69yf6n/effut4HXgbAgj+v8AsPfutU669+631737r3XduCb/AOw/r791queuvfut9eH+39+691KpaqeiqYKyllaGop5UlhdGIZHQ3BBWx9+6rSo0nrZ8/lpfNnJ7t23h9gZiKWUYGFaOGWeMsI3SwLRuS17/ANb+4n92+XbLmXlybb7xtIkUivDy6y/+6fvu82G//ujbVLCdgGp9vV5GF7OFJWJVRlknjYPFKAfSdX9D75nc4ew8tvEYdvkFwjcVFPTrsUvK091tiJciuodw6U2V7aqN3V1JhcnU/fvUELE7KP21vb0k3+nsj9u/YO9Te4ookNkjHuYGn+boFcycqpyxsE+7WNvXSMgefV/nwU+P1BsLZ+P3sk8Uk24KRJzFqR3QsNQ4sLe+vXJOwjlzl6Cy8TxSqgFuNcdcR/ernOfm3m+e3uIyn07mgOKZ6sSnUVRCXsLAAXv9Pz/vHsV5pTqJqAcOmHLYBK6naGSzqeFBI/Tbm1/euGT1vjjpG0G26XEyM8TKGQ/pU/n+h96yOqj59Ydy19SuLqRH9WiYAr9blCAP8ffj1ug4nqvzdHV2b3XlqqefJvBA73AMjLwOPe+B62aU66wPUOJ2hQ5GtnniyEhVuSwfQbX5v78a06pgYPVXuC2XhOxu+s1FLRRslFk5+TpCgCX+1ce90oM9b4mo6GDsbqaTbG6aB8XMKWmZkLeFiVQG/LBbAe013/uO35dXXGelx8d+tsTnt/5aony8RmppdTAyEEkNyfz7mDf3J5JsFI/CP8HRdEF+pY+nVrFPR/w1aehhlLxRqihgLg+gLce4jTKgDowbjXoUcNQRiKOYsFKryGvdifz7uMdV49KQzR0Q89RKIoQL3YiwAF/+Re99bPqega3j8hcRs6UClmjnljuLhQxBv9Lg/wCHvynqsi61I6pN+WGQyXa28oN0GRvDS1H3C3uFsJL8D+h94rfe42yaflD68fCkees5fuc862G37o2xSf2khIHQp/FOv27UT1OGy+GSvqK2UpFUst/F6r/U398AffC03dDb39jcGKNCNS14gjrK73ztt3igW+264MKxgkgHj1YJurC7Y/u1NRZqGIYqCBxHHIQqIdPp+vvGLxbva9we52jG4SU8MjLNn/V6dYnct7lvzb2lztzMZmYFiOJz1QV8uabbMeD3lFtqphangppvGkLAqAWa4H+x99bful2u/wC5bvt8nMCFZKAuT60HXQfaN63/AGvlJtxuoWeUxnjxrTr5+Xyomll72348hOr+Isn1JuoUj/be+5e3RrFYxRoagKOuJfuLcyXnO25XUy6Xed2I9CT0Xe3BP9Lf7z7W9Arz68D9Ra9/p/h/re/deI67H550+/deP7euPv3W+uyb24tb/effutDrr37rfXduP8f6e/dar11791vr3v3Xuve/de65XsfrcD/be/dapUenXE8n37rfXvfuvddgXP1tYXv791o9e5PPP+v791vHDrrn6/7z7917r3v3Xuve/de67J+nH/G/futdde/db697917r30+vv3Xuu/wP99b37r3XXv3Xuuwbf48fn8f4+/daI68DYEf1/wAf959+68RXrqxP0Hv3W+u+bc3t+P6e/dax11791vr3v3Xuve/de67H55tx/vh7917rwF782t791omnXXv3W+uzyf6e/deHXYNvwCffutEV6HXrbobuve1Rj8tsPa+WmmWVaigyECvEYpEN0mR1BI/wNvfh6jrWDjrcf/l99w7ok6C2t8d/nr05W/IHr3AzvLjMduEShcQ00RXXSVmhnAHAZb2P149+pXqynSCP9X59Wndebr+Cvx3otw1/xO+JmI6f7B3JQmnqN2U3+VTwzMt454llUBGXj1A34+p9+Hy68Tqwcj7Oq+919XbYzPYcvc+6aNMz2gcjJlf70TuXqTOZPJC1nvyv4597x+3rVSDny6Ojlfm18ZOwNk4Xafyr+NeI7l3DtCaGpx26clrgrpxSKI6emeVAzMqhQACfetPr1YOFPz+zogfy67+2p8rW2PtPEbFo9l9b9Y5UZXrrZdND46TbmQSVXSpp3ABLHStyffgM93Wmeoqvn0e3bfyJ+Nfc/XO2dnfMD4y0HdW4dlHH1GH3jkWeCuZKGFUhp5JI1JZbKALn/Ye9gVPXtVDj88Yr0GHePeWxe1Mn1zgKbrKjwfTPUW4qTceyOt44VWixVRRSI4MUhW7M3iTUSBe3vVBTreo1r0OMPee3/l58teud8nY1LtvKUtftrCQLDCIfGmLMcEEg1D8BBa3vVKDPXqhmBXy6Gz+ZX8t/iHtz5FxbP+RfxEwHcW4Ousfisni9w17yQVE1ZjqePxmV4A2tSQCQTzz73pJ4deLgEA8T8utO7+bL8rd6/NPetHU4uim2t0vs/LCswPUFDqhxGGx9HYLS0sNyBdVALabm3vwABz1VqkdvD/D0UP5K/PPpHuL4WdSfFrY3xqwnXG/eu8+chn+2KbwR5bdlOp0rj69Vc6FjtwNP+v8A096pnq5YeGF9OjffFv8AnSdTdYfAPI/Af5C/F3Cd+bEhfLZPaNfmTFDPs7cuRpWpzm8XMkwYuDoNmS3p/H196IJGOto4GWyPTpOfAL+dZR/D2j3J1lvLp2k7e+OWdlrhT9NZ9KaTAY1Kt20zQIZRodFNroLn3Y/Lqqk5DcPL1HRlc1/Pg+MXUe1+xcd8H/hLtn41b47K2/lsFuXsPbrQV2Sr6bMxsKuADIMDEjFvWEUE/wBfegPXrbNUDT5dEc+AP837PfDLB/IKmzmx4uxs93ttHdW0q3LV0NIxxlLuameGWdBI6epC5IsPpb6+9nNOqrgEHzNfz6K/8D/nAnwx7z3N3E+0l3T/AHipZ44sZ+w5pxU18lZe8jqFID6b3P8Are/ZGR1Xj02fIv5pJ3t8vsP8ok2qMOuJ3Dgc6+37Qfu/wavWrEOlXZTrtY+r34562vaereIf5jVb/Mz+f/Ru6TsaDYWSoa7YW3IY6VYEiK4qohooqnVE7AMdN+bf4e/cFPV61lSnl1cx/OJ/m2/FPoz5m0vRnyR+DG0e/sx1TtDYlXT7yzsjwVs9fDh4JKN9cbjyIfEwIBNiebX91FTmnWzSlGP2fLrVW/mQ/wA0Tdvz237hlhwD9e9E7bq6Wbb/AE1izHBgcRDSSBY46aGGRlLeMW13BJ+vvY41PVG4UX/V8+pHyA+fvRPavwd65+KuyPjPgth782ZnVyuW7fphTpmNyUoC6cbViORgscen0WUfXn37z6tq7AD5DqpUE3sLDjT/AIG/HvfVKdXjfy0P5xVb8Hts5HqzszrSm706PzETR5Hq7OiKTByTSxGGStXySqUksxN0AP8Ar+/Gnl1tSchsjy9R0aXt7+eb0Dhest6ddfC34hYD4xVu/wDG5fGbs3VtaRZ63M0GcX/L8bLJUyq6RMSdQVef6n3oDNT1Yt20TH+r5dFd/lj/AM4up+C+0txdS9ldZU/evRu6sxksvl+r8+Y5MJNW5QMlRWapJUZX0sQSov739nVVJoVbh5eo6Otu7+fZ8Zutdj7y258IfhNtv4y7y31i8zjdydi7UqDPlMjSZ2N46ugH3kitHHZ7MUW5/qfegD59WLkLRP8AJ0QX+Xx/N0zvwnh7yqMzshOx833NtfdO2KvIZJKeU0EG54nSadfLIh1JrJFv9v72c0+XVVIFT65/PqqHDb5lw/a1D2ZT0xElBvhN6Q0qldSmPN/xaOAAm1wbLa/v3y69qzXq+b+YR/On6O+evW3VlRub4m4TAfJTrOhw+Ii74pjTrnMzi8FSRQ4yCuiWZlvC8KOjjnge60Pn1Yt2kDj/AIOhg6o/n49I7q6Ix3R3zm+JG3/lRT4OGhhw25d1yCDKUdHjoRBBjo6unkaQrpUKuoWAt73ngOHXgfN+Py8/29Fi+a386Kh746X2x8Z/jt01Q/HH46bf3VQ56XqzAxUU2MrYKZtNSk1WXaWSSZPSxf6/4WFvDrTEk08umf5MfzkT2X8Kdj/CjoHqun6I2JtvM02dy7YNaSnkz+YEYjrMlU1NPIXEkhueeBf6e/Co60wUqVbgf8HTr0b/ADootufBrL/CP5FdQ0nfO0ZsvXZvBV+djo5J9v5WWF6emraWpmkDl0BQtcWJB4597JJrXrQAAAHAdUQ5eopavKZKroaYUVFV11XU0dGCCKWmnqGkgpxb/UKQv+w9+68K0z1AX+v1P+ptf/Y+/daPp1x9+6t1737r3Xvfuvddg83PP+v/AMb9+61T069cc8fW/wDsPfuvdde/db6//9X5/wD7917rsfUf6/v3Xj12xufyOPz791ocOuPv3W+uwbfW5H9L/n+vv3WiPTrr37rfXfFrW5v9f8PfutZr14i39D/re/db68T9LcWH+8/19+619vXvqeT/ALE+/de4cOvW/wBh/r8e/der14c/m3+v791vrr6f8a9+691737r3Xvfuvde9+691737r3XIWB5sf959+61kjHXH37rfXdxYi3P8AX37rWa9de/db65aefrx/qube/dar10TY8fgWv/X/AB9+68Pn14C/9P8AY+/db699P+Ne/de49e5t+bf7x7917FeuvfuvddgXNrgf6/v3WiadeP1+lgfp/re/deHDqdQYvIZOXw0FJPVScemJGa1+Bc+/der0+1GyN1UUayz4arRHYIraL3ZuAOPfuPWjTz6Nl8ZfinuTsfe+GG48dPRYpp4J41cWEvq1hpPxb6cf7f37jjrfHHWwLjOuKvqHC0FNi8e/2dNCiOQgCv4uQeP6+/eVB17oXeuuyJqubwVODLOB6GKf7VwRqHv3Dr1a8OhXzm76tkFPUKKGOT9MbKFGgmwtb3snrw66xO18fmITPLEJjJ6rhuDq+hv/AMU9++fXq9M+8OtcHHi5D4Arl78tyOPp711siuR0EGXp6Hb+DkFLCJHRQAg5Yi3PHv3nXr3y6DzE0dPurHZcwr9hXCkq1Dfpa5iII59++3rWKV61qfk5tep2x2rno6mRpWraqomDMPqEk08X/wBf36tc9WKsho3Reffutde9+690b/4ebKyue7Ho8vHBImKpP2pqkKdJcuCwU/4Acn201xFG4jc5PDpdBtl5dWz3USkxpxPp1sD9WbirdkzPHt9GkikYfctGGb1B73Yr7d6QagDT06ONge3q1qYLUU+qVrXDBr6j9L+/Vp1vpR1OU3FuGLyU1K6XBIdVY244+v49749e4Up0moM1vPBZONK2nlnphydaMNP+HP8AX3rrwGelJPv3cVWxgpaBwjA3AVuPxxf/AIn3s461T16Dbc8W8s5J4qemmp3tYMistj9L8e9efXugS3FufevXJNPlmllpKj9byo1gG4a+r37rVeiu/ILp3Y/eu0Kqud4KfJvG8q1K+IMjmOwI/IP+Pvf29b6oN7Z6b3B1dlqimq4pp8YsoSnrWQjWGJCs1uLH8H3rrfQO6H+ulrf10n37r3XH37r3Xvfuvdd2/P1/1vx/r+/dar1Z7/J76F2d8l/n30h0vvrCLuHbu88pNRVmKYIRUhYtdjr4FgD9fej1sDzPl0qP5v8Ai/j9sr5P7s6p6F2lQbUoOtNy5/bObgoTGfNVUMqQqZDH/iGsD73Wo61SjU6qaP149+68Ouvfut9diwPIuPfutHhjrr37rfXvfuvde9+691737r3XvfuvddgX/wB99f8AAe/daJp17kH8g/7Y+/dbweuvfuvde9+6912RYA/g+/darmnXhweeffuvHh14/wDIvfut9de/de675Fj/ALEH/W9+698uj9fCH5KYDo/dtDRZ3ExzQ5DIhv4rLIqxU7SuoWOQH6Dj6/S/sC88cuXW+2BNrKVZB8I/F1k792X3n2r2l5tSXd7KO4guWCPK/GIE01L9nW1x15vTFdi7Xx+4cQY2iqoRMywtdFDDUukj6g/194tXdpebbctbSgh18uu6nLnM+0c1bRFv2ySia1nHYw8+hf6+2Rmt47rxyYWCRnp5gXkQN6SrDj2YbLZ7lf3yLADk+Q6K/cPnPl/k/la4uuYnURsvwsRnrZB6i39uHY+xNuYWrZi1BSU8EiPe62QBl495fbLby2mzw2s571GevnV9zNzsN99wNx3faqfTzSMUpwpU9Hg2BvaDcmPikEi+dh6gv1uDawHswBI6Bvn0KzUtQ8IkQn0/2R9ef8D/AF92OPi61Wpx0Gmfjq6aVJEZmMhJYcn6G/491OD1vjnqFCy1sRjqVFxwVNgPr9bH3qleteXSAzm2opZZViYQq5uLAAm/BHvYPXgMdAvvzEfwbCVsf3FzKgKgkA2JsB/j70MnPWitM9Vp9X7cjwnYe7c8i2ZqmeQk2J5c8D+nu329bFeFOl3WjN7vz1RB4XeAuVicAm4CkLz/AL37T3QpbNX5f4errk0Xph6A2XuXD9sZ9vLMKZKltS+uzKH+h9zFv7oeSLALnt6K4wfqnrj7erbcbGWSIyAlk0fq+v6Bf3EKUp0ZGvn0JWOVmRVFwLfT/iR/h/X3vz699o6BT5Dbwq8Bt6GnoiY5ZIfUw1XvosPp72ePWwPPoiOEwmS3os09VM8jPqIGongMbcHm/PuxoB1UGpAHQc/ILrXI7P2BX5xkkVEp/Il1PF+Ta3uK/fDZbbmHkG6tLp/DUJSp6nr7rkc03uhbfTLrbXw6Lr0b3RT9f4eLKPgxkayqLPHKRzEwax9/P57v+1cnNu5vtEG4NBFAaVT8VOusPPnIdxzdILaSbwAoGpT59CF2V8mNwdjUEGNoadsREg/yggsPMCLaePYK5H9kNt5NvHv9wuGv5DhC9CUHy6D/ACd7QbRyhcNdykTMeHDHVKP8xDsufpjqbO7jhqmmqa6kkMkKSXkJAd2sl/6fT30f+7BbtvXMK2QXw1U0DU+ynRn7ue7/APrbclz3iWiXBC6VVuGQc0+XWkx2Bu+q37u3MbsrI1gmy1U8pjvchRwuo/1I+vvrFY2xs7VLYtq0ACp8+uJnM29tzFvtzvTxiM3Dlyo4CprTpFD68+1fRGeHXjYnjj/iPfuveWevEW/IP+x9+68Ouvfut9e9+691737r3XYv+Pr/AIe/dePz69z/AI39+69117917r3v3Xuve/de68Pr7917rs2vxwPfutDhnrr37rfXYvY2vb8/09+61jz669+631737r3Xvfuvddi3N/z9P8PfutHr1rW5BH+Bv/vHv3XuPXdxe9uP6e/deoaU67vxyCTb6n37rVM464D6+/dW697917r3v3Xuve/de65KQL3H1t791ognrjc+/db68Pr7917rmRc3CmwAvx/vPv3VRwpXriR+bG349+638uuvfut9LXr/AGNmuxd1YnamCppZ6zKVUVOXjieUU6O1mlfT9P6C55Pv3Wj1sh/HP+VFtLbe1ocj2QVyWQyNKs3grEhEkRnTUGCOOAL8e/dex59Zexf5VmwM4jxbdnhoOHAMSxBgdVwAU/p79k9eoOuXUf8ALW6s2JkIn7AEGSpFkBU1KxG+k88yD34Z49eoKdW59c9W9Rdf4Kk/uJgqamo4ogEmWOncFgObaf8Aeffs1x1YHGcdCW+cAiVI9CxngIsUagD6W4Hv32dV8z1w/iNLHET5FDgHV9Dx9SP9t791sdA3vXdUkKVTQm6KrDi9iLcG3v3Xj69Ftx1Fld/Zx6aNHjp45SXlX9NlPNyffutcehbh2htfAVMIqJoqmrj0uVLqSCByOffuvdDDjammq6dBTKiLZAAtgALcH36uOtgVNB1xyGKiqEkeyltB0kAGxt9fe+tdAzL2rkOltzUG68RJor8PWR18DjgmSBwyiw/qfeievDtPREPlz8z8x3F2Blt/7nhWfKV1IaN3bVqIj9CtY8/j34Yx149xB9Oqntw7orM1XZmt/wA1DPHNdLEhlcHjn36vW6Zz1XJvhIk3LkfEqorS6yF/1Tck8f19+60ONOknY2v/AK/+8fX37r1Rw69Y/wBD/tvfut9djj9QPv3Wjnh1x/P9P+I9+63148f8a9+691737r3Qr9K9t7k6S7G2x2JtioeDI7dy9FlEVDYyGjmEyAH8G4uD79jrXA6hxHQwfM35W7s+ZPclX3NvYSfx+uxONxdbJM4eWYY+LwwuzXN/SAPfqDy61VjluPRSR9Rxf/D37rZ4dd3W97cW+n+Pv3WqGnXH37q3Xvfuvde9+691yuLcCx/J/r791Whrnrj/AI+/dW697917rsGxvyP9b37rRFevEgngW9+68Kjj11791vr3v3Xuu7f4j37rXXrf7H/W9+63XrwIH1F/futGvl12pANzz7914io64n6n37rfXvfuvddkW/p/sPfuvdde/de697917r3+P49+691//9b5/wD7917rsi1rf0B/r791oZ49eJuB/UX9+68BTrr37rfXY+vPv3Wj11791vr3v3Xuve/de65L/Ui4+n+x9+60fQdcT9ffut9dkk/X37r1KdeIt9ffuvcevH8f63v3Wuuvfut9dgAnnge/daNfLrr37rfXvfuvde9+6912LX5+nv3Wjwx13YEMfz+Bx9PfuvZFB10oBPPAt7914kjh12Dfgnjk/wCx9+68fUddcX/w59+69mnXgL/T37rfDrxBH19+68DXh12D+CeD9fp791o+vXRBvb+v0/xHv3Xq46UO2trZrd2UgxGBopq2snKgJGpYICbFmI/Hv3Xs+fR++rvhBkayox9du6pMCStA7U00OmMaiCwsw5/pz7916nr1abtv4i7Gw2FoYsdg6aurXijAmjpohYXv9VF/fvmevEr5dCvVfCrG53BRKmGSOZtEqg06XGnm1gL+99ep69CJs7oLObKyGOqqLFusFAsS6BBYsY7XNwPejXrwrXo4dPBHnaGGizGLskaKjI8Y/P8Ar+90x17p1/u3tPCwCbG4hHqFTgKqhiR+fejx63mnQAb321ufdGVWpp6aako4WC/2gpVf9b/ifeyc9ex0MXX+OrqSiSnqNTPEEXUQPr+R/tveuvdO3YMMdLhpZZksACxY8c6L+/HrVeibYeobdW7ExUF6inaTQyj9Isbc29768AOB6V/bWyT11t9szj08fkhPlRLKTrsGHHurGkbN6A9O28Rnu0gHmy/4etbv5409G+9cPk4FVJqqKfzABQxLgSEtp/xHso2i5a4WSvkeh5z5sqbPNbBBTXGD0Qk88gWA+v09nHUejGD1yijMsscSi7SSJGo/qXbSB78cCvVgCcDq974p9Py7T6dqcyE01U8kE0UxjGrTJZm9RF/cPXfN1vLzQllroVNKV6z05W9n1PsvNvMYBacahjgAKnPHq5b4w9d7dzG1aqproEmqzBG8jMqNZ2PJJPPuYkOuBJPUdYG3ERt9xuLQn+zYj9nSv3psujwMzzUtNoiWRbOAAF/23v3VCOhG69qoJ8bpQqWjFiQFuSR/T37rfy6V2fxcNRSmXxB5BxewuOPqbf09649eqekTgsdEtX6goVTythYEn8+95r17yx0+5CemopNJQAluHCjj+o5/HvVetfLoPu1er6XsLbDRqEaoeNtD6VYj0cC4Hv3z69nh6dEOi6Q3XhJanF1FJM9O7lYiUfSVB4I/Hv3z69TpP5n4mQ9lMaLcWKjiiUqolnp1ZSifQfuD349eA8j1Fq/5evXlDjxSQ4yhkcIVNqWD6244C+/Hr3yHVenyU/lzVmAxtXuTa7LEYBM60tNAoDALrCuqDkD37rdPTqnfM4iuwOTq8TkoTBWUUrQzxkEWZT9Rf8H37r3UBQOPrZg1/wDXC/8AFffuqn5+XWzf/wAJvessfhe9Kz5UVcKGbqCeqNPUyKCtPNNS/tMHf0pyQSePfqV4dbB0kk+eOqQfnRuAbr+YPyI3KG8i5vs3cuU1lg5f7ys8xbV/iW96HDrznV0Uv+p9763117917r3v3Xuve/de697917rkCAD/AF5/H09+60a164+/db65GwHB+v1+nv3Whxz1x9+6314m/Pv3Xuve/de697917ru5tb8D37rVPPrr37rfXK9yATx/vvz791rhw68wA+hv7914Enrjf6D+n09+631zQkMp/CspP+wN/fjw69gGvVz/AMfP5oGF6X2ZjtqzbXyGTelpoKVptR8aLGAt1u4J/P49xVuXtyu4brJuM0lfENaUHWdvIH3w7nkr29s+S7W0Cm0DUeraiW+zHHq/H4LfzHdl75qqWWjx9PS1ktm9MsWtSy3sT9b+xhsXKu3bJ+pEtX9aDqAfdT355v8Acx/prydhB5pqalPTj1sQ7Y7ix24cFDUiZWkqIUcC4sC6Agj2JSdR1HqD0QRLoUY6Op8ZshkK+rEsjSCnL3W9yLFxb6/X3U4PVuOPLqxGnydNpMesJYG7XX6fT3fB6rkHHSNzy00jnxyCTk3KtyPrb3UivVgMdI55Yae7yWULc/W1wP8AW/r71SmOtHPQb7rzqQ3eGQGxvoFibAXN/fq9eJ6J98gN/rhdp1OVnmMfiQHTqHHPF/8Aeve8jh1vHRZ+o6zGboxlVl0nTVklZiRbklzcE/U+/N69eXhQ+XRsOtMbhqGtgWanjqCzPY2UjlbXHtPcn9A162BmoPUrYcWObsndApoUjAmIsqgH9X049ylu2v8Aqhak+n+TpIoBmNOjcUdKAV9BFyLc2uLf7z7jCP4anz6VE+fQk4qFVpgWS2tW/wBcFVNiPbgoc9a86dV4fKveFTHUCjET/b0vpkf1aSApNyB/re7Cg7mOOvUdnEcY7m4D16KL098kdqYjeFHt7KzRwQ1ExjeZiF8Y1abtq/1/Ybn5ktI7v6UnzpXqZ9v9m9+uuXjvgQ1AqF9cdHX+YOT2zkejqivoZoqnH1GMjkhlGgrNePUbW49xj95Fr6T21nfZ6mqDI/2OhL91S3vrb3cgtgpSVZCCPTPWvgOwziIEp46B0olaQU7jVpZA9ha/vmHy97W/1ti+pjux44A1rSpB+fXcg8sfWzGRpAZDTUOgm7b+Xmzem9uVGd3LPHBL4JGoaWSRVeplRC2hbkexjYfdr3fc7hYLabVkaqDgOgdzy2ych7HLvW/3KwxoDQEipPljrUl+cHzv3p8kt25Sjx2QqqHaS1DolMk8jJKsblBFGpJXTb6sOT+OPr0O9ovZ/aPb3aYwYw1xpFSVAIPmSfXriv70+8m6e4W9Sw20rLZIxCgE0YVxjh/n6rlJB/re5Nz/AI/X3N/UBgHrjY8/4fX37rfXvfuvddn6A2/rz/X37rXn11791vrsAWPP+t791o168CLG4+v0+nv3XiD10Dbke/db49d3N7/n37rVMU69ybn/AG/v3XsDHXXv3W+uwRY3H1+nv3WjXrr37rfXvfuvde9+691yDW4/H5Hv3WiK9dG1+Pp/xr37rwrTPXXv3W+u7E3/AMPr7916vXXv3Xuve/de697917rsknk+/depTh176jgfS9/979+61wOeuQ0ix+v+Hv3WsnHXAi3B9+6tx67Fub/7D37rRr17/XuP9h791v7OuUcUkriOJHkdiAqopZiT/RVv7917odevvjd252TW0VLtzaWRqYauWJWq1gkZIYnYAylQL/T8e/darmnR/cJ/KU7ZycSSVVTX0ReNH0vSKCCy3IN0Pv2OvENSo6DTtz+W12d1lhqjKRPV5NoImlaNqcKnpNtLMigj/X97NOvDVSp6rlyeMrsNXT47JU0tJW0srRzQyqVIKm1xf6j/AB96631dn/KH+PNbuHeWR7I3HjwuAho1qMVUSw6laWmZ/VqkFgS1rAfgf4+/Y8+vDjUdbCm9t9xpMiU7rJ4I46fTHpU6IhoVdK2HAHveSOtcD1J2bkKfMusiqrMzhWQjkXF/ejjqwHT9vradDmcTLRPCkU7xssFTZRocj9XH9Pe+tYJ6K71zvnK7O3fVbEzEklVj0kkSnqGL6BIW9GkHjj3rPXuGD0ZysrhDIqI1wyh1F1Juwufp78PXr1B001WTk0m1xq+pH9D9bf7173jr3+DoHd8VcppqjQ5ClGZhf/DkH37r2PLp56lgk/gU89DFepm8iGQLc3P1t78evKB0XbseXd2292/fVgn/AIdNLcyHXpUazf8Awt/X37gaHrXlU9GD6+3elbDA0MpkVkjX9VxfTb36letg+nRiaBRVQFnP1UD/AFIANxbj34daOD1XP8zHq9r4x8ok7JDLEwRU9NrPbk+9U68c8eqSdy7rn3BLLrd2CyTDUzEj/Of7z711YD06SzaY8dkpnNwlNIR/yT+PfuHXvPHRAsstVuPc88GMppauqrKw09LTwoWkmfWUUIg/r73x4dVGB0fzpT+Wx252niVy+Wp63bUErKYlkp/91umtCzSKeT/hx7959bzSo6XW9f5XO9tqUkVTHnKms1yaJFFPG3jF+WOlB78cdaFeiw9lfDvsHY1MavHQVGcii9VQqQlWRNN2ZSoA4P4PvVevZp0UqsoqnH1M1HWwvT1MDFJIZFIZXH4IPvfXj8uonv3W+ve/de65Ejiwt791oA+fXuWuTzbkkn+p/wAffuvcOvcAjSxJPH6bcn8e/de48euPv3W+vWv/ALDn37r3XYt+Rfj37rR68bjg/wCv7914UOR11791vru/0H4v791r59ctI/rx/rj6+99aqeuHvXVuux+f9b37rR669+6312CAeRf37rRr5deAvf8AwHv3Xj176C9+Twf9b37r3n11791vr3v3Xuve/de67uLWtz/X37rWa9de/db68Prz7917rn6SL3sf6e/dVyDTriLngf6/v3W8DPX/1/n/APv3Xuve/de67ABvc29+60SfLrstcWt7914CnXH37rfXdv8Abf19+61Xrs2HA55vfj/bce/deFTk9dE6j9Px7914Y69yOf6H37r3Xib82t7914Yx14Gx+l/fuvEV68DY3sD/AIfj37rxFRTro8m/v3W+ve/de697917rlc6SLcX+vv3WqZr16wte/PPHv3XqmvXV+Lf439+69TNevEAfQ34/3n37rw669+631737r3XYNjf37rRz12D6r8D6/wCt79148OvMfpwR/r+/daHXH37q3XvfuvdSaOlqK2phpKSJpqiokWKKNFLFmc2HA/3n37rR62AfgL8UsbidpQbv3HQU82Zkb7hWdbv+4LgKGH0UWAv72Otk0HVkOR6fgyNbSSkRxUsOh9AAjUFfVpOkfj3759e8s9Gb2FtvFRU8IWmjJpY0j1aQ2rSdNx9f9f3rrdOh/pIoKZE0pGlgADpFrW/p79148cdSta2u6I6txbxoBf8APvWeHXvn1Cejx0wJNPGrtwQAPr/sPe/Prx9OoX8GoIwXaIcc+o8EfXT9Pfj1qmOkbuDJ0sEX2kEaKzH9KqOb/njn3vr2PPqTt+jMNOaiQaQVMjAhbLxfn37z68cinRa+9OyFqKeTa2HUz5CaUqqopLgMAv1H9Peq+fWqYz1P6L6tO1MaNx5yMffVQM6mTlkLHUFsefe+HHr1Pn0G3y57CSLA1mOhGpPtJpRYA6SguL/63tBu9x9Lt5k9TT9vQs5C2ht65shs/wAIAY/lnrVV+S++G3du6OFmLNjTMj6vqGYhVH+2v7L9gtmgtmdvxmvQn9191h3DfI7eHhbpoP2jHRa/Z91FvSw2BikzO78DQyf5p8hBJJza6xSB7f7cC/8Ah7L91uTabfLOvEKejPZrVbzcoYG4FhX8utqTqqnw8vSeOw1HHG1WKaK4W2o6EB594L7huN3Zc7/vWQkoG4fn11X9uN3t7n25blosABEwA/2vRpPi9mWxVNl6OZ/SkkcYS4sNMlipH+t7zp2W+i3LZbe4i4FAT+fXKvmmzk2/mu/jcfFK1Pnno0u7aOmzmCq9KqSQSL2vdRcD/kXtf8x0U9Fw2bmajB7kbDTMyeWVfGpJAIJ/2r37hg9eHRo3dTRNdg10JK2ub2/Hvfz68SScnoEsZnmO71xdrJNIb/0vf8+/H1PXsDh0sd+Y+dKX7mn1HQAbLe9h9be/cT1qlMdYtibljqYvsqmQeSMABWIJ/wBfn+vvXW6+fQqS02OqVDT0sUzLYo2gcD+gsPfs+fXh8ukXuaahhHhjiiicJ6QqgfUfTj37r1fPplwVJFXyaJEUhgOSBYN/W/v3A169k9Bz2/gYEw1bHLAjQMkyNqUFTqj0/Q+/deBxU9UZ9vfDXB9iT57M4ugpoawzySNLFZZC6ggWKj3rrX+XqoztvpDdXVWRqFrqKZsZCwQVbXbSZbqNfA4+guPe+vZ8+tmf+U7LN0f/ACWf5jHyEyFDUUWcw+b25jdoM8LLV1f35jhqpKeNrHSo1G496zU/l1cZUH7etUvd25a/eG5MzujKWNfnK2avqrMW/dla7cnn8fn3s5PTYFBT06TgNjf+nv3WznHXYa1+Pr7914ivXRNzf37rwFB11791vru9gR/W3+8e/dapmvXXv3W+u7cXv/sPfutefXXv3W+ve/de67Aube/daJoK9de/db697917r3v3Xuve/de697917r3v3Xuu/wAD/G/v3WvPrr37rfXvfuvdGv8AiT3dVdQ9l4aoqq96XBVNUv3R1eiKQnhmuQLNbSf8ffutYGevof8AwYzu2e7us9tZnb2RiyMr46kkqkDAGO62swuffqeY63UHHV+nUm16baO0IqookcohZv8AHhb8H+vuvHh1v5dI7dfadfQzT+CpkjVCQw/ryfzf36uetKK8eHXfTfZGU3nl62lq2leCFmszf4G17+9g06tT06fu397ptyN6ekbXKLC45IJHNre9Hjjqvnnh0WzH9hjJCqNbUEsoa4ewtwf8fewPXq1RwHVdvya7cpdz5SHregrtUuUkEN1sVRg2nkqfr78DTh1rjjoSep+ldybc2dSUseXaOeeHUlnJ0a/UB/vPvZamB14LX7OjEdW7a3Vt7Kq2byEktMjsVZ2JUgDgrf2nuQPAYj5f4R1sVr0r+tsjSzdm7m8Ewlbym6g/7XYfT6+5V3hWXk+zDeY/ydIkI+oanHo7+MqGkc62IK/pU8WFhb/jXuLY/gz0sPHrFk97/wALrKeguRqbQbfQA8X592x175efSU35tDY+eoEq9yRU/iqlJlmkI9IKH6j+nu4XUNLYB6oZmt3E6fEvD9vVBfzI6hodm7txeV2Ev+4+qqC89bSu1oFMtyTp/wAPcU827d9LerJAtFOSfn10B+75zom/7JLYbtIGaMUVCBnHl0Pm4t4Y2k+LMcO6d0pULDhiyLNKC0YSP6WZvr+Lez2PaV5g5ZO2XLB0kFCD1DO8c4x+2fus+/WUHgsjk4HHPWsT3j/Mq2F1xlqjbbU8eUgw08gEkSxgsApHJUn2ArT2M5d5fka72lAkk3x0rk9S1yl99LmS33m7u9xdmjlJ0hsU/n1UHmt29q/zKe6KTZu0qfJUuzKetR6qZS0IoopGPiSMLe2oD6MP8fYrFrsvt3tUm63pAoK1x/q/Z1jP95z72O+84WyncZisQxHCKjxSPIDFSf5/Z0X75m/DDL/E7IY6lyFTU1MWQqRTRtUlS2owNKQdKrz6T7POUubrHm21N3YEMg8x59Yte3nuHb89wTMkLQSwHTIjijKfn/IjoiINjf2L+pJIqKdev+r/AB/x9+69Th11791vru3Njx/r+/da+Y68RY2vf/Ee/deGR11791vrsAc3NvfutHrr37rfXvfuvde9+6916x/2/v3XuvWP1t7917r3v3Xuve/de697917rkWuLW/p/vHv3WqZr1x9+631737r3Xvfuvde9+6913YWBvz/T37rXn11791vr3v3XuvD37r3XINYk29+60RUU66Bsb/X37rxFRToc+o/j32Z3JlqCk2ptyuq6GqmCzZBE/ZhiBs0nFyf8LLb2zJcQx/GwHS+02u/vKC2iZq+dMdXi9J/ycKf7Sky29MvRvJURrJLS106ao9SA6NBUWt/re/QzLONUeR17cdun2xwl0NLenR+9n/yzegttiketweFr5YGR3MYhu+kWbkLzf2/npBj8+js7E6l6162iSPaO16HGsiKocQxPfQLKRqX3qnp1vPQgZTcbUkUjSSQKFQW0U8Y/2HA9++devY4dEs7k3JU5+krMf9qlVTSRNG6iFSGUt9OB72c9aoOqVN0fAzI94dw0dRSYw4nF/er/ABBxeKOrp3n8hSU6foLnkW96HXjU8Orzds7J2j8burMbsXB01NRrj6WnWerp1S0pWP8AcXWPqP8AY+/de+zoop7apMlvg4+Or8kD1JXTwQfVyP6e9nPXvkOj7bKqMdj6CmrogiyTLGxHFySvBt/sfeutg049LfImqztOvhkaLVfSPpz+CLe99a6Dar6bSvqVyTMq5FX8gn1BW1X9N7fX37rfnXprzG2dzYFJK6ererVE+ga9lB+gHvXWjUj7Ogzx/bFBJlXweVP2UoOiKR1HqkJsQbkfn3vr1T057pp9dM7a/JHNHrXm+pWXj6f196+fXvs6HXpjHU0O2I3RAhZ2JJA+pHNvejXj1tfTrN2HszG7oopaOqhjfVG6qzAehif1X+vvdTTrR40PRXcBDTdb5sYqtH+SyzaIJCCVDFuAC3vfnjr1adGkxmfjCIgJKyLHZwAb6xxb/ePfsAY69mteq7P5gFPm/wC6UVUkck9HJERoH0WzXvYfn3rrdDxHVHNLT+SnnlB0iJ5Cb/T9RLD/AGHvXWyehc6s+Pm8PkJVR7e26ailpZz4qmthZ41EJ9JYtb6f4+/DrRpSg6tz6K/lx9LdD4zG5TeFJit0Z+nYVnn1pPPDM3PrdgTcH6j3b5HqoArXo653/hcXBFisWtPRUMSJHFDFDGoVU9Ci4t+Pfh1vpOZvKUGYp3WQwuGQsNSIdRPq+nv1et8OPRWd3baoqueqiWCFoJQwkpfGn7qkcp/sfeutEn8+qqvlb8R6DP0FTufaGJhxuWhjkmeGIaWYr6iJFUcg+/E0HWuqbMticjhK+oxuUpZaSsppHikilRlN42KFlv8AUXHBHv3Wx02+/db65Fri1v6f7x791qma9CX0thaPcnbnWm38jTrV0Gb3vtrF1dK/6ainrsrFTSwt/wAGDEe/db49bAH89j4//F/4q/3F6b6r68xe0OxP4DtPcGdqaYIamaHJ0EdZKHcKp9QJJHv1QeHWiCpFfOvWtmf+IH+9e/deHXXv3W+u7C178/09+6159de/db67Bt+L+/daIr176n+lz/tr+/de4DrxAH0N+PfuvDPXXv3W+ve/de697917ru3F/wDG3v3WvOnXK4Y8+kW/w9+61kD164H6+/dW697917r3v3Xuuf8AY/339ffuq/i64e/dW67vxb/G/v3WvOvXiALWN/fuvA1669+6312p0m/v3WiK9f/Q0ACQfoLe/daHXiCv+2v/ALf37rwz11791vr3v3Xuve/de67Btf8ANwR/rH+vv3WiOvAgfUX9+68euvp791vr1z7917r3v3XuuwQPqL+/daOeuvfut9cgtxwef6fn37rVaceuvxb83+vv3XvPrx55AsPp/sffuvDGOux+Lc3F7f8AEe/de64n6n37rfXv8ffuvdd2F7X/ANj791quOuvfut9cm/Hpt9f6e/dVH2164+/dW65W4vf/AGHv3Wq5p15jf8W9+68BTroED6i/v3XiK9co43lkSKNS8kjrGigXLO50qoH+J9+631cL8SPiJTJtuPfu9KG1Q/imghnK2ZWbUgjDD6D37rQqD1ct1vNQbUxMFLGFgpUQCKGwChfxx791ug6EybekVRH4YtLFmAAF7kE/jn3v59e6F7Ycs7IjksqsFJ4t9TyPfh17PAdCZW5NhIiltIUaLEW1H+o9+J8uvDy6VfkLYryg2ul+Pz/U+9db+zptxMpeQ6rmzWv+Bx+R735da4HqNuPJikgIWxBJsR/ZuLWFveuvH5dIPFY+TJ1f3UxLC91BNwFJ+h/3r375dbxTpLdx9ipsbCtDTMFq5I2hWO12OoWWyj/H3vquT0D3S+xKnc2Qfd+44ZJppGE9Krmwsz6vof6e6/IdbHqejV7snWDF+CEeNYgFVVtZRpsBx+fdvLrWKU6q2+UFRLKk9KQxaakmUNc/Qj8eyDmNXltBGvCtepa9nLm1sd+N3cEa9LAA/MY61oO79u1eA31kjU6rZCWWoi1fhVYKQP8ADn2Y7cytaIFPAUPQK5tinj3+5knFPEdmH2V6B8Dgm9v8Pa3oNVz0Nnx/w0uc7Mw1NErWX1sV+gPkRQCf8efYX5xuDa8vTzDiB0MORLRrzmW3iAqK562FtrV1bs3E0ADn7cUdnU8jUEtyPeG93HHuM7E/FX/L1m/tU11szBIK0oRT7R0aT4qVtTm4s9XTg6HqZJEHIB/d/HvLb22EsWxmCVtVFWny6xG95dqfb9/iuXXSZmYmv2dHTwuXDrU0chVtDsuji9ieQB7HY6io+o8+i8dj1CYTd9DlYFsE8ZNuNI4PP+w9+Pz61jgOjGbQzlNuLGQSxOptCA9ueXFr+/VPHrZGOgW30Jdq70ocsqlabyDVIR6bk3v/AMj9+4nrVaDoZzk1zmDjmicS+WBpNKcWJFz/AK3+v71Trfz6AaatkwOW1o2gtINQ5uLt9D78etefRn8DmI63Bx1RKtN4v0/k34v/AMT73Q9eJp0HWVkkrq8oy2ZnAAsbgX5Fz/vfvXWz0KWFx0eLx6SlCzSIDqNtQP8ASx9+PXuGekD3KkDbGrqs2RwbAcc3X+vv3AdaqOJ6LL0ftujy9JlTV0/mjfyWXj8k/Qn37q3lU9AP8nvjHhd64GvX+GBZJaykSMiy+kzAW4Hv3VTU9GE+cu16b4dfynqzpBguKk7n2/Q7l+xl06637eESLOFS3AtxwffqAjqx1IDXhw60nLGwP4t/xPv3VcV669+6313Yj/Y+/de669+691737r3XZH05vf37rQ69YWBv/sPfuvVz11791vr3v3Xuux/X+n19+60evc3uBa/0/wB69+698uvEEfUW9+68CDw669+631737r3XY5P9Pfuvdde/de697917rsf69vfuvddta/Hv3WhXz64/X37rfXI8D683+n5FvfutcT1tc/yNf5hWE6rrcfsneOWp6WmKU9AkVVz+iTx3u7cg/W/vw49exTT19A3qTuLY3bGyKKo2/maKq+4oyxjhkjso8fFrN9efeiMY62DQ9Ex703T/AATcsGCgdfJXMippNyQTbi39ffl9Dx6t8h0cD457bgwe3ZsvUpaaopzIWa3LeO/vRrx61UfD0WPsPfmOyPYGSxeYq46SlSqZEkdrIoN1Fy/u1CRUdezXHRZu3ohtKlq87jdzxfYSRu6aH1q3oP8AqDb3vVAB3uAftHXv0gO9qfLqp3C7jp6ztIbny+S8sVHWmWGR9RW3lvwD+PdRNaDHiD9o6prgJw4x1f11lnMXvLr6iy+JqoppYqSN20W40oB9Cfe1MTglHBp8+nC8FaIw6c6Kh3DnKiOPzvDEpb1/QWsV59pbmpt2H2f4erCmqvSF61q9rbE7C3JLuzcFLixJMdE9TLGuuzfguR7lreZGfk+ySmaf5Oi5ABcMQak9HYw/c/R4WWSXsPFX0ix8sX9Bc/r9xiltKFBI/n0ZmGauB035nsboTKTx169i4jVDdifNEtyF/oG93ED4YjrTW0+nhn1qOid9udmT12Qkptt55M7gZLLG9KwdIYwLcmMm349kVw99ZXWp1JQnqVdm5b2nmHYWjgIW4jGciteiX9m97dcbYw1XiewK2jj8o00hqWXUD+Laj+fay6tLfc4NMo49ArY+YN55H3Z/o5DqjJqOtWf+ZN/MomxVJluu+uM1G2Nk8tFTxUdyFGogairW0/1PuthYJtyeHGajr3NnNc/ON2Lu7TS+Kn16oH6q6m7V+WHY0GPxVNW5SoyFdHFka+FS6UglP9hGvduR9fx7T7zvVjsdm17fuERRUk9RxzFzHtvLFg15esAaHSle5z/CvqT1ui/AD4L7O+K2yMdJJjYf7010EUtbPOIzViqMYJmlNtWon6k++d3vV7u3fOV1JtW1PS1jJBIrRh1i/fbjf83bum+72aRKawQHjEfU+pPVL/8APmnmmyu0fIbj+POTwLXFFIB9PeTP3cI0j5T0pwoOhD7BO8vMG/SycTKB+wCnWuB7yO6yh697917r3v3XuuwLn6/7f37rRwOvEWNvfuvA1Feuvfut9dgE/T/Y/wCHv3WiQOuza1v6H6/19+68K8euPv3W+uwRzcXv9P8AD37rRr11c/7b37rfXrn37r3XK4taw+n1/wCJ9+61Q166Av7916vXj9frf/H37rfXXv3Xuve/de697917rsf69v8AH37r3XjYfQ39+60Ouvfut9d34tb/AGPv3WvPr2k/09+69UddqrOQqKzMfoqgsT/rAe/dbAJNB0ZLq34o9z9qz0D7f2dlZcXVyqr1whI0RkXLqrA/j+vsM73zdy/sEevcrqOM+hYV6HnKftxzVzhdpbbVaSOGNKhT1cb8bP5SEUNdSZrsGrqasSGF5cZVLCnjjHLxlVUD6g/UX9gO490tuvOza2DD+IcOsw+U/uX75DaruvMzBUIrpqNQ9fs6vN6t6E6m6Tx8VNtTb9HQVAgWN5CkTu5C2Pqt+faCDct23m6AUmlc06H25cm+33tzskj3aqsijt1EGpHS3zP8QrV/yT9tLWBAAAQciwA9zBtltJa2yo5yeuenO+92u971LNaqNFTSnSQkoNw0KPVmeSUKCyxn6H8gAezLy6BnE9SsdmMvWRNHLE0L/QEjST+Lg+/U68ajHTtHh5qwf7kKg2+tifqD9Bz711vyz1EOxtuVT2eON1Bu5JQ6wfx791r7Opxp9m7XglagpYKetVPVIPHqZh9DcD/Ye/ceHW6HoifyF7Tx02My1EZIxLEkihh+CAbWN/e+tEHy6pvwu+MxDv2eejleVBVN4Quo3Ov6ce9dbPr59W+dL9gZfJ4vHR5hnQlolCvccaPxf/D3vqo6PJiM9BHBAiyLay+m6gEf7370OrUrw6VZ3JDGAdQJZQALXH0/qPe/n17h0nM9X/c0crTkGJ0K6TwCv19+691Xx3TgqON5MnjT4K2EySRyK36WDkgnT9f9v79jr3TX1Nuvc+fSLF5ZpK4hvEkjAghQdI5a/v3XuHVkuxqQ4bbEKTjxyEM+hrXFwLf7f3qvW+k/mt40NDUeOd9Nybk3vfVbj3vqoA6C7d1Ptnc1K1QWj+8gBlpnDBW8p5t/X36nmOt+XQU7T7AeDPPt7JyeOSnZvFK9hqCmyKGP+29+p1oZx0KnYG08Z2htuTCZTxypJTNHESQLEjhgTf37z635U6rirPgc0ueaGLJGmx0kzu6eVNJR3ufxb6e9H5da8uPR29mbY2Z8adoxRYKKmGU8AWqrbo+safV9Pfq063StfToum/vlBDU1U9NRyipqCzIFjVydZN7ce9mvWuPQAVvbm+62pD0uKq/FqJV/HL6gTcfj3qvXvl0J+2+0NxyqkVfFLG5C3DBxb8i97fT37get+XQv4TIzZiaOV1uzkam/1vxz795da869Cg+w4c5SsJqPyhxaT6HUh+qEH+vvZ9OvUINR1WX8uPgzht24+t3BtvGnF5OkjmqjJD4w7ksWKAAcg/kHj3qnXh1QLujbWU2lnK/BZemkpayhnkiZJARrRWIWRT+QQPfut9J/37r3R7P5dPSmQ7p+U3VmPondYdub42jnK4Jxelpsys7lm5sAIyT78aUz1oNRh6dWMf8AClPfFNvz+ZXuvJUDq2OoNh7Lw1KkY0ogxmLSkl0j6XJXkj3odXZtXWv9cW/T/h/yP3vpuhrx64nn/D37q3XXv3Xuve/de697917r3v3XuuX6vov0/p/xr37qvDieuPv3Vuuwfrxe4t/rf4+/de669+6912Db6i/+H4v/AF9+60RXrwNvxf37rx66P19+631737r3Xvfuvdd3/wBt/T8e/dep14m5+n+w9+6114kE8Cw9+68KgZ68OD/X37rZ68Tf/Af0/Hv3WgOvAX/NvfuvHr//0fn/APv3Xuu/rb/be/de4deIt/T/AGHv3XuuvfuvddmxPHHv3Wh11791vrtrX4Fh791oVpnrr37rfXfFv8fx791rNeuvfut9e9+691737r3XdyOQfx791rjx669+6312CPofp9f9j791oj069fm4/rx7917yz14g/n88+/deFPLrv6cMOPrYf8V9+69xyOuPv3W+uS6f7X+H9f8AY/T37rRr5ddE3/4j37rwFOuvfut9d3Nrfj37r1PPrvgCxBv/AL63v3Vck466t/rHi/B9+63Xoy3xW63h7H7Qx1BWweWgojDVSMbaVmSoRo73/wAAxPv3XjTrZo21t2DM4/E7dwsQix+LpEp5xHGLGSD0C9h/h+ffq+XXqEZ6GbC9XzVxjikYiKH0EEG1lFve+vZ8+hOxXVNBBUw3AIQi97c2PvVfLrYGOhzxm2aLG06+PSulALC3NuR9Pfq9epnpC5qcpWhASbSqBb8Wbnj/AIj3v/L1rHE9CZQnyYAMfVZD/W/+sT71wPVuOOoeMjaISP8A2STyfp9Ob+/eVOq/PpK5yX72f7e2n1ajzcHn3vrwqep7CLC41qslUEUBck2+oF/eq162QAa9EG3tkavsvsSmgUFsbSTpHIASULB7cj6e/HPWqV6PptHG0+LwlFSwKIxBTpGbaedP1+nvfXqdM27alJImp19TyMun8Hn/AA968+rcOiV/JfYjNtiozgiPljo3CkAW1EXBB9lu9hjtrFOPQy9t5Io+coBcH9PFR+fWsz8s4mXcmKeVNMpSdGJAvYWNrj2U8ryM9o+o8D0PffSwhsuY4TAtFeOv7adFG9ijqD+j0fCnE0FXu6OqlZPuVrPGAQurSFQgAn/XuPcbe5VzJHsjwLwYdT57F7Zb3+8hpANavj9nV4GQx8c+NSA20FdN7D6Ec2/1veJ0MpSct1leYktd1/UHaDkdG8+LWCgo8PULCebsGsBzZ7i/vLD2qvlv9rmZTXQAD8usZfvOy2su/WAtRgjy+zoTN4Vc20s/FUgtHTyyrrX68v8AU3H+8e5Lz1jd5U6TfaWEmze3Y85QRMw+38pZOf0i/Nrke9de6hfHLdEc1LksfVyKJoZAgVj6gQ+k2BP/ABHvfWuPDoT+2scMpt6plSMtJG4KyiwICrwb+/EY695U6R/SO4Y6rXiaqUPJCzQKGJ/pa1m9669mlOnns7a5pKtKqJP2iSxZLG5+v++Hvfz638ulJ13UvJReF9WlUChb/XngEH37rXy6UuSplhyMLqhHqUk25vfj6+/deFTjpb5OpFJiIJSeNGq445/oR71x62MdAx2dVyZTYGRkhHlMb20KT/rXHv2evY6Dn481tJ/Dq+JwsciXDR8Agh+Rb37rSg9DLuLF0+8sli9tUAH3VXXY+yqoZm0Valxb/e/fuvV8j0Rz/hVpvaopty/DPYGBqnocRh+hsRRZiipmEaVORjplSczpH6eb/T6+9Uofy6u1GXPqetPQm1wDcXNvxce99Up59de/db67uT/sPfuvU669+691737r3XvfuvdcgAOWBsfpa3v3Wia4HXH37rfXvr7917rvkcX+v19+61g9eueP8Pp791unXZBP5BsPr/T/AA9+6qKDrqx5P9Pr791avXXv3XuuV0/of99/sffuq93XXF/8Pfut5p17i/8Ahf8A3j37rfl142/H09+699vXZ024Bv791rPn1xBtyPfut8eu+Sf8T7917gOn7be5c3tPK02ZwNdNQZCkdZIZo3dOVOoBlQi49+60aU6vm+Cn85ftfpmXGbaz2Uq6pBeIyyVVWIfHqF2BL2H+AP09++zr3y6vT6e/mh4HvPtjaAzVbBphrYlnJqS118gvq1tbj36lPPrde7rbX6v7f6+3FtLGxYfL0rvLSxXgiljFy8A5+v596OOvKc9VgfzgNo57Znxh7A7P6xq3p9001JSVFG1K37wkeceRl8R1cC59tXWsWTmP4/IdaepibT8VD/g6po+NnyWj3R0ptig7b39QvnDjY/4jS11S4qFn8Y1rIJOf6+8K/cDcOe4d9k/c0kxjrSgppH2dYbbpv3P45ivoJ3KRK/6dNXw+XHz6GLau4ukt45I4bb2ew2RyGoq0VPKJHUjkg6R7j/cN/wDc/bbVr25eZY1yT0mk33mu1kjhu7pojL8NcV/n0eDqnObj2nk8ft7FVzLQTuqfbRu+lo73IIHH+39o+SPcPnHd+cttgS9lKPMFZCe1geNfPoZcqb7vse8xWl7P4gk4ca9WJVm7pNv4ChblaupgJPqW+ox8kEfke+j92P8AEhUUagr+3rKSPESB8NTPWtj/ADRtqfKbtztfrvFdC5jNYqiqdC7gqsYtUyRvr5Mxp+AP639y9zNPBZe2SbhLgxRVFONadBzebqfb9ivNztF1ywmoX16g7Z+Gvydg2ziIMl3TVJkUgQViyLX+QSaBrV7i9wbjn3zNv/vOWNlfy2XgzsIyRWoHA+nWNL7/AO524N+8IrgQLJwioTpH26hWo+XUiq+KHfdEjQVve7UoNhZjXcfnkf19tQfeciuj/ittO9PRl/z9JJ+aefLLF9uKRg+oI/5+6Nz8dsluD48YDco37vaLf6VdLMaczNKVowVN2HlH1H49o5/fDmre90ij222kEZoCHIrX1FPLpXtvvTuvJtpLd3O5h2JwFPGpwKAmvWsf8+fmvUbu7Z3hhIcuKWkwlU70cf3UiK0XnIUCxtx9PebmzSSXOwWl5Lh5EBYehpnrKXYtxk3vYbbfrnEtyoZvXIr9vVCG+dz1W7dyZDLVU7TCaZtBMsjqVB4YeQn6/Xj2YdGFT1ar/Kl+Rmw+gM/ubK7xNH5JKyGpoRUyRxkNHTBbqXP9R7iz3U5Xvubdl/dVmxTxKhiPQ9QL728qbzzANtvdmQyPZS+IVqQGFCM09K1z1eX8O/5g2T+WnyG3pg8XRPRbZ21WrTwzI+umq43YqkkLAnj0n/X/ANbn3jj7ge0Ozcj+3IkXvuAMseNaV6h/nPlDeuV932nfbi9ZhfOEktyKBCRxH2efVdf8+oac1tFQLL/Gzx/1QyH3KP3bTq5QDHjT/L0PfYYaeYd+UcBKv/HV61wRb8/T3kh1k99nXR+pt7917r3v3Xuuwbcj37r1K9dE39+691737r3Xd7cX4Nr2/Pv3WuOevfU8e/de4DPXhweR/rj37r3EY68bX4+nv3XhXz67NrDgg8X/AKfT37rw49dG1hb6/n37r2a9de/db67vb/Y+/de669+691737r3XZFvyD/re/da669+6314i3B9+69x67t9Ppz/vuffuvdde/de6MP1L8ZO1O3paWXbu3K98VUSKoyCwNIrKwuGjVPx/ibe/cM9aGTQdH721/Kb37l0jfIZWsx+pVZ/JTImnVzY6gbW9obm+jgGMn06PbbYrqdRI3ap8zw6s2+On8rrrHYFJRZHfMVLuaeNLlJoKVptTDktI4v8AUfTj3EfOe7c6X1q9vy5EyOeDUNP5dTdyls3t9ssUd5zBMkzDJUEV/n1aztXD9a9bbe/g+0Nnw45I0AVljptWpV0ixUX94f717De7vNm4m+3q8dxXC0YCla8OsqeUvvLe2/JtvHY7NZxxqp+Ltqfz6ZDmamSpaanilgV2a4VQNK345/1veR3tx7Nbjsm2R224ykstKk/7PR7zx9+DZW2drPa7VZGYEVBqf5dRMxuE0dP5pZxI1uBfm/8AS/vIja9ls9qi0RrV/XHXOfnj3H5i573Frm7lKQEkhKtjoFcl8iKbb9VLTV0iLGvC63FyPp9D7Na5r0BQAuB0hz8pocjkjj4HQUpYaZA5sTfn36tet9LRO5qYsGSojNgDcSWsSLn6e/V68ONOmrJ/KXb2IY0tdUwqxOgu8n0AH9f8fegOvE9B/kvlRg45iaHKwsJhpCrKTbV/rH3vr3z6CDe/yLdYlmpsgJJH1WRZJGJ1fT6f8T718ut8eiy5Sr3X2rUVcGPx9RNLVAgmMStcN9Dz/t/fuvfLpQ9S/Dfd+LzcmezsdT4HkMyU8sVgrE39N/8Abe/U699vR3Mb1/msclKtHQyIICnAVhbSLX4/2Hv3n16teh/2rt3OzKGqTIpCABXBFv6/Ue99eHGg6FiiwZhi1VkuoqBYFj/vIP1/p79TrXDh0HG88s0Ec0SP6I0YabkEi/1HHvQHn1uvy6Jbv3OfdrPTPH/x09XA0nVweffuGetVoadNHTWZpMRn4xVViqjyaUBsAWLAD6+/D59e+fHqx59zwvjKd4mEoeJFXTyDx/Qe/db+3pGZnC0mWp3qqg6JSCVVjypvcX9++3rVOiybnjye3cgtVHM89IJT+2CSFUNwePe/l14cc9JfPZbF1y02SpadabI07q8017GSx5BA/wB5B96+zrX2dL/C9hNLjYnWfTIg0Fbm4K/4X9+r1uvkek9uTtowJJKJ1j8am/IBYAXJP5Hvda9bzTouO8N+bk7V+12xteKSd6io8M1TD5GKISFvx7116nRwemfhxtTb+IpMtvhlyuUnH3DxyBCys6ghSG54/wAffqY60DQ9DRnOstg46lAo8FGiqpAHjh+imw597oB1sdFi3bhdtwVUiUuPjhlUsA+lR9DYWt791qorjpK4t3xdUZIlLxLfTHbgXN+be/daxx4dDPie0KmhohSnGhmINpSCfTa3496+zq1SOkruLe65CJxURqY5ARJGVUjQx5Ua/wAe9/PrXVC38wDrzCUmQk3vjYlhnqK1ImRAoOmZ1V7hPxz7114/4eqv7H37rdethT/hOd18N/fLDsYvHqTafV9bu6V9IYQQ4UVFTLKb/SwX36vXtJYY6r4/me9rUfcPyv33umiq0rIoa+txJlSTyAHHVrwaSSb3Fvofe+tA5I9MdV56T+n6/n3rr1Rx64+/db697917r3v3Xuve/de697917rsEj6e/deIrx669+69148f8a9+691737r3XZFv6H/W9+69163F/futVzTr1uL3H+t+ffuvVzTrr37rfXL03+htb/effutZp1x9+631737r3XbWvwCP9f37rQ4Z669+631737r3Xvfuvdf/S+f8A+/de67JB+gtx791odde/db697917r3v3Xuve/de65gAjnjn6+/dVJIPXEgD6G/v3WwSePXai5t7914mg67svPP5t/wAbt791qp9OuIAP1Nvfutmvl164sRbn+v8AT37r2a9de/db69Y/09+691737r3XY45t/wAj/Hv3Wj6deJub+/deAp148i5PN7f7D37r3A0669+631yW3IYfX/eP9j791Vq+XXRAB4N/futivn16wte/P9P9j7916pr11791vrs3PJuffutCg6lUdFV10ywUcEs8z8KkSFif8OPfuvGnn1Y18ENqbrxXYKyVuHnpqWqNN45pUZWIaTS1x+Pr7917jnrZw6w2rSYOijmMahqtfK7fWxZtRXn37q3lXoxWJoqMRfVI9XqHCgkk+91p1qhPXLJ0zqB9oTqsW1D6ccnkf4e9deGDTrqgyMrI0Ev61Wx/x4sSCf8Aeve6jrVK9JitxbTzmd/+OoYf1Cg3uffjXreOJ6EKheGLHx02ofosV/1/rc+/Uznr1fTrg3jpaOcs1gS2nUPyBce9efW/KvQexFp6/wBIMt2axtfi9/qPfutefp0jO3d1DDbemoBcVM6GNFH6rOLC4HPvfWjUHoJ+odjzU1LVZmuivNVzCoh1AlrE6r8+9VNOtjJz0arD+R6Vh+m1wQBb6/Xk+9jPWjxr0iKxJq3ckFOBqj1AEHkX+lzb34Y63x6idz7XGV2RV0IjErvTyWWw4FrWH+H9fdJoxPbvF8j0/t93+791huwch0H8+tSL544FsBvqjo2BDJLVLp5sBYcf7x7DXLMZiilU+bdTL7z3w3G8sLgf74Ufy6IUPrz7E/UJHofvjhvl9ldkYmd5GFJVSpEy3I/eLDSR+Bxe/wDrD2EudNt/eWwzIoq4FR0P/bjmO45d5mt7lGITVkeX+qnWxnhZlyG3MXlH/wAxW06SBj9LuoI/2/vC+5VobySA/Epz1nIt2NxjXcuAlz+3o23RmWo9pg008qutYElXm+kM30A9zv7Hb3Aklzt0naZSB/g6x/8Afjk3dJ7eHmC2UvDFUk/l0NnbWEO5tsyVtJxUeieLSOdCC5It7yJddDFfTrFmN/EUHrH09l6HN7OymDyiCSoo4JKYK41XIUgXB90PTg9D0TDKVFd132JMKYPDRVVaSyqxAIMl/wDWt798x1o1GB0f7EeHcm1qV3GtKyk1MSR+pxpAPvYr1s0Geij76x2Q6n3rRV1DIfs6iWKaRYybDWfoffutChNejaUUse/tpRVQ0yTNEXII9SjRe3+Hv3W89BvtepqcVuMYxzZNekqSbWDe/daIp0MeYjLVVMttIexBsf6j36nXs14dZN5vJFtnj9UacMLi4tf/AIn3oYPW806QGyXgy9BVY/IWnhkjcGKQBg7G49+61Wg6AeLFVmxN41lJEhjoclVHSRwkaO1wP+Ke9468cH16Pp8Ous6zfvyl60wLsJUycklQqyXZCtNCZiWB4/F/ej8ut1Feqvv51uDwHyE7/r8BkqdaqbrKordt01RKuv7daR2gVU/pyLWHv3z69j4R1qsdwdO5nrXK1DSU7PiZZ3EEwVv2gTdQf8Px791Xh0CHv3Vuve/de67JHFv9j791odde/db697917ru5sB/T37rVM166tb37rfXgbG/v3XuPXvfuvde9+6912D+L2B+vv3WiPPr30uAeP979+69xyeuvfut9djki/A9+60eHXXv3W+ve/de674sP9jf37rXXXv3W+ve/de69z/tvfuvdci17G1iP959+60B5dco5pYW1xSPG/wDqkYq3+3Hv3XqDoUdgdvbs65neu2/X1UOR8nkgqvuZbxNe97X/AK88e99ep1a/8Z/5yndfUk8Dbo3Bm8lHShVijjqZpImVQFAFzcf0sfeuvVI6sA3V/wAKFKnsna9XszdmHyGSxNfT/bVMVYzyQupTRyknH597rTPWw1MDqq7t75P0e9Knw9WU8q7hzPjONwtJI4dmmfxWAjNgOb8n8e037v2BNVzc2yuckk9I7q22COFry/hUhBVmNBw9T6dXsfyuPinuXqnZ2T7b7kzE09fn6eDL0y100lsWZlEslMpkP0AuB7wl9+OebS/vl5U5USryMVdV/lw6xO5337aOa94+qt4fAtNsJKy07ZMEdvqB1G+RH82jaHU/bEGC2lk0n/u/kPFkzHNGNSK2h/oePp7kf2H9oLfZbccwb9EPqAA0dRlSfSvUn+1XK5mZ+Y91UMr0aD5CmOlXjf56G0tyVWPjylf9nS0xXUXq0I/1P0J/x95L3jNLCzUyaf4R1ORkVm1N0/74/njbc6dqKSfZO0I+wsnudQYaaELLNEzjgRlATf8A1vcw8yWlvL7eWf1ZARkyD5inRNcPbxiWS8qIPxen59B9U/zf/k/uNRmMf8UtypT1ymaIqtVpdJLsrjj8ix94my7F7RGVvqLKDxM6q6a8fPoHS8y+1iyFJJ4wRjiOI/Pom3yA+Zvz+7kX/fidSbm2XKVsA0k8QX/a/wAXP9OfdYNn9nbNy8FnbqfsXoqvt29m73Fy0MlPUg9Enzu6v5p1XjapMhFuaOjMUhqWM8ZAi0nXqDvb6ezGH/Wz8VRbQ26vXFAK9Fyr7ISOuiK1BqKH5+XHqpvsGo3hPuvKtvmSofcolKZL7hlMnkDFtLaOPqT9PcgwGMwqIfgAxThTqYbH6X6VBZU8IDtpwp5U6RYIAsRf/iPbvSog1x12rkX9TA2sCpt/t/fuvU62C/5DLH/SPvYG1vuKGxtzc0/9feP33i/+VJb8+scvfqn1mxf89HSz/n1H/c1tS/1/jnF/r/xb3vz7L/u2f8qgKen+Xov9hzXmLfq/79X/AI6vWuAACLk2/wB9/X3kj1k6T6dcffut9e9+691737r3XrH6+/de65HkCw+nB9+615564+/db699PfuvdeJv9ffuvde9+6912Tfj8D6e/daA8+uvfut9e9+691737r3Xvfuvde9+6914gjg+/de49e9+69139Tz/AL6w9+61wGOu2twB/t/6+/deFfPo5fxc+HfZffu6sA9Jt6rXaM9VHJV5GSJxHNTqQxVOP0sDfUffuvccDrbr6Y6B2/0lsHC7Yw0NDT1lLDA01SlPExZzDpZbkfj6e/CtKevXuBHy6HOipFSB2rZoJUUHgRxLdR7Z+lgLanFT0vbddweEW6vRR1DOa21TFkbQpBII4HA+tre1cUrw/wBjQfkOix4ElP61W/M9JzJ7uwhfxQxrb6Bh9D/re3PrLnyI/YOmzZ2tKBSB9p6D/L56ucmLGwuQ1/Ug454/Htp5Xk+LPVkt4o8oP5noCN6Qb8qkkFDTVLg8rpDcD8nj22MdPZIznquftfa3cVTkZXTD5GWPVcFfMBwfoCPeutgevQdYzbHZVDEKqooK2EwnXIpMoPH5N/8Abe99ex0ms92VvPGvJHE9XHIQVKBpBYgWIsffutEeXQYz5ffe6pfG1HV1NTK7CJdct2ksLAAe/dbx59Gv6U+E/bPYSR5fKVs+DpmIZBUvMABp1C2r34DrZ9Oj9bK+D1BhxC25stTZVoCjFWOr6Aagb+9gHrXDPRvtm9Y7E2cUXGYWniqVUAVHiQ3sLXAI4v79jz61TPQtRU8VTGElijIUWULFGLC3HAH096635dS46HHRLqNNEvBLMUX+tveutmnTbV1lLCWEAVBp0hgosCfxb3v7etVFOk1U1flGjWrAtbV/T+th73Ujrw9Ogx3htmrraSeWnbVIqM1vyxB1BfeuvY6ItuTZ+9sjmJaaDD1DozMurS1rF/rx78ONevU6FTr7425GSogyeVqFpdGiQU8hIYEm5BJ9+69xz0cGl2/R4qgip3KzeBV0WNhqA+o9769556aa6EPEx+hHAC/Q8/196r1s0rjoHt2YpZYXaQAp6iVYf7zz72eq/M9E97RqI8Dj6mtp5fFGi6iFb6gD6j37rw9ei3bS7Jyla9Y4kkWkUuIXLHSWHHvXWwcdRsVUbu7R3jS7UwyzvrqFjq5VLkJA7epmI/wt791r5dW/daddde9L4CjpJ4KWo3M9NBLNWlUc63F3B13PB4Pvx62ADjoTl3MlUVcShwxuirbSBbhQB79mlR17FepVYWrqQs6kXQgfT8j6f717917NcdFv3ttcyO0qIFfU2k2PNz+ffh8utUx0jsZteYNrmXhT/jyL2tb37rXTjkaKClha2lNKsbkAfTgfX/evfjXrf2dAZvPNUGPoKio1i0auSLjkD+o/3v37rZz1VX8hqDc/eUUG0dmYZ6+rlyCh5VDlIY1lB8hCA/04Hvw68RXPn0l6b+V92PJg48nUZV6eqkgExpGp47qdNypB5sP6+/daoetgP+Rb8Qt1fEgfK7vbdLtU0uW+Om+9pY7VGV01FfhpkiZAOLhmbkc8j3759WBK4PEkU60298z1FTvXd9RWFzVz7mzss5ckt5ZMpKz6i3Pv3WjWvSV1G9/9h791qg4dde/db65XF7249+61Q066ABPJt791418uu7X+vp44/F/futVpwz11+bX4v9f+J9+635dd8D8XH9f6H37rWT11+P8AG/v3W/Prr37rfXvfuvdetf6e/de67vxb37rVM1669+631737r3Xvfuvddkf054+vv3WgfXrr37rfXZN/r7917h10Pr7917rsgX459+60Pn10QR9ffut9f//T+f8A+/de67It+b8e/deHXXv3XuuyLfm/v3Whnrr37rfXZAvYG/8Aj791oHrxH4Bv+ffuvA+Z669+63139LWP1F/+Ne/da49de/db697917r3v3Xuve/de65K1vx791UivXVyfqSffurU669+6912Bf8ANv8AH37rRNOvEW/N/fuvA1669+631737r3XYNvqL+/daOevAXNr29+68TQdet9f6D8+/db6n4vH1WWr6XG0cbSz1UyRIiAk3drFjb8D8n37rXz6u9+IPxAws2Lx2R3Dj0rameSGU+ZlLNrGogEr9Bf6e/dep1avD0htPZE1BPiMXDQTKsba00/hri1gD7917PHozWILnFwFOCiKLA/W35t78R1v7elHT5KriIMgIVQPrcf737917h59LrE5aGddDlGY8KdQ4JH9ffj15fn1jqIkpKwEtcSXNv9fkm/v1K9bBCip6nP45VuqXIv6v9h/Qe9+fVfLrhCdL3ZjpW3HAFl5tz798uvCnSd3VuCFUWliYXa4I+pBbgH3unp16uKnrPseONTNVVmkIkLvrb9PpF+NX1/2HvVMV63x+zoovY2fG6e3KTDQP9xRiWNWRf0rpbn6cfj3rh1o0PDoz7CLBYijiUKoWNQF4W3HP+uffqdbr09YbJxRY+WZ2ADEsCT9Li9/exjrR4064baihyGVkyCspWJ2+n1Iva/19663UkZ6et2xtVUNcIlukdHUt/UelObD3ZDRW+w/4OmpF1PH8nX/D1p4/zF8j9325JAZAxhqa7gcWCsE+n+x9ku0rpRzSmT1I/PtwJpLRQ1aRD+Q6rw9nHUe9OGJqTR5THVYYp9tXUs+oGxAjnV25/wBYe6SoJI2jOQQR07BKYZllU0KkHrag6InxvY/R+JbCTx1FbjaGETpGw1K4iB9X194E85CbZubpvqBpjkY0r9vXRDktrfe+SLc2p1yog1U8jTpTbS3ktJnKfCVZ0ZCFhAiPcMdEgBIH+H9fY79ureWPfI7q3yrU6Gd9ebdde025bbuyjWp7SaYoPXq0Xb1B99tWj+4UfuY9mUNzcFLm9/eYBJMak8SOuXEqhNxuY4z2q5p6Uz0W/atUMB2LLhQfFHXVRLJ9NV2914dVFa9I/wCTe36eLJ0VXSpofyLqIAHJGq/H+9e9+fXh6+vQ39U5xF2pjYJpAhiolDA88rx7914dA93Pkqfc+5sfQU+mcARK+jn1Dk8j8j3R5Fjyx6UQ2Vzc/wBkpPRienqOXGUAhZCkKwlSrcgEL+fdTPDSurHSk7TuQOjwzXpE1rxDsBXuFBqWFlI59XHsl3bfLawiJDCvUkche1+9cz7gkckLBT/q9OhpyEsMtRSW+ioqmw5XkDn2GY+bkcYNepvuvu63tu1XjIH2/wCx1l3esc215lU3OmykD8kf1ufe15sJkEfTE3sCY7VpyMKCePQN7Jk+yrPGwuS1gObcn6exzZS/UQCb1HWLPMdgdr3R7GlAhIr9nT72nt5chiGr6OAmsgCyo6fqUINX49quOeiWh4cejs/yt81FkOxIuyS4ao63pclTz1FgPA8lG8JDD8/7Ej35iB58OrJHI4wK16q++VO0aPdXcfb24ftxJJmdzZGuWcE2PmqzJce9al01B694M3j+Hp7j5dVG/I3pqHeOzM1SU1IstdDHMIWUKrqVj4YADmxF/bJuYVwW6MU2XcpFLrEaDJNP9jqgzO4ar2/l6/DVyGOqx9RJTyqQRyh4Iv7eBBFR0WspVircR00+99a697917r3v3Xuve/de67H+29+6912QLX1XPv3VQfl1x9+6t1ysBbn62uP6f6/v3Wq1r10frx/vH+t7914cM9eAFib2/oP6+/dernrr37rfXvfuvde9+691737r3XK4+lh/S/8AxPv3WqHj16wuRcce/der1x9+6313Y/0PPv3Xq9dcj/D+vv3Xuve/de697917r3v3XuuQAsTf/Ye/daqa066AJ+n++ube/db6u+/lMfBjN9tb/oe0t2Yippdv7cyMElGlSQkVbTkK8kviIN+RYEn6f09wd70+4MPKOxvbwShZ5VIUedfLqDvdPnFUCcq7aPGe5BE2n/Q19WI4V6uY/mk/M3bfxx6uTrTZ+SggzWVonxtFT0iBzSyLAVUFYyLkWv8A6w949+xPIO4cxby3N++xFizVBbzr558uo35U5R/rJucFjaMG2+yI8ZP4z/CT6evr1pgZ/O5Tc2YyGezVXLXZTJ1MlVW1Uxu8s0jXJ/wAFgB/T3nmiLGgjQUAFB1llbW0NnAttbKEjQAKBwAHTP7t0/0cHYG4KHY+8OvN45xWrcNg/EamnmLShw4B+j3+nuT+fEef28sYI/iZcfs6C/MFlJvGxXezwsUeaoBGCKinWyps7+aZ8dKTaGAonxWIV6ajp4SG+21LogVfVzf639849z9lebb/AHOa/S8dRKSQBXGesUIvaS7223Xbf3ebjwiayMal6kmpJFa9KL/h1b46r9MZhg1+T/k5A5/4N7RH2I5uJqb1/wCfTq+2V4ooNq/1fs6bNy/zTvjxVbWzVKuNxLSz00yKqGnDm8RX0sDf629v7f7Ic12W5Q3zXjssZrT16aufai93Kzfav3eYfGpSQGhQ1FCCBWvWqF8gd7YjsPtrd27sFClPi8tkHmpoo10qq6j9B/xPvMPaoXt9vhhl+JVAP29Zccr7ZLs+wWu2TMXaGNUJOSaClT0DN+ALf7H2YdH/AFyCi6i/1Iv/AIe/dVrg9bBP8hn/AJmTvj8/5RQD/WHg/A94/feL/wCVIb8+sc/fr/czYv8Ano6W38+wA5raTcXGcsR9f+Xe/wCR7Lvu1/8AKoD7P8vSH2I/5WLfv+aq/wDHV61vwbDkXH+PvJLrJsip66H9LXJ9+698+vH6AWta/wDr8+/de+fXXv3W+uwf9iP6e/dap1788fn8f8R79177evf0FuR/tzf37r3z68Txa1j/AF9+6951669+6312Be/P/G/futE068Da9xf37rxFeuvfut9e9+691737r3XYF7fi/wCffutE9d/pb+tv959+69xHXRN/xb37rwFOuvfut9dgXIH0ube/dePRnPil0VkO9+08NtyGlFTi6aqhlyy8+qFnOlG4PHBJv+Bb37ga9apXrdH61672h8e+u8dtfblDSUtbBQw+aeOKOQnyU66lD2BFj/T36tOHW8EVHAdBhuHuCLHvULkK9YxHqKMQPweAvPv3Xq16BOo7v3VlciKTA0tTkKR20mWFHK2va/8AyL37rxrw6Fzbe1d3Z8pX1slRSRS2cxOWGm3q+h9+68M8ehex2xEiK/d/u+kHlwbn+pvb3viOtHpZUWEx9KFRadWK/wBQCTb/ABt79TrflTp1ShpQTamQG4AGlSbW/wAR71jr3E0HWKXa2DqBeux8E4f8GNPr+fx791ulcdBxu7Y20Ho6qGLBwDzRsrMAlx/T6D3vqoHVZnZ3x8+8zLVmHx3l1yuxjiIOkM/0sB9fevPrRqOhe6E+NdJDl4MlnsWESn/dSKUjhha3BX34dbzSvVlQlpsRjIqGhhipaaFFjCoiAcLpB4APveet0PQP7i3XHQS6hMrFWa4+o4NvfutV8j1lwO76TJH1SKrLci9x/tufeqnrwA8uhDpc/FHcFlFrAHjn/Gx97HWz1yqc1G8bDyqF/wBdRx9Pr791o+XQeZzcdPRaneUKGFgtwS3+PHvXHqwA6iYjcNLWKAjhnLcKPqAf62/2F/e/t6r0IEEbTRq7LoSwPNjfj/Ye/efXvKnTXXVeOo31JSxCe2nWyKSSPyOAf9jf3rPl1YAAV67p68zlSqheASv09P8AQWH/ABHvfVa16nGnFSvqFgB9CPx+Df3rrZNek9XUiR6wDYDmw/BBseT791roAuxNx0OHp3FTMgDFgAxFjzawF/fq9bI9Oq2fkFvim/hiU1LIsgrJTCFX6jUQLcHj3rhnr3QTYfDVNFtCIU0b+epZiqrfUXkUkG4/PvfXqEdWcfC/qej2zga3fucplWvniCrJMAXX9u4IDD6+99e8ukl3p2jTwZ1IaKsCzmseNlUc6Fa1uT/iPesda6EjrjdE9djqaWaQudOoMfyLcce91698+hn/AL5eNBE7qBax4Fx/Tk/63vVOt1A6a6vNUWT0Rs6ay41WtcW9769516fYMRBNSrJGykWvxb/G4sPeq9e+zovvaVRLiqeqMKNYRM2sX+g+vv3WqdVK9tdiZ6uyDbfxlXItVXSS00USB9Wt2KqAPr9fr71SvVsdWAfFbo7IbK2DT7uzlETuKqiWVWnUPL+9yJNDD/Y3v72OtHPWfsbs7em2Z677oy/aRxSFZvECgT+lrjj3YDqvkT1e98Ydx06fyz8n2PldDUm7MpXbUkqCiKJlmpSJIPx9Qf6+61Bx1elKN8utSf5Zfy/kyJz28tg41aapnyGSyjRUhBab7iUy/vIF51H/AGPv3VfOteqTdybYze0cvVYPcNBNjcjSSMksE6kH0tpDofyptwffuveXTB791vru3F7/AOw9+6159de/db675+puf9f37rWOHXZHAPHP4/pb37rw9OuPv3W+u/xb/H6+/de69z9ebe/dex117917r30+nv3Xuu7cX/x+n9ffutfLrxN/xb37rwx176C4PP04+vv3XuOOvAX/AMPfuvE069zyBf8Axt/T37r2OJ669+631z4X8Bv8f8f6e/dV4/LriRb6G/v3WwfXrr37rfXvr9ffuvdf/9T5/wD9ffuvde/Nvp/xHv3XuuyLH6g/63v3Wga9de/db697917r3v3Xuve/de69fi34+vv3Xuve/de697917r3v3Xuve/de697917r3v3XuuS2vz/T/AHn37rRrTHXrixFub8H/AA9+69nrq4tb8+/de8+vEg/QW9+68K+fXVj9fx/X37rfXduL3H+t+ffutVzTrr37rfXIrYXuD791qvl1x9+630a74pbEk3LvulyVRBroIC0aGwsZQwJuT/T6e/deAPH062d/j3s2WmxlBIIisUKRBPSoFlX0gH3sfPrVCTToVOzspJjcnSCoAWNVVdJ4P0A1E+9Gvl1voWNkzU+RpqVEjBV4Ee9vwbEW/wBv7317p33jUUmJpnLkJZdWo8X/AKfT3qtD17oPNnbjkrsneNg9OsiiwZiPrb8f8U9+JCgs3AdeRJJXEcQJYmlPt6Enc2W8NRBMJNKKuk8XA4vb/Y+yKXfrWN9FQSOpRsfavfbuzW68NtLZ4dR/9IWFx9AfuaqJXtc6j/QfQA+7R73bstembj2z3eC4EZVqH5dBfl+2leZ4saqzK1wGUluT9D7t+/LUCrEDpt/bLfdVIo2YetOHUfb0mTztaJKoSBC4Yfq4BPP1/wAPx7fj3W3mOmMgnosvOQ9329DNdoyoPUdC1uzJpgNo1LwtokWJ4zcnn0WJBHszGRU9Athpdox5dFJ6kxkeY3xLn5z5GjqiRxfjV9Bq5966qBXow3ZeTlinpKKlezSkWAH0ubWv739nWzXz663HJPg9irVvKTUuEAXm/Ki4/wBh79TyPW/n0s+lmnrMDNW1CkalL3bj6/0v70TinWgCcnoUKKKOubJ0zvZGoq0WJBuSthx/vre9+XXvmOIPWnZ/M6wKYPvV0jUqs38Qe/8AW86Ec+20SNB+njpVdXFzcOGuDWgoPs6rZP1P+v7v0mHDrLTwSVVRBTRC8tRNFBGP6ySuI0H+3PvTMFUs3AZ6sqs7BEFSTQD59bHPwNx2X6i2jBRZ6eSaly9EKiMVBJsCmrSL+8L/AHZlh5k3B2tF0mBiKjzz1n77K7Rf8vbB9VfElZ1FAfLHQwbI29kd795DJUkUgx8dVJ6lBCEGbg+5N9odorCkjiujTXoj95t9fZOWLmzDaTOSQPXHl1dNhoFo8fjqU2H2tL4ZFNvrpA95DvQEqPy6wWgYunjNhnyeil9j458F2vg8qoK0zGJ3ciyeoi5a3unTvn08944M5anxVel3gkiWXULH+zfkH3vPXmIUV6Cquz77V2r5h6ESHxBiSLXHtDd3sdtVW49Cbl/lu83hxPGCUBzT7esXS2Mm3lkpc3XXlVJy0RsTdL/0b/Ye433bfLhZCqnHWZfIXtfs81okk8eaZ4dHlhipMLQusIAd0ICixNytj9PYI3XnSayjK6qHrJnkv7vOx71cC6uYv0xT06L7kKOaq3hSzLdS8hbUoIv6v6cH3FfM3P8ALa7bJes+sjgtesj7H2p5P5OeOeFAoHEmnQ7R7cyFZCjRLIzaV+hH0B/r7hD/AF/JbN6SRUp8+jq/23lrcYjGhXP2dezGPq6XHNSVIJdhYBhwBbg/8T7mX2+91dt5qjaViEdTShPUU8x8k+LEbfbxWMg1I6CeloWosrTsW062J0kfX6+8tuTty+ttj3VAp1y5+8VygOX95UWsZJatT0KeVk04GsL8l6SZUuONRiIW3/FPY1BVmOnrGeRJYY1My6SxHHoxv8sbaFdsz4nfM3sPMrJBX0eQ1YMSggyrK82sxX54BBNvYW5i3mLatnm3FiAIyq/t6mj2w5IuOZ+e9r5ZZTS9qa/s/wA/RHd4Un8fxlTm9JNRlInqpBb1FmJLG5559xdB7l2jN4ckqivz6zl5g+6cmzO1wELNGaGlOq+a6iZcvkaaoi1RvJOjqRcAMLW9XtPe82oZ1kimBWo8+jPlb2c2N9puYLuKkgVqcONOqR/nX03FtTdUm6MNTXpq11lrpFtZVeNmuQvHB9zLsW7Q7lYxuhqadc6/c7kq75S5muonUiMv2+lDWnVd3s86jXrscG9uP+Ne/daOcdeH1F+R791s8OvH/C1j9Be9v9f37rQ669+6312Rb+n+w9+69117917r3v3Xuve/de68Pr7917rv63I/2359+6111791vr3v3Xuu/oPwb/T/AA9+6159eJFhYc/k+/deFa9de/db697917ru545PH0/w9+69Qde5J/qT7917h13cBbEer+v+8+/dVzWvl1x9+6t1737r3XYIF7i/9PfutGvS463wmO3FvjbeGy0ohx9bkII6ly2kFAdWhm/o1re0d/K8NnJLF8Sg06QbnPLbbfLPCKsoJHW/L8PqLrXrrobaFDhMjQ0NVW4iMVM+uFXdmi0hvT/T3zU90E5r5i5ule6heWGFhoFDTrDHc992S23W7uLidYryc0kBORigx5dFR7s/l8dad7byrN1by3/j8kstSaqhgmmikWlYsW9Kynjg29yjy77yb7y5tMW2W21SDwxQ0HHpDsHNCcspKNk3JIzOavqzU+o6Ld2t/Kq6IwXWu7s1R7rw/wB5jsbLUQiNaMSHQhJKEfQj+vsW7D75cw7pvUG3S7dJGkpoXINF+3oT2HuhvUN5HKdxS6GoDwVHc9fTrVY7I21R7Q3tuDbuPqhWUeMrWggnuG1JpDC7D6/X3lhZzG4tUmPFhXrLXart7/bobyRdDSKCVPEV8j0I27gG2bQqRx4Kc25FyIwR9Pcw82f8qZtv+l61DQXLn7f8PQCB3Atrcf0AY2/3v3E/Rj17ySf6t/8AkpvfuvU69rc8GRrH+rG3v3Wj11Y/T8f1/H+39+69UddW5tcfT+vHv3Xq9diwvfn+hH49+69k9X9/yJcxiMV2RvX+J18NGZJ6HxiVwusCnsSL/wBPcCfeEtbm85MaG0Qu5rgdY3/eBcQSbLdy9sUc/e3kteFeln/PdzOHyma2qMZXQVZTN3kETq+kfYSAE6faD7utpdWXKghvEKPQYP29IPYGRLnet7vbY6oZJRpbyairWn2da6F/xfge8iusoPn117917rsfm4v/AI/W3v3Wj17iw/rzz7917z669+631737r3XIEc35J+h/x9+60R6ddA83PPv3Xjwx12Tc2HAP4+g9+69w49dWI/x/1ube/der11791vr3v3Xuve/de697917r3v3Xuu7E8AG5+gt9b/T37r3Rx+i/hX2x3Q0FZS4TI0GFqAHhrhT+QSoQG1X+igg8X59++3rXHh0d7KfykNx47DiufPVJqBF5HjtHdfTcjTb377OtZHHquTtT42dg9a5+pxv8IrcjRQk+OsSIC5R9JDKPr+Pp79x4db+3q2T+S3saSLsDe1fmqJ6aoiomMfmRdSmGIhbavpyT72PTr3AdXxdtbio6Ckq3NUnlEVhHc8sLiwH+Hv3A9bAxQ9EXxnWW6e2t0eZp56PDrP6m9QjkiDaibnj6D3r5daBNano/ux+r9r7NoaeipqVZqiNUEk/oa7AfUt79T162W1Z6FdII4o1ULoiAsAAoFv8AC3+w97z14fZ1FlngQgAg2PBH4txYX+vv3y69Xz69Gh0kqD6wNLD/AHj3406159SERoxdzqI9Wr8W+lr+9dbpTqPPXghUAJI/24FuLge9gY63XpI5hndDGbkyC1iORc/j3rz61TGemjFbZp4mNXIiyM5Nx6SBf6+958uvdLLGUiLJdIxGqWN7Bbn6D6e/cOtChOemDeeSaipJXW1hHzfm1uPqPfvs69w49EF35vKqTIkrN+00jqUu1h6uD/r+/Y69n06lba3wIdLeZdQFmAY2H9SfeuvZ6GHHdhU8sYH3CO6L+m5vf+h59+69XqXNvsvEx1AD6/UgXH0HvfXvl0FWY3JlN0V8WLxkUks3k0+jWSt2tq9669Unhjo1/W3XNRh8dBVZiRpakx3KMbFS1rD1f097r1sDPQiVjrFEYk40KdKi3H49+PWq9Bnk4p3k+rM5Y83vYA+/V63Ty6c8MjoU1kk3N73tz/r+9da6WMtVFTROWspC8arckD+vveOvUB6Avd+94aJasLLGXRGa30JGr+h9+69WnHqo3uv5GPU7jyWAeEBYROYZCWALBrce9A+vWxw6JVuLfdXmqqhjqGJjiqhKou39lgOL+9H59bHVtPxg2FSdg4HEzVdEZIqeVC8tgAyql24b3avVany6sN3bQQbY2k+Jw6eKGKlAVVAGpgLc6PqffuveXVRu7Ort/b17AlqaekrIaJKxpdYVypXV9R7959br0fHYHWWYxWDp0kMjzRwqWUg3uBb8e9da8+mjdu391wU8z00Ex0hySFYqObKCB79Xr3A9FGqO2c/tjcCY3Lo8QNSULSax9D+L+9/Pr1PTo9/W+94stjqYuyy+SNGY3vyRyefp79x61wHTT21i0yuLqRTuA7wPbTa7cfT377Ot5rnquPY2wcDj+1jmN30Rq6SKfyUzS6dKSh7hvV/j+PeuHW/n1bDSb829LhqaGB4GhjpYoo4kCgBUFlBt/T3uvWs06JJ37k4s1UQYWkjWYZmcUEYjtqUzkKLW9+60aZp59XV7uwVf1h/KN6e6podcWXqey6ncNbbiU089MixIQPxz+f6+6jj9vTjHsp6ADosuB2stZgKWKtpfIZKSETh1Q6rxjVcH3s/LqtPTqr353/CLZe+NuZfc2Bpo8XuOngmrGnjSEPI6LrCgLYm5+o/PvZx1oV4nrWJz2CyO28tXYbK08tLXUNQ8LxSoVLKrFVlX+qta4PvXXq9M3v3W+ve/de67ubW/H9PfutUHHrr37rfXvfuvddkgngW9+60K+fXubW5t/vHv3XsV66sfr+P6+/db697917r1ifpz7917ru3F7/m1vz791quaddf4/j37rfXvfuvdeuR9OPfuvde9+691737r3Xvfuvdd/Q8j37r3Hh14kE8C3v3WhXz6/9XQABI+nv3XqV669+691yGmwvzf/ePfuqmvl10CR9PfurEV49eFr8+/de+zrsr+RyLXv791oHyPXH37rfXvfuvdd24v+PfuvfLrr37r3XfFv8b/AO8e/de8+uvfuvde9+691737r3XZUj6j37rVQevWFifyPfuvVz14W/PPv3XjXy669+6313c2t+PfutUFa9d2v+kfQc+/derTj10ST9T+f8PfuvDHDr1za349+69QVr11791vq1T+XviIc7OKGNA9SKyZraeeHv8AX6/7z7TF2+pWPyPRxDbx/ueW6PFT6dbO3SW2DS4FIZIl1RhVsV5GhbWA9vyOkSlpMDotsrSfcJRBbDUx8ugt+VeHnpMPDlKW6PFOqHSLMLWJ5HP09o4dxtp5NEbAnoR3/J+87XaC7uoiF9aHpbdET/xDatNXsxLw0sCsx5KtazE3/wAfa+nn0Ew1cDiOkz3rnTSU7oJSCbAKpt+o8Xt/re9MyoNTHHT0UM1xIIYRUk06auo6apnpVn8bWmZH1HUbFuQSbe495o5sg2+NgpoADXPp1lx7H+w+5c0ypdXSUFVIJHz6HXM7KzeZxzmnicyclOWv9LX/AN4494qb176cv2l+0fiCoNDnrpTsHs7YbbtP0F2ylqUGBjoDsj0ZuyYPNVSPp1X0MZPoRfTb8e37D395duZRb+IFr51x0Sf6w1rdTlvFXjjA4fs6Um3uoJcfTiWqTSVN9RuQbcG9/ZgvufYbleC3sJxIT5A9DjavZbYrWHwJNJY/0V/zdCrt7bsNOeJlQoBYEKurSeD6vx7lTlHmWNrlTdGgFOsdPvCeyd5Dsbrsqhy1SAqivD5dNe/6akrMPV0XkWRzHIbA8agn1sPc6We42+4CsBqB1yz5m5P3blSUpuiFWJ6JD1tuVttbtrsXIT+5VuVUNzbV9Ofa/gegpimrozdBDLufOQ1EoMkUbKyl/UqBTc/Xj34Y631G7Byb5bL0O1qW+hdIZSSQbcfT3rrRrwHRkdq45MDtanp9KxsIAr8D1Hjg+/DreAOp2LlMbV8rfpNJU+ogWNxwb/T3dF1VHyJ6blOkK3qwH7etV3+ZX1rvLfPbn8T2/i5shDTS1cMhQNZQxuDqAP8AS3sJDmraoLk2lzIEIPEkdSVf8j7tNtse6WcZZaCop8vs6qv3H1lvbakH3OcwdXSQX5kKMwW/N2Nvp7N7XedrvX8O1nV29AR0DLrZtzso/FuYGRfUjHWXrDaed3fvHCY/A0EtdUpkKWZljRmCrHMG5sPr/Qe6b1uNrtu3yT3bBVCniePS7lfb7ncN6gjtkL0YVoOtoLr3qvPvsfb9ZmUfHrQY2OKRZEKMGEQUhuAfeC+98yWbbxNBaHV4r19eJ66Z7Bsm4XewW1uiaRDHVsegr0bT4x9fpRS5DKTRKftZWKO0YOsNIWBDf4+8xPbfbDt+wrNKtDKikdYI++XNMm98xJYRtVbdijCvpjo4EVcv3EqlwCHsqn62v/Z9jv5nqGBQYXh0Cfd9Ik1DHlkQXpGRWa3+pPNv9b3rz691mxVbDu/ZMCi0slJTolzZypChTe492FB1V8ivp0WLt+mkptsT0gQtH9xHcD6/0N7ewRzAzC5FesmvaNYP3LIaAn/Z6Hv44YinO2aeSAKjCH1gAX1A/k/63sGXu2XcrBlFa9ZJ8s85cv7cgt5pQpBoR0YR6QSV8UEjXUkKCebXNvp9PeNfvX9bs2xyTxEo4zXrPD2x36z3XlxrmwNQvmOpe4OrK16ujrsepLGIkPpuBfn6j/H3h3tnuH48b225yFgDTJ6U8zX53jaJYY20yLw6xUNJvTFVCQpTVE8auqFVVyG559qriblzcIjIWVSfs6hJbm/21wLlyAPn0NkXX2Z3DS0tTV0MsYeNSwZGvzz/AE9gCHmscu3jjbp9NDih/wBnoQw+6mxWVi9pJKus/PoHOy+o9y7aq6XKQUpmpb6yfGwCLwbcD+nvJ72X+8U9qzWG5TFgxAqTw6ja823279xrwjdJEEgrSuk5/PpF11X9ziI4HIErT08LL+QzuF0kf8a99A+R/cTZd7smmEwY09eHWEXvB7J7u/Mgh5Zty9ujVqowQM+XVsEe3Kbqb4qDacwSk/0nYtMi6BRCZg4tcqttX1/PuC/vIc8vsHKVxt1u9JLkqyUPEV8upL+7vthTn6yvnWkm21RscCPn+XVeVZsvGRYfxKFk8ELIoAUWFv8AiffPiz503+W8VZZWCmnmf8/XSveOZmuY5Ce4t1Xl21tKl2/LXViIvmlkkZIgo9QI/wAPeS/Ku+3O4xxxsxNAKmvWOe9bzcbfcObcVZzgD59VGfLnERS9d7krs8y07CnlelhlClmQRMQFLc295oe294TBHCM1HHrFv7wexGbl5t9v10SNTBArw49UPH6m30vx7mjrAjr1za349+61QVr11791vr3v3Xuve/de697917r3v3Xuve/de697917rkRflRwPr791oGmD10Bc+/dePDrs6fpbn+vv3Ws9eIAtY3v8A4j37rYNePXlAN7m39Pp7914k+XXR+p/1/futjh117917rkQB9P8Ae7+/daFfPrwHIvwD7914nGOvGwPH1B+v+9W9+68Knj1xJv791vr3v3Xuve/de697917rPTVE1LPFVU8jRT08iSwyKSGR0bUrKR/Q+6sodSrCoPVWUMNDCoPHo1eF+avyHwGJo8Ni991VPQUMYip4gZLogAABN/8ADj2TScu7LI5d7dCT6gdAm89ueTL+6a8u7CN5X4sVBJ+09On+z2/Ja1v9IVZ/yVL/ANHe6f1Y2P8A5Rk/3kdJ/wDWv5F/6N0X+8D/ADdN2Y+bHyIzmLrMPk9+VVTQV0TRVMJaYeRGFipIb3teW9lRtSW6Aj0A6ftPbjkuxulvLSwiSVODBQCPsNOiq11bUZKrqK6slaaqqZWlmlYkl2b6kk+zqONIkEcYoo4DobIixqEQUA6HPdhU7OoCGB/agB+n4jF/6+5b5sBHJm2H+j0hh/3Ic/b/AIegCFrG/wBfx7ifowz14An6e/derTrxtxb/AGPv3XhXz67ubAA/Xi3H59+69QceuiCOD791uteHXfpt9OeP+N+/dazXoTete3t99S5KTKbFzc2Gq5tJkkjLi5UWB9BHtHeWFpfp4d2gcDyOeind9j2vfYPpt1hWZPRgCP59Suze6uw+3aqOq33nZszNC/ljaQubSaSob1k/gke9We3Wdgui0jCD5Cg6ps/L+0bDF4O0QLAp8lFP8HQTi359rejk18uuvfut9dg/i/B+vv3WiPPruy3PP+t7917PXXHP9fx791vrr37r3XIW5v8A7A+/daNfLrxAH0Pv3XgT59cffut9dj6Hm3+w+vv3Wj11791vrkQLCx5/41791oE164+/db697917pxxWKr85kKPE4qlkrMhXTLBTU0KlpJZGPAUf7yfeiQoqerIjyPpXJPV4Pwh/lkZXcstNvntanjpseqRTR4qsgBBFxJpjEouWP01e2lk8RqL0untFtY9UzAsfL062Kuvtibc2FhqXB7SxcGOxlJEsSKKanBIVAh50n+nt8+vRaKkVOOsu8MfLU0FZHFAC0kViwjT8ixvx79x4dW44PVSvfuApKKv/ANydNFVKXfVH4o721f1t78QOtAjy6GX4Y7cw2HyuRymCxy0ElXAwqNEaIWVl5/Tb6+/Dj17o4e6dmR7ozsQmF6dZB5wf0sqkcWPvXyPW6V4dCF9lhdo4yGmx0UcKRoGcqiXP4IJt/j73WvXsgZ6d9vVS5EidCAhNrcAsB71w49bpXqVuHKpQwkSXiEf6W4I+tibn3vy6rU16R1DmoqxwySBkvxc/X+v096FevGnp0IlBMhjiLsALcX/3j3vPW6Ux1MZo6hisbggcNY8H/Ae9daJGOmqoko6ZijadVibkn6jjV7316vl0jMtkII2jd3BH1/3njg+9efWzSnU/+MUlPjBPqUA3+hF1J/qT731rp22/mqPIUx8TBnuQLFfxx/t/fsnPW8Vr0FnbFfPBQyrGpVWjOprixF+bn/H3rz695dVVdtbmkxtVPJGxcoJHHJFm1f4e/da+3ovuD7knfIPTanSzEEtI9iQbE/X37rfl0YDa+/ZdazrI7g+ogOwH+tb3v5daODXoyO0qXO7uEL0NJO8EhVCQHt6uQSfp/r8+9dez0drq/qqg26oy+QjjatlVG0lV1BrfnUD79U9bp/PoWsrVrGjabDQotYKtl/J4t72OtHoKMnuKKFiXmtckclfoDx791sdML7hoZC7eZQbXJutrAf4H36nWvPrnBunEw8yVUaBQPUWUXH5PvXWzx6CzfHbWKoXlghyCFQLFvIBpP5+h/wBv799vXvs6rK7374qaatqaXDVxmkkVlLRStfl/p6T/ALb37y68B1Xvkq3Jbqz4BWSry9Y5RIlMjOzSNwo/1z711sU6Ob0X8Jt67xy+Pye51kxeNV46jw1UbaXjJDALrH9Pe+tf4Orsdl7dwHUu3INv4ijRXhiTXOqIoZguliLe90r1706DzeG/Qdcc8hVF1WBa9yG/offuvEgHoMMN2VE+WSgpdAkeTRq0qSdRtYn/AGPv3VRx6OtsxGkxdNNVW1NY3IUBlIuLi3v32db889PG4pMcKKYeBCVQ6iI1N7Hm3vVCOtn06q/+RG18BVpNmY4USamlllJCqtrG9/Tz73w69QUp0GXUvaS0FOIiWMaM0Kvra109I+h/4j377etUJGOjArv1MnFokfyAi1y17XPAF/esHrVPToKN1UlBPKagRKsnJuvpIubgm3Pvx62DQU6D+ffsmBilhEzkIrKo8jcC1h9D/S/v1PTrY+fQE4rsfL7u7v6r25SyvUNWb4xFLLEut7RzVqJyD/W/vRPaetrTWMcetvf5yYuDZOD696pnQRvR7U27mzS2CqHrMbBKX8X0BN/rb3vjjrXnTquTOb6bb1H6KZjHFFYfT+ytuLe/cOtnomnZ3d2JycFZBXIEUh1dJGBGi3IIa/vZzx6qKcOqBPmdsjFVla++8FGtpJNEghVFHjL3fUEtew5H+8e9dbx1Xj9ffuvddi359+60a+XXdgACDzxxx791rNaeXXvxqPN/p/r/AOt791v5DrjY2v8Aj37rfy67Nvx791oV8+vaja34/wBYe/deoK169zYc8f0/Pv3XvPrtSAef99z7914gkY69/iBwPr/t/futfI9eYr+L3v8AX/jXv3WxXz66/H1/P0/2H19+6959dix4PH5v/wAR79141HDrq3Nh7916vr11791vr3v3XuuwD9R/Uc+/daNPPrv6n1EfT6/8i9+615Y66IAPBv8A7z791sGvHr//1tAC/Fv8ffutedeuh9R+f8P6+/db65i1ySAL/g+/dVNaY66W1+fpb/ffX37rbVpjrr8/19+631yI9Oq/1P0/2Pv3VQc064g2/F/futkV669+6312D+P6n37rXz68Rb8349+68DXrr6/T37rfXZ/1re/daHXXv3W+ve/de65FiRb37rQAHXH37rfXYF/zb37rRNOvE3/FvfuvAU69bgm/+w/r7916uaddi4+nNwPxe3v3WjQ8euPv3Vuve/de6k0dLLW1NPRwKXmqZ44Y1UEktIwQfT/X59+60cZ6vy+BvS+Q69q8Lk6mnaNq+nFQZ3BIZpbMTYgfX2Ft03aOy3JIyep65H9tb/mfk+4vo1NFBPWwV13lIKSjESlXcodRW31/II9+5i3CNNq+oZtCkVr1r2h5Xb+un0k0fiLG9Dj06AD5LZpK3AyUCrqkeruObnkgewHyxeG4vjLG9V6yg959v2+HlHw4IQHFeA+XSj6eCbe2HeXhpKWMhP6m1z9efcxGaMINR8uud67fdSXDiFCatTov/Yr5Dd+5oqOMv4TURgqL2IDW+o9gTmnmaGzgdVNNI49ZVexvspf7/uUNxdRVVmBz/s9G127RYnZmzaWsrJIqRoqdGYsRd5EW+mxt9fp7wY9wfcPcuYd1bl/Y9T6qhnXyrinXVratv2H2s5fRHVVk04wOPTfF8kK+MvS43b808MZCLOkJKuotYr/vfuNj7JTy0uLstqbJr6nj0CTvnMW+XDXNlUoxqKeh6eY+78xkaCWSfByqQCQph0m9uOP979rbL2Hu5rgG3rp9ei/cOcN62Gsc7EP0VPsvvfe+Oln+1oquGHkaEjK3F/6D3OXKHszHs06XclNa/PoJ2vvVu+33Fbgsc+fQfbU7u3blqxWmkqo19KMjavox5uPclScvyx9tvg16G0Hu5a7kpm3KjdpwQD5dGLwmQyOcpy0gkkeRGuSWP6vqLf1/w9zZyRayW1mVlNTTrmt95vfrXet+rZAKpbyHRQ+zMfV7Q7GpKwRyRwyMjyOAberkj8fX2OvPrFZfhA6PP06TkdvTZJQWZacspIF7Bb8Hn3o9bB6CjC5hKntVRVHUYp3ADWsAJCB9ffuveZI6OFkMxDNAscbjSAAFBABA9769WvSN3hv3G7M2rW5GqnUEI8arxcBxYcH3YOI0d/6J60IWubiGBTSsif4eiJ7XocN2Pu6tlrqSCrgqJ1lGuNWJWR/6m/8AX3hB7rX81jfSXMDFSa+fXWPlnkq0h5GsXZFIljUmo+Q6EftT4kdf732jkYTgaMTrRztHJ41UBlhOn6L7h3lz3L3Tat2jIlPxD/D0xv8A7Y7HvezvHLAvwkggUzT7OiQ/AP4lbe2fv/ctTmcXDNJSbhqTQvIokEcesqioHH0FuB9Pcse8vuVdbhsUIs2KVj7vKpx1D3sf7ZWm2b3cyX0YOiTsrmmT1bX2niafx0+3aCJKeF2EhWNQoKDgXItf3EXsbYf1m5wgS+/UDN5/t6yw9xt4g5D9sbzd7ZKSdwBAGARTpcbdxlNs3bMMcemOSqgjd2WwYto1XJ99RVgW0t47JBQRjSPy64tXl9Ju+7XW6SVPjOWFfn1EpsktRIX1WYm+q/Nyfrb3XPTeOkt2XUCbbdVBIupDHKdR55UXBv8A6/vZ615dIH48ZaA4fcMFe66IprRqxBNgxFufe8daJ0ipz0kO+ctiKfbNewRNX3ahbBbkH6EW/HsHcxR65g4wB1kZ7NzpDYSW0pq0mAPtPS8+Nc8n93YViLyNOFZFU/QG1xcf7D3CfPHvTt3JNq0cq65FFAKZz1lVsX3Xtw3i2/rBNNpimOsVY+XRzKHBxwyQ1dWRI91Ii/Ia9xf/AIn3z/8Acv3X3nn+7ZAxjgz29ZZckWUPIuwttEJrXifs6H7a+AzecRB9myQKCI5OQum/H4/3j3jVvMlpYVdXq3mOgpvPufy1tFwY7mddVcj/AFHoynSvWUU25YYtw4xamjLMxeVVYXvYXuPcCe73PG5bTyzLdbHOYplpwrw8+o39wPcHYtz2kvtc48Qjy49HjquqtrzU/josXSwxlfRoUDSRwLAD3APL3PnN7Qw3u5XRYsKmvmP8/WIG43N9cTM8czV+09AF2T0lNk8VUokCTwxiRVg0fQAWUrce545A93dvjv1iuaxkmhc8K/b0FnuuY9rn+ss5mDA1wT616rM3T8bsnnN34/DYenkp5my1JM9PGSjN4JwWDC30/qPeefJPuTudnAh2eYsbghQFOCT58fLrJ72h+8BZ2ED2HM8YkfQy6nUHiPn0dL+Y0tZgdufHzb+Mjajbb2yKKiykSAgSVIRA8jafybfn3kb78bjbbjHsFpdHVILEeIa8Xwan9vRh93qKG73DmLfxwmunaL5LU46rYXI1NDhKisySNHGUYqz39VwT7xgexiuNxSC0NTXy6yYi3e2EL+LIKjooeSlpt0y5rLZiESU2NacwwuLebj0aL/X3ONnFLtqwWVke+SlSPL16B/Lca71vzXFwupYyaL69a7H8y/ObxTLU9DHS1NFt2pkCsqqwQ0vjbTe39kmwPvP/ANprW1i2OIuwaYDPrXrEz74l7zBHvS2IhaOz0r5dvDqoRbcg/kfX+n+39zB1ge1eI66PB459+6sOu9QsRpHP5/p791Whrx642Pv3Vuve/de697917r3v3Xuve/de697917rkeBYG9/r791Xianrj791brv8AFrc/1/Pv3WvOvXdtJH5/w9+69xHXH37rfXvfuvde9+691737r3XZN7f4e/dapTrr37rfXIL/AFNv6f74+/dVJ9OvWIB+tv8Ae/fut1HXH37rfXvfuvdd8WP+wt/j791rr3IH+v7917B669+631yABHJsfwPfutEkdcffut9KOv3NkK7HwY6RgIYQouBYsEFgLezO63e+vLSOyncmOL4R6dNLCiuXAyek8ALXv/sPz7LOnKmtOugSPp7914ivXXv3W+vfT37r3XZJJv8AX37rWB11791vr3v3XuuwPrza3+8+/da68OTzx791s9dH6n37r3XvfuvdchYC5sb/AI/p791U1Jp10OP8ffurdePJv9P8PfuvDrr37r3XvfuvdcxwALBr/wC3Hv3VTn5ddX+ot9bf7D37rdOGeugbfi/v3XiK9de/db697917r34v/re/de6uw/lifFzH7gyqdkb4xENRRwTQ1eIecB1WNIyyGMOv1a5Jt/h7DW57siXItFwa56mjkr29u77ZZOYZRVKGn5DrZd2/BHkYIYqSGKlxlJHGsUUUYiUKgtwF/r7EEFDCCOok3ZWTcnifND0rZKmGnjaKFlKqPqPx+LD/AGx9vAV6Q16TseVavesodNyEI1MLgX44I+vv3E463Ur0Qv5D9VZHNytLQRGpmcOUVOSCXv8ARf8AePfvPrVB5dC38bevKzZO3PPk4/HW1UOlY2/UrEf0b8e/dbpinRnaWmSjQ1MwD1DgkGxub8+69bHpXoNuxqidcRNU06sGWNmYLfix/wAPx73SnWia9e6jyr5LGKZH5Vze9z9D9L+/daz0r974aty1BMsDEBVBS31JHP8Asfe/Lr2anoqOH7CG1tyPgszdEE5RZJBwLm1wf96968s9b4noea7dRjokq6eoLUoRWEo/SRp459++XXjSvQd1fbZjlEdPWFbNywI1G3Bsb+/Vp17HWen7I++kCS1BJe2mQ2uT/T6/X3vHHrxxjp2rpqmtiSbU2kfpOogHm9hf36nXjXpO5jK10OLkgDPdFNjf6n/G349++XWj1x6p3VLPkGoXkbXG0hIJIDW+oHvXy62KHoZew4Iq3DzStCJB9tfTp/Fufe+tV/LqmPuqnaXN5SmhW6sJQiLeyksffuPXj69EkxOOrId1SUMiOZJZiIxZrk3uFt711vq2341fHur3U9JVZmkMFEhErNNcLKmkNpAI5v8A09+r17jw8urPMXtbbux6GOnw9HFCiRjUdIa7qulv+K+9/Pr3TnFnqUxFnsNPNtQHFvz71nr3QI9odjx4DHVEwJ5UBStrW5/2/wCOfez8utcOq/N0/IuClecT1hjBLBLkDTzYXueL+/cePXj0D1J8oZKnKHGwSPLqYorj86voPr711vHE9S96925jD0KNM00CzAESuSLhhcaSffgKdebOD0V/c3c1bWR1JNXJI0qkA6jwx+lyD70fl1sdF8hj3BujNBcfFNk6urk8SxqzMylzdffutdWz/En4KVclVRb+3qsccw+2qYaOrPrDEB1IRh+PdqUz1o4x1bbLiKXHU8NNHHAsdNCkSCOJEAEa6RyP8PfuPXuHDoGd6ZSOmjliFy6KSGBv+foP969+63X06KjuKGbNyssIbXd7C7X55Fh711r/AAdZtidRZKpzVJlDrUxTLK1y3IDXN/6/T+nv2eJ690fahLUNDBG66fDCiMCLWK8Em3vwxXrZOB0He79xqsckcTaVs4IBJuAeL/8AE+944nqua06r1+QmdVNvV5BKXjn9Vz6+Pp79x6tpzXosPSeLmzWOnqzfxtJN41FyUa/DW9660ccD0M0VPnMQJFjgnqNLEqyh73vxxb37rZyek5md4V1FSzy11HMrKjD1KwP1sLavfuvCnn0TreXYtflMl/DsZHLJU1UxgihQG5fVYDj3qtevUqany6to/lYfB7Kb47y2B2bvnHt9rQbmxWVFDVMxKpT1CyIwVh/h735UPA9e1UIPp1e//M53KNxfKGeoxsJNFjNpYfFQwoLpGtBRxUwVVW4+ie/DPWzXUK9Uv93b9q8FhMqZaaVLU0wQkfpYCwI/p798utHj1Stld17x3ruCtx9A9RW/c1kschQkCBGNtRP0496r1oDoA/lBXU20tl0mz3y0dTnp5EkmgLq0ywtIBL9DfgE39768eNOq7wARybf7b37rRJr17gH8Nx791vJ+XXH37rfXd+ALfT8+/dapmvXr8W/xv791v59eA5sTb/X9+60TjGevfQ/1sffut+XXibn6W4Hv3WgKdde/db67F7WA9+61jj11791vrv8AFv8AH6+/de669+6912Lg8C/v3WjTz669+6313+L/AOP09+6916/BH9ffuvU8+uvfuvde9+691//X0AAL/m3v3WjjrwNj/X/ffX37rxyOvHk+/db4deP+tb8e/daHXrH+h9+63Udde/de67Av+be/daOOuvfut9cri1rc/wBffutUNeujyLk3/wAPfuvedOvD6WA5+tx/T+nv3Xj6nrzG5/p+P9t7914CnXj/AMR791vrr37r3Xvfuvde9+6912f9a3+wt791odde/db697917rwJH0Nv9b37r3Xvfuvdd24vfn+nv3Wq5p0Z/wCJGxoN99t4vG1UQlghWOVlNv1tUoEPP+sfdJW0RM48h0t2+3W6vooH4MR/hHW15sLYNHQ4bE0dHTGObH0QjR1Av+2tvqB9fePfN97cRXxvK10kddevu88sbXc8rnlwoKzwmnDiR0Zfp/FNVmshrA0ZDuEke4sObjn3j375e8e42+2psu1dpoNTD06T8mez9rydzPdS3yVd3ZlJ9PLoB+7sPINwCiYF4RUBgSeGAe4Kn2IvYnnG4vLDwrt9bmhFeiX3A5fj3e6bbWWitUD7Tw6XOMjhTblPBBqYilWMKv1JCWHuet758hggMakArgnoOe3f3Ur273L629WsDHUK04V+zqR131pUZLMvl8pTMKaIvKGa4FkGq9yPr7xP91PdyGK0fb9vlEk8mMZpXrN7l/knYfb3bzLAoDxqRTHp0hOxaut3Jv8ApNrUsrRYCnASeGMEK5U2JJ9se0tnt1oybluqh5JO4k+vWPfMFzvXuHzR+74CfDDUoPt6HTCYHbeNxiwx0cfkjQKXIBLleAef8Pr7mTdd1hmciEgCuB8usjOSuUrzZI1ScYVaGvy65I2KlkFOkcQuQOAtr3tb2J9kklW2DV8uoc90FhfedDevTZvLqDF5vCT1qxIzmB3CDRe4FwR+f6ezyLcZBKFPUU7ny3b3NoZly1OiDy4QbW3E9LVQGCMy+jWLKQH4P09nqhZFEiceouZLi0uWt5SaUOOjXdcywuYmjKlCgIHFrk8A/wCPuQOXj+jjrEj3hjddz1H16RPyA2I+Zx0uWghJnhs4K/UBBcrx7EZ416hgfD0o/jTu+kp9n5fGzKstVTQyQFf7QIUrcg+/Yp1pRTI6Lq2RqF7YleFjCGqZD/rDyXIFveiet5rjoyVTuKupxGGcsLW5HHBBv73175joKO4sdmN1YL7GkeSRZE8zxx6hYr6iLjj3dFVwyNwIPVDJJBPHcJxV1P5A9IT4+R/YZx6GdSk8LpGyueSVe3594O++1lLaXbKwoDU9dWPafnyDmbk23slILW6KuOPp1ZLDAJsBkQTpP2k5I+h/zZ94cPLo3OOn8Q/w9TTGobbXU/wn/B0UfoNR/f3OwxLZTl5QeLXJbk8e5Y57anL0bsfwD/B1FnI7Bt8liUfj/wAvQ89pTfw7clFXvZoUj8TA3C6wbEH+vs3+7Tdw2/Nts8pp3V/I9CX7wO2Xm9e0N5abepZxXA48OnPPZWOuwNJJCBxAhNv0i62A/wBt76fTkSyGVchsjrjLCr2jvaT4kjIUj59I3D1ZklSP6aWUfW17fX2Hbne7e0k8OTj1Ku0+22671ZreWwJBFcdKXeFNFPgp45BpvBIAb/W6WF/frXeIbyXw4yOtb97dbly9Zm7u1IAznorex8flqGqydPRO/jmmJNr2J1ccD2cSSLCNbmg6j6zsp9zkEFoNTE+X29M/aGz83maF6acy+N5VZgSQvp+pt7irnTmSOAGODNfMdZ2fdz9mr2/ZbvdarTIU/b0P/wAbqSXEU9JSztbwIUsw/wBbnn3gT76gXNyJBksoPXTJLeba9hFgBRIhQfZ0cAbioqPNQHJTRxUalXYv+lipu3P+t+PePVvs009k3061c46gr3B3qex2eU2Zo9D/AIOlpu35aYjBJRYTZ9JFXzqixu9OhJOkWJut/wCnPu2ye0Pj677e2IBzQnrnzc8pc3c2bpLcMz5YkUr0MfV/y3yNOKQ1+J0zWBkJRgwufpb2BedPYTY9+SSOGQUbyBHQZ3nZuaeViVvlfw18zWnR5NkfKrCZmqp6SoVYLvpkZgQikm1j7xd5p+7Tum0xm8spWcRjCVwR6U6RWPNJroYA6vPo1uM3JiNwQj7WaGeORblVKm2ocD3jbuGx7pt8skO5wGABu1c5I8wf8nQtiv7e4VQhBJHQN752KMZlYtz4ECmyVO/l1KL6mvcEL+fc0+1vuluuzTR7TdzUZMxsOK04ZPQZ3nbBHL9TbilfTov/AGjsndXc2TxJzxmrDSJFEJDYKiIwDAAj+g9zhv3vtfxq24cw3/jtCpWPV5LTgKfZ1kJ7Oc6WPLmyzWzKFZ6mnqei9fJjq3b+1qLb21oYFilraPXLLcD1KnPI9iH2A56vudILrmN2qkchVR8geim+9wt5kvLgwvgsKD5E9VoZXrqeiqZ4Y43fHLNJ5xyFkAva595hbZzjZW13FcXFGIIx1mv93KzN5ZtuG9g62UFK/M9U4/zGdgUW7Nv5OhocUz1tNRyw0kka+qNjEfUCB9R9fec/tVuwnMF2G7JBWnQT++PsVtvPKLxwRUmUijUHwgHrWeyOPq8XW1GPrYmhqqWQxTRt9VYf195MqwZQw8+uNc0LwStDJhlND1DuLEW5/r7t0159esfrbge/dbqOvD882/4n37rXXXv3W+ve/de697917rsfni9/p/r+/daPXvof99+R7917iOuvfut9e9+691zA9I/sn+v9Qffuq1z69cPfurddjj/H37r3XXv3Xuvcn/E/7f37r3DrlYA2v791qvXH37rfXfNvzYf7Ye/dex117917r1z/AF9+6912B9ebW/3n37rXXv6e/db669+6912b8Xv/AIf8a9+61jy66H1Hv3W+u7Ek2H0/3j37rWAM9eJuP6f1/wAf9f37r1Ouvfut9di1xf6e/daPDrkSDcWA/wAffutAEZ64kEf639f6+/dbBr11791vrsX/ABf/AGHv3XsefXXv3XuuyPpze/8AvHv3Wh11791vruxH4P8AvuPfutVHXXv3W+ve/de67HH+Pv3XuvXvew+tv9hb37rXXXv3W+uwbX4B/wBf37rRFeuvfut9dgcXvYj8fn37rXnTrr37rfXvfuvdd2/pzb8j37rVfXrr37rfQ8fHbp/Ldy9l7f2zR0E1Xj5K+H+KPGDojgvfS7fi/wDvXsu3S/i260a4lYLQYr0NOQ+Ub7nPmGHabJC+phqA9K9bcWx+tMZ09sbaO0cRGtP9vFAKllA1PeHQwJFvpfj3Em2brHvO7GRzU6qfz66B87ciXnt57cm1toiqeFqPyJGejWY2qaiwECUZNyl3It9COLe5qjULEFHoOuZtzM1zdvK/Ek/4ek5TbgV6h6aaYCViUF/xf6A+7kdN9Om35I1ytbDJOl/6kr/T+h9+pmvWh6N1Py8+GpXZ6lYppyD4QQrEsD9B7916h8uo8Xm+yaqVRyt4olH6PyLAe/Dr2OHQe5LcO5Ypo1WkldFkKn0sRb+th79inXs9P0lYuWw01NWw2kmiZfEwtyRf6H3rr1Os2yNvPg6eWZVMcJ1aEPJHN+Le9569UUp0tp84ssQpg6lh9D+GI5sT70BTreBk9A7u/pLH7wE+TVhDXspZJQwBV/1Bjbke/caU68RU9FR3jltz7GaTbNXPLPTi8a1B1abD0A8ce/eXVOJp0WreG/p9tr5TUGV5D6Rck6m/1j79XrfDp/6R7Om3Xuenx1SS371lh5vdTYXB9+r1qnVh266urxe3vNS0jkwUglC2JLMRewPvfl1aor0DWy+xqDc1RNicpEtLUlmjCuulm50i1/fhXrwocdLbaeEkx29S1LE0kLPcsASAjc+/fLrXDoyu8ainpMHIkig+SmFwbfn6jn37r3Hj1Vn2HtSKu3UKqOBvDU1DI35WzSHg+9dbFeha2b8VNpVtXQbprKdPMDHN49SgsfqQVt731qnRxqnceK69w0cNHDHR01PCoZGUWNlsWJ9+69xHSBPduMz8Pho5IiUZg9vqPrf6n3rrwxnptk3qHuEnXSR9OOT/ALD37r3DovvyC3VHHtGoqTOo0QgqCf6X4Fve/s68M56p0zVVu3flfLS7cxtXkmeZolMCubHV9RYf091PW+jI9FfDfs/NZyizebgyONpIp4p5I6jUquh9ZADD+nvfnjrxIp0d3u74wybp2pBQYiJoK2igjHljIBkMa+q9hySR791oCh6p/wCwuo969d1tTBk6Gsnpo2dhPJqKBQfqDb36h6tWhz0Yz4M7fpcn2Aaqvplq41qITGrKCEYJ9fUP6+/daJ9OtkHDaEo6eKGJUjSlgCogChQqWFwP97978s9aqPTpK7qmliildLoy/wCq+l7c8e/de49FG3llKuSqZHOkFiCQP1D/AAHvXy63nrntHb3386TBdQZlHHH1/PPvfWqevRotu4FMXACCqvo/oDbjgj37r3UPP5aOKJ6cBQ1ypk/xt/X37JwOvUpnos28Mo8TSAMWAEhIF/pquBb/AGPvVOvVr0Qf5GV0+T26IaZjHJrlVx/qlPOlffut+eOhC+Iey0ye3oYJ4iGeblWPP4ueP6+/Z69XqweTrXbmNp7z0QmbTc30k/63097rXrwFOgb3b13sbIwyRT4pW8gZdAYAgE3549661QdFgboPYe3d0x7pONWohpKlaiOkZlezq1+VPv2OvVPAdX5/yo9+YDsztfc23qGigov7m7Src1JSKqqYYsbTmV5nsBwAOffj1YAZYcekB3t3XsLfXamfzL5qimkgmqaRyGXj7eoMJU3J/p79QDh1qpr9nVYnyNr8FurH19NiqmJoys6loyLWP5uPfuGOvE16ow7d73xvTU2S2ht7Csm6Cs8gz5j1RkvdUAfj83uB791qvRKtrbM7U+SO8y9JTZDcWTqJY46qqjiJSmiZiQsSqLC39P8Abn37rwHp0IfbPw07g6sjSqrNuZKqoRB56ioaDR4gBc8WAIH+v791vNKnopDo0bvG6lXRmR1PBVlOllP+sffuvddWPF+Afz791qvXfA/IN+P+N+/deyeuPv3W+vXJ+vPv3Xuu7f7b+vv3Wq9eI5t9ffut9eNxwRY/X37rQzkdeF+SL/4/7H37r2OB669+6312Rb839+60Ouvfut9e+n09+6913fi1uf6/n37rVM169+Lf4/X37rfXXv3Xuve/de67It+b+/daHX//0Pn/APv3Xuu7H62Nv6+/der10L/j/ePfuvfb12ST9T791oADrxYn6/70PfuvUA669+631737r3XYF/yB/r+/dePXXv3Xuux/iOD791o/Lrsmx9IK8f7H37rQFRnPXH37q3Xvfuvde9+691y9P9D9P959+613dcfzx/sPfut9dkk/X37rwFOHXXv3Xuve/de67Av+QP8AX9+69117917r1j/T37r3VlPwX2bXYDNR9iVUEiU3khhCyKVBRZ/SwJ/2/usi60KevTtncNb3SzjgprXraP6ZzOJz+MpJ1njLtAGKqQTc245HuIud9iVLVpB59dB/uxe6M9xvkFuD8I0+fQi5PIy4Ku82JBeAW8kaAgjV+s+m3+v7wf8AczZrWXdhHKKArx+fXQbnqzll2gcxW2XpkD+fQMdt5/EV7U1bLOkVXZQ0Z+ur8+1ntDbXm27uI0FUzn5dRRtu1JzHcpMT3KRX7ehX6RwNNuZKUNJq1Ij25F1B/Af2Xe+3ON5y6r2lrVWck6q8OslIr07DsKxpQkCnl0brNYaiweFkoqaIRu8LqCFA9TLYG494c7RuNzuu7C6nYtQ1yfn1GO6X9zfQu0rV1VHRVMZ1ey7jqMtV3/cMkgdghFvqACf9h7nm451kg21IbBqOlOHQU5MsJNg3z66UYY16bs7XUWLqKilnlEIXWqsxsOB9R/h7FFrzBve6W8UkRKkUrQ5J6mfmD3A27bLEsHGog/4Oiz7o7Dr8JPNJj4HqYo5CwmjLFbA3vce8neRObLb6BbPdaLIBSpPWGvPPM1xvV615bior5dLjrHviTPstLk4zTQq3hbyFrH/WDf4+5Hjm269uEtrSVWkfgBx6D9jzLNb2b3F4umNAak16a+6tmncKrmsaNIQNInjUeoFb3BX3KW18tkWg8Q5PWIXPfvEDvz/RDtQkYr0B2xN+123K9cXXqwMTeJWYt/ZP1APsQ7dZfRAr1D/NvNJ5jKyMM8ejbNnKPdOAelGlnmgcE2Ui7KOTf8+zPz6BAyOi+bf27ltlZzIyUccjUtR55ZNINr2J+g96+zrdM9BZsys/jHaTtVobpUygqb3v5CCR73xPWs9HE3LgD9iKmnW+mzAjjgfj+t/eqU49bx057AoqDMUtV5kDS01NPGykKTcJYnn3vgwI9R14qGBDenRQMZkaXDd70+Lp7JBUVEnktxYiQ2v+P9494w/eQ27xALiIcE/ydZkfdRlmubWS2GSX+fr1ZgscabeyE4dWjFDOdYbg/tH8++d5kZt1jiIzrGPz66CSWclvZyR0zpP+Doivx23Zjh2NuWBpozImbnQXIvbWRb3NvP22TyctxaQfgH+DqGfbpmuOYZ44xUiQ/wCXoyHdlTFkcdI1LIDJE7yhlINtA1W4/wB69hb2m+o2/fIXGDUdZT7dsqXu13VhfJqV1fBzkg5z0GfT+46ndlDk8bVB1NIdEZOo8RnTf1e+kUHNzLtkaAdyrTrmJvX3fVm50upydKNIzaaD16d6GqtuNsbCSfHMVYi+ng359hy/uWuv8Ybz6l7lHY4NkH7qUA6TTgOhY3fEh2zI5OmRIZFY/jUFvcf8b9lm3bv9HeroNc+vQ1535Bj3/lqTx46drZoPIfZ0DHSVOmXqczLJID9tUaNPBJ/c/r7lfdL9JtpExPl/k6wD5G5YnsefW25FqNdAP9t0ud9yY2m1rIEKrbUvBNwOL/4e8duYL8TSGKM1JPXYP2p5WbbbOLcbpdIVOFAK9Bnt/sLH4iuKU7ohXjTfT+P8P9t7gzn7YJNymRmHBehXzzzLb29m8dvkny6U01ZujseuigxyzU1PqVXlXXYqTYtf/H/D2AFi2rlq3MlwQW406xturTdeYZhHJGVjY/6uPRpurelsRhpKapyumpyDKp1N6vUT9QGv7hTnX3DvbtXt9uBWPqQtg5R2/YrU3LqCQKnA6P8A9U/HXHZQnMZAqtOdTKjBFUqT/wAU941c1e8u57O4srRS0nyqescPfjmnlm8hba4kUOaA4Az+XQ1ZboTauKpJa6gq0g0XbVrUeu3HKnj2R7X74b9f3KWm4W7GvyPD8+sMbbYo7ucpZnPoOkrszcuZ2Fuilpamuaqxk7Cza20hdVhf/ePYv5r5f2rnjlmW4gh8O4UGhoKg06Ss0u03whduBz0eyp3FhqzBR1lTURRq8AlV5GXkFb/X3hHDy9u8G6vZwQM88T6SRXIJ6GUu4Ws1qC7AY6BPbfcW3IdwVGGV4JnScqliLsL2B/2J/wAfcu8zey27320Q7xchiFWrJ5HHn+3oj2/e47aZkB49F8+RuDr+w+wNsS08bUuNpoAkkp+lj9WAP49zF7Dfu7kDkLcIZHDyvI7hfSvl+XRhFud1c7zGLZCwJFRn16L/ANx7Lw+EpaTH44IzzRlZ5VCfXRy3H9Tz7EHI/M+5bxfT3d5UKrdozwr10+9mb64XZopJl0lFGD1Up8leucJDRVNdVKsuqNke6oRqdCAbN76Vfds3bdeYN2itmf8ASi8s9Rp96D3ETbtpewniqWGD9oPWqv8ANfpZNjboi3Ji4y9Ll181WY1QJGCLq7BP6fQn30eRDGNB8uuSe4zpdXbXCcGz0Q33fpF13cgW/B9+61Qceuvfut9dm1uPr791rNevD+tx/wAT79175dde/db697917r319+6912Rb8g/63v3Wga9d6fpyBf8AqffuvV69z+m4sPz+P9v7917HHrzW/H+9+/deFfPrj791vrux/wBbi/PF/wDW9+61Xrwv9R/t7e/deNOB66+vv3W+uzawsDx9T791oV8+vXIuPwfr7916g669+631737r3Xvfuvde9+691ysAebH/AFj791rJGMddE3/1h9P9b37rwFOuvfut9d3Iv/vPHv3WqDrwt+fp791v7OvG1+OB7914fPrr37r3Xdj/AEPv3Xq9dk3AHPH/ABT37rQFD10Be/I4/r+f9b37r1eu1YAe/daIr10Lfn6e/dW+zr1vqeP9ieffutdde/db65En6E3/AN9f37rQA49dC1+fp791s/Lrxt+Pp79177euvfuvdd8g/wCPv3XuPXuOb/X37rWfLrkNNhf6m/0/H+v791rNeuJta35/qPp791sV669+631737r3Xvfuvdd8gcEc/wBPr791rj1MoaCsylXTY/H00tVW1UixQQRLqklkY2CqPemZVUscAdXiikmlEUY1MxAAHqetn/8Alk/EN+stp0/YG5IGTMZ6BKjwVESM8M0kK8IWW+kCwH+HP1949+6HNaLDJCG7IxU59OusH3OfZldktxzdusR8ZwQAwHAgZz/Lq1TsDaVauHjziITFRxMxAHAVfzcD3i/yZ7w2zc4x2Pwx+IAWr/PrJL7wUdvuHt9fWcMep1iIFAOkz15uymy9BNj2mV6mLUGjve1ha1vr76QWl1Fe2Md5bmqMBQj7OuCN7byWe5S2sw0sCcefHpG77osxio6nM0SOTT/ulV1G63PHHt7I6Z+Lh0QPNfKfNUe7JKCB2gqaWUrUx+SXXZW0m4HvXWh6V6FTBfIqDP5KhSuyMcUyyJ6Gka55HFmPv1Rx62OPR2tt9k43I09NFFJHK7Kn0Pqbj6i/v329erXoWqKSCpEZaEFpLDlVJUEX/wB9x791sn06h7jw0cNM1RSLolMZYEAA6hyCebD3vrQxjoNMNvr7eonxGUnVDdlQtcG54uNXv3WqevTRks1JQZHyLL5KcuSGBOmx5B/x/wBf3oHHXieHQr7Tz38XQ2OoFdLD6cAarH6+9062OHQOd5bcoKvE1lU1HqmSCVo5Aq3DfUEn6/4+/eXWgc46ppyuVos5ubI4CrN5qKSQKSefS1ub+9dez0Z7447ApMdvClyL8ytNdVH9m9jyB9ffuvdW41G24a/HqlQqvHJBGuiyj06L2A/4qPfs9bBBHQIZHoDGzZKPJ4wrR1Ky+RnGgEm9/wCzY/7f3vrRqKHoVcXgqLb9PG8wVq0KFknYAliBYEH6/wBfeut0zToJuy95CKmkg03UpYNzb62/Hvw9evHHRZ8Fl6DM7gFHWxags11fTaxLcH1e/fLrwHR59n4aI09OEZWhiWNl0/QLa9ifexnr3QMfIuMrt7KSxNoC0Y0KLg3F/wAj37jgcevGnVbPUW5q2p3BkMZIzB1lcXLMfTc/S/vWB17y6HrO5mrwMLzSyMI2T0sxawv9bge/dVAqOi2zZit7f3bT7HjyGiB6nxTEsdLoG5Ug8e/dbp1bN058Z+uOrcDQVEuLjqsm6pUvUeKmf1OnN9XPvXHrdehzlr6IKKamiijgSyhVhjT0D6cKPd/l1UZ6wGlo6hWvEhBUj9KleR+qx96oOrZ4dFu7e6l29vDHVtJW4+OY1EJSOYQw/ttY+u9r+9H161w6K50d0SvVe9J6yOz0ks+uNdIAHN/7PvfXiMdWi4uriFOsquAwhSw/JsP0gfT37H5db4Y6DffWaKQTWHKrwt+b/wBffutfM9FnNs5XFJLMyuQVsLj3r5dez0Oe08TT4uJJzYR6VIv6uQOeBz/T3vh17z6U+Z3jj8ZTPJIyr6LAlgLi39P6/n3o8OtigPQIVm/KPI1kghdZIixUW/Bvz9Pex69ewekZudYaqimqYxZkRnNv6X+p/wCI968+qnquHuzOANSwH9BrHRgLG4D29+8q9eGDXo83xWekptvUNTBDzJo1Efg/Un3vrda9G03LuSOnQBvpotwD/seSPeut1oOgEzOchqZnePgcgC5PqJ+o9++fXqjoMM/XDwz+QMCqFtVidX9SB7359aPr1ar/AClsND17hvlL3XLHonyPR+9MTjZnuESsqMa6K0Zve4/w96NaV9OrJQNnzI61x9ndkZvMbq3RRVte5q5dxZUiRppiSr1shA5P+P8Are/DrX4jX16GmagztHTzSTtLUwSRFit3PDLcXJ497611Xz2v03he1d9Y3HZSuXb6HKRNUVkojAkQtZoS7g8EfX3rz69x6u1+MfRnU3TG28cmz6Gjrc28UMs2Xp4aQxuxQArwD/X629+NOPn1sHFOp3y4ykdd11XwV9HEEmpp4Ek+3hBuxIJuB739nXuOetQ/tfbsO3d45eCmYNT1FfVzRBQAE1SamUW/xJPvXWvOnQbXNrfj37r1Bx669+631737r3XYtfnn37rRr5dev+Px/sP6+/dep59de/db67JJ+vv3XgKcOu/oLEG5PHHv3WuJr11/r/7D37r32deNhx9T/UfS3v3XhXrr37rfXvfuvde9+691yOm3A59+60K+fXRt+Bb37rwr59de/db697917r//0fn/ANvz+Pfuvdd3Nrfj62/4n37rWK167Xm/NuP95/Hv3Wm64+/dW67Isbe/daBqOuvfut9d24vf82t791quadeYWPHv3Xgajrkfpcnmw/31vfutDjjh1xubAfge/dbp59eJv9ffutgU669+691737r3XIAkcf1+nv3WqivXRIP0FvfuvAEcevAXNvfuvHA68eCQPfuvDh11791vrsix/wBh791oGvXZIt9Of6+/deoa9dKLnn+nv3XiadPW3cXU5zOYrGU8bSyVddSwWVSfTJMAxa3+F/futHAoPPrZ66H6HpKXquhxxpI6easoIKlSYgDrABuLc/UewhvW7S2VyNJx1kN7b+3lnzLsryTKNZ4Hp9wkXZ/XmVamxzVktGsnijWMuQIy3DAD2GOYOYbW4ttM3n69Tl7V+0G/bJvIudsNAG8ujm9V02/9y1VM2Q+5K1JiSSNwx/znB+vvDz3g3DarWT6iWg0gkddBVtOYrblj6S/qRp6sp2T/AC3aztzASZqoy1JRSSIJIxPdmEjfQAf6/vArmn76Oye2e+R7SIpJZSaDwxXFesR+aect75O3jTZVC1qwBpwz0G1L8Mu7OjN3Gnx0dZnMa8pjikpPI8SJquLEXsLce5K3D7w3tz7rbMlzuEqW0wWpEhAbh5g9Sftv3hbHdOXPBvV0TKPPzp0PdX8YO5d1UcFS0s9F5UEpicuCn5KkH3G9l7qe2+zztGjI4XGoUp+3qJ+Y/vAOkTRWkTErwI6TeV+OW98VUUuLyGVFPWvpUKzBWYH0/n639ijbefeWtztn3Hb0EkQrUjhjoBP95O/SMwyo4fgOkN2n8YMlhMSlXk9NRJIuu6t6iunUzXH449m3K/u1s97dNbWLU0mh+R6OOTOZuaPcDcx4ofweOSeFeifVfX9Dk6yLaVHQKkksi+WpIBUWPqVj/X3MEfMUtnbHeJ3qoGB1PX7kimkTbLdKNip/w9CDuTofa2F2XItBBDBlYo1m+6QhSzxi7Xt7DXLPulv55rjuoXZURsdCXfeRNruOWJbA0Dshz86dJzrSqx2ZxNbh826/cU5+1hSUKS4X0XAP+9++sHIfONnzdsMM8BAljUBh6nrkD7h8n3/KnMc6XKkxyOxB9BXoIeyenYzUTZLH0oQCUurItuP63Hsafb0CKADt8+khtA5PETLTuHKoxQkltPJt9D/re/deAPl0ZrELgosNWVWTeAO1JO+pgoIbxE2uffq569ny8uiUdaY+lznbVZVUKr9nDWyKxHCkeU2tb3rHHrwqT1YFn8JHLhXhp7LIEIvp/Gn37zz16lOisV+Zruv2yFaszJEY6gOqn9RK8E+9qK5/PpuSQJSv4jT9vRFMTu5sn2mufZW0Q1bNJISTYNLewPuDfdhG3m1eJF1GhA/Z10N+6Xstjy5btebkw7yGAPzPz6sdrO5cdPsqqx9BUH7h6CUMwtYHxG/PvCTa/bXcLjmZJZkoofift6z90bdPaXG4yuNIjchfyNOqoui+w6ml7W3U6SyFEz0o1aiPUXPH+8+8weauTNquOXYomjGpYwD86DrDL2b5laH3GuFmFYmnP2UqerO8zmTWYD7uScMJ4WJ1Ncgsv09wvt/Le27bciS1j0uvn10YuXtI7I3EIoHBNfy6RPVG4lxNVkY6VbvM7gso5Oon6e5Sg3Jba30zefWOW5cqzb7vRnsacTWnQlbYgmGaqsm4cvJOz3I+mo3sf+Ne63nMcQg8NeJ4dKtg9nrz96fU3GRWv7OhN3XVyNgqpWLFWgmYD+l04X2G9v3MR3niymtT1MfNvJqXPL5srRQGVCD+zorPSW63xudz1C0rRNNVsEQmwN5eCL+5M3XmUHZvBiySOsJ+RPZ1V9yV3K7Kqkb5r556HfcGArNzRStSMzSyNY2PJJFmIt7jvbrN7xzJJ5mo6zE515ht+X4ksrQdoFMdMmO6UpsJTyZfOyIPGjzMGchvQtwLH8+3N+2iGeIaskY6AnKktlv1+ZN1Hbxz8ug+2t3gaDPVuH2/EzJSSvGPEo50G31+t/cN83+3du0a3Fx8LevVt3vLXcdxa02KipFUGgH2dLXB98b9yu5UxuqqoHMv7bPqF0V7AgH/AGPv3K/sVy5zQhtYpo1kYYU0JPWOHu5z/wAwckRkFWkhOCw8sdHp258iuwcZh48WMxULoXRdXPqItfV/X2Qb59zbaYr5p7hotQ9VHUGbHyVJ7nSfvgS5k7qE8Om/cPyb30aFsdLlqx1mYEnW/Dfi59xpvX3fOWuXLkXRaKSRRhQBXrJXk/7ue2We2vdyuiyitK8ekdWfJHMK9HHXZGZDCi2d2NwVubE+0FpyFZxI/wBNCKNxA6iTfvu/2895LNJcLXUadPec+YO4MhQ02Nhz1RHFDGEAEjhWUC1rj2U2PtDs9rdPe/Srrc1JoOPSKz9ltltJFN9cKwHEV6y9afKvb22ssKrPU8ldUiZWE7O4N9V9Wr37mr2z3beNuazsZBEhFKUHp0Vcxe1/LZuFk2+RV08adHQ3B8stp76x1CMHKlNOkIRmI9a+j/VfX3jNt3sfzDyxdTC8YyxyNWnlx9OpD9s/brlm3uheX7oxUjGOiv8AYncEDqKeevDSknTIWJt6bAC/udPa/wBobje9zWJU8OOo1ft6zd5c2faLKwN8hVYUFaYHVbXyU7Roajb1VRLWCSpZ1N/oTcHmx5/w99TfZ3232zlCNZLVArAZPmeueH3xebdo3e4Nltncy0GPs6oG+bG/KKfbVFizEJ56ikWMShdWgsPyT7yCM0bS6VNT1gCNsvIrMT3ClR8/PqqAC5t7c6RE0Feuvfut9e9+691yIAAN/r791oHNOuPv3W+uxax/rxb37rXn12oHJP45t/X37rxPp10eTwLf4e/deGBnrr37rfXNrWH9bfj8f6/v3VRWvXD37q3Xvfuvde9+6913e/8AsPfuvddgm1rfX/b+/daIFa9cffut9cwQbD8fn/H37qpqKnrieCffutjh12SOCBYj37rwB8+ugeb/AF/w9+68RinXdr6j+Rz7916tKdcffut9e9+691yKi1/8B/vPv3Va5p1x9+6t1yve/Fyfz+ffutU69cWtbnjnj37r1DWvXQ4P0v8A4H37rZ68eT9Lf4e/da8uu9X0v9B+PfuvU9OugLt/S59+68cDruw9X+H0/wBt791qpx1x9+6t1z0/p/xB/p+Bf37qtePXVh6v8Ppfg+/deqcdeUA3v+Le/deJpw66Nrm309+62OGeurf7z791vr3v3XuvfT37r3XIDVcn/D6e/dVOOHXX0P8AWx9+63xHXjyePfuvDhnr34tbn37r3nXrsiwB/qPfuvA1NOuPv3W+ve/de6Vm1tkbm3pVmi23i58jOHVLRDjUzWADHi/N7e2ZZ4oRWQ06Mtt2m/3WURWcZappWmB1dF8G/wCXducbhx/YPYFCsUNAUqYaOrhIVF/URErjliOCf6fT2Ct83K/v4vo9qBBbBI6yZ9q+TOTuUdzHMHuHMjJHQpFiurywetiLAZrbu3zj8fU08dBiqHxxqnEahI0Cg6BYe8VveH2858uNjYbMGlZvjoM0PWfe0fe19sUgTa7AJb4CAigGMDA6NDS5TZG7dvT0dLPSVFI8BjZbr9NPPvnXc7PzXy3vq/URusytwzWteHQ4i5p2PmrbXu2mWSBwdVSKUp0UzB9U0+2N65jL0sqHGzlmhhVvSnquLL77Ifd+vd7v+Rbc74rIVUUDceHz65C+/dtsdrztO2xlWBY1K8KV6UO6ZKKvgmo1RPGw0OpC2YD0+5oBPE9Q4aU7eqoe2OksVit5Vufp6FZf4lKxYoD6Az3PC/S3v3VcefQXZjorMSGLK4CmmeVmDRiNnurA3ANvfuvdGL6C6w7TmzUMualqqSjovWBMzgSheQBf37j17ga9WgYnHTRJGZl9Mcaaj+SyoOR/xPvfDPXvn08VWmpjeBlBVhYH+vFrE+/CvHrdc16K72v19V1S/fYVmhqYiXbSSC3NwBb375da+fSd2hj8nXY6SjzSP9zEpVXkvdiLWsf969+68M9D31zgajFpNNOS0fqIAP4t9be/dbzSo6D3u3OiPD1iW8cfiluDYiwU8X96p14da++5c8mM7crKmK/jnqpdVrj6Pc3Hv3n1qlAergvittw7pkpMwFHiQB9ZHHoQfQj37HW/l1ZDMI6WERizAIoAJ+npsfez8+t06T9Rk6aAEtIv04vb/eL/ANPfsdVBPQV7m3jRwF0Z1Oo2UAji35H/ABQe9deJ8+gWr6UbykmpInBZQdLW/J5A/wCN+/daIJNPLpER9E7pjy1NW0UjorTq0rqWHoJ1XJHvf2dbHp0eHbGOlwOIgpJ2D1KQxh2vfUQLEE+/U69XHSL7D27SbjxNXT1EWoNCVt+Lm/HPv3n16mK9U/b/AKJuo97TZWGEpSVEt3KiwCrJY3/p7117PQkV+/8Ab+7drLVFkZfGAeRdTpsR71nrZpWg6bPid0/Ubj7eq9ywjRiqYyVEbMSUa34H+PHu3XqVNOrct5bvo8JSLFIPFFT06R3+g/bAQm591+3rx49F1btSlnr2FLVXj1FSoYHUAfpce99azTpZ0nY0TaAJ/SbEgn68X497Pr1sEUoOPTtPvGjqU0s6spHIupvcc8+9Hr3yHTfBXYh5g4ERAueFW4I+nPvfnnrQpxHSlO8KKjpmSMatKXVgbnj6cD37h1uvl59ABvfsuKWWWlDN5Dx9f8ORx718uvZ8+khsXL/d5mZmFh9SfyR9SB731qnp0PuQ3TRUONVXkAVUH0I4/PJHv3Xvt6LL2T2NTmiqVE5KjWFs30NuAPeuvfLoKOvs7U5U+QGTS07AXJZj+b/n37r1ehtrpKxKGoaQkxmGxB+hFuQf9t7914DquHtaL+J5yOkjNv8ALHsP9Sxk/H+x9+rinXh1YZ8aaGTF7WoaV21sFRw3+w+nHv32dbyM9C52BkGiicW0sIr/AF/ob/n/AHj36mKdeqAegEw009fXNG7cBiy3BsefyPfutDpT7h25NUYuqlUBmEDHUv4t+T78OtHq2nqWjTqX+V1meyaoijbd1ZmtsLUfoaoMtOUMer8jnke/ZqQOrHFG61McHTVUXadYKRyVly89QVX6lZKgtq4/1/fuHXq16tBpadajFQeWFTejiVhp+vo5uD9T72cY61qIPRPO6Ov6LLt9zj4hBWUU33LsnpY6CGvcWPvx62Mjo/Hw5rIMnsqCilvJXU0wjcO120otgw1c24969evH1PQi/IjaS7m2vVY+R1VFil0KwtpOojk/T37rR61d/lv1fU7NrKiskgBQ5NwKkA20uwWwP9D791sjojYIF7i/+wv791oivDro/U+/db697917rvi3+N/r791rz67twD/U+/dernr36Tx/T/e/fuvfEM9cffut9cySPr+r8H/W+vv3VaDy4dcSb/X37q3Drr37r3Xvfuvddg2+nv3XiK9eub3/AD7916nl117917r1r/T37r3Xvfuvde9+691//9L5/wDz7917rvn68/6/v3Xsdde/de67/F/8fp7917z669+691737r3XLUL30j6Wtx/t/p791qhpSvXH37rfXY/1r+/daPXj9Txb/D37rY4dde/de697917r3v3Xuve/de697917rs/61uPfuvDrwF/zb37rR669+631737r3Xvfuvdd34t/vP8AxHv3WqZ6FvpHL4vC9h4Csyun7cZCjUFwCoczjkg+/VwetEVZft625Ord27by21dtyYyop3RMbTo6RlVClgLCwP8AXj3GHMaytcNU46zj9m5baPZ0RCATQfz6MdjMPiaxI6mSGGVzYj0oTc/2rm/uEuYbwyTeCDwPXS32k5XSDbxuM6g6hUY6Hjq6CnizFFDDCqgSxEWH4ueePeJv3hGK2PH8J6kfmsFNtc8BQ9X2/HEsmzVUkfVfwP8AiPfDP3dRX9zYCgNaD58AeuZ3vHJ/u3by49Cdu7O02AppK2sgSdYhckop0/09ubLyyOZ9y/dcurxjkMGIxX5dY93u4NZKHU46K9v35abM2JgshW1dTTmtjgmFPSaV8jShf2+L/wBf8PeQPJH3UOb+bt3ttlsHm8J5FJKscrXK1oTToPz81RQxs7gVpxI4dUWdrfJftnfe+qzemOy9TjsXRVLtT0qI0amCN9ShFvY8fm3vvp7S/c45A5M9v49g3W2SaeaPuZskMV6h3cd+ur++DQk4bH7ehp6u+UNb3NTHA5+qWTI0FM8CrLHpkcqChNyTc+8DPfH7vVp7P7497tA/SlfXg4Ar10v+7NeWt/sfhyR6ZlAAJ8+gtbIU2M3pWvZVMFRKCOPw3Nv969qltpr7lyJDwZR1O8jw7fupY/FnoPe1u1aaio6pPuVFlkUR2Fz9AL2PsRcocnnxFfRnGeg9zDzKXUwRGpNRTojuH3tuao3UcjjHljoVkBdUBCNZ7mw/HHvOH2qtLzYF8epVDSoPn8+oE9yOTdt5o2RvGQG5IND50p0czEdsYvMY6GgyqRwyiII7SKAWb6X5/wB695PWt5HeR+KvXPTmPl+65c3BrSdTxx02ZUbapoJaynqqf9LycFb/ANbce1efMdB/UOid9i9r1ktXJt3C1DOryMrzRX0oh9LKSD/T3Xqxrw6VnUFfidqAzxyJPkagrIx/U5YtqJJ/1/eiQBU9eQMziNeJ6N9Q9gQVNCxyEiU0hQ6UlFiy2vxe3tMt9aa9LOKjoyvdqvNujWa9UxqRUE/7PRPe591UmaqWwdDUIr1ThfIvFg555/2Psl3XmjbLGNtEgalQaeXTPLkm07lzBDt11OqKxBqadT9n9HYOl2+tQwgqq+rjWYzLICwYi5HHPsLWd5tm6QyTTUYZoesxdwsuYNsNmvK8haEBaleH8j1Ebr+XAU+Wnnl1QCiqvHFrva8RI0g/737jPdbvb7e+C2wAJfrODkHl/mS+5Ue63R2/3Hcmtf4T1Wp8WaY5vuTe+MmiaSL+9NWqtyAuk35I9ifnC5e32yNweKDH5dQX933Z4t75xvYnWpSZqH0ox6tnqtr1s1HNjRK6xqjmJibBVRfpf3F1h4Mj+PLQeo6zd5li3K2tf3da1NAeH2dIDrQ0+E3LPj69wzNMyxu554exP+8fn29v0fiRiS3OBx6LfbJza3z2+5GjnhX7eh1rN24nG5iKigZBra7FRcX+t+PZBHZXE0HjsDQdS/dcx7RZ34sgwVj9n+cdKTJ7ioq/Gy061MLM8bKNJ+l154P+v7SRx65Ay5A6M5bqyurN3tXDVFDkf5+iTZha3a27UyNJrFO0waQqCVPq1E2HHP8AX3IG0vbzw+FL1irzra7htW4m+2wFTU8Ojq9ddhYsYhK6R42nSAs6ML2Yc2Iv7Wqbe2couOiCRt23uAS3ILMB8+gW7n7czmfSqxmEWSJWiePXErBQGWxPp/qP8fah5LNlrIQeiqCw39JiLUMgpx6JV1HmKza3YEkWadn++q1ZnlJYeqT+h/4r7DvPMEF7sDrEaMBgdFGx73NyzvDwbi1DKaknqybO0+Iix9JuWjSJapI1ZZECg2cXPC+8f/b7d7/aebYFLEBXp/PoPe8sMO+8nXUho5apB/Lrlj911K0iTVEhRGQuXI45F78+8+uborQ7FFucxA1pUt86dYifd05m3m15pfaHY+FG5Wnyr0IO1p8XuVGWrnjDlrLIxVRb/C/0/wAffNTnq+vk3iW7SrpUgUzjrpt9RPuMSw7fXPGnr0OW2vibU9ssgwVUs7qY+IpNJUMbcW/4p7hTmH312/kCIzb2vhRiuSMY6jLmuM7IC1++lmBwfs6PTs/+WJgocXRPnSklWIEM6tKF0uwvpNx7x3u/vt2t5NK22xOIQx0uUIDAeY9R1iFzRve+Xm4ObGbTGDjPTlmf5bGwFljZTDHpHqUTj+vF+P6e6WH31dbmERtKfVUJH7egXdbjzNEK/UftI6ah8EMRi1tt6ZCyGxC1Aa9vwB7Eg+9JY3YH71iZAeGpSOiy15r5us3rBcnjwHSZzvwDzGbA/eWNrMdTSg/UWuOOPZ5s33r9q5ek8ezJUH0HUnW3vZzrHtR215XOoU8+qr/mj8IMxsOCSZZjJKVU2E+osdBP6feZPsr96SfnMLPazExg0NR0W8v7PZ80yPuXM7h2J4E549a0HzX6yq8NihNLG7SUtN5WIJspAu1xb3n1yDzN+/6zMa1PVfe7kO02nllN02tNEGkZ8j1U97ljrDjrkAOT/Tm39ffutE+XXH6n37rfXrH+nv3Xuve/de697917r3v3Xuve/de697917rsC4Jv9P959+61164ta3P8AX37r1DWvXXv3W+ve/de67JuBxa3+8+/daA69f6f4e/dep1yPqP00/X37rQwPXrj9L2P0+n+Pv3W+vX5ueeb/AOv791vy67I+p+n+Hv3WgfLrr6WIP/GvfuvcevfW5vz9f9f37r3Drw5P9PfutnrvgH8N791rJHp1xP19+6312Be/NrD/AG/v3WiadclNgeef6f19+60RU9cSLW5vf37rYNeuyLAG/PHH9PfuvA166F+Tfn/eT7914+nXvobkf7f37r3Hh10fr/T37rfXYHBN/p/vPv3Wq9de/db697917r3v3Xuve/de65Egj6Af4+/daA64+/db697917r3v3Xuux9frb/H37rR4deINz+be/der11791vr3v3XuuyPpz/xr37rXXXv3W+plDRT5CtpsfSoZairnjghRRyzyNpW3vTMFUsfLq8MbTyrEuCxp+3rah/l8fE7ZWJ65wO6s/t6N8tLFC/klClnkSMMXYFeSTz7IEtpNwu2aXEfpw6ltt+teUNjjgsaNdn8VAadWtXoMbEtHHHDT06AIkSokYVRwAdNvZ7BBBa9tutB1Fm6bpue+yGfdJS5PoSKdIzceKxGfheGPSX5sVcAi31u349qfGYqY5ACjccdFSWkUbCSIkOvDuPTNt+OfZMMw/iOimkBZI2k1abc8Eke453b2k5C3rc13e6s0MoOon1PUh7Z7qc7bRtzbRbXTCIig44/n0idzd5Q4WXwGrRkclHb6gf4nn2O7W1tbG3FnZIEjXgB0C7i6uL2Y3V4xeRuJJ9ekBB3PSZGuFPDVJK8relVXUS172ABP9fb3TNadDxgtjUW7KOCvy1OJEkGsh+OP1D6j3o14dewc9L7F7R25jmSCDGxKsZ+hQMLj62JHvfz69UdCDS0NIip9vBHAoI5REH1+vK+9E9eHz6cZUXRoAA+ot/rcj3qp6sB1FSjV7amtb8f14/qfe6+XXumetxsdQ0iBQdQseAb24Bvf8+/Ur17I6aoNpUqsZWQI3BsABxe3vfyPWh69Z6qupMRTyxAqGVSBe3BA+v++HvXW8efRJ+8t74qqoq6g8saTCOZb/Q3sfzf3vrX29UPbuSeu7VpsdSuXetq5Y00+q5L/hh7159e4Co62UPi1s0bN6mxEtREErXgV2LcsVeIH62v/re/deyc9Lnc26HgEtnCNZuPyBewNh/xHvdOvA4z0AO4eyKaiEv3dakIBKgtwbj/AGPv3n1o44dFN7B7zxFFWRL/ABKKQeTS/J45H5De9deHQsdK9s7YyWVpmOSp/JUTKni1DUWK8gXPv3XqdWU42opJ6COeljjCtHGQdIPpZAwNx78a9WFKZ6ZKyVWYkN9dR+trf19++3rWemqogWop5Fb1ApaxPABHv3XgKdV4fJHr2LNw1SvTlrxShG4DX1X4Nvr73x61w6q9y1JujZ9bS4CCKo+zrqowJYuQNT2BBt71nrfV7/xD2XR7X62xtdMirlKqANLqs0lpgGtcc+/de6wfIymyS7fyEtEkjyCldl0f2WJ/w9+61w+ZPVVuD3vk8fWSxV87xzCd7q+rVw5ABHveOvUrw6Fx+wqyKiWoScoSg03LfW3+v71XrwHkeouN7eyHlMU9Qzfi1iLWNuOffuHWgKYHQ5bU3RX5SAVEbO6E202Yk+m/PvfW+nbM7nrqCJpGDcr+n1E3/I/3w9+HXqitOgqp6HK7py/3Wl0jdy35Fwrfj/H/AB91615dGO2nsl6Jfu2jZXZRzqJv6ebj3vrflToP+z8lPi6aUXKKFf0kn6g29761XNOiR5Gvye5MsKCCSSRHmZSuokG5+nP+2HvXz63w6PP091RNDi6SeSna7lW1Frclbm5K397HW/n0M+4dowxY+qpnT9zwNYX/AN6/1vfsnqvVdG5+sMhV7qkZaWRlFQ5QkkfV7/09+x1s46PR0ztqTFYymjqh41SJLajzqHB/p711sZ49KnsLEQ1qFoTqbSLhbcgfkW9+68RQZ6CTbmBaHIWaEhdQN9J4/wCNe99aAr0MTUlEyQY06deR0UqgEXLuQLc+/V69TFDx6Pn88qn/AEd/yjelutKAfbVz73rMrWheJHjqeQWIsbf4e9D4h1dsxkegHWqTthpIe0ozoLLK8QJsfqxBZrn8+/V6oOHp1avSUBXE02lbBqSJ9VvqCn6ffq1698x0A29MYr1E4VLNJqRmt9VNrj377OvcTQ9Zekt3VHVm8VqaqbRhq2RIkjblA7GzN711Y0r0fbe2Uxe48OK2kmjnp5oA10sV1OuvTYH3vrRIPDqpn5LdTY3fuNyuPqadJR46h478EOR6CpAvcEX9+8+tVr1QJ2LsPJbB3FW4ishkWCOVvtpiGZTGT6UZ7fUe/db4Y6QINv8Abe/daPXYAI/VY/09+68Sa8OuPv3W+u7cXv8A7D/Y+/darmnXXv3W+uwL8X/x9+60TTPXr3BFrn+v1t791759eJ4Ata359+68B59dc+/db697917rsi35/F/9b/D37rQNeuvfut9e9+691yv+Rxa30/PP+Hv3VaeR69wT+Bx/tz7917IHXRAB4N/futjPX//T0AjZje4H+v791Udop1xufpfgf7b37rdBx669+6312Rb+n+w9+69119Pfuvde9+6913xb/G/1/wAPfutZr14C4+o/2Jt7916vXgbH/e7f0/Pv3Xjkddnk8D/ff7D37rwwM9cffut9e9+691737r3Xvfuvde9+6913cWt+ffutZr163/Ivz7916vXiLf0/2Hv3W+uvfuvddgX/ACB/r+/daJp11791vrmjvFIkkbFZI2V0dTYqynUpB/wPv3XuPR/PjV8r97bcy9Lt7J5LXReOJKV2MgLaZQDGwvbi4t/X2Ht8sI5bZpQtT1L/ALV8z3NlvUdhLJpjYjJ8qEdXt9U935lsPBlshVeenkClEOo+l+RwfeNl5axXvMQ25F4nPXVTYvdi55U5VSWZgwoAtfMeR6O9133nQ09RRVMxjivLGwkckem/5/w9xP74e3pv7LUgJ7afn0oi94Z+ZoTGVopHDq47pb5rdbbX2rFR5HN4+KpMSXDS2Yn62+v598nfcD7qXMXNPNKbrZiRShxpp1iH7z7wfrvGQCgrXoJfkF/MAostSS4/adMlbCxMfliLNqJXTqJH495U+wX3D98tZxuW9lwzUIZx5VrjrEzeuakuDowCPIdV0SVG4+xcwczna2T7GRvKtF5HAt9StmPvrP7Y+zXL/t3aI5QS3SjElBj5dAq/3Y3ieCg0jrD2UKfCbakFGqppDRX4GpStvx9T7l6Yl0LMeA6LrAf4/EnmWX/D0WTqneJ697FhzlWCKCaMo2ssFLyfT/e/eGv3huV5OctrdI8uppj066ieyEP7qtYJ40NGQftPQlb33/W42ryG6PC81NVyyzREaimibleR/h7gbZOUJJLGLb9JHhgKcenU68y8r7zb2bcxPXQcgfI9F7p/452XXNUHyJSyTgFQW9Ksb/n8e535U5EMCrPcoRGnqOPURR7nt6ykSOHmPl6dGp211VS4jERv47uUXUSEJLAfn3KAljhUQx4C8OnG22WQm5lNQeHTVuHaOJejnETfa1QuUfUBY2/Te/sS7bvb2yeGo6hHm72zt99vDd3H5cOir5yg3TRVNRCM0WpQzKEBe2ki3s8TmKR6RkcfPqMLr2btoFacEHTXGOkhgdsfxXMiiiB887jXV3DWZzb/AHv2dDebaEKjMGZuo0f2z3i+eW5hhZIoiRWmD0Y7FdP1WyDTbkyE7VlIskavEWGnlw309m3jLcWxERyR0BBt0mzbwrXykIrAZ+3pg+RWfkpp8XV7ZmFNAMcryxKTp1lAQCF9wtuhvIt+SFmIjZqEDj0PPvTyxQe0x33l5lEsVvqHoGC+dM9ASaOoyWNx+Vdlkr5aVJhIf06yuoMwHPP+H+w9op4bRbmW2cEoWIPr1yK23nfmy4gtt1S4H1AVWBqQK+Vafz6ddndv5raeYjx+dkEtNeFAql/FZpNFkZwP6g+0V7ZXW3WhutpJaHIKHjgV8us+vu0/ey3Cx5it+WudmSSRRGa93hkF6UBameGPPy4dHC3DPTZja9TX0ZVop8dNM5XSQvkpiwW/9fccyzGS7jmbi7A09M9fStyxPabv7YR73t4HhXNqzCnAAqeqxfhFiI17X37LLFqlO7q1kY2NwSQOT7kXnybVt9uFP4B/k6xM+61tItecd0eccZW/483VuOUx8sJaUKSpuNIFuDwQPcaW9vcSJ2A06zY3jdNltbgidlDcM9BNWdbPksqMrQu1NMgZ25AtY3sAP9v7EG2h5H+nuB29RXzZDAIjuuzyBXUE4+zPDoAe1dzx7YyEWPopVqss7LEJ0u3jlfjSQt/YkWxh3FjY2X9moq5FPLiOuTH3t/vWXft6zWG13P8AjbEKjgtXWcUxnj034Dc24azHxDI1TQTyWJEEc3IPK+phx/Q39kW6coC3ufG2huxeIYjJHWJHst/eRc5coXKw+4ErStNIKACQppJxUsBQgca9KWgr6TIVUOOy7MXnOiOSW2ogf2lP5Hspt5ZqvEy+FInl/F1239jffHlb332X6+1eMPQEBSCDXFOhewGw6ynkT7VZDSTEEgHjR/W/059kG4c2WllceBfOFb5nrIva9gtFufBQCnn0r6zrpI42cU73ZHu1lNzbgc+y6Lnba5p/DWdSfSvQpl5esoVIjHEGvDoue8OkchJR1m56WGWOainuhCj1KrXP6faO554tLzcFsQRQinWH3vDsIa4NzZDuQ5p/sdKnD7skqdn/AGVXIBNRLHE4udQKm3qv7D0th4G7LdQ4qag9ReLv95cvvttwatSlPXHQi4t03dNgtuYpNbVMcYnkiudJBGoNb3PnuNz3bWPt1DbTyDxBEPP5dRH7V8i7lHzTPd20LIPEOaeVejZ1XVtFtPE45lDCf7fXMxP9rT9Wt7wk5L5723ed/kg3dA0Rainy66ae321Wdna6Z11SHift6l9S9+bq6R3hT11K7VmMV7y0pEjIyKxYKbXt7Gnur93jlH3b2CSG2QRhxTyrUjiOos+8LyFdb/Atxs9VYDy+zo8Gc/mQbgyFA38K2+8UrKLFRIbcW/HvFnl37gNpYSi3uJ3khTARqUp6dYaD2Z5pkTXIzDzJp0TvtD+YJ3DDMJKemnp6f0K1knFtXDcj3knyl9xzlKy2uS5jtld1qdIAqeoV5z2O75a3RLS/kIVqAsfn0MfRH8wHIUDU825YvvhLoacSFx4yx9X6/eOvu192Lbt5hk26yU2si1AIABB6k6y9pZjs0fMFtPrjcA/Lo72R/mE9cphpqyERrUJExWEsNRYrcAgf7z7xj5e+5VzrNuq2ou5bmOvClcV6D9/scqxnTpUDFeqeflb8sKrtfJyigjVKdiEiWMMV/SwsT76F+0nsNce3tilhJCyuOOoUrWnUk8le2Vzue0PuKT6gmTT5Z6o6+T2z6vd228stbTyPLV0siAgDhGQ+pf8AjXvPnkXZb3ly3S4Y1U5p05zJuVjzLy7PyTeMA8VdJ8+HWvFufCS7czuRw8uomiqHjVmFiyX9JP8AvXvIC3l8aFZf4hXrn5ulmdv3CayP+hOV/Z0w+3ukHXvp7917rskn6n37rVAOvAixBHP4Pv3Xs1699Pof9t/xPv3XuPHrr37rfXvfuvddj6i/9fz9PfutHh14/U/6/v3Wx14W5v8An6f4e/daPXiLfkH/AFvfuvA168SDawsfz/j7914V8+vWP1sf9t791uo669+691yY3t/gLe/daAp1x/Hv3W+ve/de67uT9T7916g68CB9Rf37rRr5deP9bWB+nv3Xh11791vr3v3XuuwLm3+8/j37rVeuj9ffut9e9+6912PryL/4e/daPDHXja5t9P8AH37rw4Z669+6312ST9Tf37rQAHDrr37rfXZFgDcc/j+n+v791qvXXv3W+ve/de65cEcWFvr/AI+/da4Hrj791vrsC/5A/wBc+/dar11791vrv/C/F/8AYc+/da+fXvoeef8AW59+69xHXjzcj6e/deHz69yP68/7z791vr3Fhxzzf/H+nv3Wuuvfut9e9+69139CL/4X/wBb37rXljoXOisM24e29j4mIEtV5ulRbC9iGv8AT+nvxAOG4dbVnjIdeIOOt4PpLaD7V60wGKkQmSKnhb6AW1Q/0HvyhEWiDq00ks765TU9MfakOVx9A+QotZ8YDOBcgf0uR791U8MdEqPe02Jrnhq51jk1shVi4LHVbj36vp1oBqZHQf8AZPdu4a+lYY4ukOhrSL5OeL3uP9j73xHXuHRHN8ds7nMcpleSRk1ENeQXube9db6NL8LOv96djZ+PP5cVEWLhL1EJcN43AUHSC/8AvHv1Ot46vHp8VDjsdDTQDSIIkAX8llXST73x60K8OkNX1c9NUl3vpJIv+OT/AIf6/v1fPrVOljh6mSohjfWShYfT8/jj/ifeut48ulC4vpsLf15HAP8Aj711vrgy2DG4PAP+P+39+6103oyU5JlcAm9r8m5/APv2SevY4dI/ce648XrDOlluACfqL3PI97r17icdFC7U7rpMRTzzmaOyRlnTVb6G1gfe8Dj14rIcKMdVJdx99zbhyc/8NkEdi4Yqz2Yarf7z7rqU8OrmGVaVHSN+L+za/tfuOkqJKaWVcbViWRlF/SGDN9fz70Cp4dbMcidzYB62dYqKLAbbo6BWEa0tHTqyC31WLSfp7vjh00DjPRWt75dp62ZInCkahbnnng+/Z69UV6I33Jjs9k3VKOSVNTsWZSeb/Qen34Hrw4dEz3p1nuNaWSumaeYqNdrt9T+L+9fb1v5dA315nt47W3/hZGmnSigyMZnQs9hGrc3H+9+9V9OvGoz1s2dNbtTc+zKKvgfUj09Og0km58ADDn/b+909eveXSxmP7pdm4ueDf6D37r3HHUyMPIgCg6f7Vv8Ae+PfqU69x6CLsXB0VbSVHnj1MsTWYgW5P5uPe+vfLqqnsDe2zcBud8fmaCGeWgneSCRgo0uj8G596619nR+vjl25jtxY+lipqiMReMJFCp4QKLKLf4e/Y63k46NBufFpuDHSU8g1pPEQ5NidDc/VvfuvDzr0Q7sb4zUtVVGtxhMcxd3LJpvy1/ovv3l17PQJZ/qbN42jNOY5Z2juBYXPA/oPe+tV6CHHbK3ENxw001DUCJ51QuykAKTb6H+vv3n1upPVp3VfW2KotvUpqIh5iqli2m4Pjvex/N/evl1rjnrPvzrugmpImpk1Su9jpCGwP+A/PvfDrx9B017O2EKNkLL6AT+oJcc3u3+xHvVPPrw9Oh6p9vxR04jVQboAOF9J/rx7316tTXosXcmxnraOfSGcsspGkDjk2t+ffqdbxx6Kv1/sBKHOwyVNO7sasXZgObN9efevl1onNR1aTtOnoKDB0kUUYDeNBcKvB06ebf7172QOvGvSR3ZAajUYmAYq2o/n6/U+9UHDrdOgZbC0iVBmmVZJNR1MwXgX/B/w9+695dZKzc9Jhk8SyoI1Asp4t/QD3vrXTZS7kjy8rWkWRWIHpNwOeFt71XrfT+KdYEMoshtrNx9f6ge9/Pr3SLo8tJU9l9e4vy+jI7sxtEo9R1eWoVLAfT8296+fWqVYL0e/+czuNdn7e2T0d5VE+MwGFypoz/nENZSpICyLxze/vwydXW/hqvr1rsYHbhh3RT5N49TF4W/s2Fhcf4+/deHoerEcFk4pMTEk7hv8njULb1KdPHv3yHXgOgw3PNCJ5CQJLsxv/QX+v9Pfvmeqj06DDOx0ldQSIQFliVmhYG2l/wCot70KA4630ruiOw62fIVOzcxUCeONC0DOzE6RwoUk+99exwPQmdhbMNXQVc0KHyvHI44F+VPAH59++XWqDj1T/wB1bFxWQy+ToszQK9SySrDK6oCGIIXTcf19+6tg9VWby21UbWztVjZv06mlhJFv2mc6VP8ArWt791rhjpKfT37rfXvfuvde9+6912Rb83/1vfuvDroEj6ce/de49euffuvde9+6917/AA/Hv3Xuu/x/jf37rXn11791vr3v3Xuu+Lf43+v+Hv3WvPrr37rfXZFj9Qf8R791oGvXXv3W+v/U+f8A+/de697917rn6fyCP9v791Xu8uuHv3Vuve/de67AJ+g9+61UDrr37rfXZt+PfutCvn14C/8AT/Y+/dbJp1yHpuSOD9D/ALz791U54ddW/Fxf63vxb+nv3W6+fXVuL8f8T791uvXXv3Xuve/de697917r3v3Xuu1Nj791oivXXv3W+ve/de697917r3v3Xuve/de6k0lXNQ1EFXSyPFPTyJLG6sVIZGDDkf4j3plV10twPTkE0tvKJom0spqCOr2f5a+8qvtrMUW2d2SKaOJpIW1FjG3hICuofjn+nuNOYNjsNrdt1hX9TjXrLb2g5p3HnbcINh3RyYl7aVNMdXg9kbX672ZhhSQrEa5oAIEUgElh6G4seD7i10l5pvRtzLqUnrOy+2jlr205Vk5g3ohVEbaQaZNMdF52r15nMvkZauonnhoWbywBpZVUx/qULz7l7Y/bHlrb4Ve7hDyYINBgjrlR7h+5l3zZvc0llWOEMQAfMH06U+TkrMBekpYpp2ja2sM72A/IJPsT7jvFrsSKmkIiigwB0xyH7b7nz+5Tb6s59M9ZNvdv5HG5SCjyFPIkJ0rd2cCxuAPr9fddt5gsdz/sWFT5DpRzp7R8z8kBpdziYKvmRT/J0LObq6jemKUQKfDLZ9XqYKP63N/Zrdt4dqzH0PQE5ehN1vcMY/iX/D0HybJo6+sgo6yMFEdAbKASUAtc+8cubLkoX86k9divYfZbe5t7aK4FQEXy+fQv7j2pjKvC0uIlhX7SGCM8gfRVtZj7AOzRmXdY40HxH06yq9wZrHb+RLyScDTGhC8OIHQL0GQxG3shPgMMkbVb6zHpVSdR5Ucfn/W95SwbMsttDGRgqK9cNd19zLmy3S/mjc+JHKwXJrQE9PW3+08vt6epod1QyRQOzCGaUuqWP0Ua+PZDvvK304+ohOB1KntX74/vi4/dW8YZqUJPSjxuA3B2GKvL0AlgxUZZ1f1eN478EMfeNvPHu7tHKt8m1R0eUkA0PA9ZTcu8rXvNlzqjOmI5B8qdJGs2F5KqSGWaN9D6WBsb82b6j6+xCOfbafbI7qOSjOoNK8OpX5e9mbY3wj3SjJXOB005TbNFtyamqKMJDMul30Aeqxv9be7bHzULu9Dyy1A+fR17k+33L3L/AC6YdphAD/F2r0PWEykG5sFHh6tfIHhRibAjV+m/PuU9r592rbJvDu5QK+p6wa569gL/AJxsvqdhXuBrQDNfy6JZ8hMPXYHI+FkZ8f4xeSxKohFgD/T27vG7bTvNxHfWLA0YcKZ6xD+8h7Yc+8ne095Z7wjmHwyASDTgcdN2z6KLKbex3iHAoowDyOEWx/V7C25SPDeOW82PXHOwmjhs4YxjSigj7B889ILsDazTUit49L09QlSrWsP2WDc6f9bj/b+1+23aqWVshlK/tHRxC7zXFvcwEo8EscnGlQjAnh8uHoc9CX0VvmpzWy8phq6TyVINZGiu13FNFEyfRj+LW9xvvm2Nte9GCQ1DEFfzNevp6/u5ve//AF0/aG+2a7kJ+ltmESsakaKhlFa8D6dAd8MEWo7a3s1KgMdNuurWVVH6WWQ3/wCJ9jDmeKttbxT/AIlFP5dSP7K3jR77u8tpjwpXB/a3p1bnmoBqZvolzxpHA/qVHtB4lpt9mCwFadDFLTfuZuYHVS2nV6noIux93UmytrZCuXirihMiAWDMr+kAfTj8+ypd0R2AgSrudI/Pr3vhJN7U+1lzzTfSii6oxU0yUr/gPVeWDSt3Xna7N18bSR1E4qaQnUwub3IZvY4igh2zb0jj7ZHB1+tTTr5VfdTnu859553Dcrp/Et45Q0OSR+Kp/b+zocKOjNLTszeiMqQeAvBFmF/r7K2kJcacmvUS3JjvAYpBUMCMGmDg0IyPy6CTembXHZKngxSK0lNTLPAVLmVXaazANc/W593vdsS4gXdZR+tr04AyKfLrLT7kXulzb7c+48fJ2zFjtoiWYDU5YMZMipJFDXhjI8+rVPjfmduZ3ZOPXPNHT5D+HozGUIWZx/XXz/sfeG3vdytzW27Le7dA7Rt5iuOu/u3/AHgk2m5t23JtDSgEgngT0JmRyO0MhUy4mnqaZXVxGukL6iTa4PuGYNp5i2oC6uUYGlTWvWQGxe7FhvES3EEwcMOFfXpVy9fUNbtaqoYIo5xNDKWAjRuSv6v9b2Qx83XNvvqSzsV0kcSfXpFvdrDucUkj511PVVm7trNQdpU+0Ma948lUOJwn0Rg1j6V44+n095r8jQScyWMd1IKAAGvy6gbbuX0TmID4owc/t6PP1J1pjNlTwZKqCvURoSHdQ1ibHgH2Fffnar0bOEt2OhaDB8usiLCHYNrs/C26ECRsk0HQjbgzNburJCgoyTEpKEqbgAm30/H+294ybZtUGxWn1c2G49Czl/fLGBCHpr9OmuHqbIPJ5JiHVmJI0i5F7/X2LrX3lvNtiENq9NNOJx0eTb7YzjTdR6vtz08nY9Rho1lZSEUMApF7WHA59jrkz3wvdx3iKyu8hyBX7T0G+Ydw202DpbRgGh8h6fZ0iMvtSmz1PPBV06AOxIYxrweSDc/1499IOQ2L2iXFQVcf4euUv3nbwS3hiUUODjHD7Oii9h4+u2hl6XFYtWY1SlrRMwOkfS+mw9gr3O5V5e2u4HMVxAGjHxmgpU+vQW9pOfeY9/23+pguNLNhBXy4dIOPJZ2kldMrRVKU7ceSSSbTb/Wvb2d8h23JF3bruW1xopIrgLXoo512b3I2C6ktZQ7Jq40NPy6QO5t601HVQwwC/wC6ha2o2seRc+zLmvbLae5We1UH7AOpe9g+fN12zb7jbd9fTrDAAk/5emvdGSw2axE1TkWiipo6SYM0gSwtEbA39ifbLFZdqSOQZ+zqCOceaLmw52nu7ZzoqfM/PrWc+QFRj5+0NyDGlGp4q2ZA6W0t+4SALf09n0EXgxhPTqKd0vhuV693ShY56Bkr+R9P94H+x9vdFgPr1w9+6t1737r3Xvfuvddm3Fv9j791rPXXv3W+ve/de67NvwPfutCvn11791vr3v3Xuve/de67tYcj62sf979+61xOOu7mwFxY8fjj/X9+61ivXVvryOP8fr/re/dbr14gj6+/derXr1ja/wBf9b8f6/v3Xq+XXXv3W+u+AeeR/h791riMdde/db65EggAfgf8R791oA164+/db67Fr8/T37rRr5dev9bfQ+/de669+63176c+/de65c3v9Ceb+/daxSnXX55555/3w9+695Y68bX4Fh7914V8+uvfut9cjpGnj8ci/wDh791XOevNbi35+v8Ah791sV8+uuLD+v5/4j37reeuvfuvde9+691737r3Xvfuvde9+691737r3Xdrn0g/6319+61w49eHFwfp+ffuvceHXjc/6w/3ge/de4dde/db68Tfn37r3Xvp7917p2wuEym48nTYjEUslbkKx/HBBELszW/NvoP8ffvmevAeQ6tR+Jvwn7MwvY+xewM/QVFJQ47JRVksL058fiK/2i4/APtFJfQRSCNjk9CSx5V3XcLJ72JDoQEk04dbYGBylPJgqXwAKIaaEaQBa6RWawH/ABT2uwcjoNUKsUPEdIDeGdpailkpqoKYpBpeMhfVbj8+9MO006ulDKoHn0USo+NlFvjcaZeB446PzeRksBxq1AX9l8DTNMQeHQu3RNsi25PCoZKZHn0PyfGnZ5x8dFUUsUlkCswjjPGnSfr7MTQdA5e4V6DfO/CvY2TkUNSwpEp1OXhh9VzcEG3v3Hrx6Mh1xsXB9c4SDBYKFIftkKeRI4lJH04Kj34D163UdLJs0sUpjqH1FvTbi3J+g96p69ar59J7dEMlXT+SH0pYsAOdRB+vHv3W+klT7pbDU8azuy6CLAsQpA/BHvfn14CnTmncGIWPQZYxKRYqSL3+hA5/3v37HXvLHT/j9/42rgEvkCk/6ph+OeP9t79wz16p1hBknh0H+8t+xIx+2mUFDcaZNP4/opt7LH3WzifQzZ6GVvyFv97ZreQwtpbhjokvbffdPioqmOerUS8WHlI5Jt+T/h7d/eFsVLK3SH+qu8xXKxSREZz1Vl2h3JnN0V9TRUMskkUmpDaSQggtcDj2H5txaSRhG2B1K1jy1tdjYpLuNFY+vHoAa6gycdFU1kqOZAjPZixuTzbn/X+vsuffNL+Hqz0YwbFstwQVUBfWg6t0/lsYLFY9azcFQsQrqmjYEsia1cr9bsL39ijarlZU1Oc9Rhzpt6RXPg2CdoPEDq4SpnTJQnyNrDrpFrcXHHA9m4IOR0BWVkOk8R0DW4dkxzzvKi6iTcWC/wBb/wBL+/dU6Qk/W0NbJeqg1Kp4JjQj/H6i/wDvPv3XseXUOv6z2w8DUldSRuhWxvHGeR/W/vXXqnoFc18Stv7hqZavCwRQSKNQYRxKSWNxYj37hnrZBPRsej9tPsjEjbE0gZ6e31vdSE0/n8+99ezTPQlZaBoJdQa6t/vPPP8Aj7117p5wUqMrKzL9LWuOR9bE+9/Z1qnSb3rj4qrH1Q0aSYnUMTx/r+/cOvcc9a3Pyex2VHa+RpYpHkjaecRorMoLeT6enj3rrdfPowHw5zmYwu86HE5HzRRM2gRvJLYarW4Jtb3rrxPp1fRHZaeEEEq9NEbnm4ZRY+7de6DzP11NErxWVWAblgDb1fT37rRJAr0CGXzuNadoZWiZ1Y3OlDcE+/de65YemwFbVrL4YnlDcERx2uLW/HvXXqgHPQ+4IRxxJEpCR/p08AcLbj36met/Pp+lhpJAQ5Q25uzAi4H+Pv2OtdJOuydLjZbQyIlj6kGkGw+lvfvl1vjx6mUm8YAgUODxeykE3P0B/wBf3vrX2dB5vHcEdbFKpQhNLAkgEcnk/T/b+/de8qnop+c3VDtir+8klT7cSk82Gnn63Wx964depXI6HzrDtql3FTwpT1KzKDpNpCTxxb+n+w9+698uhlytXDPTM6W1aWvf+tr+/dbIx0WPfe9Kfb8bGokCh3dRc6fofzpt72etdEZ7H7nkgrqkR1hEQuscQle7GwC839663g9DV8aM5nd1v91VU832fktGzl2BNuPr79x6169HW3Dj3ixjMLxnRxckeoDi3vfW8dB/8cto1u//AJH9fY6W80GB3hisnLe7jwQVaySMQfwAD71Wg60MuD6dCV/Oz3a28Pn1n5KOa+HpNq4Chp4FYlENLRJFYIOB9PoB72DUGvVmFCp+R6rYxVNEEjYRDUtrsBzx/j71nz6rXoS6LLzQxKgZyAttNzfjj/be/VHn14jzHTVkqqSfUx5BHNv8eP8AfD349eNePQX5qsKa18hX/Y2B5+lj78PTrfDPQZ7Q3I+3ezMXVSS2p66rhplJcqutnCgcW9++fXsV6uP3DtgNtqiymlGiqcXTTsbXuHjJPP8AT3vj1rqmv5RY6kiyc9dSKqyI/qVVVWBH/Bbf7b3qnW+qtO69nzZbEx7no6Zpp4mEbrEgZxFf1sQv+wPv3Wj0UYg/43HDA/UEH37rVadcPfurde9+6912AT9PfuvcOuvfuvdciABb6n6gg+/da49cffut9e/x9+6912Lfn37rRr5deHJ4sP6X9+695Z68bk88+/dbHXXv3XuuyCPr791qoPDrr37rfXLTxYC5/qPpb37qtc56/9X5/wD7917rw4N/6e/de67Jv791oCnXXv3W+uybn37rQwOuwSOLfX/D+vv3XqA564+/db67BAPIv791o54dde/db67JJAH9PfutUzXrr37rfXduL3/2Hv3WvPrxN7f4C3v3W+uvfuvddggHn37rRr5ddH6+/db697917r3v3Xuve/de697917r3v3XujXdG/G7IdrY2TLF3jgDFUXkAr+G4H5/1/fuq56FKt+IRxFaq1kn7KSgNdzpsjXN/9f21KxUVHS+yt4pm/VNPz6sK+PmDwPVcFNX7e8NJX00S3eCylmAuxsP8fYI5jZpoTHMcHrJr2eht7HdIrrbxR1zjo8O2MxmO48tT5SrrJXpqKWKCSJ2I1+NuTa5/p7IOULK3sLzXSrk8epZ+8JzJvXNfLos5WYQxpTSKjI8+j6U+LhOLoaelCIYKZIjpABuBYG/uXWcDvY0B65+R28sjm1iUswx02psY1WuWWJXb9TNYkWv/AEt+fcNe5l4LmLwIDX7Ouj/3LuXrjZ7r95bnHpVcjUMcfn0VztDFU1JnYqKBRHUmeFNAAU/qNzx7BfI8V/bbggyV/PrJH7zl9ypu3J8yzCMSitDQA9HH2ftynxuxaSSRVE01KjaiOT6b25/PvIy7V5duC07qHrjlsNzBY83tKDSNZAB+3y6RMNFqyXkjUHS9144P9bn8+8a+cVMLMjcST12h+7xNDf28M0XAIv8Ah6dN3VpjodKcSfbkE8goWFuT7Z5DsYrrcUZ/ir0v+9VzNfbDyXPFb1CNgkcOHQA7G2i/97hlagGaV5hIhN2st+OD7yjiTw4VQeQ64a3s/wBXuM1wTlmJPQh9o7PXdlRRY1I0Scywmy2RiusAsSPZBzpudtsvKM+43LBQobJ+zo85Itru95vggsKlyVwPt6OriqDCbD6xoccqQU5/h8KzEhSzN4/UzMbX/r74j7vdbhzXz/PdqxdBKxX0pXy67Y8rgcvcq2rSDRIIxqPnWnn0VOeuwlRkp5UmjIaQsxBH5P159zdDHuUFosfcKD160PcS48Yqr8P9Xr0i93rj8lVQwUzq7sADYiwBP0v7Ptlur6zjMjsRTog5o57uN5g+h1Vr0OHV20KdaEzPES6p6Te/JH149x9zvzTuIuF0TMKH1PQt9sNwaxUxPSjU4ivn0FnfnWlXnsNkJfCRwVDsCfSilgOB7k3299yRcwxbNI5WTBDE+f7eh17vci8u+7PIE/K1+iOWUsMCuoA0z1XVtHc2X2hma3DZKmmEDVKxRGcssSRISpkiBH0PHvLO3uI9zsE8ftZB8QFdVfU9fKt95b7r2/8Atdzle321QM8Ez1ZKkLEF80FPMcR0OlVV47cePkCSJITG9wACf0WH4/3j2nWOW2lqBjHWJ0cstlNolwwx0D/URXE7sqMUoAlkocqsKfhpZA1uP9c8e6c1Wwud2huG+Elan5ADrq5/dj+5H9VNG33EnhrIs4evDuLfy6ZvhnhdybJ7N7ArM3jpYKSv3XXVccst1Vo5CQCGYfT25zje2tzbw+C3dGAMddSPYjnjlnZeZd2i3e4VI7iZiCacNTH1+fVqB3BT5uaVYjGQmq+hiwOgaibj3G91cz3AEb1p1nDyXu/Je6TPNsNwkr18qdES+Ru6ZK7N4Pb0EpMGSSoo6hFNhqXUELn+g4/2/HsT8pbXJdbm18x/Tt6NT14dci/72b3yu+W9v/1trV20XasoCnAkaP4ia+WAeJr1F2xRY/DYuhppZIojT06oxsALg6j/AMU9iy/eW6undAaE46+eW2L2e2xwyyDUF7j6mpJ/w06jbx3UrYt6fDKlZIx8ZaNirAfpJTjkj2xGLfb28bcG0UFQD5/z6FXJXJfMnPm4RW/L8BnjZgrMCRpBwSMZI6QGzMKaipWrzAd5Et6Ko6pGBbUSSfwPwPdNv3Zd4vjcYWGM4Hl+XXZ/7uX3Rj7d7NHzFuURknKg6nzIc6qVpWgqaenRmcJuL+CpD9lO0cMY06I3sFUn8i/9Px7kza/3PeKYtzjWRTwqAehx7wcs7jfkXNkrIycKY8uhj23SpuSJ8njMh4q6FTKx8oJLLza1/wDXHuPvdLkfla92xpLCFY5KYNKdDD7sO3c07lvabVfyMVqeJ6FXbffdXt/A5PFTxy1eUj8tNANJZmJUoGH5I94MX/sdf8wb9FLawnwlYaiBjB416zB5j5rg5Q3Q8u38o8VzRR8ugK642Zl6jeGS3vuMGaomqnqaHy3vAskmuw1fSw4t7yisbEcq7Um3QChCgGny6kzkHlKwv0+smo5kyPlXox+Tz1THQVTROdYVytlt+OBz7I98thzNZHb7gfFwr1Ks/KW12du8+kAqpz+XQb9P9mtBvaSgzV4VNSERphZZATbhm9wF7kch3dtsbLFGQwBpjrF603r6bmmW2ZqoGx6Ur1Y9RSU9VFHNCyOkqh10gGwYXAv+f9h7wwuo5raRoZQQVND1KjS6xrU1B8+mvc0SChfVbgk8AXta3APsa+2okn5ptUQEkuP8PRLvF/BZWbz3J0qqnj9nQEVk8Ss8YUAgkC1ufwB/vPvuLyBYTW2w27yYqvD9nXKj3/5msd63t4bNtQXGD0WjtbDy09VT7iSAzNSlUUBdWkauWPsR8y7JbcybHLs90upJK9QjyZvk3K3M9tvEB0+GRX9vSnpsDhd+bUhCU8X3Zg/dKKodSRz9P+Ke4Q5d5NuOVnktLIEIDgVJ66Wbv7l8oc18uWb3Jj8YoNeBWtOgc3H8ZKNKCoyLNFrCl1ueRYXsSfct7Jayzx0vB+3rDj3M36x22818uuFJ/h/2Oqlvl3it8YXA5LG7PqJw6U0g0QsQGfSb/S/+29ilI1i7U4dQPdXk1+TPcmrnJ61+sumSXJVwy/lGTWof70T8ymct6y59udI/s6bgSAR/X37r1M168v1H++/Hv3Xjw68fqf8AX/1/fuvDh11791vr3v3Xuve/de697917rlbkC45/1uPfutV66tzb6+/db669+6913b+vH+v791qvp14n8f09+68Ouvfut9crA3INv8P9Ye/dVqRx66JPFxb37rYp5deB/F7A/X/Ye/dePr148H+vv3Wx117917rlwB/Un/W49+61knrocEH37rx4deJBPHA9+68OGeuRIIsBzxbgfj37rQBGT10PSQfr/h/xX37rfHHXRsSbfT37rw4ddWI+vv3W+uX6rD+l/wDevfutcM9dH629+635dde/de67HJ+tvfuvHrr37r3XvfuvddgA/U29+60SR11791vr3v3Xuve/de65EAfQ39+60CT1x9+631737r3XIHTyPqf94/23v3WiK4PXfAB5uWt/sPyffutef2dcQbX/AMffut0669+6312AD9Tb37rRNOskME1TLHBBE800rqkUUal3d2NlVVHJJPv3W+thL+W38EhHRQ9p9g4uOf7mmFVjo6mIXgbxh0iAYEcH62PJ964468raCJPTq3TdGZfHxvjMbRwwwUypFFHBTqh0oNAII5/HsGcxQizK3lfhyeslfZzcjv8ADccvygASgqKj5dLnq/NyZCGWgqi8DRrezkg3+lgD/sfr7EOz7jFuVis0ZqeoY575bn5X5knspRRQcdM3bNDNSRxy0zEKwuSD9efZpxx0ERg18x0uerIW/gNLI5tK6+q4+p0j3oUGB153aTuboQRJJHPJGx45sbmw5vYD3s+nVRUZHXOrrgKdopGuxHA/JP8Ar+/ZHDrZ9OsmMpNUBlJbVILm4/Tf8H/ePfut0HSYzWJmaYSqbtfWpU/7H6D375jrXyPU+mgnlplimYk2IFxf8c/T3rPWxToKuytuVT4yV6NhHMAOQbf429Pv329ePHqq7szfu5dk5OY1BqSsUpOu7AFdVhb/AFvbU5KIWHl0YbXEk90sEh+I06Z9m/K56ipFJU1kkao2kFzYE/psOfYB5k5sm221ZYUJNOpO2329Em4wXTtWMEE9c9+fIisiaSalllkRh6SjGx/pe3uI7ffL28nZ5CQWP7Osyoua+Vdh5fi24QozIBmg9OihZOTdPa+XqJYzVSRM7MsYaQfQkni3s8m5laxgCl8+deoK5h5gg3G9Mm3W+SfIf7HQg7B6Dz0WSWfLYqcUQfU08wYra1/qRb2T3HNV/FA1zBGSvmet7XsB5huVTcsAfhOOjU5D424vLYCeoo6JJP2LkqAedN/wPcZzc+3R3D9VytTw6lB/bm2isStsuQPXpC9I5XNdRbunxDQzR46R3hHDaFGq1/oP9v7mXk3mO6vJNDyFhTHUWXewWdm8qbggBzQnq4PZe76LJY+kczBvKim+r8lQTf8A2/uerNi9sGbiesZuZI0h3mRIfh9OlrUVNONct1a4uP6c8gjj2ox0TmtMdJetykUKO1x/rDj/AB97+XXgRTPQPbjzyiWRyxVQTYfT8/X/AGPv1fTr329P3XO8KeWqkgZgNLEeog39J55/3v3rPWq5x0J2Llimz1bVJItnAvb/ABH9B/re/cOt1r1Gz2bjSYpI40g/U/Sw/Av79TrxI4dPG28lTTJrV1s17+oH3ulOvA9cd3ytPjpo6eVbGJgAD+T/AI+9cevH16pF732k9D2JDnMpCDTmsYtIwNtPk1EnUP8Aiffj69epTB6Mb1ltva8+QxO6ccIEkBhZimleUUagR79SvWq+nR/33xGaKILKFEcCpc2IIUWHv1Ot1xU9AbvnfdNBTTTT1Sx6AzE6gAbfj37r2Oia5TtWCvycq0tRqYSMpIN7gHg/8R79T069k9GQ6kyjZi8sjm6qWF7g/wC29760Oh0n3BLQgoKi1uf1C/HA+nvXWzU9YZd8oIFDVahgTcl/rb66rH37HXugo3PvF3nZ4qgsxNrKxP8AyL3vrXWXCZ+qnCDWxYsONX4P4N/eutg+vS/rIGrqGRb2cx8X5+o5sPe+tHh0QfvZaukpqmgOsPO7xwm5DKTwD715068OHU34rJXbelEOWqmdXmdlEjXHqFxy3vw699nl1YNWZaL7d3WQaGSy+oGxI4Nve69erp6r7+SmZqIKENC7krPMeD9Be4F/6e9fb1sdV24pMlvzsPCYlGklSXKQJUJdj6Cyghrf7H3rrZwOtkfp/qfBbL2niYIKOOKVqWCaQhBwzxgNqv8A6/597PXs9JnteqakgMUAKouo3W1tIbgce9+detfLoXv5U22I99fI7sGdgkp2rsvM7gdm58C0FM0zSHji1r+6HJA62oGluiD/ADV3nS9j/JPdu5I6gVipPLRecnXzSyGPRe/4t7sBTrVa8fLoty5SCmXxAhSv+9fkW976959OmPr3nKMrFtRtYGw91IHWs9KSoMUUGqS1rFiTzbj/AA9769w6L/vXIQq7OklksbWY2v8A09+638ugcyaSyzY3JQgs+OrFqwFPrHjOoG/+w9+61Shr1br1f3/hN49UU2LyFQn8YpqKGlMUhUSCOFNH5N/6+/Enr3VWPyazEdVuGphpGDo0jBgvItq5459+62BTHQLYLbdNVY5qeup0kpa4fborpcLJNxqufp9f6e9deHVeXfnWp623lLSQRFaKt11MTC/jV3bVoUfjg3A976rSuOgLWw5PP4ta/wDsffuvGpwOuPv3VuuwSD791oivXXv3W+uwLm17e/daOOuzwLW5/r/X37rwznrq/Fv8b+/db+fXXv3Xuve/de697917rv8AH+N/futefXiSfr7914CnXgL3/wABf37rxNOvBiPp7914gHr/1tAAm/4t791oY669+6313Yn6e/der14G34v791o568eD9f8Abe/de669+63165P159+691737r3XvfuvddgA/U29+60euvfut9e9+6913YWvcX/p791queuvfut9e9+691737r3XYF/zb/X9+60TTrr37rfXvfuvde9+691mp18k0Mf0LzRrf8jU4X37rR62Pv5ffVVJWdZLPIA4bwEHgH1829+HXjgdHD3/APFuXNYyprKNGJjjmmGm1yFQsBb/AGHPv1BSp8uvKSGoDk4H59FE6o6xrqnetZgJ/I0cbTwsp5AkUkD3EXuFu6QQEQ+R6z++6h7ez7reiXcVJqCR/k6NDsjF13WuXlo3idKd6nUSwP8AqiLn8f7H2Cts5l8BVkQ1I6yZ5u9n1vppbCZDpYkD7D0drZu46bIQxkzKwsnB4A55HPs5vPcOVofBBo3QD5c+6LZDcTuLqCoNaVHQtZLcWKweFqqiWWNG+3kKKbEM4B083/Hsohlk3dg7HWSepCv9ut+QLR4ov0I0B+XDoi+1cVkuzOy5M1Uh/wCF01W6aTcoyo3Dc8e5h5b2KK0gW4kWjDI650+9HulfcwbnJtdvLqiJpivR1M9ULQYODHUv+6EEKqLcIosbW/4p7Fs0mmJm8qHqAtutjPucSk5LKf59MeAx8YppKmY6SqFgWNrX55v/AIe8cud7dpnaUjzPXYr7tm629lBBZK+Sig/t6DTdGTjaaeFpl0FggXi9r2t7DnI5urfeUKg0HUwfecg2Xdfbie3nZdfHiK1A6dtl00JqIJQV0ix1/i1+B/j7ywhYvAh86dcF9zhS33W4iT4VYj/D0HHa2/Ztr9q4CioUNSlTHCulBqBYsOOP8fcQe+0sNzyXNszuFLZpwPDqVPY+I2fN8e/FNccYz+XRzZus+y+0du45sfS11PSVNLHIFVJFUBktYce+XlrufKfKd9I1zKmsE1qRXrKXm/7w091IdutkKrESuB0EW6viR2bt7F1tePvQ0NPLUMQW4EQuTcD+n49jXafcjlfdbpLeKVDUgAVHn1G/+vNeRKWYHPyPVLnaHyg3H1fvup2++OlyctBO6zSaGYgwvYoT+fp7z29r/uxX3uhy6d5t5/pk4AU4g+fRP/wQi2M5E0fiNx6sv+D3zy6m3pXUe3exGodryziONJqtRDrmeTxhG1Ee8XfvMfdD9xuUrd7vlxZLxVBJK8BQV6kflb7zttTVJpiHl8+rtO3Oodi13XMefwGQpMhRZSkSsilgkidWilTWpupP9ffMHkDnjnGw52k2bfbZ7aS2crU1oaHywOsmfbb3ifmS/UQSBgR5eXVLW/eh6Xcu8qehxGNkqDLKkRNNbgs1i3A99afaPnhLjluR9/bWE4A8eGOob+9Lbct75Zvb3wVrl1NOFadGUq/gFmMVs6DIY+doauopTIafWPKpMV+R9fZe/uVcJuj9oeAN24zSvXKLm37vu07tGZ7BfDY+hp0HHQXwWz67orM7uhJwKKaUQtM9mZGcm3q+vHtZzB7jncjG1qvhqvHFM06FPs77c3HtzCfCc6lqASamhPS77e6s2dtCtkwmHpoky9TMmtotHkLu3PKj839sbVu19fD6iU9g9epimvdxhmaW3lKsxqc+Z6G/qz4xzxbZjyzwuDU05kPkbV+pLj6/7x7Jtz5nH1JiB4Hy6H3J/ubzvydN4+23JANCcnqrL5WdEbpwPYVHlKOGsagpaqUjwagUkYG5VlHA59yXy7zVbWFg9ylDqADA5r0B/e3km5+9HeQz30rC7jypVtL6qFajz8+s3XXQO693imk89bDFIA+iqd2LE2AF29gzmz3t23bFZY1VWGO0U6DPtp/dzbpG/wBRvckrxA1JnetR+fVgnWH8vrM11DJW5nbuQYNFJJT1BjKxyC1wwuvvHDnX303QMjuHWJ6UJVqU+3rKzb/an2w9lrYGwSOW7ShOnSaEeoHRSe4/itvbZu4qp6XH160UEkpsiNpCJ6rkgfj3JvJPuvZbhtirDMupgPtr1LO3+/G2y2S2d0EUKKCopTqr/wCSHbtV15lYtj4arAztSPXoDvLA2n1arD6A/X3nz92fkvc/cCVt2vyWtozSn59RH7te7fL8dgbfbwrzOKgjjWn+TpA9GfJ3e/X2fifdeRkqcTUs6yvLrKprH0b+g/p7yW95vYX968sSvy2hWZFqAvE08+oX9rvfzcuRd7XdJRrIbAAxQ+vVofx+3ttrsjLVmSqq6nkMtSHpVBI0qzagCD7w62t/3Bt0m0XCAXEfa9RmvDrKiwi3T3U5gj5zviaAggcAAc9HiloKaM2p1UQ/hxwCv/E+wrebTNuTF0BNc9ZU8ue4FjycEtNwkC8AK9IrOQyS1SQUyko62fSb345/2/09hqz2i4XcwpQkKepj5k9wNp/qg1ws6hphUGo4HpKV2w6KoQ5GnT7bJw3ZJAQOU5HuVoOT7Lma2NluUeocOucnud7ixcuSG7s5AXbNR0I3XHck+3MvRbb3JUa43VVimkJUJoNvr/j7wx+8B92t+X7aTeNlUshqSBmnn0L/AGV9+4+b7hNovSA2ACfPNOhx3PvOLJusVBUI8TAN+211YEcXI/w9xz92721kv+ao76+Wi27ZB41r1kR7zbZNae3dxuELUJU5/LoPZjyHB1MTY2PNxxfn318tYkt7OO3iFFUU64mbrPLc7vcNM1SG/wA/TfmaWhy+EraarUXSKV1JAPqVSRb29kHpEaEdV7dWdx5nA9sV+zqyN/4dNXyU8BZW0CJZbKQW/r7YMEDHURk9GUO67nCojjkOkDGeHRqu4u1qDbeLkoknhkqKuP8AZh/tAOtvpf8Ax9uKiKO0U6STTzXL65m1N0QHJ4OTckddkclF9xDVwTSxo4va6EggN/vHu1MdJ6+fWt38m9rR7b7PzRhXRFXVdTKI+PSUk08e/Dq2a9F3AFvr/sPfutV669+631737r3Xhz/h/r+/de65aT+Of62/Hv3Wq+uOurfW/Fvx7916vXX19+631737r3Xvfuvde9+6913yf8ffutYHXek/09+69Ude0nm/H5/1/fuvVHl1x9+6313fix5/x/I9+61TNevWP9D/AF9+69Udde/db65EggcW/wBt791oCnXH37rfXvfuvdd/S4I/41791rj13e/AFj/Ue/depTieuiCPr+effuvCnl11791vr3v3Xuve/de697917r31+vv3XuuQW/5t791omnXH37rfXvfuvddk3P0t/h791oddfT6e/db697917rv6jgfT6n/X/r791rgevf19+6313wB+Df8A249+61knrriw/wB59+6914C/5t/r+/deJp11791vrsj6c3/4j37rXXXv3W+uQsPqP6f7Ae/daOeHR2PhV0RXdqdi0GWqoZBg8HVQzVBZQsU4VwXu7fgf4fn37rfHrcU69p8Litj0W3sNFBDQ4+liVDGqi5ESqxJH+PvdaDrx7h0FOYjVM2XYCeIOSwAvxf8AI9km/WDbhYtCvmOpG9seaY+Vt/iuZ/hDVqelZtWi1ZKaqggeKIlfUeFIU3PHss5U2ufaYzHJ8PkOhF7y8ybXzRfreWBGo0LEdQu0snakKLTM4QfrAvYH63A/p7F3HqFvLpZdVzebb1IWQ6mVtK/i4FgfehTrwqTjoS6mm8cXkk5YAkc/m1/+R+/dbOMdIeeVmq11AgauL/W978+/de+3pY0WShFoRwOL3sDf/D37jjrdaCvWbIVlIket3S4H4te30H+PvwGcdePDpFVe6aWEukTKz/jTYWH0ufezg9VpjpB7j3lStRyiVkWwu5Ygj/e/fuvHHVWfygr8VkcVkJoHi8scbEOhF/qT+PaSa4jCmMnoQbXs13NNHdIDQHqtbZmCy+4sksWKpZ6ydql0Hh1HnX/re405qEMVu8r0pTh1kRsMW67hFFt9hE0jnGBno+/XvxK7B3yaWOrgraWnbQWEmrhTY3JI/p7hD95q0xSyWpPoOH59TrtXsHzVukCXO4hokOTqxj8+rSOofiDs7r7FQT5iniqKwoPI76C9yLMCLf19uRbLd304m3EkitacB1MGze13JXK1gXvUE84GMg0PQi776+2/LgqmgwVFHA/gZUZQgYMPqwsPcwbcdjTbforhBQih6x+3/lHmB+ZzuO0ArBqqFA8v+K6IVS9kVvWOcrdtbipWkx7O8UckykqwLaLAn/X9wPzpyJby3zXO3HQpNVp0Sc086b/ybdRS3alo2PcKcB59CTQbP2lvrx5LH08flmXX+2QGUvyb/wBPYL2rmbceTr+tz8Kmhr1LfKnLHL3u3BFcW0iqzUqAQKH59Oatluv8jRwVEU4xjyrHGzhtC3/N7e8tvb/3L2LmhBZJKol9K5PWMX3ifZLdvb/cP3lbK0kL0yMin5dDhFueCrpUkikV1ZV9V78kf09ywylWoesX0bUNXSMzmeKKw1LY3vYm4uOCD/vfvXVjSnQB713JKInWmkGs25U/W/8Aj9feutUqc9IDA9gZHEVkRVmDyuNQIN7HixHv3XvPo9PXmSlrsO2VqkZPNEtna4v+3e597HW8Up0lt156PWylh6SbH6XI/Pv3Wq06x7Y3P4FAWZdJNrXve4vyPfuvAmvS2yW4UkoyQ6oyrcf1N/qCD70B1ateiE/JOno8rt+srJE/fgSVkYWDA/X34da+fRKup+5q3bctLQ1NXenWqliCNc8Bv9f/AAHv1evEdWH4rsM5rEQ1FIwK+JWbSSQfTc3/AON+/fb1rPAdF97a3VWV9I1PTu8ekOHA1XPP049+68B6dE/wWZmodwuaqQhGka+ok8avx/j78AevA+fR1Ng9v43asSzvNGRIgGki/wBRb+vv3XqeY6w71+UGOpbGjaOR5CQwQarXNybD37rfy6Y8B3YM8yOakQ6iDouQSCfoR9Ofp78OFOvUzU9CdQ7hgyEsc7TJZiPTf/Gw497yOvcePQz7ZqKctEdYJLcfk3I5ufejjPWhXgeho+9pkprMw1BOWFv9ce9/MdbHRP8A5B4w19GKykjJkgkLsygWa3PNvfutDHQI7C3cKQQAgJUQS6ZPqDoX+v8AsPdTw62OPRusbvJMtRw6ZFKlVBFyLkD3sdaNQegQ7u29U5XEeYIzJdySP9q+g/2Hv1PPr2Oi7/HbY8NJ2xR1NVBf/LEbyPbSG1C59Xv1PLrdfTrYClyFLS4mkjSVTpooluCB+lBb3vr329FO7c3PDBQ1TGRXCxSsOeRp5uffj6de4cejr/yhaen2NQ/JvuKtlXw5TpPeWIx8jgFBVV1C8KlL/kAm3ujcK9WjJBofMjrXxyu7ZqzcG7qyrqRLM+5ss4lJJOg1LEKPrx7vXh1SlC329IOu3jGKmyyLIWOkWub3N7ce9db+XRgthUs+So4ZEB1Mqk/ViFPP4/Hvx60ehPyuIEWOk8vIMRAN/wC0QePe+vcB0UHd9HMaqan9TRoXIvcfU/g/7170evdMGIoHcMJOFYWZbc2+nHv329e6bJ8nldi5aavoqmSKhlW3j1NoGofgLx9fr7914GvU7r/Hw9q72niyP+UJ4wWFwPVb8k+/cePWyaCvQxb56sTb2NjehTQKZ/KF+pIQ3H0/1vfutVqcdV//ACT2RXb02w+UpoWkyNC5cj+1ogP0PH0P596+fXjnh1WJLE9NNNBKtpIZHikU/wBl420sOf8AEe99eOeHWH8+/db65D6+r+n5/wCN+/dVPDHXH37q3XZIJ4Fv999ffutAevXXv3W+ve/de697917r3v3XuuzYnjj37rQ68Db6i/v3XjnrxN/xb37rwFOvC/4v9Px/T37rxp59eBA+ov79149f/9fQB0344v8AW9+Le/dar59cffut9dgkfT/W9+60QDx669+6313xb6c/19+61mvXXv3W+vWI+vHv3Xuve/de67P4/wBb37rXXXv3W+ve/de697917r3v3Xuve/de67Av+QP9f37rx68Rb+n+w9+6914W/Pv3WjXy669+631737r3XvfuvdZ6UkVVMR9RPCR/riQe/daPDraT/lyCsn6bkmAOuJqYi1v02uALe/A469x6PFuPtbIbcx1RTywHRJHJThmH5ZCtvfmxGxrTB63H3XcSgVOpf8PQfdC7OWvyeS3PMtpamrlnU6R6RIxIH0v7x350R7p2hjzU9dffu1zRbHYwX93QdgoD0YzcvW1LmbSqh8uki4VQQQPySL/X2ArbbrmAUoespt35r2e8PiKBXh5dBvW4Ws2RTNO7tDADYar2B/HPso3JJllSKJSzsw/w9GK71b7Ly7Pvc7ARojH8wpI6iY6mzG+8fVhawtBG/C6j61P0sD7yk5V5WtbLbIZ517nUHyx1xe95Pf3fub9/urGykKQpIy0BahHDrlgqqs66MsJo2aLXeSUqfpe5Nx7HOFAVcAdY2u7ySGaU6mbJJ9enOs7bx9UpkRkcqbMgJvqP4NvdHXWhQ9O29wbScXC8R0zZfuCqpcdJ4MfIY2jIDJr/AKccj2Et65ch3BPDA49T57b+8e5cp3Auy+FwBU9B1tio3HvrIsy088FOzlySGtwfqSR7a2XlC225hKwyPkOjj3L+8JvPOFu1jG5Ct8z/AJ+h9xkFbjVShiDNItgWH1v9CeLnj8exooC0UcKjrGeV3nkaWQ1LVqeo3XHWVV2f8mtoYGWNqlGWnkeTSGCaZQzAh/8AAe8Q/vhb/ByhytPv87hfDjbBP9E9S77Yb9Ftm2vZhf1HY0P29bXWy+q8Ttvb2Ix1PTU4FLQxQsGihJJVNJPA9/NHzf7h8wcybtPewT0SR9SUZhg/n0Ok2WF5GnlBLOan7ege+UlJgtj9O7w3HkHpaWOnxFbEk8kcSqs00DLHc2+v9Le5W+7fuG/c0e7G38uwMZyZI2KqWJpq8x6Hoi5i22Kx25rilK16+ev20BuHtPedc08dVBJmKuSCRAGAR5iRy3v7hfY3lxNt5BsrdYvBLRJUEAZ0jrF3drgpO5DV1dF53fhJsWVylFNJT11LL54JYnaIK6LrVj4yCbEX9jHmTY7W7s3sb5A8b4IoP8o6pZXXiAISafLj1c9/Lk+TPfe8uv8AI7f3LXZHcuDxJNHCRJVzpSUqNpXUXvaw/F/fFn75/sryDsHOFre7XBHazTDUSFVSx/LrI72e9wLjklpWYmSlQCSaivWyL8Z+oNv7kw77xilp62upbPPAEidqaS1yjBhe9/eFmy71PZSzbK6FADgmtGHqOjnmXmS95w3E7pePqAJoKnA/PoRuyhuWjvT4ilkk0awNEZIQKvA+lvYz242zZlPQekLJ8HRWslubtCJ5aalw1arOfGGjpyn6uNR0jn2JoodtIDSyAj06Su8nCnSm6u+Iu8d/7kpd17tE6h50nCTx+pUBuo0v/wAT7TbpzZZ2Nuba0pgdeismkYM3Vktb07LgdvwY+jj1RwUxQ+ONABpWw1Ae44TeBcXJeQ0JPRqYSiUXquDvHqtck9TT1NJqqhLKy6oYibKOOSD7kPZ79JITCfh0/wCTpvbtyvtg3BN025tMiEfsrXoPfjZ1jXZvelNiKhBDS01QQv7SAWja+kgi3vE/3z3+15Y2e53dTq0Ek5Pl1l9t33hp9y5V/d12B9QF0g8Cfn1sX7Uzu0MNt3D4SrwcdSKCjSB5FpYQZCqBWNyOfcWbJ9+f2fXaLWw5o2iS4aJQD+mTX/Pw6xj3jl7ed3v5r7xyPFYmlTjoAe/+3PiV1xipsjvjrWLclXUK4GMX7anln1JZwLjUOOL295V+y3v37E+4k6y7DyfP4YpWaojUGvzYH+XQB3vaL7a+yS6q3pk/s619Plj25/Kawu44t1bx+HG448zkAKgZQ5yOGl/dXVdYxHcj82B99h/un822PNssnLXJJXa1BzGzB2NPzp/PqPN88SOIXN2TIeFQD/n6TXRnS38t35hQmHYPUOTwUs7MEp0raeo0aG5DFrHj8j3PnvP7un2MhB5q3OHuUjvIUZH7PPom26xbczW2Q8R6+XR48N/LY+PHWtJ5KLK47ZUMelllyElOrKi8hnsfwPfPCH3Q9tudN/l3S13SEmdtTUcEVJ+R6y49v/cDmnZtvG2QRGRlAAVa1IHS9x3w96D3rT/w/D/KXYmOq4vSyTVMI0H9IVuP639zTse27Y8fiWMqzI4qKdFPuL7h8zXjo9/ayWxTzJPl1zl/lmbZiDz4/wCTmwMsRyrU1XExN/p9QPZpHy7ZQyGXSKk+g6Bl57y803tlHYNM3hoKUq3Qa53+XFn43dsT2vgsqBewppY21H8WCjn2aW9nFbnVGKdATeOY73egEu2LU9a9BBuD+Wl2NVa6ymqHytbCrGCWli1Am1xpIH5tb23um2We82zWO4LricGooP8AL17l7mK65YmF3trFZQRQ1OKfZ0EW2viB8n9qZmqoJ+v92bjhd2SlakxlXKkaBuOUQ/T3GVl7Y7fyzuP1u0IERjUgUz1lhtH3lrzmHkm45X38l2YEBmJxinn0vq746/IPHf8AA3qHeFKrc3mxVYvFvxqT3LUTVhWvp1hxf9+5zyoMM2Pn0lsj072zTU8sdZ17n6dbEN5KOdfr9f1L7vXPSbSSOiT9r9H7h2flBvCHZuRiqqYu8g+3ZWL2vqLKL+9Mtc9OROY6gio6KWKTP9h7xhqM4slBBSShBT1DsDZDYAqx92GRnpksASeh/wAzj8BhduzrLVU6fb0k41M0fpAjP1N/e6enWqgDHWqh8v8AcOGzfamTixVTHVGhqqyOpeE6kjd5QVRmHGqwuR/xUe6/Lq1a5HRUeP6n/be/dbz117917rn43/1J/wBsb/19+61UddhT+Uc8/wBD/tvfutGvkeugrg2Ckf64Pv3WzTz64kG/I/P+8+/dex17kH+h9+63g9de/de697917r3v3Xujk/BP4ebq+dPyK2N8dNlZekwe5N9V60GOrq5C9LFI1yXlAN7KASbAn+g9+JoOvKpZtPyr1sFbp/4TNdWbD3Hltjb5/mk/FLbG/Nuzij3HtTI5esXJYauPpWjrEWKyyavSVvcH6ge616d0Cnn/AKvz6rI/mG/ya+1/g1hcHvTB73wXe/XOXpBWSb86/jes25RxFNf71dcBSByQwH+Hu3TZB4/8X1TJx/S3+sOD/tz791XPXuP6n/bf8b9+69nr1z9D9P8AWHv3XqefXuP6n/bf8b9+69nrloNtdiUvbV/j/T37r1fLz64+n+p/w4/437917PRn/hp8Zc98xvkp1V8bNsZiiwGe7U3HFt7GZfID/JKSpmiZ0aoJIsvp5PvR+XWwKnPSn+cXxB3D8Je/N99C7pz9BuPN7D3BW7fyGRxy2pKioo7a5adgTdTfj3vrxw1PlXovXVvX2S7R3/tjr/Ev4slujIw46jfR5D5Z/wBACXF7/wBL+9inn1Uk0qOr5vkL/Ip278XvifW/Iftn5edU4vedRiZshtnpf+IJDvbLzRU3mFMKJ1YXv6eHHPHupNOndIKkg8P2da6/H++HvfVOuSoXNkBY2JsB+B9T791r7euuP8f9t791vPXXH++Hv3Xuu+P6n/bf8b9+61no9PwI+EG5fnb3Rtzpna25cftjLblySYyjrsihaljmeMyBpje4HFuPfvKvXhltPQP/ACx+PuV+K3yF7S+PmdzFFn8z1duap21k8tjh/ktVW0sambwkEgqCeD78Pn1sih6LtY2vzYEC/wDieQPfuvde4/qf9t7917PXuP8AH/be/de679NjySfxwP8Aivv3Ws9WHfCz4Pbe+WWx+8d45vvPZHUUnT23RnKPD7rmENZvWQRK5xuBBI1znVfSeP8AH3omnVlWorXoh258RTYHPZPD0tfFk4aCqkp0rYQPHP420llsbf7b3Y9UBJFemHj+p/23/G/eut568LWN/wDYe/dez5deAv7914mnXXv3W+uwL3/w9+61WnXX05/23v3W+u/rf/W/4n37rXV93wSjoMN1k9dSBBWVFApksqBy2gc3HP8AsffsDrQrSnVunRefzGYoJ6R9ccaxuVZtVrW4/Uffsdb4dDHHh6SirzLkalPHIRqVrC4vza/vY68w1dCDJm9t46ijSneJRbk3Avx/Ue9160EoKf4T0G25ty4KroamBjG7ulgeCRb/AF/dT1vgaDpUdYVlLFj4o1YFPWyWvZf6c/0sPfuPW+B6F142qgCGOgi4Fha3+uP6+/YHWsnJ6TOUx7CTUoJMYLA8cj/Y+/db9Og7zecbGAycqVtc2t9D+P6e/Z68cDoCd09tfZtIZalEBBABYi34HCn8f63v3Xq+XRetz/IOhxchZKuN5CfUA7cgH6D37rWegvyHdNdvOlqqLDljUupUmN5CyfjnT78crjraAFwD0Xin6z7N7L3THtxI6uSmnn01MgEjqkZa+pgR9LH3FfPnOW28qWbXN2Rq8gTTrJj2r5M3Hmi+trK3UlXIBNPLHVovSXxj2N1ji6D+JUUNRmgUkllaOHWJCLXa4/r/AIe4Ln57m5otBcxH9JiR11m9s/YnYuToE3B0Ek1A1SAc04Dj0drEVWNwdDajjhiRVH0jjvpA+l1HtVs72kQ14qejXnmHebpjBZroRfQUx+zpAZ/fbTSvGGsFNuGKgc2/SfYuhuoioJNR1BF9se5EkMprX59MlHufyNpZg+oEBdV7g/S9/aHdruNIdURoehJyPy/c3W5C3ukqtfMf5+gN7l6qoN7UDV6QBKqBZJFay6tQOoWI5449t7VcPvNsbZxVl+E/PoIfeE9sdmn21y1EdlNOHQIfGlM4nY020ZHfRHK0V3LNZVbSCQeBx9fcYe5O0JBtTzXS6JE/n1zk2LmrmL2s3oxWUhMIYgUJp/Lq7rN/E0b92FUwvp+8hoRLTVKxRi0vj1Ifpc+8FbH3gl5H5xhubdyB4mRU4z8usk7r3p233C5Xk2Xf49T6KKTxBp8/n1UTkUrut95ZbYOZmLSYyWRfKxtrGvSpA/2w99ofbLnO2595Rtt4hcNI6gkDiMdYHcx7a207rIirpiJNPTJ6wZrJRywEq1gR9Oebi5t7HXRN5U6Dygxn8aySwKhlBk/QOSObi6n3v7etE06F+m+P5r6iiyJ/yWNJEkkV0X9I5JufeurUznoWc1W0m2MLHiaUrpijUFwOGKLa3++Hv3Xuiobp3pH9w0TOGcswClgSCTa2n36or17hTqZtKsraupiaPX4y9woJtxz9Pe+PWujAz4iqmxgqDHIDpBPDX5Fj791vy6IF8lc/HisNWU0zhZWWZCl+Ra4968sda6rAweLzGaydLJj6WeSM1kjkx6uAWHP+t71nrdR1bV13tHK4LYeJyVYjiOr0o2scqNNuT/xX3ulOPWj/AC6Zd97OFVQT1dNZvS72UfqJF7e/der5DqufclJk4c/NBdk0zMEaxFiCbcDj37rY6X23NlZ3MwkyZEqoXUiaiD/wUD37r3QYb4w1Zt+t0VEjsNR9TM301WP/ABX3rh1vj0lqbcGUopg1HUMdNiAC/BH0sB791rofNk7v3PVRxu7y3UX0jXzpA4I/2PveevdHR613ZkpqZGrI3R1t6iWuLDg2b3v5da+fQ8w7tkeJUYMxvyeSTz9AT70Ovfb0ybyqIq3BTvUR2RlI5F/8Tf3vr1PTonez6QZLfc2KplMkcs5QqgJtc/0Hv3njr3z6sQ2b03VxmC2vxKsUpHpsAQG+nvx63SuOufbuLxW28HIMgqGJUIIIHpIFixPvXl14gDHVY+H7Uhw/alPBh4hPBJkFj/bvZRqA/Uv9PfvPrwGK9W40u48nWbcpKsuwZqWMsLn9LJewP+x974daB9eiPd79hz4zE5YStpCUtUeWa4Kqb3JPvXWxwoerWPjjumDrP+UlV90Vk4pI955PLbThqGbQs8skNxEHUhm/Ufzb+vvwNa0622NJPpXrWpO4qutmzbxlm+4ytZN5ASQUlkLC3+wPvXWvs6z7S2/mc9nIYYIZWQTKbjUwta/59++Z68T1aZ1zsapxmGpGkjKzeCIMCLNbT/h73148On3c+KaOinXT6VjNvpwbfXj349aBpx6KNnMWJ6iX83Yi9uPr9LH8j34U630lno0pH06bcj1G1vr73149I/fuPWswsgSxskjcfW4W59X196PXugf6c3lLsXeBrJvVFLUeIC72+un8e/dePCnVi2Vy67s249XElmanJI+ttS3H+P5978uq8MdFCaCjM+bw1aFMj09R6WCsAG/wPvWOt9VB967RG2d7ZBoISlHVzPIpAAUSE3IAFrX9+49ePGnQJ+/db67+p5P+39+61w68LfkX9+68a+XXXv3W+ve/de697917rsC/5A/1+PfutE0669+631ysCv0N7/qP0/1r+/dVrnrj791br3v3Xuu72Pp4uLe/dap69de/db6//9DQBvY3Xjj88+/dapXj119ffut8OuvfuvdcgBa5BPNvfutGtaDrj791vrvkf4fn37r3HrxueT791rA68QR9R791uoPXuLfTn+vv3Ws167DW5HHHPv3XiK8euPv3W+uQ5Pq/4p791o4GOvWFif6Gw9+69U1p1x9+631yIAPNzx9Pfuq1JHXH/W9+6t1737r3XgCfpz7917h1yCk2/p/X37rRNOvaRYm/0JH9b+/darnpY7A2rkN47rw2Dx0Rmmqa+lMi2J0wLMDKxt/hwP8AE+/db8+tx34XdZ0mx+pMfSNCsclTSU0sqaQLMF9RYe9n0HW/LpP/ACAzmOWsi23jYhLWSVAZnjABFyNX09oNwn8OAquW6FfJu3w3e8JNdGkYI48K16Mh0thoaLC4ymIUS1FLFJINIB1EC5PuFt0idLhncUqeulvI19Z3G2Q21i4oigYxw6M9FjIVtqUDji4Fjz+fZGzY4dSMqsAKuTQ+vRf/AJA4XzYJ4IrRauVcWAta4tb6n2b8n7NbX+8CW5UEDyPUbfeQ9wd12T23/du3Fl1E1IPEU6DHriiqsNhxY3JCaj9QQPqb+59oqqIlFAuB9nXKZHaVmuZMtL3N9p6WG66miyuLaiZVWeUMmoqosWAF7j+n49+6v0jtudK4nGY+qy+WqYngdjVMWYcKo1Wt/j78c9e8+go3jv3aE8v8DwVMshhk8TSIFPKmxB/w96pTrx9Oh+6sq8XDikjiiRJ3huw0LfUfqbn3up68EUY6U2byuM29TVFXUKgll1pExsAHkvp4/wBt7vGuqrNgKC37M9bNBjo9X8urqpZMtW9oZ6OOepimUY2Z1u6QySf2G/1j74qf3inu+3NSvyftoZKq8bGvGgPUj8n2X0soumzQg9XrY7NQTLYH6f7EA3/w98WztaQoluf0/DAGfOg49S9DuIJNc1/y9A38neqYe7+nd0dfu4STLUcxp3JKhalImEB1D/Fvco+wvP59nvday55IJCSIshGewH06ScxW3702k2y+QJHWhV8ivh13H0Z2Nm8LUbbyObo5sjVyU9XR08kgFOZSVbUo549/a192n74ftd7rcj2l7HdR2TQxorCRgpJoKmh6xQ33l27tZ/BYFhUmoFeiqbz6e7Uq8RPUnZmWp4RdVMtPKpN1/pb3LfM3vf7eJMLeDcYZif4XU/5ekNntlxD+E0HqOtg/+RJ0JmP9EfarbywTUi1VDkDTmqp7aJlU6CrSC/vhF/eme9u1R8y8uT8vXALieNGCtkqzAHh1KXJ+2NciYupK0J/YOlj8KfmdlOn/AJq5r4z7wklqMfvbdssNJNJITBR0xnZYtKngCx9yrceyljzN7At7t7dRJLO2DsBxchamvV9t3n6bd/3VLwZqdbVFBs7Z9TK7SR01Qs0jFHKRt6L2uNX9feE8G5Xk9tHcLVdSg0+3/N0PjGmo9PsfW+xAwY46jY6r3EEJ+huOLe7/ALwvm/Ea/b1sRJxHSyo8XhaEKlLT08UaLZQkUSWH0H6f6e0kkksuXap+3pxVCj5dY8qKH7abiL9J4IUiwHq49+j1Yp1o0Geq/e66bbf3ck5eBZNTqwsg+ova/wDr+5F5f+pKlaGmk/4Oi25MYGOJ8um/4+bBopq2TcNHHEqxyv6lUDUSeSD/AMT75v8A3oefrbb94PKJbWbp6U409a/LoT7Zy/c9t/IpUUr0aTfe86XZmJlJs1c0RWmiFtblxwyg/wCPvF/kzlS65q3dbOGLTBGRqloKCnFejndL4bfb0r3twHVXHcnXG4+yKz+P52eWeNpfNSQlmtHG7X02+n0/w99Evbbm7l7k2zGw7OioVFHIAyR5/t6jC+t5ruT6iY1Jz0F3YvwV2r8lqHBYnPxQxRUFNDC7mNQfHGoTkj/Aextyt967efYPcLjmLZ5GExYlSCfPPDpNNsSbsogf4ejv/E/4G9PfF/HRPtjGq2R0t5Kokn9Y9elfeJH3nvvme8P3lbxI99uCljgNRir6fOlOHQ25b5T2zZU8Vh3eQ4/t6Vnyx66xO9Ni5Y0UUrVsNDVKiRSPG7M0Jtyv9PYV+7tzdd8q8yQ7RJO728hU1di2nPCp/wAHQktdxTat4j3O3ouk5A61ddhbSrtq9j7jx+Vmy0DPlKgU8UmQqgSqykiylvp7+k37vu5/vnlqS4Zg4RV0kfl0Efe3mob5LbR25Gk/FSnp0aWn3TlcRP8Ab0GQyMI1af8Ai4VBsfqDa/udM1r1BAVQoUcOhlwfZvZFFTpPQbjrIbcqGqZWuV+p5Jt/j79jz630sqX5R/IbA+qh3tNFGnKjyaxb8Gxv73jrVWp5dRd4fOf5pzUMcezu0ziJ4gvqEcCsyqbsNVr8+9fLr1Sc/wDFdKvY/wDNF+VmOx8eJ7A3ouangUpJPPTQPqt+QxH+HvxHoet6sZHSmy/82HL4yM/3mx9NlUH61FLAusWs30X36nXgaCp6DbM/zcekMpSzRbq6aj3BHyskKIitLfhraR+ffgD1qqfPph2j/MN+B1fVHL5v4SVtYUmHkqop5YY3JNtTaQPfu7q2uMClf5dGOy3yO/lm9g7beoqfibmMTHVQusix10hVVdNJsTz79Q9e1imM/l1VN278f/5PGWrMhuGt6dO15q2eSaetyuQfxo0nquxP9L/n3qgHXq1wAOi3yfCX+UhvqIjDdq9abMaf9BrsjTOYLnSAwZvx9T72KAdUKk8Rj7eg93L/ACPfgh2PGX2R/Mb+OmxZp7+P+L5SngMLL+JCH0kf4e9FvTq6oo9ekBL/AMJvOr6pL7Z/mn/EndM1jamxmep2nvbgFTUf7170Wp1fw68OkPm/+E3+8KBGkwvyx6i3Qo5jOHqIp/KD9CmmY3v+Pds9UYAGnQN5/wD4T7/IfHh/4Fu3GbnZSQi4umEpcgXsulz9foPe6YqeqH0FOitdifyYfnVsxWfD9J743YFI/wCLXgqqQuv5ZCgI/wBufdSVHVwkhFaY6L5Xfy1/njjQxr/ix3FTKL3ebZ2SVeDyQzJz79Ude8M+nSOr/g18ucWrtkOgOx6MRhmc1G3KyLQF5YtdeLf4+9gjqpUj16LLnMHlNt5Stwmco6jG5nG1DU2Qx9VGYqikmVQ3jlU/Q2INvfuvDppBA5Iv/T37rxqeHV5//CdPLQ4T+ap8ashOLpHuuGA35VWqw1OjH/WLXv7q/D8+rx/GfsPVl38wf+Qd8o+0vnj8nt67d3Shw++O2M9ubETVD1E05gzuTeoSNnLg6IwQEH4HH0Hu4XGeqPJVjTo7HzH6x7k/ld/ykN3fGXvuKn7Nyve2Cjqtq7i/4uUmyqOnp7CI1MupopD9CAR79k8Ot1RF1E1PD9v+bqkX+UT8T/hBujp3uD5O/NLe22JNr9XQRZCi6uq5xDuHekRnaCTH4lwbhl8Zdj+A3+HvWPh62I6jxCcD/VXq3D459U/yWP5r+0+z+qfit8TMx8fO6dtUM392dwZ7NzZXG5dY6ZniqXjlJCNKVuAPpf36hPXiy8ONflQ9VSfy3P5TmC7y+XPyJ252HDS57rv4r7omx+99tQsDVZ+momc1NPjQDcs/idQQOPr798VadUNE+LNOrI0/4YO+TG5ezfjJj/jw/wAMeytnZJsNh+0+wM/NVYnKPE32j1L0lUQq63BYf4e9AefDq5Kk6Rmv5dUv/DL4IdBdsfNvsXafYndOyML8e+nN3VEeQ3LXTKMbvHEUhDB8V6rGORfpa55/HvdRx4dV0kNoGerveo96/wDCeruD5IQ/C3b/AMO85Pn63cEuz8L3vDnqk4nL5Ig0qZKPHX0CESsrg25X/WPutaZPTgAppqK/Zj9vRNtnfB+i+Av8+v4v7LwFbQ1+wtydvU1Vs/7C6ijpGLRpTyC/0swII/x924Cp6opGvSPTogv8+7/t4z8ixzx2ZuIXN/8AjuP6+/Dh15/7T8h1x/kf9lfG/YfzF61oO8epp+ws3mtz0NPtLIo4eHEVLENrkhJN+FJvp/w9+60Mn5dbAH8/7ePwt757T64+Oey+jNw4vvDsSRcP1/u2LJVgx2Iycs4g8suOU6CLtci1iD/r+/Zrnr2Cvbx6C2p+Gf8AKy/lXdXdR43+YD1rB8gOyt+YeObI0u2sj/DZMDUSpqH37of1+oXQ+/ZY0A62NEcetjx+Veir/wA2n+Vp8fV+MGz/AJ2/BygoMB09X4r+M5vZ3m+4yuEo6qQotPkZgSdaaSVv9ef6+/Lnj1p+01/1fb1YX8bf5Yn8sXFfy5vih81u/wDbOPhh3Dg6zJ9gYKpqvtslvFqGFH8WPJN01XtqA+vvwz1Y4NQBgDquD4/fAf4l/wAx35Odpdt9C7eo+oPh58Z6w5vs6mzkvkjl2utSQAahv84zAWUg+/fI46r8X6iioPAfP/N0dbYeK/kAfMfs7OfCzo7oWfrDuKtyqbV2B3NnM1PNgty5eICklqIqOT0BJJjcC/A/1vfqfl1YOuA2cZx/l6Id/LA+PO5/jN/Ocq/jn+ubY/bFPh8ZkKfV9lVoKkxBqSX6MrJyLHge/HKmnr15BSXTxwf9R6uU+etH/I76V+f28ev/AJH9Ly9xdndw7qqqHcO5tv5xqeDYGSqJmhlrslAps7rKwZg34HvVR5darQUbB8qiteqJfnz/ACrdmfEP5ufGypx0uL3R8avlFuAZHYFHjzrZtvzR6kikC3HpEig/4r/ibeHpXrxFRWlPl1dR8jPgf/J7/l4dWbd3n8jOu6HsPP8AbuCp63ZWDw+RNLNtiSphAWTIBT6mDG5U+90JNOtAoq6j658+q1PjJ8QP5YvWVJ3d8uPk3uzbW8Oo5I5851n0LRVSxbnlpYpfJHiqdojqUlXAJtzbj6e6k+XVgp4jh0cXqP4+fye/5svx876T4gfGfI/Gjt/q7bk2To5s/l58xTZELASHjMp9LObMADx7sPTh15qceI4en2dVufyqPgt1rvvZP8yvZ/cNFPlN2fHvYdXldt1+Nq5oKT72mkWj8siRn1BgQ2hvpc/X3putxAOK0+XRof5Qn8rz4l/JLqr5K9m974zGUe3Ok8FWZ+qy2TkMVNTRR1ARUlnYjlr2Fz72cGnVFGtNY41p0Y74/dL/AMkf52f6RPid0n0HVbM7+24suM2n3DUZeSq29uvJmEgJBRMdIGsXUj6g+9UPVtSkYz69ahXy+6ErvjJ8iuzujslNFUV+wNw1eGqpIOYjJDKx/b/wC2t731qmnHRaxqFwOPp9f9v791U068SbaT9B791sAcevXAtb/Y/4+/dep10T+B9B9B/r/X37rwHXekWH9T9P9v791qufs6sw+B/aVRS5Kt2jkau6SeKGgiZuWja1l0/nSeB798+t+dOr29r7+n2VQ1EawNHIIkIsNOrUtzY+/fLr3HPQDdifIbeRnkkpKaueGO5VlL82P+Hv3Dr2PPoN6f5R55BEmUjq4wjWKO7/AOx4PvXDrxpWvSrwPam7ewMiKfBUFaY5Tp1q0hUfi/09++fXhSnVnHU2MymNwmMiyKvHVEKJg97jUgPJP9Pp7tXz69To0NHCggUfX0i7fnke6knrY4dYK5YxCxNr6CBc/wBB9Pe6nrxp0AW9MXJPBMQeGJsvP05tc+/DrXl1Wx3dHW401GhJW0q2nQWFvVbi3vZ4dbFadVub8zWTiaeZ0qAVDMo1uefJwR7rTOOtE06sf+DfVTbsx4zVbQs/3cB/dlXUPpyQT7D3MPMlpsO3y3T5ZBWnUqe3PtXvXPO4poUrCD8RGOrteseldp7UpKrJR0NI2Rni5l8MWvVb6Bvr74++9/u5vfN3M8lrFIyW6uQFqeAPXVf2s9utv5D26FVVWlQCrU86dAR2Zja/E52WpW/ilY6UXhVUP9Rb3KftNzPFLt0e1y5pw+09ZSbNvc11EqjgP8HTJT5OeWkdXZr6TySbcDke5pKtA1EOOho0dtd25eVQT9g6K12LvqTB1sqI7lmk0gB7c35Av/sfYo2+GV4tbNReoJ5v3Sw2eYkpgH06SWF7Fz9TUQKlNOHnYJSgaj5mvwFA/r71NJZyVgWUM3p0G9g5026a7H0dAynJxjoxeJ3fVrjzTZqB6eqVf3IJwQ6oy3JKv9L+5F5AtYFuvp5Vowp1j1963eN3TZU3KxmJRg1CDg06Jj2l2TUdQb4xm9sI3jjkrNdX4rLePWCQdP5Psfc+cg7dzZsslsFAdRXhxNOuYEHM9xfzadyOrUTnq07bP80HA13TlAuJYRZs0aQzr5B5CyR6WOn/AB/r75Z7992aYc6yXN8hEWokAj59ZO+2fIcO6aNwkOuKvVTnc3yEot77ok3G8TRZGWV3q5C1jIPJdQSvPvML2kS59vnSxhYmA0WnkB0n93eW9jlDW9oAskY/Mnpsx/b+My9LHF9yol4AUtzxxYX95iwTwXUInt2qCBXrEeeGS0cx3A0mpHRyvj7hcdmasZGWRGurOFax5tx9P9h7vx6r5dGj3lmKbFYxoYgI/wBrTcAAkDgc+9nrwznoi29N8zSTT0sKSu/7guLm1z71Trx+XQX7Z2dW7my5q6oPYvqCEsw5N/p791vo6GwOt0p5IQ0IABBHpAHI+t/ex1XTQ9GCyOFpKHFSpKiBYorm4AsFHNz791s46o0+W8cW49zTYfEyFpjPMjKhPHrsDYe9GnXiTSo6Gj4ifH6GOKnkzmPSpMsZI1xB7XAYMNY9+63UHqwTfmxMdj9mnF08UcMdHCZIkVAtm/2HvfXvn5dFrxu3qerxU1LMoaT9xbkavqSATf3rrVcY6ro7k2xT4rcNasapG0EskhawAte9r+/HHXhk9BZi+w1wigITK6nSAv8Ahx+Px71XrfSW3JU5De1U84o5GjUXPpNzzcf77+nv3HrQ+fS36k6Qyu6crK70ciUwVNHkUlbnm49+zXr1fTo9WD+PAwdHDUyQJfSONA4P5uPe/l1rhkdGL696kpa2O324J/SSEAB/w4/w97p6dep0L3+g2lEYkURxaObEfn6ki/vR+fVgPKvQG99bWXb+yaqGi/crZY3igCcP5Sthpt78ccOteWegm+IfROdhqpd9bqjsKiqnENPMpMgDfoazf7x78OPXjjPVmoqaHCUTONCFUAsVXkKLW/3j3vjx63w4dVyfKbdM+5IaTBYZXnqslVyUpSFjqj1tpu2n6e9YHVcnPSG6++LuN2dQYjcG4KdJ8vVzxzBXAZ4zJZlLaveqY62SBknqwzG7Ti/u9BGFSwhXSAFHAXgf09769jz6ri+SHXM+SqIsRChL5iWaiAQc3mYovH+x97FK149aNRg9WUfLXrfK7M/ko/HLobCSPS5ym7Oq9x5XxExyS01bTokaELzYWPB/r7rTux8+nGIMZHoAOqUcF0NnKOGkgehdjJHEZGKmxLKNTG/+Pu1K9NlgDTo6PU3SkGEC1VTSReUBWs0a8G1739+49a1L59GbiWmoIvFJEt1TSLqo4H0Ww976tUDI6CXeFTHIKmJDYOGvewCi3FvdetAjopGeVaSrluTpLEg/65597p1sHoLc7kCCGja+nUbAAn+v49+r1vpLVuSjq8fNCRdmR155AuPr711rosGWp58fnqKMarSVyAEcW1P9Tb3r7OreWerO9i0M8WxYWZTJM0Keq97alFhx72cY49V+Z6KVuanmod65GeoBhWdJI1YiwBP0uDx7916vn0U/u7YlPuHHVySPGlZEslUlRpBYBfUov/vuPfq9W48fLqtyWMxSyxk3Mcjxk/1KMVv/ALx791XrH791vr3v3Xuuxa4v9PfutHhjrtrX4+lv639+68K0z1x9+631737r3XvfuvdcudP+H+8+/daxXrx/pYgfWx9+68PXrq3BP9Le/dermnXiALf4gH/ivv3XuPXjb8e/deFfPr//0dAAi35uP6+/daBr14G3v3XiK9de/db65XF76R9LW9+61THHroAkj8f4+/deJoOuyfqCOfpf88e/deA8+ur8W/xv7916ma9eJv7914CnXXv3W+ve/de697917r3v3Xuve/de697917rsC/8Ah/j791onr1ubX/2Pv3XvLrwIBPAPv3XuPXgSOf8AYe/deOevXNrf19+698+uwL/m3v3XiadWF/y7dkU25e4KGpq4kmRZEijVwCF0OGY/7e3v3XsHrbO2pil21tklisFKtIyJfgBgn1H09+kkWGGSd8BVY/sHW4wZZo4EGXYD9p6CnBdTUu758nu2sUVTU1S/hfV+hbnTwR7w03X7wEO283Db7kfps2n+dOsxuU/Yncd12Fby1BBIB4+vTxS56fbMzJA5/wAkPjUIOUA/H19yjvPMFluFpHeQ0pIAesifaX223rbtVrMSSMDPQiYPtZqhkFYzFgV4cXP0/ofbuyx2W8ARxkBj037k3nMft9M13dKxhXNfLqT23nqWv2SK3hn8v1te1xexA9jnZeX5dtvfFHDrF73L93rPnLlw7aT3An8sdMPWcCZjb/lCgqqqo+v0+h/p7HpNesU0AANOHTV2HhJMNRzZCmYqkStKbtqF1Fz/ALD375dWHDPRKd395bkypfZ+HnlBYtCzopsFf0tdv9b3UkKKnq0aPI2lf2dZtl7ANFGcjkmeSomPlkZnLfuM9+QfeldHHaenJrWe3NZVIr0ZjYySQ1KLTMxjA5t9ACfz9f8Ab+7jpjj0ps/jRvLduJ2fC+qarMEgANvXe55H+9ewL7nc1rybyVdboGpIAQPz6VWcHj3Kp5dXEfGCorevsTTbcyMRjpI44kMpWyqUYD1EcH6e+FXvxYw863D7xA2qcljTzNR1I+1SNbdjCi9WH4jcdBLGhpqmNozyWVh/X6f4H3gbvnKl+sh+piOocOhZDc4DRnoRKLNQzRFJJUYabD/AEfm/sESW09tJ4c8RPp0dQ3WtKMfL+XQKdgde9Xbmmer3NhsXWSBXPkmghd+OSdTDg+5U5B589z9mh+i2KeWOPVQCpAp0R7jZ2Ejl2Ar5/b1X73B1z1JkJlwGH27jBGZQWWOlhu4HH1A95ye2/M3P0Vn+898upGanmxx0FL2GzB8OIdGS6Mw2yun+r85S42mpMcZ8dVyPFGqK0rtEdKHTb6/63vHr3qh5o90PcDbPFd3itriNq1NBpYZ8+jraZYNts5CtKupH7eteDAfEHtLsD5kVnydoKCsxe2tmZySeady0UUiQSmQOqm17gcEH32Jl+81yl7ffd6/1nLqdTfbhB4cYFCTqFP256I+VORr/AJu5mWWzGQa/s6tyovn0mDlqMVWV4jqsdI0C6zywQ21fX3DvKvKyblskNUppRQT6/PoZ807JuPK94be7Uivr0utq/Pat3NOYMdM9U6stxGCSbcH2o3flrbNoi13DAfb0H4p7h4jIBj16HCP5d5KkpVlyIlpUZfrKG/V/sQPYWt7fZbuXRC6/l1YXLsCRmnQdb0+bOTo5o6KJZQKqI+JrH1q44ZeP9sb+1slltNrbtdRsshQ5HR5y7s+4cz3gsrBSxJpjosuZ7N3dvLKLU1tRUR0c0yvpcsFszcWF/YX3vny2ttueHbAElAIqOPDrIzkT7u243e5rPvKnwxkhurWvi6G/uPAw9etgC3+JP1J98d/f28n3L3SjeU1bj/hJ6MPdLZbPl27XbbJdKqoHSV+SX8Rod4beyDCQ4eOGJag8+G4sDqUG3vIL2JXbrrlW9sVoLos2kfi6xj5qEiXcbn4aDpwqanA5bAUnjeF2an1AAL6QFBH/ABPsgtbbf9p5gm8VWChvnnpETDJFUenSd25lYcNkFjilRULW+lz9fp9fYj5l2iTe9tYzKSafZ5dMQyrBL29D/FuyEUms1QQaCbG1/pzwD7x5k5a3kXngRAlQafl+zoQJfAR0PHoA969iQxGpSSVHgaOVXBtoIYWNx9Pc/wDJXt6wjjkCESVU18+iC6vO81PVCvyDr9uR/IXA0+GeCN8lLLLUxxWJMrEk6gP94Hvuz9zafc4OT5bK9rWoAr6dRrv8hmmDE1A6wZrF1NNkZJQrWZwyG3H+8e8yjglfToO/M9Krb2UJX7ec/T9NyByfqPeuOOt1H5dKHKlGpdQIIFxawueP9f34EjrxHl0HP3DxzXuRGGIIIuAP9f6f7z7914/PpzyOF2/kMYZf2/vPHwQVDF/qAP8Ait/fut08+iidhdf5OsqtSSP9sb+kOeQf6n3vNeqfMjrPsfoemyRSpq4i0YCsdRHJ+vAt798+vUNejCZfr3HYLZVdTUVJFGY4mbUES+pV13LWuffvt6tTj0VTaHaNTNkMjtWsVlSOUwQ8ECwOm4b/AGHv1a460KdF7+ZG3cTlepNyfctUGRo5lEkFRJCyhobXBjNxa/1B9669TPWrPkkahyVdSRT1DRQVU0SkzHW6o5UM5AsT/sPfuvUHUL7mbn96f/D95/z9b+/depTpRbe3nuna1YtbgM3kcdMhBtDWTIjG9/UgYA/69vfuHXjwz0Zfa/zo+TO0PEMF2JkKXxKFS8kj20m4NzJ73XrQ+09Dvg/5vHz325o/g/dWTpfH+kqgY+k35u/+HupFet1A6GjAfz+v5p22dIw3ySzVJo5XTTUzW/P9tj79TreodC3t7/hSZ/NNSuiXe/f9furFKoBpqzEY6okAUiyr5PTa3vxWvmeraz5AHoyO2/8AhR12PX89oYN90iWlqIqlYsDiqdppZojGHZo5LcE/S1vdhQYHTZqTUjrXU787Hp+4O4+w+z6Sh/hlLvTcVRmoaExpF9sJ4kQwiKMlVAK8AH6e9dbqeggBAIJFx+R/X37rfV6n/Ccugpcn/Na+NVFWRGaCTcwkZFPJeCN5Yjb/AAYA+6twHVov7Qn5HoYf5lX83n5+defzBflntfaXfmcxuA2v3Luvb+Bx9GLU2OxmJyDRUkEQax1KDZj9OPe80z15qA0HVr/wR7o7i/mg/wArL585T5zZTPbyq+p8HiX6f7Q3Cjw0WFmqAYqjHQz1RPoAsAAbE/QW96yOrKVccAePRWP5dX8sPqTrX43d0fJL5j56m3J0NtWliyMeEpcoaVc1RTSlUhgiGsWdVIdrWuWNre7kU+zpmPuNXNPl1aB/J8+VX8s/tD5D1fXnwf8Ahjubq6rx1Nk48xvirzVVmI5Ivs5Fiy04aJAkblSyhiDb8e9DBr1dipFBx+ynVBG1PlV8kPhv/NC+X/Y3Ruyt19odcY7trLy9ubd2vj5a+hr8b5S8i18iDSA/7ykMfxcWtzUcBTj1s0qaio8+riupc/8AyZP53e+x1tQfHl/jp8ttzVZp8c9Zn56WHLZd10t4qSSCwmaa91V/rwbe9gUNetMwcBVzjOM/t6qn+Ef8ovOb4+b/AMmdi5ncP8A6P+Lm5clT9n0kmUkp1rsXQTGKQF9RDEhW0nTf6fjj3sd3WivhihNfIfPqxXo/5Q/yTuqPm9190f058Ktxbp7Uh3lFtnH9rQ7klmp5dyyMfHKlAkIZjqUP+Rxy17D3U5+XV6jyP8uld82yf+H2v5euuhlxjnf1AXx8+oVFJqdWEU2rm4Fh73QgZ6ooBkx6HqgL+feCP5jHyNuef9J24v6/Tz8WPvw4def+0/IdFI/lkEn50/HY35/vnT8/9Usn19768uOtkv5rSY2m/nK/BCpzCpLi4OxsTLXJKEMZp0yQMuoOQLW+tz721AOqoe8elD0RL/hTDhd70HzIy+VztVUybQzuaq67YdPIXFNFhmpLIaOPlQo+npPvXAZ49erV6rwp1YJ01T7p2v8A8J9PlFJ2C81fh93bH2+3W0lYTKiRQSv9x9i8tiLG97D348MdbUgCh9D0DP8AMPz2Xw/8jL+Wvi8ZVz0sGU613JLWRxSOglIlp0BYLb+yW96XBoOryAFKn1HRjf8AhNjuXqbaf8vz+ZDujdvXdf2ttLAbEx83ZnWuOqGhzm7KGWS0cFLURgugB5BjQ2HvTZP5dWjaid2DXHRbOmf5nP8AJV627g2Tvnqn+VL2Mne21twPLtDDDetVWvLuUVTeFKjHU8IlmnDDUqlbj8/ke/HrSsrYHA/KnQ3fFj5R435gfzgur+xNu9C5/wCN++8h2NS1Gb2/uKkejrvuuTFqWYBjcfS9vez8Nfn1pT+sQeIB61/v5w2F3un80n5RUU8GWO5cn23kGxCCSZquqlmmWGE0P0Kgyq9lBtcE/n3sfLqrEefy62SPnaZNu9C/yHtv9t0j1XZKbXeJzWWasonavp3SOZmOr6aiLgce6Dp56Uz6j/B1Xf8A8KNKysrqro5amqmkjoccYaaLyv4ljamtbxk2Nvx/T3c8adMD/P0LXwS/l1dAdM/DDG/Mn+YJVTb86f3ZgpNwbK2THm5KapqKSljWSWjjUiXStgQbjg/4WHvVDSvW9QB0nHr1aP8Ayn/lD/Lx7uxnyS2j8H/h3uDpKvwG3ZK/PZ6qzEmYjr8MUeNamTTGojV9JYA2P9QPfgM9WY1Hy+zoiv8AK39O6/523Ia3W2WAIvpNq+P6e9N/m69Fn9p6dP5e1VUp/K5/mkUUU0sMFb1xUrU+J2ikZVyKMAHT6c297c0P5dVh/s/zPVPP/CfyoqIfnv06kM0kccm/MQJUVyFkW30cD6/7H3s/D15B+p+R6Lv/ADqdK/zOflloAA/0k5Q2A4BP1AHvQ6248vs6qxP4N/r/AE/FuPe+qj066P1+t/8AH37rw4deBAB4uT+f6e/deIz14GwPF7/7x7914jrw/wB4/J9+630fX4DdT7l3v25iszQUFZJjcVUwGeWMOscg8oZ72HIA/N7e/da8+th7uvZmQwjUNVS0ki000dOJFHAsI1BvYf197yOPXj1w696jp95URlrKZGhYeosBwQPxce/de+Y6Ya34h4/N7oC+KNKJJULjWFXRqu1jb3qlevVHR2ti9Kdf9c46JcXjKf7+NFBmOhyTb1EXH5PvfWwPToSsdRiapaayopJKi1rAWUWA96+zr2dXSlbKU9G3hY+pQARcA/T629+p17pvnyUL3JdSv1VdX+x4/wCN+98D1o9InOvHPC92VQbf0bj/AA9662c9Fp3fs7CZVpBkIVnV7gj+t/xY+99a+zoj3yD6h2tjMH/F6ahSOCKN5Zfp9AwNrD3qh0mnE8Oq1BlXV8I49Hl+Eu7dqL13jaPFwwwVKR6JG0i50ix/5H7g/nO0mvJZbWY1DDrpf7EX+1x8owJahVfzNM/t6PhT7vq6Oqhi16qckXVQbW/BJB/p/h7wa9xvYqQLPvNgamhbT5n7Os6uVIts3PbvBDAy06Ru/ayizpPjiBfT9bHgk/X/AG/sM+1fKu62tz492pRUOK4rTqSOX9nWyUh/PoFa+mbHUpUAkvq4+n+Nh7yTLa31dCW4CwxVXh1Wz21nsjkN/wBLg8BQT57My1nigw1EjyVVTKWA8MaIpueRx7kTbbBrnaJFjFar5Zp1hh7v70lrLMJ3CLXBPWxp8M/hv1D1v1FgvkZ8vRRbPixyLlsb1luN1pM/mljh8sUUNLJZ/wBw/puB7INi5GfaL1eZeaZxHbg1jhNQ8lDjHXP+49x+aLrmiXlfkyQs9x2mZcpGSfM/Lqrz5c9o7F332tvDfvWG3jtLY9dHPBicGFAFMtO+iNgOOSB9be5f2YCW/O7WieGkhBC+g6l3nK/uxyAnJ/MtwJ7yFSTIfOo+3qjTvTsyvzM74maVnjWWRVDC/BawAN/cwxzjwRPw658T7bIm6mwALdxpT7ell1Btuqk25HUQpUSGSJzpDswBtfhbW4949e5E2q+E8dKeZA6zr9sds5gs+VkW3VgqiuPs6U2ytjVfYG96zbEVPLLViRYPGLlruf6Ef19gK4vodv2430zAKBWvp1CnP1xuEm/Krg6i2Qa+vRp+3Ph3lutOtE3NTLLTZKCEzurOS7+jXpC2+vsn9tvetr/mxdihk1xMwX7M06G3NHtFaS8g/wBadIEirU/sr0kfgX2TuXNbqyOCzAmRaMVcarIGBbxelSCePecMiaCKeYB/aOsOopNQI9CR+w9WZb4pZ62nmjViT4VKL+b2v7b6e8qHoAMP11NkMi8lVSt6nsrMeOT/AEI9+6oNXRg9m9VQY2pEzwJpI4IFrX/qf8Pe+t18j59GBoMRSY6PWAt0AJJt/T6Dn3okjgOtivQfb8zsH8JyMYddSwOosRxc2Hvwr59eIB6oLylZLku/8tT1lT5KZKiTxIxuP87xpB9+pnPWuAx1ct0nVYrHY2jjjjiWQ06KGUAG5X8+99bp59PPZm56dg9GzqAygHgWsTYc3/4j377OvA9Fayu8sVgi0ZqYIbsWdzpJA1ck2I9660cDHVWnyD7Ko67d2ShpKmOWJzINcYBDi304v78etjh0XPAVAmrUnqLvTvJc8tpPP1+nvXWx0fHp7AYXOShFp42jKLyFHLFeRqP5+vvY6qfXqwHrvaWMwzRLTUscYIQ2CgFv9e1vfuvcD0Zui21BlMfNG0Q0rASptpAYc+/db4YPUjrxaTHZWpoWKftlvra9lFve+tU8h0IO4sxQUtJNpkjBCGwDf4c8f19+68acOiR75zUG5s7QY55RNHBXKzJxbSG/Iubj3rA69QaujN4GalpKOlooBDFTxwRnSgCgtb1Hj377OvceHQZ9ubknxmCytRC5UU9HPKjji2gX4/23vdetalrSvRcPirthOztxV+69x/5bR00sjUqyG4WRW9LBWHvQyaDrxKhanh0ZPtOIrPSimULHTSoqhbKFSM2HA/HtJd3kVqB4ppXoR7By5fcwMRaoWAHEdCXs3JpWYURvYlYguq1gSF+n+w9u21wlwtIjXz6Tb1sd1sb0ugV+3oue7MTHlu1eucZNB5VyO78dRqCOGM9YiEcX/r71NcJDhsV6bsNpuNwUyxCoUVPR6/5rO9sJ0tNsXpzMJGtLjMVt/ImhcDTC1ZRJMH03Fv8Abe6zXMcS1J4dXsNlu7+cJEp0s1K/Z0U/bU2zt07XocriI6aR5KeP9CqfUEBsPYHvuerGyujbyMB1lFyr92LfOYtnXdIVLBh/q8uplXHPjsfLJBGQyqzBlH9kDgW9mFnzbY3KakcdBDmD2B33Z7gxSxMAPX7fs6DGLKVWQZxKzj1OOf8AXt+Pz7WNzBEDxHRAntFuLZKkV6RW76R0gkmBJBUsTwTYfUkD2Z2G4xXi4OegNzRyff8AL0lZEJUdE239uGlx3lknuqEMFc883/r/ALx7MuAz0DBnh0Xqo3MtT5WjmUodVxe9xf6e/Dj14+g6Tk+dMAbS/oNzf8ci1h791voMdyZUz1mPeEKahqpAh4Lag3AA96+zr3VuvRe3czlet6SrraaUj9kAsPqvjBFve/Lr3l0Evc/XoZpauGC1QmtrLqVuDe5t73TrQI6IR2NS+PB5UzRP94lNOhuOQqrbj3Xzp1vgCeqlav8A4E1X/UTN9fr/AJxve+tDqN791vr3v3Xuve/de697917rl+k8WNx791r4hnroi30N/fuvDPXgbfi/v3XiK9de/db67JJ+vv3WgKdeAv8Am3v3XiadeIt791vrr37r3X//0tADn6f7G3v3Xsceuvfuvde9+6917/H8e/de65AsPpwP949+60QOuj9frf37rw669+6312Bf+g/1+PfuvHHXXv3Xuu7cX4/4n37r3XrH+h9+69UdeJv9ffuvcOuvfuvde9+6917/AA9+691737r3Xvfuvdcm/wAOB/T/AB/rb37qo64/T37q3XK4sbi5J+vv3WqZ6tf/AJX8KDsajneQIFrWNz9AOPr791unE9bJO/8AfeLO3qPbFBXQtkKh4k0o13tIdJtb2zdos9rJak01qw/aOlNnHKLmO4jFQrKf2Hpxy+Zqep+uY7nzHKpGWa17GRb2BH5uffODnf2wv057LSKXjqWVh9taddQ/ajncw8oRhhntXP7OizY7eVRVbupcZMPLFkVWo8h/s+T1aST7lfbLK/k25LZ69goPl1kdtG8W2yb7bGYjw5UDk+WejCLgaJdNQp0syRsAGAJsPqB7kXkfab20vklkBpXqKPvQ8/8AK2+cqS7bZyoZgG4cesm7qnHttZ6KWdAE5KseTZb/AF95FCmhaenXHtlpcy5qNXWTo/d2JGEr6LWitHJpW550j+g96x14E9Bj8le2qfFYGpx1BLG1TKpiupJYazb8f7D3s9ep0WjpzZUuaEudqUL1FRIJlNgbA8gf7A+092xFsxHz6M9kQS7vFGeFR/h6Xu89yTbfr4cNpCvIwAJFjYmxPHB9hrZriV7gqxqK9Tb7kbLt9pskVxEKPpH+Doy2wFpcVtGbMVkl5GpJJYyxAs+jUAPYvC1bA4cesfK0QV8+hU+GO2o+x9+5TeeSk/awla8MJYi1gCEA1/6w94BffS58m2xYtjsGqs6ZUetR1JPLXK88u1Pu0nwoePVyW2zQT7ay+MGNd641LinrQvCqf0gEc2/1vfLTflv4uYbXcfqAsATvj9T0a27K8LxhamuD1m6+o9247Mfw3JpULRVD6o6mQN41TV6fUfpf2n52uuWr7ajf2LK0sYyopUmmet2y3McgSQEA+fR58LtGRaKEyzajLGGR7/VSPeFO+b9LcXxMChFU5B416G9vtTGIFjx4dJndfW8lekiip0h0f6N/aI/p7E3L/P37hRJZow4BHAV/b0nutnJFA3VSfyUmyXU27Imp45K8uSQRci+m6g6ffSD2YvLP3B5YFw1IwRw+3qPdyjNlORxPQZ7F7p3Rn8vR0+XxVSuEkZTLFpkVGQNYqb8W/r7G/MHIWzWFjJJYyKLgDDYqD0kiupGcax2no23a3yH2ps7pHM4bbe10gnyFPLHO6xqHaV10vKWUX/P594t8v+wvMnM/utb808x7o0yWpHhRA0UCvpXP7Opb5D53seU79LpYwtOJPVPG3cDsrO4jcG5NyZGIZSorDJTU5ls0aytytr3499Poto3LZOWNNkhL4C09B0JOfefeWublF3M6BxXH5dHy+F/WfW1X95k58pTRmIl1EsqMLF/yT7w5+8HzRzlt1vHb7bbtKz4NK+nUPbXu9jIsluXAFcfZ0YT5By9ZYrCTxR5CkeSnnhYeN4w2lTdwLf19x77Qxc+blfK99A6K6niDx8ukN7um32RJDgn06Absnf3RO4tg4usw09JR7jw2PERcyLrleKO3K/X2Pti5H90tu5kuI9zLS2U71UUPaD5dSN7We6Wycnbyl9KFYCla9EIl+U1HBWDFmNStJKI/KOVcxtYMLf1+vuY09l53Q3GlquK0+3rKu4+9VsP9rFoFfTHVlnxq+csWM2u9CmGatWJX8boX5I/tWH+294n+6/3RLbmDmCPeGuWt5V9KDB4jrHr3C94Lbm28N5ZqPn0t96/Lms36z42u2xMtIxKrKVcaLfSzH2v5R+7vZ8mML20vtco41INeobut/mvlInXHl0l8B2/K8y40N9qrEJCupgyrewWx+vsW7t7fwmM3bjWRk48+kcV2fh4dO++d9bg2NhzuGojknjeVPELNYqw4IPst5d5Z2fmO9O1xEKVBr+XTs80kUeqnQi9P9n5HsCXHUuS/3Hw1YBVmLABTa36j9PYG9x+R7LlOznvtvXxXjzQAdKbK7ed1WTAPRz6vo7DbixjxGUNLPGw8istmDD9QN/8AY8e8U9v95N52rcPFRR4aHK5qCDwOOhfHy/FcR1JyeqpvmD8Acxg5h21tCSevr8DrqHpYAHYRhNZay82HPvo590376+0W/NNvytvVIlk0kk4BqSAK+uOgdzLydPbwGaPuHCvz6qTznyH+zlnxGRpQuVxr/b1UTkrKJU4YMp/1vfam35n2fcrJN1s5VaOUahTIFegjtvJnMG6SCK2gdhwqB0hZPkTMs6GmonXkEhQ3PP19pf6z2LSCJCDXoeH2U5ljsGv5UcBRmo9OlePkoZaYRz0fjfSBqYsOf629iKKQSRiQcD1EV5btZ3TWsnFTQ9R4u7ROCgp/KJbsukngn8+99M+fXdP2DnZ6gPDFJ4NQNrPYAH8/7D3vj1r7OhLodwNmMe5qoSZIwxAP14F+Af8AH8e9fPrw+fQj9O5hq+PKU9VCYjCdMAccgX4Fj791YH06EPetRGu2MnCbO7xSKALE6jGdIt78AR1rqoY0m4MTvXKZOoSSGnFUWiLgqGQv+D731qo4dZu8aSo3d1fmqSEl5JoZHe1yb+L+vvXl14enWrrvnGSYfd+fx04OumyVShuOf13Hv3WskY6SXv3Vuve/de67It+Qf9b37rw66+nv3Xuu/wA8/wBeffuveXXjyeB/rD37rQwM9eA/NwCPwf8ADn37rxPl17k/73791vrw+v4/2P09+690f/8AlnfMyi+BXy161+StZto7rHX2TXIx4cMVFUwBVkJX/A8f4+9EVHXlOltXqKdX1dq/za/5C/evZO8O5O1P5V28ct2T2FlXz+88pQdhtS0GTzkzmeqyMNHpAjaaQl5AB/h9PegKdO6lPH/B0AfzF/ni9Ib4+NW6fir8K/j/AFnxu6h3lSU8O5NvCvauqctPSLaGeoro9Nz9DY+7ACmemyzLlKZ+XQR/y+f5xHWvTHQu/fjH8z+pa75A9Q7txkGIoMZDXTUVZi6aFtaSmeO+phqcWtzxf8+/VP29aFPs+fR0vjR/PT/l9fBLsChz/wAOfhfldj7aqoKqLemPz+XOWyO6DU05gKtXy+uOMX9K34HHv2ePWzpJoeHVc/QH83mLon5m/IfvXE9e0dV0z8iN41u4t4dV1VNFVLPjq+qed8XFUTBioRZCiN9be9AY+fXie8kcD/qr0f7Bfzff5Q/VHaEnyT6M+Au6dmfJGlrhlds7qO6ah9v7ayhs7V8eHTSkkokGpW03/BNvejUnqwIVccfs6I78Sv5y+Q6M+TvyH7g7F2rUb82D8jM5VV+/dmpI9PJmKCrkLzUjzDlC1ydR+l/dgKcOqanJ7ujhYT+bN/KP6s7U/wBmB6Z+BOcwPc0ORjzeEzGSz71+OwOWifyLVLTSga2DfQjn/H3r5nq+oAUHQGfIz+dtsnvP59fEv5rP0/8Aa1nQWbjze7cFI5Ee8aiFtaB9PKg/Tj8e99NgUp+zqsT+Y78vcJ83fk/2Z37gNpnZtJv/AHPkNxrgAxZcd98QxpkZuSFtf/Y+9DAp1tiC+ryp0C/xH7sx3x1+Q/WPdGXwzZ7HbCzqZaoxINjW+OB41h45+rA+9060DQ9Hr/mGfzLqT5a9u7B7j6u2zW9a7q2HXHIY2rjf96GZH8yEPfkavqB+Pe616rpp1Z7RfzsP5eXyP6b6w2d/MZ+F2Y787D6zxMNFj98bfzsuCra2ZIAj/fOoJkUsOQT9OPz7qR08pUY4D06Jf/Mr/nB4b5S9V7K+N/xX66qfj/8AHPauL/hD7AjqPuJ6ugp38tPFU1vGvUxYyH6/j3sdUahOekj8t/5qGxvkX8Evit8RMN1aNrZD46bPrdt1G5o2ZjuKavCtLUyqTZbFbcfXj3oAg16251JQccfy6Bb+V1/Ms3R/Lv3/ALjyUeNbc3Xe/I6ei7C2UzMaTc+KiXQ1DUILrYrexYcH3YU4+fVan4SKqf8AD1cVJ/OH/k77N7UHyX6i/l35TbPyMp6xM1gdw1ucFftrA57SPJkhhJU8bsJAXQEcX/p7rSp6vUKMf4Oq4Opf5wO49tfzDcr84extt0+7JX3jFunF7dpqRKGnQ082pIGipwoW6/Ww4PvZyKdVUkSeIeNKdWqb9/njfyl+9O+Jfk93d/LyyO4O3qWvfL0tXS5wQY+uyfqkiqayFUNyJCGN/wDH6X91oa9WJBGeP2dVF/Kv+bBmflD8xupe+dw4CWl6q6Zz38Q2N1pEzilweMS4gxkIPGkKEB0/Xk/097pXJ6rXuxw6TP8AM6/mMbT+eJ2K+29iz7Pl2pFoqRM+oVAWHxkqxP0v9B72T59V4U6O98Uv5z/x1278Y9u/Ff5j9AZLvPrzbGGbEbcgo8lLjZMGJo1SaWNxcOCR9LW9+Py6suR39GI6E/np/AX4WU24cH8PvhxkdgYbfNM9B2DPlskclXbjx7gqaOOVheNOfoffhjJ68+RpHDojPxX/AJv3X/x13f8AzA8y3UP8ZxXzI2/WYHB0jyMG2ZHVyifzIFPqKnjn+g96IJHW1IU/z6w/Gz+b1sXof4lfLD44VHVb7gr/AJHbcqMHSbkkd0l26ZJPIkqLex+n+397buNfl1pO1aHyJPRHf5bHzMwHwc+RWzO7tw7TfeNNtPP0eb/gqsVFWKZf8yWX6X/r795daB7tQ6DD57fJTGfL75ZdyfI3DbbG0sb2hueo3BT7eViy4wToAadWPJAPNz791smpp0T23F7/AOw9+6159etxfj37r1c0669+631737r3XJbn0j6tYD/Xvx791rrav/lpYnYmzes8PVCGnbO5KCKOWZkj1MSgswv7359eAPDqzLszG0ufxlNAYkeyi1gCRxxz/sffq9bHn0wYLEnbG2pBBGUYxGxNuGJsDdffuJ69084NJ1xxyDsDJIrWJtc2HHv1etE9J194SUtY8dT+kEWBJspv+PeqV62MfLp3TsWipV0s6BiSwN7BgebAn8+/Hr1fM9B9uTtCnabVHUJEbG4ubEX/AMPfutdIOr7ypsahErxylbi+o3IHFxz/AE97Hy61nruHu3FZSANJPGmsWClrEAfge/cOrA+nQZ7g7KpvuwBVRGN2sOTbk2vx71Xy61nph7AwdX2Fsito6FjM0tI6II7G+pf6+9g0Nem5E1Ag9Bh8cMfubrOt/hlb5lp4nb9erSBq+n9PYR5l2fx4jcJx6yC9mPcZtpvl2e4PZwFerW9sZ6nzNPTMHUuVUEX5/QL+4c3Oz8ZDDIMcOujvJvMpspFu431AivS/nxNK0XmQklV1MWsAOL8+wjNsEdvGWhAH2dTjs3uY93dJbOPiIHSEoOt979w7lotl9c4XIZmvr5hBPVYyJqh8YHfT55UQEgAck/T2ZbTyXd30YlUGrcMYPQZ59+8Xy7ydM9reMjNDkqTQnof957Y+Gf8AK6o03z2wcP3r8k6yCN9vbYpaunep2Hn51EkddmabksYzptGR9R7mLbttseT9s8e8AkuQK6a9o+31PXMX3V919990d6uLmzJs9urUDILj0X5fPqp3fXyp75+bfaWT3F2RueefB+Urt/b8Jejix1PEf8lgSnj0qAqgCwHuEPcDfZdzA3G7yyV0D8Kj0A6ibk7mKPljfk+hGmMsC3Ekn1r0XXt/sCfa7V+3chTuZqSFiQ2oarix4/3n3L3tlPDuPL8MzkGo/wAHUi+6O+X25RLuNoD3Dy8+iCYXZeU7X3iTj6abwCd2kCrdQGb+p9irfd5hs4vATpN7U+3N/wAz3Y3K6UkjJx1cv0p8e4cPtmCCeJtawgsCqXIKX4v+fcPbxE+5I61y4oOug3Kz2fL+2G0mjFFFKU+VOk9srrafrzvGHcNJAyRVOQhd9aAo4V7H/ePcLc9Wl7Zcvy2N4CFZSFPr9nUEb1yVYcyc1G6iFAWrT8/TqxT5Gyw7w2xiaYRp4qmKOOZAqhSpWxuvuG/uucrNfc9zNLVvAbWP29Cz3vgi5X9tVsY2ADqR/LoivV/V+F2NvGor8dSeCaokfyH0DVr5b9HvrNMcgDyA65XWwIDE+bH/AA9GvrvG8pL6WBVQPyf03PtgdKK+vUzHT0VIPJLEhH1U2HFvoT+ffuPHr1TwHSqTeWIpaZmkmhTSBxcAm34Hvda9eIA4dBXububFUglihrIdZBW3qvY+9Hj14H16Jp2p8j8VgqavjmnikeaCQIoc3LX974darXqok78er7Rm3IkqpHPUSWQXP6pL8/n3Ukdb8qHqyvrz5B0NDDRwTTRo6xxgNqPLaQQB72adepjrrtfvOnipZMitVGw8WpAGIJN7i3v1eHWqE9FGo8xvrufJLj9vUFWqVMxhNVEHKgSNwxb37r3Cg6GLFfy7d5Zid6jK5KQSyRCTVLYkGQXA9X+v79Q9br0aHaf8vnA4bZ1YMzKKnJRUztDIDFfXpOmw+vvxwM9aBqe3oHdh9WZrZG6qvGx0s4oqWRj5SvoKgkC1uPexw68ejg4XP4qgeCGreOORTGgLEC5+h4/1/euB62QKdGW25laOsxsppXSzRFV0c3PveOA62egPrM5Wbf3PVuEciXULn83NuPemIVdR69GryyiFBUsaDqbk8tNkMVWzAlWWBm+rcEj6C/59l7bnbA6aip6GMfI28SQfUKjU49EB23urI5XtyuxEUjP9pNIz21foVub39vC6j416KZNhvEqmk6h0e7GZuVqmCkXUG8aC39Tax9vmWPwy9RgdFiWN6blLYoauadJrueiqptlZ0XbnFVNj+NRj/Psj/f0Bl8IceHUot7Ubmth9YQ3CvSL+Awqv7vZLHSIwKVEreb6g+skDV+PZJzXz/s3KO3G83CRVJ4BjTp3kL2h3jnLmWLaI1bw9Q1EDyr0djIdfT7nSqLxMix69EnHrZT+L+8IPcP7xc24bkLXaH0oGyRX+XXTrZvu88v8AIOyRBAJJmUauByRnqFS7Qq9r7dlmZXIAk9RF+Bxa/wCPY29ufvGWy3q7VuYBY07q4/PrHL3Q9i7rmS51WKlVr6DoNuk8ZD2F8iOuaByJE29vPD5KqWwJSGGvSVg9/wAWHvJ+35jtOYHj+jIbVTh889RXde191yDtM9xuAIBRqEjzp0Uf+fp2/mOw/wCYvuTbmKdpMRT7fwlHRQwlrIcfi46dQoXj8X9ucw7lFZiZnNKYH5DoE8p2fhbHDOq1ctXh6sehX+C3W24qrr7GnOR1CgsSqyXuFI44f3gL7xc8mxvma1lo4HkeulPsLv8AuNpy+lvdp2V8x/sdHw3X17SUeHqh/bEDaSQt9VrC49xHyt7t80NfIonOgEYz1KvOEGybxADJCofzIA6I/wDYNQZKemnXQPPJpvYApq4sR7zY5Z57j3W3Tx2AagB+3qL772+sJtuM9kBqGaY6T+9ftYMfUglT+y9l/qdHJ559yzsu8Mk66Dg06xu9xeQo7na7j6haMqmnDqrju2od6J415HmYqqk3/V/h/T3MEMqzQiQdc6txtH2/cZLSQUoT+zoqlJka4T/ZwxPJKzcRjVq+v9D/AIe3Ok1MdPsuO3Cw0GgqFDi9tLXOrj6+/fb1rHTbTYKthz2Dlr4mEEGRhkkVxxoDi459+8+vfLq8rr7u3amF2NRYqmx0bKlNAjFVHEixBTe3H492rTrwwfs6Dncu4MZul56mnKESa9UY5I1fi3vWevefRBu5cNQxT1/KqKpJoArBRYyXW496IzXr2eqdO09ottDc1RSIzPT1V6qNuCoaRizKCP8AXvb3vrVOg09+631737r3XY+v1A/1/futHro/U/n/AB9+631737r3Xvfuvde9+691737r3Xvfuvdd8j37r3HrxN/r7917h17m3+F/959+69ivX//T0AT6T6eOP9f/AHv37qoyM9euLfTn+vv3W816434t/T/iffut9evxb/Y+/de67ubW/H++Pv3WqDj11791vr3v3Xuuxb8i/v3WjXy69xb/ABvx/re/dezXrr37rfXeo/W/+H+w9+61Qdde/db697917rsknk+/deApw68Bf37rRNOvcg/4j37r3EddfX37rfXI8i5+t7f8T791oYNB10LX5+nv3Wz8uuvfuvdWpfy5qWWbcKfZVCw1hqntc82/PHv2OtZNQerg62jyKdjYc+Z52SSnaRhIzKArgklbke09xG0inTxp0c7Jfw2lwizjt1Do43dVbFuXYWNxVIwWrhp1vY861QAG3+v7i2/snk3EtOtfKtB1nJy7u20x8qr9JKFYAN9nRWNm7azdNWJlci5kNGvjicDgBOACfZ3t/K0NRIFABzw6jjm33yv4oms0kLSINKkE4HQvYpt17iqGFIJkgi/b1Xe3A/A/4j2NYLG2t0EaKP2DrGjdeZ943i5e5uZSQx4aj5/n1B3b19vusoXmFWwhDHUlyCV+hHtXQ0p0QjiSfPj1g2ztyu21iJ5VZo5gCZGNxdgLk8e9Z8utfb0UfsGrq9w72hx08pmQ1CKy6mYE6vyPp79wz17yz0eLrqho9s4Kh1IqJ4o7lgvC2tpv7bmUvEU9elm23AtL5Lj+Eg9Af3TSvlN646uxURlplQGR4wbA2ufp7K9u276RyxzU16GnN/No36BIUxpFOoG6N+Zah25HhaF3UiFUcqzAABfUDb/evZ0bgW6vK3mCOgNZWE24XUVrDxqD0cf+XR2FDSV9bg8vIMfFXVoklqJ3KqxBJN9X+v75i/eq2G9mu33cgylQdAGesqX2e9g5bj2WwQlpQNVPXq7mPtzaOIeGgx4SaJFHmqFP7buhuXv/AL175qbvypzNuge6lJUmulfMA9S57e+xE81kt1ug0k0oCB59M/Y3yt2zhdvwyqIo5YquCEEMFJOoD8W9ofbT2T3yXmSU38jSJIjHSakDj5HoTe5fsg+1cqPum3pVkNagDh1Yp1Hvuk3v17hdxQE+OXHQsOb8lOdJ/wAfeLPuHybecpc731hftqRpCU8tI9Osb7C8DWnhuKPFhvt6zbh3PDQQTVM0oSNIpSoJPNkv7Ldm2G83y8SztFIQstR+fRbeX2mrH0PVf+W2lH3pvGRpKcy0VHPIJZjGHS2uw5a9vp7zts+YYvZ3lFFd9LsootaHh6DoENA253RKivQxnpXYO0seBkKemihpoCzz+GJf0C5u1r+4ntfeHnHmy8/3UamMjUC1Pn0vbboLdP18U49VN/LL5B9ZUcOS2ZtSkTI1SiaBpadriOa5BGlTb30C9jfbLnaYQ75vrkaqGhHAdR1zPvdgkTW0HHPDqouoq8w9TNVN9xFSyyGTxeR1Gk820j3n7DFBHaR2zAEhacOom8e4MhOs0+09Db1P3pLtGsixiTz0EMtlldqiaNTdv1cMPYT3zkHYN6QzXcIdh8gerG+v4VJhb+fRkd4b72/uLCPV1OZSoaSLUwFS7G9rkeon2S7XyjtO0ShLS30kf0QP8HSCS73KV6ysT656IRurdeOhr5qfH1MniZ2JtUS6Tz/QG3uQ4dtheL9aMH8h0qTxyMMRX5npGrmdumVGqWAYupdi55a/JJuPa2OxUAhVxQ0wP83W2S5pQMf96P8An6uT+B0/WW4sWaGseEVBLIFcq2rkcjVz75//AHkxzbtcr3W0qSoNcVGPy6lHkt7fR4dwf2nq4Oj6D2hkaJJIKKHRKnokEMXqFuCCB75xX/vrzJtl+bS4c61ORU46lSPaYZ11oKjoBeyvjUlGrV2DjMNREXdG0hQdPqHI9zPyB79Q7qwtNxaoNARXOei672hou5ePRLuws/uynzuL2lulmqMVHLBCSwOgjUFH1t7yc5W2vYpNvm3zZQFlcFsdEM8socRS8OrR+vOksZkev8NlsUq0tUuPWoglCKhCoAyk6feA3O/vLd7Tznd7XuJ8SFJAjIMmrmnDob2GxNc2SzxcaV/Z0vusuyocjPkMBLMFrcHKaNwWOqZkbRfk/wCF/YB9yvbu42mOHmOxU+DeDWwpwBz5dKtt3NlcwualMdDDuTc2CfbuSpspPTRxvR1HljqBG4b9glgVe9/9b3De1bfv9zvlsdmtn1iVD4ikrwb1Hy8uhYZUnsjrFQcU45607uwetNt7i7m7Ny0EIkpU3NWshSyppLyMuhV4H49/R37Vb3ulryFZW07HV4Caqk8aCvHroZ92j245e3PlR90vYlMoU0qoOfzHRYkx1JHu6poqamaSlpqjxseWWMhrEc3HubdkSeZ45zUHHTHuPebVZ2V3tsKKKBh8IHl0rNxYTGKEdotKAi7WAA4495G7eT9FGG9OuN/N2j+slyUGNZ/w9CBsDZuKyVN50AbxgEEAf19rOHQf4k9GQwez8YlOF8ALHi5RL8Wtz7117zx1P/hOMw0zyVCBYDqYkqoAIFxf34jrfnXqb17VU9blq2aiiKU8TkMVHDDVcnj341p1utT0ItfDHkKh4ZGAhZraTyCL2JP9OPdqY61XPRUPknt7FY84uHHRpBJURRGR9EY5vbUdP9fdfl1U4NeoeN6d/iuzYUV0m+7x8ry+lW5MRNzf34Dy621APn1qpfNPrhdgdwZ9Bwa3IT6kACi8Y/UAvHv3W/Pon4XkXtY3Pv3VScY68QASPr9bf8R791sVI66It/T/AGHPv3Xga9de/db697917rwNvp7917r3v3Xuu7ED/A/77n37rWCeuvfut9e9+6912Po3+w9+615jrr37rfXYtfn6e/daNfLrx/w+nv3Wx14cG59+60evG34+nv3W/t669+691737r3Xdyf8AkQH+9e/dap10CRyPfut8eu+Sf9f/AGH+3t791rA68f6f09+63117917r3v3XuuyQbWFv6+/daAPXXv3W+ve/de65hh9GF7fT/fD37qpB8uuHv3Vuu7/7z7916nXXv3XuuWn6fTn37rVeuJ4Nvfut9e/330B9+69176+/de697917ru5Hv3XqdeAv/Tj+vv3WiadeubEfg2v/AMR791vrwF+eLD68+/daJ68bX4+nv3XhWmeuvfut9X+fAjdzZfEbfo6CZnfHpD51SRr8EXuo/NvfutDy6vmwhlyPhmqW0wRKjsZOeAg/r/re/HrfUXcO4KOsrafD44hhrWN1jsL82sQLC3vx60OFehHxuFVcbFBpC2UHQB/VbcD/AIj3vh1rj0CnZG1JKSmlq6dD5TY+iw5t+D/xX37qxyadEJ7C3Vn9tQS1U6zLGtyrFmCgAkc/1+nvXn1rolO6++MzNWOErWjsdJBmlH9r6/X+nv1et/b0o9obgzm9gY6eqaaYmws7MbEWN7+/A461TpRZ/YfalHCJaCnrDGdJV0Mg4b/Ae/eWetjPQNjMb+O46Lb9ctQtU9SsRDNLyxNrWPvw49eJoKnq6XoHrvL02z6afKRlpZERiJFDHlNX0b2V7hu9rty1lOfTobco+3+983TAWaExk8QMdCpJ1xiqpqgmnSOpe4D+JE5P1IAHsB7hzcZqxxjB6yx5V+75DYIl3dsBKtCOA6AU7jyXWu7mo8w70+JeYpBUSsyxBWNlNxYW/HskkVLqPWoyepNsbi62O9FjLXSMA9WifFzoPsb5MV1O2LppsBseKSMV29a1G/gsdLp1yzzTuNICqCTz7a2/Yp9yn0KNKD4ieHSnm/3a2fknby0beLdsO1FPcrcOjSd7/KzpL4DbQyWwPj9T0md7gnppKLcHZ1F45aGRxEYmWhK302bkke5f2qzgsbUWlrQ/0qD+XXPTnjmTeOat8bfN5ZgHJITU1aVxXPWmZ2XvXc/cfyF3f2BvrM1Waz2Tr6itmqJKyomjkkeQutopGK8X/A/w9gD3H0W9nEg4k5+fVLfervcIxZxfDSgHRqOg3y9DvrG5JqCSOhaqjMkhUqmgLYmw9wxuvK26bxy/cXEER0ohP209Ok1xZtY3ESO3e5GPt6Tvy4p6DL79r6mkK6aymEbKoAAa4/A9qfZm43GPbGsmB/T8s9ZhbXtWxHk6D99EByBStONPn1i+Ne2I9tVIqY1jkaqBuCikg/4Ej3K1/tVzuK1etejHlXnraeT2MNu66eFcDq4DbNTBQbd+7qGSILDdrqoNtF/x7e2zlS4ZwZvhHSHnX372xLdo7E1kI4g9FGyfauIzPZMOGoqmHy0taqtptqFntbj6+wN79bEsfK0JhTWwB4Dh0DvZv3D3Pmfm0xTEhQ44k8Pz6Oplof4pQYuNmMioIWP0NgVBP19hb7tnKlrttrJuvhaLiX4iR5dSj97DeR+5LW0jkBAA4dAPkkipd4TxRqLRsLACwuB/T/Ye8tmNTXrnjGKL+fT3msslNyx0sUHHH4Fr+9V9OnDk9BJuDsWPEwSeaUCIKwJMjCw+p/P+29+8+qEmpp0T/eXZ+7N7ZA7e2PNNNVPIUL07ynxj6c6T79jrYNf9npc7P+K/d2Wx0+Vzme/d8RmjglZtbBl1KoDfn+nvXXvOnVWvyPx+8Nrbzrtv56KqYUZfTKzyBXIlAsOf9t70fl1sD06UfSXRlfvKOPOZANTUbaWjaVRbU39S3Pv3n17PRx5PjctTjlbF16JVRINNmFyyj3sjqoz+XUHa/wASd7byzNLQ5nIj+Fw1CiQSH0vF9CvPvwB6t8+rTuvOm9idQ4OkpsPj4XyapEJKj7eAgyLGAxBtzz73w69x6WdVuapNQIYxG8gsvpQKbfj9P+9e25Zo7eMySmgHSuwsLrdbpbW0UszGmOpr5nwQNNkGEMLqNTOQFC/2hY8W9l9puMd5JRDjoV77yhe8uWoku1OpgMeYr1Gp12JlY5pITTSzkfusiQ6j/qgW9mlQeHQJNV4inRcux9iYyuMs2E/aliLyAggD63FgvvQ68fXpq6s3JX4qvjwmT12jl8YlYtpsPofemkjT4zTpRb2V3d4tkLnyp0b5Nk4HOKldOFZ2AcMAp/H0N/YS5i5ht7SExQnUxHU/e0XtBuu+7gt9uiFIlNcj/P0GG/cNjcLic0YAAkVHMyngABb88e43hvbmacNq4nrMy/5W2vbtsaOKMEImTQeQ+zquX43bbfM907rzkkZeiZKrxORqXUrH6X9nu8b1+7dvqW7uon9u/bU84c4tD4NYCTmmOj30GAb+NpLHGLGULwBYAnhvp7AA9wbhh9OCanHHrLU/dL2FH/esiiiDUBReI6VHcONxNF17mXrJ445XxVSVDhfqI/8AH/ePZ1tV7JPOsoPmCegFzdy9bbVYy27IFCqQMDyx6dMnwF21Qrs2ty08Y8U9VOqPoFm9Z4uf6+8KfvVczXtxvSbZbS1VTWgJ/wAHR17HbFZWdm+5ogEhOCQOrARTQzWp6ZBHEXIJKr6+eQfeJqSSw/qznU3+DqeL+9adDJKaheg07cqpMFgJoTTE0zwn1gDRf6Gyjj2PORLU3+4ifX3A8Oow3znLarNCmtQw+zouX8tHETdhfLntSmo/3RtfZeR3MI1Gsp/DYfMWUH6fT6j30p9oFkt57d5a6I42Zvy6wq+8F7kLvmxnbLRxqDacfM9F1358ad1fJr5U7r7hlx1RPj4K2tpVkkiMt/s5TEoLyA/0/r7jv3e92to2y5mV5gNTUXP8+gl7YpZzG2s7oig0inz6s26c2FXbfwMGIjofE9ACr6YUQqIxbkKOPp75++4vM9pfbibkyVEnDNeujnK1vbWGzxdwVaCnXLsGtiVXoRE0k7KyEDix1WI49ucm2kjEXJNF6Ucx7lBY2DXMxxQmvVe/c0NdtumlyjUzqWaQqQpUrb/H3lHyVcCadIkehFOoi2P3Pin3NrK1cFa0pXokOd33W5BZo5JG+rxkFn4/2F/eXfLpgS0jkY9+M9E/PUt/ubSCMdhWvRSO05WlpFkP1Mjeom/AJJ9zDsl4WQRscdc//c3ltoro30S588dGV+M/xMbfeLp+wJZIp8eZYkaPxxuQ9rm9x9PYlpTj1DgYkfZ0ane3x4wGNhMlPSQmRIQLLBD9Qv8Arf73711oevRNN9dXUNPK4EKrNEbg6FBH9Pp78Pn1bjxx0xYWtpMPQ1VDUzFWjiLD1H+yNNgL+9069joCMx2TmsNkah8dUu9OZGXQsjngHjgf7x71mnXsHoI92b8r9ySWqNYeNw5uT+Dz+r/W9668KHHRXe9sf9/gYc0IiJEkiUsQNWhXAJ1Hn6e99aPRRPfut9e9+691737r3XKwI4+ouT/re/daqa568FJF+P8AY+/deJA64+/db697917r3v3XuuwL8fn+v+Hv3WjjrxJP19+68ABw68Bf37rfXgR9D9P+J9+60fl1/9TQAUXNr2/3n37rRNB12LAm/P1Hv3XjUjHXRH5/qTx7914enXr8W/xv791v59eBtz7914ivXtRsR/X6+/daoOPXXv3W+ve/de67P44tx/t/8ffutDrr37rfXduL/wCNvfutedOuvfut9e9+6912SCeOPfutCvn11791vr3v3Xuve/de67UgXv8A0t9L+/daIr11791vr3v3Xujx/Bnc+bwnZccVC0q0pXXrUNpWVrgi4/JHvfWvPrYB69mq6+ds9kFaSVCAruPqR+be9D59aYBiKYp0O+3sXmt2VJkIl+1jksQ+oLp1fQD2kls7ZjrkXPR/Z8xb1DH9NayHScUr0tc1BjsJkaTCOsccMqK0zabXcj1A+1CNGBpj4DoquY7kuZroHUc56ETBzY7ExRx41IvG6lmYKDqJ/wAf9j7v0n49PNXlkkpnSRUaP8goOP6+91PXqevRXu2N4Q4TG1P2siLqDqUFhyVv+P8AX9+z1r5dFF6ywtVurcsmYqI2lYVRZXbn0hrm3+w9+69wx0breclTjtrxQ05KSLZdK/UW/It/vfv3XhUZ6m7FhwNdtqoq8y0Jqo47qZeWB03Nj795daxx6DBMTh8pnXqQqHEJUiKZiLpctb1X4/23tBuI1wmGtCehLyldJYbol8+Qpz0eDrzrjaGJoKTI4gxRvIiSM0LBfX/Xj+vuKt/5CtuYAV3SMSLTzHWWeze5nLkIiZgNYoPz6HWTJJSUixQSNfRZrsWLD631H3DV97O8pxzlTaLQfLrK/l3nJdz2pJo3C4FBwx0VjvGuqpMXRzCebxfxil1oJDawZT9P6eyO69t9k2yCW522BUkVTkDyp1Ld1v1lu/KU2zT0ZmQ0+2nWwn8NtwJlOicAlMrhIaGnhfk3LKhueffC370dg9l7labgijsTTrmDvFnJtu8X0A4F2p0n/kLvCsxMcOGpZnWsq5UREW+rRK2lrD/W9j72H5Xs76I7zKg8NBxPqB1H+7TOpEVa16GzrbB4brXreXdGXqIKFZqA5GrqKgqtnEJmIJe3J/Av7hL3Q3Xefc/3BTlPYyXa3mCeGpyyE0JoK4Hr0Itst4dr2xr+6GGGDTz6oi+TPzzz2/s3uPZGwqupgigyMlKuQgdvGYlYxkq6/wBf8D762+w/3Wtl5W2+y3beoFaTwwxDDNcHqEeaecpZHkgtmxXogMOGAnlyWRkkq8rVv5aiSSQyF5W5Jsf6n3mtClvYxC0skCRrigHUVzTTXDmSYknpwG363LDSlOQpP00j6Hjg+9fUJGcnqjPpwc9Qcl1MWiM8kDmRRe6NpYH+ot7cTchq0qetrcDyHDpCV+1c5SRmmhmqPtSNPi8jkW/A59q47mNzqahPTokU93A9JgdfT1Emp4JWZ+TyTb+vt/67T2r1vxlUaus1T1N5oGYwPrtcG7f0/wCNe/Dc2TAPW/qSCKcD1E23ujsDpXNUuSwVdVQUVPUI0kMMj8oGDMCP6cfT2T73ytsnNtjJbXcSs7g5I+XS233GS3kEkZIp1tR/y+/lxtTvfZlFhK3IQwbmx0cdM9PUOpnmYLZnCkgnn/D3w2+93933ePbvmVuadviP0Tks5ANEH7OsjOROYrbcLdbOZhrPDqxjO4KlrUaN4kaMhr3As3H1sPeFW2bncxXAu9tcqTmo889SDfWSEkEUp1Xp3n1Dg8tuzGRvBGhedJlayqwCNcXI951+zfuXuMXKc1xK5JRSp+3qPN024tdiNFqSR0crrLcmGx+38fthGVJKWgNEp1KAtk02sf8AePeHPPtveJzld8wXERkjupFfVxpThjqZdt5bvoOXRc6aBQcfb0WTcNbs/rjspKgZCnp3ys/nnZpAAXd7m/495R7KvMXuF7eGGSIkQrpUUzQDHUU3FpJaXzSBTVjXotXzt7SqdubSXI7YzggaqimbVBNYMpgFx6T+b+5j+6B7cRbrzAbLmK0qkOCWHmCaZ6lTlO1juLdmuFrwI6pM6WzTboxG9szkJBJVNVVM8s0hN5HGq5B99S+XeVHkvJLG0WkUZ0rT0HWbnth7n7VyLybdC4cJpBp9tPy6S3VOOwOXqt7TVEUctRDkGCObEj1Hge5oG2/uqOONhnrH8c6NzzuN7dpJVKt5+tesfYm3rber6ymGpIWIXSP0qoufcp7NP49kK8QB1gx7k7b+7eY304EjMemjoPPR1FPW080oD07adDk8c2ABPsyFeHQJNAOjf4evi1gOyoF454uAbi1/e69e8umXecozPjw9BLqnqHFyh/SCfoSPfs+fXsUx0r8RR0OzsNHBGVjqpYlE0jWJZ7Xa5+vvWetUBwOmCTdlprJKJDci4P5v+be99bp0E/bxTO4z+IynU1HEQt+DdRdeD791o06AXq7uXcFLuAYKsqnbHCOWGNG1EBLFSpv/AIe9deqeqQf5pOMSm7SocjGoC5Gepl1AW1aoQ4Pv3lTr1c9VY3va/wCP6fX37r1KcOuv9b37rfXYFyB791omgr12bfj8fU29+68Pn14m4At9P95Pv3XqUz1x9+6312BcgfS/v3WjgV699Lj/AH3Hv3XuOeuvfut9cmN7cW9+60OuPv3W+ve/de697917r3v3XuuySbf4C3v3WgKddggA8ckWH+Hv3XiCevElvx/tr+/deFB15eLk/Tj8X9+60euPv3Vuve/de65KAQR+fwf9b8e/dVNRnrxta1uR+bD/AGPv3XhWteuPv3VuuWq17fQ+/dV0149cffurdcrji31FuffutUPn15iSAf8AX9+68MY64+/db697917r3v3Xuve/de697917r1j7917r3v3Xuuwbc+/de49de/de697917ru/AH+v791qma9cjfSOf6f71791oceuHv3Vuve/de6EPrTrTcnaO5sftvbtHNPJV1MUE9SkZeKkWQ/rkt/h9B/xHv3Wq9bMHxB+NFF0HgYHyssU2TqIYhJ6g7oSAbsT+ffs9bqKU6P7kt70mPw0sSTqC0YAt9QbW/HveetCgz0C/WW8kq97VYqZvJHHMx1Elgp1Xub+9E9eFK16PVQbjx8sStHOhsq6lvyPTz791vhx641q0GfiMLBWFwDcAg/j/Ye9/b14d3DouXbPU2GzuMqKaWjje6aUKqADbn8e9Z611UV2r8e1w9ZkZ6ajZ0JdolW97hv6Ae9EVHW/PoYviR0/XxZM19XSMKZGDGOVT6gDeyqR72B1456tGrqTCSQLjYKGm1lUS7QLcHTY8n6+/DHHr1MdApkfjRh58/DvCanptdLKKogIovpOq1vdhXJHTL0qFY4PRl9o7/25RouGZoIzCqxFVCryPSDb3DnNkdzLeGpNB10Z9gbnZ7LllFQL4hGcZOOnnKZOklq4YsWVqa+ra1DRR8y1MgPCxKOb+43vdwNo5Rckcesz+VuTIOYLI3stI41yznAUep6O91H/LzxnaeGx/a3yIWn2XsbbanLVOLzUi0dRuSliUSrR0Bfl3extYfn2MeW4LgwNum5yfT23Gr4J+Sjzr1jl7z8xbHBdpyfyHB+9dykOjxIBXwTw1PTgB0WT5afzM5drCX43fFbB1fVXU+EU46uEKqkuaRf8nnklrlAY6gT+fcr7Nue27hB9PY0EY4ep+ZPWBnuRyPzryfuIv8AmTXJdPkk1oATUAA+nVWG9d5UuXxNbJWVklRPUIzu00zTOWkUs/qfn6m/s/hiWEjRw6ia9vZ75i1yakdV8dL4Ebp72yFDOvloFqnIB51DVcgH2DObNoG7TxCTKhhjoScty21vG8zDuCkivrTq7PFdcY3HRU0FFRJB+ygB8YBuU+p9jCK1gh2ttthUBHWhx8ug1cX093uaX1walGqPTjjHVafyg2vlMJuySZo5VglJ0TEkpbX9FP8AxHuA+XYbnl/m+4s2jpE7YxjrILftzudz5NgkhYjw1rj7OpvQEmQTJUwqdbwswVGtwLWsPc/eBDQFR5DrH47ruMoIeQ8fXq0bds8FN1hlJIZVFUmNBiKNZtXjP0/x9u1oKDHSLQWbW5JPzPVJXVOfyMXeuZqcpNK4NefCWdrA+WwIB9lG7bRbb1GLe8XWo8jnoacnc1ScpXL3tthzwI6vL2luP7/GUMcjkSCKD1kfX0j/AB972zY9u2UaLBAg+XVeaeeN75wYfvOUui5APQRbjqxQ7yq5pCBGzCzfQHnj2a+fQQ4CnSF31uiNIpJYpgTDHqJBFrD8e/Vr17PVefcPc6ZGWLauImZ8pkZWpVSNvWju+hGOn3o0pnrS1LhVz1YT8PujIdnbcpdzbhpVqc7XxI5lnXyMvlGoEh/6/T3VKnPSm4VI20Dj0fCWulhVWYxr4wunQgVSFBsNI+vuw6TkdEW786f6837kZMznMbSyV7ai8hRULeq5+nvfy68T0WDJpitp0tNt/b6xwUkEtgsSgMApsLke/fLrxqR0LmyA1RHT3PqOm/1ve3PA9669w49Gm29opY0s0akBSxICm1rD1e9463kig6yZrdIpZUhSTzySnx2X1WP0B968+vHh0mtw7rx2xsbLuHLToCULiNyNV7X/AAfZTutu11EYydI6H/Ie8wbBei8aLxX+ytOiC9o/OnFViVmGoI21KDEksdl5BsTx7DMFpPZPSJsdTjunMG280wCS/hGoj04dAls/5Y1WNqJdddMi1DEaXc8X/P1/HsTWN4aaZcdQlzJy0GY3FgMeg6NRsn5L4CoRXy2Qic1KqqCQqxBf6gc+zZGV8qeo7nintyVlXSR69DLit8bNr5vv4poGlYeRNJAIP4PHsJcyyzKumHHWQnsvbbXLOHv1DU8qdKpe+6DFv9n9+saCwAD2HHHHuO5bOWQ+JNVj1l9a8x7fZoLKwAQegGeg/wBx9xNvGWo2zjVeomr4mppHjBbSsvpBP+39ll+n0SC6D005p0OeWJ25hnbZfDLmUUr9vQw9P9VU2wsR9zNFGMhVB3lkUDUVn9Xq/P8Ar+443re7ncpCWY6PTrLz269uNo5NsESKNTO2S1M58q9C/SRx04knWMFowWNxb9PPsOQ6g/ienn1Kd2FeEw+RHRJe+uyKrdecodh0LyGepqVpKiJCb+OVtPqA/HI9juy3GTadrl3C4+AqdP5dYje7Q2i4m/c6KPEOP2nq1b41dF5LafWWPooKZo4EhhrJZgp0v5Y9bDVx75o+7XuJt9/zXJLLICSWUCua16IuW57Pl20j2JGAlJqR556MhTbGnyWIqp8aR9xRxvLoX/OM0f1AP+w9xfFzDHHuUcd38Mhp8qHpLzxzknLMLpPhWHH7R0Sjtncm5Nz0tfsGj2/WVOYjimhiaKFzIH+i/QX/AMfeWPIvL+07Wse/XM6pC1CanFOueHuTz5uNzurvtsp0nyB6sE/kr/Bnc/Ve++w+8N+0Jpot0bQyO3qiCsV1mlpclTskihXF7WI595C8n+82wXD7xYbUweO0sZQJFoV1kE0BHmOoeuk3O+ZJ78k63Bz0duDrLqPrWLOYrD4ugofuK3ITsXWNn8k85ZmZmAPvh/zLzv7p8/79JcSSu8MU8irQkVVZGA/l1KGwz22x3yTFvhIJ6SWK2DtKDF5PKUUdNUTVyzRp4wh0s4IFvaLdd65wm3m3269DRpEVYk1zTrINveBpoIbaCXSsVP5dFa3J8ftckuWlClRNJNz+Rr1WsfeRvLXuUiSpt4NDQD+XRvzJ7uxb5sj2EZJbQRj16rt+U+3sbLt6ppI4o1lpzKpAUagUNjyPeXvt9fSGVJgcMB1jp7e3u7W/MTySuSC5OT5V6pM3XipsdW1PDBDUSfk8i/vMblbc0uEjtiaHHWbUG4LfbSSwqdHQCdkxJJg5ja7rFJptydRQi/vIDYI5GoT1hj7q3ltGrxggHOOje/Brv2bD7TfYlWSFFQsgYrfkLYgexzXA6xZwGJHCp6PfmM3HlaaSQKWaRSwP1uD+kf4D/D36nXq16IZ3lWSUUdVPTELKiMzKB6tI/qB71x62Oq087vjIVeTmRahoj600aiNRDf7D37rf2dLDZO2pNyRSJUJ+641BmBP6he//ABv34cK9e6Su9usMrh5payOFniuwBUG2m/F7/wC9+9deBHD16B/cm3Dndu1ePqogfBDLIQwuL6SQCPfuvH16rrq4TTVVTTngwzyxEf00OV/4j3vrQ4dYLcX/AMbe/dermnXXv3W+uwbf7a3v3XiK9e5t/hf/AHn37rWK9dobH/Ye/dabI68Bdj/t/wCv59+68TQddH6n37rY4dde/db697917r3v3XuvfT6+/de6/9X5/wDY/wBPfuvde9+6912CB9Rf37rRFeurX+nv3W+ve/de697917r3+Hv3Xuve/de675+vPH59+61jh14c/U2/1/fuvcOHXXv3W+ve/de697917rsC/wCbe/daPXXv3W+ve/de697917r3v3Xuve/de6yRozuiIuppHWNRxy7Gygf7E+/daPVzfwu6NOLxFDkcjSlMvUuKpXPB0yeoBb/i1vfuveX29XaddbLjlooYWUxxIE18W1C1yQP9b37r3l0OE2WxWzsaYaaNI7+jWQCdVuDcfT2jv9fg0j4noS8qLbHcdVzlQOkjXYePccQylZUqsjkPGb/RTyOPfrKF4o9T5J6rzLuMV7eGK1FFU06d8ckeMgWOSUSKPSvPIA/P+w9rTx6DnAU6xZPctHBHIjMulUb/AFz6fz71XreTkdEk7Ikrtx1klNS6zT+U6tNyCPoefe+q/M+XQ0dH7RgosVqjjvKpZmHDHUOebe/cOHW/t6UfYUyQulHKtnZSukg8XPPv3Xj6evSQrNs18O3Jp6GdkWRC7KGIuNOqxt/sR711ulB0gYMvRY3qzMwKQ2XGTQLIpGsEvzb2T7vVAJa8OhdyZYDcr4wMpKEHPoehq6s7PqsdT4vE5GqEbVEUToJS17HgWv7U2NzFeQaTSoFOm+aNkv8AlrcQwJ0E6h6Do3dHlPu4BM8qlWQMGvwePoP979hnmPbbeCAyqMnqcvZznnd9z3GPbpH/AE1AH8+i792ZqOPF08ETeUnJ0xZVubLrFyfcUbpoTbZpJDpXSePrTh1mva3bxzaUbih6vp+EfZ2y8b1DhcAchS/xGWjjmki1KJFcLytr++DH3o/bnmnmHnd97s4WNvG9NQGACesUOaoJxuV1dSqQpJz1M3bRrv8A7ywbRyK+Mpollm0H0WhOpuD9Pp7EvLDych+z1yrqROaha8anA6heYLd7mpBwvRFf5onzAy2G/h/RfW1etO9VF4K2emvx4o9DRsyH/X9z59x37vNpI8nujzpba7uY1BbyUsSoz6A9Bj3C5rkES7XYtpUcfmadVObJ27/CccKyp1y5OtAmrppDqaSeTlmBPPN/fTm7lQt4EQAjTCgeQHUFu7MdUhqehZ2/tx8jVRzTKWXVccfUE3/2Psqnn0CiYPTTvRft6MhiNqUdNTRy+AfpudVrjjg29kUs7FtBPSVpCxr1lrMVROjRmJbkGw0i4vcA+6KzA1PVAzA0Jr0m4OuaeveQtDw1yBYXJ/H49qfrSny62ZSBQ56yv1hSUj3EBYAH8Cx/w9+O4Owop60J1rw6g1eyaZDo8BAIIvpHB+vP+t72t5516c8Q0rx6BnfXX9NPTTIYNRKkfQXbjkW9mtnekOGQ0PTsU1Pi6LJsbtDePxg7PxO8dr109Dj6TIU8lVSlnKy6pRrug+otf3XnfkbYPc/la45e36ETNMjLWgwCOhBtG7XG23SXNuSACOtxL40/I7Ed/dZ4XdFHJEaz+HUz5JI2vpqJE9Wofjn387Hvr7L3/srzpPtaq0ltPIwgB4IozT7KdZTbDzNFv23LISAyDu+fRQPm923U9dZ/DZSlnVIYVQSjkqS4vdre8jvuucj/ANY+TrizuIy0spalPt6GPItjY71zxDb3RBixWvRf8L8qHhxlLmqWqHnnpvIVW/6iPrb3Im4fd6vNwvpLOW1d0Vv4fQ9Z2ycr8uHavoU0gUpxHQE9kdh7q7Gpq3cEVRI1ZTF2pmjLgrp9SgL7lHlXYNs5NePZNIRDQMD/AD6jy99m9qurSS7iSrDI6IJ2B2x2bvuOfa255qmGOkLQ05nDgMir49K3/FveVfLNjsGy7cTyraVmloXdBU1/LqJn2u35c8RLz9JVrSuOpfUGGfEbT3BSlyrzJU6v9qJU/n/X95ee22zqu0i8vEpKwqa8a9Y0+5XO24PMdq2yb9FjmnQLdQZCsxu+9xYOSYiPJZEsCSbWLnm/09jDctmW+cOuNPRPyX7hvyxbvFL3F+jU9i4Y0G06yOP9xWVjIRz6tFxe3+v7M7C1FpD4Y49A7mrfH5i3L6xuA4dFO6Li0ZHNueCKmQL/AK3kPBH49rDj8+g8ano0GTytfj4hLDG8lkOnTybgf4fX8X9+4darToKKDsfL7b3ImRytJKaYMziSVW0KL+9deGePSzoe05uwMhPS42YGISEERm6x/wBm3H9Pe69eGelDlVGBphU1Et9C3lN73P8Axv3rHW+HReez+38cuDqKGkdDUspXSAC3q+n59+61WvRc9rvna2U5yip5QIEckIpu17m5uPp731vHVUH8wnc+Xz+8cJHlKWSmNL5tPkUqZCIgoazf4e9Dh1rFcdV0e/db65WsAb/X37rVamnXHn6/7z791vru39De/wBQPfutV67W17k2sffuvHhjrs2NzcC1/wDXPv3WhUY64e/dW697917rsc/4e/de699Rcn6fg+/da4HrskcWFiBz/j7914A+fXH37rfXJSB9Rf37rRBPDrofX6X/ADb37rx4dd8G4PFz/tvfuvddEW/N/fuvA166uf6+/db697917r3v3Xuu/wADn/Yf09+6914i35B9+60D11791vru4Nha39T/AF9+61kVPXj/AK9/fut9eH1va4/Pv3Wj13cHiwH+Pv3WqEZ66tza4/1/x791uuK9df7z791vrsi1uQb/AO8e/da669+631737r3XrHk/0+vv3XuvXP8AX37r3XMWtbi9jyffuqmteuHv3Vuve/de697917r3v3Xuve/de697917rsgj6j37rVQer3f5Zezdp0O3q3cdVj0qMoyGojllCuVm8Z0uNXIt+Pfq5p16uK9HG3z3Fk9v5ipMsZSBiQnpYKFU24t79SuevGgx0gKvvimytJJH96gmsPSCQbjk+/eXXqZ6x9Mdo01RufI0j1CrUO7BWvc8/4n8+/efXujz4Lc2WhICyvKkgXQRexva30/1x73Xr2a9GQ2RXV04U1RIPJN7g2AvYn37gOvUx0vdzUkc1AJAVuVBAH4Nubj37rfl0TvddFh5Mk8OWSKZXkYcqLkFrfn37quQOjEdW7UwFBiDWUtPHCkygppCg3Zb2Fvx7159WBHWHdGDqIHNbjCQ4IJTk3uTwB73xPVSaCvSfx+6MpMs2NyMMix6TFrKkaR9NV/ZVfbrHY4boectciX3MiB4QafLorG5Nm9o5nsOmxvWeKye5q7JVUcC0mIjlqJopJJQEDJEDzyfYK3O5g3I6YhVj6dZLclbJuPJUaz7k/hwJkljig62A/jd0F078OtgY7ur5jbjxu5N818Am271bW1Ma53B1Txa0eso2OpRe3BHsiTlfa9sl/e/MQDUzHEpyx8i3y6lTcfe3nb3Dhi5E9rUNnD8M91Q6HXhg8Cei2fJD5z70+TeXbCUuQiwXVWNZItvbdoFkop6WOnkKxI5UgFSthyOfcY828x7ruVydR0wjCRj4FXyoB59ZuewHs/yb7e7YNydPqN9mBM8zkMrFgDivnWvRI969ZYHc9DLXJFerWNn8l1DtIbkEt9Tc/X2YcibleWt2kSt2s2R0D/vQ8nbHv3L0+4Twr48UZoQAOiK712PlqMVEUcrqgdx9bHSLgf7x7yiiNYlf1A64c7nEINxlgp8JP+HoEfjZjkx3d1YJ3Hl+4exP1ve3P+v7SXMZMgZRWnVrSbSjK2MdXmxR+VYSCqssUP45/R7XipyOkD01fnjoN+4+mML2dtqWP7YfxaCCQwSJpDmQ83H5/p7iDm/eLbZ7ptwnAVY+4k/LrOb2j5At+ceTWtJgTKy0X7SOiC7K2/Vdebjk2/nIXp3pZnWOWYW8nqsqi/8Ah/T2J+S+dts5vtRJtzq5A8j1jl7je1+6ch38iXqkJU0rw+3oye7t3QLtjIUhqFMT0QWNQeSSp/3j2NyKGh6igHVkdVXKIsTvmqy8aGN3qWcP9ASJLj6e9fZ1shSKHqyDp3suTLJTUc0i60EQR7GxNgPp7914CmOHS97pqoMNQ0WUc+OSoaMtK39oHge/daINT1VN3f8AIGuxVZVYrGS+VpE8capqYli2mwt/X3tc46q3aKDJPAevT58Sehsj2DvRewd6rUfZrNFU0sU7aUuP3fQG/H9fYE5n5rg2yQW0J1N1lT7I+wW6c7W0m97gpjt6dpPrT59Xr0GRx1BR0tNRBIYYYooVAsAAg0KbD2b7DvH7yiyKY6AHur7dDk2+YI+oVI6xZPLqYi3lUi1wR+Ra1j7Ehxw6hcGo6KT2tmdIqDGTcIxQKzWJvwB/sfeq462dPRbdubRyW5ci1RNHIkRYsrMLKefwT799vVaV4dGl2/tagwFHHPUzp5EVToLAN/QD3vrYGOpFTna2sqlpcPDJc2VpEBIt9PfuvVpjpK9hbgXr7BPmsvMpqhG0kYf/AFQF7W/w964ZPVWBICpx6qF7n+SO9d711VjYslbGGR40jGtdKX0r9D7Jr2YO2keXUncsbeI4BNKKEjorEq+p5JHaSeQlmckkXJvfn2XOxRKjoZQRxyShDgdcY6eWp0pEkhmBsNBIJb8cD2WPcupqxx0NLPaIJAFiXXXoUNobN3vkqyDxU9cKeNkcEF7W/qfaWTmRbNDRxjo5tvZ7+sdwoS3YV8/t6Pn1/wBLdm5mOB6DI1FLHYeklzbj8/717CG48924xPk9T9yj91vdliW42/8ATqMknox+A+Km7KmaOrzucMoDAsjORx/hf2HLrnuFoisKUPU1cvfdluYZhc7rNqz5Hoz+x+mtrbQq1r/t/PkQkYMpYOupRp1Af649ga/365ve1z2V4dZIcr+2mw8tET2kf6oAyfs6G9iZAFuo4Cj/AAsLDj2UO9RReh8oCdJrc+WhwWFrKmV1GiB2drDhQOWH+t7egi8WRYF4t0l3C8jtbZ7iU0Cg9Em6K2ZN2V3zmdzzU8k2Fx8fnjm5EGqIa73P549kvu1zMmwcpptYIWVzpp55PWGe6qd+52mvXBaOPIPlg9bCfU/emxV2VJtBoac19NTinLqFDftLo1X/ANh75W+4vtVzLLzDHzDFKwiLFtPkamvQW/cW47nzd+9rWSiDFPkPl0kMR2JPt/cuilXy4+rqSHjAuvjd/ofx+fYnbldL7aA83bNGtQfmOhrz3ybbc0bIYZ/7VV4/MDqwvrLprrjMtD2KmFiOXqY0llklWNlDAazZLe8Sfcf3w9wNs+p5EtrgCOGNmBGqtB5V9eue+/8AIsWx708V0CQD59WQUs8G1ep2ydLFDTwVERgHjRUDME06QFt7zl9uY5OQ/umW/Pk7MJNwBjZ2JJdnBqK8SanqOtymS53w2MYGmMeXlTrXH+dXyFruuBmKuknaGYvUODdgL3utrH2a/d09uLHfrKK5uEDB+7/eiSf8PQf3m8eOQqP29Av8Lvk9vTtDBQq8ktTE2ReI8ObAH6n2Pfev2r5X2iR7zQqME446SbdeXBXTqJ6sU7a3JunFbKSWjppXkkQ+R1jb0ggf0/wv7w95Q2nYrnmllZxRTjPWRPtNs22bxdFN0emqgAJ9eqKu+t65mevqo60ssbyTBo3BHOrm/voPyPttsLNDFmgHn1Nl9yBtWxXH1NuozwI6rK7RzdOktRGGUMAXIA5JI5595D8pQSC4janmOj7a7kRQyQnhpI/l0UHeee+6oTArA3Vhxci3I595d8u6TZhjx6wM945Lj+sLxIx05x+fT/0LXvisvG6D9zyKSR9LEXFwPZ+M9RCUKgBhSvVre3stVVmLhmZ9SmnQ2+gvb83/ANh7seqr0T75BSVcArKu5kheNlK24tb6i3vVevDqvPb21Zd27qUU6syicAqpH6i1zf8A2B9+6t5dWS7O6jgxGPpZvEUk8MQe5AsNPPPv2fLrXDoO+6KrHYahkpZFiYBCmnSCQR+Sfr79WnXqdEVnnppqnJgFEp3ppPTcc6lN+f8Ab+9cerdVm74p6am3Lk0pLeNqmZzY3GtpSWP+3976r0kzY8gAfi3/ABPv3XhUY64+/db67Nj/AIcf7c+/dayOvX4t/jf/AA9+69TNeuvfut9d2/ob/wCHv3Wq9de/db67Bt+L+/daIr11791vrkLj6gkfj8cn6Ee/daPXRv8Am9/8ffuvCnl1/9bQAJJ+vv3WqAdde/db697917rkCFPPPH49+6qcjHXH37q3XYtzf+h/2/v3Wj11791vr3v3Xuuwfx+CeR791qnn1yKji1l4+jGx/wBf37rQJ889cL8W/wBj791br3v3XuuV1t9ObfX/AB/2/v3Ws9cffut9e9+691737r3XZFvyD/re/daGeuvfut9e9+690qNlUkdfu3blHNbxT5ihRri45nB5H+w9+691ta9Y7ChwUWxXRB4ajA0MhVUQajJEDcge99VFTjo4DFsTT3pRpHi1kDgiwuR6feqdWqeHQcRvW76yH8MhlIjjl1P6tRBDWtb/AGHvRXV8XTiXDQnVDg9DI1LjcNio6KutLNToEve2oqLAce7cMDqhJYlm4npI1Mj1l/so3tyE/P8AiL+/daqKHpL1G1Mxk9S6JQCxubG3+Nwfeqjj16p6TeV2cmIx808o/cFxyq3vb+t/futVxnPSy6gMiQTpGDyz8W4Nh/U+/deFafZ0nu4i1PUR1bRFWRrljwLX/P8AxT36lOveXWbFZimr9m1DpNG7R0zoyAi9yp5Nv+I92HHPVJCaADosWOjoqrAZqBpVaqbMemLV6razb0n2EN+ml+qCj4KdZFe11nto5ekkqDcasevQp7j2pPHtWg3DShoq2ip0Mbi6gLGLken/AFvZXtl61vd6QcE9DvnflmHeuXTOw/URT5Dy6dti9u5LIUH2MkpSSlCwu2pySRxfn2JN/QTWQY+eeoT9pZ3sOZTEPwMB/PqXuKunzpSJdTsWVibkm44HB947c93ccOzvAcVbrpFypsFxukP1Y+HT0LnWXbtb1XpmqquRpqeNvCgmlUgaeBpU/wCH9PcLDki2552OfbIIxqdhU0B6hb3M2mW3uXsitAfPoaOvPnDWRVmZ3BNLokgjqqaIPLNcF0Kr9Wv7j7m37v1rFFDsV2gYMVY4HkesQ+c0PL0wkjOSD0TSbI5Ts/fGY3rn5HqpmyFRNRly0gEcr3H+cv8A195O8t7Rb8qcvw7NaABNCjFBSg+XWPu6Xkl7c+Oxz0KePoJJZUhC+niyi3H+AI9vySBRXoqZwfLowez8EEji1LYKL2Ngf9bn2TXU1cg9JZZB5DPQstTuYNCJYgafx9CL3APstD6WqfPpjVpGesNDgaipmDSG+kj6W4/H0P8AX36SVRgY6trFKjoRaPArApCj1WFzYW/p9f8AY+0bzaumC2a+XWOsxQjUgC7Mb/63+PF/dlkxjh1bWCekdl8aY42fRYAEH6C9/b8TA+XV0bPQU5rHpMpXQGLAm55/H9T7MY20kHp6KlT0SXvvryXK42V6ZD5k9QZVUcp/rexjsW4LbzVY4I6X28v4T0bb4B/I+q6e2Fn9vV0j/ca41hV5Zr2ib6AE/wCH494l/eF9lo/cffba+ePUsZJrpB49DvaOY/3NbkKfi6Snyz+S03bW6MXt+JiyTOolKyO3qA086ibexh7B+zUPI8ouDHSNPKgFa9ZE+xs1zvN7+98jSTn5V6atq+SDE0dM0jN4YlS2o/QcfT3k1Nt+3xTSSxRir18h59Zs21zMyVZqj8+jA7RysFDSmCVEZZByG/Nx9COR7xI519md53feZL2xbSrsWHy8+h5y3zRqjaCZTRcdAd3jjMfP/uWpI0jMTEssaKtyV/OkD3NPsXy1f8oO9rvf6msihOaftr1D/vfscW/7PJcWPYy1PoeHy6DzYFVFJhK9pHVSEkHj1Ek3Ui/vMq2SJYqwigbrmnuEc1teSW1wSShp0XTHEYbflTXkWP3V1N/oNX9fb/AdIDTj0bHdOc+52DUzOrSeWEuD/rx/Qn/evfuPVz0VfpCRZcrmhYjVUOwW9revm49+NfPrR+XRsJaiIRqs8QZF+o+t+b/ke/efXvs6U4w2yN2YOpoq+niiqfGwjJEYYsVsCSQP959+63UcB0UcbQyHVGXydVjY5J6OqnZkZRdUXVqAut/eutHjXpJbo3luXchNHSiREmFnAL2Btb/W9+8+vDh0GdL1pX5mvjhqVleSSQBmJBA5/oT/AMR7917yp0errXqzH4rDJRvAjvJDpfUkRsbfX377evYp0QX+Yz8QqLemx63eGGoiMjh0DxNHFEHV0huWBUX5/P8AxPv3Drw4dawuSx9Via6px1dH4aujlMM8d76XAuRf/Y+/db6h3J49+61TrwP4P0J59+68fXrs8G4I/P059+69xGeuPv3W+uz+BYg/m/v3Wh14D82Nvz791vrr37r3XYtfn6e/daPDHXiObD/Yfn37rw4Z669+6313Y2v9P9f8/wCt791qvXhb8/T37rf2devY3HHv3WqVGeuvfut9di3NwTx+Px/r+/daPXXv3W+ve/de67Fub/7D37rWeuvfut9e9+691737r3Xvfuvde9+69139Ljj/AGHP+8+/da4568LWPB/1/fuvefXXv3W+ve/de697917rsC9+QLf1/P8Are/da68Bf8gf6/v3Wz176XHv3WuOeuvfut9d8WHHPN/8ffutZ68Lfn6e/db+zrr37r3Xvfuvdd24vcf7fn37rVevcWP9eLe/dez14f1uBb/H/evfuvH067LEgA/7H37rwFD1ah/Lw7njwGak2dlKlAtVPHDSo7srPFMLAAE8kG449+69QcOr86bonCb9pxPX0zPHNEjI4WMj1Lfgtz799nXvLPQQZX4SY6ly01RSq0cEjXThDwT9CL+/deA9emqf4qUmyzLuDH3NVGPK2kILhf8Agp/2/vdOvV9OjBdU0X3cFPFVgCSMj0lQSbGw4PvXXhxr0azDY/xyhRGUWwJ+gNj/AIj37rwz0ocvAZYPtomtdQDcqbi1uL/Q+95630VfsPYNe2Qo6yN28HlBcWH+rv8A7171nrxp5dC9gJnx+DpaWI2EdrpexFh9T+ffh1o8KjpTUeRWVWWpACr9Xb9K/wBTcj6fX2WbpPLbxeJF5dDnkja7Leb4WdzQFsZ6GjqX447u+QWag2/s7EVYgrJVhqdxRQiSgxiXu1TVSoCFVRySfcZ3Mu4bteeEinj+XWcGyxco+3HLhvLuRHlC1EeNTY4AevVhO4cr0H/LP2UtFhsfjOyPkBWwmSPdNKlNWUWErlhKwyMiqxLK3Ps6DWWyReFGRLdN+KuE+wevUXRWnNXuvuJvLpWs9nQ/2RBDSL6fYR1rd/JTsXtH5H9iZLfm9Nx1FXuOorZKpvHUVEcFi+qNVpgQg0iwsFt7gbmjmy/t9zeC6JK14nz6yJ2WbauWtvj2/lm3EKIADgV+eePQKUGd33taugkyC1NRTo/qkUSBAiiylreyqLfNvv08N6V6lHlP3IvrC/SOcHSTTiejdbD7bwuTo1iyFbTwygLGyO/NyvJN/cl8i7NFdXasrVoa9Br7yHuvDZ8utHD3GaM1oekvv6q2/UyyTx1UBh5Z2AXTY3JJJ/HvIhY2hjEZ8h1yIu7r627e8PFycfn1Wz1rm1qPkzNR4uQNB/EmUFCQrAtb8f63tJuF9Btdsbu4ICjJJ9OnrCwudyuBb2q6mPkOr9oaOWJaUODzTU1zxa5jB9kOzc77BvrMlhMrFcEA1pToR3PInMFqy+NCwrnh05007RZKkXkIHUNzcEXtb/W/w946/eB3ZU5euYoOLIw/l11K+6tyTeRcvC9vQQEAIBHy6Df5I9HpvLFUm4tvlafJ0rion8KRjyqo1EXUXvx7w09ivd6+5K3o2l0xMRNMk5Fejn3+9sE542dmt1AljqeAqeqZ+0d97vwFZXYCooasGk1I0h8gFlYqeAR/T31V2XnjYN+22LcY5UXxAO2ueHXJHeeTd72Hc5dumiY+GTmnz6K4++qmsrUSoiKuJPWSSP7X1JP+29iWKWKVQ8TVB6IJbW7gYiSMjo+Hx43TianJUiyVEbSIYzoD8j+twPbunzHScsAe/HRnPlbX0Gc2FAcdXw0s9HBE7Jq5YopFh/yL34inVgynqjnC4uTc3Y1MMnJrgo8gPLI7aklQN9LN7L90uGtLRmXiR0K+Rdoj33mGKOb+zjcEnyp1cXsbfG3tt4WixdDLTxrDFEt0sttKccp7ga52i8vr83klTU9dWdo5+5Z5c5Tj5fsWVEVVJOBUgZyOhkwu/mzkwpaEtMBa7xuTf8/Qe5T5XtJLdBqFOsEvfDf7Pd7gm2kDDV5dCNOMpJT6Asmtl9P6uDb6exsadY1DzA4dBPmdqZbITn7ukmkiUn6ISLHn8+9AdepXrAMVlqCFIMdRSxhbXIS7cD82H+8+/VHHrwBJAOOpdHt3ceWqESt8sMN7F3BA96GetuNLUB6Eb7jB7EoZamoaOSWGIyuzaR9Ba9/r/vPu3DrXVWPzE72x25qKPGYeeN5FlljkWGRrlfIQOB/xT2zM9Iz0v2m3+pvlU+R6rT9SgtISWZmc6mufUb/2v949h8jW1R1MkaJFEFT06X+y+vMvvWp8NBHJYXsVUMLjkfW/tJeXcVtEdfQg2HZ7ndLxYoRWp6OP1N8YcjLlIGydNJKqSIxBRCOBzcW9xbvvMioCkZp1mj7X+z8926S3cZpUeXy6sv2t0tt7B0sN6aNHaJFKtDEDcLyAQOPcZXO7T3EhYtj7T1mnsvt9tu3qsaRgYHkvQ97ewNDhaVFooEUfQEIvP9L2FvZNJO80mpupGtLGGwg+mQAU+zpVFmlGhrafoQBb/efdT8+nQoTK9RpEiiYHVbm45/2/uhCnHTyu7LXpN7n3VQbZx8lfUsPGilma5AsB7UQQNM4jj49Fu47hb7bbNdXLAKvVc/bHyJr9+5+LYuzonq58jU/YVD0zO/hSRtLM4W/Av7Gtvs8Wz2TbrfNpCDUtfl1ixz/7unc5TsewoZC509ta9Wy/GHqSl666uelrJI5c5kqQyTSFEMo+4g4Ut9eNX9ffPj3f54uuZucIzGT4ELfOhIP+x09y9sl3YbWzXSESSjJIzkft8+gNSszfX2/KyepWZsbNUTAOWkWP1P8Akg+5QeGx5l5ejRKeIFGMV4dRlDe33L++OXzGSc+XHoeKTtmgElPIksbldLatRuGvz9fcdz8kTFGQggGv7OhsvNEEyHgdQz1Yn8c+/c9WR02GUtU0cziKJo2kYWK8AD/invGP3K9lNnuZXvwlJiKHAqc8Pz6xJ96tw2xX1xUWUjq2XuXe8m2/jRtRZQ0VXU5HyyaiVJjYXC6uPpf3lRvvLYT7oOw8i3sdJYrsPQjyyR1h0lwX3ue7U/hp1qhfzI90HeUtDhsWfLXV+RFK4jJJtKQCNPuQPu77Idh2wtNhESo/LpBukvivWvHqxX+V/wDF2p2b1hjsrmqY+SqmapUvGgJWRAVFz/t/eIv3zPeuBN4bYNsceLppxOM/LoR8r7PLKPEIqvVq+9dl0Mm3KyCpgj8S076fQh5PHBP+x94Fct80blZ7/DO0tWlYYBPUmbVFcWN7G9qSpBHy4HrVe+cS0+2NwZIwIERKysIH+Afj6f099oPZKSTdNmhdzUlF/wAHWV7zTXWyQtOamg6oq7W3wZsvVmOSykOugu31/JsPea/KOzeDAs0i/Z0D77c/3ezRE+Veg42tjcju6RoYFZtB5IGoG/1tf3J0G4S7cnhk46ifduT7Tm66a8oC3Q8bF2bV4TNxsUYoxRWGni9vrx7F207gLoUJz1j/AM98qvscmtVwKjqxjZULrjkhdhoFOjWudQNvx7PCOozUkivQEd20sU2Orqeb/jlLb6Hgrx9feqV63gHomvQ9LTQdhS0a6beUsARfm97D3ry63XNOrOciftsajL6QsKngD6AXPHu2eq0B6q9+SefmOdqEUs0QJugvY8Wvb3qterDjU9FggWWov9dFSqxfi4Lccf7f3r5nrfRI+4sCuA3fLTg3WeH7i9rcvI1/p731X1p0E/v3W+uXFxYW/r791rPXTWvx/T37rwr59de/db69Yn6c+/de65XHHHIPJ/r791qh66uCbn6f8a9+69Q0679I+vP+t7917J4dcffut9d3P+2/4j37rVB14m/19+63w6//1/n/APv3Xuu9Jtf/AJH9be/daqK0669+631yKkDn37rQIPXH37rfXvfuvddi544/r791o0GevFbcfXi/Hv3Xga9esRb/AG/v3XuPXjc8n37r2Ouv8ffut9e9+6913xb6G/v3Ws1669+6312Rb37rQNevC35+nv3W/s6703BP4/A/Pv3Wq5p10STyffutgU4dd6eL/wC8fn37rVc06c8HXNjMxisijaWosjSVN/6CGdXP+8X9+68etsr4udrbZ7TxGzhT1kEs+Pw9HROgkDFpI4wOP969761SvDqwTdGEoaDb7VU2lGmieONrgC7rpWw9+63ToNuu9lVe3Y6zJVD3NU5mhJHqKSEspB/2PvX29b+zpVVO26jN1GqQsf3LkC/A/offuGevVqcdLjG7QpqaFBHGutQurULjVe3v2OtdKJMTHFHpWNFCry2gC5HLf8a97Pr1v7ei7drTUscbwoBqYkWFgCSv9F96889aPDHUfpqmWKCSRxcs5IB44/2P9fe+veVT1H+Q1NBHtSWqVLSanVSF5+n1uPeuvH1HReuvcJlshsrK5KgMk4hjcuiEtYaTyR78Pl16lc9Fo2vLWDeFUJxIlPHkSs0bs2kv5OdQ9lu724ntS4GR0O/b7eTtu8pak9jHI6sGys9JkdpUuHjkjjmnpQoSw9QIH0HsDbdbGW/BPAHrKDnXdY7DlMvAQHdSQPPI6BXEbD/gFRPIzi8z+Tj/AGk34H49i3e+2zAXhTrHr2sD3HMReYULMDU/b0uKNYab91CjOBwCNRH+PvHfmvaRugMMp7a166we2262tvty2rJqJHSA3bt/MZWVslB5HW4iCob21iw4H9PYDs94Tk6/FvasKUqR69FHvBy1b323G8hio5p5dLdPj7mcT1Nkd1TzGGWWdJlTVZijG5F/8fYQ3D3ftd959g21xUhaH06xQ9yvZcz+20/MsgHixk09aU6UXXuNVcRDGAPKIIg7W512F2J9zZdMPDR1+Eio65ly6hdyW7cYzQ/l0Ne2sIzTqzDUytfkc/X/AB/w9lF1P2UXpsufIdGTwWNj8EQKAMVOphx+Px7JJJBUnpGXGqlOlVDBGxEZOu5sTYcH6D6e07M1KjpskZbpZ42hgp4l1RhyeNQHpUH+vtHIzHpknUa9PJjiVf2xpNrXv/Tm/uoc1o3WjUmnTJUlJG0g+tT/AL7j26KD8+r0Iz0m8qsZVo5VDAqxPFhe3H19vJUCox04vHoHslAWlkVRYXNr/wCpH4H+9ezKMnTqPTyjTSprXoM9w4ynqoZoJYVbUrKGK3FyLC9/a+OTQQVPToY1qOiQ7x/iOyNyf7jYZZY6kvqhpwVuP+Cr7Ey3tqLcNdMo+3/Z6P8AZ9put+uUsrcFmJHD7em3Z+NbM7jTM5DUXV2cRSEs0ZLfpN/6e5NsNvjm2gPbAZWoIHEdZkciPB7eSW+33Y0rMBqrinRnabMQ41dDOAoFx/hfgL7Dk0EisVZTjrKmx3exubdZbaQEMK4PXOt7So8VSuyyXkAsAG5JA/31ve47Oe4cIi1/Lpy75p2/aLVpZXAIB/l0B+b7WyueM8E9HVClaQgSuGKFfoDc/wCHsU23LboFlIFfs6gzePezb7xpbFvhyOPy6fdj09ZWCU0hdopA2pEuQoP1uPY9sVaOAI3EDrEnmqaK73ZrmD4WavSD3bijjs5H5nGqSVGK/kXYcG3+PtX5U6ISD59GaqMSKrrguT6BTXYcWv4/8ffvs63wFT0Xz4/4mCXK7jZmVjDVSBeOQuu9h7117h0bf+7hmjZyn7Z/ST+Cb3sfe+vcOk1V7Qr4meWld1uwIAJFwefx9R79jgevUxjrBV46uyFG1DV0jTkxnkx6j6R9Rcf4e/de4dBHtbA0dTlcjQ+AQ1Mc7pGkkYJYXNwAfp711oVJ6FLD7Kjpq4s0UaOr8koPrq+o97x16h6HrC0tBQ0/+62lAA0qB9fyffqV4db6Q/c9LDX9ebigkijeOWkqTpdA4JFObGx9+PXq5rw60fe9KdaXtzfdOihUjzbhQAAADTxmwA9+PHrQ4dBP711vr3v3XuvWI/2Pv3XuvDjn37r3XZNzf37rwFOvC9jb6fn/AGPv3WsVz11791vrsW5v/sPfutZ66Btz791vr3v3Xuu73sDyB+Pp791qnp17i97G3v3Xs/n12wsePof8b+/deBr1x9+6313cgW/B9+61QceurEfX37rfXZFrf4+/dar10LX5+nv3W+sgUHn8H6f4fj37qhJ4dcCLfX37q9a9de/de697917rvSSL/wDI/futVFaddsLW/wCJ9+60DXroEgW/B/5F791ug49eAJv9OPfuvE0669+631737r3Xvfuvddi35F/futGvl1424t/sffuvZ68CQCPwfr7916g49de/db67Fvz791o18uuvfut9e9+691737r3Xvfuvde9+6912AT9P99f37rRNOljsHdU2yN4YHc8Bk1YiuiqXETsjNGp0yD0/Xg/T37rQz1uLfEb5Abd391Xt7ISVcX3xp084ZwZQvjBGq/vYqB1vB6E/encWHTPYnHUtZCFacRz8rZrsBY+/dex5dC7HHiM9jlMSwyxyxLqIAdTdbkWHvXHrYNOPQHPSR7Y3SnjKxU0k66QOAAWH1A/1/fs9bqOHl0ZCgyNO8YlgdHIUEMLW5H59++R6r9nXGryChwW+pH4N+P6+98cdbzx6DfsPdNNRQUUcirZresj6c/X/AA9+639nUbBZWnrKdTG2rygeMfUMTxxb37rXR/PjL8It892V0O5911UXX3WuJBrclnM//k1Dk6QDW0VJNLYMxFwAP6+25EjlUxyCoPSi0vLvb7lbu0bS6nHRte4fl5130Vt6q6Q+LNDDhZaWk/h+6d6qiFc65XxVLUkw5UHmxB9l81klvaOlnRajJpnoYbZzNPum/wBtPzCzTBGGldR0inqOB6pJ7j7Px0lVVV+Zy7VmXqvK08lVVNUXLm/p8pNvr7iWe0mivGd/ProHtfMtjf8AL8MNmFXSBgADy6rM353FjcTl3qaGsjEqOzEah6hfkL7DfM3LcO7Wxl09y9O224WkcLmQjUB0OfTnYOH7Og/h1c9OWqFMa61UvdV45PvHXmParjZW8e2B7T5dFS81WURZpT8PU/dvSHgqa44XJLRzSIXitLp0s30YKD7OuVufN2sGV4dS5yeoX9zOZNu3uzMTShwAaCtadEu39tnubGzyYumqqqrhZivmiaQoyA2+v+t7yT5e90o7qFYLurSEceoA2PlRd6uDHbDz4dcfit0tuvD9sndO53eNRVGcvNcFgLMFXV+fYe94OcvE5WktLaut1pjrMH2D9iQu/NuO8KBBGC1WHWxljIsZuDFUMtNNCXWCJG0FfqkfH1/PvnJsXPfNXIG9SXMDPpZiSCTkV6y83fkXlHc5AkMaHTQVCjy6RuapkochFEpIYEXP+Jblhb3L0/uS/P2zXQukKlUPHzx1O/Imx22zbKLe2UBKeQp0uMrVyw4FGQgjQmsNYhgBx6T7xp2m3jfdGD8amlPLPQF5tdopnEeONeqkflDLt5P4lUHGxGvqlKeRYUBLaj/T+nvMb29g3CWOKBZCVHAVPWKfPmwbXMsl+9uGc5J0jojG29gYbIRfd1tKHlmkIC6QDYn8e8rRvdzy7tMcc0lXA9c9Rby37Y7JzFdNNfw6UJpWnQ1bW6wjwtXBk8RVpSSKytJCpIZkHNiB9P6e0+3+6UUb0uTUdCPmH7qey7xY+Ly5ImumRivU7uLE7k3DiI46KpqAoCiYam0sqix4/wBh7G8HuPs0y01gE/PqCb/7qHNtlJVhVCfyp0XLC9S1ACz04ArWcmWTSQxP1vq9lc/N1veTlGYaeh3tXsPfct7es0K1lbjQdCxtfrbdWSydPjoPuWDyxxswZyqXNrk/0t7MtqvLS7uAkRDfLoI8/bBvXL20+PdaowQaEk+XVmW0thYXq7adNVZBo5Mu8KaixBfUy3N7/j3ICRrEKKKdYlXF7cXkheZi2T516DfcnfmG287NV1sMZBNlYqODyLf0938+mOAp1D238rtnZKpWjqKyl1K1iSY/WLWH19+Pr1YZx0LZ7l2lURvLTtSkeMMCAh5Ivx7116pPQO7z+R2CoKeVEqYYjEHIIKqTyQfp731X7Oq8+2PlW+STIYyimkdZo3iDxysR6j9fevn1fIPRAq2rnyNdJXTzSO0ju/7jmT9bk8Bv9v7alQOtOlu33AtZjL59L7YWxMju/OY2BYWkppZ1ErBSVKH2Uz27ivhDqRds3u3KoLg8ePV03UnQWF2nh6Wvp6aIyLAJJh4l1D0XPPsOTbRe3xKvivUxbX7hct8q+HdJRiuT0/5zuHZuyKmShVada6NSXUhY3Ujgggf0+g9xZzDyRuKXJYV09Zxe1H3meTNw2tUOmOYUx58Oll192TBvuZWhlVoywsAwHH9kr7A17sc9r2lTXrJrlz3K2nez4izL8ujQQ0ppcdHUyAhNGoBvqL8eyUwSqTqHUgQbtZ3hBhcAHpLPu3DxzyJJVRIyXDAuARb6lr+6LFKx1UPSia8tIjo1io6A/snuTF4SCQ01QjuFYKEc3LqbaRb8+zmx2iS5eh6AHM/Ptps0RKMP+K6L7jm7G7rM9BTw1lFiJta+eUOImjbgFWPsSjbrfaoxO4q46hiPmndufrqTaoiY4TXuJx0LfWXxw231JV/3iq4YK/OSs00szaZCJG54LXsR7DPMG5Tb9D+7LiqwcKDFR1JvI3tLsHL7jdrgCaU5rxHR1NhdmUEExjyc3ii9MYUudKKEAAt/T8e8RvcvkMm+P7lhIUZrTia9HvNtzZJB4cKhTkdLPez7J3limpqaoozVOLpKugsrN9AT7AnLo5k2G98S5Ryg8s+XUPXnKz7zbmONatkg9Fno+m89LmJKeirfNT6i66HLLoPIII+nuWJud9sjshNcrpb58a9R3J7e79YXRElQvkfLo/PxvhyGzMrBjKskmKRHSZuQr24Yk/ge4K9wN4h3C1+vszQjI/I46g/3V9p726tzfsdTAV6PB3N2LuLL7Lko8vmlq8VjaY1EECv6YtIvcD/D2Tcv88c0c0tbbLuczSQxuNKFaAHgOsLNw2t9sldJk0kVr+XVUvRfQ2a+Rfe+QyGUjeba+OnnraeomUvTCSBroo1cC5t7nL3U90tp9n/bpblmAncBdI+I16I7Cwfc7zQgqOtiLq/FUmw8DQbaiWERUMax2iRVTSg0jge+OPuPvM/OXMsvNsmoxsK6TUk+eOpT2eRLGMWnqaV6LD8ofkO21K44KhJSJolLlWte8gW3H+x9y17Ge1ic1RDmO8jKEnsRvwin+HrLf239s7TfdtG4TUJrj/D1rM/P/fcGaSSvppQJJ55y9jY+o+r31/8AYDlma2gW0cUCqKfl0Pd92P8Ac1mLebgpx1Qnvat+73DNGGPrb9V/rcci3vOTaoPBtFXzGOsbeabhpL9lU4HRyPjts8rj/ujGuqUXLMtyAV/qfr7cvVDLQjprleSVZiVbB6MTUbeain+4VEUi5BVRyb8eznYATLQdAD3ZaFLMmQCuehL21uKClplWqYI6KASxABRfrx9Pp7HflnrFI0LEDhXoBe7s/BW/cSUz6kaIqQpIsP6+/dWApx6Jb1ZnIcX2s5/Qp0gEkgBj9f8Aiffut4pXqyvJbvpji445pAfLTjS1wQLrYc+9k+Z6qPQDqrv5AVhm3TJDEwdJZLXBJH1+g/p7qRTq4qc9ISjxr01HBJIoN9BB/wBiCP8Abe/da48eiOfISRm3mocWYUpN7EcGQ/X3vy61TPQD24v/AI29+635067VdQPIFv6+/daJp10RY29+62M9eBI+nv3WiK9eBt9PfuvEV66/PP8Asffut9dm34+nv3Xvt69xb/G/+8e/de67Kn/X4ubfj37rVeuPv3W+ve/de6//0NAAEg3Hv3WiK4PXrm/H9b2/417917HXjc/j37r2B14D+vHF+ffuvE+nXXv3W+uRsTwPxzx/xT37rQxx664tb83+vv3XvPrsXAuOfx9PfuvGnA9df1P5v9PfuvfLrsN+DyPfuvEeY66vxb8Xv7916ma9de/db697917rsm/9P9h791oCnXYFzb6f7D37rxNBXrieCR791vr3v3Xuu7cXv+bW/wCJ9+615068Tf37rwFOuwAQTe1vfuvE0PVkf8ubsTceH7XxuAiqZnxX3EEgi1MVjMkmlox+LG17e99erTA8+tojtGtzeS2Bh6jHJLMVFPLIka3JUEF7j37HXvPPSw2Rn6fP4egh8PjmpqaGCdTYEyABSD/trX9663ToWaXFxQldKgFgCzW/Hv1et0px6kTSx0nAIueSfpwOCCfewK8etE0HSUzW40gp5UUBX0sL8kW+nvR+XXhTz6KTu+SpyuS8alnDSXFuQL/X8e9/PqvE06FvrrEy0a08OlgXAJI+g4v9Pfut8OpfyDwBk2BUyKjTSBJm4XUR6PeuPWxUY6Jn8f8AeNZhocptyobSla7RCN0DHS7aSOf9f3sD061XyHXe/wDq2rwlU+YxdMxFbXJUyKg5OtrlgB9Perm2mktSEHHo75cgkbe4pGFFBHQinE1j/wADyjSGKOGkhE8Ra1mCi91PsG7XbTQXjRzKRU1r1kR7gGO45eguIXBCKoIHSuymKmro4GoyGLRgtptzcXvqH+HtncXu3doCCRmnVuT7Xl22tYtzR1SQBdWfPpjg2nkknQuGIb6g/QA/nj3EXMEd5HKVAPWf/tDuOxX1osmtagDz6X38KWgxD+RQ7p+6o03N1uRcn/Wv7gTmfabqe7+uQ4GCOskN55Xtd52F2WjEKSPy6XOB30u6usM5tfLaY6uKcx0URAVjEAQDc8n+vuGL3leaw54td324Egju+2vUFwcpXnOHKO5cr3aFXVZCgPmADw6L513n4abKZTE1cgV6epMEIa1yENiVP5+nvNEx+Lttu6fwCvy64Pc9bBecs83X22XyGMmZgoPnQ9Gp25kqVJY2aRD9LD08/gceyKdCKgjoKsGUcehS/vLTUqJZ1AK3P0/1/wCvsva1LivTBTPTdQ74oxWENVIgDarH+l+eSf8Ae/dntHVRivWjGQtQOlUO28PTkQNPG1zz+n8m309s/u+V8gdNiBgK+XTtF2Pjaq3hq4yP6Arbn2ybF1NGHXmhNRTp2pNw0dSwcSAsb86h9f8AW9ttbkYGOqhSeI6YNwZyJJAqyqUYWJH0Fx/X2oggDDI4dXUfhPSeWH7uzIb6vo1r2B/Pt5mCcet4rQ9J/NYgR08srkDTFK5J4vpBIA9uRsWYBPMjp1XoMcOgc682Vid5S7j3juCmD4bbv3EE0zgeJX0kKCWH1/2PuOPeXmGTa5rTZdtk/XuKEAcfKvWR33cdma/343kqagn7OPRQKOsEG491PRLpoUyUwoGHMfg1nSFt/h/T3m3yCUflqwR8t4K6vtpnqTPfC1ubG8EtChrUeXTnNl66qBW5YngEX/3wPsT3OzWs5JC0PUXbJ7jb/taLGZSVA+Z67x+zsln5FWaUqCwZQzECwPHvdrtFra5Vc9b3r3A3vd/0zIQp4jpcbg27TYLbUkVR4pKj0hSoAKm3NiOfZkBTHQGlJkfWSa/b0uum8fHS4uorHS6tDIw1f61/e+vaiRk16L/vGc5XsKOk1AwmdSQv0tr/AMPfq0x1rj0bGrnhi2KcbFp0/bEMP+Cxfj3unl16pIqeimdIZWnw+7s9jqiQRtU1zmNH41jycaQfes9br59WLY6SKemQFQUIuPxfi9wR/r+9+VOvUz08QYqOQqx06DyAVNx/h7117qWuGpVbUFT9LD9AtcrY/T/X9+r16h8+gM3xsmDak8+8qcBFibyuFawYlS1tPv329e+3oD4u7v4lVyw09DKZEkCvIiG1zxwQPe/LrQNTWvQ0bJ3D/GbyzS+Lx8lHPOq4vcH3rrfz6b+7930GH683BLNPGgWlqEDEgAXpzb8+/Hr2ePWk93ZWrke1d8VqMHSozUjqy/QjwItx/tvfutDh0F2o2t/xX37r1M1669+6312Te3+Hv3WqdeBAvcfUcf4e/dePXXv3W+uwbAj+vv3WqZr11791vr3v3Xuve/de697917r3v3XuuwSAR/X37rVKmvXXv3W+u/6+/de669+6912Tf+nv3WgKdde/db67Bsb+/deOevE3N/p/re/dapinXd+bkX9+69TFB1x9+6312CBe4vccf4e/daIPXh/r2v8AX37rx68fr9b/AOPv3Wx117917rs8f4+/de669+6912OCCRx791o54deJF724v9PfuvZpTrskH6C3v3XgCOPXuCP6W/3n37r2QeuPv3W+ve/de67Bt791oivXrjm/1P0/4n37r3XgLgm9rf7z7914nrw5/wAPfut9ePFwDf6c+/da45669+631y+g/HP+8W9+61xPXQ+hP+sPfuvefV3HwEzGbXYmYaKtmYUVDemiD8qVivYL/wAU9++3rQGK9ct69z7hotxTyzVU8M9FO5tIzDUQ9uP9t+PfurU6sg+NvyQ/vBt6ngrKsGohj0kOTc2Tnk8+/Y49aFeHQqbs7CpMnVUypKgmDKRzzctwbj/X9++zrZOehy2Dm2qKKLySF+ATd73sPoR78ePWgajpZ5KuYXdbkAcKDyPwRz78erUqAOgS3t/EtyNT4rDUc+YyzsI6THUYeSqmnLgLFGig3J/HHvdOqV8urZvht8MMJ1Tsxe+/mBmKDbm1o4lrMJ1znpxSbizJVfOkFNRv6vVwASPeq9WIIx5+foPz698jvnxmO1aGXr7q+I7M6fxzmnxG3aeMU06UsJ8f706WLEgA3I9+pTPXqgn/AC9Vp7m3iuPo6ox1L+QIzhpGLOxYHU2onn21cOBCwPRhtUEp3CIAYJ49U4fI7tLc0e4HWGtmMbSMEAZubvb6f7H6e463K5t0LMwpTrLPZRe7HtyXkjEoR/g6Suyfjx2p3PT/AMTw9NkavzJ5FWISMSGFyRYH3Em9897fs0jC9kCIfMnonuueGuZGW3eh8+jwdM/CT5GbdlpnxuEzkdTe4kEUtlLD6nge4h333L5AkLNeXsOn5sv+foHbxzRe+AY4CxZsYr0N25eou89i5Cml3stckbshkM6uq+NTY3B9lljzZyfvds42GWN9I/CQf2U6pyLyjc81X2m9loNXBj0pM9DBR4GOoSGCecINcnjDEsF55PtvlffjZ7s73IJU8B6Z6y65X9qdl2FlnYBWPEnz6KTn+yMhiMivhaOnUuR6UCFfwTx7lfeJ7TfrJQq5HHqUrrfJNjsZINuYUK07eh26p+RuVx9bSU9VXtLTGZVIJLKFt9bD6e4W5l5Esb+FmCANToFbHzvuNref4wx0k+fR34N747dCU1bT1KNN6GkIYX/rYr7h0bHebH4kEYKo4I/LrKDlH3JjW18JmqCP8nQjT7hpKrD/AGrzL5BpFzp+i8GwHsIQbPNbX/jBcdM73uH70YyQCobquD5K7diraapqqcCVoF8p03+oY/095We1F0/jqrrw6Cm68t/VbQ8rpUBa9FE2dJEzU61BAKzMGHIsBwLqfcy82zG5horZUcOgXtVhbJtzW0I0NkdG92v1rk8vB/EcPBLOrxoW8eo+ll+hAHuCb3fY7WQ287AEevUa7hzzc8l3/hzS0zwJ6GPrnqh9z5xMBnadY1kcRAyqVvza3q5vz7DO/wDNE23WP1lmxOkVx/sdHG+e+SPsIk8RSQOAA6EXdPw6qsFuFoMZjmlppUjdBF6lOv8AxUH2Hds97A1hquZ9DLXj8uhT7X89WPM1ikt1pJJPEdNSdUt1hkRUZKhNO8zIV8q3KlfTcFrD6/4e8mPu2c7Hnbf5hHJ4iRqD+fUP/fC3zbYuWbW1sQoY1BoPn0XX5E9hVGNWPw1BCMyoAhFrAcj/AB95qSUD065txL2U6q07J3DWZ2pkeSeTSLMAHIFxzzb3Xjw6cpToteRyWZxNZ9zQ1kynVwRKx0m/+396690rcb3VvOjgFM2RmY2tfW/0AsfyPe+t+eOkzm9/7izBYT10hVhdvWbkHn3rPXgKdI+SnrmRqxxKY35LsWIv+SDb37rxxnqLGz1DpFELsxA0i5LXP0HHHv3Xurbvib1ilXgKbK1FFZ0iSQvIpubcnk/7b3vStK9eMklaVx0fRt44bbeIqqWZ445EgZLNbjSCBe/vYIUUHVHDS/ESfz6pD773Xk5uycrkIJpnx85cRujP47+S3Frj6ey7dk8WEAgdC/kOc2W5MUcj5V6GToLumXaEqvVTtMgWP031fQc39x9f7TBccVB6y35Q9wdx2qg1kAUxU9G2zvzhpUoTi0WSO8Yj1+MW5/Vz/wAb9ha+5SiZNaY6nPYff66gkEMtSB0Wyu7w3du7LtDtgVUkk76T4QzDk35A9l68u29rH+uOhNN7ubxvd0V21m/Lo4HS3xy3fv8AemzW+JJTQK8UxgnYqxBXW1ww9t1t7Z6QL0axjcN4t/G3NyfkTn59WK0eAwGy8LDjsJRw00MESxvdE1syqQSzgX/1uPZPvd5FHHql49SF7ebJeXl2LewOmNePSPTHxZuqCWBjeRldtRAA+p49g2yt7rdb0JGpIJx1kPzDuu2cjctyX24SBBGmrJ4mn29Jfde3cTh4ZGSUCyMbhrFW/wBh/j/j7nXafbCwuYVbc4wxIHEdcqfdL72zTbhJBsb0VSeHyPRfZ911OMqWko8g58bmwV7iyngAE+yjmr2m5Yt7UypAufl0c+xv3m903zflsNwJZccR69GQ6Y7kxq16rlKmITOPHqfT+r6f2j/re8Dfdn27lM8i7bGVjBqAOs+N05jj3GzjmAAqAeHRpsn2LR4aiky+NlUyvCWEkRWy/wBDwfcF7byjc3dytleA6QeB6jjmVbbcNvaNqHHRek+UGb3fnV69p5Z8hVZab+HvFFqdlWZvGCbf6/ua9t9uNu2W2O+MojWAa6n5Z654+8ez7da3DfTU1kn/AA9XqfGvrbG9JdV41a+CKnzddD97Oz2+4KViiRUZhyR75j+/HOe4+7fuK22bTKXsLYlW0/DrTy/z9Rjs1qdr28zSLR2pT7OllL2AkFdOY29JuRzfj+o/p7Tx+3nj2EcUy54dMtejxdanh1VH8vd+0f8AGqyslqhHoha+sjg+QcLf/be8zfZbk17DbY7aNMV8vs6zZ9j+eIl2xbKZtOkjj9nWuH8muz4NxZSpxMFSJPHNUHUCLcH+vvo3yFtUWybct0V7iAKdDL3G512y4gMaMKr1X3W46Wt3DSSqDKHqUuRexBIFuPcwbVuwaRYpe0E+fWJe47m25XUzW+TQ06s46bpI8ZhqdHjUKype359P0v7kJ7K2lAbUDUdRxb8y8w7ZK1Y2oCc06FrLyRtTuwsFSMsD/iP8Pa2y27wJNcZ6DHNnN0m723gXoNc8egDzu5vEsyJKFdWYaeBpt+Lj2fZ8+onAoTThXoBd5bjaWhl1S6mIPIb6D6WNvfurH59FdxmT+x3itfr41LqI+pIa9j70OtnI6Mvnu3aeTF09NFKwmiRLm9hZR9L+7V6qB0WDde4jn8qtQxJHl/U1vw319162B0rpaiL+EUi2DsXRQR/xr6e/dbxx6I78jcHUQZunyrf5p4zF/t2uP95976qfi+3osvv3W+uyAPob+/daBJ49de/db67txf8Axt791quadeIt+b8e/deBr14/jm/H++Hv3W+u7i1rc+/dazXrj791vrsG3v3WiK9de/db697917r/0fn/APv3Xuuxcci4/wAffutYOOvXI/P19+69Qddsbn/YW9+68BTrq34+v+tz791uvXhf8X/x/wBb8+/daNPPrr37rfXJWt/rf8T791oivXRsTx7914V8+vW4vcf8T7916vXXv3W+ve/de697917rvi3+N/8AePfutZr165+t+fpf37r1B11791vru3F7j/Wvz791qvXXv3W+ve/de6csPj2yuUx2NQ6Xrq2npQfwPNIEv/vPv3Ws9bCnxW+MOF66qtr5gQBq+upqSs89kLEuLgsfr+ffhjr2KY8urz8RPTU23qSkqESWL7ZUdWAYBStr/wBPfsdeNekJJNR7cyKV+NYLQySDyxAWGsm9hb6Wv78Ot4Jp0YTE1grsfBUMykSRBlt9LMOLe/cD1vpHbhyIhkYLJfj6Hnn6f8j9++zrX29IGrEuRLR3JaQED+gBH1APv3XqevTfDtOnjk8sgs9+SbW4+tw3vflXrwrqoBXpW4utoqCohiV1UggG3Jte/wBR78CAK9aOotpGD0o97rFntsTQIRKHR1KWBIutuB7ozooqTjp6O1upm8NENeiAHZj7U3jDkpUaKlMqswYALYtewHtVYILi5ArjpRBast4sMgz0batzOGzdNjaSixbVTtTIGkjAe7D/AHj2PlggVAiL5dDqOOOJQIRQjpEbi2JubIwGDHUVRAsjKUPjNlUH8W9optotpW1hKN0onuL+5tTaStWM9POF2lnNtUcVTmI5ZUUIpVlIJFx9Sfp7K7jltCDIBnognstwhi027kLWvHoQoYsdVUgqI41VrXP09JtyD7grm7bnS5I01H2dZf8AsZzFFBY+DNMBIB5nPHpknp6R0aOUjRYkgkX/AFW4/wBh7ifcNgE7HQvHj1nVyx7mpZ2wt7xwyjHnw6LL2dmpto5uHIY4+OiVPHKuqysWFmcgcewvFyb4dxphQlq1rxp0cbx7hbPt99Hu1o6xxsulh61wegIyW7IIdzYXIY+QWqNU1aVa41vz6gvsbbVbXRt5Yp0IAxXrlr99XlXZ5eZ4uZ9lYaHGsgUyWXP8+jWYPcNTLQQVcT3LKrgg/S4+lx7RywIWKt5dYJAKVB8z1j3H2S2PRY5JQsgjNtRa9gOb296hsNRqDjq0VuK1PHoKR2dNV1gWKU3N/oX5v+ef6+zH6BAtCOlTRBRjPU/+8tdOplM7K1weWP5P4ufdDaoDQdUCqKUGOp9DvXI0Tg/dkAG9i7C9j+L+6vaIw0kdaMSk1p0t6buOvpAhNQNIFmuzAHjnj2ibakby6aEIOBjqa/e9HXSxU7zJ5iyr+pib3APuqbQUBYjHVPpWXJ6H7H7+p6fGUkiSpqlhDg3N+bC5v7Jns2eU18uk7R509A73N3bJgdvyGktNUSMIwqFiQsnpa/8AsPZ3sW1LNefq8ACenFt9Q7cDpyTtjBYX4j7qw+OmiTc26KuGschgJkGm7p/X3jvu3Jm5cy+9VnePVoLYMAPLJ49ZlfdxEdje6D2q2mpP256BXYm3aas2niZZdJq6qjSWqvYsZD+q/wCfeZuzSX+yt4AU6VwPkOsqfdHZ+U+bYlV3VZEoPLpXQ7PjiOoR2Ab/AJFyPY42zd7m5n0yjFesUud/b/aNm276mwkDFR5U6d/t/wCFQvMGVWjjdh9P7I/oP95PsWUPHqAcGo9OgNo9xZXfGdqsMiu8cdRpZ0NwoVgPz79x6sKnoxldkqbZW1vtUUGpWAofrfUy2HI/PvQ60fl0XLZuHrNy7u/ibBheoDKPwq6rke/de6OvU7dRsMafQVk8LAk/k6OTYe/fM9WGTpGa9Vzbxgq9kb7Wv0sifeAlzcKbt+T7oJUPn069ncoQSpoej49f72TM47GBZFMjwjUeTZjY8e9h1JwevNbzomtxQDoxtCxeKNtXCC7E/mwuT7sajphSp4dQK7dGHx8hiqKuGMjUDrYKSw4sL+/Y49bqa/Z0jMzkKfeNNLiWi8lDL6GZSLFfoG1fT375Dr1c9J/bnU+zMSKiV6TVI7Ah/QVBP+P9fezjr3y6Z99bSWgxssm0FanqiCzEfQkC5Nl9++zrQ+fVQHz67x3B1xsOs2vXpKK/JRWWc6xaR47C/wDsfr70fQ9aHCvWt/WVs+QrJ66rfy1FTIZJnP8AaYjk+/dbIxjqKfqbce/dbHDrr6fX37r3XYtfn6e/daPy68R/rc/7x/r+/deHXXv3W+ve/de699Pfuvdcuf1X5/3n37rWOHXVz9f959+69jh11791vru1rHg/6x59+61Xrxt+Pp791v7evEW/IP8Are/da68PryL/AOHv3Xjw68fqeLf4H37rw4dde/db697917ru31+nH++49+69117917rsWvzz791o/LrkdNhYcn/H37rQrXPXEWsbj/W/w9+62a+XXgpP9PfuvE068bcW/wBj7914V8+uvfut9d8cf1/Pv3XuuvfuvdcibgfXgc/09+60BQ9cffut9cgpP+B/oeD791osOuvpe45/H+H9ffuvfZ163F+B/r/n/W9+69XNOvC3Nxf+nv3XjXy669+631737r3XYtzf/Ye/daz10Of8P9f37rfXvfuvdd2P1sf9t7916o66sT9Bf37r3XvfuvdWEfEb5MYDqyOfb2ZpjqyUYpkqJNXhFxouWHA/2Pv3WhUCnRq977e2zv6GXP4XOUU9VUL50ooJEaQ39ekAG59+PWxTy6GzpHrbO7cwYyTmWOaqU+GE8NyLLYf63v3n1ono9XW/UGSq8dU7i3I0gURGWnjkupXSNQ/V/X37jnrdDx6ELbGbpsdUT0uvRDDqAYm/0P8AUe909etH5dD5sTam9e49wYrZ/XeFr87X5OdKeWoxsD1P8OAI1SziMGygG5J9+xWp69UkUH59WdV+F+MH8s3atJvDuWoxXavfddQpkNvYCiqYHfZ2VK/sS5emNzdSbhCvvWa068tKajw9eqbO/wD5j9sfLjfn9598Z95sJRTMcHi6PyUkFDGrnxRiBNK2A4At73TGOPXq1HoB0gZd/wAmPicSvqIUDgG9wtvp9faea6S2oZOjna9muN4JW1+IenQU7m7DFfHPY/VSo5P9eSB/xHsI7xvSwKXRsdT77f8AtxPuEiRXcZDIeJHRLux9lZTdOUososEr00U3kkVUvdfJrP8Ar8e4e3nmm1nLwoe71r1PXMnJs8WxraQmpUcPy62Lv5Y1Z1XFgMXh6mCjTNpFGk0cyws3pQXBVr++ev3mv62S2Uk+2MwjHpUf4OsWb3l672e/Z5lbSTxPDrZh6f2Tt/cGWohS4qiOLpfHNk3jpKc6KYKOXIX6W94D+wntbzR7ve7EFtzI8suz2L+LeqHcUjBNK5wpzWvp0qvL+1sNvZ4gPFbC1px6Ln82ulet+w58ljMFQwUyUVM6w1MccCh3P9CB/t/eQfOHOnI/tz7zXXL/ALYKf3TDoU95ZS/4qGp4efSXZ9y3jb4o9xVtMoyfKvWtR3b1pV7AzEmFl1SUTTSrG30AGqw5/wAPeWvKm+xb1ZLuEeCQD/LrKbkL3CTmu2G33LUljpX59E83j1ZjcobpGdbG+o2Ive/19zLyruzSS+FNlacOsoeWOS7XdLWk2SekRjOq6zE1IZNTRqb/AOP+8exludkt3bmSxFSBUjoO868h2exwCQLQtw4dDbtafLYKQpE7hDoA1ajptxf3Ee6W8UxMdwtGXiPPqPNqe5s5xnt6Eii3DnZapovKWRgLG7fn68eyCXbtvEQcjPWR/Ku67XJAFufip0JWG63ffYelrYpSKkBHYgEAEc3De0tz7iWHJ9iTax/qAYPUv2rbLc7ayMAQR0jt5/C9sXLDLg3Y+Yh3ACaldjq/s+y7ZPfGHd1Z7/tI9fPrHTmbYp13cttVVQnh+fRzPjtsGi2LjpKDccAm1QrHeRQbEJYG5/PuL+d+bf3rcCfbGFanrF73c9teaN3uPHgViFzgHpRb5odtYCtTPYdo4ZKZ/uNClQQ172uv+t7X8t7hc7nb/RXoqHFK9Yx7ry3zXt2q3ukcKMZr0fb4y7w623tgocnvCahhqqb0/wCUMi61iS4AMn+t7w3+8hyv7jbbcfR8hI8hnIFE4ip4joY8j8zbhsMYhJMYXNTXqn/+ZT3TjKbetRRbPSOHF0Bm8U8Q0pKEmPqBX31y+4D7T8w8g+3sW+83lje3Uaaw3EHTnjnjXqNPdHnm+5quxaTvqSMmnHqg/tHvCp3FI0EsgYwubN6iCVFieePee7Ekk9ROKDHRaqrdb1MkjSSj1X/N+D9D/wAR7rgcOt8c9JbK1kc8Y0kHUeLG5+n59+690xwQS1UqU0Kl55SERV+tyePfuPWxgV6OH098achutYJ8kkkSyoXOtQAQbH6N/tve6dar0cCH4gY2qpqXErESWYIzgRn9Q4Jv+PfvLrx/w9RG+BsWGzdPUM4elSRZHFo+B+b/AOHv1OvGtOjU0eY2r1PhBhKdoFlhp1iflRyif0HvfVft6Jh2d2nSZOrqYqVhaQlWKOQDY/T+nup6sCOoWyeosJ2vjZIJI9NbY+OQ6T+46lhy3P8AX3SVBKmg9KrG7axuRcJ5cegk3F8ftw9dZqoiqUnkomMghl0gJa/pIP8Arewvf2jwGqio6nLlbfbXdToZgrEDpY7M+OOY3jPT1MxkekdwdK2Jte/+x/p7Dd3fIMcKdTPsHK9zcyiY1ZD/AIOrHemvi/gdoSU+Rek0zrpdjKkR5t/j7Ct3PJdHSMj5dTzsW0WmxILsdh89X2dHfoqmixtGtNAEiEaKo8ahblRbgC3tONvnCeIQQB0c/wBbNrmuRaRyqZGxjpGZCpqcvVmniJWFWYvKBwQOAP8AX9g2+sLjdLxYI1JUHrIbl3mfZOReWZN4v5FWTRVQfOo6T25c/Rbaov2pYhKBdm5L3+h5X/efc68qcl7ft1olywrJx65de+/3lOa+b9+udntpdNkaqFBPDomvcHeOJxOIqmaqieoaGXTGZOdQPAFj9fcg1NPs6xEMali3EsSc/Pquar703AY58jDTTeAzS6SSxBVmsLH/AFvZFva291bGEsAepT9s33fZd5W/ihYrUZp1jwfdu8aiUV1DS1ICvqUr5B6h9Rx7hndOWNsvA0NzQk9dA9o9wN6urND4TUCgefRrtkd8dqbvoF25R4ivE0yiD7jRMwUstg3P9Pr7gffuS9h2W9a5uGWi58qdM329c3boxgsoHGrFaHHVknw46spNgbkk7E7KcVGRRI6mjjm08TBdYurc8H3AfurzRNvO0Ny7y0dOsFWYenD/AAdQxzj7c8xteW97utWEjio+09Wkv8l6vdlVDHJaOkiZaWPTqVfFH6I7D6cD+nvDSw9oLLlMySW4q0jNI1aElmyT+fU23/srZ7nyuJrcUkEYPl6dCpRbrwv2a1tRWQhJ4hy78A25F/6+6y7feyuIYUNVPkOsPN35H3Tat0ezdGopP7OqlvnHVUFTHLWY6fyRyrJq0ObAeX8H3lx7NJcRwrBcrRl/zdSnyPZ3G2xnWCp/2Otdfs7AS1e4ppqYSASTSgsST+o/4/j3mZs16IYAJMig6LOaZL28uTCrGlT+fWTYewXqc/QRVSlonmjux/xI/J973LedMRkiwVyOmeWtrkg3JGmyCRXqxOg2emHipYaUhoWhQ24vqIFvp7ILD3L3iGQ+M+oDy6ydi5R5P3LbAksQD09Bx6D/ALGzP926KQzyBQyMq3uPpx9fc4cg+4A32Q20gofn1jD7re1FnBYtf7eKAVPl0SDdO81MM08Uyl3mcWLf2eOfc161YAjrD6W2lgkaOQUKkj9nQN5bcz1ilGkW3Oog3B/oLf8AE+/V9OmSCBnoOZp0+6MqfUfnm/I+ov791sdcZqySoAbUb8qRcgcC3Pv3Xj8uog0qwYjUb3tf/Y+/de+XT/SVtbUT0NDDEzA1EIYXOpULryR73x610W35S1lTT7oosL5kkp0pFqCqjkNq0gH/AGN/futcT0VX6CxHN/8AePfuvdeBA+ov/wAV9+68QfLr3F/8L/7x791vrtiD9OBf6f4/19+6qB68euib/X37q3Drv0kWtY8c34+v19+61mteuPv3W+ve/de67It+Qf8AW9+60DXrr37rfXvfuvdf/9LQAIt791oGvXrm1vx7916ma9e4sP683/4j37r2a9de/db67F78fX37rRpTPXuQT/sQffuvdde/db697917r3v3XuuyLe/daBr11791vr3v3Xuve/de67Av791omnXXv3W+ve/de674t/j791rNevG349+68K+fXhb8j37rxr5dPG36v7DO4etF/wDJ8nRy/wBOEqFJ5/1vfuvdbXPxqztLuqDZ0nkR4oMPRxM4+gcRgC4P9PexjrWadWHZT7WjxFQglDOtJM8Z1cCyXUC35/w966t0UI7syT4qvkqmcxR5YQxszMSFMosb/wCt/j791r7ejw7Rr/FtPG1UjErJRRve9zbR/X/jfv3W8EV6RORrhksjpjN49YBF7jk3axHvdOvE+XS5xmNUxo+gs/HP9PwLe9MyoupuHV4YZLiUQxCrNw6yZrEzilYxj1MLiwF/8OR7BW9cyR2/ZAcjrJb239mbvc9N3uS0Rh5jou+51zWKlaqhjmcRvfSmoXA+v09ho86rH+m7UP29TSn3Y7i6k+rt4qpx4Dh0u+tt6UeaiNDkXWGcOq+OdrsSTY2VvdF5jkunHhPVfOh6ff2h2zZ42FzDplAIFQP83WHtjC0WVqKTE0aoaqeSMqEUBtLEAWI/3n3KnJ93HuMiiI6qUB6xZ5t5Qudl3driQaVqaCnl0e/4/wDxl8WExNdX06ky0scn70aN+of2dY9zHBaiNat03aWQZQ7efRsMl0ljKTHs/gp1McRcXghvZRf8j2oVEbCdGDWyhfl0Q/u9KPEUlTSBYrRGUAIiKfSvA9I9pJRorT06KbgBSVHp0TDbWaqp4cgg1aVmKqCfpc/QD3FW72dvczkSDifToGw8w7nsd8ZbFyKHhUjp7eKsmJHq5Gq/+Fubew2eW7XVUDoeRe9O/LCI2c4+Z6Se6+vG3dTvBUAaEgkJuBqJC3Fifb0PL1jFJrpn7B0j3L3h5j3C1+kMjaQQfiP+foseO65xAXKYuZlXKQzOaVmIJEcZuAD7rNslp9O8Ua5Py6CnNXNW7c02SwXz6ggpk18ulr11lUb7rCTsDJSSGnBJJJKtbi/uC94tJLO8ZH9cdQvPEYXKtinDoPvkFS1mJohkaTVZVtZCf62P09ubO8c0nhOOnbUg9p49Bz1hishmUiqZFclgjEMCSNTfXn2u3GSKI6R5dXnJXhjo0lHs0y04LMA4BFtKmxH+vz7DzXJDY6SeKOk9lNk1isHWT0CxsALi5t+PahLsUz1YTjyHTUm0aiZdLsQL2t9CfdzdBDkdWMlRVOg53rsDI4eEZeiZtcbq1gWDcHUfof6e1ttepKTGw6cjlDMNfQv9aZjIbjxtOk7uHoofGwJJtYhfqf8Aivsvvo44XqBx6TzoATXz6T/Z9GsFJIKttZeVUQEkj1N9bH3axlfxVSLiSB+3rcbBiqLxqOje/HD4u0fYG14pczXQNFUxQyxQSNewc34Rjb/ePeTvKPtly2sMe8Sx0uHANaDrIzk+e+2mwR7NtLMB0ZnfPw+yez8EtXt60kVNGCiQqCQiJq408exlecm2EikwqK/YOj7ctz3y6i8TxTr+09Fnoqdopp6CtQx1VK3jkRxyz8k2H+w9xtc20W3X30zLQ1x0T3Vrvt1tT3M0upV4ip6CPuyarwW2ajJUQYaH8TItwWvx+PZj+HqKjTWR59T+gNs42LAS7iKCaqq4VnYAKzKzkkgsRf25HBJN8Ar08ltLIOwV6dt04Ov3FUzJT0kpi1Wb0kgWP5/Ht5rK5Xip6u1ncjOk06VPXuxExLiaZBHIpFwygMLG97j2nIKmjCnTLKVFGHDobKyakAEV0dwNJ/21rn3V6eGa9WtiTdJpHmP8PRUO9+v4czTtV08QEoHluEBI0rqHI5/23uOrreZLa7aKtRXrMLY/bi03vYIr8oAwUHgOgh6Kzc8WcXC1BbyUs/hQMW+gb+nszst3Ct4svDoGcxe37yxtZWS9xxgdHE3z2bDszHeFo2kqJ10KoLA6iLAgD2Yw7/DPP4ceT0Dty9pNy2nbPrbptIHqOkNsfY25N8yPuHNVMkeMlP3EEDs2oozXAAY/j2IB3KG6iKdFhlaJTWhz0LzmkwqfY0vpWH0Mzfm3H197r01TqTja01ZaKLUdTLe1yot+CR73wFOtY6cMzlKLbVDJU10kROh7B7Ecp9SD791vT6dUo/PzD4PufbWbyFKkZqcXrRJNCm3jiIBF/wAe9HHHrx/hHWtVk6CXGV9Vj5mVpaWUxOy/pYgXuP8AXv791odQiCAD+D7916uaddsQTcf09+68BQdcffut9dgXv/h791qvXjawsLf19+68K+fXXv3W+ve/de697917rsH8H6fW3v3WiPPrr37rfXvfuvddgXNvfuvHHXiLce/deBr12PqLcH/H/W9+619vXrG5v+Pr/wAa9+69XGOuJt+Pfut9e9+6912bWFhz+T791rPn17jm45/Hv3Xvs669+631737r3Xdvof6+/de67uAPTx/W/Pv3VaevXH37q3XvfuvddgXNvfuvHHXR449+691yuOOOP7X+Pv3WqHrx4NxwPx/hb37rw9D14Hm59+68Rig66uCbn6e/deoadeJv/rfj37rwHXXv3W+u7c24/wCI9+61XFeuwPqPz9AfwP6+/de+fXRFuPfut8euvfuvde9+691yGqx54tb/AI1791U0r10CR9PfutkA8evcWHHP5P8AX37r2a9dsLG44/pa/FvfuvA1HQ9fHjM7k/0qbSxmPr6t4K3ICCopnnleAw+NnJ8ZJF7gD6e/daOOtoDbOOipItvwViopp0pXcFQFbVECR799vWz8+h03fvykoNvvQxvHDAkGkaQEVPRpAOm359+69WnSa+KHxk7p+V/YklHh6Cq2Z1/ipjJnN+ZqKWLbcVFq1PUS1rjQBpuf1e/dbpUenVn3enz/APjN/La2NkOofjM+N3b3q1EcTuTtfHyR1ONmqZY/DOcWV1aFDA+oG/8Aj71xPWzjDfs/z9aqPfHyS7B7f3plt8bv3FWZnceXqJZqmaSuqplkZ5CyAJIxHH0+nvflTr1KnUel/wBC9iZmQVIzFLOIggKSuz6Tf6Eav6/X37rR+fRiazKSZcNJRyFgxvpvf8G/sPcxqxsyV4gdS77OyQf1gVJ+BPSGqIJlmDMCfWdSm9iSfr7gvdLud7aWKQnh10ms9o2+1s1u7WOhIGaD06F/AvQNjFjnSO2katSKTyLG1/8Abe4SvvHFySp6BG63jidlc4PQgdWbufYO+sTLtSqkXK5OpEVPRxTSKZ5SwAjiRDYm/Hsn3raDv22va3ia14cK8egNvm27TuFo/jgVUcaDrdP6G7Di6p+Mu2dzbqrUpd6b9oRSVeMqW01lJBJEBHLIG9Sg3uPcYcx7Hbfd59qrrd+X7fVuXMsUkDoBSSNDWhJGQMnrEbdrNb/mNrW2OqG3atRwJr0XHtTsXG0m3q7JHIRylKeed5NbH/H6n8++cHt3yVuF7zCBcRsDI9SWySSakknj9vRnuDtawEOKda2XyH+SNJubfk2JMLeOGqmCygkqSJSo99WeTvb7918uqwPcFH+Doi5T3+42bmaGaJiFkYAip4V6SlJX0tdDA1hd7Nfg/UX4Ps75dgkhvX9B12a9p7uLc9rhuV/Eo/wDpb02Mp56cvpQ3AJ4BJNuPr7lvYbgQLLcPwUEmvp0c+6e2w3i2sDUAJH+Hp8x+16OphZiqq/BF1A5tzz7x05t34z77NLDlSfLqLN55OtrKBZIyD1xhwgx9QGNuSLiw1Hni/ske9+ojoOgtHDJaNVTToddq7upcDSu3jHljQFWNhbji3/Ffcd73sD7rKKnBOR0Ndu5hktbUxSE4Hqen/FduRVdWVqGSwa1jYj68BSfZXfch+FbUhFOn9v5hhectL0II35gxTtUVQGn+zoIGri/JFvYeseQOYdxuhb7UjOa0NATTo9vuYbFLXxJQp+0DopfZncW2o8nJTvNpheTSsTTP+TYLa9vfRP2W+7/ALVZbVFc80jxZiKkUGD1zn9/PcK6F+8W2RiMauNB0hqvvGPbmKWfG5Uw0wBfwxVMkR9XHIQj3kOntN7dLMtw9krOnAlFOR9o6xTl5o3mYEl8HquX5Q/IrEbkx8lLEzTVrrMvlaR3ZiTbkk3+vsdwwW1pAtrZoEiXgAAKfkOiR3llYyOak9Vbz5Crq5ZJKljaR2ZRqJJBPF7+25pfDHRjtm3m9anUdolB1C5/N7n3SGUynpTue3rZLg16yRwSTnTEpZ/qoH9o/wBB7UVzTon0nTqHR7fi98XszvnI024slH9tjadop2E8a2K/qsur3bqlT1bhRde0GJipoMOII0hjRCVSNf0DS36R795deGOlvh6NqGcyzTR6EAbkL9V4+p597NfPrXUPdG7IEpa1hKhKwEAi1/T9Db37rx6pw7t3/kW3JltFU4iDz2AlkAt/QAH3o0HXvPoBdrVlTuCu51TsXZSAzMTz+SSfevOg631Yz0XS1uEFHOqFGMkZ08i1l/P9fduqmnR1t7YLG702uEmpomrUgBRxDGWL6P6ke6SRpMvhuMHpRa3d1YTi6tmoVzSvQadPbqwu0sm21szFGtXA7BGdRzZuLX9gXdOVZLmcmE0UnrKTkr30sNn2hI9wj1Oo6OrR5BMhAGp7JE6gpxa4PPBH+9e1+18qW9qoafJ6CvOvv1uu9MYNprGn2kdY3oppGZWbSL3bkn/X/PszvtnhuIfAhFB9nQH5a9xNy2zczuW4uzniBU9Sq9KTH4eZo2jSVUYtIeCb8lg39PbO28tWFgNbLVvsHRpzp70c1810tYZikAxTU3+Dqpzvjs3csO7zgcM82RaqnWnWKleQtGXawOlT7P0VEGlBQdRNNNLO/jTnUx8+nnaPw93n2C1Lmt2VZhx8vjl+0mPr0yJ5CLP7LN0uJ4oSLcVNOhzyNtO2bjuCHc3CJqFa+nSn7J+PWz9qYmDGQYvWEOmSQQpZ2QckMB7h/cTvs0zSK1KVxU9dDuUIfbKz26Ky0qSQvdReP29e6r6s2I/ioJqONPUBqaGJuTb+o/3r3CXPfM2+7OjPGDq8skdZMctbRydBYo8MYkHEUA/zdWIdYbA602wYDFDQ/cFlbUYYA1/pyQPeHXO3MvOm8hyS2ih4E9DiOOwWKm3WwUepUf5uln3FhparFwVO220iL1MtP6boOVFk9kPt3uIiumt934tw1Hz/AD6ijnrY59xjUpHUoa8B0FmxN9JEP4VkXNPWQmzNI7XBU/WzexxzHy60p+rtRqQ+nRPsW+/TKNvuu0jBB+XQo7i3lXHCaKPIkx6CQolcc2/Fj7Cu0bLFHffrR5r6dJd/5a2PcK3zBSx+Q6I92duatykT0mVmMsSs5F3ZuC1v7R/HudeXtuhtSJLYUJp1FO5bdYWtQq06JhuvauLmYVEPjvrY3Kqfqb+5Nsr2Wmhuo+3HaraT9VaYNek3jTR4utik9AMTqbgAGy8cW9q5RJNGR69IIPBt5QxPDoQ8923QY7HK0VSnliS9tR1XUcDj2UW2xPJKdQwej685pitbeqv3D59E/wB+b53H2RI2PoYJzCGYeUFypBN/crcl7bHtl8siHPp0Bd033e+YLVrSFT4ec8R0XbdG09wYeAtVyP49RYqSdS6uT7yDh3ElQo9B1CG58osszSzr5noOWD8jUxKj8k3+v+Ps4tpywz0AN92dLfvT4R5dcIEkmbSqFiWsxFzcX+ntdXoHkUNBw6WM2AMeNFSI+QNWkizCwvc+/eXWqZ6jbYpaGWqeTIWEaAlUawLMv9mx9+p16np0MVG209p7eye+M2I4KVqaaKhWYKrLURqfE4Y/0Nve+vdVF7+3bWbz3NkczVSCRXqJkpLarLSCZjCPUT+D791rpG3ubtz/ALx/vXv3XqYx1x9+631737r3XvfuvddgX49+60TTrr829+6312Rbj37r3HruxJ+ova/v3VagDrj791brsW/P9Db/AF/x791o18uuvfut9f/T+f8A+/de68OT7917rsixte/v3Wga9dg8WN7Xvx/X37r3nUddA25Hv3XiK9evzf8Axv791umKdeJv791oCnXhb83/ANh791s18uu/r/W44H+t/j791rh1xsf6e/db697917ru3BN/futVzTr3Fvzf/ePfuvZ68bfi/wDsffuvCvn11791vr3v3Xuve/de65fp/wAT/iOLe/dV+Lrj791brkrFSrKSGVgykfgjkEf439+691eZ/Lr7Rz+4cRT7WjFTU5aKRoIaoajZIzZP94sD791tWya8OrWK6v7Ax1RFFmZ6gUksiQDXwuhzpI59ttIFbQePSyOzea2Nx5DoYNw9YQ/6P6Z6SMPWVU0Na5jsTbhibf737crXpDkVB6FbaGUoq3Z1PjFlSKroadKd4TxJrjGk2B9+63SuemWKmNPXF1Fl1jUD9Lj6+/A+vXqDh0JmPySRQhxawt+Te4/1vp7Jt+vEtduc1oQD/g6kn2n5dut+5wgiSMyJVa4+Y6m1GbMsEjhdSqtgbXsLcce8Nt89zbGz3N4JJNRUkYPXb3YfbLZbPl22DhI30AkUoa9B/T57FV+W/hmSMMTSt4wZAPo3PHsDczc7z3G1yXm2KxahpToZW268tcv2RtZnj1Ljy6ZN79Tx0FbS53bFdGksrIwggb1MzHgqo/x9hT2+9wObt1uDs9qkryuaYBPH1Pl1AHuLuHLTlt1ndAACRwHRl+lvj5n9z7jwuZ3HDJNDHFC5aRSQ1nBF72/p76f+y/Ju68v7MLnfDWWUas8RU1653+5XMdvzPvOm0FI0NOHGnDq4jGYan29iaOBKdUho6ZYtIQDSFFhz7mYuJHKqeJ6BipojC0+HHRce7e8MNs+gqKZ6mM1MkciRwlgHu3H0/wB697dxbr8+ktzdLEK9Vqb2yDbsxVZmKioAjkeRkDHjQyk2F/aGRjJE0h+fRLMTLEz16LtgTRU0tTFGyjU5uR+Tf3G9xHLLOzU4HqOLmKV7hjprnpW/xWhpz+6VAHA4+tv6/X22IJv4T019PNTK9Y6zdNDFTS+AgMY5AP6kMv1Fvevp5/4T1oW854qei8bW2fX747BWWgl+0VpHRy50o2tyPof6+zra9rMx1yig+fR3tm2NcGk3Sg7g6Y3D1DlKXctNTyS4+QpLWSQKfG5kF2YkX9x17n8neGn7xtV7RTh0Sc48vG3T6iEVXoG+wdyYncmzy0zRmo9CtGwu4Y8cA+4PsInjuKHB6jy31K3bx6xbClo8NQ0rRquuSGPSFX8/UA/8U93vEd2NenZQWPccdD3gErsiC0UT/ueoEAgaT+efZTIY4+PSWSiioHDp7q8Bk1iZjGXHNxY/j/H20s8dcmnTepPPiekDlPLjA0kg8ZVub8f6/J9rY2Vxppx6eFWU06TWf3HQV2Ilgk8fkCHgqLMVXm3t2GB45Q3l1eNOmLqTI09PQ5Z9SqRO4UHi5JIGm/t/c0cyIiiterTpRgOJ6FLK9U5rfVC9cFfwR/uILWvo9QII+vuYORvbuS4RdwvFovHPQ45f5Ue4X6uUUHH9nQgdIdk7j6z3jjMDnpposRC0EWpiyRKiS25v/Qe57tHNqFtxhEFB1JVncNassZ+FcdXa7o7w2TB19QviWpcvVVuPGpItMpRpIreof4H2crIUJcNqr5dCaW6ieMUySOqRO7M3kcLuOq3DCppYqqVZxCo08OSf0L7B297PDe3IuaUI6KJt0nitXsR8L/5egF31uXMb322uCx8MtRV1rIw0KSV1G3PsrfZ2KhU6Asm0sZdSefRtPj3sLK7J2eF3FRTVbVNMnjSxJX+o/wCNexNt1ilpD3LnoU2FmltCGccPl0v6/e+EwAno4ttyNLKSfIYmbRxa17f8T7WyNGTQr/LpU7QjBTB6BjM7xyK1UlfBG9PTrcmLToso/wBpH+Hsputtt5shadEt5t8UtSopXpN0HYvnq9Rm1LqX0kkN+rm9/wDevcdcxW1/b1itFJr6dDH255Z2m4vxcbwyhVI4/b0K/wDEcNuOgtUvCsjQsNL29RK2HH09xHuG2bxBIZ5o2Pz6zp5cvuVXsl22xmQUWgFR0UZ8Adp9jw5CjRhTS1Kys8fCnU3/ABX2Rnmiy1fQTOFk4UPRN+4ZIt8WSFdakimKjqN3VuWryO+trUUTMIKmKNpFJupNwOQf+KexXyt4cs4cGuegZ75xXtlspgCaNak8KeXR5dn11RHtfGUwsnipkjBWyjleCSPr7lzFBTrn1RhI2rjXPTTlaSWV2YC4duT/AFP5I96+fV+nbERpi6SWpYW0I7Hk2tpuDf3vrWTw6J7vPcW6+wM7WYrGGpWhp5Zo5CmvRYen+nvXAdb8+qkfnjlt+dRYybHYymqnoclpNdOQ3jRZE9Ukje/daPy6o4rKueuqZqupbXPUOZJGta7H/D37rYx1guSAvHv3WqAZ66YAGwv/ALH37rwNRXrr37rfXvfuvdcrkAj37rVATXrj791vr3v3XuuQANhzfm/9PfutEkZ66IIv+bfn37r1euvfut9dgXv/AID37rRNOvfSx/1/fuvccdeJub+/deAoKdde/db67AuCeOP9v791qtDTrxt+L/7H37r2fPrr37rfXYv9R+OffutH068Tfn8/n+n+w9+68BTHXgSDx79145HXR+p9+631737r3Xvfuvde9+691737r3XvfuvdclIF73v791UivXH/AF/furdd+n/G/wDsPfutZ67C3F/p7914mnXrC1xfj+v5v/T37r1c064+/db67ubAfgXt791rz669+631737r3Xvfuvddn/C9v8ffuvdeBsbj37rxFcddfn37r3Xv9b37r3XfFvzf8/09+69nrr37r3Q3fHXN02A7e2hX1VvEK7x3YXVWK6wSfx+n6+/DrR62R91b+VcJg8nhQ9dX1sNItDQU12nrJTGAIoVQG5/w97+fXuOOrTvhn8HqzsjbLd4/KXPUfVPUeIiGVqMRvKX+F1m6aOH99sdh1qNJeR1FgQD70SOtqpJNR+XUb5nfzMcDktrT/GT4T0MPWXUFABQZ37Nada7cVNCvgkaTKRAO2sXP15v79SvHh1oGhqpz/g6oR3D09PnJp6iCsq5a2rZpJZauqaoZndjI+lm55J9+PXgAOmvavxnycuXo5K6GSojWdCwKkhgWvYA+/dbr59Wldf8Axx25PtuOiXFrDUyxKvk8YXSxXkkj/H8+/fPr1a9AV2V09u7qqresgppqzFs/+6UZ1SNzfUzD2i3G3Fxbsp6FPJ25/uzd45QaZ6BOt3HSzxNMdMUqA+SNuHuODx7hPmDaCscoQZPXRvk7n2G72hIrpq4oP2dI3MdpQYukKx1IQqvrN9IS3pH0/HuIxs7yz0Zeo95t5mRLlvBbPVkX8obpOu797b3b3T2HQvR9b/H3HvvutyGUumOylHQt9yaeF5LKxbTawv7kTlTlWKe6NxdKBBbgSPUY7fL8+oP5s50vorP6S2estwdCiuc+n2dCX8qP5l24e2fkXvbKbEyk2P62p5/s9r4SgmKUVFFQ/wCTIkMaekfpH0HuNvcrYLXnHc5bm6QMgaka07VTgKDy6T8sTbNsdikN4oMtKuTkknia9CF0z3h2H27jqvCZd6+SjqUMPkld2XS8ZF7n/b+8eN95K2Hladdxt1RHXNABXov5s3XbL9dNmAD6jopfyI6hn2nlDllgkb1vM8oubam1Elv969y5yfzLHu23mEEcKU6i+GMw7rBK2aN/l6TWw89R5bHxwrOBUUsfK6hquvFjf2luTPtd54qDtJz8x11T9iueYbba4bWU4p/k6FCk3dFQPBDOQqarSEni39fYqlubm62aV7DBkWhHmOslN+e33uwEwNSgqvQoDeWDgxi1UVTENK67axfgfhf979wI+zXzXZjlU1r1CG97xPa6o7hsL6+nQR5XuLFCqKJWRjSx41KDYH6D8+xRbcryiPuU9Rxdc1WzSUDefWeTtqjnopCJbKqfVWAJP9L+6Ly0yTAnrZ5rs2jKs9Co9f8AZ6hbc3e2VllrDK1PTU7FjrcAMPr9fctcje1MvNcxMuIU4n+L5DqEOfPeeHl+dLW0arMaEg/Prh2F8h9t4LBtF/EIxNCrEgTAMSosePczbVyHtPLM5h2uAKx4kiv8z0KbH3KW+2Fbi5l1Gnr1WL2J8gI9zVzPQ1MwaOdmDeU24N/qPcjbas1owqaevl1BPPVxac0RudI1Dgegyr+0dy1dOIjkZ3g/1PlY+n6AD/H2MUkWRQy9Y1XthLY3DRPwHQHbmrJ8pUtNNK7DkgOxIve/Pu9aDPSVRU6V6S2lmJvx9APaZ2ic0bo8gtr6zi8ePFepNLT1FfJ9tTRtLJyionLX/AX3ppIbdajr0FluG6zBHqQTx6PD8evjhWbgmp81uKnFNj4ZYpR90pAdAtyF1W9sW87zyVHDo23fbLbarQIxBf06Oru7uLD9T0MG3tvCGmp41SKRoQE/QNJ9Q+nsyr59AsVqa9BVjvmpR0ORWhmd5A7kF9YtY/k3/wBv73WnXiD0vav5WYueI6a0JqAbiRfoeR9Pp79Xrw4dBFu35RY9KCtMU/keSGRQFkB5/pb3rrx41HDqvfd2/m3RW1c6M96lpAl2NyWPHJA96FfPq1OjW/GHYL5UQzVMALu5bUy6tV1B/PvYx1og+fVpO2evnxsMBWJVUaDwtrXW4Nx/xT3ulOtUPDoxGKxFMuOvUBVVIxbV9BYfm3+Pv2evZ6Ih3hjWw+4n3Ji5GjenqPI5jIAKK2ok2+o9+qeHVCis1TXowfRncNPuWgpqeqqFM0bCMozXa6rpJsffq9X0gYHRocnuHG0mP+486AhL/UE/T8e/V8z1sivRDe+fkXW4yjbB7eWaevqnemQU5LMrHgH035549+qetY4dKr4tdI1GZkXsrf8AQPNVVMjSQw1kWt0YjVGxR/eh69e+R6P9VC0KwQxxokYEcSxoIwAosOB/h71g8R04ryo1Y2K09D0Bm/NtvlaWRahA4XWQCATe/wBb+0rbbZyHuUZ6P4edOYbFFWCZqJwz0T3LSz7XyTpAjxESFQykADmx/PuJvcDk3bryAl4tQGRTrN77tvvBd395Ht293FRwox6XeB3lVAwzGqk1IdVhKTzbgn/invHy95S2owvZiECuMjrphtu8Wslsphoykfb0YjbHadZWQNQSOZLoEBYhha1rc+4R3b2itba4a8aVYwKkdH9jZbZfMRMoyPPpL7k2Nmcu1TmsRIYKkhpGZDpBB5IsPZdZ71Y7c42y7IcKaevWOfuXyZLbblJe7YaDJx0FEm6dy4eKTG5Kom8kZYNrJPC8exSm27ddMLmBRQ9Q4267hbAwXDGo9ei9dnb9hNOUeoCy2a9j6vrzf2Mdl2so1QMdAff96jCEue77eio53srTTSRR1ALoW/tEnj6WA9jW22urg06jPcN+ZYqRt0ENR2FWyTOfuGUEkWufYgTZuwGnQXk3xy5z0j6zLV2byVNSxzysk0iowDkg6jzwP+J9mUVnBaQF5Bny6L4JbjddyjtdRoWFerJeoOr8NHtKOono4mrSseqVlDH1JdvUf8fbO0XMaXmpTU16zxsOQdssOQUeCIGVlBJpnIrx6BT5BbIpaTFTyU0UbSAyHSigWA/40fctbbeCRwCajrFLnjYXtoHOmhqeicbV6urtxhmVNN5WBuLXAPF7/wBPY/sTE618+sTOaU3CCYgglD9vQsxdILhYGqJxG/jUufpfgfW3syoDnoFVPDpF5ynpaeilgvGNOtALC3p4t7917oDpaRHyMDxyBY4KoSzorW1RK13At/h7116vRe/kz2fLmaul2XiZWiwVHFFPNApKhqhSLXA+oNiT731rNadFFJv/AEHFuPfut9eW4PAuf9v791o0pnrtiDb/AH3Hv3WgCOujb8X/ANj791bPn117917r3+9+/de697917rsAn/ivv3Wiadde/db697917rsG1/8AEW9+69Tr1uL/AONrf7D37rVc06//1NAAgj6/6/v3Xq168Tf6C3Hv3Wh11791vru/Fv8AG/v3Xvn117917r3v3Xuve/de69a319+6912CR9PfutGnn13cgm45/N/9f37r1KjHXQF/zb/X9+68cdde/db699Pfuvddkk/X+lvfutAU669+6312ePzf37r3XXv3Xuu7E/j37r1R14j8Dn/W9+61XrwH1/Fh/vh7917q/X+T5SYd66OeogR61a6fQzKCLBub39+I4dbHnXq5Lviu+93XitqY+NFkmkhmIjUK1tYJ+ntkxfqaj0YJeFLIwJ59GFxlD4sRi6aZblMfHFJqH1ITS1x/W3Ht+np0W5rX16CfdG0q7C1T5zbbsIbmSopg12J+rHSPfuHVq1Hz69gc/RZyn8ErpT1yXWSNyQ7Pe54NuPfsUqTSnWiHYhUFSSB03bo3dHtmmMd/K7BlCj9X0/UPcc8w2e477I1lZVoQRUfPHWcPsZLy7yNt/wDWLd9JkWhCmlcZ6Dmg7J3Pmm+0x9PLHE7DkxuQQeAb/T3EO3/dPvb7cDfbhNVWbUfXJ4V6GfOH3s7++naHaQY0UkClaU6HjaHUWc3Y0GYlinNQoVhILqgIF7295Dcvfd25I2ux+jvoddRnga9Qdu3vPztvM/jwzEA/b0q927P3dtg0de3lkho5Iv27FrLEQWuP9b2J+X/Z3kHk29/eWxWgSWtSSAc9BreueOb98g+m3GclPQV4efRyNk/MLY+z9o0aZSkpTlaOCGDxEBZWZRyQPr7lFr6MoA2KeQx0QR38MUfdk+fWXcXz5pM5jmoMDt6RqmoXxxyRK721i349srcxJ3IpJ6vJuqulEGemjrbpHcfedZJuzfZqosa0geGmkLRNob1LpD/ge3dAch5vPy6pDay3J8WTA9Omrv3491+18WI9vlxiV16o1OptAW5JYf737rPAWj/S4dVurQwKdHDovO1thbGNEHq6yFMigC1ETSgSGT8kKfaKOztEFSPt6L4rO1cEsMnoVMR0tsjcCgJIJWJ0qsTqSSfra3tQljbfhHHpQm3Wz0AHQmY74f7frYda0dSdakDn63HF/bw262K5HSobXDxC8OgZ7E+Mec2KHyuzaOpp5IC0ryFTptENYII/3n209qsf9n0xJt5iPiRj9nSx6cyO3+9Nr5TrDsKkUbjRJKekeUaJpGhBUMmv6njj23LbW+62jWF6tQevLDBuVu1pcDuPVbnyg+Ie6OuhnMtjUqFwtFUPLo+qGNWuNJH+t7h7mT24h2+3k3C2HaK8Oo73rlD6JXuYRgV6LJ1jLNuDJ0WNS8hpvFFIq/htYU3HuBL6kYctinUaTL4ZI+fVufWHVEC42jnqqclTCGsOCbtYj3HG4bgNZVTnojuLhtXGvQyTdYYJo2Bpitw1yQPqebW/r7K1v5CaA9J/GdmFT0VXu7qGOjw1XX0kTKFErLa30Vbgn2JNq3ISShD0ut7jU2luqpNyZqqosnJjXZwwdkUEkXA459yDDErxeKOHR3GFpnp/68bNLUon2k5xstQjTVAQ+IesX9X09inlrl+Xc94ilkWsQ6N9n25ry/R3WqDj1et0rg9oZHYVM9TLTecUwDKSlwQvNx7ywsreCGzWGIUCinU8WcMEdusYpQDpNZ/p7r3c+RKStDG4ZiJUdUNhySD700EcnHHTbW0LtResPj2vsVEwlCJMvUACKnKSmYIp45HPvXbD2rnqrFIqIgJPTZm+iJ+z2hrshRypAyDRGF0+kH0gqePfntzOK9aa1aY1Ycelhsn4tbW23UQ1k+PZ54dJQSaSLrz9Px/hf3dLOOPy4dPx2CRNqp0YuHZuLSBYEpkKxrp0hRa1uLD2r8JCMjpYYkIx0GW5+q8XVPJLHQjW1zqKg3Frkg+0zwgdJ3t1OQOgO3P07RVVJNEtIwkKt9AAOV/3r2y8NR0me1BwBQ9E43J1JkNt5FqlKWpNOHb0hT9Ab3HtC1uq/GtekTx3ERopIHy6nYrHx1CrGkU0FSFshdiLEfW49sS7fY3Mfh3KAg/IdLLLd912yQS2MpVhwqT0y7i2rm0mWoCPMEUMCqMWABueR7hTnL2J2ve5TuezfpTDPHzHUq8qe8++bRexy7yfFRSPLyr0Ee7MHJk8zicq8TR1GMVFPkBBIU2/Sf6e4Shsua+Q95W03FGkhY/FTA6yO5l5r5f95Nvhs7ALHKqUK+Zx0c3rvJ0eUwtPTqytLSwxrIF5N/08+8h9pvDfWqzjNRX7OuffPmxR8vb1JYKNJDEdLzIY9Fpmf6kfp44H5F/Zlx6BXD4uk7VNFNh6uHyIZdJRV/NyNJHv329a+zpg2RsrH41aupWAGasJdyyguLm5IP8Asfe69e+XRVvnl0pt/e/Tm5amWmjNbFE2iXSoZLQcm596PCo68OOetN3c2H/gGfyuG1+QY+qenDm12AAYHj/X9+690yEiwsLH8n37r1DXPXH37rfXZNwB/T37rQFOvDg39+62eu/rc3AP9P8AYe/da4Y64+/db65XHHp+n1/x9+61Q5z10bEmwsP6e/deHDPXdyBb6A8+/dewTXrxNx+BpFh/jz7916lD9vXH37rfXvr7917rsC/+H+P9P9f37rVevW5tf/Y+/derivXuRwDwf9sffuvYPXXv3W+ux/T+v5/p791o+vXvpcA/8b9+69x66sT9Pfut9dg259+69x66PJJ/r7917r3v3Xuve/de697917r3v3XuuwbG/v3XjnrsHk3F7+/daI9OuPv3W+ve/de65AXAu30/Hv3Va0PDrq5Nh/T6e/dbpTrr37rfXd/oP6e/de66+vv3XuuxfkD8/wC349+60acT10Rbg+/db67vwB/T/iffuvU8+vf19+69117917r3v3Xuve/de67t9Lck/j37rVehP6k6x7K7X3piNrdU7dzu5N1VVTCtLBgqKoqqmmeQlRK3gB0i17X+v0HJ9++3r1T+fp1vVfD34edL/BfoHbfyB/mG7ixu8994zHLmNj9HpkIafd2Nrkp9dFNn8ax8iIz29BW/9fejT06voZEFeJ8/T5dU8/zAP5nHyJ+Ye6ZcLj8i21ulsIZKPZOysdG1AmGoFfQFdIdIYsv1Zhyefe6kGvVDRhpHD18z0QLbG+NxYMwI/wBxPY3Z2Z3d2J9Wpv8AH3rreOjKbX7Vr5aigNRQzBQ48hZGseB9T72Pn14j06sJ2BujAVOOpslMkKOgRnDAArZbn37r1PTh0aHaXcG1/EKdJqaLxgeu4HNv6e9569XPp1m3j2ZtvceKqsTUpS1MLxqhYorMQCQNLH/efeiNSlBwPTkMghlWU+Xp1WJ2xsGjaWtnwLmDyFmADAAEtyePYI37bZVid4xWo+3rJL205us7xht9y4QnAr0XLbPQG7u1t5bf63wBmq9wbzyNPhsekOppfuqqcJEFAvyT7g6SeeO+NuEyzBQKcSTjqSOZ+Uri3tzuzSViALE1xQCvWwX8y96Yn+Xt8JOp/h715UR0/fO7onxHectKQuQfHVMa+KhqFi9Q4b1avY43u6bZNjj2hZVFy3dOBxH8K9QTsdoeYN6k3YRkxL2wV8z5sB1T91F0Y+Fye16vP1Zajy1a9RLDM95SJiJmDE/8G/PuEdw3O9urO6lslIMC1qRjrKPa/u+S7zyfd80XwKeGmoDgfl1fXsDbW2ev9nU+YxFLHHSPTK3lZQRxF6vX7xO3ndNw5j3P6O4buB4fn1hvf2TbbfS20lewnj9tOiJ/KXvXa2QxVdiITTyVzrLFGy21o3IH+8+5m5A5TvrFhNKSF9Oi+uuRbhB8PVVnXe+8pidw1yGayM8p0m/0cm3HuZpOXBvX6MVAR69ZSe2W+XUUccoJ0gdDvW9gxSoz1FQqEIGcn0ki1zz9fZlt3Lcu0jwZcg9ZqbJzfHLtSszjA6BPcvc1dCz0WKrxIGuojV9Tf0tx7V3XJltc/wCMRJ3+eOo2505gsb2J/DcBgD59BxDmNwvVNmM3lf4Tj4m8pNY5RZFPPGo+3LHk+4u/0EjIPr1iPzJzZ+6JWk8UNnAr0Xvenzbotr7tkw1KHyeMoWVZZ6T9yOSx5UEHnj6+5L2z2t2P6JRfoTIRk9Q3fe5W/S3LPbSUQ+XVkuE7Qjz3RmL3zh6SWhhy1NfSwYNfxa9XPP8AT3IGybFZcv2wtbIUA/b0A9y3K63a5NzdEluq5t17tzO48nUfdVUvgaWZQNZF/Vb+vvd3ZLq8ZBnoY7BzLMkYsZnoB1wweIxg4kJYsL3Y/qJ4+v8AX6+yWVpa0bqTrBraVBmteuW4KaPHUwNIwsRfT9bH+ntTY3bpJpc0HRFzRsNvcQGWBKvToNZ5aqe+tDybLa/1PsRFlkTsOT1ECW1xYz6rhDQHrPjsXkchUR0yU8kjO1hpBuDe1j7Y8Dw1Mp49Hv72G4PHaRDSOHVhnxo+OdPkMtS5jckB+1VhJoksNVgD9D7JIzJe3Rh8h1I15Hact7Ct+ADI1KdH47Dixu3MD/DdtrHSxQU2keMKp1Aab3HsRxQRwJoTqGr7cbjcpzPcHj5dVeduw5eugnqZZWcI0rX549RIN/blMdJK+vRL5C5qDI0kjOshGoNzwbfX+nvXWz1M/ilfcq1RL4lHpsxueOLn/iPfutUA6VG09vS71rBjKaaXzsQBGXNyxPFh/j7917PRndqfE7cArKefJU8vhOmRC4sLMb8W978+tV6s56K6WXb8NIhhZEiNySBxYC/+v791v+fRs88aPA49JZiiRRIGBYf6hbEn37HXs16Kr2F8iMTgqdlirYPEDIhAYCwHp/s+9nHXjnh0RTtX5CJmKOanxoWp8upAUBY/7Aj3WpGevU6RfUe+d4YWpGQigqVgllJVbOAL8+9ip68adHaoOxd5bsx38Pp6eqepeMIjKH4Zhcf7b3vHWuhx6Q+MVTX5JN2dhn7xb/c09LMVLK/1X0N/T3XJ69506PnGtPQU8VDRRRw0kCrHHFGoXhRYcL79xHVhxqeupLaWa/0A9P8AsQLEe/fLr3Reu2ewMZtOjd6yeKN3DIiynSWb6AA/717uFqCx4LxPp1UmjBBktwHqeqreyO9a+bcs0P8ACZ1xxlJSsEbeB+fqHtb2Dd45o2LTJaO6uyg8PXqQOUNq5i2/dLfcbXVCCwrxoRXqHiO4oBIn+Uwx+VgoBPH0ta39fcG72YZB9XbjieHXUflLnmXatmh+tlDnSM/l0cXqDeOOqqqJ6+rgETFCHLCxJH4v7gPnhLy8jPhVxXA6GcXuTNKVe2mAB6Pfjt57RgoFhjqaeeR4lVkGnkH/AB/p7xyuuXd4luvGIZQDXo7n5vhvoAlw4cnorncIw0gqa+hkiid1ZjyAT+bAe5Y5S+sRFt7gE06iDm1LKTVcQ4J6qq7Yyss00sdIzSzFpBHEjHUzauFAHuedogVVUyYAp1jXzPOzsRHlj1y68+Ou7904Oo3Jk6aqpaaSMtFHMCpNxcEXHv27c2bbZXS2kDDV6Dquw8h7tulm1/cgqvlXoFd6bCbatdVRVrFWRnARzybHj6exJt+7PeIvh5r0E912U7ZKyS8R0jNrtAmcppHZQonUAsf6fX2a7r/uHTgeijaJPA3FZFOQQerZ9jZ+ho9mxTGsgXTTxcAgE/tj+nuJY77cLXdwsVdJOeugXtvznBebP+7t0YMgUU/Z0W7svccWXarQuskZ8oABuD+ADb3kjsKv9OkjfEQOoO91Lu2e5dIFolTToONn5nGYWlkRhEHMhKEgf649jS0mlWYBTjrGvmCxsrja3cgFs9I7ffbq0iz0kcikujrxc8E/4expGxKCvWNl1H4dywXgCf8AD0VPJbtrK6SUK5YyyMRHzqIY3HHv3TNKdP2ztpZLJJlcnkXaiooqGaVKif0xMQhYKGbj3vPXuq1uxKtKrd2VZJVqEhneAOralPjcg2PvfWuPSH9+6312Db37rRFevEEfUW9+69UHh178AAcj6n37r3nXrr37rfXYIBuRf/D37rRFR14kE8C3v3XgKcevX4t/vv6+/depmvXXv3W+ve/de697917rsgjg8e/derXh1//V0AW5P0I/1/8AX9+6qvDro2/H9P8Aeffurdde/de697917rsAfU2/1vz791r5de/P+H/Ee/de8uvG1+OB7914V8+vXufVzx/vvp7916np13ex9PHFuffuvU9euiPzcc/0/H+uPfuvD069b/Y/63Pv3Xq9dA259+631737r3XY/qQbf8T791o/LrxFh/j9eD+PfuvA1669+631737r3XYJH0/4j37rVAevAkcj37rZFePXXv3Xurpf5Tu7Y8bvKkxZkAdq6T03IJDnjge/daHn1etvyWmp+08LnKlgYxFAl2H0uQPe+vAU6NBU18f2FLPBZvJArqRcgIy3H4966sadJmLOQl3ikUSeQ6XXhgwJswt79w68Rip6ATsjFjEZeLKYFzHNIVdoAxDXP6jYf8U97WzlvHEcR6O+XjEt6DOtR/l6ZP4PWbgNM1eWeRlU2PNiTyLH2M9u2m3sYxpUa/Ph0M72/ubxfALERDyqR0ZDr3ZuOgmpYnjjVQq69SJzb+hPPs9QHzNOqW8CKoUD9vR18HnsRtugipaanjIEYF7C9/63Ue1quFGkdGqyJCNNP2dBl2nu58pQvTY+nXyTo6XsG/ULfpI9sSyYp0zcSgrQefQDbO+OtTvLIDIZNZSHIcAH08vY3UH2mitdTVbpLFaCTJHR/urfjFtjCvSVVTTB1iAYh44pAGWx4uPZlHbRxivRpDYJGQ56PRhaSmoKKGgo4lipoEVVWONEJCji+i3vUnHUejNajh0z7v2zT7mo5aGcKY3SRTcIx9a6f7XvcTqBR/PrUqLIlDw6JHnvhdLW5WbJYWr+3SWRpSrOgFz9Bb6e00kcVcefRW22l2qhoOkDW9Nb464yaSU9brEQMxVD9dIuQF+nNh7usbqKqcdNNbSxHUhwOlnsH5OybZrzit046RhDIsfmkLqnpNmP9Le9ePqUxvWvTkF8yHTN0bGDuDrneu26sisxwvGwkiYxsw1LbT6r8+7xCjCQtUdGHjxulVI6rR7Gze0usN9t2NgJImNFIzGCBlQM5BbUViI4NvbMrJHJ4y+XRNKVt5DOvHoPM58uttd0bV3BtTOYwQNPT1ipUSi95PGSpuf8faa4uI9ws5LSQcVPSae8W+t3t5PMHqq348VdJhe5s3jMhb7abLE0ev8A45Cp9GkG4+nvDXnbbZ9uklVhSpP7Osft6tHtZ3QjiT1sBbUNH/D6JoFvE0F10g20k3ufcFXIYyFm6Bz4Y0z0r5/tzAzHg/UA/Xj/AFv9f2yuSKdVWgNR0X7uSvpI9uVqVUiLCIKh7MB+Iz+SPZvtcbNONPHpXB8YIFeGetfbfdR/G+wJYMSvnYVzLpi9XBYKCo/3j3M1lbyvbrbIKu1OhTDE8iBBkmnVr3x86BzG5thR42WgaGatEciyywICCSL+oi/vKXk7YVs9mTWtHda8B1NPL20eDYKHGWHVpHVPxb/u3hqeHJVGqN4xqQMt+AOBz7GMapENDdC+KxWNBqz0ydodWYPbDf5GXjklL2JJA5H4sfd3jQjUvVZoUTh0gNo9W4xqhckEeWdiJA7gOoJb6gtf3pIQDnpqOFR3Hj0aHG0cWMoooTEhKqRwii5t9bAe1a9op0uUUGOsE8P3D2SPSW4+ij6/Sw/4j3WpBx1sEt2jp4pNt/s+RiAWudPHJI4At7uqs3Vgq6vn59NlZhQzaGtYLwbfpB/r7oRQ5HXiAcdJmt2rDJc2DX+vpQ2/oPp9PfiqsK0oemSmKDy6Cfc/X9PXqyywxvy/PjjP4t+PbJhU5PSaWEEaei957qSlpJmq6eMq6tewVFA/P09pXhFa9I3txXt4jqLR7XjeAwzQqzKLXaNeT/jqHuyqVz1sQ6hpboK959W0c6TzwxBZwslgEQLf/An/AIp7K922bb97tzBeoD6Ggr+3pTs+6bjy1uS7jtzkEeVT/k6KJitwZjrreYxtYkv2VVUW1ksE0h+OPp9PcYrtM2xTNb0/TOB9nWvcWeHmhU3WHtlXLn1NOjlxbsxlbh0nM0f7sTMpubCy8Dj8+1RzkefUPCoGluPSG2tBVV9ZkZZZAaYS3jNyVKX4sP8Ae7e98evHoV4VhxtNJNJwioTqNwNIF7m3/Ffej8utj59VZ/zGfk5tbZXV+e23QZKlly9YjCOJZiJFZotISwPP9Bx73jr3A0PWpPl8nUZnJVmUqyDUVsxmlsbjUQBwT/gPeuvDpu9+6913bi/H/E+/de669+691737r3XrH37r3Xvfuvde9+6913b/AFvfuvdeOo/W/H+Hv3WhQdde/db697917rkLXH4+tz/vXv3WjXrx4uB9Db6c+/deGePXH37rfXK/HN7c6f8AiffuteeOugL/ANP9b8+/deJp142/At/X37rwr59eFub/AOw9+69nrv8AFhf/AB/4j37r3nnroC//ABv37rZx14WH15/1vfutGvl14/Xj37rf29e+h5H+uPz791ryx142vx9Pfutj59eFrG45/Hv3Ws9de/db65W08OpBsCL8cH8+/dayeHXVja9ja9r/AIuObX9+63161ragRcXHH4P0Nj791r7Ouvfut9d2KnkEH68/4+/dawevEEWv+Rf/AGB5Hv3Xuvcf1P8Ath/xX37r2evfT6f7179177evX5vfn/WHv3Xvl142Jvcn/YD/AIr7914V65KNRsFZja9gORbk/T37rxr1x4sf6/j37reeuuP9j/vvz7917rsgjggg8GxFuCLg+/de67Ck3JBCg2ZgLgE/S/v3Wq9HJ+IHwV79+ae/sPsfqTaWUqIcnUpT1G6ZMbVT7fxCNIEeqylZHZYokF2Z3YAAe9E0yetr3HHAcfl1tE53efwz/kBdRLtjZdPhO7/nRm6GGjq98YKrocrgtoZ1aYyy1SRzatXgZgV1EAFSACSAff4evY1Anh/M9a0HZf8AMB72+QndVT2x3pu6s3LV5jKSVmW0yVcatC0haKMQLIY7ILCyxgW+gHvYpSnXmJLav+K6OHtjfPWO/wCKlro62gxgqVj1rO8aEXXm4JHv1OtVFOhGjwPXzVKJSbhxVSeCqxtEx55tYE+90690vExuDp6ZVpZYJnCgKsSKWJ/oOP6e9db6W+3sqlLTtRvriEgCkOzLweB+ffq9eI9elOgZIpDTVniJB0nyEWP4+h/23vwOOtcekhm6zetFTM+PyDylh/Zd2Nv6XBJ9+44HXqevRft2b23/AI+lnqat5XSIHXqMguNVgD70yq66GFR07a3E1ncLcwNpZc9XnfycOkn2ltntb5p95zR0mA6327Juzq4V8SBMluHH6ZYYKOSUeoqTey359hy45Z2sT/vSRKtGaqABlvKvUk/65PMW72kfLJkIWXtYkmgXzI/LokvcXeGB+T/e+8vkVvOqV6zcVfUPHSTyu6RSQyGNHWOUnTwB9B7jS/5ba43Sa8uqsZTWn+TrI/kyLYtp2y3ViCYgPT9vRZ+3u/Vo8ztyDbtSjLS1aRiKKRtRXUF4Cn+g9rrPkyG52+6tlWniLQYHQv579+5tq2Btosm7JFCkCo6uA2h3vj8p8bMZBWTLHkzRsNDM4dbwH9TN7wd3bkG82T3AcyxlUriowc+XWIV2Zd717ouddajqoLflb/FcxlK6pnLosk0kB8hK3Jv9WPvIWxhWK3VUGaDoNJLMJBbgYPHov+FzAXO1s2tWClgeeLfnlT7Eu0wukokGKnrLj2qsYzYmNxXtweoW895VUzfZ4/VLNKugRxu+o3NuLEc+x2bQXKK9MjoSXXMEm2XEliHoo6Qmc3Lt/q/bjbl3Q/nyFWrNT0pZvNDJb0XS9/rx7FG07VqIZx29Qpz7zytrG0Fu9XII4nogHZPyI3xv/wC5xzVpo8GXZaaniMiTiL6KWkVuDb8W/wBj7F8VrBB/ZKB1jhdX93fSGS5csT8+lp8RPj3mO++z8Pj4oWkwuPyMFRlpGQSLLHGwkdGMlxa3LXHtQBXpIOth7ujZuC6z63pdlYaMJjcZQI62UAeTxWa2kW+v0964daIrw6qSkovv8lO0APhM0vAAJ/X7qxAWrcOnIlZ5aRjPQn7e2a+RkgjiJuSAFFi1/p+PYb3KaJR29TNydtt+7KJM16NXtH4xvuWjSSsHJAsGVP6fm/8AxHsI3u5PGCU49ZEbByVFeOhvcKxzw4fn0I2I+F8E1Q6yU2qOMjS6xxHgi/u/L293Nze+BJ0n94PbXYdm5cF/ZDuoTwH+ToScB8RMViK2OoFJqdWvcwxEX/wsP+J9yiyAjSfPrBeGVon8ROKk0/b0u9zQpsKjjpKRBG8YRdKaUIvxzot7YitYoJC6cT0a3m931/bLb3DVVei5bv3xW1iywtqK2Ia7k3/1r+1A9eikny6JV2fvgpDPjhb1eQED688fj3on0690Vi3qYj+07MOf6nj6+9db+fT5iMBX5ieOGnjb1sFBsGuf9Y/j/H37j17o+3xj+PmRl3dQ5Gs/yZI56eRhKq/uLYMRz/X+nvdOqk9Xbx7Exrx0sYhj0rBAl1iQG6rYmwHvfXj8ulMaGgwNBKY1VGgQtcKoXgf7T/xX37rfzGOquvkr8o4qOtyW0KGN5a3wzRBonlUqxaymycD3rrXRS+tNkbg7Nkm/jTS+CVmkETFiyiQ3AGr37rZPr0Z3b3xv2pikD5NQgiGt/KqN9Dc/X34CnWiVHS3ooOscPMmOhmoZGjcL4lWLXf6HhffuvA+fRtOtMDhFhiyeLpEETaNLGONgWK39Nx731sDHRm8ZkfQiMyR2RUsBpAH+A4/3j3qlRXr2eplZWRQpq8gOk35+vH9D/wAT71Ty8+tnh0Hef3rT46GWRpUQlSSWPIt+ePd1BYhfXqvDPQS9efHLd3zC7GpsJjZJBhcdUx1dVVcmEwF7sAzf4e8WfvVfeF2T2K5Jl3C7lCTMCpFc8PQdC/knZU3bd43nFUjIPVsWa/lKdb5PacGBqIoWq46WNTVCmp7mRVtcsOebfk++Hm0f3gO4y8xy3kkMhtWdqfEaj1xWlfLrL/e9w2NtkjsLKELLGoo2BkfOnVbvcH8mHc+Kqmn2tVwvSq5eJY4o9QBPH6b+8xeRPvqcj8y2wE5MbHjqqP5HqLp+bN/s1FnI3YD5EnHRd87/AC4PkPtqgC4UVjGBWIMMJuNJsL6B7ljb/ff283OSktzGtfVh/n6NBz3eQQrHExr+fT1t34e/JHbm3hncxT5SfQSjoIZdWlfqTx/T3S690fby+v8A9321zFq/0w6E20+5tzaUa7YkfaesWU+Ova26cUsskdXSXcxMsyFTxwSb+1MPOfLm33GhWVvPHRpuPubBuDeDG4FfOp6R1J8VMVtKRM3vqSGqkgcTCEtCD6TyCrf4+1svPtzuI+m2dSK4rQ9L+WLjYdxvxJuMqmhB6Z+y/kBtzbeLGAw2PijhhX7eKKMIpPjGlbBPr7d2Pk+9ubn6++kJc5qa06lLmHnjabK1FjtwBUCgp1XhvfZ29+18tU5nHYyqjon1SqRGSrL9Ra49zDtm5bbscaxTOKjHHqEtx2beebLwzWsZVGPSz6f+I24921eqolaCaNwPCyLrBH50n3NvInLUHPELSRSBhkafMfPoAc17VuHKMoW4Q1/ipjo2PYHw87P2xsRq/EV8kq08ZdqWNCJGVV4sB/xHsz3P2Ou7CX65MgfYejzlP3BlgdbdpCOHqOqws/n81tytqsVuenmpKmCWeFzUFkJZOL2P0/1vZptGzSQN4UgI04z8uk/uRz1oh7TWvQX5reTa2ShYt/asrni/+sfp7Ey7aqvqHUGy85vPbGEnj0hZKPcW4qnXHSVFQW4VolLgfkX+vs1UUWnQDmfXIXPQl4rbm29i4lt074qYEESCX+HzaY5/2+W9L292HTRPmcdFT7q+USbloptt7CglxeHOqF5jeNii+ktEVIvf8E+/da6JU7M5aRn1O7FnLcsWJuST/j7917zp1w9+631737r3XIG55ueLC3v3WqU4ddXsTbj6j/Yf09+699vXXv3W+uwObfT/AF+PfutVxXrwBJtx/sffuvE0z119Pfut9e9+691737r3XfI/w9+69x68SSbn37rQAHDr/9bQAJJNz791oCnXiLW/xF/fuvA1669+6312Tf6/X/iPfutUp11791vr3v3Xuve/de67Nvxe3+Pv3WhXz669+6312bfj37rQr59eBI5Hv3WyK8euvfuvddgX9+60TTr1+Lf4+/de+fXhb6n6fT/Y+/dePoOuvfut9e9+6913bi/+NvfutVzTrwt+fp791v7Ouv8AW9+690b34Xdmt1r3Bia2SpMNNUNGPVKUTyrIPoP8QTf/AFvfh69apU9bEHZXZEma2thNxUIknn008muFiSqcEG49+PCvWhg0PRsuluw13vtamjaS9RTUsUTxs2ptXAP19+r69Wp0uKtGx801Q9wqB2UMb3/1h78DmgHHrXEhfXoN71OUyj5qpUmkp9UWlySrECwFj/X6+xjtNqsMHisKE+vQw2q18CISuM9DZ1vsqqzdDWZmWmKQRyWiLJZdINwAfZ7EjN3U6Po4WdfEIx0J+3Man8TSJfTpJUj6A2P9P6f09uqAWoelMa93dw6NFguvJctjTUIjOwFwQNX+t9fatIyV4dLhAKavLrkerI4phJWItgf0uoPCn6gH/iPfvCBGRnqpgGqrdCntihocOUpqaGNRZb2RfoG/FvbyhRSg6fVQpHQ00eWjp440RQCbfjggc39vErSnSpTivT/TbgcAaSQP+DW4/wB99PdTEjdW1kCvT7T5tHZFe39b/wBB7o0PmOt6qdOT5eJFKxsrXH4I+o4tf22ICcnr2sHHSGzNAubqRLIikhdPqXVxb+v+t7UBVVKHqhUNjoFN8dHYLO080r0sYncPZo41RgSOSCOfbTQLINK9JWtY5MkZ6KXlenK/aFVKmOeqWllcmSMSSfX+vp9pTb6DRekD2rxt28Ok7mupcXuagelq6adnkHqLln9ZH1PuhgDih608CutRx6LNvL43S4UT1eKiaGJY31Ki6GNvqGA/r7SvalDVOkL2TJleiRZjYlVt/edLnKeOSkmp5lV5Sui7Bwbkj8+wPzRypa7/AAsjqNZHEjoMbtscW4cRRqcerKNhdq5Hb226CfLeSalFMn+Uaj41vyDf+nvFPmf2y3axumMSFkr5DqKNz5VvbWRigqK+nS3qu+aKupC9ArVDqpA8TfX+n0/N/YQi5J3mSQIkLfs6JY9mvXaiof2dEW+TPa+8spj3x2Npq2NKtWS4LBR5BpIuPci8ue3G4ownnQiny6EO38uXfxSIadJz4k/HWbJZqDdG5YDOZBHUFZo9Z1tJrI9XvITlTkRLZ1u78A04Y6knl7lkJIJrkZHDq9LaOXxe0KKlpoaSCKOnjCLoiVQBYcED/ifcwxssQCqMLwHUmoyRRhVwB0NMXc2DNDoWVFlEf6Q30YfTTb6D3WkZbUcZ6fE6AV4mnRcd5bprd4ZL1Mxp0ey8k8E2B91dqnHSSVxI4p0LHX2Fmmp4AV9AQDkfXnjj26hFK9KI1IGOHQvnbEjr6rWFiPp/Qe3RJEOnqEjHXJNuLEwLIpIFwdP9B78HQfD1ulKevU80OlLD8A20j/YAf7H37xM9a0DpO1mPlIY6Tzzf8cfUH8+7EajUnHWvkoz0npYXQMrpx9GtwP8Abj/be6kE/D1qlOPSfrqBGUuFC6QTpsOf6e2iK46qyilOg2z+MhMcpKC39NN7+6Nppjj0wUIFegqnx4jlbQlvpxpt/vvp7YAr0lYGtT0lM/QAwm4BsCQNPLEC496daig6o6scefRI+99uwZDG1FZTQKtdTyko6RhWFgCbsOfZRuNrHdxaXGei27WsJiXz6AHb+byVVjoofumhix+iOqR3I1ernj/YewHcwNbOUYYHDqPrqB4ZjqHRzNjVGIrMRS/astwimoa4uzfW5PtmtMjpLSp6T/yP3hHsLp7O7mp2KyUcc0cZUkMbw8XP+F/eiTTqy0JoetIfvvtDcvZ/Yu48pnMlVVVPHlKtKGmkmeSKGKNyiuFJ/UwHP9Pp791XHHoEffut9cvTb6G/v3Ws166Aube/dbOOuvfuvde9+6912CQLf6/+8+/dap59eIItf8+/deBr11791vr3v3Xuuergg/U/n6e/dV05qOuHv3VuuySf9h791qlOvC35+nv3W/s69a9yPoLf7z791qvkeuvfut9e9+69176e/de697917r3v3XuuwSPp791qleuvfut9crAWJ5BF+Pr791qteHXX54/2Hv3W/t66+vv3Xuve/de67NrC17/n+nv3Ws9et9L/AEN/949+69Xq/b+R38Xfhn8t96b66f8AkljXyfYe7I4sV0+iVbU/jz00X7RlVWGtdX9k/X36nmevFvwjJ6rJ+eXQ8Pxp+Wnc/SMGNlxFDsHddTg6OmljaM+KliVHkVX/AAZA4/1x79x63pK4PVpn8kj4AdIfLbEd+dqfIjBTZDqrofD024N0Vas8Cx0MjpE0SVQICtdxb/G3v1RwPWtLcR0Z344/y/8A4Md/9CfzZe7cLtetqMJ8b9lT5/oeqp5pTHjqj7uSlgNXoOmX9FmDfT6+6moNOno9DoWX1I/Z1qsEDQp5uWcE/wCAC2t/tz7t0z59Xp/yNPh78d/l93jlNo/Inb9Xn9q0s1IHioneOqjp2i1zeB0Is31t73xHXge7Pl1Zz8hO3v8AhNR8b+6OwujMn8C++N35LrbNz7eyG5E3cKFMjkaZVM7JSu4IT1BlI4N/6+6d1enqpT/Y6afkV/Lg/l6fMX4PdtfM3+Wptmt6+xfSe04dyb82NuvJmv3DRLPOsBVG1EXVmta3u3AV6bprHb68fs69/Kq+C38s/dXwU7S+X3zR6v3bvOh6p26c5W4natTLS5DJSxxCQ0xYGw13tc/T35h59ViIIofX+Q6DKD5l/wDCZiZFkqP5dPfVM5JDQLvZJLKrWU60kIuRY8e60PT9Y/8AUOqvvkRQfDz5i/IDrXqT+XV0pn+qDv3cVJgaCg3hWtVVC1NY/jiSWoUsXBvf0/0/x92HDpkir1Xh1dHvn4cfym/5U2wdkYD597KrPkJ39V06/wB49n9cZv7Wu2/Vut1XNerSj8Eqn9L3+nvR49bSgyx48K+f+x039f8ASP8AJU/mZ7O7L2r8U+t8v8Vu6NtYc1G3j2hn1qaDclTJE3gSgkV9Ik1D6X9+Hz62wrkfy8utWTvvpvOdA9qbs6p3FkMdlcttPJS0FRkcXIZaKp8bEJJCxvcEAHgkf4+99U6tL/li/wAn7tD5q5WLs7sPIYvqH45bVlhzO598b7eXD4zP4qi/ymro8LUz6VlZ1GmyEkg8e/VqcdbpQVP+z1Zp8wP5vXRnwa2Flfh3/K723FtBqKmk2/2l2ykNLVjexMZjeLD14BkjgUHgowJ4uffq1NerV0rRaVP7B/nPWqVvXe26Owdy5Xdu8MxW5zPZiqmrK2sr6iWpkeSdi50tKTZQTwB72TU16bUClB0kza3F7/n+nvXW89ZUqKiMARzzRgfQJK6gf62k+/db6fMPuvcOCqY6rHZaugkjNwoqZjGf+DITb37rVOhaxnyQ7RxtTBURZlpBBb9uQtpYCw5N/wDD37r329Hu6s+b+yZKSix29MdVtlpQkc1WwZYNZFiBJ9Pr/X37r1T59GHx/wApesNx138Ho6qPHtDbVUTThUN/pyfx/j79XPXvOnQnU+7KDICN8NunG1SNY+JJkc/S/wDX8e/U8uvYPQ0/HXqB/kf3jsLq2KppMnRbj3BSYzPPEivHi6aaQa56wj9KgXJJ9+4deoCdJ4evVhX82b5r9R9EbZ2X/Lh6srcbjoOmJKmDc+bxc8dJFn5q8j7lpWhP7qC4A1e9Y8+HVi7RsCho+KfIdUVtHtPMUiDD73xlJEx8kkX3aoVZxqbgH+vtK9hbytrIz0ILbmrd7aMRJIaD59IvcU3WnXeOrdyZrcePz9bTxGakgp50meOUXtYXJ+o/p7digSFtSDPSG+3i73AEXDVH29Fn2n88Nz43ctfHmXrZNmSMY6GghkkLRQg6RrjH9Rzx7DvMHKOz8wsJbuIeIvBgM9PbZvd1tqmNDVD5dHCxnbmxOw8GlXRZ2gxK1UasYamoVZAWXktqN/cZXvt3uVvdVs2Bj8vl8uhRt267PIfHuQAx6n4HB7IcVBj3fijM6kyS+ZLDUb3PP+29n9rytdW0a+ItSePU88p+5HLu07c0QcK1KDpA743z1b1HQVuVr8jR7jyrxMaEUTrK8U5OlPQL3N/x7GNls5RArClOow5n9xEubmQ27atXAg9VU9o9nZnsrPT5KumlSiV2FHRlz40TUSHZPpf/AHr2Io0WNAijqHry7mvZzPMSSfU9IbCYet3BlaLD0Ch6zITLT06tcAyN9Bx7vx6SYHW0P8L+nKD499J4zclfDTDc+UozHVSmFVk/djF5NX1F/wDX9+8q9arinQcfIftSpylG+PSVmGiVZDrvqVnAsL+/Z6tjol+3I/JUkhPVI5sDcgXPtLdg+Djj0ecuqjbguvh1Yt0P03Nkno8xUxnwSqGUsgIFwCTz7jy/ll1+EoNesw+Udv22OFbyZlAUV8urLcJs/E4Cgpy4itoUmyAE2X6ezCy2Bp4fFkGT0Feavdu02vdvpLE9qeh6ly7sx+NP21HSj/Us2gXPH9fZxtnLsVlJ47fF1GvO3vHf8y2Y2uIkRjHHpzxua+8Vn8IA1ErZQQLi/sSnPUKAUqT556Bbdu0DubL1sjrqTTqAtcLp5tYfT3759erX8ui1b060jhp6xoovUkb/ANgcc/Un8f4n3vy695Y6p+7YaSk3jW0DkkRPN+Qfo1rD/iPdfPrwxw6D2kiapraamjuWmfQAvJNyOPfuPWz1ax8aOhqXOQY2tr6RSyyxszSRKQQEv9T7tw6rXqzys6Z+xxePqdsLFSVVN45ZQqLG8ixrf6j3ry68ePT1Rb+psFElLnf2J4bI8sp03KcfU/4+99eGOijfJ75GHb+Fnbb9WXlkiCqsEhvc/iy/n349eGT1Thit05Hdu/qnP56KXyTzMt5tTggtqvY/7D3XUnAHqximI16SQerOOms5icf45I6G7PFFyqADhfzb6f4e2Jry2hHew6NNu5e3fdGC20TGvCg6fuwpN3bormo8DFUU8FR6dSgqArfqNx7IZuadujk8JXBP29SrtnsTzfeWX7wkhZUpXI6KJuDq7dmzd0fxnJ1lRLTq6TSIJnNwW5UC/wDt7ezW03KK7NIzx6A/MHJt/wAvr4l4pHR3OoflTs7GQ0206kCmqF8UYad7F3VRGbX/AMR7M6V6Bak16PltXclDnYYqinlUiVFcHVxZuRY+7BK/Ca/Z1smnHpTZGelgjaSerjCBbgFh9P6/6/vQjk8x14mpx0QP5I7nqqfE5KTB5FUm+1qAqJIVJcXI0ge3IVZZA5Hw560x8vXq8T+SbSVlP0M28s9TB8lW1D0zVUsQ8jxqeB5W5t7+f7+9D5hG6e4tvssMhlQKWaMNUVApw6l72/b6OJp2GPXq5XIbojZLQDWSeQhuRzaxt75m22zSl9KKIB51AGOhtNuXiKSKnoIdzb4r6NyEpCbG+qSMMLg/4+5S5Y5C228XW8uT/Caf4OiW4vH/ABDppxnYMkzqlZQ0zqxtY08fKn634/r7NN19t4FQmyuJEb5SNx/b0zFfOmSBT7K9L2l3HgspH9jPRURp3XS0TU8Wn1H1Agj3HN3yhzLs7i9s7uYyI1QS7Hh0aDcIpk8KVRp+zPScznW+y8lSMlNiaRFd9f7VNGAGP1J0+xPy/wC5XO233v8AuwnNFFKFjXpDcWdkyUgrU9ALuj4c9Wb1WafLxRqZFIKAqgUnnlR7lSy+9NzZy+6WsEZenmATX8+nLLadyhbxLVmXPqeqZflD8MurtpbmLY+BJhFPLJZQGAs1/p+b+87/AGo969+5q2ZLi5UoWUccdZP+3vJSbnaLd7oCxFD69B5gafae2scuIpsbRRIiLHqkp47kAfg+xhfPu24z/VNIx+QPU8WTbXt0At4Y1FMVoOi/9k7gzuycrFubZdRDTRJIJKqKNQodEbUyqq8X9zn7Tc47vyzeo0MrI2oVyaEefUd8+bdt+92zrcRq+MY8+jO9a/J/H9l7cGHqfDT5WCBEnFRpbzaVs4VG4uT76c7Bv6b7taXRFe1a1zU0yesGd+tf3VuLWymhqSPLz6r4+VfWGzt9VNfWpQJjsmGlkMxjSJZCOSRb6359p9w2+2mJkRQp+Qp0FNzdr5PClqaeZNeq9MN1lhtrNlM1nYP4jj6SOVxBGupmEak2UD+vshk25lXUuegtJtxQErwHRb9y/L/Ym3Gr8btbbVbQ5Glklh1T01gssbaVN2/1gfZYwoaHorOoGh6Ib2P2zuzsvIPV52ucwCSRoqWKRlhCs111oDYkD/D3rqvE16C/37rfXOw03/P/ABv37qte6nXD37q3XvfuvddgkfT/AFvfutEA8evfW5/2Pv3XuHXXv3W+uTXvz/T37rQp5dcfp791vrsm5uffuvDGOvC35/3j37rRr5deBsbj37rfHHXuWv8A4D/evfutcOuvfut9f//X+f8A+/de697917r3v3Xuve/de67b6/7b/D8e/daHDrr37rfXf4/wv791rz68Tf37rwFOuvfut9e9+691737r3XvfuvdclBJsDbj8e/daJoK9dc2t/jf6f7D37r2K169bi/8Ajb37r1c0699SP9gOB7917h14ix+t/fuvA1669+631737r3XvfuvdTsZXzYvIUWQp3eOWjqoKhGRirftSBytx/Uce/daPWwh8Ru69s9qbCp8JlKmMZClhSjFNUEebWo0kaST+fz79wyerKrydqDI6PH0vkv7pb4TFRyEY6ocWW91uZCLD20tzbvJ4KOC58vPq/wBLdAa2QgdWXZ7rDL5XZn94YqSVYZCrpIVZbxtzf2J9t2WeQi4lFF6P9r2WeZRcSDHQI56josXhKXERhUqpaynEn0DnW9m9ighEQRdCc6IkCKKU6sDx+DxG1+ocdJGkaSVdJFLIxAuHKC5uP8fZmgC24r58OjXEdvUdAntKJqueaoVeVlVgwBItfg+2EFe7pNHqOT0evrCrvivt0CO5IVhbUf8AWt/j7MoDVadGsWkppY9CJWYD727SJZrH6DTbjjge7MpHDp5kp9nUDFbUJqraWFiPre1ieTe/uqrTubh15UHE9K2Tb8SEabm1wSDx/rEe7UQnHVtJB68KDQDYkL+Rckj+n093otKDr1KZPTRXyz0SMyqwtcc3/wB9f22R5Hqpap9OmWi3FMZNEz3Jb0g/jm319+DH4T17VTHr0uqHKLoIP1P5JF/e6as9WU4r1znqmlJQW55vwb+7hQBUHrwkA456ReaxFPWsXeNWuDq4/r+ePbb9VILCnQI5unixNQ7R0+kKx4tyf8efaZwUNR59JCoVsdA/vDNJUU08a0wb0sunQDe6/S359ts6npiVhTPVfPZ2LSreYyULR2keQMEZbMOVPA9lkyAmvRPMCWz0WvcHZG76CGLarTucS7Kii1gq/Rbv+PZdcxQ3A8KdQR9nSGYrKvhyLWny6GzYlcmBwy5SudpacxM93JK3Yekc3/PusG0bZAdYjH7OrxWdnGuoIK9T9vUeU7UzxEtMrYeGdUUNGLFS3BB9rooYS9I0AH2dKIog5ppGn7OrKeouuocPSwRxUyokUaItkAPB+tvZzDHQU4Do8todIwKAdD1kdiVU8WqGJ3DqSAt7/TgAD2+yACo6UtFXj5dIBuushHKXNNVWLC9y9hz/AE9t+FU9MmIjPQgYfr2ecwxrAVJ8ZLEH+v5Pt0RiuenVirQAcejN7U25Bi6OOEQqJFQamt+R/Qj3SVguF6WotBTpaSQIgC2FiOR9ORx9f6+0wYnPTnTfIo9RBFhwLj8f19ug46qMnqPFAHLKvAYlj+bcfn8e9lqDreSc9emx8bqTYGwNwF/H9efe1kIPVcnA49JHIUETFgABYm9gL8H+vtUjkih4daKKOPHpIZCgYG4BKhSOV4PP9f8AH35wPw9N8B0G+fol0uFBJF/03P1H6bD8+2jw7umm7hpI6DGrxpZz6SAPwRb/AG9vbTLnt6YKEcM9JTPYq1O50tq0Nbj82/1XttgfPqpoBnom/YmOL/dRFLqXl1B1JUm3B9o5ABg9Fk4XzGOiI52CpoMjVY7GBlarmu+nkXY8nj2Gt3tVlUuBkdBrdLZZo9aCh6FnY2UzO2KYU+QeRDUWaJmJW4H+v7ClCtR6dBSlO1us3fstRv7qbNbcjkMhqqeZ/HfkssPBHvZGOvE5600+39uTbW7F3Th5kdDT5ScrrFrq7ahz711odBsCRz/vv6e/deOcdcuRZv6n37rXHt64k3N/furDHXXv3XuuwbX/AMffutU669+631737r3Xvfuvdd6ja3Hv3WqZr14EWP8AX8e/dezXrr37rfXZNwB/T37rVM1669+6313fgj+v/Ee/dapmvXXv3W+uQW4J/p791omhp10ASQPpf37r1cV67a3AA+nH+v7914V64+/db67/AAP9j791rrr37rfXY4PI/wBh791rjw68fr9Lf4e/deHDrsED6i/+wH/E+/deIJ4ddX4I/r/xHv3W+uvfuvde9+690c/+X13LF8fvmN0L29NkBi6bZG9cbmKipMphi0QXOicggFS2m4Jte3v1OtBtPd1eJ/wo16R29V5TpH5obZx8VQnywpshu6XMUCJJT1MjVEiPGrRC4Jkv6Sfr70COA6swb4m8sdGrqP7vfy4f5LVA6+PbO4Pnp15TU9fA7CDJzQ4+MSMwC2ZQZtJ5vz/re/AavketZjHy/wAp6Db+SYS38nn+cSfrq6igOpuT6q+U8H68+6tx/Lp5Pg/P/J1qGg2VSPqGf/eh7v0xx62bv+EzdC+U+TeYxscixSVk9LCsrmyIZKfTqY/7H3vyz1oZbSOjzfN//hNL333T8uu5ey6D5UfG3B4Le26qvcENFufe1FRZfEQ1ekimrIS4EYUC9iCbk/4D3Qmg6eVNRxX9nQq91bR+Mv8AJf8A5Z3fPxy2R3z133P3Z8kthpt7flRsTcKZ3G46enrI6jxhVAAJdbLwb+98fz6qaxGnl/hPQw/yH+oNkd3/AMsf5Cdbdr7gw+1tsbi2TKuXzWcqEpMXSo1OATUyvcBfr9fdn6pEKLj1OOimUP8AJY/lzRYlpF+T/SVZIjzeNju2mlkk/wAoYBQWF+P02vxa3v1RXrQRqefRAv5Svx36rw/85OiwGGyOL3Hgen+xqOr2VU0NQs+MzFTSKJY/tAp0ynVyOP8Abe9eVeHVkqr04ih6IT/PA3Pmd3/zQfljk8zU1D1MvZOWP200jtHRWlY/bxQtwij/AFIHvQwOtyYz8h1Wdsbf27ettwUe5dn5irxGVpJFlikpppYkkI+izRRMNQ/pc+916oP2dY9+b23H2JufJbs3XVNWZ7KSK9bOyurSMqAKxEhJvb+p9+J69w+zqzvsj+bL31uT4X9TfDDaOd/u517sPF12OyMOOoUx9XXRVsYE5lr6dlaR2IFne9hcDn3vA69Vm+LA6qXkOrU1y2ptZZ7F2LfUl/8AE+9deBz1i9+6t1yIsAf6/wDFPfutA5p10BcE/wBPfuvE068DYEf19+69SvXY+h/1vfuvHiOugSOQSCPoRwf9uPfut9ZUnlRtSyzKTa5SRlb/AG49+61TpdbV7I3ttacDA5at1MT44JHmqrMRb0Je/H9Bx7917hk9bfP8uzac3wM+AfcPzu+Rs1Njd9d/9f5qk+PH8bmSGooN0Qoyx5Kmp5DqXSbBbH6/n3rOqq9XGgJR8+f+Ydai3bncG/O7uwM92d2HnKrNbu3HUvUZHIyyP5W5ukYcktYXP1PvfVOkHT5vL0ZJpMpkKfULN46uZQeLfQG3v3XqdR58hXVVzU1tZUFv1Gepmlv+efIT7916nUUtck2UXFrW/wB5/wBf37r1OpMdVVQqPFU1MYFjaOeRBf8A1lPv3WvOnTtQ7p3Hj1lSizGQiWZdLr9zI/B/prJt/sPfuvHHmR9nTc0uSylTHE81ZX1dQ6pGsk0tRJJI54VQ5J59+Jp1v59Hm2L8Be1d57Oot3JjMrBT1sYeONKZSASmvTcqb2/Nvcc8we5Wycv330Nyat1L3J/s3zXznY/XbXEWX5DpQdXfDHtjBdqbXqqnC14x2Myqz1U01M6gJGbKL2tY/n27t/uRsN+P02AJ9SOjO9+7/wA+2cmia2elfJT/AIerd+zey8jgoqbaUV9FDSU0bCIEICkYRuP9e/sX2m5W9ymtWFDw6A2+8jbrs85gkiYEcRQ9E13JVVuerW8upkcksrc3ub+1RuYa0B6If3Ffqpd0Ip0IvW3V1Tla+lIhPg1oW44sWubMPbpUSrTj0XQzyWk2sYI/ydW99W7bfDYijoBGqxxRIP03K+nm5HtE21WrNrIz0KE573uO2+mjkIWlOhbykQkhVBMLAfRSOCPyefa5FWJdKigHQTmle5mNxOSWbia9JSnxfnnCuC4121WFjc/1PvdeqAU6Eejxi0tMEjQ/uDTwL244v72cdaFDw6wVNJQ4qGSpqmjjMqWLOR+f9f34caHrxp5dE77q3pjMHj8hJTTwHVBIRpKkGzfge/da48OqWdw4qp7C3nVy0SFpJZpRrUXvqf8AH/FPfuPW+HRnunvh5uHMZairqundqcMJgxX0gcfX36nWyfM9XXdVdWUWzsPSUzQJ5oFTUVH0ITSBf36oGeq/I9DJNVrS+hFGkgBQefoLEEe90pk9brXoAe29uYfc2LkKSLS1kWp3kVxGCfrb37h1rHVcG9dn7XaSWmzNUtWsDFvVKHACn6Nf6+6nAz1ZQzONOaHPSGh6+64qjHU0ccMbR8lvSLkG97jn2C9x3C7t7giI46yc5M5T5f3nZUlukpIOP7Ohi2VlNoUtQuFxyCeuUImiFtTcCwFvYeu/3nfKxiBP2dTBy5a8q8vspVVqp86GtOjJJnsTtvEK9ZQiOpCB9UqEuAwtYX/H+w9x6Y7eG/KTzhZAcgnPWScPP1hPy8LRbZQtKAheim9x7tbPUE4w1C9ZUyKyxJTxs7s1+FAA9j3at+27bR4k860Hz6xR9zdkPMkP+LxfsHTH8S/gp2x392Vj8lVY7JYTGpXQSTS1qvTxeF3DcFhzx7iH3o+9Dyr7fbBLew3CiSJWJANSaDyAz1Btp7UbpICGt2IGa0PWyB2R/L2zXXew8dJs3cuOOTioKdZ4xUIZPJHGCRwfrf3z+9qv7zI81c1ybbeRzRwqx0lo6BlrQEEE/wA6HpUfZreLsUtIixHGlcftA/zdV8UPxT+VO7d0y4xZ6lsVDOEaYBvA0RP4b6Ef4j3m5uP30+Q7LZBePdKshWtCRWv2V6Au8+328bLdeDdxFafLosXe/wAbt9bO7Ao9ubnrlioxJEaySaUCNopHAkGo/wCv7FfKX3j7Xm/ll9y26r+IpAoP2dBq52zwJtD8erSOqflBgugOtNv9X7CEdY4amaolorsPMyASlin+P094F88+yi+6POc/OnNANQGCq3kK/PoSW+6NYWwtoPXiOrNOjt97y31jKTMVFPUfb1WlgxjNrnk8f737w493eUeT+V5pLGMosieVRXoSbbPdXNHOR0bqHbEOYijir6bU9gWJTT9f8feKP9Ydy226c7O1EA4mtMdCaKzE9PEGesdd1ljkjBhiAY/0IJH+w/r7UbV7nblMSbm4jenkpqf8PT1zsbRiqqaHpL1OylxoM6ziLSAWLvYAfX6H2fWfuPBukxsqeIeGB59IW2m5UAohJPQE9m90Uey6OTF46qiqcrpKKqEORIeBe3vIv2b+77zB7wb7GthauIHOXKnSOpI5S9v9zuI/3tuqFLdc93AgdVf9o/LTuTbFdUmGrqVppRJZQjWChza1j76PQf3elvtFism4RRMygN8OeH29Svtje3ctI6DUpzw8uPRNtzd/5nd8slbuSqdqiRnLLKfUATc8H6f7f2gg9rI+UJP3dbwGIJjhQY6lvbL/AG1bYW+y0008ug6rN10OThZ6eUeWxYWYDk/S3/G/ZvFtk1swWQY6S307AlaEN0Dm56qqrKeWGaYFCGUB+QP6exTYIkLB4xQj06B24TTGqMa16AmiytXtfMxT4iqaKpSbyTqsllaMHkFR7zd9k+b/AK+yO0XT/qJkfMdYpe6myXFvdDcoxgkj+fQpdh5qbfuCoKijJFaxhiqQnpYC2lmb3PMo8UY6iWVmlQBegm3513/AtjR5Ap5/uvRMVN10sPWCbfX/AA9tSQ6I+04PTc0QSLj1QZ8oMFQYjfQloIkiSphkMoAALOJL39PH5PsK3i6JqdBK6QJLT16LP7SdJ+ve/de697917r3v3XuuyQfoLe/daFfPrxJP+w9+68AB11791vr3v3Xuve/de69/vHv3XuuRv+n/AFrf7Hn37rWOPXTDSbf77+nv3Xga9eAv+be/deJp169r/wCxHv3XuPXvx/jf37rfn1//0NADi305v9f8P6e/dazXrxvxc34Fv8B/T37rw669+631737r3Xvfuvde9+6912D/AF5H1t/j791oj067FgebH/W5/wBv7917JGOuvqf6f6/v3XuvG1gPyL3Pv3Xs167UgfXkf8T7914g+XXVz9Lm39PfuvU8+uvfut9di/49+60aefXZ9RuAbfT37rwxg9cffut9e9+691737r3XYF/yB/rn37rVevEn6X/w/wBt7916g49de/db6FfqHtDM9X7qocxjZpBAaiEVMIkcLoEnMiov1I9szqzxMqGhoejDa7mK1vklnGpKio62xvgTtKT5BRYjdk8UsIjNMJPyWbyhyxLeynkXlHcZ97fddwYlEOB8vLqb98i2O4sYYrBQC4qeHHrZc3ZtDGUnUNJgaGmUfbY6OORgqs3kRAL3H+P19z6UIZkHwjh0gEUa2ixKKUHVJ/amHqsTuVJJtXgjyMdrAhRaUaeD7KZVIanQZnGlwT69Gxz+ayGW2PgMdTMRGaOnUqur6sAOfx7WtqMYXowkIaNR0Kewdgmg2VU5SpQ+URa7gf7Tf6/T25HGVjr0/FCUixw6Gf4/QNkfvp3cMkFQ62NvxwPbsJ0oWBz0otVDceHRkqsImosB+f8AAAX+l/b6Vr0vbhpHUekdRqI03J4P5F/8R7u4J6bGnh06LAzDgXVj/hzf/G/tsuB1sAnrmaAR2LLwRfn+v+w918QnrZVR0y5GjWpSSPT6QCPwD9P8fbgoRRutEBlx0FtdiTBWqY1Nr/4aQb2/HupUqKdNFAcny6V+Lxc5jF7tcC1vqSR+fd8imrq4AAqOPShXCTGxIP1NuP8AD/D3oulcdeoeo1Ti5YwfSzcfgX/23u6utanrRUsKrx6DDce2ZsgzFYmJY3PpN/6D6e2mUmtOHTbxDz6DheoMhkKtdUbhHPJ0jSoP+J9sGGprXpn6UHjnpAdq/HmKHFPLJDr/AG5T6VS9xEWsSP6n+vtuSBWGOmJ7UaCQM9VB7/2IaTc74VoSJJapViJALD1WAv7KJI9Umnz6IJIiGp0ut39Wbs25sOnlqPIaGUQMo0kWRmuADx7vLFIIx05JBIsdWHRs/j3siFcJjGSKzTRQvJZBfj+pA/PtbaxdmejG0jDKAerJdlbajRYIhGwAA1MAPqDa/wDsfZkq6VqeA6OY0CkHowlDhqWGCMeK7AfVrH2lklbVx6UKAFoB1nlwlHOpXwoL2uQENrH3QTOvWyqt5deiw9PTACKJdQ/Okc/gAW9+MzMc9eC06cYYhGPSv+t/hYc29tlq9bp1il54+tiefryfz72Mde4jPUaOmvqZxe/0/wAOfqQPdyw8utUwKddRRqspVPpfn/XJt9B78SSKnrwxjqbPEBG2kgahY/n/AAFz7opz1uvl0yy4vyxs39u978Wv/sf9b2+JSDjqtB0kq2l0BkIFgCLHjkfnj2rR9XHqhGnI6DPN0kKSMSLa72P4BI+o97dfPplz5+vQfSUXmnfSt1Uc/S5I49sUrkdUpio6T2ZxeuCRFQ3Ibg3+oH4B96b59NkEKQRU9FN3/ttj90WUtYTHgAHhPaKVanovmXNCK9VsZOmlpd5VDMLrFVgKrWNhqv8A74eymdAysvRFOpcNGeh0raKl3Bh6ergVTJRRkOsYW4KD8259gSZWSd1PQInXw52HRIe/PkNt7qjbGX+/qYFrIoqhEglcq7WjtwDb/fce2x6nprHHrV07Y3w/YW+8/ufkQZCtklpkItpivYEAgHn68+/deHQcG/F/9h7917Hl11791vrsi35B/wBY+/dar11791vr3v3Xuve/de65nSCP6fmxv791UVp10bG1uPr9ffut5HHrq31/NvyPp791uvXXv3XuuyLW/wAffutV669+631737r3Xvfuvdcrrb6c/wBffutZr1x/4n37rfXdyffutUHXXv3W+ve/de697917r309+691yLfpt9QPfuqgceuP19+6t1737r3XYtY/14t/t/futeY69cn8+/deoOuvfut9ZI303/10N/yNLA8e/daI630/5VGwtl/zjvjD1P8AGvdNq2D4oYWQUdVKv3LU9PPL52jnae/qeXhQPqTx71wavWzVkznPD7OqP/59vyeoN8dhbD+JGKhnp8X8SqrJ7Mx0TLJHGlPGr0skbRmyhtaggAX59+Gc9Wauqh6P7/It2vufef8AKT/m5ba2dt7K7o3LlunaKDE4DDUc9bk8lKtVK8iUdNTqzSMF9RVQTb8e9Hj1aIdn5nrWEpPhp8sK2COan+OfcckcjuEY7A3JExIspuklOCPpxce7VHVNDk1A/l1f5/wnM2Hv3Y/yl3Pi907R3FtnL0s0MP2eZxdbjq2OqjiaKSI0tSiuCDYA25vx735VHVDhiPl1Xl/NJ+RveW3/AJ2/Jbb6bwzdDR0+98pj0oTXZBfFSrJddUZddDAkiwFrW96BxTqzVLVqfy6qQyOZy+bmqKzLZWvyFTUTead6yrmqDI7vcsxlY839+61XNOt2f4J4nJ1n8kT5jSUkWQjSLrKomeroVlR4Yo6EXk8sXIW/1N7e/NWnXo/U/PrSSjy2URQq5TJIoICpHWVAUX54Ace/dez0fv8Ald/IWb43/On499q5DLGnwG3+xMNktyirnKRVePWUCqWodjydPA1H34gkU62rBDqP2ft6uu/n3fy7929n9yV3z7+M+Cy3Z3XPyXyk+6kxmyaCbPS4CWXUaqnro8arlZNZBAP1X6fT3qtDQ9WI1ZX0Ap/l6Df+S/8Ayc6XuXe+f7X+cG09x9YdHdeUpzGafdlBLt+XIUVOhklhoTlAitI9uB9R+be91Faf6h1Qqw7qfYPXqnn+ZVhuhtvfMzufC/Ggu3S+N3HUUuyZJJYppZMXE5SF5ZISVLabXKm3vXViQTjoiP8Aj731rr319+6912foOCDz/sffutDj11Y/7f37rfXvfuvde9+69176fX37r3Xa2vz/AI/74e/daPDHXZHPH5tZRyfp/T37rQPr1ah/KY+E+4fmD8m9oAeKHrrrvPYvcHaNWYnmag2pFL/uQnlABVIzHrDO/A+n+I99nWxxyKjo6v8AP3+aG1u0+38d8Renanz9DfGt4MHsJqOUrQzv4B95M0aEI5Y+rUAQD+b+/YOerMzVpSg/y9a7fpH1Fzb8fT37qmfLrj/vAP0v791vrr37rfXvfuvde5+g9+691737r3S068qaek3ztWoqQv28OZoWmDi66RMNVwfdXUshUeY6tHTxFrwqP8PW7r8asvR5To7aRoaGk+xNNGEIgjILaAGOog8/n3jFzfsVjPv8i3alieuvv3eYobH2zt90ttNQc8M0HS8yWOxamWNsfBqm1K7LBEpBb/EKCPYFl2CPb7vVa1HoK9Za7JvGyb/spa5iQOoz2jP8uiO9o/H58xl6nI0SMVn5tZSfqTwBz7lrZNyuoLZAx4dYa+5PJ+xX25zTQRip4YHQVYz401L1n+UxSMLj+yo4/Nx7FUe7PVTXqCLn2/ieGYsuADTHRruvOmKbbsOqSnZI40Vkuo4IFzz+Pci7XOZ7cMfl1h5zvtUe2bo0SimT1g3p2tS7G88PpjjhGkSNdVsD9b/7D2YnoH1WmB0VTN/MzGUVdJTLJHIisRw5/JsR9feh14/PoVeuPl3trOVMFBJ4Gd5IwSX/AEk88En3vr1fTo5a9qYVsfDOjwjXGrC545FxYj36uKda8+imd/8AyDpaHCSUWOkSSvkQJB45Dcub/i/v3HreOqxt41vcO6cfLXTJVigfyFUKvYxM2oAE3+o96r17A6Xnxv2PU5DKxGtp3jqmkUSa1ueW9V7+/D068T1eb1ntU4bFUsREYtCt7qoNin+H9fe8jrWehQetpKBWD1ESAcnVqv8A4mxHvQFercTnpC5DdmK8ssSTxSPGCbBgfofz7tRfXqua8cdFZ7i7Bx1BjMg8VQiPodbF7MbD1fQ+68OvEAnHVPm4d8bq3LuLKUWKo6qpjYyLG8eqS5BNhce09xcQxxnxGA6ftJIo7ldZxXPQydT9Q9m56kmqKsVlJTqNSrKji91uPr7jfc9829b5ICQdRpXqUIfcSy2C0EMZA6Mz1L0tktubkO4K+R2dHT9t/VrYH6BefYk5w3rbOS+UzvKEFpVoPtp1OH3e+Xb33k5w8KR6W0NGNPMcerFMNtDYu/EWk3BTFarwxq6pZf0r/qTb3y451515qj3Wbe7ZqrIxI48PLrpDccgcu+KNisIyVjUAn1I49LHE9DdQbfmgnXF/dVKSCSKNtDamBuAQ3sGtzp7kb6PAs1aj4qA3W7L2n2hSWeOoHrSnRxNhzbnxVMr7VxcWLoaeNLOlMsZ0xiyqHjA+g9u7h913nLnLaTvO8q0moV0NqPHP2dFG8xch8vj6S4KFzig09S8921uSWVocxkHjZLqRJK+ksv8AQE+4KuPY1uVJmjbbzCQSKhAK/mB01tEHLKJrtApDZ8upG3PlPkNpSijlqaEwFrF3jIfQR9Q5HPsmvfYnbd9YXTrJqGaBjSv2V6jD3Z5U2bctuae3QCUjFAOq5/mNncn29nEqsKjSz1N18lNqLa2kuPpz+OPeavsttlpyhsa2M5osdMH0A65pc47Rdbbuj28inBNP29G6+DHxDw2cwtNWbwoqyTKWilMlQFIAYA8GT3Cf3jvfS75TquyyIVNRQHNfy6R7Ltf1LaZcE+vV9nX3XuF2Ptuiw1FTpHHTqAjaUFh9LXHvk/7gc33fNHMB3zepTGz8CWIXP246l7adsjtrURKtT59Irt7uPb/WePfVJHLkGjYIqkXUn9I4/Pt/lPlDfPcG4O3WJ8O3U6ZJAPjU+Sn/AC9TR7ee193zVcLOVKxg9E4wPy4ykmXqKjIuj0ZJWOL1Ahb3B9zPuP3Z9lg21YtuDRzUy9c8OshN29kdvksktbcfqClTinSL358mcvna2SjxlTHSQTHRpOq51cfU29jv209gdu22W3geMzSuyjUaVJJ6X7f7K7NtG1Nf7jGXMQ1eXl0Cu5TjEpjuDJ1onrmBlLNISp/NwCff0bfdf9ptj5J9uLSS2iCzihJIFf8ABXrFT3J91pYZ5uVtvQR2ygrwoeiFd0dmYWaVop/A6xlhewBYBubX/HvIrdRDPHWUA6esbLS+mjvCIHI1E+Z8+q3+0N94+urUp8G6xyNIyMI2/F7Hge4B5p5c5e3680GMeJXyp1kDytvO9cuWi30zFkxx6ZMDksxSK0kju6CMvyb2vzf/AB9xhzP7L3P05vNs4DOn5dCC5939vvZBG9NRNOkFuvsetQupPoBKi2q+q9ufYGsPb6+WPXImkg8D0K7HmDatxUB3APQFSbznizBrJpCdfFiSV0k+5g9v+WZttvRcgFWGPlTqJ/drctuO3+BCQxFel1s7t9qHd1JR1ml8bXyR0yLzZWkNtf8AT3khHOaqD6DrFiG4HjEHAJ6Oj2Phql9kU0cP7+OqYFqEK8hS4uDc8ezGVR4Q9D0YTKdFOI61uPmRRfY7/iiW+jRMfxwfJ9PYOv1pLnj0Eb9QJsdE89oekPXYF/8Aip+nv3Wq9de/db697917r3v3Xuu7m1vx7916nn17iw/rf6+/dazXr3J/qffuvYHXXv3W+udwx5/C2/p9P6e/dVyBjrifqf8AeP8AW9+6sOHXXv3XuuyLfkH/AFvfutA1669+631737r3X//R0ABa/P09+60a+XXja/H09+68K+fXXv3W+uzb8fT37rQr59de/db65AEC/wCPz/rX9+60fTr11v8ATi3+839+69mnXH37rfXYtfn6e/daNfLrxtc2+n49+62OGeuvfuvdciVtwOffutZ8+uPv3W+uXKn+h9+61hh10CR9P9f6e/deIHn11791vr3v3Xuve/de67Fr8i49+60a+XXXv3W+ve/de6WvXm3JN1by29hkAKVmXoIZb8/tSVKhuPbsMZlkCjp2FBJKqHzPX0BvgJ1NQdQ9d7biSFF+/wATQ1clo1UBpVDkgqPcn7bbrbQLpxUV6lDbIDBCtTXHmeHVi3ZfYdPievqp4pk1lxGPVzpP+PtfK6geL0bSSAQ/MdVd9oSLnsSuT9DM+QiJZbWUeT+v+8+y2Wrdw9eiScGVdQ49GBwVNAdu4DXY/wCTUt7/AIPA+h/w9qBwHSxCpQA9HIpaanTrWZUC2amsADySUt9R7XaQ0VB0YgN4Nekj8f55MPFl6epPj81a7Rkni1zYe2YBghum7VjTPr0Yqqr0nhdgyk6hx+Tz9B7UqNOR0sBrjrjRKzul7lTa1rfk8/T3etRXqhBBp69CZSUqrEi6SSVve3+H0Psukck16dUdSpqRdPr5axt/QcW9tq7eXV6Djx6R1bTvEzKFPJIHAIsfxqPtajagD03wFOmGegSUEhAWuONIvcc3Ht4MPPrWknpSYal0qo0cAfn6g/2h7TzN1ZfTpXxUgewKaQQOSAQD+Rf/AB9oy+er4HHri2OQtfSCo+o0i/15+vvYc069iuOooxFOb3jF7kfp/SCfqfd/GYY6rpAyOnBKOnhHCIqqLcqtzf8ANz7bMhPHrwWncOPSB7ApKeqwNT5FQtaTTcC4tGfx/sfbsWqukcOqy0Mfz6ok7xoIKbt/FqmkJ93Gz8ADhxe/tJOB9QNPQYulUTg9G27iz2zKzpfFYhZKRsutJT3AVNYYL+bc39qp2RoAvDowuJIjb6V406Z/j7koRSU0OtNMKxKPoLAcEH3W2YUHy6pZsq4PHqwjauXpomQBkIa30t9P9b/X9rzRlp0ag5AHQwQ5KNtGmRTqCkW+gH9L+0RiYcelGpaVHTnDUxyE3P8Asfxx9Tf20ykdWr59ZvKjOACB+Ofz+Qbe9UNOvH0HXJiBbn8E8fm//Gves9exw6gM6hipFxf6/wBef+N+7gEivWzjh1HlqzGj2s4UWAH1t/re7BKkdV49ex+qRmdh9blR/S5sD7tJQCnVRnqZMSVIAOoGxBH1N7359trSuertkY6iPVrHER+QP9Ym/wBPdgtTXquekPlqg6XIAABb62+pF7G/swiAA6aq1aN0Deer5J5THe4vxY3/AMCOPdWy2OmWLE1HWXE4xlheWYC5B08A8cWA/HuwHW1ocdMedSGLUeFuTcm3+2Htt8dVc6ajz6LN2AYRFVN6dQEtvyT6OD7TORTpBLXST69VX7jh17vyTgekVDn/AAJAJPspJUseiRqazX59Muz9/piafdlNUsbxip8TOxsPGhtp/wBj7At9/uS1OgLfkC5b7etYH5p9s5LsHtXM0D1UrUmJrqqNkWWTxO5ksgKXtwP6j8+0Z6S9Ez9+6312STa/4+nv3WqAdde/db67Nv7P0t+ffutCvn11791vrs2sLfX8/wDFffutCtc9eItb/H37r1euvfut9e9+6913ci/+P19+69TrwFzb37r3Drxv9P6e/da69ci/+Pv3W6deFr8/T37rxr5deNr8fT37rQr59de/db697917r3v3Xuve/de67Bt/rH6/4j37rRFevG34FvfuvCvn11791vr3v3Xuu7fQ/wBffuvdeNvx791oV8+uvfut9e+vv3XuuyLG3v3XuuvfuvdHk+Gn8w/5PfAut3XkPjfviXZtVvSnjps/LGJj91CkPhEZWN1HA+hPvxHWwTSnRY+2u093d19g7m7P39kWyu793ZKXKZuubWTU1c5LSzEyFjcnnk+/dazXo5Xwk/mj/Lv+Xyc4fjPvuPZx3HGsWXD0rVcVZEjagk0DOEYX/qvvRFTXqytpBXiD1Yev/CoT+bGjq6dq7OQpfSF2JhlUahY8Bffqdb1/IdEYh/m4fMSl7ezveVFuzE0HYm46yWvyeXx+M+ySWqmbXJMtPTOqgkn34Cgp1Rqs+s9ET7i7b3l3p2LuftTsHI/xTeO78jJks5XWYfc1UgGuYlyTcn63PvdOvGpNegzU24vYH62F/pyPfutHo+uwf5kfyk616M3d8ddo7xTG9Y75w8mC3Jho45/9yOPli8TwyMJALW5tptf34568O3ohZFidJuBY3PHNr/T37r1a8evKxvybG4Ou12UjkWP1H+w9+69Tq2v4vfzsPnh8Rdm4nYPUfYdHTbVw8Xgp8ZnMf/GonisBpda53W4twQvHvxz1te3h09fJL+ed8/vlPtHJbF7Q7GoZNr5KmekkxuEx38FRIHQoVvQNGGNieSv+w9++3rZNfl9nVPbyNIXeRmkkdi7SOS0ju3LM7tyef6+/dVzXHXC5tb8e/dbp59dA259+6912ST9fx791oADh165+nv3Xqdde/db65Lb+nP4P+P49+60a9dMD9SQb/wBPfuvCnl12OOT/AMb/ANt79145x054jD5PPZPH4bFUkldlMvUw0ePpYRqmqamaTxxQxKPqWPAHv3WsD8utyTAHZv8AJV/le4jfzUKzd9/OXYGR2Zn8Skax5ra1IIw8k08ikPGZAwdCSCVU/wCt71WuD06QY8r5f4f9jrTezWYyO4cvkM1l6ypyGRyNTNV1NVVyvLNK8zl9TyMSfzYD8e98emifTpoFr8/T37rZ4Y67PNgP6D37rwxx66tzb37r1cV68Rbj37rfHrr37r3XIXJufxyfoOP8PfutHAp07YGnkrM5iqeBXaWbIUqRqnDljKLaffuvDHW7/wDBvHSU/wAaNnQ10TxyRGNbSfq9cPHJ9xLzNskt3uxniFcddAvZn3Lsdj5FTa7yTSAeBPy6MXksdAZCbKV+oPp+v9fZMeV55m8RlNfs6kYe+m07dCba3mABrXuPTDLi4zcaVbUbepVNh/h7VNsd1bx6iMDoit/c/ZN3uxH4gLOfXrnBgqOMo7JGCDdgFS5Nvaex2+8ubigU0B6N+ZOb+XNk2x5Z5FLMuB0m+wc1HjcUqUEegrq8jWsCLWvcfn3MG1WzWtuFk49c8ee95g3zeHuLb4anqq/5FbioKzCZNHe9QFLDQxvq54/r7MugWBTqo8U75PI6Nc5eepdOZHYgav6E/S3vR6t1YP0X0MZ4aPLxPI85aJ7KxLcqCNX59+61X06stx3TO4cpt+njhnMTrGoIYlTYLZTc+948utUPn0m6f4tY6oyFPWbpk+6FNL5VjGlxqHIBB49+r16h9eh2m6u2zU4dMdTUNOlNEgUKKaG5sumx4/1/esVp1v5HoEH66xuxs799Rxx06mS5AjRB9b8gD3vNOtUoehUr+7MXtrEvLU1EYdIlRQJLW0i17A+9UJ6t8h59Ft29vjs/tffs1NgZJ4sDHMNcxZzE0V7nSfp9P8fYS5p5os9jtSK99OHRXuW4RWice7o51D1oaaPyVFR5J5Y08hLXAcr6yP8AY+4dPuNuWokcPz6Czb1cFjxp5dBhvb45JutvVWIIHJ1glb6W/BB+v+v7fh9yL+Md4r+3p2Pe5acc9TNm/HjrnYsazSUMM+QVizzNBC4J08m5Hsl3DmrdNzY1OlT8z0lm3K4mICGnS3r6qho/HjcXSQRioYQqscMaE3GkAlR/T2VWlvPeXSRVJZjTjwJ6RvHPdHwSSxbAyek3k8Pl8LUJLU05EAUy6QltRtcHj2o96Yt1seVrXbb1joLDjXrrt9yzYjy9y7Le1AmaMVPnSnSw2RPT1FYciweGoe6iN2Itb/D/AB9x5yr7TrzBscV/dLrjrXgCMdZWT+5ux7DEbdnBuiSCa56Pb0t1/jNx18OTzdSgpo31BHINgF+g1fX3k77c+zW3NokMKrEh81Hl+XUQ8/fePXaLB7GycGVhxB9ejt5Xcez9r4U42hWm8PhCuyogJKi3JX/efeUtnsO12tqLR410AUppX/N1g9vvOu9bzftuU0zaqkgaj5n7eqnPlBvzF0a1k+InVJkaVkVH0kMbEAW9wR7ncgct7jGy/TA1rWir/m6lX289wL4OkV3NSmMk9Vu47eu+N15MRvPItGJyEJkkvoB4N/x7w55q5H2jlpGkt4qD7OHWWVjbLzDtgu9QYgV6OZ1eldQVlNVZBBVRo0TgyIJRcLz+u/vHfmS8iEDx27aCajjT/B1Fm/ezkfNV34sEdXrntB6t06X+RO1NtU0MFZCkDRwwi0YEYVlWzelbe8D/AHZ9reZObg67fKakk148eHRPefd5uLMrKgCH0oB0Le8vmTE1I1FtqkmqaiRbRGMtwWHBBX/b+wTyP9zHnXnfc7eDdRJPGrcNNFpj4jTPQx5d9nNvtQLjdZURBxrTy6JZvdOyOyp3zOQeeCOQ3ETvJYD6j0t76z+033DL3YNqjjcBVoMBQKfy6lS39yPbjkK1+gtWRmXiRTotu6hujZ8xmqC7wL+si4sB+ePYq59+6lfbNZrJZCp9CB0N+RPdPknm25NurqjfM+vUXGV1Vn4lyaVRVhZwgZi2pfqD7kT2U+7xZW8Md9fRBplOSQDT7Og37x+7uw8oWsmzpRtYIqDjPTBv/c2TG35KZZ5DUJGQtpXP4tY8+8/dhshtO1JYRigXy4dcrue90i3ffZb23+Fs/t6rm3jR5/KvVSVrS2Pk03diRz9bk+3J1MyMvkegIskscwkXFDXosZ2NlkzRrgXZRIW0t6uAfqL/ANfYGPKzLffVL69SWOftW0/u6bOKdDZRUzw42RqldOmBg1wBxptz7GiRhIfCcAilOHUaSMfqfqEJArUZ6K32M9JTU5ljYMTK9wp/x+gPsN3Oy2jsW0gfkOjVear62FI2II+Z6ACqkWoINiLNe9zcn/W9u29hFClIxTon3HfL3cjWck/n1HVnWuxkgNmirYmBHBBVwRyP9j7XjFOigAE16t427uSkynRcdNVlZa6GiIRzdiCE4AJ59nKsHtc8fLo5jYNb0PHrXj+Y+KRp5srUMpqxWMiCw1BGqCPr7CN/UtnoJ34GqvVfvsu6LuuWk/Tj+v8AxHv3Wqjj119Dz/Xn37rfl13a54+n+Pv3Wq0GeuiLe/deBr11791vr3v3XuuVwP08H+v+Hv3VaHz64+/dW67Fvz9PfutGvl11791vrkLfU/T6f43+v09+60a8B1x9+6313Y2v+PfutVzTrr37rfX/0tAFb34tf/H37rTUpnrr88/15t7917yx142/sg/7H/jXv3Xvt669+6312f8AAG3+Pv3Wvt669+631737r3XduL/429+698uvC35v/sPfuvZ8uvX4t+P959+61Tz669+631737r3XK4025/3x9+6rTNeuib/0/wBh791sCnXamxv/AL17914io64n6n37rfXvfuvdd34t/jf37rVM169bi4+n+P8AX37r1fI9de/db697917pa9d7ik2tvPbmZRgqUeZx00xIuPDHVKX/AOK+3YXEcgY8OnIX8KUSenW/18Qe6sF2D1RtmejrYZpKPEY+ll8MikpIkSqwNvcm2U6y26lTwHUnbdcrLArAjh59D73FQZfKbCq5MZJI5UGYqjEmyLcgW9qLjU8faOHS25UvEdPHog9NuGpm2ZkaOv1iqpcsiBZDZvRJc29oNZ0Z8uicORH35oejR4PcHk2djJrkvAlOgt/Z9Si3tSH/AEwRx6MEdSg09Hg2JlFzuz6ehZv1QR/U3ANrWN/ZjGdaCnRpE1Up1MfFJt8kwDQzcnQQLkm9/wAe96Sop1bSEHb+3pW7frpKgqkxYlrWJIP149+VieJwOnFKnA6E3GPFDIhI5HClubm/At7dkXtoOrgmuehPoplMYA4ut/6/UfQ+yqQZ6eAqar1NezW4IItyf6/U+6daHbXpO5KHyFivBHH0N+ePaqI060xA6YzRuLEEWHN+AeP9c+3tYr16vUyhcpIEP0a9z/j+T7pIKrXramhx0r6diVC/g8/7AfSwPtG2OvEHz6zvIqEFvoFAuSOfx9f6+9UJGOvAZz02T1scQsCLm/1/oPrb24sZbrzMFPr0is1u6GiDM7HSl/zwT/QfT2rS3AFW6oXA4dFv7O7fipsbURiQRemX9UlrjQbH35nRBjpHcXAUU8uqSu6OxEye+I6qkl81aKrSqxsWe5449ks8lXqvQauJA0leJ6MXs/p7e3Y+yxl3eo8607PAkrMBZFutlP8AT2qS3kljq/S2K1kmi1V6Rezd17i6k3JLt7dEVRCjzgJPIGSJUVuW1H/evbaO0LaZOHTcbtBJ4cnR8Nldx4ysaHw5CKQsosRLe13/ADe3swSdT59GkVygzXo1O2d4U+QVCtQH1KP7Qt+APp7V6wy9LUkBPQmU2Z8dtTEpb63vz/iB700NeHHp0MfPh080mZhma2qzAji4PB/r+fbDwMoqenAwrTp7FXG6AhgLcW/rf+p9saSDnq1Bw9esBlDEgEA83uf8fx/vHuwFOtN8+uaU3kIv9D9Rx/sfei1B14gnPT9TU6rH6eDf6WsDbj2nZqnrxx1xljJ54BH4P5P+HvYI68vTHVwIFLG9xfi3Nvrcf4D2+jGtOtNToPs7IBG4/tG+k24+ntaikr027UWnQZfw1mmu631NqueR/W/vYWmem6nDeXT/ACSpTQAekBQfxa9xe/HveB8+t/COgT3dm41Eq6rNZtNrfT8e2JDQ16ZehHRSexNwLFS1MzyFQqygsW45jA59opWGnounY0IB6rmzOTVqvMZI6gBM2k/XUSCOfZY7rGjTNgD/ADdE6gzymGMVZuAHRFPkl2b/AKMdi7iyqVX2eQraeaemjZ1jkmVkP6P9f6W9xdJvVtd37wQmpB8uqb17f7xtNj++NxQrG2RUeXy61stw5uq3Fm8rnK0lqrK1ctXMSdR1SNcAk/0AA9rugBQeXTL7917rvm/+Pv3XvLrr37r3Xvfuvde9+691737r3XfH4v8A4+/de669+6917j/Y/j37r3Xf+ve3v3Xuuv8AW9+6912bfi/+N/futZ8+uvfut9e9+691yYk/UWP44t/r+/dVAA4de1cWP04vb68e/dbp5jro2/F/9j7917Pn16x/offut1HXXv3XuvDj37r3XZufUfz/AMR791oYx14Xtq/p/wAT7914+nXXv3W+uwL3/wAB791omnXX0+vv3W+ve/de697917rsEi/+Pv3WqA9dqL/7D37rxNOuPv3W+uwL3/w9+61WnXhb83/2Hv3WzXy69f6gfQ2+v+Hv3Wvmevcc3v8A4f8AG/fut56644/r+f8AjXv3XuuxcG4+vv3WjSmevGx+l7/n/jXv3XuHXXv3W+u7cA/19+69Xr1uCf6e/de69cj/AGPv3WuPXvxxf/H+nv3W+vXNrfgc+/dapmvXjc3Nv+Ke/dewMdde/db69Yn6e/de674/xv791rPXXv3W+vD37r3XOwLeo6efU1iQB/Ww59+6qOGOtgb+Ql8IMZ393fuP5AdmYRx1X8XMfR9qZ+rzcMkOEyNFiX+7aigaoCxyu2kaSCRz795dbHHVX/i+iyfzg/nHmPmN8sex6/A5lE6XxW4ZR1ztbHkfwvCUNFAcfHHTQodKHSgBVQP6n375dawe4GvVSJ/41/tvfurdc7FR+OePz791XDddKeCObnj/AA9+62fX066Itx+fz/T/AGHv3XhnrwNvfuvEV669+631yC8X/H+8+/dar5dG0+HXSec7m7g2zTYyFXoMNlqKqyZKlgsUZ8g1fixsb+yDmbe4OXtln3S4+GJSehVybyvfc27/AG+y7eut5XAoOPHraybtDF9U0uN2LSV9PFR46CkukRCIjrEEkJA4+t/YY9teZYOcds/eOnJJ48cdDv3c5V3v253Jdqm1RAAY4dDJg+3NqZahSVszRyOFDOBOrEXF/ciiKIYCiv2dQ+95euatIxH29LnGbo27k4Q9NkaaX/gsi8W/pf3WW2ilTS6in2dKLLd9wsJxPaysGHzPU+fKYqKBqhqmIBLs51jlR9CB9PbcFlb241RqAfs6V7tzJvG9EC+mYqB/Eei09s9gYkYmrNFUxOAsllDAngW+ntVobiRTojaRFNK1PVKvcG6c5msjXQ0ENTNAZX1CMMw06rckD2yZErpBFelaW1yyeKUIX18ugD2vt7K1mW1rRzBxKLLoNwb8/j3cdNGnDq674q4Oop8PDJkobMoj0iRCfovHH/E+/dVr1ZBiGhXG3ESrpjvwoA97HW8HPQY7kyiq7eNgHUkkfQkfU8n6e9deGMdRMbu3F0lLI9XMqMFtZmUAgf6/vwHl1rjx6Jh2lvvM5/cE+MwKzTqjuA8IuPrYfT2W7nu1vtUeuYjPSe5uEt11OesG2ekdx73aifcFU4pNYaaJ30kxn+yR/U8+463b3EhjjZLX4vLoln3nsIjOerBeteutvbHxkWOwVFDGdI8srRqzsdNifJ7hPdd0ud0uDcXbE+gr0FbmeSeQtMa9L+qxXhiZlubAng8D/AfX/evZerqcHpPqLdo4dInJTfbh+TcAC1/p7URCvl04FAWp6QORymlDrf6Ek3v9LfX2YKoAqRw6dAIz1l6txH989709PbXFSVcD/wCqX9X0/wBa/uV/ajl9N333xJlqEII6GHJm2G/3HU4qqmuejy9udX0UWMpJVo49TxxkuIlH9gA8+xt94Pk+HfdmjtoFoV4kddLPu/8AML7fJJZA0VlC0/LolW6tsVOAeesxsgVYlMgjT9VwPpb2DPbXd9v5c5fj2HcEr4eKno49weQN93DdJN02uUgOagA+vSI278uKjZ1S+DrvuEmjJVVLGNiQdNxf3klsG8WVxYg2YAX5dYic2w7ttO8Nb7tq1r5k/wCfoSar5C7g3TQ/7j1rdVQbAhnY+scAG3s9Fy0govRB9W0i/Pp02b8ed69uyyVuaiqXpZVaZBNfkMeOD7Tz7el5C3jiuPMdLdqF+t4kgYqK19Ohex/wmrsLEKmKijA+gXx2N7/X/H3CHM3ts26CRilVeopTrLzkr3MXaLNdvmetOOejD7D+LFatEpnjWI3tZ1sbAf4+4ttfuo7FubfU7gmCeBJHQyP3hbLly4JhGo8cCvQn0PxegjmEszLpPBAfTz/Q39j7Zfut+3e2/wBtbKSPlX/D0BeZvvG7zvU3iWdVH7OhFw/VW09o2mqYBNPGAVLlXANv8fcxcse2HKfLNDtlrGtPPSOoy3z3Y5m3a2NqJWVTxoSOktvTcAi1QUKRRRxp6QsagCx/r7kGsUK6Y1UAfLqLbie4nctNK5Zv6R6LDu9qXcED0tZArEhgSEHqufr7KN02613WAw3Sg+mOluxcw7py3eC6s5mU1/i9Ogi/u/HgqOUUZA9TNZf6X/SB7Ktn2GHZqiEAD7OjHnDnW75tob52dh8+gpzKGqaRZwWJ1Cx+g/pcezV6EenQAdCBxr0C248JTyJKuiMkq1jpFrfgm59p3UaadJHXUK06Bw7agLS/txAoWY8f2Rzb/H23pBFadM4I4dIfcs1BBi8hCzokqU8lgNIJ0jj2y5AQjqjHtI4DquLduRmqqmeJnZo1mcKL8cta49lTguTU9FMr1bT0iXjaOwYaSebN/t+CfehSnVaEY64Dhlb6FGDqf6MDwfeuvceh72Z2nmBQw7YWSR0Z1iIDEoqtxfj8f4e3o5nHaT0pSbt0nqsv5sZqCDdqbdW8lQY/u5ZFI8agyXAI/wATex/wPshvpNctOg9evrmp6dER9oeknXvfuvde9+6913bi/H/E+/darmnXViPrx791vr3v3XuvC35/3j37r3XvfuvddgX/ANf/AHi3v3WiadeIt7914GvXXv3W+ve/de67DEe/daIr17kn/eePfut8Ouvfuvdf/9P5/wD7917r3v3XuuQNgLfX88e/dVpU9d3Cjggm/wDvHv3XuJz10P8AXuP6fn/be/dbPXmt+Bx/rfQ+/deHz64+/db697917rsC/wDT/Y+/de66P+39+691737r3XJrX4t9Px791ocM9cR791vrk1vqPp791oV8+uiLfkH/AFvfut9de/de697917r3v3Xuve/de65Lb8/4fj/b+/daNfLriffut9dgkEEEgg3BH1BHIIt7917q+D+Vb8wm2lWJ1huCvkZpmV4p6pzp8Qn9A1NwbCwPsTbJuBRvBkOD0Jdlvwn6ch4Up1tRdfdtYvcmMlxck8FTFU00vjS4YspWwIB+vsZxzhqjobQ3AkXR0Qvt6qfD7rqcdjo7Uk9UZ5EUcXL3LFfwPZbKAkmOHRZcgq5Veh+60zFHmdurRJKhaJYiwBBKlSAVt7URkMKDy6fiIdNC+XRxuqdxGlpGp2cAo2lAT9R/h/xT2YQOAKHozt3JwehuyGUirZIB/UR3IN/Vfm9/b7HpWSSKV6X2JxpiihqkH9hSbcWuP8f6e7ogrqPDp1V7cY6flrRG3+1Agi/+9i3veonHDr1B59LzF5pGVUmIBH6bWF1I9sSwHivTqOKUGOlOuSi+hax+oJb+v9T7TmFuvBh1Dlq1mZo/re/I/NuSf9t7sEKivVqVNSeoruEBAIHH1+lv9b/YW93A8z1UinWGiJkkLj6A2t/Uj1W97egFOvcCB0qIZgkPq4YXAN7fn/H2lYVOOrkmoPl0z5HJItwHH9ef9v7ejhPn1TX3Y4dIjLZsIrNwNCm9voOL6rn2rSMIO7qrtTK9Fi7I7IpcbBUlqhF8cchJLcXA449tyyqRjpHNMkY1Hj1Vj3D3hltw1UuLwCy1crO8P+T6mPr9ANl/1/ZPNMznSvRDdXTP2rnp3+PXxE3hvbcNFvLeFPUiiJWqRJlI9Ou4Uhv8PdrW1kZ9U3V7Sxd2Ej9XlbI64xOBw1HjKOmjRIofGV0iwFvof+N+zQyrCNKcB0JI4lRNIFOgR7w+KWG7TAeip1pckiOFmj0pqNuPX/gfaeXwpV1Pjpi5sY56aePr1Wj2z8ae3ejKeTM0VRPU4ylbkxkyehBr/s/4e0L28ijxIjjolubGe1ypqOpnR3yOqcg64vISmKupv2HWS4YyBrchvofb9tdOfy63aXhJoerJNob4OZpk8jhmNgGHIt/QezKOU/EOjlZS3HI6EelyEiSaow17888EHkcf6/t7USKnp0+vT9S5iqW+ptQPAF/wf8PdVVWahHVqsRg06UlJkNTBpORYXH1B/Nwf9j7q0VOHVwRTPSxpqqN0RlNxYG4PP+t7ROhBPTiEAU6dlmIAsSLjg/04v7ZI63gn164yTqo1FtVgQP8Aaf8AXI9+C14dbJIGek1XVvpk/rc2Nz9Le1UaEU6pxx0ha8Go+v0vqv8A4n6jj2vTAoOmTk1PDphmVIQwNuF4/pf3Q0DU62aadVOg03Rn0plZEb6Ag8m1wOfbbvQ0Xpl3CrUjPRbty5ySpMjM9lF/z/T629pdbCpPSKR8V6I73rvE0dLLi45bVE8pKAcNYqBYey+5kI/Poqun8l6KhnZXXEAINTyUsk8pAP6kXXyf6+wdzrvEGz7BJJOwSuM/MdC/2v2GTf8AnK2jC6o1y35ZP+DrXd+dHelfvndUW0I52RMEXp65ULLdVH7MTW45HJ/2HuKOTdsdEk3Kc6vFNVPyPQm+8R7gQ7zuMHLmzgR29rGEcAcSMEn7SOq9vY86xf6yHT9VsLfg/n37qgrwPXC/N/furUxTrr37rfXvfuvdcjawt9bc8W9+60K164+/db697917ru3H1H/E+/dar11791vrv6i1xx/vPv3WvPrr37rfXvfuvdclNj+LHg39+60RXrxNjcW/4j6e/deGRnrxJb+g/wB6H+v7917h11bm3H/Ee/db67NhYAg/4j37rQr14E/pH5Pv3XjTieuPv3W+uyPpyOf949+6110OT/T37rfXZFvyD/re/daGeuzawt/Tn/X9+68K+fXRtYf15vx/tvfuvefXPUD9R9P9j791XSRw6x+/dX67F/r/AE9+691yBve4uTa3v3VSOuNyP6jn/eR791ugPXXv3W+uSnhv9b/iPfuqniOuPv3Vuvf1/wB9f37r3Xvfuvdd25tcD/ffj37rVeuWqw4Fj+ePe+tUrx64g2N/eutkVFOvE3N/futjHXXv3XuuS/7f/D+vv3Wj148/kC30H049+60MddA2BH9Rb37rZFeuzawt9fz/AMj9+69mvXgTpItwbc+/deoK164+/db67Bt791oivXXv3W+uzawt9fz791oVr11791voTen+rNz92dmbL6p2bB93uXfGao8FiIQjuDVVclluqcmwH09+60B5jj1tm/Prs3GfytP5bnUnw06zy9Fjvkhv/HZHA/JBqaZIcrJhZ4mSkoahIvWmlLatX1PvVDwPVtS07R+3+Z606Sz6mZyWZzqZmJYsT6iSx+pN+fe+q4PXEkED+v59+68Aa9cffut9dg2v/j791oivXXv3W+ve/de67PJ4+n+29+60OHXR4/x9+631en/IiocZuX5R4HYmSgWaDeG4sXh5wba/DVxGE+Mn6H2Eec9sj3na/wB2zgmOUhWp6EjqbPY7mK35T5km5hkI8S2jLxV/jANOrgv52vU/XvRXcy9XdOpV0O5sUYP7z1c7kxqsoV2+n9L+yzlnZdv5T17fswPhp6+vQ09yOYdz90rODmbmVk1y1oFxwOK9VCYubszCLqx+eNbTzIA5jlLKt1F76T/r+yvmPmrdLCXVAhp9nQL23k/aJrXJBY46Nn1LuTsCnghMlZPNqJugJJ9QBJ/417Jbf3js7QiPcUqfM+nTk3spvN8hudrqUOQBno3CUPZeexRSijrLVMfjDBG/tKOR7L9y+8byhYSmBqEjpbYfdy51vkEgjcA/LoJpuge2a2sEeSyJ+1ndv8nkB1EM1ytj/h7IrT7wFpvt39JtYLA8KdT5yD9y/etxT6/e18OFBUkmmOPQ4bf+I+26bGyfxWgEuTqIPW2kMoYjkkn/AGPuYeXH3TcKXlxhDnqMveex5E5LVuWtko9xHRWYGtTmvTBB8QMFhqv+IU9Iql5CwHp4H1H+t7HflTz6xRGrJPmeh929sum2zQRQQxKjLpFlH00i349++3q1COhWSoahwrSSv40Ck+ri1/yf8Pe8U6159FD7D7Gx2MlqStTFJLHf0q+p2YcWAB9+4dbNa46Kvmuyc5vBjjdvPLDWoWWQDVcXPpuP9b2mu7qOyt2uJfIdNyyCKMs3Rkuo9rJisTHkc6sdRmJtJkdwG4PIsD+fePfM2/XW73RVWogJxw6BN/dvcyFSe0cOjD4ORGna2iMcWC2FiQSOP979g6cgL69FZDDJ6Hfa01MgtNIi/pNrj8C3sruAH+EdMOr1J6UWYrqRYWETKQVtfiwP1ufaaIOX7hjqq8APPoA9x5eCKSYBxcseeBbn629nNvGxyOHStVxpPQH7l3DDTxzMXCjQx1X/AN69mywFlI6dVKJw6ED4rb5xq7wyAWWOSWHQ5swNibfUD3kB7LR+BevXH/FdSb7eml02ro/3Ync+Jnwz0VVJEHhhJVibWYL9dX9fc1cy7RDvloYX49ZD8s85vyxfePHwxX8uq09/dtpQ1M1RHWwyQGZtSEk3Un8j3j9u/IU1lGzUqM8Op72j3pt9wnTIBFMHoDdq7Jl7s7BWuxUDsqzIJfCvpYfW9vZ77d/VQ3v7vyUHCvQG93o7bmK2G64DtStOrzugvidgqfH0NRnKInxrTkqwX6iMMTyPyfeQAto4V1efUIWdigALjqxHBbM27t2jipsVRRxKiquoxqGsP6W9tGWSlOHRyqIuEFB09yUVPJYyxRaByOAAB+Ln/D3rjjp1JpIqmM0J6gVNdQ0qeMeNbW/Tb8H6j28kbnh0y7ajrlNekTl91U9HE7h7hC1vUefz9fakQ/x9ULBVxw6BXcO/YahpfXpNiAW/TwOOfdy4XA4dMNLioPQF53c8VQzEyBvSw/1yfbBfUaDpM0wJqOgzrsjEWd9QAAJ+v+H0v7YYjj0mLamPQTZrdFP5HhV1uCRe9x+Rp9tM+Pn0wSg4HoFdz7uoqOV45CC5B9Wrj/kE/wBfbDMCKNx6TSMOA6BHMbvhLPaRdOiwBt9S3P19p2kANG6TFvIcOgyrd701Oapda6tDgHj6f0PtrxADXy6b1gVp0Ubf+9HqausihqVUNrBF7AggX49oJJCWLeXSCSbUc8OixZSaOWYsDf16if8AG9/z7Rlq4HSMsGOemqqnSVh9BpW17fXjg+9Aefl14nyHUBz6CRyQCfr+bfj3br3S923ksPsrbec3luMxxw/w+o/h7TELaqRCYyCf8fdJ5khjr59NSzLDGS3Hy6pc7L3xlewN25PP5WZZZGqJ6emKX0ikSocwfW/Ok8n2HnYu2o9ELMWbUfPpAe69a67Av/Qf4n6e/daJp14ix/4n37rwz12pH5+gubf4+/deI9Ouibn37rwFOvG1+PfuvCvn11791vr3v3XuuSkfQ/T6/S/Pv3VSDxHXVrn6ge/db4Drr37rfXfI4P8AW/8AxHv3WscevC17n6D8e/dePy67JF+P6W+nv3XgD59dAX/IH+v791vr/9TQANvx791oV8+uvfut9cgtx9QP8Cef6e/dar1x+nv3W+vfT37r3Xdza34Jv791qg67U2P+wt7914ivXRJPJ9+68KDA69axF/zY/wCw9+69XrzWvx/T37rwr59de/db697917rlxb6G/v3Ws1+XXRuOLg/nj37rwznrxFv6f7D37rwNeuvfut9dm34FvfutCvn11791vr3v3XuuwL/kD/X9+61XrxFh/jf/AGFvfuvdeFx6gfp/t/6e/deNDg9LrrbdOQ2hvLCZjHSNHN99S0zsHdP2Z6pBIbpb6WB9uwyeHIH6egk8KQP6dbh3xL3XVVuQ2RVVFVqoarCUwnkMjmMTSxjgk+x5ZuzFGPAjodWMhJV68R0Y7vTr/MUu8KPNUVPJV4uroxKZIk1r+4oN9Vv8fam6jOuq9LrmORZA/l0F+zK+u2huBKaRzHTVRV3RmPBLXsVX3RCY2Fek6Fo5Pt6PLsPNI8qTJIBrGoDkfU3uPZihr0ZxPUgDoxdBlNbREk8FDz9L/Xn2pD06WD1HRktsZmlrKCKLgsEAIuTbi309qYz0tVg606m1tOoYyobN9fz9B/QH3ZxU/b1sUHDqTj5HZDc2ZSAL/X+vHva8Kde49PglmNgC4/qSP6H+nvxA49bz090chWxcm4H1PBPH9B7TuK4HVgMdZ2DzvZNRD/U/UDm/N/dfh49b4jHTxSU6whWtY/k2+pv9efbLsW6uKAYz11VziKM3NtX0P+9297RanqpYk8ekfWM7sxBPPI/J+lrA+1aUHHqpr5cegW39uIYymqU8gD6Da5IuNJ4Nv6e/TSY0+XSWQlcnqo/5H9qtjlnp4pW1TzinsHflpn0jT7JLqTSMdEN7LnSPPo4/ws+K+DzO1l35u6A1E1ckFZSeVIn9EjiT6SXPu8SCJASKluHRntljH4fiyip6tHxu28Xh6FKKgp4YKeJNEaJFEllH0BKge1HjEno4CKooOptK0FO5HCn6kXuLD3p6sOt/LqatTH5LqbXJb82P+PuhU06sOkH2tQUOc2Vl6HIU6TwvT1DcojkkU50/q93t0q5X1HTUyB4ip61vl2ucP3DkoaC8UL5SQrGtlA/cJC2H/FPZcVK3BUcK9A/TS6KqMV6s36pq6qnigSVuA4UX/Fz7NoMDPR3A/lTo6GBpIaqBJD69Si5HPJ/1vawMNNadGQGqgJ6ViYNFW62seb8Enni4918UA8OtlTSnl1GniancKwNrXBFrD68Ee3FIbPWioAx1lpq5oObn+gH+H+AHvTx6utg5p5dO8eaYABiRwbW/AP8Aj7YMArXq3iUFKY67/ipc8N/UG5P1t9fpf37wgPLrYYnqJNK0urXfn6f73a/uwAHDr2kjh0n6+pjiR2BACgmxH1v9AD7fUhcnppyOHQXZzcCwRS2caiD6efz+PbUhr3L1RpADQ8egF3Hl2mLDUSZALm544tz7YPHUOkbyFjQjoAd3bgpaKlnZ5ApCuCSSB9D/AE9p3elR0lmfT8+iAZWlyfYe9PJEsrUdJM+qS7MjID9fVcf63stKvcTDT5dFLKXk1L0me4fsuvOrN3bsyDLGuIjmhQScFi8TBbEe8R/vC77e7lznt/KG2EmOUBnA/o04jqavandbflTbLrebmgddQUn+kCOPWoB2tuOHd3YG59w07K8ORrzNEym4KBAgsf8AWHuXdktWstqgtXFCiAEHrGnmG+O471c3pOrxJGNfXPQdm1hYf6/s16JxXz669+6312Bf/eOPyb/09+61WnXXv3W+u78W/Hv3Xvn14Wvz9PfuvH5de454/wBb/D37rWevAXNvfutnHXXv3Xuve/de697917rnxbUOLf1+v+w9+6rmtD1xsT9AT/rD37reB11791vru3+x/wAR9PfutV66ubW/Hv3W+ve/de697917rkpA5tz+PfutEE9dfW5/3x9+63117917rsWvzyPfuvH5deNvx791oV8+uvfut9d/0/r7917r1uRfj/X9+61X068bc2+g/wB5/wBb37rw+fXIMAtrG5H+wva3v3WqEmvXG/0t9ef+Ne/db+3r1j9SD/tvfuvVHDrsHhvpzb/fD37rx4jrj791vr39P99b/X9+6912Lfn6e/de+zr3544H4v791ry68b3/AMePfuvDh119ffut9di1+fp79177Ou/T6vx/S559+6rnHXG/0H9Pfurddi4PF7/j37rRp59de/db697917rsC/5A/wBf37rVevD+l/r791759eNhYf0+vv3XuPXX+t791vrsi3/FPz791qteuvfut9cgpsWtdVIB+trt9OR/re/dar5dbVv8i74ibT6s627h/mJ99CDH4TpfbUe9uoIZ4YpItwblxB108FLLMPWwZwzogNtIv79jz6steIFD8/TzPVDXz0+VWY+Z3yf7N+QeZSsjm3tl5apaesmeQQAHT+xG7MI1awIVbf4ge/fLqpJY5x/m6Jtf/ePfut9de/de697917r3v3Xuve/de697917rsafyDf37rRr5dde/db62Hf8AhPHsCvz/AMjdzdiUURdOnqOj3vWPwEgpqCOSVppWP6VGm5N/eisbEB89XSSeKphNKih+degq/m7fOzc3bXzH7R3BgKyCpo8paFqsTSyoHDmG0Vz+BHc8/n8e06WkUcjSAZY16NpeYNwnsksNdI48AZ6rQ2P8oN/7Yqooq2qSsxZc+aMLL5grG50s7sD/AK1vdJ7C0uQVmQEH5Dpm33ncbQhoZDjyz1aZ8Y/mnsPN5yk2/kIvsqyRrI9UzpqYgAugbgi/1A9xHzh7S2G7W8k22nQ5H5V6yX9nPfBdk3WGx31A0TGlTXq97Y/bGPpduRVSfbPTrAksUyqpR1ZLizEc/wC394T81exe9z7odLnJzn59db+V+ZuSN42GG9tXjCaRWgHGlT5dMS99YSqycslQ0OtW9ICg6eeSLfT3kf7P+zlvskCvOKsKVJ6xe+8R95zb+VNnl5a5c+OUFCy1Hy8uhe2/2DjcyscsDoytYm1zYfXm/wCPeV0MUdvEIIhRQKdcktxv73eL6Tc79y8kjEmpJ4mvn0/1uVx89mLqFX8Ak3FvbnDpJknHQfbl3Hi8ZSTVbVETeJGZUBOrjn/Y/wCt79175dEE7M+UGRNTUYWkZ6OmDmJ6qXyrGFv+T/S39D711qpbj1Xr3d8mNq7PxdZLU5CPJ5iaMikamneS1SwNgVQ3/wBv739vXuiH9Q/MPfmL7Rx1blqqn/g2UysVPOpMqGlpnYhNTliGvwDwPr7TXlut3AYW8x1SWNZFIbrYj2t2Tj8zjMbmcfKkmPq6amsyMxTysgLAH6e4B37Z2sb1kI4k06Bl5ZlJCB0NGH3ZAp8okW4UEKb/ANP8fYbkt9QoR0iK07eplT2a2PnV/L6CRfS7fp/IH09tjbqginWvDJoB0oF7fx9TTKBMmpgfqzaifp/a9pv3ZIrVpjqn0pqOgr3dvaKWOSeOVRY3Pqbn/C3s1tbOjU6dSPSaNnorfYvaVNHjpoVlCylGU+phY3I5sf8Ab+z2029vEqOjCGL18+kD8eu+qLYG96uuy0+uDIOka3lk0qfoPz/X3LPIdzHtm4VlwD0LuWLlLC7PiYr0Z3tfv2HMI6YWoMjzx3CxTOSS4uPqfc0z3gY6ozWvUhS3CtlTjop6UPZW666N1oshJj6mYjWElKaC34Y/8T7K7qGa7TSRUHpizuJre5E6A1B8utgH+X90D/AsFTbpyNNp8p9XljjJ1+P6Em5+vsy5f5eh26b60DuPUkNvVzudgtvN8I6uPw6wUsEaQ6VVNI0gAcqtuFH+A/r7EkxZmNemAAACOHT5LXxx/wBrTb8XNj+T7YEZPTnAVPSWyWfvqVXAC35JNhxa1vz7VxQUOemiR0F2czrhZf3LBUY2F+SDe3tRqCYHTZdR0A26N4SxQurPx6iRqcWt7TyS5p0keQV0jh0W3eG/jGkoE6qyk863suofU29pXcDPSKWTTnoDX7KLSvFJUKxuQNLv+eAbj2mMq06TfUCnSdy/cGOx8VRT1cqXaNlBLOLEn63JB908cKKk9NyT6RQdF7yvalMJ5alZ1aBXY8O5/wBYmxt7TGencc9JjIGqegF3b2vTZXJMYZQqrx+t/qP629p5J1Y16Sm4Gojz6DTMb4a+pZfxYWaT/bXHtO0xIyOmjIcsOgazu+przEyXDBhYO3J9sPK3BemGkL16A3KZqSqqpZC5u9x9SeD9OT7TVetR0mY+nSaMjPcFr83+oPvYUDPn1o5PXA2tz9PdutdT8RQSZLIQxID4IpEkqmA1BYB+tmv+PemYKtT1snSNXRTPmN2rBLVQ9Z7dnWXF0scM9TNTykIHv6oWCmxJtyD+PZFcy+I1B0SXU4lb7Oq/iLf0/wBb8+03SWteuvfut9e/w9+6913yf8eP949+69gdesf6H37r1euvfuvde9+691737r3XZBH+P4uPpf8A1/futA1669+631737r3Xvp7917rsm4ufrf6/i39PfutDHXXv3W+uXGn/AB/1/wDH37rWa9cffut9f//V+f8A+/de67BI+nv3WiK9eJvz+b3P9P8Abe/deAp13e59X9Pxx7916lBjrq3F/wDffW3v3Xq5p11791vrs2/F/wDY+/deFfPr1+Lf439+69TNevE3/wBtb37r3XuLfm/+8e/de68psb+/daIqOuvfut9cwCvq4I/4j37qpIOOuJtfi/8AsffutitM9eNvxf8A2Pv3XhXz669+631737r3Xvfuvde9+691737r3XZJP19+60BTr3Fvzf8A3j37r2eskcskUkU0Z0vA6SRsB+l0YMp/249768AB1sFfy3/krS7ow9L19n8lFBuKlNqSeaYqUSA2j03+gYDj2KdqvFlUROaEcOhRtVwsi+GT3Dh1tBfH3dOH37jf7u7qhhqGpqZoaeomRJdcSJwys349i+1cSDS/QztXMiaH6Jt8mdsxYDcM2R2yt6WCqdSkAF10Nax0+0F4lHqvDpBeJpbUnl059db5146jqdTB4Io0lUkn1/2r+7Qy9v2dbimAFT0cPaW7KPK0karUR+Qqo/WCQSLgf7x7XI4fh0YxurCo6G7bmanxjJ+4xVtLBtRtYc+1CsSPSnS2OUEUHQw0u5I6uNS5ux+pPAt+fr/sPbqv6dOK48ulNjquOb1Rm44JHBufxb28pp2jh1ZGJPDpRx1IW/H+3tYH8n/jXvekHqxPl1Liq9TAAAkcccDn8+6MlKnq1dRA8ulNjwAoNuCBf82P9T7SS16uAK08unaV1Rbkixvwp/JX8+2VBJ6uSAMdMVZN5U9R5vwFP6bnn2oQaTTqgApUdJLNVyY+med2FlVgB+S1r+3xw6o7hMnohncm7ppqmWOItqY2FmI4PHHtFct516LLiU/l1WH3FiZ8ruLFmq1GmespHkVuQSJQeAfZVMNTDoiuQWdadbBPQuQx+L6o2hQ05RFXEUqso9JNl/Ps4EOuNWXgB0LbdqRAD0HQt1O4aZIuKhRYWN3AsAPr/wAb97WHuyOndQPxdISt3lTRSFvuE4JJs/8AT/Ae3wkYNOPVXcfhx1Jwm8qaulEYmXUxsF13Y8WHP+PujRq3DrSyVNfTqfujLx/wXIQzsLPSVIsTe58PA9+SNVqw625IFSeqMIMaK3tvL1TJ/m8rJ4+Li3kNreysAeKT8+g0QGuGI9ej2bRoTBFHIoIF1Y24F7cezBAejSEYyOjLbSzn2YSCU+k6TqJBtz9Pb4oRQ9LYmWnz6GejrYatF0OmnTcgEXHH0v7aZSOnxwqOuNVFDMjB1Grmx+hB/wBf/X597QkGo69kjpgamZb2txcf7Af4+1IcHpqndjrj4iTZm/AuOB9PeiR1evkes8MJRvU3JU/7x9PdWavDrYoc9Ya2sEMTAW4+vP0ANvr72ABk9eLcT0D25dxlFdNZBXVxqt9P9b227qO49J2YAaTnoG6qtqa53OpuTwC1+P8AW9sVZj8uk5Na18ug43lXDF0ksrNyEJHP04559tOSvDpiZgB0RfdO56jd+7KTauLd3WucLKyMSFu1m1W9l7t4j6F6K5H8Rgi8OhvyW0Nvdc7Yd/2Iq56fVUzSKqkEJqY6zz7VGWHbbZry4NFVT+2nSoQrHHnHWvF/M9+T9JBt/K9bYHKrMmZQQyLRVH6Z2iZWZwv+pve594b7BtF1zF7gXXNd4vbFIyx6h+D5fb0Ht63+QWx2q3YhTxp1rzcgf8Gv/r+506BXE9cffut9dgXF/wCnv3Wq5p164FrfX839+69x49de/db67Nvxf/Y+/de+3r1uCf6W/wB59+69117917rkBxcfi97/APEe/dV+R64+/dW697917rvi35v/ALx791rPXgLkD+vv3XiaCvXYNj/Ue/deIqOuJ+p9+6312GIFvwffutUFa9de/db697917r3v3XuuQW/+t791omnXRt+L/wCx9+68K+fXXv3W+ve/de697917rsi1v8Rf37rQNeuvp791vrskn37rQFOuvfut9eHJt7917rl9Cb/Xi39P6+/da48OvFieOPfuvAAdcffut9dgXv8A4e/daJp11791vrsC/Hv3XuHXjf6H8cce/daHr11791vr3v3Xuve/de697917r3v3XuuWr6f1H+2+lvfuq064+/dW67Fvzf8A2Hv3Xs+XXrG1/futVzTrr37rfXdvof6+/de669+6912Tfn37r3DrwF7/AOHv3WiadGq+Fvxyzvyl+SXUvTWLxGSyGO3rvPEYLOVlFBM0GMx1bMI56yrqEFo1QEEkkG17e/Vp16mrtrTrYO/nifKHAfGfrHqz+Vf0JXUmMxvx1gnx3YGZwkiRDcdbloRNUGd4v8+AQqeu9uSPp78D1t14Kf8AV8utUliLmzFhcnkW5/qR791oV8+uHv3W+ve/de674/N/8PfuvdeBIBH9ffutUrnrwB+o/Hv3XifLrxN+ffut8OuvfuvdeFxyPfuvfLrb2/kn7HHxl+Dfzj+Q2fkjSq7e6E3BtXZNRIvjaGtmo5FjeCTmxXUfUOb+9EHiOrI+kZ41/l1qO5nK5DNZGsymVqpquvq6maaomnkaV2d3LsNbEmwJ4976oOPTX791vqXR1lTj6qnraCeWlrKWRJYJ4XZJI5UNw6upuPfutAsprWnz6tq6N/mDU+39k0G0N4mrlqaVRG1Q8j6HIQx3Eh4N+Db2SXOw2d1P47KK/Z1KOy+7PNGxbUNstZ3CD+keg57b+cdZNUqnX8lRHOspM8zyyCBkZrkBx9T+PZnbWsVmumEU6A++b/uPMU/1G4uWPHJr0Yb43/zKaLAfb4bey1slRUxmHzyySGnjkICi030/1vaivn0THPVibfKvA5nb7VmK3XQRSVURMETVg1RlhqUH36vWxWvHooW+/l+uyVqqrcu4Y8tTyCQRw0lUZG+vC6VJvf8AHvYoOtE9Vf8Acvy43Rvqvq4NuSyY/DzagrShlqLE/RR+OPz7114V6J9U1lVWyvNWVNRVSuxd3nleV2ZuSSzn37r32dRgSDcEgj6EcEH37rfVu/w0+SMNRt2LrzOV7RVOIgikppauc3qWQaQyFvrzwfYY5i2Zdxh8VBVx8ui69tfGUlcHqwjFdrTw2WadlVjZWaT6qf0kE/4e4uudqKPoZaEdEDWwjJUcesuY7N+4jAjmLHm5Dn6W+vtuOxK4PWhEq9JmLf06tf7mQWsQBIRb/W9umzAwB1bwx6dZ8j2XG1DIJ62zhDcM9+R+R79FZEOMdaEPRRN97wlytbNHTTuVB+qyHSRfm39fYitLbw8t0vijp8Xl0h6DE5bIsJozUehtcboXBBAuDce1U86wr4iYZevPJo7l49W4/APoim7Xz1TBu0PPHSU0UqeYmQekX+re5P8Ab2/beHME51aR0PuUpjuLGObNB1fJsn467PxlVT4eHB0zY2mjW8hokJYWuTq9y0ltGD4dMfZ1IsdlGo0qv8ujQUcOP2Tj48Lg4BS0EQ1qiosX7hFi1l9rFCxdvSxF0LpTy6e6HevgUeaTj6fqNh/rn3osjDI6sJADTpszvZdLTqB5vVfkiT/be66lXI68ZQBnpLydhU1RHfzqT+GL2t/re6mcHFaHqjTA9IrNbuidGPlBJB/tH6X/AB7rr1Vr020i0qOi57w3dEwmVptNy4uXIFifx7SOygEefSB5ABnolXa+9Y6WmlEdT+76g1pOSLcEge0Ez0HRZcOAuD0TncfbjYWkaoE5ea5IUSHV9faF5tA1A9InmAHRYNz96VeZqXMks6AFl/zrc82A49onuNeTx6SPctIKcOk2e0KqWA0yNLZv1etv9hyfdTc1FPLqiSN5nrnjqmau0zhn9bXJZjxf/E+90DZ6smTnqXlZvFGFLk2F29XHA55HvZYgaTnrTYyegnzNTA2oI5B1Nzf6+05KqemXNKHpEOS7E/m/+JHA4sPfhgdUr69cAOf6kcHi3+w97PWussNPPWVUNHTRmSWokEahebFuBe3497AxXrYwc9Ru792UvRWx5WNRFJuXK05pWo0YLUqKldIa31Fr8n2XX1xT9NT0gvZwh0DqmzI102RrqqvqXkkmq6iSod5XMkhMrl+Xb6/X2UdFPHqExuSffuvAUFOuvfut9dkW/wBtf37rQNevD6/6/H+39+68eHXIkqSB9P8Ab/4/X37rQAOT101782+n49+62KeXXRt+L2/x9+68K+fXuLfm/wDsLe/dez165tb8fX37r1M1669+631737r3Xvfuvdchc+kf6/8AxHv3WjQZ64+/db697917r3v3Xuv/1vn/APv3XuuekWuf6X4/17e/dVrmg64k8W/Avb+vPv3W6efXXv3W+uzb8fT/AB9+68Pn117917rskk3Pv3WgKdde/db674t+b/j37rXXX++P+v791vrsi3+2v791oGvXv9a/v3Xvt67Aube/deJoOujb8f7z7914V8+vAX9+68TTr3HP1vf/AGHv3Xs9de/db697917r3v3Xuu7cE+/darmnXXv3W+uyLG3v3WhkdevYW9+69TNel3132DuHrTdFBuvbFXLSV1DKpcI5VZoNV3icr/X8H8e3IpGicOp4dXilkgcOhoR1to/y7Pmtt7tXB42hq8tDSbpjjjoClRMsU7S6dDh0Jv8A76/sdbXucdwmk4bodbTuKTqAxo3VoOX6+XLyzVeYqYajH1iSVBJYOCzg3sxuPZt4dalvPo4eDWe446D7anVVDVUGaGJlSGSGWV0hYhWeKO/KD20sK5A49Nx24NRXPSJ2/ls3t7JVNRA0z01DV+GaIFiW0NZjYe6ozIccB02jPE5Hl0cnY/YmNzlJHHLMkdTZVKO1iCR+nn839rFfUMdGMUgcUHQmLn6vHSC7O8DMNLfix/x9vayo09KBI3lw6Fjam4oJxHplUa7khj9bn6W9vIwPHpRHJqz0LUVYjorf6of14JH0I9qtQ01XNOnMk56hPkpIGLf7VccX4vx7ozCtOqA0bHT/AEG5FUKOObA/S5P4NvbbIGx0+HU4HT0cj5wCn0INxc/7E8+/CPTx60SeHXF5NEbMx/rc/Wx9+GTTq5ApU9A3vvLSeOSFHshv6R/wX6296Y0+HpO7lh3dFD3VQNXzvLIjGx4JFzb+vtG6gtVui50BNG6JZ3nRLSGlq4UVXingI45GmSwI9l9wQpqo6LbkqDgdH06p7POO69wzT18a/b46EBGk9S6V+n19mVvOPCp0aw3FIASem/OfIqGn80YrhL6rWST6E/Ufn/e/fmuQBg9Ve7AzXpEUneLZWqEflcamA5c25/2Htv6kN9vTS3QY06HbaW8rtDUrVXdTHcF7/wC3/wCKe3kemeliSZ+3oRd4bwkqKJUilOqSBgVVh9TGVJt7eZwRjj0676R69Enw21Au7arIy05PnqXfVY/2mPqufaAIfEr0Vqh8bX0a7b9GqRLELWcf48f8U9qlDAU6Mkaq0p0s0ppKcqyMbD62PqH+sR7eHp1cdtT0q8VuCpplKq7argKA3LD6/ke7AmtDw6dJx0INBmvvUUSH1fm/1v8AW592ChhqXq4YK1D07FwF4BJPH+x+vutOrE0OOojeRi1voCQefyD+PbgoOqkFgT1HZ5Rc/kfm/wCPdsU6oaDh0h9xZRqWll0MQ51cauQB+SPbbNQ0PW2ag6ALI5E11UV8mq5uy34BA/w9o2qzdInfj025DI0uNp2dmAcg/W17gX/PuxNF60aAaieiZ919k+KkqoKOUtOU0hFa55H0FvaCWYpjotupqefQF9IUaUlbX7rzzXqY3aWlMpNxZtV11e2LdApLnj0ltToJd+ih/wAyP5izdfbKyuNweTWnzVWksdAwkK8tDpVQAfyfx/T2S8w3kFzYvt5Ydxz69JN4vJIYS4qD5danW8t77l35l585ubJTZCtqXLsXZvGhP18aEm31+v1/x9gS0sbaxTw7ZQtePqft6AbMXcu2SekmDY39q+qkV68Tc39+62Mdde/de697917r3v3Xuux/sbf4e/de669+6913b6fi/v3Xq9eIA4H+x9+60Knrr37rfXIgC3+tzaxt791qp64/63v3W+ve/de67A4J/At9P8ffutE+XXXv3W+uwL8/j/effutE+XXZH1I+gt9frc+/der5Hrrjm/8AsPfut5669+6912OSAffutHh161zb/X9+69XFeu2W1ufrf/ePfuvA169b0i/+Om3+839+615468CAeL2P1/w/1vfuvEE9dHkk/wBSffurDh117917r3v3Xuuzb8X/AMffutZ8+ufBX+lv9549+6rwbrH791fr3v3Xuve/de697917r3v3Xuve/de697917rkBe/8Ah/T37rRNOuP+t791vrsGxv7914569fm//IvfutUxTr1wTc3/ANh/xv37r2QMdde/db697917r3v3Xuu78Afge/dapmvXib2/w9+68BTrr37rfXvfuvdcgS1l4/1/fuq4Hd1lip5aiRYKaOWeodgscMMbSSSE/hEQEn/Ye/der5nh1uK/yy+jsT/LU+BPcHzg76w9Lgt891dc1NP8cTuWKOmrcfuyKMoa6kWWzDQLaeL3sb+9UJNR1bUiijca/wAvIdam3c/b2+O9eyNzdo9jZeTObu3PXSVWUyEpZ2mIJWIXcknSvAJJ/wBh731rj0Fnv3W+uyCCQfx791oEHPXXv3W+ve/de67Jvb/D37rVKdde/db67Btz791oivXXv3W+pmPoKzKVdPjsfTSVdbVyrFTU8K6pZpLEhEH9T791o4yetuv5d7jyPxI/kO/Cfb+IVcd2D3BVbkpd5U5Hhqo8WXK0i1Sj1WK8gEe9Any6cdRp+eOtQokkkte5Jb/Ynn3vpv7Ournj/D6cf7H37r2OvEk+/deAp1yGpQGViD/tNwQP9ce/derU064kk/U3P9Tyf9v791vrwYqQQSCOQQbEf6xHv3Xun2j3RuKgXRSZvJwpYARismaMAC1gjEgf7Ae/de6g1eSyNexNdXVlU5a5+4qZZVF/6K5Nv9h791XHHqCy2t/j/hb37rYNevajwOP6fn/b+/da0jj1xNr8fT37q329OmGzWRwGRpspiqmSlrKWRZI5I2Zb6TfS4Ui4P5Hv3y691aJ0j8kdu7yx9Ht7c832m4UhCGonbxxOygLrjc8Ef7z7D+47OlxWRAKnpBNaqxLDoyE/3kY81LU/f07KGWaAiSIqRe2tfYNuNskgYgrjorkh0HI6TlRn5onIJdCP1AmxFvafwWGD5deHDh0GmYz1dUVE1qhyjXsoc/j2YRRKAGpnq6ooQMOmrH0FdkKuJY45ZNcgD8E3B5IJt7dkZUQ1x1cnSOjydVdax5OnhhkoH1sF+sd25T/Eewfud9pVtJzQ9Fl1NpBPl1cP8ONsQ9b5OepnRaYT0yLpICs119I59y/7LGRi87cKcepF9vKhmkPAjq5TZe5qKspU0eEMYx6mC6zYf6r+v9fc/I4JqepcVwQaces+62QUnnTlzdr/AOB/AI9+kweqvSmodAJkdwPTrIpcqRe3Nr8/i3tmvmOkxlHwnoHd27nrCutJnKhrkK5+g/HHtPJI3Bek0s1CdPQey9i1FGtmd1/Fy5I449sa/wCLy6TeK1SDg9MWS7UbxkvUgG1uWP0/x9+8VjkHHXvHPr0WztDtZqakllpqu0v7gAD83B4Nh7SzTAA0PSSeZdPHoiu6eyspm3qo3mdidWn1H8j8eyx5GYUPRa8hbt49BEMHmM+ZPOJWVibfWw1fQge2NLufl0n0PJ2npnpenJ6/JQwzROBJKASDZbE839+EB1dVMFDRs9DhJ8bqWipUqVjDN4kYfmxtc+3TZgDPSgWlMjz6Ts2xThUlh0hRHq4tzx9Rfj/ePdTEVGOmzFoFPPoC921iUjSxOdJsyi31vfi3thyVXHTJoDpboFqiVzIyliyklh+SP9f2yKMKnphvir1HPP8AvN/+J92p1rrlGPK2n6W5F/7R/oP9f34YNOtrlqdPuR3Jg+qdt1W+s8VU+J0oFYXKVa8Rtp+p5t+Paa6nEQqOJ6YuZljWh/Lqo/tvtncvbe56vP7gqmlHmmWjgVnEMdNqtDeM8atI+tvzb2SO5dtTdEjszsWboLLWAb6/Q2tx7p03WuOuibm/v3Vhjrr37r3Xvfuvdd3Nre/dapmvXXv3W+u78W/xv7917zr1ysVIAsSRx791WoIqeuiOefzybfjnn37rwOOujYHj6e/dbFfPrygE88D/AH39ffuvGtMdde/db697917r3v3XuuySbf4C3Hv3WgKdde/db67AuffutE06/9f5/wD7917r3+9e/de67Nvxx791oV8+vC31P0+lvz/r+/deNeA68fr9Lf6/v3Xh11791vrlYE8cD/H37rWQM9dG1+Bb37rwr59dqbfQXP49+68RX7OuPv3W+ve/de67Btz7914ivXr83/x9+615deNvwD7914V8+vEj+zcf19+68K+fXXv3W+u+LfTnn37rWa9de/db67Nvx/Qe/daFfPrr37rfWTk8t9Ppzxz791TAwvXDm/5v/vPv3V8deA/Jt9bW/P8AtvfutV8h14C/5A/1z791446EzqjtXdfUG7sduzaldJS1VFUwSyxhnEVRHG9yrKpHNr2Pt6CZ4JBIhyOnoZngcSRmhHW0N8Y/5ig7p2RQ4Fy4zVIKelqtRk8nkLgSek8/4/T6ezuXnKysQsd6QoONRx0NrDcjexiMfEP59Wm7ezCTbXXduCqA0lPCKathR9WqRls5K+xbbzw3Vut1bMGUjiOjYaol1kUPz6ROHzECbhC1CI1LXPrqVYf25TZiL/096U0fST02CGbPn0I+R21HjoUzG3JJFJZZ3QXtwdRsF9u6CtSvHp7wwo1Iehi2dvyhzVEmOyLItVGgQ62sxf6Fufb0citg8elUcwYaX6WOPzBxFcmmcPT6wfSSeL3+vtwuBgdXDaG6MJhd2Q1NPAokUsVFhc35+nA549qkkouOlaPrjqOPSzpwatdZ/PP9QtxwPd1UOenRjHU2CidGBNhpIt/iP8be900mnl1cgEUHSkiPgUMWsNP0vx7udIGPPreqgz02V2XYK0am3P1+nF/8ffmKoMcemSSxoOkPlMUcvrJOrUQbix4P1v7bww60Qegr3htqKiACoWYJbi3PHN7e2HB4DpiVKmvRGe29rPlaaoLKdMGtlsL20H039l861x0WTqD5dFepd+5bG1abelqGWnVli4chQoOn8fm3tJ4jA6OA6ReI1dBPl0d3p/p7A74oEqq6SeSSaznSb/Vrf4+18ECycejG3tlddTZ6GfffRmzdkYV6qn8qVghLx6yq8gXtf6+1EsEaLUcelMlrDEmoD/Y6LnsvO17ZSamWRtEc4SMAnToDWv8A6/tKrNwHSNCzPxx0ajHCSuiiM+olVAIP1+tj9fasAv0vBqOlHTbYplbyeP8AV6gQBwPr7sFoc8OntAOfPpS0kMdICLgW+hvyB/T3Zfip5deppHT1DVRlNDG5I4Juf8fdjxoOnVZaU6kxzQga1vxwWA+nH9R/X34dbHSpxVWqMh1EFuPofz7dB8l63mury6W0eQVVHr1c35vz/jb/AIn3YAHq9TSp67/icdv1C3+tb/X+vvZUAZ60M9NeRzMUNO58ihtJte9xxf8AHuhK0qD1okA6R59Fz3lu+zvH5FuQTe/H09ppHqdI6TyuD29AlV7o+wLVDSK19RuOB/X6+2NdMdJHegr59AtvbsKrqRJHTudL8LYm9iLH6e2Hl9ekrTHJB6LTnV+9mFVWsWF9R1f4C/IP+29pG7vi6QNVjVugq3d2B/AcTWQ0bKgjimYaWAHpQn6X9p7u+hsLc3E50gAjPVQJZnFvbipPHrWS+bndM/Z2/JsXJI7fwStmWUanKBtAVR6vz7iqxa5ubiW7uGqrMdH2Hoq5ou4pHS2iFCo7vtHRHb/Qf09mvQRp117917rv+lwbe/da+zrs2BFuR7914Vpnrj/re/db65DVY82HPHv3VTSvXV+Le/dbpmvXXv3W+u7/AE/w9+61Trxtxa/+N/fuvCvn11791vru/BH9f+I9+6913dbfTnnn37rWa9dEk+/deAp11791vr1j/T37r3XYJ+l7A+/daNOPXY+tvqP6Dm/v3Xj69dcX/wAP6e/dezTr31PA4/3r/X9+69wGeuvfut9d8/4/4e/dex11z7917r3v3Xuve/de697917rv6WPH+t/rf19+61xx12xB+g/330Hv3XgCOur8Ef1t/vHv3W6efXuf8QD/ALAc+/dax10eP+Ne/db697917rv8H/Ye/da69Y/0P+29+63Udde/de65XXjg/wCPv3Ws9euvq4+v0/w9+69Q464+/db697917rsi35H+t+f9j791qvXYtY3/AKce/deNeuIBP0F/9b37rfXvfuvddn6AgG30J/F/9f37rX29de/db69b37r3XZtYWH+v791rPXakAgkagDypNr+/dePVjXZP8uzdHW3xL6u+WOR7V66yGB7Tp6ufGbKxuaoqnd2G+0Yq65zFxzGWnDH9BkjW/wCPeqgmnV2UquvquT8X/wBhb8/T6+99V6UG1tyV+0c9jdxYwQmvxdQKimWojEsJcAj9xD9R78MGvWiK46OT8mv5iXya+WvXHW3UfcO8EyHX/U8TxbL2/Q0v2lHjVaPxhTFGwVv9cre/P496p59bqadEWJufoB/gPp7317r3IP8AQ+/de49dgk35/wBf+p/Hv3WsdeBK82/w5Hv3XiAeuPv3W+uxbm/1/B/A/wBf37rRr13b/Ef7f37r1euja/H09+62K+fXmNzf37rQFB0aL4TbVG+Pln8f9nmH7gbl7L29hmiCh9a185p2BVuDwbke9Hh1YAMaHq9j/hRX2hjdp9j7e+GlLKrP0jSYcmGPiGnnqaRXmjjjX0rwfx73x60QQR889awnv3XuuWo2txb37rVBWvXGxP0Hv3W+u7/W30P4Pv3WqevXXv3W+u/oRcG3/Ee/da+zro2vx9Px791v7euxyfr/ALE/8V9+61wHXdv9qB/2P9f9f37r1fl1x9+631737r3XvfuvdZIpZYXWSGR4pFIKvGzI6kG4IZbEe/de6M51v8nt6bQ+yxeTnWvwcTgTBkZqkRhbAXN9R/r7Ty28cqkMOPTLxK3R7tm9n9c9tUfjxtRDiclpVXaqfxMZCtyul7H6+yC62cKdcfSaS0VqkdPEmwZqepDeUVsVywlhUyKVP5DLceyO4jmhqwGOkTq6HI6Nh0p17g62aAVMBkk1/pOkngD8H/H2DN3vp1B0efRZdSuoqOrQerus8BTvA8dLo0aGNwgA4HsAbhezMyxk/EaHonuJ3dgo4HHQtbzrP7rz0NVjV0xoQHCAj0rxbUPeYvttYRWPK8VxD8TAV6nHlWBLTaUkTDGlT0YrrLt+maioy0yI3oVlZ734BP19yPFMNQPQxguQBQ9GCyPaWMnpo43qImLKABf+o/p7VPKpOelLXClag46BbdG44Jy7xyIoIP8Arc88+07NjHTEjrpGnj0GLZAV37RfyXY82+v+JP049sl818+k+vND011+2kqkIaMhQCysoBvf83+nvzDV14xhu5ugk3Bs54hKyeX9LHn+gPA9sMlBQdJZUz0UbsHbFVKZ9byFP3OPxx/U+0M0J4jj0ilT8I6BnB9fzVOQM06O0IY/2eG/x9spHU1bpkRKrV6HLD7Ox1GjMyKoK8hgv0t9PaoRaTXy6f8ACUCo6ZK9Mdj62JggRY5Cf6Xsfw3ujBQ2odVYqaDpay7tx5xoR3TUIgti1j/QD6+7mUUoerBqih6LjvrMkxVNTG4sFYgfgj/Yf737SStXgek0j0BJ6JFunLHIVUjE86mFwTf9VuL+y6Rw0mnovLlvi6Rp55JJ45JJ9+A6r1x4VeTwDfkgf655t79k9e6Um1cM2Urvu6ommx2PIqaiWQWjeOL1MqsePpf6+/OUjTU3VWcKKnogvy87bXde6DtTb06LtjGqrPHGxbyViOUZWN7WFr/7H2Q3Mplkr5dEdxIZZCfIdEw9p+meu+fob+/dax17i3+Pv3Xs1669+631737r3XfHHHN+f9b37rWevG1+Pp791sfPr3Fv8b/7x791rz669+6313c/X+otz791rHXXv3W+u+LfTm/1/wAPfutZr11791vr3v3Xuve/de67Av8Akf7H37rRNOvEW9+63x669+691//Q+f8A+/de67tf6f05v791qtOPXja1vzzf37rw68Rb37rwNevcsf8AH/YD37r2AOuvfut9e9+6913xb/G/+8e/dazXrsG17fU8A/7H37rxHXRvfn6/X37rw4Y68Rb37rwNevcE8cf6/v3XsgZ669+6313xb/G/+8e/dazXrr37rfXvfuvde9+691737r3Xvfuvde9+6912ST9f96Hv3WgAOHXrm9/z/rD37r1BSnXXv3W+ve/de697917o2HxE7ch6r7JpqrISlcbX+KIqXdV+5MqqlgOLsCf9t7A/P2xPvexvHASJFyCOjzlu5itN4hln+AsAf29bSPw/7moa/OJgMnK1RtvORvU3aRnVHmY6b6jbjg+yD2s9yRsBGw8xvRSQgZjgUx1mLzJ7aW9/yrFzFslGquoqBnPRr+xdoVWFyRyeIdanHTP5oJYACiRseFLD6e8o2MFxGt3ZMJI3FQRw6xqnt57aQxzAqynIPUvbW+6nHUhp66GSoQoVCi7AH6c392SQjB63HJpFDnpCZ/J5w1/8S27TVKXcsVjLC5JvwB7q7EZXj1Vy5PZ0uMLvTdCUAly8VRFJEACZWcNz/r/737usj6anpxZXVKP0OXX/AGHUVpFqixRlWzOSSR/r/n27FKW4dK4ZjWi46Nft7fumBPK/p4FtRNyBySfa9JD0uWWmOhLxu8sfOVYuARzzzyfd1etR08HUZbh09yZyCdTokX1Amw5v+b+3NZAx1vByOmaSrVi5MgPP+I+n+t7byxr14MOHWaLIwwRNd7jmwvz/ALc+9gn4erVAHSXzEsVYru9iSGH0+gItzfj3px5DptjUY6LrvPbEctDkGdRaRJWHAFxpuNPtHIgH29IZkBB6qw3RgJ23bUiJGDQzSOvpA9MZJ9lUkbGTohkU+JXoQOvvnBsvpCKfE7syEVPVw1K00aTVDRt+v8C49uR38VuNLmh6UQbjHZikrUNejRb2+UG1+39r0eT2xl6etjlghUpTTlyCwAIYAmx5tz7WPeJOoMZr0vlvo50DQnj0oOrttfcmkrWU651SU2H9T+fe4kFOt28Q1Cvn0aDH0JpY2VwFOoaTawte/tYo0ig6XKFBo3Sijq0XSl+QLf1/P149uA1pU9ODTSidN1SzuWIJFybEW/rb/Ye6EmuOtGhGeo0VTJEbyFrL/tz+OPei9aDqnDh59RWz4gZ0ZrD68n+yf6e9aqHrQkrjrLDvmnpH0PIoI5T1fQD6X92EgBpw6v46/D1Kk7NpUHM6f4DX+P6ke9GWnn1rxqHj1HbtOkKG0yf0HrNj/jx7r4ucnrf1AIPSNz/a1HHDIxqNR+llkLX4+nB91eVT0y9zmvRds9v9MpUMYTcEkD1E/wCv9faYyZoPPpIZdTZ6RuSylVXIIYybW55J0i3Iv7oTXA6YYsOB6D7PV9HjI3M0geUKLre5DWuP9h7ZoFrq6aYqmWzXotu895OPIYyVFgEX8+rj6D2nldEBlkbSijJPDpFI3dpiFWJwPPot/Z+fx22NiZbcm4p1QmGcU8Ltpd1ZDyv094j+43uJec0cwpypy2SI0NJGB868B1k57W+3217bsdxzTzYwRQp0avswM9avXZ+XpM9v7dOXom1Utdk5ZYTe/pCrHa/+wPuY9phkt9tggm+JUAP29Yf8ySQS77dSWxBjMjaSOFK9IL2Y9EnXvfuvddkk/X8e/daAA4dde/db697917rkbgDng+/dVxXrq3F/fut1zTrr37rfXvfuvde9+691737r3XI6bC319+61mvXVja/9ffuvVzTrr37rfXvfuvdd3v8A7D37r3XXv3XuuwSOR7916leuvfuvddg2/wBb8+/daI68bX4+nv3XhWmevajxz9Pp7916g69xzf6/j/X9+69ny69xxx/r/wCPv3Xs9dkjnTwDa4/1vfuvAevXH37rfXvfuvdchpt6gb/g+/dVOry64+/dW67ubW/Hv3WqDj14fUX9+68eHXIqAG+n+B5/r791oGtOuPBsBwfz791vhx65eoAi/A/1vyPfutY49cPfurde9+691737r3XvfuvddggfX37rRBPXuOb/AFP0P+Pv3Xs+XXrfT/H37r1evcqSP+N+/dewevWsL+/dermnRsfhR8fab5M/Ijr3qjJZzE7aw+5M3SUGSzWbnSDG0UVRIIhLVSSEAICbsSeAPfiaZ69pLnQvHraRy/x+/wCE8Xx97oxHxE7pw3YvZnck+XpdsZPsjrbK08mxqLL1aLFFOFUMGi8ha51C4HH4vrj9nV8BaGlR6/5+qnv51n8svaHwQ3bs3dvVdUlX1B2tG2S2DMfGas40rrVawx/2gLe9/LqnE6hw8x8+qHmFiR9LEi3+sffutjh178H/AFx/xPv3WvPrZf8AkL8FussL/K3+EHftLmNwVO4u25quHcGLnraj+FU64+p8Q+1g1WAP+t78uT9vVpKLFq9Kfz6P5QfyiP5dXQ3wn6W+ZPyfqp4tsbzhyFfldv0dRBSZzL0mLkUVMOIWQ3LHUArAf7171j8+tkaW+VMn7egO+Zf8sL+X/wB8fC3KfMf+WjSbo2/tPY2IfNdgbb31NT1GdxdLDEXeRZlAujEG1h9D79UDPWjRjpBz5EcD0Cf8uP8Alm/F3DfEyo+dHz121uvenSFdiazJYXAbC0R5+pemd4mi+6KtoAZCLkc8+90NK+XWgVrTz/kB0IXyA+Ef8pf5O/FzdPyI+Au8k6jq9lbbq8vmete2M3RNu+vnpHCS0uLUlWeW59Khfp79hevU8SoXFPP/AD9aoTrZmAIJDsv+vY/X37rXDrhyD/iPfuvcR14kn6n37r1AOuvfut9e9+691737r3XZtc2+n49+60K0z12FvY/4/wDE29+69Xy6vs/kH9FY3sr5K5XsmtxyVp6JpqfsEyvGjrRQ4uX7g1EjN+lQyA3H9PfsHB62tQdQ/wBVeimfzhe3Ye9P5gfefZkFctfFuHKUTidJTNEPtqf7cpGxJ4Gn6e/Up1otX8uqxPfut9d2Nr8W/wB8PfutVzTrwJH0/Pv3XiAeuvfut9d/Q/63v3WuI68ST9ffutgU67NrC31/Pv3WhWvXQtze/wBOLf19+68a9de/db697917rsixt7914Z669+691737r3XvfuvdS6Orq6GeOqo6iamnhYSRywyPE4ZSDwyEH37rRp0a7rP5UbswGUx1HuWdKvAB44qhx5PNHGvAdySdQ/r7TS2kMqlWHTbxhxQ9Xz/HjI4jcWPwW+cQR/BspJHDHHc31kAs3+tf3D/ONgbNyUHHoNbpE0JPz9Orddi46B1V4xZXjjsLf1jFxx7hrcZio1fiHQVlYhanBGekNvXIwDOVeIqnUIUIiDfTUeOL+8xvaDcXvuWI4ZxwAp1N/Jd21ztYjl4CnQAT7orttZeXHq8scQbVG4dwguSR9D7ktnKGnQoqVNPLy6Uadk5ZwitVPdTqVvM9rDn+vt3xWPA9bEpoAc9L/Edgz19OUqpGZrKAQzN+ObX93WQnA6e8XyXpR43cjU86PqJj1XN+QVPJHt3UBjqyOK1boYqTeGKlo0BlRXbhgx/p+R/xT3cODjpQXVhQeXSL3RuHGrGbzxkFSfra4v8AS/ujFTnhTqkjrTHDonPYe7cRFI8aTRszM11BHBJt7RSuumtekErgdB3jewcLRL4qlo0BvybD6c359p/GRePTHiKccD0j91du0cEqpQzXjFiSjta1/wAge9STKRVeHVGlANPToEtxdriuntTSkfXUdbH1fX2mNxXA6YefVw6T0nYVR4tTVDto9RAlb1Afi3tvxD034tBXpC7j7KfIxPTw31WKsLnn2204IoOm3kZxRugdnlM0rS/QsxJH9Lm5sPdOmia9YCTwqLqZuAByf9Ye90690KW0thx5CmfKZ2sgxuPpojUSmq0prjUEsFLW591M6Rqa9VMiKCfTol/yP+SuOxMdTsLrdvG0eqGuyMTHxhWOlgGU8sR9B9B+fZHPcPK3HHRLcXDStjA6rcllkmkklldpZZXMkkkhLu7sdTMzNyST9SfabpP1j9+6913y359+61gddfT37rfXYF/futE069bi/wDjb37r1c068Bf/AG1/fut9eFr8/T37rRr5deFr8/T37rxr5deAv9P959+69WnHrxv9P6e/de+fXXv3W+ve/de697917r3v3Xuve/de67H9f6W9+618uvE3+vv3W+HXXv3Xuv/R+f8A+/de697917r3v3XuuVgfpf8A2P8AT/Ye/daqRx64+/db697917r3v3Xuve/de697917rkTcD+v8AvHv3WgKdceLf4/7xb37rfXvfuvde9+691737r3XJb34t/sfp791o0pnro/U/6/v3Wxw68Bf8gf6/v3Wiadde/db697917rsW/N/9hb/iffutGvl11791vr3v3XuuwLmw9+60TQV69x+b3/3j37r2euvfut9ZIpXhkjmhdo5opFkjkU2ZHQ6kZT/UHn3ogMCrZB68CQajq5r4EfJqgiyNFt7deQWKrijWJXkm0MFWUrG4Y/gge8avdnki4ETX21KQCdXb5HrMn2N90oGhHLu9yAhgFGo4A/PrZ26T3fit10NPjMjURVeHroiFlkYSBQ62jOv8Wvx7d9lffVuXrhOT+cnJDHSjvwAHkT6/4eh37p+0iXX+7zl5QUYaiFFa9Om5cTgNnbohoJHgnoKwiRXjIaNEc8Kx5t/rX95oWe+7BugEm33UcgfgFYHj1i3fbRuW1zGK8gdftHRh+vMPsR6ularp6eWnkEbElEb6vz9bexAsSYxjpu28Bu0cehb7n6w2HktnPV7XpqeOvSLUBEgAsFuRx788VIyB+VPTpVdQRmI+GM9Vw49Jtu5WRAzUxinAYOSoLA29IPsv4Y6J1OmqjHQ24neFS6xqkpY6RcqfTf8APt8NUAdKUlJyvShG98pRkFJJNIN7Am9v9h/h9fe9bLgdWMj8W6VWJ7bliYJUTN9BcsTx/t/dxNUU6dFzjHS0puxxVAFKpWD/AE9Q4938YeXT3jg0r0+0m6WqALzE/wBefzfj6e76jx60GLHp6XKo63kf0/X9V7/0PuwZtNenAdPQd7yzsTwSxBgf23RR/W4t/wAi9sliASePTMrniOiiR7GbI56rrxAGDx1ADFbi7obcjj2kCEsei8wliWpx49avX83Haeb2P23i2grK2GhrWqZbU9RNDEszRqUuqEc8Gx9gve43iuPl0C9+Vkufl08/yye1t4Nl6bbWRydbWYf+JmICqqZJ/wBqytb13PBJH1Pt7Z5pWbSTivTuzTyM+ljUDHW4D1BkqCbG0jqwJiSJRY3INvo3scw0Ix5dDy3dD0YSTTUx3Q8kE/15PNrj/Y+32qelfa3DpOTLVQtf1FL88kf7f3Qq1eqUK9O1DIJLB/z9eQLk/wBQfexQ4PVlqR04T47zoWQWK8Wtbg+76RSnV6VFD0HudwtWdTRhrrfj+0eLgg+2WBGB0y8ZGK9AnuLHZ1HMkIkBS301fQn83Fvx7ZZWrQdJCG4r0ipqPOsQ8pmuP6X5uf6e2zqpTpvvrivTfU0uaAPjkmBN7/uWBvz+PflqBU9VIY46TVVjczUsEZ5bG55JIHH+8e28npvTIx7usSYdaKNpqiSxVbkk2/1/eyrLnr3hmmr06D3cO9PtFemxhBlAZQVOq5A+tx/vPttpQRQdMtLqXSOPQB7v3T9tTzVORqh5CCQGlI9RX+h9p2VgKscdI3fT8Rz6dEK7V74o8K8s0tYkcNMLupkAZwtzcD/Yf7D3EnuJvd5NaNs2zt3yClR5V49SRyLyxFK3773eiQxEnu8wOqlvk98ttwdmldt4rIyri6dHgmaGRhHo/SEUi12t9Tfj2AeRvb632FjuN8NdwxrU5NfU9F/ub7pXG9xjl/ZnMdpHghcA0x5dENN7m97nk3+v+x9yp1BPHr1+CPwffutUzXrwF7/4e/der11/r+/db697917r3v3Xuve/de697917rvj8X/x9+69117917r3v3Xuve/de697917r3v3Xuve/de67Avx+fx/xPv3Wjjrr37rfXYFzb37rxNM9etwTzxa39P8b+/darnrxFrf4+/derXr1xax/H0/437917z669+6312CRf/H37rRFevfU/659+63w670nn6cf8j9+61UddA259+62RXr31uf8Afc+/da4Y669+631737r3XYNjf37rxFevcc3vf/D6e/daz5ddf6/v3W+ve/de697917r3v3Xuve/de697917rsW5vf/C3/E+/daz5dde/db67Nvxf/Y29+61nz669+6313f6D8D37rXXvTfm9v9hf37r2evAE/wC8f7ybe/deJp1Zn/K0+HXZny++R+2NsbByC4mix2UpFz1d949GyU055UyCwAK3Oonj8D37yr1riaVp8+tiTtPa/wDIu+GvyMpOuu2tgdudufKDDZylxlblNsbkep27TblRUp0nqImhZS4kuFZrN+AePdcnh052L8X58emb/hULkdu5roL4IZrZuIr9v7TyG06x8RhstrOTpINIKCqLgXNrc2928+vfh/P/ACdaZ09BWwxJVzUdTBS1BJgqJYJVgl/I0SkAH/Ye906bBHDz6hkWX8G5H0/1j7114Gp63MfksL/yQ/5Z4/H3ee+t72+//HvS/H1ab+xNfUdB1/Pky2Tl/ll/ywqA1tVHQwY7c8Io4p3jpJUkeeYmeBSA5DKCC39P6+9CtenH4H8uld/KUeTK/wAq3+YrisnVySY2HpVTFFUSM8FOfH+tE4Cn/H3tuPTcQov2noov8rj+bNjPjR0RL0P81um97dr/ABGy9JVY/DfYYypWgpIJ53EhpK7xMoKuzAMBYkAgkm3vxr1saQQTg+R/ydH17g/lffy1fmz8Xe5vlx/LSyO69uHrTZVXvDfuw8luuRqzGLrWSqh/hdIIkshb1alv/X34doqetFRI1IzTPlw+wjrSunheCeankBWSGaSF1P8AZaNyjAn/AFx731qvn1iItx791vj117917r3v3Xuve/de69/re/de697917rsf1/pb37rR9Otv/8Akl7Aj+NXwr+bXf24jGlX278fM1tvZ1ZMgjeGuqdMo+ze/wBQtwD9eT70QTkdWjcKM8a/yHWo1uLKZDNZvLZXKVc9dX1mQqpaipqJGklkd5mYku3+8D3vquOmT37rfXYBN/8AD37rRNOvE3t/h7916lOuvfut9e9+69176+/de65Em2k249+60AOPr11yp/xHv3XsEdd3DfW9/oP6f7G/v3WqU4ddG3+P+P8Axr37rY669+631y50fi1/9j791XGrrj791brsC/v3WiadeuSAvv3XqAGvXdiLjg8f8T+PfuvVrnq53+Xd3f8Ae0FJ1rk6rwR4ImakWaX/ADjlR4zHf6/0t7CfNW1LfWTSAVYDou3CASRVHHrYt6q3xroY1qZgk6aLhmF9IW4PP+sPeN26WBDMoHr0B7mEBjq+fQPd95mro8lDnqF30iQmTQbDSDe59zz7OcyQ28X7suTp0gAV6kHkvdY4l+kkNAOHQTS7rxm7seswniXJIAr3b13RbEE/X8+8g9Syd65r1I6sHWvSMmy2QpGAeVtKk6eR9Bx9fbIYp1XXXA8ulbgd7+N0jaXQwsGJNuBxa59uJN59eDljQdCjS7zjMQQ1CkBQV9YJJP0t7fEoK9PeIQKHqHk96SQU5mgq3QodXMukf61j70ZAo69rIFQegV3X2lknhdv4lpADgfvcCxsB7SyzEcDjpl5vI9FmzO+vuneaaqeSUNJ/uwnm/wBbe0jS1z0hZ2BqTUdBVmN41NRO/wDlEgUHjTIVHHHA9tFxSvTLSqT0marclSyaRLK3JuWe/H1/Puhdj2pjqpZq1B6T8tbK761drE3PrP8AX/W96AYDPHqgqDU9Zf4nNp0oxtyDc/W49+GoCh681T03tINRYlbt+Tfnixt/xT34KAKdbJrx6yQ09TVuIqaCSV2IA0Ank/8AFfdvLUeHW6ACp6U1V/d7r/DVO4d2VdNG8MDyQUksirKJEGoFo/rx7SXF2FWicekdxceGlAc9Vt9s/KTe+8qrI4jC5F8ftfVJT06QFo5pofoHDAjSPrbg39lDzSOTU9FLys540r0VR5HlZ5JXaSSRizySMzuzE3LFmNyT+b+2um+uHv3W+u78W/xv7916ma9dkaSP9v791oGo66+p5/P9P6+/de4DHXek3tx7916opXrj791vr3v3Xuu+Lfm/+8e/dazXrxt+L/7H37refPrr37r3Xvfuvde9+6913fi3+N/fuvUzXrwF/futE0669+631737r3Xd+CP6+/dapmvXXv3W+ve/de6//9L5/wD7917r3v3Xuu+W/wBgPx/T37rWB178W5ve/v3XvOvXXv3W+ve/de697917rkSW/H0H49+61w66/P8Ajf37rfXbX4v/AK1/6+/daFPLrtTYcC5v/vHv3WiK8evFgQb2B/Fh7914Ag464kW/21/furdde/de67Jv791oCnXXv3W+uwLm3A/1/futE067YjgD6fX/AGPv3WgDxPXViPx791uo68Te3+AA9+68BTrxFvfuvA1669+631yuSRYfQWA/2Hv3WqCnXH37rfXvfuvdd8W/x/1v+J9+61mvTphc3ktvZGnyuJqpKStpXV45I2IvpN9LgfUH8j2xcW8N3CYJ11K3EdKrS7uLG4W6tWKupqCOr4fgj859wZSKh2Nn62M5RGEEXqYMYr6UZb82PvFD3Z9qLOFm3izUhOJp5HrMv2j96b6+SPY91YO/wrXOPz8urp8fvNM5QpVZwTTH0vFKzE2tyLE/j3BW18zc18p3QGxXZAWnaxqMfb1kPPy9yzzQQN4hoW8wKcehI2t2DVmaOlxNaEMQVYxrJIUGwB/1/eZ/s5717xvrLt3NBGsUGrgOPHqPPc37uQ2rav35yoCUpUgZPCvQqzd85bbFZT4fcNdGYZ1APlaw0t/vA95YC5jKiSM6lbOOsPJpbiyma0vQVdDQ19emTc+R2tuqI5CmlQTyLrHiYKGP11G3upKvw6bdonFRx6DeHOVuJqEhhmUwqwsf6qDx7Z1FTnpoysp7ehawm4KOvRfuSpOn9y30JPFh7fUhs9KlYOuenSeixU4/alI/tG7WH+t7toBPWqJ58fl1Mx8VHCypFKeCRy/1/wBiffqAcOt44dCBRVsVPF/nBqIBHqBNhzcn27WgHSgPTrlX7onjAijfUDxdT/sOf9b3oyHTUdUaamR5dJkfxDK1IeUSMpYW+pUKfoR70Ktx6qpd2pw6W8VJTY2jaTQNek/qAHJWxP8Axr3v4RTp0YFetZP+dhi8RLXUeQaSFq1EjkhTgOG0j9IP+v7CfMIFa+fQO5iVaajxr0Xv+WJtilqMDX7gKg1dJkqhka17BLD/AIj2RWF19PKAeBPRDt1yLearcD1sg9L9huF8CTBWikRHUtyW+llB49j22mUpqXof2kwK1U4PR9tsbmSqji1ujFlBbnng/VT7Mg+odG0bgjHHoQiIqqK/BDcnkG3H+Ht7gKr0+SCK8emsiGmkBU8rwfre45sP+J90r69N1oa9PtHlIRYFlILWsfUf9b3sNqOMdXSSmD0r6WixVdEXkK3I4uVJtbm/u9PLpzSrZ4dJjP4Pb0cZOmPUVJJOk/6w59+ZRxPVGWNRjoFsvHhqZ2VViN7H8f7b/jftOwWuOkzqhGoeXQa5jL4iiWRrxj62FhpHFvbLspPSZnHnx6AzcfY+Px6TpHJDrBIH9ef6e2HlVcjPSeSbTUevRd9zdsSVQNPHOqKS1xcLfUbGx9pjMW4dIpLkD8XQWVm5p5gy4qOSWte41gawCRa59k278w7TsFq13uMqqBXBNOlW37Xum7zCKwgc1/EASOikfJ7cW4Nj7Fym5svOIZ4QzRRk6PpFr+nuAB71pzVzCNn5d/sq0LeR8sdD/cfbmflzZDve98RSgOPn1r2djd7br33UVCvUGnpJPSNJYSshuGF/wDf/AF/Y+h22ISi5m7pPXqKN3523S9tjttu+i34UGCR0Bpubk3N7kk3Nz+ST7MugTXru+oi9h+P+J9+61wGOuPv3W+u7Xv8A4e/dar11791vrkxuffutAU66Aube/dbJpnrsWDc/QXB/Pv3WjkY66Nrm309+68K0z11791vrkbECw5H149+6qK1z10Bc29+6scdde/de67t9T/T37r3XXv3Xuve/de65Bit7W5/r791oivXmINuLf1/x9+68OugCQT/T37rxNDTrtbck/QfX/Y+/daavAddG34N/9hb37rYr59de/db65Bb+/daJp1x9+631737r3XduAf6+/de67Kke/daBB64+/db697917r3v3Xuve/de697917r3v3XuuyCPqPfuvVB65AgC4tf/ABHv3VaE9dE3t/h791sCnXEAn6e/db67+h9+61x669+6313fgD+l/wDeffutUzXrr37rfXYtcX+nv3WjWmOvG1zb6e/deFaZ68t78Gxsfr79140pnrxuGN/rf37rw4dWIfy3vnduP4J95YXftNB9/s2syNI288ZFEz1ldjae4aGlZOQzAlf9jf377Ot0BPd1d32r86f5EW8u28r8rKLovvB+/Mjl6XddDja3LGr2fTbojRWerr6FiWZVkXUqkEe9fZ1vA456Lr/Oa/m5/Hz+ZTsv4o7b2b19nNnV3TNIcXvuqnj8VLmaAyqFGKgjAEYEYZT/AGiTe/A92rVqnqjCkemM5+fRI/nP3z8Cey/jv8dti/FXq/dmzO1dm0tSvce4c8zNj9z1bqwppcSh/QqAgEe658+rkppGKHqpwi50rc302/xNrEe99V+fWwn25/NY6I7E/ly/ED4gY7Y+axe/Pj3NXzbm3TJHM1FnRWz+VY6ZfoNP5P5PvyijAnrbmqFRxNOkD/Mz/mY9MfMr4ifD7oDrvZeZ2/un49plYdy5ivWQUmbXIRS6Wow/00lxf+vvWa9WLA9KD4NfzQei/jZ8J/lp8b977Lzmc3T3/wBdts7A5WiSRabCVphKJWTkfqCtZgP9v782SKdVSijPGtelv8AP5pPxL2x8aIfiL89OnZ+xuqcZS5Kk29V7Rx8NFuGhaskaSKapyoXUwXVcAt9fe69aoCNJ8ujF/wDDon8sH4d9C97dV/y5+vO5tubi+Qmya3afYWW7JyJy9BJRVhBOPw0cZIjjGkcnni3utK8enGagGjj1qnVU0k9RPUTEmapmlnkY8ajM5kLf7Ek+7dN8eo3v3W+u7f4j37rXXY4B5+otb/ivv3Wjk9cffurde9+691737r3U/HY+uytXT43G0k1bXVsqQ0tNToZJ55WayxxIPqT/AE9+60SBk9bgPzJzma+H38iz4T4vH0UuI7C7cqc7j93U1VG8EyY+njiaCnmA55jZjyOePeq5HV2A0/PHWnnKzSSPI36pGaQ/67nUfe+q9cCLG3v3Xga9de/de697917r3v3Xuve/de67JJ5I+vv3WhQY669+6312RY29+60DXPXa/kWv/vf+39+60ft64+/dW67+lwf6f7z791rjw68AT9B7914kDj14ixt7914GvXibgD+nv3XgKGvXjYk2+n49+68OGevW/Hv3W+hL6n37kuvN8YHPUFa9LDBkaY5AA2jlpvKBKH/2H591dQ6lG4Hptk1qQfPraG6r7dXc20sRvPF1C/bVcVNTEI111pBctx9b29whzZsgtrsugw1f8PQV3G0WOStKjoUMzutdyURo6xlljmTSOLlS31PPsMWqy7dMLi3JDLnHSCENayCWHDDoBc1sHP4svk9sTSBQxkkjvqGkm5Gn3MXLnuMaLb35oeHQ02zmaUMEuT0H439X01YaDOwvHKh0+R0KiwPJufcuWm5224RiWJhQ9DSC8iuFDIR0olzVPLaWlqFYmxGlhYXHC8e1pYUHr0oL0HXT7rrqHU4nbQAPTq/H49+8Q06sZWXPl0lM72jWtSNAs4DWPIPP+3HuhkbiT01LMRmvQIZbeddVoUMx+rX5t9T7ZMoI6YaYnj0hJa+pkdmMh5JN7/7D6+2StW1dNVP5dQ3Z3bUW/pf/AIn3YADrVPTrq3Nj+eOP8R9Rb3vr3XA2HFxf6Efkj8+/fb17z6zwUtZVOsdNSzPqYDWqFlHH1J97pTj1sD16VJwGLwtMa7cldBBCqlzBJKqSWAvaxPtO9ykYz0zJcJGOgL7A+TmyNi08kO0USozAUiMjTMpdDYE24/1z7L5b1yNK8Oi6S9ZlovVevY3cG8u0cg1ZuTIMIiXK0lKXjp7E3UMl7NYcc+0LMzGp6RMzNkmvQVf0916912xU/p/3q3v3VVBHHroG3v3WyK9de/db697917rux4/x49+69Xrsiwtxf68f09+6qDU164+/dW67sbX/AB791quadeAvx7914mnXrcX/AMbf8T7916uaddsQbWt9P6W9+68K+fXVza349+6959de/db69b8/j37r3XYBPA59+69WnHrr37r3XZUj/Ef19+60DXrxFrf4+/deBr13cabfn/jf9ffutZr11bi/+Nrfn37rdc06/9P5/wD7917rl6bfTn+v++Pv3Ws166BI+nv3XiAePXgSPp791sivXrm1vxe/v3WqCteuvfut9e9+6912DYcfW/1/w/p791qlT14/i3+x/wBf37rw69c+/deoOvLe/H19+68aUz1yt+P7R/P49+61Xz8uuPN7fW3H9fp791vHXgP9Yf6/v3W+uvfuvddn8f63Hv3Wh12CLfQ3/r7917NeuPv3W+uVyRyRbgW4v791WgB64+/dW697917rsWvzyPfutGvl176G44/p/re/de8uuvfut9e9+691737r3XvfuvdD98adxQbZ7d25kJ5BEjSrBySAzPKj2Nj/ALSfYQ56tJL3lu4giFSR0MuQLlbTmm1nc0Ab/L1uFdcVGD3NsLDOHQyzYtJAwvYM8QI5P5982t/jv9u3p8Ggen8+usew7C25cvQXka/EgYEf5+kbTw1+2cy88Lv4xMdJ1H6BtQ+nvJn2/wBv8TZRe00y4I6Tzc5XW2g7VP3x5Ug1Py6yb7o8vveKGsjmAqIAltDEMLcKL+8jOXOdJrSFbS9NdOK9Yv8AuN7b2+/XL7nta6WY1pQdN23afeeJRIJzLKigKpBcgqPzce5DtedttlopPULS+2XMcal1Bovy9OnrI57KUN2rIpATb6hvp9fz/rexBb7xZXv9iw6Ce4bDvG2ktcxnHy6e9sdhxa/DLIFY2/U30/wFj7MxJwCnorWZlHeKdC3QZ161lZKuMBv6OTe44v7UDV5V6eSQE1B/n0sqMVqWkEpdTY3ANgfwb/4+7DUOnVIOQelPTVdVZUeS5P0+v0PNrn3sEnj06rUGOldi44p2DSuGVbcX/J+pv/xHu4HTi0rq6V/8cxOLpizvGvjVj6r2AUcj/b+3fEUA9O+Ioy3RX+4/k1trZOHr66rrIUEMNUFBcqC6REqAL/W/tHLeIiEt0huLtEBNetP750fJ+u+QvYtYRM74zD108MTB2MUvjHjUJc/2SDfj6/T2Bdyvmu5TTgOgHud8byaowB1Y9/KNwEmc2dmYFF9VfVcD/Uj6/wDE+yzoqpU9WL7l3o2wuw6PAYqQs8hTzIrE+oN9CE/4p7Otu3FoT4b8Oj3bdxaGiOcdHu6x7ehkip4q5hFKEQXcsDw3N9RHsZW13HKOw9DK1u1kyD0bfB7/AKSqjQxTxshHIVmNj/Xk+zEOKY6MhKg6UsuZiqYyUkFzzxf/AHm/vxavTpfGOk/VZyajk16xpXm/I5Av9PdSxAxx6qWrjqK/aCUaMonsQCLFnW/9OFP19+Evkeq+OQtOg8z/AGy1QHVKsA2BuXa1rW/r7o0tcnpvxyceXQMbi7foKWORJquKSWxv+4bi30F7/wCwHtO8yrivSV7gCuei07y7byVYJkx0czaj6CpkYEWsD/re0ct2qih6QS3QGWPQVUn948xI1dkzJFTAEnXcAAC/Jb2Sz71bwMVbpbZ7fLf0YMKH/L1i2/tuTeu4o6GkDmKKURy8ixN7H2D9457t7WJ/B+IDqW+VfaC63a7haU/pMRX7OjErsfDbKoqmWXSaiFHbWya7FEuQT/X3ixzjb8x+4d39KshSKuaE9ZpXOycge2/LKMFUz6c1A4gdUBfzLu/Ycm82zsdVRTCpmNPJHA7Wi0R2kLLf8f09jv2y9sYuVSt5NmVa5PWDHvV7lxcwhtpsf7Mny6pK9zj1jT13c2t+PfutUHHrwtfn6e/dbPy67utxwbfn37rWeugRc/W39P8AevfuvZ67uvPH+t/h7917PXRIP0Fv9t/xHv3XgCOPXuRz791vj13Ytc/7f37rWBjro2/Hv3XhXz669+6313f6/wCPv3XuvD/be/de669+691yH1F/of8AYe/daPDHXRHJt9Px7914cM9dmxtYc/77/H37rwr59cffut9d3P0/p7916nXrn+p9+69Trr37r3Xvfuvddm1hYc/n/H37rWeurkfn37rfXvfuvde9+691737r3XI6gOTcf77+vv3VRSuOuPv3VuuzawsOfz/j791rPXXv3W+ve/de65C30tyfob2A9+60a8euiLfkf7D37rwNeuwbkA8jn/D8e/daIoMdeAHq/wAPp/t/fuvVOOuPv3VuuwbfT37r3Hrom/J9+69w67txfj37rVeuvfut9e9+6912Rb+nv3Xq9e/A4/r/ALH37rXn11791vr31+vv3Xuve/de67v/AK3+2Hv3XqdcgQRYi9rWso+n+JFvfuqmvl10P6kgAf04J/23v3Xj6ddH63HF/p/yP37rflnrx4444/oB/vfv3Xh69dgj8j/WsLf71b37rxHp10CLHjn8fT37rxBr16/BHHP+HP8AsD791vrsfi4uP9b/AIn37rR68eb/AO9H6+/deHXH37rfXZtYWHP59+61mvXiLAcEE88/kf1Hv3Xs1696iL2Nhxe3AP8AS/v3Xsdde/db65A/huQPoPfutEenSj2lufI7M3Hht1Yhof4nga6DIUKzxmWEzQtrXypcXF/qPfutEV6N38mP5gXyI+WWzNobA7c3BR5Da2xpRLtzH0dJJBFj2Wm+3CxR6youoANh/T37reeiQm9gSb/gf4f4e/dermnXh/U8j8j8+/de+Q66Nj9OB/j7917Pn11791vr3v3XuuSrf8j6j6n37rRNOvEg2t9B+P8AY8+/deAPn14WJN+B+OffuvGtOuJN/r791vrkt+SCB/r+/daNPPrj791vr3v3XuuwSPp9PfutUB66+vv3W+u9JsD/AF/p791qorTrxsDx/sPfuvDhnrxJJufr7916gGOuvfut9W6/Anu+GpoZet83ULBFQCM0HldirmRCkWi/F7giw/p7DnMe3fW2ZZR3KOi++h8SPAz1adgYa1a1KaQk8iVGIIBRzqU/7b6e4cuAIdSycRjoNSHT2nowGBhYKqSAOkigMth9Prex9kUtG7hinSF+0f4Okb2H1Jg9xU1RVxRLFUCMnUFRRq+o5HPsQbHzXf7W4XVUV4V6MrHd7m1ahOOiI7jx+S2ZXyRLIz06M4+pIsjW5t/xPudth5wtr5FMuGx0PNv36KVQsueoCbwgroiJGuQtioNrG1iCB7GyTxygMh6ECyrKKqcdIPNV8chZkkCmx9OoXNvwB7qxqcdNEBmzw6RxfV6mb+tj71QDh1U54dcdQIOj1kf2Ryefp9Pe6dbAPWZKarkXUtJMRb66WIP+xA9+p16nTtRbby+SYJDTywsTy8kbWHHHJHvVVpWvDrRx0+VGAw22EFXubJUwUeooHVW4/ULG3tmS5jRajj0xJOitQeXQSb++VGwNl0EuPwEazV6ho4vGDIxcDkm1+Ppzf2WyXrMKDHSCW9JwOq6OyO8949hVlQ1VWSUlDIzCOGCWZHMd+NbKwHP9APaNnZzqbpExLGp6BdmaQlndmb66nYsSf8Sfdeq8OHXD37rfXZt+PfutCvn14fX/AA/Nv6fn37rZ68bX4+nv3Xh8+uv8PfuvddkEC/v3Wq169zYci1+B+ffuvYr119ffut9e9+6913z/AI+/dex14X/F/wDYe/dax59de/db67FrfQ39+6159d/jnkfgfm/v3WvPHXH37q3XfP0/2Pv3XscevC4+n19+60aefXXv3W+u7n6X9+69Qde+v5/2/v3XuvfQ/wBf9b37rXXdx/Q2/pc/X+vv3WqHr//U+f8A+/de697917rsixt791oGor11791vr3v3Xuve/de67BI+nv3WiK8euvfut9e9+6912Bc2vb37rRNOu1v+Pr/xv37rTfPrq9jf8+/db8uuw1iT/W/+8+/deIqKdcT9T791vrs2vxe3+Pv3WhXz669+631737r3Xvfuvde9+691737r3XZW3v3Wga9e4t+b/wC8e/dez14m45+o4H9Le/depQ4669+6312Lfm/+w9+60a+XXXv3W+u/oQf9Yj37rXHqZjq6bHZCiyELMs1HVQVSFSVOqGQSWuP62sfdJEWWMxtwYU6ftpntZ0niNGQgj8ur5/h18xqauw2PwOayLRVMJp6VPNPpEAHo0MD7x75q9s9vl3QXJjqGNTjHHrpf7Me/u3XXJZ2i+YJLbppUMcmg6tlxW7MVuClSWOohqBIPIrqwYm/FwfY82jliC0sljQBRTqC+avdu5l3WUWtW7zw9OlpiIwigxMCGe/0HA+t+fZNu1iLSagOD6dS3yJzF/WHbxM4IYDNeht239m9HrqooGbkC8akn/C/+8+w/KXRwY2IH29S3ZC2MBSRFP2gdRM/gcLuJWgjpkv8ApBVALMfSPZlY73uFg2tHP5non3blrZt9iMTQj/eR0HFb0QdJloVZJSCQRcEC1+Lf09iq39w7yKmtuHz6jfcfYzbrlC8CgH7OgzzuD3Vs/WUeVvGLjSzH0qPyB9PYpsvcd30hjjqOd69lpLNS0QpTpMYbvrNY6uXHZBpAoKqWYkAfg8+xlac7200epz1F11yJuFtc+CQadDNj+zqnKqBRzGWQrqIjOri/4Ps2h5r2+TDMBX59Ip+T94j7lFVHy6b8n3RWbdkMNS80ZvYjUyi5/wAT7Ml3i2IGhga/PoL3Bns6mdSoGM46Qe5e+aiejmDVEkMTU8sxZ5SBpVdRN/bFzvCLwPRRcbzGBVete35w/KHLbpz8+09v5Wf7RTUJXPDVP6Ljx29H5J9hu53CWeorjoN3G4ST1AOOqw2ZmYszFmYkszElmJ5JJP1Psv6L+thP+UVvHG7P2TnMjkGARaytjCs1gS/pFv8Abj3utOHVfM14dG33Iz1HZ77waGWWglqDIkjAsgV2ubH/AFv8Pev8PW+PDo2ez4KTcJiloJvHNoDaA2nTcmwt7WW97NbmqnHS2C/ltyCDjoQv4rujasgQSTyRL+Vc2I/oLexHa74HWjnPQitd7BILHoRdv9w1jKVqkn1ohAJci5+lwfZwm5wUBJr0dRbnEw7T1xyvaeZr1kip45FW5seRcfjn23Lu9unA9MzbzCuK56Dyuy+562UBGmAkJP1P1PHA9lsm9xtXR0Wyb3GPh/w9dw7a3VXxku8t2uL6je1rEXHtBJvjn4ekMm9k5XqDF0fV11QKzIzyeMsGYFz9L/Qg+0LbrMT0hO6zFuPSor9vbG2tjy1WYGmhWw8gU6iovz/vXtNJeTScT0mkvZZDXoCMruKHc9U+HwNOsdI7gPJGotYGx02/HsOb1OsEJdqlupA9vbW+3vdo7YvpiBzXpf7Pp8LsKJ6maWKKrcGR5ZLAqQPoSPcWybZeX05cAlT1nBac6cvcqWKWssil0FOPy6rw+bXzdw3X+KrcXg8rHNlqgVKIKee58jRFdFh+T7H2w7FHZR65FGr7OsVPdT3UuOY7lrazlbQCR8WOtZzfG8svvzceU3Hl6iaWevqXn0SytIIw30VdXA4+th7FmKUHUAPI7tqkOonz6R/Fj9b/AI/p/jf37rWeuvfuvde9+691737r3Xvfuvde9+691737r3XZJNh/T37rQFOvXsLe/dep59de/db65cWNr/j6+/eXVc1z10Lf8U/43791s9eIsbH37rYNc9dgAgn/AG3+29+6qTmnXRN7f4e/dbp1y50i/wBP959+6154643ANx/sL+/dbpUZ69+effut9e45+v8Ah/xv37rWeuvfut9cuLG3+xv/AK/FvfuteeeurcE/0/4n37rfXrcA/wBf+I9+69Xy68LX5+nv3Xj8uvAXNh+ffutE0Feujxx791vr3v3XuuyxP19+60AB14C5t791s468Rbj37r3HrwNiD/T37rRyKdeJuSffuvAUFOuvfut9e9+691yABvb8f19+6rUjj10DY3/p791Y5x14m5v7914CnXhb83/2Hv3Xvs66/wBb37r3Xvfuvdd24B/rf/ePfutVzTrwNvfuvEV669+6313f6D+nv3XuvcWH9fz7917PXXv3Xuve/de697917rsEj6e/daIr11791vrkDewP+sP9j/X37rXDI64njj37rfXvfuvde9+691737r3XPWR9Pp+L+/dV0jz64k3N/furAU669+6912P6kXA+ova/v3Wj6dbQH8hP+Uv8evmLuPcWY+acmRwnX248LSwdY1tJWSUENVnnbyjzVepAAyFrqTbgc/X36lT1sGgJI8+q4fn18VOrfhZ/MH3h1jmMBm6/457Y3rHSU1ZTR1Mn8awEN/uFoakElnIsV9XP1H5964jHW2oGp+fVu3QWc/4TNd2b+6t6XX4/fKHFb07Jy+E2gm53y8owuM3FlTHRx1E1K8ocwvOx+gNh71npwFaeVejbfN740/8ACbj+XV2s/T3dPUPyR39ueiaimyM20szJDR/b1SLOEpjLKpLCMkkn8+/UbrRaOtMV406rs/lzfCf+Xt/MI+UXy9XZu0N67a+P/XuxMruvqbCZiqnbcMPhiL0K5iWNySRazg/U/wCHvY+fWm08V+XVCfyy692x1b3tvbY+z4pINvYWukgoI5SxcIk8kYLa7m9lH192OOmh0W/3rq3XYF/p9fr791qtOuvfut9e9+6912Rbj37r3HrwUn6e/daJA669+631737r3XZFjb37rQNRXrr37rfXZBAB/r791qvXrcA/19+69XNOuQuVtx9f+N+/daNAa9ceLfm/+8e/dW671mwHFh791XSOPXXFvzf8/wBPfurZ669+691737r3Sl2lujJbPz2Oz2Knminx9TFUaY5WiEojN9DlPx/r+9MNQK+R60RUUPWxn8Ye6aHuHaWPyC1Eb52np6VauFmvMiRoFYMPqCLe4p5t2Ixt9RCMZOOg5uVuI6uo49Hz26JKiBZUsRYAgn+n9fcZS/FT06IH4UPl085s2oZ4ybF4Sq8/QkW/HtuJaSVHWhRhq6I92fiQKWqMygk+UBtN/wBTX/1/Ys2+Z43DIejK2LLQqeiU0tfS4LK1s+RZnoYPKzerhVB5JHuT9n3y4idY5Dg46EtnuE0ThScdLXEVGwd+Un3WJytLDUljGUedbh1NiCPckw3aOM8SOhPFdJJQsaddy9d1FPJ5J8pRCivfX5Ftpv8A6/8Asfb5mVBqY9KWmiTNek/uHdfW2waYy1ldTVVSOCscimzj6/8AIvaWW8oSV4dIZr0Lwz0CeU+YOyMe0kVFGJPHwNMes8fTgA+0v1r0p0x9cSOg3zfzdqxSzx4KmkjmZGWMtCY7MRwdR9sm5Y9MtcuTjoo29e5N8b6Y/wAXylQkZZmMVPPKiEMb6W5uR7YZixqemGZm+I9BZI7yNqeR5GPJZ2Zjf88t7r1UdcPfut9e9+6912Bfj37rxNOvAX9+60TTrwNv9iLf7f37rdK9eAv/ALa/v3Wiadde/db65k3so+n05/3j6e/dVApk9cSLcG9/z791sGuevG1+Pp/j7914V8+uvfut9dj/AHvj/b+/daPXeorxxx791qgOeuPv3VuuwSPfutEV69fm/wDjf37rfy67PPPH9PfutDGOvKbG/v3XiKinXQNiSP8Afc39+68RUUPXXv3W+uxb83/2Hv3XjXy669+6912Bf/e/futE068bX4vb/H6/7x7914V8+v/V0ARc+kf6/v3WjQZ69axs3+8f6/v3Xq1GOuJ/31/fut9di35v791rPl11791vrsf73x7917rr37r3XK/ptbm9/wDeLe/da869cffut9dgE+/daJp142/F/wDY+/deFfPrxtfj6e/dbFfPrr37r3Xvfuvdd34t/jf37rVM1669+631737r3XfFv8b/AOwt7917z669+691737r3XIm4H9bn/W9+60BQ9cffut9dm/F7fQWt/T37rQ68Lfm/wDsPfutmvl14m59+60BTrxJP1FuLe/deGOHXXv3W+ve/de6fcFuTM7crIa3E19RSSRSRyaYpWVH8bagGX6f7H21LDHMumQV6XWG5Xu2S+NZOUPnTgft6sl6J+eWZ29U0GN3TVTJTr4KfWzMyMfJa5cfS4/r7Q3doxtykOKdCjYN/iXdBLuIqGOSeHHq7XqvvTb28MVR1tJkaeQzxCXQswYgkXsOfcdblBcsx8QHrMLkndtlit1exdRqFSK9GCpOxIaaDSlUgBN7CQWsRz+fZGbQk9wPUnR71FwRxnoT9jb7xk0mmSojd3ZTyym1zx7S3MDAUp0f7Pu0LOUcjPRkaOenqoEMLRvqUHUtjbUL/wBT9PZWy6TU9DaN0ZapkdJbcmz6XNo5eJDqQhrrzcfnn2/HK0ZFD0gvtvju1OsVB6KRvrpWjlnmkpoBHMA5VlUL/wAFPHs4ttwcLpJNOo43bliJiWRRXPQNYqLNde1kktXHLJArsg1AsAoJ+pP+x49mi3TsQUc/t6CT7cbQGCdAVPy6RXY28aTN11PHEQJ5WR9Cmxt+bgfn2NOX5bqbukJoPn1jX7vixsCba1AV2ocdER+ZHftJ1jgKbB42o05uvx0iQKsh1s0iAEED+gNyfYsJJNT1jwuqlSa9UQZbK1ebydZlchK01VXTPPPIxLEu5vxf8D8e9dWzTpt4t/j/AMR791vq6n+WzRHM7YqMQZGSKbITmWzWvaW9j/sD791qler5K/r/AGjD15USiWmatgQ2LOvlBVL/AJ97HCvWvkOgb6by9VDka1UkdoqafxKb8ACSw0+9DPHqw/n0fOnxMOSxkUtREsrSx6gzAE/S/PvfcDXrQNBjpmTa9DTMzmIi5IACgCx/oB7sJZqUrjq/izAYPU9MLRxxt4oVuRchlBNx/S3+HvRdzxJ68XdxUnPXB6RY2jJiQMAGtptxfm3uvVDU8enOp3LRYyl9QiRkFySAB/W3v1Bx68KAdAVvfvKHHQSxRTrGNElgDp5UfT36g68R59EV3V3HuDd+WakinnSjDtrYMwTSByffvKvWxQHPSdzvyd2X03jWNdXU0lesZEgeZC4Zhe5/4p7SXNpHdCkgr0fbNzBdbHJ4loaE9EL7m/mSnKUNZT7Xr5DVSxMsaRMxAZlsfWOB/t/dobOCJQAox01uXMO6blOZpZDU/PqojfO/9x9g5ifL7grp6mWWR5EheVnjiLm50g/n/H2pwOHRMak6mNT0iffuvde9+691737r3XYNjf37rxz165Bvb37rXlTrwBY/4m59+69gDrxFjb37rYNeuvfuvde9+6912CQLfj37rVM1669+6312Te3+Hv3Wqde4/N/8Pfut9dWP9Pfuvde9+69148G39PfuvddgXv8A4e/darTrr37rfXvfuvdcrWFz+Rxb/iffutVqcdcffut9e9+691737r3Xvfuvde9+6912Bc29+68cde0nn/D6+/daqP29de/db69/r+/de67It/rH6f63v3Wga9et9P8AH37r1eu2JvY24/p7915aUqOurcE/0t/vPv3W6+XXXv3Xuve/de65rq5At/sbj37qrU8+uH+v791brs2vx9PfuvD59de/de67FxyPx/xPHv3Wj6de5Nz/AL1791vhjrr37r3XYNjf37rxFcddfX37r3XvfuvdcgxAtx791oqDnrj791vrlYEcXuBz791Wpr1x9+6t1737r3XLkL+LN/t/fuq4J+zroW/N/wDYe/dbz5dde/db67te5/p7916vXrWt/j791qvXv9vf8+/db669+6912DYEf19+61TNeuvfut9K3Y2zsx2Bu3A7LwCJLmNyZGmxmPje9mqalwka8c/U/T37rRNBWnW4V8zH76/l+/yzvgJtTpzYe64+0qGszOS7R3NhMBlZYWjjkviqeSXHK7gmP06je1/zb36tMHqxFVquOHSe/mz9Fbx+Tv8AK0+GvzAxvX2bwPYWWqsqO2Bn8XPRZOpMEASCtllnXXo+rXcC/H096Fa/b1t6BagZHWtx/Lu0t86PipdRb/TVsYEEXF1yaDVz/jz72etLx6u5/wCFQKp/s626joGoU+39LEKW/wCLdEDZvr9PfgcEfPqjj9QfZ0rv+Euv/H+fMM/9+Myn/WiT3qvV/L8+qC/nf/2VB2X/ANrWo/P5+6m92PTY8/t6J8ASbe9dWJp14C5sPfutk068QQbH6+/daBqK9de/db67Jub+/deApjr1ja/49+61XNOu1Ngf8R/sP9j7914itOuPv3W+uybm/v3WgKDr3+3/AMffuvddf74e/db67tccfUcn+lvfutVp11791vrvi/F/9j791r7euvfut9dk3N/fuvDHXXv3XuuwSPp+ffutEV669+630az4pd71vTG+FleokXF5jxUlQHkPghYtYOVPABBtf2jvrRLy3aFxx6ZmiWVCrdbHvWnbWFzG36LMUFVFUwVUEZKxOr+twCb2/wATb3Bm+bFNaXLKinTXoIXti8MlV4dKjce/aHwlvKFNiSpYA88/77n2TxWr4oOk8cZPb0ULtTsKiqqJ6emPknYSgKh1MX1kAAD2ItutZg9SMdL4YjWnRF+08xBtrZuQy2cZqf8AiEUsMKsdEgeQWQWP+v7H202DSSCQ8B0cwxVFR1WNgN+bn2xWSVOGy1XFrmd/HJNI8Tlm4ZluOf62PscAkcOjRSRQjHQp1nyZ7SrcZ/DJclEItOjyKJQ+kf8AIf1/x92LuRQnq5lc4r0C2V3Jnc1PJU5LJ1lQ8rMSGnkEYueQIwbe6dUxw6YySTckk/1Juf8Aeffut9de/de697917r3149+6912Rb6+/daBr176mw/J4v7917gM9eItwffuvA168STzb/be/deGOuv8AX/3x9+631737r3Xf4t+L+/da8+vA29+68RXr39b/AO8f19+6314AfU/T6cfW/v3WifIdde/db674t+b/AO8e/daz163F/fuvVzTrr37rfXvfuvde9+691737r3XYt+b/AOw9+699nXXv3Xuve/de67IH1H0+nPv3Wh8+uvfut9e/3w/1/fuvde9+691//9b5/wD7917rvn6+/de+XXZN+fz+ffutAUx10ST9ffuvAU669+631737r3XZJJ59+60KeXXv1H8D/ePfuvcOuvfut9d3Nre/dapmvXVifpz791vr3v3Xuve/de697917rsWvz/vv9t791o18uuvfut9e9+6913+Pr+fp+f8AX9+6159etxe4/wBb8+/db67NrD8H/Y8/4+/daFa9ebT/AGf+Jv8A7z791oV8+uPv3Vuve/de65Np/s/7H6/X/Y+/daFfPrri3+N/949+69mvXjzzcf6359+68MY669+631737r3XvfuvdclB+o4I5Fvrcc+/daJp0Yvp75Dbr6zq6WnWsllxiMFOt5GeJCfp9eR7SXFlDcDuHQh2nmXc9oYeA50+lerVNhfLLEbgx1PJPWRLK0ahiZ7c2/xP+PsrfYoSCOhtD7pbqrKxbh0Oe2u+o/Os2NycRKsnoE2okhvoLH2S32xaFJAr1KPK/uuszCOc0PR7enfkdLXSU9DW1KG6gXLHkFv8fYNvdrKHVTh1kXyvzul2qq7Ag9HapexMfPTpaWM6o9Ra/FyOfp/r+yA27BqHqUot1idA46Rmb3FS1DGZXUg3H1/H1tz7UIlBpPRbd3SMxboAuwcpjJKCpap8QBDMpa17hf6n2a7fBJNKETPQB5n3S22ywe6noKV4/Z0TGKjirslkM9KdNNQRzspJvHZFJX6/63uWtqs/pbcBvMdYAc/cw/v/AHh5Y/hU06oQ+Xm//wC+3Z1XDr8i4R56UWJKrqYHTz+bAezPoE08h0VL/Hj6/T8+/de+XXXv3W+rSP5f/YD4CY4iGZEmlr5QgN7nyaWtb36p60KV6t73fuzelBjlBlkNDWWLAagNLDm/+Fvex14U8+ha6TkoRjnld189QYpHufVrLXJN/wCvv32de6sB27nqM4+mhZx6I7K39OOOPeiSerAADHSnSpx9UCTJFxyebgG1uLD3s9VwTTqDV11DTBryJcCwt+Tbjk/763vX29bJpjpA5/cVPTRPMZUBAJFj9P8AC5/4n3s0rTrQ49Fo7B7Mp4aaUK4LhSoVW5JPH0B96+XW8Vx0W9sbl96SPUSmSOm1XF0blf6gn/D37rRPmOgX7z3Ttvp/aFdW+aFMisMltRUSA+LUbX/x9+62PU9a8nZ/Zec7E3HX5KvrZXo3qZGpYA7CMRgkKzA/Xj37rXQYkf7YfkDj/b+/deHXXv3W+ve/de67F7H+gtf/AIj37rWK9de/db697917rlxbk3PFgPx/W/v3VfPHXQJHI9+6sRXj139QSTz/AK/Pv3VeBoOHXH37q3XrH+nv3Xuve/de69Yj68e/de67Itb6c/0P+9+/dar14C/+++n+v791vh13crwCD/t/futUBz1x9+631737r3Xvfuvdd8WP9fx791rNeuvfut9cja3A5H1PPHHv3Whxz1x9+631737r3Xvfuvde9+691737r3Xv9b37r3Xvfuvde9+691737r3XZtYW/wBj791oV8+uvfut9cmt+P8AH37rQr59dfQfX6/0P+9+/de669+631737r3XIsTb/D+nv3WgoHXH37rfXha4v9Pz/re/de+zrs2vx9PfutCtM9de/db67BIBH9fr791oiuevX+g/p791vrr37r3Xdv8Aff1/1vfuvV669+691737r3Xvfuvde9+691737r3Xvfuvdclt+R/sfqPfuqmvl10bX4+nv3WxWmeuvfut9dg2/wBj9ffutEV69f6f4e/db68SDY/n8/8AEe/daGOvAkcj37rxFeuvfut9e9+691b9/Jd+JW5Pkt8xOts9RU80+z+qd4YDdG+nipJp1psJTT+eSWeaO4jUhCCWHP8AUe9GpwOtqQpq3DrYq/mU/wDCijfvRfy17J6K6s2vsjLdO7DlpcXtjH7i2pj87BTGniETuBVRSHW1mYm1vfgAwNT14uysAq1FPXz6MR8Fv5mOW/nNfG75e/FntLEYTHbr271VV5PqOmw2EhxlBS5poXWGOioqeNVBcqbKoBuPp73wGkZ68rV7nFADT1qOtQr409Eb0+PP80L499Pb0o54tz7a772ZQz0v208dS5OXCwA0rLqDsLHSAf8AD377OvLWuerVP+FQ9LU0vzZ3bHVU1RTSLBgFZKiGSF1K0EQIKyAHj6H3scD1Vz+ov+l6WP8Awlqx+SyfYXzApcVj6zJ1snR2TWGkoaeWonmYwOAiJErG5/1vevPrZNF/PqlP5v8ARfeld8nuypIule03DZep8Zg2NuWqjljFTKRKs9NSsn+wvf8A249+NOFetBWINQRnokG5+u9+7KEbbx2Ru3aSytpjk3Ht/L4ZZGPOmM5CGPUf9b36vp16hGD0i+Rz/Xi/v3XsHrvi3+P/ABv37r2a9cbE/Qe/db67Nr8fT37rQ+fXXv3W+ux9R/rj37rx4deIsTwQL/n37rQNR14WuL/T37rxrTHXMi36SLH/AB+pH9Pfuqg+vXAkm1/xx791YCnXXv3W+uXFv8f+N+/dazXrr6n8D/ePfuvddm1h9L/m3vfWhWvXH3rq3XYYj6e/daIB669+6312CRf/ABFv9gffutEV669+630cPov5Vbi6ygpsJkZZJ8PTrohYl3ZF/sq4F+B9L+0F3t8F2O8Z6TywJL8Wej5YL5Y7T3TS662dFJQDmbSDcckAn2HpeV7eupcdIzt6fh6Qe4/kBsKgmlrNYlaLVIgaQN6l5HAv7WW+xxx0pwHTy2qgV6If3h3pX9pTihiQRYenmEkSgMhbQfQNDf7A39nsNukAonSlI1Th0Xb2/wBO9dn+vHPv3Wh6devxb/kfv3XqZr10AT9Bf/W9+631737r3XIEfQ/QX/2/v3VSPMdcffurdd8/U39+61jrr37rfXvfuvddgkfT37rRAPHru1r/AEPB+nNv8ffuvV64+/db67vxb/G/v3Xvn117917r3v3XuuRPFjf639+615164+/db67a1+Pp791oVpnrr37rfXvfuvddkEce/daBr11791vr3v3Xuvf4+/de697917r3v3Xuve/de67Nvx/xPv3WhXz669+6312SWP8AU/4e/daAAHX/1/n/APv3Xuu7m1vx791qgrXrr37rfXvfuvde9+6912bX44/4r791oV8+vE35Pv3XgKdde/db674t/j+PfutZr11791vrsEj6e/de49eBI+nv3WiK9de/db697917r3v3Xuu+LH+v49+61mvXY0/kH8XPv3WjXrj791br3v3Xuu+f99/T37r2Ouvfuvdc9Q+oHP8AiPfuq0PA8OuP1P8Aiffut8B12wAtb37rQJPHrj791br3v3Xuuyb/AO2A/wBt7917h117917r3v3XuuwSPp791qgPXiCPfuvA16mUuSr6Ig0lZU09v+OMzoP9sp9+69ToQNq9rbq2vWLUw19RUqAAUlnlP0a4IubX9+YBhpbq0byQvriND0bLq/5f5PF52jGWeWGnLKpfzPYMXuB7KLzaork1GOpC5e5+vtpospOPmerbtg/KylzFFRPHkks8Y/VO5b6fQi/sgPKwZj1LEHvmYIACSfzPQ6U3ecFRAJpZ7x6SeJG54vf23LytpFF49P2fvqzy/rVp9vQX7n7Mq9416Yyi8sdMeXkbUVFhySfZvtWxpZd75PQC5+9z7rmIfS25IQ/M9Ft+RvfmE6o2NW4OhrEmzFTEYrwysGZ5EtosDyT7EdcUHUMgcSeJ49UMZ/MVGfzGQzFVcz5Cpeoe5JPqNgCT/hb37r3TRbi/9PfuvVzTrtgAePfutKSRnoWumOw6rrje+IzCM5pTV06VCKxAAaUASWH9Pz791uvW0fsnO4PtDq3GVkLwyzvRRmQKNUgdoweb+/deNOPr0n8VDl9rTutMJPChJsuofT1fQe/deHDHSmx/dO4Y8lDh4oKlmlcKCpe3Jta/v1et8OjMY2p389BBXIs2mSMOt9f+wBA976rnj01ZXOb6igaSeKa9iB+sHkc/19663xz0DO69+ZyhoaiTJTmmiVX1PK7IAdN+fe6+fXgDwPRAd+/JHbOAq6qbI5FJ0hcEqkzMvpP9Ln/X9+HW+HQRZv8AmK7fx2MmpMK5MqoyRCMuWJtYcD/ifevs619nVa3d3yI3X3FXOa6oqIaASs4j88mqZSOFkW9gP8PfuvU8+i6+/db65X9Nuf8AiPr791qma9dcW/x9+69mvXXv3W+uwR9D9D9ffutH5deNr8Xt+L+/deHz669+6312Bc29+68TTPXX09+691737r3Xhxz7917rsMRc/wBffutUB68Tck+/dbGMdea9+ffutCnl11791vrsXsbfT8+/daNK9esTb/G/5/p7916vXXv3W+ve/de67Fr8/T8+/da8sdeNr8fT37rY+fXlNiCfp791oiop16/1t+ffuvU66ueB/T6e/db697917rsH+v0/Pv3Wj8uvXF7j6fi/v3XvLPXRNzf37rfDrvkWP9f+Ke/da4468Qfr/X37rwPl17iw/r+ffut9de/de674sf6/j/iffutefXXv3W+ve/de6745/r+PfuvddfT37r3XZN+T7914Y669+6913pNr8f8AI/futVzTrr37rfXvfuvddm3Fv9j791rPXXv3W+ve/de697917rskm1/x9PfuvUp17jj+v59+6911z/tvfuvddkWF/wCvv3Wq+XXXv3W+ve/de699ffuvdczwot+fr/tvr791UceuHv3Vuve/de697917r3v3XuuwL3/w9+61Xryi5APv3Xjw68wsSPfuvDI669+630eL4a/zBPkL8Epuwp+g83j8PN2dgRt3dEtbSyVDz4woymCEo66T6j6v6/T3qma9bBxQjosHZ3ZW5O2t5Zbfu7p0qtwZqfzV0qh2WVrltZMpYk3P5PvZz1QCmOh5+IfzZ7v+Ee96rsHozMUuH3HWJAktTVRTSJpg1FEMcToCPUbg3Hv3DrdK8eu9+fNfu3sb5U0vzH3Fl6J+6aPdmN3lBlKamkhpRncU/lpKsQhrjS9jYN71Ty6tqOqvTh8xfnZ3385t91PY3fmao87uytWnWryVPDLC1R9tGI4S6u7c2Aufe+Ap1U9x1enSk+Cn8xT5J/y6t75/sD41biodvbl3LixiMpVVtK9XHJRXuYTCHUG/+N/fs9eDAjuGOrWj/wAKoP5oMreWsyHS2RqT/nKzIdW4Srq5f+WtRNdmP+JPvVOndeKU6rI+cP8AM3+RX8wKoxdV3xFscVWKkSSmn2ltmm26PR9BJDSHSR/sPe+A6bYkmvVdlzYD8D37rVM1669+6313ytxf/X9+61g5669+631737r3XY/P9fx/h791o9eJJ/x9+69QDrr37rfXK4sP6j/Wt9b+/dVoa9cfr791bru3F/futVzTrr37rfXYtzcX/p791o18uuvfut9e9+6912LfkX9+60a+XXXv3W+ve/de68OOffuvdcgC3P8AT6/Qce/dawOuccssZ/alkjvx6XZeCfzY+/de8s9ekllcnXK73tfU7Nfjj6+/deFKdYvfut9e9+6913c2t+PfutUzXrr37rfXYJH09+60RXrr37rfXYt+eeD/ALf8e/daNfLrr37rfXIXYW44/wBh/vPv3VTQGvXXJFvwLn37reAa9de/db697917rsG1/wDEW9+69117917r3v3Xuve/de697917rskn6/X37rQoOHXuLf43/wB49+63mvXdhpv+f+N+/dVqdVOurcX/AB791uuadeNvx7914V8+u+Tzf6e/dewMddXPP+P19+69Qdde/db67uNNvze/v3WqZr12oBPPv3XiaDHXRtc2+nv3Xhwz11791vrsgj6+/daBr14W/P8AT/effut9eBtyPfutEV6//9D5/wD7917r3v3XuuwSPp+f6+/daIr13yp/H0/1x7917DDrwHFz9L/j6+/dermg64+/db67Av791omnXfKn8fT37r2GHXXHP1v/ALx7917PXrcX/r7916uadevxb/G/v3W/OvXXv3Xuuxb83/2Hv3Xs+XXXv3Xuuyb+/daAp14i3+2v791vrkpJBUW+n59+6qaA16434t/jf37q3nXrr37r3XLUbW/wt791qgrXrokWsL/W/Pv3XvmeuWg/1H+39+61qHXD37q3XvfuvddgX/43791omnXuLfm/4/p791vPXXv3Xuuwbe/daIr12AR6h+P+Re/deND29cTyb/19+631737r3Xv979+691737r3XYJBBBII5BHBB/qD7917oQ9sdnbo2u8P2lfUPFERaNpmFgDewPP8AvPv3VdJ8uje7A+V8gijps/VSRMgA9chAKgc+ogX/ANv7914/mOhH3F8yMLisTNBt6p117q6h0u73KWHqQH+v9ffvl1v+fVd++d/7g3/mJ8rnK2efySFooHkZ44gT+Abc/wCPv3Xqefn0iOLn62/Fv+N+/dezTrr37rfXvfuvddglSGUkMpBBHBBBuCD7917qxr4v/Myv61pVwW5K+QUSvGsbSO5iaJQBY3Fh/S3v3XvkerEMf83evMpFqaqp7tySZVPJ5+tv+J9+r14Cg49Kza/y46mpcgmRqZqFnje4YvGbm9/z79XrVOjKRfzIOsaSjipYqii0ogUDyR8cc3v79Xrfzr0gtyfzHeuPUonpCrIzWV4/qFNv9t798utHqsT5W/PH++MM2H2RVDx1LWlMD2VFKWZjIARe/wCPfuvfPz6qoy25M3m5pJsjkqupaU6nV5nKk8/2RYW5PHv3XumS5/PP559+6314m/v3XuHXXv3XuuSm3v3WiK9dH6+/dbHXib+/daAp12b2F7fTj/jfv3XhSp68WJ+vv3XgAOuP+v791vru17kfQf8AFbe/dar17j8X/wAffut9de/de67Nvxf/ABv791oV8+uvfut9cuQv4sf+J9+6rgn7OuPv3Vuvcf7H8e/de6744te/5/41791rPXX+v791vr3v3XuuwbG/v3WiKinXtRvf8+/depinXib8+/db4dde/de697917r3v3Xuvcc/1/H/G/fuvde9+6912Da/+Pv3WqV68Lfm/+wt7917Pl11791vrsm9v8PfutAU669+6312bfi/+xt791oV8+vE3t/h7916lOvccW+v59+6914m/Pv3W+HXib2/w9+61Tr3Fha9/z/T/AGHv3Xs9de/db697917rvj8X/wAffuvZ69x+b/4e/de69fgD+n/E+/dep59eItY/19+61WvXYNgRzz7914ip64+/db65FiRb/Y+/daApnrj791vru4tY/i9rf8T791rzx12t72FuR+b/APEe/dabhnroi1wfr+LfT37rfHrwt+b/AOw9+69ny66/1vfut9dm1+L2/wAfr79177euvfuvde9+6912CRe359+60RXrr37rfXvfuvdeH1Hv3XuuyLEj37rQyOuvfut9dg2v/j791oivXhb83/2Hv3W8+XXiOL/i9v8AH37rVc065BfSSf8AYf09+61XNOuHv3Vuu+Lfm/8AvHv3Xs9de/de697917r3v3Xuve/de697917r3v3XuvfX37r3XZBBsffutA167Bt9Pz9b+/deIr10CRyPfuvEV69YAkH/AGFv6/j37r1a5HXXv3W+uwbXt+f6+/daIrx669+631737r3XZN/+Ne/daAp11791vrsixt7914Z69wP63v8A7D37rWfy669+6313xb83/wB4t791rNeuhxz791vrly3P9B791rA64+/db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rvlT/iLf8AFffutcR14m/Pv3WwKdde/de697917ru3F/futVzTrr37rfXZt+L/AOx9+68K+fXYP0v+OR/r/wCPv3WiOuibm59+68BTHXhb83/2Hv3WzXy669+6912Lfm/+w9+69ny68Tf37rQFOuvfut9d24v/AF9+61XNOuvfut9e9+6913xb83/3j37rWa9e4t+b/wC8e/dezXrxt+L/AOx9+63nz68Bfj37rRNOuvfut9e9+691/9H5/wD7917r3v3Xuve/de697917rsnm/wDrf7x791qmKddklj+OB/vXv3XsKOuPv3W+ve/de697917r3v3Xuve/de65fpI+h9+61xHXH68+/db67Aube/daJp11791vr3v3XuuwCfp791omnXuLf43/AN49+6359de/de67AuQP6m3v3XjgV68RY29+60DUV67t6b3/ANh7916uadcffut9e9+6912AT9PfutEgdeBsb+/deIqKdde/db6yEXF/pa/1/wB49+6oMGnWP37q/XfJF7cDj37rWBjrly3PHp/5H791rAx69cPfurddrYEX+nv3Wjw69/a455uPfuveWeuj9ffut9e/1vfuvdcxbkMSD9P6/Q+/dVNeI64e/dW65Fbfn+v/ACP37rQNeuPv3W+uViwLfQD37qvDHXRtxb/Y+/dbz12QVsb/AOsR7914GuOs6VdTGulaiZF/1KSMo/2wPv3XqenWVMjXJwlbVrf8CZ+f959+61Trl/E8gSf8trCfz++//FffuvU+zrg1fWv6Wq6lr/hpWP8AvZ9+69QccdQySxJJJJ+pPJPv3VuHXuOf6/j/AIn37rXXhb8/T37rf2det+fxe1/futV8uu9RuDYcfT+n0t7917SKU66P9f6+/db669+691737r3Xvfuvde9+691737r3XZt+D/r+/daFfPrr37rfXYBPv3WiQOvAXBP9PfuvVzTrsm45/H09+69Shx14X+v9P6+/deNOHXTWJuPfuvDh163BP9Lf7z7916uadet/re/db669+691737r3XZtxY/6/v3Ws9eFvz9Pfut9eNvx9Pfuvfb117917r3v3XuuVgL3PP8Ah/rXHv3Wqk8OuPv3W+uwbc+/de49eJubn8/09+61wHXZAH0N/fuvA164+/db697917rsG3Pv3XuPXd9RueP9Ye/dapQY64+/db67H1H+uPfutHh1zvydIv8AS/8Ahb37qvlnrH791fr3v3Xuve/de697917r1v8Aeffuvde9+6912Lfk+/daNfLrr37rfXvfuvde9+691yLFha3A/p/xPv3VQAM9eJuALDj/AG/v3WwKGvXH37rfXvfuvdd34I/rb/ePfutUzXrxtxb/AGPv3Xs9de/db67Nr8fT37r329de/de697917rlcHg8Wt9P9a3vfVaU4ddW4J/p/xPvXVq+XXjewv/sPfutClevcWsfxe3+N/fuvZrjrr6+/db67t9P6n8f0/wBf37rVeu9P6uR6f95/1vfuvauHz64+/db697917r3v3Xuu+OP6/n37r3XrGwP9ffutVzTruwt+bj6j+n9ffuvVPXH37rfXYF/yB/r+/dePXrEi9uPfutVHDrr37rfXZtxb/Y+/daz17i3+Pv3Xs1669+6313e4A/pf/effutU8+uvfut9ctXJNhyLe/dap1x9+631737r3Xvfuvdetb6+/de697917r3v3Xuve/de653tyOR+b+/dV44PXE24t/sffut5669+631y1enT/AI39+61TNeuhbm/+w9+69nrxtbj6+/dez13caQPyD/vr+/dapnr2o3vYHi3v3W9OKdcffut9d3NgLcf19+6159eBINx7914ivXhb8m3v3XjXy69e1/8AHj37rfXXv3Xuu/xb/H37r3z67AH9q4/p791qp8uuPv3W+ve/de697917ru/AFhx+fz791qma9de/db67Ykm5Fv8Ab/8AE+/daAAGOvAXvyB/r+/deJp14ixte/8AiPp791sZ68Lfnge/daNfLrr37rfXO/Fvx/Wx5P8Aj791Wma+fXD37q3XvfuvddgX49+69w68Rbj37rQNeuwCRYW/r/xHv3XjjPXRWwvcEf4e/deB8uvXv9f6WH/Ee/de4cOvWFvzf+nv3Xs1+XXdwD6eePz7916lePXH37rfXZt+PfutCvn14Gxv9ffuvEV6/9L5/wD7917r3+v7917rlYH6cD/H37qtacevem/0Nrf7zf6/X37rfdTrq3F7j/iffuvV68PpcEc8W/PPv3Xuuvfut9e9+691737r3XIg2HIt/T8j88+/daFK9dAX/I/2Pv3Xq9de/db697917rv6f71/j791rrvSQbGwP+8e/deqKV64+/db697917r3v3Xuve/de67FrH66vwR791rNfl14j6cg3/xv7914dde/db67txf/AJH791quadde/db67uR9OPfutUHXXv3W+ve/de65ElvfutAAdeIJu3H+t+f9t7914EcOvX444H5H5J/J9+695569f8Lf/H37r329dC1+ffuvGvl142vx9Lf6/v3XhXz69ccEfUfW/wBL+/dez59eI/Nwb/09+68PTrsEAfTn/H6e/deIJ+zrrkn/ABPPv3Xuuvfut9dkn834/r7917HXuLHjn8e/dazXrr37rfXYt+ffutGvl14n/bfj37rfXjbi39Of9f37rQr59e+luQf9b37r3HrsEC5t6j/tvr791qh/Lrq9zc/7H37rdMUHXYIH45/B9+68QT1x9+6316x/p7917rsE20j8/wC39+61QVr11791vrly3HAt/sPfutYGeurf4j37r3XXv3W+ve/de697917r3v3Xuve/de697917rlc2+vH0/wAffutUFeuPv3W+ve/de65XsLD/AGP/ABPv3WqZqeuPv3W+ux/j9Pz/AMR791o9cvSQbD6f74e/dazXrjxYf1/P/Ee/dW68bfge/daFfPrr37rfXYNjf37rxFeuyVvwOP8Abe/daFadcffut9e9+6913z/j7917rr37r3Xhz/h/r+/de67Isbe/deGeuvfuvddkk2H9P6e/daoB11791vr3v3Xuuxa/N7f4e/daNfLrr37rfXvfuvddj6/W3+9f7H37rR68bk/1P+Hv3Xhw669+631737r3Xvfuvddk3AH9PfutU669+6313xx/X8+/de669+6913b6/T/iv+t791qvXXv3W+uYNibA2/259+6qRjPXA/Xj6fj37q3XvfuvddkW/I/2Hv3Wga9de/db697917r3v3Xuve/de697917r3v3Xuve/de67B/r9PyP969+60evG3FgR/wAT7914fPrxFrf4+/der12LjkWP+H5/23v3Xj6ddEm/P/IvfuvDh16/+v8A4/4+/dep12dP4BHv3XhXz66UXP8Ah+ffuvE0HXbAA8f09+68Pn10PzwT/wAR79149de/db65BhzcfUW49+6rQ+XXXNh/Tm3v3W/Prwt+ffuvGvl12T/Qenm1/wDivv3WqevHrj791br3v3Xuve/de697917rsED6/wCw9+60a9di5NwQD9f+J9+68aU64+/db697917rsC97f7z791qtOPXX09+631ytqubge/da4Y64+/db697917r1v+R/j37r3XZuOL8f4Hj37rQznrr37rfXZ/qBYfj37rQ+fXXv3W+ve/de697917r3v3Xuve/de697917ru/FvfutUzXrr37rfXZt+PfutCvn11791vrsi3Hv3XuPXX+Pv3XuuVwfqP9t791qhHDroW/P9Px/X37rxr5detxe4/wBb8+/dermnXuLf43/3j37r2a9eFub/ANOP9f37rx69b/kX59+69XrxN/fuvAU67uOPr/j/AIj/AG/v3XqHrqwJNuOPz7917hx669+631737r3XIn8AWH+P19+6qB5nrq3APHP+39+63XNOuvfut9di45Hv3WjQ9e5PPJ/x9+69gY69ew4uD/X/AA9+6914kfjgf0/x9+68K+fXf9CBYcf4i/v3Xvkeuib8+/db4ddi1+QTx9PfutH5ddEW/p/sPfuvA1669+6313bi/wDjb/H37r3nTr//0/n/APv3Xuve/de67J4A/pf37rVM1669+631737r3Xvfuvddkkm59+60BTA65MD9Tb37rQpwHXD37q3Xvfuvde9+691737r3XYJHI9+60RXrr37rfXZJP19+60BTrr37rfXvfuvde9+6913e55/3j37rVKDHXjb8X/2Pv3XhXz669+6312LX5vb/AA9+60a+XXib/wC9e/deAp11791vr3v3XuuyLf7a/wDt/futA1669+631yDEfT37rRAPXRJJuffuvAAcOvcW/N/949+63mvXfK88c+/dV+Lrj9ffurdeFvz9P8PfuvfZ17/W+nv3Xuuxb83/ANh791o18uuvfut9d3Nwf6e/dap11/j791vr3v3Xuu1FyAffutHA68bfi/8Asffutivn117917r3+v7917rvjn6/4f63+Pv3Ws9de/db697917rvi35v/vHv3Ws168BcE/09+69XNOuvfut9dkk+/daAp15fqPfuvHh11791vr3v3XuuyLW/xF/futA1669+631737r3Xvfuvde9+691737r3Xvfuvdd24J/p791quadd2/I+g/r/t/fuvV8j1x+vPv3W+ve/de697917rlcWsB9frf/AIj37rVD59dEWt/j7916teuvfut9d8f439+61nrxJNv8PfuvUp11791vr3v3XuvcW/x/H9P8ffuvddi3N7/4W/4n37r2evXPH+F7f7H37rVOvXvcn6+/de4dde/db697917r3v3XuuwbG/v3Xjnro88+/de697917rsC5t7914466Isbf049+691737r3XZt+L/7H37rQr59eBsb+/dbOeujzz7917rsC5t79144699DY/j6+/da4jHXiLWP4PI/w/1/fuvA9de/db697917rsC/Hv3XuHXj/T+nHv3Wh69de/db65XA+n5/r791WhPHrj791br3v3Xuve/de67K2F/6+/darmnXXv3W+u/6W+v59+69169zc/n+nv3WuAoOuv8AW9+631737r3XJb82t9Ob/wBPfuqtTz66IsSPfutg1Feuxc/n9P0/3v37r2B+fXRJP19+68BTh14Eg3H19+62RXB68Tc39+68Mdde/de656D/AIf7z791XUOuhext/Tn37rZpUdcffut9clvY2/pz/re/daNMV66txf8Ar7916uadde/db67ubAf0+n+x9+69Tz67AB+nB/N/z791qpHXVzYD8D37r1M167FyCOOB/wAb9+61gHrj791brvgnj6f4+/da8s9eItx791vj117917r3v3XuudivIP8Ah/t/9f377eq1BweuItfm/wDsPfutmtMddglSR/sD7916gPXH37rfXvfuvde9+691737r3XZPFvx/vPv3WqefXXv3W+ve/de67ubW/H19+61TNeuvfut9e9+691737r3Xvfuvde9+691737r3Xvfuvde/1/fuvde9+691737r3XI3PqP59+60KDHXV+Lf439+638+vcW/N/8AePfutZ69bi/9ffuvVzTrr37rfXvfuvddi9xb63Fv9f37rx4ddtfUbm5/r791ocOuiLG3v3Xga9dkAcc3+v8Ah7914GvXQF7/AOAv7914mnXXv3W+uyNJH+3/AN59+60DUdevqI/23v3XuA68RY29+68DXrr37rfXIEWIP0P9Prf8e/daI8x17V+Pxz/r2Pv3WtPn59dG1+L2/wAfr791sV8+vEW9+68DXrvV6bfn/jfv3WqZr1x9+6t1y1H68f6n/YfX37qtBw64+/dW67Fub/04/wBf37r2fLrr37r3X//U0ACCDY+/daBr119ffut9dkEGx9+60DXI669+631yUXP+8/7b37rTGg66Y8n/AA49+68OHXXv3W+uwbG/v3WiK9e+p/1z791vh14i3v3Wga9de/db697917rsW/N7f4e/daNfLrwt+b2/w9+69ny68DY39+68RXrxN/fuvAU669+631737r3Xvfuvde9+691737r3XYNuffuvEV69+ef949+699nXhaxve/49+61nrr37rfXvfuvdctPpv+fx/wAb9+6rXNOuPv3Vuuybm/v3Xhjrr37r3XvfuvddgEm3v3WiQOu9PFz9B/T/AG3v3Xq5p11a5sPz9PfuvcBU9dg2uD9Ofp/X6e/daIrkddW5t791uuK9euQCv+PP+w9+69xz13qB+v1H0t/X/H37rVPTrj791br3v3Xuve/de697917r3v3Xuu+Lfm/+8e/dazXrwt/j/h/xv37rx69xfn/ePfuvfZ14i3+8f7z7914GvXQ5Nv6+/db68eDb+nv3Xuu78W/5H791qma9evxb8e/depmvXXv3W+uxb8/7x791o18uuwATYfT/AI17914kgV64+/db697917r3v3XuuyCpsffutA14dde/db697917rsm9v8PfutU669+6312bfi/+Pv3Xs9e4txe/+8e/dazXrwF+Pfut8Ouvfuvdd8cWvf8AP/GvfutZ68DaxH1/x+nv3XqVweur2N/fut/Lrv8Ar/X/AHj/AB9+69117917r3v3Xuve/de697917rsC5t7914468bi4/rb37rXHPXXv3W+ve/de697917rsgi1/z791oGvXXv3W+uwL3/w9+61Xrr37rfXvfuvde9+691737r3Xvfuvde9+6912Dbn37r3Hr1+b+/dapinXX+P9ffut9e9+6912FJ+nv3WiQOvA2N/futnPXR59+691737r3XZNzf37rwx14C5t/X37rxxnrxFjb37rwNc9de/de697917rvj8X/wAffuvdde/de67BI/2Pv3WiK9de/db67AJuR+Bc+/daJp11791vru/BH9f+I9+6913qNrcfS3v3WqCteugL3P8ATn37rxPXZufVxz7914U4dcffut9e4/4r7917rsW/N/8AYe/de+zrr37r3XfHN7/4f8b9+69nrr37r3XZFjb37rwNc9dgE8i3B9+60SBjrq/N/wDEfT/D37r1MU68fqbf19+68OHXdvoT9D/T6+/der5Dro2/F/8AY+/dbz59de/de67txf37rVc0669+631737r3XIqR9bc+/daBB66Btz791vj13cHk/wCwsPfutZHDrj791vr3059+6912SSb/AJ9+60BQde4t+b/7x791vrr37r3XYNv9tb37rxFeuvfuvde9+6912L82/ob/AOt791o9de/db65WIH45t/vI9+6rgnrj/r+/dW67Nvx9P8ffuvD59de/de67BI+n59+60RXrw/5F/r+/dePXv1H/AF/fuvcB11791vrsAm/+Av791omnXXv3W+uzb8X/ANj7914V8+uvfuvdeHBv/T37r3XZNzc+/deApjrr/X9+6912bfi/+x9+60K+fXXv3W+uSi55v9P969+60TTrj791vrkSV9PHH/E/4+/dVoDnrj791br3+9+/de67Bt/tre/de669+6912CR9PfutEV669+631737r3XIciw+t73/AMLe/dVODXrj791brkDxY/T/AA+vv3WqZqOuPv3W+uzb8X/2Pv3Xvt69fgj+vv3XqefXhb83/wBh791o18uuyOLj6Xtz9b+/deBzQ9f/1dAHSb2JH0/P09+61UUr13a1iSOOOP6j37rVa4HXEm59+62BTrr37rfXZ+vp/wAPfutDhnrr/X9+631737r3Xvfuvde9+691yCki/wDvZ9+6rUDHXRBH1/1/futg167NyNX9OPfuvYGOuP1+nv3W+ve/de697917rsi1v8Rf37rQNeuvfut9dra/P09+60a0x11791vrkVsL3H/I/futVzTrj791vr3v3Xuu+LH+v+t/xPv3Ws166sR9ePfut9e9+691yKkC/Hv3Wq1x11cmw/1hYe/deoB11791vr3v3Xuuz+Pp9Px791oddWPv3W+uRYn/AGF/p/j791oADrj791vr3v3Xuuxa/P8Avj791o18uvc/Xn/X/wCN+/dexw669+631737r3XZFv6f7D37rQNevcaf8ffuvZr14i3Hv3Xga9de/db67LE/X37rQFOuvfut9e9+691737r3Xdza349+61TNeuh9ffut9d2+v+Hv3Wq9dnTbj/iffutCvn10Tf8A2At/tvfurUp14gi3+Pv3Wga9etxfj37r1c069yLf4i/v3XuPXgLm3v3Wzjrr37r3XYF+PfuvcOvG34N/futCvn11791vr3v3Xuve/de65C1jf6/j6+/daNa466v/AK3v3W+uvfuvde9+6912Rax/r791oHrr37rfXvfuvde9+6912bWFvr+ffutCvn11791vru3BP9PfutVzTrr37rfXvfuvdd2P1H+8fj37rXy699bn+nv3W/l14C/++/3r37r1evWsRfgf8R791qvp13e1wPp/sf6e/dapXJ68RwLEnj/be/dbHXQNr/4+/dep13+Bb/G9v9f8+/de8+uPv3W+uyLWP9ffutVr11791vrkrEcC3J/Pv3WiBxPXiALAfX8/6/v3XhU9cffut9e9+691yFudX+259+60a+XXiSQLjgfQ8+/deAFeuPv3W+vX9+691yJFuPz9fr791oVrnroi1v8AH37r1evW4B/r/wAR791uvl12VIAPHPv3WgQTTrofn+vFvrf/AB+nv3Xj16x5P9P6+/deqOvAE/T37rxNOvW4B/r7916uadde/db65X03HBv/AK/v3WuOevLp5v8A7D+n+8e/daNfLro2sOCD+f8AH/W9+62K9de/db69Y/7f37r3XZJIA/p791qma9de/db65C1vrY/763v3VTWvXR/pe/8ArfT37q3z67uAAR9eP9659+61nz66/Vc3A/5F+PfuvcMdeAv7914mnXXv3W+uwfxxz/X37rRHn178/wDFPfut+XXj9frf/H37rQ4dd6ja3v3XqCteuP0+vv3W+uwxX6e/daIB49dc/T/ePfut9chbkH6n6e/daNevaTe3HH/E+/der10bjg/j37rwznrr37rfXI3AF+Prb+vv3WhSvXgb/q/4n37rVKcOuPv3Vuu9Jtf8f7H37rVc0669+631737r3Xhxz7917rsm5/4p791oY68CVv8A7Y39+68QD11791vr1ifp7917rsW/P/E+/daNfLrr37rfXhx7917rskn6+/daAA66sffut9d2P9D7916o68CR9Pz791oivXYJW/H+3v7914gHroC9/wDAX9+6317n68/6/v3XscOuvfuvdcjawt9fz791rNc9cffut9d34t/jf37r3nXrtSB9f8P99z791oivXRtfj6e/deFaZ68DY39+62c9cz/tVv8AYXv731QfLrH711fr3v3Xuu7m1v8AG/v3WqZr11/re/db69/r+/de697917ru3F/8be/de+XXXv3Xuve/de67Fr8/T37rRr5dd3Lf0Fh7917C9cbf7x791vr3v3Xuve/de67vcgn8W/3j37rVKDHXj/UDj/W9+68PTrwBP09+68TTrqxPv3W+uVxptbn+tv8AH37rVDWvXH37rfX/1tAC5/qffuvUHXrn+p9+69Trr37r3XYtf1fT37rRr5dcjp/H1/2P9f8AH37rXd59dMb2/wBYX/1/futgU64+/db67II+vv3Wga9de/db67uf6n37r1B11cn68+/de697917rsG309+69x69Ynn/inv3WqgY67/V/r/7bj37rXDrj791br3v3Xuu/xf8Ax/4j37rXn11791vru5+lz/t/fuvUHXXv3Xuve/de67It/vfv3XuPXfJF/wAf7D37rWBjrj791vru5/qffuvUHXgbG/v3XjnrkLG/+q5/1vfuqmo+zrh791brs24t/Qf7f37rQ69ci/8Aj9ffut068AT9PfuvVp117917rsAm5/p791qvXXv3W+uRP4H6b3Hv3WgPM8euuTb/AHj37r2B14W5v/sPfuvGvl11cn6+/db697917r3P59+6913c2t+PfuvU8+uvfuvde9+6912NNje9/wAe/daNa4669+631737r3Xvfuvdcj9OP+QvfuteeeuPv3W+ve/de67N/wA3/wAL+/dex5ddj/avpbj/AHw9+619nXR+vH0/Hv3Xh12eDxx7914ZGeujbi31/P19+68K9de/db697917r3v3Xuve/de697917rsWsb/X8e/daNa468ouQD/vuPfuvHh12VI5/F7A+/deBrjrj791vr1z7917r3v3Xuuxa/P09+60a0x11791vr3v3Xuuxbm/1/Hv3WjXy669+6314C/v3Xuvf4e/de65E2Gn883/AOI9+6rxNeuuf8be/dWx14EC/wDX8f8AE+/daOevEk/X37rwAHDrr37rfXYueB+f+I59+60fXrwtzf6/j/X9+69ny68LWN/r+PfuvGvl163F/wCnv3W/l16/HP8AsPfutUzjrr37rfXvfuvdd8WN/r+P+J9+6111791vrw9+6914m/Pv3Xuu7k8f09+61Qdde/db67Fr8/T37rRr5detc2H+w/3x9+69wGevWJv/AIf4+/deqOuvfut9d8nj/iffutYHXhe4t9ffuvGlOvEn6E/77/Ye/deAHEdeBI+nv3XqV69+Of8AYf8AE+/de8+uvfut9e9+691737r3XJiDa34/1/futAHz64+/db67uSAP6e/dapmvXXv3W+ve/de697917rkdNuPr/sffutCvn1x9+631yFv7Xv3WjXy66+h49+639vXXv3Xuu+OP95+vv3Ws9eNr8fT37rwrTPXXv3W+uTWB4/w/2/v3WhkZ64/X37rfXvfuvddm/wBT+f8AW/3r37rQpw669+6313c/1Pv3XqDrs/4/q/Pv3Wh/Lr3Fv9q9+69mvy66JJ+vv3XgKdde/db697917ru5ta5t/T8e/deoOPXXv3XuuRHpB/J9+61XNOugCfp7916oHXh9R/r+/dbPDrs+pjb8n8+/da4DPXRNz7914cOvAkfT37rxAPXgL/7D37rxNOvG1+Pp+PfuvCtM9de/db697917rvkf71/t/futcevXIFvx/T37r1Bx669+6312b/n/AH3+29+60KeXXuLcfX8/X6e/de8+vXNtP4vf37r1BWvXXv3W+ve/de67txf8H37r1fLrr37r3XMgAf42HvfVQST1x4sP6/n3rrea9d8af9q9+69mvy66+o5+t/8AePfuvdde/db697917rsW/P8Avv8Abe/daNfLrwNjcfj37rZzjrr6+/de697917rsW/P0/wBj9f8AYe/daNfLr1v6fQ/T37r1evEEfX37r1Qeuvfut9d2K/4f6x/4p791rB69c/7f6+/db669+6912Bf6e/de4dde/de67ubW/H++Pv3WqDj11791vrsXvx/T/ffX37rRp59de/db697917r/19AEDVwPr/vFvfutE0z10QQbH37rwNevAkf7H37rxFeuvfut9e+nPv3XuuWo/wCH0t791qg64+/db697917r3v3Xuuwbf0P459+60RXrr37rfXvfuvddg2PH+t791o9eIIPPv3XhkdeIt7914GvXQt+fp/h791v7Ove/de697917r3v3XuufIU/Sx/17j37quCeuHv3Vuu78W/339ffutUzXrr37rfXvfuvdd34t/jf37r3nXroW/P8AvHv3Xuve/de65KQLnm/+8e/daIr1x9+631737r3Xvfuvde9+691yv6SPfutUzXrj791vr3v3Xuve/de67Jvb/AW9+69Tru9vp/sb29+61SvHrieTf+vv3W+uXJF+OOPz791rgaddFi1r/j37rwFOuvfut9e9+6912Ra3+Iv7917rr37r3Xvfuvdd6TcD+ov791qvn17j/G/v3Xs9de/db697917rsG3Pv3XuPXiSfr+PfutAU68Lfm/+w9+68a+XXXv3W+uyLe/daBr11791vr3v3Xuve/de697917r3v3Xuu7cX/p791quadde/db697917rsm4t791qnn11791vr3v3Xuve/de67BsLcc+/dapmvXXv3W+u7/Uf19+69117917ru3F/futVzTrr37rfXYF7/4e/dar15RcgH37rxNBXrtjc/63v3WlFB10Dbn37q3HrxN+fz+ffutDHXXv3W+ve/de697917rmGsAB+L/Uf439+6rpqeuHv3Vuve/de67+lv6/7x7917rxNzf37rwFMdeAube/deJp1zX9Lj+nv3VDxB64A2uP6+/dWpXrxt+L/wCxt7914V8+vAA2Avf/AB+n+8e/derTJ68Rbj37rfHrr37r3Xv9f37r3XZN/wDWH0/r/sffutAdde/db67Aube/deOOvFSPr791oGvDrr37rfXvfuvddi35v/sPfutZ8uuv9b37rfXvfuvddi35v/sPfutGvl11791vr3v3Xuve/de68bfj6f4+/de+3r3v3Xuvcf8AFffuvddg259+69x67JH15vx/re/daAPDrom5v/vXv3XgKCnXXv3W+ve/de67JuAP6e/dap11791vr3v3XuuQUnnj/effutE064+/db67Bt791oivXQt+fp/h791vrs2/F/8AY+/dez59de/de67JuST/ALx/X37rVKYHXXv3W+ve/de67Bt791oivXrWNj/vHv3XuIx142/F/wDY+/db+3rr37r3Xvfuvde9+691737r3Xvfuvde9+6912Bf37rRNOuvfut9e9+691737r3Xh9R7917ru92uP68X/wBfj37rXlTrs3Dc/Xj37rwpTHXRN+ffuvAU68ATf/D37rxNOvcWH1vf/Ye/dez14kk3Pv3XgKCnXYbT9Pr+b/T/AGFvfutEV49dW4v/AF9+63Xy68Lfm/8AsPfut/Z12q3/ANb/AHnj6+/daJp10bfi/wDsffut/b117917rvi35v8A7x791rNeuvfut9e9+691737r3XZA+o+n05+vv3Wvkeuvfut9dg2/21vfuvdde/de65EW4P1/w+nv3Wga566Bt/vR/wBb8+/deIr10bfj6f4+/db+3rs2/F/9j79177evX4t791qma9etwD+CffuvVzTr3p/xt/sPfuvZ69fi3+N/fut/Prs/UX+lh9P6f7H37rQ+XXV7G4/2F/fuvUqM9dcW/wAf94t791vrkVIt/j791oEHrogj6/6/v3Xga9eubW/F7+/depmvX//Q+f8A+/de67Bsb/X/AF/futEVx11791vr3v3Xuve/de67F/rb/fD37rXy68Tf37rwFOvE3N7W9+68BTrr37rfXIi/6Rx791qvr1x9+6313xb/AB/4j37rWa/Lrr37rfXvfuvde9+691yBH0I4v9fz791Ug8R17VYEAcc8n6+/depmp664t/jf/ePfut+fXXv3W+ve/de69/j7917rvi3+Pv3Ws1669+631737r3XZt+PfutCvn163F/fuvVzTrr37rfXZBFr/AJ9+60DXru4Ju309+69QgY64n68fT8e/db697917r3v3Xuve/de697917rkpI5tf37rRFcdeIJGr+p+nv3Xhg6euPv3W+uRbVb/D37rQFOugCfp791vh14m5/wBYW/23v3WgKdde/db65H6Lx/vP19+60OJ64+/db697917rsm9uPfutAU669+6313yPr7917rr37r3XIrYXv+Af9v791qtTTr2ri3+w9+61pzXrjY/09+6t1yJvbj6e/daAp1x9+6312PqLc+/daPDrwtfn6fn37rZ+XXrfUj6D37rXXgPoT9PfuvE+Q49dAX4Hv3W+uS83H4/Pv3VT69dsLWt/j7915TXrh791brkSWI4/wHv3WgNI68VI549+69WvXH37rfXLVzf37rVMU64+/db67BsCP6+/daI669+631737r3Xvfuvde9+691737r3Xvfuvde9+691yBIBAH1/2/v3WiKmvXH37rfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6913/S/wBPfuvdeNr8fT37rQrTPXjp4tf/ABv7914V8+uwR9T9fx9ffuvGvl10Tc39+68MdeUXI/3349+68cDrx4Jt/U+/de8uvW4B/rf/AHj37rfXrfQ/19+6913YWuPx9fr+ffutVzQ9cffut9d2+n+Pv3Wq9dsAPp9eL/8AGvfutCvn16x4/wAf99z791uo66Isbe/deBqK9de/db67Bsb+/daIqKdeJuSfpf37rYwKdd3IUC34Nj/sffuq0z10Bf37rZNOuvfut9cgB+fz9Pr791on066sfp/T+nPv3Xq9eIIF+OffuvVzTrr37rfXdvof6+/de68Bf6e/derTrxFuPfuvceuvfuvde9+6912bX4+nv3WhWmeuxaxve/4/1/fuvGvl1x9+631737r3Xvfuvddg2v8A4+/daIr11791vrkWuLWt791UCnXQF7/4e/dbrTrkLqP8Gt791rDH7OuHv3Vuu9JsD9ffutVFadde/db697917r3v3Xuve/de697917rsW5v/AE4/1/futGvl14Gxv791s56534JA5P1/23vfl1Snl1j966v1zsV5/qLf7f8APv3VcNjrh791br3v3XuuQvzx+Dz791o064+/db65r+f6e/dVbro6bcfX37rYr59dWNr/ANffuvVzTr3AAP5v/sLe/dez14m/+H+t9PfuvAU67HHJH+te/wBffuvHOB10Tc39+68BQU669+6312Lc3/px/r+/da669+631737r3XYF/futE068QLm3I9+68OGeuvfut9dm349+60K+fXrcX/xt7916uadeAv/AL37914mnXja/p+nv3XhXz68DYg/09+62cinXibm/v3WgKCnXXv3W+u78W/xvf8APv3WqZr119ePfut9dkW49+69x68Rb37rQNeuvfut9cmJPP49+60ABjrj791vr3v3Xuve/de697917rsAn37rRNOvcn/H37reB17m9vz9PfuvdeII4Pv3Wga8Ov/R+f8A+/de697917r1vfuvddg29+60RXrr37rfXIEgWt9f9vz791qgOeugCTb/AHv37rxNM9de/db697917rkGI/p791oivXH37rfXvfuvde9+691737r3XYNjf/e/futEV68OTz+ffuvHAx15hY+/deBqOvW9N/ze3+8e/de86dde/db697917ru/Fv8AG/v3Xvn117917r3v3Xuve/de67txf/G3v3XvOnXVza1+P6fj37r3z67Bsb+/deOeu2txb/ez791Va+fXQF+PfurE0669+691yUXvf8D37rRPXE/Xj37rfXvfuvdchaxv9fx791o1r10DYW/r7916ma9de/db697917r3v3XuuwbG4/3w9+60RUUPXXv3W+ve/de697917r3v3Xuve/de697917rkt/oLD37rRp59eIYm31I4uP8Ab+/daBAFeuPv3VuuQBP54H+Pv3WjQdcSLce/db65Akfj37rRAPXH37rfXK4tb8/1t/j791rNeuPv3W+ve/de697917rkD+CLj/D6+/dVI8+ugbc+/dW49dm4sb/Xnj37rWDjrjcn37rfXKwBF/of6X9+61WvDrj791vru5/qffuvU669+691737r3XYF+PfuvcOu7WI1fT/D37rVajHXvTY/7x9f6e/daz1x9+6t13ewI/r791qnXXv3W+uyb24HHv3WgKdeJJ+v4/33Pv3XgAOuvfut9di9+Pr791o0pnru5DE8E/T/AHj37rVAR1x+vPv3Vuuxbni//Ee/daPXha4v9PfuvGtMddlSLn8f8R/sffutBgcdcffurde9+691ysV54/p791WoOOuiDwf6/wBPfutg+XXufpb6f8T7917HXXv3W+u7cE/0/wCJ9+6912NNjf6/j6+/daNa4664sP6839+69nrwBP0/Hv3XiQOPXXJ/x9+631yFrG/4+g5+p9+61muOuPv3W+ve/de65C1jf6/j37rRrXHXXP8Ajx/vHv3W+u73FiBf8H8/X8+/dapnrxIsLXv/ALH+nv3XhWueuPv3W+uwbe/daIr14ggA/wBf+R+/dermnXXv3W+ve/de675H0v7917HXXv3Xuu7nj/D6e/dap12Cefz/AF+p9+68QOuiD9OT9P6/09+69jj14m9h/T37rwHXYsb3P+t791418uugSPp791sivXXv3Xuve/de699f979+691737r3Xvfuvdc7FvVx/vv8B/re/dVqBjrh791br3v3Xuve/de697917rsAnj37rVevcjj37r3Hr3J/rx791vHXgbf7EW9+6914Ej37rRFevA2v/jwffuvEV669+6312Rbj37r3Hr3Fv8f9j/X37rWa9de/db697917ru3Nvp/r+/de+fXiCOL/AF/pf37rQz1yBBFm/H0t/wAT791og8R1xJ/oTb/Y+/dbA66HJt/X37rfXvfuvdcvVp+vH/I/futYr16x03/H+8+/dexqp10L/j8/77n37rZ68Qbn/D/Ye/darjruxsD+P6f7H37r1c066a1+P99/t/fuvCtM9eAJ+nv3XiaddliRb37rwAHXVuL+/dermnXYUt9Lf7H37rxNOuPv3W+ve/de67AJNv8AY8+/daJp14An6H37rxp1yJA/SDzxc39+60K+fXEWvz9PfurH5de4t/jf/ePfutZr12wI4Pv3XgQeHXH37rfXvfuvde9+691737r3Xvfuvdd8W/xv/vHv3Ws1669+631y4t/tXv3Ws1+XXQBY/X8fm/09+69gddra/P8AxP1/r79141pjron8fi/Hv3XgPPrr37rfXvfuvde+vv3XuuyLG3v3XuuVm+nH9b8/6319+6rUceuINjx791sivXr83/xv7917yp12xBPH9Pz7914AgZ6//9Kgj/oG5/mE3sMZs0/+Rcf8T71Xr2j7OvD/AITdfzCh9Mbs3+n/ABdx79Xr2j5jrw/4Td/zCx/y7dmC/J/3MD36vXtHnjr3/QNz/MLt/wAWzZlvr/xdx9fe+vaPs67P/Cbn+YSF1fw3ZpP9Blx/vfv1evaPs68P+E3X8wprH+G7N/H/AC+B+fp71Xr2n7Oum/4Tc/zCj9cbsw/i/wDGB+Pfq9e009Ovf9A3P8wn/nW7M/2GXHv1evaOvf8AQNz/ADCf+dbsz/H/AHLj36vXtH2deP8Awm5/mEf863Zlv+1uB79Udb0de/6Buf5hP4xuzP8Az8L/AK/19+r1rR9nXR/4TdfzCrX/AIbsw/8AkXHv2rr2j5jrv/oG6/mFWt/DdmAG3H8YHv1evaMeXXZ/4TdfzCr843ZvH/V3Hv1evafs66/6Buf5hH4xuzP/AD7j/effq9e0de/6BuP5hR/5dmzLc/8AL4H09+r69e0Z4jrw/wCE3H8wn8YzZlv+1wv19+rXren7OvD/AITcfzCSb/w3Zv8A59x/vQ9+r1rR9nXj/wAJuf5hP/Ot2Z/59xf/AGPv1evaPs69/wBA3P8AMK/51mzB/wCRcf7D6e/V69o+zr3/AEDc/wAwm3/Ft2be/wDztxa39b+9169o+zr3/QNz/MJ/51uzL/8Aa3HvWry69o+zrv8A6Buf5hJNjjdmfTj/AHLj+l7e/V69o+zr3/QNx/MJ/ON2Zb63/i4P+8e9169p66P/AAm4/mE34xuzD+L/AMYFvr71WvW9H2de/wCgbn+YV/zrNmfj/l7j6+/V61o+zr3/AEDc/wAwn/nW7N/1/wCLj36vXtH2de/6Buf5hP8AzrdmW/7XAv79Xr2j7OvH/hNx/MJ5/wBxmzLWH/L4H59+r17Tny66/wCgbn+YT9P4Zsz6f87ce/aut6K+nXf/AEDcfzCOD/Ddmf8An4F7/wCt79XPXtH2de/6Buv5hP8Azrdmf+fdffq9a0fZ14f8Juf5hJ/5duzbf1/i6+/Vr17R9nXL/oG3/mEc2x2zD/5FwPfievaB8uuP/QNx/MKtf+G7M/8APwPfq9e0eWOvD/hNz/MIP0xuzfz/AMvdb8f63v1fPr2j7Ovf9A3P8wn6/wAN2Z9ef9zA9+qevaPs66/6Buv5hH4x2zL3/wCdwOD+Pfq+vXtHpTrv/oG6/mE35xuzf/PuPr78T17R9nXv+gbn+YT/AM63ZnP/AFeB/sPfq9e0fZ12P+E3X8wv6fwzZtiP+dutv9597rTr2n7Ovf8AQNx/MJtf+G7Mv/T+Lj3rr2jrof8ACbn+YTa/8N2Z/wCfge/Vz17R9n8+vD/hNz/MJ/51uzLfT/i8D6/09+r6de0fZ13/ANA3H8wm1/4bsz/z8D/b+/V69o64/wDQNx/MJP8Ay7NmH+n+5cX9+r17TTgR13/0Dc/zCbAfw3Ztjyf9zA+vv1et6T8uvf8AQNx/MK5JxmzOB/ztx79XrWj7OuJ/4Tc/zCT9cZsz/D/cwPfq9bC/Mddj/hNx/MKA/wCLZs38W/3Lj8+9161o+Y65f9A3X8wr6HHbNA+h/wBzAP8AvXvVevaPs68f+E3X8wo/XG7N/wBjlx9P6+/V9OvaPs66/wCgbn+YV/zrdmf+fhf9jz73Xr2j7Ovf9A3P8wqwP8N2Z/59x/xHvVevaPs67/6Buf5hItbHbMJ/7W62vf8Ar73Xr2j7Ouv+gbn+YVz/ALjNm3/7W45/r79Xr2j7Ou/+gbj+YUORjdmf4H+MD/Y+/V69pzTHXX/QNz/MK/51uzB9Prlx71Xr2j5jrw/4TcfzCef9xmzb8/8AL3H1Hv1evaTTy69/0Dc/zCv+dbsz8f8AL4X8+91PXtHrTr3/AEDcfzCRf/cZszk/87gc/wCPv1evaSfTr3/QNz/MJ/51uzP/AD7j36vXtH2de/6Buf5hVv8Ai27M/wBb+Lj/AGHvVfLr2jrr/oG5/mE3/wCLbsz/AM/C+/V8z17R9nXZ/wCE3P8AMJuP9xuzOf8Aq8D36vXtH2ddH/hNx/MJ+hxmzLfn/cuPfq+XXtPnjrw/4TcfzCfxjNmfW3/F4X36vW9NeJHXf/QNz/MJFh/DdmXP/V4X36vWggPp10f+E3P8wkf8u3Zn/n4Xj3uvXtH2ddn/AITc/wAwnj/cbs3/AB/3Ljj3qtevaPs69/0Dc/zCLf8AFu2Z/wCfce/eXXtH2de/6Buf5hV7DGbM/wAf9zCi3+39+B69o+zr3/QNz/MK+n8N2Zz/ANXcc/09+r69e0fZ17/oG5/mE/8AOs2b9bf8Xce/V69o+zrx/wCE3P8AMJF743ZnH1/3MD/ePfq9e0+eOuh/wm5/mE24xuzPyLfxgf7D36vXinnjrv8A6Buf5hP4xuzP/PuPfgevaOu/+gbn+YULj+G7M/F/9zC/T36tevaPs64j/hNz/MJP/Lt2Z/5+F/3j36uMde0fZ17/AKBuP5hN/wDi2bM+v/O4+nv1et6fmOu/+gbj+YV/zrdmf1H+5cf7z79WvWtP2de/6BuP5hX/ADrdmf8An3H9L+/V69o+Y68f+E3X8wr847Zh/wDIwP8Aiffq063o+zrw/wCE3P8AMIHK47Zlz9f9zAH+8+9169p+zr3/AEDdfzC+f9xuzP8Az8Dn36vWtH2de/6Buf5hVv8Ai2bMv9bfxgfT+o96r17R9nXv+gbj+YT9Tjdmf4f7mBb6e/V9OvaPs69/0Dc/zCf+dbsy3/a3Hv1evaPs69/0DdfzCD9Mbsz8c/xce/V69o+zro/8JuP5hJtbGbNP/kYAtc+/V69p+zrmP+E3X8wpef4dswf+Rhfp79Xr2nyx1x/6Buv5hJ/5duzb/wDa3H+w9+r59e0fZ14f8Juf5hP5xuzB/wCRce/V8uvaPWn8+vH/AITc/wAwkf8ALs2bz9P9y4549+rTr2n7Ovf9A3P8wn/nW7M+nP8AuYHHvdevaD8uvf8AQN1/MJHH8N2aL/X/AHMD6e9V69o+zr3/AEDc/wAwq/8AxbNmf+fhf9h78D5jr2j7Ovf9A3P8wr/nWbN/8+44P+PvxPXtP2ddH/hN1/MJH/Lt2Z/59x79Xr2j7Oux/wAJuf5hB/5duzf/AD7j36vW9H2ddf8AQNx/MIFz/DdmA/8Aa4Fvr79XrxU08uvH/hNz/MJ/51uzD/5GB/vPv1etaPs67/6Buf5hP/Ot2Z/5+APfuvaPs67P/Cbn+YVwP4bs3/W/i6+9169o+zrr/oG5/mEfjG7N/wDPuPr71UV69o+zrx/4Tc/zCQP+Lbsz6X/4u49+r17R9nXf/QNz/MKH/Lt2YAf+ruOR79XrwX7Ouh/wm5/mFKL/AMN2YB/2uB79Xr2nPEde/wCgbn+YSef4bs2xsSf4uPyffq9e0fZ14/8ACbn+YT/zrdmkf1/i4/2Pv1fPr2j7Ouv+gbn+YVz/ALjNmf8An4X8/T3vr2j7OvH/AITdfzCR/wAu3Zn/AJ+F/wB59+r17R9nXf8A0Dc/zCP+dbsz/D/cuPdajh17T9nXY/4TdfzCh/y7dm8cG2XHvdevaPs66/6BuP5hNr/w3Zlv+1uP6+/V69p+zr3/AEDc/wAwrj/cZs38f8vcf7x73Xr2j7P59dH/AITc/wAwni+N2bb8f7mBwPyfeq+XXtNPTrs/8Juf5hPFsbsz/wA+49+qevaOuv8AoG5/mE2v/Ddmf+fge/Vr17R9nXf/AEDc/wAwn/nW7Mtzz/Fx79Xr2j7Ouv8AoG7/AJhFyP4bsz/z8L79q69p+zrv/oG5/mEtcHG7NJ+v/F4Hv1evaaenXv8AoG4/mEiw/huzR/r5ce91PXtNfTrw/wCE3f8AMKH/AC7dmg/9rhfeq9e0/Z11/wBA3X8wm5H8M2b/AOfce/V69o67/wCgbn+YT9P4Zs3/AA/3MD+l/fqnr2j7OvH/AITc/wAwof8ALt2Z/wCfce/V69o+Y/n17/oG5/mEkkfwzZlx9f8AcuvHvdcV69o+Y/n14f8ACbn+YR9P4bs36/8AO3B9+r17T59eP/Cbn+YQDb+G7NP/AJFwP9f36vXtHXv+gbn+YT/zrdmW/wC1wPeq+XXtH2deH/Cbn+YT9f4bszjn/i8D37V17R9nXj/wm5/mFHk43Zn05/3ML+ffq9e058uvf9A3H8wkA2xmzbf9rge/V62U9adeP/Cbn+YSOf4bsz+n/F3A5/A59+1da0fZ17/oG5/mE8f7jdmf+fce/VPXtH2deH/Cbj+YSb/7jdmf+fcXP+w9+r17T9nXv+gbn+YT9P4Zs3j6/wC5ce/Vr17R9nXX/QNx/MJv/wAW3ZnIH/L4Hv1evaceXXv+gbj+YT9f4Zs0m3/O3H9Pfq+vXtOKY67/AOgbj+YVbnG7Msf+rwPz79Xr2jPl11/0DcfzCfp/Ddmf4f7mByfx9fftXXtPzHXZ/wCE3H8wgH/i27NH4JOYH+w9+r17Tjy68f8AhNx/MKH/AC7Nmf8An3Hv1evaPs68P+E3X8woXP8ADdmD/wAjC+/V8+t6Ps66/wCgbn+YV/zrNmf+fcf6/v1etaPs66H/AAm5/mEk3/hmzP8AD/cwPe69e0+WOuX/AEDc/wAwj6jG7NIB4P8AFwOf9b3ony69p9Kde/6Buv5hJJvjdm/+fdb+/VxXr2j7Ouv+gbr+YT+Mbsz/AM+49+rnr2j7Ouz/AMJuf5hTf8u3Zp/8i4/pf8f7H36vXtP2ddf9A3H8wkkXxmzf/PuPfi3WwtPMdcv+gbn+YVb/AItuzLC/H8YHv1etafs64/8AQNz/ADCf+dZsz/z8D37HHr2j7Oux/wAJuP5hRv8A7jNmf+fhffq9e008x10P+E3H8wkf8uzZg/8AIwP94v798utlfUjrsf8ACbn+YSfrjdmAf1/i4P8Are/V61o9adeH/Cbj+YUf+XZswH+n8YH497B69o+z+fXv+gbj+YVz/uN2Zx/1eF96r59e0/Z11/0Dc/zCf+dbsz/z7j37V17RT067/wCgbn+YTx/uN2Z/5+B/X36vXtP2de/6Buf5hP8AzrdmEf8Aa3Hv1a9e0fZ17/oG5/mE/nGbNA/r/Fx73Xr2j7Ou/wDoG4/mEXP+47ZlubH+Ljk/0P8AT3qvXtH2dcT/AMJuf5hJH/Ft2Z/5+APfq9e0/Z13/wBA3P8AMK+v8M2Zb+v8YHv1evaPs66/6Buv5hJ+mN2YfqP+LwPfq9e0/Z13/wBA3P8AMK/51mzOP+rwv1/pz7917R9nXv8AoG5/mFf86zZv/n3H+8e/V69o+zrv/oG5/mFDkY3Zv9OMuPfq+fXtIrTHXH/oG5/mFH/l2bM+lz/uYX/XPv1evaPmOu/+gbj+YTawxuzbfX/i8D6+/V63pzxHXX/QN1/MJH1xuzLXt/xeF/rbi3vdetaPs67/AOgbn+YSfpjNmXP/AFeB71Xr2j7Ouz/wm4/mFf8AOt2YT/T+Lj/e/fq149a048uv/9k=)
Figure 4. The Neural Reference Space for Discrete Emotion. The neural reference space (phrase coined by Edelman [1989]) is the set of brain regions consistently activated across all studies assessing the experience or perception of anger, disgust, fear, happiness and sadness (i.e. the superordinate category emotion). Brain regions in yellow exceeded the height threshold (\(p<.05\)) and regions in orange exceeded the most stringent extent-based threshold (\(p<.001\)). Regions in pink and magenta correspond to lesser extent-based thresholds and are not discussed in this article. Cortex is grey, the brainstem and nucleus accumbens are green, the amygdala is blue and the cerebellum is purple. A color version of this image can be viewed in the online version of this target article at http://www.journals.cambridge.org/bbs.
![[[@Lindquist2012-jr]](http://dx.doi.org/10.1017/S0140525X11000446)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAgEAyADIAAD/4Rd9RXhpZgAATU0AKgAAAAgABwESAAMAAAABAAEAAAEaAAUAAAABAAAAYgEbAAUAAAABAAAAagEoAAMAAAABAAIAAAExAAIAAAAbAAAAcgEyAAIAAAAUAAAAjYdpAAQAAAABAAAApAAAANAAAADIAAAAAQAAAMgAAAABQWRvYmUgUGhvdG9zaG9wIENTIFdpbmRvd3MAMjAxMjowNToxOCAxNzozOTowNAAAAAAAA6ABAAMAAAAB//8AAKACAAQAAAABAAAFeKADAAQAAAABAAAEGwAAAAAAAAAGAQMAAwAAAAEABgAAARoABQAAAAEAAAEeARsABQAAAAEAAAEmASgAAwAAAAEAAgAAAgEABAAAAAEAAAEuAgIABAAAAAEAABZHAAAAAAAAAEgAAAABAAAASAAAAAH/2P/gABBKRklGAAECAQBIAEgAAP/tAAxBZG9iZV9DTQAB/+4ADkFkb2JlAGSAAAAAAf/bAIQADAgICAkIDAkJDBELCgsRFQ8MDA8VGBMTFRMTGBEMDAwMDAwRDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAENCwsNDg0QDg4QFA4ODhQUDg4ODhQRDAwMDAwREQwMDAwMDBEMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwM/8AAEQgAeACgAwEiAAIRAQMRAf/dAAQACv/EAT8AAAEFAQEBAQEBAAAAAAAAAAMAAQIEBQYHCAkKCwEAAQUBAQEBAQEAAAAAAAAAAQACAwQFBgcICQoLEAABBAEDAgQCBQcGCAUDDDMBAAIRAwQhEjEFQVFhEyJxgTIGFJGhsUIjJBVSwWIzNHKC0UMHJZJT8OHxY3M1FqKygyZEk1RkRcKjdDYX0lXiZfKzhMPTdePzRieUpIW0lcTU5PSltcXV5fVWZnaGlqa2xtbm9jdHV2d3h5ent8fX5/cRAAICAQIEBAMEBQYHBwYFNQEAAhEDITESBEFRYXEiEwUygZEUobFCI8FS0fAzJGLhcoKSQ1MVY3M08SUGFqKygwcmNcLSRJNUoxdkRVU2dGXi8rOEw9N14/NGlKSFtJXE1OT0pbXF1eX1VmZ2hpamtsbW5vYnN0dXZ3eHl6e3x//aAAwDAQACEQMRAD8A9VQDmUhzmje4tO0ltb3CR/KYxzUdVW1Zdbnit1Za95eNwdPu1jQpKZ/bKv3bf+2rP/SajZkl7DXjksveCKjbW9rdwG7lzWrMx/qzTjXMvpJ9Vjw8OfY587d+xjmWBzNn6Z/u2/aLLP01l/r+rbZqCrJfbW60s21kuhoMkkFn5x/lJKRdKzvteN+kMZNJNeRWY3Ne0lvuDP39u9WX5FDLq6H2NbdbPp1lwDnbRufsZ9J21Uc/FdjW/tLCqe+5gPrY9RDRc36XvaQ7faz/AAft9V/816iWZWeo4NWbgudXkVj1sV0QZI1qeyz/AErfY7ckp0SAQQRIOhBWJ0nPGLgU4b63G6vJdhFgIJY47rq3WOea97G0+51jP5xXqer4jsGvLvsbQHEVvDzG22djqPdDt+9Z3W2ZLXZvua1jqq8nGfZcRFlDv0vp1PsrZWzZ6f8AN+n+k/nbf0n6NKdttzfQF1jgxobve4mAABLpJ/dUK83DucGVX12OMw1r2uJj6X0T+aljursqMe9hJ5HLXj1R/wBGxBxbm2Oa37FZjgS5rntYAHRH+Ce/a7a5JTateWNBbG4uDRPmYU0F8G6tvAbLz/1Df+rRdfikpdDtvrqLWv3EukgNa55gc6Vtd+8pyg3V3G1ltJbLQ5pD54dtP5v9RJSvtlP7tv8A21Z/6TS+2U/u2/8AbVn/AKTWVb9XDZdZe2++q255sc5mVcGhztvqenUPY3dWxtX/AAX5iCPqkNha/My3vI2mwZVrT+dsLQAdvo7/ANF/npKdyvKqssFY3B5BcA5j2SBAd/ONb+8jKvXVkG9llpZtra5oDZJO4s5Lv6isJKUkkkkp/9D1VJV2ZbrGNsZj2FrwHNPsGh1HNig7qFbSA5haToAX1A/SFX+m/wBK5tf/ABiSm2kgHKLS3fTYwOcG7jtIBdo2dj3ORXFwaS0bndgTE/NJTJVG014TrLqmxVc825A1J3mN14/zf0lf+tkrr7nUvGK1pyWiRVadvfvH/nH8tZNedl5FjuoYLLH2sa1mVgXbmwAHu24LnenRY/1W2fpXfzn+lr/m0qQRf02bt2FubkYws24+aJpDAGBjwNz/AHU7Hv8AXj1fe/8A0ibBy2ZVDLLQPtmE/wBHKkN3Ndoy7Vo2+nZ7b/Z7E+PlYuRS4UuP2fdse3h+PaDPpWM/wW1//bT/APgf5sjWfpjadrBZNOZW1oG6whjab3W/zntrb6dbf9Hf/wAElHekeDPD9RjfTe/1LAxofY4Bu97N1Nlm2sbGb9jPoJ8cdSDh9qdQ5sGRU14M6Rq9z1GlzhcwuMuMh3hMbbP+njf+CKONViDJsdTkW2WVE+pW61zgJ3N27Hn6O5E7lI2SDJqGcaHSbLG+wAEgNZyXuA217nve1u/6fpqz8FgMzKqMvM6tbYPTrY3EoYDO+0u9V7WN/e9e70/Z/wCi0YnqT8bH6W3IBz9jXZuQCN9THbnB1bPb6m51foNd/wBctQS7PPOhTNcCAQZB1BHBCyMvLb1BtmHRaWYYrByc9phvuLNtNT/b6vr0us/S0P8A0X+k9X9Gnbn72NwOhVNIqbtbkPBGPWGQ3bu+la9u78z/ANKJKdeRMTqeydUqOnY9OS/Oue+7I98W2ukMY47vSqYNtVbGN9v0PUVtj22N3N1HYwRPwSUySSSSUpJJJJT/AP/R9If099mI7CtsZbjPrNL631yHVlvpuY+Ht+mz6SqU/Vnp9BDqaq63BwslotB3gl3qu/WPdb7rP0jve/1b/wDT3LYSSU07ZFtNWRksBseHV17Q1ziz3w33O/tJs/Hzi77RhXbba2+2l8ljo923RzNvqfQ3v3/8H6aPfiY2SAL6m2QCBuEwDG4f2trVm/sUUGMS3KxmMO5jKrQ9k/vPZf73/wBXckpk/Ny6ttfU8MxLQ3Ixd1o3uMaMa0ZFP0v5xPfTidSra/f9oGO93pZNJi2p4mtzhs+l/Z/7bUXv6xjip1d9eTt3C4ZLDRukt2+nYxjmN9OHf10Brce3MbZk9LfXmlhcMrGh7Po2x+nZs/Sfznpb/wDCel/waSlV4ebbmtc6A8Abuo0hmy+tprD8XMxnn+kWe/8AN2Vf4NXaS+yA2tw31gPpt0e1p/wWQ5he3fS7+atrdb6rPU/nP59BNND8JjLmutrfXWHVZjCXvPs9NmQ5w9K67fs+n+k9X/DK9Ufs7dl3tJMm0klpJ/lu9zP3Wep/00Brr2WD1Uxso2MNj7HPc0hx2+0AS31Iaz3e5rPzkB11Rc6v7LdUSdhtcIbryd7HlyvvYHsdWeHAg/MLMdZnl+N9otqNVzw3Yyt5d7g4bTYHP2ezd731+mnakrmdmOPXqc6mXUtD67XNZ/O27qnMoeP0nqta3db61VlX8x/1uOPh1V0W1ve8236Z2TLmWudGwV1ljvUqd/o24z9lW/8AVf51X3VfpxkOtcGsYWio7dgJ19b6Pqept9n856exV8wC9hfXuZDS197TshjtHvqfq71K/wCcqsa3+2jYv+CNaLWycrFFoxdthdXNj8DHaJcy0WMdZlbttfpOc91n85/Pfno2A7qTtzrGVVUkNbRQySKwJkvt9vrOdLPZUz02bP55GrqcQ0AGwtAHq3eQgO9P6T3fy7P89Uz1To7rKbHvOTcHE0/o3PcwgWVOsbW1n6D2eqzf/hEPPRIY/tN2Sa/sOPZmPe6BfaDXQyPpP9w3fo/6nqP/ANKrlOHljLOTkZb7GgvDKGNDKw1x/R7m+59ljGfnvf8A9todXUsq97W04Fzay6PUuioBo+lZ6b/0v9SvZ+kUGM+sVserZi4wAg+mH2kn979J6O3hvtQS6izG47Bi+l9k237C31DWx0Pj+c+l7/d7lbwasurHDcy4X3Ey5waGgT+Y3aG7mt/eVhJTzpwOuem5rX4e/wDNecMiCB+c313Nc1z/APwNaLqWOoDBiD14A3tra0btPe33bmNWikkp/9L1VJZexgwIrx5zBTDPWrs2eqG+31nsY52z1P5xzFnUU/WGf1pmE4bwQKaLmQzc3e39LVd+k2fQf/6UrsoSnpU3dUXsxy6v7PQ5tgsYd3pubAB9/vcG/mK93CSnOyuk+reb6cu7EdY4etseSH6bW7W2bmVP0r+g36H+eo3YnVW2WWVW49zHOc5rLqzWWN/NZ61LnOdt/fcz/wAwt51ODdVtzgw0sIsiww2W6Bzpj6O5ZlXTfq7kCtuNjHIY1pcy1he5hDXtY5v2h7vTe52xvsdZ+kp/4NGrU2cd+XbtFwbUTY3a2q71tzA0v9bdaz2tdYPY1WqXZVlQe9npkyHVWhu4QY9z6H3Ve76XtVE4XS6hZjMx6GbgPtNQO6w02bqj6rWe9lTttn53p/o1oCyRsx27gNA46MEafS/P/sIkUAgbsAL6f5tkNHLA4Fn9lztj6/8AN9NVcXPa51hrYYnaHGNrRLne8/v7n/zasvqNr/Tc8veIL3cNYP5DP9I783d/NotQDLrGN0bDXAfez/0WmpQsPqncW+s4HSXM2j+qxjn/APS96MTe6QWMgiDLif8AviqtqusybHZZ9MuJbQxu19Tmtc9zLP0le9mU+t36avf+Z+jUrw7Grda57A0ENb73Vje47K2Dc62vdZY5lbPYhaBLrSGcQV15eRdSx2O02NtuJJrlpZY/fbY3Z7HO3IhzMKl1jH5tdJqdFg9jIc4Nt/wm7c53qNd/bQf2j9ne6jIbbTW2W12vr9Sp7WkVbi/H/mff+Zc5Spv6bnHdX9ky3Ph+hbvMxDvTc17t36Jv/baKgKSuzumtc5tmezc3VwNrGx/W2bNqd2X0ltnpWZVYsEAsfdrJ4bDn/STtxMFmv2FjNZltbDqP6nuUvS6a55Lqqw9+jt7A0u4bHvaN30WpJSY+ThWH0ce2t7gCdjHAmA7Y920H/Se1O7MxGuLXX1hzTBBe0EEduUqsPEodvporqfG3cxjWmNPbLR/JQdmW2q2lrI3us22seAQHuc9rm7m/TbvSUl+24X/cir/Pb/el9uwv+5FX+e3+9ZDejdSioWZ+W8U6t22NZuk1PeLj+lda39Bsq/0dV93qPvf+kV/HrzqcMYz2m97Wlpue+HOJn3On1P8AqklP/9P1VJJJJSln59GX6hubnOxceGja1jXEuJ27W7mu/nNzWM2/pfUWghXw9pqNXrB4gtcBsj+Xu9qSnBq/Z7neljUv6zmBrGZF1jiGbQ8PBe+79Hs9Wne1lbX/AM2rT686+kttyBi1tLmlmGQ1gbI9Efa7Wb22N/m7PQr/AO2lbNlLrziEi3IYxrjjM9rGsdubW6z+R7Hf+kUdtIb+kucHPYJB4Ywf8G383+ulfZF9mvjYQFLKRLMetoYxhncWD6Ddfc1m3/S/pf8AwupsyNrRSz3SdtDz9Egef/Bf+DKtXmu6jNjGvrwWOArdoHZDoe19PpWM3fZ9233bqv5v/Q+srAY1291wDmN1sESHFv0amD86un/z8kAbU2a621t289y48knlzlF0DIaf3mOB+RaR/wB+QqnWCz0W2t3sbvtrd7ywO/mm/SbZ+a/3v/0aY2XudjuOxu4bp1MSAyI0/wBIiRSrTvdRaX0Oc0ua0OeyRuDSTsfH0me6t+x//Bqjbkvbfj0HdZXY8+haWyxzmgvay939n9BbX+f+l/wf6xU+0EuqZvdbebzh223taPUdUH5TNcL9B/JZ639T0/U9VXMynEyafQueGU5mtDyQ0tucPo1z+e/6bWt/4dA6H9qPPRE3p97LN3SrxghpLr8SxhsYXOO/cGeoz0t82O30fzqdlONmX2U5vSxU/wBzmWvbW5rwT9L1aydlrvpvqf8A+CJ2Y+Tn1OrzA/Hy8R7mV5dctDg9u31qf+Mof+kb/gb/AOb/AEtKljdSdTaMPqYbjWjSi0vBZcP3q3uj9L/pKn+9JIvq6UAANCdMPFOklSSSSSlJJJJKf//U9VSVSr7RcHv9csAse0NDWwA1xYPpB37qy8zreZi5D8duD1HKLXlgsoqqNZhrLN2+zZtY/fs3/wAhJTvrPysrqD8l2FhY7maDfm2R6bQ4E7qmauvs3e3Z/wBuIeFl5mVa6qyrJw3NBdN7KyDD3U+19Isq/M9T+d/m1am+rIqY631G2bgQWtHA3At27UlM8LF+yYlWN6jrTU2PUeZcSqF9d/U32V5bDjdNpJFjC6H2ljp9+36OI5rd/st/Tf4Rayq5D3Br7nVvfVQC4VMbL7HN19jPztv+D/4RIKYsZArZW30yG7aGf6Ose3fB/wAJt/1/nkKq2jMzHYrK91HTy13rNeQPX97fRcxh/Sekz32er/hPzE+ZmfYqN5HqZmQQKagCSSS1oG1su9HH37rX/wDk1FtJ6T0a0lzr7WMfZY/cA59hEuLXO9rNz/oo3WyEfTcizKozc0gsbdY5lDXiH7Wfom75DXt/Tep+id9BW8nSyRxXXu/zXsd/3xBxa7aOnYlbqS615rN+wVt2ucfWvtsaPQZ/OfT9Nnqf8GrT277rGxzUB/nF6R2Hmpw8svpyc9lWM2x9F1GdTWGbnvkNqvtpZU6r9Mxw+n/hP9Gtn0twspAZvrPq0Gxu8NLpdW/ZLPoWep9F9ajVjb8qvqAeQ51HpPZ2cCRaxx/lV+//ALcRrfZbXb2nY74Ojb/4JsQKih6Xk2ZeDVbaC26NtzHNNbg9vte11R3bEbKxcfMpNGQwWVnsex43MP5rv5Sohv2DqhgBmH1DUkua0DJ/daxxY5z8pn7nqe+taiSXLdkZHStjMjdkYAHuzHGX1mTAurYz+aazZ+n/AO3FpggiQZHiEnNa5pa4AtIgg6ggqrhdPrwn2eg9wofGzGMenWZe5/o6bm+pv+ikptpJJJKUkkkkp//V9PdiYr3Fz6a3OOpcWgk/gl9jw/8AQV/5jf7kZJJSH7Fh/wCgr/zG/wByevGx6nbq6mMdEbmtAMfJFSSUwstrqY6y17a62iXPcQAB/Kc5VOnWdQvNuRlt9Cqw/q+OR+kY0S0+uR7d7/pex6Fl4WR1DMFWS308DHcy1gaQ71ntLbB6rC3211ubt9P/AK5/xemkpg6mp9jLXsa6yqfTeRq3cNrtrvzdzVn9ccHVY+GRuGXexj2gS7Y0+q9zWj3e3Y3e9v8ANfzi01Sdi3WdUZk2hvpY9bm0EGTus2+rva5vs+h7PTf/AJn+ESkXWBut6ezcWu+1MezQkEta9xrdt9zdzN6sV2PPUbqzs2iqsiHS/wClb9OqPYz9x+73/wDW1V6k71Op9OxmvLCLHXPiOGsftbtcDu3+/wDqf9tKdbR+37nFpDjisG6QWuaHvjTZ6jX1u3/4T89JVNzHH6PbP0XOb9zjH/RTZWOMjHtoLnV+q0t31u2vbI+kx/5r2orWtbMCNxk/FOkoi9HNb/ljpAPuxrHya3Bwe+uyt36Ozd9Hex7N3/QsVjpuVblYdd11Zpt1bYxwIhzSa37d35jtu5ir4VZxuq5lIB9LIDchkNIa1x9l25/51lj/ANItJJSkC51pvrqreGBzHvJiT7TW0c/8YjqFlNNserW2zbxuAMT/AFklOXb1S6rLfiuqyi2tzGuyRQPS/SbIex/+FYz1P02z+a/SK+DfXfWx9m9tgdI2gcQeykMTFHFNY/sj+5Srx8et26upjHREtaAY+SSkiSSSSn//1vVUl8qpJKfqpJfKqSSn6qSXyqkkp+isnqOdU4iig5Xse4v+1Mpi0G3ZjGl3uYzeylj7f8Gy3/g1KnOy3bxeG47mtca3VZQvDnh+yuuHNqd+lZ+l/mfoWf6RfOaSSn6qWbVQ89evyXVua1tDaq7PzTr6j9fzvp7f+s2fQ/wnzOkkp+qkl8qpJKfqpJfKqSSn6qSXyqkkp+qkl8qpJKfqpJfKqSSn/9n/7R2UUGhvdG9zaG9wIDMuMAA4QklNBAQAAAAAAAccAgAAAgACADhCSU0EJQAAAAAAEEYM8okmuFbasJwBobCnkHc4QklNA+0AAAAAABAAyAAAAAEAAQDIAAAAAQABOEJJTQQmAAAAAAAOAAAAAAAAAAAAAD+AAAA4QklNBA0AAAAAAAQAAAAeOEJJTQQZAAAAAAAEAAAAHjhCSU0D8wAAAAAACQAAAAAAAAAAAQA4QklNBAoAAAAAAAEAADhCSU0nEAAAAAAACgABAAAAAAAAAAI4QklNA/UAAAAAAEgAL2ZmAAEAbGZmAAYAAAAAAAEAL2ZmAAEAoZmaAAYAAAAAAAEAMgAAAAEAWgAAAAYAAAAAAAEANQAAAAEALQAAAAYAAAAAAAE4QklNA/gAAAAAAHAAAP////////////////////////////8D6AAAAAD/////////////////////////////A+gAAAAA/////////////////////////////wPoAAAAAP////////////////////////////8D6AAAOEJJTQQIAAAAAAAQAAAAAQAAAkAAAAJAAAAAADhCSU0EHgAAAAAABAAAAAA4QklNBBoAAAAAA2EAAAAGAAAAAAAAAAAAAAQbAAAFeAAAABYAUwAwADEANAAwADUAMgA1AFgAMQAxADAAMAAwADQANAA2AF8AZgBpAGcANQAAAAEAAAAAAAAAAAAAAAAAAAAAAAAAAQAAAAAAAAAAAAAFeAAABBsAAAAAAAAAAAAAAAAAAAAAAQAAAAAAAAAAAAAAAAAAAAAAAAAQAAAAAQAAAAAAAG51bGwAAAACAAAABmJvdW5kc09iamMAAAABAAAAAAAAUmN0MQAAAAQAAAAAVG9wIGxvbmcAAAAAAAAAAExlZnRsb25nAAAAAAAAAABCdG9tbG9uZwAABBsAAAAAUmdodGxvbmcAAAV4AAAABnNsaWNlc1ZsTHMAAAABT2JqYwAAAAEAAAAAAAVzbGljZQAAABIAAAAHc2xpY2VJRGxvbmcAAAAAAAAAB2dyb3VwSURsb25nAAAAAAAAAAZvcmlnaW5lbnVtAAAADEVTbGljZU9yaWdpbgAAAA1hdXRvR2VuZXJhdGVkAAAAAFR5cGVlbnVtAAAACkVTbGljZVR5cGUAAAAASW1nIAAAAAZib3VuZHNPYmpjAAAAAQAAAAAAAFJjdDEAAAAEAAAAAFRvcCBsb25nAAAAAAAAAABMZWZ0bG9uZwAAAAAAAAAAQnRvbWxvbmcAAAQbAAAAAFJnaHRsb25nAAAFeAAAAAN1cmxURVhUAAAAAQAAAAAAAG51bGxURVhUAAAAAQAAAAAAAE1zZ2VURVhUAAAAAQAAAAAABmFsdFRhZ1RFWFQAAAABAAAAAAAOY2VsbFRleHRJc0hUTUxib29sAQAAAAhjZWxsVGV4dFRFWFQAAAABAAAAAAAJaG9yekFsaWduZW51bQAAAA9FU2xpY2VIb3J6QWxpZ24AAAAHZGVmYXVsdAAAAAl2ZXJ0QWxpZ25lbnVtAAAAD0VTbGljZVZlcnRBbGlnbgAAAAdkZWZhdWx0AAAAC2JnQ29sb3JUeXBlZW51bQAAABFFU2xpY2VCR0NvbG9yVHlwZQAAAABOb25lAAAACXRvcE91dHNldGxvbmcAAAAAAAAACmxlZnRPdXRzZXRsb25nAAAAAAAAAAxib3R0b21PdXRzZXRsb25nAAAAAAAAAAtyaWdodE91dHNldGxvbmcAAAAAADhCSU0EKAAAAAAADAAAAAE/8AAAAAAAADhCSU0EEQAAAAAAAQEAOEJJTQQUAAAAAAAEAAAAAzhCSU0EDAAAAAAWYwAAAAEAAACgAAAAeAAAAeAAAOEAAAAWRwAYAAH/2P/gABBKRklGAAECAQBIAEgAAP/tAAxBZG9iZV9DTQAB/+4ADkFkb2JlAGSAAAAAAf/bAIQADAgICAkIDAkJDBELCgsRFQ8MDA8VGBMTFRMTGBEMDAwMDAwRDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAENCwsNDg0QDg4QFA4ODhQUDg4ODhQRDAwMDAwREQwMDAwMDBEMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwM/8AAEQgAeACgAwEiAAIRAQMRAf/dAAQACv/EAT8AAAEFAQEBAQEBAAAAAAAAAAMAAQIEBQYHCAkKCwEAAQUBAQEBAQEAAAAAAAAAAQACAwQFBgcICQoLEAABBAEDAgQCBQcGCAUDDDMBAAIRAwQhEjEFQVFhEyJxgTIGFJGhsUIjJBVSwWIzNHKC0UMHJZJT8OHxY3M1FqKygyZEk1RkRcKjdDYX0lXiZfKzhMPTdePzRieUpIW0lcTU5PSltcXV5fVWZnaGlqa2xtbm9jdHV2d3h5ent8fX5/cRAAICAQIEBAMEBQYHBwYFNQEAAhEDITESBEFRYXEiEwUygZEUobFCI8FS0fAzJGLhcoKSQ1MVY3M08SUGFqKygwcmNcLSRJNUoxdkRVU2dGXi8rOEw9N14/NGlKSFtJXE1OT0pbXF1eX1VmZ2hpamtsbW5vYnN0dXZ3eHl6e3x//aAAwDAQACEQMRAD8A9VQDmUhzmje4tO0ltb3CR/KYxzUdVW1Zdbnit1Za95eNwdPu1jQpKZ/bKv3bf+2rP/SajZkl7DXjksveCKjbW9rdwG7lzWrMx/qzTjXMvpJ9Vjw8OfY587d+xjmWBzNn6Z/u2/aLLP01l/r+rbZqCrJfbW60s21kuhoMkkFn5x/lJKRdKzvteN+kMZNJNeRWY3Ne0lvuDP39u9WX5FDLq6H2NbdbPp1lwDnbRufsZ9J21Uc/FdjW/tLCqe+5gPrY9RDRc36XvaQ7faz/AAft9V/816iWZWeo4NWbgudXkVj1sV0QZI1qeyz/AErfY7ckp0SAQQRIOhBWJ0nPGLgU4b63G6vJdhFgIJY47rq3WOea97G0+51jP5xXqer4jsGvLvsbQHEVvDzG22djqPdDt+9Z3W2ZLXZvua1jqq8nGfZcRFlDv0vp1PsrZWzZ6f8AN+n+k/nbf0n6NKdttzfQF1jgxobve4mAABLpJ/dUK83DucGVX12OMw1r2uJj6X0T+aljursqMe9hJ5HLXj1R/wBGxBxbm2Oa37FZjgS5rntYAHRH+Ce/a7a5JTateWNBbG4uDRPmYU0F8G6tvAbLz/1Df+rRdfikpdDtvrqLWv3EukgNa55gc6Vtd+8pyg3V3G1ltJbLQ5pD54dtP5v9RJSvtlP7tv8A21Z/6TS+2U/u2/8AbVn/AKTWVb9XDZdZe2++q255sc5mVcGhztvqenUPY3dWxtX/AAX5iCPqkNha/My3vI2mwZVrT+dsLQAdvo7/ANF/npKdyvKqssFY3B5BcA5j2SBAd/ONb+8jKvXVkG9llpZtra5oDZJO4s5Lv6isJKUkkkkp/9D1VJV2ZbrGNsZj2FrwHNPsGh1HNig7qFbSA5haToAX1A/SFX+m/wBK5tf/ABiSm2kgHKLS3fTYwOcG7jtIBdo2dj3ORXFwaS0bndgTE/NJTJVG014TrLqmxVc825A1J3mN14/zf0lf+tkrr7nUvGK1pyWiRVadvfvH/nH8tZNedl5FjuoYLLH2sa1mVgXbmwAHu24LnenRY/1W2fpXfzn+lr/m0qQRf02bt2FubkYws24+aJpDAGBjwNz/AHU7Hv8AXj1fe/8A0ibBy2ZVDLLQPtmE/wBHKkN3Ndoy7Vo2+nZ7b/Z7E+PlYuRS4UuP2fdse3h+PaDPpWM/wW1//bT/APgf5sjWfpjadrBZNOZW1oG6whjab3W/zntrb6dbf9Hf/wAElHekeDPD9RjfTe/1LAxofY4Bu97N1Nlm2sbGb9jPoJ8cdSDh9qdQ5sGRU14M6Rq9z1GlzhcwuMuMh3hMbbP+njf+CKONViDJsdTkW2WVE+pW61zgJ3N27Hn6O5E7lI2SDJqGcaHSbLG+wAEgNZyXuA217nve1u/6fpqz8FgMzKqMvM6tbYPTrY3EoYDO+0u9V7WN/e9e70/Z/wCi0YnqT8bH6W3IBz9jXZuQCN9THbnB1bPb6m51foNd/wBctQS7PPOhTNcCAQZB1BHBCyMvLb1BtmHRaWYYrByc9phvuLNtNT/b6vr0us/S0P8A0X+k9X9Gnbn72NwOhVNIqbtbkPBGPWGQ3bu+la9u78z/ANKJKdeRMTqeydUqOnY9OS/Oue+7I98W2ukMY47vSqYNtVbGN9v0PUVtj22N3N1HYwRPwSUySSSSUpJJJJT/AP/R9If099mI7CtsZbjPrNL631yHVlvpuY+Ht+mz6SqU/Vnp9BDqaq63BwslotB3gl3qu/WPdb7rP0jve/1b/wDT3LYSSU07ZFtNWRksBseHV17Q1ziz3w33O/tJs/Hzi77RhXbba2+2l8ljo923RzNvqfQ3v3/8H6aPfiY2SAL6m2QCBuEwDG4f2trVm/sUUGMS3KxmMO5jKrQ9k/vPZf73/wBXckpk/Ny6ttfU8MxLQ3Ixd1o3uMaMa0ZFP0v5xPfTidSra/f9oGO93pZNJi2p4mtzhs+l/Z/7bUXv6xjip1d9eTt3C4ZLDRukt2+nYxjmN9OHf10Brce3MbZk9LfXmlhcMrGh7Po2x+nZs/Sfznpb/wDCel/waSlV4ebbmtc6A8Abuo0hmy+tprD8XMxnn+kWe/8AN2Vf4NXaS+yA2tw31gPpt0e1p/wWQ5he3fS7+atrdb6rPU/nP59BNND8JjLmutrfXWHVZjCXvPs9NmQ5w9K67fs+n+k9X/DK9Ufs7dl3tJMm0klpJ/lu9zP3Wep/00Brr2WD1Uxso2MNj7HPc0hx2+0AS31Iaz3e5rPzkB11Rc6v7LdUSdhtcIbryd7HlyvvYHsdWeHAg/MLMdZnl+N9otqNVzw3Yyt5d7g4bTYHP2ezd731+mnakrmdmOPXqc6mXUtD67XNZ/O27qnMoeP0nqta3db61VlX8x/1uOPh1V0W1ve8236Z2TLmWudGwV1ljvUqd/o24z9lW/8AVf51X3VfpxkOtcGsYWio7dgJ19b6Pqept9n856exV8wC9hfXuZDS197TshjtHvqfq71K/wCcqsa3+2jYv+CNaLWycrFFoxdthdXNj8DHaJcy0WMdZlbttfpOc91n85/Pfno2A7qTtzrGVVUkNbRQySKwJkvt9vrOdLPZUz02bP55GrqcQ0AGwtAHq3eQgO9P6T3fy7P89Uz1To7rKbHvOTcHE0/o3PcwgWVOsbW1n6D2eqzf/hEPPRIY/tN2Sa/sOPZmPe6BfaDXQyPpP9w3fo/6nqP/ANKrlOHljLOTkZb7GgvDKGNDKw1x/R7m+59ljGfnvf8A9todXUsq97W04Fzay6PUuioBo+lZ6b/0v9SvZ+kUGM+sVserZi4wAg+mH2kn979J6O3hvtQS6izG47Bi+l9k237C31DWx0Pj+c+l7/d7lbwasurHDcy4X3Ey5waGgT+Y3aG7mt/eVhJTzpwOuem5rX4e/wDNecMiCB+c313Nc1z/APwNaLqWOoDBiD14A3tra0btPe33bmNWikkp/9L1VJZexgwIrx5zBTDPWrs2eqG+31nsY52z1P5xzFnUU/WGf1pmE4bwQKaLmQzc3e39LVd+k2fQf/6UrsoSnpU3dUXsxy6v7PQ5tgsYd3pubAB9/vcG/mK93CSnOyuk+reb6cu7EdY4etseSH6bW7W2bmVP0r+g36H+eo3YnVW2WWVW49zHOc5rLqzWWN/NZ61LnOdt/fcz/wAwt51ODdVtzgw0sIsiww2W6Bzpj6O5ZlXTfq7kCtuNjHIY1pcy1he5hDXtY5v2h7vTe52xvsdZ+kp/4NGrU2cd+XbtFwbUTY3a2q71tzA0v9bdaz2tdYPY1WqXZVlQe9npkyHVWhu4QY9z6H3Ve76XtVE4XS6hZjMx6GbgPtNQO6w02bqj6rWe9lTttn53p/o1oCyRsx27gNA46MEafS/P/sIkUAgbsAL6f5tkNHLA4Fn9lztj6/8AN9NVcXPa51hrYYnaHGNrRLne8/v7n/zasvqNr/Tc8veIL3cNYP5DP9I783d/NotQDLrGN0bDXAfez/0WmpQsPqncW+s4HSXM2j+qxjn/APS96MTe6QWMgiDLif8AviqtqusybHZZ9MuJbQxu19Tmtc9zLP0le9mU+t36avf+Z+jUrw7Grda57A0ENb73Vje47K2Dc62vdZY5lbPYhaBLrSGcQV15eRdSx2O02NtuJJrlpZY/fbY3Z7HO3IhzMKl1jH5tdJqdFg9jIc4Nt/wm7c53qNd/bQf2j9ne6jIbbTW2W12vr9Sp7WkVbi/H/mff+Zc5Spv6bnHdX9ky3Ph+hbvMxDvTc17t36Jv/baKgKSuzumtc5tmezc3VwNrGx/W2bNqd2X0ltnpWZVYsEAsfdrJ4bDn/STtxMFmv2FjNZltbDqP6nuUvS6a55Lqqw9+jt7A0u4bHvaN30WpJSY+ThWH0ce2t7gCdjHAmA7Y920H/Se1O7MxGuLXX1hzTBBe0EEduUqsPEodvporqfG3cxjWmNPbLR/JQdmW2q2lrI3us22seAQHuc9rm7m/TbvSUl+24X/cir/Pb/el9uwv+5FX+e3+9ZDejdSioWZ+W8U6t22NZuk1PeLj+lda39Bsq/0dV93qPvf+kV/HrzqcMYz2m97Wlpue+HOJn3On1P8AqklP/9P1VJJJJSln59GX6hubnOxceGja1jXEuJ27W7mu/nNzWM2/pfUWghXw9pqNXrB4gtcBsj+Xu9qSnBq/Z7neljUv6zmBrGZF1jiGbQ8PBe+79Hs9Wne1lbX/AM2rT686+kttyBi1tLmlmGQ1gbI9Efa7Wb22N/m7PQr/AO2lbNlLrziEi3IYxrjjM9rGsdubW6z+R7Hf+kUdtIb+kucHPYJB4Ywf8G383+ulfZF9mvjYQFLKRLMetoYxhncWD6Ddfc1m3/S/pf8AwupsyNrRSz3SdtDz9Egef/Bf+DKtXmu6jNjGvrwWOArdoHZDoe19PpWM3fZ9233bqv5v/Q+srAY1291wDmN1sESHFv0amD86un/z8kAbU2a621t289y48knlzlF0DIaf3mOB+RaR/wB+QqnWCz0W2t3sbvtrd7ywO/mm/SbZ+a/3v/0aY2XudjuOxu4bp1MSAyI0/wBIiRSrTvdRaX0Oc0ua0OeyRuDSTsfH0me6t+x//Bqjbkvbfj0HdZXY8+haWyxzmgvay939n9BbX+f+l/wf6xU+0EuqZvdbebzh223taPUdUH5TNcL9B/JZ639T0/U9VXMynEyafQueGU5mtDyQ0tucPo1z+e/6bWt/4dA6H9qPPRE3p97LN3SrxghpLr8SxhsYXOO/cGeoz0t82O30fzqdlONmX2U5vSxU/wBzmWvbW5rwT9L1aydlrvpvqf8A+CJ2Y+Tn1OrzA/Hy8R7mV5dctDg9u31qf+Mof+kb/gb/AOb/AEtKljdSdTaMPqYbjWjSi0vBZcP3q3uj9L/pKn+9JIvq6UAANCdMPFOklSSSSSlJJJJKf//U9VSVSr7RcHv9csAse0NDWwA1xYPpB37qy8zreZi5D8duD1HKLXlgsoqqNZhrLN2+zZtY/fs3/wAhJTvrPysrqD8l2FhY7maDfm2R6bQ4E7qmauvs3e3Z/wBuIeFl5mVa6qyrJw3NBdN7KyDD3U+19Isq/M9T+d/m1am+rIqY631G2bgQWtHA3At27UlM8LF+yYlWN6jrTU2PUeZcSqF9d/U32V5bDjdNpJFjC6H2ljp9+36OI5rd/st/Tf4Rayq5D3Br7nVvfVQC4VMbL7HN19jPztv+D/4RIKYsZArZW30yG7aGf6Ose3fB/wAJt/1/nkKq2jMzHYrK91HTy13rNeQPX97fRcxh/Sekz32er/hPzE+ZmfYqN5HqZmQQKagCSSS1oG1su9HH37rX/wDk1FtJ6T0a0lzr7WMfZY/cA59hEuLXO9rNz/oo3WyEfTcizKozc0gsbdY5lDXiH7Wfom75DXt/Tep+id9BW8nSyRxXXu/zXsd/3xBxa7aOnYlbqS615rN+wVt2ucfWvtsaPQZ/OfT9Nnqf8GrT277rGxzUB/nF6R2Hmpw8svpyc9lWM2x9F1GdTWGbnvkNqvtpZU6r9Mxw+n/hP9Gtn0twspAZvrPq0Gxu8NLpdW/ZLPoWep9F9ajVjb8qvqAeQ51HpPZ2cCRaxx/lV+//ALcRrfZbXb2nY74Ojb/4JsQKih6Xk2ZeDVbaC26NtzHNNbg9vte11R3bEbKxcfMpNGQwWVnsex43MP5rv5Sohv2DqhgBmH1DUkua0DJ/daxxY5z8pn7nqe+taiSXLdkZHStjMjdkYAHuzHGX1mTAurYz+aazZ+n/AO3FpggiQZHiEnNa5pa4AtIgg6ggqrhdPrwn2eg9wofGzGMenWZe5/o6bm+pv+ikptpJJJKUkkkkp//V9PdiYr3Fz6a3OOpcWgk/gl9jw/8AQV/5jf7kZJJSH7Fh/wCgr/zG/wByevGx6nbq6mMdEbmtAMfJFSSUwstrqY6y17a62iXPcQAB/Kc5VOnWdQvNuRlt9Cqw/q+OR+kY0S0+uR7d7/pex6Fl4WR1DMFWS308DHcy1gaQ71ntLbB6rC3211ubt9P/AK5/xemkpg6mp9jLXsa6yqfTeRq3cNrtrvzdzVn9ccHVY+GRuGXexj2gS7Y0+q9zWj3e3Y3e9v8ANfzi01Sdi3WdUZk2hvpY9bm0EGTus2+rva5vs+h7PTf/AJn+ESkXWBut6ezcWu+1MezQkEta9xrdt9zdzN6sV2PPUbqzs2iqsiHS/wClb9OqPYz9x+73/wDW1V6k71Op9OxmvLCLHXPiOGsftbtcDu3+/wDqf9tKdbR+37nFpDjisG6QWuaHvjTZ6jX1u3/4T89JVNzHH6PbP0XOb9zjH/RTZWOMjHtoLnV+q0t31u2vbI+kx/5r2orWtbMCNxk/FOkoi9HNb/ljpAPuxrHya3Bwe+uyt36Ozd9Hex7N3/QsVjpuVblYdd11Zpt1bYxwIhzSa37d35jtu5ir4VZxuq5lIB9LIDchkNIa1x9l25/51lj/ANItJJSkC51pvrqreGBzHvJiT7TW0c/8YjqFlNNserW2zbxuAMT/AFklOXb1S6rLfiuqyi2tzGuyRQPS/SbIex/+FYz1P02z+a/SK+DfXfWx9m9tgdI2gcQeykMTFHFNY/sj+5Srx8et26upjHREtaAY+SSkiSSSSn//1vVUl8qpJKfqpJfKqSSn6qSXyqkkp+isnqOdU4iig5Xse4v+1Mpi0G3ZjGl3uYzeylj7f8Gy3/g1KnOy3bxeG47mtca3VZQvDnh+yuuHNqd+lZ+l/mfoWf6RfOaSSn6qWbVQ89evyXVua1tDaq7PzTr6j9fzvp7f+s2fQ/wnzOkkp+qkl8qpJKfqpJfKqSSn6qSXyqkkp+qkl8qpJKfqpJfKqSSn/9kAOEJJTQQhAAAAAABTAAAAAQEAAAAPAEEAZABvAGIAZQAgAFAAaABvAHQAbwBzAGgAbwBwAAAAEgBBAGQAbwBiAGUAIABQAGgAbwB0AG8AcwBoAG8AcAAgAEMAUwAAAAEAOEJJTQ+gAAAAAAEMbWFuaUlSRlIAAAEAOEJJTUFuRHMAAADgAAAAEAAAAAEAAAAAAABudWxsAAAAAwAAAABBRlN0bG9uZwAAAAAAAAAARnJJblZsTHMAAAABT2JqYwAAAAEAAAAAAABudWxsAAAAAgAAAABGcklEbG9uZ1YP6rIAAAAARnJHQWRvdWJAPgAAAAAAAAAAAABGU3RzVmxMcwAAAAFPYmpjAAAAAQAAAAAAAG51bGwAAAAEAAAAAEZzSURsb25nAAAAAAAAAABBRnJtbG9uZwAAAAAAAAAARnNGclZsTHMAAAABbG9uZ1YP6rIAAAAATENudGxvbmcAAAAAAAA4QklNUm9sbAAAAAgAAAAAAAAAADhCSU0PoQAAAAAAHG1mcmkAAAACAAAAEAAAAAEAAAAAAAAAAQAAAAA4QklNBAYAAAAAAAcACAAAAAEBAP/hHRhodHRwOi8vbnMuYWRvYmUuY29tL3hhcC8xLjAvADw/eHBhY2tldCBiZWdpbj0n77u/JyBpZD0nVzVNME1wQ2VoaUh6cmVTek5UY3prYzlkJz8+Cjx4OnhtcG1ldGEgeG1sbnM6eD0nYWRvYmU6bnM6bWV0YS8nIHg6eG1wdGs9J1hNUCB0b29sa2l0IDMuMC0yOCwgZnJhbWV3b3JrIDEuNic+CjxyZGY6UkRGIHhtbG5zOnJkZj0naHR0cDovL3d3dy53My5vcmcvMTk5OS8wMi8yMi1yZGYtc3ludGF4LW5zIycgeG1sbnM6aVg9J2h0dHA6Ly9ucy5hZG9iZS5jb20vaVgvMS4wLyc+CgogPHJkZjpEZXNjcmlwdGlvbiByZGY6YWJvdXQ9J3V1aWQ6NDFlNWU1ZmYtYTBlMi0xMWUxLWJkMjctY2RhYTNlMDhhODM0JwogIHhtbG5zOmV4aWY9J2h0dHA6Ly9ucy5hZG9iZS5jb20vZXhpZi8xLjAvJz4KICA8ZXhpZjpDb2xvclNwYWNlPjQyOTQ5NjcyOTU8L2V4aWY6Q29sb3JTcGFjZT4KICA8ZXhpZjpQaXhlbFhEaW1lbnNpb24+MTQwMDwvZXhpZjpQaXhlbFhEaW1lbnNpb24+CiAgPGV4aWY6UGl4ZWxZRGltZW5zaW9uPjEwNTE8L2V4aWY6UGl4ZWxZRGltZW5zaW9uPgogPC9yZGY6RGVzY3JpcHRpb24+CgogPHJkZjpEZXNjcmlwdGlvbiByZGY6YWJvdXQ9J3V1aWQ6NDFlNWU1ZmYtYTBlMi0xMWUxLWJkMjctY2RhYTNlMDhhODM0JwogIHhtbG5zOnBkZj0naHR0cDovL25zLmFkb2JlLmNvbS9wZGYvMS4zLyc+CiA8L3JkZjpEZXNjcmlwdGlvbj4KCiA8cmRmOkRlc2NyaXB0aW9uIHJkZjphYm91dD0ndXVpZDo0MWU1ZTVmZi1hMGUyLTExZTEtYmQyNy1jZGFhM2UwOGE4MzQnCiAgeG1sbnM6cGhvdG9zaG9wPSdodHRwOi8vbnMuYWRvYmUuY29tL3Bob3Rvc2hvcC8xLjAvJz4KICA8cGhvdG9zaG9wOkNvbG9yTW9kZT40PC9waG90b3Nob3A6Q29sb3JNb2RlPgogIDxwaG90b3Nob3A6SUNDUHJvZmlsZT5VLlMuIFdlYiBDb2F0ZWQgKFNXT1ApIHYyPC9waG90b3Nob3A6SUNDUHJvZmlsZT4KICA8cGhvdG9zaG9wOkhpc3Rvcnk+PC9waG90b3Nob3A6SGlzdG9yeT4KIDwvcmRmOkRlc2NyaXB0aW9uPgoKIDxyZGY6RGVzY3JpcHRpb24gcmRmOmFib3V0PSd1dWlkOjQxZTVlNWZmLWEwZTItMTFlMS1iZDI3LWNkYWEzZTA4YTgzNCcKICB4bWxuczp0aWZmPSdodHRwOi8vbnMuYWRvYmUuY29tL3RpZmYvMS4wLyc+CiAgPHRpZmY6T3JpZW50YXRpb24+MTwvdGlmZjpPcmllbnRhdGlvbj4KICA8dGlmZjpYUmVzb2x1dGlvbj4yMDAvMTwvdGlmZjpYUmVzb2x1dGlvbj4KICA8dGlmZjpZUmVzb2x1dGlvbj4yMDAvMTwvdGlmZjpZUmVzb2x1dGlvbj4KICA8dGlmZjpSZXNvbHV0aW9uVW5pdD4yPC90aWZmOlJlc29sdXRpb25Vbml0PgogPC9yZGY6RGVzY3JpcHRpb24+CgogPHJkZjpEZXNjcmlwdGlvbiByZGY6YWJvdXQ9J3V1aWQ6NDFlNWU1ZmYtYTBlMi0xMWUxLWJkMjctY2RhYTNlMDhhODM0JwogIHhtbG5zOnhhcD0naHR0cDovL25zLmFkb2JlLmNvbS94YXAvMS4wLyc+CiAgPHhhcDpDcmVhdGVEYXRlPjIwMTItMDUtMThUMTc6Mzk6MDQrMDU6MzA8L3hhcDpDcmVhdGVEYXRlPgogIDx4YXA6TW9kaWZ5RGF0ZT4yMDEyLTA1LTE4VDE3OjM5OjA0KzA1OjMwPC94YXA6TW9kaWZ5RGF0ZT4KICA8eGFwOk1ldGFkYXRhRGF0ZT4yMDEyLTA1LTE4VDE3OjM5OjA0KzA1OjMwPC94YXA6TWV0YWRhdGFEYXRlPgogIDx4YXA6Q3JlYXRvclRvb2w+QWRvYmUgUGhvdG9zaG9wIENTIFdpbmRvd3M8L3hhcDpDcmVhdG9yVG9vbD4KIDwvcmRmOkRlc2NyaXB0aW9uPgoKIDxyZGY6RGVzY3JpcHRpb24gcmRmOmFib3V0PSd1dWlkOjQxZTVlNWZmLWEwZTItMTFlMS1iZDI3LWNkYWEzZTA4YTgzNCcKICB4bWxuczpzdEV2dD0naHR0cDovL25zLmFkb2JlLmNvbS94YXAvMS4wL3NUeXBlL1Jlc291cmNlRXZlbnQjJwogIHhtbG5zOnN0UmVmPSdodHRwOi8vbnMuYWRvYmUuY29tL3hhcC8xLjAvc1R5cGUvUmVzb3VyY2VSZWYjJwogIHhtbG5zOnhhcE1NPSdodHRwOi8vbnMuYWRvYmUuY29tL3hhcC8xLjAvbW0vJz4KICA8eGFwTU06SW5zdGFuY2VJRD54bXAuaWlkOjIyQTU4NkMzNUY1OEUxMTFBQzVEOUU2NUExQkQ5QThEPC94YXBNTTpJbnN0YW5jZUlEPgogIDx4YXBNTTpPcmlnaW5hbERvY3VtZW50SUQ+eG1wLmRpZDoyMUE1ODZDMzVGNThFMTExQUM1RDlFNjVBMUJEOUE4RDwveGFwTU06T3JpZ2luYWxEb2N1bWVudElEPgogIDx4YXBNTTpIaXN0b3J5PgogICA8cmRmOlNlcT4KICAgIDxyZGY6bGkgcmRmOnBhcnNlVHlwZT0nUmVzb3VyY2UnPgogICAgIDxzdEV2dDphY3Rpb24+Y3JlYXRlZDwvc3RFdnQ6YWN0aW9uPgogICAgIDxzdEV2dDppbnN0YW5jZUlEPnhtcC5paWQ6MjFBNTg2QzM1RjU4RTExMUFDNUQ5RTY1QTFCRDlBOEQ8L3N0RXZ0Omluc3RhbmNlSUQ+CiAgICAgPHN0RXZ0OndoZW4+MjAxMi0wMi0xNlQxMDo0OTozNCswNTozMDwvc3RFdnQ6d2hlbj4KICAgICA8c3RFdnQ6c29mdHdhcmVBZ2VudD5BZG9iZSBQaG90b3Nob3AgQ1M1LjEgV2luZG93czwvc3RFdnQ6c29mdHdhcmVBZ2VudD4KICAgIDwvcmRmOmxpPgogICAgPHJkZjpsaSByZGY6cGFyc2VUeXBlPSdSZXNvdXJjZSc+CiAgICAgPHN0RXZ0OmFjdGlvbj5zYXZlZDwvc3RFdnQ6YWN0aW9uPgogICAgIDxzdEV2dDppbnN0YW5jZUlEPnhtcC5paWQ6MjJBNTg2QzM1RjU4RTExMUFDNUQ5RTY1QTFCRDlBOEQ8L3N0RXZ0Omluc3RhbmNlSUQ+CiAgICAgPHN0RXZ0OndoZW4+MjAxMi0wMi0xNlQxMTowNjoyNCswNTozMDwvc3RFdnQ6d2hlbj4KICAgICA8c3RFdnQ6c29mdHdhcmVBZ2VudD5BZG9iZSBQaG90b3Nob3AgQ1M1LjEgV2luZG93czwvc3RFdnQ6c29mdHdhcmVBZ2VudD4KICAgICA8c3RFdnQ6Y2hhbmdlZD4vPC9zdEV2dDpjaGFuZ2VkPgogICAgPC9yZGY6bGk+CiAgIDwvcmRmOlNlcT4KICA8L3hhcE1NOkhpc3Rvcnk+CiAgPHhhcE1NOkRlcml2ZWRGcm9tIHJkZjpwYXJzZVR5cGU9J1Jlc291cmNlJz4KICAgPHN0UmVmOmRvY3VtZW50SUQ+eG1wLmRpZDoyMUE1ODZDMzVGNThFMTExQUM1RDlFNjVBMUJEOUE4RDwvc3RSZWY6ZG9jdW1lbnRJRD4KICA8L3hhcE1NOkRlcml2ZWRGcm9tPgogIDx4YXBNTTpEb2N1bWVudElEPmFkb2JlOmRvY2lkOnBob3Rvc2hvcDo0MWU1ZTVmZS1hMGUyLTExZTEtYmQyNy1jZGFhM2UwOGE4MzQ8L3hhcE1NOkRvY3VtZW50SUQ+CiA8L3JkZjpEZXNjcmlwdGlvbj4KCiA8cmRmOkRlc2NyaXB0aW9uIHJkZjphYm91dD0ndXVpZDo0MWU1ZTVmZi1hMGUyLTExZTEtYmQyNy1jZGFhM2UwOGE4MzQnCiAgeG1sbnM6ZGM9J2h0dHA6Ly9wdXJsLm9yZy9kYy9lbGVtZW50cy8xLjEvJz4KICA8ZGM6Zm9ybWF0PmltYWdlL2pwZWc8L2RjOmZvcm1hdD4KIDwvcmRmOkRlc2NyaXB0aW9uPgoKPC9yZGY6UkRGPgo8L3g6eG1wbWV0YT4KICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCjw/eHBhY2tldCBlbmQ9J3cnPz7/7gAOQWRvYmUAZEAAAAAB/9sAhAABAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAgEBAQEBAQICAgICAgICAgICAgICAwMDAwMDAwMDAwMDAwMDAQEBAQEBAQIBAQIDAgICAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwP/wAARCAQbBXgDAREAAhEBAxEB/90ABACv/8QBogAAAAYCAwEAAAAAAAAAAAAABwgGBQQJAwoCAQALAQAABgMBAQEAAAAAAAAAAAAGBQQDBwIIAQkACgsQAAIBAwQBAwMCAwMDAgYJdQECAwQRBRIGIQcTIgAIMRRBMiMVCVFCFmEkMxdScYEYYpElQ6Gx8CY0cgoZwdE1J+FTNoLxkqJEVHNFRjdHYyhVVlcassLS4vJkg3SThGWjs8PT4yk4ZvN1Kjk6SElKWFlaZ2hpanZ3eHl6hYaHiImKlJWWl5iZmqSlpqeoqaq0tba3uLm6xMXGx8jJytTV1tfY2drk5ebn6Onq9PX29/j5+hEAAgEDAgQEAwUEBAQGBgVtAQIDEQQhEgUxBgAiE0FRBzJhFHEIQoEjkRVSoWIWMwmxJMHRQ3LwF+GCNCWSUxhjRPGisiY1GVQ2RWQnCnODk0Z0wtLi8lVldVY3hIWjs8PT4/MpGpSktMTU5PSVpbXF1eX1KEdXZjh2hpamtsbW5vZnd4eXp7fH1+f3SFhoeIiYqLjI2Oj4OUlZaXmJmam5ydnp+So6SlpqeoqaqrrK2ur6/9oADAMBAAIRAxEAPwDf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdEB/mGdh/ILrbqbHZz480FFkNz/xORa+Ksx8GRUUaqhRo4KhWF7lvx795+nXjXSdIqetdrfv88f5JfGnJbdw/e+yah6rcGbTDY96HbdLSCaoaUQaUKQWPrP493p5qemTIwIDrTo1+Z/4UB7f6Xpaev7i6g7A+2aghr2anxrY1JIJYfKpjeSAA3/B9+o5NBmvXmdFGt+0D1B6up+D/AM1uvPnX0zje6et8NmsHgshVy0QoM4Y2rIp4YhJIGeJVBHPHHutD59OBlYAqagioPRy/eurde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/9Df49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdR6ikpatPHVU1PUx/6iohjmTn6+mQEe/de61v/wDhQRVdL7e6k6j3bVJsabeXW2/Jd10+3oIMV/ePJQUT08hpVpogJCpdG9LC5JNh9fdlB1UHTczaY9fEggj7eib/AB8/2bn+dBBhd1722HtLZnxsxggwKU+R2fi8PmYqTHwiN1hnljjklZxyg1Ef0sB78QB3A1PXqtKNLigP51/zdbQ/xk+MfWnxQ6zx/VnVtFNR7doZfuiKjxeSWseERzzWiUBQxFwvNvpc+9VJ49XAUYUUA6MT711vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6/9Hf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690Gnavb/XXSu1MhvPsrdmE2jgqCmqZzXZyujoaeaSCBplp45ZOC720qPyT791onqtj4g/zgOgvl12jmOrNvUcu1szRZOpx+JkzGSiYbhjhZliq8bGI0uklvSb/n6+7FSBUileqLLG7FUNacfl1biCCAQbggEH+oPIPuvTnXfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6iV61b0VUtCypWGGQUzv+hZtP7Za/4v8AX37r3WpV80fj71Vjfl1nO3vnPX5jfmycPVNnNrbF2dmIWyVVXwTh4aaoxzlgIma1108g+9gsBRfz6acJr1v5cK+Xz6DXuD+f/mNjvs/rj41/F3fHWuyaeuTFRVlLtv7ehjpqYinSrq5KanVLFFBZzyfqSffgATUkD5deaRgKIrEepFP9nrZ7+F3dVZ3z0Ds7sLL5Kkr83mIZGyC08oZ4JVjRzDKnBDLq54966drUAjz6Nf7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//0t/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690Tn5hfNvpj4a7ByG7+x9w4sZeOEtidoffJDm8zLJCzwfa04DNoLKAX025sDf37iadaLKo1Ngda00W0Plx/PO7HfO17ZvrP4e4LJUMWe2rmayfF1eWAkZYarAmUIZ9SpclRb+vPu57cDpkAzCpqoP7T/AJh0Pv8AMU/libE+GXTuzfkh8Q6nL7Q7J6tm29gKqvq8gpSsx0SlzkJBAqHyt42D/X8c+9AkmhNR1Z0VVLIAKDq7b+Xj8n4PlL8dNp7rm8r7j27i8Rt3d9VJI0grdw01DprKuMsAQHKFje/JP491oQSDxHTisHUOvA8Oj2e/db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuq/fkn8W/h1lN0jvH5AzU2Iq6eRJGymZzQo8SzU+myvTPGwa3puPeqeh63g8QD1Wf8xf5m/xF2b1duzqvqnqXDdpR5jbuR2vhMntDC4qrpqap8X2kVXqp6YuXUesMHDauSfdqaum2kCijHI8hnow/wDJW2d2DjvjzT7t3JR5PBbeztfkZMPt7NRzw19P5GSXziKYD0WbSD781CajryVpn/iurq/eur9e9+691737r3Xvfuvde9+690A3fW7c7s/bLZPCSNFJGl2kBZQpu31K/wCsPfuveXTJ8c+wsvvzC1lRmasVNTFZgQztpAfSb6/9f36pNa9aAA4dGU9+631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SB7K3O20tp5HLo4jkijdY2N+HMZZbEf63vxNM9aNOHRbvjt2lu7fGYyEOZqHnokqJfEdTsoU6io9XvxNT16hBr5cejn+/db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//T3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Wvz/ADt/5deQ+Uu06TuTbf8AEK7cXXtHQAYTH1Diauo6OV55GjpvoxFrG3Jv9OOLK5U8MEU6aliEqj1U1HS1/ly/zNvjVP0Ph+s+z8ztroHc/TeGxezJ8RvCthw824v4XTtDNlMfTFFLMxQNKoJJZi3591IIND1dXVxUcfMedeiJfzL/AOYdUfNPK0Hwd+I+Brd7VG581i8pUdr7aknyW34aal1R1FDMsaaVsHbUzHi3+v7uAAem2fVVB5/y+09WM9U57aP8tP4e7VpNq4ap7f3xla3AQbzwG1plqKmk3HUQ+LJSzxUmt1WMsy2IHq/w90Jqanz6dA0KFHAdW7bB3Q+9dl7X3bLjKnDSbiwePy8mKrAVqce9bAJXpJw1jqQmxuPfut+dOlf7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuqlvmr2D8aO+Mtu7409nCrzNVsHF1G59w47C5J6XJwUKQrJVqdNgoCqCSb+/Z8utYPaw49UOdafElu8d5ZDbPwC25mev8Ht6oqq6PdW9p0yOLqZlkOtVr6xdF7X9PuxP8Wem9I/Bjo/Hx0+WXyb+HPdeK6I+Wde+7Mbnq+m29gKnHQmnx1NWShaaGSkNMniPqtfjke9HgSOrBiWCt+R9cefWykja0Vx/aVW/5KF/eur9cvfuvde9+691737r3XvfuvdBN3biVzHXecpjGJJBEHj4BKkAg2v8A6/v3XqdFH+I+VbFZbLYOWQammaFYyTexlH0B9+qeHXqA9WI+/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917oo3y03IlLss4OCQx1dTKHve3pZLAC3+v+feievU66+Jm3kodoS5CRAZ5Xj0yWFzq1E8/X6e9461Q9G69+631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//1N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691weNJUaOREkjcFXR1DoysLMrK3BBHBB9+691Tl8qv5MHxF+Qu7X7My2DrtvZ9DPW5ipx+TTHYt40QyVFU9PCihfSCW/HF+PewxUY6o0aORqA6qZ3F2X0H8S91SfE3+WX1rmdz9o7lL0ue7zophuGHDZCrj8NVQROsZ8XiYks2oW/wB597zWrcP5/l1TCgKgz6DgPt6z9Y/KXen8rvc1V133zgD8nu5Oyp6LdrUeBrJK+o2vSV7Gpq1mVllVZkDEuNF1t798VTSg6t2xgK5LE18v9WB1sn/FL5QbE+VHW9PvvZclPTmLwU+XwaOWqcFWyoWbH1QIHrQgqbAC496Ip1cEHh0aD3rrfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdaqny8o6v4r/wAwLvDv/efWud3zsTuXZddsuglp5pkx+Pmy9KtFNWkpcAxFi1j72KkUHVH7TrNaUp+zqvP43fzae6/ir23uPo7q34v7739sIvU1GKzWIoKqqo1qapifItTTRtqYD6gn3s0rk9UUscKCR8ujQba+N3z8+dPyY2l3bvM5rZvXuG3FR7nxu18/SS0NRjY1Za7xAVAUnjj3vGkgefn146vFUtinl69bheOWoTH0KVXNUlHTJUmwAM6wKJiAP9qv7p08Opnv3XuqMvnN8u/lL0P2AsfXfX28N27Z+6UEYamrZYWiMrAjVACBwPfgc0r1pq0qq16r52L/AMKMOyP725XYNf8AD/fO4MthJTBXy08mU+4jdW0EyJ42C8gk8e7kGlcdNq66tNGr6U6O9sT+e51jldx4La3Z3WmV6ky2emENHT7mqp43kZnEY0I8aE8m3utD1bxFrp4H5jq7nFZrFdk7EpM5iKiKpxW4MaaujqIj5IpYmB0shP1BI96FD1c1HHqujq6uO2O862hYNHEcoV5J0sPNf6Dj+vvRr1oD08urUVYOoZTdWFwR+R+D731vrl7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de66JsCf6An/be/de6rb+U2UOZ3zjsJTvddcMbJcnVwFPH0+o9+IoK9aBzXo53TeBbAbKxtMyaDLBDL9AL3U88e/dbrXoV/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/1d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SU3tvjafXO28lvDe+dodt7axEYlyWYyLtHSUkZNg0rIGP+2Hv3XuPRbaf58fDyrhinpu/thyxTKHjdayrsyni4vD79Tr3Wf8A2e74ifT/AE8bGv8A9RlT/wBevfuvU8uvf7Pd8RL2HfOxSf6feVV/9t4vfvn17rx+d3xEAue+Ni/+dtTf/beL37rdD10Pnf8AEM/TvjY3/nZUj/e4vfuvU8uu/wDZ7/iHe3+nnYt/6feVR/3qL37yr1o4NOuj87/iGPr3xsb/AM7Koj/biL37rdOuz87/AIhj6987F/8AO2p/69e/dep17/Z7viIf+a87F/r/AMDan/r1791r5deHzv8AiGeR3zsUj+v3lV/169+OOPXhkVHXv9nv+Id7f6edi3/p95U/9evfvKvXqZp14/O74hgXPfOxR/1WVP8A169+GevZHXX+z3/EO9v9POxv/Oyqt/t/F791uhrTrv8A2e/4iXI/087FuP8Aptqf978Xv3XqHr3+z3fET/n/ABsX/wA7an/r17916h66/wBnv+IlyP8ATxsbj6/5ZU/9evfuvUPXf+z3fETj/jPOxef+m2p/69e/da69/s9/xDvb/TzsW/8A1G1P/Xr37r3nTrr/AGe/4h8/8Z52KbfW1ZU8f6/7Xv1Ovde/2fD4hf8AP+ti/wDnZU/9evfqHr3XH/Z8viD5I4f9PWxfLKVVEFZVEkswRfpD+Sbe/dbp0aPBZzE7lxGPz2CroMnh8rTrV4+vpiWgqqZyQk0TEAkGx/Hv3WiKYPTt7917poz+JjzuHyWHnNoMnRVNBUWJBanq4TBOlx9LqxHv3WiK9a43z86z2j/LJ6Xy3Yfxj2hk8HvXe3YGOnzW9Kh48jHSQ5FjJV08DupeO5Dfkce/AVIB6o1UQsAK/wCz59Ea773rsjY/8xP42fIKPMY7feO3v0Btr++GMp7SpFu3dG3jR5GOWCS4DpLJqvbg+7AEqevOVR1HqCP5inV0v8uz4l7x+MuUzO4arMNQbH7MqZ95wYiSUTBDlXespInEZshAkXgj6H3UknqygLXq49WDAMPoRcf4g/Q+/dW679+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690HfZXVOxO3MC22t/YGlz2IZzJ9vOqhlYixKS2LLf82Pv3XvOvVPvz8+RvUP8rfrraGK6e6nxlfvjtbLz7f26sFJBXVeNrGkSmSoRpEZ9bNIQoJ/Hvag9NuVUV+wUHnXqifrv5M/zUO1O+uxaKq7B3Ts6HYOKm3TU7Oq6eajqKigt9xT0cVKqr9UKqAB9PdjoAFD02rTkkOAKeVM/t62cv5dPy+yXyL2J/d3etFUUHY+2Kdm3BDWGVaqSNNESzyxy83ZiT/xHupFOnkOoV8/Pqyz3rq3USteihgkqK7wLTxDVJJOqsiL9NTFgffqde60tflx859ofDn5qbmfpfY9J3ruPsPck8eVw23KGEz4aWSsaNoJ6enjb1BvwB7uAWFfIeZ6ZZlDac6j5Dz+fRkuhv5WW/fnN2lgvlV8mKGs2ltqDMR5TB7AyKvjMrS06zrUfZrAkd1UA8M4APutaDHVvDDPqfy8v8/W05tHaGB2NtjE7P2zR/YYHCUS0GNow5k+3plvpTW3J+p+vvXDpzqtfteE7N7gx1fEugVNYsjMBYNqk/Vx7rU168KUNOrMNrVf3+3sRWX1fcUccl/63J92rXPXjjp/9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdRa6YU1FV1DfSGmmlP8AyBGWt78OPXuqtMi0u/u7oFpwzLT1aKV5Nyr/AE9XvXlp60ONT5dWjYqn+1xtBTadPgpIIitrWKRhTwP8fe+t/Z1P9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690y53cWE2zQS5PPZKmxdBAjSTVNUxWNET9TMQDwL+/de6A/wD2bX44anT/AEu7RDRu0bg1kt1dDZlP7f1B9+63SvXY+WnxwP07d2j/AOdkv/Xv37rxFOu/9mz+OP8Az93aP/nZL/0Z7916nzHXX+zafHDn/jL20OPr/lsv+9aPfuvaT11/s2vxwtf/AEvbR5/6bJf968fv3XqeXXv9m1+OH/P3do/+dkv/AF796r1vSevf7Np8cD9O3ton6fSsl/P/ACB731rSevH5a/HAfXt3aI/16yUf9Ee/deoePXv9m1+OH/P3tof+dkv/AEZ79wFevUPXf+zafHA8f6Xtoc/9Nkv/AEZ7916nXv8AZs/jj9P9L20P/O2X/oz37r1Pn10flr8cBf8A4y9tHj6gVkpP+2Efv3z69Tr3+za/HAi/+l7aFv8AqMl/6M9+69Q9dn5afHAfXt7aAv8A9Nkv/Rnv3Xqde/2bT44/T/S7tG/1t95L9P8Akj37r1D14/LT44A2/wBLu0b/APUZL/179+68Fr17/ZtPjha/+l7aFr2/4GS/X/kj37r1Ouv9m1+OH/P3tof+dkv/AEZ7916nXf8As2nxxvb/AEu7Rv8A0+8l/wCjPfuvU66Hy1+OBNh27tEn/qMl/wB78fv3W9J65f7Nn8cf+fu7R/8AOyX/AKM9+6117/Zs/jj/AM/d2h/52S/9Ge/dep1x/wBm1+OH/P3tof8AnZL/ANGe/db0nrsfLP44nkdu7R/87Jf+jPfutU69/s2nxxvb/S7tG/5/yyXj/X9HvVet6T1xPy3+N4YJ/pe2iWY2CismJPNvxH/j731qnQ47f3FhN1Yqnze3sjT5XFVYJpq6lYtDMByShIH9R+PfutdPXv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6/9bf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691TN/OOyebqenKTZEGQeDbm54o/43Qq8kYrDHVukep4yPoP6+9rx6o+BXpL/Gv+VN8Ud1fHzqbP5HaNXJmcrtqGvyVUMvNad5nKqAFBtYqfyfr71Uniet6FXAHDoaj/KE+JRtfaNbdf0/7l5/+jfe6n169oWtadeH8oP4lA3G0ay5/P8AFp+P9b0+/At5nr2lfIddf8NB/Eq5P90azk8gZef/AKN96qfM9b0geXXj/KD+JZ/5hGr/ANjl6j/o334lvI9a0qeIHXf/AA0H8Sv+eSrf/PtOR/vXv1T69eKg8QOux/KE+JdiP7o1v/n3n/6N9+z69e0j064j+UH8Sv8Ankaz/wA+89v+hffu7zPXgqjyHXf/AA0H8Sr3G0q0fj/i7z/9G+/VPCvXtK8aDrr/AIaD+JQN/wC6NZ/595x/sP0+/Vb169pHoOu/+Gg/iVe/90a2/wD2t5/9h/Z9+q3rjr2kenXv+Gg/iT/zyNZ/595/r9fpp9+z69eCgeQ67H8oX4lj6bRrP/PvP/ven3up9evaR6dcf+GgviV/zyNZ/wCfae9/9t71VvXr2lfMDrv/AIaD+JVx/v0azg3H+5ac/T/XHv2fI9e0g+Q66P8AKD+Jf1/ujV/7DKzj/eh78C/mevaFHkOu/wDhoP4lf88jWf8An3qLf7Eafe6/Pr2kenXv+Gg/iTb/AI9Cs/1/4vPf/D8e/VPr17SvoOvf8NCfEsAgbRrACLE/xee/+v8Ap96/Pr2lT5Drr/hoL4kg3G0q3/z7T/73b3ureuOvaE406Kp81/5Y3xm6p+Om9uwNqbaq6Lc2AbHTUFU+UmdERpgjAjgk6rH3rUR1rw1PEdWD/wAuXLZeu+OGxqPJVrVcOOwsMFCrFm8EPmY6NTkk8k+9nHVwaivR/Peuvde9+690EPd/SPX/AMgdgZXrrsjDRZvb+SR3EEhCtT1ohaOmrImINnjLahx/r+/fPr3lT1613d5fyxtq/EHcmc+UHefZOL3p11tAyja+yZTPFUPVJC8uBxavOi6iGRV0xt/vHu1SwpSnTYjVHMlSfkfL7Ogr6gw/8xH+ZjR7j7a2H2dk/jp17sGsn2/tfZuRlyNHBlsbSwvJjWxqxK2pWRB+4xC/Tn37AND1saiNVaA/IHo5f8uf515TZfbtZ8HvkpvtMv3RBLU1OMrstPUGuqKekgdxHqkJBSRVDITb3oqBw68rk4byH+o9bBCsrqrqQysAysPoVYXBHvXV+u/fuvde9+691737r3Xvfuvde9+691737r3XvfuvdJndO8ds7Kx0mW3Tl6TDY6IEyVVWxWNQObkgH37r3RLd8fzPfgv105g3Z8hNmYurNxFSSPWPPK9yAqKkVjyLfqHv3Xs+XRBu9f573THWG1M1vbZeCpt/7XwnhaqzVNUVcVIscr6db6bEfj/b+7BSTTpozIASfL0z1Z78M/lZtv5hdKbW7i21Rx4+i3JQ/eCiSR5RAPJ49OqTn6/191ODTp3BFR0bT37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdUf/wA434t7m7X291T3FsnB1O4sv0XuFt8T4OkHlqMsMbVx10VOkOltVyukgfj3YH16oy8CONa9VGfLzvn5Jb72xsb5T7B6e3H13vvteqG0N8YukoJI5cPj8IqUMGQqVAEixyqt7yfX35aE938uqOHC6k4+h6tb/lQV/S2Ahy2Uync+C3H3vumAUu59oqWTJ42MBJyZgBY+oW4964nHTigqKnz/AMPV7HvXVuig/ObYHZHZnxt7F2h1XvxOt93ZTFpHjd2SSvBHi3WQs8zzR8gEcce/UJ4daJp9nWqd8dd5/G/4JZ7LZrNbOxXzZ+VmPkY53NYarlql2/mnqS7T1EMiP5HVlC3t+T9D7uSzfZ02oSP4sk+dM9XkfCH+YX8iPlbveWh3z8dM91PtymmP29TV0lfDTPGJFQXlrBduDf6/63uhxwNerqSc0I+2nV0/v3Vuq7vlxiI6TO4jKBdAj8DB+P66ja3P1HvxNDnrXE0HRsOkdwpndi4gKdRpaZIy3NyCSeb+/VHl1uhHHoYffuvde9+691737r3Xvfuvde9+691wkkSKN5ZGCRxozux+ioo1Mx/1h7917oh8fz66nrvlA3xew2Qo8lvGKBJpkiaXyodIaQab6eAb+/E0+zquocOg6+UP8xjG9B9if6Nds7Gn7D3FRItZnqSgkqFlw2JADT5CcQ39KKSx+nA9++zrZNOhLrvnh1kPj/Td34eeDLmsZqKlwMJkEs2aRLyYtWB1agfTf+vvxI8uvCvE9JD4k/P/ABnyR3Jk9o7h2fJ11uOhWWSDF18s7zVUa8xMDNa2scjj34GvXga9H+qtz4KhyUWIqslTwZGdokhpXa0rtNzGFH+Pv3XqjolHyi+cm3/jjuPbu2zt+TclZl61abJeF5E/hETSaPPMI/xbnn34Z62cdLvOfLvY2OxPXuQoJIchUb+q46Oko1Lh45pAvoWx+vP59+63THRrqColqqSGeeBqaWRFZ4X+qEi5Hv32dVHUz37rfXvfuvde9+691737r3Xvfuvde9+691737r3Qc9qZ0be2bk64tpvDLDc/83IW9+4daORTojPxfw02X3/XbknOtRWO6kkHgOSPrf8Ap711unVlnvfXuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6qF/mxVO9K3rvam3dp7nbbH8XzD4+vnDyRipp6h0GhmiINh/j79ivWs0I6BHpL+VPgcx1rh8nnt0w5LL5QR5Gashq5pVIqIhI6MVBsSTex/23v1SevKiqM9CmP5SWywdX8dGo8X81T9P9YL/wAT79qbr2hOuX/DSmzeP9zy8f0mquf+TffiST17QnXX/DSezLk/x5ef+b1Vwf8AC6+9VbreleHXv+Gk9mH654G30/eqv9549+q3WgiD/V/s9cT/ACkdmEn/AHPABuD+9U/S31+n+9+/VbrwRBw65L/KS2WgsudXn+s9UD/vC+91br2haU66b+Ulsw/8v4f9Tqn/AKN96q3XtCUp1xH8pDZakEZ/+v8Au6o/6N97q1OvaE8+uQ/lI7M1Bv48Ljnmap+p/wBZffqsOvFIz1yP8pPZhIb+OqCL/wC76s/72vvVT1vSgx1w/wCGkNlG988Dfk/vVPH+F9PvdWr1oItKddj+UjssD/i+g8/QzVJFv9fT79VuvBVHDrx/lJbMP1zw4H4mqf6/4j3qrde0IDUde/4aS2YSD/HV/pxNUn/e19+q3XgqDh13/wANJbM1a/46twLWE9Va3+JK3971MOHWvDTro/ykdlkaf46LXvfzVV7/AJ4029+qet6F64j+Ufsrj/c8ODf/AD1Rz/tl9+1N17SvXI/yktmatQzwBIsQJqm1v+Sb+/VPXtCVz15f5SWzFJIzq/S3M9UB/sLL78CR17Qhz1yP8pTZv/O+H+wmqeP8eR71VuvaI/8AUOuh/KT2bz/udUn/ABmqv9uLL78CfPrehPLrif5SOzD/AMv4f9Tqn/inv1W69pTrkP5SezVBAzykW4vNVXH9foPdgzda8NOuv+Gktmck54XP/N6p/p/wX3qrdb0oOHQKfIf+VHicN1PuvMbZ3fDicxj8d5KWtlqqiNY3Di7amUc/QAX97HVWVSMdDB/KEn7AxW0d37E3pu5t1QbWVIsfKZJJUi1VCRsUeQm4sLe9GletrXTnq6D37rfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/19/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3VLn84S39xdrkmxEa8f9V7e9jB6q47K9WIfD03+MfSx/rsmgP/AFmk91HDq7fF+z/AOjJ+99a697917r3v3Xuve/de697917r3v3XuotZWU9BA9TVyrDBGLvI7KqqP8bn/AHr3sAnh1okDJ6i4rM43NwNU4ypjqoVbQzxspAb+hAN/evn14EE06cXdY1LMQAPrcqv+8sQP959+63jz6bHzmMjYq9XEpBIN5I/x9fz73QjqutD59S6aupawXp5kk/wVlJ/2w9+II68GDcOsWSylDiYDVZCdKeAG3kdlAva9gCb+9U62WA49c6DIUmTpY6yilWemlv45UIKtY2NrH/e/eyCOPXgQeHU33rrfXvfuvde9+691737r3Xvfuvde9+690Q3+ZX/2SB2Xb+mM4/r/AJV9Pfutjpo/lvG/x62n/wBqmC/+B8p49+8+qjh1YT791vr3v3XuoOTMq43INBfzrQ1Zh0/XyiBjHb/G9vfuvdavHaee3N8gewt5fEr5A9iHaznKZvd+2qLMmdTaigaopI4nf0gOqrpHvdaCo6bIDVBNB/k6Jn1x/OU7D+LVFmvhDsDqc9sdnVWXr8DturolrRkKTHVSHHxZCOCj4m0KQ8d/of8Abe7BCV19N+Kit4ABJH+D16WWy+otj/CjsPH/AMwX5g7voNz/ACf3CrPtTpzNvP8A3hSlydKfs0rIk5VU1hFDH8e/Ekii/wCr7et4Wskp44p/kHWyd8B/kP2F8mOmW7G7C67qOtquoyz0+Fw88c8a1eEMImo6+Baj1aGUgKf8PdDg06eX4QfXy9Ojw+/db697917r3v3Xuve/de697917r3v3Xuve/de6Ib85fiZvv5W7RoNsbO7QPXKU6FKt/HVulZeQvd/tQTwDb36p60QCKGo+zrXP+aP8kjB9KbI2p39vHKQ9sYTqlErt+YWOqrKKXPxGqDIFWUKdJ+hINxf25rY4HTP08YYNk06LHk92/FP5+5rpX4q/DroSm2vt/c5o8R3pLja2rrP7q1Yq0geoyc82oIg+pube9UKnPWqrOAEFAfPh+X29blvxT+NGxvih0tsnpzY1LHHj9pYePGSViFy1fIsrSvUNrAtcngW4HHtvpQejI+99e697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917pCdm02arNg7ro9vSPFmqrD1NPjpY0V2jnlXSGCsCPpcfT37r3WuZkf5Wfzs7m3Llc3XfKubY21KusqLberFrpz4HnMqxpSU8TKotxew/1/e644dV0gnLED5U6BHF/yyu9PgN8k+ru09qZKr7qbsPddNj901uHTIIMJTGeNKmproJFXSpBLhvp9R+PdtepdJx02sISTWlTXjU9bbEVzFEWFmMaah/Q6Rce6dPdAl8kdobh350pv3aW1ax6DPZvDyUmPq0/VDMXD6hb/AAFvfq0691qhfB3ur4IfDfuvu7FfJLC4jF9svWSU8e4sqJqt87Xx1xaRY4eQjOykarf193IJAPl0yrIpKmlf8nVpWB/nEUu6uyNt7G6h6Wj3VtfL1cdJ/H8YtTDFSRmQRmRYoLKLAg8j3WhpXq6umrSM9Xk7cylRmsJjspVUjUFRWU6zS0jX1QMx/Qb+9dXPRWvljt1sjtpckFLLSIoJH4ILH37Jz6darQ9c/iTmErdq1VGGu1MqXH9NL6SD/t/eh1vh0b33vr3WCWqp4ReWVU/1/wDb/j3dY3b4R1QyIOJ6hvmcXGLvWQrxe1zc/wCsLe7i3mJoFPVPHi/i6jS5+hVdUTia/wBNJ4t/X24tpITRsdUNytKr1Bk3HIpGiieQH8gn8/Tj26tmpwX6oLongvRGP5gPyZ3z0L0tUZbZOyajc2V3HNW7feOFZnbGwVCCEZAeKx41E+6vbLHRg1eti4ZlJ08Otab45b3xmyvle+9twbebJ9xZaPzpuOSadqimir01iiMTH6qCFv70Yo8CvWkkYk9uT1aX0jLtrcnyv76zm/qNBues6I3JJNDUnVIKeXFMSSk3A9P0I59tyJGnahr05G7sxJHVWvW8+epqjYOLgjqJOqD3K7Q04knakM5ygEltXpva3492KRGtPTptWlrkYqerMPl9TxYX5Q7Jm6DonpNwTx7V/jH8NLKzo0MZkXTBbi173918OIIMivTjNLrqo6Ln3xu35j0vzDwVPTUm5ExMeUw2hVFYIWTTG2qwNuf8fftCGpJ68TKGWgx59WU/Kv4p7j7B27iO1MrWNj6jFUNBkd1eWTSwiSFGqfKrc8MSD7t4cdQNXVS8ms46r32V3Tg+m9yLvPeW3V3jtPCSyVfXNLMZmpzkaZhGpgBNjytzb35oEJpqp17xGpqYdX//ABv+SUHyA2BQbxp8C+Hq6lJHqcUWkZ6Xxxh7sXubG/Htz6WOldfVTdOGpox0PsW4i/66Vo7fX9Rv/rG3vRtOND1s3NADTj1PjzuPZbyzLD/XUePbZtZfwivV1uYzxPUhMtjZP83VxOf6KTf/AHr3Q28w4rTq/jxHgw6mRzRS/wCbdW/1j7bKMvEdXDqeB6y+69W697917r3v3XuikfLDc32ezmwcTeOeqkVtXPKsnA4/1/fiQB16lesPxJ26tBtGpyE8Yaed4tMjckX1Mbe/Dh1oVrQ+XRvffut9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691Ub/ADSf+Pd6+/P+/mhv/reRffhTV1o8PzHR9/jWAOpds2/5UqYWtb/lFT37z62OFeh79+691737r3Xvfuvde9+691xdwg1H6fk3VQP9ixA9+6915WDqGH0PINwbj/XUke/de+zrl7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuuiQouf97A/3v37r1eulcMLgj/bqf+hSffuvdcvfuvdcHkWMXYgD/FlX/eWI9+69UeZ67VlcXUgj/Ag/7yt/fuvdcvfuvde9+691737r3Xvfuvde9+690Xj5TWPR++NX/KhHb/X1n+vuw4dVNa46r2/lWC1T2ib3GtP/AHMX3UjNfl1YGqn7erkPfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//9Df49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691S3/ADg7f3G2uCL/ALS/7H/L2497XjTqsnwV6sS+H3/ZMnS//hlUH/W6T3UcOrt8X7P8A6Ml731rr3v3Xuve/de697917r3v3Xug07i7IxXUfWW+exMvJGtPtLam4M/HFLe1XUYjEy5CGjFrcyNGF+o+vvw60aefWhD1p8lv5gH8/wA783jjOiuzs78YevNpbiym35MviavKUuEUUtzG9R9k8heR41Dafrz9Pdsaanj01+oZSqmij9pr/g6WPyl61/mafyPTtL5Jbn+RG6vktseu3PjdrZDa1PX5qoxkSBRNVZKqp6orwEu5v9B9ffgBUADPXnLqCxaqjjUD+XVsHzm/mIbn+Sn8oOh+T3RO7KrYmfra6bbWdrsJJKkmLzQwkU0tmjfUpVyx06uPp72qgvpbr0kjfTGWPBpUV6ob/lq/FD5xfMX4yV/dGY/miZnaFbU74yWHgwWb3Dmp66OFY9RmV1f9Oq5tYab2HAHvVACRStOthiQCWK1FcAGnW0J8bNs9rfyzPhb2v3z3d8gm+Scezdj5zcuLqZ6urqoshV4mlLik81QwcAlCTpa9vejnIFOrg5AJJB86da/nxwh/mG/z781uXvvqn5E7i+NnWcmazOHo6GCqzNPg4KjGVDNJTwwUDPd7AKbc+7ELprxJ6bUylyPhUYHCp+ZPR9/hj8ZP5pPwb+Vibe7F7I338h+r66uixzZNp8xWYKCCQCCStT7n0rp/Vz712Ba+fWx42uhNUPmcHrb/AISzQxM4s5ijLg/UMVBYH/Y+69O9Zffuvde9+691737r3Xvfuvde9+690Q3+ZX/2SB2Zzb04yx/x+59+690z/wAtw3+PW1L/AF/hUF/9fyMffjx60pqMdWFe/db697917r3v3Xuq3vmv/Lu2Z8qpIt37dykXXvb0CCli3/EtQ8/8OEAgNK8NOCTwP1f7378CQajrxAYaT/LoCejv5XXTPxF2rvbvfNYSh7X+R219mboyOE7HqFqlroJKDCTVFJBR0kulNZkAJZ0Y2HFj79XNevKAo0DIPrx6qg/lq9M7d+f3ZfZ/yl+cm4qfO5Tr7dmXoMV1hueolppDQ40vJTZplLRsaeDSPRHybf092JxpXhSp+fTKAavFc1NSB8qH/Vno5fyT/m0z0/Y+1vjh8IOv4+2IcPkcZhMrWYGmrWXG0OOKU01PRKhAEUIVgXI5AufetJpU4HVzINYRRU+foPmer7ets8+d2dtqrr51OcqsDiq3MUbTRPNQ5CppVespHVGJBjk1If8AW966sCPLpfe/db697917r3v3Xuve/de697917r3v3Xuve/de6ps/nN9m9h4744Z3pPrjZVbuXPdvYxMbBkKJZZZcakddedUjjB9RCLY/gE+9j4vl1VmZVJUVPp1WD8OaH4rfyYfjbR9hZpMTmPl531goqqo661tUZSWtFX51p6x4rvCzMF1AFWP6eefdmJdqj/UOmo1igj0k1H869W4/y2flB8ivlMN7b37k6oyPWOAk/wAo2sJ4KqnpMkhqRH46b7n9QCkng+6sADSvTiEsuoilfLz6tg966v1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddMVUFmIUAcsSAAP8SffuvdFv3H8t/j5tPcx2hn+xsPj9wibwfw+RnaTy6tGjWgK3v8A4+9nHHrQIPDof8Rl8fncdS5bFVMdZj62JZqaoiYMksbcqwt/X3rrfU9wzIwRtDEcNYNY/wCsffuvda6P8wbo/wDl1fHzeZ7O7z6kxHcHbm952ye1dpVdfU0kmTq3ncDikKkXcfm97+7AEinVG0g62Ap9lf2dEj6u/mQVPRufpNw038t7E9T9XpMry7ranys1JjaR5lQTJNWeljYA8n6+7BGrStT1UOrLqIIA+zraa6D752T3/wBXba7L2VX01bjs7jFrjBAQppWBKyQshJtpI4597EMtAWFK9e8aPIBrTPXfbBbcmz8ljGpfHE9j9yeQlkNz9D7VC2VSKt+XTJnYnC8Oi7/HWqpsDlq3A46pFRNNJ4pdJH18ljck/X8/T351gjNFNeta5W4rx6OfLiMrUMxGRkp1ubAXP549++ohUUC168YJG/Fx68m3ahgBU17Tj83B/wBb35rxa1RadeFqQtCa9OK4DGWXy0ySsBbU1/x9Pp7YN1NWqmnTi20YGRnrOmHxqfopI1/1r/0t/X3U3EzcW6cEUY8upS0dKoAWFBb6cX/x/Puhkc+fWxGi8B0w7l2ZtjeFGKDcmHpMrRgk/b1Kkx3b6myke6lmIpXqwAHDooGQ+AfRdR2/Tdt0W26OgyMKwa6WEuIzJERyilSAOOAT71U9eI9Ogp+SP8u49vdhJ2L112A/WGaq6NMTnXo4qhzlcLoEU2ObwLwrLdeSeDz70R6dWxjiOhGqvgT1qnQtD09hkgxeUxshyNFulEZpoc64vJkgunVy3qt9b/n3s1pTr328Ok/8WvgjJ0ZunJb03/vtu0Nx1KslBX1sM6tQgECEgVAuSiiw596A9etCgJp59HvyGzNs5XKwZvIYikqcpTGMwVskamaMw/5ohv8AD8X97rinXvn0me1Ni1vYm2K3a0WVOOx+Vp5KTKRgXFVTSWJjY2J/HveOtZ8uiY9tfy7evux+rdr9eY+sjwNRtgzSU2WRZHJmmkMhcKiX+p/PupqTUda0gih6M18c+hsV0F15jNmU9UmXr6RXWszRV1krAwUKpV7WAt/T3ap63QdDuaKkIIMCc/4H3fxZPXqpjQ8R1GfD42T9dJG1/wAHVb/e/dhcTDIbqvgxenUSTb1B9aaJac3v6b2/23tz6uU/Ga9Nm2jzQcemx9tVvkDw5R4QD9Br/wBt7eS8QAh1rXqjWzEUVqdZfsMrTXAqpZzbhwSSeLW9+MsD5pTrXhyp59eORy1OvNG82k/XnkXtz70Y4HagNOva5UyR1yj3FLyZ6NogqliTf+yLn3VrRfwtXraXJJo4p0Qr5K1GR3bujHwUcLmjiaBH0n03AC8/7b2xJbyoaEY9enhPFxDZ6ON0zgmwOy8fTtwZYYpCLg8gEE+2SpXj06DUdCz791vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuqjP5pBY7f6/UAj/fyxXI/I8i8e/efWjwNej9fG1dPUu2P8aGm/8AcVPfvPrfl0PPv3Xuve/de697917r3v3XutIf/hRx8+Pkam+dg9G/EjeWewG5p6lsJmDtueop6iOubIFBK8lG5YnTfgj3dVBOeFOmZJGUUXBr6VNP9Xn1Yv8A8J6vmnv3tzYm4vjd3duyu3P3F1NjpJM/W5dpJMnUyfcQq4nlqHLsVDNxa/HvTKFwOHHq0blwWcUINOjg/Mb+c/8AH34a9n4HrXsnJ43GV2cqftonq/OzltRXTGIpBcgj6W916sWA49FJ21/wpR+JU/ZkGyd6ZrC7XxGQq2gxWYdK1pq2Ifoljhkksb/0A9200FT1QShjReI6U22f+FFXxZz/AH9ienHy2Hp8fncl9lh83orFqK2EzCIVCQySBbci/vRUgV62JFLafMfs6OH8jv5svTnx57V616+zstI9D2XRU+SwmUlWbXPQVUoiiqI1SQLYn/X96yTQdW1qACfM9DF8uv5hfVfxR6b2v23uKtpKij3ZQ0eRxMEpPjqKOsACS3ikDAXP4v78Kk062WAFT0AHcH84748dJfF3H/Irfeax2GjzFFR5DFYypWb7eppa6PXBKXEmsAngW9+68TSvy6IxtP8A4Ux/FTMbWTPZrIYTE1NZWUUGKpNNf46+GucJFMksklubi3Pu5Qrx6aWZWFV/n1a9l/5jvSO1fi43yi3dnKTDbQloxUUUjKWiqJZKc1EEYUyavUP6H3WnTpP8uPWtT8wP+FInWHcnUe4sJ1XvGi2FPT5qtxVLurHCrM80rxeCli8ksnoYv+F/J93CnieHSdp1NRH8Xljo838mP5w5Xavwk7N7m+S3aFfuimp9yfd4HLZdpqmR6NIZZPt4i7lrMCDwfdWADkLw6ciZjEjSEVNanh59S8Z/wpY+PNRv+DbuRpMPQbXn3AmBj3Cxrhrnao8OlQ8oGq3Nre/FCBXrQnQvoHH+XWb+ev8ALLdp+IPVfa/xq7Or9v0G48pJ5szhpJoJJVqKeOaKnkaBgQQCeLnj35AC4U8OtTu6wu0ZowGPPpd/y5PnvT9K/wArzGd9/JTsCXP5hN1ZHG0lflzUT1eQcU8Rip+W1aVIck6uLj35h3kL1eN6wIz8aCvl0leq/wDhSN8c989j4/ZudTEbfxGVzC4bH5ofeN9xUNIUACyy2v8AT8e/FCoqetLMjtpTrY+2nuvB722/jNzbcrosjiMrSU9XSVUJujx1EKzoPzzpYXF/denelH7917r3v3Xuve/de6Lz8prf6D983/5UI7f6+s+7ClM9UPHqvT+VWCKntL+mtLf0/wCBik/7z7qeP5dXWmkgevVyPv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6/9Hf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691S5/ODBOxdsW/wCOQ/25r297WleqyfB1Yj8Pv+yZOl//AAyaD/rdJ70Ort8X7P8AAOjJe/da697917r3v3Xuve/de697917ojP8AMa2RuPf3xH7fwu2I5ZcjFsvdORaOHXqalo9uVUtQPRzbTf36tOtMCRQdafv/AAj47m2ftTKd+9AZbIRUnYGa7PzmWgxUwijnNNjca0U+lp9LkloyNKi5t9PezWlfI9UUVbSeKgdW3f8ACpbvXYHXHwVx209xZeCPMbv3mMRj6BFjqahaqrxjRUsrxITIvLGxC/1JsLe7Jh1Pz6rNR4JIxxp1S70h03vDpj/hNxuLbO9WqRkd6dz5rcODep8oc47JYRHp5IxMAdKggi3uyj9Q/M9Nuf8AFBwwuR6Z6Ll/KD/kqfLH5MfERu0ev/lXX9b7fl7GzOPg2pTVGVSLyQQI1RUAQAqCdaM/+Lcce6k0JGMevVwoaNXyNQ8v9nrZn+VHxM7W+PH8kHubond29KztXeGD2FvbM5LccrVUrCjkxDQJSB5BrILHVyPz715k9XNdKp58Pt6KB/wjy7X2o3wBy/Sv3ejfGK7U33uiuxkgiimhxtdVqsMjJIRKSdP+psP9b3qhAr1sOHPaeGPz6uv7v/m4fFTo/wCRFH8adz71gg7Hny9Lh6jFmn84gq6l1jSMzU7kA3YCzC4/IHv2KVpw69UhwpI7uA8+rS6aojqqanqojqiqYIqiM/1jmjEiH/bEe9dX6z+/de697917r3v3Xuve/de697917ohn8yz/ALJA7K/8hf8A7k+/de6af5bwH+y97UtxfEwX4tc+VuffvPrS0p1YT791vr3v3Xuve/de697917qPV0tPXUtTRVcSz0tXBLTVMLi6SwTxmKWNh/RlJB9+691rS/zUf5S/bfb9dSZ74kdnV3QeOnyQyu8Y9tVFbAtXj1UTZgTLSAWidVcurE8e91HDqjA8VJr8ug+j686k+M3xCr8F8Wft97/ItcbVUvYndGGaRctg4aekMebrFBJYBTqdm0j6E3N+PVJOetaAi6U/4v7eiCfGvtjvn4Z7K2J8sG+ROY7/ANq9i9tt17Vber56+tp8TXzvatp54XcgGPXwfe8E0UUx1rUVAZiSCQP2463g9uZT+M7dwGZkCxPlsLismyXACNX0MdUUF/6F7e69O9PSsrC6kMP6ggj/AG49+69137917r3v3Xuve/de697917r3v3Xug87C2tjM3jJMm+AjzeexVPOcGfpPT1UiFUaJrH6E3It79jietNladadXwo2R0lB82flTvj+Y/uimyU22d2zVPTOE3n/EDDj5Bl2lMdGka3ISIehAVX+p/HvZOBTqq5dq0qDj7KdWad9fziNs1Uf+iX4JbYpu1avHx0uIqvsKGenODCyCKampIPIAQpI9dj/W/PvwUEZ60z0NFFT6Cn7erp/jNvXP756X2Jmt50q4re1Thom3RhHmjapxeT8jCSmmjV2I4sRf34/Lq/A06H33rrfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdUofznfkX2Z0/1v1/111lNV4zM925Sq2iu4KHyrU4OR5oY46yGWMjSw1/X3sAHqrkqKjjWn7etajbP8tDvfrDtFan5D/KPLtXdkVK0u0sjm56+doa3JsJKeWGWQn9BcAkHj3YOAOGOmTGSwJJr68Otir+VX2D2ds7sfs/4wdgbiq94YjrDGNPit3VhqHWvYNEreKWY+oabkfn3ZYndQyitermVFJDnh69XgLnIJpxHT2lS4u4/oVvwB7f+kYLV8HplroAjSKg9arX81bcG2Olvl30N2v8iMQuS6zoMxBkzV18U00WOxkWSZ2SJLWI9LG3uyrEBxr1p2eqswoK4/Z0IPzV/mm/DXvH4r7w2F8d8Vje0q3N0VIuH23/AAiSiiDQ2EkUIRdQINhwfbHiFQAoqR59a0gsXJAr5A18v8vRcf5c3yf+ZfXWz6DZm3fjJWHb+do/to4xQ1xjwkT1K6hBISAvH1v79JM78Gp1eKIAdyZ62wdqwZHcfWmGfcmNOKzWQwiT5Oga2ukrWjbVE31+nHtoM9cnPSnSq8B1X31XWTbZ7xrKOVjFA2VIHHDp5bcE2900jVqPXjUjq09HWRFdDqVwGUj6EH6H3vh1vrl7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de699ePfuvHOOm3IQ0q0dVJLGhCU9Q5JH0tEWNvd1dx8J6oURiCw6rCgy+T3T28cTSu81GlboaMfpGmSxv9f8D7cS5lXD93TTW0ZOMdWLUu366joKQQ1rjwUyg04uBqtqKi39PalbmJyFdKdNNBIlXVq/LrNBXZiklP3FM8kIFi7ajzfj3Zo7eTCGh68HljqzVPTgm4qZGtWAU1/oTe3+xv7ZNo5wmT1ZbpT8eOnqmq6erTyU8qyLxyL/n6fX2lZGTDDpSrq3DqT7r1br3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuqi/5pDE7f6/VfqNyxEkH6/uLx78DnrXl+fR+vjYSepdsg/ihpf/AHFT3rzPW/Loeve+vde9+691737r3QV919i0PVPWO7d+5CVIafb+NarZ30aVJcItxIQP6/n3scetEVFOvmX7S+dL7l/mcd7733LsqDem19s7qqsrt+WpgmMCL968ilNIZQLHjSfbiqSNHSN3UP4jeePnjpffAj5/VvWH82rcG9KzAjr/AGv3Fv2IeWngkSCux9VVr5ILSlAFA4I96IKpniOnI5A89F+Fhw+Y6OB/OP6y6579/mX/AByq8hQ0uc2luXd1O1GJSyQ1FLNWrqFoSVtc/Tn6X/HvVaCvp1fSGcV869Bf/O4+DHxw6278+Fe3+tOusXtCSt29Ry7hnpqmolXNVb19/NKvH0AFgeLgH3tKa6+g6pPXwR82p+XQq/zT/hz8bekurvh32F1h1litq9hzbGoa7L7mpqiWaqyGQkqE1VehuUYkfg+/RHvJ61d4iFMZH7OjefzOPjkN3fy1fiv8s4cZHkN5bG65w9F/EA0rVdo6h5vIY0vwAjHk/j3pGIP246vcxqw0ngvd+zqs/dXymp/5msXxe+NW18mueqtpbMwm19yYqGMx+PJ0kyo0UxgZyXBuORf3sfp1J6bci50gY8/y6Eb+bn03hsH8gvif8bN/UcdB1Pieudm4jdW3agyigqK6n0RytM7WAa39ffo86vkOrXBH6bDzPQl/z3fgh8Huj/iR8fMv0zsDbmxc5X7Y2TWyPjq37qWtbQjpXOyyv6mP1C2HI4Fvel456tMQftH5dA386cvmn/lddN7OGQlptqVVZ19HUIFlNPOk9KsU8b6Ta0i3v78tcfLr0nBl9aV6Nl8y/wCWz8Cdj/yi+rt/7Y6z21t7fO4aPbmdzG7aeujkmmzDYhaiSOWm1n1M/q4UWI+v9NKKla+fXpCPDkVfwjGOHTT/AC6O6fjl1H/LO3rS9/TUOU29LvqDDbexWQiZ4K+QQsqUqMCLa7gX97eus9ehp9OnyGeqj/5kFBhc70rtPcnUvwxxWw9kVXZ1NWUXYePedkrEeVXiqPIzFb6f3LA+9gUI/wAHVHNVY6ceuOrS/kXWTZD+SJ0lUVVY1dUxdhLC0rX1QlMeo+15v+nge9j+3HXnX/E2PmR0NHT3fHxZ60/lB9d4j5CU2K3BV7g7Zek2ztjKI3iydZ9uqNSK6kaTIx088H3Q/Gft6cTECk/wivVGfz8w0Zqulc5sz4f47pHatT2LBU43dWPado8jFJKskUySObG6ENYH8+7jBoDXpt6sKkUAPHr6Kf8ALErqrIfDrq+orKlqqf7JYzK176Y6GnVU5J/T9PbXn0pHAdWBe/db697917r3v3Xui8fKf/mSG97f8qMdx/hqPvfl/q+XVTxPVe38qz/gV2kb3BdLHnj/ACxePej8XW1+E/b1ch791vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/S3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdUufzhL/wBxdsckDxr/AO57e9qM9Uk+DqxH4ff9kydL/wDhlUP/AFuk96HTjfF/q9OjJe/da697917r3v3Xuve/de697917qBlcdS5jF5LEVsazUWUoKzHVcLfplpa6namqI2/wZGIPv3XutKz+YB/wnS7Y2x8h/wDZo/gT2RnuucrXPU1FZtLZL5KkrfvZrPLO0tNa2pibENf/AB+o92BFAr/D0y6NlocOeJPA9Ah01/wnw+YXyu782xvH5x9v73yfW23J8Zl5tnbzlytbRVmQxrazU0/3BYed9Nhe315Pv1VHwfz68I5Wp454Gvb/AIPs62Rv5jHwBn7r+Fu1vjL05SnD0O1IqeCjgoVlHlWjxKY5ZpPECS76SxJH1PvytoIb06tLGJI2j/i6WH8nL4f7w+EnxCo+mN71FVPmY96Z3PkVYk8kUORp6eJEHlANv2r+9E6iW9erIuiNY/4RTqyfsXYuE7M2Luzr/clNHWYLeGCyGAytPKCUmo8hCYZUcDm3N7e9dW+Y49aQnyZ/4T5fLroX5Ebo7R+DXc+9dsbV3VHL4Nu7Jmy2PXEmsm1zM70hWzgFgQD9PyfdtQPx5HlTpjwnUk29Ax414HowX8uT/hOrvvb3fE/yg+YXZGV7B3gtfS5SDGbsXI1GU/iEOmZZfLUHkaxdmLXv/X6e/azTSvwnrYhBcSy5dfPrcoghSnghp4lCxwRRwxqPoqRIERR/rAe69PdZffuvde9+691737r3Xvfuvde9+690Qz+ZX/2SB2X/AIjGD/b1Pv3XvI9NP8t3/snraf8AhioR/wBZT7959aXh1YT791vr3v3Xuve/de697917r3v3XumvN445jC5fEiUwfxTGV2O8w+sQraVqYyD/ABXVf37r3Wqv2nsH5DfAztvufEbG6mk7O647R2RvDbdHnZ6SqnpRXbqx706PE1rCWMykgf1Hu3Hz6ayjagK1z+f+z0uf5ZXxw2j8f/gZubcXzv2fTY3adB2Xkuxtp7b3C7I6ZARmY1dJRSMkpe4UC402/r9PejQsadbjDogrQHJ/n0Qup/mYfNv5vfOiLpXpTYOU258WsBX0lNt3tfBU1fSJXxUAQR42qKWjCqqaABzxz9RfdAKhuqiQs48L86/5Ot0Lr3A1u29n4HFZHIzZSup8ZQCrrZ7iSWp+0QTs17/29R916epTpa+/de697917r3v3Xuve/de697917r3v3Xuqg/5gH8rLYfzHy+CyeKr6DrXKwrPPuHddLHMZ6rxl5hJJDGRqfmxYsBb+lve1YocdNyxpKNMn8sda8Px8+J+Q+TXeW5/j38XVXrbGdIZmXbHaHe+3jUQ1uTqoqt6VMxWiIh2aVo2MaixPuxIpU8T1oIdelcKvn5+tB0aLtHbXY38mnujqvsbs35Pbm7X6wySyV+7mywqi1P8A5WyiGrx008pOsJdWB5B4t71xHDPXm/TNakg4z/hr1bb8WP5m+V+YPauzqHp7ZqZfqHMQSVGc3caeeBsbHq0xlnnNhf8AH9feiCB1ZHV8+Xr1cX711fr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6wzTxU6GSZwiD6k/8U92RGc0UV6qzKgq2OiM/NL43YH5T7FgxlNUxx7o2uJ67a1cGaOahybspSaBx/aBUfn2rWzdcy46SyXSHtTPRRch8C+ye1enuqNrdkV0tbvTqTK1GUotyV9dG9bkl8uulp7glrLpAGr24rW0C/wARPl1RlmnAr20/w9GV2HvD44fH7Pbc2puiuoaDuPec8eHyJiCVNbX1bf5MkVS8bHQpsPqL3/w9sSXjN8C6R08sCsNLmp+fVg0VBRwf5qnjS3+pHtlppW+Jj06IkHAdEJ+f3Q3xk7O62k3x8paWiqeu+vqU1OQ+9jlljEMk1xGI4RqJYtwB/j7b6vTB6pZ2h8lf5aPX2OpqD4nfE3Y++spQRsuOrqikro5ah2JRn8Uv0OoD3ulPKnVFcEUBBr6Do73xt+UnyF7W2zvzE/7LrjOrcnAdOyafGSU1A+Q9YRkjWpZWYWINx72B5cet1HGlPkT1Y78X6TuCHBZ4dvY6qx2UlqF+1iqZVmJVi3l0uhZf9T9D7rmvV/w9FZ7bp5NodwY2rjHjE9WkhYX51yDn/be9efW/wgnqzLalV97tzDVerV56GGQt/UkWJ49768ePSg9+611737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690H3Z+cTAbOyte76AIniB45LxMLc+/eXXuiH/ABkwkuX7Frtxtd6da15NVjYkuWHPutetkDj1Zj7t1rrplVgVYAg/UH6e9gkGo60QCKHqFNjKGo/z1NG/9Lj6f61vbizSqaqx6bMMZ4jpin27KjmShrJKdRysCXCm3IF/alLpaaZFB+fTMkDcUPXCKry9B6JqZ5ol48jauT9Ln3Z0glFUND6dURpox39TodxUJYR1D+GUn9J+n+sT7Za1kAqueno7hWFWx0+xyJKgeNgyn6Ee0zKVND0+CGFR1z96631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3VRf80hbbf6+IJ9W5oh/1kU+/D4vy695fmOj8/Gv/mUu2f8AqBpef6/5KnvXn14cOh797691737r3XvfuvdVv/zO+iO0Pkb8a989WdYV1fQZfc+CloEqMf5fNHMZtauvi5uB9PfjTj1rurj06Il/Kz/kt9K/HbpamPf/AFlgN/dp5ZKmHNZbccVVLWSRPN5L1EJKHUb2BLG3P9fe2Nc9VjUxjSP9R6L/APzU/wCRbs7vTc3V2+/i9s6h64zmxGFRUx7cFYDWVAqWlSRY11FWC2HL8/4e9qQDnrUqFx8/XoA5v5OfyE3l2P8AFvem6J8t9x1FVxVGXnn+7LzmKqWW8xINr2/PuurB+fWwlGVvToz380D+VJ2h8ru6Pjrv/aNVXrS9ZY2kosjHF9xoZopxMzSNGpH+HPuwYrU/LrTxeIgT0Nenf+Yt/Ky7P+THWHQW19r1VatZ17taixGTWI1FxJDMHZm0Lf8A2B9+RtBr69VuIfGQJ6GvQ+fJyr6B+MX8tjaXx9+Sm7I8XNt7Y1LiWhqKd6iaasEVTLC0iML6SJbX+t7e9Dpx8g+Xl1rBf8Jf/hhU7j+X/d3e8WN82y9rb+kyO0q4x1Ecc+BfMy1FJMgkQL9GVTz+P8fdmqO1uPHpmIKx1rkAaf2dXBf8KMOg+i+1sbFUbj7Q/wBGHZjR42DF5qmp53rKeKOjUxPFLCOCSQfr9felLA48+ryqjAFjw4fb1qtfJ/427iw3T2xqPe/ys3J2lXU1RteLbO2Mk9bUrUYqGRPE0Wpn0IQALAD3ZRkgdMynALmp8utvHC/yyW+Zf8qPYnWlZp23u07fwe4duV7CoEtSaShMdIihE1A3/Tf3XVShHl08VDBkbzFPs6qFi/kRfzBOy+sq3oLfnZm+6TZOPzsdXiKutlyklGlHRL4oKeCM/RCgC8D6e/VXiOP8uqiKWhVzVflx6P3jP5BW8Kv4GVXROXyTybtxW5Vz+OM/3n3FVNRppidF06rsQCD79rIaoHW/AUx+GfT8+iW57+Sf87e2uk8X8fdyZneW29qbY3SmXx0xetNLPDScRRIoH6WAC/7H34sK1Xj1oQvoKOcfLqy7sL+UV27ur+WtsP4txTV6bp2vvNs7PIoqRLPEYhEXaw1c2vyPew36mvqzRkwGAelOgd7j/kIb17E/l/8AWPUclbLP2H1vv+bedHROKxquWWMrJBGmlbg6hcE+9Bu7UetGOsQj86DP2dE77L/ks/O35G7G632jvXL7z25QdbbhiyVBQytWtDVQ0CiOKPSB+l1UD/W971KPh6qYpCCH63AvgL0/ubor4z7H613dHNHnMCkkVUs+vyH/ACeKIMS9jyUJ91PGvT4FFA9Ojne9db697917r3v3Xui8/Kb/AJkfvf8AP+Qpf/W1G3vdKjqtaEnqvT+VWpFT2lf/AFacf9Vi3HvR4j7OrL8J+3q5H37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf//T3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdUhfzqdwYnZ3UeJ3duKaWj29h4o2yVdFBJOacPXtp9MYP197BoeqS/B0YD4hfNb45L8XekKqPfH+T1GzqaKLzU2moBglfW0qFhYEnj/AHr3r5dXrqyOjEn5ufHS9v77Kf8AWp15/wCT/fut49evf7O58dP+e2Uf4/brb/eH9+61j1HXf+zufHS3G94z/gKcX/3l/e6de889df7O58dPxvZD/rU6/X+nL+9cOvY9evf7O78dP+e1X/znX/o/37PW8eo66/2d745/89un/nOv/R/v3WsevXL/AGdz45n/AJjdP9f7cf7b9fvdOvddf7O78c/+e3T/AM51/wCj/eut0+fXv9nd+Of/AD3EZ/wEA/pf8v791rzp12Pm58dD9d7xr/iYFt/rcP79nrdB69df7O78dPxvZDbninX/AIl/fs9ax5nr3+zu/HSwP990+tv+A4/2/wCv37r3XZ+bnx0H03shH9RTrb/eX9+6911/s7vx0/57ZD/1Tr/0f79nrdPn10fm78dB9d7IL/8ATOv/AF89+z17Hr12Pm78dD/zGy/+c4/4h/fuvY9euz82/joP+Y2T/X+3HH/J/vxx1oZ4dd/7O38c7X/vxGeL2EAv/wBD+/U699vXH/Z3PjpY/wC/2T/znXn/AFvX73Q9ex69Ex/mBfLTofd/xW7EweG3gKvJ1hxsVLSRUxMzyCoD6tIJuv4Nvz70OPXiQAT8uh2/lsOs/wAc9nVMWswT4mFondGjLASG/pax9+8+tDIr1YZ791vr3v3Xuve/de697917r3v3Xuve/de6T+5cPiMxjJVzGJp8zDRh62CjqV1I1RAhePT/AENxYGx9+691ql98dYfIv+Yb/Mvyvx63pmM/1f8AHvZdBLmsf9tFVS4zIQ46nEr4pY49Ebmo0aLkge7Ciio4/wCDpoqXcBsKM0Hmetkzpr4r9G9E7Ywm3Ou+vtv7eXEUdJE9bQU8kdTWVkEISevnkdiS8ranb8c/T3Xjx6d88dCZnOytlbbydLhctnsdTZGrkSCnpTW0fmMjsEVGiMgdTc/lfe6dV1Zp0uwQwBBBBAIINwQeQQR711brv37r3Xvfuvde9+691737r3XvfuvdcJEWWN43AZJEZGBFwVddLAg/4e/de60+u38P8yP5RfyF7T3F8XOr17awXyazzbhzUk2Pq6ylxsj1sskSKthpkiaRgGB/P9D7txFK8OmiWUkgVqfX8unPH/BLtT5kVuO+Xv8AMt3XVdPdS7XRa6r62zEkj0m5aOnYz09NT4uRwQrLcLcc824v78CAKL1oxljWXy4ZwPmfXo9fUfyJ+PvZXV+7Okf5a/XVFhqja1Zj8U9fjKMYqpqaaCpKtP5ZLMdT8n1n6fgD3rINT04pDigIPofLq5voHH79xPXmDoOySq7qgooI8lGa2Csl+5APkMrQs1j9Pr70AerV6G/37r3Xvfuvde9+691737r3Xvfuvde9+691ArMnR0P/AAJlCH62tc+3Y4ZJfgHTbyonxHpgmzNfVErjqbyRk8Sc3sfpb2qFvEg1SHPSczSOKIK164x4GqrCs9bVypqN2pjcqP8AVDn343SoNKLw8+tLbNWrtX5dN2+9qVGZ2RuHB4CpfEZbIY16ajydLdammnZwwmjZeQRz9PaSSaSSuo8elSxRrkDh1qkfKb5ffzEf5amfzWQq9rV2+9vbiaqTZ2Tz7NkFyjiqMSKkTsWuBY6bX591RATQceqyuyLrcVX5cei7fy6vkvgNz/JPJ/Jr58ZTJbTr9/1U1RsXbE+MqJqSgzglWankpFlsI1DXFx+fe2UDAz69VibUxc49AfTrdd2luzB742/jd0bbrFr8NloFqKGrUWEsTDg2BPunT/SV7ixHXed673HjO1sNRZ7YlTSEZ3F5CFp6SqgT1KssakE8jjn37r3DPWqpmPmdsL+++/8AZ3wd+FXW2Wg6qqXps5k46Qx1dNGJWh82qdrt6xe/9fdqeZNOq68FaVp9gz0RLsjDfzNvll8i+nBtXEbj6swEtXAmVO2cm9NTUSmpBd9FM9wAp5/1vewEGTk9N0uD2oQo/b1uWfDrp3sjpzrOlwXZ2+svvjcLraerzE8089O6NcqHmJJBv9b/AI90NK1HTwJpQ9BN8u8UKPP4rLhbKppn12ueOT/vIPv3XqV4dGu6Nz4zuxcUwYsKamjQE2+hv/T36vXqefQy+/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6KV8sNzJRbLkwSv456ySNgwI1WZCAP8Pz79Ujr2Pz6xfEzbDYnaU1dUJ+7VLC6Ob3KuS1xf/W9+Br14jPRuvfuvde9+691737r3XvfuvddFQwswBB/B5HvYJGR1ogEUPTbUYigqFYNTxhzyJAvqB/r7dS4lQ1r020KFaAU6YpcPX0I8lFVSzWNxDyBx9OB7VC4il7ZBT59JWglT+zNes0GcqINKZKDwFb6iL/pH59+e1R8wGvTi3DLiUU6UFJW01bH5KaQSL/WxH+8H2ikjeM6XFOn0kSTKGvUr3Tpzr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6qG/mpVUNDtfYVVVFlgj3EkjuEZgqrIupuOB/sT79TPWiRQ/Lo1nx67v6wpOottO25KZBHTU0EqyTUocS/bqtgmsGwsb3968+t+XQ0Hv3qwfXc9GP9eam/6+e949etd3p17/AE+9Wf8APT0X9f8APU//AF89+x69eq3p149+9WAX/vPRn/Wmpv8AiZPfsevWxU+XXv8AT71Z9TuejA/xmpuP9e0nv2PXrXd6de/0/dWfjc9GR/UTU9v+h/fsevW+7069/p96s/56ii/1vNT3/wBb/Oe/Y9etZ9Ovf6ferbEnc1GLfT96m5/1v3PfsevW8+nXX+n7q23/AB81J/reanv/ANbPfsevXqN6dd/6ferbX/vPR8j6eamuP8P8579jr1G9OqxPnp8Lfjn/ADBJsbNvHu2m2pS4+OmgnxUklK8FVHTLYGxmUeofX8+9hyuVPTbxJINMlfy6MR8Req/ih8JuuaPrfq3N4fxpTwxZTMU1RTlso8BLJLIC/HJP9feiamp49XoFGlRgdJL5jfHj4lfNnbE2B3zubDYbOMNVJueSWn89Nop/t4kHr/s2BFzb34MQdQ8utFFcUbgeq5unP5L/AMPOvN5Y/dXYnf8AQds4/ES00uNwGUkpxFBHSzCaGnj0zPwAAoFgPe/EJweqrBGvcta/M9Xvbc7a6T2hgcTtnb2ZxuOwmCoYMbi6Gnmp/FTUdOuiGKO8n0A96FOnO7zHT0e/urAbDc1Gf9aam/23+c9+x69eofTrl/p96svYbnoj+eJqf/Y/7s9+x69aNfTro9/dWD/mJ6I/601N/wBfPfsevW814ddDv7q0/Tc1GT/TzU1/9f8Aznv2PXrWfTrv/T71Z/z09F/1Op/+vnv2PXr3d6Hrr/T91be395qP/X81Nb/eJPfsevW6N6dd/wCn3qzn/fz0XH/N6m5/1v3Pfqj169RvTrie/wDqwf8AMTUn/U6m5t+B+579j1693UrTrv8A0+9XWv8A3kpLH/m9Tf8AXz37B8+vZrSnXh391af+YmpB/rzU3/Xz37Hr16jenQAfKbvzquPo7e2vc1K+vGqyiKWmZuX/ANTrJP8ArD36vp1og14dFC/lK5WjzcPZeUxsjzUFUYmgmeNk1j7tTxfj/bH348etrTR+fV0Xv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//9Tf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690CnfnQvXvyM633B1n2Ng6HNYPP0q08q1kPmMDRyeSOWPkG6m/F/fvOvXuq7MR/KE6pwOMo8Lht55fHYfGwinx2Np6NVpqOEEkRwr5OBz78ST1oKo4dOX/DTfXX/Pf57/AM5FP/XX37PXqL6fz67/AOGnOuhb/f8A2e/85V/4iX36rdeovp/Prr/hprrnn/f/AOe5N/8AgIv5PI/zvv1WHXtK8Kfz68f5TnXRt/v/ALOi39KQf7H/AHb71nrwVR5fz69/w0311/z3+et/1Cr/ANffe6t16i+nXv8Ahprrm1hv7O/+co/6++/Vbr1FrWn8+u/+GnOuiLf3+zo/1qQf9fffqt17Sta0/n11/wANN9dcf7//AD3/AJyL/vfl967uvUWtafz64/8ADTPXP/Pf53/zkW//AFt97q3XqCn+z13/AMNNddfT+/8Anvr/AMqq/wDX336rdeovp17/AIaa66Bv/f8Az3/nIv8A199+q3XqLWtOvD+U110B/wAf/nv6f8BF/wCvvv1W69Ra1p/Pr3/DTXXV/wDj/wDPW/p9ov8Atv8AO+/VPXiqny/n1yP8pzro/wDMfZ0c82pRz/t5ffgWHXgq+n8+uv8Ahpvrr/nvs79Lf8BF4/r/ALt9+qetaVrWn8+vH+U310bf7/8Azwt/0yr/ANfffqt1vSta0/n11/w0111z/v8A/O8/9Mi/9fffs9e0r6fz68P5TfXQ/wCY/wA7/wCci/8AX336rde0r6fz67/4ab65/wCe/wA9/wCcq/8AX336rdeCqPLr0f8AKV6pkqYXyu7stl6FHjebG1VIv29QY3DgORIeOP6e/Vbr2lD5dWc7B2Jt3rfauG2jtjH0+PxWFoo6KmigTQCkdyWP+JJJ9+63x49LP37r3Xvfuvde9+691737r3Xvfuvde9+6910QCCCLgixB/IPv3XukjTbF2ZjdwT7upcBjKTcM0RhqMykQjqnhIsUklJtbn+nv1Ot16qe/mPfzcumvhdQpsPD5mnz3dO455MPtzB08UFfRw5eZlipYat43PqZmtptx+efexXjTA6ozKKAnJ4DzPVGrfCj+ZBvrPbh+e/YmQ3TtnOU9DV73p+sJauogw4xVLAMlQ1EFM76FWSIq2kj6e9hlpSnHqhWUkNq4eX+frYu/lofMzL/K/pahq+wKKmwvZ+EmnxuaxFHEi0xpMdGlPFWq8RK6mIOsf196IIwero2pajqyz3rq3Xvfuvde9+691737r3Xvfuvde9+690HnZGMoJ8BVZaTatDuvK4qmnkxFDWLqH3TodGm4P1YC4Av/AEI9+60etZrdvxK+ZHzU7Yz+8/mJvDPfHP4/dbZH7fFY1qyenwObwdPI0saxY+SRVctECNVj/T3ZSB9vz6bdCx7z2+gPH7ei2/MD5zbK6L6pzPVH8nvqXHbw7L2zV0OD3TuSlxEtHldyVMkwgrKwVashco12RgVBPv1D8R68zAdi5+zgP8HVx38nfrr5K/6IYu1fk5nc/Fv3edJjspX7UylZJVDD5Ce719NaRjpCkabe6lgxxw6ugdVpIanq6z37q3Xvfuvde9+691737r3TbXZaix/FRJpci6qBqJ/oOPb8VvLKKoMdMvPGh0k56Ts2Ty+RPioqbTC5F5RcMqH88/7D2sEFvBmU56T+JLKKLjqdSbcijlE1VO9WxALJMLqDbkDn2y92SNKAL04lsAaua9KGKGKEaYo1jX+iCw/2w9oyxbia9KQAOHWX3rrfSf3RufDbOwWR3JuCrFFh8TB9zXVTDUIIdQUuR/sffuvdaYnzV+b/AMY9+/MB9yd3dq7uresNjZ8V+z9qS0j1eKyQpa7U2PjiQlFRytjp/Hu4Bpw49NFl1VJ4evDpVfLj5qZ7u/reg3X8XPhV17ubaOIovJtnOTbbdq2hkhbmaHxqqq2gA/T/AHn3UDPGlOtu5K4Ffz6vV/lI/JDN91/GnbOD39haTa/Zm14qpNw7bpFjiix1KZUFKogB1Kbsb6h78ergggEenVo+bw2N3BjKvD5emSrx9bG0VRBILo6kfkH+n19649brTrTsyPxErPkp8x+xuqemM3N0jtaPck1Fvrcu2pXx89ZBFVMrT1CRtGZmJ+gvyfdgcdNEEtSuOhG+SH8uve/8s3acHy62l8s9+77/ANF9bQTy7PzNRLRvk3qau8X+RmqmSVBpsy2/2Fvfh3YPXmCx94qaA8TXrZW+I/cVd8gfjd1D3LkoFpch2DtGkz9XTrGIlhnmmkhkRY14Aun091PHp3pHfLHbzZHawySi5ptA+h/s3b6j/D+vvR69Xrv4kZdazadVRagWpxEQP6aGKEAf7H3sDrxx+fRvPfuvde9+691737r3Xvfuvde9+691imqIKdddRNFAp4DTSJEpP9LuQPfuvdcIquknv4Kqnm0i7eKaOSw/qdBPv3XusQyWOL+MV9EZPp4xVQF7/wBNOq/v3Xusk9ZSUoBqaqmpw30M88UQP+sZCPfuvddirpWi861NO0P08wmjMV/r/nAbf7z7917ryVlJIjyR1VO8cY1O6TxMiD+rspsB/r+/de65wzwVCeSnminjPAeGRJEJH9GQke/de6y+/de697917r3v3Xuve/de697917r3v3XuuibAn+gJ/wBt7917qtb5T5n+P7xxmIppCVSSnjZBYi6kDkDn3ug8utDPHo8HU+EXCbJwsP0aWjhkYWtbggC3vXW/PoSvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYJqann/wA9EknBX1C/B93V3T4TTqjIrfEOkzV7ekiZqjH1EkRXladbhOORwD7WJdK/bKPz6StbtGKxfs69T5mtpGMeTp/Eim2vktb6A/7H3Z7eKTMJ49aWWSMUfPShpK+lrVLU8gcL9fwR/t/aKSJ4jRx0qSVJPh6me2+nOve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xui7/ACO+Om1/kfsqfZ25KmXHo8VQtNkKePXUUkswFpohdeQRf6+/Z49exwI6q1x/8lTHYujkoKP5J9gw0zVMtQkSUrIkYdtSxqBVfRfoPfqnj1oIgwK/t6lr/JiphfV8luwzccfsScH+v/Ar36p69oTyr+3rkv8AJjpgLf7Mt2H/AK/28h/3uq9+q3y63pSlM/t67/4Zkprn/nJXsO34H28nH+w+69+qevaUp5/t69/wzJS/95K9h2/p9vJ/9Ve/Z69pX5/t66P8mOlJBHyV7DA/p9u//wBVc+/VPWtK08/29d/8MyU17/7Mr2H/AOc8n+9fde9d3W9KUoa/t67/AOGZKX/vJbsP/wA5n/8Aqr3up61pX5/t64D+THTAk/7Mt2HYn6fbv/vX3Xv1W+XW9KfP9vXZ/kx0xKn/AGZbsPi9/wDJ5Bf+nH3XvwJHWtKfP9vXm/kyUxFh8luwxf6/5O4/+Oves1r17Snz/b14fyY6UWv8lewyALf8B5P/AKq97q3y63pSlM/t66f+THTNa3yW7DFhb/MSf1/6ivfqn5da0p8/29df8MxU2kgfJbsO5tz9vIP95FV78Cfl1sqh9f29ZB/Jlpf+8lew/wDzmf8A+qves9a0r8/29eP8mWmP/cyvYX/nPJ/vf3XvdW62FQev7euv+GZaYfT5K9h/+c8n/wBVe9Z69pTzr+3rr/hmOl1av9mW7DIt9Pt5Lf7b7r3up61pT5/t6838mOmN7fJXsMc/inkHH9Lfde/Vbr2lKZr+3rs/yZKbj/nJXsP/AM55P/qr36p62FX5/t69/wAMyU3/AHkr2H/j/k8g4/8AOr36p68FTzr+3rv/AIZkpf8AvJXsL/zmk/8Aqr36p61pX5/t64D+THTXJPyW7D5/H28hA/29V79U/LrxVPn+3rn/AMMyU3/eSvYX/nM//EVXv1T17Svz/b1xb+THTErb5LdhgA8/5PIP/jrn36p69pTzr+3qBl/5J+HzlG9BkvkZv6qopV0zU01IzxSre+l1apPHvdSPTrxRDg1/b1aN8ZvjXsz4y9fY/Yu1C1Y1IhSqzE0fjq8hexvOt2/Iv9fevn1vAwOjHe/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//V3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdFX+YND3LmunM9tvpLHmt3PuSkq8RNMjiOfH0tREAKymkuLOpvYg+/efXs8Rx61c+y/5J3bHXG09r/KTcGQyHc/buG3u25tw7E3PS1NdDg8XDVfcLXQTl31FSLt6Ra62vza+vGjgD0n8EBhLxdTWp/wAA6Nb3r/MQ+VfenUU/UPS/WtNS5HOYao2l2NUGjembBYGSlFBXVtJWVH+bsobUwI44+nutAOJ6dLmlVHQUfDr+YV8VvgRsU9B0Gby28e6sTX1Wa7BrPtIK+GCkq0Ek9FT1qFmYxP8Ai/497y3VV0xjiPmPP7eth34n/NPp75b7cXJ9dZeabJ0tMsuTxVbHFT1tP6VLSmnVtQW7D6qPeiKdOKaivHo4PvXW+ve/de697917r3v3Xuve/de66IDCxAIP4Pv3XuqK/wCbh0R8m/knuDYXVHXOS3BtzpvK4/7veu5cLJUhcbU0kkstRJLHARf9kD6/X3taVz1SQMQNJpU59afLqq3C9ipthsP8Yv5b/SW1+6t34KhTC9s9i5bCq246jclExE9Qapf82qyKx1Mwv/sPfuI1GvWtWfDQA+ucD7fn0ajpH5wfL74V7z29tP5s7Di2hsHdktPN90FhlqKNZqowqaOVyRYeq66v959+NCO3r1THiWgB4EHFfTrZc2hunEb32vgd34CY1OE3JiqPM4qdgFaahroRPTSFVJAupBtf3rj06RQ06Ufv3WuoVZX0tCmuokCj+gsW4F/pf25HE8hoo6beVI/iPScqMlksgGTGw6oW4MnIZUb8+1ghhioZTnpKXllOkYHU6h2/DAfJVyNVu3JWb1BWI5AJ/p7akumOI8Dp2O3AFXyen+OKOIaY0VB/RQB7Slmb4jXpQFC8OufvXW+ve/de697917oNe2+tqHtrYG5Ng5KvqcbQ7kx70FRV0ihpoUY3DopZbkf6/v2fLr2K1PWtN/MI/lRdZ9Rdc9d792xtSm37jOv8tVZjf2Ty+MlmrZ8c1SJQZY4DICpuw5N+PdgTTpp0TgPXh0tOu/5wv8ufrno/Edf7S6/mo97UuMbHxbSp9m+LDzZGICKWeSsvqCswto0f7H34I1adb8ZCtdX+foxf8pzrHs+t7O7c+Ru4cLLtvYXaFPO21sakUtPSDXVx1CrDDIBZVQ8WH1HvVRWnp15BUV9f83V8nvXTnWvP89PhZ8ntgdqY/wCQvwlxU2e3Sar+LZ7bsdR9rTV9c0jPKKyKRwkifQ8/197Ge09UZWw6ED1r0RfMfGn53/L7du2d4fzA56LpPrDBVEH94MRS5/GwY7J0kdQHHnxcc/qJUGxI/wBb3sFRw6oUZ8Pw9BnrZ7+NNd1DR9Y7e2F0tl6fL7M2BjKbA42amkhkjNNCzFXBiP8AaYsfoPden/n0q+7MScx17mKRU1SaQ6cEkWja9re9HrX2dFH+JmVOKzGUwUzhXapMQW4U38ljx/tvfh/h60Pn1Yl731vr3v3Xuve/de66uP6j/bj3uh69UddalH1Zf9uPfqHrVR1Xp85eruwe29pS4zG7nyeyNnYNZcplty4OZ1rY6GPS1UQsJDEooJAv+PeqHVQjrepCtQePVN3xr+Utf1F3b2V1lsrsTcfY3Vi7KzFHjN07qtBkf7yR0jAJFHUksoElwOfdtDaajj1UMNVCagddUvU/aWztg7O+Y9f3Nu+pz+4uxpcfV7EkqZjiocatXqEilnKG6n6Bbcjn+lvCeumnVPEFNer8vLoXN77q3x/Mk7bq+k8Pv7N9YjYG34MnBW4CRomyM0UIkmWo+2ZWJYg8k2928JwaHrRnjoSpP5cc9R9i9udl0FHvn4Zf3ozmRTrjHVmRzO9fVLmlpoU8csyuLyGw5AJ91MMox5jrYmWtCeoPx4783zsb+9/WNXurM5za28aKsw+29y5l/HkBWynSGBqDqXSfob+9FG0462Ho1D+XV53xkw/90+rcFtvJbnh3BnqcSVGQkkyVPV1amcCRFdEYsAFP591o3n1eor0YnUv01Lf+lx79Q9eqOu7g/Qj/AG/vVOt1HXfv3Xuve/de697917r3v3Xuo9XL4KSpmvbxU80nJA/RGWHJ/wBb37rR4dVQ1VNUb37uRI2aVKeuClP1L6ZQLen/AG3vQ4deJrg9Wq4imNHi6ClI0mnpYoiB+Ci2PvY6304+/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XusMsEM4tLGkg/wBqAPuyuy/CadVZVb4h0m6vb5RnqaGaSORbssKHSjH62Iv7WxXYPbKK18+kclswFYjnruDNzUgEeVj8TLxdRcn8An3prdJO6E8erLO0dFlHSip6mGqjEkLh1P05F/8Abe0bIyGjDpUGDZHWf3Xq3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/W3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691gqaaCsp5qWqiSemqI2hnhlUNHLE40vHIp+oI4I9+691Tv/ADXcRUdYfGbcMHTOwsNgqze8OVw279zbcxcdFmcRhJoE81ZTzUSq2oliSxP44/PvwHl1pmIGriR1r09D/B7anxL+N2yfnFsaowvyDnzO58hL3Pj98wQ5HN4Da0VQfvgKGZmkc6Q/JXj0n8+7FiceXTYiC/qKMtkn16OZ/KF7T2n2X83fkH8jOudGyfj9U7SyUlNhsqItu4fHeCBfLFQxVGmMlZFJVQf9jb3srwr1pH1amXNKDh6cadbVOxd97Y7G23Q7r2llKXLYXIB/t6qkqIKhC0TaZELwMwuD+L/4+6dPDpYe/de697917r3v3Xuve/de697917qDk6NMhj66if8ATV0dXSnk/Spp2ga4H+DH37rRGOtRn4e9j4n+Tx358lcZ8mdoZpKfu/e0m5NqZrF4qWorocaKqeopxFLKtjHKsoLAMOQPduNKeQ6b+BjWncSQf8/XXyH7A7Q/m7d5bT2t1ftqGg+N2G8FRnt67jhjxVbgI8fMaiokNTMVRdXJtq5P+HvwOk1HHrTL4i5FR/h+wdbGXxO7m6Uz+yMH1H1vvOhz9V1PhcbsrI3r6GWVqnBwDHFomgkbyByhYEAccc+9KpIx06zitT0Z2szE00r0uMQSyoxRyQbAjggH2ujtlQB58DpG87O2iLh1hpcA0ziqyMjtKX1GAktGLG/09+kuwo0RDHr1tLcsNUvHpURQxQqEiRY1HFlFvp7RMzMasa9KwoHDrJ7r1vpKbs3vtTZGKrcxufP4fDUdDTS1Uz5LJ0VCTHCupxGKp1uf8B7917pNdU9v7G7m29JubYeZo8zi4qyWhkmpKumqgk8P6gzUzMBf8c+/dar0KHv3W+ve/de697917pI79Xa7bPz/APfTH02U2suPlfNUFZTrVU1TRLYyRzU78MPpwffsefXq06r06j+OX8vDtvdeWzWw+kdg1mWw5aesgqdtwxxQStOFeaFDf+2wJBNv9hx731qhrWvVkeDwWG21i6TC4DG0eIxNDGIqPHUEKU9JTRj6RwwpYKP8B711vp29+691Wh/MJ2P8st+4zb2D+N7VUVFUQldw1FHkjQTROZXW6kOt/QQePfgaHPWmBK9tK18+qh9y/wAhzub5CbTnyfa/yq7T2nuaSNZW2/BlcpXUUk0KGWOPyGfTYkafp73qzgD8+qlWAoST+dOjVfyaNg93fHXIdvdGdvTU8+3tvZBMdsXM1ldE+UzS0lWypUTI5LnUh+h/r70SK9bRWUUbOf5dXw7hhpKzE11JVOoSWmqOL86lhYjge7rGzcBjrzSKnE9Ve9Z5U7T7qqxU3iozkzpFwA6mUEHn/e/ekQs+g+fXnIC6vLqzVtw+SOOamj1RyjUhIJut+Dx7WC1Awx6TNc0OOvNkcxML0tMjXA+otY3/AMT72IbdfjPVRNOw4fy6hD+9E7HzQRoluLGxHPu4+jXz69/jJxTHWdcPkJ1vNO8TH8Brcg/4+6meJRQDrRhlc5x12NrFlGvI1er82c+9/XBfhQdb+k1DvY9Eh+XOb7v69w6VXXWJ/vFt6sf7XN09SjVET0D2apSWHm4IuLe2WvCeKjPVhaKMlj1U/hfhLvDv7tXdHY+2NvR7P2uu36iop4MZSVFFBUbjaDW9P41svqkuOPdPqDSgXpzwFrknpQ/wP5C7q602R8W8jtNoqzbW+RX1sn2k11xomCGUykXJsL/X8+2xOVYuOriNSmg9L3IdCds/Cft3N9zdfYYZeTdmBjxdjTSTIlQ8AjnQ3uOCfp7sLhiakV+R6p4C+WPn59GR+EPx4zcPYPY3yg7Tw8FPkuxsLPQ1mNnpWePwSFWqmMEnBBFxa3+9e9NO5J+fVliRV7vL/iugV3/8TN09u737QyWKxH91dobQx9TldiT4qnnhNXXJMZFiAW3J/qOfdxdMOCjrRgDZJ4dNPwC6l77xPZ+cye5MxuKqoC0sdfT5N6kQQQJaFGhM3F/pb3YXRVi2kV6oYC+Cx6vTk25IHMkVfUEhVAVm+pUAc2/r7dF4pFCoHVBaspqG6jmhzMIJhYyMOBqc2t794sDceqmKUHGesqTbmSymliZeASGUm39Rf37TZkCp68TdipA65fxbJQuqzQKQL6zp+gH+2Pvxt4WBKHq4lkUd3HqRHuOmGrzq62P9lS3Huv0bn4ePWxcjz/PqZFm8fNykjW/qVsB/r8+2WtpVNCOrrcRsaA9JPszPw4LZ+SrzMqAxPGrB1F9UTXFz/sL+2TVQSenq1pToifxpwUua7ErtyOpkgStaXWbkEGTUORx7rmvWzTqy/wB7691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdRqikp6pWWaJHuLaio1D+lj7ukjoaqeqMit8Q6S8uFq8fKsuNkkkW92jZiFW39Bf2vW5jmXTMKdJHhkjaseenKjzcbt4Ku8U9wLabLf/AFz7ZltSBrjyOrx3IJ0Scen4EMLggj+oII/3j2jII49KwQeHXfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r/19/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690jd/bE252RtTObP3Rj6fIYnO42qxlUk8KStHFVpoeSHV9GHBBBH09+z5de8qda+PZX8vzp/4MV2+e6N3dh71z/S1XRV8+R6/YTzYo0s0mqrolx/kaOQhWFhp597yeqaQDjA/wdVuSdfd8fzIcbPsr4Z9cUvUnxbpZ5qA7gwGOj2plq2F59NRNlZUMXlkYG4QE+7AqpqRX5dNkSSLSNtI9fM/Z1cL/Kr6K+ZvxI3HnOgO2aKev6D27T1c2y9017rU5DJZKqQEk1GpmK6r3F7e60rw6cUMooTXhT7Or3Rewv8AW3P+v711frv37r3Xvfuvde9+691737r3XvfuvdFh+Tez+iJNkZff/cXW+2t7R7doZPtJ8tg4MrXQS+JjTR0zurMo1DkDj37r2aUHWtHuDIfM75v5jM9QfF/qXA9V/HIVjY7cm49pYmHauVgjnX7WR5amIxO5I9RUE/T28sNFDNw/1f6qdJ3lkZqR4Prmv5enRWN3dE9rfyMeyNt5TrbsjNdlrvuqxOb3ZBmakVuSpchXTiSWkhoJWkZ4xqYawPb/AIqD4KHHmP5dJnidNMjAmpoRX18+t0HoPeFb2L051t2DlaRaTK7z2litx18XhELxVGVgFU6GOw021Di3tK0jPxPS8RhDQDoYPdOrdYameOlp56mZgkNPDJPK7EALHEhd2JP0AA+vv3XuqSPmj/Oy+P3xw/vbsXbaZ/c3aeJxmTmoYsXQwZHEQVtFFqjSoljMmu5sCNPH+PuyivTbyKoBrn08+qHegq/5l/z3s1upez90VvWXVMLZSmo6nbs8WIy9NEUP26jEUzROy2AGoiw/r72wCY49NoXlBYgoDwFcn/N0dH+XHX7t/lafLfPfBnsPeOWz/TM1JJk8Xu3cUt62s3HkqIGgVZaliWUllGlZLc396ORq6uvaQh9Mf5etsmCZKiCGoiOqOeKOaNv6pKgdDx/gfdenesvv3Xuve/de6Z8/haTcWGyOErxqo8lTPS1C2DXje2oWP+t79WnDrVOgD2b1L1p8bqXdW9KSrFBQ1VOZcrLVGGmhii8ofh2a36h+faqys7i/uUtbZSzuaAAdNTzxW0RmmNFXJPRc8N/NG+L2b322waPcROV+5NKk7VdGKSSQSeL0tqta/wCb+5PuPZvm+32794vGNNK0oa9A6Hn3YJrz6NZBq+0f4OrCsfmsdlMZT5egqoaqiqofNBNDKksci+MyWV0JBNh/X3Fs1vLBMYJAQymhBFOhksqNGJQcHogW1P5lXx9z3ae6+qMhk6nb+e2xkZsfNVZyalx+NnkhYqwhmqSnF1Njc+/JA7ioxTrTTIlamvy6Ij8s/wCcphINwVnSXxFqcdurumSrXFUU9XLi67CmtkcwNGjXdfSxFnLD+vHvZhCAF/PqgnDtpTj6Vz0p/wCXt8SPk6N21ne/yvzn8B3ZmKo5XGYDG5mKpp6mqeoEpElNDIVVNJ+g/wBt72ksUROkV+3rxjmfLkflw6u3qNuxVM888lTP+8ZT4wx0IJF0kC/u31TCMRgDA49e+mGsuT59Vm9uY5NldtYyqjXTDLUwlzptfU4PIb/Y+0pY1qOPSiikUPDqzHa7U9btzC1KxxMs2Pp5AQoPLJduf9f6+9638z1rSnkOlEqKvCqBb+gA96JJ49boOuXvXW+ve/de697917qLV0VHXwmnrqaCrgb9UNRGssTfj1I4IP8AsR791759YcdisZiITT4ugpMdAza2ho6eKmiLf6oxxAC/+NvfuvfPrAu38Gle2UTE49ckxu1etJCKxj9btUgaz/t/eqDr3y6kZDFYzKxrFk6CkyESHUkVZBHURqx/tBJQRf8Axt7317rLDQ0dPSiigpoIaRVKLTRxqkCofqojXgD/AAt7917rjDj6KnjkhgpYIopQRJHHGqo4PBDKPr79x68BTh1josTjMc8klBj6Sjeb/OvTQRwtJ+fWUAv/ALH37r3Th7917r3v3Xuve/de64NHG36kRv8AXUH3sMRwPWiAePWF6OmcEGCLn8hFH+9e7rK6moPVGiRhQjpvlwdHICFBiv8AXQB/xPt1bqQfPps2yEg8KdFF+UNTFgtrSYuOtnL15jZEd7n1LosAf9b36a4Mq0KgdbjgCNq1Hrj8W9v5XC7aqMiadSk6pIsjK2plZ78X+vH9PeoVhZe856rIZQ1V4dGu/vEkFjVRsB9CUU8G3t42gYdh6olyTXUOHTpSZeirBeKTT/g9lsf6c+2HtpY+I6eWeNjSvTkCD9CD/rG/tilOPT3Xfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6bK3F01ahBURv8AUSIAG9vxzvGfUdMyQrIPQ9J9WyODLBgZqS/DG7tYf0H9fasiC5FRhuky+NAQpyOlFRZOlrl1ROAw4KNwwN7fQ+0ckDx8elayKxp04+2enOve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//Q3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdAt3V0XszvfB0e3d8RSVeHpZ3mkoCqy0tYJLB4qmByFZTpFwfeuvY8+i57O7h6P6D33jfjRsPYqbbSnr6egj/gWIWgxstXUOI3mmESWLX+rE/wCt7uiajpHmadWUF20jpEbr/mf/AB82B3zu3oje+Qlwmf2zNHEa+ulipcfLO4uKeOacBC9iDYNf/D2Apefdvgv5bOSGTRE5jMgFU1LxHqPtOPn1CG5e/PImyc6T8j7tdCG8iCmjVUHVwGogIWIzpDaqUNM9HR2N3l1b2FQrXbd3ttmqVgpEC53GmosUEl/H5ASLH8exPabztl6uqGZPsLAH9nUp7bzBtO6w+NaTow/0w/z9CtBPBUxiWnminib9MkLrIjf6zISPZmGDCqmo6OFdXGpDUfLrL731br3v3Xuve/de64SSJEpeRgqgXJJA+nvaqWNB1VmCip6B/tHdmAxu0snX7hxkeW25TsEyNLPTioSUIpdiYyCCAPawRRQLrm49JXlkkOmLgR0Uyk+aXxL616t3lvja9RtzblJgI6tK/BYZsXR5OqykVG89PGKKJlZizgLrKki/0PtPJI74rUfYB/g6eRUTu4H1qetWzof5cddfL35Xbn+XfytoN45nA9Q7hkwW3NlpjJKrB5jE0kjpjhPRWWNtICm+jk/X3WhAp15X1kscDgP8/V+fx8/mr7X7o7gwnU2xNjbgwW25DHRYlanB/a0VJQK/hpYQVjGlUUAAA8D3Uigr04GqaCuPkersKeQywRSMLM6KxH9CRyPeurdQs3i4s5hsthaiSSKDL42uxk0sRtLFFXUzUryRn/VKGJH+Pv3Xuq1em/5T3xh6o35n+wcpiP8ASdl9wGv+4i37jaLLU0C5AkyLCkxccXsCR71ngetaVDF1FCePVRPy+6o7U/lb/Kib5gfH/bckXSGYozisrs7BUTvhKWKeBFysD4mLVGGszNE2gfQEfT24KN28PTpty8Z8Qdwpkef2joqWH3v3F/OD+Y+29z7D2jk9sdY7VqsNlKrP12KkxddDUYtEkrIqqtZUAiBSygngWHvZGlT516qpMsimhAXOR/g63QNvVeNGNxuMpMpj6+egx1HSSikraepbVS06wOxETE/VT9R7b4dPnj0oPfuvde9+6910SBySAP6kgf7379Qnh1qoHHqtf+Z9HvOv+Ne+KLZL1pqZcRH9z/DtT1AQzvbQsdySR/T3Lns8bGPmqCS/pQNjVw4DoE89C5fZZFtia6Tw60Ttg7a7DTe2Bx8T7rh3Wa6rXXU0csMs8rVxKi7DUbcW99E9zutrNlLKBGYaLwNadvWEey2e+pfQRXDTi4BkJLLTV34/YOHy6+gP8Idu73pPjvsOj7BapXKw0kklpyVqtE1MkSCZX5HFzz75qe4t5tz803Em2Aaa+XDBPWdPLFtdrtES3hNSvnx8uix/Kz+T18fflRljnsnuTdXXuZlbyVeQ2XHT009bL6j5apjIhZjq5N/YBaZ2NeHQkFvF+IV6rX2t/wAJ1NpfHfcmR3r012Vure+5MhUGrSq3aFhq6OfX5NUNSszAHUBYg+2y7EGpr1sRRA1RafPoe+tfj5/NB657O289LVY7JbCpqqM1/wDEc9S1RipzOHkWOKaVm/SLce61FOHTgRga6hTrYD28cscJjTnVRcwaVP4gsZBQVPOsKV4t791vog/y7xy0u48RlgnpT7diwXgG4uSf9f3utOtAE49OjW9G505zY2NfXrFNTwxjkMRcE24/1vevt63jy6GT37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XjwL+/de6ra+VOejz27sThqdyyxvTxtGrBjcEAnSP8AEn37rWejt9T4WPDbHwcITS8tFG0gII+l1sQffseXW8+fQhvS08gIeGI3/qi/7f3cSOvA9UMaEUp0yVO3KSe5SSWEk3HjOkA/7C3tQl26mpFemjbqVoOoD02cx7AUQEsCgXLNqY2HJtz7fEltN/aYPTWieEUTI6zQbj8REdfEyyfkxobX/wB69tG0LAtGcdbjuWrRxnpQ09ZT1KB45F5/ssQGHF/p7TPE6GhHSlJFcVHUr231fr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917ri6JIpV1VlP1DAEf7z72CVNR1ogHB6S9dgjGWqse7rOOfGDZCL3+g9rorrV2SgU6SyQso1R5PWTHZ1WYUtaGiqEOksRZSR+b+6TW3448g9binPwycelKCCAQQQeQQbg/6xHtH0q679+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/R3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690iZ+u9nVO4Y90zYDGSZ2Odahci1HE1WJUN1fzkXuDz72DTh14Eg1B600v54/S74P5SYHctAKiGp7Lq5IAlMhi+6rIazxQMHB9THyBb2/A/p7iG7iG173fM4rHIDNQjz4Gn20H+o9cvfvY8qNt/P0XMFpGHk3BDFpI+J1YeGQfU6yv2BfTpE9JfBr5V5CDIVPTO8cpQ7t2nSirrMRkczU4tZ2ZFkEOOilkZZjYgaTHz7Bm3/W8wXDuNuC6DXtcJKAOBrip+QPyz0GOVPZL3dmDXvKu9NbXNsO6FnkjjJ46YwS6sPLujofl1tGfy9qf5QY7rT+E/JeKCPOUUYFGYh+4wGiMeUnm+kG/+P+HuW+Tv36kDw7uCFBPh6qa9OKaqYr69dDfaJOfYeX0t+ffDNygpWOukn17s8OPz4Y6sN9jXqXOve/de6hVtfT0MbSSsLgXCAjWf9h7cjiaRqDpqWVYhU9JWIV+4JSZbw48Hi3pcgn8Dj8e1pMNt8OX6TaXuCC2B055jauIzmArtuV8Cy0FfTT00wZFZtNREYWkF/wC0AxsfaGWRpW1N0rSNUGkdU7Zf+Rt8aMxurL7mqN79jrHmsmcnW4Va2nOMaUvqaMRX/Sfp9Pp7rVut+HGSSRx+3o33Wf8ALo+M3VlLBR7f2RiZaaNYxPDWYuikjrnjbWJaxALOx/JPvWerUAFB0a/b/VHWW1Xim2519szCVEIUR1WM23iaOqTSbrpqYohJ/wAn+/EA9eqaU6EAADgcD+g9+691737r3Xvp7917otXyF7u+P3W2zstWdvZfYuVoKCnnqJdq5uqweQrqwRraVKfDV/kLPbixj9+Ar16pXI6pH62/mo7B7W7fynRfxr6x2n1DtHILU46t3BUYHGbSeoWS6zVIraWOKMEi+m3vdCAAeqhwWNM0/wBWOriPjl0G/WGVqdyU27W3Tjs7QLOKiPJHI0YqKmPySrBJcggM3pI4t7rk56v5dHCaRE5d1X8+pgOP9j7sATwHVSyjiek9Wbip4GMUKSSy30ghdS3vb8e1cdqTlzQdJ3uMdg6b46fN5JxJUMsdLICF0nS4U/4X/Ht0vbRABMkdMBJphqfz6nS7TwtZRz0OUo4MrS1SBKinyEYqIZFH4aOS4I5PtsX9xHKJoGMbDgVwelAtoymiQavt6qZ+WXw/6o2NvLZfdu2tqYyh/uzkv4nnKWmxiJRtElSJAzFDYA8ix9zfyPzzvO52E/L95MzGVdKEtmtOo+3/AJe26zuo91ijAMZqcY6tM6t3dj997C25uvFw09PQ5egSeCCmAEMaoTDpQD6fp9wxvdhLtm6S2UxLNGaEnj69D2wuEurRLiMUVhjoQPZV0s697917r3v3Xuve/de6KH8sNuNkdrDJRoC9MAoYre2kE/X8e9UNevcOuXxLy4qtqVdCXDNTmMWuL6kOkgAfjn3scOtk+fRuvfutde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdRqudaamnnfhYopHJ/4Khbn/be/de6qkroZ96d1KinzCGvQBR+4LrL9PevPrfyHVqOGpfssVQUhUKaemjiKgWAKix4976105+/de697917r3v3Xuo8lLBKpV4ozfi+hb/7A+7rI6moPVSisKEdMFTt1LmellkEy/pQsQh/FrfT6e1S3ZbtkGOk30oB1Keof3+axhUVsayU4tp8YDNpH159uGKCf+yND1QPPGaOMdPVDm6SsAHqib8+T0gkf0v7Ty2rxn16ejuFY6Tg9PAYMLqQw/qCCP9uPaahHHpRWvDrv37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6901ZHFQV8ZWwjkHKyIAGvbi5Ht6GdomrxHp01JEsg+fTDDW5HDSJT16h6Y8RslidINuT/sPa1o4bkao8N0m8R7chWyOlbBURVEayROrBgDwQSP8AAgey90ZDRulisrCo6ze6dW697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//S3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Wvn/ADqPjdv/ALO3D8fN89a4KbL5PaGeeasSCm8wR/vkqI5pW+mlSoJUjkX9xfz5Hcw3Ec1vE0iyxSRnTxDGhUn5VwesPvvTclcyb+Nl3TlSIS3Nncq5B4U1A1PyUgEjzFQM06GfsDoDsWi6X6w7961inxfZ+0qRM7vrDwB6ZMtHSoiNCKRWHkI08rY3Hsuvti3CPZbbe7AabmEapAMVp8vPoebtypu8fLllzRsY8O/tgJJk4B6AVWnR6Pin8osF39tOnORNNh9/Y6AR7jwkzR09QtSj+MvFTvZrc/pt7G/LPMcO+WoMlFnX4l4H9nUk8j86WvNW3q8lI7pR+rGcEMOOOje+xV0POmHKZqKiHjhBmnY2AT1BDf8Ate1UFsZDV8DpPJOFOlePTfQYmeukFdlCSTZ4Yw3psfprX29NOkQ8ODy49Nxws/dL59KxESNQqKFUfQKAB/vHtASSanpWAAKDrl711vr3v3XuuEkkcMcksrrHFEjSSSOQqoiLqd2J+gAFyfeiQBU+XWiQoLHAHRSd4/N3487J3ZHs7L9hbYXLSVCUpRc5Rr453OkRyK3INz9PYbuOa9nt7j6cyBvUqagfafl59AfcPcTlbbb9duuLqMSMaU1jieAp69GgwG4MNufF0mZwOSo8pjq2COogqqGojqIXSVdSkPGf979n8E8NzGJYGDqfMGo6GVtcwXcKz27B1YVBBr08+3un+otck0tHVR05tO9PMsJvYCUxkRkn/Xt79177etZr+YH8A8FU7o3j8lPkHvTc9PhYoMg+08Pt3Ja4Kjcyw+fF01XSq/ETMBqGnmx97BxpHVGVfibIHzx0VL4hfAf5OfLjZTb+3VS7b2h18tRkaHZtfjIKPD7jq5MaoSklqWjMcrxkFQzsCCb+9kheGeqqHkWpIAPD7Oj4fy5vkR3P1R8zd7fy8+0sg+eo9iYStylDkkmOREUkND9xCqVKhjpZdOoa/r9fbqwl11cMV6oZtDeG2eGftx1f9/CqrJlTk6qMEgSrDAzpIqPyoZTb/evb5uI4eyEft6aEDSnVIen6kxtLSKQiBybeqQKzcf4n2meZ5OPSlIlQUHTgABwOB/Qe2unOve/de6Bj5A7UXefUm8sB4hJLXYxo4joDurhwRp/3n2IuVL47fvtvc1oFbPRTvdv9Tt0kQFSR0W/4Nb2qqrbeX6vqb367BpU128mmWoC2tb6D+l/z7FvuVt0cd5HvMf8AxKyfyHRNyldO9s1jJxhx0ff3GHQv6wz1EFLE89RNHBCgLPLK6xooAuSWaw+nuyoztpQEk+Q60SFFWNB0W6v+W/ROO3SdpT9hbWXKLOKeRGzMC+OUtp0suk259i+LkXmOWz+tW2k0Ur8J4ft6In5i2xJ/pzIur7ejE43JUGXooMhjaunrqKpTyQVNLKs0MqXtqR1+vsJzQy28himUqy8QRQ9HcciyIHQgg+nQZd24k5nr/LUqoXYLrAAu3EbXt7a6seilfE7KnHZvJ4OVtLGq8QRmNyddvp7rnj1bNOrEPdutde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdIDs7Nrt/Z2UyLPoEcZS97frjbi/v3Wj6dEJ+NmGkz3Y1duR0aSJK0yq5GtF/d+ur3qvVuPHqzX3vrXXvfuvde9+691737r3Xvfuvde9+691xZFcWdVYf0YBh/vPvYJHDrRAPHporsJSVvJ1RNbjxWQX/2HtRFcvH8+k8lsjnUMHplcZfDBihEtGLgL+uS9uPrf2oBt58HB6Z/XhFeI6d6DOU9XGpkDQOTpIkGkX+n19sS2roarkdPJco+Djp7VlYXVgw/qpBH+3HtMQRx6U9d+9de697917r3v3Xuve/de697917r3v3Xuve/de697917rBUU8VTG0UqhlYWvYXH+sT7sjsh1L1oqGFD0lJqOqwkpqKQl6Un1oeSL/AF49mCPHdAJJhukDrJA/iJkdKGgyUFdGCjaXsLxsQGuRfge0c0LRHPDpXHKJB8+nH2z071737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//09/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691GqKKjrAoq6SmqghugqIIpwh/qvlBt/sPdWRH+MA/aK9UaON/jUGnqAeuf21P4TT+CH7crpMHiTwlT9VMVtNv8AC3velaaaY9Ot6V06aCnp5dVC/Kn497k6L30vyh6SWqp6Whqv4t2JgKErGlXTCfUy0FHDb68XAX3FHM+w3eyXv9ZNjBAB1TIPPP4R1AfPHKV9y1uv9eOWKrpOq5jX8a/0VHn0eLof5Hba+QOwcXntr1dPDn6qnd8ngXlD5HFPF6XWspwLoSfoCPci8s7xab7YJfKQD+JK9yn5jy6k3lTmyy5t2pL2yYByKsn4lpxBHEGuKHofcThRTD7irPmq5PVJqsyBv9p9n09xr7Y8AdCeG3C9z8elD7SdKuve/de697917r3v3XugQ+ReQ3NjOnt71W0hIc0uAyq04iuZLNjpb6Lfn6W9k+/yXEW1TNbfFpI/Ig16DvNUt5DsVxJYgmQI1KfYevmw9j5HeWS7I3Tkd21WYG4Y90ZNpjV1NQKmOoWtb/OFzcMtgBb6AC3HuPdrt7KPbI47dVKFc4BJJ+KvzrWtfPrihv8AdXLbxeSXrMbkzStKZKmRZA5rk5GmlBSlABTFOt1j+RTuvsXdXxSyj79qslWxYzcqY/bVZk3mlmnw8cMnjKSz8sqn0hrm4APs85FeL/H7e3JMUUwC+gJWrBfkCafI48uumP3S9z3ndPa2OfeHkk0yyLE8hJZ4lYhTU1JHFQTxABzWvV2vsfdZQdcWZUBZmCqPqWIAH+xPvYBJoOtEgCp6Jz8zujKP5JdQZzZNI7yZvE01Xm9vCPS8Eubih0Ucc4JsV1fW/tUluQA8mK9J2nUnQBXqlf4bfJv5L/HnfVT8bvkRSx4HZ+DyU8/94jSfw7CUWLVgNK5OJdKgrpLEN/r+9eJBG2qPJHr1pVldKSDtPD59Fk/mFfMHq345d053uH4L4vcW8/kPvXFVOKzO+MPo3DgIpqkeORKepiV9VxfVq+g9slmlfW5pXqzHwE0xJrYeXVm/8nXdH8wjtzZy9x/L7OU9Tg89HVQ4bHVCrT5W32wenlNKApCKzKCSPr7owAbGfn07GSy6mUKT5enV6nvXV+ve/de697917rBU00NXBJTVCCSGVdMiH6Mv9Pdkdo2DoaEdaZQwo2R1Wh1zUP1F8j90YttNJRb7yhpqJJgIvOrVHp0J+T/T3MW7oN95RhnHc1stTTNMdAaxYbfvskZwJjj59Wbe4b6HXRSPnHmN44L4z9jZPYS1DbppscTjRS6vOGaN1kMejm+m/sc+3MFjcc3WsW408EnNeHl0HuaXuY9jne0+MDFOvnv7jyvZdR2Jlc9kMvvOLesmRopHoTlp1V5/uwW0QX+n9Rb30ztItqG2JbRxxG3Abu0+VPXrBK/O9LvD3hmuFuSY/wBMSdnHyX09et7H+V/nux858dNvv2EtaKiGmjGPNfrM7QFyCWZ+T9PfO73ittqt+aH/AHZTPxU4V6zW5DlvZdnU3ta4pXqxPMUkddi6+mlUOslJULZhcajCQDb3EmPPocZ8uqwOrKufand9XS1ZMUE+UJRfoNBl4sP9b37zqevZAI8+rTopFmiSVDdJFDqf6qwuPfvs691k9+691737r3XvfuvdBV2n3DszqPC/xndWUo6VC+iOmkqo4p5iBciNWub8i3Hv3Xukl1d8lesu1KbIS4jM0lBU42B6uqx9dVRrVx0cal3qnjAFkAF7+/VB69npAUHzY6YyG+P7kw5yi8zVX2cWWFdE2OnqdWjwRSActf8AF/fvKvWul33N8l+tOk6HH1m58xROciVaKmhrIlnWFwCkzKQbKQeL29++fXiaCvTji/kR1rl+uk7JpM7QPhpImkEQrYWqNS8lCB+bc/S3vdPMdeGcde2b8hut98bYyu5sNnKCWLD089VW0Iq0aqhhgFy8gA4B/rb3Wvl1voQ9ib2xPYG36fceFJbH1LOkUmoOrlDZtLiwPvfXull7917r3v3Xuve/de697917r3v3XuipfKvcox+ypsKsojlr9BX1aWJN1UAAc+9GvAdeFK56a/iPt04ra9VWTxfu1SoyOVF7M92Or34EeXXiKdHB97691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XRAYWYAj+hAI/wBsffgSOHXqV49NNfhqSuX1AxsP0mM6Rf8AxA9qYrqSLHEdMSW6OdXA9MQfJ4MDy/v0pPpVfW4/ABt+far/ABe54YbpKzXENK8OlBQ5akrlJRvGwNtEhCt/vPtHLbvEc5+zpZHMr/Lp09sdO9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+6910QGBDAEH6gi4P+wPvwJGR16gPHpLZHEy08v32NJSVeSl/Tx/h/sPa+K5Vk8OfI6RyRFG8SPqdisvHWp45f26hPSwcgamH1sPbVxbmM1XIPV4Z9Xa2D0+e0vSnr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//1N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XFmVFLMQqqLkngAf4+9gEmg60SAKnpGZepps+WwT0cNZj6u8NatTCtRBJG3GkxvdSPp9R7WfTIkRaYBqjgcj8+krmK5/QlUMh+IEVx1T72j0xvX4RdtN3t1G1dVdd7jr/ALvsnCxsWRaZpryx0VPGSIwePoPcK75t+4cpbwd82cn6WQ1mT5V4AdY+b1y7f+2vMDc1bAGNlcNquoxX9qgcMccZ6tq6l7X2p3HszEb02pWwT0mUpUqJaJaiOWrx0jEg01ai2KuCPyBf8e5L2rdbXd7Rbu1aoYVpXK/I/Pqdtj3ux3/b49xsXDLIK0BqR8j6HoTfZl0cde9+691737r3XvfuvdYKmmhrKeelqY0mgqIpIJY5FDo8cqGORWVuCCCQfdWVXUqwqD1V0V1KMKgih/Pqprvf+Tp8S+4s/md91eDy2H3TVpVV5/gssNNR1uQWHVD91SwhA4ZlANzc3+vsGT8l2a+K9jNLD4gbtVu0E+YBGM+nUAc1fdt9seZ72beLuzK3UlWZo2ZNbUoNYUqG4AZzTFadQP5fnYp6hy+T+Ke9qGDDZvF1s77RUQR0KzYOkV1p0aJBZmKAENfn2Qcjbi+3XD8ubgNM2olPIkZyfUn16c9ptzXlyR+QdwURSQk+CAAo8IVC0AxgUHVtlbkKeij1yMC5W6Rg+tuL3C+5ZihaY0Xh69TxJKkS6m6TiPk84WteCiLWKsNL2t/X2t/xe2Gcv0kIluW9E6fMfh6TH6miDM7ghy5uDcci3tLLcvLg8OlaRKufPqh/+eN1b8ne0Ov+odl9A0mLpcHuLsBMfv6qx1BGuckwNW0VPKr1w9aroLEaLC/19p6gGvV2DMpXyx+zz6g0a/y/v5UfWP8Ad/F42i7Z7Z/g1LksttPLVmK3PuePJzUUbV7faTwyGmV5GfTEQWA/I+p3xOetHRGK8B/q/b0PP8t75d9g/KjdW5s7/o9zHXXVkdNU/wACwFdjHx0FNMoDQmKMIiAEEAWHvxpwHW1J8xT7eri/eurde9+691737r3XvfuvdVsfNDA1+G7E6n7MxyGmxu061KzOTxARxsi1RYGZ1/qPrx7l/wBvbmO52m+2eY1eYUQH7PLoDc0RPDe21+gosZq3R/8AZe6KPeu18NujHlWo8zSLV05RiyFCxT0sfqLg+4t3Gyk269kspfijND0MrWdbm3WdODCvT3kMdRZWkmocjSwVlJOjJLT1MSTQyK6FCHjcEHgn2nhmlgkEsLFWHAjB6ckjSVdDioPVIHzI/l4dC7Mr3+QmI2/JBVYutpqnJ0cdJSGjMrzhlIC6bAkfTTx7yK5B90eYtxj/AKs3ElQwIU1NeHUUcz8lbNBL++o4gGWlTQevr1bl0S+Mn6h6/qsRS09JQ1e26GohhpoY4I1Eikt6Ixa973P9fcG8zCYb9dJcEsyyEEk16krahH+7oTEKAqOhYcBkdT9GVgf9Yix9kXRhxx1Vn3DTSbR7kxtZGjRLLUws2kaL63B+q/X3riKdexSp4dWXbZrI6vbuHq9a6ZsfTuWJAF/GAbk+9hTw60WAOT07mspAbGohuOf84vu/hv6HrWpR59RJMvSRrqD6+bekg/n26LaQmnTJuEHUd87Tp9IpWJ+gA92Fq5zXqrXSjgK9Eg+V3WvTO8MS+7+5sxV4zAYYPkaWihq3p5q6oo2DNSQr9Cz2AA92+lGqgbr31NDQivVFWVy27Kvt/efbnSOC3dtXqPJ7fyG0SmUaqigNKlOaYzJLZVJZfUGt719OAaVFR15pWxQY6FTN9Jdc7e+HfWPaGCfcEm85uynmnyIq5WikqBUCSSA1Q+tj9Bq/P09+FtU1LCnWmmoO0GvXP48Yui+Yfyj3v113lNm22/t7aUVRjFgrJVWN6amGg+sEDgXvb3rwkHDP8uteM5JrgD8/5dR+rdnbl3B8iO6vjzs+fNzda9e7fyOVxtPJUSPJOtMOBHG3Gphxx72IEDHI6v4shWqjGOkDtXI7j6J3jlIKr+Nf3d32Z8Jk8RJUzLVYyllfwvLUwf2B/Qke/Lbxt5ivWvEcfEOrx/ip371RNg6TqnaUkq1G24GZ5Kip8grJZYxLJ4tQ/VfiwP492+jzTUOtG6pwU16OuNwR2u9LMnAJB+vIv719IfJh1b6inEdZ485SyfUOn49X+291NrIOvfUpSvUpMnRuQPMik/6o29tmCQeXV1mRvPqStVTPws8RP9A639tlHHEHpzUvr1nBB5HPuvVuPVbfyqzq7g3hh8JSSX8MkMTIGP1Vhqun+v78acevUbz6O11NhI8JsfBwKgWR6NHkIAW/JA4Hv2PLrWa56Er37rfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XFlVhZlDD+hAI/3n3sEjh1ogEUPSdrtvwyapqQtFU/2Tqso/PtXHdtTTJkdJntgcpxHUOny1ZjZDS5RWlIICzoPQBbi593MCTjXDj5dUWdo28OTpUwVMNSgeGRXBF+Dcj/AFx7RMjIaMOlYYNw6z+69W697917r3v3Xuve/de697917r3v3Xuve/de697917pP5LECVjVUv7dSpv6TpDW/wHtXDcEdj5HSWaCvcmOuGLy5eT7Kt/bqFvZ3OkMAP6n3a4twB4kWR16CUntk49KP2i6Vde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//1d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691xd1RS7sFVRcsxsAP6kn3sAk0HWiQBU9I+vr6rKzGgx91iN1lm/ssp/o3syhhjgXxZePl0jeRpGCpw6UGNxsOPgWNQC9vW55JN78H2immaVyfLpRHEsefM9RNy7bw+7sJkNvZ6iir8Vk6dqarp5URw8TG5trBsf6G3tHcW8V1C0E41KwoR1S6tYL23a1uVDI4oQfTql2uxHYH8v7u3+K42SsyfRnYGVFRko/Iz0mCpTVGyzFbiIrbj6cf4H3DzxXvIm9+LHVrK4areiivn6dY6y2+6e1HMpuIKybTePV/MQ5/kOrlNl7029v/buN3PtnIU+RxWTp0qIJqeQSqFf+yxH5H0PuXrS7gvrdbm2YMjCoI6yE27cbXdLRLyzcOjioINePSr9qel3Xvfuvde9+691737r3XF2VFLOyqoHJYgAD/En3sAk0HWiQBnqpv8AmFdRbkU4T5BdMRTQ9nbcraShnrqNVSH+BUZU1bStD6iQgYaj7j3nXl6ZWTmHbzpuIiAaY7f9Q6gz3Z5e3Fkj5t5YFNwt2C18jGCNYNM8K0+fRz/jd2Xge7+ttv7pirI6/L4mho8RnzFMZAmYp4bVflv+SwN/Yv2HfYd124SWrVpQP8mAFf5jqSOU97tOZdnivomDMihHoeDjj/PoyyqqgBQAALAAW+nszJJNT0KwAMDrv3rrfSN7BTIf3K3TNhoKabOQYHKyYU1MCVCR5IUb/Zvocfh9JHv3Xvt61KfjX1Z8fsX85ezO1PnhuMbb3hDHX1ePr945j7Ha9fWrPaCDwVAZTZOVQLybD3stgAY6bRCXJfJHD7Oj398fzits7W3FiOq/i51Rk9501ZkYcXTbs2JRefEJDE3gp/BFjYNITRZjcn/X9+K+fDrevOkCp+Xl9vV1Hx63XuTdvWG3crvVpI911tMKrI0dSjw1dMJ40ljjmhlAYEBrHj3rPn1eoOR0OPv3Xuve/de697917ov/AMndpDefS+88JHCJKqooB9u4RTLHIr3BRzyPr/X2KeTL7938w29wTRQc+nRPv1v9VtcsQGSMdBD8It+jMbA/0eS6jWddQDGVDOzO5DVBKh2P5uT7Pvcfazb7p+9R8N2dQ/Z0W8qXomsvoz8UGD0d73HHQq6A35GbPTffU249tSAMtakbEEA/5gNL/a/1vYl5Svztu+Q3Y/DX+fRTvdut1tzwN59AH8IOxW3BtHcm0aib09eVSYeJGkZikMbupsrcAXA+n9fYo9yNqFrfQ36D/coaj9uOijlS88a1e3J/sTpH2dHIn3Eq6kipZmYAi9iR9P8AD2AEtK5Zh0IXusHSOHVffynE7bgx+aaBofFJBZmXTf0g3v7YniWNqA16fiYyCpFBTo0PSVTk9x7IoZXqkeGKCKBQHYlb3JsT/re3kuIVWijI6aeF38/PoYE2tSag8jOW+psx+v8AUe9tescAdaFvmpPU9MDQJxpc/X6v/X22buU9O+AhNTnqZHjqSJgyxC4+mr1D/bH2yZXPn1tYUXgOqdP5pPwt+Wfyxwu3MP8AH7sLBbQpcRm0yVRT5WuNCksV0LqWAOr9J4Puus1zn8+ttErKQDp+dK9OPxs+IHy623s2u6u793zsvcfXsu3FoqSixFLRvWw5cUhgNSahUDEarEknn3U59f29XAIFKg/l0GmE/l9fK+lzGG6+yfY226n49YPc7bjotteSJ61JXn8j/tEfqI4+vvXzHH7et0rgnH2dDH8hfgh2lT51d6fEfcuI2Fu7KY2DG7jrMm0NOKiKKPxv4JFF7Na5BH59+p5deIqMYP2dD/8AEj4uZzozbWW3d2DU47cPdmdpJ4M9n6MRyUtTCFVo4g6jU92Fzc/4W9+z69eAoKft6Q9V8Ns3vjcnb29N5TY2TIb3wlRTbViihpljxle7lo5zHf8AbH5vb3vh17jx4dBl8M/gN2J0fv7Kbn7Jz9FmKEvPLioqGqikdJW5h8wQk2HF7j3rPWgqgep8urb5KaGXl0Ukix4HtxZHUUB6q0aMakdQ2xFC5uYzf+ur6e7i4kHn02beM8eok23cfL+HB/4Ne/t1byUGp6qbZKUXqA22zEWall0k/TWxsP6f19uG8Dijjqpt2AonWGoXPY6lnqHqYjFAkkhUXJsEJUD37XaOMjPVBHcR/CcdVf5A1O7e4w7h6loK+xjUsxOmW5sp/wBb2kKqW0r0qViBVurR8BkIKbFY6kaKSJoadY2QixQobEEe3TauBg16b+pUnIp0o1rKVzZZ49V7aSwDX/pY+2jFIvEdOrIjcD1J9t9X697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917qNU0kFVGY5kBDfmw1D/AFj7ukjxmqnqjIr/ABdJSfH1eHlaooCWp73MdyW5+ot7MEljuV0S8ekUiSwvrU46fcfl4K1SH/YlWwaOQ6ST9DYH2klt2iOM9Ko5lcZwenf2n6e697917r3v3Xuve/de697917r3v3Xuve/de697917plymJjrP3o/RUJ6lYcaiB/h7V29wYjpbKnpNPDrGpePUTHZYxv9nXnxyrwsj2VWAHA9uTW4b9SHI9OqRT6eyXHSkBBAI5BFwf6g+0HSzrv37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//9bf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XTMqqWYhVUEkk2AA+pJ97AJNB1okAVPSKyNbU5mp/h2P1LAjWnmX9Mi8cK3szhjjtk8WXj6enSGV3mfQmB0p8fQRY+nSGMcgepvqSfqeTz7QzTNM+o8OlUUQjX59T/bPTvXvfuvdITsnrna/a2zszsfeFCtfg85TGmrIgEEwW+pXhkcHSwP0I9ody2613WzexvF1RuKH1/Lot3fabLe9vk22/QPFKKMPl1UJ1nuff8A8DO5Y+rd/wAlVlOnN6V+namakmklxu0sc89oRX1CEqpA/r7inbrq/wCRd3G1bhV7OY/pt+GNa+Z6gPZrvdPa3mH9x7oWk225b9GTOmBfJXP+A/l1dJiMxjM/jaTL4aup8ljK+IT0ddSv5Kephb9MkT/kH+vuYIpY54xLCwZWyCOBHWRME8NzEs8DBkbII4EdOXtzp3r3v3XuotVWU9GheeRU4uFJsW/oB7cjieU0UdUeRYxU9JFpcln5WWAtTUKmzBhYyWP1U/0I9r18C1WrZbpGPFnPy6e6jAUE+FrsM8EclPX0VVSTJMqyq4qoDDJqDgjm/stuSbpWSXIYEfkcdKHt43gaAioYEGueIp1TNsaqyPwV+T1TsTJz1FP0pvt5cr/EJ5XFBDm8pd4Y0I9IZZJLBbe4bsWk5J5kO3uSLKbuqeGo8OseNrM3tjzo21y1G2XhLhie1ZXPAeQqTw6usoquDIUdJXUzrJT1lNDVQSLyrwzxiWNlP9CCD7mFHWRA65BFR+fWRcbrIgkXgwBH59Svdur9dEAgggEHgg8g/wCuPfuvdUm/zSP5XHXPzDhw+/s7mKbatHtOpbM7yrBVxYZKrD07rJUlqlALsEU8ueT/AIn3sEg9UeNXFD1VHU/J/YPx5w8/x9/lwdF7trN3LSvhYu4qmmTddLJmin2UtdSTNE4jUE3B1cfX3unmxp1XUQNMS1PoOH5noe/5cPyt+Z3W3dtX1x8wNq763vmdxOKODdQoq2PH0xrQHozGdBispIVuPpf8+9MFGVPW4zIcSgD0pw62kwbgG1rgGx+ouPofeunOu/fuvde9+691Dr6KDI0k1HUqHgnQrIrAMCP9ZuPbkMrQyCROI6o6B10nqsfo7J0vUff2/MBLIkUO88qVpY3PEhFQWTxKnH19zLzJA++8r210oqbdc/LHQD2qaPbt5mg4eIf29WPV2anjqno6amkZlbQZgt1BI9xJHbJ4YlkP5dDSSdixRBSnn031WGzGVppYK6qQ000Uo8TDka4igv8A7f27HcwQSB4lyOmjBLMtJTUdV2/HSig6y7s3btCZWh/vhmJpwhNhMvm1AqqcH/XPuV+bJG3rlyG/XPgKB9mOghs0SWO6SWpx4pr1aDHTQRfojUf42BPP+J9wwXZsE9DxY0U1A6Kb8rtufxPagr/GP8nI/cCgW0Lq5b/ePdOrcOoHxK3Ek+258Q8gLRGIItyTrUlf96v798+vClOjke/db697917r3v3Xuve/de697917r3v3Xuve/de697917r30+nHv3Xuve/de697917r3v3Xuve/de697917pBdl5lMFs/K5B20iOPQCfoCyMf+I96Py69SvRBvjdh5twdkZDcUo80MVcX1EBlIE1+L3/ANv78Cw68QvAjowHzK7azHRHTWY7I23tLLbvyePq4I2xGEilkq5IZEZ5ZiIASAoW5Pt1ZpEI0nptoY3qWGei+/E358dKfI7GUeJr6/H9bdleKniO0M5Wy/xmtqXPjdoKaUgtY2uP8f6e1AvGB/UyPTpgW4Ydnlx/2R1YIz53FFWmk+9p7gJ4kNyum4BX8H/X9uMbafKDSetATRcTUdOFPuKKWwlppqdvz5OBf/Dj201o3FTXq4uRqowp0+Q1dNUW8MyOT+FYX/23tO0bp8Qp08siP8J6ke6dX697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6YMlhFqT56Y+KoBuDeynm/49qobkoNL5HSaSAE6049QcflqimlNHkle6nSs5BC/4cn29Lbo6+JEfy6bjuGB0yDh0q0dJFDIwZT9CDcH2gIKmh6WAgio65e9db697917r3v3Xuve/de697917r3v3Xuve/de6Ys1h0yUQZLLURkMjDgkj6Ake1dtcmE0PDpNPAHGpeI6jYnJSI4oK46ZkGlHYWUhRYLc/n3aeEMvjRcD1qKWh0SY6U3tF0q697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv//X3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddEgAk8AC5P+A9+690jMpXz5SoGMxxIUEiolB9JQnSQCPZjDGtuvjS/l0kkk8Q6F6UWMxsOOgWJAC9rO/wBSTe559pJpTK5Y9PRxiMfPpy9s9O9e9+691737r3XvfuvdAr310ptrvbrzNbF3DTxaclEv29aERammmjuY2jqLFlF/rY+yje9ntt7297G4GG8/Mfn0HeZ+XbPmfaJdqvFBEgpXzH58eq0vjN3VvP4wdmL8Zu65qqfBV9b9l15nqqWQ0lBjkm0RRySnUGVh9Tf8fX3HPLe73nLO4/1b3gkxsaRMeCr9vz6hzkvmPceS96HJHM5Jjc0tpTWmgcAT69XJQTw1MMdRBIksMyLJFKh1JIjC6ujD6gj6H3LgIYBlyD1kIrBlDKag9NmUy8WPQhR5ZzwIluWufpwPaqC3MpzgdMyzBBQZPTHRYyoy7/eZQsYi14obkEKD6Rf/AGHtVLMluPDg4+vTMatN3ydLGONIkWONQqIAFA/oPZazFjqbj0sACig65+9db6Kd8wug8Z3z1TX4meNUyW35JNy42eIJHUvVYuA1EcCz21eooLC/sL827HHve1tE3xR949armlegNz/ytBzTsT2rijx/qKeBquRnjxHQQfAb5DVHZGzKvYG85jRb62ZWTYeHFVRkNbLhsYpghnOv8AJ9b/T2U8jcwHcrM2V52zwnSFPEquK9EXtfzY+9bY227j2XdsxjKH4tK4B+wgV6sQ9j7qVuve/de6Q/YnX23O0No5rZO66eWpwO4KCfG5KCGUwyS0tSumVFkANj/jb3oivXuqyu7t9fGP8AlBfH+tyuz9mwSVmSfJVe28NXOtVltw5eMKzUENesYlOsuAEUj8e7cT1okKK8B1rq9afzP/nD84O0t2bt2xtnNdb4fYMNfuCHZb4kx189NQyGWGONxGJGLCwX/fH3vSoOeJ6ZDu5JUYHrx62f/wCX18zW+UOw4sfufG1OD7I27TEbkxdcJUrQsZWNKmaOYAgsSb+6nB6eXIrw6sVeRIxd2Ci9rsbD3sAnA68SBx6Y6rP00LtFCpqJF+ojN7f7D2qjtHYVbHTD3AX4c9NqfxrKSB45WpIOdcbqRqB4sL/4H28fp4ARxPkemUM8rZwOq4vlZtV9j919P7+oEeLE0NVFU7hlDaY2KVFmMmmxN7fn3LXJN9+8uXb7a5TWRgQn7Ogbv9r9JulvfD4VPd1ZltzM0O48NQZ7HWNJlYFq4H+paN/SrX/2HuHry3ltLhrWX4ozQ9DqB0miWZODCvT59ePaXp7qr/5CSybD+VXVOdoomp8VNHFNXyqLRiRpdB1hPr9Pz7mflUDdOSb22kNXFQB+XQC3kmz3+3mAop4nqzPHVsWSoaWvgIaGrhSeIj6FHF1PuHZojDK0TcVNOh0ja0D+uegy7rxJzOwcrSKLsFZxa1/TC/0v7ZNaY6vjz6Jn8UK00W7q/EOxVvunXQx/oxH/ABr37ievV8urJve+vde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690Vb5TbkGO2bNiPL4nrlUpyQXLXXi3vRPXq+XTF8RduPjNtVuRqFGqrAKMQPoXJJH+tb+vvwp6dapTPRfvmr/M56q+JW78R1tlMEu+t07ix/3EG3qOo/fnjmiJEJh8bq4YcEH/ABB92AJ4daZlWlfPrWI+Y3Zqdubu/wBMvxN+Oe/+q+2oK2CvjytJU1r0SvF+5UmnpoVULqb6W/1vdl00o3Dppg1Q0Iqf2ft6tz/llfzZ957iTB9J/Lra+c2Lu+OKjoKTd+7zWUn30okMDGZapLsrEgqw/wB691IplTUdOK5PbINLenl1sbxjGZukpq+nkiq6SqhjqKWpgYNFPBKuqOWNx9VYG4P9Pd45nT4T15olY9w6aG2/LT6mx8whc3INzx/Qe1ovEcUmFekj2sgNYTTqNHlsnjHMNfC9SL/55VYqFBtyR739PDONUR09bEskR0SZ+fSjpMpS1agrIqsf7DEXv/T2jkgkj4jpQkyPwPTj7Z6d697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuoVbQw10RjlXn+yw4IP+v7dilaJqjqjoHFOkxFJXYOYRy6pqQtZSLkKvtawiuRVfi6RL4luTq4dKymqoaqMSQuGB+oH1BH1B9oJI2jNG6Wo6uKjqR7p1fr3v3Xuve/de697917r3v3Xuve/de697917plyuLFaqyRHRPGdQYcarcj/Y+1VvcGLtbIPSeaDxO5cHqJiMvrkNBVAxzxelWfjUB9Pr7vcwf6LHkHqkEpH6cnl0pfaLpX1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/0N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SUy+VllmGNoAXlYhZnQ/oR+CTb8D2vtoVVTLNj06RzSM5EcfTvjMZFj4+LNM1/JJ9dRJvwT7TzTNKc8B0/FEIxTz6dPbHTvXvfuvde9+691737r3Xvfuvde9+690Vb5SfGjC/IHaE8NMYcRvzGQA7W3P8Aolxs0TNKqM6AtpYm3+HH49hjmbl2PfLQiPsnUdj+ny/PoFc68nW3Nm3NED4dyo/Sl80YcCOiZ/Ef5V7n23uaq+NPdHno96YmqGH2rla5pdWagikMYqKcSfrQ3uCPYd5L3+4iuf3Bv1UlQ0Qt+MD0+R6j3kLni9s9xbkjmclbuM6Ynb/RFGNa+o9fn1apQYPxO09c4qqjVdHN/SL3tY/717lWW61ALENI6m5INJOs1zjpQgAcDgD6Ae0fSnr3v3Xuve/de64SRpLHJFIAySI0bqeQyOulgQf6g+9EAih60QGBU8D1TX8udkZ/4u9y7f8AlH1zTSSYvMV1FtrO4Ki1BYlqlEdXWyU8IF1ILMT/ALf3EvNVnNy1vCczWC6lchGQeVeJp1j7z7ttzyTzBFzztAJjcrFLGvoxAL0HpWv2dW07E3dit8bVwm5MRWQ1tNksbRVEjwtqEdTLTLJPA/8ARkYkEe5RsrqK9tUuYiGDAHHrTI/LqddtvoNxso7u3YMrqDUetMjpXe1XS7r3v3XuqHf543xw3P3ZsnpHeWDxlZmMb07u+fd24MfRs16jHQzU8rxyIAQwPjIsf6+9g0NeqsKr9hB/Z1WJ3/2VJvrrrbPcHwiwNRsXsjF0yp2lsrDwJLlqyixaLCwkjp11aJQrEqRbn3YDNCP2dNlmCFoqH7f59LD+Xj88OrOkd553f/c2To8V2N2RGcTnts1cslNk9u/byoTV1FPayh3TkFb2NuPbiQl2oxp9vVWmCAE8T1tJ7G3TjO2dtYvfG1s9T5TbWZhSehmpg3hddALhLk/Qm3PtQZYIe1BUjz6aEc0p1OaD06EWlw9FSsJUiBlPLOebn88H2nkuJZMMcdKFhjXgOnX2x070WL5b7M/vf0ru2CjivmIaSNsfOoXyRMst2CsRcfU/T2M+RNwNjzDAXP6ZJ1D8uiHmK0F1tkoHxUx0xfDjftPuTqrDbUdzJl9kUEWKyrksSZBIdGrV+eTf2o9wNraz3uS+H9ncksvTfLV4LjblhPxRDSejdewJ0IuiN/O3avm6iye88bGzbgwVRQCkkQL5FiaQ69LfqFv8D7kn20vdO+pt8p/SkDV+3oK82W5bbWuE+NKU6Hn49bmg3N1FsSqE4mrotu0EWRTnXFVRpokD6vySL39hnmuzaz365SlFMhK/MdG+zzi426J61OkV6FjM0kVdiq+mlUOslJUCxtw3hYKRf2HRx6Mjw6q96wqZNq931UM48cUmTYqOQCDMbWtb3qo49bz9nVqEMqzRRzL+mRFdf9Zhce/de6y+/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6rc+Vuf8A49urEYOm1KIXiiYXPNnsTx9eSffjQHr2ejo9T4dMP17h4Uj0SvQM8hsLs1iAbj6+/daA9OtMz5j9t0nXH81Xq3fvdHRW4N67G29kIRW5mV6lMRT0McpI8xYaCAvqseD7uMinn00TobW4xwH2nrY96A+a38vnerUEGy9w7E29n8mAVwNXTU8s9JJNKI0pxIY2sxY8ce288D05VTkdOHzS+Am2vl3V7X3pt/K0WE3Bi/4fNj8zCEp4ZqGBhInqp01vqAFif9692Bpw68yhqV8j1YVsDbkuz9kbS2rPUCqm27t7E4aWpBJE8mPokpXlBbn1Fb8+9DHVyamvSu9+611wdEkVkdQysCCCPwfewSpqOtEAih6ZJ8FCCZKMinlJuWNyCf68e1IumIpLkdJ3txSkeOm01WVxEgFQHroj/aTVpRbABj7UBLe5+DtPTAaa3w3d0+UeWpKuwWRUkJt4yeQfoR7SSW8keSOlMc6yccdOntjp/r3v3Xuve/de697917r3v3Xuve/de697917r3v3XusU0Mc6GOVQyN9QfdlZkNV6qyhxRukpVUtXh5vuaR2akv+5CtybE/T2vSVLkCOTj0heNoG1R8Pt6UNDkIK6MNGw8lvXHf1Kfz7RywtE1Dw6VxSrIMcep/trp3r3v3XuoeQrqbF0NZkayQQ0lDTTVdTKRcRwU8Zllcj/AAn37rxx0WTq/5b9U9w74rNlbC3BQZ6px8lRDWyUpk1QT04PkiZWP1BFj78fl1oeh49Gn9+631737r3XvfuvdMWUxiTA1UA0VMd3DLwWAHI9q7e4KHQ+QektxFqXUnHr2Hyn3a+Cf01KXurcMwU2v71cQ6f1F4HrcM2rsbj69PvtL0p697917r3v3Xuve/de697917r3v3XuuibAn+nv3Xukdj+wdm5XLy4HH7goKrMQyGKWgikJnSRTZlKkfUfn37r3lXpZe/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//R3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SezWU8AFJTHVVSnQQv6oww4c+1trAGPiyfCP59JLmRgNEfE9ZcNiVoI/NKRJWSj92W9yQ3NufdLm48VtK4UdWt4ig1NxPT57S9Keve/de697917qFkcjR4qkmr8hOlNSQLqmme+lFva5tz7916o8+slHWU2QpYK2klWamqY1lglX9Mkbfpdb/AIPv3XupPv3XuuiQASeAOT7917pLZHLSVMooMYxaR7K86fSM/kH/AF/a+G3CL403AeXSGWcu3hxH8+iY/Ln4nQ9qYJN/bGYYXura8UMu3tzwMkDwxUyl5YQsSglmsADf2AOcuXf31F9dY/p3cX9m/mAM06AHuByQm/2w3XbKRblbj9KUUquM8PXzHSY+FXywqd9wTdQ9rf7ge0NqyjDwQ5LyrW7q8DlJMhBrve55/Hst5O5qa+X907p+ndRdorWsnzHSH2656bdUbYd8Hg39udBVsGSn419QfLqx/wByF1LfXvfuvde9+691737r3SE7J2PiOwdnZzbeXo4quOtxtdFSeVVY09bLStHT1EeoGzIxBB9odxsor+0e2lFdQIHyJGD0W7tttvuthJZ3KhgykD5GmD1VX8Oew9yfHzuXcfxU7MyHlhMtdn8Hnat5GgdagGWloY5W4F1IA49xlyhuF1sO7Scrbma8XVzwzwHUH+3O6XXKnMU/IW8uSBqlikatCGJoteFRw6uOVgyhlN1YBgR9CCLgj3LfWQQznrv37r3Sbzv8CzFHVbdy0MWQo8xC+OrKJ1DpPBVDxSRuG4tz/sP9f28sDspbgB0006q2kZ61hflnjcD8efl71xt/pfCTdJ47d++cfitx7grW89FuaiqsksdVDBCwKhDcjSb8fU+3WZIVATJHTA1u5rhW6I8eoPj9vP8AmI/OTZu49nSb9zuO2jWvsKqx1WaNY8+0cbx1UcMfpNySQqj6+2JZHejE+Z6dREVmBFTRetiv+Vdtvs3ZHTNPs3fNDkMVjsSk7YjGV5kL06zSRkN+5/VLAe6Hj06tQKHy6tU966t1737r3Tbl8bBl8dVY+pXVDUxMjCym9xxw3Ht63ma3mWZOIPTcsYljMbefVbvxfqI+tO5ezdqVSkLujNMcepJ9LLUekoOAB/sPcvc5xneOX7O/T/QU7v2dAjYG+h3Oe0P4zj9vVmnuG+h30hux9sUu79n5jCVkaywTU0srIwUgtDC7IbMCPr7M9nvX2/cI7mM0IIH7SOkl9bi5tmiPmOiSfAvL10UXae28tUHVit1eDFwyM11o43kj0Rhvx9DwPcje51vGzWV5AMPHVj88dBblCVwk9vKfhegHy6sRkUNG6kXDIykf1BWxHuJuhp1Vr3HR1G0u4sfVwqUSepickDga2B5K/wDE+9UoetVFM9WYbYqhW7ew9SCD5cfTMbf1EYB/3r3vrdKdPvv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rDUTJTwSzOwVI43csfwFW/59+HWj1VTmll3t3WKWNzMkNeoAH0Fpv9j70ePXhUHPViW6N47c6i69m3Hu3IQ4zEYHHN56iYHx+VIXljhsLcsV0j/H3vhx63x4datXyD/nBdW9gdh5HZm+/gS/ZPXcORGMq+wYGqaWP7WYmm+6epiQgWQkr6vdtJNOA6aMqBjUMafLHRzuh/iJ/LY7byeytz9N4VNhdhbjo4M/DtOpzU1VUU7wTCQwqCfqCCQLe9E0x1cAFqgD7Rx6v427iI8BgsRhIm1RYrH01DG1ybpTRCNeT/gPdRw6ucnp597611737r3Xvfuvde9+691xZQ6lWAKsCCD9CD72CQajrRFRQ9MFVgIWJkoyKec39XNr/UfT/H2pW6kGHyOk8luGHZjpuhydfi3NPXo9Ql+KjkIv+xP49qHghmGqM0Pp0wkssRpIKjpUUtbTViB4JFfjkA8r/gfaGSJ4zRh0tSRXGOpXtvq/Xvfuvde9+691737r3Xvfuvde9+691737r3XFlV1KsAVPBB+h97BKmo60QCKHpLV2LloZDXY8kBTqeFb3Nzc2t/sfa6OdZB4cvSN4GjOuI0HTrjMnHWxqGIWdR60PBuDY/X2xPCY2xw6eimD4PHqDurd23dk4eqz+6MpT4jE0ccklRW1JIijWOMyNfT/gD7T9P9UrfLz+Y70H2l1/u/qjp7vbBYze08OVoC8S1STVBaheGOjVwRpMjMR9f6e909eq1Py6rG/ki937H+N+/t+9WfIigXE9h763WcptbfuamqGUUEzSokCa72ScupDAf7x72/r5dNw9o0P8WT9tT1sH/K7fHdGKh21VdQ47KZjH5Osx85qsUJZIpcfKULSgxf2SDfn8e6np4Vr9nR19pzV9Rtjb1RlEePJTYXGy18cl/IlY9IjVCvfm4e4Pv3Xvt6UHv3Xuve/de6TOUxrRyff0V0lX1SBfyAbmwHtdbzav05cjpHPEy0ePpyxmSir47XCzILPHfkW4J9szwmI1HA9OwyhxQ8R06e0/T/Xvfuvde9+691737r3TPm8/h9uUUmSzdfBjqGIEyVNQxWNABckkX9+691ryfzT/AOZ32P1NkNq9P9UbRyFNS9kxRrQdl0xqlShSSdqczxtCQAtiD9fewNWOm5XCAN6mg9Oi9bw+JvzM6u6E258sdr98T7hyz4mgz+TwGPqcichUx1svkjfURpc8WcKxI/I+vv2Py6tpYDjkfLHVuvwr+c8Pbvxvym8d30rw7u6zxEabySoaVZJ6zz+NHkVrkMR9efx79TrwaoB6N78f+/8ADd9YvMZTC0y08GJqIqd9LO2ppCwv6v8AgvvXVvKvRhvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//S3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6901ZbJJjqYueZJLpEt+TIeFHtRbwtM9PIcemJpRGuOJ6acLi5nmfJ11zNPyI3vqT8jj29czAfoxHA6bgjJ/UfielX7Q9K+ve/de697917qJX19Hi6OoyGQqIqSipImmqamZtEUMS/qkkb8Ae/deJA49afvz4+dXaX8xX5NSfBz4L9h1G3cvtDNnbe+d24KarpxhZTWtHJksrPG3phHAFha3I+vu6gadRz0w7uX8NDT1Py9B8+hu+MXzN+SX8tve+P+LnzTp8xvLZ1BWpjKfu7LPkZaOWGSoUHJ0E8nEsNtQ0/wCH+HvWGFV4+fVtZjakvA8D/n62b9i7+2h2ZtXF722LnaPce1s1B9zi8zQFzS1cI4LxmRVbg8G49149O9cMll3yExx2LbW19Msqg+g/4+zCGARL4s3SKSYyNoj6fMViosfEp03qHUGV73ux5J9p552lNPLp6KFYxXz6d/afp/qtP5l/E7I7lmg7r6bmbb3aW1zFUxzUJaOStghYy1AVIgLsQo/P9fcec3crvckbxtP6d1HShHE06h33D5EmvivMnLreBuFvkMPxAZIPrWnQqfD35QYzu3a7bczky0XYW0fFhs7Q1BkWpyFXSoUqK5Fk55K3YezLlLmSPeLX6eY0nh7WHmSOJ6PuQedbbmayNtMdN3bnRIp4lhxPR2PYx6kTr3v3Xuve/de697917quf56fHmo3ztnH9n7EAw+/NlV8WbyGZp2Mc9Zg8YqzTUkujlvSlh/h7j/nnYXvbZdxsjomhOpm8yqjh1EvulypJutgm8bZ+ld2rCTWOJRclT8iBTocPif8AIDFd69VY3cIeKnymLY4PJUnqWU1GMgWJ6gq5J9ZVjfj2ecp73Hv21rPH8Sdrf7Wgr0JeROarfmrZI7yM0dexl86rg9DvLlazKSfb0Mbxx6ipqBfSw+h5t7GiwxwDVLn5dCd5XlOmPpzocFT07JPUAT1aOHExvcMDqB/2/tia5aTC4HT8UAQVbJ6Jz82PhZtn5cbRo6SSqhwO99uh6naO52DD+E5IOJYKl/CjO2lh+Pabh0/RSKHonvwO/lPH4wdn7n7j7W3tQ9p9gbkp6iCfLgVR/wA5ZYnlFXGjMVA/J+o97JqeqqgWuak9XRRwxQgLFGkYAC+hFXhRYD0j3rq3WT37r3Xvfuvde9+691WB8kYZeufkd1JubFxPDiKypjqM3MtwiH7jQ5kCWBBtfn3M/KLjduUr6zmNZFBCD8ugHvQay3y3njFFJ7j1Zhi8lTZfH0mTo210tbCs8D/6qNv0nj3Ds8L28rQyfEpoeh0jiRQ68D1LmQSRSxnkPG6Ef1DKV9tqaMD6dbYVUjqszakn+jH5Q0W2jekpd11clW6H6SNIxZSbWA5P59zJfD98cmNd/E0IoOgLbEWW/iHgJM9Wb8H/ABHuGuh3x6r++XOJamyFDn415jkpiWHFtKj+nvRPl1oDozHRGbbN7Ex8zPr8EUMP54Ogkg3978urfZ0NPv3Wuve/de66JsCf6e/de6oB+c38wLvbYPyM2/0xsbYOSotrwbnoaWfedMlWI6+CSpSKSNnQhSOT7tgrXh02SQwFK1/YOhq+dfyc7Jocf171d1BW12O3lumlwMGRzNEZRPQVOZp41aVmjIt42JJ/2PuuOPVjXhwPQO9U/MDtb4/7m7L+NPeGdqd0bzw205M1gt9Vr1E01Xkqql89Pjondhe2oAf4/wCt795Y62D3AHoJ9odsfMHq3MbC723/ALxzmf2D2JuxsbFs2qmq3ix9H90F8rRM1guk8e/A14deIYHPQg7s+Xnb2c+Tu4ZcZueuwXXeEFJWx4tWqRSVcaIGkhQX088j3s0pjrWS2fLo6nVPenY3yd7Mjptry1O1Nt7Nkp6rKTp5Up8xBHp1RMCxuXPH+x91px6uKDqzr3vrXXvfuvde9+690g+ysumF2flq1m06YSgb+hKk3/3j344FevYr0QT45YaXO9lZHcDL5oVrfLq5Nh5QQef6X9140HVsAV6Xn8yrrPePa3xT3vtXYyVNTuGSakraeipCVmqoKSN5KmNSCD+kG/8Ar+7jPVGOPTHWppivmh0/j+1urvi72Rtyi6029SY2HD9iZOsjaR8llaQWkqaq4NmJFv8AD3sqdNem1ddfh8CBmvVi/wAUJ9rd3/zDuot+/HDAHGdQdW0BwO4q6glqDRZJ6dZKY18wb0/usQfeiCBQ9WQhm1L6EV/Prav966v1737r3XvfuvdAjl/kX07gt/R9Y5bemOot7yhTHg5dYqG1/os1tPP+v79175+vQ2KwdVdTdWUMp/qGFwePfuvdcvfuvde9+691hngiqIzHMgdD+D/xB92VipqvWmUMKHpLVmFnoyajFymMKdRgW92P9Afa6K4WQ6Z89I3iaPvTPy6kYzOtKy0tfEaaoHpGom7m9hwf6+25bWnfGajq0dwKASYPQUd6/Jvpn44YmlzHbe8aLatNkEc481SPKatlbx6USLn9XBPtJTNOlZ4V6QPQvzj+OHyOyMmD6w7CxmfzkLOsuOhSaJxp59Jl+vHvWetAilajoUtx9/8AWG1d6Y/YGZ3JTUe58lUx0lLjXUmaSaQ2VQQbc3Hv3Xs8ehq9+631737r3Xvfuvde9+691737r3SZyGHlEv3WOPjkvqMaFQWb68ayB/vPtbDcLTRNnpLJCR3R9Ui/zt+zN2bI6LumSn21tybJ0UddkWdxBLHJT2qoiYG/K3v7YaMghl7lPVxMCpVsMBnqjXaPx06O+XNJtCl+Ke1L7hpcRjs1vjtmhrZzSUeXx8Qnrqephkayl2VhYD3QMRx68VjcA/z+zqwLZXx02X8z8lgeucXBBtjtXrahpsNN2eXeISSYH0SLJDTLd2ZkuLXPuuRmvTpVXwRw/wBXHrYc+MHWXYXUfWdDsfsPeLb1yOFaOjx+UZnYLj6dNEcamT1W+hsfp7rSnVvIdGM97691737r3XvfuvddEBgQRcEWI/qD7917pIZKlkxVUMhRISrkeSNb2Fv1H2YwOJo/CfpFMPCYMmOlJQ1kdbAsycE/qX8qfr7RSxtE+k9KkcOuodTPbfV+o1ZWU9BSz1tXIsNNTRtLNIxAVEX6kk+/daJp0X7tLvcbd653Xurq3Cv2hubb1H9xBtfFF2nqnuQ0RCEMStrkLf37rxNCK9awPzW/nLZje/Wu5esuxNnDoLd1VTGmxxrf4gayWqWpMbLeZgy3tYD/AF/d9GRTpl5lVDWtfn0feg6N2v8AMT+V71lU7dylLuvtvb+1Fkxu5KcoMhBVx1UktTDL5bsVWNV0kG9/9f3Qca9PGhAX5f5Oqsvj9/NM+TWR2Vvb4IVfQuTzOd2MP7o4XdZhrpa6uaOqMJeKNvRwSSCF/wB79200Gry6bD6n8PJb+X7er9v5c3wzyfVPVW95u1qT7mr7faCszm3a1n81D4JjIaedQAVJJ+g5491rXh06o0Y8616sm6/6r2J1dR1dBsbA02Cpa2RZaqKmaQrLIl9LNrJ/qfeqefW69CF731rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//T3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdR6qpjpYXmlYKqKTz+T+APdkUuwUdVZgoqekdj4Js7WGuqw32iM3iiJ9IdfoR/sfZjLIttH4UfH16RxoZZC7jHS59lnS7r3v3Xuve/de697917rWY/nq/zAd5da5LrD4f9R1E2P3H8hJhtbIbqpPOtVtmrqMiKWGRHiYAfUXJ92pUfIZ6bZ6HT5k0H+foomP/AJOveHwA6z2t8ueg91VG8O+qeKDcXclBghVz5Tsk1LrXQ01QFQmQAWEihuPp78W8j1pYwBVKn5Hq3ToDs3qf+bJ0dltlfIrrGgwPZ+0qKZs7hKhailqdr1PkWNJxJKdaHUNUin02J4X6+9EEGo6sGDqVYY869WCdC7U2V1X1/i+k+rcxBl8ZsuneglFKyOmMEr2YeRSxYaj+fay2iVB40/2gevSeVy58OLh0Y/DYWDExMVs9RLYzS/liBb6n21cTtM3y8h07DCIhjj09+0/T3XvfuvddEBgVIuGBBB/IIsR7917jjqpH5b/G3dfWG6Kb5HfHqOTG5TEzRzbk2nifOv8AGDcy1WRZEH9AS3P+2v7ivmrl+52q4HMWwCjKRrjX8Xq3UE8+8obhs16vOPKAKTRmskK4Eg4kn59HX+L/AMiNtfILYdPlsZVxtn8NFT4/c9APTJR5aNPHURsjEtfUpvx7GXLe/wBrv1iJoWq6UDj0bzHUi8l822PNm1i6tmBkj7ZF/hcYYH5g8ejMexF0Meve/de697917pOZ+pxtTR1WHrFWpjyUEtBU03B1wVaGGZTcEcqT7eWz+oiYSDsIIPzFOH59JLlo5ENuc6+0j5HB6pJ3Bic78GvlMu+C01N0ZvuvGHx+APkkoYcpXSaZalLaVDeu/PuH7w/1F5k+qgBFlcdoXyDNSv5gnrHK7gu/a3ns7trptN4wjWPyWVie4H51/b1eFg6nHV+MoshizE1HXU0FXC8RBVo6iITJyL/hvp7loTm4RZa1DAH9vWSFs8MsSzQ5VgCPzFenf37pR1737r3XvfuvdA9233x1f0diTm+ytzU23cfoEvmm0sSmrTcJcE88ez/Y+Wd55im8DaYTK3RbuG7WG1prvJAg6TfS/wApOku/1mbqzeVJuTwK7yrEFjZFj/WSpYn8+1e/8mcw8sgHeIDEDw8+k+2b/tm71FjJrp9n+QnowvsK9HXXvfuvdFE+aGyv7ydL7kyWNhP94sXFBLjahb64wsl2VbC/+PHsee3u4G05gihlP6T11DoN80W3jbW7x/GtKHpRfFHfNLuzqHamOM/mzG28RTYzNAn1LVREqC1ze5H1v7Sc8ba9jv00wFI5mLJ9nT/L12LnbY1JqyAA/b0Zn2Duj3qsX5hY6s2r27sftlLx4/BJS01S4vpaR2AFwLD8/k+5m5BmjvdhudkOXkqQPl0AuZka33KHca0VKAnqxXZ2VGc2rt/Mq2pcniqOuDf1FTCJf+J9xLuEBtr6W3P4GI/YehvbP4lukg/EAf29F++VmFSt2FJWAXkhmtf/AGlY9QH+8H2k8qjp3PDpO/E7c9PJtufESSWaMxaFPP7gupub/wBPflVm4DrxZRgno3D5WgjYq9QgIJB+vBH1HtwQSHy6a8eKtK9QZdxY6MHTKrm9gAfr/X2+LKY8R039XHWg6wnPh+Y6fyIdIvcj9Rt/xPvf0gGGbPXjcmtAvVP/APMsoMlLP1vUbU2pKcl/fajNbWxAM0kX3ScnSpIHH9fdTbxrkv17x5CKqtei/fLJ9+dQdjdcdyVe0qmpwF9tS1dQ6yvDTRUaxvPKwtb0/U+9m3hCga+PVTPOT4mjov3Y2199fMP5Ib67z2RgKqbYuC2xTZKDOU6zRUlZVUNKNdCAAQW1DTYn37wYQxTXj168ZZmowT5dL7dPyiyndXXHTHRmH2y0u8tr7p+3zGNhhkSopKeKQRrM7AH62ueP9f376eHSG8Sh9OvC4lPaE6Pl351FtHYPUW38fFtON9/7wpaeimri5M0M80YDcBdVwxI59+WCLyfq3jysdRWnVY+5N394/G6XZOMwdZktpVtXXyeaSIT6tyJJLpWlJW1xp9P191MCVpXr31DLllp1sVfH3f8AmNz9T7Yzu8aeTHZmri0VMVRqD8Qq4c6+ebn6+/fSucJmnW2uVUAt59DbHncbJfTUDjj6f71b3prWZfLrYuYz1LSvpH/TMh/24/3v20YZBxHTnix+vRZflJuVMZsuXGq9pK9FZf8AYkhbWPts1GOnB0nPiNt1sVtqvydQNb1ah1dubAtqNiR/Qe/DHy68fLqjr+b/APIv5H72736/6G+IOaz7ZbKUcVJk6zbH3IqIchLrSppT4Tf6EKfT7sumpLdNyGaiiECtc19Oqcu/OkuxfjJhsL3F8gfh1lezslQNRrnt3Z0V8MFVXyOJXlndFX1FQQbt78KE0B60QyjVIK+tMnq//wDlNfOz4iZyj271ZtHqvDdCdgb8o6aqh25RtLUSZ2rlnvHeepJZV51AX/3r3pgynu63G6OOzy8uthz3rpzr3v3Xuk7uzOrtnbmaz7xCaPD4ytyUqG9jFRwGZ7gc/Qe/Drx4daxHeuc+P3z939mhsbvvEdB/JDE5CqpcbTNFWxVOUWjPighSoQ8OStxz9fdiSvAdN0VwaHI/aPy6Zutfmf8AOD+Xdn6Xrz5CbA3D211O7wmPufN/xOaiTEwkh3x1cG06QDfST/sPfsMcfs69rKjvyB5j/KOtgj4yfMbo75W7Yps/1RvGjz8whhGTooYpoZKGuaHy1FKVk+vjsQTf3o46spqOjVe9dW697917r3v3XumfKU9OkMteaZZqinCSoLhWcxsLLdiB7ejmdBpBx0zLCj9x4jrTj7J+QG1fkt/Mk3V0f8pGGG6s2numSj2zTV/3EcGXSPIlRRQSFtIL3+o49vOBOupO08OkusxziJzqFAaH/V5dG8/mHfBzZnxUxGz/AJKfEzckPUeP2wYszlcBi6+onfdpjdJkip21lijx2vyR9R7TFWVtD8R0r1Jp8RPtx/l6tP8AiBtbY3yk6Q6Z+R++drR/6QchDJlXr3eVJo6mjqWo1LI5P18Yf/XPuuK9O8VB9c/t6sp96691737r3Xvfuvde9+691jlljgikmlYJHEjySOfoqRqXdj/rAE+/de6ot+dvzl3VufdMHxy+MdfVV++MtKsP8YwqutbSVbRaHpY3RtX6z9QvvwyM8Oqk/hHHqpnuTMfITrmmpuqPm/tfOdg9Wbzjjev3Tub7z7fA5Csj+3hWOom4DID9L/j2tgkRh4Z4HpHLFIv6gp6Gn+XqV03/ACifkR1qaPPfEzvqpwnVfZxptxPS4Cqq/t6bH1rErR1rQra4UkEe2rgeG1Dnpy2XUCqkinH5fZ1sj/CT4a474t7GNLm8lDuzfuWnTI5ncp8hkNTIjGaONnC/UsdR5v7TnJr0rHaKDo9fv3Wuve/de697917r3v3Xuve/de64SRrKjRuAVYEEHn6+7KxRtQ6q6h10t0GlNu/bOK3DLgWy0ZrS/jFJeJSGJsLF2H+t9Pa5z9TH2jI6RjVbyUY9p6E4c8+y/pd1rafzY/mBvDGdq9ddDbd3tUda43dOciwOW3DB5UWOOTINAaqV1ZfSoNzx+Pe9JBNc06prrQDFT1WZXZ/5L/Hj5G7DxHQ3dmX7zxdDnYJt5VeCNdJTRUksiSSCrVyVK6Sb6uPe6qRkZ6qTKHGnuXzB6PP/ADX+i/j78iOv+u9y5DrCji7T3LQQnI5YymOfH1qqCZjFTgrcyXYm35/2PvwNDg8OvONSEEcelh8Wv5W3yV2Dsbo3dXXHyaqMJtenpvvszs2jmrkialqG4gN1EbkgFWU/T3qterIoQAVJ6HLsr4C9kbG+VPTHdHVjVSpjchTZXsCahHkXK1S1AmmNQscdvUAbhr+/E0wetgZ1Djnq+jHVE1XRU1TUU7Us00YeSnb9UTH6qb+6jq3U33vr3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//U3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XRIAJP0AJP+sPfuvdIetlmzmSWhhJFHDZ3kF7OQfUt/ZlGq20OtviPSCQtNJ4fkOlnBBHTxLFEoRFH0H0v+T7L2YsdR6XKAooOgD+SPyS61+MPXtfv/srNQ4igjgqlxiSAO9fXwReRKaOPUpNyVvb8e68TQdeJAUsfLqH8VPkLg/lB05he3duqiYzM12Qo4URJI1BoXVSdMpJ5DA/X36hHWwQRUefRjvfuvde9+691WV8+P5a3WXzL29HlKJKPZ3b2BSWo2jv9DUCoxeQMonSoIjDeoN9GA4497Bp1VhX8uqt/hn8vPlX8O+4qH4jfKnbOY3FtWqzDYTavaeeSuefcZkq/t4q2jqprB4it/rx/X6e/fhqPLqoqH0ycD5+v29XFdzr8efjjsTtLsXbdHt/ZO5930DNm87iiGrMpNMvlV6hXkIsQLnSB7fhiKnxGHb03JMGXQmT1rifyuP5mvU/UPZXzFyXafb9VuGpyORSp2VhamjqZ0adMmT4FdGOgFT+P6e6SlnfHDy63C6LEdRFa/wAutrPozv8A2b3dsrbO68JkKYSbjonq4KUvHExEZs4jjdy3+wIv7bI6e1DHQ9+9db697917r3v3XusNRBFVQT006CSCoikgmQ8h4pUMcin/AFwSPemAYFTwOOqsodSjcCKH8+qZPkP09vf4j9m03yD6HoaiTZE1bDJuvYGPMz0tXUznXWZOojX+nqb3EO/7RecqbiN+2Jf0SRriHAnzY9Y9c2bFufIW9DmzldCbUms9uvBiTl/tHn69Wh9JdxbW7t2Ni937Zr4aoyU1NHmKaIFWxuVaENVUUikmxRgwH+HuStn3a13myW7tmrUDUP4WpkH7Opp5d3+x5j2xNwsXDggaqfhamR+XQukhQSTYAXJP4Hs1AqaDo9JAFT0k8jm5J5GocWpllJ0u639IP6mH+t7MIrVVXxZzTpHJPIzeHEKj16k4zAxwWnrT9zUmzBnv6OPbc10zdkeF6dihC9zZPQRfJTpTA949Z5jbeXokq6+hpKzI7cdjY0ubjp70cyEC99SgC3sLcxbNb71tr20y6mAJT5NTB6D3N/LtrzLs0ljcrqIBKfJwO0j516KD8Au2ctiBlfjV2JkJqvsbZs1fXE1YYTnEjini1SG5Coot7CfIu5ywF+XNwctcQ1OeOnyHQA9rN9ubcPyZvD67y1qSTxKVx/IdWf8AuSepo697917r3v3XutOj+erN3GN/Y6CWpr12PJVwKsRSqemlomrn1qBDx9Ped33eV2P91koB9RQ14AhqfPrFf3tbeeEWIS6BsE1TV3Up8uix/wAomt7Lj+Q23aPr6qrF2sM1IucpaZKlIko/In3Al8n1H19i33rj2s8sTPuoHjaBoJpxpjoPez811+9Fj2o0tRIwIAYZqK8fnX/J1vZ++cnWZPXvfuvdMm48PT57DV+LqlDw1UEisp+hOg6fr/j7U2dw9rcLOnFT0zPEJojG3A9V1fD2pOxOye09n18pIzOfL42NjbSIp2sF1fUWY/Qe5Z5+T957RZX8Q/s07vzA6BnLR+jvri1fGpqr1Za8iRi7sFH+J9w8FLcOhuzBePRQPmjgId6dJ5rEULK2SFZSVMDrqEiClvJJYjnkW9j728uX2/mGOeTCUIP59BvmaJLza3iTLY/l1y+KfaM28+qKPHUNH56jZVJSbfnkXgvUQgoqsCT9ADzb8e688bKu376ZZGotyS4+zr3L+4Nd7aFQVMYCn7ehE7fwuU3Fsivp5FdLiaZoz9FCw8f8T7B8piVCseT0dRiUtrbFOikfFlvBvGrw8tQU0VLJpt9WVzx/sTx7TRyFD8ulMiBx8+rG0wtHdmmQTFmLci31N+fbjXLk1XHTKWsajOesv8Gxn/KpHxx+f+K+9fUz/wAR6c8GL06kx0VJELRwRqL34H/FfbZkcmpPVhGg4Dpjzezts7kNO2bw9JkTSyieAzqT4pQbiRbEc+6N3cerr2/DjqHvDr7Zu/8AC/3d3hgKHPYXR4/4fWozQaNAj02Qg/QAfX3qgPW6nqJsjq/YPXG3ZNpbI2zjtu7dmMplxVAjrTOZlKSlhIzE3BI+vv1PLr1Tx6RuB+N/SG2N1Tb2wPXWAxm6aiQyzZmnimFU8jNqZzrcrcn6+n3oqD17URw6EnP7O25uiWgnzuMgyEuNlE1E8wJMEgbUGX/Y+71610md3dOdZb8qMVV7u2diM7UYORZcVLWxOzUUiHUrw6GXm/PN/fizE1r1qg4dLWDBYimpYqOChgjpobeKFQQiWGkWF/6ce7rNKvBuqNFG3EdefB44qRHTpESDYpcc/g+3Bcyjia9VMEeMcOoTbfH9ioZP9a4t/T24Lv8AiFeqNbgtUdV1/JnI1OQ3NicIKp5hHLHF4/wQslrce2JXDtSlOrorBePHo4fUlNXYTY+IpY8aWWalYs97X4sovx7fSCFky1D1R5ZF8q9a+3z76d+TfxK+Tmyfl90BsPIdj4bDSjIZjD0happXqJUZ62lraeMlgpUsurjjkH8+6CASA6DgdVM7K4xk+VeI+XlXoC+/v5jXyH+e/WD/AB5xnxvWgn3RPj6zcSUKZCafFyUUt5KYtUtJYICxLWB/qfbWh0NTw6uJlmqkYNfOvXW3Otuosh8pPi5150XteCm7H27tPD43e26aQ1Aq8JuWhYCvhZkAF0Km5t7bB1DPTpAD0FMD+XW2ztyiq8dgcRQV9Q1XW0lBT09VUubtUTxxhZZmP9WNz79wx1fp69+690051cU2Gyv8bWJsOMdWfxQTm0Jx4hLVYkNx6SgN+ffuvVp1rgfILq7+V9332Blto9J9pbc6R+R0c1YU3HjKWviFRkuESIzSuVVy9/UhH+t72DQY6o6hjmg+yleiy5TN/ND4VtTdY/IDrPKfMz435iUpT7r3RLWVuCjxVWNEslBVRszR2U3Hq+o9+AUggY6qWYEE9w9RQEfaPPq734A9J/H3DbZou2vj7gKLZm3dxRO+R2pQvPLFjsvPT6qqnZpubx+TSD+feq+R6cAFKjqzL37rfXvfuvde9+691Tp/Nb743Z1HtXZG59ibkmoqfbGSqMxvCmoZUdJMZTTJZapVYXHpa4/p79QHqrMVUlegNqvjh8Tf5k/xi2j3rtuTbfVva2fSWGm7BjqFhqhm6Fv8sqZqZHPLNZ1IUEcg3+vuwLIar1pwrqNWfP59a8PYGx/mV8Ze/MBtP5Db03Z2Z8bsTnQku6MrLkazAV+3hVhDJTNKWTSI/oB9PatJI5hpYd/SJknglDMwMJ+Wa/PreP8Aipu7pveXSGzMt0O0H+jX7P7bBR08bRRxmJFeqQI3P62Nz/X2nkhMRxw8uliSasHj0Yz21051737r3XvfuvdcWdUVndgqqCWZiAAALkkn37r3SGoN7bT3c+WwWIy9PV1kSVFBUxxkP4pJY2hYEoT9L+/V9OvcetYT5pfCzuv4ZfI3A/OPqLKVe9cRtrIxZWu2nAlS6tJqM1SKuEAgoAG/2HPvYJI09NsoV/FXieP+x8+kV8kv5oHTX8y3p7D/AB664pKbP91/xvEy5XZlNSzxy0mXDLDV0kMspLExsTwp5t79lSGHWtYdSozUZ62Gfhf1XvDpv457K25nqmZcrDt/GFsTKC7YeWMETUpEg/A4PtXHIkzqkowK/wCDpqRWiUvH8q/6vs6N7i8mmQiv+mZLLIv0OocNYf6/tqeExN8jw6ejkDjHTr7Y6c697917r3v3Xuve/de697917oBO3/k10x0TDFP2bvCk29HM2mPWpncsG0EeOIlhY/1HvdPXrVQetVz+bF3x3dJvnq3vr4qZDNbn68k3VJW7mr8RFUx09Hi6arWRnrIoTYJov+r6+37eXw3q3A9J7hXZAy/h/mPl1sxfCH5BUXyR+PmzOyIqiKSsroBj66FdCPHV4+ljjmvECW5Nzcj3a5XU3iKKA/sr1q3Y6dLHP+Tqn3+eB/Lf318jqPaHc3Tr1P8AeTrdZ8vW4Kh85qM7ULWGpWNFhU/Xgf7H2nDUNenJYi66V41r0Rro3tzvTcfT2P6B2h8TF2L2/mY5sTu/tGgpa5NwALaminqfIp08C5I596IFa162GYjSBSg/LrYk+NXxBxOD6U2Jge7Yf7770xFNLJXZHJM7SiWVgUikL8kp9PdaZ6e4D59Hsw+Hx2AxtJiMTTJR4+hiENLTR30RRjkKt/e+tdOfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r/1d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690H8/Z2zafMpgZsrGmRdtAiOkDVe1rk3/wB49++zr3yOOl5FLHNGssTq6OAyspDAg/0I9+691k9+691737r3Xvfuvde9+690mc9lGhC0NJ66qdgjqvJRHFieP9f2ttIAx8STAHSW4l09icep+HxqY6mVb65X9buf1XcX0/7D21cTGV/kOnYowoqeJ6z5bK0GDxlfmMpUR0mOxlLNW1tVKypHT0tOhkmldmIACqCTz7T9O9aJf80D5zr81/m1gvj3Jm5cD8eup97Lk33XTUk7wZ7HQVKDJwTtAXUhgjKB/S1/biq2kt+zpJJJGZliORxb0x5dbifxs3n8bML8fMdneiavCY/p/bOFWud8cVpaejWnoEkrqiriqXBWZgl5NRAZv039t/PpXXh0V342/wAzHZ/yI7u7r2Vg6OCLrbqPb2T3HUbzKOrTUOLb9+R7OyNwCQF+vvZBHHqiurkgeXHqxPrHs/ZvcGz8fvrYWU/jG2sm8yUdd42i8jwELKpRr2sSPeur9CB7917oCu5uruuOyYMc27NqY7cW5MOJn2nXVKMarCV8n6KullUixDWYixv/AIfX2pgtzIfEbCjj0xNLpXw14nrXy+TH8qbt2n2X3h23vv5P5qfb8sLZTHbRqXyM1NQ0yzaRTgC6rbUFVQPpf3qWY10Jw6rHCrAs9Qf9X59F4/lvfHT4UfH/AKE+Q/yI+RfX20944/Z0dJXJk87dpq6N5SUjpaOV47ySShF1WawvwfbRq3y6ciAjSo86n8ui89RdPfOz5j9zN8oukMfufqPo/rzMfxDqvZe3TkKPb2/MAKxpYoqdQdDqFUK5uR70QoFPPrwMrEsPh8h/n62MP5ZPz3rPl3F2psHc+Pjxm+uksnHhNxRKoWSST7pqWRpSGYFkdbMePr70Rn7enFNVzxBoerX/AHrrfXvfuvde9+69015nDY7P4ysxGWpYqygroJIKinmF0kSVDGQR/rHg+25oo54zFKKqwoR0zcW8N1C1vOoZGFCD1SpvzA76/l/9xx9hbXjrMp0TuOsBzmMRZ2oaOvriTJrRbBWTUSGH459w/e219yRvAvrWrWUh7lzQE+vz6xz3a33P2m3/APfNiC+0TH9RBUhGP4urYtob+xPb22MRuPZWQjq8PkKSmqKiqikUPTzzRiR6V0BJuvIPuZ9qvbW4tVuwa6wCB+Vc9TvYblb75ZR3+3OHjdQcEYr0JePxlPQIPGgMpFnl/tNf6+3Jp3mOeHp0bRxCMfPpy9sdO9e9+691Ud86uptwdc7y258l+qknptyrm4E3xPRiQyvt+mKGd5QnBQqWBv7ivnXa7jb7uPmLagRLq/UI/hHmeoI9zNhvto3GHnXl8UnV1E9POEHu/wA/VjPSnauC7l69wm+tvTLNQ5CIQuRYFKunjVamNlBNrPf3IGzbpBvG3pfW5qrY/Mcepb5d3y15h2mLdLQ1SQfzHH+fQsezTo8697917oFe3vj71T3ljkx3Y+1KLcMUKhYGqCUeKzFhpYA8XJNrexDsXNG9cuymTa5jHXjTz6LNw2iw3OPw7uMP9vVXNN0Vs34Kd6bZymy9v00W3Owsl9vVTRa5ExdNJP45WJdVC2t+T7mj+sV57j8tTQ3sh8W1WoH8Rp1H42m35T3eOS3QeHO2afh+fV0ePyFHlaODIUE6VNJUoJIJ4yGSRDwGUj3j1LFJBIYpRRl4jqUUdXUMhqD1N9t9W6banKUNNqWaYAgG4HP4/r7fSCVyNI6ZeaNRk9VYd31lb1t8pesa7E05hxGekhq8jVIPT+49zrY2UHn8+5t5cji3fky8inarxVAH2dR7uks1nvtvJGMPx6sxpqGbMRJkkr3+3rCZokHKiImy2sbe4ceVbZjDpyuOh0InlUOTx6z5fbmNrcNX0lXTJVa6OrF3H1dqdlB91gvZo7hXQ0yP8PV2tYzEyt5g9V/fCqaPYW5uyNi1Z0zZzdU2Rooz/ZRJZANOqx+jfj3KPuIh3OytNzj4RxhT+wdBPlgmyuJ7JhTU5I6sZy9OtVisjTsuoS0VUlv6kwtp/wB59xAOPQ2OR1Vz1jO20+7qmGYaFkyLlVYcWMlx9be/VNevUqOrUoJBNDFKPpLGkg/1nXUPeut9Zffuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYZ5lghkma1o0Zzc2/St/fuvHqqfPLJvXu2SFWMggyP6B9E/dtYEX9+4n7evDHlQdG1+S/cifGf495zfphiqKnBUKU1NSyAMslRUxSeMAG19LW97yPt61j8utOrf/wApf5l2CzmU7Iottbx3RtLedSMztfZ9QapsPm8VIxlCwwA2KW9Ho/Ht1JCnwmnr0mZJGapAoeA8x+fR6P5eH8xvurfnc20unt3/AAs2l1Jubd8pop93Y/H1dNmJBVzinnqJJaosP0sSSB7dFwx73pjy9eqiJQTEgYEitfLrYv2D8X+peotyZDf229hY2ffOXrKmtrNxoz/dRzTnVIwDW/Vqa9vdv0bhqfBT+fW6yQDhqrxPRhqfcNI7LHUEQSkcpybN/T3Q2r6SyZA69HdBiQ4p0+xSxzIJImDqfoR7TMpU0bpUCGFR1V//ADauzd8dYfGF8jsR6mPIZvclJt2vlpQzSR43JQNHUMVTkj+tvegK9aYkKadayndfwR2n2ptXbXVfTPZUmE+Se4pMZvyt3pjjNTZqnp8hCKmbDiRiregtzb+nu1TUA9U0LQ6OPr516Nl8Dqf5rdX/ACj258OO5Idzd89N0eDpazJZXcNTVVWPVYqIvUok7lgvIsefejppUY68ok1HWa0HW2TsTYez+vMDFgtk7boNrYhn+7OKx6NHDHUTIDKWDE+r8E391A6dqTx6WnvfXuve/de6JJ8sPmv8e/jhgq7Edp9gNtTJ5elmo6KSip0r6mnnc6BI0Ub3Wx/qv+8+/UPVSwAqSB9vVDWW+BEf8wTb/a+7upvmFkt30m5MLVRxbUqmqKZ4BVStMIViDEWANvp/T3YH8uqsA2STn06IN/LE+B/bOS7x7Z+ImQ+SG5tqUvVNDlq6goofvWiatMwhdYYyVFxq1ORyFBNjb3Yngw6oisaoxIpSnrQ/Prbt2B8OdnRdIY7prvGWDuyjo4Hp3rtxLO94HjC+OBrq62IJBv8A09t5rXp6g6GnYXTuz+mtm4vZPVmEg2xtfCmQ0GEomdoITLbyaWb1G9ufa2CcEaJc9Jp4mrrj49CJjMwsxFLVftVa2XSRbVzb/b+6TW7L3pletwzhhpbiOn/2l6U9dEgckgD/AB49+691Xv8AzJe1+3+qPjXu3c3TW1juncUDQQrFTSCWojilhk87CnibX6Rze3497x59az5dVh/yIO5qHsSPuGp33vqbI9nZLMwPVbUygMLYWZqh3qoaT7ggswYBbD8f4+9sKGg4dNwszJVuNetivcO3cLuvD12A3Dj4Mph8lBJTVtDUqWhqIZYzG6OBY8gn3Xp3rWw+Rn8pDF/HHv8Awvyn+F/V1JT7gpcxT5Gu23gjVyVE9VJMJJ6uRSD/AGrn6+91oAPLqhjBbX5/sqOtiXqnK7pzvXW0clvfGPiN1VmDo5M/jpQRJTZEppnjcH8/k+/Z49Wwe0cOnHIUMuLqRXUdxEzXkQXsCf1E29mEMonHgyft6Qyo0T+JHw9OlTR1UdXAksbBrqur+qta5B9opI2jfS3SyNw61HUr231fr3v3XuuJdAdJZQx+gLC/+29+691Xl8lvnjsn4/7mg2NuSVaLK5+sTC4abwu+qunmEC3dmCj6/X8e909OtYOOtbjvbfG8cL80N+z/ACDoanPdeV1A9VtDEZaOonx1XU14E1GkDC68hgfSfe1ylR025IehyKDoV9v/AD/yuJon+P2/PjXtzZHTm+Qm28HuWCicSSvUjxGojlkJuSCL2H591KniD1bUuoagfl6dKr4qd49pfB75GZnYc22aus6G3VN9tsyulpqtqBqjI6ZEqaOQ2FxqF7fjg8e1kBWRPDc9JptcR1L+X+b/ADdbWW1M5FuPAY7LxEEVlLDK1hazSwrIRp/5C9szoEkoOHT0D+IlT1CxOwNnYPMVe4MTgKChzNdq+7yMERWoqNX6vIxNuf8AW9s9PdLD37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/1t/cso+rAf65A/3v37r3XWtP9Uv/ACUPfuvde1p/q1/5KHv3Xuutaf6tP+Sh7917r3kj/wBWn/JS+/de695E/wBWn/JQ9+6917Wn+rT/AJKHv3Xuu9af6tf+Sh7917r2tP8AVr/yUPfuvde1p/q1/wCSh7917r2tP9Wv/JQ9+6911rT/AFaf8lD37r3XvJH/AKtP+Sl9+6913rT/AFa/8lD37r3XXkj/ANWn/JQ9+6913rT/AFa/8lD37r3Xtaf6tf8Akoe/de69rT/Vr/yUPfuvde1p/q1/5KHv3Xuva0/1a/8AJQ9+6917Wn+rX/koe/de668if6tP+Sh7917r3kj/ANWn/JS+/de695I/9Wn/ACUvv3XuuQIPIII/qOffuvdd+/de697917r3v3Xuve/de697917r3v3Xuve/de66YXBF7XBF/wClxa/v3Xuq/u5vj7ur+MVO7ds11TU1Jlaq0x+X9oKpfSLcfX36gB+3rWaUp0gNhd+bx65yUeG3bFPUQqUhkaojYiNWe7G7W+g/p7950HXguK9H+2V2Ztfe9HBPiq+N5nRdcRZFKueCBz/X375deHQh+/db697917qLWVKUlPJM7Wsp0/4tb0j25EhkcKOqu2lSek3g6RqyeTLVa/ulmRFYH6XuDb2supBGohjOOktupYmRx0rvZf0s6qn/AJtUPyYzfRGK2L8bNsZPcGU7CyVftTd1Rio55KjC4LIQxRfff5PyACzm54497Gmo1dVcMUOjj0GPwM/lJ9KdL/Gql2X3bsTb3YnaOelyuQz+9c5Szy5yAZiFGjoIZ5SrRiBtX0vc/X+nvRJJqevRqEjCLw/w/M/b1rafzN+p+/fil8h8L8f/AIcdjbh31hu49xxbK3315iaidcdsLAZurSMVUlPA7ANEshvqAtpt7uDjURXplwQ4jRtNfKn8/l1Z58d83tL4nVOC/l+9N7XxPaHdOXrYMV8ht6z04lrotuZ6nj++opKwE/twayXJPBHupJYFj/q+XTygLpQCg/1ZP29bKfUey9g9NbUxvWG06vHUtHhg88dCtVSxujVZ1sRCX1c2B+n+PutfXreK0HQnZTJRY+n1lgZZARCv11N+Pp7UQQmV6eXTU0nhrUcT01YSgklY5KuCtPLZ4kLBjDc3uLfQ+1F1MB+hFgDj8+mbeLPitmvDrBv3FbVze0szi97QQ1O16ylaLLw1AZoXpr3byBATYWvwPaDpYOtF/wCYfW3e/wAy+3ewdqfGnaGVpuh+jcgaHeeBwdLXphs9i6WvYrVZaOJbSEgfVh+Pbi6QdTdJZlleqRYA418/Og6tN2j/ADcup+ifh/tb499D46oq/knsHb9HtbEbA/g0j4z7kTutQIJSOWBkuBpvcfX3UAnJ6eaQV0jjTh5/6vn0VnoX5I7T/lE7F7U7K7fhKfKj5gNRb62r1rURF445KyrkqyMhqZHjDyufQPrYcce/ULGg8utavDXW/mfzJ9B0fn44/wA335C5Ddeyl+WfSuM6k2F2BTwZDbO4qWnqIzW42onEcdWrVUzLpIIPvRA8jXq4fhqXTX5/y62GcDncZubDY3cGFqUrMTl6SKux9Wn6KilnXVFKv+DDke9dW6d/fuvdYppo6eNpZWCov1J92RGdtK8equ6oNTdBVvDaeD7ewuV2pufEU+T23WUlVCyVKlo0q3jMcFQoH9pL3U+7X+32sto1tdKH1eRFaYOR8weii+sLXfLSSx3CMPE4IoeGeB/LqpLbGU3r/Lu7iO1dyVFdlfj3urJK8G5ZopZI8dkK8/sY+MPwujyabg/j3C8N1f8AIm8fT37GSwlbEh/CTwUfZXqAbOe/9n+YPoLxi+zXDdsp/wBDdjhPszQenV2WDzWP3DiMdm8XOlTQZSipq6llR0cNBVRCWMkoSL2PI9zBDKk8SzRGquAR9h6yQtriK7gS4hNVcAg/I56dfbnT/XvfuvdJzd22cfvDbeZ21lIY56LM46px86SC48dTHoY8f0+vti5gS6t3tpMq4IP59Jb20ivbWS1lFVkUqfsPVSXQu8q74r/IzcnSG6pJMZ1XXzSx7MqyjCkqMtWSDSkbTaRybA29xXst4/LHMEmzXXZaNXwz5FieoJ5YvpuSOb5uWb39Owf+wPkXPEZ6uPUhlVhyGAIINwQRcWI9y3xz1kADUV679+691737r3RePkx1pS9jdX7ipY6RJc/SUDvg6uzeakqNX6odP5N7/T8exZydvD7TvMTFqRMe8eRHz6Jd9sRe2DoB3gdp9D0A/wAKu4pcrtrIdU52QtuPrqnlgqZp/TNVWlsqtqNyQSbG3sS+4mwCC7XfLUfpXRBAHAdE/LO7B7drGc/qQ4Pz6OZKc3lbKYmooh+l4yRqB/Jv7j9TbQAEdxPH5dCP9ebJFB06U2BhjCGpY1TAerWOCfrzf68+2XumJOjt6ejt1Ud2eih/OHZaVvUWW3fiaXXubbv2QxkqAmSKHynWI9PPA9jv243Fo98Swnb9GWur5n59B/mm017c1zEO+Omn5dDJ8b93U26+odkyGr+4ytHgKKnzCMR5IqxAUcPYm17fnn2Qc3WD2O/XA00RnJX5jo02W5W526JgasFFft6HSRdcbofoyMv/ACULewyDQ16NSK46rC3jWQ9ZfMPZ1OCIqLL0yTyj+w/mAsS7WA+vPuZrCM7zyFcNxZDQfl0A7mT6DmSJPJx1ZvHKlXSrLH6knhLJyDdXXjn3DTqUYoeI6HakMAfXqrvuilm2t3NQVcMQhV6mAswBF9TDklv9f3ShAp1skUqOrMds1S1u38NUqQfLjqRibg8+EA3t/iPdiKdaHz6fPeut9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdIfsbLrhNoZevZ/GI4SurjjUD/X/W9+zSo69506IB8dsLLn+yMluFwZovvPMGIJA/d+t/p71wz1sjoZfnx1BT95fHLd3XxykOMrMg9LU0kslTFTuz0+q6IHILfUGw97rTh1UgHj1Qr8iNq929C9R9cbxEM2f2z1FhaHCqHrkqoZgy67GCMk2/HI97FOqsKCoz0MP8vjJbu+XPfHXffv8Ao/pto4frOlSlyU1BBLTxV0zyeJJpVIsWPJPPvzg0p16Jw3cBQZ62W/rx711fptqMTRVCteBFkY38ij1A/wBb+3kmdMVx6dNtEjZpnoOd8Zil6327mt212ReOhxGKyFf4j+kyUVKahU0nghrH2pW5icaZB0maCRW1KeqJeu/lv3l89+56rBUHTdHun417ZzRxec3CaZhTUFdDLp8k80ZYa/SSAefbDQEmiZHl06s9VOrBH8/XorXz+6MzdD808VuL4iZKsr+1jt+jxR66wgdZcfTrSCneoh0G7aQCfp7aZWiOmTq4/UoyGhp1cD/Lg+L3aPV23P8ASH3pkcvN2XlI2p5qDKyTST0VJNC3likM3P6msP8AWPtutTjgOnACB3ZPVq3vfW+ve/de6TG9Kquotqbgq8YjSZCnxdVLRxrfU9Qkd41Fv6n37r3Wkfne3umNxfNj5NbY+Weamz2eym2M9jutNj5unLY6DeTTMMbHFLORpuwFtP597zSo6b1L4hV6ZpSv86dDZsLbPbn8traPx87xxeTkpM939u6rwtX1nTSCaDCYYVKxUtTJT3480bgrxwPeySxp1pUCLivHP+x1sedOfFnaVJ2BUfJOhp02/v7sChpa3cS0yOHmFVTrLNHMxsAWJJIA/I591zTp2oJr0e/37r3XvfuvdMmUxEdWpmh/aqUBKugszG9/r/X2qguWj7WyOk00AfuTB6iYnJTLJ9hXqUmU6I2a95Cv1+vu88AK+NHwPl6dUimIbw36r5/mJfLPL9C7Cqds7CpTkuydw00X8BoEjEjSLKHVyvNwR6SLe0Q406VH/V9vWsPlvmJ8u/hP2ftDe3f9LmtzbG7NSLO5Pbm4GmrsbT0ldKUenFO5IUBCQB+PbmkNhOPTJkkTMuQfTy6Ozuvp7bHdg2v8y/5cNfLTbvhWjzW/OpttCSIy5aWqU1EktPTNdl1N9GHutQvaw6coWOpDn+XWyz8dM32Lnuotl1vamAfbm92w1Ouex0odZIq9b+UOsnIP0uPdenOhy9+691737r3XCSNJUKSKGVhYg/Q+9gkGo68RUUPSRm8mErhItzRuBcc2BPHPtfUXENKdw6Qd0EvyPSqFTD9v90ZFSERmRpHYKiIouxZm4AH5JPtCylW0npcDUV6rn+Uf8zj43/GOf+Ebp3Q0mdWoeGSmo6eOshjETBZNUkUh/rx70B+fWiwpxp1Vf8nvnd2fmcX178k+gq3IZ3rjMb1o8XloYwPDQ0iSI7tPTwsdPoubt78Bx60WwDxH+rPRs/nn1p138s/iVjPkvs9MXWZrYOB/vhJXxMi1f8VoqeGaqpXSIsDJHNcWJvaxtb3rI62QGGPyPRFPir37hvmp8XKzbsHTGI7T+R/XtRkaulXJ07/eyUOITx0KLo5f6Cwv7sV08eqo+te3y4/b0/8AR3wT+Rnyk3btbI/JTrObp3B7Hzr5fG46WnqqemqBTOJIoYomU/qsF91J8h59WVWOXxT0PWxbV9F9T5WgwNDnNi7fzCbdipVxbV1J5jSS0sSxLNCSRZvSDf3759W+XSkelTbs8EVCi02MuqpTx6vFDGAAdI/wta39PZjHonh0kdw6QOGhmqvA9LOKVJo1kjbUjC6n+vtAylDpbj0uBDCo6ye69b697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//XuV7N+e/zb7b+bjfHrpDrakynXsO0KbORZmnhQVZqjjlrKpZKhTwqtqHJ97oaVPVK1egHlk9DYcl/MvU2/wBFtWbXH1J+hte4PvQp59X7h/xfXH+J/wAy8D/mVlX9f9q/4r71iuevd3+o9d/xL+Zf9f8ARZWE2/oQP9797x1rup/s9e/iX8y/g/6LKv6/7V/r/wBfejpHXu70/n17+JfzL+b9V1ZJ/Nz/ALDm/v3z69Rv9R64/wAS/mYkm/VlZb8fX/iD731vP+o9dnJfzMALjqyrvbj6/X/Ym3vx68NX+o9eGT/mYWuerKy5HqHP/EH3rHXu7y/w9d/xL+ZeP+aWVf14/UQP8fr73jHXu4f8X145L+Zeb36sq/8AbNz/ALz7914A/wCo9dHI/wAy8i3+iyr/ANhf/ivvXWqN/qPXv4l/Mu4B6sqybcfX8f7H3sU4de7h5fz67/if8y8cjqurNvoBqH+9n34kDr1GPp+3rr+JfzL+f+MWVfP0/V/vPPvwI63Q/wCo9eOS/mX/AE/0WVlyPr6h/wAT70KdaOrh/l68cn/Mv/PVlYf631f8V97xXr3d6fz68Ml/MvsB/osq/wDG+r/ivvwoM9b7j6ft68cj/MvJuerKu/0/tWH+8+9YpnrxDV/2evfxL+Zhq/5lZV2/rzwf9vf3snr2f9R67/if8zD/AJ9ZWX/r6v8AiT71jz693f6j1xOS/mYD6dV1Z5/x+n5PJ9+68K+f+Hr38S/mYX/5lZV2/rduT9bWB9+x17P+o9MO79zfzMsBs7dm4qfqqqlrMDhKvI0VOQWFRUQoSsfjJ5H5Pv2OvUfyp+3ow38q35t92fJHFZnYvfmz6Tam/wDaUNTLV09PEsL/AG0cixxiVFP11E8ke9niB1VDUHFOrkvfurde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6910QGBUi4IIIP0IPBHv3XugU7D6O2nv1C8tNFRVehx5o0a7OV0qzWPv1PTr3Dojm6Oq+w+nsg2S221XNi4pA941lZGWP9z8e/HIz1rgehk6x+VHlamxG96f7B4vHAZiio5Ymw/URf8A2PvwyMdbNOPR0sLnsXuCijr8ZVR1EEihwVdCwB+moKTb37rVembJGbMZCPHwtakQ+R5lPp1xjlSR/jcezKHRbwGVviPl0gkZppvDU9o8+ldFEsMaRoAFVQOOOQLX9lzEsanpeoCig6ye9db697917pFdjvuCPYW7n2nTmr3KmAyL4KlH1qMmtOWpIR/wZ7D37rYFTQ9aolZ8ee4/ivvfuX5n930Vfnu2+78NuHbvXOwMlFVVUGG3XM/+47JwW1BZELKyWsfe61wOmghWrNk/6v8AUeg427md6/ArqfbPyb3rhY8t8xvmXUZzYv8AdzMU5fIbcpZ5lhpMnG1RyJZSyCMryPe8E04AV61QomqlWalft8vyHQ476+JXy06H+Nua+c29+1dz0e/dgYmp3ruTratraoQxUEcsZgVklfQ4PkQGO30/HuuCaU6uFfGamn5fZ0b74F/zfcX8ifiZ2J8ge4qah20enttz7gqAkVLRLkFoiKSOOKKVgJXY2JVOT7U6hAhRDVjg9JIibg6nFAK8ejE/yx/mB3F8ysv2V2huHblJjemMiofrPNQ+CJsoYqxIZYzFEedKXJ449pjg0PHpWh1Lq4en+fq3asoqXIU0tHWwR1NLOuiWCVdUcin+yy+/dW6D/ZvT/WXXrbjbZWy8Jtv+90pn3KuMpzCmYmKNGZKxSSGOl2HFuCffuvHIoeglo/hT8T8Zu47/AKHoPrqm3l52q/7wxYVRkTUk6jN5SxGq/N9PvVOt1PWpL8lvjDtX5K/zPtoYH5W7urtipTbglouuIa6mqp6IYSGpc4yGGqYaUjPCgnj24CaVX8+k8katLpfIGR9vSo+TG3vlfmuxsV8Rdo9c0u++p6fIxUWD7VmvXV+3MPjqkPbH16EiKLQlyCfehQZ6swJ7QOPn6dWM70/nF9K/Cjcfx0+J1Xk59w5Gi2TiMPu7KpRDISUuUifwvDHNA5UaBcevn34AkdWaRVIB8+rmKT5e9NVPRsXfwzsj7HH20NVMkSNXQVdRZBA1IGvfV9PegKmg63qAWvSo6x7PwffeETc+06id9tsyeGaaJoXkEg1qGT+pA59mKtHaJXi56Rsr3DZFF6HWnpoaWJYoUVFUD9ItcgW1H2Xu7O2pj0sVQqhR0FfdPT+1O7NkZDZ+68fT10EsU0uPeoQuKLIeIinqkA/KtpN/8PZTvG02m82TWl2oYGtK+R8j0R8x8v2HMm2vt1+gdTUiorQ0wequ+jO496fDvs9+gu65qqXrzI1jy7Z3nWRyytFJUeigoEkkPEViotfj8e402bd73lHcjse8VNsx7JD5E8B9nUL8tcw7j7fb1/VPmUk2jn9Gdvme1D+0dXL0lVBW0tPWUsiy09VBFUwSowZZIZ0EkTqVuCCCCPcuoyuodTUHI/PrIRHWRA6GoIqPz6ke7dW697917oiPzj+Pq9qbFg3fhFWm3V128+5sdJAr/cV89LpljpiIxc8i/sE86bAN1shdxGklv3j1NPLqNPcnlRd+2sXsGJ7Q+KhHEsuQOnv4Wd9VfbfW9Hjd3MKPsLBealzOLkKLOlPTEQwzlL6ubc3Ht/lDe33Xbwl323CVDL8hwPT/ALc80NzBsyR33bdxVEiHiKY6On7F/UidNVdmKKgus0n7gHCKL8/gG3t+K3klyox01JKEHTKk+RzEyo0BioC3qkUkalt7VUitlqMt0k/VuOOB1Wn2Fhj8aPkLtjcWLhE2F7JyYfcNUEJFNA1Rpk8kknC29y/tdyOcOVZrSY0ktF7B6mnQLu4Bse9RTIKpOe8+nVpuNyNHlqGmyOPmSoo6qMS080ZBSSM8AqVuPcJzQyQSGGUUZeI6kFHWRA6GoPU7231fpLb027Sbq25ksLWoslPVQSa0a+klYm0g25+vtdt129jeJcx4Kn/L0nuoRcQNEfMdEC+EORl21uXtTZmZqmVzuInDQy+n9qOVl8cYfkg6hYj3KPuPCLyzstxt1xo7yPn69BDlR2gluLSU8G7erJvcQdDfqt/527YO36PFdz06aq3A1mNxyMoLOiSNxx9LcH6+5c9tL36l35fb4ZAzdArm23ESLuY4oQOjydX5gZ/rvZmYDBjkNu4upcgqf3HpVLg6eL3vf3G29W5td2uLc/gkYfz6FdjIJbOKQeag/wAuicfLjEfaZCi3EieqOenXV9foob88f19lhr0pAH8+jM9DZx87sLH1Dvq8UUMQva62juRx7914U8uhq9+631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690Vr5Tbn/AIPsqXGq4EmRjsq8XJuU+h9++Xr17zr0lviHtyTG7er8pUpd61dUZYHhTIG9N/8AD3oGpPWyKdU6fzKc12H2980+ivjyvYtb1btren2lFLlMXlYYI41nlaA1M5WRTqN76SR/T3bABJzTptiWIUGgz+fy6hb7/lg/Jvqelnk2B2DmvkTt56mLVtfOZGVqOuiVw7Cso5JXAKi6nm39D71wOR1ujNgH9v8An6ul+GG0M9svquixO5OqMP1RmVSM1+Jw6ssNTOCbvIDwSB+R/X3rNerZ8z0cH3vr3XvfuvdAP8h8Dgd69Ybv2lkM/j8VW5DA5aGhinyNHTSzVktC8VPCY5nViGZgOB79SvXq9azXww+YWP8A5VGd3/0R8nsJUbe2PujeVfuuk3NiaR8vU2mjZU8fibQ6OrIf1fji1/d9TEhlzT/D0x2qDE+AeB+X+fpWdT78q/ml/NJxPfvxvqM4vUMWIp/udyiiqMTPHJDSlZUmVQVXWVPpJP1+vtSsqvRZ8cBX/L00yMW1RHAB/wAPW0ZQ5uOFaehyMhWvSCNZiwuXlRdMjMRbk2ufentW0eJHlerR3NTpkFD0o0dXUOpurC4I9pSCDQ9KQQRUdQMtlKLCYyuy+RmWnocdSy1dXM7KqxQQrrkdmYgCw/qfeut9AN1d8oOnu6dy5bZuxs7/ABjLYmKaTIQCOMwpHCbSfuo7A8/j37r1AeqbP5mH8jbZXy43rQd79dbjbY/Ze3s4NyvS4+mnEm4qmOdZ1pPuIW9JYjm4tb3YMQCK8em3iRyrMKleHy6hdAfyxO3+x967b3R8ndzZyHFddVVFV7Y27XzVNVDLLjwkUAgDNZVOkM30491rjHVghBBY1ofLrYjpaaGjp4aWnRY4aeKOGNEFlVIkEaAD/AAD37q3Wf37r3Xvfuvde9+690UX5P8Ay66H+LmH/jvbO5JcQwiWWGPGxwVdcyvqUEU7Nf8AB/F7e1EMxj7TlfMdMSwrIKg0PVaTd3fEX+Y3nsdufr3f9fJuzZ1NIuNx2VpoMfPWCmhZxpWM6r6rfj25PGorInA+XTEUppokoaenVQGE786M3j2l2ltj5eTZHO5PrXNVGB692plMdUVmPyURlalijdj9FW4I/HtLpNAfXpSHGogmmk+fz9OrT/5Pv8vrOdFbh7A74p995yi2b2Lmny20tlwrVQYyPC1FU0/2xjlIHiUWUCxv78TUZ62qBSaYrmnWwv7r051737r3XvfuvdRFr6Fp/tVrKVqm1/txUQmYgfUiIHV+P6e/der0GXbfa/XvVG3Zs9v/ADNPjMbHrJJmpxU/trqYpDIwJsD/AE93jlaJqr1R41kFD0SDf3zB2L2v0L2HUdI5utydRRYDcMc1TLEkNTTCOkYrLEsZubWJFvaqaPxFMw/YOmIpdDiNs9U8/wAvv4S9R/N7qnsXdfbO8n3Bvo7mzmKhx+Z0tU4+KdXEdcsbuJCFa1iqm1v9b2jNQaL08oVxrbNa/wDFdFf6r3Dsb40fKLsj+W7vncAyfXeQocvRYHOgI8FDuPKQmHH1VKxN7oWA4P497arAt889aXQpEa4xj/KOjMdcfDX5/dRZrJfHTBYjM7q+PG5ZZ6ubdrTStQzY7NKJSoOo20owuPeqg5PHq4R1oART+fV3Pwt/l+dc/D9qrNbWkE2eztCFzMoidLzVEQaojV3JJAe/4H09+JJ49eVVXh1YZ711br3v3XuoVdRpWwtE31sdJ/oT7dikMbV6alj8RadM2HrGilkx04CiI2ib/Vc/T2quogyiZM149JraRlJjfy4dKb2g6Xde9+691737r3Xvfuvde9+691737r3Xvfuvde+nJ9+690mczu/b+AKrk8hDCzsqBQ6M2pvoCt7+/HHHr32dP9LUwVlPFU00iywzIHjdSCCp+n09+691n9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//9C8L4TM0P8AMYrUjZ41frmZXVHZEdTjDdWQcEe/Vx1qlWJ+Q62Nvfut9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdIPtCV4eu95yRu6Ou3slZkYowvTlTZhz9Db37r3VJf8sPTJ8m+65iLu2MqwWNyx/y5CbsffmpqFOtR/Ca+vV+Hv3W+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuolZQ0mQhanraeOpgYENFKupTcWPHv3Xuiodm/GLC7hFTk8HalrrvLHBGjD16fQFK/Xn34+vXhjoqYzfZ/RuTNDVirmpUkCqro5TwoNV/V/r+7LqVwxHDrRIKU9ejfdP997a3BSR0GYqRSZuVkJRxGgJe91NyD9fbk0xlIJ6aiiWKtOPRo4ZoqiNZYZEljcBleN1dSCLixX2z091l9+691737r3XvfuvdBn2tjNoS7Tr9xbu2ri91w7PpqnPY6jydJHViCspkDLLTCRW0udK+oC/Hv3Xq9awPyyz25/5hvYG0Ml07tykrd8fF/JT7yq9tVjR0NPS0+MyAnQ0vkIuF0Lwfx72SfPzx1SgJqp+E1z0Un5G/zUPk98790Yb4F7N2XRUWQzFdFs/5BQY2ArSY3DzVMdNPXPWRs3pDJ5HFwo/AHtQFEaFzmvDpgyCaURgFacT8qdJfsH44DvrIbK/lbfEeCSLG9MV1Tju/N54SmkjGYoa2pV6+tyFXBczQwsxVQTbj2yDQlzxPTjguohU6QKV9aen2no2Me4e6/wCTB3B8Wvi1tWrTdvW/Y+Yix4q52SpKUGQrkpKl0pJSTC4IP496JLAmuf8AD1sAKVWmDj7KZ627aKrjrqWGqi5jnjjlQ8g6ZIxJyD/S9vdenAaivUr37rfXvfuvdER+X3wG6s+WtLQ1mXlfZ+9sTGIsVvrDU2vN0EYJKrE+tOBqP59+4cOtEBhQ9VR/IjYW4/5Zfx+3HsHobcG4+++/Oy6mnTbz7soZ56nD4iqZ6bIy0hSSUK7X9Nnv/h72Kk1rnqrABaAE/ac1/wA3Wb4SfynekOrunM38i/mfT4fePYvYeOO+axt6oEqOvayop5K58Tjaiub9fBKoB/vHvY1PQDr2kJWvrU/Pou3wj7KzPyN+Q+6Nm7Aw+PyXwh2duWej7Fyu42psNi8PWU0zmkhgkqWWNtRS0dr39uRuIMnLfPpgq05Kk0XyI4k9bK/UW8ekjSy7S6tyW2qWjxb+BMdja7Hr5TGxQNBFHIWf82IH09tMxY6m4npSF0gIPLodvryOQfofeut9e9+690Wn5M/HHavyF2RU4XKQJT53H68jgcvFGDVQ5GmhLUkJlBB0M4UH+n4HsOcycv22/WRhlFHXuU+dQMD7K9A/nLk/b+btsNpdLR17kbzDD4c/b0Sj4o/JDePWG+Z/jP8AIGN8Vn8Y0v8Ad/KVY0xVGPRfHQp91KQrBlCWF/8AW9g/lbmK6268PLm/dki/CTwp5Z6jjkfnHcdl3Q8lc3dk6VMbngycFyfOlK9W1RukqJJGyukiq6MpDKyuNSsCPqCDce5TBqKjqdgQRUefXP37rfWKeCGqhlp6iNZYZkaOWNv0ujCzKwH4P596IDDS2QeqsqupRxUHj1TB3NjNw/DH5FUXcu3I5Jtldm5j+FZqiRGelxNE8wE0o8npWwN7n3EO7QXXKfMS7ta1aK6bSw8lHn/h6x65jiuvbvm5eZLUE2l64SQDggr8WcD59Ww4fetJvfBY/PbNnjyeHyca+KsjZTclQ7WKkji4v7mWxktZ4hcBqgio9KdTnDex7laJdWTao5OBH7en2gwEaP8Ac1pM8z+pkk9QU/4X9qJbwsNEYoOlMVtpy5r0pERI1CooVR9FUWA/1h7REkmp6VAAYHRefkz1hT9mdY56ggpVfcEFJqwtWFdp6WYSaz4dHNz7FnJu8vs+8RSMf0ie8eRHz6Jd+sFvrB0Hx0wegg+E/aFduDbOR6wzV/4x1ogxs881lqKpTUEBmU8nSSRe3s99xdmitbxd5t/7O77qDgMdIOV795rc2M3xwY+Z6PP7jXoVddEXBH9QR/txb37r3VXm9pYuoPllsOmjJjx+6ZVq6t19MTmaSxMkknAt/r+5p26u/wDI9yxy8IoPXHQBuR+7eYYiMLJx6s/gmjqIknhdZIpBqjdTdWW/DAj3DDqUYq2COh6rBhqHA9Av8g9gQ9k9ZZvbs6q8YU5EqwYgtQQSSrwv+PsQ8q7mdp3mO6Xj8P8AvRA6K95tReWLQkV8/wBnQDfBTfVbu3YW58TXM19m7gO36WNyAUpqcOi6U+oF19if3L26Oy3OGeMf7kJrP2mnRTyndG4s3jY/2TafyHS/+VWDjyGxDU/24ZmP0JPpS9+Pca+XQqzXpP8AxL3EtRtqbDlgxhdABxwUBF7Dn36o68AR0cf37rfXvfuvdcWZUUu5CqouzE2AA/JJ9+691Tj8lf5gnb22O5NydTfHvZmP3rXbCx9RuDe71VOlQuP29j18tdWQsh/soCxLfT340A49V1ZwOhBH8xLH574x4fuHZ9JHXbsyVdU4b+DzRQ+FcvSqFliKD6KHNhx73TrZNOpnT/z7n3V1zVS74x1Lie00FQafB0wpxRPpTVB+b3b+nvXy69k8Oj3dMbw3BvvYGJ3NuagTG5SvaYvSxroVIlt4m0/43PvQr59b6FX3vr3Xvfuvde9+691XB8sc+2c3LhcFTnmnkSF41INyJDquPr9T78adaHn0bLruOLZvU9JWeIB6fHTVDAnRqZIiUF2/Fx799nXifl1qJ434Ub0/mtfKft/e+4e8q7qvN9H7jlg2rDQ5RUlr4I61lp44o6eRSoDW9R+n592rQUHn02VDHVnBx5U6sI2b0z/NG+MW68Jtvr2kzHZWzca8dLU57IZM1v3VJJUhp5XV5GvdR+QR71inDh1ejAg1qOtiTYdXuOu2dtyr3dSLQ7mqMXTy5ujXhabIMD54lH+B96Hz6uaeXDpWEhQSxCgfUkgAf65Pv3WukVvveFHtLZ+4tyCWOY4nD5SshWJ0lLVNJRPURR2W/wCQL+9jqrHGOtOrufePZnzPo+yfkRn+1Mz11g+v94TbRo8RhsglM1ZVgGPHolDC6k6mUXbSfduHl1XuOVJABPQzfGPflHsXrKPbXzy6T232LiMzuDF0+0N4b2o2r9wth6qW1O9O1T6zG6lefwPdWoe5evKXVQrHPr1s59HdOdK9Y7TxFR0719tzY+GzeMocrDDgqP7YSQZCkWpiLsTc+hxx/vHvWOPTtCMHoYK3F0tar6o0SZrWnVR5Bb/H29HO8dBXHp0zJCsg9D69J4xZHBm8F6mmuNRkJNlJ5/417VjwbnjhukmmeA6lyOiw/Nmn7V3Z8bexsX0lS0mV3pkdo5mmpsYK2GOreWeAJDHFCTqZi34HP+HtHJGY2KHy6WIwejA9a4f8nH5TD4y9n7p6X+YONq9idr5LI5Ongnqsd4kRppLwQ1dZUaPS30BDWuQfp7o3Cq8OqxEqfDkIqf2Hrb/xuToMvRU2QxtXTVtHVwxVEFRSzxVEMkU0YljZZYSym6kHg+69PdTvfuvde9+691737r3RSvlx8sdi/FbrnK7t3LXw/wAUio/PjMYklPJU1Llyg00ztqb6fQL73Q9VLACvQG/y/wD5Q90fKPBbs7C3ztamwXX9SpfYWR+3FE2Q0TaXWQEAcKbk+/EUPXlaq14HrW0+dffmzNtfNnB4/wCTFdkqjCPn2h2ztqopfu8Hm6E1DRIlW8np0XP1v9Pz78ASOHVHID1Y4H+Hpc434x5X4/w7l+c+25cdsva1NlYcz1vs7b88RpNw0dTOpMb0iGxRQfULH29FKwIRsjpuWNVrIozTy6u56S+H/wAcvlRtbq35N7u6629jt3brof7wZfFjH6EyNYtTZpKkORY3APC25Hu00ZB1Rjtp/q/wdbjZT2yHuHDq2zC4XFbdxVDhMJRQY3FY2BKWhoaZSsFNBH+mKJTewHtN0o+3p09+690g9/dl7M60wNZuLd+cx+Lx1EjySmespYpmEcZlYRRTOpY2H0Hv2fLr2Bx6rfi/nK/COt3n/o+j3huJc3NUjHrKcJElB5pm8SkVzT6QLnhiPeyCM061qStNQ/b0zdidjdf9Cdp4D5N7o7Mytd1xm6NaKgxWOrKfIUsk2SQpTioSFiiMA1yCQR70K9eqB506qG/mr7+3xvDtzb+9KzLZui+O+ao6DI43J4eN8hDNJVU6OlNMtNdAx+jKfd1zU+fTUhYEHy8+g/65+Yu6elKvFw7S6/xVT1FuTEUe2stkqvGLFUVMNfD4aqpcSAWkKkk/n25Gc6S1OqSmoDhejF5v4sdl9d0tL8lPh/nspkdtbvgjGd29C8tFj8PUZVddVJZyE9BY8j6e/ToqvjhTr0DFwaYPp0RHuD4x7Jot9bM7nzG+chl++cru2ihyWKijkyLwSGUE6qmDUPS3HtoMaU8urtGhcOfiHD/iutyzoEbrXqjZibx8v8YjwGJQmc3lamGPjFO0l+bkfW/Pup49OjoZffut9e9+691737r3XvfuvdJXNY94pFydNqLwnW6fQGxvzb8e19rKCphfz6R3ClSJF8uPT7j6v7yljmICuw9aD+yb/T2klj8Nyo4dKI3EiBupvtvpzr3v3Xuve/de697917rxIHJNh/U+/de6SO5N8bc2rA8+Wr4Ygis2hZIjJdV1aSpYW9+690TLf/ytqaiWfGbMpWqg5aHyLErtdxZDqH/Ee/GhwOtf6boLNt7D7U7WysWZyD18NCssUjqQ6qAW5/w/HHvXXs8D1Y7sbb9TtvAUuNqqiSoljRQxkJZl0i2m597630sffuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/Ru/8AhWbfzHKkH6nruX+v0/hbH37r3AmnoP8AD1sc+/de697917r3v3Xuve/de697917r3v3Xusc00NPE89RLFBDGuqSaaRYoo1/1TyOQAP8AEn37r3QWSd5dRxZmTAydjbJTJRPoeBt04QFXPGlh57g/g3HHvdPn1XVmlD0J1JWUlfBHVUNVTVtLKNUVTSTxVEEi/wBY5oSVI/1j711brVq/nQf8KDB/Lf7f2H1psHFUe6zl6UzbmaKkx2Ukx8kFUYqyJjLfxtGtvTfUfwvuygE9xp03I7oKIupvSoGPXPV7/wAFPlXtn5mfGDqfv7btZSTf3921Dlq+jglp2loavzNDLHNBB/m72BUEDg+69OVBOOio/wAzn+a/0r/Ly27SYTcOTNZ23umhSr2dtmnjpKp6iOQukc9RSu/kZSy2CrHz9b+9hSxoOqSOsa6mP2ep+Q6oFyH/AAoR+dWwqGj7Y7K6nwlD0VVSUUwy1JtJf4iaGtm0RsDpvfTz9Pd9ApxB6Z8dvxRsOtoD4ofO7qL5O/GiH5F4jI1GJ27isCMnuqXOQRYpaCeOmaolRGlZUIcJ6BxyQPr7boeB49KNQ06wajrXq7h/4UUdj9u9n7p2X/L82dT7+x2xstW4XclVnsFTZBVraH0SiCaAMNOoHSdXI92CgjUTTpp5SrBApb5jAHQ+fy6f58W4e8fkNg/il8oNpz7S7k3JO0OKgw+3hRYu5ZgoqZABptYfk+9soWma169FL4tQVKlc0P8Ak62fkdZFV15Vhcf63uhx06DUVHXL37rfXvfuvde9+691737r3XvfuvdIDtX/AJlxvT/w3sj/ANaT799vXuqTf5YAA+THdVvp/DKv/wBzk97YUI6rH8LH59X4+9dW697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917ri7hEZ2NgoJP+w97UFjQdaJCip6CrN7Uw3Ycs8eUoYGijjeL7grrkJ4QW5A/HtdKiww6fM9I42aaXUcAdFL7N+MGRxE8ua2LNOWiDTkRqwZNK8LwT9PaCnSzFKdInZPfG+Ota5cBuOnnlo4nEUstVHdgsVw1jIOP9e/v1TTHW6Do+uwu1Nr78oYZ8bXRLUNGhkhkliVvIy3ZVF7/wCwt78M9aqB0JYIIuCCP6g3Hv3W+u/fuvdYKmmp6yCWlq4IqmmnQxzQTIskUqH6pJG1wQfyD7917qkL5SfywMhW793J2t8dty5jam4uwJJ6Pc+BwTy46jjo5pfNIyvTycxtexUgf4e3olHxScBw6YlqCBHxPHrL8VP5QGwehNp90buq6lsh3X3NtOtwlTuSSCVMjtypqFLCto6wyamn1+rUB9PoefdZXMh+XV40WNcDJyT69DD8bviV1T/LV6v7O713BW1m5N1TYCTI7+3M9MzZmuo46yORYHldmaVtem9/9v7oSSerKgUUXic56qQ+J3V3aX80H5tbl+RndFNkcf0v1Jnv7ydK1dZTT6cpFBmFNJjghKotlILW/A/PvfwjT69UA1v4g8uH7OPW2LNLT0VPJNIY4IIEZ2PpRFVF1H+gHA916c4CvRE8P/Me+Mma7PrupoNy11NubHVxx9VLkKano8VHOsvhOqvlm0hdX5IHv3lXr1RWhOR0eukrKPIU8dXQVVNW0sy6oamkniqaeVf9VHNCWVh/iD791vqT7917pJ53Y+zNx1lNlNw7ZwmXr6BQKSuyOPpqmppVU6gIJ5VLLY88H34Cpxx68TQZ4dUofzUesfl78ocrgPjl0jiZaLpbIU9Blt1buxd6esx1VjY2FRRmdCraNJKhAeSbD2q0eClXGekzMZ8RH7f9jrXa+ZVT8j37I66/l8fBTbX8M6yrNtYl+7N37djlodyZTeGJiDZ+vrEgs8mnQxuT+PbAo1WYVH+XrbaxRIzRvP5D/Oelpk63ZvxI652f3N8fO++xd89ybQzuB2dv3Y+6o5aCmp86lT4co1NBUOWni1AqG0e/caClOrN2A6STkefz63V+kO0KPdfT/Uu5N2ZjDYnc+7uv9sZ+uxlZk6Gjq2rMhjI5KpoqSZ1cr5Cwuq2vx7oOHTxwadDmrK6q6MrowDKykMrKRcMrDgg/g+/de65e/de6Jd8vfixg+9tqjNYr/cF2BtZ3zeGzuNhK5PIVFBH5abFz1EZDGN2UC3P1/p7B/NnLEG+2vix/p3EXcrKMkjIUn0x1HvPvJNrzRYiaP9O6gOtJFwxZcha+h8x0Efw1+WuW3RlKnonueAYDtfbCtTRQ1AipaaqxVDGYaeQSy6dcjhRxyfZXylzVLdSnY95Hh3cWKcAVHD8+iH2957nv535Z5iXwb+DGk4BQYDAnjWnVlgIIBBBBAIINwQeQQR7kTqYOmvI5amx6HW15D+lRzz/j7figaTPl0xLOsePPoAe7OooO+ev89tvMU0Qerx80OHmdCTSVUh4qIxfhrW59l++7Zabht0lmRV2UgE/hPqOg7zJsMHNG0S2N0B3KQv2nz+3onnwc7Tn663RuP4obumMNV1+al8XlMhpp3r2klF6dHlsX4HpA9x5yZurWV1Jyvdsa29aM3n8vn1HHtpvb7PezchbgSGsh2M2NQJx9vVq3uTOpw697917rogMLEAg/g+/AkcOtEA8eqsuyIKr43/Ina+dwytFtzsPIio3NURKViijM9nEsj8Dn3Nm0NHzdynPbXGZrVaRg8a06j+/17FvUc0QpHMe/q0DGZKjzFBTZPHzJUUVZGJqedCGWSMmwZSLj3DE0MlvK0MooymhHQ+jkWVBIhqDw6ne2ur9EF+dmzPFsX/Sri4vJuTak1DDQ6I2aRInlLMyMPp+b3HuUPbTca7j+5Jv7KcEnoH822wFp+8E+OOlOjO9Fbnj3X1VsjKGoSauk2/QfxFA6u8VX47SpJp+huL+wbzNZNY73cw0oodtPzHl0INquVurCKVTWqiv29CjkIvPQV0H/AB2o6mL/AKmQsn4/1/ZLC2iZG9CD/PpbINUbD1B6q16Gqa/q75K5DrSBTBjN1VNXmJkAMavMdUqNpb6k39zVzOkW88npu7ZeEBR9mOgDtDNYb61gvwyVb8+j/dz4c5rY2TpgpYpHLJYAm1oibkD3CB6kH7OiafFCuFBu+vxGs2NRLHpPHqUlQLHn34DrZNB1ZH731rr3v3XumXccNTUYHLQUV/u5aCojp9P18zRkJa3+Pvx4de61+ert1ba2B8p/lNj98wLT7gn6i3jBJVVKfbfeJLROBTLPLbyF7gBRe/vQp5deHxEHzp0u/gP8edt95fHjKVGQ14Xbp3FmhjlamaAQ1TSl2qI1JX6Aj1D3uprjrQpTPRct1bKl637MrNt7N+5zlTsWqkydbWz08kkNVDFMV8b1DXBWw/Jt79X18+tkZ7erMvhB8zNz9+b83X13n8DQ4ek2piozjGoYI40kNMqxurNGB+ATz+ffuHWhw+zq0D37rfXvfuvdYp5kghkmkICRIzsTwLKL+/Dr3VTm9apdxdvV1dM7yU2NrmaVUBcgCRnayp/gD70BnHXqgDppk/msfC/N5jL9C1+4N24nO43yYHJST4yKjpIagg07SRV8kliusk/5se7UIFeqkrXQWFR1Vzjf5Q9VtPsXdnyB+KHydrN1ZjfeVmzy7PfdVNQBJGk+7NNLH9xZhrKqOOfewTQA56qVGslKqfPOOj5fETc/81HbXcm2tjd6df0Z6gmrPHlNxPLDXzU1G0wvNDWKXIOn+h9+PCo6stQ1Hpn0Pn1ekTbk8Ackn8e69W6pW/mi/wAwnfPQb4/of4342Pc3yL3dDT1ODxJjp6+leiqoiPHJGLssl7cf0597AqOm2YA0OfQfPqtbon5y/Ovpvuva/TXzw2XjdobC7Xwby5WSqjpIaqlgzcRpXmp3mvp0hrgAj34gUqK9aqwIDgZ4EHz9OiafzCPgt8mvjj2lRb9+K9fjN+/Hre9bT75ZJc9RvRy18jCsSCqxplPqVja+nn8e7AilDx8uqNHLWqGgJqR0e34sbA+Xv8wLsjrjcHyM62w20+oNh4TD4iasxFEkFJLNiVtAY4oyFeVtAuf8eT7qe0UBz04gLnUwoP2mvW07hcVT4PD4rC0nFLicdR42mFrWgoadaaEW/HpUe9dOVqanpz9+690FfdHZmN6j613fvzJsgTbuAymVhjfTaaahpjMkXr45NuPfs+XWsefWnr3p88/5h+Upn+ZvWu3Ik6Tm3hkNt46ojeGOkmrsbOFSianSwdiNNxpP19q4piP0m/b0iljpWdeH8Nf59C3118tPh7/Ngx2N6j+QlF/oG+WuJiWMb425gYaCTNVHjLRRVtT+25YuABY/1I91dCO9Mn18urROkq6JMf0eBHWxD8IPjpuz4yddp1vm925Te2Ppp2qqLO5aaSWokjaERwRoZWYhAtrC/tN0rA4GvR4vfut9e9+6911zxb+vP+t7917rTT+bu/8AcmX/AJgO3sJ8yJMztXpLHbz8O0Fx1KzY7OYVcl6Vr5G9HjK/Vm934A049MtTWA3w0/n1YL3v8/KkZbrT48fy+cBicxt2uaPGVa4mmhkjp2ldVZUSEuYgARqYsLm544HutK11cOrsTwTJ/wBXHoZ/lZ/Kg2x8x+mNjZrsLH0eB742vgohJWU1MagtkUkdmjE6MCH5BBHH+PvyMVNR16SJHGlsgZ6Lh8ev5WvfVblNqbP7z3Fl06w2Jqp8Zi5q2aqoMpSLOJAk1IJCFuBxf34kE4x1vQ34jUdbB2B2DidlbRwW0tqUsdBjNuUn2WNp4FESRwXuVCg8f1+vt+3l0Eq+QemZo9VGUZHSkxGTE6mlmOmeE6Of7Vvz7tPBp71yD1qCUP2niOp2WaqTF5J6JQ1WuPrWpVP0aqWnY06n/Xa3tLjz6UGvl1pv99V3d3yd/mE7Z+MnyJ3dneq+uNxZH9nJ4qQ09MlIEZabW0jIl5QAl3cC55sPdwQF1efTZq0uhvgpXHn1cfuD+S/8bMh1XPtrC1ssW4YcY8uN33FQQSZioq4KYtSzyVEEp1KzAElCf8L+6gtXj1YxxkUpjql7qzEbn2X2hun+Xh3jk67LY7Jvk9zbVy2ZDz1FNU46lefErT/cXdVksosCPeyadw6oAT2NkU4/Lo4fwR7nrqnG73+FncvWWE372QuSy2U64xu8aKOShaioI5P4XGTOuoBgqlbMP8Tbj3o+o6uhxTzH+CvUfH/Ff5y/IDeabC7g+P2zuseq8fuNqqjyu38fFRJHj4JiINHgN2GkC3v2BnNet0Y1BpT7ePV2WV+Ie3X6Ep+j6XeGb21hWolgqclinlhnSdoAkkiJG6/Q303I9qoZCyGOlT+XSeSMRuJBgefHoCfi/wDytOmfj7nZNz1O6cz21VtVyVlOd5QR1EdJUspCSRfvSepCbi/9OfaVlYGjdKFMZ7o/8PVo8UUcMUcMSLHFDGkUUaiypHGuhEUf0AAA966t1k9+691737r3Xvfuvde9+691wkjWVGjcXVxpYf4H3sEg1HWiARQ9JKglbHZKallP7Uz6ILmw/Vx7XyJ40IccRx6RI5ilMZ4Hh0sPZf0u697917r17cnge/de6Se5N67e2rTNUZavhiAVm8ayxmThdVipItf8e9061UdEz7B+Vs80s2M2VTNUu58eoIjt610j1KD+f6e9Z698j0EeK687U7fr4snknrIKPWpliIkEei+h+LgWIH+t71ihA6tUnPRu+vfjbtXasdPVV8SV1ZZHlSWK41q2qzMT7sKCh9Oq049GNosfRY6IQUNLDSwgACOFAi8fTge9db6me/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuuLOqC7sqj+rEKP8Abn37r3WL7ql/5WYP+p0f/Fffuvde+6pv+ViD/qdH/wAV9+69177qm/5WIP8AqdH/AMV9+69177qm/wCViD/qdH/xX37r3Xvuqb/lYg/6mx/8V9+69177qm/5WIP+p0f/ABX37r3XvuqW9vuYL/080d/979+oevde+6pf+VmD/qdH/wAV9+61Ude+6pf+VmD/AKnR/wDFffut9e+6pv8AlYg/6nR/8V9+69Xr33VL/wArMH/U6P8A4r7916o6991Tf8rEH/U2P/ivv3XuvfdU3/KxB/1Oj/4r7917r33VN/ysQf8AU6P/AIr7917r33VN/wArEH/U6P8A4r7916o6991Tf8rEH/U6P/ivv3XuvfdUv/KzB/1Oj/4r7916o66+6pf+Vmn/AOp0f/FffutVHXf3VL/yswf9To/+K+/dbqOvfdU3/KxB/wBTo/8Aivv3Xqjr33VN/wArEH/U6P8A4r7917r33VL/AMrMH/U6P/ivv3Xq9e+6pf8AlZg/6nR/8V9+6914VNMzaFqICx+iiWMsf9gDf3759aqOs/v3W+v/0rvvhUL/AMxuqP8A37ub8/8AVrPv3z68fiP2D/J1sde/de697917r3v3Xuve/de697917r3v3Xuq2/5uHY3cHVH8vn5Jb56Mx/8AEux8Nsgvt6nWNpZjUS10ccjQxrySF/p/X37FOtd2rHCh6+cR8Nd2dM97LmN09+/JDvjZPyqrjkmn2vJ93itlDLxZBvtFmnrHUQpItj+bfS/tw01VC9JlroKvIa+Rr19Fr4adkU3RP8vvBdgdi7potyYzY2ArMlX5uLM09e0lKugQpWVoYorXIFi17f7D230p7qV60QfjPTdRfzPO9P5nh+T2awNXjYNxVWU6UyeXq6F6rG1E24qpYqXDVJZOBAkcmuMixa3u41fCcU6YOhh4wqa4H2ZFf8vVzP8AwlT+XDZDsb5WfDXK5en/ALt9PV8GL6wV6yBlrqXHZuoxs0VHGoBOpIZHGlm4A/J403cajgB1aE6YwpNSSf2dEF/mUVtT2h/wpE+C+zu1YVzO1YN/bZ2/Pt3PL5MJVYdM5Uq0NRTPwY5gAr3H0A96pQDrYALsPQVH2mv+XrfL7F+MXxj3P13W7I3z1b1rFsPwRQvR5HDYmhxNCsJvTsjyBEj0H9ILAe60HT2pgOPVIv8APFxm0fjF/J17Yx3xfOMw2GO8ts0Es2wZqOWJKetjqBVwGfGs62AjQFS3F/8AH3bgwJ9eqHET0Pkeiyf8JTOmOmp/iL2Xuttq7W3Hu7cW78NkNwZzI42irsxHUz0NQ8tLNJIjaAzEubEEn63sPdpR+qembJibNCfnX5562Esx8VPh7Sd97b7SG1OsNsd0YymH8NmpEweN3JWJe8dSKXWkpIHAZE5+pv7pTy6fJzk9HUiCCNBGQyBRpIIIK24II9+63SmOsnv3Xuve/de697917r3v3Xuve/de6QHapt1xvX/w3sj/ANaT7917qkz+V/f/AGZfuq/0/hlXY/8AVcnvx4jrScG6vy9+631737r3Xvfuvde9+691737r3XF3SNS7sqIouzOwVVH9Sx4Hv3Xuo8NdRVDFaerpZ2H1WGoilYf64Qn37r3Ur37r3Xvfuvde9+691737r3SZ3HXtFAtHTEmrmZdKj/jmeGP+2PtbZxqX1yYUdJLp6JpXj05YmgioKREjveRVkcn66mFyP959szymVyT5dPxKFQfPpzZQwKsLqwIIP0IPBB9sdOdBDv3pjaW+4n+6pIaSpaJ080MIGp2tZ2sRzx9R798utH16IlvHp7ffUuSfLbZmrXxkUjzRpCH0uqN6Rp5/r71SmOvVPQndY/Kiqp56fC75genEZWnErKiPqVdJJZhf6/19+z1agp0d/A7mw246SOsxdbBPHKFZUEsZlAZdQuikn3unVa9PFTOtNBJMxAEaluTa5/A93jQyOFHn1V2CKWPSZwxrMhUy5F3kSllJ8Sg6dWn02Nva658OFBCoBI6SwhpW8QmnSt9l3S3pI782XhexNpZvZe4qeOrwm4KNqHJUs0ayxVFMzBzG6NwRcA+/de6aeserNk9Q7Uxuz9i4LH4LEY2DwpDQU606yktraSQLe5J/x/1vfvn175dUpfzUv5m3Ynxw7J6v+OXRmKpsj2L3CIsbj5q+lgkgSqq6l6P7dWmU6dVwCb/63vYA4nHTbMcBRUk06pk+WHYnUnUPx97YqvlVFuXYfyayaUbYRti42OMSVpn1PJLUJa4PNyl/e1rXGR1pyqrVyB8/n5U6tG/l7/zBIfjv8Fttdh/J3L1/9znwcVb1i1TLAd07kx71JjYSU1SVkdrn9fNrEWN+PUDfD14EqtZcHzPV1fxr+WXUvyl2fi94dbZUSU+TohXLjqyemGRgjLBCk0MbEhgTyLe6/b04CDwz0N2brJamqiw9NcNPbySAH0kH6Ej6e19vGI08d+k0sgc+GnEdPNFi4aSkaBBpkliZJZlJDszrpLX/AKj8e000rTNU9OxRiNaefWuj8qvh38jPjT8qsZ8s/idtal3hBBSVM2ZpMlG0yx1lbAVrVqadWsyOWazf8T7aGBTq5Xu1qBnBB6LVL8ONv9+Q7o+bfzTnx/Rs216ypqE6/wBrUMeHwe6tx0qGrppJqTUieSVwNThSTz73WlQpx1QoG0s4yvz4fP59AN1htv5ufzDe73+Reyv4ntfpvpLFNsvbdPT1Aw+NqaLatC8mIjip7xrMXSIF2VW+ov8AUe/UVBTif8HXh4kj61OlQTg/i+fVwf8AK6/mN74783Jmugu76Kkg7L2vUVmPxFRikpjSVOJwiyB3rpoBzN44wLk3PvTDT9h6sjhyRwK8R1eb711frogEEH6EWP8ArH37r3VdHzL+JZ39TL2r1gGwXZu2XbLifGIYJ8wlDH5hTTPGRq1FSCpHPsAc38rDcE/eW3dlzF3VXBamaE9RJ7h8hnd0G+bITDfQd4K416c6T6g+nU/4h/LmXtjEHrvfqRYfuTACWkyOLm8VNFNTUcfijqVvYlzpJPp5J+vsw5L3+Tdrf6bdR4dzF8S8KgeY+fr0o9v+e/3/AG37p3WkW5Q9rxkgYGA3509Ojv47CO8hrMkfJMxP7JOuMA83Bv7Hs9ytPDh4evUjxwFm1yfs6VKqqAKqhVH0CgAD/WA9oSa5PSwADh1Vv86ekcjiK3CfIXryCWhzmya857dElBE/nydLBKJNNQy31A/Q39xrzrs0kTJvtgNLwnVIRxIB8+oV9zOXLmCWHm3ZgVmtW1yaRl1GSD69HN+N3dNF3r1Zt/fMZggyGQikTIY9XQT0s8JCv5IRyt/8R7F3L28Jve2R3y4LcR5gj5dSHyjzFFzPskO6JQNIMr5g/MeXQ+ezzoT9e9+690Xj5MdWQ9o9YZ/FU1Kj59KZWxNYqM9TTPG+thCV+lxf2LOTt7bZt5imdqRE9w8j9vRLvu3i/sHRR30wfToIPhT2lXbj2zkutM16cr1mkeHllnKpUVISYqXYGxNibX9n3uLssVpeJvFtlLzuxwGOi3lXcJLi3axmw0Hb8z0eb3GvQs6QvZG06LeuzsxgK9Fkp6imklKOpZWeCJnjBA/x9mez38m27hHdR8QaftI6R39st3atA2QR0SX4KZzI0TdmbPzs5jfGbl8WEppWAJo45JIysMZ5typ9yL7lW8Uos9wthXXH3n54Oegxyo8kXjWkx+Fu0fLqxMi4IP0It/t/cT8Ohn1Wt33SDYHyI2t2NAogpIoKaCaoF0CyzBVKtIePyfcw8rv+9OVJtqfLEkgfLoD7wotN4ivBgAAV6sGaRc7tQzsFdcnhDOP7QJqaPyIQR/rj6e4knj8Gd4v4SR+w06Gsbaow3qAeqzOtqqbZXdlTBN+2s9fIygjQLNKSvBHtmtcjq3DHVqEUytBDM7KgkjSS7MAPWoYcn/X97oTw69wGeo75XGR/5zI0CW+uusp1t/t2928OT+E/sPWtaeo/b011W7tsUYJnz+HS39cjS/8AEN7cS2nc0CH9h6ba4iQ0LD9vRBvkF8VPjL33uim3fnd8w7Tz6TiTJVmDyNJFNl4QdUlJXOXGqNvoQP6+3P3fffhjJH2dU+ts697iv29Dvteq6L656/HWW0s5i8XgVoXoxLjq2njlMssIhkqx6r620gk+3l2m+I+A1+w9NNuVmGoWH7R0hsXs743wbaym26jN46qrc5DUU9fuOoq4f4qYqh9QBmLcgX9+O0Xyj4T+w9aG4WhOGH7R1G6C6Y+Onx7zObzmyN30WQym4FaKrmqshSl1jeQSMAxY/wBAPr7a/d99xeNsfL/N04L6zp2OP2/5+jeUu+NpVZIh3DhyR+P4jS3PF/8AVe2mtbhKVRs/I9OJcwvwYft6eo8viZQDFk8fIG/SUraZgT/QaW9tmKQYKn9h6c8SM/iH7ekt2BnKfFbRy+QSohPhguCsyH6/0sT7qQw8urAq3n0QDoHAS7r7EyubngWrpXq3llR1LxlfIVfVp4+h+vvVK569XOOmv5GfylPjX8hclkc1UQVGwc3k5BLVZbaVBTw1ksoBBlkd5FuxJJJv9ffqsPhNOtFI2NXFeq2M5/Im3l07Vy7j6I+Q/Z248jFIs1DicvXzUEUZU+Xxs0dQyW1AAWP0P097qfxDrQSnwkj869WU/wAufafzP2RBvDAfKKkb+F0jmLadZLkBXTVEf3F/I7B2/sDgnn3vHWwCPioT8urKd211Vjdsbgr6OPy1VHhMvVQJ9S09PjpJoVAH9WUe9DrzGg60yfhd8s+um+eu9d5fNalztF2FtbcOag60lrcaI8MtBE0ogmqqqp0IoRdJQG4P+t7sRgU6bRlErBsEYz6fLqV8tantL+bX82MLi/jilPkOptq0ow+Qz1PXQTVGKkxyaJ5BNTkKillZ/wBVrn6+9DtGfPrzDxSNBHb/AKqdHTxX8kvtTc022W3x3VvulosLHQI+KhylXNQFKWQaohCk+m2kf09+DYwOrmM17jX8+thPoTpnC9E9c4TYOFqJauHGUsEc9ZMZPLVTxppaZxISbm/J916v0NHv3XumrN5rG7exVfmcvWU9Dj8dST1lVUVM0cEUcNPGZJCZJSB9B/X6+/E0691S58i/5iXwz7tjzvxzzW6M/j8jmxVYKHMwfaw4Jq2ttSGF67WVcc/QEe/Zpq6qStdNRX06p7+TsON+J+weregN81OXyXx6xPZ9P2Pjqrb6y1kWXetmjmlgqVjJQq6BQefexn8+qtpRRXgOjCdc/Gb4xfzD/mNH2L1Hh8h0/tDamCweap6zbe3/AOHVGVyOOpUeanqaiMxpd2F2Jb/YE8e7q7x8OB6qY45X8Q5K8OtoGkaq2vBjcTJJLV4+hx2Px0VVNdppPsqZabzStzy9tTG559qxElxHrTD1NemPFeFtDZA8+llDPHOgkiYMp/xFx/r29o3Ro20t0qSRZF1L16aaGnjeaolighjF3lmkWKNB/V3cgAf6596VWdtKAknyGerEhRU46Cpe8Or3z7bbG9trnJq/j8P8ex1y99OkLrve/s8PLW8i1+s+nk0cfgb/ADdFg3iwM3gCRa/aOi4/NX4J9QfN7rHLbN3ZFS4nL11C8GG3zjKZKrLYh3fyrNRyo6j6n6g35/P09kZDIdLCh+fHoxIVxUZHWsV8Wtt5/wDkh/Jys2n25h8tv7r/AH7VSUG2t15bGzVORp8dNWADJUc82rxSAAchv9492Yas+Y6ZQ+CNP4Sf2H0PW5R13v8A292hs3B762pPLU4DcNItbjZpoxFK8LHTd4wTY3BFr+2+lPS19+691737r3SWyuLaJ/4hSahKhBZV4uL8sbezC3nDDwZOB6Rzxsv6kfTvja+Kug4P7ieiRTwbjgkD2mniMT/I8OnopBIvz6ox/nIfFbObg2ND8guose/+lLbFfjwJaCCX7tMbRxGWomWWMkDhDcWHtoHqzrVcGhHDoT/5ff8AMe6/7O+MFZmeydx4zC7r6jx9LtvcFBkcnQUmZytXjaFleWmo5WV3ZigXUAeT9P66Ioethw6g+v8Aqz1Vz13ka35+fzLsd3D1rgMjB13t2T+F5jPV9FJHUQtRRlJFFWoClW0kAg/T3vgM9aGWqvkKdbAGQ+DPVFd3ZgO+aQ1WD3rg8VS41XxaCNKj7ePSZpJi4a7c3sPz7r5U6coPTo6cSeKKOMu8hjRELuxZ3Krp1Ox5JP1JPv3XuuM8KzxPEw4ZSAf6Ejg+7IxRtQ6q6hxQ9JShkbEV0lNUMwpn/wA2x/SGY8Ae18qrNAHTiOkSyGGbQ3A9LEG4uPoefZd0v697917r3v3Xuve/de697917r3v3Xuk/naLzQipj4lg5BA5+twR7WWsoU6G4HpLOn+iDy6mYqqWeljVmvLGtpQTyOeCfbVwmmQkcDw6chcOnz6adybz2/takerylfTxhBfxCeLyngkekm/49scOPT3Hh0TLsP5WTvLJjNl071DzERo6ormzKVI1AH8/09+oa460SBg9BBh9hdpdx161uTetp6RSDKjB1TSG0MSDbj3oDPW/n0bzr/wCNe09rxQVVfGtbXelpFkiBUMH1ckn6/wCt72fl1oDox9HQUWPiENFSw0sQAASFAi2H0vb6/wCx9+631L9+691737r3Xvfuvde9+691jWaF2KJLGzj6orqzD/XUG/v3Xusnv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de66N7G31sbf6/v3XuqDPnl8z/AJX4Dt/FdRfHXDUFcMg8MMhmigeWSZh43UF1J4Y/197FKZ6oSQQAK/n1GxmI/mn1GJxdbNtPGrVVdIktTGZacNHI34Md/Tf6+/cM9W7jjHUv+DfzTR9NpY0/1/cp+b8cWP8AT3rFa9bIJ9OuIw3808sD/dHGhbfQyU/1/r9b+9kg/wCr/Y61pNPLrv8Ag3809hZ9o4z/AAtJTfQfQ8n3quOtgMPTry4j+aeRc7RxgP0/zlOeLfX37z60Q3HrzYX+afa67Txt+LfuUw4/xuffsdboaU67/g3802+r+6WNJtb/ADsH+3sT73XrVGHCnXFcN/NQ/tbRxn0+vkpib/7f3rHXqH5de/gn800pb+6ONFj9PJTAn/YXt79jr3eM167OG/mnoAF2jjWudJHkpbAf1Fr+/V6tSprjricJ/NO1DTtLGi1rny0w4+v1B9+BHl1ogn06yHEfzTv+eSxp/P8AnIL/AO9+9g9V0twHXv4T/NP/ABtDGD+mqSn/AN7v718+t0f16x/wf+anz/v0cUL/APN2m/4r791ujevXL+C/zTgv/HpY3UR/x1pvr/sP+J9+69Q/LrtcN/NPC2O0sZc/X9ym/P8ATn37rx1YpTroYX+aaL/79LGfj/dlOP8Aej735f6v83WiCeumw3809PUm0cYfrdfLTf1/1yfeut0Py68cL/NPJFto40BuW/cpeD+PfsdeofUddnEfzULj/fp40qotbXTXP9B9ffsdaofl1xGI/mpgEDaOLIvexkph/rcX9+x1uh8qdd/wT+abcX2jjbn6/uUtueSPr79jrfd6jrl/Bf5poBttLG/9TKf/AHoG3v2OHWiGr5dBj2/2B/M76N2jD2Ll9s4mLHUmShgrG10srQwg6y0kRuVDAHSSObH+nvfHHWu5e7j1bf8ABfv3dXyH6bg3jvKnhp89TVENDXLAIxG0/hLyECIAfUe9deHAH16//9O7/wCFrk/zHKkD6f6O5h9P6Ys+/Ade/ER8h1sc+/de697917r3v3Xuve/de697917r3v3XuijfOnLdj4P4t9tZbqrbmI3ZvKh2958Xgc5SJX47IHzhZo6ikk4kAUg6SPfuvVIPXzu/k5R9N/IXpDtHZWX6v3L17818tj0TblPsTZsuL2xPuCmygZ6SeqpVDpHMmpWKi4v7dyBpWn7ektVc65FOPKh/4o9G3qsZ83/hf/JD3J8eN+4zdtRvL5CbH+120g+9y2Rxbx5n7iKGGrlDul4bRluCQASPdQAx0A06uXaFPEILA1wBUivQq/yxf+Etey+3vit1X3d2z3D2d1hv7szbM2a3HhMBDNTukrV0yUcFRFUTRmy2vcqCR6uSfeq+h6uUFe4eQpmlPlQdFbrPhP3R/Ir/AJsXxep+jJd47z6q7OzuOyXZW4avGy1klZg5c0yVEE1SpdUYR+QgXJ1Fg3JuN+VB9vWgNLiQg0+EDiPzx+w9WY/8KAPgF3vvHvr4/fzXvh1hpspuHq/D4XN1mKehkeRchQVEtdTV1bSqSykAtyFsWJBvx70tCaH8utyh1QMlK/iHmRxx8+iP/I3+eV86fmH8ZKb4r9NbMzdF35mIcDj94ZRduvQzSVVPN48hBT1wRQEZmB/H6RckX920gEsSD0yJmYBFjYAjj6dX5fA/+XF3f2B/Ko7D+OXzGaqn3l3BjqXdVHDkKg18+JzsFBJ9qPGzkoruEAFwdLX91YgtqAp05FGy2/hSNr4ip40PWtd8HPk781/+E/8Av3u74+dydf5Sq643/vKoy+1vBhZMs0QojNHjKujkdG8YaOTSY9Rtz/hbdC9WY0J6oHFuqwxoWQcCOjUfCfL/AMzX+Zp/NY64+W+66DO7f+Oe2vFSTqiPhqCHB0iNFTq+PCxqXsBqYcf1F/ppl04416vHIz5oU40B8/n1v3Y6lNFQUdIzFzTU8UBduWbxoF1Mf68c+69PCvn1N9+691737r3Xvfuvde9+691737r3SA7Ut/o43pf6f3eyX+38Bt/vPvYFT1o8OqS/5YCW+THdRv8A8uuqH+H/AAOT3psEDrUeVY/Pq/P37q3Xvfuvde9+691737r3Xvfuvda3/wA3f5n3eO7O/cR8WfgtRUe590z1zbd3dWGClrxT5Rp2p5IEnCt4FXgMxYfT8e7Bcajj/L000nf4Sip4n0A6DbsPsb+aV8LNqU/dPYOHxVdsjCmlXfP2ktFlXRqqdV0xRRGQ6CLrqta/596FDxx1Y6lFQAadXr/Dr5UbS+WHUG3ewtuu8eQqMZSyZ/HyeMSUGQkLI8bKn0vp1AED6296ODQ9WUhl1DI6Nl791vr3v3XuuLsEVmJsFBJvx9Bf3sCpoOtEgCp6SGMAy2TfJEBkp2kgsRxqHCkX/wBb2Y3H+LwiEeeekUNZZTIeA6WPst6Xde9+691737r3Uaqo6StjMNZTw1MTAqY5o1kWzfXhvfuvdFh7P+NW3d0w1Fdholo8mdbxxxRqo1sdQKtf3oA+XW69FAV+1ejMyCRW1FNHIfSysYxGh0La/A4PveRgdVNOPRndi/IrE75mpcPmphRVQkRJiZI4kZgl31cC/N/aqCVIgSwyR0xPGZSAhx0bygNH9tElDLDLAqjQ0MiSLYi99SEj2nZizFj08qhRpHU33Xq3Xvfuvde9+691Vv8AN/8Al27Z+R27tmd2YIRJ2v1k8VTtSOaFBBNUQzGeMvVa10ENY3tb/H36p60QDw49a83yQ6j+X3ePys6vwPyr6225R9EbSysFB2Fn4cLA86YiOUlqqSqAu76BdST/AE92BA4cemWXUQrgaRn8/Lj0rOq/jDL/ADTfk5BtANltpfFv4iZA7P25T0NKKOkzO2RWOtGDTkpHJLMVZz9bC5+g9+rpFRxP8ut6fFNHHYv8zT/AOjRdk70+I/8AKP7Hx2zum95b9372rvEvUUm3cfPSZbb2AheUt9hXRURdVY6P0WuAefbkaGRtUnAceqSOIV0wileAySerAfhF/NX6w+RW7qfrbeFNktr9n5mrEeLpcxTw0FNVsH8Rgo4pAjDn6EA3+nv00hI0L8I63Ci11t8RFfn1cl7Z6UdYp4IaqJ4KiKOeGQaZIpVDxuv9GRuCP9f37r3VYf8AME+A/wDs6m28J15/H8hsnaNFlqDMVj7eYU/3FRSn1rNSxsikMvpPHvwwcdaZQwz0A/zj+TnX38sz4t7b6e6rwFP/AHsrMJjNiUdNhsZTJPM1bhBjJtyVscYLvPK7eQytcg/Q2sB4DPXmcKteJ8h1VD0rvHb38sfryXfG5aepz/zA7fqqfdO0sBHAmQNHtrd1OaimOUpn1SDyCYalsOD7sSWNBwH8um6eEKnLMc08z6dHg6L/AJpvyL683rhKT5ubapdubV3hNRrt2rxOKpMcUTLrehMjhVDadSFlLA/196oPw56trOoKwpXrYoxeRpsvi8dl6N9dHlKCkyNK/Hrpq2nWpgfj+qMD711c449MtdmZJqoY/HxLM9wJvILxuhNmXj8fg+16WiiIyXGAR0ia5LOFiFc9Vl/K/wCLOe2huNvkx0ZE9JvzHS/f7rxVMojo58RTKJah6aNPq1tVwR7ibnHYLuC7/rDsNRKvxqMAqBk09T59Q3z7yXeWV7/XblZdN6mZVHBoxkj7adG0+LHyRw3f2x6PISyxUW66XXS5fDzOkdYk1MgWWb7bhgCQx5HsS8s8wQ77YiUnTKMMvnjzp0OuSOcLbmva0nPbOKh0OGBHHHRqfYl6G/TPuDBY7c2GyOBy0CVONylM9LWQSLqSWF/1KQfbM8EdzC0EoqrChHy6YuraK8t3tpxVHFCPl1TntHcmc+Gvyar9q56Kan6y7Pyoxm0FVfDQUbzVIIls91UAjkj3E1rczcocxtbXIpbXJ0x+gqePUAbfeXPt7zm23XWLG/bTD5Kref2V6ugilinjWWGSOaJwGSSJ1kR1PIKutwR/re5eBDCoNR1kKGDCqmo6ye99b66IBBB5BFiP6g/X37r3VWnZFNUfHX5GbUzmHR6fau+a9avdFRGnhpk8lT6vLK1xwQfr7mzaHXm3lOe1nzPbLSMcTw9Oo+vwdi3qO4TEcxq3p+fVnuLydHmcdSZTHzLUUVdCtRTTIQyyRP8ApYEce4Ynhkt5mglFGU0I+fQ+jkWWMSJkNkdTJEEkbofo6Mh/1mFj7bBoQfTq/VYlXUR9O/LLbWDLmmx+7JDW1JQhIHM51AuTe/1P+x9zMinmDkea4Aq0AoPXHQEdxtnMCRnAlz8urO45ElRZI2DI4DKw+jKfoR7hkgqdJ4jodAgio6IZ8+6CmqOojk8ZUUi52hzePZLVEQnNPEdUwCXv6QBf/X9yZ7YTlN98Cavhsh+yvl0EecEX92mZCNSkfs6z7Z+YvVu2Oqdu1Gaqa2tq8dgaDH19PjvFVVDVMNMIXAEZuL2/px7LN55U3Bt8liiAAkZmWuBQmvTtpzFYR2CNITVVANPs6KDl+2MTvrds2/8Ar6iyNLFE7LSxZeBqeZqv604swFwSP6ewxuO1y7XeraXRBqckHy8+jqx3CLcLVrm1Bx6jz6FfD7s+cXYFMqpgsPTYj0x080KxU7PTkaY5ZHW1zYckn2LFi5RtY1ZXYtQVr69BtpeZ7mRhoVUqaHzp09w/Hn5AbjcndOXehWUXkFDXr6dXqYLpb3U71ssApbpqp6jqx2ndpaGR6fYelFQ/BWPLxk7i33uimdgCwpquQ3JHqA9f+t+fbTc3tEf0IU/Z1ccsLLieV/yP+z11Vfy3OtK1f8p3/wBgFzfU0eQCC5+vBY/7370vPV+hqsMf7OrNyfYutHlk/I9YKX+Wj1VSm43z2BIb3u+QQ/7bn3ZufNyb/Qox+XVF5L25RTxJD9rdZ6j+Wx1XUXvvffyXAF0yCgi3+N/el573JTXwo/2dbfk3bmXTrkH+26w038tTq+lfXHv/ALELckFskpFz/hf3t+fNxddJhi/3nry8m7evCST/AHrqXUfAHA4yMzYPsDd8k6/pjqKtxf8Ap+lyPdF5ymftlhT9nVzytCi0jlfHz6T0vxd7f2/d9s7ir6t0P7YrK/0n+lwze3xzBtk2J4wPsHTTbJuEXdBIT9p6D/ez/IvZFEMPvkUv8DyQ8bSR1CyM6X0CzKTc/X2WbtPsk9sTZ1D9Ltvh3aKYC7pp+XTV1x3jlOhzV5HJ4WurcDk+RLSUTzS+IkNdWAPPHtvZdgj3SDUrgOPU9W3TeX26TKEp50HRu9hfOjprfdRFRU8uSw1Y50+HNxpQgtq0WR5rX/23t675N3e0XxCodfVc/wCDqlpzRtl3hSVPo2P8PRq8VvDbGahE2Oz2IqQf1JFkaV3U3tYqGv7DUltPExV0Ip8j0fRzxSKGVhn5jpRqyuoZGVlIuGUhlI/qCPbBBHHp7rzokqPHIiyRyKyOjgMjow0srKeCCOCPfuvdV1fMT+WR8cfmHtuqxO4MJTdf5+pjnU702RiMdj9wF5YDCpmnjEesC9z+T+T72CRw60yq/wAYr1rm4X4KfOT+S5uTNbl+JSP2V1DnM3HU7i3HuSGHJZfxPIZJYJKF3kKJIpb8c2921Bj3H8+mNBiA8NageXn+3raB+EPyF3F8j+nKHem7MVNiNywGCkzVJJRmiRa90aSUQRWA0C3HHunnTpQB2g+vRyPfuvdFH+QvzT6X+Nlbi8VvzKyT5jLV1JQU2GxL09TlVesIEcslEWDhPUDqt79nr1R0Sr+cR2NvXBfEzb+5OvmykWN3FXwSbkmoEdayn2lk8fDUVMrhP0lUfm/0PvYpXPVZCQhI8utdHYXSHQvzYwO0+hPjxufIpmdt7jp977o3jm6imp8790lp8vR/eoQRDEQ5UM3492OpcsKV6b/TcEIa0/b/AMX1br0FtPAdt971Xwq3/jMX2F1DsnakdZiN4VsEOZyRzmPx2iWNMk2oBS6/pLW490oK18+nB3Ajq93pToLrXoTa9Ptfr/buMxcEJcyV0FBTQV9Vr4AnqIxqIA9IANv8Pfvt6twwOhiqKeKpiaKVQysCL2F1uLXU/wBfd0do21L1V0VxQ9JF46nb0xeEtNQysAQ12ZebkD2YBlvBR8MOkRV7ZuzIPRQfn/2TvDZPxu3rm9kFo6yTCFzNG2mog1S2Jjb8Hg+xz7ZbRY33NUEG4ZAbgeBx0Qc1388GzySWvEr1oZYftjuGTsGHfX98N7vuI19dMmMStd6V5Y61tAsv199G5tm2Ndu/dvgQ+FRe+meHWEVvvfMku5Dd3uLjXVwIKroNGIHD5Cta163/AD4G7y3Xv34wdc7k3qsy7gq6FkqvuARMyxxp43fVzc3Pvmr7l2FltvOF1a2FPDBxThmvWc/KVxcXWwwT3NdZGa/YOhO7w+OPVHyCwbYfsXauIzEyRrHQZeqoIKrJ4sB9d6CaX9BJ+tvYC6EmPPpb9X9dYTqjY+D2Ft1pmw2ApzTUPn0iQRFi9mC8fUn6e/Drx6X/AL917rpmCqzMbKoLMT9AALk+9gEmg60TQVPVenya/mR9B/GPKQ4HeOQkqsrUGNY4qKrp1CGVdQ13DG/+HuUuUvabmTmyA3VkulB6g+XQL3/nrZOX2Ed7IFLYFSBn0+3pc/Hf5Z9V/Iugj3B13kWk4V6ijnrYpp9cknjsI0A/Psu5q5I3flaTwN1XHkQCOlGzcxbfvMf1Fg+oelQejbZvDYvdWEyGGylLT1tBk6KqoamnqIxKmiphankDIfoQCfpz/Q+4+ZSp6FaMHWvWuLuv/hP9s3PfJlexcb2VuzbPX1dXNl8rg8C/21BLUk+R6Z6QyjUGN11Efn3vUeHVPCWuoeflXHV8fRPx26v+PO0aHaXXm28ZjY6WNVqsrFQU8GTycqgj7itnjuWc3Nzf3Sma9O8BQcOh097691737r3XEOjEgMpI+oDAkf64Hv3XumjM0IqqcyKBrivJe3JCi/H+29qbeYxtQ8D0nnhVxq8x1wwdb9zTaHP7sZKkH6lQAL+/XEeltQ4HrcEmtaHiOnz2m6f697917r3v3Xuve/de68SByeB/j7917pKbk3jt7bdHLUZWvpVVRzD54vIw/wCCk+9io7h5dVND2nz6KJufvyrEGXl2WHmCjgag5IJIGmwP9fa9iJ4dSjI6RgmKUIfPoC8XsjtTuWvWryT1tPSM2oxnWqaVbk24H0PsuyMdL6Dj0bvYHxp2rtlYKzIxisr1N2WSJXVfUG/UT/h/T3v59VpnoydHj6HHoI6KkgpUAtphjWPj+ht9f9j791vqZ7917r3v3Xuve/de697917pmzW4cHtyl+9z2XxuHpSdIqMlWQ0cLN/qVknIBP+A59+61XNOqcf5h382LZ3xSpqXbO2IKvIbvzFaMVjq+FYKnGGpqwq0ciyFWBBLXuD72oLGo607aKA8TwHREM78xP5mPx/Ta3dfa+26Burd6VGNq6Z4aSnm8mBycaVsbyKFOg+Jla7AH6H34AHh59VJZTUgUHp1sO/HfvvaHyF6qwfZm1qtGx9ZTQQ5FpJYtFLlI6ZHroGkBsArN+be9efTnlUcD0ONNV0tYnlpKmnqowSpkppo50DD6jXESL+/de6ke/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuvH6e/Hh17qgXtwRr8ztrkxJcZGI6+Vf/PC/qXn3qvb14CrdXx4Fi+GxrEkk0kPJLMeFsLluffhw62ePTv731rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917oi38xBgPjhngRqVq+mupJ0m0Ev1A+vv3mOvUqp6DX+Vmqr0LlNKqo/vERZQADaA8+9nqq1oPs6//9S774UFJf5jdWwcKy9dS+n+oGLb6W9+GBTr2NR+wdbHNx/vgffqdar16/F/+IP+9e/dbr59euLX5/2x/wB697p1qo49d/X3rrfXVwP6/wC2PvdOtVHXtQ/x/wBsffqHr1R1hqKemrIJaaqgiqaaZSk0FREssMqHkrJFICGH+BHvRHXqjoLX6H6Plyf8ak6f6yfLh/J/FH2Jto5DyX1a/vDS+S9+b6r+/U63qJ8+lRmuvtg7ko6TH7h2VtXO4/HqFoaLMbexeRpKJRzppKasidIx/ggHv1OvBqcD0o8dj8diKGmxmKoaXG46iiENHQUNNHSUdLCpusVPTQKqIovwqgD3unWiwOT0nc/sDYe66+hym59mbW3Hk8bYY7IZzb+LytZQAMWAo6mvid4uST6GHPvVPPrYc0oD091eCwtdjDha3EY2rw7RiFsVU0NNNjmiH0iNFIpjK8/p029+690GeL+PnQuDybZjDdLdV4nLu2tspjev9rUWQZ731NWU1Ksl/wDHV79QU63rb1PQuxxxQRpDDGsUUaKkccaaY40UWVEVRYADgAe95PVagdBxu3prqHf9Slbvrq/r/eVXEbx1W6dnYHP1MZve6TZSnlYf7A+9UznrYYgYOOlBtnY2ytlU60Wztpbb2rSKpVaXbmDx2FplU8lRDjo41t/hb37rxNePSqJA/r/tif8Aeve6daqB14EH+v8AtiP979+p16oPXgQf6/7Y+/U69Ude1D/H/bH36h69UdeJt/yIn/eveut1p16/F/8AiD/vXv3Xq+fSB7TAbrjel2VB/d7InU/Ci0BPPvfDrXEdUmfywGU/JnuvS8cg/hlX6o2DKf8ALk/p78xqR16PgwHV+XvXW+ve/de697917r3v3Xugv7m3BktrdabrzuIWR8jQY2WSmWEEyF2Gj0gf4En37Pl1oiop1rjfyJestpv8jPmn2Nk2p87vDJblhqojlkira/AtU5iSSpfHvKCYbnSrMtj9AD9fe2pjqsY+JjxJ62Qe1NkbK7N2dluv99x42qwm4oPBPRZIQvHNpN1dYZyoYr+PeurHqvLDxfGH+VdtLeFXWbtMqbwr4MnTbUjq6WPL+JFeSOLE4lnJaM6j6k9+4nrwARacAOh3+Nfz86F+Tkgo9oZuLB5eUL9ng9x11HRZivLP4wtFQsQ8hv8A6m/vdD59a1KaaTWvR3veurdJzcVWUpvtYWYVM5XQB/qb2b2stE7vEbgOkly5p4a8T04YmhjoaRFQWaULJJf8uR7auJTLJU+XT0KaEp05+2Oneve/de697917r3v3Xuve/de6R+78Dt/NY2elzNJTyfcKyRytCrSh/wDaW+v+8+3YY2kfHlx6blcItT+XRJd+fF+rp6Q5vZryxVN5JpFRfG4RpONIU3+n597uQiyaU61AWKa249B7tHursHqqtGG3FFVSY+F9DyVKk3WP9sfrB/px7ZBHA9OEeY6Pn173BtXf9JG9DWwwVQjUywzTxqS4QFguq3+29+GcdbJoAfXoWAwYBlIZSLggggj+oI9+69137917r3v3Xugs7W6f2d29svceydy0EUdHualWmrMjSQQpk4tMgkWaGqsGDAi3J/Pv3Wx889B3sPpXYnxh6tyeG6q29Q0c9Lj6iWorEo4kyObqIKeSWObJSRcyuD9CfbkaGR9PTUjCNK9azH8mHZXW/bnyw+X3ZHyPlxmT3jsnsGpl2Jj+x5se9NHTVGQnFZUY+nzRCuIQEsig2vexAPu8jeGPCH59ViGpmlNa1oPkKeXSp+dW7Omsv/ND+OU3TUuFw8u08VGuYzOyo8dj9qivx8ktXJ/FqnFKsCOqgR3IubC/PtrgOrsSXX1Fa9WB/DL+Yp2r8wPkXVbN2FiZF6n60yNXt3f+TrKaNpKuvp52gjqaStCAujFfT6vp7qcU6spqSBwHn8+rtYtz7cmyX8Hhz2Ily12H8NjyFK9bdRdv8mVi/H+t791vp99+690T/uL4ZdR9xdkU/bW9cZ/eDPYfDmjxuGycMNbgzUUtORR1ElJL9XVwpuPes9bx6Z9etenYNDtLAfzkhmvlDTrDVYPaGTosHBuWmhbr+goKLCvDhJFGQvThol0GNSLBgPe6dtPOvTYJ8Qk8KY/b/h6sx+edF8SflF8a5N65jc+3cRtzqbLVu7qTL4SpxkNbVS7Zi8pxNJUUtriRY/SgJsbWHv1dJx1c0b4hWtOqcuov52vyl+QLx5b4w4elbpLraNdtZiHMYqiqs62I2pT/AG1TXNI8Rk5ii/zgJ+g59vrGAniPj5dJfHLv4cY1epr/AIOtlX4MfLnrz5gdT0/YG0WeHKUdQuI3BSVs6PWjMQwh62SKKysIy17cce9SzM508AMU6dijRB25rno61TTQVlPNS1UST01RG8M8Mqho5YnGl43U/UEcEe2GUMCrCoPV3VXUo4qDxHVPvyP6c3v8XuxD8iuh46g4/J1Mj74wEERXG0WIV9VS1JSi6hipPIX3E3MOz3vLW4nmLYq6W/tE/CF86DqAucOX9y5L3j+uHLAJWQ/rxj4Qg4lR/FT9vVjfQfeG1e+NgYnee3KhVNTGIazHTzRmvpqqCNROZ4BZlBa9rr7kHY96td8sEvLY8eIPEEcajqXOV+ZLDmjao9ysWqGGRXIPnUcR0N3s56EfRR/l90RRdx9cZCpoqYHee2qOWs2rUxxBp467WGUI45BvyCPYX5q2Zd129mjFZox+n9vQG585Zj5g2Z/DH+MRAtE1MhvI9Bv8F+65tz7Ul6k3dVMewuvInp8593ODUzDzBE9Dcki/9fZdyXvDXNqdquz/AIxb4ap+fRN7a8xyXtidi3Fq3ln2yVOT8+rAPY56lLr3v3Xui8/JfqmLtbrDO4WlpkOc8CyY2tSPVVQGEmQrC45F/wDD2K+Tt7bZN5jndv0q9w8jXom3zbl3GweKndTB8+gg+Ffa1ZufbWW64zThMn1o0WCBqHUVNVHFKya7NYm30/Ps+9xNkis7yPdrb4LzvxwBp0WcrbjJcQNZT4aDt+ZA6PDLLHCjSzSJFGguzyMERR/Vmbge43AJNB0KyQoqeqqv5gG6NlbJp8Z3DS5eirtx7XlpaSClxtZDNWBR+4bxxkn6e5p9robvcWfYWUiKYEmooOo/5wmt7UDctQLR0pQ56TW2vmT2/wB4bc2tF8fsFMtQmIpYMxNmqRW1VjqIWdJZUH9rm49lu78nbdy/uEy72+Cx0BT5ceHVbTma93e3j/c6ZAAYsPPHS9x/xg7m7YlQ935IU1HVMslXT4+pVgjOdMpEUbDnT9PZK3MO3bav+6kdw4E9Lxsu47h/yVGweIHQ49cfBrp/rivSvoxkc1pdJGpM0Y6ujdkbXzAxIt/h7Ktw5u3XcE0yEL8xg/t6MrPljbLJtUYJ+3I6QHyq2pittHGZXBYfG4imglgRocbRxUkTkANqkSABb/7D2GJJJJnLysWPqTXoQIiRgKgCj5Y6NL0LmlzmwcdUhEQxrHEdH50x8FvdBWtD1fFMdDV731rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuq5/lluQ5nP4fb9K3rppUjdA17nykGy/1N/evt69XgOjXdT7MwK9f4ODJYLFVsjwFn++x9NVE3sLH7hW/p+PbiSSR/2bEfYSP8HVWVH+JQftA6rW7c6s2Z378jds7aw9Mm29v7frRSbl/utTx0jrocreQUwUAn88cH3Ou0X1zy5yZPPP8AqSTLVNf+SvUbbjYW28b9HCnYkZ79OOhpzHwsz2wZzXdO7ozdTObOKfN5ItGr31lbu1jdgPx7AEPNUd4NG6RrQeajo6l5bltn1bfI32E9IeTvL5UdJ5ONO1cLHX7Vo2RJHxdLHVVLQFtYMbxqxPo/p7XDaOX93jLbc+mU/wARoK/PpN+89822Sm4JqjHmoz0bjqX5d9XdsDx0dRPt2oT0yQ7ikioGLhtAEfm0lrn6cewzuXLW4baKyDX/AKXPR9Yb/ZX2EOk/0sdGMrdz7foMe2Uqcvj0oBYfdfdQtAS36QJFJHPsnhtLmeTwokJb0pno3eeJF1swp9vXSHBbuxpWoo6LL4ubS3hraeGspJxY6X8cwZGH1tx7pPA8D+HKKEeXVo5FkGtOHUvFYTDYKBqXCYnG4imZtbU+MoaaggZ7W1GKlVVv/jb2z1foP+5+0cN0911ujfebdlpsFhMtkIwpUa56Kgepiju/HJUD6H37r3WtB8FMHiP5nPyi3t82+xNxU/8AcPrPJVWBn2vV1sL0xnxcbRUDCh4VUUIGZglha31IHvZqBTppSJO9uGafkcn7ej75L5f7T+VPyp3F8HcRhqXPdQ5LZuR2/WZyGjiyFFTFMa0E9RTVSiySxKhEYL8Hj3ulBnz6uGq3aMj/AC9EQk/kZdk/GHsbL5n4gbnq6/A7pkrFzFTna2nxuQpabL3FZBCRNyIwxAK/X34uSKHNOqLCqNqjotePV0vwY+D+3vittVK2vqZ832FlVeTNZiutPURPPHaamhqWZiVuWsb/AE904mp6dAAFB1YN7317r3v3XusFQadYZHqmiSBFLSPMyrGi/lmd+AP8T7smssAla/LqrBSO7h1U98md+5ruvdcnQnWdKuUwc8ox296nStZTJjhUHU1K6gWsP8fc5cobZb8vbf8A1l3k6JANUQ4GtPPqON7uptwu/wB07eNSk0kPEAfLpHbK/kufHDbuZxW4Z6irq56SYVklI9EAjyTgTSxPql+mon8e1F/7/wDM91A9vEoAYUrX0wPLpPae1fL0Ey3BQagSeHmcn+fVuu0dqYbZO3sbtjAUkVFicVAtPSU8KCNERRbhV49wff31xuV297dNqdzUnqSra3itIFt4RRVwOlJ7R9P9e9+691737r3SU3luTD7ZwGTr8vkKWhiSiq9BqKiOBnf7Zyqx6z9Tbj2v26zuL26SKBSxqOAr5jpLdzxwQM7mmD8vLrQy/mC4/fXYveOXzlBjt0ZLC02YtFKtDLV0zRaWRGilIsV/oR76Qe3Em37ZsCW7tGkhThWhr1hv7hWu5bluiyRJI8aSVwoIIoR/Ko6tQ/kg9Q9o4XcU2ezNPmqbb0bNIY8jTyQRMPvNahVbjge4e+8Dvm0TWP00RQynhpNTw6kj2g2fc7XTJca9A1E6hTiajraAjqmx+Vmp5CfDVTyShj+ldbauPeH5QSwBvxAdT8HKTEcBXpUAggEcg8g+0PDpYCCKjr3v3Xuve/de6h1tRDBBL5KmCmdo2WN6iRY01sNKG5/xt9PfuvdUR71+efbnQfzzl687Tkji6NyRTG4+dWSGKSvqk0U8sdRIp1WdgbX97pjHl02G7gpHHz+fp1e5Q1cOSoKOugZZKevo6erhZTqV4aqETRsp/IKsLe9A+Y6cI8j0nKsNiK1aiFT4pmCFf7PqOn2Yr/jEOjzHSCggm1evStRg6qwNwwB4/wAR7LyCDQ9LlIYVHXL3rrfXvfuvdJfcO8dv7ZpJKvJ5GmjEYv4hPH5T+OFv7917omW//lXM88uO2dC8jOSiMulyTcqQXC39+z5dbGRXoG8XsPtTt/ICqyUlfBSlyzIS6oVB1XtccWb3r7OtEdG52b8fcJtDG/cVP+VVekGojlQSqxNj+D/X2rtZCG0HgePSe4UlSwGRw6MFgIqGGjjio6SnpvCioywRqgt/jYfm3utxGEkJHn1uCRnSj8R0/e0/T/Xvfuvde9+691737r3RcO8/lP1H0BQ/c733Hj1rObYamr6b+LMoTya1o3Ooi3597p1qvp0Ujd3zu/0udY5XJfFfJ4ap39jhUVAweenomyM9LTxayabHSglzcH6D3XI62Cv29a4Py7/ms9kdnRYj44dzLnNl9k0++8c/8Vo0G3sEtG06xzJLJGiLyvIBP+Pu4WvdxoOmGm/Ae0kgdWL/AMxH4ybP3L8QOmOz9jzY3emVwOO21ujcGTp2gzcr09BjYa2sn+7hJsAwZDq/K+/A06ckSv2jh+XXcv8ANR+NfyK+EzbOqNsVmW3jicNF1xQ41Yo696XN0OMXFQZGGmVbqPQo/wACLg+9aTwPXhIpyDX7P8HRvv5XfUPZ+2vgbuTalWlVj9y7k3Pm8rt9K9TTSwUlekT091cjSNIFvejknqyghFHn1Yh8Uuv+xuueu58J2bVrWZ98tUVKSJOtQopXJMa61J/r79QDh1urHj0Z737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6914/Q+/de6oG7d/7LO2v9Cf4jFx/rzcXJ96oQtevCmrq+Hb9/4JjL/X7SO/+29+GR17p497691737r3Xvfuvde9+691737r3QV92b9k6x6t3vvqFVaXbG3MpmowwDKWoIPLYhgR/tx70T17z6pv/lv/AM1LcPzB7d3FsPcEVHTUWLkyoinWOmp0RaBXIMjoiWHp+pNvdyOB9em45A5YfwmnVydf3d07ijKuS7S2BQNAHMy1m7cJTtEENnMiyzAi35uOPdenKGlR0+7V7H2BvqA1OzN6bX3VTqCWm2/nMdlohpFzd6KRxx7916hpXpO7k706Z2fWjHbp7U2Bt6vLFPs8zuvDY+q1A20+CplVr3/qPfuHXsk0A6WOE3ltLcmPfK7e3Ngc5jI08j5DFZWhr6NUtq1tU0rsgFub39+68ccesWK3zszOVk+Pw26tv5Svprioo6DLUVXUwEfXywwuWX/Yj375de8q9Rj2HsY5CfER7u25JmKclJcWuYoWr0ksSsbUgcvqNvpp9+x17NK9UL4b+brurJ/NWu+OElLSpiqTdp28WEFMJQFrftixcR3/AB/X3anbq8umxJ+r4dM0r1efu3ufqTYcwp969mbF2pUEqBBuDdOHxMxLC6gR1kqtz+OPdcdXzSvT3tfsLYm94Fqdnbx21uinZdSz4DM0GWiK/QnyUUjj/effut8OPSx9+691737r3Xvfuvde9+691737r3XvfuvdEV/mIW/2XHO3/wCdhTf9aJPfvPr3keg3/laqR0Lk7/8APRH8/wBIW9+6qBgfZ1//1bs+5PiN82ulPlz/AKZvjBS42Xbr7XpcUKqtjppA0xoVp6qKZJiwILFuCPewRTu6bIbUGUjhkf4Oln/en+cWTqBwQVvqDBQixJuP7PvWper0b5fz66/vR/OL/stgbH+sFFf/AHlb/wC8+/ah1rur5fz65Dcv84scM2Ct+P2aIG/550+91FadaqfKn8+vHdH84leCcF9LD9mi4P8AXlffiV633EVFP59df3m/nFkX1YL/ABIhorcfX6L71qH+qnXqnhj+fXjuf+cSAOcHyeP2aT/o33vHE/5Ot1Py/n147m/nFji+D9XA/ZouD/yT71UHh1qpPp/Prl/eX+cWPqcF/wBSaL/o33uoHHrWo/L+fXX96P5xAuNWD545ho+P+TfetQ/1U63RjnH8+uhub+cUTw2Dtxf9mi/4lfftQ62SRxp/Prv+8384oajqwXp/5s0h/wBf+z72CP8AVTrRr8s/b12Nz/zjP64E/wBP2KH/AGH0X3rUOtVPnT+fXX95/wCcW19LYIMOP8xRf7exX36oOOtmo9P59dnc384vi7YK9vxBR8/8kr73UVoP8nWgT8v59cRuf+cUTpBwXP8AzZouP9ut/fqr/qp1vPHH8+uX95f5xt76sFpHFvBQ/X/H0X9+FOtVPyp+fXR3N/OMPGrBa/qLQUIGn/WCW/3j3Wq9b7/Kn8+uv7zfzjEPrbBsPx+xQ3/1zZB7tUV69RznHXabm/nFkE6sFfn6wUf/ABC296BHD/N14k/L+fXH+8384vXYNggAL28NFb/blSf959+qvHrVW+VPz65/3n/nFX0k4K/14go/+jfftS9bzSuP59dHc384wfRsEAeBeChPJ/1197JA68CTwp/Ppl3Tkf5w+f2hurb0gwDz5jC1NDTfsUAcSyqQArlBa/0P9Rx7rVT17vAqadGb/lVfEHurovBZnsL5CJTQdn7qFTT5GnpWiMYpZWWaN/22IuWB/HvZyeHW0BVdJNa8erkPfut9e9+691737r3XvfuvdRayipchSzUdbBFU0tRG0U0MyB45EddLBlbj8+/de61YO5vh583fgB3nv3vj4Y1ENX172Zl5cn2FS1C0lXPHFNVPVLSxUEpkNg3K2Xj3uooAem2BDlkFa+X+bqL0nvz+bz8u/kPsHMVpxWB6R2zPKd8R11PQYO1OvD/bRusbPI/0UILk+/MFpUGvXlLse8BRn7f+K6BboXYSfzQP5ge7Ml3jujL0WO+NO5KzaeOwP32iDOY3H1klJT04opPS9ytibGwN/eyAFqPPrwzIVbgppT1NK1PS1/mE9B9a/FL5kdD7q+P+4sriuz6DHUdfQ7Sp8gsNF+3WuxlqKODSH1quojTwPr78K5r15wAQUHHHyPR2PkV/N0y20qvqrqnqXFZDdXaOc2rS5PeOUxNOuUxeNzCQBq7HTaEbS6lWJBta9rce9Ba4rTqzNp+EV/1efQtfDj+bF1d3juSk647KSbb3bsU/2EGJydVHRVFbIkpjmmpcXIEJU/XUAfofat3MMHhqaV4+p6Sx0mm1HiB+ynVqcnf3TtPuOm2nV9i7Poc/VQGeHF1udoqWtcBipRYJ2U3Fv9b2jFD0szXhjoT8Xm8Pm6Y1uHylBlKRSVNVQVUNXTgj6jzQFl/H9ffuvdQqfdm2avLHA02exM+aVGlbFRV1O9eI0F2c0qtrsPzx7916vSg9+691737r3XibAk/Qc+/de6RVUxzWXFFqIhoyJVI+hYcm4P8ArezRB9Lb+IeLY6QsRPLpHl0s9K6dBAK2C6SLi1rWsfZWc9LRgdBlvrqXaO/qZocpQwxSabCeCCNX1ar6mta/+396p1vohm++jt59X138Y2jNWPRxuZIUgZipBktZwp/p9Qfe8g461xGelz1r8oshjKiDE77ilRVPiDSEIyqo0A3Ycc+/cc9e6PNt/deD3LRw1mLr6aZZgSsSzxtMLC9igN/offut9KP37r3XvfuvdI2StmyOaighRZaOnMsNUjjVG6yAxPcfng+zARiG31E5NCPX16Qsxkm0nI6qY+RH8ojrPfm927T6uy2Y2XvGrrJarJ0GKqI8bi6+Wa/knnMTICfV9CP8b+0BYkEnNelgQV7cdVTfPX4TbX+Cvxp3V/cCqz24uyOys1h8jkt1ZiPzTYBqif8Ay6kxeWuxjVrENY8j3tc46o5KKdI4/wCqvVi3QPZPxg/l1fAHbnaAzu1pd6b82fh8zuGlpMziF3Jlt5ZbHyaFFPGWn0QyuA+sC/1B5A96GW+fVjpRMcPXovX8t7LdoYvB9tfO/wCY26J8NtWiz0k/WUVZkVoafJ4XJGSSJFSQqruImUoNPqPvzU4deTV8fr/IdWgfGX+Z98e/k7uXIbY25LW7SraSsekpJN3VVLj4svYkQy44yBQ6yWGnn8/X34gjj1sMrfCa9WPI6SoskbpJHIoZHRg6OjC6srLcEEfQj3rq3RJvlf8AAbon5eUQj7Doa7C5YIUfc+1Wp8buGSIoIzFLkWRmK6QBY+/CoOOvEArRuHWvH/N4+AeK+Mvwz666u+NuU7BzOLy3a6zb1nrq6TI1K4F1jStE8tMqLosXBBFv8fapIyi+PJ9mfs6TSOGrFF5U4fbnqHkPjhtH4pfGqHeP8vHe2w8xujfXXqYDfG1Nw1mIk3HSZvLYgQ5owYZ3JDGRpLNp1D2y8ms0Y46cVRGCYlApw9Oj4/yD/jl3B1d0lnd5dywS4zcub3LktGNEZpqZqepg1vUJTjj9TCxt7oxBOOrICBnz4/b1sJ+9dX6as1hcbuHF1uHy1LDWUGQp5aWpgnjWWN4pV0sCr8e2poY54zFKAVYUIPTNxbxXULQTqGVhQg9U2dpbZ318F+2YOyuvYamt6p3ZX+DO4mNBJS42jaUGoP24usZW9w1h/X3Eu5wXvJe6jcLAVtpjRl4gDz+z7esfN8tN09st8G+bSpfb7hqSoOCA8WA8uraOtOydsdq7Sxe8NqVsdXjMnDrRRIjzQuthJFMqnggn8+5Q27cbbdLVbu1NVb9o+3qddn3ey3uwj3CxcNHIKj/Z6X5AYWYAg/UEXB/1wfa/o049U2/KLr3P/GbuvBfIPrVapMDuPKCr7PjiTx08dGKi7qhUkAEc+4k5lsLjlzeI992yojkas32V6x9522m55O5mh5u2gERStW6AwNPr+XVrPW3YOB7Q2Zg97bcnSbF52kWqgCyLI8XOl45CtuQR/T3J2338G52iXtsao4qOpw2ndLXedvi3G0YNHKKihr0uva3oy64tp0trsF0nUT9NNub/AOHvYrXHXjw6IBvKHqL467vr+z48tFWT1dS9XksDhayL7+oqHBkQGliLauf8P9f3Ice6bpzBtq7PIlNIorMMAfb5dBGeLb9mum3BW4mpUHJ/LoG4e4Pkp8kctUw9V0BwWxqhwakZiBaOpSkdvExWWUKeL392O2bFsUQbcTrmH8JqK9I03Ded6kIsV8OE/wAQoadCFT/y8tkboxT13Y+WzWT3NOxnlg+7FRi0qI+YiYSxDg/T6i3utr7g3+13QbbVVYx8qGnn0ofk+zuoj9axZz86j9nRlfjt1LJ1VtzI7eq8PiKSGnrwcVNRUMMEz0a6gnldeSRwRz7J+bN6j3u8S8jdiSvdU1z0b7Jt37tt2tyoArigpjox3sJ9HfXvfuvdFW+VeCTI7G+50kvFM7E/0CICOT78Tjr2fLpg+JW4/uduTYcsGMTqAoNypiUgm3+39+xSnWqEdHI9+631737r3Xvfuvde9+691737r3Xvfuvde9+691FrapKKknq5P0QRmRvxwP8AH37Hn17Pl1VTuORt9d1lQWlRK86Qt2UfvXHA9+NKY68K9WZRz0+ztqRy1elIcbSDXzpUHTcXJ+nPt+1t2uZxAmSx6bmkEUZc+XRAPhtt96/t3vDd1en3EVbljNi5JBrWNZKom8b/AOA9y17gXXg7Ft1hFgqtG/Z0CeWYPF3G7uZM1bHVlvuHuh11ArMVjMirJkMdQ1yMLMtZSwVKkWtYiZT+Pd1kkQ1RiPsNOqsiN8QB+0dFO7W+GHVnZuROc01+2srAmqlG3njx9J5o0Pj8sEIUEFrE8+xHt3NW42CeDiRTx1ZPRDf8u2N83iZRhw046qT+Us/yN6w3fg+g9v5SfK7dyzUlfCcfUtU1y0kNp3SUpdg1gRYn3PHt5b8s3u1zcyXyaJY6gahQenUa80Nv1ndR7TaNqRqVoc049WlfGj5P9c7l21t3YtRLWbcz+AxVNjMn/eeSOgM+Spx4pjFJUEF9bAkE2te3uFOZtkvhfz34AaN3JXRntOfLhTqRNk3mzlt0tiSrqADqxnh0dqmqqasiWekqIamB/wBE0EiyxN/wWRCQf9gfYOZGQ0YUPQlDKwqpr0GncXUe2O7Nj5jYG7jVDC5qjq6Kq+0dUm8dZTtTOVLAjgN7r1vB49akXzM/lwfNH+XZt3dW5P5cmWqavrXcNXka7eNPXzU75KGlrYNddEaEFtTWZ1ikVLkjj3Yd3xcfXpllKZjpTOPn8ujU/wAg7vP490fXeV2rvo0+2fkVXbnqps1k9+yUlBl6upmib7mlxVVkCsixmUsFUH1X/wAPe3wccOtxElQG+Lz62hI5ElRJYnWSORVeORGDI6ONSujLwQRyCPdOneufv3Xuve/de66JCgsxCqBckmwAH1JJ9+Argde6r6+U/fmZaro+o+q5f4huTPztisvLRMs8lDFM4jBugJT8ktce5U5K5XtxG2+70NMUQ1JXFaZ/PoHcwbxKHG3bf3SSdpp5V6GX44/HjF9PYI1uSRa/eeYiL53IThJ28kjiTxxSG54/Jv8A7a3sP83c1zb/AHPhw9tvH8CjH7ejPZNlj2uGr90rfET0ONbDWYiqNbSM0lPK376OWfSoN7AG/sNRmOdfDfB6MHUwNrWpB6UtHWRVsCTxHhxcqf1Kf6Ee0ksbRPobpXHIJFqOpftvq/XvfuvdAp293ls/qTBVWTytXDWVsS6YsVSVCNXyO6nQUhFyebXHsR7Dy1f77cCKFSqn8RGP29FW5btbbdGWkNT6A56IZtzbfbny7zP8V3lLWYPq8To1PTpejrWpQ9lTx+nUSLk/159ydd3mxciW/g7eBJekZPEV+3oJQ2+48yS67slLcHA4GnR24vjB0wu2qTbNRsnBV9PSRJGtbW42nnyEmg3V5qhhdj/ifcctznzAbxrxbh1LGukMQv7OhT+4dt+nFuYwQPMjP7ehW2jsbamxsdFi9r4LGYeliQIFoKOGmLgc3doxc/7f2R3+5325zGa9kaQn1JPRhbWlvaRiKBQoHoOnXL0Aq4C8ajzR+pT+fTza/tm2l0Pnh1ueIOtRxHWDCZBp4zTzkCaJtK/S7KByf9gfdriEqfEHA9ahkAGg9OOQyeOxNO1Xk66kx9Kt9VRWTx08K2FzqklIA/2/tJ0p6CffXdWytt7H3PurE7iw2elweKyNYlNiMlR5CZ56KAymPRTM9iLfQj36nXidJFRx61m+yvmH/MA+QmJ3l2x01msRtbqvZFfl0enz1TRY6pMmJBk8cH3CrrY6eFW/192NBk9Ngsfhp0WzanfGR/mIdFVKdjOid7dUZ6u3DJXUqotdkMZt+IzTA6ApK/t+k2PvxGk48+vI3irSlCPL0I62LP5Xvyui+THQNFPmK+Fd1bQrP7qz46eoQ5F6HEUq09NUtAQGIshBPP0+vvRwadXU6l1evVkWSphU00igXdVZk/17e3YH0PU8Omp0LpQdNuBqWMJpZr+aO7G/9L2t7euogp1rwPTVtJWqHy6xZ/d2B23Sy1WTyNJD4hcxPURrKbGxAU/8T7SUxXpX5gevRMuwvlXKZ5sfs2J5JQSgZCHYkNpNmUX+nJ96FePWzStOgaxWy+1O468VGRkrael1F2Ul1VlvrA5t+D71nh1ro3OwvjRtPb0UdXk4jVZG93DojKCQLk6r/n37iM9e4HoyNBjaHGwR09FTQU6RIEHijRCQBa5K8n/Y+99e6lyRrKjRt+lxY+9gkGo6qy6lKnz6SeNb+H5Kena4WoeyAn/G4sPa+UeNCGX8PSWM+E+n16V/sv6WdYp5o6eGaolZUigikmkdiAqRxqXdmJ+gAFz791446qn+Qn82Po7pbsTHdWUFFkN7boyU0NOBtiqjrxSSzuIwk0UCt6geCpPvYBP5dUZgKVOT5dWSbf3O+6evMfu2lp5qWTMbZOXp6apQpPBJPQmaOKZCLhgbAi3utaivTlKGnWk/8vvkl1hUfzGMTs35BQdhZKuqoZ6XHY7bs8z0RWSIw0bvSKCGANr29uAduOmSwDjWceQ6GHoPpffnS3yW2v8AJerrarDdRQVkSUOGzU5x7VlDU1GmCKahlI1XU2Pp9+JBFD15VKuSDj06Hv8AmSfGvYHzx+Reyes02im31zeIxO4KfcO0sRDTVk9RKgeGWWvgClr/ANpiefdVOerOuoDVmnRrupPgn8rei8dXdOUeQo9y/H7JbDnwlUmYqYMlk4XmomptIgkcski6voPfq568AQMnh0Tb+Xv8ENm9GfOfL9ebq2vnMht6UZHckCZTEGTANmmDz0quXultVufe2JNB6daSMKzEDjn5dbZePx+PxdLFRYyipaCjhRY4aajgjp4I0jXQqpFEAAAAB9PdaU6crXPU337r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XR+h/wBY+/de6oH7eB/2c/a35/3IRf8AW4e9Gmnrww3V8O3xbC4wf9Mkf+9e/ClMde6ePe+vde9+691737r3Xvfuvde9+690Wn5gH/nGzuQWBDdfblBBAP0oSfz70eHXhhh1p1fyjapsVuL5E5SjJp6yg2hvirp6iH9uWKWNJCHR1sQf8fbkh7R9nSe3H6jgfxdFR+KfxO73+fnfvc+Er987kh2dRYLc9bTeDPTrPG8crvYBnFr/AEX/AJF7s1AAKDPVEDOWYucGlK9Cr/Lz7L74+GXyL7Z+Om2NzZnN4vbuFyv8JjyeQmrqpsjcwrZG1H/EWPuhANGGOroHUlSdX29V5969nYLtj5d73wPzLznb23JaR6iowWSwddV43Ex5F59cf3Ej2URg/X3YfDRM+vVGJ1EyEjGM0HW2V/LaosVtn4j9u1WwezabdO3YNnSti45dyx5TNQO9ILMdDMVIUC590PE9KUrpFeiRfyiuxN9535tdwY3L7hzWQxsC5Aw01XkJ6mNSJyFIjNwOPofe3HA/LpmJmIfUa56C/bPZW+2/m1buwb7kzf8ACB2IKcUH8Sn+0EP3Z/bNObC1vx79T9Pqxc/U6K4oMdIGFXh/m2bpek8ENUOzf2JZAqRo5rB6pGtwLm5PvdP0qdaI/wAbx/COl9/NY+MGyOyewt15/u3uTJFKKrilx2K6/wB3K9bHGIzMsMtJA+pTfgi3uqEnFP5dXdaDiafs6L3/AMJ+Pk5v7G/JndfTO3s7uav62w2dOHx0O5shUVFW1A9ckZaRZr+qwH093bypjpqFqkqSSPnx639/bfSnr3v3Xuve/de697917r3v3Xuve/de6It/MQ/7Jxz3/awpvxf/AHRL715jr3keg3/la/8AMhcnz/zEZ/2H7B92PHrS/CPs6//W39Xjjk4kRHH9HUMP9sffuvdY/tab/lXg/wCpUf8AxT37r3Xvtab/AJV4P+pUf/FPfuvde+2p/wDjhD/1KT/inv3Xuvfa03/KvB/1Kj/4p7917r321N/yrw/9Sk/4p7917r321N/yrwf9So/+Ke/U69177am/5V4f+pSf8U9+p17rv7an/wCOEP8A1KT/AIp7917rr7Wm/wCVeDn/AJsx/wDFPfuvde+2pv8AlXg/6lJ/xT37r3Xvtab/AJV4P+pSf8U9+69177am/wCVeH/qUn/FPfuvde+1pv8AlXh/6lJ+f9h7917rv7an/wCOEP8A1KT/AIp79Tr3XX2tN/yrw/8AUpP+Ke/U69177am/44Q/9Sk/4p7917r321N/xwh/6lpf/b29+69177an/wCOEP8AsYkP/Ee9UHXq9d/bU/8Axwh/6lJ/xT3vr3Xvtqf/AI4Q/wDUpP8Ainv3Xuuvtqb/AJV4P+pUf/FPfuvde+2pv+VeH/qUn/FPfuvde+2pv+OEN/6+NL/7e3v3XuswAAAAsBwAPoB7917rv37r3Xvfuvde9+691737r3XvfuvdYKilpquJoKungqoW/VDURRzRN/waOQEH/be/de+fUejxWMxyuuOx1DjxICHFFSU9KH/4MIFW/wDsffuvfb1r4d//AMs/vnqr5Dt8g/hbk6HEtlaufN7so6mppqKonyk+qWZ1p9YMy6zfke91xpPDqhQa/EU0J4/PoP8Arf8Alt/JrsXuCr+XvzKzVPkt97CxuYqNoY6jqaesiqKGKiklhWrpIWNhz6V03v73XyHWgpFGJB/yV6Dj+StufqXZuf8AlvmO9K/aWF3bD2DkZ9rt2M2NjrVw6zVVRXJhWywLLH/mwViFyOBxx7siNIwC/wCr160WWJWLcan/AGOiU0PX3UvyD/m2Q/OLaebpKHqbqLFbhxOa/utXRU21jl6HHTCnSWOmIQPJKAV9JPt26kWRgR5U6Zt1K1B8/wDKen34ZfFPtX+Yh2P3F8pq/c+awmJ653ZuTbG3oanLSQR1CwRVFVjmhR2UtGqIuogcah7YJAFKdOrGzHXqIFaAfIefR7P5W3yV7j627L7D6R7m3JS57Y2Cj3NlRmYatq+joqfC0805phWAG0jqNOkH6+9NTHV0JBKny6FrY3zF+Ou38J2j8/5pNyUmG2tktz7DpaLJZeT+GVObpoJDTiJnBBuo/Rp96odVB1sMoQuTjzP2dGv/AJZ38wqH5o/GrsX5FZCCoTD7O3Jm8atCgWSqNHi6c1uuLQouGQhUNubH+nvZBBIPWlYOgkU1B4ftp0e746/IHbHyR2JLv/aeNyWMxcWZr8IYMooFQ1RjyBK4sqjSdQtx7159XxSo6GTNVy0VG3/HSe8MXNvU3F/959qLaLxZM8Bk9NTPpXHE9RcBQPT0wnqV/wArl1F2/Og8gX9uXcgZ9Efwjpu3Q01tx6UPtH0p697917rDPT09ShiqIYp4zwUmjWRTf/BgffuvdFk7R+N2394I1Zh4kosmzkkoscacm91YWt71Tr2eHRPZsd2h0XmWliNfU0MT3YhneNVZrCwU/gD377OvHo2HWfyg23uf7XF5kmjyTko800gWO4sLtcf6/vQNetmlOjG5PLUtVReLG1cNTPVACBoJFkH1BvdL+1dtHqbW3wjj0mnk0rpHE9S8HjRQUqmQf5TMA0zEclv9f3q4l8R+3gOHW4I9C1PE9PftP0/0BfyF+PXXfyY65yfWXZNA1Xgcm8crS06wrW080SssUtPNIp0kar2/Pv32deIBFGFR1rw92fyEsrv3vXZy4reWXPQOBgo3raSry8ZqwmMYSJBHjnl+rAWFlt7sHoDQfn0y8CuwqTpH4fL5dCp8xoOvPlx0Ljv5fvTNVU7a3t1yIJ8fQirSilymM2RjWE4jpYgpdpI0LNcnn/X96HEN1d+8MgNDTqtz487D2380evxtLoVcjsT5SdFbnxGzGo6+pTCxbgw+3pvDlMmI28ZZlMZZnYni/PvefMY6rg4ByDSv2f5+tkDNfObrj441XXPR3aDVtTvzH7Q27jdx5OGeNMfFlqLGRUtfK8zq2oa1Zy17e6nHTmoVGePRz9v9obR7H2/T5brvcuH3RTVaQl6jCV0VfFTedf8ANTyQ8K4NwVaxuPp7VW0QJ8R8KPXpPO5poXieoe4dtdb7yxeS6s3hLt7N1uZxlYtVgsg1DU5SOnr4SJaqlpai8ilQdSyKota/ulzN471GBw6vBF4QqePVbPXv8lX4r9cduntrD5jsGrqhXHJJtvI5lJ8GtWX1+qEKAY7n/N6f9j7ZqaU6cCIDUD/D1bxQY+gxdNHR46jpaGlhREjp6SCOnhRUQIgEcQA4AA96pTh1bqZ7917r3v3XuklvfZWB7A21ltrbiooKzG5ejlopxNBFM8aS8M8JkB0sPwR7SXtnBf2z2twoKuKH/Y6Q7jt9tudnJZXahkkBBqK4PVPeIyu+f5f3bv8ABs/HXZfoveORej2yY3LriBPPpNRWsuoRqpF7m3uJop73kLdvBuavYzmiU/Dni3p1j/BNuPtRv/09yGfarltMVM+GSfxeQX5+XVym2ty4Xd2Gos/t+vpsli6+JJYKqlkEsLakDFRIvBIvY+5et7iG6iWeBg6twIyOshbO8t763W6tWDowqCMjpMdq9e4rtHYm4dkZeGGWkz1E1HI8kaSNEC1xJHr+hH9fabc7CLc7GSxm+GQU+z59I972uHedsm26cArKuk1FePVcnw3m330Z2RvTpXd1DmKnZ8NaaTYtSInkp4E+5F/I3KhSt+Qfcf8AKJvtlv5tnugxhBpGacM9RH7efvTlndbnlrcEY26tSBvILXgerOdz7w25s/Hy5HcWXoMXTxRs4NbUxweSwJCpq55sebe5Xt7We6kEcKlifQV6mueeK3QySsFA9TTqubfPyu3z2/nJtgfH7F5GkrllNJPnayO+OqCzGJ2gqdIXTyLG/scWnLtntkH1u9MCOIUcR+XQRud9utwl+k2dTq/iPD06VHT3wlEeVXfHdmRqs9vSOpSaKmp65ajDiz+RjLEdStzxpsPabc+bCYvo9qUJF6kUb/Y6e2/loCX6zc2Mk1fXt/Z1YPj8Ti8VEsGMx1Dj4UUKsdFSQUqBR+NMKj2C3kkkOpySfma9CxURBRAB9mOnD3Tq3Xvfuvde9+691737r3QT9z4hszsbJ06oXKRTS2ChiNMJ559++XXuiZ/E7ILQbxr8PqKgzSx6Gb+16k+n+x9+A68fn1ZJ7917r3v3Xuve/de697917r3v3Xuve/de697917pB9mZJcVsrOVTOEK0hCnVpN73Nj/rD36mOvVp1X38dcFUbk7Crs2V8scFS0hawewElzdj719vXgfTo3vyb3NDtrqLdVQZhFUvRqtOdejktydX+w9izkyya836FaVAOeiXfrgQbc+aEjpGfEHbBx3XNHuV1US7lgFTI9hrktJcMW+pH4ufZhz7eibdms1OITTpPy3bmKyEx/wBEz0bb2BOhF1737r3SZ3huKh2rt7JZrISiCnpqeX9xjpVZWibxXb8XI9rdvs5L67S3iFSSP2Vz0nuZ0t4TLJwHVdXxQ2dkOy+x929t7yp0r5MLl6mi2+9dEKpDSTs6q0DTXtpX6WHuWeeNwi2faINisDpEigvpxkU40+fQL5dtnvr6XcrkVKtRa5x0O3e/w+2H2y1TuChSowG7YU8tBPiHix9NLVIvoNSkIW9yAxYn6+wBs/M13ttIXo8XmDnHy6O905etdwrKlUk8iMZ+fRXMb2X3x8SJ6PE9p0tVu3ZCyRR0D4dTWzR0Yb9bzpq0kD9V/wCnsQSWG08ygy7cRFLxOrGeiWO73XYaJfDxIhgEZPVhnVfdOyu2sLTZXb+SpY6mdEeTDzVcRyVOZBdVkg9LX/1h7Be47Vd7bKY51wPMDB6FdjuVtuEQkhb8vMfl0KVbQ0WRp5KTIUdLXUkylJaasgiqYJUYWZZIZgVII45Hss6MOqcPlH/J76b7f7bxffWwJ8jsXsClzFDV1eM281Jhtu1EEDhpZRS0ojCSHTza97/j3YMQCDkdUZASG8x59W/baxj4XbuBw8jmSTFYfG415GbUXeio0pmct+SSt7+61rnq/T37917r3v3Xuid/KT5E0XWWFj2rt/Xk957oMmLxsFBIXqMbVyMI45aqNRcLf639j/kvlSTd7g3132W8PcxPBh6DoN77vK2Uf08PdI+BTiD0w/Ff46V2zGq+zuxT/EexNzRs9V9yqVMNNTzqJYngLk6XsbGw9qedubItwC7NtPbaw8KYJIxQ/Lpnl/ZntdV9e908nH7Ojxe436FXXCSNJUaNwGVgQQf6H3tWKnUPLrTKGFD0kbSYGrZjd6WobgDkKuq5/wBb2YVF2meI6RVNs1PLpUiqgNOKppFjg0azJIQiqv8AVmPHtBobVoAqelgYFdXl0TrvL5VYTZzPtPZOrcG767TDRyYt1q4qOZ/R/lCICQQbfX3IPLfJFxfj67cf0oFyQ2CR8ug1uvMMVsfprXvkbhTNOgk6m+MO7t/bipeze8q6SsmYiuxGLSYPEFMmpErKZiQLgn6j2eb5znY7XaHZ+W10gYZv8x6L9u5fuLqb67djqJyB/n6scocfQ4ynjpMfR01FTRKESClgjgjVVFgAkYA9xJLLLM5klYsT5k16GqIqKFQUA9Opntvq3Xvfuvde9+690A/Z3aux+oKqHK7qzOMw1NLojVshWLSI80o9Eas3F2v9LezCJxNEY3PSKVPDk8RR1QF/NU+aHbcm/MP1L1pk58Rs7OUNJkVz5qHo8ZUUlbTqWKVtlB4J5v7RGMqxB8un/F1BdI48T6dBT8Ye9On/AI9bpoqDtrNZXdG3tx4ONssVzT1dE2UrYtEyA3ZSLt9L+6kAnHVwQp7j0E3YW/8AYvaXypqegMLl4dj/ABirqKXd0lQ9ZFiI6meaP7uWAygqryuPQF/N/p72MCp49aJqdK8KdBp1T8Zuyqr5z124fiNisjSdE5TD/wB3K58pTp9rUrU0ppsjMTcK4k9bKfz72TqFOtBCrgrwIz9vlTrYF+HXwJ3T8Y++MrvjH5CGPYWb246T4iOaKPx5ytpyapzRRubWkPBt7p/k6dAAJI8+rQ9wbuwG2qWapymRpYfChYwNMiysBwRp+v8Atx7917j0U7cPyApMhU167RulUkbpGFlLBnRr/Qf19mMLCaMxniOkMo8Fgy+Z6ACn2d2v3BmZpMhLXU9O7MQGZkjZSfICQfZcw0uVPHpcCGGocOjVdc/Gbb+3o46vPRiqyACsVKxuutk9ZYm/596p5nj148cdGeoMZQYyFIKGlgp40UKPFEiE2FrkqB73x691P9+691737r3XvfuvdJbcUcMBgyMlTFSmnJ5l1Krfk8qDz7W2sgAMTDB6SXK0o/p0Vb5XfNjrb4mbKTcm7Vlz2TqKSKox+AxM98jkFlRmV6eIIzEcfW3tK4o5VfXp9HBQEny6qw2j/O2693Lueh2X2NsTdm2sPvulqFxmQybTY2Gip66FqWLzTNGtwNYa9/x7rpYZ49b1oSATT8uPRB9pdebE+Of8y3qDdmZkkzuze0MjT7po8lnvFkcRBQ5CrLQOsk2pbKSCD72MrT06pQCbxP4hTrcIoZsdksZBNjngmxlZTXpXpgop5KaVSFaEKANJH0sPdR09x616v5i/8qfsbsfujGfKH4xUW3z2jgY6VaWkzVPjmivFEPJLGtURqbUCbafe9QGKY6oyk0KkAjqL1n8JPmZ8icztRPllXYrH7O24mPiqoMCaOjleejlDOIqanZQ1gObD37Hlw68ATxpX5dXgba+PvXW2MttrPUeKjmzG1sLR4LHZCeGJqj7SjXTGXe19X+x96GOr/wCXobyAwKkAgixBFwR/Qg+/de6bUwuGjrf4lHicYmRtp+/SgpVrdP8AT7oJrt/yF7917pz9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XR+h/wBb37rw49UD9uD/AJzP2vb85CI83P1mvfn3o10469Xuz1fDt/8A4suMvyftI/8Aevex17p49+691737r3Xvfuvde9+691737r3Rd/lfjavLfHjt6goKeaqrKnYW4oaangQyTSzS0RREjjH1J/A9+8jTrR4g/PrUT/lc9S9lYDI/I5czs/cVB97srfNPRtV4uWBZppoZPFHHr4JY/QD3aQ1Az0zAGDtqBGf29Dn/ACT+suwtqd1d1Vef2xuHDwT7W3dFDNXUE1JFI8sUqxojG1yTwP6+7ORih4DrUIZfE1Aip6Kt0/1T3FiP5lXaG7sVszOhP8ukoanIYh5KCSX7k+PU8nDKfeu0Lx6sA/iFiDw49ID+YTvnpze289/bI+TvQu/67cmTxJx2F3B1xtiOhaHIqdK1D1MMd2Ukc88+9LQCoNK9blFWoyk49K9D1/I++FXeu28d2ZTbUTdeJ6f3TtqtosDRbzlmjq9VSl4lmjqm5sptf+vvbFTQDy/w9UiEgDaqivAfLoruM7d7f/lnfNnsivzPXW9M1BvWvlxmCqsFiZ5II/LVlGleSIMCpt79RWUEkY693xuV0kg0NQOsnweTvPuf+Zbuntbdux91UW2svu+LKYqetxFRCIxJOspaWaQD6Bj73gJSo60A5m16SuKVPQ5b66Z7nyP8yDsLKbZ2luGF67e8s2KyjYyVKRS0hKy/c/S3N7+6hgE+fV2WQz1pigz0SvsTBV/S/wAo+wz8n9jdu7yyu4c4zbV/gUVbV4qFnY2eoHqVkt+B73XGKdUKkOdQY+h8uhn/AJN+0t3435zbn3HF1xunAbUzG7WqcZLW4OWjSGk++Vl80hA+g97YjSM163EG8Q1UqPn19Aj230o697917r3v3Xuve/de697917r3v3XuiLfzEP8AsnHO/wDawpf9v4ZPfvMdePA16Db+Vr/zIbJ/4biP/Whvez1oeVfT/P1//9ff49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XRAYEMAQfqCLg/64Pv3XuiGfzIKf5HzfFzdi/FjIxYvtGKuoZYJX0ASYKOGVstTor8FmXSAtufe+PDqrVoacfL7etSH5Mb96M7S69w/3fSvc2F+UGCoqXa++MrgUnx+39w7iqYfs67LRUECAkFiST+fr7VwFY4SW4+vSG4HiyBVrT5VrX7ej+fBT+UV2nsH+W72rsra8tTjeyu9d+7f39iZNwzxJX0m3S7GupHqZnvGzq5uhIv7SlgXLAYPSqOMrAImbIrnq3Cq2Fjf5d/wSzWF21t6qyu7s3gnxue/gkCCWbdWYxEtFNXJ9oDqEbO1nvcnn3XJNfXp2mlKLwHWt/T5Pffxj/l/b6rMrFWJ392t8gaOmxtE113HDsHPlo6sFTeQRtrAN7fX3b8Xy/zdNVYJkdxP+E/5B0K/z56R2/tH4edHfy69hGtqOzuzd3bP7w3fTUUp/if2+7cbG1XQy08f1C+Qk6j72DnV6dedRp8GnxY/KuT0dXtzcHVH8oX489TfGro6ky+R3b2PNtGq7EwkUgOTBylJHBXyVFBToVQMZHurEm1rn3WjN3Hq3ZHSNRQDA+zz6ss+Nfyo6i213ZhPh3svb1ZjsxWbPxe/K6eoDRBMjmcSuRyERp1XSCLDm/vVDSvV9WaHy/4rqwPcG6tpjcdPtrJbjw9HlUEU8WMqajRWuX5TTCfrcfS3tdC4ggMhHHHSSRXln0Dyz0IqyxBxCGHkCK3jAOoIeA2n+ntDWp6WcB1m9+691737r3Xvfuvde9+690y5rb+J3BRTUOTo4KiGcWcvFGz2/wAGYE+/de6JF2f8XDT/AHub2hMaZYQJQkbLHIqgljbQb/U/W3vwUsaDrRJUaugU2j2nvfrLKQRbjirpaChewklLlJELC1if9Y+3Q7oDHwB49UMYYiQ56sA6/wC69ob+p0alrIKOqK8008wDMwIB0XA/r+fbINenMeXQwghgGUgqRcEG4IP5B9769137917rhIiyRvG36ZEZGt9dLjSbe/de6oo+Wv8ALZ7XXv3HfJ74lZmmwW+sfiamlqaPIVkEUck9XTNT1M0UblQ+oEkgj8/n3sHyPTbLnWpp1VPn9idr/wApHG5P5ud/49cz2TmN3SYHE4zbBpkoZ6vPhmmraunpNSyfrIIZefdq6qCnVQBHqkLVNM/5OlXvrvHrc7YyPfvzJ657C3S/YOycjuPaGR29HUUkNL/HcWarGRy1fjI9Gtbgf0493hi8Q6vLqs0yRL3cR/l6S/8AI8+VuU6/667v7fztbktufHjH5DckeLo95V05nqdyQwz1GDx9JU1AN3chVCqOfblzJU+GMev29NWqVAkbNfh+Q+fR4f5WeV+SPzG+TO7vm9virrcT1hQ12e2Zi9vZKokhkqdNLJHj5qChfTqhRTGdQW3It7T4GBx6UpqJZmwDwH2Hj+fWyd7r051DyFdT4yhq8hVOI6ajgkqJ3Y2CRRLqdif8B7pJIsUZkfAUVPTcsiwxtK+Aoqfy61zflx/PR2v0529leuNkbayOfh25XNRZXJUVYgp5Jons8UfkAu1vqV4H0vf3H1xzHul9Kz7Qq+CjadRPxUwaY4DrD/3A+9JtfLfMUuybVBJc/TGkrIV0q3HRVmWrUpWmBwrXHVrHwc+a+xvmR1vTbr24j0GWpyYMriKicy1dNPHGGl8oYD6HjgkfkE/X2INg39N2V4JV8OeI6WQmvz1D5HiOp09rfc/Zfc3Yl3XaiQ3B0bDIw+JWHkQfQkHiCQQejzexJ1KHQXdvdU7Z7g2Vmdn7kooJosnRS0sNW0EUlTRu/IkppX5Q3/IPst3Xa7bdrN7O5UEOKVpkfZ6dEu/bHZb/ALdJt94oIkUitMivp1Vp052puv4W9lJ0Z2gchU9XZKvkodi5eaRjFQPPOEE1XUMGGiwsbn3Ge07lc8m7gNl3CptWNI29KniT6dQny/vd77c7yOVt7JNjI2m3cknT8mOfTq5DH5GgytJDX42spq+iqEEkFVSTJPTyoRcNHLGSD/t/ctxyJKgkjIZTwIyOsg45Y5kEkTBlPAg1B6Ln3h8hOu+l6GWuq4KHM7ksTTYnHrTvlpJWJQBVRS5N/wAXv7PNo5fn3WXtXSvmxGP29FG57pZbautgC/kBTUeiT4XrDuT5gZag3R2HWVmD6rknATCtKaLKilSS6x+I6WLaSSSR7F8t/tnLCGCxAe5H4uK16D0djuHMEgnvDot/4eDdWV9ddV7M6vwNJt7auJp6akowRFUTQwSV7j6jzVekM1rcewLfbhdbhOZ7lqsf2fs6GFpZW1lEIrdaAft/b0IvtD0r697917r3v3Xuve/de697917r3v3Xum3M06VeJyVM6hlmoapLEAi5hNuD/j79WnXj1WB1vPJszu6ogmJjSWtcov04djbge/BqZ61SooerTIH8sMMv/HSKN/8AktA3v3W+gB+QHyV6v+OG3KbcnY248Vh6atqZKSjhra+Kklqp41BaKEMDdvUOLe9gE9aJoK+nTVhPlP1xmekKrveOrSLakMdS1MjVCiavlgVWWnpCVuzPq9Nl9+0t6da1rStei1/H7+Z1013tvet2UcVk9hTU0ssFNk91VEdLQ180bFVip5HjQXe3F292KMADQ568HRq0PDpu7Z/mj9Q9Vdxf6IqjbWf3DUJVQ08+48RIr4imSVgPuJX8begXuSG9+8N6Vp1oyRhtJOT0N/c3zj6b6c2btretdlKfP47c6RtRJiaxHdWdFYxv6TypJBBA+nvRVhxB69rFKkgdKHFfLvrPK9c7Y7DhqLU+6pJIqDEmYGv8iC+nTp5vcfj37S3p1sMp8+ovfG/afIdT0WWgWahj3FAwhpp7pOp/GoWH9eOPdSCPLqwpWmD0xfEHbcuOw1flZ4zqrUJjdgPozqTZv9b3rrZA4jpD/OjNSVVLtvr6mkIrd3hYaaIN6nInK8J+f6e5X9tbcI026OO2Dj+zoF82SkolmpoZcdG46R29UbU6q2Vt6rQpU4vDx086sArCQSs51Afnn2BOY7tL7e7i6jyrtUdCXbIjBYRQtxVQOhU9knS7oP8As7sjbnVGzstvbdNXFSYjEwvLPLLIIlJWFpVTVY/XTb6ezTZtou97v026zFXc4xXz6R317DYWzXU5oq9a0/f/APOt2N2pm8t1Btqgq8bRSZihpZMj91VLBJG84iB8vjUFTqva/vLblj2Cvtktk327IdtLUFB6ela9QDu/vDs24bhJsFrJSRWUEUamT/FSnn69bDXxlj21L0zsXK7b+3eHMYCiq6qaBlYzVQBWZ5GHJOq4ufeL3OJvBzBcw3daxuQAfIeXU57H4B22J4aZUV+3ofvYW6N+mrL4XE5yjqKLLY+jrqeoglp5EqqaGoHjlQo+nyqbGxNiPbkcskTho2IIOKHpuSNJEKyAEH16rJ7K+MGf693Q3YHxp3JDRZan89flsFUZFJJAlMPJOKelBFwRcgBP8PY/sN8S6t/o9/iJQ0Aenr6noGXmzS2k312yuAw4rX/J0MfQfy9xG8KuLYnYUM21950QFNUVOakFFT19Sg0uYfMB+ph6ebc+yveeWZLZfrLE+JEc0XJA+dOjLat+juf8XugY5BjuxU9HliljmjSaJ1kikVXjkQhkdGF1ZWH1BH0PsIkEGh6EoNRUdc/euvde9+690Xf5C974Tpna8k8r/cbgykc1JhaKCUeb74gCF3QAm1z9PYs5U5ZuOYL0KMRJQuSMU8+iXet3i2u31cXbCj59Fp+M3Su5t6blre9O3E+9yObMsmJxlciyfa+oPFMYJf08Hjj8exhzjzFZ7dZry3sR0rHQMw8/UVHRJsW13FxMd13LLPwB8vTqx0AKAqgKqgBVAAAAFgAB+PcR8cnoa9d+/de697917pP7jzGAwmOnrdw19Hj6GJGZ5qyVYkAt+GPPtXZ291cyiO0Usx8hnpi4eFIy0xAHz6rG358ht/8Aa+56jrHpWnq6ameZKOXNFnfHSAOUeSOoIsFsb/X3Mu28qbVslkN55hILAVCfi+WOgDdb1e31x+7drBAONXl+3ozPQnxT291i3959yf7nN81zNNk6qpdKyjWYtqDU4l1W59g3mfne63kfRWf6dsuFAwafPoQbRy9DYHx5++U8Scj8ujfqqooVFCqOAqgKoH+AHsAkk5PQl679+691xZlRWdyFRFLMx4CqouxJ/wAB7917ogPyF/mLdF/H3OwbdymRh3LlJNCz02Gr4nkpHYXZJ10tbSOT73Q9aLKvxGnRTPkn/NW2rsvrvHdn9aV1PuHDCsxtFk8Xi6v7muo56th5EqzEnoK3sQR79xwOtMwUajw6dfmL19hvnR8M8D27siuqly9Nh6PdlfFjaxtdI1BQfcVVLUp6SrR6Ltx+ffgdLauvOiupWvRDvjDvT44/NX4qb22p3pjMnm94dO14wuLptvWn3fWUGEgYMlO5XWwOjSRf/W9qZo2KiRDwHSeB0P6TDJP+DoilZi+gvknkKXpPojofunZO5MVumGiGf3XFPNTSwUsvi1M7qoVbrc8+04qM1HT1FNVAOPlT9nV8e1P5PPx/3j1ltDHdx0mcqd3Y+LHVdRkcHkI8bVxtBAumllqEWQuL/qBP+HutW6uVQ/EK9WN7E2L1F8cti0W1dt0+NxmKwdOqQGpFK+TqPBGIl+4qlQM7gfk/1Pvxyc8et0oMcOgY398rqRRNjdqU8z1l3ijmidmJkDWXkAe9V9OvY4HoCcftTtbuPI+bKSVsMM8pY6mdI2ilBcA3Nr+/HrwqTjo1XX/xrw22I1rsmxmrkQSkWSQNIE5B5P59vQvoevTUyB1+zoxWAakiQUcNNTwvToBeKFI2YKNILaRe/wDj7cuEGrxAOPVIHLDQfLpS+03Sjr3v3Xuve/de6DrdPbvWGyKqOh3fv3a22qyY6YqbNZikx88pva0cdQylueOPfq9boTw6AT5bfK3afxy6Gz3ca1EOboKOkhnoJMdUCWGqWoJSOSCohDA2Nvp/sffhxx1UkAVOOtdHtT5I/wAzXurrVe++r83U43qiup0yGOoraZWo1lKy+IMoZuAQSB7ejIVwCcdMSCR4z4YBPHPRQPnb8iux94bZ+Pvc6xZHI1fVm2KDH9mI5kno5ssZ/Q9RDKNP55BHt6W3I/UU4PSdZCFAHEHP2dD7m+p9y/Jvo7YHyE7e7C6vg6+osRS1FHtrblRisdu+lpUmHhpJaeErIGsLcj2kBIOOljJqwx4Z/wBR6sExHxYpPmt0NtbIdaUVVtvP9WU2LwW0srnQqTV2Npn8mqHISkEmynkG3Pvww3VmAkWp6vm6h2zl9l9YbF2nnp1qszt7bWMxWSqFcSLNVUkAikkDj63t9feurnj0IxAIsRcH6g8j37rXXFESMWRFQE3sihRf+th7917rl7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuvH6H37r3VAvbbo3zP2yNVmGTiAHJPE300j3unbjrX4ur48CpXDY1Wvf7SIm6lTyt/0tz7qOHWz8unf3vr3Xvfuvde9+691737r3XvfuvdcJYo5o3imjSWKRSskcqLJG6ngq6NcEH8gj37r3TRSbb27QGU0OBwtEZ1ZZjSYuhpzMr8OsvhjGoH8g/X37r3WShwGCxjyS43C4nHyShhLJQ46jpHkDfqEjwIpN/zf37r3WGPbO24qp6+Lb+DjrpP85WR4mgSqk/4PULGHP8AsT7917pryfXmwM1MKjMbH2flqgG4nyW2cLXTA/1EtVAzf7z7917p9xuFw+GiEGIxOMxUCiyw42gpaGJR/QR0qKB/tvfuvdMmT2BsTNVKVuZ2VtLLVkbao6vJ7bw1fUo311JPVQuwP+IPv3W6nqVRbN2hjZvucdtXbdBUXv56LB4ylmv/AF8sESt/vPv3XqnqX/dzbwqzXjA4b75m1NW/wuh+7Lf6o1OjXf8Axv79QdeqemrJdf7DzFUtdl9k7Rylah1JWZHbeGrqpD/VaiphZwf9j791r59SqDZu0MVL58XtXbeNnvfzUGDxlHLf638lPEpv/sffuvVPSk9+691737r3Xvfuvde9+691737r3XvfuvdEX/mHi/xwzx1BQK+muxBsP2JfqR795jrxpQ16DL+Vkyt0LlNJBUbi/H9fC3vZ60OA6//Q3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3UStkhippHnRJIwpukiq6NcWsVbj/AA93jTxGCevVXbQpb06BPG9MdW5rcp3tWbI23JnKeVlV22/i1pibemZozGQ7fkMTe/PtVeAoVQcKf6q9J7fvqx416HKGCCmiSCnhiggiUJHDDGkUUaD6KkaAAD/AD2i6VdN2ZweJ3BR/YZmgpcjRmSOU01ZBHUQM8ZupaKUEH/be/de6I7298B+rO5e7sF2tueijFPhaOiSLEU1PSLj2qqAKYZWoiAlyVF20397BpjrRUGh9Oqw/lD8Ne7cR/M6wXzQxsMWW6t2ztDEYmkwcSRVXghwuNSlWBKJr2ChDY249+BoKeueqldTAjyFP58egj6+663B8w/5vlX21ufCVx6gxGyk/3GZCMVNFFlMRjAlOscdVdVJmQcgG3+Pv1e2h61Qs1T5Cn5k9IPtP5E9afFT+at2h2lu/JY+KTBbEyOL29hZ6sUstdX0dD9pRUNNGtvURYKoH+w92jUvw8jn7OqzOI/iOSMfkeizbu+XneOOr8v8AzEe66ytwHUO8J8ntTrLZORrqihyg3Hjk00NTBALMYR6WDWsb8e3ZHDnwE+EdUUiNTcvjH+r9vQndWfMv52b96nrfmDSd4bel2TtmStr8910Mvr3PBt7Ew+dTLjheQRPHwH/wPtkgA06urFlDYNc/l9vr1sXfy7vmTgfnN8bNu93YOjbH/cZGu29kKV5mmk/iOJjjFROxcAjyFiw496IIOenQQVDA1Bz0a/s3f+C6r2Bu3sXc1RHS4DZ2Fq85lZ5ZPFHHSUi6pC0ljb6gXt711voJvjN8mtl/KLYtP2DsKN5du1U9RT09ckwqaaaWlfRKiTqqgkX+nvVc068KEVHRk/e+vde9+690l89WXlp8YulxWHRKtr2BNufa21iNDOeC9JLh6/pr58ektu3qfau78V9jkaCETpBoimjiiW0gBszC3P1t9faWZ/FcuelEa+HGE6Ihvr4+b165qv4/tSqnkghIdI6eTgKPWylUP+Htunr1etOHSu62+UOUwc0OG3zBMxX9tWkZ1K6mAB5B+nu1a468AAKnz6PPtnem3910kNTicjTTtKuowJIGlTm1mHHvXXulX7917rogEWIBB/B5Hv3Xuib/ADV+H2xfmD1TU7C3ZDGtVSVKZXBzukDQplaWMikM6zjTpBtc/Wwt72NVRp9eqsFIOrrWO+fnxj/mctsLb/QcWdwuY2Dt/LY+PESYugpJael2XQqkZp5Z4luoEAN1J49rHljRAsfpXpF4TvISwxUV9ek5meqf9Pe+epf5aXx4xlRhdj0eP212P2NvCnSOmw9TvTF4sVu5jU1NMNNgVlQISSTxb2kBpk5J6VMlT4aYUZ/KvD8+tw3pvq3bnTnXG09gbaxtDjqTb+CxOOqTQ0sFMtbW0VBHTVNbMIAAzyOpYsb/AF+vug6d6FD3vr3Sb3hhZdx7V3BgYZPFLl8TW4+OW+nxtVQmIPf8Wv7TXsBurSS3BprUrX7RTpJf27XVlLbKaGRSv7RTrQH+dHwZ7x6u+R+9KKHaO59zY/PZurrMXlcbiqrJwz/xCraTxtPTKwGktYFrem39D7h+zuY9hD7NuHY0BOk0NHXiCCAanyI41+3rkX7q+1XOPLHOl3ZWljc38N1K0ySxxvINUjEtGxQHTpPwlqDSQK4PVr38oKsz3w9ymWg7u21uDbFFvKSOlxtbliaagpDJSppndCSouQVe/P8AsfbGx75b2vM0m4TIyQyIEDMCorU5INK4pn06nn7r0W9+3Ml7/XWzlsVvpaxCQjSBoUVoCQpJBqDQ+o62qMJnsNuTHQZbA5Ojy2NqVV4K2gnSop5AyhxpkjJF7EXH1H59zVBPDcxiaBg6ngQajroBb3EF1EJrdw6HgVNQfz6dvb3T3QAfIL4/7T772dV4DOUyR5OKCU4XKIsaT0NWx1K/ltq03+tj7It+2O23uzaCUd4HafMHoL81crWHNG3NZ3ajVQ6G81PrXj0Sus7pr/jdsTB9C7XrP78b6WN6Chr8PUPWyYaollKg5HTc8G1rn2M+R+Sbq025P3o2mGL+L8Q+R6IrCa45Y2uLYw3jyoKAg1I+3j0tei/iXmsvnou1O+6w7h3K8iZHCUa1AeCkZ5RKDWQSBgeP7NvYi3fmSOKE7dsw0R8GNMn7Ojba9iklk+v3U65DlR5DqxmGCGnQRwQxQRgACOGNI0AAsAFQAewQSWyTXoXAAcOsvvXW+ve/de697917r3v3Xuve/de697917rokAXJAA+pPA9+48OvdNVdk6KKGZGmR3MTjxowLEFbEcf6/t+OCRzWlB008qLiueqvO2mlwfclJXUULwI9VF6yCoN3Ck8f6/ujRCNqOa562jiRNSCnQkd8fOWn+OuL2wK7rTdO9/wCKNRUAm2/HKyxPIoQyylUfj8+3nSIKGB/wdMoZiSPT7f8AP0TP+YV8e9ufMzoXbvaW8oc3t/CY2ofPYzC/emiroKueNZgKhXsSo0gHj3YzxLTSMj/J14wSupVzg8egGGJqcf8ABjpjb9HPVDbcPaCY+ZvuWGqnp9MREzg2YafqD7obhBXSOrGCgqD0jP5iHXPXmzOpuvM71dUrDucfb1czYmRKOZ60IraXensXN/evqHJAHAdNvbqFJHlwz0Kfxv2B13u74Udqdk9hLDN3INn5dY2yEkb5OAU9Hejlg8oMhZm41AjTa/597N1LU06dEERoTx/y9IzpH4Y7k+Sfw56+y7ZWakq8Ln80ZZcnWqypSpNqWVmnP6QCPejcOR3de8AECvHpPYCmqdlbioNsZipmrsN0xWHK1bwVTGlzUUL6Wp6FFNmYlbWX3b6oEAOKgdN/TsG49HNrPlVtX5RT0WC2dRyYhMCqp/B5ZXNTTCnUK/lha1r/AOt7rLKrgaMevTkcboasajy+XVpPTmLrtvbIxFOYrGRDcKgBP7QI+n/Ffd0SF09D1SSSVHFMjognZ+4Kjt35U9YU9HDJJS9fZeGPNRqCYwqVILmVW/17m3ua9ntF2Hkm8dzRrlCV/MdAO9uP3lzDAoB0wtnq2ZaqlDmFZog6HSYwygg/W1h7gzRIRqIOepD1oDpBGOpPunV+iZ/PLpvcXefxz3jsLa88kGXr0M8DRP43YQ0kmpATb6/Qe5A9tN9tOXuaoNxvRVFwa/MjoL837dPumxzWduSGceXHhTrRgo/hr3tXdoz9cJsjdsNVBm8bAcy1EBFKaasVncTHmxtyffQ9ueOXY9qG6tcREMrHRXhUdYZHkXmGfcf3UIpVVGiPjdvdpYMRXj5UOOt8f4d9Z5rqnoXYG08/K8mSoMFTpUrI2qSKQsWKN75yc+7vb71zLc3lsOwuafPrNflyxk2/a4oJT3UFejSewX0fdAb373BiuntiZDPVrxvVzhsfQ0/kKSmqqoWSGRQvPDEH2JuVtgn3/c1toxRR3E+VAc9FG87nHtlmZmyTgD59AL8P9l7tyFDk+yuwpqmbMZerkkwsc0jNEmJq1Zl1Rt/VStvYm5+3GxikTZ9rAEaCjkfxDoq5btrlozfXhOpzivp0rPkZ8U9t9wQSbiwyHEb9owkmOyNNJHSQSSwpaP7gRhSTqAJYnn2Gdi5hn2tvBl7oTxBz+zpZvOxw7iPGTtlXgRjoAeofkhvPpndEXTHyAWaWrgeKlxm75JGTEx0kYKQwecgqzWKg839nW57Fa7pbHdtk4HJj/FXz6KrHebnbZRt+75atA/BaeWerMMbkqDL0UGRxtVDW0VTGssFTA4eKRHUMpVh/gR7ALo8bFHFCPI9DNGV1DKag9Bt272vt3qfatfns1WU8VQlNO2OpJZPHJW1Ma+mCH+rG/A9nWw7Hd75erbW6krUaiPIep6Q7juMG3wGWUgEA0HqfTohXSfXO5/kxvqfubs+nqo9jQVckm2duVjFKiOtgIeKd45Qf2+Af088e5P5j3az5O20cv7MR9SR+o44UPEY8+ghtNncb7dndL8ERA9in19fs6tHhhip4o4II0iiiRY4441VEVUUKqqq8AAAD3CrMzMWY1J6HwAAoOsvvXW+ve/de6DPsztjZ/VeDq8xubJ0sEkEPlp8a1QkVbXm9hFSo/wCon/W9nOz7Ff73cLBZoSCaFqVC/b0gvtxtrCMyTsBTy8z9nVdrp2t8yM7NJRtW7W6xp5f8po615KearpRMbJTMbXJB9ywG2PkC1CyATXjDBFDQ08+gXp3LmWbUhMduOIPnny6sA686a2f1ZtoYPaeOiR1gKGsqYoZat3Yes+ewP15HPuLN25gv97u/HvnNCeAJA+WOhha7ZbWMHh2ygEeZGfn0rafcNJiaOplz1bDQU1DxJVVTmONR9fU7f4c+yy4hCqJE4HpVBLqPhtxHSK2P8hukeys7kts7D7P2juvcGImMGSxGGySVNdRzK2lo5oQL3B449pOlORx6Qvy2+TWC+KPUOb7X3Bha3N0OKKxLR0b+N5JpYmZPWFbi682Hv2eA69UZJ8hXql740fP/AORnzOw/aXYOwY8hhtibKFSKzFzGeR/tamCSNY0lcDkKCeP6X92I0nS2D00kglXXHkE0HQQ/yzvjz0b8vs735n++KuuznYWE3xNQYTHjM6ZYsdUieOadYZlcyFCEUIoFvrf3o+Q6sgDVdhU1P5dF+7A+I+A6n+aEXxq2XlajJ9db5FZlKzEV1QlTWHItC8tGqU7+kFZLcW9+rUV60AAaDhQ9O3VHya+RfwiyG/fh7vHrHee86PsquyUG281RUlTU4vCYvLRyUMSB1VkWyyDUAw+lvfjQivXu9aKATnj0dX+WR/LN7d+OfZ1R35uzKUCbe3bPPuA7baWF6tKbJs7lKiiW9nCvcauR7VRzB18OnlQdMSQ+G5nqTU1I8ur6ZqrrLbkVRmxjNrYqqXyySSxYnG0ddJL9X1SxRBySTyxPPtO6Mho3SlH1rUdFl7A+V1NAZsZtWmkkq/3IY5YHZrt9FIsB/sLe6VxjHW6E4PQG4/avancteJcpLW09JPJrUM7opWY6he/HvVKnPW6mnRqeuvjJgtviOp3CorqtNDqP23/cA9Wpjf8A1/e6ClOvVzjh0aCgxlBjKeOmoqWGCKJVVRHEik6RYElQLn37r3U4gEEH6Hg+/de6SM6vi8os6i0VQQgv9ADwT7M0K3FvpbiOi51MM2sefSX7r3+etOr93b3jiedsBijXKkV9T3fSAmnm/wDreyw4qD0YDIBHn1r9/D/+ejjezfkdu/qLtbaeb2vgKeqkosRunLzTQYuOSOp8SyxtMlmU83tz/T3bTTqiyKxocenVwPzY723L1P8AGPeHb3VLnO1WOxUddiqvEk1UVQsrWWWGWG9wRyLe69XNQaU61zuifjh3/wDzO9obq7X3r2CYNxYWF58FipMrNBXUNZ5Wkhgq4XIK6iCAT7vWh0jPTQUyR6nJX7Ooe1N4dhd69M96/CbeGRmqd5dewLi9nmqqpJ3rqulrAs8UHlNm4Q2sfdaAEN1YVYFG+z+XRo/jJ89tq9W/GSv+KPZvXlYu7+rcNVbaplq6dVbNzySSShqalZeXVmtqB5HvR7c9XUhjpJp0AXwB2ft35EZ/vnpLv3A1OJ2Z2rkxJtmkyVFFHXxtFUkU/wDDfuQAukMPpx7MVIktznIH8+kNKS1Yef8ALo7/AFl/JNottbvqcTufdU9b1BHWp/DsHSZErWJj4J/JDG8P6AbAA2Hsu1EjI6WCNVODj08ur2+vOvdrdXbRw2ydn4+PHYLBUiUVFEFj8piQkgzyoq625+pHv3V+lv7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuujyCP6j37r3VIXzf+I/yBy3ZmL7X6QydNSVVE6S3bxeRJUAdvrz+r/D36tB1Wn8+kTQZX+ZjBjsdSvu0eWnplRyF+pH01G3P9Peyfn1uhI4DrP/ABn+ZjrB/vfZRwQV/p/sPegacetEOfIdefM/zM7+nd4tb8rySP8AWHv3W6H0HXNcz/Mwt/x93+3W9yP6ce/Vr59aAPoOuDZv+ZkR6d3AG9/0n8f6w9+4nrwDcKDrsZn+Zjq43dZbH6rzf/be91znr1DwoOvNmf5mNtS7tGq9uVsLf4ce/V8q9eo3mB14Zr+Zj+d23bn+ybfXjgj34YOT1o6vIDrsZr+ZlYX3cNQN76Tp+v8AQj34kUr1YA1pQddfxr+Zk2kDdwBHJ9J5H5H09+PXgDXgOuDZj+ZmHJXeA08caT/T/D36uOtUb0HXhmv5mgI1bvB5/wBSb2/23v32dbNeNB1yGb/mY3N928fVbA35H5BHv359aoRwA68M3/MyudW7gb8j0kW/w+nv3nx68a+QHXjmf5mR0hd3AC/qupuefxx71Xz68A3mB12ua/mZANfdoI/HH+2vwfe6imevZrgDrgc1/M0Nz/e8AH6eg/Qf7D37y69n0HXhmv5mmlv9/eNR+h0n8f649+rjrZqfIdeOa/mZ2W277EDm6nk2/wBb37Hr1qjeg67Ga/mZ3UHdw025AQ3P5uDb37rxr6deGa/mZgMP73i97r6Twv8AiLe/H0r16jV4Drk2a/mZkADdyg2vfR/vRt70D1uhAwOuH8Z/maabf3vW/wCTob+v+t79wzXr2fIDoOO2thfzH+79nQ9fZbc1NNSVOUhqKoyoiCWEnSwkkcchQTYE8fj34EV60QzLTA6ts+D/AENuX4+9O020N21ENRnKmeGurXgKFPMYSrj9sn8n3vrYFBTr/9Hf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691Sh/OR/mBbp+Eux+rotgfcybp7G3NLgac0XnMsDmSKGIuIgb8seD72ta/t6o7AKTxNQAPt6J/g/5n3yy6O2/Q5ftzprf+/vJBFkmWnochTiSlngE0UepUP8Aqh/vHvwI4A9eKuACVP5dW4/B75zxfMrb9VmB1buXrapooy89DnzI7XVVvYvFGRctYX966sDXo/3v3W+ve/de697917r3v3Xuve/de697917r3v3Xuve/de6TG5an/Jfs4z+9MUsL2JXVzb2ss1Hia24DpJdMwXSvn08Y2AU9HAmnSxjUv/UsRz7YmbVISOFenoV0xjqf7a6d697917r3v3Xuo9TSU1ZDLBUwxTRTRSQyLIiuGjlUo6kMDwQbe/de6D3ZfUmxOv62tyO2MHS0FdkGkapqhDAahzK2qQCZUDAH+l/egOvfPqhv5P8A8kiH5SfPKb5Gb7zMbdcCvoctBhqGvggrvu4GV5ElhY6ihKm4I59q4XSNCTxP29I54zLMnno6DL/hQd8UcpvXo34zYjZG2Kyu626w3dS/x/AbapYqaaPFY4UyyVNQlOqqRJGti1v1XJ+vtOKVrw49KXB0EAVFRUeoHRSe1tj/AB03L1HtfFfC+myGxN09n4Wi6737tLOZNJqmIy0ApK6daW4VVclgSR9eePp79U/iFeqEKQRFQE8Othb+V18OJvhL8WdudSZCaOoy75Gr3DkJYpYpIxJlII3WJTCSvp9V/wA391rXPr06F0gDjQU6JT/PU+R+U6/6x68+PWDNRPL8lsjW7EykNFPLHOlLPUw00WpouBqdjwSPp/T3sCvHh1VydNEOSaD8+nLoLubpD+VJ8Ruq+h85mqbc+/aSWsyeY2rS14/jVFVZxlqgKo6Xv6dKrz/X3rix69VUQfKgqfPqx74xfNbpr5QUNRHs/cWMp910CmTKbOes8uYxsSgEvUppW3Jt72RTj1sGvDo3ssgijeQhmCKWIUXYgf0HvyqWYKPPrzsEUsfLpJ4mlesyFTkKlG8evVSBh9Bq/F/ZhcyCOFYV4+fSO3XW5k9elh7Lul3WOWGKdGjmjSVGBVlkRXUgixBDX9+690XLsn47bZ3fE1TjIFocmxZjIPGiatJ0kcD82/Pvw68eiYVm3ezujcw1RQvWz46GS8jrI7RgKfKxup/oD70BXJ8uvY8vPozfV/ylwu4RFjdwqaOu1KjVEzsqlmfQNVwf9f3v5dbIPRuKHI0WSp1q6CpjqqdxdZYjqUi1/futV6SlfJNna9aGAsKKM/vMtxaROeT7M4gLSLxG+I8OkUgNwwC8B0pqvE0FfQzUFbSwVMFRRyUM4lijcvTzQGnlQlgeCpI9lrHUSeli9oA6AHpn4o9KdDZLLZjrva8ePymarKmurchWGGsrRNVMxlWCqaNXRPUQEDWt7r1bHkOjIe99a697917r3v3Xum+pxOKrXElZjMfVyKQyvU0dNO6sOQwaVSQf8fdGijY1ZQT8wOm2iic1dQT8wD0EfcnQXXXd22ZNt7ww0UkKRzGiqKNIqWppZ5F9LrKi3sCP03H1Psp3fYtv3m2NvdpjypQEdEPMPK+0cy2JsdyjDLQ0IwQfUHqsD7nvj4CZtUqlyO/uk6meVsXhsc8tVUYqJn8Ykq5E1FQot9fcb6t85FuavWexJ7UGSvzPULg80e1k3fqu9sr2otSyDhk+Y6tS6d7g2x3Ps6g3bt2ZESqj1VOOeVXqqF1A1LOlgRyeDb3Ju07tbbvarcwGleK1yOpt5f3+y5i29L+yIo3FfMfb0T75JfKitbNDpvqKOXKbyy0v8PmyeOeR/wCBzGXx+WqVANKji9/9f3Juw8uIYf3ruR0xJmh/F9nRZvO+P4p22wGqU4JHl9vQm/G/4rYvrBX3rvTTnuz81rmzmSmeOrpA8tmH26ShtLDnkH2X77zHLuP+KWvZbr8I4H8+luz7FHYj6m4752+I9HMAAAAAAH0A4A/1h7C3Qj679+691737r3Xvfuvde9+691737r3WOSWOFdcrqij8sbD3ZVZjRRXrRYKKnphrNwRRsY6WNqlzwGjOoA/6w9q4rQtmQ06TyTgL2Z6bvDnMkNQm8MJsCrAA2P1HtwvbRHAz0yqzycT1Ppdt0UemWoDS1INy+qwuP8Pbcl7I3amF6eW2T4nyeiNfLfELiq2h3FDFYCpiUFVAsVCkklefr7REktU9KAAq0XoxPSiYbefX2Mq8pi8bkjG0QH31DTVjI0cYKEGoVrH/AFvehXrflQ9OndfUVF2tsiXZyiGipnWZESNI4oY1lUD0RgBRyPwPez17oEKH4f4KL45HpKqaGWooK2uzGDrwIwtJl57GCdjbkC1j78KjPXjkU6Kj03/Ld3XDu2Sp+QO6aDfOz6Gd6rB4eklLPBKraoFm86kBRxewPvxNevBQBxr17tT+Xj2dlu0Jct1hvbF7a6zyRjpcxtmZjG0+L4WanSCJdBJW4F7e/VPXtIrWvVhmK6QxmyukK3qPYejFxy4eajpZ3CKsdbUxoKioPjAAuykj+l/fhTz61ny6Kzm/gtBV9OS4GnrqVOyvDOf4+/hFK8ryNIrTOwBIF/yfeuGR1avl0Q/40fFys6Y7brf47kYM3uPLVU8GUqqORJKRw8lgY0QkD6e91B6oBT8+r8PNS7W26s1QAtPjqRGk0gKBZQPoP8ePb9vA9xMsEfFuqyyLGniNwHVdvxG26+d7u713Xl4RU0lXkfJh3ZQRAHqxfQx5Hp4HuXOe7r6XlzbbGA0IXu+eOgRy7F4+63dxKKgnt6sQrdvQu5qKNmhqS1yxYkHi1/8AevcUR3bAaJMgdDFrZeMeD1ALZ3GWeeQ1UZP6Yxc8H+g/w9u0t5/gxTpv9aHHHp2pc7R1REc4FO7D9E1ub/ix9sSWzxjUuR8un0nV8NjruPbm2jUjIR4LBmqJ1CsTFUH3Or66vuQmq/8AsffjeXmjwmlenpqan7K062IIK6wq/sHT4AFACgAD6AAAD/WA9pePHp7qJkK6nxtFVV1VIsUFLBLPI7myhIYzI1z/AKw9uQxPNIsSCpYgftx1V3WNS7YA6qwlpq/5c98wT65ZOpNsTinydFHKxSTI0XCuQ/F2K/09zYHi5F5YK4+umFVJH4T/AJq9AAhuYt2x/uPGc+lR1adjsfS4qgosbRRrFSUFLBR08aqqhIaaIRRLZQBwAPcJzSvPK00hqzEk/ac9D9EEaBF4AU6m+2+rdAf3f0Rszu/bM2G3HQj72nSafE5Cn8cFRT5Ap/k8kk4XUyhgCVv7N9o3i72i4E1ucHiDkEfZ0X7jtlrucBhuB9hHEHqvbYXem9viTv1ene3qesy21JqhY8NunySDHQQykLTIs7AgkLYEH2M73abXma1/ee1UWUfEnmfXHQRtd0udhvBt+5VaIntfy+Qr0PO9+hM98guysXujMbmgy3UVOKXM4Wjop7AVT2n8RRj6rcBha3t/a+arLlnZntLSLTfNVWYjy4V6XXWzz7xerPM4NuKMoHr0fDFYuhwuPpMZjqeGlpKOnhp4YoYkiXRDGI1JWMAXIHJ9xnPPLcytNMSzMSSSa8ehbHGkSCOMUA6cPbXV+ve/de4dFN77+Uu1+p6dcTiFG6N11uqCHE4uUyVVFKW0B54kBN/6C/sdcr8lXm+N49x+jAuSzDDD5dB3eOYLfbh4cX6khwAvEHot3Xnx53339n6TsfvSvqpdsiZ6rb22zM0VZREN5Y0qon/HIHI9i3dua9s5XtTtPLagTUo8lKg/YeiSz2W73iYXu7n9PiqeY+3qyvBYHFbbxlJiMPSQ0dDRxCGGKKNE9I/LFALn+p9xBdXU95M1xcMWZjU9DmGKOCMRRCijh08e0/TnRbPlRt+fLdN7tXHeVKxqfyEwOUkZVgkT0stj/a9q4JcGJ+B/z9Jp0/0ReI60tPiZ1B8hOo97fKb5P9PbnyM9P1fn4q/cmCir6qsr6n7jIlgkNOGJsLHVYe2pAF7KdejUmsgP5H162Dvjt83fjr/NS6P3H0fu3MY7EdmY7Hsue2vPVK+Sq8tSwyRqaWnnVbMWK6kuT/Qe2yunp1HWRar0UT+WLuWr+IvyJ7P+IXY+JaHG9t7inotsPURxwGnp9ctLDrWxRldJCCD782e49eTSg8IcBn8uhA7z/lgfJ7p3u6t7f+Fm+6XaG28tWyZDP42Wuipq6WqqmJkZaeMjUtyfoOPeq1FD14rR9QNK/LB6M18Nf5e3a+B7WxnyI+T25KXdnYdA/wB3h5IJ4agxeRzIpqTyRYHge/E+Q6sq0yTn/Vw6t43LWbDw5bI7hpcD91TozpNV4+jmqxpGu0cskZYf1+vvXn1bj0VXf3yoxtP5MRtyjMpDmnT7fVcgjSpXQALf0AA92Q0avCnVSKih6Cmba+9O2aeOr8lVQ088yEq8rJdZmCtwxH9Pa24/Xh8VMUwekcVY5jGeHHoxPXnxjwOAMFXuBRXVSLHJYGNv3VH0Y2P+ufaCmM9LvOo6NDj8ZQ4unjpaGmhp4olCqI40U2UWGoqBf37rVOp/v3W+ve/de697917pmzdMZ6Uuo9cN3H+tbn2ptWCyUPA9J7hapUdAB8jg2R+PPYEIN5lwLBy3Nh5PSSD/ALH3W5TTKT5Hh+XVrdqxgenWoFDub43bs2r3NsrI09Pme59s4qufaFPgJ4abKUmZNUQPu1iAd7f0v7ayMjr1VOG6GH+Wn/NK3TiKXKfEr5dbLzWJ2VWQz4LbG4d5rULRQtJJpp5Ynq1IkQGxtqsP9v72QoJKZHn1pHkIAnGk+Xz/AD6N58Etzbu+O3zV7J23HjKncnVnbOWfGbYyeLB/gsdJNVLNT1MQhugCgi/upNBXpwK1dPAcf8/R792fy4M1jPl1tDv7q/IUGKwUWfTL7ux85pUatXymWdPALaw3IsR+ffsUp1unfrPH/D0X7+YVs3rzaneW297Yvo/du6dwVtTDWNkNvUKQ4uOoZdOqrgpks1nFyG96rnrY+ytenb4ofGnunsvs3Dd5b4RNt7e2nWiowWClpYsdWCnWYS+KSKMAsdIvcj8fX2otpNJ0+p6YnQ6SfQdXzUVdFWiQx39DWPN+D9PdJYjEQD1eKQSCo6ne2uneve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuuLIjizorj+jKGH+2Pv3Xusf29P/wAcIf8AqUn/ABT37r1T177eD/jhD/1LT/inv3Xuvfbwf8cIf+paf8U9+69177eD/jhD/wBS0/4p7916p699vB/xwh/6lp/xT37r1T177eD/AI4Q/wDUtP8Ainv3Xuvfbwf8cIf+paf8U9+69U9e+3g/44Q/9S0/4p7917r329P/AMcIf+pSf8U9+69177eD/jhD/wBS0/4p7917r328H/HGH/qWn/FPfuvde+3p/wDjhD/1KT/inv3Xq9e+3p/+OEP/AFKT/inv3Xuvfbwf8cIf+paf8U9+63U9e+3g/wCOEP8A1KT/AIp791qp699vB/xwh/6lJ/xT37r1T177eD/jhD/1LT/inv3Xqnr329P/AMcIf+pSf8U9+69177eD/jhD/wBSk/4p7916p699vB/xwh/6lp/xT37r1T177eD/AI4Q/wDUtP8Ainv3Xuvfbwf8cIf+paf8U9+69177eD/jhD/1LT/inv3XqnrsQQA3EMQI/IjS/wDvXv3XqnrL9Pp7917r/9Lf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691pCfzwN79i97fOLa3RvWUFdnc109unE7rnwuOilqahYIpIaqU+Agra1jwPd0AoSx6YkLCVQor5n8urN6b+chv/Z+V2h152b8P915yGqpcJhZsq2KmhijaKiSmnqpYKmnkjZWYatNgP6WHvRApUU6dBNdLgivRsPlJ85ds/FjCfHze20etJ6Be8s9Bjq6hp6cUYx0E8yJLrSjUXsWI9QA496A1Dr1dJofWn7erDO0/kp1z0913t/fu9c1R4mLc9DST4Oiq2eOTJVVTSpUmkgIH1GsDn3oZ6scCvl0pNk917S3P11hOx8pXUu3cNm/L9vJXyskX7Z4s5H5HPv3XuhF2/unb26qT77buVpcvR3t9zRszw3/AKayAPfuvfLoGu2fkt1f03vXr7YO89y4zE7k7JqxSbZx1ZKyVGRlaoFMopwBY3e6/wCv7917PRhPfuvde9+691737r3XvfuvdI1nTJZ+nkT/ADdJqjdfqGYc8+16gxWxB4t0h1GS5xwHSy9oOl3Xvfuvde9+691737r3XvfuvdcJG0Ru/wDqEZv+SVv72BUgdaJoK9JfDK1dWS5NrFbvED+Tp4Av/h7XXJ8OMQjpHCuuQycenTOYLE7kxVdhc1Q02RxuRpZqOrpqqCKeOSCdNEi6JQRex4NuDz7QdLeia7Y/l1/FPaG8v7+YPYD0+4hWNXpPJkZZaaOpZtZkjpGXQOf6e/Z4dexWtB0d1VSCJERQscaxxoosFVBZFAH4AHv3XutUru1N0fLz+YN29i8vt3JDanxOeq3/ALfrKi7UNWmDrhOUp1qAVvIwB9A5974D7eqfE9fIAH8z02/y/wD4/bM+bfze+R/bverndWIp6KGu2rtNMjNC2CrYsjDSxvUw3NlWMkKAoF7f63vxJoF4daRVLNIc+Q+VOut+7B2J8af5jm0aH4mVGQgrdz70o8d2liKavnq5KKjesVKullpoyNCAAgXH0+vv3Fc+XXqhZAQKVyejifIv+Zz3Vt/ujsvYuy+h99z4HrOpigkqIqPIkbkjWyu8XiTSNXJGn8e1ECEfqn8PTMzmvhgfFTo8H8v/APmG9dfNDH7n27h6KPbO/evKeP8AvdtGeeR8jjCZhTsamGYBg2s+q/59syOXkLevT0QCpT06N/TfIzo2s3e2wKbs/acu81nambba5C2UWoQEtEYGUerj6X906cp0VzLfzDOuKX5MbS+NuHx02czm5couKqcpTSOkWJqGk8dqmIqb8/4j3vyr1oEFtBOerDPeut9NOZweMz1FNj8nSx1FNMCHVkUnkEcFgf6+/de6Jp2b8WIKt58ps+T7IxkzCFZFSQ6Euo4Av6h+PeqenXq9F/wvZnZPUVf/AAfNNVHHxOsQSRpfXGGuxBB549uodJ1kVA6o4JGkYJ6O71R3Ns3dNCsa1VNj6+SzSJK7appXfTa7fn3uWd5zqbgOHVYohEKDz6MErK6q6kMrAMrD6FSLgj21071y9+691737r3RU/lP8u+q/ihs192dgZqigOtkgxjSP93UsALLGkQJuSwAFrm/Hsg3vf7fZoxjxJWwsa/EegJz57hbB7fbQ+773KqInEZqfQACpJJwAASTwHQJfEf8AmV9EfLbJ1mD2plqbE5ymUsmJr3mjrJxYspRJlXggGx/Nre0Gy81x7lObW7iNtL5K34h8iMdBP2198eTvc0umzS6JI6VjcMkgrWlVcAgGhoaUNOPVjAIYAg3BAIP9QeQfYu6mfrv37r3TRnMFitx4utw+Zo4a3H5CnemqoJo0cSQyCzKNYNv8D7angiuImhmFVYUI6YuLeG7ha3nUMrihB9OqqO1904DpncFZ1R8cKab+9O7S2Krfsp5Jjt2pnl0NUTonCoCb3IHHsS8nciWW2694n/TgGdJ/F8h1HTWtny/cNZ8uLpebBANdNeJp0af4xfF6i6jpZd3bvdM72lnVkbP5t3SphcTAOFgDglSDe5B9qN/5hfc2Fta9lsnwrwP59CrZNkG3g3Nyddw/xN0cf2F+hF1737r3Xvfuvde9+691737r3XvfuvdMeRzcFENMQFTMR/m42BK/64HtVDatICWNAOkstyEIVRWvTSKHJ5grLUymKlNrQG4bn9X/ABHtR40EAKxju9emRHNL3MelDRYqioOaeKzEcsTqN/yfaKSeSX4z0sSJEGB04+2unOve/de6Kv8AK3EJXbGSo8YYwyyMTb/UqDz735dapnpPfEvcP3O3ZMOrgiJw2kX4MasD/h71q8ut/Po5Pv3Xuve/de697917r3v3Xuve/de6QvZWSXFbKz1YXCFKNwpvbknn/ePfs9exXPVe/wAdcHPuXsSuzZImjjqZZbghrBZdTFr+9Ur+XWiR0cr5I7ki251Hu2qEojqTQaacXIN2exIK/wBPYo5QtDd77AhFVrnop3u4Fvt0jedOg++H+3TR9d0u6JADNuWLzSSWXVJolvqLDn/b+zbn+7Em6myHCHFOkPLMJWyE5/0TNejc+wF0Jeve/de6Z6/CUVc3ldCs45R1JADD6Ege347iRBp4jpmSFH+3pkEeVwp1GRqqnNrRpdiAPwb/AJ9q/wBC54dpHSas0Bocjp+x2Wp69eCI5gCWhYjXZfqQPaSaBoT6j16UxTCTHA9EL+YPceTnmoukevpHfem4pKaQz0ruXio5R45ogF/JDG/uTuQtgiRW5i3QUt4q4PqPP+XQT5k3J2I2uzNZX9PToxXx06eoOn9hUmMiT/crmI6bKZyVgpd8lNFrmu45Nix/PsJ827/Jv+5tMfgjJVB/RBx0dbLti7XaCIfE1C329D/7C3Rx1737r3XvfuvdA53T0ns3u7adRtvdePSoaNJp8VVrpjnosiY7U9QJQC1laxK+zPat2u9ouRcWrU8mHkR5jov3Hbbbc7cwXC18x8j69EB6o7O3h8Ut9N032jJLXbPmqiuC3JIz/ZoJgFpYY52FiQNII/3gexluNhacx2n7120aZfxJ54446C1nfT7FdDbb7MX4W8s8B1arQV1Lk6Onr6KZKilqoo5oZozdHSRA6kH/AFj7jx0aNijChHQ3VldQymoPXddX0eMpJ6/IVEVJR0yGSoqZm0xRIDYu7fge7RRSTyCKIFmPADietO6Rrrc0A8+q7e4/lTmd2Zqo6x6QpajK5WokekbdOMaWalopNejzOyCwVRyb+5Z5f5Jt7C2G88xsEQZ8NqAn5fb0C9z3+S6lO37UpdjgsOA+fS56J+I8G1K9989qVabs7BqpFqnrzMZqaKR7SEGKQEEg3H9PZXzNz217ENu2VfAtVxSlCelmz8ui2b6q/PiTHNfTo8SqqAKqqqj6BQAB+OAPcbkkmp6FfDh1y96691737r3SU3vRRZDa2ZpJlDpLQVV1IDAkU7ECx9+BoetNw60XviztX5Zbx+YXyT6Z6c3kmz8Nv/eVZQ5L7/8A4t01KmRdbVCuClgpJ+ntQVDxB2yRXpKzSLcGIfCaft6PVuv+Rj298YmyPfXxr35FSdwVNdFlty1mOrqiI1tfJMJZ6mipyiWBb+yB+fp7ZDADSRjp1ou/xEPd/I9XGfGT4u03ZtJ1z3N8gcPXz96bIoYIpdwFYqQVuQEnk+7mUKHdhY88fX6+61BOOHTtD58erMs9ufA7cpZJcxX09OI0LaJjy5ClhxY/X3umevVpw6KFv/5VUkPmxW2KYy1F2jjnhMhLalspBW3591r1uo6ASi2/2r3BkA+QlrIqKokDBi7qojeyFSW/wHvdSTjrVKjo1HXfxfwG3zFV7hUV1WoRguqOT1gk+okH+vvVOtgnowVVhKPD0kEeNgigp4ioKeND+k3U3sPa22cUKN0knTOsdKejqUqYVdDfSFVrfS4HtiWMxvQ9PxPrSvUv21051jlljgjeaZ1jijUs7sbKqj6kn3tVLMFUVJ60SFFTw6r170/mQdE9G7q/ulms7QVeSWZoZ4o5ZVaAo2ly9hbg+5T5d9p+YeYbL62FCikVFaZ6Bu7c7bVtM/gTMCa049Gq6Y7x2J3jtem3NsrM0eRp5lRpIoHJeEvGJACG+vHsF8w8t7ly5eG0v0KkcCfPoQbXutrutuJ7Zga+Xp0MTqHRkP0ZSv8Atxb2QA0NejJhVSOi793UbP1X2VjmNxPhFjhQH9TCRiQt/wDD2tuAJIVdf9XDpJC3huUbrXX/AJXPwb6Ez/y0707B3fsTLVW7YkmrKWtqqu+KepORTXI9MbgkAkqLW/r7RGpx0pRQKmnn1d58r/5e3RHymwOHp9x7cpsXn9qxKNsZLDQUeK8EyEaTWGmiDSCw/r/sPeiW4jregEUOR0L3SHxq2h1R1/svadTQ0eSyezxIabLeGI1DSMQFY1BTWwAAtc+/dWIyT0Zj37r3UeWkpZyDPTU8xX9JlhjkI/1tYPv3XuvPTRGFoY40iVltaNFQD+nC292RtLauqsNQp0wYZBS1NVAf7chtze5BIuPay5PiRqw8ukluSJCp6U/tD0t697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/T3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3UHJNULj6xqW/3K08pgt9fIE9Fv8AY+/de60ut6VHyE+Hf83ruX5pbn6g3L2ts7deHy2JxuNgpZZqaITwiihq4mCP4ymkFWA4/Hu2NP59NEN4lSCRQZHyPVu/VX82XoLvHLYXb3cHUtL19lsjWxUNIufjhmqo6plsFhlmiVxZvSASfeirD7OtiSNmp5/PHXv5x+39s0PSvTHZEX29LsvrfMnc9TWsUFLR4WTw1vmkYfRQCTx9PfgPLqzkFa+dR/LqtbqfeG+f5y2/+pNu7KyS43pPoHMQZaq3HHVVS4nc2JpJo1qqOlc6g8raQigfj62HvZAU9NowlFc0B/b/ALHRtvn3Xba3f2bsHpnpz5SbM652d17X0VPunYdRkamllpo47Q1glcWEhJU3FvegadXIoKE06rv68+Vvdfxh+fPSnU2yO/KPuvrnuDf+N22+29s11ZV0+2qOvrkpH++TVpBs2rke/fgJpkdVGpZVQkUNftwOjVfK2Wb5U/zYNlUdPkHnxHxJ35jDmYIKiaJKajoK2KtqRLHfS3rDA2HPvZNFr69epWUV4Ln9ox1sP9LfJ/YHee9exdnbLrYa6fruqWlyUsbMWLNKIQWU8Dk24916d+zoY8L2HsrcWZyW3sJuPHZLNYeVocljqaRnqKSRL6klUgAEWP59+69506iRdpdfT5w7bh3Vipc6swgOMSVzVCYtp8ejT9b8fX3unWq0yelrWVcFDTS1VS/jghRnkexOlVFyxA/oPe0Uu1B1pmCrU9BfsjfWw89HuLJ4LdGPy1Ph6sxZaaEMq42YsQYpr3549q7qXUFj4U+fTECkEsfPpf4XcWE3DFLNhcjT5CKB/HK9OxYI/wDqTcD2i6U9PXv3Xuve/de697917r3v3XumPP1ppKMBT6p3ENvzZxYn2qtY9cnDhnpNcvpSgOSes2FovsaFIiblz5Tb/awCPdLmQSSkjyx1eBNEYHTt7Y6e697917ri6LIrI4urCxH9R7917oJ8T0d1nhMxu/PY7bVJDld9UMmN3RV6UaTJ0cpvJDK2m9j+effuvdUT9j/y2PlJ0N3b2H3P8Se1INn4rf8ANPTfwmlmkiraCCpnMoimREAZbfkE2I/r72SKUPVNBDVQ8fLy6KR3T8au9/gV17v75N71zj7/AO3u0sdIRuijlkqn2XkZWWVq+td1DK51gh1Nr8XuCPdgNTBB1Vj4YMnE9A3tDsT5OdffEbcXzW3x8x9mZ2GDHSZSt2JNPHV7jnCzGnjjqInXWQL8kt7VTfoxqsf4uk8ZaZizkEgV4Y/1eXRY/hHkuwOpeqfkx8oNlZeqw3ZXzUwsz9abhpqiZRBVnK+TIy0MUZGksQy/4e0ppwHljp9dekvJxY1H7MdWD5z4Hdn9NfCyq+Z+7expYe+MftSl3dXZiWuqzNBV19X9sjyH/jpZ1c+q/Nvx70Grjq3h0Ouuafl/xXUn/hPX0B2Lv7dfcfyU+RdZV7yztTlKOv67ztbPLIIJ6mueWpq4vNdiQqKBzbn3uSgbSvDqltqaPxZB3En9nW2z7p0/16/++sffuvdY5ZEijd3ICqCTf6f63vagswA6qzBVJPQP5jr7bfY7Vc+dxwLw+SCF1SNQSyaFb6f8T7W3KpDGI04npLCzyuWY8OibdgfHHc2ya6TcG0p5padJGnip6eY3VYhrVSi2/P8Ah7Q0ouc9LSanHU3r75NZ/atRFgd6QTSyROlOGqPKdCBueSfqAffqjz6qMjo9m0N+bd3nRQ1OJr4JpZEV2p1J8iahe1j9fp7916vS09+631p3/wA//r/t+o7dx27pIcrWdXGhp6fHeKSX+H0WS8UUdQ8sQNrubAPb+o4vzE25a7XnCeW/romVBAx+EaV71HkGJofmMeXXOD74e2cxrzVZ726udpijKkg1RJmIAZl4ZFFVqGhqMas1AfC+r7U2V3ftLeXXeDz+SfAZahnyseLWqeEYxpbVyV0kPCxlNTeo/VfZbzJdww2gljb/ABiKjoFBLEg8KDNG4HyzXrGv21veYYedrDd+VoJLmS2lTxxEDp8BjR/EYYA0gsKnLKKVPW//ANL/ACZ6r7ZwWETE7pxUe55cfRDI7Yap1ZShqzTL5opIbXsGDC/+39yXs/MW2bpEiRSr4pUVSvcDTI67Dcvc47Hv1shtp18Wg1R17lNMgjoyH15/r7EHQt6Jh8rfkfT9X4eDaO2HXIb43Wz4rHxUruanFVEjqkdS6p+bnj2KOXNjbcZTcz9sUWSTwPy6D2+7uLCEQw90smAB5fPpq+KfxxbZcc3Z2/VOS7J3MkkuQq6oiYxxzNqClJQSrDn839u8x779YRt9ri3j4AfLpvYtnFqDeXI1TycSejyewl0Jeve/de697917r3v3Xuve/de6wzTxU8ZkmcIg/LGw/wBb3ZUZzReqswQVPSTnyVblpTTY4NDFcA1HOll/PqHtekUUA1S5PSN3kmNI8Dz6d6HCwUxE0wE1V9Wk5tf/AFj7Ty3DOaLgdPRQCMZyenv2m6Ude9+691737r3XvfuvdBT3Lhv41sjJ0+jX4oZprccaYiPeiKjr3RNPihkPsN5V+JY6F8s6BCT9SGUe9hamvXiaYA6sh9+691737r3Xvfuvde9+691737r3RY/k/uJcRsyWiEmh6+F0AuQSGfTYW/1j70a/l17hnpDfEHbT47D12WljINajeN2A/MikkG35H+Pv1c069TzPSO+c+elOP23silkKVe7LUsSgsCzGoKj6cf7f3K3tpbL4k24yfDBn+XQL5ulYxpaJxkx0arojb1RtTqfZuAqwVqcdi1hmBtfWXLE3H+v7BHM10l7vlxdR8Hao6Ee0wm326KFuKrToXPZD0Y9e9+691737r3XRAIIIuD9R79wyOvccHoDO7t54XqfZOT3lUyR0rxpNRxvIzBGnrIWSL9JBuGtb2JuW7C43vcUsVGoYJ+wHon3a4jsLVriukjA6J18Qussvm9xZjvLs5pqnLyVZm2nWVLalXGVUhEZiWXmwW1iPY/593m3trWPlnZ6BKfqAfxAedOgxyzYSyTvu+4fGT2k+Y+XVm6Osiq6EMjC6sPoR/h7hsgg0PHofAgio65e9db697917r3v3Xuve/de6A7vfpLbfdez6nCZalU5Sjjnqdv1qskT0eUKjwTNIRewYC4v7Ntn3afaboTRHtOGHqPTos3TbINztzFKMjKn0Pr0RHp35B5j42ZbN9Q99ZLRBhkqJ8LuaueQ0+Q8a6KbHUkhNieAPY6uuWm5qCbjsCVLfEg4j1Y9BSx3htjkbb92agX4WP4vkOs+Z3v3B8vM622dhpXbF2PAzfxDNT/cR0Obx/wDnTDHJ+Sy/0Hs/t9u2HkO1F5uZFzcn4UFNSNwrTpua53LmSYwWgMUPmx4MOjx9L9BbH6Yxcce38eq5iog05XJSMJZaqZ7eVlZhcKSP6/nn3G3MXNG48wzlrpv0we1fQeXQt2vaLXaotEC9x4nzPQ6+wz0a9dE2/r/sAT/vXv3Xuu/fuvde9+691CyNKK2iqaU2/ehljF/9U8RQf7379145FOqevj5/L6331H8lt2dxHM0n8EzmdlybUSik8zxyVJn0+kBr/wCx93RiBpPn1R1GouMnq2DIbzwuIjr5cvWRUiUzHRHMCGYKl/8Aex7emgMSKwzUdNwTeKSCKUOPs6Kjv/5V0cHmxW2aVpqgkok8BlJJYFVPptxf/D2lqfs6f+XQEUOC7X7hr1bIzVkWPncEFmlVVUnR6i9/x71xPXvkejR9dfGHB7ckjqtwBchOgDW1o5DhtSgmxFv6+7UHn1oV6NFjMRj8PTLS4+mip4U+ioig/wBRcqBf37gKdb869OXv3XusNRCs8MkTC4ZTb/Xtx7sjaWB6o66lI6TODlelqpsbIDqu8l/9Y8cn2uu1WRBMvSa3JRjGxr0qJpo4InmmdY4o1LyOxsqqPqSfaBVLMFXJPSskAVPRPfkR8mNj7R2ZuTD7fzFFnt3VGNq6elw1FLIaxKi4RPSoBve9vY/5U5P3K93CG4uozHAGBLEYp0Gt6321gtXihYNIQQAOPWiv8sNudnbq7oz+6t5baz1CcnPk48fFVVtXDd6ictEyoSP9gffRHk672uz2KOzsZUbTpJIUHgOsMuddr3O/30315E4Gl1QCRlrq86Cmf8/WyV/I12P2jtrbmSrN1Q5Km23Pjo0oYayaWVPN9r6SrSHnj8+8VfvD7jtF1MkVoVMoapIAGK9ZC+0FhuVnZ/45UKUFASTTA8zx62JPeLHU3dIzdWKoshF9vkYjPQVf7dZELAPGv4N/9c+11udUbJ0kuAQwcdJ7YXUfXWwq7IZ3ZuBp8VW5lSK6oiVA84LBiHsB+R/X2iZSrUPHpUrh11DoVPeut9e9+691737r3Xvfuvde9+690kasSUeYglJvEzamC/0uAbj2YxlXtivn0hkqswauOlarBlDDkMAQf8D7LyKGnS0Gor13711vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv//U3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdNGYwOIz9HNQZbH0tbTTpolSenhlupNyAZVa30/Hv3XqdFB3Z/L1+J29dxY/dW4Or6GpzmMrhkaOthq6ilMdWJPKJDHAQp5/BHv2fXr2DkgdLL5PfF3ZfyU6G3L0PuCnjTbOe25JtxYXJ0wUbQrAi67FuFUC4596NeA61SvQM/DH4EbE+DPxjm6G6Sip8FXmnyxizaHyOtbkJNccgmdQ4RB9Afof9h72evUAFB1rw9W/GhOh/mr3rvD5UdA7k7xwu8ZaqHbVbFWVkNKK2euLx18Lr9fSDxf8+7V1KAMdNKoV2ZhWvA8fy6Ab4IdVUG5/wCZT3L2jH0pl+qtj9I7h/vdgJ8u8s9K0FHW64aeKSqX1MBa2n+nv2rtCnrwQGUyAU8h+fRj/j7m5drdofzcflDmX+6gzmLqsrtWpclxTVNfmRpeEN6lspt6SPeqYA/1cerjTqc/MfyXqzP+T1tPGbJ6H7B+Umad4aTsejky9dWSStIJY8fMaiVkRzcXYKOT+ffmrXPXo6CP5nPVc/wl7U3XHvn+Yn2Jv7uaDZGKky09XsbcWUkqWp8WtVlZCqxLq9N42H6fx70RSnW1bVqI9f5U6BDu+mrtqdEdh957Z+e20zviSux1Zt+opK/ILkHSVtchiAIKkni1/dqioBGOq0IUmuf5dG63P8/O/cD/ACbd79zeHMb23rsl9vYOPdkH3Us+5I8u7wyVsct9RA5uQfbsC0kHp1SV/wBNjTNCafZ00fywO/czifgF3Z2p8gKyp6rp+wK3B1mP3VuI1ZghqshVaPCvkOoFrk+n3SYqZWAyB1q1Mr2ylxRvMdX2fEDasfWvTg3jl+waLeO3d409DurGbhgeV6MYqsjZ4JUeT6qwcHj3Qmpr0oAoNJ49Gr2pvja296eoq9rZimy9PSy+GeWmJKxyG9kbUB/Q+9de6Vfv3Xuve/de697917pI5KCbI5ZKZUZoIVjlaQW0IfqL3P8AX2vhPgwF9VCcU6Qyo0ko08B0rFGlVX/UqB/thb2hOTXpcOHXL3rr3Xvfuvde9+691737r3XVgfqAffuvdBV3F1/sXtPZGX6+7BxCZnbu5qY0VZRkRK7oG1DxySA6Tf8AIHtTbRNI9RwHH7Ok9xJoSg4nh1Tn3l/JS667B6r3T1lsmvj2/tzOY00lNh6ipdlVWn8xjeVV0ci3P090mesxK/CDjr0cI8DS2GIzTrh37/LgrNg/AXpnq/qilL76+OOFqkwUWO8clTkZq+saqqPC8K/uFWPpH1sf8PbINM9PstcDNP8AJ1Ux8oflN8zOzOieuvhBkujN40h3hiqHD7m3nJR18Uayw1uqGWqOgIoU83PuwA416acykUApXj1tBfAXpQdC/FPp3r6rpoo85hdqQwZupSONJaqskqHmdpmjA1EAgAnnjn3Xjnp7AwvDocO8u0aLprrDdXYmQg+5ptvY6oqWiuw1MsDvH+nn6qLgeyre90TZttk3CQVCDh8zw6IeZ97i5c2O43mcVSBCx+wAnrRh7l/m0fKzd3aua3JtHs3PbZ25R5hhi8DQO6Uwo4pbRLOrkk6wLsARYG34v7jaGDc7y3M97dSrJMCRoegjB4BRwxjjWp65V8w/eK90N23qXctm3A2tozlrePQh/TU416gWOviQCKA0FCK9bSv8sb5q1/zK6JrjnacU+9dlLSY3NOrSM2QqHk1fdo8jEnUCv+PPP09jjkjc7y4R7LcqNLaFVL/78VgSrn0OMj1/LrPT2E91JPdTk9ry8j8K7tmMUq8QXFCSp81YEEeYrQ5HVrNFTrTU8UYXSwRBJwAS4HqJt/j7FUja3J+fU9xoEUDqQyq6lXUMrAqQQCCCLEEH231fov8A2R8f9q71hmnpaWOhysgkb7hdKguy8HgcG/vRFevcOiOZDaHZ/S2WkqsQ9Y2Pp6guZFeR4zFCbn6ce/Y4DrXnXoz/AFZ8nsRmvt8TuiWPH1a6InqJ/IGZ/wBJvc88/wCHv1c0630NnYXWXVvf+0m27vnB4zeO2KoFjTVKJJE3lUXKswJF7fj2XbptNhvFv9NfprSvqQQfUEZB+zol33l7ZuZbFtt3y3S5gbijgMpB8iCDUdB91T8O/jv0pTZGk6364w23ocrHLFXCGCOVp45lKyKWlUnkH8ey/bOVtl2lmaziy4oSzFyR6VYk/wA+iTYPbvkzlZHi5f2+G0WT4hGiqD+QAz8+i2dwfy+cNU1z7t+PWXXqre7tNUVeSjnqHjrJS2tUVVB035HPA9hzduQ4Wf6vl9vpZzktU5+Xy6CXMHtRayytuHKcn7vuzkuM6j8x0WWo+ZfevxHgk2t3htnLbnSXy0eF3hVx1H2mRngPgjWBhwbkXHtjl7c+ZbPcV2ncLd7hAaNJmgHrXoJrz/zPyIRZ80273CHCzgHSSMD7KnowvxZ6pqe6dz1XyR7EgFVS5xvudt4KqdmbGVQ0zCpMT3sBq4H9feQG4cwwDao9v2ztqO8jz+XUqcuW374I5huciUAop/D8+rQ/YL6HXXvfuvde9+691737r3XvfuvdRaurho4jJM4WwOlSeWI/A93jjaRtK9VZlQVbpJiOu3BUapNVPQxsLxnUPKt/7J9r6x2i0GWPSOj3DVGAOlbS0kFHGIoECIP9uf8AX9oHdpG1N0rSNYxRepPunV+ve/de697917r3v3Xuve/de6as5TrVYbKU7LqEtDVJbj6mE2+v+PvY60RXqsPrqeXZ3dlTTytojmq5Si/0LMbDi3uterUqM9WmU7iSCGQG4kijcH+oZAQfe+tdZffuvde9+691737r3XvfuvdV1/LPNtmc9iNuQS6ZIpxEwBIv+9/X/bj6e9FqdeABwejd9K4VcJsDEUxQLIULO1hdvSo+o97695kdEG7xqp+0Pk51bjccTUUmys9EMzDHeyxJVXYSXuDx7m3lxE2bk68mmw1wh0H506j/AHQvfb9bpFkRN3dWoKqoAqKqqOAqgKAALAAD3CZJJqepA65e9de697917r3v3XugK7O+SXTHT8v22/8AfOK29WMjPFS1RdpZLR61Cqg/P459iXZ+Ud/31fE223aRfMjh0U3297bt503UoQ+h6rwr+xqL5y9uYbb2ycg2T6gxTRncM9P5FpZammeyySkkgksLLx7laHa39udhku9wXTfPXQDxofToFveLzVuSxWp1W6/ERwqOrXsdtbG4zbuL25TRBKLEUFPQ0aiy+OOniEaAaQP6D8e4QlvZZrx7xzVpCSfzPUhC3VYViXgooOoUVTWYKZaeqLS0f0STnSgP0X/Ycc+3nSO6HiIaH06YV3gOl+HSuhniqIxJC4dD9CP9v7LmUqaHparBhUdZfeurde9+691737r3XvfuvdEX+bfxaxHyA2XDkRGqZraKVWXpUhPhlyLwASpSl4xcsbEDUf8AD3IPIfOk3Kd1IFFVnAX101xXoJc08uQ75bBm+KLuHzpnpr+Dna+JyW0Yupq6gOD3ftL7lKrEzLapNNCAiTSk2JNlPJ/Fh7S852dy96d3ZvEjm4N5V9Ot8r3qG2/dzroki4j5evR/PYI6FnXRNhc/Qe/deOM9aav80D+aX3pTd67j6r6k3VlNiYvYOTnxdfW4maWGtydVFMWudRKiMKRzpuT9OBzFs15uG738kskjwwRMyIinTqIwWan8h+fXOD359+udbTnGflblaY2UNmQHlABd3IDaV1AgIARU0JYmmAM2NfyZvn52D8haXN9b9qZCfPbh29SNUw56oMhmrKdwhg8xYlQ/DBrf64te3tby1ut5b7w+x3jmRGjMsbsatStCrfZ5HzB6l77sfvDvvPlrc7VzO3iXVowHiAUDowqrEcA3awYCgqKgCtBsI+5F6zB6Yc3ubC7ei82XroqNLarv/SxP0/2Hv1QOPXuim9jfKPH49ajHbXCV07nTFNAH18ggcg/7178D1o8OgPo6XfHaVNUCvhqadaogxzMzqqi+j8/4H2vjYSwFPPpI6lJdR+EdGG60+MmEwRp6/Pha+UoHKFw5LXDKTcH2gK0Ok+XSsNqAZejU4zE4/D060uPpo6eBfoqKB/vI9+8qdb869OPv3Xuve/de697917rDUVENLBLU1DrFBAjSSyN+lEUXZj/re7IjSMEQVJ4daYhQWbgOigdn/K3rfZFZImAylBubci+SD+CUwkNQJVsNIK2JJ/1vY+2Xkjd9yQG7VoYeOo8OgvuG/wBjZyUtyJJPQdFkrdzfJL5GZdcZt+my3XGCqXv95UieOlkp5PXYMR/Qcexklnyjynb+NdMt3KvkKVr0TGffN7k8KENAh8z0ZXq/4c7F2vJHnd7QJu3eYeOV81JLL42cC7t4X+p1f1/p7B+9e4G5Xo+n20+Bb8NFB/h6O9v5Ys7b9W5/Vl/i6ELfvxS6H7Nq6Wu3nsLFZeqo2R6eV4o4mRo/0k+NRf8A2Psp23nfmXaIzFYXLIrcRx6W3fL+0XrB7mFWI8+hh2jszbexcPBgNrYunxOKpgohpaeNEVQqhFuVAvYD8+yC/wBxvNzuDc3rl3PEnoytrWC0jEVuoVR5DpUe0XSjpqy9M1RRyaDZkVmH1/H+t7ft5NEmfPpqZdS9RdvzaqQQtfXGTcn8+3Lte/X69N27Y0dP/tJ0p697917r3v3Xuve/de697917pPbghJpvuFIBjK/jn6/gj2ss2q/hngekt0tE1jiOnXHyCSjpzqufGob+oIH0PtPMumQj59PRMGQfZ1M9t9Ode9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdBd2d3R1j03jRmOzN34vaOMZGkWsykjxxMitpJBQMfrx9Pe+vdJrqf5MdGd5PJH1T2Lgt6PFr8qYmSZmQRrqcsJUX6D3r5de6Hb37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/V3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690xZvbOD3GtMuZx9PXCjmE9OZo1ZopRwGViL+/de6D6p6N63ei3RTUe3KGil3dTGmzVTDGqy1a3uDIwH+9e/dex0V7sT4AdX5/obtfp3ZtLBtup7Tx0VHlMw92s0Enmj1GJNRUt9bg/j37PHz68Pnw616939A/wAwnqTpHs74e9fUm8aja1TGuM2xujFJXPjqONam7T000KgaWAN+Pz73qUmh6aVJVB0iuOPWPsD+Wf3ztP4gbMkmo8lubOwYWM9p7ep2mFbuusWXyiSVY1DkqODq59+DZz1t4jp0r/q9ei1/OnZnWWS+AWI6265+GOWwPZONx+Ghzm5Fq66apFUso8qyxW4/2I497GDx6rJlKBc0xw6Mf8q62v6U/k3/AB/6AUna24Ozto7Xz+YpZlvPQTU1dxBMbfUjmzA+1UKj6cseNaj7Omrk6JNBPEAfn0q/nNHiNyfFf4GfBnGS09Rme0+o9obur4adRBJkqyIgLUSS04HqLCT6g8WHtIlANXp0/Ko/s14Nj9lK9PfyY3b3Z2j2/wDDz+Wp8ed7VvWO36XqPAYXc27KaasjoqXL42gIrlrGpmJcoEYBQhZjYWJPvwwCSPs6s5Z2AU04k+tPLpCd+7S+Vv8AKI7G627fyvedT2d1mUxdFn9qwT5P7LK5GWfxzvW0c2gi9iVJHI92Hd9vTbHws1JXzxn7R0IXyo/mJfKfdvzv6R6++O2H3Fldmdh9QYHecuMwa1kkEVblMQKyphZYWILRG9yf6e6gDSSfl1djISqpQ4Neh9+JX8zruzbnza298KvkNsrI4rL7sxyZSHPZxKtKqjhqqdqukN5WACMpUj0ng+/fhqOHWhUPpeoalfkfLrZGqchR0lBUZSqnSGgpqWWtnqXP7cdJDGZpJ2I/shAWP+HvQzw6cPz61Yfnb/Ody9B8sV+LnxTo/wDSLuSiaF6+u23FVvVO3jVqqCIwm7pGdQvpF7e1kuIAjYP8+kMbN9R21IzjyHWwP8Pt0dmbx6S25uDtfA5Dbm7K+NJanHZNXSqVGp0fWyS+oAkn6+0Xn0vrUenRo/e+tde9+691737r3Xvfuvde9+690jsoBkcrTU4XWtJIDIPqQCfZjCDFblwaaukbkvMA3lw6WAAACgAAC1vxb/W9l3Szrv37r3TFkNtYPK1cFdX42kqaqmt4ZpaeB3SxuPU6kn/Yn3up60QD09oiRqqIqoiiyqqhVUf0Cjge9db6QnZnX+G7Q2VnNkZ+JZ8XnKOalqI2+n7kLRKx4P0LX+ntDuO32+52b2VyKo4of2Y6LN42u33nbpduuhVJQQQetPrvD+Rn3Pje3K2HYNZQ1OzMrlnqYqnS8hoKWom1tHK6AKdANlBA4+vuMRYcybYy7Z4Kz0wkgJAC/h144gYNOPXOHmr7pvNEPMMsXL9wpsZZCV1IS0KsalQVIDAfgwKCgJNKmzzoL+Vb3F8cdkSZXqruyfCZiopkyGcw0DSsMnWU1pvGj06BATpChb+1d1ybzNA7XVtfhJHUa1QEA0zpHr9vU98ifd73r282Yty9vEokk/UkRqEM9AMAKAuABTPzJPQ+bL+VPyU6sqmw/ZfU+fzVBRP9vJuSqSqEdRGrlpatX4Frc39p7LmfmDbX+n3C1Z1XGsg5+fQ52/nbnHY5jbb5t8kiLjxfI+p+XRztgfNboveApcdX71w+E3VOyRttypeVaxZHbQqJwwJv9bkexfY847JdkQvMqSn8BrXqQtp9xOWdxAie5SOc/wChk937OjaUlXT11NBWUkqzU1TGssEqcrJG4urL/r+xQrK6hlyD0OI5ElQSIag8D1GymJx+ZpXosjTR1NPICGRwDwRY2J+nu3V+ib9r/F3HZA1Gb2qn206l5FpY3Kt5CBpYaQL88ce9H58OvCg6L7gd+9m9N5WOlzaVbYiKVY/DL5WTRF6b/X8X9++zr1M56PP193xtDedLAs1fT0OSkEa/avqBZ2XkC9+f8Pe6jj17NadDNW5Kjx+OqMrVzLFQ0tM9ZPOf0pTxp5HkP+AHPuyIzuEUVJwOqswRSzcB1UtunD1nzY75O1ssjT9Tdf1/8VxleIxFS10kEoMkCMA2pm0gfX3IhgsNi2ANcoHnmFCvmKjiegBdW0fNe6fTXa67WEgjAoSP8J6td29t7EbWxFHg8FRRY/F0ESw0tLCLRxoqhQAP9hz7jaONIl0oKDj0Ore3htIRBbrpReA9Onr3fp/r3v3Xuve/de697917qLV1cVJE0shAt9B+ST9Pd0RnNF6o7hBXpK08FTnar7iqBjpInBjib6Mt78e1zFLWOiZY9IlWS4er/D0sY40iRY41Coosqj6Aey9mLGp49L1UKKDrn711vr3v3Xuve/de697917r3v3Xuve/de64uodHQ/R1ZT/rMLH37r3VWnddO+2O7aSqhVo6dqxLyDgHU4t9Pz72cmg60OAPViP8AfXbm3dlUG5s/lKfG4eDFwTVFbOxEUaxQgSs2m/0/PHvRx1sZ6QXXvyc6L7VzD7f2B2JhNyZmJ2jkx9C83nR1/UpEiKOLH6H37r3lXpv3p8sPj715vSPrzePZWDwe8ZpY4Y8JVGo+5aWVtMcd40Kgkn8n36o69w6X+/e4OuOsdtwbv31urHbd25UwrPBlK1n8EkLxiVXXxhjYqwP096qK069017P756n3/suu7C2fvLGZ3Z+NhknrM1RtJ9vDFF+tmEoVuP8AW9+qOthSeHVYh7l6x7r7vydTtDdtHn6Hb9ZJU5YwRlBQ00Mt5ZZAxIIHvZznqg9B1ajt7fOzJdrT5PB5enyOLw1GktTLBwI0Ci5YfT6/09vwQtdTrBF8TmgHVJZRDGZG4Doh3xGxz7n7/wC9N6VqirxtRV+TBu4uia6sBmjB+lhexB9y9z3MLLlfbdvj7XA7/njoEcto0+73d0+VJ7erOfcNdDzr3v3Xuve/de6w1BZaedl/UIZSv/BghI92QAuAfUdabCkjrQh/mlbm7WrvknkqXd1fV0+KTMulBHUy1qK1L4iIRGFNrEWPvpP7RWmzxcrI1ioLlBWlONc9YWe6l5up3xI7p9EPieeoE4OMfl1ZF/IVyPYNDuDKYumhqpNnV0krZKa80sN/vS8RkeUf1t+fcVfePi22WzE0hHjrTSMA/D8upB9j/rIbZYVr4JLHiT+I+Zz1the8K+sk+o9VSw1cLQzIHUg2v+CRYEe7o7RtqXqjorih6R0P3e3anwuWmoH9WsatMVz+nn2YFY7qPUuH9OkhZrd9PEdLSCeKojWWJw6sByDf8Xt7LmVkOlulqkMKjrL7r1vr3v3Xuve/de66IDAggEH6gi4P+uD7917qr75X9bZvqjeuG756zpp4aqoyuve0lJqCpi4nV5GlSIAaCrEE+5B5cv4tytH2bcDVQP0x8+gTvllLY3K7rZjNe/7Ojq7T732Dn+sKXtCfNUlDtz7YvV1kjWjgkgQecFeTwb+wHvqrsEsi7ifDCZJPp0fjebFNu/eNxIEjAqSf59ED7W+ZG+u1akbN+O2067dGNr52pJd5YlJ2ipPHLZp7qeAAL+4p3Tm683E/S7DEZVc08Ra49T1EW+e4W57437u5OtzcqxIMy/CtPP8AZ6dU0/KH+SZ3xuWrl7Swe6qfc+6t710mRzlBUB2qqSeoOthI6LqAUED6H6fX2XwbPzTstspmCXSOS1F1CQFs91cHj5U6xb5/+61zhuF0/M9hfm6urx9ckUq1VTQDtZaMAAAACG+30tJ/ljfAeg+Ee1aveXZuZpBu7cVO4qIpZFBpwY0VI0hADC2k2uTyf8fYk5Z2K9S+fe90osjroVB+Ba1+2pJNeshvYD2VPtlt811fymW5um1vUABTpACqPJQAaVJNST506PN2R8pcdjfNjtrquRmYBVmhDl0PKk+kn8kex79vWSXRa1xfbnb1UJKh64Y+cg6S0wRVB4v9fwefej14dGn62+L+FwBpK/O6K6UDXJCzFixvdQbgj/X59+yePXvs6M0m28ZQUT02MpIqUAejQo4tza59vQv4b16bmQSLQ+XXLB1zTLNTTOPJTsI1B4JA/oPb11GAQ6+fTNvJWqt5cOlD7SdKuve/de6iV1dS46lmra2ZKemp0aSaaQ2VEUXZj/rD25FE8ziOMVY8B1VmVFLMaAdFM7C+X+wNvGfGbNraXeG40MkIxdJ5TIJwPQota/PsdbVyDul0BNuCm3iOdRpw6Dl5zLZwkxW58R/QdFfWL5L/ACNy5pY5st1dt+dy7PUCpSCamf8AWnF7/Q8D2M9XJ/KVvrIW8lHpSoPRCU3/AHqTSCbeM+vmOjM7C+HvWGzqeHM7jxSbm3fCyzT53yPrkkQXLqkoH1Nybj2Ddz593ncZDb2j+DAcBP8Aiuj605csLNPEkXXJ5t6/t6NXgpqVqGKnpIxDBSKtPFD6RojjGlQAth+Pr7BNykiyFpDqLZJ6PrdgUoBQDp69pun+ve/de697917r3v3XuuLqHRkP0YEf7f3sGhr1oiop0k8azUuUqYW/Q3pQHi3P9fa+YK8IYcekMLFZivSu9l/S/r3v3Xuve/de697917r3v3XuoldEs1NKjDV6SQPryBx7dhYrICOmplDRkHpm28zBamN25EvpUn6Dn6D2ovAKhh0xaGgIPSk9oulnXvfuvde9+691737r3RfPk12PnuquqNx712/jZspWYTHV+QamhDFilHSmfkLY2NvfvLrXVTPSn8wXtv5BfHNuwtrbMrxuLGdpptbIY6iWcyLjfLZ6phqJsALE39+YUwevKwYa18+rPOxPlj1L0R1nt/efb+6qbbH8Qx1ICtYjM75Fol89KQh/UrGzc+/YPW/Wnl1X1k/5z3VVFvDH0cOHp5uv8nkY8dSb3LVn280kj6TpjDWuAb297APWtS9CL8yP5m21Omettsbg6mo6fsbL73MVHiqSkWVnStqSojhCxsCHs305/wBf34CvWmbSKn7KDqnPcX83T5o9cdgbBoeyvj/unZ2A3ZuSlpErsrS5OnjagqZlKzRGbSpUq1wfftI/Cc9a1sCNamhx9nUf+Yzku8d679663xndu5vfPSuayOOmrcajVSUCY6oZKmqpneMkD9RH196H2560wbGO0cR0Yb4c9vfEPq/u2mXY24cZ1DDuGenoaPak0tTVyzVbU6xVMAd2+rPcc/197NSOvIQKhadbOlNUQ1UEVRTyLLDNHHJHIv0dJEDow/1wQfeunB1n9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//9bf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddMqupVgGVgQysAQQeCCDwffuvdBjlOmutczBkabJbVxtXBlZBLXxSwqUqHBuGYW4seePeqde+XRSvlL8Jesvlrsk7IylHBh59u/aUe3cl6h/C6SjQIkCRIrEi4v7MJlMUCjzPSJP1pzXgvRaMT/K3raP5T/G/vLO70pdyYboLaVLtjF42oaRZXp6FW+3jSnmTlLkn6j2gBoKevSxgrNqzio/b0Qj5n9Wd2/Ff+Y3sj5X9d7Bym/OusZE1bXUNIlQ9DHJVxF6ijmaJfQV1EKQeOCPdhQjPVKMjBlyCCD8vn0GXyDzXfn80DvLa+0avqHL7U6aotuS5PK1rpVfw6kymNpGlU1lXKpBcsLC/9eB70MZPHrzVc0Aovn6nozn8o3qHdNf2nuntDfOCnwlR1DSZTYmFqapSXnwlGslCggEy3KmP+0pHHI9+PACvVgKEtTj0ECYin+QH8+TbPYywfxDb22Nstil4KJFHgcM0Mcb6QATqTk3ufezha+p61TVJQeQP8z1s19gTY/F9ebylr1QYyj2nnnqUbhPs4sXJ5Izz9NPH197iUs4C9ekYKhJ9OtNv4Y/C/afZvz07M+Y+ydw0/X+2Os8TuLdOey6lpVFBgA9RWQmKzCxVSX9XC3Pt66fVNUeXSW0iGlicEn9vQxZv+bJ3n82O8M7tn4pw5Kr2NsjH19FBmttrUxxZvL4VnM0D6XtqmYGw4+tgBb2xSg1evT4k1MVH4eNf8nVln8tb+Y/u/ubfe4fjp3rtl9ldnbOxNdma6LKiWKuOOx9M0009U0rn6hb8j/W+vvR8qZr1dXrUEUp/l6N7vL+aZ8LNkb1fYOZ7dxqbippmjraWKkqJlgVWKM3mX0mxHv1MdbJ/1V6NhTd27DzfUuS7j2rnKTObMosNWZiPK05Jp5KejhE0pINiLAi496x1alfz6I10v/MP/ANOeP7frdh7T/jjdd7byOZxcVMrKctUUF704u5ve3Onn37getAhhj16OV8dO1Nw9udN7d7F3Xtt9pZnLR1r1eClBRqQU05iS4fkagLi/uyjU4X160x0pqHp0J+Biklra7IkXhqj+23FuDz/sfa26ZVjWEcR0lttTsZG8+lZ7QdLOve/de697917r3v3Xuve/de6T+brJ41WkpHCVM1ihsCdN+bXv/vXtZbRI1Xk4DpNNPJG4WPienWihMNPEGA8pRTK1gCz25Jt7Tytqc04eXT6igqeJ49RcxhcXn6KTHZejhrqOZWWSCZQyMrLpYc/1B/HtPLFHMhjlFQfLpu4t4bqMxTqGU+R6Jp2X8DOj94ioy22ttUG0d5OZHp9yUZn8sMrLZJTGC1yrci1vYRvuSNluCZraMRTHg4rg9R9vPtjy1uJNzawLBc+UqjuHRO8v058wPjFURZLa2+s93ZiGl8sGCh+7H2lLE2r7Zo2tb8gewhJtHNnLP6tvO16pNQgrgenQAn5e9wuTX8Xb7t90jrURnBAH4f8Ai+hm2P8APTMYg02P7s2NN194/FFVV1ck48fP7zspNvTfn2dWPO80YA3qD6cDiTX8+hDt3ufNbhYeZLY2ZFKluHz/AGdHe63756o7bAPX+78fuP0B3FIWVo7gnS6yAc8HgX9jLbt72zdhXb5RJ9nUj7RzNsm+jVtVwsw/omvSq3hsHbu9KOamzFDFPI8bJHMRZoyf7XH19mvR90QTsX43bj2fkJMzs6WSYBmqE8DyExjV6RpAH0Hvxz14HoLN09zdjQ7RqOucgatanI082MrKtw5MNPINDFje9h/rezDaLi3tr0T3OQmaHpDuVvPc2hghOknift6Ob8QNt7Y2V1hjNvUGSpclnDNU1eRqVIM8jVJEojuebKLi3+v7Vb7uh3W/M64WgAHpTpnaNtG22SwHJ8z616Nv7JejTr3v3Xuve/de697917rFPNHTxPNKdKILsfdlVnbSvHqrMFGpukjEk+erBM90ooHuinhZlP5/x9mDFbWPSPiPSOjTyV8h0sI40iRUjUKiiwA+g9lxJY1PS0AAUHXP3rrfXvfuvde9+691737r3Xvfuvde9+691737r3TZUZigpm0POhlJAEYvqJP+8e30tpWzSg6Ya4jHA1PVf/y9zOx9nwQb731mIds4KCcucpOnkU/bW81lUXJH59uC2XX3tQdNNOxB0ivQXVWU2J87uk8TtfpTt2mbEbWr/uN1ZGKOSjjmxKMpq6aTWblQoN/x7eZbZDXVXqqtPICCKV4U6qV712amw991mO+HUs8mW2LFLnN07lwUs5lqYcY2utkkMTcLw1/bbSQA4GOtiOX8PQ49b/HTCfJTovsD5fb4rP4rvvbu3shUULzSyrPHlsLSmR53Uk8hhdbnk/T3USwqSunHVlhkcqzHoqPVe9ewPl7hunet+wNx1dZgKDfwxORoal5o467Dx1oiNEXdvpouB72WiYDFD1oByDU9Dnltpbw6X+SHyW6S6zmq4er49qmlxu0KeaaSlx5miAeW4NtRNyTb34LA4oTTrw+ojNf5enROOhMBuXpveG7mqWmwUe8Karx1RWMZXBSpc3LX5H+v700SsaA1p1rxHhPCter4Nq909JbK+L2d2PjuwKCq3/mMHLTrTorfd1lYyoUhQk/QWNzqvc/T2JOTNumu+YIJFTUkb9x9B0X79dpFtclTQsuOjW/CPaNRheqMZnK6Ix5DMpK1SWPrb1h1LXF/pb6+zX3Hv1uN7a1jNVjpTpHyrbtFt4lbi/R0vcd9Cnr3v3Xuve/de68RcWP0PB9+690TXv34NdD/ACCqostvHalNPnYGjMWRBRCBGLDUuhieP8fY/wCWfcbmPlhDBZzHwj+H/UegzvPKm1byQ9xGNQ86D/N0KnRvx06u+PuA/gXXO3afDRTRotbKml3qZFbWXLBV/PP09k3MnNe8c0XP1G6Sl6cB6dL9q2Wx2eAQ2iBfXodvYZ6N+ve/de6wVNPFVQvDKoZWBH+INvqPd43aNgy9VdA66T0kaWSbBVppZrmjmbUJjqIUn6L7MHRbqLWvxDy6RiQwyaXwvS0VgyhlNwwBB/qCLj2WEUND0uGc9d+/de697917r3v3XumDdW3qLdm3cxtvIoHoczQy0NUhtZophZhyD/vXt62ne2nWeP4kNR01PEs8TRNwYU6p+27sml6w7czvTm/agx9S7gnmx+BxE7uaSrmq5T6InYgC5Nvp7F/M+1W/O2wpKQPESnieVVHEHqNpNqiW9ba9xNbZ6gKeBr1Z1svr3qXoHAfYbUxGP2riiNUgiuzykkEu7G55I/Fh7jnbtqsNoi8CxjEa+nQ12fYNp2CD6faoViT0XoIOxvlTh8GHpdrBMjOSV8yxlylm06gtyOP8R7MeHRwK8ei0vSdud21/3lVJW/w2Vrx8OiIitdQFUWAF/fm7s9bLMTk16M71v8YcLhBDXZ5UrJiS0kTMWZr82Nxxz/U+9D59aIqOjV4vD43C04pcZSx0sIt6Ixb6Cw597638+nP37r3XvfuvdI+phbH5aGoj/wA07AyD6A6jzf2YIRLDpPHpC6lJgw4dPmUzOOwuPlyeVqY6OihQySzSn0ooXUTx9eBfj2mhtprmYQQDUx4U6VPKkSeJIaDon3YHzJ2jjpJcZ1qId9ZxC0LUNIk11nKkIARa/PH09j7avb6+lHjbv/i0fGppw6DN5zNbodFl+s3oOi80Wz/kl8j8m0+ayOV6twDsZHo5vuUiqKduJY0QfXUBa3sWS7hyjylDpt1W9lGKilQfLomS233e3rIxtoz5evRvesvin1f1/DS1tRg6TMboi0NUZ6YyNLNKnPlVWtYk8m49gHeed963RmjSQxwngg9PToS2HL+32YDFA0nm3mT69GbjjSGNIo1CRxqERQAAqqLAAD2DSSxLHiej0AAUHXbqGVlP0ZSP9uLe/A0NevEVFOkljA1LlpaYjQhDtb6C7C4t7XzfqQCQdI4yUmMZ6V/sv6W9e9+691737r3Xvfuvde9+690kcxG1LXUtQn6ZZR5PxpF+fp7MLbS8TKRkDpDOoSUMMV6VisrqGUhlP0I9l9Kcel3HPXL37r3Xvfuvde9+691737r3XRAIsfoeLf19+FRkdaNDg9JWlf7GorpHXiOKrmF+OIUL2/2w+vtfN3Qgj5dI4u2Yjy61kPlp/ML7Jzfzo2X8ftsbxq9g7ayNXRQ1uUpjOkEZR1WXy6HsQfzdfaIfDXpSW/UCj9tOrbu6P5gvUnxv2vtDa9Zuim3j2DU4CkkWhtKZcj4qdVasZ0YG8jXI+vHvQHVifPh1B+OP8zbqXujcZ2buSoptnbrmpXq6DFSrKfuYorvI/kkY29P+HvXz68DU0Hl0Tv5j/wA0PtCj7Pn6M+K3WlZ2ruaFJ6zVhYZpq+aKliDuIUgkuQDqvZfexw1daY50gVPH8ug0+Pv8znencsG8/jV8kthv1dvnKbXzdNFTZWmnSv8AJLSOnjkM7MVN7fji/vxFeGa9aVlrpYEU6JF8f8zvjrxeyeodjZ+owNdR1W4t8yLTaiZ6OjR5wwYWuNPP09+Pz62DQkDqD8jotwfLb+XVsfdeTyktflttdzZHG5arleVXkoopB5FexJI45v72O1/9Xp1pwJISPX/Iej04f4i9D7s/la1WcxO0KSt3JtHGZDOUuVjncTR5WipwrOCLkgMR6frfn6e9VOqp6tpGgL5dVA9NZ6pxXUXUu79+RPUUG1O3ZKusrKgTSDGYrG1C/vlXvdUA+h4PvbDjTptD8JPkerNP5qH8wb4l9r9MbBq9g71xW+svi2aalgiopInx+RjRBIrNKgYMpUgqp08cHn35QS1OtzMEjLHpbdebz7o7Q/l9dOb42nsXI9mRU+4a85HbVJC0nloaREKxOqBm0lWX3r4TQnq9CyhgOIHRYNpfDLfvyk7t2Bu1vjVX9CwbT3FDkchkpxWw09WEmWZn0yr9SR79UjHr1rRVgzD4etuXb+KbCYbHYp5zUNQ0lPS+Y3BcU8Cwhuf8F96Hz6vgcOnn37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/19/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690XTePyH23s3cE+CybRxvDL4yxDkn1ab3vb3qp9OvUzx6fcT3/ANdZJIycwkckl+CosP6Hk+7Yp1rNfl0v6Dfe1ckVWiy1PMW+gU25/wAb/wCv71UdboelYkiSKGRldSAQVIIIP+I9+691jqZDDBJIP7Kk+7xrqcL1SQ0UnpO7ditJWzH6ySX/ANe5vf2ru2NFU9J7dV1Fl6VPtD0r6a8zhcZuDHVGJzFHDX46qAWopZ11RSqDcBh/xT37r3TXtrZe2Nn0E+M21h6TEUFSxealpUKxyMylSW1En6Ej6+/de644fZW2tu0uTpMDiaXFxZbztXLTIVE8s6lXkk/xOo+9U63XoButvih1p1t2DW9lYjGQruisapL1y3LhapSsi+pf6E/Q+/Z6151Hn0p/kpSZbO9Odi7Y28jz5vPbM3DjKKGO/kaprKAwwKoXm5J4t7X2Y06pW8hj88dIrsliqL656pg/l/fEbtXp3+Xx8s9sb2wmQp+x+wMP2fQ4mgqll+7qcfX4GeOmhRXUN+8z6Rb/AFx7ROdTGnSuNPDVQeP+c9UUfyyesukvjh8fOy+pe3/kLP8AE7vs9hbpzVNH/DMlV1smJncyLJJJEmtDrvxf8+/EHyGOm1IA7iASSTn5/wCboTcp1XmehuruzPl51x3vN3luLs/b25dg4zf0CV0eRiqKiFqZNaTAMraWDA39+49px1v4Kyg1qP8AiuHQZ/BHqjPY74g72373x8bv9K3YG4P7y0WG7Vy9RWLWY+eqUtSyIqtZjHe4vf3tjQkA9ejQlQzAZ/n0dr4N9p9vbC+HHy/2F2dT5HDbTruo92wdaRVglFM+5KiBkgp6PykAtpAX03PHvzgHh59bhLphxgHH2dAh1H2dvf4sfBb4Sbh2xSVEXZHyC7g3Ttjebw+U1T4T+IxUlIk6XDBXDt9fe2pU9aSqolPOlfzr1bj8l/5qmT+Idd1h0dt7Y9Ju7dmfmocdk6WqikkkppK5EummBh6izE3ZfbtuhZ9Y4Lk9NXUqqBGa1bAp0Zf45/zRNqbw7EHUfb+Fp+s9yzSRUOBpTBKBkq2YKYoCHb0k6rCy/X6+6TPrlLjgenYRpQR+fVrW5d27f2hRiv3DkqfHUpbSJJ5I1JP+CsQT/sPbXTladOWIy+PzuOpcri6mOroK2MS008TqySIeLgqT/tvfut9OXv3Xuve/de66JCi7EAD6k+9gEmg60SAKnpF0Jky+Zat5SPHu0IB/tgGwNvZjJSC28MjLdJEXxJdYNQOlr7LelnXvfuvde9+691737r3QYb56Z6z7Iini3ptLG50VCusjVSNr/cXSxDIRzb2XXu0bduK6b2ISA+vRNuXL+z7upXcbdZdXHUPXoh3Yf8u3RVy5Pojsau6faKSSrjx+ISdoqh0GqKCSRCLA/Qm359gfceQCc7FcGzoa0Ff2V6jDdfaRQ5m5WvG20g6gsYwfka4p+XQSYPt/5hdB5F8LufrnL752vjD4pt41SzyJNFTggzav9rHq+vsrj3jmvY5Pp57Zpol4yGvl59ElvvvuFyxJ4F9ZNdQJxmrxp50+fVg/W/yCxW++rcv2BXUseNOJx1VV1dBJwddLA0kkIWY8+oabe5L5Z3A8whFjXS7ECnpXqWuX+ZIt72o7gy6CBUr6U+3olPSvX2R+Su+d19vSV0mN2fXfe4zHUCqywrWQsVukdrfXnj3InMtlZ7bYptyqPHFCW86de2C7ub+8kvnJ8E1AHljpwz2xOy+n8o9fh3qnoYpHdnQSMropMY4Fr8ewYBTgOhSSadGK6t+UOMyCw4nd1qCoiAj+4kUh2KrY31EX597rXrVKdG+xWXx+apI67G1KVNNKAUkQg/UX5A+nv3HI6305e/de66JAFzwB+ffuPXiaZPSQyMkuZq1oqViKeFtNSw+huw+v+29mMSi2j8Rxk8OkUh8dvDHDpUUtOlLBHBGPTGth/jzc+0Lu0jam6VIgRdI6ke6dX697917r3v3Xuve/de697917r3v3XumarzNNAzQxMs1SL2iF78ce1Mds7ZfA9ek8lwqDtyemNY81li6u0lBHfUsgvyv9B7U6oLelBq6YpLO1a6R070mApIgGqQKqcG5lcG55v7TNdSEUXA6dW1jBq2eij/Nzqva/Z/VaYDc2Nhr8THLUvJTyEomh1Uycr/X2wztxJzXpQET4aY6qW3l0Nu3avxvn2N8NduVmD3FW19cM8duyTyVVXSS+jQ7JdgLX490wW7ut0dRSOlRw6TvwZ6x+VHWVHmeut6dA5imqN4moxO4OyK2GpFQ+JyamOqjd5lPp5JJv72dJ68gcDv8A8PQu7o+PvyX6Z3BlemusNsZjcHU+8FkpMpXUQnegp48p6qp3dRp9BJvce/Gn59eCnFOHT12h/Lu3j0/1p13nOlTVZnee0c0m5MpjaASmfI1LMtU9GoRbsAfTfj36uM9e0/w9Teo+o+xdrbm7D+S/dtHPgM/2finw0ezK/WsuPlSMRfdCKUcDgW5/PvWPLrdT5+fSQ6e+Im6e9c52zVbninw2Hixk77MrZNYjqa0kmJYdK/4i5HvdSMjrwAPEdEj7Q+FPbOJ7Y6cx1ZT1e2KTCZ1Gqsg7zgbki+5uUYEC9wLe5q9pZILSz3G6uTqNKrXy6j7neJpprSGE6RqyB58ePW0fsjBV+2NkYGgo1dngghL0wH6P8mVLG3+tf3Ge6XUV9uksz4BJz+fQps7aa2s4okNadLamz8V0irl+2mPGk3/V/jf2XSWhA1RGo6Wx3NSVkFKdKFWV1DKQVYXBH0IPtF0qBBFR1y9+631737r3Xvfuvde9+691737r3Xvfuvde9+691Cr6KKugaKQcj1Iw+quBxz7dhlaF9S9NyxrItD0n8NWS0dQ+KrGYuLvHI9/p+Fv7VXSCQeNGMdMQvpOhznpW+0HSvr3v3Xuve/de6Zs1uDE7epvu8vWR0kH+qci/9L29+691U38+t5be3ttvHZvZcq1Gf2G8uXppo0/cnmjlBjjBiuTbT+T7FXKu6raXps5vgnop+Xz6DfM23G5sxdRfHDVgPn6dBX8f+w+0/lBs3HUtXW1n8aoHaPK0ZEqPT0tNGFhZl5Nv9h7R8z7Sdr3NvBzC3wn1J49KOXtzXcrBfEP6o+IenVgfWvxaxWKMWR3GVq5GOqSmcsWa6jVcMLDm/sgpXJ6OxUY6N1iMLjMFSpRYqkipKdAAscY/AFuSfe+t06dPfuvde9+691737r3THuHceH2tjJ8vnKyOioKYXlmkI9IsW4Unnge1NpZ3F7MILZdTHgOmppooEMkpoo8+iP8AaPzDwVZTVOJ6ipU3zmFPheKGCTVHIykEKf8ABre5J2TkG5icT76fp4/t6CO48yQSKY9t/Vf5dAvtPqr5CfIqaTIb6zmW62wkDIzYWRZ/HWx6vG6LGP8AVL7EV/vnK3KaiLbYlu5D+LGOiu023et7Gu9kMCj8Pr0eHrT409X9bwUs9Dt6hqc9EqmozUgkM88yNqE1mNgb8/T3Gu8c4bzu7MskpWI8E8gPToX2Ox2FiAY0GocW9ejBjjj+nsK9HHXvfuvde9+691737r3SRzoaCqhnS6lmQFh/h+OPZjbd8ZQ9IbgFXquT0q4zeNDe90U3/wBcey9sEjpaMjrn711vr3v3Xuve/de697917pkzsDS0jMnDoGI/xuPp7U2zEPTy6T3AGip6DnsHtvaHTex5N2b3yK43G0dN9xPM+khUuQCdZUc2/r7pcaRKQOrQEtEG9eim9p/I7ee4dpbF7E6Vw8+59nbijZ58nSvF44Fik0ymQRuQLcHk+2OJ9B07TyPHo5OyN1mTrjC7p3M644yY4VVe08sYWKzHVdy1vx9L+/Cvn1skcR0Wyh/mK/ETJb9HWdH2xjJd4mqNF/DPtqlV+5DaTF9yw8f14vq9769g+Y6K18/v5o+L+Gm6to7WpMHR7hqd5Y2KuxjyBpQ61EZ8RRoZAv67A3v72AWNB5dNtIsYBauTTqsX/h1j5wda71252P2h8eMjgujN0T09ZSburqGvTFPjqqYIJaWZyI+VYkWP+t78ACMHrxYrllx61H+DrZP2FvSh7e672R2Fg51hxu/tp1GUglQeiKKupPGrC5P6SSDz+PauI64GU+VKfn0xIuiUMfPrUt+V/wAQk6u/mgdebu3FvJM1t+rhXJ1Ku0hjjepi8qwtrAFwSB9faTIBU+XT5UGQSA4Nei1fI/e++cB/Mo6u2ziuph2zS1+ChfFYuZZhHUU06kU0cbXC8cD6+7DKnqpxIuK4P5fPoym6/jv2RW9tYztbeey5vjtW/ammoMfqkvMkh8SRxRxkmzD/AHv3WtRQcOraTq1nHRPcj1j2R1X/ADJNpZjdHyBy3x22pXbYVn7AhjrpBHHW0txUKsQu2sHkD+vu3lXjnquQ9C1Kj9vyz1ZP1V1T0vRfImkq8N3xB8qez8xB5Yc7JTVsFU9NVsbWadbklTzf+nutTSox1cKCeNT0PdP8Re2sH8ut7bvodr1o2vuHq7NYmCVDL4RPk8U0Cwiy2uGJHHvVSRjrYHdU9KH4ifC7s1+j+wOiN4YiqwmPyOW3BnMZV1HkVFrqtX8LqXX63t+PeyQTXrwUhaH/AFZ6st+HHxXzHV/xSzPQ2+62StfLV+dSWeXWdNFkinjVdYvYaf6e9fPq1KAL6dAdvz4O9e9IdObuootkRdqYippMnUHAz+SFaeWpa5kGlb/2vqPfiSePWtI4Dqi/cfT3Ue8diP15s34t4+DeeXq8njoRTS1pqMJUTykR1q678j3apwa9N6QwKgU62V/5aPRO5fjp8Udmda7rjmgzFFW5DISU82vXTRV0cLRQesA+kLb6e61qa9OgAAAeQ6P57917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//0N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690XPsH467b39lZstWSiGpmcOzaWLXDFrcf6/vxr14UAp0Au4fiNU0TE4TJNIo5TxmQEf0BA+h9+Pr1occ9BxUdIdt7cZ2xk1dIoN1ZPNYWP+t71SuOrcPy6hQZvvfast6qLI1MEQvZvNbg/i/+t73wx1UZ49CDtn5Ib3FTHg8pg5CtXxJUSRMTHY8+o+1FsNUoXpm5NIiTw6E/E/KPbWNqZcdltFO8UmmRjFpa97clbf7D3SdmaQitaY69AFWOo8+hexXf/XGTCWzKxu4BAKrpAJsPzf235V6ezWnS9xm/NrZggY/KwTEmw9QXm9vz70M9bOOlYksco1RyI4P5RlYf7dffutVHXP37rfXvfuvdJJ1M25o2K3jjhFj/ALUq8ED2Yii2VPM9IiC9xqHAdKt0V0ZHAZXUqwP0KsLEH2XdLeiSdp/y6viB3JvKbf8Av7pvb+a3XVIYq3LSNURS1ULCzpKiNpN/yQB73XrRFfIY6be2/gN0ruz435v4+dd7XxXXuDeiyEu3v4YswixmaqYdMWQZnMjlgQOfeuvHhQ8OqEtm9E/zCPjttGo+JOH6szvaXXFZnMhInY0sNZLHRwZBjElRTTEDQI0OoXHuxKnJ6qqSLQAYHz6FL+YV0t2l038XPhp0ts/CVu6Nx7i7CnTsbJok8s0NHlKinJpZjGtwsQkZbt+QePegc/6uPXmB0UHGor9nn0lex/jju7L9z/H/AKlXb8o2T0Tu/A7okrFScwU8c80VZVuX02+oP1A96rQHr3EinAGvTX8UtqbP+WP85b5RQ75wEW6No9Uu+d2zJWJNHDTVOPqYqSjaIgWb9wrdQRx7VxExW5Y/iH+x0lcGS6oPwUz9oqen/wCYOI2B2Z/MP69m2HhqXb56W3rjqvflVRRteogoaxRLNMsf49FrN7SLWlKcelTYkHqP8o6B/C4f5D/zbfnp8gtk0ncG4Omuq+gsuarC1GN+6en3PjRkEx8EEdJTyBSz6g7EkAAH3s48s9aALk5OkU4etOg16r7I+XXxT/mS7S+LcO9tw9n9e7i3/Tbe/idRUVM9HR0D1YhkqoULnRZFYke7UBSvn1UMwuPCJ7aVB9OrXezf5vu68r352l078aev6Ps0dJbgqMH2FUPBIs2Palm+2qJDeQEgODyoHuoFc9XLUNKcOrBfhJ86+ufmNjd10G3akQ756/mjpd74JKaWGPEVMkpgEaSyM2v1gi49+PVlIYVHR0t0ZOlw+Dr8jWzimpqSCSaWYlVCJEhlYkuQPop+p93iZVfUxAAya9MXU8NvA0s5og4nqozef85D4n9b7zk2TNusTT01clFlKiKgEqwVZcJLqlp30nSf1Enj829gy952gluCbGJ5YlJBfSadvGnqPSnHrH/dfvHe3Wybm+1z3qaomCSUBOhj/EVqBT8RJovnTq0XqbtzY3dOz8bvbYGagzeDyVPFPHUwtGxXyA+lxEzAG4NufZ/tu52m62wurRtSnj6g+hHkepv2Lftr5j29Ny2mVZonAIZSCDX0IJHQm+zDo5697917r3v3Xuve/de697917qJXUNJk6Segr6eOqo6qMxVFPKNUcsZ+qOP6e6uiyIUcVB4jqkkaTIY5BVTxHVX3zSzmMwVDtbp3qTH09HnsxuCOLcmMxkRXxYevKLLJLoYtyCxNx7GvI+zWVkZdykQRxqtVI82HQH34wW0abXtSBHdu4KPI8T0fDpDrDF9R9eYbaGJN6eGNa2U2IvV1cCPUnn/agfYe3a/l3K9e6l4nH5Dh0LNtsksLRbaPgP8AD59CbkcZQZWnakyFNHVU7/WOQEjn/W9lvS/on3avxdx2Z+4y+2CKWsuzJTR6wbltQICjn3rzr17ot+L3Z2d0lklpcmlXUY+KU6Y3SVl0BtA+vH09+PXvzr0d/rn5AbT3nTQxVdVHQ5VrK1OQFW4UFvqbjk/63vfXgDToYcxkVFLHHTOrvWArEyMG+o1D6e1drFqYu3BektxJQBF4nrNhcd9jT6pL/cTDVNf6k3uL+63M/itQcBw6vBFoFfM9PXtN0/1737r3Xvfuvde9+691737r3WGeoipozJM4RB+T/wAU93RGkOlBU9Ud1QVbpKVFbW5iX7eg1w05NjULcWsfyfa6OOO3GuXJ6QtNJM+lMDp2oMJTUgDSgVE4AvK454HtPLcu+BgdKY7dVNWyensCwsOAPabpRwx1737r3QT9z4Vs1sfJwqmswwTSW/w8dvfqV61WnRPPiZk3xe7sthwTHHO8sUkYIswW5W+rnhrHj36mevVpjqxv6+/db697917r3v3Xuq7vltuCTNZbE7XSQ2p6rT4gFtqaSxJtzzb6n3U+vWhU8ejcdJ4psP15hqJ1Ksis1jzwyqQRwPdvLrdKHoivfmZynY/yI6x2/hp5JaTZu4YnzEEVtCxLUXcyeO/1/wAfc08sW8O08p3l1OAGnQ6T86dADdne83y3iXPhtkfLq0JVCgBQAB9ABYcCw9wuSTk9D8CnDpryGIpK9SWRVm+qygeoH+vt+K4eLHEenTMkKv8ALpPWyeCcAl6ukvYub2jU8mw/4j2qHg3QP4T0xSSE4yOlXR1kNbEJYmvwNQ/Kn2hkjaNtJ6VI4cVHUv231fr3v3Xuve/de697917r3v3Xuve/de697917pP5zHGoiFTAumphIfWv6iiC9v949rLSYIdEnwnpHdRFgHTiOpGFyQyNLqZdEsZMbp+fTxqt7buIvDfHA9PQuWWh4jrrM5/E4CmarytZFSwqGJZmW/p+tlvf2n6e6Kr2N8psJh45qTawXJ1bCSP8ARqKlW0GwVvfj8uvfb0WBh2v3JXF4xWpi5nbTEBMqIrHyD6fge/Vr1qh6MdsD4pYamozNuphWtVBBU0cmo60IBkUlhxc/197UmNhIpyOHy60RrBVhg8R69Fq2hR0XxW+SuZjFIKTa3YVYMViKYEmKMvNpR1ZtNjcA8e5Enrv/AC+mk1ktxVj646A8RTZN8eoolwaD7eraFIZQw5DAEf6xFx7jroddd+/de697917pMbq3jt7ZmLnzG4chFQ0NOLyyMysyi1x+3e5+ntbY7fd7jMLe0Qsx6T3F1Bap4k7aR0SPe/zOXLvPiuksMd8ZBv2tP28qvGzXjawU24Nre5G232++nAn5hk+nUfPj0F7rmUSkx7YvjE/LoM9vdA95d7V38c7P3NmNjY1CjybYP3Bhq1VwPGY7gWK3ufZzd808t8tRfT7NCly5/wBExjpBDs27bs/i7jIYlH4PXo8PXvx96x66paYYXbGOTKRoPuMqEf7mpl1ajKxJsObH6e433Xmred2c+PM2g8F8h0KrLZrCyWkUYr6+fQkU9ZUwZX7KWV5IZSxjD6f21BsqggX9lLRK8BlAoR0pSQrLoJx5DpTe0PSzr3v3Xuve/de64l0H1ZR+OWA5/wBj7917rl7917pmzUJkpldVu0bX/wBhbnn2ptW0vQ+fTE61Wo6zYmQyUaszXsSLk/QD8H3W4XTJQdWhNUHUXcucg25gctnJyvhxdFLWS3P9iL6m3tjp3om3QPzi67737T3R1Xg6uL+PbXo6qtrIdHjIgpeZHuzk2A5PHv329eGeHl0cen3Pg6uiqshS5CCopaONpah4XSQxovBYqhJ9+690E21vkp1DvHObp27gtzw1OU2bSvWZ+AhFNHTxtpeQ6WJNv9b3unXuq3O7f5uvXmzNxZzb/XlBDu87aDvnJJ6eVVpY45jEzAo4B+l+fdk+IV6bkI0kDP8Aq+fRVPmH8g/+HBfhb2LR9KSSPurE7aL53H0KhZcYI5vI0yJCzNYA2592lAWjDz49Uiq60Br0SP4h7O712L/L17Gope08/Jmdq7cqXGJ804bFSy1Au8ak6gSBb8e2206q9Wj1hKE5p0Ifc/yc7c3J8IemercVuPKY/L5TDSw7h3VEXFZGwqSqPNIh1D0t9T72PTy6szHTqBz0se1f5ZW1cH8Edtd37Y3KkXcSbZoMtV7xilkWsrqmqmklaRJb8yLpAIHP1/p70GNc8OvGNR3Lxpx6IpnZpe5/jZHvztKhbc2d+PtDjMMNwZETPVVPmrB4mZudVjf6n3sUDUHTbVePUc0/w9XefLnAHuv+U31iMRA9alZtLaVbDCoa4joqZ4NJSNSfRo0gcf4+6glen5Bqr8x0Yv8Aldb4rc/8XMRtiuV0k65wkG34YnjZDAh1SMq6+TyPb9v/AGypXiR0nlI8Iy04D/N0AH80D4x9gdj5/bPY/W2CqMzlaA4iDx0omZ/Cqosz2jU/Tm/Ptt6+I1fXpxRqjXT9vWf5K/ysq/5LbL6l7F2Nvqp6Z7o2xsLAUVVmqM1KVS1cNErzxpPTrqVwxI/HP590BI4Z6syhjq4EenXL43/yvu28LunA7u+Q3yBz3a1HgPtqePbmZmrqlKsUkmryyyVN7A2Fvz73jyHXgDSpJJ6Pj8l/gh0R8mMRj6Xd208auYxMMVJjsyEk88FHEgRYGCfqAAGn6W96B08OvFQ2G6i/HP4CdB/HWrps9t3aONqd40ixxU25XWUVcEES2SOJSbC39effjnB6sABw6O/pW4OkXH0Nhcf63v3Xuu9Kg3AFz+bc+/de679+691Hq6Wnrqaakq4lnpqhDHNC/KSI31VgPx7917oOcX0z1dhcv/HsVsrCUOY8nm/iEEDrUeW99eosRf8A2Hv3W6k9Cd791rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//0d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6903VWJxtaCKujhnDfXWt78W9+OePXhjoOdybD2iIiKfC0sNbNfxzJcMtxYkA39rbJaP4lMDpJdkFNBPHoLsx8Xdm7iRKqUrT1EyK8jxoxLOy2JJBHtLNmViPXpRGAIlHQQ5/4h1FExbB5OSUclAnluOPyP9f3Tq/2dB9L0b2xtos2Oqa/QpuNHlIOn1AH/Y+9AE9brTj1BizHem0Kj/KIshPGvNm8huFGo3Hvx9OqjhXpX4r5T75wLChyuCeQXDPPJAzEc3YXYX+nv1fTrxHQs4L5bYKsCJlIkpJG0p/myPWxtexNuPewakAjz68aAVJ6E3DdxbPFSKrIZHxNOt0JCWAcccEg+19yVWMJ59I4Vbxi3l0J+M37tbL6fscpDLq02OpFHq+n1PtAKHh0tOOPStWaJxdJY3Fr3V1YW/rcH37r3WQEH6G/+t7917r3v3Xuk3n9n7Z3RJQS7gw1FlZMXOKnHvVxl2pJ1IYSwkEWNwD7917qLW7F2tWzZGsfD0a5DJ05p6quWO1RIgWya3vzb3qlePXvKnRcejvit1Z0r2n2D2Ns7F0EG6d9weLcNfTl/uZ0NSKm0ysLC7qDwfr7WyhhbpXHSWJwZ2CmteifYv4O7M6h7z+R/wAnO093mLZe/oqmsyAkM1sJTVNQZDKAysp0swHA9pBwp6dKGALahxNP5dU7fG/cXfHwb+RnfHePV/Xi9mdDd4Vkqbf3LWSyCkbHQZBqunqonhZfG6FBddVj9CD72aGgrnqvehOKqc441pnoIv5aPce+vkD81vmF8ou5tttidufGqsn3vjphA/28ivX+FIKaNAVawY2F/wAe9mtNJ+3qikMTIPI6fsxnpU9rdRUm2twdx/zFvgTnm33PV5Mbw7b64RTQxVFdkaszVVJlaSUr5otRNwAfetRA0ny6sQC5mXNeI8sf5erkf5OfbnT3eOxd6di7P6o291V2rnGpqvtvF4Cmkp4ZcvU1BZA2vi2pGayn6n349vaePVkYSLrXh1YJ8xdu7q3t0LvrZ2xJ54d25fFVMeMamLCZWSkkZyNHP0/p7JOY7S/udjnFgD4gGKefqPzHQO9wLK/3TlK92/aG03MkbKhH4WKmjfl186PsbrbfmzuwNw7Y3fjK2n3NSZeWGWKqin88kzS38kZcBnEhNwfzf+vuPrDcLN7FWRggQEMpIBUrxBHlT+fXFXerK75b3CXY95VkuoGKyg1JdjnUCRVg9ag0qa+uOtmT+VnL83vjr0znBB1Dks3sncslJlsDPlUmEmPgWT/KGgia1ka7MFN/qbj6e0O3bnvdnczXmxWjSW04VtTAhdQwWQehHHGTnrOz7ukHuzyVylLBPtmqynfxbcM36iK2WDLnSC1WArXuNQOrYaL5zbyxbRR752N/A3QAVRWCZtDA+rj/AAHPsRR85XkY/wAdh0H8+sj4vcm/gFN1tTCRxxXoTMb/ADDPj1EkcW5d0S4msbSpjagkK62Ngv1B9rhz/sCdt1IUb7OjSL3Y5RSi31wImPkR0PWy/kj1F2AkT7X3PDXCcxiPWq0+ryGyH91h7PbPmHar+n00mqv5f4ehVt/N2w7pT6KcPXh0N6VVO4VlngYMoYaZY2BBF7gg8/6/s61KfPoRh1IqCOsodW/Syt/rEH/evfqjrdR0zbly6YDb2bzj6SmIxVdkSGIAYUdM04W545029vW8fjTJEPxED9p6pNJ4UTSfwgn9nVW3xaxNX3h8id4fIXMRebDS09bhKHHSo4p6epgBiilQfpJAH9fr7kPmKSPaNki2OL48MT59AfYkbdN3l3iQduVA+Y6tjACgACwAAAH4A4A9xv0POu/fuvde9+690j907G29u6knp8tj4J5JUCrO6kuhBuGFvfuvdEL7G+Nee2rWT5vZks0kSfu3i8t01uWIsOOB+PetNTQZPWidI1V6T/XffW4tlZRaDd0UtTHG7J5qqFm8F/QGQt/rH2+ZnVfD4evz6aVEJ8TiT1YHszsrbG96dZMTXRvLpu8bMim4Avbn/H6e2enuHHoQffuvde9+691737r3XvfuvdQ62up6CBqiofSi8f1JJ+nHtyKJ5n0IOm5JFjXU3SThiq9xTfcSlqakQjQgvaVb8Ej/AB9rneO1Xw0yf8HSYK05qeHSxp6aGljWOFFRQFB0i1yBa59l7OzmrGvStUVB2jrP7r1br3v3Xuve/de6as7Tirw2UpiL+agqo7f1JhNh/t/fq0691WB15LLtHuqrpi5VZaqQafoLM5H9q3096Bp1ojFerT4HEkEMgNw8Ubg/1DICD731vrL7917qPV1KUdNNVSfogjMjfjgf4+/de6qw3VO++O9gsdpqdMjwn1XiUkWK+/EinXgGOR5dGM+TXy+6h+Huy8bXb9yq0NTV0lOcXRL4T9wxPiIbyOtgWBHHsd8lcg71zrdmHbUqinuOcfsHQZ5i5o23l228e+kCE0pUgDPDJPVXX8v35t9X95fJrtLNZbJ00L7lmmXa8TpTiJqhagN400n6lSbW/PuaPc32/wB32DlC0trZCRAB4nGtKdR7yXzftm873PKXBLkhaUOR6dbCHvFzqaOve/de64uiSKUdQ6t9VYXB97BKmo60QDg9JHI4+rx8orMaWMYJMlOP02vf2uimWQeHKM+vSF4XiJaM4PT5jMlHkIbj0zJYTJ9NLH8c+2J4DCfkeHT8M3iCjYI6dPafpR1737r3Xvfuvde9+691737r3XvfuvdM+ZzmLwdHJV5SqjggVWLanQMQB6rKSPfvn1o+nRV90d7Y+jqq5Nl2yEiQzExhUJDIP1AqT9fZiv8AjMNAe4dJCfBk0nh69Fcn/wBLXcmRaaNK2DHyzPF4QJQiozXP0/wHstyG0npZ5ah0ZHrf4r4vF+HKbjcVFUdDy00gcktpu4Or6c+9+QI61nh0bnD7ew+AgWnxNDDRxKoUCIEXAFuSf9b37rfTz7917oinzh6sl3Vs3H9hULGOu6xE2ejEYYvOIpVcJpQc2P8AX+vsX8pbgLe5Nk3C47fs6DHMtj49uLtfig7h0Mnxe7LbtPp3a+56yZTlZ4JYshTsyCaGSnk8Q1Rg3AsBa49lXMFgNu3SS2UdoOPzz0Y7LeG+2+Odj3EZ/wAHQm727H2h17jnyu6crFQUaLqZgVkkA558YIP4Ptjbdov91l8CyQs3Su6vbayj8W4YKOiT7t+Ym4t1SzY/ofbLbykcFBI9NIrqCTGxFrj3I1hyBaWSibmWbwAPKvQVueZZrmse0J4pPn0kNrfF3tbtyuXdHbe7MztuJpFkn2heV6aZQ2pUZLgabXB9r73nTY9hj+j2KBZT/v3FR0mttg3HcW8bc5Cg/g8j0enYXSnXXXVNTrtzbONoq2FR5MhDERUzPe5dnJ/rz7jTdOY923ZybqZmU/hJwOhdabXZWSgQxgEefn0LHsi6MOve/de6S2ZU09XDVqLaQPV/Q6vZhbNqjKHpFOoV9Y6UcEnlhikHOpFP+8e0LjSxHSpG1ID1zkbQjufoiMx/5BF/der9UL/NH+b5mejPkKnxp6n2jRbt7Jq6P7mgxtVTPIZ/JCJISp1C9+eLe9gE56oZAtARUnovVB8i/wCb13FuzCiX4xVu3doVdbRVEmZoqCekjFBLLaSXWzAEWub29+7fWvXv1MUAH59bDHXGbrMds7bNDvmqgxu7HxlJ/FKKsqoI2irmjHliBla978W+vvVa9W4de7W7i2D1HiIMjvnMLi6TIt9vSuBExldzoXT5CB9T/j7vH8YI6rJlCOqr/lb/ADFt8dcYfH4D477Th7A3Xmqx6Ompqml1afu5AtKY7EAlgeCfaq8UatXDpPASBoOeq6qH+aT80une1aLrn5b9I0Gydn79qaPCQVFZTRNJLFlHAQprdrD1Dn2kp21GentWlgpHHANcdR+vEXpT5+d5ZPAacZid79eVEGEracaRHVbhoRJH4WfgkF7WX3riAOrDDGnnTo8f8s7s3cma7o7j6T3dnKvMNjdvTzlK5lLIk0yi6Kx1Gwbjj/H3v5jqwqa14joivR2Xpesv5iHyb23u/cs2L2nunH5XBpMhWRIFqqjSkiqx/H+HupJoKdVFNTfl0z5LKLmN/wDbfSvxf6A273VlpKNqXdW66qn05SnojIfLVrdgQBe5J97ycnz68fVeA456AH+UnnMhsP5M/KfpTdLTYbOblwtft0bVRStNQ1ktQAYI5W4NjdePdnJKqfLpmMadSEUNa9GE6Y35uXZHaPyX+Ne8sTUUyb5jl29tyOSknZZag1FoikrLwCObj3Q5Grp8EBynn5dGM2v8AOz+xOhO5thZXF1GC3JiqCnh2DOPuFnqJPL5mFPcA8i3v3mPTr2gBSPWvRb8TjP5om6upaX4bbg6LzON2Fg4Y8LQ79ajq46qop0qDapar+hFmY/T8+99vEdVAlpQgVpStere+jP5Z2z9t/GLJdP70rHq8nvmhpJd1VDrK7R1tPN5UB1WJK2sfeqmtR1cABdPR/uvOkNp7H6n291BUU0Od2vt7HrjaSlrIz4vtlJYJpvcck/n8+/cRQ9b4Hp9w3XGzevsLkcfsjb1Bt6myMiSVkVAjKkzgn1yB2P+t7UWxAmB9OmZ/wCzp5dLTESCehSNyWMYCMP8LcD365WkhI8+tW7VT7OncAAADgDgD2n6f679+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf//S3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdI7OaZM1iIGYgSE8D88nn2vtsQOwOR0jnLGZVoCD0r0UIqoPoosPaEmpr0rAoKDrl711vr319+6903VeIxlcSauigqLix8iavfuvfLpIZLq/ZGTSQT4GjEjg/uBWuCRb8n37rw+fQO5v4wbIrXkq0K0hS8iqsZCal5AYg/8AEe7IpZxTqrMiIS3QSbi+N9fuGJqnE1k0S0xenjSLyAMsZAQ2H+HtXf4kCkeQ6T2p/TJGc9BxN0N2ztxWkx1VXmNeU0eb+yfSD/r+0IBPl0pJ9em6HJd57SnKVMVdMkIv6xKQwQ25JHv1M9bHDPSrxnyl7AwLLSZTBM0SWDvJTE8KLMQSP6+/Ajz69SmOhZwXy62/UJH/ABiP7VyQHtFpI454JH5978+vY6F7EfIbrbLgCHKlGIH6hH+Vuf7Q9+NOtZ8+lzSdh7WycMjUOSikYISFLRBif6D1H3aOjOB1p6hT1N28IZZajIpNE33Qtp8qMwA5F1B49rbtu1Y/TpNbKKlh0lO8eq8d3f1PvbqrKV8uNx+9cQ+IqchTp5JqVGmSbyxJdbkFB+R7QZHSvHn1r8bp/lx/KbqLAZnorrbcm4eyesd0xfZwZmrmq0j2jDPN5HnoadidDhT6tHF/fiRWpHVQlF0qcfPj1Z18G/5efX/xV6v3DtbJpT7tyfZWLjp+w/4jTySQZjU3kNLVrMbuoBIYEDm9veyamvVlARdK9FC7o/kg7b39ujK5HrPv7efS20M7UrNmNibUpZhhq+ISmT7efRUx6lsbDUp9+BINeqGNSCASK8aGnVnfxR+JnW3xL6+pdl7Fo4pa54lXP7leIxZHcM6NqSorrs3Km9gD+ffiScnqwAUUXh0OGMEmRzNRVMTaikeIc8EMNDD/AG3swmPh24SlNXSSKjSswPD/AIroLd0/E346713FHuvc/Um0szuBJVn/AIpV0bNUmVW1LIxDWJB5HsHz8r7Bcymae1RmbiSDn7c56Ib3kjlTcbsX17YQyyg11MgJr5GvQ8Y3EY3EYymw2No4aPF0dOKWmooVtBDTgECFFP45Ps7ihigiEMShUUUAHAD06EsMEUEQghUKiigA4AenSayPW+w8uWOT2nha4vfWamjjkLXFjct7TybfZTf2sSt9o6Sy7Xt0/wDbQo32ivQP7n+JHQ25/I9R15t+GaQMTLFSWKuV0hlW/Fv9f2U3PKuxXOXt0r606D99yLyxfV8W0jqfPT0XTdX8ujaOWWRdrb8z+xwxYxjCR6BDcWXRaRf0/UeyC75BtplK207Q/wClHD+fQRvfaTbZwRZXMlrXh4eKfz6CCf4M929XKa/ZXd29N+SoWmgoMrUTg3j5jgYamWx/1/ZOOSd52v8AUtL2ScjIBJ6If9bXmTYxr23dLi79Fkav5dQsb2B/MP2VW/ZJ09DmMHSWDZKYmWZ4ovpIw5PP191j3Hn+zfwls9cY/EePTcW5+7e2yaP3cksQ4tqzQfLoNPkz/MF7O6367hx/Y+zE23PuKqbA1Qp6RpWdqm0DxFDewu39kexnyvzNvP7wrfWoHhjVwPlk/LpNzL7tbry7tnib7amHWQh0qWyxp5fOnR7f5f1diano+hNDT/az11a+VmiMTws/30QlV9DgfTn/AG/sVz81Rc1zm+i/D28PTqUuQ9yt9x2NJ4BTV3UpTj8uj1+6dDbr3v3Xuve/de697917pLZ+rLmHHQ+v7pjFOo/sqSByR7W2qAVlby4dJZ2Jog8+g13v0RtLeGNFOaSCirQljVIrFpH+t2P+v7SzN4shfh0/GuhaHPRHty9WdkdQ1/8AEcA1W+Nik1XiWUq667m4A9t9Xqehw6w+UlPN48VvFDRyxFk8rosbtyAvLkX978x17OadHLwu4MTuCkWtxVXFUwN9CroWH/BlUm3v3Xunn37r3WGonjpommlYKi/k/wBfwPdkRnbSvVWYKKnpHxwT7gq/NUArQwmypYhZV+oNva8utrHpT4j59IgrTsS3DpZRRRwxrHEoRFFlUfQey8ksanj0uVQooOsnvXW+ve/de697917r3v3XuuLKHVkYXVlKsP6gixHv3Xuque7KSTa3d1PX06+Knkq4/WL8qz39V/8AA+/YHXq1x1ZRtbJRZXA4uribUHoqYN/gwhW4449+690oPfuvdIrsPJRYvZ2eqXcIRQShORe5IFxf/C/v3Xuq8PjvhJdzdm1GauZkhr5JpCb2CRyFm5H+A91ZaqacaHrYbu+zqtr+e/8AGTtftOm2dubZ+NqM7hMO1NNkYTDUzwU8MNcZHQpADYW/r7y9+7lzds+zma1vHEcjVAyASStBx6x297eU9y5h25Y7RQwDxuQ1aUV6kY9R1WF/LR+KXdeW762znaPbdRg8Hhs01VVzU9JWQoKZGXUzF1A/HuXPdTnLYbflyaCWbxJJEoASDnPQH9t+U93i3hJjEsccbthQRjFCfLy63oYIzFBDEWLGOKOMseSxRApJP+NvfOpzqYsPMnrL9RRQOsvuvW+ve/de68QCCDyDwR7917pI5DHz46f+IY5SQDqlhH6WP9SB7Moplnj8KXj5dIJY2hbxI+HSioqtKyBZFI16R5UH9h/yPaGSMxtQ9K45BItR1M9t9Ode9+691737r3TRmM7i8FSvWZKrip4YwS2qSMPYC5srEX9+61X06Kp2J8p8JhhNR7VAyNZaSMXRXs4X6gKT/sPe8Vp14V49Fikbtrumu1wLXwY+dypjRJlQLMbEED8ce6+VD1vh8PRlOsvjPFtynOVzNZI+QeMmSmcPYgLdla/0v7ft5DGw+fTMyawSfLPRmdnYvFYugWnoaGCjljLCRYxy1uNXP+8+73MYWQsnA9VgkDLpPEdLL2m6Ude9+691737r3THubBUm5sDlcBXKHo8rSSUdSjfpaKS2pW/w49uwzvbSrPH8S5H29NTRLNE0TcGFD1SLs3s7troXuvsjrDYO21zlJXPJR7bx0qMKeOqkmshp9Vrm9jx7nteWdj33YbbfNzn8MgAuQc0+fUWQ7luO0bxPtdomsHCjyB/1Ho0uzPiRvnsqrj3p3PufLUFXUyl6vZhMktChBEgiKsQAnJHHsh3DnvbtoQ7fy/CrKBiXgft+3o/tuW7u9YXO6yMD/BxHR8NmdW7E2DBFFtfbeMxMkaBWqKWHRNI1gGZnv+bfT3Ge473ue6MTeTM4PkTjoWWm32lkoW3QLTzHQg+ynpb1737r3Xvfuvde9+690z5yLy4+QAG4ZW4FyLe1NqxWUU6T3Kao+vYWfzUgQ/WGyf7D6j366TTJX163bmqU9OnWRdaOn11Ky/8AJQt7TdP9ab/84PMJ8Uvmzt75K4XZGMzWapnxuPoqmqpmmdmeJPECSLfk/n3ZRUEdMSsU0mlfLo71B8+v5onYmN2JU9OfGfH5PbuawOErJcnT4mWf/JayFSKjXJdUso4CgAe9UHr07qbgF/aelD8zOv8A5bZTqDbHd2XqM1tXeuJzFFmN0baxkk/jpaGiRaiud44f7Kjix496FBjrxDHPAj+fRcPlx8ytt/K3orpcbcyzw5/C70wO1c7SjwLUT1cDx01ZO0CMX9TAk8W/p72Kg6utMVYU9T0ov5lfc1T8Vdl9ObP2nsPHPnt2022aar3lHSSPlcemQpoStfBOR6XQvcH8e1kxD24Pn0kDlZ6EYrTqs/5k9V7f2VP1Bvip+QO7u8d9ZTPYjMZLbGejrshBt2OpVKlY4pnLg+MnSLWtb2lBrXpQ9FIJJJrjo6PdUm5YMl8d+0cNi6mpO7d07awuXm+0qWdaGDxRMJECkAab+69OGuoH1P8Ah6F3ufov5ufFT5ebj+Rvxd61quwsL2fiMbQzwilqayjoKerpY5JgUUWB/wAfe1IIz+XVXVq4pmnnTh1j6l/lpfI3s75BJ8je2qOt2vU7wycNXn9uOKpaeiiciWUiO1gNVxb3qtBp6to7tR6HXcn8qztnr7v7e/YPS3YefweL7IRKTK1GFnraR8dTSgeYloyLgf0v70Tj7OvBaE0xXox3x/8A5TfX3RnaOO7drOxK7cPYU1elfl66sWUz5WoJEhE0rsWY6r3JX/Y+1IDPHQA0Hn5dJtKxy6icn1PVndf0V1Bk91Jveu6/27UbsjnWpTOvRn78TqbiXyKwF/8AG3tNjpZU9CuqKpJVQC1iT/WwsP8AePfutdcvfuvde9+691737r3USuTXTSL/AFHt2E0cdNy00GvTNt0OsdUr/UTcc3uOfp7UXmSpApjpPacGHz6UntF0s697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6/9Pf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690i6pmqNwUT2uIGCg/0sb/AJ/xPszQBbRgPPpDq13ALeXl0tPZZ0u697917r3v3Xuve/de697917onvZ3zI6i2D3JjPj5lMsydjZqipq6mxySUzf5NVxCWLXEWMnKsD+ke37cVl4cOmbg0j8s9GmwQSHEwVDuEimiFWWdgqRpIgkLMx4AtySfe7lw8lfTr0Ion29S6LK4rJxSS4/JY+vhjYpJLRVlPVxIw+qu8DMAR+QT7Tj1HTvEZ6xyY3D5INJJTUdYGurP6JQf6qXUn37rflTpLZTrLZWVikSfA0IeQEeRYzcXNybX9+8qde869A5m/itsXLlmQmjLXIEMXAJNx/a96z17Hp0D24viHLSiT+72QmZmuV8fkWxv6QffiOvD59IKl6O7K2hXUk8lVWfaQzh39UpWRAbFWP5v7ftl/VUnhXpuYnQwHULMbn7X2/nKqekirGxwAECqkhTj0/Qe73jVuWpw6btQBEK5PShxfyq39gwlLlcITAn1len1tb6fUjj2mqT0oxw6FnCfLvAS+IZiI09/84REEI5H9SP8AH34Z49aJoehkw3f3X+aZBT15QSGytI0YH+NwSPeiQOtqK+fQlUG7tvZK32uTpWJvYNPCt7f67e9jPXjjpwyFfBDQT1EdRCwRCQySoR/sCD7egXVIAfPpqZqIT037cgCU8lSCD90Vk4N7/wBb+3rtiX0ny6btgNNR0pPaPpT1737r3Xvfuvde9+691737r3XvfuvdePI59+691Ud8jMRtr5CfJ7EfHev2xi6qn22tJu2arlhEhdo4kq51kZASL25v7kLZbSw2rl2TebmBZTLVO7546j7e7ax3rel2eW3STwwHJYV6tL2ztPAbUx1Fj8FiaLEU9LR09KKehhWGILBEI1Fl+tgLXPJ9xvFa21uW+mQIGJNAKDPQ4t7O1tFCW0YjAFKAU6U3t7pT1737r3XvfuvdcJHEaNI30UEn3sDUadaY0BPp0kMXA9Zl6yslH7QOqH/A6rD2YTuEt1jXj59I4aPMWOell7LulvUSsoaPIQmnrqeKqgb6xTKHQ8W+h9+690U7tT4x4bcXmy23UWjr1IdKWFGAci540/X8fX3qma9e9B0VWg3D2Z0llminStqKeKUXheJ2iW8lzfULfQe/YB68cjHR2+sPkFt7e8EMNfNHRZJl/cRjHGitq02sSPz7t50HWgfXoVq6Rs9VxUtJKj0QJE8kUiuAVP50E+zCIC3j8Q8ekEhaeXQMAdK+ngjpoY4YwAqKFFha9ha59oHYuxY+fS5VCig6ze69W697917r3v3Xuve/de697917r3v3Xuq+vl7ifsqnH5+JdUv3UZ+l9IUrzf6fi3vVPPrRzw6M30FmBmuu8ZUlrutkccXW0ShQbe99bqKY6Gr37r3Ravk1uEYbZM1MJNMlfDLGguBck6VIv/j70RXrYxnoNfh/t2ShxlZmJo2D1qyWZlI4Yg2v9PfvPr3l0crMYPEbgopcdmsfTZKhnUpNS1UfkhkU/UMvtRb3NxayCW2cow8xx6alhimXRKoYfPpj2x19srZhkO1dt4vBGXV5P4fB4tWrlgeT9fam93XcdxoL2ZpafxGvTUFna2tfp0CV9Olj7L+lPXvfuvde9+691737r3XRAYEEXBFiPfgaGo60RUUPSImSXAZETqzNQ1DapjzZGc/T2ZDTdx0PxDpEQ0Enb8PS1jkWWNJUN0dQyn+oIuPZcwKkqfLpaDUV6hV+UoMZBJU1tVBBHGpZvJLGpOkXIAYi59662M8Oir9lfJ/C7caWg26RXZCzooskg1W9LDST+ffuvUPRTanJ9q9z5M+Fa6lpJ5LeNElWPRJ6SfTwfegPLj17HkejKdbfFfHY9IcnudzLX3R3gkR2LMOW1aiLX9+GRnr38+jd4XbuH2/TR02KoYKRI0VLxLZm0j6sfe+tAUFOnoi4IP0Isf8AY+/cM9bORTpGGR8ZnX4/yaZLLf6ayPp/j9fZlQS2nzHSGoinp5dLQci/9fZb0u697917r3v3Xuve/de6qw+U1PF1V3j1Vu+ipk8m5dxRmqqkDB0tVANrkPA9yFy+8+57Rc2bSELGuF8ugRvgWw3K3uVXLtQnq0LH19Pk6OnrqV1kgqYklRlZWFnUNbUvHF7e4/dSjFG4joaowdQw8+pvuvVuve/de697917ri7rGrO5CqiszEkABVGokk/0HvRNMnrRNBU9VhfJz+ar8bvjTuiDZ2589PVZx/EZ6fFLT15p0lNg86RFig/xaw9hG95st4bprS0jeZkFWKgED5V9flx6hLnf355D5Hv12zdLoCdqdi97AH8TKoYqv9IgD59Gg+NXyr6p+U+0juTrjMxZGnWO9RDJNTedG1+IhoYjcEH6gi49meycwWe8hjBVXjNGVhRgfs49Dbkrn3l3n7bP3jy/cLPHkEqQcg0I9QQeIOR0OeDZqSqqqOT9Tzvp5+liePYruaPGrj06FcAMblPU9Kz2g6W9a4f8APg66mfYuB7Ej26meTG7ix008L0s9UBT0MKvI7JECOBxz72Pip69NSntr6UPQf9f/AM7PF0vU+y9ldT9f33Ntna2F22cc+GmSjNbQUCUilVAH1KEkAfU+96SOPXvFUioBr/Pqwb4d/IL5DfLqh3Ttfvzrah27sXcG3a+ijraegajkb72FkQAcXNha1/8AX914Ggz04BqFSKU6qQqv5OXdG0fnd/ePatPkKzoSnzaZ6mKx1aYoTibz3aMenWG921dtDx6b8IiTWvDjT59bCHyV+H/V3yT2xtfAb/pKOCsw2MoqOiyrws1XSy08SJG0RW3Kkcaj/vXtRAC0RAFaf5um5aCQEmlekL05/Lb6G61vLubFU3Z1Qlmo6ndVJ5moyo0x+IBzcKPpc/09paZz0pFB8PRvJOkuqZqHG4yfY2CmoMPUJVYqllpS0VBPGLJJTLf0kfj375de+fQnRQpDDHAg0xxIscagkaUQaUUW/AAA9+691l9+691737r3SUzo0V2Plt9HBLfmwP0v/T2vtjWJl6RT1Eqnj0qVYOoZTcEXB9oSKGh6WA165e9db697917r3v3Xuve/de6xyqWjZR9SLD3ZTRgeqsuoU6TeEe1ROmok6m1Xv9Qfx7XXWYwekkA0vQ9Kj2X9Leve/de697917r3v3Xuve/de697917r3v3XuuOtB9XX/AJKHv3XuuLTQqLtLGoH1LOoH+8n37r1R1gavoUF2rKVR/jUQj/ez7917qRHJHKgkidZEN7MjBlNuDYj37r3XP37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/U3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdI7F6qjLVpIBFPMV5/A/1j/xHsymOi3A9R0Xx/qTk+nSx9lvRh1737r3Xvfuvde9+691gqpvt6aon4/Ygmm54H7cZfk/7D37r3WkR8gPk31lhf8AhQZtXcfY2bziYTF4Cjp5cfjKcVmMSOkxQgi/eW6ix03ub+1todEur/Vw6Q3NSoPnSn8+tlPur559DxfHbsLM7a3jHS1lX15uim295qqioqqPLPh5YMdFaaS+rWVNwD/re0jElzXjXpYKKoPl1VV/Jt7V7P3R8Ae4+y+w9y1qZ+u7hy+3ts1WcyUdJFJFOpFKtPU1RVW1M40qpubG3vTAAmgpw68hLIrk1rX/AA9Xn/ELF782z09AvalWqZqqzVfWxVFVVIyPjqlUakZZ3OkqedNjb3UdXOejU09ZSVa6qWqp6lR9Wp54plH+xjJ97610WnuL5Y9V9J7j29tfeOQaHK7jycWLoIYpoQxqZnEaCRH5AJI9+698vXozUMqzwxTpyk0aSof6rIodf94Pv3XukxuiYxx0cegSLNPoZCbAg/Qke11ktSx9B0kujSmadTI8DhqimjE+Ko3uguHhVvqOeTz7SSZkJ6fjACADpPZTrDZWUiaOXA0KlhbWsX0/rwT7rXq9OgjzPxZ2FlvIVBpnfkeOBQFPNvo3vQrWvWzQjoG8/wDESogUrgMnUKyX8ZiLqCf8bnj6e9HjXrYwKdBvkejO3tsxiWmrsi8cf0ZGlbUDcm5BPPHvdPPqpOdPXDbWS7ZxU0sOaFdJSR3UeQOQwHNyD/re1dmg8XWfLpPcsRHQ8D0pKf5Rb523VyY2qwzNQwSaI5GpdR0H83K/8T7ZnfXKT07CtIx6dCpg/l3gJWhGbienRiokKxKpFyASL+2c9OV6GTB9/wCwM5pFPXGIv9PI8QH1t+SPfqjrdCPn0JWP3ft7Ji9NlKO3H66iFCb/AEAu3vfWvKvT8tVSvYJUwMT9As0bX/1rH37r3We9+RyPfuvde9+690x7mr2xe3M/k1Nmx+Gydap+lmpaJ5x9f8V9vW6eJOkf8TAftPTczaImb0BP8uqwfhhSVHaPcG7u+shGz1pfJ7eNRIjF9ChoVXyL6T9Pcgc1EbftkW0Jww1P8vQJ5are38u5kZNVr1a17jnodde9+691737r3XvfuvdM2cqhT0MygXeZGRB/iR7U20euQN5DpieTQlPXrrARtHjKfyC0hBL3+v1/x9+umDTHTw61bD9PURQnp69pulHXvfuvde9+690k9ybL2/uikqKXKY+nlacczmMGVWtbUDxf6+/dap0Qztj45ZHa038U2TVVCh7SFYFcEaQWI0i/5H4Pu6IzA04dVZwuPPpOdd977s61mTE7opZpKaRkSWoqIvXzIObuL/T3UuxFD5dbEYB1Dz6P9sfsvbe+KBKrHVsCyaAXilmiRidWk6VJ/wBb3UZ6tw49CGCCAQQQfoQbg/6x9769137917r3v3Xuve/de697917r3v3XuirfKzDiv2TFUCIO1O8rE6STZdLDke918utUpnpN/ErcP3eBlxIkv4GcslxwY0I4HugPl1amOjm+7da6ry+W+eky2Vxe24ZOYKgR6EIJb9y9tI59686de8qjo1nRuFGE69xFO0QjmZS7mxDsCi6b397rXr3DoYffuvde9+691737r3Xvfuvde9+691737r3XvfuvdMeeqsVFj5/4lVU8MIUteSaONrqL+nUfbkUhicOPLqkiF10jj0WTe3yApto42ogxFq2aPVFAVCTWJS62IuPr7U3aY8VOk9s9CY2yeinT5Ttrt3IOIBkKaiqZVICK6LpkOi1lt7RD59KyM0HRkOtvixRUaJkd3SPPkAyuUmRpGJDaiG1MOPp7117jx6Nxhtr4LAwQw4zG0tL4UVQ8UYVyVH6i3vfXvKnSg9+691737r3XvfuvdJ3cUBemjmRfVDIJGYDkKvq9rbNwHKt5jpJdR6lDDiOnLGVgrqOOccX9J/11AufaaVDHIVPT0LmRNR6cPbfTvXvfuvde9+690S/5tbFhzvVeT3oEV6/YdHJlaH0s0iuJQS0QH5HH19inlS7aLcktfwzGh6DvMtssm3vcfiiGodLr4k7sl3d0BsPPV816uooJlq2lZVZJYp2Wz/09On6+0XMtutpvM0QwAelOwXP1W0wzsckZr0P1TuHBUaM9TmcVAqAk+XI0kZsB+Az/AF9h4zwrxdR9pHRk91bxirSKP9sOgQ3T8oOpdoNKmVzZdoTZxRiKqF+T6TG/P09k11zHtdoaSvX7M9B295x2Ow/t5eHpQ9Fl3j/Mb6sxKzDBxZqtkSwS2NeQObWvwp/PsO3nuDtcGIQzH7OgXuPu5sVsp8DWx+Sk/wCAdApk/mb8i+z8Rk6PpzYUdfLU0tVFBNWULQyhHpnjYi6/UEg3Hsnm5t5h3CEjZoAxIPEeVOg/N7g83b1aOvLdnrZgQC40+R+XWnp8uet+99tdu7hqu6cHlqLdGbyMmQkkqKaoelmWoctFHQzFbFEWyhByLcj8+0Ozz29vEbSasc61aRXoGLHJY5yCfMYHDrmDz/tvOG0c13dxz5CyXt05fWKmJgfhWJiAKIoC6fiAGRmpvZ/kJdUdt43P5zedfHmKHY0hTTT1cFTT01QyyhHmRZgAQxsOP6X9reXwb/md7uw/solCyMD2s35YNOH/ABXWW33O9i5hikut4kDxWMj9sbKy1KgBnFQMMcCmDpr59bSMi+HcCSEkK5vz+SR9fc1DutPmOs9zi6r0rfZd0u6SG8tg7M7DxbYTe+28VufEsWLY/LU4qaZiw0teM2vcce/deGDUdBdg/ip8bttP5cF0j1vjJfIJfNS7XxqzeQfSTysha/8Ajf37reo9DPidu4LAxLBhcTQ4uFVCrFQ08dPGqgWACRgDj36lOtdPPv3XukzuCEv4Gv8ApkU/7Yg29mFm2COkVxhwelBBIskSMv00gf7ED2hcaWI6VqaqD1m916t1737r3Xvfuvde9+690nNwovjhka40EkMADb2stDQkdJLpdQFOnXGuJKKnYc3T/ifbEwIkNen4vgHU721051737r3XvfuvdcDJGv6pEX/XZR/vfv3XusRq6Vb6qmnW173mjFrfW9z7916vSPp8pi4MnJ5a+iT1va1XCQQebmx9r5WBgGekEWoT5Fa9ONTvHbtLfy5Sj4v+mphP0/5C9oOl/DpiqO0tnUwJfKQ2FwbSw/gX+mr37hx6950HSUr+/tg0AYy1hYKSPS8V+PzYE8e/Ch61npI13yr64pFNp5mezWIKEAj/AA97x16jdIis+X23LsKGKSTk6bxA3/p/t/ej8ut0xnpIV/y+ydycdjGkvcKv2qkfXj6j3Wp8+t0r0l6j5Y9k1ZaOjwJVSGAZaVQT/Q3C+7VNetY6T9T3X29mf81jKhGNydMJFr88ce9cOPn1uteHTLUbi7vyagxQ18YZiDpEg0g8i4A/4j37HHr3A9ScftzuzJVEbyz5IRsSSD5OLj0j34da4nqxvrChymO2fjKbMM7V6reYyX1X0gc397AoM9e1aj0IPv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//V3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdI7boX+I5cgkkzfkfm/sxuxSBPs6RQU8Vj8+lj7LulvXvfuvde9+691737r3SQ7ByYwuw965c6v9xm09xV4Ci7FqTETTqFH5JKgAe/de8+tRz+WX8auvvm73x2n3b2vhTLmdvbpzWBo8hX4xzWaJHkSBIZmAUWCjTf2qQFYda8R0jcI0xD1ySB1C/njfy7upfj98ddq7+wPae+MTWZjtTHYSHDxOIqWpiyitM8EQic2VCoVgfqGH9D7TgktnpRKqCMlcUp5/l0g/mLt/O9Mfyvvit0N1/XV+3t0ZnubaG9cw2JeSkyFfhclTQ+Kqq4h+4b6y2o8c+/VJavHj16hSPQDQ1Ufzz0bb5F/Lj5O747d67/l3fFKNs/mcH1nsnNbky9U8IyUqHBRVeSmkr39SpEC5N2sPflppq3VW1atKeea14Z9Oi8d8/Oj+Yv8AyyOxepOuM3gMdnNpb/3nt7E5GbKQU2WmkiyU8UNT4JpA5UDyEKyn/W9+pUFq08+tl2WRUI1AmhIPD8uuP80nv/PVnzh2ZtbaqVeVz2Kp9lbn/hVJD99DS5LKUUFfJE8CBiqq5tz+Pe1FVJ8uvO2mVQBU+nVrfxV/m3Um/O/ch8d+58VJtfdePxmPipnjooqSklyDUqnS0kuks0jfgHgngfj3UjFRw6vqGrScHq6fcIE8uKC3YtOrqtr3DEc+11mdKuT6dJ7kamC9JrfXcPXvW8cC7n3NhqKolcRLRyZSgjrARYanp5JAwH+JHtCcmvSkYFOlBtHf+0N8UcVZtncWFy/kTyPTY/K0NbVU4H1WogpnZkI/IYe9HHWx0sffuvde9+691wdEkUpIquh+qsAQf9cH37r3SNzWOxMlVSUbY+kvOSS3iUE82tx7XWq0jZ+klxl1p1GyXWezMnTtDPg6EOwsZlhu59Om5ufaImpr0qUECnQS5n4ubByytZWpmN7eOFbA29PIb8e60zXr3l0DG4/iNUwyE7cyVQoXhShZL8f6/wDX3s8et08ug1yXRXbW2l8lNX5BkWxVleQhgvq+tz73T061w6ZYc53TtiQSyJXzinOn1CRtWg6vpb/Ye64Br1s56W+M+VHY+Jljp8xh3NPGFRnelDMFHDG7L735Y690K+D+X233MYzkEsFyobREqEXPN7+9V9evdddy/JvZEHV+XqqGu8T5ihrcYFkmhRh9zCYiBe973I+ns42O3N1uCqB8JB6Lt2m+nsWcmlQR+0dMfwJhwWI6cq6iOpoqd8juKqq5dc8MZIn9UJIYj63NvZnzfdePuoUmulAOizle2EO26gPiYno9KVlJJbx1VNJfkaJ4muP8NJ9hXoScOPUkEHkc/wCt7917r3v3Xuve/de6SGddqmtoKeKzeGpBlH+xB5HtfbgJEznzHSGdy8ojHl0rgAosoAH4AFh7QdLhQcOu/fuvde9+691737r3XibAn+nPv3XukhEi5fKNJIBLBSsUaF+Yvpa+n6H2YPSG30ji3SKNjJNq9OkXv/pXaW+qWRJ6OCjqSjaJYYQLyaCqk2It/sB7LwM16Wnoh28Op+wuqciK3ATVz49GWSKODWY3jRtbAqOB/j71Qk14de8qHoaOrflHxT4Xe8T0s8OmnEjqkbkl9KkswubD+p9+BHn1rPR0cNuLEZ6miqsZXU9QkqhlVZo2kAP0uik+7UPW6jp796691737r3Xvfuvde9+690FHcuEbObJycKpr8FPNKRYnjSB9B799nWicU6JZ8WsumG3jX42oYxhpp4wlwvIuhHP+Pvwpx68SeA/1cOrIqmpSmppap/0RRGVv+CgX+vv3Wz1VlvOsk3l3kPHaWmTID0X1hQsl+Lc+/A+fWiM9Wh4ijjocZRUsS6FipoRp+lm8Q1ce/fZ14dOXv3W+ve/de697917r3v3Xuve/de6gV+Ux+MgkqK6rp6eONSzeWaNCbC9gGIuffuvdFZ7G+UOA26ZaHb5+9yQuiACOVdRU2ItcfW39fegQevUPn0VCbM9sdxZFo4RX0tLUSAkxo6L6joIFrf19+NT14UGOjJbL6EbB4Rzu1fu2Z4nc1CtLIGElwDf6cfX2vtmMiG3Pn0jnUIwl8ujXYDbuCxFBSR43H0kKJDHpkjhVXY2vqLHn6+0bjSxHp0qUgrUefSk916t1737r3Xvfuvde9+691737r3WCpjEtPNGRfVE4sbfUrx9fdkOlwequNSkdJjbk5hkmxrLYxF5P6WF+P949rrxAyiYefSW3qrGPy6V3sv6Wde9+691737r3Qf8Aae3k3XsDc+3pEEkeVxslK6EFgwdgbEDn8e1m3zfTXkc/DSa9Jb2Hx7V4f4hTqtzoqXfdfi+yuotlERPtKneClVpBCIZJJ/EDFexHJF/a/wBxLS9urGK7sfinpU+fD16AcUe4zWk+2bdgxig+RPp02Yv+XPuPfFR/Gexu197YevJ1vQYrITGnJL6rABwtvcGj28mv38a9upIz6KTT/D0DYvaO63NvH3ncLhG40RyB/h6MVtP4EdabaWNavP5/cRT6tl9E2s8G7Xc/09iO15I2+3UB5Gkp/F0LbD2w2Wypqkean8ZrX+fRisB0L1RgKfwxbI25WPf/AD9ZiaWokI44tMGH4+tvZ9Dse1xLTwEJ9SoP+HoX23Lmy2qaVtoz8yoP+HoQcZtLbGF/4tG38RjOCP8AIKCmpeD9R+yq+18VrbQf2Mar9gA/wdGcVnaQf2Mar9gA/wAHQfb76A6c7LqIqze3Xm2NxVsDI0VXksck0yaTcWa4/wB59orvZNpvn8S7gR2pSpGaeleizceW9j3Vg9/axysDUFlByPPpcbT2RtLYuOTFbQ29i9vY9FCilxdKlNFYfS4Xk/7E+1VpY2linh2caxr6KKdGFnt9lt8fhWUSxL6KKdc8zGBVQy3sQAL/AEHB9nNsaxFevT1EgYcOlHEbxRn+qIf9uvtE2GPStfhHWT3XrfXvfuvde9+691737r3TNmkDU6EsFCv+SB+L/n2qtTRz0luh2g9ZsfPEKSMvND/riROB+L3P19tz/wBoadOQn9ME9SjW0Y+tXTD/AF54h/vZ9s9PdRJc3h4P87lKBbfg1cF/9sD791omnTTUb121TX8uWoha97VMJ+n9Df37rfScqu3Nk0l9eVhYj8LLCSebfhvfqjrxx0gNzd+bFipWQVZdrW9MkTDk2+gv7et5RG+ojpqVPEXT0lE+V3XuOoY6cvUNURrpsNB/24t71M6vKWHn1uJCkYWvScq/l9hNRNFBI6/i8Kn6/wBOPbPTn29I7IfL7OWb7DFM1+FIpFJP+PIPv3n1qvp0mqj5Vdn1oApsIyxkcFKQKxB+lyF92r6deGemKq7i7kzJumPqE1AadMLjj6ckD+h/r71wz1vHDpmqMp3flWKiOuiSQDUyiQEc2twPdRnr1QB1wpdndxZCZVaevWZrWB1i9zYk+3aNp1eXTepCdI6VUPx77cymmWqrcimoBrF5FHq+p5PuhFMr1dSeB6d6b4p75kkD1uWrSLnUPMxvf6/2v6e9Z639nSwx/wARpJCr5HJVJa/KsS62J5/P+t78OvH06WtJ8PdlhVNZXTyNcMQsZNj9SPUw928+tU6VtF8XOvqEqUR5CpU+uFebfX+1+fdadb8qDpY0fRmxaPTpxsEmm36qdbm3+N/e6Z695U6VlL1vsulXSmAoG4tdouf94PvfDrRqTXp3i2ltqBQsWEx6BTcf5OpP9fq1z711unn1OXCYhP0Y2iX/AFqeMf8AEe/de6kpQUUX+bpYEt9NMSj/AHr37r3UoAAAAWA+gHv3Xuu/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691/9bf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcJDZGN7ek8/wCw92XLDqrmi9JfboU1OTdfzKASfqfa28wqDpNbLQknpV+0HSvr3v3Xuve/de697917pvy2NpszjMhiaxQ9JkqKpoalCAweCqhMMqlT9bqSLe/de6LT0/8AHrrX49VuRxfXWJpccm4sm+bySw0q07GqkkLlyyGxvqPtfAv+KsW4dI5hquAV4jorH8z34JV/zw2JsDYjVr0mL2lvjG7tfS9hI1PInlUoXW/pU29oASD0rKqwz6j+WR0WT5X/AAG7W7m+RPWMWFxNM/Tuw9lbK29DUSxlKeKq27RQ0j1TqrABrx3Lc+9hqCnWnUswYfbTogm691n4CfzWcr3XvbFVO4+tanZFLtOPK7cpzXVjtLjEx0iGoS4HjY8o/wDS3vYyKDqrHQ1T5j+Y6CDtf5m7A/mOfzD4OmdtbcylZjutY8fvWhXcFC0zQw4wJXSPEmkBSqobLfj37gKHz60Tqbs/DQnoy3wgwdL2z/Ob723ZuTa8We2xjuuXXEy7gxS1dLSVlBRw0VPLSGoUxh1IITjgcjm3vXBQD8+rqSzs3oB06954TbXc/wDNErNgbU27itv5Lr7P4+vyGQ2/QwQVlcmOdGm+9anVTY6TqB4Hv3BevcZV86dbNGYrKbGtSV1dPDS4/GRUtRW1c7rHFTRainlmlfhVH1JPtXFQW7A+dadMSg+MCPl1q7/Jmp+K3afyw3xVbs7S7RaliyWp6XZsz5jFQhalvIKcwHSAD/QW9o80+3p9ipPzA4f5+i+fymu2azD/AMyPtnYfWG7917g63OSlx9ZDuyqkP2GGknWP7l6eW4jcKBz7sRqAHn1RSF1GpK/4D6fZ0YfJfzyO4t2/N75OfGrq3bAyuH6Nz9VhoalMPHVCZKStNF51qEiuwci4Nz78Kaak5z153KyFApIoDX7erOdy/wA27pHqfYWwZuwv4pnOwNwUgGfwO2BT1VRhay/1r4YrtHqv+gqLW+p9+pn16uWA+XRv/jH80+m/lTR1cnX+U+2ydAt6rBZWopocuhBAfTSXDta/Nh7qcGh62pDCoz0usJ3FsDfm/wCs2vtjOUOVzm1q16LO0dFXU9XLQTRvpaKpp4iTGQRzq9r42MdsyngekkgBuF6G85nELWLj2yuOWvc6UomraYVbn+i05bWf9gPaDpZ05e/de697917rg8cco0yIrr/RgGH+2Pv3XumypxGEeNjVUGP8f1YzQwheRblmHv1OvVp0kK7rvYueik0YvFurFkeWmSKWzEcqSCR/sPfvl17oLc38XNgZgswVqZjcr44EsGP+s3vw68eq1Pnl0EdjbB29Lh8lOseS3RHjoo9ZjVvLIqp6T/W/sZckQGbcpWI+FK9BTm+4MG3pnDOB+3oUNmfHbtTYnXuGgoKqqRaihoMkqU7sReaAShWUE2bnkH2Gdyma53F5j5Ej9h6P7CIQWCReoBx9nThT5jujbMgkeOvm8fBDK7AmPjn/AAPtDjpdTpY475R9k4aVabM4uQwx6VLNT6jpHF/Up/3j3sk9VHqOhdwfy+25ZEz0E0Uj8WRFjIfTe1iLfX3uvqOtfZ0MuD7+2HmlUx1ggZremSWK/Iv+SPfuJp16uK9KXD7kwdfmamq/iFKkUgBh81TGoLWvxc29mEhpaKB0kjXXOzHoQEraOT/N1dNJ9P0TxN9fp+k+y/jw6WVHUoEHkcj+o9+691737r3XvfuvdNmYnNNj6iRTZtNl/rf/AA9v26F5QB0zO4jjJPULb1GYKX7lz66sCVgfqCT+fd7p6vo/h6rAgC6x59KD2l6UdRKugoq+MxVtNDUxkFdM0auLMLEC/v1evdFa7S+NOE3P5MngEWjyl2aOKKNY0DeM6SGB/r70RjHXqZr0Uqmq+0ej8wGZa6ppaeZVKurtGqJ63+tx73mmfLr2PLz6OX1j8jdubwWnx+Vmjo8tIFDa3SOLUTYggge/HrQ4Z49GUgqaeqjEtNPFPGwurwyLIpB/IKk+/db6ze/de697917ppzsC1WGylOwuJqGpjt/XVEQPfuPXuqsdsrNtPuZ6ZyYoZcg9lPpPql4vf/X91p59b49WWb3ykeO2TlazWFLYo+IltNy8YP1P+B928utefVdnQODm3T2XW5VrytHVzSXN2QKrXvquPx70B5der5nq0pRZQP6AD/ePe+vdd+/de697917r3v3Xum+uymOxsTy1tbTUyICT5p44zx+AGN/fuvHor3Yfyi23t0y0ODJrMit1UjxyRszCy2FiPr79WnXqV6KZX5ztnuLJMkYr6SlqGspjWRUZb6TyLD8+9D19evfL06MR1x8VaKlMeR3fJJPWoAVSRA7Mdd/Vc8cD8+9jrVBXo3mF2vg8DTQ02Ox1JAIhYSJAiyMb31Fvrf3smvXqdOeQp/uqWSEAEtY2P0Nvd4m0SBuqSoHQqemzA1KyRywE+qF9FvrbT6SB7UXaUIYefTFqx0lW8ulB7R9K+ve/de697917r3v3Xuve/de699ePfuvdJCEfa5+drf51AP8ADni/swb9S2FeA6Qr+nNQHPSv9l/S7r3v3Xuve/de6xyxrNG8T8q6lT/rH3sGhr1VlDKVPn1XD0pMuyvkf2ZSm0aZyqaDTfSJT57r9frYgEexzuoN1sUDfwDoJ7cRa7xPEPx9WRewL0Luve/de66JAFyQB/Umw/3n37r3WJqinT9U8K/8GlRf97Pv3XusTZCgXhq2kU/41EI/3s+/U61UdQ5dwYSC/ly2PW31vVwm3F+bH3sCvXq+XSI3NvrbFNThmydG5RgbpVRXHNyLe1VqRqox6TzqWFOog7i2TTwReTJwi0a8CeFjwv0HIv7YkADnPTyV0jHDpL13yQ6+oSytUtIyXuFlj/H9Pr7b6tXpH1vy569p7xRioaext6lZbj/AL78aDrwqRnpGV3y/xUmoYymma17aogf9b8c+9VPHrZGMdJKv+XO5XBGPxTsRyo+2TkA2HOn8+91PXiKZ6SNd8lu0Myhi/hMscRPp0U+n/D1WUe7KzA9nVSBTPn0zjsvuPJKRTUdSqH/UxtY3F+LD3ViSdTefXlWg0r02yzd3ZQ61FdGWNiAJf63uPesU6tnpwo+vu5MsNUlTkAzEagxkH6ufqT791oHp/i+Ofa+Wsaqvro42vZvK683vY3PvdPIdbFRnp0ofiVvPy6qzK1DK3B/yj/bAjV71mvWznJ6V0PxKEETT1VdUysF1MrMX+nJFgT7cjQO1Hx1R2KqWHSxwvxL2RNTCWsqZnlb9SiLVpP0+pYe7Sp4bU6biYuKnpdUHxl6/olVfCZNP0vBGB/seT7Z869PY6V9D0lsahtpxlPKB+JaeMi97+9jr3nXpVQde7OgC6cDj/T9P2Bb6W+nvwPXj07w7X27B/mcNjk/1qaM/72Pe69ap1MGIxagBcfRqB/SniH+P4HvVOt9Mc8NJBmqWNaOnS63BSOxPI5449ro0Btmp0jdqTgnpWjgWH09oeln2de9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/9ff49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdR6ptMEp/2k+7x/GOqP8J6YsABqq2A/Uyk/j8n2puxTT0zb+fSl9o+lPXvfuvde9+691737r3XvfuvdI2sQtuinvbT4FP8Aje3N/ZjEf8TNPn0hlH+Mg9LEfQf63su6Wjh005+gfKYPM42J3ilyOKr6GOWNijxvVUrwK6MLWILXB9++XW+BqOtabtXpj5pdDdobtaXY21exelswmRbbuT3FjaTceUTLVDeSmBMpkeMqT+fx7920+fWiGr6/4elt/LV/l69lYf5Gbo+Wvduztubfrd2YLIYmipsZj6ehqZEqqZoY5DFEQQgBH9mxtb3sngB1VFpqJPGn8urxOvPjr1l1ju7L732vgqOj3Hm6Y0mQyKU0UdTNCzairSryQf6e69XwBQdF668+AvXewvlhvv5XQ5KryG697Q1UdTj6qHVTU71QIeWOQtybm/K+7VNKdap3VApw/l0YzuraNX2P1z2T17j6mSiyO79uSYWgqYZPFJBIwN5UkuNNh+b+1RSlur+lf58Okwf9cp54/l1rA9N9IdrfAruDsTF9ibJ/vy+8KepoNj1+RxH8aiNZWVB+2qZZ2DjTY3vf2lND8qdPKrJUVrq/1U6CD4AdQ9z9Od4fzI/k52ntV8Hj8XsGvy+2fsMY1G38Tq8ghp/4eBxwpP0HA97OKAeXWlUsG1YJI/YB0dr+U7sHGba2R8rPmjU9fYnK7n31inylHNnMDFNX1tdT1bVbRtUSqS4aQKXKgE2+vvT0B+zq0VRGT5segS/k09Tdb95/K75idl9rLj9yZiTKrX0e0M+aaqxmFkrcsVnahoJ+IlQHSAALcf4+9sNNAOqp+oGduNafYB0FO95I/id/NP2DnOgdwS5LYeT3zJN2VjcfVxthcVTyVMgqKKaGHUscYXgBgPezTT1WhEwORUf8Ueje/wAjfbOdyfy4/mKdpZvMZTKUOc7Enk23HW1Mk1FS0+SzVRUOaS/FgihRb8H245ZYwvr/AMX1qMrJI7nNG/ZQAY6Y+rPkvv7sr5+b030+WyeQ616S3blKTdePxdYamKOClmkVVaGMAKCV/I49s07R09q/UNOAx+fXo/5xPyD7K3R3BvjqXH4ifp7qDdgw2fw0tPRVO6JKWWrMEJo6dYzIxCqbkA2/PvxGR8+tazk0qB1db8dPm5snur4/ZXuuoxmawlJtGkQblpspTCmqGrNJbRSowW5Nvpb36nVga5HRXsZ/Od+OVVuKtxFft3eWIoqTJR4x8zXRLHj/ADSyeNX8siKtj9fr9PfqEdVDCuTjoT/5gXy0p+sPiBF3V1vmaOtjz+WwlHi6+krYZI1GSjkbxvLHqAYWAK2+vuuSQOrE0Qt6dCf/AC+dzb93n0PjN19gO0mS3CaTLUcjPr10VZTmVGDWFxz79ipp1sfCK8ej1e99e6qn/mItNu2XYmyaaWQy0m68XkTDE7agvlRiSi/1/r7kTkilr4143mhHQF5v/wAZ8G1HEOp/n1Z1tynVdtbfp5UV/HhMVGyuAwulBGpuD/rewBMazOfVj/h6G0QAjUegH+DqZNh8VUKVmx1FIp5Iami5/wBiB7a6c6SmV612fl1Inw1EhIsWSnS/1v8An36vWugnzPxd6/yztLokp3N7BIUsDe/FiLD34Y69ToF838Rp4KxZ8DkKhYo3LAeQJdQbjjV/T3tfjHp1pwCp6Ru5+pOzMfTwQYapqiKb1F0c62/s/UH/AHv2ruyqqFXj59J7YKxLg9I6nzXdO1ZBrjrpghHLK73/ALI5IP09osdKqA9LPHfKHsbDyJT5jGytEgsWNPqawsBYlfe69aA6FXB/MDboKx5ykqUdjbhRHybf7T/r+/Djnrxr0MuE+QWwc2EMVaIC/wBBNNEP6fk2/r71Xq1OlNWbuwuZqKGGiyNK9PLfyuahPGov/aI9r7Wixu58ukV0NTqh8+l7SVVAIo4Ya2ll0qFHjqInv/rBT7RsSx1dKlAVQo6cPr9Pderde9+691737r3Sb3DtTCbmo5qPJ0NPKswYGUwoZQWGkkMeffuvdEf7Q+MFZi3nzWw5Zo/FepZYwFlVlAIC6TewI/p7916tOPQe7H7w3x1lWJiNzw1b4+ORI5ZahTyqGznUwP8AX3rNMdeoej6bF7a2pvmkimoa6CGdkTVDNPGGLtxpW9vdvs699vQohgwDKQykXBBBBH9QR7117rplV1KsAVYEMD9CD9QffuvdVcd3Y1tvd1wZJLxUclchH9kAGW4596J9evDPDoznd286FOqaN6StgaatoI4GjjmUuCIFXkD/ABB97NOvcM9Iz4h7clo8fV5qWI/5V5tMpX/VkfRv9b37z60OPR4Pfut9e9+6903ZDLY7FwvPX1tNSogufNMkZ/wABN+f9b37r3RXuxPlDtrb3kx+FY1eRDaUdGSWMm+k2AB4v+ffqjrxB6KXX53truHIvFGtfTUk7jQ0QdVI1WJJFh9D71TPWwQfy6MT1z8VKOnEVfvKR56tAHRSodibhhru3vY+fWjU8eje4XbGEwNJDSY/H0sKwrZZFgjEh5vcsBf/AHn34568MdKD37r3XvfuvddH6H/W9+690isehx2bajN/8pJlN+Ba1+P9j7M5SJbbWPLpDH2zaW49Lb2WdLuve/de697917r3v3XuuiQvLEAf1JA/3v37r3WFqqmT9dRAtvrqljW3+3Pv3XukllshjqathqDkaFdbIraqlNQ5/wAL+10DgxlDjpHMn6gfp4l3LgIFDS5fHqLD/lJiP+9H2i8+lfljplqew9pUpIkzNDxf6VMX4/1z7117pNVndOx6JmV8lDJp5JSeE/8AE+/dez0lK75K9fUJIknkcA8lJYzYf1t71UdbOOPREMr2FjIe4f7+4tZBjTXiqqAT6mjV9ek2HP8AtvYkG9wfuz6H8QFB0RNtE37wF7XFc9GLyny+o3ZjiKKoKMf2rxBvxyCdPPsNDV59HxA6RVb8tN5sLY7FSsxA4alB5I/Hp97rnPWvs6Y2+R/b2U9MeNnSNx9Eh0/T8cKPez6darTPTJVdidz5hiI6WsQW4tGwPPHFh71k8et0z02GHu3JyjmvRSDfTrW34A/H+v79jrVaHp3pesO48kR5KquUvYgGRweRa3+x97Gry60aE9On+y29o1i+Wrra3R/aDSMB/ttXuyozNpHn15mVVqenvGfErdMy+SuydSNTcKZwRY8f6r3plKGnW1cMtV6XNB8QqNihyFdO3PrOsNcfk2v7r59bPS5pfiL1/HoaeaokcWJtEv1+trs3vZ49ap0raH42dfUIAWmMmm1i0MY4H4PJ9+OevDj0rqLpvZFCVKYunk0/6uCP+nvQFDXrZoelBJsXaNPTSFcHjxoiaxFOtwQOD7diFZAKdNyGik1657ewGAipB4cVQghydRpkJJH59V/p7eulAkqBQdNW7MV0txHSmGNxy8LQUaj/AApoR/vS+0lB0p6ypSUsf+bp4U/4JGi/9Cj3sY4deOePWcAAWHA9+69137917qFkf+Ac/JHoP0+tvzb27D/ajpuXCHqBg7/bkXJA+hP1J/x9vXYGr59NW5Jr6dPntJ0p697917r3v3Xuve/de697917pH5EuM/S2A0hOT+ebezGGv0pp0Xyj/GQD0sB9B7LujAcOve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/0N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691DrzakmJvwv49uw/2g6bl/sz0zbeVgKksQQWBX+tiSbH2pvOI6ZtiCDTpS+0PSrr3v3Xuve/de697917r3v3XuklUBW3TALm/wBtf8W4AIHswTFkT8+kj5uQOlb7L+lfXvfuvdRauhoq9BFXUlNWRq2pY6qCKoRW/wBUElBF/wDH37r3WWGGGnjWGCKOGJBZIokWONB/RUWwH+w9+691l9+691737r3SRF/47MzE8P6Sp5A/P/G/ZhwtekfGfHGvTvXbfwWUlSoyeFxWRnisY5q7H0lXLGV+hSSoRiLfix9l9B0sBI4dJjdfWezd3bX3LtHIYPGQYrdlC2Pza0mPo4HrqZmDlKgxoNfIH67+/dbr69NPXHTWwurtiRddbXwdFTbYSOWKSgFNEkVSkvDLPGgs3v3nXrXlTrX47K/lT/Jr4/d5didw/A7cdDSVPa+RkrN10m4MhSY+npUnqDNJT0UNQ+nQNXFhfj3sH16oUJqVIFePzp0O/wAWf5S1ZsvaPcW8u5skua7y7ZSWsmqHmir6DCZWS8/mxtVrITVIbEr9B70fl5dbCmuc16rd+Ome+Y3wC3t8i+uJdpvWZvsTMtHsSvgxQq6ZWieXxVlTKAw1aXBU3uPamdSArNio6T27dzoorQ1/2OpH8rba+/8A4X9T/wAwX5SfMDaWYzmP3XumkFJicfjfPlq2XNVc7VJo0e+getWZwOLe2DTVRfSnTyglD4nGpP8Aq/LovPcewKT449Y5752fBHcmD/u3unNYvL726533WYyXPRVuVmM8sD7bnY+dFcsNRj4HvYphW/1f7HVDgNJHT1I8j/mPVgW8/wCZ/wBLbR/l/wCJy++tkZXA703nR4iu3TQ7eoIMPRVucryV89LTRoA8PPKgWH+x90ANaUr04zpp1E0HVevzP7lrqf4S4zcOT2ridubY3nl9vZjB5eDEQYzPTUtdHanL1yoGb6gnn3Zf29Vc9pHT18hcpufbf8rfr7Yc+Srchl97bx2PuvC0tTVNUVf8GnS6PFC1zpIa9wffhltXWmGlCvHgP59bd3xQwMW3fjZ0ZjkhEMydV7Hlql0aGNVUYCGqqNa/1DyMPdR0+RQkdGE9+611Td8rOw8Ptj5W4WlztPVV+Pix2PqDS0oMjq6Rq+vQPp9fr7mDk/Y5t05fcwsFJJyeo43/AHGO135FlBIoMdH+2z8l+tspS4+COqahvR0oC1kscPjtEF0ENzwB9fYOu+Td3t2YldVCeAr0LLffLKZQAaY8+heo+xdjV0Mc0O68B+4AdDZWjWRSf7JRmBv/ALD2QybRucbFWgfH9E9GS3lqyhhIufmOnao3TtyloKjJz5zFR0FLCZ56tq2AQRRL9XeQE2H+PthLG8eUQrExZjQChr1driFUL6hQfPovuG+YPQ2c3S20qPsHa7ZJZ2p1tmacpJKradEfHJ/w9iqfkLmS3s/rXtpNNK/Ca9EycybbJP8ATrIuqtOPRjqqrimxklVSzpLDJCHimhcOjo4uGV0+oIPsJRxss2iQUI4g9HMjVi1JmvULBRRS0IkeNXLsQTIgYkfW1m/x93u6eJQdUthRSfPqfPh8VUqVnx1FKD9Q9NEfzf629pelPSRyfWez8oLT4ijX/FKeO/8Atz78cinWgKdBbmfjB19l5GlMTwOSSuiJLKTf/Uke/AUFOt5xTy6BTP8AxCm1y/3fyE0KsRpPmCWu1yeWB96qa9ePy6S1X1B2BtDF1WPpKuomrGX9txIXIuv1DAm3049mAIS2On8XSQ1aYBvLpB0mW7p2kQzpXzGMXBYSOP68k/61vaAEjpURTpa4/wCTvZOHKRZnHVDRrYMWp7kC/wCC6n8e/GvXsVAHQq4P5gbeULHnaOoSV7DgCPSSf9a3096rTj16mehjwnyC2HmlRo6tYC9uJZ4xYE2vyB73WnHrfnjoSaLe21q7T9vmsexZbgfcx3/1j73TrVen5K7H1QKR1VJOGupRZYpAw/IKgm49668DXoN9/dR7U35j5aesoaennKSlKiCBFYyOosWIt/T8e91695Y6IdvTpTfvWFacztySrkoI5GkjSmJZfHFYqCEvzz70Pn1vy6Evq35PVNG8OG31HNE8WinDyAIylVsSWYX449+rXh16lOja1naG06bb0m4FydJLCsBmWAToZDxwpt791o4NOqve3uxx2LuWevphopYJpPAw/WGST02P9Le9U8z1YHoPchnc5k6OOgrauaSjiv40aYsAbW/T/sPfsE56rQ8OjqfHHubC4mgptmZACOTyIkUp9BLlP9bn/b+91NKdeAz0eSrzGLoac1NXX0sEIUPrkmRQQwutgTfn/W9+63w6LD2L8n9ubbWehwt63JC6xPEyzIXDaf0ge/daOM9FKrs/293FkRAFr6ekqHKq0auq21aluQB+CPeqjj1ag8ujC9d/FGkpSlbvCWSoqP1qrkSvqPNiGPHP9ffs9ayePRv8HtbB7fo4aPHY+liSEWEggj8jH6XZrX97690oCQouSFA/JNh7917qM1dRp+uqp0/4NMi/72fewCevVA6jvmMTH+vJUS2/1VTCP+J9+II49aqD1Bl3VtyEEyZrHLb8Gpjv/tveut9M9V2Ns+kYLJm6AkgnioT6D/X97p1UtTy6DTNdybJpcvT1q1sM3gjKMUqU9XPHAv7VRSAQtEfPpNKhMokHl1Arfk9sGgUmVncgE2inRvoL2/T7SYrQdKq4r0ja35h7HGqOjpalpLmzFtQ/w4Cj6+/Y4deoRk9JCu+Xqy3/AIbQVBHOkmFW/wBh+n3okeXW6HpIVvyx3xNcYvFTFubaqVTwPr/Z9768emSb5Edv5a/ix9SisNICQMi3P+0oPftRPDqtKHPHplqt9d15Ualpq1dfPEclgW5NyOL+9E9W48OmSoou5siyGp++VHICAl/1Hkm3u6AtUJ1VwBQydKKj6n7hyzKJKuuVnCkDyOB6uQBq91pU9bPqOn0fFzs6vbVU5Ooj1/q1VOm3/JR9+Pp1qp6f8b8Q9xxE/f5WWQEc2qVb/X/tEe9Gp6tQdLfFfETE+VDlayoaM28lm18fmw1e/DhXz60ePqOiydv7Fx3XHZWytr0mpsbn8glPUGQWJjaUIdIPsWbVs1re2Ety3xIOg9uG6XFpexQL8Dno/GJ+PXXsFNTSJSGU+ON1Z447epAbaf8AjfsKstGNehEGqtellSdR7LpNOjFUzW/1UEXP+vx7114Yz0pYdlbXg06MJjhpNx/kyD/Ye99aIzXpyiwGEhOqLE0CEfkUsX/Ej36vW+HUtcfQL+mipF/1qeEf8R7rQde6zLTwJ+mGJf8AgsaD/eh7317qLkwxo5Av14P1twP9b29AaSA9MzU8Mg9QsDK0tNLqYtplsLk8H8gX9uXQo46pbAhDXp99pelPXvfuvde9+691737r3UWt/wCAlR/yyb/evbkP9oOm5f7M9Mm2X1UTDiwka39eTzc+1V8P1K9M23A9KX2h6Vde9+691737r3XvfuvdRK4A0k9/wl/95/PtyL+0HVJPgPTVg3JEq/hQLf7E/Ue1N2KU6T2prXpQe0XSvr3v3Xuve/de697917r3v3Xukflj/uYpQP06Rr/re4I0/wC8ezG3P+LkdIZiBOD0rk/Sv+sP969l549LV+Hrl711vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//0d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691CyIvRzj+qW/2/tyGviCnVJPgPTPt1dKTgnkMAef8T9Pai7FCD69MWzBq9KX2j6Vde9+691737r3Xvfuvde9+690kZOd0RE/UQm3+wX+nsxH+4R6RE/4x0rvZd0t697917r3v3Xuve/de697917r3v3XukhAzf3hq14t9R9PyfZiR/igPSQ18bHSv9l3Svr3v3Xuve/de697917ri7aUY/SwPPvaipA6q5opPSHxOLwuUq66WvwGDqKmCclKyXGUstU1z9XmnVmuPwQfay8jChGBOR5+X2dMWpND1M3BsTaO59v5Pa+Z27hqzBZeNkyGNlxtI9HVakKB5qcpoZhe4LA2PtF0p6qU3n/JP+M2794ybgkzG8KPD1tYldXbYgqkOE8kD+SKGKjDKgQni2nge9hiOqMiscjHS/8Al7/K26f+QHT+39i7cxVLhctsrHUVBtsR09JDQVMdAn7AyCHSCdQBLEn+nvwNDXrbKGFD1Vn8pP5Vnzn+SPSm3Old6V+xJtnbGqsJQ7XpMLNiYqqPCYdgIRIsMi3bSD9Tf35DQ1PVZYy/BqVI8vs6MP2T/LQ7X3ruL477aSCmk6+6w6/2rtzO08sULRtksJGiyyGItZidP6vfhjB6swDEMPWvV/mz8Ku29qba2+qqi4PBYrEqigKqrj6JKVVVRwAAn0HuowOrnJr0o/e+tdVEbnweH3n/ADMDtjcECVmOXr0VS08iq6iWPGeRSA3A5A59yjZ391t3IYuLM6WMlCflXPUfTxW91zr9POK0jr+dOjl534fdUZwu7JlqOR9Q1UtUqKobmyooFv8Ab+ya25/3y2pTSwHqOhFLy3tsopQj7D0B24P5f+2fuDWbd3HuKOdGMkSzZKRIgwPpBXX9PYktvdG606LuFCPOi/7HRQ/J0FS0UjA+WcdEd+dPUHyr6t6Kzk2wc1LW4BaCppcslFVebILReQFb6CSCVPB9yH7eb/ybvXMEa38YWWoK1GK9BPmvbOYtv2pzZvqFDwOetSrrkdk03Z23chjK7sF8+N2VhnpZcnUNpkExv5IQf0X/AE+8zt0/db7RLFKkAj8NaEL9nA+vr1ids6byu/w3CSXhYTSakeSq+Yyv8PDTT5Hr6FHxTrN05D43bMqd6CcZ58PD9wKnV59K0sXi8mrm/wBffMrnWOzi5vnSwp4YbFOHE16zw2B532GJriuor58ejH7b/wCLTTn+pf8A3v2Eb3/cg9Hlt/Yg9PvtL0/1737r3Xvfuvde9+690k6ZI5spNriWUeU38qh7A/n1X/P09r5DSADhjpFGtJifXp9mxeNqFKzY+jlB+oemhb/eSPaDh0tpXpIZTrTaGWJ+4xNItx/uuCJfxb8j3smvXhgU6CzMfGHr3LM8phkhls2jRHGFBI4+hHvQ60eNegWzXxBnZ5Dg8hJApYlAZ1j/ABxxqHN/emr5dbFKd3QZ5PoLtPaMUklDX1crBv2ys5e6KOLWJH1/p7tQUz16pJ6Y6LK927RfW8ddMY2+pDEaQb/X6e6gmtB14gDpZ0HyZ7MxDx0+ZoKl4ltrdobkBf1AMV97NRw6rjj5dCliPlxtidPs9y4ySQSKqyeVF06nFmFnUi3+w9+r1vyp0jd8UnUPY9NJk9vywYnKOHls0scUYZiDZTx70fl1sVzXopuZmymJnlwSZaWpo42YqYqtpYyoPp4vb36p49ewemGNRGtl5JJJJ/Nz71x6tTrJqa1/wfx71x69QcOstNW1ePqoK+jcxz0kgmQq2m5UcXI9760QB0qch23vXeJ/hFZk5I40URsUqnU6VAC88e7UA60cU6W+yaLYOAdMlu6aTIODrC+USm5X8k/0PvQ869bNKDoyVB8pOr9v0qUWB2+8bxgIspiXUWAtqL6b/j6A+/fI9aFfI9M9d8up3c/YY6pP10gRXFvyP0+919OtEVHSQrPlP2DUlzjcbVC3KgQG1iPoRp/3n34VHHr1fLpsbvvuLLx6RR1iBxwPGy/mwFrD377OvVINOmaXdXdWU1nw1yx8hV0vc/6wt70cdey2B01pgu7cpMDLJXxo97cvwD9B71jy6tnp8pemO3Msy+atrFZrWHlYDj/C/u2eq9KMfFHsTIlDU5aaK4uddUF+v+u3H+t78R1upr04v8TM5j4Fmq8jJOdahilQr8E2JNm49uQQmV9PTc0ixgN0IOI+JGFMUUlfUTM0kas93VrE8/Qn3QrkqfLq9agEZr0uaT4o9dQIPIs7ycEnRHa/1I5P096Py68Pn0sKH4/7BoVVUotYS1i0UQPH+vf37zqet+VB0qqTqrZlGQ0WKpiR9NcER/29gPevPr3ShpdnbapECR4bH2FjdqaIngW5uPdq9ap69OqYjFRACPG0KAfTTSQC3/JvvXXqdM+fgpYIaUx0VGb1K3vTx8AC5tpA9qrUDUfs6TXRbSoB8+n6lWFoY3WGJSVH6I0ABA/Fvad10sR0+h1KD1L916v1737r3XvfuvdVZfMupMHcfTpS4Y5aLUb2J/fsbH/H3IXK4LbZdBvJf8nQI5gam4Ww/pdWabfk8uIoZP8AVQR/7wgHsAyjTIR0M4q6BXp59t9Ode9+691737r3Xvfuvde9+691FrBemlH+F/bkX9oOmpv7M9M23CDBVW/E5H+9+1N78a/Z01aiin7elH7RdKuve/de697917r3v3Xuotb/AMBKn/li/wDvXtyH+1X7em5f7M9Me2UtSuf+bjfni9/au+P6g6TWeVJPHpTe0HS3r3v3Xuve/de697917qNWDVTTKPyh/wCK+3Iv7QdNy/AembBAhqgH6jSL/wCx9qbv8PSe18+lF7RdLOve/de697917r3v3Xuve/de6SmV4ylORa+kf4/j629rrcExHpBcMRMAOlSnKKf6qP8AevaJuJ6XL8I65e9db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/0t/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691DrwWpJwLXKG1/6/7H27DiQdNy/2Z6aNvlytQH+oYcgcHn2ovAag9M2xFDTpR+0XSrr3v3Xuve/de697917r3v3XuklMB/emL6/8Bx9P9b/kfswWn0RPSNq/U/LpW+y/pZ1737r3Xvfuvde9+691737r3XvfuvdJKnIGfqr2ueP9549mBzajpGcTdK32X9LOve/de697917r3v3XusNRbwyX/wBSfp/vHu8ddYp03LTQa9JrbhUzZEAEHyjVf+vtXeEkLXpPailT0q/aHpZ1737r3Xvfuvde9+6911YXvYX/AK25/wBv7917rv37r3XvfuvdVIUqv/w7DU3N0/0XyMo/of4Kb+5KNP8AW3H/ADW/5+6AQI/r6f8Amj/k6tv9xr0Peve/de6Z8/gcVubEV+DzVHT12NyNPJTVVNUwxzRvHILG6SAj/Ee1FrdT2VwtzbsVdDUEGnTU0MdxEYZRVWwR0RTb38tb4z7a7Ck7Gx22I/4w9S1WKeWko3o45mfWWSPTxz/h7ki693ObbvbP3XJL2UpUE16CcPI2wwXf1kcY11rwHR5a6ip6HCNRUUMVNTU1OkUMMMaxxRxxIERVRBYWAH49x1DI0tz4khLFjkn59CiZQkBVMADrvbgK4imBIJ9XI+n196vMXBHW7U1hB6ffaXpR1737r3Xvfuvde9+60eHSXxZc5Cqt+nycn+vswnFIRXpFEwM2OlR7L+l3Xvfuvde9+691737r3XFkRxZ0Vx/RlDD/AHn37r3UObGY6oUpPQ0kqn6h6eJv97Hv3Xjnj0k8p1ztLLK61OJpRrvcxwRL9f6ce/HPWgKDoKM58Yuv8uJJBA8U7XKlUjAufoLi3veBw69TPQH7n+HtVPHMu3si1ONLGNTOqX/1I4b/AF/evPrZ+XROdzdbZ/rfO1GO3DWpIE1hSz3N0awFwT79ny69gY6alWRUHBt+LD6g8j3Wvn1agrQ9dWPHB5/17e/U6t1grZ4qCkqKlyqmOMsdR4NvqBf/AGHvYq2OqHzPTl1TR7a3RXz1eRnSGOQsuppVUDSPre/vxIHXlqR5dG/wXUHU+ReL7rMwMth5AKlX4IubLf37B49e8vs6GXH9A9MKsTU9ZBMBYgaomJP+t/xr3anl1Xz1dL7HdHdcwkNTUsc4HNgIG/H9oge9Upx63x6WFJ1ls+j/AM3iqY/T9UMJ+n/IPvVOtg0NR0oINq7ep10x4jH2/wAaWL+t/wCnu1cU6rmta9OMeKxkItFj6KMf7TSwj/on3rrfnXrOKSlH0pqcf60MY/4j36nXusoijX9MaL/rIo/3r37r3XP37r3SX3aSMXwbfvR/7Hn6e11h/an7Okd58A+3p+obmjpbm58EXP8AyAPaSX+0b7T0pT4B9nUr3Tq/Xvfuvde9+691737r3XvfuvdJ/cNvt6f/ABnt/vHPtZZ11Gnp0luvhH29OWOUrSRA2uRfi/5/1/bE2ZDTp6L4M9TvbXTnXvfuvde9+691VF825IY+3+npJZUhEeVibXKdK8VFgfch8rsg225DmmP8nQD5kkWK/tySANWa9WZbVyWPkwOMKV9LNqpo+UkW19IuoBsePcfzyRmZgrA56GcEsRiUhga/PpR/dU17eeK/9Na+29Q6f1p6jrIJY2/S6n/WPv1Qet6h69ZPe+t9e9+691737r3UKvqaalpZZqqZYIVVtcj30qNJJJt72HCHUcU6blZFQmQhR6npJbMzeEySVseNydHWyCpfWlPIzMlr/qDAe3rm5hncCNg1B5Gv+DpJZXNrMCIJFc18j0uvbHS/oPezOzNrdUbYrd27ur4MdiaGGeeWeomWFNNPH5H9bA/j37rVeq6KD+b38X8q1a2LyP8AEqahrJKKaqo8pSywCaI2ILqlh/re9kY61qFejFdFfOHqTv8Az67d2dJJ984UprqoplOoEgWRRzx79TrYNeHRxazmkqLfmJuR/rfX3eLEg+3qkv8AZnpk2zGUomLMCS7HgMLXJP5Fv9t7VX5rIOk9rwPy6UJljDhC4Dn6L+T+eB7Q9K6jh0GnZ3bezepMSua3lk6bGULK7CapnWBLJwfUwP8AvXv3W+njrzf+3ezdqY7eO1q2DIYTJ+X7WqppRPDJ4n0PpkUC9j9ffuveWelv7917qNWC9NMC2kaCSSC1gOTwtz7vGaOD1SQVQ9NOFCAz6JVkHAsqOtub8lgB7UXRrTpPbefTbn+wNmbWrqTG7g3HjMVX1xC0lJVzFJqgn6CNQDf2kqOHSvpC1/fGxaDdmD2c1dHJlNwSrFj1EoXzMzaRoQrc/wC39+690NLMqgsxsB9SfoP9f37r3XYIYXBuD9D/AF9+69x68SFFybAfU/gf6/v3XuHSTyYLZOnkjOpdAv6WIBBH5It7MIB+jnovnI8YEdKtP0r/AKw/3r2gbiel68OumljVgjOqs36VJ5P+sPeut9NWb3BhduUbZDO5KlxdEl9VRVyeOMaRc8+/de6DDN989dYfbX97EzdJkML999gaulmHiFRewXWR795V61X1x0vdnbwxO98NDnMNIJaKY2Rw2sH0hv1WH9ffuvA16VIYG9je31/w9+69UHA68GBJAPI+o/p791uvXfv3Xuve/de6L9L8lurYt+zdcNn6D+80M/270P3aecSa9GkxWvf/AAv78cda48OjA+/db697917rg0iKyqzAM3Cg/Vj/AIe/deqOufv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r/09/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691FrADTSg3tp/H+v7dh/tB01NXwzTpl2+TerH4Dj/AHs/X2pvPw9M2owelJ7Q9K+ve/de697917r3v3Xuve/de6SM7KN1QLzqNOv+tyvHteoP0ZP29JHNLgAdK72g6V9e9+691737r3USorqSkF6qojgH+qlJRf8Ako8e/UPWqjrDSZbG1zFaOshqWHJWFtZA/wAQPfuvVHTj791vpJ+F487LLpIEhADEEAhjZbE/6/tfqragDpISBN3dKz2g6V9e9+6916/+8f4H/e/fuvV697917rDN6oXsCfS3FiDwP6H3aM9wPVJBVSD0ntvIyyV7kHS0oseSL/WwP5P+t7WXlKJ0mteLD06UpYKpc8KoLEkEWAFyTf2h6WdEe2H86+qd/wDeMvRuIyFDJuKKorqRjHUO7ipo0ZvFpKgXJW1r+90NK9aqK08+s3x6+T+U7c7A3/s2v2vmMQNo7hqsPFVV8E6RVSU87RGaFnUDTxfj3quOveZHRv8AK7o2/g3SLL5ajx0kltCVUnjL3+mkEc+/db65ZXcmEwuJOcyeQgpMSqCQ10rEQBGBZXLf0sCffutfZ0W7ePzL6F2fUQwVXYO2pxJPHTPIMgI1jmkNhGQwvfke/VHXjWlejEbU3ftze2Hps7tfL0eZxlVFHJHV0MnlhPkXUF1WHP19+OOt9VeiNof5qhqSPRL1uYQ34u+H0Af7c+5HJJ9utI8pf+fugGKDnwsfOH/J1bP7jjoede9+691737r3XvfuvdNmYBOOqQCB+2fr/wAR7UW1PGHTFxmM9RduAjE04P1Gr/e/drzE569b/wBkOn32l6f697917r3v3Xuve/de6SeOl05SpisVHlP1/JuRwD7MZhWAN0hiRVl7elZ7Lul3Xvfuvde9+691737r3Xvfuvde9+691737r3UKuyNFjYWqK6ojpolDMXkNhZRc29+690Wvsr5Ibe2tTzUuKZa+tbXFG0EhZlkvYEBR79inXiDXHl1VL8gtzdg7sqv7zxiWOhjmepZbMC0IfWRqP5I9+4jrQ+fQfbL7Wh3A32s9PJHPTqsUus83QWZrHn3qg8+t18h0rpt94UVi0BqoPuGYBYhJd9R/HvQAPWzjoW9n9c5TsStix328wopiqyPpXQ0ci6+G/PHuw4CnXic06N/jvhlhaDFxwUMhp5WRWJLohVmXkXQ+9EHrWD0wVXxR3TiNc2KycjBdRIFVqAF+CF1X97pjPWmzk9Iuq2B3Jt6RmoqyqkjQnTZ2awH0vz/h7qT5dbAByeuot891bdt5DVFlsGP7n0I4/wCN+7KadeNOHT7j/knvzHkpmaatkZCbm8gFxwDf3pmNetUA4dCThfl7jAUpsnjZ/KbDyOZAL3sQeP8Ae/ex1s+nQtYn5HbNyWkSTQUt/r5J7EH6fke/daJp0I+N7O2bkwDDmaMXIHMt/qbD6D3vr1elVBnsNUgGDI00gb6aX+v+3966305xyxyi8bq4/qpuP9v7917pNbqUyUKxAFiZUawBN7N7X2FQ5bpFekFQvn0oKIFaSmVhYiCMEc/UIP6+0clDIaevStMIPs6k+6dW697917r3v3Xuve/de697917pM7mkCQ0gIJ1VAAsPybC3tdYglmp6dI7yukU9enyjVlpog31Cj/bEXHtLLlzTpTH8A6le2+r9e9+691737r3VN/8AMe21uHdO/OqcZtesWiy1RVwxUlQzBVjl855Y34/HsTWFhuF5s9ytk4VtPrTNOop9w9vv9yeC329/DkLYJ+3p+xvxx+blFg8dDiOy8JTvHGGVZapbhXQFSefcF3HL/Oiu3gXKg/b8+iP+qHuRFAqQX6Aj1/4vqIenv5guGZp6zsXD5CIm4+3qI3H1vawN/oP6e042nn2LMlwrfYemRy/7qW/dNeRv9g/2euxkPmTt665HIvXFT6vArNqt/S3u+vmy3oZWJp6dOeLz/aCsza6enXN/kV8idtgfxPbe4MiI+HFNS1D3sPxYH3r+sG+24/URm/I9VPNfNtsCZIJGp6A9Rf8AZ9+zMW4grOp98VTfl0oKgj6/8EPug56v4u2S1kb8j1Ue5u9W7aZ9unY/JCelVQfzBNwMq/fdW7wpm8ZY+ainUAj/AAKD2qj59kPx2zj7a9K4vdK6P9rYzL9qnqsb+ZD/ADWt/wC3tqjZnXuKzO0czkiS+VqGmh8FKacLI0aWGp7kaQePyf8AEru+Z73mK/O02ga2SMK8jV7iDwVR86Gp8vt6x+99fvCbts9tFsPL8bQXd3WjuDpRQBqb0ZqsNK8PM4FDVv8ADL+Zj8itk9ybYp8/vHJ7kwWfzmOpMjRVMz6gKyrWHyxmPgm7epSDxyLW9tXpvNjQbnZTvSI1dGNQ6+fHII4j9nUB+3vv5z1y7zPaR7rcm7tbqWOFwwAZGdgqupUCuSNSkcMgimd7PZebbcm0tuZ91KNmMPQ5Blb6qaqASkG/+v7lyyn+qs4rgfjUN+0V66vbfcG6sYrluMiK37RXqtv+bhHSyfFzMrXSVKUQTKmr+1meCVqcUQ8qh0I+o9qxXy6VsO09azv8vPM/y1qTpHeFLvvZu/shuJ97TtNVpnbxqyOTPpElyLn8H3Zg1adNI0WkaqV+fV3nxjqPiVBtreW6vi5tzdGK3rt/a2Uy9K+UyArF10dM0kOgILg/q9tkGnTw08Fp+XQq/Bf525rtjr3svF9k1Eqbx2hS53IKlVJOahcbi1csSrDgAD6+3FOlwR1T4wV4dAn8f/np2r2Bvntuvx8eRymxcFt3Oz4CCi+4nC5Oh1eEJwTq4+nty4Ic6umbfAOMg/t6I7vT5PfzB6rCZLvSj3+229i42qyVQ+3slPNTZBqWhlJEawSAObgWHtnHDp5tdNQA6EHtX5U1HzG+DmE3DnJa6LcuOOSpMjUJU1Eck/2y3dyxAJufoffqkGh68xDLjzHSw6/+Wu7/AIz/AA3+IuE2tVVKLvTfWWoq+Z5p3qZKYVSp4XmFyVuxOm9veuLHr1SFUHIwOrN/mr8pN4dK/HzbvYeAqZIMjksRDVyujyo/kZAxbVHc/n8+9gHFOtkjSSfLoofanzs7ryfxG6G3D17RZg747bavoZc9TR1UsNDPTTmINUyqvA/pq92Qam08OqOx01pWtOiDbd+e3zC+LvbXX2N753wm7cJvLJBHx9FVzSGmppJtIFSIgQCAfow93lGaenTUR0HupQ9B9/NL3/8ALrdXyP8AjDurqneLYzZe/JKepp6JKyVH8P3VtBFwbi9ufbahc9Ouz40+vRttodn5vbHyd+M+3+1Zayt3FPXIv3a1Mq04cSBkMoHBFwfr70Rio6up7hq869KX5k/Pf5Fdr9/jor4i1mRxtRjMi+JyGRpXlenNTTArM9Q1gigke/AClW60WJOhBkcenD4o/wAx3uHq3tNfjx8p5K2v3vmfvKPEZStmqI4kqI1LRyIttLA2sCPfuORkda1fgOD/AJOi85j+YN8x++e5t79V9JVuYoTidwZCipK5TUmljp6JmRgXYaQDp9+0igNeva2JK0rTz6N3/Lr+bvcGf7on6C+QdTWZLeE9fLQUNXVPPp8lPUlZCqt6SCBwfa1GCQGnHy6SU8SYI+CM49Or2uzt3jYexdzboELzth8Lk66OOO+oyUtE9QhH+xX2hrmvS7yp1qfbV+enzI+UPYO79x9Tb7XbW29l7kmxVTtutrJfvqqOllYf5PCwu1wpAA97K4z02HLEhaUBp0PPzG7x+VPanwbly22azJ7V3thtzviK+syRqKNq+FYNE00DS2JViAR72DwB62+rSSvGmOi5/G7cvd2z/g1Wbj74zlTuDFz9gUVPBHTVcjVBnbh2EiksF5HvRoGNOvID4QL8Rx6Fvvz+ZB2F8behNnw9YUuXlk3Ln6LDU60rVUlTTvVhVglYqL6bHnnn34Vrx62zYqvHh0Cfan8y75rfDrL7Z3V3JPns/t3senxlNt/GU5qFlgnzMKTUtQwjB/T5Be497AqMfb1UuymhA9Pz6G+h+fPy76b3xsXsztuuycvWPYmSxNFi8LUtURSomV0Sq7ADVZQ9yT70KEU8+rVbV3DFadWLb/8AnDu+m732dtXCVcibd3A+3leNJJ9P+5GJJJAdP/Bvfq4JHVsagD0t/wCYJ8tt4/Hzc/WeI25USxU+7chiaasEbzKHjnlUSf5sfnV+T71T168TQ9VK0VVLkfnXU7tlrK81M+Vx9cYvu5hHrqGWVrx3/wAfp735U6qR319B1av8PPmVvPuz5p/ITpjNzyNguvcLTTYqAmQRRNDLDTsRGwC3OokkfX3qhAFerA1qMYp/g6A/f/8AMO33sf5L7g2NNLU1e3cPl/txRxyVA1xLMyaQoFvx73Xz61UVp0RDcXze+YXfPdXasHTu/YttYPrZzVvgKqslWrqqaOcDw0sLX1Pawso9+8vt6rUljppj16vp+CPd2/u3OuvF2PjcnT7lw8YFZkK+GeNK5iVQGEzgXtyePeuBp1cZGej4e/db697917r3v3XuuLtpVm/1Klv9sL+/de6p8+RX81HBdRdoJ1fgdiZrdWVSQwzPifPK3lCkldCIR9Rbj36hIqOtMQppxJ6Ctv5tG51bS3SO+lI4I+0qbgj63Ux+9Hq2KefXEfzatyk/8yS319Of8kqP+vfvWfXr3lwPWT/h2jcv1/0Jb5+tv+AtT/vI8fveOtV+39g64D+bTuaxH+hPfN/6mln/AN78fvXlxz1vHGh66/4dp3N/z5PfX/nLP/1796z69eNPQ/s65/8ADtW5P+fI76vx/wAotT/vXj92/PrXlwPXX/DtG5l/V0lvm3/ULUH/AF/91+9Djk9bNOABr17/AIdp3Kfp0nvn8j/gLUf9e/fj9vWvtB67/wCHadzBf+ZI75v/ANQtRf8A61+/D5nr1RwAPXX/AA7XuSxP+hLfJ/6pKj/r37358evEH59cR/Nq3Lc/8YS3yBb/AJVKjj/rH71nyPW/Lgeuf/DtO5bAnpLfNj9LUtRz/wBY/fs149a/I9cf+Hadzcn/AEJb6t/T7SoH/XP37zwet44Z65D+bVuS3PSW+b24/wAkqPr/ANS/fs9ar6g9df8ADtO5eR/oS3z/AK/2lR/vfj978uPXv29eX+bTuYcnpLfJt/0y1H9Pz+37rn163Ueh67P82nctzbpLfP8A5yVH+8/t+7eXHrX5H9nXh/Nq3IQR/oS3xe9r/aVI/wCufv3Xv29cf+Hatynj/QjvkW/6ZKn/AHvx+9eeD17yrQ/y67P82rcwHPSO+b8f8olT/vfj9+8uPXvnQ/y67/4dp3Lx/wAYS3zyPzS1A/3kx+/fn1rj69cW/m07mH/NEt834+lLUH8/8s/fvLj1YfMHryfzadzcE9Jb6sTaxpZ/qRx9I/fsjz69itKHqBJ/OJq8RkxTZ3pzeNDSmIsKiqhqYYjIB+jV4/r73Q0x1okfZ+XX/9Te+3F2XsXalYMduHcuNxNayhxT1cjLIUK6g1gPpb3vr3TN/pt6pvb+/OCva/8AwJP0/wBt7rXrdD14d2dUk6f78YO9r/8AAhv+jfe+tcc9d/6bOqrkf34wfH5+4a39eDp9+69Q9dHuzqoC/wDfjB8/S1Qx/wBv6ffuvde/029UgC++cFybf8CG/wCKe/EjrwqRXrzd29Ur9d84L/YVJP8A0T71XrdD10e7uqAFJ3zg7Ne1qhvx/X0+/V61Q9dr3b1S17b5wXHPNQw/6J9763Q9dHu3qkED+/GC5+lqkn/iPfq9eoeux3b1Sb/7/nBcf9NJ/wCKe/dap17/AE29U3A/vzguf+mhv959Pvxx14Z68O7eqTcf34wfH/TQ3/Rvvwz144683dnVQUsN8YI2/H3DE/7a3v2K568KkY683dvVKgX3zguf6VDf9G+/V69Q9eXuzqlvpvnBcG3NSw/6J9+r1uh68vdvVLfTfOC/2NSR/wAR7916h69/pt6pvb+/OCv/ANRJ/wCKe/dep15u7eqV+u+cF/sKkn/on37yr1rrod3dUk2/vxg/6/8AAhv+jffvl144z10e7uqh9N8YO/8A1EMf96Hv2PPrxB8uvf6b+qb/APH8YO3/AC3b/inv3Hh17Neuf+mvqq1/784I2F+Kkn/iPeut0PXR7u6oCNId84IKqliTUNwB9T+n3sZ61Tpc4HcWE3Rj48rgMjT5THSkiOrpWLRMQLkAkD37r3y6evfuvde9+691737r3Xvfuvde9+691hnF4ZB9LqR/t/d4/jHTcvwHpgwAs9aLg2kF7f1uR7VXY+E9M2zVB+XSl9oulXXvfuvde9+691737r3XvfuvdIqrbTuyn45+3Q3P0+lvz7MY82RHz6QyVFyG6Wo9l3S4cOve/de6bcxUvR4uvqo+Xp6WaVbfW6Jq49+691pvZjvj+YN8rf5q3bfxW6g7cGzdn7VweUzNHTZGrqYMfBS0MP3DD6H1HgDTzc+7YAqfM9N1Zm0rQAAVPnU9K/5Vbn/mL/y3anqzsfs/uGm3ftbd27lxFTjMNkKmoIWlMZqI6nxgWDI4I+o/2I9+ABNBxPWnJjGskEDiKeR9Or9a/wDmF9TbT+MOxfkDu2vpMTHvKKnx2Hw1bUyJU5TMw0qfeQ0xClm9Z44/I5v7rw48enAaivl69FU6q/m9dY7q7IxW3t94gbBpN0ZdMRt6vz1TVQwV1S8miNaTyCzE3Fh7WBv0NPSY6TKPPoyfy0/mXdLfFHJYvC7nyGNr8nuBIRgKP756eXJ1EwBEFOoUlj6hwPaPiMdKCwU54dFu7w/m9bS6+6jwG4Y9sVNLvDfdPJHtjAtPUpXy1IayrToo1MTddP8Ar+/AE4PXmYAVrXovPUPz5+UmM3X1rmuwdh7zrtrdn5FoYKOSmrVGHh8g0vU3X6BSDzb3s48+HXhq4kUr0JHzv/m0UnWe++vujeo6Cry28+wpYqOTIY2WpaTb9XK+gw1iQra9zax9+FSD6deJWoB4nFOgw+N381veOyPkRtL43/IPG5CXLb/yyYfE5nJS1UCU0s0ipFNGkg0kfg+/UoNXp1UvVvDbjn8x0kvll/N53j1L8tMT8VevNq5rMZbfmdOJw+axT1jwQyeY06kRxqVOo2/P59q7vKqeH+z0ntmXWVHE+X2dCl0p/Mp7L2P8hNqfGjvvA5eiy/aorKfb+fzP3cFPjzPG0FNPpYWKaxyb/j2jpjUOlRYltLYJrT5jz6q4+L3Qvcuzf5wuKafecDYbLbyz2UhU1jPFVUtM8tTqiRSbqQPpbn24TWMD0PTSqRcs5OCuB6dWR9KfN7ee1PkR3lHvSankxG3N0bsx8EKRGHyTxmZaVy0Y5swB9tjgOnKmpr5Y6AL470vzk/mBQd3dm4rs5Nu4DZW88pjsFQZWqq4nqI4Ipqqlho7gm2mILc2HI597ODTj69bQlgXrQVNKeg6Yfj984O5+zvjH3v8AHztjKVtT2ngN15LE7UzFVPUxzRY6iilidY3e10vYg/7b3ugrUdV1MQUbiCRjz65fy+PgzF3x8Q/kHvjvHP5HPb829m9z1+zchR5V2jojhcRPkYoqksSf1Rqtrj+v49+J7zTrap+mpOa16N1/wnY7b352P8fe6sXvzNvmptm9rS4TCO88s5pcVB54Epw0pP0st7e9GgP7OvISQAfKv8j0cnsaqpNn/PbGbvq5oqSGTbdJSGefhCWpwpW68259yRt8M19ya1nCpYhiaD7egReFYOaVuXNAVpXqzGHee2HgglOaoQJYIpQRJZbSAW/HHJ9gM7de1I8NsHoZC5goO4dVs/Mb+aP1N8VK0YmZKXdWVVpBPQ0U9SZoFiYKzOIR/j/j7lrkX2c3nnGPx6mFDwJAz+3oCc1e4W1csA+OdVONKn+Qr0KXwv8Anx1z8wMdJ/d1qbGZyCLzS4N5ZmrEh8RmMjLKPpYeybn/ANtNz5Gk1XFXiJpqxStaeXRhypzjY8zwh7c91K08/Xh0f33GPQz6hZFFkoqgN9PGx9uwsVkBHTUy1jPUDAsooliU30E/0+l7e3brMmrqlue2nT57S9KOve/de697917r3v3XukgdUWaUG4E01+ePzYWJ/r7MSA1vX0HSIALMSfPpX+y7pb1737r3Xvfuvde9+691737r3USsrqTHwtUVk6U8K3u7mw4Fz9PfuvdF97E+Q+2NnxTQ0csWRqdLoghdiyyW9Ngvv2OtHonGX7C7T7Ur2pcOayLHTysioDJxHKfwD/Qe/FjXPXqLxHQvbH+N0ONpW3Tviq8kKQiqmjncSMSP1+k/S/8Aj79TyrjrxJpnj0BXZ1Tit4Z1dlbLoB9n5mpWdY0e/Om9z7rTzHV6imehm2T8JNnUOE+/r6Y/xuthjkJCwADygFtVvpa5PPu3y6qDTI6IHvj47022u9oaGYv9r/EFXQJF0hfJYekG309+qSOvU7s9XX9edfbe23hcY9HSxtK1JSuXMaCzCAAjj6+9kDqq+fQpe9dW697917rG0UTCzRxsD9QyKQf9gffuvdM9dtvC5AEVOPpm1CxIhiB/6F9+r1oCnDpI1nU2yq1WE2LjufoyrECD/X9PvVOrY6QWX+N+xK4M8FM0Up+nEYH+3AHveBw6rT16CjN/Eajq9TY+s8L/ANn94L9PoPejk9bPDHQd1/xt3xt5WOMyMxCHgrPq1ELcEEH3unn1rHAdJF9u937fk/bqKt0Q2BVmICjkWI91rU46tiles57V7fwJCTR12hLeT/OWJH1PHvZPr1UE+XT1B8ldxq8TZajrG8ekSKxku2k3b/YW9vxymMdNSQrIfLoZMD8tsDVRw09VjJIXVEQySNIouOCWJv7ZJNanz6dFKADy6FXEfIDY+SjVpchS07EgWM35PH0I96r1sDpf47sPamSAMOWpjcAj1Eg3/wAQPduvdKWDMYypAMFbBID9LNa/+3966904ggi4Nwfoffuvdd+/de6S25mIGPVRc/dKfpf/AA+ntdZVGsj06RXdDpB9elOv6V/4KP8AevaI8elo4dcveuvde9+691737r3VXvymQZXvDqyC1/tcxECDb6LP/Q+5B5fIi2i4LfiHQL3weJuVuPQ9WbY+HwUVNCABogiWw/qIxf2AHNWJ6GKDStOpZVT9VB/1wD7rQHj1eg64+KP/AI5p/wAkL71Qda0r6dcTTwH6wxH/AF40P/Ee/aV9OvaV9OuH2lIfrS0//UmP/invWhfQda0J6D9nTRX7X2/k0KVeIx0gIYFvsqbWQwsRrK39tSW0EvxoD+Q6Yls7aYUkRT+Q/wA3VaPzy/ls7G+VWwaqg27S0mE3pFL9xjsk0dLHHG8cHjRS7BbAkAEX5BI9hLfOVVu5Fv8AaqRXSHBOFYeatTyP+z5dQj7veyWz+5Oym0iAhulOqKUAVjYcGHD7CK9wJB6rD+H/API13J1/2Zjt79qZnFV+M29kYZqKho/tX8s9NOJPMy6nJIA9P0A+v1tYk/qpzHf3CRb4I4rcEMVjcsX+RrTHy/b1A3tv90m42PmOLfeZ7xbxLc1jj0BVDg4c1ZqsPw8KHPGlNnfDYunwmKx+IpBppcbSQUdOtgLRQII0Fh/gPclwxJBEsMfwqKD7B1nVbwrbwJAnBAAPy6rT/muR003xrr465NdDI+TSsFwv+StSqJeW4/Tf27546dOUIPVQn8tD4+fy09wfH3PZDfFfQUef/vnUnLw1mdFK+uRf2TTxxI9wW16/6ce/EEHJ6qmjQCFB6ut+N/XHwk2NW1+L6Hy2JnyWWonxs8CZJshJLBVqYSojljUG9yefeuHVsHIFOtc/5970zvwQ+WW88piqaoi2Z2jR5PZdBSQP9tEWzX+TtJ46fj+0fqPbqAsdI6TzP4dDwr/h8urAuosNkPiZ/LzyW/qPEu+9N5xV+Toq6SCKR6SHJU3nhmlkF2K2YcH3qUd+nq8JATUfLqvbF4DuXtv4h7z7f3/3Jsz+ElM9BS7TNbTQZU6C0mkUi2PP0Bt7pQaqdWqWSpIz0GfQVcrfDfIU0Q1tFLnL2kBVrcEgfTke7Nx60hGivQw9wvUYz4cfB/cNRSuuNxm/81U11Q5UQ08UeQS7SPfgWB91A7j1ZmpGp9KdG3/mSfK7q/c3xX6xw22MtjMxV5jEpRNHRVTSPDJGgRw4PFwfx7sKseHVJGVYzU8eidb3+TvZW2/jn8L+mNhtFtWp3NlK2mm3DlIkNLBNWVYjE7SyCyoAQSR/T3eNdTVPl16STSigcTQfy6L784+ud09Zb+6jXe/ZO2OwM3VSU8tXV7eyENTHC0k3k0lozwR/h7vLUtQ9MjSE1Vpno8vypzMOHf4C56uWGLFwY9JqitqfEYooxUrd3kb8fm9/ada5p0panbX1/wAnU3ufL0nYPzK+MVRtWogr6epq4hHPjnUp9bXUr/T/AA96rx68SKqvS1+DXZO0fjj81+6cN3AKLC1G7d25CLD5LcKRsYA9SzCelklBKkg3BHvZ8qdeWgYqeg0+deS278g/5i/U2a6n8GYosIi0VZmsBoWjEtKhEs1W0drsfqxPvYNASetGuoAeVejV/wAqCijx3cvdNNLj6Gaspc5mYpaiopKepfWsj6ykrDUCefofejQnPWwCOHSB2i+v+a/sxlp44f8AczMT9skcEZvKblo47f717UgqYuHTWn/GK/LrY47NbEjY+5FzdMarGS4fJQ1cYtfwSUTrKef9pv7SGlOlHmOtR/e/x0WJ9/dmfEftjaHXNTh9wZGuy2BzeWpIMlVz0amd1p4PqxZgwAt7tXybqhTu1Lg/tB6UfVnye7S79+I28dtdmY+SuyWz9zNRNnkpljpKhKZGNRUFrBSGtcH346QcdaUsy5+z/iunKq3xgNx/y8JsVh6yjq8hQdqwxzU1K6GZNDhbkf7D3U/H1cf2WT0hd10tLWbO6pWux8VWE3ltthHPFDOikTLyUlFvduHVK5FfXo5f85jFYvJUfxlhq8TSTRQ1OynVYqSnQglIiByL6QOAv4HHvw49WbyPzHXv5yOhOmfiZDR0FPAPu9o2FNTw07Ainp1BPjA449+FAfy6qxrUH+IdAP2Vuyj607R6X3Hu2VKDGVmU2nGlZWShYiiwxBmLjmwv78PQdWNFILevSv8A5t3yU2DuztroPC7LyVDuVVzm3Ypp8bUeTxtJLFIQ4/Gm5H+w97pUHqrMAwHmT0h8PHKPmXHTxwMxqZ8CRoINzJCjWt7qAQOrFu4joUvg12zszqb+Z38u17DytDtalrcM1NRV2Vn8EVTUJURTCGA83LaCB7tQ06qMah9n+Dos2H7c2n2Z/MI3xB9muT2/R7maSWalkEkdTTLVf7rLf1Bv71Qha9eqpk0jhTpW/ID49QNv7uHsv4q9r7R6/wA9ihFW7hxGZylLBX1BkPkljhgJ9XPGm3vwP8XWyhrqU0P7f29WF/yXPk/2p3TB2X1/2ir5DJdeU0Ij3JFD46LKO9bHTN4GUWbg6gQfp78QBw63G5danFDTq+f3rq3Xvfuvde9+691jm/zUv/LN/wDoU+/dbHHrXp6NwWDrvn9US5KihrpF3HXFVq4YqqMHzsFuswI4NvfiRQDrVO9q9X6vs3aUjmR9r7ed2JZnbDY8sxP1JPj+vv3WqdcBsnZwOr+6u3L/ANf4Ljv+vfv2OvEVFOuX9y9ofja23P8AzyY7/r379g9eAp10dlbPIsdrbdt/2pcd/wBe/fuvAU68dlbPIsdq7c/88uO/69+/deAoajrv+5e0P+eW25/55Md/179+68BTro7K2e312tt0/T/ly46/H/Tv377OvAU66/uTs61jtXbtv+1Lj/8Ar37916ma9djZOzwLf3V25bn/AJcuO/P1/wB1+/depmvXQ2Rs4XttXbnPH/Flx/8A179+FB14568Nk7OF7bV25zx/xZcf/vP7fv3y69SvXv7k7PsB/dXbtl+g/guP4/1v2/fqDr2euzsnZ5IJ2rty45H+4XH/AF/6l+/deApw68dlbQN/9+ttznj/AIsuO/69+/Y69SnXhsrZ6/Tau3B/5Bccf+ufv3Xqddf3J2fz/v1du+r6/wC4XHf9e/fuvUr10NkbOBuNq7c/88uO/wCvfv3WznrsbI2eDf8Aurty/wD2pcd+f+nfv2OveVOvf3J2dYj+6u3LE3/4suP/AOvfv2OtUzXrl/cvaFrf3W25/wCeTHf9e/fuvU64nZOzj/zCu3P/ADy4/wD4iP37rwFBTr39ydn20/3V27Ym5/3C4/8A69+/Y6317+5Ozx9Nq7cHN/8Aiy46/wDt/H78Mdap59V6fzD9q7Xg61xL0+CxNJMcumqWkxVFTSEF0FjLEgJ92wePWuA6/9W2ehxPdXy3+e822Jd9fw3bUGxUrnoJ6qpjCmmxgdnWK5uSy/gX597BNPn02RVqVpTo/B/ltdkf8/Gpbfgfc1X1/wCSfegfXq2nNa9e/wCG2uytIH+kelvf6/cVX0/5J9+qet6R17/htrsokk9j0vI+n3NV9f630+91PA9a0/Profy2uyh9Ox6X/W+5q/8Ae9Pv1c8Ot6R59cv+G2+yjf8A4yNSf+dNX/0b78TjHWtA9euv+G2eyfqexqX/AGFTVf8AEr79Ude0+VevH+W12Sfr2NS/4f5TVf8ARnv1c8OvBacD17/htrskf81Gpf8Azpqv+jfe6j069o+fXR/ls9kH/mo1Le//ACs1X/RvvRPWwCPProfy2eyR/wA1Hpf/ADpq+P8Ak334H5deK1xXrl/w2z2Sfr2NTf8AnTVf9G+/A+vWgtPPr3/DbXZViP8ASPSj/Wqar8fT+z79qpw69oFa167H8tvssC3+kekP+vU1Y/29l96qevaB69eX+W32SDz2NSkf9RNUf9a3p97Bx14qK9df8Ntdk/8APxqX/wA6asf9E+/Bj14oD59e/wCG2uyb3/0jUv0/5War/etPv1c169oxSvXX/DbPZH/PxqX/AB/yqq5/5N96r1uh9euz/La7JP17Gpf/ADpqh/h/qfe6ivDrWn59e/4ba7JtYdjUo5/5War/AHvT79X063pHmeux/Lb7JH07Gpf9jU1Z/wBt6fficY61pzk9dH+W12SeD2NSf+dNX/0b79Xr2mnA9eH8trsgC3+kamvzz9zVDj8f2feq/LrZFek5vD+XR2fjdl7srKXsal+9p8FXTUjfd1CBZYojIxLMABxf8+/V+XWitfPph/lM797Mpt4b+6Y3xuB87TbSpaqrp5hNNPD5EnWnUI8hIPF/p72woR1pCTWvV7XvXV+ve/de697917r3v3Xuve/de64supSv9Rb3sGhr1VhqFOkvt9rVWTjH0Wb+v59rrsHQjHz6S2x7ivSq9oOlnXvfuvde9+691737r3XvfuvdI7MII8tBUkfRUQHn629mNuf8XI6RTlvExw6V6G6If6qp/wB4/wAPZeeJ6WL8I65e9db6g5NBJj6yNraZKeVDf6AMukk+/daOB1oc5/s3u7p/+ev3tVdPYLL5OvOByVJT/wAJppJ5GimotExKwg3Gkm/u4AK59emu4SDSKkqK/kej6dgfF/53fzFd6dd4jfuYrNu7A2nn6XMV1FudJIFTHvItRWpCtSp1MVUiwF/eqgDGT1soznu7VrkevRUv59218r0nnPiB0ntilyZ6+2/vbGJQ01PUzRJNUyTU8VfOVhsrB2BNrfT3sUpXzp1p/INwLDp3+YfxczHZVV8cclEZdjw7dzWIyOBqMgxxqVdaY4plKSDSZPUB7fiICNp4kZ6ZlU6lJ8mx0oflz8f9vdmd7fCug7nqKvPZLF78wy1FbS5Bljq4hUwKFu5Ib6cg+0wI0npSF/UWuQTXo0f86/o/b3W3dPws3rtPa9TTbP2ZuBKzOvAw/hdHBR5ZGH3iWsR41F9Q5Hva+Y+XWnqoDD+L/J1bNvj+Yh8dcZsPrrKYqkwO/azcqQig29jIYJKiilip/E8dMmk6GB4Gn8ce9CoJHVuIp1qg91bO7QxHz+647FyAqdg7f7E3jHk8NJuNm+2wtLLkdSPIJ/oqgj3ev6ekdNUIuQ5wCKdWfdgdMbR3h82ugM1urfWA7Tz9DmYpMRkdpVFOkVG6OJIxOE5JU/X223Ak9OKo1KOiwfI7sjYXx2/nNfFul3ylJStld40lVBU5HQy0kLZDS8r6geVHqvbj6+1t2f00B9AeklqB47P5dw6OP/Ma3vtHv/8AmH/FvDdSy0+7K7EUtNjW3Ht91MNHUy18krxzzIAx0+QAX9pBgGvSpu5lp5VP8qdJjtxN5fGb+bP8UJsxgq6v2/LhBPnM1CNFGjVsEsUhnlQcnm9z9fesAU62PiDU4hh1kwHTW5Oz/mfvPbFHiaukwfYGT3VuOnrrKlO10lnpgHPB1GwF/ej2gdeALOT+fTf8Bfl5uL4V0fyP6c7f2XlxlcnvTN/3OnmU0odokqKalLKVOuNtanixPHPu3E1HWg2j9NsZNPn1H+B3xb7V7nw/d/eu5sHX4epO5ssNv0FXCkEmQo8l5FjmpoXALqOPUB701Bw8utoGJLNgny6tt/lv9Eb36x+NXdux940VRRZPdOY3aaCKpjCM0GUxFTRxMq2+hLj3rByOrrVVAPl/n6C3+Td8R+wfidtvt/Abuhlp4d1dh1+fpUcKA8DTSlHGgW5DD34kHh1UJTJ9T/h6XH8xfbNdT7n2nunG1a46rrcxj8WtWWMd1kKJp1LY+5k9tN2t7OOaK6QyLoJoPz6jrnO2lkljeFtJLAV/PowlN8ZuyqzC42Sk3rAq1OBxwW8svolkgSYtf/C/4v7QHnTZhMxe2OHJ4eXRqNh3AxALNxUft61fP5mvw37v2X25kt2ZGHLbrp64VccVRQ1LvSyLJKCG0ngG30495d+1PPPL+5bIlpCVhpTBGRQdY5+4/J+8Q7n9fHrmIVl0hqKakZocV/wdHV/kZ/Gfs7Z3bOY7TztJlcVgavEtTmCvmd4GZse0Xiij+l7/AO8+wB94fmvaL3YU2eBleQN5DPxVqehZ7Ncsbjtt/PuU+tRNpNGaoFF00X0+dPPra2H0F/r7wu6ySHDrBVKHppkIuGjYf7ce7oaOD1V/hPSewP7c9RD9Aq3C/wCOq1/979q7odoY8T0lt8uc9Kj2h6Wde9+691737r3XvfuvdJTNEw5LHz29AN2/p6T9P969mFsQ0LIekc8f6gkGKdKlHWRVdTdWFwf6j2gIIND0rUhhUdcveut9e9+691gqaqnpImmqZUhiQEs7mwAAuffuvdAN2F3/ALV2dBKlLUw5CsGtBFGzFg2i4IA/p791qvRMs72v2N2jXNQYFauCinkICqZbaZho/r79U+fXgKZ6E/r34x1+SlizO7qh5dbJI0ckmpma92AR7+/D59br6dHI27sbbGz6cNjqKKEQQ+uZ44yVVASX4Xji/wCffvPHWqevROfkb3HUVtSdk7Wl8kju0MklOZBrvwwNrD37r1RWnHpX/HPphMVSpujPwGSsqdE0cdQqO/kddWolgTb3rjnrfRzAAAAAAALADgAD6AD3vr3VafybxhxnY2PzwUJ5K5CXsBwHB+o96rXHWqeY6Ph1pkf4ps/FVerVrhRbg3HpjXj3utc9b8h0vffuvde9+691737r3Xvfuvde9+691737r3XvfuvdcW0gFmtYAkki9gBc+/Hr3DPSHTfuw6zINhUzmLnyOsxvQka5dd7FShW3tEu42Ty+CkgL+nn0XDdtueX6cSqX9PPpxrdnbcyQJqcbTuHF/TFEvBH9NPtb5U6MQfMdBZU9R7FzGX0jHERKHRrJDa4HP0H+9+1rRhLercekSSO89V+Hphzfxm2dXa/sovAXJIJ0KRcf1Ue0X2dLKdBJmPiDLJIZMZkPEq3ZUE+k6gOABx78B5de8s9IDKdF9l7eJjxeQqHEZOnTKzXCfT6e9EevVhw6YzF3bto6pJa6WOP9IAl/H5A/w97BHHrRrTHUyn7u7Wwsqx1kdc6LYNzML2Fvpf3ok+fXqA9L3EfLHIYtUTLYqpmII1B/Lz+Cef6n3sGpz1oig6XFN8nsBm6inkqKVKONXVtMrSccX/J49qEm0JpHTLQ6m1H/AIrobcT3fsTJBFGYpI30qWUSXsdNyLW/HtgUbPT2RjpcUm9ds1yhqfK07g2/1X5+l7D3rrfT/BX0dSLwVEUoP5Vr+90PWqjqX711vqsfssfx75G7YpWGv+H5tB9L6QtRxcex/YHwdilPqvQLu2M+8xr6HqzcAAAD6AAD/WHsAdDTrv37r3Xvfuvde9+691737r3XvfuvdQci6pSyMwBFvoRe/wDsPb0ArIOm5SQhoadN23/+A81lRQZQbKoXkjkG39Pbl18Q6bt2LKa9P/tL0o6CLunpraXeWyshsjeNO1Tiq6nqoGRNGofdQ+JyDICBx+ffuvdVJYH+Sr1Fs2PJ0O062uocXksjLkZKeSvOrzSMWv8AtqB/vHv1T5561pHlgdGk+PH8vja3Q+7INz43IT1E8LQsqvVySgGK5B0kD+vv1evU9OhK+VnwO6I+Wr7cru0sJLVV+18nDl6CppDSRE1NOwdfM0ykkXUfn27CaP8Ab03KAVrQY6GfMdHbB3n1RD1XlsYkm2KfDx4SCBI4AY6anhEMZC2K3sPr7tcDvp1qH4SfXqveg/lMdVQPW7eqZqh9gVUs8n8HjqFSZfuW1THSF0X9sVPn06VFKDocNsfy2fj3s7aUuxsDhqmDbcsUimmn+1kl1zIVlYsFA5/1vfs+fXgKcB0ud5/BXpLefRuK6NyWGZtubfWqkwjKKdJqSqqTqeQHSQL/AOHv3nXr1KinRQtq/wAnToeOjpsRvWKvzGIxDzS4OCGrjdqeWV9V5RMpBH5sB79U160EXgc/b0PXcH8uDpDsbrzZ20KfFmiquvEvtSqtSRCnmJBLysE45F+D7vGxVh1WRQUpjoGsR/KI6IzkWOr+zIK7O5zF3ahqaeqhkiSTVqRm86m4H9Afe5HLOacOtKg0+R6Mn3D8Bul+3uvtn7IzOMl0bFo1pNuVCGBJKZR9QXKkgf63tsYyOnDVhpPTJ1V/L36i66z+zN0mieqzexm14WocwTLG+sNdy63/ANt79XPW6YFOPTN8pP5cXUHyJ3JTb9ko5MTvaml8i5Clkhpad259cixqGJ5/r79U+XVSoPHpQ/HP+X3030TBUZGPFnKborDK1Rk6p4anQZh62gaRCwvf+o9+63Ty6GbqT4tdadN7k3NubadA1PX7orKityDMsKh5akkyWMYv9Sfr79QdbqTx6SdL8SesMb3hju5aahI3XQztNDUf5Pw7eotptq+v9PapQPBJ6SszCbFOjX5jEUOdxtbicjF5qLIUs9JUx3ALQ1ERilAJv9QT+PaXpV1VPvb+VF1DkNzVWZ2d93iKTJ1xr8pST1gZZpZTeYIqqBY88Ee/fZ1WlTU9Gp2n8Jekto9U5HqvHbfRMTmYJP4nL46XzzVU0eiSUMqf48Xv79Tu1Hq3lQdAH1v/ACweltg4fJ7Vipqio2tksy+ckopJYZX+8LFlYKy6ePp9Pfjk9aAAFOhpqfgn0jV0uKo5sTIafEV1NkKNfFS3SelcPETxb8fj37B69ny6XPdnxS6v71j2qm88e9UdoyUEmLKrC2j+HBRTg+QfjSL2978qdepmvWLuz4m9X974bZuC3vQPV0OyJKGTDpGkF4zQACK/kBAHpH09+wTnr1PTzz0EfyO/l69LfILaGA29mcbLBV7WeCXDVcDwwlJKcKIjIwS9hpH0PvXDh16lcHh0Cmx/5SfQVDkMXn980dbns/hauCqx061MMkKPT/5ksJ0J4sPp/sLe/ZPHrdFBqBXo09D8K+n6Hf8AB2LFjJBn4JqaRZjHTabUgCwj6X4A9+oPLrWTx6L337/K96b7d7AqOzcbTz4fdeUmWXM1MU0UEdUUbUpPhUMb/m5Pv3yHWtIPHpRdQ/yw/jr1XnP730WGq5911jiTKVsslNJHUMBbSl01D6Xvc+919evAU8ukB2X/ACrOp90b0ze7NsvV4tt1SB9xQT1YZKhS+pliRVA/2496qa+vW6A18ujs/Hv4z9a/HLb5xGw8StFUVkMaZWs0wiWrdSG9TRKpIBAtcn3qnW+HRife+vde9+691737r3WOX/NSf8s3/wChT7917rX76F/7L8qFsOdx1x1f0/yhiPfvIDr1SHJ62CPfuvde9+691737r3Xvfuvde9+6903ZbJ0mFxldlq6RYaPHUs1XUyMbLHDAmuR2P9APfutE9VN9jfzmfh511vqt69yfZu0f7y495Y6rGSZNoqyJoW0PqiI/r79x4darTj0L/Sn8zv4td1Z+LbOC7I20MzN4xDRQ1ck8zmRNaDSBxf34462DXI6Mz3Z8kOruhduUG6uwdy47B4bIp5aWsrneOOaMgFTHYX5v/T37rZ6d+lO9uve+trR7u68ztDn8NLM8MdZQSNJAzINTKGYA3Atf37r3Ubvr5Adc/HTZNdvzsvcFBt3A0MMk0tbkZTDTqkQ9bO4+gFxf3qoHXqGlR1WPiv55vwrzkJrcR2fs6uxyNKslbT5cyQoIXKM2oD+o92A6rrH+o9WDdKfL3o7vnZtXvTr/AHxhs5i6CneprZKCWSWOniSMOWaQi1v8ffqdbrTj0SvfX85n4f7I3nk9kVfZW0pM3i6g01VQtk2WphlDmNkkQD6gg+/ceHWiSvHoROlv5onx67v3RHtXam5cNVZKSbwiKCtaR2YMFIAI/wAR78RTrwYHgejP9/fKrpz447cbcHZO78TgI2jSWnjyEskSzLJEZUOsCwuB7150PW64qOiHbE/nRfEXeW6aLbT9gbXoHyEqx0s7ZJ282uURqyLbn6j3uh6qrqW016th2tuzb29MNR5/bOUpsvia6Py0tbStqilS9tS3966v0ovfuvde9+691737r3Xvfuvde9+691XT/MTH/GM8Sb3H8XjGn8j91Ofe/LrTAU+3r//Wu/8AhUB/w43VH/v3ctr/AOOLPv3XvM/YOtjn37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcWYKLkE/6wJ/3r37r3XatqF7Ef64I/3v37r3XEuFNiGv+LKSD/rEe/der1z9+691HaqgSQRM4VyQApBuSeB9PfutVHUj37rfXB3WNdTmyj6m3A/1/fuvcOPXGKaOYao21L/qgDb/AFrn8+/deqDw6y+/de697917r3v3Xuve/de697917r3v3XukB2r/AMy43p/4b2Q/60n37r32dUm/ywD/AM5L91AcD+GVX+t/wOT35uIr1WP4W+3q/H37q3Xvfuvde9+691737r3Xvfuvde9+690jNvsEyeWUkeqewFx/Wwt7MblawIfl0hgY+MwpjpZ+y7pd1737r3Xvfuvde9+691737r3SP3UGhjgqr+kTRpb/ABLcezCycZTpFcjSQ46VcB1QQt/qooz/ALdAfaFviPSxfhHWX3XrfWGeMTQyREXEg0H/AFmNj7917qgXp34n9lbJ/m8dkfIGTEz/ANxs9hq+hhrpKSGSncVdG1PpjmZSwPNwQb+96hTqmg11fIfy6v8AEjQWIjRTYfpUC3HvXV+q9vnz8C9o/M/amBSo+1xu+NlVM2S2jmp44jHSZEyLNE8xZGJAZf68e/DBqOtEav8AV6dV5bQ/lj/K7P7o6+X5DdxYjeWz9h52LIYzHw1UWpaSBx44IQ4B/SALW9qbfSA+OI+Z6TXFQyd3n9nRz/kT/LwpO2+1uot97ayNJhcV1tmMflDRSGPyTfaGNn8ZC8lil/aalOlArWvp0efubofrfvvaNVszsnCR5nE1NM1Pe0SVMGqxMlPM6sVa4/Hv3VvLOeiQ9T/ysug+t90Y3Ntjamvo9tVT1e1qR6wyChldw/74kQhh/gB79k8evAUyOPRh/kn8LelPkzj8X/fnbcUmb23TiLbWUpBBSyY9k/zYcol2UfS1xb8e/daIBFKdBt0N/Lt6Q6azeM3ecO+V3dhJjLiclNUtLHSMTcuqOty3Fhzb37JFOvDtNeqDP5jPwqPfv83n4j5/O7er8hg48tjxkMvSssMNFBJXkVBc24shP+v7V3BLqregA6RwJ+qQOFSfz8utg7pL+X70L0pvar3/AIPAPU7kjyc1Xha6snFS2OhZw0egyKW18fhrD2lJr0rUaR0YnsDofq7s/NYvcO9Ns0uXzGGjEWOrpAqT06AltKSqNX5P591pXq3TrhOoev8Ab2Xx+dxOApaXKYyBqajq1RPLFC4sy69Nz/t/e+vY49B32b8TOiO39xUu6997HosrnKNo2hrIz9oxaI3VpRABrPHJa5P59+68MdDrgNvYfbGJosHg6Cmx+Mx9PFS0tNTwxxokMI0opEYAJ/qSLn6+/de6eQqqLKoUf0AAH+2Hv3XuulVV/Sqrf66QB/vXv3Xuq3f5km3WyvXexa2BLS0W+cfLI62v40kR/Ufr9L+x5yLcGO9lQnBjOOgfzhAZLOJ14rID/Po+fX+TpstszbFVTNqT+A4hHH5WRKFEdT/sR7Bt7G0V1IrfxH/D0KbZxJAjD0H+Drju/r7aG/KVKPdODoctBGxZRUU8EjAta/rdSfx7f2/dtw2tzJZSFCfQnqlzZWt4um5QOPmOp+19o7e2ZjIsPtvF0uLoISSkNNBFELnj1GNVv/sfbV7uF3uMxuLxy7HzJPVre1gtU8OBQq/LpSe0fSjrxF+D791ogEUPSSo5DT5+tRjaKQBUBsB9fx/sfZhINdqrefSGOkdyVA6Vvsv6X9e9+691737r3XvfuvdMuepfuMfMUH70a3ja3IPtRbPolFeB6YuFLx0XrvCVKzUUcQN5KdRHIf8AEG3v1yhV9Xk3XrdqppPEdOk00VPG0sziONASzt9AALkm3tgCvT/QK797z2nsyJx95T1lUFa0ALhi+i62t/sPfsdazXoke6e4uw+yMlJS7aSqgopZNOiMy20yegaR/S/ulR59bp0uevfjVmc7UxZrdtRKxJjkkjmkNyS1msrf4e90z1ojz6Optfrbam0o4hi8dEksaqDKyoxZhzcC3HPu1fTr1AePS94A/AA/px711voqPyG7kj2rjZcHhqkNk6lJYX8ZfUC6BdPp/pf34EV69SvQM/HnqWq3Hlzu7dEUtRC8n3MZlIYamBZbGS/v1AOPWqnyx1YhBBFTQxwQoscUSKiKoAAVRYcD37rfWX37r3REvmRij/D8PkIuXWcu1uCAHC345/HvRFPz68D0OfxzyQr+t8YpfU8B0n/YoP8AH+oPvdKDrVc06Hr37rfXvfuvde9+691737r3Xvfuvde9+691737r3VeX8zrvLdnx++KO99/7LaWPPUksFFSTR6x4jWQtGHJjIPDMDa/sJ84XNzBtYjtZPBaWRE1+Y1GmPn6dRH7381X/ACd7dbhvu2ELPCh0E10hiKAmhGASCc8OtHTD/Nn5L4vfh3rH2huOWvauhq5aRqyVoHKTCUxGC9gp+h02Nvz7BMmxweARFJIk1D+qHOqvqfI58iKdcsIfdb3Btb798x7pKZywZtWkxNQ106KUVTwOmjU/FXPW6x8QvntsnsbovY2a3jmaKg3GdvUhyX3EkvknrC+h2YSc8X5v/T2JeWua7aaxS23CQePF2u38TA0J66ge3XuxtHMvKdpuc0ih3iRjk8SBXjnBqOjpbL7X66zKy1NFuXGzSTvqQIWuoY/QfX/W9jiXebCbSkUoOOpMsd62qeskcq56F2ly2OrUElLVxTIfoyE25+n1HuyyIwqp6OUnhkFUYHpwBBAINwfofd+nuvWB+oB/2A9+691HqKOlqo2jqIIpVYWOuNGIH+BYe/de6StZsDalcSanFwuTe9lUfX6/j37zr17yp0jsr0XsHJI18YI5PqGHjI+obn0j+nv1BXPWjgdA1kvjTgMpkaunoWSnjVD/ALsUfU3HCj6/7D2plhVYg/r0xHIzSFfLpBZH4j5Og1VeKyRDA3Ma1Fza/Fhx/vHtNTz6fqfI06QuQ6l7bwzMcXX1XiivZVZzexK/1/pb3XA6tSvTXFl+6tuIVlauZRwCfJb66eBz7sMZPVSM0HT3jPkB2NhpETKUddOIGvc+W3HHI96yTU+fW6AY6DHBdgRp2bJvfM07VB+6FQsD3DRNqvzfn2eneNNh9GF4inRONsLXpuieHR78V8m9m1/NRJDR/Swkd9V+L/763sir69G/Hh0JeM7c2NkhH4s3S63+qgsQp/Av/wAa92+fXs9LKm3HhKu32+Rp5L2tYnm/9CR7114GvTvHNFKLxurj+qn37r1R1k9+631737r3TNnnMePkYC51KB/rkH2otf7Xpi4IEeRXrrBJpo7/AOqYE/48fX3u6I8Sg69b10dPXtN0/wBe9+691737r3XvfuvdMuckC0yqVvraw/wP4PtVairk9JrgkAU6kYmMx0aAm9yT/t/x7pcGsp6vAKR56cvbHT3Xvfuvde9+691737r3TFn9X2h0mx/p/W5+ntVaf2lOk9zTw+pWHBGOp7/XSSfzzf3S5/tjXq1uKRCvTn7Y6e697917r3v3Xuve/de697917pKxIGzCuCGAZr8fS3Aufa56iDGOkSms+elV7Q9Leve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6xy/5qT/lm/8A0Kffuvda/PQ//ZfdRY/Xclbf/YVJ9+Pl1YCrN1sFe/dV697917r3v3Xuve/de697917oK+8QP9D3Zh5Ftlbg5BINhj3PBH0/1/ewadeoCevn5fDb4O/H35UfzP8Ac0XcO38xmo66sy0UxpczNTN4/O7lB5GKj8829uGlB0kRTreuaHFeHR3/AOcJ/L52L8Bsr1d2p8Up8nsRslmqdq01eZnq52ionSN4lMZQ2tcA6fdVJJ0+vV5VCjxAKEenSs+c/a+d70/ll9U7m3VX1NfmKGSsxzVq1FTE8kdPSqLF9VzyL+/JiQfLrUwrbk+uerT/APhPKgj+G+NUPK187kjeWaSZuIVAs0hJ91bLHp2IARr9g6a/+FH1HT1nwe3HHW+Y0X8Jzgqlhnlpy0TRx6vXEQfx78OPXpcp1qYfyxYP5QlP8Ntxx9311K/ac2IzLUFNNuurpas5Np2OiWl5P5Fvp7v5UB6bBAqXA+dejjfym6ntSbe/f1H1ZNlKPoxcTVHB0y1dVUUn8O8liFkJIJMduffnrUDqsJqrMvAnquXadZ8QsH87u3JPkHtOv3CG3FMVC5yrpSD94SbWbj8jj34Ale3rTFUlJkz9vW19/Lz27/Ld3HvLJ7l6X61yeCz+Keepjlm3DNXRF1pw4bRJ+NYUm3urBq56eV4ytYwOqqOy9ybw/mEfzC26N3znqyXrPCb1O3q7Cfe1NOsmPgrTSBYi7aQdBPIHuwwgYcT1QktcGI/CBX8+rzOw/wDhPL8O5MfhKvqLEZfam7sGIhBlslnZ6mncxMJNYiRLglh+f9v7pqb16cMaEUYD8sdW9fGDpTLdD9a4fYWTzKZr+EUq08NQjSMLX1MbyAH3rq44dGR9+631737r3Xvfuvde9+691737r3VdH8xQf8YzxJHB/i8fP1+kqfj3vy6rxJB6/9e774Vhl/mOVI5t/o7mv+P+XW1vevTr3mfsHWx17317r3v3Xuve/de697917r3v3Xuve/de6QfZPZO0Optn5jfW+cxT4PbmDp/uMhkKm/igjJ0qWC82v/T37r3yHVZOH/nTfB3LbzxWzk7e24KrN1RpcaympMlTIJPFZEawvf8Ax97OB1UEk4pj59Vof8KQf5iHanxB6R6o3v0TmqyOi3zi3yNNmMRPUxxVMUlSft5DNTsuldBUkn6D35VBYhvIdamdkjVo6VLAZ4Z6D/8A4TH/AM0/s/53YbuTavdNfPPuHYlBSVlNXVtXUVEdUGqYzIIaiqYg2jfURa/B/p78dNaKPLryFwhMxFa+XCnVEPzF/n/fKOl/mKbp6m6/ymYx2xtn9pVuz5aenrcgslSKTJGJ3FGj2MYW1zf+v492QK1PXPTVw00RLLQqKYPE9bU3z3/m403wb+C/Sm98jSHcfc/bfX9DnMLtwPUDKVK1TeL7uJYmD6iz8E3+n0591UE8Mnp2V0jy2Af8Pp1Spi9y/wA3DufrGm+TG2M/2NisFu2ipd3YjakDZXyJjZHMhgp4+SVspHH9Pdv0qitemv8AG80A+Q9ft6vv/lC/N7ufu/q3f9L8jNmZrZmQ6soplqdwbhSrhbLNjQ8tU5+7tz41LWH196NK0GenULaQzihOKenVLffH83r5E/zBfkjuHoj4P0ufoMBtTN1m1shu/a7ZFoabIUOqBvv2iZlV2ZWb8fS1vewoC1fz9OqGR3do4h8ODq4fl09dE/Jb+Zf8Q/lJsfZ/dWG392hszMigOTrq1MtNjEjqJwHBkPpDWP596rHSi8evKbgN3gFT5jj1ugbVzMm4ttYHPS0rUUuZxNBk3o3vrpWraZagwNfm66rG/uvT3T/7917r3v3Xuve/de697917r3v3XukF2lb/AEc70v8AT+7uS+v9ftzb36letNUDHVJf8sD/ALKX7q/7VlV/7nJ72wyOtR/C3V+PvXVuve/de697917r3v3Xuve/de697917pD0dqfNyqF/zs1yfpcEkD2aOC9qCfIdIFNJ6fPpceyvpf1737r3Xvfuvde9+691737r3SX3bE0mMXSCStTEbD/A39rLL+2p8uk10Kx9P9G6vS05Ugjwxjj+oQA+00ilXIPr09GQUBHUn3Tq/Xvfuvde9+691737r3XvfuvdJXdCM6UJBAEdRrJIB+g/r+Pa+xNC3zFOkl18Ir0/0UglpYnH0Kgf7YW9pJhpkI6ehNYx1L9t9O9e9+691737r3XvfuvdBXltm7Qqt6YnOZHCpU52kkV8bkfFAWpSDq4dl1Dn+h9r9LS25f+H7ekWoJcBfXoVPaDpb1737r3Xvfuvde9+691737r3Xvfuvde9+690V75abTk3X1fPHFGZGxVScncAEqKeLV+f629n/AC5dC13EE/ix+3ol363NxYFV/Dn9nWD4gbkbcvU1PUu+p6PI1GOa9+PtUVFHJP4935mg8DcitKVAP7eqcvTePtwYmtCR+zo1HsO9HvXvfuvde9+691737r3SPz0Rp62iq42Ch5h5r/kC1/8AePZjaESRtG3kMdILoMjq6+vStR1kVXU3VgCD/r+y8gg0PS4EEVHXP3rrfXvfuvde9+691imeNInaZgsYU6i30At72K1x1okUz0Xzc3cW2uv6iphkqoJZJyWSIhgxbSSv0P8AX2tndJI1Na6ek0CspbV59FL3h3pvnsGukx21oamkpZZghePyhCGHj/r/AI+0JJbPDpVpC8OlBsX44bh3dJHl931ModSslppHGoav7KsD9R/h70M9er0dXaXV+1No09OlDj4jURKuqZgramB1AgEe99e6EcAAWAsB9APoPfuvde9+690CvdPZ1BsDbVVIKmP+JTxyxwwcmQXjuGsLf19761WuOiO9VbKzncO9G3BnjI2OWoEwaRjo8bEtbS4P4HuoGetn14jqzfC4ehwWOp8bj4Vhp6eNUCqALlVCljYD+nvfXunX37r3XvfuvdFl+TuITIbMafRqenjma/8AgpDD/e/ficdaxXpE/EXMtVYGfHM+r7Yy+nkEabAfX3qprp68Ojoe99b697917r3v3Xuve/de697917rokAgEgE/QX596rmnXqjrv3vr3QQd49MbR7568zfXO9aMVuDzUUizwnT/nDA0Mb+oH9Nwfp+PZVvO0Wm92LWF4DoJBFDQhhlSD5EHNeg/zPy5t3NWzTbJukYkhnUqysKgggggg/I9a9e3/APhPtj6DtQZ/K72pKzYEeVWsjwivatekWcSCjnn8d2W11+lz+SefYBHKfM8jC0nuY/p65ZQwlK+leANPMDrDC0+5XtUW/C6ub+SXb1fUtsaUpWoRnpqZPKmCRhic1uAqfgl0Dh9tYnaO19sx4iroqSOnjqY6nxmTQBd3UWUEHkBVHHsW23JGywWwEURGkUrUkn1J9eso09seUrSxTbtut0h0KANNFGBjAAH5DpGZD4L5mhgI2bvX+DSqlo1aSc2IW6jUotwfZPNyfOCTazaPTjjpBJ7bTRj/ABG5MZ8ugly/xt+W+z6wVeJ7XlyWPgs4oIJZS0oT1aQB/X2Tzcv802r+Il1qUeQPRFcco89WEniW1+XQfh9euk76+UHVzJS5DrPcu80itE9TDDVMjBOWkuotbn3U77zJtY0PbPMB50PVf6z87bNSOWxkuacSteHQl7b+e1ZTKqb82RVbVe6+Q5FZ1MX/AB0vqI+n59mVvztKo/x6Ax/bXHRxbe57xKBuls0B89QOOh32180On9wrFo3Fjo2fSr2d7K5+qm5+o9ntvzdtM/CQDoS2XuFsF4BpmWp+fQ44fuHrzOIjY/clDOZACFRif1fT2bw7tYT/ANnID0JYN+2q4p4coNeluMxjaqkkmp6yKRDExDo1/qvHsxidXYac1PRiJ4ZEqrCh6btu0wZHr2N3mLrc25sRz/tva+8fIj9OqwL+IcOlR7Q9KevEA8Hkf0Pv3Xum2txGNyCBKqkgkANxeNAb/wCuB78c9eGOHQcb02Zsuh29lcpW42COKkp2nklso0hTz9R9Ofb1tEJZ1j9T01NJ4URf06IV8athYzsbfXaMuZpGmwMSN/AmuFjDvMI1Zbg/S9zb2J+YrK3tLS3EQo/4v2dEOyXc11cTlzVPw9GBzfxLw1ff7KdKYj9DGVh/vS+wjSpqehGOg0yfxc3Pg9T4rKSMRYp453YD8jkf639Pe+H2da6Rk2wu7MFJqpq6taJD9AZSP6/X3U+o63616wNvjuTbmpag10p4I5muLcke7V+fWqeXSmw/yT3li3U5TH1coT9RfykX/qSf8Peut06FHFfLqilkjgrcSsQJAaRllFhqtc8+99ax69Ll/kNs/MUvhlrqWndmFlIe/Jt/qvai2cI2o46anXUtB0LG3+ydm11HAsGXpg1gLc2/pfi/ukzBnJB63GCq0I6W8GZxdSAYK2CQN9NLH/ifbXyHTlenFXRwCjBgRcWP4/r791v5dcvfuvde9+690mdwTWFPH/WUD6/1/HtdaAAFj0juSxYKMdP9NGIoI1HHpB/2JHtJI2pyelKAqoB6z+6dX697917r3v3Xuve/de6Tefc2giHIcm4/qL/T2usx8THpHdZIHTzQp46WFLWsnP1+t/8AH2llOqQnpTGKIB1L9t9X697917r3v3Xuve/de66JsCf6A+9jj1o8Okvj115F3Hps7mx5vY/Ue182IQD0jizLXpU+y/pb1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3WOb/NS/8ALN/+hT7917rX76FAPz8qQTa+462xP9fuW+nvXEA9WrRz1sEe99V697917r3v3Xuve/de697917oKu87np3sxQOW2Xn1H+u9A6D/e/e6dernrQG+Gfy26s+MP8zbdGU7WzWLwOMxtZlZpKjJM5jZfO6EWS39b+3GB0gjy6SRMBI9fM46NF/OR/mGbE+cG7eq+rfjtNSb+gx2dpY6ufBfcPMsFVMjNJLrv9BqPA+nuqDT3nq0zKx8Bcluhr/mA9T5Ton+WB1djMrQyQ1Qgqc1V00rDXHFUUi3ZtIBte/vyfGDXr02ICvpQdKH+SN/NR+KHSfxcpNo9h9g7b27k0yuSqJI6kzmohMSaGjBT+pXnj3pgdR+Z6cjYaQPQU6Hv+dd8rumvlB8DshV9Sbtxm7aXKYHMyVC0DSXjEoVPGdf9At/9j70BU469KaIa9VcfyLv5HXw4+UPx3xvZPb+yKrJZtzlI6uSPOywVc0/mCQllAN1UEW44A/p72cdaUau40NfXrbQxfw16G+HPxp7B270ptJMFTQbXnjM87xVVW9lWIgzaFNiL/Xn/AB91z69O44U608v5f/w16K+U3z47gou59nS5un/j9RJG8NctI7BclYhGKnn1e7nAAGOk69zseNMZ63R+mP5d3xY+PrZCbqvYcmAnr0dZJZMjLVlS8Xj9HkUf4Hn8/wCHutT0/pFKDHWnZ31j9z/y/P5j1H2nunBzxbKze9jna/L1Qc0MNFJkTOHkMYHBUX+vuw7o9A4jPSd/0rgzHgcfn1ah87f5/nVtD1bgf9lnzmP3tvjMR000uGw0lY2Qp5piI5IEKEX0sbDg+/BCervOiAE8T1b7/LE707P+QPx92/2B2dgclgcrlsdT1K02SWYSlncq4/e54tz7p08DXPVlHv3W+ve/de697917r3v3Xuve/de6rp/mKD/jGWJ/7W6f9bU92Hw9VPE/l1//0Lv/AIXMT/MeqB9V/wBHUvIAtf8Ahbe/D/P1vzP2D/D1sc+/da697917r3v3Xuve/de697917r3v3Xuqo/5u24ursP8AGLc1D2t2TQde7dytA8Mz14fx16RzaiupDcBT9T7159eOR18+X+Zdh+ltp/HHqnsX42bBliymMoZqpe48Pkaqanr5P4ifJX06E6VKqCf6f4e3QzEk8ekbxR6AtKZ6sf8A5lO5NzdpfyRf5VGT3/lX3Lujc+xNzpm81MD5cglPlGpqdHaUkkRpZRc+9RdxNfn07eEKi04Ar/g6JD1v3TTfya+iutt34OGekz3yG21LmaKOmM01XU0THTJNeAodA/tFifwL3t79iucU60FkAoc1z0XreXSeHnym2flxkceEy/erR7ux9U7AtI9RWLLLLGji63J5N/8AX93QDgOmJy1NRPGnVl/88x8xF25/KA3pvNZX6sp+gNrPn5ZnkFEGOROlZY76T9R9fdI9Ws09OnbvT4S6/wCMU+3HW/R8UNz7Ub4l9J7nxtVSw7Q/0YYGsoqpViipUxUNCfGyonp0hFtx9fzzf3QZ4dK3NDVuiI9r/Jn46/Iv4+/I3bnxS3NiMrvnbsuaod10eFpPtKuOugx1QtdJIYByGAYXB/Hv2a46qfhziv8Ah61mv+EgWd2vi+0/mbgq5oot113bk0dAskcU00kMbzxzhJJ7uthqBKm9+fr736kcOqDiobiOtzb5MfJ74v8AR8uLxnfeew+Nqa2alONgyNHHUSB6lwKdllb1KCf6e9UJ6vUV6NFtbMYbcG2sDnduyxz4HL4jH5HDzRC0UuNq6ZZ6OSMf0MbKR791s1rnp+9+691737r3Xvfuvde9+691737r3SA7Ut/o43pcX/372R/2/hNvex1oivVJn8r8j/Zl+6hcE/wyrJ/rzXJ9R701ajPXo/hbq/L37rfXvfuvde9+691737r3Xvfuvde9+690jcl+xuLFqgss5u4/Btxf2YxEtaPXy6QyDTdL8+ll7Lul3Xvfuvde9+691737r3XvfuvdNmWj8tFIB9R6hxe1h/T2otm0yjpi4/s+oO25jNQsTf0zMvJv9AB7veCkvpUdVtSDH0ofaTpT1737r3Xvfuvde9+691737r3TRm4RJjqlrXaKGR1H5JC/T2otnKygDz6YuE1JX06wbadpMPSs9wxDgg/UWa3vd2KTkdbtzWIHp+9punuve/de697917r3v3XuktmUC5PHVN7GMnj+vNuPZhbEmB06RzhRIG8+lOrBlVh9GAP+39oCKGnSsHUKjrl711vr3v3Xuve/de6i1FZTUoLVEyRgKWOogWUC5PtxIpJPgFeqGRFNCekBmO2tiYMSfxDOU8LRh9Qb+qC5H1Hs1t9h3O5p4UZNekcu5WkXxOOgD3J8yOusU81NjK+kyNYpdIoFMhaRxwqgA/n8exRZ+326zAPOpRfXoouOZrKM0jYMei67/wDkf272DtndGK291ZlJ8fUYPKJDkqeGbxgSwFYZL8j/AFifYq23lHYNruYp7y8UMHWqkj8+ie73rcruKSG3gJBU93UD+WFubO0Wy9xbE3k8tLuUZ2sygxtSD54qdi5bl+eBb8e03u5Dtkm5xXe1UaPQASvCvTXIMt0lo9peYcMTT5dWve4d6kPr3v3Xuve/de697917pqzFMtRQzem8iIxQ/lTb6j2otn0yj0PTFwuqM04jrBt+Utj4opCTLECGv+oi/BI92u1pKWHA9atmJjockdPntL0o64syoCzEKB9STYf7z72ATw60SBk9BfvXtramzaWZ6vIQmrQDxwH6OSOALEfnj3qvWx0SDfHyD3dvWpGP2tTTwRtZA8AkCuCdFxY/4+/Gvl1UU4EdRtsdB7v3l/uY3bLO/qEieV2GlVYX+vu8Ka20nqsraRqpw6O1sLq/Z+3sdAlLQQSVkKhaiS6v676h+P8AD3aaIxnSetQyCQVHQvqoUBVAVVAAAFgAPoAPbXTvXfv3Xuve/de6Rm+t4Y/ZeAq8tXyrGFhmEGr6NKsZK/Tn629+60a9VnQf3i7838fIJnxiVl1ILqhi1Wv6rj6e/fb16tOHy6sx2TszF7Mw1LjsfAsbrBEs7ALdnUc8gD3uvXuln711vr3v3Xuve/de6CfufG/xHYuYUJqMVJOw/wAOB/xT37rx4dFG+IuRFDnsrjZTYu9RGLmw5Pp/3kD34Z61kfPqxH37rfXvfuvde9+691737r3XvfuvdUy/zdO++4vjvsjBdhdaLUtj8WaZcwIjMsAM9S6Bp2h+igWv7jnnafdI7y1hspDEkmC/kDmnpx4dY3feL5x5q5J5fTe+W4/E8Nl8SpIAVm0liR5KDU/LqpnqT+fF2li8dj6fcXXkedqgmiWWKqlfyMWALKJHU/T2UjeuZdqQjVDOF85GZSftwesZdq++ndbTEsG+7e8kgwTE6sCfkGKn8uravh5/NcwfyW3tj9iZDbMe3MzXtZaZ0mWUXnES2JYqb/4E/wCPsy2Tnua+v49v3CFYnkwCp1KeHA/5x1kV7T/eW5d9z9zG0WMbRTgVKOpVwK0B4lSD8ifnTq5GeVYInlc2Cgn/AGNuB7kxFLMFHWT7sFWp6SeGgkr6yXIVBJMMhWJWBvY/kX9rrhzFGIl8+kcSCV/E9Olj7L+l3XvfuvdeIBFjyDwQfoR7917oKd29I9Yb5aRt07Ux+VMpYv5VKXL/AKv81p9ll3s+3XtfqYw1eiW95e2fcSTewLJX16BvMfCnoqalePA7RpMHUsWImppZtIY/29Lajf8A2Psofk7ZKfoRBD8ieg/ce3nLTL/itusTeo6BrMfBrNwpIdob+bDMC3gQGosgPC8gfj2VS8myrU2s2j9vRDP7bzLU2N0Yz5ccdF+3f8cvlZsuoSXDdq12XpUkNqCmkmJKBtIHB/I59l8fLPNFvKZUuiyrkAE9BO/5T5629w1rfl0B+Hp0x3dnyq6mp46Oq6s3FvGKOyzVSQ1TKqKugysV/qefbUm/8z7dJSW2ebyrQ+XSmPmLnnZAEksJLkebD/D0K23PnnXY9VXsPZsu1gbeR61Z1KEL6r3I/PHPtfb87SKP8fh8L1Jr0b2vuc8AH72tzB66q9Dptf5p9O7kaNY9yUCGQi4Hk9Btdhyfx9PZ5b837PccJB0J7L3D2C9I0TLnodcT3B17mlU4/cdJUavppJ5JGoD/AG3s4h3awnNIpAehLBv21XGIpQT0Fvyl3xi8V0VvuSjro3yNdgpI8XEhJeaZ3Urot/gD7FnLEQu92h05UNnqm9XcS7VKyNkrjpO/CHA/ZdC7QzdZCY8xm6aoqK4tbVbz6ERri/8AZvyfavmyXVvMsS/ChAHTXLUZXaYpHFGYVPRvpJEiRpJGCoouzEgAD/XPsM9CCtOPTdS5rGVsrQ0tXFNKhIZEdbi31+p9+61UevTmVVhYgMD+CAQf9v791vB49MNbtjB5Bi9Vj4JGJufQo5ta9gPeiK8etgkcOkvXdUbHyCss2GiGrklNPJ/x1A+/AAGvWjnpAZr45bIyCkUdGtMxW1yR9bcfpX/W97OeqgU6BDcXxPpRNqoqxELXK2mI0/heLD/evb8UWpCem5JtLBekXk/jrvzBqFwuSqGGnWpikkYCwuACvtg8aHj06uRX16TLbb7x26+s1WQeGK9v876tIvYe98OPXhkEDrqPtHtjCyD7qKtkEZCsn7vqC3vyf6+9AdeJPDpc4n5P7ixPjbIYuWQKRrVxLZj/AGr8+/ZGOvDoUsP8scPW6PvaSKkvYMHEqkE/6597qOvZ6VQ702ZmqumH8RplPkQ6QrC1+fqT/h7VwyokZrx6TzRs7AdDXQb72xkEQ02ThfUq8AH6kf4e0ZIrTpTQgDpRwZGiqf8AM1Eb/wCsbfi/59+p1qo6m3B5B4/r791vr3v3Xuve/de6SuYYtkKFB6gZACPqAL35HtfAdMLGnSGapmA6VQAAsOB7QdLuve/de697917r3v3Xuve/de6w1DhIZGJtZT9f62493jFXA6pJXQadMOETyPPMx1FXsD/r/UD2sujpUL0mt1qdXSk9oOlnXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdY5f8ANS/8s3/6FPv3XutfzoVC3z6qSBe24643/p/lLX/Pvx8ut17z1sD+/da697917r3v3Xuve/de697917przWJpM9icjhq9PJRZOkmoqqPj1wTpokXn+oNvfuvHqkvuT+Qz8L+5N+5HsLcGykfPZOSd6mpFdJCzed9bACGMfn36pHDqukVrQfs6F342/wAmP4T/ABu3Cu79q9bRvuiJojFXz1z1UKmJbA+GdL3/AKWI97qTx68FC4AFOjk/JL4odX/JvYi9e9gYiKu29HQnHxUg0RLHT2C6FOk2FgBwPeurUHVRlP8A8Jz/AIPUEc8OO2alNHUSTyOq5GqUBp2LubKgHJJ97DNwqadVKKwowH7OjZ7T/lD/ABj2v1lJ1bDtpZNvSU0lM0JqnkJSVdLeqZG/p/T3qp8+t0XgBjo5Xxn+LPWXxZ2bDsrrPEricRCZCsKsr3MpDOSyqv1IH49++3rfQ+7kwGP3RhMjgMpH5qDJ07U1TFxZ425Km/8Are/Z8uvdEz6e+AXRfSvYmZ7J2bt9aHcGaneoq6nXGzM7yeZiAIxb1f4+/Anz60BQ46PP791vopHyk+FPQvy625UYHt/aMOdLU6w01Usi01RF4wQgMyoW0+o35v8A09+4daIr1Xj01/IR+DXVO8KXd3+j5chV42XyY1GyM0iQlZBLH5EnQhhcc/n3up8+qhADUAfs6uj2rtXA7KwVBtvbWPhxmGxsfho6OBVWOGO99IsB711fpRe/de697917r3v3Xuve/de697917qu3+YnDI3WGJYISpy6ci3/HROPexTh1U4JP2df/0bwPhWbfzG6oAFgeu5vUPoD/AAs+/D59aNdR+wf4etjj37rfXvfuvde9+691737r3Xvfuvde9+691r6f8KBPgD2j86fj3iMN1pWVMs+1KbIyV+26R51nz0dXMjCKMQg3KhT/ALf3ZDpavTc8fixeGMGoP7OtY7sD+XL8vu7vgfsf4pbY+PW6+uarpzDV1JU5dqOtEm7RPVtKFkM8d2tciwPI92XSp44Py6adJpEHaNQpxIpj/P0fv5Q/yz/k9vz+Vv8AAP4+YfZm4KveHRm2M5j87TxxS/eQyZDJGrRZ0CXvz+QPdYiEbPDPVrqKSaPSnHHn6dBF/OC/kt/IX5GfFD+XfSda7ZyNXv7qDqSbZ++aJKaSeSCoqcj5mWpiiQHUE0WZrEEW+nvSaSauaDpybxlWkChjUYJpinr0MvzE/lI95Vnw9/lwdbdb7PzOQ3L1d1mmI7KSkV2nhywrlnU1YjjuCI7A6vyD7tEwU5Pl01dxPIgWOlaivVn/AM9P5R2Q+dX8s/pbp+tqF2z3V1X11jsXisvValq6T7RjJJQKQjNq1Iukccn3RSQMdPyqGOcgUp9vWvhg+uv5yOxep6D4Q4bbnba7a2pBBtXb2+4KbNfYU2PH+TSyU1Uq6PGwJY2Nvbg8KlMjpNS9BrRSPWuethL+RV/KV3l8E9g9l7l7m3Eu6939yJUTZWkq3nlqaWateQVzVa1C8M6OV/2PurMSa/l06kQRQDUmpJr6nqnX5VfyjvlT/Lw+W2Y+TnwXh3FmNnbjzNburL7I2XFXytV5Wvkaq8lVDSoCGSRiLc3+v09+1KQFbgOqvHIHaaLJbjXgPs6TPTn8uT+Yf/Mg+Vmzu5/lNkt6bA6/xs1HLltp7xjytPBUGjlD+VEnW+sKoCgDn3Yuq18Pz6p4MstDcimk1FD59b3HX+0qbYWx9o7Jo2D0m09u4jb1M66rPBiaFKKJhq55CA8+2+lRya9K/wB+691737r3Xvfuvde9+691737r3SA7U/5lxvT/AMN7I/8AWk+9jj17qkz+WAtvkx3Ufz/DKu//AJ3J783EdVj+Fvt6vy966t1737r3Xvfuvde9+691737r3XvfuvdJHPgx1+PqgLGE/q/pc/X2YWmYmQ+fSS4Olgw4jpUwSeWGOT/VqG/2/tC40sV9OlKGqA9Zfderde9+691737r3XvfuvdYahA8Eqn8xvz/T0/X3eNirgj16pIoZCD0nNrjxU08DN6xO7Bfza5HtXeks4enl0ntaKpUdKn2h6V9e9+691737r3Xvfuvde9+691gqo/LTzR/6uNl/24/x93jNHB+fVJBVCOk3t+cpPU4+/FNdtP8AS7W49rLtaqJDknpNbNkqOHSr9oOlnXvfuvde9+691737r3TDnqcvT/cr+qAAjkA/X68+1lo9G0evSW5Qmj+nU2mqYoqCCWeVEAiDMzOvA+p/P49sujvKQo8+nVdFQZx0hs523sPbwkOTzcMPiuHIGoKQuog8j8ezO22HdLungxk16TTbjZwf2jgdAFuT5mdaYxpabF5Gnr6wXEUQWQlmK3UWU/k8exPZ+3u7zAPMpVfPonn5nsIzpVgT0CeX+T/de6JmpNodYZGsoKi4iyVLFNYRyDQJBp/oefYkg5N5dsl1314quPwkjopffN1nfTbW5ZT5+XTDT9S/JDsOsjrq7deU2tFOQz0crzKIlc6WiKniw9qX33lHaojFHCsxHmKZ+fVBtu93koeSQxg/b0M2D+IOQYQy7s3e2Zk9Pnik85D/AIcagPyOPYeuefohVbG38P0OOjKLlok6riXX69DVi/jB0tjlp5ZNk42rrodDNWTmZpGlTnWLMB9f8PYdn5z5hmJAuGVT5CnRrFsW1xj+yBPr0NmKwmLwlEuOxdHDSUSJ41p41GgJp06efqLf19hy4uJrqQyzsWY+fRpHFHEuiMUHp1VpVVI6h+e1fn6++N2Nn8CtDDputP8AxKupQpCDhb+Rvp7H6Im48liGPM0bVPrQH/N0CC37v5qMjYidafmerXIZVmiimTlJY0lU/wC0yKHX/eD7joihp0OgaivWT3rrfXvfuvde9+691xZQ6sp+jAg/n6+/DHWiKinSUx5ahytTFM2mKSyx3IC/W9+f8fZjIBLbArxHSJG8OYg4HTNu3tDau0IZWyNfGk6AaIjyHLcAAg+y7FCT0t+zoj/YPyO3Duyt/he06eSNAdCTwJIoPrIve59+NfLrw4GvTZtDoze/YVXHX7sqJzDI10ad5FVAG1A8/wCH49+4nr3AdHY2V0vtTaNNEoo4qmqiFvMeQRweQwv9ffqDh16nQvCNFjESqoQLpCgCwFrWt78DQ1HXiARTpGfu4TJlrEUdQ5eRudIJPszotxFj4h0iBML0OB0s43WRFkQ6lcBlI/IPstIKmh6WghhUdc/eut9NeZy9Fg8dUZGvmWGCCN2LN+WVCwUf7b37rR9Oqx+0N+Zvt/eI2/iPJLjEqliaBA6owJCawRcfj8+9A9b8+jy9O9X4/YGApQIUOQngieST9TR6l1FdRH1597pQ9az59DP791vr3v3Xuve/de697917pMbzpxVbWzsJGrXj57D/ABA1W/3j348OvdVo9I102I7XrqNrxK1XIlvp9XsL/wCPuvz69xFOrVF/SP8AWH+9e7de679+691737r3Xvfuvde9+690UH5y9O0nd/xv7A2LNAkk2Sx+qJjpDBoY3IsxB/rfj2G+a7GW/wBlkitjpkBDKfQrkdR/7n8uRc1cl32yyD+3jZPnRlI8/t61Iv5f/wAetv763h3VtzI4qLNbx65q1osLgQR5ci9NWvDU+BWB/UYz9Bf3D1xYtvssfjVc+EHWMGgZ9VG+2nof2dc2PYD25sL3dt7g3RFudysJBEqEZIVnBKg1pqZOPGgpXrYe+ItL8V4NzUuOruuMVsDuDbE5pAtQ5FVU1SzjWVsB6tQv/T2MOV4eWBdrE9uIbuM0FfM/5+s8OQo+SIrwQGyjtNwg7eADcfUCvz6tQyk02QqoqGlJMLf5yRTcBlIvyt/cywIsUZkfj6dTXKWkcKOH+HpTU1OlNEkSADSoBI/tEfk+0TuXap6UogRaDrP7p1fr3v3Xuve/de697917r3v3XuumIVWY/RQSf9YC/vYyadeOBXpJ0k1RW5yWUSFaSOOypZSCQLH6j+vtc6LDb0pk+fSVZXeXTWg6VhAIIIuCLEHkEH6gj2g6VdBbujpPq7eYYbn2fi8uHLFvuUk9RZtTX8bL9T7LLrZ9tva/UxB6+vRPd8v7Pfil5brJX1HQPZv4YdHVVOV2/tDH7dqTe01G04QE86tLFjf/AGPspl5Q2XTS3iEZ+VeiC45A5cdNNrbrEfl0DOX+DWYjEkm1uw5sQwN4okFQAP6DVa3spl5MlWptJ9B/PoOze2kq1axuzGfKlcdVrd/9Nd/bU7i6b25W9kVu5duZLcCU2ZxheV4jStUBND8kC3+I9iDlPlTmbbZpL83LMi5FDj54+zqO905V52seYtuVNwM1r4hEqEcVIxnyoeth/aG3qHau3MVgcdD4KTH0kMUcQtZT4wZLFQB+q/sTXM73MzTSGpbrJa2iWCFYVFAoA6Ix/M27pyfSfxR7F3Tt+sej3FS0dM+OeIXkHkmMLlbEEHkH2n4mnT5wCfTrXY+K3Tn8xbsLrkfInB9k7wzmD3JBPkcRgI6quby+GUu8EUZYj8W493bQDw6ajWYrUkV6se+Cf8zDfmS7apvjj39tifbO7K/JDFYSoyUVQK3IVJnEK3Mjcfn8W9+pio6shq2luP8AL8uthT3Xq/Xvfuvde9+690mHleXMRwnlPyeDa3NvZgoCWpYcekDgvcBT0pxxx7L+l4wKdYpoYp42imRZEYWKsAR/vPv3Xuk5UbM21VEmfFU8hb9Vwefxzb37r3SUyPTmxcgra8RGrHUbgiwLD6/T34U690GuZ+Mm0q8SNSxw07G5W5cc/i5A/HvRFT16uOgSqviy7ZCdcZWiMxo7oySMbEfpt7UPblIxIT0ysoZ9I6SGR6O7WwzM+KyNa6rfQsZltpT6AW/wv7Tmlenhwp0y+HuzbgBnlyDlLm58puAdP0/1vexjPWjkUHUmh7m7LxE9q6lrJo09Jv5bWAtcj3oDOOrVxQ9CNi/ljkaC0Vfi2awsTJGx5+lwf9f3YcM56p/LoU8L8p9tVoU5Dw0hNrgrIpXi9+T711YCox0rMX3RsrM5FJDlIQoe6kLybG9hc/0/x9rRLGsOlT0kMTmXU3lx6GKj3ZgK8aqbIROv11XAHtFilelVc06fIamCoXVDKjr/AFVgfe6Hr1R1n96631737r3XvfuvdQcjII6SViL8W/2/t2AVkHTcnwHpq26jLDUFvo8oZf8AAc3Ht+8cMwA8umLUEBq+vSj9o+lfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdY5b+KS318b2/POn+nv3XutZrsHO9tfGf5jt2DB15XZzAPlp6z7pknaneJ2M1j4x/T/H37iOq8HqeB6Nu/8zvfkkjyp1fojd2ZIzBOwRCbqpvcmw/J97ofLqw6x/8ADnHYJ+nWP54tTTD/AFrC3v3DrVR/q/4vrx/mb9gj6dZG3F708x5v9Bx79x69UcP9X+Hrpf5nHYP/AD7In6/8osv/ABI96008+vV+X+r9vXNf5nPYH0PWAbm/NNN/tjb34jr1eu2/mb9gH0/6MNNhfimlX/X59+HCvWzTz64D+Zx2B9f9GFwf600x/P4Fve6dar/q/wBR65/8OcdgfQ9XKP8AH7Wb6+9U69Uedeuv+HOewPoesASf60sp/P0+nv3y63UeX+r+fXZ/mb7/AAST1iLH8CmlIX/G1vfqEinXsfP/AFfn17/hzjf44/0Xhh/U00p/2Nx71Q9aqB69d/8ADnHYFiP9F6jg8ilm/Hv1OvVHz6xj+Zv2Fwf9GNxxe9NN9P8Abe7eVD1quf8AV/n67/4c47As3/GMAb3/AOUWb02/AuPeqHq1R10P5nHYAsT1je/4NPLY/wCwtx78fTrVfT/V/Prtf5nPYHqP+i8Ef40svH+A9+49br9v+r8+ux/M37A+p6xuBaw+2lsfzyLe/UPXq9d/8Ocdg/T/AEXKL/1pZube/fn1qor59df8Oc9gcf8AGLl+n/KrN79SnW6j1PXTfzOOwDc/6MbWv9KaYD/bW9+oT59eqOuv+HNuwbXPWRBH0/yeX6f4i3v3Dh1qvqOu/wDhznsC4P8AowFrW0/azW/re1vfqdbr1yH8zfsG1h1jclgSPBMbDV/W3v3nnrRPp0VL5T/Nbt3uzE0Wz9u9UVVQ1PUCuvSU9SHAVlcj0g/0N/futE54df/SuX+HW9dsYf8AmXz4bKZSKkrpuuXEUctkUs+IMiBnkta4/p72Mjh1qo1EfIdbEB7S2QrFf43TcEi4eO1wbW/V79Tr1cVp17/SjscGxzlKp/xkj/6O9+pivWtXyPXX+lPY/wDzvKb/AFtcd/8Aob36nXtXyPXQ7U2Of+X3TD/AvGP+ivfqYrXrxNDSh67PaeyALnN03+t5Irj/AFxq96qPXq1D6dcB2rsYm38bpx/rvH/0d79jrXzp1zXtPY7EgZun4/2uMA/6129+x69er8uuz2lscAk5ylFhexkjuf8AWs3v3XqmtKdcV7U2M30zdN/yXH/xDe9jPXiaeXXL/Sjse1znKUf4GSME/wCt6vfqda1YrTr3+lLY3/O9pP8AqZH/ANHe/U69X5Hrpu0tjryc5TH+lnj5/wBb1e/UxWvW6/Lrte0djsLjOUv+trjvz/yF7916p9OuB7U2MP8Al903+weP/iW9+p8+vA18uuQ7S2Qf+X3TD/XeMH/bavehQ+fWzUeXXj2lsccfxumJ/oHjv/0N73ila9aqa0p1w/0rbG/53dP/AMlx/wDR3vWPXrfXIdqbHP8Ay+6YD+peP/o734UPXjUCtOvf6VNjWv8Axym+pFtcd+PzbV791qvXf+lPY3/O8pf+S4/+jve6fPrWr5Hrw7S2QWK/xumuBf8AXHb/AKG96x69b67/ANKWx/p/HKa//B4/+jve6da1ZpQ9Ibs/s7Y56135O+bpvFS7ZyMs13j1BGhKKVBax5/x9+4ZHW+Jp1Th/KnzuO3D8jO5sliZ/uaGbFVfjmCkKxFcn5+n0/x96NcV68lNLU9etgn37rfXvfuvde9+691737r3Xvfuvde9+690m90J/uLmkX/OLbSR9fa2xP6wU8Okl2tY6jj05YdtWMoyTc+BdX+v/j7YuBSZvt6egNYh05e2eneve/de697917r3v3XuuLC6sP6qR/txb3scevHIp0kcZeDMSU5Frq7D+hA/oP8AYezGfut9fSSPSJdI6WHst6V9e9+691737r3Xvfuvde9+691737r3HpJUEP2udrnFmE/ptqUEWN72PswlPiWq1xTpDHSOcqvnjpW+y/pd0S/5jfMbY/xO2a+Z3DWRLm6qBJMRQSojrVszsmmzMv8AqT7kHkLkO/50vfCgB8JT3EeXQW5m5mteXbbxZyATwB6Ib8V/5x3Xndu42wG71pduSTSSR0TxwKvnZH0gf50/1/A9yTzj7E7hsNr9Tt5MtOIJ4fy6B/LfudY71IY5aJmg+dOrAd1/MnrDDSyUeMysNfWkhII/FITIxW4tpNvcaWPt9vNwPEnTSvn0M7jmewhOlTU9AHuT5Jd5byhmotm9Z1eQx8p9OQp4JfSv01en/XB9imz5R5a29hJuF4EYeRI6J5963a6Ui0tywPn1i23s3v8A7DgCZOvym1VA0yozTKEJOi1h/gT7tebhyttTVhVZvTh1q2t95u0pISnr0t4PhNmK+rhrNydiVWRjBVp6NhO6S+q5DMRb6cceyx/ca3iQx2doE9DjpWvKjyHXczFq8R0YLbnxd6dwtNAKnZ+LyOQisXrqhZGdmVtSlRcWt/rewrec67/cuTHOyKfIU6OrfYdthUBowxHmehzw+CxOApFocPRQ0NKgAWGFdKgD6Dnn2Gbi5nun8S4Ys3qejaOKOFdEYoOnb2x051737r3Xvfuvde9+691Xd/MB6+qc7tHZ25cIhhyWE3ZSVlfUxkh2ooWSQhtPJAsfz7G3Jt6sV1JbzZDoQPt6CPNVi01uk8XFHBP+Ho4/Um86DfOxMFmce4kjShpKGZrj/gTS0iRz/Qn+17C+42r2l28T+pP5EnoR2VwlzbLIvoOhK9oelfXvfuvde9+690g91djbY2lTTTZPIRJJGtxECGLG9rfX3unWq9EV7G+SWU3DWrR7Yo2tC2lamGJwX0sb+oH27HKQpUefl020aswYnh0w7Y6e372lWLktw1FQlFM2pRM0ioo/UPwf9h7Z4dOEqTjo5ew+hNqbPjjaWmirahAT5CW4Ygf1A/p70Pn1s1rg9DpDDFTxpDCixxoNKooAAA/HHvfXusvv3Xuve/de6hZCijr6Z4H4vyrW5BHtyKQxNqHTcieItOk/jK2XHStj68FEVtMMrX5UcA/63tXPEJVEsWfXpPHL4Z8N+lRLPFDC9RI4WKONpWe4sEVdRN/9b2gp5dK64r1Xn8gO46ndFcdkbbLOjVAhkkjU8M6hSCym9yf6+/Gnl14ZPQu/HrpaHa9BT7izEevJVKRyoj3JBPqDkMP9b37r3nXo2nv3W+ve/de697917r3v3Xuve/de6g5OA1WPrKcC5mp5IwP6llsPfvs69jz6qzEDbc70emY+IGvuG5N9T/0PvVBwPWzjq1SmcSU8EgOoPDE2r+upAb+9nj1UVpnrN791vr3v3Xuve/de697917qPVU0VZTy006h4po3jdWAIKuhQ3B/wPvRAYUPVXRZFKOKg9Vn9K/y5tvdJfIncfe22M/8Aby7myr5LJYyN5mE2uQyGPS6BBe5vb/X9gLbOTH27eP3ks5ZAWKp/CGNSBjzPz6hTlf2Y2rlXnm+52sHIlv2DSrUkErwoDgcTWnEmpz0pflN8ScZ2FXNv3rGqi2x2PRO1XLNQF0qMpURlpGYiME6jwCfz7X79yet7IdytP0ZlGqo/Ef8AP0c858hwbyTu2yuIL1DWq8WIzkefQe/Gn5Z5PamZj6g73oZNtblppFosbkK6OQT5kmTSKgySEX1XBPsp2bnKZLgbRvgMbrhSRlvQ9E/KPPc9ncjYOaU8CcHSpP4/mOrTqapgrKeGqppFmgnRZIpUIZXRhcMCPchqyuoZTUHqakdZFDoag5HWf3bq3Xvfuvde9+691737r3XvfuvdNGarRR0n1sZj4V/rdhbj2pto/Ek+zPTFw+lKDz6x4Kkamox5R+7I7SFj9Sr+oW/2/vd1LrkoOA6rbpRdTcT09+0vSnr3v3Xuve/de6w1FRFSwSVEzBIol1yMSAFUfUkn3tVLHSPPrRIUVPVXfXk83d3yk3stVOZMZsKtNbREgBF0z2RVIuDc2HB9yJevLtPLsQU0Mwof2dAy0kbcd7lB+GE1H7erSvcddDTrWd/4UD9mZxNnbe6h2u0s+b3rj6eKkx8AZnqpjVM4VShve9h9PdkoTny6anqE7fPHRc/i38pv5j/xi+MPWOw9vfGrcW4cNi8ZUQ0lfNh66cN531s6vo/sltQ/x9+OnjXrY10FV8vI9Kz48dN9kVPZ2S+d/wAsduV3W0PX+T/j2Nos1FNTtVVLOaqCCngnCAgkcc+9HAAHDrSq2vWePAfZ69P8P83P5Fd8713/AJrpbqyoy+ythZTxxzY+Cdo5cckukS1DLI1/SLk39700+3rZcMCVFQPP1+zq2X4Q/wAwra/yK2Ru2u3g9LtrcOwGFPnceUEZjaO4kUFnILC35t/r+9HjTq6kMuroa9lfP34z9gbjr9p7Z3zHW5vH1bUNRTeKNQtQH8egOJCD6uPflyR141A6MhjM9gnroJJcjEs1YDJSI81PaRSdN76yR9fa+5rHEE4VHSKEFn1ny6EJWVgGUhgRcEEEEf4Eey/pd164/qP9v7917rv37r3XvfuvdR6p9FPKwNjoa3+vb25ENTgdNymiHNOmLb6yOss73vqK3/qPaq7IFE6YgQai46UvtD0r6hVeOoq5dNVTxzCxHrW/19+690mazr/aVcpWfEU/N+VFjz7917oO8z0BsrJsxjoooNRub3PB+o4Hvxzw699vQZ5n4n7fqVlekqI0LL+lmdQPp+be9qK9eY0GOgom+LuVgmnjxNfJH4SdLRPITe9jyoPtx4tAqfPppW1kkdJWs6g7kwTM1Dkci8EViQDNYqTz9P8AW9tCvE8OnK5p59cI9zdxbXS81PXTmO55EvqN/wDWt70a8OvA049PuO+Rm/McAtfh6hW+hLJJYWPH1P8Ar+/A/PreCel1ivl0+tYMjj1g0Eq7vER9D9Sb+9160OOehWwnya2XXFRXVcVNqsBZSDcm3AY/7H3uoPXhXpa1XbuycpR6KLKpIzkH6J/W1hz7UWxUPU9MXBbR29LvCZzCzUUTU1ZEdShm1MoNz/rH23K2pz05EpVRXpTJLHKoaN0cH6FWB/3r21051k9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3ST3Jsbam71VNx4WjyqqukLUpq9NrWuOffjkU68MZ6Rg6E6iAsNj4aw/5tP/ANHe9aR1bU3Xv9AvUY+myMOP9aJ/+K+/aR17Uevf6Beo/wDniMP/ANSn/wCK+/aR17Ueu/8AQL1GPpsjD/8AUp/+jvftI69qY9df6Beo+f8Afj4fn/m03/FfftI69qPDr3+gXqP/AJ4jD/8AUp/+K+/U69qPXv8AQL1H/wA8Ph/+pT/9He/aR17Ueu/9A3Un/PE4f/qU3/FffqDr2o9e/wBAvUf/ADxGH/6lP/0d71pHXtR69/oF6jP12Rh/+pT/APFfe6Dr2o9e/wBAvUf/ADxGH/6lP/xX37SOvFieuv8AQL1H/wA8Rh+fr+03P+8+/aR17Ueu/wDQN1H/AM8Rh/pb/NP/AMV9+oOvajx69/oF6j+n9yMOB/TxP/xX36g69qPXX+gXqP8A54fDf9Sn/wCK+/aR17U3Xf8AoF6j/wCeIw9v6eJ/+K+/aR17W3Xv9AvUf42Rhx/06b/ivv1B17Uevf6BupP+eIw//Up/+K+/aR1rUeuv9AvUf/PEYf8A6lN/xX37SOt6j13/AKBeo/8Anh8N/wBSn/6O960jr2tuuv8AQL1H/wA8Rh/+pTf8V97oPPr2o9eHQnUQ5Gx8OD/XxPf/AKG9+oOvam67/wBAvUfP+/Iw/P1/af8A4r79Qde1HqbjelusMRVGtxu0MVSVRQoZoo21lSCLXYn+p9+Ap1qtev/T2VO6f5S6b3+Qp+QXX/Zkmx8v/A6bDmggSqR9NPSLS60miGkagLm59+BoKdVZVJDmteH5dQ3/AJZHb55HyByVyfp5qwW/Nxz79nr1B8+uk/ljdvKoB+QORJ/r5q3/AF/fvs61jjTrJ/w2P27/AN5A5H/qbWf05PvwqerY9OuJ/lj9ug3HyByP+sJqzj/effhXquAeB67H8sjt76n5A5L/AFvNWf7zz798ut+Xn17/AIbH7dt/2UDkb/8ALasHvZqOHXhQ+XXj/LG7e+g+QOSB+nE1Zf3ry63j0PXR/lj9vAH/AJyByRNv+O9Z9f8AY+/Z4dexTz64/wDDZHb7Kw/0/wCRBtwfLV3v/r39+z17trjrw/lkdvgID8gMiTax/frLD/ef969+6929cz/LH7dt/wBlAZG4/wCb1Z/xX36pr1rHGnXQ/lkdvG9/kBkbD6DzVn+x+h97A69j0PXX/DZPb+oKPkBkgD9WE1Z/vPPvRFevCnz65f8ADY/bp+vyAyP9L+at/wBvwffhXh14U9OvD+WR29z/AM5BZIW4H71Z9B9Pz78a9eH59cf+Gyu37E/6f8lqBIt5qy/+wN/fvl1vHz67X+WR28Rz3/kRz9PNWfT/ABsfezXj1rHXR/lj9vL9PkDkT/h5qy3+2J9668adcv8AhsbtwD/soHJX/wCW1Z/vPvZBHXhT064t/LJ7eB47/wAl+ORNVjn/AGJ9+pTr2KZHXQ/lk9wfQ/IDJf6/nrOf8L39669gddj+WP28QD/swOSU3/47Vn9f6g+/HreCOoOZ/lddtZvB5jb83yAyCw5nHzY+eVpK1k0Si4Lgcm1/x79nj1qg4Z6N78FvgrhPhttWfGruAbr3JXmb+JZ4pMjTpLpPjHnAawIvz78ak1PW1UKKDqwH37rfXvfuvde9+691737r3Xvfuvde9+69025aITUE8ZFwyj8X+hufb1uaSimOmpvgr1DwMganaMNcREKB/Qf63t27HcD69N29dPT97SdKeve/de697917r3v3Xuve/de6SFU4p9xxSMNKNCFLfgkr7MYxrtCK9IpBpnDDpXA3AI+hAP8At/Zcelo4dd+/de697917rq4/qPfuvdITsDsrZ/WOGfPbyysOKxyBj5ZHiDNptfQjspP1H09+690HfU3yZ6g7orqvGbD3RDk6+iF5qaUwQynn6RJrux/wAv7317qkf5B/zlKyn+VG4/iz0Vtmj3t2F1/nZsXuvHR0TGqpHhqjTOPKDduQPoLe1kD1jYD9nSOceHKuoZOQf8/V9nUGd3duXrzbeb31hX29ufIUSz5LESKUakkcalQq3PI59oulnVGX88j4q9g934HZu8NowVWQodnQUz19HF9wVkMdTJIPIIAePpf3lH93jnLbNgnnsLwhGnJAJp5gDFeoN95OU7rmOwiMFSIXV6AkfAa0NPLqkj4DfBntPsnvHaNdW4Go25iMRkZGyVQorI4hCKlbs5KgfQE+8gPcb3A2faOXp1SUSu69oNK1ocdRLyJyRul3vcN1cxCER6gQpalCRk163TdsfGXqPA0dPHV7RxOYrqc8ZGthd5msoUNa4seL+8B73nLfbqQtHO0an8I4dZZ2+xbdCgDRhiPMjobMRgcRgKf7TD0EGPp+P2addKC30sDf2HLi6uLp9dwxc+p6NY4o4l0xgAfLpnxdZUQZOrpKl3cTzloy9vTyRxb2omhDQLIvkOk0LBJGStanpX+y/pb1737r3XvfuvdRquspqGB6mrnip4YwS0k0iRoLC/6pCB/vPu8cbysEjBJPp1V3VBqbA6YMTvLbmcnemxuUpKmeNirRpU0zNcf0VHJP+29q7jbby1QPMhAPyP8Am6YivLeZ9ETAkdTMvuTDYOBqnJV0MEShix8iEgL9eL+24LO5uW0QqServPFHlzSnRcN7fLjqzajS0i5gT5BQ6xweIuHmAGmO8ZP19i3beRN6vqOY6L558uiS75jsLYU11PRSux+3u6+7MDn8JtnryaqwEmLrWpcnDFIl2ljIhe4F7kexvtvL3LfL80c9/dASahVSf29B2+3Pdd1jeK0i7KGjfPp3/l1b4zGDw2W6U3tHJSbwxlbWZhqOfUs0VOykup8hufoPx7DvuRbW0+5Lu21kNbMoWo4V8ulPJlxPFbttl7iZDXPp1aT7jPoddI7c2+tu7UgkmytdHEUW5QMhb625uR79w49aJ6Jjv/5RVlfUTYraNKatG1IksKG91bk6h/sPfq468eg1wXVnYvadelfnJKqHH1DH0yCUKqt6+fr71kjHVsAdHE2N8edpbUhT7mnirp9ILMb8OUAPLAfn3YYbV1RhqWnQp46SPC1ZxiR+GkL6IE/sj6WKn2ueMSxBkGR0ljcpIVbh0svaDpZ1737r3Xvfuvde9+691737r3TTlcbHXwG5EcqDUkv006efr7UQTmE5yOmJoBLngeiWd592V2Bhk2djGZ6qdjCs6AF1Bj0lbg3tc+/XBjY6o/PrUBkppcU6Zvjx03Lkq8by3BH5S7pUaZS6l3Z9YOkj6+0wqDQ9KOIr1YAiJGiRxqERFCIo4CqosoA/w9769w65e/de697917r3v3Xuve/de697917r3v3Xuqu+8qV8P3ZDkgCI5K5SP6f53kD3rz69Sor1ZBtKuGR29jKofRqaJbgg30Rhb8e9g1690pPfuvde9+691737r3Xvfuvde9+690m8nmjE4paNDLO5tdfxc24PtdBbD45cDpHLcV7IznqRisWKUtVzeqqmFySPUmrlhz7buJzJ2L8I6vbw+H3Hiei4fIv4r7O71xktXoiwe9qfTJjN0wqwrKZ4UPjjBTmxNrn2D9/5ZtN6j10CTj4X8x8uglzbyTt3M8BZh4dwuVkHxA9Es6v+RfYfxn3ZSdU97UNVFthJo6LGbtrEmkaaAy6Y50ZiOGH9fYQ23f7/AJeuhtm8qREMBz6evUd7JzZu3J24LsPNCkQ1ASY+Y8urZsDnsXubE0Obw1XHW43IwLU0lRGykSQv+lrKTa/9D7lCGeK4iWaE1VhUH5dTnbXMN3CtxA2pGFQenj270/1737r3XvfuvdcXYIjOfoqkm/8AgL+9gVNB1okAVPSQjvmskS63o4uVHNgycf7yfa9qW8NFPcekmkTSVPAdLAAKABwAAAP6AcD2X8c9KwKCnXfv3W+ve/de6xTzRU8Uk88iRRRqXd3ZUVVH5LOQB/sT70SAKnqrMqKWY0A6KJ8jfkFsnb3XO7MRidw0sm68hi6ilw9JBPTO8lZqChBaS4Nx+B7X8uS2V9u8cJcUVu7I6DG68x7bHbyQRSgykdoBHH9vTD8Iutn2/wBc0e/sxE0W7N5wzy5nXcuUWp1Qhiw/wv8A7b2f8233i35sozWKKmn9nTvLVoYrIXMgpJJk9He9hLoSdU5fMn4Fbl+R3yM6g7LFbJHhtiZKCerhLShJYY6vz2BVSP8ADn36tMDqpUNTq3TDYanxGFoMLEL0tBSw0kaNpZfHEgUIOBwPxx73XNR1sDFOqwv5x2zt3b4+EHYu3to/cGtnmxriOnVy5SLya2AhGrhSffh/l60fP7D1rv8Awm6s2nsP4/blmxvetZtLcsWPSHd+BpUmjaeqlYoI5tdi3+PB92c1ap6bijCxAD/UehQg6oi+NPxc7k7g23vJ8zU7qy1Gs7RpMrzfxFipkZXtc3b8e9VqerABVNPmfz6ndB/DnH/G/wCLm5PmRvfsdUyW9KilzO3sTIJjOHr7zRKFksP1EDg/Ue7oe7PTbRgIaceJ6Gb419T/AC0+Tezdyd0Ue+Nw4Sn27VJJtjCRy1SR5uiOqaNoVQ6bFV97mfW2TWnXrdXCZ/F0ami/mPbz+KHxv3lkfkLh5MNvjCZ8bf2vTZOO8uUWUERVDrKwvfi17j8+2Rk46eY0XPRYNm/zRvlrj6em7V3D1UYth5LK0Qx9fNCftJ8VWvqWpj1NY3Q349+Iqa160HNKUx1e7tz5tdNVXSmG7i3FuGPG4qpp6OLK6VhH2mUnTVLSqHe3pP8Aj79jq9D099e/NT4+9mVcNDtXeSVlRUSRxRLNHBCrSSm0ahg9ufx795de6X3dHe3XvTOGx2Q3rmosbBmZ46ehcGncyvIQEAEjDg3FiPb9vp8TuNOmJz2UAr1yi7q2BhdoYvdOQyqwYrMVEUVFOqoRNJUgNEALgcj3qZ9b18urRIUSg6FvDZiiz2PgyePk8lLUKHifjkMoYHj/AAPtnp3p19+691737r3XvfuvdQcjOIKSZi2k6SBb68+3oE1yDpqZtKY8+oGD8jQNKxJWQjSeObC9/wDefbl1QNpHl1SAYJ6ffr9faXpR001OCxNYSaqhgmv9da/8U9+oOvdJbKdZbPyasJMRTxufoyC1uLfQ39+690GmS+OGzMgxcU8UTH6kKxv+efeiM4699vQc5f4kYeYO9FVxxkA+MAyKRYccEe9gdaPz6CTcPxx3Ph5lhxVdMzHTp8ZlPFubD/X9vJGxQuKinTbMAdHHpPHrvujbrJOlRkXgQXEZ81mAGr2z5knp2mKDj1zj7H7b20RFUY+smVbC5WQ+lTf3r59aJxQ9KzGfJzeGMZBX4mSy21a4mI4PIs3+HuwI690IGJ+W9LUuoyFJHTJ9HPhIsQefqfegc568a0r0LGF+R+wMgi/d5Fad29IAUW1f8hH/AIn37HW8gDoR8Z2ZtDLKGpcnGQ1tJYoAQfoeG9+86de6V1NlcdVoJIKuB1P0PlQcf7E+90691OV0b9LK3/BWB/3r3rr3XL37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//U3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdY5YxLG0bfRhbn3ZW0sGHl1VlDKVPSU25IRVZGE2ASVgo/4K34/3v2vvFrGj9JbdiHKdK/2XdLOve/de697917r3v3Xuve/de6TufpTLHBJFoE33MKEs6oSjNpNi3tbaSUJRuFCekd0hoHXjUdED+XP8wHYXxP3dtzbW8K2OjjzdbQ0EcjpTMA1VoUFnnZbH1e0dSST0qPaAK9H82nubFbx25hdy4WrirMdmsXQZOmnieORWhrqZamO5iJF7NyAfeqefVqipA8uqp/5jP80/r/4GzYTE7lqIY85u6uXBbZp56VZYqrNzSpHBTs7kfqLj+vvdOq1px6IxL/Mp+ZHT+f2Fv3uvpz+7fWXYOboYKTKz0Mqwy4XIOrpV08kllF4nDA3t9D73QHhx6pqIywxX9nRJv5lHzd253J8iNlbW3P2bk+vuk2zeLXBZzHQioizRqZI5KyilELaW0MSvBPvw4EjrzGraK49egxzuyO6+qvlT1Hv34jbgzO/evsrvHBS56anWajgp8Z5I3qHqEkIBBQsW9+r254+XWgrCQaT25r0Yztzr/YXxY/mWYT5GVO26KmyfyH3TjBVVUcTvK89S8RrZC7jTcSkn0n2/buEqh/Fj9vTc6s7Kx4L/AJOHWwf8s/kP2h1BnunqXrjaKbnxO+MmIc7WGAz/AMOpHZDHKCoNrq17+07goxU+WOlKkMofyPRw8aIN3bYpxnqCGeHJUyGsoZkJie6gsjpx+fbkM8tvIJoGKsOBHEdaeNJF0OKg+R6x7d2Hs7aLySba29jcM81xK1DCYi+rk6uTf2qu903C/AW8laQD1NemYbS2t/7BAv2DpXe0HSjr3v3Xukhl4zT5eirQLRx/5wc2e/p9mVuxe3aI+fSKZVSUSDp/q8pQ4+iOQrqiKlpAFLTzOsca6zZdTOQB7LmGlip8ulYNRXqRS1dNWwx1FJPFUQSqrpLDIkqMrC4IaMkf7z711bqR7917qjH+dR8muweken3xuyVq6Z65J2kyNHCzzRiSkB0qy/0PP195G+wPKO2b9vInvqGhA0nhx6h/3Z5lvdi2SaS0BJRGYUFTgda2HxL+ZHyM2j2thxi90Z7edbnavGOcbU08pjgWuYalIJPIv9PeVnOXJHK99s8glhSBYw2a8adQBybzlzMm6Ks0zXHi6CAE0BAwHH1z656259q9AdqdvYLA5veO68ptj+K4ejyVTRKZSqNWJ5WhCccgW494V3vNOybDcS2+3wLNoYqDjy8+spINn3Dco0e6kMYYA4+fRiOvfiR11tKMtuKjpt6VoIdKvLQszLIPq+nVz/t/YT3bnvdr8/4oxt19FPR1Y8uWVqKSjxT6t0ZjD4LD4CmFHhsfTY6lAUCCmQpGAgsoCkn6ewbcXNxdv4ly5dvU9HscUcK6IgFHoOquvklhsl0N35h+/wDbLLUndVXTYLJ0ETCQQU8mmCqnZPot7k29jbZ7213DY32W8OkRAsD8+I6CO52txY7sm62wr4lFI/y9Dn2Z8nTH9vjNnIa9qjG0kskyRBnFTNGHlS6k/T6ewCGqWpwBI6GZGmlfMdAvh9g9ldu1YrsnLWQ0M7kyQssoRI39YNh795VHWuGD0bzYHxz2ztXw1NZFDWTjS7KdZ9RQBgxIH597GDXrVMU6MXSUdLQwrT0kKU8K/pjjFlHFvz791vqT7917plzNAtTTtMgtUQKXjYAXv/r+1VtKVbQeB6TXEWpdQNCOuGDyBqqfwSn/ACmmGmUHg/Xg8/7D3q5hEbVXgetwS6xpPEdPvtN0o697917r3v3Xuve/de6L73j27R7AxE1HBIDlKqJkRVKMR5IzpAF735F+PfqVx1ommRnoovUPWeV7Z3BJunPCQUizidXlEi2tJwPpb35aDh1pqnB6sCoIH2pDT46nhH8PhQKX/BA51Xt/sPZgsUc6VXDdJDK8Jo/DpX01dTVagwyAmwOk8H/Ye0ckTxmjDpUkiuKr1L9t9X697917r3v3Xuve/de697917r3v3Xuq7/l1jjj8th8yi6TJVBiRe5AltY+/U8+vZ6Nt0lkP4n17hqrVq1Jp+v00xr70OHXq16Fr3vr3Xvfuvde9+691gnqYaddUrhb/AEH5P+w93SNnNF6qzhRnpL1OVq66YU2PiZoidMkoButzYm/+HtckEcK65TnpK0jyYQdO+PxMVJ+7Laac2OthcqfzYn2nnuDJhcDpyGAINTZPTx7TdKOve/de6Brufo/ZXde2qrBboxlLNUtGRQZR4y1TQShToeJgfoCbkeyndtntN3tzDcqK+R8x0H+YeXNu5jsmtb1ASeDEZB6rDx26u4Pg/u5MNudK7cXUNTUxxJuOeOaePHQFtSwQB/oApIv7jeO73Xku88G5rJZk01ny+Q6hqO+3/wBt74W98DNtxP8AanOj5fZ1a31n2ftTtXbdJuXauQjraOoijkcAoJImk+iugJI/2PuT9v3G23O3FzbNVT1OG0bxZb1ZreWThlYA/t6Eb2v6Neve/de6TGdyEilcfSjXPLbWB/ZQ/n/Yfn2stogR4rcB0lmkz4Y6dcXQLj6VYgbs3rcnk6iORf2zPL4r6unYkCL05e2eneve/de697917qpH+b98jN9fH348VNZsSCT+IbkR8Y9eiM5xuuoRBU3X6W1f8R7AXPF/cRRQbXA3hi8YozjBUAVND5E8B86dY7feT593fkHkSTcdnWsspWMNSojLsFDt8lrWnnw60lMJ3/2xUb8we7shvTcOVyGOy338dJkMhVVFLKwn8ssMsBbQUflSNPH4t7JFs02+1ZbBjE4Ao9STVRgkk5+dePn1zGXnzm3ab6PfWv5Z5rdvE/UlYo1TVlZK6dLZFAMV7aUHX0Nfhhvyv7M+NnWG9MnjP4RW5rA0809BoKeBlQJYqf62v7HHK+63W9bLFuN5/aPUGnA6TSo+RpXrsZ7c71NzFyXt+9XEZie5hRyh4rqUGh+Y4dGk9iDobdEq+Xfzo6U+Hu2zmOys8KSpmhjmpKanWGqZlkcxgyRiQMOR/T3sCpx1VmCjJp0uPix8pevflZ15Qb/6/rmrMfVReVy0QhKevRbRqYjn+vvXVh0YfN4LEbkxtRiM5QU+SxtUpSopKlS0UgKlfUBb8E+/de6qd3v/ACj+q9x79qt3bd3LV7SxWRrVrMntuhp3kpK8rJ5NMz6h9f8AgvvxJ4daKqelb8pvgfjuzOnsV1BsyFcVhaZKD70w+RVrpaV7mWUIDY8X9+GOtFdQp0gvnL8SNw7u+A21eg9lU8lbl9n43HIkMPleWRMVTmeUjxjV9R/T2ohSrVbgOmp3wQOJFKdV5fFL5/7s+MnTtR0bX7Tkl7GwdTQYfAYmahnbziM+F52dgCSRcDUD/X2y1dR/l04hAUA8fTosn833Yfa/yR6n27vTsHbs+yKPK5fCVpeKCoKmwXyS6Y1sLWv78hz03IvbUinTruPqHZ0vQ3U20sV3plcrjv7u7ap8hjNMzQ0WQ8CRGnES+oW+n091qa5HDp0haYOD0j/nL1fX7D+PnX3x32/vSohqNxbpwGclq4mKO1LX6E9SuQ9rHkW97Q17uqPgeHwDY6Mv8kfgMvxS+PXV3YW0u2K/+8MybZrqyOENEaqp/h8FZIKZg13Qs5AYgf7z70CSc9WKKgqlcdER/mA9rd9d4dedNJhZsjPlKDdu3sXJRRq2uanjMcX3LIlz6/ySPbyjQK08umWkLsFHqOrlOz+5cR1J1L0T1P2hpxufnOy5Z4pESSRPuaWJHlfWQw5a5v8AT2yRU16UA0x8+hs7r/mkdS9A57GdQbIqv7xZ3D4bG1+UApEqUgglpVeXRJCWBsP6i/4v72BXrRYKaHo1/wASfnt1J8pp59u7dy1t5UNN91W4uSGOl0wrF5HKI76iR/QL70cYPWwaio4dHTfdW3Uq5aF8zjVqoE1ywmupBIi/1ZS9x/tvfuvV6d6WspK2JZqSpgqYnF1eCWOVSP8Ag0ZI9+630zbhdjClOgu01wP63P0t7W2YyX9OklywwK9OWKheCgp4nFmVLG/+vwfbE7a5Sw6fiFEA6cPbPTnXvfuvde9+691737r3XvfuvdJZpnnzMNmIWMlbA8EA/U+zAKEtTXz6RfFPX06UzxpKpSRQ6NwVYXBH+I9l/S3plqNsYCq5qMVSS/W+tDzf63sffuvdIzMdRbLyyt/uKggZr3KA25+tgffvn1r5dBjkfjDtCr1mHRCzEmwV1Fz/AK3+9+9U8ut9B3lfiNRANJQ1yBvUyqjSX1f2SFNve8A9a49Bvlfjh2FjtT4XJVvovoSMS24HB+nvWfLr2PPpPjb3c+1lOqSvm8V20sJP7PH0t/vHvdKihx14VHd1Iou4O1MRUQw1GLqSPJpdij8hB9Tx71wPW6jqwLrPO1+49qUOVyURhqZwNcbAqRZAfof9f3vrQzkdCB791vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/9Xf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SMt/D8/BCCAKxmkP+Nz/h7MifFtCf4ekPwThfXpZ+y3pd1737r3XTMqKzMwVVBZmYgBQBckk+/de6bVzGNkpKusgrKapgoo5pahqaohn8awIZJA/iJsbA8H37r3Rfdq/KrqjfCbzi23mRUVuysTnctk6aUwLKIcDStU1njiVyxsFJHH+x9+8utedPnTqhrcP82T5E977lzNd8YNiUW7dgbS3FVYnI5GopqeKenqMfKfJrXVe1lPJ9vRv4bavUU6Zkq66QK0Pr6dFT/mxfHfs/8AmCfDvr35Dy10uzt4bU7Jpm3VTYYsZqfHYsJJPIY6S+lF0WJP+Pv0i+HIB5Hqq1liJIzinV+H8pbP4jP/AA+2Y2H3bVbxiwzx7dqclWOrTw1mIx0EM9KQpJAW4tf20RQ9Pr8APGvVVv8AwpK+N+C37sD45dovg4K59jdrrmdw1MkDymDG0UtLVrUkqCBpswu3H097Ge37eqSAKPEPkQehd+Yny++NXyH/AJeOEqttVMOZzFdt6DbGz6LRT1D4rduPw0VCfDHTMxBVo10GwsOLe/AeRx1ssCuoZHVYNH/K57R7f/l6fHXs6n2bLvHuHr/svObpyW2q6nqzPkMFHVR1GOhkiIuVIHII+h9+1Cp+fVfDPhqRxFDnh0bjbvV/zW+YGZ6w21lemYvjHt/r3JUpyVZsujq8TDk6KjVIi9eE/WzKn5PN/eiR5Z6tRyaUpmvHq5Lvn+XfszvPG9HQ5vOyfxXpmvhrqbL1SytUZR0KOyyaL2OpLgk+/KSrBh5dWdQ656Onhdv4usSDC57H02VlwCwpQ1NWmto/FEsI8YP+C3B9rbtNarOPxcf8/Sa3bSzQ+nDoSIoo4I1iiRY40ACoosqgfgD2h6V9ZPfuvde9+691737r3TFuCFpsbUNCuuaJdSgH+nLDj/D2qs2CzANwPSe4WqahxHVYf8zbuHDY34vbq2Zt3d5xO/q3+GzUMeLqoJK2OKgjarqA3hfUt1ABFgb+251pMQeBPVonrFUYI/w9AX/I6+acnyt6f7C29k8nLkc51BmcZtqrlqvEtTKrLMjTOqnU3qQAsR+Rz7bYUNOrRMXjDkUrX+XV5vuvTnRWvlP8V9j/ACl2LUbN3XDDC0glMGQKuZIWeHxL+jmw49jTkznXceTdwF7aZGKr+deg7zBy7acwWhtrnzrn8qdV0fGv+Tb110p2FHvjIZqPMtQVkM1FTf5USBTPrht5ABb6fn3KfNfvvuW+7Z9BBHo1ggnHnx6BPL/thtmz3huhmhHr5cOPl1dpSU0dHS09JCLQ00MUEYJ+kcSBEF/9Ye8e5HMjmRuLEn9vUrqAqhR5dNOb3Lh8BTvU5GriiSNWdlDx67J+r0kj3T7evdFB7F+UtNSSVGP2mprHYvCpEas2tSPoyk/6/v3Xsn59F1rNr9jdy0+QrslQz1VPTU81bSUk4lWNJm9cbqBxe/u0PiGVVU6dRofs606oYzq7tIqAfXp7+LWL2tuDd2T2bupYF3nglepqcWzIH+2iUqAFvq4Fibj6ez3ddnWxtlniaqt8+iTbt3S7unspMSpmmOHVntBjKHFwinoKaKlhAVdEYsCEFlv7IOj3qd7917r3v3Xuve/de697917pJZSCXGVAyNKn7ZbXUBRz+rkG319mELrPH4L8fLpFIpifWvShoa2KugSaI3uoLL+QfaOSJom0t0qRw61HUz231fr3v3Xug27L7Bx2wcFUZCplT7nxt4YiU1XKEq2kn+v049+Jp14CvVdmDw+4e/N9LX1wm/hUdTGwZkkCKgkAtdf8Pe/t61gGo6s02jtXH7Qw9NicfGiLFGqyMlwJGUmzWP8Ar+9db6UkkUcy6ZUV1/owuPdlZlNVNOqsqt8Qr0mpcLJSyPVUTsWvqEI4Ucfi319rRciRfDk/b0kaBoxWP9nXGDNVkL6K+n8ajgkfq/1x729tGy6oz1WOeRTpkHT7T5GkqheGUH6XB4Iv/h7RmJ14jpWJUIrXj1O9t9Ode9+691737r3XvfuvdE6+XWH++23QVWn00od9QBJ1LJq/H+B96PXgDkjpSfFzPLXbHp8cDf7Ysw+lxZQpHv1c060AejQ+99b6jy1UEIJkkUW/2Pu6xu3AdUMiLxPTBNnmlZosfGZXFvqP9uP6e1i2ipmY06SvcMfgGD1jhxdXXsJq53hIIIj+oIve3vzzxxgpGK9aSKR8uelHT0sFKumGNU4AJAALf4n2iZ2f4jXpYqBBQdSPderde9+691737r3XvfuvdI/fGxNsdiYGr25uzE0mXxtVFKvgq0LpFLJGY1mQAj1Le49pL2ytr+A290gdT5H/AA9INx22z3W1a0vkEiMDg/4eql969edsfCfdDbw60+93T1tVVaVGQoZUlenxkV/JJCIRcAIL2t7i+7sN05PufqtvrLATkeQHpTqDNx2rfPbi9+v2YNPYsasv8A4mg6sa6G+QWy+9Nr0+XwFen8Rhihjy1DMY4JKeuYESwxxs2ogEH8e5A2TfbPe7YTW7dwpqHCh9Opb5Z5p27mWyFzZvUj4gcEH0p0N2RyEVBAZHI1PdYx/ViODx+PYhhiaVqDoQzSiNfn014ehLu2Qqhqnf0rf/AFNvrz/r+3riWlI0wB0xbxE/qPnpSe0fS3r3v3Xuve/de697917okXz32d11vnoPeW3d8x0s1VkcLVU+346ixc12oNeJfrf6fT2xPylFzen7ukOitKP5oa1DA+vQF9weX9o5k5duNr3dQ6SoVAPnUGvVFXw3/kjYbczbc7S37uKR8S2TqK6HBzU8sUc8FPLqp0kXQCfx9Wsbci3sHcwch7pHurbU96rWqadRCUZ/UavIHzoOHn1iXyR90TZ47y33XfLmS4SJy6xtQIwB7AwC1bTji1CckUx1tFbM2hhdibaxW1dv0qUeJw9LFS0lPGLJGkcYTgf42ufYvtLWCyt1trZdKIKADrOHb7GDbbNLK2GmOMUAHkOiE/zBv5hXW/wn61yuYyeRSp3nLRiXB4unSnrfJI7tH+/FqJFiPyvtUASaDpU7BRU8OtY6tx9R8+urezvkv3fuo4nDYCgGU2tt6u0xU+Y01J00kVOx9IF/6e7fBhT00AJ01SDHkOr8P5M+HxWG6KrajD01NQYOWJHpUi0x08UaS3ezHgAWJJvYe6E91OnlACVHVt+F35tPcFXX0WKzmNqqjGvoq0jr6OQxte3KxyMf9uPe6dbB6V6srgMrBlIuGUggj+oI966312SACT9Byf8AWHv3XuHSRgdq/LGQEvAgkhlS/pKONDBh/Qj2YSDw7fQRx6QoS0+rpB1nxy6LyGeXdFb1Zs+q3CsglXMTYuN64Sg3EgmJve/59l1Ael+o9T+0elOvu29mzbI3dgKKvw/20kNFDIraKF2iMUcsKg/2OCB/h731o5/Pqurrv+U91j11vWo3lWb1y24MRBUGvg2zX02jHUy07eaOMN5G4QD66ffqnrQVVyOqKP5iGwuzvkd/MYxXWXTlBXVQ2/iKOoxOJo45hC7YakSdIRJGLAeg3J93RtINfPpp0Z2TSfhyejgHqP58fL/IbO6y7x69r9g7F2LBiqOPMCnqYUnp8Mq0xaVhfUWVAPehmmnq5BqSaU+3oGd+YvC7R+fmD6byqywbK2vQ4SRalYJvDJlMfCjNIZWH1Ljn2/IGij0Nx6YRlklqMYr+fTV8zK7K9+/MDfG0MVBPkYdidbPnsXI8EjmI4fHLUQSo8QPI0i3tMKgZ6fahag9Oh0/k+bG6Z7ZzG+N5d1YHbu5d9pj6nGN/euODVTUNG4EkrQ1JAb0KVF/p9fe2Hl5dejagLHj0BVKuO2L/ADEe4Mx0NGm39uYegr6s4zCrbH0mPhb97xzR+jSFB/P+HveNND1rOssB5Cv7epHRnXHyg+U/ym7o3Lhd6bmx2yqfb1RUYmWGonWiaoFkSCNozouT9OffjQACmetgOzFq4FMf4ejt/wArP5M9yZX5Uds/FrsMT1uL6/xNdUUuWqgHqJZ6LTGA0xJJv70RT8+tqSSwPl/l6v2yTfc5OnhjYkwsha1+L+r/AHr6+10IKQEnpJLSSUU8ulUBYW/p7QdLeHXvfuvde9+691737r3XvfuvdcJGCIzE2spP+2HvairAdVY0UnpM4hVnq55hz43P1+oJPtdcnRGF9ekkCkvqPSp9oOlvXvfuvde9+691737r3Xvfuvde9+691FqKKlqv+BEEc3FvWL8f09+690yz7Q21U/57DUTkHVfxkNf/AFwffuvHPTzRUNLjoFpqOFYIE/TGl9I4txf/AFvfuvdS/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//1t/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYpZ4YRqmmiiUfVpZEjA/2LEe/de6aV3Jt9pzTjOYgzf8AHMZKiLnm1guu/v3WqjoD+8vkhsrotsRHuiWZXzIVqQwx+RCrSGMan+g5Hv2SaDrxwK9CFSZzH7j2/t3fdHLElFXUMVbDLNKkKeGYEpd3IA/PtdZuGVoT59JLxWUq48ulfj9wYfI+OOmymOnqHW/ggr6SaW4/VaOJyTb/AFvaR10OV9OlSElAT59Ex+WPz16c+KEkGI3pkZ23HW0v3dHjaJIqqdojF5VZqZW18i3491Ar14kDJPVdHT386Prr5DfJ/FfFTb7zY/O752nnJsMa6gjoHet/hUklIvmlsRdhx78RTy6qrhyQPLJ+zrD/ACvvkHv6XtvuP4+dzVdR/HszuPcLYCmyE0aVAx1NJOjvTxyHWwKDi1x70aUx59XQ1Ygnh/g6KbTbgofj7/OZqvjhkMxLDsXsjZOYTIcolP8AabuwjEtKJOdQEnIB+v097Pw0HVakSV9RX8wemSr+BVF0N2VvfrzqPsWtosdnv45vd4aaplxdPOah3mWJhKy6uDb36pJqc068EQVK4qa/t6m/AjevcfYvZnbXws3Vimq+uc51/vKChzE6uyw56TGTClq0ql+ra+fr/T29IGeHxBkdMRMBO0We2n2dDV/KS258sfiR2huT4e7s2Q79UNurObwi3hXUs6VaS1fp8UFRN/ut1jj4N/px7aqCajz/AMnTqhgpQ8F4fZXq4f8AmAdJ4vvX4nd1bTr6cy5P/RxuufAOqu702YXGGWnljWP1E3TSAP6+9VI6coGp6da6v8jn+W9smu6Wp6DuDemczO7Nn9h5fNRbGy9DVrRtRiS6Tlpyg+vHAPH49+Y56bjjAStPXrbpwuDxO3cbTYjCUFPjMbSRrFT0dKnihiREEahVH+AFz71078uptPSU1KGFPCkIc6nCCwZv6ke/de6ke/de6R2RH2GVp50uv3cyiQi4uAb2PszhIltynoOkMilZdXl0qmqaZGRXqIUaXiNXlRWkP9EVjc/7D2W0NadLQwIqDx65yzRQRtLPLHDEgu8krrHGo/qzuQB/sT711vpL0O+dpZKvkxlFuDET1sTFWgTI0TOxHBEarIS3+wHv3yPWq+fRQfl786esfisuOwGfq5pd7biphUbfxkEMVQs6kNpaVCdViRwAvvwFTjrRNOPn0QT41fziNu9id+4HoXs9I8FuLf338ezovt6WkjqJEp2kpVnZ9LAMQBfn6/T3bh3dUDBqp5+nVWfwE6yyf8wP5X/Kym+QXYma2nUda79zGE2bt1ZESLPURr56Fkp45XjDCONQfRcn8D2++p4w48hnppTGJmjJrnA+wDrH1JQ7d/lCfzaOvfipgNzy1ux/kRPSbr3DW6BCiQ5WpdqZKtJmJDRn6gH/ABHtgnWM8R08oWNqcAQT+fW5XRVtPkaSnrqSQS0tVEs0Eo+kkTi6uP8AAj3XpzqV7917ppymbxmGgeoyFXDBGisxDSIGIQXNlYj37r3lXopHZPyjxeMafG7ZJq6m7whgiOxkt6bWv+ffqjy60Kk9Fwp6XtXuPJgMK2loqiQXCLIq+KU8nj/efej8h1ugySejP7E+M23tvJDlNxzRvLFommWoOkXXmQyPIQB9ffiQvcxwOtMVUVY0H29ILvb5dbB6Xp22tsanhyW7qi9JRUNDAlTC9ShEcUUrRauCbi59g3fub7XbQYbc6pjhQBXPUa82e4u37GPo7M+LcthVGat5cK9VwUm1vkB1Z2pjfmLnsZU4uXf2TXF5bbkSSGiocLL9azxt6V1I35HsZe2KbxzC855mqihSY1rxJGOgLsO381Wu9nnfdFMf1NFMQ+FVzRvtPV9Wxd54bfu2sduTBVa1lDWwRkyqUOmoEStPGwQkXUm3tVd2stnO0EwoR/g6n22uY7qETRGoPSv9pulHXvfuvde9+691737r3WOWJJo2ikUMjizA/ke9qxU6h1plDCh6SFp8DVkogaknYXP4RQfp/h7Muy6TPEdIKPBJjgelbBNHURrJGwKsL8H6f6/suZGQ0bpcrBuHTRuLP4/bWKqstkZlhp6dGJLMqktoLKBqP+HuuPPrZr5dVhb13RuDvTfCYmgExx0dRHHEEjvHLGGEdzo/w9+NDgdbrQCvVg3VXXGP6/wFPSwwqtbJEPuX0kFWvcqL+/dax0Knv3W+ve/de697917rDLTwTf52JH/4MoJ92DsuAeqlFPEdMNRgFLeWnmeNgdQReB/rXB+ntZHdgDSwFOkrWxGUPWIQ5eD9AZxb/VE/QfT3YvbuaNjqnhzR5XPWFMrlI2tLTgC/JIvbn3toIDwPXhLIo6lncCJ/nI7Hi55t/vXtv6QHIPV/qH8x1x/vJT/UKdJ4BIIu39Pe/ompWvW/qlIwOgQ78mGZ2NXRrFcx00z3sTa7W/4j3R7bQuonraT6np0BnxFzNXAMjjUQSNGKgKrf1BuLW5/HtqGONz3mnTk0jr8IrTo7ejM1Sh5EaMsASqtpA/1h7VVt4zQZ6ZPjvkcOsy4BpWWSoqXuDfxjkf15ufejeAYUdbW2/iPT7BRU1NYxQorW5fSNR/xJ9pHkdz3HpQsaqKAdSvbfV+ve/de697917r3v3Xuve/de697917r3v3Xum3LYjGZ2gnxmXoqfIUFUjRz0tTGJIZFZShDK3+BNvbcsUc6GOVQyniD01PBDcxmGdQynBB4dVPd8/HHe3Qe4n7m+P5qJo45nqsjs2mWRcesMQEs0zQi4Nube403blq/2i6/e/L9TTLRiukDzPUH8zcpbnyvdnmLlIE0y0Iwhpxx6+nRhvi98n8D33QJQblm/he8cZ4qSrxM4ipFNTHfzBFexY3FhYexfy/zZa7lbiCoWcYYHGfOnQt5L51suabbwrnsuEwyGgIYcePR60ChVCW0gALb6Wtxa3sQVrnqRxQCg65e/db697917r3v3XusM88NLDJUVEixQwoXkkdgqIo+rMx4HvaqWOleJ60SFFT1VJ2bXZD5Td+UPXeJqZVwHXeSGQq5YBemrKWOYPJHJLDwQQLcn3Iu3oOXNma/lHfOKD1B6A94z75ui2aYSE1PoerT8RiqHB42kxWNp46WiooY4IIYhZEVFC8A/1+vuPZpZJ5DLKaseJPQ3RFjQIuAMDqbM0ixO0SeSRVJRL2DEDgX9tHq3VD25v5VGQ+SHyI3hv/v/ADeRqdkU+Rar29gKuOWppK+Jpi0dOFLqqot7nn/Ye7A0FB1Row7VbPp0Sb+ah/LTouiPjzvHffT+985jdu0VHTrNsukSeHH0ifcBbQxRuy6GH9Re4PHva0rnqkq9lFqDTpX9Ed8ZT46/yk8ZlMJMw7A3Bt+T+HoWQVcvkmanbTE/rP4PHv3nT59WJ0gGuKdFc6I6V+ZnTvQu8/lzNuHcWRj7HpqbPUWLylXKkFIs1WznxxVDWAOlgLAA+/EqDQDryrIF7jUnPV+P8sn5a7o+SnUuXrN+w02OyuyTBRV87+GnjbWWJkaVrAqoU3Ymw/Pup+LT1ZCGTVw6sX/vXg8jBMmJzOLrJ1Ogxw5Ckka97cLG5J/2Ht6FA70PTU7ER1HThhaJqaKSR7a52Dm1jz+bEe3LqTUwX061bLRK+vT37S9KOve/de6w1EIqKeenY2WeGSFj9bLKhRv94Pv3XuitbT+JHXW0u6Ze86NGk3jLR1FCahkKlYaiIxOofUeLH6W96zw68KCpHn0ZrI1H2tM7BmDOdKm/Kk/lb+1ECeI4Hp0zMwSM18+iob6+HfU3ZmZG88riKam3ZLMZnz0MDNXOuoMEZiw/p7tdOWkNOtQKCobpi2B8Gusdj9o7l7RdVzOY3Ltqo2vVirhcEUNTD4JPVrIN149phwp0951HVc/an8pHN4LsHMb16R35mMFSbnFZBV4TGGqgioIaxyZG1K1vofp78SeHXtK11DHS1wv8utvjz0tv3N4PIT707U3dtWvw9TWVkdQa1Wq4WVlRmJNxfn/H6X9749apQ46Xnwh6o3h8dPiNvPcG5qOpx296iHKOSqzCp8DNeIaJOeCQVt79UnryinHovv8AK064y03yc7u7mzdDPFUbjxtZDHV1EE8TyNLWpr9UgAPpv78fTryg5Y+vV7uIX7jIVVYRcOBp/otuB/vHtdcHTEsfSWGplJ8h0qfaDpZ1737r3Xvfuvde9+691737r3TdlZhBRSyH6Dj/AG/t+2XVKB0xcGkZp1BwFO8VPJI4sJ2DqR+R/X27eOGfSPLqtshVKnz6f/aPpT1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/19/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdan/zB/mTfLfK/Myk+KfV+Fo0oMtulNtY1nhp6apqS9W1KrQzOAxJuOQfdgpC6iemWk/X8FR5Vr0u+1/ij/MM2LtXLdm7MOYze88Oi1keBSteqgkEb+SciFWIZVAJNh71jh1ciSlajpR9c/wAwL/Zk/hl3NV7x2ziMt8gPjcuNoM1jq6lp5GhrJqrwVSU8CjUxRlJsRx78aeX5daXgT6Yp8+PUD5K/zKd9YT4N4Sr67xsA3VDicTSZxaOKCQY55yY3pwkd/HpuTYgH/be3ICRKKHqk7UhLEdIz4j9o9hUG9eluxdx73kbb+6dsxZXdcGQzVLBBj62oP+Y0zMAp/wAD71MulqjierRMXWjHgMdMPZK7e3z/ADjekH7Rmo9w7Jy+MQxY3MNT1u1amknpmSkkq55LwlV9JsTY2APB90Hw9XB7xX0P7egc/nE4DpL4Z/MPqj5ZdWYfbu1tx4TH4LBmk2lR0kFMZqlvA8zCmvEjGMgMAB/X6+9pmq/n03MdJVx54P59D38luovkXsPvT48fzDfilt/+9OJ3R1FtjMZbHrC9bQJms5iVlro66FbodZma/wDXn+nvVVIz1dg0bDTmlQR5/l04bL+BfyK+ZfyM2582u6cc+xN9Y1MRQChx9PPSxSUFGqxhIUUhVVY1UWH+x9+qVFBmvXtGshzxHVsXyH/lq7T+Q258DvSs7L3jsbM43btDg6mLbzWhqUgplimeQ+VOWIN/ehXq5CnP+x0MPx8+JPXPxPwsS7c8u5M/UTxxTbszMZ/jToVKSRmQMw0sGN+fa62BnjMHpnpJKRA4kHA46OFHicU9auaFBTfxKSJF++8YNR4yvCeX62t+PaIjSSOlQNR1Nq6Wnrqaejq4UqKWqikgqIJRqjmhlXRJG6/kEGx96630kNr9b7F2VNNUbU2visDNUFzO+Pg8JlMhu5fk3v8An37r3S39+691737r3XvfuvdNGVokqFinf/lGYyf6wAvf2qtpNDFfXpPcKStR+fVEf8xT5N9/fHH5H/GFNsUaydbbsy9NLuytkVDDT0RyPimEk0nCfti/PtudR4rFetxOAi1GD/gpXoSf5lvzbz2wsJ0/1x1JNDW13yFo1/g2TgeBgqSVHgVI6rmxuRfSfx7apk+g6dZqAUGSeq6vkB8bvk/8QOjF+YWF3lnczubZ+Opc3uPb1XVyTUNO1bVLCP2pCVkS7gHi3PvwoTnrTalXUM4yOk/2b2xsfsXq/wCPHzo7KrVz/bGHwcOSbrzJU0VTgK+oFQY4oqmP9Ijva62+nvwqcDrRZBRz5cPz6Ip89Nob52jF1z/NB3BtTEdeVu2Y8Zktmbf2bTr9vXQV1RoDLRw3CgAn8fT3ZRXHl1STsOqmR+3Pl0efAdBd2Vey+g/n98OMNV5PN722rBu7fWBSnmjifcVZIfumrqY+lvWb2t+fb0DFh4YHHqksYRvFHlx+YPS/2/8AyxO+/nb8gtl/Mb5Ufc9e9i7FjoKTCUdNS1CLVUVHUBo44tJCoqre3HthgUYp06qq6K5zStKHrZywNPQbG2lhcJW5GJlwWJpaAzTTIsk4o4RHrOs3ubX91ANOnajouvY3yexGCmmx+3v8qqgHiDBUk/d08WIuBz71Xr1OivvX9sdyZEwRx1tNS1My6dCSKDFIdBB0ge9CnXv9N0ZXr/4t4nHLBXbibz1iiN3jlQuzSfViSxHu3WqdD5mMzsPqjAT5Gulx+LpMfSSzOFkgSolSnS7hEdgSefp7S3d7b2UJmmYAKPXpDfbjabdA09y4UKCeI8uqw97fJvtn5KbpqthdAYiom2u0s1DkcvPTmCSCnbiWdJwPqoBIN/cY3nMe58xTmy2JCYyaEkUNPWvUI3/Om9c5XrbTypGTCCVeQilB5kHz6NF0H8KtmdZ1C7p3ZO+99018K1FS2fgFVHj6yVdUn23mJ5UkgEj/ABHsS7JyfZ7cfHuj48hz3iuk+dK9Dblf2723ZJPrrom5ncVJkzQnjSvRq9+7FxG+NmZfaNbSU7UtbjZ6OkVk9NHI0WiGWEL9CnFrex7Y3T2Nwk0RppI/YPL7Oh9c2sVxbNbMO0ilP8HVefxr39kOj+z878e92TvDt/Hz1E2GyNUqxrLVTt6I1kmsSDwOD/T2Nd/tE3exTe7Yd7AagPQdBLZro7ZevtFwaAfCerRVYMAykMrAMpBuCCLggj2AOht137917r3v3Xuve/de697917rBUU8dTE0UgBU/Q25B/qPdkdkNV6o6B10npFT1LbTd6irkvjDrZ5XYAqLFgBq49mDFLmP0YdJQHik+XRDe8e1Mh2FnYdsbZeV6AukU4h/cUgHSdQj+t7+y1lIbS3l0sB7cefRmeg+oaTZWHiyVfSxtkqqNHjkdWEiAnXq9X0+nHvfXhnPRlfeut9e9+691737r3Xvfuvde9+691737r3XvfuvdcGjjb9SKf9cA+9hiOB61pHWI0lM36qeI/wCui+7eI/qetaE9B1x+xo+P8mh4Nx6F+vvfiyfxHrQjQGtB0h+ysXS1Wys+gp49S0D6dK6bC4vwv+v71rYjJ63pUGoHRD/jRllxG+slRrYM880RQ24ubGwP+v7bB09epq6syH0Fvp731br3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917qLV1kNFC0szAADgcXJ/HHtyONpG0r1R3CDPHpKQ00m4pZHr0L41leFqZiVimRxZldB9QR7WS+HBF4S01Hj59JwjTE+LlD5eXRDvkb8O5ZMpJ2z0m8u394Y0NVNgcUppqGvMKeRpCsZsXJBuCOb+4t5g5TYzHddn7JhnQMA9RbzdyCxuDv8Ay4TFdJnQuFamcj16nfGD5fTZ+rXqzuGE7f7ExzpRxpOkdLTTwQKU8jySabseD7c5b5qa4k/dm7Dw7hcegoOr8mc/m9f9zb+PBvE7aHANPOp49WKRSxTIssMiSxuoZJI2V0ZWFwysvBB9j0EEVHUsqysNSmoPWT3vrfXvfuvdEG+ePyPi6U62rMfjJWnzO4aKpo0ShEdTVUupggZoluyn6/j2JNi28Osu5XApHbLrNfOnQN5z5hGw7TLcKNRCk0GT+zpF/wAuas2RmOtYt0msVuxczNV/xxK1lhr/AAMVliQwvZjxc/T8eyG454s+bHP0T/pISAMcRg4/2Oir263rbd52hb2Jv13rqDYb7KdWW+2epH697917r3v3Xui8fJfoDEfI/rTM9b5yukosbmYViqGXy6SEYupZYuTz79TzHXsefVTW7v5a+76zL9Odc45JKjqzZS/Z5RmSoNNPTGo8jvMv1/x597r1XTw6cv5o25shs7pjaXxM6ypa2BsrjMZRUK4+OV9UVKDphJS7AC5IA+l/fuJr5deOF+fVf02+N/dUbf6O+HnS0H2m+u38HSxbzq6UCGvky003hEcsjAOCL8+/ep61kEKnz/lx6T/yO3V8pP5Yabb33VZfIbipqlKCXLwV84rkjnqJgXjSGUsCAQeRx7VQr+mXGKdJJ3dXVCNQPHq+7o/51bLzvx42B2z2jlcdhcru3AwZr+GpV0MNXItQ1lVKJmRhxz9PaSuo16WU048h0azqzvPrvt+hNbs/OUVVYKRTNW0hq2D8DTBG5Y/7Ae/HGOrDPDoYffuvde9+691737r3SRzEj1lbFQxMbIyu1uf8SD7MbcCKMynpFL+o+ny6VcS6I0W1tKKLf6y29l7GrE9LFFFA9OufvXW+ve/de6xSwxToY5o0kQ/VXAI/3n37r3UGtw2MyOOnxNXRwzY6oQxzUpX9p0P1BUe/DHWiOkbQbN2lsWknG2MHQ4aSqVkk+zVkMgI9RNzzz7UW6a3qRUDpmckLQHj0rsLEEokbSAzXuQALgHj365aslPTrVuulK+vTv7T9KOve/de697917r3v3Xuve/de6TG4XaUR0S/7tsTbknn+ntdaArWX06R3LA0Tz6f6SMxU0ER+qRqp/wBgPaSQ6nLevSmMUQDqR7p1fr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/Q3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691qd/wA6PYEvSXzn+GnyW2ZjBi8Rt7K0uc3tXUqPTxT1dBn2fyVdSPQNcai+r3dRUFR01IwjIlOaGn5EdbOfU/ZmF7V6w2p2TQ1FGMNu7ELlaaTzx/bfby6g6eUmxC6WBN/wfdOnT1qJ/AvZOP3T/ME+T3W+z4xkNmb/AOwNwf3ujEZmxgihrJ5otci8MoP0N/dz8I6az4zgcMH8+lx8PNrYncafPPpHuHBTtt+m3xPS7XqZMZNLNj6XHV8yhsettVhHYix/A91NRQjrYAOpG9f5EdEn7k6r+NO5utN4bB6e7W7r/wBL2O3LjqPDYd8XX0+KFNFLokgp3jOpRqtYW+ntVMAQrjI8+k0VAHQEhhw49WyU/wAJ6nH/AATwXZ/dtdlsH2ttfFYbH7Rz1NHK24DjgdMf3E2rUhYKTz+B9fp7S1FccOlWkle456rz+aPSOy96fGPEVu79157dXaybl28mAxT+TL+fEWGmepERYq99P197FdVRw6pIFK5yRSn29bTH8v3Yu59v/E/q3bfYVMKqOPaW33xlBXr5Ep8eKH9iJqaW+gqNPpI9049PdHlpaSmooUp6SCKmgj4SGFAkaD+iovA97691I9+6901ZiiSso2D/AO6CZ14v6oxcce1FtKYpKjzx0zOmtPs6jbdrJKzHhpbB4pGisD/ZTgX92u4/Dl+0V69Aax/Z0/e0vT3SVn3ptymzDYGbJ0iZRdN6QzxCcauR+1fV/vHvwz1416VIIP0IvYG35sfobe/de6StVvfatFlf4LV53F02S1pGaWorqeGYO50quiRgb349+691Ud8p/wCa/snqzfmb6y6/SqzO6toSum5ftoaevp4tEpjbSFViALfW/vY6qT5dDr8GP5inV3zInz+ysNPNDv3adG9VuCgqWpkvEGVWMUEViNOrm4+nv3cprw68CsimhrTB6Sn81zpuTsv4kdt5nC437vfmxsBTz7KkjieSpim+7QzCDRzqKsSOPx7UvTSrDiak/wAuky6gWRsDgP2Hql7a+2N2fLH4G9VZ3aA/jHyO+G2Cq5a/Fyus1eXlyDVMd4o7OpjAvZvaetD/AKuHT3xoD/qqBw6m9g/zC+0Pkb8RJ/iRUbOy1d3ruDDUWAzUNRiJvs556euBFKXA5+ifX+g5t78QAdR4de16gUAyR0dTYn8qGt3d8L9kbd3jNVYrtal29GJtvNGy46kqFnLRUoDMCpsAx4t/j71Ug16toGnSel/1h/LAzvYe3NvbN+SlZUVeztmUcVFidryTHJYmvhhlBjWWkaQoABci/vRJ4dW047ura+suq9i9F7Ch2Jt1qeg2njKZoKDFO8dJR0lMqk/b08CNZQR/T3aPUHDA0p1V9JUg5r/PoJd2/JfDbYiq8ZjFV6qnYwUqRusy20EoVJv+fb1wVNGXpq3RlBVuHl0Wmr3N2n3BkWpqZa2mo6iQWkjRlJR/QbsvtN516U4pTowfXvxboaZIq3c8jT1TFJmWeMu5bVcizHjj34iuetVPRssPtvA7ao44qGjpKRKdOZ9CRmy8lmc/T/b+9kgZPWiQoyaDorXyG+X+xemqGegp6lsvuWoElLj6XGeGuArnUCnEixauCxsfYT33muy2mMop1yHApQ58q9AHmvnzbOXozGG1zNhQvd3Hhw6J1szpDvL5a5CDdPdtbkdpbMlqxkMZBjHMEldRP+5HBLT6luGFgb8ewlabPvPNTi53kmGKtRp8x6U6j2x5a5l58nF9zIzW9sTVVQkFhxow+fn1aT151bsvrHC0eG2rhMfjxSwJDLWwUkUNXWMi6DLUSr6iW/Ivb3JVjttnt0SxWyBaClQKE/M9TZtWy7ds1stvYxqgUUqAAT9p6ET2v6Neve/de6JT8t+go9/YGPfG2lOP3ftDyZiKehRxVZR6ciRIJmTlgLf7b2LOWt5+jlNpcd0cmM8BXoN79tX1cf1MGJI8gjiaeXTv8Ue+Zey9rQ7e3a60m/sIskOSoZDHG5p6YCKJxGbNew5FvbPMez/QXBntswvwP29ObFupvYPBuMTLxHRv/YZ6EHXvfuvde9+691737r3XTMqgsxCgckkgAD/En37r3RFvkh3I/Gztuv55akrFM8JWQq5urH08/m3197DEYHVSK0J6buhen6nFJHvDcFOZSV8yCWNrufIGtY8e34IllqG4+XTM0rRkEDHR7KCupqyENT2VVFgnAKgcCw90lieI0bpyORXGOp/trpzr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917pj3LTmrwOVpgLmajlS3+v799nXuqwdih9ud0VNM37ayVrD/U3/AHbH6+9UxTr3z6tYQ6kRh9Cqkf7EX97691y9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3TRksvBQrpvqlbhVXnm35t7UQwNI2eHTMkwQYyemanoZ8zIKmtLxxCxVBwr/4W9vyukA0RcemY42kOuTpWxxpEixxqFVQAAAB9Bb8e0JJJqelYAAoOuRAIIIuCCCP6g/Ue9db6JP8AJT4hbb7dhfdG23bbO+MarVdFXYmLwTV9VEoMMU8sRBsxFj7B/MHKttuqm4g/TmXIK41EeR6jrnDkKz35frbQ+DdJlWTBJ8q0/n0Xjo35V7s6q3YnSvyBpZ8ZkaSVKHGZN4gsDwxDwwtLUSCxLcE+r2QbJzPd7ZdfubfhpZcA/Lyz8+gny1zxf7LuI5b5rXw3XCPTDAcM+vqOrU8fkaLK0dPX0FTDVUtTDFPFLBKkqMkqB0OqMkfQ+5MjkSVBJGQQfTqbYpY5kEkZDAiuDXj0H3bXZGK6w2Zl9x5GpiimpqKeSggd41epqY11KkaP+r/WF/Zlt1jLf3SwRgmpFfl0mv7yOyt2lkNKDHVe3x462yfyM3hnO5Oz8WMntHJyVMOGxeVhLRCaOTUGjgawA5BJA9jDmG5g2uxGx21NVBrpwIPkegltdid7ke9vl1RNgBh/k9OuHeXxf3l09uCp7l+PklUamO9Rl9rRkx42npojYmlgUkEFfrx7xx3jlq62idt22GuSSyDAH2DoF8zcm7jsFy3MPKVdYy8X4aD0HQ//ABi+XOC7gpTtndDLg+wMWHjytHW+KigkaBArGDyabksDwByfZ9y3zTDuyfTXPZcL8QNAMenQp5M58teYI/pLz9K7TDq3bkelejtBgwDKQykXBBBBB+hBHsY9SLWvDrv37r3Xvfuvde9+690WzuXrzqqCWLtXsCgjrZNsK8lNLLB5ZEdom0JG45H0vyPexTz601Tw49a1/ee46vbnyi2p8tOraaHLbP6qq1qswXKSzUMkeqcwqi/QECwv7si6sHpt28MeIPKtei7bh+Zee/mk/IrE7NXBtV7DxJqoM5Qz0RehofskZoQqRD6Bh9W5J+t/aiYiNPDTgePSWAmd9T8BmvRO8Ye2PkN8i5pM3WZrb3Q3QOSn2Vkqbb6SeKDH4+Zok8dHB9TpUkXHtP2hfn0/3tKQTRV/mej+fGPujbPVvy72dTdHbt3nn9m0sEkm4YdyI9LTCWJzLLE0b+nSAOPes0z04GGqi+mej9bv/np7U238rNvdOR43J5DbFWFp8rU43HwVyRVjOEZFqYlNgpNrXv79Q0qetGQBgnn69XzdQdq4XuDaFHu/BRVMVDVqrItVEYpgHuVDIbWNh7rUeXTo6E6olEMMkhNtKMRb63txb3eNS7hR1SRtCk9JvCQGoqJMi97lnjAN/r9Pz7WXT6VEK8OktsHcl24dKr2g6W9e9+691737r3Xvfuvde9+690kctIKuvp6Zf90yrqsL/kXv7MYF8OEufPpFKS8mkcOlWiLGiogCqosAOB7LySxqelgAUUHXP3rrfXvfuvde9+691737r3XvfuvdJNS1ZmY5DyIG0/4WBvc/7b2YEBLanr0g+K4B6Vnsv6X9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//0d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdVQ/zhfjRk/kT8QOwKHaOPNd2Bi6KlO2jFC8tQn+UmScQ+Mgg3I97UkGo6o6B1KngeqgPjjkP5mO5Pi91/8bdlbYanXYGEfEVVdUU6UOQaEzsCJayUKdKiR7Xbi/vfbq+R60DIV+fDq3b+Wd/Lro/h9Rbu7B3ZOMl2r2Y/3m5TPH5f4VUPN55kpKkswOs8Ej/b+6k1PyHVlQKPmcno+OA+OfVe2905vdmO2xio8juKpnrMuDQwlK6pnJLy1Ia4c8n6r70RUU6t516TzfG7ojb+8ot3UPVmxqbITsZqmoTAUwlaqLahVFyCuq/IuLe18I8WBo/Py6Ry1jnEgPHoS+yOsNsdrbErNiZyERYDIwohipo0UQosbJH4EFgNOq62+ntEQVNPTpWDUV9eiY9R/wAszofqndrbrafL71YVAqIcTuxIshjKeRG1ReKB2YDQbEAi3H096z5nr2lfIdWJ0tLTUVPBR0cEVNS00SQ09PAixQwwxjSkUUaWCqBwABx791vrP7917r3v3XuuLLqVlP0ZSp/2It72DQ160RUU6ALtXtbBdE41NwbgiqpcbVyyRJHSrqPkWxJYAH+vtfKfqIQyjIx0jiDQyFWwOihfJv8AmX9U/HXq/qbtXcDVEOC7M3odqUavJBC0csc0ccsk7zrYKA4JNuPaACuOlZKjJNBj+fQD/LftzqDpndvU/wAoqbeeWzGN7dyuIxtPSYLJQ12HpRLHC8plqIWZAAHAa6ggg8e9Z4DrdQMt9nXt1fzQl6t+ROBwW/KWdunN9VOKw+0s1jo4ZaQVFWiR/cz1xUKQrODJZv8AX9+9etE0IHCvRT/mBnNxZf5jQxbK3jVVeN3Hl8SmC/g2UWoo5Zq91ljRGhJH9r6fj6e9inXmapBHVTXxp7Y7J29/M0+YXT1VtTAZ/dNXQ1eCT/SDRUwxqVktSEElBPXLp8n5BB974gH0r1Qkh2A4mh6Pj8PeiR8MPmbWb43PlG/vt3VkBjclt/a08VVh8X/EJkl0MKUkKt/xYce6tkZ8urIoDGnE/wCTrar3dhcXm8bJj81Tx1OGqyFytPKpeKenVGASVB9Rcj2rtiTVAKmmPz6YuBpIb16qL+BPwN3p8XPkr8k+xZAavrrumqmOPxVQ16KCikr/AL2IR0jswsBdbMtrH2nkUq5Q+XTyHt1D8Rr/ACp1arSdMdR0GXOfous9jUmbLmT+LU22MPDkPITcv93HEHvf83908qdOVPQhVFXSUMeuqqIKaNR9ZZEjUAC9hqI/Hv3Wqjou3YPyL2ttZJ6Ojn+4r7lImhkjkUkqbEAX/Pv1R1o8c9FDzO+uyu3a846gSup6KoawliVlIUjRYlR71knq4pToRdp/GOtpJKfLbo1yqhjZ/MjSO3ruC4Jv9P6+34YhKdBOek88mjvXgOjt7W2lt/b+OpocZQUiWRSZlhUOSOeCeRY/T2266GKny6dRtShh59T9xbowe1sdVZPN5KioKelhlnb7mphgZxEmsqgkIuSPoPaa4uYLWMyzsFA9TTh0nury2s4mmuHChRXJA4dVc9lfL7e3cWbrOtPj3i62pyE88uOqK6em/wAlMMq+FpI6pVtxyRz7jfcOar3dZTt+wqSxqCTwofn1DO7c+7jv10+z8poXeuksR2+ldXQpfHb4R4zaddF2D2tPLuffFVIaioxmUCV+MpJWGolElLAkEi1hbj2Y7BybFauL7cj4kxyVbKg9HPKXtzDt8o3beyZ7pskN3KD8gerDaamp6OCOmpIIqanhUJFBAixxRoBYKkaWAH+AHseqqqNKigHl1KqIkahEFAPIdZ/e+rde9+691737r3XCSNJUaORFkjdSro4DKyngqyngg/ke9gkGo691V78hOptz9Ldgr3v1ksxoaqpNRuzH0sTLBTUKyXl8UYuCSCfx7kHY9xt91sv3NfU1AUQnzPQI3exn268/etl5/GB6dHm6Y7i213LtCh3NgahVeVRFU0M0sf3kM8SDytJALMAWvbj2D902y42q6NtOOHmOFOhPt24Q7jbiaM/l59C/7LejDr3v3Xuve/de6LX332/SbJxMmLoahWylWoRRDKhdS900EDn/AF/fiD5dewcHos/R/U+T39nX3buRJHpjMZW+5QnTaTUoQn6+9KRTHWiDw6sgpKCloqOKggiRaaGMRrHYadI/w92BINR14gMKHpOVuKq6GY1WLuQfU8Z+gt9eB7MIp0lXRNx6RvEyGqcOnDH5uOe0NQDFOvpbUNKk/wCufbUtqy9yZHV0uATRun4EMLqQR/UG4/3j2kII49KgQeHXfvXXuve/de697917r3v3Xuve/de697917r3v3Xuve/de6xyxrNG8T8q6lWH+B+vv3Xuqrd/Qtt/vtVYlImyVlYDTe8vBuffseXXhk9Wj4yZajH0cqsGDU0BuDcX8Qvz72etDh1P96631737r3Xvfuvde9+691737r3XvfuvdcWdUBLMFAFzcgf7372ASaDrRIAqek5V5nzFqahBaQnSWIuCCObEe1kdtp75OHSR7lXGiPj11QYVi/wBzX+qW/pS4YWPJv73LdY0R8OvRW+db9KVVVQFUBVHAA4A9oiSTU9KwAMDrv3rrfXvfuvde9+690W/5BfG7ZXeu3KijyVJFj9wQJPPjc7RQRpkFqwl4UkqBZiuoD88ew/vvL9pvVuVcaZBWjAZr6E+nQS5p5R23mayaG4XTIKlXAowPlnjx6rt2p3/vr4O1/wDcjvgVVTsVahv4RlgVkkaAftw/5VICAtiv9r2CNr3Tc+WLxdr3NGaJ2opArT0NfTqJ7LmrcfbaQbbzWxNrXtkPp5VPT/Juur+d/Y+Px236yeHrPa80OXjmidGSpVrStBO0VgwNgpBPPvJfbbyw2XZhfW5V5J1pxqQSOPyI6H0F/HzjcK9m9bdaNUGoNc0x8urZ9u7exO1sPRYTC0VPQUFFBFFHBTRiKK8cYjaTSPy1rk+wJNNJcSGWUlmPmepDhijgjEcYoB08yxRzRvFNGksUgKvHIodHU/VWVuCP8D7ZIBFDkdXIDDSwqD1XT8n/AIezbmrYux+nb4DfONmasFDi0FHTZGYSeS0wiIBvzcW/2HsA8ycqG5b94bT2Trmi41HqJ+c+QZL2Ubxy8fCukNaLgN559eoXx0+X9W+UbqruiN8LvjEsaWaoqlSlopDFaKNYppAoa5H1v7rsHNbGT92bx2Tpgk4HTXKnPzeP+4uYh4V0mDXCnywTx6sgp6iCqhSemminhkUMksMiSxsGFwVdCQePcgKysKqajqXFZXXUhqD5jrN731br3v3XukjvnZuL39trIbYzGr7DIxmOUoAzC6MgIB/4N7917rXW7V/l0fJzrndG7cL0TjqbeOxd+1y1GZTOFTDBE7EPGIp3tYAkf63tTEwVdWMfZ0lmVnYJ5Hjx6OJ/Lz/lk7e+MuB3xuDd2CxMW/N7/cySCkpgoxj1Ub6vBKrEcFh7YZi5qenkjWJdC8OPRHaj4c/JL4edtbz3F1RsbB7w697FzlTldwHLUArYDJWM0jK8MhcFhqPvRIoAethSGJXzz0j+4uuG6L6Z3R3VubZNFge0s1mRR4agwWH8EKUmXQo8gWMWGkNwLe9A1wOtkEDUejn/AMqf4C7B2v1jkO1e3uvsBurem/snT7lweR3PhIqvIYzHVOqf/JpaoG13Itb+nPuxIPWlUrj9vVyWWl2p1JsvcOZpaShwuGwmLyGXelp1ipadVoqZqgxxJ9PxwLe6+denOI09a9uE/n69dZjvKv66kwe56jb1Dk58dPXUOOilgDQSEOxqoo+Bx/X6e1UK8HYY6QzSBmMaGtMfLq9n49d99f8AyA2VHu7YVYktDrSOop3qYpaqGZ1JImjSxU8H8e2ZGLMSelMS6UA+XQ9h0LFQ6lh9VDAkf649t9Odcvfuvde9+691737r3WKaQQxSSn+wjN/rkDge7Kupgvr1VjRSek1hUapramvdOJBZSbcEHgj/AGB9rbk6IxEOksIrIWPSq9oOlnXvfuvde9+691737r3XvfuvdRK6YQU0shNrCwP+J9uRJrkC9NytoQt0y7fPl+4mK/VhZiP8f6+1V2QAEHl0nthWrHj0pfaHpZ1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf//S3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691hqKeCrhenqYY6iCQWkimRZI3H1syNwffuvdQKDBYXFO8mMxWPx7yfreipIKZn/AODmFRf/AGPvQAHW6np197611737r3TFuGm+4xswUfuAqVP5+v0v7VWcmiYV4dMToGSp8uuWCrBVUSJ/ap1WFr/UlRa5/wBt71dRlJCf4s9et3LJny6e/abp/r3v3Xuve/de697917oIO0O8uuOpcHXZrde5sNS/YwzSnHvk6WKvmMI9UcUDEnVewAI/Pv3A0PXvs6I1j/mT8ePllgd37TxVTSxbo27icjkqDDZatppaqeSnW6mGitqOq3Bt7V2rVJjPA/4eks44OvHz+zrXk+RuG3B/MjxO/PikuvEVfxvwW8e1sXSxBKer8OMiNZMaWmUK2srGum3J9pyPAkA+3pwUuo2QYrT/AGOh2+DvUu1vkX/KT3ztKry2TzfdXTX989xYLG5qf73KUr4mjUY9oqKT1qjeNgGX6Nb+vvTYavW0GuNVJqaUJ+fQzfy/do7V+eHwXX459x08VF371RmNwZbAZd6aAZSCNJguOM1S4WRQGsh1Na1iPoQfHtavr15CJE0niOP2jopHxS6J+WeyvnjH1h21iK6t2b1puZMng8vUU0lStStLVA0patNwyFQCpuffiQRjrUaSB+/Pz6uA+cv8qrHdz9kHvvpYU+3O1stWwVm4Zafw4wZCSAKUmkrFZLlj+oHm4v70DQZ6uyamqMEefQq/DX+XRB1HnantPtrK1e5+x619RpK+YZKhxsqquiopJ3dgGFjbT9PejQ8OrAU45+fVr1TTpVQSU8n6JFKn3eNzG4ceXVXUOpXpKY/Jx4qoqKHI1UUFPFpWlaeQrZRza7+1lyqSIJU4+fSWBmRzG3QXb/8AkDtPaCTU8c6VVYPSngmRxcg2sFB549l4I6WEE/Lom2f7V7L7WrJMbhoayKgmIRZEVlIUHQeVt/X3s563SnQl7A+LeQqp4Mlu+WSQtpa041nTq1KdLG/HPvXd59eND0c7bGw9u7UpVp8dQU+pbfvNCnk+t+Dzbn3vyp17zr0qayCOop5IZW0IVN2vYKAL3J/oPd45DG2seXVJFDIVboo3bHyq6/6Lx9dSZmsGWzAkkhoqHFVMVVUeRov8nvAoLWueePZNzJzHYbVD47NralNKmpr0COYOd9o5Zgb6h9T8AqmpJ8qDoksOxu//AJn5eDLbiqKrbPUk1XDMvjkNFkpICQCixXUt6RyB7jf6LfOcJg9wTHZmhrwP7PPqNf3ZzT7izCe7YwbeSDQdrn5dWe9TdIbE6ewFJhNs4qlaWnWPyZaopIP4nUSIttclRYt9bm1/z+fck7Xs1ltUAht1FR+Igaj+fUzbFy5tmwWi2tkg7fxEDUfnXoYPZr0fde9+691737r3Xvfuvde9+691737r3TbmMRj87javE5Smiq6GtheCogmQOjo4sbqfbkUrwyCSM0I4dUkjSVDG4qD1UZulc58Te4shuPaNXHkdj5eoYZHEUUqS/wAOpHlHlLUiahGwH5sPY9stx2zmq0O2ysPqoh65r1HtxK2w7oXtCGjb4lBrQfZ1aR112LtzszbWP3Lt2simpq2PUaczRNUwOoGtJolNwR/iPYKvrG4sLhre4FCv7Oh3Z3kN7CJoTUHpe+0fSroKu1OzMZ15g56qeZBXSRH7WMuoOo3Aaxv/AE9+4da4mnVf+zNrbj703rLmsiJnxqVHkIkUvGI1lB4Y/j3r7et8OHDqzbbe3qDbOKpsVQRRxxQJpLRppMjflm/r7316g8un/wB+691737r3TTXYinrbHmJ1HDR+m5/F7e1EVw8Zrx6Ty26yDGD0yxnKYq4e0kJPBPqOkce1b+BcDtwek6+NB8XDp7octT1ajnxuByHIUXH1te3tJLbSRn16UR3CuM46dAQ3III/qCD/AL17T9KOPXfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuq0/kvjxiewsbmlGkz1wZnI0/wC7b31H3o049epinR8eta5chs7EVSsX1wgEk35VR+fe+vdLv37r3Xvfuvde9+691737r3XFnVeWZVH+1ED/AHv3sAnh1okDiemStzkFOfHEDLIfoV5Xn2qitWcamwB0mluVTAyemtKfI5diZ2aKAm1xdTYD8e3y8MC0GT00EmlbUeHSjosfT0UYSNQxHJdgNRP9b+0UkzyGp6UxwpHw8+p3trp7r3v3Xuve/de697917r3v3XumvMZjHYHHVWUylVBR0dHDJPLLPKkS6I11MAXI5/w9uRxPM4jjBJPp03JIkSF3NAOqe/ke9T86dwHqLZ+Ipp9u4uScVWekpkmAgNvMyVnABAXgBvYvk5Z2m12l5t7UeJItFx3Bjwp1GHNW12vP8MmxvGGiIILEVwfn0a34efFab4vUFTtujliqcC9Mjx1kxX737jxWeORtROgG9r+4o2Ta77a5ZIZJWkhLErqNePDpXyDyUeSLb912xrbKKLU1Ip5Z6SPzM/mU9FfDmHFw7qrRuTLZKd4Xxe3q+lnrceY3CE1sA1FRc/n2JaE4HHqRWcIupjQfPoxvxi+SuwvlH1jieytjVkP2mRaSOXFyVcE2RonRQ4FVBHYrcMCOLe9Z8+rAgioNejGe/db6J98l/iftXu3DtkcZGuA3tjUM2JyeNSKkeqqxJrVa+ZNJYf4k+wpzFyxbbzF4kX6cyZUjFT8+gHzjyPY8y22uP9K4TKOuDX5kU6K30z8kN39A5ym6a74p6v8AySZqHFZiwEL6m8cTzVTggrx9b+w5tPMFzscw2regcYU/5z6dAzl7m2/5YuV5f5prVcK/kfSp6tQw+axeeoYcjia2mrqSdFdJqWZJkswBtqT+l/clRSxzIJImDA+Y6mi3uYbqMSwMGU+YNenX250/1ByFYtFTSTE+oD0L+Sfb0MZlcL5dNSyaFr59NmFjqJddZUO5EvMQLvYAn/U3t7fuXUDw1Ax0zboalz59KH2i6V9R6mkpqyPxVdPDUxXv45o1kS/9dLXHv3Xugq7I6S2D2nRQY7dmGpK6igkilFLJTRSQs8JvGxjbgEfQW96ORTrwwehGwOEx+28NjcDiaeOlxuKpIqOjp4lCRxQRCyIiD6D/AA9768BTqmP+c/8ANXbnxm6MjxM1dImU3Llo8DVU9HVxx1YpMpTeMloiCdNmN/8AYe7KCzUHTcrqqkk56AT4UdHfFnrn4Rb17P3JitgZnc29sNkNz0eTzNLjptx0kuTx7TU9PSzyXkDBnuNFuRz7VXL6VCJ59J7ZQxLGoz/P/iuq4/i/8/8AA/CfZu9N6VdXlW27mt45TDYegqqhIkORkkZaQiKUEGPkfQe0pDN0o1KlanFejgbB/mafJXErjO1t8vSVHW+4MvDFSxU8kMlStBWN5otQVTwEI5t71TzPWw2cDHWxP0j3LtnuzrvD7/29Ov8ADq6ngWZnlRhHVGBXmQuLD6n3r5dX8q9C/HLFMuqKRJF/1SMGH+3X37r3WT37r3Sfz9QY4Y4Y2tJKxUD+t+OR7WWiVYseA6S3LqBo9enDFwPT0cUcihX5LAf4nj6f4e2Z31yE9OQJojA6cPbPT3Xvfuvde9+691737r3XvfuvdJzcFReEUScyTjgf4Xt7W2iZ8Q8B0luGr+n07Y+AQUkKaQraAXsALt+b29p5n1yE9PRIEQDqb7a6c697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/9Pf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcHVXRla2kjm/0/wBj72pKmo60wDCh6R2FyGKpMhUYyPKUE1TUSs608c4M99XChP8AY+1tx+rGJADj7OkkTlZPDNOlmzBQWYhVUFmJNgABckn2h6WdICXtXriDJrhpt7bZiybP4xQyZalSq8l7aPEx+t/xf37r3QFfLX5fdb/ErYcO8t6zrWvXTrTY/E0dXGtfUPIAY5Y4gGJU3FuOffuJoOPWiQBU9Uc72/n34vaPduC2hmdm7rw+0dzVONx9DkK+l+zo1kysqxwzCreIX4a4sfdwppUeXTbOFIDY1YFR5nom/wAtKXufdv8AN625193XuefFfDfee0drbsw2Qx9eY3eo3HjYsjTxK9wGvIwBU+9AYJ+fXnqGUedPyrXoQP5kXxVo/wCXj291L8uugstuGrwvYGdwe0cljGl14+noXSJqiSRE4KzQuH1Mgvc/0Pu0TaHDV6rKmKgYwD9n+x0re3qGl6T+X+3vlTsPH1dXsr5AbY25sXMJgYC9Dr3BjIabPU87Qkhr621KVF/ay9jVkE6cT0xAxWUxHhw/2ehNwXxV+TXxI+bmW3X1Zj1k+PvaGIo6B6KGFTQyUG4KZWr4amhVjZlLMHBA59oCRpoelQUh+3geI+zz6ug+Pvwk2B0L3DuzuDaDzU1RvjB09NXYpIlgoqWonVZ6toYlY2u4v9B70ePTgAFaDj0dFtvYB69so2FxJyclvJkf4fSffPb6a6rR5D/sW9+6908e/de6xSzQwIZJ5Y4UH1eR1RR/sW9+690CW+u9tobPhqVNZFV1Ma+kQTK3qN/7IF/x79w69x6I9u/tze/Z1bo29DWR0qMAZEDAlQ1rcW9upLJp8IcG6ZlSMDWfw56EDYPxnzGeanzm56iSRJ2LukzayLEEXVz/AEJ9tlCjaW8unFbUmoefR2NqddbZ2jTiHHY+mL2s0zwR6vrc6frb3rq3S6AAAAAAHAA4A/1h7917ppzWexG3qKXIZnI0mOpIUd2mrJ44EtGhcgNIR+B7alnigQvMwUDzJp0xcXMFrGZZ2Cgepp8+qze3fmxlNz7hfrLo/G19fuCtf7JcvFF9xjFE6mFi1Qi6RyeOfcdbrzhLcXH7t2ZS0hxq4jqHN+9xZ7y8/cvLKNJM2NYFUFcZPWLqL4M1DZkdr9618+Y3sk8FVTYtJFrcUFLeTTPAWIv9Ba3tby7yezXAv97JeYmumvb+fVuXvbRBefv7mlzLcjIAyo/2terNcEmMhoIqfFUlLRU0CrEtNSQpBFGFFlUIgA+nsftbi27FAUfIUHUwW/gBNFuoVRwAFOnn3XpR1737r3Xvfuvde9+691737r3Xvfuvde9+690Sf5SfLHBdPYxdvbek/jW98yWo8ZS4yVaiairC+hTUwxhiBf6k+wfzLzRDtEYgt++d8ADND8x1HXOvPNry/D9JbfqXMmFVckH5joLfjJ8fN37iG6+w+7L5Cq7EoJI1xtSRKKKnlIdHihkJ0NybG3tnkuy3exvW328YiSSppXGfPoq5E5c3QyTb5v7anux8J4KPLHl0GGYwXZHw17FqtyYk1WT6nzFS6CmjtMaWmM1n1xjUIyBYjj3kDHPY802Qt5aLcoOPqadH8sd5y5dmeLut34j06sYwvdeztwbAj39ja6A0U1N5hRyVEf3cbD9SSRr6hb+um3sAX1lNt9w1vcChX9h6GVpdRXsImiNQeiB57L7h783vFR0X3L4uKoKra8iaFl/tfT8H2iJIOM9KQMDqxXrXr7HbCwkFDTRItU0dqiRVUXPHp4v/AE559761TPQke/db697917r3v3Xuve/de66IDCzAEf0IBH+8+/AkcOtEA8emStwkFReSItFKLlQp0rci349qo7p17WyOmGt1rVePTSq5rH8Eq0VzYg6vSPoPb9LeQdvH59MkSRCvn04Q55AwSojdSbDVpsP9f229riqHry3RHxjp3hraacApKtyL6SwB/wB59pmideI6VrIjYB6khlP0ZT/rEe6UPVqjrl711vr3v3Xuve/de697917oiXzLw2uhwWRgQh0kLSuo+lpf6/j3vr1T0Ovx4y0Vf13i4QxaSANfnVwVUcn/AGHvWOvZOT0PHv3XuuJZR9WUf65A97oetVHUKfI0sBKtIrMP7KkE/wCtx7dSCRs06baZFr8umWXcPkcx00MmoEjUykjj6c2t7Uiz0ish6Z+oLfAOsLUmWyB1SsEjuQADpNr+7eJDDhR02qSymrcOnmjxFLSEPbySWFy4DAH/AAv/ALx7SyTu/wAh0qSFE+f29OoAAsAAP6AWHtjjx6d4cOu/fuvde9+691737r3Xvfuvde9+6901ZrN43b+OqcplauCjpKWGSaSSeVYlKxrqIUt9T7ciikmcRxipPp1SSRIlLuaAdVY9xdl7++Tu8B1P1GZ4MBFUsuZyusJSy0F7VBWqFlvpBsL+5D2rb7LYLb957nQuR2r51+zoEX17db3cfQWHwD4m8qfb0fTpLpbaXRWy6fA4ZULqi1eTy1asQq5Kp4h9yZKo8iMNqsNVrewZu263G63JnnOPIDgB5Y6FO2bbBttsIIh9p8yfPqnr+Z//ADasf8fsLnusOkIqzc3a9VTZDGU8+BMeRGOyOoQ0zSRQqxUavqT+OPZaor8ulsj6Bgaj6DqpX+WD8ecd86uzd/7i+c+4sinZNfjqiqptvVVfBSrKtS4mhaCnq/TG6IxawXm1vr78xKmin8+qxqJATIPy9OjTbK3Dm/5XHzXm2FSZebM9LdkZeHbG08dHWK7wCqnSmiqaijGpVZLjkD6i/v2SM8R1ZaI2kcG4f7HW1nDKk0cciMCJI0kH9dMihlNv9j7r8unOsvv3XugC70+Puze79v1GOzVLHTZIRH7TK00ES1scgOpf3hZvr+b+yPedis95gMcwo3kw4/t6C/MvKu3cyWphulo1MMBmv29V0bV7B7S+Fm6Itn9jQ5DNdeVsrRYKth1VDQwPKFWSqmOoCwH5PsA2l9ufJ1x9Hfgvbse08cV416iaz3Tefbm9Fju4aWzY0Rhmg/pcerXNh9h7Y7FwdHndt5KkrIKuLyeGKpilngtbUsyIbgi/5HuTrG+t9wgE9uwIPzyOpw23dLPdbZbm0cMGFcGp6kVryZPJJTxq3hhYpLwbcNcEj2fRqIINR4npxj4zlD5dKyKNYo0jXgILD2XMdRr0rUaRTrJ711br3v3Xuve/de6xTFxDKY/1iNyn/BwpK/7z78OtHh1rT/NL4A9l/Oj5iUOI7BNXD08KX7oTwRLLHT1tNCFhKxsygWKjn8D2sthpBlPADpJMFdwhzTy+fTnS/wApDvPG5rE7CoN1helceKaFov4rEtY1PTNpCfaeW9ivFvaWR9TaqdKFjIWlafLogH8034IbQ68zGx9lVdFmx1PDncRV1NdiqG0hyqlDVTylWtpsb3P9ffgTx8+tOoxjzz0u+5Og911WJ652N8Zc9tzcuxKvE7bhqqGSvpKvK09VPSIkvnpNZZHXm/HB96HHq5XFBgdGC+S/yE7H/l2fGLr7pDrmtar7W3JnqN8jT0siz1VDNWIiJG1OFYqhZh+bcD35aE1OOqsxVKAVI4dHn/l59u/L7N1tHg/kNSzeOrp6XJQzfb6F+3raXzwhyoHPI49+PHHW1J01Iz1dHqATU3A03P8Ahxf36lTQdWrQVPSQCHKZdiT+zA2tfyAVP/E8ezAt4EFBxPSJf1ps8Olj7Lul3Xvfuvde9+691737r3Xvfuvde9+690ktD12YSQcxwOQSOQAD/T2Y18G30jz6QYlmB9Olb7Lul/Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//1N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddEgAk/QAk/6w59+691S78+/5kFR1P2Vt/wCMvTUM+V7c3lSxvjcnjZfuaaklq4miSlnjRSNYcj+179QnPl1UsBQcSeA6r13Lj/5rXUOZwPdu4a1a3as9ZjzU0lC8U9ZonnXyJJTRXYXF/wAe1UTs6GJTx6TSIU/UoD1Yv/MU+dG8fiZ/L82z3pPDVQb03XUYHbs7DXDJS1uegl8jv6TYgBeLe01M6T69Pl+wso4D/N/n6rlk+CnzI7E6RwfygwO9oJN97j27iO0MbTrmQsY23X4pdwrE8TMP3vCQdFrn6fX34EH4h1ZkYcGz1W52p8mewvlLBtzfHaH8aye3uut34HYuZhgeSWnfIY2pWnqJ3RwF0kIb8e9gU/Z00TqYfI9XFfzbdm/G3sj4KdXbmwkey2z+1KXa248XUbXpsONwBMThInnoqueiCyrplX1LLzquR+b6X4gOnZfhY+mfzBr0RaXB74/mR/y/uru5+uaaqXubqXt/b+yclI/OWl2XgqdEoWEzaXKKsdjxb3Y4YjiBjpsd8aycCaH+eetj3d/x+wXyz+Luxth9kY/RmMDt/Bwl6mljleHPY7Dw0U1RGJyNJYoCWB+vtuvmOnSNQofPqR1H8RNjdd9Lbb6x3xRU2dG3t0HO4uurKWGuqaeVCopljlkJ0qP6A8e1kBllqgyAPPpNP4cNH8z6dHdShx00FKGpaWoSniijp2mgilMaRqFQIXBta349pWUqdLdKVaoqp6cALcDgDgAfj3rrfWOWWOFDJK6xovLO5CqP9cn37r3QNb87v2lsuCQvWQVs6g/twTqSpBseAPfuHWq5p0TDd/e2++xKhsftKnqkpGYoSl7AA8/QD8H6296+3rZHp09bI+NG4dxyQ5rc9ROVlYtJHPIpHJ1kaXPNr+/Z4jrxpw6OntLqrae0aXwUWPglZ0s7ywR31EDUVA92BIbUPLrVKih8+nnGyvisnJQSgJT1EgFKo9Ki/wDQezCZRNCJF4jpDHIY5SjeZ6WX+Psu6X9F87s+Rmwul8BWZPL5CmyGRiGiDC0VVG2RmkeNjGY4BcnkC9h7It45gsNotzNKwZh+EHuP2DoL8w817Xy9atPcOGYcEB7j9g6riP8Ap++bmRU0ZrNq9TeWM18NaWoa94tWginvoJJF+B7ANd65yk7KxWuKg4P5dRIG5n9yHHharewr3Bu1yPl6dWSdKfHTrvpDCLi9tY2OqqZEiNTlMnBDU5CSVfUxWeQFlBPNgfx9fcgbPsFhs0PhW61PmxAJ/b1LnLnKe08tWv09ileFWbLE/aeh6ZFdSjAFWBUj8WIt7PASDUdCYgEU6Q6PJt/J+N7/AGVSxcsPoCx/r/vQ9mB03ENV+IdIlVoHzwPS4R1kRXU3VgGB/qCL+y7h0uBqK9cvfuvde9+691737r3XvfuvddEhQWYgAckngAf1J9+68TTJ6IT8qPlzj+tY02PsbXnd9ZpmoKSPFSCpkx9XI/jjepjiBKqD9SfYI5l5ri21fo7PvnfA050n59Rfztz7DstNs24GW6l7VC5o3zpwHSM+MfxMr2zL90d2E5nfOZP3sFBVhaiipUmAlicQyEgEX/p7Scucry+Md43rvnfNDkD8ukXJ3I0wuTzFzIfEu5MgHKqDnAPDqydI0iRY40SONBZURQiKP6Kq8D3IQAAoOpcAAFBgdJXe+G21nNt5PH7rgppcPPTNHVNURJJ40Yj1RFwbNe1iPb1vcS20omiJBXpueGOeMxSiobqmLcdDU7M3BuXH7IqMhV7TcssNLHMZAsZlOoJCOBx7NLzcZN6McU1A48+kFlYptCSPFlPQ9Hs+JddsaoxtR9pPRR7iC2qaSoeP71GOnVaNjcH8e015td1YgNIDpPA0x05abjb3hIQio8ujvey7ow697917r3v3Xuve/de697917r3v3Xuve/de66IB+oB/1xf36pHDr1AePUWooqepUrIg54uoAP8At/bqSunA9NtEj8R0zS7ehRGNI8iyn6anuvtQt43Bxjpg2wXKceoiY7MQgnyqQDwFbm3+t7uJbbgB1VkmAp5dZfvMlTm0oJUAWsNX14+vuxjgc9vVFeVV6yDNTA6WiYng3CECx/HttrdBkHq4nk8x147hVT6qeUj/AAVj/vIHvYswwwevfUsG4de/vLDqI+2qOAP7J/4p719E1K6h1v6rNKdFt+SrnMbReQQOEpqdpLlOQS5+v9PbMsGgUrXpyOUtkjpMfEzcUk+EqcahLClime178qOLj/ePdI41YVbpyV2QdvRtY6vK1BcINIP044HHHtUFtxx6S65m4dY2xuZnYNLLGE+unVz/ALx7t41sOAyOvCKYju8+psW3qYMsszyNN9TZuA35t7ba8fgox06tsoHdx6eYqWCIeiJB/jpBP+3PtK0jtxPTyRogoo6kfT3Tq/Dh1Gq6yloKeSqraiKlpogWknmcRxoALkszcD3V3SNdTmgHmeqSSJEpeQ0A8z0gsP251pn8o+Fw299u5LKI/jahpMhFLUB76dPjH5vx7QQ7vtlxL4EE6O/oGBPRdDve1XE308M6M/oDnoRvZj0ade9+691737r3XvfuvdJDeW+NubFw9XmNwZKkooaaCWVY550iedo11eKPV+T+Pam1tJruURQqST6dJ7i5itozJKQKdVj7r3t2f8u90SbV69jq8NsOlqnWvq6o+GOWlBtMUnsqtqA45/I9j62tdv5bthdX3fMRgD16BU9xfcwzGGyqkIOSfT7erB+m+mdq9N7YpsDgKYPOVjmrshURxPWz1TRgTXqANWm97C/sFbnulzulwZ5z9gHAD7Ohdt+32+3QCCAfafMnoR9w4pc5g8th2do1yVBU0ZkQlXTzRlNSsPof6ey3pf1W50l/K+6R6v7x3L35mIX3nuncMVTC2K3TRUeYxFI1Q2o1VPHV+RQ6/i6+/VJ49aChSaDj5+fRTvnN/La7NHYbfIH4Y1Ee3exa+US57DwvT4zFxQQOpT+H06MAbqOUC/63vYPkeqMv4lNCP9Wf8/VYe0Om+3ar5Ebf3b829zYfLZWmzNOdo4rG5CmmysGYpZAsBqaQG6DyKL8c+/VA4fn14LkFyD6U62Xvj9me6sv2PvOTfq+PZiYrHx7YiMZjZQkUaoWFhyU5J/JufdKgmo6dpQZ6OX7t1rr3v3XukLv/AK62r2Tgavb+6MZTVtLVx+MzPBE9TCPw0EzglSL/AIPtFfWFtuEBguVBB8/MfYei3dNqst3tWtLxAyt8hX8uqjd5bL7c+FG5qvc+yanIZjrOrm1VVDG7z1UdNJJqZEjXUVNh7jptt3TlK4N7akvak5HE0r1BN/t2/wDtxeG+2wtLYk1KZLAfLqyToXvjZPce26TMYirgpctUx+StxdRUIa+FwQpV4jYjn8ex7tnMFpvUIkgYAj8NcjqXuWuZ9s5gtVuLZgHYVZSe6v2dGI9mvQq697917r3v3Xuve/de6jVdQlLA8znhRx/ix+g93jQyOFHVHbStek/iIEqqp8g1JSK6kos4gC1P1J/zn1t9L+1M5CL4a9MQrrbxD0qfaPpV0D/cnR3X3em16nae/cNTZHH1CSoJvBCayAyqEZ6edwSrcCx9+61QefRJ+sf5Z3x5+OGWzPZOzZN4ZHMUlLU11NSZjKff0cdTEpkgWGlVB9DYAfj37J8+vUUcBx6rs69+O+8O9fnpujt7urETy9SbaoKrLUcGUpBNSLNio9dMscdQQASygX9+HDqunu1HyFB1eV1F3J152dUTUe0KKiifGRmmYxR02uNaNfEIx41BCgCy8+9YHDpzPn0NeYqhT0pF/VNeMW+tyPqPam2TW9T5dJrhmCUXz6x4SmEVIsjA+WQsXJ+tieB73cuS+nyHW7dQE1evT17TdP8AXvfuvde9+691737r3XvfuvdN2UqFpqOaRr3CmwBsfpY29vQRmSQL01M1E6g4GNhA9QRYT2Zbjm3t67Yagg8umrVKAsfPp/8AaPpV1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691/9Xf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3WGo1GCcLfV4ZdNvrq0G1vfuvdagOza5et/5y/X+I7kXVk9w7knqNs1mYZHo6Wjqp2FG481wPqNJ+gPux+HHr00ppOdXmMdbdmQpsTW4uVqmjx1fQR08lQkdTTQ1VGREhkWQROCpHF7gf63vStpNRj7OnGFQR1rr/M3sjZP8zfa+f+EGzcfFJujEbqk3HQ1dM0cuPWfbNPJ4KVYJdKpyXsAf9Ye37igkVk4U6TwmqPCxz/s9AH1R/Mb7z6K6rr/iTvzrXd2Z3ft3Gy9Z7VzdPj5/s6PDw0LYOmSTSh1CNLBCG4AFj7Y0jy6eDsMMDX/D0br4Cfy7aSn+MHZ9J8gdvxyTb+zmQ3tg4oaWmir6ETUs1QkzLZjrDaLKbE/1HvRPn14LQUOc/wCXqtek6m2N1/kew9sdtVG4NwbMEe46Xb+24cgajIpkQjpjmFGzEBA2m4A92NfPrS8SG/n1Z9/JG6g3FtHqvduZqsLNhdpZHc9WuOwuXo1gkeIh3p6yKmYWuAQC9vz701Cetxgqorjq+iOKKFdEMccSfXTGiot/+CqAPeur9RMlRJXUskTC7hWMZ/o+nj27DIY3B6bkUMuemPA1hp1lxtY2ialBcu5spjv/AKo8e1N3HWky8D0zbvjQfLpIb37h2psylaaaupquZfrBHNZxZtJGkC/tCcdKuPRLd6fIbeG+Z5MXsqCqiiclGClyNIb/AIp70fQ9ewO4dctkfHTde75lzG6Z6grM2tkmf0jV+4OHP9fevkOt/Po7GyOptq7KhQ0VDBJVFP3ZJYYWGsqAdIIP9P6+7Upw61x49CgqKg0oqoo+iqoUf7Ye/de65e/de6Se7JcbjqCTNZCsgoEoEL/c1DiOIWBazP7fhukgBWUhVPEngOkl2YYk8eZgoXzPVdHbnzo+9yUPW/UFBVZndddppv4vjWasoqWRx4WMrIPSAxB5PuOt/wCbtNz+79oBkkY0DrkD9nUS777lJJcfuXl5DLO2NS1ZV8sny6jdOfCrc+7txw9l/I7KyZvNRlKzC42mqlemiUyB40radrj9P4I9otp5NuLq5G5cwNrcZVQcfmOmeX/be6vbwb1zfJ40wyig9o+0Zz1Z3i8RjMLRw0GKoaSgpYI1jjhpKeKnjCr/ALTEAP8AE+5HjijiUJGoUD0FOpmhgit4xFCoVRigFOnH25071737r3UDI0MVfTtFIOfqpAF7jkC/t6GUxPUdMzReIvzHTLhcgyO+PqjaRGtGW4BW3AHtTdQgqJo+Hn0zbytXw5OPSp9oOlnXvfuvde9+691xd1jVndgqKLsxNgAPqSffuGetEgCp6rs+VPy/p9nVP+jDrhJs7vrNM+NR8S5qGx00zCOOWdYwSFHOo39gHmfmoWf+6/b++Z+3tzpPUUc8c+rtzDZdmBmu5TpGjOknzNOAHn1K+LnxIfa1c/bPbZGf7MzYFVKKvx1lDTRVEflj0RSagrrf+g925b5V+mf967r+pcvk1yBX/L05yVyKbCU77vv617Lkk5ABzQA1pTqwpVVFCqoVVACqoAVQOAAB9PY84dSoABgddO6RqzuwRFF2ZjYKB9ST7917ogHyJ7sqcjVtsfasjvMz+CqaCR/3P3dNwAPyPfjTrQDV6WfQ3RNJTYj+M7opxUy16uyRVCJI/qsbtrv9Ln8e7IxQ6l4jrTqGBRsg8egk7f8AjLuzrzcT9o9G101HKsn3mfxskw/dAk1MKSFfqOF+g9jjbeYLe9gG27utRwU0/wAPQRv9kuLWb67ajQ8WHr9nU/Z3zw23jMJkKXsmmkwu48OArx5GRqSTJyF/GPtY3X1c/wBPayPkC+3GcHbO+M+YFQPt6onN1rDGwvgUdfI4r0enYO8KLfu1MTuvHDTRZenFRApJJVL2sWIF/YJ3Swk2y+exl+KM0PQps7gXdutwvBsjpY+y/pV1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddFVP1AP+uAffq9eoOuJjQ/VEP8ArqPe6n161QddeKL/AI5x/wDJC/8AFPe9Tep61pX0HXEU8AJPijufr6F978R6UqeteGla06CvubER5HYWcVYo9UdG1vStwCf9b+vvWpiOPW9Kg1A6KN8SMimNz2WxrkWczxkGxIvISCLf4j3ocM9er6dWJBQPooH+sAP96966t137917r3v3Xuve/de697917qnD+cd8i+wujPjxlz1/LVUORyktHSnJ05ZDSR1YaCVw6/lQdQF/qPYF5wvZgYdtRiizMAzD0pw+0/wCXrHX7x3Oe8co8j3Euz9sr6VD/AMGtgpb/AGoOoDzI608ek/lb3r172hiN6YDemdmy75GlnqoqrJ1tUteXqlZoZo3Yi7fQEC4/HHHsHXey2Fhbm8sB9PLCCwkXBwK9x/ED5165j7Hz5zfylvScwbZfzzXGsallkeRJQT8JDMaE8FK0K1xjB+h90FvDJ796b653fmo5I8tuDbFDkMgst9QqpFtKSW55PI9yly9ey7lsdrfz4eaNWP2nrtNy3ezbhsVre3Ao8kalvtpnoX/Zz0eddEgAkmwAuSfoAPqffuvdFy7s+R2zupMVMXqYstm3EkVPiqKcNWCXRdG8Sgk8n8ezzadiutzlAA0p5sRjop3Hd7awjJY1byA49Erwmwe1flzmoNybvqKzC9YxVUUi4uWQ01dISbkRo5UsNI/pb2K573bOW4TbWoD3FPi4gdByOzv9/k8a4OiAHCnBPRodvdpdTdM7/wAd0LjqGPETRUcMn8UqgkP3MzRhQrSKvqJ/HPuItw5vW43w2F8aO3A1x68Onf6y7PtO8py2BobTWpxU/wCXo36OsiJIhDJIqurD6FWGpSP9h7NQQRUdDMEEVHn1y97631737r3VS/8AMm+du6PjNhcPs/rjbGazG+t7zTYbDZHH07zw4vItMsVPNUKin0m/0uPewPy6oxIFRnqrLYHUGO6py8Hys/mC7sjy+7ctPJl9obLxGXjpspPk3H3FEtTQ3JUayoKaffqg8OHWlFO9uPl6Dq4D4v8A8y7p/wCQGcO2ZKNNgVqj7XHxZ6r8UuQeCMLFDAGQaiQAAL+9fb1dWBGDXqzQEEAjkEAg/wBQeR791vrv37r3UOtq0pIWckaregf1Pt2GIyNTy6ZmkEa/M8OklJgKHddLVQ7goaWvoqldPhqoUmX9JX0iS4/P9Pbt6kLRfTkBgQQQcjpIbdLyNo7pQyn1Feq0e3fjNvTojctT2z0VVVUeMSZKzOYZX5fU/klWmpozwvH4HuJd05cu9kuDumyE6eLDz/IenUPb7yZuHLd62/csEha1dPXzNB0Zj45/Lja/blMuDzbDAbtoV8NZS5KUQy1dT5AgEEBUHm/sScv8z2+6r4M36cq8QTSv2dDLlLnqy39Pp5/0p0wVbBJ+zo54N+R7FnUgde9+691737r3SSylU1dWR0ENyhKliP6q3PsxgjEUZkfpFMxkbSnSmpoFp4UiUW0qL/4taxPtC7F2LdKkUKoHWf3Tq/XvfuvdcWVXUo6q6MCGVgGVgfqCp4Pv3Xuk5l9o4DM4nJYaox1LBS5Wlmo6t6KngpZzDOLSBZY1uCf6+/de6Crqj469b9K1VfkNl02Qp5a4zPUmsq1qVPmuZNICLb6+/AFiB14sFU9CYT/GMkY9X7FOVcc8cHkf6/sxINtDXzPSBS00ppw6VqqFAVQAAAAB9ABx7Lyamp6XAUFOu/eut9e9+691737r3Xvfuvde9+690lMo7VldBSKf2wwWUX/qbXI9mEC+FEZPPpBK5kkCAY6U0MSwRRwoLLGoUf7D2hZi7Fj59LVUKukdZfderde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/W3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3VJX80P8Alx7h+Q2aw3yK6SqYsT3tsCipkw1UGgpUejx4aZ38hKsZBYWI597Bp1VkD/Ijz6Jdt35p/Nij2nQdC1O290Pv2kxn916jc70FQ1JNLVRfazzmZksQNR0vfj8e9CnWtTniuf5dHQ/lq/y9M78ZshuDubuWrjyXYe7q1stjpIZYZhj48i7PNHUlSSrsr3A/2/09qo0WaLw1Hd/k6TvWGXxWNa8cefVw0+wtjZGuizNVs/bFVkeJVrp8FjJapmf1eRp3iLFv6MefaYihIPSoHUK9KdqSnNK9EsMcdM8LwGGJFjjETpoZVRQABY/ge9db6KDnPgn8e9w74XsHKbbqZtwCcVF/uk+zeXVqu9KYyp/2Pv2evY9OjZ4bCYnb+PpsXhsfR42gpYo4YaaipoaWJViQIpMcCqL2HJt7917pzZlUFmIVRySTYAf4n37r3QY717X2vsykaepr6WeddV6cS2cBTY3sPr79w690RzfXyBz26sjUR7Ihng8peNnRnKlQ11tb/Ae3jK7RiPpvQFbWD1w2b0PvPsWVM3uGqnVJGEj+WSyEMNdgJD/U+2CpGOrgg59ejpbF6X2ps2GCSOiinrlCmSSSONk1BbHixv8A7f3sYz16lcHoYURIwFjRUUCwVFCgAfQAL791vrl7917r3v3Xugb7c7v2R1Dt+tzG4MtRCrp4g9NiWqBHWVjMDpWFCDfkW9lO6bzZ7VAZZ2GocFrk/Z0H995k23YbVri7kFRwWuT9nVY9Tvnvv5p5GXC7HhrtmbEU/wC5GTJ64I6yCOTTI1PK+m91PFvcdve73zdKYrEGGAca4qPkeocbcuaPcWQwbZqtbXzLimoedPy6Oz0j8Xut/jxT0lZi8c1fuCvjLZzJ16RVgeeRgzSU7yAlLEcWP+PuQ+XeWLHa4WEK6pGGWbJrxNPTqSeWOS9m5ShBtUrK9S7HNSfOp6N9DJHNEkkRBRlBXTawBF7cezhlKHSfLocggjHWX3rrfXvfuvde9+691737r3SczWMMv+V04AmjsxtZSQP1E2+vHtbbT6R4TcD0knjaviR/n1LxOSjrYQpNpo/Qyn6nTwSB7buIDE1RwPV4JhItDxHTx7TdKOsc0sUEUk0zrHFEpeSRzZURRdmY/gD8+9EhRqOAOtMwUFmwB1WX8nfmFUpl5On+mY5s7vHKNJjZcjiZGqYqJ5yIxI4jHpC/2j/vPuO+ZOaiJP3VtXfK2KrmnUO858/Otz/V7l0GW6kqtVyFr5n0A6X/AMVPiOnXLt2X2g0e4e2M0rS1tbO0VXSUtPURh0jjjk1WkXUbnggj2s5Y5V/d5/eW5nxLp+J4gA/L16M+R+Qk2au77ufGv5csxyAD5D0pXo/AAAAAAAAAAFgAOAAB7HPUocOu/fuvdFJ+QfdsG16Cfb2Hl8mSql8LNE7B0fyafqo4Hv3y6qa+XQS9A9OVu48gu89zI8heVpgZ1WTUWIdba/8AX+nvXHHW+Br1YXDDFTxJDCixxRqFREAVVAFgAB731vptzuYx2BxVZlctLHDj6SMyVEktvGqD/VXB49v21vLdzrBAKs3AdNTSpDGZJOA6pI7J6Pj+dPaeTyW1E/u/RbFqkQ10eikx1eI5iEKBdOu5t/X3kNtu/t7a7CsU9HkuRgcWGPPqKtw2WPnHcS8dVWA8eAPQ3ba7Q7R+J1TQbC37j6zcO2qQimxtXQIzUdNA7Ah5JYwQOB/X3Hd7Z2XNTvuNswSZ8kHiT0f217e7D/il2peNcAjgB1Yd172/snsfGU1dg8zQSVMyFpcetSslTTEG2mQADn/D2B73bLuxkKTKaDzpg/Z0LrW/truMPEwPy6FL2XdLeve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917pKb4pvu9p52C1y9DJYfXkEH37r3VafR1VLh+16yhkYp5Kx0F+Bbyc2H+v9Pda+Y6tQY6tX926r1737r3Xvfuvde9+691737r3Rc/k18bdj/JvrTNdd71pRLSZOO8U6pGZYZ0jZInR3sVsTe4PFvZLvmy2+92ZtpSUYEMrr8SsOBB+3oH87cmbRzxsU+xbxGJIplKkH/PxBHEEGoOR1rE4/wDlQYf4+/NvqTbu8q6LOdZZ+sizCzSeGR4ft6oeOmqgDZgrLcEgcj8/X3FG4Wu8RbtbcvbrRoZnUtKp+KLOpGHkSaA+RFesDrP7tsHJ/u3ttrvE31mzyapUV8ukqkaAx4OoywJAIYCtePW3PtvE4rA4LFYbBxpFiMbRxUmOjjt40pYhaJU0gC1vyB7mu3hht4FggFEQUUDyA4ddGbSCG2tkt7fCIAF+zqJuXd+3do0M1fn8rR42CGJ5NVVKIw2hdWkE/wBfa6C1nuWCQqWJ9OtzXMNupaVqU6r27F+XGd3znX6+6eoKyWvqWanXOUvknowsq+MM8qrpAvf2M7HlyC0h+s3NgAM6Tx6Cl3v7XUptNuBZvUcOlT1H8RK+rzK7+7xrzuDdWqKooooZ1kpYVJ1BJ4muLgW4t7Y3HmVUh+i2keHHwPqenrHYGeT6rczrfiPQdH8o6Kkx9PFSUVPDS08EaRxwwRJDGqIulQEjAHsGszOxZjUnoVKqoAqigHVefzm+PFZvHAQdm7EE9Jvja1YuYrauGQKajF49BNJDxY3AU25PuPudNgku7f8AeNhiaM6ifVR5fy6in3K5Sfc7Rd42uqXcBD6h5quSP2dDT8RO+6PvXrCiyrqKTM4WRcFkqGZiKozY+AQPUFHsSGKk3H+x9nHKm+De9tErDS6HQQeNQOP59CDkLmiLmjZEufhkTsZTxquD/MHo1vsT9Djr3v3Xugi3711svIip3fl9u0Gay2HQ5CgFfQU2QEdXCweIxRTo1jqte3+v791oivWrp0919RfOL+Z13ntn5OZSpg676zirtwbLwkOSTFUy1lDWRpBS+OX9tY1UlyoXkLp97rQV6oo1OdQ4UoOjZ/zCPhZidq7t+OfY3xyoq3E01BuyKfdOTx8xONOPiqovt55paZVAGkMGBv8AS/vw8+rMASCuCD1F7C/nobVxncmC+Nuytr1m5t602To8FmczhKueZVqF0UjvJTIhsNdyefe9Ladfl1TxUEng17vTrYD6+y2Ur9kYbK7hinpcjNRLPWRVKsk0bONYWRX5BsQOfelUu2kcenCdIqep1pc1WhgR9pEfp+Cv9B7McW0JDcT0gNbiXHAdKuONIkWNBZVFgPZcSSanpeAAKDrqWKKeNopo0ljcFXjkRXRlIsQytcHj3UgEUOevEBhRhUdVw/I/4gGWrn7T6enO3t54pXyMojlSnpZPtkaeRvHHpH9n6W9gHf8AlTW53PaeyZKn0GM9RNzdyLqlO/bCfBuY+70BpnI6p33t/PQzXV1bNsHJbZyWYz216t8NlslBWGKOqraaXxVboH5spBH0/wBb2RbdzLzJf2SyRxqBkamfLaSRWlMVIx1jPun3sBsl7NsT2ss89m5jkddIUspo9NTAnTw4Z8q9W3/y/v5iGzvmRhq2CmgbFbixgRaigqp3epd2cD6NwQQQwK+xVy5zLLuM8m27jF4M8VDxqHB4FT6f4COsg/aD3l2X3RsXkstSSxMUdHFHVhQ0IyCCCCCCQQfWoFkmXrzR05EYLTSEKqqCWsxtcex3bReI9W4DqZ55Cq6V49RMTTQQMpmcfezqZlidSsiIRydJ/wB7HvdxNU+GnAdehjC9x49KL2l6Ude9+691737r3Xvfuvde9+690z5mtFLT6P7c94x/Xng/737UW0et9R8uk9xJpXSOJ644WiWmp9ekiSQknV+oA8/n3a5l1vQcB1uCMqtW4np69pen+ve/de697917r3v3Xuve/de6j1U600EkzWsi35/17e7xoXcKPPpuR9CFuk/hI2qZ56yRfTqvEfwbm9x7WXTBFES/n0mt6yMXPSo9oOlvXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/X3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XTKrAqwDKRYqwBBH9CD7917pt/guHEwqRicZ9wPpP9hS+Yf60ujV/vPv3XusWZoFraBoFAXQQ6AAKBoBsBb6e1FtN4Emr1x0xcR+ImOPWDAVzVVKySEa6d/CAf1aUFh/vXu93GEfUvA561buSlGPT/7SdKOuiQASTYAXJ/oB7917oPd39l7Z2fSyz1tfTtLGrEwayG9NuL/7H34kDrXRK99/JvN7imkxmz4J0ErvCGhMuk+r0H0+/Anr1Ac9JravSO/eyK0ZLc1VULDKfIVlk0r45Br+jn3r7OHVj/Po4eyOhNp7VoVWSlSev0EGZljcK+jSH5HJ9uI2hgR1Rl1AjoTMDKtBM+DZEQUq/tMqKist+B6fr+Pau5jDoLhOB6SwOVYxN5dK32h6Wde9+691Gq6yloIHqqyeOmp4heSaU6Y0H9Wb8e6u6oupzQDqkkiRKXkNAPM9V7d9fN7Cbarf7kdaUk28d0Vr/brPg3eoOOl1GMmZIxcWNvr7AW+c5w20n0W3DxpGxVfw/b1FfM3uRbWU37s2dTcztjszp+306CrrP4g9hdx5+j7D+Rucly2IdzVYbb8dQyVFJEr64o6mM8fXggj/AGHsu27lW/3a4G4cwPqXiq1yPt6Jdo5A3Tfrtd25vl8ROKRfw+lerSNt7Ywm08TR4TA4+noMfQxCGnihhijIQfTW0YGo/wBSfck29vDaxCGBQqrgU6mi1tLeyhW3tlCqooAB06VtJHW07wSC4cW/1vayKQxOHHTk0QlTSek1iKl8bUtjaon1vpp2P4UGw/2HtXcKJl8ZPz6Twy6W8JvLpYey/pb1737r3Xvfuvde9+6910QCCDyCLEf1B+vv3XuPSQyVDLi6g5WjJWGNbzpyQqDmRyB+LezCOdJYzFN+3pG8XhHxE4DJ6C/LfKDorA5U4LN9lbbxeXQfvUNXUPHNGwIEilSOLX+vsKXvMmwWF0bO7vI0kH4Sc9E0vN3L8Evgz3SK3oT+39nRLPkL8tspv/OjpnoCSTN5DLr9tW7mxLyS0SUlWgUnyACwAvz7B2+c1ncJ/wB0bEfEL8ZFyKHqOeauerjc7z+rnKv6kkmGkXKgHzr+3offi18SMD0nTPurcIjznZGXVpMnmZmWpSJaiMM0MPlBIYEkFgf9b2ect8qwbR/jdx33DcW4gfZ0KOSuRLTlqM3tx+reS5eQ54+Qr0dYAAWHAHAA+gHsYdSH1737r3QIdy9r47r3BVAWaNslURPHCiuweJ9QFyF/2PvdMdarmh6JN1fsPPdwbxl3DmxI9Ak7ylpSGiaMNrsvk4/PvRBI63jqzTD4iiwePp8bQRLDT08aoqqqrchQCx0/1t7917p09+691XB8pO0c1vzceK6N6+mlnk3BULj87U0juPsW82gipI4UD/H3LvJeyW+12b8ybqKeENSA+ePLoD79uEt5Om02WfEwxHl0bzpLqPE9Q7Lxm36RI5MmlKq5bIIF1VlTrLs5cAEj6fX2AuY99n37cHunNEr2r6DoTbXt0W22qwJxpk+p6Xm6tn7f3niqjD7gx0FbR1I0yB4ojKBY/olZSR9fx7Jre5mtpBLC1COlssMU6aJRUHqv7fHxD3dsLLzbm6CzrYKjUfdZLG1dUWmqNH7si04sB9R9Pr7GlpzNbXkYt95TWeAIHDoJ3GwT2khn2p9A4kevWDaXy93JsXKU+1OzdtZX7i6xT5yqE0dGsjOB5dbC1rXPv0/LVvdxG52+QU8l8+vW+/T27+DfIRQ/EeHR7dm9o7K3zRxVOAz2PrndA0kFPN5GiOrTpYj/AB9hK62+6tGpMhA+fQmtr23uV1ROD9nQhe0PSvr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917qDkqX72gqqX6+eJo/8Ab/T37r3VWgjbbferQlwobIWUCw5aXm+n3rrxby9erVIHEsEMg+jxRuP+QkDe99aBqOsvv3W+sU08VPE807iOKMandr2UD6k292VGdtKCpPWiQoqegiXv3qGTONtuPfWFkzSSeFsckkhqRLewTTb6/j2fHlbfltvrDbP4fHVinRcN324zfTiVdfpXoXopY540liYPHIoZGF7Mp+hF/ZAylTpbBHRiCCKjrJ711vqq7+ZhT47a2xqftmOsp6XN7XqKGGmldhFNFGW8rhZR6h6eePcfc/WNx9ENztlJeMqoI+Z6hP3lRLDZ/wCsKkK9tQgnFAOOepmM+bdJUdY9dUWyMXW7ozeR2tjv4hV4p5Z1paxqZYruVF/13JJPuYOUuXHvdjt9wvJAoKKSDxr51r0K9p5riuthtJrIGV3iUkjIyOvYzo/u3vutps52JuF6TY9XaSTBTTNFXENwyhTyCF/r7EL7vtWzIYrFNUw4N5dLI9r3Pc28S+ekRHw+fQbZ+JviH8mdn0mEtTdSZDGQU+WfIIkrvkKkWUpPYkNduPcD8081b+vN0U97LSyoKgEgZ6Ae7Xt1ydzpblDTb5Eo1f4zwpx6uBxtbDksfQ5GnN6evpKesgP9YamITRH/AJJI9jWNxJGsi8GAI/PqbYpBLEsq8GAI+wivU33fpzqHkaKHJUFZj6hFenrqWekmR1DK0VRGYpFKtweCfr7pIiyIY24MKHpuWNZY2ibgwIP59U3Z6ryfwy+S0eZEU0XUm7alcfBT3MNKMhXnS0njh9NwW/p7iaeSTlHmLxci0lNAPKrf8X1j/dST+3vOIuG7dvuSFHkA7H0HrXq47GZCnyuPosjSSpNT1tLT1UTxm6lJ4VmWx/1mHuWY3WRBIuQwB/b1kBDKk0SyoahgCPzFep/u/TnXF0SRSjqrowsysAysP6EH6+/de6pw+Wf8tLc2998z9p/GbdGO6739uKaX+92QrpPtI6yGWXW/ikpIyxuCbhh79WnAdaKg+dD1Wz2b8i/ksa6l/l/bQygz/aOIyRwG5N0UTPX06TtJ4FkEjAiNVP1YW4+vvYGNR6ozd/hrxpk+g6B3dHxx+PH8uDd+3t9Z+WLtL5d7gqUrN20uJrIa+DHZCodZok8A1FZAx5H1PvYq7UHWqxxDUR+fmetoT49dhbl7p6V2XvbP4qqwOVzqO+RxlVA1PNTosYKB4yBYG/HH59qotEDan40qP29NSa5QBHw/2OjFUVHFRQrDELAfU/1PtPLK0raj09HGEHz6l+2+nOve/de6bcvRnIYvIUQCk1VFV041AMNU1M0Smx/xI90kUshUeYI/aOmZ4/FhaP8AiBH7R1oefzBP5fnevX/eu5cvjtr5bceF3ZmqjL0NZh6M1fiauk8hp50gvpAJ4b6f1t+YUjuH5ZZ9u3hSoDMyOAWVlZi1O0GjCtKefl1yb95vaHnfYeeLzctvsZL+23CQyKYVLNG7UqrqOC+YfhTDUpU3D/yWfhL2J01HX9t7+p6zBnIRRyQ4arZUkVFl0oGiBPqK3Zv9gPZ/ypYXW8bwd8ZDHCoCxVqGYVyzA5FfIHgOOT1kj91v2r3/AJUtpuZOYEa3muCCYSQQgWqqMEgsRliKjgK4qbivlR8mNlfGfrjO9lb5yFJRU9L93Q4+mrpJYDLUy0ZeiZHX6XawH9T7lx2EUQReJ/w9ZnAeJIZT8I61oti/Oj529gdi5P5O7f8A71VnQm08wtDVbbgFc8VfQytdJ4iRcxlOSw4t7SaQMVz09rc9wA0+Xr1sZ/Dv509ZfKbbUH2eQocHvOlWnpq7a9VVlsk9S10do4mFzyBcX/P9PdTWuenFIZajo93v3W+ve/de697917rhI4jR5G+iKWP+wF/ewCxoOtMQqlj5dJGON8vkjK5vSRcqPqNSnj6+15It4qD4j0hUNNIeliBYAf049l/S/r3v3Xuve/de697917r3v3Xuve/de6S+ZqhUTR46I6mlOmTT/Z5+p9r7aPQpmbpFcNqYL0/UdOKWmigFv21sSPyf6+0kr+I5b16VIgRaDqV7b6v1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//Q3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+69176+/de6RsqnFZqOT1JRyglza6hnNr8ezFf17Y+o6RECGap4Hrrcu/NtbVgeTKZOmgkEZkSF3szAC9v8AC/svFPPpYSfLomfY/wAoKuvq5MJs+nlJcPCtRA8hDMf0kkf8U96+zrw4VboMNv8AVnZPaGQSq3DNVJTVEgkvJIwTxyc6fVxx/j71ivW+PR0Ov+gtqbQp4XqaVKuvjCEuQjJrUfqJIN/fqdeOeh5hghp0WOCJIkRVRVRFUBVFgPT7317rL7917pJbhppIXgrqVW1JJrnK/lFNze3+9ezC0kDKYX4Ux0jnTS4kXpQ0NZFXUyVERuG4I/IYcEH2jljaJ9LdKUYOtR0gOx+2dk9W4tslu3OUOLBXVBDVS+J6g6tOmO49le4bnabZF4l04X0B8/s6K923zbtmhM19IEp6mleqtN9dwd5fK3dI2R1DBkNqbW85gqNwuZlxtbEknMkc5sLEH8H3Gt5um9c0XP0W1AxRg0LeTf6h1Cu6b/zJzxdfuzl4NbwVo0prpI+R6Ot8fviBsDpqGnz1XRrmewJwz5jO1EgqYqidyCzQxTLwLi9/Yu2HlSx2j/GGXXO3xMcg/YOpE5V5B2nl1BcMokujl5Dmp/MdG/AAFgAB/QcD2LOh7137917r3v3XumDOY9qmNaqC4qYB6CPyPqPay0mCHw2+E9JbmMlda8R1zw2TFXF4ZvTUQ+hgxN3I+pF/ermDw21LwPVoJNQ0txHT57SdKOve/de697917r3v3XugO+R+5cztPpfsTN4FHbJ0W087NTNGCWjljoGZJBp54P5Hso3y5ltdsmmh+II2fTHH8ug/zRdXFnsdzPbCriNqfs6+cD2F2x2XvLsDcO5Nx7gyk2aq83knlknrqoyRhqlh4afU3pRR6VC/j3HFvt23z2qzSosrSgMzMoJYkVqSRn5dcSeZNxu973i53bdZ5JLh3ck62pGdR7Ez2qvCgp1uG/yHcBR5X415HfWaxs8u6v7x1GMiy1cxqZZMaplEX28k12UW4Nj9Pp7POSNvs4Jb2eBRiWgIAoBpFQPSh49dJfuh2MLe2UW4zKWnMkq63JYlBI4WhNTSgp8uHV8XuQusseve/de6Rm+d5Y3ZeDqspXTJG8cLPAj3/cYG1hb37r3VZpTcXfG/neMTtivupBYOxjC+Qkmzf4e9Cox14muerMtjbPodl4GkxNJFGGiQeWRY0VmYqAQWUXP09760Oln791vosvyY7qpOq9n1VJRVA/vZmaYxYGlVmWeWZn0gw6OSePYy5O5dk3q/WWQfoRnvPl+fRDvu6pYWpVcyMO0fPoK/iB05lcVSZDtXfCfcbl3mDWJT1gEs+PDSh7nyAlSRe359nfP3MEM8ibJt2IrfBI4Nj5dF3LW1SRI24XeXlzQ+XR7vcZdC/r3v3XuvfX6+/de6Lz8gI+n6ba89b2fQ01TSppSOGMRxVcskt447TAX4ueSfYj5dtd3vbsQ7WSD5nyx0T7w1glsWvQCP59E5q/i/vHH46k318fNynbeAr6JcqMRU1UklRUwhTLaNhxyRwL+xFJvlutw2373H4jodOoDgeiD9yTJELraX0KwrT+fXtnfJjtzYVX/CuwtqZ3KJRutPLkHjqPt5AG9Uob6EAe6XWxbbep4ljKq18sV6ta7xuFr+nexsaefRr9p/KrrPPLDFks1j8LXSlFFHUTkTBnfSF0Hm/sOXHLt/ECyIWUefR3BvlnKdLMFPoeh9od1bfyMUU1HlKaeOZQ8TKWs6n6EEj2UfS3ArVDjo0FxCchgen8EEAjkEXB/qD9Pafhg9Pcc9d+/de697917r3v3Xuve/de697917r3v3Xuve/de6q57somw/clLlEUxo2RBv/X93n6f196PGnXhXTnqyHaNb/ENvY2qvcSU8dv8AAKgUD3upOT17pS+/de6JF/MG7A3d1p8Yt/7q2WKhc5QU8RpJKfWHTXFIG0tHz+L+5F9rtssd15ut7W/oYyTUH8ugrzje3VlssstoO8A0/YetEDEfJXuul7AXfUu5qxNwS1MVTPSNkq8zRzGv5DIW4NvfRiflfYn2/wDdyxAxAEA6RQjT1hXb818wLeDcJmAlbSWj1Nir04fYfTj1vF/F35ZbVrvjl17nd9bmof72VWFkerpZ52FRNJG14wddySb2987eeeVZrPmu5trKMiLVjGOszdh3+CXZopZ3BfTnPHps3t8vN15ky0OwtnZerLERQ1lCk8kbhxo1gqP6+y215agh7ryVV+Rp1q636eQ6LSJmr6dF77L+LHyA+T/X2Uk3luUQYWulWd9u1kjJVyLAml0Cy8D08c2v+PZVzbPs37mk26zi1SkghgMVHA/t6C/NPKW584bDLY3r0RxlD5imRno0fwa+Om3uodlVOMqsDLDl8XPFRLNkRHOHijLMrQhwbAEC1j7A+wblvL2LWt8xCqaAcMdGvtzsCbDs/wBAyEGKiiucDhTqwRVVRZVCj+igAf7YezTqReiD/wAwjq6Xf/S8s+IpyM3iMxR5RauEKk609GvkkXy/qAsv4P8AX2BufdsbcNn/AEcOrBq+dB8+ox91Njfd+W2FviVGVgw49ueP5dCV8Oe3Iu2uoMXUi33O1UpNr1puxLT42mEOpi31JCjn2Z8qbou57UpHGIBD9oFOjnkLfU37YI5l4xUjP2rg/wCDo1/sTdDXr3v3Xui2fKPpXE909ZZPFVsAkyGEp6vM4R0VBMuTpofJT6ZCLjlfwfYc5n2aLedtaKQVZAWX/TAY6CHOvLsHMeyvbSirRgunqGAxQ9F1+CHb2TnwVT052FXl+wNu1lZ44Z5HadsVAuinVg/9FXix9kfJO6StAdo3Bq3EZOPPSOH+DoKe2O/XEtmdg3Zv8bgJFPMqMD+XVjvsf9S11737r3XRF/8Abg/7Y39+691S98zfjri/jBs/5H/MvqKkkj7Nl21U5aSZo4KiRKpqm7S0g060a7CxU3v798h1WlK1/wBVOq7v5XvUnUPcm05fnD8uN6YzI7ty9bkK2PEZmveSTBzUM2v+IZWnlDsEFjoGmx4/HtWsRRNS4ofPpIHEpBYVJHEcB6AdHMm/mS5fsTvvD7B6O23WV/XeIykdNmN0bfiqDhXx0LiBqqRIRoRQgBP09sSSMzVP5dKIkVQEH5+nV2e0d+bV3klRFt3O0eZnxywrkBSl7wSOgNpA30JJ/r7bqDw6dpQdLT37r3Xvfuvde9+6902V2IxGQs+RxmNrSn0etoaapKj6cGZTb/Ye23hjl+NQ32gH/D01JDBJmVFb7QD/AIeg93ruvAbQwORytaaLF4PF0dbLLMI0p6cPTUjVCp+0ABe3AH+v7MIo0tI9dKHyA6TkKzeFEAF+Qp1qFfMDcXb384nvCp6Z6Myc0HXW3K1Ist9pVVEeNWtw8ZjV6iWQ6AzkEAH6+05OdTZJ4dX0agY1NAOJ/wAnRzPhb8j8p8NzD8N/kJsF02tUpHt+r3RlaFI6HxnVSeRpGXRIGjb0liebMPdD/F08vb2N+XR+dr/y6tmbL7lwvyF6A3XBtTbOWWDNvSVlUxinNW3lvAIQE0tf0j6/j3rNM9OaVBrwJ9PPq3nHrKlBRrPKs8600ImnU3WaXxjySqf6Mbke/Dr3Uz37r3XvfuvdJ7NV2hBTRAvI7hXVfqFY6f8Aifa21iz4jdI7hw36Y6cMZRrRUqRr9W9bX+oLcke2Z5TLIT0/DGET59OPtjp3r3v3Xuve/de697917r3v3XusFTMKeCSUkDQpbn/inu8a63C+vVXbSpPSaw1MZ6qetkF9Tala9wb/AFAHtbcvoQRDpJCplbWfLpWey/pb1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/9Hf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SV3FvLAbXjZ8vXxUzBGcI55IC6vfuvdE87C+SFJka/+C4CD7os2lKmAvpUmwGoj/iv+w9vW8vhv3cOHTU8fix6V4jz6DCv65352hn6OSuqp2oZ44mADuiqjAKQb/63u11EFkBTgR1W3kLx54jo1nXPx32ztSmjlylOtbXrofUSrBZFH1LEH/ePafyz0/546MRTUtPSQxwU8SRRRqFRUVVACiw+nv3XupHv3Xuve/de697917rBVGJaeYzsEh0HyO30VTwSffg2g6/TqrFQpLcOq+e7/mRtfqaorNr7KWLeG5ptUUeMxzytWU0wkKkFEvz7De/8228cRhsD4844qOI6i/mj3C23Y62m3j6i5OAi/FX7K9AL1r8bO0fkvnk7B77r66LZb1Blx2zquWWGrp43YTIhRuQLH6n2DNt5f3TmKYX++sRDXEZqGHn0Fdo5P3vnG4G7c0uRbk1WA4IHHPVrGytibX69wdLtzamLhxmKpP8AMwoqs44A9UttTfT8+5Ns7O2sYBb2q6UHAdTdt+22e12wtLJAiLwA6V/tV0u697917r3v3Xuve/de697917pJZegkpKmPK0YIEVvNEt7uSfqAPZhbyiVDA/nwPSOdTGfEXpQUNbDXQrJGylrDWoNyrW/PtJLE0TaT0/FIJFr59TfbXTvXCWRIY5JZG0xxI8kjH6KiLqZjb+gHv3XuihV/zt+L+N32vXdX2tt6Lc5qRRtjC8n3K1bNoEBX+t+L/wC8e/Y61mtOjRZKgxO8Nu1ePqQlbhtw4qWnksAVqKDI09tShx9GVgRce2Z4Y7mF4JRVXUqfsIoemriCO5he3lFVcEH8+qHuzv5EvUe8+1m3zhMu+FwFTk3yNfgNa6KlpZvNMoVYyFBJNwpAP5v7jeTkW/t2+k2q7MdpwCsauq+it5eg9Bwp1iLzB90Xk/eOZTvlu7QRSSGSWBWIjkYmp1LTAJqSEKg1Na9W6dO9MbW+NuyMTsbYGOFDt2ihpVkgjSJFaogj8ckhEIUWNyfp7knZNosdusRZWopTJJNSzEZJPmSesluWuXNq5M2aLZNmhEMMYUBVACinoBgDowVLVw1cQkiYMCOQDyp/ofaiSNozRh0J0dXGD1Gy+Vo8Lj6nI10qw09NE8jMxtfSL6R/ifdB1YmnVYHae9893FvP+7+CkkbFLUvCqxvIFAEn1+v9Bf37zPWyKgdHq6c6wodgbfpVaBf4nNGjzSlULDVGL2IF7/X37HEdaz59DP791vpKb13di9jbbym5sxKsVFi6c1EpYkXANrAj2u26wn3O7SztxVnNB0murmO0haaQ0C9Vq9U7KzfyT7hrezt3Geo2RgK01uz1Z2NM2icMEiD3DfX3L++bjbco7Auz2FBcSikvqMcT0CNus5d73I39zmKM1T0PVqUcaRIscaLGigBURQqgAWAAHuEySxqxqeh+AAKDrn711vr3v3XusU00cEUk0rBEiR3ZmNgAilj/ALwPdlUuwVeJ60zBRU9VWbw+9+VXfWKxGNklk692lWR0O6aaGV1iklgl06nvx9b+5tsBFyTyy881PqpwTGSM5HUf3WvmHdhCn9jEe7q0HBYSg25h8fgsZGYsdi6ZKSkiY6ikEf6FJ4vb3DF1cy3dw91Mau5qT8+h7FEsMSxJwUUHXDNbexG4KOSgytFDVU8gIZWjTV6gVOlyCRwfx7pFNJC+uM0PXpIo5RSQV6Kzvr4a9Q56CsyWMxD4rPLFLLT1y1biKOVIyUd0A4sRf2IbPmfc4nWORtSVFRTj0R3nL23zgui6Xpx6qxx21PkfD8gUwGzewzl9m4mGYPTUU8zxRmkXUIGA4vf3Os1zyrb8nm4u7XRcPShIya+fUdR2m/HfPAtZ9UK8R9nRt8V3/wB8bVqRRZzZO4q6noXMDVhhqfFMsLEF1a1rEe4nfaNougXhmVS2aYxXoWpue6WzaJomYDz6GbC/MnyOlLm9q1GJZVGuarEwH+1XJI+nssl5XNC0Ugb7OjGLmEHEqFft6EzG/LbqeaINlNxY3GSatJjlmIIP9CD7QPy3uQP6cZbpYu/WBNHcL9vS5xfyG6kyyr9nvDGyu1vSjk/X6EH/AB9pJdi3aE1eFgvr0oj3ewlFVlXpZ0fZOyq//gNnqN/z+o83Fx9PaV7C6TDIelS3luxoGr07x7s27KAUy1KQfp6yP94I9tG2mHFT04J4zwPWR90bfjNmytIDa/6/94+nvQt5m4KevGaIefSLy/dPWeDrKfH5PdeOpqyqkEUELs2t5CbaQLfX2lkkSKYW74cmgHr0gn3nbbeRYZZQGY0A8+iP/KDP7fymdxFdgshT1c8dQklQIf1IC+oFv94PtZc2dxaU8dStejCOaOVisZqfToa9lfJTrPbey8XQZXcuOTMU0fjmonm0yhtI06r/ANfoPa6z2W/vFDoh0+vRfc7xZW0hjdgCPLpmy3zKxEDMuGwhywNgjU5le4/JFv8AYfj2bx8rSnMjaft6L5eYIkr4a6vSnRcO2u6Owe4du5PaSdcZurw2VVUlijgkkjZdLRg6XBH9q/sSbFa2ewX0e4GdQydEe5315uds1usDEMPTqu/rX+S3Xbw3TXb4zk0OCoa+oE8FDWfbeanAm1+MwqpP5/p7me++8Jb29mtjDGzsoyRWn7eo/sfadp7xr2UiPV5Gn5eXVwvWfwG6v2riqDH7opnzjYuIR0JhqGhjjAYH1KFt+PoB7x+3rnjdd0vnukbQGNeFepW27lSws4VjkGsr59HG2n19tPZVMlLt7FQ0cSJ41JVJHC31WEhF/YSub25u2LTtUnoRQ2sFuKRLTpZHSqkmyqAS3AAAAuSfaXpR0D+8++uoOu0nfdu9MNg/CGeVahmVrImtiQq2PHP19+p1oU8uiX74/mwfDva4qKbF9tbXzuUpw98fFPMkupI9ZXSB/sPr72BXh1piVFTw6fvjX85unPmtjN17X2rW4yprUo8jRrRQM0zSFoXiDqZrgEWP49s3MAmgeJxWqn/B0nuYY722eE0IZSP5dBf8PMo/Tfcu6+hKuTwS5zJ5DcENNISXsC7qEt6RwfcZ8pSfujdZNkbBkYtT7OoY9v5jsG/3HK7nTrZpAD6Z6tk9yj1OPXvfuvdcWVXUqwDKwIKsAQQfqCDx79x68QCKHqnv5hde7l6P7cwfyP2FHMyZTKQUW4KWjJVKXHoV+5mlihAsmkk3PuKObLG52TdU5gsAe9qPTyHngdQFz/tl5yxv0XOO1AkOwWVRw0VySB6cerRer+wsB2fsvD7u25WxV1BXU8SvLCbqlUkCGphv/VWNvck7bf2+5WaXds2pWHl6+Y6mnZt1td52+O/s2DI4GR60z0IPtd0addEgAkmwHJJ9+AJwOvEgZPQcb8wW3Ox8Dk9g7gohlcBuCH7DMU40MstI5BkRhICByByR7WRwFFMsg4DAPSR5i7eGnrx61jfmH/Kk70xfYuFo+lOw6Tr/AOP1bkj/AH4o6rIfa0se36ir1z/dinAQKFJHP59sSza2OPs6tHB4Yopp0Eu89zZnaeDPxM+AmDyEm6Z42w+7u1MGz5CCslZvHPUCqRWMcQNyTf6e26V7m6d1EAJGM9Gz/l9bA+Y3xJ7AxmB7qrc3utN8zfbZnLAVdTSwq8iSRzyu/At9effjQmo6sldOTXrZeR0kUOjBkblWBuGH9Qfeurdc/fuvde9+690mMvkGkvQUnMsh0kgn6/0Fva2CHSPFfpHNKrHwx0nN59a4LsDZGV2duOBqijy9JVxyhG8bLNVUrU6yarfi9/bE8nivXyHT0Mfhr8+tZHcPxy7m/lN9v13afT9BWbq6s3Pmky+4cRQRmr85lmMzpVMi60CjVe9vp/T20CDx49WKFDqXIPl6dXC9dZb4yfzJNhYrO7t23RVO5MTFBNksZSzLS5DG1QYskTTspdihWxDHj/W+msjq9QaefVe/y17T7LpvlFsj4rdG5mrw2NxkeIjpaaGSoLx46jCs8Umk2NgDfj3sUpnrRJ1AL+fV08HyI6o2ONnbA3Zv3ER73kwGHgqsc0h+6krY6NKaoLr/AGWaRWNjc+9AeXWyTxAx0Y+CeOpghqIW1Q1EUc0Tf6qOVA6N/sQQffut9Ya2qSkgeRjyFOkfkseB7cijMjgdNyOEX59J7CwyVkz11SnPIAbnn8EX9q7lhGPDTpNCuttbcOlZ7QdLeve/de697917r3v3Xuve/de697917pKZur808OOh5aQlZCL+m5sAfZhapoUytw6Rztr7V49KGiplpKeOEclVsx/qfaOWTxHLeXSmNNCgHj1K9t9X697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/9Lf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdJzPbrwW24zJl66Ol9BcB73IAv791qvRROyPlBTRNJitrR/eTPqRJqfyFiXSy3ZT/X3onGOt0Pn0AdJtftLt2tWpyktYtFK6WUtKto3bQQef6e/avQdeoaUJ6Nv1v8btu7ZjhqcxClbUFEcqzBiHB1DUWB/wB4PvdMUPW/l0NLUcW2aqJ6OFUx+hYtAVeCx/LAfj2Zx6bmDwz8Q6L3DQzaxw6W8ciyxpIpurqGH+xF/ZaRpJB8ulykMKjrn711vr3v3Xuve/de6Q2/uxdp9bYWbObty9NiaKNGZZKgkByv1C29or6/tdvhM10wVR69Fu57tY7Rbm5vnCKPM9Vb9l/I7tf5FZobG6Kx1fQYSonehn3lRCf7LR5eJ3kBsAB+T7jXcOYNz3+Y2GyqVQ48QcOoW3fmzfebrj92crqyRt2mYV0j5/PoyXRvwl2h1wg3Tvpl3p2QzGpn3Azs6+Zk1MFjnXkhyefp/r+xRyzynBs7/VXREtyxy54fs6F/LPtztuygX25f41ecWlPEn8+joYfJCb/I5gsdRAoQjSE1BfStlXgex1cwiP8AUXgepEhm19pFCOn/ANpOlHXvfuvde9+691737r3Xvfuvde9+691xdVdSjAFWFiD72CVNR1ogMKHpGz00+DqxU02pqSR9UsYublvre3sxR0uYijfEOkLhoGBGR0rIqqKWnWpDDxldTG4sv+qB/wBb2XuhRtJ6WqwZdXVVv82b5hbg+KXxoqt47Ex1Rm8zmMyNs1C0flvRUeQpGjqKh2iIPpDe9AVNOtsaIz+g6qA6G/lq4P5c/G7I/ILY3ZFJN3hkMi256fKRVlUf4ZUikOQG3qgJqKzaxoBYgX/PvdSDTpsRqw1eZyD0cf8AlnfN3t7Bb5PxE+SuEytPvDE1Zx+D3Fm2qY5v4fQoyRxnz8NGw0lD/wAR701OI4dbjZjVJBQj+fWwgjrIiuhDI4DKw+hUi4I/1/eunOuMsaTI0cihlYEEH/Hjj3ZWKHUvVWUMKHpH1MEu3Wlr43ZqBPXNGLkIuoa3b/b+zASx3KaHwekRikhcMnRG/kB3PV7qq49mbWdrNI8Ezw+QeQlrMCQf8PZdJRGKcelqfqKG4dDP8eum6XbWLiz+Wp9WRqgJUSUK7IXW+o3F/wA+6jhXq3A9Gx97691jlljgjeaV1jijUu7sbKqj6kn3tVLMFUVJ60SAKnqrTvDfG4fkL2rRdP7K8w29iK77XdlRTNIsdVQ+ezmRrlbWJ/Hua+W9utOVdkbftx/tZFrGDSoNPLoAbvcT71uK7bZ5jU0cj06sb6+2RieutpYjaGEjEeOxMJihUAcljqdiQBck/k+4k3Xcp92v5L+4NXkNT0NrO1js7ZbaPAUdLT2XdKuve/de697917olny/7hn2hto7F23K3999yrCMRDEXE7JITG/j0fnn3InIWwLfXn7zux/i0NdRPDoL8y7kbe2NpAf1X4Dz6V/xV6gTrHYseRq0K7h3fDBls+ZDqlFY5LFHYgHi/Iv7L+dt+O87kYYz+lASqelOlWwbaLC0DN/aSZb7ejSewX0fde9+690C3fnZNH1d11l9wVmgJPFLjImfVZZq2BooyNJHNzx7EXK+0PvO7R2qeRDfkD0V7veixsmmbhw/b0Xj4M9dV23Nn7i3TuGn8uU3PnpcrjauYB5FxlUGeMRMwuAbj8+xX7k7tHdX8VlamiQppYD+IevRNypZGG1e5lHfIxNfkej0VNLT1cLwVMMc0UilWSRFYEH68MD7jRWKnUvHoWMqsKMOg3zHTfXme1/xPb9PUeQMGN9B9X1tp9rYtzvYf7NyOkslhaS/GgPQS5f4adCZcyS1W1pBI121LWFQrXvqsV/4n2ZRc0bzFQLJ/Loum5e2qWpaPPVbuB+PONyvy83N1vtjK/wALwGIw0+Qjg+5lcL4kOgALe/NhwPcx3m9Rwcgw7ncxapHYLWnQCt9lEnMkllC2lEWvRs6/4Y79EmrCdlR45LWCkVZtb6X0p7jKPmm0H9tb6v2dCxuXLgD9KenTJN8Ne8r3p+7Iol/Cla42tx/qPbo5o2qubT/B02eXdx/DdU/b16L4bd7B1abu+GUB1YgpX/QG/wDqPdv607QKUtKfs68vLu4g/wC5X8j1W33T8be1aP5F7Fwdf2lFkI/7xwLIq1NQqlXmHDKxH+xBH+v7hHmfmaGTmq3e1i0BZASPlXqB+beX91bnywT6w6EkqVB419c9WMds/H6Tr/Efx6XKLk5GhiRyCxIaGMIX0uAeTz/xPuTt73obsV0pooOsmbKxNpKZdVSaD9gp0+dDfHjp7sPFtndxbfavywBaSQVRRNQYC5QXP1Ptq35h3O2i+mheij5dNz7Jt1zKbmZNTHo4GI6C6swaquP21BFp/TqcuRxa3I9tSbzuMoo8h6cj2yyi+BOHQo4zDY3D0602PpIaeJfoEjQN9LcsAD7LnkeRtTmp6WqioKKKdOVgPoAP9h7p1frv37r3XvfuvdMm4qOvyGFydFjKn7StqqGrp6ef6eKaenaKKS/40sQffutHrXk3V/KF+UXZO993bk7E+R2PyO3cpU5ObG4N6nKvJFTzwMlPCSIjGoFwOW497DEYA6q0YY6ixHy9Oqpvg7/Lw6f2l866zon5JUbb2ye4avMVGIWDLTJ+3GjyUw13/QQBewvb6e7F2IHl03HEquRx88+nR5OrOuNh/CT+cTsrorq7GVG2+vt04qSvkgqa16pGOVpHnp9brpXjWB6l496qSOrqAslAOI8vkerMPmLjazpPv/aHyepSRjC2O2/OqMfHJUTlIncrHYm973v7ifm6N9l3yPmX8OEp8+oK9woH5b5nt+dVNFGmIj/TECuPn1a/tnKx53bmBzcTB48vh8dklYfQitpEqf8Aoq3uT7aUT26TD8ag/tFepxtJhcWsc4zrVW/aAenz2/0o697917pE9h7Mx+/doZ7bGQgimjy+LrKBGkVCYnqY9AkRnBsQbc+0d/aJfWclrIKh1I/b0Xbrt8O52EtnMARIpX9vVWHxT3HmvjT3RuT4+7xq5KLZBmqanb9ZUvJJTy5Gpc+KKJz6bn6WA9xpyxNNy9vEmwXR0wZKk8NR9OoT5Eubjk3f5uUr9itsKmIk1BJPDq4LyJ4/KWATSH1H6aSL39yuO7h59T5qAGo8OkrkMlLXOaPH3YXs8i3/ANY+zGGJYR4kvSCWR5W0JwHTtisVFj49RAaofmSTkk/0Av7TTztKaeQ6VxxhBnj0xdibMoewtk7k2Xkgv2O5MZNjKq4H+amIvzY2PHBH09p+neiWUPV/QX8uzqDenaGAwNPRVFHQioz+RqpfNNXPJLfSjhVdEJH4P+v791qlOHz61aN0fzCfmT80PkJkcZs/PZXp7r+DLSLgNwyrU0mIzNDHOUVqSVfqNIH0Pu9FC0PHpnXIW7eHzHVvnwR+a3Zuze4aT4+dyZGr3dUbgyS4nb+5qlql4qySSZESopXdiCv190NOI6dUktpPHj1sRe/dW6Ysvk/t18EFnqJOLA/S/H49qreHWdbcB0nmlCjSOPXWJxhiAqqn11EnrF73jve/J97uJw36acB1SCCh8RuJ6fvaTpX0w7l2zhd34au2/uChgyGLyMLwVVNPHHIro66G0+RWsbG1wL+/de6Lls341dS/HLBb53B1PtmXD5Csw+cyVRGlS0/3NcKR5YjGtl0+r6Ae9ig49eNagjrUfXsz5Nbq+eWS3ltrrfc82ZhqslhaPNwwVMqUsc+qnSo9QNgl7/X3shNIr0zql8Q6Vx0dqh6QpOqcnU9i/IXt/Hby7oyDT5bB7MatqGzNG8zGSkpZkY8MCV4/w91r5dOYrXj/AKvLoVuqv5lHeHTW5MUnemys7QbAqq6nw2GzOZhrI6H7KRilMIHYgGykWI92oCaV6rqK5YU6vzouy9q7n2RjOxo8nCm0qjEUmWqMiAzU8KzxeZ0YE39I9rEpBFV+J6SuGml7eA6UXW3aGxO0sPLmNhZyjzuLpqhqSWpoxpjSoQkNGVP54PtESW7ieloFO0DoRveut9e9+691737r3Xvfuvde9+691Gq6hKaB5GNiFOkfkn8ce3I0LuFHTcjaUJ6YMNSeaeXITLcyEtETc25/x9q7mTSvhLjpLbRHV4jGtelR7QdLuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/T3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdMmZ3FhsBH5crWxUiWLDWeSACfp/sPfvKvWjjopfYnyix1B9xj9uRismuY45Ig5ZtS2DXB/r/T3quade+fp0XGPC9rdvVQqKyWsjoZ5EdELSqEjJ0Ff9t79mvXh8+jU9bfGfB4BaWvzapWVOhXZdWpgwYsNWoEf0/PvZFOPW/n0aTHYqgxMApqCmjp4h9FRQP9ufr798uvfPpw9+691Cr6RK2mkhcXBGoD/alFx7dhkMUgYdNyprQr0ybfqXj14+oJMyM7Lf66B9OP6e1N2imkqcD01b1Xs49Kj2h6U9dEhQWYgAAkk/QAfU+/deOBXonve/y62V1YlTg8PVU+f3mUkjp8FDraoNQCFWMBfyfYV3rmmz2zVDGfEmH4egDzNz3tuyKbaFhJcGulBxJ6JbtDpfu75b7ifdXbVXkdp7CSf7iDaFcZ0+7ppW8ipGvPDAjn6ewdZ7RvPNFz9ZujGKD/fZrw+XUd7fy7zJz5dm+5hLW9oMiE17gfXq1LrjqzZHVGEjwGyMLBhseqxh44ru8romkySSNySfqfx/h7kzb9ts9rg+nskCL1Nu07Lt2yWwtdujEaADA+XQh+1/Rr0ks3QSQTR5OkBDI4ecLxqXUCSSP9v7MLaRJF8GX8ukVwrRnXGOn/H10VfTpNGwLFfWv5VvpyPaSaIxOVPSmJw6A+fU721051737r3Xvfuvde9+691737r3XCWRYo5JXIVIkeR2P0Coupif9gPfuvdAd1J331z3jUbuxuzcxTZafZ+TfFZmKDVennRzGQ+v83FuB72rEHUOtMoYUPVFv8yz5kdwJ8vtl/y/+iZa3D7v7GxWPydJuHHNULVU0uQpNQgVomAAXm5tf2tBFyndgr59Iz/i8gC51V/IDoj/AMmOoflf8Psbjsl8mslk/kR1duAU1Fkqd6mqq8bh8plYzGnmlluFljFz/sPaTBJUcR0+S4Go5HngV6XPxhHfvwb7U2XuLq6LK9ifHjsulwu4svjsSaqbEYibNHyDEVrRelJqcOV55496NGFT1saozRcr/g/2Otms9HdT9h7n213dnNm0w39LgsbJBldTx1VJDPSJKtM4WwYoDpuwvx7r8+na4p0P8caxRpEg0pGiog/oqDSo/wBt7917rkSACSbAC5J/AH1Pv3Xuib/Inu2mwtHNtXBy+XJVPkhl8YYsLkL/AGT9PfqkcD144z0F/RPTNdXyRbxzETya5zVKJDfXqXyEAPf+o9vQqjP3+fTEzMialz8uj/YzJ00yJShfBJCiRiI8cIukW/23Hu81u0ZqMjrcUodc4PT37TdPdEn+WXef908MnX+1pFqt1bs1YuIQazNQzSSBEN0PBv8A4e5G5G5a+tuP3reikEHdngegtzFuogi+jtzWSTFPTpX/ABa6Z/0b7OgzOegZt856OSTPVktmmkDyB0jcsNQIt/X2X868wfvfcDb2x/xaL4B5dKeX9s+gtfElH6smW6NT7BPQg697917r3v3Xuk1u7dGM2bt/IbizE609Bj4jJLK36VJB0X/1z7WWFlNuF0lpbirOcDpi5njtoWmlNAOq3+itvV3yE7jyna+6I3bGbIygG2nm1+OspzMfF4o3BBWwv/T3LvM13HyrsCbHZnvuF/Up5GmegTtEDbxuTblN8MZ7fn1aIAAAAAAPoALAf6wHuFuh7137917r3v3Xuq4/l5k37F3Vg+kKaTymvqqHKNGpI5jsSNX04/1vcuchwDarGTmNxTSCvQJ5jc3lwm1A/EQadH12Xhxt/aW3MIF0/wAKw9BQkWAs1PTqjDj/ABB9xhuNwbq/muT+N2P7T0L7aMRW6Rj8KgfsHSn9oun+ve/de6ZtxVX2WBzFZe32uNrJ7/08UDP/AMR7UWieJdRx/wATAftPTU7aYWb0B6rd+LlANzfIPd3ZapqjqKGvxfmB4LAFdNrf4f19y9zrJ9HyrBtPmCrU6BPL6mbeprwGoII6s79wz0O+ve/de697917qm3tGL+LfMOhpra/4fnoZbf6kLUXv7ibcR4vNQUfhbqA95Hi+4CKPwtX+fVjHyExYyHXuUkC6pIaRtP8Ahf1D/evcsitK+nU+ADoBfhpkbQ5nHzOFdEl8aE/lZVLC/wBPoCfeqefXs8Oj6e99e64u6opZjZQCSf8ABVLH/eAffuvdVld1fzU/jH0/uOv2fVb0xlZubHymnqMYyVIkjqFBJjJj4uNJ49+FT5daYhcMadV01/8APlwvZNduDbnQO0KTsTNbeqWpMrTYgVj1NDMoK+OQAkg6uOB73Q9U1qeFSf8AD09/BT+bP2n3P8h6DpHuPrKs6+yO5Mp9lgkyUVYs1UDU+MNEKoj8Xtx78VpmvVlcO2mhHn1sZe9dW6xzGMRSGUhYvG/kLEABNJ1XJ/w9+691WRnfjJ0rkfmhtH5OUe/cPjtxbYo/sH2zeJaqqqPH4Q0arYkN/U+/H+XVRQGvnw6Ffs74V7K7N+RG1vki08cG6tvY6joqScc64adAsZBRTfj/AGr37PAdWoKdLf5adUU/Z3Stdt6phFS2Gi/jEXIB8+MpTIrqxBIuRfj2Gea9rXddqaFhXR3/AO89Arn7ZRvvL8lswrp7h9qio/wdBj/L37Kym+Oo63HbgqCMntfOTYGmpZi3lSgo1KQadfJHpPst5E3GS92pknPdExUfYMDom9q94n3Tl4pdmkkLmOnyXA6P37HHUnde9+691737r3Vd3z36WrN3bRxXYe1WFBn9g1zbjrZoT45cjS0TLMKZig1MSQR9fYE512Se/tkvbLteA6zTiQM06ib3R5cl3Db491sOya0bxSR+ILmhp69Lz4pfISP5Edb46V3XG7jx0v2OWx5DrMkNFF4WmcOb+or/AE9iLlTe4dz21Ll6CVRpZPMUpn8+jnkTmuHm3YkmPbMDpZSMjSKGv59HFoMdT0CaYlGsj1v+Sfz7PpZWlOeHQ8SMIPn04e2unOve/de6Jp88ekMl3/8AG3sHrrEFzkc5jUgp40YozskhkNmAP+HvXXiAR1rpbG2ucd1IfinkdgSbV7J2vDNjNqb6qFCSyVTzsdSuFBN/wb+7cak9NDFOm/ovpTsX4b7iyPfPyg3E2d/0dVEWR2XBkXkP3k3lE0BgHBK2H4PvbUPW1quSeH+Dq6H4x/zSdid+52jweToabadVuOS21YHEzPlAXEQEbyNybn6+3oogzd3DqjyVAEZoT69Wi0GJZKqeqqmMjmT9sNzbgG/Pu8k4CaI8A9USEltUnl0ovaTpV1737r3XvfuvdYp4Y6mGWnmUPFNG8UiH6NHIul1P+uD7917oJqjq/a21MZujMbKwNDjNzVeGyi01fDTQtOK2WmYxSgsv112PvVOvdaomf6a3DTfLvcu7PkzvT+6+Op4sxX0W5My84hpTChekZYbWOiwNgPds6aDpsj9Wrfl0Sfc+4u7fl/8AJPE/HvanYdZ3F1LgMxHmKfduOjkixdFR0UmuSnJsNIRAQb2+nt+NFC6mwemJWZ38MGo4nq1b5U9+bhrMNsT4VfHaumydbiaDEU+78niPuBIIY6VYMik8iNYrFZtR90lcyGpOB06o0di5LZ+wdXQfAjZXXvT3VmN6z29vCi3TuOeCm3BnnpWZzBkKilBqqV3JPqibUD7ax+fTqjGOA6P3791vr3v3Xuve/de697917r3v3XukhXvJk69KWE/twyDXb6aQeb/j2ZQgQxGRuJ6QSsZJdK5HSqhiWCJIk4VBYey9mLMWPn0uVdIoOsvuvW+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/9Tf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdFN7X+R9FtCqqMNjIlqchE5iZFV2cOQRbg/1t78a8B14UPHy6Km0favcdcZG+8p6Kd1Ij/dUKl9H+9H37y6159Ga66+L2Iw5pcjuAitnsHeJmJf9WqxuPeqU630bHF4bHYaBafHUsdNEo0hUUXt/Qn3v5dep05+/de697917r3v3Xuve/de6ri+c/zd2D8RafbtZk6uCTcmbzFLjosYYfNK0VRp1ShY2DWAOom3HsOb9zemyhbKKPxpjUlfRBksc4p1Dnun7r7F7Z28E24OPEnkWNUAqzFj5AZoB3MeAAqejZbX7q21X9VYfs/cFZBiMTkMNS5VpJCAPFPAJQUjJv8An2YjcoFsFv7g6FZdX7epDtOYLOTZ03a6YRoyhjX5ivVfPbHyt7G7or5tj/HvA1mWxtTPLjajdmMjqLUgkewqGKkgaQCfcf7pzPf7u5s9hQujY1r5fP8ALqK99533bmCX928oxmVWJUyrWi/P8uhn+P3whxO0J4t59t16dgb3qvHXpW1xlY0E8qazGwe1yhNvr9R7N9h5OjtKXW6t9ROc1NcdCDlb23t9vkG476/1dy2dTfhJ8h9nVg8caRRpFGoSONFjRQLBURdKqAPwAPY6AAFB1KYAUBRwHXP3vrfXvfuvdcXRZEZGF1cFWH9QRY+9gkGo60QCKHpFMsuByHkUE0M5sQL6Yxf/AG3szGm6hp+IdIMwPnA6WcUqTxrLGwZHF1YfkeyxlKmh6XqwYah1k96631737r3Xvfuvde9+690Sz5h/L7anxJ2um79/JT020JVWnr8pUpJJDA1TqTxssZH1A+nvYpTqpr5dVEb5732pJ0Xu3vn+XHgaje0dflKLJ9iU+3IqlngmqAZqyaohhYuI1Ibn6e9AZz17V26lp+Xn+fVf/b/ZuT7/AO6erP5gXQ8Jy/dvSu38BtfePX+KhlOQgzVDD4K+qeA3d7erWR+fd1LICR59UIVqEDIqP2/5ejQ/Kb509jfMPpXbfQWL6wqJ905jI4Cr3SKaCo+5hyML6GiKsG0GMyPqsBf8+1AjjdNQND6dMGV0bwyKg+fpXq974f8AQsfW3xy2hsPdSpklrcZispXY2oXUKGs8Cu9MzkAkoyr/AKxFuOfaVq1oelSUpUdHGghipoYqeFBHDBGkUSD6JHGulFH+sB711brL7917oBO6+3cZsDCVlKkiSZWpp5oY4fq8ZZQFcWP+Pv1ada6Jn051dl+0t0Sbrz4k+0NU0yzSM+hozeTQFYH/AFre/EZ69XH29WbYrGUmIx9NjaOJYqamiWNEH04UAk/6/v3Dh1vy6b8lhRMzVNG/gqQC2pf7RHIAA9rIbrSNEmR0mkh/EnlnoLOzu3qHqbaFdmNwuIqyOkklxiSrqNZOrW8Wm4Ps62XYZd93Bbe1FVrRvkPXovvt0Swt2kn7cEjoonxn6yru1t75LvvfETVeOyM8s+2sfOWCUNTHLqWZI3v9L3FvY85x3mLY9tTljbTpdBSQjzHp0HthsDuN0283WVbKA+Xzp1Zf7h3odde9+691737r3XvfuvdVp/KvfmT7K3jgOitmyvPSbhmhps7WwawuPlWWx8rITawNuR7mHkjbIdn2+XmXcBRogSg9RToC8x3b390mzW3B8MfTo8nVXXmO6z2Vg9rUUUYlxtBDTVlUnLVk8dy87tYXvf8Ap7jXfN2l3ncZb2Q4diQPQenQt2+yjsLRLZPwihPr0JHso6W9e9+691AytVHRYzIVcriNKaiqpmYkADxQM/1P+t7dgRpJljXJJA/n1SRgqFm4AHqrv47UmR7l+QGX7beRp8XtCvqcHp50eZdaQkq1xxYn3NHNkkWwcrJsYw84D/livQD2VH3Pen3M/DGSvVqfuEuh/wBe9+691737r3SC7Rr4sd15vOolcIV2zmhFf6tKaB1jC/43I9mmyxNNu1ug/wB+J+zUOke4SCKzlb+i3+DonH8u+hefqHJ56uiIr6zdmXiV3HqEET8Wv/XV/X2P/daam+paxntWJT+f+odBrkuL/dYZWHcXb/V/PqwT3FnQx697917r3v3XuqeaoHJ/ODctMqlvsq1ZCPrYCXnj3FXbJzhKvo3UDyfq+5Mqfw06tG7OpBW7Fz0OkMTRXFwOLEc+5VHDqeB5dEH+NWWGL31X4w3RzPKv/Brtb/if6e/VAFOtUNa9Wce/db6xyxiWN42+jo6X/prUoT/tj795168eFOq28z/Kr+Iu8N+ZrsLffXVPuTO5arkrDNJX1MUaSyqwZmjW1z6jbn6+916rpFa0H7OqGPjF1J8cv5dP8w3dmz9zbMpNt4zunddRVYSorZGq/DR1FWWp5yZABbixI/Hvxzj06ooCvX14f5ehH+X2/ev6/wDnG/DSu6Wr8fkqTH1GFiza4ZW+2mr/AOIES6WUC90Khi4+t/x78CdJ/LrZp4g04NG62xkYstyuk3YWPP0a3vXTnVG383n5V9m9f4ei6J6fp6yLfO8aWOpo8ljRN99DBVUrRDxNGwtpY34HvYArU+XVHZlFF4nh1rt9d/Eb5uQd8bb2/vz5WZrC7w3Xhm3pjsDkq+uFVHR0kRrTSAu3GoDSLce9nT+EY6oBLWrn+XWzP/KK+RW9e3+u9/7Q7BrajJZ3rXcn93IsrVFzJkY6WaSneUtITckgEe6kUOOnFYsO4+dOresnSJX46voXUMlZR1NKymxBE8JjI5/1/dJE8RCh8wR+3qsyCSJoz+IEftHVSfUeR/0JfMuu6nnlFLhs7RVOVXUSYTU1Ss8RLiy3uf6e4t2uT9z83naRhHBP7f8AZ6gvYJ25d9xJNhc6YplMg9Kn+Xn1bwrK6q6m6sAyn+oIuD7lXj1PAIIqOuXv3Xumivy8FIGRCJKgDiIfX6fn2pitnky2B0xJOqfDk9JuXFT7ooq/H5hCcZkaZ6app3UESwufUg1Di49u3K26x+GO6uD+Yz0wY/qonhmHa4ofmOqnMjjZfhT8lf70RBouvuxa4Yihom1yQU71MvjeUKNIBBa/HuFpw3KfMxuzi3uDpUeleoJmQ+3POP1lKWd8wQDyDE8flXq5LH11Lk6KmrqOZKimqYY5YpUIKusiBwRa/wDX3LKOrqHU1Bz1kBFIk0YkjNQRWvUz3bpzr3v3Xuve/de6K53v1hsKk29neyIdiUeZ3tiYHrMXVxLoqhVi7CSw9JA5JFvfscT148Kdav3cve1V351H8n8f2tn2lzGxRRxbW21VI0cnpqjE0MJNidI/p7diQs3y6YeQaSD8+hN+Evx4m78xHQO/9gYFtuVHUtCqZyop5JQayoFaHRnFuSf9f29NKqjw06ZhiLkTN5cOtr7b0VZBhcdFkHaStSmQVLtfU0v5Jv7SfM9Lfs6effuvde9+691737r3Xvfuvde9+690Vb5K/FvqT5D7Wy+F3vgaOqy2Rx9VRUWTaT7eanephEId3UXIAt/j7eijZzUcOmZnVaA8eqZcl8Nav+XF0ruDH9I7aO59/b+zGSx65vFs1VNicZll8CMBoDqRr/P+v79M9e0DqkMOmrA5HRKkxcvw16/nlrCdz/Kbs/J1WMknJabKYHD7jjKBmVxqDqZLXt7arXHl06KItRxP8+riPhH1NV/CL49Zftr5Bbtat3HmIZtwI1XK8k9NQ1tL9ytG4lKtca+bfS/+tfxzkdWXtwTnz6PD8WvlHtz5Q7brd27R8M2Epa6egFVCrovmiNwjCUk3t/T3rrfn0a/37rfXvfuvde9+690yZnI/ZwiNLGac6EH5v/hb2qtofEbU3AdJ55NI0jieuWIohBF53W08o9RP1t9R79cy6joHAdegj0jWeJ6efaXpR1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/9Xf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdF/znx/2vnt0jclWsesz+eWGzfuHVq54t79kde48ehmw+38RgYRBi6OOlQLp9A5Ivfk+/de6evfuvde9+691737r3Xvfuvde9+6903ZfI0+HxWSy1U4jpsZQVeQqHNrJDRwNUSMb/4KfbcsiwxNK+AoJP2AVPTU8qwwtM3BQT+wV60cvnB8hNpfKj5+AZPJL/o225kIMZTmQNHTSVWI9c9Op1EfvSxRw6788+4H3q/a5t7zmWIFRMVVTQkiFTQkDjnJr6UPl1yh94eb9o9w/e+1+ruBHYWEvgmporSISzJWtKO6xxavWoHEHq2vY243+Q2b2lgtxbzbq7qzF43E4SmxJbTRZJaeLwxrYMf84AB9PZla3B5thgaaX6WBVVQPJgB/l6yw2q+PO11DBdXBsLFAq6OAf/YPV6vT3UnXfVO2qPHbAxdFSUk9LTvPXUxDtkGMYJqGf+jn1D3K21bVYbVbiKxQKCBUjz+f59ZK7BsW07JZrDtcaqtBkefz/PoXvZp0fde9+691737r3Xvfuvde9+691FraSOtp5IJACHUgE/2T+CLe3I5GibUvVHjWQUYdJXHVM+HqTQ1d/tixWBzew54tf2vljW4j8SPiOPSRJDC+h/PpZqwYBlNweQR+fZaQQaHpcDUVHXfvXXuve/de64u6xqzuwVEUszMQAqqNTEk/0Hv3Xuqe/nrv/wCGfyu2Vnvid2R2FjcLnM3UAU9bV0wMVDkIIZIqdTVRseCzjj/ePexxB6qcgoeBFOPr1SB8Vdr9q/yPOxcrs7PR/wCkj4x9wTR1mU3wqT1mDxmJmLCOngLXVX0ORY/UEi1vd2Yu2o8emYo0t4xGldArx4/n0Ns/UGwNz/Jfa/fHwD3a1VtbcbQ5beXVWFWdTka6tmLZCepj0kaSGa4I4/HuhPrinTtC2Qak+nDrZL6u+PXUe2oMJvPH9bYfb29K7G0FdlatY3aujyk1OstWJWY6dQkLfRB70vadQ49XPcug8OhdP8QxFU0ju09EWLC/6Yw1yRwPZl+lcpQYbpAWa3NTkdKimqoauMSQuGB+tvwfyPaB42jbS3SxHWRdS9IfsXfmK2HgKvJV9QsUxgmFKvpJMoTgkEj3UdWNeq1cPh9zd7dg/fVCSyYyOtL6yJAphvq+huLWHutDQ9br1aJtLbGP2nhaTE4+JYo4YohJptZpVQKxFve+vdKb37r3UHJ5GkxFBVZOvlENHRQtPUSm1o4k/Uxvb27DDJcSrDEKsxoB8+qSSLEhkbgOqqN4Q535g9zR7boTJS7B2bW/dLkvXHBk4IZbyQMy3Bva3ubrBrbkHYPrJO65nFNPmpPn1H10snM+4/TjEMRrX16skxmEi2NjaDB7fpPDhaGKJIaeMemMqgSQ3t+SLn3ENxcvuc7XV01ZGrU/4OhlDEbBBDCOwUp8v9R6VtFlaasuquFlUXdPwP62PtDLbvHniOlcc6uM9Ontjp/r3v3XugR777Uo+qdg5fOmRGyaQAUFJYM87SNoJC3v7EnK+ySb3ukdtTsr3H06Kt33FNutGmJ7hw6LN8Nes8lPJn+3d3QGes3hMMlhBUay1FFJNqJiV/p9P6+xj7g7xEixbDYGiwDS9PPHRFyzYu5fc7kd0uV+Q6sH9xV0Muve/de64u6xo8jkKiKzuxIAVVGpiSf6D3sAsQo4nrRIAqfLqub5ffLnrvbGx8ptPb25oZd51WSpccuPj8LP45rxygMslxfUPctcici7rc7kl9eQ0twpap9Rw8vl0COZOY7KK1a2gk/UJpQfPHr0P/xR62ouv+r8dV0unz7yho9zV2ltX+UVkHkOpv62b+p9hbnjd5N03p434W9Yx9gPR1y7YrZ7epXPidx/PoznsG9H3Xvfuvde9+690U75g7qG1+sg2vQcrVyYwf7V54wum1x9b+xzyDZfWbwf6A1fs6D3Mlx9PYV/iNP29TviHt07b6ioKQoI/ua6orgo/wCmlFe/tvn26F3vzODXSoX9nVuW4jDtaqfMk/t6NH7BXR/1737r3XvfuvdVCbCj/iXz87Xpyb/ZBpR/gQ5t7iiwBl57uk9K9QNt363upfIfwAfz6te3HS/eYHI03N5aVlsPz9Db3K3l1PPVYGxFO3e7ZYm4WTIMtv8ADyC/v1KinWgQMdWtKwZVYfRlDD/WIv731vrl7917pvyuVoMLQ1GSydRHS0VMjSTTysFRFVSxJLW/A9+GetE061kP5mXbv8vb5Ob0x2Iru7sbsDuXZxTEYndDY+V5aKUSsqrHJTXLAM1rg/63uwNOqsKjNMfPPRjv5a38t/pTaNZgO95u3cZ3/ujFtHV4XPQPIHw1QJvKjtFMC+q4sNVh701Tx68iKgwSSfM9X4+9dX6qd/mZ/FXdHa20Y+1OrUqW7Q2maU0LUTFatcVSRmWqaIgG9gpJHvwwa9aapGOqC/kBJ2Z8n+7OrtyddZ+txnbvXOzcfs3M0FERDXzvQ0wpshUloTydIbVc392FAD8+mm1sRpNCP8vV2/8AKsx/VfT238v1hLvKmzvbu5cgczn6RlvUwVsRkarp/I9jqViS3HNvdTxz08BRdPVz/v3Xuqov5ieyavaKbY7z2pTs+6KXcOLxtXLFr8seNVl8jegXsFBvz7jDn+zktVj3mzFZdaqSP4eoP92tvm29YeaNvXVPHIin/SVFf5dWL9Z73w28tgbc3Hj66Opglw2MFXKpB8daKJBUQtYn1B78e5C2q4TcLOKSE6iVFaetM9S5tW4299tsV5E1VKrX5Ggr/Pp8qstU1X7VBEzhrqXXkqDxq9niwJENUnH06eaVpW0pw6k47CJD+9WEVM72bU1/T+QOf6e25blj2x4HTyQiuph0oAAAAAAALAD6AD2k49KOi1fKTpTGdzdb5ShnpxLnMNRVddtqW+l4MqFBhZGAvckC3+NvYe5k2ePeNvaMiroCU+TeXQP515ch5i2eSBxWSMFo/k/kR88dEJ+K3zi2D1ZRP0F3luiPEb52i9Z5KiuZDLPTiTRDG+tweNNgbewPy1znZbVAdn3yQrLCSCxU6a0+HVwrTy49RXyT7o7NsjDk3mq7jiv7dSxVmAdkqQG0k1IxSvVsOzd67c37hKbcG18jFksXVAGGojZDqBUOLqhNuCPcn2d7b38AubVtSNwPU7bfuNpudsLqycOjcD0q/arpd1737r3THlq2gMT0VRGtV51aJ6e1wwYabfn+vtRFbvJk4HTDzxr8yOqWfkd/Jv233dvSXd+0t/f6OqfM1CVO5MTTw1FQuWYSmVtbQqFF7/Q397eUIdEfAdUWASVeQnPp1Zr8YvjltT4zda4zYG2gk70yD+I5JVs+RnX9MrhlBFueP8faWmanpVgCg4DoxvvfXuve/de697917r3v3Xuve/de6jVdTHSQtNIbAfT/ABNuB7vGhkbSOqSOEXUektRU82Xrfv5iRTofTGbhTblSAf8AYfj2ueRYI9CcekiK0pq46fMjgsVlvGchRQ1JiKmMyKDpZTqUj/WPsu6WgUFOtcH+Yb8Zuzdo/Jj/AGaTD4Cp3rtyhq6eqfEqsrRUMNAyMZXjjXTpA5/2HvdRw9eqFKEN6dFD7r+VndX8xHtHAdN4nCTbM6z2hHST5/MUiVEFJJj6SNRlUrHJ02spuPpx/T3umkVPWqiQ0FR69HH3p3fiOkuvdq9PfA7ANvTIY2akrNznCozzS5RYlirmdo2uQW1X91GT1fNKDNPLo0/wt/mN5DevYlR0P3hh02TvqggRjT1cbCoNQ0NxFM2skXaw5HvdPTr2r1werB+0Plt0t1PnsTtbdG6qem3BmamnpaOgULIWkqSBErlWuCbj8e9HhXrYyaDj0YehykNbi6fKpYQVNOlTFyPVHKodCCf6g+7qhZgo8+tMwVSemKihfKV71Ew/ZicvH9bX+gHPtfK4ghCLxPSONPFfUelj7Lel3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//W3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691FqK2lpbfcVEEOogDzTRRXubDmVh7917qSrBgGUhlYAqykEEHkEEe/de6Ld8rcHvzc3TW6dv9dw1E2fzePrMUq0xYSCCupmgka6c2F/ZBzJDeXG1SQWQJdwRQehGegnzrb7pd7BNb7QKzOpA/Mdaq23P5HfyFrcjX5XNZT7GoqMlNUxN9tO04inkMv7ryD9dzyf6+44j23mia3W2jtkiVVC0apBoKVoKYp5fz650r9zzm3c3kfdbvQruSAsdWIJJqzMTV/U0456Npsn+TV3zFktv1OS7uz1FjcLkMfXjFS1VeYdNG+vxRgekXtpsVta4/x9sWvIG8yPokNvCG/EqsGWprUd1AR5GnUjbH91PmayurSWTmK/8ABtZEkETSEqdBrp8hpPAgg9tR51Gxj1Lh6jZ+0sLs7KZF8jksPj6SjeplvrnFPAItZLf1tf3NsNg1laRxhtYVQCfmB1nHstu9jYx2craygAr60FOhW9+6Oeve/de697917r3v3Xuve/de697917prymPSup3Fv3lX9pvyD/gfai3mMT/Lz6YmiEgrTI6acRkngcY6u/bkSyws17yc2H19v3EOv9aPh0zbyFT4b/8AFdKr2g6W9Yp54aaNpp5Y4YkBLPK6RoAOeWcgf7z7917qnz+c/wDLbdnxo+E++989TxjL7uSuxFBF9jNFUSw0uQ1pUaEpnLX0kXNuPewK46oz6QT6Anquv4WfAnY3ze+JmH7kreyPtu4N64umzWRqWDvkNn5YqzpRTRhtSqzAXYjjn/W9+rQ060oR1rx+fUz4DZ6p7ByfaHwG+XWKl7Fx1VnqnGbKrc87ytRx0EjxLV0U5uwUgAqoNve2pSo/1HraltRjbJHnTiOrkfh18AuvPh/WbjrNp1QrXzFTI1CBG6fwuhdyfsYzJe62NuLe6HUTUnqyoqCi9H/97631wkjSVGjkUMjCxBF/e1JU1HVWUONLdB3uDIQ7FpKrNTz/AORrFKRGxAHkUarWYj/ePa7xknjpJxHSMRyQsNHA9Vzb43ZuDvTe6YnHJK2NSqERhQMECE2LXW/4H9PaChPDpbUD7erDequu6DYO3qWliij+9lghaeUXLAmO5S5F/wA8+/de+fQp+/db697917qu/wCV/cWSzGTp+mOvjLW5/KTHHbhigXUaKknbQZy0dyABzz7ljkfYIreE8wbpRY0GpCfMjy6BXMG5vM422yy7Yb5D16M58fuoKPp7YONwGpKrKurVOQyPqMsz1IWUxMzAGym49g3mrfpN/wB0e5GIxhV9KYr+fR/s23LttksJy3mfUnoc2VWBVgCCLEH6Eewzwz0bccdJ6swMcj+aikNJIOWCDh/zY+1kV0VGmQaq9JpIARVMdRIcpWUDGGtiPjUgCVv7X+IPt57eKYaoznpOk0sXa46flyVGaWSseeJIIlZ5HaRFCBRc6iTYf4X9ozBIHEYBJPSxZVKljinVXu6psj8qO+MZiMa8h2h13XrBuGnQMtPWwrUcmRkuDcni59zPZLFyTyy881PHuhVD5g06AtyG5h3ZY4z+lCe70PVoGJxVDg8bR4nGQLS0FBAtPS06fpihT9Kj3C888tzM08x1O5qT6noeRxpEgjQUAwOnH211fr3v3XuiifOLd+7dj/Hfeu4NlRzy56mpJkgSAM0miSjlEjBU5Nvr7HftxY2O481QW24U8MmufUEU6DfNlzcWuyyzWvxgY/YetAfcPaHaea7TyuXze5MlV5QbmoJI8fLTVOuJjOrNFqP15/w99KLbadoh2dIbeIKvhtVqjOONOsILjed7l36Rppy1JY6JoI0jFRXzr6+XW9d/La3vvPe/xy21V7zjqErKGkpaKjNQrqz0kYcIyh/wLAe+dvu3t9ht/NUi2FKPUtT1x1mlyRd3V3siPdijDA+zqwX3F3Qx697917r3v3Xuq9P5gsn8Q2Ps3D0z/wCUNu6jkkQfXQXRbH3KvtavhbjcXD/D4R/y9A3m99drHCvEuOjndZYxcTsfbVKqaP8AcPj3Yf7WaZdR9x/vU/1G5zP/AEm/wnoS2EXg2qL8h/g6Xvsq6W9e9+691737r3VRfx/dcj/MX+Q8bepcfj3kt/Qs+hP95PuKdgbX7g7gD+EH/D1A/K9J/d3eB/Ai/wA69W5MgZCjDUCNJv8AkWsfcrdTvTFOqq97BsB3tC2krHJkSLkcEeYgH8f09+z59eHVomHqRV42jnBuHgj5H+CAe/db4Y6cvfuvdU//AM53fHYG1PiRvSn2HV12Lrq2OgBytD5BNTap3idVMXIup597FDjqjErVh6da42R/l79T7WX4U/I7srdtDuLEbx2t/eDtJaiR3kpK6OZmSKu5sGIA4ex/3v3Yu3wjpvwIwwbz+fVln8n3eORxnf8A2tgOttw1W7eqc3uOY00QV0o9u4v+Ik08UIJKgAWFx703DI6vHSpFaivW0b7r050W/wCV1ZvSi6U3ZJsOoqqbPSUNTCJqMkTR0ctHItU4Kg8BeT7959er5dajeE6mXqvqjs35I7M7vqMh2rh9/wBNT1u3YJAmQhopZjJlXYObsoF1IA+nu+Wx00QFq9c/6sdCdsX5KdZbu+cfRfZPx/3JJXZ+k2rgaDfe16ekkhSr3XIgiy8sxT0sWdmuT+fegCB3dbLAt2muM/Z1uG4bIPVYPFZGtUU81XjqSqqIzx4pZ4Fkkjt/tJJHvSqXwuerMwQd3QPd74Og371xuXBinFa64jJVECEEaamKlIhcEA/Qn2j3fahd7fJHNiikj7adB3mO0i3XaZramrtY0+YGOiRfy5MxX0OzNzdU7oqpTm6TctfX00Utw60McrMECudVrKB9PYO9vrw2lq+3TGsqOxHzBFP5U6jf2iuJYdpm2C/OqVJGI/0uokD9nVp1PSQUqhYUC8AEj6n/ABPsfySNIdTdTRHGsYovUn3Tpzr3v3XukxvSsrMftXO12PjMtbS0Es1LEv6nmQgoo/1/aa8eSK1eSIVYAkfb0i3GSSKylkiyyqSPt6+c987t2b0znyr7bymYNXi687jq08TB0cLGbRrfjg3Jv/j7ifaEhureWa4QF5JXLg/xVpn8gOuNfu/c/vX3I3S43aOsqyKgBwQgRaftqT1sLf8ACf8A7a7R3ZBvTZ26KzI5TauFx6SYuqrfI6003ijX7ZJm4awAt9SL+1vKZa05on261JNuYfEK1qEctSg9KjNPLrLb7m3M+/7vb7lte4zNPbWbKsJbOldOU1fi00FK1IrQnh1s5e5U6zr6TWUyzhvtKJfLKx0ORfgkfQH/AGPtdBbimuXA6SyTVNE654zDiM/c1l5ah/Vpe58f+HPvU9zXsjwB1uOEHuccfLpRe0XSnr3v3Xuve/de697917r3v3Xuve/de6xyypDG0jmyqLk+7KpZtI6qzBRU9JVpJs1UAIp+zRrEj9Nx/X/efZgAlsnd8XSFtc744dKmCGOnjWKNQqqAOPybWv7L2Ys2o9LlGladZfderdNuYw+Nz+MrcNmKSKvxmRgemraScaoqiCQWeNx/Q+/de6q4+YvxS29tHpndT/HrZVNtDcGVxmXhy1fg/uBLJT1aXk8qnWeSx+hH9PfgB1o1PDHWu5t+l2L8ZugMtlcj33kdnd9xZfMVdDhaKlqjW5WWFy0GPlmYDTrb0/05924nppaInpX/AA/Z1g+EuG3jPkt5/NP5H1ldtv7ygyC7YyGSEhmzeTo01UaQpNp1awAbj3tjnSvWlDEFpPPh9nT38dfjN8jfkZ8psh8y+wq/MV/QhzX3VHFkpJo8Vjkx8gfXSxzEJwqj6e9HC6R1sBy4kbgOH+frYoxvfXZXZ3a2OwnWG1Jsl1fRHH0TZmmUmFYKKnWCokLRErxY/m3+w9qI6wr4jefDqpHitoGOrOoII6eJI41ACqovbk2H1Pth3aRtTdPIgQUHWb3Tq3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//19/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3TTnsl/BsNlMsVDpjKCsr5Ab/5qkp2qJDYfWwX6e/dePy613/lX8pM98x+k99z/GPc+Vj7P6z3oaar2tjQaOpqsNh1aWsqY4w3kkAKnVx79+LPDqp1GoQ5/wAvR6P5Xnzkx3yv6jXB7geHH9jdbLQ7S3BRTaKaoyFVjac009WkLtqZrxgNZRc8/W/vZFD1pXDCvn5/b1ad711fr3v3Xuve/de6Sefx7o4ydLcTxka9P5jA5v7MLSUH9F+B6STRlW8UH8unjFZKPI0yyLYOPS6XuQVHPtNPCYmp5dPxyaxnj06e2OnOve/de697917r3v3Xuve/de697917pizOKFagni9NVCNUbD6kj6D/AHv2rtrjwjobKnpNNDr7xgj+fWPCZJ6hWpaoBKmC62Y2aTTzex5+nv11AE/UTIPW4ZtZ0HiOqCP5ovzF36vyK6i+FfWVecRP3BHTwVO5KaSGCpx9XM7xFY6wuNIH59pQOJ6eY4A4E1/l1ULjMh251z8j8d0r2R993z0Ecu2K7fzu4JJ58dtmcSGD/gTIzIDGLMhJ59+NKY6bQvXJqKfz8un75NZj5Wfyra6ftH4gYrI9u9Fb+lgr5JoIaupw22aXJMWixR8PoBiDcG9jb3ZQDx49Vcug1KDQ5I8x8x1b7/KA6Qpt/wCB/wBmz39X/wAR7G3fPHnExlQxabbUuQZ5p6WND+lR+kD3Q+nTykE6h5/4Or9/furde9+691hqKiKlglqJ3WOKGN5XZiFAWNSzctb8D37r3VbPfvbFdvjPrs3bckklD9yaZ2jXUpZgENzHf6n348OvZr0O/SnS7bKw1NuCWO+XmSKo8Xq1206rkMP9j7U2zp8EmPn0xOjDvTPRmMbmkqhoqAIZwbFT9L2/r7vPamPKZHVYrgPhvLp99o+lPQEfILtzF9S7GrcjUz6Mtk4Z6LBRLpZ5a/SBGNF7/U2Fh7E/Kuwzb7uSxKP00ILn0Xoo3ncY9utC7HubCj1PRdfiN1LWZCpru897wk7r3M1RHHSVCSaqeAsHjqLSD6kHi3+v+PYs5732OFF5b20/ow0qR5n06JeW9udi26XY/Uk8vl1YD7izoY9e9+691737r3SA7P3tgeuNk53e25GSPE4CjerqpXVWCIOL2bg8/wBfZrsu33O7bjHt1p8cpoOkO4XMVnatczcFHWqh8gv5127K7dWe2bsXDQna08ogjy0FLSq6wrUtD5Cyt9bH8e8zuWfYWwhtIr6/f9YfhJOTSvp1jjv/ALtlLx7C0jZk/jUCi5pk1/wdXlfy68rtrOdRQ78xkq1Wb3VBJWZ9lA8iyQaZR5Av+vYe8ePdeO7h3wbbKNMcRon546mHkyWFtvN0mWkFWPVi9PkKWoC6JBqIB0nix/I59xQ8MicR0NUnjfAPU321091737r3TFuLb2I3ViazB5qlircfXQTQTU8vKss0TRMbf1AY29qbO7nsZ1ubZtLqQQR8jXpmeGO4jMUoqDjqqDJ/yfui8j2Qd/CcxK+Qjr5cUI38LPHIJAuof639Pc2Q++m/xbX+7ilSFpqr8uo8f212l703laEkGn2dWn7J2VgNgbcxm2duUMNBjsZSRUsUcIsGEQPrb+pJJPuGdy3G53S7e8u2LM5Jz1INtbRWkKwQigUU6VvtB0o697917r3v3Xuq2PldNJnN/wC3NuJ6/BlqKoKjmyhlYm319zDyOottplvD5qR0BuYCJb5IBxqD1Yrh4xDiMVCPpFjqGMf8gUyr/wAR7iS4bVO7erH/AAnobRgLGoHkB05e2er9e9+691737r3VOXxYnaf+ZR8th/umLEweL+h1Vyg29xJyya+4W5n5H/COse+RZGf3o5lH4Vjgp+da/wCTq433LfWQnVafyWx64fsDD5jTp81asjPaxA8x554/23vRyevUNOj5dbVy5HZ2IqkbWHhtf/FQOPexw6959Lr37r3QJ/IHqLaPdfWO4tjbyaGnxGTprzVso5pfAGdJAf8AA829+60RXrT37/w/WeB6+7p+OFd3Jk637nJ09LtLJGjqpI9t01IxYwUbAXAcCxsB7sK8R1RioBVj5dCd/LG+ZXWXxL3vsboHF4ukyUm9IqTGZbeklCyVlTI1YpFU0lRpI5/r722o9x6bjaMUiXyr1uMY3I0uWoKXJUUglpKyJZ6eQWIeNv0sLXHP+v7p0o6wZyiXI4bK0LRCb7zG19KI2Fw5qKV4dP8Asb29+zw69QVr1qI9i/yk61+6d1bm7A+RNX1NsTM5quyNVt9ZaiSmq4ZSXfXCrC50nm6n3YMahV6adEqSxIr6HoWPj7tj+W1/L/7Ax+Vpuw6LsffE1fAsGR/hxVRUzVGuJy89gCbgH2oWNnFW4dNF0gosdAT8+PWyrsrNr2PtrC7ox8oiwubx0GSxjwsjRvRVN2iC+IkDji3t8TQwL+mAT034cs0lXwOhBjxFIlLLTFAwmhkhkcixZJBZh7RzStNhuHT6wKqFP4hTqofKZKbon54DITRtS7GzmMFIJzdYjWV0Gkx63st9Tfg+4ikd9n538UikDLSvzI6gmaVuWPc7xCKWsyUr/TP29XD08y1FPBUIbrPDFMp/qsqBweP8D7ldTqUMPPqe1bUoYeYr1m976t1737r3XCSOOZGjlUPG4syNyGH1sR70QCKHrTKGGlsg9VFfK7+UD0d8nd7/AN/Z6+o2hnJ5WkyMuNjZv4grSmUpMPp9SbG1x+D7A248mvPdNdbZcm1Mhq4Chgx9aHgaYqOscvcr7tXJfuRuy71etJbTgULQtpLrUnS3EEZNDSorg9HV+MXxW60+KuyKfaeyaGCHwxhKnJ6Ss9UqxqhMl/6lbn2d8v8ALttskJSHvlc1dz8TE+Z6lXkfkPYfb/aE2rZoxGiACo4kAefqTxJOScnPQ7V+SqK6QUeNBIY6ZJADdRf6+xfDCsQMk3lw6FjytKfDj4dO+NxiUcatJaSotdpDyQTzx7TzXBkNBgdPRQLHk5PTt7T9P9e9+691737r3Xvfuvde9+691737r3XCSRIkZ3YKqi5J97AJNB1pmCip6R9TWVOYqPtqWMmlV1u/NiV/J9mMaRwJrY56Rs5nOleHSqpaaOkhWKNdIAF7flvyfaGWQyNqPSmNBGunqT7b6c697917r3v3XusFTTQVkEtLUxrNTzoY5onF0kRv1Iw/offuvdVjfKP+Vn0R8ks/hNx1GNoduV+LyseTqmpoJGFc3mEsyHQfText9feweqsobjnqqH+YT8eOxRvHpzp7AYqrw/S+1N3Y9J5acVL0FVQiojgqJJiq2AZAb3/r78BTqpBYj0rnrj88Ple8FDtD4TfEkzUu3MPTY/Gb0lxEIiWjSoiVK2seeKzFVZmY6ja9z7djjDd54f5emppGX9NfiP8AIdKHpb+YVsP4S7Ix/RGIy7b93Ng6hslu3KVFE1QYlqowXiSoQ3YRn/H6+9PqLaa1A6spRFDNx/1ceryPip8ueuvlFtaPKbVyccuZgh82Sx1o4pKddKksIg5YAFrcj22RQ9OhgeHRt/eurde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//0N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Rae8flH0z0hGKDsbNwxDIx/bPSQtTVBeKqQxvFPGXBAKnkEfQ+/daJ8x1ro/NDrKp+O3Z+D/mHfC40+T6vjkpMXunY+DZpsRV1uUg05eoylHTswAcM5If/b+9qM0PVHx+oM+v+r5dAp2NmKvqHdWwf5kfxAW2yh/AqHtLrzGxs2Kod9ZePyZ6OsoUPqGt3Kll92pXtOOqsSoEq5Xz+Y9fy62q/jF35t75G9RbU7FwcxaoyOIxr56lMJh/h+ampQ9bSBP6K4a1vxb3T5dPYIqOB6MJ7917r3v3XuuLKrqVYBlYEEH6EH3sEg1HWiARQ9Iupp5cFXCsgB+0lbS0Y/SmogE29mKMtzHob4h0ik1wuCOHSxhmjqI1libUrfn/AB/IPsvdSjaW6WKwYVHWX3Xq3Xvfuvde9+691737r3XvfuvddXA+pAv9Ofr7917qjT5u/wAx3PbQ7Kx3UfxsxMW8uxKeqGPyWPFLDORkJJGikp0MRZri4HP59qYbhkUo2VPr0lmjBcFfi+XVQH81P4qfJbujr3ZPyvjx9fsDuvrLFY9WxOLM9LWLXJJ5pGjVLENYljb35owv6nEHj14sz0RDRlyD/k6RnVnc2Y7Z+JB+Km39u1GQ707RXEVW792tQ1IzUOXgq188Jqwuo6j+o39sUpny6urBqqONOtob4y/E+iwHw/2r0V3DE27XrNtR0+fpMt5iKWtKMieMPZlePhgf8PevLp4gfb8+vfCb4SL8Om7EpqLsDJbvw288sK/F4qsSZINu0qTNJDR0olZhYA6ePfsHrQFMDh0fj3rq3XTMFUsxAVQWYk2AAFyST7917okHyO7nFMjbR25Us1bPrikeDS7XdArD0Em1zb3414dV456ifHDpQoq7w3LD5aqZ450inEgLM131WYfjj34Up14aqU6PSqKiqigKiqFVR9AoFgAPfurdM1dhoqi8kB8Eo59AtqYfT/jftVFcsva+R0mktw3cmD0msjuyDZ+KyWQ3JIKahxtHJVieR1UvHCPWqlyLn2sisG3CdIrMVZzTHz6TtdC0jJnwFFeq3dt4fKfLnvGs3JkXqh1Rt2oNVhWMb+CorqVr+AAG3JXk3/x9y5eTwcictraRAfXSijeoB8+gZbpJzJuxnkr9NHlfmerWKKipcdSw0dFBHT00CLHFDGNKIqKFAA/1h7hCSR5XMkhqT59SGqhRpUUA6le6db697917r3v3XugK+SvWdT3B0tvfrqkfxz7mxjUSSDV6CTqB9PP1A9iXlHd02LmC33SThE1T0Vb1Yncdsls14uKdaQ28P5ZPe2G7Xqtj0+1at8XPWNTR5NaevJdWrC+vWFt76D2Puty9cbOu5NMNYFaVX06xAvPbHdYtyawhQeEcV7tXxE8fTPW4J8CPjpW/HvpPC7ZzErTZGWnb7mKUSaoQwQhbS882PvBz3M5qi5n5ge6th2KcH149ZTcobK+z7WlvKO6mfl0cypwcch1QStAR9FQWH+tcewEl2R8Yr0I2tRTtNOmvzZjHSeqJpadTbWxuCo+v09vUgnHoemlaaI1bh0CHye7Z3d1p0tubeuxMSMxuPFhvt6DQZQ4Smad20fW3A/HtO9sytpH+rPT6XCstW61yemv58vYNZ3rQbQ7Z23Hh9vUtWtBlXpcTaSP16JDewNwL/m/tp4GWhr16O5iaTQQRTzPV1cv80/4uptN90Dcdd4Y6hKVompY1k80r6I/2y2rn8+2yCOI6UVHkR0dDprtrCdy7Rpt47fLPiq5I5aSVkMZkhlBKNpP+t7qK+fW8dC57317r3v3XuuiQASfoPfuvdVsbnH97PmL/AHeUiVKSgjn0XBVTCgJbj/b+5hsv8Q5A+r4ajT9vQFnpc8z+CfwivVkVPH4oIIv+OcMcf/JCBf8AiPcQOdTE+p6HIwAOs3uvW+ve/de6i1s329LNN9PGur/bMPdWOla9UkbQhb06qH+IUHl+eHycyukf5VjIx5ObsRWofr7iflVQeedyk9R/lHUD8gIG9z9+n82WMV+zq4L3LXU99ER+ZmJM1Pt7IRLpNOQ8hA+tpiOQP+J9+8x1r16Hf49ZePIdfYyEG7wLz/W2hV59+630O/v3XuoWQoYclR1FFUDVFURtG4P0swsffuvdV+dmfHb4W9Srl+xO2Nl7Tr6mtqDVSnJiF6iQ2aQtHRySIW+hF/e/Lr1c8eqvu3Yv5bvyQ3ltrH9cti+rd44uWCGjzmLxQx5Yxzjx6WVigINhfVz78NQ4DqhKvgn9nV//AFfTY3anWWy8LR5j+PU2KwVJQwZXWsr16066DUO6Ei5P+PtyOB3+XVZJ0UgDz6UtRlayup54qCE6pYpYg631IZU0LIp/qt7/AOw9qhbxoKyHpP40pagGOqne9f5WGb7+7Ii3/n/kBuzEUN7ttmFKmppdFtBQkyoouB/qT7YeRVNEA6eEWpaSE9F/+R/8kzZm5Oqnptl7gqK/fWOyNHX0mSqKWVaupWm9QhQxMxuWA/4r7a8SSvHHp1swRU4VPVlXwA697N6t6Oo9j9oQVcGT2/PBjcYKvXrfGUsJjjdfJzY+26Dy6eFQM9Hl97691XF/MW6tqd2db4Lcm3oWp85tncdNmaqtgWQzPj6EpPMj6PxZfz7j/n/a3vdvjmt+143DEjjQZ6iP3a2OTcNmju7PtmglWQsONEIJHRm/jL21Rdx9U4Pc1GAPtIYMLVWK3arxtKkE7MFJtci/sRcubpHu22JPH+Gin7VAB6GvKO+RcwbJFfxcKaT9qih6MF7PuhP1737r3XvfuvdN1fkIqGO7ep7XVB/h/W3t+GFpT8umZZljwePScSPIZybySFqalBGnTcagPrf/AIp7V6orYUXJ6YIe4+Q6VFJRQUaaY1GrnVIR6mub8n2hkkaQ93SmOJYxjqZ7b6c697917r3v3Xuve/de697917r3v3XuuLsqKXY2VRck/wBB72AWNB1pmCip6SNbVzZWqFDS6hTn9co4II+vP+PtekSwx+K/H06QyOZ3CLw6UlFRRUUIjjAvb1sBYsf6+0ckhkap6VxxiMUHUz23051737r3Xvfuvde9+691737r3XvfuvdBr2fsvb29ds5LD5iip5566klpqOqkB8tHKwulREw5BU2PA59uxRNK1B5dNSSCMfb1Qb8o/hPhPjH1p2d3HtytbcG8N3bbykUuRWGq+9xfmk/bqYjY3K3AH05Ht6V1T9IeXTUaVrIePSo/lt9WfE6l+HNJ273NtvaG8d95Wpza7oye5BTTZs0lPLop6UQTyKygjUS1r/7b2kIoelCkhceY6J/8GOwMT1Z8yPkp21iqyTbvRWHocrW46gg0QUFNQCo/ZipUnKo50iwC+9kig6qoqzEcOr8Pjt8/OiPkllK3G7Gzx1U+rxPkfBRpJ4x+4A8jgHn6Ee9fI9WBByDX7OjxI6SKHjdZEYXV0YMpH9Qy8e/dW65e/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6/9Hf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3UOvyFFjKWWtyFXTUVLAjPLUVU8VPCiqpY6pZiFHA/J9+690VTMfLnrKv/j+3tpZRcruqkpchDS0itA8ctVHSMYTG8TtqBfSOB7317PWsZlMBP2Bge/O2PlhvjMbT3Vg9012H682fJGznNQ1oeCknipJCGKIdFmUH37NccOmyBkv5H+XRZv5fHyH7B+A/dub+KnzJoctubpH5EvJuHD0+eglqo6bEbgR4sRkqCKo5XSsgYaf6e7EKRqX8/8AP1RGZGCSHB+E/wCQ9XZ/FP4Gbv6e7yqNuY7HDsr4fdtiq3zOuVWpXEUslcr1GMgkoyeJI9SAfTj3VjWnTqLoqOAPl8zx6uz6d6Z2l0lga/bezKSKhxFbk5sklJDH44qcyk6YYwSeFBsPdQKdXPQu+99a697917r3v3XusNRBHUxPDKoZHBBB/Fx9R7sjsjal6q6h10npIU08uDrjSTljRyG0cn1Bc/Qc/T2ZSKt1EJE+IdIotUDlX+HpaAhgCCCCLgj2WEEGh6XAgio6796631737r3Xvfuvde9+691QL8of5p24sH8zep/jT0vRw5CWs3NFgt3nIpTQCOtmqzRsKeaYgkC625/HvdKjUOm9Q8QRHjx/LojW7afdP8vX56UXYnbG1YtxbX7b3Q+brc5NSNXvgaOfIGR5cdU2KB1s1rN73xWg68SEk1PwOK+hp0Z3+YT8qNr/ACFzHWmxuhc3l81Wbnjh8eOgpnhpctI9RfTVpEWIIDBDc/j+nt2OSg0v8PTcyFh2fF1b18a/jx1ZsXr/AK2ymQ6n2htvsyl2/RpmMjT4uJcsmWiuamWSucCQszWbn6Xt9APemhJBdcgU6slwMIxyejke2enuve/de697917ouXfPb9FsXB1GNoJ1kzlZG8KQo0bELJFb+pN+f6e/VA69kjHRWOjOpspv7cTbw3IsstG061OudHFixLBB+P8AW96pXI6rXGnqyujo6egpoaSljWKCCNY40UWACrpv/r+99W6k+/de64u6RqzuyoiAszuwVVUclmY8Af4+9gEmgyevdVffKLsDN91b5oegOtzJUVdLXrLuGrpP3I/4a5Van96K/AUEke5n5M2q35d21+Z93wGHYDx1eWD0Ad+vJN2ul2ixzQ9xHp0dDq3q2k6V2Rjtp7ZRqiniaOsralwUd6gxgTgqSbgm5F/9t7jzed5k5i3N7+8NCcKPl5dCay29drtVt7fgKEn5+fQsUWZhqCEk/bkNhzwL/nk+ySS2dBUZ6MIpw+GwenoEEXBBH9Rz7S8OPT4IOR13791vr3v3Xuve/de6Zp9vYSpqxX1GMpZqxWDLUvHeUMPoQ3tSt3cpH4SOQvp5dNGCFm1lQT69PP049puneve/de66KhgVYAg8EHkH3sEjI60QCKHoPOys3szZ+zcxn9509I+36GmmkrIqlIzDKft3IicSWHrClbn+vu3iPxr1Uovp1Qj8WNodF/NXuXsLN0PQWxsZtXaGXq4a7NRULJM8bSPFFO0kYs7G2of19qEuQq6XUE+vSc25YVVuk9W/E/4qdl/IZOnoN3HbuSky5ej27FRmKkrPtZtSIqOQSWtYfjn24ZIChznqgil1UI4dX87B23t/qjaWD2Pt2ggp8dt+igxitBFo8/2t08rAfn6/Un3VbUMuqvHpwTnXopw49CBFnKRwNRZSf8Ppx+R7o1rIOHW/qF6nx1tNLwkg+l+bL/vftkxOPLp1ZUYVB64V9VHT0NXUF1tBTTTH1DgIhNz73EheVUpxIHXpHAQt6A9VidPTVO4fm9ubcifu44bdyFOsg5QTCBksCvF/cycwKtp7dQ2hw2tTT5V6A21frczyTrlQpFerSfcLdD3r3v3Xuve/de6Ydzy+Db+WmBsY6R3v/wAFIPtqc6YWJ9Okt6dNpI3oOqsPhZGJ/lR3xk/qamisW+t7Vq8X9xjygteZ72b+If5eoX9ulLc6btOfxAfyPVtvuU+py6K78pMWK3ZUtUAGakiLWvyBrLA/8j97Ck9aOkgjz6R/xDzYqtvV1LK4X7aNyASBZVcEnn3UKS1BnrZKgV6N+2XoUvql+n+H+9e3/p5aVp00Zox59RBnqZ5fHFdgRIdRB40Rllv/AK5AHt36VtGrpv6gVq2B1qy/zEId/wDfX8wz499Yby3JlNi9ZZSpxuNyktNrix0tE9aY5KupY6UuVJuWPuuiOI95z02TJMQvBcn7fl0Zn5w/y4+jerPjXn997Q3VU4HMbefHx4/OY6BXqq5ml/T5KWR7ubXNuP8AH6e9G4OmhA6dNuGyppTo+v8ALHrN2ZT48bfp93y1eRgocdSwYbI5BXFRXUbMzNO+vm97C/urTPgLjqyxKRqcVPVkiRQU6koiRKoJYiygKouSSfwPbbOzfEenAqjA6Qmb7T2Bt5Jny268BSCDX5RLmMbGyFF1sCskoN7fj3Xzp1bjw6Kv2J/MM+OfXVPU1OT3K9ZHSCYytjXo6pLwprYB4pHB/wAPeyKY61UUr0pvip81eoPlzisvlescjNURYetNFVRVngiqBJdgNMKMSR6Tz79SnHrQavAg16OH711bpG9g4Cm3LszcuHqIllFdhMlTRggkq81MyqRb/G3tJfQC4tJYT+JWH7R0g3O2W6sJYGFdSsP2jqt34K5qTrXeW6fj9Wt4pqetyW4IIGCx2DFmIRJLPyLfT3H/ACXMdsupNhbjUt1EftpcHZr+flN8aGZwPtJP29Wqe5L6mvrHJLHCpaRgoH9T+f6Ae7KrMaKOqswUVPSbq8zJO/22PUs97OfoRfiw9rEtgg1y8OkzzFxpj8+slDh3L/c1zM7tyYXOpRf36a5WmiIY9etR25J1SdKFVSNQqhUQfQCwA9oiScnpWAAKDoMOxu5euuq8VPl95bnxOMp6dA7xSZKgSrKkkDRTzSqx+nvXXiaGnVG3Y389TrXCdzYjYW1Y6us2/NlPtMlkZ8XAyJB9x4mkiq/02A/N/e9LUJ6oZEDBT/Lq+XrrfWF7K2Zgd67eqFqsRnqKOspJlkjk1Kws1zFcXBuCPx71050tvfuvde9+691737r3XCSRIkLyMFVRck+9gFjQdaJAFT0kqmumy9SKKjuKXUBJMBb/AHn2ujRIU8R+PSN2eY6Bw6UNBj4aGPQgBY31Pbk39pZJWkOeHSmOMRig6n+2unOve/de697917r3v3Xuve/de697917rDPPHTxtJIbKAf9jxew92RC7aR1R3CLqPSTjWbN1vlYslNC2pCL2YD6L7MGK28WkcT0jANw4ZusPYHX22+ydqZTaO5aCCsxeUpGpJUljD6I2IJ0/T+n9fZacnUePS8YFB1Q/2F/JT1bxye4Nv99bj23sGpnkqJtq0kdb9jj6aefVKojR7GwP9Pe6nqmlQc1p/LqvvefVee7k7Qxvwy6Bo6yXAbZyX8K7D3nBRVUM9djhOIKmtrnUXZFF2tf35eGo9VNWOgYHn0K/zY+KvXvwL612Fh+ouxcm3ZlPE0tNR49Ujrc3WyyBHWsihd2VC19APJH4F/dgdRz1pgIoz4ePTrYC+A+5d9bv+KHUm4Oyaaal3jX4SZstFUK6Tao6ySKB5FfkFowp904cOn/IfZ0cK4va4ufoL8/7b37rVR137917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//0t/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3TRlc/hMHEZsxl8bjEC6ga+upqTUv8AVfOy3/2Hv3Xq06pY/nVd/wC7NifFjKxda5WJlymQw6VeZwlbDUT08VQ7I0QmpywUMrA/X3sDIB6o7EIWXj1O+EPws2Bh+qekvk1kt+ZjLZqp64xm7c/j66WGTHVNXLSGuqo5JjJ9VKmNgy/UEe9Vrx6uFVeGfn1UN85t75H5M/KjE/IT477cn3D1T0r9ntTeu3XoBJha3cFNI3+UVtJSDxnU40gkXNr392FAM+fTbl2oV8iaj/V6dHm6K+Iu4/5gPZGzfkN8j9l0G2IOvcTQYLbtLDQzRrNR40/5DTwQvZQFUD8j3qpFQvn17SHpqHw1/b1sT7awNJtfAYjbtALUWGoYMfSC1tNPTJoiUD8WFgPeunOnz37r3Xvfuvde9+691737r3XvfuvdNuTx0eQp2jYASKC0b25D249vQymJwfLpqWMSCnn0x4evqKSVsbkBpKEiORvzc+kX/wBb2quIhL+tFmvTMT+F2P0rvZf0r697917rHLNFTxvNPLHDDGNUksrrHGij6s7uQAP8SffuvdU1/JP+cX0d0Zu/M7Zgevyx2nOIdyT0dNT1lJGRKYmMVRGSByPqW97Ck5HVC6qDXqrf5GfH3Ynzmocd8/8A4WZjIVPa3U/j3TkNpJB4JsrlIqv7p55FpTZgCPoQePewTQg/s61pUnxVy3r/AJOrG/hj8gOv/wCZP1Vkuo/kRsukm7t6+xssG6qvJ4mJHxFYJxT6sXKwVtYJJYFub888+/HBr69eRtS5/wBR6OV0P/Lx6U6M3LHumhpzuXKUbl8RNlqVQ2IYtrU0RV2tY+6mp49OAKPhx0d/L4tqxRLTsUqI7FAvpBP55/r7V29wI+x8qek00GvuTj1DxWXcMaOv9EyEIrEWBt6eT7cuLdSPEi4HpuKcqdD+XSoBB5BuP6j2g6W9Bf2f2Pjev8JPV1EqGreKUQQ611hvHdGKN/ifeiadeAr1XXszbG4u8d8vla81EmOWtWZXlRyixB72DLxbj3v5HrR446tG23t+g21iaTF0EMcMcEMaPoBHkdRy5B/2Pv3W+n737r3XvfuvdFJ+VveUPV+0P4RipVqNxboeTB00FMyS1VK1cggSbwglh+rg29jvkjlpt5v/AKicUiho5J4GmaV6DvMG7LYW3hoavJ2gDjnpn+JHQj9cbdO7d1xmr31uB5K2bIVIdquOirYxLHCzub8huR+Pajnvmcbtd/QWJpbRYoOBIxX+XTPLeztYwfUXOZXyT8j0c0gEEEXB4IP0I9x50KOOD0yVmEgqWMkbGB7fSPgXAtcW9qo7plGlsjpNLbh6lcHppWur8VJ9vOhamHCSkEsT/sPx7f8ACimGpTnpOJJIe09Pi5rHrTvU1FTDSwwqGmlqJEhijB/LySEAD/X9pJIXjOeHS1JFcD16AiH5V9KT7uqNlpvHFfximmMD3yOOFMXDaDpm839f8PbRFMdXBqejC0tXS1sKVFHU09XBILxzU00c8Tgi4KyREg/7A+9db6ke/de697917r3v3Xuve/de6LR8uOuJO1+jN4bKgytPiKiupWqoqmeripBeip5JTGJJCL6vpb36tBXrXE9a33w4/mG7O+DLdodSdqbeiospUZ/Thq7b1JBWfxAU/kVZa2ZLhtQa/J97oTkdUDBOxsU6E/pLcWY+b3z96/8AkB1vt+owWxtq/bxZupgxz0CsYZdDTTGIBNTWvf37gPn15W1Go4Aft62f1o6cKA0SORf1OoLNc3ux/J92DuBQE9W0Lxpx6iy4mkkuVTxkm91H0/1h7dW5kHHPTZgQmvUN8BEQQtRKh/qODb/YH24t4w4ivTTWoPA06Su7MQ+O2numqWsmYw7ey0o1N9GipWkHJ/wHtdYXKy38KaRmRf5npi5hZLdzXgpP8uiG/BLE1+fXcvYmjzlc9kcWJpD62VSwKj8f6/uTfcy4itvB2nhVFanQR5SimlSS+GTqI6sNeTciMwjp0dS5Kkn6Am9vcU/4nQZ6GYFwcnqTFPuAqNdNHq5vc2+p90ZbOvaetVuh5dcHqs8GIFMpFgRyef6/X3YR2hHxdeDz6qHpK79rc5FsTc87U8avHiZnUg2IseTY/wCw9odwW1W1kIbgp6S7k8o26dm8lx1Wz8Dky03cXamXaBbVdLUBmN+WFQCgv+fUB7jnk1bc7rcyA8R/l6ib2v8AFl36/nkFA1f9jq0pY85LYTRstgPo3H059yqWth8PUxFZwuK9Bj3JgKyu2LmjNqbTTAablgTckH/bn23JNH4ZVOnIY5FbU/n0Vr4qAU2YyeMmqGjEhlQIDb9TWtb2mhl8Lyr0/JEX4nHVhP8ABKU2uzG3+A593+ofpsW6jz6mxUFJEAFgjuL8lRfng/X3QyyHz6dEaDy6rR/mL/GvpTf+wJe0Owdzy9eVuyaVfs9zYvTDWq0eueOGN9S+o24/1vrf3QknJz1agIp6da+G6f5zXV8FJtz4dTy57eGHmFNQ0OSzOGWqqMxHDMD91JU2LP8AmxB920kcemvGUnB62Pf5ffyt6d+QHW0W2eto1xtZ15Rw4bLY+VKelfyQzGLXFAtmI1Hm491Ip04G1ZHA9Hr3XSV1ft7L0WNLCsq8fV00DKbMsk9O0aMD/UEi3vX2dW61Dv5h/wDL0+Q2J2PvLtrL9n71w9JLuyKOno8ZVV0wWjq1Nn0QsbKFU/UW92BXgR03IJDUhiOhx+Af8vD4/br+Pddl+0e+a3K5SeHVkY83lKJKigRqdmlZoqyYO3NgAqk8Wt9PejUtjryhNHcK9OnwH2xh/j5/MDTpfpDM1W6+ospRZCtymb8BFMa1KOWT/OpdeHNhzz78anJz16MKhKKKf8X1tG+9dOddEBgVYAgixB5BB/BHv3XuPHqmz5KQZD4/fKnCd7L5KTCbqq6Lbl410xNLUaYZNRf0Dlj7iTmBZNg5nTfTUJKQg+00HWPvOCPypzxFzSSVinKwmnCrH9nVtke4YJcXjqqldZ3rsfR1cZUiRClTAsiHUvBvf3MNrCZ41kOAQP8ABXqd/qVMKOhrqUH9vUeKgyGUZZaxmgjJDAKeCByOPasyxQqVQVPTfhSTAasdKOloKek5jQF/y5A1f7f2jeV3FCcdKkiVOHU321070QT5+9+9sdF9ax13TuDXO7xyyTR4umel+5Q1MbgKjLY8EH3sdaPDhXrVE+W/xL/mI9/deU/yZ7tl3VsPbW2YZ8/mKLDy1SRyY/7ovEWx0LfoJNgStufdlKLx6YkScqaEAen+z1az018d/iT25/LPodxY3EbKPaX93Klq7duUelpd40+Rhl0OzRu4kF1AIGn63591PToCkaRwI6MR/Je7wzm6tvb+6LyNVU1+O6fjEWMr5yrLPHU5BYWCSD6j6kc+9EEGvr16MhgfkadXq+/dX697917rHLKkKNJI2lVFz/xQe7KpY0HVWYKKnpJzyT5ufwwFlpQdLsLiw+l/ZgqpbpV+PSIu00lF4dKOioYKGIRxKNVvU9gGb/be0MkhkNT0rjjCDqb7b6c697917r3v3Xuve/de697917r3v3XuuLusaM7GwUEm/H0F/e1BY0HWmIUVPSPlmqMxVmCO4gj+pH0AvySPZiojtk1HiekRZp3oOlVTU0VLEsUSgBfza1yfqfaB3ZzU9LFRV4dSPdOrdQsjRR5KiqaGVmWOpjMTlfrpP19+60RXovDdWdf9F43e3ZGyNnYqPeeRpHkrMpDSla3IOTfRKVJ1cm5/r78B14jqjbor4j9w/Lj5c7n7U+QsWRodh7KyzZfalJVQz/Z5bRXXgorXC6SPr/hf37AFF6qAS2tvLgP8vWxbmcjgeu9qVNalNTY3DYamJipKVY6eGJACVjhj+n4vb88+/Dq/WvnV/wA6uok7i3ntjF4+So2hsWvaDPTrQ0zzxQJU+FnjdV/3s+9gevTfiCtOryeju/8AYPemwMfv3a2Wp/4dUU/mqVq6imilpQtg71ADAIL/ANbe9Z6uOHQp4bdOA3DLVQ4bKUWSajOmoNFVQVKIb6bFoWP5/r71XrfSg97691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//T3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvddEgC5IA/qTYe/de6KR8gvmd038eKGWfdWbp8jVxohfH4atpKusiaQkKssMTMynj6Ee/DrR/Z04/Gz5d9TfKLE12Q66yiS1GMjDV2NqKqmavgP0OqmQ6xb83Xj37z62KEVBr1rh9u/J7sn57d5dudf4rdQ2ZsXo/PVuDzMtPlKTFV7CilkhiengBRpC7qb8n3vhn16bJJLKOA6Ll8XKrufvwds/Fr5CTS0nTFTnhBhN6ZlWSvhix87JjKiCrnsWDnSCA30974UPn1UBjqUio/w9Gv2ptT+aT0Xgsx0FsLZ4z/AFtkphiNg5WeNKr7zaBkMKVFNOxYhGjbUOfz792kV63Rl7cEeX+z1cL/AC7vhBT/ABu6s3VR7+xlDkNydoZSl3Nu3EVtIs9LSZJGkdqfxTalNme9wB70TXpxRpFB9v7erKsXh8XhKVaHD4+kxtGhutLRQpTwKbWuscYAH+2966305e/de697917r3v3Xuve/de697917r3v3Xuve/de6Zcxi1r4tcZKVEfqRl4LECwUn2qtp/CbS3A9MTR6hqHEdQ8RlAsMtPkJEp5aT/OyTOscYUmwJd7D/AHn3e6hA/VTIPVIJanQ3EdVYfMv+bN0v8YMnJt2CSv3LnKKRkyI2/FT5eGC03iKHw6rEWPPtGBXh0oZgoz0mOsv5hey/nx8cu7ds9K1ldQduYraVQcfia77ejr5pyVkMlPTIQ6soBGm1zf68e/EZz1pXU1INafy61oeiPkR11nt+9vfDHe+1KLM90dj1L7S3Hmt6YyKEYTKivKpLjMhVgFWDfUg+9sKjUOHTcbFT4T/Ean7erUOnRlP5dXdvQvx+6hyNNvU9l10WJ7dx9OYshiMeK2qWNqQNDrACo173HvR1Elj1ZFVaIo9eti/Z3xv6s6+3rnd87J2zi9s5fclSKrMPiaNKY10urW5qGU83bn3WnTvnXoe/e+vde9+69005LFxVsR0ARTjlZFsGJ/oT7UQXDRNnI6YmgWTPn0isrvKm2VjKiXcEqxiFZPAWkVWfTHqUnV/X29PHG48WM8emopJA3ht1XLuLNbm7538lJR+d8dFVpGiqpkieLVoLegW+ntD59LOPVj/WuwMZsPb9JQUlOiVRhQ1UoSzeS3KAnm3Pv1OvDoRvfuvde9+690h+wt74nYG18puDK1UNMKajqnpFldE89VHCXihTX9STb2ZbTts+6XqWsKltRFaeQJ49JL26js7dppDSgNOq6vj5sjP/ACF7UyXfHYVI8u3qeeeixGGq4mWk89P/AMB54YTYcAKbgf737lnmrcrXlTZU5a2pqTEAsw40PEE9AvZrSXeb87veD9MVCqfX16tKREiRI41CpGioij6KiDSqj/WHuFSSxJPE9D4AAUHXP3rrfXvfuvdY5IY5RpkRXFreoA2/1r+7KzKaqeqlVbiOqsf5qvecfQXxm3q+Mraihz+5MDUU2FlpZFjljqoJeGDEcA3tf/D2pW4LVRhx6TSWyrWYGlPLqlX4n/yyX+UfxY2x8jcN3huYd0Z+TLSvtmDIxvTCpp6omOCaRJg6vp9ZBjtpP19+IjYEL5fs+zqqLIqgv8WP8HGnp0dP4Z93/Nzo/tTE9Ad/7cnTZq5D+G4XLS0gnq6gMy08Uk9XpJK6Re5Pthoyg6eik1nScn162KfbfT/Xvfuvde9+6910eAT/AEHv3Xutdf8Amh9/9vby7/2F8Uupsk2Kxu98epyNeKmPH1IqWgdZVSZrMF/HDc297GePAcOqsTjTxPn1WX8dvjhuvHYXtzPZ/bu3d/5jYu5Vo6yu3H48p4DSVLGcrUyFgLhT+efeywJxw6oqFRnPWxx/LX7N6y7P6szEuxdkYHaOS2jkosHuL+C0MVJHVV0gkkLXjAuBoPPutM9OA1FerJffut9e9+691737r3QKd/boTbHW+ekdggyWPrscCWVTeopig0lv9e3sR8rWRvd3iA/Ayt+w9FW8T+BZOfUEftHQGfAjbz7d6YqqeQENWboylfdgQSKjSwNj/h7EnuddC65gVl/DGo/Z0VcoQmHatJ82J6O97jjoVde9+691737r3QedszeDrXesx48eArn+tvonHPtBuh07dM3op6K96bRtNw3oh6r6/l+Q+bMb0yo5+686XvfVapB+vsDcjgNNLL6/5+or9rKPNczD8RP+Hq0j3JHU0dJHfdP91tLNwadRko2AH+OoW9+60eq1+jqmbEdr1VLI7JE9awZL2ABmufr+OfejSnb1sVOW6tW97691737r3VOX86jrze3YHxG3fjtqTZKJDNjfuP4Ykss9o3d3Iji5tpvfj3YcRXh1RwxBC8adUp7vg+NnUXTvxb+ROP6q2rm890TsWjw3Z9RW7fjqMhlNxSl1iqsgHFy/IHq/p71SuOvE07h5fyx0ab+TvS5ntHt/dnd21KWPbexc1lps3VYlEXG07w1lbqSGOmbSHALC1h72409vWoiG7/I9bSfuvTnSD7J68292ltDL7N3PSx1eJy1JUU8scq61jeaBoUqAtx6k1Fl5+vv3Xj1qjfKj+XfsL4ybly07/Ifc1Bt7M5GXJ1GJpMqtPUUcDr5mp1oWlLFVBtwLe7avI8em9FDqUkfnjo+n8r7sj4W7AiO19mbvn3LvevqIEm3duuSnglp5pWZGpIauVrKGLWte5tb3o1JqR15SoFFNer+1qIHhSoSaN4JI1ljmR1aOSNl1K6OvBBHIIPPvwUsaDPViwUVPTNVZ6nh1CINIwBsdPF/9fn2qjtHYVOOmXuEU06Jl81up6vuXqF5TBpfZz1e6aZ1R2lM9AgmRQi/TlR7CnOW1W99tZPFof1B+Q6j33G2A8x8vkEZgrID81yP8HUz4E9g1fZPROOyOdYS5jF5Oqws0dQUaoSDHBYaa6fUCwNvaTkvep922YSzYZGKU+Q4dX9sN2feeVori5/tFJUg8RpwOjwAACw4A4HsV9SP1737r3XvfuvdM+V2/g86IRmcTQZMU7aoBW00VR4m/rH5Abf7D37r1emrd2yNub12jlNj57HU9ZtzMUJxtbjZEvTSUZI/ZMYIuosLL9Pfuvdau3ff8oDv7b3aO7E6Z3fPj+pd71bxVFF/GVxlFhKCSo8pWkpZZB9FNuPdq44V6bKt+FqD06u0+Anwx2Z8R+vRQ4bLR57dOco4xufLLKk8lRKsiyrHJMGYsVYcn3omp6sgVVoB1YF711bqJVVsFGheZgLD9II1f7b25HE0hovVHkVOPHpNRrWZmUu10pP7JHpuL35HtcTHbAact0ipNM1D8PSopqaKljEcSgAAAmwu1vybe0MkjSNqbpZHGsS6V6ke2+nOve/de697917r3v3Xuve/de697917rokKCSQAOSTwAPfuPXuknXVxyNQaGlLEBgrEXAIvYm/sxhiESeJJ0ildnbQmelDRUcdHCqKBqsNbfkn/X9o5pTK1fLpRFGI1p59TPbXTvXvfuvde9+691gqKanq4Xp6qGOeCQWeKVA6MP6FW9+691HocXjsYjR46ip6JHN2SniWJWP9Sq+/de6p5/mMfKLfvT/YXWWw6Sgd9k70qIE3FVeAPHDTNM8JZpWBC2Bvcn3vJ4eXVaqvHzNOqOJo9pdJfJ/HbNpcDt7ce1PlDmFpa7IGGCuqaGkqasAsXTUUZSb8W97PCp8utKaNTyb/J0IXze7C7D+B2E3B098Xs5W7r3J23HSDG4+C1XDjFkb92ljhXUYzewtx78oDnPl03I2hQVFa8Pl1Yl/LL3tvPo34ybm7K7xyOTk7Dz9FDVV2366TVUU1cs+opR0k3qIsb+ke6+eOnVBC0Jr1c90f2unbm1Ytxx0s1KkqoypPA1PJZr/VCB/vA96+3q3Q1e99e697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/U3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdVNfzZfm9vH4YdJzbi2FQVFTunK0jNjahKQVFPTOJzTv9w7qVW45F/ewKnT03I2hS5FQPLqvP4G/DzZnyH6uwvzT+UvZzibsVarNz7brcxTDG0hjnYSfdidtEfAbTGqk/Tj6X9/pevUrmT0H5dEV/lodr9edY/wAyTuOh623rlF6wO+snRZl9w18NBhIcbNWSU8JoHlIjaMXBBB92/CD1QU8VlXhg/n6dA/8AzG/hHvnoH5twd6dRbhq8l8eewtwtuvsWs2TXjIMqVNWaxoJFx7srsAbH6j/H34MunTx608beJ4oJFeI9fQ9Hgpd7VPzs3H050z8etnZTaOx8Vj6Wj3vumTES47NVNTTVKmStrKoWubLc+r3rA6cGpuGOtpnrTZEWyNhbI2nUzPk6raW38fho8lWkzVkv2UAhMrTOS12tzz7qMdOE1z0IXv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuqjv5xPd3cHRPxa3ZuLpehnm3DW4avjnnoqYzVUIpyrwumhTY3ZufamGbT2NwP8uk08ZK60FSPIefVZP8lvbHxv3l0Bmvk78nM1tncncuZ/i8m8Nn9ith6+LCUFPMGhraPEZe+uSQaxylhx9Pr7pMlO5eHy6tbuGXPE5P2+nSK/l29a0m8/5nvyC7S6Rx9RhusBlazKU2KpKM0O2P4WK5QlLRmICJlYEBVX+tvbZ+EdXXukZ/LAP2jqxn5s/yYenvkN2E/eOwjHsLs85FMpUT7ex0NA+RrQ5d55aundDqY2JJH1+n9PegSBQcOttGrsGPEefp0L/wn/ll7U+POYqexd95rIb+33XSmeJtxaq04aoBUrPR1EsjENxcH3oknraqF+316tk9+6t1737r3XvfuvdQcjkaTFUc1dWzJDTwIzu7sFHAvYFvz7917qsnuzsnI9obqi23t3yPQJOlOzQESDn9sn0Dn6m/v2phjrVFJ4ZHRoemusY+tMTSZCrpI5KuqgjkeRo2EyMTqPP4NuefayKOOaMgcekzu6PU9GYpa2CrRXiYXIB0k+oXH0t7TMjKSPTp9XVuHUv3Tq/UerqoaGlqKypdY4KWGSeZ2IVVjiQu7En/AAHu8aNI4jQVLGg+09aZgqljwHVU/bG4878ru3qTqnaVTUrsbb9VHk63JQf5mWWks1TE80fFjpIsW9zfsVpbckbC2934H1Mo0hTxzwx1Hu5TS8w7oNttj+ihBLDhjy6s+2ptvGbTwOMweJo6eipqGjpoDHTRiNHlihVJJmA+rMQSSfcMX15NfXT3M7FixJz6E8Oh7BDHbxLDGKBQBjpRe0nT3Xvfuvde9+690gO0Oxtv9TbD3N2FuiYQ4Pa2NfJ5FvIsZ+3jYKdLPwDz+ffvt691qa/Mj5PdkfPnGbpx+1du1tZ07iaarjosjT441dR4I6ljNJDUUyAM9rXt/h7tSh6ZY6x2io6Jv8JexPlR0BmKjGfHjIZPK7d261RVbixGclYT0VI9knNPjp7srW4Fhf3tqE+h60gIHqPnxH2dbl3x6z9P3V1TtTsLdu2MeN41UJ+6mrseorKaqiQaZY3kXUh/ppI9uLKajUcD5DrbxCh0cT9vQ6CozGPP+UKGpUPDfrc/65HtSy283wcek+qWH4sgdOlNnaWo+odLmw1Lbn+h9svasvDPTq3C/ixXp2SaKT9Eit/UBgSP9ce05Vl4jp8OrcD1Uv8AL7+ZBg/jH8peoOmcvNGmD3rSwz5aqaSmighSoZheSolU6CAP6+9AZ68WAp8/8nVePzs6yl+bW6v9mC+F+5MVk+zesaYUEeLgzdGksrxQmN50WCTgk/g3v78CBWvVWBNKGlPX/B1W/wDGHY/8yzbW1u0+iqjaRn3B2luSjrspLLC80okFWTUmGqa9gVZr2b3Y6fi6bTxANFQT69bY3wX+M0fxo6cx+DrYlh3buWGky+840jCJHnEV1kRWuS1g5ufdMnJ6UUoNI8ujq+/de697917r3v3XuiHfzAs1VYbqfBmkLCSt3PS0ZCn9SS6VYEf7H3JvtbAk++Sa+Cxk9BLnGQptqgebAdGK6D2//d7rXb1P4xH93j6OvKgW9VRTqWJH+v7CfNF19XvEr8dLFf2Ho72mHwbFF9QD0M/sPdGXXvfuvde9+690E/esog6e7FmJsI9rZN7/ANNMV/z7Kt7bTtFw3oh6JeYm0bFdMPKNv8HRHf5cS/cbPyWSAulRUVqhzbkrPyLj2Dfb/vtGm9Seo19o6Sbc84/EW/w9Wde5H6mTqFkab7yiqaawPmiZLH6H8+/de6qunI213a4j1RF68g/jny296wOtcfy6tOxVT95j6Sp/46wob3vf02Jv/r+9nJ68OHTh791vqp35y/zDuufjrVpsbeG0ctuWPNU7lqaDFmupGRqVyplup5B597Ar1R2C5PWtH1v8mMp8xM72p1N8cthTZ3CZvPf7+PD7hwmumo6iOQrDFBCFsqh/8fe6Uweqaq1Cefn0OnWXZfzj+FvffXmD7h2PhuvOlaOeio8oMNRxYuGfHvVKQ0jgWLaeRf3vFOtZVgGpT1+fW5Bsfc1BvPaG3d2YuTy43cOKpctQyFxJrpaxPNA2teDdSOR7pxz0/SmOnnMSSRYjKSxf52PHV0kdvr5EpmZLf7G3vYyadaPDrTz7P+OlP8yvmHuep7r7J3Hs44ugz8dBikrmioqinoaVvtrUjut9QUcge3FRhinHpqQqWo/AfP16Ie+yerelfj/uPb+xt4bmm7ype8qOnxFNC5jqp9uUlcwEyRg+QhjY3HFvb6xEdzUp59JTIFXSnxA4+eet234xz7y3T8f+ra3eiPTZZ9jbXAe5SeoCY9EeaqDWOtgov/xv3tZY4T2Z/wCK6d0NMatwHRjKXE0lMbhRIdIHrFwPySAfbDTyMKcOnUhVPn1h3BiosxgMvh3RTFkcdV0TR29JWohMZBA/HPtJcRieFomzqBH7eq3MImtngp8SkftHVVXxuzy9QfKbd/S9TKaDCPS1lbSwMfBS/dzXdCiP9WPA+vuMuXrgbXzNLsx7VoSPIV6hTlG7XYudbjlxzoQqWA4CpJ/n1bcpDAMPowBH+sRce5S6nMZFeu/fuvde9+691737r3QQd9dtYjorqPe/bOfv/B9k4eTL1+kqGMCSLFZS/F7sPr78evdaUfbXzn/mA/LTubH5LY01djOj89mJIdpV0NS1BF9hDVsv79RFoQ2X6n3egUVbpPqaVwIxQevV1HwJ+Tfc/W3Y1J093pWQ5Kj3JULj9qVcFbDWtLUSlBG8s66iQDfm/utF4jp0FuDcer/shkkoysa+qVxcD6244vb/AG/t+GHxBqbh03NKUHbk9M1NiqmunFXXsRGSSsd73U/T6+1Mk8ca6IuI6YiidzqbgelWkaRqEjUIo+gUWHsvJLGp6WgBRQdc/eut9e9+691737r3Xvfuvde9+691737r3XTMFUsxsALk/wCA97AJNB1okAVPSTr8hJXT/Z0lyt9LMt+b8GxHswiiWFdcnHpHJKZDoTp6x2OjooxdVaZrlntyL/gH2lmmMhoOHT8UWgVPHpz9sdPde9+691737r3Xvfuvde9+691737r3QA9//HPr/wCQ20qzbO8MfAtRNCIqLOR06yZLGkXs1JIStv1X+vv3l1oivVJHZX8rbr74obbz/wAgKfdu499ZvY0sVbtehydLPWGlnmZn1U4LOF0kDm3vxJoR1UBQakcPnXoD/gDidoZvNdz/AC0+U8dPXQbIkkzOz8XucJaSrkYtDFSxVXAAIFgFI/2F/e2oKU61Gags3E/yHSr6i773j8ku+Nwd5Z3D4fbHx46sy33GQoJVgx2HqsYJi0MSLIFjlLIP8fejSuerKScjhwHz/wBjq1P40/zIvjZ3vubdGy9hrT7Zg2xkPslqqs4/GYytXzeFJqbxBU0k8i3497Ipx68GBrQ16shgqIKqGOoppoqiCZQ8U0LrLFIh+jRyISCP8QfeurdZvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//V3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdAb8hugNhfJLrDcvV+/8AFUFdi9xUJpPvKihhqqvHuG8kU9HI9mRg3+pYfU/X37Hn17NMdaI3bG2/l38Zvl1t34Sb4zGY238Yeyd1rtrB5qOqko4aHbs+U+1+/SVzaIaCWb1D3c5XV59JhqWURnKEY+R9OthLK/yGfjzP13CerewtwDNVNMJP7zYyWAtm2trSokrqWYiQ6x+q/wDvPvWo1z04I0pUCh9c9KX4HfA75KdFb2yu2u5aPB7+6iNRJT0CbqqafPSPQtMLS/azPJZtI4BFveiV4qM9WRXUkM1fQ9XebY6y662U7y7Q2NtPbEkjFnkwWAxmLkYnk+ujjQ2/wvb3o5yer1PS49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Sa3Xs7bG+MLX7e3Zg8Zn8RkaWWkqqHKUcNZBJDMLMuiYG34PFvfqde60hP5xf8svvz4s79g73+KdXn5Ok91ZCrm7gxGNeWKn2/tv7kTT02NoYXIBKs1rL+L/T2ohl0kKRUefSG5gZgZEJVvQefWwz/ACaeyfj3vT4u7Soeo6WGm3Hj6OpG55MnDTJuurkSVEaoykh/eKE2CqfSLX+p9+uIwrll4Hp+CQug1/EOPVw/tP0/1737r3Xvfuvde9+691xd1jRndgqIpZmYgBVAuSSffuvdEH+R3cs9dJ/cra8zyfcPHDUtTsrSa2GlgNHP1P8AX37r2R0q/jr0guFpINz7hgEtbUKs8Kzxkk+u92ufr73Wp61pHRy3hikj8TorJa2kgWAtYWH49+VipqvXmUOKN0lKzFVlFN91jWOkHW6MeAALkBfZhHPFKmibj0jeN4m1Jw6dMbmYa0+FwUqEH7gIAW4+pHtia2aPuGR08kwbB6JP8ve9qvbtLT9ZbId6veOemjo6qClYSvFj65RE5MaAm4DH3IvIXLMd053jcu2CMVBOO4dBbmTdpIQLCzzK+MZwehi+NPSON6d2PBT6Eqc5mzHl8nXzxKa1J6yESSUpma50qT9AbX+v09h7nDmOXf8AciwNIo6qqg4oDxp0a7HtSbZaBTl2yT59GR9hHo7697917r3v3XugA7576wXRmOwFfm6arqlzmSFDEtJGZGQ8DU1geLn37Pl148Oi0fNCbH/Ij4f9q7f2NnqCLc+d2bVQUOGNdEuSapnKmOH7QEOSfqPT79qpw61SuT1Ux/K1+YvSvx76Xpfit3ptGDH796+rs3UZjN5XFUCw5CnrqsOkUn38d5GBAABexH497IBNR59UR9K6ScgdIT4o4Sj7v/mS9rb16n2+9LsZMvUZWuxbUZj21JixVrI8MKJpiZWBHpA/2HvxPb9vWxmTUeAx1tW0VBQ42BabHUdLQ0y8rT0dPFTQLf8A1MUICj/YD3rq/UplVhZgGH9CAR/tj72CRw60QDx6bajE0tRzp8X5/bAUX/1vbyXEifPpp4Ec1PTVFh56Kp80MrtGqT8O1x6oWC8fX629vtcpImlhTh/h6aELowK9aR/847ddMPkLSp37Rbhx+MiYRYDP4SkdZaXHmN0pyK6wC82/PumlGciPh1SVnVQSMV/Z0Xn+WL0p3LjsV2zuvo/sec1NdlBkNqU+7tzx0C1tGKrUq6KiQB7mwt+fbbxZpT7erQOzKWB/b1fz8Tu8/mxh+6NrbH7t25smpx9ZIkZzGBp6GqqVjNQNcr5GjuSdPHLe2mQqtRnpQrliaAdbEMT+SNXtbUL2/p/T3QZFenOsnvfXuve/de697917qun5v1ZzibY2r/nFTP0FR4+GIJkS5Cf8T7lr24jNt416MVRh0Cuaiswjtz/ED0fXaVKKLa23aUC3gwmMjIHFiKNL8f6/uML9/Evpn9Xb/CehfbLot0X0Uf4OlD7SdPde9+691737r3QKfI+f7bojtWe9vFs3Lvf/AFofZLzEabHdH/hbdEHNTaeXL1v+FP8A4Oie/wAsVRVdD0GUAv8AcZHJpq/rplva/sKe23dsSy+rHqPPZdS3KsU38Rb/AA9WW+5F6mLr3v3Xuqru8KM4btemyCqUE9eHVvoOZz9T71Q1z1o+VOrINg1P3e0sNPfUXpRc3vfk/ke99brXpY+/de6DTdPT/WG86oZDdewdqblroUlEE+cwtHknjLxGOyfdKwHBP4/1re/cetU869avGI7Bzv8AKa+Q3bOa3F07jDtLtDcE2Vopdt7XQy0tBPUtV04pJ1VtNgfwR7tpJGPLpv4TU5B6xdt999i/zXN2bf2Hsnacm1+tJ6nHyZbM5rHGirqA0s/6xXOFstuSL+3BCxp6nh1V3VhjA4162W+mjtTrnq7YnWmN3Nic5WbH2zidsSfZZCGrleXF0y0rOwjJNyR7tHavXQ2OvPcq3cv/ABXQo68vkUdQqLTzxvGwNgdEg0sOf8PanTaxDuyR0lDXMjVHDrX7/me9JdRdadk0Pfhy24sVuqOmWmOG28ssFHPF9sPK8mg8lhyeOfadrgnCjp/6YUqxyPP7eqLd47+6A253xt/v/C7c7Fydbh6GnSowNZhp6jAz18MWp62ZSNBJPJJHtsl2+I06tSNWrQn50PW3H/LP+YdF8wOjX3VFTRY6fbVbS4AUAWKGSOnjgZYP2EAtpCBbW449tEFTQ9PqysoZeB6se9+631737r3VP/z32rW9V7+2H3rtuOUZHM7qocRmJaZCGSiEia2mk5ABU839xRzxbNtV9BvVtXVI4VqeQ6gP3QsJNk3W05qtAdbzIj0/gJFSfkOrW9n7hx+6Nt4jNYypjq6asoKSTyxOJFErUyPKhYflSbH3KFrMlxbpNGQQwBx9nU42N1FeWqXEJqGAz+XSl9v9K+ve/de697917ouXy36eq+/fjp2r0/QsiVe+ttTYaB3sFWR50mUksQP7H5Pv3XutXqjgk6x6Vzfwxz22BgN79bJW0e0NzrQNjXqMhU1Ta/NkTa4+tjqI92ycjpsUUBTggY6Iv1B8tc/8WN8Nmu+I8luKt65qfPtmXCOmTmadqgFDJKNV7Afk+1qQxKutukLTtXRXu9etvT4J/MvZPzL2a298MJaWveFamTF18sS1VOissPqpVAK8m9re2HnJGiPC9K44hTU5q3n6dWDAAcDgf4e0/T/Xvfuvde9+691737r3Xvfuvde9+691737r3XiQASTYDkk/QD37r3SSyeSlqphj6G7Fra3HIt+eR/vXtdFD4S+LJ0jkm1nQnTxjMZHQRD+1K3LMfqCfqAT7YmmMhp5dPRRBBU8enX2x091737r3Xvfuvde9+691737r3Xvfuvde9+691737r3TPnsBh9zYyow2dx9LlMZVDTUUVZEJqeUAWAkjP1+vv3WiK9UvfMz+XnvbtLcWF2x1UsG3uq8wkJ3tFjStEYhATIzR04cauBYW9+GD1ogkdVqd6ba3R2lm+u/gJ8d8VkMRspI6fb/YefoqNqKtyVfBPpmrq2pisWAAP6mt73UcT1Uip0r6fsHSL/mK/Hzqb+Xt8fdvbZ6Y3Pmsj39FSYxchDjZLVFVMCtRJLKkRLuT+Cb8W/rb34EscnrTBEQ6celPXo/n8vz+a/i6/YvTnVXduNzeO3jW7ZoKJsrlYhQRPVtOI1eommQE2DW5PvdP2dWD1+LB4062D8dkKTKUNJkaKZJ6StgSoppUYMssUg1KyMOCLfke69X+3qb7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//W3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691Wl/Mi+A+2vmX1ZV/wukgoe2tt0mrY2diSGCeOoap8skEtUSpUG5ZX1cH+t+Ng0NequgddJx8/Q9C58Euse1+l+gdl9WdrzrWZnaGMNClYZxVSzq9QZQJKkFtWkEgc+/Hj1sVpk16Oj711vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuktvTZm3OwNs5faW68VRZnB5uimoa6irqeOphkhnWzftygi44IPv3XuiH/GX+Xf1v8AEjtHf3YvVWRyfh3xSS077eqVSHF4pZ5BLIlHDGxXgjg2H+t7URSVOmQ46YeIKCYxQno+uNzYklajrB46iOwLEaVPAHt2a1oPEj4HqsU1Rpfy6UntF0p697917r3v3Xuiq/IfuWDaOKkwWGqVkzNaBGhglBZfIhBRgL+/fZ175Hz6A749dQV+6cqd67ojkkj8omtUpqLOZNQCajzcD37rXy6sWggipoY4II1iiiUJGiCyqo+gA9+631l9+6914i/B5B9+690BHenYO3uqNnZLc9dKkVdLT1FBjYUmWJ5K6oiPgYJ9TZrexRyztd3vm4JZxiqggsafhBz0S7vdQ7fatcMaHgB8/LonPxU6w3Fubc+R+QHbsL1GdknenwVPUQian/htSrLTTgPxqVdJBtb2Pedt4tLKyXljYjSKlXINDqHEdBzYbS4mnO77kKuTRcVx5dWaUtTBUxK8DKVsLKLekDgCw9w66MjUbodowYVHUn3Tq3Xvfuvde9+691QX/NU+c6fE/KUlb2Ps/Obm2P5nlwww2NNU0E1PMvkeSTQwH+xPvYWp09UZ9KazwHVdHWG9+6Plvhpfkd8UdzUeJmV5amfZ2683S01cGpv3BEmJqCNYtwFC+/Upg9aJ1ZU/5j1K2Ln/AI4fJztXHdV/LPBbs2T3k+TGNy+e2hi0xmKyFQto4VlZVXXqci5F+Pp73lRjh16quQHGfTrZw+O/xb6p+NeAGG67xKJJNEqVGaq4omy9YmlQRUVSAFgdINj7r055U6Mj7917r3v3Xuve/de697917qtT+Y78bPip3n1DmaH5AT4PbimGEU24NFFT5qPxElUgqJdJAtfkkD3urClDSnVWCnDCoPl1Uj1/8C/gFl9tYPCdL9+1OKzeIoY6WKCfdFJQpVzr/mywilF7uB7c8aRRnPTYhipQLQfI9WC/DX4rd19T7tWqyWb2zu3Y8M/jhyceSp8tkEp1n8lknLswaw+nuySD4nHHr3ghQVU46tuqIs1DKTSlDBrY2vchCTYAf63tQjWrLRuPSfTcBqnhnh1GfO1VN6ZYWZlNmsn9Bz72LVHNQaDrazSDjw6mx7ghNtcUi3AJuDwf9t7ba1OrSD1YXB8x1MizNHKpYMVA+ur/AHn20beQGnTnjp1XF3dVx7o+ROL2zHIssUApKkoCGGuMKxOn/Ye5e5bRrLlN7wihNR+3oFbswut5SFTgUPVkePMUGPoIjIg0UdNGAWFzohVeB/sPcQShnldgOJP+HocqVCCvoOpokjP0dT/yEPbWlvTq2oevXLUp5DA/7Ee/UPXqjr1wPqQP9iPeut9AT8ndL/H3t1NQBOyMwPqONUVh7I+Zgf3Bd/8ANJv8HQc5u/5Vi+/5ov8A4OisfyuYRB8W9vgkav4xlSSSL6fJ7DXtkhHK0RP8TdAT2SGnkW2B9W/w9WNmRByXQf8AIQ/H19yFpPp1LuoevXA1EA/3bH/yUPdvDf06qZEHn1Xb8vqWGjzG3chTBSZ2jd3A+l5b8n3p0K4PW1cMO3o13TW46ao6/wANrcvLDAQ4U6m0gA3t7dWBmFR020qpx6EOXcSerwwytx6SV4/oCePb62nDUemjcENSmOsP32YrPRTIqFgxBZQLAf1PvZjgjyc06qZJparH1UT/ADMPm9198a5sH19vPqvF7+3Vu6hhq8RUz4CDNSQCRWjXQsyObh1tx+Px7prgDEoD+fDrdJlQBzSvp1VfnN7/AMwXuPrSqj+O/X+0tk4fMRCqo3ptu0W364U3jMZV5tCG9/8AivujT0fXHxHXhAxUhsfMnodv5VHS/wA69r7hzu+/kdm8XS7fwmXgiyD5DKwCAk1nrCEsbkj/AA59tvNJIanB6ehhEa8QetnSkqaespoaqklinp50EkM0DB4pEb9Lo68EH+vts9OjopHyf6U6J3rQw7t7pmlhxlC8ECFmQwPIFsiFH45A5uR70MHrZFR1z666F+Me6uu6rFbW692RlcHXY6pxJyNVtvGVVYYqunaJZfPKrEMAbgqwNx9feyQ2OtAMOkv8T/hPsz4l1O4qbYlVO2H3BlanLT0zIKeCGacsVjigDEWXVYe/EknrygAUAp0eX3rrfXvfuvdAF8k+vKDsLqndVHVU6VFZjcNkq7Eq8Ql0ZCODVE6AnhuOCPZHzDYpfbXKrCrKpK/aBjoMc3bTDu+yTQyCpVWK4r3Uweip/wAujsyprOvKjqzdFS8m8ts5LJzSxTSapVx+sLHqU8gDSLf6/sMe3+4O+3nbro1ljZj+XQG9o95km2Y7LfNW5t2ateNK4/l1ZT7kPqYOve/de697917purcnTUSgyNqJNtKepv8AbD2/FbvLkdMyzpFg9Vk/N74mbz+S8WHTYOJ23hlYKMzlxBS0WXqD5i5Z6xijMQDxz7ckSKAaRlumo2lmOo4HQQdXfydPjF15tr+MdozT5/O1MJmztVmXpZsdDISWKI0+vVa1739pjIzDJ6ULHGvkK+vn0yfE/b3xN6L743Vg+pd+0UtRk8iaWfFLl6aOjpFFQAY4qKInSAVHFvdWr5dWSlcCnV36OkiLJG6ujgMroQysD9CCPfut9cvfuvde9+691737r3Xvfuvde9+6910SALk2H9T79x60SBk9JjI189TMtHQgm7aXcfQgj1C/tfDEkaeLL0jklaRvDj6dMdi4KBSVGqV+XduTc/WxPtPLO0mPIdPRwqvcePTp7Y6f697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6wVNOtVBLTuWVJkaNijFWAYaTYj/D37r3RZavprZHUVPujf8As3blHNvGu888WQmo45ayKpmhca4ZV5BvY39+4Z68RU9UzdAfDvePevf+9vkl8w2P92euMvUVWAoMuiSYvJ0cDtNFDVRVBAICAfUH+nvdcDqunur+zoo/yj3/ALd+VPyjwON6o2/gdm9V9cscZl8xhqGjxcUX8Ncv5pKulRFUvpFre/fZ1pjqIHp1Zz8F/mL2N3/2HiNgbUo6letuqoztbO5Coj8hr/tJTTQ1Rqgvr9K8Ek2HvVBgdbLVJby/y9XTQ5nE1NU1FT5Kinq01a6aKoikmXR+rUiEkW/Pv3Hq3WCDceBqa7+G0+Xx8+QF70cVVFJUDT+q8Sm4t79175dPXv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6/9ff49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3TLk8THVgTRAJUx8owsuojkXt7VQXBj7XyvSeWHUKpx6gY7LvFL9hXgrKraFkP6T9Pz/sfb09upXxYuHTcMzKdEnSoBBFwQQfoRyPZf0s6BvuHtCh692/VSrURLlZItNLGZAGVnBANv68e/daqOiIdW7Hz3c28huHcIqJMZ5xM5kBkQBZARp1e9+WePXq+nVn+Fw1DgcfT43HwpDT06BFCoqlrfQtp966306+/de697917pvyuSpcRjq3J1s0UFNRU09TLJLIsaaYIjKRqb8kDj27BC9xKsMYJLEDGePVHdY0LuaAdVTRNmfmX3YzhayHpvbFZ9vXRai4lyVDwCl7Ahiv19zewt+QOXaYN/MKg/0T1Hv6nMm7Yr9NGc/aOrUaDBY7G4ejwlJBHFQ0VLT0kMaIqWjpkEcZIUWvYe4TlupprhrmQ1ZiSfz49SAsKLGIlFABTppmxdZjnMuMc+P6srG55HPHtQk0c40zcek7pJE2pPh6lUeeR28NUpjkFlLEaVLfn3SW0IGqPI6vHcA4fpRK6uAyMGUi4INwR7RkEGh6Ugg8OupJEiRpJHWONFLO7kKqqPqzMfoPeut9UhfPH5s/FityeX6O7H2EnYkdIZY8pU4/H0uXqqKPyaJmhEkZ0Xt9b/AI974jHVNQrSvVeWwviX1/vnHybx/l67vnwG/caJMrUbK3Hl6egeJyfOYkxisBYLxYi3vVTWh68FFKrQdGO+G3w0+SvZ/elVvP5mYjDUUm0JFyeByG38dQ0lTV19MVFMtTU01tS3HqLX97PCg68Aa6jTHCn2dbHqKEREHIRVUX/oosPeur9cvfuvde9+691737r3XR4BP9Affuvda1v83umzPavy0+P/AMe83lcvieqt84ynXc9TjalqV0aSWUGVTcC4B/PvY4EjqjZIU/OvVZGw/wCWptyu7Q3Jgen+xs7Qz7br66DHSZ7cBgSpioAZEcEOAblRa3vxf1HVRFkFSR+fV0P8j3I9q1O3/kDguzcvU5Z9lb0g25ipZa2SsTxiaoMxjd2PH7QsR78aVx1dAdHdxBI6vp966t1wMcZvdEN/rdVN/wDX971N69aoPTqLPj6WotrjAIFroAv+9e3EmdOHVHiV+PTXNgqGKGok/cH7Et/UAALar8f0t7fW6lZwPmOm2hRUPVZe2aeDcPznqMc5eajpMHUVXqOoXp6cyEXP0PHuY72R7X23EowzOB+006AlvEsvNhTOkLX8+rMpcI5KskvMaCNBfjQCbX/2FvcOLcqPLj0OTC1KV6xpiq5b2kXn/avdzcxHy6p4EnXFcZlF48q/kj1D/Ye9meD068YJaYP8+uzQ5dvSZV0jkeofW1vx70JbcZp1tY5+BPQC/J6lykXQ3Zy+VLS7Sr43Gr68EgAf7H2QczywfuC7oMlD0HOcVlHLV4teMTD+R6An+W1h66m+M23G1gK+UytgTa/q0nj/AFz7D3ty0acrRLT8R/ydBP2ct5IeR7Uehbo+TYircKDIPrc2c/63sf8A1MYPaOpO8CQk1PXJMDzqeU3NrgEn/efejd+g6v4BI0k9FO+Vu1oWwFLkVDO9NGDc+oKyubn/AA9p5ZjIc9OomgBelF8XZaXKbSszF5IItLqWuLOQpNj7qs8g7fLrbwxtk+vRrI6aCJdKRJb/AIKvvzSM3E9bVFXgOswVV+igf6wA/wB690qT1agHDqon+Zdg9sbbfbndbbKqd5b32ZDHDgqRcImYowrBpFapjdWuFbm1veqiuevGvl1UVnvlV87t9/wmm2TltgbAxVdHAyY7JRY/bzU8cxCPHJDME0/W9vdqDPr1UljSgH59WxfH/o/vLsn475vBdg702vk9yZnI0OQSs2lloZoUeKo1SLUT0zablb/n3rzqOrDhpPHq1nr7b0+1Nlbb25VStPU4fFU9FPMza2kkiFmYt+ffut9JLuvpja/euzZNkbtmroMVJVxVjPj5BFUeSEEKAx/HPv3XupnUfUu3Om9rRbS2xLWy42EoytXSCWa6LoHqHv3XuhS9+691737r3XvfuvdRa2kir6SpopxqhqoXglUi4Mci6WFv9b3V1DoUPA9UkjWVDG/Aih6qJ21Q03x6+Z25MlKRSbc3ZCcZSLL+xSSTVTaR4lF7nUQfcV26LsPN0kj4jl7R6Z6gq0jTlX3Cml+GK5AUeQr8urfY3Ekccg+jorj/AFmXUPcqg1Fep3BqAesc9TFTrqkcD/C4v/tvbiRs5oo6qzqgqT0l6nM1FXOaaijcAsF1kcG9vz/r+1yWyRLrmPSVp2c0i6m0mFBtJWkySA6lFwRc88+25LqnbFgdWS3qS0nTjkqj7DHVU8ScwwsUVABY2sCB/vPtGSSanPSoLQUXHWvXvH5Ydt9u7w7y6zkzsOFxG2JWgo1r6xqaSZYZHiC0ha1yR/T3oZGevcCR6dVMdCdHbdpq75HdtbCzO9cj2xsaZK6KNKqepx5qpa4eSRAhNgD/AIe91OB1QqMsOPW4F8TNwbo3T8deqc/vTyf3oyW2YZ8x5g6y/dfcSJ6w4BvpC+9dOdGK9+691737r3Xvfuvde9+691jkkSJC7sqgAm7G1/8AD3ZVLGg6qzBRU9JaWtq8rOYKMFIVbS5bgED9RDD+t/ZgsUdumuTJ6QvI8r+GPPp+oaGOjjAA1SHlnNib/mx9o5pjKfl0phhEYqcnqf7Z6f697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuuEkccqlJUSRD9UkVXU/66tx7917oDu++qpe0etMzsbDumKbLMpllo9FGWAjaOzGPSDwx+vv3AY69x48OtYn5vdCV3w72zWdU9fx10sfZVVi8puXcUsKpLFK7B5oYMihNlFzf1c/n3sVOT5dNt2jSo4n/UerAqHv747fy9/hvtqq2LkcJmOzd+bXxtVlqTHV1LJuCXN19F+5UyRwgvZJXsAx/Gr8j35cnqzUQGmAOiwdL/LjuLpXqXevyG73y8s2S3LWP/o82+1Y0OXlpMxG32lQtK3qKLdbm3vfyHXgcVOD0a3+V9ifkZv7Mbg7/wC+M9HSYbK5CSq2pj8hUtFU1GKyTSNAyQH6hVKi9vr7qQK0HW1ZiKnz8ur2VYMAykFWAZSOQQRcEH37rfXfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/Q3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3TXkcXDXJcgLMnMbjj1DkXI9vxTNGfl0zLCHFRx6Q2Z3cdlY2qkzDLI0SH7VRfU/oJDcA/ke1MkSTKHi49MJK0R0ydV1zJuvvXsU08/3P8LWrXxgsWjEccuk69XH0PtAaq2kjpWCCuoefVlexdk4vY+Fp8XQQxo6oPPKqKC7/nkfj3UCnVqDpbe99e697917rokKCxNgAST/AEAFyffuOOvdVsfK7tzcG890Y3oPq+o82cystNPXV9HLqhipXGmop52Xi4BNwT7l/kjYrXbrJ+Z94FI0BAUjNfIjoEcw7jNdTjZrH42ySPTz6OR0p1Tg+ptlY/B4qlENZU09NWZqZkQSz5WSAfduSovbUSBz/j7j/mPfLnfdxe4nNVUkIPRa46Eu17dDttqsMYycsfU+fQweyDoy697917psrcTSVvqkTS4UhWSy8/gm3t+O4kjFBw6Ykt0kOo8emEx5XFOXDCSjW+lF9TGwsARz7Vg28604HpORJBnoKvkZvyv2r8fO2d1YairqjN4fYmbrMdSUkUr1Uld4DDTrAIwTq1EMPaOWBo+GR0oiuEdtJwetYb+Vz2Z8eO3e0t+5D5LY2uwPYueOZx8tTveUQ4+SMynx+Ja1SUkP0U2+vujr88dbhkqDqFGrSnn9vTz8t9k7R+LPyq6L3d8Jd41GZrexN90NB2FisbkXrKTH4upr40en+3hsNDRluSo490GV49OadMgx5/y623KSKNYopVhijllhjaRkjVGZigJ1FRc8/wBffurdSvfuvde9+691737r3Xvfuvdcda3035/oAT/t7e/de6K/3f8AErqzvndG3N5bzpKhtwbVgeHEVlN41eEHUVu7DVwWvwffuvYpQ9UyfKj+Vz8oKnt3b+6fizufF4jayinbPrksrTUFZLKW/wArOmSQFw3NuPdgQK1HTZRzQq1DXOPLq4P4dfGPFfGjrhcSAH3hufwZbfVWhQw1eeGsyPE0ZIIGsjVc39149OAUx0br37r3Xvfuvde9+690056Y0+Ey9QPrBjK6b+n+bpmc/wC9e1Fquu5jT1ZR/PpuY6YmPoD1Wt8WKEbv7w3H2SoMq05yWJM/6wCUeK3kPP8AsLe5f52k+g5ch2nhXS1P2HoD8uhrndJL4+VVr1Z/7hjoede9+691737r3XvfuvdFn+Wc+jpPfMF7fcYCtS17X9Nhf2HuZ2ps06+qHoIc7vp5euV9Y2/wdB3/AC+KU0nxp2tGfzX5Nvpb6yL7LuQ00cuxgerdFPtZGI+TrZR8+juexl1IvXvfuvdAL8h8SMl19lWtd4YfTxzckn36gpXrw406An4bV7RQ5vGTPZkUhFY8+iQfQf7f3viOtfPo+HvXW+ve/de6Kh80u4JOiOgd39k0mFoM1X4xIqSkpshRRVsKyVqPGsjRSA30kA+/de+z06oG+MPwX7V+cOK3h2x3TurG0mOyletVtSm2/NFSSUSThjFBUQU5UIFAF1PPu1aYHTaqGy3n0r/jbv3sn4NfzBOu/hLld0Lu3bHZ4hqYpaHIy1sNFT1Beenjqb6gsiWs63HvxyK8OHXkw+itQQSPy62efdenOve/de697917r3v3Xuve/de697917r3v3Xuqwv5jWxMlPgNhb920hp67au50zGaqogEd8bSvHK/klHNhz7jrn/brieG3u7TDQvqY/wBEU6hv3a2yZ7O23Wx7Xt5RI5Hmi0JFfs6OJ0v29j+1uqsBvXCWlSrSLGs0RaQfd0sKxzsWI/rc+xxsN3FuljHdqaqR/MUB/n1IPLm+Q77scW6W5qHotR6gZPQnU+MrK0mTIP8AtvyAD6rfUC3+8ez17iOLEQ4dGiQSOaycOlFTUkFKgjiQAD8kAt/t/aN5XkNWPSxI1QUUdSfbfV+uEkaSo0cihkdSrKfoQRY+/de6o2+c/wDLd7Q7A3Ad6fGvKYvBZWuYVO4Ur6mmx/3Eis8kja2ZdYtb6+/efDqrAkVBoeqi+su3qn4p5Te/T20sBWb57o36xxW78nhYv4riaCu83KTxxh1NmH5PveD3Hy6oO06BmuerIvix/ME7X2BvnbHVfyAw+RoaTNS/a4ionppMfj6GJ5lSNmATSFsTwPfq4xnq41MaEUPWwlj8hR5Sjp8hj6iOroquNZqaphbVFNE36XRvyPeurfb1M9+691737r3USrrYKNC8zAf0X8n25HE0hovVHkCcek08lRm5VSMPHThhe9wNP1uD+fa4KlstTk9JTrlbHDpTUlJFSRLFEoFh6mtyx/JPtDJI0jVPSpI1QUHUr231fr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6L/358butvkRter21vzGLPHPGwhroYovvqaQRGONopnF7Dg2v79U9exnqhLff8pH+4PadN3J2DnarN9TbLpppIcJDXRVNQ0dHAZaGF6N3I9RUDge96qCnVPD7q1wPLoB+v8Aa0vzB7NyXcPadO+0vjh0cJcGMTWKKOOrixStJjIIqY6Vd5NABsp9+4fb14dxq3AcP8/Ri/iP3D3l8tfk9iz1tTT7Z+NXWRO26qiaP7alehxzsIJ/SFDFgl7j34qAKdbVtTE+Q6vkTv8A6sot80fVy7uw025kpwr0CVuqrUwrpN6fSWP05IPutKAdWzXoaqPIUWQTyUVVDVR/6qFg4H45t73175dTPfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//R3+Pfuvde9+691737r3Xr/wC+sffuvde9+6916/8AvrH37r3Xv999D7917rq/+v8A7Y+/de67v/vrH37r3Xvfuvde/wB99PfuvV697917r3v3Xuur/wCv/tj791qvXf8Avvp791uvXv8AffQ+/de697917rq/+v8A7Y+/de69f/X/ANsT791qvXf+++nv3W69e/3309+69Xr3+++h9+6911f/AF/9sffqder137917r3v3Wq9e/3309+63Xr3v3Xuur/6/wDtj7917rv/AH309+69Xr3+++nv3Xq9e/330Pv3XuvX/wB9Yj/e/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6902ZfLUeEx9Rkq6QR01Mhd2Y2B/oL/AOPv3Xuqzd/9obo7Q30mF26zNQwVCxoIyxQp5Cp4Isfe4pWibUOqyRpIulx0cvYWw4thY6gr5KZHrqlPJWMsKa1csG+o5HterQ3Y4UbpGwlgxxHQ3UdbDWRrJGwuRcrfke0ksTRGh6VRyBxjqZ7a6c697917os/yZ7zx/Tmy55oZUqNw5IiiocbGzCrP3UZjSeNV5sCRyPYx5O5bl3/cAHFIkyW8seXRFvu7JtlqSMu2APPPQV/EfouowVHP2nvmM1279zTfxTFzzskr0ONrVLrHdrsG5Btxb2dc98yrcyDZNtOmCEaWAxVh0g5c2loUO4XeZZMg+gPl0ez3GnQs697917r3v3Xuve/de6w1E8FNBLUVMiRU8KGSaSQgJHGvLO5P0AHJ9+rTr3Wul83v5lG96ftTsDqLoXAZOvk69xNbltyZukhnr8RksfQN5KhDCFMeng3/AB7URzFfj4dJZo1OEFW6In1Dj+kPm/tYb23Pjqrq/t3PZCfF4XcUrx4DbuQzavdE9AQFy/BHtSvhTGnA9MgOB4g48Ogk2Z8ffk//AC/PlRT9i96YTI9qbMzuQiTbEODb+Nw0mtgKSsMi6xH6LMDx7altjWi0oPTz69BOVqWYsT/Lrc06n3/D2bsTBbzgx9Tiky1LFI1DVgieCQQqZUcEA/qJt7RkEYIp0vBqK1r0I3v3W+ve/de697917r3v3Xutej+Yt8kPmB8U95r2bg6vI1vVOIljqMnQY5ZmMkUjNIIXaIGxIFh72BnScHqjHGoDHQH/ABY/4UL4jts5Ci3L0tvPFjGHxy5fJSVMdLUEyiPyxloxwBc/X3sq3CnVVlRsqT+zq6r46fPXor5JV38G2TujDz5+NmiqcNDkUqa2nmWQReOSJVBBJP0PvXnTpwGoqM9Hd96631737r3Xvfuvde9+690iuxMrBiNk7oq53CAYLLJH/tUr0Lqii39SfZltEDXG5Qov8a1+yo6SX0oitXJ/hP8Ag6JR/Lqopf8ARduzJ10QNZV75ywWVwC/gRyVCt9bHV7kP3YlH75ghjPasK4+fQX5LjI2+SR+Jkbqwz3FXQz697917r3v3Xuve/de6KX8xJ/F1Ln478T4yrQj8kW9hnmokbVJTzB6BHPjU2OZR5qeo3wapxTfHfaaKLKajIMB9Pq6349tclrp2CIfM9Ne2yFOUrcH59G99ivoede9+690hux8cuT2hlqZhe8Nx9D/AIfQ/wCv79mhHWjgjohPxzyZxfZGWxAYgNVNGqC4tebSRYcH8e9GoFR14eh6sv8Ae+t9e9+690F/cfV+C7h6+z2xNxRiTHZWmk9LIjgVEcLimYh+LBmBPv3Xvs609u2N2/Nb4Nb83dsDZHb238VtXN180eOoKXKQgwY+RDojljiJCsAeQfdu1hnpkllNEoR8+sHwe7W2dsD5DYTvT5TbyxW+e2qfJU8m0Kmgy0dTJRQ1dT41E5BZgQrcKB/vHvx4UA68jANqalaU/wCK63RdtZ6j3Rt/D7ix5Joc3jqXJUhPJNPVxCaK/wBPwR+PdRnPT5FD09+/da697917r3v3Xuve/de6jy1UEIJkkRbfUFhf/be7rG7cB1RpEQZPSeqs8zkw0UTs97GQDUtiPqPapLbSayfs6TNc1OlOg87U2NLv7rjd+ByipMuT29W0lKrIryJLKpsyhvof6H2g3mKC426a1Val1I/ljop3nbf3ntU9tMKh0IH59EG/l57oqNkbi3h8basSr/dB6zLQCpZndjLJysd7gC3PB9xryNfyWtxJy2wxBqYfmc9Rb7S3km23Fxya/C0qwr6Hq2X3JnU6de9+691737r3XvfuvdJbetBW5TbGXoMexSrqaSWONgbEegk2PH1tb3o1IoOtHh1qy/Dbf+xfhT8svkTku+9tztPv7Lz/AN18xm4KaWGiY5A1IlpJatXCK4XSWUXA4Hu2SKdUqqklsVz09fJH5J0vzX+RGxtp9a7WkxOBw9XBj59001KftDHHUkvVzV8SKAoB+vFgPfsAdbbOAfz62Q+kK/bNFsTbmysRunF7jyO1MTBi8k+PqfuGWop/84Zb8jlrXPuuOHThJOT0M3vfWumjI5aCiUqpEspB0qhBII/qB7UQ27SGpwOmJZhGuMnpnpKGpyrioryRCCNCfQ2H0tf/AHv2oeZYBpiGemEjaXubh0q4oo4UVI1CqoAAAA+n9be0LMzmrdLFVUFF6ye69W697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917pKby2Zgt+YSbb+4qdqrG1DBpYVYLqspWxJBH0J/HvRFRTrYNOqdf5pXx03FifibW7R+PWIbGLVblpspnabFQRRVdRTU0N52kkplUuCOTce9rQZ9Om2BIIGMdEQ6/wD5ifVHww+JDde7W2fUUvbGQhgw+ZjLCDK1GcrKc00mQCwguyoxJFzxc+9irGvWtcaLSvRJ87iu9OuevH+Z+fzuTqeydybhipds00NZP56TEZZdUVRPAOQqA2Jt72KcOtEuAX8/To6Xxp+Xnyi6Srdr7z7c3OvYGwdzVuJpxR4irkqPtJck1/t6h0/S66rMp/Pvx41HWwW/FT8utpLb2WTPYHC5yNDHHmMTj8okbcmNK+kSqVD/AIgNb3XpylMdPHv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6/9Lf49+690Sr53/KiD4k9IZPsI4qfL5CtaoxOHpqZpFlGReDXC4MQJ/Pv3E09evEhULny6pv6M+ZXzM7j64bsGkw26Kamqs3V0lNTSwVhkFPq1xMmsAlQD/T3vAwT1Uam7qcehV/09fNUgf7ityW/wCoap/2PvVF4163qk4Fevf6fPmsAP8AcVuQ/wDVLVcfn36i1rXrYZ+AX+fXv9P3zVuT/Cty3PBP29VwP8be91X1613+a9djv35riyrity2HP/Aeqt79VfXrXfxK9d/6f/mv9P4XuWwA/wCUeq/pYn3uo9etnX/D10e/vmtxfF7lP9P8mqj70aHz68DIBhade/0+/NdgP9xO5AF5t9vVfkfj3rt4V69V1Pw566Hf3zWGofwnctj/ANM9UAffu3hXrdZDnT14d+/NUA2xO5Bf8Gnquf8Abe99tMHrR8TGOuh3781zcDE7kt+R9tVe9UUefHrZLjivXf8Ap8+bAOo4vcvpsF/yaq4H/Ee91XAr1UFifh66/wBP3zWNwcTuWx5P+TVXPvXbwr1s+Ic6evDv75rLc/wncv8Ah/k9V9Pp+Pe+3162TIfLrke//muf+XTuW39Ptqo/n37tIyevDX5L1y/0/fNe/wDxaty2/P8Ak9Xf+nv1R69Uq9Kaesf+nz5rEcYnctwb3+3qr+9UTzPVy0vGnXL/AE/fNf6NityG/Nvt6r+th792nz60NfFV69/p++ayjjFbl+v/ACr1XH+w97qvAHr1ZCaleuJ79+a35xO5Cf6/bVXvXafPrYL0wvXY7++bAA04rcoX6W+3quD72Co4HrXfXI68e/Pmsx/4tW5eOSftqr/ePeu3jXr1XAyvXY7++aw9IxW5bAfmnqgLn3vt9etd/HT10O/vmuFt/Cty/ngU9V7920pXrdZK109cf9PnzVFiMVuW5PH+TVXvVF9et1fPb1zHf3zYvb+Fblv/AIU9Vb3uq+vWqtxC9e/2YD5sA84vctxb6U9Vx+efe6j169RjwXrw7++ay3P8K3Jc8n/JqsH3rt9etd9aBevf6ffmwxucVuUH6j/J6r/X96OmvHrepwKBegpqf5k3yD6O+R/UXXfbG19z1+A31LTNV1MiVkdLTwVFUIdUhsR6efr73jqlSGoRQHrZL25nKXc2DxmeoQwpMrSpV04Y3bxSEhbnj+n9PeunOnr37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYppoqeJ5p3WOKNS0kjGyqo+pJ9+691Xf8ge6KzceTXZe1Hd0LiCp8DuDPZyrMPoPz78acOtCp49DF8eulYdr46HcGbgWbI1K+WnEqxM8ZLBrtwT/X8+/U69x6NfJGkqNG6hkYWI/wCKe9qxU1HXmUMKHpMVeKqqLVUYyTQqku8fJYj8gf7x7Xx3Ecg0zCp6RvC8Z1JwHUvFZuOsBiqB4J04YSHTqYGxsD7antjH3JkHpyGbUKPx6x7s3PidoYGvzuYrIqOjpIJX80rFUaURM8ceofliLD3uwsp7+6W1t1LMxGB6Vz1a4njt4TNIaADqsbqPa2f+Vnbdf2lvWOZNmbQyL42gxlQT4sisRYU0sEcosV9IJIHuY99vrXknYl2XbiPqJ11Fh+H1r+3oDbdBJv8AuZ3C5H6cZoAeB9OrVKOkgoaWmoqZFip6WGOngjUAKkUSBEUAWH0HuE5HaVzI5qWNT9p6H6gKAo4DqT7p1vr3v3Xuve/de697917oF+5uzOvdl7ayOK3juvE7fn3BRz4mhTIVJpzLUV6fbR6XH0sWB+t/fq568Pl1pR/J75DdsfCD5P7wo6PbNfuDr/to1u05d0LRCtxb4jMy+J6gVjKwKhXuST9PdqAjHEdMFnRwKYPE+nRit35HY3YnW3xk6N6PxzZfeG1+w4dyblzO2ZF8Z/jEsdUy1Rp/xHc3uePfqUyfPq1Q1FX1/Z1t57Y2VRnZW38LunHYjM1dFhsdSyzVeNpapg8NCkV71KsdakEavr7sk0iEUJx88deeGNxQjj5/5unCnwdTt+nWkw2iHHwkukEaLGFuOQqILD/Ye1iSQS/2gyek0iSplDgdOFPnnX9upgkXT+qVvSCPpx7q9sCf0zXraTsBR+n2Gtpp1BjmRr/jUL+0rRSL8Q6UiRDwPUr2305x697917qqb+cTj+wK74XdiR7FxiZScvjDPSw0EVXXOiyOZGiZwSoC/Uge9jiOqtXSdPp0Q/4e7p/l11/xd2FgO3KPBbb7DptpLT79p6haSgr5sygkR21INQZl06f8fr72dVeqgoBQDoBP5X/xz6twvzF3fvzpDH7jXZtPumerqKyryL1lFLE1aWhZLcabkfj3piaAdejRdbOOtsT3rpzr3v3Xuve/de697917opfzJ3O+1uqGqlm8P3mSix7HUV1CpQpp4/rf2O/b6yF7vegiuldX7Og5zNc/Tbfq9TT9vTt8TNtLtnqykhSLxDI1AyhGlV1GriDlvT/X/b+2Oerv6ve2Na6Bp/Z07y5AYNuFfxGv7ejPewX0fde9+691737r3XvfuvdE0+alR4Os6hP+O8FRF/t7cewlze1NuI9a9R/7hGmzsD5g9Kn4cQfb9BbQjtbmtb6W/VIOePanlNdOyRD7ejDkNQnLFuB6Ho0XsSdDDr3v3XumzMUorMZWUxF/LCwA+vP197HWjTz6q+2dKNs96VnJVHyNwv048oPIHuvHj141GB1ahDKk8SSobq6hlP8Agfe+t9ZPfuvdcJF1o6f6pGX/AJKFvfuvdU+91fy3+id97/qt99z7khoqCrqJKjwNVxRytHpNwofn6XF7cf4+9gn7OqlR6V6qd+e/xL/lz9M9cTb16M3DU1vceLzuJhx9BHuRatWiMgkdxTCOMizAWsT/AMSdhiTQdUdFC1IA/ka/t62EPgHvHc+8fjZ13WbngqIail21iaWlaoU6pqaKJkRw7fqsABf/AFvdT078z0df37r3XFmVblmAA5Nzb3sAnh1okDj03T5algbSW1G31U8X/A9urBIwrTplrhBWmadMb5WvrZDDSRPFckB2Fl+l/r7V/TxRjU5r0x40j4A6zQ4OeX1V8okJIJ0k3/xB90a6QGkYoOt/TMxqx6faWhpqNdEEYA/qfUf9ufaV5XkNWPSlI0jFFHUp0SRWR1DIwsysLgg/UEe2+PVyARQ9VCdyEfHn5RYvsSEChoexMvDhVY/sxv5ZhEwHi5P1/PuKt2P9XuZF3IUC3DaR+Zz1A/MBblPnWPdk7EvWEfpxOOreKeVZoIZkN1lijkU/1WRAwP8Atj7lRTVQep3UhlDDzHWX3vq3Xvfuvde9+691737r3Vd/zo+Nfxn3vsTJ9kdx7HymZfa9Oppo9qNT47IVE+hmhWRkS7D66mJv9PeqeY692kUYA/b1rz7PwPd3e+VyHWvwg693H0/jKZjT1Ga3XAH+6RbpJN/FZUXSDbVwfd6jic9N0LCi4H+ry6Gv40ZH5I/y6PkLgtj/ACZ3vS7vl7CqoZalaLJS1MCx1FSrCXWjFeV/r79TWe0Z60tVFGao9SOtoek3THnsJhstiEdqfOU71FNICGAiuAt2A+pv/T2pgiFSZPKmOm5ZCaBPPp1oMMotPWXeZubHkA/1N/dprn8EXDqkVvnXJ0oAAAAAABwAOAPaIknJ6WcOu/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdRayipMhTzUlbTQVVPPFJDLDURRzRPHKuh0ZJAQQQbEEe/de6rP7a/lUfFnsnd9d2ZXbdyce646eqq6WGnqKX+FNkUh10rmgMQWwdRxfn+vv1T69aIWtSo6pT7Sye7em+6c5sTv3CVOU6oo9tZug23EIUgx1POlGRiWUAePUpK/Tn3sA06oSdVW4dKj+UNszJdsbh3Vsnsbb9fkutF3BV7iwskqCaCCop5JJcbGtRMG0rwvC+9sRj169HqNfTy62ssdQU+LoKLG0ilKWgpYKOmQm5SCmjEUS3/wAFAHuvTnUz37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/09/j37r3VP8A/OXUH4/bIJsdPYdGQDypPhXhlPBH+v7959aNdJp0cP4RNHU/HrZyyUtEgip41AgpIIFa9LESzrGAC3+1Hn37iSetgaQAOjb/AG1P/wAcIf8AqUn/ABT36g69119tTf8AKvB/1Kj/AOKe/deqevfbU3/KvB/1Kj/4p7916p699tTf8q8H/UqP/inv1B16vXf21P8AX7eH/qVH/wAU9+69177an/5V4f8AqUn/ABT36g69119tTf8AKvB/1Kj/AOKe/deqevfbU3/KvB/1Kj/4p791up67+2p/+VeH/qVH/wAU9+61119tTf8AKvB/1Kj/AOKe/deqeu/tqf8A44Q/9Sk/4p79Qde66+2pv+VeD/qVH/xT37rdT177Wm/5V4P+pUf/ABT37r1T177Wm/5V4P8AqVH/AMU9+69U9d/bU/8Axwh/6lJ/xT36g61119tTf8q8H/UqP/inv1OvVPXvtqb/AJV4P+pUf/FPfut1PXf21P8A8q8P/UpP+Ke/UHWuuvtqb/lXg/6lR/8AFPfuvVPXvtqb/lXg/wCpUf8AxT37rdT139tT/wDKvD/1KT/inv1B1rr321P/AMq8P/UpP+Ke/UHXuuvtqb/lXg/6lR/8U9+69U9e+2pv+VeD/qVH/wAU9+oOvV699tTf8q8H/UqP/inv3Xqnr321N/yrwf8AUqP/AIp791up67+2p/8AjhD/ANSk/wCKe/UHWuvfbU//ABwh/wCpSf8AFPfqDr3VFf8ANbdP9LHSMAp6YBKcFJVpoVnF8g5I+4A1WH4F+Pe1406q9NPVw/SIt1LsAf8AZuUX15P0P591HVzx6FP3vrXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdE4+RndUWDoZNr4Gby19UPFMYi4dSbgrdf6H36vl1U8ePSD+O/S0+SqY95blid21mSMT6XLFmDi2u596+Y63U8D1YCiJGoRFVEUWVVAVQB+AB731vrl7917r3v3XumTJYqmmSSpFoZo0eTyi9gFUsSQPaqG4dSEOQek8kCkFhg9VTd59i7q7+7SxvSWxqmWba/3VNHnslSvIKSnqadhG61Ln0r+RY+5v5c2mx5X2V+YtxFJqEoDSpB4U6jvcr653a/G1QZjqNRHy6s4652hh9i7WxO2sWlOrY6ip6askhWNXqKqGPRNLIY/qS1/r7hrd7+43O9e8nr3kkfIHgOpAsoIrW3WCLGkU/MdLz2V9LOve/de697917r3v3Xum3M1v8NxOSyABJoqGpqVAUsWaGEuqgD8ki3v3Wjw61a8Xk8382P5mW7upu1N1vi+u9r0VfmMNgZqyaB0rKCPyxRwxOyrd2Uf8R7tTFfXqgJJNfLo3P83LbHWHWfQHQuzM7tOHceDqt6phKqaloaX+JTUSLAzyGvKFwSHst2/HvQ41HW3A092eGPz6On8JPhv8U+qtuYXs7pjYtfichnsXSSzy7hqY8lPDPLRL5/AkilV5YlWU3HvVa8erhQowAK+nVjX04HAHv3Xuve/de6jVNJBVp450DL/tj/t/biSvGaoem3iSTDjplmwWjihfwrxZWJI/1r+1AutVDJk9MNbfw9QpP43QsP3TNGOSqi4+v0v7c028uRjquqSI0PU+jzTyuIp6d4ms3qb0j0Lc/X/Afj23JbAZQ16utxnuHHqp3+YL/M062+NtMmyqTaMXbWUy0Kx1+36DyTzRGVWDQ+MKy6ltzdTY+2PCkU0I6uZo8ajTPlx6qY+PG3/gB80Zd3b83BsPOdLTYioiqt4U+TzcsNMKl6kRBYoHC6CWJ9Kj22Q6nScdWBjcalz5cKHrZP8AjD8fOkOitj0kXSWPjTb+epYaxMt5UqZcnAzGSOcT6QdJPNr/ANP6e9cc9OAACgFOjN+/db697917r3v3Xuve/de6r5/mCTDJ9c4Lb8JYTPujHTyAEi6iRQv0/wBjf3KntYhh3aW6bh4bDoGc5kSWCRLx1jH5jo5vWmPixuwto00UYj0bdxGsAAXc0asSdPH59x9vMpm3Sdya/qN/h6E9gnh2ka/0R/g6XPss6Wde9+691737r3XvfuvdEZ+c9WKfYOPjP0qJpYwP6kkcewZzn/uEq+pPUbe5TEbWqjzx0LfxUiMHSO0oyLWjnNrf1YezTllNGzxL9vQg5MUJy7bqPToxfs/6FPXvfuvddEXBB/Pv3XjnqqLsNH293Yk36Y5cgtyOL/v8fp+vvxNMdeAOD1aBtioFVgcZUD6S0ysL/U8nn3ocOvdP3vfXuve/de6rJ+b/AMRu6/kZurblZ15vyj2tt6holp8rR1NTJC1Q9ikjKIwSbg+/V61pr506A7rj+Th03BkaDPdw1FXuzK0Yinl+2yDCknq4ZRIjSLOp9PHPHvYJJoOqsqjP+Hq3HbdBtnYuAxG09v08dFhcFRRY/HUsWhUhp4v83GNIH9frb2+trKwr1Qzx1NOpUmdqJ3MVLTSAW4lClh9L3/4j2+ttGg1SH8umDcO4qgp11FQ5epu01RpRieG+tj72ZoEFAK9aEMrefHpyiwlNoH3AMkgN7g2A9stdOMR4HTq2sYy2T08JGkahEUKqiwAH9PaYkk1PSgADA65+9db697917r3v3XuiI/PPqQb/AOs4N2xRlqzrJqjdFN4yiyF6bTIoDHm1xfj2Ced9qF9twux8VtVx+VOoz9zth/e2yi8UVazJlHrVcjoQvhn2rP250TtXc2SnDZZjVUNXAzMZo/sCsEesPz+m3Ptfyhux3jZIrt/iyCPPGOjX29308wcr29/Ie9gQw8wRjo1nsT9Dfr3v3Xuve/de697917pO7o2thd4YiowefpFrcbU289O9tLgf1DAj/ePfvKnXuGeqhfm/888f8M6Wg6y6g6y/i+aylGkFHNh8eEqITOpRjppFuzhv7TC/+t7usTnAx008qKtTn5dVj9K9B94fIjsDH/KP5S7yixXX226qHIDD56Q0tfS0qVCyR0QSosw9PtQAIRqXj00DJKQrYHp1szdJdl9T7/25BRdWZzH5nEbdhFHHHRyGY0cAbSqu5vyT9efad5GdixPHp5IxGAtOHQ3e6dOde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690BHdfxt6h+QWJhw3Z216fM0cEomR4fHSVfkFhdqtELkWFrE+/HPXqDzFen3qPpDrbo/Af3a6429TYPF3RiirG87GMFV11AUMfr+ffuvY6Fr37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//9Tf49+691T/APzl/wDsn3ZP/iQqP/rSvvXmOtN8J6N/8Gf+yeto/wDLCL/Yf5JFx72eJ+3rY+EfYOjg+/de697917r3v3Xuve/de697917r3v3Xuve/de6Kv81/kxh/h98Ze0/kVnqNK/F9bYWLK1NHIXVJxPWx0MaM0ZBHqkH0Pv3DrY86+QJ/Z1rH9N/8Kes731R5jLdc/G7LZ/GYUBqiqxcmUlhVPJ4yWHJAFvdip86ft6TrKhyNX5jI6tv+Bv8AOS6f+WW6v9Gm8o8d1h2ZV1H2mD2pXy1Rr8xUmURJDEJL+s3NhYe/MpXj1aKRZK6M0414jovX84v+fLtT+Vd2ZtDrXLbAG8cpunEUOajYSVqPHR1N/I4+2IFksb3H+H1496Ffl+fV28uOfT5dWM/Cr+YPsL5efFap+TeMhpcLg8TtuXN5imDytFRyQUElY1OzSEkk6Ao/xI/r79Qg0PXgysmteHVIPxX/AOFOux/kj86aH4jUPVgx9Nld21+1qDdSTVzxSzUFYaSeQI7FRpP1uAL3+tj799tOvGmokE4r9hp0Yj+Zv/PsoPgX8y9qfEDE9U1HYG69ybXxe41mpJK0TpHl6I1dLGqU50m4V7cfj34A1Ip1p2QKHJND6fs6BHE/8KPH2puzBQ919G13WeycxUUFG27NxSZGDH09RXz+KKPXJpGr8jn3YKfl1RpFBHGp+WM9WZ/zCf5tvVvws+IGy/lnjY8dvrau963G0GMWnnn8BqcnQLWQRloSHDeogA/W3utPIcT1djQFjWgHlk9VUbL/AOFGPZe/tvwbl278V9zV+Jq1ikgrKQZWSCSOeLyRurgHgj6e9lTwNK9UWVWFV1H8urtPiL89qfuv417k+QXa+1puo8dttaqaqo86Z4y8FJSvUSyoZxqIGkA2HH1914YPl04DUA5z69U79qf8KLvuN3Zaj6L6Xruz9nYSWtgfdO3pshUUVRPQyaZYneEEX/r9P9b3fQafb034yVoK1HHHVin8ub+btsb5vZQ7JzG3qfrzsGCnmml25WVM/wB6VggMrF0qObki3491KsuG6skscg7DWnGvHq5wEEAg3BAIP9QeQfeunOu/fuvde9+691737r3Xvfuvde9+691737r3XvfuvdUP/wA1247f6TIBI+2W9vp/wPf6+9qKt1pz2Z6uN6R56l2B+P8AfuUXB/1j7qOrNx6FP3vrXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Reu9+26br/BvT0k6/xOsTTFoLa4iSV+qn37hk9arXHRP+mutsz2hug7q3AJJqVZnmbyPeN18ge4Ev8Ar+/AVJ62CKdWcY/H0uLo4aGiiWGmgQJGigAAf1Nvz79177epvv3Xuve/de6LBvT5TdfbL7f2l0/X5OkG4d2FBSU7F/MS7aQEsbf7cexnt/JW53+wz77Gp8OHj6dEM/MFnb7jHtrnvk4dIr5fdzZDZ+349g7MqWXsDdAhXFwwM/3IppCY5tCp/UH2ZchcvRX90d03Af4rDXUTwqMjpHzLuj20P0Vqf1pKU9elL8W+iqDrDZa5PJ06z7p3ZHBls5NUKks0VXMDJIiyMNQNzc2PtHzrzNJvO4eBCaQQVVAOBA6f2DZ0sLXXKKySZY/PowdbjaujlaroHOkvqeMXJJ+vPsLxTRSr4coz0aPHJEdSZHUqhzscjCGrXwTfT18avx9PbMtsRmPI6ejmwA3HpQqwYBlIIIuCOQQfyPaUgg0PSgEEVHXfvXW+ve/de6oT/nbfObcXxd2V1dhNj5lsNmNxb8paHPTpLMjNiJGh9DCEj0m5JB92UV6o7hR5eXQd9qfBLMfIvrrZfye+IPY+PwvdmUw+Jye68zST/btWmeiSpnooXiGryfX0k8/4+9Cg45HXiC1fI+R49BF198LPnT8jN7bX278guwKuu2TsTLU+RMOZmkdJIqRgJUpVqFNy5A+g9+qKdvXgr1q/D5DrZn2ftXFbL27i9uYanFNQ42jpqaONTcXhgWFn/wBjpv711fpTe/de697917r3v3Xuve/de697917pEVm+NkwZaPD1mboIMs7vHFSyFhM76GVgotzxf3dXYCgOOmyiFtRGetPX5r5jH/FT5K7q3v21tOuz1PuLNS5HaOXrWl+xxlDUzOY3Eb3UgD8e30nkJq2acOk0kcS54Vx+fQPvvXpTNfFnvug2TWUeW7f7ly+Gy20KfF1TRVdJBHXq9VDFFBb63+nPuwkT8QyetFCUIGeNOts/4Nwbq2v8Ovj3jN00tW+4KPr+jp8nFUamqIZY5XKLOzc6gDbn3qOKOQkV4U/y9OPJIgBp/qx/s9Gxhz8ZQNPEYWsLhuP9uD9Pe2tCMqa9eFyK0YdOEWUo5Rfyqv8AgT/t/bTW8g4CvV1njOK9ShU05NhNGT/TUL+2/Df06c8RPXrhVVtLR0s9ZUzJFTU8Mk80pPpSKJdUjm39B72kUkjiNBViaAfM9eaRFXUxx1Tp8m/k51Du3tzDdf0e7cXVTU2RoklhQSFhURsqspubE3v+Pc+8nco73YbHJuMsLDUpp9nUa75ve33O5JapIMEftHVu21ZaWbbO35KF1kpHw+ONO6/paL7RApHuCL5ZFvJVlFG1tX9p6kiBlaFCnCg6f/aXp3r3v3Xuve/de697917qvv5+TNHtDakamwlyrg/0NmTj2Bud2paxD1PUX+57ldtiA82/zdGR+OMQh6h2sgGm1O3H+uF9n/Ly6dqiHQs5TTRsUC/Loc/Z10JOve/de697917qs75PUMWH31h8kVIM1Ssl+BcmU88e/aWGD14EHIOOj09YZaCr2Pg53mS5pbWJ5ULzyPr+fd1idh2jptpUU9xp0sHzdEhYeRSR/Q8n2+LWQ06a+pXqFLn2BAhpXe9rEAm9+Ba3txbRaVZh1T6ksaKOohnzFYCIvJBqII1KbAfX8+3NNvHxz023js1B1Ijw9bJY1VQGNubk3vfkce6G5iX4R1c27k4NB1OXBUJUCaMyMCDe9uR9PbRu5a9poOnVtoxk8enSGCKnQRxIFUf7f/Yn2nd2c1bp5VCii9Zvderde9+691737r3Xvfuvde9+691737r3Sb3hg4dzbXzm36hBLBl8fPQyxkKweOcaWWzce2LqBbm3eB8hwQfz6SX1uLuzktjwdSP29VVfDuuyHV/yU7S6iyFV9ntWjgmfAUsrt41qmmDtoVfSCRccD3GPKbybbzHc7U/bEAdI+df2dQj7fPLsnOF/sEp0wLmMfM5Py6t59yr1PPXvfuvde9+691weRI11SMFX+pNvewpbA60WC8ekzWZp5nNNQqzN9DIvPB/1va6O2CDXL0klnLdsfVTv80ba+A6+6Pzva+3MDM/Z+K+wOIz5SKaCgaYMkjFHBN9XPtp5qYTGetiGne+cdUy7Sy/fuzOuNt9i/Jvfh3r0vvfGplMtj4pZIYMXHKbRR1UkNtDAXsPac5NR0+oZVGuh/LoVPhf3thdpfMfq/aHx73Gud6r7Eq4ps/i8ZWVFWtFDPUD/ACaqubakuPr70R+I9eUhW0rwIJ62xqivo6WRIqiojhkk5RXJBYf1HvXV6jqUrBgGUggi4I+hHv3W+u/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//V3+PfuvdU/wD85f8A7J92T/4kKj/60r79ivWmroPRwPg1f/Zeto3Fv2Iv/cSL37zP29bHwj7Ojge/de697917r3v3Xuve/de697917r3v3Xuve/de6pH/AOFF9/8AhnH5oWLKf7j4UhlZlYW3bQfQrY+6tw6unE/YeqQf+EiPUfXO6fjN2nVZ3a9LkavIU+Ojq6isihmEiVVfK04h8ikgtYX/AMP8fbjijCnoOk8PdGxb+Nv5Gg6JH/OP2Htv4h/z8vgkOiKSr2dTZyl2vuXL4+mq5FgyGUq83MlTI8VMVBV1VQoK8Ee9VJx1agHeooTj/V+3pJf8KUeuZPlH/NO+C3WWW8scvZfQ+Kqcn45TTvd5Z6ho1P0BYwckC9jb3v8AH1qreCPUk9Bp8Gvn3H8QP5dvzx+Me4q5cfueLs6l2ZsqmqZKp6lcVT1UlHI1M6uBpa63sLAD3s9z6vs6aWiQeEOIr+3y6Iz8I+hK34+fzq/hvS1Zkan7ZfB9mUyvJqKru2hqqycKy8lWliLAN/U8/T3puOOrxagml/If5OrFv+FAvaeK6H/4UOdQdrZnbbbxxmB6n6uln24rkPWFsFXUSooANrNIG+n4J/HveTIadeNBApbgOgj/AJr/AM18l/Mu6Z6z+OPS/wAQ8ptbdcG9ds5NN1YZZpjJSxO1I0c8QjW49XJuf6gXHvxRlFXx1RZonekVSfn5dHi/ni/Hrdnx5/kHfGLYfYNdLLuJex9hZr7Wpmkeeg8G3hTti5iT+rS9yP6/4+6E1oen41Kqw+zP+XoIv5ff8zDf+xfjzsLZVB8Mty57HBtr445k0dRIksMcAhNSjCFvQ49Quf8AX93YEmvDpmNgqgUJpio4dWzfzjfkturGfyvuncrtXZtd1m/Ym9a3bO4sFplpZHx2VoUNR5BAEJVdRX1D35PiAOa9bmLCGRhxAFP8HR4P5EXwe+PW1vgft6pXaVHmMhu3O5bK5nIVZjqmkevpoJpKaOSQMQAWYsL3ufdWFHPV4gDClcmn7erDetf5cvxM6f8AkFke5uv8OuA7ByFIwqaBaxHjRZV9UkEI06QwJ4IJ/wBf344FergipFOrHEUIioPoqqo/1lFh711vrl7917r3v3Xuve/de697917r3v3Xuve/de697917qiP+a4D/AKXekyGAH263FvqPv2/Pva/F1V/hp1cT0l/zKXYP/huUX+9H3UcOrnj0KfvfWuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917oO+yewcb17gZ8pWyIJtBNNEwJ8hBsQAvPv3XuPVcmCx+4u/t9vW1CzDEJUOyoWYIiCXX6g9xbke/D0618+rNto7Uxu0MPS4rHxIghjUSSBVDO9gCbgDjj34db+zpU+/de697917rpv0t/rH/AHr34cevdao3yr7FxGB/mG7A33mpNcOzsmkQjMkwU6JCFugYA88+81eT9qmuPbKfbYMGda+XWPe+XsUXOkV5N/oRIAz59Hi278iOqabeFV3x3Lu7GrmMLkT/AHMwleXMs9FUVAWIQxyE8KCLH2ALzlfeWsF5a2GFvDkX9Vx5EDz6FVvvFgkx3fcpBqQ9gPofTq4zqnsvD9rbUod24IocbkIUnpmQkgxyXtyfz/X3AW+bNPsd61jc/GpoepL26/i3G3FzD8J6Ez2TdL+mmvxFNWgvoCT8FZRxpP8AsPb8c7pjy6YkhDA6cH16ZGqMjhmEbo9XDx6luVVQPpc+1QEVyfQ9MEyQ4HT/AEeUpaxQFkVZTw0ZPIP9PaR4JEzTHr0pSZW+R6TnYG/dudbbWy27N0ZGDG4zFUNVWSTVBIV/tovJ4xb8nj2z071q3/Nv5jfDH5cT0u3+19k0FDjpM9Jt/bXZc1ZNDRR5xplp4JWWUEFk0rxx7tUjj00ShOnjX16nbA6y+X/wSqNnbn6o3bX93dWbiq8dUY/b+1JKnI0lFjauPVRvUxR6gumNv6W49+qD8uvBWTA7h/g62Zund6Zjf+wcHufP4KbbuXrqSA1uNqIzHNFUGnR5iyNyLsTx7r070KPv3Xuve/de697917r3v3Xuve/de697917rXd/mt9A98bHwee+S+xe6Ydr4naRp6p8AtXVU9ZVNNIw0wIhAbkgEA+9ivpUdVbAqDQ8eqZMJP8oPmD1g9f3v0jv3s3AJQCXa2eFHViGspIiyLJTVciHWoCt9GNre7MVU0U9NJ4rpWVR8ujBfymdifHncPyAg2ju7pnIYPcu1cn9hhIq+qLGnnWrCoKimnGl9LC5Fvp70wOKnq0ZAJAGfPrcypaSnoqaGjpoY4aaCMRQwxoqRoi/RVRQAB/rD3UY4dOnPHrFNjaOo/wA7CG/PHp/3r24szrwPTbQxt8Q6hTYKjZQIk8TC/ILfkW9vJdyDDZ6aNsn4cdNv936oG61YWxFuCfofofb31cfmvVfpjShPRSfnTkN+bZ+Ou6ajY81Q2aWjyaM9L5A4p5KMk/o5sLH2OfbWLbrzmqJdyA8Oq8fUHoM82m9ttjkNmavn9hHWhE24+yZ+267L1NVVNvBN4qsMhrKwyeIS3sVY/W/vo6Lba12RYUUeB4WcDj1hV9VvDcxSSTEeP4wCjU1NFPThXre6/l3bm7Bzfxm29X77jrHzUEFDDSip8jySUmmyldfNre+dnura7Xb83SR7dTwzUmnr1mjyXNfybAj3tdeB+XR3HzVcov8AYTXv9NBvb+vPuPBbQk/EOhQZpR5dcVz9d+cZMefra3vxtYqV19a+olHlXrIc7VDj+Hy3/Nv+N+/fSR/x9bW5cipFOuv49VA/8AJCOPp719IlPi68LhtXVef8wHOVJ2516v2Mq/c5ojSb8kzKtv8AeB7j7nuBVhgUN+P/ADdRb7qTstja4rVx/h/2Ojb9JV1ZR9VbZUUEpkMaoQB+lWiVi/8AxH19inY7eL6GNdWKdDrl55E2KEgZwP5dCj/Fso17UUq/gHSf6fW3s6NvAPxDo5WSVhU165LWZRwB45VLccrb6n3URwcajrRlmrSh68Isu7KGkfSfoCfoCLXsfdtVuOA60BMw+zolPy5wNTow+SkYv9ukTahxpsxBJt/j7STurnsx0oijZFoeht+Plsrsig1zaxHT2sCeCxC+3EudCBAOHVJbcStny6HpcDjhy8OtvySfz/h7qbqWtQadXEKAZz05RUtPAFEcarpFgbXI/wBifbRdm4nj04EUcB1I906t1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3VRXzNoKjpzuHrLszDxMx3bumGlzMkQ0+KAVCh/L4rEizD68e4q5wDbRu1tuEAr4zgMR5Z/zdQN7iI/L3MFjvcH+jyhXI9CfP7OrX8FmKPPYqhytBKs9LWU8M0ci30nXGHIF/wCl7e5QhlWaMSIaggHqcra4juoFniNVYA9OpIUEk2A+pPt0CuB0/wAOmWqzMMReOBfNIOAAeCSbce1UdqzDU+B0nknVcLnptWmyOVGuSRoIwf0Pfkf4W9vM8UFVUVPTKiSbLY6fqTH09IAUUGQCxc/U+0jzSSYY46UpEqZHHoOO5uqdvdybDzGyNx0y1FDkoWOkrGT5Y4m8P+cBFtRHtr5dOEefWov3d8TPlDvDfOZ6G3HuSv250HU5NaZqqvYxYXHYync6GeYAILJzZT7tVRw49UKyMafh9fPpY9TVPxs/l1vPSdK02M+QfbdIwpKCXbtTLVSY+pc+lowgLalfj0+9GrGp60AsI0Dj5evVg/xMpfmv8j+0cb21v2u3BsDZgqBWz7UzX3lP54HqrPHDHPa4Avz78aHhx6sNQyw49X/UkLU9NBA7a2ijVGfn1Efnn3rq/Uj37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/W3+PfuvdU/wD85e/+y+bK/wDEhUf/AFpX3rzHVX+A9HB+DYt8e9ogHV+xFz/1Rxe98SetgUUfZ0b/AN+631737r3Xvfuvde9+691737r3Xvfuvde9+691Tr/P12VnOw/5TPy92jtyklrcxltlYtKOmh/zsrQbmo6hwnB/soT9Pej1ZTSp+R/wdabn8gT+Zjuf+Xx1B2dsXffT1ZUVVVSJHhf4gaqmmrKmmqGlpXAj/sXYqyfm1wfbxjdjnyFOkKXMESkVqCxb9v8As9D30n1D8hv50H81zpD5e7x2Lk+u9gfH3ceMhyMdbDUvS5fC43KPU04o56lTpQhyOTzwLj3R6D7enomZ2rTsoaH59Dz/ADuus975L+fx/L/y21trVtTtPA9d7fxUuQp0vRUKfdVsUkEhsbELIlvre5va3Oskk9XJCqqfb/g6pZ/mt/y++ycf/M96h6f2rt/MUu3+2o9u7xytTRNKMa9dWZNqnIS1vhUJ5AiIEDgkljb8+3OOBwHTBAHdxZ+A+zj/AC6s0+S/Re8dj/z/AH+WNhMVs+vXA7P6F6fxmQycUJNIlXQYqviyZqJY1Cq6tNCACbtqNvofbYH8ulDOuB5sCQOlr/Ne6Kqu1/8AhTn8b6TeOwMhurrHIdb9awZXxhloZo6PbtYlUj1LKVVkmlisv1N7j6H3uncSOHTZIEaIeOcH5dblXU3wW+KvWcmC3LsTqTD4PK0lHSSUdXKDUVkBCiVfL5LoXDHkhbX+g91qeNenSARQgda+v/Cv7Ym79+fATp7E7NwVVnKun76xU1VR0KnXHQx0OjXoAtpDab/0H+29+FajrUjBY2J4U6s3/lC9D7EPwn6vXcmyKWPLjDbekqvvIYWkE0GHiK6CqAjS5a9/z72xJPWo1AQADpY/zdPhPF8qvh7uLZe18fCuW2Di8/ufbVDAI45JMhHj7ww0w0m7lluLD35Tpz6dekTxEMfk2OtWH+WN/Nr74/l3fH3evxa7l6hyGV3hgtyblym3ctmDkoarTpWmp6GOL0ggiNNNxwfp7uUZ6sOHSfx0hAhkqCBThx6Mv/Kn7e/mEfNf+YnufvffKbx2J0bk6OpWm23knyS4amZKcyQPGs3oOq3Fh+fdG04rk9ORNK1SRRRSnqfXrdwgVkhhVzqdYo1ZueWCAMef8fevt6f+zrL7917r3v3Xuve/de697917r3v3Xuve/de697917qiH+a4QO3+k/wAf5Mn14H/A9/e1+KnWn+Dq4vpH/mUuwP8Aw3KL/ej7qOHVjx6FP3vrXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdQ8hVfY0VVV6DJ9vC8ugfVyo4X/AGPv3Xuqv+yMzuvuPfo21HDUQ0NLVePSC6rEolJBN7j6e/VxXrWK9H46q63x/XuBgpIokNe8Y+5nGkk3UXGoD+o9+6sehU9+611737r3XvfuvdcW5Vv+Cn/evexx691pr/J34w96/IT5e5+baeDysWMx+5Hhaug8ghCCYxmRjY8fn3nnypzZy9y1yVEL2RdTRjB+zrGHmHlveN95oMlqGVI3yQePEdWYdLfyg6/JJict3xu1N0U9MIpKTEfcSmWlSKcSJEzBCt7C3uJuYPfGCAvBy7D4RPFqYOOPHof7X7bl1WTdpPEp+E+VDjq8Lr3r3bPWW2MZtLalCKDEYunWnp4dWohF55bi/wBf6e8dt13W83m8e+vW1O5qepUs7OCxgW3txRVHS49lvSvr3v3XuuLKrqUYBlYWIP0I97BINR1ogEUPQDdu9t9TdJY9Mzv7ddDtKCWRVjmqUaXyOxFgFUg35H59qku2ppfI6TNbAvqTj1VB/NU7Lyfb3xCwG7OjctPunb1Fu6Sp3fX4pZYov7tRIpqfuliYkR6L3vx78yRNV1IHDH5Z60zyIugivz/Py6oz61yPw0+de9cD8Zdr4DEbTwPXkEW+Mvu01biF9w4eEVWZ+4lkPOpkYWHN/acq47unFkSQlR+GnVv38uXtfemc+b28NhbK3LLuD48YPr+TFYpqZZGxIr8NQiKnaNpLgPrT6ggn/W90wB8+nVYkn0x+3z62JwAosoAH9AAB/vHv3Vuu/fuvde9+691737r3Xvfuvde9+691737r3Ven8y34+bo+RHxh35s3Zzytnp6Omko6WH/Oz/bTmVwnB5t9PewSM+nTbJqr8+qfvjz/ADNe4PjB1Dtj4+796Jr4KzqjC/3ZoctX01VE+eiWaT7ciNNKHSWJ1AeofW/v1KnHn1tWx3gjoVf5Z/UG6u2O+N6/I/cWya3Y9Mc5Lm8elTGYIckZa3Xopl0C/wBeLe9MfwjraA11nFcdbG/v3Vuve/de697917r3v3XumnN4TG7ixlXiMtTJV0NdBLTVELqrBo5ozG9tQIvY/wBPai2uZrSZZ4G0spBB+zpqWGOdDHIKg9VsVf8AKz+PtR2ON/Jhokf78ZBqW0PqmEmu/wDmrf7C/uWk95eZF2v93a/w6a54ft6BJ5C2c3v1mgVrXgOP206si29t7E7WxFDg8LSx0eOx9NFS00MaqoWKFNCX0gC/9ePcTXd3PfTtc3LanYkk/b0NYII7eMRRCijp79punuve/daoOurD+nv1T17SOugiDkKB/sPeyzHietBVHAdVwfzBZV+z6vgty+41I4H5mVR7j3nqQ6bZfVuol9020xWSDzlH+Ho7XU8Xh2HgI7abUkXH/TpfYy2xdNkg+X+TqSNmULt0YHoP8HQj+1/Rp1737r3XvfuvdFg+UeINdsWrq1HNLD/Q/wBWb36mK9a4HpI/D3LGt2zXUjNc0q8A/UDzWt7951HW/KnRzffuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdFX+XnWlJ2B09uqp8YfMbcw1bksG1hrSujAKlGIJBPFrew3zVt63u0StSrxqSn29ArnzZo925fnqKyRIWT5MOHQR/y/u2E3R01i9mZmqMu9tprXx5ilkLmZYkkvAzaySf6ey7kO9a42dLS5P60ddX2Vx0Qe1O9vuHLcdhcn/GbcESDNag9HTEmTypIXXTxuPSfoOD+fcjgQQCpyepG/VnFOFOnjH4eGkGuS00/5kNzz9fz7TzXLSYXA6djt1Q1OT08+0vSnr3v3Xuve/de6Il81/jJ2Z8k9oPtDYu/oNkUtUIfu5ZI9Ik0XSUM8CNIdSm3vw4560cj06Db4nfyv+hvjlSYzP5TBUm7Oz4FR6/dc0s8kEtSjiXzwU0yrY6h/aB4/Hv2T1oKBgf7PViNXuvbmMq4sbVZGmpqprCOnJAI/oAF4Hv3W6imOlGrK6q6EMrKGUjkEEXBB9+631y9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//X3+PfuvdU+/zmCf8AZftkAG1+w6P/AGJ8Kke/efWm+E9HH+D8Dw/HvZwa/qpomW9uR9pEPx795nrfkB6dG79+691737r3Xvfuvde9+691737r3Xvfuvde9+690md4bP25v3bmU2luzGQZnb2Zg+2yeNqhqgq4NQfxyAfi4Hv3XgadEyr/AOWX8I8nPHUVfRO13mi0hGT7mK2htS8RuL2P9ffs+vW8eQH7OjSdX9NdbdM4ibBda7Wx21sVUOkk1LQR6Vd0BClna7H6/k+/da6ad4/H7qLf29sJ2Lu3ZeLzO8tuRpFhc5VRlqugjQ3RYW/wv791uuKdQ93fHDpffe/MB2ZuvYmHzG99sU0dJg89Uwhqugp4gBHHE39FsLX+nv2fXrQoBSnUjcHx56e3T2Rgu287snFZDsHbVLFRYTck0V63H00H+aigI4AW3H9PfuvcBTrrcfx46d3Z2Xie39wbJxWS7FwdNHR4vc08WqvpKeFdMccb/wBAPp791uuKdDQiLGixoAqIoVVH0CgWAHv3Wug17R6d647owdPtvsva+P3VhaWsWvgoMiheGOrT9MygEci3v3XulLtHZu29i4am2/tXF0+Hw9HGkVNRUw0xRRxroRVH9AOPegKde6UxAYFWAZWBBBAIIIsQQfe+vdE+7I+BPxQ7b3SN6b/6h2/n9y+UzHJzCaGVpC2sl1p2VTz/AFHv2evCg8h0Yfr7rTZHVuDj25sPb9Dt3DRafHRUUYRF0roX1fqPA/JPv3XqCtel37917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuqJf5rkf/GWelJWtp+3QAm3/ADsGB97HHqr/AA46uH6Rt/ol2Bb6f3corf7Y+6jh1c8ehT97611737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XCSNJY3ikUMjqVZT9Cp+oPv3XukfQ7B21jctNmqKhWGtnJaRxpIJP1tcX/r+ffsU6159LP37rfXvfuvde9+691737r3XvfuvdJ/E7WwODqq2txeNpqSqyMhmrJ441Ek0hNyzPa/8AvPtXPfXVyixzuWVBQCvAdMx28MTF41ALcelB7SdPde9+691737r3XvfuvdInsTfGG652ZuTeOcq4qSiwOEyuVLTXtK+PoXq1hUD6lilgLi9/fuvdaonXPXfcf85vvff3bS73m2X0VtGpyO2ZMMz1Yx0WVghMFFJT00bPqkbx6z/hck8e7ZUY4npmglOahRX8z59N3wwk378eP5pO4/5a25s43cHTG4tj5WTMZD96TFCCrwT10lOdbEI0YPjkA5BBAN+ffm4ahx62g0kx1qoAIr5Zp0bzsL/hP1tHGdgZPenxn7Apum4M5XVsuXxt8jUNU0dfIWqafzQRk2YErYm1vew5GOtmJTU1IJ9OHVr/AMQPiFtH4bdfwbXwlLFm87O/nyu5Yw2qqdowk/8AnFVwGJJsfahUSfNdJHl0wC1v2cR69HSpMxR1K38qo4/UhJuPdHtpEOBUdOJcI3HHTmrq6hkYMp+hHtggjB6f65e9de697917r3v3Xuve/de697917rXX+b/zg+ReX78wfTnxw2lmcvJQV4xuZrsJHNJLTVImaN/KI2Isf6ke/D1PVSSTpUVPRZd3d6fIfZm4KWn7q+G+a7EenljSXceUpKuLx6pAVk1ItjYEnn34AevXjqXiv7KdbF/xX3zBvvqbbuUp9ojZETUgK7fCBPsQTbx8AX/1z78DXq1PTh0ZP37r3Xvfuvde9+691737r3TJn9xYfbFD/Ec3WxUNGZUgE0pAUyyfoXn+vvwFetE06ktkoJMVJlaN1qYPs5KyBkIKyosRkWxH9be/db6Jv1r8pslvnvvIdQy4AUtJR0NZV/xPxuCzU0LOE1k25K+9nqq0z0dr3rq3Xvfuvde9+691737r3VYf8wKqDZPqSl/7OaFbf6q9SLe4457cGW0T+n/l6hr3SYmawQf79H+Hqw/Y0Ig2th4wukCipjp/pqp0Pse2aaLZF+Q/wdSvto02UY+Q/wAHSt9qul3Xvfuvde9+690Ffc+PTJdfZunZNRMYI/rwCPr/ALH37y61TPRQvifl0xObzGFUAGR3hYXN+ZuP9559+B63xI6sR9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XCSRIl1SMEX+pPvYBY0HWiQBU9NNVmqaG6Qss0tiQoJ/HtTHau2WwOk73KjCZ6YK2jq9z4+sxtXG9PSVsYhk1AWMRYFrXv7vNHAq6G7q8emHje6haF+DY6qp6SxqfHv5edi0eTu+J3tMcfigWuomknBUoOFHIF7D3D+2u2zc3XPiGkc/ao/PHUJ8uR/wBU+fr2OX4LwgL9vy6uMRVRQqgKoHAH49yoSSanqewABjrl711vr3v3Xuve/de697917r3v3XuqjP5q3zT3Z8Y+razHdb4SbPb7yKQ/Y0NKspqWiqqdxrj8TA+kkH3ug6qWIrjrW56bqPnJ2r2ttvNdh9l7n2FUbspRuTE7dydRXRGSkh/fMUEcjAAECw97OgYXptTMT30H2dbJX8rT5Ybi74252Dsnegf+N9aZw4OGtnDiXJpDUvTPJqdmubqDx7104DXI+zq2/wB66t1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//Q3+PfuvdV5/zJ+iNyd89CPhtqQy1Wb21kJdyUVHCW8lbPSQDx0qBQblj9B795168RUEdVYfH75D/OfrTrCHZr/H/cLnF5SWCCWWkrld6anXwrIDYcNYG/vZ9R1UBqd46F/wD2cP503sfj7nf8L0tfbj+gt79kefXqA+XXH/ZxfnT+fj5nb/8AULXH3qp9T16lfLrmPmJ86ACT8e86SeP+AtcSCPqT79mlamnXgPkOuj8xPnVpF/j5n/8Azlr7f7x79njXrwBJwOvD5i/OgXB+PmeNx/yq1xt79k5B63+XXR+YnzoJU/7LznWsbkmlrrj36p9evAU8uuX+zifOg8/7L7nOR6VNJX8f7ce9AnyJ60y1FaDrivzE+dJLf84+51QOCPta7n+v0Huw1Hz69pAA7ePXL/ZxfnTyF+PWePNr/a1wt/t/fs8K9aA08R1x/wBnF+dX5+Pme/8AOavP+P496yOJ62B8uu/9nE+dIIv8fM6wP4FLXn/eD72K+R69Typ14/MT50m9vj3nQv5vS13B9+BYnj1qn9HrofMX51D6/HzPAc2H2td9PoD79kmletkV4Drx+Ynzo/7x7zv+xpa76+/V+fXqfLrl/s4vzoBufj5nWFrf8Ba8/wC8H3rNKg9b86U64/7OH85x+n495y1vxSVwF/6/T3sEngT1qnkR17/ZxPnRpF/j5nSb/wDKrX/T3qvoevU+XXf+zh/Ohjc/HjOi34+0rf8Ab+91J869e0+gA68PmH86b/8AZPmdH0v/AJJX/wC2HvVfn16grw67b5ifOe9x8e87e31FJXA8D+vvdG9evefDrj/s4nzo/wC8fM7c8f8AAWu5/wBhb36p9T16ny67PzF+dI9I+PWd/wAT9tW8j+nvR1A8evcR8PXv9nF+c4Fh8es7e3NqWu5/23v1Wpk9ep6Dr3+zifOm3/ZPmdH+0/a13P8AhwPfiT5nPXgBTh10vzE+dP0/2XzPAf0+1ruB72NR8+veXDrkfmJ857G/x7zrXHH+S15sAObg+/DV5H7evU9R1wHzD+dFrD4950C3/KrXfT/Ye9d3qevUA8uPXv8AZxPnSAT/ALL3nfz9KWu/p/T36tfM9bAofh6LF2NivmL8wPkz0hJvDqTP7Q2nhDRUeYybU9XFR00MdUDLNM8ikfQk8+9ilK+fVGV9QHl9vWzts/b0e09sYTbcUvnjw1BFQpNyPIsV7MQf9f3r7enOlL7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xui5fKjpWv786c3T1zjM0cDV5fH1sUVaNfLy0bwrExj5AJbk+/daIr1qebc3F8zf5MO2uzevtt9U5Lf/XO86zOZWXdU1JWz4uGskpnWOsgmVQA0YkJAJ93w3nnpg1jXgSo/b+fRkP5E+7+re7t9bs+SHZO44I/kFUZXIYWixNYviq1oMtFJDKitPY6UViv19+YUNPT+fVon1DWM6uPy62vkZXVXQhldQysDcFSLggj3Tp7rzKrqVYAqRYg/ke9gkGo60QCKHpkq8DTTL/k1qV78sgPIvf8AHtTHdOoo2R0ne2Umq46b/tsljARCZKlF9QA5B/1x7fDwTfFRem2E0ZqM9SKfcEfArY/tiTbn8H6c39ttaV/sjXqy3FDR+nyKrp5gDHKjBvpzY+0zROvEdPiRG4HqT7b6c697917ppzmYocBjKvLZGYU9HRxPLNKRcKApIuPfqgcevdacXbPa/wAjehvl9n+3+j9n5PfmHrdyPk68UkTTUyxyVTM6TqCbCw92FCADjpttQfWBUdWndEfzX4+2Ny4rr/vPraj21VbinFPFi5acmzM6xRoDJqINyTc+609etqwJoa/n1eFtfH4CixVNJt2jho8dUxLJDHEulfGeRYXP+9+/Dq/z6Ufv3Xuve/de697917r3v3Xuqlf5zOK7GyfxErJOsc3W4PcGL3PQZaSegeVJ56Oip5Hlprw+qzG3uy0rnqrkhDp40x1Wr8Lf5w+R2h1VBs7u3b0dLkdvCkw4yNbBVPNWQ0sPiqZp11erWB+eOfeyuajptJBp78Hq3L4h/K745/JHdM2S68wGNo97CmlapyFJDJDI8JiYygq/+0g/n3Qj1HTqsGHaerIvfut9e9+691737r3XvfuvdVTfPqcyb06cpAb6t3UwJ/qDVLZePcYc9P8A41Zj+n/l6hP3PNdx25PWZR/Pq0TCQrT4jGRqNIWgo7j+jfbLq+v+PuS4hSNfsH+DqZrdAkCKPID/AAdOvtzp7r3v3Xuve/de6Te7aUVu38lTldWuBrD/ABt9ffqVx17qtjpmpOE7prqJjpSbIlCD/ZvMBcXt70fIdepjq0v3vr3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691EmrqaBS0kqgC/0N/p7dWGR+A6aaZF4npinzs0hK0FMZ19QEgB+o44HtStsgFXP5dMPOSdK9Y0x+QrxeqkeIXvpbV+fwB7u0sEWEFeqLHNIe446dKTDUlOqmRBNKPrI31/2HtM9zI2AaDp9LdF456dwAoAAAA4AH09pyScnp8AAUHVVX8wfaOTwuc6v7Q27HJDHtvODI7hnjv/wGhnEheQIBcW/qfcZc+Wckc9ruNvgRtVz8geoT91rC4t7ix3yz/wCI8mqT/S1z1YX1X2DjuxevcDviknjNDlaNZhMSqJeMCOUnkgeoH8+x7tt/HuFil7Ge1h/q/n1K2y7pFuu1Rbkh7ZFrXoRYJ4amNZoJEljb9Loysp/2Kkj2vBDCo4Ho1V1calNR1m976t1737r3Xvfuvde9+691Uv8AzNPjLuftHaydibIoZstubby0yU+Lpixnmjp4yxKoFN/pb3sEDJ6qwLDT1Udv/Yvyd+VGzdutg9j5Xr3f/V2JptvUWRpkmpqyooKePx1coOkHhQWY39+BA61TUvbg9GT+A29tofEPdcHSVdX027+4OwnparLSPG01TTZNqnyToZV/t/qBvzfn3ule7rw7BorkdbHdBPLU0VLUTxeCaaCOSWE/7rkZbsnP9D7r1fqX7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//R3+Pfuvde9+691737r3XVh/Qf7b36p6916w/oP9t7917rv37rVB11Yf0Hv3W+vWH9B7917r1h/Qe/de69Yf0Hv3XuvWH9B7917rv37rVB11Yf0H+29+6313Yf09+69Qdet7917rqw/oPfuvdesP6D37r3XrD+g/23v3XuvWH9B7917r1h/Qf7Ye/de679+6911Yf0H+29+r17ru3v3XqDrqw/oPfuvdesP6D/AG3v1evdesP6D37r3XrD+g9+6916w/oPfuvdd2Hv3XqDrqw/oPfuvdd2Hv3XqDr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6R2+ev9ndlYCq2vvnAUO5MBWKy1ONyCM9PIHQxsSEIN7E8g+/de6pE7i/kvYil7Lo+z/jNv1+m4aXNU2Uq9rYv75YqhVlDzCGeO/wBTf0sfp+T73qxTqhQFg1aH/D1eDsfHV+H2jtzEZOeSqr8VhMZjqyrlJL1dTR0iwTVTX/LsCxv715dW6Vfv3W+ve/de697917qFNjqKouZaeNmP9q3IP9R7cWWRDVT02YozxHTDLgZo28tNUsApuIgDz/h7Vrdqw0uOk5tytSvn/LrgarMUdrwPKn5LAkAL/Qn3bRbyeeeqAzJ28adTINwQcLVgQseB9Tcn6ce2TaSZKjA6dW5Xg3HpJ9m0m3907Rym2MtnkwUGapTCMgWRPBq5ViZCB/vPtMyFfi6fDqwqD1RBv3+Up21SV2e3j1X8nMpm6bKytULtmmqWjV4xqmZI5I2dGFhawN/8Pfqk+XWioHmf9noq38u7+O5T5UZbrfvLpGmyVXtPPPj8ZuzJeT7oSRVYj+6DfQ/TV78evLqBII62z6amgo6eKlpYlhp4EEcUSXCRoPoq3916c6z+/de697917r3v3Xuve/de6IB/MK+T/X3xu6WyGe3fhMfuyesnfH0O3K9BJHVVUtMWiRwxFgxKrce9jJp1okAEny618/j38oNndt9r1e1O1fg3tChircfV7joIQlXTSVuHEJqVrwbg6XQAkj3sr8+qLJUldJFM5oeryP5eO8vir2NhMtuHpnqrbnWW6cVlazEZHH4syvO6xGRS6s7ngAMrAj/Y8+6njnq6kkVHVofv3W+ve/de697917r3v3XuqhvnrVAdp9J0wa2ve1Gtxf6GsUW59xTz2a7jZKM/qD/D1BHua9N82xCeM64/Pq3GkXRS0yf6inhX/kmMD3KiCigfLqdUFEA+Q6ke7dW697917r3v3XusU0SzRSRN9JEZD/yEun3sGnXuqp6txt3vOZrlC2RBA/rebmx/w+vv3z60GHVpeIqmrcbR1LG7SwhmP9Tex9649b4dOXv3Xuve/de697917r3v3XuvEgcnge/derTj1CmyFJBfySgW+tufbixO/AdNtKi8emSfcOq6UUXma4AN+eeOPaoWhUan6TNck4QdY4/4tWAalkgDX9V7Dnm1/diYIuGevHxnwMDrNT4Aqb1NQ0qn9SG9iPyOfdGu/wCAU6uLfUQzdP0FLT0yhYYljA/1PtIzsxqx6fEaA1A6ke69X697917rg7rGpZjYC1/9ibfj37PAdaYhRU9Vq9yfLX499qVnafxfl3PTJ2CmJnxiUUiRRPBVyNo1JNK1+Db9I9x9u/Mmy7pHc7AzMJDWMErRdY4gMcVHmBnqHeYfcDk7eLq+5EkuoxfeH/ZahrAbAala0+dPTqrvtr5eZ745fF/ePRWJyLUm5NoYeojxGSjMUkz1FbU+OEq2oE8kHj2BU3uWw25eUI3IkLBQw40JzQ/IVPy6gbnT3Nm9vvbu72G3l0XNpCRE5oaux0x4JFe4jq1r+VrvLe/YXw+603lvurqK7L5mCslarqCRJURpIFSUqfwebe5M5JaZtkAlYvpkkVWPEqrECp8+HU9exG57xvXtlte7742ue4hV2alNVQDWnlXj6enVi/sX9TF1737r3Xvfuvde9+6910QCCDyCLEH6W/N/fuvdUhfzWvnvjfhfsqrwHX2Ihg37nHTzVdPT+Sb7bJUxWd1YciyvyL+9gFjTpuR1Ranz6rY+DvyD+JnS8Q777s3dHvXujc5p8/hUr6KZ6jbs1ZJ5JKYPJYXFza3097KngOvKw+In7etkH4vfKDaPyc2zX7m2pLG1LSzIoVRobQ7MoJUsx/Huvy6uCONejSe/db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//9Lf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690Uv5I/NHo74uRUB7Q3E2PnyYvSQ0ccdZJ+op+7HG2peRbke/UPWiQK5pT16c/j58vOlvkpSyTda7jGRmhRnlp6lYqWb0gFhHCz6m+t/0+/fb14EHINegk3H8uMvh/kFg+n4cSJaPJ5dcfNWCmuY1aYRhvKBb6fm/vfEV63gNp8+j+e9de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de66IBBBFwfqD79w69SvHqDNjKKceunj1fhgPUD/AK/t5Z5V4HpowxnNOiG/OLoibtjYwosf2rP1rUwU0cMU1PVpTyTsmptQUurG/A492a4JFWHVDDQdpz1SrjtmfODo6jGP6f3BuDttV1JHVpWmZiJTocqmstwPba6CSxx1ZvEVRp7iPXo6/wDLZ+Lfde1d5b17l+QcOTxe489USZGhxWUaYvDUtOJSqrILDi4Fj7fMMbKBGak9MrJIg1S4pX/YHV3EWfoHLKz6Cpta1/r9Pfms5Rw6utyjfZ06R1dPKoZJVIIv9Rf/AFvbDROpoR06sqNwPWZXRv0sDb+h90II49XBB65e9db697917qi3+dD09u7P9W0HYO3NvjeceEy0FVV7fdJZIooKCBZGq2SMH8Kffhx49UfINOI/n1Q71T/Mj2fj/ljg917h2dQUKYLq6XYH8IXHlqdskMSaCOEhh+XNr+3Ch09NrMmrIp1df/Jw6z3xlZ95d4Zzbn9yMLl87kVxmAgjlhpa+myBmeKvSOUAWAINx7ofT06dThX162Bfeurde9+691737r3XvfuvdUxfPSpCd2dGxlrg79oRb8AGuW9/cQ89yf7t7JfPxB/h6x89z5AOZNoQcfqV/wAPVzUf6E/4Iv8AvXuXRw6yDHDrn7317r3v3Xuve/de697917qrHvTHnD9q46tIK+erR3k/6efn3onj14fMdWRbGnFTtTCzBtYejU6v+Qj78Mjr3A06VTMqi5IA92AJ4daLAceoslfSRgs0y2X6259uCGQ+XVDLGOJ6bZM/SfSnbyub6R/X/Hj28to9NT4HTL3Sr8Ir02Nl8rNIEjpNKk+lhf8A21vr7fW3t1WrHPTJmlc8MdZxSZOqF5XkivxpuQOfqfdRLDHwFerFJpOOOs8G3kR9c1Q89+WDD82+g90a8J+FadOC282NeniKgpISDHAisP7VufaZpZH+I9PiNFyB1L9t9X697917r3v3Xuve/de697917rplDCxFxcH/AGKm49+60QDx61/v5i/8rTK77zOV+Q/QuWqcL2bSynKVuOxpqop8xPHOZVKtBchtJ06h/rEFfcXcx8pywtNuO3r4ySktJDmpY8XjP4X+w5p5cesOPfX7usvM94eeuTJ2tN6hoVZSVDaTUBqZ4YDD7GDLjrVw7T3F3JubsTHbF7MpcjR7pgy0eBrY8hTPBWz1MtUKdDVqB6+bWYC1uR7CO3bdtdjb3G62jNK4GQ5qyMoyprkEHiD1zs5q/rnv+9jZub3ZtyhdYQjoEOt2ADMFoHHAqyihXIrXrfe+DeyB118WuotnhNBw224IWWzD1OxkY2fn8/n3K/Jts9ry7bxSmrEMx+1mLH+Z67P+2O1LsfIW17OooLa3jjH2KoHRs/Yo6HnXvfuvde9+691737r3XvfuvdE++RPwr6T+SVdDluydtY/K1VJGumoqhIGRIo9PLJ+AB791qnWuX/Mf7h+Jvw9zdH1D1L0FsrtTslKemjx1NkaSSrshg1HxlD/ZN7D6+9opOB5dNySrGKsK19OPTL8O/nt3v1ZiqHetX0HiOvut6qvxtFljj6Ux0tNPV1GhodMrcEckD6+9kUPr15G1LUinW2r1rvjHdi7J21vDGyrJBnsPQ5PSgsIzVw+Qxn/WNx9fdceXTvS69+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYjPCDpM0QYfgyID/tiffuvddfcQf8AHeH/AKmJ/wAV9+691354P+O0X/UxP+K+/de699xB/wAdov8AqYn/ABX37r3XX3EH088N/wCnkT/ivv3Xuu/PB/x2i/6mJ/xX36nXqjrr7iD/AI7w/wDUxP8AivvdD16o6788H180Vv6+RP8AivvXXuveeD6+aK39fIn/ABX36h69177iD/jtFx/zcT/ivv3Xqjr3ng+vmi/6mJ/xX37r3XvPB/x2i5+n7if8V9+6917zwf8AHaL/AKmJ/wAV9+6917zw/wDHaL/qYn/Fffuvdd+aH/jrH/1MX/ivv3Xq9deeD/jtF/1MT/ivv3XuvGeAfWaIf68if8V9+oevddGopx9Z4Rf+sqf8V97oevVHWVWVhdWDD+qkEf7ce9de679+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/09/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690Cvc/b+M6u2fnstDLS1W46KgabDYWeWON8nWFwkVPGrsC1+SdPPHv3WqjrVH+E3YG1/nB8+u/dt/LydKahwktbU7W23nU+zxQr0yAWKip5KjSAArMQAebe7kaQAPPplTrZtWKHh/l6G/wCYPxnl+CHyT6W7R6K3xkKbb+8Nx/dZLZOOk0UdFSw1a/tLCjtrhkQ2DH62PHuoNRpbj1dlCkOmPIj162ROvMTsfsjB7X7WrNl4aDcmWo4siK56NfvqeoDFC6ynkG6391pXj07Xob/e+tde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Wrd8mKbs/+YR8ucn0Zjezsp1LgeuczJh5Mnjqk0iVdPBVNH5pRCyl2N7D/Ye7KPPj1Rvi01I9af4OmT5A/Dzvj+WRsOt+RexfkFu7tPEbLanqK3E5aqlgZZKmdVCT0MksgZOOCCbj+nvQIJH29aeqqdNTQcD1sHfDnuXJ/Iz4ydRdybgoo6TK792xFma6kWNY0hqPupaZgqLYD/N3/wBj72GKtVcdXIBFDkdGNlxVDICPAik/2lFjx9Pbi3EoNa16aMEZ4CnTU23QZNa1kqAf2BfSf6XF/b4vPVemzaitQadYWxlfT6vDLJIb3vf/AGFuf8PbnjwsO4dUMUtcYp12JM1EtvCzkfjnn/Y+9AW7GpNOtfrKadeGUygv5KcKB9WseP8AYD3v6eClQetGaWtF6rk/mN/NXF/GnrSrx0e36Dde6cw8tBHtzIQxz0tTHUQAwF42uSGLEcD2n8BWHYerm4dFq4yPLrXApt/9ibdz1H3Rvn4C9e0O3Kyvp8t/EXwdakE2PdvuJMiARpuE9V/p739K4Woz1Y3S6u4Y/n1tUfBj5R9QfJfpej3V1VjqTA4vAvR4XMYWijiipsblmiKy08McX9kMpAJ9teBJ6dO+NGV1Vx0csZahN7Tcj6jSfdvppfTrQnjIrXrIMlRkXEwte17H6+9eBIPLrfjJ69dnIUgFzKP+N/W3vXgyDy62JUPDrmK2kOm9TCpY2VXlRWJP4Ck+6FGBoR1sSIcVHVKHzmmjyPyG6ZpEe5pN8UEh0m9rVi83HuHed4XbfLSnlIP8PWO3uT+vzftUY4rcKf2dXXmqpolVZJolcIh0F0D2I4Oi9/cxIjMBpHWRLSIg7j1FOXoQSPISR/tJ/wBtz7eFtITTqhnQdYHz+PS95DcX/sn6j3b6SXqguUPDrF/eGkZSYzqItxpYfU+7C0bi3DrTXFB2jPWFs3O66oYVIF/re/uwtUByemzcPUADok3yKxdKcni8rlj9o4MUsYtYuNVwf9497MFuGrq62ZZiQFGOjLdVV1TX7Oxi0bGSGOnsjA82Y8Hj/b+6g244dWZZiKjz6EJMTWSkmWodAT/Uk2H+HuxuY1wor1U27sQWPXKLbiIWL1UkoY3KsPpfmw59tm6J8unDbKTXp2ixlFCF008ZK/Ritz7aaeRuJ6cWFBxFepoVVtZQLfSw+n+t7aqTx6cCgcOuXvXW+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuq5/kX/Lb6T787G2b2dLi6Pb+59r5RMnVV1IJ0ly0iTedRU+Pg882bi/PsEbvyTZ7hdi7tW8AuQZQvCWhqNXzHr1DvOfsrylzhv1jzHcwrHdWL61dagnNaMQe5fPS1QDmlej97awcG28HjsJTW8GPp46eO17aY1Ci1/8AW9jG3hW3iEKcFx1LNpbJaW6W6cFFOn3290p697917r3v3Xuve/de697917qHkYXqMfXU8ZIeejqoUI+oeWBkUj/Yn37r3WqL3R1xsjoP53YzfXyI2VBuPZdSla65yuo6meajetpmFGImClfQSpHPvYrTqmA9fXoqHYOO70yHaa9EdbbVn3F0R2PuGl3nDuWpWZExlFLVfcQRRyOAFVFax97oOJPXjq/COPW178XcrsPZ2w9j9WY/c9LktzUm26E1lCldRztTz08ZSSnRYnLAi30IPuo4dXJz0bj37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdJLfmdk2zs7cuehGqbE4XIV8K2B1SU0BkVeeOT72OtH0612tnd9fMH5I9ubjfYW26yXa+PqshSrNSKyRJ9uSF1GLgfT3o9aFTwHQ5jZnzzUtba2XPLAW81rXsD71j59XAb5deGzvnsOP7rZf8A289r/wBePfvlnr1PMU66OzfnqLMNrZi9/wCs3I/x9+x6nrxr5067XZ3z1/O1suCfrzP/AF97NPn14ahwp147M+ev1/uvl/8AEXm/4j37Hz61Qk+XXhsz56C/+/XzBuPpeW/+9e/Y+fXu4enXv7nfPUCw2tmD+BczfX3o0+fW+6lMdcTs356kc7WzA455msLf7f37FPPrdD8uvDZnz0AH+/Wy9/63m+v49+wfXrVD8uuxsz56C3+/WzH+3mt/vHvePn1o6vl13/c356jk7Wy/H05lP1496PDz6351FOvf3N+ev1G1svz9Ree/+w96p9vWxX5de/ub89Rz/dbMEgXsDN9T9fr738s9azxx11/c357fX+62X5/2qfj/AGHvf2169k+nXv7mfPUf8wvlzf8AxmH+3t70c+vXhUDy65f3M+en/PLZf/YtMR799vWu75dI7sGn+eOyNqT7nbauVc0EyvOh8jBII/WzMp4tb+vvY681eOOj2/y+vkLvjuLZ1dTb+oBRZrG1KUzIQoOqMN5Cbfnj6e/E+XWhU56si9+631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//1N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdIbsjf+C6v2ZnN87klaHDbfpDW1zxgNIIUYBiiH6n37r32daiH81r+Z51ll+0PirvXrbc24v7v0m7Fl3rQyUaRUr0tJlFQpPp9OkqGF2/2PuwUnA9OmWkCgNUUr/LoW/liPiV8q9t4Tuf4vZc9cdyBaesFDtulEDbzySxCZ4quala1wbcj6/n3oGhyOrnvFQc+o6rl+MPaHyR3B8rdlbS/mAYzcG0dn4bcMFP1lVVVPVS/wB57VKRQx1ZkBATTzf+nux0mtOmk16gsvHP2HrfO2/Fh4MPQxYCKGDEJAgoYqdQkCQaRoEaj6D8+6dKOnn37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdB/2lvin622HuPe1WoenwFC1bKrWsVVgtjcj+v9ffvs69jz61EsTSdpfLb5A9m9ufFkyf3v67rZM9mcHQ1cNGctoqyW8sUbDyeoqCvPu1KcemviJZD8uhyynUH8yD5hZXGdcd0bIyW2+t8lPBT7qmjEy0Ip4ZV0SVCxE345HHvWF4Vr1Yq7ChpTz62XuhupsV0X1FsXqbCOsuL2RhYsPRyqGAkjSVpi9n55Zz9fdRXz6vjy6F33vr3Xvfuvde9+691737r3XEohvdQb/Xj3vUetaVrWnWtX/Owrs/1rurbPbWI2vDvFaCqxkM2FrKCWtohFSwxu9Q6KpFwPe1JrQHpuWijWVrTpA9g/znemm+M8O3s71fR1mYm2DNtiTGVOCklo6GvqsUKZpKZCDpKMbiw+vu1Zj26uqM0IGvTn7M9AF/Jf/mA9L9H7B3ZtXfC5bC5Xe/Y0dXjcTS4VkpBBXTusDu0igixYWt9B7szuBSvVIREyDFMnFKcetuTEQ47PYrHZyidvtcvQ0+RpiVAP29bEKmK4/HpYce3FunAp1drdCcY6mtt+FrgTSKD+AB/S3u31jenVBagZ1HrEduIVANVKSDe9v6H+t/e/rD/COr/TrWoPVJ381Pv7ur4sVHWW7diUtRWbXq9zqdxVZR5Ep8bBUDUL/RWf9Klja5F/r7jjm3e9zsb6CO3IjV9RBIwzLkJXy1Z6xd+8V7gcx+2VpY77tkJltfqEFwRXtirkinmTRVJxqIB49F/7H+RfWfZ8vXHyETcMZwGOymMrcjJLUUqSQS0umSvVkJABVwQQRwfYG3nma23GaDdZVZPDcBkI7ww4jTxr6eop0Rbrz7y5vcNhzyl0htUKyeIWUKAPi1EkAFSCCDwIINOhw+J3zn378uvl5v7bO0qB5+qMDQQvjcv4wFlSji8bsZIwF9RUlfVc/XgEXFWxc373um/rCYwtvKrsqaaOioQFL/OTJC0qKetaGvtn70XXup7kbpt2yxF9tsVjC3ANVkY6gwFBSgpVTU6hmgBWt5pwtMSrcg2QH6H9KgH/AG/uURcv1lV4CE18upC4uhAAMCsR9S31J/x91NxKfPq4gjHl1lGPoh9KaIf8gj3XxZPXq3hpwp1lWmp1FlhQD6cKPetb+vW9C+nRPvlthxVbep69RZqOJbWHAAdiB7oT1sCuB5dKb4tZk5PZYhJuaSNFtxxdv6D3oDPVj0aL3vrXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYZqinpwDUTwwA/QzSpED/rFyPfuvdRxk8afpkKE/61XTn/AKK9+6913/Esd/zsKL/zqg/6O9+691LR0kUNG6up5DIwZSP8CPfuvdcvfuvde9+691737r3XvfuvdF87/wCseqN5bSyeU7E2XhdzNjqSWppZMlTNNJHNSwGWEJpuT+kC3vx4de4cOteLsPenyd+QecyPWvxy6apaba+JllwCbnxlBNRVuMpGT7ayVAtpVPqPfsU6bOomi8fU9F42n0D8g/5YnaG3e9Oxt/Zvc6ZPL0SVOIydeatIJa6TVPS+KodtSqG/p7tUFaKKdaCujVJLVx/xXW3R1dvan7F6/wBpb0pyAu48Fj8syAafE1ZAJWj0j+huPdAa9PHHS/8Ae+tde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdBj3KL9X75Fr32zluP6/wCTH37yr17qor+VlPLHuPetPFK8cMmXyokgR2ET2Z2GtBweRf36uB15Rx6vD9+691737r3Xvfuvde9+691737r3Xvfuvde9+6910SB9SB/rm3v3Xuval/1Q/wBuPfqEcetVHr13791vr3v3Xuve/de697917r3v3Xuve/de697917oBvklNLD1NuTxSvFrpZlfQxXWop3bQ1vqLj6e/fZ17Hn0QL+W6dTbnJWzfxipufqTw3JP+PvR49bHVu3vfWuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//1d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3TFubcWL2jgMtubN1C0uIwlFNkMhUuyKsNLANUsjM5AFh/Uj37r3Wvh8mP5uewctJvnb+Fo2zXWWBoqibcFTLRU9VT1dDTylJvFKoZZPpwoPvxWvVdYpUft+zouHYnxz+G3z3/l79l91dM4+kpd3bd2hk6rAUE2NSkqUysMomcU9OtnLNd9JjU8jmwPvdSO4eXVSqyCh8x5f6vXpc/yifgZ1bvn4vdVVe6txZzH9n7EymUqM3jRQy07ywGoUUTSPMyEi3HF7e/MSSadbjUBQacB/Pq9jtD4edMdqwbPbP7Yx38W2VJFLh82KUS5CKSNFXXrZhckrquTf8fT3XPl1fHDoxe2MDDtjBY7BQTy1MOOgEEc83+ddV+hfk/737317p/9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Rfvkydn1/Um69t7zzVPhMVnsc1JPVz1MFMI4y4bWHnIH1AHv3WiR5+XWrdsX+X73/8fN+bn7f+H2+U3lTb3lllraSj3HSxRmnepMskEsUc1mUWUH/X97LVGem1j0V0YB8j59XHfCHtX5oV+5Ztt9+7QpcPjxMyLURSJOXIKorXUkHi/IPvR9R06tODD9nVwnv3Xuve/de697917r3v3Xuve/de6xySxQoXmkjiQfV5HVEH+uzWHv3Xugs3ZgunOxaj+7u8aPY27J3TUMTlajGV87La3/AN3LfT/D6e/fb14Hy6DLKfEv4iz0pTL9K9Smj1XIrMLio4NQ/JMhC39+oet6jXj0x0/wAJ/iPWT0mV270913SSUNQk0NVhcfB40nhbVGymBioKn6W/23v2OHWqk5r0bWgoqfG0VJj6SNYqWip4aWniQWWOGCMRxooH0AAAHv3Xupfv3Xuve/de6Kr8xuh8J8gOi99bMyONp8hkanAV0eEM0buYa6XToZBHzfi4t+fYf5l2tNz2uRVXVKg1R/JhQj+Y6A3uHynY848q3ez30ayiWNlAYVGRT/V8+tCLNbJ7i252XUfFuoGUiqqPdr4kYgpOsElZVVIijrBCBqKMtpPzYc2vf3EjxbckTcw3UNLpQFKk58QdumnDUTQA0qRTy64yX/KnN9rvn+tlcSyCVroRpb1IiZ2IpLQCpQrSQ8QMmlQet3n+XP8ADvBfFPo/b2NqcXSDfuWpI6/cOYWJ46ud6ynSQxSlzfgk8H6cD3J3KmyNt1qby9UfVTks59AeC/Yox11w9k/bKx9tOTbba0jUXJUNM4FC8hA1MfmT68BQeXVh3sW9TJ1737r3Xvfuvde9+690A/yKxYyPW2YKpqljQaDYkgaSb8e9461noB/hvkxBSZjFzudYuArEA6lkFgB+T9feuOevCuB0fD37rfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdV5/P7bPyY3JtHCw/G/FDK5qMv/EIvPHCVXy3WxkYD6e/Yrnrx1FaLT8+tXrvHvD+bz0N2T1/sPP9eyUNNvbIrSLU1RRR43qDCrx1DErb/G/u+Dw6ZPiVyAfz6V3yO+T/APMe+PGOwpkwlNWZvcMMUmMpaespsn5mcrcNHTMxBFz/AI+9gAmg607sqaiK/Z1sk/y0Owu/ezvj7t3eHfmCGD3DlqUyJB4jCSyuoIMbWI9PI90PTy8M9WNe9db697917r3v3Xuve/de6i1lFSZCB6Wtp4aqnkBEkM6LJG4IsQytwffuvdVpfMf5OYH4fbarKLrHZNDR7rzLsi/wzFRiKaqqoQaaSVY0tcOwJLX9+4Y60TjUfLqkjeHVPyS+TGVj7m+WWTfaXWVFCNy0lM+TiUVNNRR/cqIMcXB1FVHGn3sH06qanLfkOjD/AB8/nSbU35vrbPxs6ZxdZVy7Y+z2rTGqxMUazUuPY0wmWQLflVvf/Y+/FSor1pXBYqMkdbJe2q6rye38NkK+MRVlbjqWpqYgLBJpYg7rb/XPvXTnT57917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6DHuU26w3wf6bayx/1/wDJj+PfvLr3DPVQ/wDK353VvMgC38Xy30+oN5Px78fI9eXgerxvfuvde9+691737r3Xvfuvde9+690V35ffJbavxT6O3x29umrWmpdqYd8mFJhLyKkgjYrHMbNb+lvfq069xx1ST/Lf/wCFA3Uvzh7fyHW8daacCskosdK9HR45JJvIEiQOwXyXuCNJ5B492I9OHTSsW44Pp0ZL+YP/ADJuyviRu04vD4BqvGQSQGedccKlPHMDItpGQjlf8ffgpY0HXmfSKkE19Oqi+sP+FNFZ29v3K9b7NxstbuXCzvT5Cl/gcC+ORJvA2llT1WYFSR+ePeyteq+MB5HPVsHan82aToz405zu3sijOMqcPQ01UYpKKGNP33VNTRzqBb1e6DOD06TQV6sa+Dvylw3zA+PWxO7cIwaj3hjDkI2VI4wVExh4SL0jkH6e9Z63UE46OB731vr3v3Xuve/de697917r3v3XugA+TH/Mpdxf9Q83/uO/09+60eiEfy3NX+/mv9P4xU8/n6P9fej8XW1pmnVu3vfXuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//W3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdFh+aW1dxb4+KPfm0dpGYbk3D1tn8Zhvt9Xn++qIAIvFp51fW1vfutg0NT1pGfF3uPF7kxG1/wCXPujZ+JpuxdqbzycfcW7dz42LH1Mu3sjX6JYny1UFLrGpuq6v9793YfiXpNGTiOQfb/n/AD6t+6727iti9+ba+KPxk053rjamZpm3vLRRK2KrMdWIjZG8sFwyC5H191OASePTopqCjAHWyxszrXZXX0csOz8Bj8FBMiI8VDD4lYKoBLG5JuRc/wCPvXV69Lv37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Wt5/PN3vmN6Z3pb4uYXP5PbMfdEseDqszj5TSvRy1WR8KS+e4Itcc3t73TOfLqhJI7eJNOijZ3+U78qvg78fsnuvp/uTcu/NwbYxpqTR5Dccy07PUESOLtKVYDjn36uanrQU6aAn8z0iPgT/M6+a+081XJ8k9r0EGztuPbKZSGCOWpWnadYQ61bggki9rn3thTgeqIxPxLSnnXrb76239hO0djbb3/tt3kwe6MdHk8a8hUu1NIxVSxXi9wfp7rWvSggjB6XHv3Wuve/de697917r3v3Xui1fLDDdr57p3P47pmnSp3vPHKtBE8qwhtVO4FnYgX1WA9++fXs+XWpNtTpz+bbtv5g0e4stt/IRtTRsKXH1Fb/AJHPAF9L+EtYqfwfdiUI006ZVZlYsXBFOHRxuzsn/Mz3dDRdeZDEUGNyVRl4mligr6anrvtJZtLsI7h7WPHvQK1z1ch6UFPt6v8APh11pvXqzpvDbf3/AFdRWblmSCuyLVMrTPFUzQDzQq7c2U+9efV+AA49Gs9+691737r3XvfuvdcW02Ou2n86rW/2N/fuvfb0QLN/y+OnM98iKj5FVeNoZN0VNRHUqpgmfRKja/IrhtIJP5t/h7Bc3JdlPvLbpKxIZg/hmujWODfb1Et17Qcr3fPI5+khU3oGlXoahfQeXmc8aEjh0PbfJDryHser6rEtR/eLHSLTyINHiDg6AoH14+nsacB1LVBwHRgwbgEfkX/2/v3Xuu/fuvde9+691737r3SE7JoRkdn5amIvqhvaxP0BH0H+v795HrXA16IP8bcj/Dexsph76NWQ0Bb2BUS8gj/H3qh6sD69WZ+99a697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XutYf+eZ31h2n2vsTrR6/J91wQrTbVgpFedYMt946CJvASUNzfkc/X3sA1z1Rz2449cv5Zf8vbszcG29pd5fOTNz0uYiJrMHtfcOWHiqZ/Mst5YayRdMej+n1v8AT+m2IPDHVURhmQ6j1sw4Slw1HjKWnwENDBiY4wKOLGiJaNYxx+yIfTb/AFvdenft6dffuvde9+691737r3Xvfuvde9+690WL5SDb+E613Duqo2FgN5Z2ixtc2Mjy2OSsdKuKjZ6aRbKWJVgtufe+vcD1rN9MdO90fOzNbv3z8lt6ZHqbrnZ+bqMdTYjF1jUNPNi4WK/b02ILrqXQADZffjjAyem6F+5sD0/z9Qdq9nfFX4h/MPC4npfZ9FuSsxdNBDU5mswMX3FQ0SXnrjOQTqIJLMT79QkZ62GVTRfPj1tT9Ed1bU722HQb02lOktI6xU9ZGjRMKatEd5oLRHix+gNj711foaPfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3QYdzf8yu31/4bGX/9xj72OvdVCfytP+Po3le9/wCMZW39P92e6+XWx5jq8r3vrXXvfuvde9+691737r3Xvfuvdakf/Cjn5kbK27S9c/GvI5qKkh7WqJ9p5KNayl0iaevEdp0fkfW3uyjOemJSdPbxr/LrXnz2w+rv5em4/hrneoMriY6+q3ilfvysx9RRwVDw/exzMKlrk6SCbE+7ZNU6oNKFZfStet+zdmX6h+TXxJl7grdk7S3dHuDa8FTDV5OipMhJHVwKsBmWrjXUGT6gA25/p7bIz0pUnTx609/5LXXfVtf/ADOvkTTV/WWza+ki3VU+GmqcXE0UQGUAUoCv44tf/ivuzcOmovjI+fVwP/ClnbW3cN8L930eGwGJxsJw9ArxUVFBBHIP4j6QYowF4At9PflwR1ub4T69Hk/kEwJB/LZ+PQSOOIf3TlskahFAGQYcAe9eXVx/kHV1fvXVuve/de697917r3v3Xuve/de6AD5Mc9S7iH5+3m/3inc+/daPRB/5bhN9zg/jL1Atzx+r3onPV6Y6t3976r1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/19/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691XH/ADCvn5sT4O4Drqo3NVouW7I3K238LTrPTI5lUxoXeOYEkapAOB+PfqE8OtEgDJpw/n0cbp3ddb2H1xgty5mBTLnKSOraJ0Gh6ephSeK6WsQQ39PfuPW8Ux1W38sv5OPx0+Tm767sKjqqzqjeuTmNRltwbJx0NNXZSQtrY1k0Mkbtc8n6/wCA97BZeHVWRXy3HowXw3/l/dYfDylrp9vZfKby3Jlaf7ev3HuKFGr3T0+lGZ3IFltyf9t71Unj1YBR8PR+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691Td/M++MvWPyMptsbgouwNs4DuHrgA7Po6nO0VBO9X9w1QizO7qYyGsAfrf3utDnz6owDKBn5EdVi4nZn81rsja9R0lXLTZTZ2QjNFUZekzMNZ54IjpWX76GQlgEFyb+99o4cOq/qECpFenrcvxk7K2W/Xnxgwe1lyVV2EIcb2bnKvHTT1NBMalWDQ1cpNlX63B96+Z6uBjSfPj1sr9E9axdPdR7F6zglM8OzsJDiI5m+sixSM+s/X6lj+fdR1c/LoWve+tde9+691737r3XvfuvdRqyf7Wjq6n6/b0089v6+KIyf8R7917rWm7L+YHa2W/mPPt5anGYfrHA7MzVTJNWV8WPnNbi8SaiKONpCDdpFACj6+907a9VqC+kDy6rhr+9vmz3bLuT5sbfkal2VsTtJOvHNTXCjp2iiqykISFiBJqRb6gpv7tRV48em+5u5cKDw+VetvX4s9o5LtvpvZ+6sxDJDlpsLjI8kXW3lrRSDzyqT+oEi9/z7pWvT3Ri/fuvde9+691737r3RU/mP2juTpzpDePYe3FDVG2MLX5aUekHTTKugjVcfU+99aPp0Sr4IfzQOr+9OlcZnOwNy0WI3pFk8nRV9PXV9DAGhpW/alHkZNIFiumx/wBf3qhBoetK6uoZejGbN238Zewu28h2dtbekGS3nkpFmqsfT5Clqo0lPrJTwk8H6jnj34+nW6itacOj6CwAA+gAt/re/db679+691737r3XvfuvdNuXpvvMbV0/B8kEg5/4IT79x60eHVXmyHj273lUFmMYbJ8LewNpRb6/63vwr59bJpw8+rUIZVniSVDdXUMD/gffuvDh1l9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3WtJ/MDm6I+LHyAxHZGYhq96dob2yf8S2xhslQitxtBXGqayoJLkDXYDSPe6E9Nkqvcfs+3phw+yP5mPy4yuH3TX42m2r1nUShqBKerjxENPRxFSFp6V3U8pYcDn34FR8+tgOfRR1sPdK7fq9kdd7Z2bmstTZHN4WhWmrXWsjqZjIGufIQSb3NufeqenVyaGh6Fv37r3Xvfuvde9+691737r3XvfuvdQshjaDK00lHkqOmrqWVGSSnqokmidXXSwKuD9Rx7917rWQ/mL4Cs6a+Q8O5Zhm9rdOTwWqaLaiTw0E1bJCrvrhU6CT/aJ9+oTnqpbTknosPxrp8P3d80qPb2x9o4/N4LM7Dr6eLMZbGxySQ1dTinWKeaqAAR1b6kni3uxwK9URqvQenWwd/Ll+Ou9vjb1RuvaW+DKK7Kb0r8vRxOToio3Z0RYVJNl5FvdenRgU6sM9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdBj3Lz1jvcf121lh/6rH37yr17qob+Vv/AMfVvO/B/jGWsBb6AyW96+3rYrnq8f3vrXXvfuvde9+691737r3Sb3fnV2xtvL59kaVcXSmqMaAszhXC6Qo+v197FPPrRrTHXzqfk98Re9f5vH8yzdmydy0W5cNsTr7dVXX7ZzNJjapJEgGQMjsZlCgEgDm9+ODx7sKBc5NemKM8vbUADPz6MR89P+E1e6NjdOZ7s7aXYG9t65nr/DplcVQ1ElVUtDXqFU+KnilYsARYm1voT79roanrzQBhQClQfn1aX/JJ7f7S3f8ABzsH469j4rLx53rTa+QnpXyFDLFI83KCCMuAWuVU6f8AD34jSf59WjYyJU1FMZxw6ID/ACaupuxdsfzKvkDnM/t7J0OLyG5KmWlqZ8bU08Tr/FA1/K/pPHvzGgx1qMEua+R6tA/4Ux7U3nuX4W76XZWIq8tlo8LRrTQU1HNVlplrGlA0RA/i/uoND1eQAjIr1TD/AC3f56+R+I/xB6v6b3bsvd38V2jtyPH1cCbZd0grZ6u7OvkS/F+PbmkcAemhKfiZSOHkfLrcC+APzKh+ZHXP994cZk8aopo6gpkqL7KQ65RGNKBVH+v7b6fGRXqwH3rq3Xvfuvde9+691737r3QAfJg6eptxH8imm/8AcZ/fuvdEI/lucnc545zFQOPwbMffjx62D1bt791rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv//Q3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690XX5D/JvrX42bXG5t+ZGIJIZFgx0FXBHXztHYERxNdvq1hx79SvWiQOPWsL/MR2h19/N3m2Xl+j87k6bs3pbL1O9o9n5bIRK1Q9JUpPTpBREo1iIxcaTf3YGhzw6bK61JQioIPVi/8ALt/mcT7hy9H8XPkNiavbvb20qeDE/fPTxY3BtSY6lFJS08IdVDSNoH0PJN/fiMVHW0IJpwPp1fYrBlVhyGAYH/Ai4916c679+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3QZdybzquvest4b0oo/LVbexMlfDFp1eR0kVAukg3+v9PfuvUr1pf9xdX5z5T/AOnTuHf/AGzl9uZvD0U2R2HgsPn/AA1VVVpWErTTUcbqyi34A93qAeHTIRnUkkj0oaU6Ol1F3z3v8QNt/AbaMWWXLY7u2plTc0lfWtV5c0s2RWhMUwlLMpCAEDj3UitT04Dp0hhWuP5dbUCYXA1VXHmf4Tj5MgdEseQlooDWoSgKslQ661NrfQ+9fLrYNRUdPfv3W+ve/de697917r3v3Xuve/de6w1IiNPUCcgQmGUTE/QRFCJCf9hf37r3Ws187/hH1b3P8gqfsLYnZ+L2xmEnhiy1DNuGjxl4lVErQUklXggG4I97DUx02yVI+XS87D+IfX+8OrdlfHHp7tDZkOx0zOE3RuqGg3RQxV1duinsKyR6GOW8hLFhq5v7159WpQUA/wCK6vw6t2Viuvev9obQxEcK0uC27h8Z5YFVVqXo6JInqWI+pdgWuT78KUx1alOl/wC/de697917r3v3Xugc762LsnsTqveG1uxK7+G7QyeGqafOVutUEFA9vK5LED+n59+49erpzx61C63ob+Vvt3tjI9X7X7Z7kp60ZKpRqzELEuCkqJqgiaOjkjc6rMbe7sZDnj0yq26nSBQ+lTx6uV+BvxR+JmwexsrU9Vdt763Ju3G00NVWYTc9WqFYZIQSVp2cngEE+6E1FT06oA4D+fV4IFgAPoAB/tvfut9d+/de697917r3v3XuuLKGVlP0ZSp/1iLe/dePDHVUPYEUu3+6YKlRpWSvUggaRYy25v8A196r5HrQFOrRNszfc4HFTmxMtIjkjkck/n34cOt9PnvfXuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6oX/m7dF1WN/up8n8fhjuGPqd4cxX496E16s61RkCLHzwWANre/CtKeXVWoW9Tx6Ipt3+bp8he7NkUWF6pw1PsjIQ0/ib+I00eDo1WG0JjjeREAU/0v7sVpg9UV9QBCkfb1Zf/Ly3P3Xlsvkcn3pvzZdXV1pkego6Dc9DV1MkiurKoggYk8Ei1v8AD3U+nTlDQE9XSe/db697917r3v3Xuve/de697917r3v3XugK72+PHXPyF2pPtLfuKhqaOYuwqkgiarjaSPxkpI1vxa1/fgacOvEA8egi+MnwU6V+LE9ZW7CoqisylXI7jK5ZI5K6njcFfBTzIfSliRb3rJOevDSBRRTo6XvfXuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xugw7mt/ov3zf6f3Yy5J+lgKUkn37y691UF/KylpX3TvS1XTiRcvlj4mmAlY+selPz/t/fs060rZNerxfuqX/lZg/p/no/r/T6+/db66+7pbX+5p7f180dv979+69139zTf8rEH/U2P/ivvVR16h6991Tf8rEH/U6P/ivvdR16nWGoOPq4JKeqeknp5lKSwzPE8ciH6q6sbEe9VHXqdIvB9d9Zbby9Tn9vbS2lhs3WEmryuMx2PpK+pJ+pnqoAHa/5ufez8+vccdK3IwYfLUdRjsmlBX0NUhjqaSqMM0E0Z5KSxObEf4H36vXqdJLbfXfWWzzXttXaW0tvnKKyZI4jG4+hNarfqWqNOo13/Or36vXiK8eu8N171nt7J1OZwO1NqYnLVba6rI4/H4+mrKh/rqmqIgGY/wCJPvxPr1vJx05bo2rsje2PlxG78Jt7cuMnGmagzVLRZCmlUfRXhqQwP+xHv3Wsg9Au/wASfic6aH6D6YdOLhtkbZI4YMLkw/1596oK9WLPTiehi2ls3YOw6EYzZW3ttbVxwUKtFgaKhxlKFBuFENIFW3+Fvfsdayelb91Tf8rEH/U2P/ivv1R16h66NVSj61NOP9eaP/ivvfHh1rr33VL9fuae39fNH/xX37r3Xf3VL/yswf8AU6P/AIr7917oBPkrPTHqXcTGqplC081i8q2Y/bPdVt+feuOB14/PohX8txlY7nKMrg5ipN0IK8hubj36hB62DUV6t397611737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/0d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3TPmdwYPb0MNTnctj8RT1EyU0M2QqoqSKWeQgJCkkxALEkWF/fuvdamX8zHtHb1N8ztxUvyGzGYi6JwVImS2gcPWCOmrspTIkopkdvQ4aQWYC/uyA8B01KVBBfhT8ugq6A6UwWE3L2J/Mh2dn6nB9ezbdr6TaO3amu+yFfnMdR3p6RaZCBJqsCQF5v78eOn06sq/jrxHVwvxT6L6w+ffUHWnyk7A25Js3suDcde1TW7bo6fET5J8FURpTmvZFVpFcG5N+fdaUJHVwBRWIzT9nV2EEK08ENOhJSCKOFSxuxWJAikn+th7917rL7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuq2/5j3ytwHxr62p6fcEaPR7vhkppPI6IniSbRIrs4PH0497AJNB1V2VF1twHWtzuf8AlrfJrt/D7W+Ufxcz2PzW1t/1NVW5TD/xhJEpIoqotIj0mu2n8cD3YMKUIr028bE60fT8vI9WD/FD+Xj8oOyOx+st8fKtaai2/wBMZGOs2HSU3jI/ZnSZYVhDDSmoEk2PupYeQ6uoqAWINOtmP3rq/Xvfuvde9+691737r3Xvfuvde9+691GrIop6SqgnfRDNTTxTPcLoikiKSPqP0sCTf37r3VIPeX8rv4cd077yW68n3vunbu4aqeoaoocbuqmWljqZVCsvgLoVsfxf3sEjB60yg+X+H/J1UF8j/gjtH4Z9tbT7C2B3PuTdWDGfxNJHQxbmM1cx86sTJTQyNwSTc/T/AB97rih6aKEMGBI+Vf8AP1uF9YVQruuNh1oMpFXs/btTeY6pj5sTFJeVj9W55PuvT549Ln37rXXvfuvde9+690Q7+Yq+6F+M/YUe3DVrFNtvJx5VqEuKpaOyHVCycg+9+XWqgH59ajGxcd8atubH6CzFRn6iLsB+1Z23n/G6yGOtjxYyKg+ZpDq0gA/q492NTjplBHg/P/P1ZZ07ubCZL+YTmKz43VucyGPqnxi7ilE/3GE/hpp0FX9tJF6bAe6mmnpxcyVH59bVSfoS/wBdK/71711frl7917r3v3Xuve/de697917qtL5P4o4PfeEr7ALPNE5cccNLq+p9+IHl1oHz9Oj39YVYrdj4GfVq1Ug5vfi9x79Q063WuOl97917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XukvvHB7b3Dt/IY3dmKos1g5YmNbj6+mWrpZ4wP0yU7cN/sffq9ap5+fWrfubqXcHzM+Re5el+usLs/rvrzYuTko8zNh6KlwVY9DBUFGkg8AUsxHNr3PvfzHVSSTRjgdNHyM+I20v5buX2/2ZsDuHM5bIY0w11XjszuDx6neTyFY6d5G1Alfpb6W9740r16mirD/AA46tN+O/wDNc627ap+rdqriszkN4brpxHlqumZKmihqRKItWuNf7RP5+nutCOPWwytTSerf0fWitYrqANmFiLi/I9+6t8+ufv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuknvrAPujZ25dvRHTLmcJkMdESQAJKqAohN/8be/de6109nfEP55fHTu7O5zrSOhOyMhUV1TT1CyQurtOSyhwC1jz72eHVAKManB8uh4FN/Mpd5PTQlSWcD9sDlr3/4j3rFOrHXwx10KX+ZRYDTQ2v8A82/r9Peu3reeGOuzSfzK9RAWh0gXHMdv9h79RT1oF/OnXjSfzK2AIWgt/X9r6D6j36i9bqQfLrj9r/MsJBtQ6SbfWIfjkWt792Up1o6+Ap1y+z/mVhvQtDa4Df5vkf1v732kdeqw9OvGj/mVXfStDqA/5tcH8G5HvQ09eqwFcddCk/mVgcihu31/zX59+7Ot1bzp14Uv8ygXGmh/w4j9+On1613cRTrzUf8AMrsWtQf1+sX+sPfu3gevVbyp12lJ/MrI9YoOf038X0/wsPfjo69qfhjr3238ygcaaCwuP91/Qf63v3b1urfLrtKT+ZU2rig+th/mxwPp9PfjpHXqt8uvGj/mU8eihufof27e/dvWiTXy64Gj/mVqf00Gi3P+a5v9OLX9+7Tjr1W4Y65Ck/mU24WgsPzeP/Ye/dvr1upA8ukvvXZf8x3fm3jtSujonpqyoCzBNGowzWSX6W+g9+FPLrRJrmg6sG+Cnxs3f0TtCrTffiXP5GWOpmWIqQsjg+VTpY/197NK9eFQOrAvfut9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691/9Lf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdRqyqioaSqrZzaGkp5qmU/0igjMrn/bA+/de61kvnz373981O4pviv8AE1qmkz+yamqz8eQWoFNR1c1GFIeeqOhAE0n9Tcc+9inE56bYlgVQ0Pr1Vv8AJHC5zdWE2x8ef5istbt7sfCZZf7s70gaKgxk1a+mOCmr8kAEmjDBSwDe7DjVTw6owFPDfOr9lf8AJ1YV8fv5ZPy83Zt3YOw91b12fkfjFh8jSbjwH92s5jqkV1HPEGQPDBJ+4WjIDqb2P+PvRIrWmerqh06S2B5U/l1s69Y9bbX6l2ZiNjbPoIMbg8RCqU9NTwxwR+TxKk03jiAF3K6j/vfuvTnlTpf+/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuqrv5pHxl60+S/WNBt3fm98Ls+ejV1xsuWyMGN8nklMjsk07KLXsPr78CQajqrqHTSRXqmnprpz50/HHblFs34+dvbC3T19hfuDi8Zjd1Y/M1RWd7uGo45HIa31sPe61rXrWhlGSPz62FPhlu7ufcuzJo+6KcQbjpI08gSFoEZvQtwhA/wAfp7rWpx1ccM9HT97691737r3Xvfuvde9+691737r3XvfuvdNuZRZMPlo2fxK+NrkaQnSI1aldS5b8W+t/exx60eHWq/2t0nU5vtDc390vkFsvGZCfL18q4/I79o6Wohcm6o9PNKpGk/UW9+qK9VKsD59Z+jPh3tPN9w4fc/yP+QPXubweNnpp4sXQb5oclLNU0z6ol+2WVvqbAnT70HAFOttGagtU9bQGyq/a9btzFRbQydDlcFjqGkx1DUUFQlTAKekgEECCSPjhVHv3Vq16Vfv3Xuve/de697917pE9jR7Rm2RuWPfbUqbTkxVRHnXq9AhXHtYTFjJwPx/sffuvdaPXzG6m/l+5juzM1OxE7RyNNBX1c9XNt2XyYKEirLSGg8PCkG/Puw1evTDLDWpGetg3+UvtD4b4LaUtZ0rWVlV2DVUQizlPuuaGfcENOsEZbQGBYDgg8/7D3o1OT06oQDt4dXXe9dW697917r3v3Xuve/de697917ojvy42vkMy+Fq6CmmqGhWMN4Y9ZGmQ/wBD7917FOh26HatTY2MpayKaKSmp410TLpYH6Ec/wCt70M569nz6Gz3vr3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdY5YkmjaKRQyOCrKfoQRYg+/de61fv5gPxP+aWxO9sRvb4PFMfNuit+53XUSCGmDTSVDF7lzYrpIJJHuykDj03IKmq0H29JLJ/yz+9u7KPB7t/mGdqYbA4aJFaqFJuOjhqvAhDxqacSAcjVbj36tBjHWtOsd3cPTo8HTPX3wW+LW1qjO9Bvld/5XZ+lFqCsWVlvI6+unkhBK2IH0X3XPHj1cBRgCnVvXTvZSdpbPxu5lx9XjWrYBM1PWRNDKmo20lGAP+8e/VHWxXiehY9+631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XB443/Wiv/gwDD/bH37r1OuH21P8A8cIf+pafj/Ye/daoOvfbU/8Axwh/6lp/xT37r1B177an/wCOEP8A1LT8/wCw9+69Qde+3p/+OEP/AFLT/inv3W6de+3p/wDjhD/1LT/inv3Wqde+2p/+OEP/AFLT/inv3XqDr321P/xwh/6lp/xT36vXqDr321P/AMcIf+paf8U9+69Qde+3p/8AjhD/ANS0/wCKe/dep177en/44Q/9S0/4p791unXvtqf/AI4Q/wBP82n/ABT37rVB177en/44Q/8AUtP+Ke/deoOvfb0//HCH/qUn/FPfut0699vT/wDHCH/qWn/FPfutUA699tT/APHCH/qWn/FPe6nr1B177an/AOOEP/UtP+Ke9dbp14U8AIIhiBH0IjUEf6xA9+69QdZvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//09/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdY5ZY4IpJpXWOKJGkkkY2VI0XU7sf6Acn37r3VZ3en8yforrne+R6nyNTDlXrYJsXWZahyKijpBVp9tUmVwvGgM1zq4I9+49VLAYJoeqS4u9ct8O/mnkO8thUUfZPQm7qVqKGbbkYyk6TZRdM0dRkacMVMbMbgn8e90xT06rUqxPkePy6aP5nGS2n/NqXr/AKo2Zs/L7XzOxa+PdE25YIhTFzWiKaSOuyKBCI4wnIZjY397XtIPp1qQB0KVpWmejOfycOxO5OtfkNvn4Ybmzc24+vdg7SfLYrJpWPkaIV9LTRweOGrOoc3F1DWuP8PejQ9w6tGCupGzSlD9vr1sye9dX697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de61LO6aff/wDMR+e/ZXxo3PvKfbewer8xV00EZyr0MlRQUlcYzFRISoeQ34A97BotfPpthWTSTQD9pr0qPk18C96fy3Npy99/Hzf1VU7d2dEmR3BQbjyrTV9YrSKrxUlJIx1gkfheB72DXj1pgRlTkZoeHV+3wb7uyHyO+L/VXc+YooaLL70wTVWQjigWEtPSVb0RZgoF7+MG/uv29O/Z0bX37r3Xvfuvde9+691737r3Xvfuvde9+690ieyatqHr3fVUgdnp9nbmmQJw+uPESlSpH0IPPv3Xs1x1pmfy5/5cvT/zu3v3Z3D2n2LuXb2S212Dk8JDg49wNDXVIqZJTHUQ000ikxjRYkD6+7k8B0wqAs0hzk+ZxToNvkX/AC1aTo75cQbW627JymYxskKy0VKdyNLVJJKdULT06yfRS3qNv6+/BvKnXjGdQYMR8q9XDfy2u0+6Oo/k/D8Oewc0m4MPPtCo3TT1cFV97DTGnxclfBCZubNawazfUc+6H5dOpxIOaDrYs9+6t1737r3XvfuvdEK/mN1GYp/jF2NJj6p6XHrtnKtmpIpjBL9iFS4jluLH/H37r2K18+qX/wCWnl/5b9R8eyOw87g03xLk8zHuA7lydFNWNTNKAogkqCWAvq5t9fdirVz02jxkUTP+fosXTXfHx76i/mDbtn6E3jDjNmNlXiyE2Tz0aYl8S82mdIVVtIGn6H341K160GVZKD7T1t6bE7M2V2TQLktl7gxm4KAxRSLV4yrjq4CskayACSMn6Xsfdenel97917r3v3Xuve/de697917qHV4+hrgBWUsFSF/SJo1ktzfjV7917rNBTwU0Yip4Y4Y14CRIqKP9gvv3Xus3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917oufyizO+8D1LuOv6zo6efea04GLkmpVqGic3DNHcGzfSx9++fXs/l1reZb4+/KbuF6rOfL/uvbO3Nh1LGSDEzbnhxeXeBHMix09IXU6tFwBb+nvdR5dUCluPD0HQK5b5p/HH4J7swfXvSWz99b9m3BojyWYnmfL46eR5gPSqq6kAi/vwWpz1Quqig/Zx62aPhL3JmO7OsoN4ZPb9Vt2KuRZaOhqaQ0bJFr0i0ZVfxb8e9Upjy6e+3j0dH37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/9Tf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3RIf5gnbe8+mvjfvbdOxsTksvmpMPl6FIsXDJNUwJNjnBqB4gSun8Ee/Y4HrRLAVUVI617v5U/V/xf+aHWu/aT5I5DI4zvzKbwy9FQYnJZtqbONiauJtNVTU1SjFir/XUy/Qf14s2Djy6bQKyAsMnjXyPS0yvxZ+Q/wO7toto4vHSdgfFHdedjpqPFCigzeXT72URGqmZgzRFNX1496JHlx62qsDQZH+Dqyr5K/B3s6baG2h8NXxGza7ckES73kzf2MFXJi8hGssiRVMgRkIV2BRfp9AfegfXq7LxoaHo0Xwt+Fu2/jFtyPJV6Q5Hs3KQSJuTcCeKXz+dFMsEVRYuy67nlv9h79x62AFGM16Pf7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XusNTUQ0kElTUSLFBEuqSRv0ot7Fjb37rRNOqsPjh8/f8ATp81O5PjfQ4XIRY3rqCvnjy7tI9FUrRP47xa1AFyRax9+FaV8j14ldRA4in8+iT/ADO+FOa3f3juHtj4Yb6wlF3Hh6n+MdhYxK+CmySTGdppT47hm/Fx79qAGny6qyljrrQ8Oi0YX45fzJ/lnurF7L7v3OarYME6U26Yq+ZFinxcEig+OOVgrkC9ve9QHw8etFWfDUA/w9bNfR3UuB6N6u2n1btlBHhNp48UNEoRI7KzmaQ6Y+OXZjx7qK+fTnQs+99e697917r3v3Xuve/de697917r3v3XumDdE+FgwGXXcNVBR4epx1bSZCeobTEtJU0rxVGo/wDBC3v3Xs+XWrh3l8Ns5sHL7v78+DXdez9obDjy9dHuapyW4qbH00eZkTXNG0aMATqLWut/e9WKHPVDHmqmhOSPI/Pp4/l+fFmfund2f7a7U7d2rv7tmix9bjcZBhc3DlGNVNC8dPVTIWNlVrG/4t70c4HXlUjJz5dWl/CT4N7q6W3ruDtruDIUOa7JqKmvocJXUbwVKwbfqTJCsEk6ksD42AC+9f5Or0oTmterS/e+vde9+691737r3Qb9sdW7Y7k2HuLrzeEUs2A3NjZ8XkkgZVmNNUW1hGcEA8D8e/de869U6UP/AAn3+D2KmrJcfN2NS/fSzzTRw5yBIQ1RIZHCJHELcnj3Ysx8+mvCjX4VH+r8+sWH/wCE+HwawWRrMvj27ATI10bJUVMuYpnchhY2KxA3/wBifftR9et+HGOCj9nVofxv+LfXnxg2um0+vpsvLjUXSGy9SKmciwXl7D+g918+rjhToyvv3W+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917pszBmTG1j00MU1QkEjQpNGJYzIEOnUh+vv3WiSBjj1prfIzqvtLJ909o7v8AlpmcjUdf4zLvNsfbuBr2oK6qoWqWaGmoqaIgH0cHj3cHGOmytW1Hh0Gmzd79dbWzOM3Ngfj52BWbWw8sclHUZzDfxHVTmcHU1VNGf6E+9UqaVz17gaqpA9ett74j979d949W4XJbG/h1FNiqCCky+Bo0jhmw0yftrBVQIq6GNrkf1v7r07Wvz6NT7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/9Xf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYamQw088ygs0UMsgUfViiFgB/r29+691q0fLb+dJ2h0R8+Y/i5j+vN411CaBcgKqlSsNHKrwLOq2iQi/Nhz7sATXptpApWvnXHRu9r/znOoYsNTYzv8A2suwqXJtHQVY3nI0FPVQVBMM4eKtj0MCp/SR+fevPPWw3kOPRJfkn0D0h2dvrH/JX+Xz2/tTbXZ880WQye3qXOQUgko5EEkseOoImALPZtIsb+/V8uPWvDr3DH+X7er+/hXuXsPfXQ+3K3uCgd9349/4ZVSZKlBnqBR08YWpvMCSGYkq3vVc46cIIoG6OAAAAAAALWAHAt9LD37r3Xfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917oCPk7uiq2X0F2nuehilnrMPtStq6aKEsJXmVlCBCvN+fx791sCp61v8A+Wx8uui+tN298dwdoblwG1t4ZTCZErDm61Ya6omhqhMsWphrOtlAa/vZBB+XTUbBlPrXob/5UG4qLtv5kfI/uqLdFLmMFuTH1k+KgWtlrKZ3qa5CwhWUkApGWP0+n096NBg9bXzb5/4OtkGKKmFnhihX+jJGinn/ABAHv3V69Z/fuvde9+691737r3Xvfuvde9+691737r3XvfuvdVz/AM0PL77wvxdztV1+ta2XfJ00NX9iZBMMVJBIK43j5A0/U/j3759ePCg61gtzVOL7V6ppPir0bPlKjcO6M/jt1b2qospPUGOpI05aFheyhQTcX93GDqPTDHWPDX/ZHr0eX4c9NbN6B+c3WO0Pj9k8hksU2ycT/ft5Mi1dT/3mFHpzSPECVsr6uCOPdSfXpxUCk6MY/wAHDrargLtDEZbeQxoZLfTXp9Vv9j711frL7917r3v3Xuve/de6ow/mR/zLt8/FfK1+2tqbA3JlZ6aOfTlMctSIZGWUIpRolIPH+PvYFWx02zaV7gT0XjoP+c9vrcPWONzO5ent6V+UaudZqkx1ra4VW5U3Tj6e/EH16sGFOB6Ef42/Nj5K/Jb5HVOLo9s7o2tsCkqhO0eSp6yOk+1QeRo1eVVBNuOPfjgdeQsW4YHn69bBy3KqT9bC/wDr2966t137917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917qgf+a/17l/9JHWfZmSxMuS2PtZaaTN0+NhSGOaNahpGjqbfqJAIu1/9t78BTI6qccfXp337/Mc+PuI+OFFt3ZG08PnMzU4KLGptmho6Serog0fjMhp1jJE+q13v/U/n36hr14uKdpr05fyXOm+xdjYbunsbeNBlMVhu083SZfbmOySyI1NAk8sjJHG/wCkAN9APez8uvICFIbjWvV5/vXVuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6/9bf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691r7fzlfipuRMdB8vunaGEdj7MqKCTcEsNBTS1Mm28VTCWrmL6S59ETBjf3YGvaeHVGB+ICpHD/L1UTtTr7r/wDnu9p7aiptm1e0Nmdf4nE4feFVQztR0NRuHC04GWqvKq2DytGxCAE3PvxqmD1SiT5Umg/1U6uh2P8AyWdjdL9o7R3j05navE4Lb9JiEqMdma9qyWapoIwsrBAukqxv9fetTdOCNa14dXp0FP8AaUVLTlYleGngik8KLHGZI4gjFEXgC44H9Pehw6t1L9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690x7k27it2YPJbczlOKvE5amakrqZrWmgcgshuCObfke/de6pv76/kc/EftSLN5HAYbN7e3Fm1damSDKxpQMZZWlkfxJGpHJ+nPveogdUMaMcgdV0n+Tx81fixS5DK/FLuTFYLEQQyvNjoq+WLJzQu2lkB0DX6bC17+96gfiHVfD0/A3HyI6vR+BdP3zQ9XUuP79ytTmN3UyutRWzmRjJYqq8v+eD7p50HT2aZ6Pj731rr3v3Xuve/de697917r3v3Xuve/de6hZKoekx2Qqo0aSSmoqqojjXlpHhgaREUD8kiw9++zr329VRbk+Um7891R2nkN5dVbhrsbiMtlNvxY2ooJpnyFFLC0TzweRfyp4I+nv1SR8+tHHVM/RHaXR3Qu+Ny782V8aN90+5sq+RgqJ8qstRTp/EUZZmgjZPT9Tpt9Pe2rTJ6oulSWAoT8uPVmX8swdQ5rfGe3Vh+ss/tfe+Vq6yvbJZeTyxxx1jyPNFF5RqW9yABa3upr59OLTJHn1eh7317r3v3Xuve/de697917om3zGxnS2C6o3jv3s7a9Jn2wmCyFZRUX29O1XkKmFQy08LOhJYki/P09+rnrfEU6pFrPnP0zsT4k4zduK6alwWU3tk8vgNm4qsoI/uZMnTzGEPTOEBYtdbC3vdDq6bqAurP+foR/5ZPz6zOW3pV9Zdu9fVWx67IySRYqvzGMGPqJ5XAelijmZFJDCy/wCN/ezppjPXkLE9wIP+rh1sdg3AP9QD/t/der9d+/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6Dzs/Y2yuwNoZXb2/cfBX7eqoWNakiRmQRohv45HVtJsT9PfqV691redpZj+X70JvbNS9S9T7j37vTEVRR6Kmyi1scNSCzkGjQECzgCwHvdT59U7QaCgPy6QXSf8ANq7sq+9dpbX3f1zuHrTq2LIRUjplqOqoMdFRyVSxF30oikW5v/h73QUqD1oO2ujKQKcetqfbW4cZuzA4rceFqY6zFZikjraGqiN45qeW+h1P+w91+zpzPn0+e/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//19/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3TBuvbuO3dtncO18tBHU43cWEymDroZUSRHpMrRPRTqVkBH6XP1HvR68OPWtj8MvjVv34F/zGMn0v15gcqnx/3cmQ3JkKxIRLQSZLK0sjwztURqACjOPSbEW592wRQ8em1DKxNMH/AA+fWzYDcA/S4Bt/S4966c679+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6910QGFiAR/QgEf7Y+/de66VVX9Kqv/BQB/vXv3XuuXv3Xuve/de697917r3v3Xuve/de697917ppz2YosBhspmchIkVHjMfWV9QzmwMVFTNVSD/H0oePfhxx1o08+tU3vv+ZP3l8r+ytw9W/FnYWYO1sFlZ8NlNz4GinnoY6yMGAtXsq6QzMpsW/p73QKNTnj02WdyRGK0qPl+XQN1u8vl38dpKXd/ZXV+5exsEk0LZKkp8WdEVPEddTPM8cZsoW9yffqr5Y6t+qB3Cv2dbA38u/vfor5KdcLv7rfb1FtrcOJmjxW4cTGQJ6SueNmdGW17ghgf8fejxz1tGDLUefVk3v3Vuve/de697917r3v3Xuij/NHqLIdu9I7txOId/4vRYavqMbDHcvUVJVdMSr9CSB9D78eHXqVOeHWrL1f8W/kv3lvDYHXO99u53C7Z6H3jPvGjqclRNBRV0cNatS1PAJBokL6AAoHPu3bQ08+mxrJAbGk1+3q2Pq3obsXvv5nbj31vnC1mA2Ns+LG122tVBHjKaaqx0KQIsXhVdWplvx/sfdaY6uMmvp1fcosoH9AB/thb37rfXfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuk9uipwUGHrBuKoipsXJG0dQ8jFRpcFfqtz791o5x0TDb/w1+InV+Wy/dVPt6GCfLynLVucydYaui1OxnMkcDR2ANjwb+/ceHXqDzHVMn8wXvXafy73njuiPjxs6PP5VaujoG3Xtygig/hssVRpkSpenQH6m178Cw92pTPTbd3YPz62AvihsDdPV3x96w2HvKpeo3BtvbdPQV5kuZUkVy4jkZuSVv8An3Xyp06cmvRiffuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/0N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdRmo6R5vuGpadqgDSKgwxGcD+glI1f7z7917qT7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917om/wA8N1VGzvjfvbMU85pz9rUUksupltBU4+dJrlOfp72OtGlM9Ul/yRPkH8X+mOqu9qHK9hbewudzfZa5mvgqWMtXOziWEKkoBZrO9yhYAfX35vi6rEf0x61PV2G7vlv8a9y7B3nSL2Jt+vp6vae4YpIHjLrKsmLki8fjkDKSS3APvXzHVzg9Uk/ySFw1F2h2nT7CqPLs+t3Zl6qqEMzvC9U0szRPoJIH+sPe2PDqiZqRw62gveur9e9+691737r3XvfuvdBD3jh97Z3rnceO6/yTYrc9Rj5kxlYihjDUm3jfSfr+fexWhp1o04nrXrz3xn/mpVAyTw9v5ZlM1S1PHDH4ndJJz4UVokBPpt70AB1UaqcQPy6Sv8v7u/5Y7I+Wm4+oPkp2NkZ8PRNLS4+nzCyhKmut4o44XaxJLce7HSBUdVXxCSrU+VPTraaU3VSfqVB/3j3Xp3rl7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917orPy+6u332z0xuba3XWeO3tz1cKNQ1wdoyhiDMw1ryL8D37NOtUzXrWX752f/ADKs31jVdEYWXer/AGUUVEmfgWvakqDHqVpRMosQV4+v5977Bx6owm4AA/PpJfFLrj5WfEXFz1WK6b3llewclEGzG5vsqisFTVeUSeaPUjFTfnjn34kE1J69GjogFM/b1eN8IO6flD2Dul4e5dn7mwtJNO7tU5anqYIhecDhZQB9Df6e9Eigp1cBvxCnVuvv3W+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv//R3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdBp231ht/t/Y+Y2LuWAVGKy8EsU0Zta7wPCGJIP0Dn8e/fZ14dUbVX8hnrbE5PN1uwNzwbYgzdfJkKmDXWPeZzqBPjjtwfe9RpQdUMUZNTj7OkpmP5HfYEtNLSYPvekx8EyyRyIy5Szwygq6ELGfqCfe9RAxTrXhLwqerCf5eH8vPH/BzbGbxLbjg3NlM5XCvqa+BHVRNrZmI8yK3Or3okk1PVkQRqEU1A9erNveurde9+691737r3XvfuvdA73vuHdu1+s9zZnZGMqsvuOix002OoKNC9RU1CW0QxqPqT70a9eqK0PWu5WfOn+Y3TVFTHH0L2Aniq6gU8gxFdYrBORFpsLG4A93IX16pql4af59K34UfF35Cd5/JDL/IT5DLl9s0gnGWxuCzULU1QK7iWNI4mAP6jyD/r+/HIoOtKTr1Pg9bK6jSAP6AD/bC3uvTnXfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r/0t/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690Tv52diZvrX41dn7g2vlZcTuem2tkp8FUQg+b7+AK0YiI5vzx70RXNetV0nhjrWD+I+yf5qvyQ67ynZ1J3DvnH4jI1mXp8RFLPULFNLTz8R0/B5AZR/tvdz4YNAOmo/qCKuR8sdF32L2x/MY2v352jsbfXYe9MpW9X0dZkocdWy1bNk6imOkUaIDySfx79+nxHW6z1IYj5Y62of5cPyvzvyJ6upsdvnEy4Xf+3oSM3Q1AkWoEKKkUUsqyEm7Ekn/X91ODTpxeHVk/v3W+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r/09/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdEr+UXza6s+MVZjsHu3LQwbiytOKqix8kZkMsDC6v6GBH0/p79140A49FfX+a91Y0UMnkpiJY1cWp6j8j/AIN73Q9V1r69dn+a71Z/x0pj/rU9R/0d71Q9eLIPPrj/AMOvdXX/AF0trc2p6i9/+Sve6Hr2pPXrx/mu9XWIEtMx+t/t5wB/h+r3rPWyyjz69/w691aVBWWlvfm9PUcD/kr36h4da1KOJ67P813q0N/naXTb6/b1P1/5K9+zSvXtS9eP817qsWvJTc/n7eot/wBDe/UPXtSngeuv+HXOriT+9Shbcf5NUXv/AE+vv1D1vUvDrw/mu9XBfVJTXP0/yeo/J4/te99aLL1x/wCHXesAeZaW31P+T1F7fn+179Q9e1r1z/4df6rtfyU1v+oeo/2N/V71Q9b1L69e/wCHX+rCOHpz/wBU8/8A0f73Q9a1KOJ66P8ANf6ssbSU1x/Wnn4/5O96oeval9evL/Ne6tIA8lMSP1EU9Rb/AGHq9763UevUOu/m39QYuE1WQqaWCkj0meb7eotGrMFvy3+Pv1D1rUPXqzfqPtDbXcWwNvdgbUro6/D7go/uqeaMafo2l1KXNrcfn3rq3n0JXv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6Lf8qumm7w6Z3nsuln+2ylfgchBi5wdJjq5UUowb8fp9+61/g61qOv/AJQfPz4D7fyXRWK6Uzm9Nq7frstWYjcs9DLU08rVM5FoKgDSQbA2v7tQcSeqd6ilKj5dCB8eOh/kt3Y/ePys3ZjspRbs37tnIDDbTlIFSMrInnQQwqC3B4A96b0GetrUmpwerZf5aXx43j1d19PvvsmiqcVv7dpqIcvja0OtXBBFIrQtKrgWvbj3o8cdXAKih49Whe/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/1N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvda4vzw2vt7e3813437a3jjIc/teu29iI6/CVJKwVKuzKwYj+o978j1U/Ev59XJR/Cv4utDEq9ObVREUqgEM9wur6EiT3rq1ajrsfCj4uj/AJo9tb/qRP8A9H+/de67/wBkp+LvP/GHtq8/nw1F/wDrZ7917rpfhP8AFxfp07tX63/zNR9f+pnv3XuvL8KPi4vA6d2r/sYaj8/9PPfut169/sk/xc/PTu1T/h4aj/iJPfuvDHXv9ko+Ln56d2qf8DDUW/3iT37rXXY+FPxdF/8AjD21iD+PDUW/3iT37r3Xv9ko+LhFj07tU/0vDUcf61pPeqde69/slPxdtY9O7VP158NRfn/p579TrfXX+yT/ABdtb/Q9tax4/wAzP/0f7316vn14fCf4uC1untrcfjwz/wC2/X71Tr1evf7JP8XLsf8AQ7tX1fX9mo/6+e99er15fhP8XFvbp7a3P/Nmo/3j9z3qnXq9FP8Am/8AET427c+MXZ2YxXU23KPJUWNp56SspknjqIZhUKilWZyLc3Itz73U+XWiAcHh1x/lQvUxdHU+PM7fw6jpUFBR3JjpEM9mVL3+tvfvPrw+Hq1n37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdE++bvbuY6i6I3ll9vxyDN12EyFNjKqIMZKOr0qEmSxHI1e/ZJ60SB1rQdr/ACr7Yh+KvT228turJ5TfPde8crs16+dhJV7eiqq0QUtWhQllI13F/ewMn5dU1dqg8Sf8PQ/fB7Ld9/Cvvqh6n7B7GzHa1JuCpp6U0eQlqKiHGLmIlKTRiQ3UoGH4/HvRpSox1tdQOhjWvW1Apuqk/UqCf9iPfur9cvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/1d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvda8XzKv/w7t8Zv6fwDC/8AWxr+9jrxpUfn1sMQ/wCaT/W/4n/D3o8etDh0HXaPbOyOnNuf3q35lVxOGNUlIKlghHnk/Sv7jKP9597p16oHTvsTf+1+yMDT7k2jkocpialUeKohkhkFpF1KG8LMAbfgn349eBr0G2+Pkv1D13vSm6/3RuaCh3VVwxTwY3XTtI0cyhkJDSBhcEfVfeqGlevVFadOFF8hOrK/e9P17S7ip5N01VJHWw48SU+toJV1o1hJq5H+0+99er0NakMAw5DAEH+oIuPeut9Fq7T+W3R3Tedbbe/N2QYnLpGsjUhNOZArAEHRJKrfn+nv3Wqgcekht/53/G3cuTpMRjN7xNW1tQlNAk4poFaWT9I1PLb/AGPvwz16oA49Dx2D2/sLrDb0G6d35unx+FqUEkFZ5ado5UZQwaN2dVINx9GPv3W6gcei0D+Yn8WmBZd8hlBcalip2UlDZgCJrf7z79kdex5dGT6z7h2L23iTm9lZVcjj1UO8zeNAqkBrnSzD6H37r2egT7K+cnx16pzlRt3du9IafKUsnjmhpTS1KK2vx/rWX8H/AA9+615Vr0LvVXevW3c1E9fsLP0+WhjUs6iSnEwUAEkxRyMfoQfp79k569UdDB791vomf8wAkfE/ta30/hVPc88D7teeP8ffuvf6v5dFz/lTW/0MKb3/AMmj/wDcj37z68tNGOrWPfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYaieOlgmqJTaOGNpHI+oVBc+/de6JR3lvXpzvDYvZvVdbmVpcrQbbrpZ614iWxADDXVRspvcBf6e/ccjrXn1rSdI/HD4/wCzey8juDsr5C1O7NvbXyNXmNmYmqp6h4qTLwVRki8CvcCzAfS3vbMaYHVFQA1rWnCvVp3ww2f03vf5Pbx7iyfZsm+8rmaURYjbGSglC4ieGILDPGw4uqj02/PvWB1cZNfPq+kWAAH0sLf63v3W+u/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/9bf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3WvF8zNP/AA7n8ZrsQf4DhT9L/SVve/LrR4j8+thiHiJP9b8/4m/vXXhw6qs/nGfHbc3yP+F++Nq7S3HkNs5jbq1u8Y6/GrMZ2hw2Jnnkh/Y5AJsb+91yD5DqrgGMr5tgfn1RV/wlM+Z+5989B/JTqLt7dE+S3j13vLL1G2HysqR1FRhNuYyc18YjnbWGQgFgRb0n6e9niWAoOtIaIsLGpGCeq39jZ7uL+Zh/woLra3bXYGex/VeLwGYpqqOlqag4Kin2thpUNOIaMtDqeSJTrPIIt+T72RQBvJumwwkZ4/xR9OfWG/u0ab/hSTidgHsbcNRtLC7OFA2DepkOOmbHY1qTWaYtp1F116rX/Hup/wAvTqmo+YHX0LMYScbjyTcmhpCSfqSYFufeur9fNA/4URZfe2R/nB0W05+5twdfbMrc5s+jENDVTQUsSVEFJJUWSKRFbUNQIcEcni/u6jsZq8OmZmHjRxkVDDj6Z49GD/nF9JdHfG/p/oPfnQ/zC3JlN8yvg6mfCYaMtJksqlBBNHR1DUrE6ZHZgQfe17mCtwPVJlWONniJ1DoaPmR3V8gO0v5HfS28ewc7mtg7rGaz2NmzETSx5CbCU9Iv2Vc5kIb8av6c+/Chk0nh16UsLYMDk0/b0Wr+V/tD4o71+I20q/tv5c7rXfFduDMQzyS46etYQ6DoU1DuzEKbgamJ45/r713Ux1sJGaVND55p1fj8kO5MZ/L2/lu7j3f0Jvuq7DyG+NoV521mp4hBUJkIV0rFGs59LMovx7qAWOOPTkhWJdXkP83HohH8qT+V12B/MH+N+L+XfdveG5Nv5vs6qzdO+1WhqK5sTUUdTcSxyGRUsSw4U/T3YtQ4x1RYVYVkJYnPGnVmXwa/lUfIn4ZfJjcW6E7b3HvrrfJ1hSngnlq48fHRtKtykEzEfpFiLe6u1Rjj1dI9OAcfPrZL966c6Jr8/hf4odq8XH8Lp7j/AA+6Uf73b3o9ez5f6sdFx/lT/wDMmzxb/J4+P+n/AL35068Ph6tY9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691Hq6daqmnpn/TPG0bf6zix9+691S98wO+Pir8Pcruas3ZuXVu/fNDPg67GUdHBXyQCokCOJokYutr/ke/AHrTMoHGnVHtJ3t8ZnzlTk8400G03nnqVrBhJ2ZY5HMpcR6f6e70NemdadXyfy99rfFPsCm/0g9H7vXM5FY45azHzQJS1VOoiCSH7a+sAEjkj/H3U1r06KEVHVw4FgB/Tj3rq3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//X3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691rxfMs2/m6fGYaQb4HC2vb/AI6H3vy60wypHz62GIf80n+t/wAT7114cOmXdeIiz+19yYKaMTRZrAZjEyxEXEkeSx8lG8ZH9GD29+6318un5Tdobz/knfzQ9/4bEUdZidnbz2hup6zHUNJJLTSVO9cfJDDVKlSLOq+ZSxS5AYH6X931BgE9Ok2hkZ5QK6s088dbC3/CVv4rZ7ZfQndnyp7Rwr0+/wDd269xZXaGRqlqPu325uCiqK9pQZgAFIIsFHGq34B96NQ2k8B05EFZBKooXyft6qI2z2HtbYn/AApYrdybtrXoMXT7erWqamTRCoaSB5DaSeyn6+/EVp1qtKn1636dt/MX4/5ibA4XGb4x01bkocbT0kJrMfq8tUFhhicrNy1/rZfeqUHTmqvXzv8A/hRv1ovdv84Gn68bLVm3Vr8xtWhGapqWaaSmFbSUqtUROn0ZCbow+luPd1/s2Poa9NSt/jEafxgj+fWxJ07/AMJieu8fnNh707J+Rm5+2sHQY/buYXbe48ZVCmjrBj0klFM71L+i5GhiFJA5APvQdv8AUOrGCMNXNR8+sX/Ckrr3avWXwe6l6h2fjY8LtSnOTw0dNQQtpWKnpoIzO4W5LMSxJJ/w/HvcfdIK9UuiEtmPoQf59F7/AJL/APJa6W7g+CvXm/8AJ71yMFfXbhzhMEVA8hiWEIXWVpZEOpml1CwIt7qWcVUeR6ssMTASMKkiv7ejtfzo/hVkdj/y3MhtnrdazP0XUm1slkPJ9vMZpJtWmL9mEubNe3JPvynSdXXpU1x+EBxGPy6RP/Cdv579Rbd/l77I6v7Qz67Y3rsWsz9Zm6KtNPTCnhqZgyxpFUukjG6G3H196KkHPn1ZXDLUeQA/Z0KvTf8APQi7++cub+NPW1A2R21hNyriKusbGU7MlK1UIFmWshBuLEG+r36lFqevCTVKEA8uPl1ss+9dOdE1+f1/9lQ7Wt+cVTgn+g+7Tn37r2fL5/4Oi5fypz/xhkD+lNHf+v8AwIPvx+Lrw+Hq1f37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3TDunJyYXbuZy0QvLjsfUVaD+rQprAN/9b37r3WqH8WOldt/K7+bT3zuPvL7bfGycfg8rlcXsjcEiy4xK+OYRwGCnDC4Tl2AtwDz7seA/PppVqxr+GlOtjKp+IHxFr8a+Fn6T6rkx8sQpjSpiKBLpbQqI8TBweLAg391p59O6q4r1r0dA7ej+JH80/tTCdeZXxdfbhyFTg8VtKhmWTEYumrJtCpDFcgFBxwPdq9temwKSaRwOfz62vUN0U/1VTz9fp7r051y9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/Q3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691ryfMq3/DunxmP5/gGF/wCtpt735daPxD8+theH/NJ/rf8AE+9deHDrJ791vqjj+Zv/ACPuh/5lG+8D2Pv7IwYTc2EOLjFb9jUVMs9PjY1hSNpIXBsVQKRa1vr72DQGnn1V1V6a64rwNOPr69Wi/H3477N+O/TW3+m9nwpHhsJt6lwbzKrr90aeiNG1U6uSbsDzz719vW6UwMdUBfL7/hNj1Z8nfkVlPkXQ9zZnr7d+TilhaXFUNZJURxTADQs8cgFhY2FvdlZq4P8ALqjRxkZBNPQ06SfVX/CbB+tewNtb4Hy/3/mRgMxjMomNqIa9IpxjZhKISTIQNVvyPfqngafs6r4a1BAbH9Lo8Pye/km9U/Jj5LbX+R+4t3S0W5Nvw7bSWH7aqklr5tvwRwrPJMjWBk8YJPvwYqpUefVmiR5EmYZThnq7zGUSYzG4/GxnVHj6Gkokb/VJSU6wKef6hfdenDk16IV/MA+BOzfnlsTA7K3hmXwtPgauqqoKhIpZtRqgocaYSD/Z97DFTqHVZESWMxuKg9C18Nvi7t34fdG7f6S2vkXymKwVVWVcdW6PGZJKzQJLJISR+ge9Z8+tigUAcAKdGC3ls/bm/wDbGY2fu3FUub27nqR6LK4utUvS1lM5DGKZVIJFwDb37rxr5Y61jfkN/wAJpevexuzM3vbqvvXcvUmE3HVNJUbU29RVUdDQJJMZmEZhm9S3NrG3uwZqUPl008UddWk1PGhp1Zl8Bv5RXx8+DStm8ZR0G++wqmn8VbvnLYsR5aaUlS0/kd3JY6fqfp9R70x1ceroixrRMDq2f3rq/RM/5gBP+yn9rD/VYqAE/wBAKpW4/wBt7917/V/Louf8qbjphR/0zR8/k/5R71ivXh8PVrHvfXuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XumHdOMfNbdzOJj/AF5DH1FKv+vKmn37r32dalO8PgZ/MQ6Z+YfZXdPQu0KzLYLdVNX0dHURTt4vt6uUjhGa1yvuwKlaHj02UlDllIof8PSvPXH848KSnXdcG4ZSJlvqU6lIu39fezppQ+XW9Mo816VHwm/l+/MpflDW93fIvbVTh456v70tUzM4aawYFAGIHq90ZhSijrypIG1OQfs9OtptAQqg/UKAf9cC3v3V+uXv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r/9Hf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Wsn/ADNd+VvS/wDMo6C7gyWMyM+0tvbexD1tXSUMtQqyQEu6CUAoDb8H3viKdNv2spPDPVilP/Nc6Onx2Nq1p9wa6umE8gahTgs3HGnjj3rq9fn10f5rnSAP/AfPf+cKf8U9+r8uvfn11/w690dbV9tn7f0+xXn/AGNvfvy61+fXL/h17ozSCafP67/8qQt/ttPv2a9bp8+ubfzW+jOAtPn9Rt/yhA8WueCvvWQc9e/Pr3/DrfRqj10+fuTa32Q/pwf0+9569Sua9cf+HWukb6ft89e1/wDgCv0/23v3Xsca9dL/ADXejbkNT5/6cWoV+v440+/GvXuJ49d/8Ot9H6Linz5bkn/IVtYfi2n37PXvz68f5rvRojV/ts/cmx/yJbD6/T0+/Zr1r8+ul/mu9HMQPts8L2JtRL9L/wCt78a+Q635Vr1yP81zo0Pp+2z+m4H/AACH5/x0+/Z69Q8a9eP81zo0Fl+2z4P0T/Il+p+l7r/re/Z69+fXFf5rfR+oBoM8ebEfYqPxf6hffj178+is/Nf+Zt1JvP439ibW21R5+bNZajo6elQ0OsEeYNICirzzb34Ak0OOqsxQV446Nh/KdxGYT447f3PkqSakh3FQ+amSeF4JLRzgnUj2P5v7959WXAp1ad791vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6/9Lf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Vb/zx8f8LpNX+y+X8Sf8zU+5/vD/AJs/8W3xcaP6X9+8urDquek0fZ0n/ZOf+ZT/ADX3Hh/T/ui39n37y6oePXN/HY/9k4/X/ppv/vHvw4efWxx68ujn/snH6/8ATRf37rx49e/bt/3Lj+P0/c+/daHHroaLn/snT6fj7jV9Pfur+XWRtFx/2Tp9B+v7i3+w96HVPw9dnx6z/wBk6/Qf8rN/p7314/D1wOi3/cuX0/6aP+J9668OPXaeO3/cuv0/H3On34dWPXL0WH/ZOX6v+mjT/sPez1UddejV/wBy4/j9H3H+8W/Hvw623HrptGof9k6fUfX7i/8Avv6e/eXWv2dcW0X/AO5c/qv1+4v71+3qw4eXXaePV/3Lp9R/ys6/fv29ab8uodV49a3/ANlXtqF/7x/c/wAM/wA8v/ArRz/tvz78fh6sPLq9/oXT/op2hp/uTb+H8f6PfJ/dT9f/AC6vL6tP9b/n3peA683E8ehh926r1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/2Q==)
Figure 5. Logistic Regression Findings. Selected results from the logistic regressions are presented (for additional findings, see Table S6 in supplementary materials). Circles with positive values represent a 100% increase in the odds that a variable predicted an increase in activity in that brain area. Circles with negative values represent a 100% increase in the odds that a variable predicted there would not be an increase in activity in that brain area. Legend: Blue lines: left hemisphere; Green lines: right hemisphere. Arrowheads: % change in odds is greater than values represented in this figure. Abbreviations: OFC: orbitofrontal cortex; DLPFC: dorsolateral prefrontal cortex; ATL: anterior temporal lobe; VLPFC: ventrolateral prefrontal cortex; DMPFC: dorsomedial prefrontal cortex; aMCC: anterior mid-cingulate cortex; sAAC: subgenual ACC. A color version of this image can be viewed in the online version of this target article at http://www.journals.cambridge.org/bbs.
![[[@Lindquist2012-jr]](http://dx.doi.org/10.1017/S0140525X11000446)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAgEAyADIAAD/4RqdRXhpZgAATU0AKgAAAAgABwESAAMAAAABAAEAAAEaAAUAAAABAAAAYgEbAAUAAAABAAAAagEoAAMAAAABAAIAAAExAAIAAAAbAAAAcgEyAAIAAAAUAAAAjYdpAAQAAAABAAAApAAAANAAAADIAAAAAQAAAMgAAAABQWRvYmUgUGhvdG9zaG9wIENTIFdpbmRvd3MAMjAxMjowNToxOCAxNzozOTowOAAAAAAAA6ABAAMAAAAB//8AAKACAAQAAAABAAAFeKADAAQAAAABAAADnwAAAAAAAAAGAQMAAwAAAAEABgAAARoABQAAAAEAAAEeARsABQAAAAEAAAEmASgAAwAAAAEAAgAAAgEABAAAAAEAAAEuAgIABAAAAAEAABlnAAAAAAAAAEgAAAABAAAASAAAAAH/2P/gABBKRklGAAECAQBIAEgAAP/tAAxBZG9iZV9DTQAB/+4ADkFkb2JlAGSAAAAAAf/bAIQADAgICAkIDAkJDBELCgsRFQ8MDA8VGBMTFRMTGBEMDAwMDAwRDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAENCwsNDg0QDg4QFA4ODhQUDg4ODhQRDAwMDAwREQwMDAwMDBEMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwM/8AAEQgAagCgAwEiAAIRAQMRAf/dAAQACv/EAT8AAAEFAQEBAQEBAAAAAAAAAAMAAQIEBQYHCAkKCwEAAQUBAQEBAQEAAAAAAAAAAQACAwQFBgcICQoLEAABBAEDAgQCBQcGCAUDDDMBAAIRAwQhEjEFQVFhEyJxgTIGFJGhsUIjJBVSwWIzNHKC0UMHJZJT8OHxY3M1FqKygyZEk1RkRcKjdDYX0lXiZfKzhMPTdePzRieUpIW0lcTU5PSltcXV5fVWZnaGlqa2xtbm9jdHV2d3h5ent8fX5/cRAAICAQIEBAMEBQYHBwYFNQEAAhEDITESBEFRYXEiEwUygZEUobFCI8FS0fAzJGLhcoKSQ1MVY3M08SUGFqKygwcmNcLSRJNUoxdkRVU2dGXi8rOEw9N14/NGlKSFtJXE1OT0pbXF1eX1VmZ2hpamtsbW5vYnN0dXZ3eHl6e3x//aAAwDAQACEQMRAD8A9VSVTDdTDW0XuyGTZLnP3mQWy3d/watpKUqLhhjJvOTW0uLmlpcyZbsaNHbfo7t6NmZ+Hg1tty7W01ucGNc7guIJDf8AoqwjRq60KLF1ezzj7eqModYMLAst26UM3SHa/wCEcwMf/U/8ESLuqaObi4Gsu2ulp1DWtqdtZb+dvtsfu/4JdGkgposGIcqk4zGhwLtxazbpt7u2/vbVyed/jf8Aq3g5uRhXYuabcW19Ly1lRaXVuNbi3dkN9vtXdLyv6q9M6Q7rvW+pZuOcjJHVcimp5AihrHPv9dsvbte5zv5zZ7NjPT/nEkuvZ/ji+r9U+r0/qNe0w7dVWIJEwd2StX6vf4wOlfWL1f2fj5DPQsprs9cMb/Pl7GOZ6dt27Z6XuQvUpv8At7OoVV5VDrHTU15yQS5sN/QOZW30G1bHV1bn/pvZX/R965v6jYGN03r3X8HFJOPRnYLap5Dd97tv9idqVjodtD4KfUUlUyBifaAX1bsn03hluwmG/ns9bbtb/U3K036I+CSl1RtGKMyw5LGuljNpczdoDZw7aVeSSU41hefVFVGG2LB6DngkGraNzrWtr9tvrbv0f+jQg7M31k4uCGAtFsE6gvZ6jmfov8Fji7b/AKSz0lvJJKaAGEcjHOPW0PDySWsiG7LBq7b+9tXF/WD/ABtu6L1zK6QOk/aXYzxWLBkbS6Q13839ms2/S/fXoa8lOV0HD/xh9ayeo3CvNbawYYe0huraw/Zczf6dz5/cZ7P8N+lSU9Hh/Xr6z5bPUH1YNDCJacjMbSSZ27Gsuxm2b3fu7Fn53+NnqfTc2vC6j9XXY11u3aHZQIIcdoe1zMVzHt/quW1ifaPTi+urADPa59jpc0vLq9hfZDat297a6v0387/pFy31/wDsVmDRqX24+bWKHMnaC/8Andxez3V+nWz6Fn86kp//0PTaXXuc03VNpd+k9rXb9AW7HbtrPpt9ynlZWPiUOyMmwVUsjc92gEna3/pOQMR1LiDTechs2y9zt0Hc3czT/R/RVL63mPq7lH/i/wDz7WnY4iU4xP6UhH7Vs5cMJSH6IJ+xqfXSxlnR8W2shzH5FbmuHBBZaWldIuR+tDo+q/TD42Uf+erFp9ez8zF6l0mnHtNdeTcWXNAadzd1I2+9rnN+m76CmMDKGOI75N/6rEJ8M5yPbHt/WdtZVHVciz6w5HSixgoppFrXid5J9LR35u39Kmyeq5NX1ixOltaz7PkVOse4g75AtPsdu2f4L9xUsM/9nOcP+6rf/ddNhjoSMhvj44/43CulOzERO0+CX+LxPRrzln1D+uWH1LqeT07qWIzG6lkWXOx7Wvsb7rDdW7Y6pzW3N9n0F6MoVXU3Amqxtgadri0h0H907VCyvnfTvqJ9dul2WPwep4jX3lrrbXh73lxH6x9KnZ+kfv8AT9v6Kv2K39Xvqp1T6v5mZl9Sya8p3VMzFcw1lxILH3Pd6nqNZ+bYzZtXdqh1fjD/APDdP5SgAAr7ftbN5yd8Maz0Njt7i47w78zawN27P3v0iM36I+CrZHpes0uvLLPTfto3gBwj3Wen9J3p/vKy36I+CKl0kkklKSSSSUpeYfWf/FR1frXX83qlObj1V5Vgexjg/cAGtb7trdv5q9PSSU+XX/4r/rVk42PjZnU6MyvFe97GXvvcw7wGsBYNv8z73sf/AMJ6f80rGf8A4vPrV1KquvNz8V5rtZbuBtiWGwucynb6VW9tv0K/0X/Frtf2vb/zh/ZPpt9P0PW9WTumfo7I2rURljqr6+sarRwyur9Jrr8wf//R9EzsvJx8HIyjW2q2im57Gk7x7AHMc7bs+ksbrGddnfUV+Zdt9W1tZdsEN/nmN0BLlu5mNW+m2q8WZNVrLA+oGC5r9rXUs2+l7f7ap2dP6aekDBdhXPwdrWjBBdv+mXif0od9Nu/+eUuOcY8JI1E4yv8Aqx6Mc4SlxAHQwMa/rFx/rW6Pqp0o+NmP/wCebFb+tTo6z0AeOSf+rx1fz+ndOvwq8fKxLcnHocw1Y7HHdWWs2s+jYx3ta7Z9OxEzMHCysvFsycezJtx7HPx7WuIbUQ5mr/0lf7rf9J9BOjmA4dD6fc/8cC04ieLUa8H/ADHN6g7/ALOumN8cd/8A1OSmwj/2fZ4/7qN/91lo24eE/qWPl2Y1tmYxjmVZgJ2MBFvtfD2t/f8A8D/hEmYOG3qF+czHsZmupDH5hcdr2xWdjfft/NZ/gf8ABoe6Kqj/ADft/Xi4k+2bvT+c4/pw8KHonUsvL6v1bGvs31YljW0t2gbQTb+c0bnfQ/OWd9VC09G6sQAIvumP+LatjEw8LHzMu7Hxrca6+xjr7nE7bTueBsmyxv7/AOZX9NRwMDAxsW1mLiXY1V73uupcTusLme5w3WWbd/0fpVpHJGpACuLg/wCZ8yBjlcSTfDx/8/5XL+r2a/C+pJzKWtL6Ra9rXfRkPdzthX3Zdmb0zpWXYA1999D3BswCZ+juUqun9Oq6UcGvCuZgua9rsKXF53Pbuj9KX/Sd/pk2RRVjY/T8eit1NFd+OK6XGSwS/wBurn/9WhknGXEQPVKZlf8AVPRdjhKPCL0jARr+sHSvc0WbfRc55Y6LQBDQB9AvJ/OVLF6vbb17K6Sa2ivFqa9tgJ3EkVOhzY2/4VaF4tI9jmhkO3ggknT27fc385ZWPg4lXXMrPqyt+dfW1luMdpa0N9Fm5rG7bPzWf4RNgY1LiH6Pp/vWulxXHh/e9X910MTqWHmXZFGO8vsxH+neC1zdrpc2Je1u/wCg76CF0zrOH1LEszKt1VNLnMebYbG0Ne52jnN2bXKr0nCw8XLz78PKOQ/Kv3ZDXQW1uDrd1bNmzbtf6jff6n0EHpnTun4fR8rExs03YdptORkO2lzN1bd/uYGVt2V/pPexE+36qv8AR4f/AFItHuaXX6XF/wBw6Gf1anH6Tb1PGLMqtglmx42u93pn9K0WfRcrGDk/a8LHyi3Z69bLNszG9oft3e3dysgdKwW/V79lfbHfYC0j7Sdu/W3f9OPS/nHbP5pRz+m9Oy+hY2BdmlmCxtP2fIbt3PDWubX7nB9b/UY3d7K0qx7WR6z6q/yfT0q/Wb6fKPTf+U83oElUsy68ctZfeyrc4Mr3CNx9g/e+lusVLKwsXI6xh5V2Wa8vG3+jjtgNeCXs3PYdz/8AppgA6mtCvJPQW0t3/Z7t/wC6c/itTE6xXldUy+mipzXYYaXWEgh24fmt+kqn2PD/AG5+1PtX+UPs/p/Zvbs2x6m7Z/O/+CqeFg4lHV87Ox8o3ZGQWMyKTBayC1sN2gPa7+u9SylCQ13EBGP98FjjGQOmxmZH+6//0vQz13ofqNd+0cWACCfXr77f5aE3r+F9o93UenjH11F7S+JdtEbtn0di0KPXhjcltfqEOLvTnboW7Y3/ANZG2t8AkpzH9d6QG2mnqOH6hH6MvvZt3R7d+1+7buQ8f6wYRsP2nqHT21wY9PIaXEyNn0y3b7Vr7W+AVZrLLbrgLXVtY4Na1oZH0WP/AD63u/OSU1Hdd6SKCKepYQt7b7mFvPfZZ+6nHXukOxyLeo4fqkEEMvZtn+Tuel1DJdgmsE5N3q7ta2VEDbt9hPpfztu79Xr/AMPb+iUsC5+a17t91Br2h1dgqDwXNFnuZ6Xt+kkpDldfwgR9k6j09w7i29ojzmtz0Ude6MRUX9Qww8a2bb6yAdpnbLm+3crJZZXfSDa6xry4Oa4MjRpd+ZWx35q8z+tGZ9Ycr/GHd0bB6zb0vFNdTpD37Gj02Od6VNf07bHu9rP8I9JT6E7r/TPtDNvUcH7PHvJvZvmD9D37PpbFU6n1no9j8Ysz8VwGTQTF1fYv3fn+a5yzoPUDZfRV9aOpVvqZWWZL7vUYXO9r2nGYyp/b/T/o/wDCLJ+ofU+tZXXc/B6rn2dQGDdSxhfYbGb2ZApN1O/95v5/7iSn0h/XehuY4DqOKSQQP09f/k0K/wCsXR62NDMmjIMatrvoBBHH89fV9JaDn4zbBU5zBYRuDCQHED87ap7Gfuj7klOd+3ui2MY452MwyC5pvqJHx22Oag3fWfpbLHBllVwZ9CxuRjAOkD/SZNb2/wCYtfYz90fcq5rfZk2MFhrYxrIa0M5O/d9Nj0lIP270TeHftDFjbB/T18/56HV9Y+juue1+Vj1gSBacigtcB9Hbtt9T878+tP1DLrwHMre/JuttY99ddFTLHO9PYHDa2r/hmJYOT9ssLGWWsAYLAXioEgucwez0d+5uz9Nu/mLP0P8AOJKXd1/ozA9zc7GeS4FrRfUCRDW/nWBqbF+sXSbQ7fdVjAHT1L8c7p1dt9DIv+j/ACladU+u2j9K57XvLXNc1kRssf8AmVsd9Ji8q+uln1uyvrxm4HRMzLox6m0F4qvsqoqD66/fZse2tm53v/0liSn0x3W+kMx4+149rg0A1NuqDj+a4fpLWM/6akOudIsqaTmY7Hna5zDdUS3UOcDteW+1eXN6R9ZmNm36zZ9rw0bmVXWgbvzvTtsv9zP5T66v+LWV1P8A579Iux7resZuRg3WtY25uTdEk/zd1brP0bnt/sPSU//T9Kwn4721uxrjfWWv22OcXz7mT73K2gUvue5jrqxU/a+WB27uyPdDfpI6SlIL8Wix5e5vudEkEiY8dpCB1PquN0yuqzID3C6wVN2CfcQT7tW+32qOf1evCzcPDdW57s5xa17YhsbR79x3fn/mpwhI1Q3uv8H5lpnEXZ2q/wDC+VsDDoHAcJ8Hv/8AJJHDoPIcY497v/JKnd1ayvr9HSRU013Um027juBHqe3Zt/4L95aaUomNX+kOIeShIG6/RPCfNCzForeHtb7myASSYnmNxK4F+XgV/wCM/qeBlGLc+nFrxy7RpLGMt9PeP0jLNza7Ktv+i2fuL0Rch9Yv8XPQOsdTu65nZeXj3bWuc6mytjWCpoaHt30WWN2tZu+mmrl63ZVvUsnHupZ9kfWwF0OaNsO3e/cNvtdd9HZ/wr1gfV7qHTeo/XbqeR007qdmFW+zSHvZbVW97dvtc32+nvZ7H7N626/qN03qeEXV9e6tk4uS0se85DHeoAdjvVc/G3WfR9L+p7FLC+pXTPq3mDNwrr7rs7IpruFxr2gB/q/o68ajHaz3MS1BoiqQDeuleb098+sIx949N36xLfb/AMHr+k9/8lWG/RHwVXJdUMlrXZLmWGqzbjAtAeB9K3bt9TdX/JerTfoj4JJXQrMWix+97fcQASCRIE7Z2kfvIqSSkAw6BwHCeYe7/wAknOJSedx+L3/+SRkklIWYmOx7Xtb7mztJJMSNum4/urzb6x/WDo/S/rZ1TGziarLH49hdsL63tGPS1otNJbe3Z7/o/wCkXp6wOq/UP6qdYzrOodRwfXyrQ0Ps9W5k7Witnsqurr+g391JTxHSOq0dYqz/ANmZ17L8et1te8sBduD2fZKMRwqpfW2v/Bsr/wBH+k9RD6n1bCwejnCuuHUK85lbMZjnj1fdtdXZcxrXt249rfV9Vn+FZ/wi7Wj/ABdfVDGdW7Hw30upf6tRZk5ILbNB6rP1j6ftUXf4tfqW+0XP6e59oMh7sjIJka98hLXwVq//1PSW2U41Ystyd9VbLHPvsc2AAWbtzmBjPYq3W+sHC6K7qWCa7hLPTcZcwhzg3d+jc1LqmJZk9OyMPDpFVltd1dbXQxs7mjf7N3ss+n9FZ+b0bqWR9Va+jVtZ9trZVuJfFftdudD4L/zP9EpcYhcTI/pgSH9RjyGdSER+gaP9dh9b7i/pvTLXQHWZNTiBxLmOKJ9Y3R1/oI8bX/lpUuu9J6h1HAwcbGY0W4d1T7t7obDGQ70y0P3fTROsdNzs/q3SszHY0U4Fr3X73AOIJr/mmt37v5t30nMUkJxHBqNBl/50fSxyhI8em5x/80+pDlu/7OsJv/dQ/wDuwukWDf07Nt+s+L1lrG/Y6cY1vBcPU3H1j7WfQ2/pWf4VXKrOst6nfZe1h6WWN+ztaR6oefT3ep+Zt3er/hFHOpCNEemA/wCl8rJAGJlYPqn+zd0lV6oY6ZmH/gLP+ocqnTm9Zpy81/UHCzHttH2JrS0ljJs9r/bV+b6X59qHj19dt6VlYvUxW/MvFrKHVEBga5m2v1DDHfT3/mJoiAb4gaMfx/71JkSK4SLB/D/vmp0XqBwPqZVnBnqmkWHYTtn9M9v0of4/uq7k5Jy8DpeUW7Dffj2bJmN437d0N3cqjV0bqTPqj+xC1n20sfBD/wBHrb6v85G/6D/9ErJpuxumdJxbwBbj24tb4MtLmjY7Y793c1PymB4iPmOSX+ItxiY4QflEI/47q3HI3QxrTTsdvcXEODvzA1m3/vyIx7D7QRuAEtnVByRULGuc+wPLHtYxpdsOm529rPbu/d3LGwqMmv629QzrKHsxLaWNryC3RxDaG7f9J+a781RxiCJWaqNj+sf3V8pEcNC7NH+r4vRKvn3Powcm6sw+qp72E6iWtLmqNOc22y1jqbqvTdtDrGENfz7qtu7c32qHU3G3p2VTU1zrbKbG1sDXSXFjtrdQkBUhfcJJ9JrsUX1ezcjP6Pj5eS4Ous37nNG0e172D2/1WrSWN9VmXYnQsXGy6n05DBZvrc0kibHuH0Nzfouag/W9mZldJNPT6rbbxYxxZWC122H6y7008xjLMYgiMTIi/wBERtYJSjiBIMpCI0/SJp2cjMxMZ1bci5lRudsqD3Bu5x/MZu+k5GXOfWSjLyrulHFosuFGUx2QWj6DQa3Fzt21S6nTl3fWXpeTRTY/FpFgvtAIa0+5vubo7/ooCESI+qrEif8AA+Uf4SjOQJ9N0Ygf4XzH/BegkTHdOuc9DI/54ftD0H/Yvsm37Tt9u76W3/Sf9BH6c/qY+sHUXZQuGA7YMUvk1z7Q702/me5I4xViQ+UT+p/QSJm6MT8xj9P3n//V9PvqZc4VWTtc107SWnln57C16q7el3EYIyA9wMek293qSwuc7c5tnrv93096un+eb/Vd+ViZv8876P0Rx9Ll30v5H7v9tJTXyKMQ1vdkuNdVJncLH1wNrG+59b2Jqn9Nyshjqsht91e57G12yAN0Pc6qt/pu2vds97FaZ9Kz+sP+pamp+ifo/Sd9Dj6Tuf5f+k/4RJTWsqxGNZm3vNZraG7zY5rBumpu+vf6Lv53/CMU214xDsul3qG7b7w8uaQC3+bbudU3j/Bo9P8ANN+ATW/zR+X5UlNV7+mYNr/VyG0WWkPcLrTBkvd+jbe/az3ep/NKdeLjNpbTUXOptDjPqOeSHN5bc57rPo/uvVi3gfR5/P4+X8pMfpV8d+OOElNZ1WAI6e60hzwSKfVd6pBPqOcH7/tG32/vIPVGtYMNjfotyaAO+gLvFaXft/FZ/V+cT/w1R+VySm/Z/Nu+B/Igtos3mz1nuY4CKXBuxv0folrG2/m/n2I1n8274H8iyPrD/Rq/6N9B/wDSv7H81/6N/wCDSU6LabK3EvvfdvfLQ8MAYCXHYz0mV/R3bf0nqJm41wrcw5Nj3HdttcGb2S3YNmytlXtd+k/SVpsf+i0fzfb+Y/m/+tfyVh9W/wCUbv8Ak3g/z/0voU/03/0V/ISU7xpftFXr2b9p/Tezf9IO/wBH6P8AJ/mlH7Ncams+1WBzQ0OuaK97y0Oa7fNbqm73H/B1ow/nG8fQ7cchZ+D/AMq5X9D/ADv5n+f5Z/P/APo3/rKSm3ZTZY8ll76NrwSGBhDtK/a/1WWeG32bFIUWeq2z1ntY2ZpaG7HSXOl5cx1v535lihlf0e/+a+k3+f8A5v8Awf0/++fy1m/VjjI/on+D/ofwP87/AN8/kJKdJuPa4B4vftdXHowzZJbtmfT9b/wVS9J7HOe659ge9paxwaGtG5vtZsYx/wD24+xBz/8Akt3819Bv8/8AzfLf5xExf6Bj/wA39Gv+Z/m+W/zP/B/uJKf/2f/tILRQaG90b3Nob3AgMy4wADhCSU0EBAAAAAAABxwCAAACAAIAOEJJTQQlAAAAAAAQRgzyiSa4VtqwnAGhsKeQdzhCSU0D7QAAAAAAEADIAAAAAQABAMgAAAABAAE4QklNBCYAAAAAAA4AAAAAAAAAAAAAP4AAADhCSU0EDQAAAAAABAAAAB44QklNBBkAAAAAAAQAAAAeOEJJTQPzAAAAAAAJAAAAAAAAAAABADhCSU0ECgAAAAAAAQAAOEJJTScQAAAAAAAKAAEAAAAAAAAAAjhCSU0D9QAAAAAASAAvZmYAAQBsZmYABgAAAAAAAQAvZmYAAQChmZoABgAAAAAAAQAyAAAAAQBaAAAABgAAAAAAAQA1AAAAAQAtAAAABgAAAAAAAThCSU0D+AAAAAAAcAAA/////////////////////////////wPoAAAAAP////////////////////////////8D6AAAAAD/////////////////////////////A+gAAAAA/////////////////////////////wPoAAA4QklNBAgAAAAAABAAAAABAAACQAAAAkAAAAAAOEJJTQQeAAAAAAAEAAAAADhCSU0EGgAAAAADYQAAAAYAAAAAAAAAAAAAA58AAAV4AAAAFgBTADAAMQA0ADAANQAyADUAWAAxADEAMAAwADAANAA0ADYAXwBmAGkAZwA2AAAAAQAAAAAAAAAAAAAAAAAAAAAAAAABAAAAAAAAAAAAAAV4AAADnwAAAAAAAAAAAAAAAAAAAAABAAAAAAAAAAAAAAAAAAAAAAAAABAAAAABAAAAAAAAbnVsbAAAAAIAAAAGYm91bmRzT2JqYwAAAAEAAAAAAABSY3QxAAAABAAAAABUb3AgbG9uZwAAAAAAAAAATGVmdGxvbmcAAAAAAAAAAEJ0b21sb25nAAADnwAAAABSZ2h0bG9uZwAABXgAAAAGc2xpY2VzVmxMcwAAAAFPYmpjAAAAAQAAAAAABXNsaWNlAAAAEgAAAAdzbGljZUlEbG9uZwAAAAAAAAAHZ3JvdXBJRGxvbmcAAAAAAAAABm9yaWdpbmVudW0AAAAMRVNsaWNlT3JpZ2luAAAADWF1dG9HZW5lcmF0ZWQAAAAAVHlwZWVudW0AAAAKRVNsaWNlVHlwZQAAAABJbWcgAAAABmJvdW5kc09iamMAAAABAAAAAAAAUmN0MQAAAAQAAAAAVG9wIGxvbmcAAAAAAAAAAExlZnRsb25nAAAAAAAAAABCdG9tbG9uZwAAA58AAAAAUmdodGxvbmcAAAV4AAAAA3VybFRFWFQAAAABAAAAAAAAbnVsbFRFWFQAAAABAAAAAAAATXNnZVRFWFQAAAABAAAAAAAGYWx0VGFnVEVYVAAAAAEAAAAAAA5jZWxsVGV4dElzSFRNTGJvb2wBAAAACGNlbGxUZXh0VEVYVAAAAAEAAAAAAAlob3J6QWxpZ25lbnVtAAAAD0VTbGljZUhvcnpBbGlnbgAAAAdkZWZhdWx0AAAACXZlcnRBbGlnbmVudW0AAAAPRVNsaWNlVmVydEFsaWduAAAAB2RlZmF1bHQAAAALYmdDb2xvclR5cGVlbnVtAAAAEUVTbGljZUJHQ29sb3JUeXBlAAAAAE5vbmUAAAAJdG9wT3V0c2V0bG9uZwAAAAAAAAAKbGVmdE91dHNldGxvbmcAAAAAAAAADGJvdHRvbU91dHNldGxvbmcAAAAAAAAAC3JpZ2h0T3V0c2V0bG9uZwAAAAAAOEJJTQQoAAAAAAAMAAAAAT/wAAAAAAAAOEJJTQQRAAAAAAABAQA4QklNBBQAAAAAAAQAAAAJOEJJTQQMAAAAABmDAAAAAQAAAKAAAABqAAAB4AAAxsAAABlnABgAAf/Y/+AAEEpGSUYAAQIBAEgASAAA/+0ADEFkb2JlX0NNAAH/7gAOQWRvYmUAZIAAAAAB/9sAhAAMCAgICQgMCQkMEQsKCxEVDwwMDxUYExMVExMYEQwMDAwMDBEMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMAQ0LCw0ODRAODhAUDg4OFBQODg4OFBEMDAwMDBERDAwMDAwMEQwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAz/wAARCABqAKADASIAAhEBAxEB/90ABAAK/8QBPwAAAQUBAQEBAQEAAAAAAAAAAwABAgQFBgcICQoLAQABBQEBAQEBAQAAAAAAAAABAAIDBAUGBwgJCgsQAAEEAQMCBAIFBwYIBQMMMwEAAhEDBCESMQVBUWETInGBMgYUkaGxQiMkFVLBYjM0coLRQwclklPw4fFjczUWorKDJkSTVGRFwqN0NhfSVeJl8rOEw9N14/NGJ5SkhbSVxNTk9KW1xdXl9VZmdoaWprbG1ub2N0dXZ3eHl6e3x9fn9xEAAgIBAgQEAwQFBgcHBgU1AQACEQMhMRIEQVFhcSITBTKBkRShsUIjwVLR8DMkYuFygpJDUxVjczTxJQYWorKDByY1wtJEk1SjF2RFVTZ0ZeLys4TD03Xj80aUpIW0lcTU5PSltcXV5fVWZnaGlqa2xtbm9ic3R1dnd4eXp7fH/9oADAMBAAIRAxEAPwD1VJVMN1MNbRe7IZNkuc/eZBbLd3/Bq2kpSouGGMm85NbS4uaWlzJluxo0dt+ju3o2Zn4eDW23LtbTW5wY1zuC4gkN/wCirCNGrrQosXV7POPt6oyh1gwsCy3bpQzdIdr/AIRzAx/9T/wRIu6po5uLgay7a6WnUNa2p21lv52+2x+7/gl0aSCmiwYhyqTjMaHAu3FrNum3u7b+9tXJ53+N/wCreDm5GFdi5ptxbX0vLWVFpdW41uLd2Q32+1d0vK/qr0zpDuu9b6lm45yMkdVyKankCKGsc+/12y9u17nO/nNns2M9P+cSS69n+OL6v1T6vT+o17TDt1VYgkTB3ZK1fq9/jA6V9YvV/Z+PkM9Cymuz1wxv8+XsY5np23btnpe5C9Sm/wC3s6hVXlUOsdNTXnJBLmw39A5lbfQbVsdXVuf+m9lf9H3rm/qNgY3TevdfwcUk49GdgtqnkN33u2/2J2pWOh20Pgp9RSVTIGJ9oBfVuyfTeGW7CYb+ez1tu1v9TcrTfoj4JKXVG0YozLDksa6WM2lzN2gNnDtpV5JJTjWF59UVUYbYsHoOeCQato3Ota2v22+tu/R/6NCDszfWTi4IYC0WwTqC9nqOZ+i/wWOLtv8ApLPSW8kkpoAYRyMc49bQ8PJJayIbssGrtv721cX9YP8AG27ovXMrpA6T9pdjPFYsGRtLpDXfzf2azb9L99ehryU5XQcP/GH1rJ6jcK81trBhh7SG6trD9lzN/p3Pn9xns/w36VJT0eH9evrPls9QfVg0MIlpyMxtJJnbsay7GbZvd+7sWfnf42ep9Nza8LqP1ddjXW7dodlAghx2h7XMxXMe3+q5bWJ9o9OL66sAM9rn2OlzS8ur2F9kNq3b3trq/Tfzv+kXLfX/AOxWYNGpfbj5tYocydoL/wCd3F7PdX6dbPoWfzqSn//Q9Npde5zTdU2l36T2tdv0Bbsdu2s+m33KeVlY+JQ7IybBVSyNz3aASdrf+k5AxHUuINN5yGzbL3O3QdzdzNP9H9FUvreY+ruUf+L/APPtadjiJTjE/pSEftWzlwwlIfogn7Gp9dLGWdHxbayHMfkVua4cEFlpaV0i5H60Oj6r9MPjZR/56sWn17PzMXqXSace0115NxZc0Bp3N3Ujb72uc36bvoKYwMoY4jvk3/qsQnwznI9se39Z21lUdVyLPrDkdKLGCimkWteJ3kn0tHfm7f0qbJ6rk1fWLE6W1rPs+RU6x7iDvkC0+x27Z/gv3FSwz/2c5w/7qt/9102GOhIyG+Pjj/jcK6U7MRE7T4Jf4vE9GvOWfUP65YfUup5PTupYjMbqWRZc7Hta+xvusN1btjqnNbc32fQXoyhVdTcCarG2Bp2uLSHQf3TtULK+d9O+on126XZY/B6niNfeWutteHveXEfrH0qdn6R+/wBP2/oq/Yrf1e+qnVPq/mZmX1LJryndUzMVzDWXEgsfc93qeo1n5tjNm1d2qHV+MP8A8N0/lKAACvt+1s3nJ3wxrPQ2O3uLjvDvzNrA3bs/e/SIzfoj4Ktkel6zS68ss9N+2jeAHCPdZ6f0nen+8rLfoj4IqXSSSSUpJJJJSl5h9Z/8VHV+tdfzeqU5uPVXlWB7GOD9wAa1vu2t2/mr09JJT5df/iv+tWTjY+NmdTozK8V73sZe+9zDvAawFg2/zPvex/8Awnp/zSsZ/wDi8+tXUqq683PxXmu1lu4G2JYbC5zKdvpVb22/Qr/Rf8Wu1/a9v/OH9k+m30/Q9b1ZO6Z+jsjatRGWOqvr6xqtHDK6v0muvzB//9H0TOy8nHwcjKNbaraKbnsaTvHsAcxztuz6SxusZ12d9RX5l231bW1l2wQ3+eY3QEuW7mY1b6barxZk1WssD6gYLmv2tdSzb6Xt/tqnZ0/pp6QMF2Fc/B2taMEF2/6ZeJ/Sh3027/55S45xjwkjUTjK/wCrHoxzhKXEAdDAxr+sXH+tbo+qnSj42Y//AJ5sVv61OjrPQB45J/6vHV/P6d06/Crx8rEtycehzDVjscd1Zazaz6NjHe1rtn07ETMwcLKy8WzJx7Mm3Hsc/Hta4htRDmav/SV/ut/0n0E6OYDh0Pp9z/xwLTiJ4tRrwf8AMc3qDv8As66Y3xx3/wDU5KbCP/Z9nj/uo3/3WWjbh4T+pY+XZjW2ZjGOZVmAnYwEW+18Pa39/wDwP+ESZg4beoX5zMexma6kMfmFx2vbFZ2N9+381n+B/wAGh7oqqP8AN+39eLiT7Zu9P5zj+nDwoeidSy8vq/Vsa+zfViWNbS3aBtBNv5zRud9D85Z31ULT0bqxAAi+6Y/4tq2MTDwsfMy7sfGtxrr7GOvucTttO54GybLG/v8A5lf01HAwMDGxbWYuJdjVXve66lxO6wuZ7nDdZZt3/R+lWkckakAK4uD/AJnzIGOVxJN8PH/z/lcv6vZr8L6knMpa0vpFr2td9GQ93O2Ffdl2ZvTOlZdgDX330PcGzAJn6O5Sq6f06rpRwa8K5mC5r2uwpcXnc9u6P0pf9J3+mTZFFWNj9Px6K3U0V344rpcZLBL/AG6uf/1aGScZcRA9UpmV/wBU9F2OEo8IvSMBGv6wdK9zRZt9FznljotAENAH0C8n85UsXq9tvXsrpJraK8Wpr22AncSRU6HNjb/hVoXi0j2OaGQ7eCCSdPbt9zfzllY+DiVdcys+rK3519bWW4x2lrQ30Wbmsbts/NZ/hE2BjUuIfo+n+9a6XFceH971f3XQxOpYeZdkUY7y+zEf6d4LXN2ulzYl7W7/AKDvoIXTOs4fUsSzMq3VU0ucx5thsbQ17naOc3ZtcqvScLDxcvPvw8o5D8q/dkNdBbW4Ot3Vs2bNu1/qN9/qfQQemdO6fh9HysTGzTdh2m05GQ7aXM3Vt3+5gZW3ZX+k97ET7fqq/wBHh/8AUi0e5pdfpcX/AHDoZ/VqcfpNvU8Ysyq2CWbHja73emf0rRZ9FysYOT9rwsfKLdnr1ss2zMb2h+3d7d3KyB0rBb9Xv2V9sd9gLSPtJ279bd/049L+cds/mlHP6b07L6FjYF2aWYLG0/Z8hu3c8Na5tfucH1v9Rjd3srSrHtZHrPqr/J9PSr9Zvp8o9N/5TzegSVSzLrxy1l97KtzgyvcI3H2D976W6xUsrCxcjrGHlXZZry8bf6OO2A14Jezc9h3P/wCmmADqa0K8k9BbS3f9nu3/ALpz+K1MTrFeV1TL6aKnNdhhpdYSCHbh+a36SqfY8P8Abn7U+1f5Q+z+n9m9uzbHqbtn87/4Kp4WDiUdXzs7HyjdkZBYzIpMFrILWw3aA9rv671LKUJDXcQEY/3wWOMZA6bGZkf7r//S9DPXeh+o137RxYAIJ9evvt/loTev4X2j3dR6eMfXUXtL4l20Ru2fR2LQo9eGNyW1+oQ4u9Oduhbtjf8A1kba3wCSnMf13pAbaaeo4fqEfoy+9m3dHt37X7tu5Dx/rBhGw/aeodPbXBj08hpcTI2fTLdvtWvtb4BVmsstuuAtdW1jg1rWhkfRY/8APre785JTUd13pIoIp6lhC3tvuYW899ln7qcde6Q7HIt6jh+qQQQy9m2f5O56XUMl2CawTk3eru1rZUQNu32E+l/O27v1ev8Aw9v6JSwLn5rXu33UGvaHV2CoPBc0We5npe36SSkOV1/CBH2TqPT3DuLb2iPOa3PRR17oxFRf1DDDxrZtvrIB2mdsub7dysllld9INrrGvLg5rgyNGl35lbHfmrzP60Zn1hyv8Yd3RsHrNvS8U11OkPfsaPTY53pU1/Ttse72s/wj0lPoTuv9M+0M29Rwfs8e8m9m+YP0Pfs+lsVTqfWej2PxizPxXAZNBMXV9i/d+f5rnLOg9QNl9FX1o6lW+plZZkvu9Rhc72vacZjKn9v9P+j/AMIsn6h9T61lddz8HqufZ1AYN1LGF9hsZvZkCk3U7/3m/n/uJKfSH9d6G5jgOo4pJBA/T1/+TQr/AKxdHrY0MyaMgxq2u+gEEcfz19X0loOfjNsFTnMFhG4MJAcQPztqnsZ+6PuSU537e6LYxjjnYzDILmm+okfHbY5qDd9Z+lsscGWVXBn0LG5GMA6QP9Jk1vb/AJi19jP3R9yrmt9mTYwWGtjGshrQzk79302PSUg/bvRN4d+0MWNsH9PXz/nodX1j6O657X5WPWBIFpyKC1wH0du231Pzvz60/UMuvAcyt78m621j3110VMsc709gcNrav+GYlg5P2ywsZZawBgsBeKgSC5zB7PR37m7P027+Ys/Q/wA4kpd3X+jMD3NzsZ5LgWtF9QJENb+dYGpsX6xdJtDt91WMAdPUvxzunV230Mi/6P8AKVp1T67aP0rnte8tc1zWRGyx/wCZWx30mLyr66WfW7K+vGbgdEzMujHqbQXiq+yqioPrr99mx7a2bne//SWJKfTHdb6QzHj7Xj2uDQDU26oOP5rh+ktYz/pqQ650iyppOZjsedrnMN1RLdQ5wO15b7V5c3pH1mY2bfrNn2vDRuZVdaBu/O9O2y/3M/lPrq/4tZXU/wDnv0i7Hut6xm5GDda1jbm5N0ST/N3Vus/Rue3+w9JT/9P0rCfjvbW7GuN9Za/bY5xfPuZPvcraBS+57mOurFT9r5YHbu7I90N+kjpKUgvxaLHl7m+50SQSJjx2kIHU+q43TK6rMgPcLrBU3YJ9xBPu1b7fao5/V68LNw8N1bnuznFrXtiGxtHv3Hd+f+anCEjVDe6/wfmWmcRdnar/AML5WwMOgcBwnwe//wAkkcOg8hxjj3u/8kqd3VrK+v0dJFTTXdSbTbuO4Eep7dm3/gv3lppSiY1f6Q4h5KEgbr9E8J80LMWit4e1vubIBJJieY3ErgX5eBX/AIz+p4GUYtz6cWvHLtGksYy3094/SMs3Nrsq2/6LZ+4vRFyH1i/xc9A6x1O7rmdl5ePdta5zqbK2NYKmhoe3fRZY3a1m76aauXrdlW9Syce6ln2R9bAXQ5o2w7d79w2+1130dn/CvWB9XuodN6j9dup5HTTup2YVb7NIe9ltVb3t2+1zfb6e9nsfs3rbr+o3Tep4RdX17q2Ti5LSx7zkMd6gB2O9Vz8bdZ9H0v6nsUsL6ldM+reYM3Cuvuuzsimu4XGvaAH+r+jrxqMdrPcxLUGiKpAN66V5vT3z6wjH3j03frEt9v8Awev6T3/yVYb9EfBVcl1QyWtdkuZYarNuMC0B4H0rdu31N1f8l6tN+iPgkldCsxaLH73t9xABIJEgTtnaR+8ipJKQDDoHAcJ5h7v/ACSc4lJ53H4vf/5JGSSUhZiY7Hte1vubO0kkxI26bj+6vNvrH9YOj9L+tnVMbOJqssfj2F2wvre0Y9LWi00lt7dnv+j/AKRenrA6r9Q/qp1jOs6h1HB9fKtDQ+z1bmTtaK2eyq6uv6Df3UlPEdI6rR1irP8A2ZnXsvx63W17ywF24PZ9koxHCql9ba/8Gyv/AEf6T1EPqfVsLB6OcK64dQrzmVsxmOePV9211dlzGte3bj2t9X1Wf4Vn/CLtaP8AF19UMZ1bsfDfS6l/q1FmTkgts0Hqs/WPp+1Rd/i1+pb7Rc/p7n2gyHuyMgmRr3yEtfBWr//U9JbZTjViy3J31Vssc++xzYABZu3OYGM9irdb6wcLorupYJruEs9NxlzCHODd36NzUuqYlmT07Iw8OkVWW13V1tdDGzuaN/s3eyz6f0Vn5vRupZH1Vr6NW1n22tlW4l8V+1250Pgv/M/0SlxiFxMj+mBIf1GPIZ1IRH6Bo/12H1vuL+m9MtdAdZk1OIHEuY4on1jdHX+gjxtf+WlS670nqHUcDBxsZjRbh3VPu3uhsMZDvTLQ/d9NE6x03Oz+rdKzMdjRTgWvdfvcA4gmv+aa3fu/m3fScxSQnEcGo0GX/nR9LHKEjx6bnH/zT6kOW7/s6wm/91D/AO7C6RYN/Ts236z4vWWsb9jpxjW8Fw9TcfWPtZ9Db+lZ/hVcqs6y3qd9l7WHpZY37O1pHqh59Pd6n5m3d6v+EUc6kI0R6YD/AKXyskAYmVg+qf7N3SVXqhjpmYf+As/6hyqdOb1mnLzX9QcLMe20fYmtLSWMmz2v9tX5vpfn2oePX123pWVi9TFb8y8WsodUQGBrmba/UMMd9Pf+YmiIBviBox/H/vUmRIrhIsH8P++anReoHA+plWcGeqaRYdhO2f0z2/Sh/j+6ruTknLwOl5RbsN9+PZsmY3jft3Q3dyqNXRupM+qP7ELWfbSx8EP/AEetvq/zkb/oP/0Ssmm7G6Z0nFvAFuPbi1vgy0uaNjtjv3dzU/KYHiI+Y5Jf4i3GJjhB+UQj/jurccjdDGtNOx29xcQ4O/MDWbf+/IjHsPtBG4AS2dUHJFQsa5z7A8se1jGl2w6bnb2s9u793csbCoya/rb1DOsoezEtpY2vILdHENobt/0n5rvzVHGIIlZqo2P6x/dXykRw0Ls0f6vi9Eq+fc+jBybqzD6qnvYTqJa0uao05zbbLWOpuq9N20OsYQ1/Puq27tzfaodTcbenZVNTXOtspsbWwNdJcWO2t1CQFSF9wkn0muxRfV7NyM/o+Pl5Lg66zfuc0bR7XvYPb/VatJY31WZdidCxcbLqfTkMFm+tzSSJse4fQ3N+i5qD9b2ZmV0k09PqttvFjHFlYLXbYfrLvTTzGMsxiCIxMiL/AERG1glKOIEgykIjT9ImnZyMzExnVtyLmVG52yoPcG7nH8xm76TkZc59ZKMvKu6UcWiy4UZTHZBaPoNBrcXO3bVLqdOXd9Zel5NFNj8WkWC+0AhrT7m+5ujv+igIRIj6qsSJ/wAD5R/hKM5An03RiB/hfMf8F6CRMd065z0Mj/nh+0PQf9i+ybftO327vpbf9J/0Efpz+pj6wdRdlC4YDtgxS+TXPtDvTb+Z7kjjFWJD5RP6n9BImboxPzGP0/ef/9X0++plzhVZO1zXTtJaeWfnsLXqrt6XcRgjID3Ax6Tb3epLC5ztzm2eu/3fT3q6f55v9V35WJm/zzvo/RHH0uXfS/kfu/20lNfIoxDW92S411UmdwsfXA2sb7n1vYmqf03KyGOqyG33V7nsbXbIA3Q9zqq3+m7a92z3sVpn0rP6w/6lqan6J+j9J30OPpO5/l/6T/hElNayrEY1mbe81mtobvNjmsG6am769/ou/nf8IxTbXjEOy6XeobtvvDy5pALf5tu51TeP8Gj0/wA034BNb/NH5flSU1Xv6Zg2v9XIbRZaQ9wutMGS936Nt79rPd6n80p14uM2ltNRc6m0OM+o55Ic3ltznus+j+69WLeB9Hn8/j5fykx+lXx3444SU1nVYAjp7rSHPBIp9V3qkE+o5wfv+0bfb+8g9Ua1gw2N+i3JoA76Au8Vpd+38Vn9X5xP/DVH5XJKb9n8274H8iC2izebPWe5jgIpcG7G/R+iWsbb+b+fYjWfzbvgfyLI+sP9Gr/o30H/ANK/sfzX/o3/AINJTotpsrcS+99298tDwwBgJcdjPSZX9Hdt/SeombjXCtzDk2Pcd221wZvZLdg2bK2Ve136T9JWmx/6LR/N9v5j+b/61/JWH1b/AJRu/wCTeD/P/S+hT/Tf/RX8hJTvGl+0VevZv2n9N7N/0g7/AEfo/wAn+aUfs1xqaz7VYHNDQ65or3vLQ5rt81uqbvcf8HWjD+cbx9DtxyFn4P8Ayrlf0P8AO/mf5/ln8/8A+jf+spKbdlNljyWXvo2vBIYGEO0r9r/VZZ4bfZsUhRZ6rbPWe1jZmlobsdJc6XlzHW/nfmWKGV/R7/5r6Tf5/wDm/wDB/T/75/LWb9WOMj+if4P+h/A/zv8A3z+Qkp0m49rgHi9+11cejDNklu2Z9P1v/BVL0nsc57rn2B72lrHBoa0bm+1mxjH/APbj7EHP/wCS3fzX0G/z/wDN8t/nETF/oGP/ADf0a/5n+b5b/M/8H+4kp//ZADhCSU0EIQAAAAAAUwAAAAEBAAAADwBBAGQAbwBiAGUAIABQAGgAbwB0AG8AcwBoAG8AcAAAABIAQQBkAG8AYgBlACAAUABoAG8AdABvAHMAaABvAHAAIABDAFMAAAABADhCSU0PoAAAAAABDG1hbmlJUkZSAAABADhCSU1BbkRzAAAA4AAAABAAAAABAAAAAAAAbnVsbAAAAAMAAAAAQUZTdGxvbmcAAAAAAAAAAEZySW5WbExzAAAAAU9iamMAAAABAAAAAAAAbnVsbAAAAAIAAAAARnJJRGxvbmddoGeLAAAAAEZyR0Fkb3ViQD4AAAAAAAAAAAAARlN0c1ZsTHMAAAABT2JqYwAAAAEAAAAAAABudWxsAAAABAAAAABGc0lEbG9uZwAAAAAAAAAAQUZybWxvbmcAAAAAAAAAAEZzRnJWbExzAAAAAWxvbmddoGeLAAAAAExDbnRsb25nAAAAAAAAOEJJTVJvbGwAAAAIAAAAAAAAAAA4QklND6EAAAAAABxtZnJpAAAAAgAAABAAAAABAAAAAAAAAAEAAAAAOEJJTQQGAAAAAAAHAAgAAAABAQD/4R0XaHR0cDovL25zLmFkb2JlLmNvbS94YXAvMS4wLwA8P3hwYWNrZXQgYmVnaW49J++7vycgaWQ9J1c1TTBNcENlaGlIenJlU3pOVGN6a2M5ZCc/Pgo8eDp4bXBtZXRhIHhtbG5zOng9J2Fkb2JlOm5zOm1ldGEvJyB4OnhtcHRrPSdYTVAgdG9vbGtpdCAzLjAtMjgsIGZyYW1ld29yayAxLjYnPgo8cmRmOlJERiB4bWxuczpyZGY9J2h0dHA6Ly93d3cudzMub3JnLzE5OTkvMDIvMjItcmRmLXN5bnRheC1ucyMnIHhtbG5zOmlYPSdodHRwOi8vbnMuYWRvYmUuY29tL2lYLzEuMC8nPgoKIDxyZGY6RGVzY3JpcHRpb24gcmRmOmFib3V0PSd1dWlkOjQ1NDAzYjhmLWEwZTItMTFlMS1iZDI3LWNkYWEzZTA4YTgzNCcKICB4bWxuczpleGlmPSdodHRwOi8vbnMuYWRvYmUuY29tL2V4aWYvMS4wLyc+CiAgPGV4aWY6Q29sb3JTcGFjZT40Mjk0OTY3Mjk1PC9leGlmOkNvbG9yU3BhY2U+CiAgPGV4aWY6UGl4ZWxYRGltZW5zaW9uPjE0MDA8L2V4aWY6UGl4ZWxYRGltZW5zaW9uPgogIDxleGlmOlBpeGVsWURpbWVuc2lvbj45Mjc8L2V4aWY6UGl4ZWxZRGltZW5zaW9uPgogPC9yZGY6RGVzY3JpcHRpb24+CgogPHJkZjpEZXNjcmlwdGlvbiByZGY6YWJvdXQ9J3V1aWQ6NDU0MDNiOGYtYTBlMi0xMWUxLWJkMjctY2RhYTNlMDhhODM0JwogIHhtbG5zOnBkZj0naHR0cDovL25zLmFkb2JlLmNvbS9wZGYvMS4zLyc+CiA8L3JkZjpEZXNjcmlwdGlvbj4KCiA8cmRmOkRlc2NyaXB0aW9uIHJkZjphYm91dD0ndXVpZDo0NTQwM2I4Zi1hMGUyLTExZTEtYmQyNy1jZGFhM2UwOGE4MzQnCiAgeG1sbnM6cGhvdG9zaG9wPSdodHRwOi8vbnMuYWRvYmUuY29tL3Bob3Rvc2hvcC8xLjAvJz4KICA8cGhvdG9zaG9wOkNvbG9yTW9kZT40PC9waG90b3Nob3A6Q29sb3JNb2RlPgogIDxwaG90b3Nob3A6SUNDUHJvZmlsZT5VLlMuIFdlYiBDb2F0ZWQgKFNXT1ApIHYyPC9waG90b3Nob3A6SUNDUHJvZmlsZT4KICA8cGhvdG9zaG9wOkhpc3Rvcnk+PC9waG90b3Nob3A6SGlzdG9yeT4KIDwvcmRmOkRlc2NyaXB0aW9uPgoKIDxyZGY6RGVzY3JpcHRpb24gcmRmOmFib3V0PSd1dWlkOjQ1NDAzYjhmLWEwZTItMTFlMS1iZDI3LWNkYWEzZTA4YTgzNCcKICB4bWxuczp0aWZmPSdodHRwOi8vbnMuYWRvYmUuY29tL3RpZmYvMS4wLyc+CiAgPHRpZmY6T3JpZW50YXRpb24+MTwvdGlmZjpPcmllbnRhdGlvbj4KICA8dGlmZjpYUmVzb2x1dGlvbj4yMDAvMTwvdGlmZjpYUmVzb2x1dGlvbj4KICA8dGlmZjpZUmVzb2x1dGlvbj4yMDAvMTwvdGlmZjpZUmVzb2x1dGlvbj4KICA8dGlmZjpSZXNvbHV0aW9uVW5pdD4yPC90aWZmOlJlc29sdXRpb25Vbml0PgogPC9yZGY6RGVzY3JpcHRpb24+CgogPHJkZjpEZXNjcmlwdGlvbiByZGY6YWJvdXQ9J3V1aWQ6NDU0MDNiOGYtYTBlMi0xMWUxLWJkMjctY2RhYTNlMDhhODM0JwogIHhtbG5zOnhhcD0naHR0cDovL25zLmFkb2JlLmNvbS94YXAvMS4wLyc+CiAgPHhhcDpDcmVhdGVEYXRlPjIwMTItMDUtMThUMTc6Mzk6MDgrMDU6MzA8L3hhcDpDcmVhdGVEYXRlPgogIDx4YXA6TW9kaWZ5RGF0ZT4yMDEyLTA1LTE4VDE3OjM5OjA4KzA1OjMwPC94YXA6TW9kaWZ5RGF0ZT4KICA8eGFwOk1ldGFkYXRhRGF0ZT4yMDEyLTA1LTE4VDE3OjM5OjA4KzA1OjMwPC94YXA6TWV0YWRhdGFEYXRlPgogIDx4YXA6Q3JlYXRvclRvb2w+QWRvYmUgUGhvdG9zaG9wIENTIFdpbmRvd3M8L3hhcDpDcmVhdG9yVG9vbD4KIDwvcmRmOkRlc2NyaXB0aW9uPgoKIDxyZGY6RGVzY3JpcHRpb24gcmRmOmFib3V0PSd1dWlkOjQ1NDAzYjhmLWEwZTItMTFlMS1iZDI3LWNkYWEzZTA4YTgzNCcKICB4bWxuczpzdEV2dD0naHR0cDovL25zLmFkb2JlLmNvbS94YXAvMS4wL3NUeXBlL1Jlc291cmNlRXZlbnQjJwogIHhtbG5zOnN0UmVmPSdodHRwOi8vbnMuYWRvYmUuY29tL3hhcC8xLjAvc1R5cGUvUmVzb3VyY2VSZWYjJwogIHhtbG5zOnhhcE1NPSdodHRwOi8vbnMuYWRvYmUuY29tL3hhcC8xLjAvbW0vJz4KICA8eGFwTU06SW5zdGFuY2VJRD54bXAuaWlkOjI0QTU4NkMzNUY1OEUxMTFBQzVEOUU2NUExQkQ5QThEPC94YXBNTTpJbnN0YW5jZUlEPgogIDx4YXBNTTpPcmlnaW5hbERvY3VtZW50SUQ+eG1wLmRpZDoyM0E1ODZDMzVGNThFMTExQUM1RDlFNjVBMUJEOUE4RDwveGFwTU06T3JpZ2luYWxEb2N1bWVudElEPgogIDx4YXBNTTpIaXN0b3J5PgogICA8cmRmOlNlcT4KICAgIDxyZGY6bGkgcmRmOnBhcnNlVHlwZT0nUmVzb3VyY2UnPgogICAgIDxzdEV2dDphY3Rpb24+Y3JlYXRlZDwvc3RFdnQ6YWN0aW9uPgogICAgIDxzdEV2dDppbnN0YW5jZUlEPnhtcC5paWQ6MjNBNTg2QzM1RjU4RTExMUFDNUQ5RTY1QTFCRDlBOEQ8L3N0RXZ0Omluc3RhbmNlSUQ+CiAgICAgPHN0RXZ0OndoZW4+MjAxMi0wMi0xNlQxMDo0OTozNSswNTozMDwvc3RFdnQ6d2hlbj4KICAgICA8c3RFdnQ6c29mdHdhcmVBZ2VudD5BZG9iZSBQaG90b3Nob3AgQ1M1LjEgV2luZG93czwvc3RFdnQ6c29mdHdhcmVBZ2VudD4KICAgIDwvcmRmOmxpPgogICAgPHJkZjpsaSByZGY6cGFyc2VUeXBlPSdSZXNvdXJjZSc+CiAgICAgPHN0RXZ0OmFjdGlvbj5zYXZlZDwvc3RFdnQ6YWN0aW9uPgogICAgIDxzdEV2dDppbnN0YW5jZUlEPnhtcC5paWQ6MjRBNTg2QzM1RjU4RTExMUFDNUQ5RTY1QTFCRDlBOEQ8L3N0RXZ0Omluc3RhbmNlSUQ+CiAgICAgPHN0RXZ0OndoZW4+MjAxMi0wMi0xNlQxMToxMDoxNSswNTozMDwvc3RFdnQ6d2hlbj4KICAgICA8c3RFdnQ6c29mdHdhcmVBZ2VudD5BZG9iZSBQaG90b3Nob3AgQ1M1LjEgV2luZG93czwvc3RFdnQ6c29mdHdhcmVBZ2VudD4KICAgICA8c3RFdnQ6Y2hhbmdlZD4vPC9zdEV2dDpjaGFuZ2VkPgogICAgPC9yZGY6bGk+CiAgIDwvcmRmOlNlcT4KICA8L3hhcE1NOkhpc3Rvcnk+CiAgPHhhcE1NOkRlcml2ZWRGcm9tIHJkZjpwYXJzZVR5cGU9J1Jlc291cmNlJz4KICAgPHN0UmVmOmRvY3VtZW50SUQ+eG1wLmRpZDoyM0E1ODZDMzVGNThFMTExQUM1RDlFNjVBMUJEOUE4RDwvc3RSZWY6ZG9jdW1lbnRJRD4KICA8L3hhcE1NOkRlcml2ZWRGcm9tPgogIDx4YXBNTTpEb2N1bWVudElEPmFkb2JlOmRvY2lkOnBob3Rvc2hvcDo0NTQwM2I4ZS1hMGUyLTExZTEtYmQyNy1jZGFhM2UwOGE4MzQ8L3hhcE1NOkRvY3VtZW50SUQ+CiA8L3JkZjpEZXNjcmlwdGlvbj4KCiA8cmRmOkRlc2NyaXB0aW9uIHJkZjphYm91dD0ndXVpZDo0NTQwM2I4Zi1hMGUyLTExZTEtYmQyNy1jZGFhM2UwOGE4MzQnCiAgeG1sbnM6ZGM9J2h0dHA6Ly9wdXJsLm9yZy9kYy9lbGVtZW50cy8xLjEvJz4KICA8ZGM6Zm9ybWF0PmltYWdlL2pwZWc8L2RjOmZvcm1hdD4KIDwvcmRmOkRlc2NyaXB0aW9uPgoKPC9yZGY6UkRGPgo8L3g6eG1wbWV0YT4KICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCjw/eHBhY2tldCBlbmQ9J3cnPz7/7gAOQWRvYmUAZEAAAAAB/9sAhAABAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAgEBAQEBAQICAgICAgICAgICAgICAwMDAwMDAwMDAwMDAwMDAQEBAQEBAQIBAQIDAgICAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwP/wAARCAOfBXgDAREAAhEBAxEB/90ABACv/8QBogAAAAYCAwEAAAAAAAAAAAAABwgGBQQJAwoCAQALAQAABgMBAQEAAAAAAAAAAAAGBQQDBwIIAQkACgsQAAIBAwQBAwMCAwMDAgYJdQECAwQRBRIGIQcTIgAIMRRBMiMVCVFCFmEkMxdScYEYYpElQ6Gx8CY0cgoZwdE1J+FTNoLxkqJEVHNFRjdHYyhVVlcassLS4vJkg3SThGWjs8PT4yk4ZvN1Kjk6SElKWFlaZ2hpanZ3eHl6hYaHiImKlJWWl5iZmqSlpqeoqaq0tba3uLm6xMXGx8jJytTV1tfY2drk5ebn6Onq9PX29/j5+hEAAgEDAgQEAwUEBAQGBgVtAQIDEQQhEgUxBgAiE0FRBzJhFHEIQoEjkRVSoWIWMwmxJMHRQ3LwF+GCNCWSUxhjRPGisiY1GVQ2RWQnCnODk0Z0wtLi8lVldVY3hIWjs8PT4/MpGpSktMTU5PSVpbXF1eX1KEdXZjh2hpamtsbW5vZnd4eXp7fH1+f3SFhoeIiYqLjI2Oj4OUlZaXmJmam5ydnp+So6SlpqeoqaqrrK2ur6/9oADAMBAAIRAxEAPwDf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Wnv0d8Qe/v5l/wDMK+c/bHeXypfGdcbDqqLrvrTrLbWx6Bp9pJjsrJR4sipqGiVqeOGlqDI7SSSyyTXYjSPZk7fTqKZU+Xp+fQZhiO5TSGQ6ZI2I1DgVxQEeVK/6uHVga/yHdvLf/nJneNj+P7kYTj+v/KV7b+rX+HpX+6G/36f2dcR/IcwA/wC5m94f639x8J/vf3Xv31a/w9aOzsf9FP7Oux/IdwFv+ymt4/8AoEYT8/8AVV799Wv8PWv3M3DxT+zrr/hhzb/4+TW8P9f+5GF/+qvfvq1/h63+52/36evf8MOYAm5+Te8D/wCSPhP/AKq9++rX+Hrf7nJ4yHr3/DDmAsR/sze8f/QIwn/1V799Wv8AD1r9zGuJT+zrv/hh3b/F/k1vH/H/AH5GE+n/AJ1e/fVr/D1r9zN/v09eP8hzb5I/5ya3jYfT/fkYU2/9Wvfvq1/h63+52/36evf8MO4D8fJveI/r/vx8J/8AVXv31a/w9a/cx/36euv+GHNv8/8AOTW8Ofr/AL8fC/8A1V799Wv8PWzs7H/RT+zrsfyHcAAP+cm948H/AJ4jCfT+n/Ar376tf4evfudq18U/s69/ww7t8H0/JreIH9P7kYQn/b/de/fVr/D179ztT+1P7Ovf8MO7fH6fk1vFf6j+5GFNz/sar376tP4evfudv9+nr3/DDu3yCD8md4cn/niML/8AVXv31afw9e/c7f79P7Ouj/IdwBFv9mb3hb/wx8L/APVXv31a/wAPXv3OaU8Q9e/4YdwH/eTe8f8A0B8J/sf+Ur376tf4evfuc/79PXj/ACHMATz8m94m30vsfCD/AOOvfvq1/h68dnalPFP7OvD+Q5t8fT5Nbw/9AjC8/wBP+Ur376tf4evHZ2PGU/s69/ww5t/k/wCzNbwuf+zIwthxb/la9++rT+Hr37nb/fp/Z13/AMMO4D8fJreIt/2ZGF/2P/KV799WvmvWv3Mw/wBFPXX/AAw5t+1v9mb3h/6A+E5/9Wvfvq1/h62NnI/0U/s67H8hzb44Pyb3iR+R/cjCC/8ATn7r376tf4evfuc+Up/Z14/yHcAeP9ma3iB/T+5GEP8A8de/fVr/AA9eGzsD/an9nXj/ACHdvn6/JreFv6f3Hwn/ANVe/fVr/D179zt/v0/s66/4Yc2//wB5Nbx/w/35GF4/9W/fvq1/h61+5m/36f2ddj+Q7gB/3M3vH/0B8J/9Ve/fVr/D1v8Ac7DAlP7OvH+Q5t82/wCcmt4C3/ZkYQ8/+dXv31a/w9e/c7cPFPRcfmJ/I7zW2Pix3puTq75a7g2z2NtzY1buPamezGxsW+KoKnAOuVrFrFpZ5JR5oInhjkjRijMGsbe7Lc62CIKE+fp1STaljUySOWVcleFaeVRkV+XVr/8AKA7S7c7b+AnSef7x3hSdgdk4TH1uyc7valxcOG/vR/dGVcTS5eqoKYLGs0kaDyMiLqI1EaiSU90gjl0j0H7el21TyXNms0nEk0HoPIfOg8/PqzX2n6Meve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917pObwytTgto7pzlEImrMNtzN5WkWdS8LVOOxktZAJkUglS6DUARcfn3ZQCwB9equSEJXjQ9af/8ALw/l5d5fzAqr5WfKH5MfL6uzeZ373hXwbNwu29hYuFNnYvHo8s2LqzK8CeNYJqWmpo4QwVYS7MWcj2YSSfTnQcjiPkPToN2tudyQ3BOhwSrUyGIJz8vsHVjo/kObe/PyZ3gT/X+5OFH/AMde2/q0/h6U/uZuHimn2de/4YdwH4+Te8fp9P7kYT6/+dXv31an8PXhszD/AEU9e/4YdwH/AHk3vG35/wB+PhLnn/qK9++rWnw9b/c7f79PXv8Ahh3A/j5N7xA/A/uPhP8A6q9++rX+Hrw2dgKeKf2deP8AId2/aw+TW8R/X/fkYX/6q9++rX+Hr37mP+/T+zrofyHNvAf9lNbxvxz/AHIwn/1V799WvDT179zsTUyn9nXZ/kObd/HyZ3iB+R/cnCm//q1799Wn8PXjs7f79P7OvN/Id2+TcfJreI/pfZGE/wBj/wApXv31a/w9b/dDf78PXX/DDm373/2ZveH/AKA+F/P/AFV+/fVr/D1r9zkcJD1y/wCGHtv3uPk1vEf4f3IwnH+sfuvfvq1/h68dnbylP7Ouv+GHcBx/zk1vC4vc/wByMJz/AOrXv31a/wAPXjs7eUp/Z11/ww5gD9fk3vAj+n9x8IP96qvfvq0/h69+52/36euR/kPbfP0+TW8VH9P7kYQj/H61Xv31a/w9e/c7HjKeuJ/kN7fNr/JreF//AAyMLyP6f8Cvfvq1/h69+52/36f2Drkf5Du3iP8AspneN7W/48nC/wBP+or376taU09eOzsTXxT1x/4Yb27Yf85M7wve5P8AcjC8/wCFvuvfvq0/h63+6G/36eu/+GHNu3v/ALMzvG1vp/cjC8/+rfv31afw9aOzseMp68f5Dm3yb/7M1vH6Wt/cjCHj+n/Ar376tP4evfuduHin9nXj/Ic28bf85M7xAH4/uRhDz/W/3Xv31a/w9a/c7Up4p/Z13/ww9gLcfJveP/oEYT6/+dXv31a/w9e/c7f79P7Ouv8Ahh3Afn5N7xP5H+/HwgsfweKr376tf4et/udv9+nro/yHNvkD/nJreFx/2ZGE/wCIqvfvq1/h69+52Br4p/Z13/ww5t/6n5NbxJ/H+/Iwv+2/4F+/fVr/AA9aOzMf9FP7OvD+Q7t+1j8mt4Hm9/7kYT/evuvfvq0/h62dnan9qf2ddj+Q9gP+8mt4fn/mCML/APVXv31a/wAPXhs7A18U/s64n+Q5t8/X5NbwP9f9+Rhfr/51e/fVr/D1r9zNWvinrv8A4Yd2/wDn5NbxI/8ADIwgt/6te/C7UD4evfuY+cp6Ij/Mk/lAdk/Hv4qZ3vD49fMTNbM7Q6q3rsvedHmc3sTFyUk+KoMskMuOpft5ZylQZ5IZkZ4ijBCjWDX92Sbxm8NBpr58emLiwWyiN1ITJoqQtaVNMEkenWy78Nuxd39tfFjofsff9bSZLe27uuNv5XdGSoaNMfS5LMtT+CtyEVDGSsXndDKY09ILEKALAJJ1VZWVOHR1ZSSTWkcspqzCp6Mv7a6Vde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/0N/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdUBfydhf5E/wAwh/ye14h/628l7MLv+yT/AFevQf2ev1Vx/pj/AJOr/fZf0IOve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917otXzJv/sqXyGt9f9EW+Lf+eKb27D/ar9vSe7/3GevoeiYfyUf+yAuuv/Do3z/7uj7evf7c9Idk/wCSdH1bJ7SdG3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690iuyf+Zdb+/H+/K3T/wC6Of3ZPjH2jqkn9m32H/B1Td/IZUD4x9o2/Pee57n+tqGnHtXff2g+zon2H/cRj/Tb/CerxvaLo7697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuq0P5vn/bv3vf8A6gdu/wDvTUvtVZ/24/1eY6K95/5J0lPQ/wCA9DJ/Lyt/skfxntwP9FmC/wB7k9t3H9s329KNv/3Cj+zo5ftnpZ1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691/9Hf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3VAX8nUf85EfzCG/r2vCPz/zu8n7MLv8Ask/1evQf2iv1Vx/pj/z71f77L+hB1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690Ube/zq+LnXfyy6m+D+7+0aDFfJju7ame3p1z102NzFRNmMDt2lqa6tmqMzTwNQ0kskNFWyUtPU1CSTrBKY1bTzbQdOo+fTfiKXKDiOPyrw6Nz7r051737r3RRfll87fir8HcXsbM/KDtrC9WY/sfcke1NnzZamyVWctmHlihaNY8ZDM0ccbTxeSaQBF1C5+vtxI2cFhwHTMs6RMFatW4U+XE/YPPo2lPUQVdPBVU0qT01TDFUU80bBo5oJkEkUsbD6qykEH+ntvp7rN7917r3v3Xuve/de697917r3v3Xuve/de6LV8yf+yUvkPb/n0W+P/dFN7dh/tV+3pNeU+lkr/CeiXfyT7f7IF13p/wCeo3zf6/X+Mn+vt69/tz0i2On7tjp1bN7SdG3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690iuyRfrrfw/rsrdQ/wBvgp/dk+MfaOqSf2bfYf8AB1Tf/Ia4+MXaA/p3luj/ANwaf2rvv7QfYOifYf8AcRq/xt/hPV4vtF0d9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdBn2l3N1R0lhKPcnbnYW0+usFkcnTYagye7czR4elrcrVtpp6CkeqYGSRv9SgNhybD3dI3kNEFadMzXENuA07BQTQV9ehHgnhqYYamnljnp6iKOeCeF1kimhlQSRSxSJcMrKQVINiOR7pwwenQQRUdZffut9e9+691737r3Xvfuvde9+691737r3RPuq/nt8Tu6/kb2n8Tesu38FunvzpjFyZnsTYNHT5KOtwmOp8lFh6yoWsqIUp5/BVTRQzCCVirML8c+3GiZAC3n0zHPHKxVPLHyxxp9nn0cH23091737r3Xvfuvde9+691737r3Xvfuvde9+691737r3VaP8AN7Nv5fve5tf/ACHbv/vS0vPtVZ/2w/1eY6K95xt8h+R/wHoY/wCXlf8A2SP4z3+v+izBf73J7buP7ZulG3/7hR/Z0cv2z0s697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//S3+Pfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691QH/J2H/ORP8wk25/0rxD68f8XvJH6ezC7/ALJP9Xr0HtnNbm4/0x/596v89l/Qh697917r3v3Xuve/de697917qkL5Sf8ACgj+XV8Ye5NyfHOs3/uPszvjZ1VNSbq6+6/2luCsTAT0aiTI01fuitpo8c08CHU8FNNKwF9Wn2qitWlfQSFNK56LL3dIbOHx9LSKG0nSCaH0wCf5dGO/l9/zb/g//Mwg3VR/F/tI5ze2w4Eqt79b7jw2U2zvTb9E9SKFcqMflIkSsoWnIiWso5JI9RCsVYge2Xj0E0NQMV6WQTiZAxBUsK0PGnUr5AfzbPgZ8WvkBuj41d+d34zrfsrZvR9R8gdwQ5rGZSTE0uwqeoki8UFdQRTNPk5I4nqIMbBG88kQ1IrEge/LEWWqnPp8vXrUlykUgSQUBoAfIseC/aeiV/HH/hS3/Kj+S3eOO6J2v3XlNn5/dNbTUHX+4+yNp5vZuzt51tY3ipaGPO5WJI8dUTP6YYsmYNfALKxC+7GEUGlgSfLptLss7iSNkVDTUeB/1fs6ZO7v+FPP8pforvPMdG7i7l3JuCt2hujK7L37vTZuwd1Z/Z209zYeqahr6Fa+mpteTihmjkjmq8Wk8AKnxvJ78IhoLMwBHl1trlhKqRxsytnUKUHVq3ZXzt+MPV/xJ/2eTNdkUmY+M0uzMR2Bj+xtp0VZn6PL7Uzsay4zJ42kp1WZhIHAMbqjowKOquCvuiRl20jFM9OzTpCmtqmpAFONTwHQsfHjv/qv5T9Jda/IbpHci7u6p7a2xSbt2VuAUlVj5K7FVbvAy1WPrlSanqIJo5aepglQPHKjowup91IoadOg1FeHReOv/wCZR8Oe0Pkf3t8Vtldt47L9t/GnDZnOd2Uho6qj29sSl26yJuGPKbkrFSm10BY/dlGKRlJAz3RgHPCbw/EBHp889MfUJ4/05BBpWtMEDjT7Oqsd/wD/AAqa/lR7MzOZXDdhb77E2NtrNxbbz/ZWytgbgq9sw5upDtSU+Ijr4YKiuilWN2SpijEbBSVLLYl1bYGMyM4FDQ9JJdx8O6FosTMzDUDTFBxzwH5mvVkX8vz+ah8P/wCZnht8Zr4qbxz+5YOvK6lody024tq5TbFZTGtQPTTxRZBbOjXA/Vf6cW59tPFpXUp1DpVDc+K5iZSjDND6Y/z9FG+YH/Ci3+V/8L+6818eux+38zuTtbZ2Yjw3YeD2Bs7P7joNg1Kwipr6XO7gihWiespoyDNQ0k0sqN+24SQFfe44g5IZtOOtT3LRAFEMgJoaeXVDmU+bHxf+f3/Cnb+Vp3l8Uexafs/YI6I7DwuSyi4TLYCtxOdx+w99zTYzJYnPw09ZBPFHKhKzQIRf03Uhjt8Ig+R/wnqkR1TynI7l4j+j1tC/Pv8Am4/B7+Wvktj7Y+UXZ1ftzfvZ2KyWY682Hgdpbm3Nndy0GNrVxktUajE0slFRQtVOtOktdVRBn1BA2k2pGmtgpNK9PzzeBE0uktpFaDjTouXxA/4UDfy8fl/3rhPjBt/fO4uuvkDuqtqaDaHXnYO2MzjU3VVU+ObLJQYjc8UDY77yWnSSSGjnnjkk06YhI5C+7zQeE2nUCfl0msr9b2HxwjIpNAWFK+XDiM/LqpT/AIV+wQzdYfBvyxRSf8Z3mX9yJJPS2Qw+pQWH0P5Ht22/sn/LrV1i6ip/DJ/x3rY3+WH8wX4m/wAuvorZPa3yy7Ph622pmqLFYTbkcGCz+5s3uPLxYiKd8bhcHtumqZ5HVCpZ3VI1BBZ1HtMEZqkcBxPp0sklSNgh+Jq0HmaZx1W51l/wpp/lhb37I2j1xu/fm7un5ux6mmi673T2BtLL020txU9fULS4ysr8zj4pUxkdS7qkclbpjDGzyLb2oltTFTuBLcOi6z3VLxnAjZRGaMSCAD+wY+YqOtg+CeCqghqaaaOopqiKOennhdZYZ4JkEkU0UiEhlZSGVgbEG49pOGOjXjkdZffuvde9+691737r3XvfuvdFq+ZP/ZKXyG/8RFvj/wB0U3t2H+1X7ek15/uLJ/pT0S/+Seb/AAC66/8ADo3yP/W0fb17/uQekOxmu2x9Wy+0nRv1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdIrsn/mXW/v/AAyt0/8Auin92T4x9o6pJ/Zt9h6ps/kMW/2WTtIWII7z3Pe5JufsKf2rvvjX7OifYf8AcV/9O3+E9Xke0XR31737r3Xvfuvde9+691737r3XvfuvdVKfOP8AnYfAf4B9i0HSPc3ZeSyPfGZx9JkMN1Js3a24c9nJosiuvG/xPMU1McbQ/cD/ADS1FUJG/EZ9vxQNIyqTp1cK+fSK6voraJ5KF/DpqCipFf8AV5V6afgz/PH+Afz37SyPQXVnY2Q2z8gMXDWzzdR7+weS2/m8nHjITU5T+7OUqIhRZF6VFLVEME3lQAkx6QT79LAYmKghtPGnl160vku4lm0sgf4dQoT/AJvsNOrgfbHS3r3v3XuqV/53O0P5YO7uhOsYP5o2Vz+I6vo+zaaXYFTtnc269s5X+980EcVQrVG1ZonkpvF4/uBLcKLFbNz7eiEjBghoMV6S3LwoyGQFjU0oK+Wfypx+XR8+/PlF8d/g78Scp8kOzNwzbe+PfVGyNnTRZbG01duCtk25kpKHbezqXE0oL1NZNUtVUcUILF31amJNz7bVdTUJp8+nncompRX0A6q4+Qv/AApi/lNfHrK7fwOR7yynYWazO29p7yrKPrfZu4twwYHbG8aGPJYuqyOWECUb1Ygmiklx9PPJPHqAkVDx7dWHJDsFoK/b0la+GhXgQyajTHkRxr9nVr/xH+ZHxy+c/SeG+Qvxh7Ix3ZXVmarMhihm6akyOIq8TnMQEOXwG4MNmoqeqoq2lEkZmgniU6XR1LRurFkgjpYGBFeH+TqpLsv/AIUv/wAsTYe7t24LAb93j2htrrPM1eD7V7C2LsncNVtXZNZRZI4mYxS5CmglyaeZWCTUKPFIBeGSQW9qY7YyRmTUAF49Ft1uiWt1HaGNmMnAgEj9tKD8yOrjMJ8oOj9x/HCl+WmF3xR13QlZ1zJ2tTb6FLXU9M+yYsa2Ulyb0NXHHUIyxqwaGSJXDDSRf2x4b6wlMnpf40fhGYHArX8uPTJ8S/l70N83eoKTvP45bxXe/XVZns1tlct9pNQTwZzb8qRZSgqaSf1I6eSNh+CrKffnQxtpP8uvQyrMniKCPkRQ/s6RW3vn38Zd0fNjev8AL6w295an5M7A6toe3txbSOMqkx0O1K2ppYClPmyPBJWQpXUc89Kp1pHKrH8+9tGVQMTk+Xy8j1pJg8hRQaDFfKvmPy61gf5SMEEf/ClP+aIVp4UlHV+8/wBxY0VgH7XxBcagL8n688+1NwP0x9g6R2JJLf6eT/CvV8Xzo/nX/An+X52fjehu7uxszUd+ZzbeL3Vg+p9p7R3Jm8vVYfN1U1JhqiuzsFL/AAuk+5annMUc1X5WCFhHYqSniiMrBagV9en7u7W0haYqWCcaCp/z9M3wO/nj/Ab+YL2lX9BdQ9g5fA/IDF4zMZiq6l3xtvNYPL1mPwDgZmXA5qeAY+uemRhJPTxVAnVNT+IojMNyw+GxUHVTjTrVrdi5hWYqUDcNWK+n2fYerg/bPSzr3v3Xuve/de697917r3v3Xuve/de6rQ/m+f8Abv3vf/qB27/701L7VWn9sP8AV5jor3n/AJJ0n2H/AAHoZP5eX/ZEfxn/AD/xizBf73J7buP7Zun9u/3Cj+zo5ftnpb1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691/9Pf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3VAP8nW/+zE/zCRqBH+leL0/n/i+ZPn2YXf8AZJ/q9eg/s/8AuVcf6Y/5Or/vZf0IOve/de697917r3v3Xuve/de61wPkR/Nb+DXV/wAnewOjfhl8K8b8/fmfgZtwVnbe3Og9k7Kotz4Wuoq8U+46TcPZGUxNQauohnYrVwxSyLG/oJ13UKlieRdRbI8q5p0WSXcVtMsKRMQ5PcB2g+df9iv2dUq/yzu3N0dk/wDCpPd+68v8L85/Lzyu7/iPuqk3Z8cc5FQ0WUrp6bbGLyh3tl6fCY/F0TyZWeKKtV4aSzeMM0jymRvbZBNdXkP8vT+pUKmP8T544qMjP7ehg+YnxS6Y+ZP/AArf6p6t76wM26Ng7Z+JGx+0ztiKSCHGbmz3XOOym4MDid1wyxv93jJZyBW0ht541ELnxsyny8K+in/D1uXJ0HIZwDXP4a/4QOnr/hYD8ZOhMF8YPhPuHZ3U/X3X25cj8tdv9WT7k2FsvbG1c2Nj5/YmXqanBLWYeliLU8U1LBPTwSBo43W6qLm+kJaiHgSOrTgRo0y/EqNT04VyPy6tr+UH8vH4hdO/yMfkF1HsTo7rmHF7O+CXYGdxG6c1sza2W35U7uwvU9Ruai31l94S0f3lRmhXItSMi0vlVwNDKqqBuQ6ZWUcASPy4detwHt0Y4JAY/acn/V6dE7/4T/dcP80P+E+e4fi32RXtkdr732l3R0liJch/lS4HD7sxkwxr0onVwgoK2r+8p9KnxyKGUah7clCgRHhUZ6S2hd/qFJrSRqV/aOi7f8J8/m1nviH8LP5oXxX7zzdFktyfyo9zd1ZTaW3qip8WTzOzNm0e4MjnKDH01Q6h41zeIm/zJUAVsZt6tR0w10IFVXGPMfP8unYmEdUZtMkg1UJrQnBpXyrwHRUvgZ8Zu3uwf5LP85n+ZNumuo9r9s/zAqDubt/aGap0MOdoutttVOcr967dgzJ0SrDmquqyVF447JIIIRZr8OI4RiF4OCaenp/g6TSxNLCC1Q8RA1HGrhqp8iSekX8Hf5tn8nfr/wDkF13w/wC3YtoL8m4fj73vsDK9TVvRubzub7E7U3bUZ19kbgpd4Y/B1OOeeeSvxrw5Crr1mpTELEGGO6YDhRuPl0YM5AYshweOMj1rXA9erkv5I/Tndnwd/wCE/PY29t57NpesO2G6V+S3eu1MbPt6nw+88fDRbSzG4NkZTeM0SpUzTzNTQ1tPFUnXDA8a+m+kOOaaI+BXj9vSe2BPiTk6lc1U/wBGlOgb/wCEnvxE6W3h8GN//MjtPaW1O4O+vk529vf/AEgb17FwOB3pnoaLaeaqKSPGU1Tm6eZ6aOtmqJq2sjQqJ5CrPcJGF1L8KvmrZP7erWpOuSPGlCAoHkKefzJ6LV2X8ROiPiL/AMK2fg3J8ftn0fX2I75657G7V3ps/CQY+g2hjN4ZDrDeeCylRtHAY6KKLG09VHRQ1EtJGojEzSPGqhyPemUaVbzIP8jTpyNj4rx+SkU/MVPWwX/M2+e/8v8A+Im+eo9u9/dW7P8AkD8tt047IVvxz6bg63wW9e08lTS1v2tXUbf3JmsdWRYKConieNJHnjaeSNhFHJocruGJpSI66QeFfOnTN9dxWcZuWQuVoCFFSNRoMfb1pj/zwPmF27218kvgf2nuL+Vt2f8Ay8t97D7Bw42N3hvOtx9Pkt64fJ7uoDFSCbaGJoKd4aIAzU8VVVySwmeXQqiQ6nEBjYkAtQ0r5D/V8+qystwgBOkFCSpGTwII/wBKfSvVx3/Ct+SSp6X+AlRNKJpZu5aWaSbk+aSWbCu8tzzyTfn3u3+B/wAut3eLmIf0ZP8AjvV8H8xL5gfy+vhv0F1p2D88KTrfcsFRQJB1F1/vLZ2I39uLeO6osHTmto9n7dytLVorqrQfcVjxrHCChZw2kFmMMxKatKnien7pkjXx2QuyVIAFT86dahX/AAoB+cW+fk3/AC/Mjh8p/Jd7M+N/XK756uyPWfzH3pRbPhw218NLlxXUNDgE2/g4KmjXcFOBQJTtXpCVlOpHkWIe7SK0VUJr8/8AN01bzR3QWbSUOcEUPA4Pl8+PW9R8HMrX5z4X/EzLZSrevyNf8b+lKitr5HeSWuqG65xwlrZZJLszykeR2PJJJ9tygK5A6UW7F4VY9Gl9t9Pde9+691737r3XvfuvdFp+ZX/ZKPyH/wDERb4/90c3t2H+1X7ek93/ALjP/pT0TD+Sf/2QF11wR/v6N8nn/tdHn27ef256QbH/AMk2Pq2X2l6N+ve/de697917rh5IxwXQH/gy+/de695Y/wDjon/Ja+/de695Y/8Ajon/ACWvv3XuveSP/jon/JS+/de695Y/+Oif8lr7917r3kj/AOOif8lL79Ude695I/8Ajon/ACUvv3XuveSP/jon/Ja+/de695Y/+Oif8lr7917r3kj/AOOif8lL7917r3lj/wCOif8AJa+/VHXuveWP/jon/Ja+/de695Y/+Oif8lr7917r3lj/AOOif8lr7917r3lj/wCOif8AJa+/de695I/+Oif8lL7917r3kj/46J/yWvv1R17r3lj/AOOif8lr7917r3lj/wCOif8AJa+/de695Y/+Oif8lr7917r3lj/46J/yWvv1R17rkGVv0kH/AFiD/vXv3XuvEhQWJsACSf6AC5Pv3XuiB/Hb+aB8IPlb3l2B8b+h+78NvzuPrBMrJvTaFFjczS1GKjwdf/DMq61ddBHDMIJz4nMUjWbj3qterFSOj++99V697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XukV2T/AMy639b6/wByt02/88c/uyfGPtHVJPgb7D1Td/Iav/ssfaN/r/pz3Pf/AAP2NP7V339oPs6Jth/3Ff8A07f4T1eN7RdHnXvfuvde9+691737r3Xvfuvde9+691Q18+v5lH8vv4w/JbEdQ474+7U+X/8AMN3JFjxhOnOvtjbUzfbsCfwc1mFp8lvnM46rGMklpAHgphL5DF62QLpJVRRvKApfTThXouubiO0fWsTOXIDFRWnpXrWh7l+R3Y3dn/Cgv+UFv7en8tveX8tjeh31LjMnHvVMNDn+9sdl8tUY+fN1dXtvFYuGpjooGlxxeRp5T9yyu4jES+6gEsEbhnpx2VUaZOPbjPr6GnGvX0Q/afpb1737r3WoR/wsQijk+Fvx18kUcoHe0v8AnEVwAcdSgj1f1+h9rLX4X/2v+HovvCRPCQaf2n/VpujS/wDChBhH/wAJ3e4QqR2PVvxYjClFKop7G2iP2x+CPwR9PaXzPS05A6H/APksfy5/hv0//LH+NGPxHR+wt51XdXTW0ez+z9x9mbP2nvXc+7txdlbXpszuChyWWytE7nGQtK1PQY4HwwQiyrraRmvISraR5Af4K9NW4DR62AqxNaCnnT/J1SL/AMJgNj5Ss27/ADcvhxiN0ZbbGypO0N/YPatbhqqWNdo1FfWZPr+DKYimUqscqU0NMZBHpLCNVuLAhQaRRoxFaMf2UHSMoZ55Y1YrrjX8jqda/wAugM+CXyA7F/4T34ztP4bfzOv5f2X3F8XN+/IfdW7qb5v4Ta+L3psjMYncNNRYXHLmsTLi6tMxAPtY54aVq6Orp/uJYxSuoVi3oLVKnHGvln1HHp1ZfBRY3XupQL5mnEgk0PrTj1tg/LLtPpnvH+U38hu0OgNwbY3d0zvT4q7+y+wsvtCmjg2/VbfqNpzNRxUWMjjiNN4x6HpXgjeJgUZFII96jDiddXGo/Z1u5dHsZTFw0tw8jQ1/OvVb3/CT1YV/lR0nhEIv8iu5fJ4fH+sPjQfIE+jWtcHn3qcUYfZ0/ASQ9f4j/k6Kp0FFGv8AwsL+VzBItR+DKyak0E63x+yFZiV+jW4P5/B9+m8v9KvTdt+P/mo3SE/lLf8AcSr/ADRv/EX7y/8AfrYj2/cf2Y+wdJ7Hi3+nk/wr1bz/ADCf5jvwL+MXyQ271ZU/Hrb3y3/mFVu0sfVbU6m2X1ttzN9u0WyK95KmhiqOx81i6pcfFL654MctSZWQmYxJGyuzcMLzduqlBXPW729hsqSFGcuwTtFaE+v5cT1qa/K/5Rdqdi/z4P5cHa24P5dm/wD+Wp2TWdhdTbFydbu+upost2ltfJ7zalmyORfbGMxtBUMaeplx3k1zS+FVhkbSgVfJVATQk5FflTh1eTTK9CRQaDppkHUO77KY/Pr6S/tL0v697917r3v3Xuve/de697917r3v3Xuq0P5vZI/l+97kf8qO3f8A3pqX2qs/7cf6vMdFe8f8k+T7D/gPQyfy8rf7JH8Z7fT/AEWYL/e5Pbdx/bN9vSjb6fRR09Ojl+2elnXvfuvde9+691qS/wDCmv8Am2/Lb+WVnvi3Q/GXceGwNL2bQ7urd2jKYOhzD1Yw1SsFMkDVqsI7BgePrb3U1J6tWi1ABNfPrVeP/Csn+aj+N/7XH/kmYH/r173T5nrWtv4V/n12f+FZH81G3/H/AG1/9f8AuZgf+vXvWk+vXvEP8K/z64n/AIVk/wA1G/G/9rf+gZgf+vXvdPmet6z/AAr/AD67/wCgsj+alpH+/wD9r/6/9y8D/wBevfqfM9a1n+Ff59dH/hWR/NR/G/8Aa9vz/vzMD/169+p8z17Wf4V/n13/ANBZP81C1v7/AO1/9f8AuZgf+vXvVPn1vWf4R/Pr3/QWR/NR4/3/APtf/X/uZgef+sXvdPmeta2/hX+fSkxn/Cq3+alX42rrz2BthPAkhsNk4NheNS3pKxW/HsWbLt+33Oz3U9yrNLEpKkGnSWa4kSVUCrRjTz6TS/8ACsn+alb1b/2tew/5gzAnn/X8XsJU+Z6V6z/Cv8+vD/hWR/NSH13/ALX/AMP9+Zgf+vXvdPmetaz/AAr/AD67/wCgsn+ajb/j/wDa9/6/3MwH/Xr36nzPXtbfwr/Pr3/QWT/NR4/4yBtfi1/9+XgP9jz4vfqfM9b1t/Cv8+vf9BZH81E/Tf8Atf8A2GzMD9P+pXv1Pn1rWf4V/n12f+FZH81G3/H/AG1x/wCSZgf+vXv2n5nr3iH+Ff59cf8AoLI/mpf8/A2v/wCgXgP+vXv1Pmeva2/hX+fXv+gsj+aj/wA/A2t/6BmB/wCvXv1Pmeva2/hX+fXv+gsj+ajx/wAZA2v/AI/78zAf9evfqfM9b1n+Efz67/6CyP5qNv8AmYG1yf8Awy8B9Pz/ALq9+p8+teIf4V/n14f8Kyf5qP8Az3217/4bMwP/AF69+0/M9e1n+Ff59df9BZH81Li+/wDa/wDj/vzMCP8Arl79T5nr2s/wr/Prx/4VkfzUfxv/AGv/AOgZgf8Ar179T5nres/wj+fXf/QWT/NRvf8Av/te39P7mYH/AHvxe/UxxPXtbV+Ff59e/wCgsj+aiTxv/a/+t/cvAn/rl79T5nrWs/wr/Pqz7+TP/wAKIf5hnzQ/mRfHj46dwbz25kusuwsjmqHcmOpNrYmhq5/tcRLXU7xVlNGrppMY+h5v71SnmerBtVagDHl1/9Tf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3VAX8nVQPkT/MJP5/0rwj/YfxvJn6ezC7/sk/1evQf2c1urj/AEx/ydX++y/oQde9+691737r3Xvfuvde9+691od/GjZP8z/+Qz81vmrjdn/yz+wP5ivUXyc7DzXaGze6ul6WuoNxwxZ3O1eWx+Jy266LCZ6emZPudOUxFVHGpnX7mF3Vr+3mK6NIzU1r/kI6SIjmfxWwANNOIwahgfnWmejNfBf4t/zTt+f8KC6X+Yd8yvjzV9f9bb8+KGZXEZLFy4VdsdRYnN7fp8XtLpDNT08zVFRuHGSedMgHQvIzvUMyoQiWOha0NRp/n6dVXxpNJdaEPU/YAaH9mPt6OBmfh58nan/hUXtj5iw9O7lf4wUXwlk2dU90A0Y2fFukYOuw3922qPJ5P4j91NGopPHqMR8/+buRRWAU1/hI/n07IjM4oODg/lpp13/wp++H/wAnfmJ8a/iDtP4v9Obn7m3Hsn5obG3lu3EbV+zeswG1l2fmcS24sjHVyRhKGKpqIY6mpuVhDq72W5Go6BgT6jrdwrNA6qKkqwH2kY6t7+XvVHYe/wD+Wv8AInpTZ23JM72nuv4b9jdcbd2nS1lKs2V3tlOoanb9BgaWtmZYS81YywRuzhCxB1AG/v0hBlYjgSetwKywIjYIUA/s6r4/4Te/Fr5CfET+W9tXqr5K9Y5jqTsNd6bizB2juCooJM1T4ytSEUs9fT4+WVYHYqw8bPqFufbk7KVRQa0Wh6TWUUkbzFxQNISPmD1qqf8AClb4w7r6H/m7dU1XxjqMxtSX+Y1sLGbd7PwO2Dppt47n3hv6fr3e0OcxdIpE8ddTTUtQ8ssZZW8jK40k+7JLJ26RgYPz+38vPpuezikEgZjqPcpr8JwMfImlV4H7c9bK/wDNZ6X+PnwG/kE5/wCMs/Z03W23eresNudZdNVRqcgtb2T2/BFU5nb+zpaekEktT/Ha9a6pqo5fQEDyzEIjH36CQLOWQYzSv8uvX0DSbeYpWJONRHmAe7h8q/bw6qN+A3Rv85vbPw36J7D/ANkM+C2/GwO0ZM115id5fG/p2g7pqcRRVb1e16vNZpGppY8lUoEnp6oCKQh0eUl7t7UItqQTcEh/OmM9Fs0m8qyJtaI0AAAL1JI8z/mqRXq7n+Vl/M9P84LrT5l/FP5FfHvNfG7vbpDG5Lo35A9eT5Vq+lrMf2BicvsfO1OJilhhnoJ6aWlrKWqopXlEbGN4p5Ef0oWUAllBoD59H0bMUCSEFyK1HD0rT9nVKvw5wv8AN6/4T7dj9+/E3rf4A9u/zG/iLvHea7r+P+9+sKqpxw2w1bJM1RNl63EYrLGCWpiliXI0NbHAqzw+SGo0OwVxyhAAyvlkVFeIPTEKSozMQAx+LBoxAww9McRTj+3pZdF/CT+bxv8A/n2fCb+Yj8xOqnret92bJ7Kz1VT7YeijxHxP23J1tuDBYHqLsIqQqZNKjIUphaKaoNW07esPDMkfpihNENVAx/q8+tWazaC9wumRjVqEkGmBT0x0K383H4gfOT45/wA4b4xfzefjH8ad4/PLY2zMHW4TeXSO1R9xu3ZlbDtau2mi4WOKmyNTBBLDX/f0OQpcfMYKqNkkjCtrPmcPGo4aRT/Z6vHC8U8j1r4hBGOFBSn+UdEy/medefzrf5y83x37Kov5d3YXxp6Z6S3zgKrIdA763Hhf9JW7sjkN1UFVk94tkczTYyoNJT00Cp4pKSlEaK8hSU2JcQosVFehJ7gfP7Okz/UPeESRVQKQjg8Cw7qg0+WerX/+FIfwc+WXzC6n+GGA+MnS+X7XynXfaNDkd6UGFyuGppduUIfEhKusbJTRBqdft5vJMhIUJcjke6wMqowJArTpRco7XEbqCQA4x6sABX5ep6R3/Chz+X98xO3Nx/AH5x/FDrCg+RW8PgvlZZ92/GOpxSbjq97UmVy2Fy65TCYOQSRZEQS416aupUheYxNHNCrmIoaRMqnUfLyPnXHVrqNpV8JQRrxqH4aZGPMHh0Q7+aF2j/Oq/nHfBHfPSew/5Vfa/wATNmbIznWW4uyNjb8q1y2+++J8fnY6jH4Xqek3DjMBLT0mEroafKVkUUEzvEip9x6WieoClWNc/Pz6szSiVFC1TNSPI0xj063JPh1sjc/WnxN+NHXm9MS+B3fsfojqnam58JJUQ1cmHz2A2RRYzKYyWqpy0cjQTRPEzIxUlTYkc+9SkM5I6vbqyQhWwc/4ejIe2+nuve/de697917r3v3Xui1fMm3+ypfIa/0/0Rb4/wDdHN7dh/tV+3pPd/7jP9h6Jh/JRt/sgXXVvp/ejfNv/P0fb17/AG56Q7J/yTo+rZPaTo2697917rHKxSKRx9Ujdh/rqt/fuvdfM++U/wDwpp+WXVPyZ7662oGro8VsftHdW2MSsbRACgxGTeji9DsCL6Lj+osffsEdeJIJAA6AY/8ACqn5dAW8uRPP+qgHH9OZPfqDrVWPADrr/oKp+XV7+XJf8lU//Xz36g69U+g67/6Cqfl1biXI/wDJUH1/5L9+oOvVPoP59d/9BVPy6I/zuRv/AMGg/wCvnv1B16pr8I/n11/0FVfLr/jrkv8AkqD/AK+e/Y69U+g/n14f8Kqfl0eTLkr/ANA0H/Xz36g69U/wjr3/AEFU/Lu3+dyN/wDg0H/Xz36nWgT6Drx/4VU/Lk/7tyV/+DQf9fPfqDrdT6D+fXQ/4VUfLvj93Jf4+qDn/bSe9469U1rQde/6Cqfl1/x0yP8AyXB/1896p8+vVPmB17/oKp+Xf/HXI/8AJUH/AF89+oKdeqfQfz67/wCgqr5dWP7mSv8Aj10/P/WT34jHXqn+Efz6cqn/AIVN/MCDH0VW75Tx1RYLcIoH1J9bOFP+Fib+zy/25bfZrO9UUM2qp/0p6ZjmLzPHQdtP59Nv/QVT8ur/AOdyX/JcA/3jyeyMADp6p9B/Prr/AKCqfl3/AMdcj/yVB/189+oOvVPoOvD/AIVU/Lu3+dyJt/tUH5/H6/fqde1HhQdeP/Cqn5d/XzZHn8aoLD/rJ7914k+g67/6Cqvl3+ZMj/j6oP8Ar579Qdeq3mB14/8ACqn5dfiXIj/kOD/r579QdeqfQfz66/6Cqfl3+Jcj/wAlQf8AXz36g69U/wAI629P+E7/APMm7Z/mH9P713f2kJvvcLm62hpzM2ptFM4ThgSCDfgj37gKDrwzk9bHuTlMOPrZB9UpKph/rpTO4/3r3sdeJp186r/hNVIqfz4/nSyxxqZcT3URZFGkNv153CkfQFgCbfX3X06ueB6+jEpuAf6gH/bj3vqvXfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6RfZP/Mut/f+GXun/wB0c/uyfGPtHVJP7NvsP+Dqm3+Q0D/ssnaV/wDn+e57f1/4A0/19q77+0H2dE2wgi0av8bf4T1eN7RdHnXvfuvde9+691737r3Xvfuvde9+691pKfJLoL+Yn/K3/nbd6fzH+hvg/uD+Yv0p8t8c89TiuucXUzdhdUVKYmgxtXgky9NjsxU4iojkpf8AJayKianqqRhC5R0Kl4MojI46v2in+fpKUdpwxFAlfsbV/gIp0x7+6h/m9/zBP5tX8qP5390fCrdPRnx36335X1OP6qr6vGVG6Pjrt3D5gVm5833Jl6xKKrepz3gpKuh10EJKRLTpTxvfXZNClWr61+XTcgnkEiFfNdJrxFa/lTPV6/8ANH71/mudT/Ib4K7b/l79F0Xa/Tm+957qpflPmqrbOOzkG3cLS1uHhwVNlcxXVELYWlaknytUtdEAZJYVi1XsjUiCFx4nDz6fuWmWJjAKv5D1PV1cDTNBC1QixTtFG08SNrWOYoDIiv8AkBrgH8+2jSuOnhWmePWtV/wpv+Gnyi+aPxX6R2T8Wuosz3BunbfcP8cz+Gwdfi6OsxuKmooIYK+RcpLCrRa0YSMpOgC5HI9qrd1VX1Glaf4ekd0jNJGwBIGutM01IVH5VOeh3/ncfGfvvvr+SH2z8eemes852R3bP158fqGm652saev3BkazZW+Nt5LdNLh4Q6rVy0sFHVSiOJi0ixnxhjYFPxJ6VngOrHv5enXe9upPgp8QusOycBPtXf8AsL469SbV3jtmqnp6mqwG48NsujostiKqalZozLTzI0UmhiAwIube9yEF6j5f4OqQKUiCtxz/AIT1r2/ySvhL88fh1uj+a7vve3RU2xN5dkb433uH49/3tymHrcLv7JvubMZzBNTLjaglqWcPSESyNGt3AJHPtU7RsEViKVz+wdIik6PJJCp1eGAK8CwZzjj/ABDP8ugD+Z38xH+bt80fi73T8Hcv/IR7yxnYPbWDn60Ttjcs82f6kwdZJXx08+/0w2c21/D7RqjVNDL/AB8xQuUkWeQIAzHasn6fD58KfPp9fEktwLgd5A1Ba1Bpmh9a+ePXq8j+Wh/LMrvil/KywXwV7g3bLufN7z2Tu2PsmpobS47bWZ7LxCxZrb221kYo9LjXOmKwVHcNpAUgmzSqJldchafnTqq27yW0kctFMuokDgNXl8/mfPrXE+AOb/nNfyP0+QHwW2j/ACxO0PmT1xuzuzdm/uk+8Ni1lfBsfE/3mpKbCpla+soaCrpXo5Fo6atqaKtraGSKTyjyaGDnZERcFsitag+R8qfLqoN2IiEpqC0AYcWGNVRxBFPLoxX8p/8Al/fzRenv573cvy2+du0zvDF9tfFHcuTqu89sGji68xmf3puHblRhepadYxHoyuIhoamjnookdVSHzrLJHIjtWZgzEg1HAfYOr2kbpEviLpcnU2ajURkg+ny/b69Gi/lv/Bn5bdO/z3f5hHyf7P6Zy20ug+0ev9y4rYHZNVlMPUYjdWQynYuNzGOpsVBSzPO7PSwSyyao18emzcke3J5FZAAfIdUtInjZtQI7nPyyRSnrgZ6Jf8qPjf8AzA/5Z388bev8znoX4Zdk/wAxrp/vHq/c+3psL1/JIN6ddZbdUtOlXh3rKPH5mroTRNRxLSVMePeGejmlhZom490kcSUYYxSn2f5OrQQvAXUmoZiwJHkfL7R69BR8hug/5yP8yv8AmK/AX5vdk/CDdnRfx86k7m6ipR0NuLcO34999eYDB7z/AI9uXf28a2qWknqIWDPMS8MLIqqgplLGzvYIgEcUzUHjWnr0nj+pe6czxU4BHBqNIIahByCSPn/n3wfaLo1697917r3v3Xuve/de697917r3v3Xuq0f5vdv+G/u9r/T7Hbv+x/38tLwPaq0/th/q8x0V7yK7fJ9h/wAB6GP+XlY/CP4z24H+izBf73J7buP7ZulG3/7hR/Z0cv2z0s697917r3v3Xuvnz/8AC3n/AI+T4Pf9qPsT/wBz4/evPq34Pz60H/e+q9dj+l7e/dePXv6e/de69+Dz9PoPfutdevxb37rdPPrwtfn6e/dePy65Ajn8C30/r/re/dVz0Jm2ADtrI3v/AJuq/r/qD7GnL/8AyRb7/SdIrihuI/t/ydBh7BfS/rv8f43+v+Hv3WuvW4/2P0/4n37r1evKrMQqgsx4CqCST/QAe/db6EbC9S9i7ghjnxG08vWRTANHIlLKoZT9DZgP6e/darnqJN1lvynyrYWbbGWiySIZGp3pZQdI+p120/7z7916vSq250P2RuHJnFrt3IUki0z1Aknp3VGC/RFNvqffuvdBzubbOY2ll6rC5ukkpK2lco6OrANb8oxAv/sPfuvDpP8Av3W+uybgf4e/daA66+vv3W+uyCPr791oEeXXuOP6/n37rfXYP4vb+h/3v37rR64+/db6vf8A+E1H/b5X4h/9r/cf/vN1PvR6svn9nX//1d/j37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdUAfydCf8AZiv5hI/A7Wit/wCf3J+zC7/sl/1evQf2cD6q4P8ASP8Ak6v/APZf0IOve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6oO7F/lIdwd2/ztOrP5lncvbPXG7OgOiuq6vZ/U/SK4PODdOG3DHT5BsHWVjVEZxzpDkcnPlpavzmV5oYIhEEAKPeIqgeHUGlD/l6S+A8jN49GGoMtPIDgD+eejpfzUv5bnV/8074lbl+MnZWdzG0KtM3Qb+613zhGDVWzOzNvY2sx+3c3V0L+msovHXVNPW0hKl4pGKOkiowaUgHPShgSKKafz/b1Rt1d/L3/wCFRfSHV+2uhuuv5m/w2k642Rgl2jtLN7i60mzO8qHbVMhpcfDLnctsSqrmmgh0rDJLVSyRgACY6VPtwtqOtjU/Zx/ydNrGEXwkFFpjPD7P8nVsH8oj+U9hP5aWyu191bz7IyPd/wAr/k9uSj338me4qpqiLD7k3VSVddXw0Oz8XWIs9PQRz5KsneSoJmnlkLvoVY409JJ4hrSleP29Vt4BAgStaYFeIHpXz+Z6uI9tdKOve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917otPzK4+KPyH/wDERb4/90c3tyH+1X7ek93/ALjSf6U9Ew/koG/wC66/8OjfH/u5Pt+9/tz0g2M122Pq2X2k6N+ve/de6w1P/Aef/ljL/wBCH37r3Xw1/wCYf/2XP8rv/E5b/wD/AHeSe9Dh1ZsNTom31+p/H9P94976p9nXgPz+L8+/der5dde/db68PqPfuvddn6/09+60OHXv6W+v++t7917rr6fX37rfXZt+Pp79177evXvYH6D37rVKcOvC359+62fl17jj+v59+69176n37rXDpe5y/wDdDb3/AAZf+tTexhvH/Kr7b9sn+HpBB/udN9i9IIjgG39ef6+wf0v8+vC1+fp791s/Lrr37r3Xdjz/AIfX37rVR14W5v8A7D37r2euvfut9d8XFuP9f37rWaZ6+kf/AMI1R/zjZ2Uf+zoyv/W8e9efVgcdbquZ/wCLXkP+oGt/9xH92HVG4dfOr/4TV/8Ab+L5x/1/hPdFvzY/32k918x04eB6+jMPoP8AWH+9e99V679+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SL7J/5l1v7/AMMvdP8A7o5/dk+MfaOqSf2bfYf8HVNn8hkn/ZZO0gfx3nucAfkf5DT/AF9q77+0H2dEuwV+kYH+Nv8ACerx/aLo9697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuq0P5vn/AG7973/6gduf+9NS+1Vn/bD/AFeY6K95/wCSfJ9h/wAB6GP+XiQfhH8Z7fT/AEWYIf7ZpB7buP7ZulG3illGP6PRzPbPSzr3v3Xuve/de6+fR/wt5/4+X4O/9qTsT/3Pi968+rH4Pz60Hve+q9e9+6912bfj6e/de+3r34uR9fof979+6159etxf37r1c067/N1H0HPv3Xvkeuj9frf/AHj37rw6E7bBLbaySKrM/jqVVURmYkofoFBPsa8vBjs19pFew16QXJpPGD/F0H1Jja+uqqehpKOonq6qVIaeBIpDJLI5sqItvYK6X9WsdB/ylPkB25gsfu6uxFbicHVLHLZ6OTmKRSwYzH8cfge/Y68QeljP/K7r9u9gLgdwZO+NcCIDSw/f1WKhl5vz79nr1OjrbB/kyfddl9bV236OoyuBmyeOkzK6JJohC76Z9YkuOBf3v59bpnrcH6R/lRdFbT2rjom2XiaidKdGZ5aCjZgR/Z1OPr7qadeHHoHe0P5P/W+d31DuXCbOxdGtgkrCipPD47+q6L79wx17ia9AP8mPgF0Z1l1/UVv8OxePz9PF+1LR00EUryAAhCYxcj/D3sZ62RXqhDd38q2o+TdbU1uHxb4+rp5nFPXiFolljLH1aksTe35976r0Q75E/wAm3tTqtqaPBmWsqTHeSnSF5/Ixta35H+w9663x4dV7dk/DTuvrLG/xLObYyBiDfoWklRzH+ZBq/p/T36vWuiuV+PrsZMaavppqWZODHMhRhbj8+/dex1CNuP6/n37rw669+6312LcXH9b/APEe/daz12PpyOD79149Xuf8Jqf+3yvxCt/z0G47/wDoNVPvR6uvn9h6/9bf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Wslguuv5jfwA+V3yRn6G+Pb/ACS6z7prk3tQbzgghx+Pp6uvyM1bBipaeKoSWOrovNJFMpYo49YuCCDXXazxL4jUI8v8vQVEO62F7L9NH4qSd2qoFDw00/nXodx81/5wNv8At3vHf/ltL/vX3vuohsv4v5/7PT5vN8B/3H/mP83Xv9nX/nAk/wDbvdP9bzSj/e633oRWR/F/P/Z699ZvgH+4/wDMf5uvf7Ot/OBJ4/l8Rj8f56X/AG9zW+/CKyP4v5/7PW/rN8Azb/zH+brv/Z1v5wPH/YviO/5/el5/9Xfe/Bsv4x+3/Z619bvn/KP/ADH+brr/AGdf+cD/AN6+I/r/AMdpbW/p/wADffvBsuGv+f8As9e+s3ylfp/5j/N1yPzX/m/kDT/L2QH8kzy/7x/lvvQhsv4/5/7PWzeb2P8AiPX8x/m64H5r/wA4If8AdPiM/wCtNL/9W+/GKy/i/n/s9bF3vZ/4j/zH+brkPmt/OB4v/L4j+ov+/KOPyB/lvvfg2VPjH7f9nqv1m+V/3H/mP83Xj81/5wFzb+XvGBbj9+X6/wDnb794Nl/H/P8A2etfWb5/yj/zH+brr/Z1/wCcDz/2L3j4FuZpRz/Uf5b78IbL+L+f+z1s3m+f8o/8x/m69/s6/wDOBsf+xe6XvYfvS/7f/gb714Vl5t/q/b1v6vfK/wBh/Mf5uvf7Ov8AzgR9f5e6H8cTS/7f/gb794Nl/GP2/wCz1r6zfDwt/wCY/wA3Xf8As6384DT/ANu+I9V+D55fp/rfe+/eDZfxj9v+z1763e/+Uf8AmP8AN11/s6/84EXv/L3Q/wCtNL/9W+/eFZfxfz/2evG83zFIP5j/ADde/wBnW/nA8f8AYviP/H96Xn/1d978Gy/jH7f9nr31u+f8o/8AMf5uvH5r/wA4G4t/L4Tn/m9JYf6/+W+9GGy/j/n/ALPXheb5TNv/ADH+brr/AGdj+cDxf+Xuv+P7svP9P+U33rwrP+L+f+z1Y3e+f74/mP8AN13/ALOv/OAP0/l7oP8AXml/+rfdvBsf4v5/7PVfrN9HGD+Y/wA3Xv8AZ1v5wNz/ANi+Ix/0+l/+rfevBsv4x+3/AGet/Wb3TNv/ADH+brw+a/8AOB/P8veM/wDT+Uf/AB772IbL+L+f+z1o3m+eVv8AzH+bro/Nf+cD9R/L3S39PNLf/wBzfevCsuOr+f8As9bF5vfA2/8AMf5uux81/wCcBa5/l7p/1Olv9f8AqN978Kxp8X8+tfWb5Wgt/wCY/wA3Xv8AZ1/5wN/+3e6W/p55P97+9968Gyr8Y/b/ALPXvrN8p/uPn7R/m67/ANnX/nA/T/hveO/9RPLb/wBzffvBsv4/5/7PXvrN84/T/wAx/m68Pmx/N/vc/wAvZCP6eaW4/wAf+Bvv3hWVPi/n/s9bF3vdaGD+Y/zddH5rfzgvqP5fEdr8fvS3t/52+/eDZU+L+f8As9eF5vZNPp/5j/N13/s6384G/wD274jt/Tzy/wD1b734Nj/H/P8A2eq/Wb5T/cf+Y/zdAD8q/kh/O27h+N3cXVPWnwbptmb/AOwNpzbX27u2oSCvp8H/ABWdabKTyUdfUzQOXpWljVpInCE69Jt714dqprG4BHCpx176nd37Li3JRsNQitPMjhnq3r+WR8aeyfib8Lunene49xYnc3a2Ow7Z7sOuwNH9nhabdm4tOQzGKxq/7sjppCYvLpUMwYqoW10dxIJZCw6N9vtjaWqwE1pX+fl+XR+fbPS3r3v3XusNTxTz/wDLGX/oQ+/de6+Gv/MP/wCy5/ld/wCJy3+f9vnJT70OHVm+LomvvfVeuwbe/daIr14W5v8A04/1/fut9de/de675PH1/Pv3WsDr1vp/j791vrx+vPv3Xvs6fcbtjP5mGapxeHyFbTwW8slPTSyIl/p6wLH/AGHv3Ws9N9RjMjSMy1VBW07ISGE9LPEQR9b61Hv3W+ofFvpyP+K+/da8+sghncalhlYH8rG5H+wIHv3XhTrIaGtRdbUlUqf6pqeYLz/tRW3v3Xullm2/36eAUq4IYA6kcAWjYfVhbn8exjvAP9V9tJHnJ/h6QW/+5s3+16Qt/qB9D/X/AIj2Dul9PM9cffut9e9+6913+Lk8+/da86desb2/Pv3Xqjr30PP+xt7917iOu/62sBb6H37r329fSP8A+EaxP+y2dk/4bnytv6/58e9efWxw63U8z/xa8h/1BVv/ALhye7DqrcMdfOr/AOE1gt/Ph+cf+GI7o/8Ae2k91HEdOHgevozL+lf+Cj/eve+q9cvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690x7mwx3FtvcO3xVGiOdweWwwrFiE7UhydBJRCqELFQ5j169JYXta497U0YH06qwLKVHmOtY345Yz+aX/L3r+6Oiuufin/p02IexKncG2Oy4wlLi9xQ18QL5LEww1EUio8YjWSGYExupHBvc1f6OcK7NQ04en8+grbjeNvaS3ih1rqqHqBqrnh5U4fPozP8As6/84D/vXun/AFPl/wDq33TwbL+Ift/2elH1m+/8o/8AMf5uuv8AZ1/5wNh/2L3S5H/HaT/eR97794Nl/F/P/Z639Zvlf9x8faP83XY+a/8AOBH1/l7xn/p/L/tv+Bvv3g2P8f8AP/Z68bzfKVEH8x/m66/2df8AnAi4/wCG+Iz/AEIml/8Aq33rwrL+L+f+z1sXm9n/AIj/AMx/m69/s6384L/vXxH/ANT5P/q33vwbL+P+f+z1r63e/wDlH/mP83XZ+a384D8fy+I/+p8v/wBW+/eDY/x/z/2etC833zt/5j/N14fNf+cBzf8Al7of6fvS2H9f+U33rwbKtNX8/wDZ6t9Xvmmv0+ftH+brr/Z1v5wNv+3fEfP0/el4/r/ym+9+DZfxj9v+z1r6zfP+Uf8AmP8AN13/ALOv/OA4/wCxfEY/r+/L/wDVvvfg2P8AF/P/AGetfWb5/wAo/wDMf5uvD5r/AM4DkH+Xulx+fNKAf/V33rwbH+L+f+z143m+eVv/ADH+brofNj+cBf8A7d7J/h+9ILf65+996EVlXLfz/wBnrZu98p/YfzH+brr/AGdb+cF9f+G+I/8AW80n/wBW+/CGy/j/AJ/7PW/rN7H/ABH/AJj/ADddn5rfzgbD/sXxH/j+/J/9W+9+DY/x/wA/9nrQvN8r/uP/ADH+br3+zr/zgLf9u9054/z0vH+J/wAt96EVl/F/P/Z639Xvg/0D+Y/zde/2df8AnA25/l7pf6f56X/b/wDA337wbKnxfz/2etG83yv+4/8AMf5uvf7Ot/OB/P8AL4jPH/HeX6/+dvvfg2P8f8/9nrxvd8/5R/5j/N14fNb+cD+f5fEf+t5pf/q33oQ2R/F/P/Z62bzfBwt/5j/N17/Z1/5wIP8A273Q3+hE0th/r/5b734Nl/GP2/7PWvrN8/5R/wCY/wA3Xh81/wCcDex/l7xkfk+aUD6/j/LfevBsv4/5/wCz1v6ze/8AlH/mP83Xf+zr/wA4C7X/AJe6WNwtppfT/Qn/AC337wbL+P8An/s9e+s3s/8AEf8AmP8AN1x/2dj+cD/3r3X6W/z0n1/r/wADffhFZVy38/8AZ699XvlK+B/Mf5uvf7Ot/OCv/wBu+I/+p0vP+t/lvvwisq01fz/2evG83ulfp/5j/N1y/wBnX/nAD6/y90Iv+J5f/q334xWXk38/9nrQvN8P+gfzH+bro/Nf+cDfj+Xulrf8dpf+JrffvBsq01/z/wBnrf1m98fp/wCY/wA3Xv8AZ1v5wX/evmP/AKnSf/Vvvfg2X8Y/b/s9e+s3v/lH/mP83Xj81/5wNv8At3xHf+vml/3r7334w2QHx/z/ANnrQvN8J/3H/mP83XX+zr/zgr2/4b4T/X80lv8Ab/e+9eDZV+P+f+z1v6ze6V+n/mP83RO/nhv3+eD8xPjzP8dus/iDiOoc32DvzZtFuTf2bip63E47ZEWTWbMxVaZCapSFAyxzSzpEXCIyoVLXG9FtH3wuNQ9f59NvJul2Db3cDCN6glSKg0wc+VeP+XrZH+MnUlf0N8fuounctm6fcmW682NhNt5XO0lIaGkymTo6fVkKqkpGLMkJmZxEGN9IBIBJAQSuHkLjgej61haC2SFjUqKV6HT230o697917r3v3Xuvnz/8Lef+Pk+D3P8Ay4+xP9f/AIHx8+9efVvwfn1oQcmw976pwz11791vr3v3Xuve/de67I/IHH/E+/daHoeu1vewGq/Fhe5/w9+68RXo+/xT/l99wfKhKms23SzYvHQJ5IKiekdzVKDdwt7W4va49+68K9XB7D/lh7T6y3BtnbWWx1VV7pyAp6avp5ryQSzSJpd/FIfyT7lPk06uTN0JAroah8+kNwB9RHSvVkVH/JrwOzYcN2XNt6lmXHVSZWSmNHGQkYNyrqPx/X3FERrGD8ujAjq+f4w7d2nuXrOPbWMxdHRSY2kWjanjgWPT44z+kD/W93r17jk9FYofiNtHM9+Zhdw071KxQS1dHTNYp5w11AVv+I974de49WB9RT7D2puKk2RX7Ygo56eRIKOYQIj8GyMWX6n3o5yOvdCB8tvkJmOmduY6l2gAtbWPHAluTpdAef8Ab+/U9evD16Kjh/l327tjF0Wb3uskmIyCq4lWNwqRufoSB/j9ffsDr3HrLtoY75VZfIrXI+SolWWSlgHrW6i6go/1+nvfDPWskZ6M51d0bhurcHmanMYmloKaCOU0waFImNhcFT/tveq+Y62eimU/XGO7k7TlSahSqoaOqeOH0aotAc6Q1zYnj3vr329Qvkr8IevMtgslTZja1EySUUqxzNTQ6IWMQswBP49661jrSS/mD/B6LZGbzddgsUzUsUtRLSy0kNlAAZh6kv7314jqifIYytxlTLSVtNNTzQuyMsiMt9LFbi/+t791odQPfut9cgbX+n+x/wCI9+60RXrof0J4/wAPfuvfPq97/hNSLfzlviGD/wA9BuP/AN5qp96PV0Nan5Hr/9ff49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691hqf+A8/wDyxl/6EPv3XuvhsfzEP+y5/ld/4nLf/wD7vJfehw623xdE1Jvb/AW976qOvW/3u3v3W+u/62+tje//ABHv3Wvt64+/db68Db37r3Sh2ztfcG883Q7e2zi6rL5nIzJDSUNHG0ksjuQF4H0H+J9+61wx1cZ8UP5PXcHZvYu06DtrG1W0ds5QwT1SV8E9KwR2DaZJyQLFf6Wv79jr3W5T0r/JG6C2VsvFUFBisRmoo6WmNXM0UFQ8pEasxLXv/X36o8+tkZx07drfyWPjjvbbtdQYrZVBj6yWGSB6mCkgj0M8ZTWGB/BN/eq9b0gnrW1+TP8AIKTrnsbDUe069psHkq0yVFN9zI7uklQboi6jYD+g9761To1u1P5TXW3W1BhqXdOCienSGnaSeanRgSUViNTH/X9+698+h9zvwN+Mldg48RS7ToEmmSOITpSQX1EaSSQf8fdWOD9nW6CnRf8AIfyWuv8AtSfdGExOMljp8fBBNRmNdKxu0RMZBQ8ckfQ+5D35U/qDssi01EyV6Rxj/G5K/LrXg+cH8uvs34s7+lw9PhqzIYeolmFL4o3YxRwlrya3PIsOR7j7h0q6raqKWoo5pIKuCWnmido3jljaN1dDpZSGH4Pv3Xvs6j+/db697917rvk8/wBP8ffutY669+631737r3X0kf8AhGqP+cbOyj/2dGVH+H/Age9efXhw63U81/xa6/8A6gq3/wBw5Pdh1V+HXzrP+E1f/b+H5yf1OJ7osP6/7/aX3UeXTh8+vozL+lfxwOP6ce9nj1Xrv37r3Xvfuvde9+691737r3VffyJ+au5OuvkJsn4m9N9K7o7b7m3715mOxUzMFTjcf13sPCYvIfw5H3pkp545llnYF4YUCgqUOslgvtTFAGTxZDRf8PRfc3jRyi2gUtIQT8gB6/bXHTj8nvmblfix1j1VVZzqnO9sd+dqbk2psrCdK9W1VLXZmTL5yoWizO5z9wzOuExLsstbUKHIUhVubsvlhEkh8M9g8+rPdNDCvij9Vvw/OtK/Z5nrF8Y/mrW9u9udo/HLt/qvcHR/eXWE8dRR4fcM0DYTtjaRgSWXfnWtQCXqMfE8iQzBixVvUrMurTqaEKPEjNV69a3bSOYJhpkH7D8x0fj2n6XdIzJdh7KxGQ/heT3HjqPIeRYvtZ5SsgkY2VCLWuffq9bp0sI5I5kWWJ0kjdQyOjBkZSLhlZeCPfutdc/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6918+f/hbz/x8vwe/7UfYn/ufH7159W/B+fWg/wC99V697917ru/0H9Pfuvdevxb37rVM16EPr/qvfPZ1a9Bs3B1eXlhaMTmnRmWESMBqNgb2BvYe/db6v2+AX8p7B9i53Gp2hTVH8QUR1IVldFEj2IiaJ+ODx791rraw+KPwRn6S3RFjtuYhKXbSwqgbw+NLfmR7G309+r1vIyekB83MBTdb91bK3LSUsA8dfSrIY1FyYmAJut/cpclZ5K3Nj5I3SK5FLlKevVxXRiYjuXqampq/xxmrokVkcfh0LWN7cf4e4oixEoHp0uPGvXLA7L62+P5yBmzFDFVVTSTCnV0W2oEAab/4+3M+XXiAeikvvqlzPdsFRt2I1S1Eqo9RDZ1AZvoxW/8Avfvfl17z6Fjem4MdsjdMW4c9i2QwhZBVmI6dS83LW9+69jj0UL5Nbsynd5xFTtOOWrWjrIZCIg7hVRQLWA/w96+XXqGlR0evq7oSn7R6ZxOP3DQR/eUlNFHJHPEOXUfQBre/VPXs+fWD479RZXqPsXMucZ9pg6L7giQx6IRGt+eOPz78aU690195dp7n7F3LPszaOoRM7QM8YLIben+yP8PeuA69SvTltulpOh8LjpsgIZt05Joy8YA8odzcki5P59749e49cvkxv6obranyFSwWsrsdI4hW4cu0XpAUf19+FevdVObh+P8Atrurq3MT7lxanJVENSKUzJ6yWBC8Nbj3vrxz1rdfLn+Wxm+v8FntyRbd+5xFYKmoFTBTeqEqGZB5Fvb37qp61w9xYerwGYr8TVwtBJTVM0YRx6vGspCH/ePfutDpj9+6t13fm9h/rfj37rXV73/Cak/9jlviF/juDcYN/wDw2qn3o9XXz+w9f//Q38qmpp6Onmq6ueGlpaaJ56ipqZUgp4IYl1yTTTSkKqqASzMQAOT72ASaDrRIUVY0A6i4vLYvOY+myuFyNDlsXWIZKTI42rgrqGqjDFC9PVUzMjgMCCVY8gj35lZTRhQ9aV1ddSGo9eudBksdlacVeLr6PJUhkmhFTQVUFXTmankMM8Qmp2ZdSOCrre4IIPPvzKymjCh68rq41IQR8upvvXVuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XusFT/wGqP+WEv/AFrPv3XuvhsfzD/+y5vldf8A5/lv/wD93knvQ4dbbj0TYW/N/wDYe99a+zr3+x/P++Pv3XuuzYfkk/7x/sffutceugL8C5JNgB+ffut9Hr6V/l9d6d2ba/vTgMVJSY1l8kTz0ksnkhI1LIGUj8fUe/fPrWfLrZg/lU/ystq9ORU/a/ZlJS5zc0E8FTBFVIkqReI6tLQsfSFP0Fv8fr799vWx6jrarbqLZvdXXMtZtqlpMTuLD0QSE0KiBw8MQVSuk3+o/p711sUJr0z/AAybtPbnYlfsjdgqqvC08z06T1Adk0LJpHqYfS1vfvn1404Hoefld3VSdW4+sxm31jbIiF2mVByG8Zawtf37j16tOqAtq9+7w7g+TeKw+ZLvj6CuZPA99AHluLf7f3vy68OOervN8dB4ztPYdLFRxwpX/ZR6SukOJFh9H5/r71WnXsHoufVnwN3TLuFf7ycY+mqkcFrgGNZQRyf8PemOCR14caHowNVluu+iN3dn46aCnaWjoMdFGpAuZEh0nT/sR7Ge8GQ8n7WG4VcjppMzN1R58j9hVfy97twSjbyJtXHZCWOpnkgLLPDUzaSytwOAfp7Bw6c6Kb/MV/kQ7ZynWMW7+sKGHHbipse1dEsC6FedqdpXMixn1f6x9+49epTrTH7m6R330buqr2rvfFz0VTBPLDBUvE8cNV4jZigf6H82v9PfutdBB791vrsG1/8AH37rVOuv8ffut9e9+6919JD/AIRrC3xs7J+v/H0ZU/4czjge9efXhwPW6pmf+LXX/wDUFW2/845Pr7sOqtw6+dX/AMJqx/2Pi+cf/ap7o/2H+/1l591HEdOHgevozr+kfngc/wCw976r137917r3v3Xuve/de697917rVD+XvVfxrov5r/YOW+afyu7R+Mn98Oj85urpndWxO3KzqqjrNo0VPFictTT5mmjkBalEVQRCVBZ49IuWAJojM1uvhgMRTFOg1NFEm4u107RBgdLBiv7CP8B6RvU3yC3v1d318f8A5f8AyU3jXZj4FdPUrfH349/J3eFQ1TJ2Dt7fEsuNwe+91ZmQGprJpTGuqslU6xG3JZW93Ii8Jo4/jatQPL0HTCy3RvFubofoR6Qj1qWrSrHGKf7Jx1ZX1vu/ZHzE/my7T+R/x13niOy+nfjj8eN7dOdjb12vXRVu2R2BvPJ/xbF4OmreBPNHC5JMOpbKxvYXKZ1MVr4b4JNadGcTpc7n9RB3IqlSwyK1B49Xh+0PRz0WPujo/ZmdwG+d3VpyUeYh23nK+CemqRH4Kumx0k0E0YAvdWUW59+62M46gfCiuyGR+NnXVVk6+qyVY1LkkkrK2Vp6mZYspNFH5ZW5JCgD3odePRq/e+tde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6918+f8A4W8/8fL8Hv8AtR9if+58fvXn1b8H59aEH9fe+q9dG34+n+Pv3Xvt67vzwB/sR791qmM9GQ6K+MfZHeWUpI9vYWsbEvPEJcgIWaNozKFkCW/wJ9Xv3W+ty/8Aly/CjofoDZcGT3bRQSZuekiNcuQAkYSEBpLFjc8/197691e/0rtfoKWqO4NuRYumaiUyMadUTlPVzY+654db6TPenzFz+Kzy7S6zwsmQ+0AilkoIS8gUcEsQDx79149V7bs3XubvjtvaeG3viKvGImQpw33qaCWLC5GoD8+5T5LH/IK3P00N+3pDck/UR/I9X8dO4jY3W2zYMI2apaOsemWGHVKq6RoKr9D9fcTw5iX7B0vOST0V/v74m7m7WqKjcWL3TWvSBGkjFHVsAVJDDgH6ce3agdaHp0F/xq2gnWW7Ttjc1LLX5AVQNPXTAyELchQztzfke90qOtdHK+WldtbD9drV5TH0zl4LrI0V2I0j3rJPXhjonXx47I6fwxhmzFJGyVEoUI6gJqYnkLx78eOOt16t7wmaw8uy2zeyqVWh8Pkggp141Wut1HvVOtnqvruHvTuPG/e4vHbSqpFr5GgeaCkJfRJxqLW974DrX+Dp26D663FicLmew9xUJhrTTzVkS1KEMsltSgA/4n+nvx4deHy6LDhdzZruTu2akzVWYKfGZPwJHI2mPSkpCgA2/p72T1qnp0cPu7rza9LTYWr3JlIZMVQUiM9Gr3dwqA8D6fT3oH069T16JXu/dFJmcjjcL1vh3fEpJHBKYIr3GvS2q1/e+t+WOvfJPqak3F0xk9vZDFa5sjhaqSzx3eOX7UgaeeOT795de+3HXzg/nL0xkere3dwCchKWavmiigYESJ+9IwN/z791Xz6JD791vrsc/m3v3Xj1e7/wmo/7fLfEP/w4Nx/+81U+9Hqy+f2Hr//Rut/mn/IXLZP57/FfoCHrj5S9rdbbDoNw7y7V67+NNXUCq3pNuHCCbD027sVBLT/eUVJHEjiIzKbySBXQsD7MbdAsJfALcCfLoPbhcNJfJbUcogJYJxOMV+Xy6m9yfKDrb5O9RfFr4M9AVfZ/xjb5J7yyOzu1toVE9fsv5DdGbT2vmIqyWlrtE8/2cuVnjdWf7hzJCdNwHdTZYmLNNLnSMU4deluogkVra1TxCag4YD58aenQx1uzh8A/5kXxi2j1Luzf2a65+bmJz2wd9dfbx3HV5/a21sn1Fs9a/G7y2pSOB9pX1zWavl/3a5kv6WCq3qE8DO4yvSjSbPcIoYidMtag+VATj7er2/aDo56LX2rvrt3bmclp9lbaXLYmGmWoec0Zna0cXmqFDAfgA291Na9bp0pfj/3NRd6bBTedHi6vDtBlsjgq2jrABIK/EuIap1UAWUseB72OvEU6G/3vrXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3WGp/4Dz/APLGX/oQ+/de6+Gv/MPH/Oc/yu5/5rlv/n/yOS+9Dh1tuPRNvxf/AB+nvfWuuxb6n/be/daNeA6FDqHp7fXeG88dsbr/AA1RmMzkJooylPGZFpYpX0fcTKvOkH+nv3W/s625fhx/wm6psphtrb57YyUozEEVPXVWM+6YQvMp1ENSu1uQfpb3rr1Or4+v/jLtrpWKHYOMwkNJhlhFLG0cIVdIUJc2/wBb37j17h0r6jrHO9eV7GmjafbWTQtZAzeHX9Ppe319748Ot/b0MPxxz+6tldixUcEU9Zt3KzKrKAzBFdrEG449+PDPXvs6t8TD4DGQVGdoqCKCuahepaRUAkMnhEnBH+PvXl1rqjr5L1eQ3VvbKBqjXPWTyoI3N7ISVP8AvHvfAdbz5dVjbm2bnOse0MHmtmUv3WcqpRJOwTXbVJdrjn34Z49eIp1ZTtL5M9obGixM+XoZWpEWm89kNhdVLD9PHvXn1o08+rm+k+z4+0dk4/ckUPgkqYlEiEaTqVFB4t/ifdWPafs63QdU4/KOKpyvyQ31gEaV5KuqoowgPDgkgi3+t7H2+LTkbZj5nX/h6Sx5uJPTo7Px9+NuLiwdBW1tBHTiJYagu6BXkayyHnj8+wHXHSrz6GruHacWfx4xEMCmjip/txGo4cLGY/x9b+9fPrePPrV//mefyuNtd57by2TgxEVLnKVKmqoquFFjkEyI7hpJL3uDa1vdq9VI60Ru+OlNz9D9g5jYu5qaaOahncUtS8ZVKiEOQLN9Cwtzb/X9+6159Av791vrsH+v0/p791r7Ovf71791vr6SH/CNX/smvsrj/mKMrY/9Pxx7159eHn1uqZn/AIteQ/6ga3/3Dk97HVW4dfOq/wCE1Z/7Hw/OP/HE90f7D/f7SfX3r06cPn19GZf0r/wUf71731Xrl7917r3v3Xuve/de697917oKOxeh+ke36rH13a3UHWXZVdiaKsxuLrd97G2zuusx2OyJDV9BQ1Wcpp3ihmIBliRgrEXYH3ZXZfhNOqPHHJTxFBp6ivU7NdN9R7j2FRdV7g6u69zfWWNhxtPjuvMts3b2Q2Tj4MMwfEQ0O1qqneihWlKqacRwDxkXS3vQZgdQOetlFK6SBT08up2werutOqsdW4frHr7ZXXeKydfJlcjjdkbXwu1qHIZOWNYpMjW0uEggSWcoqqZZFLWAF7Ae9szMasa9eVEQaUAA+WOl37r1bpCdof8AMtt+2/54/cf/ALqZffutjj0AvwdFvjL1x/1D5X6/9rif+vv3Xj0bX37rXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdfPn/AOFvP/HyfB7/ALUfYn/ufH7159W/B+fWhARbj3vqvHrr37r3QtdPdVbj7W3fisJhKR5YXr6Va2fQXiii86F1a35ZTx791rrel+F3X3RXx+6m2xR5fGYps3JQ0pryYAXEzwgMGI/2r37rY6O7u3H7U3Jtp8jtjCzS/cR3hShi4YP9Da3v3W/Lqb030R2JU7eyUW1ZqqnrawNJ4JyQUVxcLYe/daApg9HP+MXxO3LhchX5XfVEk2Rk8heokUEGO1wFZrG9/fh140p0T75a9c7ypvkJtaDZdEKNf4hTosqxlVYhh+R+fcp8lf8AKmbmf+Ft0iuB/jKdKbuDbHZ+GpsBJV5t6arikheojEzKzgWuLD3FEP8AZD7Olx49We/F7dtUeso13JUPIscSIZ5WPqspB5/p7v55690NuM6/2ZuSrG4qKijapikB84UXZh+dXv3XulDvnqjavYWEXHbjpPPT0y+lCqsjWtwwJ596Bzjr1KdV677+PGFqMzBgdn7YaliinAM6U6ILj+0rD/Y+9jh1ro9fX9TgumNkUGG3JWxU8vjSwqHsAf6EC/8AX3rj1anRZ/kx8qNsbFxdLW7eocVlqmaVBbw+VyT+F+vHvfHrXHpU9Hd4/wCm3YeSosjQ/wAIL0ExYBPCgS1iQLD6W96Pr1qleHVZ24MBNiu4ZKPYVbNLWTZItUvHI9xL5Tc8fj3vrfDo6/bJx2N6kNNvDKxTbiloGWnDyXkBMAAXn8/09768TTou/wATqiqoZMnW1OEaopqGSSRJpYdQZFctf3rj17/B04d4fJXbLtl6bLUTUkFLDNGrBLRqgUhgPfsjh16o60Vf5uO3JuwN8Vu9tnULz4aGrlqJnhUEMv7l+f68+9gdV456ojKlWKMCrKSrA8FWU2IPv3W+uPv3Xur3/wDhNR/2+W+If/a/3H/7zVT70erL5/Yev//S2nPmf8Uflnn/AJJdIfKb4Qbv6Y2fv7amM3TtTt/Edx0W5avC772vX4kUu21pv7uqziamkMiS3aM6PGVYlTdVFMmjwZvh+XRbdWkpnF3aEBxghuB/1fLoq2W/lRfIDdPXG9PklnexOr6L+atuaopc/jOzcHFuGl6BwOUw2fjqMPhKXbjwGuNOaCMRzVDIzGW3oKBmZ0XlKRD4B/xXSU7Mru14xAuGFCw4ca0p6V6NL8dPin8vN3fITHfIv+YFu/pLeO4+p8FTYn4/be6PoNy4nDbZyWZxTUW+tx7hGeRWmnqWYinRXZVBuQqoi+25Jolj8KCtDxr0pgtLhp/qb0qWXChagAfmfPq2D2k6M+oeRJGPryCQRR1RBH1BEDEEe/de6JV8Ax/xhjOG5N+zd8Ek8k3yA5PvQ49Wbo8PvfVeve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XusFTzTVA/5sS/8AWs+/de6+Gx/MP/7Lm+V3/ict/wD/ALvJPehw6s3xHomv+v731Xr3+t7917ram/4TmdFpTdgZfu3eu3j/AHapYavHUtZV04AeSaIilmjZubBgSOPfqefXhwPz4db1e1403DDSTbcycX2uhAI1f8WsBb3XrfnTpa1WxKSuljGWoxJKrD98Jck/1DX9+x175HoT6TrPbuUw4oKyCOWHRpTyICyi1vz79w69054HrLZ+1FRqSgikqUYNGxRbpY6hY/j36tcdeOOhKPiqKKWOSwWaF4dNxwHXQBb37y68OHVFHyk2LvfD9rVFXgcdV1dCZ3YNEhdQGYt6be948uvfPor9e2RxW98DXZihkSq8kcTrOlmBkfT+ffq9e8q9We7U6Ek3thcZkKunjkoK6OGQq4B0rpDEWJ/x9+4Z610fTrTa2M2DhKTb+MRI4IVX0qoAVjbVex/w91bgadbr0XCj6d2zvX5Fdjbjy6hqvGtj5oBwbseT+fY13eVpOTtrT+DVTphVHjMfXo69FS0lFjI6GljEKRIIwq8elF0j6f63sGeWenx8ukLuE4yjhmq6uVI40QlmkNtOkE8/7b3o9eHp1Wh8ie4tly1cm2aGopJ6ypJgdSdSoGul2Fvzf+vvY618utc/+aP/AC0cT3P1rlO19tUtLPm6Kmlr2FGg8+tYnkDtY34/p7316lc9aQG5MFW7Yz2W2/kIylZia6ooZw66TrgkKFrf4/X37rXHpk4IA/P9ffuvedevH+g+nv3Xh8+vpH/8I1f+ybOyuf8AmKMrfnj/AD4/HvXn1scD1uqZn/i11/8A1BVv/uHJ7sOqtw6+dX/wmr/7fw/OM8cYnug/+vtL7qOI6dPA9fRnX9K/6w+n0+n4976p137917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XukL2h/zLffv/hn7i/8AdTL791scegG+D1/9lm65uQf8nyvI+n/F3m9+68ejZ+/da697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r58//C3n/j5Pg9/2o+xP/c+P3rz6t+D8+tCD+p/3v6+99U6n4rG1OWyVFjKVS1RW1MVPGP6GVwmo/wCte/v3Xutp/wDledA9XdaUGJyvYcMU+RycdPIXkUcs6K2sfXn37r3V+mQ626WyaLksVlYpJPHG1NjDICGb6qqx/wDEe/D06sc56HDZO+6vrPbiyVux5KzFwFUhZKO7NFfQpBIP49+p14Hq3P4ybg2vuHZJ3jSYMYu0BnnjqYRG6gRmQ+9cOvHJ6L32r85aHGb6TZ20qJ6mojqxBVeBSyi8gU39P+P9ffhg9eNOhDzdXit1bz61y+Tw8K19XPSTuXhGol47n6/m/uQuWWdeUL3SaAhq9J5RWYfLoC/kts/cWf7Io6KjpC1EKhbRhbqIvILC3+t7juPEY6UefRv9n9UVH9yMViEH2d4YnmH6ATbn3v5Hr1aZHUXtDt3D9EbdoMDTPF/GKplij1/V3IAB5B+t/fqder69Jan+TYwO14MtuDxyT1YDxRR/Qq3IBFve6V68ajpWbB7+wG5IajLyUkNGIQ0qu66XYC/ANvevLr3RGvlP8jMFvw5DA4h6hMrRxuKbxE8yIAEPA/w97HXscOgh+O/Qu5O0ainym8pmq6CE+QQTtrjVVJIJVza/v3lnr2R1aJt/ZWw9oYStwmBq6Klyk1JJS6UYIAzrptx/j711vPHy6C/qXofGbb3Zmd15hI6qp80s6SMAy2JLXXn6e9nrWCOiPfLutm3vu6Lb+GqpFlhyEaokTlQEQ6dIA/1vfuvcR0bf4/YJOs9nmr3m1FDjqmjsRJxM4dOSTzfj37rw6KL8hP8ARfvafI7a2/Q00tZnxNDTymMM4eYGLUp/wJ9769jgOqbvkd/LD303X+dqNdLNRvT1FVDGGDEq0bSgMNX+Pv1etUpw602/kX0xXda7tzeMkpvDPiquZKwBdKNeZ+VAvyLe/da+Y6K9791vq9//AITUc/zlviH/ANr/AHH/AO81U+9Hqy+f2Hr/09/j37r3Xvfuvde9+691737r3UHJ/wDFtyH/AFA1f/WhvfuvdEs+AZv0xnOCD/pN3xe4t/y8B70OrN0eH3vqvXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3WGp/4D1H/ACxl/wCtZ9+6918Nf+Yf/wBlzfK78/8AGct//wDu8k96HDqzfF0TYWtf/H6f4e99Uz0ZX4l9FVvyH7z2R1xTxGShyubx8eX0i5XHzVAiktyP688/T37rY4gdfTN+IfxE2t0N1njuoKDEUdNh0x8Ky5NYEWaWoVOJXkQ8nn+vvxNOvDIz0KMVFnek868VHX1NfjJagGnRJC4Ck3AFri3v329b+zo//W2brty4alyFfTsEnVCokX1C/wCb2918+vdDFC32oNraVFtJ/P8AS3veOtdep380nkkPBBIHFwRwAffseXW+kXvjf+2tjURyW6cvTYfHowQTVMvhj1H9IJ5PvR+XXhnoOsluzau+9uTZLasmF3FWsjGlkh0zyPxzz+effh17qrrufpbt7d+88Zkcft2OnhiqRJIEg08JKWB4P9Pdsdez1YB1JmN3bUwkFDvGWlwtBjaRVVqo+IFViUOf94PvR+XXvLpb7X7/AOrdxbjfbWD3fi8nuBJlSahp6jXKGEgBAW3upGD17p+2UxbuHs+a4QvFjCQeCLgH6+xhuhP9UtuHzbplf7UjowSk6D42H9D+SefYP6e6A7uHZmd3TipKPEVDwGRCsjKxUjUCAT9P6+9+det/Z0QmT4UQNXVWd3JkZhWOTI0sziygeoMGLe91B60OgY3ntCmoM3B11jckcxQZqOooKyl1+VFjMDRrdPx9f6+/cevGlc9aEf8AOW+LrfHn5M5iahoxRYTPSloYhGEvVSl6l2Gkn8XB9761wOfPqnz/AF7+/de+zrsn6n8nj/C3v3WqdfSP/wCEa3/ZNnZPHH96Mrz/AI+ce9efWxwPW6nmgDishf8A5Uq3/wBw5Pdh1VqUz186v/hNYf8AsfD84/8AtUd0/wDvaye6+nTh4N19GZP0r/wUf71731Xrl7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XukL2f/zLffv/AIZ+4/8A3Uy+/Hh1scegG+DwA+M3XNuR9vlf/dvN70OvHo2fvfWuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6+fP8A8Lef+Pk+D3/aj7E/9z4/evPq34Pz60ICb8+99V4dDt8fdl7p3V2FhKnb+363ORY2tjkqlpIfKI7WIDC/+N/futfIdbrP8uzpmg7HFBHvXCTY80lJSxRUs0GiVJFjVCNJ/wAffjwz1sAHqxXcHxxfrntaDLvj8tVbWV6eWnhgi1JpUhipW/049+49bp59WLbS7B6rzWKodv5jaFZHBFHFEz1FMNB0i1249+oQK9arno5e2YdnNsevxm0Y0pYJqORVgWyctEwtYe69eNeHVJ+78XH1J25mNxbnxompaioZopzHqC6pQQdXPuwp1v5jo3OD7mxW4c9sbckismOoJ6fwxpwWSNLrYW9yLyumrk6+c4ADVPl0mlxOvz6Gk/JnYVduiqny+JCNTSusLzoNbBSdJB0j3GkEkMkYMDq4/o9KjUGhx0p8H8ocduHdX8KxcJix0UIKtay8f2eR7dIx1rgeiJ/KTfuP7H39R4vHSGXJ0NSgiiBvpZSoDEcf09+6959J/dvRvcW8cRhK7HzKtNQrExp9ZVWCWuGQH375de+3pL1tf2FtOrodlfYzivq0jEksCN4QXOkgn3vHXujQdefGHb9PTLvTf00UVRMglcTabtfkizH3rj14+g6EXNd1bJ2Th6rbOwKNnyBjen8lOo+pFtQKj36nr17rh091vuzfC1W5MxX1ePGtpo2mkKKAbm9yR7916vr0JnYXau2+uto1+IOWSrzSU0sfkSUNyFsbmx59+A68fl1Xd05l8Fv3tB8lmp4wyZBXT7g8NeUkcH6+9/Z14Zwejk9p9Q7s3bmaSuOfgx+xaaJJCiT+NGhRASqrf+nvVaY69nos3YuO6rwMkFXtvzV2bxFrulmHkj9Tabf4j34fPrfHoY8Pk4+4epshQvCYK37CaNxMNLBVi0L/AE/HvYx1o9aIf81Pov8A0e9z5masaN6TNVFbO8Y+o0NLYEAn3r59a+R61987FHT5euhhXTHHO6ov9Bf3vrQGOryP+E1H/b5X4h/9r/cf/vN1PvR6uvn9h6//1N/j37r3Xvfuvde9+691737r3ULJ8Y3If9QVX/1ob37r3RKvgEQemM4Rf/mZu+Pr9f8Ai4D3odWbo8XvfVeve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XusFT/wABqj/lhL/1rPv3XuvhsfzDjf5zfK4/9/y7A/8Ad7L70OHW249E2/w4+v197618+r8P5AHXOWzvytqN0T7eqMhgaHByxQZDweWmiycMjugD/hh/j79XPWsGvX0ceueua3cGNhyNdmRQxLovBLLoYIFtwt/eqV6tWgp0IGW231FjVVtw5zH1MtLa6zSq2l15sfr78TjrX2dPmB7Y6rXx4jC5THnwDSkMEo5C8X029+p69e+wdL6HceByystHWRtMOBGHGpuP7PvXXqU66FQaZo9dwmoE3Bva/vXDrfHPVdv8wrrrM9zdfZDbGHr63Fs0Ek9LWULmNlnigKxgsCD9Tf3Ydez1qOz/ACZ+Y/wx7Artk0tZuLMUdLVypj5KmSeZJoRKW9P6gfoPp73nqterWOjv5nHyT3j0pvzN5PaOTkz+3KRXgqHoZG0kxarhtF/9j78B1snFeqyt8fPD5yd+UtRi6NMrjaaprZaINT+eCZEeYwX9IH0B9+oBw618x1bH/LW+K/Z2wszj+zt85vPZXcWYnp6uWGuleWKIM0craSzE2+vFvem4U6sMZPWwFsXJvVdtdjFl0SCkxCyLa12EQB9i/dQRyjtprxLdNLTxW6MTDl8Zj9JyFVFTu2ohJX03H9bewfTz6d8qHrnDuHBZGVqehyVNUOtiY0lDM1/xp/PPv32de6J38parfq04o9tQ1AgnGhZIAwJ1XBuR/S/vYNetcOgj6P6Cy2PxeR39n45cluIQSTU8M6+TxMULKUueD79w631po/8ACjTrreGW3Bjd75KiEENJkZnI0EFI4tUbj/A2PvfVSM161M7jg8n+t/fuvZ68SPwP+K+/deAPn19I/wD4Rqg/7LZ2Ufwd0Zb/AHicX968+tjget1XM/8AFryH/UFW/wDuJJ7sOqtw6+dX/wAJq/8At/F84uOP4T3Rx/5O0nFvdR5dOHz6+jMPoP8AWHvZ49V679+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SF7Q56337zb/fn7j5/8hMvv3Wxx6AX4O/8AZMvXPIP+T5Xkfn/cvN70OvHo2nvfWuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6+fp/wALbKGWu3V8GYomVTJiewIAW+gaWvQg/wCw0n2ustunv2PgkCnGpp1p3CJU+vWiOdkV4BP3NPwL/Rv+K+zL+rt3/Gn7emPqV9D1s1fyWOvdhw7fhzedwkWQyE+pJJvtwwMiqvqLH/X9mo5E3bTVnQH0r1oXKeVT1sObV35tzqPsGmbH4iSGmr549McUYUAeQHiwPvR5D3Uf6In+r8+ti6jr1cvg+5uvtzdXrn67asuRq6alJCvTCRrpGTzx7bPI+6B9HiJnrf1CkVPDonsPy/6Woq6roc9tlsbNFPKiBokjNlYgADT7v/UXdf8Afif6vz619TFUAA9LjEfNLqyjpJv4NFU08ZUiMEqB9Lcen35eRd04GROt/VITwPRF/kv8nNm7qo5MbBQTVuZycvixq2D+SbWDp4W54/p7pLyTuMETTyyoFQVJ+X7etC5Umig16btgfJPorrCLbb9l7kpKfK4mGKsn221QElv4iBEYGB5vbj3HfNtn7nb37d7py77exjVOrJ44BIWv4qj06V27WcV5HNdnh5dFX7F+b+H7L3rmcztDB1OOwkFVNDQRQR6TOiSftyDSovceyr2Y9j+d+S+VIbPmjcVvbplDO5J+IjIpU0p07uW5W1xcFoE0r0tdmfKncuLxlVX0m2co9V4GWF/tW1s1uCCF9y4eQ91/37HX7f8AZ6QfVJ5DoA9j/KTc2M7jO6N4YLIn76r+0p6aWFl5d/TIFZf9b8e9nkTdRjxY+qi6TzB62Oui++6TKbWhyWbw1XS440wmVpItK6efyV9+PIu6E4kT9v8As9b+qQ8R1Cq/kN0XlN6U/lwBqK+KdY1mSFSbg8WOn3o8ibqGp4idb+pQjh0l+9d5ZbemSx2L2tQ5amw06Rj9mMiNFaw5tb3sciboBUyJ+3rQulPAHpY9V9X7FwBp8luKjr6yvCqz+SLVqJ/rf6/Xn2mfk/cVxrQ/YeriZaV6F7sTtGmocEdubOwtfSPKviV4YdNkKgXDAe3Y+SNzlFVkSnzPVTcIMEH8uiz47rnaEiVGZ7Jrq6NJ3aWVao8LquSBq/HPvX9StzLFVdD8x1vx1Aqem1Kb4y4HIw12Dr2p54pVMjIyAMQ3PPu/9Rt2piROtfUpwHUHv75HbZG0aXbu08jUyRlVi8qSXBUrpIuB72ORN2JHenz619VHmlei34euw2P2yM5W4+uq5apw80zx6gLnnST7ueQ91pq8ROvfVIPLoTtt90bD2nt3I1onkpUkpJ9cRIRVbR9CLf4e6/1D3Y/jTq31ScAOtKj+a93lj+yu68vBQzh6LH1FVDHIbuWDvIBpYAD3b+oe7BDIXQKvz6p9WleB6ogym0a2syFXVJUwaJpndLqQbE8Ai/sqPLd4p0l0NPn14XKeh6u1/wCE3mDqMX/OX+IIlkR75ncc91FvT/AKiHT9f6m/tJd7LdWsRmkZSo9Dn9nT0UquSB6df//V3+Pfuvde9+691737r3XvfuvdQcn/AMW3If8AUDV/9aG9+690Sv4BDT0xnBzb/Sbvf9X1/wCLgvvQ6s3l0eL3vqvXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3WGp/wCA8/8Ayxl/6EPv3Xuvhr/zEP8Asuf5Xf8Aict//wDu8k96HDrbceia+99a62TP5B3y4xHUG6NydbVeGiqKysFRnEyDwhmRDr0xrJY/U3BF/fuvYGT1t9bS+S+8d3LUVGKFZT0bU7aEp+F5HFgB791up49AtuneW4aivrancWXydNA8jyMJJWUKt+eD/h791r5jpq683siZuoy2GyFTVpRak0q5cSkG4uP8feut/Lo8Hxp3pvbem/ajI5A1FFhMaWLI4ZEfQLi4P+t791759HzO+WyefWhtaGVlVLfSw9N/p719vXq+fSu3HiaCrxzJVwJURqoJuuo/S/09+HH5dez+fVfPbnxg6f7Iyy5jM7cx8mTog4gmanDFixPLNce91PWxnoq25U2p05ic51Xhdr0DVG/JIYRUJTj9mJCYSGbk/T/H3s+vWvt6GfpL4r9a7Zw1BUZLA0M+QkcVUhMCsuqdxMObn6X96Jp14D06sq2Rt7CY2kpocfj4IUiEaR2jC+NRYDT7q3wnrfl0D1JuA4HuzsWmhYN5f4dqdeQALf09jPdVI5O2sn+n0wv9uw6T/wAlcju2swlBndrzTK1MhNSkBOoCxPqC/wCHsHDp4A9FX6r7rq8buKGmyeSnp8gJo1eOeQqrNrGoWPv3Xq1werJM129thtvY+XK/byu8KENIQbnRclTb3rPXukTR/JTauG10tIqSROjKyKVIAKkWsB9Pe6U68etLf/hSh37h8zHi9tYeONmyNXJHMq6bx/c6jq+n4AJ+vvY6qa8B1pscf7H/AHj37r3XXv3W+vpJf8I1f+ya+yf/AA6Mt/1vHvXn14efW6nmf+LXkP8AqCrf/cOT3YdVbh186r/hNX/2/i+cf+GJ7p/97aT3UeXTh4Hr6M6/pX/WH+9e99V679+691737r3Xvfuvde9+691Gqa2jokElZV01IhvZ6meKBDpF2s0pA4/PuwVm+EE9VZ0T4iB9pp1mEkZjEodDEUEgkDAxmMjUHDji1ub+60NadbqKV8uuME8NTDHUU00VRBMgkingkSWGVGF1eOSMlWB/BB97IIND14EMKg1HWX3rrfSF7PF+t9+i1/8Afn7j4H/apl9+PWxx6AX4O2/2Wbrm30+3yv1/7W8/vQ68ejae99a697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r59f8Awt1mlh3P8G3ikaNkw3YcileCHWvj0tf/AA5/2/uySyRnsYj7OtkApn160LxmMpyfvZuLH9Q/4p7e+ruv9+N+09N6E9OrwP5Xvz9210ZkKDZ+9KWn+0DoFqJ1RRKLIryM5Uj6jke3Tue6E1+of9vWwqAUp1sOYf5udQdp9lbUFFLhxRmpp9arIhsGYEk2Ue9Dcd0H/El/29e0x8KdbI/QnaXUOS2VQ4Omo8RNHVUirKRpYESQ6STx/j7qdx3Q8bl/29b0pTA6AbtfpTo3P72WeKjxxklmEjpEPTqd7kEXHvf7z3UCn1L/ALet6Upw6y5f4/dNYrDq8dJj6dPEGZyApU2uSbn34bnuv/KS/wC3rWhPTohPd+L6OwWMrJsX9hPujDlp8TJGwbx1JbSTb+tvdZbzcLqJrae5cpIKNny68AqnUFFeqyewevert5vRZjPx10+9cjUhFLJqgMZIKjVce3eQzzrtu1bjFYTMtkqnzzTrV19NJIjMO/06PX0H8VNu5GjxMOPxtNUSNDC7QmMsQ2ki7c+2Y903Uxgm5fPz6sUQGtB1YLTfF6sw8VLDT4PGldKfttHyAR9LX92/eW5/8pD/ALeteHHxoOp7fCrZX8Rod676xmNpaXGSpWCMx6FcxHVaxP8Ah73+8dzp/uS/7evaErTT0KW7O0NtZbCLsPrbbFK0cEH2Qlp4QS1hb6qP+J9+G47oDX6l/wBvW9CU4DoRvjZ8XsUPLujfuFpIZGlM0Rq4yoUknTYk+/fvPdOP1Mn7etaI+FB0fuDauwKSOFYMbhZTEoSPQqsRb6f196O47ka/4w/7et6VHkOuDYTANMSMNQaNVgfFxb8Wt7r9fuI/0d/29e0r6Drs4DbUck0zYLHySRo7RExA/TkD3b947lT/AHIcD7etaV8x0VPtDq/K9m5RKKKgpqLGglXEKlFK3tcjj35Nz3SP4bhx+fWyiEUp0C+d+LPXGMpZMdPkKFMs6EhTJyHtyP1f192/em7f8pL/ALeq+GlfhHRetwfEPLTCSejanqaOG8kGhiytp5XjV/vXvf703Xibl/29bMaV+EdQ9pzNgqhOv934CmalmIhhnaG6qL6L6j71+890/wCUl/29e0RgfCOge+UHX21NtbK3FNRPBSxRY2pn1KdKqPAZOOeR73+9N2p/uS/7evGNBig6+el83t7Ty9tbjo8VVAwNXzgzIFa6LNIulGI/1vfv3nudCrXDkHjnqulDgDojwzGU+n3s/wDX6g/4/wBPbf1d1/vxv29e0J6dXqf8JsKyqqv5y/xBaonklIz240u5v6P7uVLaf9a9j7aknnkFHcn7T04igVoPLr//1t/OWaGnTyTyxQxg2LyyJGgP9NTkD3sAnhnrRZVFWNOuUcsc0aSwyJLFIoaOSN1eN1P0ZHW4I/xB9+IIND14EEVGR1xhmhqE8sEsU8ZZ1EkMiSIWjcxyLrQkXVgVI/BBB9+IIwevAhhVTXrL711vqFk7/wAOyFvr9lV2/wBfwNb37r3RKvgEb9L5z9XHZu+AdX1v/EF496HVm6PF731Xr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917qJX1tPjaGsyNW4jpaClqKypkP0SCmiM0zm/9FUn3eKNppViT4mIA+0mg/n1SR1jQyNwUEn7BnonZ+fHxrDzJ/fqgJgmlgktPDxLDIYpF/V+CCPch/61fOWkN9K2QDwPA9Ab/XI5TqR9SuCQcjiMHr3+z7/Gz/nuaD/qfD/0d73/AK1fOX/KK37D1v8A1x+U/wDlJX9o68vz3+NjSRRDfVBqmmigjHnh5kmcRov6vyT70faznIAn6VsAngfLrw9yOUyQv1K1JAGRxPRwsbkKXLY+iydFIJqPIUsFZTSqbrJBURiWJwR/UEe49mieCVoZBRkJB+0Gh6HEciyxrKnBgCPsPU3231fr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rBU/8AAao/5YS/9az7917r4bH8w/8A7Ln+V1uf+M5b/wD94zknvQ4dbbj0TUf7b3vrXV1P8jLAYrcXywyFBl445aU7WkbRJ9CweS1h/X37r3X0JOgesNt0lMqDHU9Lj49IV5V0Bv8AC7e9ZHW61HRCv5lYy216CDH7OwjocnVR0v31JC2gJKtmfyAe9jrR+XSE+KnW2ZwODwc2QYVs2QaGWuMxY6VZ7uCf9b34nrw6tq2lV4bDU9dSY2nioneaMMyLoMg0DUf8fej8+rChFB0MG3aOnkUZMMTLGbRgfi/N7e/A9aPp0MWAqlyFHPT1JBZhZSeeLafUfevs690CG/MGcZVyziQ/bklmsfSLXPB9+8+vD59UK/Lf5Bwba+TPW2yqaMSPlJ7GUBTbRUlSQfx7tTy60T59XZdYgZ+kxkEDEkY+kqZ3+oVFpllex/1veqGhPkASfsAqT1uoqB5sQo+0mg6Mfi94bWlwtdJiq+EDDskOQmmkUCOS+ixYfTn+vsg2zmbZN5huJ7OddFtiUk/D5Z6EO7cpb/sk1tbXkDF7sViCjLDj0VGgz236ns3s7ITZakDBKF45DMlmtz6T+fY43rmjllOS9pd76EKxcKdYz0ii5Z5ka8khSylZlpUBDUdK7I70opsARA0NfSViywq6ESRuQpWxYceyC2uLe8hFzZuJYjwZTVT9h6R3VpdWFwbW/jMUo4owoR+XVX3fWy8hFVzbloq2HBtjmlrVkSTweUAmbS7cf0sPb/nnpOaEdF+wXfXcG+qGlXB42tzWIw87UktRBC9T5PA3iYgqPfutVrw6sY6J2dmd64ZspnsRNjpI4C7ioieMkiO5uDb/AG3v3Dh14jzPWov/AMKMNoUeH3pg3pnTytkYUYXsFA1Lzf8A1/e+tfPrVg966312Lfn37rRr5dfSQ/4Rrf8AZNnZP9P70ZUf6584PvXn1YcD1uqZn/i1ZD/qBrf/AHDk97HVG4dfOq/4TV/9v4fnHf8AOJ7ov/6G0vvQ4jp08D19Gdf0rb6WH+9e99U679+691737r3Xvfuvde9+691rZ/zQN5Uuwf5iXSW4e9Nt/NTcfxeT45bziyEvxy2dl9yde7f3R/F5TU5nfFbih/k0kUQVndmDhNDWKX9mNs1ItMZGsnzx0Rbih+o8acP4Kqa6BU1x5AE4z0NvfvZ1B8osp8Gfhn8ae8t99ddY9ybOxXcuZ7T2Xn2TsbI9adbCN1663F5gJIVy8aSQZNZSGZ0KadKEN5UZC88o7lP8+vPcJKILS1bskANfOgI4/bXh0s+jqvdnxA/mX1Xw6oOxd8dk9O/JXqrdnyA2Zgd+ZQ5NelK7ZlUuGrNqbLqAARj6oq8rxPb1MGtqW7NSFZYPGI7gadOweJa7h9GprGylhXypQUH7errPaPo46Jp3xR/JCXHb9baT7SXZj7czgYV1SVyBof4fIZwkduH03t715Z6359O/wcBHxj61B5cUeR8n9PL/ABSbyW/w1Xt7315ujae/da697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r58/wDwt5/4+T4Pf9qPsT/3Pj968+rfg/PrQg/A+n5/1/8AY+99U8+u0d42DozI6kFWUlWBH0II9+630Y/499yZjYfZe2srlc5XfwenqoxPCZdS/rUJ9f8AW9+611tPdP8A8xLE7ax2EnxO4yqPBTxyRy1KjQNAButvx791vhw6sb25/MH6sp8P/H8luOCtzD06yeH7mJ2WQJqsFt799nXgRw6bsJ/MEi7lkqMDiA6x1cr0MMoIutjpDggD+vv3l16tKU6MPtP41y7nohla2SWvqagfdlZtTAq/qte/Pvw63x4dTn+O9PW9i7TwlRtyOMPVwRLOImH9BcN/h7lPkyTTyVuYJpVG6Q3P+5CUHDqzbB9abb+P2Gqs7XTxeaOB2ijYgeKwIF9V7Dn3E8R/TA+Q6XGtei7UPc/Y3Z27aiHZzVU1LBMyIYF1R8H6AqPbnz698uhJ3rR97biw8eBr8fVss5EOrxSXUMANRPv3Xq9HJ+Lvx1oNlYFMxuejhmyksYldZ1JdGNzchv8AYe/efXukR8le29zzVqbN6/gljaK0bGlW3rAAI9A/r/j79nrYp0HvV22+85qyjqMzVVyU2tHkSV3ACg8ix9+OetD06NRk90Z7b+QoaSUNUROE8tjqKk8cj3rHXs06WlZvSijpg33CCcwlmjZ7G9rEAe/Dr32dEt3x8ltzbUztTSwY6ZqO7os0cRKaf66rf63uwoevdEG7i79rmycu4Xy8lFJEWl8UkuhWAGoiw/PvXljrXn01bC+c1RUinop8t/k8RVXLyJpKq1mtx73THW+hP3H8o+sMyYayeSmbIU2nU4KFmf63vb+vvXl177eqhP5inyR3tvLaOS271vRVTLVUssU08SHiNomRhdfxa3vdOtZ8utJr5AYrNYrdEsO4NRyhqal6hpL6y0srPY3/AKc+/de8+gB5H+Fv9geffutdXvf8JqP+3yvxD/7X+4//AHm6n3o9XXz+w9f/177P5xm7cpsv5JfBLNbzx/y3y/x1iyPZn+k3HfGDZmW3fD9+uKi/h1XvdcQjvBTxKQ4kaxEYkKXII9rrU0RgtNR4V4dE25ozSxswbwlBLaRU/LGT88Dpz7q79212j8Yfht8Zfhb3d2DsXD/MDdOT6/w3dMWTMXdPW+3NqZZMnuMVlBWos9NWzytLQSmRUKRBkW4cMLiJjK0k4ppH5ft6Za7i+lihsmJEpIB/EPWoPD9nSwpaLdfwE/mE/Hbqzb3a/YvZPT/zaxWa2jL11vvKLkMR1tvLqvaS5Sv3vtWZFBWozLny1yMAGLsObIwoSLiBpGGV6dUNY30cCMWSWuDTBArjzz1eT7Q9HXRau2YvkO+VrP8ARrDtZ9vtj5VZsxViKp1tTMswVP68nSf6296+XXqV6Cr+XstUnRVetcEFevYW8BXCM3jFZ94v3AjP5XV9D78OrN0er3vqvXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3QX9z1r0PV+/HS95No7jjuDa2rEyg8j/An2t2w/7tbT/mvF/wAfHSW/FbC4/wCaUn/HT1ouGkpRVZf9k85zMtbXL6i2SkJP6vfUcSP4UWf9DTyH8I65jCCLxp+2n603mf8AfjfPrIaOlI4Q2t/x0k4/P9ffhK/r/g6s1vGfI/tPXGGhpxX4Y+NvTnMQy+uX6rXowvdvfnlbwJTX/Q39P4T1oW6eNACOE0R4n+MfPreM6Ar3r+pNjPISTFtzDQg3uNKY6KwBPvl/vh/3dXX/ADVk/wCPHrpjtAptdv8A800/46Ohl9lfRj1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691gqb/bVFvr4Jbf6/jPv3Xjw6+G9/MJp6ib5y/K4xQSyBe8t/aiiMwF83KQCR/Ue7Kj04H9nW3Iqeid/YVt/+Ak/448T/wDFPe/Dk/hP7OqVFKV6tJ/lE5nM7X+Wu26ukWppopvs4a5wpUfaNVHyByfxp1e7CC4K6kjZvsHXgQSM9b+3YfyV2/Q4Ci2ztTL09JkdUCM6TojB/GPqR/j+PejbXPHwn/Z1YMOjAUeAwXdnS1NQbhr8PU7jpqcNBLUTxmRpAOGueb+/eBdf76f9nXqrXojOH6z7k2VvFMXTSwyYN6sCCRJtUYh8h029+Fvc0qIn/YevVAOT1aljuv6Gl2DS5XJ5Cngyq0gMreUC8njBv714F0T/AGT/ALOvalHn0BtR3TS7RP8ACzkaWpdJCjFZgw4a1z734FyP9Cf9nW6rxqOhBxHym2Lg46Y5vI09P5bB28qABm/Davevp7rh4T/s61UVqD0ot4d3ddblwSyUG4Mc4qkCIfuU1Xk4HAH4v/X3r6e5H+hN+zretPM9Uy9y/Gqk7A+UPXW+jkKF8Zh46qeWYz2S3kM17+7eDc/76f8AZ1qqkVJ6tj6n7p6p2hO+GrNxYta2koTBKDUxg6IoAjqLj+gI92SC5bXWJ9IRi2Pwgdw/Z02zDVHQ9zOoX/TE0U/t6DPZIfvXsXedLsbMLi9gLXQitWjm0SV0gk1EgD68n3w4+9r7/wDM/I3NV7yZ7aW8qPu0hSKNAVqy6iS7eQ/w9dOLO22Tkj252ffedFS73QxkxFqEKCB/k6ds18bKiLeHYIx+bqI4qemox66ghvVFYlrH3jfzx7p++Pt57d8r3XuBb3caXLuKrqMak5wfn/xXQT5D90+XpeZ7uS6t4W8Qr5CgFR1n+Om3Jqo9h7EyeTgqU2jTrUYyeaYF5Jai5K3/AMCPfWX7iPuhv3uLyjc7TfxuY7ONZULVL95pQ19Oo/8AvccvbAku1c97QqxvuhKOi/CoQEg0+fRdd+dJb87Wz2Y29U5JKPCmZopKkTBUjhVyt73H4J954GC6GPCf9h6w31Cnl0ar4z9b9N/GzaFZhMpmcTmJz5ZZElnhkPmflwRyfqPejb3XHwn/AGdeBUcT0m+1flBt6ihyNDsz+H0yqkqJHTOgvcEAHSPexbXXHwn/AGdeJX160ov55Dbi31V0W58hUvMRWeaKGJtYtEx4t/re9+BcAEmNgPWnVSR1rUigrSAwpZyrAEHxPYg/T8e6+HJx0n9nW9S+vXvsK3/lUqP+pT/8U9+8OT+E/s69qX16+kN/wjWDD42dk3Ugf3nypBsebzjn3QqwNSOtjget1TM/8Wuv/wCoKt/9w5Pex1VuHXzq/wDhNX/2/i+cYPF8R3R/r/8AH7Scj3UcR04eB6+jMv6R/rD/AHr3s8eqjrv37r3Xvfuvde9+691737r3VbnzHqv5lOUrdybK+KnWPxd3N1vunZWYwM24u2d7bwxO56DJZjEtQTyNh8XD9s8atJIFW7Aixb8j2piFsAGcmo+XRfcncDqSBEIIIBJI8vQD/N0Q2X+Vz8ieh+ivj/2f8WMh11U/PDquXaq7pp98bkzMHS2bwdZXvU7623hzDTGoiiRJPHTsyr5VDklCV9vm7R2KuOw/t6RrtUkMSvCw8VacfhPmR6iv59HM+KHxw+TW4fkLu75efOTHdV4fuHD4iu6x6W2v0xm8xl9qbZ6uyKpV5WfL1OajWaWuqanVYa2CrqJNyoVmWWMR+DDkeZPr0qtba4M5u7ygfIAWpAGPXzx1aT7S9GXSE7Q56238P67O3GOPrziZffutjj0A3wdBHxl64DfX7fK3/P8Ay95/fuvHo2nv3Wuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6+fR/wt5/4+T4Pc/wDLk7E4/wCq+Pn3rz6t+D8+tB/iw/r+fe+q5669+691yHBvcgixH45HI9+619nSsoN9boxqJHR5WoijQBVUMSAB9LXPv3XqdKih7n7CoWBXO1MgX6K7kcfS3Hv3XujxfBv5abw2z3htTF56tepw+XydPAELeiF9QLEhvrqsTf8A2Hv3Xj8+voTdMZvJ5fY2EyWHOp63F01RHb/UyR6wPp/T3ulet1pjpN733t2XtjsPa0kmId9FZE0UgiPN/pZrfX3J/JyK3Je5E+SN0juGpcJ9vQibvn7T7inTEfw2s8NcwhkYRPpVWa5vb3FcWIwT6dLmOadG46/2btv4mdeDIZyhjm3JkI/uYRMl2V3U2vrvxyPdzU461QdFz3t8ne6s5VnI7awKfw1ZDolWDgJ+GFh79/h695dCt078nd/19dSYjcFJUSS1MqQygx8IrmxA4+nvxFOHXuPRmd9b86q6np495bkgpvvagCZzUWAR2AYk6wR+ffgOtZ6CQ/zFOia5/s6fP4emqHjKKgqaddLfQcBfx79Trwqep9F8iOu95ANjszQ1NSwKoyzIxJP0AsPp79Tz68M4PQN7+3PNT1orUziJFqB8STWAX6/T/W978s9b4cOgk3d3DtafGNTTrFVVUcZUspVy5tyW4+vvVM9e6rI7z2huHtWXxbbjlpKZp1aRxqS0X0Zbg297r17iMdI/CdEUGCoYqfIZbRX6OY45buXPNiL/AF9+61g9Sl6QyqTLUUxnqIHbWS+sqbG4vz79Wn29e6C7ufL47rTZudqs1jo9cVJMqSSoLMfEeQWv/T37rwqOtNL5a7kXenYuVz1FBajesnuyL6AfM4/1vfutcD0U2/1/x/Pv3Xur3v8AhNR/2+V+If8Ajn9x/wDvNVPvR6uvn9nX/9DcV+Vu6fn/AILK4aj+IPUfx77E29XYypj3FX9vb93HtfKYzIyRyRwmhoMVF4poVPjYhpCWGoen2oiFvQGRiD9nSK5a/DUtkRh/SJH+AHqq/BfypfkNsz4qbf7K2lVbCoP5jWydy5jsjaOMi3TkI+iaPcWX3b/FarbH3D03nWJ6W5NUAB5P2yNPrChrtddAO3+fRdHtMgg1swE2Tj4QTnHn/Po3fQPx8+ZXdPyW218jf5gG1emdmVXQuEjo/j7tPpHdOW3BRtuHdGHNDvzcW6qjLxl/qSlLEkg+q/RUs7TyxJF4UNTq416Uw2tzLci6vdIKYUKSftJJ9f8AB1bx7SdGvUHJ/wDFtyH4/wAhq+f6fsN7917olXwBuels2T/z83e/+xAyA96Hn1ZvLo8fvfVeve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xugg73v8A6Kd82+v91s+bj8WxkntZtn/JVtP+a8X/AB8dJr7/AJJ9x/zSf/jp60dySarKXA1fxrL2uR9P4hJ9PfURQPCj/wCaaf8AHR1zN4TTD/h0v/Hz1yNj9LaedV7C/wDT37qwPXFNQrMV6r/7msT+Qf8AlOT6+/PTwZf+ab/8dPWgT48Ff9+xf8fHW7p8cAB09sgD/nQ4n/3XRe+YW9/8lm7/AOar/wDHj10p2j/kl2//ADTT/jo6HP2WdGPXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3WGo/4Dz/8ALGX/AKEPv3Xuvh7fPvOy4z5xfLOJYElEnem/DdzYgpmpU/A9mabrMEC6VoMdNvEC1eikf3vmuLUcP0/1X5/r9Pdv3tN/CvVfCHr1sT/yGKjobcu+crjOxvs4d8GpnkxlRMyh46NndkjUnk3uLezK35ouLaPw1iWh694IPn1uSzfEn4910cGdqtwSR1NSEmjBY6VkIFtN/bh5tuP98r17wB5HoS9rfFGhrXp12t2JUU9NqUoi1WhdA4AAv7t/W6euYE694A8iejs0PQuE21syWszVY+UmwuPkramrAEjPHTgFmDC9+PfjzhdUosKAda8AevVAnye/mJbSqt6VfUOyspLRVlLVyY6VmdImWVHMRFh/rcX97HN9yB/Yp14wr5HpSbB2FW7g2/QZyvydVkKqtjWZirNKQZPUv0P+PvX9brjh4C9e8AevXLs3aexcBgZX3XmRRzNC0yrNL43XxrcAXP1/p70Obrn/AHyvXjAp8z1R53j2b2Fgc3kK7Yu7cgdrU1UDSgVTKpVWOkAA/Tj3Yc3XA/0BOvCAevQLzfMjuuqqaCCPO1wmp4npRL9wwJ8o8ZIPv39cLkDECde8AV4npT9Z5ntjPdg4muy28MiKGvnVsnL925WFJZAdLNf+h93Tm6Yto8BKONJ+xsHqjwAUepqhDD7VNR/g62i/gbLtXaWcTGUm51rVqUoqmcedWDSmNDKHYfm/49/P7/eBX197X+9EPOFlt4ntbWeOVqD8BJL6fWg6zt2qPcPdL2dtb25astqhUAZYeXDq0LBT7Fyu9+10fIPLUSUuNDRXUpHrQA6j/T3Kvv19832E58+69sn7ysovq5YqRoY8q6rTsGmuqvHzHWOu0ci7/ZcwyIupVU8c5z0Uzc2L2tsPvTZ8mGyUsFHumump89Q0jBvuoydMQZR9bE+4j/u3/fHmi993m2vbIPC2m6lWMA1GpAtRSoFKHy6yI5u5fk3L2U3K+35Wkl29AYGYfCTg/wAurGqj49bNj29VVctXPSR5KhiqpZkS0ixVERkFyR/Q+/oHn5suhIQIloOsFY4VKDNcDoju5OguhIZ8hDVb2nWodpLtJLYh2JFr39tjm654+CvThgUnop26vibso1FXk9tbvFfTvrJQzgsByQQPdv623AFBCvWhAK1r1q6/zhOrKaiqzt+PN0wloxJMglqAgYyKzW590k5ruJYWhMKAN1owAkUPWqzV7lnoauooTTQyfZzy02tXuH8LmPXe3N7X9lX72l/gXrfhDrB/fCe3/AOID8EMfr/gbe9/vaX+Ede8EevX0ZP+EbUzS/GvsksAP9/Pkxcf7RNb/eb+0NzdPckFwAR6dOIukGnW6bmTbF1/+NFWj/1Tk9sDrzcOvnV/8Jq/+38PzjP/AFae6f8Aed7Se6jiOnG4Hr6M6/pX/WH+9e7Hj1UcOu/euvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690hO0OOtt/W5P8Ac/cdv/PTL791scegG+Dt/wDZZeub3v8Ab5X6/X/i7zf196HXj0bT3vrXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdfPn/4W8/8fJ8Hv+1H2J/7nx+9efVvwfn1oPi35/3j3vqv2dcr/UW/1v8AD37rVOuuSf8AH37r3Ade0nj/AB9+69Xrx/2P+x9+68Olj19uT+5+89u7kKMwxOTpqpgv1CK9mbn+guffuvHPX0Tf5afyz2r2z03tSooa+mFRi6Clo50eRQ7PTw8m1vp791vjkdWO7j+QvXdd2VsjH5VcbOaSsp1nF4zfTYXe49yXylG0nJm4aDTSrE/Z0knNLhCfM9GV3v8ALjq/YNL5tq4WkqassQvhhjkKyfkji/uL4sxgnpaePRUexO891fIWWhpoMTWKdaJH+yyoqXFrFbC3Ht3z60OFOjj9Y5DYextj022t6U9O2aqY0IDqheMMCObgn8j3onNevZp0H3Zfdvx/6OxtTujK1mOpp41eaLW0akOouL6h79kjPXqZ61Ff5qX86XFbxiy2y9g5A+ZZJYKeWkb8BQoKMnHH9fe+tdawdR8w++HyhyNNvnKwnzeUR+Rj6A19BbV9PfutdHx+Mv8ANW7k2TubH0ecytdXRytHT095nmaWZyQqqv4Pv3W6kdbPPxy7/wCyu7sVQ5HcMWRpafIJG1OZY5E1K44BB9+PWx6dHITbOYYPSrjJp6gnUszxvYi1/wBXvXy61noQcL1du2roAWkGMjYXZlZQwX6c397qadb+zpV4fofalO65bcGWepqIDqGtrq5BvZvfutdP2bxeFp4FgoGpIoyAI7MLkKLWt79w68Pn1Vt/MgptpUPS2VlyNZS02Qakl0IXCs/7LBbe/dePWjj2NmcbCczS3EklXV1X2pYfgTsL3P8At/fj1Uny6LufqffurdXv/wDCaj/t8t8Qr/jP7j/95qp96PVl8/sPX//R3+Pfuvde9+691737r3XvfuvdQcmL43ID+tDVj/rA3v3XuiV/AIW6YzgH0/0m73t/58B70BTqzdHi976r1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690EXexA6p31fn/fq58fS/wDy7JPa3bP+Sraf814v+PjpLf8A/JPuP+aT/wDHT1o6XP3WU49QzWXtxxY5CS/vqGP7KL/mmn/HR1zOAHjzZ/0WX/j7de9N/wBNwfrb6j/Ye9163TPXcZ/yzF+nTfM4mwA/V/lyf191f+xl/wCab/8AHT1of28H/NWL/j463cvjcf8AjDuyf+1Difzz/wAW6L3zE3z/AJLV3/zVf/jx66VbR/yS7f8A5pp/x0dDr7K+jHr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rDU/wDAef8A5Yy/9CH37r3Xw1/5h/Pzm+Vx/wC/5b//APd5J70OHW2+Lom1uL/429761506EvqXtbd/TW88XvjZmRnx+UxtRFIwhlaJaqKNw5glK82P9ffutefW518BP5ic3yi2Lj8ZnMp9puLDpHTVtO8zJM7xcOQp5NgPqPe89b+fVwW0uw96JW4vHberMgRUyRIjxl2I1H+gP09669w49W+ZftodWdJYxt7P91U5egWlrVmN3eCdR5FN/wDD3rjWnVvkeqJO1P5XuyO597VfyG68y32tJPPLl8hQUz6mjqS5m8bLcfUnjj34V6qc9GY6m3hszq7alXhdyKgrcPA1JTGoAXyNEoVW9Q+vHvxr1vFOqmvmDlM93NkslDg8hLRRiSU0y0zsA0AuOdP4/qfex1o8Oqb+wtyPts/3Jy+ZgE9FL4ZkE4ZpJGbjVfm/v3AdaAHSPxmPjqshSxRVlP5JGDRL5BrJJuCF9+630bHYnW2/J2cUNbpjrEU6/JYxgKLMDfj3sYNetYOOrMPitPletZlx2X3Y75esmj8VS9T6oPWv9q5ta3uD/ev2J5T96Nkay3iMLcgHTJQE8Dg1xTPn1K/th7q7v7d7go/trA/HCfh/IdWIYHs7cuE3Fv1sJnZczk8nBjY2kEnkDkqApBB5t7xF91f7tDZ7j2v5dm2orLNbSOwQ0CkE0OB6D06m7Y/vB8nbhzi8252YitWFcDgaV9PM9WB/HjqqhyuCoOy+xWlrdxYl2rKAVOoorSESAAm/09zx7G/dR5N9ols97hUfvKChNBQA0HCmMcOgF7le/W481w3vLmzxCHbLkaaDFQtfL59H3h7DTcW0snSRSw1EiUj06xI4cqiRGONQB/T8D3lizF6lvPrHhV0UC8Oqf+zuvN15LcmQnpqatWmnqJjZRN44wXJDDnj37y69Whoeq3vlV2Bv3434v+MwbhlggZJWNNNUsikBTdSrHm1vfh6der6dabf8xb5Zbk7t7HP+5qp8kGmSrFPKTEVsQiawf6/j349a88dVflmZi5JLMSxJ5JJ5JPv3XsU69c8g83/r+Cfz7916nmOvpH/8I1if9lr7JH4/vTlj/wBZx7159bHA9bqeaF8XX/8AUFW/+4cnuw6q3Dr51n/Caz/t/D85D/1ae6P/AHtpPdRxHTh4Hr6Mq/pX/go/3r3vqvXL37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdIXtC/wDo337b6/3P3Hb/AF/4TL791scegF+Dt/8AZZuub/X7fK3/APPvP70OvHo2nvfWuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6+fP/AMLef+Pk+D3/AGo+xP8A3Pj968+rfg/PrQg/wt731Trr37rfXZ+p4t/xHv3Wh143tc/n37r3y68ST9ffut0p165/P5/4j37rVB0d742/OHsr45UsGL2tU1MWPViHjiqGVQr/AKm0k/4+/deyB1YF1V/Me3ZvDfOJzW4cxVU0YqlaaUzMBGb3N2Y29yfybJGnJ+6QMe/w2P7ekU4LXCEYp1s//GT5N9D7527jsnmt2Y6tqgkfmSoq42LTaSzXHP8Asb+4siGmMKfTpcanPVrnVnyJ6QoIW/hCYWYxR+moEkTBWVTyWt7coT144HVa3zX+cmzOusnktxUW4aSpq4EmdKGKpVhGi6SNCL78OGevH0HWpN83v5ou5+8ZcjgcRlKmmp4mkiHgeUKWAC6W5A/Hv3WqnqlPI5Kuy1XLW5ColqamZi8kkrs5JP1tqJ9+631B/wCJ49+690d/+X1tfZ+6/khtPHb1p4ajFpKkqJMAVFQJgqEA3559+PXvPrfw2psin21h9uSbOxVH/BKSjp2iSJVB4VWXhQPr9ffuHXvPoYq7u2j2Tt3L7g3PiYqHHYPH1NfVVssWiNYqWPW4LsLXt+PfsjrZp5dE/wCi/wCaz1D8ld/ZXq7YlO09ZjTViauigU0oakYpJGZ4/wC1x9PYV5y5x2nkbZX33d9RjUgaVFWNfQdEm/cw2PLll9beHUSQFUcTXzp0w/L759ddfG7DUybsyCUtdXrJJQ0aOvlnWIEsBG/JI+nHtDyLz/tXuFYSbhtKOiRNpOtdNSfMeo6Q8rc3bfzdHO9iNJt2CuPmc9Av8a/lrkPldTV2b25SZPD4jDyBEqcjSy0UVUGN1eFzwy25v7Sc6e5/LXIYVd0LvK/BUXV0i5k5/wCX+WZltLqQNOQTpGQoH8Xp+fVYH82ug+R02Bqs/RwS5XYlLGfK+NkmqZlFmACpGbcfU/4e2eUvdTlvnCQwWxMUvkjDSftI49E/Lfu3yrzLuH7rhcxz0qtRRH/0rnBPyGetWfLZOsydVJNWM2sOw8bXHjOo6gVP0N/r7knqTB69NluCf6e/dernq97/AITUk/8ADy3xDv8AX+8G4/8A3mqn3o9XXzp6Hr//0t/j37r3Xvfuvde9+691737r3ULJf8W6v/6gqr/rQ3v3XuiWfAQg9MZwj6f6Td8f6/8AxcB70OPVj0eD3vqvXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3QR96/8AMqt9fj/fq5+3+v8AwyS3tZtn/JVtP+a8X/Hh0lvyf3fcf80n/wCOnrRz1MKzKBrXOZzH9Lf8XCQfX31EUDwY6f77T/jo65nA/rTf81Zf+Pt12DY/U2/r/gR791cKB16M3rMXfUbZnE6Tb+tcn9Pen/sZT/wt/wDjp60NInhH/DYv+Pjrdz+N4t09sk/k4HE3/wDPdF75h75/yWrv/mq//Hj10o2j/kl2/wDzTT/jo6HT2WdGPXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3WGp/4Dz/APLGX/oQ+/de6+Gv/MP/AOy5vld/4nLf/wD7vJPehw6s3xHomwF/e+qE0669+630az4h/ILLdA9sYHPw1k0WFqK6mgysQlZYkgaT1ylL25Bsffutdb5Xxm+enQm5Np4jJ4PLYifcUWNSpNJNPAJVaNBq9H1vce/cet1A6Plmu36f5F9Q5SWSpiinoUZaBUk4Kqlk0W+v+w96Aoa9bLAjoO/jP3ZS7Cwu5tsZjJRyBfPDNDUS8a7ELcP/AE97zx69w6I/8tuxWmw+Yy+BptYgE85ko9b3jUF2diluAPz731XrXF+Qn8x2p2TA+K23MtXuNKWsoZIRL+5C0paPyOR+nSf6+9der1SruTvHsHdO4avcWVzM1RV1NYauzs9ktIXVbg82Btf37rxr0PeA+Q1Sse2qmPMSpnfu6OKpjZ7CJDMFk5J+gF+ffuvVxnq97pjd26t24nA1GBzVIKeoo0NRUNVImkrGuos1+PexUmg63Qk46EuhqdzPveKhwu7KHI11LMn8TiiySOaZmb08Rtxz72Y3IOD1bw5fJT+zq2H4rZp8JmM9UblqqaolhhoGlllnEiqbKfqb/wBPch7+JI+QNkYgjukpUdJERhcyeICK06tYPyl2tTbOlxNNlaCMRUroFhqI7AiIrc2/p7jmnSr+XVZW7v5rO1/ic+692bvzcWTwuOkMkdB5vPJUK8pBVaccta30HvZFevVxjoburP5tGI+RnVZ7K2btfF0O2cvFULT5Ovhej0PGCjMzOLCzG3uM+Z/dPaeV9z/czQtNOQKKFqc9RRzZ7x8scnbnJtF+GMsShmIUkAHhmnWsT/OU7y7vy1NBuL+8VANrZCtkgoI8bkRU6Q1y6vGCLek3Hsx5X56HM87RfTmADhVSD+der8i+7vL3uJeT2WyVrbU14pTUKj+XWsVW11XkamWsrZ5KmpmbVJLKxZmP+ufY76lPqIDbke/de49e9+6919JH/hGt/wBk2dk/+HRlv9j++D7159eHA9bqmZ/4teQ/6gq3/wBw5Pdh1VuHXzqv+E1f/b+L5x/1OI7pA/pc72k/PuvmOnTwPX0Z1/St/rYf71731Trv37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdIXtD/mW+/f8Awz9x/wDupl9+62OPQCfB3n4y9c83/wAnyvP9f9y8/vQ68eja+99a697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r58//AAt5/wCPk+D3/aj7E/8Ac+P3rz6t+D8+tCC9735J/P8AT3vqlPTrr37rfXIXNyT/AK/9ffuqmg4dcffurde9+6912bcAfQe/daHqevWsAfz7917zp0KO1yV21kWR2jcJUFWQlSDoYBgw5v8A7H2NOXyRs18VNKoa9IbgAzx/b/k69tDujsvY1OKXbO6snjqcOXES1ErKGb+l2+n+HsF9LqdXg/y+4/kX8ldh7hlwHcuTo9xY5q0w4r+I+DyRwMyx6o9eqzWtcD8+45559wrfkUxS30dYZCBqoTQn1pwHUS89+513yTu8G3x2JuIpE1M/AChA0r/E/mFHl0FWyfi18sflT3vvzrve+f3JgcNtVquiq87O89LBWCB9MrxVchYEXUi/5v7Y5h90tg2DZId5lcOJ9NFXJOr+H1P2de5l92dq2fZI9x21VurmUKfADAFA3m/8AHmTw6rd+V3RVF8fOz67YdJuA7iekaoWqq2limdaiB1V1d4gOTqP1/p7F3Le9Hf9rTc9HhrJlRmtDwrXoW8lcyzc1bIm6zQGBmNNPEfaD5g+R8+gK2bteu3tuvb+0sa0aV+4cnSYqkeU2jWaqk8aFz/T2dTyrBC0zcEBJ/LoR313Ft9nLezfBErO32KKn+XVrdD/ACeu8a2ipqyLN7bdKiNZFK5SnAsf8Cbj3Fc/uzs9vM0MkbgqaHtbrHJfvUe2Eg1RXWpfIgHOadLrYf8AK1+QfVG6sTv6j3FgYF25ULkKzx5eBWkpIPXKlkIJ/BA/w9+h93dmeVY1jdixoBpbz6UWv3ofbeW4SCOYsznSBpPE9bPPxn+RtPlNm4PFNl4q2pxcVPjss4mWRI62wSxf/Ye5StJhdWkd4ooJRqA9B1kRZ3KXtjFfphZlDDyweiNfzYPmXmt24TH/ABJ6GmiynZW+KdqiufHzajDSFRBVU1RJALoLfW/5tx7dd0iieaQgLGCxPlQcennZIonnlNFjUsxPkBk9LD+Vr8G6X4pdepuHeZWo7K3kafJ1bVenzURkS09PRfkJ9OTyfqfz758+/fuW3OF0dk2YlrOCokYcC3lU9Yj8z81f1r5mW+kPhW1uSlsNX9spNGdl+RqB+3qtj+dbBjqH5N/HrMb1Bk2LBXu+YEhcwCjWvWWUSD9IBVWuT+Pc7/dklt35LdIx3q/d9unHQ59io5o5d9ebIa6BX7PDFB+3q03srcEMvwlrNx/B/GY1slUY/FfYR41xHJHTrRaatpDTjVrUgG359xVdILn3xlg57alp3mEP8HlSlcevUf79Fy5PzjeLzEzRqZkFwygknhoWlD2EcSPLoLPiZle490/ETtcfLmgphTUmKyP8CrMx6ZKiT7KSzypVi6ESWC8+789Q7XY+5W0t7fsPGdh4oj80qK6gPl0Xcy23I9juujlyR/p454AFQELHIWXSIyBkMfipUevWmdu1acbr3MtGQaUbgzIpiLafAMjJ4tNvxptb/D3m9b6zAmv4tIr9tOsyrYt9LGX46Vr+wdJ7jn+v49u9KOr3v+E1Bv8AzlviHf8A53+4/wD3mqn3o9WUcfsPX//T3+Pfuvde9+691737r3XvfuvdQcn/AMW3If8AUDV/+47e/de6JV8AL/6Fs3f6/wCk3fH/ALsF96HVm+XR4/e+q9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdBD3tYdU76JF/9+tnv6/8AOsk549rds/5Ktp/zXi/48Okt9/yT7j/mk/8Ax09aObW+7yht/wAvrMXH4P8AuQf8++oa/wBlH/zTT/jo65nf6LN/zVl/4+eufptbUbfnj/be/Zr07Sq565R3+9xPJ/4vOJtYD6fep7o5/Ql/5pv/AMdPWwo8eD/mrH/x8dbuXxwN+ntk/wDahxX/ALrovfMXe/8Aks3X/NV/+PHrpJtP/JMt/wDmmn/HR0Ofss6Meve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XumifcGCpZnp6nM4unnjNpIZq+ljlQ/0dHYEf7Ee1CWd3IodI2IPmFNP8HTDXNujFWdQR5EjrF/efbn/O+w/wD58qT/AKP92+hvf99P/vJ/zda+rtf9+L/vQ69/efbf1/j2Ht/X+JUn/R/v30N7/vp/95P+br31dr/vxf8Aeh06UtXS1sK1FHUQVUD30TU8qTRNb66ZIyQf9v7YkjkibRICpHkRQ9PI6SLqQgj1HUj3Tq3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYKnmmqB/WCX/AK1n37r3Xw2P5h3/AGXN8rvz/wAZy7A/2/8AHJfehw623HomvvfWuvfU/wCJ9+69w67HB5Nrf0/qPfutfZ0Iuxe2OwOuK85HaO5cljKlozEQtTM8RjP1URlrD6/i3v3W+rRerP5xvevW21qfbAiqK6OKJY5aj71tUpX6nQ7D6/6/veOvAnr24P5tXZVclRWY6KrpK/IOZKsQzlDrb63Oq3+2PvQ61U16h0X83HtCPb+UwWTwr5VMlj6qgMtTUIxjWqj8ZcamP09+63nqrmfccG5uwIdx7jAFFktx0tblUYmRUoHrlkqk5vcCPVx7pJq8M6eNDT7adMXHi/TyeF8eltP20NP59XUfMP4RdcZX43fHzvroLFxvitz1GHxm5/4NQEVUz5qvTH+eSKMerxm7hv8AA8W9wnyZ7gX91zfuvK2+gRm0OqMscsgWpP8AsdQDyJzhzBtfMFxyzzpdfUXUwmmjwFjhVD2w6vUqRSuTk9Jz+Yv8W+kvib0T0ntjGUMJ7S3JhTWZLJpTr97LNGBIxqpkFwQp+rEfX3f21513nnffNxuSKWFtJ4cR8nxk/t6M/bTmDmbmjer3cr6b/FwxU22P0CK6aEcdY7jXhXqvX49d79hYDde2trDdWTpNtTVcNNUQ0yvNMIWlUMFsbn0gj3Le7SXMdhI9odMgGDw6mDdpbmHbpZLQgSBSVJwK9X57WyHSe1Ms+48blN5fxTJ09FNXyfwKs/dlESuxuw/r/T3Dc/MPOUilFm0cchxX/B1inHzt71rI6tPalQ7AfqeVTSueNOPSJ+U3z7oOpMRtiDr6bPRz5aZqfcFRX0U9A0ixg+LxlrazYG4F/Y95O3/nTcrVNr5hufqLW3qYgSCU1ceHHqU/ajfed95vLz+tpiIQgL4T6xSg/Znoo2b/AJo26qilpsfs6TM5jJVkRiejpY6l6mSZ4zcRx3ubE/j2NyyJVnNAPM9TS0kcYLyHSo8zjqv3tzfHyC7eyYqt8YPetPgKjIU7Sx1WJy8VJFDUVQAaondAukBr3JsPbH1VucLItf8ATD/P0j/eNi1RHMhbNAHUn9letnjqan6w2n/L52f16O0KPZb1SzB62graNqyOSq0nRoZgw0n3hxuVlv1x7w3W4R2huY1jUKzVotK5GKE9YTc5b5uG48xJuEe3TXdxBNIGthGSsqLUKXalApGQa9VR/wAw/wCI/aG1uodt9qYLtjNdqdZSyPUVK1b08lHj444NZqFlpS4DA8aWbn3NHt9zrtO6btPsE1sLa+g+IUyR6ioHUp+zvOm13l49lebPHs1zOaRgU1zFR3BlopGn51x1ReD/AI/S9v8AY+5n6yTI66sef8Pr/vfv3Xqjrr37rfX0kv8AhGt/2TX2T/4dGW/63j3rz68PPrdTzP8Axa8h/wBQNb/7iP7sOqtw6+dX/wAJq+P58Xzj/wC1T3Rb/wBDWX3UcR04eB6+jMv6R/rD/evezx6r137917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XukL2gL9b79B5vs/cXH/AJCZffutjj0AvweFvjN1zxb/ACfK8f0/3Lze9Drx6Np731rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuvnz/8AC3n/AI+T4Pf9qPsT/wBz4/evPq34Pz60IfTccm35976pnrj791vrlp+v+HPv3Wq9dWPH+Pv3Xq9e5HH9bXH+8+/dewc9eAvf/D37rdeuvfuvdCjtn/j2clx/uipP1tf0t7Gewf8AJGvv9IekFx/bx/6b/J0F4F/9tf2DOlxNOrBP5bHbnZ/V/wAm9jU/XUddWxbny9Di9wUFPHNNCcazsXlmRfQq3NizD+ljcewH7jbDtG/crXVvu5CxhGOo0wQMZ6jX3V2var3lOe/3ADxLJWmhPn4qCqgeZJ4U8+tw75TZ3OdX/HTfPYPW+2aeLfGWwFfPkZKKJRVUUr05eSoIQAllPABP194I+3tvFzHz1FsO+XRazs2HgBjhtJwBXj1ids227PLNt13cw6H3eREu65IVhUh6/CPL5V60GN87p3JvTdmd3Lu2oqancOWyE9Vk3qvIsq1LGzo0cputrfT30htLaG0tkt7cAIoAFPTrO7bbGz22wisbBQsMShUA4aQMUp089R4+qyvZuxsdRZU4OrrdyY2npsupVTjppJwEqgz8DQeeffrxlS1kdhqAUkj1xw6a3qVIdpuZZI/FVY3JT+IAHt/Ph1s7YX4x91TYigmX5m5mkEtOjLSitwNoFsPR+v8A3v3jrd84cvxXLxttOsg/EEbP/GesCo/cjlqUav8AW/VDU4KRgjPmAv59R9y/GTuin2zuOom+ZmarIYcRWSzUv3mCIqY1S7QsFe9m+np596tuceX5LiONNnoWYAHQ2D/vPS7bfcLluXdLeMciJETIo16UqlfxAaakj5dUobe+Y/bXx9ym79j4nNTZiOLMS3yn3bq8s0Zus3puLi/1B95FWMniWUTKugaRRfIfLrOzbpRPt8MiLoUqCF/hFOHR9v5P4r+6flrne4exMxS12RWkrY1OVnVxTXUzAwipJH5+v9efcM+/XMG6bNyTPBtSt4k4C1SuqhNCBTyPn1Gnuvvz7dtCbbHKYDcMuqUmgCA9y1PDUMdXz78x3d+4flLszN7X3HRUnVW36OSiq8VS5GEjJeSUMk7QK39kXFre8X9j/qxtvtzPBuMDG8nBZmaM1D0wKkcOsVo+ZNgeW62mfRNuAuENq1R2QduoAj8/z6rT/nafHrePYlb1vviHLw022KDO4vbVXR+VRUl89UiBKiOA/q0EmxII/qPcq/ds5n2+DZ5tmVCJBqkY0NO358OHl1LvtTzSuxb7uuw33c1wDdQ/6SNAGU/n/h6MRuDf+2P5YPwi2rFj6STcOdymIpdENR5HWaeSMKgeZ/SCWKqPxf2HV2i796Pcq4luT4drtraBTBb/AC9Aba4Lz3K5kleCVrdt6Uyse0mCOI6KIPMnjXyPUf4XfLrDfzFuney+r93bej29k6Gjq1NPCxeNpPtWlppTNFb/AFQINh9D717gclTe0/Mdnzps58SIusbBj5MQDQdKeauSZOQLiz2MXskscrGeKVgPErBR2RsZBp+w9anvyQ6ubpvube2wGnapXD5WoMcj2v4qiZpY1uAOALAH+nvMrZb7957XBff79QN+0dZX8n78nM3LdpvcQ0idAR/g6Aw/7b2adCbq97/hNR/2+V+IfF/9/BuP/Yf79qp596PVl8/sPX//1N+XM5rDbcxWQzu4ctjMFg8VSy1uUzGZrqXGYrG0cK6pquvyFayRQxIOWeRwo/J97AJNB1okKKtgdJXDdp9Z7h2LF2hg+wdl5braelmrot/UG5sNUbNeipqlqOoqxuWOY0fiSZWiZ/NpDAqTf3sqwbSRnqqujLrBx69YNg9t9ZdpxZCXrzfO2d3nEyiHK0+Ey1JWVuMd2IgOQoEbzwpKFLQPJGFlX1Rll597eN4zRx1WKaKddURqOhE906d6hZEE4+vABJNFVAAckkwNYD37r3RKvgELdL5wC/8AzM7fH1FiD/EFuLe9DqzdHi976r1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690EHe9z1Tvq3/PLZ+//nsk9rds/wCSraf814v+PDpLf0/d9xX/AH0//HT1o6n/AIF5S1iRmcx/ha9fJ/t/fUJT+jHX/faf8dHXNJVAmmpn9WX/AI+evcciw/1r/X8+/fPq5FMU49dxMRW4q/8AzucTx+D/AJcnvUg/Rlp/vt/+OnqqsTcQFv8Afsf/AB8dbufxv/5k/soX+mBxPH9P9x0X598xN7/5LV1/zVf/AI8euku0f8ku3/5pr/x0dDr7LOjHr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6xyyCKNpG/Sgu35sB9T7qx0qSPLrYFTTrUE+ZndnalF8rO5aPC75y+OxFNnI0ocfBNKsNNGC6lUAcD8f099APbHYtkuvb7bJ7m2R5GQlmIFTw+XWCfuXzBv9p7g7ja2l2yRrooorita+fRbT3v3QAD/pEzh/ofPN/wBH+xweWeXTxtE/YP8AN0C/61czKf8Actqfn/n6wz9791LDLp7EzeoRSEf5RN9dBN/1+7Jyzy7qA+kSlR5D/N1WXmvmcRt/jbVoc59Pt62ff5X29M9vD4t7RqNy5OfMZdZKxp66pZ2mkBdQoJYngf6/vB33ptLWy9wLuCzQRxgLRRwHHrNP2fvLu+5Csrm9cySMtWY+Z6sa9xT1J3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYan/gPP8Aj9mX/oQ+/de6+Gx/MPFvnP8AK4fX/jOXYH/u8l96HDrbceiae99a67t+fx791qvXrj6f4/q9+69Tz674vcn/AGNvfuveWOur83+l/wDY+/depinXr2+h4/1vfuvUrx68APybe/deNevfQ+/db49ba/8AJY7VxfdPx63X1B2FQU2QwvVVXi5sPFXBJooo6CP+JCoBblQtr2P+PvDb7xO27jsG/WXNexN4UtzqhkK8W1gKK+vHrFb3S2Lb9s5rMut/H3YGVQCag24FQvpUUqBx8+qYv5r3ftb3R8pd3UFLko6/aOz6lMdtoQsDFDF4Ak6RhCVAFgOP6e579o+VzytyVaWcv9q6h3/0zZOeJ49S17VbTa2vLi7wkJhnv6SShhQ6h2io9aDqv/rbd8Wwt54XdM+PGTjxNVHUtRFlUT+Nw4W7cfj3IO4Wn11o9rq0avPqQNzsv3lYSWeop4gIqOIr1btH/NtwsccCf6DqRjDBFDdq2nOoxxhCfr/hwPccH24lNf8AHDn5f7HWMB+67YmR5P3zd97Mx/UP4iTT8q46Kd8wPlknyS21tJYuvqHaEGMrJKmGqgqI5p6kzRNqjk0f0/HuQto5EPK+1wbq1y0xu9QKkUC6fT7epK9rPbGL24mvoIb2W7W4ZW/VYsV7QKAnyx0UrqHsCPrDf+C3rNi0zMeHnMrY+TRpnBtx+5x+Pz7d3G0+us3tdWnWKV9OpO33a/31tU22eIYvGUrqXBFfMfPqznef8znEdgbVn2OeoaPFrmlosXNlHrIHFJC8qxT1CxgHlVuy/wBPp7ANvyDLbzeOLommaU/Z1j3y593Kz5Y39OY4d2vJ5ItbCOSZ2Qls5Umnlj06MX3F8BM3270X1nuToDsWPM0ktSlRmcJW5kU1LQvURjyFEick6Wa6Bl+o/wAeAntPPcez8zXW073bsrKBpkCk6hnzp0UbT707Xypvl6nuDAu3VkaKF2+JwhopIIwH8qV49Dt8ln2/8SP5ZdH8dN+bsj3D2DuymylPSRvVpXVFNPXItRHHr1MwVdJUG1gL/wBPYW5Rjl5q93rjm3b4jFZwoEBoV1sKg1BA9emuVdxPuR7lWvM2zwhILB2aT00spVGRhglq1YcQOtXr3lB1lt1737r3Xdib/wCHv3Xq9fSP/wCEatz8bOyv8N0ZW3/U8e9efXhwPW6rmf8Ai15D/qBrf/cOT3YdVbh186v/AITVEj+fD84/+1T3R/sP9/tJ7qOI6cbz6+jMOQP9Yf71731UcOu/fuvde9+691737r3XvfuvdVo/zQvnFvf4U/HLfm9ei+vcJ3d8gsJszOb+2/1DX5SppJZNibUgep3jvzI02NvUtQYyNQZCmkMxtq9Le34YfEqx4D+Z9Okd3deBpRfiY+laCuWp8uiz7T/mg94dqfCP4F9p9RdKbT3X8s/m5tfZm56PqKr3LPjNtbP2lWTCHsHsE5OVdctNjImjqaelkZS+oq7ARsDaOEOxZsKvH/N03cXfhKkSHVJJw/wV/n0Zn42fLXuV/ktv/wCHXyw2btjaPZ2PxlRvXo7d21Mp9/jO6urcciRZfc1Vj1AGPrKWoYxvAAoKq11R1s1poU0eNF8PVbW5l8b6S5HeBUH1Hr/OnVlvtJ0ZdEm7/wDlx1Rsej3115nafejZ59vZrHK+N2rX12PNRVY6SGImvi9AW55b6D3rq1KZ6UvwcIb4x9ayA3WWiyMyE8HRLlJpF1D8Gx5HvfWj5dG09+611737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xz6P+FvP/AB8vwd/r/BOxP/c+L3rz6sfg/PrQf/B/2HvfVOvX4At/sffut9de/de67Bt/rfn37rRHXX09+631737r3XvfuvdCftkodtZMEgHxVN+Rx6Db2M+Xz/umvv8ASHpBcf28f+m6DEf1vz+PYM6XdbBP8pGX449L7M3N3/2bnccu8qSnr1o8bXPTyfbwUEr/AG8sccp1Bj9RpH+v7gP3ksebuYo4uXeXwVglIEzDBCnjTrGX3l3HmPcOZLTlWwsZZ4CnjRuobwzcKQFSVxhVoa5wejIfGn+bFtvsrvjszr7t6vpW6o3PLX0m26rIvFLSVFLUSsiRpA9ggC2tyb8fT2D+b/ZUWHLllecrJ4W5WhQ6lBDOQM6j8/n0R8w+1O7bFsg3q08S5mn0/UwA4hBHe0OK6lPAefl1S/8AzE9g9VbM71yVf1LmqfK7f3Q9ZlXjpniaGikLoscUaxEgCxP+v7nnkW53q42GIb8um4QAN9tM9TJ7R75uu98rK2620luYGMSeKpV3RCQrkHI1AVz0Q2mqqijqIKqllkgqKd1lgmiYpJFIhuroy8gj2MmUOpVhUHqUWRXUq2QePQoxd5drQxpFHvbNhEXQq/dy+kD8Xv7Kjse0k1MC/s6KjsO0MxY26VP9Edek7x7Vmikhk3vm2jmjaORDVy2dGHKnn3sbJtSkMIFqPl15di2hWDLboCDjA6CyaeapmknqJHmnlcySyyuWd2PJLM3JJ9mgAUUHAdGoAUUXA6EvrTuTsbqDITZPrzctZt2tqUKTTUpF2UixFm49or3bbHcAFvYlkA4ahXon3rl7ZuYrb6Te7dLiP+FxUdWFfEH5593DvvZA7H7YyR2s9fSw1kVT4Ege9Uh9Tm39kte7Wt+PYF5y5M2a65cure0s0MjowWg8yDTqKedvafk/9wvPs+2RR3MLLIrItH7DUgEZNeFOjTfzgvmHJu/sLrzEdY76GU2li3ps5lcTRyQtS1WTxksVRQ/cSpqIAJa4H5/1rewV7H8if1a5fkTdIAl0xIJ86Hog9m+V2vDufMG+WhjlndordpFpIluVClaHgCwJ+eOjvdYfIf40/P74uYLqfuLPY7E7zxOJipqxqtoElpa6GMaEDTMAbcMGH559gbcOUucfbnne45j5XQ3FpdGskfClfMdR5zLtPMPtjva3W0W0nhQVFtJCjSmSNjqeORQO2pwD0/8AVmR+In8tbqDeWU29vXG5vde46SsYVCyU0tS1Q9O0NNxA7Em+lQB/re0vMO2c9+6+8WtjfwG1sLdwzHNXIIPCnSGDd+cfdLeoJ7nb5UnTtWOaNo0gjeiykOR3MVzTrVX7r7OyPcPZu6ewMqAtTncjPMLHUft1lb7cn/EqRcfj3ljtdim27fFYpwjUL+wdZjcvbLb8vbNBs9oP04FCjoKfa/o66vf/AOE1H/b5X4h82/3P7j/2P+/aqePej1ZfP7D1/9XZU/mF9i7Q+SnZUn8u3JZ3C4/qPcPWHY25fl7uSszibYy+zNs0uzTneuMbgayuaJamStrUWarSAtoiVdTAawFsEZjjM9K+n+XopvJkmnWy16Rxb/n39v8Aqr1Qv/LXTYva/wDKK/lc/DjM5eHP9F9+/KP5D7J7oxUWaeGtyW2Nj9yZLMbdx9bkKaUVFHDNM9LMI1Kq14vxb3eBBV5FGAMfb6dMXkjskMEjCrsQ/wBnkSPKo6u17X622f8AEn+aT8Ksl8dtqwbH/wBmqxG8ut++6LCy1cuK3DtDq3Zay7FmrcVUSPFC9M6qq1MahnaMMxLlmaqsZbVy+SvDq8qLb7nCsPaJAdQ9aA06vX9oejroqfc/TXcm981U5nZPyE3D1phloWV8FjcTTVlOWjgPldpJWB9f544/HvX29bFOg/8A5edPUUnRFbSVdU1fV0nYO8Keqr3AV6+ohrFSWsdR9DIfUR791sinR7Pe+q9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdBD3tf/RTvqxsf7q5/wD91cntZtn/ACVbT/mvF/x8dJb7/kn3H/NJ/wDjp60bBJqq8pcW/wBzWY/2/wDEJPfUUCkUQ/4Wn/HR1zRQnxJ6f7+l/wCPt17UrH68/j8e9UIHThyeu4jauxNuAc1iQR/j98nvz18CXP8Aob/8dPVMePAaf6NF/wAfHW738b/+ZPbKGnSRgcTfm9/9x0XvmFvf/JZuv+ar/wDHj10m2j/kl2//ADTT/jo6HX2WdGPXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691ByX/AAAqrG37LG/+tz7q/wAB+w9bHEfaOtLX5jaW+VHcdyQf42n9f9VJ76K+1dR7c7X/AKQ/5Ouf3ulT/XH3MH+h/wA/dFw5HrHF/ov+8H2Pvl0BeOfPrFObRTKwuPFJz9P91k+9r8QI9R1WU/psKeR/wdbTv8pck/GXbn9P8q4/5DT3gV74/wDTxbwf6X/L1nP7L/8ATvbE/wBHq0/3EXUq9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691jlTyRSJ/q43T/kpSPfuvdfOS+UH/CVX5OdwfI/vDtLH7lqxit/9k7n3XjFWiVtNHmsi9bGmto21adVh/hx71gDrxya1HQFf9Ah3ym/56at/wDOGP8A68+91FK9aofUdd/9Ahvym4/381d/j/kMf/Xr37HXqN6jr3/QIb8pv+emrv8Azhj/AOvPveOvUPqOvf8AQIb8pv8Anpq3/wA4Y7/9afesVp16jeo69/0CG/Kb/npq3/zhTn/rD79Udeo3qOvf9Ah3ym/56Wtv/wBQMf8A159+x59eofUdd/8AQIb8pv8Anpq3/wA4Uv8A9afe8deofIjrof8ACQ35Tll/381bYuNV6KP9H9rkxe9E9pI4gY+3rR1BSRQmh6tb+L38jPtz4x9Tbz612HgJ031urFz0OY3GgeKWepqKFqEzlmFiQrfj/Ye8XeYNk56569wLaz3SyCbXZSatTEFXowIIHGuOsc9+5F5s5x5uS9vrrwktpFMBVcxxhgZErShElKH5dVYZH/hI/wDLTL5CvymS3bkamsr62qq5ppaTySMaidpV1vLGxJAIBN+f8PeUKxxRIscYooAA/IdZFRxLDGsUVAqgD+XUP/oEN+U3/PS1v/nDHz/1h92x1eh9R17/AKBDvlN/z0tb/wCcMf8A1691qKV69RvUdOFR/wAJHvlbUUVJRvujIGKkv41NKGW544QxED68W9nt9uSXGzWlgOMGqvz1HpiOErM0tR3U/l03D/hIb8p/+elrf/OGP/rz7I6jp+jeo67/AOgQ75Tf89NW3/woo+P+sXveOvUPqOh066/4TXfzKepaJ8RsDu7cuFwrggY1qJKqCO/1EImT0j+gHA/HspvNg2Xcn8W8gV39adBPfuQeTOZ5VuOYLCG6kXgXUGn7QekZ2D/wlo+enbeU/jPZXau4N05BBaFqymAggX8eGmEZRTb6kC/tTZ7Zt22posYhGDxoOjTZeXNj5btRY7Dbx20Q/CgoOg+H/CQ35Tfnctb/AIf5DH/159rAQejijeo66/6BDflN/wA9NW/+cMf/AF596qOvUPqOux/wkN+U3/PTVot/0wpf/rV73jr1D6jrbB/kB/yue0f5aXUu8tl9lZSTI1mdzVZkKcyQmAolS4fSI7AAC3A9+8+vDhnrYQzP/Frr/wDqCrf/AHDk97HWm4dfOs/4TV/9v4vnEB/zqe6Lf+htJ7qOI6cJwevozD6C/wDQe9nj1UcOu/fuvde9+691737r3Xvfuvda6f8AMF6K+dG1sz89/kpszrDYPyAOe+IXdHVHTORl3LLid4dZ9a7g2jNX5zam39l0kX+5OsedZJwCDJUSKAHJbQVwe3MIStDx4cT0TPFfrePPpV0OBnIU0rT5+fVe/wALt09v/E74LfyifnF8huqtyYXrLoDpkdDZvbW28HuHcPZkO3+xdy1OIwe9M3sZKZZ6Rfs4oJArEsLqz6PKg92iKGIxV7m4/sx/s9UuhMlyl2VrHHgUyaEipp8vL5dW1fHDeG3fnz/Mbo/mx1Djt24zp7419Q7u+PUmX31tfO7JyG8d37xqlzlVJgcLmYkknpaVJCGqGC2soIBcD3WRfBt/CY9xNenLdjd3/wBZGCI1UpUgitaZoR8uryvaHo46DztCioj17v2dqOkeb+6G4z5ZKaGRyf4TKeWZST/X3ojHWwc9Ad8Hbf7LL1zY3H2+V/Fv+XxP+Pfh149G097611737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Wtx/Pu/ku7t/mw5j495LbG8m2snUlNuWjyEQjicVsGbmE4IMgNiCo5HPvXn1ao00+fWvT/0Bsdk3/wCZsSaf6eOG/wDvXv1etUHXf/QGx2Rz/wAZXl/6lw/8U9+6917/AKA2OyP+fry/9S4f+Ke/Z69Qddf9AbHZN/8AmbEtvz+3Dz/vHv1T16nXj/wjY7J4t2vL/j+3F/xT36p69Qdd/wDQGx2T/wA/Xl/6lw/8U9+qevU66/6A2Oybc9ryX/5Zw/8AFPfqnr1B080P/COztSko6imi7g0QyoweN4k1PqBB06R7EO2bzDYbdPZSRa2mBGr06Ykh1yB/Tpm/6A2Oyf8An7Ev/UuH/insPV6fp1Ij/wCEcnakcTQJ3FWpC1w0SOFiYH6gxrwf9t71T5daKITUgfy6xR/8I3OzImEkfbU8cim6PGsaOv8AQh1Fx/sPezny68QDg8OuU3/CODtGpcPUdvVU72trmKSv/rapATb34Y68FVRRRT9nWP8A6A2OyOf+MsS/6/jh/wB6t79XrdOuv+gNjsnj/jLEv+P7cP8Asfx79U9ep17/AKA2Oyf+fsSf9S4f+Ke/VPXqde/6A2Oyf+fry/8AUuH/AIp79U9eoOvf9AbHZN/+ZsSAW/45w8/7x79U9eoOuS/8I2+y0YOnbM6OpBV0WJWUj6EMB9ffutEAinWSb/hHB2hUNrqO3qud7W1zaJWt/TU4J9+GPLrwVRwFOstJ/wAI6O2KBjJQdy5CikP1kpJftmP+u0Nj/vPvRVWw4qPn1po0cUcA/bTrLWf8I7e3q8KuQ7symQRf0rV1D1AW3+pExb/ePegqL8Cgfl1pYYkyigfYAOm//oDY7K/5+vJ/h+3F/wAU92z1enXj/wAI2Oyfx2xJ/sY4v+Ke/Z69QdH+/lc/8Jkt/fBT50dKfKLKdjvl8P1lV5atqsUYof8AK5q3GyUMXrADAASH6G3v3HrYx+zr/9bdL73+EXxd+SdXlst2/wBRbb3RuXL7brdqzbpf76gz8WLrMfJjVCVuOmh1vBHK3gaUNoIFuAB7eSeWMaVOOkstlazN4kiAt69VidI/yGemPjv/AC/Jfhf1X21urBdj47dW5977M+VDbdxUvY2z9zZ3dv8AeGjnxuJEwpxBTUqRY9oUmTyBTOSJDxZLhozReHp1SaxjnU6z3H8Q4/L5dG9+LHwL7B6l7azPeHyU+T+6vl72PBgMPtrrPK722ZgNr0/VuOpqBqPP1G3qTDSSIarJliaichW5f9TOW97kuAyaI10jz+fVbexMUhmncyv5EgCg+VOrK/abpf1CyX/FuyH/AFBVX/WhvfuvdEq+AVz0vnCfr/pN3x/vGQHvQ6s3l0eL3vqvXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3QRd7EDqjfZP/PK5/wD91cntZtv/ACVLT/mvF/x8dJr7/cC4/wCaT/8AHT1oxl/8rydrX/jWZt/Q/wC5CT31HX+yi/5pp/x0dczlqJJv+a0v/Vxuvav9t+T9APfqdO1/l13GT97iOOf41ibD/quT3V/7GX/mm/8Ax09aArPD/wA1Yv8Aj463hPjeSenNkX+v8AxP/uui98wd7/5LN3/zVf8A48euk2042y3/AOaaf8dHQ6eyzow697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuoWR/4A1Vvr4X/wB690k+BvsPWxxH2jrS1+YpJ+VHcg1WYZxf6fTXJf30W9q8e3W1/wCkP+Trn97on/mI+51/4X/z90XCzfqB9P0vb/eh7HvQHp59YZx+1Mp5Ijk4/wAdB/PuyHuH29UlHY32H/B1tN/ylr/7LNt0nn/gTY/TjWn494Fe+P8A08W8H+l/y9Zz+y//AE7yw/0vVqPuIupU697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuuAjjB1CNA3+qCrf/b+9UANR1oADI65+99b697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XumvNf8WvIf40VaP9b/ACOT3sdVbh186v8A4TVg/wDD8Xzj/wC1T3R/72svuo4jpw+fX0Zx9B/rD/evdjx6qOHXfvXXuve/de697917r3v3Xuve/de64PFHIhikjR4iApjdFZCo+gKHi3v3Xuuo4ooV0QxxxJctpjRUXUfqdKgC59+48evdZPfuvdITtEX6238P67O3H/7qZffjw62OI6AX4Of9kydcf9Q+V/8AdvP70Ottx6Nr731Xr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//X3+Pfuvde9+691737r3XvfuvdQsn/AMW7Ifj/ACKr/wCtDe/de6JX8AjfpfOG9/8AjJu+Pxb/AJeA96HVm6PD731Xr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6rM7j/mb9QdN7/zXX+cp55cnhaqWlqGQEqXi/Vaze5p5e9ld/5i2iLd7ZgElAI+w9RFzB7w8vcvbrJtN5iSM0PQY/8ADw/RP/KnVf7FW/6O9nX/AAPfM38Q/wBX5dE/+v1yp5H+fXGT+cT0RGju1HVWRSx4b6KLn+17sv3eeZmIUOM/6vTrTe/fKiqWJ4fPqwf489/7Y+ROyl3ttVHTHPIsYVxZrsur6En3EnN3Kt5yjuR2y+NXHUpcscy2fNO3DcrH4D0PnsK9CTr3v3Xuve/de6B/vggdUb6v9P7qbg/3jFye1m2/8lW0/wCa8X/Hx0mvv+Sfcf8ANJ/+OnrRd8g+7ypH6f41meP/ACISe+pKj9KL/mmn/HR1zNV/1Z6D/Rpa/wDORuuRcAEA8Hn/AG39PeqdO5OR16ORvvcRbg/xvE/7EGuT3px+hL/zTf8A46eqj+3gHrNF/wAfHW8V8bTfpvY/9RgMTf8A2OOi98v96/5LN3/zVk/48eulO1Cm2W//ADTT/jo6Hb2WdL+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917qFkeaGq/P7L8f7D3V/gP2HrY4j7etLP5kBT8p+4yAb/xxPyfprkHvor7VE/63O11/gP8Ak65/+6Q/5iPuePNP+fui4cAmwuP6X9j35dAfgMdYprCGbj1eKTm/+0H3tKlh9o6rLhG+w/4Otp7+Ut/2TJtwj6f5V/vLp7wL98f+ni3n+16zk9l/+ne2P+l6tP8AcRdSr1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6902Zn/i15D/qBrf8A3Ef3sdabr51X/Cay/wDw/F847f8AOo7p4/r/AL/aS3Puo4jq54Hr6M6/pH+sP9697PHqvXfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6QvaFh1vv2/A/ufuO5/oP4TL78eHWxx6AX4O2/2WXrmxuPt8rz/wCRef3odePRtPe+tde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/0N/j37r3Xvfuvde9+691737r3UHJ/wDFtyH/AFA1f/WhvfuvdEq+AIt0vnPr/wAzN3x/7sF96HVj0eP3vqvXvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcJG0qT/rf7z7bmNImPyPWxlgOtMP5tTGp+S/ZEroGtuCuBZgCf9599Ffawn+otgP+Fr1gN7qoDzxeEgHvPRVdMZtqjS39n0j/AG/09yDU+R6jsqvmvWOeKPwzgog/afnSvPpPvasdQNfPpuSNSjCnkfLraE/lH1MjdIV1K3EcNTAUA4A9NgLf7H3gt77knnEfYes4fZUAcoIR6j/B1bT7hXqX+ve/de697917oIe9if8ARRvzjj+6W4rn+n+4qT2r27/kqWn/ADXi/wCPjpLff7gT/wDNJ/8Ajp60UTJeryptYLm8yL8f87GT31LUUiiH/C4/+OjrmehPjTsOHjS/9XG65XNyQv8Aja/1/pz79XpyhwOuMTuK7Ef4ZvEX4/6bksPfnA8CX/mm/wDx09UqTPB8pov+PjreV+Nlz05sgn6Hb+It/wCe6K/vl5vX/JZu/wDmrJ/x49dKtqzttv8A800/46Oh39lnS/r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6g5L/gBV82/Zfn/Ye6v8B+w9bX4h9o60tPmN/2VN3Hz9c4nFv9qf30U9q/+ndbX/pD/k6wD90R/wAxF3P/AGn+Xot4te5F/wDfcex8egJxNeuE5H28thz4pP8AYek+9p8Y+0dVl/sWp6H/AAdbTf8AKWK/7LNt0D6gVP8A1sT3gV74/wDTxrz/AGv+XrOX2Y/6d7Y/6Xq1H3EfUp9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdNea/4tdfzb/Iq23/nHJ72Oqtw6+dX/AMJrP+38Pzj/AO1R3Rf/ANDWT3UcR04eDdfRmX9K/wDBR/vXvfVeuXv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6QnaH/Mtt+/n/fn7j4/8hMvv3Wxx6AX4OkH4y9c24H2+V4+v/L3m/Pv3Xj0bX37rXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//9Hf49+691737r3Xvfuvde9+691Byf8Axbch/wBQNX/1ob37r3RLfgHf/QxnL/8APzd78/8AkQX3odWPR4Pe+q9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691im/zZ/wBcfm3tqf8AsX+w9bX4h9vWmD80ST8kuyrGwG4a78X/AD/X30U9rDXkaw/5pjrAj3TxzzefNj/g6K6foCW+o4Fj9Pcg1HUeUJ6jVFjDJY/SKQfn6aT7uh7x9vTcinwz9h62ff5RgX/QtkyOD9zDdf6en+vvBT32/wCVxH2HrOD2X/5U9PtH+Dq3H3C/Uu9e9+691737r3QQ973/ANE+/T/TaW4uP6/7ipPavbhXdbT/AJrxf8fHSa9/3AuP+aT/APHT1oklr1mVIHBzeasAfqP4jJf6++po/sov+aaf8dHXM1PjuP8AmtL/ANXG67LC/H+x/wBgOPeunvt67iZjWYgEWvmsRwD9f8vT8+9SU8GX/mm//HT1UD9eCv8Av6L/AI+Ot5z42X/0NbG/8N/EW/8APdF75d71/wAlm7/5qyf8ePXSjav+SZb/APNNP+OjodvZb0v697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuoeR4oao/0hf/AHr3V/gP2H/B1scR9o60rvmQf+cqe5CPqc4v5+nrk/Hvor7V1/1udr/0h/ydYBe6NB7j7n/tP+fui2iSxH4H0t/X/Y+x9Soz0Ba0yOuE8g8Uoa7ftSXsf9oJHvaDuH2jqstPDaucH/B1tQfylCT8ZduE/S1Vb/qYgPHvAn3x/wCnjXn+1/y9ZyezAp7e2Nf4erUvcR9Sp1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6902Zn/i1ZD/qBrbf4H7R+fex1VuHXzqv+E1n/AG/h+cf/AGqe6f8A3tpfdRxHTjefX0Zl/Sv/AAUf71731Xrl7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XukJ2j/wAy239f/njtx/8Aupl9+62OI6AX4N/9kydcf9Q+V/8AdvP70Ottx6Nr731Xr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//S3Y/kN8oth/HOmww3NjN0bizWehyeQoMDtXC1uTqxhcDTGu3Bm62pjQwwQUlOryuZHBbSQB9SH4YGlBatAOkd1eJa0BBZj5DoIdq/zM/hTvP4dy/PTb/dWKq/i/BLX0c/YL4nPQvDmMZuI7TrMA235KYV7Vq5EfbCBKc6riRSYj5PbaozNpXPT7TRpF4zmgpXPSt+PHzo6D+Se+959U7JzmQxna2wcNg9z7l693Ni6zD56n2ruiAVm29x0RqEENTT1UDxyHwSM8WsCRVuCXJYGizWo9embe8juCUHaw8j0cb2x0r6actW0UVFXQzVlJDK1FVWSaphiaxgbkq5Bt+b+/de6Jn8AmDdL5wggj/SbvixBDKw/iAsysOCD70OrN0eL3vqvXvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYpv82bC/I/3v/H21P8A2L/Yetr8Q+3rTA+aV/8AZkuyNJ4/vDW6h/W599E/asg8i2JP++16wJ91FP8AXi7pw1norZP4t/wXn6e5BHUfMCOPWOYn7eYW48Umo8c+nk+7r8Y+3pmShiYH0PWzz/KLt/oXydvp9xDb/be8Fffb/lcR9h6zg9l6f1PSnqP8HVufuF+pc697917r3v3Xugg74/5lNv78/wC/S3Fxe30xUntZtv8AyVbT/mvF/wAfHSa+/wBwLj/mk/8Ax09aIfkJq8pyP+L3mr/+fGT6e+pgH6UX/NNP+OjrmerASzn/AIdN/wBXG67LXsB/sTb6f09+A6uSaddxuxrsOSeRm8RpFvr/AJenvUmIJf8Amm//AB09aGZ4DX/Rov8Aj463o/jZ/wAya2Lf6/3fxH/uti98ut6/5LN3/wA1ZP8Ajx66T7T/AMkyD/mmn/HR0O3st6MOve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917qHkP+AVV/wAsW/3r3V/gb7D/AIOtjiPtHWlL8ynUfK3ucA2tnVH/ACdJ+ffRb2qH/MONrP8Aws/5OsAvdGh9x9zX/Sf8/dFrLXOkcf0Psf8AQFoDkcesc5tDMb3vFJx/yAfz72uWH2jqkgKxso8gf8HW1T/KQv8A7LFtw3uD91b+nEie8CPfI/8AMRrwfJf8vWcvsx/076x/0v8Am6tV9xF1KnXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3TZmf+LXX/40VaP/AFTk97HVW4dfOq/4TWf9v4vnH/2qe6f9h/v9pPdR5dOHgevozL+lf+Cj/eve+q9cvfuvde9+691737r3XvfuvdE8+Tnzb6g+KazSdiQ7prYMPtfKb/3jVbe2/kcnR7L67wMTT57eOcrIozEIaZFZjCjmRgOAOLqI7cyIZCaDpFcXqQSCIKWJ408umHOfzG/h9g/i51l8x5u26Gr6E7oqNs47qndVFh89LV74ze8a6TF7fwOKwLUy1qVctVDNTyR1EEYhdHExQLf22kbSNpXp+aeOCPxJDT09SfTpc/G75jdJfKOu7G271vnapN99P52DbPaWwM7QVWJ3PszNVNOKqCGspapFWeF0IKVNMzx39LFW9Pu0sLRHOR69N213Hcg6cMOI8x0aj2z0q6DvtGtoR15v2neto45v7obiBjkqYEcXxUosVZrj+n09+62BnoD/AIO2/wBll65sLD7fK/8Au3m9+68ejae/da697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/9Pc9+aWxPk12B07mMN8WO39jdO79bH50VmY33sOj3zj8xiZsPJE+Hp0rZFSid24NUY5AoOorZbF6Exg/qAkY4GnSW6S5Zf8WdUOfiFetXj4c7h65wHxd/lgdF5Hq+Xq3rvq75GdwT/J7aWaaHM9e5HPV29JTQbhzNeNWNnhr6yZ6uGKZj4UOkC3sxEYVnZcVHaOg79Q0kUEcxLaGOtvWmP2VGPlQdXPfKqgWq/mk/y6avqpccuWpIO0pe5Z9rRwLWy9fnZZi2lFuuoxILNSLIH+0SrOgKR4/SR7TR6vpX1fz6M7mn70t/D9DWnClDSvV0ntD0c9Fv7X+OWzey8jW7kzlfn6esTHyqseNytTRwXhpmEZMcRA+tr/ANfeqdbqegm/l70qUHRdfQRljFQ9hbwo4mclnaOnrVjQux5JsOSffh1s8B0en3vqvXvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYpuI2N7Wt/vftqf+xb7D1tfjH29aX3zRYJ8kuybcltw1pA/pf30S9q88jWJ/wCFr1gX7pgf13vKj8R/wdFc1av9hx/re5D6jzrFNcwzW4Hif/Y+k+7J8Qr69UlBKNTAAPWz5/KM/wCZL5Pj/lJh/wBhx7wV99v+VxH2HrN32Y/5VBPtH+Dq3L3C/Uude9+691737r3QP982/wBE2/vrcbR3ERYn/nVSe1e2/wDJVs/+a8X/AB8dJr3/AHAuP+aT/wDHT1odhl+7y3PP8czN/wA/8vGT6++pwP6Ufyjj/wCODrmeuZp8f6NN/wBXG6561/Bv73T16ua+XXo5L1+IOr6ZzD8f9V6D3qTEEo/4W/8Ax09UUHxoa/7+i/4+Ot6r41/8yZ2Lzf8A372I5/8AIdF75c73/wAlm7/5qyf8ePXSnaf+SZb/APNNP+Ojod/ZZ0Yde9+691737r3RFvn/AN2776O6Qym6OucrTYjdChhRVdVSpVxIVH1aGS4PuS/avYNp5k5oi27eYzJC3EA0P7eo99y993Xl3lmbcdnZUnQEqWFRWmKj0618oP5ofziaCB37G2+XaGNpCu26W2pkBYi6/S/vLZ/ZL2yDkCzkpU/6IesUIvez3RaJWknt9RArSM0r5+fWX/h0H5vXH/GR8AOf+ebpfp/r6fdf9ZP20/5Q5P8AnIer/wCvX7mUzcW//OM9GB+K/wDMP+WHYPf3XOzN878w2Q2vntwU1DlaODBU9NLNTSD1okyqCv8Arj2Eue/abkLZuUr3dNttXSaGMspLkgH7OhhyH7r8973zZZ7Vuk8LQTPRgsdCRT1PDrZw94WKwZQw8+svuve99e697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6hZL/gDVWF/2W/w90k/s2+w9bX4h9o60nfmY1vlb3OB9f48l+P9rk99Gfan/p3G1f8ANM/5OsAfdL/p4+50/of8/dFrL/Qt9P8AYex/59AWh4efWKokvDMoPrEUpHHH6Cfe1HcPtHVZMRtXjQ/4OtrD+UYdXxg22w/rVg/6/kT3gN75f9PHvPsX/L1nL7MH/mH1iD/D/m6tY9xF1KnXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3TXmv8Ai11//UFW/wDuHJ72Oqvw6+dZ/wAJq/8At/F84h/XFd0D/wBfWX3UcQenDwPX0Zh9B/rD3Y8eqjh137117r3v3Xuve/de697917qsj+ap1b8p+0vi32th/jf2tsHYtA/UvZlL2btDeexcfuSp7O2jU7ZmXK7Qw+48hKExElbSfc0a1JhYK8qsWW1wogMQNJVJz5HpBfJeNGTaOqHSfiFeqafhlvrqrL1f8oDYtRsCbrTonq3oXJbby/Xu9tGV2ftvvCorzDgUbJ5ENj6quNdEauKrDGRWmAHIPtaY2RJCuCeFPSnROtwJ57bUSVAGonNTUf7NSePVl+Zx80n87frbI9cpRx7Zp/ih2DH3e23IY46ap3lPmk/us+8JcaNElasHhEZrD5QugDgD2n7vozq9R+zpfj98gpw0HV6aqin50r1c97RdHHROu+PjRsnduK35vbIZDcMWUXbebrkipMvVU9H5qfHyTIrU6HTp4sRbn3qnn1upOOnH4OH/AJxj61XgCOjyMS/8FjysyKSfybDn34dbbo2nvfVeve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//U3HflP/L7+O3zFzu2dx9zU/Y0mT2nQ1mNxbbK7T3xsSkkoq5JI6mGvx+26yCCfUsjgs8d7Hkm3t+O4kjXStP2dJJ7GC4fXJWvyYj/AAEdNdP/AC2fiNS/Das+BsPX2QX46V9JUwVWBO69xNuaSrq81/eCXNf31aoOSFd91ZlqRPqCAR/5v0+6+NJr8SuerCzthB9OEGj06cviZ/L0+MnwrzG9dxdG7b3PR7h3/Q4HFbizu8d8bm31lJMVtun+2xeNoKrclROaaBQFMiQhdZVS19K23JPJKKMcdVtrK2tSWhWhPEkkn9pr0d32z0r6hZP/AItuQ/6gqv8A60N7917olXwCFul84LEW7N3xwTc/8XAe9DqzdHi976r1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3WKYAxkH21P/Yv9h62vxA/MdaW3zUcr8k+yVA4/vBXf6/199E/arPI1l/zTXrAv3Up/Xi7/wBMeiua+ASLcAD/AB/xPuQ/l1H2KdYZ31QTC5X9qT/G9lv7unxj7R0zLiMkeh62g/5RJv0tkz/00wXF+f0j3gn76/8AK4/kes3/AGY/5U9PtH+Dq3T3C/Ut9e9+691737r3QQd8EDqXf/8AX+6G47D+p/hUntZttP3raf8ANeL/AI+Ok19/uBP/AM03/wCOnrQx1XqsrqXn+N5r6f8Aaxk99StZWKL/AJpx/wDHB1zSVKyz0/39N/1cbrssAAbaf63N/wDW961k8OnNOevRuwrsQQOf45h7D/qvQfX35iPAlH/C3/46eqAfrwV/39F/x8db2nxp/wCZMbE5v/v3sR/sP9xsXHvl7vX/ACWbv/mrJ/x49dJdq/5JkH/NNP8Ajo6Hj2WdGHXvfuvde9+691VH/NlY/wCgl1B+vkNr8WOr8e5h9jj/AMjWMfZ1FHvIAeTJz8j/AIOtW6n1CnhA5tDFc/19A45955P8Zr6nrBiH+yUj0H+DrMT9LD/XFySB7rw6cpXoyXw7IX5NdRkG3+/qo7f1J/1/YF9zc8g7n/zRboce2QI5+23/AJqf5Ot1GIkopIsebj/Y++cVv/YJ9g66BsAGIHWT291rr3v3Xuve/de6rQ+bHz/f4lbpwm3Itiz7tbLYtK9pIKjw+AvJpCEEj/X9zN7a+1K8/wBhLe/ViDw300IrXqI/cb3PXkO5itmtXn8VdVVpjPRIT/O4qbf8yWyFx9f8tH+9avcl/wDA1LTG5L/vPUbH7x0QNDt0v8v8/XX/AA9zUAj/AIwxXn/qsBv/AIfq96/4Gkf9HJf2de/4I6H/AKN8v8uj0/CX58j5bZXOYuXZE+0pMQ0qqZ5xKZ/GGNwLn/U+4x9yva1eQIYpfqhP4lOApSvUme3PuYvPrShbZ7fwyR3edOrJfcO9Sz1737r3XvfuvdQ8h/wCqf8Alk3ukn9m32H/AAdbHxD7R1pK/NBiPlf3QNQt/HV9P5uGk/Pvo17Uf9O32r/mmf8AJ1gD7pj/AJiRuf8Azb/5+6LPqP8Atv8Aif8AW9yB0BqYr1ilcCCZWF7Qy/6/6CeD78o7x9o61LUQt9h/wdbXP8oog/F7bVrj/gXcf9PE94C++X/Tx7z7F/y9ZyezP/TvrH/S/wCbq1r3EXUp9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdNeascXkL/wDKlW/+4cnvY6q3Dr51f/Caz/t/D84/+1T3Tz/5Osvuo4jpw8D19GZf0r/wUf71731Xrl7917r3v3Xuve/de697917qvf5R/wAsb4sfMLej787ooez6vOT7cn2pVw7U7e39s7C1uFqYBTTU1VgsFWxUrEoACwiFzybkk+1CXMiLpFMfLpFLt9vO5eStTx7iB+wHp97H/lvfErtP4q7G+Ge6dgZEdFdbvtefZeHw269xYbcWDrdozGoxWRp930NQuQadnZ2qZJJ28xdvJfi1BNIH1g56ca0tnhEDINI4D0+zpbfFH4SfH74X4ne2K6L29nMa3Ym4xujd+X3TuzPb0z+WycdMKSBHzG4pp5kgjQHRBGwQMzMQSb+/SzPKav5det7SC1BEK0qanJJP5mp6Nr7a6U9ITtD/AJltv23/ADx+4v8A3Uy+/dbHHoBvg9b/AGWbrm30+3yv/u3mv70OvHo2nvfWuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rC9RTxNpknhja19LyojWP0NmPuwR2FVBP5daLKME064feUf/ACtU3/U+L/ivvfhSfwn9h61rT1H7evfeUf8AytU3/U+L/ivv3hSfwn9h69rT1H7esscsUoLRSRyAGxMbq4B/oSt/dSrLhhTrYIPDrJ711vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/9Xf49+691737r3Xvfuvde9+691CyX/FuyH/AFBVX/WhvfuvdEq+ANv9C+ct9P8ASbvj83/5eA96HVm+fR4ve+q9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691jl/Qbi/tqb+xb7D1tfiH29aV3zYe3yU7K9X03FXC3+x/Pvol7Uf8qNZf8016wL91aDne7+bHoq+s2sWHP6fqLf6w9yNTqPfLHWCdz4ZeeRE/+N/Tf3ZPjH29UkxE1T5HraM/lCtq6Uyf/URAT/yT7wS99f8AlcvyPWbnsxX+qCfaP8HVvHuF+pc697917r3v3Xugf75YL1Jv/i5/uhuOw/8AITJ7V7b/AMlWz/5rxf8AHx0lv/8AcC4/5pv/AMdPWhTrX7vKmxH+5vNW/pzkpOffUgt+lF/zTj/44OuayL+rP/zWm/6uN10Sf9USB9bfTn6W91r07Q1r13GzGtxF72Ocw1uf+rgnHHvZP6Ex/wCFyf8AHT1WlbiD/mtF/wBXB1ve/GkAdMbEt+du4f8Ax/5dsXvl/vX/ACWbv/mrJ/x49dINq/5Jlv8A801/wDoefZb0Yde9+691737r3VUH82Y26Lbi4tJf/D9XPPuYfY7/AJXaP8uop94/+VMn+w/4OtXGnA8EHp+kMZJv+NAt7zxf4z9vWDUX9kv2DrJcXsFP+3P1/wBj7r05muOjJ/D0L/szHUdv+erpOPyP8fYG9zD/AMgLc/8Amiehx7Zgjn3bf+an+TrdSi/za83PNz/jqPvnFb/2CfYOugLfEesnt7qvXvfuvddH6H/WPvw49ePWtB/OPlll7n2xHqsi7bpLEngXINre8wPu11PLl0Tn9ZusT/vEL/u2s6Y/T/y9U8AP9f1WNrf1/wAfeSNR1jpQ9d3HAIt/U/4/ge9dWpU06uR/k6SyR9k5+NWOmRarUB9LWf6+8Z/vHk/u+3P2f5OskPu8iktz9rf5etlX3iP1lT1737r3XvfuvdQsj/wBqv8Alk3+3/Hukn9m32H/AAdbX4h9o60i/mkyj5Zd0m51fx5L83H6pPx76Oe1Ff8AW32r/mmf8nWAXulT/XI3Mn/hf/P3RaNdvUDx/qfyPx7H/QF/1U6wzNannub/ALMn+ubISOfd0+MfaOm5aeC32H/B1tgfyhTq+Le2D/X7z/rYnvAL30/6eReD/S/5es5vZjPt5YH+j/kHVr3uIepU697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6bMz/AMWvIf8AUFW/+4cnvY6q3Dr51f8AwmrH/Y+H5xj8/wAJ7oA/9DaT3UeXTh4Hr6My/pH+sP8Aeve+q9d+/de697917r3v3Xuve/de697917r3v3Xuve/de697917pC9of8y337/4Z+4//AHUy+/dbHHoBfg9/2TN1zzf/ACfK8/T/AJe83vQ68ejae99a697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917romwJP4BP+259+691rOfzOfkF2rs/5Q/wPZe7a7EYSPalK/wBnT1M8KGoAQNJpjIHPPvMn2M5b2PduRTd7jAsknjMKkA4z1iP72cz7/s3OsVnts5jiaAkipHdUU4dV+H5T9/3BO/8AK2/p99Wf1/4N7mAclcrcPpE/3kdRQeeubRn6pv2t11/s1Hf9/wDj/ssf8Pvqz+v/AAb37+pPK3/KKn+8jrS89821zdN/vTdXw/yh+2t59jdYb4bfObqc5koN2SRUk1RNLM8VOusaNUpJtx7xT+8FtG3bNzJaQ7bEIkaGpAAFTj06ye9ht53PfOWJ7rdJDJIJnFSScAkDj1cP7gLqcuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//9bf49+691737r3Xvfuvde9+691Byf8Axbch/wBQNX/1ob37r3RK/gDb/Qvm7Cw/0m74/qf+XgP6+9DqzeXR4ve+q9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691jlNk/2I/3v21P/AGL/AGHrY+Ifb1pR/Nl7fJjswD6f3jrrn/G/099E/aXPI1n/AKResC/djHO90T/EeireS9xzz9Cf7P8Ah7kinn1HWrrDUNaCW5HEUl7/AFPpPuyDvHVJTSI/YetpT+UEdXSWSP8A00Qc/k+n3gb76/8AK5n7D1nB7M/8qhH+X+Dq3v3DPUt9e9+691737r3QPd9j/jEfYBuBbZ+5Pr/2qZPavbf+SrZ/814v+PjpNfU+guB/wt/+OnrQj1MKzKn8/wAczXH9f9yUlyPfUaoEcf8AzTj/AOODrmwoPiz0/wB/Tf8AVxuvFrEC4sfr/h/S/utTx6coeHXo5f8ALsR/UZzD2/x/3IR/j3pv7GWv++5P+OnqoUGeD/mtF/x8db4nxoA/0L7DIvzt3Dnn/tWxe+Ye8/8AJYu/+asn/Hj10f2r/kmwf801/wAA6Hn2W9L+ve/de697917qqH+bNz0U4/wkv+bj1H3MHscSOdo6fLqKPeQV5Ln+xv8AB1q3UxtTwC/0gitz9PQOD/X3nk5PiGvqesHIlUQqK+Q6zFgebi7cf8a9049OkUX7ejJfDzT/ALMv1F9b/wB66P8AP+t7A/uaf+QFuf8AzRboce2Q/wCR7tv/ADU/ydbqUNvGtjf68/19XvnHB/YJ9g6z+b4j9vWX271rr3v3XuvH6H37rx4dazP84wE907bHLH+7dJYC449P1HvL77tZ/wCQ7df81m6xR+8QP92tpT/ff+bqn64H1DX/ANc/X/Ye8kusdyfTrrUv+oP+PPv1Oq9w8urjf5PFj2fnCDf01P04sNL8H3jJ94//AHAt/wAv8I6yS+73XxLn7W/y9bLfvErrKbr3v3Xuve/de6h5D/gFU8X/AGm90k/s2+w9bX4h9vWkH81H/wCcsu6xf1fx5OP+Q5PfR32o/wCncbV/pD/k6wB90xX3I3P/AJt/8/dFkaQ/hhpt/t/cgYGT0BtPn1inkbwS3tbwy/0+pQ297SmsD5jqki0iamMH/B1tj/ygWLfFjbBJF/8AK+P+niW94B++h/5iTej/AEv+XrOX2X/6d5YH+h/kHVsPuIOpU697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6bMz/wAWvIf9QNb/AO4cnvY603Dr51X/AAms/wC38Xzj/wC1R3T/AO9tJ7qOI6ueB6+jOv6V/wBYf71731Xrv37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdIXtD/mW+/eL/wC/P3Hx/X/cTLx791scegF+DpB+MvXJAsDT5Xj/AMi8/vQ68ejae99a697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rhJzHIP6o3/Qvv3Xhx61Kf5pQv8sKgs1iNswCxP09Sf095x/d8/wCnfH/mu3+XrCv39/5X2Ej/AHwf8I6rxJ/IPH9Oef8AG/ubuoa0knrvUBb83/3j3o8OrKADnrYI/ksf8eNvgAWtueoP+w/c94efeTP/ACKLP/miP8nWWn3dRTlO4p/v9/8Ajx6ve945dZB9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//X3+Pfuvde9+691737r3XvfuvdQcn/AMW3If8AUDV/9aG9+690Sv4Ai3S+cF7/APGTd8c/+RBfeh1Zujxe99V697917r3v3Xuve/de697917r3v3Xuve/de697917rHL+g/wCuP979tT/2L/Yetj4h9vWk583mH+zMdmC9r7irri/15/w99E/aSv8AUez/ANIvWBnuuB/Xa6r/ABHoqJflQP8AU/4cH/H3JXz6jqgr1gqHbwzW5tFJq+nPpP0HuyfEPt6rKP03+w/4Otp7+T7z0fkj/wBNNPf/AA9PvAz31/5XM/Yes3/Zof8AIQj/AC/wdW/e4Y6lrr3v3Xuve/de6B3vz/mUXYP/AIZ+4/8A3VSe1e2n/drZ/wDNeL/j46TXv+4Fx/zTf/jp60GjIBWZb13/ANzmaB/wvkZPfULV+nF/zTT/AI6OubS/20//ADWm/wCrjdcrjj1Cx/P+t9Pr70a9XoP2dejkP3+HPBtnMPxb6/7kEHHvx/sZR/wt/wDjp60cT2//ADWi/wCPjrfL+M4YdLbDv/zzmGt/h/uMi98xd5/5LF3/AM1ZP+PHro7tf/JNg/5pr/gHQ9ey3pf1737r3XvfuvdVOfzam09FNza4l/6K9zB7H/8AK7xfl1FXvEK8mXH2H/B1q0U8h+3hH/NmP/b6B9feekg7z9vWDcIpEn2D/B1nD8jnn88fQe606uTU9GT+HTg/JrqNNX03XRt9f949gP3PxyDuR/4Uehz7YVPPu3ef6n+Trdci/wA2vFvrx/sffOO3/sE+wdZ/t8R6ye3uq9e9+6914+/dePDrWb/nG2/01bcW9j/dukItxf8AT7y8+7Zjl66H/Dm6xT+8MA26Wlf99/5uqe9f5NuLi/8AQ/4+8lM9Y69vXRP4JHNjYcWPv3XtPAdXG/yeSP8ASdmrcempv/Q+lveMn3jv9wbf8uskvu9/2lyfm3+XrZc94ldZSde9+691737r3UPIcUVT/wAsj7pJ/Zt9h/wdWXLD7etHv5rsp+Wvdgt6hnl/22uT30c9qjp9t9q/0h/ydYCe6NP9cfcyOP6f/P3RYtSj1f2fwD/tvz7HhNO3oD8Vx1wnkAgmW1j4ZLX/AOCE+7xt+oD8x01KKxuCMaT/AID1tm/yfmDfFXbBtzer5/r+4l7e8BffT/p5N6fkv+XrOH2XqPbywr/B/kHVsfuIepV697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6bMz/xa6//AKgq3/3Dk97HVW4dfOr/AOE1nP8APh+chv8A8unuj/3tpfdR5dOHgevoyp+lf+Cj/eve+q9cvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690hez/wDmW+/f/DP3H/7qZffutjj0Avwe/wCyZuueb/5Plef/ACLze/dePRtPfutde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691xf9D/8Fb/evfuvdalH80kn/ZsKqxFxtqEhSLk+pPz7zj+75/074/8ANZv8vWFfv7/yvsJ/5dz/AIV6rw1EccH6Em30J9zb1DlfL167DHVYj/Cw/I/w9+PWgATjrYJ/kr/8eLvj8f7+iosP6j9z3h395P8A5Wiz/wCaI/ydZa/d1AHKdx/zXf8A48er3veOfWQfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf/Q3QvlP3V3n1LjcT/oS6Cy/cVdU0GbzecyseRoqHBbcxu3qJ8jPRVETzR1E1ZWpG0VGkfBcqPUTp9qIY4mBMjU+XSK6muYyFt01V4nyH+z0Q7p/wDnO7F7e/l1bb+d+M+PnatDujeW7t0debN+L/3WJrO1t4b32pvRtn5DHbckVUjnpwF++efwAxxehkMmkNpYHZ9PDrcl7FHCJKhieAGakcR+Rx0Y3on5z7o3T8g8l8YfkL0luDofsWs2tid49ZZ7MVcE2ze2KLJUP8Uyu3tqVBLOcphY7x5GJnILpIF0sug3lgUL4kRqvTVveu0v09yulzw9CP2nqxr2l6MekDuzsDZm3UrsZnNxY/F1rUNSRDUuyvZqdmUgAc3H0F/fq9bp0Vj+X7Ik3SWYmicSQzdlb1lhkX9MkT16sjr/AIEe9DrbdHl976r1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3WOX9H+xHtub+xf7D1sfEPt60kPnCdPyc7Mvfncdf9P9ce+iXtGa8j2fyResDvdbHOt1/pj/k6KeXN/wA8/wBLXsfcl4rnqOqMOsc7jwT2P0hk/P50G3uyfGPtHTUtfCb7D1tSfyeCT0bkb/8AKzB9f6afeBfvtnnRqeh6zg9mscoR1+X+Dq4P3DHUtde9+691737r3QPd+EDqLsK5+uzty2/89MvtZtv/ACVbT/mvF/x8dJr3/cGf/mm//HT1oGav8qy5Iu38dzVuPx/EpPr76f58KP8A5px/8dHXNxSBNP8A81pv+rjdZi1uQD/tQ/3oe656cx11G5+9xFiLfxzDcEc/8XBPp78x/Sl/5pv/AMdPVQKzwD/h0X/Hx1vqfGb/AJkrsIWPG28LyfzfGRH3zI3n/ksXf/NWT/jx66ObZ/yToP8Ammn/AB0dD37Lel3Xvfuvde9+691Uz/Nva3Q7c2t5Tf8A2/uYPZD/AJXeIfZ1FXvD/wAqZcH5H/B1qu08z+Cn5uBDHYj8+gce89nHeftPWDkf9kp86D/B1m8puf6ng/4D8H3WnV6E9GX+Gshb5O9Qggf8fZR/6/49gH3RFOQ9x/5pHod+2H/K97fT/fn+Trdni/QP9j/0MffOO3/sE+wdZ+N8R6ye3uq9e9+6910fof8AW9+691rKfzlGCd07ave/92qS1jyP0+8vfu1n/dBdL/w1usU/vDiu6Wn/ADT/AMvVOomJH1Fv8bcX/r7yVp1jtXzPXYl+guP6fi9iebH36nWvUdXKfycn1dmZzn6LUgD/AGD+8Y/vHmllb/l/k6yQ+70KyXJ+bf5etmH3iT1lN1737r3XvfuvdQ8hf7Kpt9fGfdJK+G1PQ9bX4hX1HWjl82mZPlv3dYg3z6jT+f1yX99Fvao/8w32r/SH/J1gR7n1/wBcbcqf8L/5+6LCZLcEg/4Dm/8ArD2PdWc9Aih4jrDO/wCxNwb+GXkfQ3Q/n3ZG7l+0f4empgRG9PQ/4Ottb+T0dXxU2s39fvP+tkfvAr3z/wCnkXp/0v8Al6zg9mRT28sP9IP8A6tn9xF1KXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737rVR1737r1R1737rfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdNmZ/4teQ/wCoGt/9w5Pex1VuHXzq/wDhNXb/AIfi+cf9Die6P9t/faT3UeXTh8+vozLbSLfSwt/rW976r137917r3v3Xuve/de697917ppyufweCFEc3mMXiBkq2HG485OvpaEV2QqDpgoqT7ll8krnhY0ux/A92VGb4QTTqjyRx01sBXhU9OUssUEUk88kcMMMbyzTSuscUUUal5JJJHsFVQCSSbAc+9AEmg6sSAKnAHUXGZTG5mhpsniK+jymNrI/LSV+PqYayjqY721wVNOWRhcEXU+/MrKaMKHrSOki6kNQfMdTveurdIXs//mW+/f8Awz9x/wDupl9+62OPQC/B63+yzdc2Fh9vlfr/ANreb3odePRtPe+tde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691xf9Df8Fb/AHr37r3WpJ/NLYD5Y1QPBO2YbD8kak+nvOP7vf8A0700/wB/t/l6wt9+6HnyH/mgf8K9V46wpH5Fuf8AjfubRnqGyFH5deV7kG1uRa5/H49+6sKHJ62Df5LB1bG3xb6DdE/H+wk94d/eT/5Wiz/5oj/J1ll93Yf8hOcj/f7/APHj1e97xz6yC697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r/0dk3+bn/ADR+s/hdQ9efGmg7R2T1x8k/lNBX4Prnc3YVBkMjsDrnbq1sOLz2+t8HGfuRIIppYcchFnqBqYFE0upghEjAvwPD5/Z0gvrw28Z8P4hxPklfNvl/xfDohk3XXxc+FfxM/lzdpfHDtjG9zfEH4a90733v3X3JRbqg3Yq1/aWThr9w7lzFVTF5IqdsrVTeGmCfsx+CJtRszKYQwLq9VLYFei68eMpDLFR0jNWK8Mjjj59G63T2v1R8+v5jnws3V8Uux8B23tb4lwb73z3PuXaFaKrB4LH9k7S/h+1MZWVk6oHnnZVdYY9TgsQwBV9NArQ2zrJgtwHTjSpeblFJbEOsddRGaVBoOr5PaDo86BvsDpnZW9563NZ+mqJqpaCdNUUgUBY6ZlQgWP0+tveqde6L3/L5hjpej8pSw38NL2NvOmhDfqEUNcqID/sB78OrN5dHo976r1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3WOQXQ/7D23N/Yt9h68PiH29aRHzkk/5yd7NuQCNxV//AEN76H+0hpyPZn+gvWCXusP+RrdV/iP+DopwkuDyP6f0/wBj7ksMKnqOtOOsFQ94JfUP81J/0Kfdo2PiL9o/w9NzL+k1PQ/4Otq3+Tpf/QZkr/8AK1Af+TfeBvvqa86N9h6ze9nP+VQj/L/B1cN7hnqWOve/de697917oHu/Lf6Iew7/AEGzdy/+6mX6e1m2/wDJUtP+a8X/AB8dJr3/AHBn/wCab/8AHT1oC+QfeZYm+kZ3Ng8/X/clJ9PfT2v6cf8AzTj/AOOjrm6g/VnJ/wB/Tf8AVxuu9QFrkgH6m/8AT3WpPToAp13Ef8uxJ5t/HcLf/wA+CfT35jWGX/mm/wDx09V0kTwH/h0X/H1633/jOQelthW/G28Lx/S+Li98yt5/5LF3/wA1ZP8Ajx66NbZ/yToP9Iv/AB0dD17Lel3Xvfuvde9+691Up/NzJHQr2+tpbf8AJ3uX/Y//AJXiH7R1FfvDT+pk/wBjf4OtVOnc/bw3P6YIif8AC6e8+3Heaep6wXizEtf4R/g6yCUWPDc8f4n/AFj7rTpwaqA+vRm/hi4/2Z/qAWP/AB9tHYX5/H19x97pj/kBbiR/vo9D32w/5X3b68df+Trd1jN0H+u3/Qx984rf+wT7B1n43E9c/b3Wuve/de66P0P+t7917rWL/nMvp7s2yt/+YZozz/yD9D7y9+7V/wAkK6/5qt1ip94b/kpWh/4X/l6pvM1vx/sP+J95MU9OsdK0x10Jh/rfXg/4/j36nWwteHn1c3/JrYN2bneebVJ/1uH494wfeSxZ232D/J1kh93gANdU9W62a/eJPWUvXvfuvde9+691ErzajqDYn9s8D6+6Sf2bfYf8HWxkjrRo+bbJ/s3fd/qIP8fX+v01yD30U9q6/wCtxtX+kP8Ak6wJ9zgP9cXcv+bf/P3RX9aqSARb8X/1vY74cegUePWKZx4JkJ48Up4/Foyfr7svxAfMdNy/2TA+h/wdbb38ndifilte5B5rPoLf2094Ge+JP+uPej5L/l6zd9mv+ne2H+k/yDq2n3EnUo9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690QH+Yh8gN/wDx56ex26+uKmipc/V5xaLXXxeWH7fQpYabHm5/p7lP2i5W2fm/mR9s3lWaJY9XbjPUa+6fM268qct/vPaNPi6wvdwp1RYn81r5kFATmtrkkXuKI2/6E95QH2K9uvKOX9v+z1jOPfL3EoCRB+1v83XIfzWvmPb/AIvW1yef+UL/AInR79/rE+3Z/wBDk/b/ALPXj75+4P8ADDX7W/zdGX+IX8xn5Ldqd5YPZW/Mnt+bbdekPnSkpfHOZJJChCsVFuP8fYJ9w/aTkrl7laXc9sSQTLWlTUcPt6Gnt57sc5cxczptm5iIQkA9ta1JPqOtjcEMAR9CAR/rHn3iCDUV6yu6797691737r3XvfuvdBRv/vLqPqyvocX2Hv7b20shk4HqaCkzFWaeargQkPJCoU3AIN/Z9tPK/MG+xNPtFpJcIhoxQVAPoeifcuYNk2eRYt0uo4GfKh2oTT06Qf8As4Pxk/5/Tsj/AM+Tf9GezX/W852/6Ns3+8/7PRb/AF45R/6OEP8AvY67/wBnB+Mn/P6dkf8Anyb/AKM9+/1vOdv+jbN/vP8As9e/rxyj/wBHCH/ex0I/X3cfV/a38R/0db2wW7ziSi5IYaqNQaNpP0Ce4Fr+ybd+Xd82HR++LZ7fxPh1ilaenRrtm+bRvIY7VcJPo+LQa0+3oS/ZL0a9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6902ZkgYvIX/5Uq3/3Dk97HVW4Z6+dX/wmr/7fw/OMj/nU90H/AGH99pPdRxHTh4Hr6My/Qf6w/wB6976r137917r3v3Xuve/de697917qof8Am89Q/Has+MHefdHc+8d3bX3Tj+kd/dedStiN/wCT28Z+0Nz7eq4+v8ZsrCwt45dw12TENPQmNCzEkkWUuqmGSSnhoMfz6QXVvbk/UTGhHCpxXyH5/wCHqsvozYPd/Yvxe/k+/wAur5F7l7r62ym8umqbunumur935rGd1VeS6wzs2exHXO9M7K5kainpZoqeopJiXKKkcikxAB6JUCPMPw4z9n+qvSS4kmeWG0cgCQVJWo4EYz5EcAfsPR7en8O/w4/mry/GHrbObwy3T3yv6d3h8g8ttfduertwYrrjeWyqtcGlFsIVBIoKKpjRmkpr2Ytcliqt7q5EtuZW+IEDq0Ia23EWqH9NlLUJrQigx6cerv8A2i6OOiZ987q7vo8dvyg23gYKrbI25nEerej1utG2PdZ5Fkt9QpPPuvWxx6d/g3z8Y+tSf1GjyRf/AIOcpMX/AN5v72Ott0bX3vqvXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcJP83Jb66H/wChffuvdajv809iPllUA2v/AHXh/wB7T6H3nJ93of8AMPT/AM12/wAvWF3v4B/X6E/8u5/wr1XaWN7kE2HIHFx7m6mOoaJpj1695SALAm9iPyR/Qe9069xI62FP5KLa9hb5JBBG6ai9/qD+57w5+8p/ytNn/wA0R/k6yz+7qP8AkJXH/Nd/+PHq+P3jl1kH1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//0t5Dsr44fHvubO7c3R290Z1D2luTZ4qBtTcHYXXG0N5Zrba1a6apcHk9w0dRNSiQfrELqD+fewzDgeHVdCmpIGeOOPT1RdKdOY3YWR6rx/U/W1D1ll4poMt13R7I21TbHycNQyvPFkNqw0woZldlVmEkBuQCeQPey7E1JOOtCONQQqgA+g6kbA6f6m6oGTXq/rHr7rkZtqRsyNi7O29tP+LNQRGChOS/gVPB5/ChKxeXVoBIWwJ9+Zmb4jXraoiCiAD7BToRvderdQsl/wAW7If9QVV/1ob37r3RKPgB/wAyWzn/AIk7fH/uwX3odWbo8nvfVeve/de697917r3v3Xuve/de697917r3v3Xuve/de64SGyn23N/ZN9h68BUj7etHz50G3ye7LI/Su4q+9/qBfke+hftKf+QPaD+gvWCvuuK87XJ/pH/AOimGQf61/wBP+P8Ar+5Ir1HRHWCoc+Cbj/dUnH+sv493jP6i1PmOqTA+E32Hra2/k4Nq6KyP+FTT/Xkn0+8EffSn9dG+w9Ztezlf6pJX5f4OrivcN9Sx1737r3XvfuvdA735/wAyh7DH/Zm7l/1uMTL9fazbc7raf814v+PjpNeitjP/AM03/wCOnrQALf5XmAbAnO5uw/Fv4nJ+PfTokeHEB/vtP+Ojrm9HXxbiv+/pv+rjdd6rC17j8m3A/p71qp05QmvXcUn+XYf/AAzuF4/rfIx+9MT4Mv8AzTf/AI6evBa3EA/4dF/x8db8vxnt/oV2DYfXbWFP/rMi/PvmZvH/ACV7o/8ADZP+PHroxtn/ACToP9Iv+AdD17Lul3Xvfuvde9+691Uh/N4bT0G5/wAJRf8Ap+r3L/sf/wArzD9o6iz3gFeTLj7G/wAHWqTTy/5NCLg/sRH/ABPoHB95+SU8Q/af8PWDMIPgp9g/wdZBKTb/ABNv6WH9R71jq9Wr0Z74YSE/KPp5QeBuyiJP9RxwPcd+6oP9Qtx/5pN0Pfa8/wDI8sPm/wDk63f4/wBA/wBdv+hvfOO3/sE+wdZ9Hiesnt7rXXvfuvdeP0Pv3Xjw61gv50Dae7dtD8/3Yojf6X/T7y8+7T/yRLn/AJqt/l6xV+8MtdxtT/Q6poMgHPPBta/+395OUr1jnWnDrgZVP9R+Tz79Q9bHV0f8mNg3Z2fsRa1SeeDwHNveL33kx/itsD8v8I6yR+70e65+1utnT3iR1lGOHXvfut9e9+691DyBtRVJ/wCbTe6Sf2bfYf8AB1sZI60Yfm61vl53h6h/x8Cj/W9cnvoj7WMf9bnax/QP+TrAv3PH/MRdy/5t/wDP3RXS5AYcWFuT/ifY9qSK9AkAdYJpP2Zr2/zMvA+o/bPuyV1j7R/h6pMaRN/pT/g625v5OTaviftU8fWs/H+1p7wO98T/AMxHvfsX/L1m37N/9O9sP9IP8A6tt9xH1KPXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdU/8A84dm/wBBm2kB4O5Wvc/1CfT3OP3ej/yN56/75H+XqGPfYD+pSn/ho/ydaysWnSoDGwH9fz9OfecLcesK1oVHWQk/S45F9Q+n+tz7qOrmg6Nf8HnI+R20iNQ/fpBx+R5z7jX3d/5Umb8/8HUke0Q/5Gsf2L/hPW5lB/mIf+WUf/Qg989k+AdZ4n4j1l93611737r3XvfuvdayH85qvOR7y6yR2KLRbbroVRbflmJJv/ib/X3mN92pyOXdy/5qr1iH95CIPvu1V8kk/wAHVQun86z/ALDTf/W+nvI3WesfPAXj17S/FmP+IOn/AGH4961db8JVyOHV4n8k/IS0Wb7hpNYZK6qoAw+o4ePngD3i195k1O1fY3+XrJz7tiBIN0p/v3y+wdbEvvFLrKLr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XumvNX/hdfb8UVaf8A1Tk97HVW4dfOr/4TWf8Ab+H5yH6WxHdPH+H99ZePdR5dOn8XX0Zk/Qv/AAVf96976p1y9+691737r3Xvfuvde9+691Ql/MI+DH8zv5F/NvpjunoDv/4y4j40dQ7VWsxfRveW1d07oxs3bNNPLV02+Mrt6gx9TS1tSkrQ/Z1n3kL00cWlY2LavaqGZIxppgjPr+R6LbuzmuTrVwGUgqDUrQfxDzPz8vLzHQ9/I34efMjuvqno/szb/aXROx/5g3SuextVju0Mft/eLdNV+2ajJLPuzbcu3Kv7itZauJQUdo9IbUhUK2pdrPHHVFB0njXj1qaznnCys4EqHtIHb9hBz0JHxN+J3yAwXcu9/k381t+dYdo98VNLNsnq2r6mw2d27tHr/qyojSWswq0GZCSTVlTUBmklYNZb+pmclayzIUEUQovz49XtrSVZjdXTBpMgaQQAPTOerKvabow6QvZ5I6337YkH+5+4+QbEf7iZfz791scegF+Dot8Zeufz/k+W+n/a4n96HXj0bT3vrXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcX/Q/wDwVv8Aevfuvdaif81YhflvVAGx/uxCfr+bp7zl+7zn29an+/2/y9YXe/oH9fID/wAu5/wr1XMZG5Ibgfqve59zh9vUNcRjriJv6m9zcW/pf3sgdaBIPWw7/JJYPsHfR543VUA/6/7nvDb7y2OarP8A5oD/ACdZa/d1oeUbg/8AD3/48er7PeOXWQXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf//T3+Pfuvde9+691737r3XvfuvdQcn/AMW3If8AUDV/9aG9+690Sv4Bc9L5z6f8zN3x9P8ADIL70OrN5dHi976r1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3WOT9P+xH+x9tzf2LfYevD4h9vWjp862t8ouzhew/vHX3/wBv76Fe01P6j2n+kXrBf3V/5XW5/wBMeikGTnj+oI/qP8PcjVr1HnWKokfwzW+vikv9bH0+7JTWv2jqkobw2HyPW19/Jsa/RGS+n/Aqn4/tfp/PvBT3y/5XNvsPWa/s7/yqMf5f4OrjfcOdSv1737r3XvfuvdA734CeoOxObAbL3MT/AOeiX2r27/kq2n/NeL/j46TXv+4M/wDzTf8A46evn7l1+8y9mP8Axfc3e5/6uUnvpzX9OL/mmn/HR1zhWhlnp/v6b/q43XLULWBXn6/4W+lvdanj06Big69G7fxDDfp4zuG+v5vkI/e2A8GWn++3/wCOnqmRPbn/AIdF/wAfHW/Z8ZDfpTYP0421hbkfTnGRfT3zO3n/AJLF1/zVk/48eui+2Z26D/SL/gHQ+ey3pd1737r3XvfuvdVHfzfCB8fpj+Qsp/wt6vcv+x9P68w/aOot938cm3H2N/g61OaaX/JoDYj9iL68c6Bxc+8+Za+IftP+HrBuGvgof6I/wdSfL/T8/X/AH6ke26kYrw6doMdGg+Frf85SdOi4/wCPuoub/Xn6H3H/ALptXkLcP+aTdDv2xUDnuw/0/wDk63i4/wBI/wBdv+hj75y2/wDYJ9g6z3PHrn7e611737r3XR+h/wBY+/Dj148OtXr+dM9u7drgfU7XoiP6fRfeXn3aTTZrn/mq3WK33hhW+tf9J/l6pf8AKT/Qf1v/AF95PUHDrHCvXEvcWJFyb3HvwoTjq2errf5Lvq7OzxF/01X1+psH594ufeT/ANx7b8v8nWSX3eTm5HlVutnv3iR1lJ1737r3XvfuvdRK/wD4B1P/ACyb3ST4G+w/4OtjiPtHWir84Tb5f95gm/8Av4VAtcf25P6++h/td/07nax/QP8Ak6wO9zgv+uLuX/Nv/n7orZNjzc/1N+P8Lex3XoEkUNT1jlb9ma3BMM36vrbxn3dD3AfMdNzAeE4Hof8AB1t2/wAm83+Ju1SPoWrbH/p5H7wP98P+njXn2L/l6za9nP8Ap31h/pP83VuHuJepQ697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917qn3+cQbdG7ZYiw/vMw5/4LHz7nD7vmOd5v+aI/y9Q176Z5KX5Sj/J1rKxuojU8c2/3n8X95wsDXrC1CukHrJqXV+Ppe3/Gvdc06uPTo1/weYH5G7SII5qaP/Wt5z7jT3dxyTN+f+DqSPaOv9dEPyX/AAnrc0g/zEP/ACyj/wChR758J8A6ztPHrL7v1rr3v3Xuve/de61fv5w3HeuwQwFzgq3m305P1v7zD+7WD/Vzcf8AmqvWJX3i6fv3a6/wSf4OqmSFuLg8Di1v9595GVPWP+nPXG/0Iv8A7V/xHvfVaenV2P8AJcJG6e0uB/wMorf8lx/T3i195jjtR/ot/l6yZ+7gD4O6f81R/gHWxx7xU6ye697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917przX/Frr/8AqCrf/cOT3sdVbr51f/Cas2/nxfOL/tU90n/bb1l91Hl04eB6+jOvKqf8B/vXvZ6r137917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XukL2h/zLffv/hn7j/8AdTL78etjiOgF+Dv/AGTL1zfj/J8r/wC7ef3odePRtPe+tde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691wf8AQ/49Dc/7D37r3WoV/NbYJ8uam7D/AI9eGw/5Itf3nP8Ad4z7eH/mu3+XrC33+P8AyPYP+ec/4V6riM2n6W5/P4v7nIDqGCDUZ68JT9G0jn+nJ/4L70erKKHJ62Jf5IbA9f78tz/v66gcG9v859feGv3l/wDla7L/AJoD/J1lr93Sn9ULgjP67/8AHj1fh7xx6yC697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//U3+Pfuvde9+691737r3XvfuvdQcn/AMW3If8AUDV/9aG9+690Sr4A/wDMl84f+/m74/8AdgvvQ6s3R4/e+q9MWX3Pt7AFBmsxQYwyGyfeVCQ6j/RdXtVb2V3dV+mjZ6caCvSea6trenjuEr6mnTN/pI2F/wA9bg//ADvi/wCK+1H7m3X/AJR3/wB5PTH7027/AH8n+9Dr3+knYX/PW4L/AM74f+K+9/ubdf8AlHf/AHk9e/em3f7+T/eh07Yjde289LJBhc3jsnNEuuSOjqUmdEvbUwX8e09xYXlqoe4iZAfMinT0N5a3B0wSK5Hoa9KD2k6U9e9+691737r3XCTlf9iPbcv9k32Hr3mPt60afnc9vlJ2hwTbceQ/P05/HvoN7T/8qTaf6ResGPdRf+RpckfxHoogmFh9b/QWP4t9fcj1PUfU8+sNRJeCcajxFJfn86T7tGTrX7R1SUfpP9h62yf5M5v0LkTa3+U0/wDt9P594Le+Gecm+w9Zq+z4I5Sjr8v8HVyPuHepW697917r3v3Xugc7+v8A6IOxLH6bL3Of/WPL7Vbf/wAlS0/5rxf8fHSe8H+Iz/8ANN/+Onr59uv/ACvLlvxns5wL/wDOzl+vvptWscf/ADTT/jo65yJTxZ/+a03/AFcbrlqvYDV6vqR9Rb37yr1fPXo5H+/w/pN/49heeef9yEfvZI8GX/mm/wDx09UP9vB/zWi/6uDrf3+MnPSewDYC+2cJ9P8AtVxfX3zQ3n/ksXX/ADVf/jx66K7Z/wAk6D/SL/gHQ+ey3pd1737r3XvfuvdVFfzgDb4+zf60pv8A7E8H3Lvsif8AkcQ/b1F3u7/yptwP6Lf4OtTKll/yanBIJEERNv7XoHHvPOVv1Wr6nrB+Bf0EH9Ef4Os5m/VweRYj+gHutfXp4AeXRn/hPKrfKfpsX4/vdRcfn6gD6+wD7pZ5D3Cn++m6HPtkD/XuwP8ATP8Ag63mY/0D/Xb/AKGPvnVB/Yr9g6zzPHrn7d611737r3XR+h/1j78OvHh1q4fzrjp7u2wb8na1F+f+Cj3lx92sn9zXP/NVusWfvBiu4Wo/4X/l6paEvPNuP8eL/wCPvJnUesdio8+u/IALhhctf/YW+nv2rrXV2X8laTV2buC7c6aqwB/wf3jF95I1trU/Jf8AJ1kd93vjc/6Zv8vW0F7xL6yi697917r3v3XuoeQ/4BVP1/zZ+n1/2Hukn9m32Hra/EPt60TPnG//ADmF3oB+NwqQBy3Lye+h3tfn262r/SH/ACdYHe5p/wCYjblX0jx/vXRWxIbc3P8AX6/n2OwfPoFEVHWKeT9mcG9xDL+f9oP09+X4x9o/w9VmI8Jx/RP+Drb1/k12PxL2o1+S1bYXHA8ie8E/e819xrw/Jf8AL1mv7OCnt9Yf6Qf4B1bn7iXqUOve/de697917oO+1921mxuv9z7px6wPXYjGVFVSrUi8DTRxlkEg/pcezTZbOHcN0hs5yQkjAGnGh6L91upLLbpbqEAsikivCo61tK/+cR8tKfKZeki271qYKLLV1HTE0UpZqenmKRNJx9SPr7zKh+73yC9vHIZrirorHPmR9vWIE33gefI7qaFbO20pIyqdTVIBoCe3H2Z6jf8ADxny2+n93utL/i1BJ/xT3f8A4HnkH/f1x/vXVB94Tns/8Q7b/em/6B6l4z+cR8sanK46kn271sKepq4IJitDKHVJJljZl4/oTb23L93vkJIXdZriqgkZ9B09D94DnmS4jja0ttLsATqNRUjyp1su9f5+o3VsfaW5KwRLV5zb+KydUsAtCtTWUaTTrED/AGQxIHvDDc7eK03K4tISSsUjoK8aKxAr1mDYTPc2UNxIKNIisftIB6WHtD0r697917r3v3XuqeP5x509FbZ5+u5mN/6cR/X3N33fjTneb/mkP8vUN++QryWKf79H+TrWISUBQOeFv9Rb3nMRnrC1aaR139xyLnm3+x0/1t71p63UdG3+DEuv5IbSUH0mek/17mc8+4x94BTkib7T/g6kn2iqedEr/CP8PW6BT/5iH/llH/0IPfPeP4B1nafiPWb3frXXvfuvde9+691q7/zjpvH3v1+th6sDXH/eW95i/dqFeXNy/wCaq9Yk/eMqN92un8Mn+DqpP7m/5At+b3vf+nvI3T1AOo8adcPubWN73/s/kf69/ftI60GPl1dz/JXfy7p7UBNtNZQ/6w9afn3ix95gUO1fY3+XrJj7uJrBuf8AzV/yDrY+94qdZO9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6902Zn/AItdf/1BVv8A7hye9jqrcOvnVf8ACav/ALfw/OP/ALVPdP8Atv77Se6jiOnDwPX0Zl/Sv/BR/vXvfVeuXv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6QvaHHW+/f8Awz9x/wDupl9+62OPQC/B7/smbrn6/wDAfK/X6/8AF3m9+HXj0bT37rXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcX/AEP/AMFb/evfuvdagH82JgPl3Ui4v/dWG1v9dLD3nP8Ad3qPbtj/AMPb/L1hd7+g/wBfLev/ACjn/CvVbge354/N/wAH3OWr16hvQOI678l/qeSePfgRT7OvUoetiz+R2wbrzfvNyN11F7fk3k+vvDX7zFP62Wf/ADQH+TrLT7ug/wCQhcf813/48er9/eOHWQPXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf//V3+Pfuvde9+691737r3XvfuvdQcn/AMW3If8AUDV/9aG9+690Sv4BEHpfOEcD/Sbvjj/yID3odWbo8XvfVetev+cRvfcuMz+ysbgs/ksKIaxDL/DpzCZQQt1kt7yo+7rZ2d0t211EsnbjUK9Yy/eCu761htPopmiOsVK+Y8xnqlodh9iG4/v7uQW/6bT7ya/dW0eVrH+zrG39775/ymSftHXX+kPsb8b73IbHn/LT9P8AC/v37p2j/lFj/Z1YbvvY/wCJkh/Mf5urcv5PO9N1ZfvDemPz25MpmKZtou8UNdUGWOORJdWpQfzx9fePf3i7OxsuWLSW1hWMmcAlRQmo6nv7v99uF5v18l5O0oWJSA3lnrZI94g9ZX9e9+691737r3XCT9P+xH0/1/bc39k32Hrw4jrRf+eD/wDOUvaHNv8AfyZDg/k6v6e+gntST/Ui0/0i9YN+6f8Ayulz/pj/AIOiharckgE/S/8AxHuRQTwHUfEDrDUsft5yCotC9/pcnSefbkbfqL9vTUq/ot9h/wAHW2f/ACYiT0Jk+Sf8qpjz+PT9PeC/vef+Rk1fQ/4es1faD/lU4/y/wdXL+4e6lXr3v3Xuve/de6B3v4kdP9jEfX+5W5+f6f7h5farb/8AkqWn/NeL/j46T3n+4U//ADTf/jp6+fF5VNXl/qFGezl7/Un+KSj301zoi/5px/8AHR1zljzJPX/f03/Vxusge35P+3+n491PTvXUUv8Al2I+v/F+wv8A7sY/eiT4Uv8AzTf/AI6eqkDx7f8A5rQ/9XB1v/8Axj/5kl19/T+7OEt/X/i1xXv75qbx/wAle6/5qyf8ePXRHbP+SdB/pF/wDoffZd0u697917r3v3XuqhP5w7Ffj1Of9pk/3tuB7lz2TNOeIPt6i/3dFeTbgf0W/wAHWpPSSA0tPyRaCL6m39ge88ZT+q32n/D1hDAoFug/oj/B1nMn1vqv/sPberp4qOjSfCYj/ZqemQfqd30P9P6j6+wH7oEnkTcP+aR6G/tmKc9bfX+M/wCDrelT9I/12/6GPvnbB/Yr9g6zyPHrn7d611737r3XR+h/1j78OPWjw61Z/wCdo5XvHa9rX/utRfX/AJB+vvLb7tpps1z/AM1G6xb+8H/ufan+h/l6pV8r2JsvH+vf3kvqHWPP5dcRN/gPr9b/AEPvVevaQT1dx/JQcN2fuG9ibVVrfjh/r7xl+8ea21r9i/5OsjPu/DNzT+Juto33ib1k/wBe9+691737r3USu/4B1F/p4z7pJ/Zt9h/wdbHEdaIfzmYf7OJ3tY2P941Bt+PXJ76Fe15I9utqH/Cz/k6wQ9zB/wAxH3I/KP8A5+6K15Atxq/pbn2Os8Ogb5dYJ5P2Z1/Phl5/P+bPuyfGpr5j/D01OP0np6H/AAdbfn8mc6viTtNrW9VaP+skfvBX3ux7iXg+S/5es1vZ7/p39hT+Af4B1bv7ibqT+ve/de697917ou/yqkaPovfZUkH+FzC4uP8AdTH2e8rmnMVp/px/hHRLzD/yRbn/AEh/wdaVeR5y+cNyT/GMgSL/ANZz9PfSy2/3Eh/5pr/g6503Q/x24H/DX/w9Q/p+R6v8fp+fb/Sc0DdS8fJbKYw6T6a6l5H/AFEp9PdJgfp3r/Cf8B6ctyv1UQ/pL/x4dbv/AEGxfpvrZiSSdnYT68/8oot75hbz/wAl6/8A+a8n+HrpLtP/ACSrb/mlH/x0dC97L+l/Xvfuvde9+691Tj/OXfR0Ntji99zPf/D0x8+5t+79jniX/mkP8vUPe+IryWP+ag/ydavSSnSnN7IOb3HB/p7zrIyR1hQoGkV66Mw/xBva9/7P9Pfqdb869G++CUwb5KbRVQbeej/PH+fPNvcW+8YpyRKfmf8AB1JftD/yuiD+iv8AhPW6jB/mYf8AllH/ANCD3z3T4R1naeJ6y+7da697917r3v3XutWj+cvJp7469uf+XBX8n/Xbg+8x/uz/APKt7lT/AH6vWJf3jAP39tf+lk/wdVEmbgHi4vx+P9ce8j6dY/Uz10aj6fTn6fT/AHv3vT1WuBTz6vD/AJJknk3R2sL/AErKHn6D9cfB94q/ebFG2r/St/l6yZ+7ef8AF90/5q/5B1sl+8Uusnuve/de697917rDUSiCnnnNrQwyym/0tGhc3/23uyjUwX1PWiaAnqjjuv8AmyZnq/tDcmw6PatJW0+DmaJapqeRzLpkKE6lH+HvJnlf2Js9+2GDdpJypl8gRjrHXmb3vfYN+m2fwNXh+dD606C4fzodw3sdl0X/AJzS/wDFPZ9/wOdj/wApLftHRH/wQzjBthX7D1xb+dHuBQL7Lov1AcU0v0LAfke9j7uVj/ykt+0dab7w0g/4jD9h6uL+J/ek3yI6fxPZNRRx0E+QrKumeliUoqfblbHSf63948c+csx8o8xSbLG2sIAa/b1PvJnMTc07DFvLLo8QnH2dGV9gzoV9e9+691737r3TZmf+LXkP+oGt/wDcOT3sdVbh186v/hNWB/w/D84/+1T3Rbn/ALPaT3UeXTh4Hr6My8Ko/oB/vXvfVeu/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690he0Lf6N9+3+n9z9x3/8APTL7914cegG+D1v9lm65t9Pt8ra30/4u83v3Wz0bP37rXQRb8766e6wr6fF7/wCwNvbVyFWC1NS5ar8EswX9RRbG9vz7Ptr5X5h3qIzbVaSTovEoKgdE248xbJtMgi3K6jhZuAZqE9IX/ZxvjF/z+jZJ/wBbJHj/AF/T7M/9b7nan/JMm/3n/Z6L/wCu3Kfnfw/70Ouz8xfjEP8AmtGyf/Pkf+jfe/8AW951/wCjbN/vP+z17+u3KZ/4nw/70OhL6+7i6w7W+/8A9He9cHu7+FiM5D+D1X3H2ol4jMvAtf8AHsn3bl3e9i0Hd7Z7fxK6dYpWnp0abbve07wGO13CT6Pi0GtPt6Ev2S9GvXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691xf9D/APBW/wB69+691p5/zZ3KfMGp/P8Av1ICDf8AqU95y/d5ant4w/4ef8vWGPv4K8+2/wDzzn/CvVbRmPBP0vc88W+vI9zdqHA9Q8ENa8KdeEwvc8/0F/x79q8gevaaGp62N/5GzB+u9/EC3+/tqOD/ANPPeHP3ljXmuy/5oD/J1lh93YaeUJ/nO/8Ax49X+e8cup/697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//W23f5ln8x/Y/8s7pvBdzb/wCm+6e38Jnd1UG1ftuodsxZqPAz188cKV+6MnUyJFRQt5LQalYyupRRf27HEZASPL8z+zpPPcLAQGHxeZwB9p8vl6nHQtfIv5pdafGv4z0nyZ3Vgt67jw2awu08ntPr7aGGXKdk7uyG84aefEYDCbdaRGkqkSpElWNVoUSRjcgBvJEXfQPLz69LcxxQiZ/MYHmfs6D34+/PXE9rd37j+N3ZXVe+uju3sVtfB7327R70oPtdt9g7c3BRfxSOPZmZmK/dV9BAyjJUgS8bB7E6WCuSwaRqjNV6Yt73xH8GZdD+XoR8urA/abpf0EG/u49gbKlrsLuPOU2Pr2x9QwglkAc+SmZowB/tX0HvVevdF3/l9TQ1PR+Uqqc6oKrsbedRC/11xS1ysjA/4j34dWbo8/vfVetbb+cgwTfG1yfzVRfX6fRfeV/3b66Lv/S9Yv8A3iP7G0/046pd1pa35JuT9T/X8e8oaE9Y0do695R+rmw4sOBYe/UPVgBXq3H+TjIH+QG7FUWH9z5j/vDe8c/vLf8AKp2df+UgdZAfd2A/rBfkf75H+XrZ6H+P1/PvDvrLXr3v3Xuve/de64Sfp/2I/wB79ty/2TfZ14cR1orfPSUL8p+0hwbblyH1/wBf30A9qTTkq0/0i9YP+6Irzlcn+keifeW5UWP04P1AP4JPuRq9R6RXrFUO3hntYFYpL/4+n6j3aNu8faOm5kPhtT0PW2v/ACXiW6DyRNiPuqaxH59P9feDfvcf+Ri35/4es0vaEf8AITj/AC/wdXN+4f6lTr3v3Xuve/de6B3v4X6e7H/p/crc9/8Azzy29qtvr+9LT/mvF/x8dJ7yn0U9f99v/wAdPXz3GYfe5cW4/jucvYfU/wATl99M/wDQ4/8Ammn/AB0dc6FI8af/AJrTf9XG67ubHm/9Lf8AE+616vTjTrlG3+XYXi189hb24J/3Ix29+NfClH/C3/46etEHxrc0/wBGi/4+OvoDfGPjpHr4f9mxhD/sDi4vfNXeP+Svdf8ANWT/AI8euh+2f8k6D/SL/gHQ+ey7pd1737r3XvfuvdU//wA4zj48TH/lr/0V9P8AH3Lfsr/yu0H29Rj7tf8AKn3A/on/AAHrUepZCKan5FlgiP15/QPr7zulP6jfaesIrdf0Ur/CP8HWbzH6k/Xg/wCH+t7pXp2nRpvhC+v5WdMAEkjeFD+fxccewF7nn/kCbh/zSPQ59tAf682BP8Z/wdb2SfpH+u3/AEMffPGH+yX7B1ncePXP271rr3v3Xuuj9D/re/de61X/AOd21u8drAk3/utQ3t/yD+PeWf3b6/ue5P8AwxusXvvA/wC51r/pP8vVJZmtyNX9LE8/7D3krqPWPFM468ZhYXBIJuT79XHW6Vx1eB/JJkB7P3F6b6hVc/0ur+8ZvvGmtva19B/k6yL+7+KfU/6Zv8vW0r7xQ6ycHDr3v3Xuve/de6h5DijqP+WZ/wB7Hukn9m32H/B1sfEOtDv50OB8x+9wOD/eNf8AX/XJ9be+hPtgf+Yd7XX+A/5OsE/csH/XG3L7I/8An7oq5fg2HH9T9f8Ab+xzUdA2nWGd/wBme59Qglvb9X+bP497jbvX7R/h6anH6L0/hP8AgPW4N/JkbV8RtpHj9VbyPp/nI/eC/vaa+4t59i/5es1PZ/8A5UCw/wBIP8A6t49xP1J3Xvfuvde9+690XP5X/wDMiN9m9iMZKR/1Jf2e8sUHMNqf6Y/wjom5gFdluB/QP+DrSgyVURl83yCTmcgCQQbfvkc++l1qo+jh/wCaaf4OudVyT9bcD/hr/wCHqF91b6j/AAHP+9e39PTFQfLqXjam+VxnNwa6lBB/H+Up9b+2rgabaQ+it/gPTtvQ3UX+mX/jw63jOgDfpnrQ8c7Nwn/uL75hbya77f8A/NeT/D10h2r/AJJdt/zST/jo6GH2X9L+ve/de697917qmv8AnPsF6C2ufpfc7i/9Bpj9zZ7AGnO8p/4UP8J6h73vBPJuP9+D/J1qzpKQgCudJX8nm5952s2a9YWKq6QM9chMQP1XP454/wBb3rV69e0+mOjh/AyYN8mNngX/AM/R3At/x3PuLfeU/wDIGlp6n/B1JXtB/wArqvrpX/Cet2KD/MQ/8so/+hB757p8I6zqPE9Zfduvde9+691737r3Wq5/OjkCd9ddXNidv11wOLm7WPvMj7so/wCQ5uX/ADVXrEv7xhpve1/6WT/jvVP3mJA5+n1B595I4r1j9kjrvzMD+OOLfT6/S1/e8daoePV5n8kJ9W6O2Pp6ayi/xB/cjHPvFT7zvxbT9jf5esmfu3rS23T/AJq/5F62V/eKHWTvXvfuvde9+690055imEy7D6rjK8i39RSv79+Jf9Mv+Edab4T9h/wdaVPyjkkfvzfzsQwNfPe55N6pre+jft1p/qZaU9P8g65+e41f663X+rzPQCE/6ofX88X9jWh6BgOaDrgxBU3sCGXj/DUPx72BQ9aapXPW1z/Knldvi1h0vdVzuRt+bAopNrf7D3gN74GvuNcf80k/w9Z0ezoP9QbT/TN1Zj7ifqUOve/de697917pszP/ABa6/wD6gq3/ANw5Pex1VuHXzqv+E1n/AG/i+cf/AGqe6f8A3tZfdRxHTh4Hr6M6/pX/AFh/vXvZ49VHXfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6QvaH/Mt9+/n/fn7j/91Mvv3Wxx6AX4Om/xm65NrXp8rwP+1vN70OvN0bT3vrXWrR/N7lgy3eeAjqqdJ/sRVJFrBJQaXHBHvNL7ussqcuXOliKkf5OsOvvDWsFxvlmZkDaSSK+R0nqqAYzGgD/I4ufrwfr+Pz+PeQn1E38R6gL6C1pTQPn102Lxx4+1h+h/Bt/vfvwnlH4j1o2VtX+zH8+r4v5JDQ0OV7ho4YViWekxjsFB+sbxkE3PvFT7zczldqLGtWcf4espPu3QRRQ7msYp3If5DrYV94qdZP8AXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde+nPv3Xutd7+ZL8qu4+r++6ja+wt1ZDDYqmx2Odqamq3hTyyR3lYhf6n3lx7L8mct79yku4blbrLIWYVIrgHHWLXvFzrzHsHMgstrmMcehTQGmT1Xx/s+HycNx/pAzP1/GQktx/sPcuf62XJf/AChp/vI6if8A1z+duH1Tf711x/2fL5OWBPYGatc/8p8v/Ej37/Wx5L/5Q0/3kdbHuhzpxN0/+9dXT/ysO/ew+48DuNewc9W5qupmZ6d6udpiia/oC3+HvGn3z5b2bl2+hTaohErcaCnWRnsvzHu/MW2SXG6yGRh6mtM9XB+4B6nDr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuuL/of/AIK3+9e/de606v5tzKnzDqhe19pwW/A5MfPvOH7vhI9vW/5rn/L1hn78mvPlvU/8R2/wr1WnrK/2r/69ufc1gnqIPPr3m5/1/wA/kA/ge98R1vz62QP5FzBuuewOL/7+2oAJ/wCnnPvD37yn/K12X/NAf5Osrfu8Z5QnP/D3/wCPHrYD946dT71737r3XvfuvdcJZY4Y3mldY4okZ5JHNlRFF2ZifwB72qliFXJPWiQo1HgOgTr/AJK9A4uuqsZke3th0WQopDFV0dRuCijnp5F4KSxs1wR/Q+xLFyZzXPEs0O3zsjZBEZoR8uiOTmfl2KQxSXsKsvEF1BH2ivUP/ZpPjr/z+fr3/wBCOh/6O9uf1H5v/wCjdP8A8426p/Wvlr/lOh/5yL/n67/2aP46/T/TP17cmw/38dDyT+B6vev6kc3/APRun/5xt1v+tXLf/KdB/wA5F/z9DJg87hty4ukzWAyVHl8TXR+WjyNBMlRSVMd7a4Zk4Yf4j2Hrq1ubKdra7QxyLgqwoR9o6OYLiC6iWe3YOjZDA1BHyPTt7T9Pde9+691737r3X//X2ff55HcPVfUv8u7uI9m9kbO69bdk20cLtj+9WcocTPnsrFvLH109DhqWoby1MscEbyyLCjFFF2sLe1NpiYMcAcT5dF+6d1k8a5ZhgDJP2Dify6Ll8qu3etq3YP8ALA+X23uwNq75+MnSG68JX9sdi7WzdBmtnYHH5XaGJw2Pymar6WR1hRKmN1KyoGVrKwUsAVMIzIpwWrT/AGOi68dfCt5F7kQjVTyxTPpn16X3b29ti/Ln+ZX8Dsx8a+wNsdr4n43w9jb37pzPX+cx248Ntnbu+toii2nT5zJ42Rokeol0vFFrZryAaQ1x7bQNFauHxq4dKJWjuNyheGjeGDUjNKjH+fq8r2h6Oeg53h1psDc4rstuLaWFzOQWhqB91XUolm0x07aVDn6W/B/Hv3Xuiwfy/YooOksvBBGIoIOyd6wwRKLLFFHXqqRp/gBwPeh1Zujy+99V61p/5zkoj3ttUtfmqiHH44W3vK/7tlSLwf0esYPvE4trQ/8ADF6pO+8BHDN9SL+8p9NOsZxTh177rjlmt/X/AB/r79p68cCtOrf/AOTJL5PkJu0An/jzpzZvp/a5HvG37zB/5C1mvpOOsg/u7U/f1/8A80R1tFe8O+stOve/de697917rg/6fx9R9fp9fdJP7NvsPXvMfb1ol/Phgnyq7THHO5cgOf8AX/HvP32pIHJVr/pB1hB7of8AK5XIH8R6J5rBFvzbj/jf+PuRNWOgBTND1hne1PUfX/NSA3P+0m/u0ZOtR8+m5cwvT0P+Drbh/ktH/jAWTt9Puqaw/p6PeD3vbX+uLfYesz/aLHKkf5f4Orn/AHEHUp9e9+691737r3QO9/X/AND3Y/0sNk7oLX+v/Fnltb2qsP8Akp2n/NeL/jw6T3lPopq/77f/AI6evnq3H3eX9XJz2csSRcf7k5eAffS8t+nH/wA00/46Ouc0aDxJ/wDmvN/1cbrlrIsABb/eP9if+Ne61x0/kDh/q+fXGN3NfhiNI/3P4W1/+1jHe3uxI8KSv++3/wCOnrR1fUQU4eND/wAfHX0D/jESekevr/UbYwf/ALq4vp75r7x/yV7r/mrJ/wAePXQ3bf8Aknw/6Rf8A6H32XdLuve/de697917qnz+cg1vjtMPp/nDf+n6vcs+y5P9dYPt6jH3bFeT7in8J/wdaiVK5NNAQfpTxXueT6Pedch/UP2nrCaBf0E/0o/wdZNf+PJ4t/T/ABv71q6dp0an4Otf5X9Kgnn++NDbnnkjg39gP3OP/IF3D/mkehv7a/8AK9WH+nP+DrfAT9P1vy3+w5+nvnpD/Yr9g6zrPHrn7c691737r3Xj9D79148OtVT+eESO89qhSAf7q0J5/wCQeB7yx+7iQNoua/78brGD7wAre2oHHR/l6pF8p/JU2sPrwPeSlfTrHnSR17WTxqHJve/+9e/aqdWArjq8f+SG9+0Nxj62FUOeQLh+R7xn+8Wa21sfkP8AJ1kT7AY+pH9Jutp73il1k11737r3XvfuvdQ6+32dRf6eM/737pJ/Zt9h/wAHWxxHWhn862H+zld8AXB/vEt/+S5PfQb2wI/1vNr/ANIf8nWCvuWD/ribkPlH9n4uiq6rAgX0j+vsc16BgWvDrDMy+Gc83aCa9/8Almfe0PePtH+HpucUhbPkf8HW4Z/JgP8AziHtK3A11xt/08j94Me9hP8Ari3n2L/l6zT9nwB7f2A/oD/AOrfPcU9Sb1737r3XvfuvdFw+WJt0Pv0/kYua39P8y/19nfLVP6wWtf4x/hHRPv8A/wAke4/0h/wdaQmSqb5rOAEcZvIhv6cTn302tB/iUH/NNKfs65z3Wn664p5SvX9vUM1J/Bv/AE9v0Hn0npmo6mYyo/3LYzTyTX0mq/8AT7lPz7T3gpZykfwn/Aen7UH6uIH+Jf8Ajw63qPj9z0t1kfpfZmDP/qqPfL/ef+S5fH/h8n+HrpFtX/JMtv8Amkn/AB0dDF7L+l/Xvfuvde9+691TD/OnJHQO12uB/v6H/wBsFj+vuavYI052l/5pD/L1EHvaP+QZn/fg/wAnWq3HP6Fsfx7zmc9x6wvQHSD1kMxFv0/S5AIvf+vPuuoU6uV6OJ8BJS3yb2fyDeeiB5/HnPuMfeB68jSj5n/AOpH9pF/5Gan+iv8AhPW7pT8QQj/m1H/0IPfP1PhHWcx4nrN7t1rr3v3Xuve/de61Tv51kvj7+65W19W3q7njg3b8e8w/u1H/AJDe5V/36vWJ/wB4kA73tX+lk/471Tt5xf6G4Fr/ANb/ANT7yM1dQFpI4ddeblbaj+SP8fe9ZpXrWnPV7X8jhzLurt/68VmP+v0H7kZ94s/eZNTtP2N/gPWSv3c8W26f81f8i9bMPvFXrJnr3v3Xuve/de6Z9wf8WLMnjjF5D6/T/gK/196/Ev8Apl/w9aPwn7D/AIOtJX5S1Gnv3f1m4/iE/wCf+mp/fRz22FeS7P7P8g65/wDuP/yul0P9XE9AF95bgk2/wHscaegVjy64vVg25P6lsf6erm/vYXrTEdbY/wDKeYP8V8OwJ5z+RBv/AIRj3gD73U/1xbj/AJpr/h6zp9oK/wBQ7X7W/wAnVnXuKepO697917r3v3XumvNf8Wuv/wCoKt/9w5Pex1VuGOvnWf8ACav/ALfxfOL+v8J7osf6f7/aTn3UeXTh4Hr6Mw+g/wBYf71731Xrv37r3Xvfuvde9+691737r3VZP8wr+aF1f/Lpy3Q2K7K6i7w7Bi767BwvX+J3D1rs8Zfae1KvMZNMaKndWemkSOGRPIJUpEVpZEBK29uxxF11eVaep/Z6fPpNPdJAwRvMVqTQfZX1PkOJ6Hr5f/MjYPw+2JtTde5tt707Az/YO9tudfbC6/66wzZ7du4c9uarWkpqw45WV4sbSFkevrdLCFWX0szAe/RxGRqHAHE9enuUgQNxJ4DzNf8AJ0m/jB83Ns/ILsDtXpTcext4dP8Ad/TeThody7H3xQNj49y42amScbs6+rptJyeMUyJHLMiDQxA9QOr3eWDR3oar69N214JiYpBpccQcfsr0eD2n6W9Fx7n7n68wO1t/bZy2epKTL/3Wz9KtJJKqyyVEuNkjjiVfrck+9HrY4jpL/Bs3+MfWzD6PS5N1/wAVfLTMp/2x9+HXm6Nt72eHWutVz+bKwHfFALgeqp5/rw/vM37u/wDyrtx9o/ydYi/eA/5Ldp9p/wAB6q5Ein/U2/x+vPvIPqBAvCvWMtwD6bgG/wDj78KdeIPHq8/+Si2vcfbZ/wCmGh/176o/6e8VPvN4XaR/Sb/L1k793L+x3M+pT/AOth73i11kx1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SN7D3viettj7o37nUqJMPtPDVmbyMdKnkqXpaKPySrAn5Yj6D2YbTts28bnBtdsQJLhwi14VbAr0i3G+h2ywm3C4rohUu1Mmiipp1VTTfzrfi1VwRVMWC7D8Uql1JwhDWDFeVP+t7naT7tnPMblGlt6j+n1BkH3kfby5hWeIzlWyP0nH+EdZ/+Hpfi7/zo+wv/ADy8/wC290/4HDnf/fsH+99Pj7xPIBNKzf8AOJv83Tjhf5yfxizu4MBtylw2/krdxZekw1E0uG0wrVVjaYjK34Ufkn2zc/d251tbOa9kkg0woXbvzQcadPW33guQ7q+g2+MzeJcyCNP0moWNSATSgwDk0HVtFJUx1lLT1cV/FUwQ1Ed/romjEi3H9bHn3AjqUcoeIJH7DTqcUYOoceYr1I916t1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691wkvoe310m3+29+60eHWqH/ADVyP9mizhZufsceAAfr6fz7zd+72f8AkCIP6Tf4esNvfsf8i9T/AMLTqtX6j9Vmv+Dx7nOvUJBa/aOu9f1JKkHgC/5vybe/fZ16ma9X0fyZWBG6LG48Mtv6f2veI33j/wDkoW1fl1lb93zOyzH7f8PWwB7xo6yK697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rhJ/m3/wCCN/vXv3XutOH+bm9vmJUj0ljtOnNmP0/zf595vfd9JHt63/Nc/wCXrDb33APPlviv+Lt/hXqs4zWA4X8XAP59zXq6iErjrwmJYcAX/wB9x79XHXtNc9bJH8ihtXW/YF7C27qgAA/8tPr7xA+8lnmqzP8Awgf5Osqvu845RnH/AA9/+PHrYK9469T71737r3XvfuvdJLftUaPZe56kcmHC17D/AF/AR/xPtRZtpvIf9Ov+Hpi6zbSf6U/4OtH3uikx9d2/2LWS0aNLUZ6Z3cl7k+RuCQffTHlW4nXlmyGrAjHp1zf5ssbSTmy/kKAsXFTnPH59Br/CMWePso1P5N3B/wB79n31U/8AEf5dEH7ss+IjH7T/AJ+uv4RjAYm+0S6yKw5e40g2/Pvf1M5r3dbO3Wi0/TGPt/z9blvwBqhUfFTqxVXSsWKlhUDkAJLqHP8AyF75ye6DMfcDclbycfzH+x10P9ugq8lbcq/76HRzfYC6G3Xvfuvde9+691//0N2z5GfET4w/Lzb2I2p8neiutO89uYCtlyWExHZG2MfuSkxNdOixzVWPWtUmJ3CKGKEXsL/T3YMwBUHB6o0aMwdhUrwPp17B/EL4u7a6RynxswHQfVeJ6DzdEcdmOoqLZ2Hh2Hk6ItG/grtvrH4JQGhiZS6kgopBBAPvZkckEnh1oQxBSoUUPHHHpz6V+Lnx0+ODbgfobpXrjqSTdn8OG5ZdibXxm35s4uIhanxaZOaiRXlWnRmWJWJCgmw97eWST4zXqsNvBbgiFAoPGgp0PPtvp7qDk/8Ai25D/qBq/wDrQ3v3XuiV/AG/+hfOX5P+k3fH/uwX3odWby6PF731U9ay/wDOrm8W9to25vWRAj/YL7yv+7URqvK8NHWMX3h829oB/vxeqPjVcH+gP9Pr7yoDr1jToNevLV/W5/HHuuvrwj9OrjP5K03l+RO71uT/AL8yoJH+344942feYNeWbMj/AH+vWQX3dwRvl/Xh4Q/w9bTv0494e9ZY9e9+691737r3XB/0/wCxH+9+25f7Jvs695jrRB+fcgHys7TuL23Nkf8AYer68+8+/axqcl2v+kXrCH3QFOcrig4seieeQE8G1vz+SR/T3IYPy6ANKmnWCok/YmIP0hkuP8dJ93RqOPtHVJV/Sevof8HW3T/JXa/QOS/p91Tf9C/n3hB711/rg32H/D1md7SADlSP8v8AB1dF7iHqUeve/de697917oG/kB/zJzsg3PGyN0cD8/7h5farb/8AkqWn/NeL/j46T3mbKcf8Lf8A46evnl6v8sy/HAz2cFrfg5ST+vvpaW/Tj/5pp/x0dc6oqCWeg/0eb/q43XLXpIAuL/7AC3PvVfPp6hpQ9dxTXr8MLcDP4T68c/xKPgX9+J/Tl/5pv/x09aAHjW4Jp+tD/wBXB19Bj4xG/SHXp/P918H/AF/51UP4Pvmzu/8AyVrr/mrJ/wAePXQvbRTb4R/QX/AOh99l/S3r3v3Xuve/de6p4/nKsV+OdQR/tfJ/5C9yx7MGnOsP29Rl7sAHlC4r/C3+A9ag1LMftacccQRc/wBbxj3nTI36jfaesKoFAt08+0f4Os/k+l7+o2P+H5vz7oWGenKdGt+DbA/LLpWxBH98aD68cgj+nsCe5jV5Gv8A/mkehr7bJ/yObA+jn/B1vlJ+n/Yn/e/fPiH+yX7B1nSePXL2517r3v3Xuuj9D/rH34cevdapn88ltPem1eSCdqUIFv8AkH3lf93Q02e5r/vxusYff1Sb+1I46P8AL1R55CB9Ppx9OD/t/eSFesftJ65Bvx+SL/UWH+A9+r1rFK9XnfyO2LdobjDcALVkWP1srkXv7xr+8TQ29t9g/wAnWQ/sDgXH+mb/AA9bU/vFTrJnr3v3Xuve/de6iV4vR1A/5tn/AHv3ST+zb7D1sYI60KfnZIw+ZnfXHA3Eg/5Pl/HvoJ7ZUHt5tf8ApD/k6wW9y6n3F3IfKP8A5+6Kr5Ba5Jv/AEHI/wBgPY3rQ56BlBWvWGpkC08314hm54/CH3aMguPtH+Hqky0hYf0T/g63E/5LrBviDtEj8vXG5+v6094N+9ZH+uHef7X/AC9Zo+0A/wCQBYH+gP8AAOrfvcU9Sb1737r3XvfuvdFv+WjaehN/f44qYf8AWJj7OuW/+S/a/wCnH+Hoo37/AJI1x/pD1oy5Ootm88Bb/i+ZO9/p/wACD/X30ztpKWUFP99p/g652XMdb64/5rSf8e6iCq4J51fj+n9Pb3iHpjw6dTcZVp/F8ULuL19H+Byfuk459pLyRvo5f9Kf8B6U2yf43FT+Jf8Ajw63xPj2b9J9Xn+uysF/7iD3zH3n/kuX3/NeT/D10Z2sU2y3H/Ck/wCOjoZPZf0v697917r3v3XuqXP51x0/H3a3P13U5PP40x+5o9hTTnaQ/wDCh/l6iL3rr/U2g/34P8nWqWsn7a88WuP9f/D3nCznUT1hminSOuvuPz9WI54H0/J91r5V6vQjI6Ob/L+nB+T+z1F9Pmojf/H7g/T3Gfu+f+QRKR6n/B1I3tLX+uaj+iv+E9bw0H+Yh/5ZR/8AQg94BL8I6zhPHrL7t1rr3v3Xuve/de61QP5283h+QHWqi/O3K48/jlrc+8v/ALthI5c3I/8ADR/k6xS+8MK73tY/ov8A8dPVNn3Tf1AtfV/jf+nvInWeoF8OufTrj90/Fvqfe9Zr1rR1fL/Isl8u6O4dX9mrx9v9fyR+8XfvLmp2n7G/y9ZI/d0FLbdP+av+Retm/wB4sdZL9e9+691737r3TRn/APix5n8j+FZC/wD5yP79+JftH+EdVatD9h/wdaO/yqmt8g+wLtYCvqOP6f5Uw99H/bSn9R7M/L/IOsAfcgf8ja6pxoP+PHovxqgTa9ifp+Ppz7HIoDToEkNpoB1xapGn9X0KAj6f2h72CDj7evMCMnrbf/lJtq+KOGa9/wDc/kh/toh759e9Zr7iXB/4Wv8Ah6zq9oQRyHa19W/ydWh+4r6kzr3v3Xuve/de6a8z/wAWuvuL/wCRVv8A7hye9jqrcOvnWf8ACasD/h+L5xi//Lp7ot/T/j9pP6+6jiOnDwPX0Zl/Sv8ArD/eve+q9d+/de697917r3v3Xuve/de6o6/nw92dPdWfGfqXE9l9r7A66yWf+TPTFfh6Td248biK7IYzDZ2WbNZKhoalvPJT0iEPUzRoVjH6mF7e1lniQscChFf8nRXuwD2wjGW1KaedAQSacaDzPl138se2OuNl/M/4FfLrcG+9q1PxaGz93dfSdtU+exlZ1xQby3tOi7TrKnccMr0sZl1K0bkg2RuQVIDkQJhkQYYn8+k13Ii3VvOTVAKE+VSRTPTlg81gPkR/OI2H3D0buvC9idY9G/GXffWvau69lZShz+1cfvrdGX+/wODrM1i5Hp3q1hcERqzkAN9NJtVlaOz0vgkggf6v29XRo5948WHIRGUkZFaj/B1dl7Q9HXQC9w9VdcZjaG/NwZbZ2FyGZ/utn6j+IVFOXqvNHjJXSQSX4IIBBA96p1sHPSG+DwA+M3XCjgLS5NVH0sq5aZVX/YDj37rx6Nn731rrVM/m4ymPvrG6SPU9Vcf04f3mf93MV5euR8x1iJ94Go3q0IHmf8B6qsNQLgauObf4/wCvb3kNTqBKimeuPnPPqF7HVb6D+n197p59aIBx1e9/JGctuPt43uBQUFjf86oveJ33nCSdqHozf5esn/u6Yi3MfNP8A62KveLXWTHXvfuvde9+691hqJPDBNNx+1FJJz9PQhbn/be9imoA+Z60cCo613e4P5u/fOwuyt07PwXXOxK/F4TKVlDS1lXV1i1EqU05iV5VU2uR/T3l3y/7Acq7rssG43F5OryqrEACmRXHWKvMXvzzLs++zbXbbfFJHEzKGMhBwacKdBuP50vyOJseruux/wBVld/0d7Nz93Hk7/lOuP2Don/4IrmzP+6uHH/DT/m6xTfzp/khFE8n+i3rolSth97W8guFP9r3Zfu4cnM1Prrj/eR1SX7xnNqJqXa4T/zdPr9nV8XxU7nzXe/UeF39uCgoMZlq8AVVHjXd6WJigayNJz9b/X3i1zzsFnyzzBLtNk7SInAtxPWT3J++XPMOxxbpdoI3k4qpqB0ZP2D+hR1737r3XvfuvdFd+ZVTJT/HDtlUayz7My8b/wCIKL+fYl5Lr/XLa/8AnoTog5rAPK+4evgP/g60rcWXFBSr+fGT/rDyHj30wn/tmPz65uWVPpUHy/ynqbr/AKj+g/2HtrpURQjpTbEbx9hdfSL/AGN44dwf6MJrg+y7ej/uivv+aD/4OjDZVb+sO3Hj/jCf5et43rypkq9m4CeRizPjqTk/UgU6298xZ/7d/wDTN/hPXSOD+xT7B/g6Wntrp3r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rhIbI55/Sfp9fp791o4HWpr/NhnKfKXNjgWosfb+p9P595ufd5zyOo/pN/h6w39+6jm1SP4E6rNFSf6r/re54p1COny66NQdI/Rxc359+0563qFer9v5LTeRN0sLECGS/8AvI94g/eQr+9LcfZ1lf8Ad9A/ckv2n/D1sFe8aesh+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de64v+h/+Ct/vXv3XutNX+byw/2ciq5sf7o0/B+n+6/p/X3m5934/wDMPmB/3+3+XrDf33H/ACOren++G/wr1WVrA/P1H+Puaanz6iIr12JF1D8njTf8f096rTj1YDI62T/5ELautuwzwT/fGe//AFl94h/eQP8AyKbMf8IH+TrKf7vdRylN/wA15P8Ajx62EfeO/U+de9+691737r3SH7L/AOPB3dxf/cFX8f8ATk+37X/cyH/Tr/h6Yuf9xpP9K3+DrSH7Ym/4ynvwE2/3NT/UfkSN76WcrZ5asj/wsdc7eaQBzTfemv8Az9IE1AP9Prz/AFv/AE9nuk9EZI4Drg09io/IYf70bj3ulKk9aJrQdbi38vRtXxU6yI+hoak/4cOo985/c819wtyP9IddA/b0f8gzb6/77HR2/YG6GfXvfuvde9+691//0d/j37r3Xvfuvde9+691737r3UHJ/wDFtyH/AFA1f/WhvfuvdEs+AYt0xnOSf+Mm745Jv/y8B70OrN0eH3vqvWsH/O7kWPe2zzcr/lkV7fT6L7yr+7eaG9H9H/P1jP8AeDFbe0P/AAxeqMfuh/X/AGHP+395P6usbtGPLr33X9r+nFrC/wDsPe/EHWtDA16uZ/kjTmX5G7xB4I2VUH/e/eOH3lGryzaD/h46n77voI32+rn9Jf8AD1tY+8Q+srOve/de697917rg/wCn/Yj/AHv23N/ZN9h695jrQ4+fr2+VvaYJ4/vNkR/ydxe3vPj2qP8AyC7Uf0F6wk90f+VzuD/SP+DonOsi49P+Fz9P9a3uQq1x0AaHj1gqpG+3n5UWhkuR9T6Cb+7x08RT8x03LmJvsP8Ag627/wCSgS3x+ybEfWrpv+hP6e8I/eo/8i9vsP8Ah6zM9pK/1Vjr8v8AB1dR7iLqUeve/de697917oHPkAP+MN9lf1Gx908/n/izy/T2r27/AJKdp/zXi/48OmLv/cOb/mm//HT1877yf5VmCGPGfzlweD/xdJf6e+lZI0Rj/hcf/HR1ztQAzT1r/bz/APVxuspfgEm5P0H5/wBb3Sv8unKGlT59eikb77DWFx/H8Jz+R/uSj96P9lL/AM03/wCOnr1D49vXP60P/VxevoQfGAH/AEIdenjna+C+n/aqi983d3Nd2uj/AMNf/jx66Ebdiwh/0i/4B0P/ALL+lvXvfuvde9+691Tp/OauPjdUm/H7n9f6t7lf2aNOdIPt6jT3WFeUbgf0W/wdafVI4FPT3Y+mnhJ/pYxj3nLKxLn7T/h6wrtqCFP9KP8AB1nMo55PP0J+oJ590qenTSuOjW/BdrfLTpO5JvvPH3H9SSPYF9zP+VH3D/mkehr7aj/kcWFP4z/g632k/SP9j/vfvn3D/Yr9g6zmPHrl7d611737r3XR+h/1vfuvdaon89AlO9dqEf8APKUH1/5B95Wfd2NNpuf+ajdYye/eb61/0nVG/kYcH8/T/er295H1r1j/AIHHPXQkB4JPP+8H34setaRTq9T+RmQe09zC/Gmrt/yS9veNn3hzWG1r6DrIb2EXSLg/0m/w9bWPvFfrJbr3v3Xuve/de6i1v/ASf/lmf9791f4D9h/wde8+tB/54E/7Ob33Zv8AmJF4J4/XL9D7z/8AbJh/re7WB/Af8nWDXuUv/MRNyb5Rf8/dFQ1kC5P1/wBv/T2OK1OOgaFIHp1iqZR9vMGuSIZeR+f2z/X3aOocfaP8PVJwDC/+lP8Ag63Hf5LJ1fD7Z7CwBeuAH54kj94O+9QP+uHeE+i/5esz/aGn9QLCn8A/wDq4H3FXUl9e9+691737r3RbPlvcdA9gHjjEzHk2/wB0v7OuXMb9an+mP8I6Kd8/5JNx/pD1olZOa2a3ACxt/HcnqvzYfcH6e+lNu/8AicA/4Wn+DrnlcITf3JP+/ZP+PdRfuWHHHPAv9eP6+3fENOm9A8+puKqtWYxIv9MhSGw/r90nti7J+jl/0p/wHp61Cm7iA/jX/jw633fjvz0h1af67JwPA+n/AAEHvmdvH/Jbvf8AmvJ/h66J7Z/yTrf/AJpJ/wAdHQ0ey/pd1737r3XvfuvdUp/zuH8fx52qebndUnH/ACDH7mf2H/5XST/mmP8AL1EnvT/yp/8AzcH+TrU9SoJjU3N7Wt+B/j7zbY93WHKjtAPXYlP9ef6/4fn37VjrVBXo6H8viUP8o9nr9AZqMcD8/cHn3Gnu6R/UmX7T/g6kb2lFOclH9Ff8J63mIP8AMQ/n9qPn/kAe8Bl+EdZunj1l92611737r3XvfuvdamH88OUp8g+tFP8AzzVd/j+W95efdup/Vzca/wC/R/k6xW+8L/yWtqp5rJ/x3qmH7g8f8V/3r3kNrHUDaW8uu/uCfyAPx/Ue/FjTqwGakdX4fyIZNe6e5QbHTWY4fX+skfPvGD7ymTtX2N/gPWRn3dv9xtz/AOa3+Retnz3i51kp1737r3XvfuvdNOe/4seZ/wC1VkP/AHEf378S/aP8PVW4H7D1os/LWqKfIrsRB/yvzfX9P/At7++i3tq9OSLOv+rA6wK9xlH9dbo0/CP+PHovH3VzyQP6n6nj2N/EpjoF6AeuLVN+SRq1R2/p+se9iTzHoeqOlf5dbfP8ohg/xMwrA3/38GSufx/ml+nvAP3oz7gzn/ha/wCHrOT2kqORrUfNurTPcW9SV1737r3XvfuvdNea/wCLXX/9QVb/AO4cnvY6q3Dr51f/AAms/wC38Pzj/wC1T3R/72svuo4jpw8D19Gdf0r/AKw/3r3vqvXfv3Xuve/de697917r3v3XuilfJr4HfDj5mS7an+U/xy6t71qNnLNHtep7B25BmanBxVEvmnhoKglXSN2uzR6tJubjk3trYLorjj1Qxxl/EI7qUr8vTpSZv4efFvcnSVB8bs90J1hlOhsXUY2sxvVFXtXHSbLoKzEVQrcZWUmH0hI5oJRrjlWzg35sSPe/Ek1a659eqfTw+GYtI0nypjpUdJ/HPon434XNbd6H6o2R1PhNyZl9w5/HbJwdJhafMZt4Fpjk8j9sAZZfGqorOTYcC3PvzyPIaua9bighgXRCoUegFOhp906d6QvZ9/8ARvv231/ufuO3+v8AwmW3v3WxxHQDfB6/+yzdc3tf7fK3t/2t5vfuvHo2fvR4da61Of5vkxi7+xR4F2q/r9Po/wBPeZ/3cgTy9d19V/ydYkfeCr++bKnq3/HT1U8ai1iB7yKDL59QEVbiOu/O1rfm3qP/ABT3rWvHrenyPV9n8jmQybj7jX+yuPx9v+S4ifeJf3m2q+1/6Zv8B6ye+7qoW33If0l/wDrY694udZK9e9+691737r3TVmyRiMkQbH7Cu/3ikcj23JwH2r/hHXutILvqoU9x7+Lksf7xZb6/9RZ99L+Tc8r2dP8Afaf4Oud/Og/5FV5Qf6I/+HoJPOo4NiT+R7E1OgxSo+zrBUTRmJh9P0cf8hjk+7IDq6pKP0z+X+Hrbh/llyGT45Yu5JC1Shb/ANPEePfPX3bP/I2n+w/4es+vbLHKNvX/AFY6sT9xt1IHXvfuvde9+690VH5rkj429rMP7Oz8sf8ArGB7EvJX/K57X/z0J0Q81f8AKsbh/wA0H/wdaUOMqiKCl9RP7ZJ/w9Zvz76aTr+s329c3rOn0iH5f5T1ONWPrcn6C4/x9s6T0pqtK9KPY9UD2D1+vPO8MR9T+fN7Lt7U/uG+P/CH/wAHRhsjD+sW3Af8pEf+Xree6yv/AHG27f8AONpf/cdPfMKf+2f/AEzf4T10hh/sU/0o/wAHS99tdO9e9+691737r3RKfmr8q6j4r7Ep920mHpc1PPOkX21S7qArEgt6Ofx7kf235Ii553c7bLIYwBWo6j/3C50bknaDuixiSnkajqr5f50u4SL/AOj3EfQc+WptyL+5wP3cbAH/AHMf9g6hn/gh5sUs1z8z13/w9LuC4B69w/5v+9U/7D37/gcbD/lMf9g63/wQ0/lZr+09GP8Aip/M2ynyG7jwPWFZtHHYmLMJVMauCSZpEMEWtQofjk+wdz57L2vKHLc2+x3DSeFTBApk9C3kb3ifm/mBNka3EesMagny+3q4f3jz1PHXvfuvde9+691xf9D/APBT/vXv3Xj1qQfzbJQvypza35+xx/8AsPR9febv3dz/AMglR/Sb/D1hx79qTzUh/oJ1WAahT+WB/I/1vc+fLqEdJ49d+dT+TYf7b6e9VFK9VIPWwR/JNk8sW7COLQSC1zzyR7w++8jjd7cfZ1lf933Oxy/n/h62F/eNHWRHXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdcJP82//BG/3r37rx4daZP83/UPmXVC3oG0Kbn+1/uv3mz7AGnt83/Nc/5esO/fUV56tz/y7t/hXqsUONQuTp44/r/Q+5oLeXURUFeuw6i5ufrf88f63vRavWwM9bLf8hltfWnYhA4G8JySeD/u23vEX7x2eabP/mgP8nWUv3fK/wBUZq/7/k/4+ethf3jz1PfXvfuvde9+690huzP+PA3f/wBqHIf9aT7ftf8AcuH/AE6/4embn/ceT/Sn/B1o2duVR/0r79B+i5ufn8n9xre+l/KQryvYn/hY65282f8AK1X3+nH+XoPfu1tf6H8i/sQ6eiCor1xNTqK82Gof6/0PvRFFI+XWiOB+fW5f/LsYv8TusG/rQVP1+v8AnFtx75w+5n/K/wC5f6Zf8B66De3/APypu3j/AIWOjw+wP0Muve/de697917r/9Lf49+691737r3Xvfuvde9+691Byf8Axbch/wBQNX/1ob37r3RKvgCLdL5wWI/4ydvjg/8AawHvQ6s3R4/e+q9auf8APJmEW9tm35vWR8f7BPr7yl+7o2l73/S/5+sa/f8AANva1/34vVDv3Itpuwuf98PeTur59Y56cY6993b8t/T/AA4961+fXjH1dT/I4mEvyR3mAef7kVX9fqL3N/eO33jzXlq0r/v8dTx7AADfL4j/AH0v+HrbFH0H5/x94j9ZUde9+691737r3XB/oP8AXF/9v7pJ/Zt9h615jrQt/mAyqvyv7WX6j+8+Rvf8c+89Paw/8gy1p/CvWFPucAecbmnmT/g6Jt5Qb/W44A5sf9c+5B1HoA6PLz6w1Mg8E5/5syA3/B0m1vdkPetfUdNyp+k32H/B1t8/yTDf4+ZKxJH3VN/t9HvCf3pNeb3P2/4esyvabHKsY+z/AAdXV+4j6k/r3v3Xuve/de6Bv5A/8ya7L+v/AB426v8A3TS+1e3f8lS0/wCa8X/Hh0xd/wC4c3/NN/8Ajp6+dxqArcuQOP49nbn6m/8AFJffSc/BHX/faf8AHR1zvQjxp6f7+m/6uN1z1fiy6vqLEWHPutRWvT1fLr0cl67DfUWz+D/2P+5KP3s1EUo8/Df/AI6eqZM8Hl+tD/x8dfQp+MB/4wd13/4a2C/3nFRe+b27f8lW6/5qyf8AHj10H27/AHAh/wBIv+AdD/7L+lvXvfuvde9+691Tf/OfOn421RuRxJb/AJO/PuVfZokc6wfb1Gvur/yqFx/pW/wdaeFNKDS0wJ58EX+x9A/r7zikP6jfaesL4BWBPmo/wdZvKOSTc/S39P8AEe66ger6CMdGy+Cjhvlv0iLkk70oOf8AYjj2Bvcxv+QRf0/30ehp7bJ/yOrCue8/4Ot+RL6ef6n/AHv3z/i/sl+wdZx+fXP2517r3v3Xuuj9D/rH34cevdanf89Z2Xvfaaji+06D/on3lT93k02m5J/343WM/vytb21/0n+XqjAzNbljYG178e8iwa9QCUp59cfuAQR/jb82/wBv72CetEDq9v8AkXMD2puX/Fav/oV/eN/3hTWC2/L/ACdZCewwotwP6Tf4etrv3i11kn1737r3XvfuvdRK42pJz/tB/wB790k/s2+w/wCDrw4jrQY+eTAfM/vsEm/95F4+g/XL7z99sjT292v/AEh/ydYPe5Q/5iLuVcYi/wCfuioauAF+n9fz/X2N9VcdA7T1gqHIp57gcQzWt9bmM+7o3ePtHTM4PhOafhP+Drcj/kqm/wAPdnm5Prr/AK/8tI/eD/vR/wBPCu/sX/L1md7R/wDKhWH/ADTH+AdXC+4r6krr3v3Xuve/de6LV8u/+yf+wubf7iJ/+tL/AE9nHL3/ACXLX/Tj/COire6/um4/0h60LsrUL/G9wjUf+L9lL2/6iT76RW8n+Jwf800/wdc+blf8euf+a0n+HqF9yf8AVf8ABfbviDpvQepuKqQMziRe5bIUY4P/AE1J9fbN0/8AiktD+E/4D05boPq4a/xr/wAeHW/78deejeq//DH2/wD+4Y981d4Nd6vT/wAOk/w9dDdsxt1v/wA00/46Ohp9oOl3Xvfuvde9+691SZ/PANvjttU3t/v6pLEfUeiPn3MnsSac5yH/AIWP8vUSe9ALcn0/4YP8nWpZHKAq+s20fX/H3myzd3WHiqAB1k89hYNc/U/635911eXVio4Do6v8vOUN8ptmgcjzUQI/H/Ag8+4193G/5BUv2n/B1IvtOB/XEN/RX/Cet6iDiGIf0ij/AOhB7wNX4R1mucnrL7317r3v3Xuve/de61If55jhfkP1iNZB/uzXG/5PLe8uPu5NTl3cf+ao6xZ+8Gtd52oj0k/471S4Jb3N7/S3P0/r9feQNeGeoKA66E/+sP8AW921efWwPI9X9/yF317p7n+nFZjv8frJEPeMv3kTU7V9jf5esifu9KBbbnT/AH9/kXraH94vdZI9e9+691737r3TRn/+LFmv+1Vkf/cN/fvNftH+HrR4H7D1oe/LupZfkd2KCLoMhOLfn/gW9rW99DvbhwOSbT/V5DrA73FQ/wBdLk/0f+fj0XP7oNwf8LsL/wCvb2NdfQO0Dh1wkqhp/A9SDkn/AFY92V8/t6064/Z1uJfyfWDfETBkG/8Av4snc/8ATpfeBXvKdXuBcH/ha/4T1m77TinI9r9rf5OrVvcX9SP1737r3XvfuvdNmZ/4tdf/ANQVb/sP8jk97HVW4dfOr/4TV/8Ab+L5xn/q090c/S3+/wBpPdR5dOHz6+jMv6R/rD/eve+q9d+/de697917r3v3Xuve/de697917r3v3Xuve/de697917pC9of8y337+P8Afn7i5/8AITL791scegG+D5v8ZuuT+DT5X/3bze/dePRs/futdak384qUR/IHE83uarj/AFPDni/vMr7uzEcvXYHqv+EdYl+/1DvNn55P/HT1UoZxYXci35N7H/Y+8g9WKdQPQAZ699x9f3SeObn6+9avTrxFag9X+fyKpRJuHuX1E2x+Pvcc/ri94p/eWOqTa6/xN/l6ya+7wNNvuX+mX/AOtkj3i/1kl1737r3XvfuvdNWc/wCLRk/+1fXf+4j+6ScB9o/wjr3WjD3/AFYHc3YALHjceWuOf+Vw8++mHI4rytZ/801/wdc8OeARzXeU/jb/AA9BD96ACvNvyTf/AIn2K9Nc9BapXrBU1q+EnngoPrYH9wfX3tFz+3puUkr+z/D1t9/ywHD/ABtxTAk3qlPP/LI/T3zt92f+V2nr8/8AD1n77af8qlb/AOry6sd9xx0P+ve/de697917op3zcYL8Z+3D+RsvL2B+n+bX6+xLyUf+RntX/PQnRDzUK8sbgP8AhD/4OtHzFVp+wpVOkHxsb8/6s3HvpvcMvjMD6/5Oub23ofo4/sP+E9TvvjyRa17WueOfx7Z1rw6WaM16VOxa4t2L10FIN944b6n6AzWt7Lt6IOw348/Ak/wdL9lT/kRbb6fUJ/gPW+B1jf8AuNt0k3JxtIT/AIf5OnHvl9MKTP8A6Zv8J66Pw/2KfYP8HS99t9O9e9+691737r3VIP8AOVqWj66wsZYlGaMlebE3fke8gfu8n/kUuPkf8nUFe/Qryka9a5a1SlVFv7IuPrfj3mgVNa9YfrTSB6de+5iufx/X/D+nv2k9arQmg6Pn/LSqgPl510Yzy0tUOObgoARb3EXvr2+296P9L/h6lf2SFef7c+qv1t+obop/qPz7wKX4R9g/wdZv9cve+vde9+691wk/Q/59J+v+t791o8OtQ3+bxP4/ldmlA5NDjr/9S/x7zW+721OSx/pm/wAPWIHvuteaU/0i9VamqvxqPB9zxqPUJaafb177pf8AVmx/23v2vPXtI62Gf5IEokg3bY6h4Zfr+PUfeIv3j31bvB+X+DrKr2AAGxyU+f8Ah62JveNfWQfXvfuvde9+6910SACT9ALn3omgqevdVX7t/m7/ABg2bu/dGyspBvFsttLM1ODyZgxSvAaylNnML6uVP1B9zpt/sBzpuW3QbnA0Xh3CB1q2aH16hXcPfnkXbd0udouJH8a0fRIApw1K+noQemH/AIeY+Kd/+A29v/PQv1/p+r2q/wCB056r8UP+9dJv+CD5BpXXJ/vB/wA3WKo/nPfFCmgmqJafewjhRpHtiFJ0qLkj1e7J93LnuRwitDU4+Lqk33hvb6CNppJJAqipOg8P2dWX9T9m7d7i6+2z2RtX7kYHdWPTI45ayPxVSwOSAJo/wePp7hffdmuuX92n2e9IMsDaWpkV+XUx7Nu1rvm2Q7rZV8KdQy1waHh0Ivso6M+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6Tu4d3bW2lDFUbo3FhtvQTsywzZnI0uOjlZRdljeqZQSP8Pay02++v2K2MLzEcQilqfsB6TXF5aWihrqRYweBZgv+HpIf6bunf+fobD/9CnDf9fvZh/VrmH/lBn/5xP8A5ukn772b/lKi/wCci/5+vf6bunf+fobD/wDQpw3/AF+9+/q1zD/ygz/84n/zde/fmzf8pUX/ADkX/P09YLsvrzc9WaDbm99rZ2uABNJic5jq+oAP0JhppGb/AHj2mutl3eyj8W8tpYl9WRlH7SOnoNz266fw7aeORvRXUn9gPS39lnS7r3v3Xuve/de64v8Aof8A4K3+9e/daPDrTB/nCvp+aFV6iLbPpjb8H/N/7z7zW9gz/wAw/b/muf8AL1h776D/AJHNv/zzt/hXqsDy241XH1N72HHuZS3UShamg695VuAxsTbSR9be9hjXreny62Yf5Cjq3WfYgBv/AL/Cf/rr7xH+8aa802f/ADRH+TrKX7vy6OUJh/w+T/j562HvePfU79e9+691737r3SF7N56/3f8A9qHIf9aD7etv9yoj/TX/AA9M3H+48n+lb/B1om9x1Fu3OwFvZRnZwbf8tG99LuUnA5Wsf+af+brnlzYv/Iqvj/SH+XoO/uuOGJP5/oP8PYgMo6IQlMfz64mp5j9V1Li1/wDWI96aXtb7OveHwPz63S/5cpv8SerGJuWx9Uf+sqi3vnF7lZ593E/0h/g66A8ggDk+wHpGOjz+wT0MOve/de697917r//T3+Pfuvde9+691737r3XvfuvdQsl/xbsh/wBQVV/1ob37r3RK/gH/AMyYznN/+Mm7454/52C/096HHqzdHh976r1qxfz15lh3vsq5IvWx8DkH0p7yg+7wTrvKfw/5+scffsD6a1J/34vVB5quCAxve9/8P6e8l9fWOpXy6x/dEAEOb3t9fz7trHVDG3V3H8iqcyfJTeove2xKo/7yefePP3ijq5btP+a46nf2EXTvd9/zSX/D1ts+8TOspOve/de697917rg/0H/Bh7rJ/Zt9nXvTrQi/mCuF+WHa/H/MTZLn/kLk+88Pa015Ntf9KvWFPudT+uNxj8R6Jj5uABfmwFvqf9f3IVR0BKE8OsVSxNPPdr2hkuP6eg+7Ie8U9R01NURMfketwX+SQQfj1kSPzV03/QvvCf3m/wCVub7D/h6zH9pj/wAhWP8A2v8Ag6uu9xL1J/Xvfuvde9+690DfyB/5k12Wf6bG3Vx/5BpfaqwqNztD/wAPi/48OmLqn0c1f99v/wAdPXzrvIRWZj6W/j+dufpp/wBykth76SlqpF/zTT/jo654RKBLcUP+jzf9XG65hiCQVW39QebfW/uury6eABHHrmkgFfhDZT/ufwnF/r/uSjHuzH9KX/mm/wDx09VIAntz/wAOh/4+OvoY/GD/AJkd11/4auB4/wDIVEffODdv+Stdf81ZP+PHroJtv/JPh/0i/wCAdD97L+l3Xvfuvde9+691TX/Ole3xpqv8BKR/ycPcq+zhP9c4Pt6jX3V/5VG4/wBK3+A9acVPKPtac83NPD9f+WY/p7zdkb9RvtPWGUIBgSn8I/wdSA/AJtdidP8Agf6+6agOndJ4no2/wRJPy46RBI/4/THng/4j2B/ckj+pF/8A80z0NPbgMOeLD/Tn/B1v4L+kf7H/AHv3gJF/ZL9g6ze8+uXtzr3XvfuvddH6H/WP+9e9jj148OtTL+e/Jo732kfpfadBx/hxx7ym+72f91VyP+GN1jR77/7m2v8ApP8AL1RKZyFJ/F7/AF/3k+8igc9QGc8OveUkD1AAnVfjg/i3v2sda0ngOr4/5E7hu1dyi9yFq+f6+l/eOP3gyTBbfYP8nWQXsOCq3AP8Tf4etsX3i51kl1737r3XvfuvdRa3/gJP/wAE/P8Ar+6Sf2bfYf8AB1scR1oFfPd7fNHv4ksP9/Ko/r9ZJfp7z89tD/zD7bP9If8AJ1g77lf9PF3H7Iv+fuin6iLjUbcf61jz9fY18+gfnh1hme8NRe5/YlHP0/zRv7snxqPmP8PTc39g5P8ACf8AAety3+ShYfDjZwvf9zIWP/TyO/vCL3n/AOnhXn+1/wAvWZftGf8AkBWH/NMf4B1cR7izqSuve/de697917os/wAviB8fOxT/AEw05H+v4X9nHL//ACW7b/Tj/D0Vb1nargf0D1oJZaoAzu4eB/xf8qPz/wArJ5Pvo1bPWzg/5pp/g65/3S/7sLkHH6z/AOHqD91caTe4+n++Pt3VTPTejy6n4eqH8axAH/Owo73/AKfdJf2nu3P0cp/on/Aen7da3UQ8wy/8eHX0Gfjlz0X1QR/zwu3v/cIe+b+7/wDJZvP+a0n+HroLtv8AyT4P+aaf8dHQ1+0HS3r3v3Xuve/de6pC/nlsU+OW02Bt/v7Jb/62iP6+5i9jSRzjJ/zTH+XqJveXPKH/ADcHWo2k50J9LBQfrc/7b3mizkMRXrEJU7R69cvuWB4C3/330PvWokV62VFcdHb/AJdc2r5WbMW/JlouB9P+BB9xv7tMTyXIP6R/wdSJ7VAf1wX/AEq/4T1vfwf5mL/COPn/AJAHvBRfhHWaPn1l97691737r3Xvfuvdaiv89aQJ8ierwfT/AL9eutb8ct9PeWn3dCRy7uP/ADVH+TrFz7wK13navsk/46eqT/Mv4LcfkX9R/wAfeQFfXqC9IIx1w+4YfS/q+o/pb3vVXj1sDrYI/kEys+6u67gECsx3B+g/di+nvGf7xxBbav8ASt/l6yH+72CLXc/Txj/gXraV94xdZH9e9+691737r3TTn+MHmf8AtVZH/wBxH9+8x9o/wjqrcD9h60H/AJh1Oj5K9kLq4GRqOfr/AMpbe+g3tw1OS7Qf6uHWDHuIv/I0uP8AS/8APx6Lb9zz+o/61/Y0L9A4R4r1jeqBAsxvqQXv/tY497D0bqjJUdblv8nJg3w+wR/7OTK/9al94Je8WefZz/QX/D1mv7U/8qTa/a3+Tq1/3GPUjde9+691737r3TZmf+LXkP8AqBrb/wDnHJ72Oqtw6+dX/wAJq/8At/F84z9R/CO6Lj/ydpD7qPLpw8D19GZf0rb6WH+9e99V679+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SE7Q/5ltv3/wAM/cf/ALqZffjw62OPQC/Bz/smXrj/AKh8r/7t5/euvHo2vvfWutRP+ctKY/kJiLWFzV355+j/AE95ifd6amwXQPqP8nWJ/v2hO8WdP4j/AMdPVQ5qRwCAbXsP97t7n8M3UGFBwPXTVDWsNI455+v9Pe9WevacUPWwV/IclMm5e6r/AEGNx1v+pkXvFf7yZBba/wDTN/l6yV+73/Ybl/pl/wAC9bLfvGPrI7r3v3Xuve/de6as5/xaMl/hj67/ANw5PdHyB9o/wjr3Wh58hawL3b2Gb8LuTLi34v8AeH6++lPJEtOVLQf0F/wdc+ud4x/Wu7b+m3+HoG/vuLFvrzf6exT4p6C/h5oeHWGprlEL3Ygegf8AJ459uRSUcH7f8HTMqDwiBxx/hHW45/K0cSfGbEMDf/KU/wCtR/p755e651c63Br6/wCHrPf23FOVLcfLqyb3HPQ8697917r3v3XuikfOPj4w9wP9NGyMwb3+noXn2I+S/wDlc9r/AOehOiPmj/lWb8f8Jf8AwdaLeJrB/D6UklgY25J+v7h499Lrhj4zD5/5OuctgoNmh+X+U9T/ALlmFxYAkD6/7z7YLjpYI+lVsKo/4yP1utxzvXC35PP7x59oN4b/AHR3w/4Q/S7ZgRzDtuP+JKf4D1vy9Ym+xtuH/q20n/uOnvmRN/bP/pm/wnroxD/ZL9g/wdL323051737r3XvfuvdUV/zqZhB1xhWLafVFb+oF3vf3kD93cgc2sD6H/J1Bnv0K8otTrWxXILoS0rWAHN/6j3mrqTj1h8I2pSnXYyC83k+tr/0/wBf37UvDr2g+fR/f5YtWsvzG61QOSWnqrC/B9IPuHPfg/8AMOrsA+n+HqWfZIU59gJ8lf8AydbkKfpH++/PvAtfhH2D/B1m31z97691737r3XB/0P8A8FP+9e/daPDrT2/nBzlPlnm1LWtQ43SP8fH/AIe80fu/GnJgp/E3+HrET31FeaIz/QXqqz7liD6lJH15/wAbe511nqFtBJp1390Pr6dP4F/yPr7rr62Ux1sU/wAjOQSUu7iLXEMv+2u1veJf3ijq3eD8v8HWUfsH/wAkOQ/b/hPWxr7xv6yB697917r3v3XumnOTy02LrJoTZ0hcgj8emxPuknwHrw4jrRn7ryKt3d28zWLNvrKkseTqMnJPvpvympblLbD/AMu6dc5ubKDnLdsf8SD/AMdHQbmujJtxe1+fZ9oPRHWuCOoVfWxNQVg9P/AeX88/p9uwoRMp+Y6YuyptpBT8J63HP5d1bLVfF7rSNmvHDt+mEY/Cgu1wPfOT3O/5Xzch/wANPXQn29/5Uzb/APmkv+Do8/sB9DPr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuqMv502S0bO60x5eSNJazJSM0cskRJ/zdrxlf9T7yS+7dJXeL8UHaqjgD8/PrHr7waa9itKkgazwJHl8utdZFhKL/AJTUm6gf8Dav8D/g/vMMu9eA/wB5H+brEb6eLTxb/e2/z9ctMX/Hap9PPNbV8/0/t+9a24Y/3lf83Xhbx/F3f72/+fqyL+VlkxjvkjH43mb7paWFkkqKiQWsv0EjEe4T9+2J5PWtME+QH+AdTR7DQiPmuYrXOnizHy+ZPW2l7wWBqK9Zrde97691737r3XCT9D/8Eb/evfuvdaXH84x9HzSqRx/x59Lzfm9ovx7zS9g8cgHP+jn/AC9Yge+n/K8W/wDzzt/hXqrsyXHAUj6fW3+w9zLXqJB8uuvJdhcDiwuPx/re/V62MnrZr/kGG/WHY5A4/vjPz/1N+vvEr7xf/K0WZ/4T/m6yk9gMcozD/h8n/H262JPePvU69e9+691737r3SF7N/wCZfbw/7UGQ/wCtB9vW3+5UX+nX/D0zcf2D/wClP+DrQu7nqtPb3Yik2tnpxe/58j++knKjU5Xsf+afXPzmsE803wp+Mf5eg2+6AJGokk/1/wCI9nwfoh0U48OujUEOhLenWOAf8PfjJ2N9nXincPt63aP5brF/iH1Qx5vjqrn/AKer752e5Gee9xP9If4Os+OQxTlCwH/Cx0e32Cuhf1737r3Xvfuvdf/U3+PfuvdEZ+ePyq3j8Z+tMHR9MbLwfaPyT7X3BS7N6K6v3DlKjCYbdW4DVwHMT5bLU9jDT0VJK0zetS76EBALFX4IfFOeA49I7y6FsoAyzYA+fQI9T/K/5IdafKzA/Fv5kbX2LjaHuLbcOY+O/Z2yci1VJu/deOxP8Z7A2LuHEJZKYYsloqOpCAyqqltayK4ckiR0MsOAOI6TQXM0NwLS7yXqVPr5kfl1ap7SdGvRXe5PlP171Hlara25cPvmur5aF2Eu3tsVWVows9OSgNVEQB/tRtx9ffuvUPRF/iX8vesusescpg8zhOwK6qqd97oy0cmE2nWZOjWmyVV5oY5KuIhRKoFnj+o964dWOejQH+YH06AT/djtc8X42JX/ANP+De91HWqHz6oz/mkf3j+Xm4tsZTpfZG+KqmxNRHLXtmdt11C9lChjGig/045Puavabn3l7kw3B3xnXxRRdIrnqH/dPkbf+b4YY9kCVjdWJckYHHqqT/ZOfkkFP/GOs6f/ACFZDgf8k+5l/wBff2+/37L/ALx1Ef8ArJ898dMNf9MeuI+HnyT4t1znAb6b/wALyH1/wGn37/X49vf9+y/7x17/AFkuezjTD/vR6s0/lcYfeXxH7k3RvruXY286TBZPa8+Io3w+3K+vnatc8JJCQPT/AIg+4o92/cblnnTZ4LHY3dnjkDHUKY+3qUPaz2+5i5Q3K5ud5CBJUAUoSTUevV+v/DgfTnH+/Y7WuVBt/cSv/P45b3A9R1Nmk9R6r+Yf0hj4TUZHCdmY+mW96is2XWQQ3H41u4F/eq9epTrLF/MH6YqIYqin252nPBOokhmi2NXPFNGfpJG4axB/qPfq9bKkdeP8wTpw2B2x2v8ArANtiV/HP1/V701CpA8x1rT69asfy16E7i7n76332Hsbr7dE+2dw5qrrcbLX4WvgqjBUH9syRBbA/wBR7yi5I93OTOX+XYds3KSQSooBAWox1jdzr7V827/zDLuW3rH4TkkFiQei3n4afI9SAvXWdDDi/wDCshz/AI/p9iz/AF9fb/8A37L/ALx0Fx7J89Dyh/3o9Y5/hn8kWilReus5qljdecTkLFmW1+F93T329vVYEyy0B/h6bk9kefHQqqw1IP4j1sI/y1e3sd8VeoavZfa+zOw6XcE1RDIqYraldXU5SNdL3b0kH8/X3jb7i8ybXzPv7bltJLRGuWwesiOQ+X9x5d2JNv3MASrSunIwKdWRD+YH04QD/djtcX/rsSv/AOj/AGAa9DUKT1Dn/mK9E0tXTY+qxXZFLkKxDJR0FRs2rirKyNfrJTQM93A/qPe+vEU6m/8ADgXTv/PMdrAcc/3Er/z9ONXvVR17Seg97T+cHVm8Ovd6bXxO1O1JMrntr5zEY9JNk10MRrMjjnpqcSyEtZS7C5t7ftZEhvIJ3+GORHP2KwJ/l01cRtJbyRrxZWA+0ig61D/9kx+SBqMjM3XOb0VOWylZGFxWQ9UFZXPUU5a6/XSwv/j7zIPvt7eaEUSy4RVPb5qoB/mOsRo/ZHn1WlZxB3yyuO4/C7llr86HPz66/wBkz+SLWI66zgPP/LqyH/Er71/r7e31P7WX/eOnD7J89V4Q0/0x67T4Z/JJanHTN1zmylHlsZWyn+FZAkQUtYk8xUafrpU29+Pvt7e6HUyy1ZGA7PMqQP5nrR9kufDJE4EICSxue5vhVwWp86DHlXrbe6W+avWGw+s9m7Tzm1O04svhsDi8dkFh2VXTwiqo6BKebxyArcalNrj3h5fzx3N/PcRfDI7sPsLEjrLOzgkgtI4ZPiVQD9oFOhU/4cC6evb+6/a3+w2LXn/or2kr0o0nqHD/ADFOi6mtqsbTYjsqoydCiyV+Og2ZVyVtFG/6JKqnV7op/BPv1evUPl1MP8wPpwAn+7Ha3H4/uJX/APR3v1R16h6rs/mPd1Yr5R9KT7G6r2Z2LVbklaTxpldp11FS2NwoLcm/N/Y69vOYts5Z5ji3PdiVhTiVyegdzzsN/wAwbBNt22gGVwQNWBUjz613af4bfJOOngV+t83rSJEe2LyFrquk29P9feSz++3t4zswllyf4Osc4vZHnxY1RhCSAB8R8h1m/wBk3+SFrnrjOKPrxisgfqb/AOp90Pvp7ff79l/3jpweynPfpD/vR6Hv4s/HnunqL5BdZ9k706+3NBtbae46TKZeSiwtfNVJSQkF2hiIszf4ew1zl7wcl75y1dbVt8khmmQqoK0FT0I+UfajnDZOZrXdr8ReDExLaSa8OtrNf5gXTlv+PZ7X+pt/vxK+5F+D+r3iqmEAPkOsl9OTTrqT+YL0zFG80u2+04oY11SSybGrkjjUfVnYvwP8T7tUcOtUPUai/mI9HZKFp8dhezK+FJDE81HsusniWQfVDIj2v/h79Xr1MVHUn/hwPpwg22x2v+R/x4lf9R9edXvdQM9eoeqGP5pON3Z8uez9vbu6a2NvWsxONwFJj6x8ztyuoZfuoSNSrEAfTx9b+5v9pvcDlvk2ymt97Z0Z3JGkVweob9z+ROYObbmCXZwhWNaHWSOquD8NvkgQVPXGbub8fwrIW/p9dPuXv9fb2+85Zf8AeeoqPsnz0c6Yf96PXv8AZNfkeq2PXOc+v/OqyH/Rvv3+vr7ef78l/wB561/rJ89+kP8AvR6tO/ldUW6viTvjN5/uPY29qOgrlqBSthtuV9a95QwTXGwH+qF+R7h/3Y5/5c5yihXY2dvDpXUKcOpX9ruRt/5Q8Ub0EBcsQUJODw6vi/4cC6dtf+7Ha3/oCV//AEd7hGo6mPSeold/MT6LxVOKvLYjsnF0rSpTpU1+zKymgeok/wA3AssjgF2/C/n34GvXqU49Sh/ME6aZVddtdrMrAFWXYleQwPIKkP8AT3vr2k06xVPz/wCnHieM7W7XYOLWGxa4E8j8lj/vXurZUgeYPXgPXrVO+Vfx47r7h+SPa/Z+yuvtyzbP3hmRXYOStwtfBWSU4d2JqIwtlPqHHvKvkz3h5I2HlOy2XcJJBPApDgLUVxwPWM/OPtRzjvnN15vdgIvp5xGEqTqquqtfKhqKdAKPhp8kPp/o7zoA+o/hWQvz9P7PsR/6+vt8BiWX/eeg8fZTnutNMNP9MeuMnwz+SUkEqL11nNbxyIrHF5AcspAuNPuy++3t6GBMstAf4Om5fZLntomULDUggdx4kU62Kv5bfduI+LHx1271l2ts7sam3djHqjVx4raVdW0QWZlZdEnB50/1PvGn3G5i23mjm243raSTBLTSWwcV8usiuQNg3HlvlS12fdAvjwoFbTXTUAVp50+3qwf/AIcE6d1af7r9r/8AoC1/9P8Ag3sDV6GdD1Dl/mLdFU9XS4+pxPZNNkK8suPoajZtZFV17J+paOBn1SEfnT7316h8+pv/AA4H07cg7X7XUrwb7Er7g/0tq96r1vT0DXyA+ZXW/ZHU+8Nl7c2n2hLms3jZqXHrUbLrqeBp3iZV8shLWHP9PZhtV1DZ7jDdT/AjAmnpXpDuVtNc2MsEIq7KQPn1qh13w/8AkjUZTM1kXXec8Fblq+tpr4rIamgqZi8Wsafrb3l7D77e3yQRRtLLVEVT2eYFD1ijN7J89yXM82mGkkjOvceBNRX59YB8OvkkBqPXWdJ/H+4rIW4/5B97/wBff2+/37L/ALz1v/WT56/hh/3o9Scd8P8A5G0+Sx1XN15nhDT1lPNMRichqEcU6yOQNPPAPuk3vpyBJbvEsstWBA7fUHq0fsrz0lxHIyw0VgT3HyIP+TrbX6j+bfV+x+sth7RzO1e00y23drYfE5JYdk100SVtFSiGdY5ARqGocG3vD/cJ47ncbi5i+CSRmH2E1HWWFnA8FpFDJ8SIoP2gUPQkj+YF06f07X7XJP0H9xK/kf1/V7SV6U6T1Dp/5inRVXPLS0eJ7Iq6uAFqilptm1ctRAF+pljV7rb/AB9+Jp1qnl1M/wCHA+nLXO2O1v6/8eJXnj+v6vfq9e0nqtD+Z32xR/LPpvB7H6g2b2FV7ix+ebJTx5fadfQUwpWVBqWT1Xa6njj3IXtnzNtXKfMLbnu5ZYmTTVcmv2dAP3E5c3PmfYP3dtQUy6we6oFB1r+p8Nvkmqqv+jnOXA03OKyH6v6/p95HN77e3p4Sy/7x1AC+yXPaj4Yf96PXL/ZNvkmBpPXWc1Wtf+FV/wBPz/Z96/19vb0/6LL/ALx1v/WS57HlD/vR6M18Peju4ek+99tdhb76/wB0QbZxUlMa2WhwlfPVgRSl3CRleePp7BvPnu1ybzFy4+17ZJIZWNQCtB0LuR/a7mzl7mAbnuQj8KgHaSTUEk9bSUf8wHpxUVf7sdr3VVWx2JXfhQDzq94wg4FesjdPXpf5g3TMEUtRPtztKCnp42mqKibY9dHDTwoLvLNIWsqj8k+91HWqH/VXrBR/zEOj8jTx1mNwvZmRopgWgraHZdZU0syg2JjmjexHv1evUJ4dST/MD6cFv9+x2vz/ANmJX/8AR3vdR16h6oC/mj4Hefy47f2NvbpjYu9KzA7fwdTj8o+b29XUdSKqUHQIY0FrXP159zz7Re43LPJO0XllvzurzuGTSKig9eoR91/b7mTnLcbC62MR6LfX4mskHuUgBafPjXy6rSPw7+SOkEddZ3/EnFZD/W/1PuVR77e3vnLL/vPUZ/6yXPdO0Qj/AGx68Ph38kT/AM07zvP/AFashx/yb79/r7e3o/0WX/eOvf6yXPfHTD/vR6tp/lSSbg+Hma7Or+7dj74pIt0T0kmF/ge266tZhC6M5qFccW0n6Hn3DXu/z7y7zubH9ws7fTgh9Ypxrw6ln2l5G3/kyC8j33RWeUumip7SAM188Hq8EfzA+nCob+7Ha3Iv/wAeJX8f8ne4VqOphoeodd/MV6KxdP8AeZbE9lYqj1LGavIbMrKanWRzpjjMsjgXY8Afn3vr1Opf/DgnThCMu2O1mSRRJG42LX6XjYakkU6uVYcg/n3qo69Q9Qsp8++nqvHV9Gm1+1jJV0VXTxg7FrlBeaBo1BYsbcn6297FKg/MdaKkqfs61TfkX8d+8+zO6N5772n17uSbbmcrJZse9Xhq+KoaN6hpF8kQXjgj3lbyj7yck7Jy5Btd9JIJo+NFxw9esZOa/aTnHeuY5d0shF4Lig1E1rUny6Bb/ZPvkqdRHXObvYWJxWQ/3rT7ER99/b6lPFl/3noP/wCsjz15iH/ej10/w++Strf6OM3f0G38KyH0DXPOn37/AF+Pb6tfFl/3jrTeyHPTcBDj+ketjn+Xb33g/jJ8dsN1n2ps/sel3ZS5iurZ4sXtGurqRaeoRViIm9PPHI94ye4O/wC3cy80S7vthJhdVAJwaj5dZG8i7Hf8vctQ7XuIAmjJrpyM/Po+J/mB9OC9tsdrEjk/78SvHH/JXsFV6GGk+fS064+Z3UfZm7oNkYim3his9UhDBT7i27PiVl8n6QjTNz/tvfq9ep0bb3vrXTZmTbF5D/Girf8A3Dk97HVW4dfOq/4TV/8Ab+H5yf8Aap7ov/h/v9pfdRxHTh4Hr6My/pX/AIKP96976r1y9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SF7Q/wCZb79/8M/cf/upl9+62OPQC/B63+yzdc2+n2+Vt/595vfuvHo2nv3WutQD+dHKY/kPhf8AE1d+bfQP7zA+741Nguq+o/ydYqe/A/3b2f8Apm/46eqfTOOCW/5OHPue9R6g7SKA066NQfza9r/q+nH1Hv2r59a0dbDX8g+TXuTuz8gY7Hc/1/ci494u/ePOptsr/E3+XrJL7v4pBuX+mX/AvWzR7xm6yL697917r3v3XumrOf8AFnyf/avrv/cOT3V+A+0f4R17rQT+R9bo7w7GUc23Ll/Tc2P+Wn6++jnJL/8AIXtAP4F/wdYCc6oTzVdn+m3+HoFfviWAI4/Jv9P9b2KNeMdBjw+sU9Z+y5PFmQcn6/uD6e7xyd46anSsR/L/AAjrc+/lUPr+MGHbg3q1/P48J98/fdKh5yuD9v8Ah6zt9uqf1WtwPT/J1Zl7jzodde9+691737r3RQ/nUxX4tdysLenYuZPP/BFt7EfJppzjtZ/5eE6I+ZxXly+H/CX/AMHWhzia8jG0g0jmMn9X/NxvfSK5lJnY/Prnjt6qLOMU8j/hPU/778kf4WuPp/X214opjpVoqa9Kzr6t/wCMldbf0O9sIv1/Jn9l+8Sf7pL4f8If/B0u2ZP+RBt3/PQn+A9fQI6v/wCPF25xb/cZSf7H/JkN/fNKX+1f/TN/hPXQ6L+yX7B/g6X/ALp051737r3XvfuvdUK/zvpfH1lhSPy0P+9v7nz7vjaebCfkf8nUI++iluUn61kFryFX6elRf1fU295l+JnrEcJih8uuxXXPJAB/2r6e9+JQdeEdeOOrDP5WVT5Pml1egYcz1d+fr6FP09w/75SA+3t2P9L/AIepV9mI6c9wn0Rv8nW6THfQL/Xn/e/eCy/CPsH+DrNAV8+ufvfW+ve/de64v+h/+Cn/AHr37rx604P5yEuj5cZwH/lQxdjzcft/4e8yvYJ6cngD+Jv8PWJPvkleZ0P9BeqoDUEcgD/XvcH/AG/ucg56hjRnHXvuOLaV/wCSj71qHW9HWx5/IlfXSbv+h/Zl5/P6jx7xQ+8Ma7tD9g/wdZP+w4psj/n/AIetkT3jn1PvXvfuvde9+690x7jt/Bchc2/yd/8AevbcppGetr8Q60Le7ciqd49xLqPp35lVHP51++nHKkoHKO1gf8o6dc6ua4wec93p/wApB/46Og3/AIqoABc3/P8AgP6ez3xh0S+HXPl1ByGUH2NbZj/wGl/P+0/Xn29BMPFQfMdJruH/ABWSv8J63SP5bDiX4q9ZyXuW2/Sn/Ya25984/c2h593M/wDDT/k66DcgCnJ+3j/hS/4Oj+ewJ0Meve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6oH/AJ3tUKbb3VhblWnyP+28re8i/u3tp3y/+xf8HUA+/wAgbYbX5Of8HWuaMsukEEgFVsL/AE4HNveZRkStesTBHjHDrkMsOPV9CdP+P+v70ZV8uvCPqxz+VpXmo+TGPjvcmWk+hvb9P49wf7+sDyelPU9TL7FJp5omJ/o/4OtwsfT3g0vwjrMzrv3vr3XvfuvdcJP0P/wRv969+691pUfzkSV+a1VY/wDMHUp/N/pF7zR9hm08gEf8OP8Al6xB98hXnm3/AOedv8K9VcmTkfU/kn/H3MWuuOomK468J/8AA3BFgfz/ALH36vW6VIPWzt/IEcv1d2T/AE/vnPfj83l4v7xM+8QT/Wez/wCaI/wDrKH2BxynN/zXk/4+3WxV7x/6nXr3v3Xuve/de6QnZ9/9Hu8bfX+7+R/60H29bf7kx/6Zf8PTNx/YP/pT/g60Ee66kjuPsdb/AKc/Uf8AWxx+ffRvlV6csWNf99j/ACdYB81JXmi9Ncah/l6DP7ksDe1zaxvyPZ8HAz0RFPI9Y2qmDxnV/bHAN/wfetfaQPTrxQKQT69bw38tQlvh51Mx/OOqj/1mHvnp7i/8rzuH+mH+DrPHkUAcpWI/4WOj5+wZ0Luve/de697917r/1d/j37r3VQP82LF7h66ovjp82cVgM1vLA/DTsWt35vrZ22MXX5zdGW2ruQUeFqqvB4XHo71L0zIGkUcqrBx6VYhZaSKA0bfi6KdzilYx3EY1eEakfl0B2y+1tqfzPPnh8b+4Ol9tdl7b65+EFJuHde7Nx9n7D3D1+24c921tQ4/Gbc27SZeIGqenRVao5CoQ5vbSWuy/TwMjnLenTSP9ffRzxAhIq1JBWpI8qgVp1ff7QdHfTXloIWx+QkaCF5BQVdneGNz/AMB24OoG4/w9+698uiN/y86BU6PzsdXTwOx7T33JZoo3FjkF0nkfgX/1veh1Zuj2/YUP/KnSf+c8P/FPe+tVPXYoqIfSkpR/rU8Q/wCI96oDx69U9d/Z0f8AyqU3/UiL/inv2lfTr1T177Oj/wCVWm/6kRf8U9+oPTr1T10aKiP1pKU/69PEf+I9+oOvVPXX2FD/AMqdJ/5zw/8AFPe+vVPRVfmBiFq+qJ6ekgpomasHIp4foVF7Erx71ivXskdDB03jaeLqrYUM9NTvJDtugRi0ETeoJ6rXHv3XjWvQlfYUP/KlSf8AnPD/AMU9769U9cvsqMfSkph/04i/4p71QenXqnr32dH/AMqtN/1Ii/4p79QenXqnr32dH/yq03/UiL/inv1B6deqeujQ0R+tHSn/AF6eL/inv1B16p66+wof+VKk/wDOeH/invdB16p6IT3hg3l+Ynx4yUEVOtPRbbzsMqNBC6HVVPpvG4INh9OOPeuvHh0fYUNEQL0dKTYc/bw/0/1ve6Dr1T139jQ/8qdL/wCc8P8AxT37r1T139nR/wDKrTf9SIv+Ke9UHp16p699nR/8qtN/1Ii/4p79QenXqnr32dH/AMqtN/1Ii/4p79QenXqnrr7Gi/5U6X/znh/4p73Tr1T177Gh/wCVOl/854f+Ke/deqeiDdMY51+cHycqJ4IDTTbU2jHCBHFYgNGGBQD8j6k/X8+9efW/w9H4+wof+VKk/wDOeH/invfWqnrsUNEPpR0o/wBaniH/ABHvVAevVPXf2dH/AMqtN/1Ii/4p79QenXqnr32dH/yq03/UiL/inv1B6deqevfZUZ+tJTH/AKcRf8U9+oOvVPXH7Ch/5U6T/wA54f8AinvfXqnpDdm4+mfr/d0cNLTJI+FqlUinh/Vp4vx70fl16pPRffhnh1o+t8zFVw08rPuetk5ghtbTxxp/H4978+vVxQdG7+wof+VKk/8AOeH/AIp7916p67FFRD6UlKP9aniH/Ee9UB69U9d/Z0f/ACq03/UiL/inv1B16p699nR/8qtN/wBSIv8Ainv1B6deqevfZUZ+tJTH/pxF/wAU97oOvVPXH7Ch/wCVOk/854f+Ke/deqeiN/P/AAJyfTO36ahip4JI+zNnVJIijRdMdW2u9h+Rb3U9bB416OZt2jpXwOHaWlpHc4yh1MKWBQT9soNlVbD/AGHu1Oqgnp4+wof+VOk/854f+Ke/dbqeuX2VGP8AlEpv+pEX/FPeqDr1T177Oj/5Vab/AKkRf8U9+oPTr1T177Oj/wCVWm/6kRf8U9+oPTr1T10aGiP1o6U/69PEf+I97oOvVPXX2FD/AMqVJ/5zw/8AFPfuvVPRBvkNinl+W/xHnp6emFLSV+4nqAI4ka7xEA8C5+nA/HvR62Oj9GhoiSTR0pJJJJp4iST9STb3vrVT177Gh/5U6X/znh/4p79Qdeqeu/s6P/lUpv8AqRF/xT3rSvp16p699nR/8qtN/wBSIv8Ainv1B6deqevfZ0f/ACq03/UiL/inv1B6deqeuvsaH/lTpf8Aznh/4p73QdeqeuvsqJQStHSggGxFPED9Pwbe/deqeiFfHnCSU3ffZ1RPHTtBPBVCNfDD9Gql4sF/p7qKderx6Pr9hQ/8qdJ/5zw/8U9269U9dihoh9KOlH+tTxf8U96oOvVPXf2dH/yqU3/UiL/inv1B16p699nR/wDKrTf9SIv+Ke/UHp16p699lRn60lMf+nEX/FPfqDr1T1x+wof+VOk/854f+Ke99eqegw7sxtPN0/2dBBTUySVGx9yQqVhjXl8ZIFBIH9bH3o8MdeBNegi+E2IFH8aOtKOupqV56fGVEbEwQk2+7dhdrcnn6nn3ug61U9Gs+wof+VOk/wDOeH/inv3W6nrsUVEPpSUo/wBaniH/ABHvVAevVPXf2dH/AMqtN/1Ii/4p79QenXqnr32dH/yqU3/UiL/inv2lfTr1T140VGfrSUx/14Iv+Ke/UHXqnrj9hQ/8qVJ/5zw/8U9769U9Ek/mB4f73465KmoKWlWV917TLWhhQlFyQYgOBcf7D37y68Ca9G/2vQ0p2xtoS0tMzpt/DIS0ETEaMdGLAkfQe9Dh14k16fPsKH/lTpP/ADnh/wCKe99eqeuX2VH/AMqlN/1Ii/4p71QdeqevfZ0f/KrTf9SIv+Ke/UHp16p699nR/wDKrTf9SIv+Ke/UHp16p66NDRH60dKf9eni/wCKe90HXqnrr7Ch/wCVKk/854f+Ke/deqeiE9k4OSX5X7Ir41ijgh/hvpjREJYKL6tABP8AsfdTTy63U06sB926r02Zk2xeQ/6gq3/3Dk97HVW4dfOr/wCE1n/b+H5yf9qjun/3tpPdRxHTh4Hr6Mq/pX/go/3r3vqvXL37r3Xvfuvde9+691737r3XvfuvdJzde8NqbEwlVuTem5MHtTAURjWqzO4cnR4jHRSTNoggNXXOiGSRrJFGCWdiFUFiB72qljRcnqrMqKWY0A6Z+v8As/r3tXES5zrveGA3fjaWoNHXTYTI09ZLjK4Lragy1Ih8tLOFsxhqER7EHTYg+7OjRtpbqsU0cya4zUdLv3TpzpC9ngt1vv0AEk7P3HwBcn/cTL+B791scegF+Dv/AGTN1zxb/J8tx/5GJ/eh149G097611p2/wA7KoWl+ReCWU+Nn+7KiSyXGl/oGt7y79gFY7DdaQTw/wAI6xW992Rd2sy5AqxpUj+E+vVNRyFP9PIvH+1p/sPz7nmjeYP7OoQDx8NQ/aOvGuj5bzLe3PrT+nvfefwn9nXtUQGXH7R1sWf8J/ZvuNx94mM60TG425WzBSZYgLkf194wfeLDatsqPxN/gPWR/wB39kNvuWkg968D8l62e/eNPWRPXvfuvde9+6901ZsE4jJgAk/w+u4AJP8AwDcfQe6sCaU9R/h6918+r5K5ERd7dkxlkGnc+ZB1OoYH748EE++iXJWv+rFrQfgXy+XWBXOjxrzRdjV+NvMevQIfxSO4BeL+otIn/FfYo76cD+w9BjXET8Q/aOsFXlIvt3PkTkoB61t/nAOT7vH4msDSf2dNTPCIiSw8vMeo63Zf5ThL/FjByW9LVSWbnSSIebH8/X3gJ7n55wuDSnH/AA9Z1e3v/Kr2/wBn+Tqzv3H/AEN+ve/de697917ooPzvV/8AZUe7pFW6x7BzJZgD6fSpuSOAP9f2IeTxXnDa/wDnoXom5kP/ACHb4f8ACX/wdaB2GyMBxlGRPG14ySRIliBIw+t/fR25EnjtVTx9D6dc8dvlgexjKupx6j1PTicjASf3l/p+tPbGmSnA/s6V+JDXDCv2jpVdd18J7O6xXzIde+sEB61JLefhRY8n2h3ZX/cl9g/2D+R9Ol+zvD/WHbRUZuUHEcaN19DjrJHTYu2g4YE4qiYBgQdJpUseffNmX+1f/TN/hPXQmL+yX7B/g6XnunTnXvfuvde9+691QF/PVl+26mwsz/tp5YQJG9Kmxe6hjx/vPud/YEN/Ww0FcHqFffEqOUnLGgA9adatKZaIqP3Iz6RwZF54/wBf3mIwkrwPWJavBgah+0dcv4nFx+5ENX0/dU2t9fz79+oPI4+XW9UOe4H0yOrHP5UFYs/zc6tRHWRvuKs6VYMxAQE3A9xJ72lj7f3QIPl1KPs1pHPcVGHwP5/Z1u5pcKL/AF5/3v3g8PhH2D/B1mZ1y97691737r3XF+UYDn0nj/Ye/de60yf50FQsPy9zaSP4v9x2KYaiF1BorgjV/vHvMf2FD/1RoBkM3+HrEr3wZBzOmsgdi+fVSX8QiBt50sfodam3+2Pub6P/AAn9nUOa4/Jh+0de/iEJvadbj8l0/wBa459+o/of2da1xcdQ/aP8/Wyj/IUl+5od5SL6gsMoJX1L+sgEsvAv7xT+8EGO7w6vQf4OsnvYllOxOVINa8PtPp1sqe8dep7697917r3v3XumfPoZMRXIqFyaeT0qrMTZSTwvPtuXEZ62vxdfPm73zCR9+91xmoiBi7CzCFGkUFCJPyD9D/r++lPKzP8A1T2ygP8AuOnl1z05qMX9c92BIqJ8iox2A9Be2ZiNgKiK4PJ8sfP+vz7O9cnof2HomHhcdQ/aOoOSzcK4+vZp4gBSzf7sSw9P9b+3rdpPHQUPEdJb1oRZTEsKBT5j/P1vEfyyLyfEbqmoAJSTblNZ7GznWx9LfQ/7A++ePuSa89bkf+GnrP8A5DFOUbD/AJpL/g6sH9gfoXde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691rz/z4KsUW1uppHbxoZ8mNb+gEiUmwLfn8+8hfu76l3u9IHEL1AvvzT9w2xOO8/4OtZ1M7CVW1TFyot+6l/p+feXxd68D1imPDoO4ftHXL+OxC5+5iGn/AJuJ+f6C/vWtj5H+fVqR8dQ/aOrOP5SuRWt+UuPjikWUl6M6UIZudJ+i39wt78MTyilQeJ6mH2P0nmaYgg001/Z8ut0D3hKOHWYfXve+vde9+691wk/Q/F/Q3H9ePp7917rSe/nNyCD5uVMctoZP7mUh0SERvYeK40tY/wC8e8zvYYM3IDUBP65/y9Yh++TxLzzbamAP07UqRnuTqrAVSHUfLGBzwJFP/E+5j0Ofwn9nUTmSOlQw/aOvfdQhgTMhsLga04t/rH37w5PQ/sPXvGjHFh+0dbQX/Cfl/P1T2ZJHeVBvSbU6epVOqUcstwP9ifeJ/wB4lWHM9nqFP0Bx+wdZO/d/kSXlGV0IYePLkGvB2Hl1sY+8fup3697917r3v3XukL2cC3Xu8goJP93skbKCTxTkkgD29bf7kx/6Yf4embj+wf8A0p/wdfPo7yyUUfdPZcZkQEbhqLgutx+6/wBQfp76Kcrav6sWRAPwen2dYDc1vGOab1dQrqGKj0PQYfxSnIF5EJ/N5EuP959nnf5A0+zojLQkjuFftHXA5On1paWMXcAHyL/Q8D37vocHh6deLRLkMOPqOt6r+WYS3w16icjhsZVFWtwymYAFT/Tj3z79wwf673/+mH+DrO7kan9UrGn++x0fn2Dehb1737r3Xvfuvdf/1t/j37r3XRAIIIBBFiDyCD9QR7917rpURBZFVB/RVCj+n0Hv1a8evdcvfuvdQcn/AMW3IW+v2NX/ANaG9+690Sr4AcdLZz/xJu+P/dgPeh1Zujx+99V697917r3v3Xuve/de697917r3v3Xuiz/Kof8AGNJv+otf+hfej1sZ6FXqb/mWuyv+1BQ/9Ce/Dh15uPQh+99a697917r3v3Xuve/de697917r3v3XuiS9yf8AZVHRf/ahzP8A7kv711by6OyPoP8AWH+9e99V679+691737r3Xvfuvde9+691737r3XvfuvdEX6gH/OZ3yP8A/DZ2nx/yFHz7159b8uj0e99a697917r3v3Xuve/de697917r3v3XukZ2J/x4+5+L/wC4mp4/2A9+PXugK+Jf/HhZf/w4qvj+np9+690aj37r3Xvfuvde9+691737r3Xvfuvde9+690Tj5u/8yowv5/4yBtXg/wDUU1/evPrYwD0a7btv4FiLCw/h1F/7jL7seqjp596631737r3Xvfuvde9+691737r3XvfuvdEd74/7Km+Lpt/yl54E3+n7bW966sOHR4ve+q9e9+691737r3Xvfuvde9+691737r3XR+h/1j/vXv3XuiS9Ek/6bexLn/dNTxb/AKahz70OvdHb97691737r3Xvfuvde9+691737r3XvfuvdBz2/wD8ys7C/wDDQz3/ALrpPejw62vHoLviD/2T9sH/AKgqj/3Ib3vrR49GX9+691737r3Xvfuvde9+691737r3XvfuvdE8+c1/9AuQt/z0+2Pxf/l4f09+68OPRqNtf8e5t/8A7UmK/wDcCP37r3T37917r3v3Xuve/de697917r3v3Xuve/de6JJ2B/2UztD/AKoP+hRx70evdHb9769015r/AItWQ4v/AJFW/wDuHJ72Oqvw6+dZ/wAJq/8At/D85P8AtUd0f+9tJ7qOI6cbgevozL+lf9Yf7172ePVR137917r3v3Xuve/de697917r3v3XuqXvn5tzb/yF+b/ws+I/cOOjzvQWepM/3dlduTVE+Pps32V1tkmm2UlXW0bpJKsLr6aYtpbW35PtbbgLC8g4j/N0UX5L3kNuxorZI9SCKdMG1NoYL40/zlcT1v0bgItndefKD49b37h7xwWJkmfB5PsraeRGI2/nfsJXaOjmanjACwKqkM9gFYj3tmMlprfJB49VjVbfdhDFhXQsQPUUAr+XV4HtD0c9Eg+QnSvbWfod97rwvyF3TtTbsO3c3WvtCix0EuOlpqfHyTS0EkzOGCSKChIW/PvR4dbBFei0/Fbozuje/Q2xdx7b+Su79k4mviybU22Mbi6Wahxqx5GSFhDOZAza2VnNxxfj8+/dWNOB6MCfi78hjcf7OBvwXtb/AHEU1xb/AKe+/Z60NPp0WrtP+Upg+8c1BuPt3uTcO987TBhBkchiqYTIHvqsS7f19iPZ+b+aOX4Wt9ku2t0b4gPP+fRBuvK3Le+yrNvNotwyfCWAx9mMdBp/wxb0b+N45If+Qum/4r7N/wDXO9wv+jm/7P8AZ6K/9bvkX/o3R/sH+brr/hi3o61v755O/wDX+FUv+9X96/1zvcL/AKOT/s/2evf63fIvD93R/sH+bobOnv5X0nx8lys/Sneu5tgzZ1EizD43E0pNdHEQY1k9a/Qgf7b2Rb1zNzBzH4Z325Nx4WUr5V6Oto5e2Ll9XXZbZbcSfFpp3fb0O3+yvfIX/vL/AH5/56Kb/r77I89HNV9OgG752N8i+mcLi8rT/KneuYkyFYlK0FTjYYlTVKqakaN2+gN+ffh9vXjSmB0NGH+NvyFyuHxOT/2b3fsZyONoq9ov4TSkRPWUy1BjU+XkDVYE+/Z6928KdTpPiz8gpUeOT5fb7aORHjkU4ims0cqGORT+7+QSPfqHr1V9Oib57+SX1PujMV24Nwdg5fJ5nJzy1WQrpsVTeSqqJ31yyP6j9W59jK19xOebG3W1s9wZI0AAUDgB+fQSuOQ+Tbydrm6sEeRzUsaVJPHy6af+GLej+SN55S9uP9xdLwf6/X2//rne4X/Ryf8AZ/s9Nf63fI3/AEbo/wBg/wA3XR/kWdHMmh955NwSCQcVS24Nx+fex7n+4Q4bm/7P9nrR9uuRGFDtsf7B/m6NL178Dex+psBBtTrb5Oby2rtqlbXT4mixNMYI2A0gj9wfjj2Eb7cL7dLk3m4SGSVuLHz6FNnZWW3QC1sk8OMcAPLpeH4u/IYm/wDs4G/f8f8AcRS8/wDWX2jz0qqvp0XKv2v8i6X5JU/Qsfym3qtDNs2PdI3BNjYvu/PISDRiGN2XTxwS3v3WjQZHRjf9lc+Q1/8AssLfx5/OIpRcf4gS+/Z632+nSb3b8Le4d+bfyu094fKze+b21naOXHZjE1GJpvBXUM4tNTy/uHhh7ftri4s7hLy1bRLEdSN5qw4HpmeGC5he3nXUkgow9QfLooEf8inoqFFih3hk0jQaUUYulAUE3sOf6n2M290PcNzqbc3J+z/Z6CCe3HIcahI9tjUDyAFP8HWUfyL+jhf/AH+WT5Fv+LXS/T/b+9f653uF/wBHJ/2f7PVv9bvkX/o3R/sH+brPRfyOel8bX0GUod75SnyOLq4q/H1K4ul1U1bTnVBOnP1U+6Se5XP80TQS7k5RwVYU4qeI49Wj9vuSIpUni2+MPGQymgqrDgRjj0cyh+J/feMpqeioPl1vumo6SJYaenXEUxSOJFCogvL+AAB7BRJOTxPQwUIoCgYHUxfi78hdd2+YO/bGwNsRSkgfmwMv+29663Vei79LbS+RXau+e7dnVHym3rjk6l3XT7aoa6mxtPKcws0byNU1aO66CNBAAv79XOOvUFM9GI/2V75C/wDeX+/P/PRTf9fffs9eqvp0DvbH8trO9842PDdx/IXde+cVAwaGiyOJphHGR9LfuH+vs22jfd55fuPq9lnMEv8AEOizdNn2nfLc2u7wieM/halP8HQBD+Rf0cP+YyyfH0/3F03H4/r7Ef8Arne4X/Ryf9n+z0Hx7d8ij/lnR/sH+brh/wAMW9H3/wCP0ylvwP4XTf8AR3vf+ud7g/8ARyf9n+z17/W75FpT93R/sH+bpc9c/wAnnZfUG6sfvrrPtbO7U3hii5x2bo8VTGelZ+HKDWPqP8fZdunPHN+92bbfu1600D8VI4/z6X7byfyts90L7a7NIZhgMtK0PEcOjTf7K38hrAD5g784P1/hFNf63/46+wsMCnQkJU9Ivsjo/wCQ2wtl5rdSfLXfNdLioBLHSy4qnSKS9xZ2WQkf7b37PHr2DwHTJ0z1F8ie09g43eVR8sd8Y2or6mrhakhxcEkMa07hAVd5A1z/AK3v2evdvmOhSPxe+QxBH+zgb85/6tFL/sP92+/UPXqr0WTtD+UZtvu3cku8e2e3s/vPdE6RxzZeuxVMszxxC0aEa2+g+nPsSbTzlzVsFt9Hst40EX8IH+z0H905U5a3u4+q3a0WeSlKtTgPy6Dj/hizo3nTvLJrf/q10x/4n2a/653uF/0cn/Z/s9Fn+t5yN/0bo/2D/N12P5FnRgH/AB+OT/8APXTf9He/f653uF/0cn/Z/s9a/wBbrkXh+7o/2D/N0OHUn8sas6DjqoumO+907CjrVK1a43FUzeYN+oMda+w/vHMW+8wyCXe7g3DLwJx0e7TsOy7DEYdnt1gQ+S8OhxPxe+QxFv8AZwN+Xta/8Ipf9j/u32TZ6NwRToufyB2t8jekajqqmpvlLvXNHsXfEO0auSqxsESYuGZVYVsPidixGrkED377eveWOjFJ8YPkI6o6/MHfhDKGv/CKaxvzcXl9+z16q+nXCb4sfIKdGjk+X+/GjcFXX+EUwDAgggkS/n34iooeHXqgcOiZ5j+SH0/uDM5XcWa35lq7N5ytlyWXyD4umElbXTm8tRJ6jyT/AI+xrb+5HPtrbpaW+4usUY0otOAHlx6CE/IPJV1cyXlxt8byynU7ECrHhU49Om7/AIYs6P5tvTKC5/51dN9P6fq9u/65/uF/0c3/AGf7PVP9bvkX/o3R/sH+brpv5FfRkkbRSbyyjxupV0OMprMrCxH19+Huh7hqdS7m4I+X+z1R/bnkORSj7bGQcEUFCP2dGv2J8Ge0erttY3ZvXvyj3ptnauHhWmxeGpMTTGCkp0N1jS8g/wB69g+9vbzcrp7+/fxJpDVmPEn16Ftpa2ljbpaWaaIkFFUcAB5dLT/ZXvkL/wB5gb8/89FN/wBffabPSiq+nRdsrtP5FYr5K7V6D/2afes9FuTZeS3ZPn/4bTiuo5aAsFo46dn0Mrab31e9de+3oxH+yu/Ia1v9nB35+Of4RS/j/p779nr3b6dcv9lf+Qx+vy/35/56KX/r779nr1V9Ouh8XvkKP+5v9+f+eim/6++/Z69VfTr3+yu/Ib/vMDflvwP4RTcf9Zffs9eqvp14fF75Dfn5gb8P+P8ACKb/AK++/Z692+nXX+yu/Ia4I+YG/Ba/H8IprG/9f3ffs9eOk9Yqn4yfISmpqupb5f78YQ0004UYel5aGIyKp/d+htz79nrw016ALpPZ/wAi+28rvHG1Pym3pihtaoWnhkpsbDKarVL49UvkdbfT8X9+rXr2KZ6MJ/sr/wAhf+8v9+fm/wDuIpef+svv2evdvp0C3b38tHKfICmxtF3R3/unfdJhmkbFw5HFUyrSNMbymM+Q/U/X2b7PzBvfL0rT7JcG3d/iI86dFW67Hs2+xrDvEAnRMgNwB6AQfyK+jQT/AL/LKWNhb+GU3AH+Or2Iv9c73C/6Ob/s/wBnoh/1u+RRw26P9g/zdd/8MWdHX/4/PKf4f7jKfj+vOr37/XO9wf8Ao5P+z/Z69/rd8i8f3dH+wf5uhA63/lA7S6d3Cm7OsO2c/tHckZUplaLF0zTqU/SR6x9PZZuvOnNm+2/0m8XjTxjgpH+z0YbZyjyxss5udqtFgkPEqBmnrjo0X+yvfIbn/nMDfnJJ/wCLRTfU8n/dvsM0pgdCPt8+gr7v6n+RHUPVG9eyaf5Xb5y9VtLFDJQYyXF00dPWkVCwNFNIJCR+u4sD79nr2PLp+606P+Q/YGwNob2f5a73opNz4HH5mSjhxVO8VLJWwCV4Ed5ATpJtcj37PWscD0tj8XfkKfp8v9+D+v8AuIpuR/T/ADvv2et9vp0VztD+UFtTuzdJ3x2v23n947uamWibN1+JpvuTSJbTD+s8cD8/j2J9p505s2C0/d+y3jQQV1aBwr68eg5unKPLG93Qvt3s0nmUaQzAVCniOHyHSAH8izo4C398smf/ACF0o/Fv6+zL/XO9wR/y0n/Z/s9F/wDrd8jf9G6P+X+bro/yK+jDYrvHJhh+f4XTH/or37/XO9wv+jm/7P8AZ69/rd8i/wDRuj/YP83Q8dPfyz8r8e8bX4fpP5Abp2BjMpVGuyNJjMTTaKqra5aV/wBxfrc+w9vPMO+cxzrdb7cG4kQUVjxA9Oj3aNi2Xl+3NpslutvExLFVAA1HJP2k8ehr/wBle+Qv/eX+/L/1/hFN/wBffZPno27fTou3yK2n8j+isVsbI0fyk3lnZN475xGz5YKvHQQ/aw5OQJJWwsrsCyX+htf8e/cOPXgAeHRi2+L/AMgiQYvl/v7QQpGrEUgb9IvcLLb6/T37PXhpHl1ByHxP77ytHU4+v+Xe+56KshenqoGxNMFmglGmSNrS/Qjj3tSysGXiMjrTBGBVhg4PROct/JB6dz2Vr85mN95Wuy+UnapyFbJi6YSVM7ElpHGo83J9jOD3G57tYEtbbcWSNBRVpwH7eglPyDyVdTtc3O3xvI+WYgVP506bv+GK+jb3G88p9Of9xdLyf63v7d/1zfcH/o5P+z/Z6a/1u+Ra1/dsdfsH+brzfyKujCF/3+OTupDA/wALpfqOL2v79/rne4X/AEcn/Z/s9e/1u+Rf+jdH+wf5ujabH+D/AGp1ntjGbK2B8pN6ba2phkaLF4akxNMYKSNyCyRnyD629g+8vLvcbt76+fxJpMsx8z0K7S1tbG2WztE0RIKKo4AdK4fF35C3/wCywd+gEj6YiluBfm37vtNnpRVekV07k+39j/I2r6x3h25nuysLplKvmKVKZoSkJdbKjEfUfg+/dex5dWU+99V6/9ff49+691737r3Xvfuvde9+691CyX/FuyH/AFBVX/WhvfuvdEr+AZJ6Yzl+D/pN3xx/5EF96HVm6PD731Xr3v3Xuve/de697917r3v3Xuve/de6LR8qbf6NJif+Vpbf7Ye9dbHQqdTf8y12V/2oKH/oT34cOvNx6EP3vrXXvfuvde9+691737r3Xvfuvde9+690SXuO3+zUdGf1/gOa/wDch/eureXR2R9B/rD/AHr3vqvXfv3Xuve/de697917r3v3Xuve/de697917oi/UH/ZZ3yPH/ZsbTP/ACdGPehx6sfh6PR731Xr3v3Xuve/de697917r3v3Xuve/de6RnYnOx9z/n/cTU/70Pejw68OgL+Jd/7hZcEWtuOs/wBtYe/A162RTo0/vfWuve/de697917r3v3Xuve/de697917onHzd46owp/7+BtX/wBym968x1scDXo123v+LFiPx/uOov8A3GX3Y9VHTz711vr3v3Xuve/de697917r3v3Xuve/de6I93x/2VN8Xf8AqLz3/WtvdTx6sOHR4fduq9e9+691737r3Xvfuvde9+691737r3XR+h/1j/vXv3XuiTdFf8zt7D4/3TUc/wDVSPdRx690dr3br3Xvfuvde9+691737r3Xvfuvde9+690HPb//ADKzsL/w0M9/7rpPejw62OPQW/EH/sn3YP8AU0M9/wDX+4b3vrx49GY9+611737r3Xvfuvde9+691737r3XvfuvdE9+cv/Mhch/T+8+2L8/T/ch+PfvLHWxxz0afbX/Hubf/AO1Jiv8A3Aj9+HDrR6e/fuvde9+691737r3Xvfuvde9+691737r3RJOwD/zkxtD/AMh/+9fT3rr3n0dv3vr3TXmjbF5D/qCrf/cOT3sdVbA6+dZ/wmr/AO38Pzk/7VHdH/vbSe6jiOnDwPX0Zl/Sv+sP9697PHqo4dd+/de697917r3v3Xuve/de697917oinzq+FTfMPZW0KfZ3bm4vjt3J11vDC7s6/wC9NkYPG5nd224sfWCpyu344MhJCslHXhVEqGUaWVWAPqDPRTNFjiD5dI7qzjugCSVdeDDiOm74ifCbPfH7dvYfavdHfW6PlT3ZvmuWnx3Z+/Ns4bb+U2TsmOnSNNjbYosTJLHBSPIgllKFdRCrpAB1Wln1roQaVHl1W1shAxllcySH8RAH5Cnl0fz2n6XdIPtIButd/qRdW2duRWH9QcRKD7914cegD+DMUUHxi63ihXRGtPltK8m18xOT9f8AH37rZ6Nx791rr3v3Xuve/de697917r3v3Xuve/de6J38wKaCp2zgFniEqrk4zY2+vnT3o9e6NNtYBds7eUCwGDxQA/oBQxgD34cOtnj0/e99a697917r3v3Xuve/de697917r3v3XuiNZekpv9neoavwqakdb08fm/taNR9PvXWycdHl97611737r3Xvfuvde9+691737r3Xvfuvde9+690Q/wCJ1LT0/cfy1eGMI8/ZlE8zXN3b7ebkg+9efVj8PR8Pe+q9e9+691737r3Xvfuvde9+691737r3QQ96xRzdX7mjkUMjUvqU/kc+9Hr3SY+L8MVP1BhIoUCRrWZGyj+plF/fhnr3Rhfe+vde9+691737r3Xvfuvde9+691737r3RIPmXR01VWdCGohEpg7UpJYSf7EgiSzD3o8erA46OxTG9PCf6xIf9uvvfVes/v3Xuve/de697917r3v3Xuve/de697917oiW56OlPz368rWivVL1TnY0lubqhLgrb3o8erDh0e33vqvXvfuvde9+691737r3Xvfuvde9+691CyIBx9eD9DRVQP+t4G9+690TX4p0tNTbi7LMEYjL1yF7fk+cnn3odbzTo6/vfWuve/de697917r3v3Xuve/de697917ovHyvghqfjz2jBPGJYZNusskZ+jD7uI2PvRr5dbHSq6Dijh6Z62iiUJGm08UqIOQqiAAD3vrXn0Lvv3Xuve/de697917r3v3Xuve/de697917ojfzio6WswHUK1UQlEXbe25YwTbTIsy2Ye9HrY6PEn6V/4KP9697611y9+691737r3Xvfuvde9+691737r3XvfuvdEcpKWn/2bOoqBEBNpnvJ+T+wR7r59W6PH7t1Xr//0N/j37r3Xvfuvde9+691737r3ULJf8W6v/6gqr/rQ3v3XuiWfAT/AJkznP8AxJm9/wD3YL70OrN0eD3vqvVdfyD+ddVtTujJfFX4ybN2d398rdrbPw/aW/eo8/2bh+rV2h1Tl6j7al3TU5vPRvHUSytpMVLApIQh5CqkXfiiRxWRqD5Cp6R3NzLF2wJrb0J0imK5PmPT7PXoKsH/ADlviHlep+ke66rN5HHda9s/IXN/Fbce9KiIJtXqbu3bVJI+cw2+M/KFpYcXHPE8UWZMq08iFJQbOB72IAagNkcB/q4dae80aWZe1jQn0J+VMj59LjLfzRukMD8as/8AKnK7c3knV+M7t3B0zgqqjpYaur3ocLWy0VJvXa6eharGVxhdqaSFnDAEqzAE+3BaFpBGGzpr0nk3WOK2N06kKHK/b8+FcnhjqJ8cv5g27Oxu/P8AQD370LWfHncO9tmUvZfRWRyG8aDc1B2ZsiulP2azSU0EC0OVMQMj452aRSrIQGAB09sBGZI21U4/Lq0W4Mbr6a4j8OoBU1qG/wAx+R6s89pOjPos/wAqf+ZaS/8AUUP96HvR62OhV6m/5ltsv/tQUP8A0J78vDrzcehD97611RXV/wA5+feO8Pl+nQPxuznaPU3wE7UzvXnyq7IyO88ftGXa1BsumqqzfW4tqbeyFO0uUWgioK90hik1SLCW9Ksp9qFiiKgl8+lPPpBJc3SSlRCSo86jh6/7HHoyW8f5nXWW68B8fMN8RYttd/8AeXyv6j/08/H7qvcO76XqpN29RUgU5jd1fndyRFaX7dmMK0njaV5EkAGmMsdRxKzHW1APTP8ALpy4uJI46wJrc8ATpH2EnAPy+3ovPbH89L49dPfETsn5Ebi21lMt2L0z8htufEXtLpja+Rjzcm2vkbuCD7uPalPvCkhakrMctOstQMlToyssbR6fIAD7wkEulmovrT/J1Vrmb6YzRx6nH4agfbQmgP8An6tm6J3n2hv7rrE7n7e6tTp3eWQeR59kruei3Y9HRMiSUlRPkqGOJEkcMQ8DLqQrZuTYVlREaiNq6et5JZU1SpoPpWvQxe2un+iS9x/9lUdGf9qHNf8AuQ/vVc9W8ujsj6D/AFh/vXvfVegv7n7m65+P/XG4+1+19y0O09kbXghlyeYyDlIVmrKhaLH0UWkG8tRPJHDEPpqYXIUEi8cbSOEXiemZ547eIzSmijqu/pr+ZnUbg+QfV3x7776qwvTW5fkxtveW9PinUYjs7AdhL2ntrZFKcjlTm4sNGn8GqXpv3YEnLByGT9Q9vyW6oDpatOOP9VektveSSsBImkNXSdQNaevpX06cNp/zUOr907txvx+TaeTpPmpW5PMY6q+Ls9a8edxEOGyjxVWYy26pqZaKCjOPT+ILJJZivpC29fuxtKNUt2evTa7mrjw1X9X+H/Z+zNOP5dIbtb+adumHuzvfpX4x/GjcfyCy3xLO1Zvk3kTuWn2fDsqm3PTvX+La0FZTSnNTU1NDUTTLTHgRudOlS3vyW0ZoHehPDHH061LuEyk+DCXCkasiorx/Z1Zb0T3l1t8kOq9p9x9TbgptzbH3hRy1GNyNPqBiqqOoehymMq0YApPSVMctPOv0DobErYlNJG0blG8ul8E8dzEJojVW4dC77p090RfqD/ss75H/APhs7T/6Gj4968+t+XR6Pe+tdEV+a3zw6/8AhRV9BYjd22c9vDcXyK7Ok6s2DgdvSxR19Xno8WcixVJEkL8tFHoVb+u9+LF+CETEgmlB0ivbz6NUOktrbTj59A90V/M02/uHsj5I9HfKTYB+MPcnxj2Fiu6N+YXJbih3ftI9I7nqTT7Y3tS7xx0EdO08rBY6nHoGkjkYJy1wNtADTwjqrjrUd4Rr+qXw9Irxrg1z/LpH7S/mp0Y7l6O2p2z1hh+vunPlz2DXdY/EvtbGdmYDeOS7N3XS0zV1HTZ/ZWKQVOIiq4lHilmchZGVG+t/d3tlRTRqsBWlP8vVIL6SZgTHRCaBtQP508h0tcB/Mby3a3zAq/il0D0Rm+wU613tW4T5Ddk5jctDtjbnWu06SRqJdy4ZJYZf4rPJVKYo8ekiSN+PyR57URwiV24jA+fVIdyNxdtbQxkhDRmrgenlmvoCaefVo3tJ0adIzsS39x9z3+n8Jqb/AO2Hv3Xh0BXxL/48LL8WH946y3+tpFvfut+XRqPfutdVy/KT57HqbuGg+LnQ+x9v94fKmp61rO7a7qnN9hYbrOkxnUOKyL4vJbmk3FnEeKSo8yEQUSLqZQXYhfaiGFXy50g44V6Q3d1JDVbdPEcCpBOnH2noP8h/N6+MW1Oo9j99b/rKzbPTuZ31iOmOwt/0vlz2G6p7xysaOuxtyjGxM0lJH+4ZcxDeBVCki7EK41oy9te7yHqPXpmLdIpV8RcoMMwNdLDiv5efp0kfkJ/OH6n+Ovxvq/ktuvr3c2S2buz5P4T4zdAfwirWpj7syG5ohNgd84erSAimxM6RVziZlkBEGlWZ2sGzCiuEdqVGfOh6dW7lkgaaGMtQ4BxUeufl0K/QP8wmq3Z8m5fhf8l+qj8dvk3uDZGW7f6r2HHuin37jew+n8PWHG5Dc8G4sRBHT0lZT1CSpNQTHXZHKklGHussaJlG1Dp23nllJWaMxnyzWo+3/J1Zj7Z6VdE3+b3/ADKjC/8Ah/7V/wDcpvevPreaGnRr9u/8WLEf9q6i/wDcZfdj1RenOpqIqSnqKqZtMNNDLUSt/qYoUMkjc/0APvwFTTrZNASfLqkvO/ziqjIUOT7N6l+M28ewPjt1nvmh2F312hUbjx+EyexMrk8suMpKjb21jDNJl6dFdJaiaGRVjDqH03BK36NahS+SKjHRL+9y0ZmiiLIraWNfhzSuRkD5dGb7/wD5gmI2jlut+v8A417c2j8iO3eztu53fGE2o/Z+3NiYej2VtjH/AMTzWUye4Mp5UjqRFqEVGyCQlSGtx7bS2NCZTpA+VelU24AMsdsokZgTTVpwPt6CrsP+cR8eesvgJUfOzcWNy9ZicflsPsjLdc7bnj3Bnsb2dnMqdv4/as1Xj0ZWgesAArkiKeNg2m9wNtasj0c0X1p03HukctsJYwC/8NRx4cftx9vVgnxy392l2h07szfncfVkPTG/Ny48ZTI9eQ7ppN4HCUdW3nxX3GZoooU80lO0bzQFNUTko3IPtPIqK1EOoevS+B5ZIw0q6D6Vr/PocPdOnuiPd7/9lTfF0/8ATXnv+tbe6nqw4dHh926r0DXyD7/6o+LfTe/O/e8d10myOrOtcRHmt3bmrUkkhx9JPXQ4ukURRAs8k1TUQQRKPq7i5AuRZV1Gg6q7hFLGpp6ZPRBOuf5mbJ3R051J8jetNu9Nf7N1U5B/hXmsB2dg+yx3TjMfgTulnztLt+IHByy494JoTUFo2d/Fq1jl94Y1pRvtxw/z9I4bqaQsHjp/DRgaj1P8P59OvTf82n46d87v6u6664oNzZbsnfXdPZ3Su7+vZIY4N49R5TqlasZ/c+/MCQammxM70yrQ1ssUcU4e6MShB81tpDNqwBUfP8q9VS/Dukeg6mYqRmi0rWpp8ui57m/nW0P+jbt7v7rj44bq3z8ePjD2nvHYfya7IXdeOo6nZWC2ZnDgcnuTaGBFM0mblSQeSWip31RpZmax1e7m2Raan+Lhjphdzlk1+HEaRkhiTQADiRjPV03Xm/NtdpbC2V2XsytbJbR7B2pt/em2K94Xp5KzA7mxUWZxNRLTS+qN2gmQvG3KtdTyD7SEFTQ9GqsHUMvA9LA/Q/6x/wB6966t0SXom/8Aps7E/A8NR/7kj3ode6O37317oiPzl+dez/hVi+nsfkdo57sDsv5EdlUHT/Sux8Leli3J2BlAn2FDl8yUlShpiZFDVDxlR+bfX2/DEJKljQDpHdXX05VFXUzmgFadB50z/MawNZie9cL8stlQ/F7uT457J3F2z2V1tkdxwbsVOnMBTmqbsfb+aoooo62kYKUIiB9TJpLB/TeS2K0aM6lOK9NW+4CTWlwvhyIKla1NPUetekR0h/M+h7A7E6noezOttvdY9N/KWkxVb8Re0aPs/A7zrO2Uz0inAUWc2ph4xPh6isjdSizMypIRG5Buy7a1Hhl4zUrxx6dNpuVLhYbhQgkpoOoHUT5UwQeufxa/meT/ADJ+VfyQ6L6J6Ny+Z6v+KHb+X6O7j7ozm6KPAQY/feDkqKXIRYLbFXT+athFRSzxp45dRC6zpVlvTw4vC168+lPPp/x7r6owmGiClG1DIPnTiKeh6tl9p+l3Qc9v/wDMrOwv/DQz3/uuk96PDrY49Bb8Qf8Asn7YP/UFP/7kN73148ejMe/da6rN+RX8wWt2BvneXXvx86+2n3zuHpLDZPd3yRpq/tTA9dP1TtPEY0ZmskSPORO2QrftBJMKeAWFlUtqYgKo7fUniOaA8MV6LZr8rP4Fuodl+LuA0+fn/qPSfp/5uPxnn2b8Y+6KirqcJ8a/kv1R2B2Zi++dxTrh9obRq9hxqJdl516hNJyVXOlTS00CyhpJYwI1fVx5bUsrUIqpAp6163JuCRPGJFIV1JJphaeR9P8AVw6m9l/zXeg+uPhDtL521WA3zV9W9o5/BbY6iw0uFqqLde9MtuzMzbf2savEvG0mPgq6mCQiSVXKxaZLEMPfhbgvo1cOOOHXnv8ATAJ9OGI05414HhjJH+rPSn+N/wA6d1757nrvjd8numD8Ze9MtjKndfVe0Kvd1JvKh7Q2RQUn3WWy+DzNBDFEJ6IA/c0xuVs4JDoVPpbcKniRtqHn8uvW9+0k5tbhPCkyQK11AUyP29WPe0vRj0T35y/8yGr/AOv959sWP9P9yHv3Xhx6NPtr/j3MB/2pMV/7gx+/dbPHp2nmjp4ZqiVtMUEUk0rf6mOJC7t/sAD791rqhuu/nh4Wr6UzHy02d8ct3bh+InVnZGb2H3926Ny0K5PY1Ljt0x7Rpc9tjaMVMZ82v3M0JqoqdwYQ6hm5B9rDbICAWyeGOihdylcM6xdqEhiTSlDSoxn/AFHo63efzukwe59s9VfF/Zu1fkD3hmtrYfsjLdf5nsjD9XR7Z6yzWHGdx25sllNwxsplqaZllgpFXXpOp7Ai9I7fUpdzQfZXp64v/DkEFuokfiRqC0HrU/6vLj0Dz/zk/jM/w32V8yMXQ7j3HtXd3YGe6kl2vgEhrcxi+y9p1dTitzbf8yjTURU9ZT+NKmBCskckcoAB0+7JaF5CgNKZ6pc7olrbrPIvxHTT0PD0Na1FPt6Pj8X+yu2+3um9t9hd19Px9F703HLka2Pr0brpN4z0O23qi22clX5OjhgWKorKQxzzUbJrgYlGNwQE8iqj6VNadLoHkkiDyroY+Va/z6MH7p090STsD/spnaB/r9gOP66R70a9bpkHo7fvfWumzM/8WvIf4UNaf/VOT3vqrcOvnV/8Jq7f8PxfOP8A7VHdHP8A5O0nuo4jpw8D19GZfoP9Yf717sePVRw6796691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SF7Q/5lvv3i/wDvz9x8fS/+4mXi/v3Wxx6AX4O8fGbrm4t/k+V4Bvb/AHLze/dePRtPfutde9+691737r3Xvfuvde9+691737r3RQvlzc7bwAH5ySX/AMbTJ70evdGh2x/x7e3/APtS4v8A9wU97HDrx49Pnv3Xuve/de697917r3v3Xuve/de697917okeXJHzUobC/wDxjuD/AGHJ9+8uvHo7nv3Xuve/de697917r3v3Xuve/de697917r3v3XuiLfFP/mcHyw5uP9JVHb8f8o83v3n1Y0p0en37qvXvfuvde9+691737r3Xvfuvde9+690EveH/ADLLc3/UKP8Ae/ej17pM/GT/AJlHhf8AqLyH/W0e/DrZ6MB731rr3v3Xuve/de697917r3v3Xuve/de6JR8xP+BfRP8A4lGkv/h+0nvR6sOHR0KX/gNB/wAso/8AoX3s8eqDh1n9+631737r3Xvfuvde9+691737r3XvfuvdEa3Nf/Z7evuPr1bnP97f3o9W8ujy+99V697917r3v3Xuve/de697917r3v3XuoeR/wCLfXf9QdT/ANaG9+690Tz4tf8AHxdk/wBfvkv/AIfvn3odb8ujn+99a697917r3v3Xuve/de697917r3v3Xui+/Kn/ALJ+7Pt9f7vN/wC5cXvR62OPSo6H/wCZOdcf+Gpi/wDrQPe+tHj0LXv3Xuve/de697917r3v3Xuve/de697917okfzaBOC6k/p/pX27f6f8AHZbfX3o8Otjo7S/pX/go/wB697611y9+691737r3Xvfuvde9+691737r3XvfuvdEkpP+yraj/gk3/uOfdet46O37t1rr/9Hf49+691737r3Xvfuvde9+691Byf8Axbch+P8AIavn62/Yb37r3RK/gD/zJfOf+JN3x/sf9yA/Huo8+rN0eL3bqvWtl8xf5ZO96f8Ami7m/mH7c+M2P+be2e6ej9p9G7i6mre2qfp6t6jr9tLSUU2/6TJVLouUgq6KjjgNIp1Rs8rCxKEKoCq516D9nRdexySMAYhKg4ZoQeB+3HSi7w/lkbs7/wDhXsD4E9P/ABr2j8G/jV2f3Vkd1/J/a+F33g+ws9T7XmqV3BWZrbOSctLJkq/IwU7u4mDqI44yyw6h7cDQKWaRtdRgZHSaWK+lSKK3XwArgk1DEKBwAP5DNcdAb3d118q/jd/K76f6K3j1Nt3Idk/Hj5S4zqz4u7SxGax2cpez+m9g0lTjeocruR6fyRQVmRpQEnjqCZRpDTAOzFnITE0pMeAQMfPpi9S8is1SUh3Dmh4ArXFfQ06N90bs/wCe3y++WPTPbHzF+KuA+KfV/wAfNgYndW0v4D2btzem4N5drVdSErMDWUmLH3FFj4FLTGGRQqqgj1s76g00kUcbIh1E/wCXpVHBdXFwk86+GEFAK1Jp+XV7/tF0b9Fn+VX/ADLSb/qLX/oUe9HrfQq9Tf8AMttlf9qCh/6E9+HDrx49CH731rrRu+KdT8+t39h/8KBvjD0F8RMb2rsH5G/OLvjrbM915XsrbnWsPV9X2XhMjsnK5VsBmYvNnocfiZaLIhaN1YO+lmbyABSqAqDK2mnDHEdIJJHV2W1QSVrqOrg3oRn9gp/Po7PZf8lrsTpfcn8sjuPbvVmO+clT8Ivh1SfD/efSk/YNP0yu4J3lqquk7Wwu5at1Qpj5clXIaB21SosLfqBtuIorEh9J8jTy6rdLM8ahohIOJFaUb5fLJ/1cRL+Un8srfPZvwMwnVHxS+EWwPi/v/N/zCOgvkR2f1nW9pYfeFNuvb2xtxU8+6exq/d9dJIrVIp1jC0GoyCOB/GrNJpb0jr4ldWrHGn+r8utwxOYAmjwqHADeQ+fofMeletm32l6MOve/de6JL3JY/KfowH/nQ5n/ANyX9+8+tnh0dkfQf6w/3r37rXRGP5kHw8h+d3xE7M+N8m4Krbk+6mwGax9VTSeCOvye0c3DuOgwVfUggxU1dJTilmmW5jV9YDabF2FxHIGIr0mu4DcQNCDSv+rz6rr+CnxQfbncHUe/Ox/5R2zPjv2D1jsjeGMwneK/I+h7PrNq5KlpP4XQ4/B4VpZWAzyB2edF/wAn8hLX5uomfUmrxK18qdILK38KbR9OEAHxaq8OGP8AVToue1f5b/zdxHzmxP8AOVpNtbfwfyG3i2Y2t2/8G13dhKrA1Wy0on6927k8P267yUgrosVDT5SWJYFUyELqLa4/fg8OnwqkCnH16dZLsMJyisQSQOBFRQ58xQA0416SW3u7v5juwPnl/M+238Yvgttz5B7b3fmOrxHuTNdrbc65h23ncxiZaDN0mSky6gZKOhWqqX8UTIziHUr6WI9uyhNKljQLw+f+bpHbSTCaVYF1Fz3VwENPz1f5PnXq9j+Xb8RYPg78S+tvjwmbm3BXbdk3JuLO18jB4Y9xb43DU7szmOoJL+qmpairengc2Logey6rBFNIJJCw6NrOA21usLGpFf5mvR3fbXSnojHUH/ZZvyP/APDZ2n/0NH791s8Ojz+/da611f56u4Ozdo97/wAoPdvSvXMPbvbWA+ZW4qrZPW1ZWx4jH7qrW2Gi1VLW52o/aoVSIGQVMh0xkaz+n2qtqHWGwKZPpnos3FnUwtGKtrwD5nScfL7euVP8Ivlr89t//OrdPy+6xxvxO2n8g/jdszoPrnAba3/gextzYjJ7eyxy9bufI5rb6xJNBI6KhhcKNDnQNQ1e3vGjgjVIzqzU46SC0ub2aWW5TwqqFXOr1qTinn/l6YvhR8HqzrzsT464btT+Ujsva2X+P28azF7a+Tb/ACTot6nBja+Iahw/c+J2RUTMTVZd40kNGsYkgZ72UoAPSy60L+JnyFP8vHq9ramCVY/pwABTXq/yfP0+VerSPhD8e+yOjdxfJ7I9h4rbVJF2V3fuHd2zMjiK2nyOUym1a2rnqKOfLzxoskRPkUrTyOSp1cD8s3EiOihTkdKbK3lhkkaQAAnHn0f32l6MukZ2J/x4+5+L/wC4mp4/2A96PDr3QF/Ez/jwstxb/fxVnH/II97690af37r3Wuv89/5eG8d2/wAyHavziwfxFxPzG2zVfHSo6czmyKzuSDqqu27mYcs8tJlImqnRamFqdgGj5U6n4DgH2vt5IxFpLaGB408uiW+t7k3XirGJoiuV1aaMOBp54qPl138o/wCWn2H8rfibjPgB1D0JtX4HdB9mbhxnaPd278VvPB9m19Nk0k1ZvZmJwiN9xPWVeiEnINUKiiNFBRdWrzSqWMjyFzSnDNOvRWpjjW3t7dYkDB+ONVa1IFPPP29VdfzResPlP8Zv5TXxR+POQ6UxxzHxE/mi/Gfqv4s02O3HR7um+RPU2xKPIT9Yb23DFF5WwtVlcgy0NVT1Dh0A1MEBJZjtMlYuBGfOnr0tOpIaXJFQe01pqPl6cTinA8M9W5/Hn4xfMj5F/wA0zaP8yj5T9L4n4nY/or43br+Neyun6Lsnb3beR7DO8c1Nn63e1TuHbqRQ0VPC1VKi07R+RikXJs5OpFhVf021H7OtwPdSSVmjCAD1qa/5s9X8+2OlnROPm7/zKjC/+H/tb/3Kb37rY6Ndt3/ixYj/ALV1Hb/znX3s9VHn11uUI23c+srFImwuVEjqCWVDQyB2AH5A597T4x9o6rJ/Zt9h/wAHWpL8bN/fzQdzfGPs748dCfBraG8epN6dqPhtu98bh7I2xtGN9mZrcaQb0zFXtjLBZK2eiiivFIGLLyArsE0mchiSQSE0IHD1r0G7P6yaBrdUFGY91aAUPpSteh67p/lKZTprvDpHfmwPi+fnXseLb28MZv7Ebr7jpeqMlsTLVlCPtpMFIJYTV0dbI7qIiG0AFZBYgmiXKOCtdH8+nptsaKRZlTxqVBBOkivpx/Po4/Y/wczu5/gbsjqvoT4rdffG/eNX2RsDeO6+kpt643cmIwtFgtxiqyRfdzCWCsmWNUqF/tfVRdhzpbgBysj6h608+rybeTbp9LCsLAiqg4AqD5Y/2erq6SJoaWmhfRrip4Yn8f8Am9UcYVtFwOLjjj2Xnjjo9HAV6ke9db6I53yL/Kf4uH8irz3/AFrb3onqw4dHj976r1X9/NK+GVf/ADBPgZ8hfiRiN0pszNdsbbwsOC3DMhlpKPNbV3dj96YuCujXkwVE2OWmlI+iyFvx7smnVR8DpuXxBGTEAW8q9Vr/AAm+FTbA3J8dMv2B/KP2r152b8ZsDFtHYXeFV8msTviXFVuE2sNrzb5xGDlkkVGy6o8vjMRkg8pt6lHtY7KyUMtR6U6K4I5IpC6WwU+uvj0jfi9/Lh+Z/wAcv5gGK/mEZDGbO3v2z8v+yuw9k/NLGR5nBYzb3Vfx/gjXIdbZjZMcLL97kTPjMalUlLGZGsgckSTkake2KlUGaChzkjqtrb7okqyzSAguxKUFFU8KHiSPOvzp1Xz8Lch/MF7O+DPzc+LXxn+IW3t59YdqfLH5QbZ238gdydl7d2j/AAaXcXYZbctRk9i5xDPkUolQRwzxuqknSQ+m3t2kKuryNQgDH5dJa3r201vbxBw7OA2oChLGtfWn8+HW4F8feuqvqPozqHrDITxVOT2H1ztDa+WqYG108+XxGDhpMtNTNYftNULKY+B6bcD2hlfXIW9ej63i8GBYvQdC8fof9Y/717b6e6JL0V/zO3sT/ljUc/8AVSvvXn17y6O37317qg7+d5vHv7YO6v5de6vjj09Rd49gYr5YYuoj2HXZOmwseRpEgpiyw5aqBWByeBJ/ZHqsQPa22zE48jxPp8+ibcjpuIXGWBwP4jnHy+3y9D0gab4KfJr5z7m+X3yf+WvSuM+MvbHYvxV7J+GPU/SWH7PxO/sZmdnZTzZTG793du7AAQK9XVTfbxwCMaEu7qqouvYmijURLkVyfl1t7S4uHa5fsfTpUVBoeNSft/Lrl/Lc+BEXTZ+OmyO2v5X+1eu9w/HnAYOkw3yMf5B0O/Hj3RtyB/t91YraAmaRXmks6xWIid7qNKgC8so0MI5e0/hp5enSe0tGaSN7y1USpQ69Wru82BIrnPzz0bH+Vf8AErvP4w9hfzMNxd0bb2zgaH5IfPztHvLqms29lcdkp9x9bbjx1JFhs5mYqABqaokZHU09R+6pVtQFwSjkZWC6fIdHUMbIXLGupif8H8urfPbXT3Qc9v8A/MrOwv8Aw0M9/wC66T3o8Otrx6C34gi3x+2D/wBQM/8A7kN73SnXjx6Mx791rrWp79/lrZ3YXzU+QvdmO+EFB8+Ni/MNpt17tyGd70j6irem8rh8KMOdh02EeaMZKhyy6WFSt9AGlgLEMvimHhiMPopxxWvRJdWR8d7gxeOW4Z00FOHz/PpRfMP+WVvn5rfGT4qfy+sL8eNq/E74eQbH3F2T2Nj9rb1xW5M30R3Vs2CWfqjY+EMEqvlqOprMhWSV9ZCGBVpDIdSqC2HhVmZ+81x5Y6UPDdSRokNIRQ6uDEH0zxBzU/5egc+d+F+b3SH8oLoHYe+ut9sdnfJjpjujpfaeHoMVlsK2B7Ox2287NR4PIiOgRYqCVqJaeJ4yASyl7hmICmPwy7NGdVfLhTj59F04nS1ijuV0FCKEZDEEeWDn18vy6Ob0F0x8v/ll85+s/mj8yPjni/iUnxO2Tu/Y/Tez8H2hhOz67tWTtvALDunN56pwoRaCnxzXSGnlQSa7L6vW4Ss8SRGJDqqePRnHHcTXCXUyiPSCNNQTmnnTq8r2l6Meie/OYD/QLkeBcbn2va/9f4iPfutjo0+2v+Pc2/8A9qTFf+4Efv3WupOZCnD5UMdKnG1wZgLkKaVrkD34cevdaTPwSj/mK93fys+wfiP0B8RtsZXqnsTvTt7Abc+R+4+y9tbcFLhD3U2c3Lk8j1vlk+6qXilhlp4agOEYKRpZkHs0bwVmEkjUIAx+XQZT6ySya3tow6szUfVSlWNajzpkD1p1Zb8i/wCWJn9g/LfE/J7avxSpfnNju3esNh9SdkbIy/c56jHS1R19tiDFnee26iaWL+J0+W8JiemB1Qlm9IUqfdEuSw06tGeNOPSibbFSXxxH4uAAtaU9c+eejE0fwe3LP0J8MNrdLfEzrz4sYrrDvzNdi9g9Ayb6xm8MLtjGZeumbJZ0bgVZY8rVVpiiqXjOqRfKE/scVSWNJGLNq4UNPt6cmtZpoIgkYQg5Wtafn58OrwQAAAAAAAAALAAcAAD2g6Ouu/fuvdEk7A/7KY2h+f8AgB/hbge9db6O3731rprzRti6/wD6gq2/+t9nJ72Oqtw6+dX/AMJq7/8AD8Xzitz/ALiO6P8AW/4/aT+vuo8unDwPX0Z1+g/HA/3r3vqvXfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6QvZ/8AzLffv5/35+4//dTL791scegG+D1/9ln65uLH7fK8Xv8A8veb3odePRs/e+tdQ6vI4+g8Arq6jojVTLT0wq6mGm+4nb9MEHmZdbn8Ktz/AIe90J4deqBx6me9de69/j7917qLSV1FXxmahrKWthWR4mlpKiKpjWWM2kjMkJYBl/Ivce/de6le/de6KB8urf3b2/8A9rJf+tye9Hr2ejRbX/49rb//AGpMX/7gp731s8en337rXUR8hQRVUVFJW0kdZPq8FI9TClVNoXU/igZtbWHJsOB73Q0r1qorSvXq3IUGNh+5yNbSUFOXSIT1tTDSw+SQ2SPyzsq6m/Avc+9UJ4dbqBx6zSzwQQtUTTRQwKodppZEjhVT9GaRyAB/jf3uhOB1okDJ65RyxzRpLDIksUiho5I3V43U/Rkdbgj/ABB964cetgg5HXP37r3RIsubfNWhP/fuoP8Aez71Trx4dHd9769176cngD37r3USlyFBXRST0NbSVkMTvFLNS1MNRFHJH/nI5JIWIDL+QTcfn3uhHWqjqSHQqrh1KPpKMGBVw/6NLfQ3/Fvr71Q9bqOos2QoKaop6SorqOnqqs2paaaphiqKk/0p4XYM5/4KD73Q8etVHUz3rrfXvfuvdEW+KRH+l75XgC1uyqP/ANx5vevPqx4dHp976r10zKiszsFVQWZmIVVVRcsxP0AH1Pv3XqgcesNNVU1bBHU0dRBV00oJiqKaaOeCQA6SY5YiVIuCOD7914EHh1jpq+hrXqI6OtpKt6STw1SU1TDO9NN/xyqFiYlG4Ppax97oRx61UHh1zerpI6iGjkqaeOrqVkenpXmjWoqEhF5nhgY6nCAgsVBt+feqHj1vqR7917oJO8v+ZY7mt/yqj/e/ej17pM/GP/mUeE/6i8h/1tHvw630YH3vrXUSryFBQCJq6tpKJZ5VggarqYaYTTv+mGIzMupz+FHPv1CeHWiQOPUv/D8+/db6xNPCsbStNEsSag8jSII10HS2pybCxBBufe6Hh1rUtK164UtZSV0K1NDVU9ZTuWVZ6WeKohZkbQ4WWElSQQQbHg+9cOPW+pHv3XuiUfMTis6IN/8AmqNJ/sbRJ791YcOjoUxvTwH/AJtJ/wBC+/dV6z+/de6itXUSVcdA9ZSpXSxtNFRNURLVyQqSGljpidZUWN2At73Q8evVHXqquoqFY3rqylo0lkWGJ6qoip1klf8ATFG0xALH8KOffqE8OtEgces0ssUEUk80scMMSNJLNK6xxRxqNTPJI5AAA5JJt718ut9ehmhqIo56eWOeCZFkimhdZYpY2F1eORCQwI5BBt7917rJ7917ojW5rn53dff0Xq3Oc/65f3rrY4dHl97611737r3UOiyOPyKSSY+uo6+OGVoJZKKpgqkinT9cMjQMwVx+VPI97oRx69UHqZ7117qM9ZRxVMFHJVU0dXUrI9NSvPElTUJELyvBAx1OF/tFQbfn3uh49aqOpPvXW+oeR/4t9d/1B1P/AFpb37r3RO/iz/x8XZPP/Kcn/W8+9DrZ4dHQ97611hqKinpYZKiqnhpqeJdUs9RKkMMS/TVJLIQqj/En37r3Dj16nqKerhjqaWeGpp5lDw1FPKk0MqH6PHLGSrD/ABB9+691ipchQVxnFFW0lYaWUwVIpamGoNPOv6oZxEx0OPyrWPvZBHHrQIPDrmtXSvUPSpU071USh5KZZo2qI1P0Z4QdQBv9SPfqGlade1CtK56ke9db6L78qf8Asn7s/wDP+/eb/wBy4vz70etjj0qOh/8AmTvXP9f7q4u//Uge99ePHoWvfutdRJ6+hpZqamqa2kp6isYpSU89TDFNVOPqlNFIwZyP6KD73Q9aqOpWpdWm41W1abjVpva9v6e9dbqK064ySRxI8srpHHGrPJJIwRERRdnd2sAAOST7917rDSVtHkIFqaCrpq2mYsq1FJPFUwMyHS6rLCWUkHgi/Hv1COPXupPv3XuiRfNsXwXUn147X299P+Wy+/Hqy8ejtp+hf+Cr/vXv3VeuXv3Xuo7VlIlTHRPVU61k0bzRUjTxLUywxmzyxwE62VTwWAsPfqHj1qo64VVfQ0Ph++raSj+4lEFP91Uw0/nmb9MMPmYamP4Vbn3sAnh1uoHHqS7pGjO7KiIrO7uwVERRdmZjwABySfeuvdY6epp6yCKppJ4aqmmUPDUU8qTwSofo8UsRKsP8QffuvdZvfuvdEjpP+yrqgf7RP/vNOfdT1sf5Ojue7da6/9Lf49+691737r3Xvfuvde9+691Byn/FsyP/AFA1f/uO3v3XuiV/AI36XzZ/7+bvf/eMgo+vvQ6s3R4ve+q9e9+691737r3SU3ZsXZ++ocTTbw27itxwYHN0W48PFlKZalMdnMbq+xydKG/TLHrbSf8AHkH3dHdDVDTpuSKOYaZBUDpV+6dOde9+690Wf5Vf8yzm/wCotf8AYiw96695dCr1N/zLXZX/AGoKH/oT34cOttx6EP3vrXQZ9fdOdZdVZHsLL9e7PxO1cj2rvWu7E7Bq8ZHKku6N6ZKnjpa7P5EyMw80iRICECrxcKCT7u0jOAGPDA6ajhjiLGMU1Gp+Z9ehM906d697917r3v3Xuve/de6JL3Hb/ZqOjL2H+4DNWv8A9RD+/efVs06OyPoP9Yf71791Xrv37r3Xvfuvde9+690jsB19sra24N27r27trFYfce/Kuhr94ZiipxFW7hrMbTmkoajIyX9TRxsyggD6km5593Z3YBWOBw6bSGONmdBQtx+fSx906c697917oi/UH/ZZvyP/APDZ2n/0NH7159b8uj0e99a6DnenUnXHYm4uu92712jitxbj6m3FU7s66y2Qjkeq2puKsxz4mpyuMZGUCR6d2jOsMPobXAPuyuyghfPB6beKORlZxUoaj5HoRvdenOve/de697917r3v3XukX2L/AMeNuj/tU1P+9D349e6Av4l2/uDlrf8APRVn+9e9DreaZ6NR731rr3v3Xuve/de6DTs/p3rPujF7dw3aOz8TvPF7T3ntzsLbtHl45XixW8tpVRrdu56m8TIfNTSMzJclTchgR7sjshqppXHTckUcwCyCoBBH2jgehL916c697917om/zdNuqML/4kDav/uU3vR49bHn0a/bv/FixH/auo/8A3HX3Y/Lqo8+naSNJY3ilRZIpUaOSNwGR0ddLo6nggg2IPvXDPWyKih6TWztk7S6+wVPtnZO38Ztnb9LNU1FPicTTrS0UU9ZMaiqlSJfy7sWY+7u7yGrmvTccUcS6YxQcelR7p051737r3Xvfuvde9+690Rzvg/8AOU/xcFv+UvPc/wDTtvej1ZeB6PH731Xr3v3Xuve/de697917oOusOpOuOmNv1u1er9pYrZm3sjuTcG763FYiOSOlqNybpyDZTP5V1lZz5Kmdi72NhwAAAB7u7tIauanpuKGOFSsQoCSfzPHoRfdOnOuj9D/rH/evfuvdEk6JBHdnYdx/ump5v/01Lx7159brUdHc97610j90df7K3rX7Uym69tYrPZHY+cj3LtGsyFP5p8BnoozDHk8c9xolCm1+R9Da4FrK7KCFNK8em3hjkZWcVK5Hy6WHuvTnXvfuvde9+691737r3Qc9v/8AMrOwvz/v0M9/7rpPejw62OPQW/ED/sn3YNxY/Yz/APuQ3vfXjx6Mx791rr3v3Xuve/de6R29+vtldk4qlwe+9tYvdOIosvjM/SUGWg+4p6fM4ao+6xmRiUEESQyepTf+oNwSPd0dkNUNOm5Io5gBIK0z0sfdOnOve/de6J585r/6Bchb6/3n2vz/AE/3Ie/deHHo1G2v+Pc2/wD9qTFf+4Efvw4dePTy6LIjRuqujqyOjAMrKwsysp+oI4I9+690HvVvU3XXSmz6XYPVe08XsrZ9FkMxlabA4dJI6KLIZ/Jy5nL1SiZnbVPUzSStdvqbCwAAu7tI2psnpuKKOFPDiFB6fb0InunTnXvfuvde9+691737r3RJOwP+ymNocjn7D/oUe9db8+jt+99a6bMz/wAWvIf4UNbf/W+zk97HVW4dfOr/AOE1fH8+L5x/9qjui3/obSe6jy6cPA9fRmXlQf6gf7172ePVR137917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XukJ2hb/Rvv2/I/ufuLj/yEy+/dbHHoBvg9/wBkzdc8W/yfK8Xv/wAveb37rx6Np791rrUe3T2j8lvnP/Mq/md/G7d/xYp/kNsv4Vr16nxWebtifopetN37s2cK6i3K8onp1z7Vtei1hlbX4o4ljJCui+1kMgRaV0fOla+v2dFV3C00uFE2nIGrToPlX1r6Hpn+YvyZ/mefAD4Zfy5u6Oz96UHYn8w7CZDtXYvbXxbwbJuDCfIPaG56qqTGZzJZTaoaGCp2bjv4fVvWQBllme7uTFITeMNIrqi6lFD6dNXckVrJC8shR21LWmokUJAoK/yFP5dGl7Xzu9qH4Z/y39m7F+X2SSi7r3vlH7G+QFRl/HDkYt0MctunatZUQz2SOinr6vEp5ZNSGnu2hrqrsTHxXJWhpw9OkVyv+KW4ilqC1S/8VTUimPsr5dD78aNlbc+H/wDMZzfx1+OG4Kzc/RXbXXlNvTe/X/8AeLK7xj6j3fjMXLUJuabL19RUvTnLuocxSOFInAUCyWbkPi22txkfl59KoEFruRhgJ0OKkHOacR6V9OHV6/sv6Peig/LkkbawHF/9ySX/AOpye9Hr3Rodr/8AHtbf/wC1Ji//AHBT3vrZ49Pvv3WutCTp/ce49w/ybvn5/M03X3ZuPe3zl+Ovyl7/AKnpXtZd0ZOnrtiVW2ewMRhdvYmLaCTrRVEVRT1dXURU1VRuPBJGousTXVieXTowwp6fy6LDY22szZRga/FwrSp4kfZ5D08uj/7x+QPyL+aH8w3avw67X+N0Hyf6Owf8t74+fKRtnPv6ToAU3de+cbQ1ef3z/eKGam+78lTVyUEWKN/tvHIygBX97gcRg50/Olfy/Z1q9hNwQhXxcV0atNPRq8eP+D7egh+T22/5jWL+Ov8AKh+MvzL7TTrTdPen81SfrLeu0tn7nosvV5f4suZ6zrzr7Lbw28W+7losdHNTh2m8rCSmeWQvCVNUciRpIzw4/Prc1uk1ulvcA1YilD8J48QeHEfPAPW3/wBada7P6h2TguvNhYyXEbU23TNSYmgnyGQys0ETymZ/JX5WWaeQlmJJeQ/0Fh7YkkaRtTdL4okhTw04dLv3TpzokWXsPmrRc/8ANOoOP9ifeuvGtOju+99e6qQ/nMfJTvL41/F/Z2Q6B2xPujd/a/efX/S+UpYY/H9vtXfUVbT5qVsuxC44OIkh+/Zl8WokMp5Cq0VWkq/AZ6Ld1lmitf0PiYgcONfLHCvrwHnjoqnw0+Kny865+W+4KdOjcL8af5fm/fi/nNrdsbIT5A0XbWc3b3rk5yrdj4PIrVVFTj2ekkmhMkRWNUJYktYByaVTwOog1GKfl+zpmztZI6gp4aspB7q1qMn5Gv5dEz/l7/LTtPuT+aV2z/Lg7h7CytZ8dPixQ5nLfHHHNTPRt2vQdeZ+mpNqbqquxcaUfKLDHMzpSmpkVxHwCqH25I+lTIq0c/nj/V/s9NwRO8gjeUPEtccCCpFPnkEmv7OnvavWGP772n/My+UndvyAyO7PkH8bfkB2LlfjjlqTdtRtZunqTrzbH8c2V1um3aaqSnq0rKtDAqzwF5CyiPkENdpGjdYlFVYcP9WB0mjhjuopLqRiJImOk1OCAPLz/wAvWwd8A+3+x+/vhf8AGvuft7Evg+zOx+qNs7n3pjHoDi3p85W05FSzY8hfF5Qqy6AoHq4AFh7L51CSso4A9HlnI8trHJJ8TKCfLNM9G99tdKeiLfFI/wDGX/lh/wCJKo//AHHm96HV24dHp976p1QR/MXzG7t6/wA03+Wv8aavtDJbA6W7Q2D8h832PiaHMTYQb1bA42mEG2BXRyxAS1sd6aO13GslAWt7WW7FInYCtKdFF/GJruCMsVB1E/OgGP516IvnO++6f5eOx/5t+zfihncn2p130f3D8edv9E9aKlZuhPj+na+Nij35jIM5kpqqpqo555WkWSeVhSPZmswv7UKiy6JJePpw/wBVOkU08lmk8NouFpTiaEgY8zn+XRlPh/8AHf5t7A+XHxm7d6/6AwXx7+OGZ6u7CyvzPrar5B0XbFb3fvLObdOS2NmcVt96qploZ6avaNwtOqqmp0kYKvLc8iMChao8scOlFjBNERL4eg0JY666q/Lypin+ToY/5Xnx+q+4+wd5/ODvTsbO9q9gbf7A3/tPpHE1mczseN6hwGQnmpt0UH8NjnFNVy1YmVImnR1iQNpAYgj13KQixqKA8fy6ptEGqSS4lYswY09FB8vnw4nPV9Hsv6P+gk7xNustyn/plA/25t791sdJr4yf8ykwv/UXkP8AraPeh1rowHvfXutVP5IZX5P/AC6/mi/M/wCM2b+NSd99P9DdJdWbi+O9PN3L/oVh6z7Ez1PBUZDf33cVRCcxJWVDm8DK2hI0S6qbAxgIhRXLaQflWvqPlTogu1N5PJAqGRkoaatIT+Fq+dc4p5edeg2/mS/Kn+Yd/Lo+I3w++Qma3pSf7O9mO1cnsHevR+36GPf+B7n6vpkqP7ubSyCY1J6enmxkBiqpMpBF5ZJZTqkaQEe7EwysyxiqDh5Ek9aRb2CCN7p1SdqhiDVVABIoDx4AU4k8OufzbzO4cHtX+Qz0/gO/c9hOsfmT8xKjOfJWM7nWdt41e+MXQbu3JsWs3JRyH7anxlfk8jhkpkmTRO8bNaSIqE7SuJSw7fKnGvS1bWJ7VY2JcV1aq0oa/l9lM/Z6HL+CG6s10Z/Of+aP8vjqjL5Gn+IPXnxW6s702rsCsqchuim2h25vXcVDS7sqqHd2XkqKuIVaVMvkoZakqSoKoGiY+25XaQanHSi2gjtyY4jjjQmv+znP7Otif2x0r6JP8xRes6IH9O0KU/8AWJPfutjh0dGm5p4P+WSf9C+/Hj1ocOgu75zeU210j29uHCSyQZjC9ab3ymLniLCWCvott1NRSTxFLEMjqrKQeCPbkX9ov29M3BIgcj0PWsjhOrcePiJ1L88sx8m6vdHzJ232xhptr7w/vjk55q7b8eSNHB0HPs/7swTyGLVLIzU5kKnklD7NGkPjGMjtK/b0GY4lFlHdFyZFfBBp+VOBFP8APx6Zeyuy/ln/ADCO/c/s7cvxFbfNBtb4+ZiTYGxq/uKr6LXYPbFTUj+HdmSQ5KphnyASTTZGDLpCqLKPdQI4EFDpznFcdWd7q/mIMeuiYGrRRvXHl0OnzL2D83dwfDP4D/G7u3t2Dpnem+vkZ1h1r3jLszNfx+q7H6kj8VHmtkVm4qFxpqKiBhG1ZE5JIVjdrk0AhYtJbnPE1H7P2+fT7C7EUVtuAFGoO0/tzXy8vPJ62MusuttndPbA2n1h19imwmytkYalwO28S9fkMm9DjKQHwwvkMrLNUSm5JLyysxJ+vsuZi7Fm4noQRxrEgjTgOl17r1fojW5r/wCz3df2+n+i3N3/ANu9vevPrfl/q+XR5fe+tdUnfz+/l1398Lf5f1V2t8bqMVvYWf7x6c6vqojQGsL7U33nJqPcNHHMpDUklbHEmPjrlYNC9QHUhre3IjRq0r0zcVMRAbTXz/wD8zjoBvjr8b/mJ1V8sdu1Wx+jcL8OP5fW6fjp2dkPk5jT35iu38pn+68vt6WXau/MJLWVFRV0MuNaQOz07JCY0kaRjdB7UySgkMraiPlSnRfa2uhWR4zGpyRq1VJ4k+fHh5ceibfyn/mD8j/kJ3V2NsXurubIZ7BfCfqbvTbfxp33LTZHAVHzywORqchST9s5HE5F1BfbdNRUCIKQP/nkLENDMxcmDopqlKkZr0lsJre6kUJOX0q4C0oONK186eoNP5dAHtvDZ/NfyY+of5htf8lM1h/mttvtLF1Wwu133HkspJsaiyfeJ23lerRs41Zp6lshRLL51qoHkAkVlCoranC7NN4IFBTOOPSeONY9t+tYlnrUZ4d3DHH0/Z1um7PyNfmNpbXy2Vh+3ymU27hMjkoNAj8FfW4yKprIdA/TpkZhb8W9ljABiB5HoTISyBjgkDp2yP8Axb67/qDqv+tDe69W6J38WTfcXZJ/6bk/p/x3PvQ62Rjo6HvfWuqKf5l2HyHbHzt/l6/HXeHcE+0fj12TL2fWdsdSx5OfBRdxTYnBTT4HEyZqjqKeeEwTRoyiB9bXIXkj2ttyUgaReIP+bomvgst9FbSV0sD5kev/ABfVd3b/AMkfkj8KOnf5uHxv+IrZfcfXPxY6u6l3b8Y9/wBVWjdJ2dmu1Nx09JvjZv8Aeipd/unoo6ieOGN5fJBKpk9LavbxQTBZSKOfL1/zdJVlay8a11EwoBRskqDWtT+Ly8x0c34FdGfL/YPe/RvZ+x/jbt/469Bdr9W4ncHyiiyPfNP23W773jX7IOUx25sVh6ionmxtTNk5kk0U9tKM6yNpLD3WaWFoSjtqYcMUp/xXV7W2vYr5ZoYxHC4JkqxYk0FCo4Z8/wDB0gf+E6PXuT7R6R7h+Y3eO/s73F8hNyfLH5KbIi33ks5mPs8ds/am6l27SYeiwKyrSIrSJUzJqhYxxSRxqR47liWaUJ4RIpT08vTpfbWlt4jXKBgSTgk8eFQCTQHiPKhrTrZY9pejHovvyp/7J+7O/wDDeb/3Ki496PWxx6VHRH/Mneuf/DVxf/Wge99a8+ha9+691qBbuyXzd/mDZ/53bj2z8WIN4909D9odidK/EDtDId2wdNp0/lNr5iQbe3cuw6urpxlIxZag1U8bLK/DMdRsa6o4YlAbSTQkUrX8+g34dzc3crOniBSVVg2nT64PEj9h6d/5rXya+bfwE6y+L3dO3N7w575d9kfGfa/x8+VmCwtQu7MH0tiIKdMju75X4fbmND+aWkyxlhSV4Vp/J6kcqCoajHiLpCVXVWvD/VTp+7YW7eM0xSQx0001Co4E+lfU9G0+b2U3BnPjb/Ko6+2/8rKv/Rt35291BjO6+z5JTiZ/kTtDPYCir85T1Va80b485mSeWfQsgaEsEtdLDcZYtI3muB9nXrnQiwRD4ZDmh4kjoVPiQlT8Wf5nnYHwX6JkyGV+JWY+O9T3/NSVmYrN5RdbdtVG76fAVm2afc1bLPNTippyagUc8xbSyEhioY0nbxbcSuKNWn5dOWcQtL5rSFqxaNQBqaMWzmv8ur3vaHo56JH82rfwLqT+v+ljbtj/ANPl96PDrY6O0v6V/wCCj/eve+tHoGPkjnsvtb47d97n2/UvR57bnS3aWewlXHq8lLl8RsauyGNqU0EG6TRowsb3HuyfGPtHVJP7NvsP+DrUk2xtrL0n8rD4DfP2l+SWaxvzPyvb3S2Ng7Rl3DkM1bam9Oz5cTurpZtnvVvSzJV0ilKh56dpY2JPpjFiZtI8kxh4Ch8vl0G4YY4NvjvCS8lVoa8NTDH+TozXb27vlF86Pl183Op838QG7XwXxaNRtH46xZHuiPpIbP3xntrLWYHf4V6inOeVKwJVSSgOgQKoKqAPegIoYxpahPnStf8Ai/5dbY3V5eOHi1qlKCpWhp/OlehF7H2J86d3dJ/yxPiD8l+72687737hN0Yr5Lbo2lmfvabd9FS16UMm2v4nhXWOeeTDtHTyVEZZfMzOHJJY6hdR4jgV/wBjPV7+OUvb25YqK58yRWlK+WKVI4eXWwx0z0/sHoHq7ZXTnV+ImwWwev8ADRYHbOKqclkcxU0tDHK9Q33GUy0s1RNI8skkjvLITdjay2ALndnYs3E9H8UaQxiNOAwPPoTfdenOiSUn/ZVtQf8AaZx/1gPvXVujt+99V6//09/j37r3Xvfuvde9+691737r3UPIgHH14JsDR1QJ/oPA3Pv3XuibfBCCjp+nc2lFWiuiPZO9XMoUoFkavXXHY/096HWz5dHV97611737r3Xvfuvde9+691737r3XvfuvdAl3zsvM762U+GwcXlrGqBIEuB6QPrc+9Hr1adL3YGIq8DsrbWGr10VmNxNLS1K3B0yxLZhce/Dh1s8elh731rr3v3Xuve/de697917r3v3Xuve/de6JZ3FFA3yf6QkeoCzLg8wI6exvKPunudX4t78Ot+XR0h9B/rD/AHr37rXXfv3Xuve/de697917r3v3Xuve/de697917ornXnWO5dv/ACR7k7Gr4dG3t44TAUWIm1A+SagKGcaRyLWP1968+t+XRo/e+tde9+691737r3Xvfuvde9+691737r3SM7DAOyNzhm0A4mpu31twOffjw690CHxQjij2HlRFMJlO4KslrWs2nlT7117o0XvfXuve/de697917r3v3Xuve/de697917ouXyc67z/ZewsZgtuQmeupt24HKyKGC2paGcvM1z/QH3rrY6HzEUz0eLx9LJzJT0dNFJ/weOFUYf7ce99VAp04+/db697917r3v3Xuve/de697917r3v3XuiTd5wUsnyg+MkstX4J4qrOGCn0E/c3RrjV+Le9Hqw4Ho7PvfVeve/de697917r3v3Xuve/de697917ro8gj/A+/de6LJ1X1puPa3Zu8Ny5OEpjsvFMtI+oG5ecOOB/h7159bPRnPe+tde9+691737r3Xvfuvde9+691737r3QedtKjdYb/WR/EjbSzoeS19CnHuC1v8Pejw62OPQYfEhIoug9iRwTfcRLRT6JradY+4bmx9749aPHoyXv3Xuve/de697917r3v3Xuve/de697917ouvyi6/z/ZnVNZtbbUBqMpNm8HWrGCFvDQ1nmmN2/oPfuvDj0O+EppKPDYijmFpqXGUFNKP6SQUqROP9uD79148enP37r3Xvfuvde9+691737r3Xvfuvde9+690Srf8cJ+Sm0WM+mQfw+0Wn6jSPqfdaEnrfn0dX3brXTZmf+LXkP8AqBrb/wCt9nJ72Oqtw6+dV/wms/7fxfOI2+mJ7p4/w/vrJ7qPLp0+fX0Z1/StuBYWH+w976p137917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XukJ2h/zLbfv/AIZ+4/8A3Uy+/dbHHoBvg6b/ABl64Nrf5PleP/IvP/X3odePRtPe+tdEc+QH8ub4j/Jrsvb/AHD2r1tVVXZG3KVqGn3Xtbdu69kZHKUXi8MNNuI7TrKRMisS+mE1auyCwUgAAPJcSRjSpx0kmsbad9cgz6gkf4OhD2v8NPjftHLdf5/Gdb0lXnertvbk2tsbM7gy+f3NksFg93QGl3DRU9Tn6qoJFTEzxszXYKzBSLn35riVgVJwevR2VtG4kVe5eBOaV40r6+fVf3yS/k/9G9ndR/Gr4n9f7LTa3xU637G3runfWzMdvDN0OXFFvCrOdrqjEZ6vlqa1pJK6Sc6Fm9AZVWyCwehuNOpnOT/k6SXm3LMI441GhTUitKV4n/iujhfEX+XX8TPg3k+wM78cuuqvaef7QjwEG9s7l92bo3dlsvT7ZpjTYmmFXuarqTBGgYu8cARXazMDpWzEkzy/Eel1vawWo0win8z+056O/wC2ulHRQvlz/wAe3gBewOST/rclvej1706NBtj/AI9rb/8A2pMX/wC4Ke9jh148en337r3Ws3/LX/4T0fHHpPa2594fKzq5t7d21Xyk7c7awzy9h7irdnSbUye4m/0fwZLZ+ErIsRVKtGqzSRVlLI+s+sgegKTMYxohOOPDz8+i5bQXBM14nfUgUJppB7cevmfnw6uZ+SnwB+K/yzy+x9xdz9cNlNyddNEm1Nx7Z3FuLY+doaCElosNUZLaNTRyVFCrMWSjnLxKSSqi5u2k0keFPT89nb3FDIvD0JH+DrlU/AL4p5HA/HzbmY6zGex3xa7Vj7r6PfP7l3Tma/ZnZUS1Ij3FFkchWSTVJBqpSIKp5IQdJEfoHvTSu7FjxPV4reKGMRpwGeNejk+2+n+ve/de6JHlv+y1aEf9+7g4t/ifeuvHh0dz3vr3SC7R6w2L3P19u3q3srb9JujY2+MJX7e3JhKxpo0rMbkadqacQ1NMyTQSqG1Q1EEiSRuA6MrAH3tWKmq9VdFddLio6J70h/LH+IHx5xO8MJ1jsjdOOxm++p8p0luSly/ZvYW4UquusyjJksPT/wAZyM3glkViBWQ6ahf7Ei+3nuZZAA3lnh0lhsLa3ZmjBqwock49MnpcD4BfE1Ouel+rYepMVS7X+Pp24eqJqPI5ij3LtobXIfGo+7KaoXI1aMw11MdXUyLM5LSBm596W4lRiwPHqz2VvJGIiuF4U/1f4eqvOmf5BvxRyHcvyA72+TPWVRvLsDsL5QTd3bYnouzN4Uu3p6fFOlXt6ryW2sDV0tK2uUyGopKuOQMAAVAJBfmuBSkPA5P29I7awqxe7FSpoucU+wfPOc9bAlPT09JTwUlJBDS0tNDFT01NTxJDT09PCgjhgghjAVERQFVVAAAAAt7Rccno2AAFB1m9+690Rb4pW/0wfLDi3/GSaP8A9x5veurHh0en3vqvVL3z/wD5Y9J87/m/8Luxe0cQ24fjh0lsruaDsHD0O663a2Yn3ZuGmpm2UYanDSwZBoTNHeT7WZbaTqZR9VMMqxRsQe7FOi28tWuriNXWsQDas0Ncafn5f4Ojv/Hn4F/Fb4t7A7A6y6c6tocJtHtWokqexqTL5TM7orN4ySY9sXbOZPcNRUVEoWB3RB5Bp1Ej1c+6PPJIQWPDh0/b2NtaqyQrQOanzqfz6QPxv/li/D74n7ywm++ktjbn27n9u47c+Jwv8T7L7B3NjKGg3hKZc5Trh9wZGopnD6isZliYxj9GkgH3t7iR00Nw61FYW0EpmjFCajiaZ444dGy6p6a6z6Q29V7U6r2jjdnYCvzWT3FWY7GmoaOozOYn+4yFbI9U8jlnb8atIHCgD228jyU1mtOnoYIoAREKVNT0J3unT3QSd4/8yy3Nb6/aj/e/ej17pNfGS/8Aojwtzc/d5Hn/AKej34db8h0YD3vrXRF+/v5cHxF+S/ZUPcHa3XFdXdkR4vG4Sfde3d57w2fkcjisPJ5MbQ5Q7ZraZKhIuFXyKTpAF7AWfS5ljXQpx6dIZtutJ5vqJE76AagSDQGoGPn0J+0vh78ddlb8XszEddUdXvWLA0O2qPN7kymb3bLjcVQU4pYY8VBuepq4qaRoxoknhRZHBIZiCb+a4lYaSf2Y6cSyt431hc/PP+Hqm35z/wAjHqPu5f5e/TXT+xGwHxm6M+ZHY/yC7x2rDvvN0uSqcX2XG24dyy4bM5SqlyEYqMpGqrTUEyeGNgsIjADL6N1y7nuAx8z1SeJ9KwQqPDYnXmlFNSafMn+Vfl1bp8T/AIM/GL4S4HcWA+OfW8OzIt25SXLbky9fms7uzc+ZneZ54aav3Rumpq6+SnhZ28FO0+hL3Avz7rJK8lA3l07BawW9TEKV+ZP+Ho2/trpR0Sj5h/8AAzoj/wAShSf7xEnv3Vhw6OhS/wDAaD/lkn/QvvZ49UHDpB9ubZy29Oq+yNn4CaCnze6di7r29iJqpgtNHkczg58fRtO5BsnkkXUbGw592jIVwTwr03MrPEyrxIPVOfxV/kOfCXqSj6p7G7E6vym4++dmbyx3bNdlKzsneGR21R9m0MheDI0W36Sqhx0kMZCMIZKd0LDnUOPama6diVQ46LbHareBQ8iAOPnUfs4V6PZ8jv5c3xM+VvY+E7c7l2BlMv2Ht/aFXsPH7lwG9t4bPrDtasrGrpcbVptmtpY5rSO+iWRC6hiobTYBmK4kiFE/n0rubG2uiGmXI8wSD+0dDFkPi30VmdqdQbLzexKXOYDojLYHO9Ww5nJZnJ1m2cvtqNY8RXpk6uoaoqJIwqlvupJAxA1A2HvwnkBYg01cerm0gYIGWujI+XRgfbPSnr3v3XuiNbmv/s93X39P9Fub/wB7f3rrY4f6vl0eX3vrXSA7Q6t6/wC6dibj6z7R2ti957H3XQPjs7t/MQ+akq4GOpHRkKvFLGwEkM0TLJG4DIwIB92VmQ6l6pJGkqGOQVB6J901/LE+HXQ9JUUHX2wdzQ0NVhcvt2oos92h2RuSklwmciaDJY6WmzGUlR43R2QB1OlbAWsPbxuZTThj5dJE261SukHP9I/5+lVmvgd8caE9bbo2F1fhNudhfH7p/tbqboDL0NblKOm2Pg+08C+Mz9AKNZ/BUioch2lrY5XRnkkVg7sTQyu5Oo8Tnp5baCJFWNANAIWnlXy6q2/l+fyDfi11L8fumW+T3Uzby+RO281ld6b4q6nsLdGS2tUbtO8KnMYSddu4qtTEOlPAKUoqU59WoOzXICiS7cMRCe37Oi622mAxK10lZAanJpUHBpw/l1sOqqoqoiqiIoVEUBVVVFlVVHAAHAA9oujnqLkf+LfXf9QdT/1ob37r3RO/iz/x8XZP/Ucn/W8+9Drfl0dD3vrXVPv8wv8Alb7B/mA/Jr4g7/7cwE24ur+im3zUbhpMfvLKbQy0VXmKAnFinkwzxVM0c0wRJBDKrKATqW9/aqGVY4j/ABVx/LouuraSe4T/AH2RRvKua/bxpw6Nh1F/L3+JHR/xl3B8QevuqKOi6F3ZHnI907Syuaz+frNxtuGr+9yM+Z3LmKmbIzyeTSYXapvEFUR6be2vGk1iSuRw6UfSW5ga3KDQwoR6j59d/HD+X/8AGH4obgq9zdJ7Q3Dt7K1lA+MJyfYG+Ny0NNj5EWM0lHi89X1FNGgVQEAi9I/Tb3aS5lkXS1P2dN29hbWpBhBFOGSQMU/wdCr8cPjD0d8Sev67q34+7EoevNi5Le28ew6zBY+ryVbDNu7fuYfO7oypmyk08oNRUPcRhwiKFRFVVA9slixqelaqFFB0PnvXVui+/Kn/ALJ+7P8A/Deb/wBy4vp791ZePSo6I/5k71z/AOGri/8ArT795dV8+ha9+691X/3v/LE+G/yO7Zr+7+zeucxP2Tltpx7KzOd2xv3e2zEzODiqfuohlMbtiupaaaoU+n7tovMVAUuQBZ+O4ljFF/n0jnsLW5OqVc/Ikf4OhXPwp+NEu/q3syu6zoMtvDJfHyj+LeSyObyWazMOQ6Roaj7iLaFZQZKolgkLHiWtaM1Mg4eU8+6+NIOB86/n06beE1qoNV05/h9OiEfOz+Ub098outPiZ8f8BstMb0R0p3vht/7g2nj95ZrBVGM27RwhJnwORlllqAVAZRTwyD9QKgEEhQlzqBMpyOGPP/V69IJtvClEthRDhhX8PoPTBIx0eT4ifBT4x/Bjbe8NrfGzr59lY/fu6qzeW66rI7i3Hu7N5fNVqqjefO7rqqyrFPGF/ZpllEaEsQupiSmeV5PiPRhDbw266YhT/V69G89t9PdEk+bP/Fi6k5t/xlfbv/W5fej1sdHZT9K/8FH+9e99a6DHu/aeY350t29sbbrxR7g3n1fv/aeCkndI4EzG4tqVeHxjzSSelUE8yFi3AH1492WgYE8K9UepQheNDTqlL+X1/If+J3RPRfxtru9OpX3L8j+uKTGbr3hka7sHdWa21/pLpcvJl4cpT7epa0Yg/asYhEIabT6bMX5JVSXUgYrGe3gMeXRXbbVB4cb3KVkFCcmmoedOH2Y6se+SH8uv4n/K7sTa3a/cmwMlk+wdn7fy21sTuXbe893bLrZsFmV0VVDl12tWUqVugXEElSrvECVRgtgGY55Ivg6WXFlb3X9qP2Ej/B0K+C+KfQ23qPpKjpNh01WfjrjZcV1BXZnKZrNZXalJPBHTzacnk6iWaqdlijOureQggEW9+8eWrGvxcerfR29EBWvh/Dxx0Yj2z0p697917okdJ/2VbUf8Fn/60H3o9b6O5731rr//1N/j37r3Xvfuvde9+691737r3UHJ/wDFtyH/AFA1f/uO3v3XuiVfAD/mS+c/8Sbvj/D/AJeC/j3UdWbo8fu3Veve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6JJ3Lx8qOiza/+4HM/wC81Lj3rz63jTno7Q+g/wBYf71731rrv37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SL7G/48bdH/AGqKn/eh791scegM+Jdv7g5X/wAOKrv/AK+ke/de6NP791rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuiO98W/2ab4uA2v93nrX/wCWbe9Hqw4dHi976r1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Qc9v/wDMrOwv/DQz3/uuk96PDra8R0FvxBI/2X7YNv8AlRqP/cg39760ejMe/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917okfYA/5yY2gfz/kH/Qo/PvWet+Y6O5731rprzV/4XX2/wCVKt/9w5Pex1VuHXzq/wDhNXx/Pi+cf9RiO6P/AHtpL+6g8OnDwNOvozr+lf8AWH+9e9nj1Udd+/de697917r3v3Xuve/de697917r3v3Xuve/de697917pC9n/wDMt9+/+GfuP8X/AOXTL+Pfutjj0A3wev8A7LN1ze9/t8r9QVP/ABd5vwffuvHo2fv3Wuve/de697917r3v3Xuve/de697917ooHy6/49vAGxNskn0BP+7k/p70evdGi2v/AMe1t/8A7UmL/wDcFPe+tnj0++/da697917r3v3Xuve/de697917r3v3XuiR5aw+atESD/zLqD1WOn9R/PvXXjw6O57317r3v3Xuve/de697917r3v3Xuve/de697917oivxRFu3/ljw1v8ASVR2JUqD/k830J+vvXn1auOj1e99V697917r3v3Xuve/de697917r3v3Xugm7wF+stzcX/yUf7370evdJj4yf8yjwv8A1GZH8W/3cPfh1s9GA97611737r3Xvfuvde9+691737r3XvfuvdEo+YnFZ0QbMf8AjKFJewLWHiTk296PW/Lo6FL/AMBoP+WSf9C+99aHDrP7917r3v3Xuve/de697917r3v3Xuve/de6I3uYE/O3r7hrDq3OXNjY8v8AU/T3rz6sPh6PJ731Xr3v3Xuve/de697917r3v3Xuve/de6h5H/i313/UHVf9aG9+690Tr4sG+4OyCQwJrk/UCP8Ad5+hIF/evPrZ6Oj731rr3v3Xuve/de697917r3v3Xuve/de6L78qf+yfuz+Cf9+83Cgkn/K4vwPejw62OPSo6I/5k71yORbauLFj9R+x+b+99a8+ha9+691737r3Xvfuvde9+691737r3XvfuvdEj+bZIwPUlgT/AMZY24OAWt+8v1t70eHW149HaT9K/wDBR/vXvfWuuXv3Xuve/de697917r3v3Xuve/de697917oklLc/K2e4PCT2Njb/ADB/PutM9Wr5dHb926r1/9Xf49+691737r3Xvfuvde9+691Cyf8Axbch/wBQVX/1ob37r3RKvgCCOl84CLf8ZN3x/wC7Ef096HVm8ujxe99V697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917okvcht8p+jDYn/AHA5kcf9RL+9efWye3o7I+g/1h/vXvfWuu/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdIvsUX2Puj/tUVP+9D3o9bHHoDPiWCNhZf/wAOOs/6FHveevHo0/v3Wuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6I53wf+cp/i6Lf8ped5/p+23vR6sOHR4/e+q9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690HPb/8AzKzsL/w0M9/7rpPejw62vHoLviD/ANk/bB/6gqj/ANyG9760ePRl/fuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690STsC3+zM7Q/H/ABb/AK/n0j3qtOt+fR2/e+tdNmZ/4tWR/wCoGt/9w397HWm4dfOr/wCE1XH8+H5x/wCGJ7oJ/wBhvaT3XzHVzgHr6Mym4B/qAf8Abj3vqvXfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6RHZjtF11vqRLa49o7hdbi41LipWW4P1F/futjj0BfwprKiv+N3XlVVaPPLBlC/jQRpdctMoso/wHvQ68fLo1fvfWuve/de697917r3v3Xuve/de697917pK7p2Zt/eVPBS5+j+7hp5BJEurTpcENe9j+QPfuvdKSmp4qSngpYF0Q00McES/6mOJBGi/7AAe/de6ze/de697917r3v3Xuve/de697917r3v3XuiU5iqm/2cyhpCUEH+j6CQegay9yf1/W3vVPPrfl0db3vrXXvfuvde9+691737r3Xvfuvde9+691737r3SM21sHbO0crubM4KhFJX7uyCZPOShgfu6xFKrLawtwx9+63XpZ+/da697917r3v3Xuve/de697917r3v3Xugn7vdo+tNyOttQpltcXF9X9PfuvdJv40zPP1NhZH06jVV99IAHEo/A9+690Pnv3Xuve/de697917r3v3Xuve/de697917pIbr2LtzerYV9wUX3jbfyS5XFnVp+3rUACy/Q3+g96p1uvSuVQqhQLBQAB/gPe+tdd+/de697917r3v3Xuve/de697917r3v3XuiTbkrahPnFsKhVo/tpOsM3K6lAZNalwLP9QP6j3o9WHDo7PvfVeve/de697917r3v3Xuve/de697917rhIiyxvG4ukiMjj+quNLD/be/de6Sm2tj7d2lPkKnCUf2suTfyVjatXlbVqv9B+feqAdbr0rve+tde9+691737r3Xvfuvde9+691737r3QBfKKeSm6D7Mni0+SLb7MutQy3+7iHKn3o9bHHpS9Fu0vUHXkr21y7XxjvpFl1NACdI/p731rz6Fj37r3Xvfuvde9+691737r3Xvfuvde9+690jt47D21vuDF0+5KL72LD5OnzFAurT4a+mN4ZfofoffuPXgaGvSxAsAB+OPfuvde9+691737r3Xvfuvde9+691737r3XvfuvdEppJ5P9mpqIbr49Ex/SL8QE/q91630db3brXX/1t/j37r3Xvfuvde9+691737r3UHJ/wDFtyH/AFA1f/WhvfuvdEr+AJJ6Xzl/r/pN3wPx+Mgv9Peh1Zujxe99V697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917okncv/ZU/RljY/wABzP8Aj/ykv78OPWzw6O0PoP8AWH+9e/da679+691737r3XvfuvdJ/cu69s7MxozO7M9iduYo1dJQDI5mup8fSGtr5hT0VKJ6llUySuQqLe5Pu6I8hogr03JLHENUhoDjqdT5jFVeSyOHpcjR1GVxEdFNlMdFPG9Zj4skjyY+SrgU6kEyxuYyw9QU2+nvRVgoYjB6sHQsUByOI+3py916t1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SM7E/wCPH3R/2qKn/eh78etjj0BXxLFthZcHn/fx1n+9D37rx6NR791rr3v3Xuve/de697917r3v3XugI69+TvQfa/ZfZHTvXXZ+3N3dl9Q1YoOyNp4l6ySv2pWa1jamr5pIVgZlZ1VxDK+ljZrEEe3DFIq6yMdMJc28kpgRwXXiB5dDv7b6f697917r3v3Xuve/de697917r3v3Xuve/de697917ojvfP8A2VN8XPx/lee/2P7bce9Hqw6PF731Xr3v3Xuve/de697917pL4/e20MrujO7Kxu5MPXbu2xSY+u3DtylroJsvhqPLBmxtTkKJCXjSYKShYc/7b3YowUMRg9NrLGzmNSCw4j06VHuvTnXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690HPb/APzKzsL/AMNDPf8Auuk96PDrY49Bb8Qf+yftg/8AUDP/AO5De9j59ePHozHv3Wuve/de697917r3v3Xuve/de697917r3v3Xuve/de6TuJ3dtfPZXP4PC7gxGVzO1amno9yYygr6eqrsHVVcRnpqfKU8TFoXdAWVXAJsf6e7MjqAzCgPDptJY3YqhqV4/LpRe69Ode9+691737r3Xvfuvde9+690SPsDj5NbQ/8AIf8A70Pesdb8+jue99a6a81/xash/wBQVbb/AF/s5Pex1VuHXzq/+E1n/b+H5yD+uI7o/wDe1k91Hl04eBp19GZP0L/wVf8Aeve+q9cvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690hO0P+Zbb9/8ADP3H/wC6mX37rw49AN8HRb4y9cD6f5Plf/dvP70Otno2nvfWuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6JJliP9nVoR/37uDj/AJCPvR4dePR2/e+vde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690EveIv1juYf9Mn/E+9Hrw6THxksOo8Lb6feZH/AK3D34dbOOjA+99a697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917oje5h/znb18T+Orc5b/bvf3rz635f6vl0eT3vrXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdF9+VH/ZP3Z3/hvt/7lxe9HrY6VHRH/Mneuf8Aw1cX/wBaPe+tefQte/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917oklISflbUf0CTW/wBf7c+6+fVq46O37t1Xr//X3+Pfuvde9+691737r3XvfuvdQsl/xbq//qCqv+tDe/de6JV8AhbpfOCxH/GTd8fX/tYD3odWby6PF731Xr3v3Xuve/de697917rX8+U/8xHs34ifMDqnbm6/lX0R2F112z35Q9Uj42bb69li7N2fgsvH+zkq/etNLL/lVKSnmSo0KzkBdQ1BTBII3Q1Uqaca/wCT/V9nRFPe3EFwoEiOpahQDuA9a/Lzx+fQt/JD5XfN/fnzM7A+Gvw/wG0ev06w+O9F35Vd3dgbfG7Nv72ytdkIqCk6uxeM5+3qpGkBFQYyBySQB7qsEcag3CnJpg9OS3d1O7pYso0rqGoEg9Jrsf8AmjbopviR8eN34p9v9X91d7/JvZvw2yW6d54h8nsvYXa+UiKZzc0mHhe8tFqTXDHMyhNZV7hAxqII1kYNUgCoHn1Zr6Z7ZHSisWCsTwGOP2dBt8Wf5mvdG2Ogvnz3X8ut/wDX3YeF+H3eG7/jxt3I9fbFqtowb/3xtDQFy1KSQjUta1TTIqxK2lVkcMbANcwRO6xxAgnJr5fLplL67t7ea5vSpVCVXSKVpwb/AGK9cPhX/NX3V2H8iut+r+w+4+vO+9t/ILrKk7D263U/XGX21V9F5ytq9MOw96VkylK0ol45Z2OtWGu1uGu1mGUlBpINMnj0zFvBWZVkYSKyau1T28ME8K/s88dGS+B3yq+YnzB+Tnymz279t7f6a+Lfx67T3R0NtbYOT28uS3t2tn8LClTD2dQbvWQGloNLxj7bSwcsVCqVLBPMkUa6NJ1evr0Y2st1PKZC6mMYoAag/af9Xz8urh/aXoy697917r3v3Xuve/de6JL3Hb/ZqOjP+1Dmv/ch/fvn1by6OyPoP9Yf71791Xrv37r3XvfuvdBl3L2ttjpDq/enau8Jmi2/svCVOWq0jt56uZbQ0OOpr/7sqZ3jgS/ALAngH3eNDI4UefTU8qwRGVvLrWZ+VvY38x7sbqLoTtnumXatH8fu9e3ts1X+jDam2P4PuTpvEQ7pWTadbvHcj6nyH8QpfXDZ/STyLkezKJYEZ1WoIHH1+Xy6DV1LuEkUM0gVkdsgYKivGvn+XRw+1P5j+8Onu5/5xdL1101nu0t3/CHqD4q7l25s7HaBWb4m3/tmpmraunaMNI8GKSaSuqktqaGB1UgkEJlHiLHGeBJ/z9G0r/TNcTj8Kr8/L0/LpZ0X8xjcHRv8sjuP+YTvnt7YfzLwm1cHhNy4Ch6a2lTbGO3q/OZSi25V7G3LTmomcTYqvyEf3jVEUU4jhe6BnUjTxoZVjClfWvVoZ5fpmnaRZBxBAoPz6YPh78mvm72J82z1Hv3vbpLtbp6h6ixPdO5qfZHWj4DMYDFdhYBMrs/a38bgd42qqKWogSZzMS6X1qHuquyQwCDWFKtwyf8AV9vSWC6vm3D6d3R4wKkhSDnIFakf5a+XUHaf827eu5/5RvyM/mJDYuOpd1dPb67i2piNrGnkehrYNhdwL1/iaiogE12JppkkktILspJHJHtrwU+oEflT/J0pN1N9A1xjUC1PTBI/ydO3ZX8wruHcPzc6p+KOF3tsT4n7b3d0Z1f3Fs3tLtfaSbrwHyT3T2BAtRleo+v56uamgx9ViY7mQyVHnla4jVvSPe44k8MuVLZPA9anuZBcrbrKqMVrQg59aHh/Mn5efRa+m9y/O7N/8KOPlB1ifkjgm+PXXvxP6T3xuzq+t2xVVWGy+E3A1LR4yj2hhnnYYvKLkXrpqnJrVNqifSyy6tC1Z4ynDt8h5g/b1dI5hMO/vxqNDpK+QA4A18/28RTZo9pejHr3v3Xuve/de6RvYdhsjc9/p/Can/eh7917oCviZ/x4WX/odx1lj/yCPeh1s0p0af3vrXXvfuvde9+691737r3QM/I07wX4/d4P19n4dq76j6l7Dl2huWogephwO44tp1cmHy8kEZVmFPOEl9Jvxce7xisgHz6amNImPyPWsH8Wvlr3L/Ky/lVbf+R/y87B607Z3z8gu9Ns9dbG3htTY0OJyu2X37lqnFneHaOVpY0qs5Fi2iatlgZdegKiyfuEe10qhpAr5p6cDXgKfP8A1fIms5GitjLEAASeJqwAFSS2K0GeH+zZv8Ff5ge6Oze/N7/EjfXbuzfkpvGk2jleyNq/IHqrr+r2b11RYSBIY/7tbjxc+lGqYJJ00zQTEOQyEm4YantlSMTKNIrShNSet2W5tLdPYuwdwpYMqkKBWgqTUV/Pyrw6C3pH+Yl2Ntf+YdsH4edrfKfo75MUvcW0t55rHUXUnXsu1a/q3MYCuKY7HZfMwySQVscqq0TapPIOWKAgFtNbq0RZVKkHFTWvXo9wlW7WOSRZEINdKkFTXFfXj8vPodvij8qvmH8rPnV8ptsy7c29058TfiVvdupzjM3t4Zrd3fW4crjJ6/H7w27uTWn8OpKLxoZo11BtaJpLFyrUqRRLpKnV5GuOldvNc3ExdXXwxgrQ1r9v2fb/AJOrh/aXoy697917r3v3Xuve/de697917ojvfF/9mn+Llh/yl565/FvG3vR6sOjxe99V697917r3v3Xuve/de61it17g+d9Z/NQ/mEwfE19r7Ix/WvRXW27sjn+w8Cm6sH2lXeGOqg2VtulUaqOthjiqAJ2YKT6T+oezMGExxpMCa0oQfl0HGW9W5nms3UFRkMCa5J/ZT/VwrZLlvlhvXuz4TbM72637t62+J25avOLgN77w7g2rDuzb2LzmA8+O3PhcPiJZ4lkearhMlLrJbxAg2YElgQqspRlLeYpjpebt5LVZ0cRVNCWFR+z/AIronfTv84PdGb/lXfMz5mGn253t2N8O27MxUuR2diqvaW0ezqnas4gwe4qTGVK+WkpWDtUVSrHcQxEqpY29+nhjjkXTUg+Xn9nWrK7muLZ2crqUkBqUUjNGp5D8+mjrr+Y98vOpch/LV7O+U39zd6dXfzR9xdUdXbF612NtSHbG7+g9+dpbRTemFzm5M7OzLlaJYXWCrgWzpyycrzSQQEERggjp63+uDAzsrKQOAoa+tf8AAPTrFkv5hnzz7e65+c/d/Q+29gbb2N8C+/e3dnVO183teXN5z5C4LqC75nY+3q6abTjquWJQ/wDEABZ5UQAC5V0x26hQQSW6SC4v5DMylVSJiASK1A8vWvl59Gh6l/mtwby76wexexOsn6u6irP5fOyvmfuftLO5OOPG7M3Xm66Mbl6lr6mZliqKugp3Zx4fU7JYAlre6vbECig1r/q/4vp6PcFLFpGAUJWnz8+P+D/N0dH4Wdz9ofIDqzN9sdhbex239vbt39uTI9MLQxtDUZbpmXwvsrN5ONmb96pQyvr41rZgLEEtTokbaU/PpRZTTTxGWUUqTT7PLo33tjpZ1737r3XvfuvdBz2//wAys7C/8NDPf+66T3o8Otjj0FvxA/7J92D+f8hn/wDchve+vHj0Zj37rXXvfuvde9+691737r3XvfuvdU47I+aHaPZX8z/tf4sTdw9b9SbP6bhwBxnS+59jTVnYPeeNyWGGSyW5tq7xrJYIoYodQZYqYyOFKkxMA7BaYY1txJpLEjiDgfaOidLyaXcXtdaoEpRSMsPMg+nljz9Og1/mnfObtr4PZ6TtLD/K3obbGyMFP15TD4y7j69lz/Ze9zuLLx43Kyw7mpJGmoxKGaWnbxKiopuytp17t4UkXuU+ea46ruF5PbyExSrjT2FSWNeND6/t/LoX+6Pl5353h2V1R8ePgxldp7T3tunaM2/+1e1N+7Zk3Xt7q3Dvt6PM4XBzYM2E1TWvIIRJZgrFAP7XvQgjjRnlz6U6s95cTypBaUUmuosKgUHCla8egn/k+ZnuXO9m/OCr+QdLPRdvU3ZG18ZuuF0SGCrnxtPkKT+LUdLGAsUNUVMsSAcIQPd70KFTRw6Z2Vp2eY3C0Ian2j1/Pj1ej7L+j7r3v3Xuve/de697917r3v3XuiSdgW/2ZnaA/P8AuP8A+hR7159b8+jt+99a6a80bYuv/wCoKt/9w5Pex1VuHXzrP+E1f/b+L5x34/3E90cf1H99pPdRmnThyD19GYcAAfSwt/tve+q9d+/de697917r3v3Xuve/de697917r3v3Xuve/de697917pCdof8AMtt+/wDhn7j/APdTL791scegG+DpJ+MvXJP1+3yv/u3n9+68ejae/da697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917okeWt/s6tD/AF/0dQf72fej148Ojue99e697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917oJe8bf6MdzX+n2o/3v3o9e6THxjFuo8IPp/lmR/x/wB2j34dbPRgfe+tde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690Rrc//AGXb17/4i3Ofj/an968+t+XR5fe+tde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690X35U/wDZP3Z//hvN/wC5cXvR62vHpUdEf8yd65/8NXF/9aB7t1rz6Fr3rr3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdEjpP+yraj/gk39f+OB91r5db6O57t1rr//Q3+Pfuvde9+691737r3XvfuvdQsn/AMW7If8AUFV/9aG9+690Sr4BEHpfOW+n+k3fH/uwHvQ6s3z6PF731Xr3v3Xuve/de68eRb+v9CQf9uPfuvccda2GK/k5/NiLLNtfcPy46M3T1HD8zNw/JjHVOd6Mkyfb9BtPL1bTUmx4N61UutZoUIQyCXQTze1lC4XSfGQdXD5U/wA/RO22SEiNXAjB1cKsT6V9OrhtmfFXIbR+W2/PkoN+vX4rePTG3+q6bZr4lIqjGVeFykde+cbKrJpkDrEFWPxggsbmwF6zXXjKMUNa9O2u3m2kZtdQV08PnWvWvp/Mz+IfY/TPS3TfSeP712r1hgd2/NxO/s98pexNoz5HYOzsfV070lPsabb1D9zJTZfUFqIa8BVkUFRJH6yqmORpe6Ly4imT/wAX0WXNvHax+FdMQGIIcGgU1xUfI/t4dG9+L22+yfmd0j8jfiX2lujqHvnrTHrWZjqD5e9Q7CG0Nhf6Q4FWPCUG7Nql4Rl8xQytHVV9VDqWdFkSV2LxyikpETeOMMeKn06ftUN3EbOUiSNeDqKCo+VT0ev4S/G35kdFSbbwff3afxm7A2TtjYNNtijj6p6Ol6+3bU5yjcJTZWozIlWIQ+IBZIhAS7XY+olvaeWWJhWMMGPqcdL7a3u4+y4ZGQDFFoftPEdDZ8RPi/k/jHR90UmS33/fkdqdy7n7RoZP4SMU2Coc9DDFDhZR5JPM8fjJaQWBuLD3W4mExBApQdXsrVrVGVm1amr/ALHRwPafpb1737r3Xvfuvde9+690SbuP/sqjoz/tQ5r/ANyX5966t5Do7A+g/wBYf71731Xrv37r3XvfuvdBp3D1VtTu7rLeXVe9qNa3bm88LU4mtQ69dPK1pqHIQFGU+SmnWOeP1WLIAbqSDdHMbah03LEk8ZikyD1Rnhf5Y/8AMp3X171L0z3388Oq939WdUb8ptwUdBtfqGvxWezm2sLnhW7ZwmYyM8qGaSjox9ujSMwBP1YAH2rW5hWraSSePRNLtd3JojEwEcZBA05I9Cf8w6NhjPgD3Z1v8qv5kHy66b7t2TjexPmZ1/0NtbqzE722JV57bPV2Z6g23NgMnkd3UdNUIctT1wlJhij0FATrDWsU6utFDAkA9GjxPWR4iAzAAVFRjhUef2dVW/M/+W73r0Z8Ff5ufbW6u09v9hdwfNLY3S2NyO2Ottp1+0ul+tU67ztNSZPdeG2EzTJHLPHLJUV1THD5XKB5C7XPtQpjllVYaqR5nOei2ZZ7ayme9pKD5INOPP1P58R0tv5cPcfyG6u7d+NfSGxe2eofnz052bsqi273lu7prYNLtHdXxaG2Njwtgsv2VugyGWtoamqvQUtPVaJwwcLCjFSXLijxkupVl4ZwemtuEkcqrDKsyN8VBRlxjNT+eB6nrur/AJG/zA258VO+/gP1X8yOsNufEbuHsvd+/qDDZvqnJZXsjE02+N9pvfO4Kv3Ms4WWFpo0K6TfUOGCEj20s9uvcFOr1r0oksr+WsbzKYyeGmh0+lQejrfPD+XJ8ofl31anxe21391BsP4zydSdcbBNRX9W1+R72wG5dlVNO2R3bs/f8NQUoHq4aSBEECo8Y1rqIdiaRSQKp1gkn0OOn7mC9kceA6Kop8S1NQc5rwI8ulnSfy5+2+tf5gfX3zQ6L7w23isPk/j/ANY/G35J7U7F2rX7q3L2dsLqtmlwWUwG4Y51NHlpZBFJNVSMLsGL+QHT7qJIfDKuprWoP+fpxobv6oSxyAJpoVI8/Uen+x1bz7T9Leve/de697917pF9i/8AHjbo/wC1RU/70Pfj1scegL+JYtsHLf8AhxVn/Qo9+68ejUe/da697917r3v3Xuve/de6Su+tuPvHZO8dox1v8Nk3VtXcO3I8j4RUfYPnMTNjFrft2ID+Iy69BI1WtcX92U6WDeh6pIutCnqCP29VO9r/AMq+t7H+APWnxEfsrbFXv7qPfu1uzdndjbn2NDmNsjdO1dwy5miiyW05ZH8lK8cvikVmbVpF1I49q1ul8bWwxSlP9X29FL7XIbIW6SUdWDBqVGCDkeYNM9GB6w+PPyuqere59jd/9q9E1m4uwuutz7I2jvHovqev67ym1KzO4aXFUuXrKiWqLVCU7Okwp0C3ZQA4HHttpYhQxg1Brk16VpBcsGS4ZdJBHaCDn8/29V8fEn+VB8xup+xfiPuLv75OdEdjba+LuyNzbWafYvSLba7F3xVZnJmuoqrL7xqnEjLGmlJDJqJILaS7Mxe+rRTrVTU5Pp+XST92TNRWcBVBAAGaH1J4n/UerVvjJ8Ysl8fd7fJPdddvkbvp+9+0oewsZQfwgYx9q0kOK+wOKmmWRxUOzEsZAFFgOLk2YuJ/HoaUp0rsrM2msFtWog8KUoKf7PRvPafpd1737r3Xvfuvde9+691737r3RHu9/wDsqX4u/wDUXnv+tbe9GvVhw6PD731Xr3v3Xuve/de697917ol2xPihk9nfJf5M99y9gff47v8A2dtba9BtZMMsE+0p8DRy01RkGyHlIqNZcNGuhfzqPtSZwURafCekCWbJNNLq/tRQD04/5+q8+wv5VPyOo+rPi7tXoX5PbN2zvDoT5Rbx773TVdgdeVW49i9h7e3ZBNTrtCs2nFO6CSmBiZZnvqZpSGQ6D7cN0pbI7SKfP9vSddtkSMaXq6sWFa6c/wBHhw/yenRc/k18Du9Pjr/Jp/mt7O3zvzZHafafeeyezt+U0PUewD15tqgirpfvY8HjsFCWLO8byLKVRVtwAQSfdXk8d1SMU+3/AD9OQwfQRSTztr/EaD8yAP8AB1E+Cv8ALF+YfcvX/wDK+3789vkP1vv/AKq+GezOmu7eh+qOvevsjtHe2F7NwuxYKTZydjbzmdVykWLoXpYplSCMTSxNrQkmRqOYlBXSdX2+fr05Cl2zLIZF8PiAFzpPAE/5QB0XT4jfEb+Y38iNo/zMtl9VfIXrf4+/Hnu35wfJjb2X2VvbrTI7o341PmMrFT7r3LtfckJhFEtfA8MUIAf9LOpjYAlQXiiKGVSWABFOi5Iby5E62soRGdwQy1NeBI4Ur5cfXoZf52nwz39U/Eb+W98QOksgRU7j7/6S+MXZ3YENBLFuDKdSz7clodyy11bSq8oppqmnWskikkIDGwNyxOopZZHZge0ZI+Xy6fu7a1igjhcdxogNPOnFh5gcc/IHrZT6f6x2/wBK9UdbdQbUMx211jsfa+w8E9Sb1EmL2rhocLRzVBuf3HSEO/J5J59ombUxb16No0EcYQeQA6Ef3Xq/Xvfuvde9+690HPb/APzKzsL/AMNDPf8Auuk96PDra8egt+IP/ZPuweb/AORVH4t/ykN731o8ejMe/de697917r3v3Xuve/de697917qoH5U/Av5SfLHuen/vb3/1Tsz42UG59ubixuO2B1dVYP5F0ke3K8V0GLg7eFQZoopLGOXwSIjA8x8W9rI54Y0wpJ+Zx+zopubK7uZRWRVQfwrRuP8AFXh8v59Fs+TX8qb5o9s9w/L/AD/XPym6RoOnvlBUdMzYvaPbXTNT2HvHrdetKOmp8tHg9zVUjMjVbQs8Ukbg+oFgrDUdi6UgaxkcKYHXpdtkJZYXor01EjU2PSvDoe+3P5ePya252N0F278JfkP170pv/am3KvanyIq9/wDX9dvDEd0Y7+A0+OxdXR4mKV4qGWCojkkEenSF8dm1IS2vqVclZV7fl1b93vEA9s4WQeZFRn5dD18Bvhx3D8Ych3hvPvbu7F93dj93bnxW48xm8NtmXbVBQvjoqgSQxQTyOzB3qDpUBVRVAH14rcTpMAEWlOrbfZTWhkaaTxC5rwpT5dWOe0vRl1737r3Xvfuvde9+691737r3RJOwDb5M7R/x/h//AEKPej1vz6O3731rprzRAxdff/lSrf8A3Dk97HVW4Z6+dX/wms/7fxfOPmw/hHdPP9P9/tJz7r6dOnz6+jOv6V5vwOf68fX3vqnXfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6QvZ5t1vv0/9mfuL/wB1Mvv3Xhx6Af4P/wDZM/XP/UPlf/dvN791s9Gy9+611737r3Xvfuvde9+690WTr75nfFftXs3K9Ndd96df7s7Rwr5GPI7JxWX15uJ8QSuTSGGZEWYwaWMggd9IBJ4BPt1oZVXWVx0mjvLWWTwY3BYeVc9Ovbvyz+NfQmWw+B7j7q2B17ms/UQUuKxe4s5BT11RNVSiGnElNFraFXYgK84RT+D78sEriqqT16W8tYTplkVTwyfM+X29K7tjvXp7ovZX+kbt3sbauwNkEwLHuTcGTipsfUtUxeeBaIx63nLJ6wIUY6fV9OfdVjdzpUVPTks8MK65WCg+Z6Yervk78fO69mZzsTqnt/Y2+tjbYg+63FunA5unqMPhKcUjV5lylXJpEAEKPIfJawU3+nvbQyoQrKQT1SO6tpUMkThlHEg46zdLfJXoX5FUmeruj+1dn9l02164Y3cL7YyYq5MRWMWCR1sEipIgbQ+iTToax0sbe/PFJHTWKV63DdW9xXwXDaeNPLrNs/5G9Fdg9mbv6b2P2pszdfZ+wcdTZbeOzMFl4cjltv4+qmFPDUZAU2qMfuMqMqyFlJswB9+aKRF1MpA68lzbyuY43BYcQOhq9t9P9e9+691737r3XvfuvdEiy/HzVoSBc/6O4P8Aez71149Hd97691737r3XvfuvdYaiogpIJ6qqmipqamhkqKmonkSGCnghQyTTTSyEKqIoLMzEAAXPvwBJoOtEgCp6q57O/mjdFDsLqPrT4+9h7C7h3Tu7uXbfXe98dhKusyJ27i8rVilq5oaun8cTTgm6lGkUW59rI7VtLNKCKcOiqfco/FjjtnVtRofP8v8AD0cnb/fW1/L37kd5bo2pgNq9I7zbAZnOVFaKGkwlBFtihzU7bgqqtyizLJUsBptcFVClrXbeIUQRipYdKo7jMpmICo1K9PfX3yK6O7U64yPb3X3Z209z9Z4f+JfxXelDkPHg8b/CIhPkvvqmsWIxeFCGbWo4IIvce22ikVgpGT07HcwSoZI2BUcT0HvTXzh+I/yFz+S2t0r3/wBddi7iw+Kr83lMNt/MGSvoMRi6gUuRyFTBVJEVihdgJGP0vf6c+7NBMgqykDqkV7aTsUhkViMkA1I+306Glu1+s12ZiexG37tNdiZ6fG02F3e2cx67eytRma8YvFQUGVL+GV6ipYQRKrEs/pHN/dBG5bQBkdPGaIIJCw0ngekTv75PfH3q3fm3OsOwu3NlbT7A3ZEJ9v7Ty2WSHMV8LNojmNOgYQq5/Q05QN/Zv7skMrrqRajpuS6t4X0SOAfTop/x9/ms/ET5IfL/AOQ/wj2Jvbx93fHnLUGLyWMyaiCg3ysuMFdmqzY9ct46tMZL/k9WuoNqs6BkuRtoWVa8SOI9Pt6rHdxvJo4V+Eng48yPs6sn9s9Kuve/de697917oJu8P+ZZbm/6hP8Aifej17pMfGQW6jwv/UZkf+to9+HWz0YD3vrXXvfuvde9+691737r3QG/Jf5B9e/FPoXtT5Fdq10mP6+6j2jkN37mqYV1zfZUWmKKCFT/AG5ppI4lP4LXsbe7IutgvDpuaTwoy4FSOA9eq/v5c/zfre//AI19gfMTt/5MdG7y6Ryufy+Y2fU7L21W7Pj6l2rRzyyHa++sjlH8lXkIIjCrFYSzkEqzFtIUSxJUJCpJPnxr0htbmRlea6dQoPClCv8Apvnx/wA2OrEOuvkJ0n231hN3R1t2XtXeHVlNTZasqt74jICTCUdPgomnzElZNMqND9uil5VkRSF5tYi7LRSI+hhQ+nSuK5gmi8eJwyeo4dJvo35Z/G35LT5+l6H7k2T2fV7W8Z3DRbZyTT1uJSaUwxTVdHUJHKI2caRKEKXsL3I9+eKSMVcUr1qG7trglYHDEcaHh0+4T5G9Fbl7czHQ23u1dmZvuHb+E/vHm+v8Xl4K7cGLwwlWBq2sgptSJpdlDxl9a39Sj34xSKutlIHW0ubeSQwo4LDiAehq9t9P9e9+691737r3Xvfuvde9+690Rnc1v9nu6+55HVuc4/2L/X3rz6t+Ho83vfVeve/de697917r3v3Xuqsetf5nPT/+zC/IzozvjsPr3qrLdddn4bZnWNDma6bG1u6sZk0aiEk9VUF4XnarVFUAx8P9NIuFslqwRWiFcZ6KLfdIWnkiuHVSCAoOOPDo7XeXyZ6D+NOGwu4O+O1NqdYYbcVbJjsHX7mrJIIsnWQxrNNDSLTpIzaFZWZtOkAi559pkikk+AVp0YTXMFuA0zhQfXpV03cXVVZ1nD3NT9g7Sk6pqMOm4IewTm6KPaj4Z38a1/8AF5HEQQv+3ywOv0W1ce9eG+vw6Z9Ot/UQ+D4+oaONfKnSO6f+UXx77/y+7MD0x25svsbNbFekTdmM21lBV1mE++XVSS1MLqpaN/oJY9SXsC1yB728UiCriletRXME5KxOGI408umfJ/MP4tYfd2zth5HvrrGm3b2BlMphNn4X+9OOlnzmXwspgyeNp5oGaFJopFKFJpEJb0rc8e7fTzfwnqn1tpUDxBU46H6lzmFrsjksRRZfGVeWwy0j5jGU1dSz5DFLXxGehbI0cTGSETIC8RkUawCVuB7bKsBUjj0+JEYlVIJHH5dNm0N67S3/AIWPceydx4fdWBlrMhj48vgq6DI4963E1r47JUq1NMWXXBPHJFIt7hlI9+ZWQ0YUPXkdJF1IajpT+69X697917r3v3Xui+/Kj/sn7s7/AMN9v/cuL3o8OtjpUdEf8yd65/8ADVxf/Wge99aPHoWvfuvde9+691737r3Xvfuvde9+690BeN+TXQOY7jyPx9xXbOzcj3RiKWasynXVHlFqNxUEFPD9xOKqCIFEkRPU8Jk8ij6qLe3fBlCeJp7fXpOLq3MpgDguPLz6TGR+Z3xXxHcsfx6ynevX+P7pmrqfFxdeVmX+3zr5OrjEtNjgJUEPnkDLph8uskgWvx78IJSmsKadaN7arL4DSAOcUrmvT/3n8pPj38aKLD13e/bWzusodwzPT4GLcmRMNbmJo2VHTHY+nWSeWzMqllj0gmxPvUcUkvwCvW57q3t/7ZwteFTx6CD4SfK+f5abf7a3VHQYunwGzu181s3aeRxQqBFmtu0lHDVUGQn+4d7ysJDqKkDkcD25cRpGwCV4efTFhcvcxs7kGhoKdHb9p+l/Xvfuvde9+691737r3XvfuvdEkpbH5W1H+CTf+45v7r1avR2/duq9f//R3+Pfuvde9+691737r3XvfuvdQcn/AMW3If8AUDV/+47e/de6JX8Abf6F85bj/jJu+OD/ANrAe9DqzdHi976r1737r3Xvfuvde9+691737r3XvfuvdMuf21t3deOfD7pwGF3LiJJYppMXn8VQ5nHSTQNqhlehyMckRZDyrFbg/T3sEqaqadaZVYUYVHz69gdt7e2rjkxG18Dhdt4mOSSaPF4HF0OHxyTTG80qUWPjjiDOeWYLc/n34kk1Jr14Kqiiig6eveut9e9+691737r3Xvfuvde9+691737r3RJe4/8Asqfozj/lw5n/ANyX96635dHZH0H+sP8Aeve+tdd+/de697917r3v3Xuve/de697917qPV0lJX0tRRV1NT1tHVxSU9VSVcMdTS1MEq6JYaiCYMjowJDKwII4I97BINR14gEUPSb2xsLY2yfvzszZe09o/xSSObJ/3Y27h8B/EZogVilr/AOFQxeZlBIVpLkXNveyzN8RJ6qqIuVAH2DpWe69W697917r3v3Xuve/de697917r3v3XukZ2IbbH3Of+rTU/X6fQf096PDr3QF/Ev/jwsv8A+HHWf9Cj34U8ut+VejT+99a697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917ojvfFx8pvi5/jV57/rW3vR6sOB6PF731Xr3v3Xuve/de697917r3v3Xuve/de6wVVLTV1NPR1tPBV0lVE8FTS1UMdRTVEEq6JIZ4JQVdGBIZWBBHB9+BIyOvEAih6yRRRwxxwwxpFDEiRRRRIscccca6UjjRbAKAAAALAe/de4dYaaio6ITLR0lNSLUVE1XOtNBFAJ6uobVPUzCIDVI55d2uSfqT72STx60ABwFOvVFFR1hp2q6SmqmpKhKukNRBFOaWriBWOppzKDokUEhXWxFzY8+/AkcPPrxAOSOHUn3rrfXvfuvde9+691737r3Qc9v/wDMrOwv/DQz3/uuk96PDrY49Bb8QTf4/bB/r9jP/wC5B9768ePRmPfutde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690STf5t8mdo/8AkPH/ACb7117z6O37317pszP/ABa8h/1A1v8A7iSe9jrTcOvnV/8ACasf9j4vnHc2/wBxPdH4/rvaT3UcR1c8D19GZeFUf0A/3r3s8eq9d+/de697917r3v3Xuve/de697917r3v3Xuve/de697917pCdoi/W2/h/2Z+4/wD3Uy+/deHHoBvg6LfGXrgG/FPlfr9f+LvN70Otno2nvfWuve/de697917pn3Dl029gM5n5aaorY8Hh8nl5KOkUPVVaY2ietemplP1kkCaUH9SPe1Gpgvr1V20qWpWgr1pj7d+cOM+XXy6/lzfILqvuf437Mw+U7v7hwbfHvr/raqwPdlN/DcjJiK1N7bjWP/Ko5lXWw0pG5dmCuzOQbogFVIJU0yTUH7OgrJOzSRuSquNRChaMuK9xHlSh6su2H8X+lu2fmP8Azgdy9kYbKb0ym4tmdZ0DU26FkzGN2dFiNqZLKUVVsGkyMTDH1Inp4JHkpCrOUAJsSC3IPCaMRnzp9vl0ph03SXBnWuAcjgaE4qPXqurvT5a03VPXv8lXsLuPfOJl2Vi+8vkZtHcu3ez8TU7ox/aG2cfjRtLCQLjpIpCBh1nhMclRHZdKqhJsPdtALMlaFqY8+mfqDHbRXBUssbGpOQAQRU+dAaft6EfP9aZvpj+ShnKL4w/JnqHsbZO5++OwuxO9u/uuMOmM2htHYG6d5VGYg25l8LDetNNhjLQ0NXDLGjKq6QgiILaRv8YPjVqBQV/Zj/VXr08enax9CQFZtTlRUAE1PD0qfl/Lo4vwH68k3n3fvT5D7U+dHx37Zki+PO1tvbu6l+Nuwodn4/8Ag9Ht5lxmb3DQrUGRp5pCriokpg6vpAVLKo8XC0R4zSuCfmerxQtLI1zFcIaIAVQUAAFTXJJJHr0Pf8iTozrPqT4hblzey6HJ1uf35373Znt0bw3YRkt65WabeDRQYytz9VGtTJR0iRqtLA7FE9TD1MxKS6BSTRU0oPPo02wrJb+NQAknIFK0Pngf7PHq7D2l6Muve/de697917r3v3XuiR5a/wDs6tFb/n3cF/8Abn37h14/Po7nv3Xuve/de697917ogP8AM8quxKL4Xdv1fXslTTx0+LSfsSroJxT5Oh6mhV5ewq3GSj1iWOhVmbx+rxeQjgH2otSolBbpDuKyNasIs+tPTz6okqezf5aPZXaf8qDJ/Dbd+39y79wG6uucJEu0aGeneo2w1fSx1s/YZjpYTUZUVa1N55yz3LnVoKEmA8YLIW+Fj/L/AFf5eg/M1h9RaJFTxEwAK1GMg/8AQ3QfYXr7ZPyIwv8APr2B3V8ln+Oe19mfzY9kbs2V2zvLJ1Eeztt7x23icPV7S2tmqCqkgjrcPVVUEFPUY0SeOVXUkEDlGhOpMFscBj9nRzcqohl71i7gdTAEVxxBNPKnl5etekN82/5gm6O7v5T3duRyq4frbL/D/wDmCfHHqzefcPUmJqdo9IfKHG4vedHkMhufr7CRfv8A8HyUMkX31BOZEJRLMUOhNhNE4D1UEYrnqjXHiWLSQaZGQiunAJBz+zjSp8uPVj/w+n6C7fT+Yh8i+kuyukd7dm9ldNbwotq9UdPbVj2lu7rPFJ13Ki4yqgqEinkqK2vWn1T08bIZSGZ9ThfbtzrRFioaVGT/AKuPSXa/pp55b1GQsw4AAED5+f7fXqmmX5i9Y5L/AIT/AHwL+O9H2yz/ACq2z3/8e9ubx6rirMnU9j4XLYL5FVmXr6DccBgDxxtRtAQ0npcOqKWs1qrFL4zSMukUP8x1Z7mzazjtoZA5DKKD+i1Tg8MCtD1Zp8t+6dify3Pn73N8l+v+zth/J3IfIjs/48bK79+EdXjhuz5C7CzmbxNLtDam9On81NLKmJx1NStHkq7GTQLDIGB1LIEcVRWeEBVao8wcdPySRw3x1yx0cqNDDuBp5H5+Qpx6Xf8AL6o+oulf55n83zaG/KTY+zu4O9909Fb36LXI46lot0b82VF1acnv2TaNfJENccVbKklfBDKGkcGRlbSWDTo/grIOB4n/AD9KIp7f62S3LDWuminyBH4fkTxp59bLftN0Y9e9+691737r3QSd5EjrHcxH/KqP9tf3o9eHSZ+Mf/Mo8L+P8syP/W4e/DrZ6MD731rr3v3Xuve/de697917omX8xHG4fL/Bz5SY3P4yizOHqunN3LXYzI0SZKiq40ovKqT0Tq4kAdVYAqeQD+Pb1v8A2y09ekt6QLSQn+E9a6/zj23uzrT4i/yVct0FuvrP409Q7e782huPtPI5zacqdVmCs21+1U9j7Wx0QStoZitS86Tr625LAKpVdpMksiipoK0H2eXoePRIZVt7S3c0UM1AxrQEk0LeoNKZp8+jj9Lbgw3xV60+e/z/AKr5SdQ/Knq2j6oq8vk+oOgNrUW3+r9sV+Lneary9TtKkqamELLFqSoYIWkgE5LPc+9SESeHGymOh4nif9X+z1a3jMBuLqOYThgKRqAFXjkCuanj+zorXwM7/qu1f5qG0e0dpd+/Hffu1O1vhdt7ODq/499fVuyxs2jybfxKnot7RyIfuK2lkA1u7XWyqFQKq+3lQMvhuCFJPH/COk5nZbkyxsrPoAOkUAzkN8+P2enn1Yh/JB6E6y6w60+Um99s0mXy+8+wfmH3Zkdzby3jI+W3W8a1dH9vt6izNfGKiPGU5LNBSK/jVnY8n6IbsaGCKTQiv59G+10kjaVlGrURWgBoOHl/Pz6vD9pOjTr3v3Xuve/de697917r3v3XuiN7mH/OdvX5/wC/W5v/AGHL+9efVhw/1fLo8nvfVeve/de697917r3v3XutZvqT4o9C7y3j/PC3vuna024t17k7Xo81Nm8/SnJ121cls/a2QzODk2LNWxNJQKtTEks32jDXYA3HHs0YsskYU4IPQbSOKeG5MiglWFDTI4edPLyPTXvT5947avxs/l99F9hbx6k2T2N8keqO6p8F3j8n9kz792Th6fraiqcUgjpnXyLV1UX28atNpUoqKwkDAHXhDxGNak0wDQ/6vPqy3LfTxLhRQ97gEfzpx4eXCnVVXyQ3Rkar/hKl3Q24e0p9/RYbv3ZuMpN9daiq25istjZfkBgp3pcNTTImjHfuzHwFdPIFgePbd1QTB2xjy49P7XWSxaNACK41DtOc49PTo/e3O2+ke/8A+az/ACdcl/Lv3bgd8dcdWdKd7n5b5LpydMZi8JtjK9J0lH1zB3Z9lDTLV1L12rww1uudZzcqrMp9tOs+g+JWnSuKSyMyrbldVDwH+GmAftz1XF133D8Fcr/Jb+QXT+++xqTM/Mfc/wAgt9v/AHcoq6oHe9PuaD5ARz7ZXYOW8Lz0lKMdEGkkpJUUq8ySNdlBWaJXmX+AD8uHn0R+PZW9jKtQszMcHjUtjSDnPHHR9Pnh3H2n/K97T3d3TtLLZ/sH5AfzUOr+gOlun9gLkMhKuxNw9QbUodv7jq6XBTq8f3xpMjd6pf1zOHPqBHttPAdxxqp4ep6W3BvbeNiACHAzwC+WTxzgAZyftI2ePiz0ttv4/wDQ/XfWe2cbU4uDG4OnyuZgrahqqtm3VuFRmt01dVM1vXJXTTGwACiyj6X9oZXaRyzdHFtAlvCI4+H+fowftvp/r3v3Xuve/de6L78qf+yfuz//AA3m/wDcuL6+9HrY49Kjoj/mTvXH+G1MX/1p976159C17917r3v3Xuve/de697917r3v3XutaDqTvDa3ws+ePZ2x9g9tdXfMbHfJ/wCQ833PVHWO1oqz5K9D53ddSsNZld1bx8jU1XgqGzpPTyzK6rqdAh1gmbxtNECQUKjzOD+XQbhnjsr1o1cTCVqhQKupI8z6elc+Xp0Sn+Yp83dtfInsHe+29i9x/HHo7J/Gr+YT171buXZOb63qH+RG+8ptmqiiqM1i95QqSsM7eRT4wfSgR5fTGrWiTR8FTjJrj/V6dVuptbfq6U7wQpXvOeIPpxqft4dHY3J3J8bNq/Ozen/Dn25tubc3NvH4+5+H46YjftNFkNsZPqCqqrZ3J4Wgjpp4ocubSiOXSswcuARLYe/MDo02/wDEOHVo2jS48e/4FCAW+dMZxkftp0Z/+RWNgp8fe6IOqarI13WUHfe5U2RWZVaj7+pwjY+nemkqJahVLnSV5HFre2L/AFeKNXGnT+weCLRhb/AGNOPrnjmleHV33tD0e9e9+691737r3Xvfuvde9+690SOk/wCyrqj/AIJN/X/jgfdePVqdHc926r1//9Lf49+691737r3Xvfuvde9+691HrIWqKSqp1IVp6eeFWP0VpYigJt/S/v3Xui+/GTp3N9JbAyO0s9laLMVlZu7cO4UqqBHjhSmzFSJ4YGWQA6kAsxt70Pn1YmvRive+q9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690XrfvUWb3X3N1z2PR5WipcVs7G19FXUEyOaqqkqpmkRoXUWAAPNz79148KdGEHAA/w9+69137917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917pg3TiZs9t3MYenlSCbI0UtLFLICUjeT6MwH49+690HHSXW+U6y21XYXK5ClyM9VlZ69JqVXRFjlFlQhwOR70Otnh0M3vfWuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6Lv2T0/m96dydO9jUOVoqTF9cz5KXJ0E8btVV4rVKoKV1BAtfm5HvR62CM9GI97611737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SV3zgajdGzd0bcpJo6aqzmCyWLp6iYFooZq2laCOSQDkgFrn3o8OtjBr0jejOvsl1d1jtnZGXrqbJZDCU8sM9ZSIyQTF5S4MasARwfe+tHj0Lnv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6L1ufqHN5zt3BdhU+VoocbivtfLQyI5qZPCAG0MBYf7f3rPW8dGF976102Zn/i15D/qBrf/AHDk97HVW4dfOr/4TWD/ALHxfOO//Op7o/3nesvuo4jpw8D19GZf0r+eBz/sPe+q9d+/de697917r3v3Xuve/de697917r3v3Xuve/de697917pC9oW/0b79v9P7n7i/w/5dMvv3Wxx6Ab4P/wDZM/XP/UPlf/dvN79149Gz9+611737r3XvfuvddEAgggEEEEEXBB+oI9+690FOA6G6N2rl6PcG1+mOqNt57HVVbW4/N4DrraGHy9DWZJzJkaujyWOo45opahmJnkRwzkksTf3bW9KVNPt6p4cddWkV+wdCTDi8bT1NdWwY6hgrMp4RkquGkp46nIinjMVOK6dFDzaEJVPITpBIFh71U4zw6tpUVoOPH59VifMH4E7t7X7V6k71+P8AkOlMF2H1pU5KMbf716+PYPWkVHk6NqefMbV25TIRi8u5ciesplRpRpZmDrdlMU6BNEoJ9COPRfcWcrSCW1YIRxDCqkfYKdCb8QPhFiehdr9kVHZKbF3ZvzvGpNX23gNl7Yi2/wBF1KxyTrBj9sdX1aGkgieKdxVvLCWmJswCjms0/iUC4A4ev7er2dn9OGZyC7caYX8h0cLZ/V3WfXslZLsHrvYux5chFDT18u0NpYDbUldBTcU0FW+Gp4TKkf8AYVyQv4A9sl3bDEnpWscaGqqBX0AHSsx+MxuIphR4rH0OMpBLNMKXH0kFFTCaokM1RKIKZVXU7ku7WuxJJuT70SSak16sFVRRRTqd711vr3v3Xuve/de697917okWXv8A7OrQgG3/ABjuD/ez7914jFT0d337r3Xvfuvde9+691Hq6SkyFJVUFfS09bQ1tPNSVlHVwx1NJV0lTGYailqqeYMkkciMUdHBDAkEEH37r3QZ7X6L6S2PUUNXsvp3qvaFVjCDjana/Xu0tv1GPKkkGhmxNHC0ViSR4yPqfdtbUpU9UEcYNQo/YOnnK9XdZ53HbhxGb662LmcTu3Kw53dWMyu0dv5DH7mzdOsaU+Z3BRVdO8dbVxrDCEqKlXkUIgDDStvBmBqDTrbIjCjAEH1HRMPnb8Ddr/LD41L0Vsin2H1xNg+xdl9p7XpKrZtBUdez7q2VkRXUMe6Np4uOOKpp5Bw48bG6rcEC3t6GfQ+qWrYp8+kV7Zme38K3IjIIIxjBrkDjw6DH45fy/wDe2E+Su3vmL8ldw9Vz95dfbD3J1RsTF/GnbGU6x6xrdkboKvkq7sPbtbqfMZRCoWikkYR0yj9scKE9JKhTRHUDiamvW7e2lWXx7gqXAIBUUwfI1Py6sAh6F6Npso2bp+mOqIM02RhzDZeHrraEWUbLU8v3FPlDXpRiU1EcnrSfXrVvUGB59teI54sf29KhFEDUKP2Dp2qupOqa7dUm+q3rLr6s3tNJRyzbxqtmbcqN0yy45BHj5ZNwTUxq2aBVVYWM10AAWwHvQdwNIJp9vWzFEW1FRX1oK9OlRsDYdZu2i3/V7J2lVb7xlHLjsdvWo23hp924/HzoY5qGi3HJCayKF1JVoo5gpBIIsfftTU01NOt6E1a6CvrTPSu916t1737r3XvfuvdBJ3jb/RluW/8Ayqj/AG9+Pejw62Ok18ZP+ZR4X6/8C8h9f+Wo9+HXj0YD3vrXXvfuvde9+691737r3UeqpKWvpp6Kupqeso6qJ4KqkqoY6imqYJV0SQzwTAq6MCQysCCOD78CQajrRAIocjpgzuydmbowabY3NtLbG4ttR+Dx7ezuBxWXwcf2y6KbRichFJTjxgkJaP0jgW92DMDUE160UUjSQKfZ014nq3rLA7eym0cH11sXDbUzlNPRZvbGK2jgMdt7MUdVC1PU0mVwtJTpTVEUkbukkc0bKykqQQSPfmd2+Ik9aWONPgUCvoAOom0Oneo+vq85TYXVnXOyMm2PixLZHaGyNs7ar2xUJ1Q4w1eGpYZDToeVh1aB+B78WY8ST1sIi/CAPy6XNBjMbi45YcZj6LHRVFTPWzxUFJBSRzVlS2upq5Up1UNLIeXkILMeST70STx62AFwBTqb711vr3v3Xuve/de697917r3v3XuiNbm/7Lt6++v/ADK3OW/oeX96PHrY4f6vl0eX3vrXXvfuvde9+691737r3TdDh8RTPkpKfF46B8xIZsu8NDTRPlZTF4DLkmRQZ2KejVLqOnj6e96jjPDrWlRWg48fn0jtx9SdU7xiwVPu7rLr3dUG1xVDbMO49l7bzkW3RXRCGtGCjydNKKQTIqpL9uE1qAGuB72GYGoJ60UQjSQKelOibfP/APl99e/Nf4Vb7+GWMg2z1ds3d+T2TkqeDBbbpMft7GHaW+KHeLxRYPBrTxqJzSMh8aj1Nc3592jdQ+qQah01cRyPCY4G0MaZ/Mf5MdGq676E6h6l29JgesuttgdfGp25jttV+Q2Tsvbu16zI0WLxi4ugOQnwsEMk/iRF8YmdtNhY+6l2biT04qIuVAB+Q6J98EP5a3SHw06J251VXbN617I3bht4753lV9k5br3b9RuSvye7921O4oZ2y2VgnrBLTRSwwK/mFvGNNgB7dlmLtWOoFB59JrW0WGIJIAxBJrQeZr0o/lD8D9q/Jv5SfB75JbiyePhn+GO9N/71w+DrMUa2TNZHeG3IcRRNTVOoLCaWeniqLup5UFfUB71HIiIwI7jwPp1aeGSWWNg1EUksv8WMfsOft6P/AO2elXXvfuvde9+691737r3RfflQL/H7s7/w3m/9y4vp70etjpUdD/8AMnOuf/DVxY/6we99aPHoWvfuvde9+691737r3Xvfuvde9+690g8T1X1hgNy1288F1xsPC7wybSNkt14naG38duTINMCJWrc7R06VUpe51GSU3/PuxdyKEn9vVBHGDUKAfsHTJV9D9HZDPVm6a/pnqit3PkMxFuGv3HV9d7Qqc9XZ+BQsOcrMvNRtUS1iBVC1LyGQACzce/amAoCetlEJ1ECvrTp13d1H1T2Bk8Xmt+dZde72zGEpamhwuW3dsvbe5MliKKsYPV0mLrszTTS08UrKpkjidVYgFgbe9BmXINOvMisKMAR8x0qMBtvbu1MemI2vgMLtvFRu0keMwGKocPj0ke2uRKLHRxxhjYXIW5t78STxNethVX4RTp696631737r3Xvfuvde9+691737r3RI6P8A7KtqP+Cz/wDWg+64r1vo7nu3Wuv/0zFf9Bs3xU/7xF7n/wDQs2t/1696z1aievXv+g2b4qf94i90f+hZtb/r179nrdE9T13/ANBsvxTv/wBkjdz/AOv/AHs2t/169+z16ievXv8AoNm+Kn/eIvdH/oWbW/69e/Z61RPXr3/QbN8VP+8Re6P/AELNrf8AXr37PXqJ69e/6DZfip/3iN3P/wChZtb/AK9e/Z69RPU9eH/C2b4qf94i9zj/AMmza3/Xr37PXqJ69e/6DZfin/3iN3P/AOhZtb/r179nr1E9T17/AKDZvin/AN4i90f+hZtb/r179nr1E9evf9Bs3xU/7xF7o/8AQs2t/wBevfs9bonr11/0GzfFT/vEXuj/ANCza3/Xr37PWqJ69d/9Bs3xT/7xG7oH/k2bW/69e/Z69RPXr3/QbN8VP+8Re6P/AELNrf8AXr37PW6J6nr3/QbN8VP+8Re6P/Qs2t/169+z1qievXX/AEGzfFT/ALxF7o/9Cza3/Xr37PXqJ69d/wDQbN8U/wDvEbuf/wBCza3/AF69+z1uievXh/wtl+Kf/eI3c4/8mza3/Xr37PWqJ69e/wCg2b4p/wDeI3dH/oWbW/69e/Z692ep66/6DZvip/3iL3R/6Fm1v+vXv2evUT167/6DZvipb/skbue/9P72bW/69e/Z63RPXr3/AEGzfFT/ALxF7o/9Cza3/Xr37PWqJ6nr3/QbL8U/+8Ru5/8A0LNrf9evfs9bonqevf8AQbN8VP8AvEXuj/0LNrf9evfs9aonr17/AKDZvip/3iN3R/6Fm1v+vXv2evUT1PXv+g2b4qf94i9z/wDoWbW/69e/Z69RPXrr/oNm+Kn/AHiL3R/6Fm1v+vXv2et0T167/wCg2b4qf94i90f+hZtb/r179nrVE9evf9BsvxT/AO8Ru6P/AELNrf8AXr37PW6J69df9Bs3xU/7xF7o/wDQs2t/169+z16iep69/wBBs3xU/wC8Re6P/Qs2t/169+z16ievXf8A0GzfFPj/AJxG7o/xH97Nrf8AXr37PWqJ6/y69/0GzfFP/vEbuj/0LNrf9evfs9eonr17/oNm+Kn/AHiN3Qf/ACbNrf8AXr37PXqJ6nrof8LZvip+fiL3OP8AH+9u1v8Ar179nr1E9evf9Bs3xU/7xF7n/wDQs2t/169+z16ievXv+g2b4qf94i90f+hZtb/r179nrdE9evf9Bs3xU/7xF7o/9Cza3/Xr37PWqJ69dn/hbN8VP+8Ru5z/AOTZtb/r179nr1E9euv+g2b4qf8AeIvc/wD6Fm1v+vXv2evUT169/wBBs3xU/wC8Re5//Qt2t/169+z16ievXv8AoNm+Kn4+Ivc//oWbW/69e/Z69RPU9e/6DZvip/3iL3R/6Fm1v+vXv2evUT165f8AQbL8U+P+cRu6P/Qs2t/169+z16iep/Z11/0GzfFP/vEbuj/0LNrf9evfs9bonqevf9Bs3xU/7xF7o/8AQs2t/wBevfs9aonqevf9BsvxT/7xG7nH/k2bW/4iL37PW6J69df9Bs3xU/7xF7n/APQs2t/169+z1qievXf/AEGzfFT/ALxF7o/9Cza3/Xr37PW6J6/y66/6DZvip/3iL3P/AOhZtb/r179nr1E9evD/AIWzfFT8/EXuf/0LNrf9evfs9eonr12P+FsvxT/7xG7n/wDQs2t/169+z16iep66/wCg2b4qf94i90f+hZtb/r179nrVE9euz/wtl+Kn4+I3c5/8mza3/Xr37PW6J69e/wCg2b4p/wDeI3dH/oWbW/69e/Z61RPU9df9Bs3xU/7xF7o/9Cza3/Xr37PXqJ69e/6DZvip/wB4i9z/APoW7W/69e/Z69RPXr3/AEGzfFT/ALxF7o/9Cza3/Xr37PXqJ69d/wDQbN8U/wDvEXuj/wBCza3/AF69+z16ievXv+g2X4p/94jdz/6/97Nrf9evfs9eonr/AC69/wBBsvxT/wC8Ru5//Qs2t/169+z16iep66/6DZvip/3iL3P/AOhZtb/r179nr1E9evf9Bs3xU/7xF7n/APQs2t/169+z1uievXf/AEGzfFT/ALxG7o/9Cza3/Xr37PXqJ69e/wCg2b4qf94i90f+hZtb/r179nr1E9evf9Bs3xT/AO8Re6P/AELNrf8AXr37PWqJ69e/6DZvip/3iN3R/wChZtb/AK9e/Z69RPXr3/QbN8VP+8Ru6P8A0LNrf9evfs9eonqevf8AQbN8VP8AvEXuj/0LNrf9evfs9bonr17/AKDZvip/3iL3R/6Fm1v+vXv2etUT169/0GzfFT/vEbuj/wBCza3/AF69+z1uievXX/QbN8VP+8Re6P8A0LNrf9evfs9eonqeu/8AoNl+Kf8A3iN3P/6Fm1v+vXv1T6deonqevf8AQbN8U/8AvEbuj/0LNrf9evfs9eonr11/0GzfFT/vEXuf/wBCza3/AF69+z1qievXY/4Wy/FT8/Ebucf+TZtb/r179nrdE9evf9Bs3xT/AO8Ru6P/AELNrf8AXr37PWqJ69e/6DZvin/3iL3R/wChZtb/AK9e/Z69RPXrr/oNm+Kn/eIvdH/oWbW/69e/Z63RPXr3/QbN8VP+8Re6P/Qs2t/169+z1qiep67/AOg2b4qf94i90f8AoWbW/wCvXv2et0T169/0GzfFT/vEXuj/ANCza3/Xr37PWqJ69e/6DZvin/3iL3R/6Fm1v+vXv2evUT1PXX/QbN8VP+8Re6P/AELNrf8AXr37PW6J69d/9BsvxT/7xG7n/wDQs2t/169+z1qievWGf/ha98T6qGWnm+IvdPjmhmibTuza1x5YWj/Ef+PPv1T14hD59a0f8pj+b10t8C/5jnyM+ZfYvX+79xbL7jx+/afb+2sHUwJmsZLuvcL5ilFbOySowiUiOT083JH49+yB14EEkHHW0iP+FpPwtAA/2XrtvgDgZOj4/wAP+Afv3d17s9T+zrv/AKDSfhb/AN499t/+fOi/+o/fu7r3Z6n9n+z17/oNJ+Fv/ePfbf8A586P/wCo/fu7r36fqf2de/6DSfhb/wB499tf+fOj/wDqT37PXv0/U/s69/0Gk/C3/vHvtv8A8+dH/wDUfv3d6de/T9T+zr3/AEGk/C3/ALx77b/8+dF/9R+/Vb0692ep/Z17/oNJ+Fv/AHj321/586P/AOo/fu7r36fqf2de/wCg0n4W/wDePfbX/nzo/wD6k9+z6de/T9T+zr3/AEGk/C3/ALx77b/8+dF/9R+/d3p17s9T+zrw/wCFiXw17Mv1zSdCdq0VZvxTtCirajJUjU1JV7iH8Jpp6hVpQxRXlUkAj37PW+z1P7OsMf8Awqb+KvwheX4w706T7L3HuXrGRqLKZzEV9JDi66XJn+LIKSKSmdxoSZQxY2P4/PvWRwHXuwjJz9nWb/oNJ+Fv/ePfbX/nzo//AKj977vTr36fqf2de/6DSfhb/wB499t/+fOi/wDqP37u612ep/Z/s9e/6DSfhb/3j321/r/xOj/+o/fqn069+n6n9nXv+g0n4Wf94+dtf+fOj/8AqP37u69+n6n9nXv+g0n4W/8AePfbX/nzo/8A6j9+z6fz69+n6n9nXv8AoNJ+Fv8A3j323/586L/6j9+qfTr3Z6n9nXv+g0n4W/8AePfbX/nzo/8A6j9+7uvfp+p/Z17/AKDSfhb/AN499t/+fOi/+o/fu7r3Z6n9nXj/AMLSfhZ+Pj521/586P8A+o/fu70692ep/Z17/oNJ+Fv/AHj321/586P/AOo/fqt6de7PU/s69/0Gk/Cz/vHvtr/z50f/ANR+9d3p179P1P7Ovf8AQaT8Lf8AvHvtr/z50f8A9Se91b063+n6n9nXX/QaT8Lf+8e+2/8Az50X/wBR+/VPp1r9P1P7Ou/+g0n4W/8AePfbf/nzov8A6k9+z6fz69+n6n9nThQf8KafivnKSt+b79N9iU+zMUD1o226nIQjMfxeGPzitjngp9BjKkWUp/r8+/Z68dFaVx9nTcP+FpXwtIB/2Xrttf8AA5OiuP8AbUfvVW9Ot0j9T+zrv/oNJ+Fv/ePfbX/nzo//AKj977vTrX6fqf2de/6DSfhb/wB499tf+fOj4/8AVP36renXv0/U/s69/wBBpPwt/wC8e+2//PnR/wD1H793enXv0/U/s69/0Gk/C3/vHvtv/wA+dF/9R+/Vb069+n6n9nXv+g0n4W/9499tf+fOj5/9U/fu7069+n6n9nXv+g0n4W/949dt/wDnzov/AKj96q3p17s9T+z/AGevf9BpPwt/7x77b/8APnRf/Ufveevdnqf2de/6DSfhb/3j323/AOfOj4/9U/fs+nXv0/U/s69/0Gk/C23/AGT321+OP4nR/wCx/wCUP3qrenXuz1P7Ovf9BpPwt/7x77b/APPnR/8A1H73VvTr3Z6n9nXv+g0n4W2/7J77b/8APnR//Ufv1W9Ovfp+p/Z17/oNJ+Fv/ePfbX/nzo//AKj9+7uvfp+p/Z17/oNJ+Fn/AHj323/586P/AOo/eu7r3Z6n9n+z1KoP+Fdvw+73rqbqnE9C9pUGQ3axoaWurclSmkgltqHlWKmVjxf6H8f63veTg9aOgCoJr9nXLJf8Kw/iT8Wq+o6Y3D0Z2dl8rt0ipqMjjsjTJRS/xAeeNI1lpme4W1yTb37I62DGRWv8uof/AEGk/C3/ALx77b/8+dF/9R+9d3p179P1P7Ovf9BpPwt/7x77a/8APnR8f+qfvfd6de/T9T+zr3/QaT8LP+8e+2h/r5Oj/wDqP37Pp179P1P7Ovf9BpPwt/7x77a/8+dH/wDUnv3d179P1P7Ovf8AQaT8Lf8AvHvtv/z50f8A9Se/d3Xv0/U/s69/0Gk/C3/vHvtr/wA+dH/9R+9d3p179P1P7Ovf9BpPwt/7x77b/wBb+J0f/wBR+/Vb0692ep/Z17/oNJ+Fv/ePfbX/AJ86P/6j977uvfp+p/Z17/oNJ+Fv/ePfbX/nzo/9t/wD9+7vTr3Z6n9nXv8AoNJ+Fv8A3j321/586P8A+o/fu7063+n6n9nXv+g0n4W/9499tf6/8To//qP37u9Otfp+p/Z17/oNJ+Fv/ePfbf8A586P/wCo/fqt6de/T9T+z/Z69/0Gk/C3/vHvtv8A8+dH/wDUnvVW9P59b/T9T+zrr/oNK+Fov/zj122f8Bk6Ln/1T977utfp+p/Z1zqf+FNXxb3BBB88aPp7sUbJ6/depsntX76n/jtXm9whpaWrinenEaxoDyCnP9ffsnrdU4V/l1x/6DSfhb/3j323/wCfOj/+o/fu70612ep/Z17/AKDSfhb/AN499tf+fOj/APqP36p9Ovfp+p/Z17/oNJ+Fv/ePfbf/AJ86L/6j9+z6de/T9T+zr3/QaT8Lf+8e+2x/5E6L/wCo/fu70692ep/Z/s9e/wCg0n4W/wDePfbX/nzo/wD6j9+7vTr36fqf2de/6DSfhb/3j323/wCfOi/+o/eqt6de7PU/s69/0Gk/C3/vHvtr/wA+dH/9R+993Xuz1P7Ovf8AQaT8Lf8AvHvtr/z50f8A9Se/Z63+n6n9nXv+g0n4W/8AePfbX/nzo/8A6j9+7vT+fWv0/U/s69/0Gk/C2/8A2T321/r/AMTo/wD6j9+q3p179P1P7Ovf9BpPwt/7x77b/wDPnRf/AFH793enW/0/U/s69/0Gk/C3/vHvtr/z50f/ANR+/Z9Otfp+p/Z17/oNJ+Fv/ePfbX/nzo//AKj9+7vTr36fqf2de/6DSfhb/wB4+dtf+fOj/wDqP37u69+n6n9nXX/QWT8SvlRLD8edsdG9n4XP9rSLtnFZjIZCklx9BVysKlJKuJKZHZSIyPSf9f37Pn1vsHmf2dS6z/hXd8O+hZv9DeU6K7PymU65QbUyVfQZOmFJUV2I/wAlqJYlmpWYB2BYAsbX+vvxr5daGg8Sf2dRf+g0n4W/9499t/8Anzov/qP37u692ep/Z17/AKDSfhb/AN499t/+fOi/+o/fu70692ep/Z/s9e/6DSfhb/3j323/AOfOi/8AqP37u9Ovdnqf2de/6DSfhb/3j323/wCfOi/+o/fu7069+n6n9nXv+g0n4W/9499t/wDnzov/AKj9+7uvfp+p/Z17/oNJ+Fv/AHj321/586P/AOpPfqt6db/T9T+zr3/QaT8Lf+8e+2//AD50X/1H793enWuz1P7OvH/haT8Lfx8e+2j/AOROj/8AqP3qrenXv0/U/s69/wBBpPwt/wC8e+2//PnRf/Ufvfd6de/T9T+zrw/4Wk/C3i/x77bH9f8AcnRf/Ufv3d6de7PU/s68f+FpPws/Hx77b/8APnR//UfvXd6de7PU/s69/wBBpPwt/wC8e+2//PnR/wD1H733de7PU/s/2evf9BpPwt/7x77b/wDPnRf/AFJ79VvTr36fqf2ddH/haV8LQL/7L122fqbDJ0Vzb8C9Hbn3qrenXuz1P7Onh/8AhTJ8WqLCr8wH6Z7EbbNVIKZMAuQhGXE0x+2AMhg8X6j9dFv9h7tQ0r1XUmqlTT7Omcf8LSvhabH/AGXrtsXH0/idFcf4f8BPeu7q/wCn6n9nX//U2bP+GpPiNfn4j/H/AJ/79/jf959+ofTreoD8X+Drs/yo/iJ9B8RugL/+GBjLD3qh9OtV/pddH+VF8RgL/wCyj/H+xvz/AHAxvFvr73Q04dernj14fyo/iNb/ALJH+P5/8p/jPfqH069X+l1yP8qH4igAf7KR8ftXN/8AfgY23+HHvek+nWtQ9evH+VH8RbW/2Un4/wD+x2BjLH/Ae9aTXI63qrmvXv8AhqL4jaT/AM4kfH/gcf78DG8e/aT17UK0r11/w1H8Rfx8R/j/AP4f78DG+/UPp16voeuj/Kk+I1v+yR+gL/U/8Y/xtvfqH069Wnn17/hqT4ikf9kj/H/Ve/8Ax4GN9+ofTr1R/F/g67P8qP4ij6/Ef4/3P0P9wMbYXPF/e9J9Ovah69dn+VF8RQL/AOykfH882JGwMZ/sRz71Q+nWtQPE064n+VJ8RedPxH+P5t/37/Ge/UPp1vV/S67/AOGoviPa/wDspHx/INv+YAxv+8+/FSPLrQYeR68P5UXxGPJ+JHx/t9OdgY3/AIr79pb068WA8+uh/Ki+It7f7KR8fx+f+PAxo/23vwUny62WA8+vf8NRfEa5/wCcSPj/AP152BjOffgDTh14kClW67/4aj+I1gR8SPj8L2/5gDG/149+0t6daLD1/wAHXj/Kj+I34+I/x/PHJ/uBjDz9bj37SR5dbDfPr3/DUfxGtqPxI+P/ANfz1/jfz79Q04de1CvHrr/hqT4jfn4k/H7/AAP9wMb/AFtYX9+Cn069X+l/g67P8qL4igkf7KN0Bxa19gY3/effiCPLrwPnXrr/AIaj+I//AHiR0Bx/2YGN/wBf36h9Ovf7brs/yoviLxb4j9AE83vsDGf7C/v1Gpw68CK01cOuP/DUfxGHJ+I/x/t/4j/G/wC8+/UJ8ut1H8XXIfyo/iKbk/Ej4/6Ra9tgYz37SfTquofxf4OuP/DUnxH+n+yjdAEj8/6P8b79T5dbr5V65H+VH8RLX/2UfoCx+l+v8bcH6G9vftLenWtQ/i66/wCGo/iKbH/ZR+gP8f8AfgY3n/Wv79Qjy69Wo49d/wDDUfxFLAf7KN0Bb/xH+Nv79Q+nWyaCteuv+GpPiLzf4jdAXH/fv8b79pPp16vqeuz/ACo/iNpB/wBlH6AH1/5gDG/1/Hv1D6der8+vf8NQ/EU8j4kfH+3+OwMbf/Y+96T6da1D169/w1F8RRyfiR8fgPoP9+BjOT71Q+nW9Q9euv8AhqP4jEcfEf4//wDov8ba3v1DwI69WvBuu/8AhqP4iC1/iN8f7cXvsDGW/wAefes+nXq+deu/+Go/iJex+I/x/JbkW2BjBYH3uh9Ovavn1x/4aj+I1z/ziP8AH4WP/Pv8b+D73pb061qH8XXh/Kj+I17f7KP8f7Wv/wAeBjR/sPetJPl1uoB49eH8qP4i/n4j9Af+i/xv+3Pv1D6der/S67/4ak+Ihtb4kfH/APp/x4GN5P5/2Hv2k+nXi1fPrw/lR/Ea5B+JHx/4/P8AcDG/X+nv2luFOtasV1ddf8NS/EO3PxH6AJ+gtsDGf7D36h4U62WHk3XZ/lR/EYED/ZR/j+bjj/fgYz36h9OtBqnj14fyoviMbj/ZR/j+G/s/78DG/wCxPv1CfLrZNOJ66X+VJ8RDe/xH+P5/1tgY23+x9+0n069X+l14/wAqT4jC4/2Uf4/kD622BjfftLenXtQpx66H8qP4jXH/ADiR8fufx/o/xvv1D6deqPJuuX/DUfxFsf8AnEj4/wDFuf7g42w1f1Hv2k+nWgw4168f5UXxGBIHxH+P5t+P7gYy/wDh79Q+nWwwIwevf8NR/Ea1z8R/j/x9b7Axvv2k+nXtVOJ66P8AKk+Iw+vxH+P5/p/xj/G/7H36h9OvVHmf8HXY/lRfEWwP+yj/AB/5+h/uBjLD/X9+ofTr2oevDrv/AIaj+IhDf84j/H/UP+zAxthzxYe/AH069qFaauuv+Go/iKOP9lI+P5PP/MAYwj34q3p1oN/S69/w1H8RLG/xH+P9x/XYGMt78AT5db1Afi65D+VH8RGAH+yjdA/152BjP9j79pJ8uvaqDj10P5UPxFIF/iP8f/rb/jwMZ/th72FJ8utFgDx66b+VF8Rh/wByj/H/APrz1/jfp71QjFOtggjj17/hqP4im3/OI/x/t/jsDG+9UNaU69Xz1ddf8NRfEYAn/ZR/j9bmwHX+NH097ofTr1fn14fyo/iL9P8AZR/j/wA8j/jH+Nt9fr79RvTr1fPV14fyo/iLcj/ZR/j+T/4YGNsCfoT79Q+nXgw/i67/AOGo/iIp/wCyR/j+SP6bAxt7/i/v2k+nWtQ9euP/AA1J8ReP+cRugL8f80/xtvfqH063qB8+uQ/lRfEc8/7KP8frD/v3+NB9+ANOHXtQ4V68f5UfxGFv+cSOgOeR/wAY/wAbcD/G3v2k04deBHDV/g67H8qL4in/ALlH+P8Afgk/6P8AG/7Hj36h9OvV+fXQ/lRfEZrkfEjoC1/z1/jbnj37SfTrWofxddj+VH8ROf8AnEf4/wD+t/cDGAj/AF/fqHjTr2rNK9dD+VH8RPp/so3QP+1f78DG/X8W9+ofTrZ40B66H8qP4jf94j/H/wD9F/jfeqH063UVw3Xl/lRfEY8/7KP8f7C3/NP8aPr/AK/vdD6dVLDybrs/yoviNcH/AGUf4/2P/fv8bx/r+/UNOHXq/Prtf5UXxFYX/wBlH+P4/r/vwMZb/C3v1D6de1fPrx/lRfEZRz8R+gCT9LbAxv0/x9+0n068GrwPXQ/lRfEb6n4jdAWFuBsDG39+0n069XyJ68f5UXxHvz8R/j+f/Kf43/e/fqHhTrdQc6uvD+VH8Rvr/spHx/AH1P8AcDGgj8e/aTStOvFhwr1x/wCGo/iNcX+I3QFv6f6P8b79T5der8+uR/lR/EVSAfiP0B/rDYGM4v8AT36h9Otavn11/wANR/EX6/7KP0Bbm4/0f436X/r79Q+Y63x8+ux/Kj+IzHn4j9Af0BOwMb78FPp1ot8+ux/Ki+I3/eI/x/te3/HgY336h9Ot6gPPpozX8qn4l09BVyQfEvoOKaOmqnSRdgYu6Fad21DVcX4uOPfgM560xOmoPWkB/I6+LnV/ff8AN9+YHU29Ou9o7q2rt6j7QqsZtnNYSkyGGx5w+6JYIBRUMw0R+NBpTT9B70OtkUBPp1urJ/J7+IpRT/ssPTnNj/x5OMuB9efe6H0615ceuY/k9fEPkf7LD05z9D/cjGXHv1D6der8+vf8M9fEP/vGHpz6W42RjLH/AB9+0/Lr1T69dj+T18QgCP8AZYenDxx/vyMbce/UPp1r8+uP/DPPxD/Hxh6bA/8ADIxn+9+96T6dbr6nrv8A4Z5+If8A3jD059LW/uRjPfqfLr1T6/4OvD+Tz8QwLH4w9OEH/syMZ71Q+nXvz67/AOGeviGb/wDOMPTnJBH+/IxnAH4970/Lr2PXro/yeviEb/8AOMPTg/pbZGM/23v2k+nXq/PrLS/ygfiVR1FPW0nxo6hpa2jniqqOrg2VjUnpKqB9cNRBIOVdGF1I+h9+Kn068D5g9OGd/lL/ABg3Rl63cG6Pj31ZuPP5ExnIZzL7Rx9Zk64wroiNTVSeptK+lb/j36h9Ot1r59NX/DPXxD/7xh6c/wBhsjGW960/LrVfn10f5PHxEtz8YenL/g/3Ixn+x970n068Dnj/AIOuz/J5+IfFvjF07/sdkYw+/U+XXqjzPXj/ACefiHbj4w9OC/4/uTjB/vPvVDwp1qvz69/wzz8Qw1v9lh6dF/rfZGM/2/vek+nXqj166/4Z5+IY4/2WPpz/AGGyMZ/vHvVD6dbr8/8AB1y/4Z6+IR/7lh6cH/kkYz+nv1D6de/Prv8A4Z6+IXA/2WHpz/0CMZz73p+XWq+p64/8M8/EM/T4xdOaf6HZOM9+ofTr359eH8nr4hWN/jD06ef+eIxn0/BHvVPl1utc168P5PXxDB5+MXTp/wAP7kYzn+nven5dar8+uz/J7+IX/eMHTv8A6BGM/p71T5db/Prw/k9fEIAj/ZYem/p/zxGN9+0/LrVfn/g66/4Z5+If0Hxi6cH/AJJGM/1vfqH063X1PT7D/Ki+NVPgH2lB0F1nDtN6tq9ttx7UoVwz5BhZ640I9HlP5b6+/aT6der8+mL/AIZ4+IQXSPjD02B/hsjGD36hrw69X59dj+Tz8Qx/3LD04fwW/uRjPr73pNOHWq/Prw/k8fEQ8j4w9O8fn+5GM/4n3rSfTrxYDFeu/wDhnn4h2/7Jh6cP542RjOOPe9Py69WmK9cf+GePiGTf/ZYunP6H/fkY38/196ofTrdfn13/AMM8/EIcf7LF05/sNkY3j3uhrw61X59e/wCGeviH9B8Yemz/AE/35GN/2PvVD6dbqKceuv8Ahnr4hH6/GHpwf4f3Ixn+8+909B1qvz/wde/4Z6+If5+MPTh/19kYz36ny6358eu/+GeviEAR/ssXTh/xGycZ+efeqZ4der8+uv8Ahnr4h2APxg6cP+P9yMZ/r8n36h9OvV9D13/wz18QuP8AnGHpzj+myMZz79Q+Q61+fXj/ACeviEfp8Yemx/T/AH5GM49+0n069X5/4Ouv+GefiF/3jD05/wCgRjL/AO397p8ut1Pr/g6mY7+UX8VMPWwZLEfG/qTGZGlbVTV9Fs3HQVdOxNyYpk5X/Ye9UPp1sEDz6z5j+Up8WtxZGXMbg+OvVWays6qk+SyW0MfVVkyJ+hZJ5OSB+B79pPp1qvz/AMHTb/wzz8Qzb/nGHpz/AGGyMYD/AIe909R1qp9euv8Ahnn4iH6fGHpw2v8A8wRjPetJPl16vqeuh/J5+In9r4w9Of8AoEYz3uny699h67/4Z6+IZv8A84xdN2P4/uRjeP8AW96ofTrYNPPrv/hnj4hgXHxg6ct9L/3Ixl7+9kH061UV49cR/J4+IZP/AGTF05b/AA2Rjb8/Tke9UPp1uo9f8HXZ/k9fEM3A+MPTn/oEYz36h9OvfOvXh/J5+If/AHjD04eLH/fkYz6f0Hv1D6der8+vf8M9fEL8fGHpz/Y7IxnP+297ofTr359ch/J4+In/AHjD059NRvsjGWt/h79pPp1osPI9df8ADPXxEP0+MPTh/wDJIxtv9t71Q+nW6jzPXv8Ahnr4hWsfjB05/X/jyMZ/tj79Q+nXgfn11/wz18Rbj/nGHpr/ANAjGf7H36ny69UevXv+GeviHz/zjD05b+n9yMZf/W9+ofTr1fn0+J/Kh+NMe3arZ0Px/wCso9oV9bFkq7a67UoBhK3Iwf5mvqKEelpU/DHke90NcdaqBxPTIf5PXxCsf+cYem//AECMZ/vHvQU+nW6/Prr/AIZ4+IZ/7li6cv8A0/uTjOP9iffqH069qp59eH8nr4ic3+MPTf8A6BGMv/T3uh9OvVFOPXX/AAzz8RAOfjB05Y/k7Ixn+w9+oR5der6Hrv8A4Z5+IX4+MPTgB/psjGX96ofTr359dn+T18QyOfjD05/sNkYy3+x9+ofTrxPoeuv+GefiGeT8YenP9hsjGW970n069qzQHro/yefiH+PjD05/rf3IxnP496p8utV+fXZ/k8fEQ2A+MPToPNx/cjGf6/0Hvek+nXgw4k466/4Z5+IYHHxh6cuP+zIxlj71pPp1sn59dj+Tz8Qxz/ssPTd7cf78jG/7z73Q9er8/wDB17/hnr4hkC/xh6d+v42RjP8AYe/U9R1r8+u/+GeviGQQPjD04P8AH+5GM4/1/etJ9Ot1p59dD+Tz8QwLH4xdOnn6/wByMZY+/U+XXvz/AMHThif5SHxZ2/laHO4D46dU4bN4yU1GMy+O2fj6bIY+cjSZqSoT1I1uLj36h8h16vz64ZD+UV8VMvXVOTyvxt6lyeSrppKisyNbs3HT1dXUzNrlmnmblnY3JJ+vvek8ada1VrU9Q/8Ahnn4h3/7Ji6c45t/cjGe9EH068D8+vH+T18Qjb/nGHp3/wBAjGW/x9+p8ut/n14fyeviHf8A7Jh6c5/7MjG+90Pp1rhmvXv+GeviGT/2TB07x+BsjGfj/D3rST5der8+uv8Ahnn4hn/uWLpz/YbIxn+9e/Up5dbr8+vf8M8/EO9/9lh6cP8A5JGM9+p8utfn12P5PXxDuT/ssPTn54/uRjLAe/aT6db/AD68f5PPxCP/AHLF059f+eIxpt/j79Q+nXq549df8M8/EMAj/ZYenLX+o2Rjf949+ofTr1a5r/g69/wz38Qx/wByw9OEfQH+5GMv/r+909B14n59dj+T18RPz8Yemzf/ALMjGf7D36h9OvV9D1x/4Z5+InF/jF059b/8eTjffqfLr359d/8ADPPxDub/ABh6cP8AT/fkYz/Y+9U9B178+u2/k8/EJhY/GDpyxBBH9yMZYgjm/v1Pl16vz6f2/lS/Gx8ANqv0J1m+2Vto2+dq0JxKEfS1GfT/AK3v1D6deqPXpPD+Tz8Qxwvxh6cH0+uyMX/vA9+ofTrdfU9f/9Xf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdFw+U/yX2R8T+pM92/v9tO3sBTzVFUfJ4vRDH5HJazfQf4e/dep1Q7T/APCo74K1ESSpkKXS4JU/xiK3Bsf91/4e/UPVar/F/LrMP+FQ/wAGDz/EaX/W/jEXH+H6PfqHrdV9f5ddf9BQ/wAGL/8AFxpSP+1xF/vH7fv1D16q0+L+XXf/AEFD/Bix/wByNLx9P9zEXP8AyZ79Q9aqv8X8uuh/wqI+DBt/uQph/icxF/X/AAj9+oet1X+L+XXf/QUN8GPxkaX/AM/EX/Rnv2evVX1/l10P+FQ/wZC3OQpf9b+MRf1t/qPfqHr1V9f5dd/9BQ/wX/52NL/rfxiL/r379Q9aqv8AF/Lrj/0FEfBg8DI0tz/XMRcW/wCQPfqHrdV/i/l13/0FEfBe9v4jS8/T/cxF+Pr/AGPfqHrVR6/y66/6CiPgz/zsKX/W/jEfP+t+379Q9eqvm38uuR/4VD/Bi4H8Rpeb/wDL4i/6M97oevVX+L+XXX/QUR8GDe2QpR/r5eLj/kz3rPWwV9f5deP/AAqH+DFjbIUtxbn+MRc/8me/d3Xqr6/y6zt/wqA+DaRiRq+nCkA6v4tHY/6xMftXJZyRW63LHtbgOqiRCSoPD5dYT/wqH+C4F/4jS/8An4i/6M9pM9Wqvr/Lrj/0FEfBkgEZClsDyP4xFyL/APBPfqHr1V9f5ddn/hUP8GPVbIUo4BH+5iL+n/BPfs9aqvr/AC69/wBBQ/wYFv8AchS8/X/cxFx/yZ79Q9eqv8X8uhG6B/4UX/Eb5B/IXr7ojadbSjJ9jZvHbcwM38TjmMuVyEgjihEYjBJLEC1/fsgVPWxRjQHP2dbCWa/4tdd/1B1v/uHJ72OtNw6+dX/wms/7fxfOP/tUd024+h/vtJ7qOI6cPA9fRnX9K/6w/wB697PHqo679+691737r3Xvfuvde9+69Tr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r59H833/hSx/MT+EX8wfvn43dRRdQDrfrvKUFBtyLP7Jgy2YaGelE8k9bkZHUsxJI0aeLXvz70RnqwIoKjqtL/AKDC/wCazz+x0T/6Lql/6P8Afqde1D067H/Cwv8Ams/8cOif/RdUo/6L9+p17UPTrr/oML/msn6QdFf+i6pf+j/fqHr2oenTji/+Ffn813J10NHFT9EFpA5s3X1LELIuonyFjb6f09mezWI3Hc4bJjQSNQ9NTzLDE0hHAdR6z/hYD/NdoquopZoOiNcErRsF67pWUW+gDhxf/XsPaS6gNvcyQE/AxH7D1dJFdQ1OPUf/AKDCv5rP/HDoq3/iOqX/AKP9p6Hq2pfTr3/QYX/NZ/44dE/+i6pf+j/fqHr2oenXX/QYX/NZ/wCOHRX/AKLql/6P9+69UenXv+gwv+az/wAcOiv/AEXVL/0f79Tr2oenXv8AoML/AJrP/HDon/0XVL/0f79Q9e1D069/0GF/zWf+OHRX/ouqX/o/36nXtQ9Ovf8AQYX/ADWf+OHRX/ouqX/o/wB+p17UPTrv/oML/ms/8cOiv/RdUv8A0f79Q9e1L6ddf9Bhf81n/jj0V/6LqlP/AEX79Q9e1D069/0GF/zWf+OHRX/ouqX/AKP9+p17UPTr3/QYX/NZ/wCOHRX/AKLml/6P9+p17UPTr3/QYX/NZ4/Y6K/9F1S8/wDJ/v1Ovah6dd/9Bhf81n8wdFH/AMp1S/8AR/v1D17UPTr3/QYX/NZP0g6K/wDRdUp/6L9+oeval9Ouv+gwv+az/wAcOif/AEXVL/0f79Q9e1D067/6DC/5rP4g6Kv/AOI6pT/0X79Q9e1D069/0GF/zWP+OHRX/ouqX/o/36h9evah6dWafyd/+FKf8xH5yfzE/j98Z+4I+oj1p2RlMxj9zR7f2TBicykNHh5q+GooclGzFGDxqpTTyD9eLe9ZHWxRsU6//9bf49+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3VTXdv86f4J9H997y+NOV39ld6duddYuiye/tvde4uHcq7R++0GPH5iqjnjRKmMSRGeFbmLWivpY29qEtpJBUU+zz6LbndLa1aklaeoBIJHEVGKj06Qv/AA+x8Kbgfa9uD+p/uPHYf+rftz6KX5dJ/wB+2NK93+8n/N14/wA9j4U/8qvbh/x/uPH/APVfv30UtfLr37+sT/F/vJ67/wCH1/hTa/23bfAvb+46X/233fvZsZfl179/WP8AS/Yeuz/PW+FQtel7b5/7MePgf1P+V+9fRS/Lr37+sBip/Yf83Xf/AA+r8KrXFL22R/4Y8f8AT/qL9++il+XXv39Yj+L9h64n+ev8KgbfaduHi/Gx4+P8P+Bfv30UvlTrw36xPmf2Hrw/nr/Cki/2vbY/wOx0uf8A1b9++il+XXjv1iPM/sPXj/PX+FI4+17cJ/IGx04/1/8AK/fvoZfl179/WP8AS/Yf83XIfz1vhSbAU3bdz+P7jp/9V+9/Qy/Lr37+saVz+w9dH+et8KR/yjdtnmxA2Olx/wCrfvX0Uvy69+/rHhn9h/zdeP8APW+FQv8A5J25w2n/AI8eP/b/APAv376KX5de/f1h6n9h68f563wpAv8Abdtn/AbHQ/8Ax3799FL14b9YHzb9h66H89f4Un/lF7bH+vseP/6r9++il+XXv39Y/wBL/eT11/w+v8Kf+VXtz/0B4/8A6r9++il+XXv39Y1p3f7yeuz/AD1/hSP+Ubts82sNjpf/AHmr9++il+XXv37Y/P8AYf8AN12P563wpP8Ayjdtf6x2Qn/1X799FL1r9/2Hqf2H/N13/wAPq/CsGxpe2x/j/ceO3+3+797+hl+XXv39Y+p/YeuJ/nrfCoC/2nbhH9RsdP8A6r96+il+XW/39Y1p3fsPXh/PX+FP/Kt22P8AX2OnP/q3739DL8uvDfrE/wAX7D/m66H89j4Uk/8AAXtwf4nY8f8A9V+9fRS/Lrf79sfn+w/5uvH+ex8KR/yiduc/9mPH/wDVfv30Uvy60N+sT/F/vJ69/wAPr/Cm9hS9uH/EbHS3/uX799FL8uvHfrECtT+w9cj/AD1vhSLf5N22f/JHTj/b1fv30Uvy60d+sQK937D/AJuuv+H1/hTa/wBt23z+P7jpf/3L97+il9R179/2P9L/AHk/5uvf8Pr/AApuP8l7bsf7X9x0t/7l+9fRS/Lrf79sfn+w/wCbrv8A4fW+FVgfte27EkX/ALjx2Fv6/wCV+/fQy/Lrw36x+f7D14/z1vhUP+UXtw3+h/uPHz/6t+9/Qy04jrX7/sa07v2H/N0w7n/n/fAjZW38luvd1b2ht/bOGSGXMZvIbKWOixsE9QtKk1SwqibF3UAKCSeACbD3U2cgFSQAOrLvtm5CoGJJoAFNST+XVu3UvbXXPevW+ze3epN3YXffXO/8FQbk2nuvb9bDX4vL4nJQCop54Z4SQGAOmSNrMjAowDAj2mZSpoejZHWRdSmv+rh9vQi+69X697917r3v3Xuve/de6o6/4UHxxy/y4e31kQOpwWaBUgEEHGm/Hv3n1o00mvXxzPfut9di1jf6/j37rWfLrr37rfXvfuvde9+691737r3Xf4/H/E+/da8+vG3Fv9j7917PXXv3W+u/95/x5/3j37r3XuOf9hb/AIn37rWeuvfut9e9+6912B/vY9+60eltlqGki29QzxU8azEoWkUHWdQJOo+xBfADZbf5E/z49MIT4rdIj2H+lHXduP6fX/Y/63v3WvPr3+wH+8+/db69xb/H37r2erbv5Gu3KHPfzPviNNW3Jw/cWz8hSqPoahMkNDEf4e9Hh1ZD+oB8j/g6+zrmv+LXX/40VaB/h/kcnPuw6bbh186v/hNX/wBv4fnH/wBqjuj6/X/j9ZfdRxHTjefX0Zk/Sv8AwUf71731Xrl7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuuLukaNJIyoiKzu7sFREUamZmbgADkk+/de4ZPVNu4v57n8vvF9ldm9Xbb35uDsnN9Q7jbaO+cl15g4dxbfxm4oWeKpx0WVjqEWbxSxywtJGChkjdVZtJ9qktHddQI+fy6Kp94tbeTw5dQ8wdJoR6g0z1FP89f4Vf8AKp24f9bZEf8At/8AgX7t9FL8umv39Yep/Yf83Xj/AD1vhULf5J25Y/n+48dv/cv342UvqOvfv6x/pfsPXZ/nq/CpTY0vbf8Ar/3HSw/1z93799FL179/WHqf2H/N14fz1vhV/wAq3bY/19jp/wDVfvf0Mvy60N/sDnu/Yf8AN13/AMPrfCn/AJVu2v8A0B0/+q/fvoZet/v6x8yf2Hrr/h9b4U/8q3bQP9P7jpf/ANy/fvoZevDfrE+Z/Yeuv+H1/hV/yqduf639x4//AKr96+il+XXv39Y1/F/vJ/zdeH89f4Un/lF7cH+vsdP/AKr9++il+XXv37Y1p3f7yf8AN12P563wqJsKTtz+v/Hjx8f6/wDlfv30UvqOtDf7A/xfsPXj/PW+FQbSaXtz/X/uPHb/ANy/fvopfl1v9/WHmT+w/wCbrofz1/hTz/kvbnH/AGY8fP8A6t+/fRTfLrx36xHmf2H/ADdeP89f4Ui3+S9t8/8AZjx8f6/+V+9/Qy1pjrQ36xIxq/3k/wCbrkP56vwqP/KL23x/2Y8f/wBV+/fQy+o68N/sD/F+w/5uvf8AD6vwqvb7Xtu/H/MDx/kf9Rfv30UvGo63+/bH5/sPXj/PV+FQAP23bfJsP9+RH/8AVfv30Mvy69+/rH1P7D/m67/4fU+FX/Kt239L/wDHjp9Pp/yt+9fRS9e/ftj8/wBh64/8PrfCm9jTdtj83Ox05/8AVv376KX5de/f1if4v2Hrofz1/hSb/wCS9t8f9mPHz/S3+V+/fRS/Lrx36xHr+w/5uvf8Pr/Cr/lU7cv/AE/uPH/9V+/fRS/Lr37+sf6X+8nrw/nr/Coi4pO3PrYj+48fH+P/AAL9++im+XXv39Yep/Yf83Xf/D63wp+n23bd7X/48eP/AG3/AAL97+ilpWo61+/rH5/sP+brr/h9f4VAavtO3LatP/Hjx/X6/wDK3719FL8ut/v2x/pfsPXv+H1/hTYf5L24L/j+48f/ANV+/fRS/Lr37+sf6X7D10f56/wpBt9r25/6A8f/ANV+/fRTfLr37+sfU/sPXf8Aw+v8Krf8Be3Pra39x05/x/4F+/fRS/Lr37+saVqf2H/N12f563wqH/KL23/6A6c/63+V+9mxl9R14b9Ynhq/YevD+et8Kjc/a9tgC3J2PHzf+n+V+/fQy+o69+/rGlc/sPTHnf8AhQF/L12amGruwN0b76/27mdxYnbI3Xuvaa0G38fkMxP4aVsjVpUuyRrZpJXVG0orMRZT7q1nIgqxAHVot6tJnEcQYk+in9v2DzPV0uDzmG3NhsVuLbuUoM5gc5j6TK4bMYqqhrsblMZXwLU0VfQVlOWjliljZXR0Ygggg+0pBBoejdWVhqU1B6dPeut9e9+691737r3XvfuvdfHc/wCFJHH84X5X/wDa+xX/ALrx7914jA6oq9+6917/AF/fuvde9+690qdm8Z+kv/qJ/r+LxHn2IuVP+Vgtv9N/kPSS9/3Ef7Om/cItm8n/ANRUnss3X/kpT/6dv8J6fh/sl+wdNHJ/oLfj2g6vgddA2/AP+v791s5669+691737r3Xvfuvde9+691737r3XL8G1rX/ANj791rz66txf/G3v3XvOnXXv3W+u/zx/vPv3Xuuib8+/de697917r3v3Xuu/r9L3/Pv3Xvt69+eP949+611631HH+uffuvV8+r3v+E1Bt/OV+If+Of3GP8A12qn3o9XXz+w9f/X3Zuy/lz8feqd2R9f7n7Gwc/YZqMRDUdfbfqqfPb0x0WcI/htdl9vUDtPSU8ikOklQqalIZAw9vRwSS5Xh8+ktxeQ2xAc1J8hk/b0o+4fkf0p0LS7bm7U7B2/tWt3pNW02ycDW1sI3JvOqxtKtfkqba+B1CprXpoGWao8KERoQzlbi+khd20Dy6vLcxQxiVzg8PU9KvqvtvrPu/ZeN7F6j3vtzsLZGXkq4KDcm18nT5TGS1WPqGpMhRPNTk+OeCVWjnhkCujCzKD7oysh0tg9ORyJKmuM1B6ET3Xq/Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvda0n8or4+9D535d/zFex830715muwsl2CcDkt5ZnbdBl81W4Wo3BWzTY2WTJJKgjZ6eBmKIGbQoLEAD2Y3RKorrgnzHnx6Du0gPPPE+VDGgOQOGR6V+XV/3+y/9E/8+Z6s/wDQB2t/9S+0XjS/xHo8+ng/hHXv9AHRX/PmerP/AEAdrf8A1L7940v8R699PB/COvf7L/0V/wA+Z6s/9AHa3/1L7940v8R699PB/COu/wDQB0Wfr011b/6AO1v/AKl9+8WT1PXvp4P4R14dA9Fj6dNdWj/W2Dtb/wCpffvFk9T176eD+Ede/wBAHRV7/wChrq2//hg7W/8AqX37xpf4uvfTwfwjrw6A6KHI6Z6sH/kg7W/+pffvGk/iPXvp4P4R17/QB0V/z5nqz/0Adrf/AFL7940vr176eD+Ede/0AdFXv/oZ6sv/AF/uDtb/AOpffvGk9evfTwfwjr3+gDor/nzPVn9f+PB2t9f6/wDAX37xZPU9e+ng/hHXv9APRf8Az5rq3/0Adrf/AFL7940v8XWvpoP4B17/AEAdFf8APmerP/QB2t/9S+/eNL/Eet/Twfwjrr/QB0V/z5nqz/0ANrf/AFL7940n8R699PB/COvf6AOiv+fM9Wf+gDtb/wCpfe/Gl/i699PB/COvf7L/ANE/8+Z6s/8AQB2t/wDUvvXjSevXvp4P4R13/oA6K/58z1Z/6AO1v/qX37xZP4j176eD+Edd/wCgHov/AJ811b/6AO1v/qX37xpPXr308H8A66/0AdFWt/oa6tt/T+4O1v8A6l9+8aX+Lr308H8I66/2X/on/nzPVn/oA7W/+pffvFk9T176eH+Ede/2X/on/nzPVn/oA7W/+pffvGl/iPXvp4f4R14/H/oo/Xpnqz/0Adrf/Uvv3jS+vXvp4P4R13/oA6K/58z1Z/6AO1v/AKl9+8aX+Lr308B/COuv9AHRX/PmerP/AEAdrf8A1L794sn8R699PB/COvf6AOiv+fM9Wf8AoA7W/wDqX37xZPU9e+ng/hHXf+gDoq1v9DPVlh9B/cHa3/1L7940v8R699PB/COvf6AOirW/0NdW2/p/cHa3/wBS+/eNJ69e+ng/hHXh0B0ULAdNdW8fT/fg7W/+pffvFk9evfTwfwjoq3zk+Mnxw3P8O/kjt7cvQ/U+awNf1TumorsVUbHwEENVPiqB8ljpWmooI5VeGojjmjeORWDKCD7cid3kCsag+XTNzFHHbu6AAgVBHEfZ0B38jnD4rb/8u/q3CYLG0mGw2L3BvSixeKoI/FRY+hp8v46elpoh+lEUWUe7Xn9v+XSXZP8AknI3rUn7Tx6t29pejbr3v3Xuve/de697917qj3/hQX/27k7e/wC1Hmv/AHWn37rx4dfHKt/vX5/1/futV669+6313xxa/wDj/wAa9+69nrr37r3XYt+ffutGvl1zjjeaSOKJC8krrHHGguzOx0qqj+pP09+690b/AKe+EPefblfjY6Pa2QxGNyMkXhyFdSv4pYpGALxspt+fqT7917Pl1dV1x/IioqzC0dbvjcbU1ZPAksiR18iKrMoYjSsi2+v9Pfut06VuM/kF43cu4P4fg8hkZsehbyVSVU5QAfU6hKffuvUPr0E+X/kw7Wwfc2I62qctUNDW1CwTua2bXGdeg3/c4/2/v3WqfPq0bGf8JethZfC0uXoNx108U9NHNZKyRiDJGHIsJvxf349b4noB+2v+E2mF2biqypxefqvuIoJpIhLXTKdcaEqCDKfyPfvLr3E061wvkT8Iu2uid3ZLAyYaqzlHT1U0cM1DC0jJEsjKmpiSG4H1B9+619vRQ8tgsvgKpaTNY6rxlT+rw1cRjfTf6gH37r3HpV5kgbYx5HNzHz/VdPB9iC+/5Itv9p6TJ/bN0HvsP9Kuve/de697917rsi3/ABUfT37rVerjf5Dd/wDhzj4t2t/zNXaP+v8A8XIe9N8P7Orp/aD7D/g6+yvmf+LXX/8AUFW3/wDOOT3YdNtw6+dX/wAJrOf58Pzk/qcT3QAf6f7/AGk91Hl04fPr6Mq8Ko/oo/3r3vqvXL37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdIrsrnrnf4/rsrdX/uin92T4x9o6pJ/Zt9h/wdUC/wAhX43/AB4i6A723TB0h1kNzbz70zcm8c7VbTxOTyW45sdTLPQy5GoyUc3+beondVj0rqdmILG/tbdsUkGjFRn59EmyKstqfEGrSzAVzQVOBXgPl1en/oA6K/58z1Z/6AO1v/qX2k8aT16OPp4P4R17/QB0V9P9DPVlv6f3B2t/9S+/eNJ69e+ngP4R149AdFH69NdWn/yQdrf/AFL794snr176aD+Ede/0AdF8f8Ya6t4+n+/B2tx/6q+/eLJ69e+ng/hHXX+y/wDRP/PmerOf+zB2t/8AUvv3jS/xHr308H8I69/sv/RX1/0M9WX+v/Hg7W/+pffvFk9evfTwfwjrv/QB0Ub36a6s5+v+/B2tz/r/AOS+/eNL/EevfTwfwjrr/Zf+if8AnzPVn/oA7W/+pffvGl/iPXvp4f4R13/oA6KHI6a6sv8A+GDtb/6l978aX+I9e+ng/hHXf+gHov8A5811b/6AO1v/AKl968aX1699NB/AOuP+y/8ARX/PmerP/QB2t/8AUvv3jS/xde+ng/hHXf8AoA6K/wCfM9Wf+gDtb/6l9+8aX+I9e+ng/hHXv9APRf8Az5rq3/0Adrf/AFL794sn8R619NB/AOvf6AOir3/0NdW3/r/cHa1//cX37xpPXrf08H8I68egOiiLHprq23/hg7W/+pffvGk9evfTwfwjr3+gDoo/Xprqz6W/48Ha30/p/wABffvFk9evfTwfwjrr/QB0V/z5nqz/ANAHa3/1L7940v8AEevfTwfwjrv/AEAdFf8APmerP/QB2t/9S+/eLJ69e+ng/hHXX+y/9E/8+Z6s/wDQB2t/9S+/eNL/ABHr308H8I67HQPRY+nTXVo/1tg7W/8AqX37xpf4j176aD+Addf6AOivr/oZ6sv9f+PB2t/9S+/eLJ6nr308H8I67/0AdFWt/oZ6ste9v7g7W+v9f+Avv3jSevXvp4P4R149AdFH69M9WH/X2Dtb/wCpffvGl/i699PB/COuv9l/6K/58z1Z/wCgDtb/AOpffvGl/iPXvp4P4R13/oA6KP16a6sNvpfYO1j/APGvv3jSevXvp4P4R149AdFH69M9WH/X2Dtb/wCpffvGk9evfTwfwjr3+gDor/nzXVn/AKAO1v8A6l9+8WT1699PB/COquv5y3xi+N25P5fHcUG4uhuqcvBg6za248PHJszC0TY3P0udhpKbLUlRjYoZFlSKWVP16SrEMCD7ftmaSYBzUfP7ei/dESCwkeIaTQ5GDwPmM9HR/l2QRU3we+MdPAixQQdUbfhhiQWSKKPyJHGg/AUAAD+ntq4zM3SnbgBYxAcNI6Oh7Z6W9e9+691737r3XvfuvdfHb/4Uj/8Ab4T5X/8Aa+xX5v8A8oA96Hn1s8B9nVFfvfWuve/de67tx+b/AO8e/da6VGzyTn6S9v0Tj/YeI/T2IuU/+Vhtv9N/kPSS+FLR6enTfuL/AIveT/6in/4j2Wbr/wAlKf8A5qN/hPSiH+yX7B0z8n6D/Xt/xPtB1fHXrf63v3W+vD+pv/sPfutfZ17+v5/1/fut9eH+9c+/dePXvr/W5PH+N/futddsjobOrIfrZlKm39bH37rfUykxuQr3SOioqmpeRgqLDC7lmP0UWHv3Wq9KjcHXO+tq4+lye49q5jD46sCGlq62laKCYSfoKtz9fe+vCnl0iSCOCLEfg+9db67Av/T/AGPv3Xuuvfuvde/1/fuvde9+6912Pyb2/wBj7917r3HH1/xt791rPXXv3W+r3/8AhNR/2+W+IX/a/wBx3/8AQaqfej1ZfP7D1//Qsf8AjBlP5iG4/kL8zPmF1D1D8QN7V2al63XeU/bcmdxfZ1JjNr4w1OQxu35cGgjg/wBx0Ehp0lZEZ0j+pUD2ay6NKxElRmlOFegxamcyy3SqkhOnVqrqAHpinqejafDfvHqH+ZL/ADDewe5t1bSize2ujdh0WI+OuC7BoqSrq9nbvemiwva2U25SOPGfOzzK0jKxZCG4sAtZo2trcAYPE/YerWN3Due4PKDqRKqoIxqHxUr5D+fQ8fy9sFD1X8+f5jHQ/W2Ho9pfHnZlV1LuXZGy8BEtNtXB703fiaiv33Ji6JPTDLNVajUKlhcAW9IszcZgjY8T0usOy9uIk+AEUHkDmvV1bgsjqpsxVgp/oSLA+0XRv1Xn3une3Tm2tzduYrdFLX0m26iCshwtZUvJTzwVVetOUkitY6Qw496GBnrfHo9OzMvU7g2htfO1iJHV5nb2HylUkXEaVFfj46qZYx/QMxA9+HDrx49KX3vrXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdUBfydiD8if5hHFie1oePxYZzJj2YXf9ko/1efQf2cD6q4P9I/8+9X++y/oQde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3RavmT/2Sj8h/wDxEW+P/dFN7dh/tV+3pNd/7iyfYeiW/wAk0k/ADrq9+N075HP/AGuj7evf9yD0h2P/AJJsfVs/tJ0b9e9+691737r3XvfuvdUe/wDCgv8A7dydvf8AajzX/uuPv3XjwPXxyuLf4+/dez117917rv8Axtx/gPfutdeIH4Nx/vP+x9+68Pn1YH8OP5dndvzCylJ/c3GmkwZq4UqK6qidBLTMfXJBJcAcfQm/+t7917/B1sJYT/hNfkev6XaXY2bzlTkafG1FDlMlQisEsWqmdZ3Ro1YEj8WPv1R16h9a9bKHx4+LnT27NgbZ2jhqDHYbLYOjhpC8SLFI0saLET9frdf6+/eVR1bjx6OLj/5dtGTDWZXLu9FTqJSPICjRpzb9X0t79nrWOgE7Z3z1v0FXybTwMFJ94AaaSdVGosbxkk8/ke9cc9b4DPVW1TVbRy/yPxW4clVwq9XWeZS7AadT6vT/ALE+7da4nrYI6G7b64w8dPt7JZmjvUQQRU6SSrYFo1AAuPdfPrdOs3yK6G3Bv+nOU2hUiakqondRA10ZZFJW1v8AXHv3A9e6qtyv8ujCw1eY3h2/QUs9BIJZAlat1Frk2u355/HvZ+fWvPqo/vT+VV0h8j+wqvC7Sx0NBTI0ifd0cVlViCqsHDA8E/j3o8MdepnPVG3zH/kz9vdL5HJ4zr+ObcOFxDPVSMzTVc3gVGfSLM1rAexru1nHHyzaXQNCxOPLpLHUTsOqMNxbdy+1cxXYHOUU1Bk8fO8FTTzKVZWRitwD+Dbg+wX0pBr0ye/db67t/re/de68RYfUH/AH37rQ6uF/kRVdNTfzO/islRKkbVHa+0Y4VY21v/EgNKj/AGPvTfD1ZP7QfYf8HX2X8z/xa8h/1A1v+x/yOT3bqjcOvnVf8Jq/+38Pzj/7VPdP+w/3+0vuo8unDwPX0Zk/Sv8AwUf71731Xrl7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XukV2T/zLrf3/hlbp/8AdFP7snxD7eqSf2bfYf8AB1TZ/IYAHxk7TIFr957n/H/TDT+1d9/aD7OiXYK/SvX+Nv8ACeryPaLo9697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuq0f5vZA/l+973/5Udu/+9LS/X2qs/7Yf6vMdFe8Cu3yD5H/AAHoYv5eBB+EfxnI/wCfWYL/AKGk9t3H9s3Sjb8WUf8Apejme2elnXvfuvde9+691737r3Xx3P8AhSMCf5wnyvHA/wBz2K/9wB795nrxwB9g6oq9+691737r3XfNr/0/4n37rWOlRs0Xz9Lc/wBic/7HxH2IuU/+Vgtv9N/kPSW+/wBxH+zqBuHnOZP6D/Kn/wCI9lm6/wDJSn/07f4T0/F/ZL9g6ZgbH+v+9H2g6vx6fNvbbzu7MnT4XbeKq8xlaprQ0VDC0s73NvoPxz9SffuvefVvXxr/AJPfc3b+Omy+6o2wNNLSpLRU1zBOkpUsUnMn1/H0Hv3XsnoY+wf5Evc+Mxn3GzspR1dYnqkjrKhZRpH1UIhB59+63T59FfqP5VXdWzKkf3+jpYoZHEaLTeRAp/LFy1z9R791rjxx0f8A+LX8pvbzSLufPxtnZoiJoqSb/KYRpuVQrq4/x4v79140I6Nhur4G9bUuXSvynXeMipKaPxyR09Dqi0LwzHUfr73nr1BTPS82z1H8cdvU1NjMH1ZtbIV8breSuomM8Ui/VgFP+v79g9eB6fextsdS7jx9Jht2bCwNdTUCq1NSpS646dI7FURf6e9dbJ6Ij3J8NPjL2Xi8g+Jxz7T3DJBImPSkiFLR/dMtojLICCBf37rWOqGu9PjL2B0bkp0zVKuQwvlkNPmKENJSiHX+15n5F7Ecg2/1vfqdb6LiDb+n+x9+69117917ru/Fv98P9b37r1Ouvfuvde9+691737r3V7//AAmpt/w8t8Qrf87/AHH9f/Daqfej1ZPOvoev/9HZ37u/k+7G7a7q7g7k2t8pvlT0SO9Y9lRdj9f9O76xe29jZeLZqRw+KHGPQyNGteiFaxS7Btb2sGt7VC6YJpYaiOBPEdFz7bE0xljZow1NQXAanr0I/f38rLpzt6fo3I9a9idrfFXP9EyZoYTcXx1y+I2Vl91UWdxa42spN7VP2khr2upnE7kSF3cuzBgBpLp1+LuB8j1abboZAPDJjZfNcHoy3xF+KOyviB1PSdY7U3Hu/f8AkZMjks1uns3sivpM32NvnMZKtkq2r9056nhhM5hV/DToVtGg49TMS3LK0rVOPl09bWsdrH4cdT6kmpP2no0ntrpT0Uz5wG3xn7G+n/AbGfX/ALWsP09+62vHoeer/wDmW2wP/DN21/7p4fehw68ePS697611737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3VAP8nX/soj+YRwP+Zrxc/wBr/i+ZP/ePa+7/ALNf9Xr0Htn/ANyrj/TH/n3q/wC9oOhD1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdVT/wA5v50dn/y7fgvvn5LdQbW2ru3fGB3FtzBY7G7zNacFFFmTO1RWVENBJFJIyCEKqeQD1En6e3oEWRiG8gT0muZHjCLGaF3VakVpU0rTz6M18L/khkPkD8I/j58p+0YNs7ByHZfRe0+2N+JDkhSbR2q+S24uaz9T/FMtIBBQ0yiSVpaiW0cYJdrKT7bYdxA6eRqxhm9OqC+7f+FU/QkfZW/+qvhN8TPk983Kvr7NSYLJ9o9SbHkz3VFbWQu0bVOEqdvyVWSqqSVkkFNVy0tPHMF8kReMhjdFjr+oT+XTMzz6f8WVSf6RI/yf6vToUvhX/wAKbPjB8h+/9o/FX5H9Hd4/Bzuze8dPS7Tf5BYSg2rsXcuerp/t8RgqfJ5SeCvoZ8i/7VAa6hWGWW0Qn8jKp8yJX9Ov5463HJIEBnAr56cgf6vXq9L5b/J7r34a/HbtL5KdpRZip2R1Ttms3JmKPAUf32Xr1pk/ZoqGEkKHlchdbkKoux+ljqKMyNpGPPrdzcJbReI+cgD7Tw61a9sf8K3P78bXpN67H/lYfNTeWz8rLlIsJu3a22Zc/tbLviqlqWrShz+JppqWVoHASpWOVjG91axFvblLf+l/LpnVf/wp+09bK/wZ+Xv+znfGTaHyPy3TXZ/x1TckGQmrthd1YZ9sblw0OMP7+TmSsEd6KRQZIqllUFQT9BctyIFoVNQf2/n09BI7giVdJU0PofmPl1Rx8nv+FS3xp2F23v3or4d/HX5B/O7eXXlQuP3Fvfona0e5+qKXIrIYaiPH5jCS1NfkYY5Q1P8Ad09GlNJIreGeVVudoqV/UJ/LrUskug+AAT5aiRXowf8ALc/4UE9RfO/vnC/FDf3xn+RnxV+ROe21m9ybfwHb2zhi9r7tpdt0jVuaXB5GeWOtikSKOR1jqqIK2lgJSwt780YILR1oPXr0criiTgBz/Caj+dD/AMV1sH+2ulHXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690Wr5km3xR+Q//iIt8f8Auim9uw/2q/b0nu82sn+lPRL/AOShY/ALrq3H+/o3z/t/40fb17XxzXpBsdDtsdOrZfaTo3697917r3v3Xuve/de6o+/4UF8/y4+3gTb/AHB5o3/1scePfvPrR+Hr45PH5/p+P9f37r2fLrwIF/rzcf7D37r2evXP0v8Aj8/8R7917HVj/wAE/wCXf2R8zM1KMNR1cGBpyyPUojRl2AvrWUn6H8cc+/db63OPhf8ABjur4xbFxm1tsUE1PHR00SNVADzyGJQF1MSb+/HOOvAU6uv+PFT2HnVm697Dxc09NWxNTipmiJCahouWPvXAU63k9Cz118Odz7D7Rnz9JXSLt+eqFTFCknpRC/k06Ln37rXVhu4qSaHb8lEkrD/ImiaQ+nSfFoYn3sinXq+nVFHyK6P2PLubI7i3HnvJMJZJRF5hcWYtaxPvQ638+qvM5s3bWa7Qpqjb7TyihlZUewIJVrAgi3vfWvs6m7vyO8sJunHy0pyVOaSWDxtGWAbQ3pub/wCHv3Xutlb4Cbx3LvTrKF9zRyO9JSQiNqkcuFjQfU+/Dj1s8Ohv+RfT79ubMqsDRuaWbxMAYzpB1A8Ai39feznPVRjj0Tfpj4dYzqSmyOczyB/CJZXlNi7WXX+Sfrb3RuHVh0UbtPc+wKreW9cHTbbbJtUUclIiyUwkJaSNotQFv8fY/wB7BHKVkScVPSWP+3avWiv/ADdPgTvPY/YGf7lw+Dah2/kpZ6iSmSIqiwiR5dWm/B+vsAjh0p6oD97691yHAv8AU/71791rz664/wBj/vH+w9+69nq2j+R1t3+OfzQfiDUfctB/CO5Nn12lTbzacko8bf4f1HvTcOrxn9QD5H/B19nzM/8AFqyH/UDW/wCt/wAA5Pr7sOm24dfOq/4TWf8Ab+L5x8/8unun/Y/7/aT3UcR04eB6+jOv6V/1h/vXvZ49VHXfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6RfZP/ADLrf3/hl7p/90c/uyfGPtHVJP7NvsP+Dqmz+QyQfjH2lY/81z3Rx+B/kNP9Pau++Nfs6Jtg/wBxX/07f4T1eP7RdHnXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdR6uqpqGlqa2smjpqSjp5qqqqJmCRU9NTxmaeaVzwFRQWY/gD37jgdaJAFT5da738uf8A4UgfE/8AmK/MPsD4kbN2LvnrbK4z+Mnqfde9Z8Q9F2uu3K6SlyiUlHjZJHoZpYU+8pIJyWaK4YiQFQ94alCUrVeNf8nSbxpEmCSgAP8ACRU5Hr+XWxT7Z6Vda4Xzm/4Uq/E/4s95bg+LvRnVHc/zg772rSVX97MJ8cMLj937U2Xl6SY09fg89nKGeWonrKJwBXwY+jmWnYiKWWOXUiuKq176/l0zI70PhAE+VTQV/wAvST+EP/ClHrX5G9/dW/Fz5A/Dv5R/E7tjuTcY2l15mextiT0GwNwbjnBaiw71OWemyNLLJ6UDGkljDMup0BHuxjVq+FXA8x02ksqgC5Cgk0Gk1+XnTz8x1eD8x/mh8dfgX0Zuj5CfJnsHF7C2BtuCRKZJ5oJdxbvzxp3nx+z9kYJnSbJ5Ws0MtPSQc2DSSNHEjyK0BXpSTQV61mJ/+FZkW46/F7t6p/lb/OPe/R1SPO2+l2BNJU5nGM5VMpgqvBfe4oRmx0iStcG1ta/h0LDp7tWr7BTpKz3ZkHhhCnnVjX+Q/wCK+fWxB/Ls/mFdJ/zLvjzTfIno7F7523g4t05vY+5dn9kYWmwO8tpbv26sMmUw2XoqKoqoG0rPE8csM7Kwb8EEBt0KcfPp+KVZQSvlg/4ePn0e/wB16c697917r3v3Xuve/de697917r3v3Xuve/de697917qtD+b5/wBu/e9/z/kO3b/+hNS+1Vn/AG4/1eY6LN4/5J8n2H/Aehj/AJeIA+EfxnANx/otwfP/ACFJ/T23cf2zdP2H+4cf+l6OZ7Z6Wde9+691737r3XvfuvdfHc/4Ukf9vhflf/2vsT/7rx70PPr3l1RV7317rscA3H1+nv3Wuuvfut9KnZt/4/S2/wBRPf8A1vEfYi5U/wCVgtv9N/kPSS+/3Ek+zpSYPr/dvZm/5Nq7LwldnMzkMiIYqaigeYp5HCeSUoOFH/IvZZuv/JSn/wBO3+E9PRH9JfsHVmNL/JQ+ZdXHgJ0wOOSnzqU8iyTGaI0iz2JFQpvYqPrcj2hp05nh1sx/y7/5KvXfx0wdDvrsjx53fdRSxVE0Uuiqp6afx3PjYE2sfoALe9dbHy6stp9z7R2pviLZ9NjKanhkYRKTEqAMTa62Hv329exw6QXcfcOD2ZvDDbdo0i15OpihJAX6ykcfT/H37HHrefPp77N6twPbey2po6RBlGpNUM+gDTIV4IYW5uPfutGtOq69ubg7B+N2YrsTm9tVtfhkmcRVVPTGb9oHgsxvYc+/dayOjVbf7N2T27tevp6enaLJTQurU0ip5lkItbTYEfT37PW+q0J8fWbM7jfGVVOft6uqLxahxoZ+B791r7elJ3HTU2N3DhWCfb01dJAszH0rZ7XF/e+vdD/S/HzrjdWAxtXWVQo/JCkpqEFix0g/W/vVet0Hn1E3f8POmN77WqNvZqUZXHvA6CapjD/b3XTrj5+o/HvXXgPXqmftP+U/0TJuGpx2zsruR62oqWl1reKhiQMdSKVaw/23vfWvs6rm+YX8vvLfHzBUu6dtyVuWxCqDkI2LVM8YYX1qASQFHLfXj37r2eqxj/rfT/fc+/db69+D9f8AiP8AY+/de669+691737r3V7/APwmoH/Y5b4h3H/L/wBx/wDvNVPvR6snn9h6/9Lf49+691737r3Xvfuvde9+690Ur5x/9ky9jfkfb4u9v+1rD70eHVl49D31f/zLbYH/AIZm2f8A3TQ+/Dh1o8el1731rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuqAv5OygfIn+YSQSb9rw8H8H+N5M+zC7/sl/wBXr0H9nINzcf6Y/wDPvV/vsv6EHXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691rrf8ACpL/ALdIdqf+H3sb6/T6VntTa/E3+lPSO84w/wDNVP8AD1VZ/NC+TvYPx5/4S6/BzZ/W9d/C8p8o+vOgfjluCvUkVEexd29XZPO7so6fTZgayDGrRuVYeiVwbglTQV8RhStcfZXz6tJQW6sSQEAY08worT7OtjL+TP8ADnrj4Sfy7Pjb1VsTbuKxmay/Xm3999m7kpcdBR5fffYW7qBc1mtx7gqU1STSDzLTUyySMIaeOKFLKnvUy6HMf8PV7aQTQLNShcAn7f8AY6qy/wCFZ/xS2b3B/Lef5D02GxtB3P8AFrsHbO9tm79goUG46XadV9xS7p2n/E4ikv2dRI1LXLE7MkdRTxyKoYsT6JS9RXgK/s6rcSLEFcjiQv8AvRp/I9NXevyN3h8u/wDhK/XfIPflTLkd89jfE2obdVdIyyTZHcO2c/V7MyldI6qupppsc8pYi5LXNySfb8DapmYClVOP2dItwUxWSIzaqMuT5/b0zfyCP5pv8ufoX+UH8Oere6vmf8eurOx9kbc7Lxm7Njb47HwO3904Gsm7k3FnKePI4WukWeMS0dZTVELlNLpIpUnmybSSKjoxMqK2k4P2HoxX/Cgn+YVSddfyWew/kF8Suytq772/8h8ntfpTavZ2xs7RZ7AVe0uzxkMTu3IYLM4xpI3k+yo62k9DhkdmVtJUj3taxtUiv+rj1px4yURiuRkccHI+Veh4/wCE9Hwq67+Gn8sT4/Ue3Nt4mj7H7j2zH2x3FvGHHxU+c3jufctbUVeJTKVx1SyU+Mxz09BQQs+iONGZVDyyFvSqY3Mfp1W2kE8QnH48/l6dXGZbrTr3Pb32t2Vmtk7XyvYOx6LMY3Z29a/CY+q3PtnH7hiEGcosJmpozUU0VUihZ0icBx9RyfdQzBdIOD08UQsHIyPPpb+69W697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xui1fMn/slL5D/APiIt8fX/tRTe3Yf7Vft6TXn+4sn+lPRLv5J4I+APXV7f8fRvk8f9rk+3r3/AHIPSHYxTbYx1bN7SdG/Xvfuvde9+691737r3VH3/Cgsf9i4+3j+Bg8yP9vjjb37rx4dfHI9+6913bi/H/E+/de6eNu41MxncTipGKJkMhTUjsDYqs0gRiD/ALH37rRwK9fSS/k07B6l6A6B2hmauChSqylDSVFVOwTyM2kAjkH8fX3qvr1ulOB62desc915vygimw8FL4GRSX0JoACi/IX34FevEU4dClS4br/GVfniqMZFVg3sxCyX/wAAB7328etGvQhQ1dHLSh6WSOZOLabHj/X92JFOq6T0mM7NRVVJJRyukMk0bIFY2J1C3491J6sBQ0PVRnyT+KG5NzZiszFHkKg42UvIYw5C6eSeCfpY/wBPeqjz6t8h1WBvHZ+M6Zy8ddOEkeGYiR+CxYMeD9fyDf36vn1r7Oj2/G/r7avyBpaerrcTAxQxnziIMDo0/wBq3+PvxwMdbwD8urkeudl4Xrbb9PhsRHDBojCSRKNLEKoAsP8AYe94HHrRzw6W9XnKOlhMtVMlOjfqLm2m355vb36o69TpKVubwm4qSqoRU0tXC0bB40YNqshBFgPdWI01OOtjjjqtfctf1fsjtHcuRzG10li0RKkgp9QEha1ySP6+z/c+ZNiuNlg22K7jMkVSRqGPWvTSQyK5Yqc/Lqt/+ZZsnYfaXSO68jW4kQYg4yolpFlgVVuYZGUXI9x7tPOXKm/Xcu3bPfRTzQmjqrgkEfLpVJbTxAPIhAPXzKu3MLR7f7G3bjKCwo6fNVy06rwEj8xsgt+B+PYl6YHp0HI4BP8AXj3vrRyeuI+o966t1cb/ACHP+3nPxb4H/M1do/8AuzA96b4f2dbT+0H2H/B19lfNf8Wuv/6gq2/+I+zk492HVGrTHXzq/wDhNYP+x8Xzj/7VHdIH+v8A32k91HkOnDwbr6My8Ko/oo/3r3vqvXL37r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdIrsn/mXW/v/DK3T/7o5/dk+MfaOqSf2bfYf8HVNn8hjj4ydp8Af8Zz3P8AT/qBp/ay++Nfs6Jdg/3Ff/Tt/h6vI9oej3r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuktvjO1O19lbw3NR08NVV7d2tuDO0tLUSGKCpqcRiZshBTzyjlUdowrMPoCT72oqwB6q5opI8h1o/8AwO/nJfz1/wCbT1Xk+vviD0z8fdsby633NuCPvX5H7+XLbd2dT4HcVdJNsHavXbJPLD/GaWATJWlKSqkMUcMxSAS6vagNDqEjL9o8vy6L2S8YNbq/lVXxXJ4EeVPIjj0eL+W1/OC+c+wv5imc/lO/zfdq9XbZ7uyuCxlX0L2j1nSVy4jsTKVWNXO0WJqsmKmWkyEORxxklo62KmpZo6iCWmqoRIT46MA9WwvmKcD9nz6URlogsRJk8iTSo9CaeR9ej4f8KMvmPW/D3+Vx3lkdmbk/gXcvcEeC6l6mgptUmXrchuTcdFDu+oxVMgJkamwhrmIA4Z04N7e9RBql04r1u6aPQIZa0kqPsFCSa+WBx61ZPmf8CqD+Tv8AH7+Sz/Mo6d68yce5ur+w9pdr/MCphLrXS7g7HxG3t00+HrYpGDpAEbMYN4S7epwLKjEKoZxKcCirxp88E9IY4Gs1prLySYXUTTFSB8sYH+qu4n/Mo+cGV6V/lC97/Oro6qjXKyfHPafY/VuRn5FJJ23JiMXtbKNqW+unXNxTgaQdSD9J5CVSFetK06M3UyRFa6SRSo8vs6rG/wCEnnw/2V07/LzHyfyeDxuS75+WW9Nx723l2LV0Sybln2Uk0A2xtCPKTtJKtHHOtVkJo0ZVlqZ2kkDFE02kUpQHzAP7emreRJlLr+ElR/tTT+Z62Yd29ade79yezczvbZW1915brvcMe7diZPcGFoMrXbP3PFA1LHntuVVYjvSVQjZkE0JVrfngWoGZQQppXj0+yIxBYVI4fLrSQ/nMjO/zFP8AhQj8Hf5Zm/1jrvjX0uvXPbu8tpsHkp92Puxotx72pMkkRQhKrF01Jiy5cssUs2nTfl2Mal04wC3208ukszeFKrEk6yEA8hWpJ+2g63jdu7dwO0cBhdq7Ww2L25trbmLocJgMBhKGmxmHwuHxlMtHjsXi8dRqkUEEESJHFFGoVVAAAA9sE1yelYAAoOmLZHWvX3WlPnqTrzZW19k0u6Ny5XeW4qba2Ex+Dgze684yvmNw5OLHxxrNWVJRDNO4LNYXPHuzMzfEa9aVEQUQU6W3uvVuve/de697917r3v3Xuve/de697917r3v3Xuve/de6rQ/m+G38v3vb/qB27/tv7zUvtVZ/24/1eY6K95/5J0n2H/Aehj/l4i3wj+M4/wC/WYL/AKGk9t3H9s3Sjb/9wo/s6OZ7Z6Wde9+691737r3XvfuvdfHc/wCFJH/b4X5X/wDa+xP/ALgD3oefXvIdUVjnj3vrXDPXh/rXA/H49+68es1NS1FbURUtJDJUVM7hIYIVLySOfoqKOSffut9GI6t+PfcO6cia7DbJzkkFHFI8rNQTFysiaV0RCzck/X2JuT4nm5ktYo6ai3n9h6TXal7d1HmOPWzD/Js+Km5+qdwz757T6tCVtbVvJR5OvxcsdQpMgaGS8l/ovAPsr3pGi3e4jelQ7fZx6dhFI1+wdbTXY+x8ru/asc+2s3U4OZ4QYYqfxxIthqVb2Fv6eyzp3qvybvXuPpLcUe2d4Y6XK4yrm+1p8iY3nupOkO0vI/PvfWvt6UfbFJW1mJx/ZWMo3atCJVladGLaTyvA96+3rZoRjy6BTdGzJO1sXj99rJLBmdv+Oskhk9DM0PJFv68e9/LrVAehk68+V+w9v4dcTuMTw5LGoIJmIXS7x8Ekke/dbr5dC1R57Gd40rNQ4WnqcRMtvPJEpaRT9LsB9OPfutdJig6p646brK3deSehx8TBz9s7hFMjcgaGvx/jb37rfDj0W3dmC2X2huqPdeNioqanoKhGerjICaIjc2YD6ce9dawc9F77R2Rju2dyJiMFWyMuI9XkoSHXywfRWbj+nvfXuhx6oxNSaaPZO5klhNIgipKlgQZEQWGo3/Nvfvn175dLTNUWPpq9NtUNWx038ixG7sB+oEf09++fW6+XSdyVMmEWSKh2RJkRpPkr1o3lmNxcurD37PXuie7321VdsZuo2ZldnVgwtZFNTyNWURRAXXRdCfzb6e/dV8+qUfmZ/K7put8RuDeXX8GdlyDStWriJYBHQxrpMsywr9QD+P8AH3unXvPqjesoqvG1dRQ19NLSVlLI0NRTVCGOWKVDZldG/wB9/T3rr3HqJ791vr3v3Xur3/8AhNSf+xy3xC+v/Hwbj/8Aeaqfej1ZfP7D1//T3+Pfuvde9+691737r3XvfuvdFM+cNj8ZuxwTYfa4z/3aw+/HrY6Hnq//AJltsD/wzNs/+6aH3ocOvHiel1731rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuqBP5Oxv8AIj+YR9L/AOleH6f9rvJ+193/AGS/6vXoP7P/ALk3H+mP/PvV/ftB0IOve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rXV/4VJkj+Uh2oR/z3mxQb/0P3l/am1+Jv8ASnpHecYf+aqf4eqmf5q3xw7F74/4S7/A3efW2LbMZD4ybL+Pnf8AuikiV3nh2Jt3qzK7e3LlI1T6rRLkoaqUf8c0Y/i/ulD4hpTBr+zq7lPpwrgkMAuPLUKV+z1Pl1si/wAmz5ldYfNX+Xv8bOxtjbw2zmt0Yfq3am1O0dpYnO43JZ/r7fG3aAYTK4Dc+MpX89HIzUxnpvuIk8sLpImpWv71KSzGSmD1u3VYolgrUoKfP7fsPVRf/CsT5ibR2R8HMT8MNhbm21uP5I/KPtLY2zMf1ZQ5GmyO86XYdV922S3BNt6kdqiJKisFDjqSSZAHlnugfSR7tGHQFgPiFB+fmOqXBhlIjY/CQxpw7TUAngKny6X/AMrfjDuD4Vf8Jidy/GXc8sTbt6h+KRxO5pKZw8Ue6Mrk59z59I3W4PjrK6ZCVJFxwSLe3YABMwQ1AU/5Okm4apLJTKKEstR0Xb+RD/Ji/lffJn+Uv8Qu6++fhz1h2b2t2Ft3sfKbx3zuGo3aM7nKyi7g3Bt2iepqMbkqdVWGioqaCFI0VUCXUBixKSvRr0Z7/hQL/Lnw9Z/I+3n8cPhd1Pgdl7P+OO5tp907f6v2fSywUVNtDYtVksvvpsekryTTVIp8jW5GaWaR5JWV2dySWFlBc6a+X+z01IyQrrIJyK0FeOK9Ge/4T1/OTrH5kfy0/jvBhd57Xq+3uptlr1z2519R5ugm3RtTMbWydRisdkMlgg4qYaTI0SU1ZRzvHodJNIcsrAXlLSHxfX+Xl03arHBGLVTlMZxXzqPX8ure8v3h0zgOz9r9J5ztfrrEdxb2xuTzGz+rMlvPb1F2DujFYWA1OWyOB2hUVC19VDTxhpJpIYGVVDMTZSQ1paladKNaE6QRXoUveurde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3RafmV/2Sj8h/wDxEW+Pp/2opvbsP9qv29J7vFrJ/pT0S/8Aknkn4Bdd3txujfI4/wC1z7evf7c9IdjNdtjPVs3tJ0bde9+691737r3XvfuvdUhf8KB1Vv5c3bqsdKnCZgE/4fw839uwojyBZDQdVYkDHXx98fhcPPRwTT1ZEsiMWXyBbEOR9PxwPZilpYlATL00XkBpTqZ/d/BfX7xrW5/eX6293+jsP9+da8ST06UmzNt46bde31oqovOMpSso8qkcMSCbf0Iv7U2u3bbNLpkm0ileva5PTred+C2/MTkOqdpbIylfDBUYvHQxuxm0LIFANw3sx/c2wAf7kfz62Xl4U6s/ovmnHsOih62648dXmtHgeeJy7eS2n6r/AIj+vvR2bYeIuP59b8SX068vaHf0jrmsjW5DySSLMI45ZGTSfWFtf+h97/dGwkf2/wDPqviS8SOrKfjz3n2VX7eilzeMk8ESL5J6hXF9Nhq1G/uv7n2KmLj9p694knp1M7s7v3WKWlrsBSa3gIeYwBj+g3YGwH9Pfl2fY+L3H8x1svIPw8OoO0fk/Xb02xVYLI49EyopnhHkSzFimi9yPdjs2w/7/wAfb17xJDSg6oW+Zu6d21vYcOzaekZZshkFKFFIupna44/r/r+9HZ9h/wB//wA+teJL5jq3r4g5yXovqvG1GboolrnpI5VRwdbtJCjA2N/fm2fYvK44/PrZkkGKdGc2L8htx7x3Bkax8YIqCAgRM6sIiL8c2t70dn2RR23H8+t+K54r0AHzA+QHY+ExP22CoZIVqkdI6mBXCj6i+of7D3tdn2Ir3T/zHVTJL6dEX6G7U+SqbqXKTTVVZhTMrGKWWRxodwXuoP0t7ib3P9w/b722tfB3q60NKripIoO00ofXows7S7uyGjWtOrK4M/t7dEWbqN4YmgarShp5qgKpaTVbUTa3JB/x94N/vPZfdX2Rn3f213KWLeJprlVmbCgamC59OHQmEcljuYjvVBiAGPPqn/8AmhfJnZUHUeW6n2hRpUZuso6iIER6ftlWncaWAB/1/r7jv7kP3XPcnkb3M3LnTnfdmktrohiNbHW1BqJqxFKgnAHS/mjmGyvLKO0tYgpXz/1Dr5z/AGTi6Cr3zuSbJT+KsfK1vmjWUALIJbH6++ud9t+1xXGiCbUtOPUdB5KVI6RP938F9DWt+OPKvpv7SfR2P+/eveI9eHXEYDBFuKtgB+fMtz719HYf7968ZJPTq2z+RnRY2h/mc/FzwVakt2xs9FRpASb16tYW/JPur2dkInZZakDA+fV4nYyrj1/wdfY5zQvish/1A1v/ALhyeyrq7cOvnWf8Jq7/APD8Xzi+lxie6Pr9P+P2kHuo8unDwPX0Zh9Bf62F7fT6e99V679+691737r3Xvfuvde9+691jlligjeaeSOGGJS8ksrrHHGii7O7uQAB+ST72ASaDrRIAqcDrtJI5I1mjkSSJ0EiSo6vG8bDUrq68EEcgg29+oQaHrwIIqOHXkkSVFkidJI3UMkiMHR1IuGVl4IP9R70QQaHrwIIqM9c/fut9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdIrsn/mXW/v8Awyt0/wDujn92T4x9o6pJ/Zt9h/wdU2fyGL/7LJ2kTa57z3P9P+oGn9rL7+0X7OiXYP8AcV/9O3+E9Xke0PR71737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Qb9xqz9Q9qIgLM/W++FVRyWZtsVQAA/x92X4h9vVJPgb7D1qi/8ACNvufrDK/wAvfsPoei3jgH7d2R35v3eW49hHI0Sbno9rbnocZTYfPtiSRUNSvLTSw+ezIHGm4Jt7cYfpKR5Vr8s9J4mpcyKRSumnzx5etPPoAP5k+6ts93/8Kmf5YHXXU8abq3p0DnNq5Tud9uA5J9r0dPp3kw3HLRgim+1x4UzCZ/QJEVgC6qbsf0UXiRq/IHh+3puMUvZZK0DBAPmRWoH2ef29Bj/woE3p8lP5hP8AN0+L3wF+DvXeK7K7X+D+PfurcWL3fk8bjOvanc+9sfg93E7qqMvUUsH2lFjIaGGoBkDH7l40YuxVfRdqiSP4lya8KeXWrn9VmtpsJIAq0rUnJNfQCg881p0Jfzs6S/4VIfOb4jdm/E7uj4pfBSk6y3lQbbnyeQ6+33hKTetLBsLNU+66CHa7ZDdNZTx1FRLQwwXFFqKFo08fkJDerUTQAFvt9elHhlFBYlgnlQeQp1G/l7b27a/mp/8ACY/5Y/DSjq6XP/JHovB5XorB7alqdWVrsH1jksF2H1rRzRLdg9RBQz4iJlLKZIBbghPdiGkIY0Fe2vlUdNxtHAjxglyndTiaGpAH2cOj0f8ACUH5p9fdufy9sT8Ws/uvbmG+QXxo3puvZGa6wrsxRU28Z9k05gqtv7qpcBOyVEtNqepoqiSJGWOendXIut9SapAGp8Iofy8+vWwigrEp+IlhXHxGtB60/b1sqb57v6a6y3LsPZvY3a3XWxN3dpZ2Pa/Wu1937y2/t3cG/txzAtDg9oYjK1EVRkap7WWCljdybAC5A9taWpWmOlWpa6a59OtJb+cbXbp/l4f8KLvhT/Mb3/BDjfjT3VTdXdWbm3vUsYcdtxdtGPam/myNS11T7Cgnosq1/rAsvAsCXoyVXVijAr+3z6SXAWSRUINYyJK0waVBH20PDreR2pvjZm+9p4nfmyt2bc3bsnO4yLM4Xdu3M1j8ztzKYmaLzxZKhzNBJJTywlPV5EkK2+p9tFGDaSM9KlkjZdakU9ek31f3T1B3djM7mum+0ev+1cPtjc2U2ZuPKdebvwO8cfgt3YXR/F9tZaswE9RHT11N5IzNTSsJFDKSoBHvRBGD1YEEVHQm+9db697917r3v3Xuve/de697917r3v3Xuve/de697917qtD+b5/27973/wCoHbv/AL01L7VWf9sKf6sjor3n/knSfYf8B6GP+Xj/ANkR/Ge/H/GLMH/0NJ/X23cf2zdKNv8A9wo6fw9HM9s9LOve/de697917r3v3Xuvjuf8KRz/ANjhPleRf/i/Yr6/X/gAPevM9eNKD7OqKve+vdLbr/r7dvZ258dtDZeGrM1mspUQU8VPRwvMYxNKIvLLo+ign/inv3WuJHW6h/K3/kZbH2TtCk7Q+RdFSZndeWooaqhwddEk1JRyPEJIzpuWjPNiD/sb+/eVet+fz6Ptur417c+P+5cxmcFgqGm27ViGOPwU8ZjiiEwIQXHAH0HsW8hAnnCyA82P+Dpi6zbt5no3eU39XY7bu3arBbZp6/GLjaN38NMrEnxck+NRz7KeYRTf7sHP6r/4enIK+EtPQdMlTvrsvsCCGDAY0YKkpiPMkoamBUHmxb2T/PpzpI96VdAdpYen3FJRT5eCVVLRujzl1sDbi/vefLr1PPoQNuZDDUvW1FNloI2oY6GJisoHKaPpY3597rXr3DovFPnabc9ZW4jaWHegxssjrU1RiKIyk+og/wC396z17pr3/wBJbHo9sO9NhKHJ5qrJM8/LS62tc+n839+60R04dabvy21cPTbQweJWgaOIeWoaMoEUAghXPvx+XXumbJbXrOyc3VUe4slVVWNQOZo3ZnhUj6hefeh1s54dQX+N2L+0qKTC7sr8ZROChpaRhosf6+r6+9469Q9Ytu9ebW6dpKhoakVldVTa5aua33Dt+b6r/W/v3XuIp0273ocxuGhpa3bJWkrpCoSW4iex/wBqFuPeutV9OlR1t1JX4gSbm3FWy5HMBWLQO3kQAi5sbn3vy699nRmeu56iWqkhqqCD7RjZVljAsn0sQR78Ot+XQ5f3B2RV6a6ox+OinUq6sEQWb6hidP1v79jrXQQd39LbR3ftSvlrKageGKmlDvpRl8Yj9QY2+lvfh1uvWih/NI6Cxe1d8Vu6Ni4Glp8RQVUseZqqKMXkMjuEdyv1C2uf8Pfuq+fVOvv3W+u/8Lc/63+9e/da+fV73/Caj/t8t8Q//Dg3H/7zVT70err5/Yev/9Tf49+691737r3Xvfuvde9+690Uv5xED4zdjX/5VsZx/X/crD9fej1sdD11f/zLbYH/AIZm2f8A3TQ+/Dh148el1731rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuqAf5OylfkX/MJ+lv8AStEL/wDkcyZ9mF3/AGS/6vXoP7Qa3Vx/pj/k6v8AvZf0IOve/de697917r3v3Xuve/de697917r3v3Xuve/de697917ojH8xT4H9ffzHfjJub4w9l7v3bsbbe48vh82dx7LNCczR1uGMvgUQZJHhlicSuHRgObEHjlyOTwyTStRTpmaHxdOaFGDDzyOGD0LHRfxg646P+LHXHxFihm7A6s6/6jxXTM9Nv2nocxJvHaNDgP7u11PumjaMU861tOZFqYTHoKuUsR7qzVYt04q6VC8ada7vY/8Awlj6ywPcG8ezvgX86vkx/LxwW/J1qs/1j0pPlchtWKdGZ4osPWQ7gxFdDTIXcRU1RPULGpCx6VFjfxTSi4HnQ8T69MfTIW1ydxzQkZAPlXzH29GY+Bf/AAnU+NPxL7qb5Od99s9k/PP5I4+qpKrZvaXyHhFY2xpqIlqatwWCqK7IiSsjZnaKrrKmZoy7GMI1iNmdyKHj6+YHp1VbOFH1LheJXyJ/iPmT9p6t1+afxU2f83PjD298Xt97h3DtTbPbu1K3bFfuPar0y5zDGpX9mvo46xWilMbgFopBpcXUkXuKxyGMkjzBH7enLi3W5QRuSKEHHy6TP8vr4a7X/l9fDzpL4fbM3jnuwNvdL4PN4mj3luWjo8dmc/PuHd2Q3nkq2px2PZ4adfuslMkMMbtojVAXdgWLZ6f6OBXUNFk6KsxuSo6XIY7IUtRQ19BWwRVVFXUVXEaeqo6ulnDJJFLGzJJG6lWUkEEH37r3Wrb33/wlm6FzXeGb7x+Dnyv72/ly5LdSlty7R6DSsn2pUVLSNUPLhYafNYiqx8RlZ3FJHVPTx6isUUa2Ht3xSAAop608+kxtUZy8ndmq1/D6gEUND6Ho1H8vL+QR0x8LO9IflX3H3/3D84PlBgaeuoOve4O9aqqWt65x+Wxs2Ky67cxRyORMlRUQ1NTGamtqpmjSWQRBCxb35pWZdLZ+Z4genXo7WOKQyR9teIHAn1PnXq/T210p697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xui1fMn/ALJS+Q//AIiLfH/uim9uQ/2q/b0mu/8AcWT7D0S7+SeCPgD13cAH+9G+Tx/2uT7fvf7c9Itj/wCSbH1bN7SdG3Xvfuvde9+691737r3VHv8AwoM/7dydvf0/geavb6/8W4+/dePA9fHK5te/+88+/da69zb6/wCwvz79Tr2OjufDL4Y90/Kre1HT9a4+eCmxtSlRUZiVZYadVgOqVYqgEDUPpyfeiAcHr3WwVvnqfsf4v7e27iqjcVXT5qGgSGolinOqSYqPq2r+vHvWhPTrfViXwq6X3fuOmxXZVdma2pyVXoqGFQS2rnkKWP8Aj79oU+XXgTXq/wA6qwYq6GKHctOJIoVX1SICG0gDm4960J6db6EzfG/YsPhht7bMf2lOE0yyQqF/s2/AHvQRT5deyBU8egk2Nuupnyj4/KNLW0NSxR/IofQXNri9/ezGnmOtgnpHdo4vKbJzS5/ArUjHOyu3hjPOs6rED37QlOtZr1W92Nmnz/fW3s1lIppYYpg3qQEAo1xr/wBj794aenW69WD7eqs72PXUeOoJJRQU0UCIv0jVERQbhbAce/aE9OvGoGOhmzuardo0MG28MzRVCqorKhOCzAfgj68+/BFPEdaJz1NxmYod54x9ubvRaxkUCCeUK7AsP8R714ajgOt9BjuXD5/Yj1qbQp1lgqoZFjGkBYfQdDAge4f95PZnl/3j2SPat2GhomrrUkMRXIr9nRht+4TbdIZI/P8Al1WJvz5B9qdL1e9stl5cjna+ri8NLQJ5KiCHyMVQNa9gPbfMP3b7Ll72t2flT28VLKKNz4so7XYef2knrcO8GW9knvKsxGB0RncXVndHfWMzXZ2RoRSjKwVPgp2drwq0LHUEY8Gx/p7kLlPl9uXNji2mZzKyKAXPEnpFcSePKZBgHy61EPmL0pn+l+49w4jMR1LjIVctf9zKp8ZnqJSzwo5+tuCPYmHp0xXoqFz/AFP+39+63165t+b/ANbn37rVOrZP5H2Drsx/ND+H89FUfbpjO5doVdUA5UzQjIreE/1Btc+9MKj9nV4/jA+3/B19n/Nf8Wuv/wCoKt/9w5Pdh023Dr51n/Cav/t/D85Pz/uI7o4/8naT3UeXThwD19GZf0j8cDj/AGHvfVeu/fuvde9+691737r3XvfuvdayPz6+b+1j/Mfw3RPYu8vlXSfHLrDp3cc/ZHX/AMd9j5TdmL3rvptGYhq93yYGGWqjjpad1gjQXF1+i62cGUMRSHWlCzUpXHQcvLsS3xt5S4iQGoVS1T5VoCR50x0YHuPvvE/K+u+GXw1+IHeW8+odn9s7dxXbe7OwNsVTwdpYPrXYcwqKjqndeEyempx02XiWWnr4qkpKAoUDxm76VCNdxMMjh6cPLp6SYExWNqx0uKk+dKjjX1r6fs6EnonMb5+I38yGv+GNT2jvbtrqX5GdYbp7/wCt8XvurSqn6P8A7mVS4fIbL2vVoLy4+qZXmZXC2cq1gQdTUumaHxuDA06et9drffRA1RlLCvlSgoP2/wCrHV0ntH0b9FT7N713psPMZZKbZUmTwGGjlq6zIxw1LsKCmQy1MqlOLhRce618ut0+XQ0dU9lYLt3YWA7B22lRHiNwU8k1MlWnjnQwzNBKrofpZ1IHu3XiKdCJ791rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917pFdk/8y639/wCGVun/AN0U/uyfGPtHVJP7NvsPVN38hr/smLtD6Af6ct0Wt/1A0/tXff2g+wdE2w1+kav8bf4T1eN7RdHnXvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdBv3JN9v1D2rUaxH4Ot98zeQkAJ4tsVT6yT+Ba/uy/EPt6q/wH7D188v+QZ/JYwXzx+CafKPqf5e94/C35JYfvTs/YWb7Q6Qqq2XJ7i2lR0GOraDCZmmp8ripI40NUZE+1qo78mQPcaXteiIBcVrX8uHSIQiW6Z3zoA0g8BUZ/bTrbQ/lZ/ySekP5aG5ux+46zs/ffyj+UfbBlp96/I3t6nT++VRiZpVmmxWJgmq8hJTLM0cf3U71ss0wRVaTQNHujSkrp9eJ8z6D7B08lrHHJrHAfCMUX1I+Z8yel78RP5R3W/xP+eXyx/mAxdzdm9s9q/KvDwYDJ4zf8GEkodj4gZaky1XSYPJUSConDnH0NPD5dAhgi8aqQ3GnfWagUrT+XVoYfCWhOrJNTxyan/V6dW3+2+n+qZfgH/Jd6b/l1fL75R/J/o7tjsQ7S+TeOkirfj5kqfGxdf7JzlZuhN05DcGJrKV/LUyiRZIKJJIIzTwzTR65FZBG476uAoPTyr69NRxaMsdRzk8aVrT7B5dFD+an/CZr4x/Ibvif5N/FzuztD+Xv3XnKzJZLeu4vj1StHh925fM1jV+WzUmCp8li2oKqpmPlqGx9VFFI95HhLkn3sSlVoMH1HmPTqj26SPqk7higPAH1Hn05/Dv/AITj9XdG/IXZnyb+V3y37+/mC9ndU5XHbl6dk75qq+DCddbvxdamQod10dHLmMrUVdVBNFBLBHUVfgSSKOTxMyi2zMxXSc+lfLrQtY1lEqdp86fi9K/IdXF/NP4N/Gj+YD0tlOiflB1th+wtm1U0mTwVRVo9PuDZW5xQy0NFuzZ+bpWSooq6BJnAeKQLIpMcqvGxUtq2lqkV+XT0iF0KglSfMcR1rc4X/hKbvLY+MqdhdW/zifmx110oamVMf1PhKSePD4/CySFo8Peh3NS0MqorFA7YoKR9YzzdzxnBqpIpwzw6Y+jgKlWUHV8WOJ8yaUGfPHWw3/L9/l7/AB2/ltdEw9EfHXAVdDi8hmp93783Zm6uXI7r7H37XUMGPym8dz1srFTPLFTwxJDCqQxRoqRoOS1HfWa0p07DF4SaKk/b/IfYOjx+6dO9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691Wh/N8/7d+97f9QO3f/empfaqz/tx/q8x0Vb1/wAk6T7P8h6GX+Xn/wBkSfGj/wARZgv97k9t3H9s3Snb/wDcKP7Ojle2elnXvfuvde9+691737r3Xx3P+FI/H84X5YW/53+K/wDdePfuveQ+zqjOkhWoqqWnZtInqIYWfiyrLIELC/8AS/v3Ws9b6X8k/wDl+dE7V6lw3bFXi6DOb4rIIaw5GvhjaojYwpMjIeQSCeOOPfifLrYp+3q0v5H9vPs3J4PaG3a9KCsqqkU1kkWI6FcKiKotxz78evdNvcu5IYelnxu4ZBU5HIUVJJDWy21I8hDH1n/X/r7FvIGecrInyLf4OmLqvgN1n2rvXG7D6nxC5Cqjr6yopIXp4I9E7lHi/bUggkeynmH/AJL12Dw8V/8AD05ED4an5dAhX707N3HHKKMpgcRPI2msRvBMkTG4a3H4/Pso6cNa9cqTae2qXwZTeu6n3I6WlSEzCe0g50mxP59+695Z6fa7LvutExdCFx216dQraz4w8SfgX9668AOk1m9147b1G2D2wYUZ1Eck0LC4P0Mhf+vveevV6j7dyqU8JatzAqZ2/cKSSq2lj9RY39+r59e6m4vKw1ldXyzVlNDEkLgSM8aKW/pf+nv3XqGnSh2lUYaaiyqjMY6KRpHAcVUd/wDYE+/Z68OuFaP4RQSSY/JJWTTX0FJFdNR/OoH37gOtDpJPiaWpWKq3DK07y+soeUBvewPv3W6+XSU3dkMylPFBtbHB46V1EZdSF0Lza49+Hz68B0JuyMbuvK4yGqzGU/hJnGlYo5AEDNwByf8Ae/ej14cadCLR9Sb0kmWooN31ml/0okll5/1JB97p16vr0rh0zvujEeUy++6+mx9OvllieeyyKg1MCC3v3y695dSIcfnOzo5dm7bmqhiowafJZQggyf2HKNwCD7917oGe/wD+WB112L1BuXAfaRVOay1PJK+SlgvIswiI162/oT791qnn1oafOf4M7m+JW8a2gaSpymE+9qQKwqrLBGZ30F3X6AWA559+611Xxc3uL39+698vLq97/hNR/wBvlviHf/nf7j/95qp96PV18/sPX//V39mdEtqdV1EBdTAXJNgBf3sAngOtFlGCadeZlRS7sqKouzMQqgf1JPHvVK4HXiQBU9cvfut9e9+690Uv5xW/2WXsgkXApcYSLX/5esPPvR62OPQ9dX/8y22B/wCGZtn/AN00Pvw4dePHpde99a697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rXv8A5PObw0fyN/mC08uXxcdRJ2j9xHTvXU6TPBT57Ix1E6xswJRGdA7DgEi/19mN0p8JaD/VnoObO6/VXGR8R8/s6v8AP45hB9cxi/8Az4Un/R/tBpb0PQg1p6j9vXv45hf+dxi//PhSf9H+/aW9D17xE/iH7evfxzC/87jF/wDnwpP+j/ftDeh694ifxD9vXX8dwn/O4xX/AJ8KT/o/37S3oet609R+3rv+OYT/AJ3GL/8APhSf9H+/aW9D1rWnqP29e/jmE/53GL/8+FJ/0f79pb0PXvET1H7euv47hP8AncYr/wA+FJ/0f79pb0PW9aeo/b17+O4T/ncYr/z4Un/R/v2lvQ9e1p6j9vXv47hP+dxiv/PhSf8AR/v2lvQ9e1p6j9vXv47hP+dziv8Az4Un/R/v2lvQ9e1p6j9vXv47hP8AncYr/wA+FJ/0f79pb0PXtaeo/b17+O4T/nc4r/z4Un/R/v2lvQ9e1p6j9vXv47hP+dxiv/PhSf8AR/v2lvQ9e1p6j9vXv47hP+dziv8Az4Un/R/v2lvQ9e1p6j9vXv45hP8AncYr/wA+FJ/0f79pb0PXtaeo/b13/HML/wA7jF/+fCk/6P8AftDeh614kf8AEP29dfx3Cf8AO4xX/nwpP+j/AH7S3oet609R+3rv+OYT/ncYv/z4Un/R/v2lvQ9a8RPUft66/juE/wCdxiv/AD4Un/R/v2lvQ9b1p6j9vXv47hP+dxiv/PhSf9H+/aW9D17WnqP29d/xzCf87jF8f9XCk/6P9+0t6HrXiJ/EP29dfx3Cf87jFf8AnwpP+j/ftLeh63rT1H7evfx3Cf8AO4xX/nwpP+j/AH7S3oeva09R+3r38dwn/O4xX/nwpP8Ao/37S3oeva09R+3r38cwn/O4xX/nwpP+j/ftLeh69rT1H7eu/wCOYQ/8vjF/+fCk/wCj/ftLeh614ifxD9vXv45hf+dxi/8Az4Un/R/v2hvQ9e8RP4h+3orfzd3nszAfED5IZvcW8Nr7ewdB1HvL+IZzN53HY3EY8VOIkp6dq2vqZFjjEkjLGmo+piFFyfbkKsJASKdJ7tla2dVNSQcDj0Un+SJWUOQ/l89bVmNrqXJUNRuXe8tNXUNRFVUdVC+Y1xzU1RCSrowIKsDyOfbt7XxyT0j2Ig7bHT59W3e0nRv1737r3Xvfuvde9+691R7/AMKDP+3cfbx/pg81/wC64+/efXj8J6+OUBf/AGA9+61WnXQF/wDjf09+631v4/yNdp4Dafx8osym1qCR66Kmmnq44keaYMlmfURe/wDsffjx68DjPVk/yh+KWwO9tl5PdGIoi2YxVPJMaVoiGSWKPUAFH+N/fh6deOcjosXxm7CzW3cTF13jdt6K7bc38PkqXiZWcrIRwR/gPexjr3l1bdtDdFeu0wcjEIq2ZFsgFiLgf191IJOOt1PA9O+OxUuei9YJDnk/Xgn6n/b+/efW6Yr0tsbs2kxCqKWlEtS7KVcKSb+/GvXhjpS7mxUjbdcZahheNUB0yKOE083/ANh79XrQp1Vxv/t3oPbnbuE2Tk8PSSZnKTNGKxgv7EmvRYN+Ofe+HWs16sq6w2vi4cTNUbepYEWogikhkhszMskQcc2/pb3XrZp1nyeyXrJHashIqrufI4I1fW17+99bqOg+l29UYmqceJldTeOYAi+nng+9Vx14UOesGd3LJ/durjig82UiikAVwCWAUgc296PDHy68SPPj1Sr3F2zVx9qS7Gym16KQ56rhpnqWj1Op8oAPIsDz7kHfCTyjZAYyekUZH1Brknq1HqHpihqNm4rGTxRrFX0sb+EIttM0X00/7H2AARQDpZTPVHX87b+WNg8n1Tk+xtuYaP8AvDQwzVZlip21EjUY2Zl+lvfq9aIrjrQwy2MqsNkq7FVqeOrx9TLS1Cc+mWFtLDn3vr3UC9hxfn6/0t791qmerjP5Df8A285+LXP/ADVXaP8A7sh70fh/Z1ZP7QfYf8HX2WMz/wAWuv8A8KKtP/qnJ7sOqNw6+dX/AMJq/wDt/F84/wDtU90f+9tJ7qOI6cPA9fRmH0H+sP8Aevezx6qOHXfv3Xuve/de697917r3v3XuqWvkb0T88OnvmnW/Jj4JdY9I9n7Y7P6kzeE7Z2v21vyr2XUxdjUdVG+28ji6inQu1NMkMJn0vwRIpCgq3tYk0bxeFKSKcKdFM1rcxXRurMBiwoQxp+dc8PSnRZz/ACzPlr0/gsT8z+iafrzKfzFd1752zvLtDrLPbtfGdC0GJylY7b42ZtHMrF5RHDC4VJGKrL6woFkLufWK48Jx25/n0mXaGhka8gP6zaSQSdPb5DOB+X29Hs+Knx9+Te9fkhuj5i/N7anXOxO1dt4Gv6p6T2R1Puqo3Vt3DdaZQJW5jKZzKViB3ramp1hYw1gCzEAaFDMssYi8GHI8yelttbTtcG8u6B8gBTUAfbj/AAdWo+0vRl0hOzwo64361lv/AHP3FzYG/wDuJl4N/fuvDj0A3weFvjN1zcAf5PleB9P+LvN9Pfutno2nv3Wuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XukT2Yyp1x2AzkBV2TutmLEABRgqgkkn8e7J8Y+0dUk/s2+w/wCDqlr+Q3ncIPjR2rAczihPD3buGeaE19MssMNTRwpTyyIzAhXKMEb6EggfT2tvVYuCAeHRJsLr9KwJHxt5/M9Xj/xzC/8AO4xf/nwpP+j/AGi0N6Ho78SP+Ift66/juE/53GK/8+FJ/wBH+/aW9D1vWnqP29e/juE/53GK/wDPhSf9H+/aW9D17WnqP29e/juE/wCdxiv/AD4Un/R/v2lvQ9e1p6j9vXv47hP+dxiv/PhSf9H+/aW9D17WnqP29d/xzCf87jF/+fCk/wCj/ftLeh614ieo/b11/HcJ/wA7jFf+fCk/6P8AftLeh63rT1H7evfx3Cf87jFf+fCk/wCj/ftLeh69rT1H7eotfXbYytDW4zI12ErsdkaSpoK+iqayimpqyiq4TT1VLUROxV0kRmR1IsQSD79pf0PWi8ZwSP29AF8Y/jX8Wvhn1p/od+MOwdi9O9a/3ize7G2ptevc0Um4dxTioy+UmnydTUTu8hVEUPKVjjRIowkaKo2Q54j+XWl8JakEZ+fRh/47hP8AncYr/wA+FJ/0f71pb0PVtaeo/b17+OYQ/TMYr/z4Un/R/v2lvQ9e1p6j9vXv47hP+dxiv/PhSf8AR/v2lvQ9e1p6j9vXf8cwv/O4xf8A58KT/o/37S3oeteIn8Q/b11/HcJ/zucV/wCfCk/6P9+0t6HretPUft69/HcJ/wA7nFf+fCk/6P8AftLeh69rT1H7evfx3Cf87jFf+fCk/wCj/ftLeh69rT1H7evfxzCf87jFf+fCk/6P9+0t6Hr2tPUft69/HcJ/zuMV/wCfCk/6P9+0t6Hr2tPUft69/HcJ/wA7jFf+fCk/6P8AftLeh69rT1H7evfxzCf87jFf+fCk/wCj/ftLeh69rT1H7evfx3Cf87jFf+fCk/6P9+0t6Hr2tPUft69/HcJ/zuMV/wCfCk/6P9+0t6Hr2tPUft67/jmF/wCdxi//AD4Un/R/v2lvQ9a8SP8AiH7evfxzCf8AO4xf/nwpP+j/AH7S3oeveInqP29e/jmE/wCdxi//AD4Un/R/v2lvQ9e8RPUft66/juE/53GK/wDPhSf9H+/aW9D1vWnqP29Vffzkd8bG29/L17srNyb32jtuhrm2viaGuz24MZjKOsy1TuOmelxVLPUyKslTKFYxwqSzAGw49qbUFZgTjos3dlawkCmpoeGfI9GJ/l3yRy/CH4zSxOksb9V4FkkjYOjqTJZkZeCD+CPbVx/bN0p28g2UZGe3o5vtnpZ1737r3Xvfuvde9+6918d3/hSKjN/OG+VqIGdn3BiAigEszNQgBQP8T9PevXr3kPs6Dv8Al2/yxd1/IfcGG332RR5TA9cUdXTVYZKORpKxYnWa86sBxaxCj/Y/0974da4n5dbyPQWzeuOjdhUuD2hm0iosVjYqaGlqJEgeWWKm8QLRE3B4Hv2OtnB6Cum6Xl7b39Pv3dOTb7HF1b1VHCjmRFVJNQ/NgLD37j16lOsPyRze39w7OqdoYOUVNbjo4YPLEQzQLG+kG6/8T7FvIRpzlYn+k3+Dpm5zbuvn1I2lgds7T2ThKveeQFTURY+mkiSpY6QBDdU9R/Hso5h/5L13T/fr/wCHpyL+zX7OgY3f2NPvmvG1dlwq1O0njaqhA9Ed/wCy0f8AT2U9XyOm6SnpNpTUOCqlkytfO6+bVqcpI31VefeutdD7tzYW8t5xwYvD7eygjlCrGVo5lgOr6a5VHvf29bpmvRidj/AOvMq5jf8Am4Nu01xM9O8hVWj5JLlzx/j79XFeqgjoVT038Qtp+Wk3BvjbIq4gfI02Ugia4+pOo+9dWFSK06SM21/hXVyS0NLv3bv7nBWHK0rE/wBbWa59+p59aqSeHS+2R8TvjdnI2qMHn4a+mqG12hlaRbG44KE+/Hj1vyz0OdP8TOpoce+PpqMDUhWnm8bm9/o/PvVadbrXHVY3yL61PUO5lpaqV5cNM3kp5JBpSJSfSgPvfHrVKY6Lhm97OaVaHb8cM7TMAZPSQobg+u3197APHrWOHS0i3HT0mBxVJlK2KGuMkV1MoVixNwV9+r17PR0thVEKYejr6iYNDDTCTyA31ADUCffuvdI3I5/P9p7nO28UlRHgaVyK6pRWt40NmDFf6j3444dbHz6NTsKp672tjJ8FhchQw5OmhK1KSyxLPJNpu3pPP1/x96p1sHpx2v2JtncrZrZdbmKWkympoaeOSeJCwcWIW/PPv1OvV6qj/mR/EDqrdvV+fhyW38dmtx1lJUS00wi807ytG7Jot9eTf3sdaPXz1O/umtz9N77zOFzOErcZQtkKo4+SoppIYzGah9EKF/6KOP8AD37qvVtP/Can/t8t8Qvx/uf3H/7zVT9Pej5dXTz+w9f/1rXO0fkH1vs/+aV8qtofL7ur5a7A2fS5DpIdJPgNhbopuj8HFLFTzMo3xQUdTSqKmZkEuvSpvKxkDLpJrEW8JRCATTIJ/wBX+rj0Grnw47qSS9dkQlQpAxXhxzTNBnHp1YR2zlN/fMv57x/G3aHdW6urekfjNsag3927htiV2Pqo++qjszCQjbGHyuViBalgxMkkdTEiu2pwzspuoVhB4EXjkZJ6WSs15dGzB7FAJ8614V/PPl0+/wAuPtPtLa/c3yn+B3bnZm4e9M98Wq/aW4MB3Lu2OGm3Rn9n9pwzZvB7dzNPAWVji4glPDLqJKggsQFtS4RdCzD8XT1hJKJZbSQ18KlCfMHh1bq+oI5QAvpbQD9C1vSD/sfaToz6q++X1d8i5uo+w4M7t3bsGwvJQGprYsjFJkFo1yqeGRKcG/Pp1D+tveuPWxg46sL6y0/6ONhaf0/3O23p/wBb+Dw29+HDrx49Lj3vrXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdUz96/yXOmO3O9N4d47P7a7O6Mrd+UMCbm2j1mcdj9tV2ZEplyG4FhbSyTVbWedB6C92AF7Bal86RhCA1PM8eiSfYraa5NyrvGSKEKaKfmR6/PoNT/ACK9nH/ub75H/wDnwxv/AEd7t9e38A6p+4IfKaT/AHrrx/kWbQP/AHN/8kP6f8XDHfT8f2vfvr38lHXhsMHnLIf9t14fyLNoD/ub/wCR/wD58Md/0f78L9x+EdeOwQH/AEWT/euuj/Iq2hY/85f/ACPBN7n+IY7/AKP9++vf+Ede/cMP+/ZP9668P5Fe0AAD8wPkebG//A/G/wDR/v317fwjrw2CEf6NJ/vXXj/Ir2iSD/s4HyP4N/8Agfjj/wBF+9/Xv/COvfuCH/fsn+9dcv8AhizZ4vb5f/I8E/U/xDG/9He9fXt/COvfuGH/AH9JT/Tddf8ADFez/wDvL75H/wCP+5DHc/8AJ3v317/wjr37gh/37J/vXXR/kVbQ4t8v/keB+R9/juf+T/fvr2/hHXv3BD/v6T/euu/+GK9nf95ffI//AM+GO/6O9++vb+Ade/cMX+/pP29eP8ivZ5/7m++R/wD58Mb/ANHe/fXt/COvDYYR/osn+9ddf8MVbO/7y++R/wD58Md/0d799e38I69+4Yv9/Sf7111/wxVtDm/zA+R5/wCq/G8f8n+9G/c/hHWxsMA/0WT/AHrrv/hirZ97/wCzf/I+39Pv8d/vev3v69v4B1r9wRf7+k/3rrv/AIYr2f8A95ffI/j6f7kMd/0d799e38A69+4If9+yf7117/hizaFz/wA5gfI/n6f5fjeP+T/e/wB4P/COtf1fh/37J/vXXQ/kVbP/AD8vvkeT/X+IY7/o/wB6+vb+Adb/AHBF/v6T9vXf/DFmz/8AvL75H3P/AE343/o73v69v4R1r9wQ/wC/pP8Aeuuh/Iq2fzf5f/I83/6uGN4/5O96+vf+Edb/AHDD5Syf7117/hirZ/P/ADl/8j7G3H3+O4t/jr9++valNI68NghrUyyf711y/wCGLNoXJ/2b/wCR/P1/y/Hc2/5D97/eDfwjr37gh4eNJ/vXXX/DFezj9fl98j//AD4Y7/o73o37H8I69+4IRwmk/wB669/wxXs+1v8AZvvkfb/tYY3/AKO97/eD/wAI61+4IePjSf7110P5Fezwf+yv/kfb+n3+N/6P96+vb+Edb/cEJ/0WT/euu/8AhivZ/wCPl98jxb/q4Y7/AKO9++vfyUde/cEPnLJ/vXXY/kWbQH0+X/yP/wDPhjf+jve/r3/hHXv3BD/v6T/euuP/AAxXtGxH+zgfI+5P1+/xvH/J/v37wf8AhHWv6vw/79k/3rpGdj/8J6uo+2thbk6y3/8AKn5E7g2Tu+mpqPceDqchQCnydLS1aVscUpRwRZ0VlYcqeRz70b4sNLICD1ZNjSNxJHPICpqMg/yIIP2Hq6/489AdWfFvpjr3oTpfbFJtHrfrPbeO21tvD0gLP9tQU6wvXV9S93nqqhgZqieQlndieBYBE7l21Ho5iiSFNCf8WfM/n0M/uvTnXvfuvde9+691737r3VHv/Cgv/t3J29/2o81/7rj9ffuvHgevjlWFh/j/AL1791roSen9oRb87N2VtGoV2pc7n6GgqdCliIZZLNcD+v09+68eHX1Hv5dHxy2L090Zsfb8lRDR0qYmladnATzGwb1a/wDXt79U8OrY49Wj0/U+zqvEZV9tvTVEWRgkjljjZXBaRbXIHvRPp1quOqMe46Zvjj2Zlp6jFS09LkK55kmhp7htTkhizC359762aHo3nSu55OytuvlYoneKNAwMiqPov0sOPeuvAV6N319j70UkssBSwspC/gcH37h149CXjBRRuSADMD6NQ+hHH09+z1oHHTTvSCoyeOlppf20aNh6f7QIsL2/1/esUx1sVrnrWW+V2yJIfmFseCnpp/FLX8yIJPq1QeTb6c+7DPWjjrY26cwlRtvbuHSKokkL4+hLI4Pp/wAmTjm/vXz62ehtr0pKxV+7tFJ+GUWLH3rh17jw6Qe8sZSx4aSoijLSQ20+jk3PN/fq169Sh6Kn2LOdtYepzzRyCAwOWkC3+kZvx70RXH2dbOOqRdzdd9p96/ICk3FsTFz1eNw+Tgnm0xN+4nmW+vT9Le5C3wj+qFl9rDpFHQXDfZ1sb7No6DrLrDDZPeMdNBuKkxsA+1eQBkkig+mlv8R7j70p0r+3qur5G9k5/wCStBuLrLE4xmx9ZST0z1AhDKoEbAWIB/JHvfDr32dfPL/mM/FTcXxs7qz1NlInSjyuQnmRpFdSZpZGIKahaxUfj3vrX+Xqu0m/9P8AYe/db6uO/kOaf+HOfi3dkH/GVdo21Gxb/ckOAPem+Hrcf9oK+h/wdfZWzP8Axa6//GirR/6pye7DqjcOvnV/8Jq7f8PxfOT8D+E90f7Af32k91Hl043n19GZbWFvpYW/1re99V679+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SF7Pt/o337f6f3P3Hf8A89Mvv3Wxx6Ab4Pf9kzdc83/yfK8/X/l7ze9Drx6Nn731rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917rDU01PWU9RR1cENVSVUMtNVU1RGk0FRTzxmKaCeKQFWR1JVlYEEEg+/AkGo60QCKHqjbdv8irprJdi9h716+757l6hwO/s7/Hh13sifH021tuTOuqejwyOwcQGQs6I9yl9IJA9rxfyaQpUGnn0RNsFsZXljkdNZB0hqKPsHlXz9emz/hizaBIP+zgfI+wH0+/xv1/r+v3769uOkda/cEP+/ZP9669/wAMV7Qvf/Zv/kf/AOd+O/6P9++vetdI63+4IaU8WT/euuv+GKtn25+X/wAjyf6/xDHf9H+/fXt/COvDYYR/osn+9ddj+RXs8f8Ac3/yPNvp/uQx3H/J3v317/wjrx2GE/6LJ/vXXv8AhivZ/wD3l/8AI8/1/wByGO5/p/b9+F+38I69+4If9/Sf7117/hizZ97/AOzf/I//AB/3IY3n/k73769610jr37ghpTxZP9666H8ivaFgD8v/AJHm3/Tfjh9f+Q/fvr2/gHXv3DF/v6T/AHrrv/hivZ/0/wBm++R9v6fxDHf9He/fXt/COvfuCHj40n+9de/4Ys2eOB8vvkf/AIf5fjf+jve/3g/8I61+4ITxmk/3rr3/AAxXs4/X5ffI/wD8+GO/6O96+vf+Edb/AHBCOE0n+9de/wCGK9nH6/L75H/+fDHf9He/fXt/AOtfuCIf6NJ/vXXQ/kVbPBJHy/8Akf8AQWH3+O4/5P8Afvr3/hHW/wBwQnjNJ/vXXj/Ir2eTf/Zv/kf/AOd+N/6O9+N+5/COtjYYB/osn+9deP8AIr2gbf8AOYHyPFvr/l+O5H9P1+9/vBx+EdVPL8H+/ZP9669/wxVs7/vL75H/APnwx3/R/vX17+ajrZ2GHylk/wB669/wxVs7/vL75H/+fDHf9He/fXt/COvfuGH/AH7J/vXXY/kV7PH/AHN98j//AD4Y3/o73769v4R1r9wQnjNJ/vXXh/Ir2h+fl/8AI883H+X43j/D9fvf7wf+Ede/cEP+/pP9666H8ivaAJP+zf8AyP5/6b8b/wBH+9fXv/COrfuGD/fsn+9deH8irZ/N/l/8jzzf/gfjh/0X799e38I61+4YfKWT/euu/wDhivZ9rf7N/wDI/wCv/Owxv/R3v3170oFHXv3DDWplk/3rrr/hirZ5/wC5v/kf/wCd+N+n/JXv317/AMI63+4Ya1Esn+9deP8AIr2faw+X/wAjx/5EMd/0f799e/DSOtDYYa1Msn+9deH8ivaIBH+zgfI/63/4H47j/Aev37696U0jr37hhrXxZP8Aeuvf8MV7QBuvzA+R44P/AC8Md9SLX/X799e/8I69+4Ia18aT/euux/Ir2gPr8v8A5Hk/1+/x3/R/vf17fwjrX7gh/wB+yf7110f5Fezz/wBzffI+/wDX7/G/9H+9G/f+EdbGwwj/AEWT/eukF2P/AMJ0vj53VhcHs7un5Dd99ldfYreOA3lW7JzWUpYcZmKzATmSKnqJ6ZxIiyozwu6c6GYfn3pr0uNLoCOrx7KkTiSOaQEVrkEEHBBqPT8x5dX+7P2htnYG1dvbI2ZhaDbm09qYegwG3cFi4FpsficRi6ZaShoqWFfoqRqBc3JNyxJJJRsxdizZJ6OERIkEcYoBwHSj916v1737r3Xvfuvde9+6918fL/hRSgk/nQ/JaNjZX3pthCf6Bo4wTz70OPXjw/LraD+HvW+4tsfG3r2t27U0mRppsLQVdVQBYVaSOXHo7KdAu3vZHXgcDocaldmbmB/jbVGzqyiB+4RUeKGdk/VzIQOTf37rfDPShwG+sTQYqpw20xU5GMK8ElXKmlXBHjLLIL3/AK+/fLr3Qb1+zsXtjam690ZOoMtXXR+RUb1MC0urRpP9P6+xdyAC3OViqivc3+Dpi6I8BiTTpM5HD1PYWMoJqmWSnxcFHTpDTKSGqCEtpVQR7KOYDXfrv/mq/wDh6tF/Zr9nQ59E/EjeuYhkn2nt6SBKp2H31WksLIjDmSMyA34549k5Pn07wwT0ZvOdcfGb4m4eTenyI3NiMzuNA1RDiHrKc1aSL9FEMpBPNvfutfZ1Xl3J/Pk6l2pT1u3Oj9k0kLQa6enq5KKFWup0oyyr/vd/fqevW649eqrN6/zKPld35mZaHG7iyuJiyLmCCnxtQ2kRyEaV0Rn3vrVfPoausfiT2xv+nTdXavZmVo6aqQVEkU1XKZ2R7ll8JcH8e/E9e0149CDWY7469VVDULQ7t3Pl6EEPMtDWGCSVPyHVj/X3vr1B5dAd2R/MW3719GmJ6swGT29TU7WSZo6iBmRfyzSH6/X3rHWgT5dbIX8tj5A7h7v6FpNz74yUNXuCOaGNo3nV6hIyCTcC5496IB6tWnHoF/5oG59vHa9EkFdAMiI0JEMiNKqgD9YHI/x97HWjQ9a6GN+TkGzsrJiamtasUVOn1lW0HVwOffqevWgejq7W3HguwsTR5ypyLGQBJ4Y4zq8ZHIvpPFv8ffut1z0ZrF94w4fak+BoaqTJV6xFIYYLTTkBLBDGvP8Ah9PeqHrY6VPTfyifDZOPaldtetxmSzgdGrZaF4g7u2hSXkH5/wAPe89eHQ65XoTc9V93vegz1dDlcvUR1FJRQCQoqSeohrHj6j8e/dep6dCjt342KmFTceQzkidgNaeCNZX1OY7NZuRa/vXn175DpQYHqbdO8q+eq3zRGqp8WjpBFPraKVVFl+oIPv3Wz6DrWb/njfFGmzeDqtx4LbUFBXU0cxiqIafQquruQ5ZFHvfl1UjqrP8A4ThYeswP86f4lYvIIqVNNuPcqOFOpbrtyqW4P+wPvR62nn9nX//X2QvnNjP5nvyH2p2D8ddk/EroKbqnfT7Yxp7Mz3e8RylBio8zBX5qvTbbU0MgqIBFqgIW55AViRZdEbaJ/EDk0+XRNeDcrmI26xKA1KktwFc4AzjhnpI7l+GHy5+FnbnWXcPwF2btLvip3pgqjaHyX2x3B2PDs5DSYvCQLt3N7WyUkaM6CrjIWAl3iCi+pXOnfjxzAxydo8jx619DcWjia0o58wxpX8/l9nRxvgP8c+2tkVfa3yY+T2D23tf5UfJDI46fszamyM5HuHZe0sFtGSbH7OwOFyyLeoZaRleaQyMASqA3ViWZ5VYCOP4V4dK7K2li1TT08STLAcB8gerHvabpf0Uv5xf9kzdj8XvTYwW/H/F1h+vv3Wx0PXV//Mttgf8AhmbZ/wDdND70OHXjxPS697611737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdUff8KCz/2Lk7eH9cFmv/dcffuvHh18ckW/Pv3Wvs6MN8VK2soO/OtpqKlSsmO5ceoif/lsGBH+Nxb37rfn19LPpHsw5bZG2cNnYJ8LLT4ynVkCOqkCMXYMbe/de8urM+hN57OoJI6EbikqZZraad3VrsfoLMf6+658+tnh0I3dPx02F3JBDJuLFQTs2mSOVl0sBa4IZR73Wg699nWDr349bL67wxw+EhjSBxYrzcAf2fp+feqkde8ul5NhKDAUZigxv7cSkllQkML3Jvb37rRHQY5KqomilnpAIJEuWsCCpvyLD3s9eHHpAT7gqKqQK9UJooWKsgN+L/0Hv3W+PVVvf+16uv8AkjtDcdPiDU0VNVK0k3jusVpif1Wt72OHXjk46tAoN3Gnoce1HojWOjpkdSQq3WFQf9a3uvDHXuhB2/uOmrg9dkZUlhhtwj6gp/Fre98OHXiM06FnC1WF3LE9MlCJ42XTcqbWIsLEce9fl148MdQ9zdMbb3PgZ8HkkQQTh9KEcLqBNhx70wFPzHXh0Vvb3WNJ0tkd2x7Cw8FbkzTKQ5iVnj1C4ZdS3/p7HW9SluWbWHyUk9Jo1pMWPReN35fcOdrJjvrJVVOfK96RroiDV9LXH+9ewOAKdKa56Fb4+YTZdVlZ8fRYqKSpqkZRUhAzAlf1kkfn37r2adakv/Cmvoo0G46Ld1BTrFFBUCSXTGFA+3uHuQPyCfex6dVPWmv791vq2P8Akf4nK5L+aH8PZcdO0EVB3Ns+prwrEeamGSW8RUfW/wBfem4dWT4x+f8Ag6+0Dmv+LXX/APUFW/7D/I5Ofdh023Dr51n/AAmr/wC38Xzj/p/Ce6Ln/D++0nPuo8unD59fRmH0FvpYf7172eqjh137917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XukL2h/zLfft/p/c/cd7f0/hMvv3Wxx6AX4PWPxm650/T7fK2/8+83v3Xj0bT37rXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6918nL+cp1ZT9t/z/e8tmZJC1DkN44OebxiRhppYEtrEfNtQ5vx/X2c2HL267ouqyUEHNSQP9R6bklRKaz6dbDOF25uDq3Ze0aTYG55ZaDC4XGxtii2mPVDRqki2Y/nTax9mR5H5jAoVjP8At+qC6h4Z/Z0xbp762vujET4bsHET4bLRlEOTpadvFIytYl5BYc+9f1I5jpUrH/vfXvqYjwJ6GfYm4dkzbA+2wctTHLFC0kdSkAbzWQsCz/8AG/exyPzGcaY/9762bmIVPQebM3pjuys5ndg541y0cagCrgp3lluHAA0Lf2Bedk90+QvpN+5MsYr26RzWMyUqvA5+zpVbCyu9UVwxVfWnVoPx76D6/iFDlt4rmZcJjVjOPp5MZMpqJIRwXBH0Nvr7KNm5v3DmO8S25kt0sN3uW7rYNUAnOqvHJ6cktljWsJ1IOB6F75j/ACxw3x16QzWX60wOQqa+loZoaCnhxhUU7KCiy64wDe3uSo+TuYJa+GsZp/S6RGeJeNf2daCPyU+VHZnyH35l812JubLOrVszU+Lq5Z4lpwX1KoTV+P6W9vf1I5lHFI/97HTf1UJya9AFi5KGaoRBVXJYC3qctc2tc396PJHMlMJH/vY6sLmE+v7Org/gv1jgGrYNz51J3hpys0bGlLIGUMVsWFvezyRzIBQpGP8Ab9e+pgOanHy6tyFftOvqCZ6yr8IfxRQpCUCR/ghVsP8AePe/6jcyDisf+99a+ph4Z6U2Q2v1hPj/ACUh/wAteP1PJSRk6iPqSwPvX9R+ZAPhj/3vqwuYfxV/Z1XV8mNq7FTE1sni+9qYldm0UagqoA/1Cj3r+pHMfALH/vfVTcwnuz+zon/U3zf7D+OFLXU2yZcj/CfWophHIsKEE2/IFxc+9nknmQCgSM/7cde+qh8yegw3v86d7dtZLIZLeGRqpBKskcdK7OwTV9BpJ4t78OR+ZfJI/wDex1s3MVOJ6IxuHdS12cnrYJ5RJNKzBFV2JZjwAP6+/HkjmX+CP/ex1oXUJ4E/s6Mr1T33vbYFPGldQ5U4mYrGjy0lQsZQ/QqzcH37+pHMnmkf+9jrf1ER4Hq0L499xdfxxVO6aqd3z0jLKkUqhvDxcjxvcfX/AA9+/qRzJSpWMf7frwuYfKv7OrZNjbt6t7U2hjshXFIdxUaL9rXQUsXnjdT6LiMf7372eR+Y/SP/AHvrYuIvn0vB3BuTY9Madqatz1Oo000rUrsVQfpso/wHvX9SOYznRH/vfXvqogPPqJT/ACKfbzLvHdNDlIqIctHFSSs0RJ4UIePdTyRzLTVpjp/p+t/Uw14mvU6m+Y+d3bkIajbGPyibdV1VnageN3jBs2oD/D3b+o3MgHwxn/b9VNzDxyOgm+dNb153J0BnqOojqE3IMTUtDE9IBIJxTOQNNr/qPv39R+ZP4Y8f0+vG5gpWp/Z1qs/yJ9m5raH8+T42Y7LIysm8N3TwGRJI2+1OHq4o1AkANwT+OLe0G4cubttcImvUAU+YYHpyKVHJC+h6/9Df49+691737r3Xvfuvde9+690Uz5wi/wAZuxv+obGcf1/3Kw8e/HrY6Hnq/wD5ltsD/wAMzbX/ALp4ffhw68eJ6XXv3Wuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917qj7/AIUFn/sXJ29xc/wPNc/0tjjf37rR4dfHJ4t/j7917Nej/wD8tXrr/SP8o9kY4J5Hx2QoK+NbXu8dWLD6H6gH34io62KVHX0Ma/EblxeNxVGNsSSRUtNBTvVU8bkxoY1u3oH19+63Q9Kza+/9u9aypmKjE5OsyFLeQK1NUAeRRq+l+ffj16nn1aj8eu567uLBQ19XiZqCmjjCwGaJ4iUA0j9f+t71SnXuJ6f+9d1QbE2/BkoMjFQTOVb1yqjMB9Qoci/vXE9e4dAftz5NV+bpYMRSYc5V5FMT1PjZtVzpJ1C/HvZx17oMe6h2l/dnI5La9A2PeWJ5FRWKtzGWBAJHv1B16tOqoz2/8iOqMrLuvcyVOR27BM7V1NUu/j8KuS1lv/QH3vB60K9B9u7+ar1JLuWhiqMNTxmN1SuPjDPHIjaZLFiT9b+/cMderXh1y3r/ADEIu1psZtTpOApVzCKOolF4mDNx6fVz9R7116tOHR4vjme/vtBWbljnrsRUJFMVkkLEBlVmAW/vZ+fVgDx6PFQ/IVtgTw0dVgZIYtOmaoMUhIbTpuL/AOPvVPIda+3pZdad0Hfu93plzKtTOwMNEZEWRNXOkRg3/PvR+fy699nQ8bKWln7M3TT1SK8ZpIRd1DCxsPo3Hsa7yKcu23TKH9Q9Bd3P8a4t+ZwV1LXwYukMup9TpCCNXNgRb2Chwz099nUCLBbW+NuIiyENJ/HskYdJqYkMyp6CLllAA9+r5dap5nrUv/4UNdl0HY3VDz0+NMdav380zNGQUjcnSLn+nPuwxnrx4EdaJwtz/X8e/daNerj/AOQ2T/w5x8XLGwPau0bj8f8AFyHPvTfCerR/2g+w/wCDr7Kma/4tdf8A9QVb/wC4cnuw6o3Dr51n/Cav/t/F84/+1T3R/wC9tJ7qOI6cPA9fRmX6D/WH+9e9nj1Udd+/de697917r3v3Xuve/de697917r3v3Xuve/de697917pC9n/8y337/wCGfuL/AN1Mvv3Wxx6Ab4PWHxn65ta32+V+n/a3m9+68ejZ+/da697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuvla/wA0LKbhpP8AhQ731R7fIFfk9yYyljkKhpKYLSxzCSIn83ve31v/AIe1MN7eW4K20hQH06qyqQNXV5XX3VO4cpQ4sZ/sKWgmnpqV5oqgrGE1oCyjWfbh3TdP9/t14Rx+Q6F7ujojBf6K5KfHSU25apljZ8jA0ZqFKtdmAhF/9597O7br5zk9e8NPJesXSMmx9s7Nhw2WKJOsXglWrAiK/t6LEyWPH9ffv3tuvnOevGKM5p1Pl3Fjuk8Pu3ePXuxcDvnMV8IMNZW1io2Oby6w0aIRqPFufZny2m8bzzZaW73rRoxaooDXt+fVZWSKBmC1PSw25/Nv7e2hLjI9/wDQ+DjwcNJTRP8AYieYPGqi8nijY8kX947XPsbue2+89x7kX3Mt1dSJIwFsyqIgoOFFADj7ejhd2R9tWyWBQKfF51+3o5W3/wCY58Ru+NszYHfmx0wn30BhqqKpxyxwkshBKtUXP1Pudm3XdCSVmIB/l0UmNCc9VI/Ir+Wl8TO7M9kN19TbgjxctY8lSKBIVVEaRtWkMq2t70d23egAuD/q/Prfhx+Y/Z1XjT/yuG2/vOnijyhrMdBUqzni2gNzzp92/e268TOetaEIpTq6XAfGzanW/wAf6r7H7Siy8NEft3VoxNUSqvHpIB/23vw3bdq18c9eKIMgdEyw26srTzyU7Y9qiop2MP6SS+gWL/6/vx3XdWwZ2p17w4/TpWQTdk7hLxYXbtS3pNiscv1/BFvfv3rumR47de8NONOkovRvaG4stNT7hwMkdFVKyyecEKVY83Lg+9Dddz/3+3Xii+Y6Lt398VtxbL2HnKrEbRpq4l5ajyx6ZGjjCjn0r/U/T37967r5Tt1rw468OqO/7vZWpzFZj48fIK/7h4pYI4maSJ7kFdCj3v8Ae27D/iQ2evGOM8R1YP0D8OaHcuKGczNK9RkkIqFpZ4zHIukXvoKn6e9fvbdqf7kN/q/PrwjTyHVk+R6N23kOvcXRTbVoKimw9IErHMcUbh4wDqZgt/oPev3pula+O3VtCemeiaYf427m/vHnc5g45MbhFkllo6aMsYZIFvcFvoPdv3tupGZ26r4a8KdDj0V2jUbB3P8Aw2Wsgq1pKpIshj2nVvHpfkBb396G6bqP9Hbr2iMilOrxtgdnbG3jg6OSgixi1phUPTzNBr8hUXsj3J5v72N23av+5Dde8OP06fd44/GZDaeRGUwlG1OJogPRGFIYcEWHv3723alDOx62Ej9Op+0araWzdr4lItvY7w1bBNZSKyeRgL3I/wBj70N23av+5Ddb8OLAI6YO7OudrSYOp3LNTUn8PqaCeabQ6mNQYS54At78N23X/lIbrXhpwA61jPghVbfb/hRh8UabbUkJo4c7vJJlgVAoLYyqKX0W/K/n+ntqa/vp08O4lLivA9WRFDEgeXX/0d/j37r3Xvfuvde9+691737r3RS/nF/2TL2R/wBQuM/92sPvR62Oh66v/wCZbbA/8MzbP/umh9+HDrx4npde99a697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuqPf+FBf/buTt7/tRZr/AN1x9+68Rjr45VuL+/darmnVkv8AKs7Hw/Wnyw2blszPFT09XVUNGjylQutqsf6q34N/fuveY6+nHsDsvbcOGx2Xr3oa3B5Smgn1MkEgKtGvAJv70QerClPs6EDKdpfGR4kOSwVLKx5ciniIJt6v7PvVOtmvRkuoN+dW7kx0dFsRaekghHppIkijYAfghAPe/PrXQZ/LPpTIdqYrFVFLkJaKixrRvUQxMx1pGdTagv8AUe/Vp17j0BO3KXI7FpKSn2liKetegQJK7OjSuy8ElW5+o9+rU9epTqfuPvKvrcfWY3c2Iio5oYJNCsAoJRDYX/2HvxHmevV9OqS+++zt99sZzcWwsDCmPoojPDEYVUrIpVk5f/G/vfWvt6qiyHwy3bX59xXYd2mqKwPPWmVgoQzFmP0t9L/T37HWs8ejVYrpPbPTD7YrtvzIM+Jac1awuJiXLKGDkDgi3vw62ePWwb1bvWrxXTeMzMtXHBU/YxHQyRgystOptY2/PuvnQ9bI6Dau7/8A7xu2Ny+2466FXaMVnj08XsTqX/X92Ap16uKdC/8AG/ps7h7TpexcDkJaPGQMk1Ri1LmIhQCVJ+nurCuPPHW/t6sO2dQLJ2dulgQpjpYFs3C3Uj6k+xnvDMeXrYdJ46CRqdBp8jcL2PVRtU7YqZTTwglYqWVjcrcm/jP+HsGDp+majoluJ7j3xtbyYbsfC1FbipSYnnqYpJDGpBXUuu/veOHW6HrW6/n+b06xg6jaLE5GnjymXiqlioQYxIhkuY10/UfX3sdVauetG/37r3Vx38hsn/hzj4t8X/4yrtEn/wA+Y96b4f2dbT+0H2H/AAdfZXzP/FryH/UDW/8AuJJ7sOqNw6+dX/wmrP8A2Pj+cQ/rie6OR+P9/tJ7r6dOHz6+jMPoPzwOf68e99V679+691737r3Xvfuvde9+690FXYPePUHVNdjcX2N2LtTZ2UzFHV5DF4zN5WCmyNfQUJ01dbS0NzK8UZ9LSBNN+L3HtxIpJMoK9My3EMJpI1P29Kmr33svH7Uh3zkN04HH7OqaGlycG5q/KUlFhpaGujEtFUR19UyRkShl8YvdiQACTb3rQ+vRTPp1fxE8Pxa9tK1+XWLZPYOx+yMR/Hth7qwe7MSspp5azCZCnrVpapRqakrooj5KeYCxMMyq4BBK2I96dGRtLih61HLHMuuM1HSx916c6QfZ7IeuN+prRT/c/cd7sBpH8JlFz791scegI+DwA+M3XIH0+3yv/u3m96HXj0bP3vrXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6918kn+d72pkOmP583ffYOKpI6+rxO9dvqtPNIYkZKmCOKQ6x9CB9P959+Hn148B1tqdB47rjvbobavYlZDF9/UYDGz5JqKoZ1p5fsElkB8NxwxP59+z177MdBPvrd1DtIVFB1xlKnM1UDmOTCOjyw+ltJGpifx78Ot1x0jKbdeIzUMR3xtGoxU9TZJnp0nVdVv12W359++zrwI4Hpo3lsKobamXyuw91yChWKORsU72d7vfxsGN/949i3kH/lcrIVodTf4OmLr/cdqdZKHcW5aHGYzH5/aUNbIaKnRag0izhk8dlYtYj2U8w0/f13/wA1X/w9XhB8Ja+g6QOZ2i+79w00aYgYhAyyB4kNOoJ5+igf09k/l1c06VP+kHbvUky4ytq6mOYDQZBrdWINjdifzf3vPW+m3cXyY2lh4FzprC6izSalB1IP9j9ffqdeJA4dCXt7t+r7Xw9HlaOqq2wEQjUxSLJHAvF7/Wx49+61U9DPjcL1ZszbNV2LuGuxjU0IZmpTPTs7yKLkeM8+/de+zo1Pxr7R6r7Ax81ZtyloIihKoskUCgjmzAsOfp78fTrYoeh73PT7dyMbwmnphKykBooolJ/A9SD3XI6tT16Jz2FgxiY62Ouj+72/UxvHU0rQCRAr/VvUD9Pdh1U8adUdNsjppfkwsW16GAJPO81fFUokafcFz5CquLEfX37qp9Oj07168ymzdw4TcWzaaI4usjhiqaeLQkAWQDXfSAPoffq9WPRncFsWm3RtespYIFhlqoD91CttIZo7Oykfge/de6LR3bRVGxuv67bu14qeOsjoKuOWsVozMjlb2P0N7+/cMdax0QPoH47VTw7i7C3RXVD5GtFTWU+syETMrEgKPp79w699vSl2tu3fu09wVWQkjrsbiaKovSVq+bRJDG/1IuAOB7317Ix0fDBfLuHcm01wNXPHUzsYkaRXVn9FkJIBJv71Tr2OhvruxcJl+rmoaTIxplooDLTszqpicDUCTfi3v3XvPqlP5j/zMtwdNbNynX1duQZGqlhqqaK1QH8alXjCiRX/AKD349bBPHqtL+Qt2PV9p/z1fizu2uLNLWbk3NZmcyFg2CrJgwJJ/wBX/X3o9eQ1r9h6/9Lf49+691737r3Xvfuvde9+690Ur5xm3xl7H/xpsYP/AFqw+9Hra8eh76v/AOZbbA/8MzbP/umh9+HDrx4npde99a697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuqPf+FBht/Lj7eH9cHmf/AHXH37rR4dfHK/H1/wBh791vp1wWbyW28xj87h6l6PJ4uqjrKKpjJDRTxG6MCpB/3n37r3W8L/KR+Y24fkb1Njtq7yzDzVm2YI6TU05kmqWgPrLAsSLWt7914Enj1cQk1Nm644fHUbM6Eq0zMwubfWx9+HVvLqz74n9LTbYgTc9RWMwqAHFMHJA1/jT/ALH3o5PWsdHbz+OatoqiBG8iSQMGSx4Gix4968uvHHRH6XbUVJvWpjpswKRlqDro5NNnu5OkB/8Ainvw6959JD5B9QV+Xw1TlYKiSntSysWghJ1t4jY3Qe7A9aI9OtfnP5PcnWvZlfS1uNlrqWvqXT7qeN4Si+QrqBNvfuvGvQuU2aiainevyyx09RCWRFijeSIOtyA314v71x638ugCrs3smny0FDHW1GVyc9YhUSRPeI+a5Cg+9560aeXVjOLz2X3LtvZu0cPR1OiZIUlRVkT0FVF2t/hz711uvRl957XwOytm4nHCCJs3VRw+YhEaVXkABvxf6+/Y69Ty6Pd8V9pSYHalJMkbCWti8jEqRYPGH/PvTEfzHXs8OkX2p2k+xd27wpsZVJFlJqSJUfWFKN+bcj2Nt4jI5btpPJumEb9XSeirbe+W28sZl/4dlqt69Z6gqFm9SFWb6EsSPp7BQGOn8dLbvv5HdZY7rerzW66bE0X29FLPJM7U8ZL+Jm+ptzcce90p17FcdfN0/m1/Kai797xyVDtyqaXbeLqG8IiqTLBrhkMaroViv+P09+6r8uqjffut9Ww/yQH3An80L4eDCKzQN3Ps9cta2kUP8SXUSfre/wBP9j703Dq0dPEH2H/B19oPNf8AFryH/UFW/wCw/wAjk592HTbcOvnV/wDCav8A7fxfOL+v8J7oH+x/vrJ7qPLpw8D19GZRZVH9FA/3j3vqvXL37r3Xvfuvde9+691737r3Wvn/ADHOjfkFifnb1l8vdk/CfaPzC6c2P8ct57U35Bn+3MXsrI7Omhr58rVTYrauZLx5CVqa7x6aWXVdgNMiqQtt5F0CLVpJPGleie9t5PGN2I/FCqQFrQ+XCuPLz/aOkTvvt3or+Y725/L9+NgxQoPjpltnwd37z6npaqrwrbY7H66k17E2Vk8rjjAKiOklhljFPENEinUbM9leEbQrJI2W8j+XHpKbhLySCGPtjIqy+hqKCvQ/bJ2fhPiz/OMoOqOjcK+1etflV0BvXunufb9HU1dXgZeydn5AYfBZmioal3joHkgjtogCqwZwAFawaZjJaF34g8elUcYt918KLCshYj5ig6vE9oejjomvfHTG/wDP47fm5Mb25k8DiP7u5ypOAhoRJTtBFj5JZKVpdY4cC1wv+w96PWxTp1+DgC/GPrVRzoo8khb/AFTJlZlZ/wDYkX9+HXj0bX3vrXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+6918d7/hSG7RfziPlZJGSske4cO6ML3V0oQykH/Age9ca9e8h9nWP+WH/NJ746o7O2V07uveMlX1DnKtcbkKarm8ceMpWiEbSMZGsykCx54P4/punWuFOttrBdr/ABi2xkot2w7521VS5WGKskE2Wx8giedfK48bOeRf8+/Up1voXqL5H/Fvfs8OEfdO05KiQ6Io0rcYjs7i3pIP9T72R17y6Zex+qNsVO18xlNj5+GDywxyRR09QJo2DSAgjQbexZyFjnGyb+kf8HTFyP8AF2r0MPWvX+6E2ljRlzR5ALQwFJpUgvo8fpu7An/efZTzD/yXbv8A5qP/AIenIsRLX06D7snb2PokeWevxOLqYySkr1dLCCR+CxtYeygnpwCvDojfa9T1VVQR0ea3Tgq3N1L+JY4slRtpJ/SdSsT9ffhTj14n16C1+qujNpYSbeHcvZ1FBt2OE1FBtijkgrZasAXRGMbFhfi/Hv32daHz6KX2n8+6eHGT7F6I2uMHtClDQQZWnL+acJ6VkkFwRcX9+PXq9FAPd/ce66Oqx1bunI1uIZnnqKNppljAJ9QHrI9+HXur4/5bHWvcm68NQZxnq8NtiLxJHUM76Kgeogeo839+68B5dXvQbAylJAkkk/3ckcYJYkEtYfX3U9b4Y6Qm58PFX0dbj66jVg8LrdlFyx4vyPe+tnI613e5870j1N8gxDvBHw9fNM7xViq3AZzpY/QW/wBf3Y9V6sj663l1n2Rt2DEYreGKrUqI43o3nraVJom02jXSzXuPeuOOt4pXo2m2NsSbT2xKYTHkJRTPHFJC4lWUMLizJf63H59++XWqY6IB2L1xvze24ayhgw70OPqppJaid5jp0gkFQG/r/h7917z6Fei6kTbXXtOlYESPH07a44QHeUAa3UhBc3t798utj59Ei7Rym9924mv2ztHYawYuDyQiv8RimkUqVLci5J96610UDa/XnaWyq2XK1WAqocdDLeaV3lYMrvqZtLf09760PXoMvlj80Nv9JbCqKWhyfh3DX05R4hOQ8JKlHVV1A3uPe+vedetXDvnu3Ldv7glr6urqJo2nllLSSyMJNbswB1Mf6+9der69Wt/8JqP+3y3xC/8ADg3H/wC81U+9Hq6+f2Hr/9Pf49+691737r3Xvfuvde9+690Uv5xf9ky9jj+tNjAP9f8AisPvR62Oh66v/wCZbbA/8MzbP/umh9+GB148T0uve+tde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3VH3/Cgv/t3J29/2os1/7rj7917yPXxybcX/AB79175detb/AGPv3Xuj9/Ar5obl+JfZeNyUc7ttStq4xlKbyFIo0kb96aQlgNNvqLfX37rXz63x/jf3btD5F7KxPYXW2YomnmpIZa2ljnhaSGRowW1gEn+tr+/dWGerd/jT2TmsfT1a52pVsfRKdbvIAllF7Lc2/HvR+XXvt49Hy6735hN9rWSY+0kcIMbgG63PF7gn377evZ6r2+T229zbU3xT7uwtdJFSJP5ZKaNiokAYsb2t+Pfvl17h094L5B12V2/T02dwJrMasCxySWMhJChOQL/X36mevdV4/L7bGI3Zi593YLAihbHBprLAEeVQGduQt/e+GOtceqtKbuzaSUWRo63BVEGUx6yQgMswWR1BUMAePqPe+tY6f/jpg6PfW9Tu/MU4joqSoMkdKI/NrVXGkWAPPPvXWx8ur+/j5Q7WzTT5BtvxUFJiaYBMhNGsarpiA1aZFFuR70eHW/PoNt6ZCi3F2vS4ihrBlqda+NLQWlRFWewWyXHHv3AV69506ty2NQxYbauPigjELLjlC+n+39t9Ctv6+6ngK+o69TyHVN3yCxO+s52zu809JPIiaGjKO1nTUSPp+Le5B3yg5Rsqepz0kjr47V6K1undGH6+xNbk9+QnDNj4ZJfvZgQt442b9TWA/T/X2ABwHSs1PWq5/Mw/maVe9lznXvXu4ZZ8XGZaSX7WrZbqrlCT43/F/e+qk061uaysqa+pmrKyaSoqaiRpZpZXaR3dzdizOSf959+63TqN7917q47+Q2SP5nHxcsQL9qbR+oB/5eQ96Pw/s62n9oPsP+Dr7K2Z/wCLXX/9QVb/AO4cnuw6o3Dr51f/AAms/wC38Xzj/wAcT3T/AO9tJ7oPLpw8D19GZeFX/go/3r3bqvXL37r3Xvfuvde9+691737r3VdPy2+K/wAwO+M7lV6a+dmX+PPX2e2xW7byux8b01s7eEtsjQfY1ldS7kydRDVJI4Z2BFiuqykAA+1McsCga0qR8+i+4tryVj4U2hWqKaQaY8ifPoDd9fylcTL8aenur+ke68n0j8iOnshtDLY75W4XZOKye9dwVmDrDW7gizODkqYYTT5JmP7InIh0qBqBcNYXbFyWFVPl02driEAjjYq607/MkeZHD7ejCfEb4Vby6P3x2D3J8ge+sl8p+8941X8OwXZG4tl4jZ02wthLAi/3I21i8TNNFHBJKnmmkTRq4UKBqLUlnDqEjGlR5dPW1l4LmeZvEkP4iAKD0AHl1YJ7T9LukL2gL9b79H/Zn7j/APdTL78eHWxx6AT4Of8AZMvXH/UPlf8A3cT+9Drx6Nr731rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6+O5/wpHW384T5Xgf877FH/1QHv3XjgD7OqMYKialkSopp5qaojOqOaCR4ZY2/qkkZBB/xB9+61mvSn/v9vnxrGd47m8aiyqc5kiABwAP3PfuvdOWA7U7E21l6TOYneO4YchRSLLBJJl8hMgZSCNUTyWP0Hv3W+rNunP5t3yd2zgqvZuV3BXZ/wC9pRS0dWah1NIkA1oxjkf/AAsSD/sPYn5Ll+n5otLildLHH2jpNdnTbOaVx0PE/wDPj+T+HwDbUo67JQVFGWpjWfcoLqi6ANOsm3sr3qTxt3uZeGqRv8PTsRPhrgcOiS9l/wA0P5bdlSVC5HsKvhpp2c+NZJjIFb8BlcAf7b2WDq/zPRaD8me5JK5MhU7uyNTVxy+VZJaqrNnvySPJb37HXurPPiV8tOid0Yuvh+W+9MrQ1lDFJ/DNH31ZHUsgJhj8OpgAePr7916uOhRT5+/E7am559nYHrem3dtrPSvQR7pqJpKRsVHKdKVfhawJ54Hv3W6joXMRTYalydHm8NJHWbRzrJU00KFSI4Z2usbMv9B791o563K/grT4/wD2WzbZxMESR+OlYJCFDC6tyzLz71Tq3RxKTKVUOlGkOlRZlJ1ekfj3r7OtkDpo3BHS1jxSIoDGxZuANP5v795168OFD1qxfznunqrGbkxe+cZTyTRSxxRzTQqy6AzAuSyD8X/r7t8+qnOOqtPj7s/vHeOfok64z+YoFhmRnKz1ZRVRjcgare/U61Xy62lfjBvffm2trY3Z+9szPuDOLTJ5JahWLRuigaCWJ9+49bFfPodtxrld0pVRY+tXG5CAEkJGAWa30BH9ffut16Akbt3TtTJij3LLLVY5AwmLqzxub2ViDx9Peh1ofPqVm93UmfpozhYqNJEW7CKCFC2n66gg59762fQdF77/AOxMdtDp7cWcr6mipJqanljcNHBHqlMRKAE2549+qD1ry60TvmL2rlOyuxq6eoqpZqGCrqWpR5HMJBmdeFuR/vHv3WvOnRP/AH7rfV7/APwmnF/5y3xD/wAM/uM/+u1U+9Hqy+f2Hr//1Lav59Pzm+Vuz+nO4sp8B+56zqfIfB7fHS9Z8oM1FiqCSTcFD3JmKSg2ptvAZKuimEkVN5BJko4SrMHMbWCg+1scQWOrfE3Co4dFNxcO8xVK+HHTWQcmvD8h5/6qn67J3n398qvl3sP4xdbd013TfX/S3Um2+0fk3NhsLR1Gb7bl7M2zA+0cLt7JEiXGw0NU61UmiRfJqdW1KqD36MLDF9QRUk0HW7h3ubv6FTpCirH1rw/n0tf5dneXcq9m/JP4SfIfsE9z9sfFvJ7cyx7kXFU2Bfd2yey1mzG08fksTB9KrH04SCScsxk/tEldTNzxqFWVfxeXV7GeVpZLSXJipnhUHh1a5IWWN2RdbqjFE+mpgt1W/wDiePaboy6qn+XXbXyEyfWHYO18z8e5MPseV6Onn3024oJ0pKJMtH46449RqOqy8X4vz7159WoPLqyPrEBeuNhANrA2dtsBrW1D+Dw829+HDrR49Lj3vrXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691R9/woL4/lx9vG9v9wmZH0ve+OPHv3Xjw6+OR/xP/Fffuvdevb/evfuvddgE/T37r1adWr/y1fnHvr499mYPZ9TuerptlbgyNPSzxTVErRxNNJoKMZHChQD6f9t7916tOvo39AYCj7D6x2/uLbW8qObEbgoaeqnkgmik1eSNSwZ0Y/kn8+9fb14EdWT9MYDbew8QlJjshDWVL2aqZJFkLSX5JCm/v3nU9bPRUflfj9753P002KQthF1GdAf7P5JX6/T37HXs9ReoajacOGqMIaMy5ho7COWnLqJgo+hYW+vv3n17iMdA127sDeORnX7+spY8DJJofErFB64pGIswA/offgQOvcc9Vx92fGjG1G6sfTbeoKelgr0c1pSnT9RBuWIX3v7OtdK7pz437s69+6yG1FhyDBoz9h6CWLEFrBh/xHv329e6sIq8nuTaHTtXT5ilXHZvPximihiQRSqWXx3Hjsfqfx799vW6enSz+L/SFLsXA1vYe8KaTI5CQPXQy1GuRlDgTLpWQE/T3qvl16mOjG0vyc2TPFVUalaWWlE0JjchDdQUFlY/1/w91NafmOtcPs6rL7T75kxnYe5s1T5Cmix0aCR2laIARJdzcuf6D3IO+LXlCyI9TXpLD/uQ2etab+cX/NE23ktl5Dq7YtfSvnquOWkqqqheLyLIQyXHge9hfn2ARgdKa1607aytq8jU1NbW1EtRU1Urz1Esrs7SSSNqZmJP9ffuvYHUP37rfXvfuvdXHfyHAT/M5+LZANv9Ku0f6n/l5j6n3pvh62ldY+w/4Ovsr5n/AItWQ/6ga3/3Dk92HVG4dfOq/wCE1f8A2/h+cdv+dT3QP/X2l/r7qOI6dPBuvozr+lf9Yf717sePVBw6796691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SE7Q/5lvv3/AMM/cf8A7qZffutjj0A3wdN/jN1yT/yr5X/3bze9Drx6Np731rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6+O5/wpH/7fCfK+5v/ALn8V/7gD37zPW/IfZ1RV791rrs/X6W9+60Ouvr791vpU7N/4v8ASH6WSc/9Yj7EXKn/ACsFt/pv8h6SX3+4j/Z1A3Dxm8kfx91J/wAR7LN1/wCSlP8A81G/wnp6H+yWnoOmb6nj2g6d4dde/de697917rmjsjq6elkYMp44YG4PPv3WqevVsHwj7gz+5YK/Zu5q5Z6Ogh1Y6pqpUiEcnPiVS7DlePoPfuvdbqf8q75PYE7Z/wBF258nT080UYp6BpZU0MVDBXVi1vr78Rjqw+fVwOQiSnn8tJPHWxPcq8MiyqQfyfGSPdf8HVhnppy9VIlJ444XaQrywVjYfkce/Vr1rhnohnzU6ax3bHT2aStohNW0VPLLE8kfrQIg+moE8e7CvWiOqJfjVV0ew90TbfjEdCmMyH29XLpVWfRIdRLcW49+49VpQ9H03l83/jT0fvvaO0c/uqlXem8DTJicfRBK+WV5yIwJBTuShueQw90lkigjaac6UUVJ9AOtPIkSGWUhVXJJxjo48nZGOM9Jl0yEFKlTDG2qaVYdYnAKag5H1v7BWxe4/KHM2+Ny9slyZrlATTSQtF493A06Dm1848v71uZ2fbZ/EnoTpp5LxI6D3uHtnZuE2lmK3N5HFLJFRzyI33dI0rMsWoBV1XJPscdCbqnPbnznpsPuXLSSzePbtPNMUmlkCo0UbnUV1Nb6D349V8uqrP5h/wDMyp+z6Go6667qZGx8j6cnPTSssYniDLd9D2vzaw9+699nVFmaztbnKjzVjagGZlB/UCxuxLnk8k+/dep0zAkc/Ufn37rxFer3f+E1PH85b4hf9r/cf/vNVPvR6uh4/Yev/9W0P+Z5/Je+X26P5fHefx7+Hff2R7Dffe8ertyzdQbqwe29v5Pe0O2N3UtbX/xHs+uqWnL0gRK2OGchHWDRfVoBWSXCyrSlD69FdvYPbNUOWXzX1/4o56OFubpn52fDLuXp75CdOdXVPzY3Z2h1btbqj5RYuLc+1etqnbjdf7Lo6PbuXwVRXnx1FO1XA6LII2clSZAPID7sJYZU8Ju0Dgf8v59NvbXVtN9VF+sTgitDTyycY/Lo3f8AL76J7Wx+4e6fmH8jdix9UfIX5RVuFXdvU1JlsduGi6/2vsRp8TtHFtuDHs61U8tMVlks1kGkfqLBWriRCBHHkL59PWFvMjPc3GHlpVeNKcBXqzj2m6Muim/OA2+M3ZHNgaXGA8kXH8Vh4Nvej1sdDx1h/wAy22B/4Zm2f9t/Bobe/Dh148el1731rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6o+/4UFj/sXH29/wBqPNf+64+/dePwnr45Nja/49+69161v9j7917r1yP9j7916nXOKWWCVJoJHhmiYPHLE7JJG6m6ujrYgj8Ee/de62sf5LH8y3sStyeC+Nu693vRUaLBSYuqrarTGKVToU+WdxybAH/H37rw631+ldpzbT21BuvJ54bgjqaWOqUQT+cMHjWTjQSOb/196Pr1vI6RO98tuzsDPqu2MPUR0scumYPrCsmrSbBuPfvLrw6U2VpqbZO36b7THxQbpq1VZSI1Yo7Dnm39fevl1vz6Bjdsu29tUUu4uys94Z54pJ4IJZTGiOq60UKxA/2w9+zx60SOHRItt9oR9idg5SLHY9cnhcfK0VJOsqgPGzabg3P4Pu3XuPRq9sbByUFbR5fbdeKaWSVGqsZ5fObagSCL/wDEe9GnWxWlT0YLeXWWZ3zXbVmqXSOhxKCWvgsul2jUPci39f8AD36vWsdM+4+4cxjasbHwWPElNTwijeJLN5AieL6D/inv1MdaNK9Ew3Lt/clX2HLLXYebD4iTVNPXNIY6fTbySFuAB+b8+/GvWx8utb3+b18vqPqXO7l2j1zuKOSvmgalqWpalXKsIHUqTG/9f8Pcg70CeUbNR6k9JI/7dyetPXcWa3b2Fm8jn8iMpm62eaSeokjiqq1acSNq0+gNpH+v7j2oHE06fMiLTWQK+p6SDI6O0boySKSrI6lWUj6hlbkH/X9248OrVBFR1x+l7j/jXv3Xuvcc82/4n37rfVsX8j/LZbG/zQvh7DjIDPFX9zbPpsgQpPhpjklvKW/Fvp703DqyD9QH7f8AB19oLM/8WvIf40NaP/VOT3YdNtw6+dV/wmrNv58Xzj/qMT3QR/sN7Se6jiB04eB6+jOvKi/1sL/7b3vqvXfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6QnaP/ADLbf3/hnbj/APdTL791scegG+DoI+MvXIP1+3yt/wDz7z+9Drx6Np731rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6+O5/wpH5/nCfK+3N89irf+cA9+Hn144A+zqiv62AHPv3WuHXXv3W+ve/de6VGz7fx+kB5BWf/AA/3UfYi5U/5WC2/03+Q9JL7/cR/s6g7i/4vmT/6in/3oeyzdf8AkpT/AOnb/Cen4f7JfsHTML2Nv9j7QdXNPPrr37rfXvfuvdLPr7B4Dcu9NvYLc+cXbmByeQipclm3UMuPgkB/eKtx9bDn+vtPdyyQWzyxLrZQSB69IdyuLm12+a5tI/GlRSVQcWI4D8+rkewv5T/afX9B1TuroLsit3zt3svJ4qmGWx1OYExkWQUMahxAp1FAfo3B/r7iDYfeXZr+e/sd5T6KewDMyOR3Iv4h8j1D21+8tu7zxb3ZtaPEuBUNrlFf0RT8Q+eOhi723rvf+W5U7Iope5Bvjf01LQz5LalNTiirKKmuBUSSVCsNRS/J4BPH59n/ACBzvc87rNfxwNHZqxEbH8YHBh8uj72/5/3Lne4nkexaC1jwHLA/qA5joPMeZ6Ezrv8A4UEfIKsyuC2RtLH18Vbma6KhStq6mnKReQXLMZ5DYCxJPuQrmcW8DTkatIrT1+XUj316u32kl7INSxqWIAqcenr1d31/8lvnLnsNjszT7tocqctRpWCBa/CXpxJz4ipe/H9fcN7h78ct7VO9te2zq8Z0miSEfkQtOoKg9/Ly8meKx5ZupUUnS5GkOB50JqPzHR++ruzuxN+7HyW1u2MZUVe4cpFJBFXQLFNBDG4sVd6W6c2+t/ZNtv3hNu3jmO32Db9sd47ghfGGsaCfVSox8+hHsPuze7zMttf7HLYuzhRrNQQTTVUVH5da4vzd2h3F09unckXV9LKM1mMhP9k4R1jWSVv25Gkta1z7yGddD6D5dS866H0notnxm696B6g7H2z2B81d+0u4+9N418dRtnD18lRK9BJNPqpKShQakRQ7AH+vF/cIe63+uBfWMu28qxaIypLSkihXzAB+XUD+6Q9wbvcYrDY7bxdq8NmnKtpcsKaaH+HjX8urZvn5vfObf6Uxm4tkV8mMknlxZpZINVxTM40i6f4W9wd92a1a253uo7jMsayAn5+fQF9qPEX3UtldfDpaS9voajifM9UNdj96bllimrN/b6qJ6aKJnWiNXIocolwjKH/I/qPebnWV/CvVaHZHyg3VuOesxWEf7LAa5IowGImlUXXWJEP0P1HvfXs+XRWKieSqnlqJmZ5ZpGkkdjdmZjckn37rY6wj/H37r3XvfuvdXv8A/Caj/t8t8Q/+1/uP/wB5qp96PVl8/sPX/9bf49+691737r3Xvfuvde9+690Uv5w2/wBll7IuL/5LjOAbf8vWH8+9HrY49D11f/zLbYH/AIZm2f8A3TQ+/Dh148T0uve+tde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3VHv8AwoM/7dydvf8AajzX/utPv3XvI9fHKAv9PfuvVpx69b8/j37rXy69wT/Qf7f37r2evE/j+nv3Xh0qtlb23J17uPG7r2pkqjFZrFVEdRS1UEjxnVG2oK/jKkrcfS/v3XutsP8Alyf8KJew9jybc647ryclZhImo6Oatr2UwvGCIysZeS54H0Pv329e+3ra427/ADIek+5sRt6Tq7emGxmeyS0plpgaaImSULqBYyf1PvVPPq3n69HH3RvfFbe6xbsLdtbi81JiqT7ycwZCiLsscP3Bssbk/T+nv3EE+QFf2depUkjy6o9+Tfybo/k1BDHtCiTGYegkaAsuViieYozRAiNnuefx7iXfPeTZthvzYPZvNT8QqBjj+zqId997uVuXdyk2i8hdpIiAxANBXhnhnpcfEXZ9LQyz0GWy0eEr8kqx0P3c6apXcqqsuoi/J/HtzlP3l5Y5y3f9yWEbRzngDXNOOej3l/3K2jma7SxtITE0mVzWo/ydZO5/mLmvhn3HieuazZuQ7CzWeZpMfPBUVUNIYwdVvJF6ALD+v59yRuW52ez25u9xbRGua/LoYbvu+3bDZNuG6uYoUyzUrTowGxv5lW8tw7jwmHy/Rk+3cTkxoyOXfMySpSRSJYSlGYD6G/uP5/eb25t0aSa+UKvGgYn9g6BUPu1yFcyiK3vNTHgNJFf29Su3Plb0n0lkZuxcxvLFVSTwSVTYg1EDTJKI2kMZPkvwRb2K+WebuX+cbM3/AC9KZoh5lStPyOehltm87fvKl7B9VOP/ABfWt786f+FDctVFuLY/V1Mj+cVdFFWUXjaWnVw8SuX13HBB+vsRkVwOjIGh61WOze1t597bhbI5jJVud3NujLqBAn3E8o+4mtFF4l1G6lrG3+9exDuu9QS7NBaV0rDUmvl656TOyQF55DpUCpJPp1tKfA/4EbB6f+NeWyHYGHxmT7D3HgpMnkGniiNQlLUwGpowYZgWQBTp/rYcn3gn7ie6W6XfPFrt+wMVtFfSzipViDQj0x1g77o86XPOFxeXlpdy2sG3sqwRozRuZUfukZlI1I3ADgRX161SPkDQ0uN7m7Eo6KNaemh3Pk0jiRQEjUVBAVVX6Dj6e8udlkaXbIXc1JUdZn8tyyT7DaSy5YxISfnpHQN25sPZn0e9de/de6uP/kNEf8OcfFwFVa/am0rFlBIP8SHKk+9N8PW0/tB9h/wdfZWzP/FryH/UFW/+4cnuw6o3Dr51X/Caw/8AY+L5x/44jun/AN7aT3UcR04eB6+jMv6V/wCCj/eve+q9cvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690hO0P8AmW2/v/DO3H/7qZffjw62OI6AX4Of9ky9cf8AUPlv/dxP70OvHo2vvfWuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r47n/Ckf/t8J8rja3+57E/8AuAPeuveQ+zqir3vr3Xd/oP6e/de669+690qNnf8AF/pf+CT/APWo+xFyn/ysFt/pv8h6SX3+4j09OoO4TbOZM/8ATVJ7Ld1/5KU//NRv8J6fizEv2Dpm+vN/r9ePp7L+r9d2J/3r/be/deqB1x/w9+631nggnqJ46eljlnqJXVIooEeSV5CbKsaJck/0t70SFFW4dVLBV1NgD163d/5X2G7F6p+Lm2aLvDdNJBPuqsibYlNmfG1RF93CTjVhWqsUKggWAH9feAPvh+7uYecmPLls0slopN08ZIBjB7lNOP2dYcc8btYXPMt22zyCOEhotPh6v8ar/a18lAxXh8+tZL+ZxsTuDZ3yj3xP2rVVeRjzeTrcjtStkaSamXASzD7eCGYjQLcekMfeXvthuOy7jyhaPsiiONUVSvmGAzXz49ZAe1m5bNf8rxptNNcJ8OegpqmX42p51Oa8OiR9dNt5d7bdbdVTNR7fXIRnJ1ME0tPLDThSS6TQkMpvbkEex1dGYW7G3FXpivr0PNwN0LKU2YDS6TpByCfKvWyt8eu2Pg1iptrba2r3K+Nz+Vip6N0rdybhq2+8luPHonfQD/rNb3FV3ce6CvI9jZRz0rpUpDUj7T1imNy+99JuzQWtntiWpY0YJVwnkdOKmnHPW0v8Wdu7x23tWjpsBlcVubZ+SeOqmyqyx1s5Rl48chZ3HFv8PYO2q99+rrma3bcdqjtNvDDxaxQHt8yHXuB6lfl2P3phdBzT4M6lwWZIVTStcin2eda9F7/mPdeYRet8nuOloAMtArypVCPTKGCBrhrX+v8Aj7yRk0Fj4eR1MkmnUdJqOtIfu7NZfI/IvqGpydXJUS0e4KJKd5maQxKuQQhf3L/kD2h3BdVhKp/gb/B0Xbqf91s//NNv8HV7v8zrsnPbL+D2w8ziJ0GTraHARmeSPyKPK6RFiv8Ahf8Ar7xT9jbZIfcfeZkFNDMAPtp1ix7WXDL7qQ+dLOY59fEA61K9z743Ju2tqa3M5CeY1T62hWR1gW34WMk8e8t+steJ6SPHHP8Ar/4e/db669+6913xz+P6D6/7z791rPXuOOf+Ne/db6ve/wCE1H/b5b4h/wDa/wBx/wDvNVPvR6svn9h6/9ff49+691737r3Xvfuvde9+690Uz5w2/wBll7IubD7XGX/8+sPv3Xuh56v/AOZbbAt9P7m7at+eP4PDb34cOtnj0uvfutde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3VH3/AAoLP/YuTt4f1wWa/wB4xx9+68eB6+OR7917r3v3XuuwP9t/X37rVeuzfj8j8f4+/deHXrf9K/8AG/futV68rtG6yRsyOjBkZSQysOQysPz791b7ehv2H8jO3uuJqefbG8MpTPSOjweSqqH8eg3VQQ4Nhb37rX2dHtxP84D5ZQbYbamW3fXZHGSwmmlg+5qWWSMp49OmWQ/UcW9+zmnn1up6sB+De2u4e2I6vvzuPMZnavVWCjlytBi0lrNOVVQKuGaSOMAi5BGn3AHuVum0Wl2u1bJCkm4XHaR2+eC3ypXrFb3v5p2UE8r7BCkm4zsqyS6VIiJ+DWf6RwPTj0CPyU/m39t0XduOr+oGymM2RsasajjmejraUVopXMTCN5dIaxUEu/B+g/r7EXtv7a7XypFHuUihrtu4titTx/zdD72j9v35ZsU3Xe2El/KoOmoIiBA7V6E/B/zwu4e7uwdrYne9VhcLQU8CQTbkzOHx1VVQrCqx6Uq2JdSQSfr7HXNlnLuGzywRjUSOFAf2V6kHnT6xuXp/orcXMgU0jP4j6Zx1YVL86tk1tGaSo+QO2kiqKZFkMePSN0aWKzBZEIIsT7xmfkmYk12+ufRc5+zrENo/cQhxHy2qkk0IdQfyx1r5fPbvLN1nY9bQ7O7YqN27erVLtNR1dR4l8mpnCIWsBc2+nvIHkKyNjtmj6b6etMUAr+zrJn2qg3SHliL982ptLgjuQkNQ/aOgg+IHWfSPama3J/pv3nT7dEL07UU1dPIrVRlF5pCVNyb8ezHmjcd126BJdrjMhzUADpz3I33m/Ytvil5PshezM1GUsFCrTjnj9nVp/wAc+h/gtsDvzBbmh7PwuQptry/eQQTmSWGWWRAFJWa4Yg/Qkcc29xjzZvnPlzyrKLS0IluAVXAH25+zqDd45v8AdPmHZjse77YbZLgMtw8b9yKRjTShyePVy3UT4LsTtTt3dGC7pxO69uZPatBi6LbNDA0Q21BSJoSpe/pIZRbge8d+ZJr7Ydi2u2n2toZEmLFiQdbNxzxyc56iS/2hn2UbLuML2s8JJN0wosyipVaHiy4qT1qFfO/rnB9c/ITd1Bgt1U264MpX1mTnqaZBGtLPNVEPTMB+R9P9h7zU5Kvbi/2CCa4iMTaRg/Z1mp7Y71cb5yhbXdzA1uVXQA3FguA32NSo+R6JmeOB/sT7FnUg8cnro/778+/dbHVxv8hzj+Zz8WvyP9K20Of/ACJDj3pvh/Z1tD+oD8j/AIOvsr5n/i15D/qCrf8A3Dk92HVH4dfOr/4TWf8Ab+H5yf8Aao7o/wDe2k91Hl04eB6+jKv6V/4KP96976r1y9+691737r3Xvfuvde9+691Xr/Ms+dh+AXxn3t3Xt/rOu7w35t7EVmaw/U+Dy8WNzGQwmLiaoz+6axhHPMmNxsYD1cqQn6hQQTw/DCZanyH+qnSS6ultwBjUxxXh9p+Q8+i8Yj+avmt6fCX4Y/IrrT4+5vfnePzTxGycjsj4+YncFFBlcBj9wTim3TvHI5CoUmXEYUMk8jBVeVHQBlIcr6OHWxJwo49VnuxCir8TvwA4H55PDownxm+Ze9t895dl/Fn5F9WP033XshTmtizDK02R253h17DCn3u+9mCAs0MdPM/hmgkdjcMwI0sotLCoXxYsr1S1u3aQ21wNMgz9o9f9X+athvtN0YdE/wC+PlT0VsvE762FufeiY3c77czePGN/hmTnY1dTjpIoYRNDEY7sxAuXt/j711sD16mfB06vjJ1rIP0y0eRlj/xjlykzxt/sQQffh15ujae99a697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuvj8/8KONr7myv84L5XyY7b2bromzuKdJaTF11RG8f2IGtZIoyCL8XB964dbOAPs6o6/uFvf8A55Dc34/5ceTvz/069761n069/cLe/wDzyG5v/PHk/wDr179Udez6dd/3B3x/zyG5f/PHkv8Ar379178ulDtXZW86XOUkk20txommdSz4XJhReMgXYRH/AGHsR8pivMFsfRv8h6TXas9s6KMkdQs3sfek+YyMke0tyMrVMhVlwmTKsPwQTF7LN1/5Kc/+nb/D07FURqPkOmr+4W+P+eP3N/r/AMDyf/Xv2X9Odd/3D3wf+YQ3NwP+dHkwP+tXv1etU+XXX9w97/X+6G5v/PHk/wDeP2/fqjrefQ/s6EnqAbr6y7H2pvvIdc7kz1JtzJLXy4psHkCKvQjIEKyRWP1vz7R39s15aPbI2guKV9Oiff8Aa5N62a52mKVoGuI2TWvxLqFKj5jy+fR5Pk389/kx33mNgS7d2ruzY+3euKrH5PBYqmwOQp5XyWNsIGqzRoFeIAWKD63/AB7BXK/txy/yxHdeAgd7wsZWbNdXEZqQOo45J9pNu5Z26SDeZTuV1NF4DzutC0WaDSMBs5YZPHpv+WHzK7M+WHWe1dm7t6i3FT7m2/Fjlm3L/d+qRqh6KxmKyJEZCr2+je3OU+RNv5Pup5drYiO4YsUqSAT6eQA+XV+SPa5uS+Yrrd7e9eW2lQRx21KJEoJNfVnNcsTWlB1XENhb45/36G5f8f8AcHk/+vXse16lv8j1IpNm9g0NTBW0W191U1VSyLNBPDhcpHLDKh1LIjCIEEf19+BIyMdbBINRUdXE/D7+bX81PipSviGwu7914BKYw0dHLhMm8kPBUa2qFINv6j/be7+I5FCxp9vVtcpFKk/aT0PXeP8APD+Ufde06ra2U6q3NTwVUbRs7YCqA0sAPqiE/j3THVBq4UH8+qVN3bs7d3fvTFb1q9m7gjrcHWpW0EK4LKaVkjnE41kRf1H49tyIJY2jPBgR+3pueFbiFoJBhwVP2EU6OJ8kPm33N8iui9p9K5frLc1HS7cgoYp8g+Dr/wDKHoXV0ZCsZNrrxcD2AeWvb3beWN7ud7sm77okv8yeot5W9rbXlrmeXmTxzKxRo4kpQRRsalR61Oanqts7C3xa/wDdHc1v+1Hk/wDr17kLqVvy68dhb44vtDc3+H+4PJ/T/YRe/dep8j17+4W+P+eQ3N/548l/169+6313/cLfBsP7obm/P/LjyX/Xr36o69+XXQ2Hvj/nj9y8/wDVjyf/AF69+r1r9vV53/Cbja+5sV/OS+IkuR29mqGFc9uKSSWrxddTxJH/AHcqE8jSSxgAXsLk2596PV18/sPX/9Df49+691737r3Xvfuvde9+690Uv5xf9ky9kf8AULjPr/2tYfej1sdD11f/AMy22B/4Zm2f/dND78OHXjxPS697611737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdUdf8KEJRF/Lh7ePjnkJweZAWCCWoc/7jje0cKs3+8e/efWjw6+O0uNyLqGSgrWRuQy0k7KebcMF9+63TPXIYrJ2P+46vv9LfZ1H0/wCSffutde/heT/519eP+qOo5P8AyT791vrr+F5T6fw6v4/6ZKjj/k336vXqdd/wvKfX+HV/+v8AaVP/AEb7916nXX8Kyf8Azrq//wA46j/o337r3Xv4Vk/+ddX/APnHUf8ARvv3XupVDSZWhraStjxlY70dTBVIr0VQyM8EolVWXTyCRYj3VgGUr6in7equutCh8wR+3q7Xq7+bpvvr/rHD9cHY7SUGNpFpZEj29G8FQukKwkV/1Dj+0PcMbp7L7Fum9HfJ3fxycHWwp9lOHWP2/exv72vLma3vTElydTDTVgfUMc48vToAPkd8727x2JWbQp+sKPDyVcUkbVlLteloZFMga7iSmW97m9zc+xbsXJn7luhMk7svmGdm/wAJx0s9vvZebknd23a53e7vyQQqTSsyKDTAXhimCc06rBGMyikFaCvB+oYUlSDcf0On2OzTz6naleI6k+LcVv8AN5m3/LOu/H+w910R/wAI/YOteGleA/Z1Hkx2XlbVJRZKRj9Xelqmb/YllJ92FAKDrYFBgU67joMvCweKiyUTC3qjpqpGH+xVffiFbBz14rUZFelrV1OXixFCKD+8ENdGR5J0pshA78EMGqFQav8AD1ezu9Nu+zW6BRUE/h/y06SiCMyNVFofkOjafEH5k9jfFiffc+Pos1l33pi4cbJ97BV1ywCK9nQVBupuRyD+PcYc3clbbzbHAl2oHgPrX7fy6AnP3t3t/PVpBZ3DGFYXDnR2lgPwkgcD59FG7D3BuXsHeWf3dlKLJvU5rI1NbpkpqlvCs8hkEaXXgc3sP6+xVZWkdjapaxCgQAdDjbbCHbLGKxgFEjUKPyFOkX/C8n/zrq//AB/yOo/3n0+1fS7rv+FZPm+Nr/8AD/I6j/o337rWerff5FBlx38zv4qJWUlfCantjaSRE0FWyFhklJ8jqhCfUcsQPej8P7OrIP1B9h/wdfZXzP8Axa6//qCrf/cOT3YdUbh186z/AITV/wDb+L5x/wDap7o/97aT3UcR04eB6+jMPoP9Yf717sePVRw6796691737r3Xvfuvde9+691rX/zF9vfL/Ym6/nv8kcn8cMh3lgMb8Ku9eoOis9tHduNxuH2R1pufak2U3FV7h2ZNI9Rk8isvllmMcXkKKyx2jsAvUwmAKrUPE/M9EsgvRetI8YZMqp9FPEnog/wM7qynx6+F/wDJn+b3yI2jkOtPjz010TU/H3Mbwrpny0tXUb83FVYDbW75NvY5Hraamlp6ZJ4nljIk9TJdSl9xUaJo69zeX5dUuiUuY7lk/SjqK8RQkZxwA6t66T35sD50/wA0bB/LH447nj3z0p8aOi959E7x3pS09fjcRk+wN514zdNjMXBl4YZqnwQvdpEjCgAkmxW9JFMNt4b4JINOrwOt3uX1cHciKyavIk0P+Tq872i6Ougj7Y2ftGt2NvvJ121duVuQG09wyffVmEx1RV+RcVKyyfcyxl9QPIbVf3ojHWwTw6Cj4Pi3xm64UcBabKKo/AUZeYAD/Afj3vrx6Nn791rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6Bbc/xw+P+9M5W7n3f0r1dufcWR0/f5zPbH27lcrWaBZPua+tp3ke341Mffsdeq3kemH/AGUf4t/948dL/wDot9p//Uvv2B5deq3qf2n/AD9e/wBlH+LZ/wC5d+l//Rb7TH/xr79QenXqt6n9p/z9e/2Uf4t/948dL/8Aot9p/wC9fa+/Y69VvU/tP+fr3+yj/Fu9/wDZd+l7j/v2+0//AKl92RmjbVGSp9QaH9o63VvM9e/2Uj4t/wDeO/S//ot9p/8A1L70SWNWyfU9eq3qf59e/wBlI+Ln/ePHS/8A6Lfaf/1L71j061VvU/tP+fr3+yj/ABb/AO8d+l//AEW+0/8A6l9+x6deq3qf2n/P17/ZSPi3/wB479L/APottp//AFL79jr1W9T+0/5+vf7KP8W/+8d+l/8A0W+0/wD6l9+x16rep/af8/Xv9lH+LZ/7l46X/wDRb7TH+9Uvv2OvVb1P7T/n69/so/xbH/cvHS//AKLfaZ/+NffsenW6t6n9p/z9e/2Uj4t/948dL/8Aot9p/wD1L79j061VvU/tP+fr3+yj/Fv/ALx36X/r/wAy32n/APUvv2OvVb1P7T/n69/spHxc/wC8eOl//Rb7T/8AqX37HXqt6n9p/wA/Xv8AZSPi5/3jx0v/AOi32n/9S+/Y69VvU/tP+fr3+yj/ABb/AO8d+l//AEW+0/8A6l9+x16rep/af8/Xv9lH+Lf/AHjv0v8A+i32n/8AUvv2PTr1W9T+0/5+vf7KR8W/+8d+l/8A0W20/wD6l9+x16rep/af8/Xv9lI+Ln/ePHS//ot9p/8A1L79j069VvU/tP8An69/so/xb+n+y8dL2/8AEb7T/wDqX37Hp1urep/af8/Xv9lH+Lf0/wBl36X/APRb7T/+pffqDrVW9T+0/wCfr3+yj/Fv/vHfpf8A9FttP/6l9+x16rep/af8/T9tf44dAbJztFufZ/S3V+19x43X/D87gdj7dxWWovINMn2uQoqdJY9Q4Olhf36g9OvVbzP+Hr//0d/j37r3Xvfuvde9+691737r3RTPnCL/ABm7HBF/8lxnH/kVh96PWxjoeer/APmW2wP/AAzNs/8Aumh49+HDrx4npde99a697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917olO/Pn/wDGnrjdGT2funfNLj85iKiSlraR0ctFNEbOtx7kna/ajnPd7FNwsrUtFIKqfUHqPtz9zeUdpvn2+9u0SWM0ZScg9I//AIc3+JX/AD8Wj+tv81J7X/6y/PtK/Rt+0dIf9d3ketPrY/29dP8AzOfiQiNI3Y1GFQEt+3JwB+fex7Lc/E0Fm2fmOtN7vcjqCTex0Hz6N31V23snuXbEG7th5VMvhKgqI6qMFVJYalFjz9PYA33YNy5dvTt+6J4cg8uh1s29WG+2YvtucSRtwI4dCZ7Jejbr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6T+5tp7X3piqjBbv27hN0YWqRo6rE7gxdFl8bUI4s6TUVekkbAjggr79SvWwSMjoDF+GvxIRQifGToRVH0VeptjAD88AUPvVB1vU3r13/sm/xJ/7xl6F/wDRTbG/+offqDr2pvXr3+ycfEr/ALxl6F/9FPsb/wCoffqDrWpvXr3+yb/Er/vGXoX/ANFPsb/6h9+oOt6m9eu/9k4+JX/eMvQv/optjf8A1D79Qde1t69df7Jx8Sv+8ZehePp/xifY3H/qj79Qde1N6nr3+yb/ABK/7xl6F/8ART7G/wDqH36g69qb169/sm/xK/7xl6F/9FNsb/6h9+oOvam9evf7Jt8Sf+8ZOhP/AEU2xv8A6h9+oOva29evD4bfEkfT4ydCj/yk2xv/AKh9+oOvam9evf7Jv8Sf+8Zehf8A0U2xv/qH36g69qb169/sm/xK/wC8Zehf/RTbG/8AqH36g69qb169/snHxK/7xl6F/wDRT7G/+offqDr2tvXr3+yb/En/ALxl6F/9FNsb/wCoffqDr2pvXrl/snPxLtb/AGWfoa39P9FGx7f7b7H3YkkaScDy69qYefXH/ZN/iV/3jL0L/wCim2N/9Q+60HXtTevXf+ycfEr/ALxl6G/9FPsb/wCoffqDr2pvXrr/AGTf4lf94y9C/wDop9jf/UPv1B17U3r17/ZOPiV/3jL0L/6KbY3/ANQ+/UHXtTevTzt74tfGnaOYodw7W+P/AE1tzPYydKrHZrCdbbQxeUoKmM3Sejr6KkSWNx+GRgR79Qde1N69DNmf+LXkP+oGt/8AcOT6e7DptuHXzqv+E1n/AG/i+cfHP8I7p/8Ae2k91Hl04eB6+jMv6V/4KP8Aeve+q9cvfuvde9+691737r3XvfuvdYp4IamGamqYYqinqIpIKinnjSWGeGVDHLDNFICrKykqysCCDY+/de6T1fsrZuU29FtHJ7S2zkdqQR00UG2K/A4qr29DFRMHo44sLURNTKsJAMQEVlIBW1ve6kGo49aIBFDw6z7e2ptbaNJNQbT21gNsUNTUvWVFHt7D47C0k9ZIoSSrmp8bHEjSMFUNIwLEAXPHvxJbJNevBVUUUU6f/eut9ITtAX6336P67P3H/wC6mX37rY49AN8HRb4zdcj+lPlf/dvN79149G09+611737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvdf//Su/8A50Hym+QfxewW6Nx/Gf5gbFo/lpNnersb8e/ilkMBjap9wYfcm5qXHbx/jWHrJ5I8hNNSPLUQ1FXGiRohCsrNH7XRgvFoij4+Z+X5dEtxKlvceNczgEEAIME1wABXz/Po3fZ3a3ys78+UvXfxQ6f7A270zSdX9YYTsT5kZk4NsjuDKVW+Nr0821tudY14k1Uq02RcvUSFlaRGKFtKeusaJFH47ZzQdOzzS3Fx9DEdNBVj8qYp+fS6/l8/Izujcu9u/wD4h/J/cm39/fIX4w5bC1G4Ox9n4n+Cbb3hsjfvmyWxqg4tlVoq2CkRI6piB5CdRAYMWpPGqqsqcG8unLG4kkeS2ly0VBX1rw6tCdtCO9i2lWbSPq2kXsP9f2m6Meqv/l93vuPMdR9hbNquod2Y2gneho/701EkZxcCJlYytS6hAbNp9PP5966tgHqwrrIaeuNhKDe2ztti4+h/3Dw8j34cOtHj0uPe+tde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3QEdtfJjpLo2ooaXtLfeL2nPkmCUUdf5dU7EXAXQD7FGw8mcy8zIz7HatOE46aY6Dm982cvcuFRvV0lvrNBqNKnoH/8AhxH4d/8AP6tt/wDqz/0Z7EP+tF7hf9G2T+X+foi/10uQv+jlD/vQ67/4cQ+Hn/P6dt/+rP8A0Z71/rR+4X/Rtk/l/n63/ro8h8P3lD/vQ6FzqT5N9Id55DI4vqzfmL3bXYqnWqr6eg8uungdtCyP5FHBPHsh3/krmbliFJ98tWt0kNFLUyejzZObeXuY5Hi2W6S4aMVYIa0Hz6Hr2FehH1737r3XvfuvdcJG0IzD6gG3+vb23MxWJmHkOtjLAdaVHzEMNf8AI3sqeop4JXO5MiGkaJGPL83JF/fSH22d15KsNLEfpL5/Lrnv7mwwyc73rOgNJDxGfLotIpKL6faU4/1JMMfP9Px7G/iS/wAR/b0Bfp7byjX9g6xVVDRNTzr9rTH9pwf2YueP9b3ZJpdYOo8fXpuW1gMTDQvA/hHW1T/Kgqmk6AFKFVI6eenCIqhQo0MPoPeBfvgzHnR6muD/AIes7/ZwAcnRAClKf4OrTPcO9St1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdNeaF8XX82/yKt/8AcOT3sdVbh186z/hNX/2/h+cf/ap7oH+P/H7Sfn3UeXTh8+voyr+lbfTSLf7b3vqvXL37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdITtDjrbf3/AIZ+4/8A3Uy+/dbHHoBfg5/2TL1x/wBQ+V/928/vQ683Ho2vvfWuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//Tus/mz5Hsj5VbE7D+PuC/lXfJPee+f4t14evfkjt6i2LiZMDWUu5KbIvmsFvmhnfJU8VCiyJUwrOEZGbVa3tfEFiaplBUeQ/zdEd6ZbqMxLbHWaUY6aAg4NeOPs6Ebcmz/mD8Dfkbsf5F7f6R7M+cVR8hNiYnrnvij6tpcXQ5/reXYG2KRNsV9XJkpXWqheWLwipITyMsmrSzKWsZIZo/BrpA4V68ILu0uPq9JlLYYLxp5cacOjZ/y7+n+yMrvrv75yd19a57o/tP5WVW26Kr6T3OKebcWwNqdcCfCbbiz1dTNpepqYdMugIulNJPLWVm4dNKwx5C+fSqwhm1yXU40tLTt8xThWnVqftJ0ZdFK+cAVfjN2ORGpJpsZ/ZF7/xWL1f64/r70eHWxx6HvrD/AJltsD/wzNs/+6aH34cOvHj0uve+tde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdFn+WnyJo/i701m+2q3BVO44sRV0VL/CqNtNROatmGpPr9Av09jLkLlN+duZYuXo5RC0oY6jwFPXoIc881Rcl8tXHMU0bSpAASqirH7B5nqqgfzucCVVh09n/UqsB9weNXNj6fc7H7tV1Wn7wj/Z1CQ+8dt1AfoZs/0evD+dvgbj/jD+fsf+mg8W/5B9+/4Gu6/wCjhH+zr3/BHbd52U3+89C50L/NpwfdfcO0Oqo+s8zhX3VNLEuVqJi0FL47AtICB/X+vsO82ew9zyzy7cb+16kgtwKqBk16EHKnvlY8zcxwcvR2ksbThjqYUUBaefrnh1ckCGAZTcEAg/1B5B9489T5137917r3v3Xuve/de61qP5ycqV3YO1KSqhinSmqImiEkaSaPT9RrHH+w95efdueRbG6ZSRUHgesT/vFwxTS2iyqGAdTkAjz9eqZDQ4/6mipQLfmni/3jj3kz4038Z/aesbTaWnnEv+8j/N1y/h9AeBRUgP15p4vz9Px7940vmx/aevfR2hOI1r/pR/m6ua/kuvDR9vdg08MEcKz7XTV440Qnxvr/ALH1+nvGr7y8r/uCwZiT+t6+o6yO+7lCse67gI1Cjw04ADz+XWyt7w/6y0697917r3v3XusU/wDmn/4Kf9hx7ZuP7Bvs6svxD7etKn5bf9lDdlngD+8eRuCPr6x+PfR/23/5Uqw/5pL1z89yqDna9+bnounAW115+n+F/Y36A/HqPUqPt5bNyI3/ANj6fr7uh7x+XTcoPht9h62lv5Tf/Miqjm/79Nf/AJJb3gX73/8AK6N9h6zo9nj/AMg+L8v8HVrHuIOpV697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XumvNG2LyH/UFW/+4cnvY6q3Dr51f/Cav/t/D8478H+E90f63/H7SXv7qPLpw+fX0Zl/Sv59I/3r3vqvXL37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdIXtD/mW+/fz/vz9x/+6mX37rY49AJ8HP8AsmXrj8/5Plfp9P8Ai7zfT37rx6Nr791rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//U3+Pfuvde9+691737r3XvfuvdFM+cAJ+M3Y9v+VXGf+7WH3o9bHQ89X/8y22B/wCGZtn/AG/8Ghv78OHXjx6XXvfWuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuqrP5u1RIPiluGmU+mTLYxz/QEK4ufcw+w2Pcy0/wCab9RJ74rX21v/APa9atMQbxpf8IliPz6fr7zxY9x6weUdg+wf4Oud/wDbH6/7Tb6X966sQejN/Cx2i+VHVcsbgMtZMFsfVcun59x77sH/AJh3uA+Q/wAvQ/8AakU9xbD00v8A4R1ueYtmbHULP+pqWAt/rmMe+eHWffU/37r3Xvfuvde9+691rQfzhT/xk3bwuP8APxfXn+z7y5+7gP8AELoH0PWK33h8y2n+nH+A9U8EA29SfX/X/wB795L1PWOAC164m9h6hcG978/7E+9460agdXF/yZh/xmbfVz/zCt7XB+t+R7xq+8znl6w/5r9ZE/d2/wCSvuAP++062XfeIfWWHXvfuvde9+691hqDaJv9Y3/1tJ9s3H9g/wBnVkBLgD160qfluSvyI7KAGpTuPIX4vzrHA99Hvbah5JsD/wALXrn/AO5VRzte/wCnP+Toulx+QRb6fT/k72OPl0B6Yr1gqDeCayn/ADbg/wBB6fx7unxj7emJamNh8j1tLfynP+ZFVFrf8CKUf6/pbn3gX73f8ro32N1nT7PY5Pi/L/B1ax7iDqVOve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917otW5vmF8Ztnbhy209zdx7Nw24sFUGky+JrK90q6CpH1hnQIbHj+vsZWXt7zpuNpHf2W3TSQyiqMFww9RnoK3fO/KVhdPY3m4QxzR4ZGcBl+0eXTJ/s83xK/5/rsX/AM+L/wDRntV/rY8+/wDRrn/3n/Z6T/64XJP/AEc7f/nIOum+c/xJVSzd7bFCqCxJyMlgo5JPo9+Hthz6cDa5/wDef9nrx9wuSQKnc7eg/wCGDowmx997R7J23Qbv2NnqDcu2soHbH5jGSGWjqljbS5icgXsf8PYS3Pa9w2a8fb9ziaGZPiRhQj7ehNYbhZbparfbdKs0L5V1NVI+R6VvtB0s697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917pszP/Frr/8AqCrf/cOT3sdVbh186r/hNZ/2/i+cf/ap7p/97aT3UcR04eB6+jOv6V/1h/vXvZ49VHDrv37r3Xvfuvde9+691737r3TRltwYLA/Y/wAbzOLw/wDE62LG47+J19LQ/f5Cc2hoqP7ll8kr/wBmNLsf6e7KjvXSCadUeSOOmsgV4V6dJZY4I5JppEihhR5ZZZXWOOKONdbySO1gqqASSTYD3UAk0HViQBU9Rcdksfl6KnyWKrqPJ4+rTy0tdQVMNZR1MZNtcFTTsyOLgi6k+9lSpowoetK6uupDUHqb711bpCdoXPW2/gPr/c/cf/upl9+PDrY49AL8G/8AsmTrj/qHyv8A7uJ/eh15uja+99a697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv//V3+Pfuvde9+691737r3XvfuvdFM+cJA+M3Y5JsPtsZ/7tYfej1sceh56v/wCZbbA/8MzbP/umh9+HDrx4npde99a697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6qm/m7i/xZz554ymN+n1+j/7x7mH2H/6eXa/8036iX3w/wCnbXwHy61boyAicPwiauf9p4t7zwPE9YQLUIPsHXfp5/Vz+Lj37rfHI6Mz8L7f7NJ1Wfz99NYD6/rT6+4992K/63W4/YP8vQ99qv8Ap4thU50v/hHW6Div+LbQf9QkH/Wse+eA6z76n+/de697917r3v3XutaH+cL/AMzN2+QR/n4r3/4L9PeXH3cf9wbr7D1ix94UVltP9MvVOot9CL2P4+t7/wBfeTFesb9OOurCxt9f8bfT/Ye9Vp1sKT9vVxf8mcD/AEz765uBtXkXufzx7xs+8xX+r1h/zX6yH+7tjdtxI/32v+HrZe94h9ZX9e9+691737r3WKf/ADT/APBT/vXtm4/sW+zqyfGPt60pvlwyj5DdlX/56TI/699Y99HPbUf8gqx/5pr1gF7lkf11vf8ATn/J0XPWOP8AH+n1v/r+x1T06AvHj1hqCfBP9f8ANuf+Tefdk+MdNyUEbH5HraY/lN2/0E1BA5+4pfr/AE0t7wM97v8AldG+xus5vaCg5QiH+l/wdWse4g6lPr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuo1ZM1PSzzqATEhcA/Tj6+6v8ACaenWxxz1pNfLynocj8qu8K6ajpJKiq3Q8ksrQo0jlWcDUxFz/sffSP20nnX282pA5oIvX7OudnuXY2knuPu07xqXZkBJAqaA0qflXHp0Xj+FY611oKMn6X8Ef54P49jY3E/DWf29AsWFkMiJP8AeR1hq8VjjTVK/YUWk08y808fOqMix492juJg6kO3Eefz6pPYWTQurRIRpYfCPQ/LrbV/lW1n/OJPX2LSNIoKKGt8UcahEjvKvpVRwB75+e9jM/uRfsxqSV4/Yes+vZyNIvbrbY4wFCxgAAUAGOrJvcU9Sf1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3TZmf+LXkP+oKt/wDcOT3sdVbh186r/hNZ/wBv4fnJ/wBqnun/AGP+/wBpfdR5Dpw8D19GZP0L/wAFX/eve+q9cvfuvde9+691737r3XvfuvdUf/zrI9jbV6B7H3znOl/kh2dunL9H9m7L2zvnqCsrZNo9GV2QwskVF2XumJK6BKA4+aVaxq5KaVvFE4texVdbGRl8NWVRwNeOeifcjDCfqJI3kIFRpyMdEa6Qwe4u6PjD/J+/l+7l+Rm8d8bX390liu9eyu8tgb2y8O7OwoutspJmcTs2s3NUSyzzYuTXJjamKokd5RTqr6vGPdkiEQeQ5K4Hzx03LctcvBCp0iQamAPAVGK/n8q9Hn6exUvw4/mqy/F/rjP7wzHTvyu6e3h8g8ltbd2drdwYvrfeGyqtcGtDsH7j/gDRVSRlpKYkhi1yWZVb3RyJbYysO4GnTkKta7j9NGf02UtQmtCKDH7ervvaLo46Jl3yvyZXHb8k2jW7Mj2QNuZtmiyFGsmSFB/D3FQqSAXL6CdPvVD59bqOA6efg5/2TH1r+W+zyOs/gyHKTGS3+Gq9vfh1tujae99V697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/W3+Pfuvde9+691737r3XvfuvdFK+cX/ZMvZH/AFC4z/X/AOLrD9Pfj1tcHoe+r/8AmW2wP/DM2z/7pofehw68eJ6XXvfWuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuqpf5u5v8WdwKDY/wAUxd/6kaX+nuYPYf8A6eXa/wDNN/8AJ1E3vgK+2t9/tetW5GPjT+mhByPr6R9feeJ4nrB8A6R8wOuZuf6D/eLf63vVcU6uUJNa9GX+GNl+UnVfNz97Nz/yGn19x77sH/mHe4n5D/L0Pvatae4lh/pX/wAnW6DiTfGUB5/4CQfX6/5se+eA4dZ8dOHvfXuve/de68fp7917rWe/nDHT2dt+/wBDPGefp+g/T3lv93D/AHBugPTrFj7wppLaH+mOqdSRbj+p+n1sf6+8mfn1jgQTjrsG30/17/Q/4/T37HXsjq4z+TP/AMzm33aw/wB+qf1fW/P9PeNX3l/+VfsP+a/WRX3eP+StuFP99p/h62XveInWVnXvfuvde9+691im/wA0/wDrH/evbU/9i32dWX4h9vWk78unK/InsoD1A7kyNzxx6/8AiPfRv2zzyVZf801/wdYBe5f/ACut5/pz/k6LgX/qSPwvP1H9fY8A6AhJ6w1Ln7eYBiCInuL8n0/Qe7oKuPt6bmNImr6Hrag/lLMT0PODx/lFKeSb/pYW94Ee93/K6v8A7brOf2gH/IOi/L/B1a57iHqU+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6bcv8A8Wyt+o/Yb6fX3V/gP2dbHEdaUvytsfk13J6DzuaTleP7T/X30d9tT/zD/a/+aX+brnv7kL/zELdP9Ov+A9ALYgcKyk/7b2NajoHKp4nqPU6hTVFgTaGX6/4xm3u6U1j7R03Kv6L/AOlb/B1tdfyqyT8XdnXH+6Kv/rcnAv7wA96f+njX/wBq9Z4e0H/Tvtv/ANJ1Zp7ivqTOve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6a81/wAWvIf9QVb9P+oOT3sdVbh186v/AITV8fz4fnJ/hiO6L/1/4/WX3UcR04eB6+jMv6V/4KP96976r1y9+691737r3Xvfuvde9+691Wl80uvP5k/Y9bufaHxd3r8TMJ0/vLrzcG0sxQdy7U3zmd2x5HP4lsXVVJkxDPQy05WWVfE8RBQ2ZGPtTG1uoBYHUOkFwl+5KwMgU+oNR/Pomw/lLdydR/GL46p8YOzOuNn/ADd6LO0tfYm5qPcdT01X4uPIvXb22pg9oUsbyUlBN5CtIohBYBvKB5PQ79ZqYhx2npIu0+FCvgv+olKMeGPl0cb4l/FL5C4nufevyg+be9Orexe+JaSfYvVUvUOLzuC2hsLquoiSWtxLUWaVZJq2pqAzSSsXAW/qLOdLUsyFPChFF+fSm2tJRMbq6IaQ1A01AA9PPqzD2m6MekL2h/zLbfv/AIZ+4/8A3Uy+/Hh1scegE+Dhv8ZeuP8AqHyv/u4n96HXm6Nr731rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//X3+Pfuvde9+691737r3XvfuvdFL+cP/ZMvY/BP+S4zgfU/wC5WH6e9Hra8eh66v8A+ZbbA/8ADM2z/wC6aH34cOvHiel1731rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917qqL+b0QPivuBrG4ymMF/p9Q9zf3MHsN/wBPMtR/wt/8nUUe9/8A07W+/LrVqikHjj550La5uP0i9/eeTDu6wgQnQPsHXMvf+2Be17n6e9U63xHRmfhg+r5TdVi//KbNbm/9tPr7jz3Zx7d7jT0X/L0Pvar/AKeHY/6V/wDJ1ujYr/i2Y/8A6g6f/rUPfPHrPfpw9+691737r3Xvfuvday3841ynZ23edV6iLj+g0ce8uPu3D/Erv7D1iv8AeGP6tmafjX/L1TeZv8bXv/r/AOsT7yap1jnU1695j+Wa/wDr+/U69Xy6uS/kxNr7n31cn/j1eLnj6H8e8aPvMf8AJBsB/wAO6yI+7vX96bgf+Fr1sz+8ROsrOve/de697917rFP/AJpv9Y/717an/sW+zqy/EOtJL5eSFfkX2Xzb/fy5Li/58g+vvo57Y/8AKlWX/NNf8HWAPuav/I1vCP4z0W7yk3FxxwPpYH8ex7SnQEJr1HqXbwS+oAiJvz9ePbifGKevTc5/Tavoetq3+UoS3Q05PP8AlFNf/X0t9feAvvd/yur/AO26zp9oP+VOi/L/AAdWve4h6lLr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xum/Lf8W2st/xwf3V/gNOvDj1pRfKxrfJnua1wf7zyX/p+p7299HfbYf8gDaz/wAK/wA3XPv3HNfcLdARjWv+A9AKb2JBY/7E8exp59A+mK9YKk2p6gXPEE3N+P8ANn3dPjH2j/D0zMF8J1/on/B1tc/yqufi7s3/AJYVf/W5Pr7wA96f+njX/wBq9Z3e0P8A077b6fwdWa+4s6kzr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XumzM/wDFrr/+oKt/9w5Pex1VuHXzq/8AhNZ/2/i+cf8A2qe6OP6/7/aT3UcR04eB6+jMv6Vv9bD/AHr3vqo4dd+/de697917r3v3Xuve/de697917r3v3Xuve/de697917pC9of8y337/wCGfuP6f9qmX37rY49AN8HgB8ZuuQPp9vlf/dvMfeuvHo2fvfWuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//Q2Cf5yvyG/mBfCHoHt35n9L9ndG0ezuqMp1tj9rdab0wuWfFZrF7w3dRbW3HUbzqnkh81UHqkFEtPUJpuWUqwsytTAU8NR3HzP+fy6LJBepJ47tVF/Ctan7B5/n0PPYfyS+UXeff/AFH8aPjXW7F2HNiusMH2f8ut47gpMlPl9n47fGzqXM7IxXVl20zF66cJUzSozeMhQVKux3HEka+NLkeQ49anuJZpvpLftNASeFBxx0J3wH+Ufb3Zmf7q+NPyhh2evyg+N+Yxyb6yXXcVSmytx7P3g81bsPO0K1R1R1UlGifdxWC6zdVXlQ3PEFUSLwbpyyuXlZ7eX448EjgfT/V/qNkzsEVnP0VSxt/RRc+0/Rh1W58xu+9r5rpjsPZUGMy6ZKf7CjhqJIgKQSx5WPU7v/T08D8396r1sdHs6xBXrjYKn6jZu2gbf1/g8Pvw4dePHpc+99a697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6qf/AJwDafijuAj6/wAWxnJ+ltL+5h9hf+nnWn/NN/8AD1Evviae2l//ALXrVdikbxp+mwRP9j6fx7z0YCp6wiQ9g+wf4Osglt9bG3191p1uuKHoznwqkv8AKnqm9rNXTabcn9SX9x57t/8ATutxp6L/AJeh/wC1P/TxbAH+F/8ACOt1PFf8W2g/P+SU/wBP+WQ987hw6z36n+99e697917r3v3XutY3+ck5Xs7bpvYfcxA/1/Sfp7y4+7bm0ux8j/h6xZ+8N8dp/p1/wHqm0uOBf6/QX95O9Y3EV64F/TfUt7n6fn/D37q2mp6uY/ktEt3Rvsk/Tav0J5tzyAPeMv3mf+SHY0/391kX93Yn947hX+Betm/3iJ1lV1737r3XvfuvdYpv82/+sf8AevbU/wDYt9nW1+Ifb1pD/MByPkZ2YPqBuXJG55+j/T30c9r6Hkqy/wCaa/4OsBPcwkc53g/pnotnkHB+rH9P0/3n3IFOgEcEAdR6l7U8tiOI3uSfzp/Hu6fGPt6alH6TA+h62tP5RzE9CT35vUUpv/X0N7wD97v+V2kH+m/w9Z1e0IA5Php/R/wdWye4h6lLr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xum/K/8AFurP+WD+6v8AAfs691pO/K+Vf9mb7mH9Nzyc/j9T3t76O+2g/wCYf7Uf+Fdc+/cj/p4O6f6df8B6AFpT+LW/N+T7GwA4noGtqBoOHWGpkvTVC8C8E3q/BtGbWPuyCki/aOm5a+C/+lP+Dra//lTnV8WtmNfnwVdx/wBPk+nvAD3qA/1x7/7V/wAvWd3tD/077bv9J/m6s39xX1JnXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdNea/wCLXX/T/gFW/X/qDk97HVW4dfOr/wCE1h/7HxfOM/8AVo7p/wAef77SfT3UeXTh4Hr6M68qt/6D/eve+q9d+/de697917r3v3Xuve/de697917r3v3Xuve/de697917pC9of8y337/4Z+4//dTL791scR0Avwdv/ss3XNyD/k+V5H0P+5if3odePRtPe+tde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X//Rsu+Wfy/3n8k/5g+Y6K+S38vf507t+F3xiye3M9trA7E6hz+4uvPkPvTOU1FUY3P9r4HIRx4/JY7C1bGox1JT1cnjaGRpgGYgro0QIyBgG9Tw/LonuZZ2ljlaNjFU4Ud1fVh6fZ0ejubsHsP4c/NXA/MHAfHLv3uzqj5g9bbb2Duja/VvXNXmN/dL1XX+3KetwR3ht2N9NJHOFESRmRAG8kerUljsaHi+nqAVPGuOqsZYbv68IzI4AIAqwpwx9v8ALodv5cnXW799do/JH5/766+3f0zlvllPtLE4Xp/sPD1GG35tTavV0c+AxVbuijm0iKWtUCSOMRg6RrPDLdq4dQqwqa6fPp+wjlMkl3KNPi07TxAHCvVtZAIIPIIsR/gfaXoz6Jd82dq7apPjl2PkIMJjoq4QY2QVcdOi1CyfxWL1rL9b8n8+9YHWwc16Mx1h/wAy22Bfn/fmbZ/900Pvw4dePE9Lr3vrXXvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdYKmpp6Onmq6uaOnpqeNpp55WCRRRINTySOeAAOSfdkR5HEcYqxwAPM9VdlRS7mgGSegqk786WhkeKXs/ZiSRsVkRs5RhkYfVWGr6+z5eVOZWAZbGYg/0D0UHmLYlOlruIEf01/z9Y/9mA6S/wCfo7K/8/tH/wBHe7f1S5m/5QZv94PWv6x7D/ymRf72v+frknfvSsskcUfaGy3lldY40XOUZZ3Y2VVGrkn+nvR5T5lUFjYzADj2Hrw5i2EkAXkVT/TX/P0K1JV0tfTQ1lFPFU0tQgkgnhYPFLGfo6MOCD7IZI3icxyAqw4g8R0cI6SKHQ1B4EdSPdOrde9+691737r3VTf84U2+J24jf6ZfFjj68o/uYPYWv+ufaf8ANN+on98B/wAw0v8A8utVKN/REQTbxrYn6fpHvPg8T9vWDyAaF+wf4Osgf6/Wx/I5Nv8AE+69WIr0Z/4TN/zlX1PyDeunA5/OtPcd+7g/5hzuP2L/AJeh97Uf9PGsR/Rf/COt1/FW/hlBb/lUp/8ArUPfOwcOs+ep/vfXuve/de68fp7917rWF/nNuV7O20LAn7iL6D6jSbe8uvu2ZtLsfI9YsfeHHdaE/wAa/wCXqmQyXH5H+P8AQj63PvJ7rG/Az1x8n4t/r3+gH4Pv3WwadXRfyVST3VvwX+u0/qfpe3vGH7zFP3LYj/hw6yM+7wf92N//AM0162dfeIvWVXXvfuvde9+691jl/wA23+sf969tT/2LfZ1sfEPt60e/mJIR8juzluB/v5ckCB9Td/fR32uA/qVZn/ha/wCDrAb3N/5XS7/05/ydFo8t9PJuOAR/xPuQaU6AOeHWKpk/YnH1/ae5P9bfj3ZB3j7empv7Fh8j/g62vf5RZ/4wFMPwJ6W3/JLe8Afe7/ld5P8Abf4es6vaHHJsNfRf8HVs/uIupR697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917puywvjawc/5hvp9eOfdX+A/Z14cR1pHfK9x/sz/dAuf+Pmk4v9PU/wBB76Qe2v8A077av+aX+brn37j0HuFup89af4D0AXk+mkgj/EfW/sbU8z0DCfwjrBVSsKeoB0keGX/Yftn3aMDWv2j/AA9Ul/sXJ46T/g62zP5Ufq+LGy2+l4Kv/rcnIHvn7701/wBcjcAfVf8AL1nf7Q/9O+27/Sf5urOvcV9SX1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3TZmf8Ai1ZD/qBrf/cOT3sdVbh186r/AITV/wDb+H5xj/q090cn8f7/AFl+vuo4jpw5B6+jOv0H+sP9697PHqo679+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SF7QNut9+n622fuM/8ArJl9+PWxxHQC/B03+MvXJ+n+T5X/AN28/vQ68ejae99a697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuqpv5p/cW9urOq8fPsLcOQ2/mJ6mNjU4+oamlMZLXGtP8AW9zd7HbLte98xtb7pCsyAHDCor1DfvPve6bHy2brapWik8ipp1QgvzO+TNh/xlPdP6V/5e031t/re8sP9bzkuv8AuBF/vA6xc/1xedqV+uk/3rr3+zn/ACXHH+lLdnH1/wBy01+f6e/f63nJn/KBF/vA6t/rjc6Uqb6T/eujt/y9vlF3Zvr5ObM2vvXfefzeDyK1yTUNfXyTwSN4bRlo2HNib+4x94OT+Wdn5Gutx2+0jikj00ZVoRnqSvaXnLmbeOc4dv3K6eWJlc6Sag0HWzaOefeFwNRXrLvr3vfXuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv/S3+Pfuvde9+691737r3XvfuvdFL+cQJ+M3Y4Fr/bYz6/9rWH37rYpXPQ9dX/8y22B/wCGbtr/AN08PvQ4dePE9Lr3vrXXvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdBL3vWvj+neyKqNtMkW0cwUYfUM1MUBH+39m/LxA5gsVPBpkH8+ive6/ue6pikT/8AHT1pJ57L5WXP5mQ5WsvJkJiQJWAHrP0HvphaxQ/Sx0QfCPLrnFdyTm9lpI3xHz6bP4ll7grlKs8EWMrfT+v19v8AhwfwDpkvc/78P7ep+Hy2Yiz2Ak/idURHmKJjeVvVpf6fX2nu4ofo5+wfA3Si0lnN9bjWcSL1ur/GzJS5XpTYNXMS0jYSmVifqdKfU++a/NJA5iu0HlIeui3L1TslsT5oOhz9kPRz1737r3XvfuvdVLfziiR8TNyH6Wy2L/1ydL3t7mL2D/6efaf803/ydRN74Z9tL/7B/l61So5WEMYFjpjTj+t1H1958Edx6wgXCD7B/g67EtxwbX/H096p1bBz0aT4RSFvlh1IvFvvqix/r609x17uf9O43H7F/wAvQ99qTT3Gscfhf/COt2rFf8W2g/6hKf8A61D3zqHDrPnqf7317r3v3Xuve/de61gf5zrBezttm/1qIhz9f082I95c/dr/ANxbz7D/AIesWfvDCrWn+nX/AAHqlwyCw5NiT9PeUNOsb6GuOuvJwbHn/iPx71jr1D6dXTfyTn191b+vwBtO/wDjf6X94w/eZH+6ax/5rDrIv7u//JSv/wDSL1s+e8ROsq+ve/de697917rHN/m2/wBY/wDQp9tT/wBi32dbX4h9vWjf8xn/AOckuzgTz/ebJcj8Dyf199HPa4kck2ZH++1/wdYD+5oB51ux/TP+TotQdf8AY/j/AB/xv7H5k8ugGEOeo9TJemm9XPjfVb6/p9ux/GtPXpqYDwmr6HrbG/lDG/QE3+E9KOfr+hvr7wA97f8AldpP9t/h6zo9oscnw19B/g6tr9xH1KHXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdN2WNsbWW5/Zb/ex7pJ8B+zrYyR1pA/LVz/s0Xdf4tuh+f8AkJ+PfSP2zFfb3af+aX+brn17kGnuHuop+JP8B6L/AOSwJ9Vx+PqPY3p0DQSc9R6pz9rUjm3gmJv9P82fd4x3r9o/w9NTEeC9f4W/wHrbY/lOtr+KWx2/IgrAb/X/ADye+fXvX/08ncB816zu9n/+ne7cRw8Pqz/3FXUm9e9+691737r3TFuXcWN2pg8huDLy+HH4ynepqZP9THGupjz/AID2qs7SW+uUtIBV3NB9vSe6uYrO3a5mNFQVP2dEik/mQ/GmOWeFtzNrp55aeT/N8SQvocf7A+5NX2b5zZQwh+IVH59R6fdfk9WKmfKkg/aOuB/mSfGgf8xK5/1vHx/r+/f6zfOf++R17/XX5P8A9/8AUii/mN/G2vq6aig3I5nqpUhiB8fLyOEUf7cj3WT2e5xijMjRCgFT+XVo/dTlKRxGs+Tgfnjo9OOrqfKUFFkqVtdNX0lPWU7/AF1Q1MQmjPH+BHuLpomhlaF/iQkH7QaHqRY3WWNZF4MAR9hz1M9t9X697917r3v3XumvNf8AFrr/AK/8Aq36f9QcnvY6q3Dr51f/AAmr/wC38Pzj/wC1R3QLf1/3+0nuo4jpw8D19Gdf0r/rD/eve+q9d+/de697917r3v3Xuve/de6IT8zvmnuL4m4jN7ixnx27W7b21sfYO4u1ewtybTxTNg8LsraVJJX5umx9dZlnyhijZoqdtKj6s1rlVEcKMmtmp6Dzr0huLqWOURxRlvMnyp8v8/QSt/Nn6Lr/AIXfHH5mbW2D25ujFfKmo2pjupeocXtqkq+2KnJ7qzMuALZvA0FTPHDQ4+phf7/IQSzRLGY3j1iRfdY4WdqcAOJ6cmu44ow3EngBmuaeXlXz6G34v/NzbvyB7C7X6S3PsHePTndvTeThotw7K3zRfYQ7qxc1Mk43d15XS6TksYpkSOWZUGhiB6gdXvc0ITuQ1X16pbXfjMYpBpkHEf5q9Hj9sdLegk7X3Ztek2LvzGVufxVJXHaW4Y/tZ6yKOfW2LlVUEbG5JJAAA96qOtgZ6Cf4P2/2Wbrm30NNlCLfQg5aYgj34dePRs/e+tde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3VIP85L/AI8DDeqwvH9Pxy/vIH7vH/K0P9h/ydQR7+U/qka8P9nrXVX6LZ+dI0i/+HN7e80Dx6w9FKDrsMwtcrcfXn6/63vWOt06Pb/LZYH5ddc2Zr+Wq+p/wX3Envl/07W+/wBr/h6lX2S/6eHbU/gf/J1t7xm6KbW4/wB6NveBK/CPsH+DrOIf5+ufvfW+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuv//T3+Pfuvde9+691737r3XvfuvdFL+cVz8ZexwCAftsZzyLf7lYfoR78etjjnoeur/+ZbbA/wDDM2z/AO6aH3ocOvHj0uve+tde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690CXyMP/GE+zAbW/uhlv9f/ADQ9muwf8rFt/wDzXTos3qv7mu/+aL/4D1pKZZlOZynBv95LyRz+s8H300tQRax/6Udc47qn1soHHUf8PTfwOQTyeQOP9ce3q9MU6m4prZzC/lRlKW1/+Dfge2Lz/cGY/wBBunrP/ko24/4YvW6r8Vrf6COvrG/+4aK/+vYce+anNNP6y3v/ADUPXRfl7/kiWv8AzTHRhvZF0c9e9+691737r3VSX85AlfiRuNrn/i8Yr6fX9L/S3uYvYPHuhaf803/wjqJ/fD/p2d/9g61P0ktGp/GhP9j6R+r3nsXGog+vWEKKSg+wf4OuQdbD6An6/gj/AFvfia4B6tp9ejT/AAekH+zZdSAHj76e1vr+pL2t7jz3b/6dvuJ+S/5eh57VD/mI1iP6L/4V63dMT/xbKCxJ/wAkg5P1/wA2PfOkcOs+OnD3vr3Xvfuvde9+691q8fzpZAOztsAg3+5i/HH6be8t/u2NS2vK/wAJ6xb+8IO6z/06/wCXqlfWvBJ+hP8Ar395O6vIdY5gHrj5Tbj/ABv/AK3496qOt0xjh1dV/JHYt3fv8c2/uiDze31P494yfeYauy2J4frdZD/d5UDcr8/0F62h/eI/WVHXvfuvde9+691jl/Q3+sf969tTf2LfZ1sfEPt60ZPmU9vkp2gPx/eXJ/T6W8n199GPbBqclWY/oL/g6wK9ylU853h89Z6LKXPBuBYWHPsek56A+kU6wVMhFPN+P2nuf6ce3IzSRa+vTEy/pNT0PW2l/KDN/j/MeSDUUvJ/Pob3gN71/wDK7S/7brOT2kxyhD9i/wCDq273EnUn9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+69035bjG1n1P7LfT3ST+zP2Hrw4j7R1o5/LiQj5Sd2AEi26JOD/gz++kvtl/07zaa/76659e5P8A08TdQP4k/wAB6L4ZW0kgk3Iv/Uf7b2N6dA7h1HqpdVPUi5/zEx5+n+bPtyMUdftHTM/9g4/on/B1tyfylyW+KGxyf+OFYP8ArMnvnv71/wDTytw+1f8AL1nj7Q/9O820/wDC/wDIOrQ/cVdSX1737r3XvfuvdFq+XFY9H0Lvt0NteLqI2+vKmne44/2HsScnGnNFmP6Y/wAI6IOaB/yH7o/0D/gPWlZVUtOMjl28XL5fIva78lqljf6++lkMjm3iz+Bf8HXOOa3iF1NUf6I58/Nuo/21KSbp9PqNTWP9L2PtzxH416b+niGKcenfbtPBFuLCSqp1pk6Ijlv+VuO359p7x2NnKK/hb/AelVhCov4WpkOv/Hh1vKdRzmo6x2FKTcttPBEn6/SgRBz/ALD3zA3M13W7+U0n/Hj10q2//cCD/mmn/HR0IvtF0r697917r3v3XumzM/8AFryH/UFW/wDuHJ72Oqtw6+dX/wAJq/8At/F84/8AtU90WP8AT/f7S+6jiOnDwPX0Zl/SP9Yf71731Xrv37r3Xvfuvde9+691737r3VH/APNh+fHW2zaqg/lzbP7e6l2H8kPlVs7I4HM5PtTcw2xtrrDo/eFJV7a3tvmozQkRI8wtEamTC0kzASMjSt6VXUpgh1nUfLIHmaeg6L7278ECJOLEAk/CtfU+RI4Dz6Lu2wek/wCXzvv+VXkaHtKj3Z8QOientwfH5PkFn9yYLKbeyG8NzVQ/u9mNxbmxcj0CPUVEktRGI20opZVYhCfaiIM0UimoY+vHh/l6RXPhxXMEgp4a4JFKA1FOGBT0+fRlsDnNv/Iz+cNsPuPovdmF7F6v6L+M2++tO1d2bLylBn9q0G+905f+IYHB1OaxkjwPVLC9xGjObBvoFNm2Vo7TS+CTUDpyNo5928aHuCIykjIqSp/l1dl7RdHPRVO8ehdpbmwO+915Gqr1rl21naxUia0SyQY6SVLi/I4sRYe9UoOtj06i/Bz/ALJj62X8JSZOMf6yZWZB/vA9+HXj0bX3vrXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691R//OT46/xFxwWi5/H1f8+8gPu8V/rS4+R/ydQV79mnKZPWuon0XgE6R9foePeaBPWHoA8vPrlcciwP+9j37reK9Hr/AJbFh8uuuLXv5qo2P+sPp7iP3zP/ADDW+/2v+HqWPZJR/rhWx/ov/k62+4/0L+P8P6c+8Cl+EfYP8HWb3Drn7317r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r//U3+Pfuvde9+691737r3XvfuvdFL+cVz8ZuxgPzTYwf+tWH3o8Otjj0PXV/wDzLbYH/hmbZH+2w0Pvw4dePHpde99a697917r3v3Xuve/de697917r3v3Xuve/de697917oEfkcP8AjCHZx4/48/Lcn6j9scj2abD/AMrDYH/h6dFu8/8AJHux/wAKf/AetIzLzD+MZQX5FbLc/wBfWffTW0qbSM/0R/g65yXlBeS046j03eYar6vwT9f+I/p7UU6YrnqfiZV/jWEN7/7laUf4C7H2mvB/iU/+kb/B0oszW/t6Z/UXrdb+Klv9A/X1r2/g0X1+v0HvmnzPnmS8/wCah66J8vCmyWv/ADTHRiPZH0c9e9+691737r3VRv8AOWJX4jbjYG3+5fF83sB6X9zB7C49z7Q/8Lf/AAjqJ/e4E+21+B6DrU1jmJjQ/jxpf839I954s+T9v+XrClF7Fr6D/B1y8q/Wwufof6f6/vWvPWyB5dGo+DTn/Zt+oxwL10/+uLul/ce+7LA+3G4j5L/l6HXtYKe4lif6D/4V63g8WLY2gA/5VIP+tY988Bw6zz6n+99e697917r3v3XutW7+dS5XtDbNr/8AAmP/AFh6f6e8tfu30+lvP9L/AJesXvvB/HaV/jX/AAHqlLy/gnj6/W3P5+vvJmvWO1D1x8tuQRz9ef8AbW9+6ppx1dl/JCbV3fv8X5/uiCbn+twPeNH3lv8Aki2P/NbrIb7vONyv/wDSL1tHe8SOsp+ve/de697917rHL+hv8AT/ALx7an/sW+zrY+Ifb1otfMuXT8lO0QBf/fz5IE/kHyfQW99FPbE/8guz/wBIvWBvuZ/yud3/AKY9FjMgI+nP0H9D/j7HtfXoC/Z1HqHP28wsD+29/wDD0293iNJB9o6bnH6TfYettv8Ak+tf49yj6j7ilIP9fQ3vAv3q/wCV1k/P/COs4faUD+qMP2L/AIOrcvcSdSd1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Tflf+LdV82/Zbn/AGPuknwH7OvDiOtGb5eSD/ZqO7rsb/3qkF/xfU/6T76R+2ZA9vNq/wCaX+brn97kD/mIm60/iT/Aei8+YAk3t/vf0449jYvjHQOCkGh6wVUo+0qQfzBMQf6nxk+9xuRKv2j/AA9NzqTA5/ot/gPW3h/KTYt8TNjH/VQVhNz/AEnT3z896jX3J3D7V/y9Z2e0Qp7e7d/zTH+AdWje4r6krr3v3Xuve/de6K/8xf8Asn/ff/aul5/6cP7EfKH/ACs9n/px/hHRFzR/yr91/pD/AID1peVLgV+V5BIytfa//Lcj30ph/wBxov8ASL/gHXOqen1U2M+I/wDh6xF14sE/x55P+v7cANemyKDHTlgTbPYY8f8AFyorc/8ATWntPeGlnKR/C3+A9KLGpvof9Ov/AB4dbxHTP/Mq9gHjnaWE/wDcMe+YW5f8la8/5ryf8e66R2H+4MH/ADSj/wCOjoTvaTpX1737r3XvfuvdNeZ/4tdf/wBQVbb/AF/s5Pex1VuHXzq/+E1n/b+H5yf9qjun/wB7aT3UcR04eB6+jMv6V/4KP96976r1y9+691737r3Xvfuvde9+690TLuv+Xd8Hfkd2fQ90d6/Fzp7tPtTG4CTa9HvneG06TKZ6PASI0f8ADZKl7CRFV3WMyqzIrMEZQSDdZJFIKmhHDpp4IZFKuoIbiCKg/aOl/mviD8X9xdK0fxyznQ/WWU6Kx1Tj62g6rq9rY6XZtHW4qrFdjq2mxBXQk0Mo1pKtnBvzYn34yOW11z14QQiMxaBpPEUx+zpUdLfHfo3454XM7c6K6q2T1ThNxZmTcOdxuysHSYWny+bkgWmbJ5EUwBll8aKgZybAWFvfnkeQ1c169FBDAuiFQo9AKdDN7p070he0P+Zb79/8M/cf/upl9+PWxx6AX4O2/wBll65t9Pt8r/7t5vfuvHo2nv3Wuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917qjz+cw2nrvCgkC7RWN/ry9/eQP3eP+Vqf7D/k6gn37H/ISPWuUstlHP4Wxv8A4fj3mkRnrD4UoOu/OF/2H1/x/p/tvfqdbx0fH+WrMG+XvW4PP79Vb/Yhfp7iD31FPba9/wBr1LHskK+4NvTyR/8AJ1uBp+gf778+8CV+EfYP8HWb3XP3br3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3X/9Xf49+691737r3Xvfuvde9+690Ur5x3/wBll7IsSP8AJcZyP+1rD791sdD31f8A8y22B/4Zm2f/AHTQ+9Dh148el1731roMs53P1NtrIy4jcHYm0MNlIReWgyOcoaWrjH9XglcMP9t7Orblzfr2EXFpaSyIeDKhI/aB0Vz73tFtJ4Nxcxo48mcA/sJ6af8AZhejf+fsbD5/7OTG/wDR/tR/VHmf/lAn/wCcbf5umv6xbD/ymRf72v8An68fkL0aPr2xsMf+TJjf+j/fv6o8z/8AKBP/AM42/wA3Xv6xbF/ymRf72v8An6ELbW7ds7yx/wDFtqZ7F7hxglaD77EVkNdS+ZP1x+aAlbj8i/spvdvvdul8C/iaF6V0uCpp60PRha3lrex+NaSLKnCqkEV+0dKH2j6U9e9+691737r3QI/I7/mR/Z5/ps7Lf9ax7NNh/wCVgsP+a6dFm8/8ke6/5pP/AID1o7ZmcDNZYW+lbKCf+Qz76cWIJsoj/RHXOS9IF9N/pj03Cosfpf8AHP1/1varT0lBA6cMRN/uawek2vlqPj8WLHj2lvR/iM9f99t/g6VWQJv7bSf9EXrdv+KXPQ3Xp/6ssX+9D3zO5m/5WK8P/DD10X2D/ki23+kHRifZJ0b9e9+691737r3VRP8AObNviHuP/tc4r6f8Ff6+5f8AYb/p5tr/AM03/wAI6ir3sp/rb31fQdalUcgEcR9RCxpcfjlR7ztY5NPXrClB2j7B/g678qkH9QP4t9Prc+9Z63QV6Nb8FnB+XXT45J+/qOf9V+4nB9x97sn/AJhzuP2L/l6HftYB/riWP+kf/CvW8dir/wANoLgA/aQcD/lmPfPQcOs7+p/vfXuve/de6979148OtWj+djJp7Q2vYC/3MP8A0La/vLL7uJ/xW8/0p6xg+8EKm0/06/5eqSfLb6j/AB/1z+feTFR1jqMEk9eMg/1PNzx+Le/V9D1sU9Ort/5HpJ7x7AuQf9+gOR9Rz9PeNP3lTXZLH/mt1kF93sEbnuH+kTraW94l9ZTde9+691737r3WOX9Df6x/3r23N/ZN9nWxxH29aKPzQkC/JbtEE/Xc2T4/x8nvof7Ykjkuz/0i9YIe5a15yu/9Mf8AJ0V4yggWJuBb+ov7HoPQGIHAdR6uVvtZrf8AHN7/AI/s/wBPbkZHiD7empwwgb7D1tzfyejf48vYcCel5/5Bb6+8DPek151k/PrOD2nFOUYqfL/B1br7ibqTOve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6b8rzjqz/li3+9+6Sf2Z+zrYNCD1or/L6Qf7NV3gtxf+9cllvbSNT++jvtt/073af+aX+brAL3GAPuHup/pJ/gPReDKAT9Df6G9ja3saV6B4A6j1cxFNUjiwp5rc8/5s+3IsSqfmP8PTNwP0JP9K3+A9bfv8o83+JWxD/WCt/63J7wA96c+5O4fav+XrOn2j/6d9tw/wCFj/AOrSvcWdST1737r3XvfuvdFe+Y/wD2T9vv/tWynn6f5h/Yi5R/5Wa0/wBOP8I6IuZ/+SBdf6Q/4D1pVVVRfIZUkcjK5D/Y/wCUEAn30uhX/Fov9Iv+DrnRMf8AGZq/78f/AA9YWmH9T/jz9OOPblD1T8unPAzH+PYYX4/iNF/sf8rT6e0l/ixlP9Fv8B6V7fUbhCB/Gv8Ax4dbzPS//Mquvj/XaOD4/P8AwDHvmDuP/JVu/wDmvJ/x7rpDY/7hQf8ANNP+OjoUPaTpX1737r3XvfuvdNea/wCLXX/9QVb/AO4cnvY6q/Dr51f/AAmr5/nw/OMfW+J7o/x/5jWT3UcR04fPr6M6/pH+sP8Aeve+q9d+/de697917r3v3Xuve/de697917r3v3Xuve/de697917pC9of8y337/4Z+4//AHUy+/dbHEdAL8HRb4y9cjj/AID5X6G4/wCLvP70OvHo2nvfWuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917qjT+c8bdeYY8fqi5/p+v3kB93j/lbGHyP+TqC/frPKLU61vRI2kf6wAsTxx+Pea5GesPBlc9cvKDY6mv+R/xr3odeZc9H3/lnSBvmB1ta/8An6r/AF/0r+PcOe+//TubwH+j1Lfsiv8AyP7en8L/AOTrcRj/AED/AH3594Fr8I+wf4Os3OufvfXuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917pvymVxuFop8ll66lx1BTKXqKysmSCnhQC5aSRyAB7dggmuZBDbqXc8ABUnpuWaKBDLMwVRxJNAOgrHyH6LN7ds7D4JB/38eO4INiD6/Z7/VHmfj9BP8A842/zdFX9Yth/wCUyH/e1/z9e/2Ybozn/jLGw+Pr/v48d/0f79/VHmf/AJQJ/wDnG3+br39Yti/5TIv97X/P1npu/elKyohpKTtLY9RU1DrFBBFuHHvLLI7aUSNA9ySeAB7q/KvMsaGR7GYKMkmNsfy6svMGxuwRbuIk4A1rn+fQto6yIrowZHVXRlN1ZWF1YH+hHsgIIND5dG4NRUdcveuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//9bbP7V/mlfHvqH55dKfy9N0ba7dPcvecFRPtXctNsKtHWMJTFPlaaGbdtQ6LMziNopGponjheyyOCQPbvgto1+R/wAnr6dJTdIJvB8xQZxx4U9fnTh0Kfyb+b2wfjj2F1J0uNq727I7n7xbcX9wNk7IwdTlIoqXa+P/AInk8nu7LQBkxlM8YdKV5UYzSKyKBpLC0MJlNThfXrVzdCDsQanPAf5+ld8R/l91V8y+tZ+xeso9x4Z8Rncrtfdeyt8YeXbm9to7gw9Y9FVUebwVSTJHHK0bPSVFtE0fqWxDKtZYmiOeB4Hq9tcpcpqXBHEHiOjTswVSzGwUFif6AC5PtrpR0Qf5o9r7CynQXYu2qTcFFNmpUxtNFQRzRtO8y5SMuAgJ/TpNx711ZePRwOsBbrfYIP1GzdtA/wCv/Bofexw60eJ6WdQ+inncfVYZWH+uqE+/VpnrR4daafz6aDM/KHeNdOsrSeJ4Qy1FSoAWcDgI4H499CvZ+Z05Et0FPi9B6fZ1gN7xWkM3Pksjg18MjDMPxDyBA6J5/DaIj9M44t/wLqx/sf1+5M8eT5fsH+bqNPoYBmjf70/+fri2MoypIE9uLj7qrv8AX+mv34TyV8v2D/N1v6OCnA/703+fraK/k6TovxoyVCmsJT7xr2UPJJIQZU59UhJ/s/194O/eGkY8/Kp84FPCnA/L7es1/YOMR8goFGBM/mT5+pz1bb7gvqbOve/de697917oEPkgSOje0SPxs3Ln/rEPZnsX/KwWH/NdOi3eP+SRdf8ANJ/8B60X81UH+N5fn/lPlv8Ak/rP099O9vp9BCf6I65yX1fr5h5Bj02/cm/0v+P9f/E+1lAekoFOnDE1DDN4EenjL0fP5Prt7S3wH0Fx/wA02/wdKrLG42wH+/F/y9bwvxPJPQXXZP5wkJ/3ge+ZXMv/ACsN3/pz10W2D/kjW3+kHRjPZJ0b9e9+691737r3VQv86FtPw93MRwf41ieR9R6Htb3L3sOae5tr/wA03/wjqKvewE+21+Pl1qPxzHxxjV9I0uCfr6ePedZOT1hWqnQB8h/g65GYf1tqFv6Wt71qPVqDj0a/4IyD/ZvOnrG9q+o+v+Lp9Pcfe7DV9utxHyX/AC9Dr2sH/MRLE/0H/wAK9bzeL/4t1D/1CQf9ax757jh1nb1O97691737r3Xvfuvdar387clO0drkW/4ExH/k33lh93PFreD5dYxfeAFTaf6deqQzPcDkjnn6f195L+fWO5APXhISTyf8AP8AePdc8evEDgOrvv5GpJ7x7C/p/dC/+JN/eNv3kc7LZV/371kF930Abjf/AOkXraf94mdZR9e9+691737r3WOX9Df6x/2PHtub+yb7OvD4h9vWiH805APkz2lf8boynP4H7vvod7ZH/kGWn+kX/B1gl7mf8rndU/iP+Tor5lAt+Cbc8c/j2O69AcDrBUv/AJPUeoWEb35H10+7RnvX7R01OP0W+w9bd/8AJ2a/x3kH1/yil/22hvr7wQ95z/yNJfz/AMnWbvtOP+QlF+X+Dq3j3E/Um9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+69035XjHVf/ACxb6+6SfAfsPXhxHWiT8wpB/s1veSgXI3ZISf8AkOQ++jXtuae320k/76/zdYDe41P9cLdafxJ/gPRdPJ/aF+foPr/h9PYzJHl0DwD6U6wVUlqapFibU8/JPBvGfd4m/UX7R0zcD9ByP4W/wHrcK/lEHV8RNhtYf5mtB/r/AJ5Pp7wE96P+nj7h9q/5es5vaT/p3+3V/wB9j/AOrTvcW9ST1737r3XvfuvdFa+Zf/ZPW/f6/wAOlt/sIJPYh5RNOZrT/Tj/AAjoi5mFdhuR/QP+DrSXqqlf4jlxfj+LZAHn+lSffTKBf8Wh/wBIv/HR1zplp9VOKf6K/wDx49YPuOfrx/j9fbunqhww6dcFUf7+DCfXjJUNh/X/ACyPn2j3EadvmP8ARb/AelO3924Q18nX/jw63sOl/wDmVPXp/rtDA/8AuEp98vtx/wCSrd/81pP+PddIrD/cCD/mmn/HR0J/tJ0r697917r3v3XumvMm2LyH/UDW/wDuHJ72Oqtw6+dX/wAJq+P58Xzit/zqe6Lf1v8A32k91Hl04fM9fRnW+lb/AFsL/wCvb3vqvXfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6QvaH/ADLbfv8A4Z+4/wD3Uy+/dbHHoBfg7x8ZuuQP+VfK/X/tbz+9Drx6Np731rpAbp7U642RVQ0O7967c25V1AJgpstk6ejmlC/UokpBNvZrZbHvG5RmXb7aSZRxKKSB+zouut22yxcR3k6RMeAZgD/PpM/7MR0Yf+ar7G/8/wDQ/wDR3tb/AFR5n/5QJ/8AnGekv9ZNg/5TIv8Aex11/sxPRf8Az9fY3/n/AKL/AKO9+/qlzP8A8oE//OM9b/rHsP8AymRf72OlntPsXYu/Puv7m7rwe5vstP3f8Hr4K37bX+jzeEnTf8X9l1/tG6bXp/eNu8GvhrUrX7K9LbTcrC/1fRTLLp46SDT7adLP2XdLeve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuqLP507aeusNdioLQ8i3+qfn3kD93f/lbWHyP+TqC/fr/lUWPWtosjBR67iy/71+be82cV6w8zTroTG172t+q3196oB1vJp/Po/v8ALGl1/MTrVb/qnqrg/wCCr7hn35H/ADDq7/L/AA9S37IGvP0HyVv8nW4/H+hf9j/vfvAtfhH2D/B1m31z97691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdE8+dOQNB8dd7Dm1TQzQGzFTZoW+hUg+xt7dsRzfaU/iB/mOgjz1Q8r3QP8ACf8AB1pjw4yiCyftz81VYeaqr5vUub8v76RNcSmhqOC+Q9B8uuc0NhbqCAD8T/jfzY/PrN/DqK17T8fj7qq/3rX7r48vy/YP83Tn0EHw5/3tv+gulr1lSU1J2PsmpUT6otzYRgfuaki65FDYgvb/AG/st3qWRtmulNP7J/Ifwn5dGuwWsCb7ZyANUTR07m/iHz63otrT/c7awE/5lwuKkP8ArvQxt/xPvl9K5eeQn+Nx+xj10riFIkH9Ff8AAOn73Tq/Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdR5KumhbRLUQxta+l5FU2/rYn3YI7CoBPVGdFNGNOsf8Qof+Vym/6nJ/xX3bwpf4T1rxY/4h17+IUP8Ayt0//U5P+K+/eFJ/CeveLH/EOs8U8M4LQypKAbExsGAP+uPdWRl+IU6srK2VNesvuvVuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6//X2LP5jHcPTuy/5iP8rjC7y7V6y2dn6TsPsWtqsbund23sLloMfkcJR0+MkmhyEqSxRVUwMVO0mlJHuASQfa62/sJB6jHz+zon3A/45bEcFbPyFPP06UuB35tLpz+bd3Nnu5904HaW2+8ekev6PoTc+78vjsVt3OVO28VBPumg2xmsrKtO0miOTyLBIC1iDfUL7ILWgVeIOR1RWVN3Z5DhlAU+VaZp1L+BFNLvf5/fzE+/9kZCDcPQ3YMnUO3Ni7twkqVe0dwbn2jh56PeU+CyVKxpqkrUczSw35YFmOoXrc1EEaNxHVtv0ve3E0fwsRQ+RpWtOrm2UMpVhdWBUj+oIsR7RdHPRAfmd031jiOhuxd1UG0cbS7gRMfURZVDP9xFUSZWLVKupyObm/Hv3DrfE1PRxesCT1vsEk3J2btok/1P8Hh59+HDrx49LCt/4B1X/UPP/wBaj70etdaZPzgmC/JTeKXAXVN9f+W499BvaEE8i2/2/wCTrBH3coOeZK/wf5eioedP6rxwOfwP6D3JlOo4FOuJnUDgi/Fuf8ffqZ69ilOtnX+Tg4f47Zthz/v8Km//AFLf3g794X/p4Mf/ADzj/COs0PYjHIQH/Dn/AMPVvnuEOpl697917r3v3XugN+Sn/Miu07G3+/LzB/20Q9mWxj/d/Y/8106Lt3/5JN1/zSf/AAHrRKzU4GdzCm4Irpbm3JPkPvpnYH/EYf8ASjrnZeqPrpjxOo9N5qP9cf63Nvx7Vh6Z6TafLqbhp757ADm38Yo+SbH9X49sXkh+guB5eG3T1mo+vtj/AMNX/L1vO/E0k9Adc3/50kP+9D3zQ5lP/Ihu/wDTnrohsP8AyRrb/SDoxvsl6N+ve/de697917qoH+dOdPw53MxJH+5vE2sBxaOS/uXfYnHuZa/803/wjqKvesV9t7/7OtRGOT0Jx9UTi319I+vvOdjk9YYoBoX7B/g6yaraRxc/Q3Pp9+r/ALPXiCB8+jafBBv+cvunFuCRkJyTxz+4n1HuP/dYg+3e4/Yv+Xoc+1wK+4diP6D/AOFet6XF/wDFuof+oSD/AK1j3z6HDrOrqf7317r3v3Xuve/de61Vv54Dae0Npgc/5VEf8f0m/vK/7ueLa8/0vWMfv/Qm0/5qL/l6o+8q3A/P1v8Aix/HvJOvl1jz8XXvI31DWN7X/NvwPftXn16h4eXV4X8i5y/eXYRueNnkc/Tg/X3jd95Gp2ayH/Desgfu/UG43+PwJ1tUe8Tusoeve/de697917rHL+g/4gg/61jf23N/ZN9nW1+Ifb1ocfNNv+cm+1P8N05MW/r+5a9vfQr20IHJlmP6C9YLe5P/ACud2f6R/wAnRXtd/wChsPT9Db2Oq9Abzp1Gq2YU0trf5p7/AOPp93jaki/b03OoMLfYf8HW3z/JyN/jtIbc/cUv+xGhveCfvMa85yfn1m17U45Ti/L/AAdW+e4o6kvr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuoGU/wCLfVfn9o/72PdJP7NvsPXhx60O/mM//OWXe4BsRuyW5/5DkFvfRb23J/1vtq/5pf5usCPcXHuFuh+cf+Bui5+Q21avpx/j/tvYxqSadBIY6w1kg+2qFJvanmv/AEJ8R/Pu8RPiLX1HTVyC1vIB/C3+A9biX8oM6viFsJr3/Zrv+tye8Cfeb/p4+4H5r/l6zi9pv+VA27/mmP8AAOrU/cXdSP1737r3XvfuvdFY+Z1v9l438b2Ixs1j/wBU8n09iDlPHMtof6Y/wjok5k/5IVz/AKQ/4OtH2qnX+IZa/wBDmMl+fr/lJ99MonpbQn/haf4Oudcy/wCNTg5/Vk/48esf3H6efof+RD25qHr1TSR067enZtw4X1f8vOh/p9PvI/aLcmB26b/Sn/Aele3r/uxgr/Gv/Hh1vj9Km/U/Xh/7M7A/+4K++YO4/wDJVu/+a0n/AB7ro7Y4sYB/wtP+OjoUfaTpV1737r3XvfuvdNeZ/wCLXkP8aKtH/qnJ72Oqtw6+dZ/wmr4/nw/OP+oxPdB/229pPx7r6dOHgevoyrfSt/rpF/8AXt731Xrl7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XukL2gbdb79P8ATZ+4/wD3Uy+/Hh1scegE+DhJ+MvXJPJNPlf/AHbz+/dePRtffutdatn83upTKd5behnM5+xFUkfjnniC+lx9IWAP+x95pfd1kdeW7mlM04gH09R1h794aCOXfbLXXBbgxH4T6EdVQfY0ptc1X+P+W1n+w/3Z7yD8Z/l/vI/zdY//AEcfHu/3tv8AP10aCCxAeq4Bv/ltZc3H/LT3vxn+X7B/m699LEMgt/vb/wCfq+P+STMtNk+4qNWmKzUuMktLPLLzG8f08rG3B/HvFP7zUjadq4ZZxgAevp1lJ92+JUh3PTX4kOST5D1J62EPeKvWT3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3VEv87BivWuEYW5aK1/r9X95Afd3NObj9h/ydQb781PKDU61p1nIVbi9lW/8Ajce81TJVsdYgBBQA+fXITj+gt/vvr7r4jdb8OmPTo/8A/K/mDfMjrRCLjz1XII49I+nuHffdtXt1d1+X+HqWPZNdPPsBHmj/AOTrcuj/AED/AGP+9+8DV+EfYP8AB1mx1z97691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+69176c+/de6rr3P8AzQ/irtLc24dpZfcOcTMbZytRhspFFh5JI462mOmVUkDcj+h9y/Y+yHPW4WMO4W8SGOdA6EvxB4eXUVXvvLyJYX822XNyVmt20ONJw3pXpl/4dh+I3P8Av49wcf8AVjl5/wCTvan/AFhuf/8AfMf+9j/N0n/17/b/AP5Sj/vB64S/zZfiHDE80m5NwBI0Z2P8DlNgvJ/te9r7Ce4DMFEMdT/T/wBjqr++Xt7Ghke6YACpOg9H5637C252psrAb+2nPNUbe3JRLX4yaeIwzSU7mwLxG+k3H09xTvG03ex7lNtV8AJYG0sAaiv29Sfte52m8WEW5WJ1RTKGU8Kg9Lj2WdGHXvfuvde9+690Sf5+kD46bsJFyIpP68Dwm5v7Gvt3/wArfaf6Yf4eglzx/wAqzdf6U/4OtOiGQaJCTc/c1Vjf8GoYWPvo4w4fYP8AB1z1StCf6Tf8ePWYsLjgfn8/7D3qvW9BPSr6+c/392aPr/v48NYH/tYJ9fZbvP8AyRroj/fT/wDHT0abEGG+2gJ/0aP/AI8Ot5bZRvtPbfAH+4HD/T8/7jozz75gt/ay/wDNST/jx66QIKRp/pV/wDpUe/dW697917r3v3XuiXfMb5c474oYHbuar8QmXGeqKmBYWeRSn26g3Hj/ANf3I/t1yDJz5dzWscvh+CATw8+gDz7zzb8j2UV5cJrEhIp9nVea/wA6Tb7AH+40AuAf89V/nn3Lp+7jcD/iUf2DqKx94bbTwt/5nrs/zo9vj/mB6f8A1/NV8/7z71/wONx/ylH9g63/AMEJtw/4jj9p6Mn8Wv5keF+SPYkmwqXbcOKmSOKQVCyTksZLHTaQ/wCPsGc8+zsvJu0DdHnL1JxQdC7kr3atOcd0bbYItJWmc+Yr1aT7g/qY+ve/de697917romwJ/oCf9t7917rWR/mfd9dwbT+UpwmxOxc5trAxbTpWbGUEjJTmqGgNNYMOTzf3mV7F8t8u7ryK13utmk0vjEam40zjrEP3u5m5l2jnaG02m8aCFoCSgFQWqtDWv29V8H5P/JD6DuLdBP/AC3f/o/3MI5J5OGf3dF+z/Y6iU87871xuT/7z/s9ePyf+SOoD/TDuf8A6ntwbf8AB/ev6kcm8f3dF+z/AGOrDnjnbz3F/wDef9nq+r+UB2pvzsLq7fTdh7qyO68pBuyWOiq8g+uSGlUuBClybL9PeKf3g9p2rZ+ZLSDaYFgRoQSF8zjPWT/sHu+77zyvPc7xObiUTuNRFMajQAZ4Dq4z3APU69e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691//9Db0+QH8qv+Xp8p+3KDvj5BfFfrjtHt3GLi0o98bgO4kyqphNP8KSaLGV1PTzCDQvjE0LWsPdxI4pQ8OHTRghYsSvx0r86cK9Cv3T8G/iR8isB1ptfuzobYnYmB6cadur8dnKKrEeyhU41MRUJhJaGaGREemijidGdlIVSRqAPuyzSq2tTk9UktLaWMQyICo4DoW+mOkup/jx13gupektiYDrbrjbIrP4FtHbVM9Nise1fVvX1skays8jPLNI8jvI7MSeT9PdHdnbU5qenIoo4UEcShVHADoUvdenOil/OIA/GXsfV9BTYw/wDrVh9+62Oh66v/AOZbbA/8MzbNv9b+DQ296HDrx49LCt/4B1X/AFDT/wDWo+/GtMda60q/nZOF+Te80JvYz8f9PwRx76E+zYryHB/pv8nWCXu//wAr1IfLw/8AL0UVqn6ceq1gB/vfuUNPUbVHn12aiymxuSAb8+n34DPWiB5dbR/8mF/J8bs0w/57Gquf+nb/AF94KfeE/wCngp/zQH+EdZpexII5CQH/AH6/+Hq4j3CXUy9e9+691737r3QGfJXjortU/wDZkZnn/p0PZnsf/Jesf+a6dFu8f8kq5/5pP/gPWhtm6kfx3MWPIrpif9YSH30tsSfoYf8ASjrnjej/AB6bH4j02mc/qB4Nvra/PtVq6S0ByOp2Gn/38GBF72zNH/hxr9sXh/xGf/mm3T9iK7hbH/hq/wCXrew+JR1fH/rg/wBcHD/vQ981eY/+S/dn+meuhuw/8ka2/wBIOjIeybo2697917r3v3Xuqff51bW+G+6Dblc3iLf7GOT6+5b9izT3Mtf+abf4R1FnvUK+3F/9n+frUJin9Ed1/sJcf8gfX3nKx7j1himEFPID/B1z8qm5uQD+R9R71qoet6QR9vRs/gWyf7OD05YnjIT8/wBf3EPP+t7APuo3/MPdx+wf5eh17XIB7g2VPJH/AMK9b2mL/wCLdQ/9QsH/AFrHvn6OHWc3U73vr3Xvfuvde9+691qo/wA8VyvaG0/pc1UX+20m3vKz7uxpb3nrpPWMvv8ACv0n/NReqNfIeb8/77kgn3knWnWPdPPr3ksLcf74+61x1umeryv5Fblu8uxB+Bs48fg8+8cPvGkfuey/5q9T/wCwIP7wv/8ASr1tXD6e8Uesnuve/de697917rHL+hv9Y/717bm/sm+zrY+Ifb1oWfNeTR8ne1V/runKjn6/53k++g/tof8AkGWn+kX/AAdYL+5Irzndf6Y9FaElrcf8b4/p7HdegQAOo9VIWp5/zaJ+D+Dp92jPev2jpqZT4LfYetwT+Ta1/jpL/wBRNJ/0Iw94Le8przlJ+fWbPtVjlOL8v8HVwHuKOpK697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917qBlP+LfV/8ALI/737o/wH7D17zH5daFvzJm0/LPvcckjd0gN+P7cl/p76Je3J/5AG1U/wB9f5usC/cT/p4O6f6aP/Aei3mUhuP9v9PYxrjoJUFesFXKxpKnn/lHn/61H3eM0kWnqP8AD01cKTBJ/pW/wHrci/k/MW+HuwG/Bhrr/wCuJkt7wM95f+ni3/2r/l6zg9p/+VB2/wD5pj/AOrVvcX9SN1737r3XvfuvdFW+aPHx27AP9MbLx/X/ACeT2IOUzTmS0J/jH+EdEnMdf3Hc0/gP+DrRsrJwMjmORxl8jb8/8pJv76UxNW2hp/vtP8HXPGVCLmf/AJqv/wAe6wedTbkD+ov9fd9XWgtcU6dNvSk7jwnIA/ilDYCx/wCUyP629pNxb/dfN/pW/wCOnpVYqw3CD/Tr/wAeHW+90p/zKbro/k7N2/8A7b7BSPfMncf+Spd/81pP+PddFrH/AHCh/wCaaf8AHR0KPtJ0q697917r3v3XumvNC+Lr/wDqCrf/AHDk97HVW4dfOs/4TVm38+H5yf1/hPdFv9hvaT3UeXTh4Hr6My/pX88D/eve+q9d+/de697917r3v3Xuve/de6qA/mu/NT5m/Cyk+PW6vjd8feue3utt89wbN2B3HuneW8KnD5XZVBubNpRQjC4KneHymaASlasvKEcBTDyCVMESyg0yR5cMev8AsdIby6a2ozYQ4qMnUTQCnofM+X8wPHzb+V/aHTc3UXUvxr2FtvtP5Kd3bnxlLt7Zm6szNg8Rt/rmCqWHfPYldWxgLIcXE4eCmLgSsG1eldLaih1VZvhXj165uvD0xR/2knD+Wfyr0lPjb8sO7IPk3vz4bfLPau1Nu9j02Jqd9dDb02fkXrqHuTqvGKkOY3BlceAFx9dS1DGNoAqhlRrqrj1WmiQp40XCvTdtdSCf6K5+OlQa8QKVP8+rMvaXoy6Jr3v8iKDa2N37s6ba+frZjtzOUSVtJjamajDT4+SISvOqldIuSf8Ae/eq9boQc9OvwcOr4x9avYDyUeRksDe3kykz2P8Ajz78Ottjo2vvZ4dV61Wv5s0vj75x3F7vU/k8fr95nfd3zy7cfl/k6xE+8B/yW7T7T/gPVW/lGq1hYD/exz7yDpjHUC1XVnh1wM4vcW/4n34L14tXI49Xo/yUJPLuLtz/AAoaE8/8Git7xU+82CF2n/TN/l6yd+7l/Y7n9qf4B1sP+8WesmOve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuqHv53DAdY4UngBouR9eS/uf/u8mnNp+w/5OoO99gDyi3Ws0kvoU3PCi3+PHvM6tCesRAO0fLrsTC178n9X/ABv3qp63xGOrBf5XUgb5mdZAfTz1dh9dXpUn3EHvpn27u6/L/D1K3sr/AMr7BT+B/wDJ1uex/oH+x/3v3givwj7B/g6zV65+99e697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6a8zUtR4yrqUNnihdl+n10n+vtuX4D1ZaFs9aOXdLpN3d2/O5YPLvrKyNcnli9/fTflMn+qO2f886dc4ObFU857vQf8SD/AMdHQb6VbkMfTa/J4t9bez6p6IwvlTqHkYw1BV3Y6TTyW9R5uvF/bkBImX7emLtQbZx/RPW4n/Lzrpan4vdZ08jErSbfpo4x9QFLtyD75ze53/K+bl/zVPXQz28FOTdv/wCaS/4OjxewH0NOve/de697917okn8wBivxx3cw+ggkv/1JNvY09va/1utP9MP8PQT53zy1cj+if8HWmfDUNaQlgP8AKaoAcc/5Q31HvpEVFB9i/wCAdc80NNWPxN/x49Z/OfpruB/jyPzYe9U6tUeXSt68qGbsDZovYf3kwvH/AJEE+vsq36i7JdH/AIU//HT0abBU79aH/hsf/Hh1vXbJv/dLbdxb/cDhv/dbF75fn+1l/wCakn/Hj10gT4F/0q/4B0qffurde9+691737r3VFH86Odzt/rGG/oNRkDpt9TqIJ/3j3kj921v92+4g+idY9/eDUHY7P/TnrXuvYAaRwq3t/QLxY+8vz59YlA0A66Gkn6W/p/h795dXFCeHVh/8seV4PkhQmNtId6VWP+qHp4PuFPfg/wDIOH59TL7FqRzZKT/R/wAHW217wZX4R1mj173vr3XvfuvdcH/Q9vrob/evfuvdak380yWNflhVBh6v7sQ8/wBOU95xfd7qfb01/wB/t/l6wt9/B/yPYf8Amgf8K9V2+ZPqL/8AFT7m+nUNhacevCcX5FwbX4+h/Pv1DTrYArnrYO/kqSatib5sPT/eecC/4/znvDv7yeOabP8A5oj/ACdZZfd1A/qlcEf7/f8A48er4PeOXWQfXvfuvde9+691DyNbHjqCsr5f81R001TJ/wAEhQu3+9e3IYzLKsQ4sQP29UkcRxtIfIV6qY3h/N06U2fuvP7TrcdknrMBWvRVLpC5QyKSLqf6ce55232C5i3Hb4twiddMy6hnqEdx99eWNt3CXbZ6iSE0OD0nh/OV6MN/9xeU/wCpL8e1n/A7czfxr+3pJ/wQHKnz/Yevf8PKdGakBxmT9bBB+y/1b6e/f8DvzMR8a/t68fvAcqCnHPyPVpXUPZWJ7e69252Fg1dMZuOj+7pkkBDqmorZgfzx7g/mDZp+X93m2e5NZIDQ06mfZN1h3va4d0t/gmXUPs6Er2TdGvXvfuvde9+691//0d/j37r3Xvfuvde9+691737r3RS/nDb/AGWXse5sPtcZf88fxWH3o9bHQ9dX/wDMttgf+GZtn/bfwaG3vw4dePHpY1v/AADqv+oaf/rUfezw611pJfPaoCfKHeik2IMx4/5bj30E9nHI5Cg/03+TrBj3cSvPLk8fDP8Ax4dE9+7v+k2P+H59ybr8z1HBjFMdcDVEg8+njV/t/p72JCDjqrJjHW1N/JSbX8ac6xP/ADGlVb/W8bce8GPvAmvuAh/4QP8ACOszfYwU5DSn+/X6uV9wp1MfXvfuvde9+690BfyXNuh+1z/TZGZ/1v8AND2Z7J/yXbIj/fyf4ei7d/8AklXP/NN/8B60Ic5Mv8ezB/tCvmv/AK+s++k1i3+IQ/6Udc9rxf8AH5/9Mf8AL02ef63AI/V/jf2q1U4dJtJPHqfhpv8Ac9t/+zfM0f0N+NfIt7Yu3/xGev8AvtunrNCNwtqf79XrfD+I3Hx763+nODgI/wBbSPfNzmLO/XR/pnroRsf/ACR7cf0B0ZP2TdGvXvfuvde9+691Tz/OxOn4ZboNyCc5iB9L/wC63HuWvY0D/XKta/77b/COos96an25vwPT/P1p/wAUoEaeo3CJc2t9VHvOFq1PWGiE6B9g/wAHXZe4J1ct+P8AY/Ue/V8ut1FKno2/wGP/ADmL00bkj+IT/m1/WnHsA+6hJ9vdw+wf5ehz7XY9wbJv6En+Fet8DGf8W6h/6hIP+tQ94Ajh1nGOHU73vrfXvfuvde9+68etUj+eXIF7Q2l+T91F/sPSfeVX3dz/AIvef6U/4esZvfzDWh/4Yv8Al6otMwI5+n+A/wBj7yQr1j6RinXvLwPTe30/4j6e/aqHrWmoqOr0v5EchfvXsUH/AJ43/bc+8cfvGGuzWQ/4b1P/ALAj/dhfn+gnW1v7xT6yd697917r3v3Xuscn6D/rH/YcHn23L/ZN9nWx8Q+3rQl+bb2+T3av053Rlf8AXH73PHvoH7asf6m2g/oL/g6wa9xxXnG6P9I/5Oiq+T6C9jbj6exzXoEkU4dYKqQ/b1B+v7T6v+Sebf19uRn9QfaOm5tQhbzwetwv+TQdXxxl/p9zSEf8kN7wZ94z/wAjGT8+s1Pav/lVYvsH+Dq4T3FPUlde9+691737r3XCWWOGOSaaRIookaSWWRlSOONBqd3drAADkkn3tVZmCqKk8B1okAVOAOkZ/pK67/57vZ/BIP8Av5MPwQbEf578H6+zL9zbv/yizf8AON/83SP947f/AL/j/wB7X/P17/SX11/z3mzv/Qlw/wD1+9+/cu8f8os3/ON/83Xv3jt/+/4/97X/AD9djsnrxmVV33s8s7KiKNyYclnY2VVHm5JPAHvX7m3cCv0suP8Ahb/5uvfvLb/9/wAf+9r/AJ+lmjpKiSRuskciq6OjBkdGGpWVl4II5BHsuIKmhwR0sBBFR1y96631737r3XvfuvdVufzMO2N89VdIz5Lr3c1ZtXcExbw5Kit5UANha5H5HuW/ZvaNp3rmtLTd4RPF5qeHUX+7W67rtHKst3s8xgmAOlqVofs611Ifm58vPDCzd77lZjFGS3+qbQNVxq95gv7a+32s02qPz/1cOsR4/cj3GMalt0NaCvYPT7euf+zt/Lzn/jOu5iwF78fn/kL3r/W19vv+jXH/AKvy6c/1x/cOn/JUP+8f7PRo/hd8uvknu35Jdb7c3t27ns/tzK52npsji6qwgqoWteOT1Hjn+nsDe5PInJe18l31/t23pFNHGSrjiD0N/bbnjnXdOc7Sw3S/M0MjUK6aVx9vW1R9feDikMAw8+szeve99e697917r3v3Xugo353b1p1pWwY7em6MdhKyoiWeGCsqIYXeJjYOBIw49n21cs71vUZm22BpVXiQCf8AAOibcd/2naZBFfzLGxFQCRw6QX+zc9Af8/Dwf/nbS/8AXz2af1A5r/5RH/3k/wCbov8A66ct/wDKUn7R/n69/s3PQH/Pw8H/AOdtL/1896/qDzX/AMoj/wC8n/N17+unLf8AylJ+0f5+hG2D3F152c1Smydx0GcNJcVH2c8UvjK/UN42Nvp7KN15e3bZdP7yhaLVwqCOjTbt723dq/QSiSnGhHQneyXo1697917r3v3XuoGU/wCLfVf8sj/vY90k+BvsPXhxH2jrQh+ZjgfLfvlb8/3um+v9NcnPvob7dN/yAdq/5pf5usD/AHDC/wCuDun2x/4G6Lb5LXH1/H+PP9PYx1eY6CYAA0jI6jVcp+1qhbj7ab8/82iefbkTd6/aP8PTU4/QcH+Fv8HW5Z/J3Yt8Oevz9LxV9h+OJ0594H+8hJ9xL+vqv+XrNz2pH/IC27/mmP8AAOrXPcYdSL1737r3XvfuvdFT+alv9l07BP8ATFzc/wBP8nk+ns/5V/5WO0/04/wjol5i/wCSJc1/gP8Ag60Uque+TzB+mnM5IAc8/wCUsPfSOJv8Xi/5pp/g658S5upyf9+yf8e6j+UXt9fyb/j+gFvd9Q6bANc9PO2pb7jwY4F8pQ2+vP8AlkftHuLH93zf6U/4D0q29f8AdhDX+Nf+PDrf26S/5lJ1z/X+5e3v9f8A4t6++aG4f8lS6/5rSf8AHj10Rsf9woaf77T/AI6OhS9pelXXvfuvde9+69015q38LyF/+VKtt/r/AGcnvY6q3Dr51f8AwmsP/Y+L5xf44jun/wB7WT3UcR04eB6+jMv6V/4KP96976r1y9+691737r3Xvfuvde9+691rtf8ACh/5idVfHro3o/r7ee3e2dzbk3f3r1vvyhoutevM3u2Gl2xsTNvNnarJZalVaWCZtYSmpjKZZG+iWsSrtSI28V+HD8+ivcw88YtohVqhvQUUiuTivy/2en75X/IrauE3L8LP5s21drdm7p6C2jiK/q3eG2afrrdKdpYTG9o160lNnp9hzQLUaKZmdJpFBRXCjWQ6n27GAFeFiKtkenSa4ZmkhvolLLHgimckZpx/1evQg/G3dWG+e/8AMTpfm51dg97bd6h+N3Um7fjzHX9ibSzmx8tvbde8KpM5WVOBwuXjV5qSlSQhp3tayggM4UUlXwbfwmPcTXp23Y3d/wDVopCIpSpBBNaHgaYx1eN7RdHHSC7Phh/0db9cwxFztDcd2aJGJ/3Ey/Ukc+9HrY4joCPg8NPxm65H9KfK/wDu3m9768ejae/Hh1rrVH/m5zePvzGAflqr/YWD/n3mf93MV5euvtH+TrEX7wWN5tPtb/AeqqzV2vz9f8P6e8iNPUBVpnrxqgB/ib/7D3qnXqDq97+SHJr3H3ACQSKCg/N7XeL3id95z4tqH9Jv8vWT/wB3Mfobl9q/4B1sXe8WusmOve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuqFf5376essLx/ai/3t/c+fd8xzafsP+TqEffWh5SavWsasrWXnT6QOPxx7zLJyR1iNpNK9cxIOCSb/ANrj/bD3rVTHVgtcnqwr+Vk4b5o9YD6AVFXYcnjQOb+4i982r7eXf5f4epT9lwBz5D/pG/ydbpiW0i305/3v3gqvwj7B/g6zRBqOufvfW+ve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XumLcwBwmQv8A8cH/ANvb3ST4Cetrk9aMvdFQw7q7bGoXG+Mpx/rP76b8oqDyjtn/ADzp1zj5sNOct2H/AC8H/jo6Dn7hj+Rzz/T/AF/Z/pHRICQK9RMjVstDV83H20n+9e3YF/WX7R0nuz/ir/6U9bi38uZtXxh66JXk4Gm5/wCQ24984vc7/lfNyH/DT10L9vf+VN2//mkv+Do+XsCdDPr3v3Xuve/de6I9/MHbT8bt3kf8cJSf9hAfYz9vj/yLrP8A0w/wjoK86ivLN0P6J/wdaW8NTfycXIqau555/wAobk399Jj5D+iv+Adc8o1Oa/xP/wAePWQVPHHAP6vr7rqUefVtPnTpZdczu3YWzP6ncuE/P/VxQeyfmJlGxXVP99v/AMdPRvy+n+760r5Sx/8AHut8rZP/AB6O2f8AHb+FP+t/uNj98wD/AGkv/NST/jx66OJ8C/6Vf8A6VPv3Vuve/de697917qhj+dXMsWA6vbm7T5Hkf8tW+vvI77tuN63EeoX/AAdY+/eBBbYrMj+M9a8616hV9RvYf144/r7zGKmvWJagUHXjXRkkazz/ALzb3rSet+VerD/5YdQsvySxyhr/ALtLx/sV/PuEffsU5PU/M9TN7FknmqUemn/B1t4D3gyvwjrM/r3vfXuve/de64Sf5t/+CN/vXv3XutQ/+azOIvltUqT9drwkD/arpyT7zm+7wNXt4x/4e3+XrC33+J/r7APL6c/4V6rm+6b6BuLXPucNI6h0k0x14VRFvV9D9Px79p6rmo62H/5I0nm2Bvtv9TuqoB+ot/nPp7w3+8sKc12X/NAf5OstPu6/8qjcf89D/wDHj1fb7xx6yC697917r3v3XukZ2JK0Oxd1yqSrJg8gQR9R+wR7UWhpeQ/6df8AD0xdf7jSf6U/4OtIrtyYS9q7+lcAtJnJmLH638jW99LOVs8tWVP99jrnZzUKc033+n6D3VGBYqNX5aw/1/Z5nokpXriSt0NluHFh/sD72CequtQPt63GP5fdQ03xV6w1W9OOqEFv6LKD/wAT750e55J9wdyH9Mf4Ougnt7Qcmbf/AM0x0dT2Behn1737r3Xvfuvdf//S3+Pfuvde9+691737r3XvfuvdFM+cP/ZM3Y9/p9tjL/8An1h9+62Oh56v/wCZbbA/8MzbP/umht70OHXjxPSwrTajqr/8q0/+P+6iffjw611o6/zAalV+VG9VJ9X73PPH74959+0DAchwf6b/ACHrB/3aQ/13f/mmf+PDonH3QN9LW/oOfr/X3JesV6jrTmtOunqfQ1m+lj+f1X5HuytnqpU0x1tefyRnMvxizrE3tveqP/WNveEHv6f+R8tP98D/AAjrMf2QFORUr/v1/wDD1dB7hfqYOve/de697917oCfk02noXtk/02Pmvx/zaA9mWy/8luyP/Dk6L92/5Jdz/wA03/wHrQNzst8/mrNf/L5iw/p+4bge+kFg3+IQ/wClHXPy9Um+m/0x/wAvTV5HFz9V4tz/ALz7V6x0kKHiOp+Fntn9v8X/ANzVCCSeblz+Pae8atlP/wA02/wdP2Ypf23/ADVX/L1vtfEPn489an/qxQ/7yB75x8xf8l26/wCah66BbH/ySLf/AEg6Mp7JujXr3v3Xuve/de6p1/nbNp+F+6LHn+PYe/8AraH/AD7ln2NP/MSrX/mm/wDhHUXe84r7dX/+l609I5LpHwLeNPz+r0/Uj3m8xyesOEFFUDyA/wAHXPyn/D/D+g/1veh8+rUFCR0br4ByX+ZHTAJ+uRnB/HHkQC3sBe6h/wCYebhT0H+Xobe2C/8AMQrP10P/AIV63ycULY2gub/5JBz/ANOx7wFHDrODqf7317r3v3Xuve/de61Qv56Bt2jtG4BvVR8/6yn3lN93ltNvef6U9Y1e/a6vpP8Amov+XqiUzcA/Tkgn6ce8j+OOsfq0yeu/IxHBuf6fQWPv1RTrwJrUdXrfyHn1d69jgDj+5puT/W/vHP7xP/JJsv8Amr1PnsJ/ufuHpoXra/H0H+sPeKx6ya679+691737r3WOX9Df6x/3r23N/ZN9nWxxHWgv83pAPlD2qpvcboynP/T0c++gHtsf+QdaU/gX/B1g97jgHnG6P9I9FTMguB9DxY/4+x1XHQJoK46w1ch+2qPUeInuf+Qfxf3aNv1FHz6bnp4Ln5HrcV/kxkn43yEm96mkP+3RveDfvD/yuMtfn/hHWaPtYa8qQ/l/g6uJ9xX1JPXvfuvde9+690E3euS/hfUPYtQGKN/c7cCI6kqVZ8a6agy8iwJ9meyGm92Xzni/48OkG6/8ku5/5pP/AMdPWjTBkawmsP8AG8sf9ymVN/4vkCRqyEhtzL76fsiEJ+mnwJ+Bf4R8uuaEHiDxAJpf7SX/AER/9+N/S6kfxGsFx/Gcpc/n+L1/+v8AXye66I/99p/vC/5uniZP9+yY/wCGP/0F1Jx+UrBm9tn+MZT07lwTEfxevsdORQ2I8nIP5B490nWMWk58Nf7KT8C/wn5dbiEv1tqfEk/t4v8ARX/jH9LI+XA9by/UWSbLda7Kq25Y7bw0Za5Jbx42IaiT+T75gboKbncf81ZP+PHrpVtxrYQ1/gX/AI6OhH9oelvXvfuvde9+691T1/OEm8PSNM1+LSfm1zrbj3NXsP8A8rrH1EPvSP8AkGzkeQPWrpBkYzBFY8+GP8/Q6B/T3nc8feft6wjib9JccQP8HWQ5Ffrxx/iefddHTgB8ujZfBivEvyn6lVT+rctKOCf9Uv19xr7ujTyBuHzjPUi+02efLL5N1urRcIB/i3/Qx987of7FfsH+DrPI8T1k9u9a697917rx4F/6e/de61hv5yuUMne236RpahEp9s0ShYameEE6lbkRMPeZP3b31csTmgJ8VuIB/wAPWJP3hU/3eWtCR+mOBI8x6dU+/cx3/wCBNaB/X7+r/wCJf3kXTHwr/vI/zdY+aSfxNX/Tt/n65fcqDb7qs/BF6+rvp/5L9+/2q/7yP83XvDP8T/723+fq8L+S7lmXcm7aBJp5I5zVM3mqJp+dT/8AHVj7xa+8kSBa0AGF4AD09Osmfu7IFt7mpJ7m4knzPqetjf3ih1lB1737r3XvfuvdQMnzQVQ/5tH/AHsce6SfA32H/B14cR1oJ/M+RV+Xffqji28JQT+f1yce+hft0QOQdq/5pf5usEvcFa+4W6fbH/gbotnmA5BNj+PqPYuJHQUAAXqLVzqKSq4/5R5vr+f2z7cjP6q/aP8AD0xcf7jyClaK3+A9bnX8nQ3+GvXp/Bhr7f8AU5L+8EveL/p4l+fmv+XrNv2q/wCVB27/AJpj/AOrXvcZdSJ1737r3XvfuvdFR+a7AfHHsPmx/hU9vr/yrSf09nvK2OYrT/Tj/COibmH/AJIlz/pD1ofVc4GVzB4BGZyYtfg/5S3199H4m/xaHP8Aoaf4Oufcq/43PTylk/491HM45IN/6/4e7aqY6rpPTxtuc/3lwPPLZSgtz/02R29pNweu3zU8lP8AgPSqwSm4QV83X/jw6+gX0hz1F1ufydlbd/8AdevvmvuB/wB2d1/zWk/48euhdl/uHD/zTT/B0KntJ0q697917r3v3XumzM/8WvIf9QVb/wC4cnvY6q3Dr51X/Cav/t/F84v+1T3T/wC9rL7qPLpw8D19GZf0rb6aRb/be99V65e/de697917r3v3Xuve/de6xyQxTALNFHKoNwJEVwCPoQGB59+691zKqV0lQVtbSQCtv6WPv3XuulVUFkVVH1soCi5+vA9+691y9+690hez/wDmW+/bfX+5+4//AHUy+/dbHHoBvg9f/ZZuub/X7fK/+7eb3odePRs/e/LrXWpn/OBn8ff2LUHktVH/AAHD+8zvu5GnL919q/5OsSPvAAtvVl5UJ/46eqnPux+Wtb+l7f7H3kR4g6gTQOuvumt9fwdX14/p794o634dcDq/D+RnJ5Nydxi9wMfQc8/8dIveJf3miC+1n+k3+XrJ37uy6bbcqfxL/gHWyB7xd6yU697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6oO/njyaOrcKb2tJCB/hy/9Pc8/d/NObD9h/ydQn75V/qk9OtYETPpBB/C2F/rxz7zI1Z6xK01Axx65CZrEEjn9XP0t9Pfiw6tpIxTh1Yl/Ksl1fNXq8E3vUVdh+P0L+fcRe+DA+3t1+X+HqUfZkf8jyH/AEj/AOTrdYj/AED8/X/e/eDS/CPsH+DrM0dc/e+t9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdMe5D/uFr/wDGBx/tx7bl+A9WX4h1og92Vqju/t8cgLvvKr/tn99PeUtI5Q2vP/EdOucnNoc86btUY+oP/HR0G5rk+oc82/J+n9fZ9qWmeiTQ3UPIVi/Y1o1H/gLKQSf9p49uwMvjJ9o6TXiN9LJX+E9bmv8ALdfX8WetmB4OApT/ALHW3Pvm97nf8r9uY/4aeuhft8P+Qbt//NJf8HR+fYE6GXXvfuvde9+690Rv+YYbfGnebWvanlN7/T9lvYx5AxzbaEfxD/COgtzmK8uXI/on/B1pOQ1QtLzY/c1fH/VS499H2c4+xf8AAOufMajNP4n/AOPHrL9x6eH5v/X3Uvnq4Wgr0tOtal/9IuyV1n/j5cJ/7sY/z7J9/b/dFdf803/46ejXYVJ360WnCVP+PDrfl2SLbR2x/jt3CE/+e2P3zJP9rKP+GP8A8ePXRVP7Nf8ASr/g6VPv3Vuve/de697917qgf+d1IY9u9WWa2qbJWNvraRufeRn3b/8Aku3/APtf8HWP/wB4DOwWgHHWf8HWugtUwH1BIUXv9bW495llxXrEpUwOuP3Nze9tXA5tbT+ffta8OtlDxHVjv8rSXV8mMcur/dtIbk/X9PuDff8Azyen2nqZvYkU5pm/2v8Ag63DB7wZX4R1md173vr3XvfuvdcJP82//BG/6F9+691p7/zZZjH8vKldRv8A3VgsD9Pqn195y/d5YL7dsPPx2/y9YY+/iFufLf8A55z/AIV6rb+6A083+h/5F7nDXivUN+FTrr7y5A4Fz9fevF634ZHDrY0/kbyGXrvfx/1O7ajn+v8AnPeHP3lzXmyyI/3wP8nWWH3dQf6oT1/3+/8Ax49X+e8cesgeve/de697917pD9l/8eBu76f8WHIfX6f5g+3rX/cuH/Tr/h6Yuf8AceT/AEp/wdaNnblWV7X38jObDOTi39PW3099MeU1ryvZH/hY65282Y5rvh/TH+XoP/vTYjVyLcm/Ps/0dEOePmOumqz6DqP6hxzYcH3pgNJ+zr3EivW5X/LtYv8AFDrBi1/8gqf9j61984vcw15/3I/0l/y9dBfb7HJu3/8ANMdHh9gfoZde9+691737r3X/09/j37r3Xvfuvde9+691737r3RSvnGL/ABk7IH1/yXGf+7WH3o9bHHPQ99X/APMttgf+GZtn/wB00Pvw4dePHpY1v/AOq/6hp/8ArUfe+tdaLX8wmoC/K3fCEgj976mx/wCBA956e0TH+osFf4v8nWEnuyp/rs3/ADTP/Hh0S8VPHPH4Bv8A8R7kjX59R7oIz1xaqIUgEDn+pN+fr78Hz17R69ban8jmTX8Xc839d81YFv8Alk/vCj36zz4h/wCED/COsv8A2SFORk/5qv8A4errPcNdS91737r3XvfuvdAR8nP+ZBdtH6H+42a5+v8Aun2ZbLX992X/ADWT/D0X7t/yS7j/AJpv/gPXz989UL/eDNgfUV89/ryNZ499GrEn6CGv8I65/Xn+581OOo/5emr7j6HUbfgfgD2qr0yQfPpxw1R/v4NvAer/AHNUOn/kv639sXh/xGcD/fbdPWigX9sTn9Vf8vW/Z8QCT8dusyb3/gMHH+wHvnTzDX9+3Vf4z1n5sf8AySLf/SDoy/sm6Neve/de697917qm7+d+dPwq3Q17AZ7D8/8ATuT3LPsf/wBPItf+abf4R1F/vLT/AFutwr/D/kPWnTFMTFFz9Y0/2PpH195ss2SPn1h2p7B8wOuYkuPqAD9Obke9VPl1scBXo3vwAJPzM6XF/pkp7j+v7iewH7pH/mHu4D5D/L0N/bEH/XCsz5aJP8K9b7GKv/DaC4t/kkHF7/7rH594Ejh1m4Op/vfXuve/de68ffuvdam/89dxH2jtA/g1UfH9DpPPvKb7vZrb3Y/onrGv36NPpP8Amov+XqiMyL9L3tz7yMr59Y/mgGOuQkI/tD/b/wC8W91J6sKDHV7f8hZi3e3Y5Y3H9zT/AIf2vp7x3+8R/wAkiy/5q9Tz7Cg/vG/Na9qdbY/vFfrJnr3v3Xuve/de6xyi6H/AE/8AJp9tzGkTfZ14fEPt60DfnBKR8ou1wLAf3pypuf6eX3n57bEf1PtPXQvWEPuMCecLqn8R/wAnRUtZYc2IHAv/AE/r7HNfLoFUPUeqcimmII/zMnN7/wBn6e7Rn9RftHTcwIiYgeR63Hf5LxJ+NspPN6qk/I/1De8H/eCn9cJPz/w9Zoe1pJ5ViPyH+Dq473FnUj9e9+691737r3QD/JdtPSXYzH6LtHPH/Wtj39meyV/fdlT/AH/F/wAe6Q7oK7Zc/wDNJ/8AjvWhxBlADWWb/l6ZUW1EXtkJPfUB58J/pE/46Oua9vCaSH/hsv8A1cbrN/FRcL5DyCb3Nhb8e6ifHSgwCtOs+Py1sztzkkncuCBNyLXyKD3WaYNaTg/76k/46etxQgX1rT/f8P8Ax8db5/QX/MpdkH/Vbew7f7fGxH3y93LO53P/ADVk/wCPHrpDYYsYR/QX/AOhj9oulnXvfuvde9+691TN/OZm8HREL302D83/ANrb6e5l9i20c6xnqJveRdXJ04+R/wAHWqLBlWMEIEg/zMZHPH6Bf3nPJOdbfaf8PWFcMX6afYP8HWcZawsHuf7XP0H+v7qJT59OmL06N98BciJfln0/GX4fdFGByST6l4I9xv7tyFuQr4f8LP8Al6kL2qipz1Zn+kf8A63i0/SP9dv+hj758w/2S09B1nMePXP2517r3v3Xuuj9D/rH/evfuvHh1qpfzpK00vyIw6g2/wB+zREc2+ujn3mF925wnLs4P+/W/wAvWKH3gl1bxbEf77H+EdU6HK/7Ve/+1fS/vJDxx6dY/wDhY69/FCDp1fkc6voD+PfvHHXhEaU+fV6H8kysM+9t0C+oBasDn/an594sfeScN9LT0X/J1kt93tNNvcf6Z/8ACetmn3ip1kz1737r3XvfuvdQcl/wAqv+WR/3se6v8B+w9e8x9o60AvmnMB8v+/1v9N4SgD6WGuT8++gvt4B/ULa/+af+brBX3Cx7hboPnH/gbotJmH+qP49jCuKdBMhuo9XLelqhe9qae3+H7R92jIEin5j/AA9NzitvIf6Lf4D1ujfybiW+GPXb3JBhyA5FufOn094Le8FP9cO/+1f8vWbHtZ/yoe3V/wB9j/AOrY/cZ9SF1737r3XvfuvdFO+bZA+N3Yv9f4TUW/2FNIT7PeV/+Vhtf9OP8I6J+YP+SLcf6Q9aEVZUf7lMzf8A53WTH1+tqpvfReNv8Xi/5pp/g6wCmVTdz4x4r1/3rrEJxckNcf2v6C/u2qhpXqlCamnTztqcjc237k85Wgt/h/lsftJuDVsJv9Kf8B6UWK0v4P8ATr/x4dfQl6Pv/oi62N/rsnbn/uuX3ze3D/kpXX/NaT/j3XQay/3Dh/5pp/gHQq+0vSrr3v3Xuve/de6a81/xa6/i/wDkVb/7hye9jqrcOvnV/wDCav8A7fxfOP8A7VHdH+w/3+0nuo4jpw8G6+jOv6V/1h/vXux49VHDrv3rr3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdIXtD/mW+/f/DP3H/7qZfejw62OPQCfBz/smXrj/qHyv/u4n9+HXj0bU+99a61IP5x9QIvkFhxfgmq/3p/eY/3eGA5fugfl/k6xM9/RXeLOg8z/AMdPVRhqQV4cC/8AQ/Xm/HvIHXmnUFaRSvXRqj9dR+nI5/1h9fe9Rr17SKE9bAX8iSbXuTugXBC47HG9/wDa4r+8VfvKsS21/wCmb/L1kx93oAW+5EfxL/gXrZR94wdZIde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdUD/zz2CdU4Um3MkP14tYv/X3O/sEac1n7D1CvvgT/VJwPTrV1WWyKbckLbk88W/HvMUvmnWJoXtB678xvbTz/r8+9aut6adWK/ypnv8ANnqzgH/KKv8AP+0L7iX3uavt9dD0p/h6lD2bWnPcR/oP/k63aYxZF5v9ef8AY+8HR8I+wf4Osyh1z97631737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690ybj/wCLLkLmw+3f/evbcv8AZnra8etB3vCpA707jXUB/v8A3LAf9TPoPfTPlRz/AFR2welunXO7mr/lct2r/v8A/wCfR0GX3IBtq+n1HPA/2Hs811FeiYL1BydX/kFaByPtpbW/J0+3oHpMlT5jpLeLW1kp/Cet1r+WoS3xQ6xcn9W36Xi309be+dHuYSefNzr/AL9P+DroHyDjlCwH/Cl/wdH+9gXoYde9+691737r3RFv5iRt8Zt6G/P203H9f2G/PsX8h45rtD/TH+EdBnnDPLtz/pT/AIOtIKGpFpSf1Cqqx/rj7l/fRhjWhr5L/gHXPuNKBvXU/wDx49ZWqF+n1H55PPuuo16d0DgR0tOtKm/YmyPV6v70YMf6/wDuSj9lO+kHZLof8Lf/AI6ejXYqjfbQ/wDDY/8Ajw6+gLsm/wDdDawP/POYM/7fGRH3zQP9rJ/zUf8A48euhyZjU/0R/gHSo9+6t1737r3Xvfuvda/X88qQQ7b6pNwB5sl/1tb6H3kR93I03u/9e3qA/ftQdita/wAZ/wAHWt0tXdAdYuQPz+r/AF/eYhbPWKCp21HXlqhYcjkn8/T8ce9ax59b0Z6sn/lVS6/k7j1JDDy0ZHqv9dPuE/fpq8noPmepi9jBTmibFPh/wDrcj94PjgOsyOve99e697917rhJ+h/+CN/vXv3XutOD+bpOE+YtTHfn+6VNYXte/j5t7zf+76ae3jD/AIe3+XrDX34H/I7gNP8AiO3+FeqzvuSSDwdJ/r/T8H3NWs9RDoHXf3RFyQPr9b/T/W96DE9b09bI/wDInm8nW/YFgB/v76i//WX3iD95I15psyf98D/J1lV93kBeUJ/nPJ/x49bBfvHXqfeve/de697917pC9m/8y/3f/wBqHIf9aT7etv8AcqL/AE6/4embnNu/+lP+DrRI7jq2Xt3sL9zUBnZxp4FryNz76WcpSEcrWNP99/5uuenNkY/rVfeZ1j/L0Hf3rW5Zf8Tq+vs/Eh6IdA66+9LFPUP1AWv/AIe/GVtJ+w9aMamh+fW6d/LjbX8SOq24ucfVE/n/AHavvnJ7k/8AK+7j/ph/g66AcggDk+wA/wB9jo9HsE9DDr3v3Xuve/de6//U3+PfuvdFA+SvzZ6Z+MG7eq+td5TZ7Odrd3z7hg6s682rha/L5jch2rQDJZypqKmmjaGigihuVmqWUSMCkYYhtL0MLSn0Hr0lubpLYCuWPAevSy+Lnyr6a+YXWEPbPSWfq8ztr+MZbbuTpMtjKvB7gwO4MHVtQ5PE5rC16rNDIkiHQxBSRbMjMPpSSNo2oer29xHcx+JGft+R6MaSACSbAckngAD6kn3Tp/omvzb3BgKj449j0MGfwz132+NApI8pQvVsy5WLUi06uX1fXi349662Pn0OHWu7tqQddbDim3Pt6KVNm7bDxy5rGpIrLiIVYMjygggggi3B49+Bx14g16VlXvPZzUtSP727ZF4JgCc9i7XMRHP7vvdR1qh60Yv5hs8lT8sN8y4+KXIUzCZkrKBGq6V7zj9FRBdT/sD7zm9p7yzi5IhWWeNDq4M6g8PQnrDD3TtL6XnRmit5JF8M9yozD4hioFOiXkZCwJoMj+P+UObn/ePciHcLClPqIv8AnIv+foAiw3If8Rpv+cbf5uuDGu08Y/I/g/8AASf8n/W9+G4WFf8AciL/AJyL/n68237kf+I03/ONv83W2p/I9zeGxXxazkeZy2Mw9Q++qx1psrkKWgqmTxNZxDVurWN+Db3hr75yxTc7q8Lq4EIypBHEeY6y19l4poeSkjmRoz4j4YEEZ9Dnq6X++mzvp/ezbX/n9xf/AF99w/UdSzQ9d/3y2gfpurbfH1/3OYv/AK++/deoeuv757P/AOes21/5/cX/ANfffuvUPQGfJfde1aroXtmCm3Rt2aol2Nm0hhizeNkmlcwXVY40kJYm1gAPr7MdmIG82jMQAJV4/b0X7qrHbLigJ/Tb/AetAXOrV/3hzRFBkCBXznUtHMVYeQ8qwHPvobY31h9BCPqIh2j8a/5+sDLyxv8A66altMasc+G1PP5dNemuANqDIAE3H+RzX/1uR7U/X2Ff9yIv+ci/5+k/0G5Cv+KzU/5pt/m6nYQVy7h27/kGQ0/xqiLlqSYLGNf6iSPoPbN3f2Bspx9REexv9EX/AD9PWljuIv7b/Fpv7Ra1jannk4/n1vx/EjdW16P49dZwVe5tvU88e36cSQz5nHQzRHSPRJHJICpH5BHvnrzAwbe7kgggueHWeGyKRtNuCCKIB0ZD++mzv+es21/5/cX/ANffZR0a0PXZ3ltBQC26ttgN+knOYsA/6xMvv3XqHrr++ez/APnq9tcfX/c7i+P+svv3XqHqnz+dpnsLl/hVumnwuZxOarTn8Qy47FZKjr651EcgLJTUrsxH+w9yr7JyxRe41q8rKg0NljQDI4k46jD3jjmk9u9wSFGkYrhVBZic4AGSetPZIa8RRn+FZEHQmv8AySe6nSP8PeaJvrDUf8Yi4/78X/P1iEtjuOhf8Wm4D/Qm9Ps6yeGvF/8AcbkW/wBT/kk/P9fx799bYf8AKRF/zkX/AD9W+h3A/wDEab/nE3+bo3XwD8tL8yOmaiugqKClTIzGasrYmpaOEeRLeaomsq/4XPsC+513ZvyBfrHNGzEDAdSTx8gehn7aWt9Hz/ZtLBKiiN+5o2CjK4JIpX0H29b4OO3js9KCjjbdm2dS00KkDPYs8iMXt+77wSHDrNahp1NG89nn6br20f8AWzuL/wCvvvfW6Hrs7y2gDY7r22D/AEOcxYP+28vv1evUPXX989nc/wC/s21/j/udxf8A199+qOtUPWqf/PPr6XKdm7NmwtRT5qMVURZ8PNFkkT0G/kakLAEfm595P+wE9tFDd+PKkdVPxMBX7K9Y4e+0FzKLU28TykSLhFLU45NOA6or8Fb9Djch/Uf5HP8A6/8AT3kN9bYf8pEX/ORf8/UDfQ7h/wAo03/ONv8AN1wEFfb/AIt2Qvc/Wjn/ANhfj3762x4/URf85F/z9a+iv+ItZv8AnG/+bq9T+RBXU+K7z7HqM5UwYWE7OCxy5eWLGxyEuBaF6soGI/IB+nvHv7wM9vPtVmsMqPSX8LA/4D1O/sVDcQ39940Lx1RPjUrX7Kjray/vps7/AJ6zbP8A5/sX/wBffeL3WSdD13/fPZ5+m69tH/yO4v8A6++/deoevf302d/z1m2v/P7i/wDr779UdeofTrg+89nFSP72bZF7jnPYsckWt/nfdJKGNh8uvAGox1oRfOBpZ/lH2rLSU09dSyboyhjqqKN6mlkUy8PFNDdWB/BB955e3N3Zx8oWqyTxqdC4LqD+yvWFPuHZ3z83XLx28rqWOVRiP2gdFWENbz/uOyNibgfZz8j+n09jX62w/wCUiL/nIv8An6Bhsdwrm2m/5xt/m6j1UVf9tMoxmRsYpOBRzkg6eABb25He2HiD/GIuI/0Rf8/TU9juJhYC2m4H/Qm/zdbiH8mrPYTE/G94cvmsTiqg1VIBTZLI0dDUkBHufDUurW/HvCb3ckjl5ukeNwwzkEEftHWZXtjHJHyvErqVNBgggjHoc9XA/wB89nj/AJizbX/n9xf/AF99xh1IlD13/fLaFtX969t6RwW/jmLtf+l/L7916h66/vps7/nrNtc/T/c7i+f+svv1R16h6Ar5K7r2rVdIdmx026ttyTjZe4dEEebxsk8ztj2CRwxJISzH8AC/sx2Yhd6smOAJ48+nd0i3IE7dcClaxv8A4OtBWmq8hesvQZEf7lcsQGpJhcfxCSxHH0P1HvpNJudh2UuIvgT/AERf4R8+ueFttm5aZP8AFZh+rLxjb/fjZ4cDxHy6kfc15P8AwByPH5+1m4/2NvbX7zsf+UmL/nIv+fpUdt3Ef8Rpv+cbf5upGPq64ZzbV6HIAf3nwJZ/tJ9KKMjHdnNuFH1JP091m3Ow+lnH1MX9lJ/oi/wn59Vi27cPrrVvpZqePDX9NsDWMnGAPM9b+HQ+7Nq0vUuxIajdO24pRtrClonzmNSSNv4ZEGSRHkBBB+oPvm1uJB3G4oa/qyf8ePXQqyUi0iH9Bf8AAOhf/vps7/nrNtf+f3F/9ffaOo6VUPXZ3jtAAE7q22A36Sc5jLH/AFj5fe+vUPXX989n/wDPV7a/8/uL/wCvvv1R16h6pj/nQ7gwuR+PyfwjM4rKyrrU02LyFLX1PDt/uildmtz/AE9y37LTRQ85RPK6oPUkAftPUXe7kMsvKUyxIzmhoFBJOPIDrUtgqa5aeEfw/JA+CLg0c9/0Dg8fX3mq+52Gs/4xFxP+iL6/b1h5Ft24iJf8Wm4D/Qm9Ps6yfdV3/OuyH/nJOT/vXuv7y2//AJSYv+ci/wCfq/7t3Gv+4s3/ADjb/N0cn+X1XPD8u+nJa+OWgpU3VRmWqr0akpIUDrd5qieyqP8AEn3H3unfWcvI97HFPG5KHAdSTx8geh37YWN9FzxaSS28iKCcsjADA8yOt59N57PC2/vZtk2Zr2zuL49RNv8AO/j3gelAgHyHWaxBr1y/vns//nq9tf8An9xf/X33eo69Q9dneW0B9d17bH+vnMX/ANfffuvUPXFt57O0n/f2bZHBFzncXYcfn9336o69Q9aof87XL01Z8icHUYeohzVOdrUA82ImTIwK10ujy0hZQ39QT7yw+73d21rsE6Tyoh8RviYD19T1i978Wl1cbvbGCGSSkYrpQt5j0HVMn3VeWA+wyA4vf7Sfj8W+nvIP96bfx+oi/wCci/5+oLG2bjw+mm/5xt/m64msrwbfw/I/0A+1n5/xv73+87Cv+5MX/ORf8/Xhtm40/wBxZv8AnG3+bq+j+R3mKSh3lu2TNVlLhowlYVbMVMWODnU+lYjVstyfxb3jP94W6t7hrYQyo+B8LA+np1kT7D2txb284nieMlmwylTxPkfXrZx/vps7/nrNtf8An9xf/X33jHUdZF0PXIby2gfpurbZ/PGcxZ4/rxL79Udeoeuv76bO+v8AezbVv6/x3F/9fffq9boeodfvHZ70dQg3bthS0ZAL57Fqv+xPl90f4D9h/wAHXqHrQY+Z4lqPl139UUlNU1tNLvCVoKulgeopKhDJIdVPURXVx/iD7z75Au7JORNsSSeNWEeQXUEcOI8usHefrW+k5/3R47aVlPh0YRsVOGrQgUNPP06LW0NeORjciQPp/kc/N/8AYexd9bYcPqIv+ci/5+gsLLcSM203/ONv83UargyBpqnTjsiGFNP9KKck/tGwAt9fd47ywMi1uIuI/wBEX1+3pqay3H6eSlrMe1qDwm9D8uPW5z/J4z+Dxnwz67iyucw+Lqkjr1koclk6KhrIi0yEeSmqHVh9PyPeD/u68b+4F80bBwStCpBB4+Y6zP8Aa5JE5F29ZEKMIxVWBDDAwQcg9Wr/AN89n/8APV7a/wDP7i/+vvuNupBoeuxvLaBBI3XtsgfUjOYsgf658vv1R16h66/vns//AJ6vbX/n9xf/AF99+69Q9FV+ae6NsV3xw7Hp6Pcm36qpOHnKU9PmcfNPIft3GmKKKQsx5+gHs85ZZV3+1ZiBRx/hHRPzArHZrgAE1U+XWhhWJXfxPNEY6vI/jOTKkUk9mBqmsym3IP8AUe+hMd9YCCKtxF8C/wCiL6fb1gdLZbh9VPS1mzLJ/obZ7uPDh1g8dcBzjcgCfx9pNz/vHu311h53EX/ORf8AP1T6HcK1+lmx/wALb/N077ZSuG59vl6HIKv8WoCWakmCoBWRkliRwLfn2mv76xNhMBcRHtP+iL6H59KLKw3IX8Ja2lA1rxjanxD5dfQY6T3ZtSm6k63gm3TtyOWPZO3FeJ83jVkRhjk1K6NICCDwQR9b++dm4EHcrkgg/qycP9Mes+7NSLOGo/Av+AdCj/fTZ3/PWba/8/uL/wCvvtJUdKaHp4ocljspB9zjK+iyNMWKCooaqCrg1L+pfLTsy3H5F/futUpx6m+/de6a8z/xa6//AKgq3/3Dk97HVW4dfOs/4TV8fz4vnGB+cR3Rb/0NpOPdRxHThyD19GZfoP8AWH+9e7Hj1UcOu/euvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690he0P8AmW+/eL/78/cfH9f9xMvv3Xhx6AX4PC3xm6540/5PleL3t/uXm9+62ejae/da61CP5zs/i+Q2GFrgmruAf8H95gfd8bTsF0B6j/J1in78rXd7M/0j/wAdPVQS1RFxb6W+p/B/Huf9Y49QZooOuP3L2BB+gP54Pv3iZ62Y69bCn8hSTybk7sN/pjcaQP8ADyRe8WvvInU22f6Zv8vWSX3fVCwblT+Jf8C9bMXvGXrIzr3v3Xuve/de697917rAamnUkNUQKR9QZYwR/rgn3bQ58j+zqutfUddfd0v/ACs0/wD1Oj/4r734b+h/Z17WnqP29e+7pf8AlZp/+p0f/FfetD+h/Z17WnqP29ZlZXF1ZWH9VII/2496II49bBB4dcveut9e9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691r/8A89c6eqMJz/uyHj6AWZ/z7nX2Er/Ws09D/k6hf3uAPKj19OtWkS2VfVYBVtz9OPx7zBrUnrE8KaDrl5x/U/4/i/8AT3qvW9NTSvVjn8p+TV83eqx9f8orLWI5/bH9fcS+9n/KgXQHy6lD2d/5XmGv8D/5Ot3WPhB/sf8Ae/eEY+EV9B/g6zGHXP3vrfXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3TLuIXw1eP8Apnf/AKF9tzf2Z6svxDr5/HetcF757mXTbTv/ADA5P/Nz30o5Wk/5Cm2/886dc9uak/5GO7Y/0f8A59HQY/fD6/73/T/X9nnidEoj1V+XUDKV1sdW2IAFNKRzyfR7dgk/XWvqOk17FSzlI/hPW7z/ACy2aT4ldWub2bbtMebfXW3vnl7kmvPW5f8ANU9Z98hjTyjYD/hS/wCDqwX2B+hf1737r3XvfuvdES/mMNp+MG9/+oWX/eIG9i7kU05qtP8ATj/COgzzeK8v3I/on/B1ozQVLWlH/TVWfn/pqf8APvoi7cM+S/4B1gHFwP8Apn/48es33APJIP8AQ39t6j69O6RStOlt1lOW7J2MCbX3Rg+L8/8AFxj9le/Mf3Lc5/0N/wDjp6NNiUfvy0x/oqf8eHX0Idk8bP2t/wCG5g+f63xkR982D/ayf81H/wCPHroOmUU/If4OlT791br3v3Xuve/de615/wCe7IY9s9TWP1myYt9L/ut9feQf3eDTer4n0X/B1A/vuK7Ha/6c/wCDrWiWrYILn+yL8+8vC+esVwmMdcvuxxyR/Xn6D3rV1vTnqzP+U1Nr+UmPGokF6Pm97/p49wx77MTykg+Z6mD2RA/rNMf9L/g63QPeE44DrL/r3vfXuve/de64P+h/+CN/vXv3XutMf+cDMF+ZlUp4b+6NNz+Vv47W95s+wDU9vm/5rn/L1h177Kf69W9BUG3b/CvVYv3IFiJDx+L/AF/x9zNqp1Emnz9OvfeW4JLajf8A1r/j3oSenVtFMdbLv8heXX1n2Ibf8xfOL/4Xk94j/eOavNNn/wA0R/k6yk+74tOUZj/w+T/jx62GPePPU9de9+691737r3SF7O/5l9vH/tQZH/rQfb1t/uTF/pl/w9M3H9g/+lP+DrQc7qrFXuLsVS59Ofn+n/LRvfSDlRx/Viy/5p/5uuf/ADXGf6033+nH+XoNvvVJ/VwPzf6n+tr+z3xBTojMVc9dCsF4zrIu44vb8c+9FxRvsPVGj4AevW7t/LYIf4f9UODe+Pquf+no988fcbPPO4H+kP8AB1nvyJ/yqNgP+Fjo+PsF9C7r3v3Xuve/de6//9Xf49+691TRsjL43a/84XvgdtVdDj13V0tsJOgavdTwQ08k1BjIDu+n2dV5QhEmKLOJ1pCGYCRWvyPa9hWzGjjXPRKhpvD+J5qNP2+fWP4JUVXkP5if8yPe2znp6no7Ny9OY3aeQwGg7Prd54rDVEW9ZcTPQf5FLN9zrM8kJLEm7Hnmk9RAgPHq1gAb24dPgJFPSvnTq5l0WRHjblXVkYfS6sNJ59o+jjqrX5q/Frpna3SvZPZuIwGRh3hTrR1cGQObyU0cdRV5WOOdzRyOYypDsNOnji1veqenVq149ChsD4NfHPO7F2Vm8htTKPkMptPb1fWypuTMRrLVVmLiqaiTxiSy3dibAAD6e/eWetHB4dKw/AL4zsLHaWWIve3958z9f+pnvfXq9YW/l6/E+TmfrKkqnJuZqurqKmdv+DzTXY/7E+96ph8MrqPQOwH7AetaYjlo0J+agnrr/hvL4lWt/oqxlv8AlrJ/xT3rXcf7+k/5yN/n69ph/wB9p/vA/wA3Xj/Ly+JR/wCaVYv6Af5x/oPp+Pftdx/v6T/nI3+fr2mH/fSf7yP83WWP+X98XYBoo9iVGPi/444/N5Gihv8A6oxU7Kt/8be/HW2XZmPqxJP7T1saRhVCj5Cg/l1zPwE+NBIJ2ll7gWH+/nzP0/6me/der0T7vL4r9QbS7X6427t3AZqkw+byWPgy0Cbiyzx1EM0umVSWckXH5B96z1vJI6OG3wF+NDMzf3Ryq6gQQu581ax+oAMvvfy61XrG3wA+Mj8SbOycq20lJdyZeSN1/wBS8byEMP8AAj37PkevV8iK9Yv+G8/iX+eqsWT+SZXJP+JJHv2u4/39J/zkb/P1XRD/AL7T/eR/m64/8N4/Enn/AIxTi+f+bj/7xx73ruP9/Sf85G/z9e0w/wC+0/3kf5uux/Ly+JS/TqrFg/UESuGB/qpA4Pv2u44eNJ/vbf5+t6Yf99p/vI/zdZh8APjIo0x7OyUMf4ih3Hl4okA+gSNJAAP8B7rnzPW6+VOua/AT4zqVP90csdJHpbc+aKkXvY/u/T3vr1eib9I/FjqHdvfvyA2duXA5yr25s7cFPSbTo/7x5aOHH0jpeSMsH1Pc/m/vQ68Sa9HI/wBkF+NHP+/Ty/qtf/fz5n8fT/dnvfXq9Y2/l+/F2W61WxamvjPJgr85kq2nYj6FoKh2Un/Ej34FlyjFT6gkH9oz1o6Ww6hh6EVH7D1iP8vP4lN9eqcXx/SR/wDinveu4/39J/zkb/P1rTD/AL7T/eR/m67/AOG9PiX/AM+qxf8A1Mf/AIp71ruP9/Sf85G/z9e0w/77T/eR/m67H8vb4nodUPWFFTSj9M9LVT086H+sc0VmX/YH34tMcNLIw9C7EfsJ63piBqI0B9QoB/l1m/2QP4z3v/dLLAj+m58yPr/rSe9dbr03Zf4HfGuhw+Wq4dpZcSU2Mr6iM/3nzN1khpHkRlvJ9QQCL/n3ulevA06LD8MPih0x2r0uN074wWXymdO8t2Y3719xZeGU0GOyHgooCiSBbInF7c+9cT1sno15+AnxoJudpZe//hz5n/r5731qvWJv5fPxWlN6vrpcgQbhsjk62uZT/tLVLMR/sPftUg+B2X/SsR/g60QjfGit9oB/w9cf+G9PiV/z6nFf9TH/AOKe967j/f0v/ORv8/WtEP8AvtP95H+br3/DefxK/wCfVYv/AKmP/wAU9613H+/pP+cjf5+vaYf99p/vI/zdc1/l9fFaE6qPrtcc5FmfG5OsoHcf6l3pSpI/wPvxMrYeR3/0zFv8PWwEHwIq/YAP8HWT/ZA/jORb+6WXt/T+8+Z/6+e/dbr0W35QfELpPr7YNNldobdzVFkpMgIGnj3Hl5G8WkGzLJIR70evVqKdDF178GvjtnNjbTy+S2plpMhX4Shq6yX+8uZVpamWINJIw8lrk/0Hvw62T5dLA/AP40N9dpZc83/4+fM/X+v+c9+p1rV1gP8AL0+JznXN1hRVMhOppqmqnnmdv6vLLdif9c+7a5h8Msg+QdgP2A9a0xHJjQn5qD17/hvP4lf8+pxX/Ux/+Ke9a7j/AH9J/wA5G/z9a0Qj/Q0/3kf5uvH+Xn8Sm4PVOK/6mP8A8U9+13H+/pP+cjf5+vaIf99p/vI/zdc1/l+/F2IaaXYtTQx/8cqDOZKjh/6lU7qP949+7jl2LH1Jqf2nqw0j4QB9gp/g6y/7IL8af+eTy/8A6E+Z/wCvnv3Xq9E37G+K/UeF+W/UXXONwGZh2JuPZ2Wr85jYtxZbTU5GnqHipp3meQkEAC4B5/w9662TivRxx8BPjPwRtLLiw4/38+Z44/5ae99ar1xPwA+MbE+TZ2TlVuHSXcmYkjcfQq6NJYg/kH37Pkader6ivWH/AIby+JX/AD6vGf8AU2T/AGJ/2P59+13H+/pP+cjf5+taYf8AfSf7wv8Am68P5efxLAt/orxv+xlk/wCKe/ap/wDf0n/ORv8AP17TF/vtP94X/N10f5ePxKNj/osxoI5BE0gIP4INvqPwfe9dx/v6T/nI3+fr2mL/AH2n+8L/AJupA+AXxlFtOz8ogXhVj3LmERR9AqosgAA/A9648et1pwHXL/ZBPjRcE7Sy5I/rufM/n/p57916vRPen/i509uj5P8AyJ67zODzNVtLY9Ht59s4+XcOXVcdLW6fu3hlSQFtRvbVew96p1vgK9HC/wBkF+NP/PJ5f/0J8z/189761XrC38vv4uSgrVbEqK6M/WKvzmSrYv8AqXUOw/3j34FxlGKn1BIP7R146SKMoYfMV/w9cf8AhvP4l8f8YqxfH0/cfj/Ace967j/f0n/ORv8AP1rTD/vtP95H+brr/hvP4lXv/oqxd/8Alo//ABT37Xcf7+k/5yN/n69ph/32n+8j/N1yX+Xv8UI/VB1lR0sg/TPS1c9NURn+sc0VmB/xB961THDSuw9C7EfsJ69piGRGg+xQOsy/AT40KLLtLLgXv/x8+Z+v+xk96p1uvSK7J+Dvx4wGxN05jF7Wy8eRoMXNUUko3NmC0cyEBWUGS35/I9+62G6Cr4w/EDpHsDrubM7u27ma3KLm6ylWok3Hl43+3iCmNNKSAW5/p7916tOjFn4CfGhhY7Ty5H/hz5n/AK+e/U61XrBJ/L3+Ks/NX10le4+kuQyVZXSqPoFWSpLED/D3sGRfgkdP9KxX/B1ohGNWRWPqVB/w9cf+G8/iWBb/AEV4z/X8r3/29ve9dx/v6T/nI3+frWmH/faf7wv+brw/l5/EsAj/AEV4zn+sr3/3r3rXcf7+k/5yN/n63ph4+Gn+8L/m65J/L4+K8HNH16ceb3Jx2VraEsf6saZlv79qkPxuzf6Zi3+HrwCD4UVfsAH+DrMfgH8ZybnaWXva3/Hz5n6f9TPfut16KJ8zPin1B1b17s7LbDwOYxuRyPZO2MJX1EW4cvNLLh8hKy1lKQ8hADACxtce9Hj1sZHRsaL4E/GmooaKZ9pZjVLS08ratz5nVqkhDnURJ9efe+tVNM9Sj8A/jOfrtLLHgj/j58z9D9ePJ7916vUY/wAvP4mudUvV9DPKf1z1FTNPPIf9VLNJdmP+JPveuf8ADLIPkHYAfYK061piOTGhPzUE9cv+G9PiX/z6rF/9TH/4p79ruP8Af0n/ADkb/P1rTD/vtP8AeR/m69/w3n8Sv+fVYv8A6mP/AMU9+13H+/pP+cjf5+vaYf8Afaf7yP8AN1kT+X98X4Ropdj1dFF+KehzuTpKdf8AgkEDqo/2A967iasxY+pNT+056tUDCgAegFB1kHwE+NAAA2ll7D6D+8+Z/wCvnv3XgadE77X+LfT22flP8fevMLgc1DtLedHuCfc+PO4su8Fc9HFI9Jqfyal0lRex5/w96PWxno4jfAX40M2o7Sy35sBufM2F/wAD9z37rVadcW+APxjcFZdm5GdG4aKfceWmicf0eN5CD/sR73kGoJHXia4IB+3rD/w3p8S/+fVYv/E+R7k/kk25Pv2u4/39J/zkb/P1XTD/AL7T/eR/m66/4by+JVrf6KsXb/lq/wDxT37Xcf7+k/5yN/n63ph/32n+8j/N17/hvP4l3uOq8YG/DLLIpH9CpA4Pveu4/wB/Sf723+fr2mH/AH2n+8D/ADdSB8AvjMvCbQyiKBYIm5cwiKo+iqiyWAH4A91p1uvXm+A/xpRWb+6OWOlWNjufNW/Sf6S+/db1Hp9+He08TsrrbN7fwdPV02Mot8biSlhrKuetlSITKqKJ6gliLAWF/fh1o+nRsve+tdNmZ/4teQ/6ga3/ANw5Pex1VuHXzq/+E1f/AG/i+cV/r/Ce5+fyP9/rJz7qOI6cPA9fRmH0H54HP+w976qOHXfv3Xuve/de697917r3v3Xuve/de6bMzmsPt3G1WYz2Ux+FxNFH5azJZSsgoaGmj+mqapqWVFueBc8ngc+9gFjpXJPWmYKNTYA6a9pb12jv3ER57Ze48PufESOYhX4WvgroY5lAL08/hJMUqgjVFIFYXFwPe3RkOlxQ9VjljlXXGajpT+69X6QvZ9/9HG/bC5/ufuOwH1P+4mX37rY49AL8Hjf4zdcm1v8AJ8rx/T/cvN79149G09+611p3/wA7GpNN8isEjCxb7si50kjS/wBAfeXn3fiRsV19o/ydYre/B/3bWWfxH/jp6px/iSt+QQfp6x+Of6+551N6dQjROBI66bIoARdRceoa1v8ATj37U3p1UqnqP29bFH8gKo+53J3eV9QXGY0Ejkf52Hi494wfeMqW20gU7m/wHrI/2ACi33Eg17l/wL1s7+8aesiuve/de697917qNWSmGkqph9YqeeQW/qkRYf7172DQg/MdaPA9aenyD+dPyCwXc2+sPhuzKrHYuhzmSpaWjXyWhihqiiILSDgD3nzyjyFyXe8t211c2YZ2RST6mn2dYQc389c52PMlzaWd2FjV2oKZAr9o6B4/zAPkrx/xlesb+v8AnLW/6mexH/rc8i/8oS/6vy6Dn+uFz0eF6P2H/P1Hqf5gXyXjhdl7Wq7jRY/uC13AI/znuye2/IbMAbFf9X5dUk9xOe0QsL0Yp5H1/wBN1tS/AXsncHaPx827uTc2Sky2WkISorpAbzExA/kn/e/eDvuTZWW3c1z2lhH4ca8APLPWafIV5eX3LUFzfPrkYZPr0dj2Auhn1737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Wvx/Pgbx9SYRyNI8sPJNgTd/oTxx7nP2GBPNRA9D/k6hj3tYLym5Pp1qsCpBWxYXCr/AGhxx/j7y+IJPDrFLUoABIyOuxUC4BcW5ubj/Yfn3qjenWxo8yP29WS/ylZkk+cXVqK2o/cVdrWJsIxfge4p96w/9Qbmvy6kz2dKf15ioa9jef2dbwqAhQD9ef8AD8+8JB8I+wdZj9c/e+vde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdM+fUtiK8AXvTycAEm2k34HtuXETdbX4h1887vmtA7+7tQkAJ2HmR+oD/dnNxf30h5YJ/qrtv/ADQTrn9zRT+uG7KSMXH/AD4OguNav4cD/kIc+zkE8KdE1F9R+0dN+UrdOOrTrXimkP6gben/AF/b1uSZ0FPMdJr3QtlKxIwp8+t53+WCzP8AELqh+Sr7cpiGtwSHa9j758+49f68bjX/AH6es9uRqf1Tsaf76X/B1Yb7BPQs697917r3v3XuiHfzHAf9ld301jZKSZibGwH27fU/j2LeRv8AlaLT/TD/AAjoNc3f8kC4/wBKf8HWiRDXxWlPkWwqqwfqU3/ypxa499CXDYqDwX/AOsCImSjKCPifzH8R6ymsS1hIAPzyD7qK+nTpZaVJHS56vrFbszYoDgsd04G3qH5yUYtYH2V76T+5rkkf6G//AB09Gex6P35aaSP7VPP+kOvohbJBGz9q6r3/ALt4K9xbn+FxX9832FJZP9O//Hj10CT+zX7B/g6VHv3Vuve/de697917rXX/AJ90vh2t1Ex9IM+UGo8D/OHjn3kD93yo3m9p6L/g6gn30YDZLYNjvPWsYtamlf3h9Bf1L9Le8tyzV4dYtjSRgj9vXvv0JN5B/r6l/wCK+69/p14lPXq0L+UXUCb5UY9FOo+SjtpOo8afpb3DXvmT/VNAfU9S97J6TzLKVNfh/wAHW6z7wtHDrLzr3vfXuve/de64v+h/+Ct/vXv3XutK/wDnI1KQfNSridvHIdnUxCMQp/3VYkNY+80vYSv9QWPEeOf8vWIHvmVHPNshIBNu3+FeqtjXJwRIv4B9S/X8/n3MtWFcHqJiU8iKfl1779AReVfrx6l/2H596JY+XVdSA0qPlnrZ0/kCzGfqzsl0u4G851Zh6go/d4JF7e8S/vFk/wBabQf8IH+AdZRfd+IblCYg/wCjyfydutif3j71PHXvfuvde9+690hOz7/6Pd5WBP8Av3skbAXPFOT+Pb1t/uTH/ph/h6ZuP7B/9Kf8HXz8+768J3P2SLKANw1A/UBz5H+oPvopyq5/qzZUH+h9YEc1Bf6z3tSPiHp0GH8SUmzaR+T61H0Hs+q1KgdEXYxyR10cjHeIghrOBbUot9fzf34s1GweHWiFFDUH8x1vP/yzn8nw36icAgNjaoj+hHmHIPvnz7iV/rvfk/xD/B1ndyLT+qdjT/fY6Px7BvQt697917r3v3Xuv//W3+PfuvdE7+VnwK+LfzVqOuq75E9cneeS6oyWWymxMrRbi3FtjKYSozlItFlIhXbbqqV5oZURCYpiyhhqUAkkuxzSRfAek9xawXS6ZlrT8j+3j0LHx6+O/UPxX6p290r0btKHZfXW2Hr5cXhY67I5Sb7nKVj1+QrK3J5aWapqJpZXJaSaVjYKosqqBV3aRtTmp6vBBFbxiGFdKjgOhs906d6KZ84Rf4zdjggMPtcZwfof9ysP9ffutjoeer/+ZbbA/H+/M2zx/T/cNDx70OHXjx6XXvfWuve/de697917r3v3Xuve/de697917okffwH+mDrNrKT/ABTHAXAvfyj8n3rqw8ujue99V697917r3v3Xuve/de697917r3v3Xuve/de6JJ0OB/sw/wAifSqn+8MBuANTeg8sfevTrZ6O3731rr3v3Xuve/de697917r3v3Xuve/de6aNwf8AFhzf/aoyX/uE/v3XuiifAuw6IsFCj+/O8uAABzk7/j3odWby6Ol731Xr3v3Xuve/de697917r3v3Xuve/de6K38sQG6+pbgH/cj+QCAdK/g+9efW/LobOtONgbQHH/FioPp9P8yPp78OHXjx6XHvfWuve/de697917r3v3Xuve/de697917okPZwB+YHTnpBP90ct6rDUP8AK34De9VIPWz8PR3R9B/rD3vrR49d+/de697917r3v3Xuve/de697917r3v3XuiOdIqB8tfk2Qqgml21dgAGP6bXP1968+rHh0eP3vqvXvfuvde9+691737r3Xvfuvde9+690ge0v+Zfbr/7VM3+P5Hvx690EHxQsOtp7AD/c7W/QBR+lfwPeh1s9Gd97611737r3Xvfuvde9+691737r3XvfuvdEt+cAv11s3hT/AMZN2qfUoYf55vwfevPrY4dHFxv/ABbsf/1BUv8A1oX3s8etDh1N9+691737r3Xvfuvde9+691737r3XvfuvdEg7jC/7Nz8cCVUt/D9yWYgFhaKT6H37rY6O/wC/da697917r3v3Xuve/de697917r3v3XuuEn+bf/gjf9C+/de6AzoAW2xnxYD/AH+Od+gtf94e9Dqx6Hb3vqvTXmv+LVkP8aGtB/8AOOT3sdVfA6+dX/wms/7fxfOLj/l0d0/+9rJ7qOI6cPBuvozL+lf+Cj/evezx6r1y9+691737r3Xvfuvde9+691737r3VMH8wfbuI+SPzG+HHwt7Q+/fozdUeY733Fj8XlK7AVOf3r1Zk2qdpYqsylBIjyUwdDrpPpJrN+dNltuAsLy+Y/wA3RRfOz3kNoT2PUn8iOk119tHDfE7+cDTdM9IY2u2/1f8ALHojeneXbe3DkK/J4Gm7I2ZXrhMPk8LSVkjpjfLBHYxQ2VtT2AUgDzsZbUyPkggV61Gott0EEWEdCxHzFB/g6vE9oujjoj/yC6v+QeVpN87j2v30m19nQ7dzdVJtQ7fiqvJRQY+SWoozWtyPIgK6hyL+9ZpTqwpXotPxY6p+Se7eiNkZ3ZfyJXZ+3q2PJNQbd/uxBV/w5I8jJFIv3hOp9bqz2I4v/j79x63gcejA/wCgT5gG/wDzlgn1BFtpQi1vfs9eqv8AqHRSu6P5S2U+RO4oN2dyd0/3rz9KGFNWnBinMYYEN6Rf8E+xXsPPPNfLEDW2xXPgo/xCla9BffOTeWOZJUm3q2Ezx5Uny+zoHh/IW63/AD2ECR9P9xNv+I9nn+u97jf8pw/3nomPtX7fn/iAP5deP8hfrc2/4yELfkfwr6/7x79/rv8AuN/ynD/eetf61fIB42A/l0YXoX+WLvb4wS5ufo3vZdnzbkjjizL/AN31qvu44WDRCzcCxUew1zBzdzHzV4Z3+fxvC+HypXoRbDyty/ywJF2OAQCXLAeZ9ejJDoT5f2APyvQnm5/ulDz/AE9h3PR/29Ah3fjPlx0zh8blpPksuaOQq1pfAdsw04jLSLHqUi97ar/j3occ9eJHl0L+L6T+XuWxOLykfytEIyWOo6/xNtGA+I1lOs/jBH1C6rX97z16oHUuX4//AC+milhk+WClJY3iYDaUIJWRSj8j/Am3v2evVHREdx/yR9ubvzmR3LuLtM1+cy1TNWZCs/hLL56md/JK+n/Fufcg2fut7gbdarZWV6EiQAKKcAOgLee2nI24XTXt7ZB5XNWY8ST0yD+RLsW4/wCMlGwP0/hJ4/1j7U/68fuV/wAp4/3npN/rUe3ta/QDrz/yI9hSIUfsjUpPI/hR/BuPfh7x+5Smovx/vPWm9pvbxhRtvU1+zo5PWHwq7+6X2vTbL60+Sv8AANs0jB6fHttlKjQ2nTcO/wDUfX2Bd03XcN6vG3Dcn8SZuJ4V6G23bdY7VaiysU0RrwHp0Iw6F+X4vf5Xob3t/v0YeB+PZfQ9Lqr0XutHy2pvkRT9CL8k11TbRj3V/eKXbMKcvcfZCmXVzccG/v3Hr1adGE/0CfL7/vK9f9jtKH/eh71Q9bqvXR6E+YH4+V6Dnn/fpQ/T+nveetVXrr/QJ8wP+8sEvqv/AMejDbT/AE9+z17Ff9jrv/QJ8v8A/vK9P/QSiP8AvfvVD1uq9df6BPmBdf8AnLBOLXH90YTf3vPWqr5ddt0J8vz9Pleg5v8A8elD9P6ce/Z69Udcv9Avy+JF/leoF+bbRhuB+bfS/wDsffs9eqvQBdSQfLbtfencezYvkoMTJ1Juin20K5NsU86ZppkeQ1LqbGOwQi1jz7959exTofP9Avy//wC8r0/9BGG/+396oet1XoB+7/5dHaHySw9Nt/uv5B/3uxFKQ0NL/d1aYIwJIIK8fnnj2ebFzJvfLN19bskvgy+vHom3rYNm5itDY7zF40R4qeB+316LAf5C/XFxp7CAAFh/uJ/H4/HsWf67/uP/AMpw/wB5/wBnoLj2s5AH/EEfy64/8MLdcc/8ZDFzb/l0/wCP+t79/rv+43nfD/eevf61nIFf9wB/LoRuqv5OUXR29Mb2L1Z28Nubyw5kOOy38F8/gMo0v+2Rzccc+yzePcTnLmCxbbN4uhLA/FaU4fPox2jkPlHYr0bjtNqIplBAYU4HiOjpL0J8vwLH5Xqx/qdpQ+wXnoX1B6RnYfWvy62Ds/L7qk+Ui138Ki8wpP7qwxrKP6GT8f7b3rI49eqPLpo6k2X8uu1di4/eUXyfXFGvqKmH7P8AurBMsYpnC6vJwSTf6W97z17HQlf6BPl/z/zlenP4/ujD79Q9bqvXf+gX5fWt/s1yf6/90offs9aqteuLdCfMA/T5YIP/ACUoffs9eqvXj0H8v+LfLBRb6/79KLn37PXqj/UOvf6A/mBcH/ZsEt/T+6UP9Pes9eqvXL/QJ8v/APvK9P8A0EovfqHrdV/1Dov3e4+W/SNR1lTP8kf44/ZG84dn+RtsQU8eI8yK4r9XJe2rkW9+z17HEdD8vQvy+ZUYfLFGuoNxtKEq3H1H+Hv1D16q064T/H/5e1EbxP8ALABJFKOE2nGhKsLEXX34iuD1qo8uiCZ3+Rpszc+ezO6M72UKzPbiyE2VzVaMRo+7r5zeWfTbi/8AT3Idt7r+4Vnax2VtehYoRpQaeCjy6Adz7Y8iXl5Lf3NiGmnbVI2Ks1KVP5Y6af8Ahhnrrm3YgFyf+XT/AGT/AGf+N+3v9d/3H/5Th/vPTX+tX7f/APKAP5dcJP5CnWs0bwy9gh4pVKSKcVwyMLMPp72vvD7kowZb8Aj+j1ST2n9vJUMcm3qVYUINKEHy4dHa62+GnyI6f2jidhddfJv+BbSwUC0uJxZ2wk4paZORGGf/AHv2Atx3C93a9k3LcG1zSmrtwqehzY2VnttqllZLoijAVV9AOA6X56E+YBI/5ywQDi4/ujDe1/6+0eeldR/qHQC5SL5aYv5D7Y6Df5KCV9x7PyG7DuBNrwF6MY8sDRmlNgdem4Or6e/cOvYp0PR6E+X9rD5XoOb/APHpQ3/2/v2evVXy6Q/YPw8+R3au2shs3fnydGZ2xlUMdfjRthIRMhXSV1p/gT7V2V9d7bcreWTaJENVPofs6TXdpbX8DWt0uqNsEevRHl/kKdaKCF7AAu7yf8Wr+3I5dz9PyST7Hp94Pcg8b4f7z0Bl9qfb1RRbBRkny4k1J/M9c1/kMdcAc9hAt/X+E/8AGvev9d/3H874f7z1b/Ws5A/5QB/Lqbiv5FeyMJk6HM43slYMjjKmCtoZ/wCD38NVTSiaCS3+DAH21P7se4VzC1vPego4II08QcHq8HthyHbzpcQ2IV0IKkUwRkdWA0fx6+XFBSU1DTfK1UpqSngpaeM7TiJSCmjEMKajzwoAJ/PuPTqLFjxJJ4+Z6Ho0AAAYHXOp6L+XlNT1FS3ytUinp5p9I2lD6vFGZNJ+n1ta/v2etgr0BnTtL8t+3Mlu7GxfJVcQ216hacSjbEE4qy0vj1MDbT9Pxf37z61joev9Avy+tb/Zrkufz/dKH8f09+oevVWvRfO9P5bvZPyYpcTQd39/jd1FgmlfFQjbq0v27z/51vR9b+z7YeaN+5Xla42KbwXf4ic8OHRJvnLmx8yQrb73D4yIagcM9FrP8hjriwC9hBbf9Wm/HsUn3f8Acc/8Th/vPQZHtT7fDhYD+XXv+GGuuja/YYNv+rT/AMa97/13/cj/AJTh/vPXj7Ve3/8AygD+XQmdTfyfpeid0JvXqjuP+7W54ymjIjB+bT4+F9Bt9LeyfevcDnDmS1FlvVyJYhwFKdG2zcj8p8vXJu9ntRDI3Ejzpwr0ckdCfL/8/K9D9fptKEfX2D6enQrqP9Q6DDujZvy76h6v3l2VJ8njmP7o4sZIYpNqQRCuAqEgaJpr3X9YINj/ALD3rPn1uo8un3rrrX5d9gbE2nvaP5SR0K7owdBmRRDasMopfvYRMafy/nRex97z1qo8+lkehPl+Rb/Zr0H+ttGEH37PXqr0TjuL+T/L8gd6P2J293F/eneT0a49su2E8LfZpbTFoH/BRz7GOx+4HOHLVj+7NjuvBg1atNK93r0E965F5S5ivl3LerUTTopVWNKhTSoH20Ffs6C4fyFet73PYV/6f7ij/vPHs2/13/cf/lOH+89Ff+tX7f8A/KAP5deH8hbrcG/+kK5vx/uKPH+8e/f67/uP/wApw/3nrX+tV7f1r9AP5dGX6H/ludk/GHE5TBdGfID+5uJzNW2QyVKNvipFTWsSTM2rj8ni3sLcwc0b9zVcpeb/ADeNJGNKmlKL6dCbYuXNj5YtTY7FCIImYsVFKaiak/mSSejAjoX5fgAH5XoTYc/3Rh+v59kOejuq9F++QEXy66Jxuy8lL8k0z398d64nZ6wf3ahp2o/4pJ4zXJ9Q3jvcr79w49e4nH+Dowf+gX5ef2PlgGUqCC+0IFa5Xm4UkfX/AB9+z5de7eoGQ+Ony1ytDVY2v+Vgmoq6mlpKqIbUiQywTLokTUvIuDa/valkYOvEGo604R1KtwOD0QnM/wAjLZu4szk9w5rswVeZzFQ1Xkar+EW887kln0n6fX3IFr7re4FlbJZ2t6FjjFFGngOgLc+2fIt5cveXNkHkk+Jj59Ng/kM9dXv/AKQ1+vH+4g8e3v8AXf8Acj/lOH+89M/61Xt8P+IA/l1yH8hrrvknsQE3BB/hH0t9Px79/rv+4/D64f7z14+1Xt8eNgP5dHc64+HfyL6j2hiNg9ffJpcJtHARNBiMWdsRz/bRO2pl8j8nnkewJuG4Xu63j7hfvrmkyx9T9nQ3sbK0220Sxsl0RRiij0Hp0uB0J8v7+r5YLpuPptGG4H59o89Kqr/qHSV6e3b3ntT5CVXVfY3aA7FxhSUxzDEx40wmOEyC4T/W/r7917qx73vqvX//19/j37r3Xvfuvde9+691737r3RS/nF/2TL2OfoRTYwg/+RWH3o9bHQ9dX/8AMttgf+GZtn/3TQ+/Dh148T0uve+tde9+691737r3Xvfuvde9+691737r3RIu/v8AmcPWY/H8Sx5P/U4e6+Z6sOHR3fduq9e9+691737r3Xvfuvde9+691737r3XvfuvdEj6Ia3yI+RA/ruCn/wBh6Peh1tvXo7nvfWuve/de697917r3v3Xuve/de697917po3B/xYc3/wBqjJf+4b+/de6KL8DP+ZEWH43xvL/3Z+9DqzeXR0fe+q9e9+691737r3Xvfuvde9+691737r3RWfll/wAy/pP+1l/0SvvXn17y6G/rX/jwdo/9qHH/APWke/Dh1s8elv731rr3v3Xuve/de697917r3v3Xuve/de6JD2d/2WD059f+PRy3P/VW/uvmT1s/D0d0fQf6w9268ePXfv3Wuve/de697917r3v3Xuve/de697917ojnSJ/5y1+TYt9KXbX+x+nvXn1Y/D0eP3vqvXvfuvde9+691737r3Xvfuvde9+690gO0v8AmX26/wDtUzf72PfutjoIPieb9bTn/q+1v/Qq+/dePRnvfutde9+691737r3Xvfuvde9+691737r3RLvnB/zLrZv/AIkzav8A1ub3rz62OB6OJjP+Ldj7m/8AkVLz9L/sL7sePWh1N96691737r3Xvfuvde9+691737r3XvfuvdEi7iH/ADlx8cjf6Y7cfH+vFKPfutjo7vv3Wuve/de697917r3v3Xuve/de697917rhJ/m5P+CN/wBC+/de6AvoD/j2M/8A+Hjnf+tw96HWzw6Hf3vrXTXmv+LXX/8AUFW/+4cnvY6q3Dr51n/Cav8A7fw/OMfT/cT3QP8AAf7/AGl91Hl04fPr6Mq/pX8+kc/7D3vqvXL37r3Xvfuvde9+691737r3XvfuvdEC+efwx3P8q9s7GzXTnbh+O/yH6t3dh9xde920e2IN1V2IxUFaKjP7XqcVPPTrLS16quq7nQy/pIZgX4ZjFVTlTxHSO6s1uaMDpdfhYZp+Xp02/EX4Ydj9Q9g9gd5/JvvKD5O997qnbD7W38+x6HY9P1712YE17L27h8fNLEiTTKZZ5VC34UDly1pZw6iOMaV9OqWtm0Lme4fxJD50pQegHVh/tN0v6QfaSh+td/oRcNs3cikf1DYiYEe/dbHHoAvgxBHTfGLreGJdKJT5bSt72vmJz9feh149G597611737r3Xvfuvde9+691737r3XvfuvdE5+YNHTVm2MAlTH5FXJxlR/QidPeut1pjo1G1lCbZ26q/pXB4pR/rChjA97HXjx6fvfutde9+691737r3Xvfuvde9+691737r3RF8vQ0p+cFDXeIfd/6N6eIS/nxgk6R7114nHR6Pe+vde9+691737r3Xvfuvde9+691737r3XvfuvdEP+JtLBB3H8tZYkCvUdmUTysP7Tfbzc+9UoerHh0fD3vqvXvfuvde9+691737r3Xvfuvde9+690EPe0Mc/V26IpV1xtSDUt7X596PXukt8XIIabp7BxQJojFZkiF+trzC/vw62ejD+99a697917r3v3Xuve/de697917r3v3XuiPfM2jpaus6DNTF5TT9qUksJ49EgiT1c+9efVhWmOjt0xvTwn+sSf4f2fezx6qOHWb37r3Xvfuvde9+691737r3Xvfuvde9+690RHc9FSt89+vK5o/8AKo+qc7Ekl/7DF7i3vR49bpjo93vfWuve/de697917r3v3Xuve/de697917qFkgDjq8H6Giqgf9YwMPfuvdEz+KNHTUu4uyzBHoaSuTyG/wCr98n3oU635dHZ97611737r3Xvfuvde9+691737r3XvfuvdF3+WNNDV/HjtKnqE8kMu3WWRP8AVD7yI296PWx0qugoY6fpjraCFdMUW08Uka/6lRBwPe+tefQve/de697917r3v3Xuve/de697917r3v3XuiMfOWgpa7AdPiqjMgg7c23NHY20yLOulj70erDo8qfoX/gq/wC9e99V65e/de697917r3v3Xuve/de697917r3v3XuiNUlFS/7NrUVXjtU6Z7vxe325HutTXq3l0eX3bqvX/9Df49+691737r3Xvfuvde9+690Uz5w3/wBlm7HsLn7bGf8Au1h9+PWx0PPV/wDzLbYH/hmbZ/8AdND70OHXm4npde99a6LRvP5i/Gjr3ufbHx83n25trAdvbxijlwG0Kxcl5anz3+3iqsrDA1DSySW/biqqmN2uLD1Ld0QysniAYHSZ7y2jmFu7gO3AfZ0LmT7P66wsu7YMtvfbGOqNh4NNzbzp6zM0MFTtnb0kDVEWbzNO7h4KVkR2Wd1CGxAJPHvXgyUBp8XDq/1EFWGodnH5dJDsP5G9F9UbQfffYXaWz9sbXG2Yd4w5Guy0Ly1+2alQ9NmMTjqXyVVXFIGHjNNDIW/A97EMrcBwNOtPdW6AFmAqK/l69OnS/ePU/wAh9hY3s7pbfGF7B2LlpainotwYSSc05qqRgtVR1FPVpFNBNGSNcU0SuAQbWIJq8bxnS4oetwXENzH4sDB1PmOHQre6dPdEj7+I/wBMPWQ/P8Uxx/2HmHvXVh0dz3vqvULJZLH4bHV+Xy1bS43F4qiqsjksjXTx01FQUFFA1TWVtXUzEJHFFGrPI7EBVBJNh72ASaDrRIUFjwHRWOs/nX8R+4+yIepOse9dlbx3/VYaXP0GCxU9eRlMVBL4ZqjEZOpgjpKtlPJip53fSCwUqCQ60EqDUy0A6TRXtrOwSJwSc/l0/dw/ML419A722R1z2/2zt7ZG8+xamOl2nhclFlJ5K6SacU0L1lTj6eaCijdzZZa2WJDydVgT70kMsilkFQOrS3dtA6xyuFZsAep6cu6Pld8dPjxLtWm7n7c2jsOs3uZTtHH5SsmqMluGKAqJqrGY3GRzzywoXQNOI/GLj1e/JDJJUoK049amu7e3KrM4Utw+f2dDri8nQZvGY7M4qqjrcZlqGlyWOrIdXiq6GugWppKmPWAdLxsrC4B55HtsgqaHiOn1ZXUMuQep/vXVuiS9EAH5D/Ij/DcFPb/kk+9V62ePR2ve+tdIHs/tHYHTOx892R2fufHbO2TtmlFZms/lDMaajhLiNAIaVJJZXZiFSOKNnY/QH3ZEZzpQVPTcsscKGSU6VHn0Fu2Pl58a95dPT9+7c7e2rkeoqSrpcfX7zEtZT0WMr63IRYukosrRVUKVdLK888KKk9Ohs6t+g6vdzDKH0EZ6bW6t3i8ZWBX1+3oXcl2HsfEbi2ztLI7ow9LuTeVPkKva+HerRqzM0mLpPvq+rpEjuPFHF6/IxCkfpJPHuoikYEgcOrtPEjBWYAnh0Ae2/nL8S94dzx/Hza/eeys727OK8U20sbU1lS1ZLi1LV9LQ5lIf4fPNEFbVDFVM9wQATx7ubeZU1lcdMrfWjzC3WQFzkDzx0a/2z0r6aNwf8WHN/wDaoyX/ALhv7917oovwMt/oI4/57nef/uz96HVj5dHR976r0XbuH5afHDoJNw/6Xe3tobKqdq4Kj3LnsXX1k1Vm8fgshUGlosk+Dxkc9W8cjghTHCxtzbTz7dSGVxVRUdJ5bu3hYpK4BArT5dLbH93dS5Xqgd54zf8Atuv6kO3pN1f38o65ajAjAwxeWataeIFgUsVeEp5A/wC2UD+n3UxuH0Uz1cTwmLxgw0+vSS6S+U/x9+Rmz8zvzpjtLbe9tq7dmqYdwZKkkq8a2ENJE080uWoc1FTVFPF41Z0mliEbKrFWIU220MiEKwyeqRXdvNGZYnBUcT11138qvjz232fu7pvrPtba2+Oxtibdxe6917f25PU5JcRgMzU/Z46uly8EZoZNcllMcVS8ik+pR788MkYq4oOtxXVvOxSJgxHH8+HRgfbfT/RW/lgSOv6Xi/8AuR5/5JXn3rz635dDZ1r/AMeBtHm/+4Kg/wCtI9+HDrx49Lj3vrXRad7fMT40dc9xbT6C3p29tjA9ub3lp6fbezqk5CWqq6msYpR0lVX0sElJSTTEftxVdRGzAggWYEurBK6a1FQOk0l5bRSiGRwGbgPXoWX7R67ji3tM+8sAqdbw19RvsNkIRLtamxdF/EK+py8F9cccUN3aTSV4IBLAgV8NxTHxcOnBPEdVGHZx+XQNdk/NT4r9Q7Y2nvHsXu3Zm2sDvvazb32bPUVNXV1m5tpLBHUnP4jEY6GarlptE0Z8ggtdrfqBAssErEgDhx6akvbWJQ0jgBhUfMDiehn607N2D3Hsfb3ZHWO6cVvPZG6qCLJYHcOGmaWjrqWZdSnTIFkikX6SQzIsiH0uqkW90ZWQ6WFD0/HIkqCSM1B8+l37r1fokXZv/ZYPTh/P90csB/51v7169bJ7ejuD6D/WHvfWumrOZ7CbYxdVnNx5fG4LDUKo1blcvW0+Px9IssqwxtUVdUyxoC7KoLMOSB7sqs50qKnqjyJGuuQ0HReevPmZ8X+1+29ydFdd9z7P3X2vtOCaqzWz8ZU1TVi01Moepnx1ZLElLWpECDI1FPKFFyeASLtBKi62GOmEvbWWXwEcF6Vp50677F+Zfxg6l7Q270z2L3LtLavZe6pKKHD7XyMtaZ3myTiPHRV9bTQyUtG05I8S1c8RYEEcEH3tYJnXUq461LfWkMnhSyANwp0oe9vk/wBA/GbD4nO969pbX65oM9kqTE4NMzUzS5HMV9bULSwQ4zD46OernGtlVnjgKpf1MPdUikkroFadXmure3p4zha8Pz4ft6HGirKXI0dJkKKZaijrqaCspKhL6J6WqiE9PMmoA2ZGDC4/PuhBBoenwQwDDgepPvXW+iOdIm/y0+Tf/UNtsf7bT7159WPw9Hj976r0heyuzNidPbIz/Y3ZW5cftLZe16J8hm87kjMaejp049MNMkk0sjHhIoY3djwqn3ZVZ20qKnqkkscKGSU0UcT0HnXHyk+PvbPVkXdmw+1dq5fq6Sb7eXd9RVyYXH0FQar7JYMtHnUppqNzIQqrVRR3uCLgg+7+BKH0ac9MreWzRCZXBU8D616W2a7f6t25JuuHO7/2pi5di7Zod57wjrMzRxPtvamTMi47cGWUteGlnMUnilbhrcXuL6EUjUoOJp+fV2uIUJDMBpFT8h69BXsn5mfF3sftao6R2P3XsrcfaNPgaPcy7Vx9dMaiuwtfH5qerxNdLGtLWnR63ipJ5JEW5ZQAbWaCZBqZcDqkd5bSsEjcEny6M37Z6U9IHtL/AJl9uu3/ADqZ/wDex78evDoIPigLdbT/APa9rb/8kr70OvdGd97690Wvvz5hfGj4vw0cve/cG1OvZK8K9LQZGarrss8DMFFW+Hw0VTVJBc288kSx/wC1e3Y4JZRVFr0lnvbS1/3IcJT18vt9OhIyXcvVuI6uHdWS3zgKXqttu0+7I97vWBsLPgKumFXS11NLGC8hlRh44o0MjMdAQv6fdfDcv4dM9OmaIR+KWGn16SPTfyf6E7/65yfbXUfZu3d3deYOTKR5zcsT1eLpcG2FgNVlDmYM3FTTUqwxK0rPPEq6AWBIBI28UkbaHFCeqw3UFxH40LBlHmPl1H6Q+VXx6+SMmej6M7T232V/dkxjMy7dNdLS0qyymCOWOrqYY4pkZ1Kh4HdT/X3t4ZIxVxTqsF5bXOIHDfZ0YL210p6Jd84P+ZdbN/8AEmbW/wCtze9efWxwPRxMb/xbqD/qCpf+tC+9nqo4dTffut9FmqPmR8Y6TvP/AGWyq7h2rTd1/tKNj1D18FUamaMSw48ZSWAUP3TqRopfuvKT6Qhbj274EpTXTHSb6u28XwNY1+nQo47uHq3K13YGNod/bXlreqsrQ4PsaF8tTU42bl8nRpkMdj89LUlEgkmikRowzeq+keq4918N8Y48OnPHiOruHZg/LpFd0fKT4/fHja1DvPuXtTa+xdv5WLz4moydTPUVmVgCeRp8ZiMbHNVzxqvLSRQMqj9RFx7sIJWYqBkdNyXltEgeRwARX8vXpf8AV/aPX/dGxNudm9Xbpxm9NibsoRkdv7jxDyPRZClLmJmVZ1SRGV1ZXjlRXUghlB90dGjbS4oR07FLHPGJYWDKeBHA9L73XpzokXcRI+XHxxH4OP3J/wBapPfuvA9Hd9+690mt47y2r19tnMbz3vn8Xtbam36Q12b3BmquKhxeMpBIsXnq6qYhUUuyqL/UkAcke7KrO2lRU9UkkSJDJIaAcT0CPXnzB+M3bHaGf6Y677k2durs3bWLpM1lNp46rnFa2LrYRUQ1mMqKmNKeuURkPJ9lLKYxy4UA+7tDKi6mGOmY7u2lfw43BbjTPWHffzI+MnWPbu3uid/dw7V2v2rumOlkw+1cm1fHJIK9tNCtZlEgaipWmNhEtVUxlriw5F/LDK661GOvPeW0cvgu4DenT53l8pfj38a8FW7j7x7Y2j15jaClpK2dMxXNLlGpK+s+wpKilwWOSetnV5roDDTtyDfgE+/JDK4qor1ua7toDSZwv+zw6G/F5OhzWMx2YxdQlXjMtQ0mTx1XGHVKqhr6daqkqEWQKwDxurAMAeeQD7aIoaHpQDUVHUuT/Nyf8Eb/AKF9+690BnQP/Hs7g/8ADxzv/W4e9Dqx+fQ7e99V6as1/wAWqv8A+oKtt/5xye9jqj5HXzrP+E1f/b+H5x/44jui/wD6Gsnuo4jp08G6+jMv6V/4KP8Aevezx6r1y9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SF7P/5lvv3/AMM/cf8A7qZffutjj0A3weFvjN1yLW/yfK8Xv/y95vz79149Gz9+611737r3Xvfuvde9+691737r3XvfuvdFB+XVv7t4C5/5eSWH9f3k96PXujQ7X/49rb//AGpMX/7gp731s8en337rXXvfuvde9+691737r3Xvfuvde9+690SPLf8AZa1D9P8AmXcH+v8AU+9Hrx4dHc97691737r3Xvfuvde9+691737r3Xvfuvde9+690Rb4pLp7g+WH/iSaP/3Hm9+8+rHh0en37qvXvfuvde9+691737r3Xvfuvde9+690EneX/Mstzc2/yUfT/X96PXuHSa+Mgt1HhR/02ZH/AK2j3sde6MB7917r3v3Xuve/de697917r3v3Xuve/de6JP8AMUA1fRF/x2jSH/rEnv3Wxw6OjTf8B4P+WSf9C+/daHDrP7917r3v3Xuve/de697917r3v3Xuve/de6I1uYf853dfm3/NLc3z/wAhP9feureXR5fe+q9e9+691737r3Xvfuvde9+691737r3UPI/8W+u/6g6r/rQ3v3XuidfFgW3D2SPrauTn6f7vPvQya9b8ujo+99a697917r3v3Xuve/de697917r3v3Xui+/Kn/sn7s7/AMN9v/cuL37qy8elR0R/zJ3rn/w1cX/1p9+6r59C17917r3v3Xuve/de697917r3v3Xuve/de6JH82gP4D1Kf6dsbdt9fzMvvR6svR2k/Sv/AAUf71731Xrl7917r3v3Xuve/de697917r3v3Xuve/de6JJSf9lW1Fv9RNf/AM5z7r8+t/5ujt+7da6//9Hf49+691737r3Xvfuvde9+690Uv5xf9ky9j3BNqbGGw/7WsPv3Wx0PXV//ADLbYH/hmbZ+v/amh96HDrx49Lr3vrXWhl8f9mdw71+O3z1+C3zT/mGbQ+KXcHZ/ye7grTsnsvqup3n29iOs8zuSmr8Vu/bHZ0k0M8GPycsbfw6OlnYww8K8PlKFeqvo7lJxxBpx6I2kgExCyhBqqVZak0Oc8aH/AFenQ0/zOtkbtofkF1VnfjTmex891P8AGv4a9RbZ/mcdu0eZqxgvkb8QdrVtC67RoMW6zo2bqIKetqp6ugkinBmBMhWlfW5GrKgYtoOo0Bz5f6s9J7yWOScpHC0yiPuINBSvD+kaVx5fmOj4fKzvz4P9cfNz+X3253luGbafxwn+OWysr0WKyjzM+AxP8YgqJ9kQ7twEUdVNLBHStBqjnjkKlQW1EEn0aymBwOJJ/b165ltU3GGWQ0VVxXgMeY4ft4eXRpv5S+79idudxfMXun4tVsr/AA03d2RlqDbURpq3FYvI9u4+vjl3fltsYKuihamo/FIRZY1UB41tcWVq5b9JUb4h/m6V7ah+olmiBEbEkehNeIHz+XV53tD0c9El79P/ABmDrT8f7k8dz/09HHvXn1b06O1731XoiH80bwj+Wz89zUNXpAvw/wDkQ0rYuqkosgEXqnKMTR1cJDRvxww+nvY4560wJUgcetO7qbujo75Hfy1v5F3xb+F+dO7vl/1h8nvivvbt/ZezKPM4jsjaOwNjbsyb9y5fdG61pKeM4paiSN3X7xo3gEdkZYtIU6ZiurOmmKny6LxNaJIIyV8TVQgDzNfLy44qT+3o2csnbuE+WP8AOf6C+T3zh2X8Tcd8ke6dyzfH/C9udWSdlZrN9K7i2vU4yk3v1duWodf4VR0mOmo6VaemctHURvKEjdVlLkSuYxVdXpQ0/b0muZYUnYeMIieIZa18yRnApjFP29Dj198Ytk9c/wA5H+S/1PlN3bh+QeI6T/lN9lY/b3am46WoyOA37UYKtqqDDbwqqetNTEk0kNfJPH5ZmkF6PUxeKMlmrKGYVU1p0uokjohCslNQ+3yoPSlfsx69bbSqqKqIqoiKFRFAVVVRZVVRwABwAPafpZwwOuXv3XuiR9DkD5D/ACJH0J3DBYfn9PvQ6s3R3Pe+q9a9/wDOnn7q2h3J8I+3KbubF9A/FHYG5d5HvftPc+26rfuy9u7jyooo+v03TsKljlFbHUzRyQQvLGUViwLLc3XWjBVbFT6efRNuscjvEytoVSSzcVH2jz/ycegx+OGK+L3Texf5nXy+7i+dO0PlL1v3vkut8f2rgtrda5Da/XvW26I8Ou29g4zCbOo2qCK7I1FZRiGSnhX9wLJJJrVnXb+IHRVXQfKprjz69C9u8EkjyCUYDUFM+X58Bjoo/wDJ6o+/fjd/s7PSv8wnCb02n392xtXc2Z+N2696ZLJZWPsjruPZuVmrqTrqstOmLakhNN5YfNE3FioaMj262uQrpyAckGmP9jpPGsFuHVqhnUaVapIalCQanBOa4/KnRXeo/kD8Oe3v5WPRHw5+FO8lrv5h1d2vmZOm8CMPuaTtbH7nw/cUmc39lchv6qoxoojiTPqeSvKlVjGm0V12CVmLg/pkfl+XTLKktikFP8YWnDDDNckeoz8wet6DbsOSp9v4OnzMnlzEGHxkOVlD+USZKKiRK6QSf2ryhjq/P19lZpXHQmFaCvXe4P8Aiw5v/tUZL/3Cf3rrfRRPgZ/zIj/yed5f+7P3odWbo6XvfVetV7sH5DfEv47f8KBPlTuj5ib4xm1MHWfDvo6k6qfd9Dlc9tGPKyU4TcynE0FPVolTLAGSFpYLEs9iHK+1qLK1sFi41NeiSZ7WHdDLdUHYoUnhxNR/lHRGMBtnuCg/lv8AS/Znx63xmujviNi/5hffnavc28t6UWd3PBtDpzL7mcYfeeR2nKjVFbt8VDMYcekBjiLLdRq9qgY4pNJFWp9tei6T6m7tVmRtMfiMQaUooJyRTII/Pz6P/wBObM2Ruzc38xD5h7A+fG0/lLvbfPwlwPXVb1l1N1lUdabV2ZtTHQSZDB7yXbdK5WStqoYpokeKASAO4dgNK+2TqDIChUE4qa9Lo3jkglKTLIQudK0oPKv7DT7fPq3D+Vl0X1j0r8POr5OvduVmJrd+4o7z3blM7G8u5svnctOzVH8QyFWi1DU8WnTSQSErGhOkDUbsXjs0tGPp0r2mGKGzURinHjx4+pz+3qxn2l6M+itfLG3+j+kuL/7kb3va3pX3rz635dDb1p/x4G0fz/uCoP8AD/dI9+HDrx49Lj3vrXWkV0XtHe2d3h/MO+M3yG/mH7I6I3l2p84N+Z7auzd79ST7o7GjosjLGcVkdn7/AMqYZKGjfRGlNFSSFYghs0ayEOcKPDIkCa6j1oKeh6Ccr/VrLaGf6dtRHCrVpXUpI8wcUwKevUj+ZXsvtLO/Jb4p7y+F2S3tvrqb4cdFYzYv8wj5EYetyQ2RuPpjaGS++7M2purbqWTJZirxkNXVvNB5ZRrCCQiEsGgZQCzdpY4r6eg6WFLdmVErIka0IHm1RVmOPKvkan8uhF+Qvyk+DnWf8+T+XR8iewt54javxU3Z/KXy1b03ujO0OXqeu6TD7g3Fm6nZAba5pamWmV8WlRHeSnBWYU/ks0asqRfEZiq1154H9vRnI1vHGkkhURAClR58B/m4fn62BfyBcdvHK5z+aF3JBQZ2D46d7/PDePYXxVy1a1dS7Z3J1TVYzxwZrYmBrVjNDjZL04SNYIgWBXQPHc0lDqQH49XtXhkUtB8NSOFPP/MRnrYm9tdKuiRdnEf7OB06D/zyGWt/51N70OPXjw6O4PoP9Ye99ePHqtP+bvLFB8Be7JqupqqTFRRbZfOz0Uz09UuDXctKcn4JY/UG8d7W59qrT+2x0WbxT6B9WB50+w9Uk0PyG+GXyJm+A3T38tHdb5T5b7M2Juir2jkqShy+Bye3dvYrERje0PY25cjSwxVTvUxTyBpZZFClmv8AuW9q+5WdpfhJxXh+zoqBinSBLLMiLmlarw4nH8/kegPosRuPd3Y/zK+PPyt/mIdc9Adjb17Z6rrd97M3V1B/fPeceUo2hqsCdn9hzCOOngqrRqhpJrWYMNKFibVACvGmqgPA4/Z0zod2ltrq48JnKmhWrfKhPr/hPVqfafxm2Xlf5pf8sifsqu3V23n+lOhu1m2tuzLJWNtjKVhwc0EGYzuHXzUM1RFpE1NLM5aNijK2oKfbTkNAZFGkj0Pz6MY1aO+S3dtakEior5YoTw4+R6v/AAAAAAAAAAALAAcAAD2W9HvXfv3XuiN9If8AZWvyc/wptt/8Rf3rz6sfh6PJ731XrWr/AJ9S/Iza/f38rHuPYHZ8nS/xx6r7631kPkn2tlMJUbv2BsPH5TD4uHbeZ7F2RFePJ0MkMWTpYI5kKrNIrao7F1UQaq9or6j16QX5jVAXbSa4biAeNSOBFPXoBKzrf48L13/NY+TnYHzBpfnJt75U9E9e7Gq+iOj9r1/VVFgl25rotn5HamMxVTIaeunq56Wd6yBAQsbvM0rMAVIjkaVQF0gHzNekD3MEdlK7uJiQcKtCfQAD+VKf5eiVdXxd0dafyx/5tXTXzBz26st/MN238WOv907g3tk6ioZoPirk6aloOk9m0WTpY6emWrxcEddDVkRedmljSWSR4Gb34hg6UYMAx4Yz00rBra41wmOQxitTqqKGmfkfLoxVJ3X8JPkJl/5GXWfxGztRvDuXqHtTYW4dwttWh3Jic5siKm2DTR70ru08nNSwivpp6tagvBLUyxtYsLJb3bQ9ZJJ+FMenWvqLfTbWu3sNWrIpkADNSRjOPnw4dbjXss6EnSA7S/5l9uuxt/uJn5/2I9+690EPxQ/5ltP/ANr2t/6FX3odbPRnfe+tdatXyC+T/wDL9+Mf877v7cnzQ3jj9tVOX+D22sZQPvnFZ/duxn26uSpayvo6bb1DSVsEdZNDHKnkEAd1JTV6vZgAxgURfEM49OiItGl/K1yP02AUVyCantHz/wBXl0Qnt3afyix/wM6h7Mw3b1X1J8d6/wDm1bb7z6L3hm8Lmtx43b3xUyglqtlV+5torG8kmDHqnTGVMel+W02cEvUWV6DLBc+Vfl9vSMs9nBreoiaXtJqdIJwxAHwj04dH73B1ptHe/wAZf5s/yP2v83ML8o6LsT4Ld2bJzGy+mdhf6MsFtuuGychkYNyUWExUsgauZaZoI5Y42dg0uqRixBanbUiRMmgA8a5+zpVYxabia8juPG1LhAAFBBrUfb59WafyPNibY2H/ACnfgZQ7bwYw5yPxv69zWVlqcfHQ5jI5TNY85jIVeYbxxyPI008hXyD0ppVbKAPaKQnVpqSBw6OIFXww+kBmFTQeZyf59Wue2+n+iW/OD/mXWzP/ABJm1f8Arc3vXWxw6OLjf+LdQf8AUFS/9aF92PHrQ4dJLtGl3PXdZ9iUWySw3nWbF3dS7RK1H2jDc9RgKiLAlaq48R+6MVpLjT9b8e9r8Q+3qr5Qgeh60supuv6vvHoHr/4j/Kn+Z7sTozvfYnyd2Bunt3rJum65u6tv9oYDfH8R2zsiftaoNNNUff6YiJFmeGzAOxTUpNJCzd0aVoONcfPHQZtQIgIp5wpLfCRVqngNWOHl9mR0qv5plDuDe3z7ynYPRW3OxN3fDj4/929F4r+aZ1rsetyO1a7sTuIV0Vb15vRJqdWmqKXF4+SjNYtJJFASj60czBxSLWyKpYHBovnT7enLwwxzvKkbCjprcVoTjiBxHDhWlM9Ht+d3yU+D/UP83j4IdyfJfe9Hidi5P4t9u/3AzeWps1muuxBlKWo11OX2zQU9TG0njfTE81KXjawsCthQI4g0D4/8lenpJ4hfiZyPBCkEngDgAfnw6NJ/Jymrd673+cXe3VWRkrfg13Z23tTcfw9EVPkMTg5cBjsFUY/f1ftnbmTSKWjopMoNMd4UDaQLXQhWrlqqgY1YDPSnbU0vMyAiNmBWvClBwHkK+XV5ntJ0a9Eh7iP/ADlz8chb647cf/WqX349bHR3vfutdU4fz/Z62l/lJfLypoausoWp9tbLmqqqgqXpKqLHR9mYZskY5oyGF4dYIB5HH59v22J18ukO5qXsJUArVaU6rDxfdPwm+Sfyg/kzbZ+FucG6+xulqLflfmcptfG7nxMeyMXV9OQUU8fbUs9LB/EfNVxzloppZlEheReHF1gR9LyXANPt/wAHRX9TbtNbwbewrmuOAp+IkdE8p9n7y3t058svhp85f5k3Xnxi7p392vWZLsnZu5uoq/eHaG39l1m+osxsLNbZ7U/aZYMpGIvtvs6ltEZ+qKzXuxMgHhoWoOINB+zpPArQl0uZhHrbKsKmhNAK4OT+z59XW0vx16/35/NR63xfZVPn+x6ToX4u9d1fXeVzMM9TtvN5DF4YYr+8W46eVJKOqnml8lWquzaZyG5I9sh3W1amM/5eljQRSbwjSAsUXBNacOPoTjq+AAKAqgKqgAAAAAAWAAHsv6PuuMn+bk/4I3/Qvv3XugL6A/49ncH/AIeOd/63D3odbPDod/e+tdNea/4tWR/6ga3/ANw5P6+9jqrcOvnV/wDCas/9j4fnGP64juj/AHje0nuo8unDwPX0Zl/Sv/BR/vXvfVeuXv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6QvZ/wDzLffv/hn7j/8AdTL791scegF+D1v9lm65tyPt8r/7t5veh149G097611xLKttTKtyFFyBcn6AX/Pv3XuuXv3Xuve/de66BDC6kEXIuCCLjgjj37r3Xfv3Xuig/Ln/AI9vAf8AayS31/47J70evdGh2v8A8e1t/wD7UmL/APcFPe+tnj0++/da66uAQCQCb2F+Tb62Hv3XuvMyqLswUcC7EAXPAFz7917rv37r3Xvfuvde9+690SHMEj5q0P0567g/3s+/deI6O97917r3v3XuugwYXUgi5FwQRccEce/de679+6910WUEAkAtfSCQCbcmw9+69137917r3v3XuiL/ABTN+4Plefp/xkmivzf/AJR5vz791Y1Ap0ej37qvXvfuvddAggEEEH6EG4P+sR7917rwYG9iDY2NiDY/0NvfuvdeuLgXFzewvybfWw9+69137917oJe8BfrLcv8A1C+9Hr3SZ+Mhv1HhSP8AlbyP/W0e99bPRgPfutddFlW2ohbkAXIFyfoBf37r3Xfv3Xuve/de66BDC6kEf1BuOOPqPfuvdd+/de6JR8xP+BnRP/iUKT/rUnv3Vhw6OhS/8B4P+WSf9C+9nj1UcOs/vXXuuri4W41EXAuLkD6kD37r3XiQPqQLkAXNrk/Qe/de67JtyeAOST7917rwN+RyDyCPfuvde9+690Rrcx/5zu6+H/frc5/vb+9efWxw6PL731rr3v3XuulZWuVYMASCVINiPqDb37r3Xfv3Xuuri4Fxc3sL8m31sPfuvdd+/de6h5H/AIt9d/1B1P8A1ob37r3RO/izY7i7J/6jk/63n3odb4Do6HvfWuvEgC5NgOST9B7917roEEAggg8gg3BH+B9+6914MrXsQbGxsQbH+ht7917rv37r3XvfuvdF9+VP/ZP3Z3/hvt/sf8ri96PWx0qOhyT051zcW/36uL/N/wDdHvfWvPoWvfuvddFgCASAW4UEgE25Nh7917rv37r3XvfuvddAhhdSGB+hBBH9PqPfuvdd+/de6JH827/wHqS3/P2Nu3/6nL70erL0dpP0r/wUf71731Xrl7917rq4uBcXIJAvyQPqQPfuvdeJAtcgXNhcgXJ+gF/fuvdd+/de66BBAIIIPII5BH+B9+69137917oklJ/2VbUf8Em5/wCnB91I8+rdHb926r1//9Lf49+691737r3Xvfuvde9+690Uz5w/9kzdji9r02MH+3ysPvx62Oh56v8A+ZbbA/8ADM2z/wC6aH3ocOvHj0uve+tdJPJbD2Nmcr/HcvszaeVznhSm/jOS27h67K/bxm8cH8RqoWm0KeVXXYfge7B2AoCeqlEOSAfy6eJsHhaimrKOoxGLnpMhR/w6vpZsfSS01djxG0X2NZA6FZYdDMvicFbEi1ifeixPE9bCqOAp0Vzdnw36n3z8i9u/IPd+Hwe5qna3XqbEwuy9w7Yw2a29jjT1z1VHmsfFkEdIJoUkMcYSL02BVh9Pbwm0xGMDPrXpI9oHuhcE4ApSn+X+fRn8Ft3b+16BcVtnBYfbuLWaaoXG4LGUWIoFqKl/JUTrR49I4w8jep203Y8kk+2SScnpWAAKDHTx711vokffpt3D1nfkfxLHfn6HzDm3uvn1YdHc926r0Eff/UuN776L7j6PzNWaDE9v9Yb660yNcIRUmiot77aqduVFWKcldfjWoL6NQva1x7spAYE5A6q6lkKqaEgivp8+gj+GPw86v+Gfx66T6M2Vgdrz1XTvW+H6+j3tRbVxGHzmcjoY9eSrqqrp0NRerqGeeZXnYu7FnJYk+7SPqYkYB8vTpuGLw41VzqYChbzJ9T8z59GPzWx9lbkrafJbi2ftbP5GjhenpMhmtv4nKVtLTyG7wU9VXQySIjHkqrAH+nuodhgEjpwojfEAfy6dkwuHjq6OvjxOMSvx1DJjMfWpQUq1dBjZShlx9HUhNcUDGNC0SMFOlbjge9VJ6tQDA6c/euvde9+690STodQfkR8iD+RuCD/oT3odbPR2/e+tdNWaweE3JjanDbiw+Kz+HrAi1eKzWPpMpjapY5BLGKmhrkkicKyhhqU2IBHI97BIyOtEAih6aKfYWxqTGVuFpNl7TpcNkp4KrI4mn27h4cZX1NK6SU1RW0EcIileNo0ZHkQlSqkEWHvZdiakk9aCIoooAH2ddbx2Zgt7YTKYfL0NHJJkMLmsLT5OSipqjIYqLOY58bVz42omUtE+h+dDC9gDx73HI0bAj/i+qyxLKhVvMcfT59F8+Knw/wCpfin1dsrr7aW2No1mW2cM9JHveDZmAw24ayr3HkpshlKo1lHEZkaVZVhlImJdUGokcC80viPUYHTVrbi3iEeCR506Nh7Z6U9NG4P+LBnPz/uIyX/uE/v3XuiifAz/AJkR/wCTxvL/AN2fvQ6s3R0ve+q9ENwfwK60o/nH3d819xR4TeOf7j6h646n/uxuLbGNytPtuj2FK7zV2Prcj5QPvFEIkRIl/SdRIt7e8X9IRrgg1r0lFsPqnuGNQyhaHypX/DXo7H929u/wSTbP8Bwv925aWShl2/8Awuh/gklFNfzUcmK8fgMT3OqMx6Tc3HtqpJqTnpTQAUpjqFg9lbN2wlTHtraW2NvR1sMVNWR4PA4rEpV08CGOGCpSgijEiIpKqr3ABIAsfeyzN8RJ6qsaLXSoFfQdKSOOOGNIoo0iijVUjjjVUjjRRZURFsAAOAAPda1yergAYHXP37r3RW/lif8AjH9LyB/uRH1/4KvHvR635dDZ1r/x4G0f+1FQf9aR78OHXjx6XHvfWukrWbF2RkMsuer9nbVrs4jpImarNvYipyySIAqSLkZoWmDAAAEPce7B3AoCf29UMcZNSor9nTqmCwkdJkKCPDYpKHLfc/xWjTH0i0mT+8jMNX/EKdU0TeVCUk8gOocNce9FieJ6sFVfhFOiDbz/AJdfUm+vnt1X84c9RbdyNZ1P8atxfHTbvXmS2ni8hgqTGZvcb5iDLY9KoNBTfb081RRxxR04tG5VSFYr7cEgEZUDuJ4/LphoWa4EpbtCkafIn1+0cPsJ6sAxmLxmFoKTFYbHUOJxdBClNQ43GUlPQUFFTxi0cFJR0qpHGi/hUUAf09tkkmp6fAAFBjqd711vokXZ3/ZYPTn/AIaOW/8Act/z70KV62fh6O4PoP8AWHvfWjx6AT5MdB4L5M9P7h6f3HlKnD4jcVVhKmpraWmirWthstDlBTy0k7Kskcvi8bqW+hv+Le3YZPCfURXpNdQfUwmKtOldsDprqzrCgw1HsbrzY+15cJiIsNSV+3tpYHB1opBEiVCfc42COQCdkEkw1+tuWuefdXdnJJ/Z07FEsSBB5ClfXpQ5DYGxMtlGzmV2VtLJ5p2pmfMZDbeGrco7UfFGzZCphaUmK37ZL+n+zb3oO6igJH59bMaMasoJHy6UzUtM00VS1PA1RTpJHBUNFGZoY5beWOKUjUoaw1AGxsL+9VPDq1BWtM9Z/eut9e9+690RzpEf85a/Jv8Axpdt/j/W968+rH4ejx+99V6bstiMTnsfU4nOYvHZrFVqeKsxmWoqbI4+rjuG8dTRVivHItwDZlI97BIyOtEA4Ir014zZmz8LTSUeH2ptrE0cyxpNSYzBYugppUiIMSSQUsSKwWw0gjj8e9639T+3qvhxj8I/YOi9/Mj4s7Z+XXxs70+PmUrKfah7w2KNj5jeFHi6eoysFDBXx5GiWodDHJPFE6MFiaWyh302JN7RuEkDtkDpu5hM9u8KnSWBFfSvQjdP9Fdb9MbK2FtHaWz9o46XYeytt7Los3itrYXEZGoo9vYWLDiZp6KISL5hGZHXyHljcm5J00jMTk0PlXq0cSIoFBUACtPToY/dOnekB2l/zL7ddv8AnUzf72PfutjoIfih/wAy2nv/AM72t/6FX3odePRnfe+tdE6m+FPUeT+VW8flRujB7c3fuPdvWOK61OF3PtHBZuHGUmPq0mqK2kyGSjlcCeKNYnhCAWLXJBt7fM1YRGBQjz6RraBbprkmuocKcOjXVG38BV4iPAVeExFVgooIKWLCVGNopsRFS0qhKWmjxsiGFY41VRGgSygAAC3tkEg1HSsgEUI6iYvaG08HQ1WLwm19u4fGV0ckVdjsXhMbj6GsilQxyx1VJSRJHIrKzKwdSCCQeCffizNxNetBEX4QB+XT1R0dJj6WmoaClp6Gho4Iqako6OCKmpaWmgQRw09NTwhUREUBVRQAALAW966t1I9+690S75wf8y52b/4kzav/AFub3o8R1sefRxMb/wAW6g/6gqX/AK0L72ePWh1N9+690jp+u+v6rMTbhqdi7OqM/UVNNW1Gcn2zhJsxPWUahaOrmyckBnaWIACORnLKB6SPdtbAUBNOqlEJ1ECv2dPiYLBxtlHjw2KR83NHU5p0x1IrZeoiiWCKfKMqXqHWNERWl1EKoANgPeqnrZAOD0Vbf/w06t7J+TnWXyW3Zi8Fncp1jsLcuyMRtjO7Ww2bxltwOdOVpZsij/byQo7qFjj549Qt7fE58ExHj69ImslN2tz5AEUp5+o6NZh8NiNvYyjwuAxWNweGx0Ip8ficPQ0uMxlBTglhBR0FEqRRICSQqIBz9Pafpdw6cvfuvdEi7it/s3Hxx/qMfuS3+P7Unv3Wx0d337rXRSvnP8U8L82/i32n8ZNwZ+bbGJ7NosFSVeahpPvmpFwm5qLcaq9Hrj8iSGkETLrHDX5+hchcRyB2FadJ7uA3Nu0AbTqFK9DZ191J151nhtvYraOzdp4SXb23cTtumyOG21hsPWyUGJx8eOhjM9BCjhSkYumoj8e9M7NWpNPt6dWNVAoBUfLp5yvXmwM7lHzec2Ns/M5qSKngky+V2zhcjlJIKN/JSQvkKuB5SkTeqNS9lPK296DuoopI/PrZRGNWAJHy6VS09OsxqFghWoMSwGdYkExgRtSQmUDVoBJIW9gfeqnh1ugrWmes3vXW+uEn+bk/4I3/AEL7917oDOgTfbOf/wDDwzv/AFuHuq9Wbodvduq9NmZ/4teQ/wCoKt/9w5Pex1VuHXzq/wDhNX/2/i+cY/6tPdH/AL20nuo8unDwPX0Zl4UD62A5/rx731Xrv37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdITtH/mW2/v/AAz9x/8Aupl9+8utjj0AvwcFvjL1wPp/k+V/9283v3Xj0bX37rXWpN2F8iu/vnD/ADFv5knxhznxo7S7l2r8CZ9gSfGyq6i7UHTI2vvjdO0RlKTJb4lqq6gXNPX16LUQHVP4qeFh4TGwPtbA6ohFQvqSK1+zoovYXllB0mXSagK2nSRw1ZyD6H8xw6j/AC0+cn8zr4GfCn+Xh8ou95sLn/mIuR7N697/APhvi1jqa3vLC7krqqm2dujHS7SWsgpKzadCtBX5CaGJxLJKodwRL78g8QMqLqAocf5+tXUq2rxSTSaGaq588E8BXPn59G07T3n8gcJ8Q/5dmzdjfLHz7n+QG8sgewe9pcgExtdi93A5rK7XNTDLJY4/+IS4uJmcMklMTpjYFFejoZn1L5cPTpFda1trdYZahmqW/iBORQH8vl0NPxu2jV/Cj+YRlfi11jvPd3YnSHcuxIt+Vm1d27pzO99wdSbwxmMmq6/N1WYybyPBT5Z4yywOQumVLfpQ+25CJrYOwow9OlMAa13I28bFkcVoSTQ08ia8fTyzTq8T2g6POig/Ln/j29v/ANP4klx9L/vJ70fTr1adGh2x/wAe3t//ALUmL/8AcFPe+vHj0++/de60UOpuxe6t0fyivnX/ADX97fILfG7PmL8YPk18gZelt04/dWUg2Ttqm2d2Bh9t4vAZnr6IrQV1HLHXVJip6iEr9u1OGY2csrNw+nRQEU9OiwWEfieNVwdVaavWlf8AYB4dHf7F+Wnd3zS+ee2PhN2P8fewe9un8H/Lx+P3y7q8T0pv5ukM9Tdv9g4uir8ruXJbiqK2hFTjHlrTj6HG+e8UquwjkKuvvcDBK0IU+pz+XWr6JpqBlZ1x2q2kg8a1BGPKn+EV6Cb5Gbt/mf1Hxp/la/H/AOU28q34/wC9vkN/NGqekd9bfwucxr70zfxTgknn69wGa3Xt6Sfy10eOimjmqUlWad/tJpSD5FbSsVkaSOhA448utzxCW3jgm1KWYAUOQaVyR5cR9lOtuvrDrfbnUexsB17tJ81LgNt0rUmPk3DnMluLLvG0rTM1Zl8rJJNK12NtTWA4AA9p5HMjam6XQwrAnhpUj556X3unTvRI8t/2WrQ/+I7g/wB7Pv3l148Ojue/de6qe/nEfLHtn4mfGXZ+b6X2jW7y3n273dsLo0Y7GRS/xODE7/hrYMlU4uuQhaOpKwrFDVyG0TPqFmAYKbVVaTv4Dou3SaaG1rB8RIH218h6E+XlXj0T74V9H/M7ZPy63DsGh6G7U6F+AvYPxm3Djuzx2j3Zi+zN7z/IzKVZp4917HysWQq6yhDUcrw6YkVL/ulUKqivTSIRWoYggigp+XSezt5Vquho1ZSDqbUanz4kjPl0WP4A/NXuTvn+Zx25/LD7V7FrF6j+HVHlsn1jNSrWYzfPc1P1pnaahwG58vvuicvX0SpMHmpvLaVU9WqxI27IiGULRjTj8+qwxXEkogeQNEK5XDAqRQHNc1rw4cOnbbvXXY/yM25/Md+XfbXyK3XX9xfE7vbsbJ/GyHZO6cltbbHXe2esNtDce39gbm2pTyCkr2rp1anZpUBcslrnUGcZjCywhQQw9M/l0lSP6uKS7LsrxsaHUaYAwQOI+XD06vw+BfeW9Pkv8Nvjh312LhE25vntTqzbm7d04aOjkx8dFmK+ErViOil5iVynlCfQBuOLe0EyhJWVcAHo9tJGmtY5X4soJ+0jPRuPbXSjoi3xS47f+WH9f9JNH/7jze9efWzwz0en3vrXVEf8wjdvbO8P5nX8ur4n7c7SzHXHUncWwPkHuHtGmwmVq8PX7oo9sYynvg6Oqp3QCoqIPJBA3LIXLqLj2st20Ru9K0p0U38bS3cESuVrqJ+YAGPtz0RfNfKLvb+WvsL+apsTp7cOX7w6++O/bXx9218dNn5aTK7w3T07D3Jjozu3Ebj3Pk5JKmu11MzS0RnZlgaxY8sC+sYm0SSUz5D/AFeXSOWeSxjnhtgTopQsScmmPPj/AKvkOfw+65+cnX/zC+OO9dhfHTtPqL4276617D3F82833D3Xi+yot5b1yG3P4r1/ktjYWTIVFRj5Yq8qQlPTx+hzFIoVGd6TtGwKagfSg4dPWMc8ZEpjZSRVqtUGvpk0pjAFPs6HT+Wj092F372bur5tfIHuHde/8xs7fW/9ndC7Jo9052g27sHb+TlnpdzQ7mwNPIlNkJ3EsaUfnV1iUFguoIRq7cBQiilR/g6rtUMjSyXE7liCdIzQA+Rzk/M/lTq932g6Pugm7w/5llub/Ck/4170evdJf4yf8yjwv/UZkf8ArcPfh1s9GB97611qsfKDs75JfK3+Zj8vvipkPjr27211R8a+oer93fH+bqrtWHpw7W7M3DSR1WQ3XuvJz1dKmWjqp2Ahp2EpSKLQsfqLKYQUjUEkKDxJFa/L5U6Ib3VcyPEis7JSgVimn0JNRUNnHyzxHSO/mHfO3+YP/Ln+IHxJ+WG/shh8X8ntzdo5PrTsr43VdKm7qLtnriiWoO2sTiYMKZ4KLMUcRjrazJUwMshlCk39BswhdmSPuRcjGSSeqxNfxwRzXWmOd6hu6qqoBPyrw+dTwHU75o7x7n2vtf8AkY9KbY+Qu68Fs/52/LmbL/Imrnzsibj3Bi9/4mi3xWdeUG4KVpHpKbFyZWtxccUUqkVS05NgrR+2DKRKWAAoKU6WLah7ZI2dmq2rUDTifP5Z4dG2+Ce/94dD/wA4X5g/y0Nm7ozuZ+LXWXxb6w+RGycFvXI5PeG7ds9jb/z9DTbpSHe+WZ6lqCoSsZ0oZXKoQpj5WS7Mkhk7iAD8ulVvAtv+mhJHzNfT/P8A4OthT210q6JR8xP+BfRP9P8ASjSf9ak+vvXz6sOBHR0KbmngP9Yk/wChfe+q9Bl3nuHJ7S6X7Z3RhZHhzG3uuN6ZnFzx3EkGQx23qiqo54yvOpJFVhb8j25EAZBXpm4JWBiPQ9a2mK667Nf4p9UfzGtzfKLcG4flHtjsrEHEVkW9Mu+ys314mRNCnTGR2Qsn2tRk5E1SSylPIykg3HPs0ZgZjEFGnTXh0Go4n+jju2lJkV+IJCkehWucef2EdIftn5E/JX+YN3Tm9pxfFPujdO2tldD5bK7B2t1r203UP91O6ZqsDFbx3PU5SppHraVZNEaRSFl0hVAYk30qxwIKHTnNRXrzzXN9KwMZeiYCtoo37f556Hf5fL/MF3V8K/gt0V2Jv5Pjt2Z2n8gOtep+/K3G5qPL7k3n1HP46HcGEg3DhJJPt8pPCwDVUb8uA55JJppi1NLbmpGcj/Vx6UFrowx224ArqoO1qZ88+dPLhx4+fWxD1X1rtnp3rrZ/V2zf4udr7HwlJgMGc9msjuLMmgowRE2RzeWeSoqZTclpJXJ/AsAAC12LsWPn0II4xEgjWtB65PS/916v0Rvc1v8AZ7evv6nq3Of9DP7159WHD/V8ujye99V6pg/ny/Nnuj4E/Aqp7o6GxEGZ37n+7OouphSy0klTP/AewczNRZyPGSRc09bUww/ZUVVYmKadHUFwvtyIgOCwr0zcBzERGaH/AFY8uPr0Wv4ydW/NHrD5c7f2p1v0L2Z8Y/gXv74/9n7l+SG4e5O5MB2xuGg71yGCeXZW5Nm5OXK1dZjhRSyIKmCBo45FWR5hEIwoVzSAkHUGI9B0XWkBCshjeNTk1etSa186jNKAY49Fn/lW/O/5R/KLuXtPr7t/tqmylD8BusO6tr7Q31iFyGMxvz4myM9dSYrtrEYzIrB44tuU9DSK5pTU3aeF2ZHMxXcwaNTVKVIz/P8An0xZTR3MqBZw+hWGkeedNSfUU8v83QC7eqO9c9/J96d/mb1vyh3Nifl3trsfH1Owd15Pc+byOxsDtbMd3f3QzfXWf2V5mp8hPXUizAmqQsheMLZQ2p0vqm8BQAKZ+fSZI/DsPrpGZmBqKGlKtwNOPp/s9bkm0crV53ae2M3Xwfa12Y29hcrWU2kp9vV5DGxVdRBoN7aHcrb8W9ljDSxA8uhKjFkDEUqB065H/i313/UHU/8AWhvderdE7+LNv7xdkj+lcgP+P7596HW+joe99a6pA/mRUnYvbHzZ+BPxcHcNbsH499ut2ZW917GwWSrtvbn7LjwODmq8DjMPuXGyRz0xhmjVmCNdgTYXt7W2/ZA0opUHz/LoovT4l7FasSFcGtCR68P8/Ve3bvy9+SvwG6l/mufGH4/LubsWh+G3VnVG9fjd2Duh592ZrDTdu7igotxbSzm4Kkn7yfGx1UgptfrimXVZRwHWUTBZgO704V/zdJI5nsjLZs5MaAUc1JUGtanieHlno0nwE2V8v9ld/wDTW79k/Gjs3p7oDuvrih3X8na7ufurHdoGt3tV7NbMUGd2bjHr6mqxkkuSmRTCgBKMweONfSvpmgeAh2BccKD/AA9btY76K/UwwlYXqXLPUg0xpXNK/kP29Jv/AIT1bd7F7y6j7g+aHyN7b3f2v3nnvlP8jevaCv8A70ZWHZmK2Xs7c67dosZR7TgaOj9My1TU5eNjHAYFv5FZinlmkCeE1Mj06MLe1gMrXKFviOCTSowaA+R4jy62SPaXox6L78qf+yfuzv8Aw32/w/5S4vej1telR0R/zJ3rn/w1cX/1o976159C17917rUG7D7Z+Znzx3B82d79bfFzuPd3b3xm7P7E6W+Ie9tjdtQdXbE2du/aGWkXD53dOzsrX0gy92CVE8sqypIP2yFDKyGY8OKJaEKSMgitfz6Dn691dyF0ZwhKqVagH2gkVI/OvrXpZfzRvml84v5evVfxY+REmZpch8j+7vjntP49d/dcpOu4Nm9AbxemjzG6PlM+Awhqknp6HIvNTsphWm1jR52S6K3HpddASo1Vr/q9On7tpLdvH8YI5SmhuBI88efpno2/zX3J3Jkfjz/K/wCtNn/KWmkwnyT7a6mwPf3beMkqsdUd4bM3LgqLI7kG0sgkvlxi5Z6h5qURuPEhRSt1I97jqxkbFVGMenW7qiiCKpAkarZNTUcP83QhfE2t3J8Sv5lm9v5eOwspuzeXxvzXx5qfknhsjvfNZHeGe6+3vWbsp9vVu0huiuZpRBURlqiOnnN9BRhc3ZqTsJYBMy0atMcOr2aG0vWskcsmnUKklgS3Cp8vTq872i6OOiSfNoD+A9S3+n+ljbv9f+Oy/wBPej1tePR2U/Qv/BV/3r3vrXQO/Irc2X2V8fe9d5bfnNNntpdN9nbmwlSuq9Pl8DsmuymNnGjn0TRI3HPHHuyfGPtHVJMRt9h61R9tU3dlN/LH+B/8yeL5O7npPlvn+0OnMN/efOblzOY2HWdcdg9my4Lc3VNbsiSZ6Wrq6umRl+4nXyRuSo0qtyYljJMYQABQ+XQdhj8GwjvZGZ2qKUNB3HgfL/VTHHodu8O6PkT84PlV8zOpYvih3X2Hg/h7VTbX6OPW/a8fTsOG7Oy+2Vr8Bubdz1dbRjLo1Yiy+l3RIwAEZbg7VY4YhRgCfMitfs/ydeeS4vLx1eJnWOlAraaGnBq0rT9n+DoUN/z/AMxXf3Q38tH4yd5doDpD5K9v4Dc2N+UGVwWUhK5PH09euLlw6ZDb7vGchJiGQtLDIwWpdm1aifdYTGNcgFf+Kr1e9S5cwQaymc+pFaU44PDNTTrYE6U6g2j0F1VsfpzYb56XaPX+EiwWDl3Rn8lujcE1LHM9S82Wz+XeSoqZnkkdmd2sL6VCoFUF7uXYsfPo+ijWKMRrWg9TU/t6FL3XpzoklJ/2VbUf8Em/60H3r/P1an+Do7fvfVev/9Pf49+691737r3Xvfuvde9+690VL5s0tTW/GzsSnpIzLUPT4zxxi12IysJI59+62OPQ69ZI8fXOwo5BpdNnbbV1P1V1w8IYH/WPvQ4dePE9Lj3vrXXvfuvde9+691737r3Xvfuvde9+690Szvihrant7raWnppZYoclj2ldVJVFEouxIH491+fVq+XR0/duq9e9+691737r3Xvfuvde9+691737r3XvfuvdEq6Lgnh+QnyEkkiKxz5+AxNfhhoNz70M9bPR1fe+tde9+691737r3Xvfuvde9+691737r3TTnlLYPNKoJZsTkVUDkkmjcAAe/de6Kb8F6Ksx/R329dTS0k/99t4P4ZlKv43yd0exA4PvQ6s3RyPe+q9e9+691737r3Xvfuvde9+691737r3RX/lZFJNsGlSNC7fxHgcDnSv9fevPrY4dDV1urLsLaSuLMuCoAw/oRCLj34cOvHjjpbe99a697917r3v3Xuve/de697917r3v3XuiWdlUNdN8uOoauGllkpYdp5VJqhVbxxu1U5CswFr/AOx9+HHPWzXT0dMfQe/da697917r3v3Xuve/de697917r3v3Xuve/de6JH0pSVUXyv8AktUyRMtPUU23BBISCHKWDWHvXn1b8PR3Pe+q9e9+691737r3Xvfuvde9+691737r3SD7Pjkl2DumOJGeR8VMqItyzMSOBb3o9bHQS/FilqaTrqeKqgeCX+N1raJAVaxVbH1Ae/Drxr0Zb3vrXXvfuvde9+691737r3Xvfuvde9+690TT5s01RU9ebOSniMrL2Vtd2AIGlBM2puf6e9HrY6N9jb/w7H3+v2VLf/X8C+9nj1qlMdTffuvde9+691737r3Xvfuvde9+691737r3RLO3qKsm+WHx3q4aWWSlp8fuIVFQqkxwlopAodgLC/v3Xh0dP37r3Xvfuvde9+691737r3Xvfuvde9+691wk/wA2/wDwRv8AoX37r3QHdBIybZz4YEE7wzhF/wChmFj70OtnodPe+tdNmZ/4tWQ/6ga3/Yf5HJ731VuHXzqv+E1fH8+H5x/9qjuj/wB7aT3UcR04eB6+jMvKr/wUf7172ePVeuXv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6QvZ//ADLffv8A4Z+4/wD3Uy+/dbHHoBfg9/2TN1zzf/J8rz/5F5veh149G097610QLvz+Wp8YvkP25ju89z0fYuyuzqajioMxuXqHsnc3WM29KWmgFNQRb5g2zLFHlDSxgJTvUqWQAAGwFn0uJEAUUIHqOkc1jBM/iNqBP8LEV+2h6ELDfBv47YzP9Y7qye2MzvbcnUG291bV2Jmuw917g3xX4rEb0pmo9wI0m4Z5xJJLEzospXUgY6CDYjxuZSpUYB9MdeSwtlkWUjUy1oWzSvGn29V4/JT+Tr1ZvzpD4ufDrq6g3bt34ydc9l753NvdKTsHIJvnE0G8K5tw1NXh9z5WR6q/3ktQsUcN/GhVQth7fhuB3PIaNT9vSK+28SiKGFexSTQGlK/6vLo6Pw3/AJbXxn+DG4uyt4dJ0nYVbuztim27Rbw3J2V2JuDsPLz0O16Y02NocdVZ12+2iNw8yRAa2VSeFUBNJM8uG/l0vt7OC1/swanzJJP7T0fj210q6KB8uv8Aj28B/jkk/wCtye/de6NFtj/j2tv/APakxf8A7gp791s8en337rXWrJ/LQ/4TydO9dbD3puD5fYTsvL9h5X5XdwdoQdfR9r5mn6hz+zp9yNFsD+83XWCn/hmQWWlQVEv3atIxK3KoAntU03hdkRBHGtPP/Y6LVtfqiZrsFWqRQMaaQcEjhniers/kn/Lq+M/yj3RsPfO+cRvLaG+OuqKmw+A3j0/vnO9Wbin2zRMZKPaOayO1XhNZioWYtFRTAohPpsOPbaTumMHzyK9PzWcM7Bm1AgUqrEY/Lj+fXPN/y7vjPurC/GbC7xw2896D4j9xw979KZbdu/8Ac+dz+F7Fp0qUhyOWzFbO01fTr905WkqCYhpjsoC2OmlZmLGmfl05HbpEgRSSAa5JJ6PL7a6f697917okmWH/ADmpQ/8AiO4P+hiPejw68ejt+99e6DLuLqDYPfPW27ep+zcKud2bvPEVmHy1Kk8tFX0yVdO0CZLDZSmKz0dbTlvJS1dO6yROAykH3ZWKmq9VdFkXSwqOig9F/wAs749fH3Cby29svc3fuXxe+eocr0lmKffPee+93/b7LzMbRVk+HOSqLUmTCuRFk4QJ4/7DL7de4dwAQMZwOksNjDAzMhY6hShYkU+VTjjx49PVH/LZ+JON2N0hsjE7ArMM/wAfRt2Pr3fWHztfjOz/AAbbYSQ0u5+waQpkctDVONdbFXTSJMxJIF/fluJFYt69eext3QJQinoaH8/X869VddNfyC+htz9y/JHuz5KU3bMm4+xvlVP3Lg9vbW7nz+C2BuLGYeRKvCz5zae3ZUhqKepkZ1qKWrNyg02QHl6WcKAIjWuT/q8vs6SW1iXdnugag0ArQEepA4n5mp8uGOtirH4+gxNBR4vF0VLjcbjqWCioMfQ08VJRUVHTRiGmpaSmgCpHHGgCoiAAAAAW9oySTU8ejZVCjSuAOpnvXW+iLfFLnt/5YG/J7Ko//ceb3rqx4dHp976r1R//ADE/5amU+dnzv+D29d5xbog+O/TmyO6z2Fl9j7wl2Tuqi3Zl6Wml2XT0ubxkkeQiimnQBzSWNla7L9faqCQRRswI1YoD59Fl7bG6uIkZSUAapBoQcUzx/wBXp0dv45/y6fiz8X+vO1+tOu9n5jLYDvGs++7WrOw9z5bfe4N6VKY9sZTyZfOZxnmZoYnYwupDI5EitrCkUkuJJCGP4eFMdP29hb2qskYJ1mrEkkk/aekt8Zf5ZXx2+Ju78DvHq3cffVbV7bxW6MNicLv7vDfO+NqxUG7ZTLkop9u5ud6aYx300hlVvCANHIB9+a4d00Gn7M9aisYIZfFQtXIoWJGeOCadGv6W6J6w+Pm167ZvVG3F21gMjuDLbnrKQVtbXtNmM3UGpr6kz1zyOAW4VAQqgWA90kkeShfy6egt4rZSsQoCa9C/7b6f6CXvHjrLc1+P8lHN7fn3o9e6THxj/wCZR4T/AKi8h/1tHvw630YH3vrXVf8A35/LT+M3yJ7efvbd6dn7W7PqsJiNu5bcnV/aW6+vJc/h8E+rF0WcgwEyJULGAEGocqBfkX9qEuZI00ChA9RXpDNt1tNMbhqq5ABKsVqAagGnH8+hKwvwh+O+M7Bo+zsxtTI9g7txeDosBhKntLcWX7Hodv0tFTCkFXgsXu2SqgpquWPiaqjQSNcm4ufemuJGFOH2dXWygV9Zqx+ZqP2dUt/PP+Rpsvtqn/lv9E9K43f9H8delfmr2X3v3RKOyMlLvLb23uyQd0Zptsbqy8zVtFAMlEsFFDQ800ZQRgOA/vcbAkyOaFRUfM+nVZo2VVt4Uqjkhs00g1NR86/6q9XD/ED4A/G/4Q0m7/8AQlgNyzbi37Xfd7v3/wBi7vzXYnYudginaposRX7z3K8ta9FTM7Gnpmk0r9TqIBFJJnl+Kn5CnTkFpDbEtHWp8ySTj7ejp+2ulPRKPmJ/wL6J/wDEoUnHPP7Se9efWxw6OhS/8BoP+WUf/QvvZ49VHDoP+4tu5nd/U3Zu1NuiBs/uXYO7sDhlqWRKd8nlsDPQ0KTPJ6VUySKCzcD6nj3eMgOCeHTc6s0LKvEg9Um/FP8AkHfFTrjHdSdjdqxdwZzt/aG/Mb3LktuVncm4Krreh7GopC8KJtDHOKGakSyFoGLKxGknTdSqnumLFUIp60z0V2O1QRRq8qnUPIsSPtpWlej/APyP/lt/G75Q9s4bu7fz9p7d7HwuyKrr6LN9X9pbp66Ffturq2rDFlaXb0saTzIzMkc7DWEslyqqA1HcyxjSKEfPPSu42+2uHEjAqwFKqSpofs6GfcHxM6V3btbpPaW7cHmN04/4/Z3b25etq3cG587k85Q5vbMax43IZbMzzmevf0IZfumfWQL8ce6ieQFiMauPTrWkDhA4r4eQSc9GT9s9Keve/de6I1ucj/Z7uvR+R1bnP97f37rfl0eX37rXQW90dLdZ/ITrbcvUvbu1cfvHYu66VabK4evUgrJDIJ6LI4+qjtJTVdNKqzU1TCyvG4DKfdkdkbUvTcsSTIY5Mg9FD6b/AJZPx16RxtXhtt7j7+3DiK7b2Z2rV4zfnfXYm7aGowWdgelrqGamrqsK6+NzHHrB0KAB9Pb/ANVJ6D9nSVdvgXgWP2sT/h6m1f8ALn+Nm1puq96dXbBfa/YHxy6W7g6l6PkxedyNDiqDF9r4CTHZqHclAriLKSySkyRz12sxvJI45a4qZ5HJ1n4iK9XSytoEQRIB4YIWnkD5dVW/y+P5AnQezPjX0v8A7Nfg+z8z3Ridw5Lf++9jzdt589YjdsO9KnLYFv7jYib+FlIqdaZxoGolm1ub2D8l0yOViIK/Z0XW20wyRJJeKfErqI1HTUHHaMfy62UI40ijSKJFjjjRY440AVERF0oiqOAABYD2h6POo2R/4t9d/wBQdT/1ob37r3RO/izzuLsk/S9cnH/T8+9DqxFB0dD3vqvVMP8AMe/lf4X5/fKb4Zb432d5U3WfR779q9yZTYe/ajYm4cdWZPH68YsNbj2WqZZ5RHGWprOo1epb39qoZFSImvcDgevD9vRZd2zz3KAg6CKEjBHE8eI8uFD5cK9Gp6h/lr/FLpX4sby+Hm19p7iy3T/YaZob6l3lvDNbp33uyXN1v38tTnN95B/v5pKd9P2ZMloQo0AXYs2J5BIJAcjI6UtZW7wNbMtUcUI9R08/Gb4B9IfE/OVGd6uzvc1bLLi/4NDi9+dwb03vt/H40RrFFS4/BZmoamjWNVAi/bOj+zb3uS4eRSrAZ9B1S3sIbVg0ZbHAFiQMU8+hG+KfxH6O+FnWOQ6g+Pu2avamxcp2Bvrs2sxlbmcjnZn3b2LnH3BuWrWtyjvIsbzvphhB0xxqqL9LlpmLGp6VqoQUHRlvderdF9+VH/ZP3Z//AIbzf+5cX9fej1scelR0R/zJ3rn/AMNXF/8AWge99a8+ha9+691XR3f/ACuvi/3x3Dmu8twTdv7K31uXaybU3Iepu3d3da4HPUyVP3P8YyuA23LHTSZM8I9fpErqBrLN6vb6XEiCgoftFekc9jBO2ptQP9Fiv+A/7PQm5P4E/GjcHYmS7P3Vs7Iby3PmPjbS/FLKjd+4ctuTFZDqKlqTVNTVWLyskkTZSZjafL2+5kH1fUSxqJpB8Jpmv59Xa0gepdakroznt9OiB/PX+UV198j+pvhv8b9lY7d2I6Q6P77wO8sxRYTsHIYncO1tq0FP4zJgM5kZZJ0jhAdUp4LlSVKKCL+1MdwGBMtKjI8q/b6/Z8z0gm2/RojtwdBwc1oOGK8MeYyKChr0fL4a/AP47fBTC74xPRuN3jUVnYm6Kzde6t1dkb2znY2866qqwunFx7n3I8lTHj4SuqGjRggYljqaxCWSV5Pi6MYbaKAUjH5nJ/ac9HU9t9P9Ej+bRIwPUtgT/wAZY27e34/eX3o9WXj0dpP0r/wUf71731XoLO9tsZve3SHcmzNtJHLuPd3VXYe2MBHMYxDJm8/tGsxWKSUzEIFM8sYYsbW+vHuy01Cvr1SSvhtpyaGnVE/8vf8AkK/Hfq349/GOv+ReD7QzfeXXy4ff27toZXtzcNd1pj+zqDNSZilqqXY1BMcWi0x8QRYV5sdbMSfayS6dWKRkEcK0zTontdphaKOW5VtYoxXUdIYfLh9nVm/yQ/lv/G/5Qdpbd7m3y3Z+1Owtvbdym1zm+puztz9af3hxWUjEJG7KXbckceRmp1GmkqKhTJEvpDaQoCeOd48Ch+0V6MJ7OG4NXqD6qxX5eXQs4n4hdIYyLoI1WFzu5Ml8asbUYzqvcO7N15/cG4qCKqgip5583l6+dpcjMRDG3kqy5BHFhx78J5BqpjVx68bKBtBYEmPgSan8z59Gd9s9K+ve/de6JHSf9lW1H/BJ7/8AUg/T3U9W/wA3R3Pduq9f/9Tf49+691737r3Xvfuvde9+690U75vSSRfGjsZ4neNxTYy0kbFXT/crDyGH09+rTrY6HjrEk9b7BLEsx2btoljyWP8AB4eSf8ffhw68ePS59+611737r3Xvfuvde9+691737r3XvfuvdRJqChqJUnqKSmmmjIMcssMbyIR9CjsLj/Ye/de6l+/de697917r3v3Xuve/de697917r3v3Xuve/de6JP0Q7t8hfkQrO7Km4KfQrMSqeg8KPdfs62ejse7da697917r3v3Xuve/de697917r3v3XuuiAwKsAysCGUi4IIsQQffuvdRqOio8fD9vQ0tPRwa2k8NNCkEWtzd30RgC5P1Pv3XupXv3Xuve/de697917r3v3Xuve/de697917orvyvYrsCkIYof4kLFTY/pX6Ee9efW/Loa+tiTsHaJJJJwVAST9T+yPfhw68ePS397611737r3Xvfuvde9+691737r3XvfuvdRZKGilqoq2Skp5KyBSkNU8MbVESE3KxykagD/AEB9+691K9+691737r3Xvfuvde9+691737r3Xvfuvde9+690R3pGSRvlp8mkaSRo0ptt6EZiUQm2rSp+n+w968+rfh6PF731Xr3v3Xuve/de697917r3v3Xuve/de64SRxzI0UqLJG4s6OoZGH9GU8H37r3WKmpKajTxUlPDTRkljHBGkSaj9W0oAL/4+/de6ke/de697917r3v3Xuve/de697917r3v3XuiYfN55E662aY5HjJ7M2sGKMVJXzNcXHv3n1scOjhY3/i30H/UFS/9aF9+611N9+691737r3Xvfuvde9+691737r3XvfuvdRJaChnqYK2akppaulDCmqZIY3ngDfqEMrDUt/zY+/de6l+/de697917r3v3Xuve/de697917r3v3XuuEn+bf/gjf9C+/de6A3oEk7Yz92Lf7/DO/Uk2/eHHPvQ62eh1976102Zk/wC4uv8A8aKtH/qnJ72Oqtw6+dV/wmsH/Y+L5x/9qnun/wB7WX3UeXTh4Hr6My/pX/go/wB6976r1y9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SF7Qv/AKN9+2+v9z9x2/P/AC6Zfx791scegF+Dt/8AZZeub/X7fK34t/y95/x79149G09+611737r3Xvfuvde9+691737r3XvfuvdFB+XRA21gb/8AOyS3F/8Adyf096PXujQ7X/49rb//AGpMX/7gp72OHWzx6fffutde9+691737r3Xvfuvde9+691737r3RIstz81qIc/8AMuoD/h+o/n37rZ+Ho7vv3Wuve/de697917r3v3Xuve/de697917r3v3XuiLfFIg9wfLGwtbsqiBFiP8AlHm+l/evPqx+Ho9PvfVeve/de697917r3v3Xuve/de697917oJO8eOstzH+lKP8Ae/ej1vpNfGT/AJlHhf8AqLyH/W0e99a6MB7917r3v3Xuve/de697917r3v3Xuve/de6JP8xTar6IP/f0aQfQn/dSWv7959b8jXo6NN/wHh/5ZJ/0L78ePWh1n9+691737r3Xvfuvde9+691737r3XvfuvdEa3MP+c7uviP8An1mcv9eeX968+rDh0eX3vqvXvfuvde9+691737r3Xvfuvde9+691DyP/ABb67/qDqf8ArQ3v3Xuid/Fmx3F2Tb/leX8Ef7vP9fehx63Wo6Oh731rr3v3Xuve/de697917r3v3Xuve/de6L78qTb4/dn/APhvN9Bf/lLi/HvR62OlR0Ob9Odc/X/j1cX9f+WP9Pe/LrR49C17917r3v3Xuve/de697917r3v3Xuve/de6JH822C4HqT/Htjbo+hI/z6/W3vR62OPR2k/Sv/BR/vXvfWuuXv3Xuve/de697917r3v3Xuve/de697917oklIf8AnK2ov9Sk1v8AW+3PuvVujt+7dV6//9Xf49+691737r3Xvfuvde9+690Uz5wm3xm7H5A/ybGfX6f8XWH37Hn1sVrjoeesP+ZbbA/8MzbP/umh96HDrx49Lr3vrXXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdEl6HI/2Yf5EfT/i/0/8At9J96p1s9Ha97611737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3RXPlhf/AEf0trf8XH/olfevPr3l0NnWv/Hg7R/7UVB/1pHvw4dbPS397611737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3RHOkL/7Nr8m+R/wG23wP9h7159W/D0eP3vqvXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdEt+cIv11sz/xJm1T/ANZm968+rDgeji43/i3UH/UFS/8AWhfezx6qOpvv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuuEn+bf/gjf9C+/de6AvoD/j2Nwf8Ah453/rcPeh1s9Dv731rpszP/ABa6/wD6gq3/ANw5Pex1VuHXzqv+E1n/AG/i+cf1/wCLT3R/72snuo4jpw8G6+jMv6V/4KP96976r1y9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SH7Mdo+ut9SIAWj2luF1DC6llxUrAMP6f19+62OPQG/Cqslr/AI3deVU6RRySU+U1JCgjjGnLTKNKD3odePRqve+tde9+691737r3Xvfuvde9+691737r3SN3jsTAb5pqalz0Dzw0somiVGC2cEML3B/I9+690q6Wmio6anpIQVhpYIqeIHkiOFBGgJ/1gPfuvdZ/fuvde9+691737r3Xvfuvde9+691737r3RKsvUyf7OZQ0uiPx/wCj2B9YUeXVqPGv+nvXWzwp0dX3vrXXvfuvde9+691737r3Xvfuvde9+691737r3SC2l1xtnZWZ3dncHTPDkN65NMtnZHcMJ6yNWVXUACwsx96p1smop0vfe+tde9+691737r3Xvfuvde9+691737r3QT93OU603IwAYimFgRcHn8j+nv3Xuk58apWm6nwrsEUmqrxZBZRaUfj3ode6Hv3vr3Xvfuvde9+691737r3Xvfuvde9+690ht6de7c34+AfcEEk7bbyqZnGaHCeOtQBVdrg3HHv3WweluqhFVVFgoAA/wAsPfutdcvfuvde9+691737r3Xvfuvde9+691737r3RJtx10yfOHYdCI4TDL1jmpWkKXmDKWACP+B/Ue9dbHCvR2fe+tde9+691737r3Xvfuvde9+691737r3XCWNZY5In/TKjxt/wAFddJ/3g+/de6Rm1Ngbd2ZUZOqwlO8M2WcSVjO4bWwbXcWA/J96Ap1smvS297611737r3Xvfuvde9+691737r3XvfuvdAH8op2pug+zJ0CM8e32ZRINSE/dRizL+fej1scelL0ZI03UPXkzBQ0m18W5CDSoJgBso/p731roV/fuvde9+691737r3Xvfuvde9+691737r3SF3113tvsOnw9NuOnkqI8Hl6bN0AjYLorqVg0Tm4NwCPejnrYNOl0BYAf0497611737r3Xvfuvde9+691737r3Xvfuvde9+690SulqH/2aeog0po0zHVpGsfsH8+9db6Op731rr//1t/j37r3Xvfuvde9+691737r3RTPnCAfjP2Nf/lWxn+3/isPv3Wxx6HnrD/mW2wP/DN21/7p4ffhw68ePS69+611737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3RJOh/8Asof5E2/G4Ke9/wDgt+Peutno7fvfWuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6K18sgT19S2NrZH/olfeuveXQ29aC2wNoD/qxUH/Wke/Dh1s8elx731rr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XuiOdIf8AZWvyc/6htt/8R7159W/D0eP3vqvXvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdEu+b/8AzLrZv/iTNq/9bm968+tjgejiY03x1Af60VKf+sC+99aHU337r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XCT/Nv/wAEb/oX37r3QGdAf8exn/8Aw8c7/wBbh70Otnodve+tdNmZ/wCLXkP+oKt/9w5Pex1VuHXzq/8AhNZ/2/h+cf8A2qe6P/e1l91HEdOHgevoyr+lf+Cj/evezx6r1y9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3SE7Q/5ltv3/wAM/cf0/wC1TL78eHWxx6AX4Of9ky9cf9Q+V+v1/wCLxP70OvHo2vvfWuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6JJlv+y1aH/xHcH+9n3ry60ejt+99b697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917oJO8uOsty/X/gMPp/r+/Hr3Sa+MnPUmF/6i8h/wBbR70OvdGA97691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3RG9zH/AJzt6/H/AH63N/7H1P8AT3rz62OH+r5dHk97611737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3RfflT/2T92d/4bzf+5cXvR62OlR0Pf8A0Odc3/55XF/i3+6B+Pe+tHj0LXv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6JHSH/nK2oH+0Tf7zTt7r1vo7nu3Wuv/X3+Pfuvde9+691737r3XvfuvdFL+cX/ZMvY5H1FNjCP6X/isP196PWx0PXV//ADLbYH/hmba/908Pvw4dePE9Lr3vrXXvfuvde9+690x7nzT7b25ntwR4nK56TB4fJZZMHgqdavNZd8fSPVLjMTSsyiSonK+OFCwu5AuPe1GpgK06q7aEL0rT06JP8afnhhO/u1N09I7q6S7j+PPau3drU2+KPZ/cuGxmIrtzbPqa37Bc5gmx9RMJEWS2tGAIF78qwD8luUTxFIYfLpFBfpNKYXRo2ArRhSo9R0BZ/nFfGjIdlw7V2jt/sje3WFF2vUdH7y+RO38HTzdO7D7Qozprtr7lzcsyyxSU5K+c+IaQdQBHtxbNmUkMKjy6afdYUkVSjaWNNVMA/P06Fz5efzF+t/idVfwleuOz+8c/QddVncW6sN07isfn5tm9SUFT9rV9g56eeeNFoFIZlePVdQWuARduO3aQVJC5pn16dudwjtzQKz0XUdIrj1+f5dOlJ/MH6xrvi7158nqbZHY70XcM+OoOoerWxOPXs7srJ5uUJgqPa+E+5KSirQ+aOUyhRHZj+pQ2zbMJClRjifIdaXcI3t1nCt3/AArTJr50PTx1b8+el999K9t9z7zg3F0lT/H8bh/06bL7Qo6fF7x6v/uzSGuyUm4MdRyTDx+IFo5EYgkEGxHvT27IwAIIPA+XV4b6OZGahUpgqfi/Z09fCr5l7T+cfWVb3N1x1/2JtXq+sy7wddbx3xi6PG4vtjbN3Wn3vsdqaaVpsdI8bIJJAvPAvzaksQjpRg1fTpy2uDcAkoyUNO4Ur0cb210p697917r3v3Xuve/de6JJ0Px8iPkQADzuCC5H0HpPvXWz0dv3vrXXvfuvde9+690BfyP+Q/XPxb6k3J3H2hW1VPtvb/2lPFQ4yBavNZ7M5KoFJicBg6JmQTVVTKwWNCwFrsSAD7diiaZ9C9J7q5jtITNLwHl5n5Dqqjpb+YvvH5IfzButeoaXY/aHQuD271pv7Ndg9U9qU0GIz1bH/BXyu3NyZLE0+tER0MMtM3lb0835I9q3gEVsSaGp4/Popivzc7osA1IQtSpxj1pwz/k6GHZf8zrpjB/HDoXftLktz9279+Ru/O1tk9E9cbMloM32F23muvd35GDc9HtwVUkEDx4ikgVqmSSQCOMIOfw08aySmhCgAdK4bhobZSwaRmJAA4mlfXoyva/zc2X031l1hujeOwewKbtzumOtpOqvjFHRYt+8d77lxNEclm9sYbb61JhkqKGlH3NU4nKJGVJOpgvtpYdTlQwoPPy6VNc6IhIUap4L+LoA+qv5svS2/eg/kP8AIvevWncHSewvjOcbH2G3ZWAx2OrDWZCq/h0mPxQgqWjknp6rRTzxu66Wdeefbr2jI6pUHV0kg3WKeGSYI6iLiGFD+XRwd5/KrqDYm7PjVszO5ipTM/LDKZHE9PpDTxvFl6jGbQG9qh6tmkBjX7JkYFVf1ML2AJ9sLEWVmr8PSx7hEkjjINZOHyoK56Azqn+YRsrvrem88V0Z1L25251jszObs2bU987LxWDresMpvzY80lFuraOKr6mthnkno6qM0skni0mThb3F3Rbigq6gny6ZN62ohInYA0qAKGmOin/yXP5p3ZP8zrrvunc3YXx13Z1HJ1b3Lv7YGL3kKT/jHm6sfgcusVDhaCurJBUHN4+CZIsrAsTRq6k+QE6PdJEQDUmPKh4/b9nTkEsrMY5VzxqOGeA+0efV2ftnpV1737r3XvfuvdFb+WAv1/S/X/i4/jj+yvHv3n1vobOtf+PA2h/2oaD/AK0j3ocOvHj0uPe+tde9+691737r3XvfuvdVN/zn/mR2H8JfhXneyOsusN+9nbk3nvLbfU/2vW6v/enbWP3wlRSV+5MTNEGaKrijjMdHKFOiZ0fgqD7ft0DPkVp5dIr+V4oKowUk8TWn8gcnyx0juivkvsv4k/FLoHZON2J8re3e9uy+vW7VxHxz33mo9+/KGXbrsn96c/lYstUIkWNoJCAqhwRrVQhctZ+SMzScVUcK/wCQ9JIZxZ2o7ZJHpqKnLH1Irx/b0dTEfMfaW9Pjxke+eq9ib97XyWHWkoc10zsmixtf2xgt1TyxwVez87gHqFWlraVpA06ySWCDyLqXn2wYGWTw2NPn5HpWL1Ht/HiUv6qPiB9D0nfj988es+7Ngdz7z3NtXfHQ+V+PE+STujYvbdBR4ndOxaPGYZ8/NlchTY+adWpjSxySCRD/AGSLcqT57dkK0IIbgfLr0N9HKrkqUaM0KkZ/1Gv2fPrP8JfnP1/88dpbq7N6e2T2FQ9QY7OPitg9q7pxVHj9o9v0FPNLSV2f2DPDPJJNS09RC8ErSItnFr3BArJEIwDqBr6dOW9wZ61RkoadwpX7Ojue2ulPXvfuvde9+691737r3XvfuvdEb6Q/7K1+Tlr/APAbbf8ArXuDx7159W/D0eT3vqvXvfuvde9+690z7hq5qDAZyup20VFFh8nVwOQCEmpqJ5omsf6MAfdkALgH1HTcpIiYrxoafs617/5dP833b+8NnbZ2p29i+190Yzcff/ZfVOO+T1ZR01d0/wD3gx2Zknh21k91VLwvTyUZdaUoY2VAQdQUWC+e2DkvGRgcOiWz3EwhYbpHGpmAYjHyNfQ+Xr1Zz8sPnC/xSrqWqyXxv777Q6/psBFufeXanW238Vk9j7Gw0lYaWSpzdfV1UTkxKPNKsaelCGvY3CaKDxMagD6Hj0vub4W2TGzqBUsoqAP9XpXrh8lf5jfxp+LnXHQ/Y2/NwZHKU/yb3VtvZXRW3NtUkWQ3H2HubdONXK4vF4ijkkjTX43jR9Ug0ySJHy7ge6rCTIY2IBHr07JdKkKzIpcNTgK0r69MPVv8yjpDe+A7yff2F3l0R2X8dNlZfs/tbo/tShosT2ngOrMfHJLQdjJgYZmEuJrvGyU9SHCiSyPp1KW20BVgtQa+fVYrxZImkKsukVIPGnr0gtlfzbfjpvHsXpDa023Oytp9e/JObD7e6L713ZgKfGdXdj9l5pPNSdXYLKJPJI2WKAkLo0MQQGtz7u9qyVqwqPLpmDcopwGVW0twamOjI4P5y/HHdXUffXdO0t7xbl2b8bt6dg9cdptjIg2Qw2/utCke5Npx0sjLrq/JNDHCmoa2kUcc2bELFwgoa9KGu41hadgQFqM8cenRjOvt64rsnYmzOwsFFXQYXfG18FuzEwZKn+1yEOO3BjIspRxV1NdvHKscqiRdRs1xc/X22y6WKny6fjcSRiQcGFelh7r1fr3v3Xuve/de6Jd84BfrnZv+HZu1fp/y2b3rz62PPo4mN/4t1B/1BUv/AFoX3Y8etdTfeuvde9+691737r3XvfuvdQMrk6HCYvJZnKVCUeMxFBWZPI1cpIipaGgp2qquokI/spGjMf8AAe9gFiFHn1pmCqWbgM9FC+JXzIxfy8pdzbj2f092zs7rnG1VVDs7svfOJxeN2r2XTUWQfGVVbtFqaqlndFlR+ZIgCFb1XBHt6WHwuLAn09Okdrei7BdUZVzQkYNPT5Hy9eg76/8A5hWH3L8h8J8d+wfj73v0ZmN8Tbop+r94dpbexeJ2n2PNtTVJXxYCopaqWUPJGvkhWSO7Bk+moE2a2IQurBqcQOqJuCtOsLxsmqtCRg09P9mnQO/MP+atsLoHMdtddbQ617T7Jy3V23YF7N7M2ViqWfrrpnL7nonTbv8AfLPO7PFKJSh0JAbP6Sbg2cgtWZlZ6ZOAfPpPe7pHCsiIGOgdzLkLXzJ6M/8Ay8eyd39vfDzpjsffWbqdx7k3XhcnkqvM1snmq62B9wVcdA9RLZdTLAsaXI+gHtm5GmYilOlO2yGaySSpOoVz8+jp+2Ol/Xvfuvde9+691737r3XCT/Nv/wAEb/oX37rw6AvoC392dwW/57DO/wDW4e9DrZ6Hf3vrXTZmf+LXkP8AqBrbf+ccnvY6q3Dr51X/AAmr/wC38XziP9MT3R/72svuo4jpw8D19GdfoP8AWH+9e99VHDrv37r3Xvfuvde9+691737r3Xvfuvde9+691737r3XvfuvdIXtD/mW+/f8Awz9x/wDupl9+62OI6AX4Oi3xm65HH/AfLfT6f8Xif3odePRtPe+tde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690STLW/2dSh/wDEdwH/AHk+9HrR6O3731vr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xugl7xt/oy3Le3/AUfX/X96PXukz8ZP8AmUmF/wCovIf9bR731vowHv3Wuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6Izua3+z3dffW56tzf+t9X+nvXVhw6PN731Xr3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xui+/Kn/sn7s/8/wC/ebj/AKq4vej1scelR0P/AMyc64/H+/Uxf/Wge99a8+ha9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3RJKS3+zW1Fv9RNf6f8q591630dv3brXX//0N/j37r3Xvfuvde9+691737r3RTPnAbfGfsfi/8Ak2M4+v8Ay9Yffutjoeer/wDmW2wP/DN21/7p4fehw68ePS697611737r3XvfuvdFa+be7u69hfEvv/efxz2lkd9927b613BleutoYanFXm8/nqaAMtBhKU/5ytMXlakTktKEABNgXYdHiAycOk90JjAwt6ayMV619f5f/V/cW9/5i2yPkrmegvnbsfN5f4jbexXbu7/lXlWq9pUG/KuBjmdq7Okq3HiRJTaGnVQyi50qL+zJZY0P6jCleA9PnTz6Ifpp5JC0SPXSBVyOIPBanC/6uPRs+u/iR2Nt3+W/8yeqYPj1NhN/76+QHbm/dsbFpaDF0mU3TUZXcGOnx286BUcRyT1EcMjwzM2pwnFxa7LPH9QCCKaePShYJ2250ZDq1kgeeT1Vh8lct2Q3zv2xtnC/Hn5O98YSr/lRYT46fK6H43sQm19r1jSruzYlTNIrxU+6KZwqlWKtqZVRSNRDgEdSrYzUE8K/6uHSaU3I0yRDXRdDKo7wvzrQGnA5zXFejNfKHprrzvT42fy9+4/jBtX5S7/6C+K1bsfqCt6N6frpqP5LdbVeB+2pFzG4ah5nZMniY6aCLJSgsAzJIp0P6KROYi0clAWzU8OnbuBboRXNvqZIqDSuGB9fLhX16Nl1n8Seq8z8Qfm5uDpj47/KzFdmd1YHe2B3Htf5Y11TlN6dqZLI0YNRUUdJkqueOpp6kFlUyEMzDSL3Hu3jurrHIylSPLy619DCYJZ7WOQSEk95yxOCck+Xp5U6te+E/X9X1V8Rvjn1vX7Oi69rtl9RbK2/WbIhpYKKPa9Tj8PHFPhhSU10jMLXVkX6G4+vtBNpMraeFeju1Di2QSYbSK19ejQ+2ulHXvfuvde9+691737r3RJeiGt8h/kQD+dwQWt/wU+9U8+tnB6O1731rr3v3Xuve/de6qT/AJt+yOyn6r6074676s3T8iG+O/YOM3rnfjjtHEjOZvs/G1uQpaFpMVjGNpajG6fuApRiIy7geggq7WREJDYr0W7jbyzBHizoNaev2dF06m3b3F8pf5iO0O93+E3cnQGysD0Hvja+a3p2rt2i2/l9w5jLbVlhxWIyckbsG8U1QtJTrqPCFjYCyuyBI7XQHDGvAfb0kheefdBM0DImk9zU4+mCT1Xd8b/iZXdcfyoPhn1x8p/iD8x07/6876+Tu6Onsn8a6E4nuL4657Pdi5rOYLdmWzlPVQvjKDNxNSrfxyiaLTqXTa9YzqkYIVAIFdX+TpyePTbRmVZGZGJHhmhBzxzw8vP9nQYfLLf/AM7oMb/IL7N+R/Tu6d5/PHA95fJL+J7M6tx32u9jsh8OuD2/kM7RVOuOnylVhEpZchKQsTTeV2AuR7tEFV3BoV8z+EdN3U07W8Lx1WStdBprYUoRxpXzqTTHGvRnN87e2r8rf5Uf8xv4CfF7rvt/ZHzLxWDx+9e0eiPkLNDlO2J9wbw3rQ75gyQeKR4p6XJU2PqExpjAuxjFmLhjuQt4ytqGk4B8vz61ahPo5YvDcSLllOGJOcHI/ZXpRHN/JX5afKL+S2tD8J/kZ1Ftz4g7l3FlO6N99q7SpMBgMLLB03BsquioJ4ZpNVPPNSMKWZtDSeRFEYPttkWOJhqBr6dPpLLdXUT+E6BKk6gPMEUwfnXpcdIba7q6l/mI7Go/gZ8Z/k10f8b+yu9e4pPnNtXv7GzY74/y4yKmmq8d2p8fcFFVTpjcplcov3Bqo3VaqJ443isWA3IKw1YqSAOHH/i+q258O7McUcoVixJJqlflU4B8qDj0On8j3Yfcfxbw3yl+GPcHSHam1KraPyd727r2l3JlsHDF1N2HsvtrfX8T27QbWzwlLyV8EAEk8IjZSpL6wfT7anUUDhgcD7elVnK5d4niZKMaE8CPUH58aeXV9vtN0v697917r3v3Xuit/LG3+j+lubf7kf8AWv6V96635dDZ1rf+4G0b/X+BUH/Wke/Dh148elx731rr3v3Xuve/de697917qu/+Zz132V2b8cMTt7qnZ2Z3xuim7l6pzkmFwRi++TD4nPmbJZE+ZlHigUq0hB4HP0BPtTaMqyEsaY6L9zR3tqRrqOoY6qs/mQ/FTd6fzG/ij8zcv178v+xdibR+Me9Ontxx/EjJvR732zuPJ1LTLSZCpoZ4JhQ1S1DtMsUqiRtRudNvaiJkYMVKgg4rwIJr0huVnSSJXWRoyvdopUMAABxBoc1A8+PSw7J2PvX4Z/y9snvL+Xv8avlnLuXuL5DbCyvc+w8/JVV3yeh2fnMjHjd5bp267SVBgaCFIws4JKK8jkBdTDzSlpgZyGIFBSlPz/1cOvRWawWbCwRow7lnDE6jwJocmp4Dyr8ugu+CXxq7Aizv80/NYbob5ZYHaXeeLylJgIvltXxZHefZmVr9peGqpsXNXSuaiF5XkiALFdJWNmJ9vCWFQBIQTTy4V/4o9JRb3TMzwowUmtWPcRQcfzAoPl8urrv5c3VmQ6S+EPxw6ry2wE6tyWzOv4sZXdfx0dPjxtipky9VXS0LUVL+3GzGbyuq/wBpzfm/sunKmZinDo+slkW0jWWoYDNePR1vbPSrr3v3Xuve/de697917r3v3XuiN9IW/wBm1+Tf1/4Dbb+v0+o+nvXn1b8PR5Pe+q9e9+691737r3TLuSmnrNu56kpYjUVNVhcpTU8AZVM089DJFDEGewGpiBcm3PPuyEBwTwqOm5QTEwXiQafs61+ds/EXtPD/AMozeXSuO+PNfhe0H7e3xvPD9bUOPxtJnDVZLsaWvj3TRU8bBPNUQO0uu+po2JsQfa/xIxdagQBQdEngTnaxGyEsGJp50qf+L6LV/Mqn+WHZGb7c+LPYPxx+cG8ejofhJ1lL1nnviZUTY/a26e6alIod27X37NCwWsNNURlJafU1okFkIdXFo2iBqhAoSST/AJOtXUV06ESqzAqAoSoz/Srinr+eKdFp/mr1m6fjL8cP+E6+R3J0XvPeW7+pPmd1hWVXT/8AD0y/aNVV4bbxqoNmUFKutXysyiJ1iY2+4hj16SpISudc50njTPpjj0YQq0VmhcEFBXSPOh4fn/m6EjuDr/uPvrf384H+ZrvjpLs34y9Lbt/lY7p+JO1ti9+7bp9q9q5ndW162TcWT3W+Fo5qiOHFWh8MT+dzKZE5AUgbCCN1GoNny/1fPrTSvPDKSjRjQctQZI9Py9epXSnZXb/zK6A/lBdF9dfATu/r3C9I756C7G3D3F2DsbHUPVWE2tsPaT46p37sHPiQytPkGeWshmaCJgXNvIXB9qAqRs8rMDUcPPou8Sa4igtUhdaMKtjTQA5BrU18sfb019zdZ732r/N/6m/lY/GHEVOI+P2/N90v8yX5RvlaOnmx26I83uKbG7xxuYyOlWqlnkpECpI3qkkjUhnUE6SdTGX00IFK/wCQdOyWbpOkIYshbVp/bUt8h8uJPkB1uDU1NTUVNT0dHTw0lJSQRU1LS00SQU9NTQRiKCnp4IgFREUBURQAAAALey7jk9HgAAoOs/v3W+ve/de697917ol3zgt/o62Z/wCJN2r/ANbn968+tjgejiY3/i3UH/UFS/8AWhfdjx6qOHU33rrfXvfuvde9+691737r3Se3bHRTbV3NDksRWbgx0u38zFX4HHQioyGbopMdIlViaGnLoJJqmMtDGmtdTMBcXv7svxCmOqvTQaiuOHr1r5/y+8L391N3Dnuu/ip0F8nOq/hxjsT2Rnt47X+aVTNPW0fYvhqKnatF8fo0nkekoKytaIVFPNK8bJqdrMob2YS+Ey652Bb+jxP29ENmLiGUwWcbJFn+0OAfLSM4PlX/AD9V+dQ7Q+VXym+TfwV7l7k+Mv8AMF2f8gdtbi7xx3Z+e7Tq5I/jz1vj8hkKjH4DLYPETMq0cTQMsqKFGoKoXX6T7v4kYGoEAeXr+fTD29y8ioUdmFdRJ7M/w1P7cfy6NpS7v71+KtP80/hrmPgB3t8kqresNBl9v9+7U2TT5DrXt9d5Ykx08OdytU/kkkwrSHUvraORXQheGNQ0crrIH06fXj060dxaQSQeEZDIMFaU/OtP9XHq5v8Alv7E3t1t8L+kNodh7TyWxd3Y7BZKXK7Ry6xx5LAGvz9VXU2OqoYyRGyRSJZPqosD7SXJDTGhr8+jTbUeOyRZF0GnD0+XR4fafpd1737r3Xvfuvde9+691wk/zb/8Eb/oX37r3QF9Af8AHs7g/wDDxzv/AFuHvQ62eHQ7+99a6bMz/wAWrIf9QNb9Pr/wDk+nvY6q3Dr51f8Awmr/AO38Pzj/AO1R3R9fx/v9pOfdRxHTh8+vozL+kf6w/wB697PHqvXfv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6QvaAv1vv0f12fuMcfXnEy+/dbHHoBfg6APjN1yAbgU+VsTyf+LvN79149G09+611737r3Xvfuvde9+690Bmy/k38eOxt9ZbrHYfdfWW7+w8E1YuW2Xt7eOEym46FsexWvE2KpZml/ZIPlsp02Oq1vbjRSKuplIHTCXNvI/ho4LegOenTsf5A9HdP1+HxfanbfXvXuS3BURUuFoN3bsw2CrMnPPIIYUpKavlR21OQoNrE8X9+WKRxVVJHXpLm3iYLI4BOMnz6UHYfavWnUu1Zt8dnb82nsLZ8BhWTcu687j8Lhg1QuuBEr66RI2ZxdlVSSRcjj3pY3c0UVPV5JooV1SsFHzPTZsHvDpztPbmS3h1v2jsPfO1cNCKjMbh2vunD5nD4qA0rVvmyVfRSvHAvhVpCZWUaVJ+gPvzRyKaMCCetJPDIpZHBAzUHrvrDu7p/uujy2Q6i7N2P2TRYGuOMzVTsvcmL3BHiq8Fh9rXnGySeJzpbSHtextex9+eN0+MUr16OeGavhMGpxoes2B7m6l3Tv7cnVm2uyNlZ/sjZ9BTZTdWx8RuPF5Dc+38fVyCGnrMtiKWVpoEZyFu6ixIBsSPfmjdRqYEDryTwuxRGBI4gHoS/dOneve/de697917r3v3XuiR5b/stWh/8R3Bcf8hH8e9dePDo7nvfXuve/de697917rhJJHFG8srpFFEjSSSSMqRxxoup3d2sAAASSTYD37r3DJ6rn7b/AJj/AEltrfPVHXnUO+evO4d1b27f271vuvE7X3PBmqjadFmKoUtTWTvhmkjWoRm9Ecj/AIOpfp7VR29VZpARTh0Wz7giyJHAytqNDn9n+X9nRsNs9yYPIVfdUm4svtrBYDp7drYHK5qfKLSUeOoIds0WdqavcNXXFIqd4zUuG9QUKF/P1beKgTRUlh0oiuNTS+JQKjUr8vn0/wCyO5+pOydl1fY2wOyNl7w2FQCvNfvDb+4sZktu0IxcXnyJrMrTSNDF4E9cvkYaV5PHuhjdW0kZPTqTwuhkRgVHnXHSO6w+VXxr7qytZgupO9erOxc1j6Gryldidob0wWbyNJjqGoFLW11RR0UzSLFFIQsjlbKSLn3toZUFWUjqsd1bTHTE6seOCOhV/vvsz+79Duw7s22NrZOSiixu4zm8aMHXy5GqFDj46PKmTwStPMyxRBHJZyFW54910Pq00yOr+LHoEmoaTwNekhvHvXpjrzde39i777U2FtDeW60aTbm2Nxbpw+IzeZiV/GZaDH1sqSOur0hgtiQQDwfdlikZdSgkdVe4gjbRI4B9CeivdHfzLfiJ8hPlH378POu+zsVV95/HfKYzGbs2vXVFJSf3hatxwr66q2RMZSMnFQMfBXGEXjfmxT1e9tEwXV+0eY+3qqXMbyaOHoTwb/Snzp0fj210o697917r3v3Xugk7y46x3Lf6faj/AHv3o9bHSa+MnPUeFt/yuZD/AK2j34dePRgPe+tde9+691737r3XvfuvdA98ge9OvPjN0t2T352xmEwPXfVe1shu3deVkKgU2NoQFCprIGuSR44kBNtTD3ZF1sF4dNyyeFGXpWnl69EZ/l7/ADN3N8j+g9+fKztbuT405bozI5zLZjr3M9X5HJUsewtj0U8jCg7UzGem8EeQhhMPlChfVdvoyr7USxICEiBJP7D9nSG0uZZEea5KqoOBkEf6avA9H+2V3H1T2PsI9p7C7F2du7rdafIVb74wG4MbktsRU2JVnyk0+Yp5DDGKdVZptbDQBdrD2w0bq2hhQ+nS1JoZY/GjYMvqDj9vTJ1P8iOie9v4wOme3evez329Ikeci2TurEbgmxLSOY4/v4cfK7xBmUqrMoBIsD780ciCrinWoriCYkQuGp6GvTzQdzdS5XsfIdP4zsjZWQ7TxOIGfyfX1FuPF1O7sfhTIIf4lWYOGU1EcWplBZkFri/19+McgXWQQOtrPC7+GrAt6A9CX7p071737r3Xvfuvde9+691737r3RG9zf9l29f8A/iLc5/0M/vR49WHA9Hk976r1737r3Xvfuvde9+691XD1T/MU6p3F3v8AIfpLtjePXPVOa6s7IxOy9g0W49z02Fy2+aHIxtTfdxR5iSNJZjVqsax05v6x6eL+1ktqyKpQE149FlvuMckzxTMqkEUFaGh4cf8AVx6ON2x3v0x0RjMZme5u0dj9YYrNVjY/E1+99x43b1Pkq1FDyU1FJkZE8jKrKWCXsCL2uPaZI3f4BXpdLPDCAZWC19T0qqXsDYtbs2HsWk3jtmo2DUYtc3BvSLOY5try4hx6ckmcEn23hP08nktfi9+PetD6tFM+nW/Fi8PxdQ0+tcdJTrfvnpbuHIblxXVXamw+w8ns6elp900Gz9z4nPVWAmrUMlIuTgx8rtEJACUZhY2IBv720boKsKV61HNDKSsbBiOND1Fr/kP0Pi90bV2VkO4+tKXdu+K7I4zaG3Zd6YD+LbjyOJYx5KgxNGs5eaaFgVkjUagw02vx734Uv8J6qbm3BALrn5joWYq6inqaqjgrKWasofD97SxVEUlTR/coZKf7qBCWj8igsmsDUORce6EECpHHp0MpJUEEjiPTqBgdx4DdWOTMbZzeK3BipKispEyWGr6XJULVWPqnoq6nWqo2dC8M0bxSLqurKVNiPfiCpocdeVlcalNR08+9dW697917r3v3Xui+/Kn/ALJ+7O/8N5v/AHLi96PVl49Kjoj/AJk71z/4auLt/wBSPduq+fQte9de697917r3v3Xuve/de697917oKqTvTpmv7Mq+mKLtHYlX2zQUb19b11TbnxM28KWkjj80k1Rgo5TUJpT1sClwvJAHtzwpNGuhp69Mi4gMnghxqHlXPTFVfJr48UXaCdKVndfWVL23JPDSx9c1G8sHDu96uojEsFIuEkmExmdSCsWnWbiw59+EUhXWFNOtG6thJ4JddXpXPTt2z350r0Rj8flO5e0tjdZ0OWnamxMu8dxY7CPlalSqtBjKerdZKhwWUaYUY3IFrke9LG7/AACvVpJ4Yf7Vgv2noE/hz8rYPlfhe1txUGJx9DhdidpZnY2AyONqKmop9wYehpYqqjy7NUgWeRZPVo9P0sB7cniWIgLX8+k9lcvcozPQUNMdHI9sdLeve/de697917r3v3Xuve/de6JJSf8AZVtR/wAFnH/qufderU8+jt+7dV6//9Hf49+691737r3Xvfuvde9+690Uv5xX/wBll7HsbEU2M/8AdrD791sdD11f/wAy22B/4Zm2f/dND70OHXjx6XXvfWuve/de697917r3v3Xuve/de697917ogvfP8vHqvvHs2btuj7I7z6U3jlMDJt7ddT0T2JP15R71pHl8gqt00lHTyLU1IX9szkhmUDVci/tTHcuiaKAj59IJ9uhmk8UMyEgg6WIrX1A6Nn1T1RsbpbZOH2B19hYMNgsTBGruEibJZnIeJY6zPbgyCqr1lfVFRJU1Ut3dv6AABl3aRtTdK4okhTRGKD/Vx6Ef3Tpzr3v3Xuve/de697917r3v3Xuve/de6JJ0OD/sxHyI+lv7wQfX6/p96Ax1s9Hb97611737r3Xvfuvde9+691737r3XvfuvdFI+Uvw06x+V56+yW8M92HsXenVeXr831/2J1TuuXZu9tt1eUpVpMglHmIYpf25UVdSFPqLg8kF+Kd4QVFCDxB6R3VlFdFXclWQ1BU0I/Pp++OvxR6u+NlLnK3bBz+8uxt4R0EfYPdPZGRi3V292HHiA6YWDd+9JIop6qGiR2jo6ewjiX9K3JJrJKZDWgA9Bw6dgt1t00glj6sak/aejM+2un+ve/de697917r3v3Xuve/de697917orPyz/AOZf0n/ay/6JX37z695dDd1r/wAeBtH8f7gqD/rSPehw690uPe+vde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+690RzpD/ALK1+TfB/wCA22+T9ObfT3rz6t+Ho8fvfVeve/de697917r3v3Xuve/de697917ov/dfxl6l+QO5ejd29l4aryma+O/aVD3D1jPS170SYze2Ox82MpqusRFPmiEczXiutyBza4LiSMgYD8Qp0xLbxzMjvxjbUPtpT/L0uO4eqNmd6dW7+6c7EoZsnsbsra2X2fuqgpqhqSoqsLmqVqStigqVBKPpb0tpNj9QRx7qjFGDjiOryxrNG0T8GBB+w9Pewdk7f602Ns3rradNJR7X2HtbAbO27STTNUTU2D21i4sPioJZ2sXZYIUDMQLnmw96ZizFj59WRBGgjXgoA/Z0GlP8buqqb5H5D5WR4er/ANMmS6tpOnajNvXyNQjZFHnjuOKjixxGlZTUm7S6uV403ufd/FbwvC8q1+fTX08f1H1OdVNPHFK14dDx7b6f697917r3v3Xuve/de6Jd84P+ZdbN/wDEmbV/63t7159bHA9HExv/ABbqD/qCpf8ArQvvfVRw6m+/db697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de64Sf5uT/gjf9C+/de6AvoD/j2M/wD+Hjnf+tw96HWz0O/vfWumzM/8WrIf9QNb/wC4cnvY603Dr51X/Cav/t/D84/8cT3R/sP9/rL7qOI6ueB6+jMv6V/4KPp9Pp72ePVeuXv3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6QvaH/Mt9+/T/jz9x/X6f8AFpl+vv3Wxx6Ab4PX/wBlm65va/2+V+n0/wCLvN79149Gz9+611737r3XvfuvdNOfzFPt7BZrP1cc81Jg8TkcxUw0sflqZafGUb1s0dPFcapGVCEW/JsPe1GpgPXqrHSpb0HWm/hvlzt75E/Lz+XZ8lvj/uP4XdabC3T3R3Ht2t636xx38N+SeZOOyEmKyUXZXjjVJkm9TgW0kuxFw3BuqVDIakGmTw/LoKvN3xTAIjDUQAKOMV7vlwP7Oj97d+KXS/fvzH/m6bk7dmyfYdVJs7rSn2/id0vFlsX1Q2J2vkMzDlOvIKlNWOqXqKWCSeSFgz6NJOlmBo+qBo1QnjT/ACdPxrFepcPOoNAGFRwOTj8/9WOiM97fLGj211x/Jj3h3xvPZu4erx3b8g9k9i7d7iihyeF7S27hMT/dPay0VHPDLHUth/NAxeoiYKFVfVwDsJ3soNCw8uPTbXFLeKeQF1jY1J+EAg5PoAaD06W2Z6o7I+L/APJazGJ+Pfc/x/3pjeye9t+bw7d726mnifq7aXVm6t3VFfhNt5zIA/cLT4uGSixdcjIvhGqIKFfVJ5WLXB8ap0igB/Yafy63PCYdrAsCqhyWYj4QCa/sGaeXRz/gh05vtu7N4/ILa3ye+FeU2pW/H3aWF3j058QqZoqOSLH7aZqLdu5aAOQlXUS/u/dvGreS4AH0OjIgpHIjccE+vlThw6vFbzmRrqKaOmgAqg9Bkk5qSMeVPXoWf5EPx/606t+LG8t/YCfJb07A7K747oye7e0d6/bZbsHKUg3i6Y7bFTuORBUHHUCIq01Lr0KSWILc+0l1qR/DqSKDoy2zw5YPqAihiSCQKVpjPz9fXq8P2l6M+ve/de697917r3v3XuiRZf8A7LVoeCf+Mdwf72feuvHh0d33vr3Xvfuvde9+690QX+ZtuDsXbXwz7ayXXT1lI38Oipd95jGyJDktvdX1Kum/M9QSuQUenotTO68rGXbgAkKLXT4tW8ukG5GQWjeHXPGnGnVEtf8A8NsVfa/8qbcXw+7J2LubflPuzrnFVcGxazGz1G78PNkKZavcXZIxyB5cwKz7pddVeX6j9CofZgvjFZA3AnH2dEE67es9oYiBIuABxrTII8/PjwPSL2/sGp+VeH/nzdVbv+UmO6ApNjfzWtk7h2Z2J2NuWPC9c0FftrFYeswnWm8ZaiWBZsDkp6WOjq6BXPlDKNLc3RRtR0NC2KUHH8v8PR1cxEwS6SsZLBtTfCaEcc8DSlMfz6RfzO/mCz9m/wAqjvH+62I2b8f90/Eb59/HnpnvTdPRtBBtv459900e8qOp3HkOqqmmAlrdvZimeJaynqU12jClnjdfdwpE4D1oQaV49NtMpsWkhCOVIqE4Ghzw9M4PpnB6sG+IWyuit2R/zEPlX0TuH445TPb56Z3rj+vuu/j5T0tNu/rjFjryWpnps5jokh8FZXV8cBXw04RpSeV1BS7cl0RYskVGT0k2uO1luJb6LRVxSigVp86cfP8Ab1UfP8wdiV3/AAnj+AnWlH3djpPkjQd8/Hzb+6Ov03HSVPaNDksL8javI5TFbkwif5TTg0XgYmeJA6MgBYkj3QJL47SMCAQePzHTrz2psoreN1Zgy0Cn+Fs4ORQZz1ZF8uOydk/y4vnj3H8styb16d+WGxfkJ2b8ethdifGHLtQb5+WXQG5M3iafaO1c90lgquSSKjwBXRk8pRTLEZFdXjOvS5qgZoRpDAr5jh05KyRXzGRoir6e1vjBp8zSh9KcfXp4/ly7J6O6o/nl/wA4zG7twfW+0u5uw939Gbq6RGWixNDvHc2ycp1j/GN+1HX0VXaRofvZYnya0IDM41SKVBIadZPCDitDxPr9vSqKSD6t4CRqSmlcYBHl9p9Ps62bPabow697917r3v3Xugl7w/5lluYcG9J+f9f3o9e6TPxk46jwv/UZkf8ArcPfh1s9GA97611737r3Xvfuvde9+690Sn+Y5g9ubl+Cfypwe7sTjM5tyv6b3bHlMVmKZKzGVkMdIJ40q6aQEMBIiOLjggH6j29BmZft6S3tBaSE/wAJ611Pl/tTcvSXxC/kt4f4qnpDojpfK/IDaWX7lot7UKYrpLc1JXbeDGn7YoIEtV46pf7iSdZiSxAsVCLZdpLSuAT20wPSg4dEjSJDawM1BragZjipJ+P1BGKeZPR6Ol2qPhz1z87fmZ2B3t8Z+4/jVTdX5DOSdA/FWmjl6i2jlKapYZSvq8EHlp4I6mFjBUlSS0JmY6goHusjCVkTSVIPFvPpy3iktBPOZFljYDTHGuF41+2v+wOisfBHuOp3N/NN2zv/AGNvn4dQdYdy/DHbe5Mf1j8UqOXHvi6XJMMpQy9iUJQA5KB1XzMQCNKji3L4jEi+EwNKnj/k+XSQTeFcmZCmrQMJjFfxfMcOj0/yQPj11tsDZnyp7Zo63J7/AOz+wPl13QM32RvdqfNbxx+Egq6NqHY2Hzs8f3EGIpWd3hpEcJqclgbLZFd1VhGCaUrTo22sRyI0+katRFQM4/zcOr1/aPo2697917r3v3Xuve/de697917ojW5if9nt6/H/AH63N/X/AF3+nvXn1by/1fLo8vvfVeve/de697917r3v3XutXvrP4cfH7srsL+dv2Vvmhm3fvKp7boM5iM3nVjyNX1fkNm7Zr85i/wC4Msy66BZKmFHqBEw1hbG449mrM6SIoJowPQaWGCeG4d1UlWFDTIIpTPy8ulDvX5u7Pyfxu+A/SfZ0vxxy3dXyM6r7ordn9mfMnELkussanW+PqsRU5GGrCFmrqhEp40YhQ2gaizMD7qI6OzVOaYU56v8AUGSCJSo4HvcVX04/Ph/IdVVfIzfW7l/4SudyvuHtuLI1+2u9Nnbbh3r0tkamkwNRhZ+/sHJJhdvTxmN/4bpnmvEGAKm1rekt3VBMrGoqPLj09tYD2DRIFcA4B+E5yOHD06PpiN+dL5/+a3/J1o/5em8Npbo6/wD9Cner/L+q6DyuCq8TV7YTpWkfret7/G3fQ1U+QDy0/wDEgKkVFjYNo9tSeMVPiA06WQ/RCVVtiuog1p5/bT/Ln06rr2LvP4X5n+Sz31Tdgdy4Wf5m1XyD33NimotxYlfkXtDctN3+ke38f1pDMDkKSnmx0ZaeSjURyRySrIxKqArpK8ygV0U/yefRGr2kG3yuxUS6mwxyCWxpBznyp0fT5qfIntz+Vz2f2d8g23HuHsXtT+Zf1T8d+qfjt1bHlKiopdl7x6w2lR4Hd2WTbtTG18i1NkTPNKlled1Yk2K+6qIJCKE1U8D59LJ3vbaN2Cj9QDuGAPtP5gDFa8K8Rsq/ErpPAfH74/dddb4CHIwrS4ZM/m/4rUGor5t1bqP94NzzVD/RSa2omCoOFUAXJBJQTOXkLHo4tYFt4FiTh/n6Mf7b6Ude9+691737r3RfflR/2T92d/4bzf8AuXF70etjpUdEf8yc64/8NXF/9aPe+tefQte/de697917r3v3Xuve/de697917rWu6r7P2v8ABT539nYnL70+PvypovlV8iJqfa9L07R027fmD0xmt4TrSpj+wfG7KNvU2pknCVCunLgfqjJm6tPCKAppHnhT0GopY7C9dWZJRK1RpFZAT5GnkPInomv8xP5Z7L747B3bVdL7i+GXUOe+N38wTrvqzsPIbsxpx/yw3Zn8DNFHkKzBZCJATRylWQjUeIwGOpebRKUGkEtj1qv+r061duJH1uEQaxSopIc8QfnkH8x0cjcm+/jdu/5w74pv5nG69lbTy1V0JnoPjrs3tCfD0e1sr1lWVOvM772xQ1yGL+LRjytBOgEwfV4/3EUe9sGVNNuPMcOrJ4clx4u4eSEAtSmfTgOHn59GV/kWRbEo/j/3Vi+sM7U7m68xnfe5afaOerZGmrMliDjqdqaoqJG+pK6bf4W9p7/V4oLcadP7B4ItGW3OpAxoa1885+3q772h6Peve/de697917r3v3Xuve/de6JJSD/nK2oP40Tf7f7c+6nj1b59Hb926r1//9Lf49+691737r3Xvfuvde9+690Uv5xEj4zdj2F/8mxl/wDW/isPvR62Oh66v/5ltsD/AMMzbP8A7pofexw68eJ6XXv3Wuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6JH0Rc/Ij5EWJAG4Ke/8AQ+n3qvr1tujue99a697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917orXyxNuv6Ti98kP9h6V968+tjobutf+PB2j/2oqD/rSPfhw68ePS397611737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3RHOkB/zlr8mzz/wG239f9cfT3rz6t+Ho8fvfVeve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de697917r3v3Xuve/de6Jd84D/xjnZv1/5mbtX6f8tm968+tjo4mN/4t1B/1BUv/Whfdjx60OHU33rr3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3Xvfuvde9+691737r3XCT/Nv/AMEb/oX37r3QF9Af8exn/wDw8M7/ANbh70Otnh0O/vfWumzM/wDFqyH/AFA1v/uI/vY6q3Dr51f/AAmr/wC38Pzk/wC1T3R/rf8AH7S+6jiOnDwPX0ZV/Sv/AAUf71731Xrl7917r3v3Xuve/de697917r3v3Xuve/de697917r3v3XukJ2h/wAy237/AOGfuP8A91Mvv3Wxx6AX4OX/ANll64ve/wBvlfr9f+LvP70Ottx6Nr731Xr3v3Xuve/de6xy+LxSefx+Hxv5vLp8Xi0nyeTXxpte9+LfX37r3VeHU2M/lR0/YG1p+kJPgY3Z4z24m2UvWmW6Iqt6/wB5Zahzun+69PtyZ6z70S+T70Ui+RWvrsb+3qz6R8VPz6TAWms00avPhX8+jt4wdb/x/epw39yf70FMf/pE/hpwRz3jFG/8L/vmKX/KAv2/k8H33Hj1aPTf3Q+JQVr8v9jpweDVqU/pcP59Uk/zAOl+gch2t8X+5Nkdr/y+qTPYTcW6sd1n1H8vt4bGp+pM3mJaVodyZfpXbq1EYbOozf7k1x8btcI8gDA3VxyY0zK1fIjj0W3EJ1eLaOgH4lb4f5dDN8FOlvjBsLorvneW6+1vijvfZHZVTW1ff20unt6bIy3w36/p6aWo+6x1Ph6momx2PWSORjkZMhIgYgAqAlzW4kZiFVSKcKjPVtvgjjR3Z1Yt8QUjQPy6Op8dKH4OUdfuP/ZUX+MT5OTE47+9n+gjIdZ11eMEoAxX8d/uPLJItJ9PD57RnjTfj21IbggeLqp5Vr0qgFiC302ivnp0/wA6dGC2IOvhgFHWX9z/AO638Qyej+4xwpwP8UNa5zGk4H/J/P8Aca/ubevyatfqv7afXXvrX59KE8PT+nSny6WXuvV+ve/de697917r3v3XuiR5Y/8AOatDx/zTuD/ez795dePR3Pfuvde9+691737r3THucbbO2twjeX8F/ugcHlhur+8hoV27/ds0Egzn8fbJ/wCTCi+18v3RqP2vFq8npv78K1x17oinReM/laUeb2mfjfJ8GX3GssI2N/ofy/R9dmvP5G8H91htOaScvr1aPtedV7c+3ibjT3aqfnTpKostfbo1V8qVr/n6ErfVH8EJ9pdzRdjv8Vl2RP2PjX+QJ3dXdV0+217bWCkGIHbU2VkWnj3D41ovt1yzLVlfFoFit6oZQw8OtfKnV5vp/Dbx9Onz1Up+dcft6IV/MR6M+F3d3wQqNpbJ7h+GfTXUuB7s643nj9xbt3n1vtn45ZzsjbuWSpw20N75jHVcGPkqK8mNI6YzGeRhGoRgRZRG8iyVmBOPTP5dIbqGKW1ItHRAGB4jSaEYanr9vSP+F/Q/Wm4/mTifkbvTuH4Wba782F1TunYGH6A+Ce/tt0+yN6bB3AY5a7f/AG7slJmrq+ro7xpQziFYYFKEyHSi+/TN+npQNTzLevXrSNPqDJIYvEAIpGfwk+Y4/n0cvHYz+U+u8jPipPgQd+HeGMl00WW6Gfc39/BkQcOBTxTGoGU+7t9sgXzeWwQavbTG5p3aqfn0oQbfr7PD1V8tNa/5+hI3LQfy+m74yFZvB/iT/syv8Q2z/FU3Fkepl7k/iqU8Y2h/EKHIyjL/AHHi8P8AD/JHrK6PFcafflNzo7NWn5Vp1txYeN+p4fiY46dXy+f2dL7MUvxJb5F7bq883x/HyvjwFVFtNMrWbATvk7aajb7s4KiqnGdek+31a3hjKCO/IX3QeLoxXT+dOnT9P42dPiflqp/h6Mj7b6f697917r3v3Xugl7x/5llub/qFH+8G/vR68Okz8ZP+ZR4X6/8AAvIfX/lqPfh1s9GA97611737r3Xvfuvde9+690x7mG2jt7MjeP8ABP7qnHVQ3D/eU0I2/wDwnxH73+MHJ/5P9vov5fN6NN9XHva6tQ08eqvp0nXSnnXh0DXasHxbqOqcXD3S3RS9JMcZ/BD2PUbEpus9QgY4f+Ez7iZMYD4tX23gb9F9Hpv7cBm1nTXV58a9MuLXwh4mnR5VpT/N0hdiUXwTg6c7Eputn+LLdBvgskvazbPr+rajqsbaehlXLf3+q8TI2LWi+28/mOQcRiPWWsur3tzOaeJX5Vr16EWlD9Ppp56af5Ok58c8f/Lno94hvijJ8PH3+doUkaf6Ccn1BX7u/uIpH2ZjXY8slYMXe2hlHgPFifem8anfWnzr16P6Wv6Wjj5U4/l59Gr2QOvRj8n/AKN/7m/wr+8GWOZ/uScKcf8A3pMwOc/iX8D/AG/v9en7oS/u3tr5t7q+uv6la/PpyPwqfpUpXy9fy6WfunTnXvfuvde9+691737r3XvfuvdEa3N/2Xb199f+ZWZz/W/U/wBfevPqw+Ho8vvfVeve/de697917r3v3Xug9xI6qE/YBwf+j/7k1sh7T/hR26Zv4h9gRN/f4UnqE32urV/EPV4r39N/dz4mNVflx/l00PBo2mn9Lh/Potfc1D/Lxq8d1H/p9f4gpiYafcR6L/0pZDqGix5pGpkXdf8Aou/vLLHHJF4vH9+MXdQuky2493Bn1nTXV+deqOLXwgH06PKtKfzx0Sj+Y98e/hH8lP5V3YvS22e7/il8evixn8919VQdrxb7662P8esHWYHs7G5s0ku8cPV02GhmrqmAUaaakPJUSJHyzAe/IQJAZwSOqyqzwMtmwVjTIpTy/wAnRtfiVt3+XdicVnsN8Ksp8RJ81HsrBYneOV+OGZ6my+ZfC0mEiocLkdzVXX1RNUGFohHLFLVyWk9L6mJ1e/OZvx6qfOvW4vpa/oaK/wBGn58Oq7/5d3x7/lPfHjpza/XGa7q/l6d+dxY3szsPN1vZNP2B0bvDdmU3Xn98VOUgxq5HIV1RXmsoVenpGpgQY5E0hA1yX5XmZqwhgtB5HpDaRWkUYS4aN5KnNVrk1HzqOHRkvnj8ePjf2t81/wCWR2v3V3p0Z1vubofszsrP9S9Y9m742ztnd3c+5M/tuix1JiOsdv5usp5MzU0FUlFUy01JDMyXibTcqCzEwCsKEseH+XpXcxu7xtqAjU1YHgfT9hoftp1bt7Y6Wde9+691737r3XvfuvdF9+VP/ZPvZ/1/495vp9f+BcX096PWxx6VHQ//ADJzrj/w1MX/ANaB731roWvfuvde9+691737r3Xvfuvde9+690T3YFF8Cou8Nw1XWD/FM/Iyapq/7zrsmv6qm7fFZZvvv4vR4WRsskttfl8iBv1avz7fY3Gnu1U/OnSRBY+J+no1fLTX/P0F25MZ/KkbtXc1Vu6T4Gf6aW7JpJd4ruHLdEL2L/pcQItGNx0mRmFeNwcJoSpT7omxsSfdQZtPbWn506u30vid+nV86V/z9CH8i8f/AC+qzfWxZPldJ8Sk7Jh25lh1uO98n1RQ7zO0nqV/jX9z4t8yx1kmO8wH3BpAYQ/6rE+9IZQf061+XW5fptI8fTTy1Up+Veh36Yj6Nh2bHH8fT1adgrWTeI9RS7Wn2p/ENC/cWl2gWpTNp0a+dVtN+Le9Prr31/Pq0Xg0/RpT5U/ydCz7p071737r3Xvfuvde9+691737r3RI6S/+zW1H1tom/r/yrm3upp/Pq2ejue7dV6//2Q==)
Figure 6. Proportion of Study Contrasts with Increased Activation in Four Key Brain Areas. The y-axes plot the proportion of study contrasts in our database that had increased activation within 10mm of that brain area. The x-axes denote the contrast type separated by experience (exp) and perception (per). All brain regions depicted are in the right hemisphere. See Figures S2 and S3 in supplementary materials, available at http://www.journals.cambridge.org/bbs2012008, for additional regions. A color version of this image can be viewed in the online version of this target article at http://www.journals.cambridge.org/bbs.
![[[@Lindquist2012-jr]](http://dx.doi.org/10.1017/S0140525X11000446)](data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAByoAAATkCAYAAAAzVYcPAAAMa2lDQ1BJQ0MgUHJvZmlsZQAASImVVwdYU8kWnltSSWiBCEgJvQnSq5QQWgQBqYKNkAQSSowJQcWOLCq4dhHFiq6KKLYVkEVF7GVR7K5lURcVZV0sKIrKm5CArvvK9873zZ0/Z878p9yZ3BkAtHp5Umkuqg1AniRfFh8RwhqXmsYidQANQABkYAsceXy5lB0XFw2gDPZ/l3c3AaLsrzkpuf45/l9FVyCU8wFAJkCcIZDz8yBuBgDfwJfK8gEgKvWW0/KlSjwPYj0ZDBDi1UqcpcK7lDhDhZsGbBLjORBfAYBM4/FkWQBo3oN6VgE/C/JofoLYRSIQSwDQGgFxIF/EE0CsjH1EXt4UJa6A2A7aSyGG8QCfjG84s/7GnzHEz+NlDWFVXgNCDhXLpbm8Gf9naf635OUqBn3YwEYTySLjlfnDGt7OmRKlxDSIuyQZMbHKWkPcKxao6g4AShUpIpNU9qgxX86B9QNMiF0EvNAoiI0hDpfkxkSr9RmZ4nAuxHC1oNPF+dxEiA0gXiSUhyWobbbIpsSrfaF1mTIOW60/x5MN+FX6eqDISWKr+V+LhFw1P6ZZKEpMgZgKsVWBODkGYk2IneU5CVFqm1GFIk7MoI1MEa+M3wrieKEkIkTFjxVkysLj1falefLBfLEtIjE3Ro0P5osSI1X1wU7xeQPxw1ywK0IJO2mQRygfFz2Yi0AYGqbKHXsmlCQlqHl6pfkh8aq5OFWaG6e2xy2EuRFKvQXEHvKCBPVcPDkfLk4VP54pzY9LVMWJF2bzRsep4sGXg2jAAaGABRSwZYApIBuIW7vqu+Av1Ug44AEZyAJC4KTWDM5IGRiRwGcCKAR/QiQE8qF5IQOjQlAA9Z+HtKqnE8gcGC0YmJEDnkCcB6JALvytGJglGfKWDP6AGvE/vPNg48N4c2FTjv97/aD2q4YNNdFqjWLQI0tr0JIYRgwlRhLDifa4ER6I++PR8BkMmxvug/sO5vHVnvCE0EZ4RLhBaCfcmSwukn0X5RjQDvnD1bXI+LYWuA3k9MRD8ADIDplxJm4EnHAP6IeNB0HPnlDLUcetrArrO+6/ZfDN21DbUVwoKGUYJZhi9/1MTQdNzyEWZa2/rY8q1oyhenOGRr73z/mm+gLYR31viS3CDmFnsRPYeawJqwcs7DjWgF3Cjirx0Or6Y2B1DXqLH4gnB/KI/+GPp/aprKTcpcal0+WTaixfOD1fufE4U6QzZOIsUT6LDb8OQhZXwncewXJzcXMFQPmtUf19vWEOfEMQ5oWvuqIDAASE9ff3N33VRW8F4HAt3P69X3W2xQDQ6QCcm8FXyApUOlz5IMB/CS240wyBKbAEdjAfN+AF/EEwCAOjQSxIBKlgEqyyCK5zGZgGZoH5oASUgeVgDVgPNoNtYBfYCw6CetAEToAz4CK4Am6Au3D1dIAXoBu8A30IgpAQOsJADBEzxBpxRNwQHyQQCUOikXgkFUlHshAJokBmIQuQMmQlsh7ZilQjB5AjyAnkPNKG3EEeIp3Ia+QjiqE0VA81QW3QkagPykaj0ER0IpqFTkUL0WJ0KVqBVqF70Dr0BHoRvYG2oy/QHgxgGhgTM8ecMB+Mg8ViaVgmJsPmYKVYOVaF1WKN8D1fw9qxLuwDTsQZOAt3gis4Ek/C+fhUfA6+BF+P78Lr8FP4Nfwh3o1/IdAJxgRHgh+BSxhHyCJMI5QQygk7CIcJp+Fe6iC8IxKJTKIt0RvuxVRiNnEmcQlxI3EfsZnYRnxM7CGRSIYkR1IAKZbEI+WTSkjrSHtIx0lXSR2kXrIG2YzsRg4np5El5CJyOXk3+Rj5KvkpuY+iTbGm+FFiKQLKDMoyynZKI+UypYPSR9Wh2lIDqInUbOp8agW1lnqaeo/6RkNDw0LDV2OshlhjnkaFxn6NcxoPNT7QdGkONA5tAk1BW0rbSWum3aG9odPpNvRgeho9n76UXk0/SX9A79VkaDprcjUFmnM1KzXrNK9qvtSiaFlrsbUmaRVqlWsd0rqs1aVN0bbR5mjztOdoV2of0b6l3aPD0HHVidXJ01mis1vnvM4zXZKujW6YrkC3WHeb7kndxwyMYcngMPiMBYztjNOMDj2inq0eVy9br0xvr16rXre+rr6HfrL+dP1K/aP67UyMacPkMnOZy5gHmTeZH4eZDGMPEw5bPKx22NVh7w2GGwQbCA1KDfYZ3DD4aMgyDDPMMVxhWG943wg3cjAaazTNaJPRaaOu4XrD/Yfzh5cOPzj8N2PU2ME43nim8TbjS8Y9JqYmESZSk3UmJ026TJmmwabZpqtNj5l2mjHMAs3EZqvNjps9Z+mz2KxcVgXrFKvb3Ng80lxhvtW81bzPwtYiyaLIYp/FfUuqpY9lpuVqyxbLbiszqzFWs6xqrH6zplj7WIus11qftX5vY2uTYrPQpt7mma2BLde20LbG9p4d3S7Ibqpdld11e6K9j32O/Ub7Kw6og6eDyKHS4bIj6ujlKHbc6Ng2gjDCd4RkRNWIW040J7ZTgVON00NnpnO0c5FzvfPLkVYj00auGHl25BcXT5dcl+0ud111XUe7Frk2ur52c3Dju1W6XXenu4e7z3VvcH/l4egh9NjkcduT4TnGc6Fni+dnL28vmVetV6e3lXe69wbvWz56PnE+S3zO+RJ8Q3zn+jb5fvDz8sv3O+j3l7+Tf47/bv9no2xHCUdtH/U4wCKAF7A1oD2QFZgeuCWwPcg8iBdUFfQo2DJYELwj+Cnbnp3N3sN+GeISIgs5HPKe48eZzWkOxUIjQktDW8N0w5LC1oc9CLcIzwqvCe+O8IyYGdEcSYiMilwReYtrwuVzq7ndo71Hzx59KooWlRC1PupRtEO0LLpxDDpm9JhVY+7FWMdIYupjQSw3dlXs/TjbuKlxv4wljo0bWzn2Sbxr/Kz4swmMhMkJuxPeJYYkLku8m2SXpEhqSdZKnpBcnfw+JTRlZUr7uJHjZo+7mGqUKk5tSCOlJaftSOsZHzZ+zfiOCZ4TSibcnGg7cfrE85OMJuVOOjpZazJv8qF0QnpK+u70T7xYXhWvJ4ObsSGjm8/hr+W/EAQLVgs6hQHClcKnmQGZKzOfZQVkrcrqFAWJykVdYo54vfhVdmT25uz3ObE5O3P6c1Ny9+WR89Lzjkh0JTmSU1NMp0yf0iZ1lJZI26f6TV0ztVsWJdshR+QT5Q35evBQf0lhp/hB8bAgsKCyoHda8rRD03WmS6ZfmuEwY/GMp4XhhT/NxGfyZ7bMMp81f9bD2ezZW+cgczLmtMy1nFs8t2NexLxd86nzc+b/WuRStLLo7YKUBY3FJsXzih//EPFDTYlmiazk1kL/hZsX4YvEi1oXuy9et/hLqaD0QplLWXnZpyX8JRd+dP2x4sf+pZlLW5d5Ldu0nLhcsvzmiqAVu1bqrCxc+XjVmFV1q1mrS1e/XTN5zflyj/LNa6lrFWvbK6IrGtZZrVu+7tN60foblSGV+zYYb1i84f1Gwcarm4I31W422Vy2+eMW8ZbbWyO21lXZVJVvI24r2PZke/L2sz/5/FS9w2hH2Y7POyU723fF7zpV7V1dvdt497IatEZR07lnwp4re0P3NtQ61W7dx9xXth/sV+x/fiD9wM2DUQdbDvkcqv3Z+ucNhxmHS+uQuhl13fWi+vaG1Ia2I6OPtDT6Nx7+xfmXnU3mTZVH9Y8uO0Y9Vnys/3jh8Z5maXPXiawTj1smt9w9Oe7k9VNjT7Wejjp97kz4mZNn2WePnws413Te7/yRCz4X6i96Xay75Hnp8K+evx5u9Wqtu+x9ueGK75XGtlFtx64GXT1xLfTamevc6xdvxNxou5l08/atCbfabwtuP7uTe+fVbwW/9d2dd49wr/S+9v3yB8YPqn63/31fu1f70YehDy89Snh09zH/8Ys/5H986ih+Qn9S/tTsafUzt2dNneGdV56Pf97xQvqir6vkT50/N7y0e/nzX8F/Xeoe193xSvaq//WSN4Zvdr71eNvSE9fz4F3eu773pb2Gvbs++Hw4+zHl49O+aZ9Inyo+239u/BL15V5/Xn+/lCfjDRwFMNjQzEwAXu+E54RUABjw3kYdr7oLDgiiur8OIPCfsOq+OCBeAMAjCFAe4znNAOyHzWYePLrDpjzCJwYD1N19qKlFnunupuKiwZsQobe//40JAKRGAD7L+vv7Nvb3f94Og70DQPNU1R1UKUR4Z9gSqEQ3DATzwHeiup9+k+P3PVBG4AG+7/8FX1+PrPDTZoMAAACKZVhJZk1NACoAAAAIAAQBGgAFAAAAAQAAAD4BGwAFAAAAAQAAAEYBKAADAAAAAQACAACHaQAEAAAAAQAAAE4AAAAAAAAAkAAAAAEAAACQAAAAAQADkoYABwAAABIAAAB4oAIABAAAAAEAAAcqoAMABAAAAAEAAATkAAAAAEFTQ0lJAAAAU2NyZWVuc2hvdPXD450AAAAJcEhZcwAAFiUAABYlAUlSJPAAAAHYaVRYdFhNTDpjb20uYWRvYmUueG1wAAAAAAA8eDp4bXBtZXRhIHhtbG5zOng9ImFkb2JlOm5zOm1ldGEvIiB4OnhtcHRrPSJYTVAgQ29yZSA2LjAuMCI+CiAgIDxyZGY6UkRGIHhtbG5zOnJkZj0iaHR0cDovL3d3dy53My5vcmcvMTk5OS8wMi8yMi1yZGYtc3ludGF4LW5zIyI+CiAgICAgIDxyZGY6RGVzY3JpcHRpb24gcmRmOmFib3V0PSIiCiAgICAgICAgICAgIHhtbG5zOmV4aWY9Imh0dHA6Ly9ucy5hZG9iZS5jb20vZXhpZi8xLjAvIj4KICAgICAgICAgPGV4aWY6UGl4ZWxZRGltZW5zaW9uPjEyNTI8L2V4aWY6UGl4ZWxZRGltZW5zaW9uPgogICAgICAgICA8ZXhpZjpQaXhlbFhEaW1lbnNpb24+MTgzNDwvZXhpZjpQaXhlbFhEaW1lbnNpb24+CiAgICAgICAgIDxleGlmOlVzZXJDb21tZW50PlNjcmVlbnNob3Q8L2V4aWY6VXNlckNvbW1lbnQ+CiAgICAgIDwvcmRmOkRlc2NyaXB0aW9uPgogICA8L3JkZjpSREY+CjwveDp4bXBtZXRhPgp5iVYKAAAAHGlET1QAAAACAAAAAAAAAnIAAAAoAAACcgAAAnIABLnoBvR8jwAAQABJREFUeAHs3Ye7pElVBvDGhIhIVEBRLooIiJJRMTAkcUkKCgKKLAjILvCAyh9iwABKRlHiAou67K6MkSSoGABJA5gAExgIoti/gnct2753+t6Ze6dn5q3n6em+3fVVnfOe94Sq+rrnap9ZtkVbESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCReAIEbhaDyqPEO1OVQSKQBEoAkWgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKwECgB5UlQhEoAkWgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkeOQA8qjxzyTlgEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEikAPKsuBIlAEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEjhyBHlQeOeSdsAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEikARKAJFoAgUgR5UlgNFoAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEikARKAJFoAgcOQI9qDxyyDthESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCPagsB4pAESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEikARKAJFoAgUgSJQBIpAEThyBHpQeeSQd8IiUASKQBEoAkWgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUAR6UFkOFIEiUASKQBEoAkWgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCRaAIFIEicOQI9KDyyCHvhEWgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCPSgshwoAkWgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkXgyBHoQeWRQ94Ji0ARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEikAR6EFlOVAEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEisCRI9CDyiOHvBMWgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEikARKAJFoAgUgSLQg8pyoAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEikARKAJFoAgUgSNHoAeVRw55JywCRaAIFIEiUASKQBEoAkWgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCRaAHleVAESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCR45ADyqPHPJOWASKQBEoAkWgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKQA8qy4EiUASKQBEoAkWgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASOHIEeVB455J2wCBSBIlAEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBHlSWA0WgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBw5Aj2oPHLIO2ERKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEikARKAI9qCwHikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEikAROHIEelB55JB3wiJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEikARKAJFoAgUgSJQBHpQWQ4UgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEikARKAJFoAgUgSJw5Aj0oPLIIe+ERaAIFIEiUASKQBEoAkWgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCRaAI9KCyHCgCRaAIFIEiUASKQBEoAkWgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCReDIEehB5ZFD3gmLQBEoAkWgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKQBHoQWU5UASKQBEoAkWgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKwJEj0IPKI4e8ExaBIlAEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBItCDynKgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBI0egB5VHDnknLAJFoAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEikARKAJFoAeV5UARKAJFoAhsNQKf+cxn1sp3tatdbe37fXMzBIJrcdwMr/YqAucqAokF9Gs8OFetfH7rNXO8PD+/uVDti0ARKAJFoAgUgSJQBIpAEdhOBHpQuZ12qVRFoAgUgfMegWws/vd///cir4FiI31+nPdA7ROAYJnnHEzkeZ/DtXsRKAJnMQLiQB7U+LzP+7yhTePBWWzUin4VAslzec4HqSHyd5+LQBEoAkWgCBSBIlAEikARKAJF4Mwi0IPKM4t/Zy8CRaAIFIE1CDic/K//+q/FJz/5ycW///u/j2d/20T/4i/+4vG4xjWusfjCL/zCfgNoDX67vQXXT3/60wPPj3/846Pbl3zJlyyufvWrLz7/8z//qkOK3a7v+0WgCJwbCDi4EVM/9alPjXjwiU98Yvj/l37pl474Ktb2sPLcsPX5qkXqiP/8z/8cPPc3Tn/BF3zB4Ljncvx8ZUf1LgJFoAgUgSJQBIpAESgCRWDbEOhB5bZZpPIUgSJQBM5zBGyg21h0kPZP//RPiw9+8IPj2d9f9EVftPjyL//yxVd8xVcsbnjDGy6uda1rjQO2bjaenDTB9T/+4z8Gnn/zN38zNmm/6qu+anG9611v4cDSwW9bESgCZx6BfAPsMGKbsT3E1I9+9KOLf/zHf1x8+MMfHvH1Zje72eL617/+eO3mhbYicLYikDri3/7t3xb//M//PA4rcdph/A1ucIPFNa95zXE4fxg+drZiVrmLQBEoAkWgCBSBIlAEikARKAJnCoEzflBpo8Td3P/6r/961Tdm3PGaDZp1wFhk2qz2rRrfqHHXdxajvnnj2yK5W9Ymtm+KWIR2IboOzdP3Xu5cZk8b4b4J5cEeuYvZJjjbsRv7sc1udzTnWz/u8vdYHct14YENdq+P6htB+Ekemx+efSthlbf4hpuRk6503k3f02eJjnTYCLD//G0/G2Deu/a1rz02vti53/Q7mBXyDZ9/+Zd/WThIy8PfYosNxpvc5CaLr/marxmPTTfU2cfDGDbnE1fkDnPyV/GD3dgvD3HlXMgf0T+Hv+9///sXf/ZnfzaM9M3f/M2LnZ2dcVgpNm9TkwfYTJzNw3tspsVm4mseyQXbpEdlKQL7RYDP4rpYxW/VVeoMMYqfqm8Tnw4yNh/62Mc+NmKsePCud71r3KzwLd/yLQuHlUdx40LifXQUa82bWqnf6tyvZdt/RgCvHMQ7hHfDkzUin3FI+bVf+7XjQF5Njmen2virHGU9a16+aux5vSNfbWNLfUB+6xqxhg7kv+51rzvq2uDER8+VxkZiEF6o49WD+JH4qqY4F2MQe9NbXpED2J2edM836mPvM2nryJn9APbxmh+Rk7yr9nENu6obcRifNTclyCtsuq1+uF+s4YG79Mxahv5pfBU+9GVPeMFADeG9bfXl+CV+Zu1HVn5pnU2HVbtH5214nusa9vE3vHFvrtu2Ff9twLAyFIEiUASKwPmNwBk9qEwx+Q//8A+Ld7/73WMhaXGkuPRZHqsmUqhYZPo2jW+CKLgsRP/u7/5u8Z73vGcUpQqBG9/4xotb3OIW49s321Bwr+pxrv2dgtk3oP76r/968aEPfWjBthYJbMoG7MJ2X/mVX7m40Y1uNF5bbKwWzGyvuLO5YBy2dce/BZV5jGXR4VtAxvnqr/7qseFwFBtrZFN00g3fyBjeKq59rvici1ILfd8CyzfAfN4C9ez0APZlZzbHSRx461vfOjjxTd/0TWPzi52/7Mu+7P/x+uzU+GiltuC2MLVp/vrXv35x4sSJET9IYZHN5x1S3vSmN13sLA/XfLNSLPDZXo3NxA5ji09iCt+VOyyGNbmFr7KfuCJOXec61xk5xqL4bG7hrRj6t3/7t4s//dM/XVx55ZVDpXvf+96L2972tkNfvD1TsYmMWub3Nz8Taz/ykY+MR/IAnuhnw8LGRWoCcZYN2fJsa6v6n23yV97Tj4BDD/4qZnmopfBdbFLf4j0/iM9sKoF4aMOXX/3VX/3VuGnhzW9+88hb3/d93zfigXlsrB1mE3v5tDqPfuqmxHX1ovkPot9hytyxzx4ErCNw633ve9/iL//yL8dhlNxgzXDHO95x3PSkdsC7U2litwd/esc73jHm5KtqE76qplCrWKOc6bYuz6Q+ynr8Ax/4wKiR1ANw4pNn+wb7qt7+VhOqMdyo8Ud/9EeDH25+czOc+Mpmp4MfZ9rmq/OL/WKv3PIXf/EXIwZbV8snDvDVv7H36rVH+XcOKNWtuKl+59P8yM005J3tw6YeWUfQ773vfe94T3/7Quy7DX54OnCEh9j293//91etZfgyDORNcQ0+Yp69Fusnsci6Ru3M5tvYsp/kRtW3ve1tw/b2+tjPjZWec9i6jfLzLTc+WGeeWK5hw1nx5OY3v/ngLezP9nXlNmJfmYpAESgCReDcQOCMHlRmYSSJ/8Ef/MEoJhWXufMomxOeFV2aZwsniwgb1RK+YksBqxj9kz/5k1Gs2ahUbFtgWZBuc0FztlMpd47ZULPxr7BkC4VzFhWKTgWzRbuNZLbLN6IsNOYDxtyRbMFv48p4FhtsbA6Lf8WdDSxcyIad8RSB3jtMe9OXPuGtRQLe4jO5PHBWEUoOCwQbfhYH9CajBaD3W6ieefaLKR4WxLiVB9tY2GWjNJLqiwPzgc8VV1wx3vuO7/iOsYiyqWNBWPsGtc2eYZuN+be85S2L17zmNSMGiBnw5NvihRtUPOQBf++1ocIv2ctmlENJscQhqLhig9x8/Fdjawt4MUU8kUMsiM0bHpAxuUuswhvjhy82BbaxhbcWzzblHAK/4hWvGLHqwQ9+8OLbvu3bFnhL/+Teo9aDjJETthb38oBNU/nETTByCt9jM3ImD9h8srmWb9v622fsot/Z0OiOT3STY5JT6CGXnA/xJBtUcGA3dUPs6PX5trGD826GEbPe+c53js1l8VDtq74VB4PJfuJS+tpIsxH4xje+cfE7v/M745D/wgsvXHzrt37riLnqlMNsYjL/pt+f//mfD47TK5t5akZ2P1t8+DCx6tj7R0CusI7gOw6i5BN1HX5953d+58jxiS/7H/2zV8w5y9rn93//98fNt2oO+fSWt7zl4hu+4RsWX//1Xz8OSQ46z36ui0zq2dQpfChrEs+zT6lh9FUX/fEf//Hi7W9/+zgAUXd993d/9+Ibv/Ebr7oByHXztfuR6zD70llckzvonLrOHkFsnPUZOfTXTw3oEPuyyy4bB8xqPoeUd7rTnUYduM0HOgfFk63dRCy3/OEf/uGwO4zUvm5a21nWgttwwxc51YH2Fqy11e9yIl7e+c53HmtqOQqfNTxmU/sF+udBN3rxQfa1ZjgXmtgmd7vZHy44r05MreQ1bOjvATd7YmrlrKm8r/82NTYUu91E9Xu/93sjhltbfd3Xfd3iu77ru8bzNsotpnhYp6hr3LRiX5JtcE7esdaytpx5u03YV5YiUASKQBEoAtuAwBk9qMxiws/PvfzlLx8LI4sfxYeFpMVBii2JX/Ps0EehJdFbTEj2Fp8KbgssBYICW0GgkLXhrFDbtkJsGwhwOmTInWMWuIplCwPFs8WFwpJNLR492C+HlTaUFZ1s6RCPzTWHB4pT4xjTgsNYCnCLELyxmWkB4z18YGtcsBlgPPbHn8No5qezzT0b/TZAcMt8M2/NHXld47DDQSXe0ttCQf9tPdg4DOy2ccxs2OKYQxwLCs/iikWtgzD2zcYMDrNrDirFHBscbHzs2LHF7W53u7HId1CS+LWNem+bTHD14P82rd28wr9sIvkpwmzywZV/e3jNr/Za8OXgIxsd2fRn42zWmTcxij8a02YGX3UXthwiziSWZREtNuGBeOAafm3xv40tvMVvGFj8v/jFLx46/eAP/uDYtI389Azfj1oX9sq3KE8sb2Ky8SsfyOvJ4XwtfIl8bCmeyiu3uc1tht0c6Hhv3pxM/219zkE9feU49ZAYZMP7fMgX9HaQzq/YW72wiZ9vqz1PVS7+mkNKtQbfcLOTeKi+xXf8Fr/2E5eS99RqfhFAvHXDjfj1hCc8YcQDuB/2N0/EUJt58igZ+PG97nWvkUcdwoq7zaOnyqLz9/ochDsEf8Mb3jAOPMRRh4f3uMc9hh+dykFU8pAaQOyynnWDlWd/8yEHXne5y11GHcNfj6JlnUIG3ygVN+R0/mQd4nlualq518/juknMwZ06Qe654IILFn4tJOuqM1kfzDLPr2MHMVAOVUc61NIcvtFDjSafprkmMdO3Cn/zN39z3BCl9sMPdSesXOPA51xqOaiUW9SC1tr0FHNvf/vbj/pJLlBbn8lmjc+H8ZIP5yZo62cH6Le61a3+z42K2Y+wF+EAzz4C7uvvgEj/c+WgEn/VBK9+9avHfoSaSS2gVlQ3zfte4oE1Lhvbb7FPYt9EPFJPHHae3y+H+CW7088NVOxvbUbmu9/97uOZLlkT7Hf8w+qfusqNlfgqlrpxRQyWd8TRBzzgAaO+2Qb/OiwcOm4RKAJFoAgUgVNF4IweVGZD8k1vetPi2c9+9lgYWUhI3opKCymFiMIrixAKuytp/saLRaZNZ3cvWWxYpGQxpihQlCrYjNN2+hGAfTaT4W/jy52abMdO7GVhmzsjSaAotonsEGBneefmvHC2YMIJm9MOoC08LBJzZ6BC0Hs2uNjcAluh7aDSnfi3vvWtx3iK9cNo0cMiCG9tsuGshXB4i2/ktECCj0MSf9u0cBhg44K8+p8rd3YeBtZHMaYFnEWRDWF8skl+YnlAYlPm2PLgkb1wL/FDLGJLiz52def57/7u746DShsbFsIW+/hnEXWmDnyOArvTOQdc+XIWeDC95JJLBu7f//3fP3ybn8kRNk/EEA/x3wJ2twUrO+VGFj93avPNHOwpN7DT7K/44DPvW8Q7rHQDBP9myyz4LTxthvhGkgW1xb9vaIhl29jC2xwEW/z/2q/92hD1h37ohxZ3u9vdBtfFqDO5ESl3wFZOsdlrI01OwQ0Ypy5gbzqJx3I+H04fd87baGQLnNE3/ruNtpllElNscNBZ/rC5IQaJKTi4bRtKs+yn47X87ydI+ai4i498MLmVv59PLQe3Nlz5hI1bmKib1LdwOUhcksPmgwmb1a997WuHvzzxiU8c8eAo+MbXxWXf8L788stHLPfTs3IpHc82/z2fuHk26Co3q+nkEgfh+KbmVqc5EM/NrmLNQRo/knf4aQ79HXjl5wrVkW4o+PZv//ZxMCqWzwcIB5lzk2tyAEcmaxT1rRwoj1h/WJvNjR5qG3HXOk7eVQ+TX20Apxz0bWNNm/pG/SCHipXyqCZOJofIp2muEQPViPRVE6kVrSf1z8/h71VfZqyz7Tl68w3fNOYX9FRj8Q0HWWqtM51vc/Cobnfo4+BKjYDHD3rQg8aBDzn5FP5mL4j/WUPggnyJvw4qPdt7ONvrKNz18G29F7zgBeNGH7Ww2Kbulbutk/g8XMRBaxU1JTvzf35hv8T6Bkbb1OjCl9U99lrEJTamW24gOIo4ul9MIje/cgOYvSx5x00TfAnmD3nIQ4YOOIy7bLSNMXW/urd/ESgCRaAIFIHTicAZPahMQne30c/93M+Ng6n83J67pmzAKCYtIC2iFGWaZJ+DIQsKB2KKLwsxxYCNHAWawksh0AOD00mZ/z+WRYNCUsG8elDsmy3saKOYXfLtSIWzO/58ZkFkQ4qdNAWeTTMLEwtI9ld0+zyHeilgbWhaiOMHW9sMsCngjlCHEIfRsgGAt09/+tOHnDYfcNa3HCz0cJSONi/gY6NAoW3RRU4Lf5sFFoQWTS1SD8NSm41pwc4u7GRjw4aWjVPcvHD5E3h3uMMdxgJpPugQixK/LP7Y12aVzQ28s7kjBrFrbbuZHeDHFuwglogBv/VbvzWwtHHOt8WM3EUrpuSx10JPXhBLjGnBaHwLdXayEeUw0rhiCH916CXuGFue8blDr/gpu1v02wizwWODyyYP2z/84Q8fC9HNND76Xng7H1T+6q/+6uDnfFBJ5zPFW/LZWJJH+CB82Qsv5HObKvKFXIAH+rMV/OUWm1RsaeMlD3azoTH779Ejv/mMbpKwYSqm4CN7yBU4KHfYLD6XW74hwGfZmM5isBzLlux7PjW1LZ/Fb5voqZ3URLCx0cVf1SX7iUviHb9SP+WufweF+HbxxRePn1fjc4e9oSt/2tAT7x3wiM028sR7MZUMasCzxX/PJ26eDbryCQduajs/cZmDSvH0nve85zi4UK8f9KBS3aImyAGfbwbbnBbHzMV/rINsrptPzS9/mfMwm7ghZpBD3cPP1TTy4v3vf/+h9zy/XOqRXwohu7ijjnWNevhUcJrnOozXZBfT1AAON9jht3/7t8dUcJdDxBO1+dxcI6aqO9x0SH/rTPE1tQbczrX4g7dyBhtbc+cXDNQXamN5gO3VTmeyWZulZuVXbMSf2cYvgbg5OYc9anc8t4aLH5Ldt5nZ3yGR69QQ8szZ3PCdDe2BPOMZzxjrG/6pJrCusZeGx3hr74U/iwWwU0f4zP/1aL/Ef1mi/za18JOs/Fkc54fqAX6Mn9vol7nR0roQB61j8FGdI844KP/e7/3exV3vetexX6SmxUW6tBWBIlAEikARKAL/i8AZP6hUWNqU++mf/umx8ev/wVBMKqAUlO5+VChbTCjMNH9b/Ck2FVsSvA0XCzPFtmIgfRSw6w4MjJWFTcbO+DlYyHPg8reiz3M+yzjztfk8fXL93Hev/nO/XJvnfBaZyZNH5tV3tZ/+c0vfWZ9Veef+e732rSI/falgtrlowW9D2eI8/+k5G1kUOTRQyCmcvWfBbkFoYZQ7XRV0Cjwb1IpV9lPMWWDqQ2ab0+ayuaUI9LMguOD/VVJ0OwjcWR5+HkbLhiDe/tRP/dRY5Pm5Tw/6WijgJ8zxkR4WV35KyYM+vnnl7k6LBDzHV3jEbvTWYh/vGy92ZyufuSa21D/X5zn9fZaW/hk7f+fzPGcMz2n6at7L2KufZ9w859p1z/Mc85hz38iX8fL33MfreazdPiOzNo8ldohDDghsmDocsSB2QPXkJz952CmHY7OubCTWuNaGkM8sovAwC/zgNcsTPVfx8350y/Ms5zyG1/rnEb3Ch4yd51xrXGPuNW76bvocGTxnPq/nlnmjV57TR392sClhIW3j3DdsXve6143Y8BM/8RNj49xCnK/k+jxnnHXPNhAtFt1Mcfz48eGTbioQn2xyuEOXv5KBv3qwKV3YkU31kUu0bJw4SHKzgkM1Mctm3uMf//gxpn5kW8XZHB7GXve56+aWvvprs+3mfnk9jx9b+MxcebaRJ3aKX7sdVBpHy7zhVcYZH07/pH/mz9+r+k+X/L+XfEle4IdszxdhK97yRQdVux1U8j8PG1pkdbOIuGpDQ37JQUfkCjb+jo4RiI557CX/qq7z9fNn8/jGzZh5dl3k8myDw08H+kYAfeQ+hzbqop1lTpMHZ/lc7+/VOTOXz8Uqn5NlvhZW2uq1+Xt8uPwn12TMWfb0yXOuzXx5zuerY/hbSz/5/EUvetE4qBV36SyvO1jILzTon3FyfcbIOPOzz7RcQ/69dPhs7//911irD2PFL/6352dfrfbN55lzltln+mvhpdf6ePALta2ax4Mt4SJm5cBe/xxobhqXzGls9Qlfc5iRb1RedNFFo0bBNTE3+phHix55XtXns702+9fNheK9ePQbv/EbY9Nu/inqHFSaI/iQx9+Z3/t5ZFafp0/65bPdnqNnxvL33DLe/Ozz+TqvzZfm74znPZ/hzTqZ5nFc4++06JPnddenb8bJc8babTxjbTrePFbGCx4ZIzJGnjyvymOsueW6k42TazJ/ZMqzz40FZ/nc+kAukVdyUClfn46DSj6kJnDjkoNQtb6aA6/VBZo1gXpD3Z/DktQTo8Me/6xilr9zyTrM9FHDpPa58sorR16VB61RHvawh43/c3J1LGPKt9ZoYo2YYhNdDlLX5jA3OK/OHZlWnzNPnn3u2tg573vO2HkvY839c5330lKPO3D0azx+5vfSSy8d8ziYdTDgkFg9kDlyrbno61CE/mKeteZu+weui3yrMmdMz+tknj/P64wVuaKfz9eNb9x57Pyd8TZ5NhfM6K0mVHuZl+751ZLVdWnknOfLe5Ezc0e+6OLvg7TkNXURm1pDO1h1sObGQGtodnKzgFwm/+Wbl/wPd+9973uPg0rXOOCKXpEnOgR/z3OLvrMu0ce1qa/Sb36ex8nrzJdn77sm46ffXs+xn3WNG6bVTfK1gzD1ojUOXMRAtuUXMBQH7a9Yb+mrtvqe7/me8Tp5KXIFj+hDHp95PxiROddF3lyXvp7TMtasb95LH8/GCD/VweGnukdN4Nl16TtefO4f72fOVRkyV7DO3/P1p/LaHpc9MTx0c4xDVthbn/AzaxP/xybeZp9zr4Nz8uexKlfen21Bn1k318z9Ypt5rNVrdsPEOFrG8HdeZ7xcu27M9MnzPF7Giaz6ZIxZH++1FYEiUASKwPmBwFYcVPp5DgeVFpOSt4dvEOzs7IxiRFGpJamlMLJo8pC4LBYVXhYZEp7CyXU2dDzPyS3J1TWKW9cpiFynJTl6znWejZk5s2DLtZFNnzzIOTfjmyePzBVZ018/sumXcX0Wuc1Jbn/TzULSM5kyhs+Cif5eRz99PGZd5mtnmTd5beHwkpe8ZBzwKMYUYjYAfKtxZ2nDbKzSh9yRBaaxkcVRMLUwMY7FfzBnR3rq4zpjuHPSoZJC3QaEsRV+ivT73ve+4261TeTfbx8cc3cf3v7Mz/zMWBw5dHTXpoMPiyEHHGSHvb4WUA5y/Z97ilWbFXhOVt/CtDCkX2xvDg0+bAU7OtPRaxjov3ogxu7pq7+Hv3ezvfHJaTyPubkm/M7n6ZM58rnrfBZOhY8Zex43r8m6X3mNb0yYRJaMN8vrvcicOcgKP+/HZ4zlPTax0WTD1iLXs01x/1cX21oUmdscwdIcGRvOxtIPVyOnPnNLf7J4uC5j+oxsxgmO5PR3dMlY+mYsY0QvfgQb7+V9tsrYxmUb43q9Om7G3/Q5cmS+PGdO45Bn1inYeF/LGPzdN8hyN7zDYht/7uTPNyptHkXHjJlxxmBr/hEnxAcb8XzW/P5vmxx++Fa+98gRm3jWzAGr2f7ZABT3yOfZ5qdF/+Me97ixIWIs18I6Pua92W/IHa6ss8PcP/Eg+cy1rpmb/rjEBjO/9NE/19i4dRjskHXdQaXDD3J6GMc8kTNjrM6buXOdZ3134+98fV5no9BGigNquMoDbmLxE33uEvfa5gtMja/RFz55+FsTG21K2gRgCy1yhqf0IyvcfGZMD3yYH95bxTtYu961Wvr52/vGN5e+rs+Y5Pd6VS798o1C3wAWl2zKiEE2unMneWQMDsY2Z/TyHLt5HzaRxfuuYxt2NRZZZ3kjc/QipweZXevhb2PNTX8PYxnDnJnX+/q7LmMYL5hlfjcV+EliG/7kk0/lVgeVDqzZ3zWRJ3IYPzbJnGQw7urcdN9Nh1kfr13rEZ08m4cMYlHmz3Xr+ntP/3X+kP7GJLfn9CcjzIKlOO+zYJj5XcOvHUxsEpdmuztQWT2olPfUJuoSMsx8IE94sE6f4LDps81kOVdszkHlQx/60HGog+85qKR37Br86UEeuHhETu/BOzzbxNbGN64xMpbX3tdmzhlv1X/15Wf6h9euy/ueNdeKTbAzJlm1vebXxyOcn+fP9WOQz41jrPDVvB7+9pj1IWdsuRtG+me84J9n789yrfrVLFvGIYvrMwbMNVh4RKY8e29dy3iuz1ie6ajBCs7yinzHL6wVHEapI07XQSWu8D3j+wUI9YYc6n2+5dCPDnJXbma0TrFOOlmjYzgZO3r2XrA3dvjgGeYwUEuRRTyVTx3uwNTcj3jEI8ZNP/AK/pHFuK7P+8bMRnrGxnOfx/ZzHMg4eY4OxpxlZ5/EIWPl89hyliHzhBOZj+4+M4frrB3FEzmUrdnD5w5ixDPrLjUB2xg/TR9/e99r45NNfDWn91YbmWOTyOw9D41skTf2ofPqWGT3MFbih/76asYjlzkis8+MTU7PGXd17DHALv/ELsZVe5vf9caj+yyDvubW1+vMF/l8Fix8rulDvugeW40P9/HPfFBpTaA+5M85qLRXpCZwQOQALms4PsnH3Px+7NixUZ/zezLNspDXg/z08/B6tuNsy9jUe66jO7t5hh+9g52/V23iGmMHM6+95zq4e96kZQw8//mf//kRf6yV1In+D0f1UvYVzCUOOjCDkQN8scEhrpjk/0yEU9Y5wQPvzEOmYGYsh4Yw0uDhOjrTNdcGT/09vB88gmG4kfFnvfX3IAMOsIvr2W+ez9g+I2fGjxzByOf6aT4zbx7mznXz/Ad9nXpG/WpPAe7k9XxieROmdrPlf3kEb3Wt/R81jly1rtFhVf7oF91hpMFm9jn9NGPEHsbyGrYau8IgNvG8lz0yVsYjg8ds34xHlk3GI1MexjIH2eN3/GIeJ3oNBfpPESgCRaAInLMIbMVBpW/FOai0qLMhZwPZz+RI5hZICoq9mgSn0LYgVJxKoK5xrQWjAiBFlmSqv6LBAZKNUAs6fyfZ6+v6FDCePRS57sZz8KYAlEwVxzauFaqu08fD55Lr3PTR14OsrtdH/7lQUQTaoCQTWTXykJ2edHSHmevp51qLL8V65jSGfhblxjKf+TUJHybmdX0e3jtIAeBOPhveDuJgaQHhDlYHcV6Taz+NXmyokRWu65rDAd86cQBhQUpPmw8WpH5aQ6F+GA2OOaj82Z/92bEwnr8hubPzfw/Y2UJ/cj7zmc8cxatvBznI9c3P3OWrqNMXpxxsKtgcouR9Y7AnDuADm8/fGIKb69laXw8cUOSH265TNLs7ml3Y3ussyGe8XMc/POMFHyBL5gkPfY6bPuNzxsVJzzjm/XWNjWGJ53hjPDJHXmOSFy/JyKf4njG957O5uY5fRJ7IDEfvGR9HvA87Y/IXmMHVogKfbDi5I9sizv+VZXMDZvRPQZ55jYWfPjOehSJM18nnWjqzD1zjl2Tj53DFd7agI11nOc1hPrh4sCns8MUGtc/5m+vFGJj6LHHE2BkXr9gndjfufhsZ6BR+s505zT1vfLB/OAcb83rA3rzRg+3w3t2o7GBDgi1g4P+iscFGfjxIHM44e8meQzkb4R5s8+AHP3hshIsR7LyJ/tET1ieWC04LUZsiDlbZ0rcuxR0HlmzJF2xOktVr78FIX7rCxTW5w3uOc7BNnmJDuZGMDmr0X+VX+kdGGxJswfbJh66BHzl85nDg13/91wdvfviHf3jEIvFEP3OSUz9yOrTHx3V8MR5Z8Zh++M3+7Msf2IivzvqtsxefsMGYQxNcwFkLev9HqYNK8u8WT2AQHMii4Z25YUfOcE0soBt5YSQOuTb+lziGr/jm79V4A2s4xX/NQWd6wjy+CQ/z8s+My4aJvd6HH5nEIXdh+wYMLPgS/GDgDnn2T0xwfeIhuenAZnl4zzd5yGXzhKxkMZ/58R4/yUzWxEfP+hmPf7se5ngR3yMDW8x2hR+M55iOhzBOPICNccjtQQ420uiK5w5o5XN+ZV6bjfwUN/HQNckHuZ5OMA8H+Rf7GpOdyAUPc8FN/DO/cXbj0xBq+Q8M2M/YGRNexvKtXeOZH06a/jCAIcz1JZv+/AGGM27xH/3JDStNf3mUfczvfbYxPpnhb27zwlg9BLOTxSXXsTv9jY1zYkG+UWlMN1y46YpN4Wde85sbjnBzPdvQhzwzBkOBDf/Jxp7YnJ9+dVDplzFyUIk35CArXdkCj/DbZ3SHNTlhDxOxii7kjM+QnZxzo6PHHMMyD9t5X2Mz2PFHvPMMB+OxD26IWeb3mfnJYQw64qJGnvyMMfzJNM9vHLEwscn8xjG/+BHszQF7Y8wt/IsvGCdjwRAXjcdmOGYcPIOVsY3n8zTj0Y/8+Cm+xAbGgj9djYMP9IOTcXxmLPpFLjaCr3ESZ3wOB+OE17EdO8/yrMoVP4tcdDSX6+il4caJZc5WV+gHRzHFDZU2is1r/oM0fgEXN02+7GUvG+P7mUl40NODPY0v38tjm/ziSzAzPpnxwoO+dBQ3cI+e5oI7fWHOVuoUNa18Ym2Gg2RQn9znPvcZ62vcMs5qgze+wcW47Iob3mc/hxzkMB4OyiWe/W3+ucWv9McbsmuJQ+SnI5ljQ6/x17VwCMfIgBeZL9zQz9h0ZGe5M7+iQWZ7CnBna9fjM47MTT+P8Ibe+maO9CUPzMgcm8x24RMa/OJf4g8b4R2/y1z6xb/oDle6sKO+8DSP+EY/r83Pt4wHh3X+ZtyTNTqQVXzCX3YJzpGVDtqcz8gHE5/Rw2c4wWZs7HO8pDvZxEr6+Nv7rtlPozP/sS5wKGdNkINK/2WB9Zkx3ejuxkH7EfIg/HLzMNvLvfSb54+PRYe94hLuwSVxiW+4nl3MHX/QL/yE0cn8wdzGcZ28zC82aZEdJv7rJN/gkw8dPLqxz74CeZMfzMNGcr39Gj4CI/hZt1hfkQHn2BCn5TO8IBN96YMv6jQ2JwMbJ08nF8LEY/aL+FvyjniBF/BMHltnG/Pgv3mNT0a2JJP5va/OxBOy5xEfZRdy+Nx75ocLuc1vPBi57nQ03Du+/OUecVcMhpt9KY2NcFmTC3yzEv7sRo51TRzkn57pCyM4wNN78BE7NPp4pN6ha3BgD/yOTcRAY7Ep3zQ/TGDjQe65mQ8vzLmXfTPeqn2NN9s3cY99yC9Hk834c41CltQVsdVB4sisS18XgSJQBIrA2YHAVh1UKoocVDqkzEGlYkSy3aspVBQyilfFrMQnSUr+fipOklOESJKSdgqfbIj7GTwFhSLO5/pK3OaVED0rzI3nbjULPYeoknaKZklWgtbHYYHPFWBzk4QtosiZBYk++tuItHmmKWQUkYpBumhwkLx95nqfS/QKRAtuxenOzs5YFChmFDYKJpt+fu7DfPSmn4JBIWPTM/+vokLKe6uF/Jj8JP8oyGx4+1kWdlBEzt9YMu5+Gvk9NDabi5uM43NFl005ixN34rOBhSgOPfCBDxy4rLs2Yxz0WcGvULOxloNKm2rmdTirAGUvfNDYyTXk9BMtilUbnBYGx5Z3erI9jsKefenkDmjX2PBQPNKNPd0N6fW8wMBJ/MRHdmdrNtffXdRwwiOYuY49wlFcJouFFc7PjU/gD5uSDW/oRR8yGBv/LRK85zMLLdzPxrqx+eK6ZuFgHH7PF22qGNN85MVVeim8jYPr7Et2eNFlbq6zSCU3eSKzhRYZEx/4N/lsehvD52Tgmw4o+R37+sx8sHGN8WZuZm4cEyfIhQcWiHwZHuGf6ywMxCqLWnKyEZnYPPqKNRb2fNPcOIJP4gQsMh79+DO52YDvudZPHMHLHDDFJZs3sGY/vsnm7kK2WWouetItY0evkz2TAUfxC9fE0cQaPDSnceEADzYUbzwytznpQV7XOpwkM9npZgGDl7BgczrABi5wNg7/2KsZO9+m9H8WsZU77N1c4GYBMXuThQ878V3ykRNX6E1XtmUjcnrmi+T0c2O4Zg7vwcj17M8vfPPb53RMvAjH5CO2M5ebeWB5j3vcY/SHp2vgpz9b4BYZ+ZHDPnjyBfzGHX7PNz3DEca+4Y03DiqPLWMRH6OLmE5XfSwOLajlGXzJvOFLeG0u/cUfnNBXTDQmjplnr8b36WnDF6eNy6fYSD6Bbfxwt3GCXTYL9SenB67KwZETTuznb/7uGrjgE3vtLPMpX/EsVqxuJIQP4gXscYjtzYn7bMfWyeP0x2E2UJfQh2+zDVvDDk9tJLG5eEYmmyh8Gp5k48Ou40e4I+Yagw7qAnPCD+bsRh7xgT3JZDz5Xs1gA5fM3oeF68XBxEA2oBeb4415cy1/NgfewhcHcRzGfJd/yB/GFefxUD/j0B2fyC++aub185/0dy3fx3lz0N0DDskx9BcD4EEGmLMlu/IvePJ98iSXmNt18io96GAjZK+GN/FFY+IpfPigeIsjbAsnjZ5sYW521Nd7bA/v+EP8HWb4w28cpMhV9NbfJhb5wlu60RMPyI5H+sbu/HaTuORmLtfitdwDc9+w9tOveP6oRz1qxFayy1HmxXdYsIfYBj+HmfhMxuQnNt5Pwz3zrzuonOM7f3D4kHodFxz4wNH1OM828gkZ8AXWeCZfwEsuYqtZRtzgZ2yMP8ZnBzrPtQhc4GVM2OGRMcUM17LB8eUGJVvBhHxko59v9LOvRh71Kf9jR9eTYXX+6Ikb5CU7ToiJsDe/eYwxt/AvvkAPY5GDbD6PX+GwMfghHqfW8Hkwynh4gM/05NNiPD6IL3KZcdS/fFL8JBf9jSMu4604wM/ZEU9hwlfEDjjwT9fSLTFCfJjlia7mTu1Arpn3fAon8ceY5IAvHWyE4jAMT8dBpXHFO7H7BS94wcCEffEEdnyIfLDnIw5N3HgDp71a6ivxAOb0E3+MB0c6BjNzsR870pV95P/kcJjDiq3wkq3EfXOwzdz8nXipr3HFi51lHhTj+Jg6iu3kBXzMT+gnH8zjiS/8SDzERbKbg+8Yl5/iEtvQUR+vxRu2IiOdyMv/8ILv0DncwKHwKhwlJ87jn+tw3VzyH/085kYmcxlT3OXjbOS6mX/6wJ5dyAtndsEBNsZzc8Ii/mWNhovkwEt2MKY5k1/YyroPXubV1xiwEd/4sFqT3MZgDzgkj9ENTsbdpNGBrOzCH+HtevrD2Pz8mozJZ2Is+fQhGz19Bms2gwGbpdbBHbqkToF98uQmMupDRrLBeD6oFKvUrXjAJ/iIb1x6bQ4xRN0Ie7Eo+Snz0stjv3GJTjDnG2zBLm4sg6M5+IP6gn1gBIu5rfqDOocc/MF6wfMmzTW4OB9Uspn4koNKPiNGaGo5sQqX8tP65MN18UqOTI0r5+C2egznxGT6ykF8Waxjb/rjiuv5C/uKMzDhx4lXdBQLZ7+AD1643mv8nf1MTuYbYpn52BU/xS7rbDjDAOesd/gj/PPAb7KLU3wUh+CF0+ZLvMQjurnuVBt58PBVr3rV8CnxQH3qG66whWkebIOfciau0n9dg/cm+xqwS03NlvAxJxyMwSb8PDbBQ41N+bP+bOEhVpGHvdKSb43FJsbzSD6a7Svnwhgn4MxmxuOXGTPj8RtxBS5kE++MRR+yudZ+hTjCN/jyXG9Hvj4XgSJQBIrAuYfAVh1USng2Nvf7jcoUygp9BzmKA0lRwjWWQkRiU6Qo1CRaCVFitKmn0FGYSY6KI4ldUae/ppizQSDhKiqy6HCNDQjzGtMcCg7Fgj4WKXOTgC2i9Fc4mU8f/ecCFQ42a/UxroRuQagoVAQqyCT3FOPmIpfCQGKnI/1cT0eFGkyy6HOdokxxJvEruvMzrYo4hW2KiVn+3V4rzF796lePzVDFqWJVoW5MD/iT3/tzIbrbeJu8ryCEgw0lNrjiiivGgokuOOQAgL32o8cm8+rDHji37qAyP1k8L1DYDaccVPqpWJxTFComFbCKOTzQz8LaprKf0rWgh6MiEldsPLkWBxSgPvPNAwsNNmXjFI/sj9cWCPjgepjhKFtY6NpsVPylMDUPG1lEaxafFirGdH0KR5jiEj6fWG6AZNMkvLKowUmLEA+LJ3zLuPwKhhYOKXhxFU/JSz+LR3hEXrw0LvuyK5lhhqsZ18LJgobuxjAPfWAPG/LyLbLAjP78hSzepy854KyIzyaMwtgc4RIcNeOLE7CAMf9zZ6qY47WFiGvooZ9FkznIx4582HvmIoPxYEw+c/JNsYHO4kr01Udf1xoLX/yfdt5zAAdvn9EJvmKIWKXRl2w2ltgoOMKJ/TZp9DEXudkeB/ihuWDsPXjQWTO2ecUbsrEf7ooLeMzWcBFHjGPRgnPsYBx48BfXszdcyM+G5Pf+Xg0OFue4YUMBFnQXm3JwwFbhKHzXNfFG/HYYYOEotuIczsobOEpOuuKc1/ybn+Munlgo+naDAzly/+iP/uiQwbVw0vALxnCEqfjmQJF9/N9tyRWuwS9zGVvskFs8YMif4MdWuExHecx1+Cw+wMT7j3zkI6/6RqWxbH7IQfKpa30DA+a4I7fCKP7AfuEi3dgSxvzLQY4YM+u3DlvvZYFvXrKZ10acmCqms/umLT5KRvrjIjuxlxxlrvifWEFe/eCCB+R1IGbBjq90yIGgzRINr8iKB3wsHGBH9hNzYMEn2IFMdDKO8fAXBy3q+T9ftgFsTPK5jkxioHidR+QSG8ItfqE/nrOdXEMXG/FyEc7iEg5nc4o9cYlMdDC/PrAXm/k3ncjN3q5LXeVGMvOThQ/jJozpPG888GNz4gieamIbXF0PA2OQ1dwOyuh/YhlT5Fjz0o3ueGp+MSR1Fv3FRTLyFzrgvjhCDrjDlj3gSFYbm+QXA7IhMwTb5R/X0wt3+C8sxQFy+0YHfvL5+C/c8EEf+ZNMfMpc8wEVnmlyq/HJLT7hFYzoiPf0FgthKe4Ynx/DT/yCj2vJRe/9xCU+hfv8XEzKTyVecMEFw39hShcyea0vW5MJjskjdBPP9rNRHrjFLbxdPah0I4lxzcV+fImMuQkDJ2zIsil/wR9j4Q2bwZeN5Aj5hs+JX8ZL7Ar+rsF743iIocaib/KY8ZKX1WxqG/wTK8QWOrz0pS8dcReu6ql8hjdwNC/eqdvwhu/D0/zmU3/gC1v7W2xK7QZbPsj2cFEXpI7xPn5rOE8XPmQ8z3IBn/ZZ9DEvzsKILtFJrsJn4/ErsR2ngs0c0xIzxRD5nE7GsrmYepLt8F8cyOa18VLv+cw45IExH89BOLuxGRzxHH7GS01HT7aCvbHZQawhNx3ohifGTp7yGa7S91QOKo1DbviY20G/GzbFMgcoDgzIKw7Js/hJV/PKt2K/vmSbG/2Mqy+fwwX6wQwnZh3ZCCfhJV7gFgz1Y3fYuD61gPnwn435jbnokeY138E7DYb08IsWuMYG5MBzfMBJcSC/DMPmuDi38IcsYiKu6yN+qQ/Ij/N5iLPinZhIFjkIjnR1HZ5Z7+KZub0HK7yUO8RIssEd510rR+AVHhln1pvO/ubr+pPHN/HEVg/zsZFcpG/8iz3Nw/aJj3wVN403+yvbiEN8lq3Iwk7Gi53Jbt1HdzWHfuIDubJeYHuYkEUcYG84eIg5ZGejTZo4wM7sKU/hCGyMI64nppMRLnRVV/BjOPJ5/ensvTzkJw13HNLgu4MlOpGP3uRnl00avzU/fswHlTjsl1HYlmz00Af2O8uYgbdu1PJ63Xypm/cTl2DDhnw3tb452Q2/2Vzsk7vozUZ7+YO8jeezP8B9k8Yu4sTqQaWcJK6ZH95ivH6pS+R6+zX8Aw/1t1+iP9voSyb9fEOcX8EYF/k3zvhMjIG166xL5FfXshVb8AucSv2LtxpfIhNuuMb1eIzvYhJOaeyD++qaV77ylSOOuU4c8ktH4gcMxDd1k/k01+OmxrfIQ1e+g6tslPolcQRH5IT4+bh4n/+IU8YX4y655JJRk/Exvq/ugx0ZxXJ1Pv47qFSD0x8e3lv1C/Fwk30Nfg9P9qQXPOlDb3VA/EOsEoPJmjgFLzaGA1yNgeuzPfCHzfHBeOIfXNfZlw1wf7ZvxmNDLfU6ufjOiWW+UiOJr3jEZ/GRn4lHiffkpKtc1lYEikARKALnNgJbdVApCUraHjZoJE1J7WQJSVElaSoQbJRKzIoOCVeBbDEjcUqASbKKH/MpJCRpRa9nf3s/n0vmxpK4PRQUWYQrgGycmNc1ZFXESqiKZIXH3CRhiyj9JWYFeAp5Cz0LIs3i252B2XQyjwYHCVpRq4BQhCoeFQAWAd5TTCicFTZk0t+iIgujuWDNpoUCim4KRuP4W4GzWjANIdb8o1gxn4LZJpKFKVmy0aawZEs4wsjYp6OZ1yZ+HgpbeuCODSx2P4ym4FbozQeVFiYWtRa47MhGCmItCyIY+YkW9oGN4uvY8ltMeMqW+rG9fn6aRVGbxQGbGJNNjIureMhXFG7k0R9vPFtM4DxeK8pxR1FKdvbhBwpr/GNzspBpXlgpHN3ha5MN1riVucmLV2RiT+MqWM3Nh8hr3Gxo21zGCc3iQV84WJx7JpOxFdeR1yLEWD5TrMMHh8ipEDc++S2INTrxR/J6DZPgFb8hszHoySdcCwc+Rl++aeGjmCeHO77hHN7CkFweYoPYYy6+atwf+IEfGDGHjP5WbMNGTOLzx5ffuDA+vYypD5myaQFjY5LH+LC1cLXwFHv0pws/hqEi3wbZy1/+8rFRABMYRjdzxOfhKCb4G6/4u7gjZtFV300aO1hAGsv8YqVvTrGrWERG+uBc5qYPDMQc2OOtGMlHcZmtYG4BZNPBxgxesLuNRnEcpviGW3wMt2f77yY7vY2JaxaIcgX7wYnPksMmUsbiN+sa2ecNYPa00UBudnM9vYxLbzjIBf7GfQszi1QHy3hAD/8XHN3oyK4a2VxPblzB6Re/+MWDCzZAbb7Qn83w29w2t+QWC3b64V2wMjed8BzOeGMDgf3kJPyycSoWsQe+iuNyqZiO8/xNjLBBA3e8NIcWXPgAfI2JX+KauAhbsuyG6xhk+Q8eudObncgprvE/m99yiL8P0uRZ3BOb8BQX6A87fOUrkY1f8j+fuw43d5a5Sxyz6GZL/qrNeRpf2QsmrhEf2VRfWPFln+MzfH1mPAfZfDY+gFNkxJETy3jkOtiRAe58xwMWYqpFPB6Ylz1hJ3/ggRzAt/Ey/siW5hbTjIlH9GRvHMI9Y/Hd2JgMuIs3bEtWsYhNPMhhfDyAMR6S3zV8AL7wCAeNq7/3HCToAye44xBZXE8fcuhHT/NEd/rDw/vilmvh5Xpxgy7yCW67nq3ZVtwS870f/eGxV0v+xgm5RY7lk/S68MILh8/P+Rl35Uv9bFjJKeYWb/mvGEGP5C0ywZa/2fDEE7jyNfWEfnJMuJFYYzz+BT/60l88xsWTxSXX4j7uwC4HlX6ZQqwWB3yOO/ok5/Aj8tnchanPyYrH/Jxd4b6fxs7i4l4HleKhuGZDXb2nv3hGDz7JR8JxfCYf/uEk2fHcTQ/HljEO98nuoekn7+COeMfOGtzZCTdxN7HBZzjFt8RuXDQG3/N/q8KeD8GNbPgVX/KMs2pU+czY7Ckm4QkZjEUf17GtucUMvIIV/WDMH+QucTY8Jxv74QC88EJe1p8veWhzfjEefXaW8cB49IIX2cQD2OCFMcnB38R38sGcrnwEzt7n03Qku77GlyPh4qCDDt43Ph8yhrjED9iNnrA21rwu8Tf5faaPWCrWyBseZGBbvphcam78NifeiomwhZd85hcKxF/yBJsB0Ab/4J/xjQ0b/i7/0cs3kuUt45LTjZRiBxz4lnwrp+qrT7hoWnoYV6yFPR3FBrzAKQ+4RUc28hCXcdHnXsMTB3ALPniBfzhpbnaig2vT8MI84jhb4A5++T8t6UNen6k3ySU3iAHqKLaCK9nmJk+Sn/7iE1vDQH+xEH98fmIZv8xHhvCVvP6GB3nUQ5rr5U8+jWv0ZUU6sLUAAEAASURBVFu6msOzeOmhiRNkkzP4Nb3JQXfj45SYLfd437h0OraMF64jDxv5DKbWRfxV3DaWMfkDP+KvbEgeNsRVf7ONmE5vdSGemp+dcFPsET/IzLb8hx969jcszC8Gw8p13qc/28jJYsoq/gOANf/wZTLiLn7iKS6aU97hF3xOwx364jdOsge7qS3pjXOJBal1YMM3xFAH0rnpgD7wpM8mzVzml9f5mDhCVnOqhfkzHWANZ7xmOxjDRr/ZvzKn3LCfuAQrmM1xiW+QzUEaf2ATdlYvsrOcSN+5zXVj/EHc0l+9K+9u0sJdvPF/VMJATjDOfFAprpGbrmzID+VPduJHOInnXrOJvjimj3UHvXCcreGJ4/qxv4Yv8qD3zZEHXzZ3HvriL66LIT4XT4zJ32ApB7GXxiflRvI6MIUVvstRbvTh0zAQEy677LLRT4wgv358kkzkxk192VAfPsxPYWXu3DiLS7h5kMb2OA8vN9yp2ciBhw4q8VJtoX5xQwv+4a9agL+xu7mDa2Qwzib7GvxeTGEPOZwvy/9wU5uQTZxMXeFz/iI+6Ud+c4vnMBZfE0/4D7sZC3Y4rD952RcfEh/lD318zgbsmzoFz9lE489ujMNbfY3Dt8jHDmQhHzm9Ty4Y8Sf2PKidgmufi0ARKAJFYPsR2JqDSt80U3wqmvKwoFJMpKBVaGjek8gkK0ksG2QWY4o2iVJik2htxCvA9JOoFT0eNgQUSuawEFFMSPSKI0WUTROLHQWbxGhhRy79stiRrI8vN5zNqzhTlCi2LBj0VZjMTTFgbv0V21nk6W+jR5GmKWbooVC0aek6BZ4ErgjRz7OiJIUHfSV1GJqDXMZXiClaXON6GCo4FAmKCUWQ9xQANpxsfCkmFAHBfQi1xz8KdYsrmFpIKBxtDigQ4WV+BTvcUjiSm03mgnePKf7PR+S1uLRgUugowsyJDxZsij7FH9seRlMIK+7mg0oLDIsTPGEb+qXQUhzjXhaiCmV2Ied8AGNBxvYOnvyElAIOzy0+cJldFGvsyAfYHLdxQwGpQMYZc8GCTfHQM27qp/Bnd7LgrOLS+HAjk+IfrzT9LExtIrAvm7KZhSd7kkcRqhjlNyeWi2dFOA6yD/4blwy45ToNX+iGL+TFbzzAD/2NSX5NAYujZLCRYMEEA5tMil/jWhhq+tnIxAcLMjw3Ls7hv4ex4cY+eZA1i3/+5lq+b1x37JpDX4U0DPOAJVvSw8IExhZQOfwzD66yP7vSwZ2s9MVN+MER/myqwVhcYU+chincfFMTv+iQvuzMLnhog9scxsUJccj4xmYzsvocR+AoZurnzlSYmwPXNmlsYlNADGE/toQ9HtggyAYHPGDL3nzVg178QsyzONSfzHDiU2TUD6ZkFSvx0U+l4ZGYYnFrMevZ3/Tbq9E1m4k2StmXvLgBf/KKUbhhLmPDgvxzHOT3xqK76+lNf/lCzuH3vnkAexwxBh8gp7nYHQfIYKMLv574xCeOxao5zaclvgU39rWBZYwf+ZEfuSqn4YF5Es8d7uAXWchALwtNPHQtfPEJt+HLT+lD9xxUyoV8jG427Rx64SQb8Tc3gBhzxsV8uEA38RjX2MqDn4o//Jn/7Nbo7LDkec973phXLmMPGy54Qq5NN+BW5yAfX8Il9qI3W+I8n+InuIuXicFkYWOYiR9kiD7J6/xUHBVvxA0c0/SHEf8yNjvBlN/ByLjiJb7A00aJ2MdGbI73Ng7ZAJau5y+4CkscxXvvhzd4QB7j44t4hBfmgJ38QW5xzHXGSC4W+9QM0dXnHrEZf5cr+CU9ySQGyxU2WshFHjjjFR6qP/i+vEh2ttQHDmzrQV+4RxefmQf2Dkb4lTyXueghnqnHXMevcMJYsBUL2ddr9sXZY8sNODFTnsuGJ64Zg/9tEvfoYQ72xSE+4Rvs8HHwqGZiF7agEzz5Q24wcI0x5BH+CzN4wJhcfJDM8v6v/MqvjJyHFzbxcYOvGU/cgk3wh6tcAxO1rFzEv9WWm8Ql88NXHjEueenFBrgCbzLjsjnoxsb8h07wgLNY85CHPGT4iDFPFo9X/dM45MXb+f+ohCsZ4Ao/PixuwgnH2FOM8yADG7ArW8kj+CPW4TWZxC/f/KBTYhHO6cuH+bL6BZY4nQeeJIcmZooV5mMjc7K52tcNZnyEf/OvbPaxt79dh7Nig7hMDvq7NrWHPuxOL3PII3CWP5Jr2Uy+gI+6bd7gTn1FL7UHu5kLjvHpxCM6i0fWCuTCKePBit5wlwee/exnD/+K7OSLT5MXZsbEZ/7MrzzTD2fYgY3FBXZjg6xLyKUlTqs78VhMExfJhAswYwfxiL/AAm7iEu6zAxzwVXxgA7naWPqyHWx8Jnapv+UXdjZuOLHKz93+hllyKv/hE/TkOw9/+MNHvmA7sl5++eVDVp/Dxg1tcg+/Esfg5qHBBxbJqcbFffKRm354kXgKezka/jiBZ8YV58wtbvAXfo0LcPfLG/DkK/qZG97snRhMVjkFxy5c3pChVjM2XvFTfMVJOYU9kx/NnWZsfujGGRjhGvxhr7/ryC1vGYuOMMMz/diFTGwOA7GXv+gjt5KJj2VOn+EP2clHb42f4jRb45s5jUs+GMBc3KQXm1q3Glscdo0YrD+s5UrfKMUpeMql4TI5+Bju8VEyk0VffkIGa0Q83VnmJfqaW1+yih9yKLn4LN3MT1f8JgcOkwE/6CvmGtNDLR8sYoPdnsUUutJbvcBP4E0uutOJbrhBPnUJHuOjv8nOP9UYOIn3uGxcvME78VS/Y8s8zObZP8EZumzS4ENn+vL31O/GJaNx8JxNyZu6hEx4IsfNLTyng/XIQeOSOGnfhq1yQy/fgCEZ8JvOMEqLP6jv+IOYtOoP4u4mzVg4CZNf+IVfGPaBP73Zj9/6G+/EKrLhFv+gu/hBB3LiDt4YM/FHTrTugI+aw1jJZziSfIaXOMoXcIgfi0fmxWH8zPqZvOQgs778mj8m75BFvafxIbURnByYspX4JveIr64hL17wWzFErOGjZMVHcuGn13ggHxpHHiIjvxKDxAdcogtZD9LU21m70k9NgI/G94s08NVHDHvOc54zcEo8sF8lHuPCKl/FzE33NcQeeJsXtnyFzdkQ71NXwMRcYp8cqXY2D9vxJ/bFDXlXXzGA7LBjP/kp9mW/2b58H3fwjP3Iw++NZx3FFuzGrtZ74g8/VU+wP5u4Bp/Ykozm14dvsG3qmIPYqdcUgSJQBIrA2YPAVhxU2njwf/0p5m0uSVhZyEvaEpbE5qFJjJKZAtCiT0EjKSqkLbwl1BT5NnIUbIpHCdRmjCQqgUt2FgySt+JO4SWZK6AUBgpy10jCig39JFLzKpIVmQpU81owKKJsmOirn4Q8N0WABYb+krPiUVGm/7qDSoWXAkMhAQe4KGgUi8aGg6JCwQAbi3UyK0xsLkjo97///Ycs2ZDTT4GmKLGAIo+FqWIgG06KJ3jRcZOmmLCQYAPzs6OCxjxsp+ggr8U1u+0siyC6sBFsN12wRBYLEsV0Clh40gt34MNOMDL+YbQs8Gys4S2OWBgo9hVidKOXotxCUGGnEGQXNoWJwpisFjLkZkOLAX35w/Of//xRgCs4Fds2/OkEQ3bXH3+9Nn/u2Pba+HDWP3zFHxjhHH5bFOOg+dgZ/82Bh67V2NMGg6Kb/OyskNeXruTCq+jKHxzC4IDXZMy4Fvx8eh43CzVywQz/LLTpmGId1saCWxaoilbFM3ndqcu/tRT0YgCs8Y8PmBfONv74mwUajPAOz/FJX4scfkMufoGzj3nMY8aGkqIeTuJDHmwLb/PZuNByUAl7my38it/zCZi789N8x5YLd7aBTzB0PYz1t/gSp+BPVgsdnBEryEUWiwU2FKvMb/FpQZBFn7EtjsQl41rM0ysP/kFeMRCe/HSTlgWLcegk9sERzg7eLfjEUvPCKhuQ7CeeiVPsazHvTlPyWoTAShyJHfTFPTZ73OMeN/RnO3yjP/sZBz57tSx4xTk8zuKN3OTjRxZAsBKbYOHZvNHD+PqSMXZnU3w3Jp34w6Mf/egRf3CabPQiK5vzNTx2zfHlQplv56Bynse1YgcsLHzZ1x249MyBIqxhkU0htjCuOMDv/JQTH+XLxvYeufkSuWHrmc/wPQcuOMmXzON9fmxRLV7AEKfxRXxlW3pp8hN/0xe/zSXviG/Goydb7Wan6EsmC3i6JHbwb34iJvDlgzQ6k83GpbwkBvI58SYHlThgTvbl02IAnugPY1wQ2x/wgAeMOCIGsSff56fG5n9wgad4Ay+cIrf+5GBL4+Ke2M2njcsXxFZzscull156VQwTR4zHt9ldv3DffMbGL/6NX/KSDUX5hG/J5+bAa5zBST4U37GZQDY2JhN5xTs+bexsFKoX+DtuwIkseCYmuQ5f4SF/iVswdYMBHpjbuGxNVnryJ5wwD5nMJU7BEt/NR0a2F+vZK/kP93zmWrzmJ/gqfie+6E8+fqnRz0YV34IbDMhknL0amckaf1RDusPfe24gET/lFjGEXcRHNmZr3zxiT7qxsQMKmz/6i7/0t6E1+7n5HKjlENjfYgz98JJd5QC2Vd/iG1ngIMbJCZvEJbIaGxeNm4NK4ySH4I48kpwjT55YxgP9yUMu9YtvkOEyX4frfpr5+f4mB5V4xZfxXOMX4hHfgAPfwC85FH+MK5fyTXnpsY997LiG/WGG+7CnuzHpxEcc5PAZfON/+MYPxDrXuBbnYYPnchzuO2g2hlhinaCe4R/6kU3TX0w2Jr47zDB/4oJYE7/SD8fNRwc6sa++bGEsNuJn+KWRTwzzOc7jdw5+vMY5dk8+Mh6bkplPG08eZ1d+jktuJIUTXxKPcTO6J27q64FXagn6md88fJpvipViizhqHLryfS35iY7600GcYDcH4eSiL39R8xtPfcDfzMen+Ba5cBAP2EUdK6fwQ1zgi2KD/HgqB5X8MJu8iYv0lfPYDyfJyyfFRLLyG+/5Jifuspm4AUNyzTjAAN/FWzZjjzmvwA3eeCmuwcoY8DWmOAdDufn4st4wDpvgY36q0fX6uQ4nYKZf8iVb8Ct1sDiOj/CmL13w0Lz8hD39lGHqcePhrRjhG2f6u1ZOjH/RyfxqPjrwS3qZB070NgZc9RFTjceu4ieZEqPonRhIbnirhzR1hDWauMYP6W1cMhrbpj4+yTviEc7BWt2N8+Tgqz4zrnqbXbPOkp/oHX81PllSS6iL+K88CCe8w0H5B+ZylkO9F77whYOnMNDXuDhibHKzHwz5LDzFHHPBgH7zPsJQfI9/2I2MxiKfGCDW7CxrX7jCQDzHDbLzuRxUkoHs5iUfjNiVDYxLb3g6QGLXHIiJqTDjr2y9SUv9wXdhL76KmewhVos3bAkfspAJbtmn4Vtzg5cx1Ut8DI4HiUvqOwdmMIINPzMeLsGCnf2EunikxR/EofgDHpMTz2DDH+C6STMe35G3nvGMZwxscNn14ji5YMx+3mczNlb/wl8fnGEb9oOfMWFDLuM6OBc7cVSsTj7Djeyv8FP5hQ3EQ5zCAXYxJk7MeYffwFvsEJs1nGM3eQXvNTLLZezjoJJP00e+z0GlfvLSfFBJDjjKB/riG1nJQ3fz+j8kYQErvmUdkBoCNgdp8py4iJ9ilWZuMYc/WXPgLFytc3CYnDgbf9Mn+TAyiMfiHl/fa1+DLuKDBgM+GJzNBQPrMjizCyzZO3Vtakx/G4fc8q5nPoTX4p/8iyOxLx+c7asP34czHHBUDjHvseUak329Rx+/MMYf+AAeil+4BQMxl6/m4W9cIre5/d1WBIpAESgC5zYCW3NQ+fSnP30URIoESdTGiNcKZ0lSQs1DMpfUFCIpahQ8ikWFgoJMUlZIzgeVEr5CW4KUlM1jo8mCxDgpUGzaWVgq0IwrIVpUmFNfRajiT6Fz2AeVEr6iURFCHwsuBZUFkcU4bBRfOWixMaNQUtTD0IJUAayYgCW5FX+KCZuwiiZYKB4Vqw4EFG0KBcXAJk0RSwYFDoyNSwbYKVwVrYoLNoGzwhCeXltQ081cqwuK1bnZXwFGfgu7LBgViAodBZ/CG050p/NhtCzwbKzhLR7ghg0AtiGL4h0ubIdPFoOw8bcFhA0VRT+Z4UIvCzrFpU0rd5rRyyLH2Ov0YksFpYIwd+IaJ5vq5IEFjHEYfmQyB25bAOCLhQVfsLizkUomTZFpo0Sxi0+uVXCm8CZbfNTYOISveShojUtPBS9Z9COvDQGLOryhv0UdPeFH/hShClpc5duRl38rWPH6YQ972JAn/qigz2JWYW0s8iqQFcA2F/kBOfAN7yyIYEBfeNCXX7Djk570pLFpF3+Abx6uYVv28g1YstLTXDCEDRnY3bjGFJ/wUj82tVDhywp9TVGOIxbRNgjd0WoeCxn9ba7tLBd15OHzsKSzjRNz4ZU+dNYPtsbGWfbR34aIhR2fhF82xC024XiyZhz6wJk+bMi2FoU5+MniFM54jcsWJGKvZ39bxPk/3lxDTvbQV9wwvrFtmIm3T3nKU4acWRQZl6x57CUze8HV2DawLdpwT5ziC+KJccQhc5ELVyzks/kNwywC4Syu8elsMMMEBhdffPHAP/zCY/PDH1dwi13lDX6510GlzSGLcHFm3UGl+E92eLGFB47xOQeaDg7wP3GQ3OKmMeNL5IH9fFCJF2I5fHCF78FMvM7BfWK2mMCW0QsX8X51fjjsxi1+Yxw2980di3E+ygY2PvHqVOK5DQ4bQ+IS/eFh0Wxcz/w8+Ycs+siJYiQ8cYQ99XUg4zr6eB9HjUtm/BJf5DY+hQ/J0/rzaXyBPRvgkxgt9vEBcsDI3OTFUbGAfRyGmT8bluE9nwm/8QsHclBIF/HA2GTGZXEjmwrGoJdNB7HEONkIIJux87lYLgfwX3jiv81p3zgXh3FOXPY5HeUNNvSNbbErm0WpQcjmoZnH+5kP7uKpWOX9nWUcE/vkBnPiF7nI7xoY4Sqe4hDZ2JMdHBh4prP+5sy1dAwWe8WPfCYOi+X818YZ33OgoyZkG7HceHyH7dhBnJVXyCNf2qBmD/GZb3pff/US3Hwjml9fuPwGEw7hhNgh1tDPuPyYPxhDfQsfONgE209cope4iMc4aX7y8kUbRvSCOd7IDThsbjwjB+4fXx6A4Li4QF7xU2zYT5Pf2e1kB5X4h1fwNz8u2+Alp9oVJuZmY5zgY/ryT/jB6aKLLhq+JAbQXYyFPT3wTiwT55KX8No8dE+dAGdz4KbPtXAfb/kHjlqf4Ie6xphig2YsPJE3Z3uRF6f4uINH9jWH/rjLR8V6fsHPT3xu0xn31O3qN/0ip/H5NI7RwXMaH5AL6YxXMMAzfBOPrDPIDCMYuiGPj+Ugk63pR8/VAyVzyCF4xGfkDvKaCzZ4r+4gL3vRkdwO/shkHnFXHBQz4TH/X84wI5Mx8YYtYCwO8S18Nab38dUYdDQ/nsNFPNKfP9JD/+T3YHSy59jDuHzd3/BjN/F6Z8k34+KYeeVI8RHO6kM44wec2ZgPa+IB3fmXb5bjFn5nHUBevAvGxsNJdmdXc7KLv/k2+ax7Tyz5Yh7zys9yDxvhCPw9w0y84g/whRufhr96gq/DlHx0Uh+wh7whx+CtmEEXY8ktuOWGDc9kootfntCfT5iXjj4ju8fMKeOQU+w1n1yLUzgNZzwypxgFC/Lgmb6+Ia7ZoI8vWn9Fb3iZF/dw7rnPfe6IbzgnrjhQJS98yIib7CJOwlytbX3jwY5iNx6R2efyJo76xRP+je/ykRs34S9mGVvNgdPiB91grn6AOa7gFZuyX3iHS25oIpdaSY50MBZ5h+J7/AMr/ik27nVQaYjESTxie3GKHwdTXIcruxmXPvRWx6iN5Ub68M/wF1abNBziE+J/OOk1HOAhluKMOXA7D9iua/yfXeDM5geNS3D+sR/7saGPuIXfchif45vishv7xANyxh/w1/85L47jHjziD2xMl00a7hqTDr/0S7801krsiX9sY6zEfHzHcTjyTdjgDP7yHXE9OZ795E9Yu/EGP+ijj3qS33otFmixgTnUwHvlHf1hBYPkHX5CHuPK5znYhQ2eiTMnO6gUJ/FNrMcHPgZ3MZ5fyvdwkc/oA3/rULHH3GIIfww3ybmfxhb82y/KsCus4WNM+JIHvjCiu3U9OciEt1mzizv6ze3EMm5vsq/BRhp/SawSA8VC/stH7TvII/w03MAZNR15cIlfeI/c+E22YBf74p33xGB4z0385Qvs6wELa132FZ/o6Hrxy0GlefEVVupOcUJeIKM52AgXNDJ7D+fEzbYiUASKQBE4txHYmoNKCUtBIvFJUopeRb0kqLBSCOShsFCAKOAVTBKWBbykaBEhSdvck/Dmg0p9FOQSpOTtwFPiVjwbMxu6Ck19jKf4MG8OjPRVWGgKiIMeVCqSFBRzglZIaDZL6GGDhQwKOf0UGDYQFFNJ2PorBCwGLCCim2LB4sZBLBwkdg9YplC38FOkKxQswOho41wxumnBHJsoJoyrwCGLhQTsPPvb+/pY0MJa4eahWM4i7GQFuuJGEW1jSyGkqFLAKWIUmnkoTNcVULA6HS0LPBt7DirhbnPEIxuyikbYWNRbHCig4YPTOKv4o7sFKUxgAyPcM66DSjy2yYtzFs3GnvUyvsLXBsgzn/nMIYeCkF9YQHrGceOnqHONBR7b2+zz06547HOF5IXLDVILPX/bsHBHsn7kxzk2U1BamPMD7+GVcS3++Bg/Zhfj4pvxfNMMf9mQvM961rNGQQ8PfLbxZWEBD344ywsXPqHA5284BUty+IYEWcxDp+PLDRZ+g8/mgrUFo/HhN8trDtexpwUR2cmGV/yIfk996lMHlima6emhwVGBT1f2sgiaDyrhzob8wHg4axFgs91GKA7wM1whhxY/MrZrjGsOi2J64IPXNqr4vTHp7Bs+mo0ittcHFxI/YUEWOlpM+78AzUVeMZJdxVyYBPsx4Jp/4ExniyB42XCwwMRTnDavxUR0YisLHHxgGwtg14rReCHeiAlk5S/6GVcfvoATP/mTPznkzEZQxDqZrLGVZ/Yhi02UxCc8gS9sfIYfFoq4Qhec9Zqs5jafeMynxXBYitFinDgrfvLVNP0Tt8xDLxtSbAbv/R5U+unIY8uDcDKJlxag+CVf4AL77izzCFzJQeYsJMUKOYesFtMOgS3w8Q8fjSsH8D92YGeLf2PbiIVBDiTEFVjpZ0421VesYK9sLBvL+LvZiV3gI4a7/rnLjUJ4ugb+eIX3+MGf9tuML27YACQbDOQH32gR73Lwhatk1F8ssJkiL9IbrjDjIw46bAjCVB6CjXHlAHEkG9XijdijXxbVxqQjebJJRZbEJ3bT1CYOrIyLl3jCR/SzyLcBEjzDL9zGLzGDXfFLzEo8cB37pQZwXfSFv+vpDgd/8wU2gRe+i8FyALn5DB8VH23E4Zmx+Xg2r8VpY6jTxHUxHtbiFk7M8YEcM/7w8c0Sc+E4XGzY2jiBr5yR5lqy4ar+eEg2MuubjQ8+7Tr+gLfRP/NmvL2exVt1FR+20eU1PxQjyEdO9hZf4CCPiGHsjs/8AhbyFu6RL/0TF9kORvzMmOQ2Ly7yC5zgc/zBWPNBZTZ0N41L7M3ucit5xVo/AYg3vrHgECCHlOSEmf70NgffgAMbqRvIor/6Xd9N234OKnOjofnVxOSUd8QjvGLb5FA8gL8HP9Xf/wnMl/gQfuqDb+K43AxDdZMbEnCXbvFf49Kff8RX4MLv+al5/BIGLu4sucDGYqq4YT4c0GCDd+olNZb5Xct/jc1Xs6FnTo0MOI1b/A+35DAbyvjnRh61Af21yOm1MeLj9PMZ+cksd4hdYpzx8YremV9fMcs3QHCQ//BjsSg+Ld/DHhbmin54yrZZl3hW3+K9dQmcyRt82SO1KUzMJxbyMXGX3fiDWKwuTc4ztwMlPFD3JM7RUfzKIYe1Dv+COz35Yb5RyTZi0n6a3CvmsAXf0choHaIWgAs704mdzS/3Jx7wX78oMeNgDJ/zc5yUn9mAnOyLU+IueeFGR5jT02sPc8KVPWFlLH4jT9PRGG5Mon+uYTOv8UQ/6wL4klncSjzCY33IyEbGxSG5Qnx2AIo/5ucXsDcGPWDAn+idg3j9zOt6epDds789jJHX5oQ3XoSLYpT8g4/sDis+gjewlkfpBmd1MRnl0+gNb+PjqtzhRikxQe42Ll6picgES7Hfg83xXX5Rx7O3GMQmdDA+m5BH3HZQ6dnf7EcWdjA2THAJN9X75sEJ4+KTPln30YVd4Upedby8rI4g7/3ud78Rb+JT9NutwVbuhOWmB5VuFBEn4cEvHbjCFPZkNC8d1QL0dZCKT+KG/nKaeAgr/TdpyWviHZ2NSwZ1WvK5cdiVLNabYgGc4bXaXHc64hKOqN+tP9laXJ7/zz025A/iAX/FM9zkDw664eJ9/uCGL8/8AX82afFXXLT+x3e64aX4Jg8bDwbsxd5qCdfBTR+28MBffs4m/Fte4f/qMFhnn4q96SUOqxM048dv4eD6+Fd8WWzHC+/LO2yZvEMmtmO3/GKJceFFH7XJ6k+/5gZl44k/4ot4xC+sqfmWvEhWXEu9SR798V3Ow3/9xUL4exa399PIYFy8dACJB/DFDzEbvsb0nr7wVGeJy3gIc/GLzDBgl7mJR8c33NdwXeph+rEhnokZO8uaxA0b8jY5YrfZv+R6sVOe5ks//uM/PmKVGKU/G67a13tzXYE7dDQOTMQCfKGbvG9+Y9BfvGM39sE/GFj3qSNhpmbH58gbmYOPv9uKQBEoAkXg3EVgaw4qHfhIahZCEpTFbBb3KfwlK48UpAp4xagiQTJ2vcQsUdrsUaTYPFEwSLQ2ECys9FGcS54KIwsC40iKEp+FguLPosi4kqhiQxGj6EwhoYDIQaVFlDkUfPpJuOkX+uijSCWnjRbJXR8FkgXPzrKQ0Gwc2biQyCV7SV3iNubcb3Re/mMhTM4UJopA7ynQji03Sow7J3rFpMLFhpOiTX8PGyQ2BGyyBvvMsduzcTW4sUMWKQowCygYwZ1OinRyKZ4VJTYV6ARbMiqs17XYHV6uVwgqytnHuK7LBkEOSrJgXDfeqb6XBZ7NCQfsCi14Wawp6MmjiCcD2VPIKUgt1uCsWGP7bPCwsYJuPqhUpDuYjm58IgU3HTIurpDDRoBxLZzwxOuMP+vM9hYL5Fbgw9F7eOvOfNeTHadsCOmHK+RXvFoAKKzpOrds5OK4QtxChM30f/zjHz9sbfFhM/0Xf/EXhx35nQI2CyA+yI/mZgybS/htgUEeC3v+bePRAh2n8E1Bz29wDlYwwDELBnOta+wJa/oq1C2Gyah/CvXVjSx2tSCwuWhOG3nsMR9U4gCf5+tsxNfwF+8f8YhHDL3pSs658U86kyU/AYxbuG2hL8bAns1gYn4HlQr6C5cbxnzexsXsT5GXvzhktCFj8WiBLEayq7gr1u5W/McPLULEUTqxhXhs4Ui2xNFZH9yGsbiqvw1ud7rjkwM18opvFnJ0EjNyoGezDu+f9rSnDTlhuorXPNfqazJr0cnfNqXEIliwOa7gDltZ6JIXD8mEm/wii03jsA3/YdfVg0rfwMXHubEn/VcPKuHtG5ir8ZaMZCALzrBXvlE5H1TCQZwlB74m7uO8fnivTzY+Yj/jiqHGtbD2+XxQiTdkNjZ+iXM2nPALb3MIxyey2c1nyCF+ybd4OG/0z3jMr8mUvJENcfnRXPTINyrlC7FsP824cJzHJTP/yU8gidvZcMnYbCUWw+j40rdwkW5i95Of/ORxgIOrYmJ8gE+YTx/xLoc8GdMzzsGTfh7iJQ7gl41ouGnmXT2ohDl76kPmudFR7MFjPJAb8VJ9M98IsA6/4G8MsRm3+Qf9+aJxPYuP/EOOic84sMAHm4L0cC2+OnTDGf35kTzHlnI8XxYvvA9DdpZr4p/0EiPmg0qYii1ilOtX9SAfbM2tbgsPjc8W6kT8UQ+R09zmjW/Mc8+4rr6mnzzEdjYk4SDvGJ/9PBuTLHKQvMonYYlj5GY7+V/NKb6QA07ZgKUDHR0kqDUdzsszxjKv3Mo+qRtXDyr3E5fid+Igm4nL4jr/l69xbt5QTvyIz4sfNsrFDxuyZGErOsJ0U1zFGT6KszZwYeIAXN0DK/LgJ2zIx8Z8RI5zo5IbB2CbuoGc+ovpan16HV/6sTWBbwgYVz7FG/U1W+knP7Gn2iYHOascmP8OHsaBCb/LQYOanl+ow/CCbHg+txx0mZ/9zW9MvBA/8IBPes+12billzjC3611xA9xiY+Yhz0S++LT/Fe8wR3xDT5qEbbn03xGX36i/hMfya2fOs5P5fFLXGQfeR7++ogD/Co3XcGQvPOGqbjEZnDCKbjgihaupJ4VV8VT9naNmBe7sSGZ1a1s5nO1EL7gwbr4EDnIj2P8U2xUP/Jb9dhqfTfbafV1+MVecgPdcEy88c0ovk0OcYZucMdxOJJ5ztM52HMt3Iwth+IDWcUZePulE/6oDhE/2fhk/sXmZEw9Ll6JT/KIfC82zs3ceIMPOajETbFmPqhkWzqJSfzV+LjMV/kN/XGBXxjLGPKBPvyOf2XD3PzBk/1xMPnHsxrVA2fVLWpD+PFb9SNM+Jk6DQfVLezrc/6It5oN8vmgcrz5uX8yJzu6gdIcOEePHNgb1/v0oA9+inPWLeI03VfzcuZg90suuWQcuPA53ONjZBYn6AEbB5XPXd6oRX5z8+c5vsfe5IU/jog3ZOG35LCfob7BJRzZq8GUb+SgUjwRP+Rp6yU5G6bsQz4xBy/Sj9/Q33xz/E3NIKZYr/FjvBIzxDV+h1Ob3ngm77I9u9OZHHglNhmTf4iDuM3v1AnmYRPYwmJubMCW+Huqccn6k534L/8Sl+RSfgdH9ZH1gLhFD7biL+IFneIPDpjhvZ8Wv4GJn36VF+GMh2zDduIaHuiLN3xJPaKOwSfzi8N8UizHIzjCU6yy/pRv+APOqsetQ/FEv7mZYzXv4Gke+OZznEveyR6BnGC94Bc4jK+RVwxzUOmgn0/TR/xad1CJF+aCNVnlRTj4e27shMf6yzVqM3GVL/LJ1X2N+drV17EBXNUw8Uf8xkPxCa5zfMcNtsITNR1M+ASf1x9v5oar9hnxhu/BHVfgsG5fA69gpT9uuAaWfMX48jUf1XDDa7jJO2IB3yIjmdwAJW5m3U1fsdA1+BzbeoZB7GssOLOvOM4G4pP8CGvjkNE+gHiqLpBX8I/+MFMHyXXqP/OnrkgcJP/82t9tRaAIFIEicG4hsDUHlf4PFIWo4lJxIZlJ9ik0FaWSm6bAVdhIflkQSYqSvsJzt4NKxZGFhgJM8SNJW+Qo0rIAlCwlWYsciVQClSglWQs7RZTiTksBYV6bTYo3RbsCSRFB/rnpo3hQICkIJHZF0epBpcLQBqhx6SUZKyaNu64woa/FgIU43RTDikELZWMrWLUkdcWEQkMRaFHkoWBTHNjgNodiV3Gwn8Y+Hsamm4WqIsX4ih9Fi6JEEcSebAgnBdo6vTI3eRVH9FR0sZ1Ck8w4oGizMGFDY+IHXaNvxjldzwpuxT4uOWAnjwLaYo082YTFXQ9/w9MiCrcVY3PxRU54GBP3jGvBCj8LTxtrcMKV6AXnyIGr7qjEGxyxMDq2PPxhz3UbMFmg4qGFFZ5ZvOGujW32wGVY2xjUD/ZwVXx7mCe8Cq7ZsDOeBSqO00l/B6D8R3HLR3/5l395+E8WQBa8fCsFacb0TF5ctfDNnYhwgqWNSj7sOu/lxgHcoIMxyWoBsuqPmSM48jWLDcU9GRX0CnX4Gz8Ls/Ccj+Wgkr3wfj6oNL+YAwf+zD64TKb834z6ZNzIg+v8hz/7SVebcewoBvATCwhcgiXbHP/cQaU+vsnG9hYleJdG5vDLBpgNGfayMLR4ESf4jvi3m9+QHU/FGnaAEZ/GZxtuNgrFRpyfx3ANbNiEDfHbN3FsMMwHb/glXpxYLsxsAMKMrHTNNyrX8SM6bvoMXzZPfMJt3EkMNb/FnsUlW/EH/ORPbMU2PqcL+fBdTOM/NoxP90GlORwoss2Ml79xCr8sfL2mG5lzhzZ5s2GFA2xIX7nWQYOfjGarHFTOG276ySU2VWwUwMxGhVgrZ8IHbjjAXl6bAw7mx1Nzz1xYZ6PIxfccoMsRriOLGzVsYPBFuX4/DRZ8xLg2XMjJJ8QDG6WecQ4P54aruAEj2IsH8jYZbEzRnyw275KncYaeydNiEh+cm/5sxWdxhj/atBQf4SoWGoO/rx5UJgfM9snY8S/xmk/KB3xMfCavjQbz8J25wd3DJga/kwPIaBNLvIUBLLIhoRZiY3WMnE4WvOH35jIWHyc/zPgHTrKlQw0PGPJnMSObEHg080Tc8w1YG4li2s5y42I+qJzjGn0SL83F1vhjDDrIx+K+TY88/A0Pthd/+dEmLXkIL2BMPngYT57mdxps2Jc8ZOBzsPfsb3GWPvTCPbEDj+QfmOOB8fDCdexhLmPiIVuZU8yeN7LZaT9xiTzsK0fNB5WwyQ1r8GMfLXxJXBA//DSchgdkIbvrZ3uODnv8c5CDSrGarzlQxb/kBf5DTrrBHw/FL/WiGkwdwpfwFY7BVA1HL9wS2+RF9eReLXiwGd/ACXUAH1BvyNUO5GCyDo+sB8yNt+zAl+EnhsKeHuahl9c4iA/6kpfv4oFvgNqchYO+7Eo/ffUjn3iDI8bgM/4mKw6LHa7bWXJS/sI/9ZW58VN851diF47xGfKJAa7h0zipRhUb+S2uyx9Zl7AZeWCcesOcmufEMZue4hDZcMMcqS/Mya5unGFbPiPO5pu16g98zbjGTp1GdjYyLhnY9yAHlbCDoZhvPL4r7tJb/SdfZf0EJ5jD3xqIrcVF1+IFPson1gNiEbzpZx0nhuOtOOXAA2/F0P9h795+bT2r+oHP3QOUUgQLWCyt3dRDAZEWI9F4ookXxgtjCAhy1zsvTIwm/h9CNJ5jGqskXMiNiYe70pASBRGp8VBQtmg5FUShLT3str/387K/5WH+5pprzrXm3nvtNb9PMteca873fZ4xvuM7xjOeZ7zvnI7V76gjPZcb/NmO3eRrsJIPWEsoKK4qVLIBm8mjySDesOtYqBSzHMee8nzcoBuf4jfmPnEe//Ca/c17OEl+40YXMid+4yNu4au5CA/gnIf36IP78m5+4psR+BmdjG8M8dfanW1WFSodlwZvPmQsNswdlTjHDxQq2Um/4r47CsVi9tSPmCde0Oug/MR5Cs6wJBt7yj2cJ05YN+K7QqULHmEAQ7yQm+s7tiYvWcQcOZ94w178Tl/yGrFjk3kNrnyDbdhRDHAef4bpQYVK8UKOLz6Qz3H8ybka+dieDeCFJ8Yy59NJXug1vTdpmdfgyOfJS1bnm0fxURzn1/CBF7vg2aq8Eb9g5hwc30Vc4pO46cJedpED4acCJFviED34Cn+Qi8IJ1vEH+ck2ja78EG9/93d/d47Nxsx+CA6LhbkAgg3EBL5K73NTDLKWwWP7W/jDj/AYRo6RN/M3sZIe8lTroFXxJ/Gb7fmqPnB5ed4hAz/IvEMPdrLOOmqhUsFLXKWPvJIu8icYwGRsuCN3MR+wGd7CzJwkdot3mzRye7ArXdkBXvrHQT6ePI8N+LDjxQ6cEw/EKOcbXx6TgnX8nRzitnhMXjmDuVWuqX96LssLW8eK+/hGHrjAge+ZI+USmnG8FgMTX9mMjRyb/SD+bu5I7EleEfuKR/gllse+sBWn8YKM4qlCpfgEB3O7+RAOo/+Sk934RtYKcmY+RvdceDBiNCvTP0WgCBSBInDqEDgRhUoLMoVKE6rkXbLkIXGTjEpEs2BnAROVBMyk5TOJkOTLZCdZMIla0Jpos3kiGTHJS3Qd4+E8i1wbHiZ+CY7Jz6LBRO/ZBgM5UiAw4SbBluilMKJvY0joDkogsjGhb8m25OGgQqUExnEmekmhiVsiZTPC5D02yYJFgQk/i3fJosWDpCB65RxJQjY9JBiO9ZBcWZgZZ1x45LyjPktcJKV0tqi30GNrelnswNYCiw1WJR9ZUMECJhJ9CY5jk2DDXLIroUoic1R5DzuPPLiDt7kTWHIl4ccPr/HTghSO+czijIw2McZEnz0kc/qEk34tQPHDlcAWgxYzkrU050hwcR1XFJ4khvzGggLvcYZ/GGtsklI2YQeFSnhKtOF/z3QnVDa84K1Q6Tjj4D2fsgDwGBf8+k/CzkYWvs6T/DpWQZGNbAjgqA0B/UvQUwjCBT4kIR4befHTQoQ8bG8zh34Kc/Rld9jxR+PyXZt2dDK+uDLiN/Yfe1oQSOwtNMUHsWEsVIZXsPegGx0sethLEj8WKmFPTnjgPft4j0x+Oy0bCsv6ZuPGeX7jxaKXDcc4JAbgh2Puv1CopC+c754KlT5PnKIrefHL4oC8+AJTC0ObRnxeLBp5OWLktZhBR/LY+GJHCy6cVkCz0MoG3ejHOS8bSPitAEtvG5DkZUv+gh9iAxvAzLF0UajkB1l4L8t2nP+z0WwOsWGfjTU+YtHOH1zBy7fwEx/xCx8VstiAbdnzUhYqzU2wElfZgm29h18K4eTBWe9pOMAWfN1GDdmXCw3sYO5kv8Rtfbtj1zzD/8VcG07sBTPj80m2E+PEqvGOmU1sQzYLa3GBDfDMfGQj2XjkEke3aYkbfNodetk8Il8KQav4xP/EMhjhH/3ph4cKI9lYxoFxPoOZDSm+ZFHu+LGFM+KTXIU/4pf4ZAOIjvogpw1W87lNDRsLfBT/VuGQDSJcELfMB4oz4p2vJlOoNA7uji3niWM2o8RPD3JpeCMeBA+8wXMbgrhAFhve/F6OgWtiCh7SwWYc3zB3iM+w9qCPmMG+Z6fNCPPIOG/zPxs+eEVm+ZkcAU5isjl1bLjjASubmfwSvmTFATroH39gYmx2Yp8UNsb+DnqNk3gBK7axCY8b5PFVhnI/WIlzdIcrfcVluSocvE+W5KUwgzl74Yf4qZ9x/qYXLOhkXBjLM/DMcTBkK7KFY5vEpcRl+vA5G+XiOv8fC5XkT4Nz4od5hF95T6FSfA4/180j6SvP+AJLMm96RyU744U77cz/ZDSf8B/y0I1e5hHY2kCGrXyBnHggvsHTA7b+xwe5gjzYhuy6ZhwP8wd/EAvunfIA9jdn2KS8e5rbyLmq0RvuYouH//WHp/Iiz/TwnubZ/+Z9BQZ5C7vLm3xFpbjPX/AUZ6w35B/kwUXvyz+coy88whd+4jjYkRWe4iMZNGP5nE/DSpzRn4b7YotnvkkGfs2nyZeCAGy8hrFjsy4hS1r047NZlzhe3sMe/N+556Z1VwqV8i+f45+Yuyr/SKFS3CUHPPggPztKoTL4wwO3xFz2w0H9wS0bqvwA7vRgDxiQn9xikI1gfMy6EgawTgznEzZoFaLNOynwbOJfxhRbzKvy5sMKlbhlTjCnrypUisEjvvQQA8yvsKW/eUo+LzbJBxSqzAF4BnP5g8+Sj9OXnLhPVrqLL7jjs+TcMEz+Gr6KUwqV/MzciHOOMz+xjXjmgh9NPkQ2HBrXLcbANzKwo0IhvcSxsVDpGH2Kj2I435Evm1uz9uQHqxo88800OAAn/IeBwghOw2csVMKaXrjBn9LIS0exXmxzB5dYKJ4pWih06FscWF5XpI88w3jbQiVesBOZxUf6iz/+T35BPra3NnC8Z/6M4ylUmnvH9UlkWvVMV/F1LFTyH/izq7HFUXsgZDPnwhYe4rC4xF8Sa8RGMskz2Pw4cUncxWc+GhvyB33Dny+QQQ5Lj1z4Sx/5Mz7ijwcbbtNwgb/SwTcq0R+u9HaBHx7AWMzXcBhvHecCCDFLLOdjjsV3F2SyEx9UqMw3BPBbeV4KlavkpJ/z+B8/9iznwYfMO87LvMPn2VETw41vv0Nc1uKX5DzsjkqFSn7pHLmoeEBmfixmjY3O8noxC1/oj8PsyE7036TBX8ykIz3MLda1+mdLc6k+vTa3as6hvzkI7+jGD/kSfLMuGf03hUry4hh+m2dxJ3F0lBfu5mlxX15FRzaAg/PETXJ4RKbkFeKg+Q2vYGfexQ0xy3t0jX3FMv/DfNm++M2+eKBv8xb7uoBVf5p4Trb4orjLNx3Pp/PAaXLjCNvyG74Vf547658iUASKQBE4lQicqEKlSc1kJrGT4J2dNmBMSkm0MrGaoCSeST4tmk36FkSSNpOnyViikg0hE7/JUwIpwZeomBhNvsYwGVpIpUhgwpYQkMHGpgRb4mHCzgLA+RJxmxwmbWNIsiQnq67Mckw2RWymSewkRY63sWssDQ6SHv1KgOi7bgOUXpK06GaDRmKibzjSTaLkMSYoEgL65j1JgCRCYkUX+O6iZaNT4kJvyZkkWLIGe/a20SDpMmaSuozNnpI19nWeRJB+FnAS6yT5EptwIudejOcs8HDot37rt2Z5cMSDLeGNO0k28dfGhYf3/T/qCH88TOKmXxte+JFC5XJC6hyLP9jY1LDBbwEvQbfRlE3DVXYkP37D08Yg3vAh3L3nQqHSebiXhZVxxkIlPccFP5zJi4v6taCXWNPJsQponscNAf1bIOAoO3oNo/hXbJcFC+6QRwKO45J7V6SLGfxS0ssfjS+h1xedktBb2K5q8CDnxShU0pE8/Jl8bE+mFCpX6Rt/cZ6vjLJB4jx8t4HBR3EM3o65/0KhUl9joZI/pOELHfngUQuVOMrvsmEjjlpk2PjPHQ8WQzg+8jvn8XeLZPy2oKPTeIegWHA5CpXkwwGLbLEFb80jdMMj9oIr7MnIPniOj7BkA77HLocVKi0c+Sue8p/jfPUrjCOvggkZzBXksIFj/rMIz4IucYae5ssUKvWTO6IsTs2d3hNfLBzplyvu2cxGrA0L/IIBvMxV2Zgx9yUmjzwIFw96NjcqfOCVOMFfxTHxYVW8Oagf79MVV/EJ5suFShfG6BNXsxma/pxHdxjhKmzJZsMrd1TSHZ/5NXzYAc5yBLqLSXx0bLg04sUfbXCx064LleQ2j+OXDdoUKkd7iKvwEffwnWzig0Z2/JQTsLnNC5wXz+Qn4u/ZKV/BmxQqxeDELrrBBy6Ox03neM3f5CY2H+DEv7IBb2zz0TaFSuewd/TBWXqIt8YjMznIRH+6GddcaW6g4yaN3MZgR5um/FhBWc7x7ne/e9YHTvT2ufwKRvwp86kNa818Z/7CPTLyRfEcd+QRZIOJz+F+KQuVCmRjoZKPpME58WO5UJlc2/mr8rj0sfzMVkctVLpwIIVKWLEvGT3wje+OhUpxnD/wDTFGLopvfJjP47xcy0avWLiuZRznsZFYcO+FQqUNT4WG5PWj36XP6C2fEV/wFk/EGZvNZMQhjzRj+l+M0iffdBwfwmO4Z27CGQ98dI4YID4pxsanrUnwz1yH3+Zy8XYsVCYeygFtUsav4Muv+YP80fjmSLrzK2PwO9gmbzfPiuv8j57j3EQ3jaxk8ew98mZdYo7hW8uFyvErFsWhEW/FGHKYp9gIHuZxmB2lUIk3MNCf4pIiidhJNz4LZ3p5kCP24t9ikI1qx8p3XShmbel17AdX/eKtB3+SlzuOreXh6Tu8WPUcOcdCJX6ILfI1cWZswX3TQiX+ptCOv3QS0/TPx8wtCgywIa+4i1vmO7ksnNiXvuYfPDEX8Sc6sjuOmGsdBzvHnpvW3Tjg/bunCwFwjr/wXVzF0YtVqJQH0YcNzWEpVNpEp8+qJo+wbhFncE885R/wJzdd+dBYqDSnsjccYRU+w4GOuHQ5CpUKXHye/+BsCpV0F7s08rG9mLKqUEn3cb5fhdn4Hl3FNDbHEzHNnC5WuYAQR3DC+zBxLPnMpeLYmOfBUZzFDzmr+UFfR4lL5nVxSTyju3lRn3zBvJIclp3NJ/TwW4v4ww8Ta0d/GPU+7DUuiJMwUai0B8THzF/yC7Kxi3Whxr/lZ+wnJvBd867Ybd1mH8aFtvaa6DIWKvU1FirDx1FGuJOBfmKjuYV8ZDDviG9wkiPxUbizKb7wJeOLh+KylnU/LLcpVIobYhBczeHi59jMBfJjMorj8gZcxpNtCpXBUxxkc3bAdzFMMw8Z23Mam7EBvolnuMg+5KQ/nuAtrJJ36W/c19AfjPgRWy/va2SfkY09YI1vuBH9yOGh5XX08Z7jzTXsLq763/xv/R770pvt2Bdn4E6X5MDyCvalqz7oNxYqE8fwT1zES5xgE6/JDR9+JsdjU/Lgyqp1G7nbikARKAJF4HQhcGIKle973/vmCcqiw6TkIbmRcFrwrmsm0MMKlfowGUrOXPEmQTURmlhN/IqUJmNJhCTM8SZqm7ISHsmD/8cmObZZIzk1sTrH8Y6VEEjGxyaByMaIpNqELnlwfDY0HG+CHzdAyZNCJVycM7Yx8ZaoW8xJEOEn8XFuEsskJc7Pe/S2AQADSYEkYJOF8CjDJq9zZTMbKGRJYshpsearhWBGDmNrElULQvpkM8li1vsWbxYAFkqSPDZ07qVoEi+62FxUqJSwWVDirg0Ysm2TSLEJHCSukjRJtA0vCxsFBxuAdB0TUufgDzlw5Q//8A9n3ki2bQY4R/INF4n42PTLZ/BWoRIXLSzxPHZwDn6nUIlj4xXR5Fn2B7biU2zlPEmtTQf8TqHSMRZA7uiTgOMmu1tY2bg2xrK89JTMWljkymTjON5GJX0tgCS5EnrH8XW6OwYmqxL6YJJFA3kSGyw6LIB/7dd+bbbtiGN8iEzGFAPYS+I93lFJJtzFWZvajrUYIJOvQCGX+LPM2yTwFhkWtvxEQSNXXVqk4AJs9X3/UKg87KtfcZW8R72jUny0AcF/YcTGFg4pWFiQiNmJLTDGbRgbG9dsvtngh6k7Cmw22ejxP37gHTvgNV8Q7y7mHZXhATzxPMVUi3hcEkf99iQftzlGn2wGw5IN+C3/cZwYPbboL67r04aUDR8LuxQqxwUtjjjH5pUNFXEGD/BkLOyKkzgFU7bwGnfwAw/5HV9KPE2cyaaPfvMbc6sKlWxms8Xcir+wwGfcc6WtzQ1jwwtHzR0K8Pw5G4wjD0ZMVr22APaVwHSBp1igLzHVpp2NnU0bXXFVrIO5wpdNFPbjd/nqV5uE/HRs0RvP8S88ODsVndhLAYJs5vNt5mn2zCJf7BXv+bUNhxRG4MXm/IP/46NNC/OchboYsLzxgivsbgMGdmxFbvFZwQlvs2E12gPfyWSTxTzAhj7HS3mMeYytx80lx+TBLxUIUqiEiZb4yJfJLzey+Q1/OonNeIkjNi/M/7ia+cT8ka9+NTbcza1wwgE2XNeSN9BNLGFHuOMxjMhpzsAr/dKTPPKgdS3+Y87NPO2OA3yxASNOsCn9+AluGUNskxuSwVc4m2vFPMcbU19sRgbzIM6bF9nZ5/yVn2UTFK5iLh6b5+HDx3E9HOPbh8UlcQZvyConE5fEdeNuU6iEGR5cjkKlC5XwL/Mz/oZ/9FpXqGQH3Mc3D7FODJMrbHtHJZ6z4b0XCpUu1hCzktePfheOiR/8wYMcNodxk4+7QANv+DY7jS19mQ/wynxLbrkE3XGe/a01+Cp+mpf1Z27gZ47FF77pGFzBK/6Ofzb45SljS15CTpyVL/Fp8YqP4aCNROeTXxzhC44RJ8mlD/4m3tHTOfShI9m16OczOkY/ObVjjOcOGrGODOKICwXMu3SDYfrQnzyN3/MfNiKTeEjWoxQqsyHPZvgFP/k0W4lNbKIt6+Nzuay4G6zNJWIEvoht4p08KbmP/MIcYd43B1hTGiO4zQMd8GcsVPJrWMHTPLKLQiXOsztczZFwJT+OubMPv1wgJO7iH7z5BF6JMY7FB7HNfBcfxE1cxRMxUp6hseO5iUPyDWM6Ds/Y3RzC9vojh2PEM/1qLvg0D5pDyJLGRimI4NNBd1Tql67WH3yF7voR8+DUA1DMAABAAElEQVQpBrPTqha+0g9PcM851lDmPXwVt3dVqJSHwvaw+UxcMJfBPxeiOY8u5kU5AB1hRD56y1v5vrmN/8LUOpNOztVgJbbSO4VKvBdb2J8NdlGoxA/f6IBPuG48/iLesI+4gIdyDL7FPvyGDRLzUqgk87Zxid3EXc/6heW5iZ/meesX+T5uwUcBzmv+IBeCBTvBD844bg7bprGLmIK3KVTC1fwlrumfzvxkbOIX/zDnyxPZU+4hBrlzmzxjodLn+uJnP/uzPzvbfYyv5PCw/tCfeYct+L/cNfOOnIWOmXeyZ4crcDR+fsOQvPHLbQqVOG1M/kXmi1mohD3e0APPxUAxAnbwgSnsxdxgRK9gxydwEXfIjMf8g/3GPF/cXi5UOvagfQ17I3xVXgE7vmtOgkXykcy35EqLXOLGmFewmePwWYzSJx8zj5njR/uao+llDot9s84Ro/3WsnlgbNZpzoGbfIQP4WcexsZJuMj1vDZ/bOsv45h9XQSKQBEoAlcGAqemUCn5s3iRtJn8JT4SlbunDSGbPpIFi1VfRSOhcLwFn0RRAiV5knRomaRNhhIIfa1KJJOYGdckboKXYFh8WHwuFxRNwsa2wJVISMQkxyZgclogaPraZgOUvpIZfUq86Wnil8xYwFsYSEJM+ElQkpR49iA7jOjuOZ/PAu3oj4RUEiWBsjEq2ZFY0l8hywYHOSRuGr0kOxZSkmp400syboEK5yxAnHcxZF6lehZ4NoNSqLSJYHFnA/TsZEeLF4nqJo1dJL3bFCr1i694a/Pld37nd2asLIok/BJS9h8XkJGFHbJJZoEqudYXOyhUOh+WOKXgiN/sJgnFJQtsCwGLrbFZqOEu+/IzPoZvjlWQch7sFCp/7/d+b17M8BHj2qDV96qFlU0d45PDRoGFIKwk6ilU4owEd5uEPrKTSX/0VejnQ2TErV//9V+fY4SkOPZkL4/DCpXOSXGOjSzecJqNLLDh7Bj+Nja2oDM5snGCU/RNoi4msSOfv/8SFSqjN5/ECzqJZRYr73rXu+ZFlkULzo2+GJ6GG86zcSamsp/NHjFY7KGT+HU5CpXxa5sYFmTmEosyOrlT0lxhzuCrNkLEJRsjeGkxbwH2q7/6qysLlTBIQWC8w8cdetnU5V9w4zMWbzbccN3if9UdqHhjTiGH+SKLZH5/z3QHhniKs4mn+iW7hSsuil8KoGLnqkJlCn3GoCPfIAs72bDAQViJy/zTpqIr3I1rczkbVyO3173mv7gMd7wS682PNkjGK63X9ZHPcDX6mjv4kY1CsrOTDSxxB3fhPjYxlX3NTzASz8QyC2Q8sLkMVzbfZp6GO3sq+MIzC3hx76BCpU0lOUgKlWKHvGRs2TDhN7DjX+xk43ZdoTIb+HT44Ac/OMtj/hJn5DF8Mpvi/JK+uJZNEHIpULGPDU1zXhr8swFhQ8P5+E9v3CMrucV/80PuKHU+O6VQyV5i31ioFF/WtfgP/piT6Mme+jXfsQNemY/4Hn9ZNe+sGoNe+CGO82O/DWWDxfwFbz7LZt4zfyiI27jnq3jkt5nNvWK/PEZ/fCdxlP/AEy+zAUpeuMPOceYGtmGn5UKlsR2Lt45fF5fgNMYlMW+fCpV4yR9hKi+Hs1iAF6s21Jb5wHYe+sEvfnfvFoVK44kFxjff+Z/N2RVv+DobeRhHy7zqWVwXu8UifuI9Psf+Lvhgf7LxTZzCw/g0Ptrc54eOl2vQwZzMHxV2+MXYyMFnrVv0ywfMDeYAOpjfcdM8oNBpY9ScKnbiluN8Lo5al9BznJvoOOoXHek3Fl7McwqVcjSbmfzOVy+LEck/Rrn5v9yOjPwCzvxL3D1KoVIcM7Z5BcZim1goH84FOsYfbcZOsCALnPAu6z9848dwEW/5sDhOVv6Ik+Ks/EMsFP83We+wL/vIYfTDDs6zYaxQaf4bW+zrOHk0HWGGM9YF8M06mG3Sv3kSf9hCLGMD9tCf38CWA+OeghE98QJfcQl28grxno3ETLy3tsMjOYYc2bFwMXeI4+Ys3BkLvasKlX46QUuhEuf4QxobpSAC8+TbZLSGd8GC/QN+RT75Hjxx2Zxh3qLXqnk5Y/BxeRYszUl4Akv4O08xAyf5iPHpKc9XtGBzc1X8grzJp+G26qtf7XuM/hI5lp+T7/L/K7FQiWf8QrwSI3CdDcVyHIEVe8tp4IhPjpN3iXvZL2HXo8YlPBZL2UdMZDvxiT/o31qJvc0n5MEDOQIbkYs/4Jd++OY2TX/8gs4pVMqnxBP5iDx3VV4jzvPVFCrFM7jISY5SqEzcENP4G+xhLCaO8468jZ7ihjVD5h2YKXzzh/yGIRzil/xt0zsqcdq4OHEpCpXGoy+70odN6MIO5IA/X2Qnn2m4gjPs4BzxUVwQ69iOHcQUfWj4uc2+BjzFZHKJEfJN45nvso5iM49RJmORLXmFc8juPfLjjK/SFsfkt+QVp/BMLhv7iunmOLoZH9/FOPblB+wytvB4zCvMI3xHzsxX8cncap3EtuYJc2VbESgCRaAInG4ETmyh0iJBAmfDU3KzrlkwW1xZNEra/G8SNSHefaFQKRGzuLDAzdVuFmAWMBJYyZMFgGTRwsgkaOL1sHmwKvG3CWQ8CYEFlPNNphZZElD9muQtciSxNmIdT07JrM8sVGzsOv7stAjVTPLbbICOC0b9SygkBxZbCiKSHotdiYfkZGxJWCQi5EnC7PVhzTmSizz0BT9YS0w8px/JiEW6BFXy4cpMeGgSGAmyBbQkCV4WDnCQpFrowdcGoUWmhItdYcdWxkkzXh7eG8eXVMbOxqErObfRWZ9Z4NlIGAuVkraxUHkYb/WlJVGTjOGUTeZ7pw0v+h50R6Xz4I1zksE/+IM/mDc/bGLYYLLItUjLxgkcjOMci2IcYQeba5JCfgb/bCDpn08pDLIBu7EtzLP4s1BMoU2/OGdx5ng8zAJE4q1fCS3OWEi6o8+xeImnFpKSdH1K7kd5JfT8RYLML2CEyxJW/Haetm1CP580/Qkv8FFssEFj00DyrfBENvEgSTscRz7bvGAvthjvqBQ3JPRkz6aP1xYg5LaIYy8Y4KMGH4k+LOlqA4e9+HAe4gp72YTnG/dfokLlLOD0x8IB93EH9+Dit4LoI/ZZmPPJ6JQFvPjreLyzqIXvPVNBzaaNeM2mdGcHHKE/X4DXUe6oxEmxJPHJa3YzDt6Ofu99csY3jE1OcQfeCj58ik3ZCCfpwPZ4CRN+545KccAY9MdjcjjHRoo+6eS3YdndnRL4NfKevGyrT33jjoUqDo53VMKdvGR0jHkIxmenecSFAeQYeRue82VzEB0VnHEJH7NpgpPxP5zGYb7F3gqm4pS++anY4X/Ymm/f8573zP4Y/cOZTZ5xnlx8UBEoGyjivQ0M84RNgMSc8Au2MKOfB7zJDx+6iXU2TG3M2BDAp7un+UO8k2PgnuOjs/wBB8UnuIqD5g1xLxfUOJZ8MHQcW3lPf47jC9loiO5shTN8lp5wgyH9xkKlGOGrDcVR+DoG3uR1rE12fNWMSV92IoO4y06bFCoTV3H4Ax/4wGxD+ZDNX1y28QtDfTvWxgEM8YwO/DyFSpsy7OJY8mjsY84Iruxr8wKn+Tde4ac4Pn4lFGz8dqpn55rD6C+2wBcey/qzPx44PjmI13hJJp8b00Ywf0mhQtxiL//zx02a/szNuPH7v//7c57CLs636SSGwYA/uICD3LBgT998wEez0UJmvo5LdOO3cl99iU90CG7wgJ3caFWhEre3iUvGpov5lF33rVBJd3ENrvyY/4gf8hT5Cl4mj8WL2D1cwzN2wjFc1oc8gL3kPPzIBWPmD3ZcbuJMxuez8l3N+Ao9eMP/jBO++9xr/PLwGreSa5MRz9y5Qy+fKYC5MEOeJLbzU/KIR/zYnCRuyM/5AdnlnXI9HMnDOTipTw1WuCsuZP4Ru7IxT37H47Z4ZCxzIDwUyuQz4jPZEzPmjqc/0c/YGTexJHlpxjQnKziKEfzQ+GTUBxnFUBu39HSOuCummE+2KVQG94xvnvIantZwinGKSuJTWuxD9vhn8kG4aC4KURAjjzhqrkme8Nd//dezHuYH+olV8ia81Ccd8Y+ebO9/cy9MvJcLEvUj7mjGgT9+B1MYOxfe7CnfwAnx5qBCJX1if3kdPMgtpiu061uBVKxka/LLoeQ6Ws4317twURzCPfHPsdYF+MGWjoUXbotVxqOnC3jMtebGFCrxGsb6dQEOncgjz6I7feirsQ+cxGz6joVKazl2IQdscJeOjsMj49895RHGZxf9Ju/VL3uI1TAxv8pD+LP+nMc/5Aj0I/NJKVTisJxcDoCPGvnkOSfpjkpzqHwTt+zT8GsxBj/EYnOx+COesr/4xzfZUoxmD3aUk+0iLsXecke5Bp6I62TwW5b4cv+0VnMczsQf6HGUhmO4bZwUKsUGeopr9HZhAc45lt5iBVzkcPEl8xBcxDD5ijzEfAYbhXB+qS/yrrqjMv3iuTxb3+QS1+DO5+TY5kp+Z18A5sbPBTLG58NXSqESnvIAcYMO5n1zOd+hN7zYFf5iNTzgpPF3XDAv4SoscEafzhcDxwv39LtNoTJrR/7KD5wvfpqf5DXsgRNkMGZaOEJOD3LiLpuRn119Iwg76w9P5AnmEfY15zgnOajjYCN2WLc5Lr/lrr889J+46XVwNbfioDnI/C13kceYH+QQsG0rAkWgCBSB043AiStUSoY8MhmZ4EyW65pEy+LKxCxp87+J00aRzVcLA304xgaxyU+SahHkc8mBlgTCwtO4kicLd8/Z8B3lkGiYRJPwZeNNomjDT7IiGbDwtDmcxMGka/I2vqRB0jrKsW2h0kJLkm4hKjGRJEsqLDLGO53oMDaJgqTAQlLCZWE5XjEraVjXnAdrektOJOASIHjZLDCeBETzWTbEgpeECr4S5FxpZUwbJhI3iw4LUhtIxmJTeNmMsIhybhbakZMNvefhdXSgK8xtUupf8khXckZnttqkwZvsY6HSAtzi7lIWKiWWHnjlij5JLztK6CSFEn/YWrzAQfJJdgkg2XFXEgsnPiJBtnErCdR85s5IySZbsIHNAAkvfdlB3xJUCyD+df+0EGNfr40pabeQdIWnfslrs/rP//zPZ3lxxyKTz5AXfjaR2I+8eIMLNh0kvjju/W0SenLCg61XNT5gQwHP+I7FJT3wLXf8KZBn00QfZMAjvrqqUMn3+YHiF99nG3EH7uKL+ABDm4H65iewgbGFpBhBDvHK+75S0/Fwogd8LPhgaXx44vTF/OrXYEe+LDLJKPbYfOKbuZN3jNtig40j9rMZY1OAvjBy54UNG7ECX/CXvdlBLDtOoZJdxV52IgMbiwPsaIOIn4hXsIQxPMUHsRPnycF++HPPVFDFY8fjurjHRrBnW/pZuP3yL//yHAeM4djEH/2bL/iqc9zxQpZsoLGrjS6yZD4jBwxsOPNFnFeoNFfwJX4HL5+RNxto/MkGjk28LOjIAQfHZmOe/5MbFqsKlexNbriJnWx377Qg59tkzSY93plr+Bh9bIwbL7qHN4c908XGKvlcIEE+XBGf3fkmlulbzAm/yJd5iP08LKLxi0z8S4yxUYtTfNxncgPxTjEBrt4jL5vwaVjiq7iHA+Ke+GRhHv22nadhuFyoZCv8ysYEGeDrDofIC9/EcvER9uTUwl1xkm7k5pObFirlQfhoA4K/8EdY8+XwkX/TlV3ELzGHLuaMXHAhVpLFsfqhB/4nb4p/pVCJr2xFf7wZN6hgbj6DP+6Jj/R2HKzM+3xQM2eb0/g3u5vLxFecMb5xxWoPY7oDkgz6EHfgbpOZ/M7bpMGejvjpq5PZlI3IQSdN/+KFC1fIDA/4KcDya00/9CCbBnMFBLamF120TQuVxt8mLhkXb1IA2LZQaY6kj6ZgLS6JgeIqLtB5k2Y+wXXc8k0bcnV5q8ICjplHYSVWubBCTigPENsO+upXutHLsXIH8QQnXGigX/5MPvZiPz7ALngu/zC+vDyxxrG4rU9cE3PE+LNnz862lw9mY1ecT2xZV6jUh/HxUZ5lbuBX4pENPZwnp3HGxma47sHmuEI3uOETLBVc+A+55WvyDTLhJZ7D01wgTsCI7vplP3qLA/Dlf+Q0lnHYwvm4CWPvswXbZR4kswsPjIcLdPK52CRv8F7WJXQVA7TwJf5lXOMbaxzXOgLW+iM7GcRk8wO85Zx8P/6INwpM/BXWYgJM4GtD37mOp9+6Ri4Y4Ys7S+hCNxxRiOC/dDcvRxdjeU0H8uCJ+E529paLBy+429xmQ3HZMeYtHMEzOULuAI+99SnHwUn8hIW50fH44H16u0BXvuJ48U6cYR9xi95kJKuxybRJoTJ24hM4J37wM2OIfXiiL/w1zyaGJw8nKzz43m//9m/PeTIbWkfdPRXyzN1yDDyjG/kVyvAMhnjDfo4XM80T5IeXcfm9eQSPrD/kQ/YVYCw+aXQmL57Be7lQya74xH7yCFynp9+OJ7s5iV5kEKvMd3DXb/It8w6+yG2MTVaFZ/lv1k54pF/jkx8X5PvszUdGPuEgnXD/YtxRaTz6XAmFSt9AgGu4gI9iL3+HufmWH4k37AhTvu5YNhf/5NeO3UVcYnMypBAotvI7PJAD8Aef8V1zEC7yaf5/lJbx8FahUlziW/Ia63g5rrH4OL8Iz+FDNviY7+S+YzyU9+F65rNNC5Uw9BML/NN4sJbX8Wm5OP35sjw1847YJA7gHPuYp8iiZf2m35N0RyXcPcQMfivOxG/5DX3ZQB7Bv+FLb+domQ/EZvORuVf89L/jxQfzozjtWDxdVajEKfEn8XTufPojNsgDx7zGHoR1nn02OCevyTme2YwtnO85eQX+4LV8wgVQZLUmEhfN8/xP7BWr6WgsmIhP7OtYYxsXH5znPeN4iMV8Ej8Sl8U4MdzcYDyxnx/bB8neMG63FYEiUASKwOlG4EQUKi1i/UalBU8KlSY1i+ZsSK4zQzZ2JV6SK//beJKoWbhaQFg8nJs2nvOVhdkskihKKEyyWbQ51kPSYAI1IerPYpk8FrUSiIxrMpbwSsBM9hb4EmOJviTFBqyEUNIs6TKRe0+/Frd0tclzdlpcao612UgfMlskwULy41jyjk0SIQnNglHiTR4ySoglM9lA0pcmoch5EkIP+lpI0XmTjSaJmk00iSzdLSaNabEmEYJXNgv1Tz7JjkRZwi4RgpUF81ggc5zkW6Ij6TWGfhRDJCkWsJIWiU10kQiS2XHetzimj9d0TuKDa8ZmR8mTpBie2VCdOzzkD6wtUsNb/VkEjwV2Y+PQJi22gB8s9XvvVBCQxOWOSjxZTkjTt0Q2C3jJHQ7SCf8kzfSEDYz06Rj8ch67WETony/gmKRTS798ir9YcGUTRL/61zdd9aNfCxCbCLhhU4xtJd5jQo0rEtBsbDnXYs3CxqIfB5Pcw5oP4LP++Q3bkzeP4ELeccOP7cko4SdDjgtueU6Cjnd8GTauBCeDhRNdYWJcPKGvWEEWvkwXGwb0sMDgyzYhxAw+hr8WDXRQgJCk6xPe5KO7/tjHZ4ozEnOLGJtZ4gQeZDOEXzk2G9jGV6ik72GFSn3jl68gNBb9xAh2h88mfm8Ry8Y4QScY0JPd9JXNADrhtgWYAiB9xDR482H2o5MYQHY6wZS9+T7ekRXHfuM3fmPGVZxml02axRbO2rwQUy0qbeoZi6zsoz86k98GkRhNVjEKh8VCcsKfTY1Nfv3YtMqCm174YeGGa3ji4Xx8EZfEDP1mowuOcPfgH2Km/h2H6/AyXxiHDhZ4NhjvnjbxYKZ/nGPT2AL/6cIO5IAzPemoX9jiFt+jq3ijr1WFSrFcMwa7mFfuneJSNiL0STcbG7DBaePmjolNbDQeI0bjZDa+cQD/jW/+witzeuYomGrOwTEPmNIf7jY/2A6vcrcHO4mzPoeNmH12mnf1lXkCV4zNVjhDx8QQ8QkftW3naX2xqUIce0UOfbtSXZ4C82zkWKCzEf3pDGPzn9hu7uRfyY/ozA/5I37hpZj1K7/yK/NmJ30dH5uSP/mLTQUbQeSRX4jZZEo8wHWxUQwzL/N9x5o7Fc6dg2P8wmasfseNDv5m3MQN/kUvc1HmnuhPLp/b4MMzcyvbsBX9xzil3/gA+cZxx8KGPuHD9gqycIKh+CvvoAd8wifHr2v4pD86uJCEj5IT/+Ai9yCvzXnxVYzXHO8303CR34lNOC8G8SE4jhdsRQZxPpwxP4lLjmejxIJwd5O4RNfwwXlkhzUeyI/FMXeQm8cSIyNL4jkOmHcUKr3nq6NtquOw8zeZR9Kn+IXrYj085U2u/BdLxDy2hDd/NH/GN8WtXBiCC2InnpGHjXBCzOILKVQqbJKTb7C32EEXurOLeKMvuosdZ6fYACtN/MQdPCOPedsx5POZ+cqGpVgtFrK/Qgo5R7+L3vyFD9Ar+ZDYhz9kZN/EupxPN5zhX541MQCfPZMLR9iFTennMzyxsZkNY2PThcyO53MarpJboVIcpxe/xUF987fMwZFFHIQzPcyh4qMNRdgYD8/ZTGyCsyZfzroEx/AwtjOOmCf38yz+0oEd8IqfZW4Uj8QcnME9dsMZNnQueeSF0ZGseEJPsVQBi6+KjWLqupa8X+zmx+Kh9/SlsJx5f1UcoRvMzaNkEDPIjptk9ZV8MNGX8zOn26CWC8AErvIl86Dcms7GF3NggksabrM1vMQDsYNfkdtxOOWCIsfAFm/kmJkfzk05gvUEe1mvyUsU+PHCceQd+Yjv/Ib/+upHspNVv5p5mG7JR8QHjezkk2+7M50f8hW8Ny97TT6+7Dj8wh9jiZ+4peCnoKdvesMJ3zI3i/c4gGseOIlLOKxvvkYXfsIevumF/riEH9bx8MYNx9AV182X5mlzJL6ZM8lLNziSGdZkhr/Ywv7yFzkE/xZb8JndUvD44z/+41le+sMMn+E34s3vncOnxBtjkIMM7GoMHNL3uhY+k09BnN7Oowee4SJMNf4Pd/kl7NmW34z5fnjP78xRYgoekQ92eMRWeDfyY52MPnNusGQj8opdeJw7KsXgYIl/ZIWPuEpPtocPm8Kd7fHCvCMG8kPtOHFJDNPISndcdUev+CNuGhM38IX9cRbP+NRRmviL7zDx0y/8m8/rk/0yd4oTuAcH8Yc8zsFftjKPma/Iwz/0Iebzf/m+eVTupU/+wO7hI7kjR/IrXBeT5Cj23/izMfgcOfim8cVkORFfIXPypeVCpXksdyTDj5zsLhc0dvLFxGNxwTH8AAaJN8GY3mJV4oj5gI+TEw54uq6Rlx4w4jPs7II1WLrjLxznk3wCL8g5NvjBiCxwwFNzv3mdf8irFQCdDy9+RF5+KP7yTfLS0zhjI5t+zBvWAjjuXNjhnbzX3I53sSP5xBUcoQcd+Tgfw13/68OFH+yL42zKvsnJ8Rsfk3eZO6w1vcfn2Feh0lpuzCvIKw7TK3ilHzxhX3obz3zNlyPXqHdfF4EiUASKwOlD4EQUKu+frsz+zd/8zXmCkwx5SNQlLyZLida6lg0wE76k0yQqWTHhj1fYmkAlpBIwk7dk2oRtopfMmGiTVJi0Td6SX5OohN2CQYKg7yTFFhgmYwmBBNoYEgJjS9QkQfqRONjgkOST10JDQiiRkhyNhTpJuORH4ikRIhMsJBiwsdAam/4lEhJviwI4WPyRTQJvI8AkL+Egj/4kAh7Ogy+cLR4kpI6ln+PWNQsIY0i26C6hskjUkphn8UJ/n9Hfed6HO5wklB5w1eDpjiMYeC2hk8SM8lk4Rj46RA/jspVERsJpDPYwPjwtMCVw7A17PJOkLm/IzYIc8CcLPLxVYLcwssBhG5uNFno4cxhv0z37sYXkDT6SaAtQOCmQ3D1tMq1KSHM+u8NJIik5tPEo+UuiSVd4Gwfv+AeOOQZWuKV/eLA92bUU/iz49I+/dNIfv7FZ4lhcgQk5bDbAGp76szDEKbZjGy0LOZzBVT7jfP6nX+eOvOEv5JVAGxu+fMaCiY/RU2NXG5IWTDYULIyyuU0/x65quAMLyTA9+Y6NcuPiZWTHJTpJur0HT/5pkaFQKWbYaLa5MPIZf9mVbSz0+Qk+8014e8ZRjc0dD0vxyRjGEx/4ynhHBB1t2Bmfv+gjhcrlRF4/+sVVxytU6t9GusWLxQZ82DKLl1VYRUZ+D+8sRnBFEyNxKnEbr40DW/yxUObnNmsstHCPvcUlNrAhpS+cYwcxANYKlfwARxy7SYuf8gdxkbywx0+xF3dhJo5kbHLyQ1xLfIaNRSM5HIsvPhf3LKTMOeYU74nR2QgTf/APJnR2HvxtUtpwtzDVLBhxmm3JE8zwj+y4T3a+MRYq8YZdHedzC3r8d47PjMmmXvNb4+MXO9jcxV240MsdUfBlDz6GA+EBeejGr3z1K7s4N/HDnDgWuNj/KI18xgqvxWtY8R1j8Wf6kC88cE427jxr7Ib/fJXPiCUw5Nv6wy825oN8Kz5FX3rCR0wSK9kD7uxvA0W/+tf0uc08jVv8FY50IwPb8ANfMUdeMtBfXORbbIozcCEnfcjMZ2CBW/rgt7hNHjkOHxev/KameIRjdBkbX9O3jQcb7/hMN/ZLHA4fxQ5y0dk54pP4e890p7F5jzzwwif4Oo7/ifn6oBffx0345jN8o5NNRPpo9EiOZo7Ab33Qhw0iG//iN3wgY5LNOPxUjpB5j8/7TH+O148ND/M/DB13WKEi2DmfPehITjFAvIrO5BTf5ARjXgdfc7t4If8Mr+FNFhs5Nt5t5IwNb8IZ/BVP6S4uyW9hwP58YZO4xN/NT+Gy+RgH8CaFSgVuvME5fpdGd8eLXTiWOyptHDoeh3EB/my+SWOXbCorqLCFQqV5CcfwG942rNxxmc0rOJETbnzZvGBMMnrgEazJmUKlwiY5YU4vmOEkf+Nr5MBPejsGJ5MP4bfPNDLRVdxzrPfFxfwGqXhhE9/GGjlXYREe0Z9dycDO/FKM068Ybi4NN8VBMjuXzo5jRzHY8d4PTjiGk5o5CWfkG2InPJOXwIn/sBlZ+bO8gJ/xPbzlN+b80a/oxI8do4/4NFzYxDOczDnigngmPuGz9/EExmKE+Unf5KKfZ/akO76aRx3rPfyHGT3paLNbvNYHv6YjbMjHH+hpviM/WWGpTz7H32AnvgTjGbAVf8RrWGSeha1xbO67c5o/ko8ey40s9DJH0R8esHCBGdzc2Y0v8lp2pOO5aY7ll/J6ebXz2YR+MDOW/nDCMx1gdHaaH8wp8PW+sVx4h9v8VqwQc2HAB/AneQtusCV/saalq+P4jXhmHgm20RGm8g/HK8woGOAhG4tN+rZ5L+bgYPJ1cvMpvLAuw33n0S16mo/YRfwWd8gGdw/6pVBp/hB3YQQ785OCogIHncnCl+jiODiJ+3jlM3KIKwqmfCG+reCOU7Amr77N4Qp79MRtvNMfefQlfuEvPzZnWvfq3+e4giewcI6mT9wUf301uNf8R/y7e8rJYJgGHzaNvOINP6CTmMNvk/Ow87qGdziNX+IuvcluPPaKz+mDHuyDF3yNDa2bych/2An/ND5HJ37pePLBie1xHDfZgZ03ac5lT/2I5+TlhzjiAhlrTLizA9zhTUa8Mv4Y28yX/J7tcYVfJS7RUT/sxDbbxqX4vXhjfP7mTtrkenwLr4wffxjnl02wGI/BBTrguTsq8df/5gQ20bcx+Y/3YYM72Yfx2rFyMNxhTzZyPF8znync8zmcwkmFuOVCJZnIws+sa8Qssct4Yos+8ZMs5BjnHcc5lz3kYuNPAOARH9JfvrlC7OHrLqwUV5yLZ3DmlzjNdo4hM1npODZjygnwmZ34Nrz4pbyNvOta/E/+hos4KW/Cf+t/cZKd8QgnxH5yjs17Yhpb6EN8FmvJw/8UKuEtZpm77BuQF0dhwDfJu2pfw1iwd56+8VvfxoKF+QVGfJjumuM9nEsPfsB2Yh8ZNL5CT/bwWjxMXhGuOT/rCjEa1mzuOPaFjxxDvuRzeYX50Hl4Z2yYwdi84hixTg7mPH2wKR3GvHQWsH+KQBEoAkXg1CFw2QuVNpUscFwRZvEpUZB4SpwsuExQEtB1zaQnGTXpZaNBkmCSlUib+E2C+rex5BiTd5JnSZTjTXxZ3JLLhCup0iStErlsxphQTa4ScpOpBbONBosX50jcJRQSAf063wQsOcnD58aWUJFTsqZJViSd9NE3mejguHWJFFkkMhJGiYnFo4RTwkdeOMJBf5IHz/6XlEgkJACSPzJ5P4n3LNSKP/Sw0CGjZNyCWFJLf31bKCURknhIbCRDbGo8CbKE6+xk52ygGEZyJTG1kLcgoResLJIlkc5PUsUGScLpKIHJxo1kEW6Ol+zAxG8owIe9JaeSyiRjmyY+kmE64q3fVpEQSqDw1kaDBZQxo/sK6L7treigT5tmFgg2AMlsA9uiRlIKo1UNjy1u2Ruv2UE/3rfwZPf4EDvAy5iSVjaHAwwkmzCI3PqTINtEk3DyB/05JhsIsMAv/XrGCZ/BHl8tyuAhKY4M5MJLPjvyhp1HeXFIv2xNZvaXrJJX37hg8RN5yWgBYsECD5zhszYF8Pog/GDhIZHHZYm4DVHcwyky8QeLawuDbDTQx/F4kK9OtFFggUFGXORDMNK35F7ssbDip+xLdv1Eh+jKznDEZTayweE17PmV4+iYwqrftCOrQha+kDUbQvqiH/vhKnl9RU/4RV4YwUffcF/X2IQN6UAnMU/ss+BwPqxgRnfjktWDnnRiQ76Sohg7OVa/OJDNQAtwi0B29tuP9Br5uU5Gn0VOOtvk8IAX3MhFT7h7FpvyIDvfEI9tqsAf9mR3rHPxUb8Wj/giVsED5vShq/Nhqw+v9YsH4o95SJw7N3E2PpNz6ThuMPBlfo1PNvDhoD9xmzwwG7mAX/wL5vrRH254Tlyig/lQTORXNlr1K176X79psSG5LajpisfGJQdu2gAKR+F0nJaClvjDl43Lz7yPW2yW+YlsGv7AHSa4z+/x6+w0t3jfPGVugjkO6Fu8Zac8wkG4sa852zylL/FjOY5tO0/bbDM2fzG3Gx9W5kB3XvELuOMIH6A7bjmHrXyGh+SAO14613xNBzzBKw8+bjPCV1ezq3FwYGz8X+5EHnHTs80BuuMiruCPvmHjfbHMObAkhwKVnMR8yj5k9qAfDjo3MSX+zXb8y/nijrnC6/AGTjhsbqA/W8GE/0R3PGUX8wA+8CM8cR4cyEJe5+DIOE/ZeGFPfgk/OEXGEZ+DXutP33yOruJU7nxyDs7we0UseWw2vsQHOjmeD5FZP/jq+OS9OT7jGyecEW/4HZvDTv6C4+yj6e+wuOR45znfnA8rczzeuHKf/7uDCm/Ylf+k0R1HzN04psCguaMs8YNtyTPGkJy/6pm8xtef+MJmNiwVzPgEn+aT7GvjWa7tNT3ICTcy4pUxyehhIxF/9Hv/dFGZfvwGKKxxALfjb2wBY33TDd/0kc3q9El+POV/uIP79HW8ucq8ah6UJ5DfeoGc67CIv+Mu+4p14hWd6QUP/NTMUV7zZfMBOZLj4LEmVojPWY+QBy+cw5701gcbecgLYEVHXLTJK+/EY74un7QegQvukStYk4dfe8CXLHIf2CSflBfiMJuZb+QMWZck9mYeJiesyKc/Mohzci79Oc5YkZn/ien8iqziVnIE+tKVrPolB/n1L7aR8+6pECT+ONZx61riElzZmh+wvRw3X3m+7vxwSNxnI3m+9Qi5cUW8wCc2IKP+YU8/PBY/yAgzOIQTXuOJmAArDzqRzZhitbH0QXZ8w2vY0NvxLqhgM75vHP5CR7jqN795jfvGGvnMFmIu2yp4GQNWjrUe8hAHzcf6N6bGJmyJD8ZznjlVbklH8unD8Ykn9IEXLtJB4Yv/yz3krz5nZzLxJRfgeNanz/TFB/BK/BcH4W1+gM+90x1k5l5xh9wKoTgNY428ZHQs7okb7MQn2AP3wjfvkcWYfNOcJebqG5fpRyaywUv8FT+8xgWcoJ85L83xfM7cLVY5Hn76F3P4rbXcGDNy7vIznXFR3BNX6e08MQWm9BYnjZm5Tp7A1vyHPcVSXGML52ow4mf83fGeyYtHdEps4KubNOeKF/SEO3lxmg0VPsSr8ER/8mkcl++L/TjMTuIJnvMxMsB/7Pe4cQlHtdgT7xR2zPdkwQ142SsgA/zIFH+YT97iD7t4mDtd2Iw/5g085AtsMsYJPjv6EcySR+IlP4AJ3vIxct93333z3IBTOMkfyD36f0QW08QAfiz/M+/oi35iEc6QLTLIO5wjDpAXf93Jj3ca33GMtY8LJGGoL75uHST3o78Y6cIB86dzzIuOITNfE3PGhjt4QUZchhEOw4AMeLquRS7+QrY82FLewndgCf91DTZ8kPz2WbI3wJfxmq/I99li3NeAAYzIu25fg2/TFW5sQl5+IX6I4WJs5jyysCkfFh/DCzGSbXwGK1zjT+YkeZu+nANz3NHfaF/n6NvnsLU/om9y8Dccy1pMX843VuZr+OCpOYrO/FZ+jEvkbysCRaAIFIHTjcBlLVRKaCWhkhuFKYlNNpBMkCZ7E2oWCQeZwoRsoS0hHjcAs9jIxo8FRhZfjtVMfhYNSbQlPkl0TbImUUm65MTC1oauBYykxARtEk6ya0NRQmCjgUz0k5iZpCVCkn5JhjHordHPxL1qQ8tETgYTPywkiJKzbEjMHQx/soCx0Dk3JeYSFAkFbCwc6CABSCO/xQJ86CaxkwjoPwlljl31nETLwpbuxvOQfEqYJRljQitRh4GFl7Ho47XEiCxJmvTlN0AkbpIhib/P8EByAg8Yemh0IgssYX122hiCE52yiGJT3NCvhE3yih9wd4xE2vmbNDjCE2/z1UKSsBSw4Kmv6HNYn/Sggz5xTTJogU3mu6eNFAsxiRqcVrXYgf/gjMSXHfDdIlo/MPNgV4mfRR792QFebI4Lo91x30ILh8hlHH5CN7YgrzFwnQ7L+JNZ37CFhfG1yIunNhz4IptLqvkSefEmMhsTpqO88dlRXrzDGXJL7nGKb+EZPz8IPzKRn0/Cy4LH1c1kIo/FEl82pkUIztickizjpw0DV3TyPws5XKA3jo1Jt75xj22yIQk7+rI/feFKX35CX/HQBsdoH33CUH/nJj/HQxvM9LUQEJ8cb+GQRr8soMnLD8Iv8sIIPmQYfTbnj8/xN/aHDxtavLCjeMX3YWFMfeEFLNgQJ/gbe9CRP7Oh4zIfsKPNCBjxBce5gpac2/gVOcmBC2JyNv3YAPfgQcY84CeuszMZPcQQC9dsfgYH59ATZ21Y2cjle7Gnz9lNAY//wpddxcT4Kb9iC32IKXAwDts5F158JxuHsLIhqj+f8dcRN1zE3WzesoXx2DSxFy/NJTAlB97wTRtceJMF9sgBusAyG3QW+QozFpsw4g/uHLJwZmdz3HEam+Emu+ECftGJDXGePzqGTGSDC2wtqOkHa3aDkff5FG6FB+ykPzwQ1/BAozOs+DWbix3iI1uIl3Qb4xi74P2m87Tx6SI+OYf/GIuf27wyhkZ32PJt/fMD8noP//HTPG2OM+ckT8Jp+QefERNwx1dR8xtcgdPYYCjm4Yn+zY8eeJTcBW/4hHlVf17Diy/531eBydW8Tx/8w2sy4DVdjKPBjo/htzjARtl4EKsin/7ZRTyxMUkmuJGVPW3e4BrdxUfn0gGmsQfekCexBU9g4FxjkxnuZGGDke8jRqte68uDr8ut6Gsjib7GYw++ZAz+ASdNvMA5x8tD+RMOO97GpeNxF8/Ghjc4Aw+4w1SsdqxYgA+J2+TaJC4pjOMFjrM1+9twIxeMV20Ak2nUW+xyQY/38CDxQ34Bz00xxVvxHmcVVPirO8zIBxt2M9/Bl3zw85p/kxMP8Aq/0sgkvoWL4jO9fPWYfmHMl8goRsaW5nwP8jjfuIkz4hrc5ax4i78e+ANzvidvMyf63GZpCk+Ra9Wz2K+wwu9wWH7Aj/yP8/EfOml8EqdwWXwiw5jf0AU+uCJ24A2e8is4OR+m5gAxB2fpigfe5x8unBQDyUUmdoGJmOc9uGgwT4zgTziA89ko1zf8cJadz11Yl/Bp/BfLV61LYMq+dKMjPPlqbExWmNMLVrgoRngv647ECrLAS3zxmXPJz2/Yhz3hQtZ1jfy4YTwPOpGJjDaVE7/X9cGGZBDL+Zs7KsU68WyM5zAlJy7Bn37syU7w95lGJ/5GF/EA59iBbZP/kZu8fMGY5tHMofCE7/JX9dpAFkudpy9fTSvW4gesRt+GgxiFZ/yTbcU5NoRvigZk5HP8W4OFB/1xXgwiI/ngw0aO14/5hRw4QXdj0Z1MsGMDc5SGb3wGH9zBnlwblmSFLZuLA+LM2Sln0Kfx5cXmDrZkD3YVI+FEZ/LC33wjJtOVffzP7+IXuCReZK2lL36R9a7PHKM/MtFX/PDVubCklzVG4vus2PTH8cbAIWOLN3xdXGJHhU35CowP43PWsfRgN3rDlJ2MT29x0phkoqf5mL1gKP6Q0TMbOVdLXsGOjicfHdln5Kc+NmnOzZxGZ3aFN79mP7EBLzN+8OEreIXH9NPkWWR2DszIKqfUJ67r/6hxyRyssQ15cVTx0NgebJ4Lj+GQvC0xbT55yz9sQ248ECPgYs6ARTiLt2TDCTGBTWGH42KGZ//DEGf4Ao7DzboWjjiFEwqt7L2q4T/s6J15h+3lYXRnb2PgAd3JJzYZj5+bd+6e9jv4o+Z9uoi5LmAyd4g9fF2+wZ/oj2cu/GJD9hQrHJO83TwwNvOq/BSfcZkc/BIWfChxZDxnfJ35mm7Rk+84TyFcPDLf0GldI7tYBbPEW7ryJTjgKR80b7MteXEVBt4n7zjvL48V/zZn83HzhxgXjsBXI4fGRriBD3DwSFzFIZww37KvOM23YGAucW5iNRz0gW9iLjnIzF7wYXucIBP/kFfAD3/YT2MT/Ymf+CCvwA85ifd8Hn+bT+ifIlAEikAROJUIXNZCZRJKCYbCgOTOpCRp8DBpmuAOS7glMyZEE7Kkxf8SMotIizbJkcn+3JTomBhNsBIUCU0WqibFLBwkmiZlfVk8ON5riY87liRrSexMlsaTcJm0TeKeJXfe93k2lZwjCfGe5Jv+XktqJB2eNThY6EmoLGAkCT4zSUsSlxOvMFPC4WFcCQ2ZJQHk0ScMJAKOMa7kAjb6pZvkR/9w8blx17WMp2/JlnFgTG7jk4OO+oEtuS2CjMEuFp2wWU7o9MNOsKQ/e5DHeJHf6zSvjYMrZDdG9JEw4YKkDKY2rNiGHegteXI8HLLQSb8HPZOBTBItCTKujOPhrb4O4236D476lKzpV8FNgpcN1ciYc1Y9S/7oxhZ8yrP/JZISYhjCCOZsfXbyNc82ACwisoGQvsmhUGnxxsaSQ5jhIkzxKdzSNwzZlO+yr4QUFsv2Tf/O5zeSX2PpKxthkReO+tAnWeHMr42/bC+ck8zrAxbkpRuZN8GPXXFWIs9vYCiBxkG2oLcNhyTwOJQFPHvhoMXFuFGE+x5szOfZl54WDPyTzGQ1Ngz5JA6TmX0sqMUNdqOPpi/4sKs4BjuLSp+76lUskcw7J805dAi/+MHIL+PRJ/LmvHXPZKYTjpHh3OT70cn7ZOQDbJXFC9n4ZGw49g8/PgBTnOCv+hU33JkC1002YMY+6b3Ms9EvxAXHwJ4P8A0cxjOcw2G8XtXoiBv6s1lA9/gbXXBOEQ9f0g9MyGO+ghe7OS8+yldgZXw44a8xLGQ1/cEBF3El3DKePoyPt/gFw/Tr2LFf9tYvHpDJQjBzEBn0mxa+8SuxQGz2FU9iAn7inMKHRfPI05x/1Gf8FFvNJ+aC4IvDeEJnzZj4jlPmFnbz4Ed8gm019sIt/o1X+hsXyHTGL+exlz5gDSscHP1Jf/TfZp5OfIYjnejALvzbIhz3tPgBjrClmGaTQaxkC/GQrfKgN9lghVPOoR9/lqvQwTiJH/Mg0x99GQueZApvYEJWvHAePIyBix6ZS31msxfmfBy+mXtsuOgH/9hRE6/hSF+yi29iuv7HuJM4DCO5F/3pRT86kMG8nQe/xWXj4QmbsLGxNX2LQ3wGFh7G1s+qeW8+6ZA/sKMv7PCIH2fTi5+RLXHOGJq5Bf/Ef/g4XmN3G0Jw9DrHzx9Of9ginKEXPMe8DR9G/DaJS/JecUmMogu74Qy8+X94s4yPY5NfsosNNM2GOlzJn/hBpk0aDJMDwIWNs6HNZvrDRfwnH/y89pmNZxzit/FzY5KTfzqWXvyHLmKV2Jr8gYz6xjnHw5cfkMNrsdpnmvNxhl3xl70yN7MR3skD8NQxOOY4cq5rZCWD89hZP+YTfPYeeybW4TE52PygeWqMm/EdHKUffcWK+CHs+LHP+DUc6WSz1RjGZhsbk3AhH3vFp/WFi3Chr/yIXPDVl/HoF96E/+YntuG3wdgxbJg+yYFTZIS1OEH/8Er8JDf52Atu5r/ELjqyg/gtftGTLelJL5+zD39d5s8qexnLGOwCBzZjW3OFfuC1SSMD2eHpoh+Y8EOy0jVxiZ6OMxbeO97YjueD8IIVXOgIf3HE69Fv4QEXHOAHfN3cog94mufcDW3+pg97iKPnpphrPDi5QI9sfJF/xgb0hae+jMHXYGQMsskP2JB+5uhVOCcfSlwkJ/9mI8fThVz4QFZj61/DC7glv5rfvPBHH3hBJn2Lt7irT33BSvxwLv9yjDwLvuKYvmHCHiPv2I9s7AJPOOGg87zPLjDiA+lHrCUnjsAwzbF8CQb6ET/MW7BmT9iNvHJ84hV5Hc+W5np68FtcJ+8YDzPe+Bw92A2/yE9u48GZ3nTQyERf8QSGbEs3MvIfvuVcDZY4AW/Hk8//4oTjyYdT+tikORc+7Eln8oob+GSegOs4Pow88N45ZHARDF9iz8QUesLJ+9kv2UVcYk948QPzo3xZwRyWvnnAXbzhFcwOs9NhGPFR4+AizI0dnuRcY8i/yMCXxGh88fB/4oXz4I3H5FdoFvfieylcpd/xOfMOnvA58UofztfYm93TFz8gq/PI533zB35oeMQ29HOBBd34LtnlneIceePneIHTfMEx+kk+Mnd44Q/uZC4jGw7gsP7gcdB6L32Ej+Q5N/mscekhDtuv0UfwzDnrnvkdjpJJ7KOjeCFPETeN53O4Ggdn+CZ5fX6QvOEAPyArW4iHYpW8QsyGseNgwB500KexxSC6sJmYy076Gu1Lf3hq7EsWuDsXBuQlv+Z9F5WxCbvKr5JXyPX1430NV/WVGIMXZGFbn43zz3xC/xSBIlAEisCpROCyFiqXJ1JJhsktDxOSBOawSclkaxKVXElo/W/ilbya2JLsKD5IGiW7kn2LS1e4SRodn3GcTxaLIQsBDwsYiW2+ek8in0RXX8YwtonZM1n0kwSRHGMy7Rz6a/TMBOz/JEL6oBe5HON8yYRE5qCmz+BBDgmJZ3160CuJiYRHX/qUwAZ37wWLg8bJ+/rSr6THOB5ekztjOpb8Ehk6GMd4xvU+7MfmfMlnkhxYpRnPY7l5D9bL+hiDnvogmwUJXL0XfuQY52/SjAXjLIRgGvyi0ya8HcfSJxlhpl9yek+SGKzIvK6RAw/hpw/6+j92Z1P4xAaxA7uwwbL+FhspVEqmyWFDnd8sJ6LOD0fTb7BYtm90gOGyvGwzyhs7hZtkIK/3l+XFOZzRJyzoG87Fxhl71XPsCj9+E18mj8/0xR4SaM/08pnjYi+JPl8OpqMf0dfx+rfYj33IyvaOpVdsRFfjrOqLPLE3W0vy4WHRaXx9LOMz8svmgf8t9o2DE8Ye5V2F0fhe8IJ3sKITG5LN5zAiCx3YgGye/W+8sTmeTOwII33p1/nZNKHTNjLqnyzpU3/6JrNHbGsM/PUgH0xwbh1vRnviCzvo23hkdK45Al+8NsaIGf3YzXPkgMnon3AiO35p7Eu25bipX+PCnhz4RRb9auygL+eSxRg+C2/JmTlo2V9jFwtUC0vFTXdikttGkc3/fMWhc7e1zyzgij+4EJ2ysB7jA7nwYeRY4oRneJMl8oz2SnzU3xgv4B+u6iO+oa9lf4IfDPTBRsZJDIy9R7XoYizHe9AvdsER52rB27Hk9KC/c+jguMShMdZEHsfjOV1swNDhILsYy3n6dl54Qx9NH9EJLh7kpjc88IaujiMbzkVez3TwPmzSV3Al12inecDpD5mcQ6YxDpPJZ2QQt2BGf7rRwdiJQ147HuYa+5HTmHnox7nLdp1P2OAPGYO5DS+60lO/5KJbxtCd48lENjg7XnNMYsR4/Pzh9GfkjXPhTx8+TJfwPMdHLseui0s4pA+NPWM3mIy8WcYnesBaXqDhAVlw5SCuzQeu+BPbZXwYmkfH/tgddvQhq9fGSjx0jsfYYpv0i3853uvopW8Px49+kHF85thgHh6xL/x8BhPjwEM/PvNITB3lWvXaGIl1xqUn++nL++ExuclB91GO+KC+x7jJRuRa1gXPwh/yG4cMMPQ+bhiDXuTAb/3AB/aO1yKL/tiLX5Jl2a/0HV6SJfmPfqKjY5wXHSOffvVJntHG9HQu+dIf+YIV2Zynn/iVMZznYZzEz1X8mRUc/hiL7MYzjjbawetNWrCApzwMnmQMZyIrmWAmTox+HPz1Q4fI4Pz49OiD+iAvueM/ictsD18bwpmDHB/eOAeO/Eb/y3alLywj5xh/9U2e2E8/q3DOuaMdI5/jl3U0pmM19iWXZ8eNbcSN3WDAhuQKx+nlXFjqUz6U2KJfmLAHOdJiP/2NODkv3DNGfCP9GIetYDg2+DmPjMb32jlw81jmlfE9crxx6a5/eEfeUeZxvLyOHsFJP5Hb+PoJpmSiL84GQ5/nuJFv+qWTfh3v2Xv6oo/zYGOsTdrILzaKHfVhnqD3OL4+R93YSJ6tH/4eXMNH79MdnokjdPSgt770T37nHBaX9GVMhSEFUntHD053pJPVV+W7KMD+kVi5yh82wWQ8BibmHWPCOjKPx3hNB4/EC7iNnIw9gjc8Mp/F1pkXlvv2v/NGnrC9Bzx8No6NN/53vM/gYAyYeNbCI/qliEVG+Ms3PDtG/8mn/M9OPvNY5W/sSq7wKGM7Hh7h/CzEij/BZ+Q3PXDjoPlvRTcvvEX+YAVz8iRuksl4kdc428gLD+eQVd/LeQUstHCbDjDgI2zktfc0cqSvxD196ttnsS+ZnT/a1/nehw+bOJ5d2U1fbKEf8pDZmI7DBX0FV3KGp/psKwJFoAgUgdONwGUtVF4qaCU5rlZy1bsrWBUqTaKu0nGFm6ugTIwmQEmASdSkKdH0lR2Km54lmr/0S780J5qu4vO/fpyz3Ey22qrPlo+9VP/Ty6JQo2v0vRjj0994Eg8YBN+LMVb7XI8AW8TuB/F1VQ/LhUqLdj6Th81MNtZ2yaWjyrtKh+O8Rw4PDYcvhi+n/8h5McZI35fqmU4Xgxe7lj/xSb+7iE9H1dt54cGueTb2e1T89CF+uHDHPOjqbd80wOfdiekuz/wuy1HH2Oa8XeMV/ciwTXzcRuZdHHup5DTOLvxXH3ijPw1fjoPvtvrv2r93YcPL0ceu7Hk5ZL+cY4a/ZAhvL/X8zHYeNgi142zU7YIH6WOMD8fN/YKzvkecj4K1PrSjnDufeAX8iQ08B/uD9F2Fh/fG9w8693JCMcpIDjLuQs5R713rN8q8K3l3LeO2/V1MvLaV5VIff9S4pOhiz8ldyfJl+07W0u5U81XlcmV3lynA7EPDoXG+2IUf7wNuF1tHNjlJeQWeRB7z2mFz28XGp/0XgSJQBIrA5UdgLwqVrgZWdLS56re1JJCuYnInpR+/9jUSrthxxY+FskTTla6+fklh09dOOMdVcPfcc8/8uxWuQHXF0ZW0ILnUC6kmqJffwSMBW2jbLBKWC5Wusx1a3QAAQABJREFURvXVQgqVivsWW0fpNzKte75Y/a4bc9VnJ0WOVbKd5PeuBNzIuOsF9FH1Pup5l4ID2bDxtUF+C8fGi6tgXahg08Vdlb6qyP+Xqu0ar133d7FwuFRy7mIcfaQfeOwiV0p/m8xjjt21f18su17sfrfB7WLLcqX0D7MRt004dzF026Ucoz5HlXWX8pBhuT/vXS6sjX0ltNiRrEfBKucf5dxLhU9kPKqOl0rOcZzIfJJxHeXt64MRYMtle25iV7mxr7KUJ/tGIntP7g6z3+Q3Na2fc0fgwaOfrk+WcTxd2l2Z2izzexNuH6TpLuy7LI+xjiPTQbL2/SJQBIpAEbhyENiLQqWvTfC1dX5/wG9h2nD11aKKjX6Lx1dZultModIVy4qYvvLCHSSKlZJOX3Hgd4dcEWdj1tVw7sLpRLqe7LtIYNaP0E8vFgJ+F8GP2PtdOv6QQqUipd8aUKhsKwJXMgKNT4dbz13xLt5xJ+V99903/xaOudPXQPvaV79D5RsGfF1RWxE4SQjUv0+SNSpLESgCRaAIFIHTiYB8w1rZRb6+8vUv//Iv5/9d4OuCeL+J+rrhN5JPJwrVqggUgSJQBIpAESgCx0dgLwqVNlp9H7qvfHV3pIKlHx53p+X4nfSKjgqV7qrMVw94rYjpN9JszPqqO5uyOe74JmgPReBkIuAu4r/5m7+Zf5fOb6fkq18VKT1uuummkyl4pSoCRWAnCNh4cZEO//ftAn/0R3+0+NznPre466675rnQHZUu4Nm3K8R3Am47KQJFoAgUgSJQBIpAEbiiEZAr+waH3E354Q9/ePFXf/VXs04/93M/N38Tl4Lla17zmvm39+wttRWBIlAEikARKAJFoAisRmAvCpVJIG22+q1KBRh3VSpc+i0BPzDtLkpJpuQxPxxu81VSeeutt85Xwfnq19e+9rXznSOKmr2bcjWp+u7pQOCzn/3sfAfyww8/PBf13UWsWO8rbO644475juTToWm1KAJFYBUC5k7fSPDoo48uPvrRjy4+8IEPzFeIK1D+8A//8OLNb37zPCeOF/ys6qfvFYEiUASKQBEoAkWgCBSB04aAXNlvcvv2roceemjOl32Dl2/q+vmf//n5t9xd5P6KV7zihYvhTxsG1acIFIEiUASKQBEoArtCYC8KlcCSRLqrUmFSIvnII4/Mz74C1m8K+Gq7FCr9noDfn/S7lb7eUrHy5ptvXtx44439ytddMa/9nHgE/E6rIqUrRPmOAr67KPmDh6+CbSsCReD0ImDeNDd+7WtfW/gq6L/927+d/3exgq+wchFPN15Or/2rWREoAkWgCBSBIlAEisDBCMiV7SHZY3IRvK9/tX62bn7rW9+6uP322+c9JRf19UL3g3HsJ0WgCBSBIlAEikARgMDeFCop62q3p556ar5DRDLpYQPWHSO+HlaS6Stffa2r5FIhRrHSRqy7KyWYPu+dlNBsO+0I+MpHhXx3HPMPfsEnfAWsZz7SVgSKwOlFwOZLfqPShQs2YMyjLuDxO5XmR1eMd048vRyoZkWgCBSBIlAEikARKAKrEZAre+SCeN/gZf187bXXLm677bb5QndrZt/a1Xx5NYZ9twgUgSJQBIpAESgCQWCvCpW54u38+fPz5quve7UJa+NVkTItv0+pMCPJdIel5yaYQajP+4BACvv8hX9YXPGJPFq03wcWVMd9R4DviwXmSxcvmEcVJ8d5cd8xqv5FoAgUgSJQBIpAESgC+4mA3FiubF9JvuzCeOtk39AlX+4e0n7yoloXgSJQBIpAESgC2yOwV4XK7eHpGUWgCKxCwIJM65Whq9Dpe0WgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKwCYItFC5CUo9pggUgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEikARKAJFoAgUgZ0i0ELlTuFsZ0WgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCGyCQAuVm6DUY4pAESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEikARKAJFoAgUgSJQBIpAEdgpAi1U7hTOdlYEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEisAmCLRQuQlKPaYIFIEiUASKQBEoAkWgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCRaAIFIGdItBC5U7hbGdFoAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEikARKAJFoAhsgkALlZug1GOKQBEoAkWgCBSBIlAEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKQBHYKQItVO4UznZWBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBIlAEikARKAJFoAgUgSJQBIrAJgi0ULkJSj2mCBSBIlAEikARKAJFoAgUgSJQBIpAESgCRaAIFIEiUASKQBEoAkWgCBSBnSLQQuVO4WxnRaAIFIEiUASKQBG4shA4f/784umnn154fv755+fHmTNnriwlToy0wS3Pzw+Sja+Ht/tyrxHga1ddddXi6quvXlxzzTXzs/fqg3tNiypfBIpAESgCRaAIFIEiUASKQBHYKwRaqNwrc1fZIlAEikARKAJFoAh8CwGFySeeeGLx5S9/efHYY48tnn322RcKlS2UfAung18pPqYo6dnjquE9n+fx3PRaG8/55jv9u18I8Ls0BcoXvehFi5e85CWL7/iO75ifvad42VYEikARKAJFoAgUgSJQBIpAESgCRWAfEGihch+sXB2LQBEoAkWgCBSBIrCEgGLJc889t/jCF76w+MQnPrF45JFH5jsrvdc7upbAOvDfFB1ToLx6OnJ6nLnmwhlTcfL5Z6fX56eH5xQtU9y8cFif9gqBFCo9v/jFL54LlDfddNPi9ttvX7zqVa9aXHfddfPdlXsFSpUtAkWgCBSBIlAEikARKAJFoAgUgb1FoIXKvTV9FS8CRaAIFIEiUAT2GQEFSXdQfvKTn1y8//3vX3z84x9fPP744/NXwLZQuSkzUqh099tUnDzzoun5xReevffMVJt8enp845uvF+6qzDnTy7a9RGAsVN5www2Lm2++efGmN71p8ba3vW1xxx13zIVLBcze1byX9KjSRaAIFIEiUASKQBEoAkWgCBSBvUOghcq9M3kVLgJFoAgUgSJQBIrAYr6b0u9SfuQjH1m8733vWzzwwAOLr3/964tnnnmmBZKtCeJOSoXK6y48XjL9r1CpSPnk9Hh8ekyvXyhUTi/b9hoBxUqPl7/85Yvbbrtt8aM/+qOLX/iFX1jceeedixtvvHH+CtgWKveaIlW+CBSBIlAEikARKAJFoAgUgSKwNwi0ULk3pq6iRaAIFIEiUASKQBH4FgLuqFSofPDBB18oVPqdyqeffrqFym/BtOErX+Xq614VKqci5Znrp9cKlU9NBcrpbsr5jsoWKidA2i4gkLsq/S7lrbfeOhcq3/72ty/uuuuuFirLkiJQBIpAESgCRaAIFIEiUASKQBHYKwRaqNwrc1fZIlAEikARKAJFoAh8E4GxUPne9753vqPSV78qVLZti8CFQuWZ6WtfFwqVL50eU6Hy+RYqt0Vy345XqLzlllteKFS+5S1vaaFy30hQfYtAESgCRaAIFIEiUASKQBEoAnuOQAuVe06Aql8EikARKAJFoAjsJwItVO7a7uMdlWOh0le/PjEN1jsqd434aeivhcrTYMXqUASKQBEoAkWgCBSBIlAEikARKALHQaCFyuOg13OLQBEoAkWgCBSBInCFItBC5a4Nl0Ll9LWv/98dlS1U7hrt09JfC5WnxZLVowgUgSJQBIpAESgCRaAIFIEiUASOikALlUdFrucVgSJQBIpAESgCReAKRqCFyl0ab/zq16lQedV0R6XfqJy/+tUdlY9P//eOyl0iflr6aqHytFiyehSBIlAEikARKAJFoAgUgSJQBIrAURFoofKoyPW8IlAEikARKAJFoAhcwQi0ULlL4w2FyjMX7qhsoXKXAJ/avlqoPLWmrWJFoAgUgSJQBIpAESgCRaAIFIEisCECLVRuCFQPKwJFoAgUgSJQBIrAaUKghcpdWrOFyl2iuU99tVC5T9aurkWgCBSBIlAEikARKAJFoAgUgSKwCoEWKleh0veKQBG4YhB4/vnnF+PjihG8ghaBInBiEThz5sxifJxYQY8pWAuVxwTw205vofLb4Og/GyPQQuXGUPXAIlAETiAC4zrM67YiUASKwC4QsBa76qrpZxSm5nXbbhGwDnzmmWcW58+fXzz77LML/xfno2AcbuZZH5kL83yUfnvOaUFAbiSWXX311Ytrrrlmce21174Q206LjrvUo4XKXaLZvopAEbjkCCTBklxloXzJheiARaAInCoEkkhKJj1O66Kthcpd0raFyl2iuU99tVC5T9aurkXgdCGQtZd1mM1ueUWLlafLxtWmCFwOBFKkzIa+/0/reuxy4GvMp556avH1r3998dhjjy0ef/zxOYYX522tYf3noaCe14qTz00Pz3lML9v2EoHkSQqUL3nJSxY33HDD4uUvf/nixS9+8V7isYnSLVRuglKPKQJF4EQiIOh/4xvfWPzP//zP/JxiZZPYE2muClUErggExBXFSYnkS1/60jmZfNGLXnRFyL6tkC1UbovYuuNbqFyHTj87GIGLXag8qGjQXOlgm/STIlAENkPA2stm9xNPPLH42te+Nr9ubNkMux5VBIrAwQjIXa677rp5Q//666+f70CyPmvbHQL/+7//u/jMZz6z+O///u/FF7/4xblYmTtYdzfKae4phcmJl2eunRTFTwXLqUj5/DPT8/lvvp6LltPLtr1FwJ6LvaVXv/rVi1tuuWXxvd/7vYsbb7xxb/E4TPEWKg9DqJ8XgSJwIhGQvFocP/roo4uHH3548aUvfenbvrKii+QTabYKVQRONAKSSHHFwviVr3zl4qabblrceuutC4WE09haqNylVVuo3CWa+9TXxSxUypXWFSqbK+0T06prEdg9Ak8++eTi//7v/+ZN7v/8z/+cX5/mb6LYPYLtsQgUgWUE5C3WKO46et3rXjevx172spf1DqRloI75v5j9wAMPLD72sY8t/vmf/3nx5S9/ee6xueGmwFr7KUxOFzSfuf6bz3OxcipQPv/E9P9T00PB0t2VbfuKQNZhCpPf933ft/iRH/mRxc/8zM8sbr/99n2F5FC9W6g8FKIeUASKwElCIImrhbErd//jP/5j8Xd/93fzAllxwS307n6ySM6kcJLkryxFoAicTAQsyp5++ul5k83r7/zO71x8z/d8z+JNb3rT4rWvfe18FZyvHzpNrYXKXVqzhcpdorlPfV2sQmXudPK1Xl/5ylfmK+X5vCt6X/WqV80XYMibfBVRWxEoAkVgGwTEF79t5ltt/uu//mtej/3rv/7rHGdsxokzbUWgCBSBbRGwf2OfR2xRnHzjG984b+hbi73iFa/onZXbArrm+H/7t39b/MVf/MXiQx/60OLv//7vF5///OfXHN2P/n8EUqi8bipU3jA9zHvTXZXupnz+69Prb0wPhcpnp0fbviPwXd/1XfO+0t133714xzveMce2fcfkIP1bqDwImb5fBIrAiUQgG2/upFSk/Id/+Ic5uXI1ryvuvvu7v3tOYhUrbch5tBWBIlAE1iGQ36T0FTif/vSnF1/96lfnzfvbbrtt8ZM/+ZOLH/zBH5y/puO03VnZQuU6Vmz7WQuV2yLW47+JwMUqVPpqfLHsU5/61OLBBx9cnDt3bi4s3HzzzYuf+ImfWLz+9a9fWDT7iuu2IlAEisA2CCgkiC/iirXYP/3TPy1servw4c4775zvgHLRVy4azfM2Y/TYIlAE9gOB8Q4+axNfQ/rJT35y3sexBnPRqLhiXeYuSxdZtR0fgRQq77///sXHP/7xFiq3hjSFyum3Bs+8bDobL339qzsqW6jcGs5TfoI1l3imUPnOd76zhco19m6hcg04/agIFIGTh4Crd/0Oiu/Sf+ihh+arvz7ykY/MP/4tgVWsdKeAK3nPnz//wgL55GlSiYpAETgpCChU2lzzlTcWxo888sj8u7cufPjxH//xxVve8pZ5U19sOU2thcpdWrOFyl2iuU99XaxC5WOPPTZvOn30ox9dfPCDH5wLCXKo7//+71+8/e1vX/zYj/3YvOnnDoW2IlAEisA2CIgvX/jCFxbuovTNNr420J2Viggu8PKNFCk+tEi5DbI9tgjsJwK5sEG88JWkH/7whxe+EcLejs39t771rYsf+IEfmC+C6AVWu+FIC5XHxXFVodK3lLiLcipUPt87Ko+L8Gk6P3dUvu1tb2uh8hDDtlB5CED9uAgUgZOFgK9mfPzxxxef/exnF//4j/+4+Jd/+Zf5R8B95atigu/6VkxwpZ1CZe+oPFn2qzRF4CQi4KuiFSp9PaK7AiyQvb7++usXb3jDG+YF8g/90A/Ni+OTKP9RZWqh8qjIrTqvhcpVqPS9wxHYZaEyBQEbfr4eX+HAZt+f/umfzhdh+FYKG37vete7Fj/1Uz81/1aKr2lsKwJFoAhsg0Diizu2XeDlAlLrM7/t7QIvv+89tsSm8b2+LgJFoAhAQM6SQqW1idzFN0F86Utfmr/+1YUPb37zm+cLrXwFrK+EbTs+Ag8//PD81a/uqOxXvx4FzwuFyjO++jV3VF4oVOaOyuefnjruV78eBd3Tdk4Klf3q18Mt20Ll4Rj1iCJQBE4QAgqVrq5LoVJBwdc1uiNAofLs2bPzb8spXNqQ68L4BBmvohSBE4qAxbFCpd9DsWj7zGc+s/jc5z43S2tB7A6ku+66a/Ga17zmhGpwNLFaqDwabqvPaqFyNS599zAELlahUm4kR7IBde+9984Xd8mJfIXae97znvmrh9ydcNruFD8M735eBIrA8RFIfPEzHPIm+ZOLvm655Zb5zie5U1sRKAJFYFMEUqiUp7jwwbdBePb/K1/5yvmbbVyQ7iIId263HR+BFCr9RuXHPvaxfvXr1pBa+01f9ToXKsffqMxXvz4x3VXZQuXWsJ7SExQqXfieOypdDN+2GoEWKlfj0neLQBE4oQikUClxdQWvrx1SkBT4XWlngezrQPxGpcTWo60IFIEisA6BXMnrt25dBJHHU089NS+GXQDRQuU6BPvZtEqdQJiuoj3jd0qunx5+9++qaYH61PR4cno8Pv1vsep3kzsvTSC0XUDgYhYqXXShUPknf/Ini0984hPziAqV7373u+dC5R133PFthUqxsK0IFIEicBgCKVS6GMLvVPpN3BtuuGEuIliPubOyrQgUgSKwKQLJP+zd2N/xzVnurHSntr0dRUq/UdlC5aaIHn5cC5WHY7T+iBYq1+PTT0cEWqgc0Vj/uoXK9fj00yJQBE4YAmOh0lc0Pvroowu/L+dOJ5tvCpV+n/Laa689YZJXnCJQBE46Ar7KzEUQFsY23/wersWxRXELlSfdepdbvhYqL7cFrtTxL1ah8qtf/erC3U4Klffdd9+86QcjX/2qUOmK3te//vVzoTIbhHm+UrGs3EWgCFwaBFKodGGXnMn6zLfb+IpG6zEbcm1FoAgUgaMg8MUvfnH+KQ7xRazxkz6KlOJLC5VHQXT1OS1UrsZl83dbqNwcqx7ZQuXmHGihcnOsemQRKAInAIGxUPnQQw/Nv11ALFfuupVeodIVve6obCsCRaAIbINA7qhUpLQ4doeA30GxOL7zzjv71a/bgLl3x7ZQuXcm35HCuyhULn97hIKjr2L893//97lQ+Wd/9mfzt1A4TqHyF3/xF18oVL761a9+4feholILlkGiz0WgCKxCIIVK+ZJC5ZNPPjl/A4V8qXdUrkKs7xWBIrAJArmj0jdnWYu5iNRF6L7dpoXKTRDc/JgWKjfHavWRLVSuxqXvrkKghcpVqKx+r4XK1bj03SJQBE4oAsuFSlfcSWgT+F1lZ9OvhcoTasCKVQROMALjxpvFsY038aSFyhNstBMjWguVJ8YUV5gguyxUpmCp0OiOyk9/+tMvFCpd3OXzN77xjXOh8qd/+qfnOyoVKn0zhXNSoMzzFQZlxS0CReASIbBv+dIlgrXDFIG9R0Ce8vnPf37+FogUKq+//vrF6173uhe++lXe1HZ8BFqoPC6GLVQeF8F9Oj/71f2NysOt3kLl4Rj1iCJQBE4QAi1UniBjVJQicMoQ2LeNt+eee25x/vz5xYMPPrh473vfu3jggQfm34IRZ9u2RaCFym0R6/HfROBiFio/9alPzYXK97///fMdlUZUqHznO9+5SKHSwrmFyrKxCBSBbRDYt3xpG2x6bBEoAkdHoIXKo2O37ZktVG6L2PLxLVQuI9L/D0aghcqDsVn+pIXKZUT6fxEoAicagRYqT7R5KlwRuKIR2LeNtxYqd0nXFYXKM1cvFs8/NT2+MT2e+ObrxXPToM/vcuD2dYUjcLEKlb76VaHyQx/60MJXv7qjUlOofMc73jEXKt/whjcs3FF59dVX947KK5xHFb8IXEoE9i1fupTYdqwisM8ItFB56azfQuVxsW6h8rgI7tP5LVRubu0WKjfHqkcWgSJwAhBoofIEGKEiFIFTisC+bby1ULlLIl8oVC5evFicuX6xuOql0/NU/FmMhcqnp6/efHYatIXKXSJ/pffVQuWVbsHKXwT2D4F9y5f2z8LVuAhcHgRaqLx0uLdQeVysW6g8LoL7dH4LlZtbu4XKzbHqkUWgCJwABFqoPAFGqAhF4JQisG8bby1U7pLI316oPHP1y6av07xmcfVVzyyuOvPkNJA7Kp9ePPfcs1Ox0l2V395szGie88j/8wdH/DP+7uByFxlz1e8RjjIsn9f/d4vArguVsflXvvKVlXdUuoty+atfe0flbm3a3orAaUdg3/Kl027P6lcETgoC8s/+RuWlsUYLlcfFuYXK4yK4T+e3ULm5tVuo3ByrHlkEisAJQKCFyhNghIpQBE4pAvu28dZC5a6JfM3U4XWLM9PdlGeufvni2mv/H3t3/m3ZUZYPfPd4u293pzMwJCjagAbQyIwyBUIYIoEgIaCLtRz4Afx7WC4FEYEAGg2ESRlNSAIic4AQQbNEaUVdznwhY6fT3d/61Omnu/rk3u7Tfadzz6laq84+Z+8a3nqr6q33qWfvfXYOuxaODdu3FbJy64ioPHLkcCErjx4nI8tDl1u2nCAmnU88cuRIk+7cnsBUtv8ezP8Pam3IyfGWt2SlNOrvZOW4ltbmdycq10avvdSuga6BtdPAvPlLa6fJXnLXQNdAq4FOVLbaWNvvnahcqX47UblSDc5T/k5UTt7bnaicXFc9ZddA18AUaKATlVPQCV2EroEZ1cC8bbx1onL1BvLoKbYdhRRcHLZt3zds33nhsGvX7mHP4rZCVh4pZGV5BWx5DeyRIw83BORJolJfIAcffvjh4fDhw4O1ztHvlrRcTuIQjSM5RgSlp+S2by9PdR7//8E2bwjL5ENmCs6TJXX7fjaEZcrNsa2zf19aA52oXFov/WzXQNfA9Gpg3vyl6e2JLlnXwGxpgP/Yn6hcnz7tROVK9dyJypVqcJ7yd6Jy8t7uROXkuuopuwa6BqZAA52onIJO6CJ0DcyoBuZt460TlaszkEfk4NZCCO4adu48rxCUFwy7Fh817N2zbzjvvJ3D4uKWcv5weQ0skvLhQvwh/5CCJ4lKZGQIygcffHB44IEHarz//vuHQ4cOVeIyTzmOS92SkwhH5OSOHTtKnTuHhYWF+juEZJs3+RxDVIakTJ3W3JYoXaqclGlzSf6zJTeTf16Pnaic157v7e4a2LwamDd/afP2VJe8a2BzaaATlevXX52oXKmuO1G5Ug3OU/5OVE7e252onFxXPWXXQNfAFGigE5VT0AldhK6BGdXAvG28daJyZQO58Hv11a3btm0vZCBScG8hJS8c9uwtJOXexxSScv+wf/+u8n3bsHtXeapyu/+mHD2hqOYQlfohRGUIynvvvXe45557hp/85CfDfffdNyAsQxraxBGFkIwtQYmcFHft2lUj0nKcYPQ7Ud4QleRQDzkQpiICdTmStApx/CPtyFOgyRNZ27T9+0kNdKLypC76t66BroHNoYF585c2R690KbsGNr8GOlG5fn3YicqV6roTlSvV4Dzl70Tl5L3dicrJddVTdg10DUyBBjpROQWd0EXoGphRDczbxlsnKlc2kLdu3VKfVkQKLi7uK4TkBYWcRFA+dthX4v79iMrdw76928v1Y+UJx2OFEEQuqtdH/VKfQETuhSBETCIojUfxxz/+cf0dslJafSeEoMzTkyEnd+/evSxRGYLS0WthleG7zSHkoqcp1RWyklxtnbXisY/kzVOhyvBdjKxjWfrP4xroRGUfCl0DXQObTQPz5i9ttv7p8nYNbFYNdKJy/XquE5Ur1XUnKleqwXnK34nKyXu7E5WT66qn7BroGpgCDXSicgo6oYvQNTCjGpi3jbdOVE4+kBF5Qo6+e1Jx9+6FQlDuG84//6Lh/AseM1xw/sWFnHzssGdfISz3nVdIy12FqNw27NkzlKccEYMnn2QcJyqRe8hBRCVyUvzRj350gqz0lKXr0iEObeYgGZGUiMnFxcUaW5LSNa+CFaRPkK+NISpbwjRPVIaobPOnHOcSQ1LK1xKdzidN8vXjSQ3ME1G51Dgw9tp5Fc20aXM9x6Tpx66BroGN0cC8+Usbo+Vea9fA/GnA2t//o3J9+r0TlSvVcycqV6rBecrficrJe7sTlZPrqqfsGugamAINdKJyCjqhi9A1MKMamLeNt05UnnkghxhxzJOH+e7pxfPP3z9ceNFFw2Mfc/HwqEddMlxw4SXDvkJSLuwqr4Bd3FtIzB3lvypHROWuhaESlcoZtpRYAu7w6NEjlXi0viEhEZVe+yrmaUpPWPqNrEQCSqf/lOWJzr179xYydE89IioRlIhUJCV5pR0nfsaJSmk8UYlYVD55QlLmNa7jGkuZyReSsn11rXMhVqXv4VQNzAtRqe+Nk4xFWhgfg9FMO658ly7zThrfe+ga6BrYOA3Mm7+0cZruNXcNzJcGrPmdqFyfPu9E5Ur13InKlWpwnvJ3onLy3u5E5eS66im7BroGpkADnaicgk7oInQNzKgG5m3jDWGAQPrSl740vO1tbxu+8IUvVJKMnZ33gAgR82pUhF++O/qNGHzUox41PPaxjx0uedzjKlF5fnmicvfiReX6eYVA3F3IykJSLm4taY8Nu/JEZXllbKFoKklZqMpK3oToo3tPTSIjEZYi0i8RWemc6whFBA7CFFG5b9++KhOiEknpmmDTRz/r76VIopA+0pFD2sSWWHI9aWvBx8tOueTJE6F5de3//d//Vdk7WRmNPfI4D0SlsZOx7Wh8GZ/5P9X2yV/jybhLet/NN2M66czB8bH4SM32M10DXQNrpYF585fWSo+93K6BroFTNcBf6ETlqTpZq1+dqFypZjtRuVINzlP+TlRO3tudqJxcVz1l10DXwBRowMaVjdp//dd/He66667hP//zP+smbAz/4x//+PKqvfPqZtYUiNtF6BroGthEGpi3jTeEAMKgE5WnDlIECBIl5AgyBUESksTROWuNtefiix87PPbixw0XXviYQhg+qhAq+wv9uFjy7xwWd2+pEVG5sLPQk9tGT4OVfZhKVIbkQ8aIyD7rnCcaE5F87ZOKIS2lE/JEZZ6q9BuRIyhTH0sb8lGdCS3ZY3PItfbouyDdeEwZaYM6yIlI9SQokvJ//ud/6utrEZeupfzkTfn5PY/HWScq9bFxaNz+93//d/2/VWPB/Dr//POP/5fr/kq46/8Q3ny9//3f/62/kfFuDKArrzg2BzPG53HM9DZ3DWy0BubNX9pofff6uwbmRQN8hk5Urk9vd6JypXqePqIyuG6t8VXqiQbXur7Us5mP2a9+yUteMrzhDW8YnvrUp27m5qyp7J2oXFP19sK7BroGVlsDNkI7UbnaWu3lrbYGOGvZkF9uc3+161ReW2/KVz/SZdyhzPV+PKmBedt4M0Y7UXmy/zNXESjIvpAjiJG8TtW1kJZIk0c/+lHDowtZ+ZhHP7YQLhcNu3ZfUObb3kLM7Bq2bd/xSKLSQ45b2AevfB3ZiRB96Y8Qio5Im5CXedIS4WMdRGSa8+TJa19D4pjzykvekJ5+I41ca0FlvucYXShHRArlu2NrT+SJ7ONE5X/9139VshJp6SlQbZK2jfKn3pO9MT/fZpWoDPGOuGZbkdWiccyXM8cuKq9NFi+88MI634wPadyEhuw2npQjmIPSXXDBBTUa6+NjcX5GTW9p18DGamDe/KWN1XavfSM1EB+F35LAB+rrT7Sxukf67kTl6up0udI6UbmcZiY9P71E5VItWE2s1eLA1LWa5afMWTp2onLy3uxE5eS66im7BroGpkADnaicgk7oIpxWA5y0bNACtYCsTX4Eh+9rFdSrvpaI4ESqVyTDUk7lWsmzGcudt4034wUx0J+oHI1W89NcQYDs37+/RqRInuLK61RPpttdnwgbEScXDXv2nj/s3LGv/P3knuNEZSEQ62tftw779pYnH8sTlf6acjRX2YmTr2LVF5nD7EcbQ1giG9tXq4aoJBeiktyi3+a6MqyZ0iF8RN+dS/lart42aJ8Yu6W81o7kemtPlJH6EFPIJuQkwslTdJ6MQ7Cqm41qo3zjMoz/buWbte+zSlQabz/60Y/qGzDuvvvuOiY89esGAOPJmDV3PFXpaLwZOz/84Q+HO++8s36/+OKL63lP5hoTiMqf+qmfGp74xCeWmwQeXclO+XroGugaWF8NzJu/tL7a7bVNiwbi2/BTcqMV38e6wzdybH2haZF7M8tB552oXJ8e7ETlSvU8nUQlm5QYPOWY7+fS6vG8KT9l5XqOOd+PJzXQicqTujjTt05UnklD/XrXQNfAVGmgE5VT1R1dmDENALI24D3tZGPVePXUiP+Os8HqiZC1CJxCMRvDCAGEhM1gdee/64DqHpbXwLxtvHWicjQWArY8mWiOIimRIIl+hwCkM8F8M5/27FkscW955eu+QsDsLXMOUblYbhrwP5EL5XyZg3sLUbnPf0kCjqM6RyTlyScqM4cd1cGWtJFdYU/McWROnqhUGjlCUpLfb22SP3nkyxOZLVkZglQ58owTlHTChjkql01picrojtyxf+oJUemJSkSlGJnJRIbIoW1pc9offZBr1sN6E5W/8Au/MFx33XXDi1/84vraIeM8m636U8jxXHSfvtPf//Ef/zH84z/+YyUe9feBAwfqvDJOjVlrkyPy0jhAbv/gBz+o6Y0nr0Uy/pCXxpV8/hP20ksvrYQlklPeswkZqxn7fidkDrRjPNf6sWuga+CkBubNXzrZ8tn4FjuddduRTWT7rPdZ61eyFsyCppCT7Q1i/BfBjTb+o9yRjuZdT6vZ18ZmJypXU6PLl9WJyuV1M9mVjSUqY3vGj3z6xLQjNr/1edvv0rW/x7/7Hb9Z2nGbl2vStXml7WGkgU5UTj4SOlE5ua56yq6BroEp0ACAYPOr/0flFHRGF+EUDXDKbMQan//yL/8yfOc736mvrvOkyM/8zM8Mv/iLv1j/z+6UTKv0Q902GTy19A//8A/Dv//7v9dX7dng/dmf/dnhp3/6p8v/6F1cSJPyWFcPy2pg3jbegIr+RGX538iyMReyz3xF3DzucY+rcwaoCBkC9NGXiFQx75yTd2FhVznuLr/LplV5orJsY9Vz5523MOw/b3shP7cXQsVTiiMiaPTa15OgT1mJ+sV8Vk82EUNUhmx0M4Jz2VRE9iBxkDYhKpUjDbvktasiklMZzrmmfPUCnMrSHhuUISiVl5jNy3YDswWqkVX55PM0nRs2kJQIS+SlesVxstJv8ohpN/nnIcwaURm7kqcpEY/f//7367h8xjOeUQlGYyoEuD5GpLO/NieRm6LxbN2U1nkkprHlt/lpXbO+uZHgbILxpT5jzpgVyWwsG9vmjzoczYl2jJ9NPT1t18Asa2De/KVZ68vYaX6B12yzrdZfdtmNlfACG8gnmOdAP7CVNQi2oysBpnr6059ej3TU14nVGyV80k5Urp4+T1dSJypPp51Jrm0cUcnm8FGD3dihfG/92NinYMzljtaE9lrbetf4yi0+kzbB9UTp2mtJ049D3Qf8pV/6paH/R+WZR0MnKs+so56ia6BrYIo00InKKeqMLsopGuCUAbQ25b/73e8Ot956a/0OzNps5ZQcKE+TrEWIA4nA/8Y3vlHJShu97vS97LLLhic/+cnDk570pPp6vbWof1bKnLeNN+MG6JjXV78G5AXQITwQk5dccskJct9v5xEX0tvIC8l2+PCI6DD3t24pG1Vbd5Y05anpLYvl+95h967F4fz9uwrRuVDm3s7y5JjXMAOVWwqgyytgTwLDgDx90kb1pU4ECztjUxEYRKzYWMwTlS3BQq7IKw+SEoGY/MpUj3TaBsyGpFSO2BKg6snGZYCvfIIyyKO+EJU29MaJSnVKo962XchLbUuMbHSi7FkOs0ZU6ltjQN8fPHiwbu5am6xHz372sytRaRwZT8hI6Y0VrwluN4I9reLJSWMwZLfNS2PDk5iPf/zjB0+HmqNnE+R3QxEZ23GWOYCQJ1eIf3Oih66BroFTNTBv/tKprd/8v9hd660bQDyxnhtBrEdere3JdbaXrZ7n4Aarf/u3f6tvBoDt3HQl0NHLXvayiq1CEMyznlaz7Xy+TlSupkaXL6sTlcvrZrIryxCVw8MFGN1T4v0legp79F/rk5W5fCp+ashJx9xc59hG/jWfO5iQjRLMLTF40xF2S8x5adUVjOc87BZMKb1zKc9va0qi36lHWWcT5JvVAK90onKy3u1E5WR66qm6BroGpkQD2WDqT1ROSYd0MU5ogGNl8xO4+vrXvz589KMfrd9tpj73uc8drrnmmrrpGqfvRMZV+MJZ5Bz+0z/90/D5z39+uOuuu+pTx+6I/uVf/uV6168NXU+K9bC8BuZt4y3jZh6JSvMQcLMJh5AwVy666KL6pJantUQbdcgSpAiwJ09AGpLDJt+hQ8iO8mTiw8De1uHI0fJHlMPuEVG5uGe48ILFEneVsneVOrxG9eQTCrCYPkgE7FogmLryBGJIvBzZnJA95A+piGgJKGUXrJvkDVHpKCpXHeqPPvIkpbJEQDekTYhKYBhAlkcMGFWX8tSFEEU+eRIByYSEYh/T1jZPSEoEqnwhU7VTmQG8y8/czX1lrYnK22+/fbjhhhvqukBTa/3qV/2p7/PaV/6aMWA8uWnHOqT//Ta/jCW2V3pPXxpDbvDxX5TeRmD8ZW21qWZMGXeeqHzOc55T0006AtRrXJHPOPM9c8A15Rrf5oE55ckicjrfQ9dA18BJDcybv3Sy5bPxLXg6/wnsyM6yz8973vNO3NzI95nn4M0A1qXvfe979UZQb6wRnvKUpwyvf/3r63oan2ie9bSabbcWd6JyNTW6fFmdqFxeN5NdWYKo3FJubjt2uMTVJSr5obAdewO78lOD2fLdeZHdzt/+BLsFt5lfcFUwp7WAHywGb6lL+kTYTTr+fYsdU06wZtK05ckb/5pOlT3uU7s+HifT/+ZK1YnKyfurE5WT66qn7BroGpgCDQRYdaJyCjqji3CKBjhYufP2y1/+8nDjjTfWu3APHDgwPP/5z6+A1v9tjTtnpxRyjj84gZxCr9f73Oc+N3zrW9+qT6ZwUl/wghfUp1jcwXW2T56cozibNtu8bbwZN8DFLBOVmW/t0feQlMAcQsLrXW3QeZpSRFL67VWwrgN6AFuAHYIDIXf//feWYyEsHwTgCpA7LM1CfapydyUqy/8YXYSQWSxPZu4qpEt5KnH76D8k3TOqD5QZkBcQ2B7Vld/5bi0kT8jEAFK/yQrIaqfypZWvJQIRleR3TRppQ9AghlKutoeoDBCmO+lFIW1gg8gZUtR8Qip5ss5TCOqXR/6UwW7KR76Qm+woIikyuh7wLP2shVkjKvWzfuenWZMcPa1jrj3hCU8YtNe4Q4IjG41XfY/M/Od//uc6NjxJiaREWBp/xpUNYq9Ud5TfzQQvfOELa7pJx4TxY6wZm+NEZTZcjGeBnG408kR1O94nraun6xqYZQ3Mm780a33JDlpn/YfwV7/61Xr0m/9z5ZVX1ptK3LzFF5jnYO2iI2sPPVnPBDfd/MZv/MbwtKc9rfpO/LEeVkcD1ulOVK6OLs9USicqz6ShM10/DVF5tBCVw8qeqIzvGewUEpI/HWzmOz86GE0a1/iwudk0uDB2ahzLWg/41c4L0ikHLhTNyRaHBpfBrmJwHPyXqEzn+dTya4Ny06Za0fEP14MlyeB78rkmzkLoROXkvdiJysl11VN2DXQNTIEGLKLu+OxE5RR0RhfhERoIUQnM3nTTTZWo9B9av/IrvzK87nWvq3fgctBWO3DmOIrAtFfOfvvb366vckJeuDP6Wc96VgXVnag8vebnbeMt42ZWicqAoRYYhSQL2LMJh4iwIefprhCUfiMvA/QAQWBNAKKsRUAYMu3++0u8rzyxeP9D5TticFt5srIAx/Lq1wsvXBwefdGecpPA3kJ6Lg579ywOOxd2FrA2eg1PwGJbXgvyfFfPUgCRPOQzz3NsX/Wj3enjlkAkM5JG2c6Tga6UB8wCuMpJpCPtB4KliT4DHsfboFzrtE1PhFWiuqJ3dfiuP+TXRjKZgzYGPcXgO5vaypk6Tz+TN9dVfYews05ce+21wzOf+cz6JB8dTbpeBMQ7ZtzTu/8svn2dn6g0vvIa17vvvvsEUamvrUHGETLTuDpQbuQxfq1f+tpTldrtph5rp/TGns0OBOWdd95ZyzMmPHF5+eWX13Rn0+PGobrIYN6p2zxRh3KdNxbZBU/NuGEhY/5s6ulpuwZmWQPz5i/NWl9ac63Rbib5yle+UvEDu+gGkE5Unuxtvghs9bd/+7fD1772tYrrXPVmgje+8Y317zXiF53M1b+tRAP8mE5UrkSDk+ftROXkulo65RJE5VDw3bHm1a/D0q9+Hffv48ennvjyfGc2hu/MP3YDKXzmGHwWjCYNbNWmS5qQlcqFu/jC/N34w37zhV1XRkt+OsdHlkb0XRkhKZ0b4eGTb8bhRzsvncCPDgb3vQ3BqsoV+eb5Lj/djOunzb9ZvneicvKe6kTl5LrqKbsGugamQAMWrnknKs+0UHMmJgkpZ7n0uZ6ylkuX62ebPvnW+jgu1+nqO1Mbk3e8zOTj7NlMRVR+5CMfOUFUev3qr/3ar9X/ikzalNUex8ttr50uXxw8r3697bbbKlGJzOfE2vxGVJ7pv7zOte5Wxs3+fd423jJuZo2oNFdEQChkWMBRwF4IOXMEUYSY9JrHlqB0LUAQ6AtRZ5wDZw89NAJ5I2LjvrI2PVji4QL8jpUnK71+Z2E4/4Ldlai8+OJ9pfy9hZQZ/WfItm3uUEV6PlyBXEhP61vAnnIRJyHqAhAD2rQBmYpQcUT4kJesaTdZpbd2yh8iRh2+q9d1+qIbYFcEgoFbUXkBuPQosBfGj7wBlcpXHrm1gz0M4WhuSau8AOzoVFnkA2qlR1SG3PRdOeQN4FXv6exVFXATfcwaUakfbTL6v0lEKdLSmDCGgHTjTJ8ac8ateWUMGTvy0gei8kAhMZGESxGV0tpQPxeiUl2ZU+axcWk8GV/GrbHqKWDzyZsIPNXZicpNNKG6qOuigXnzl1ZDqadbt9jD9QzWXOuq/1/034uwi99u2IIZ2F++Bbs9HtKOpWTOtTbPUuna6+Pflyojac62rOSb5Jh62zr4JLAVotLfetCXkFeoX3bZZSdu4FqujpS71PW2rqWuz+M5+upE5fr0fCcqV6pndrvgoi3lFdlb9pa4e/T7xKtfHyi/R0TlyMSP7Lx5P07WGfexFa6LwazwUrAT31T02/mQlHAff1p0LhguOE7aYEN+Lz+azwtn8b2tCfCVOtl9+ZQjklWeEfY9eYNfiEo+tfUjPjQfX1SHNmmLupUdGZwTXFc233wcp8rvnGuzgP06UVm7fKKPTlROpKaeqGuga2BaNGARtQjO4xOVcV7Gj+mbLPiO+Z5r40dlpJykzTHnkybnx48pczy989Iul348X37nmHz5PX5MfePnx/MlXdqR9OPpct6xvdZ+b9OkvJQvXdIam+dKVI6XmzpT9vgx1x05b5zFcyEq047xY8pP+1J/zs/icd423jJuZomozHhFqAFrgBnABRiJvgd4BcQBfDbkECQBfy2wSzr55Ae0zBdz7tCh0Stg773XXan3lycUHhx+cs/DhZgBQj3xuFBf/XrJJchQTz4iEneVu0pHRGVL0ACKnnBwFJE7ISsBtRasAY3a5tW0idoQolLb6UC6yJq6AD/lOlpTAUQh4FQ7lZ2orABLZRk3AZXKVAbZlCcCq+RnD0XfnScL+aJvIJs+9ZlyyKT9NgYRlV4FKvpubqYceieHOAth1ohKfW6TkZ/mVa76zRiyAf7EJz6xzr+Q0Y76Vh79mnQ///M/X1/p6gln49EYs7baLHY03vLqV69nPZuQ8euYceSoTOPPmFOH8W8TmgzG7jysgWejx552vjUwb/7SSno7a9X4MWXGb1lPGxP/zwazDWVH6zobzKewPlv32b6Ecfkjb46uJ8qT8475nrKWO7b5U1/SppzlyhpP3+bL96WOyZdjyndcKVGpzJSbY2Q4U3uSbt6O9NSJyvXp9U5UrlTPZyAqhweGLSeIypEd3LLl5H8/tva19UmdDyaDm+CE3JjqO/vcYlJpYT42mw2HrcTguOBY16RTF1zG53VjHh+cny0kn3pDhipfMDf56pE1WFBZfH1+Ab+e3VS2Mtk5+SOv8v1O25WhTGnhQLIEC1qb/LY+SSPtuB2tgm2Sj05UTt5RnaicXFc9ZddA18AUaMAiZsGaN6LSohxQSQeijV3nhDg0HBCOAAcgAGi82+JkcCrklzZRnjgLAa0pW7nZrI5zoSxOQ5wLeQXlcUTIMw50XY9jIz0Z1MtxihwpX9o2pL44NfI6J28cH+mdpx/R5mPqcU1a5Uc/8vvd1t+mkSchbVWuNkd2cmsrnfovNnfdfvjDH66bnf5n60xPVEYfymzljV7oMPqMbJHJkRzaeLZEpXypW73a5TedjNetfnXPcpi3jbeMm1khKs1j0XxhrwAzpJij+WkMJ7ouAnGuA2SOzmWuxZ4mjeu+O28umCcPPzx6Euy++xCVDxRw9kABWGWdutfm1JZS//byJOXu4ZLyRGWISnUgKo8eHZEjAWYBZ47IPjGEItvCPpjnsXkApCdAlyMqY3vN5fR1bJf53s75zPe0vdUD+0avQuxFbFXKcSRjIrmtIWLkJo/+ALJboK1s5crbAl3g2dN4LVlJN9JphzbRxWYPdDFLr37Vh17hiuzjqxkL5o1XtXoKxVxDPrvmlXrS2ZTQl3SB0KSPRHmVId3f//3f1/Gg7xGVz33uc2u5qzEGjCtrgDH3wx/+sNoB/0PWicrV0G4vY9Y0MG/+0rn2H1vVrpvWrmCSrLt8ivgo1tv1COytGN8gvn98KD4O+URBWmnIzlYK/ATrujTKsdYnSq+M+GNJl/JqAWMf8tCP8pWTdZ5Myop/Eh+szd7K1+o3+Ez+8SBP2z/qI596yKte4/xcnqhU9nh7WrnIk353JOfpdDMu+yz/prdOVK5PD3eicqV6bojKrftOPFG5pT5ReW8p/IFh65aHii0Z7TWZ5/Cfo8jGZN6zRca+EPsAd7oJFc7LG3/gqNhe6eQXlaVMtiu2RbpgWEe2TZpgLkSgfStHNleQR7388WBj+SJr5CQrm8ZuwnrwGYISZnNUpmtZA1IWOcinPGVkfYQj4V54ANkJA8IKylI2HKC8rFXRVRV6k3x0onLyjupE5eS66im7BroGpkADFlGLlQ2uu+66q24oWahi+N1Zb2G1AM5SyCJuAQeaLOIcAIu6oL0cAHdbcWR8jwPU6oGuOBjK4ZhY8G12i/IIqYOjwfHgBLlm85CzxMHgdAgcFDJwIoAKG5QcD2nIIT2HioPTBnm0QV36VB5OkaiulN/m8Z0e5CVX8mpP6nPkrCmTLHF2ktY1stAX/fhNJ9oor/pbB5BcCdJFbuWKfkd27fVd/3zzm9+sRKVXBE1CVKZceemS7PqGfBzGbN5G/5E9stGBvjhbolIddGksxan0m56jE3UbV/qfjmY50L8ngESvLbRZov3+K+3pT396fQXgLLU/42YWiEpzxbwwt9mzjFvgjk0xjwLQ2BdpHaV3vr3umvGf8lxTJvvg6Ld827b5D0g3anga0Ct0HijzXzxc5tSR8j+Vx0r6rWX+LBSyY+9w0fFXv6rPHbUPPzwi59gn9sTaJpr/zoXsMw6zcWeeC2TXLnYnRKU2k888JV/aSTcCG2ZuB1g6is5Lo83yRC/az6a5Jk3yxQazFSLZnGN3RXZFVJcor0Cm2Hh21ne6UK+xKI82a7+5CKhahxKBVufYK+nUlbJzrBVtsg/9NktEpb7RV3wChJ+xbQxdcsklwzOe8Yy6nuhH66NXwyI1jXdp6CIREXnppZfWcSI9ovIHP/hBnSPGqOtezYpIXI1gHFsH+UadqFwNjfYyZlkD8+YvnWtfWjetk9b2+NnsnfPWPv4Em2cd52dbF9cjWHOtz+yeNdfRb7Y1WMP34CDXrLnawb4LXs3ND9EO14yJ4CPts74rS7us+W154220hpOJLGwwXamLX8AXoZf4OjAJvbVBfdpg/RHlIzt/SD7prTEJ6hPJrX/US3Zt0a7I69y5EJXxabSBD0M36ffIRTfaQj5yqruVMbLO21G/dKJyfXq9E5Ur1XOIyrL/tHV/eXLAK2DL/hKi8ug9haT0NpmHi+0bYaAR3jz5ph+2LcG4N/9FtpLNYl/ZCH9NMr4PJ508bI0osC3siPyjuh75hhzXpWcv4Sn2nJ3zW3nysbfsEzsYDN3a79TZ2l1lBLfFfiuPbVOOdljr4FR1kMN1ZbDDcGTssPzsJtkclescGxp8Ka+4mUL2q1/ykpcMb3jDG+rfXGwm+ddT1k5Urqe2e11dA10DK9ZAQNK8EJUWY4tyiCQLtbuLAB/AyuJukeaYcCA4AZ5asBACPpyLOAOUz7FQpicG/u7v/q6WIx1nhPOgPNc4BhwCdcijHJuCNgQDTDlIHApgQh7A0m9gllPCoSKHfBws5duoFpJPHnVpAzmkJ7+0SwXykIsO5OVUabfNeq+U41hFVxyb3IkFtEZXyo2+AgoDZMmsjS1oJJt6gEyy2lRVLofMeAyApntOpfQ2Xz/1qU9V5wrJ5X8i2/+olIae9K0+Tb+qg/zpW7Iqn1NHh3Tv/7J811b61A/6SPvORFTKr27OoDFEl9qkfu0hjzYpL44y3dAvwjXgXTvpjh5nKdB/JypHY2Cz9KtxGmBmXJor2Rgzbs0VAMk1tiJzvs1njplLjrmu/bETAW3mHFvoaF4or1R/fLMPwCo3YPy/Yit+fHj4cSUqy00Uu7YUeXYMj3n0YrHPiM7Fmk/5hw97ReyImDP/zElzUGQDRLYnR3PTPBfUTRY2i71if7QzsmmPNNqkHdormv+iOS6yQ0L0IX30kPmddOqPPGTMhqBzkU15ym/LVE50SW9iCNWsT/Jom7KUSxcBvWwUe8+mB7Caq7HrbZ214k32Yb2bJaLS+mId02fsqTVTf5mXT3va0+oxa56+Nb4F4854MBaMA3pBVBov1lx5lGOsmteIzyc84Ql1/K9Gl6vTmkjuTlSuhkZ7GbOsgXnzlybtS/ZLzKYrPcXP9j1+dtZe6zT/JH42HBSfJev3pHWfTTr1k5FMcJwjG8iv4O+TJ36Ocq3x1uU82fLAbkUAAEAASURBVM4v+Lmf+7lqz+FKNt/6zIZKZz3nT7H7cCDswl9h17XZNUE58sOF9KQMNtj6nzWe/0AWvg6MRj5rgPWCjqwJZNeG4EPrinqkO3DgQM0rnZh66UB/yJMba/hOnvwnM3+KXJMSlfqdnsgdWZQdnZAxPpx66FqbYCuRfpyLz1QFncMPeuxE5fp0fCcqJ9dzbEfsiOPWrW4uKDhr22IlKrdsdSw2aXi4kJX3Dtu2PlTs0NFiq3JD/Oim+IWFk5jUeE/Z5j6bxj6weewBGwrjBc+6xn6ynbGfLQ5yLf60tUVUFnsZ2ytffHU2N3tAypGG7VOvyGaPk4upVxktZsu+kvXAee3RBvJbU4JVyURObWcTlcNms7fyKic4gQ21NoiuScfOklWMHpQ17cH67gbLTlSeuac6UXlmHfUUXQNdA1OkgQCleSEqAZ2DBw/W16NxJjnuAKDFmNPAEbFAW7Sl5YR4qhRp99SnPrU+CQb4cDhaZ+Duu+8ebr755kowAkccB46Esl3jHHAgAj45LTYFkW5PecpTKqBynTxf/epX65MRhon0QB+HC9AG9KR/8pOfXCNZBOV75Zu6vvvd71aAqXzpvMpNvqUCx0U+8fvf/37Np0xg+VnPelbVh1fDIQrpTT0BhXRl/DgHLMYZ5Cxx/rTNpqeyyEIn2i2/dn7nO9+pr51TLqdJW5VB53TBWaJnYFxffOUrX6n94r+2nv/85w+vfe1ra/u0S5lkUNbXvva12hZlkk+dZFW2MlugTi5PpNATeQFc6dK3ZyIq6YrcNgHoKJFOhDi2+o8sZFR/NozTl+RoNzBq5hn40G+dqNw8RKVxKpozxiObY04AQ8CA8W5us22AnfFtrojmlqNg/rrmqDwh18yvlG9+KytxtEk2AlpIxwfuR1A+VIBUiMpjBSAOw3nl9a8XXeipCf8XEnsBJD5U7SW7CyQCaGy5eQe0ib6358xfgbzk0EZtBib9JmPAKLm1KVFbxLRROWmnc65Jm3SuW1/YK/XGHpBTZOvJRs7oUznKiB2LXp3z3fkAZ8cAbnWpJ21WdgtY2cdsYLJfAGuANVulHWmLsjZTYF9niag0FowXfWZNss64McpYetKTnlT735ojnbZno9a41f9ufLKOK4NenLf2y2+M26jm41iH5DWOViMY3+rpROVqaLOXMesamDd/adL+zJppjYqP7ei3NcpayGZZK7OuWuv4L/AC/56vzb7BedbItQjWTessPP29732vHsnBf3rOc55T8RC/gqzkhgXYRv8T/LnPfa76A/AaG+5mlNywGgxoPecfsN9wkbbBaWx3a7fZfD6QtQKe5IM7p07+gkBPgt/wIRykHPiVjvgsyvCU/sGCq+BD/oG6c7OotaT1f6w/dGCdkscadccdd9TyXvnKV9bNZHJaFzzJr93wmjoEZOZ1111X/8eYb6M/9b01RHnw7Z133ll1Fv9HX2qX9qlbu/iu1jQ35dA7eek8ZdbK5uyDjjpRuT6d3onKyfRsfouxIaM5Deu4KdR8LXZo2/nHCcuCvbYeHbZteWDYuePIsFBuWN29282lozfzLC6O7LqyBONd8JuNYyfYLnagxZ1sbfZeYm/ibzuKKYf9SBnK8V0kd/KyQ+0NJr6zSeRQj30wNtAxdctPXunY9+A0edlc0XfrBRsnfevnw+RZV9qygi/ZcTHl5ciustX8DnUqn/wiWaw3YnRZFTGFH52onLxTOlE5ua56yq6BroEp0IBFzwI2L0QlUIjIAyJFYNCCzNmwQc2RAIws3HlVJQcA4fbsZz+7AijgkGMQx8Li/q1vfWv44Ac/WMFcnBCOiAX/YAFYHAa/LfrqpPeQcMoDOtVt49hrTm06qlfgpKiDnOoFuJ/5zGcOL3zhC4cD5a5Wjgl5AXdyfOELX6jOR8DnK17xigpopYsTl6EHuACSSEP64JSRx/9Iaa86lUdnkTub9xw1chk7uSuLs0Zu5C5iVyQvQMuxcp2TxJG//fbba7nAOD1pv6hcetImMgPO6iGf/GR70YteNFxzzTUViGoLfXK0ANlbbrnlxNOt0uuPbE5oj3I5Z2TWJ/7vEth3RxadkYGeyHAmotIGBNnpQL/Rk34wp/SvjRJRedqgX+lRUFfaAlArS92zFOi6E5XTT1SaJ6L5BtQZhwFC7KI5KBqj5oyxDfjJw16yG+ZgC+xib4x9tlI6UQD45A9gzDxR78LC6H82jh6x+XSkzKXD5f8pjww/uVc9hahcGAo43VbmdQGpewBQG1o23o4UkOf/OO6t84/dtMFnformMxl9Z49DCLIJ5CMvedhYbdR+ti6AlF7ILV2O49+jxxy1XfRbUA8dqTMykDFAkg1zPjLJS08B2Y6RwzVyiM5Zw5LWNSH9okyRDVIfGxw7aE0U2WE2EXiVjr70F5k3W9B3s0RU0r9+MKb5JdbPb3/723XNsyFrDOhPY8CTMdZf7Td2jSk+iPVJGuNbeuPAeLEO2ch1Q5E5no2X1ehzY64TlauhyV7GPGhg3vylSfvUOmRN5GfDOPxsNpA9ZOP493AHu2ZdZ3OQX2wmbIUAe8ELXlCxCBsn/VoEvgWZ3PiJIHTUpwjSK6+8svr7fCh+Btmsw+R0E+ZNN91U0yLW+B/WYnnZb+3XTjqAIfgx2spmK9ffKLD7wYzyhJT67Gc/W/VmTXRd2fwWelKOsukIpoRDrAfSSkOP2uAmFxgRbiI74heuVL908W/4DGw+jIOk/MY3vjF8/vOfrxjoTW960/C85z2vEojqnYSo1Ed8GP6J8qx5iE1yxEdr8RXdw9D0xHeDPV/+8pdX3B69xzdbi/6f5jKNt4wJPoSxpy/tbVj/8zc/09yGzSJbJyof2VOZd47wiTiOn9i0hQWYDgHoTTmehB4Rldu2uTG2vOp1x+FiC736FF5bKPN89BdHu3btrjbROGfT2CJBHcFHfFt2NNH4txY4r26yyRuM1mKg4C1pE4PHgrektwbAc+wQW80mOads+JbdsifFHrGzZJBfnWynvMFnjmw5X901dlMdZGX3lGM949Oz7cpXljaoM6SnMlIObCe6pi42noxtGvW5rk7lqJNexWkMnaicvFc6UTm5rnrKroGugSnQgIXPYjVPRKX/4nR3qFeRaTsnhrNiQ54DY5EHhKQBIi3kHAJkFhDptaM2AQX6s8B//etfH66//voK5jgknIb2laucEY6J9ECCTWHlWviVHceFk6E8MmUDUjrAD+HDYQiofNWrXlWBL9k5FfqQHH/xF39R0yrTXbJekYqEI4Ny2wAs3nbbbRVQAqMcMMSd9O6uBZY/+clPVl2QTZlADV1Jy6GhR7qyeUAO7QZ46QsRB8xyJOhWeu0AOG+99dbhYNlA5UjSF6CkfM6fcugIUOdUkYPeOGIAtVc8vPrVr6710GHGMVD9xS9+8YSupOfIceq0HYBVpnpteHAOkan05E5msuonstL1pEQlHShP/WQ1hjihicaEftUmaTixnEAg8Vd/9VcrKdxuNLR9tJm/03UnKqebqDQ2ze0RSFw4sekH/JgL5rrou3lqA8i8CrALqANozENzzPhPucoWpHM+1+Q3z5RnfporbJmyzVX5S1Fl/nuV2LHhnhKRlwsLW0q6YmPP89ofYHf0+tVDh07ejcqui2wHmczlRL/JGhDmN5kE8mgjWchEHufYJPKKAdf5TdZE10RtJn90UAsvH3RADnWqn3zsAlmB0gBS8ihDueqnE0c217nU0coS0EyuVufj7VWvetRpbbHWhahkn3wHXNlgeacZoEav40f9N2tEpTYaLzY/rPU2w/ShdV3Qr75rt81ma64xIY9XC9roZY+NI+ND3xpX/Anps0GdsVsLXeEHmYwxY6q/+nWFyuzZZ14D8+YvTdqh1kN+RYhKeCOEVHwH67b1kL2Ln+3oN98asebmS0+gwxlrEdg7feiGyb/+67+uBB/7BxNdffXVlVBkl/kV1lXrLP9Y2j/90z+t7ctTjWw3W82mJ/JbYEF6gIf4aIg4BOBll11Wbb5yrRFumIQH3bjJx4DF4A318yHohbzWAZvnsA8M5jqdWgf4BtriyUekI2xDFnW5URQRSOf8HIFvw2+AmxGbX/7yl2v9+ubNb37z8OIXv7iSttJNSlTqd32tPdYwspAd+av9/EcyaId1RjqEHB/HWvjSl7609nuwXSvvWoyBaS3TuOhE5fr0TicqT9VzsJC5B5+wPyH7HINddu503pOPyMTFknZveeXrecUW+e58ISd3Hys2aGuJ/mPXG3WCi0Y3zrItbIZozLNj6kw9Oca+trhKWmtN1pvgNa3RhracyBx/WZ5gO345Wxmikp2Vn7/NFgZbwynOuca2s9PWD9HawIY5ry2Ri0z0aA2RHy63nilXW5QVOxycp1w2M2UpT5rUKZ00Ihwqhrx0jfyR4dSenY5f1qz+6tfJ+qITlZPpqafqGugamBINcO4tqoAPAg+ws7jH8OcuO4vyLATAixNp485ibNG3uFv0LfTaySEALgEp5B1STTqEFpISMAR6OCgWb/pz9+y73/3uCuYs/oAfEAcYAqnIMuXTN10D2vQNdKoPMMwGo41DJKU+4BhxHsjiTlJAwzmyICCRgcrmnHCKgMkPfehDFdAlnVekJp12CnGqgNBPf/rTNR/yjtMD0JMdAETUffzjH6/1AvvaDfCSkbMZQtHreNy5SwZAm9OA8HQncwg4MmqvJzvIqT10p93yIDcRIuRWbjY3AU9yAPTkRxZfccUVpxCV+oGeAGRPXurnOHOcOPmUyzmTTru9akkd9OcOYWBf28irP6SdhKjUp+YNEO0on7qUoV7AXtu1Sfu1x7hC7JIN4M/d3tovrTgLYd423jKvvvSlLw1ve9vb6tPIHH1jZFqCsRVwZY74HuBovLJdAVLsgegc+xWSEtiTV1nGu/kHzIjsn3OCsqUT6IYdFQMg2VvzQ7mnkoPK31HybC1llleklb/cK2ocjhQ+0ROVi4vlVdh7txTwCUCOnlJ88EH/Y+S1OffUOW6eB2ClXkey6g9yOorkJZ95G3kyd60P5HRNW9qj82LALz2K0U10rf3arH51qZM9CEBkB61HAZPSpk/IA8yK6kn50W1kIleielNf2uiozdqvbnWp11oHVLOF7JPoN3mkk55ulCduhjCrRKW+0G/6TD8ZP/pfvxhXxoj1zPylA+NBn0tvbTQnjBtBnxrb0prfot+rGcw/dWctN36tsfwbchinPXQNdA2MNDBv/tKk/c5WsW/xs9k+tpDts06LfAg2hc1hb0JswVls4VVXXVX9bNiE/WF7Vtv+xN7BAreXN8Y4wkRw9Gte85qKw9RNXjZbfx8suEva97///fUmRgSc9AcOHKjYCNaKbWbL6QB28KSiMrxdxl9hwFuwFB1IA5PBWEhQenrZy15WcZn62Xmyxh/w27oRXy+b53CUNigL6Ykw5I/4X+TXve51Fd8pO3rUR3wGN6XAzX/zN39TcSH53/KWtwxXFNwGD0t3NkSlNQRuTuRHaYdyyUqGjI88bat8sroBVYQb6UfarIGTjr9ZSGesdKJyfXqyE5UjYi821pzji8JKbI05a26eirEWCr4ZEZWeqNy507z25PmeQla6edXTkF71Wv6fcQ+7780/sNcIm/lfS+tEcI4jm0AG9QerOcJQkYdPGvwmnZD1JrjR3FFO8sgfHJYR1WI7fjbbyb7DUWRhc7Qdjma3+N3WLPKoj18vbchN2Ix9FtIG36VVFpnlV55ov0lZZJUv2JIswZX04XqC39JaC6SRFkFJbjY38js/jgVTxjQcs1/d/6PyzL3Ricoz66in6BroGpgiDVhALYjzQlRacIE4izinY9zx4ABYyC3aHA1PySHqbPIBWO7IBTgRhRwFzokyAcL3vve9FczJa+EEHKX3Wh5PK3Bs6Fu57gr9zGc+U+86zV2qyMG80tVdv5w4jhNnguP7V3/1VzU9sAhwuUNXHchQQFzZQLlXCAGJnB4yu6MUSPNKV0BUSL8DdYhKd79yTDwtCYA6cpiU5zpHBhjWHiAVeOY8kY0DkydK3eWLcNRmxF/IRw6aOhGE2k2vyGLOlTYoM2mVaxNCH9ls8OpZxCbATIfawiHRD9oUOchoLHOuyNU6oPpZOn3rGjk8KYowVBfQC8ySQ9/Sk3RnIioRi8YAXds84fTpN3VnfOlDdUtHpwhXr3r62Mc+VuvQjzYbPNVpk0L6WQHS+qI/UTk9RKVxaHwZm+YSYGOsmp/GLTti7CcCVM4BRICldAF6yhFacGgOsgfmuvOpL3bVOfNANBeVoUxlq4c9EP0GVstLYsv83Fbsz5bhvvtLXQVjLezcUu6qHYq85TVAOxFoNjGRb4eq3WY3Es3H1Je6zWvRvCenNPmuPfRCJ/QBUKfNdEbe6M9vunA9UT7nXNNm7W/nMllSF12R0/rBfrBd1g7X5UvZdBHdK598kaM9Rr7UG1unvNTZfle/+tTL9luHrI02s2z0WqfI1fYnHSp32oOxZI10Y9G1115b11UbA9mAnUT+tNNRf4h0Yk26vWws33DDDXV9VFb+W8sTI9YPm776Jvmk8X0lgRyiMWTc6j/flZtr6swYNE5c02fS60fjPnLII33mc8bVSmQcz0tGY6sTleOa6b+7Bh6pgXnzlx6pgaXPxO61frY1ka3LuufItrGJNluRZW4g/fCHP1xtJaKOnx3CKmv00jWe29nYO7gCbnHk81uLvAEGvoMN+RbalP5GOl5f3sjjBlbYChZ086InF2ExNzRa19lxbYO3PIEJc1h74DJtgwXZc+dhUrjJjaHWvje+8Y0V41ibrIP0ZG0QlS2fSKdZu/gEsBJ82BKVyF43ylrrkpbGrC/WGfsJyM0Qldbjt771rcMVhaiEhaWblKhMmfGP6FiddMjvSv30SV7rM3m1n3/lxlN61/9utLXOtT7ZufX05stFP52oXJ9+m3eikh02x8xNdpZNMVfZbFgSxgu55vvu3SNb7olKf+WxfTuMg3RbGB4+Agf6v8qFgvt2DOftLzeB790xLO4pb+QpxObIjvOBR/anJd7gndg36WLjfBfZgvjL5HMu/rF8bKRo7jifPNoVu2NEJW3wFUzHtluvYCxlSK8OtjDR79h19o1t5y/7zs6pNzaZnNKqSyCL/OxgsKo6yCCvMsihfjphR4X0i7TKdz552Esyk926RRbR79hfaaOTWuAUfHSicvJO6ETl5LrqKbsGugamQAOAjwVoXojKtNcCbZHnBFi4BedylM6iDej9yZ/8SQU/wB4yyROKAGTAZktUIggF4NKmJRIOaYcIEyzyHAdE3Y033liBNFDJcQGg/e8kYtEThhyRyMbx/cQnPlEJLq8OBVw9+YgMBFSBT0E6BBy59SnHMK+sJQviUiAz0ALQeTWQMjlAAPJ1111X5QX4gFNPHnLS1Ee+EJXKiYMGFL797W+v5SBZpSEbYEhv2kKfQLMNXgSpc9p5RQGv9ApkI0ZSLl0BswC/17ki95xT9uWXX37iPyrJRg7XOF2cKHqLY6df07fK9t2GAAIWmLaJylmmJ4CWLBwfZZ2JqKR3dXMEtU9QlnGVelO3o7mGoNWm97znPdUpfOUrX1nbY7PBndRrsWFcBduAj2zEICuR2BxoY938MC9sysxSMBaMm2l7otIcCdAyNluAE6CTuzORlOyLuRiC0lwSzVkRyBEz97S5BYgBRsa89Oaj6HfmqHkamcgTAHveeZ5E9npZ/92xswDVcoPHoa3DfQ8Ao1uGXeXh/rLPVmybpw5ttiH/EDGIt9FTimysDTM2QT0BVo6xWeRg50XjMkQOGc1BMok29eisbbe2+00nSRMy0e/M4ehIO5Wb8ZE62Qy2mF0AEMlBrtgvZSk3ZatP2eOySJ+6fBfUp72xi+3RdzLoJ7pSN0Bqo896hLB0QwywGrAtPdm0QdnTHNiYWSQq6dxY2izBeDauOlG5WXqsy7mRGpg3f2lSXVtvROuV9VI4nZ9tXWVzPE3oTTduMIHH4Be45EB5WtGabs1czRB7h9wLUcn+5cbSYItgx/R3iErkqpsvYZErCi5CCMJs2ipYt/kpsNC73vWuiktdlw4mkpcfZe2WBn4LUfnrv/7rte18butj/LKsJ1nT/c65EJVuWIUp80Ql/IuohBfjd5CPfxCiErZEVLoxVX2/+7u/W28wPRuikhx8GOWKZOTfxN9xbOWma5gNDocZ+THqhhm9uQaxGt+MvPMU6KkTlevT4/NIVMZmOLIJLT5i79glczFPFMKYfrNt8I0bU0fYJlhxa5nz24rNL2/WOTYiLj1BuX//jpLHm2+QjAWHlicpBbiEfQyWswYET7lOHuW3WC6/g+OyJmiD+aLM4J20S9tam+c6GxWsRQaYKsQe3KQsdcFzwdtwpXLkZzORlInkVp480tOP9NoQedg+v5WZNqlH3rZ+a6ayyJ888qlbUB6ZpSMHvQUPsqcwYGLbJmWqbxpCJyon74VOVE6uq56ya6BrYAo0YBG1+MwLUWlRbgGPhdaCK7oWJyAOD8Dz53/+5xUQ2rgHeNwZCxhysiz+gHNe/cpBRV55vdmVV1554lU7nDRB+epCkCFAgT9OAfAmvaf6QjwqWxQQiTfffHOtx126nJcQocBvCB9kEHB4xx131Feg6l93lAK+/gsRcSgEgGofUgWo50BqHwDqOxkRkMA+BweB6qlKaVKf9tCVdiDeEICAMwCLGPVKVU4Ex8g4I9sf/uEfVjBJFrLbPAAgpdMuQb+I5KRToPtTn/pU/Y0IJUf+I4WOkj79lzLavk0ax4PldUs2EjzZGkIXcYaoRBhqn3FyJqKSzMpT73LjKtcdjRVgn87f9773VcdUv9OBevUVx5O+ZiFkI6YTlRv36lfzw/wFTIytAEY3ELBpIrBo7jnmd8gxYzHARjkpyzH2yTxrwQ6wJJoXbV7pnZPefPFdkEZ9ZNu/P09w7i3nd5c0hag8XF4H/aBNqXJvbUNUbt/OZo9ItxCVISkBL3WoSz3tPPTbtVZmcxNQc46cwGsArGP0kPb7DSDSKdlbQCk9kEnnAYTambazywGU6mUbRefIK0/qDkhVT8pVdqvXti/SJ9qrrOhae6MPbfS9lSPglM1lo6wJvtvkBaDJGZ1Gn9o0jcHaPGtE5TTq+UwyGc+dqDyTlvr1roGRBubNXzqbfs/6bd3y3ZonZE3LdUc+ALvjPxK96cYa5glFuAFR6abIbPyejQxnSht7d65EpXUWHiAnLOKGPj4ZP0Owllu7kY/wAywIq8FSMATc5S0vykFSwk1wDn/BDZHwpbRwC7+CP0GPS/kP6gtO3Ciiklxtv/ouxI+iD9/5PNJqN8zmRljtd9OVtsPj3qyAYI1fVguaow+660Tl+nT4PBGVwRuZg+ZhcBFMFHIS3kRO5shuBXfmZlC4Z+tWGNENrUcLHjpW3qTjzV1uikX07Rj2n7d92FuJSjeKFsKt1CewA2wjDBeizncYx9hXdjBbbIAjWYO1gq+0IUFeUfvSxrTZ+dgi65D6g6mCJV1Xd3QCx/nunGvSw1bWKxgQUciukY1e6AkuznoVechHTuVEHvhM/kS/I7u2KzNtl1f9wYHSii0uZU/JZf1ssaDytVV+5W90sJ7Zk+2vfj1zT3Si8sw66im6BroGpkgDFkmL47wQlVG9xdmCbMGNcxCnJgs33XA4vXIV4LEYIrMQfhZFG/oWe04GQOjOXSDpQLlbF8j0X5aAIyeDY9IGJOAHPvCBSiiqVx7lhqzinLTBk4W33XZbBV9AGIcK4EYGyoPoFGwuI98QkICa3xwccnv1D6DGsfEqW+SqJzuVp60hNL0ih4NJRu269dZbqzMDPKc+ZCTHh67oD1GpPQgp4BowlNYdvohbeuLwuHP4937v92r95PdqPhF41mZOXBuU7QlEhKlX2vruqUNl547eOGnyxfHSt/olkQNG1sQ8TUqvHDDONDJXBObPhqiMvBw29RhTqVffOsepU7ffNqMAf0++Oo8AtomSvtdfnMlZCPO28ZbxNy1PVAZYBSQBhoCipybNyzw9GcAYws0YDJhSRuZYvjsGIPlu7ANX7Ii5JxrrziVt0hvX0ZM5IQ37wKap3xOVe/eOnqrcudN/lOwaDj+8fXjwkFfVeKLyWLFpx0paBKeNqhHh5mnKEVk5etUNWZQfMEXGROfUa/6RFbDNk4N+C+YgmUS6aIlBbRGlYbPoi81cjqykAyF6Ui+7EEDNzonOkU3Zyk3Zyvc9sqg38ig75ddKjtejLmWlrdqbGDtIjvRZZAkwRVaGsARUnWfX6Ed+5U9r6ETldPSMsWLc8EM8LWT89v+onI6+6VJMnwbmzV86lx6w7lizWj/b2uWcaI3je1hPYSFvovGbjw1zhKjka6y2nx17d7ZE5e23316JR/n99YanI92w6k088cPoKms5bObNNP4Ow/UDBT9qlzywGT/CNbjJG3GMK1jMTZ5IWmn4f3xBa2VIS35HG6aBqCSPPhfjN/FD+GvGgLbqc8E5exlwuDcC2Wi35sDrb3jDG+pRn4+3s2ae8Q/660Tl+nTyvBCVwR7wW4gw8w1Wse8FVyInRTdQwJtsTsg3eAm2kWc0LxGEsKS3VNkvOTrccy9777ybN7eXvF796sbWcqNoQ1SyAeyDtYBdCJ6LfSBrZAx+cmQ/g7McW3trtJg3CWlvfrvWYqrgqaxFwXLq1UZlq5O+5JMuuIuNJrdzrtML/eXGYbLJK7Qy+Z2yrHPjdlHdYupm+7RDGXCc+iJ3cKFzymJn2VD4z9Pp1gNRHa7Lp43j8pBpPUMnKifXdicqJ9dVT9k10DUwBRqwIAF080RUWlQ5BzbPOO420BCRWYCBRZFubNQCPPIg07ye1Z2pNts4XxwKi3lLVCLqEF4IP0QlZ4OT0AavxQlRSRZ5EJXKRxiOE5UHyxOASD7kIwDKYZEWqSaGqORAaI/yPQlJdvJ5YvF3fud3KhnG4VGeV58iNLWdU+lJSU84kp1T5SlOhIv/UuSoaHNeT0sXnExOIPJQOv+ZQoch+5CJXu2q/RwputSGd7zjHdXp8UQocK1M4HlcT3RubIao9F8zCFa6ClHpiU1OV4I+4+yRw5iWVx/bLHWec0VmTqFrylcP8pMsykUYni1RqQxtVK56ySlqcxxQ9RpX+pvOPX2qn40Td0PTm77noMYhTbs261Hb+39Ubsx/VJoX7JOxZK4CPMAip94GmCPgaAy2gNHcB2zkFQJEjHExYM31ROkyB8xB4zxAxnkBQEp655QLGAE7ua7uvXsRfnsrWblz555S367y+tedlagcjhVC8zhRuae8+tUTlUfKE5WHy6tf779/9OoadZtryjUnI7861Dtet/RsA1tgbvotDb2JZHKM/NGrYwAgmxqSly31PZt/0mRjrK2bjKlX3ey0c+SVR5lsQSI5EsnT9lHalqN6hLTdMZFOxIDSgNRWD+at1/2wXwCqtdJ358gaHdVKpvCjE5XT0SnGSScqp6MvuhTTr4F585fOtkeyfto8jY/taH2iu6yh7A7/w9oFA/Fv4A1kJR//QCH2nLOOrmaIvVsJUekGzBCVcAjsGP/BGm7ddqPjhz70ofrWHD5Vbt6E0bRNu+hEOkTl9773vWqH+Sx8Pm8bCHEJo8GP8Cxfow1wlLZs5BOV5Em79S+ZYLeDBcPqX2OB38Z3cj03VRkPfCR+LozsL00QlvHl2nbOw3dzpxOV69PT80RUsk3mFLzCpvK9RTgItoQ72RYEZUhKez1sDTwTAs1cZZ8KSqv45ARReU+5AfyhQqwVonLXMkSlsQ3TwJ5sATtgL8yRTWYzpWEr1UNeR3WTgSxk1wbnIos8ie3IGck5wpNsk7rhqGAp9TkvnbrEtlzp2StrFBlhKnbMOfVJ2+rPd7IpRxivv223crRZHdornzbKq6+UH13Jp05yk9c1wXfnyEc2uK/FgXz6cbKyZtygj05UTq74TlROrquesmuga2AKNGChshDNC1FpUbaQW3S9JlW7kUZAjUXZAi8NvViogR5P3VnwATsg1381nomo9ERliEpOG8ejDcsRlUgyRNw4UYnsQfJ5chFRyZGSNmSlO2QF8muLNJ7ARETmKcff/u3frk8vKhsJ6QlFdxxrp/xAPLIScOXQ0I1XJ3n17cECCoFmOkCokZFjR1fAj/qQlRwopBtgqCzlcpKko3NteOc731nB5hVXXFFfcaS8A8c3Dlo9cZqMzUmIyjhWHCjtRdbKB9RyqDiv2pm+zRhwPkA/mxhnQ1S6K1ndCAdjRZ3u5nUkC/nbMaV+44xcdMpx70RlOzM293djAUjZyCcqzUHRuDb3EGbAorGKoLzkkkvq5o3fzodYM/cCquQVtAeoEX0PkHFNGpGtSJ1pP/tp3Du2IDFyOQYwmZfyOaf+xcXdJY7+02REVO6uROWhQ9sLWC2vZEVU7jpW7I8nKkevflWGpylHcfS6H+fIrZ5W7tQbsGY+Btz6Lp800QW9sLfRibYnOOdawC5dA+F0KgLAAYhps7LphC1QL/vETojOpW55RWUCmwGc6afonSzytDHncsw1ehbTp+QQ9VP6LOSpdYRNs+FpjRSRlc6RWz5lTWPoROV09Aob0InK6eiLLsX0a4DNnacbuybtkaxf1h2bpq2fbT1ays9me/jZ8BvMs1mIyte+9rWnEJV8C2u9YL217sJtbtxEQApwFszljTnwG9+DLwNjuLkV3kQ40h0/xHV4DsEpfSKfkJ+RjXr6OxeiEnmsTn/14YZY6/G5/kelvtcW6wisqT+VT7b0e/xMvot08B0CgO+lnXB7Jyo7UVknyzp8zDpRyYaI7AQ8EtzDzrZkpHkPDznCmvkdTBMso5zYnEcQleWJykOH4EVPVG4rtnxbfaJy9251j25+ZSOC54LlQlSyHexDsApbKrINLW6znwVvkSnYKljJUR1C2u4oJA3cmEiWVj/qSvukJw85RTaLrRLZdpiSXPRGnyLZnFOGEBmUJQ/cmHK0W/nSqJeu5fVdSPp2X0o50gfPS6cN0pArPnxuWGV7nQtmDWaWbyNCJyon13onKifXVU/ZNdA1MAUasKBZbOaFqLTQetUp8OWJtgA3ToFNfEcLNqfE4mvTAFkH+ACDCMirrrqqAsLTPVG5FkSl/xpBVAKnHJaliErOBbkRkYhKT3p6TRDn0atvgFntPFgA7J/92Z/VV9sCrZ78RJgBuu645bjYmJb/xhtvrMCYTjhMgF/0pD7OlmscPAQmwk85ng7knHL6pAlR6T8qOTpXFKJS2hCf5FBGgjKNTeB0/NWvXhebV79Kl3EMINCT9nOyOGieIgshQ2/aZnNDWmVz7LQZ6RviVxs5dEjHEL7mCBnVTWavWZJP3a55ja7/jEGSchw5mOoVOcBC5DT28t+f/pOGHtTdn6hM72/OY4DDRhGVATDmnLlkvJr7NqAQlMa1+W/cGp+u25iSVgRm5A0ppz3muOi7ueYotMBG+jZPwJDxzh7JL68gnbxCys81dZMnBJ1Xv5Z/pSz5d5b/qATSyv+fLBwnKssTlTsQlcfYPK+qGd0EEGCq3sic8h1F58lIPnZCHkDPb9fJSBcim5G2Ja/8opC0bA37yDaK9Ask0qsysq4oI/phF0NUOrIbgnqVJb9IH+mnyERXgvIiV37XC2MfbTqy6xNyJGq7+umDLtheYDRPVbKVISuth9LRsXKnLdgQ6f9RufG9Yoxkk8O6aI64yYsNau3AxkvaJega2HgNdKJy6T7Ieo2gip8Nm1m3rbV87DwRaJ21Lllb+dnwgDXKDZS5GfEJ5Q0u1ldr6WqG2Dv4Ur3wAPtnLbr66qvrG2tsqqqbTOnv25tXv64GUcn3sK7zKazb9Mb++p6bjdRNr3xARCXdePNOcDD7DCdpC9LxjjvuqHn5I/Bd8Bc/JP6cOulgnKjUR+dCVOobfUlmcuRvTfgmfNqQIvTJzzIe4ExEthuRyaJ93hTUX/3aicrVnOunK2vWicpgHpgkWAemNB9F/nfwC+wSPDeOZx6Jadwgi3RE5tlb8upXr4EdEZU7F7YO+/dtK2VvK2UWjLujYMlip9hSeAaGCZZjI+AY6wBcwzaNYzb1k1NkJ8jJjmhfymR/2rzBx7F54+mkF6SzvvB5gyHVH8xJLrImktF1dbdYki7pTVmRq1ZQPqTXbnmVJ2ozeVO3vNpEXumlVae0ZHHONWnkIauQcunRWsEG27dL9BsOVJ8ypaeLjQidqJxc652onFxXPWXXQNfAFGjAQmUhmnWi0gIqIp68Cgd5B0xZpG3eI9guvfTS+qSRxVqw8AKa/uMEyOOIeXXMq171qnqcRqIyQ4q87mZFmPgvSk6TP5r2lCMyDJBzR650XvsDyHlS1KtUOW+cD9eA08985jOV1OP8cGI4dcqL88Z54pz6/xNA1/9hHihPSNKPsgROEcfGE5Vvf/vb61M6L3rRi+oTlQg6GwccXg5agv4yNvWTNpAX0CZ/iEryJp1rNjE+9alPVZCqPOSyjVF3DkceThkwK63XQsmnTXntLTIXqaN940SlMtWNiFauDQF3nCGPP/vZz9ajtkrn6VPjyiaFdPRkvtk8ULf2SHvFccKWHqTnLGcMRheb9ZiNGJtK2TjgeGuncWLuzVJgT4ybjSQqW6BjXtKxaJ63JKUxb74FnDia1wFV+sXc0iax/e6aeSTKo05HvwW2E2ATjXk6CZBJmhyVKwrbthUisgAmcu3evVhk2VWuLZT8OypR6T8qK1FZ/qNyT+EwK1FZ/qNSPTal8kQl4ORc5FZ26kk7yEQ2aUd5T/6nJh0EuKVdbT5tEZUvSENmoND4Fm2e+u18ypBWOanb/A9RCfSRhV7UrX8S2QTlOE825dG5kDZGlnqy+Wj1nPTSitpAFjF9RYaAfTK5mccNPuxknqy0gcnu0VvKaqrc8K/034nKDe+GOj46Ubnx/dAl2BwamDd/adJesT5Zk7y5xRN6/G3YwPqIZONPtjdGSm998kYZfrY19vnPf379e4f8R6W8q+1nWw/Zu40mKmEdfkb8IroiF9wD63sqkYzWc1iPfwiPuQHTTas2fvkZaYtXv3oykx/AD4GDr7322nqzJj8kPkb6SR0wNgwKF1mPz4WoVB75DpYba92kCzcpl/8Dp8HsfFrt5WsZI/Ai3GaM8FPIh1h94xvf2F/9Wnw4+oPFzAk6g73NHxhZP/Wwcg3MKlFpnpt77CZMYrzAmPbG8h+U5mJIP7ZCWrgl0blE+0P5PsKfI1ty9Cjb5b+Gjw4/Of5E5dGjBRvu2FL2VrYW7OeVpt5mU54CLDJBj3BI7B1cxf47sgkwTYtDtYN9U7+9GvJaD7L3wmYEG8mrXLaITdX+RCNlKVyo/LR31K7RK1eVEazZytWWn7xkIRP5yEledQV7qjsYLu1WtigdvSrDHCeDEPusbrqRT4guyKxtQmSlRzjQWgALivbyrCW+u6Y8ZZFnI0InKifXeicqJ9dVT9k10DUwBRqweM8DUWkBtfACXB/84Afr0cLKuQJigDP/u8jhsmgL8riTNE8echqQU69+9as3hKic5NWvGVKcCIAEuEOa6GNgBKjXThsit9xySz0i1Dyd+dKXvnQ4UAhGTg5ASleePnUXq/w23hFunFOOHOckDhG9BewgB6VtnR5pOTra8Pu///sVTCLmkH42EMgkT4jNtCMbN57s9F+ZgDVgZbPBHb2AKseN4wSYAtO3l7uTgTDEqb7VNrJx2MhEdmBWmdoG1DovLQLSa4ImJSqNCWBYvZ48RcgB/eoG5h39js44vRw7T+m++93vro5eXknVicr0+uY9xs5sJFHJfhnrISlt4CEpjUPz1BwOeJHWfAjwytE5cTyY72KCNAGvyeO3NOytyM6K5in9JL90gmO+q3/HjvIfImU+LuxYKDLsLHl2Dg8d3j7c/0C5y3Xw6tehADBE5eg/Ko8dG9WRV78CYAGX6hRSp6MYsEeugFhH1+gEsAP0fA9wI7t8aZej3/Joe4ChuS6yZ+xD7GDamPqtvWRlW9krkQzSyRPwrAyAU/nkSp9JpywxevU9IfXl2OpB+rSnbROZ6CRgmmxssLtorSnsL8KSvWXPbSLKEz2k7o0+0n8nKje6F0avojdO+BNufDKG+xOVG98vXYLp1ED83Xm5sWvSXrAe8ZvhGXiMzw67wQL8dvgB7oifbW22wcrH52dbu2CN4A35rKvZwD2dHFlT23V0ufTkZO/Wk6gkF/8Occe2whzIAiFrPH1Yp+mEfMYX7ENOuuRTwMCISnjsQMGBfBA6dzMnHAzjuMFVfbDNm970pqr7Vi98B1gPEaavEJW33nprledciEpl6Ts3DLtZFR61jiAn4XA33oYYgR35U65r1ze/+c2K87TdDbSdqOxPVC43b1f7/KwSlewEH47thCPhSU82i+yxucgGZ6+FfW0xZb47z74oy7x1FEd4C67ZUjBFeeXzg+Vm9UJUFrM6HD2CXCyvrS5/+VHgUEk/+h37w07HzgW/sB++i8F5bJS0ZFF3S1T6nTUBpmEzk9/35Es74D7nxBYDua49ykp5kY1MInmcE5bLD/fBgXSlneqQL20I7lOOc2R0JJc+CNkZGVxjIyOD8qRVvrbTv3qUqyzpYDzrBixoPQgehAX59chK18mlvI0InaicXOudqJxcVz1l10DXwBRowGJkAZr1JyotoNoKPAGuQBpnC6jzSiBgF5nF0bJwW9At0MDODTfcUIEPh2GzEJWcB68/BTCRYvpX0D6gXvvoQNAmZB6ACvDSE7DndavuYE5ed1wiO5TB8eHECBwim/LAI2eVA8tB4vDEieTE2Ij/4he/OHj1K3BM956qo3+vUVU2HQv6i4xAKtAJpLozF8hGTnpNkVckIVmlJSMgTWbtdS6vxkVqImvIHCfsYLlDF2mKzHV3MUcN6AXUz+aJSu3ksGnX9ddfX78rB+GpfrqmE+k4oxw6zp3XvhqH2oOopHv1GoPSxrGsytjEH/O28cbBBxo2iqhku4wdYNF8MmfdnGATjzOfuWm8S2s+GJeOfostCMu5pG2HYvLk2KaVLptk5jGdiAFWqVPe5Hf0ROW27eWpzm3+s6TM12FHybd9OPRQeX10ISrLjbYFABagWv+j8virX48/Udm++hW4ZMfUKagvR9/ZB9F16cgoCvQX4Ow7fQjj7ZFeGc5LIy1bCPjS/zghXAspH6lffjY0INA6TBZ6UFZLVAKR+mwE5E/+J6iyxFavfkev6vS9DcnjGD04RgeO5KBDayBwyk6xWyErfUdesunSRRcpu61vI77TfycqN0Lzp9ZpbGTsdKLyVN30X10D4xqYN39pvP3L/ba5ar1BfPGb+fswA/zgBj84gH8D01mDpHcDIb9ceuuVGxBhAb52iErrqfRZi62D1nLnrcHZuG3X0+VkdD72bqOISjhW2/ge2qRtrc/A3+FzuFET6Qf/wLjWeT6it8q8/OUvrxiLD8QngY9gRaQv3KSNdP+bv/mbFefQE/3wQZTN3nvlLiyWyBd961vfWt/qA2OSI+VKQx5Buf5LUjvIrf6QqnxqpKny9TeiFF7T5yEF+CpwL3mVa5zQgXI7UdmJyjrI1uFjVojKYAdH+A4GgUvsAdnXYHPFvIaZ320+Bq+wpcGO42W5pjxpxczhHTvgrYL1ysN5Dx5CVLKrSMjy5F+5d3ax3KC6q+C/krXYCHLBNyOMw34H09lzYqtyZJv8hm2kI5d6tSdYy292RwgOCrEnb9aHrBGOaRc7I+Sa9ihLPWxjMJXyyEROQRplyC9KKw+7qgyY0nfXlJH80okCudoovXz6QvRbUKd2JCqTvK5nT0CZbTp6E9VrnYD5rK1uWIUHrcvsrjTkSztqhev0YQy6ecZb47zi240pPSytgU5ULq2XfrZroGtgSjVgYZkXopJzAGz9wR/8QQVpHC2AyH8zWuQ8RcdhESy8QA6w40k+d4hKDxhv1Ktfz+aJSpvfCDSAGYEHlAKZnBDkBYdMv3vqCqmmXXTgDjl6khbh5ylFeuBMIRZDViLTODMcKt/jFNmkF53jALkucPo4OWR5//vfX4EkGYBqrxwCkL3GVf0cHY6UTRuEptfPyoesVA5ZL7/88uGaa66pRCA5gFmbGNJIr24EqI0JBCAQTpbI4Q7dm2++uRK5nC0OdohN6QFr5aq//Y9KbWtf/aoNQLb/o7Eh4kkjJGru3kYSGVPkMddcB7ZtoHzkIx+pzt0VV1xRZe1PVNahsqk/4uRvBFEJ7BhnAId5ba6aX3nawDmgJeCJos01MjvKL5onyhGlzXdH19p0fudcvjsmKBuAMpcc/U590kTmlIGoHJWjrvK6nKPbypzdWgDr1uE+T1RWotJ/VA7Hn6j0ROOIZAtRCVAFiKlX2wTHxFYuNkE6kTz0ByCa274HSCZPmz5tIrO07GCAb+wgEEh3CWRQlnLISd6AQOeEAE3liQHxyon+x8tL23JeOqE9SiMkbdqkHWkXOyVaB8hlnWCLgVP2i710BFCdZ9e1Q/7oI/XUyjbgoxOVG6D0Jao0LjpRuYRi+qmugSU00InKJZRSTlljkI387He+8511k5SfjnxEPMImyDnrpnXI2oSwggm8+tUaJp0IE/CL+ELWU+s+vMePt9maNVx50ljX2/V7aQlHZ2PvWqLSTaNumvEEIJzTEqrp79tX4T8qYQ43nR44cKD6LtpkfeeP8GXoRrBG0w8c4tW4dGQ9h73cZAnferpSHus//Ac/woNwk36g79/6rd+q+qcnPhKfgY8QYhF29lYeGIq+EZVXFKyTt9VMQlTSj35EqPKp5dFHcPub3/zmSlRqV3xOvombc70iFmbULv1J7/0/KjtROZqla/85C0RlcIa5xf6Z4+YSHImYtEfi5nT2zDk+t+v2ftjMYCf5ldViDj3gnHKDtRwXyutydpQ36sB+XvV66KGh3Mg5erLy8OHy5potR4fFXUeHhVOIypOYVB3BctaB4Bg2I5GNDtZTZ/Aa2f3WTuXIL23wmfzytfqQ1u+0MXpik7TfeUFZqd/Rb3XIz86mTufYZ0FZyshemrrJYy1ThnTSC45sfX6r39qVqF1kkYZO1B8Z1CO212E/dTlKn6he+rCm2V9kb60HfHxyRbetLFXANf7oROXkCu5E5eS66im7BroGpkADFiALDDACVFh4LHYx/Da7OSAWzM0cLOoWZkTlO97xjgpmACzkmLtwQlRa2DkEAA5n0xOY/tMSsAM2kXqAHKCUpy+Rgsr94z/+4wrKPBXnTk8EKMC3lP6AqQ984AO1fAu/PMpFlvlOtjYAfyEqkYccEXUgt4B1RFwbOBiAqnwBbYAeIIlw5ViSH5GmDHecApM2140JxCygSU7glO48lWg8qEs6gYPDCTI+yMTZ83SlNtNlHD+OCycG4P3Lv/zLehcvB0c7tQOQ1AecXw6sjQljUR8gHgHVtt1XFMD7mte85gRRSV5AGlFJboFetE10F69yOVqcLHLoV21zhxidhNRsiUogGVGpfk+EaLd0xoFNAU6kO8tsoLznPe+p311Lv4SoJI9xEtBPxtvL5gSZrrzyykpUpu+VSZezELIRYxzqI2PA2DDGEc76e5aCcc5+bBRRGWBjPJvPxp+7z41/cw0oMuYEsrKLou+Ca2LACwCVmHMATfvd7/GYOgKgUpejcwFU0rV5R79HMrir9kh53c9DhaR88FCxCfcXsDuUV78uICrbV7+ObPuDDyLWRkDKPBfZrbbO1O1cZJKGDgJC6ZCeRPbLXNReQR7ppRVTtjYkX8ChY0Co6wnRifxsrTmRSA6BztUtf4C/c205bXlpV/Saa47pixzbtNFD2qNtAaXWEDpki9lIc9n6YS0UfQdQXWvJyuillWG9v7Mx/YnK9db6I+szrjtR+Ui99DNdA0tpYN78paV0sNQ5GAkm8yaSP/qjP6q+OLwAO/DHQ1RaH6VFOsIL/GyvHrWuuiEyNxHyjazP1kTrHN8UscVWwUb8UnjHhryNbGv7JCH2LkSlN9LAG3DT1VdfXXHD6YhK662/tHAjJkxGDn5A639YXz0xeNNNN9U2kstaRw/y+C7QgbZZC7UhuEKbtRO2yRt3jDt10ekrXvGK6jOqV344DQ6DceEgBCeC4vWvf33FOdpDP9oezAZfwZ3q0G9w8OmISrhIID9CMU9UwrCITnLyqT2pCUfB7Z7ohBvTN3wXbfaEKAKWrMF28FgnKjtRWQfZOnxsRqIy+MBRDMaDf0TzzP4Hm2j+u+Fg/C1a8Ip0iTCLclJ2MFdwR3CT9AsL8JajGw/8p2MhKg+VfZP7ClH5gKcJj5Zyjhbsd3TYVTDgzvJmnTxRqZwtW0avYU0d7GTwDLtqXRB9d01QL9toLQhRSV4YKdgMthGTL7qhk2DjtNOxPa8OdbGjqVs5ZFSP+tXtKER2311XHjusTvIog010tKYth/fIoD3KFoNDlZM6gjXTN37TFztORke/ye+aGJ1YL6wLidY4crU4UD3BmtqzliH71f2JyjNruROVZ9ZRT9E10DUwRRqw8MwDUWnBtNACT9dff30lvzgPnC7EklcFhFSySLtbCMhE3iLKpHUdiXXVVVc9gqhEkr3rXe+qwAwIBqC9Qmc5ohKIClFpcT9Q7oJFVAJUpyMqgWkAnAMjrUgm5GEcDsNLW8msHUCjJ/g+8YlPVNDHKUJcAKUh1ZAZdAHMck4ATvUg/hB1yDhEhzTqQkbGOYtDxTniyHJegV4R6OcoSaNcGwLK1A4kMIeLvgDPEJXKoRP/hwKk6geAV17OF0KPQ+KJSnJrK/noNPICqJ4kQybqC+0lr/rSt4AvAo1c2pQnMOmU7Bw09droULb6gf6WqOTk2rTX/+973/sqkJaXXOo1ZtyprG84c+rTHzYaRPkR2urWLpsj2VCYIjNxzqLM28Yb59y4WW+i0vgC1oCazEHzygaesWhMOm/us4UtWCFvQIWOTlnmbOb2+NFcyvxXr+uOicoQA1SWOrZ1jdIDm+ofDbciZtGljcRyg8GDJRaiskDV0sZCVPqPyhK3bweGkIwAFiB3EpCyIdY39kFMG1tZci1gjPx0FILQkU7ZJDIKSeuYsrS/zWcOB7S7lryjlp0EpMpgfxLJKMijTmU6imRry/E99eeY8pc6Jm/SOqpPJAddkcNRtH4Aq2wmXbKpwKhNTpE9czTHXbOBaO2UN7peSo71ONeJyvXQ8pnrMH6MERsa/dWvZ9ZXTzHfGpg3f2nS3mZH6AZZ9t73vre+KYZfzb+BYfj6Niutm9Lx1eNnI7msxfkveE8eHih4Cy6xBlq34MIPfvCD9UZCPjg8ggRTvnrgjklC7J0bIN28GKISeejGSmXCR6mbrAfL22vchAo/WD/9pQWiEnbht/E/tEvIWg07fOhDHzpBVCJCPS1KbnqwZsNv1msYw3oIs9EDP8DajUxBqJJVgFXJRz90wOfgF1jb4bC82SZvrEH8khG24u+ok52XFqGofBjO/gLicfw/KqXTDmRyS1R6RWuIyuBAfcmnll5aOqRPmFHb+EbSuqa/tZ0MwXba1F/92onKOtDX4WOzEZVsQiJb0+IPczt4JntAbAyi0g2xbnhnY4JTcoRdRDjR/BTYWzYluMs513fsQFCO0o8Tlf6jMkTl1q1HGqLSX2SMcC+SUhSOHTv1JlR4hF1mH9gokV0QyMcWiy1Ryc7Kxx5bH+Txneytfkayj/CZdud3sJp2ppyU4bcgPb3SXcjIYDLXlZF+IA88pg3WjMgiffpNHsFvZbP1ytc2R3WkL6SRN5E+lK/cVk7tlUZw1B66ZNNz06pjsKC1wjVplCk92ZO/flmDj05UTq7UTlROrquesmuga2AKNGDRnAeikqottBzIT37yk5UkA2ospMg3ThcwyWmgE4u1RVcaZKVPpvySAABAAElEQVRF90ABt8gkd5wCaIBfABIA7U5f4Eu6lqgMQdB2NzAVolJd8uSJSiCRQ9gGBBfgi9wDwjgi6ghRCSxyPhK0lYPAgQDYEJWAeNqCDPUknzuMleM3h4YjQyfIN/kQcJ6ABKYBQroCijmnnChBXRwaepOfDsmDINEu6elAuZwar+9xxysC1IYCZ1F50iE2/SY7p4yTaHwiF/Wd8oFYIDlEpbQ2Q8nobtpbbrmllss5Iy/ChkNNZ2RQLkBNDu3koAG+nsBE+rZEpf4cf6IS6LU5YgzQiXGiPz/+8Y/XzRH9SZd0aqNB28jNCdQWfRLCUrqXvvSldZNBP9ABuck6C4FTra2ivubA0hnd9CcqV6+HA2qALXaJPUOSJ3LkAybNF9FcSPTbPE5QXhvN9dRhLI/HgLekiS0KIFJuys+xTTP6PrJfW7YEQHllzlDm51DmztZKVh49dipRuWMHuUdPOR4q7wlCVJrfiS1g0lbtbGUZ1wP52Z+APHNRdE6bySlP8qUt2i8NMCiv7+awPNFjrfj4h3wpg+3Md+fVET3KH906L6bOfM9vx3xv6xr/nnRtO6xv7Pd4DFlJj+xa7DGbZ26zZXmy0m/X5VHeJLKMy7Zav9kY6zlbfe2111Z7bW3Rl/Q2SYj8jtG9tnqi5PbyNLz/rraeCp4E8d9aNsPd9GTd0W/JJ82k9Uo7K8G46UTlrPRmb8daa2De/KVJ9Wk9saazt/xs/ra1xlrLZ+ZnIxStu2wOP5vd4XMit6TLjYj8/AMFl/CVrIHIPK8/DX5DevJNYSO2HDbgO00SYu/yRCUS0BppLTrdq19DVMqPqIRvrCn8OG2ylgjk5S9ok1fawoLWFeUjKi+99NKqB+MIFoJfrcfyw2DZEKcfN3daz+iR7uAexB/cBi/Joz4ySasuJCVdaRPZ6Eba6FJdylO/JynhNjIoM/9RKZ825NWvblr1JKR1NutoiEr+iH50wyjsCzciLQX4S99b6/layuSXkFW9brbl81n34Xbrs/ZJG33Wgubkg35zQ695QVf6DT6Gx6LLOVHHmjZzsxGVLd4IjuErGx9sn30K39kQ84m94OPa2zD/pG0xoTmmHDHzjZ0yBs1T89LRb3NR3qTduRNmGj1RWV6SU+xJS1SWJyp3e6rSE5WFkKv/UTm6gfMkUTnCQWxXcA0bBr8kBp+QLW3TPjLQhbxsmT0b9szRb+fTTnnHo7bEtmijetQtf8rQbnVYk6JTdtk5gU4Ev+nMUR75rYHmre9kkTbpaqbjH86RLXWkL9WjjWRUdnSkbcqmH0cyk9116RL9Tlo2nm12bKP11PqS9tKDmHa1cq7W905UTq7JTlROrquesmuga2AKNAAIWFRCyHHuLSgx/HFeLW6zEBBUAJcn5IAuAAhY4whwGrKQA2CAkAVWWs4BxwyAQvC5+xQhYMG3sLsr1KtfgS9OPyJLOne4cvTG9Qdw2+gEJjkH8nhSM2QV568NgAWyUXrgl7OkDumBMMTgeNCP+tZdrQjHG2+8sebXlgMFqAPENlb9Lwmgr0xBmwFRJJ0nFD2JCeCow3gAbLRdujhiHBVgE/Gn/IBD4DevpiUPJ4fjF/KUHg4WgpHDozz61Ae5cw8AR7ZwfhC10iFylYmoBMxTrrbaIAixSB6ycNq0Tbkcbf0Y0E4egFrf61O6aIlK/WkTgZzmiL5EZrrz2CYGR10ZrrkrW9RHdK49cRg5w3RGz3SvPfpTsIGiTJsj2sup5GTOQjAuOlE5erJvLfvTvDFmjE8bR54QNk+NNXPWmA8QI4c5A3S0sZUPMBHGj8Zz5pM5pU7HRHKIATZtmfk+XmZ+k2kUAjT9TwiicksBTp6qLACyEpXD8Ve/Hi31y/NIopKNYQ/YVjZKZOO1N/Wot9VDO1/ZCvpKNCfZcO0Uks9ROdE/fSRddJVjzdh8tLpvZVKePO3Rd7ENyTN+bNPkuzRtTN2x4fTDF2DLWrIyumuvBzCzYdZOayob6rtzrisj9UWG9Tx2onI9tb18XTY8rNnGSH+icnk99StdAzQwb/7SpL2edYpPj6yKn83vttYIWbP59sgwT0PyA/jZ8ucmxLy5xNruvDUrRCV/P0QlvxwmOFuiUh8iKtTrGKISvuPj88XgAUHd/GPY5v3vf3/dIIZrEJUhSfkT/AvBus2PgWE/+tGPnngaEm5wUw5CUPlkQADCrog963P8h/gSjnCWvAhCRKf8ztngTno6gqPoXll05UlRNt1azyeiS2uufQP4DNbhw2gbPAYPvuUtb6l4M0Ql3Kcd5ISXBPp2YxF5lMuH4HvYl4Dt9Dssq1znXZeOPmExspODvuLPkFG7vKoWfuejRZ+10jn5oKtOVK5PZ28GojLzm0bMIfMCzjGX4EjzSDSn/A4W8p2Ndc3Rb/ZCGW1kt4KHzDf2wBiMDWNX/Ha+zbdjB/yILPQ2Ha9+daNqeRXr4fK61K3Hihz++qM8DblQnkosRGXsWUGrtXOVmXrUETsQkpI9iO0gn3Zpd/ZdyCMfLDRO3LlmnUnb5Be1TySLutno1MsHVpbovOvaGxKRDn1Pfo1o+0Z6ZdlnTPTbeenIlPTOkT1lkC31pA/9dj7tJJ92WiuV7ztZU450idEnHcLXYrC279azlrx0Le3W7+Rbi5D96v7q1zNrtxOVZ9ZRT9E10DUwRRqw4Flg5oWotAgj05BwQA/SD2AC7CyiFnCOmk1+wM+iDiDRkYUeoZgnEP22gFvoAS6EHsCFjEKgBRBzgDgmbUCQ3nzzzRXIWsjlQb7JZ9ENkE0eQM1dxBxgII1TI62IDEVOLBW0F0h1N6r/NEGiaav2AW4AsfzukBM4Ipy4bAjkvz6MEwAeaXigkB+cK+mc57TYpCaXp03ogl6Qee5K9gpceekgjhyQi/yTln6NP06OMjmC5AF4tQ+4pCMAWVuAXjKTXRohDhr961eAmiyAGWdJu5QrL+IGWAbo5VMmeZ2jFxsE0skjPx3oL+n0pevaQw+ceOnIrm4AHmmNiLVBYWwI9KWP6Q8Brp3SOWofgK5MfU9O/TsLwVgzLsT+ROXa9ajxYtwYjzbWjHHj09G4M54BMgDLHEwkkTkgBuzk6FyutyDDdfMl4JLNzHdHsriecnJUVr475nutpHxEjvLNrwJUTxKV9dWvhaj0vyWlmY8gKgHA9olK9trmGvvHRpln0rTtaOt1XhS0gS7N9YD2gNkAPLJHP75HH9ouf9qfa2lrjmlrjup1rU2ftDlK04bU71y+5zieLvVoI3vlSB/0Qj8iW+XI1kZfSSt/dORagCq7n6cX2HQgle6VkbytLOv13SZKNm/7E5XrpfVH1mNMdaLykXrpZ7oGltLAvPlLS+lgqXNZf6zpfHK4DcHFx4fnrPOC9Zpfz8+GP6xB0lmPYIZEPpI13poGH8BWH/vYx2rZ1g3ElpsW4T2+E19gkmD9VJ6bMpTpSGYywZPIU4QaXClIC9vBGJ/+9Kfr+gvXqPtA8d/UHX9C+qzfBwtm9X+d/GpB+cg9/p7ylQtbwVjIyhC69CFouzXSzacwlrzqhIHiv9WExz/oTzvUp11wDoxD9/wI5QWzwTLK4TPBkYhKfqcbY/WJdOTQjwdLO/SlcgT69tpbbW/l4FNID//CsMF2zgv6XZ/ydbVJ2/hN5Lb+uGYT27ho9Vkzz8mHOUSHxkN/onJtO33aiUpzQ4RTzDPz03y1pwVDutGAHXEUnWezzHMYyJFNTHQt5J35JY2Yc+1cjg2LLRrJMSL6tm2DK2HHlqh0k2oh/w4XUnPbcaKyYMCdO8vTmGVbbcuJrhx9M84FR3WwT8E3/FHYxTnXyUr2RL8F15MWeRc8ox1pc9rW2hNtg4+kl19Ud/CUOv8/e/f25ElWlY0/q/o8A/i+oL7IsWeG04AwEBoSGgj3xntDxHjnH+WVd3rJjRFe+AcYeKVhoIAcIggGplF/iPrCnPtcVb/1yaxnandS1V09XdVTXbV2966duQ9rr/3s/GauJ9fOTJjDhpxgrv9gBA9BXfKii3M63ck0LnIStRnHqzx9mSv9uAflPGmb7tqS7dppjM6ltoOPvskd9SI3HFHd1M82GWwY977cL7NNNp31RSa9jjq0o/LwiLaj8vBYdc1GoBE4AQi46Li4nBVHZS76SJcLqRutLqoutC7CjA4Xc0Ya0uMijejAiXGBALkouujbF7Qjg/PTRVl7xh1ypF4u9ON0c2whC2Rrr43+tGPARHba0A+ZU998MTLUpafI0NkvMDqMERH2ClevDKIjhxziaJUxcss4FRhU+lLPK5aQUYYO8ue1OZxpIaGwTCRTP0gswu2mNcLPqWslMSJsjAwkhgq9YCZqZz7kZQ4YU8aHqCO2+uEIVIc+xkwn+CaQSw84iZHLSFIG08wNwxvO8o2XQabM/AbPUZ75Us/xoZxu6tJFPceHvs2PG/f6ZljKJ9v8aCOGqKtnvJzMkUknx4s2pyGctRtvIRa+p/Pnf/7n80p5x4Xj4DiD49Ix5jjyNGUclX6rjjfHqmMr5MoxluhYE51Tsk1Xx7XoGE0M0VAv7aX6H+NIoNQVxjTbc0H90Y+wpHFUIoxTkRxxeaLybUelb1TWylqvfq1WpR9HJQfa8i0S50jnlGDvvGYM0X/ubPePPuUrt013OMHLOcE5xvnCOWk/QmksonawCI5kpSxp+lU2xrQf09Q9KFU3IbLs205IvvFljMYJD8ek6PwoOrdK5Tnfqh99jMn82idTe+c352/nMTfApK6pzpVwV4eMUZ/oddyp470dlceN8oPlO55cE9kD/UTlg/HqGmcbgbNmLz3MbLuOuDaxw51TOPjY2a71bgwL7HFcBm9gV7v+uC65nsljH4mu5a7VuZbhC+Fvrvex022T6dp3mKA/1z06RTc6sxvwO7qN8tQ1HtfNa+W0c21mv9E9/CL2hP7pK7Jt8CGpazJbxfiiL7muzcaVa7J+yCePfQMDfYRjsRPlkSeOQTvjcM0nM9zNdR628GErGR85tuXBwb0FfXIScqjCwhi0pb9jnu0hjz6jozF6BCd1XUtgy85wfRFgauyJxqct3dSBD3uA/BHPcYynfRu+7ah8PLN8kh2V+X1L3RMJz/HbcN7L+SDnS/nOC+r67QjaOn/6nfk9J/odylN3jOpqIzgOnSdFYdRncxOPcv6xKNVCA9+erTfq1On97t1dR2Vxv0uXlrfpbG6WjJJX/yssafqZc3b7ch5w/kp0PqNHzoV0Fu3Ld75xTnLOdJ5yfaGvMY1ccMQk5/60k2qnbzIFODgvwkkM9rblB1916agtGWRFD3mC+uSJgv4zTvrbhkX6cW6M7hmreuQ6r4v6kadvQXs6Ra9cB+g0Rrjah5dzuuuD63OuFWQrpxMsgsfcyRH8aUfl4UFsR+XhseqajUAjcAIQcIEJmbD60sXFRSQnfje8GSoubKch5CLpgumCbPyITC7sLvqMrhgSxqyMEeBizTiAhXoxiMhUx0XehVwZA2Zdb8RP//rNhVsb9WP4RHbakBsdtVGuLn3WBk7aSBkhiKqnDDkqkXFjs4rWtzY5H801YinkePDK17/6q7+arhV5fqacmp76syJYO/UZrnSCi/EzUBgnVi97xax2HLVW0P7Jn/zJ299zzHEUo8pYGDAZ24gzPY1R1Id6+sw8KIfbGCI38yolW8i8RC455JKZ8tRJGXmjbuoF8xF3clJXn3Q1v2SbK3UZpcZvO+ORZoyRt577Wbkn9M9Zu/GW4+9xOiodL45pv2E3uJyzRU8buzGEoDj21HH85TyTY9Exn+i4J090bIqOYdFvVWqMgjrqp63UMZx9ZaM89dNu3hj+pK+lfCGc1XX99pZvVHJUXq/vVCKvdXqeX/3z1FMLWd3Y4GTcrnOX74EsZAkxcl0L8fIbju5jX8YiZoy26ZnfK0xd/0LyXBsyxnE8tsdoHPoRgoFtdZIfPaTy13ip/6CgnRBZ2U4qP+OWmkNY5BwFn0Tn8FwLgwOdcn7K8SKPHO38vtkMbga7ceomInIK+5GY0udxBnPWjsrHifj+fTme2lG5Pzad2wisEThr9tJ6/A/adz1zbXL9yTUsPErbXLdj46jvOiTNdUydXJPli66JsdeVi+qzZXJdf5BuyiPPtXa0OfQ3Xj/H67a6xqJ/7dkY+qbDfn2ro034kH5H20tf6uRar56Y8SknO/Zg+JA8ZfuFjAv2sR/onDFqR2djFIMbHfRrHGxQddKHMYjqSAXt6JP20UX/+837uh356SM6q3OQ3Mg/Cyk82lH5eGb6pDoq/Q7z+/c78ZtkK4uclBZxcFAmysN9nJP8hhxD+b3azu9euXtC4Zk5t+jDeSW/Z/1r57cs2hYFqfJz5xYuVWeDOi+eq3tLm3X+Ki5Yjspz57ZLl/o+5UUPFdS9lfrsx/Z2OR3r7Tvbu3IyRrqlP7Kjd1J5wWLU0fnCeQ2/wYlwSec69Z3bOPvwwpzP0kfaqO++m7byxn60h00i3MjJOTM6w4YeuS5El+gR3HPOzvj051wvFemmL32Ew8bprEyd6Ds6QvUvqBOd9CE/sqWwTLRPT3KyYB83xAntR75xRf7cyRH8yf3qfvXrg8FsR+WDMeoajUAjcIIQcOE7S45K0Lvg5qLrgpkoLxdmhpULtDBeVOXlwj0X7v5RxwV4lLFfvbRRL/UP00YdfUhFIfLpLO4XrG7yJMM//uM/Tn/zN38zGw1ea5TXso7fJNGekeN48H2Vv/iLv5ifqOSoVJ8R4LWlHJAMteghTT++H/LXf/3X8yt/vMbH62z/7//9v/MTlYwyhpWQcUiD/344Z4zaZPy25Y9l8oTIVTcGkTwhOI1Gs/x1eeqlLP2mXvod64111U//2qRejin7QuSOY0nZXOEU/DlrN97MKcP9cTkqHS9+U4gIkum36XfHSWNlOqKJqCCMjj91kSLR71EaMqlsPEbJdvyKfkuJOW4dnuqIjuG0lWY7v5Wk+x3S6SNlJW4OfrbFfeqctHyj8vqNoqW+UVlrZq7MT1TGURkCdS+xCzEKMc35gP6J8pSHfGVs9IdpHJWIPIIqLzhR0tgTxu3kSYPRunwcd+rcD6dRZuQmHWVlW2o86zGO5DfEEqkWnf/VF2Dg2MjxkmOFjuSSg+R6ugEh5ajksERM/e7hr867EdpR+W6g/ut9ukndjspfx6VzGoH9EDhr9tJ+GDwoL9e3XNukouDaFNsj11tl2ijL9TVl2kSe655tZWO9sa76DwqRN6ZrmaMM9TIW+QfpuW4TfeWP8lMvciM7OChPH2NKxv3GSp5Azhjlp//Iixxloz2hfCyLjpGtzPylztzh7h91Up/McTzqj32nfdrcT+7Yx2nehkU7Kh/PDJ9ER2V+I2x63A+fydPneKJoP47LbHNw4ZDa+82NjjDHlN+dcg4wEWciX9SXmHNy0CfHb1gMPyNL4KhczgEXquz8vED19u1yqt31sEA5KjkpL1iwXuRwh6OszgW754NSctZH+/A02wnjeSt543lDHp3wljgH8SL79MOBRkelPuBiDHDBeTzpjRdpL5987YIJDmkbZonRVV390FNb/bKhRfLolnNZcLUfnVM/TkTy9GVe1nobq3rhfuSH/+l/vxDdMn8jniMG7Bg8EB90zvGwhDy40JG+me/9+nnYvHZUHh6xdlQeHquu2Qg0AicAAReNs+aoPAGwH7sKMSQYCIxmjhOvcmWIcDh6OvKFF16YHRpZYUWpHA8cm3/5l385f4uEA8QrX/NEJWcIo4eBFMOFceZVtl79+vd///ezQeLpS6+W9eSm9owxhlOHs4PAWbvx5nfHYH8cjkq/P78pZAe59PSkV71yVHJSMt79Tv3m1BVsaxPipG0cUNKRMI2/Vb9z5CLEZE0yyE8/0nVUnjge/eQkJn+p5wbi8upXjsob5aR8i6Oy+NPsqKxVtZ6o9J2SjY1Fxt0isiF3znNiSFF0H1N1U1899e0bI3KL4CHdCDuyLsZRmbHSdb8w4pNxJ0391JGmbExT736p+gnBcUyN15jiiA0uiC+Cipwi11L76mlvfI4FZNhxkWNEnjJ1yNXGb9xTlEhpIuelxSv6Ufdxh3ZUPm7E9+/P/Lejcn9sOrcRWCNw1uyl9fh7vxFoBI4HAXZYOyqPB9u11JPmqAwfY8fjMOzjvOLVq5rd44kja3Q4youjkgx8IhwifMnY8YS008a2fvDLkVPiK45DcsJJcC/beBc6c/48JyPn5sVpa/tCcRNPVnIGKt8qLlKvmj7nLVnekrP3/UfOSgJwN/roWyrKC1fS/5qTpIwOdKNTnIO4kX1t4Jfx4YfGJuBC6nHEuRfmnqp98tShCy4ZjOKopBuZ9INv8NEuvG3EW756GaNtesFPPTpLw+PUMw/6Tf+Zk7Sjp3aicRq/fpQnnQc5/AleyYq+2uOEuF84oe8Ux1kJG+XBM+0fNW1H5eERbEfl4bHqmo1AI3ACEHDBaEflCZiII1aBocIg8DSl175ynPzd3/3dbLT86Z/+6fSVr3xleu6552bnBmOJwSNox3Dxjcq//du/nR2PjA7Gjae0OEI4RBhrjCDGTI4hTlHGiVS518V6EtN3KjlOGDdrA+eIh93iThgCZ+3GWwz243ZU+h35zSI8fmvIpt+m35nXvvoeEiKqPIQjBERb5AlB8rsOabI/Eju/7/TjsIocqbgO+W1L004aOckf2+4nq5rM7XVx967vlEx1TipH5fXFUck5eWXXUXnhAqfanm5eAWucIXm2M+5sj+XOXSFpUuc/5XQOucwKYzjDSpkIf2Nah3F8yjLuMU2bsW7K0yZ1kqavdZuUS4On1LFoLMZknCPhDTmVZlsduOnH2HIsOFbEjDl9qEsmR2ecUXkNrOuAV4GTrd6o86jvcW23o/K4kH04uX5TOTb6G5UPh13XPnsInDV76ezNcI+4EXh3EGCDtaPy8WB/khyV7PnwvSy8xBdxRPdybLOXcRs2v7pS+xxcWaDJ/ncMhUfEsYVnhIum/ugUI0vbkb/gJJGDI+AekXPx4sJNL1y4XHkXpxs3z1f54qicOCrP367+1LewdFmQegd3Ka6TseJuHHThtvvp4EgYeYntkTON+oXDkGNsZOPLsNJOOTziqMziT/KCpXbBRXv5IhlrLhk9yA1/k+orPEwq4Hjwg2N0lhcOZ07irIQHvZXpg/y0kdoXtRejR3AKvvpez6l29AgO7H58kN3vO8XOPXih+83qkB2580Ae4U87Kg8PXjsqD49V12wEGoETgIALSzsqT8BEHLEKjA6rl15++eX5u5H//M//PH3rW9+av2H3Z3/2Z9Mf/dEfzY4NRuhoJMXo8XTkP/3TP02+W/rSSy/NT80wqBg8jFopQ4Wh4RiKgSKPTE5Nr4n1atmrV6/OrxU54iG2uCcAgbN24y3G/3E7Kv3OEA7Eh0PSU86clIm+VelJQHWiE6KDTNgP4VAeMofc2Q+B0kdiSKZDLuQi6X6Hofqic0vOLwfJiJyUV7O5re9R3uuo9NqxWtVazknfKck3KqsLWs1lyskzxjE6r4WEJXXOcp5E8BLlKTdueCB5znfOabCWF0ySRu/gkPHYV5bybCcd6491kp/26/1RfsrSXlniOO8ZK4IYYjpur48NY3O+F22bQ4HsyIWTduSwITgmve4n3yUJKdU3/B9naEfl40T74L4cG+2oPBifLmkERgTOmr00jr23G4FG4PgQYLu1o/L48B0lv9uOyvABdjv7HW+x0BInxA3xxQ9/+MOzo9IrX/Ec9bRj30vxQPd54qjEDeWz+3EItl34kvxwyXAlMvVLTjgg2bnHFM7FqUeO4/PcOa+RXRbQXrhwpfIuVl8Xptt39FsrV8tReaEclbV8teTcKP6Bu92rh/5w2jgF6UG3OATNk77WabhNuCKdEo1ZCCYZF8y00wYmxoILeUgAPsarX/rQIzrZJys6wU8UyIsuI9fSh/zwMfWV42D0jKMyWJJNT3Mo6jM4pK1xqU9GOKC8YJBt/Qj6pveou3zl2pABB+N3/xEfdC9R5LDEDfEBupJtPEcR2lF5eBTbUXl4rLpmI9AInAAEXKTaUXkCJuKIVWAIMAg4Kr/zne/Mr3D96U9/OjsnX3zxxfkpx6yii4FEBYYDg4ORYRXUj3/84/mpSnLceCZX/RjA2qjPKGKIearLK1+96vXZZ5+dDWKGKwOpw9lD4KzdeAuxOE5Hpd8fooD0eGUPI93vDvG0StY+8qlcvfymnetDSvxmhZAohCZRG7/XkBHkRBzPE9qu99ekQ7kYkpr6qSfN9ihvaVc5bzsqN6brN6cigc419brX2VG5XeebqfRcXv2q/SJvSff2710lG0IWLJAq5zTESmofgaIzYheiHoIJF1jALfgcNL6MP+Me95NHz/uFsd64rU2wS740mEpzLBqz8RqbmLEHizUZNR7RODO2yHPc5BhKSg7sPHmf71UiqKLfP2zVib73G+9RlbWj8qiQfDQ57ah8NPy69dlC4KzZS2drdnu0jcC7hwD7qx2Vjwf/d9NRiQeIbPfwO05KnNATlDgivih6mlIZLhNnE7uena89ThjHIy5EnuMIZwiPiMMJX1AfVxLxJ/vhEdqRiwuwCzn1EslSvui7y0UvlqNyulSc5UJ9n9JrZ3FOjkqvfb1Z+9erjKNS6nWny9tgjMW9KHobm226kG1MCcY7Bv0HA3oaF12l8mFKxsiNIy/1My4pnbQzfm1GhyFcyFKW+VrrYj86RS95+tRGCJ760icc6atcn/pJtD9ikPlQ/0FRXSFjGbmv/OgHLzoEB29Xu3bt2tvx5z//+cwRzXtw1f5RQzsqD49gOyoPj1XXbAQagROAgItKOypPwEQcsQoMBc5GxMQTkTEQXNC/+tWvzk7EGEvrrhkljgvGhPY/+tGPpp/97GfzaigylcVwi9HEELRazxNdHJUf+9jHZocJYzF11v30/ulH4KzdePO7YIAfl6MypMbvjZMS8eSgzJOU9vO9EXUQCm3ohUQgNAupW15xqkydkdTYTgwhCdkMMdNOSGo7ZMa2oGwdU0ea7aV26kcm+VbRTqVvOSpvcFQiujtFVLeLeG4XGbbCdXn1a/VeY7xXB33rI3PivDXGYOFcydE2Oiq1hR9yGSdl8ET24OL8KR3JpnYJ49jlpWydn/oHpWk3yrAd/FIuDa4Zd4hsCKTxh3w7TpWnTfrP+TpzLT9EXFt4BTvylEldL/zekdO8Apbjkn2hzbqf9HccaTsqjwPVh5dp3vuJyofHrVucTQTOmr10Nme5R90IPH4E2F/tqHw8uL8bjsrwCnY7ToKn4CyjkzJv3slrX92zwXG0CT/Eg2zjTXig9ok4D9mOpTi31LWvf/XJ068YZxzUwyFwB4sX8QKvSs1CRnXIvnx5ca5drFe/zo7K2xerL31aQGmh6p3qy4JS3zq8PrdfOO2egw5nwwFGvXG1cBr6Gt8Y5CWfronqyR9xpac4ygsPCjcKhurAIQ7D3HdL+/C3UZdxO3pJhXWf+hHD7eirjrkw5qR0SFty1mPVLnOUNHOsjJ50Due1LW+Uk/owYPvHUelBBw9K+F6lPPNOX3KPIrSj8vAotqPy8Fh1zUagETgBCLhYtKPyBEzEEavAYGAsuHnsSRfGIOODEclIZcDF0NivawYEGYwNxwc5tkVylTFQyGAIiQww8skeV7E9yBDbr//OOx0InLUbb/ndHKej0m/Ob4xTkoPSogDOSiTUa2CRNL/DkaA4mvxmne/zG14TqZAav+P8nqUITggnoiOOv+lxO2RKf/LF1E+ZNFE9IXWXNE69xVF58+biqHyTo/Kub36Uo/LSTo2zXgNbD2pvbCBwiCe5s7i3/+jHec9YQyBhYDt5wSS4wInOcEC2Q7xhETykwdh85FyacVAg29LsJ13nzRV2/6QseKUs+UmTv06DbcYe0mm8xiY6TsXUjUxptslVrt6IIZxcT9zQgF2OeZi6VnjynqPS4hhPVXJU5fpD3uMI7ah8HCg/uA/HSjsqH4xT12gEIHDW7KWe9UagEXg8CLC92lH5eLB+3I7K2O14C66Gn+AuFopbuOp1r+77JNrPK1/xPoEtz6bPvR48gSx8khw2te04qkZugAPQIX3rP/Xkh0PgCPrgqHJfSmpfX8L58+4nlXPtAt55adreuVSvffX5Egtu8dBpunTR055e/eoJQjxkeaKSjI2N5U049OSEzb2oOAfhI9A9cc7Y/ZM8+orZV6ytiOtJjSvylKsbnkQXUZ562sADPmLaKxPHsN5PWWSN+8E1HC86k5++Rn6atknJzFjHlLwxpu/omzTtUzf6hFtbqOohhzgqvf4VJzT3jjftjiK0o/LwKLaj8vBYdc1GoBE4AQi4WLSj8gRMxBGrwICI8RBjIAaL9CBj6H5qRKZjJkYYgyhGYIyX+8nosrOFwFm78cZQ99s4Lkel35vfLxLGOfnRj350euaZZ+Ztxrp8JDFEETkS/TadB0IgRqec37VArrp+z2OMLMQn547xt74+l5CXPCmdE5SNUX5k3ZvOJYVlveSnXvtafrGJo/Lu8ETlU08t36vkqCSTo3Id5I/jRpSdv0RYJI756tPFuINDMAie8t0IkA8TY4z+s+bVPvvS++WlbK5Uf1Kf7mNIftKxbNxOO8figsviaDQuMYQ09dKW3FF26knhBDM4cTomyk8fjns3OXyrkqPS6lk3xqygZWMoJ+txhHfLUfm1r31t+sxnPjMvIhh/K8Y8Yvs4MDgJfbSj8iTMQuvwpCBw1uylJ2VeWs9G4ElHgJ3WjsrHM4vvhqMSB8FP8BKOOk46Tkqvd8UNLWSVWuCaT4Ooy051bIxOxCxCZLNakMlRqQ2Z2uCCymL7S+3TQRk9wosgPnIwHAEf8HYujkr94gba+kblhQt4Jmee19H6PuWl6gfPwmXLEXlpq7Y98Wmx6Y154e2dO5xey/cg6UvPOFbD06Izfegr7hdSti4Px0tqvImRo82aYwUX7RLlJYzb8tb7qbdfWXQNV4vOY59reersl0fGWp4849kvP32au/WYM9/sGQtWfaPSE5Uclc5B8kcH9TjGd7LdjsrDo9aOysNj1TUbgUbgBCDg5mM7Kk/ARByDCqNxQXyMl7WR8jBdkzkaLmStb8g+jLyue7oROGs33hjvDPfjdFQigZ6cjJPyueeemz7ykY/MJBRB83tMDGn0O81vN065pJxNftOCdnHQIXgip5w8MWSPvJxHxlQfYxjr5XykfKyXOvem6uSJSo7KnXJUIrzLE5WXLm1PV+r1r56urC9k7BKppefIsTeSqbWD0jwlxmEZwkWGsYrGDfNgmTy4yA9xzZiS6j9lY5ptfQgHpaOc1FvXlb+uN+YpSwwW9m2P9ead+hP5qQMPGMHH8TI+TclZqUyb1Ec+/eatmuWoFBHVvDI81470d1zpu+Go9O1nr1XnqEScD5rn4xrzSZTbjsqTOCut00lF4KzZSyd1HlqvRuC0IcBGa0fl45nVx+moZH+zNXE3Tjq2L6cijihyTHqCksPSfp40DK/DYRwb+BF7jfMwT1XKVw+vHB2V4T3QDJewTQ/8KPwzfILdTz5+QH4clfqRT4a2S3uLPjdLJ9+lPF/coxyVE0clHuqJyXJInvMEpqcqLThdnJT6x9XipKWzN3vRn05kRx91jU0Y8+aM3T8pH+uou45jue20SzqWp6+kyh4mpG9tRhn6StxPHnxTHhlrPNJOvdSXiuZPxANHHh3+rCxt0o88HNHCVecdT1ZyWOKE3rrjGCDrKEI7Kg+PYjsqD49V12wEGoETgIALxeiodIPRhcaJ/wtf+MJ8M9wFnwHQ4clEwHwKo2HzKCOJvMg4KrmR1+npQYCDIgYqIxVRsTrz4x//+PTCCy/Mr6I5PaNdSBvj/bgclQgg4oV0Xr16df7W7Cc+8Yn5PI2MImkjEVmI37L6Fc4hG3E8cT6JdEY0tHWu1wfSy1GZVD7CF/n53Scl3/Z4fkhdeclPqr6QOntp5ExFjPJEZTkq3/REpVWzHJR3Ss8707lNei+v2HGay3iloqA/4xZHYmU7+WOqPl0iK8Q7qTmwvSbrowxYCqMMbRKjX3TUX0K2R5ySt07TZqybvDFVvo4pT9sxtZ3x5FhxE2N0VNo2TnhkHI4lr/VhR/zHf/zHHK2i9Qog9clKP+n/ONJ301H5/PPPzzeHYGK+Dpqz4xj3SZNpzvvVrydtVlqfk4pAOypP6sy0Xo3Ak40Au+sXv/jF9N3vfnd2GLjvw7a/WjzC5yMsfOTE6vDoCDwuR2Xsy/AR9+pwQ1wwzkmpJys5GnFvHNG843M4TO7tsffZa5yHIicT+55sDr/xCUV5sWvVcWyNuoQjxQYm273G0VGJJ9hXpn14xLlzOCQujf+dq3b16tcdT2haGLoxP1F5/jxH5e2qtyw2NWPa46vGCAfjtG98yqJfZnfUOXnrNGNMfvZHWWPeul72pfp71LDuN31Hvj7GaG4yP1JBG/MSvDNHc2H9GevbxpPjoMTvzFm4nPzcOxj7JUtbc+5Ywv84KMMJ8UO2To6xR8WmHZWZvQen7ah8MEZdoxFoBE4QAvs5KqnnNRFxVLrgM2g6NAKNQCPwMAggIxwVIoclwxSJQIq/+MUvtqPyIcBEMBBEJBP5dIPh2WefnTgqPVEpD6FMCKkZyQzygBgiGGIclSEc6jrXx0FJnmif83IkfKNcfWY/6ahHSIy8NSlRfy/uyim/HV5XKk43b0xFdhZH5e07y2t/NjdvTpsbiDTn162ZFFmFCx96hoSP+qZvGIwRHqK8Uc+QuZHQyROi74hnsJTCkyxtYScGx+hG1zVJnIWv/uyHZ/KSrjFdiZh3U+d+aTCQhqAaC2KKoIp+wxmj/o3HWIw19oTXvY6/e8TUSmrl+o8O++l5FHntqDwKFB9dRjsqHx3DlnB2EBgdlc6ffj/OZVnYhZd1aAQagUbgnSDglfzf+973ZkelRaTsUueWdlS+EzQPbvM4HZU4BBscv+Bo9gRlIqclB6V89/HUwY/CkZKy3dnk7HM2fhyVbH82vnp4O+6JH6Z++E/sebrgAuLIC5TjE65nZOMCIh6hTDsyRRSrsqo+fTamGzfP1zZHo37rCU8LVOvVrxyVtfS26noac3kTUDCIrvQONwlXWs+a/PuV3a9+2iVVdy0v2ETOej/5kZHypMk/KNVe3URzknnJtlQgY8R6D/O9Rb1pY75Ex8Sa/9l3bIy8mXw6aB9dlI+cMJ8EkeapSseAeo8S2lF5ePTaUXl4rLpmI9AInAAEchFBihmwLiAuMgydz372s/O3z1z8GSfyOzQCjUAj8CAEGMSiG2/Xrl2bibFzDOPW62eeqe8qtqPyQSjeWw7PEEaG+dVyVMJR/NCHPjSvmkUkx/P0uE2afRExiCMKUQgp0YdzvX6c9xPd0EA8ERt1xDFkP+m6LP3KX+s0yrONqBalqnqLo/LGzZ3pejkqX3/DCk0raT3J99a0dfe12n+jiPVbdVzJX5xmdIZDdA6xXkjwva//CSmLfklpICz67D0ZpzzkTOp4hh/ixoEnIvry1dW3Fb6iG84h0HSLXsFM/YNC6qTcfmLypKOMdZuxbKwrXwwW0hwb4/jiqBzHB9Pg7IaAdlbqI6F+7xYnvPzyy7Nd4cYYnEJkR72PersdlUeN6DuTZ777icp3hl23OnsIxF7KWyica107LET63Oc+N/Oys4dKj7gRaATeKQLswNh+XsH47W9/e36yKecWn464Wlyin6h8pwj/ervH5aiMYxDfYfNyTOZblHFSyh+fLsQ7RHwOz4vz0XGC04TLZGEhe52dr14cf+rKD4+0LUQf8tUVbUe2+4040toRSr56589zVC7c8s4d382sRarXN4vnLa+C3dzwNp1bpQ8nJWfl4uDyXUs85Omn3/O2Uza8JLyPfmOgk7BOxzpjefLVP6jNWJb6+e0dtJ/8yEz9pMlfp2knVVc0D5kXcyPaV6b9fk7KEZ+0xePMleh4cK4Iv5XKU0d9IbrZTn/RSaq+ReteA+upbveZpbgBvohDpx0ZDxvaUXl4xNpReXisumYj0AicAATiqPRaRq8EYci6uFmF5WkdFwBGjpuQufCcALVbhUagETjBCISweOXHD3/4w9lZwTBFRp4px5rvuH3pS1/qJyoPOYeIgMgJ5zU+v/M7vzOvgrYa2g0GC0sQUuQz5+kxTft0l7IQzRAa8xaSiejF4RfCOZKayJKGqCQdy2ynv2yP5WObRc9F3viNyrfe2p4dlTduWH15vQgUp9f/TG+9+av6/uGrMxEqPjSPnzPQNWu9gng9hqWvvZs49qOnNMRpzIMT553r5uigRLwRrpA5JE7IfHHOu3FgZbMbzyH98Mz49SMknXd2/6RO8uybK+lYtl/btEmaOtJEYxVzPNA/TsqME1lFOO2rR3fHW8YSR6UbHKOj8qc//en8nUqkFD7BNfocR9qOyuNA9eFlOl7aUfnwuHWLs4kAR6WFHddqcZfoXOoa4uYzh4LrSM73OY+fTaR61I1AI/AgBGIfOlewuywe+4d/+IfZHmPnf/CDH5x+93d/d34rCx7Bburw6Ag8DkeluWWD4zU4hTfquF9nTvHBvOrV9UMd9rmojZTtjuON5Y4T3Iad7trDfrMvqK8d3iE4nsITYtPTSR39jdxRnwJOQWYcX3hEOCf5Fy8uzspigMUxOLh2pjfe3JgdlihV5Zb8W9WmPJj1ROVGOS716Vg2DpzPMTyOKTxpVqD+qC+s0zlznz/qpe663Zifpsk76Pp8UP7YbqyT/KTpZ0zVNwciTMXsSwXtzUPmXwobqTIytAvvM0/jXJmzRMeEuddGW3JEIbqnjHxy8cYsYuWo/PnPfz47K90jcqzpV720n4Ud8k87Kg8JVFVrR+XhseqajUAjcAIQcMFxk9W7w3/wgx/MqX2GBoPHTXEGB+MjF7wToHar0Ag0AicYAUarcwYj9Pvf//5MkD1RxVHz+c9/fibHnhBgYJ6m4BzJgD/qb1QiA/DkhHPT0hOUnJQf/vCHZ6el8zRylvM0PRj8MfpDJsbUtvLUNQ/mDelDGl0DEuWRPRKScd7Iul8YdYlO+9Unh6hF3kZhuTHdtKq2nqh87bW7RaC9cubNIky/qrxfTK+9+t/TL3/1/8ph+daM+/nzF952UsZhCRfXsDjUMhYEKmRtHBc8ECbzKNpOHElcyDaSlSgPuTNGst1AyGuYpOYJkUaoYQvTYDdidBA2yV9wutdZGVyTpu6Ypix9SdfjNWZ2wTjWOCuTr//c6AiuxqstLLz6lU3hySCOSt8ncS5gW7xTMjqO40Hb7ah8EEKPp9xvoR2Vjwfr7uXJR4CNND6J7rfjXJvzmbRDI9AINAKHRYBtGzvPgnSOSrYYx9YztWjUm20++clPzjyCzdzh0RE4bkela4J5ZYPjElm86p6daFEk7oH7qDNyN+3Y6rHftc/Th0bO7sdjOJXY62w4HCEcaTye2PLqKxfpFdnhXFKcS5k65IVPaE835QuP4KzEiTaLJ9S7c25O5ajcKX3KgVpPWE47d6bz5/YclZubi6PMWIzVWBzD6ZMu9F0HuogPCmO9sf6Yv5aReuFa6/KD8rVTtl95ZCaNzNSVZg5gmu1RXuYP3sFFGpna4G+ZG87qOCbDazN35jw8LnIjKzrRUZ65VSdzbzGWT4Gwc/BCDkt2jj7wy1HnjPNBaTsqH4TQXnk7Kvew6K1GoBF4AhBwYXKBcOF46aWX5lc0unC4YCDFLv5uqLrQjBegJ2BorWIj0Ai8SwgwfhnEDFCkzRNWDFUOti9/+cvza8yQZKs+T1OIsX/UjkrnX2QMXpyTVj/7rownK91wQEqVqxcCEYPfXMgXEYfE5MF/rBPiiGAkjm1CbMZ52y9Pea4Z0WXMG9uP2/gjecgqR+WtWxvTm/VEJUflW2/dqP3Xy2H5yyLS/1nH139O/1PXrtdff62uY8Vqa82t6xWiirSOUV4IOazEjC+pfuOoCymLcw45s43IuWYicoi8aFtU7hiArf7MV17FZJ5GRyVdYK2uEIyC2Zy5+2fBY49YZ1/bbEdG2kVO0jE/fYXUGluiMWScUjgkL8cWvY0vEX50US+rZkdHJZuC8zKOShgdZ8iNfeear3/96/PT255Goi+8DhOCmzQYu8H34x//ePrmN785feMb35hfl0+W1+S/+OKL01e/+tXp+eefn53T67k5bL+H0e1JqePYaUflkzJbree7jYDzIy7mBp4FHt5C4ZVpziVuPuNjHRqBRqAROCwCsTvYXHn1KxvWE9pslRdeeGF2WMY+OqzcrncwAsftqHQ9YHO7HuAYHDVeD45f287CVVwId1NfyLEgD//BQUZHpXI8AJdhx1t0aFteQmTYd0yFQ9iOrYwfkM3ezj1EfSoPl2Ab2lc3nO3KlcvVjs4ejFgcla+Xo/L69frsx61ywO3cnh2V5zZv15ju1niW19LCIn2N/elTGHWeM3bz9stPeVJ1Ui+psmwnTf2kxjaG9f5YNm6rN9aN/KRjXdupC/8xJl+qrWMAHqLt5Ekzj3FSht+aI1F+5s2xgCOHC2ofubbTb/LNrfkxz9o6rnBBjkoLJ0Q2j0Va+o3c9Tjvt9+Oyvuhc29ZOyrvxaP3GoFG4IQj4KLg4uEiwYj1BIQbcVa95EauC4wLWy5AJ3xIrV4j0Ai8ywjESEV03HRDdhAJTwH+wR/8wbyCl+PmtN14Y/Az4o/aURky52YlDDkppZxgIaXIAtxDVnIIOHeHpJCTGHKR8qTy1Um5NGXki/uFdX6uF9JE7ZIfGeP+InrpY1lVW09Ulv9xz1F5/W1H5Ztv/tf06iu/KJKzfOvizXqq8vbtO7OuiNFIlENikSZR2bidfeN0PUTO4oBEnuzLF0PmHNPKRHnKjIUM8qzsdfPHHDnWbbupkCc91aGn+pm3EYtsB1fpuJ12Y/6Ib9onXZe59otxzBrDGBFT+9IQUzIcD2wDNzjgCkfHi37Ud2MdEfW7R0Kv1SsMbctzPtCnY/Q4QzsqjxPdw8v2u2lH5eHx6ppnGwHnTzfy3LjDxSz2cO6Un8VIZxuhHn0j0Ag8DAKxGdlnvhFnAQR7zeIqjkqf4bDgkT3HHu3w6Agct6My/Aaf8KYWDspx4Sr7F78wz3hCQrhAbHi2exx7ZCrHY0ZHJR6EA7DbhZF32NdGjLNLPXXiCMURwnXUxyXCo+xnLE899XTpYvHjovfOzmZd96b65MdOfafSq0N9E3FxVJ4/d6d4iH6WN9dEhn7SFx3GsetrDGveNJZle6yT39FYlu0HpcH9MPXWddNv0pSP+/IS4Z9t/akXLMLl5SWqo03mJLx2dFAqUydzZzv9aB/5ZKZv25kXx5njwTHieLLgMwtZvereQlaL2fFDfT0sP2xHpVk4XGhH5eFw6lqNQCNwQhBwUXFRYIi4QFnl4luV3iEuuAC5uLnodGgEGoFG4LAIOGcwOhmmziEcNcgUguz1NIxX+acpOJcy5o/SUQlHhBPhQ0qfqSdRr169+raj0ut0kbORlDmvCyEQcCZDRB6yLx3r2NZf8rItTbzffKkjpP+QlqRj2VxxqGt/t/lyvfH6H69+3XVUvvpqPVF5/cZ0p56ovHmrvkt543/KKfY/RXD+e74B8+qrr9WxtvdUo7EZq+MMPiFL2ZbC1A0aZF1q3LkWcri5aez45Yx0LIvmV5048Wybd2PXH1mckW4iOOY5l/MkZV7HNN4cMCfaBqNgF3yCqfSgmLpjGjmRm7Lsh3jSHyl1/ZeGoBpfxpzx0RXhDG5SY6YXeWRY5IR0Ip95jaFFUHFUwo+84wztqDxOdA8v27HUjsrD49U1zzYCzovOua49OJiFHhaO2neudf7t0Ag0Ao3AO0EgCyHYR3gYLoGL2Q8XeCdyu829CByno5KtjcuYM3zQk5R5w04WruIy7HRzqr4w2tzylYtkxYZXxzHCjuc0Em3LY98LOU5ci8IT5YdPqEuOeuS6bqUP9dUTXefoRofLl68UZ1p42FNPcVReLIl7jsrr5ai8ebMcWPXq1wvnb1e8W3W2Ky5P8+kn47EdTpWx0y8hvChl6zT1kj6oPPUelKbfw9RLXX2n//u1C6eTwn6c68xR5s3+WmY47cgD8UJzFHmRLU99czj2Q6Y68qX2zUm4rm35uLQFE/gh+4ajUurtER6YGY+1+415LGtH5YjG/bfbUXl/fLq0EWgETigCuQi5mfijH/3o7Vc1UteFrUMj0Ag0Ag+LQM4riApnDYPSq0s5bE5jYLgz4o/aURnCB79nn312vsFwtZyViCnCGkdlSAiSMBIUJAV5S5rt1EkaAjOm621zul9IvZSlnnQdU0eaeskr1Yvk2KunG+8ur/95883t6dXdb1TevV2rLu++XiTqlen6W69Mv3rll9Mrv/plkZ9Xiui8PpPrkB06GasIw0THI8ziVORYFNVD0DgnOdzcIEbWkSvzKppjZCzb9uFHJnnmg/M4T1DaHh2UiHucpcibPoPdGgsopEy6X1TnoBDcx/Lk0R/pRE6Nz5jXzsoQVf3S0xjpb5xuhtiXTybcYAU3dgTnpKeBOCtHR2Vkjjod9bY5cAOnX/161Mg+nLx2VD4cXl27EYCA65fzKGelJystmHHtcp3p0Ag0Ao3AO0WAvco+8kYWjq5+UvudInlwu+NyVDr/43BsbwsgPQl7tXigBcCclfkMSBa1qJsQbsGWl+96wnYP/3BchA/gAVmgyYaLzZ622oRLpQ/tcYAs6tQffUcHorrhHziU/fAmY8IrlicqPdm7OCrfGJ6o3Nm5W/LuTBcvbFX/OxWNZe8tQRnP/a6TwQEuxpMwbicvqbJ1eeSs09Rbp5F1vzTYpM5+/aZsTLWDvxgHojztYSzCRgqb6Jb+tGFzhAuOTkr9pH6OkZELy0v/5Ij2M/fmFd91vMh3fLBtOCZxwzgqcUUOzBxv6h42tKPysEjVXBawh0f28HK7ZiPQCDQCx46A05eLFVIs7dAINAKNwKMi4LzCQGaoMlhDoh5V7klsz2hnxB+1oxLJgNt+jkpP7ikbiWfIyX4pAiEf+QhpCRGB6dqMTVnSh8WdvP3iQXI2NpjRC4G8c2djunFzpxxgyzcqr9+olb133ypC9lbVeauuU+VIfOO1Ij6vTL8sR+UrFZGg0bmoH7qLwQNWcbohUm7WuHkjHylfOypdD82tOAb4jYSMDA55cyLaNzei/hz/cVLaz5zRawyZg+itLNtjOrY5aDuyxvIcp7mxYLyJblIgk8qQTsHxN+IFM2NyE4I+6iGYcEc2PVGJiHJQesWPbc5LtgV8I3fU6Si34d6OyqNE9J3Jckz0E5XvDLtudXYRcH7OTWPnZTaF60yHRqARaAQeFQH2HBuOTWe7zy2Piui97Y/LURk7HLfwJCzn5DP1VGxe+4p7hGtkXtnnCbbDWVKujD0+OqrY/+EArkM4hLZ4Cs4isv3DXyIDZ3C9Iss1Szv96UsM7yRL1J6+Wfi48CNPgtbix+1dR6VvVN7gBN2qNlvT5UtbddxuV/94CS6Lg+19e1F/ZCeE/yRN/n6pdqmXNLpGZvKl4YS2k69eMB7bpn3S9J929sft1Eua+us0fZtDmIdf0U1buGfOMg/yxbGtuTNvIhnaC8FTqg9lifpTT5q+lZGrL/MZR6V97R1XnpzMt7iv1adBPFHJUemVsHmCl9wRj/W4x/12VI5o3H+7HZX3x6dLG4FGoBFoBBqBRqAROJUIxGg/akclooHMxVF59ery6ldk1RN7yEBWro4EEjkIaUoa4pN0nIgQl/0IQuqvU+2TN8oat0e52R7L97bvXetXnKeITX2jkqPy9bvltKzXrG7dKDfmjWnz3I3ZaXn9xpvzk4+clJ6q5CjjrOQUQ4qQo5AuesIBLnAKRfe4rQAAQABJREFUkeKoRP7lIVrIUmQg3kgcvbUXEW5yQtyR7cjIk5ScZebMzaBxbsb9EH2y9sMw/cEn26mXNHO1Tvcw3WsbOfCAi3HByBjhJR0dlerRjZ7BypOny8rnxTlOJpJKDhkcUxyViChHpVf8SO0jqPpU/zhDOyqPE93Dy25H5eGx6pqNQCPQCDQCjUAj8GQjcNSOytj+uAPb1pOTed0rLugtRXmaUh32ejhK2kKULT9GnGHkAWx/kd0WO10dbXAmcslPDH8hG1fQZmxPtqB99Mm2fbwId0q8dGlxfnI87mxvlNNsmt4oR6XFqnfu4CLbVXe7nJU7pQMnHD62jGkcpz7t032M8sf9bCc/6ZgfufSNTGONc05qnNooz/jglTiOXR0hqXYHhdQ5qFy+9vqnB+4aJ7E8upgjOOe4oBN9EtI+zkdp5i3jIce2fOXmOQ5N28mT0kN9fYYz2tZeGZ7oTUWcknihpypFC1rxxv14e3Q9KG1H5UHI/Hp+Oyp/HZPOaQQagUagEWgEGoFG4NQjEAJzVI5Kxr2IZHBGjo5KRNWrX+OoDHlMuiarAR8xGaN8+0nHsuQriy7SEJcxbxaw+0e+kNT2KNf+GFJWtd7OplLxnvoWZT1R+dbO9Nobteq3vlMy7ZSjcuPmdP5cPeVY27duLd9Tee01TrJXy1FZr4ItZ5noCb+RcNMHSRNDuuHKyWg1MuyQMWRqdFQiYGPbkD0pORx35oGzkxw3E7KSVHkiwiZmP3LIHiMQ7K/T5MnP3AQ7x56Y/bTVxnyNZFmdkE43JdxcgFUclblRoZ629EU6jTOOSvvyBfjAjAy4cRZ7gtKTlFbKxlmpjGz1yT6u0I7K40L24eSa636i8uEw69qNQCPQCDQCjUAj8GQicNSOytjvbG8ckGPSq3s9SenNIV7hy+bl8GOTh+NoJ47cItuQHTlAbH92fJxdo/2PG7H595yKywJMfZEZ7svm037kXfoKD6EP3iPiQiNfvXBBPnmbJW9xVBalmB2Vd+96M9PO9NSVndJjqna+lxnehA/pZS+EX0jDi6TGPKa29ytP+2APV9vy8ZeMMw47+cYID+OCVTAL5zPmcT5oq81B4X5l2mRso5My/MqY9BV8pfQYOWf6VZeMYJOxZ86k6cvYjdlxIupPNN/aC7DCFWEgwkSZOuGJFq3iiPghJyVnpcWsOCI+qh96HSa0o/IwKC112lF5eKy6ZiPQCDQCjUAj0Ag0AqcGgRj8R+WoDElCDjnCfJPkmd1X/SCrIahIwUj6QkhCigAcokFHcU1KUiep+iEs8oToM6ZjH0ut5S9ykzjmZ1uZkH6SevVrdT3H4iqzo/Kt6zvT6/Wtktu3y2E43So9bhXhquW2O0WQ7izfWHzjjTfnb1TGSWnFppWbIT3kR29kDZlC3mCH5HMwIlWwQab2c1SmXcjeKMP8xFnphoI5Mw9Imnpj1F6kT/ALVmMKn+zbFoKTlK5SczlG+ULkr/UmUzvkFHlENN2oEI095JscMjJOY+KAlcJKvhDizlGJgJoDJNQ31hDRRA5M86HPjGMWcMR/2lF5xIC+Q3GOq3ZUvkPwulkj0Ag0Ao1AI9AIPFEIHLWjMhzCQsg4KZ999tn5qUqcMK98Dd8YuUU4QHhGgGTbr+1/tjmbLU4ibcgil73P7seXpLgT+59u4RNxRmkvkh+Oot/okvGMvMT28hpXvHBxVBZNqMWPxQGL6r3tqHxqmp+oLJVKt2U0RWXu4RO4hSDVvxh+FL2yL13XWetsnMFWWRx18MKXyNQXHEauBKssWA1fMvbMxZrb0Vne/ULKw5/or3+cKnpFH/2YJ33HUTnOWfoZcUpedBv705c51U8c2zAY+aLxhVdL7ZNPJ/UcXzDTHh/MYlaOSk9Y4u1ZYKy/w4R2VB4GpaVOOyoPj1XXbAQagUagEWgEGoFG4NQggMQw5I/KUcnIR5A88ZeVtByVVtF67avXjHKKhQSF+IUQIhkhNNIQDTraHglZCInJUHdMbSsnl07Ijr7E9DU3qD9paz9l2qZ9+kmqq0XHfJNC3wjm7hOVN6dybhVhracq79zxOpvb07mKs6OynJZ3ylF56xZH2/KNRM5JTkaEh9PMPhIn0D0YhXzHUfmBD3xgxhEuyBcHi7YIlfb01Va7kD0p7PNUJgeZubKPpKVe+pTSQVxjQ/5BcVa+/sDJnGXupKL5RATFzK02+ojOa6KqjDz1jQ95TLRPlr7Uo6/2opsUcVSSLZARZyfMYJfvVMZJiYjKQ0IR1vHYm4Uc4Z92VB4hmI8gqh2VjwBeN20EGoFGoBFoBBqBJwqB43BUsr3xPU9R4oCf+MQnZkelt+qwd/GSkV+MPCN8Y49zLTyCjc/WZ6fFSWkfpxC0I1ffeE6i/bE/csNNwj/CTUY7X73wnuiUdMlXjud4QnOzdNso3leOynr169uOynqi8lK9+rXUqroLV9zjkHuLaxdOubeIM2ONUy36SekojtvaBwM8zpjpqB4Zo6POmAXlqRsnJX4eZyUZmaNgkTlJSs64bX8M0Usduujb2OIENJfy1IOtPkVzN3LXUabt4GWb7ET7gnL4kK0PXDGO2vGYMX796E//2tFPG/US6es1rxa0euPOz372szm1uBVvVx5cFw0O/tuOyoOxWZe0o3KNSO83Ao1AI9AINAKNQCNwBhAIcTgqRyVSw+C3YtZKWiT16tW9b5JYYYsQIQYhfCE5IR5SeokhGogDEiBPCClBtA4K6uiDTuJIVNNu7JMc+dqMUV76U2fRD8Gky0g8F0dl8aEiRdP01nXfyNguWXfqta93ihDWMttyVN69i/xYqen7KMtKzTjLOCxDekb96R4yxemG6HNUInQwQsDi6ORYgxe9jVsdZMy2VHtzgJCGlMpLHfWCm5ScEYNgIU0Y89RNyPFl7sYY8kdP0Rjgqi09QlSlYuZOP+qpT0bIru0cH+pE/7RPKl/QXlu4BfvRUenVPv/2b/82k1LlSKux6Ps4wml1VMIrMXM8Ypi5Go+v4Ktejh/b6owx9Y4yNc/9ROVRItqyGoFGoBFoBBqBRuCkInDUjsrwDt+h9MpXT1M+99xzbzsqLY5UR2TTsctjs2d7tAljC+IKB9n96pMXrhSb3z5OIZKtXkLksjPHKD+BjTrqMm6jQGL9rfa7r359a6P4AmcXm3W7Xv26PV26uD2dv1D7G9XPYBNrOfZFB3ayceInicYsPza0etrZH/WOrsZq3HRVrn2cdGTiSrG9M1fhhVm4ah+GwQ12kT+PeBn4nDfur8eU8WlLF33Th61NF6k8QR/6dA9BGt6XvudK+/whW1yHYKk/jsr0CV96wcf4c3zIC+bqj7iTBcM8VXnt2rX5FbCeqsQZlEXuWo/1fjsq14gcvN+OyoOx6ZJGoBFoBBqBRqARaAROLQIhDkflqGTwIzhe8cpJiaRevXp1+tCHPjSvruWQQUCQgwQ6IAFr0pV8xj8iI5UXsoS8iCGOkSdNnRCREFjpSHrIQ04S1Q9pSZr6ZKZe0sVRqUdOlcVReevWRpGijel6xa3tnenC+a2Kd0vurdIL4Vycc14Le+vWQqCsduUQE0dH5Vp/2MEXmbRa2T5sOCc5Kjk6bcujN6I3Ej77SCAZVs1KRfnGCx99aivaNu7EYDumtoXU0UaAEXzNm4j0JbWdSFdzrz4Z9Iie0VUqL/qRr51ITrb1J5Cjrqhdxhfd9KcdcmmVbJ6oRDrz/RGOSt8giaMyOs4dHPGf0+qoNB9wM+9uAEj3fjvLjYnMrXnP/KSOOXI8k5F5zHFgjo86uJHhN2SVtCdq/XY+97nPzd/WzW/hqPtseY1AI9AINAKNQCPQCLwbCByXo9Jiyvs5KnEMNnr4Rmz27I82V2xJNmF4BLs/Nr+62oe3hc9EpvJRXnCOrUmObWm21QmvWadLmXJcZ6o2936j8vbtervLxla99pXtulVj3Kq69zoq9TfasfoNp4kTj01qvOEfqR+95Wsnjb7wM351R5nkkK8tLIKzuuzq8MJwQ/ZvcFR3jWF0STorMPzRzxjokrmL09A46U6GPsIH1n3v10fyko592c7Y9Rn+ESy1ER0faR9OmDf1qDuOgQx8ED/wRCWO4A08FrnikcozL2tdxv12VI5o3H+7HZX3x6dLG4FGoBFoBBqBRqAROJUIxJA/Skelp/U4KhHUOCrzXRJP8CFECA+DPv0jTyEQyVeGwChLlBeCFQKK3IR0hHBIQ6qUh8BK5Qtk6UsfUkFd+iFJIUzy1m1SX5vqag5EFK8psmJF7eZ0veJ2OSovXtgusrdVq2pvlxyEM47XxYFj3IgRZyWnjIjwJOh/HAO94OipVbpqjyQhS5yc5BgbgjmOI2Q0eUipbTFkNJglXeMancY0WKSutraDLfInGlNi8jKv5iBzm7mKno4nuhq3vMwh+drkGIkMumX+g5s24zyqSwe4x1HplT4clRyUiaOjMsfLOPaj2j6NjkrzY37dkHBMI/jmP78l5Y5f30nNjRFzBGfHtHZxJJPjGM2xH+e6+u806H8d9Dk6KvXXjso1Sr3fCDQCjUAj0Ag0AqcBgaN2VLLL2OoclXmrjle/+gQIJw17l00+Rm3G/dFmD59gG4YzSO2La3ufrMiThpOoN4bYgGTYxgvCI5KnTeSPKTkRx5TE9cp8nF/9euOGJ/PK+TbdLRu33qZTC1U5KqdyXKqXfqWjTvo0rvClOPPkpa7xpM2oc/A4qF7GQ+/wuzEl15yFb4Ufhhuu5yM6JCVXyD49ErJNB7a98WVstuVrpw86iLiBvsf5i7z0YT/b6SPjV2Y7mMJnjZG2YvSiEz6Id9hWP3XoQXdluGI4ooWteCLegE/m+NH/QaEdlQch8+v57aj8dUw6pxFoBBqBRqARaAQagVOPQIz4o3RUIjgI6kc/+tG3n6ocHZXIB4LEoA95QABGEhGCkTpSUdAWoSEnRCbkLaQiqbrKEu0Lo3xyQ27I45wwhhA1fZGXNjATlj6k816V1ytF71pVuzndrHhjdlRORbi253j50p3SA7leniC8excpNa69V+EgOghSyJs+9/paVpzSEZn0Gl26qYtceaIyT/9pk3FIQ/qkIYG2k09OMIRR8JOm/3mj/kSn4CEVtUsMXrClHwdVxmZbnjnPvI5zQA/6RNcQZ2nGony/uYwudM04yIteGUP0ohMnWl73mW+PWC2bJyrjODbv5B9HOG2OysyDYxmBR+x/8YtfzMeAOTQn8DSnvlnk6eC8DizHszY5nnPDwG+Sg96xLyUrx+jDzEv002acU/rmWLBSmvx2VD4Msl23EWgEGoFGoBFoBJ4UBI7DUcl+Z6PhgXmqcnRUhr+x5UX7+23H1g+nYDeKbPjRJlceO18au1+6thFHm2+Ux84MFx05SeSRFbnmdpcezTakb1IWtZneeHOnnFm+C4lXclLemj/9sVncr9yZ99ibZKx1iQ7s4PAkOhqDvuFEn+ChvqhutslULo66B2dybCsnW7AtDz9k94rhiPITyRPTZt7YbZ/tdUofUV/wpWt4oO3oa2zhpzke6JqxkDtu29c2abbtqzfmZ55TZyynEy5oke/oqAzuxmv89sMR8uYdT1XiNvg3rmg8jp37hXZU3g+de8vaUXkvHr3XCDQCjUAj0Ag0Ao3AmUCA4Y3cHJWjMkSHI+GDH/zgvILWilqOSs5LTwKGaOmXQT86rEIigB9iEUJqXwjJWjvZkImQD2n2x1T7yE3/IRXRnfMkUR/yx3bRI31sbi6EsKhRjWdjuh1H5Y16onJno8jeTpGvrenK5WVV7c7OQrA5Kn3Dcnt7cdiGwElHTPQHF6k+jR+JhKWxIUZxtiFa2qrjKbXRuQd3MWMKETS+kMFgZbz6ErMtpUd0CY7BQ9vEtMv8IndWosZZSefgn/GRqx0Z9KFfSDMHlbFIQ6DVUXetFzlC9I8u0ug/6gU73x2xKpZz6lp9eySOSs4ymIZMR/bcwRH+OY2OShgj/Ug8Mg9XNyecB8yheXcM+5atJ7CdM4S0+clPfjIf1+Yc7o4fx6qbX5yb2mnjOMlxcNgp0Tf9xBwT2sax6ulkT9c65tpReVhUu14j0Ag0Ao1AI9AIPEkIHKWjkp3NNscz2GfstDxV6RMgbD2L0pSvuQhbbh3ZfGK4BVzZb+EN2Q7eqRd+IA0HUCf2XtLYguFfcaDZV0YeGfSKLvYF9IgcT0niczfLUfnm245Ki1Jvl5PyVrW/U3LuVlw4LK4YnZb2exyPTYobxeFlX4gOOFHGpC09xeiddsroTmc4h0vFEZgxkB8M1U/d1MsckaONqF70n5WrP+v95EvpknFmbPQU7eufXH3oN1iPfUW+NNuj7PQhTZ2xXuomT6qu8cMOD8Q9pLiGPHqpFxzJoK8FlPgBvojb2MYfvdEIxzUfZB8U2lF5EDK/nt+Oyl/HpHMagUagEWgEGoFGoBE49QgwxBneR+WoZNAjGZxknpLiULCilqMSQeWQQUQY//pGEsQY9fJFQZ6o3lgeQhNCGwKVtknJiKykGa8xIxMhaWQiZJwiSHReRakPY4oeqU92SBRH5eZGEbeqt71djsM756ZbXv1ajkpcZc9RuVVEzFizIhhJWhyVIxbBhI7rGByMmbPHuDhX8mQg4qeOsXACZRz2g1vwSmp8YnALVut9Yx5xoBu9RXUjJ6n6yuBMRw4/BND2SFDVSYic6Ap/x4uxxlFpjuRF/+hLt8TIk8pLqi/7MKY/XeLk9d0Rq2SvlUNNRECtkqXzg4jn3MEj/DltjsrMO/xg+dJLL01uhsl/9tlnZ2ejeRgdlV4B67hA+n/84x9P3/72t2fcP/WpT83nFDcFzJffpxten/70p+fzCuwc3w8TzH1u7OQ3TR95bkLkm6X0a0flwyDbdRuBRqARaAQagUbgSUHgqByV4QxsczYZRyVbjbMSD7R4NQtW45BSj52vTThdUmUplxfOxVZjS4psuXAmeNMh/CEcASdJ0HZsH1uVbYlHjRxFmbbkZIFkZC7yFll43OyovLlTr37dKBmeqOTovD2d27xZ+twpGZxxe2+e2SjOKMT+HNOMiZ6C8Rg/zMJ9jDO6s1vpL9rGV5RpBz+cCX8Kh5KnTEhf+ifT+IL7GnttRJioKySddw74M2KuPzEc0LZyctN3ME4/+kg/47buMpdjqk7aJo1qkWMfRvqHW5yU+KDjAI7BhAw6SbXBZfOWGJzRm3jyVCU52tJHSDrv7P5pR+WIxv2321F5f3y6tBFoBBqBRqARaAQagVOJQAz1o3JUIgEIBzLlJv9v/uZvziQ1jkrOCGXq6FtgyKcdIhBiIT9RPXVSbyQ0IRDaHRTI0R/igZggciFz2iCBiCidOT44+NaEMESSHHpE13N0rvHMY9rxHYtyVNZTldevIzUcldsla7vI4la9BujXHZVxXJKb8eoruiJ00Tc60Fn/2ihPHe3pYSzGgJiGYKuvLGm27Qfbg1L97YdhdL4Hj90+5CkPEYyjMiRQvvGQmzDqRj/zgiybC2Q7hNt+blzoRwh26zQ6ShODL0JJL46pOCpffvnl+bWvCKgnLZFYuo56Rt+jSk+boxK+sPVk4k9/+tPZUclZac6+8IUvzN8pgp35tIDB7848I/nXyrHpaUo3z9T/4he/ONfzlKvXyJoHT1U+99xz80p95xbtHybQTV+OxdwwcWzYdqNKGYel81U7Kh8G2a7bCDQCjUAj0Ag0Ak8KAkfhqAwHwMdir3NUcspwULLT8MF8k3w/Z5i2bL5EcsYoHy8Q2JjhciOXwCFGOeqnjXbsx5EHaMseZPeJ6yfqyKMrHkUXssnb4xm4mgWZXgu6U3xioz77UYveblvIWQ7KeqLSNyovXNipuOh2/vzCw1Af/a8j/cI39GXc+qaDKA/eMGCz0p8tG2dluK166uOBbOQ4KjMOeKRvsshMf5kDacasbD++SM79QrAyLv2IdNS3bYFs/aQv/aSvyKZforzIlZIT3NRJe3LTJmnaakMH2LH5zf3aUUl2dIssOOcTEZyUifgOjuI4Ijf6Rf+k7agMEg9O21H5YIy6RiPQCDQCjUAj0Ag0AqcOAYY9g/ooHZXIALKBIHEoIKierAxJRZKQHwGZYPyL8qQjOWHoJ4R8pO6a1ChP0C6kJdshSXFMGLdAHp0QOU9rIdIcKNEn7UcipN1ChBZiR8a5c8Z0ftdRea6eqOSo4wQtR+XFevXr7KgsAjothGpr694nKvWTSNfMTZyUSek9bmcc9KFzCK0x2BZhFVylwXkZwx4hTJ0QuqTGO+qjT1GekHoLDntzaDxwG28EcAwieuZBmToJ0Y2+xpKYcRhTxqUs+o642Q5+STN32ZfKi2MKueSo9NQeR6WnKZFPZDTHS3Q8jvS0OSodG+aco9e3PoOncf7+7//+vHgBjvntmQs3CLw+6Vo5KrUzR57K9uSkOXcTQLl5cvz7nVqp//zzz8/OzsPOi+PDjQTyOCNtm2P9kWub7lL9t6PysMh2vUagEWgEGoFGoBF4khB4VEcl+58tziZnq1kkiUtxVOJ97ChPUmYRqDrqxlkZO38/u1/dRPXZjAIbk73GVrPNhhQiS13b4T5zYf0JB1A/dio+gpfEUcUmlKeuccVRKY28hWfgcuRwvHmKcprfpFNN6806pc8OB+Wd4n/bxYc3i4stTyvSyzjIoANbM5zIvn6FYIr/jItO6aBtOAwMtBeDBxn6gJ22ifYzDvOmHjlS+9qIwU5fyQvfCt9LOiu7+kO/hGzrI1GfieSQPfaTviIjqfwxkJ0xkJe+IivpWlf11Ic9R2XmPo5KWJoL9fQZPGw77tTzxhhPUnoLDL6In+AunJ5pP+qa7XZUBokHp+2ofDBGXaMRaAQagUagEWgEGoFThwADnzF+VI5KAIV0MOw5E6ymZZgjrIhqiBICgQSFUI5EgAwBSQjxsC8/7bQVEYeRvIQIJdUu24iJ8YYMkqdfOiHW9JOOq1b1n/bS6JNxnj+/rJJdHJUXisScL7Lo1a+Do7KeqHzqcr369QLyWsSwyO3iqFwIYuRHdvqkL1JE3zhRQqyQaXnawgGZNQ6R/nBNhBl9ozO85AW7pCmXCmlDn2Cnz+gUfdNulBsZ9FMfcaNziOBIqOfOdvvL/JqXxDgqjUu0n7mnQ/DSFz0Ts5809ZLCkk4clUgmR6Un9zxNiYD65gjdyTvO4Lj7yEc+Mn35y1+evv71r09f+tKX5ps75jI4Pqh/YxKkmTdE2mtUv/nNb07f+MY3pu9973tznc9+9rPTiy++OH31q19929Fn7tJOpcP2Owtc/YGX+UXa4clRCVPj8YSkxQvmz1y6qeX4piv8PYFpXpwzvDbs6tWrczvk380A4yGXfhyVHJ/qHTbAx40J/XFU2s4c0yM3faQWWrSj8rDIdr1GoBFoBBqBRqAReJIQeFRHZex+9h3OJ3JSJrJvLQRl64WbsO3xE7a8lD2YOJbFwSZVFz9gw7HZ2InsNNtsfHqE80Rm7FptRPXCB7TDS9iAa0cluexY7WOr0ss225MMb8PZ2lqcpHfueIqynqa8UW/UqdSrYKtWOSnv1ph3yoY9X3F5c090yzj0RYdxLPrQlzHDzPildKBTxqDN2B4m7FghegdD8yOSSY4+govUPtnhYFJRXiK56onr7Tlj90/kJi/70sxBttWJ/KSRP7ZPvymLzPA9qTwhuhun7bRVln7hFA4YR6XjwFyMjkY6Ze5hqh918EOvgLV4krNSah+fJI/s6KjfhHZUBokHp+2ofDBGXaMRaAQagUagEWgEGoFThwDCwFg/SkclkBj2DHoElaPS6x3zRGUIVwjqSFxDLsiIgR9SIxUQjtST6kfeSD4QidSfG9WfyAuZsa89AkInDsqQabqtyVDkSxOWce69smZn50LheX66WY7KG7uOyov1yh+vfn3qKQ5FRGrR7U6tuB2/UUnfyJaKdBXNEVKNkI4OP0RIQHzzqldjCRGFjWicdA1Zs528pPLGOhmjNnQb9aCLffmjzMhISkbGoU1uCGTFMv0jJ/1pSyd6G5f5MR7HiVSUr86oG13gFD2lYwym0Un9kFSEk6OSI8wTgEinfc4xeqt7nOG0OSrhBTdPLXqN649+9KPpBz/4wYz3xz72sfl84DhF2J955pl5bjkNOSo5Is2Vb1mqywlp7h335uf73//+7EQ21xyVf/iHfzh//+hh5odujsHxxpA+7bvB4HjgDHVOaEflwyDbdRuBRqARaAQagUbgSUHgURyVbHA2OzudvYTreXpS5LDMK0fD82LTS8XY86MjyLaoDTsxMXZ/7Ev2msgWFHCCUR7d2HXqjzGcgx0YRyV7kJMqjipy1SODXH2HT+lrkYsDLTzu7t2deopys+RZrLpZi1Hx4J3pcjkqr1zZqDGcr3ix4uIoPH/+wiwDB9HX2h4NpjDgXIRBxq9/ukV/YxDJkWesQjBM+zgqMxfGJRhj0nC38DBp5jj1Une/VB5sxDFkP2Vjqh7Z+/Uzyki95EUGLML9jJ0cYzNfMLCdcWirjvrBL05qqXnAU8NN1deenMy/fpWrjyfgORa2clSKFkHiEOToJ3qSJbSjcsHhMH/bUXkYlLpOI9AINAKNQCPQCDQCpwyBGOzH4ahk3HPAeO0rw5yj0hNKyGxW1oaEjSQQoRAY9wjIGOkrXwipSX1lISypF8IykhTt5adM3whcCLVtpCQhJGNMU7a5adWrVaflEKy4M7/6dXmi8saNWvVaql66OBXhtqK2vlFZr4HlqFz0RJaWbWR3u/TK2GqjyNRCsD19GVKFiN68sbym6I03Xp/JknEg53GyIrT2Q6pC0mAQHKTyU5ZUvkDm27rUPjzF6CENAQuWSdNP+iIvcxNCjsCFDEZW5kx9skIyzY/xOFakovmJrjk+ImdMU7aWrQ/jC9nME5UclHmaEtlEREfir91xhNPoqIS9V+deq1e5uhHGwcjx6xzgGDW/bmp5tasbWm4QIPkcm8q80vVqPU3p3GHuzQNH5ne/+905dWOGo/KP//iPZ4fmw8xLjmU6jr9rx6cbDG48OBbo2Y7Kh0G26zYCjUAj0Ag0Ao3Ak4LAozoq2Wvsck9QWliWT32wa+NgUyeR/c6uF2PXyxNTJ3a/9nGwycMN2G9sd5HNZl+IHuEHsfNwAvYje09eovyZU5UtGU4iZVuSHfsQ18CRyB95jScqRVytRFcf9TTlrYvTna3igtucZfXK10s++7FRnPd8xeWtN5cueTLS61vrFbG7Y4ijkk5sUv0ZRziy1H7GbzzaxkkZncNX6EnftM9C1mBpvpSTlzHZHvfX+Tme1UnIdlL5o02dfHkpmzeGevbVS92kqTemYxmZ5tI8wS3zqk7mC2bht/JTB07ho7C3QFEKR/mZh/1k0Ue5Y8UCS85KC1tFvMFTlfgjmaOsjKMdlUHiwWk7Kh+MUddoBBqBRqARaAQagUbg1CHAaGdwH4ejEgmKo3J0ViKznJUhoAgTAqq+iBwJIwFBKkS6SkfCqS7CIi/Rfoga2Wuyogx5EUdCjNTZl59A1jqmbOmjCGw5KTerzc7OuV2yeq5IzGY5LsuJWI7KK1d2arzlxqwnKmtks56eqIyjkjPSePUjSGyHVGXcSA9yxIE2P+13+9ZM7mAYBzBc7RsDPKX0RLgy7nU61pkVmHXYc5xGjxBC+tiOvmQL6YP89Ckv+Jm73AgwFhHZJi8xcrWnv/kwL4kh2MrJHXUajxP54ohr9KOrtvpHJjnUQjLzrRF5cKYXGccZTpujErYwgx/SDlOvgIW1eTRPyL25vVrOSMeuPA5jddXhIPz4xz8+OzPtmwdl3/nOd2ZnsuMojkr1HibkeJQKSd2kyNO1HNbRw/krx/PD9NN1G4FGoBFoBBqBRqAROKkIPKqjEr/iAPMUZd6CwRnDro29Hts7PG9t19snR7nUvrbkssPCaWDIrme7x95nv5EfvsBWY3+yGeN4Ut++trH/1CEjzkJ1RPvyR/4Q/ckWBXkbG3gSjrNZ8muR6u2L1a6elizud+5cLVAdHJWeqDSeixcXJyEZt2/rf3lTTriQfJwsGBm7bXl0zvjpmvFJRzzIgCPscEKOysRgqjzjWcayOAvX22QlKBvDel9Z8LWd8tjYSZUlyEu95K33kz+m2sEjmEgjC1aiMUojTx3HAaxxCM7JRBhmDtQTIifHrX0hc4DjZHFjOKSnK22HQ5JJL1FoR+UMw6H+PFGOShO830nDweeHloMwI8+Bq13Kk6ZOp41AI9AINAKNQCPQCJxFBGK0H5ej0pNSXv0q+i4dA/3973//TGCRJoQpJAwRQALYaUJsuP2IZAhliKc09WPzheymD/vpY0zliwhd6kQHepC3jvLZnCEx0Xt7e7OIzmaR3XJU3kRoNqcrlzdqnNP01NPbJZ8HcnGgxVF5d/cbJ/Tf62fpd3mqshyaVQcOt27WK4JmR+Wbi6OyiCo9YMjZk6cq7WeM0S22cnQ2xmyPdfZ0WMYdvaTBmC0uqrsOsbOl2U6dEMTMX24KIHKZZ2lka7/fDQvzZDz6J1PULmn0o2/COH552iKmnGecZp7mQzA5w/KdEQSWzFFO5B1letoclcHXvHKoW3VshTG8zSmHpG9Rmn9PWvuNmg94q+fc8JnPfGZ2VDpnKCfL3Pzrv/7rnJJl9f5XvvKVh36i8qC5IzNP13oFsN9RP1F5EFqd3wg0Ao1AI9AINAJPMgKP6qhko3OGsdW8st/31i3uiqMSv2DfscFt4yZsK5FtF0ckOcrY97ZFZdK1zR/OwD4X2JXaigI+wJ4bnVDhFuz58AdcIbxhTG2PPCIcQD9jPHdu8UHgelvb56vfXUdlOS75tC7XZz88UXnlyrmKu+O5UE7HevVr9LxzZ3kyNH3SbY2Dffl0Mo5wKGMMjwoHiq7axFGZhax5cxDsYTpyP/ITbCfKy/yNdVJ3TNXbLzxs/oP60QeZovEm2tfWuHKsmS95qRv8RkdlMDQHwU+7yEkqb5QDfxyHUzKLMj1V6e0v3szCiamOedNOaEflDMOh/jxRjkoHTw4kE+6gy8krP7aM2sHgQMzBkR+8ejlgU7fTRqARaAQagUagEWgEzhoCDHK21XE4KtlbHJWcCZ58EvNKIPmcESMJZachA2w0gR1HN4SU7ceeC7GwilGUp4y9ZyzaaB+CiwSnnxAzeon6G6O89B+SFGIhDXmpTmb70xOU+hrbbG1tzI5K36i8WY7KjQ3k9FyR+PpGydM708V69WueqLx7h3NtcUKyabdFhKvk62/Hq1/nfsspCIciszc5KsuZw7mGHN3edVTCERGNs9J+8Ix+dE2Ut1+5cc867GK53g4OY/6sa+mZQIaor6Qp085c5SaDuYtdn+3sm3vBvKxvaMgjX9/kqZsUliMpjA7qG7cojy76yqt7OCnFvLqHw6odlZm5d5bCOHMDa/uC1cYcjoh8fnfmxBz6XfvNXq0nLa3Od9PL79hvn6Pyhz/84exUJte55ctf/vJc551peG8r5xPzzmn97//+7+2ovBee3msEGoFGoBFoBBqBU4TAozoq2edsNgtSP/GJT8x2m21vz1HG5marxxbHPUaOhq+oJ2+8r882tK8++11g27P9cAgpmzJyY9+rw95kS+JKUra8PGVpo134S/KVJaav2LDy6bG5ufCnCxc8scdBilvgI+dKpwvVx/mSQefyU1zaqbFtlMOSc7aeEq2nKWebtz4ZInirzt27yxuD9EefcBXjVjfjD98JhzIeOCTSU4wc7WBrbsINR0elcpjpDxbrIG+dP+6P23Q7KKzLxv1sJ43MpAfJTL52orkZZQTDjI085bCBH+zY+44Lqbzgpp52ieGM9oX0l+OCjDxViT96Gwv+YDufECGbjkI7KmcYDvXnxDsqx4PBQeXGjAPKj9KB48cnWsnhByc4GJQjtU5QDiQ/dD9WZNfJ0H4OuEMh1ZUagUagEWgEGoFGoBE4RQgwnNlIR+2oZJ+xsxBVDkrOho9+9KPzU5W+S8dRmVeUqiey4US2GVIR3UZCEeLJFkQM2HhsPSRDfW1DgGMfStl+8tl/sQHTpzb0Td/7TS9bNJFumxXP7eq6WQR1JlX1+p968LEI0MZ0s56qvHnLCthyUnJUPr05vacclRcuInNFhMtOnQlqOStnYrlLLrdLwHZtG8uOuLWQ5rtbe98jefOt69MbNe5XOSrL1t0o/S9fvjJjagWzyN41PuMSjTExecabbek8BtpVv2KI1azLMH74JC9pbHVl5CTqM9tpF3I3ku3M8ZgaG/lkZE7H+ZOv38iTitFJv+pknNJsK1NPf6OjErEUOdI4rBxvkUn/4wqn9YnKHBf57QQ/DuF/+Zd/mR3D4zxlThy7XiPGEfnMM8/Mv1/zpJ0nMf32zaXyF154Yb5BFtmPkuKXeQ1wOyofBclu2wg0Ao1AI9AINAInHYGjdFR+8pOfnN+Egff5HjnuxZ4TRltPHns+9+VxFna+fCn7znb4S2x29jhuMjqWlKUem18dziP2O7sxzso4pNiZ0UUqjiH2qnpk4SK4gn7T14V6KvLSpXPlgKxXtJYj0hCVbW2dKx54vt6C460vxSsvKKs6xlVRu/CQejvsLJNcMf0aw8hdbAvRJ9xJOka6iuqJMIEvDhxH5ciH6ZG+xvFnO7gklZ/tpKlL9weFdZ3sZ9zaR27SB8lUPsqxr61xSccYXIKf4yE+peCvvZD2SZfce5/izLw5Nhxj3syDP+IO165dmx2WuKRj0LyoL7SjMmg+OD3RjkoHnoPKAeCEk0drHQxOGk5gXiHmRJhVG+o76NxgcGDk4PBjdFNMXTcErChwIuzQCDQCjUAj0Ag0Ao3AWUSAzcSAPg5HJRuLzRUnpe/Ief2rVz1yVMZ5yJZjo4WcIlcC3RAKNiBnJNLJDhTZeEgBG0++eggJWSMxCylLX8jw6Owaya1tMvYLIVIhPcjLTPKqjVWzQqk71cLY0ncqJ6XXv+46Kp+yqG6j7M6pxlgV65smC+4L9lul+xLLMVffKtm6dXPaqbydIjV5ytIcsXtvlH37Ro33VaTo9TemG+Xo3Jjt26en/132MFtYNN7gStdEY0ykvxgMjCF2N/2QKqm4JoLBI2VjvWAYgpd98tUjN+TamHITwDyz35PaNm6Bjo6PRGMjX1CHPCnZdFOmjXpjm2BCJ7roy/Fk1WuepvTKHuTS8TU6KoPB3OkR/zltjkpYwdecZF4yN7D3xOL3v//9+YlK85MbVtpl4QGIOSs/9alPzcezOfIEptcrkQkzjko3xnDBowiOuXZUHgWSLaMRaAQagUagEWgETjoCj+qoZMPhWt6Y89xzz82LyyxMxfU4yNh34SDsP7wjtnn4WOx0aez0cBNtRpuSLSmyMQXl6koFfCCcEUfkN2Dny8M32I/qhieMqe11f+Ek2lVvpf/CR65ctgi1Xut6eacWh25N5zbriT08sByVt+sVsDvbF6fNc5ytxXHPLa97PT/vF2+sBa5CxiUVhegWzObM+jPa1HRJhIXtpOqJMMF5+T/MQ/iwvPC/jDd9RAf79EjM/n7p2GbcHuvaXpfZT1Q+Bv3uFw7KH+vup7NyfQVDx0j4J+zkJ6z7iI7q7Led4w13wG18NsKCSinHpfsV+jI/2rejMkg/OD3RjkoHjpOKiXcDwQraTLTJ9kNDYkU3w5zcnEzcXPjZz34238DyA3RQaKccmbWaP+/OzknwwVB1jUagEWgEGoFGoBFoBE4PAjHaj8tRyeZCWEWOSo4FNhtHZZyGI2kNeYIw3RAAzgOL1ZDNLEBj5+X7D8rYiyFlFqKJiFme2gwZlqZfZDjkNoRQOgb24xgQGHW0O3ducfJtVJP5Ocl6VWupO926UY7Km/VU5c0ivPXdkqcuT9PT9erX9763viVZ36vc8IqgknN3a3kNzd1qtFW26/bN69P29Tem7ddenbZvvFVsu8hTjcuTlcYHi+tV7/XrN6Zf1lOV//VmvdaoZGxcKiL63vdNH6ibAu9nD5ejEiHN+KJv0mCcNBgYK8wT2dnZhsEai3V9dcc6+hsJX+qHTBuPyG4PYcx2bgzE5teWvBwrGRu9go2UDoIxsfkz35nz3AShF5n6cQPD8YRneLUoRyWy6fiKE5zOwji+OeOI/pw2RyW8YJtFpnDE58wR7BH3l19+ef5tOx84T3CwKxsdx+bZjS/zyFFJnjnw+3YusfABn7N/FKEdlUeBYstoBBqBRqARaAQagScBgUd1VLLT2Njur1+9evXtmNe/jnyEjcc+jy3PJh8jWaLykUOEi6x5CXzVEwX24cgb2ZOJ7NAsgFRPm/Q18qHI0idZeAj79U5xMoGj8vJl3+XcLF63Pb3nqbu1mO5Wjauch8XJ7tw+V4tVr5Rj6kp5+67UG3gul2OynJabFljWuOpJy41a4FpQvM0p1twCTolzp/WHPsYfDJKO+SknD87mBQ8enZTwViYaq34SRj3Sf1J1Ujdp6q/TyEu9pMmXapOY/ZTvV1/ZQfkpS3nSyJPqC1Z4SHij+YXZqEfqpr42iet6+tEe3+Gnwh29+vUnP/nJ7IvCJ/FLx52+tG9H5Tgr998+0Y5KhNSqZmTWt0zcpDK5ucHlR2dbdENKcJBcq8dtvVLIQeGjvk5Cbj44QNT1OPqnP/3p+VVBOXneH6YubQQagUagEWgEGoFG4HQhEKP9KB2VDHcEiO21dlRyLHBMcMx46o8DIs6jpNoKdGO3seUQTGSTo5KTI09XsfmUq0sWmWRn9SiSRo/EOK7GvugbnZG2kFQ6IBWCcmEuR/DqGyUclfNrX992VFrJy1G5M928LlaDu9vTlQt3p6ev7JRO9YrWp4uo1ndKiulOW0VWt2r57VZ9e3LrRjkp33xt2n7lv6fpF/8x7bzyK97OerKSo7LIVRGhO/Utk+u370yv1Xcq/7tI0b/drO0ivTtX3jM9VYv1fuv/fHBewQzz95R9HOJN55DS5ElhEqJqbCFlI/mF65qYqZugPHFN9oKrNLLVDUkciaJ5djNAKn90XCKA8rUVxjHYj7x1HXPveGDnI+q2czzAhE76wS0QSQQTqfTaHk5LjkrHHLkZm/60O+pw2hyV5sxv1dOPSHvwdLPHMafc79jv9Pnnn5+eeeaZ2fGoDO4vvfTS9K1vfWvmdG5+mXNz4XfLQXm1bobhcZyU5lT5UQTnkn6i8iiQbBmNQCPQCDQCjUAjcNIReBRHpbGx29hmHhpyj923xdl03qYz3rfHOfCBcJLwkPCz8LLY+OqFe4VDSBP1HZ6hrhCOwbZnz8VJKWXr8y0oI0Ob9Cld98vuZ/9H1uKo9ERlLUB96vz8lpzfeN/t6b3vuVH714sXWnS5VU7KC9XP08XZ+CbeWzo+XfFyOSe9AteY6HovjxjHWYX3hJTJNL6MX5r98LZwFWXGwz4OF5JmnJmD4Ke+MPaV/dRJmTT1pYnqJ9+2MLYd2y+lS/20T5qytJcqi6yUH6avsa768Mq8hoMGs2ApTT1lYsoijy7BkFzHiGMsnMcTlR6aw39wmvG4w2k+//nPT1/72temF198ceZAkdvpvQicSEelCXdQuAHlpoET6He/+935ZgRCi6S66ZDoxpOTJALsAOHY/MEPfjAfQL5d4oeZJyzV8yi6VwVZ4e8E6ibGw4Tx4N3vwNWH6ABe/6gepp+u2wg0Ao1AI9AINAKNwHEhwIZhrD8ORyXyGkfl+mlH5GkkUMYb3ZBN9h0SEEclxxJ7zz6nh8AWzAI2T1jFEcoeCyHWh3rrvthrIR2x3Wah+VOkBLnkmJxtPNtzLPJUhNO/7XIoFpTTTY7Kt+ozBG/YuTVd2bk+PX3x7vTe952fLr338nTuPUVaLz81bdW3SupLKeWorCcB6wnKrddfmab/+vdp4+UfTtN//2LauH5zmm6XDDZxyb5TTs3r5ax8pZyV/1kO0J9W0a8uXJ623lPf+/zAb07/53c+NK9o/sDKURmbNBi4KRBCrizkMbZ3SFnsW/nifkG+evuRudi/0lG2uo65pKOjMdvS3BxA8OKsJGcch7lKm/0clY4Hx0KclfiA8WsnkMsJnicqcY68rsfx5ZijB131nbgfFo+Sd1odlQg6ou71RyHrjrn8tjnV8bqr5Xi0+l6ZubAamaNSG1zPfJkr8+jJbItQPWmp/VGGdlQeJZotqxFoBBqBRqARaAROMgKP6qhkn7Hd8Dr31Tko2WdsNY5Li1PZbnFU4gRpo50Yuz5p8tULR1ljOHKM1AnXwAfwRvY9nsinwJ4fHUb6uHjxUnHCvac69Z8+2f1xVLI/cQ1U6OKFclQ+vTm97713p//1Gzdq3G9OT115o9TzzUPfx7w4vXX9ffX61/9Vef+7WF698WOznq6cOCrxoV0H3bxXf6S7DzXuxzHGcSofQ+qHryXVxlhwnsv11p1Lly+9vWDXuJVnnMFu1qP08z8hfc9lu0qmz6Tpc61b2uonMXnkpf06Td9jn8kjRxjbpIzsjEmefSF60TO807wmLvO6cNnUUS/b0oT0oZ8ct8pHRyUeyRfF92ThKx7jOHQMqduOyqD54PREOiodMCYTWTXBiU5yv/d7vzefAHOTyUGivlUSIcRW2js5eXoSAXazKq9xUuaARW6dSH37xAn0YYID2wGH0NrOwezgdVJwEwyxjo75UT1MH123EWgEGoFGoBFoBBqB40SA0cyGOkpHJX0XAri8bj+vfuWo5Ixgf8VRiUSxmUQkNgSKDLYa3dhZbEIEE9FEONmH7Ln9HJWcPuxFstlhIRMhwLHNxr7W5MO+fzN7tI0s7xIT7ep5vGmziMzGdr3Kcqseo6yUvuVzLF2Lrr55d7r5Sm28+fr01I1fTk/VStv3vPfCdOk3np7Ovb9szvf9xvwk5HZ9r8RTlds3b0zbb7w6bfx/P5nO/fCfps3//LdyVN6eNmp1bnU1bZNd0atef1nxP7Y3p5d2Lkz/79LT09b73j9d+a3fnh2Vv/3bvzV9oFY1G3/GS1/YrqPykDq6OxZENq0obx3NC2wS0i5tIiPlM45VX73UkaZe+jPPicpDIHOjQYoICrGz2fbGFhlxKKaO+WeLrx2VGbd22uSJSk/9capdq7eyeAtLHJV7Nyj28JgVOcI/p81RaQ5hm98sjPEm8zoeP+YHB8PXbCuDt6ctzYf24VCOIb9dr4hVXzTHRxnaUXmUaLasRqARaAQagUagETjJCDyqo5Ldxk5jk7PLOCfdX7egjLOS85IdjueNIbagtiJ7nn0u4itSeWLqpK8xjRyyw0nYmuy5cYGrbbZo7FDyL19e3uwTZ6U8fZGJI4SLsEu3torgFSG7eKE+P1DPOL3vfbfKUflmje2NcgbWG3G2faKkfBJvXar4gXJUfqBkfaBkva+IE1v1fNUhd+FbxSh2fYJxPu7PMcax02scb8Zs3PV/1s9fmF04X5yzHJSXyhl74eIenhlfZL2dzpxX6wR9ZRu2tve4InwSl/4zjqWNfjKn4xxGYuaKDMH+fiHj/f/Ze9PnupL0zC9xF1zsBLivVWQv6lIvI5W8dChCaumbx+NxzPRI/XdZiolwOLx87X+gNTERtj90S+PSWNYydivkUJVU6m6yWNxA7MC9F3eZ5/fmeXAPUSAJkmA1wM5Tlciz5PrkAZjPefJ90+10WtfpfDyv961elutiPOEmjCcxAe7pftRjl085HgO/k465D9/hXeN7BJ5ALVSy6PWwUEnaIlTWR+bF56dSqOSl4Q8Jg80HAwacDwb88fvwww9jRT4vI3/EILa8YN7HkvS8LKRl5f6dO3cijUkvprh85CIvf0hd3othevapX8a6WMmLTZv4RYRI88eaD0UErnlWjoLAWUSAfzb4Yx1+12XF0ZdFR39fHzP1D+0X/k074Q5Sd4PfK/lybzWn0rT8wrflcoFz7pejIFAQKAgUBF4fAeYuzKHehlDJPAtXQCwKs1hpi0qEGeZvzJUgrxYPmSuZHDimjSYXCBdeHYtQCfFkTkZaz70sglr8NKEgrgcTGtCjXoLJSFzrHzhi/aie8W+RiJduNUcSKfd7qdHvSkyUj9chroSEpXTF3r7Elh2ESm1WubmeZrcfpLnxnuaDWsi2vJgaFy6mdO58Gs8vp5EsIkdjEVcpnKPdnTR1/9PU+rv/OzXu/7wmVMrqUm3s6x/ETbXpybiR7o5bIVSudhbSSGXNSai8ikXlEUIl4+AARpwbB/cb/MAZEkXg3PeIORwHJnEn3zucz3lJ4rQuy2mdxvctUjqujzd8AKGSZ5THuPLe8OGD/nDQZp4TczAXJx1CFvNw3rW6cE05pIWs8g7xLtWFSvMOnpGGsmmrQ1Rygj/eNaHS4+pxAT/GlHHnAH9+13gP/bvPmHGQhvTgTh7K4iAPafwuk5cyTvLgb0lx/XqSiJayCgIFgYJAQaAgUBA4rQhMhMofy5PFX4UHjFdta56bT8d8G8+HH3zwQfra174usfJOLFDluzxzcs3kqjndZD59eI7HPM8cJc/34G4W2ixeZnGTegn1g8WfgwEWlV3pCXlx69raurjjxsGCOeaOlI1QCTdAqOSaeprynEOZ5iu4fM3zVxbaDST8DSRU7suiEqOozTQ/t6m8m0qzpzoGaWt7TkLlZS1cvaw56kUtcsX9a0cSnxZWaqEp5Y7EIVmAqpvCI7d+wi+y6Ei3aId5qePor+67184f6XWXdE2+l6o/h0VK8uIdyGnzXpkZwyixKjSaVGsXbYbjxsJatd0xomsevyoxI6zCow21cfR83XW7r3CEfJ4xoF2Hj9zm3EbeH0BzfsfkqafzOfdJw0FdFieZ63fl8ajfz/yOZ7mPk7LJE56UgqtM3km/m/SR9vCu8W0C7uAtRLCoRKiES8Iv4bFwGnB7Vqj8AxnVfZOqynEEAqdSqOTlYbU8K5o/+eSTGGhES8gspuT8seOF4mMUH7/4YMELwsvBi8ELyQoOniFG+jnl/d3f/V0IoOTnD+lv//Zvx0e0I7A58hZl0z6ET146znnp8x+d/GGHjLy8fHjgoxztrP/CHFlwuVkQOKUI8A/pQH9Yd+TOYG27n9a2emlVYU+WHtIP491+W03n9w1Rcna6mZbm5P9+oZPOzU+nhdm2RMuT/UD1tvpQyi0IFAQKAqcVAeYuiAInLVQyB4IkMV9jHkZgTsa8i0k6AiYCEvMzCxXk4W8+7TFhqM+duMd8y2IlpAAhCRGLOZ3FKYtSkM9MPDOxhVyYABNTH+VzPBtXpI1/33gmUtRQu5oEOWttiqw1BxIpuzsKW6mxK/lwsKdUEsvEh/pDuanc0wrLTYmYWxshVOL+dW5W+6Hg9nVZFpWLK2k0i1A5n8YNuYCV+AjhS6ufp+Y//L+p8fB+mtrppbH+nVW1sqacSl21dUP7Wj7SvpSfSaj8VPWst2fT1OJyWrh4SRaV1wJbW1TSdwcwJvjapNGYg7vnsSZsHgOecTg2Vr7n9B435+O507r8esw5gfwEPgQQLFIxv2ZBoAke5dJ+xpd5NXNs3h/6wFGvl3ukpc98HCFwzrjTJtJSD+8OwjdEEvekrH79mRY8whfw0sL7xTtH+9xe4pM+3jWhEnzqeHls6tgxDgTGilB/V0hXx5zynN55fM2zkzp454pQeVJolnIKAgWBgkBBoCBQEDjNCJyMUCnHpjE/n9W376sSKn89rCq/+rWvifddSyvLKyEIZlYFGhPOgWiHgIMQxvSauV3mahj/5Hk8LlrtpjWLluZwee6oLAcHZTG/Zz63qTk8Bkpr0hU2NjaDUyAsMeeEE8zM4ho18wPan7nRZEsM5q6Ul+ejfX2T7IpP7EmolMvXhW1xkQ3l31J7sxC1sTGSULkgofK6hMor2i7kourSAtUGbl/hKVr0qECMwcdohFcYxFuC582kZE9GsJi4Ta3z1l6Z8bwAAEAASURBVPr8N86Vw3PpjJ3E3eBAFnhzn0h7MAaUz3+6F/d5VNEbsbPcHvU/Y5C93gRHqzjbGM4aGTyPd2x3voh7uf6WhNNGY4Irc3zKHTHmlENR0V/FnFYD6vZmLCZtn/CL/M5kvCZ9cT7uR5cqTtHX1imIk7wbWajEonLiHdMcxbjn7wZY+NYXOk/6Qh8QIBHED/NI+OT9+5+HpgWX7fXgkhYqv609Kn9fe1QWoTKP+NE/T6VQyUcohEkGmD+exAw+v3h85OLDBC8Fq6lv374dHyy4RtzEbRN/eFjJwTPMzflYxYcGPjyw1yXqNukp63d/93dj09+j4fniXV5g8vJHzxaV/jjDLy/1OPDh4Wv6A41bI35h/Ev3xVLLnYLA6UWAf0j7+8O0vqPfodXd9NnqTrr7ZCdt7Q3C0hGx8m0c8Q+2/nlBkFySMHlxaSbduDifrizPpvOLsqjotN5GtaXMgkBBoCDwK4MAk2zmMG9DqGTOxjzoypUrMd/C9SuCJe5fmRfxzGIiBIuDtjCHIma+xX2IQiYLzbjHHIz5l4VK5ozMvzio08KUy3bew7FJneuOAuIH0iScSURRJGpK7n6aIjKt/X4IlK1hT+dySdkTOQ2hckPWlbKenJJQqXx9iY69vvao3NHq2125BJLr11lZVM5OixiLFE/NL6XxzGIaTS+lUXMujZuyqmx2Iox3t9PUg5+lqbWnabwnK00JlfFvsEjsrgjrhsKjKQmVIra/6IkQNyU+LiylxfPao1K4IgKfDxF4Lj4aWKxjXkwwEQcn+u15aRDGipDWzydkMKjeBKLqzGkh8Z4Lc69+uI56WaThmpi8jF89MMaIiIwtY+3xZWwRKRHAcQGKaEmf6A/lcXi+zT1C/f3hOen8nlE+83lESYRKOAT7isAX4B0IlaS1aOb8xCd5vItC5Uni82WVVYTKLwvpUk9BoCBQECgIFAQKAr9sBLJQ+SfpJz+pW1S+2sc95t3MtWdnO8HxPvjgG+nrX/+1hFB5Q9/bWZzK93tb1FE6c/FhxQGYYztwn/KwbGR+j7VjtnyUB55wZTp9wAlzeXxjB8XcZngF83bmc8zh1zXHX9fCVguV8An4D+5QZ6ryMjdCVMsiVPDCaGQ0NAS1hrbwmG7vKs+2rCh3tNh2K6wpp9vsT7mr+vZVhzzfbC1pj8qbMvKohMrmQvRlShwxjeUBaKRFqGNEKzjvtNLhEhbBksW69IMjn9COZ0PF3fTxFYzwLufYnKfZRKS0sJZ5EGkIUbKKruS7uAa36lG+5rkaAo7Gcl/8F9zgZgO5DkJg5GiIk7reOscGx2hHjb8HpjFGiKA5RDl0NXdX1qeZw1F2bm9um9vuMea5yyCOAqo+Rl+rd4H78ZT+SCSkD3xjoB8Ergm49aWvpFaN+r6t9kfbWfD7LJYeD9qA8Mh7lnnkY2lM90O3+vnPfxZ88sGDh/GtYmdnN+qlnolFpYXKX6eochyBwKkUKlk9jQsmTGaxqOSce3yg4IMXH1v4w8MvxsWLF+NDFy+dP1zxEeOb3/xmun379sFzXkILlXyI4AMIH8wQKrG+fJWDuvi4wYuZX+5sLuw2sDobi0s+PNAOxFJe6skv2avUVtIWBH65CGBN2ZNQ+WSjm/7p4Xb69MFm+sfP9Q//dk+/g/kfybfRwviHRSJpR9aU52VFee3CXPrK1cV08+JCurIi18ozeXXS26i7lFkQKAgUBH4VEDAJOWmhkvkOczSEJERJvEvY0wWuYJmoIzTVLeIsWDG3Yj4F+WDuVBceKZN0zOGYZzEXZO6XyVMWN0kD6USoZL74LHky4ZjEpPf8DL5DvXgSIJ4a9MO9a3NvO7VlOdmWMNne35ZQuZ2aQ81D+7Kq7CnIwlIbVoZQKVvL1JN7n25PLFT96Ax20mzaT3NtEWMRn6mW3JUiTCZZUqZOGjZ0LsvK0YwsLLHZ7O6mkVZe4g52KDfrA5lp9vRvoRzDpnW177GE0Af9Qbq/0017qmtmfiGdWzkvofJKzHn5ILAga1UwINRFSq7pb51o0c/oc0VKTU6DtOkZh9MQ+/A9f1xw7PukA9d6Xdxz+aTnY4KJomPGnvFkjHkXiElLu3lfmFvz7hBYsGhBmrocXHc9pl3UTVnURdnmDQiVcAQWMrI4Eg8tCJU8p43k46j3P26c0I8iVJ4QkG9YDO9Esah8QxBL9oJAQaAgUBAoCBQEzgQCH3/8SfrRj/5dJVT+P1q090DtxhX/8T2XMfeGazEfv3LlskTKr8tY56uxTyWGQVmonNM8fiK8IQjBKJhfM892YI4+1j3ckiJ4YUk5o7k/8//Zak9Ji0fZ8g0BLotwOtE8nfLY2kFCpfSD+C6/thExc/q+RDfqzmV7AWde1IjVn9s4hUgYHEZcVPyu1eqpf9tqw7a47W7EMzPZmnI83Et78jy3sdFUncsHQmVTFpXN1qyEQ3nkkdDZmELU3FEbu+Iiooj72sJuf1ZipbhgCJaTfkSvhIE5VD2etC3jGduSiCNlcTCLbLkftqQEmXwEvxW+5mpmda45cyWEvczRwMu8DG7G+IhshZjHmJtr5tjcOou+mW8e5py0I/NO4ji0GFdFxjDyLnG8LHb7SRvUVNmCA3Ij3gfqiJriDkLlEM6phcf7WFaKB8IF83snsVKWvTQnthw7ECltkZpxjpLVPt47EvPu4mJ4Y2NdRnYIlfcqofIXwSkxvONbBTwWS87hsK/vHxfTd77zzyqLyu/L9WsRKmOAjvhxKoVKPjzZBdOnn34aHwt4EfgggbjIRxcGHULJC8kLxjm/RLxw58+fD4tKb+DLH02eUaYtKklPWb/zO7/zShaVYMgvqD+KcU7b+GXhnl1I8ZEDt2bf+c53YmUJf1z8C3fEOJRbBYFTiwBCZVdWHY839tI/fL6ZPvlsM338mdwoyP1rCJVvyaSSf1z4x3S2LaFSFpQ3ZU359etL6f3Li+nq+blw/3pqQSsNKwgUBAoCZwABE8S3IVQy52H+hfUbi8xu374dgbkZ18zVeM6R28FqzbwQjDkb9yxOIXh6z2/S5/nWhoTK7RCTWDzmeRnPmXMhcEKcIFImU46PEu/Ilwma5nnsH6KVj6kny8mdzdTaepqmNx+n6d21NL0vwXK4o70q9WzUDSvLKbnxgbQMRJR66ndXulZXAuNY5XQkUs5OjdOcvANMi9xM4eZnKPejA7l7HeJMtp2GrYU0nL+QhjPn0mBmPg3kDlZbQou8jkWq9G+wSPeO/i1eV1iVePlYxOjhhtySSsCc1VxzRaLdZWF6UaIwAh5YmTzSV5/TfwuVnpPmPueVs2BO8D3i5wXwcnrPhYmdnueMA/W5Tudh3s58ncC4ef7ue1z7PucctN1CJe9UCLLiBdyjj9RR75P74Db6IwgxZUMc4RsIU48fPw6OgFhJMLnkvSJ9vU/RmBP+UYTKEwb0NYsrQuVrAleyFQQKAgWBgkBBoCBw5hD4+ON/kFD57yVU/iT95V/9Rfr8PkIl+78fU6gU50G4QTxkPn5ZC1G/8pWvpDt3bscCVbzoeGGqOYjjKQmDHHCHoXjOQCISc25zCcQ2+EtHXLEuVOIS1rwii0ZRTPxgzg+H62uev6M5PMZOm+JLBwtbJRjhbhS+EJZ/WP/VBMosCEpcU90IgK3WWItMxeU6XbVBC09n2coD0XJX3GNbwtau2i6hUlt+bGy2VN+5tLt3Q324IpHygkRKXMsO5DZ2V+WtiRdtqp1aBCke2Ostiv8siuctqN9atBpWlcITxQxcESoV05Zol3AmRqikbbmtnGeeFXwrrBlzmizYAUsoduIyz3I9L8r1c1JyL0Q9cWA42QEfEz9HtOQ5gigLb82lg2fiphfOrcCzg/GhnXBf9YNDLTn4yZ18N/+sLnLfI131Ayw4PYir+7lbLtSFRVo1M3M311jxW96t4IPEWhDMOeIl95UhsG1XLmt59wJ71Ut5vFtUBv4cvEd73b2wqIRHstj13t17YWyH7vT48ZPQhuCb+/uI091KqPyN9Hu//730h3/wryVUfiPKKj++iMCpFCr5g8LgMthYVbL6gQ9OrMDHlSofrDCx5UMCaXgxEAh5efgjRro7d+6EAMkqDj6EQT4p82//9m/DQpOXkhX+r7pHJRDWP35w7nt8+Ki3i3q//e1vxwe5+KNS/YJGhvKjIHBGELBFJULlp7KkRKz8WGLlmiwq2T+y+Zbea/4p4B/DGYRK7U15QxaVX5NQeevSQrq6UoTKM/L6lGYWBAoCpxgB5k3Mh05aqKTLkBLIC3M2PEswL8vk9U64grVQGQRVhIi9I1gBizjEOfctTrFQDTGHeRXzKcgTe0IwX9ze3jkQKyFU9IeDvJAMYp+bVFGOA/NLntPeICIicqy6HKgt4+2N1Nh4lNrrD1Jn7X7q7K7KQlJWlQ0JlNPap7mpIKkRUjmSGEnNPbGprqaGeyIw/NcRyZ3Vv5Vzave07k/19UMGmMNd5ZEGN5K6OdKK28HshbS/fCX1L72X9hfOp32JlexN2ReR6sqCckfkerPbT08lUq5KoH0igY1/n7GeXJYVJR8HwBQRD8zr4mQdBxPbA9KoTvMe1Oe2XPte3BcmEDLvJ8I9B6dzHl9TPnXV6+Ye42Piy9ycuTPBIiXP8zuRXUFRHvkohzHDa4rdv/JeWKj0PNv9cBmUx8cP3imTbuqFW8AdECqxqMR7C8HWlMznSUd+2vA2j1+GUPmDH/wgfe9734uFnfAm4+f3wvHb7PdpK5vxLhaVp21USnsKAgWBgkBBoCBQEHgbCHz88T+mH/3J/55+8uM/q4TKhyIqCJXH22IJoZCAi9Z5cQ/mk++//166detmui6jILwfLmnezly9zsXabXm8YZFhiD6TveODf2nOHvNu8SlEsbpnnbwItRLCVK8qF9PSUfGSEVxFQiXz/cwp97RnpHhixRXhG3CC4BbKxlwXQZA4BLWwYsxWgC0WmIq4zUxrwanEyXntTTk319c5wmVf21PtKZ+8vshzzoFQubOk82sSSy/JvexKJebtS7zbUv8fZ7FyarMSKlfkfWdF24WsqE3zytNSN7Jop+ZUbUPoy3zK/K0RbeQ+IqtiLdSMZxY09ZwjsIDYgk3gAreDW4lnPcPzIrl+gB17xIs3iXtma0osEL2XI5au8Lq8nQiuc82r6yIlAjBtyu9Gxja3SFXECfc45b98L8aQcw7am8+e+RljxR0yHxz1c25mfkoB9CWXxD0e5Wchxqr/0c/gpPmbA/wRHP3dIPqhd4z0cEowgQeDOW2hSN6nra3N4JG4fv3snjzz3LubHsgyeXX1qVwBbwaXRKgcyfL20qXzMmT7TQmVvyuh8l9KqPw1Gl2OIxA4lUIlHw8QIREW7969Gx8T+CVAWPzWt74VHyiwqMSFK65hWf3MKgkOyD4favgohkh5+/bt+MPIcz5AkJ69LHkJef7hhx9GuUdg88q3ILi0i7YjsPKHlPZiOUB9vNDlKAicNQQmQqVcv1ZuXz++L3/v22wqzT/qb+e95o+/hcqVSqj86jUJlbh+PT+bForr17P2KpX2FgQKAqcMAYgaQszbEiqZ7CMuQVQRKQlf/epXD4RK5knUnwWkLFjhRoVr2kZ+RDeEKeZ3nHMPwsBckYBY6fPsXiVbYzLvitWlIkuIXCZTtAfCTFnEBJ57hST1DvsSqCSEjtYfp8aTz1J77V6aWf8szfSeps5UN7U62rdyQf/+dSCPGlT9MyhDSdlOokHK7atIzZ7+EZOX1tSRy9fZloRKEUhpm2lK5pbjHRHFbVlthlgpApSEg1y/9s/fSt1bv576K9dTvz2n8pohVPZEEnexAJSIuyaRcm19I61qLktblyTWsVqZjwPEYEUfTbQsFIIF98Dl8JzU4h7lRf+FL/Ez96uVuJBd7hN8+LoeM+elTnCnDQSO/PGgF4I040WwUHlYFAxSWpVDfvrFuGExikjJOfdYyQ0h5nD7KYuPEfndwkJzsi8JdfLOWKjEzStzdxY+EhCr4A2k412jzLd5FKHybaJ7/LKLUHl8rErKgkBBoCBQECgIFATONgIff/yphMr/Q0Llf8hC5ecIlXi7Oa5QKW6huT5c6ty5JX2Dv5Ru3byp7+tX0yV9j8f7ydzcvObqeVEonCALjzNhKcncHotAjjx/Z+7OAkE8tGTPLFjpYUXZkRtY0pvL8AmSNFmcRHgyd8nnlIPY1u1mLyp74lBwTIuVTh+VVz8oEz4Bf2nLWAKhcm5G23dIoFxY6CseKKQ0Ix7YbuPOFYtKCZXao3JzsyErzgWJj5fVrgtpuoNQyT6b+7I2lFDZfKRyV5VnQ22V953eBYWLkX5/f0ni4bTygQUWn7kdB1aV5m6VSEkbEVYnomWlNSgfB4IaeygScKVrboS3IPodnoMqngeGweh0QrrMn/ICYngU6eF31An+0+LuCNMdjaU5njlnWKkKu8wzc5vqGgj9onPxH32EQOt/2kAr6jwyn8fd3MCclAIqfHQj8hPrUCGT/PRZ1wpxVEnMKyMdfY2F0gix7mN2Yxx7VKofHGCyj3ArLs57aQ5NWXBM+CI88j6GdhIq792TUKn9KZ8+zVwyv297Wai8fCF959tYVP6OhMr/TkLl16OO8uOLCJxKoZIPA3wkQFhE8ENY5CXgI8xv/dZvxWpxXgaEzJ///OfxnBeAl4YPF/5Ag1j5jW98Q39M5uLlYZU0oiZElI8cCJ8ffPBBfED7IjSvfsdCJfUgsBah8tUxLDlOHwJFqDx9Y1JaVBAoCBQETgIBE5K3IVTSPogLJAah8s6dO88IlZBXnjG/2xeRREiypR2kCAIAIWIOZ4tKiDB5IBjMubC+tFBJXBe9SEMZFujIx/yQMijTIcoU6SId6YPQaU/KwdMnaSSRsvHwn0Ko7OzIqnKs/dJnJaAuSqBcFhGbUxCXH4s4ImXJSajcvlYBwqT7nWntI6lVufMSK9shVIpIS6QcbuQYsXIwbKeB3L/2Vt5Le+/9RupKsOx35A5oqhXkCRdGXc2N6eO6LP1YFLeGUKk6ELlw/cpeoJzbRS5zYvrk/tfPeWaixTiZ2GVSO7FoxP0P70gEC5X0qwrg5XI4d1nEXHv8/WGB+4wxY2dx2W57eQ+oh4O2Og9lcE5grxrm1nzw8LvARwD2leEgP+8OJDu/V/nd8nt18LFiL783kMt1ib5wDngFq1+fPl0NfHlGO/MHkyJUBsDv+A/Gu1hUvuODXLpXECgIFAQKAgWBgkAgkIXK/1NCJRaV7FFZCZXiHy8/4AAS8+TedHFRhgSXL6ZrV6/IkvJ6fLfPWzSweBKBURaUwQm16LAjrzaz8LE5zelngtfhYpMD96S4gWU+zzlCVFMWei1ZVmYRDGvKvEIULkK6fRYmSkhi3m9RLQqD04VgJ1ewcA/xKHgigfke9xCgsiiayyJfcBvVwSJI6XFy9TqUtehAXHQgjjXW+ZTariWm011ZDmrPyRF8Acu6pEWls7KQPK8yVtTPZYmxCLNDWVRuqw9PhNdTPVuTUNgQ70WkvCKR87rasax7M2ovCy+jFbkd4pEspA1BshImSZA5V45JrrMQGwOTqs/h2hRRkiCMIpYwx/6T2aKy4nNUp9xhaVilRbiD/8QYqALwzwJz5mBwangZFq9wtvr+nvkaITW3nbbm9oJtVKYf3JucR/0HC2HNPfOYMMa0LToYWTK/BxfE04NylASBNt4d8vAO1YRKc1/zVooCE94Dcz3a6T7l9yyn4TnvDOlzf+DY2cMTfJbFw2hT2f3rRKjE6xP7Uw4G8tI0kkXlgVCJReW/KEKlX4Ej4lMpVPKyMOCsakaIxHIS8Y+PSuz5yAcrPiiwEpoPHPxC8ovDLwsvHi8Rz/mIgatYPkzh1omPDpTNNVaOCJU3btyI1edHYPPKtwrBfWXISoYzgEARKs/AIJUmFgQKAgWB10CASTyT77clVFpwQkS7c+dOBKwqmX9h/ce8jXmZAyQzEyNETlaydkQKswUdloLM65jvwUpwRcO8a3d3TwHrSlbKZvKJ8JnLyWTGghl5KSOLlPOKs0Ulq0Nb2pMiyI5WS462tNr10Wdp/OBnqfHgH1JL1pSdvvapbHfT9IoI8wURswuacy5I3JTFJFu5hEWlCFFP+0oiVmq7EgmVuEOSRWV7Ks0rtCFMsqgcbYswritIrMSyctBrys3rXOou3UzbN34z7Umw3J9eSoPmdBCtgXDpaRXw7g57rWykDQmViJXMf3H1CpYE5sf0EZIFo4NMZWKGYOkVwnlvFxNJ0kAY84rkTEzBLlwDiZCFWCniF26DVB911svOhK4ufFJnrtfjD7aMJ3UwNoyThUrivNo07wXpPPSD8UeIzPnzKmo+HLQUeHfaKpP0tD/Iud5nPjrYkpL3ye8W7wohf6TYrUTuneAGuHldW5P4K8Fybe1pYLu5mS0qwYKPGG/zKBaVbxPd45ddeNzxsSopCwIFgYJAQaAgUBA42wg86/rVQqV4ljy6PP+AB4g7iD8gYM10ZuXRcCW9d+u6vq1fn7h8rTzhMF83D5iWYNmRcJkXjuIVRbxOohfzeqwH4Re2AAyLOFUVVoXiU01EKYQ71QsVYW7OHL3fy/vdM4fDQo62wX2oF44AzcGC0LwRvgh3JH0WkbIVJyJUiFsqPOdHnGvIolJWlXMSKOdHlVDJ9b7avhMCZEpoDD0tJtW2H90ZCafnVO+S+Ka2p+hggThSuj2ViVfHdbVuQ+1sStC8pDZcVZ6bsvy8KJFtTv2Ci2YR70DoU1+y+9fc9zwuWZgECPgPgiJ4ZTEy45L5kPqGFaWExwmvy/0MfEP9q0ROY1/hABa0xXwc/sy4MV6BLRxMQnWIhRXvAzf4ZnDPEBHVZt1jDOKgrdUpUeag+UbmcVVf1AYWzyKYRp/g5upfbEGiMijO9QZeuqbcwKG2yDY4q9LTAO/lyTeB3MZmpMfqlvdoMEAYz2Of+5F/BygTLHElnN2/Tvg1PDMLlTKiu5+Fyrt370q7ehQWlTzLArr8LY172qblovQsLCqL61eB/cLjVAqV/nDGB4Of/exn6e///u/T3/zN38THhVu3bsUvBlaW/ILgvhWx8ebNm/GLw2pz8vz1X/91fHxgA1/SIWryUmJl+f7776df+7VfSzzjYxXPT+IoBPckUCxlnDYEilB52kaktKcgUBAoCJwMAp5vZaHyj9Of/umfasK9HeIORO9NDxM9XPIz93r//dvpzp07Wix2NVwEMS+D3CEmZZEyu16BWLJaE1JkoRLXQSFUiuCyPwQEhjyIXJAH3PkwD2NVK2VmYpDLczuyVWUWJzPhgnRJDOtkl0SxnldkaLyxmkb3/jGlz/8xNR98klpbD1Jb1pTtBVlTXhPhviySdmk6NRblSlUWk2O5QYd67YtEhVCpfSf3Brqv/UNCqNSK3PnOWGWIBPUnQuUAsXJdYtqOVtb2p9Nu53rauPTttLd4M4TKUQv3S3nvlmEl8G1JQGM/DOa1PMPVEkLX4mLewxNMObLABunKB6SMZ8x5Jyth8ypYUkDmMilELM6CcSbt6hmrUvmvYpeQQpM4yoPETlagZlG0IVNTxpFnpHU/ECoZp4m4vHsgVFJubl/+iIFQ6fJZrWtSmoXQTOSD5KphmaBPXBYx/tQ1eUdklVq5f8rvCe8KIne2VAVPxF8WOsI/sLRkMST5s1Bpav3mvxd5RFxe3rbi5s1b6bvf/a/T97///diWgt8Z3tHoX87wwp9BxpWCmDwE+sKWFz/+8Y/TD3/4w/TTn/40yvjmN7+Zyh6VX4Sz8LgvYlLuFAQKAgWBgkBBoCDwbiLw8cefpB/96N+nn/zkJ7Ko/MsQW1JYUwYjek6n8/yVOTnC45w8FV7RN/avfvUr6fbt9+O7/CUJMgviJbMSthAh4R+IleYguHHthOjFgsQsVOa9/yqxCVFKgTkttWVhatKc4K8Ssyw+5kWrec6uCbCEQbYeYXGr+JoWPDInhtP05b1nV15V9mKR60SsRKza1172IeopHelxaTs9TTlt9WNKVqCyrsSyUm5g5+d3ZFm5If64JU4qLzwSunZ2EB9n1O5F9VmLbOfEY2fY55KtTGR9qb0pU5KamfaUvi3ug+vXa2m3q8Wp+5clwi3oWSeEPQuT8B1zn4xB5iDwMTAgxF6LCHshtsmyVPGBSKnzECjrQmWV7wBb9RXM+A+hjv/jlrgb3M5ud2fESYI/mvPpOfwuW3xynrnhwZ6ZlFkFFQyLzIteg1Nyp6qnsoqkT9Gm6FPuj/uS+8P4sIA2c/vcZpUBRtH2nN+4IDBSHtaYPIeTstiVd5Z3oq33kTIYu8wZZZGrugNnlZlF7lwy706Pbwu8I8KSOsAIb1AYwz0R37p//7MD168PtZ3IuhbABo9U+aORBPTxIFwjZ6Hy92RR+a9kUflBBqL8/AICp1KodCsZdExoIdkIj3xAYOU4vxA8Y+U4FpNf0er827dv6w9RJ9J8+umn6S/+4i/CipIV+KSHfOIO67333ov0X//618NNVv2Xx/W+blwI7usiV/KdZgSKUHmaR6e0rSBQECgIvD4CTLSZoH/00X9Mf/TH/1ZC5Z+lHe3N2NdEPGbg1cT/+DWIZXBUUZACkQHc/9ySEMNiM+ZhuPJfWFgMEmBRcV/iWHbRksWqWdwCaTHZfLhpRVCsVnIiVIbFYCZpQTCwyhTJZFWtLehsVUkf6UYmKOxxIpdDKiMIslwO5ZW82VovaPlQ7Vh7nMZ3P05T9z9JzYcSKncfpXZzL7WWZel5S6LmNYVLaseSrCo7WnEpERAatK89R3oSKbv7zbQn4TGEymm5CJoZprlOP003hOu+CNauVm5uiBBJpNx/KrFV3LW7rUV1U5fT08VvpJ3ZG2kgi8pxezb6CmFFLAQrVgJnd7eQ3anwCjK/IOtQuVDiAwBHkHHEX2GK+CimFgSSDwW2VIS8s+9LJmLqOWSueh+C9AYRyytYYzj1/IAUilQyt7bwCfELS8fqQwTjwzPKxp0TNC/Iosq3cLjHh4LKBRNjRZvjo4c+WEQbNacPkVJlxSprVU87gsQi+PolU7vyPQuteWW0SSex34ksaiNsI2Da1bA+WkiQ3NJCx42N9fRUZJPFkE+frscq2WibxOuD+tSXkznoUPQohOaJUPmvJFT+ZqxOL0LlySB93FIKjzsuUiVdQaAgUBAoCBQECgJnHYGPP/5YQuW/k1D54/SXfymhUt/ew03MC+e6zF2hBHiNmY5v8levXpMh0NfTnTt3tCj1vTAOYgEl89iJUCmuEB5WFGuunwUj4szBWFDJt3nm22E9J86QXZTahSfzZv6f8BXm98zhYx6/tR1ze8rA1Szf/jFKgk+aH8EZ96QLwD92wxMPe1ZmwZI5YCycVRrm53AY2gYn6XQkcHWS+NxA5UlXmNtW+euyJtXeky3tUyme0N1ju45pcRa4ZUu4wP/GEveGKqunfu2pXLkAHWPB15HwdV4axfW017stDntFvVpS/2fEr1jgieiXhb9n9Qo4T+a/8Ka6yDbxJpPFSp5lS0q4HJaJCGbZ8hLexEHZISwGrxNbq2I4Hvs0IvjGWMHNBABjmS0TxQOV9lmRUvmVj7wRVDb/qVC1OXO1XH8WEF2/rWSVMHAP960HfVNf9E0iOD5cP4RCxNeJpxsVH/2IDumHBW6d1c6xqBRvVftjb80Y047ama0qER8ZezCL7wYaDfoQ/VEF1FfnjXBL0vEebWxsSnd6HFsMss0gAW+eLIDlnaJsRFPag9EcHkJ/7/d+P/3hH/4bCZW/7maX+BACp1qo5GVBkGRF8L179+KDQf2PDKbHDDYuxXB5xUcRXgbSs7clH3K4xy8gLxfpWaFMesKrfAA4hNuRl4XgHglLuXnGEShC5RkfwNL8gkBBoCDwHASYZDOB/ujP/0JC5f8kofL/Sjuad0EIkqwBgzQ8J+/RtzPx0fw+DuZgCIIryyvhAeP69Wvh9vX8yvkgj5AAk8IgHSIy3IMUMUfDNWuQTJWR9ycUoUVgU7kQJMiPSQ/5s8CWharYm0N94z6klrkgLoomhIsVlQoizZAxiFrsNCnykdazUNmQNWXzsYTKroRKue1prUiTfE9E7bpWCF9SOCeSI/KqAkRCmlrp2U5dCZTdfift9nHhAyEai6h2JSRuSuzcTY2RhNCuCM629qyQWLm/JgvDp1pju95K6/0L6WHrTtpqX0vDtgjr9PyBtWezmssy14QY7YiYcyDkstcLq1zBhNXAYMpeLAhzrEKFIEX/1c9w2xMELedpBulEouXjQCa/B26XILIVmQVrjiC1QWQRKid7STI/z2IlHxzyquksVIIrxWSXRBYOQzSU8IpoCLmmTsbe1p6IqLFnTTQh5419VTSeMea1ttUJMKttwyK0X62QVdlggGiLC12vMua9CIKv+3y0wD0PbnWfap9K9hp5+nQteEQIldU7pF6AADCcwFFhOx5KqFzUCnQsKv9LWVT+SwmV/0x8ZfmVeIo/OhAzRoRiUflqw1R43KvhVVIXBAoCBYGCQEGgIHB2EchC5Z9UQuVfSai8r84cb57LPBMRD0EQT4UTA6L345rv8/PzCzGvD7FSPAtewGJGYrgCfC4WJYaVHp5Tct2Zn2ZBKgQ3cYS8mJV5eOYU3IfvIFIyh8fbDJwiC6h530wMm7wtBpwFEYt5PR5WLHCGaFmJnV156MFKk/oRseCI9LEt4bHVgqOwN2VPXGpXvG5L53L/qm1BEKH6/cybW628h+XSYl9pBno+UFlwMXGdCFgLVkKlLCr3el8RH7kqhrEsYWwusIEPNeWth74Yk8x1bEWJcEb/WYyZvQiBRRYrxYMP3L2SPvMmuKBFPMrMdTAOk8Wlwdui7nzfYiVpOEektIBK/ixsIkxWoqoF1mfancfLfJ3YnIV2UGf0Mcii08LRJkIsffNiU87pO+VkXgY35a0AL/Lnd2hqKoujsWA3P1UfsscmxpQF0FiL0gfKyuViMamyxc2qEiPmeeavPXFwe3CCO27H1iEIkw8ePJDLV8Kj8MzDglzKNOa09Vmh8g+KUBnoHv3jVAuVfiEgjgiWDHT94I9aXiWR9yziBefDg9PzMtd/CfgF48MXgZeT65M8CsE9STRLWacFgSJUnpaRKO0oCBQECgIniwDzrCxU/qWEyv8l/emf/bmEyg3Nt7Co1BxJk/fjH5k8Qg9MXFh9OS+SyN4lV+Xu9fLlS+nixYthQcYz5m3Z6i+TKOqC+DC/Q1Aj4DYo5mwhhGUBDLKZic2EwOV2ZlICmY1VuJoTMi/E7QvsJVZTiiAjgmVirHMRpBApVW+4OBXhG68/Sunex6nxUPtTrir0JFROS6g8j1ApgfL6bGpfFplcmpNQOSdNd0b0g31JZuTGhzAr4jmvOShukdjHZFtkdVXnm6kx1ora/W4a7WqV5abIpYTK3dVx2nrcTI+3z6XPBjfTerqUBq0luZWdlwhpVzsaD/UDosT8dk9uS+kW2EC0II/MefvxHNequFSFcEPmshsjhERbpgZBwyWSsGA+nF2rMnqTjwAZUxO+LH4xZiak+UNDXnFLGVEW2LayWx3umYDmMRHprERDVsgy9l6VStsZH8qASPIe0BJWA0O4ERnjYwUrhGUlSnr6z4EQzTXvs8XqKB9hUukjn98FpfG+mxYr+TCBpermZhYqESlXK6GSDwCUSflxBBwZk3zjdX6qrCgOa1cJlYsLlVD5X0io/G8lVH67sqicid+R49Tg9hEzRoQiVB4HuUmawuMmWJSzgkBBoCBQECgIFATebQSyUPmj7Pr1wKLyeH1mnsmcne/xeMq5c+eOwu10+/YdLU69FoZB3pYieEqIlMzv8xw/e195ViijTI7MT/P8HZ4QXE7zdx+e88OHslC5HRZs6AVwUPgj22KwENCWnbSBA06AoGmhMufficWJiJZwJ3gGXAcxFZ5FP7OwhxtXechp93R/L+J2u6+FovAPRDeMo3oSR/fS8pLETLmJbbe0WLSRBUb6RTcG4ov9/oo42zVZVt4Rz7kmnrpSCZVwpyz+AQf0w/3NlpEWKe0ZBqGybm0IbvBqC5QTvpThzaIiHO2Au1Xn5m1wt2dDtpI0v2ec6gFtm+vgh5VI6efPtt/8LXOqnCdz+cxjzOfM6bJYybjCxxg3xvzwO0G/KEs/Y4xz3Rk33h3zRsaQPjKmaEJ4WGKMOfJC1sxLQ6zUQNEmAmXQBngCorhD8EYtboVvPZK7VwTLVS143ZZ3Ktpq/ggGHBOhUq5f//APi1CZYTny56kWKusvhl+Wei94Af3LxEvHQR7+APjDRz19/uPiP4yTFRv1NG9yXgjum6BX8p5WBIpQeVpHprSrIFAQKAi8GQKZCGJRiVD5v1VC5ebrCZWahGdikOdmWMaxinVlZTnEycsisXi1wIU/5ID5G0cmU/VVkeQXcZXbHlzuQCy96hYB64A4VUTXbmZiD8YDEpxXeWpSGHPCECpVF+07yO+0irmP69YRIpgI6mjtYUq4fX0kkfLpp2l6/3Fqi5C2Lozl+lVtvy43q5fkunZpWUIl6uWyxNAl9WUhRMputyMhsaPqR+pDV0RoQ0LlI52viszKz+tgK4200nK4JTdDEip3ngzTxsOp9PnThfTp9qW0ui/y2tDWBZVQOSMXtTMSFXEBG2RJhLSn1cD0i1XKzG9FGSsiBZFjxeezFpXkZU+OWVzoCn8v2gsCHhhPVsqCRz4yEYX4eWxx0RPzadVJbNEYa1XG1CLwgWWkCCDpOBCCQ3QM8bASkUWkmbtTZZSncqiDe7wbmZyKoKrPrBQeDkQcgzyqPJVJS0UjIz3vM6QQDpBdBWFFKcIeYnVFOL3njcoHS9JmoRKLys20BuEMoXI9PlpkoZJV0NQWL7niEzhcnvYsWVqyUPlbEir/m/ThbyJUFovKE0D5lYooPO6V4CqJCwIFgYJAQaAgUBA4wwi8iVBJty36sCD1xo0b2uLjvcQ+lZxfkufDsKqcmw9hyHwBoTJzsbwHor/nE5svZH46ESonWgB8QQsZNYcmDRwBoRFPilhUstCWMhAql5fPVWLlUoip8BLyWnSyUInolPNvqaydEKPgH3WhkraRd0qC5NTUQO0fSoAcqP+ymJQQ2ZAQCUeY1qLWhfmNdG5pK51fxj1sX7wo809RH/EY6pf3m8GswooWU17VjiDvqz9XVa4sKptYVFa8TgSHfhJoMxg4IEwihFnAyy5RWdBpboUgan4FhwNreFvGnv4QEGAn3nDy4tI6TwZLcMgcMAuK5oOZF03aSP95lvNMhE3/ejBe7g9xlKOHxDCsZ4VV+p3LNrejj+53FipZiFtxs6qczFepkTZTJlxyghtPaB/fFrJQmQ3YuOd6KLuep85FeWdw6eqwvr4W24UgVBLYqhADO6wu4aO5z9SajyJUGomXx6daqHx5809XikJwT9d4lNacDAJFqDwZHEspBYGCQEHgtCHABJoJeBYq/1e5fpVF5TZED9evCIm4sXnRAUGYHJkINeXutSOiJktKiZRMyi9dvpguyZLy3Dm5AdLK2yCLIgUc9Uk8pIIjiA5CotJAmJ4hPboX1yJdvo+AiXseiAd7T+DSNQiv0iAWBjGqyqWNzhfkS+XxfAiZYcWmSOrw6YM09blEyicSKTd+ljqDJ2la7lvbF0TK35NQeW1BFpVLEiovyT2rVsE2L6uMiyKcSxK95kRy5QJ2T6RHLn6m2zsSGVclVD5Q+x5JbFwVU92Q+9cd6ZV7qf+0n7YfD9La5+N079FM+vuny+nB3mLqpfmU2nLrirAowj0zoz4JC49ZFtBEjmt7QA5Fxhi7HqtOJWQi7mGRKLoXWII7+34euNSVYBl7dYKZyGpgJjyCPDIYCvEfsUPgn61Tg/ByrTaY2Jr44qqXcYg9QPn4oPxBREW22cslr27NY8OwHwjOjIeuQ0QUKUVERHilXxDHeF+CmCpR7UC0heQGuRTJzOS9WhmrOuNNhfXqYLwhr5BI0oUr3dijcjMTzppFZaysVr3KUautelFrd17tNLdDiquyIVTi+vWmXL9iUfnPi1D5amCeWOrC404MylJQQaAgUBAoCBQECgKnHIE3FSrhU/CHxcVFWVVejO093nvvvdju48qVK7FAlUWrcBnzDPgCXMEx577mXghX8LKaOMc5838OnnNwzRzeFpEIRCxohJsgQrEw9ty5pbCohPewbyV5nc8uXxEpCYhPiJbcR6wiLe1iQSftUs3BH8RQ9GwkXiX6p9stuWglbjRHWgy6K5FyVSLluixKN7VgVxaVbdrbVFvbKretNk5rAeW8+rcsPnRJXPGmnotPNuRJpzEbfAjuBe+grXUcaBchc5wsVCLeZWtAC2NwnIpfqWrwMMaOPRaO831EzCyS0l/G1lgDeT7PfDDzqEn7cn15mxGLoZTh8VQHK+wmMfd8sLVH3cMSdG1Sd8Y9L0TNi1Fj4Wq8E/CpilMJs8MH7arjBZYc9Du8NmnrFN5N+k8fSc93EcrP+TLW8AMC7wliJGFtbU0Ba8qncc49nnfl8Yi8E5evk1YVoXKCxcvOilD5MoRe4XkhuK8AVkl6ZhAoQuWZGarS0IJAQaAg8EoIeEL+0UfsUfk/5z0qt+X6VaRH03iFL076X1QB5ASrPfYruSDrySsSKW/cuJ6uXNF+4rh8FZGFGEAIXDb0AjKCkMURoiJCEiyluiauHnMW6UnNnowIXJSHCIcLl7xCMruLhXBNCEzUlPNGfWKYVfcGEBlEMZHTPa3IHTz9PDXl9rW9+k9pdutumh2tyQpRouMFkVGEyusIlcsSKq/IovJ98c+baq9WxWp1LG5fu92WiC4ksSuhci3Ndh5LqLyv8wdq82P5l1mXRaWEyg2JcKsSKh/20+png/Tzh6300ycL6bPtubQ7mkkjEVZEX0jyjPqHIMsB0WJlKcQOYBAiw7VqtWoUgkSwsAe+5J1ROYzNwsJ87BszNz8XFpYHVpUmpkFOqQmqTBUV5sR6diD0Cn/E5HyPVbSZlLI6lw8C0W6NN2MDKWSMGdfYJyUE5DwEMf61crGYxGUrRJx9ZLKbHwmV6i/P8kGbqtPqfcHdb5BLYdMPcVMxOFT5qjcqYyVRE4woe1duX/nAAcmEcEaQZSX3eE6ZtPnEj2eEyveqPSr/hVy/fqdy/aoPFgedfHHt/N5wEHu8iuvXF2N2+GnhcYcRKdcFgYJAQaAgUBAoCLyrCLypUMl8EzEKbseiuwsXLobb16tXr4n7ZaESF6z24gIXOCpkoSyLlxbI4Kj1UJ/fcs6zg8WGlTtOxCzKmp2dC2tKBNRFba9A++CEtBduxPyfOV9d5ESoRGjiHs+pg77BZWgT166X98F9zx5l4KJTWgi6J5Hyabp4fk3C7brq7okXi6yMp7WIdFbtXYgwGC6KDy2qFLl7nbosHiVryql5hWnVIcFM/IC+xOLLqr1uN22rB6cBD9pXP2g/eIC5uZiv65hj7WoOZ45XL4dzcwvOqaveHgvJB5ho8TB12oqW+0cdtDeEWPiYFtnSL67pBu2g/YSMP5hk4TbXh3hNf93nLGh6nIhpI+8IXC5zSQREiczVuPLdILZCgWPrHkduE/kyR6y/J7wjcMWnlbtXeBZi5fr6Rrw7WFLCGY8aC8ouQiUoHO8oQuXxcDpWqkJwjwVTSXTGEChC5RkbsNLcgkBBoCBwTAQy0ZBF5Uf/UULl/yih8j/ULCqZsGerxxcWd0CKMlmdlotSVrBeFUG9fv16eu+9W+na1asSKi9IIFsMwgc5yoSnJnrJtQwHpCMs40RUICJheVe5r8nkY9IaeA9iZVPlWRRj1azduSCYIYVlKS/ni3rjtFqhqbIRsyAXuyKoW3LjMlz9PLUe/0OaXf9FWtj5XLaNW2lufpg6F0Wib7FHpYTKq0upde5KmurcwcxSxOaGxLXzEirn5fa1qfIQ1bZlUbgqgfBBmp+9F0Ll1PhJSv2NNNzeS4M1CZWPe2nzc4TKXvqnB430n57Mprs7nbQ96KTBlPbOgNRL/MUNLhaVHHncIHNZPEOo5B57L2bMMnbc48jui/I+7ezVkvdsWQzREjIfQqWwwi1Qdm1UEwGVPxNMRMosKIcwacvUEIvz2GX3QNkdrIVKC8e4r8VN7YSsilhqAPmPfEGMq7IQXS1U2rURxC+T10xIJ+MIRc0fD3iOq1eEdghpXnGcXSGBha1rOSdtfODQytcdjfuG3L5CPJ88fpKe4MInhMptWaZqn0+9H3ahRJtP5IjfG/qiPSrDohKh8r+SReV/L6HyN4pQeSIgv1ohhce9Gl4ldUGgIFAQKAgUBAoCZxeBkxAqmY/D67JYuRTbfbBnJWFlZSWsLdnHkufwDQKimWMLlxbOiOEEE76Q8Y15P7xBAT7IXJ55PHM3rCAJ3KM8uAciJdacnFNfFruyMJb5QfaogjCJQLmxsRELFLGqhENQFnkoj5jrHGAdFf+U+NlqyYNMC28+bXkTGqYLK5vCYCNdubSm+b0WrHbgKR0toFyQGLeksCIOsiQOtKC+aJuP5jlxWXnRUZrRKG/jAe+AyxBbiAyOI+HN1xb1uE+7wMQY0V6LfBYoicE2C4hZFHYax+QnL0fm3Bnn+n2emUOZa9Em2kA6tmuhDrwcEVM2ZfKsfngMo6/VODIulMUz8tXbS37nqcec+/B92kIbKZs2EnhPKJ/7tCe3UZ6GqveDa/ed/M5bt7yti5R5P0qEyrXK3au8JEncNNd0m+pxESrraLz4vAiVL8bnlZ4WgvtKcJXEZwSBIlSekYEqzSwIFAQKAq+IABNxCMFHH/15+qM/+mMJlX8mtzfbMdFG4HvVA0LRkVC5wl4lEilv3bqVbr//frgCunDhQlqUIANRRcQKUqQVnOTJJEbuflShyUVYxkHIJDxlEmYilskTtI8DYY0Vm7ganVhUZneplD0hRhMiQz7EMCztICK7Equ2tyRSbqynDYlUw9X7aUbWlItb99NKbzUtNXdFPqdS55IIl4TK9g25ELq6kJorF1Nj9paMT2+qNbckrl0UEVqUUNlOuyFUbkmofCiLys8kVN5N061HaWq0EdaUw3WtHH0i8vR5V0JlN63e66VPH6X0N+ud9PO9TtoctFN/3NZ+JSJ4EhDb6iPWi9F2rbQ1KczX8VPkaEIsIWsmbwiQYI41JfvFEHCJVCfwIVJCJJ8Z92cxA8scssDMOSIj70qkVJ3wUIge2DMmCKF8oFiQ9SZuZvN4Z8I6EUUprxIrFdMPxpw+Mj68C+6v+0Sd1BU9p68aT5NSyL0/QkAayZvz53colz2M1bt7sd/IVqyIffJkNT169CgRh1CplbMHxFaugFjhTNtO9hhXQiWuX78rofJfS6j8sAiVJwvysUorPO5YMJVEBYGCQEGgIFAQKAi8Awi8qVAJBOYGcDlEHywo4RiIlJzDNSxU8hwe6NhipcU0xCICXMEBTkHIXHEieHnOz9zNghntoSzKzy5n7WGnGZzIvNf8gnwIUYiTFirzHoPZfSfl0Q4O8rIgFM8w0A+4GaLc9DS8lm1NEEZbEipH2u5kR16F1oTDrnixFo1OidOFNeWyuMdFuXyVOKn9KBsN9qScVWktlS2HO/vD2PKCPhHgQPSTug+LcPX7tC9zsowTbTaWjrkX3Fvj5Ni4HsY2+qo6XYfLJqYd4GfcwTBzrImwyxh4TF0XeTnM41wOfI2yGAeXxTPy0fZ6OfUySOPgcrmmzQTaRNmU6bZaqHT5lG3xnLqMg8sB/8NCJQIl4fHjx7HAFfGS96dednT0iB9FqDwClOfcKkLlc4B5ndv8AqCo85Hj7t278dJ/61vfCrN3/gj4F+t1yi55CgK/LASKUPnLQr7UWxAoCBQE3i4CJmwfffSRhMr/oRIq82T7dWpGMGTCDznF5Sv7lNy+fTtdv3Y9LCqxtJxY8GEpKCJTEVJczsBhaJMJBiSDkIkSRG1CSnQWRBGxy2QGkbQjyz3iLIYiVOaeQDoQN4khmgO51MFaDwKypZW061pJuyH3LZsSqcZy/bqwfjct7z1Kl0eb6Xy7r5W5IuCVUNm6IVJ6fS61zi+nxtxlCZVXVfJ1tfVS2uudU5mdECpHoy25bH0gofJ+mp+5l9qNp9qScDeNd+Qa5onEtEcSKu9LqFRYVfh0dSr9p52Z9LPudNoYtlNvJFdD6gDzxxDyqs5EHyorU3Uoi4QHwl1OT6/d94zPdJD28+cvxPggVtodEquA40NAZKgTyYwXZWX8/EHCQmMWCwPZamwYP3BmXCB9fJCY136l585hwZndL0EMeVfymFNGFiqzpWaun7HOZJPx9zuQiTpD6b5xnvHweyNhMqwqJ66RLHSy78jBPpY6h7zuak9SrClx3wPpfPDgoc6faD6/HitkWWnNPpW4Bg4XsuBNh0/w4EPOzZs3KqHy31RC5fn40HJc7uDxISYPobh+fbVBKjzu1fAqqQsCBYGCQEGgIFAQOLsInIRQ6d4z70QMY96PMMniSAKioYVJP6vfgxM4WJg6SsCEy1hIok74BkISApG5Is8thFJH5hs1oVF5nBaOYSHLQiWuPREq89w/W1V6Hp65aOWphr62EdKwEFU9nSUtTF3OPGtlWhaV++nypU3tV7mn9gzkkaYpbjKnBa1LspS8ILFT+1FKoGw0sPRsipeODzgpbaF+5qS0lYM2wNM46DdzfQc/Mz5g6MB41AXK4HqV8Mt9ro0p55RpHm6RlH57bIlJwzOLf3VxkTLAvF6/y3det5t66B9jQFkEjyX9JB9trJdH+T5cjmPuU6bL9btB+zzO7hN53Ce/c8TGkLJIQ/vIbzEbUdIiJZoPnngQuHlO2dT9oqMIlS9C59lnRah8Fo83uuKXqwiVbwRhyXwKEShC5SkclNKkgkBBoCBwAgh4Mp+Fyj+SUPmnB6sCX6d4JviIhKykvXLlaogvt29LqLx+o3IBtHxAWrOgmF0A4aK1LlTSLhNCzk1CckzLslgkvhQki7wIXxNCNnFjGqlFNmJfxIrAmGBBBvN+ExtplT0mcPep0Nh4nFa2H6bL+2vpemM3XZ4Zqk9ynXoZi0q5+AmhUntVXtBK2HntMTK9ohZdEgG9oH0uz8lCc1ZCpVop16/TbSwqH6V5hXbaTKkrtzObItWPJFQ+0CpPCZRbn/fSqq7/abOZ/j+5jv2F3L5uDOU+FqGStqsTlS0l3dG9+HmAC1cQLgicQ7ZWzESLcYHo4XoXy9bzsnhFqPRqY9zLRj4ArY5nMZ8QYj9XjaozX5HWwqJFQcaNMnO9CyGOnju3HKur52VdyX32i0GAPRAqJVjSD8qlj4yZ3wPHk3ZRd26X3xfSHAQJkYjRiJMQTYIFT9Jwzf6XWajcCCtKSOeDBw9EPFfjQ8WWrGyxMPYHA6wzyUsbTvLIQqUtKr9fhMqTBPcVyio87hXAKkkLAgWBgkBBoCBQEDjTCJykUAkQzPstMFmsshDEvL8uVCJm1gVJnjtYbCQ+bPFG+RzMxT2f59ycoy6CwgvraeEL9WAhCj6IQAknJHDNnJDnlO08nGeBC17VVttm1Kd5xefkNeZiml9YTivLC+JZzXTpQk+cp6/tP/YlSkpEG7ZkSTkrbnJOTcqWlFNTEilVZk98ZHs714/wZaGU+sAEDC0sRof0I/OlzP14ZrxJbz4MJk5H7GuPEek45xkHeMKRCRYg6Tv5KJ+Yaz+3CEg+Dqer1+866UsdS48dIiLlWaSkfLe13i/aWW9rfUw490Ed7gdlunzqYDxJ67bQNmPh8g/X4bYhRjI2CJXwRQuVvC9FqDT6JxcXofLksIw/ZkWoPEFAS1GnAoEiVJ6KYSiNKAgUBAoCJ44Ak3Um7ScpVEJkEMAQxK5du5bel+vXGzduhHeJ8+fPx54hXmFVIMInAABAAElEQVRrAmryQwfrBAIiwVEnWb52bDIDsYBwOJh0mYxQLsQFwgKh2BYhhXDg4tNhUysld3WvtbOeLnTX0tXxVrrV6qWrc6N0frmZFiRUtm5qb5UbEiuvy6rywkxqLHS0T+V8GqUlYbkkt68KWFTu0pc9Ecynaba9lubaG6k93JM1pQSzNZGyB3JH81CrNCVSbjwepMdrw/TznXb6/wfz6d5gOm1KpNzTXiXGQxN2uhzkKk6qH8bI/YbU+RwMeA42EFfGhTGou2PiA4DxJ5+xdl11/Hxej0lHGwl14sq56/b7wDvBnjUIc3ygOKpej5vLZcwcPIau3/X6Pul8jzS+79jPiXnv+QjBxwivkJ0IlU9jvxr2rCGQhneGd8dl0L6TOopQeVJIvlk5Rah8M/xK7oJAQaAgUBAoCBQEzg4CJy1U0nPP44nNRyw4wUXgHfA/AtcW1uAEvuYZPMGio7mKOQ5l17kA9ZCXfHBMi6Ck91HnB84LF0BsQxhkvo9YaYtK5oRwGc/7yc9BXSywZWHuzAzWoniLWU7TM5dU94oWtiJWdsS3xmlxYahn+6nVRKRj8WlbwuScQOpoO49p8ZRxcAvqZpEkWgKchPZQL/ytjgH9qePK8zq2YMg19z0O7r/z8YxAWmL6w0F9FuXAxEIlWJEOfEkLDof5Hnk5eE79BM5dNs+O4mXgT17HpKGdBLfT/XFb3S+PJzH56ofbSDsJLt953JZ6fW4vsdvgdH5PECXxvgNffPgQDzx5YWsRKuvon8x5ESpPBscopRDcEwSzFHVqEChC5akZitKQgkBBoCBwoggwYWfyfVJCJZN7iASkCvEFUYp9KhEqES25zi5HFyONSSokpE483Enuucw66TGZMJFwXL9PGRAMB/pqkRKigbsWVkU+FNmAaKyxx4QIal+CVKe3m84PtL/I1F66Nd1L1+eH6fJyI527JJdGEik717NQ2bogq8AF7SM5M53GjY4sKjuyqOxI1GqlHQmV42FfxG4nzbS6abbRS20JXeN1ESa5fd2XNeWerCi3H++np+vj9PlOM/1CLl8/HcymB1p1uz2ckuvXSrhV29WRgIX+HHXUcaqTOdJzDcmHvGPtytggHprI14ltPa/HhDJM7iCUPifmmUkm+EKuIWwE3i3KoF5EyqtXr8a7cPHixRCsGf/6mLk++udyTTLrRJP6/Jw2cF1vl/M7Jm09kIe2MW/ngwQfBUw8sajkQ4FFSguV9VXDlHWSRxEqTxLN1y+r8LjXx67kLAgUBAoCBYGCQEHgbCHwNoRKI+A5PbHn+nAM+BzBQhn3HLiHIAZngaMQ4Apck4d0lOeyqYtznlmkXFxcjHxwUfMb6nda0jOPr3MBOIvn++YxcBq4gjmG5/6UhVA5M9MRl0VMXUwzsxIqOxdU74r4zbJcvs5KrJwS10pq+1jtkwCHbxxZUE5pv0o5NlVoBC/Fe8uavPo8esSeh6vBSeActJN+0x8v8AUbY2UMuUcwnrSPtporcW78nYeYYFxIC8+yQEnMNfc5qJO20CZwq6d1OurxWJCesrnm4Bn5CJTpc8fOSx73z7HbTlkOh8skf/2gXMbO48c1aRy4ph31trgMt9t1cZ909BPOaJHSQqVdv1IX5b7oKK5fX4TOs8+KUPksHm90VQjuG8FXMp9SBIpQeUoHpjSrIFAQKAi8IQKeyJ+UUMmkngk+hAmCiPXe9evXQ6QkvnLlSghWCDMmXSaR7kp9kk9ZJlNOZ/JzmEi4bp6bXFCmSQkkA+KFdRxCFATjgcL9+/dDsDTRGIqYdob76dxY+4s0+ulWp5tuzA3T9XNjtV1WlVe1EviqCOG1mdS6qNWoixJZZ1tprHoHsoLsduWydXdKQqUImerstPdTpzFIs7K5bO3JhdCqhMrH2Zpy98kgbawP02OJlHf7M+nefkehnVYHjbSr/Ur6CuAR5AkSVhExkyljVu87eJnccZ+0XEPgGRMLlCb/JrfgWw/G2WXQDpM6YhMy3+eaeTAE3wGSz0G9iJO2sOU9QDDlfr0e94c2Uy5lUoaJsz8auG63x20gPoyNyySmLxwu20IlojVCJe8EgevnfbB4WR1RwSv+KELlKwL2lpIXHveWgC3FFgQKAgWBgkBBoCBw6hB4m0Ll4c4yByfA3+rB94jhBHAR80iLlOaA5n4u23l5DqeAWyLsmesczm/OQX44BOITcz+ESmIH8w3zDLiF+QVtyBaVWaicm1uQ29dl3VsJwXJhngWhs2lZW4bMzzck8CU9s8iGeDcREne0qtUchIWSLJxlMS3tgs/B1eBL7hP9MW8jdqD/9A08zInMleircTW2Tl/HwVwLDNx/yvK4UBfnxo00hLpQCTbUZZxJb+wo63CoY+p8blu9DMrxcbhMyqBcl0W6ej2u32U4Pf0wzyTmmnwcpPW7xjn36SdjY6GyPl68P/X8UcgRP4pQeQQoz7lVhMrnAPM6t/mlLq5fXwe5kuc0I1CEytM8OqVtBYGCQEHg9RFg4s3k/KSESlrChB5yBdFAgMGKEmEKoRKLOq7tAhYCBvGBjHCYPMSFfpjwUBbhsAhHXQ4mOKRxOp5xUC4EgnkaqyEhgp9//nn67LPP0r1790Kk2tT97l5X+0qOUlt7Iy40JEw2B+lGu5duze6n9+W+54pWxy5fEBm+rP0/JFY2LyqIiDbmJVS2G2kwnmILShHeUdrdFuEZjlJH7n46U+M0q/PWjiwBsaaUFeWewpZEyqc7KT3od9LPx0vpM7l8fdRLaXN/nLpKz7+/QbAgTuqD+xIntR/0E6wI7ruvScY9CLwJLqtiwRPsjS3nPDep55p8xpB3xaSOd6ZO6rgPgYPkeiUyMfc4qNuugG/fvh3vwYuESr+X5GfMIIAEyjd5NiEk9jn5Aq8Kq6i8+uH3BFzcF8r0+1AnnhYqXSftcH8p/6SPIlSeNKKvV17hca+HW8lVECgIFAQKAgWBgsDZQ+DLFCpBx5zCc/Kj7sHnzE0cv0i0oizSwV/glRb1ECsRLr0w1hwIbsNcnrk9nIK5n0U3z/XhCQ6e9xNTl7lWdv0Kb1oQn1pMrTZWlefUhkW5fLVQyUJRhM2J8CVCF5yiq30p8egSC2clUiJ8wT9YUAuvoU+0ne064EwE+nQUV6NNtI02m6MR02aeURa8Dv5HML8jPTjAr8x5ODcOHh9zbOpw+RYpuaYcDmNDetISOGgHgXQ+J+YgDelpk8Ph8XaeyKAf9bIo08HpXLbLpw5jxD3S0273nT6b6zmv0xOTnucsavZ4EfM9gYWtvEOMmfNSx1FHESqPQuXoe0WoPBqX17pbCO5rwVYynXIEilB5ygeoNK8gUBAoCLwmAp6on7RQadKBQIWrV6zpECkJiJZccx8CBuGCmHAwwadNPiAHJkeQLJ8TU4cD6SjDRMyx01EuBIJ5moUpLCkRKn/xi1/E6khWSUJUtF1IaqnsueZUWmmO0jWEyun9dHu2n64tSLxcaqT5C7Ja1H6VjfNq0xJCpYTWaQmV4mNdiYx73WHa1V6Uo8E4TascLaZNMzpvSrwcPxEhXB2k3aeDtL41Tk/6rXR/OJt+0ViWy9fptNYdpJ19CX+DvLKTtgfxcVzhZIyITcDqeHFO4Bl4MBaQVGPotMYK4grBrxN6nrkMxgUMTVCJua7fh7Sa6Jq00T7qRpxm7N97770EUUOc477bSTofrofxsHWmBdA6gXb9tIHg98eYBW4qlNjvCjEH7aeNdgMM4XTwhwKek65evsuMQk7oRxEqTwjINyyG8S4LTt8QxJK9IFAQKAgUBAoCBYEzgcCXLVS+DBTP1Q9zFbgCh+fwLsfpzQHhlBYoiREtLVyac8JtyIfwVBcozR3q9Ryuz5yl1WLPRsQ/3NLOSqRc0J6T51JbgiV7Vi4szEhYbKUFLWTNQuVkv8axFsPCZRC4ELrgoiyehYMgXMKjaAu8jD4gVDrAF8zT4HTuC22Gq8BZLLhxzQE2dZGScsGXOkhPW5j/wrM4BxP4Fc/pP30mPTH3zNFcTz0t6aiP2GNDTD4O4vq5y6ds8jkv187v9OZ5LsPczLHbwXOX69jt59rtoO/0lb673/TJ9bh+YsomLeODoExggav5IvndrqjgOT+KUPkcYI64XYTKI0B53VuF4L4uciXfaUagCJWneXRK2woCBYGCwOsjwGScifpJCpW0xpN7CBSEymIlQhWBiToWdhAuW1WaPNAmH9yDrBBMMg6THz+jLogY4pdXm9YJHCQDIsFKVYgFJAOx8u7du0EOs+vXrhy0ykWO3PLMtJtpWULlleZ+utHqp/e1z+S1mf10cW6c5pe0+vO8BMolubdZEBmbRajUKlPx6J4Eyb3+UEQTS79xajem0rTK7Ay00nRHYt9TWQmuDSWQjdLaXiM91L6Wn0/Np/vNc+nJqJ22tcK225+IgAfEp0byjI9j422sDsfgAC4QQONMXmMHaeU5Y2USDIbg6Ty0g7EBR94ZYq4duD6K9JOfsinX7l8RLamLemlDvU3U47JMnBkzgkk09ZCGYHxoh9voNtVjnnFQl+ugfIRri1O2quSauiClLp+8LoPzkzzA5ubNm+m73/1u+v73v58+/PDDEHbBrY7Ni+p024jJQ+ADyCeffJJ+/OMfpx/+8Ifppz/9aRTxzW9+M/3gBz9I3/ve99IHH3wQVs4eB9fn+EV1vmvPCo9710a09KcgUBAoCBQECgIFgechcNqEStrp+SdxPdT74DS+Zz4D34G/mNMg9DHHtmBZ5x7mEeYPlEk5FsvMO90Gz5NzGlt9IhbOiKPOpamGrCpbsnicnZdA2dE+lbh+baotpIXrILZmjoMYiegF78CzD0IlW1DASeCq1Ek/aDccGt4Eb7YbWJ6Z4zLvN2+CH1lsAxvaSjr4loPz0W/S0hZCnWPxzHgYB8qiLp7BA+tcsJ6W8s3VjZnHqR6bt5CGUK+H8vyc2PUejmkLfXfsPJRXH0eXTbkcpCMfPA+8CZybX1IeaRzoKxjBD/l+gKjM2PHtANzI57rrfTx8XoTKw4g8/7oIlc/H5pWfFIL7ypCVDGcAgSJUnoFBKk0sCBQECgKvgYDJxkkLlW4KxACxC2LIalCEKibpBFzAcg8SBuEyqalP9CEUDiY7jn2ffJAR6jE5Rfy0AEp6DhMSiAZWdN6TkJWskI0Qp+T6dSRi0hCZnNZqWemQ6WJjP11t9NLNqT2Jlr10QYrj3OxIe1NK0JyTYNlRG6VEjuXWB4vKnsRJrCp3exLSpHqqmCR5MLVRQPfkakjuXne3RmljO6VVWVM+HM+mx4359FjkdkN7XO5JqOz384pOxkfM54D81LGJTlU/jAV9deCez01Swcmki5g03IO8gpcJPTEk30IlGHOQx2SQ2IH74EuAzHGfe9RPGZTFOEOwIdoec5Nl2uHD5UMYmVcjUHq/SMaOezyjLtJyUJeD89fb43OnJyZQFmXzseDJkyfxTph48gzi+WUcRaj8MlB+eR2MuUVrFjDwe/Gtb30rFlfwLtff05eXVlIUBAoCBYGCQEGgIFAQOL0InEah8nXQYn5GgK/ALcwJ4TZwDgJzbfiN+aHzEDPHc17yEyx0mUvVYywq2aey1UKUY9HlrCTIBV2zCHM2zSFULjYroRLRDH6Ga9ZsxYi4xaJZBC+ESriHrfPgUdQFd6K9CJXwZQuVdbEVLkN6+IqFNq7NwcyPwYM5Lf0ypyMdeeBWDgh29fykNQ7gxEGddV5Vr4u09fT0o44z+UnPQTkcfm5846Z+mNcRk9bXnNeD+WC9PMryGLo9bgvlOz+40WdiB/rv/hFzTRr4ohc6M14sBuV7AtyBNMc5ilB5HJRymiJUHh+rl6YsBPelEJUEZxCBIlSewUErTS4IFAQKAsdAgIk6k+u3JVRCPiA5ECSTLcRKREpC3QUsaUhrInQ4pjsmM44hHeSBjEDAvIIWEkegTNJw1MkGBJFVkJAMSCIiFQLFju4PRSKhYm2VO699KpfTIF0Y99Ll8W66kLppudVLc+391JQ72GZrKOvLoYRNCWXaixKa0hPv2pNV5a72mpRmmRpttVvklP+G2r+yvz1K27tTab3bTKuDdnqSZtNaYyZtNTppV3l7saKzEvsqEmdSdzg2Do7pq0P9HiStjoXLIQ3PwA68GCMHC5UmtZRL+jpRrJ+b9HGPdKQnL+XwUYDgOhCUTRxJ64O8fichhcyrIYaMF4IlRJr7h4VK5ycvZXisebdNPMnDdZ18cs/vAuST94F3wSuaSUt5b/soQuXbRvh45RcedzycSqqCQEGgIFAQKAgUBM4+Au+KUMlIwCcc4IYE+A38A25jsZIYPgIXgRvBR+ArBK4JnHOfAJ9xeT5HqCQ0mwh/06p3Rgtd53Q+p7zwnnZaCqGyoTbAh+A6kz0k4TTwDhbNYk0J90D0gufAO6jPQiUcwWIl/fDiXvMl5q7wmTo3Aod6v9w/yuUgL/yIPNRJGQTuwX04SOsyOKfvHOZq5kjUxXPXYcy4Tx6PCTF5HWgDwYfLr9fhtPXY+eqxn5OXctxu2uS2H1U+vPCoUOeOFikZH8YJi8qD7wYaR+PufrwoLkLli9B59lkRKp/F442uCsF9I/hK5lOKQBEqT+nAlGYVBAoCBYE3RIBJPhP0tyVU0jwTGIthkC1WhSJUEtvCzsJYnVCY3NS76XsuFzICqbQgBvl04D7p6CeEir5CPiAdCFQIUljTEcJ9i8jaQHtDQniayteRI9i50SAtjnppedhNS+NuWpR15cxUV/tYKkjAbO3LRc8Qly/CUvm6wykJlVNpt99I+0ielbXlSMRpX8963ZS2e1Npo9dMa8NWWlOCLdlc9mTF2ZNIui9rygECmdpMO3z43DH3jQXki3AUdnWcSMO183Jt/OpiIuceDzAkDWUTyE8bCHWSyLnbRjrIIWSasXDZ9Y8CbnM0pvpBfsapPkZ2+Vpf7WsiTZ2HD7erPtaMtwVOk0+eQy7r7wFitVc0m3i6T4frOcnrIlSeJJqvX1bhca+PXclZECgIFAQKAgWBgsDZQuBdEirryJsfwV8sVjLXxrMLblSJuYafmJtYoHRM3jr/Mc+C4+RnOW40YoMPLXRluwYWYnZUZlvCaCPNz4pLyvNOs4WIyqLZzLvhHnBPeAdiJefwHbgHB+WbQ1lktUhJOzjMlZi7ko9r8yLzO7hYvR/kI405EPmc37yHNM5PXot9xtR8z/zI/NB8kTzkr/NNn5ujUQZ8z2VRp8snzVGBNPX75HV+7nNQhjko7Sa4LfU2uBy3g7Y4mCf6WwHjwjcCC8sscGZhK98QGEewpJzjHEWoPA5KOU0RKo+P1UtTFoL7UohKgjOIQBEqz+CglSYXBAoCBYFjIMDEmgn22xQqaQYkgWCyaKLoPTcQL1nhanHMxAKyYeJyOKY8yBDEyOVaoLTQRjmQEYtfkA/66wAJYe6GCAbZ4Hxfz0mPUNgYDlJL122ZQs4MFMb9NCdXsB0Jl9PD3dTqbafW3laa6mkF7EBuY9ifUhtV7u43006/mfqpmUYzIqbTjTSYbulaYqTMLnf2p9KWvIpuSdDcQthEoGQsIErUr3Pqrx/030TM97kHDiaJxCaI3PdRT+M89XtgSIAUMwbEdeLOM7CkfJfLu+P3B7w4p32UT1qPCcTaYiXlHB5Tt5HYYwVZ9phAEC1SmkwzfvU6yUu9BA6Xw3iTx2NMORYsTc65h9UmK2Uhonws4F0gb7wHFfmNgt/SjyJUviVgX7FY3pPi+vUVQSvJCwIFgYJAQaAgUBA4kwi8q0Ilg2G+Ay+C2zDXhndeuXIlFstyzj34DpzFXIf0DuZX5kzc9712G86FEAbX7Ihbanmr4oasLDudZlqcn0qz4oAhVMqQESnNvAkuAr9B7IJ/wEOYg8I7aDf10CbaBqflHG7FM9LAUcxhic1ZDvfbbTU/on7yw4EI5kTwKp6RjjzURXB/6X+9DPNR7pPmcHqnpT1OS0wdbgPt4NzPyeN83Kvfp5z64eeOeUZe97fennqZpKvn4dztIfb3AfNQxgVeiDCJu1dEZeLwwqTxAz/6QTnHOYpQeRyUcpoiVB4fq5emLAT3pRCVBGcQgSJUnsFBK00uCBQECgLHQMCT8rctVNIUEwiIIKQLcohACVEkrouVJmR1ggQZMlEiNpEiLQSUMi1Ucs190pnQMUeDeJiM0SaTFaeB6PEcsXKAULXfTyPlEQNMUxIiW+NB6sjV6/Son9r7e6nd3U6NnY001d2NdP3+KO3JkhKRcqcnK0kJkyFUyu3PcKaZ9puN1B9NyepSYqXS7vQHaVuhG4SRfUG0GrYiPLQNzOr95rpOhoypSaJjsCGf++h0xs1lmtAZS4gduDFGxD7nmkD5LtfEDrzAj8DhshgPgl0scU5+2nL48D36RjkmiBYrLSLXCTXpaINxogza5rJ4zngy7uS3C1muHVy+hUo+FvDRgOd+T+p4H273SV0XofKkkHyzchj3IlS+GYYld0GgIFAQKAgUBAoCZwOBd1moZATgBXAbOAiLZPHmc+PGjQOxknvmjHAUB7iMOQ/nlEPsMHH9Ci9qi4+0tXXHtBaaSkyUl5y2FqguzI1DqBR9CotK2gOnGI3gJ+J+mnPCQwicw1ngNfAY6jH3IuaavHATuBDpCeZF5kTkrfMhcyLqJo25ETzLgXuUTR302dwP3LhXLyO3P7cRTEhfFwW5R/Dh9I7pn/mjORzP3O56XS7jec/Ix+HnHiO3yePHc9K6DT4nr+/V22WMbUlpkRKXrxYr4YuMm8eMco5zFKHyOCjlNEWoPD5WL03JH4tCcF8KU0lwxhAoQuUZG7DS3IJAQaAgcEwEmJgzIf8yhEqaZAID6WKVKNZ2iDQrKysHgqWFLRMliIYDhKlORFwOJNNWe8TkJS0HJAIiZ7HLpKJejss1YSFNtyKOPc3tBiJ0I+EkM0e5hB2HYNmWqNjs76WGxMpxlabfG6ZduXbd6UqsVOiNJcpNixzJ7U+abaSx9igRvZML2FHq9vppZ6+bNrUiEzK0s43Ln14QONoNsaJdEED6X2+jCRF94BlpTGidnmeU4bK4Phx4Xr/n+igDDD0GlM09t4EyTYot6HFNebSHfHwUYCw8now3zzhov/vANfncVu57zCwkmsQzjjyjTtdP+no/XA7vNmnJC76QSmLeA+75PsIkAWtKYp5D3k2gad/bPopQ+bYRPl75hccdD6eSqiBQECgIFAQKAgWBs4/Auy5UmpMwz2arkatXr6abN29GfPHixeCgcEhzJ3jO4WCeZA6Un8O/4FVwzZY84rQkHLa00BVBkWdTcvtaCZUdXLlmvsUbA28xR4GnwDmIzWvMaWi76zQ3Yp5qDsO5ORHlcZCX4Dq4TzBvqtdHvdTJQV3gAFezcEvdlEV+0hHMuXgGZuaH5Acn1006B7fhcMxzDrfZ+X1NXH/u+3Gz+uF7eUzy2Jmv0iae19vBOe04HLtt7ifYwgnRdnD1ikjJXqIIlbiAZYGrOanLr7freedFqHweMl+8X4TKL2Ly2ncKwX1t6ErGU4xAESpP8eCUphUECgIFgTdAgIn5lylU0lRIA4QCAgEhQsxiRSuE0daViFsIXTyHBCF8QTwIEBnymiCZVFEO5Ip8PKMeCAdEDFIHqSC2CEU5pCO/RTnaxYE1JWl3CRKuMhnBJavEOPog3JpyC9sY9NK4300jCWiImT2Eyr2koD0qJVTKaFLsTy5kOxI35yTUSqiUkWVg3u32ok24lCFsKDCPhPBwGKN637nn56QxFqSp98M48dwHeHBdjzmvB57X67X4aSLKM5dpkmdyxzXPqZv2MB4OjAn3eM6YkMeE1/2gXLeP55BpcAcTAueMHcGEm3TUe7gP3PPYM36QTTAmtljJfd4J7iFSEnhGPZRPGV/WUYTKLwvpF9dTeNyL8SlPCwIFgYJAQaAgUBB4dxB414VKOAkcBK89uHy9fv16unXrVpzDO+Gb8CfSmeOYjzg2b6pfN+Uhp9mEQ2kR6lgecyRQ7u1py499edKR55yW6OSchEoVLf4zJS6b+ZV5jnkKfON5nMbt4W0jDfwEbgpXMTdy3sOcxRwIzgSfqgdzKe5xUA/8GSy8XQecjbZyuA7K5CA9PNzc0JwTnGiHeSFpzdN8j+cOPDe2R8XcczBuXPuIZw1xW40BbXKgbZy7/a6b9h/VHj93TDpwZoHrkydPQqBEpCQc3k+UtPX+uG3Pi4tQ+Txkvni/CJVfxOS17xSC+9rQlYynGIEiVJ7iwSlNKwgUBAoCb4AAk3IIyJdlUemmQi4gEBa1IEesdHVAuOSexUev7jSRNEGCSEEwCaRxOggKxMHEzmIVMXM17rt+8kBiqctiGu5X95TOhJDzniwgyTeSVeVYFpHaTDL2pRxp70qEzYFcuPb6kJukvA3Vo30plWyqqX0up+UudmakWNcIlUq/K0UTcWxVJAgiRKB9tJu20QfjQ7vcNmNITDqIIv2nD8aAfGB0mNCRJ4hdjfz5nmPKNL51odLEz+TPZdNeDufzmBzGlTZx1Mky7x+H81KH2025EEDS10m1hcoYi0qojEJqPyiXfIw1hB6cWQGLWIk4SagLlXb5yniTj3q/zKMIlV8m2s+vq/C452NTnhQECgIFgYJAQaAg8G4h8K4LlfAY+B388tq1awdCJZaVLI6Fa8Jb4ChwEYJ50uGR9n3ihgSyrJkhzk2FQJmFyqa28GBBLkLlKITKmZmG6njWQw5lm4PDZ+AdXMN9cvnPCpukYY4KT4G/EFuEdD6XyTWiptORzzzKMVyHMukv/UegxLsR/Jtz7vHMXIyYAAeEp5lnmhNSN8+pu94e5+P54cN4HsbdY+DYeNSvnXeqEipdhmOec1A/2Dq4fW7j4TZzn7TgC2fEgvKzzz4LkRKrStzAwinB1uNGGcc9ilB5XKT0vULAHh/Z45f7K5myENxfyWF/5ztdhMp3fohLB99BBPiXfcTkTBOuwUCCDROvEZPML6ezmjempia4rDZss+owJvSe1H85bSi1vBwBJuRMtL9soZKWmXhAdhDZIEgQSVtWQpYQcLy6E+ELwgkJIQ/nFsUcc89k030zWcNyzgIVBI3DdVMHZJU6KIupMWTOhBDLSiwg+xA7CZTDgUgPxDKInleqDpQHl67ag7Ina0rtUTkcinA2+iJ12tuyI8Gyzb4euDbtqy07aX1tPT2WSxlWaD6uhEraBTaHhUr6xr06+XIa2m2cLLhCJMHq8OH8xD53Gt8zCTWe4ETgvoPJKXk4J9THhDYRvFKZtjAmjIcD7x5YHx5TjyHtMmmEOJpYmxxyj+cj/rZFzN85vdMal57Gb1ci5ebWZuCMEIwLnxAlt7OLV0RMC5e2pqTML5saFaHSb+AvNy487peLf6m9IFAQKAgUBAoCBYEvD4FfBaHSi2GxpmR/SltUwjstVMJt4DHmRfAAAhzD5x6VzJXgatzBNWoS/2tokaosKhUPZVGpTx9ptjNMM7OKESpn8ODTCZ5EXZQB34DPmNuYe8CJzKvcJtLCneClDlyT1zzJMfcQ0uA7cBzSkxZe6zykoUzKh6vBQ+DgYMI592gnbXG7zPeMldvGfWPkuI5VoKQ0GbcvxtThZz4/Ks7isNLyH+UdfFuKOwxFPoj1vYu20Efj7PP6mLq9xBw8Y0zghCxwxe3rvXv3QqhEtIRH8j0BvmAO67xRwEt+FKHyJQDVHhehsgbGm54WgvumCJb8pxGBIlSexlEpbSoIvBgBRMl9iSl7vUHa3N1P212txJPF10D3q3n1iwt4w6eIk7PTrTQ/00qLc9M610pC3WOSWY7Tg4An5L8MoRIUIBoQHcQsiBEECaHSlpVc160rEcIgSASLYsQOJk+Ua8JmwRFiQbAgxXPSQ8YQ+ajHFpwQJAgIeesrUnvaQ7K/L1LZlzipZ30Jjtzjmvv9PgKnRMo+7n+m1QZ+33oirH21sa94X9wpk82tzS2RoLUQKS2iMY80LrQBbCwS1sW7SKQfpOE+fYBsO3ANJuQ/fECowIfDsdNwXa8XfCifshwba64J1FFPZ9GYMeXc7aBeiDF9hDQTgy/jQL2kIw9tdz7KdRvJT1qPqwkn4zTYl4gZMeOiOiDzIulbiJRaEbv6dDU9efwkW1VuZhewjKvH1iSe9lEP4cs8ilD5ZaL9/LoKj3s+NuVJQaAgUBAoCBQECgLvFgK/SkIlFpUIlexRiRvYulAJn4H/wDnMN8wzzD2IfWS+hBiGuDUlfoMnnZa4H/wRfjYSlxlKrNQ+lXOIgfoWMgsvyluakB/OQYALWfTifp1XwYNoF3WTti5Wck4gr8vimrksAqVFNbiOORfPqY++USZ1mYObf9vStM4laQMHsXHi2ngR+5rY6ZyWmLrq105P3no4nJdnWaSsBMlKkXSdlGPuhtejCPBFeT+in8ET4YgKMabaxoX0lHZw6JRrMIHX871g9clq7E2JUPngAW5fnxxYUxrDV+WLRag8QPylJ0WofClEx09QCO7xsSopzw4CRag8O2NVWloQMAL7mpwhUq5v99KDtb20utVLGzsSViReNphM1uZmznOS8Uy7mZbnO+nCYiddXp5N5+YlQkisbLHEsBynBgEm5Ezcf5lCJUQDkQqrSsRCiBLh0qVLz1hXIiYiYNVFMYtl3IMAEUx26kST+RmkA+JGjDBFv0kPEaNsxCKEPsRKyiM/xM+i2t6eiF5F8HAD21fo9iS6qexM/BAryaO9SvbbEs4kVMqiMo27alM3tRryCSvRcjhErMPta943EZHS+ydCfDjcBxO7et8iQfWD57QVkknb6QOB/nCPZy866iSPdFybIJLXweIkZdYD90nDODiNhUqLjR4TyCH9s0BIzDXjQL2UQdngbze2Lp82ua28swSTz0zys0CZCbrcHWkl7JasJtn3kxWxT1blWlcE8+ma3L9uZPevJu7koQwT2FclnS/C97jPilB5XKTebrrC494uvqX0gkBBoCBQECgIFARODwK/CkIlvALxDXevtqpEMLIgB/eoC5XmGeYF8A3fgyPAR/S/vqcwjhK35PoV7odFZU8edfBkNTU1TJ3pgXiNXMBKqJyb6wS3oS64DQccyMIhdXGY15GGNsGx4FEcpMk8Jy/4hJ96oSXzVwLX8Bu4rr3ImO9Qn4P7RNnwNTgkeJh/cw4nh4/RFtLRd3Mkx7QLPHheD2439+iTn5ljZgwnH6N8TWzOV79HPXGwjhRqTVT9GIc4yWJWWU9W/HDIucYN70fgNgmMJUJlnWszngRZwyrtnjj6+vpGeiwLys/l7vUzCZVYVuL29T+z96bfcVzntfeDngfMIADOIjVSky3FsfUmuZGc+zEfsuIk9r+V5GP+AWflQ9bKWlley/5w31cZfH2vE8uyKNmmKIoz5qnRA9DoRuPdv9M4ZBMCgQYJkA3iKaq6uqtOVZ3aVQ2d3ft59gOuOPY0m08W2OpCZfs2dvPqQmU3KHXZxglul0B5s2OFgAuVx+p2eWcdgYBAXRlG5bUNm5VI+fVM2e4tVG2+pAGsxEtsWINYeQRYMWZkIpNyUgLlubGivTTRH8TKYl5iRsqFyjZCvfEK8WLw/ryFSggNRCgSJURKBvPMvI9Rr4iKELdI3thvJ+npJDa8h0xBzMikhGAwR5GM7VEci0IlIl8kkZCch6SQiFRFr0L0iFjV3CaE2OpIrFzX+m2hsimRstnMiiRBnGqaq7rhFREgxLn1BySyVGpn/MUsT84XJ/rGtPN64va4jeuHZNJv8BseHg5LiHm8jkgo9zoWbeJ2jhlJZbw34ARh7Zw5b9zOuZijQMl7tnEcjh0JNtfKHIkzIiETbeO94Fo4D5/jOSJx7RQpOebGdnYr94VxeK3Wvs+Iv0GklBBMHdDFxaVA2kurJatihaS2PBccg2PSxziHDj3DFxcqnyHYe5zKedwe4PgmR8ARcAQcAUfAEXihEHjRhUr4InwC0Q1OiVhJZiUZlYhxjL8jt4SvwIOiKBmELvGynTyhzZUQFcnBg2fKSUeZlG2hUvUqVQYEoTKTacj2Vb+JFOlDNgRiwmvoE+dgzBk5JufiuPQh8imWtI39og28JYqRBN9GThV5VfyM7Su1FOG8tOdc7AvnYo58k3NyDjAAIxyNwAh8wIt1bKMPu+EBN4s8MHK22G/WRx7I/sy0j3wOzhUn+hHXx3Vxe+BmlPiAp2mJW1GYESjhb5pZhv5FgTIuhdmmSrZwD5tyPmoqYD/so1PzW1joT+yXPje2S7PAHxEpp6em7f7U/SBaLq8sB+7K8cjajP2L/e1mCabvvvuuffTRR/Y3f/M39uabb3az24lscyyEyvgQxGW8UzzQcep8H9eFh3n7CxC3x2Vsc5hLJ7iHiaYfq1cQcKGyV+6E98MR6B6BdQmVpWrd7i/W7Mt7Jbs1W7bppZpVJVSmECrbYYDdH3C/lg/HmqFlv0TJc6OFIFK+cnbIzuj9QCFtWWVa+tQ7CDBYZ/D+vIRKkGBcBnmBzEAmEdqI6IQkRaIEcYJMRvEtkh8IBtPO8V48XiRGXCPkLWZUIpJBDpkiiUXki2If4hj9Ah/aMUP0WEL0Hq6r6Zht4XNdgQH1uqx5msqobKaFa1ZL4duo6P2q5hXZklZEFtuWo+VyW7CjX1Ew2znODR3c44U+MndeA/iBFTNkke1x4n0kinHfiF1c0ja2iziDR6eIGLMeWc+5mTuJdSSpnItjxecM3CKJZhmJM9u5lxyPe8Dxo1gZz8F2+si9DEQbgZJMSBHGhiJbm5sSKiUCV9cqhhi5tLwYbHsW5rScX7KlhWVlWELaJVJy/3eIlPST6aD3IOz0lC/cK6yoPvjgA/vBD35g77//fvgBhR8HYr/2O0XsN0v2YSb69/r16/bxxx/bj3/8Y7t69Wo4zFtvvWU//OEP7cMPP7QrV66EYADwjfvRqNvz7tevx22P/d3rXLSJ7WJ/4vJxx32a9c7jngY939cRcAQcAUfAEXAEjhMCL7pQCY+BR3QGwiJUIhhFbgn3pA1t4xgTXhJnBDDed45HoQw7hcraGhmVZEpGobJpBdWo7O/PiNe0HWM4D9wILhMDNhl7Rh4U+xt5FPyKdUz0Ax7FfvBZhEgEScRIPncKlzEwt5NrBc6k83JuzsfE9TL+53zgQGBwzDwl+xSMWE8fIv/q3J/18EPaRN7G59h/tkcuHjkh54tTJ6as43PEnfME7OUSFvF/IFAGkbKdQdnavj+btJMQ2eRzXBfuI5mVCM6IltzL9o9WQaiU01cq2Q56BgsCkeGQuB1NSaScmp7Sckp8asFWhXNdTkrs3+73jh+/4kXtsXShcg9wdmzqeaGShyA+sO0H4uEV8DDtnOPWuB8PI1NsF78YfD7syQnuYSPqx+sFBFyo7IW74H1wBA6GABmVWL3eX6zaNYTKuUoQKmvrjWC/epRCpf6vbQNBqCzaRWVTvnJm0IXKg92+Z9aagT9E4HkKlVwsYzNmyE0kk1GohDBBlBDgIEGR/EB4mCKh6RzvReJEW2baIDRC2CByUahkPcfpFOE4P0SScSLHjMJkFNUgeqxjbpPFdt3LWg3bHdVKpEZlQ3VKlFW5obqw9fqqsi2XNS9oe0n9qIa+hCxM2cdG4rhzjBsurosX+tl5DeBEBCwz1xXHvyzBuJM0cvg4xuZamWM/OGYUIcEDEsoMPp1CJceL7eK9icQ6jrnjcwZm8T6AHTO4xnvH/pHsdhJezkG/eFZpv64MVo4DdqK0tqVaMK2UsG9JqNwo22qFbErqUmqeWbal2RVbmReZX1EmZxWR+tFMyi5gPtImJ0mojM8XzwRT5/PZCTLtmOOzEdvFZ4rPhz05jztsRP14joAj4Ag4Ao6AI9CrCLzoQiVjRjhJ5C9kUcIvY3kRuBJ8A75EO7hPHGdyzxiHMl6NY9I4FmUI2o75JhiTLMqE1dZUp1JCZUPuOtSozOU2rVhIiDeRTdkW8zgHx4APwUdjEGzko2yHc7FkjpyN88OBaM9+iJRYu+IeEzMnYyAobeBXUaRE3IQvMZ7mPPF6uL44lua6ORccDzENMZcASnACH7ZH4ZAlE+3pK9vZL/J0sI7XwH4RU3CN52PJNXVO9Itjg00ICBaYG8pwJBMSS1d+X7I+Zl2H/m1uIVS2RUkcjLZkubupAOEWcxA32zwiXG/IwtR6zskc7h/uQfr9YfuewyrgASvKnJybn7f79+8HoXJGmZVgXVMJmA14J0IlYqf+O+jkQmX3iPW0UBkfVr5YnT9kcHk82PHLEb/I8Y8K+/FFjD+I8Jm2tONHEN53flG6h2vvlk5w98bHtx5PBFyoPJ73zXt9shGIQuUUQuX91UczKpMSLI7gR944YGMgGYVKbF8RKk97RmVPPpBxnPW8hUoICzPjNAgOZCfav0IoybAkyhMyBPlhLAfxYYpjvkicGN/F43AsyCcTYzTIG8QNAgcJYh/OGwWymMnHZ9YzlmQMGgjTNtFjH9a1x5jtjMpKhWNyfIRJnUs1Sur1lGyAlIlZW9G8qG3zmkvqB+de1zE6LWiegO2Eq2q/gAV9pv+QbvBjJiMvjndZxnbgw3uuMeLHNXFtURhiexw3gyFYMnMPWHJs7gVj6jjG7lzynuMzxecMzLgPkUCz5J5wbtqwD8eO94H3UWylb7SNUcIVRbY2VO8zlZX4qh8CUgPiBhkJyH0So9ex1F22xbllW7hbskX9DVy6r2hjWWHXlfnaUDYm59tJkttoPvvXkyJUxmctfofAn2csPo88o6xjdh737J9DP6Mj4Ag4Ao6AI+AInBwEXnShEh4SuQ+cgvE2wa9xjqVF2BbFNbgI41Fmpjgu5X3kNdAbGA7sDaGSINWaalQiWIpeKCic2pRkUyZ1TrIpc4HP0BfGt/DKaMvKe8bHkXft7AfnjPvAmxAnEc6wJ2WJUBmDcOFYcea48C44FvtH3sP17DZxHrgdYi6C2rlz5wL/hlfSJybasH/ElPaIlJF78j5iCY60A0f2i/POc3M8+kh/Az8Up65UFVSswFKuBdecrT7hk9W9zOrcOfU/tR2kilgpUXJT4nBrQ2Luuo5V0+c6oqX4hHLWgqjI/UpKQNZ9SShBtY/fwqQ0J7nP23xYXRR/XxeeKzYvoXJ6ejrUppxXduXqalmuPcKzLq68LsFT52sfW1ezO5w7LzN8dqFyV1h2XdnTQiUPbPxRgi8hD2qc+KLx40X848KPGhDd+KDTllRovsAch7a04Y8TX55IiuPxDmPJOennnAqv3r17N3yh33777RC1Eb+gh3EeP4Yj8CwRcKHyWaLt53IEDgeBXYXKZWUwrcv69YiEygfjNL1BqDwbrV9dqDycm3oER2EshWjwvIVKLg0CE4kPYhjjOwRKBvUQyjje6xTIImHiOiBhTIy34piPcR/tWcdYkHEa48o4c+2MGyFTtGOOY0SOzTHZLxI93rNPFFo4TrVKRiWRqxyXceuWiNaWaZWII5GviyJbyqZcWxQJW1U0Zl3HhTS2BRnO/7QTuEEi6XsUKSGNXA/b4szYF2yi0Ms1xmeg8xrpD/vQnuOyT8SGc/C+k0iD386Z/Tk+UzxH5z2AVHeKxmDNPvE8kF3e01f2B2vI+ML8QsiWhKhLMrbCiATa8Yz1n1Fk75iwLIpIbgl3Wbwuy/J67kZJc8UWbqh25ZyiimV/vdlApNQvCWquuxD6+DxfToJQyXPO9yb+EMEPNKzjux6/p9z/2M553PN8Iv3cjoAj4Ag4Ao6AI/CiI/CiC5XcP7gIHAVOA4dhzA1XQpCDW8KXYhAm3AbeEbWCyGM6n4M2b4M7iEuISgShUmU/6hsZcTtlTEoJy2UlUA4kJeKlg1CZz7cdfjgOXAhOE4VK+Bc8hzFw7CdLPjNxPtrAmaK+QWkHdA64EMdhW6cw2clXuxEpw4n00hn0GrNOO2t5wgfBhnaRd4JdFCvjeJ7t9D9qIBHHncvIDxEo0VGWlxRkqmvjPde6puDeZqtpyfyWFUbFETXnRnU/ixI+MwJfGZZkTzYlTDaqW1Yvqfbnosq2lBAsJVZuCD81S6YUDF1oz5miglslevZpHWJlsPDVM9InuOHo5TLBrm2xcmlJGasrq+L0a7oH+h2h0rDairI+yxJIdc6tRvv43VJJFyrjk7b/sieFSr6MzHyBo/BH6i2f+dGCh56Hmi9CtATjDwxfaL6gPNSzs7PhBw2Ibvzxij9M8Q/Szkjz/aHavwXn4qHm3C5U7o+XtzgeCLhQeTzuk/fSEehEYDehckZCZdWFyk6YTvz7SBB6RaiEwDDGgwh1EknGbozbWMc4kBmCxLgvkh6uhalz/0iYWMdYEKLHWI2ZMSWfWR/FzSjIQcKYIj6R8NG2U6jkOLVau9ZkpYJYWZcw2bRyZVNEZ1P1LBQVWl5QmyVZvxI4V9X5FBmqMEwNcw9t4voghYxzGQ8zgxefuZZIfGkTxb94jVxTFBBZ8hlM06ltIprLPhArwYd5p0jJ+SOp5j0zmMYJHON5GKeDfRQqWYIjuHLeTuIb7zH9iuR8dmY21A4hY3Ir07Thc3kbeUkRwK8oqveccBgVaU2JSEokXpmu2sy1VZv5bdlmvhDBn5JIrNoxLUXZxhvgQuXR16iE0/EM8GMKHIlIZX5g4ZmhBg7EnR+LeD55PhCkCfpkybPBs8PkPC5+o3zpCDgCjoAj4Ag4Ao7A0yFwEoRKEIr8MvKgKKxFvgS/jGJl5DmR10SeGceyjGcJdmzPHJ0syoxqI1JXPm/pDIGjWRseStvQoAIpVaMyl4OLtTUMyk8gzO0UKkMflfKXVDt4FJ/JBiSjEN5UVlbfMm4xiJSL29mUqqeISMlYOXJV+BR97JyjvkJv95o4L/wQ/gyPJGA4WuXGgOHIBVnCKRmb056Z95EjwgOjUBnPGbGMS3g4WMAJqAc5OzOngNT5cJ0EAtdba2bppmWHzQbPKTD1tJx3TqUsM4DYqPsqcZFrawuVCmpd2rTKjOp/zun9IuKltjfbwmRuWBmjmnMSO9MSOhMSKvsCVUWsJNOyT/cQrHFLaovCZS3Xqvxu0FDWbNOqKw0r3VeQ8rwyQEv6TUDByWRWSrPuanKhsiuYQqOeFCr5UkFKEf0QKG/dumXXr18PPyjhl8yXgDb8gcE7mS8PXxweUoRN2l+9ejV8adnGF4QvNBM/dkGKX3755fDF44vEF/IwJv5A0GfI9Z07d8KX1DMqDwNZP8bzRMCFyueJvp/bEXgyBFyofDLcTtpejKUgNL0gVII9xIUZkgT5YbwHeYwz475IhiCUkCTaMo5jDMi+EEvWR9LJe9bFa41iJaQvRp9ybsgrbeMxORbHZDzKHMem8ThR3As2NbKoKSuDb3UVSx6Elg1bLjW0TiKmMirXVaOy2VjVcdZ0TETKLhkNHetiYpy7k3xDMCNGbAMnxrxR5OUz18h1RKJIHRCmiCHtsxIq4/5xyb60YQZ75k5C2klMwTBiF88F9hBrREpmPtMHpthPyC7nY4rj68WFRZHYWVtaXpQYXLbUYMvGXy/axJV+m3xTWbiX5J4yoetKb4Wo1+X7VZv6vGT3f6P5k1Ur3d0WKoPudYhKcejlk7/wrMJnPvjgA/vBD35g77//fuAr3CvuUTcTODPF7wH7wX3gTx9//LH9+Mc/DtyINm+99Zb98Ic/tA8//NDefPPNYLNM+zjTptvz0na/ifvPvYfTffLJJ6FPCJZ8n//gD/7AXn/9dbtw4UK49/xYcUs87vPPP38uPA5xlL45j9vvrvp2R8ARcAQcAUfAETjOCJwkoZJxLXylk2d0CpaMxfkM/2D8DQeB58BpmCL/gzfDC+FzGi1re1pzQRaigzr2gOXyRR1HCVLDGXFRRD8CP9t6A/utr7VLYDzgPxIum5tt/sV5wvm2h/6UCamrjEhVPBOdAa0jiJQSLBmvwqXW1tslNGK/6CdcYOfMsfebwIjzw//Ag6xKxufoJySIobmAD1yZNmAU3wfOuL0u4hax6+QU8T39o/8LslaFH3z99dc2NTWl61tRPciKbfaplkp+w1IjGxIoEzYsjtd/Wpmcg3LQKSpANiMOui02tuRS1FAwfk1C4ur0hpXvaXlbmZWLcg6q654rg7IwqazMyT4dS0Gtg7Kj1b7RvxfBkmMRvArmZE8iKLNsNiVINhTUq0DX8uy6zV6TYHxbAbezqqWpDM6tbRvY/bBluwuV3aDUbtOTQiVfYAgtBPuWyOpXX30VSC1dfu2114IFGA82P1YhXCI+8p4fnHi4af/ZZ5+FPyAQcL5A9+7dC2o9XyzES0gxvstEUfDlOsgUf3DZ+UeA8xPxTaQw/eAPnAuVB0HW2/YiAi5U9uJd8T45Ansj4ELl3vj41jYCkXT1ilBJryJJikSSsVQUJ+MyEks+R2ERMhTFM8Z6ECy2Q5xYz7iR62V8yXgNcQwBjJlttImEi3N3EqnO8R7vGadCCDnOuggiZHNFhHF5uSyCVbXFJZUCWBFRKlODcUXkpiRrmorOUxcFgthKVIIfdWhQ6kJYzaZdJ9rGfXi/3Z79EmJY9J9r5ZojPiwDRtlo1SMb17zIpaJ9uUYOAh5ggH3tpq4ppUxKtheKIqK0zSJyMiNMSqBkmYQctgXKZPLRiNmIG9cArhH3+Kw9PF9bqCSiGLGXPtCG+xjvAz8osI57tbK8EiJuIbSr1ZJtSPTNTyTszLsDduadQTvz9qCNXi5av2xgE6mErZcbtnS3avc/W7F7n5Tszv9dsZU7OscBCCXX8CymF1WojPefe849vnnzpv3iF7+w3/72t8F5BjEdsfTb3/62Xb58OfyABF87ah7H9xdhPP6oE/vJ95nvMmLpzMxM+DHGedyz+Ab4ORwBR8ARcAQcAUfgWSNwUoRKcIWfRI4ZOR8cE67EOBxdgPcPuJM0ArgSbZmiRsGYlvEjQmVCGZCplNpl+tV2VL//D1uxX8caLNqoSlNg/ZrPq410ypY4UUPCF5ymHeQqkVHvESIbOh58h/EoEwGtmy2NVcm+VGmRVexIsUaVQAkf4nMQOsVB43iW/eMx4nHax3ocsQyn2vUlI94Hf5wYn7BLGp9fvHhRAZXngg4Thcp0pu28A0YRp8738LmIeTxJ/Ez/wBOxlXE/mg+BlTMKRl2ryEmlr2HpgZalRxuWPVO3wrk+G7wgTjoiJyXZtZKdmknqvBIr07LZJbOyKR/W9aq497w0mXtNW70hoXJOgcw1/Q6QVibleFOzygdOyma3X78JiEsHZNQX+hVFT9ZtwbPRpzVzvlZTx6opke5eze79Rraw16sh+HVtWc8BFrAhADZe5eOXLlQ+HpudW3pWqIQsRqGSB/f27dtB+CPKGHGSh5sfZPiDwheCB50IAyJgedghmFEo5EcshEO280MI64kKIHqZLx3HOMjEH6fwR0V97CS5/JEIP1Yp2oG+80fOCe5BkPW2vYiAC5W9eFe8T47A3gi4ULk3Pr61jQCEhnFMLwmV9CwQhu1MPcZ4MUqT8VwkldGCJghxWo+4FQUuSBRjMJasQ/Bi4nqjSME4jjEhM+toAxnlHCwjwYpkr3MZySDjwXp9vS1uaIy5sCBXjVnVUVys2eIy9UQU5bpR1vHlPdOqihDVdW3KpkyIBqlWbNtyRh3Txy1Fg6JhBo4KS+qcECghTLoMIkAROENSpg61FWxMFQGrBmBF/6NYGeuGsA4rVwTHjERICGjARCcjWnRN0b1E69IH2oJbsV/1PQuxFuW2UBnIaMyklEApkRIC96jg+jCKNxJmlkwR/ygIIVIS4BcignU/wJOJvnHfeA64N2uqDUI7iDkzNSgTxZYNXcja+feHJFQO2ekrAzZ8oRBqVtKfmjJal25LqBShvPurFbvzfyRU3t4WKg83oTX0+WlenpdQ+dFHH9mVwpRv5wAAQABJREFUK23r1/jjAUumuHya6+J+8/eFH2QQ/+BoiJTXrl0LP0jwHf3+979v7733nr300kvh+bhx40Zod1Q8ju8x33k4Ic8f73nu6Gv7+9wOYqDPCKnO457mCfB9HQFHwBFwBBwBR6BXEThJQiX3II5t4XiRa8B7Il/qFCwfCHLiPkzwkThO3AwZkFs6BkGi2J0OWTZ/SvxrVPxzWLavRRtRRmV/v7bLHKZPvI/xMPynti1U1rBsFf9aR6jUOHRTmXuMRSNXYh1ctSLHnpI4ECJlaXW7JqXKjhAoi5DZqUd0ctXQ6Sd8gTPmdV0T45P26iuv2EuXLgXtZGxMYqy4YSYXeWGbE8KbO2ewjTw6diFiz5J+0m80EzIpv775td26eUsibEllJ5XR2a/6nmdSlj/btPSFmmVOS2CU5WtLXLimQOCtdZUK2VLwrQJxiwMKvu3XhoIE5C2EXc2yfq3clJXrnIJXqwPSG3W/i+tm/XI2GlDdy7Qw1/1syVo3TrAf7F+TGQX/FpKWlcicUeZmWkIzJUNqSxu28HXFbv9q2WZ/X24LlUsuVEb8DnvZ00IlUdOIlBBbZn54wSKITEgefP6oQHJ50PkBA6seHnTIJ+vGxsbsjTfeCD9qcSy2Q3z50vNHiDRm7I9IaT7IxB8YzgeJ7Yxg4D3rECshvwigTnAPgqy37UUEXKjsxbvifXIE9kbAhcq98fGtbQSikNBrQiW9g8gw1otkEgIUxUgIJWMsxnKMDSGTCJjMQWTTOtrwGfEuiHI6JmNDiGYki4znolDJ+SJp7SRY7PO4CaIag9QWVS9kbp5MrBUJltWQUVlbqyuTErtX1djYquqaNmRLqkhOkdZkVtcmMgQpUpitba5LRF1v19lAfAyn1SYEykCcZFuTZM4hDmqXjXbbZk2kVvpen8I/k4rsJcKV8THXHgVLPkOmiTzNhAxJMiN1kJBRCXGWDZEyKjlnAQyVTdkvoTKn9xkFBSJsgiNzW/iVkKtOSKJs91P7BZxY6l8k2nHZiSHrImaMlYmmZdwcx9S0jfcC3MO9CrZHijyuK/K4KZGy0JJ9T8rGXinY+feG7fSbg3bq5X4bnFS0rSyBEHEhsku32hmVd/9bQuUvZJmkjMpg0eNCpf3oRz8K2YxHKVRy7/iRJdSeUcAojjNwNLjdp59+Gp5TxNJ33nkncDu+j7///e+DDRTPwVHwOI6LME6ZDvrCe57H+HcBjki/manNA0+MZUR4Ln1yBBwBR8ARcAQcAUfgRUDgpAmV8Z4xnotcY2eQZ+SXkUfGwMk4Noxjxr4+3HjgXFihjliuMCHuNCZuKqFyWOUoJFQWCwSRQo7EtTTWxKa1ukYmpRx36rXAvzYaGyHTEnvRzYYC/DZw/VAgrLhPqJdYK1u5qkDNivhSrbId3LludXHMRl1jVlmStjYfZmPGa3xkCZ/c5pQPAkwfM6SFC8K58xmEygl7RSXzXrr4kp2VBjM6OtbmhgomxWkHd53gsrMd7At/jvNuQmUcR8MFGW8vLC4E/YaktKl7UwpMVVKZ8OwfVY3PixnLn29Y3zmJl6Pib/kt1YuU66aCUJslWbm2ZNVbUJDqKQXWnpIwOqZg1FxdjjuyZF0xq99LW2u+YKnKiD4kTBUmbd1kl2uLVtuUm484B2P/8CyIz8JgU+Lm8Mj8mETmCXFp3UNsZpt1ZX9OtW1f73wivn9N5V7ucx5ZAB/AqcczKh95Kvf80LNCJX8AolAJoUWA5AsDmSWjkuh6REpILBNqPN7G2AWxL6QSQZOMSX6s4UeQ6elp448xbfnisP273/1uyK7cE6WOjRDcGInLMXdG4kLI4+yRuB3A+dtji4ALlcf21nnHTzACLlSe4Jt/gEuHKEC8elGo5DIikYxLyA9jwRj9ihAXhUqWkMrOmbEi7RnzxYlxXCRIkKRIOFkfRdFOcsV6JpaxH2xnbh8HNw3RHo0t5+YVZSmhcnGppsw/BMCG9qP2IkJlTYLjhiVFtDKK/MwPSzQckHiYTwRhrb4qy5qVpq0vKmJX4iNiG5YzyYxqdhYUOTqk6E7qcmimHxtVtS1tWm2haY2q2qu8CcJhxIhrj4Il75MSJtNZ7V/UefPMEhqDFZH2b+rcIrzsn1NNSgTKAtmU1KdMYxMrohYyVqlHomP1yQZJYa1bDWU8Kqq1pXOH/gqj8E9LsIEAsozYgSMTmMegPmquIFhGoZL2TODLfkQYc5/qG3Ix0b++7GYgpIiUE2/021llU46/IpHydJtQpguKuJVFT00ZrUu3a23r15hRGa1fXah8JkIlHIn7i6sNmZJ8RvxDsPyP//iP8Gz88R//cSjHAZ9DNPzd734XeNRR8rgoVNK3+Nx1CpU8b8wuVIavor84Ao6AI+AIOAKOwAuIwEkVKrmVnZwOsRKuFAM8CYRlJvgVoRJOAl+OY8UtcRWEv1TYD6FyWJmGE3KjGVMg7UjIqByWyFXIy6Y0SZaksiMlUtYkUJZrq7a2oSA58cKGSoKI5YTSG826AjnF/zYqqmOprEBEO8TMtabaN6U7tFQmY0NuQOJraxUF2pYllFXEjNbEtySW7TUR4Iojj9xSJcYpEFg0jusX7XtkEu0K3AsBNp/L27jG7IiU6CanJ8/ouiTIZvsti92tDgbHJhA2ipU7RUpwY4pYx5O1+bOcb5aX7O6duyGZjNqbcL5+WecOnsrb4Hlx5NNy2zk1Z/VcSRis29Js1aZ/V7LmQsqGE6dtbOiUjZ8ftv5zypg8J2egIWVLimtvrimQdq5gfUtDllkds+Zq0ipkpZbmbW71ri2XVUZEom9TY336H4JvlfUKP6cO5uC5vIRSufRIsES8XBdHX7hRsZnflQOvXLpZs9q8gpUr4rjYvnbJK12ojE/A/sueFCrjg8uPPqjrkFtsgvjDgDUQZJY/JNh+kRXJHxaidaNQyReBGpS0pegrQiVRuhBlaley5EcS7F//5E/+JIiZ+0PVbsGPJpF48yMLx4XMxj5HkRISTP88ErdbZL1dryLgQmWv3hnvlyPweARcqHw8Nr7lIQKMXXpZqHzY0/Y7xneQnhj9ihDH3JlFSRAbYiXrgkAncTMSJJZx4trj9cf3bIttGO/Fme1xgoTRB8gZ25sS+RjzLS3K+nWxZPMLZYkzsvapUf9R7EVZlKYYToTKZLYhcXLLCrKvGTwjqyHVUyRak+NUF+tWnqrbyi1lN8pKhizJhGpqZPr7FNmZsoFJEWjaq+YJU432MxuquyjCGtqLXargRsSIfkbiTV1JBE9ESgTS/LAigAexs0EJlagoltVoSATUv8x21iSZl7yHrLYtY/nMnJVzrcTAuix1KhJMFdXarIkQAhE8WRBTW4VrYtzOzPt4D+g74+YoVJJRiVDJ+DmOp2N7lmBPnRbmRE5lH4YTNnQxZ6ffGrTJNwZs8jVl155VNu2Q+iarnqRqlRBZvCbr1+W7NZu6qvonLlTaj3/8Y7t69SrwB25ylBmV3Ddm7jFuNrji8GMY35l333033O+f/vSn4Z4TMPqyorX5QYjnAGtY7vnz4nHwOsRLvtPujBMeF39xBBwBR8ARcAQcgRcQgZMsVMbbGXlT5Ewx8JVxKToCXJI28JmHfFHBrVoXOFKGNkMK8hwX/xy1QYl5Q1i/DlGfErcbuFB7bElGZHmtZOtbFdvMyHFHvDCZEx/VPxxy1lcVgLooN5KSnBrLypqUi8xmpqZZGYUZBZa2JGBibSrXmKpqMa4tK5BzVTypvrdQmUCgzMEpk4H/pcT/EpQgeUiLAxwaumPyI+FR7cSvR0eGpZmcNgS2sVPjNiRr23xy0HKJfsv0qVRKQkKeDt4pUEaeHHlf5zJiDo78/sC4PzqcwBkoDtlfFIcXNxacEh5Vi7N431ZbC7ayKo59v2TTv121rZWcTeQu2eT4aTt9ecQGLokzXyiZjVatla2LaEp8XBm05NKIpZbHbH2+zxan5HqkxLV707dsfnHGVpWlCvdNp3QNCuZNZODnSRu+nLORS3nNchbSPYTPlOfqNvPbks1KqJz/UtmU4uv1snCXE9ID/hsvbo+lC5V7gLNjU08KlTwM/CHAkgehksKqX3zxRbCHgjTypeGBx7IVyyIyFyGVPOQx8xKB8NKlS+FLRXt+/ECg/M1vfhNqWPKDCJEBf/qnfxoEzR247PkxRoJzjM5IfN5DbPnC8cMLf9xcqNwTSt94DBBwofIY3CTvoiOwAwEXKncA4h93RSAShV7NqNyt04z/EAqjWBhFyyhWQjCZGfu1bUof1smI+0XSFI/PuDOSz53LTmLKfpGMIbowIVRSQ3GltCqBsqpZ9R5rm8oATGosC3GMQmXVUvmG5Uf7FCWasXEJbCMXi0Gw5Djl2XVb/KpqU79etdK9ddnMtC1oChMpGzqftbGXizakCM/iqaxEzJat3FuzxRtVm/+9xp0SLCGpZDbSR6ZOnMimRKjMSaQcPJOxgdNyJZnIBsESstonsoq4h9AIRsltfMO1yk42JWsfLGXTIWpWbHcjYSq9afVFCabTCWusKvuxqeOIYJKlyYEiphG/2C+2xvFyFCsZO7MO0sp+zHEK7wnITSnSdTBhg2czdurVgp391rCNv9pvw2cV8SrhNVwj16nI1y2x7I21TVu5v2bTn5fs/icrdlvWryueURlghZscpVAZ/67A4xAp4XI3b94MvAjeBWf7l3/5l5Bt+Z3vfCfwMAIMKKuBUMlE6YxLR8TjeCbhhTE6Pj5zPIM8kwS/4sLDD1RewiPcDn9xBBwBR8ARcAQcgRcMARcq2zc0cqYoVjL+i1ySdXAjxraRn0C1Egk4IRmFcp7JDEqsbNeoHBwcklBZ0KzA0JziQSVUNht1cUOViFuThevaim2kKpYYqVtmVC4xo+KpopS46axJpFy937DygsTIkqxMEw1Lj0lMGxEHGlIf9K8qkRLOuHxH/G9awiXOOgqOfeykviJSEuhZPJUOvDOnviXTCuTVNTzIquQQ8GEtk8mEOLSCZPsLNnpqSAlYuqbBEevPjlpx65Tlt4Yt2+o3SbFtoVIXELij9otYghnvd870M3JtEsAYd0ddBR5KcKz0T9vKKaM0uWhLm3dtsTptcwvztjAt1yK55aTrA3Zh7BXZ0Z6x068NWf8lBcSeU/m/EYTKhiU2Fby6OmqJpWFlVg5a5d6mTd9YsHs374uT3NKx5qxSLYuntyyTkluQAl0zwy0bvJCy8TcLNvKyMjoVBAunDIGvd+TQ8xuVdrmueqH3NoIDUuDdZFPuAT3X2jm5UNmJxt7ve1ao5I8ADy2RuPfu3QtZlYh//KGAXPKeH6VeffXVIFRCNrHwoS0/TkWCi5i5m1DJl4KMyicRKjkXEbcQ2vgDDP1lHSIlxBzbWv64OcHd+wH0rb2PgAuVvX+PvIeOwE4EXKjciYh/3g2BKCgcJ6Gy8zogP4hpkVgy3ovRr0TARnIZBcoY5RnEuO3MSNYxRdIUxQvGd53vIzkN4p326RQq20RLdUcU5RpEStXC2GypNkZIxJRQuUVGZVUWrg0rjPcpSlMZgbIsnVBG4Nil/pA5WZ5R7QtFiX79n4sSIGUHJCuftIjT8AWJlLI5pQ7j8AWJcqOZYPs692VZ7cs2c7UsUitbVEV1IlTuNuHUSn3Lwlha4qjqflzISfQsBNGT2htJ1coMfdVYVpRV0/ar8CViOCHiiViZlAopiViWOsoRXVbU77SiTG8pq3IpCpXaKmsh7RLIPJjGcTJHBUNmxvGQUgQr5k53kogz7eNEZmmq2GfFiXTAIli+vjtsI+dltyT73GRK/ZcozLnTeRFvCa+bsn8FUyJvHwiVt0X6qSXyMEE2nuK5LglsPH/+vH3wwQf2gx/8wN5//33VgVHUsn4o4RnvZoq4sYw/CuBMQ7Dnxx9//EwzKqPgRwDpLZXvmJmZCdwIK9U/+qM/CoLkP//zP4d177333gOhEg5FjUr6T6mPy5cvh6DUw+Zxj8OT5xB+CfdEXOVviPO4x6Hl6x0BR8ARcAQcAUfgOCPgQuXDu8fYE34IvyPQlbFnDHhlWxxnt8fYMSCUmowZ8RBlGGYk4OWHQ1DewEBOZeoSskndaguVTQmPQahUvcn6sm3ma5Y7u2lFWZYSjIpQtineRzblyt26VeYQKlWSI9NSm6T4jzIcR9MhALY8JwtUCWcL15UcdVfBsbMKvlMJkF0nKIRmLE2L40lZmuZs7LICehX0iqUpvKlzEoUIEyJsSkJmPp+2gcG89Q+IfyrTsZgcsWJjwvINXWtz2DKtoqX6dP3ih+3al21hMoqUkX+3MXsoWoIlcwgcVD3O5qachBCCpZKyfkMZqNWNki2vzdns6m2bXZqy2blZKy2XlcnYtGJuyF46d9nOXpqwCQWvFi6o45Ml2xpYs1Z605INOS6VlZI5P6halVlb+nrd7l6btrtfT9nd2xI+FZCIjS7nQqjMjySt/3yfjb6atYm3FRgsnpxTECz1P5cVGDwvgXL6M/Xntqx3l4S37tVBMikjxi5URiT2X/akUEm3eWiiIMiXmmhrftjgIUcEJOKWyFvIPUSS9tGSFYHwjTfeCMSXGif8kYE0Ex2L7RGZlVGoPKj1a+xb/OGF88Z1EFzEUmqvcA6PxA3Q+MsxR8CFymN+A737JxIBFypP5G0/8EUfd6GSC2ZcGIVHREQIZuccSdJOkZI2UXTkGEwRjyhS7iZUxnOxbI9VqXOpWfUotdBYVWRTIuWWokyD7Kf6I51CZX5MROjlnJ19b9jOSKycvKJxrOoqYv06/cWqffm/Zm32mkoLqEZJtl+RnSJgk1cG7KyEuWFlVCJeVhc37N6vl21KpGnmc5FeRdRiP7OnUKlo2oIiaUclko5clKWNRE+IanaAbMRt61ZACMNavbAMsGwTS5HWPq3jHI3qpiJ4FdF7r2mlrzT+XlBb2ezoTgTSC8EFG2aw7BwzR4wZlzNuZgnOsT1d2DmlZZ2UH0va0Es5m5RgO/mmLF/fUO0Y2eCSXRrquiiDEsKdl01PECvVh4ownf3dqt37pGS3//eyrbhQGaA96oxKBEfKcTATQArnwo6Z4FHKciBcklGJKIhQeenSpWCzCtf76quvwvcZxxzWHwWP2/l8xc+RRyJU3r1714XKCIwvHQFHwBFwBBwBR+CFQ8CFykdvaeSUcDx4YuSQrIenMLU5I+8VICnXmb4+8SgJlan0mHSHISsWJOgV01q2LCuhsW39So1KlfaoV62mmpN9AxvKAkzY6CtZG39dVqcj4o3KzltfkVApN5iqOFa9LFtYud4Mi/tQ+gPb1oYcexaVSbnwVcXmxG+WxWtq83sLlTjdZFXuY+CsOODlQgiSHTytGpM5BZ+K/z2Y2pcXuF8UKjNqkyuoXEg+ozGx7FhtyAprk1aoS6ysn7LsZr8qiEio1L+Ay/bhOnEEw87Pbfw4jTDlXweuQf9Zr9tqWTavS/M2Oz9l92dUw3J2WqVVFiRgrsuONWVjZ4bswqtnbPKysj1fUv8mJdQOyR43p5J8+pdcl7PS8mlrThds5UbTZq7LWeeaeMltCZ4a41eqspnVuRFXUxlho7qU42/rXrypupxv9Itzil9KNI2Wr3O/r9j8tapV5GDUDgwWWBGvBwDu/8aFyv0xii16VqiMHdxtCfH95S9/GcTA+MMTDzwPNhHaCJUXL14M9SuJUCbzEvKLUEmkLhG+/GhCRuX3vve9YAG723kOug6CSzYlx3eCe1D0vH2vIuBCZa/eGe+XI/B4BFyofDw2vuUhAlE0Oq4ZlQ+vJBLH9hJSFMeHuxGkKFDGZSRNncJaFNg6RTbaMcdj0v6hmEn2YFKES341qtux1VdQ9+RZ2iFUpkSgsiMmUiWh8ttDsi/VLAESwQ3yOfflql37fxU9KvJZKzWDUHlahOmM6jFidTowoePqqCuqvXjzFwt2/9OSImoVzDfXsM0NkT0saHaZIKkJZVQSMUrk7tDZXLC0ITszI5E0CJUceHuGOIZpe9EWLLVZpG1TGYmNqsbbS7LJnJZF0U3Z3C5pW1MkFaGyo+YJ+PCMdc7hGKo3GXFrNqlh+fhIYPqeGyIKuG35ekZ4BdtcCa3YvUIiseVp1JohM3RgUlm1EitTItfUcGkLlSKo/1vWr6r/2Wrq6h5zul2geyarXrSMSoI2yUiEC8HZCDQlQ5RSHVwr4uXPfvaz4JxDzcrLly/b2NhYeCbYh+/YJYmUcDnncc/kEfSTOAKOgCPgCDgCjsAJQ8CFym/e8MgJI+ejRVz3sHUkTeI9sj1NJIoKlhxVVuWAsihVs1LBobmMbFtTTW0jW7AZglrrTdWd3KxZcmjTBl5K2anX8nbm7SHZscpCVRmOm8rgqykYNfCatWbgMliQZuUeo6FxCFTFUWde85yCWkv3VCtRDjeIZ7tO2gcelZFQOXhGrjqqvYibD1yJoM7kdkZl2Du8wHPFGXGokYCXzaXaQmUOoTJruS0Fia4hUo5rHrPM5oAqc8iqNQiVCmYVwYq8OfSH8+uAiT7hxLzNz5PJlLBp17ZEFE0o0BhODjdERIwlGKam2gGPM9MzhktMK90IrkQTryiA98qojSs7dFAZp9kxna0oEpwS5xTPS1QLlpmftPU7GfHAqt3/csHufHXPpu/PBr2mXl/X+SRGZ2X5WpQYfClr5/6w3ybeVFmWl8QvdS+q4rmLt6oKDF5pY327LiFZnJWyLo+Be9d70LHShcoOMPZ5eyyFSgTHX//610Go5EeQ+MDzQwjR2Vh9QYapQfn6668HoRIBkf2oYRktZBEqv/Wtb4Vo3X1w6mqzC5VdweSNjhkCLlQesxvm3XUEhIALlf4YdIPAiyRUxuuNxDIuWR8JZlzHMoqNcVvcn3FlFNji+7iMbeNx2usR4thHvGVLQqVldcK85qIO+ahQmchsyMJ0S1GtGTv91oAyKmX5KQFySAQ1IfvSlfs1+/rnCzYrAloRUcUK6LSyKbF9hcjmBtO2vtpQxmXZvvp4zqaurlr5vqJuSxAnhMR4FY8uxQ0DUU2LjOUkVuZHVTdFljZkbKZyiItkh2qf8NLBv3YQMURGLFab65uyn5VAif3rnDIaFZi6tSk2So1K6kl2TBE7lhA7li3NiJNt3MKJO/Z4+DZY1iqbEsvXU7K/hVyfeXtQGaEFiZGywJU4Oa+o4sp8PZB7rmf0paL1q/4mmaLrpQ2bkfXrPWpU/nw5RB7vhdPDMz/bdy+aUBkzKhEkqVGJAw7fN/gZNlrwMYIj4E1vvvlmECrhZGxH5GQaGRlxHvdsH0M/myPgCDgCjoAj4AicIARcqHz8zY6c7/EtIr9UoGZCPC4xLD5VUGalMgyTLdVuXJM4J/vWvoY4z6b4kxxmFKi52ScBUlwwP5mUw05eAatDwY61f1wWsAqyhGdt4hYj0ZLgT/gMfGlteUN1KWty31G24ZcS88imVH1KHHW2JM7tOomaEWyaLiTkqiOxUvxzRDypoIzBlMp+wP8eTBwiCIvS+xQIms2KKxayCjCUVawSwQr5ouX7hmX7qszR5ojlNocss7Vt/Sqhcku2OwiNjO3RZBpNBdHqmuF9ffBDEcSkRN2M8MnnVAO0UJQQKgvabSckOAAWsOVK2ZaWFwNXmJ5WRqWcKudmZm1BQuVWbsPGXs/aqddVEkWZqKMSXvtPK6NS3DYhvggHDVis5KxvatTKNxJ2//MVCZXzqk85bQtzS6FUX1NWvEmVNMkOJFWSJaksyoJd/H9G7NSrukYFD+PUg70ufJvalEtf1+QktJ25+hioH+C4xxsXKvcAZ8emnhQq+eGMh5oIXB50bKF4z3r+YGD9+rvf/S6QWexVmbEUYqKuJfvQHrHytddeC9tR4FHmOwnwmTNn7OWXXw5keAcuT/SR83pG5RNB5zv1MAIuVPbwzfGuOQKPQcCFyscA46sfQeBFFCrjBe5FMPfaFvdnGcS1zhW7vm8zli2RMFE7kTyESmVT7iJU9iU3tHlLFj6Kon01LwFywM6/N6I6lSKAym5cW2nYXVm6zt+QnasyBbHlmXh9wCZek23ma4paFaksTa/Z1G9Kdu1/KfNSNSqpFdmstYXSx0Z4QjwVUZtShG9mSORTkbW5QUWzSgjlmNR0DPvqUh7LvxAXJYS2NttWq2SA1ldbtr4IcdM2sjnZGRh2TNoa1oSo2bTkW1kNIcyG7MsOjhwOQB80Q9QRKlOqqzJ4RjVDREgRKieFB2RehEAC5bpNfV5SLRdqgCqYdlTkVVgh/OaUVRlEXWWntmtUtoVKMk/1O0G7rzv6+bw+vmhCJSU74F3YKyFKwr3gZXyfECzJmvz5z38eXHAQKuFq8DEccAgmZX/ncc/rafTzOgKOgCPgCDgCjsBJQMCFyqe9ywhwIliWE7kZlFAmgUxWsH0SI8VSNK9pbguVkJvAhiToqZn4WF+ohUjQKk4xYwRait9ki8rQVEYjnKtds9GCQwwi5bzqUk5/vmpLN2tWnlXJkbJqJSJShgM/5lrEy+B/uZGEAjnTykCUfqJgVVxp4GFMYXde9BGOnBZHy2bT0ljyNjw0ZENyQ+kvDFo+NRxEyuzmoOpT9ltaZrDphDIqt6NU15WpyDgeTaZaqwbBEnGWMyCKZjJpCZ551b0ckAYzbMX+YliXlniJUAnHrFbXrLSyKlFxweY1z83N29zsXHCtbKSrssw1G76ctrFXKGOiY53OBuvctKxxg5Wt+GOrJNHzbtGWv9y0u79ZsvvXJHzembeVhVVxj3VpTRKLdf2URBl5OWun3+m3i9+T84uOhx0sJVZmf7+qYNc23qX7dQXl6rjikE8zuVDZPXo9KVRCTrFqhdhS7xHxj/cIlhBcHnyshHiPLRCZk6dPnw6fETGJ4L1+/XpQ9C9cuBCidzkGP8hBgnlALl9W8VVF7xKxi9B5GJMLlYeBoh+j1xBwobLX7oj3xxHYHwEXKvfHyFtIDNK4iOhHspv+7u/+zv793/89jL8Yb73I0+EKlREpyN7eQqX1SahUlG1uOGFDFxHfikGopN4iVjPY1WDps6T6I2WJbwmRKETM0YvFQMYQ7xZuyIZGGYLX/795RdRWrSmREDvWPUmqxEDEyHRR9q+j7YxK6qFk+7FIjRmV+xxDm4nobTVb1lA9yHq5abUlWfTMiChLsAxCJRmdbc4bQQnLKFSmVWuFa8+qvgg2Rm3bIQmlRMDSEoVS/xFJvKFzcJ8yinYdPJMLQu2pl4WFiHxGdkV12c8uywJ36rOVQCixTBqUldHE64PB1hYhtl5pyq5nNQi7t3+5HIg9oupmXT2ir+GkoYvP9eVFEyrhcYiN8DV+sKAsR7SC4p7C0X7yk5+EbVi/vvPOO/b2228HjhbrQzqPe66PpJ/cEXAEHAFHwBFwBF5wBFyofNobDOmRUKmSHyo8Kd7CUp+3GqJD2M2saXtDM2JdnNqBmAmJh/0TKmvxWkF1KosGF4TzUT+SLEoNl0Nm35rKWCzfW7MZCWdzv8f2tWKrEs5w0wmWr11wmSBUDvcpe1BCpaxkcxIqEepCoOp2t+JhJBBZSjasCJUDyqQclaA4NKTSDf1DEiqVRdnUrNqUqaZESgm0qW2hEq6HhkMZPDSZpaVliZUVa2zWBVFLnE/C4IAyNEd1zFPDNjF5ygaHBpRVqTIkGTIrqdOZUCmPLVtXfc7qyrpVlmu2uqz6kBIs0XhKGwvWGi5bdqJlw+dyNny2oFlJaxJ44bcpOQcl0ipRUpZEfFcJbtfW7favl2z699KV7qza6pICIdebopvKeNX1D6ocytn3++3se4N27tvKFlU2JeeO/HL2txVb+FIlVuaVNCeuHbhuvI1PsHShsnvQelKo5AcybIKIxp1Sqi+klWxIiC7iZNxOFiXWrpcuXQpiJdsQNrF3xRoWckzNE9KJIcvYDZFFibhJ9O74+HjwWmb7YUwuVB4Gin6MXkPAhcpeuyPeH0dgfwRcqNwfI29xcoXKo7n3kNV9hEqTANzXsky/yOlkStGgBdWqHJYF7FCwd6WuYmlqzVZn1qy6ILsgRXVSt2RAVqbUsUR4C1amv1qxm/+5JOFNAtAetSnjdSIE9imLMTOgaFpZ5BBR2696l9RyRCwM1j9E+kaWGnfcsaQuRxMRkRqVyv6szGzYimp2rC2L9ElkDeLfjn3CR6DRTCbn4DmdezKjrFLVOpGVLQIjEb0he5XoYfWhWd+0NVncIiQSWUwtlVOvqG6ILF+HTivaVe3BB1uemd9KCFONStpB7sdfHdgm+RIq1c+FG5Vt+9dlW7xRU+2XpjUqcm6BcPaIWPmiCZUxAALBEvsngiFYx8z01Vdf2U9/+tMQhPrWW2/ZlStXwkzgKJzvxo0bzuMCUv7iCDgCjoAj4Ag4Ao7A0SDgQuXT4grBiUKlHB4RKlUGJIiTW+XHCJXarN36lDWZl2Xp8EvZIFSeeWc4uMaMqe5icSwbMh7hWyvihXPKpLwnC9LZ3ymYVTakNQlnm2sEj3bRf50rLVvUkFEp/jV0XhmVcvFBqCNzM06BAvIiIpaUrpJRhmN/UULgsGpoDqiuZb/qbyprNFWXw8+GanLWJVK2JDAGoZLSJ1vBVhUnFcTK+fkFK1dXrbFVt0R2U0434qC6rpHTyh49PWTjp0clVPZLj2lnUyaTEmebEivXMrZVU3ZlJR0ce9YrG7Ywv2S3b9+22ZX7VmrN2FZ+XcfK2dBkwUbP9gfeV5SVbnasz1IDkoVlm1uZVnkQBfTe+pVKAIorLt6uWmVhQ8Gq4iKJLQW9JoL17qX/oVIT76k/ci5KbjsXkbl671OJm7+vBJ5bpzZlwCai9WRLFyq7x60nhUqILcIiWZQ85NG2FbJLJC4/ZkB6IfaXL18O4uPo6GgQMamLgtreaQ3LPhwTAkzmJZmUZGGyP0Vb2X4YkwuVh4GiH6PXEHChstfuiPfHEdgfARcq98fIW7hQebjPAGPJLoRKUxSnjDyopzHyUk52M+0ozvPvjwT7V+xKmRECmRAoqVVJdmVFdrD3Pl2WlWnJ7v+qZFjREN35WIEwHEFkWJw5kRFJVW3K4QtZkVSRO0WgFlWjBIEP+x/shRAGd5viajI+mxvtbMqKhMKVO+uKVhXxm1OkKYJpW4f65iEEDX0ojKmGJFY9uu6hc4VwveHaOi2OdAyyKYkg5nxkT/afErFVVunQ2ZwV9Z5+llWXsqSanos3q6FWZU7Rx9SmpB3tEWDJ/FxR1uX8V+WQeTn/VVVRyKrpglipzMoQHcvFxQv8Zs+fyZoXTaiEpzFHcTJ+jmDiivPf//3fIfI6OuPEGpUEqjqPi0j50hFwBBwBR8ARcAQcgaNBwIXKp8UV7rdDqOTzlhTEB0IlLkWdGZXb59SuBHD2n6UcSNHOfWvYTr81qMxKiYIK0Exlk8rua4rnVCS0rdqd/16SHWnZSnc3VPZDAaLdZPjBvzSnVUaDsiNDyiAcfblfPElCpY7fKVSGXokPMWbHyjWlGo45ZTti/1osUm4vJ7vXgvWVlTlaKVpfVXU5G+qn8ioTwf5WZfiqlRBwOC+7VjSc6nrFNlX2JDUgZ8sJBaueVo1MWc+OnJZT0Hi/FfolyOo8iYQ0GVMNy6aOtyZL2fqQZddHrU9iKAG5SwvLdvPmTbszdcumFm/berNiuaKyM0eUnSnRE37Yf17XeFZc96yuIblpNWFEsOqt/1pSmRBZycqRqDqHUKk7pjIkOPxMvNlvr/7ZKTujOqEEBuPoEwJcKRsioXLxK4nCs00F6IqcHgJXdKFy+9nvYtGTQiUiJKIkYiWElZn3ZFIiKsaZoq6Tk5OhFiXvWc9+ZF9CchEtESKZIMu5XM4QNJmxfOXzYU4uVB4mmn6sXkHAhcpeuRPeD0egewRcqOweq5PckrHRSbR+PZp7LibYpVCZzEAalS15TvavV2Q58+0he+l7Y7KxaZcigJRt1CC5aidrVibEu6XbNbsD4fpNSVGesqKRQCg+uS95CkJlVpG7EgoRCUcuqubIhUIQ9LL9D4XKcKxwtvYLV8QUuVkQKpXtiJBakVC4fKtms19IqFRmZchQ3IWHhwME61nVkFQtkFNv5EWSFaF7vn3+TGG7FotOgrUsYmdD145QyTEzyjKlfmew9iELVHZF9LO6KJ4gsbQ8uy7BsRVskgoSXsm4zMvWNi0C3ljfDNsRM8m8nL9escWvZaur/lJfc3N9W1yNFxg6++xfXjShcj8ECUQlMhrOBicbHh62AUVrw9mcx+2Hnm93BBwBR8ARcAQcAUfg6RFwofJpMYQpdQqV8DiESlxhYkbl44XK4HRzVtzo1cK2UDkUhEpKWeCygw3pgjjM9BelwP9mleG3erdu62T4RWeYvS5B3cNVh/MMyFFn+KV8qIc5ID7F8ZMdGZXxMAiV6Cq4VabTKtWR16zakpmM+NpG1raWVKpkJW9bqxIq1yU0bkmo1DXjArS2vhYESsb5qytlq2+qRmd+0zIjW9Z/JmkD5xQwK7F0QJa3/UPKpBRXawc2Ci5xSITPrETKfEO2sFtnLLOl9MithBLYSnbr5i3NNyVYfm2rpdVQG7NIxuf4cOCIAxfFq1/Wdb7aZ8nBltXFJRdvKaPyl4t2/zNxwGsqSTG7YVu6HdjQ9p9O2el3B+z1/zlhE7LdzShwFzcf+OL0F6s2fbWkbMp1q6+0+WLE52mWLlR2j15PCpU8rNQyYSYTkjnaBnFpfHGYsGzNZrPB0rVTkETQxCaWfWJbjkkb7F+ZKdYa9wkHO4QXFyoPAUQ/RM8h4EJlz90S75AjsC8CLlTuC5E3EAIuVB7mY8DYtLuMykRKY1LZ8BRlwTomcnpWkZwIlacU5ZpT/cZkKhFqQSr5UoNeCxamZWVTzqk+yc1fLEgcVO0/ImpVn+SBirjHpTwQKrczKocvtDMaOzMqEQl3PVZ7yB22bcr6FTudUDtEQuXKHdkR/U5C5ew+tTuCUKnaKBJKx17dP6MSgRExFKvZtIg04iTRxcXRtl1swEQ2uFjh1ivtzFME17xILxmXOdW/xFK2KXsfLGKXVfNz9lo51P+ktktJWZVry6rtojoood6IC5WBL8GZIm+Kyz0eqyfeBK+jjg1/fyIng9NxTtY5j3tiaH1HR8ARcAQcAUfAEXAEukLAhcquYNqjESRph1DZJy6IUNmSUGk1vd9FqNRuCIjZ4aQNXZT162vFUAZk8sqguGDbGSaVUfAe1q/3FBR6bdXufrIcrF8Rz2oLTWUGbnOYPXoHX4IDZgckEkoQHb2EUKmMTZXKgF99I6Oy41iMySnNkUxSokOzPm/VVENyPmXNxYy1liVa1iRUtqSr6CQJZUY2m+0a9etrEvjWJQpSm1JaY35cJU/OSRw8qwzSMwlZz0oEFQ/WFVh9rWEbCoJtiLP1bajqZaPfBmzchtPnrJAZsqQCbcsVuWbeuWe3rt+1G1/csqXpstpKC0rlbHB4MFi/Dl2Sje6VpI19W5mgE322KV67fLdqN/+vhMpPS0a9ySBUKg44NyjB9FJWlq+D9uqHE6G0CIGylF6Z+rwtVCIKl6fqthkceDqAeYq3LlR2D15PCpXdd/9hS4RIpqMk1g/Ptvs7Fyp3x8XXHm8EXKg83vfPe38yEXCh8mTe94NetQuVB0Vsr/aQ1e6Eyr6kolVlxZofFam6KPvXtwbspe+O2WkR1CFlVUKgJBmFDMOmhEHENqx/phQR+vXPFxQVWrW1JQltqk+yq7i4o5uQVGpUZgeJqFUUq2qUEE2LABhqVGK9yjB6H8GuhVDZpEalrFmXN6w8LRHwZttKdb8aldSVzA2JkF/Quc9kg01rjhqZiqalf5wfkhiCFXXNDZ0DAguRBg/qteSH21a1HAsREose5qSuDevXvGxyB2LtS9UZoU1lYT1kokLy578sh6xKLHOJRnah8sNQG3J8fPyZCpU7Hs/w0Xncbqj4OkfAEXAEHAFHwBFwBI4GARcqnxbXPYTKkFG5i1CpXeR0qrqNCuAcV8DqKwWbeF1OkeKA1KekNEde/AhxkMBNHGRw1Jn+QmXxZP26cL0W+NdGheDRfXggQiUZlbJ+7Z9MB+vXMQmhBHWSUYkQudskTbI9sYwfdKqmRLuNOTnfLIpjraiW5JpEypYCbCXOppR9iTBKsllrE0KnepHieDmJsYVT4nESKYuTsmeViJgZhJfqOA1xyRU5BJUVQFpTOZONhOVaRRtMj9rY4FkbGBq0rGpbrjcl1k7N2d3rM3bj0/u2cKtsjZKOL5G0IGdNMlCHLift1DsZO/2HCmw9l1Ssr4TK+2t2S0LlvV+rvufnEipl/colFWV9O35FdrvvD9nLfzRm/eM5Wys1bEkZmPc/W7aZ31VkAUstUFnsdiMIb8O138KFyv0Qerj9hREqH17S83vnQuXzw97PfHQIuFB5dNj6kR2Bo0LAhcqjQvbFOq4LlYd5P6E+3QmVlpBQSYQrtUnOpIINz8U/GFGNjGGbfF2RrhIRExLfsFpdV9YgtRhnlU1591crEiqXbOnGWrAtpcZiV5O6xvmSOcTKZJgR/9KqfZmUYApRbQuVex+PNoiVCIAIiWR0rimqt1ETHcT2dY/d4bkpWe1kR0RChxUFq+jecH7Vx4REx/MHwRJBVNfGPgipIVtSoirLtCyIYn+DuKVz0obsUHCj9mVRVrFY5lKjsjStOppfq76LbJOwfl0W2adGyUZZ54B8krW6R7+7wvcpG50069enhOvIdnced2TQ+oEdAUfAEXAEHAFHoMcQcKHyaW8I3E8Eq08l5fr6NW9bv1qsUblDqISPaYaPZRQ8Oigb1DNvD0qoHAgiJYJbVoGXuMLATeBccEHEygUFrM7LHWZG9RaXbq1ZVSJaQ7UT2/xpj+vQ+VI6HzUZCyrBQbAogaJJ8a/HCZWPHE0nCNxM/WmK79WXxQFLel9OSMST2LklkTIhfiZ7WMRKXCtTSa1TIchcniDTbOBlRYmyOOuEbMpCm/vhgrk0v2QryyWV+1M9SwmvWWE4MDBo45OnZOmq7FKJjlvphpWWyjb99ZLd/GTe5r8qq3akBNF18Vo5bA5IeBx5JWOT78pC93sDytzMSDCVUKls1Nv/hVBJRmVVmagbwe52SKVXzr43FITKi++PBNvXFYmac9fLoTYl7jsrsRYo/BaueAiTC5Xdg+hCZfdY7dvSCe6+EHmDY4iAC5XH8KZ5l088Ai5UnvhHoCsAXKjsCqYuG0FWuxQq+yTCiYOmCtSNTMqKR8RKdSrPfXvEzr+nWhuq30gUKtmC1SVF0irCc+rqioTKkt3+PyuyfVXUqYjrgQQ2yLG6h4UOBBkSHEXKEF7azcE4pcgapBky2VSNx01ldSKY7keUQSecX1Y/EOYk55cYS3bkg/Pr+EyBEFOrBIzUDiESUZPsypQyJfsUZRwDfNk3K6vXobOqvSkcx1+R0Cuin1F7Mj+XbmP7uhqI58JX2NRimdsM2ahb/I7AObfPG07+HF5cqHwOoO9ySudxu4DiqxwBR8ARcAQcAUfghUTAhcqnva2wm12EykAwvmn9CveTpieRUgGWqhk59rLKf3xrONi9DirYkjqJbQ5EwGYrCImsC3yQzMrAB0vKqqyq/IYcbZa6s4BNKHsxJf6VUZAoJUao0RhEStWVfNwUtsD7QofafKkdqNoKjjQqPykuqmDXLQW9IkxKpKS8HqUcMmmV29Ocy+esMJC3wpCyHEeyWmYsPyg3nZDNmVAW5ZotzC3Y0qLEylJJFrCNIHD2jxTs1MW2De7Y6yr1N9wXrGSX7lft7mdLCjytBltWMjypo0mw6ugrOZtQluQZ2bnmR9PBNndRHPDOfy/Z9Gdy1bm2Jv7XED/ss9HLBbv4waidf3/Yzrw1KE7eF0RK6lPe/3TFFr5SPctpWdKWuxCCHwfgLutdqNwFlMescqHyMcA8yWonuE+Cmu/T6wi4UNnrd8j75wh8EwEXKr+Jia/5JgIuVH4TkydfA6XrUqhEGVNzRMPsQJ+seHI2+faAamUM2yXVqhy9VLSMxDkIYWlWGYGKHL3/m5VQY2P6U9XMmFa9kycR10SQyayEkGE7xJJ+PBD99rn4IOqpzZbqfgSbVkpkSrTsVuyDoIfzS5wMS07ccf54/NCN7esL9khqTx0VRM2E+kyZhweXrzd51d6k9uWkLHTPifCPXCyGOpXU0pyVTdKUsLv9yxVb+rpmGxXZxQpXMkC77fc+sDz1ZhcqnxrCQzmA87hDgdEP4gg4Ao6AI+AIOALHAAEXKp/2JonEHECoRKRMSjDsn8gE3jJxZSDUphyW3SviHW4ytSUFVFYawRWGQM1hOcXgKAP3qsyv24x4zZwyK+evVax0T7UgVyQcKnB0rynyL9x62pmUol9wwANMcKat7QzPFoGeZBrK3pV/CZ0g1LFUhGlieyarMqNsxzxiZVFiZf/2XFTWaC4bRE1q0i9LpFxeXrHl0rKt19espWDe/Eg6iLeTb/Yr65GyKDkF15pVVzZC8OmiBFvqRzZqmwpglbUsQuVL+RCwSvAvnLE0s25zKvlx79fU9lSG5M26cNoMmaUTb/Tb5Q9PiXeP2PjL/UEIJiD4vkqsTH26bItfr8n2VXUzlUH6kHAeAKzHNHWh8jHA7LLahcpdQHnSVU5wnxQ536+XEXChspfvjvfNEdgdARcqd8fF1z6KgAuVj+LxdJ8gfAcQKtU6RrhSM3L8DWplDNsrfzJup17pDxmBWJcu3ZHtqzICIVozn5dtURGhtUUxxCedDsZL9z/L3tz4m/sf8Pyheec+8f32eSHOWAmNvZ6Xde6Anf8D1TWR0Iu17fpqw2Z+SzZlye4oE3VZ0cctZYIGcv3Nnj23NS5UPjfoHzmx87hH4PAPjoAj4Ag4Ao6AI/ACI+BC5dPeXEjJLhmVu1m/qikBqhkCVCW8EaA6KaFy8o1BK6hkRUsZlGsrDVu+q2y+hXrICMQxZvzVARs6kw9i5UataYh0CHAzKmmxfHPNKjPNEIS5r6C2zZ/CYvv9ga8esTLutP2G4FGmziXvESzJrsxmssGeNZfLWT6Xt5xm7Fozyr6knmW5rADcctlWy6u2Vq8JuWaoSzn8UtYQKi/8IRmn/VaQeElmaWl6zcqzdYm2Eh7rm0F4pabnwOl8qD3Je/jzwtdtR53pz0uhZAoZkoi9hVPJcNyXvz9hZ98dshG5GG1Um3ZXAuXUb0o2fVW43lb2pSxum3INOszJhcru0XShsnus9m3pBHdfiLzBMUTAhcpjeNO8yyceARcqT/wj0BUALlR2BVOXjSBqBxMqQ91I1YgsqG7H6Ct5RXYO2asfjduESCnRs/Vy0+aUTQnJuvur5RA9W77fsPrqIRXL6PLKnnuzvQi1OGRuJGUjL2eD5Q8WSiMXFH0scg9+8yLz06rncv8TkeB79WBbG+pSPveLetgBFyofYvE83zmPe57o+7kdAUfAEXAEHAFH4Fki4ELl06INQdkhVFLnYquhWdavW/ij1qXibSqDUSxRtqO5kaSNKPuP2pTjrw3IBaagbMpEKEuxKhFu/kbFWNYrTQVdpuUWM6gAzH4bPE0dTLOyMgURK7EpXbxRU1blhrIqN4PTzUMVcZ/r2otX7bXrHrpdFCrj7lGsfFCzUqJlOiXb12AP217SlqzKer1ujME3GhvWVLpmqmBWnFBplJfzbWvcV/tVr1JZp8owbSgrsrEmcVblPajhSfYk2ajw5oTqbrIOEXNOQb5z1yu2qLIfq8q+rJdkpat7UBxP2vgVZVT+j3EJlioXMp4TX2zYPQmV01dXgxPP6v26bF9lv1vf44LjhR5g6UJl92C5UNk9Vvu2dIK7L0Te4Bgi4ELlMbxp3uUTj4ALlSf+EegKABcqu4Kpy0awvm2h0sSwEkV9FiOCoAaiWtNnWbYaIqOIj5o/ECpPpSW05UKdylc/nFD0bH8gY2vLGzb9hUQ2WZciVGJFU1/ePPQIT3XoWE9Z1XoZOCcML+dt4o0BGzwjWyHVdCFClnou1HKZ+60ilOcUTSsC60LlW/ajH/3IPvzwQ7ty5YqNj4+HSGh+VIg/NMTlsX4wDth553EHBMybOwKOgCPgCDgCjsCxRcCFyqe9dXC/nUKlPlOjstVRoxKhUhQxVUhYfkx2rheUUXll0EYlUvaPZ6GEVlUNypV7qtl4Q7amEiMR4rJyh5l4vV+1LCXUKQgzlUXQbFhpSoLm9bIt3WwLleslCZWY7Ryurva04DzCLYI9bLCFpaZlUrUowU2lPGSNQ2Zls6Fcyk3V3GxtWiKzpTqefeJzWTv1WlE1JYshCLUwkpEFbCKIvtEdh7IgwdpWx9rcUFbqNj5zwmf5Vs0QHdeWdVxlRyZlfQv+Y68W7PwfjgRcC8rAxIFnWhmqc7LTBdOq+CLZlC3pzYc5uVDZPZouVHaP1b4tneDuC5E3OIYIuFB5DG+ad/nEI+BC5Yl/BLoCwIXKrmA6QCOxUCPiVUJln4RKmNPWumYiaqNQiUi5BTcTEesL0bXFCWVUXi4oanTIXv7jU6FGZVLEq6z6lHdl+Xrvk5Ld/3XJVu7WbVN1SKgL6dNDBIhQhnj2n87Y8HnVQFGtEqJrqUUC2SeStnRH0bpEHMfalA93f+7vyKg8d+6cffDBB/ZXf/VX9v7779vo6KjquuQfCIf7dRKiz8QyCo6Li4t2/fp1+/jjj+3HP/6xXb16NbR56y0XKgMQO16cx+0AxD86Ao6AI+AIOAKOwAuLgAuVT3trdwqV4oAIlyGjstLmfwSpRqFS9SmzwwquFF+hliLZfBkFVraaLavK7rU8I76iupO1pUbI5ssUkzZ4PmtD53OhVmW6IG4ju1PqWK7ckwgnflOeaSj7r9WTQiXoxsBHalmG/7aDIiNXoQ0UZktRpC1Zs7K0hERF2eTmhtpYDUiwHDona1eJuli7kllJrc1YZ5MsSkRKBEeyKRFyV+7KFndW2ZoScak1iZBLjc7skATQc7jwtANbwZ+MygVlsq6oREh5Vs5F2ucoSoW4UMnd7m5yobI7nLpq5QS3K5i80TFDwIXKY3bDvLuOgBBwodIfg24Q6BQq//Zv/9b+/d//3Wq1WrBh6WZ/b7MTgShU5kXGtjMqDaGSuarGhGZKSErKqkZkicjanAgrAhuE9bTsfS5+ZzTY+1CHY0V1Su7815JEylXVp6yIcLUzAnstYnYnCs/6M4Jvur9NaPOj6WD7CoEFQyKP1xVJu7awTVTR83pM53Wh8lk/Mbufz3nc7rj4WkfAEXAEHAFHwBF48RCIQiUBbb/61a9senr6xbvII70ihEoFpfYRpDqg91nNEirhe8H6VfyP99vWr4mcalSKr8BVBiZUu1EZk4mU+Aoim1x0aktNqy02rFGRaCfxjQzKvKxiC3LeKao9QiV1Fsm2rCkDs7YofrOkbESEuG3DHp3weExApykKmeFDECz1gqYpWFPCKzsgrqz6lMXxjHDQrHqeOZX3AJuEgnoROcFvQ3UpqfFZmatbdV5YLkikXBU2IcBXt0PtEiGrVSVXRlMSgFUzU8clQ7NRU2Dr1LrEYnFGWcS296EzoVeH9oJQ+c4779j3v/99++u//msjcNSn3RFwoXJ3XJ5orRPcJ4LNd+pxBFyo7PEb5N1zBHZBwIXKXUDxVd9AIAqVP//5z61TqGw0Dtnr5BtnflFXIFRCUhEqlVUJeQ12r2RUbguVyqZ8IKyptmL/pKJEz+ZCjZLxV/rt9JuDoc7GWmkjRHfe/uWSzVyVvc+NdZFRhYMeMmlSB4/9FCx/VKOE6FvEX5ZYAUHmyUBtrrXCstU4fNJ5GOBFofJ73/teIK6eUXkYqB78GM7jDo6Z7+EIOAKOgCPgCDgCxxOBa9eu2b/+678G541PPvnEhcoD30bUNmZxvyBUklEJ90OoVEYlwaohSKONfZgAAEAASURBVFUqIuKbaGLkKpStQCTb0nr4SVPiZFOCGSJZ5CtwGdqnxW3SA2ov4Y6J7Q2JlWQKNmtqv9Gb/CZ0dr+X9iW1W3VyXK1PSPOlBmVSNTzTEiwzwiA/lFJ5D1nHSqh8wPXAQ0JlvSyxUiVSNipt8balgNUHTjo6NnyxLVbqWMrWTOUldur4m8JvY6UpTMUXlQDbwrmosy/7XUOX2xEq33777SBU/s3f/I0LlXvg5kLlHuAcdJMT3IMi5u2PAwIuVB6Hu+R9dAQeRcCFykfx8E+7IxCFyl/84hf293//9/af//mfVq1WDaGy05Jl97197TcR6BQqJVY+ECpjRuVGuy6lrEpzowkrTqa37XzyNnQmL1ufvI3IuhQrm1VZls5/WZXt64rNX1O9knuyopG1T+BznaTum504uWtEQPkRwETssdYNHBOrV0Ulm5ZHQTqfBuxo1zowMGDnz5+37373u/aXf/mX9t5779nIyIhbvz4NuE+wr/O4JwDNd3EEHAFHwBFwBByBY4kAGZU/+clP7N/+7d8eZFRSP/CRLLdjeWVH3emoYkHImAlSVUZlH0uREaxfjQBVhEqKR5LuqElN+yjNKFedZEafxfdaSvULYprEslBncmdmJPuI2/QRhMkyEBy11SkQLB/sE07wgr7A74QXQm1G4mJawmVSAiN8GS6FsNhUVmVDdSURbhEeA+TxNnXCso0nQcOBM3K7dIta9aPDkj5ShzNmVH700Ueh1Mebb77Z2TN/34GAC5UdYDztWye4T4ug79+LCLhQ2Yt3xfvkCOyNgAuVe+PjW9sIIFQycP7lL39p//AP/2BkVpbL5SBUOlE96FMipmNpmKSWRNaSVQl5JapWRLVVE6HakOXrlqWHzAqT1B1J2/BLeRs8m7WBU1kryoImN5gOGYDLt9dt4XrNZr+o2cot2djMtaNDiQZtq5Va+vRNBICcKS4jSY3L9taeeA3kWt9BhMoLFy7Yd77zHfvzP/9ze/fdd4NQmcvluv6xKIqeLGOQgdeoPNhtdh53MLy8tSPgCDgCjoAj4AgcXwRu3LhhP/vZz0Lpj//6r/+ymZkZy2QylkoR9efT4xGIpAKygdUrqqNKfkShEnFyq6Z1HRmV+hSmyE/gc0w6FEfr29Yyw7pdXsi8DEJn3J+d4rxL+xdu1fb1Y/cKvY71KblOuA/WtzjpdG2B28mnwXEf/J8Gz2azGcrqjI+P27e//W370z/9U/uLv/gLe/3115/msC/0vi5UHuLtdYJ7iGD6oXoGARcqe+ZWeEccga4RcKGya6hOdEMG9giVn332mf3TP/2T/frXvw5CJQNqhMpEIrKoEw3TPhcf6KXagBXEHrFShDWQVVgVQiU+MusiVU2tbll6cMtyk6K0ZxM2cCGluiMpy6tOSSavmhvJlDVWzVbvNKx0qyGRsmHV2abqbOheKdozROJGkqqj+7SNALdBkx7pR6YAFS89hlkg1dvfv/7+fjt79mywA/rwww8DccUONpvNulD5yN082g/O444WXz+6I+AIOAKOgCPgCPQOAnfu3AluOoiUn376qS0sLISxpwuV3dyjyP8QKsX9QtkPlhAO7FzqWor/wQM7VTDtFrgKS21hilwlxLc+WBE2hUYHar+92wu16MQM0CJwcbnN8wJ+er8rjhGQ7X06MX0E//AhNj6cJb+r1Ot1Gx0dDXavOOj82Z/9mV2+fPlwTvACHsWFykO8qU5wDxFMP1TPIOBCZc/cCu+II9A1Ai5Udg3ViW6IWEJW5e3bt0M07ddff21ra2tBvPSMym4fjch4YDaIlRBWvHlYsg7PUQiromsTWiZl31qQEDzSsNRow9LjTUsNSbzMywZIxThSmzlrldO2MZOyjYWE1Zf6VDOjbWODNVAkYTqoT7shEElr57YjIJ2dh3+S91Go5PtH5iRWrxcvXrR33nkniJbFYtHS6bQLlU8C7hPu4zzuCYHz3RwBR8ARcAQcAUfg2CEwPz9vX3zxhWEBCwdcXV0NY08PVO3iVgZuoZeQ6hjFyu00vT7ICJav8D/NnUJlPPROvrIfVzlo+3ieF2m5E4Pdrm0/HDv36TzeQfbrPEYX7+F6lNWJDjpkUpJZOTmpqGWfdkXAhcpdYXmylU5wnww336u3EXChsrfvj/fOEdgNARcqd0PF1+1EIIolRNBCUiGsGxsbwUIl2kfu3Mc/70Rgm+VET54gTsJ2YjYq29sFR7ZEXLckVm5lGrZVqNtWUZZAA2sKwlVN0FRLEmfKko28JWo5s1LetippJWImpXHqWBKVif58ECW6sxv++VghEL97LIlcR5gcGxuzc+fOBdGSbMqDBAtwHCaW8bvr1q8HeyScxx0ML2/tCDgCjoAj4Ag4AscXAYTJ+/fvB8vXubm5EKzqjjoHvZ/ifIEDbouU8MAgVLa5364i5UFP4e2PNQKx1A6BqXC9M2fOhJIfQ0OqBePTrgi4ULkrLE+20gnuk+Hme/U2Ai5U9vb98d45Arsh4ELlbqj4ut0QQNio1Wq2tLRk1Wo1ZFhGsWO39r5uPwRiSGZcbguZ24VEECs3Ew3bTNWtyZxes1ZS4rAETITKdCtvqabmjZwlGrKQ3VSUbiDA8Tj7nd+3HxcE+J4x86MQ2ZP5vOqVyvIVInvQH4o4DlP87iJWulB5sCfBedzB8PLWjoAj4Ag4Ao6AI3B8EcCOErGyUqkEDohFZQx2O75X9Rx6/iBYdfvckQJ+w6P0OfTNT/ncEYh8j8BUuB6ZlYiUBKX6tDsCLlTujssTrXWC+0Sw+U49joALlT1+g7x7jsAuCLhQuQsovmpXBBg8U6eSTEoIahxMQ1R9ehoEIn6dAqOEKQmVLVkBNVWzpKHaJQ1bt1afMiz7WsrBlGC1lbWUZpYJCZd9W0nF5upYITr3afrj+/YiAlFYxGYrCpYsD/pDEcdhisdzofLgd9t53MEx8z0cAUfAEXAEHAFH4HgiAP/DkpI5ckCuxDlgN/dzm98FkXKX9pEG7rLJV50sBCJH43uFWElwKjN8z6fdEXChcndcnmitE9wngs136nEEXKjs8Rvk3XMEdkHAhcpdQPFVj0WAATS2JMw+HTUCCJVtsbKluiVIlgiXkjCDIBnsX7GARaSUVOk/Fhz1/eiN43Ofd84H6VkkwS5UHgS1R9s6j3sUD//kCDgCjoAj4Ag4Ai8uApH/xeWLe6V+ZY5AbyBAYCp8Ly57o1e91wsXKg/xnjjBPUQw/VA9g4ALlT1zK7wjjkDXCLhQ2TVU3tAReOYIIEryo0D7n2TL7fdkTkZxMoFIqX/855MjsB8CPENMLKPg6dav+6H26HbncY/i4Z8cAUfAEXAEHAFHwBFwBBwBR8AReJYIuFB5iGg7wT1EMP1QPYOAC5U9cyu8I45A1wi4UNk1VN7QEXguCCBStidEy4ddiOJkWD5c7e8cgT0RcKFyT3i62ug8riuYvJEj4Ag4Ao6AI+AIOAKOgCPgCDgCR4KAC5WHCKsT3EME0w/VMwi4UNkzt8I74gh0jYALlV1D5Q0dgZ5DoFO4fC6d80TO5wL705zUhcqnQa+9r/O4p8fQj+AIOAKOgCPgCDgCjoAj4Ag4Ao7AkyLgQuWTIrfLfk5wdwHFVx17BFyoPPa30C/gBCLgQuUJvOl+yS8EAghOCJVhfg5XpLIZsg5lbnvOtl+fQ0f8lAdC4HFC5Zdffmkff/yx/eM//qNdvXo1HPOtt96yH/3oR/bhhx/alStXbHx8/IFd7IP7vn3/D9SJY97Yedwxv4HefUfAEXAEHAFHwBFwBBwBR8ARONYIuFB5iLfPCe4hgumH6hkEXKjsmVvhHXEEukbAhcquofKGjkDPINCSOtlqbdnm5pbx/95NPnfYwh51RxMSp1LJPksmE5ZKqFqmPp9AveqoYT6S4x9EqHz77bfthz/8oQuVO+6E87gdgPhHR8ARcAQcAUfAEXAEHAFHwBFwBJ4hAi5UHiLYTnAPEUw/VM8g4EJlz9wK74gj0DUCLlR2DZU3dAR6AgH0yE2pkhuNTatvbNraRtM2mhIrW60gVh51ZiOCZEoCZS6Tas/ppKUlWpJh52JlTzwie3biSYTKjz76yN544w3PqNxG1nncno+Yb3QEHAFHwBFwBBwBR8ARcAQcAUfgSBFwofIQ4XWCe4hg+qF6BgEXKnvmVnhHHIGuEXChsmuovKEj0BMIIFQiUlbXG1aqbthSpW6VNYmVWodYGQTDQ+5pTNZEiOT4uUzShotZG+7P2lAhY4VsypLKrHSh8pCBP4LDuVD59KA6j3t6DP0IjoAj4Ag4Ao6AI+AIOAKOgCPgCDwpAi5UPilyu+znBHcXUHzVsUfAhcpjfwv9Ak4gAi5UnsCb7pd8rBFAaKrVm7a4um7TSzW7PVe1eb2vrjWsIRvYBIKh/h32pIqYQYhEkBwqZuz8WNHOjhXszGjRhiVWplOJcO7DPq8f73AReBKh0mtUPnoPnMc9iod/cgQcAUfAEXAEHAFHwBFwBBwBR+BZInCshEpIOHNLkeWbm5sBp0SCH1DaM9HgTLEdbTY2NkL7ZDKpmjtJS6VSoX1sG3Y4pBcnuIcEpB+mpxBwobKnbod3xhHoCgEXKruCyRs5Aj2DAGPXikTJ2eU1uzNXsS+nSjYlwXK11ggWsO3MxkMWKpVS2SlUjg3m7OXJfrs0OWAvTQzY6EDWMrKA5dw+9TYCPD9MLEP2rTjR4uKiffnll/bxxx/bP/7jP9rVq1dDm7feest+9KMfeY3KgMbDl6Pkcdwe7k1THHaj0VLmdDMEIMSzH/U3jKeDc/BsZNMJK+bS+m4n2s9K7IQvHQFHwBFwBBwBR8ARcAQcAUfAEXiOCBwboRJyx4z4WK/XbW1tLQiQuVzOstmspdPpIESCJUJms9m0Wq1mS0tL4T3tCoWCFYtFy2QyRyJWHiXBfY7PiJ/6hCPgQuUJfwD88o8lAi5UHsvb5p0+wQgwxi0jVIZsyopdu1+y+4sIlRsPhMrEIXuwIl5wXg5LfcoxCZOvnBm0ywiVmvnsQuXxeCi5j0zt+4ldrwuVB71zR8XjuDXcl0ZzM2RNL5TW7ZaCEVZk8czE94/v9lFkTHN8ghFaOj/nSPM9JyDhTDsQge99Ow7hqKVSeuKTI+AIOAKOgCPgCDgCjoAj4Ag4Ao9H4FgIlZA7ZgjkysqKlUqlMJNJOTY2ZsPDwzYwMBCyJaNASZvV1VWrVCrWaDSCkIlQGdsiWCJuxqjjx0PU/ZbYv9nZWbt7924QUN9++22bnJw8EmG0+555S0fgyRFwofLJsfM9HYHnhYALlc8LeT+vI/BkCDDOjUIlIsa1exIql6qPZFQedmKjTvlAwAhC5aCEytMDdlnzpclBFyqf7FY+l714fphYRm7jGZUHuxVHJ1QSaIu1c8OWVXv27nzVvri7YvMSLLlt3K9EH9mNB+tvt63b3/NWyIzOKYvy/KmiffvlUTsni+dsyJhuZ1Z2ezxv5wg4Ao6AI+AIOAKOgCPgCDgCjsBRIHBshEoyKefn54OFESLgwsKC5fN5e+ONN+zixYtBDER4RJicmZmxGzduWLlcNjIpmciuxPb11KlTNjExYWfPnrWhoaGQhYngeRgTBBeBFKHyzp07LlQeBqh+jOeOgAuVz/0WeAccgQMj4ELlgSHzHRyB54rAA6FS1q+3Z8sPMyrXNpSJ1RYZEDQOdZKC0doWSh4RKmX/+pILlYcK9VEfzIXKp0f4qITKTX3JyKYsVes2o4zpr2cqEirFF0t1ff/6lO9IRiVctPP73RaeH1338FPcGq+6c8+4jmW7nV63Wsqm7LP+bEK2zkX7zqtjWvZbIZeyTCr5QNzu3NffOwKOgCPgCDgCjoAj4Ag4Ao6AI/AsETgWQiVZkti9IlB+8sknQaycmpqywcFB++53v2vUWvn/2XsX5zaS64z3kAD4At8UJVGkqPdjtbvel10bO17ZN//D7r8olyvlOKm4kqpUxVpfO7mJvdmVtS/Jq/eTlMT3SyRA3vOdwQFBECAxmAE5gL5RjWamp7un5+smZrp/c04DVgJETk9Py7179+Sbb74xC8zx8XELB+QE7ESasbExOXv2rAFLWFkCcIZZMBgA97JYfWAA6VFGgEpc6/HjxwZJaVEZRlnGTaICBJVJrBWWiQrsrQBB5d768CwVSJoCu0Dl0wV5+sotKvOSVnPKuEElIAaui3zd9etZWFQSVCateexbHu+PeH2iTmlRua9sOyI0ClTae/R6Tl4uBPPP3lFQ+f3TJZle2NA5K2HVjL/t8o9mHUVuI8jtvaDYu2PsuB07QJw2/a9NNqUzLdLf1S6nj/YoqBxWq+le6e/pkK6OND3/7JaOIVSAClABKkAFqAAVoAJUgAocsAKJB5XocKPjCDeuAJB/+ctfDELev3/f3Lj+8pe/lA8++EDOnTtnAy2I88MPP8itW7es0wVQiDksAQ5hYZlKpWR4eNhA5cTEhIyOjtq8lWF093kyAVCxjzJiBajENTAvJqwq4V6WoDKMsoybRAUIKpNYKywTFdhbAYLKvfXhWSqQNAXwHmmuX92ikqAyaVWU6PKg/WDBFpCSoDJ8dcUJKr0+YAQNi+iVtXWZnluVu88W5O7Ustx7+VpeLuVkY7Nd8goqg8W3jiD9Hjx826ISZ0pjbcfwNMF5NAvYbKZ07dbvcgfU0dCpI13yozODZlk52NulVpWZwMMQCsuFClABKkAFqAAVoAJUgApQASpwSAokGlTCYhHzSwL+wVISFpWAkLdv35Zvv/3W5qX8h3/4BwOVp0+fNmgIQAm3q3AN29vbK++++67AavLJkyf2ZTHy6ujokOPHj5sVJiwrh4aGQsm/vr5uLmZXV1etfA4rEb68rF+/K1TFXJqw3iSoDCUtIydQAYLKBFYKi0QF9lGAoHIfgXiaCiRMAYLKhFVIkxXHwRhBZf0V1xBQqYBwXd2+Lq2sy9TsivzwdF7uTa/Ko9l1mVnRc/k2yZn7152Q0I9KYSTuzMP9Lv18eXjxPEClnjRQmd6SoW6RyZEOeXuyXyZHszLc362gskMy6v41rqlQ/NrcUgEqQAWoABWgAlSAClABKkAFwiiQaFDp4A+uVAEfAf/QAYd1JCwrYR35ySefyFtvvWVuXAEOv//+ewOFgJSAkbC0xFyWSIu5K2FxiXksYWUJt7Dvv/++xatVNFwf14E7JUBPh5UIB1RFJ9dXAFCCylqVZbykKkBQmdSaYbmoQHUFCCqra8MzVCCJCuA9khaVSayZ5igT2g8WbGlRWV+dxQ8qUScKI9Xt66KBymUFlQty/8WqPJ7NycwqQGVgURkYVW7jRt9zEBn6jrw9KNo0i0r1/9oNUNm1JSeHO+TKRK9M6lyVw/09ku0GqKT719AaMwEVoAJUgApQASpABagAFaACsSqQaFC5srJiQBBgEpaUsFzEXJSzs7Py+9//3sDg3/3d3xmMHBwcNHD43XffCVyynjlzxuICRgJUIi/Ma/n111+bW1bEwbmf/vSnAhewtS4YAHBQCctJdGoBVBEOUAn3rwSVtarJeM2gAEFlM9QSy0gFdipAULlTDx5RgaQrgPdIgsqk11Jyy4f2gwVbgsr66qkhoFLr4/W6egdaeS3PZ5bljrp+vf9iTZ7M5WVWQeXrIqh0NBmU3Y8aASonhjNyZTwrJ82iskd6ezoNVKZS7dZ2dttt1qcnU1EBKkAFqAAVoAJUgApQASpABcIokGhQCRAISHn37l0BgARw/PnPf26g8F/+5V8MTP74xz8WuH2Fm9X5+XlzCYtOOqwsT506JceOHZOuri6DiXD/+te//tWAJYAiQCXyA/wMsyAtrDIBLAEpAT1xTYBKuH6FpaW7fn3nnXesDHCng4EDLlSg2RQgqGy2GmN5qYAIQSVbARVoLgXwHklQ2Vx1lqTSov1gwZagsr6aiR1Ual1sbW0GoHJ5TZ4ZqFyUBy8VVM4DVEqJRWXQR9yvp1gOLj1+pXCEoVm4RWVPelPnqFSLyqGMXFZQOXmkR4YHsgYqOzJp81TkkJJ91vraEFNRASpABagAFaACVIAKUAEqUL8CiQSVAH+AgXCv+uDBA5ubEsByeHhYfvGLX5h15D/+4z+aZeUHH3xgoLKvr68IKgEFAQgBMEdHRw1UAiICVN64ccO2gIwAlXAdC6AZZvHyAVLCyhNzaWJBmQFXYfEJd7XZbNbKAVhKUBlGYcZNkgIElUmqDZaFCtSmAEFlbToxFhVIigIElUmpieYsB0Fl9HprGKh8vSELy6tloHJT5tYOBlS2q/vZlLl+DUDlxGBaLp9QUDkagMq+ni7JFEClA0rfRleVOVABKkAFqAAVoAJUgApQASpABWpTIJGgElaJ09PT8uzZM7OohIUi5qM8evSoXL582SDgb37zG7NaLAWViHfr1i2Dgpgb0kElLDEBKuH61UElOqMnTpwwUBnWohJg0gElBgZ8cAB5wpLy5cuXdq2enh7OUVlbO2SsBCtAUJngymHRqEAVBQgqqwjDYCqQUAUIKhNaMU1SLO+LYEuLyvoqrSGgcjMva+r6NQCVK+b69cHL1/J0fjOwqNxsk/xWuxoyum1koewwhcRSFl4WSxFksOwKL6TX1iA7QGXnlowbqOwx169HBt2iMmN9bQeUvi1kzw0VoAJUgApQASpABagAFaACVKDhCiQSVMIq8dGjR0VLSlguwrXryMiIuVEFcPzd735n1ovvvfeeAcmhoSGbG/L+/fsGKi9cuGCWkmNjY+YyFp1PgE/MUfn8+XMDlwCVYeeo9BrBQIAPCnhYaQcXFqBwOQtgSotKV4jbZlSAoLIZa41lftMVIKh801sA77/ZFMA7JV2/NlutJae83ifBNkmgEuXBB57wRuMfeaJ8mUwAxqp5nEE6xEcfEPv4YNVXv7+41S/tx6Ef2tnZWXc/DmWG39WtAqicX1KLylnMUbkoD83165bMwfWrg0rcTBFKIm3h7nYQyDbFjjuXitE0il0f2yqg8tKJHrOoHFHXr33ZLuko1IcDSt/uvBqPqAAVoAJUgApQASpABagAFaACjVMg8aDy4cOHNu8joB8sI+FOFaDyD3/4g7mAdctJwEAscLmKTi8g5MmTJw1iwrIRc0oiHSwu4VIWHTDE+eijjwQws57FO4GeFh1cWFTCGjRqB9fz5JYKHLYCBJWHXQO8PhUIrwBBZXjNmIIKHKYCeKckqDzMGmjua3ufBFsHeejv3L59W65fvy7Xrl2Tmzdv2k1euXJFPvvsM7l69ap5qsE0GZ7GAZVvo6iCsgBSAjaiH4ZpNzBNBoDj4OCgoH/mwLL0Op5uZWXF+lWAnN4PRF8Qaby8pemi7h8kqHw6r6Cy4Po1txXgR0BHB5HOLFVCXbZxZHm9eL2Xh3sqpN9lUTmUlktjBJXQiAsVoAJUgApQASpABagAFaACyVEgkaASnVlYPQIsYl5JuHRFB8y/pEXYH//4R+vwvvXWW3L+/HmDkvjyFfNDws0r9tHxPnfunAHO+fl5g5jIE51kAE8ASqQ/cuRILDXioHJqaoqgMhZFmUkSFCCoTEItsAxUIJwCBJXh9GJsKnDYCgA4EFQedi007/UdWGHrEO8wQSXKAcDokBL9M/Tv0AfDB6Xoh/X19ZnHHADLjo4OKzesKBEHcdH/g5cd5AM4iXjwsNPb22vg0oFlXLUWZz/O6kM1qGZRWQoq8woqgSKxboNK7O323hMASY+1fX5n+LYimwSV22JwjwpQASpABagAFaACVIAKUIFEK5BIUImOIsDizMyMzfeIjio6qfgqFx2x++re9T//8z+tEwuLSsxbefHiRevAIg065rCsBKw8e/asdWYRhi9zkQ86uoCUWMfHx62jHEctxfklbhzlYR5UIA4FCCrjUJF5UIGDVYCg8mD15tWoQFQFADYIKqOq+OamNzCmt4/tYYNKlAHAERaUAI2AlC9fvjQPOQjHB6VYASsnJiZsigxM4ZFOp62vhj4bPNMgLe4F/T//CHV4eNg+MD169Kj136q5jq2nJXj/Ex+cwqNPI12/Oqjc2GwXWFTCRSsWR5BB+bdBZHAM77CIoatqjEVj2LYYbkfb/21qPJ0BU1JtW9Kd3pQBzFFJi8ptgbhHBagAFaACVIAKUAEqQAWoQGIUSCSoBExEZ3F5edm+pkVHFx1UdG6x3L17V/7jP/7DzsF9EUDlpUuXiqASnVu4eMUXuQCR+EoXLlnRccaXuHATe+bMGdsODAxYRzSOGiGojENF5pE0BQgqk1YjLA8V2F8Bgsr9NWIMKpAkBfCOSlCZpBpprrKg/WDB9rBBpcNI9OP8A1LAR/Tn4B0HH6A+ePDArCTh2QYflWK6DlhIIj76cd9//71BS3i9AYwE7MQCl7HHjx+XU6dOGbCES1gAzjALNMIKAIrFtUO/0T3woAzIGx/Eot8YFohanrhOlTkqawOVXragbgEojVNaoa3oBUwJgBkcB//7gd6jJgUGBajsSW8ZqJwogMqToz3COSpLdeM+FaACVIAKUAEqQAWoABWgAoepQCJBJTp36OQCWGLFPjqT3pG8d++e/P73v7cvbWFJCfeup0+fNuCIjizOf/HFF9bZBYhEpxjWlLCkxJe76NwCVKLzi04xOp9xLASVcajIPJKmAEFl0mqE5aEC+ytAULm/RoxBBZKkAN5xCSqTVCPNVRbvI2F72KASfTdAPwBJAEp8LAoAiL4c+mKYhuPzzz83t7AfffSRvPvuu/bRKT4sffz4sfzwww/y9ddfW78PIBOWjQCHAJ/YhzXlhQsX7GNUWFhi3sowi1toopyl/UsHlSgzpiBBWRsNKtfz7WKuXwuk0RHjzvtxUAlwiTN+vA0ud4UXMtDes4JKtajUbbeCysGuLSGoLIjDDRWgAlSAClABKkAFqAAVoAKJUiCRoHI/hdB5/PLLLw0+wmISX9ai04ovajGnCeaw/Pbbbw1U+pwn6IzCxdCJEyeKK+ZGiXMhqIxTTeaVFAUIKpNSEywHFahdAYLK2rViTCqQBAUIKpNQC81bhiSBSnxgCuiHj0ThvhV9M1hTYgH8u3PnjvzzP/+zgcwf//jHBirhGQcfluJjU1hbAlgCQL733nu2BaiEtSXyQX8OFpiTk5P28Sk+Sg2zYN5MQE/029xjD/Tz+TRRZsBV9BMPDlTiDhQ87nkj1VzBuuXldmLPKQCValG5A1Rm5NJYj9Ciclsv7lEBKkAFqAAVoAJUgApQASpw+Ao0JahExxIdSMBHdGKx4gtbfEGMMHRi0clEhzOYsyMQGiATbnw8TVhXQftVF0HlfgrxfDMqQFDZjLXGMr/pChBUvuktgPffbAoQVDZbjSWrvEkClSgLLBXRJytdoRi82GB6jn/6p38yd66wpoRnnLGxMYt7//59A5j40HR0dNQsJ9FvA6TEh6oAlqVecpAerllrXVA2pMecmbDyxD76i15e7AOyYgtrzcaByk2ZU3a7rnNUwqKyzHdrqdHkjlvzevZA7+dWC8cclbtdvxJUun5vyra8fRzcfQfovWAwfHCX5ZWoABWgAlSAClABKkAFmlKBpgSVYZT2F3PvyIVJGzYuQWVYxRi/GRQgqGyGWmIZqcBOBQgqd+rBIyqQdAXwvkrXr0mvpeSWz/s72KLPgxUuTG/fvi3Xr1+Xa9euyc2bN+0Grly5Ip999plcvXrVXK4CCHoa7y/5NsodA/65dSUsGPEhKSAgrCb/67/+y/bfeecdm5YDVpKIc/fuXUsDi0lM14EtQCXOwWUsPOa8ePHCwCK85Hz88ccWr9ZyQh/ASeQBUIl8ASURDutK7GPF/sjIiECrxsxReTCg0tqDWlNijkq4fh3ohOtXgspa20uzx9NmbW07r3+Lm5vBnKVql3sgt9Wuv0H4MCGF3yOdZQdWvrrLhQpQASpABagAFaACVIAKVFWg5UFl1TtvwAmCygaIyiwPXQGCykOvAhaACoRWgKAytGRMQAUOVQEABYLKQ62Cpr442g8WA1MAA7oeNqgE7HMoCEvJZ8+eWZkACDF/JUDghx9+aFN4AGp6eXEfFy9eNLeugIQAlehjAVQCtmILmIjpP/7+7//eXMAiTa0LYCnc0aJsKCOsPqEb8kQYyoYywqVsQ0HlmlpUFuaojJ3gFFgUoJRbVBZB5aCCyhN0/Vpre2nGeNu/ByJBPy4n6xubspHXjwcUWDaaFwJIplLt0pVJSSatq+63twe/S4SVzdiiWGYqQAWoABWgAlSAChyMAgSVMepMUBmjmMwqMQoQVCamKlgQKlCzAgSVNUvFiFQgEQpgYJmgMhFV0ZSF2AYTybGoBPiDxSJctmJeSrhtnZ6eNhAIQAgI+dFHH8nRo0cNFsLK8W9/+5vNVQmXq6dPnzb3r5jeA/EBKG/cuCFPnjwx60yAyk8++cSAZphKcytP5Il9QFIsAJiAlHAzi7JgPs2Gun4lqAxTbYwbQgED77lNWV7bkKXVnG3X1nMGKmFZaaCywbQypdaUnQoqe7rSMtDTIdmujHTocUqBJRcqQAWoABWgAlSAClABKlBJAYLKSqrUGUZQWadwTJZoBQgqE109LBwVqKgAQWVFWRhIBRKrAEFlYqumKQqWRFAJS0XAP1gnTk1N2byQs7OzZlkJ96+YhxJzTB4/ftxcRCIeLC8RXgoqu7q6iqDyr3/9qzx+/NhAJVy/1gMqoZUDSoeUqGT04+bm5gxSwvoTlpyNBpUbOkdlDnNUwkWnlgHuMW1xllOwjAwC8X95QJWIBbM13Osui0q6ft2Ws+X2ttvH3NK63J9ekuczqzK3/FrW1hXKa3uwNqHto9BEIingV9vRCj1Qt2jaQ70dcvZ4nxwf6pHBbKd0daQiXZOJqQAVoAJUgApQASpABVpXAYLKGOv2IEAl3v3z6rbltbpvAUBCZ8P7AzHeyq6s9DLWoYErqXSqzVy5wKWLd0x2JWBAyyhAUNkyVckbeYMUIKh8gyqbt9oSCuB9jhaVLVGVh3ITaD9YtiHE4bt+BQQErET/CK5WscLCEjDyiy++MNh47tw5OXLkiMBqEnNYAhACVF66dMksJQExASphnemuXxEHxwCVP/vZz2wey7CiW/+poJmnLe3HAYbiugcBKvNKc1AU1CBBpdcGt/UpEPwOoEFNza3Jzfuzcn9qSV4uvpaV1+ri2DLVuSNBKb0TX0hS3/U0ledTyMD+rDTPzS3Mi7kpowOd8qPTQwYrjw72mGVl3ddiQipABagAFaACVIAKUIGWVoCgMsbqLe3gwr0ROt31dnArFQv9CHSsl9dy8nJBO/2rOq+Kwkq4cPE+RllfoVI2ocP8ugGkbJe+7owcHezWjkba5sBpxDVDF5IJGqYAQWXDpGXGVKBhChBUNkxaZkwFGqIA3u8IKhsi7RuRKdoPFmzxvp6EOSpRFqywXgSwxIp9WFP+8Y9/NMtFzFPZ19dn1os4D4tLgMqTJ0/aOjk5aS5YATgBKL/77jtLh3sdGxuTn/zkJwYscRx2cc08HfpxsKiEe9qo/TjLG/e/mVdLtg2ZX1IIO7ssd54tysOXOt/m/KYoRxJYVAagcmdvyq3dNIvalp3J0RAsHf6nRWVtErZCrKBNa61r/T+dWZG//O2V/KBtbnp+XZbW8pIHVSz8PqDJhG1eHr+8uZVqV2yzCik3N3MyNtQpPz4/LBfHB2RsJGvjCB4Hv1NcqAAVoAJUgApQASpABaiAK0BQ6UrEsG04qNTeQV7f7Gf0q8i7zxbk5fxq0bIyhuJXzQKdEnR88PUl5pYYHejWzka/jPR3WRj7GFWla4kTBJUtUY28iTdMAYLKN6zCebtNrwDeswgqm74aD+0GAkARvK8fNqhEWbACSvoKyyqEoWyAgH/605/MHSwgZW9vr2SzWdMOVpdYcIw5LM+ePWugEm5hHSACWsItK0AlPgjFHJdxLA4q4aaWoJIAKY42dbB5BH939venHzE/ebUsf779Um49XZTncxuy+FotnGG9K+oRaVfnvRYEWfvd6ACT/hhtSvvWhowDVF4YlssnB2T8SFb6db7KLfiE1WV3OWq/BmNSASpABagAFaACVIAKtJ4CBJUx1mmjQSXmlchrx+PpqxX56s4refRCXSipG5eN3Gbtn0TWcb/a7bHrmsvXjrScOtorP7l4VCa0s5Fqj2eOizqKxSQHpABB5QEJzctQgRgVIKiMUUxmRQUOQAEMLhNUHoDQLXoJtB8sDgMBAF69eiW3b9+W69evy7Vr1+TmzZsW58qVK/LZZ5/J1atX5fLlyzI6OmrAAGkcHPjWEoT8D1ByY2PDXL6urKzYFnNVIjyVShlwxFyTgI8DAwMyODgoQ0NDdg6gcnFx0VYAzPPnz5sL1pmZGYsPSJnJZKzMAJWnTp2yPEIWsWL0OPtxVh9aJ/taVObb9CPUdu1p7QSDzpEK1bpd3p3Rtvt/u8IL7UFT0qJyW76W3kN70xV/Z1ifKqj84gf9DXi2LM/n1ap3bSuw4NUWgX+NXKzNyaakJScTwx3y4dlBuTTRVwCVnYX2vv1708iyMG8qQAWoABWgAlSAClCB5lGAoDLGuoqzg1upWACVufyWPFRA+f99Ny13ny+o+9ecrOfy2jHZThF310PZqM0zASjZ05mW8ycG5Oq7Y3LmWB9B5bbsLbtHUNmyVcsba2EFCCpbuHJ5ay2pAAaYCSpbsmoP5KYMjOmVsHXgeFigEi5cMd/kwsKCADDCnSvgI+AlICPCMU8l4gE2Yh5KWEWm02mL9/z5c7l7965ZY8L1K6bSQB6Ij3kj4S4W4Ug3PDxs1pVxiBxnP87qQ+uiNlAJrIPe23YPjqAyjhp9s/JAm8O6aZbMOfuw+cu7M/K3ZyvyfCEnc6vboDLA143RJ2jJW5Jq25TOtpyMD2bkgzMD6o2pV8ZHYVHZqZdPWXuP8kFEY0rPXKkAFaACVIAKUAEqQAUOUwGCyhjVj7ODW6lYDiofTC/Jf383pfOcAFRumPtX7ZrYV7U2OFEpcYQwt+SERSVA5QWdY+KXAJXH+wkqI+jaLEkJKpulplhOKrCtAEHlthbcowLNoAAGmAkqm6GmkllGA2NaNGwPG1Sur68bcHzx4oU8fPhQnj59am5eAS8BHXEe4BEWk7DuhHvXiYkJg5gIB6T885//bJDTLS1hmQlIiXinT5+WCxcumFUl5rOElWYcS5z9OKsPgKO95qhcFVnfVItKXbXS9BZgYRbciddnOcjxcL9fP18pHN+wIpwWla5Wa29R12ZNaXPCbpjr16/uzskPUysypaBy3udEVdevWEq+cY5PGC0DFrTjtFpUZtrU9auCyvdP98vFE70yAVCZ7TJQ2dYWlCOIX2j4lpr/UQEqQAWoABWgAlSACrypChBUxljzcXZwKxWrFFT+17cAlfM2qLW+ofO+FLobjXDlgo4PXM6mUu3S05XWjsaA/D8/IqisVEetGEZQ2Yq1yntqdQUIKlu9hnl/raYA3rUIKlutVg/uftB+sGCbBFAJF62wpnzy5IlBSswviX4SrCaxwBUsXL5evHjRrCMxHyXOwdoScPPGjRvy8uVLg5O4H8BNgM2TJ09afGzhNjbOJc5+nNWH1kVFUPliTZ4ubKqFm4JKc/2qkKYBoBLaBO0C1m1b0p3ekoHOLZkYysilsR45OdojIwNZ6VNw1KGWrgC+Dj59G6e+zKuxCqCuASpheZzbWDdQeeMeQOWqTC3mZQGuX7dS6iUJroaDxbeOCf247pLi90cToznrlQqgMi3vneqTC2MAlb0ykFWLyva0xtmeK5PtrW7FmZAKUAEqQAWoABWgAi2lAEFljNUZZwe3UrFKQaVZVD4NQOXr0jkq0detlDhC2LZFpYLKzoy6bumnRWUEPZstKUFls9UYy0sFdBB4Iy/zy+s2R9GtJ+pmb2pRns+uyPJaTmAd326DovEqVRzg0p2+7oycGO6xOY3PjfXLcd3v68lIZyYey5d4S87cqMDhK4BBZoLKw6+HZi1BAKSSASodlgAuworSVwAUXwAmYF3Z19cn2WzW9tvb2809LKwn5+bmDGx6fGzhNhZxe3p6bMVxnEuc/TirjxCgckv1iLP/5uAnaBcElXG2kyTmhXrGir+9vP6drSuoxByVAJV3HFS+lgKodFfDtd+Jv995G/Xj8hz8vM9R2aEWlScG0vKjU70KKrMycaTXLCrbUwoq2wnGy/XjMRWgAlSAClABKkAF3nQFCCpjbAFxdnArFasiqFzb0M6IWlQWegwNGHu2jk9OLSrTalGZVVC57fqVc1RWqqdWCyOobLUa5f28CQoQVL4Jtcx7bCUFMMhMUNlKNXqw9xIAqWSAytI7LwUoDlJwHtaTAJMO1ErTYN/T5dWNJRaPi/jV0ljECP/F2Y+z+gA8quT6tapFZYTClyWFRugaohx0/VomTgse+t+LfySwsf46sKi8P2+gctosKgug0loERHCsuL8gDiY9hR+XpwzOB1aVsKh0UPnuZDawqDySlYHeLp2iMqN/0wSV5frxmApQASpABagAFaACb7oCBJUxtoA4O7iVikVQWUkVhjVaAYLKRivM/KlA/AoQVMavKXOkAo1UAAPNBJWNVLi18zYwprdoYKoA8169eiW3b9+W69evy7Vr1+TmzZsmAuaF/Oyzz+Tq1aty+fJlm+vRAaBDQN/GoVpp2Tw/v54fV9oinaf18vi2UvyoYXH246zcNYDKjYLrV1hUYnEQFPVeHCRhS1AZVc3kp/e/FQeV6+pa2SwqH+wGlfktx4loHIUWp23VFj8uu2Vvl96u/Nijebj+wVobRjY+RyUsKgEqz8P1awFUthNUunTcUgEqQAWoABWgAlSACpQoQFBZIkbU3Tg7uJXKQlBZSRWGNVoBgspGK8z8qUD8ChBURtN0U70IbOQ31YXupqyt63xPeR2G04G38sG5aFepnhoeDDrShVXd5abaD+rK1cvEM41VAAPNBJWN1biVc3egh61DwKSAymbRPc5+nNWH1kVFi8qXOkflfDBH5cZmm+R1bRio1EcXZiTEHJVdGc5R2SxtMWw50d6w7gSVS3LjwULRonJ+TSSnc1QCVGpsvUTJS42TxoqvGttuiatF8/BqoPIds6iE61dYVHYLQWXYGmZ8KkAFqAAVoAJUgAq8GQoQVMZYz3F2cCsVi6CykioMa7QCBJWNVpj5U4H4FSCojKYp9FtY2ZCXCzqg/GpFltTNervCwkZa82CADwvmD+3pTMtwX6etI7rt1mMura0ABpkJKlu7jht5dwbG9ALYElTWp3Sc/TirD62LPUGlgqOiRWXZZzBu2FZ4LBRvyDlSEQwVzni4R/TzSF8ElWkFlV1bMjGYkUsneuTkaI+MDGSlL9slHTrfZypFV5yuX7Nt0d6w1g4qd97hzvbmrcfjbL/7WLvW4L3fhQKrylKLSoJK15JbKkAFqAAVoAJUgApQgb0UIKjcS52Q5+Ls4Fa6NEFlJVUY1mgFCCobrTDzpwLxK0BQGU3T5dc5mZ5blYcvluTWo3mZWXytg7htBhGj5Vw5dTAsiMG94BoD2Q61POgx6wNYIPT3dFROyNCWUQADwASVLVOdB34jDhCwJaisT/44+3FWH4BHleaodIvKwwCVnQoqhwgq62shyU2F9oZ1L1Cp313JRsGiEnfi7x3YD8CjWlpqHn4G4cHioDK4BsI8fiHCjg2gubkb1jkqM20bAtevBJU7JOIBFaACVIAKUAEqQAWoQBUFCCqrCFNPcJwd3ErXJ6ispArDGq0AQWWjFWb+VCB+BQgqo2m6sLohT14uy+0n8/LlnVcGLdMAlY1ywVpiwAA3r7CmPDfWX1xxzKW1FcAAMUFla9dxI+8uAAy0qIyicZz9OKsPwKOqoDIvcwYq2wuuOHfaRO60cNu+K49V8siwkx6+HTPATeBOOywqCSpLJWqZfbQ3rLWAyk3DiGgxlcBj0LL89wQfT9liDbJS/DIJtQxIgegpgsoycXhIBagAFaACVIAKUAEqsJ8CBJX7KRTifJwd3EqXJaispArDGq0AQWWjFWb+VCB+BQgqo2k6v7Iuj9Sa8rtHc/LnWy/k+eyqNBRUorg6PojBPbh+PdLfJRfGB+TixIBc0u2IHnNpbQUwMExQ2dp13Mi7c7CALS0q61M6zn6c1YfWxWGCSqigRQis2zBHJVy/ElTW1zgSngrtDWt4UBncmHFIQ4wOKgvhft8WAdcoj+8RCluNQFBZpgkPqQAVoAJUgApQASpABWpWgKCyZqn2jxhnB7fS1QgqK6nCsEYrQFDZaIWZPxWIXwGCymiaLgBUqkXl94/n5Iu/vZSpAqhMBaN50TKvkLow9mcDfLDaxLyU508MKKzsN2A50kdQWUG2lgrCIDNBZUtV6YHejIExvSK2BJX1SR9nP87qQ+tiT1C5qq44N9WiclPRTqOeLQSV9TWGJkuF9oa1FlCZ30J7q3aDVU5o3rb4aX9pqZANopjr1za4fs3R9WsFjRhEBagAFaACVIAKUAEqUFkBgsrKutQVGmcHt1IBCCorqcKwRitAUNlohZk/FYhfAYLKaJourGzI41c6P+Xj+QBU6nyVmQbOUYnSYhwQY9WwqBzpV1Cprl/PnwCoHDRXsNHuiKmTrgAGmQkqk15LyS2fgTEtHrYElfXVU5z9OKsPrYuKoPLFmjxdUNevAJX5wPUrQWV9dcZUgQJob1j3ApXz6mo453NUOnAsEksnj8UTJdLqufLTflwSq3QXoDJNUFkqCfepABWgAlSAClABKkAFalCAoLIGkWqNEmcHt9I1CSorqcKwRitAUNlohZk/FYhfAYLKaJoSVEbTj6nDK4BBZoLK8LoxRaCAgTHdxZagsr5WEWc/zuoD8KjSHJUVQOVWwaKyEiaq7268Xbh1G12/RtEx6WnR3rDWDCpD3pDmbil8zko/Ls8Gvz1YdIBJUgoqO2hRWS4Rj6kAFaACVIAKUAEqQAX2UICgcg9xwp6Ks4Nb6doElZVUYVijFSCobLTCzJ8KxK8AQWU0TQkqo+nH1OEVwCAzQWV43ZgiUMDAmO5iS1BZX6uIsx9n9aF1QVBZX10wVTgF0N6wNgpUVjKprGRUSVAZrt4YmwpQASpABagAFaACVGCnAgSVO/WIdBRnB7dSQQgqK6nCsEYrQFDZaIWZPxWIXwGCymiaElRG04+pwyuAQWaCyvC6MUWggIEx3cWWoLK+VhFnP87qA/BoX4vKNslvtau9WsESzU0qNa0tBQu1+u4I7YEWlfVq10zp0N6w1gwqC+2s2NwKN+vH5ffuUNLP+3F5PGtwGohmmxbOUblLHwZQASpABagAFaACVIAK7KkAQeWe8oQ7GWcHt9KVCSorqcKwRitAUNlohZk/FYhfAYLKaJoSVEbTj6nDK4BBZoLK8LoxRaCAgTHdxZagsr5WEWc/zuoD8KgmUIkZ/YCA1LFmkQRFB5XIQYugucIN55Z0p7dkoFNkYigtl070yMnRHhkZyEpftks6MhlJpVLWdqCeW8Zhn0vyFUB7w7o/qMScqO1oFLYUm1vhFv24/I4dTPp5Py6PhwaHOGjHKYLKXfIwgApQASpABagAFaACVGBvBQgq99Yn1Nk4O7iVLkxQWUkVhjVaAYLKRivM/KlA/AoQVEbTlKAymn5MHV4BDDITVIbXjSkCBQyM6S62BJX1tYo4+3FWH1oX4UDldrn3BULbUffc0yIE0EjnC+xOSwFUZhRUdhNU7qlcc51Ee8O6N6jcktxWSkGlt67479EBtw4w2RyVGc5RGb/IzJEKUAEqQAWoABWgAi2sQOJBpb945/N5wYoX8Pb2dkmn07b1znh5HZW+rGMfaUrX8vhxHMfZwa1UHoLKSqowrNEKEFQ2WmHmTwXiV4CgMpqmBJXR9GPq8ArgXZWgMrxuTBEogPaDBVvvG7169Upu374t169fl2vXrsnNmzctzpUrV+Szzz6Tq1evyuXLl2V0dLSYpggaiqZ9luSN+C/OfpzVh9ZFfKDSbdjCQSY0i6JFZQYWlW1qUUlQ2WoNGu0Na2VQuSLTi3mZXxOCylareN4PFaACVIAKUAEqQAVaTIFEg0p/6d7Y2JDV1VV5/fq1rK+vG6Ts7e2Vrq4uc1MDAFm6eDrEX1xcNMDZ0dEhnZ2dtgJyeke8NF3U/Tg7uJXKQlBZSRWGNVoBgspGK8z8qUD8ChBURtOUoDKafkwdXgG8uxJUhteNKQIF0H6wYEtQGWgS9v84+3FWH1oXtYPKsKXdGb9Q/TsD9QitAr3kouvX7gKoHKNF5S6xmjgA7Q3rYYNKYHGg9ACOc47KJm5SLDoVoAJUgApQASpABQ5FgcSCSrxsw4ISkHJlZUUWFhZsC/iIOTSy2awAVvb09BiwdPiIF3TATIDN5eVlA5W5XE4yOvdGd3d3MQ3AZelcHHGojw7u3NycTE1NyaNHjwyKvv3223Ls2LGi9WeU6xBURlGPaetVgKCyXuWYjgocngIEldG0J6iMph9Th1cA770EleF1Y4pAAbQfLNgSVAaahP2/dUGlz1Gprl8JKsM2i6aIj797rEkClWl1N5ym69emaD8sJBWgAlSAClABKkAFkqJAIkGlv2wDSi4tLcn8/Ly8fPnS9gEu8RIOyAhQefz4cRkaGpK+vj4LQxrEBywE3AS0BPBEngCViIt1eHjYjr0zH0eFOKicnp6Whw8fElTGISrzOHQFCCoPvQpYACoQWgGCytCS7UhAULlDDh4cgAJ4TyWoPAChW/QSaD9YsPW+DV2/hqvswwCV6/k2yW9qOau42i119Go1rP9hWwwv7BSqv3jDyA7xEN6ue4FFJUFlUaAW28HfPdaaQaU3IDQSXbz5lbej4Gz1/8uysYgIw0pQWV03nqECVIAKUAEqQAWoABWorEAiQaW/ZMOScnZ2VtDRBqjEMRZYS+IYrl/Pnz8vk5OTMjY2Zi5hASmfPXsmP/zwg4FNWF5igXUl4CYg5dGjR2ViYsL23bLSIoX4zzsEpUnQwXVICotKlI8WlaUKcb8ZFSCobMZaY5nfdAUIKqO1AILKaPoxdXgF8F5JUBleN6YIFED7wYItQWWgSdj/Dw5UrsrT+U2ZW90SA5VbinWM+Nh/u4rtoQ4eEcHDfKccMBmo1ARIQ1C5S9KWC/BxCR9DWdcPt5++WpIbDxbkzlSFOSq9AQU/Gw0BlSm1qMzQorLl2hpviApQASpABagAFaACjVQgkaASrlphGQmLyBcvXsjMzIwBS4TD1SssFv/nf/7HLCU//PBDeffdd+Xy5csC6PjkyRO5ffu2fPXVV4L4ly5dsvDHjx8LOqAAl4CaFy9elPHxcenv7zfLxzAiozOAjgBWHxhAeuQP168oM8oBC06CyjDKMm4SFSCoTGKtsExUYG8FCCr31me/swSV+ynE83ErgPdJgsq4VX1z8vP+CLYElfXV+8GAygV5+GJNQWVeQaUUQCWAopOj3WX3MwWmtDvCHiEAmO2aAS0q9xCpBU7h7x5rVVC5kJf51yK5zXbJW1sLWlW5JaUfl0uCdoTFz/txELr9v7fV7Tkq83JiIC3vTGblwlhWJo5kZaC3W9pTGZ0WJ2W/VUG+nnI7L+5RASpABagAFaACVIAKvHkKJBpUwu0rwB+AJSwisQA03rt3T373u9+ZxeRPfvITee+99+Stt94yi0qcu3v3rllUAly+//77BgwfPHhgsBMv8AMDA3L27Fk5efKkwUq4jQ2zAIDCqhNuZbHvwBLHi4uLRStQuKYlqAyjLOMmUQGCyiTWCstEBfZWgKByb332O0tQuZ9CPB+3AhhkJqiMW9U3Jz+0HyzYElTWV+8HDyq3LSr3ApX13U2QCs2iaFGZcdevHXJprFtOjvbIyEBW+rJd0pHJmOchtB0svg1y4f9JVwB/91j3ApULayIbWwrpaShfAABAAElEQVQqzYLX6zm4M7QTLA4ig6Pt/8vP+/F2jGDPcSNBZbkyPKYCVIAKUAEqQAWoABWoRYFEgkq8aGNeSUBAh4HYR6cJ7ltv3bolv/nNb8zSEtaUcP8K6IiX8/v37xvcRB4jIyNmUQnLRriKhSXm06dPBfNcAk7C/euVK1dkdHS0Fq0sDvIFpIQ7Wrh5dWDpHQNYgqKjiy3mwSSorFlaRkyoAgSVCa0YFosK7KEAQeUe4tRwiqCyBpEYJVYF8H5JUBmrpG9UZmg/WLAlqKyv6g8VVCo8KtSgFd5Boder35GH+7GfLw/fPk9Q6Vq08hbtAKuPR+xy/aoWlQ4qN9HWyoikA8YCr9wlVfl5P/aIns6z1QEmteKF61daVLpG3FIBKkAFqAAVoAJUgArsr0AiQaUXGy/bAJaAlQCC6EACMt65c0c+//xzm7PynXfeMUgJK0nEgTUl4hw/ftysJU+dOmUWlbB0xNyVgJyAlujQwfUrLDJPnDjhl9x3i04A5sqEe1dYe2IfULLYMdCyorwow5EjRwyEHjt2TN2btNs1973AHhE29dq5/JY8mF6S//5uSu48nZfFtQ1Z34AL2iChdxD2yCb0KdxzbnNL0ql2yXZm5ML4gPzy3TE5c7xPUupPqBHXDF1IJmiYAgSVDZOWGVOBhilAUBlNWoLKaPoxdXgF8K5FUBleN6YIFED7wYIt+jhY8VElpsO4fv26XLt2TW7evGlx8JHmZ599JlevXrWpM/DBpqfBFotv7eAN+a/VQGWhSaijT0CjLelWi8rBbu3/DgUWlZO0qGyZlo2/e6zF8Qgdm9gxRyVApbp+3VDXr40GlfgFcVCZJqhsmTbGG6ECVIAKUAEqQAWowEEokGhQCdgH+Ig5Kh89emQWkbOzs3YMUAg4+cEHH9ickwCB3iHHizqsLCcnJ+0cLCrR+YQ1JTrpz58/N5AIUPmzn/3MQGcYsQEmYU3pkNLdv6K8CIObWoDRwcFBgsowwjJuIhUgqExktbBQVGBPBQgq95Rn35MElftKxAgxK4B3V4LKmEV9g7JD+8GCrUNH7xcRVNbWEA4NVG66NaWWs2CqplzRFnjpLF0K1VwM8o9Fy8M9AsLbNbOU5gNQOaCgcoKg0uVpmS3+7rFWBZWLblGZMlDZyBs3UKkXgEVlui3HOSobKTbzpgJUgApQASpABahAiymQeFAJ6Dc1NWWWkk+ePDFLRkBCwEJ8Afzhhx+a9SRgISwl8eUwoCW+Fj59+rQcPXpUurq6DEwi/Y0bNwRbdEZhSfnJJ58IrC7DLLiWu3zFPqw+saBMAJSwtMTgAOeoDKMq4yZVAYLKpNYMy0UFqitAUFldm1rOEFTWohLjxKkABpkJKuNU9M3KC+0HC7YElfXV/aGAylybzRlY4JL1gUpNXExfdutBexAFRgSVZdK01CHqGStBZUtVK2+GClABKkAFqAAVoAJvnAKJBpWAgOg0wqIScBFzTAIAYgs3rtlsVt577z2Ba1V0ygEI4RY2k8kIXMICVAJmAlQiL1hUfvXVV/L48eNIoNI7AQCU2EfHAItbWgKYonw9PT2co/KN+5NqvRsmqGy9OuUdtb4CBJXR6pigMpp+TB1eAbxLElSG140pAgW8L4ItQWV9reLwQGUAGtGbhDVa6VINQJbGsf3yiIWM0EXVLrKCSrh+baNF5S7hWiMAf/dYfYyi4hyVJa5frVE06NbR9AJ3w7Co5ByVDZKZ2VIBKkAFqAAVoAJUoCUVSDSoBAiEO1VYVcLlK0AkLBYBGr/55ht7Gb948aLByHQ6bfFwrrOz0+ZcgaUkICZcv2LeSIDKv/71r7YFVIRF5d///d+bi9gwteudAWyxYItBgdIOLsAqAOnbb79tZeAclWEUZtwkKUBQmaTaYFmoQG0KEFTWplO1WASV1ZRheKMUwLskQWWj1G39fMv7JOiX0PVruHov7cdhyhH0J+vtx1l96N/01mZe1tY3ZH5pVZ7NLsudZwvy8MWqPJ3Py9zKlqzn2ySn/l2dM8YKKjVT5Is8MUdlTwdBZbgW0Tyx0d6w7gkq13SOyq1gjkqCyuapW5aUClABKkAFqAAVoAJvkgKJBpX+wg1gCdDo67179+RPf/qTWVoCRGIuSHfvik55R0eHnD592gDkxMSEWTZi7kiAym+//dYsMvEiPzY2Jh9//LFgrsqwi3VAyxKhgwuYCle16OASVJYJxMOmVICgsimrjYV+wxUgqIzWAAgqo+nH1OEVwHslQWV43ZgiUMD7JdjSorK+VnGQoPJJCajMbypQVLBc0+JE0yN7svLwwnltDsakCCpdsNbc4u8e696gcktBZWGOylrbm7ejwnaf5mZQHHFoUdma7Yx3RQWoABWgAlSAClCBRiuQWFDpL9t44faXb9/ev39f/vCHP8jz58+lv79f+vr6zA0szsPiEtaLQ0NDBiLPnj1rFpULCwsGEAE5sQ/3sACV7jo2DqHLQWWUL3ErlWdT7y+X35IH00vy399NyZ2n87K4tiHrG9AoSBGy31HpMrvCoGtuc0vSqXbJdmbkwviA/PLdMTlzvE9S7W2N/ChzV1kYcPAKEFQevOa8IhWIqgBBZTQFCSqj6cfU4RWwd9jVDZmaXZUHU4ty6+mCPH21LGiL67m8pO19y4eJw+dfKQVeHXFdgC284430dcpZfbc7c6xXTh3rt+OOTMre9SqlT1qY3w/eiYP34sLL8QEVNACEGKQ/+Hdj1CMWr09aVIav9FYDlcHfg4j+dNCiMnxzaKoU+LvHWjuoxO3V/jzxX1JP4cflIgXn9ZmitryKRCUD16+DOiXPZFYujGVl4khWBnq7pT2V0fGalD17rCSNGMAoLxyPqQAVoAJUgApQASpABRKvQCJBJV603e0rXL9ixbF3vgEov/jiC8FckJinEiuAJTrlsJyEW1fEB6yEa1i4foWlJVbMd4mXeFhhAlSeO3dOhoeHY6moODu4lQpEUFlJFYY1WgGCykYrzPypQPwKEFRG05SgMpp+TB1eAbzj0qIyvG6lKfw9Oa8f121iVU19YL00XiP2Mc7erkQorR9LYtuuAQc59o72g8X7SgSV4Ws5zn6c1YfWSTXXr0/m1PXrauD61SwqDe1UQEflDbicEPn58nC0BWsP26CyW12/Dna3ycRQh1wa65bJ0R4ZGchKX7ZLOvQD3lSK4Ch8q0lGCrQ3rPuCys0y16/+I1X4/dj+wdzZsIqnC/GtfVe4dc9OB5gUjhNUVpCIQVSAClABKkAFqAAVoAJ7KJBIUAnICNiIuSnn59VqUK0ksY9wzEWJ+Sp/+OEHWV1dlSNHjsjIyIht0cFaWloyIIk5IhH3zJkzNscIICU6oOi4DwwM2PyUAJVHjx410LmHRjWfirODW+miPgBDi8pK6jCsUQoQVDZKWeZLBRqnAEFlNG0JKqPpx9ThFcDAL0FleN1KU2zkNmX5dU5Wsa7ntN9QAJU+5l4aOcZ99C0AJjsy8DySlq6OtGTS7QdqiergAFuUh6AyfAXH2Y+z+tC62BdU5qRkjkqtt/DFrprCQaU2B0nrHJXdmQKoHCaorCpak55Ae8MaHlQWbthBd5UGqFnbgraExY+Do+3//TxB5bYm3KMCVIAKUAEqQAWoABWoXYFEgkrMRQn3rC9evLC5HjG3JOZ9BJiEO1VATJzr6emRK1eumFXk5OSkuXOFxSQg5p///GebLxIgE8ASsBNpMR8l4CUsLTG/JcJwPo4lzg5upfIQVFZShWGNVoCgstEKM38qEL8CBJXRNCWojKYfU4dXAIPMBJXhdfMUGDhfUUA5PbcqLxfWZGZxTdbW82rVE0A7jxf3VvGAWVBm1HVuX3dGjg52y1Bvp2R1v0Nh5UEtBsb0YtgSVNanepz9OKsPrYu9QeWmunVuU1CJettJiBz4GB8qQKIwd4U2gGT4u0BeAagUtahslwmCyjBSNkVctDes+4LKrcCistzW3Fsffs8qLY7Q/azH97geri3OYDvO06LS1eGWClABKkAFqAAVoAJUoFYFEgsqARbh2vXx48cCUAl3r+hAYm5JvIQDWsJ961tvvSWYhxIAEudgbXn37l1zDeswE501wM3eXp1z59QpA5VIA9ewcS5xdnArlYugspIqDGu0AgSVjVaY+VOB+BUgqIymKUFlNP2YOrwCGGQmqAyvm6cAkJlfXtd53Bfl8ctlm99zeS2nVo2BK1aPF8+2dFheB+TV1Sug5Ehfl5zW+T2PDfUYrOzqSMVzuRpyMTCm8bAlqKxBsApR4uzHWX1oXUQGldtNrUKJqwcZqNS0SG7zBapFZY+5fiWorK5a855Be8MaHlR6A3P06Mc7tdgGlcF5P/ZY26kIKl0TbqkAFaACVIAKUAEqQAXCK5BIUAkXr5iXEh1GuHLFCtevCAs6XsHLeFdXl80vCVeumKeyXQcjACRhjTk9PW3zVSIMC17eOzo6bC5LzGeJFdaUcS5xdnArlYugspIqDGu0AgSVjVaY+VOB+BUgqIymKUFlNP2YOrwCeE8lqAyvm6eAfq8WX8vtJ/Ny59mCPJhaFPwdp9XSEW5ZG7mkUm3q7jUlx9Wa8uL4gJwc7ZUjA13So25gD2rB/WPBlqCyPtXj7MdZfWhdxA0qy1uyA6Jq4WgWbQoptYlug0rMUXmCc1TW10qSmQrtDeu+oFLnqMwXrHfRdoo/jdUaUuF2vX3tE60ojsPxTFteTgxm5J3JrFwYy8rEkawM9HZLeyqj4zacE7UoGHeoABWgAlSAClABKkAFTIFEgsrSl21AS6y5XM5evq3jp0UHgMSclLCihOvWUiDpoNPT4E7RaUd8xMWKfYTFucTZwa1ULoLKSqowrNEKEFQ2WmHmTwXiV4CgMpqmBJXR9GPq8Arg/ZagMrxungL6vZhfk28fzhZh5fxyACph8Rj/omCgkClgaLeCyhPDOiXF5KBaVfaZC9hsVyb+y1bJ0ftH2BJUVhFpn+A4+3FWH1oXe4LKFXX9mt/H9as3skLZy1uyn64WHrQHuOFUUGlzVBYsKgkq92kNzXUa9Yx1L1A5v7YlGwoqNxVUFttNoeFoUlvCDI2Ut7ltxYLM4G6YoHJbFe5RASpABahAayhgz9wDvBU8b+NmFwdYfF6KCoRWIJGgEneBP37f+ou3H2PrnXAASt/38/bDUXhZR5gvHs/TeHhc2zg7uJXKRFBZSRWGNVoBgspGK8z8qUD8ChBURtOUoDKafkwdXgG8uxJUhtfNU0C/6QKo/P7xnNx5uiBz6goWc0c2BFRqN6Uwtq9Wm21mPWmg8tSQnFFQeWyoWwgqvXaaYxtnPw7tUTuz+4LK1woq85vbbcnhj7ctP3YFkS2WcqDk4cHZ4DyiIhxxASq7FVQOdbcFc1Se6JHJ0R4ZGchKX7ZLOvTD39KPeDkg5ko2x7Z07AMfaq+rh6mnr5bkxv0FuTO1ItMLeTFQqZDSLSqj3ll52yzNzy0qO9o3aVFZKgz3qQAVoAJUoKkVwPMW4/Kbm8EHi+Wu0Btxc/Yepx9d8t2sEeoyzyQqkFhQmUSx9itTnB3cStciqKykCsMarQBBZaMVZv5UIH4FCCqjaUpQGU0/pg6vADq+BJXhdfMU0A+g8psHM/L943n5QUEl5qzMpNWjSjnV8UQRt3pJW9I6P2UPLCpHsvI2QWVEVQ8veZz9OLTHxIBKldRApc5RSVB5eO2rkVdGe8O6y6JyB6jclI2tbdevjSoPACZBZaPUZb5UgApQASpw0AoASm7oV2UbOaw6TZ3u5/LBc/cg4CGu0a6gEh9fdmZS0qH9Dkw70eipLQ5aZ16PCrgCBJWuRAzbODu4lYpDUFlJlXBhPqi0naowyrQd0NC9g3iQxX0DBJVxK8r8qEDjFSCojKYxQWU0/Zg6vAIYZCaoDK+bp4B+BiofzsitUlDZKItKvbC/U8Kislvno4RFJUGl10jzbePsx6E97gcqZ831qwQWlYV5AzVRReG8/2D5VoyxMxDxkROKAXCUUjec3QYq3fUrLSp3KtbcR2gXWKuDylxgUbnZFrh+LTOH9MPKrW/7t86/+UC7qrQEkNKteLeEFpWVVGIYFaACVIAKNI8CW/J6Y1MWdd77hZV1mdd15fWGrGsYxufhtcXf0Rp1T3jm4rGb7UrL6EC3DPV2SLYzox9jtjfqksyXChyqAgSVMcofZwe3UrEIKiupUnsY9Mvrly95/SIGK76MQdhBLf4VDL5+wdf9eKB5h++gylDPdQgq61GNaajA4SpAUBlNf4LKaPoxdXgFMMhMUBleN08B/QLXr4FFpbt+7VBQifevRi14jcQXzl2dwRyVBJW7lXaAgi1WvP/6NBx7De4AumDFErwz+7tzY+ozzn4c7rMaqHzwYlWezOVlDqAyJ5JDV8S6I7gv28Et71hcJ8t3x5nKB4hvRdDTyJWgsrJOrRLqf1vVQOXUQk4WbI7KAqjEjZf8Gflu5dZnTdmk8n4r2lalBfnYqv+hzRFUVlKJYVSAClABKtA8CmzJ0uqGTM2uytTcqkzPrRmsXNvI6zuqPuu0jxFHN6P8sVr6XMYzN7+1KYPZTjk31ifj6sFluK9TP0BLN4+MLCkVCKEAQWUIsfaLGmcHt9K1CCorqVJ7GEz0V1/nZHU9b9t1Nd0HhAOw9I5X7bnVFtMfOHjQdKiZfm93xuYx6tL9NAbP9MKNunZtJdw/FkHl/hoxBhVImgIEldFqhKAymn5MHV4BDDQTVIbXzVNAP4JKAIUABAJUvXr1Sm7fvi3Xr1+Xa9euyc2bN02uK1euyGeffSZXr16Vy5cvy+jo6A4QiEgOxlzfercoTz6fF8yb51tAys7OTkmn08XrluePdEjzWufawz7mT0R8bB1ylqeJehxnPw5l3hNUzuZkbnVTtFtSmKMyGJLygamo9+LpUQz0M7Zdv9Ki0rVppS3aG9Z9QSXmREXn1Duf3uC8w+qieLgf+3kP92M/X7JFlKDNEVSWyMJdKkAFqAAVaDoFgmfrzOJruft8UR5MLcnjVyvyanHdxpR1KPlAXLDas10vdnSgU94/OyTnT/TL2LDOMa5jy/rEbTpVWWAqsJ8CBJX7KRTifJwd3EqXJaispErtYWsKKGeWXsssVn3YYEAQg/mwsgz6a3H/yO/sxQFSHhvqlhH9+qW/p8PchAVf4MR93do1qSUmQWUtKtUWZ2eLqC1NXLGS3criukvm4woQVLoS9W0JKuvTjanqVwCDzASV0fQjqKwNVL799tvy6aefNhxUYmAFcHJ9Xd1krawI+knYB2zs6emR7u5u6ejoMADp8BF/B0izsbEhq6ursry8bMeZTMbgpqdxYFl/i9mdMs5+HO5jX1CpFpWv840DlVYEvU30cXSqVrp+3V3lLROC9oZ1L1A5v6Zzaymo3ISbYesU2H+VNSg/5R0YD/fjyqkLbY6gsoo8DKYCVIAKUIEmUCB4tm7Ki/lV+f7RvNx5prBSvWK8UFC5sq5GL2ZR2V4YS477hkoeuGpNuaXvxmNDnfLj88Ny+eSAnDzSK/3ZDr024nncuMvA/KjA4ShAUBmj7nF2cCsVi6Cykiq1h8Gv+JNX+hXMyxVdl+XlwpqZ8cPS0iwba8+qppjlfbjh/k65qF+/TI72KrDsMVgJv+KAlUleCCqj1463BRu4ip5dXTnYK0xA5OtKz0TNpQBBZbT6IqiMph9Th1cAzweCyvC6eQroR1AZDlT+4he/kEuXLjXMohIWkegbLS0tyczMjCwuLhqwBJTs7++XgYEBW7PZrABEIhyQEmkWFhZsnZ+fN2gJoIl4SNPX12egE2niXOLsx9n7HuDRpt7P+obML63Ks9llHeRasEGuJ7Co3AEqgzvZv0fgMfzN0hXw8ODY3ze1CAE00mmMenSOysHulEwMd8ilE5yj0pVrhS3qG2t1UKltEK5f3aLSBzW92Xhz8uOCKH6IrUfBtjQcUYvndAddjQCOl4LKXrkwlpWJI/o33Nst7Sn8vacKA6yI7zkiNy5UgApQASpABQ5XAX+ubulHd9Pq8vW7R7P6DrcsD1+9lpdLOVneENFpKu35pT7ydKfG8voD0+P7cbXkGi8lm5KWDTkx2CEfnBmUi+N9+jztlQGASn13Dq7vGVbLiOFUoHkUIKiMsa7i7OBWKhZBZSVVag+bW15Xc/1FuYdVTfefq5/xRZ0MGS5gMX9R3D/t5c+co4Nd8s7kkPoV75dx7agN9XaaO1iCysp1CP3Q5uEmF1qNj/SYdvh66Nhwt/R2xTtAVbkU0UNxH3jRwbyoOW1red0/kKVwGfT9AeLRzszdcMLB+IFo8wZchKAyWiUTVEbTj6nDK4DnBEFleN08BfQjqEwGqLR3nhLgCEj54sULA5WwlMQKINnb2yuTk5Ny7NgxGRoaMstKWF7CZe3jx48NVCIvhy9wFzs8PCxHjhwxuIr0cVpWxtmPQ7nDWVQGLTmuvohdXrPEFq99+l0kQaX/WLTgFu2t9G9lXV0mP9WPc2/cX5A7UysytRANVLpk3oOp1k7R3tDvwPl0O0Gl68YtFaACVIAKNJcC9lwteAYxUPlQQeXzZXk0uy6vljZlOdcm65t42hWeiIVNtbv003hO2lIMqJaiEE3jpSUvHW05GR/MyHun+uXCiV4bTx7QOSvbMSVCm77kWX6e6d558iwVSLoCBJUx1lCcHdxKxSKorKRK7WHzAJXTi3JfQeV99S+OCZEXVgugUntV6FjFvZQ+iI4OdMuVyUE5d1xBpU6APNjbQVC5h+B4hrcEqNRGAEgJcLS0mpN13fr7yR63H8spvGABwsNytyuTlmxX2vZjyZyZJFoBgspo1UNQGU0/pg6vAH6vCSrD6+YpoB9BZXhQiTkqAf5g0eQrNI1i4eRzS8Iacnp62laASsw3Cdg4Ozsr3333nUHG999/XzBn5oULF6Srq8vi/vDDD/Lll1+a69fTp08bwHz+/LnNWQmgOT4+LhcvXjTACVewcVlWoh83NzcnU1NT8ujRIysr3OQCpLprWm9v+23RHkOBSsQvWVx/y6ckvLbdoEODHJEeoBJzVPZ0tMtQDy0qa9OwuWKhnrE61N8NKnNqUamuX3VQNQ/Xr754s/MgP/bzYbeaD7LCmmrbks7UplqAZPRDXVpUhpWS8akAFaACVOCwFCg8U9XzHt5pp2ZX1KJyTu5OLcuT2Q15tbwpK7n2AFSGHUT29z1P58dlt+qPYzxP022bCio3DFS+q8/T82MKKkd6dTy5U1I6fzveUQukMtL7e1kReEgFDk0BgsoYpSeojFHMBmTloBKTIN9XYBmAyg21qMxrZyoYoIn7stZx1Ewx4DA60CVXTg7J2bE+GdfJjwkq91YbD+dWAJWAlBtqSQmL3uczKzYQrUNHjYeVKiD0y6TapUetTwfVNcTRwW6DlXsrz7OtoABBZbRaJKiMph9Th1cA7wsEleF18xTQj6AyOaASfSK4bwWghIUk4CQgCuamfPr0qfz+97+3+So/+ugjee+99+Stt94yMPjkyRMBqPz666+tan/0ox8ZwHzw4IFZZMIF7OjoqIHNkydPGmRFnmEWlAMWnVjtPV3bDrYAqYCrL1++lGfPntkcmg0HlTmRHF549fqlSzRQiZza7D0T97UNKtsKoLKTrl9LxW6BfW/HSQCVQevDwCpBZQs0Ld4CFaACVOCNU6D4TNX3xI3chkzpGF4AKlfk6dyGzKxsGajExz9bDhwLKgEsYtn5VheEBScKZzxd2fufx/T0Dio7FVSODablnZMKKo9ni6AybVMn0JW668ZtayhAUBljPRJUxihmA7I6HFBZuBF9whBUhqtUPJxbAVRiDtSV1zl5qnOjfvNg1gZS8fLT6AUodFMhaVdH2lznYl6Yi+MDMqJzpXJpfQUIKqPVMUFlNP2YOrwCeC4QVIbXzVNAP4LK3aDy1q1b8vnnn8u1a9fk5s2bJhfg26effiqYo7IRFpUOSwD+lpeXzTIS+6gjgMZ79+7Jv/3bv5n1Iq5//vx5cwGLL8IBCAEKATkxFyUAJqwmYeWIcFg8Ih6sQCcmJiwt3MGGWeB6FuVCmdwNLcrm5QWsBFjF9RsKKvWL/Nd5nRZA5zjyt0If4ApzP9XiIk+9LYLKagK1UDjaL1b/26tqUVmco9Jv3lvc3i0QeWOpCtAL2Xhu2ML1a2dqixaVphz/owJUgApQgWZRwJ6phY/a8J44pXOMf/dwXu5NK6iczxmoXDXXr+1FUOnPv7jusfSpnFLXrwYqB9Ly9smseujTOZ/NQ1+XAFSmUuni89mf03GVg/lQgcNQgKAyRtUJKmMUswFZEVTWJ2pOQRugx4v5NZ3bU+c6ebYot5/Oy9zSus552KYDIHE/loNy4uHcCqASc6Bi8Bnzov7P99Py5NWyDRw12qYSYwoAlT2dabOkxNyo758bkeND3fU1BKZqKgUIKqNVF0FlNP2YOrwC6BQTVIbXzVNAP4LKyqDy+vXr8qtf/erAQKUN8Gh9wGIRLrPcehGDJ5hTEvD0t7/9rVlbwoUrLCPhXhWQBRaVq6urks1m5ejRo3Lu3DkDlQCUcP96//59WVpaMutLgEpYYx4/ftybQU1bt/YshZUo8/r6ul0b4YuLizI4OBgvqJxZ1nfoBXnwYlVdh+VkboWgsqYKY6R9FfC/uZ2gclnnqJwvzFFZcP2al52uX81Jqw+H+mXK+3WAoMG5oMunFiQeUJbEU2ILi8qO9Ja5qmsm16+4Ndwf+qDwioO+FPYRfhALNE6pGXRKP8jAFscceD4I5XkNKkAFqECggD1T9Z0U77BFUPlIQaXO+fx0Pi+zq7CobDN36oFFpf5WxyyeP3KQb0pdvwJUHu9PydsTPSWgMpj+AO5f/Tnh25iLw+yowIEqQFAZo9wElTGK2YCsCCrrE5Wgsj7dPBWAEQaf//ZkQf707ZQ8nF7SL+fR4fXXD48Z7xbZo4ONeSnHhnrk4sSA/PjCqJwYCeciLd5SMbeDUoCgMprSBJXR9GPq8ArgmUBQGV43TwH9CCqTASpRJzbIo3VSukU4BlC+/fZb+fWvf22gEvNTTk5OCqwiAQoBIgFbAC8BIrFi7sqVlRWztkRauJNFHMxV+fHHH1sc5F3LgvKgvwbwCeCJfQxCIT9sAUmxAlZiPsxYLSp3gMqNAqhU16+wqCy8EgYgqORO/IQHlY+E+atkebjGxykkD1y/tukclXT96jK22tb/zqqCSrUAmV9TF3YFi8qtXUOq5Q3JG5SH+9bDXcHd4YjRppAyrTvNCCoBJuENZ31jU9Y2cvaxLvbz+hux6+/TZYhjq1JCTXwE3KPecLr1Q9POTErn6G23j4I5+ByHyMyDClABKrC/Anim4nmaV1CJd1PMUfk9QGWJRSXmqNywOZ+D56I/H/y1zY+3r7b/83M7Lt7fgucrfvt3gMpxgMoec/061Nel87R32DyV/ozwbWle3KcCzaYAQWWMNUZQGaOYDciKoLI+UQkq69PNUwEYAXr87cm8/L9fP5cHCirty1zrjnqs+Ld4t0Fn20Clzol6WUHlTy6N6ktNNv6LMcfEKUBQGa1KCCqj6cfU4RVAh5SgMrxungL6EVQGAxsYpMCKuSFhvXjQFpVeJ6Vb1A++TAdwBGz83e9+Z/NBYg5KQMne3l4Dh5ifEq5d4RL21KlTZmkJUImBIsxteePGDbO6hJtWgMqf//znBjpLr7XXPsqBtHDvChiJvhvKhQEpt6hEGQExG2tRSVC5Vz3xXDgF0K6xVgeVGwoqFcbXDCrLrx8MmCqCLDtRORygMgOLSnX9Oj7UIc1kUQlPOMv6gSmexwsr67K0tiGv19U6XPtUdvflEpQpUu+hVp/VYYeCyf6ejK4dtma7MtKhwBLWlVyoABWgAlSg8QqUPk83DFQuK6hcMFD5pOj6VUGlzlHpDwYHk0iLZTcwLP8Nr/z8DO4Oz/RgD/nsBJXdZlGJMb2hPlhUElQGSvH/VlKAoDLG2iSojFHMBmRFUFmfqASV9enmqbYtKuflj99MGaj0AQW82JS/sni6erf+yoOXGweVx4eycgmg8iItKuvVtdnSEVRGqzGCymj6MXV4BfBcIKgMr5ungH4ElckElf7Og3knHz9+bPD0yy+/tKr74IMP5MSJE2o1lJaZmRn5/vvvDVS+8847cubMGZuLEqAS1o4OKpEHrB6R7urVqwY0vR3UsgWQdEiJfOGaFmX0OSpRTsxR2d/ffwAWlYU5Kn1Aym9ge8TLQ4Jt+Uujv/RVCMcprIFFpaiVVrsM9aRkYrhTLp3okcnRHhkZyEpftktBCOY4ShUH1nYPsO0sBo+SpYD/jdUOKr383nC8IdUbvjPdDlA5qKDyVK9cGNM5tY5kZaC3W9pTGf07T2Z7W1YwOT23KlO6Ts+tybzCytcbgRtY/7P0u91vG0Zd67fpf50KKgcUVB7p77TpOob7uwxYwrqSCxWgAlSACjReATxT/XnqoPK7AqgsnaMyp6AycP1aAiarPU6LxS5/MvixRwgyQBkwUlgOKq+MK6g8FjxPCSpdM25bTQGCyhhrlKAyRjEbkBVBZX2iElTWp5unKoJKndfzT19P69xEizYghnePsB1ez3O/LV5rghcsWFRmtKMbuH79CV2/7iddy5wnqIxWlQSV0fRj6vAK4DeboDK8bp4C+hFUJhNUwmIRcBBzTMKa8tGjRwIYODAwIO+++66MjIwYJMR5B5VwuXr69Gmbp7IcVGIeS/S5ACo/+eSTUBaVaC8Ak4CSDikxGIX2gzDMTQlIOT09bfNkNt71K0Gl/w1zW78CaL9YfWB1Xdvy01clc1TOl1tU+rV8gDQYGPVQDI4GS63hnjJI18ygcnbptdx7vqBTdSzLo1crMre0oVYz+vFnHf22MCpq9Wkdqrtcdf3a152SowOdcvpo1jzhHB3qtv6cq8wtFaACVIAKNE6B0udpJVA5u7Ilq5ijsuD6FU/K4gdeOChd/EFQDPMAj+jHHiEIRxkCUOlzVOZsjkqCSteJ21ZWgKAyxtolqIxRzAZkRVBZn6gElfXp5qkIKl0Jbg9SAYLKaGoTVEbTj6nDK4AOKUFleN08BfQjqEweqES9oH+EOSHv3r0rf/7zn82aEXNPAkSePXtWOjs75eXLlwYwb9++bYM9Fy9etPNjY2PS3d1tecCi8uuvvzbgCfAJUPnTn/7UXMd6O6hlizIB6DigxDEWdwmLOTABQwFIGwsqFZjmCnNUFgpePlxVCN5/U55Qbwl3hVszi8p2WlTuL2LzxkAb9nZtHwbUCCq92QR/Adt4slyJ8vN+XB7Pj9s046Lr18GMWlT2Jdqi0n8DUG5YU968PyO3nyzKfYWVcyt5FUatGdv0jwiLixYc7fm/R92lVyEA18MS/AQFrv4wt2dXektBZYdcHO+Vs8f75PSxfhns7SjE0yJ4wiA5/6cCVIAKUIEYFSh9npaDymcF169r6kodrl8Di0r94a74u16hUOUPBj+uENWDdrp+xRyVwUcstKh0hbhtNQUIKmOsUYLKGMVsQFYElfWJSlBZn26eiqDSleD2IBUgqIymNkFlNP2YOrwC6BQTVIbXzVNAP4JKDHjrPGo6iI31sOeoBDAB/IOFIty1PnjwQO7duycdHR2CuSkBKY8dO2ZlhjXlw4cPBXNUIh1A5uTkpLl1BaiEpeOzZ88EIBMuYrEAVH744Ye29XYQZetAFdaUKAsAamNA5Yo8mS3MURkXqKxw44AfGDcD01BvknT9WkGjVgnC3z3WPS0qV3WOSrUMzKsFSGE8tcjcyo/LdSk/78fl8fwYba75QCXuakuez6zIl3deybfq5u/u9KqByrb2jBL/Aqj0m6xh6+PPu/QqBEAnLPhbDf5a9W9Vy5Bp35TjAxmdtqNXLo33yfkTAzLc16VnggQEldCLCxWgAlSgMQqUPk8rgUqzqDxIUCmb0tm2oc+FlLw9TlDZmFpnrklSgKAyxtogqIxRzAZkRVBZn6gElfXp5qkIKl0Jbg9SAYLKaGoTVEbTj6nDK4BOMUFleN08BfQjqMSAd3JA5crKisFSAEpYQsLd6+DgoIHFc+fOyfHjx829KsCkWzHC6hLzR/b19Vm8S5cuSU9Pj+UDUAngiXwBL2FteeXKFRkdHfVmEGnroHJqasqsOw8DVPoN7Ac4PF6RNCFgFw0JAAhgSEoZSxZzVGY5R2VRuxbawd891r1BZV7WdWA1X6GdxCZFoeFik9E215HakvEmsajc2lKKq+vU7Ip8dXdGvlOLynvTazK7otbXAJVuURlGLK0TW3YSyT1yUEjZpvNUGqhM61yyOrenrjtApeZFULmHhDxFBagAFYioQOnztFZQWf7e5sflRUHeWPx33I/L45UeBxaV6vqVoLJUFu63sAIElTFWLkFljGI2ICuCyvpEJaisTzdPRVDpSnB7kAoQVEZTm6Aymn5MHV4BdFQJKsPr5imgH0FlMkAlYAnmgYTl4/37921eyi+++MLmqQR4PHXqlFlSDg8PG6hMpVJ2Du5h4d4VlqCwoOzt7RW4gIULVriGnZ+fl9XVVclkMjZ3JUDlyZMnpb+/35tBpG2c/TgbeNI2ubWZl7V1nR9waVWezSzLnWcLOlf5qllUzqpLSbh+zYOPlJXcB7jKw2uPGOSJ8bAdrl8VVJ4c7lQA0iOToz0yMpCVvmyXdKimqAcfOPNtWbF4mFAF0N6wVgSVz1dkakHb4OoBg0oFbh3qwnR8sCPRrl9dOwOVmzl5rqDyr/fn5PsnS3LvhVqEqyVqvi2jf3rtxe8C/O+y6t+ptxP8AWKpAVQGecGacku6U3k51p+Wi2NZdZnbK+dO9AcWlQpL22z1KwfZ838qQAWoABWIT4HS52klUDmjH7Cs5dt3uH71X+Xy50N5qZA3Fn/P8uPyeKXHBJWlanD/TVCAoDLGWo6zg1upWJv6o5bTzyAfTC/Jf383JXeezsvi2oasqx+X8vfgSunrDcOPZ05nkE/r57jZzoxcGB+QX747Jmd0zoSU9n793bve/A8qHUFlfUoTVNanm6ciqHQluD1IBQgqo6lNUBlNP6YOrwDetQgqw+vmKaAfQWUyQOXGxoZZRcL68ebNm/LVV1/JX/7yF3MDe+HCBYOUgI+wroTr16NHj8qRI0cknU6b1SUsMG/cuGF5wL0rwCQgZbu6XkQ8wEnkA4tMwEy4ko1jibMfZwNP2ib3BJXLPkelD2vFcRfbeSBXLUIBVLYFrl8JKrcFaqE9tDes4UGlt73yIdZo4mBsIKNr0kGl/Z3qrUI3/K1ulYDKW0+X5f5LgErVFaASFpUuVzR5qqQOHLtm2jalC6CyLyXnx3rkwnGAyj4Fld1q1KkfE2BtlsGXKnfKYCpABahAkhUofZ7uBpUbMrOi86/ndI7KrXZ9LPjzM9478scNcieojFdb5pZ8BQgqY6yjODu4lYpFUFlJldrDCCpr16o0JkFlqRrh9wkqw2vGFNEVIKiMpiFBZTT9mDq8AugUE1SG181TQD+CymSAyvX1dVlaWjI3rd9++61ZVH7zzTcGKjHv5MDAgFUbICNg4/j4uM1JiWOkxfyQ//u//2tWlIgLQAlLSriARfozZ84YqAS0LLUC9LZQ7zbOfpwBEMCjShaVOu/dk7kNmVVQuZbbMovKesu8VzoMcmkRSiwqU+b6lRaVe6nWnOfQ3rDuDyqDOSq377J0KBShfrwdo549cLS0rp1w/TqUXItK1y3QLi+b+Q1z/QqLytvPVg4RVObkKEDl8R5ds3JuTC0q+7v0tzCjoBJWlXT/Wk+7ZBoqQAWoQC0KlD5PK4FKm6MSoHJT53y2D0fih5V4GnuuBJW11BrjtJICBJUx1macHdxKxSKorKRK7WEElbVrVRqToLJUjfD7BJXhNWOK6AoQVEbTkKAymn5MHV4BdIoJKsPr5imgH0FlMkAl5pxEnwiuXDGv5PPnz21FOKwoYU2J+oIlJEAkwkZGRmzeSYAWuIzFXJWY0xLWlBiUR1rMSwk4iRVWmNlsNlbLojj7cbg/UMLaQCUgk7fkvbc2HlYSxdNVC0e2getXt6hM0/VriX6tsov2hrU6qFw316+v83A17EOfuz2Slrcj18fasx64JZ8f+3k9Ybuesw7dSloNEDtT0jygMp8zUPlcXTTffLggsKh88GrdLCrzbWm9I70hLPpH5TpV+/sLIuJ/V8T/wP14Owb2oCc0Q76wqOxUi0oDlce6C6BSLSr7u6U9lVFYGVhUel3szIlHVIAKUAEqEFWB0udpraCy/LngZfBwP8YzxJbC46D4HCkE++nCoT1FACo72oI5Kq+M98g5/YBlYiQrQ2ppn8l0SEo9kvgzwbeenlsq0IwKEFTGWGtxdnArFYugspIqtYcRVNauVWlMgspSNcLvE1SG14wpoitAUBlNQ4LKaPoxdXgF0CkmqAyvm6eAfgSVyQCVDkvQL4JlJdbl5WUbjO/s7DQrSMSBNSSOASBhLekuXGE9CViJrQ+4oH5xHnASK+IDYsa5xNmPQ3lBHyuDypVdFpWIXstSPuDl6aqFI1uCylqUbe44aG9Y/W9v/fVrefpqWW7cn5c7NkelgkrMiQpQaW0tGCH1dlOtHbkq1p71oPTv0c/ZtpCRY7hSUHlCLSrfPdWncy3qwOqRrAz07gRuSO/57sjzAA5cN2zzAJW5dXFQCYvKXaCyIJSXt1yX3UV2RfwP3I93xkQ+u0BlLywqu+XcMVhU9spIf08AKgtzyXoZdubEIypABagAFYiqAH6T/Xm6F6jMFSwq8Qte/jz1Mni4HxNUFpXgDhWoqgBBZVVpwp+Is4Nb6eoElZVUqT2MoLJ2rUpjElSWqhF+n6AyvGZMEV0BgspoGhJURtOPqcMrgE4xQWV43TwF9COoTAaoRF1gzefzZgkJa0jsI8xdtWIfA+1w64owzE+JfSyICxew2JYupXGRxuOXxomyH2c/DvcXD6h0wOF3Vhl0lA+E4fJYsDVQmXKLylTBojIrk6M9MjKQlb5sl3Qo9PW6QTpCEKjQPAvaG1YfWC0Hlc8XAlC5XgSVwb15PVt7LbndsOHbSXWwVg/a2rYtKpsBVEK3TQWVeQOVK/K1WlTefg6LSnXRrHNUFi0qC39YwV1u3/WuveKfqe/s/XcM/QPd3KJSXb+WgcphBZUpWFQSVO6SmwFUgApQgTgVKH2e1goq7UdcC6GPP1uKv/r7PQaKEavfQanrV1pUVteJZ1pHgaYAlfihKF0hPzqneIn2F+lKVeI/MNh6XN9Wih81LM4ObqWyEFRWUqX2MILK2rUqjUlQWapG+P1tULkgf/p6Sh68WCz8nqEjHz6/WlLgfSf4/duSbFdGjg/1yMWJAfnJhVE5MdJTSxaM0+QKEFRGq0CCymj6MXV4BfCbTVAZXjdPAf0IKpMBKr1Omm0bZz8O7VFfxGKwqCwfwar84lj+PonLY8GWoDLQopX/R3vDmgRQCZ0BKjP63UFHwfVr0i0qi6ByQy0qZwEq522Oygcz5aAyaEWV/wpLWlgxgu/s/XeMukNM/B0XXb/2tu+wqCSoLNGXu1SAClCBBipQ+jwlqGyg0MyaClRRIPGgsvRHAl/WYgVshPsffH1bDTwinX+Ri5dPfCXqX+xWS1NFo5qD4+zgVrooQWUlVWoPI6isXavSmASVpWqE3yeoDK8ZU0RXgKAymoYEldH0Y+rwCuC9laAyvG6eAvoRVBJUenuoZxtnPw7tMRyo3Aky0FfFYvnsuJnawAeSB0XQmfX0IKW0sqezXYaytKjcIWeLHKCdYA0LKht1+2h/rQUqM+qtTz+SN8Hwt11QrvB3WgzwcBe2/M/Vj/18cYsTCis1fbp9U7owR2UZqKTr16JY3KECVIAKNFSB0udpzaCyUCL/mS9/HEQpMC0qo6jHtM2oQKJBJX4gABtf6zwLmKcEW7gCgjUl5ibp6lJXNQVg6RaWnmZjY0PQ4VxZWbE8MI8J4nqapLsMqtSYCCorqVJ7GEFl7VqVxiSoLFUj/D5BZXjNmCK6AgSV0TQkqIymn6eOs5Pmeday9U5iLXGTEgfvrwSV9dcG9COoBJza9iLz6tUruXXrlly/fl1+9atfyc2bN03gt99+Wz799FP5xS9+IZcvX5YjR44UP/x0QObb+muk+VK2EqgM1Ae8Cqy0ACqzat5moHKkUy6doOvX5muh1UuMv3usVUHlvLp+XVWXymWuX6vnGO0MnsFptajsdIvK08meo3J/i8pSUKk3hz8sLLGBykJ2mu82qOQclYEq/J8KUAEqcLAKlD5PCSoPVntejQpAgUSDSoeUi4uLMjMzI9gCWAJKDgwMSH9/v62AlgCRCC9Ns7CwIHNzczY/CiBlNqsTuGu63t5e6e7utjRxNoM4O7iVykVQWUmV2sMIKmvXqjQmQWWpGuH3CSrDa8YU0RUgqIymIUFlNP2Q2gZOo2dTVw4YJG020AK9CCrrqm5LBP0IKgkq629BYh+4zs7OyvT0tDx69Eg6OzsFUPfYsWPFKUdqzR/tETBjazMva+sbMr+0Ks9mluXOswV5ML0iT+bUpeSyntvQj3I3NV6NGSPb0sU5yV7h+D00UGkWlWk5SVBZKmFL7NvzVhvB/qAS7S3+Wy5tlsHz10Flm4wPdci7TQ4qN9vSZlHpyuEesZTeN449HPtY7HdAt/4+4sd2ssJ/SE9QWUEYBlEBKkAFDlAB/Fb787RWUFn++x9ncWFR2dGWk+MDKeEclXEqy7ySqkAiQSV+GAAcAf4AJ73TCPAIS0mcw9rX1ycTExPWgRwaGjLXrgCZgJqPHz8WxLcv5NT1ay6XM2vK4eFhGRkZkdHRUQOWsKz0l8eolYTyAoxOTU1F7uBWKgtBZSVVag8jqKxdq9KYBJWlaoTf3waV8/JHnaPy4YulYABfe7c+wBQ+171ToOMcvGBxjsq9lWrdswSV0eqWoLJ+/TBgvqkD73h2YAAe7y7lg3n1575/SpuPTT9cS6fabXC+Ub+z+5ckXAz8ZhNUhtOsNDb0I6gMnv3o12ClRWVpC9l/P84PTtEe9wSVsxsyo6DydU5/K/V3stYF2ZYu/vtWKTwoAly/OqiERSVBZal+rbKP9obVB1bX1QvV01fLcuP+vNx5viLPYVG5AotKHWMpa0NxaFCapQ3W6n9w/dqZagFQuaK6AlS26Q0VFh+QLr1vnPJwj2e/Awgv/KH6sZ8v37bp29JOUNkj545l5dxYr9D1a7laPKYCVIAKNEaB0ufpNqicl3v6odmz+ZzMrGzKWq5dNjb1V9t+3/W9O1JR9n6aEFRGEpeJm1CBRIJKQEW4eZ2fn5cXL17Yl634uhVh+LoV4BKujAAZ33vvPbly5YpcuHDBziH+nTt35P/+7/8MdJ46dcrmsnz+/LnBzcHBQRkfH7f4x48fN8tKzHUZx+KgEmV9+PBhpC9xK5WHoLKSKrWHEVTWrlVpTILKUjXC7wMYLaxuyN+ezsufCCrDC8gUdSlAUFmXbMVEBJVFKULtYGAc7yrr+ru38lrf5XSLZ4jBykIfzAfWQ2W8T2RkjWtjQD6tkLKzIy3ZrrR0ZlI2P1sjrrlPkUKfRqeYoDK0bMUE0I+gkqCy2CDq2DlIUPlYQSUsKl9v6Me0ISwqw96W/y5uW1RmaFEZVsQmiI/fv9KB1d2gUsdVDFSKgcrC4zh4cOr91QrS9pPC89GM9VncJl3p1gCV+baU3pHPUbmfCvWfJ6isXzumpAJUgArEpUDp87QcVD6FRwz9gGUt16agsr0AKrcNEPDehaVa37P8fHBcfCoHiRV7enqctzkq2/O0qCyow03rK5BYUInOIkDly5cv7YtgwEkATLhvffr0qXz++ec2X+VHH31ksPKtt94yMPjkyRP54YcfivOw/OhHPzJLygcPHsjS0pK5ez169KiBSlhjYl4WuI4Ns8CaE3NlwrrTLTbxYwaQimvAohNWlci3XpdBlcpDUFlJldrDCCpr16o0JkFlqRrh9wkqw2vGFNEVIKiMpiFBZX364T0lr+YaS/pxxsuFNVlYWVfXhgGsDDpihXyjfXa6q3DWoURHDgOjmbQM9HTI6GCX9HVnJKOWle0gmAlfcA8ElfVXEvQjqCSorL8FHazrVweVawoqw7h+DXt/+N3Fzx9BZVjlmis+fv/sOVjwInW4oDIYcMUclQGo7Gxu1686IA1QqX9JDW8UBJUNl5gXoAJUgArsq0Dp87QaqFxVUJkrsagsz9RBY3k48g4W75v68c6Y/uEP4m+DyjRdv+6UiUctqkAiQSXgHyAgwN/y8rLNSwlwiT9SzDV57949+fd//3dzswpAef78eYHlJCwsATFhVQnICdewOI/5KGFRCRdICMdclnD9ClB57tw5gTvYWheUwd3Lwi0tyuXuaAFScYwVceCOlqCyVmUbH4+gsj6NCSrr081TARhh8BkWlXT96qpw22gFCCqjKUxQWZ9+GHBfz+XllULKe88X5fnsqv3+ra3n7R0OXTHrlnnfrL7L7EoFz4lwNwso2atwcmy4Ry6c6JfRgS6zqkzpe1+1DuOuzA4pAO+XBJX1iw/9CCoJKutvQckAlf7T6MNWfuz35eF+7OfLw/28/lkEoFJdcGY72tX1Ky0qXZtW2uL3D2t1169uURm4fkW7CLd4gmotzsODXNvaSi0qmx9U+hyVAIm2FG63XMdd7xllsnl8V8tPe10g/0z7prrMzcvR3pScP07Xr64Nt1SAClCBg1Kg9HlaC6j033L/bY+7nKWg8u1xfS4cz8r4SFaG+rrVEKtDUuoh0sGmb+MuA/OjAgepQCJBpf8wwHIR8M/npIQwgJG3b9+W3/72twYkL168aJDy2LFj9nIOi8qVlRWDk7CcBMQEqIRFJmAlXLLC6rGrq8tcwMJ1LFzA1rqgbICncO+K+ShxLVhXelnd0hLlxlyYBJW1Ktv4eASV9WlMUFmfbp5qG1QumOvXBy8WgwF7faPZ1aH1RBG3eFkKfkc5R2VEKZs2OUFltKojqKxPP39ePJ9dkW8fzskjnZN3dmnd3MDqN2g6BBd05eL+7UPegKQdOinWYLZTTh3tlR+dGbJOXLe6gcV8lXFfsz6FqqfCbzZBZXV99jsD/QgqCSr3ayd7nceHpugvoo/36NGjSFN4oD3qi5hsbeZlbX1D5pdW5dnMstx5tiAPdI6jvSwqMdAV/FIWPuwoKbSHe1BpXA8rblEEPcBvH9xwZjsJKovatNgO2htWgspwFVuq22Zex5w21vUDqxX5+uG83H62Ig9mAhd/tYBKG6C2/0rK4H+whXD8LGDxaH7aw9yikqDSZOJ/VIAKUIFDUaD0eVozqPTfd/+BL5S8+LtfY3ilGyaorKQKw1pZgUSCSgjuL46lW4TjC4Fvv/1Wfv3rXxuoxPyUk5OTBgVhgXn//n17SYe1pK8AlQCKsLb87rvvLB1e5DFX5ccff2zxkHctC8oDa0l0ZGFRiX13AYstLSprUfFw4hBU1qe7Dzy/mIeFzIIOsizKbbUOnNPB57R+od3eoNFfPOvhRrBL5xgb6u3UAWf9emisX04e6ZVjw93S25Wp74YOOBVB5QELzsuZAgSV0RoCQWV9+uF5AevJJ6+W5ev7M3J/alFeLap3jDUdAFSQiN9166eVddbqu9p2qi0FlTl9r8OclHhenB3rkw/PjcikAsvujsD9a4MeVduFiLiH90uCyvpFhH4ElUH/CX0lrPAkc+vWLbl+/br86le/Kk6LgY8oP/30U/nFL34hly9ftmkwPA22WHxbf400X8pDBZXafou/j3VIVxgfK6b0n1jN1upSv/NVUJmSYVpUFjVqpR0fL6kNVAJqFmB4oeGUPx+3tvx3YGfLQjos5fGDUPz+BOdwfnuOylawqKxtjkrXxXVyXfbbQlYoTlC5n1I8TwWoj+6FoAAAQABJREFUABVovAJ4pvrzdAeonFKmMB98wFJ0/Wq/3CjTzudl2FLufn5sP4dT6qWg0+aohOvXrFpU9sgELSrDSsz4TaRAYkFluYb4sYCVIsAgQOW//uu/mhtXzEF58uRJ6e3tNUtJzE8J167oeMMdLCwtYT0JS0eAyhs3bgisLgE1ASp//vOfG+gsv161Y5QDecEqEx1at6b0cFhbLiwsWNn6+/tpUVlNyEMIJ6isT3SCyvp081QEla4EtwepAEFlNLUJKuvTD8+LVYDKl8vy17uv5J6CSsxVufy6ACoLg5j15V49FQYFcW2AyuG+Tvuo5ccXjphlZXcnQWU15dClxvsroBSsTkdUu7PH++TMsV45dazfjjtUU8xv9/+z9ybMcSNH+neSfZLdJCWRkqhzqJE0Gns8tvf1bnjXfx/7JXb3K/pj2A7HOtYRdng8mnt0jUa3RIlXk+yDfPPJQqKrQTSJi2R3T0JqAihUoVEPqgpd9UNmjfqCfBio7N9P3FMDlelK7WmByi5e4kAjFixZals/tTsJzuHqt+f6lS0qHais053LDbp+fpYWFxo016izNXpFvBYpoNa1XpOtR1sBlB98dGA1fo7KLrV5DumueDcI8hMUHB0g1VzmAZU4B85X4ZdZJ2WOyj2eo3Kf56g8qm6qjl51VkkPXeM24NwCKqcC169z5vr1UNHsoClgCpgCx6SA/zwVUMkeMb54vEYP2COGA5Xc39U5KqXl1l9d2S/o4PPDPXHgSt1AZXZdLeV4KjAWoFJ/fMOCEZARbwf/7W9/kx/k//Iv/yLAscx+mVdXV+nLL78UUPmTn/yEbty4IW8JA1TC2lFB5ffffy/A8/Lly/Tb3/5WgGaa26duXrFGhwDXhwUdXMyBiYEBuJmdnZ01UJlG2GOOa6Aym8AGKrPppqkMVKoStj5JBQxU5lPbQGU2/fC8EItKgMoHzqLy9QaDSraohIU8fi6hI+a6Xtm+Iy4Vj/OHoPIsQCXDtl8wqLx+ni0qDVTGSSZh+PWK37AGKodKdOQBlGnMjVpna7XLPDfqR++dZdA7RxfPzlDjBD0/aF9E76eByiNv3YEIJw8qu7TT2WNwFFi4SV10l8U2sbLBRw5cZ5IApHb1G6ByiiGkc/1qoDKJeuMXB/Uen6NA5S6DSn5My7PYz+XBAVL/qHtuIwTtHRaN7/b64bqP4z9EUKn5z7rmgTm2RDVQmVU/S2cKmAKmQBEK+M/TZKCyiG8dfo4yLCp57uLlhTJ9FFhU2hyVw/WyI+OvwFiASgBBWC4C/sF1K+YNgetVWCzCohJzQepxBZVwa7SyskKYpzIKKgE7YZkJi8rf/OY3qSwq9Zaj8You2sF98eIFAYbie22OyqhKp7dvoDKb9gYqs+mmqfqgco3nqHxJNkelKmPr41TAQGU+dQ1UZtPPB5WfMqiEReWbzV1q7XSoh99N/P84LHUAQbs8AFvjOSoBKt9fnqd/vcWuXwNQaXNUxt9P/JLF71kDlfH6JAlFsTZQmUSp0Y+j/biTm6MyAJXcdqEeoj4qACoWVFIAKtX1q1lUjn5pTHeFUn64DBmozK7b4XNUJrOoTPft0dj8LOZ7WJ52A9IXzKIyKpDtmwKmgClwIgrgmarP08NAZWcPdvDuxbLjvDCxqGRQeclA5XHKbOceIQVGHlSikYCb1nfv3tH9+/fpr3/9q7hdxfyTcO168+ZNAYKvX78WgPn111/LgMsHH3wgoHJ5eZkwRyXOAYvKu3fv0rNnz8TCEhaV//Ef/yGuY4u4J4CfuE6ASsBUA5VFqFrcOQxUZtPSQGU23TSVgUpVwtYnqYCBynxqG6jMpp+Bymy6IRV+79oclfn0M9evffBsFpXpy9LpgMqevGTB1d8twXiXDntpsOZGw3Vfjw8LR7sCi0rMKY85Ks/KHJUGKlW/SVnjPuOjA6uh69dHa3TveYuevdultVaXdruwqNRSkyb3WsKiaTV88FwA7hV2GV6vTNGVsz+cOSoHVUi7Z6AyrWIW3xQwBUyB41DAf54OBZWdKVJQ6Z6M7q++kItz5Fn0xTWcxr3AsmegMo+glnasFBhpUAlLSgBGWE/CQvHRo0d07949qlar9PHHHwukxByUWGBt+d133xHmqMRclgCZ169fF5gJUIk5JQEoATIBNbEAVP7iF7+QtQTk/FNkBzfuUtRi4NHLTfrLFy/o3tM12mArhTa7DdJ2UBu0uPRZw9DIwi0RLBIa7ELt9pUF+s+PL9ENdq2GeYuO4zuzXuth6QxUHqbO8GMGKodrk+SIgcokKlmcohUwUJlPUQOV2fQzUJlNN6TCby0Dlfn0M1BpoDJ7CXJTeKDPeVIWlatbDI46JwQqub/WqANUVunaYs3mqMxTUEYwLZ4f+MSDyq0IqDz+DABfiuvXEFTO0+1LDbq61KCF5gxNlyo8VU9JXi7H1ejA7vFf2eA3+LrFWlS+6dDb7T3amyqz1czRc1QOnn3YXnTwWmGvgkq4ft0jZ1E5Qzd5zuibl5q0OD/rdCs53U5Ls2G5snBTwBQwBSZFAf95GgcqV1s8R2UIKgdz3W/RB8PT7vnnca5fGVSeMdevaXW0+OOpwEiDSlgoYr5HAMrPP/9c5n+Eu9dLly7RrVu3CNaSjUZDwOSrV68EZsLqstVq0dzcnLh2vXPnjswVqfNGAmbiOKwdcR64Zj1//nwhd89AZSEyHttJDFRmk9ZAZTbdNJWBSlXC1iepgIHKfGobqMymn4HKbLohFTrFBirz6Weg0kBl9hJ02qAy/k38KIxAO+Evejwarp7IEH2aE5TkZVN2/doEqIRF5Sy7xp6lxYUGzTXqVK1UOM7pgyM/b7adXAHcf3yGgcrnAxaV3E4Ep9ZyM7QcJboEHU51kbGHl5jLJaIZdscOi8qfrjhQeWXsQGWb3rZY1ynn+lXl0Je0I9XxwMvbenwwvqofni1M51y/6hyVZbq1bKBSVbK1KWAKmAInpYD/PD0AKt91yIFKCiwq3TNV2/kjr1EfAfro1P1oQu+4s6jcN1AZ1cj2J1aBkQSV+JENa8rV1VWBlJiX8m9/+5vMQwmXrnD5CkvKc+fOCahExwpzVOItWMw/CSgJC0rASsSHRSWsKOGWFZCywp0xwEmAymvXrtHCwkIhN9hAZSEyHttJDFRmk9ZAZTbdNJWBSlXC1iepgIHKfGobqMymn4HKbLohFTrFBirz6Weg0kBl9hI0GqASxAJjU1GApPnScN0fCpjkJO48cP1aEtevZQOVKtyErVEu8EkCKrt7/cxreRpajvpRU21hwBauXwVUnht3UKkWlTpqfLgUOlitkPLw2O6opnGgUueoZFB5kUHlsllUJtHQ4pgCpoApUJQC/vN0EFRu0dMQVE5Rm5+nOkdlsidEiivUEzLINFCZQjeLOhEKjCSo7HQ6tLW1JdDx008/pX/84x8CKgECb9++LZAS8PHMmTOyDei4tLTELkSmaX19XVzA/vOf/xQoCRgJV7Fra2vc95uixcVFcQuL88AiEzATx4tYDFQWoeLxncNAZTZtDVRm001TGahUJWx9kgoYqMyntoHKbPoZqMymG1KhU2ygMp9+BipdOUJ/Bx+8uPnVV1/RH/7wB/r9739P6FNhgTeZ//qv/6Lf/e539OGHH0ofStNgjUXXsvMD+VNkP04AEODRXo922h1a29ymZ6tbdO/ZOj162aLv37bJuX7dC+ao5LjQvSCtcR6cD9dhoLIgUUf4NLjP+AwDlc/e+nNUomw4Ew4tb25vePmLHtf9YZKgGZkMUMlTELFFZY9gUalqDcu1C9dYR2mkZ5H4aLMRwPcQA9L1co9dvxqoVI1sbQqYAqbASSrgP0+Tgkr3qwvPUfcUkOds9EHAh9xRjd3fj+ZPkyJ+mV1j1EpmURnVyPYnV4GRBJWwjoRFJKwjP/vsM3H7evfuXbGahAUkACU60M1mU2Aj5prEnJRwA4u0cO/617/+VTrosJaExSUsKWdnZ8WC8saNGwI8ATh9Nzd5b3ORHdy4a7E5KuNUSR5moDK5Vn5MA5W+Gum3DVSm18xS5FfAQGU+DQ1UZtPPQGU23ZAKnWIDlfn0M1DpypFCRwOV6cpTkf041GdAh0NB5SbmqNxj12EOMuFqdQAr3ZUfjI3zyCUwXjFQeVCfSQtBefMHVtu7u/T0zRZ98miN7j3nOSo9UOksKnUI9KAScWVQY+sx3T+Y2oVMFqhkT18CKuFE+XgW6IVlioUtT+9RHXNUzhuodKrYX1PAFDAFTlYB/3l6NKgMri14MGp7jt9gcUv0uO5H42pyPB76FpUV+uhKgy3tZ+nKYoPOzs2wt8gqlcrl8AXDH+KLhlHtbH/8FRhJUNntdkk7i8+ePSP9wNLy7NmzMr8kGo9arSZuWxEGS0lYWeJNQnTM7927J9aVcPOKyopz4jgsLy9cuCAfgM4iK7Je88uXL+nx48dyfXhrGW5qYe2Z97sMVOarcAYqs+lnoDKbbprKQKUqYeuTVMBAZT61DVRm089AZTbdkAq/aw1U5tPPQKWByuwl6GRdvz5e5SlLto4PVAYGc9KuTLELzrK5fs1TNEY+LZ4f/sDq4aAScZGlvo2gPyCKIyGQ1A0EYolGdKHhX5wXg674OIvKEl0Za9evsKhUUBkVI8x2ARvOBmeKBRaLSgOVBWhqpzAFTAFTIJsC/vN0OKgk5/p1X58N0Qek7us1uHgKJvEdWIYyAk3OyQxUqoa2/qEoMJKgErARUBLgb2NjQ6wrsUZ4vV6nMr8xgG1YQ2IfABLWlOrCFdaTgJXb29thxUdDgOOAk/ggPiBmkYuByiLVLP5cBiqzaWqgMptumspApSph65NUwEBlPrUNVGbTz0BlNt2QCr9TDVTm089ApYHK7CXoFEFlj8FRQIB0uCtPPiQtD3DJGBe3KxgEE1BZtzkqc+s6oifA8wMfjI/g5ew4UPmuxWC8y25MYyx4vfHQwRxGC+TQiC4ZX4JATgGVDMdnKgYqBwUdvgep+6By3ywqh0tlR0wBU8AUOFYF/OdpMlCpD0e9LLTo0TAcw9QMWOsLQwCVLhx/wyWSVEBlGa5fzaIy1Mg2JlqBkQSV+mO71+vJj2384Aa4xALLRHwQBx0vwEr9IBwL0sEFLNb+guOAnPggjcb34+TZNlCZR73jT2ugMpvGBiqz6aapDFSqErY+SQUMVOZT20BlNv0MVGbTDanwu9ZAZT79DFS6coT+ET6HuX797//+b5mj8s6dOzZHZVDsiuzHoT5zpU7g+rVHHYBKGZTqj0zh/mGR8wTXd+gqiK9x4ELSnVL7y1PUYFC52KzQtcU63bncoOvnZ2lxoUFzjTpV+eVd9I31e3Wt57P1aCuAcoLPoaASFrwKKsMBVC1zrrz1c9m3tpSwSHnUObj6ppcuJc6GMyF6ZaxB5Tp9/WyLHr1Ri0oee3I5cxkd+neYnkMSaHRohm9g3ZxFpYHKIYpZsClgCpgCx66A/zw9GlS631tow90SeZ66H3hKJL3H5mD8SKrgd6FLJnNUAlQuMKi8aq5fj70A2BecugIjCSpPXZWMF1BkBzfuEsz1a5wqycMMVCbXyo9poNJXI/22gcr0mlmK/AoYqMynoYHKbPoZqMymG1KhU2ygMp9+BipdOQJkwsdAZbryVGQ/DvUZo0xHz1GpoFIHrPSadcgqGq7Ho2uOr0n4kIJKXAfKglpUGqiM6jYZ+7jP+MSBym95jsrnPEflu5SgEsqERQoEjRcp1xIeHNEIQTHFCkGIbqCShThsGajaARiGbtP7PEelgcrDpLNjpoApYAocpwL+8/RQUMl2UWjK3bNPG3V9MAZXyM9mWYLnqB7lp3YQwYVoOALliHe4zD/qagwqL7NF5Y8ZVN5abtgclYF6tppMBQxUFnhfi+zgxl2Wgco4VZKHGahMrpUf00Clr0b6bQOV6TWzFPkVMFCZT0MDldn0M1CZTTekQqfYQGU+/QxUunJkoDJbOSqyHydAB/Bor0c77Q6tbW7Ts9UtuvdsnR69bFF/jso+qByAQDpixQNV/cEsB4Bce+HnUV2J9cOQHGNcGB/D2FipNE3NWoktKqvOovKKWVT21Rr/LZQdfA4DlWtw/drZpy5cv7rSEWZcLST9shYe5A2vOEqw7vtx/G20QeMNKtfYorLVt6jcdxaV0MmNHTulojq4Y74Sh2wHkeUc/EfPBYvKGR6QvjBfplsXZ+jmcpNuXmrS4vwsTZcq/HGWz9DYFlPAFDAFTIHiFfCfp/GgsketzhR7xHC/s9Cc69NBfnThkvADLMeiqdHSq0XlZbaoDEHlUoPOzs3wdHZVKrHHSH0m6DrHV1tSU+DUFTBQWeAtKLKDG3dZBirjVEkeZqAyuVZ+TAOVvhrptw1UptfMUuRXwEBlPg0NVGbTz0BlNt2QCp1iA5X59DNQ6coRBinwMYvKdOWpyH4c6jMGqQ4FlZtd2uFRLrSbiK+DUrhqHWiS83jZUDaB0/uLxtcwDGy5S3AWlSV2w9kU168GKlWjSVpL+eEbfjSo5DksQ1DZV6BwUMlDtpXyhMxRudWjHgFUuimGoJpWvygq1HBRdmCHQzRyNDw4pHXbQKWoZ39MAVPAFDg1BfBM1edpLKjk5wJAZXvP/dbChR4vqGSLSra0dxaVTWdRaaDy1MqHffHxK2CgskCNi+zgxl2Wgco4VZKHGahMrpUf00Clr0b6bQOV6TWzFPkVMFCZT0MDldn0M1CZTTekQqfYQGU+/QxUunJkoDJbOSqyHyeAkev0MFD5/eourQqoZHDkgUqFFZxUFt3XHGm47utxPxw8BOEIwwfbZbaobMCicq5mFpUq3gStk4DKd1sdnqNyj3oCKl3mo+xM96PSSHnmQAXiuh+Nh31AT5S5Cpe5mWqJrpyr0U9X5un2JXZVxwOrC80ZZxk4ffpzovq67fW61Ou02U1ui+5+B4tKnqPyNeaoZFDJFpV7HqiMy3dcmNbLsJ4iUgRUiuYcwWm/z65fiWYqgUXlcmBRyVaVZlEZp7CFmQKmgClQvAJ4NhwNKonagetXXMGw5+fBq9OHgKbQ/cGYgUNwPu9+36ISrl+vGKgcVMr2JlEBA5UF3tUiO7hxl2WgMk6V5GEGKpNr5cc0UOmrkX7bQGV6zSxFfgUMVObT0EBlNv0MVGbTDanQKTZQmU8/A5WuHBmozFaOiuzHCcjhOn00qHSuX0EUMVQVAo1g3Er3NUccbWDR47HhHBfXgfIgc1TWygYqB9SbnB3cZ3x0YLW9u0tP32zRJ4/W6FsGbs/e7dJaRlDpypYWvMMHVlVRlDlx/cqg8uq5+oSASuRd8685PXzttDtYr6OptB7j7DJHJYPKi/MVtpoxUBnVyvZNAVPAFDhuBfzn6XCLSoBK99tNryfdE0JTxa/9p27o+tVAZbxYFjpxChioLPCWFtnBjbssA5VxqiQPM1CZXCs/poFKX4302wYq02tmKfIrYKAyn4YGKrPpZ6Aym25IhU6xgcp8+hmo7IMpgAJz/ZquPBXZj0N9Fvg4ZI7KvkWlDyp1WArXrcNdfpgfrnnT4xp/MByXMS2gEhaVAJXm+lUVmqQ1yps/sHoQVO44UNkZtKhMooGUsKCYKVBDueICfiA52h0s+CsWlWzFOzGgch+5cvnToWm1g0SesWi420v3V87Of2BRWRdQyXNULs+6OSrNojKdmBbbFDAFTIEcCvjP0yiofPK2Tatsab/dCUAlPwrxNHRPh6O+1HkcQCz3HOV0QxL6xw1UHqWrHZ80BQxUFnhHi+zgxl2Wgco4VZKHGahMrpUf00Clr0b6bQOV6TWzFPkVMFCZT0MDldn0M1CZTTekQqfYQGU+/QxUunIEWGCgMn1ZKrIfh/qcGFSyO86DgENHrqIwSMM1f3o8PhyXYaBStZrcNcqbP7CqoPIfbFF5TywqI6AyWmyi+4FUGqwjqgoipXzHyKnHURonBVSuYo5Kdv2qbviQbdUlvtbFCJMwCAPW1en9AFTCotJAZULpLJopYAqYAoUp4D9Ph4LK9n44RyWeCdHnQZqL0WdnPw2e6W4Pz4Xy9BTVysEcleb6tS+TbU2sAgYqC7y1RXZw4y7LQGWcKsnDDFQm18qPaaDSVyP9toHK9JpZivwKGKjMp6GBymz6GajMphtSoVNsoDKffgYqDVRmL0FERfbjBOQAHg2xqHwsc1TynIHtPerEzFE5FQxQiRFXjkzhOgZBpc1RmUPOkU2K+4xP1PVrH1RuD1pUhqQtGFrltLIkMe1ARI3vUh34i0HXsQaVj90clQ9f8RyVDCq7XBH3/cqoI9KhjgckkAC1uOS7E0RwCTW5pgqP8gEDlaqKrU0BU8AUOB0F/OfpMFDZYlDZwZzP3IBLGx405PoYHfaYVCiJ78AizwNNFGY3OFkQAxaV9TLRZbh+vWpzVIYy2cbEKmCgssBbW2QHN+6yDFTGqZI8zEBlcq38mAYqfTXSbxuoTK+ZpcivgIHKfBoaqMymn4HKbLohFTqsBirz6Weg0kBl9hJ0kqByixyo7DKodK5fUf8xLKVjVYWASj4hzmqgMk+pGI+0Un64DCUBlV0eWHWjql6BCwZMwwLoZ1vGS+UPErhRVY3vx/O2JwlUOovKKdpjUBkCRt3wZPGyH25qisSgklNWS2ZRGQpoG6aAKWAKnIICeKbq8zQWVG51qdVhi8pe/3Gqz1X9HTfsMXno40MPRvIMi0oHKqsMKhtsbd+gK0sNOjs3Q5VKlUrlsnhSQTIFoZFT2K4pMFYKGKgs8HYZqCxQzGM4lYHKbKIaqMymm6YyUKlK2PokFTBQmU9tA5XZ9DNQmU03pDJQmV071c9ApStHGKTAx+aoTFemiuzHoT7jNft4i8ooqGTXr4jO/3SMStfgIMpC0uWmHxtloVyaomatwnNUmkVlX5nJ2UoLKocNoA5VRAuhFkyNqCcKRmb1MHadRWV5POeo9CwqY0FlWCs1x4Eg+oZBWJNVqCFr1VWT8+mq4RyVnuvXS01anJ+l6VKFPyVp320weoimFmwKmAKmQE4FDgeVu4TnQggqg3Y88jQYegXa7Gt83XcJBvcQhngCKitTbFHJoPJaACoXDVQ6zezvJCpgoLLAu1pkBzfussyiMk6V5GEGKpNr5cc0UOmrkX7bQGV6zSxFfgUMVObT0EBlNv0MVGbTDakMVGbXTvUzUOnKkYHKbGWpyH5celDpbK504ErXGLI6OGyVPH/gSM6icooadYDKKl1fnKE7Vxp0/fwsLS40aK5Rp2qlQqUAgODsBkGSazwKMZOCyp3OHvUCV3WprjsCJMO0MeEou/hU2FfdTJVB5WKdfvrePN2+7CxAFpozDrhNO+CGc51WefN12+t1qddp0/O3LboLUPl0ix6+5gHpTcxRyRaVnCutl/1a2Q9BPigNqIyp2KIbW1TOVIguzvugco5BZaCbgUqR2v6YAqaAKXBcCuDZMNyiMieoDJ6b+tyTR0H/j8tS5NFSkRdYACprBiqP66bbeUdKAQOVBd6OIju4cZdloDJOleRhBiqTa+XHNFDpq5F+20Bles0sRX4FDFTm09BAZTb9DFRm0w2p0Ck216/59DNQaaAyewk6Wdev38sclV3aEdevsKhkUMkDVYFhWghEoiwjuq9jWdFw1QHnxWAYrNsUVF5jUPmhgUqVaCLWrvz0B1bbu7v09M0WReeo7INKV2JQ5rCEA6Ya4IJT/XVlMYB5vIMyN1ubJFDJOqVS5CDOPFBPvQCcWyxReUAaoPICg8rblxp0k1383bxkoDKl9BbdFDAFTIHMCuCZmgxUut9ueKNMn6MHW353GTinv2h8DdfD+jtQY+PZUC7huWCg0tfPtidbAQOVBd5fA5UFinkMpzJQmU1UA5XZdNNUBipVCVufpAIGKvOpbaAym34GKrPphlToqBqozKefgUoDldlL0CmCyi6DSoxyYVQqICEKRHSgys+XhmkcHNMwP56Ec7tioDKqyuTt4/mBjw6sDgOVu2xRiTkqDw6MBqUJ58khj9ocCnCbGFDZpS5bVKpjZr/eHSWVaqlpdD9MFwmYYotM5/p1SiwqDVSGStmGKWAKmAInpoD/PD04RyUsKnmOynYwR6XXjitk7F+otv7u93k//OAWP35l0XPoaXEGdlDgQOVZs6g8qJyFTKICBioLvKsGKgsU8xhOZaAym6gGKrPppqkMVKoStj5JBQxU5lPbQGU2/QxUZtMNqdApNlCZTz8Dla4cAUzhY3NUpitPRfbjBAQB+uz12GqyQ2ub2/RsdYvuPVunRy8jc1SGoNJBxcOuWgeuNA4GsBAWDdfjuA4DlarG5K5xn/E5GlT2AlDpCo2WGx0Y1YKk4VkUQ5nUMjfuFpUPXu/QW3b9ClDJfFffI8giS6I0uA8OVML1a9UsKhOpZpFMAVPAFChWAf95Ggcq3wSgstPDsxe/vY+2qNQrxLmx4DmJRfdlZ+CPO45feBWeZ1wsKg1UDihkO5OrgIHKAu9tkR3cuMsy169xqiQPM1CZXCs/poFKX4302wYq02tmKfIrYKAyn4YGKrPpZ6Aym25IhY6qgcp8+hmodOUIgx/4GKhMV56K7MfJwBPX6cNBZUdcv3YBKoOBK0dCdHDKXb/uuaGt+DzFHcM5cdrpaXX9WuY5Kmtkrl/jNRznUHevDwGVb7fp3VaH2l2Gbjqw6mVYy5gGxZUnPXbUWs7F7U91AiwqASpXN3nuSgGVTqWoVkfpkeY4xq37oBKuX5vm+jWNgBbXFDAFTIECFMAzVV/86YPKdXrwcpOevN2lN/xcaHXYorLrfmfhKxU8Aiy6Jf5pob/3NL7uB4m8laY3UOmJYps/EAUMVBZ4o4vs4MZdloHKOFWShxmoTK6VH9NApa9G+m0Dlek1sxT5FTBQmU9DA5XZ9DNQmU03pEJH1UBlPv0MVBqozF6CTsj169PAopIHulY3GVTudqnTA6iUViC4fB2ccru6p0NfSfLo4gagkumHzFHJE98ZqEyi3vjFwfMDHx1YPeD6NQCVOx0HKmUgFYVEC9cRWXblEwOxLqLuR5PhuLp/rbKvutlaha4u1umn783T7csNurLUoIXmDE2XKgzQS+HArg7YRs933Pu+bns9BpKdNj1/26K7j9fo66dbpKCyuzdoUamyReukhqe9brkV0I4FNFCZVj2LbwqYAqZAsQr4z9M4UAnXr1tw/ToAKt016PNRn5dZr8w/T5lfODOLyqxKWrpxVMBAZYF3zUBlgWIew6kMVGYT1UBlNt00lYFKVcLWJ6mAgcp8ahuozKafgcpsuiEVOsUGKvPpZ6DSQGX2EnQaoLJN27sAR3vsVpJRBf/XgS0FIAfBRzQkPiZjK5ECp53mkwqorDOonDeLyjxlZFTT+sCt2+2SDyq/fbbF8G2bXZh2CL8N/Tkqw/IWFCPdP5DPSDHTAdRoPKR3JVQtKhlULhmojOoU3Ye80E1AZYmIq2owR6VZVEa1sn1TwBQwBY5bgSygMnpN/vN02DMzSRqcx81ROU2XzfVrVDLbn1AFDFQWeGMNVBYo5jGcykBlNlENVGbTTVMZqFQlbH2SChiozKe2gcps+hmozKYbUhmozK6d6meg0kBlnlJUZD8O9Rlmkgdcvw5YVLbZorInFpUKKvX6dYArOril4RpPj0fDpU7wH1yHA5UlahioVNkmbo37jE+cReVBUBm14B0uh4OOg8cVqg2GBntcEDVNtVRyFpUTACp7gUUlcon8Sf3GdqTiaTjiYdHjGq777ujBvzhdVUDllIHKg/JYiClgCpgCJ6KA/zw92qISz193WZFHQnitejwMGLIxLL2bo9JA5RDZLHgCFTBQWeBNLbKDG3dZ5vo1TpXkYQYqk2vlxzRQ6auRfttAZXrNLEV+BQxU5tPQQGU2/QxUZtMNqdApNovKfPoZqHTlCIPh+IzaHJWAKL1eT24yrm96elquM27wHvUBcTudjtQNxC0x+MBH85e9tMSnLLIfJ2CC83CqoFJYKYNKdhlWZt2aBirjb/wEhKK84ZMUVGJUNRhXPTT3Ch11jTT46L6uw3MFo6wId6CyzBaVM+Pr+vUVz1HJLv7Y6Jmtnl1up1gBlk8Wl92+ChrujgJUui0N13097taa3sU3UDmoju2ZAqaAKXDSCvjP0+Ggci90/aptPK5TWnRt1vlZET4fw0xoiEbCAQ3Dc8APd8fKJbh+LdEVs6gMVbSNyVbAQGWB97fIDm7cZRmojFMleZiByuRa+TENVPpqpN82UJleM0uRXwEDlfk0NFCZTT8Dldl0Qyp0ig1U5tPPQKUrRwryRg1UAjpub28LTKlUKlQul+WjwFLvvg4QIe67d+8Irizr9bp8ZmZmCGk1j5qmiDX6cfi+Fy9e0OPHj6lWq9FHH31EFy9eDKFq0u9BHgQG7fVop92htc1tera6RfeGWFQiPpKE41M6ZhUZr0IcWYJwPRyGRy4Q5zVQGRFlAndd+TkcVL4LXb/CotIVpEhx8oZKnUhavnSN+PhgX8MQU88THuOCPBmgcpvnku2yu9zBOSr7OfZV8JXAdtLFnQN1H1shqFyo0u1L5vo1qYoWzxQwBUyBohTAM1Jf/IkDlW/4edrCHJU999tNnqn6IBz2WDg0fFhilyOxqOSHg4HKou6wnWfUFTBQWeAdMlBZoJjHcCoDldlENVCZTTdNZaBSlbD1SSpgoDKf2gYqs+lnoDKbbkiFTq6Bynz6Gah05Ugh3mmDSpRptYrc3d2lzc1NWltbk5u8sLBAzWaTfPCo8RF3a2uLNjY25ANQCTiJuPPz89RoNARaKrDMUmrwXfj4i4LKly9fCqgEHD0RUNl14Eggjw5k6aXpfnCh4SUH4Xo4DPczxNvI4xRbVFYCi8pzPEfl9cUZ+vBKg66fn6XFhQbNNepUZX3VWhWnOPhGf+TEtjtSCmh51oFVf47Ke8826dlbhvCb7XCOSi4YAhejxUz3NXNavrDGMT2OfT2GuBqOtRybOFDJFpVsVYklfJnA7cnf/h9Voh8ybEvrrH++aY5cKRNbzkzTRQGVDbq5zLDy0hzPLztD06UKf47PqnzYtVq4KWAKmAI/JAXwTNXn6QCofLFJ37/b5RdYAlDJv9/2tNnXtf9whGgD4boTDY+qyyfRqLxpoDKqj+1PugIGKgu8wwYqCxTzGE5loDKbqAYqs+mmqQxUqhK2PkkFDFTmU9tAZTb9DFRm0w2p0Ck2UJlPPwOVrhyNCqgEYETfCHDy1atXBACIDwDjjRs3aHl5mc6ePUuzs7NisYj4sKIEYP3+++9pfX1dgBkGi2CNCQvHc+fO0eLiIl24cEFAZ9QaM0kJQl0DQMV5sWAfi4JKXOvTp08FjB43qNzmOSq7IahkqDiAf/iiIgNewaXK9eKPHpYc6KCWdwB5Q3molKapOVOhc/N1us6uOD+8bKAyFHECNnCf8dGB1YOgkq2TQ4tKLiiIL/9c5rXcOXw5KIgrY+6vHtf4YUw+Hxb81fZnkiwqO+yxei8YjXb5k+xCxoGlDx0DPSLH/cia1qVxik5N7bNFJbv4q/IclQs1tqg0UOlrZtumgClgCpyEAv7zdBio3Npl169iUclPRm7r9fdYuBHb/scGxmQJTsbdgvMKqKzBorJOP77WoFvLDbqy2KCzc/AyUqUSeyjBswmLrmXH/pgCY6qAgcoCb5yBygLFPIZTGajMJqqBymy6aSoDlaqErU9SAQOV+dQ2UJlNPwOV2XRDKnSKDVTm089ApStHCgpO26ISlpGwinz+/Dndv3+fHj16RE+ePBGLyJ/97Gd069YtgZWwksQ1az8Kble/+uorarVaAiVxTC0xz5w5I65Y33vvPTkGq0e4kE2zAFK2221xKQuwo8AS3w84Ct0AVGG5eRKgstPtSf1HHg4AoHDky+VQ4YbmVw9Hw3WgDO0K9DNQqYpN5hr32R9YPRRU8oSLAI46CApFtNwNhva1ih7X/TBGUABxTpRJlLlquUSztUowR+Uc3b7cpCtLDVpoBpaB084yEOdA/NNYfN32ejwXZadNz9+26O7jNfr66RY9eMWuXze61OHB6NBq5ojrdVnB/eCIgS6cwYHs4Xvd4sJxGJoaqByQyXZMAVPgGBVw7Z97weQYv2bg1NLiSXt3eu3+wAUN2fGfp31QuUb3X2zRE/ZQ8IbnLhZQGbxohtO41tzb0GY+PBIGDPnWwWCNjfPiN9yMgcpBgWxvohUwUFng7dUOtroMyjO3Sdxl2RyVcaokDzNQmVwrP6aBSl+N9NsGKtNrZinyK2CgMp+GBiqz6WegMptuSIVOsYHKfPoZqBwtUKnWkQ8fPqR//vOf9PXXX9N3331HcPv661//mj7++GNaWVkhwEcsgIQAmd9++y19+umnUid+9KMfiSUl4CXAJcAkrClv375NV65cEYtMuIRNs8A6E25oAVKxDXCJ+qfuaXEdb9++FTezxw8qGYLoQBdGpcKRrkNypKNXiKLx/TAvHDwEEERcv4pFJbt+XZo1i8pD5B3HQ27A+WiLyh02DcRzWoCkD8sGytGQwpRQGJxK4DiDysa4g8qXbo5KByqhixMKdQpLKKHbjfJIaVdwSEEs7tPBBYDSnRmwslJi16/VaVo+gzkqzaLyoF4WYgqYAnkVQFvU47cv8HHNUlzblPdboumDto4bPPZITyX+o21jNOZp70Mf9VAwCCo3GVTuEuao9C0qIaLmRZUMHhP9rMiB/utAYXx3A/rxdEtPwOkq5Wl+8UctKptmUaka2XpiFZh4UOn/INTG4LjupoHK41K2mPMaqMymo4HKbLppKgOVqoStT1IBA5X51DZQmU0/A5XZdEMq/F41UJlPPwOVowMqUZ7RL3r37p3M9/j555+LleQ333wjloq/+tWv6Kc//SndvHlTgCAsHGHJ+ODBA7G8BNAEgPz5z38ua+yvrq4KTMTcltevX5fPtWvXBHymKTl6XZgHE3ASLmcxIIVrAFwFEAXIBEA9SVCJsSoFIEPzoyNgGsEbyNIgWSOc4+I+hHNUhqASrl9ZQ5ujckCycd7BfcZHB1Z9i8pveY7K52+36S3mqGwzqOSyLvE5w1p8woKHQhi7aEw9rvsHI+MIxlwwsOpA5Sz99L0xtagUUNnxLCpdvrWeRuXScFUFOmPRMSjd1+O61uM4exWWM+z6FaDy1iU3IG1zVKpStjYFTIG8CqAdQn+txa7n2/zyioOV3FYNb9bzfiWnx2sYxIByiufDds+HerVEZW7v0P4d61dnuHpopM/TQ0Fl1z17gR+RCzwDhj4lg+dBeDx4YAx7LoSicAJYVAqoPAfXrwYqM9xSSzJmCowdqIxWZP1hF9Vd4+kax6URDBqEaPwi9g1UFqHi8Z3DQGU2bQ1UZtNNUxmoVCVsfZIKGKjMp7aBymz6GajMphtS4feqgcp8+hmoDMAUBn34c1quX1GW8VEXq7BQhLeZe/fu0d///ncZ/PnJT35Cd+7cIbhwBZAE0FQXsYhfrVbp/PnzYjmJ44CUOI65KwETMa/l1atXxSoTFpZJF1wX0r9+/VosOAElASh1QArgUj+YD3PkQGXijLqIyK+ByqSijW88rXNajg+AytUAVHoWlRgsTTo4zMVIFh1G0f2oYjiug7WV0KJyHEHlOrt+3aQHB0BloEOQcR1wVh2ieupxDdd9je+vVTc3R6WzqDRQ6Stk26aAKZBXAX1WtHa69Hp9hzZaHdpl9/OYg9e179pa5f2mg+lx/vL0NFUr/BJLvUxnmjWarVfEshIAc5QW6KTP00SgEr+1jjELBipHqXTYtZyEAmMFKtHhRecRDQfmIymVSjTNjV0UQOI44sKdj8ZHXKSpVLgx5O1omiLENlBZhIrHdw4Dldm0NVCZTTdNZaBSlbD1SSpgoDKf2gYqs+lnoDKbbkiF364GKvPpZ6BytECllmuAQFgvYp7Kv/zlL2Kx+MEHH9D7778voBJ9M502AzATfThASFhL4gNQCQvHp0+f0hdffEGvXr2SASS4fv3lL38pLmCTlhzUM+2v4ZzYRn8RA1JYY1+tKo/donKDLTjbfdevfGnFDXTxuQBFkN9pdq9W5r5vky0ql+ZrdG3JLCqTlpdxiYf7jI8OrJ4mqIRmGGfBHJXja1Hpg8q2Z1EpuSumWCi1DAa3scKnypao6vr1Frt+vbXcJLOoLEZyO4sp8ENWAM8IuJrf4d8dqxu79OTNlqzhEhxWlVNTPK5+jAIB5MHdK9q4s82qzFl8bq5OsKwEiMNzY1QW/3l6OKgMPBQU+fstRgQDlTGiWNBEKzBWoBIdR8wZgg4s3qTFPCV449YHj2hUcByd4o2NDXmbGB1PdHIbjQbNzc1JOkBLQM4iF+34amfb5qgsUt385zJQmU1DA5XZdNNUBipVCVufpAIGKvOpbaAym34GKrPphlT4/WqgMp9+BipHA1Rqeda7iX4Y+nCAkH/+858FOgJUwu3rysqK9OMAIR89eiTzUyIdjsPacnl5Wfpw6GMhDuauxBr9PIDK//f//p+4gNXvSrJGPxEvsuK6sA24gwVhsOaE9SZgKPqNx2ZRubpLq+yKc3sXbiV5oIsvwbkOOzwHyjY0lg7rxYUjjJsVcbVW5oFBByrrPEflDN0x168q4USs8fzwB1aPBJWI7+V8WDnSKNHjuq/HB87FBzHe7EBlla7ynKjj5/r1IKiUgXzJcDT3qoJb+1oMHonsaUToxYfkw3/cgDQsKmvs+tVAZUQ12zUFTIEMCsgYOcPIze0OvXrXoqdvWvTw5Sa9ZmC509ljl+D4rcBj42i88cOBF2xiCXbdDv/VcA3Q49Hw8Li0bnxO/j89tU/s+ZUucPv2o2tn+MWpBp1p1GimhrF55w1E053m2n+eHgSVO/05KkPXr64NL/KaoavcAv4jLsExR6W5fi1SYjvXCCsw0qDSf7sVb70CUr548YIbR/bbzx3XxcVFAY8AgoCOGh+AEi591tbW5K1dzD2C44CbcOODN2Tn5+dJ0xX19gY60XBdhGt8/PixnD9rBzeuzOxxa9Xt7dMjfqj85YsXdO/pGm3sdNi3ON7kcCmGPSDizpc0DA11lx9s8CGONyNvX1mg//z4Et1YngsmQU56ptONZ6Aym/4GKrPppqkMVKoStj5JBQxU5lPbQGU2/QxUZtMNqfBby0BlPv0MVLpyhH4NPqfl+jV6F9EPU1D5pz/96QCoRB/tyZMnMj8l5rDEC6joP62srIj7V7yYCqgIQPnJJ5+I+1f0uQAqf/Ob3wjQjH7nYfuoa/pBPGxDL/+FU1wPvjdrPw7nROdsf6/H1gsdWtvcpmerW9x3W+d+3BY99kEljxC66zk4ABjNR9DdC4NlEAt5CEPcBsLdJXDeePAPbjgNVEZEmqBdLc/DLCqfsevXd5vs1lhcvwbl38v/sHKkUaLHdV+P++WPq5IMZE8MqGTL5zaPv2AcRvOt+dX9OB0kLGFEaIYF7RBcv2IusnEElSiHPGTEY3JOL6xPYsG3cDMnoKPEGo4S8DiJ/Nt3mAJHKYC62OUXs94ymHz8alPGk++/2KRX6/zCVAdjvdz+8G8xuKB2y/C6KzG8aBpzIFzPsq8REYA2lMeUp/bo0tk6/XzljIwnn1+Ykd8n+O2HujsKi7Rl/BIbfr+eJqiEFng+mEXlKJQKu4aTVGCkQaW+2frs2TN5CxfwD/OTNJtN+tnPfiZv4l68eFFgJRo2xAcoRLzPP/9c3AwBTOJHH8KxBty8dOkSraysCLRUV7BFiI7vBxwFqMRbwWZRWYSqxZ3DQGU2LQ1UZtNNUxmoVCVsfZIKGKjMp7aBymz6GajMphtSoVNsoDKffgYqXTlCfwefUQKVgICwqPzjH/8YCyox9+TDhw/p66+/lqk6MIcl+mqYp1JBJfqD//jHP0JQefny5UygUutbtLRFXzg9eVCpw31uYMpd5+BVRofwNMWwcMBKN0dlYFHJA4Lm+nVQ00nYGwoqH67Rt882GZIzqNxSUOlecB60qRymgitZWr40TX8wOyYdR+bmZ0IsKlvsGpFfCmer5wFQGVQ85NNfUN+yLq7ddgPSjTEElW5gn11LslZtBuLyYfoxACsjeqXWaoi+CC4z4KizVVatUpJPiV+wxzVhgba2mAI/XAXY4yCDyg7Xyzfr2/ToxTo9eLHFc/C26CW3b602UWePW3UGlVxbAlR5sLIhJKxJuhGNpuEqNoNKF8W96FGe3qMafy6frbKl/Rl6/2KTzp+ZofnZGk/RpqAyehI92cmtXXuWAlRKJl1ODyoYFSllPrj9gkUlXmAxi8qU2ln0sVVgpEEl3r6F+x1Av7t374o7IHRkYQ35q1/9St5yvX79Oi0sLMgPEFhdAmSiIwzXQHj79tatWzIvJd7CBUiEGx9YY96+fZvQwYUrWADFNItabqq7IOxjwflxDRgYwPfB3WzWN3HjrscsKuNUSR5moDK5Vn5MA5W+Gum3DVSm18xS5FfAQGU+DQ1UZtPPQGU23ZAKnWIDlfn0M1A5mqBSXa0qqITnmzt37sgLp3DvqhaVAJWwqMSg8ocffkgrKyvSZwMwhKtX9K3QvwOwRJ8LFpXoD2IeyyKWKKjM88KpDJBznY6zqHzIFpXfw6JS5qhk16/dntR/H3KwBLL4YQiIDt/p8NewcFyHA5XOonKRQSVcv35orl+dwBPyF/cZnwMWlR6ofKsWlTxuIeUzUd5dydLylQhUopxygioPOjdq4+76tUVvuJ52ohaVQcXTeqpSRuurhidZQ2O0fRV209yowfVrfaxcv+rvvy329rXO5KO106U2t219l7lQgXOphSmJKNE4MQLjVqA8w+KoUa+IZVZztsrzfPK8d6zlNGtqoDIqpO3/sBTA1Gh7bFHfpTdr23T/2ZoDla926NVGj7Y6DlTyj4Wgrugvi4hKGhytw8PCg+RabeH2tTK9T7NlBpVnKvSTa3N0g0HlhbOztMDuX6uVMnvUcLA08s0nvus/T2MtKhnwbu3ySxl4GUPyryLopapI0XA9nm7tngtlA5XpZLPYY6zASINK7TACTn711Vfylu23334rLlz//d//nT7++GNaWVkRcAkoCaj53XffhR/MX4k3ctHRhAsfWDsiHuAkOrX4oJOL/aQLGi10ljGHSavVkm2YhCu8BFwFrMRxAFUDlUmVPf54BiqzaWygMptumspApSph65NUwEBlPrUNVGbTTweqnrzeok8frHJHmOcK58HRFg9c9dBT5f/HMWCkL3LVKtN0dq5G7y/P07/eWqTr55s870lFXNfzWNVIL/h9aaAy+y2CfgYqRxNUKjwBqPzDH/5APqhEXwxecfCiKV5MBagE2ATAxAcvpOLFz62tLQGUX3zxhcwfifuNF07/7d/+TdbZS04/JfqdmGbk5cuXuafwwPXx6PlwUPkGcxxhjsquuGMTqyOkwaA6X1JSIKRXL9+nO1gHDR7CHfwwUOnLM2nbuM/4aF0L56gMQWWL3nJ5w0A1ntNc0iT+4Y9F76huIiEW3Xd7KLDhgqKHMjdRoFIGo2ER5DLuZTfMd9wGqjSWoDr2ZYqcQDSTeKwbw7XZsQOV+7Td7tHqOoOPtR224G3ROs+FB0iJQfxocRFRCvqjZR+gslkv00KzRkvzdTrHvwUXGlWqV8sFfZOdxhQYTwVQR+SFsXaXXq+1QlD58E1bQGWrM0Vttqh0DZW2cpxXrbiR9qqvgh4YHtHFwPF9cc9cY1DZqABUlulHV5r03oVZuni2IaCyVq3I78Hj6Cf2rznZlv88PQAqV3WOSp7vXOeoDBp7VULFw2+5sP1XuY64BD2HRsc+3Pc3uH0zi8ojxLPDE6PAyIJKNA6AiuiYYr5JAEi4A8KbtOjQ/uIXv6Af//jHdOPGDbGSBByEy9X79+9LBxadTcxFiTjo4OLYq1ev5IO7B5ew6Bzjjd6lpaXENxTXBUAJq0mAT4BJgEvtGOANX/3AzSy+H+5p8bZw3kZXB+JsjsrEt2sgooHKATkS7xioTCxVbEQDlbGyWOAxK2CgMp/ABiqz6WegMptuSIXflwYq8+lnoDKADzwqgj7Habt+1YEe9Oeic1TCs837778vIBIvlGKKDlhMoh+Hfha85WCqDvTzZmdnpc8FgIj+IPqGsLLEcbyQeuHChewFx0t5kqDyMYPKVQGVgxaVuG9uSE+H93TIyrvQmE1oPbDweRCCcFgUVbjvjDkqF9nFmllUDig1ETu4z1rf8AJ1PKj0Xb86FH506QpiaEQtZ1ymBhYpfkEZ5EMoc7COmWWLymtLs+zib45usxXvlSUekG7O0HSpwmMjpXBsJO8YycC1pNjxddvrdanXadPzty26+3idvnq6SQ/ZNeJrWFQGLkyldkayftjXReUKa2m44VJDTpwWOowfqOTM8P/17TY9ebNF37HF+IPnm/R2q037bKGFrIaSab7DgEA9DQ92+wk0YPha7iHT0ArP7dmslwRQXj43Q5fOzdKlxQbNzVSHJ7YjpsCkKxA8GzA/5S7Pl/3qnQOVaNseAVRu7rFF5ZS4fuUGiNXwKqfsc5A2ZAe00oqraXR/MKJ7xaMPKpsMKi8xqPzw8iy9d36WLSrxXKjzSwX8YikDOfc80HMOnuuk9vznaRyofC0Wlezimp8N0gZ5F6ZXrmqIjLyj+17U2M0D6TmWWFSyxbiByljJLHACFRhJUDnwo5Hdk6Bzizdc8SbuX//6VwGBP/rRj8St68rKCsFyEp1xWF4iDkChPxclQCXe3oWrIFhkojMM2AlrSgBPvJGbdMG1oYMM8IlrwjZAJd5Skcl2uTOONfYBQA1UJlX2+OMZqMymsYHKbLppKgOVqoStT1IBA5X51DZQmU0/A5XZdEMq/L40UJlPPwOVrhwJ7OKRkdMGlegLAVKiD4aXRfHC6f/93/9J3+nGjRsCKrGGBxrERf8MfTlcN2AlPN5gCg/04+A1B8fxYmq5XJY+FkAlrC7xYmoRy8mCSp7eJLCo9F2/6gCV5kfHCXU/Ol6oxxHuD4KhDGAf7YoDlZijskpLAai8Y65fVdKJWPtjJ4eDysCi0i8sCRTQ6Fo+dX8gaVA4pf3h+QKrYgFSo6sMKn82rqDyxSCodAP5nOthQmi4CqNCabju6/FgjXqMKNBurFy/Bvd8f3+P27NdttTaoG+ertMX36+zy9wOTZXKwbx3QUaH5D8ix/Bd1dGPwdewz+OF7FCDZqtTbE3p4PjKxTm6eXlBwGVwmQEA8RPbtikw4QqgfnD9xO+MnV2ASn6RgF2/CqhcZQvLrT2CRSXmqHTtm1fJdPNAvY0GDI0o4qL+uTZun2qlwKJyoUwfXJoRUCkWlfwCS51BXIV/37nW0LWHp3V38ExVQ6R4UNlm168OVMJqHPGRx0OXQDaNxtN3ysIecWMXDUY0Z1HJoHKxTj++1qRbyw3ebrAXoRme5q5KJdYNzw8supYd+2MKjKkCIw0qoSkqGjqvgJV4y/bPf/6zdFLRccWbuOigqssgvGULEIkFx3EMnVh0cNH5hPtXndsEHecsc5ugEcK1oMMM9644r29RiX2AUsQ5e/asuX6VuzEafwxUZrsPBiqz6aapDFSqErY+SQUMVOZT20BlNv0MVGbTDanw+9JAZT79DFTqYMloWFSif6Qvd6IPB1B59+5d6TvhJVG4db1586Z4nmk0GvKiJyAl+muYrxJ1An05vJCKl0PRd4P1JV5GhVec5eVl2UY/r4jlxEElW2q1eOCw672RHw5gBRlyg3tuB4NWLEm4SNwggR+OCOg/w2YOGgJUlvkFXVgWLfG8d9cZHBmoDGWciA3cZ3x0YHW4ReUxgUqvAEqZ5T+wqGzUJwdUylyLyByWaEV1of1w3df6Oix+EC88LXRjy8BGrUTLZ8dgjkqv3L1ht6/fPF2jL7/foHJtUuIAAEAASURBVM8er9HLjS6Dygqb12LOuUAa1SPId+qV6sgJw1Oh7AWgkj290vm5iriTvHlpjj68dpYW2Q2s3hgbwE+tuCUYdwWkjjKoZLff27tsQQmLSq6nD1+16PFbH1RyPZWGqF/JtF3ymvdAjbD2BfuaJhruDiM9zoVfJQoqL82XBkDlGQZudbbA9y0qT7O++s/TI0Elk0rkXFRQKQJlBlaBPBrlIKhU/VwMfy+0qFycMVA5IKrtTKoCIwkqITYaByxooPBmIDqPsJb805/+JKASLoPQuV1ZWRG3qtqpxdwmcLOKuSFxDO6A4B4InWW4FPrkk0+kA4zzAVT+5je/kY6yfFnCPwCnOB86zNhGpwALACXeGgbEhHsiuCqyOSoTinoC0QxUZhPZQGU23TSVgUpVwtYnqYCBynxqG6jMpp+Bymy6IRV+9xqozKefgUpXjtB3wue0LSphFQlLSkBK9L/QR0N/DX0ovMyJl0nRlwOMBHgErMSLnpirEh50cP2YqgMvpAJ4AkhevXpV+nfoB6KPB4iJ40UspwMqGRyxpQPqPz5822SBqzQszi5SNuXPUeEaE+dBTxrnNFCpqkzuWsvPcFC5xXNURly/cgHpDwTrkKhqFBTEYBfnx6LxdT84HK4kFf9BvCpbeDRmauPt+jW0qOzxfIvIV5jVQjfUpSzOX+W5Fh2orNGtS7CcaRKg2+J84DKX2zvoq/ei0AtJeTIpdzwO1mO3uQCV99ii8mt2mfv5kw16sc6aTZXF/SsrF7RoKb/gkOiuRPJftJv8qbKlVpNB5YX5Mq0szdD7rN0HV844UMkuaEdFs0OyZIdMgeIV4LoBi8o2QOUOQOXWAKh802IjHLGoZDfNQQMXtnP6WIi0e8HjILxWjR8NDyMEG1NsOujmqOzRJa6nHyzzi1Ps+vXiuSadYdevM3WASrUMPN02Dm2bPk8PBZUddf3qwcqgvXMh7rcYJFAZdT1E3lA2/3jfotJAZSiQbUy0AiMLKn3VASoBAWEt+cc//lFA5QcffDAAKuEq6MGDB9IJRocVc5bcuHGDzp8/L6ASUBGg8h//+EcIKvE2L0AlOshFLOjgwi0R3MI+fvxY3vo1UFmEssWcw0BlNh0NVGbTTVMZqFQlbH2SChiozKe2gcps+hmozKYbUqFTbKAyn34GKl050gHZUQCVuAZ4vPnyyy+l/7W2tiaDP3iJFLASfTHAR3zg6hUDQ5iq4+9//7tATsRDftCPazabAjRhiYm+G+axLHKZOFDJo1xoV6bZDadZVBZZUkbvXAKMvIFVtaj85OEaffNsk56tOlC50+apapi4IT4WHWCOz1F/ODWIHsbX/bh0SIU6KxaVEwEqd2WOSrGoRIYPFy1OkkRhoW5l36JydEGllKGwzHUEVN5/zq5fn23RF0+32KKSXX/vl2mP1FIrkQwJIunwPUeVTQcqYanVrO3ThbkyvbdUp5vsGvH2ZQaVC3iGMNxlQwaUS1tMgR+SAu7Z4CwqWzu79Ootg8pn6/QosKjsg0p+LUrqBwBhoJBWNd3XYA0P9jX+Yc8FRJ1mUFmdhutXgMoS3V7mObMBKnmOSlhUzrBFJVy/Sj3lk55mfYVuyUAlGy1xXMl7oFNELtdMcf41XNcqo+4HcoYr/7iBylAW2/iBKDA2oBKdR1hURkElOqoAk1GLSswNubLSt6hUUPnPf/5TgCXOh87xr3/969QWlcPKRpEd3LjvQCPY7e3To5eb9JcvXtA9Ntvf2OnwGzLocLgU+qCIS581DA11l03ay/KGX4VuX1mg//z4Et1YnqMSd36P4zuzXuth6QxUHqbO8GMGKodrk+SIgcokKlmcohUwUJlPUQOV2fQzUJlNN6TCby0Dlfn0M1A5WqASXmfwoimsITFdBqwl0R9DWUffDdaQsJLEB15oKhV2E8iLTrGBfpW/4DisLvWj8f04ebaL7Mchj+ic7e/xvFDtDq1tbgssusfztz18uUWP3/A0IgOuXx08KnJgDl1DXIZYVPJ8geb6NU/pGO20KG/46MCqDyq/ZVD55M0WvWOLyp124PpVshMMHgzNWn/4VLc0he5rUg2XAscDAyhzkwEqt+g119N2Bx60OJeS8WjuVYV8a4yniG5ltaiE69fRBJXSvnF2sUY73+V2/fX6Nj14sSmg8qtnLQGVXQpApUqjBSUqoYZrvEPXcZGdS8lmZZ9dv5bovcUavS+gcoFB5SxN8/NmetpZoh56ajtoCkyYAu7ZEIDK7V16CYvKAFR+z65fASqjc1Rq9dSapvtppdH0WOMcAJVqUbkMUHkxsKhUUMmuwscKVO6wMRW77scYvXv2BQppxiPC6S6PnktEtbjU/SB1uPJPY6AylMU2fiAKjAWo1I6ugkq4V71z587AHJV4+xZv7MKtEH6kqyshuHdFBxidXlhUfv755+KWFVaaAJW//OUv5S3eIu53kR3cuOsxUBmnSvIwA5XJtfJjGqj01Ui/baAyvWaWIr8CBirzaWigMpt+Biqz6YZUGEwwUJlPPwOVrhwBduFz2haVWq79ddwd9uEctt3AmoMu6NNhH9N66Afn8NPEnTNLWJH9OFwzBq6SgUrf9asOZWXJwWAaDHKJdqwp5n1yoJItGGyOykGhJmDPrzMY44gDleL6VUElyibnm4uGLCiuWHTf7fl/tVwGEXmgNRqC2LgOnGMQVDbop+/NsXVbk64sNWihGbgw9cDRcdRn/+qHbfu67bH70l6nTc/ftuju43X6il2YPnzRB5U9bouKuU5VTq/KaYpQBypLNFvnOSp5PtkPWDN1/XpuhFy/Qjdp33jdZVDZYd3erG3TfQaV3z5v0dfPtunVJodnBpX9cqYqubWG+6FwiB24fgWobJboOoPKGxfZopJfrl9a0PJm7l991Wz7h6GAa+MC168KKtnY5dHrbTopUKlKlzyLyosBqHxvwKKSQSXPbSztLD9Iimlv9dvTraGbvvgz1PUrQCUbDIlFJbdB4fNTm6lIUx/ZHbC07D9R3XXqKfSqq/wbrlGv0BWbo1IlsfWEKzAWoFIbCYDKP/zhDzIPJEAlYCTmNIEva8yBAlAJ97DoaF68eFGO3bp1S97ShUtWwEwcxzbSYG6Un//857S8vFzIbS6ygxt3QQYq41RJHmagMrlWfkwDlb4a6bcNVKbXzFLkV8BAZT4NDVRm089AZTbdkAqdYgOV+fQzUKmg4GhQ+T//8z/029/+lj788ENaWloS4TEopANDus5+RwZTysD2YFC4F/ddGl/XiOxfX5i4wI0i+3Fy3Vyn04JKzY5q4udfjyVZIz0GupAe8MNAZRLVxjcO7jM+OmYSBZVPQ9evmBPVwX+UD1dKsBEMofI5Di7R4dXBGH4KPc2kgcoOW1QCVKpOWi+1nqoiGq778etBPfUeaPtWFYvKMi2fDUAlW1XeZC9WIwsqAcbbu4FF5RbdA6h8DlDJVlz7JXH9GuY43AiU0cITCQ+LYSQ8HNkfEHYQVC4BVJ6rOlB5eZ6WzsyyBX9FrCpV44HktmMKTLAC7tkQDyofs0XlamBR2d3jlkgacK50QQXU9jysjxGdtHoOqcZ4wMiyv4/flu55UyvtsevXPRpnUPn96g69YUv7rSioDBuoqDIR4WJ2DwOV0M6ByqqByhjtLGgyFRhpUIkf23ARhI4jXAYBMv7v//6vzFEJAHnjxg2ZowQugGAxCVj58OFDwvwn+CGyuLhICirxVvHq6iq9fv1afsTPz8+LRSWOnzt3rpC7W2QHN+6CDFTGqZI8zEBlcq38mAYqfTXSbxuoTK+ZpcivgIHKfBoaqMymn4HKbLohFQYTDFTm089ApStHUBH9IPR7vvrqK3nJ8/e//z19+umnIvBHH31EAJW/+93vxEPNSYDK7Hf25FIW2Y8TYMF1Ogmo7HSdRaUODiLHuH9YkoEPiTrwB+ndACUP4rM1KtyGiUUlWxdhTqg7bKmF9eJCg+YadXbTWRF3vPq9uh44qe2MrAJ6rxOBymCOSgw+KySTUWTkbsiINFAQFo2v+xLo/QmKrZTfKr8U3sQclefH16LyAVtUvtnY9Vy/DtbLaD3JUl9xRlUX56tW4Pp1vEAlXHy/Xm+x61cGlS9a9M3zndMDlY0SXVtkUHlhlq142aLyzAyVymjfnKVW9J55xdc2TYGJU8A9G4aDyjdbe7TdneZpvtC6u5bIiaD4MZ0k/hmQUs8p7VxoUblHywt916/L7Pp1geeonK2Nh0XlIKjsz1E5xflzi6qg+0FwypWmlueCWFRW6apZVKZU0aKPqwIjDSrxowduXl+8eEH379+nL7/8kjDHJKDk1atXBVLevn1brCcBKwE1ASvh4vXRo0dyT65fv041bvTevn0rxzEfCsAkwmFRef78ebG4LOIGFtnBjbseA5VxqiQPM1CZXCs/poFKX4302wYq02tmKfIrYKAyn4YGKrPpZ6Aym25IhcEEA5X59DNQ2QdbGNjAS5pff/21gcqExarIfpwAizSgMnBxq6BH0QWG+ZIsPl9y53ADZbiOqSkHKpuzVTpvoDKJnGMXxw1GD7eofPKmReL6lcdK8JzW8pk0o1oKhw2/aricjwsgyqADlXW6yqDyZ2Pq+hXg7TWDSmdRqSokVY3jacXsV2yXWMN5D9o5eAbdGFTCorI+rqByUywqv3mhoJLnqNyfhhCS0YFy4kL5b7+1kx3+04cbg5preBhPTos2Tl2/7tEiQOU5dv3KoPKDK2xRyXNUlsIXMZz7V01va1Ng0hVwz4ZhoLJDb7b2abtD1GFQiZqIOubq6WDd83Xyj0TrtB8P21pnceZpjlwt7bNF5f5Yg8onbFGJuYu3duH6VUGl5jSqQMy+ChgVLxLu74pFJb/4Y6AyRk8LmkgFRhpUAkgCMMKl62effSbzTz548IAAMPH2L+afVPevcPVa4R8hm5ubEv+TTz6R7QsXLoibVwBPuHsFmATkRDocm52dlXRF3N0iO7hx12OgMk6V5GEGKpNr5cc0UOmrkX7bQGV6zSxFfgUMVObT0EBlNv0MVGbTDakwmGCgMp9+BipdOYKKBirTl6Ui+3EKgg63qNyl1m5HIMj+PuAR7ptet27oUJWGx6897iERcP/dACXPe8ejg7CobM7Wxg5UIvf7ezwPHn96DNh6WHNm/fyqUvHKFBdaYh2rlRKVSw4ondT3JsmBu9cJQaVa8CY5cRBHS6HmWff1FBoe7gO48VxjjZm6WFSOJ6jcEAvB1+sMKlkzlD23RHOruY5Za0HtV2wXScODJKivOKsDlZiLbAxB5RosKhlUskXl1wwqX2/A9WuZesRwkCEFlmEyHAiX2Bw/WOtK70C4zwFOtz2qTjsAstScZlBZdxaVDCrPG6hUuWz9A1TAPRuioHKdHr1q0eO3AJV71PJA5TCJ/Doq9dCrjKiDuqvraN1FRcUrCwCVzSpAZZluX6zT2M9Ryc8GjNFD5wN5HibmMJEi4f6ugkrMUfnRNcxd3GA3sA06y5aolUqVrcaDuT35O/EcscUUGHcFRh5Uwm3R999/L66LHj9+LG8HY5L4ZrMprl0BK/GBdaRaVcKiEpaXSIuwaXZ5A7g5MzMj7l4BKmFReebMGXFzg+NFLEV2cOOux0BlnCrJwwxUJtfKj2mg0lcj/baByvSaWYr8ChiozKehgcps+hmozKYbUqGTa6Ayn34GKg1UZi9BJFON4AXZly9fEvqc8MgDN7l4GRZ9xTSDP6jPoGnpQCUPdA0dYHJDVpjryV80Or4ubsF1TMOikgGbs6icpetL7Pr1yui7fkWWcP0dnlOxxXNBAeq2dnu0G0Aj5Bn5H1QkToUUYapjzEln2B3n4lyd3ZmWGVZOy9yfKc58rFGhEz7DXL8+ebPlLCrbPEdlz7kaRvzh5c1drsqhF6+yRMNR1nXBOfGZJFDZ7nQFVKqtkeY11xqaBRUYuoa6iYu/MQeVPEfla5mjUkGlKqXlREtSNFz3060BQtVSKwoqYVFZFotKc/2aTlWLPQkKuGfDcFD5eqtHrXbfohJ51to5UFs50A/3mvzB8EA0jRtqyAF9UEl0CaByeTxB5fdvtt0clWJRuceg0nkpOJDnMPPRDY05oHAIe/XFjv5R95JUY8bNUfnRtTkDlVFJbX/iFBhpUKmuX+HOFbASLowAA/EjHNaTgJVnz54VYAlLSYBILACUcBX77t07saJEWI9/lOM4rCgRFxaZsKYscjFQWaSaxZ/LQGU2TQ1UZtNNUxmoVCVsfZIKGKjMp7aBymz6GajMphtSYTDBQGU+/QxUGqjMXoJOE1R2pf4fDo7ckFU6UAlQGlhUAlSy2zBYF8kclVcavG6M9ByVGAjFS7pbO2z1sYaBwV16s7nL+z2BlzgunEfH/PLcfE3bHxnUEFnjK842q3Tz0jxdYPe59SosK4t50XngizLuuMHoo0HlTptdvzL4RXxuLQSOHfaVKofGUamj4RMJKp+wReVLdv3KFpUOVO7xgDwUUBVUlQxr0R+ncudyZ1XAOwEWlQOgMinejZSqyO4Blb3bIKASFpVsqbXUVNevDboVWFQaqDygngX8QBRwz4Y4ULklFpV4oQCgss2/FaRt4zbpICgLWj3UOa6Xw6pmNFyrqIbjicnv+7BF5fiDSnX92obrV7jud+rx3yRLVBm3338sOMVUN9wReMUAqLy6BItKA5VJVLY4463ASINKVHrMOwlgCTew7XZbgCMaXLzZCleumHMSH7z1WiqV5G4gHlzAIq1aSyINjiOefjR+UbfQQGVRSh7PeQxUZtPVQGU23TSVgUpVwtYnqYCBynxqG6jMpp+Bymy6IRV+pxqozKefgUpXjqAirHNsjsp05anIfhzqM+CNWlSub27Ts9Ut+vbpOj1k+PGY38hfZfDmXL92eaALQ1I6LOXuH65ezjOQjegAlx7UcN1354LbVPSFB12/NtiicvRBpfMktEdv13fo4YsN+u7VFj1Z5WlhNttsVclWDAGE1Rwfz9rpiIHbq4uz9G8fnKf3l+dojuf7rFUw7BrV/Xiu4qizopzgc5hFJcqbgEp2oYuy6Ze3o86f5nhoGTjurl+HgspAPhYl4IxOzhiRtHRozdb9mKjSZsP6ucrlqlGv0PLZOn1wmV38XWrSTS5z5+ZnaLpU4U9J4kLn01rC9o3LETydYazute/6NQSVJXbVnPQ6VaUgV5FdzasGa/b17OL6lUHleYDKRTdH5a0rC/JyxriCSqmmmvFjXKuWx/gVdupTUsA9G4aAytUOwaJyqz1FzNtonwsC6pOWBy1/uh9mgSuh1kOE6XGNr/G0bmpczCVb4+H6udrU2FtUYu7iLfb0gPEWPHeTL6rKUSlC1SQi3M43MEelgcqjhLPjE6LASIPKcdO4yA5uXN7N9WucKsnDDFQm18qPaaDSVyP9toHK9JpZivwKGKjMp6GBymz6GajMphtSYTDBQGU+/QxUunIEFQ1Upi9LRfbjwoH8vZ7AoThQCQvBbZmjEqAyMkflsJG/EIzpIJbmMzr45Y6PM6js4YVldvP68u02ffn4Hd17vkGPX7Nl5WaHdjqYq5KtRaGTaBXVQ3XJuoaefE5ul6d5cLXEn5ULs/SbH1+kO1cX6EyzRjNVNg0R2aPaZ/3O7OncYHQ8qPzm6SY9ZUh+cqDStT9V9n411nNUngqoDFz8TQCofLXBls/7JbaKTlo/Dq/D4VHdCE6rZ6+JRSU5i0oGle9fnGWLyvEAla7+uuc3sqfAR9fZW4YEKVlANKE8Ba+0p6cJwBNcrUVJqYArW4eAyk0GlZijssev40hlYliJMsHfo+VPHrHe97rwfkXU44PhwTlwHv7gfIg3/qByg+D6VSwq2duDA5XuRaHkdQdqYOlr6Pb1rwvv66+uXw1UqkK2nnwFDFQWeI+L7ODGXZaByjhVkocZqEyulR/TQKWvRvptA5XpNbMU+RUwUJlPQwOV2fQzUJlNN6TCYIKBynz6Gah05QgqGqhMX5aK7MehPmOU7zCLyjfsUrK122YPQElcv6bPD1IMgspq4Pp1HCwqGUSy5R9+yzxnyHb30Tu6/3yTnrxt07vWHrH3V7HUSj4wmE0/WFKWxQqkRyvn6/TvHyzR7cts3cZzVWLOyim2gBsFWOkGo08TVOqAq2t7cF8mCVTuch3dgyVqysXZJ3E9DAakdXg6KDQDZ4NmAO9iOVPXOSrZxd+YWlQCVLb3pgsDlRBLSpkWtUBMrFBPq1wV4VJyaQ4WlXUGlePh+hV1F2N8PX7zAr+he2wFj33+H5SagWJSyI6WQ3wH4FGJKWW55NxLYvu429VCMmEnSaSAezYcDipbDCrh+tVVLW21jjp9pCKG0TWcA4JNrFDO8KlxORtvi0ofVPYtKuU3X6CB1h8/LJSHWyvogAX1D4vuu73+X1US0d2czwYq++rY1qQrYKCywDtcZAc37rIMVMapkjzMQGVyrfyYBip9NdJvG6hMr5mlyK+Agcp8GhqozKafgcpsuiEVOrQGKvPpZ6DSlSOoiIESc/2arjwV2Y+TASqu07Gg8sUmPV7doTfs0lRdvyI+PuEAV3DpwXhWuowEsfl0cs5pHniu8HQpcFfq5qgcD1DZZWvK3XaXnrzZok8fvqX7L7bo+VqP1nYYYLL1RxfmHzLCl0cliKXDgYMy46z4VKb3aKbEoHKpRv926yyDIwcqZ9nqbYrd6rp7lvcaBr877Z6WnzjXrwctKuHjLz7Pg9+bJM5gCrfnQEetGrWonKMrSw1aaNadC9Np58IUabTcx53tOMN83fZ6Xep12vT8bYvuPl6nr9Sicm2XgTkfYwvftHdZ46uSuu/yFNnjXehQg4u/0KJynEFlV0Cl8t3B3B5+V+P1GlZTUU+dS8kGg8rzc+WxApXud3OXNrc79G6rzRb4XD95CauoCqeiHC5doqM4JeA5PI7j+VAtw91wmc40ajTLL2Bg/l2E2zL+Crg2bhiobNNrtqgUUInHQphdhZUaghrmL4N77kmJ4/1wLT1+OUaRqpUVVFbo9nKd3js/SxfPNujM3Ay//FOjCrsMl+cBt4Wn9VyQnPCF6/O0w9PKvXi7RV88XpMXppxF5aDrV+isi163H6bHsJafLbzWJG5fFevH1DPiSN/166zNUdmXyLYmWAEDlQXe3CI7uHGXZaAyTpXkYQYqk2vlxzRQ6auRfttAZXrNLEV+BQxU5tPQQGU2/QxUZtMNqdChNVCZTz8Dla4cQUUMlBioTFeeiuzHyQAV12kFlWvBHJX3MEflIaBSB/yiA1npctKPjb6js9IaJ1DpoC1A5Q67xlVQ+eBli16sA1TuC6iUue9CoYI861ifjvD1pThk62BkhOBU+AioLPfoPXYn+a83z8h8gYvzdZqtVxm4AVRirkq36ACl7p/UGuUNHx1YbfOcgU8Z8H7ycI36oHJHwG+315O4GFTmJLKEMg5IMbCTKCuaf6xhUdmcrdO18w362XsLbInaHG9QycRNixe0C5SL6KLhkeDY3f7ZcBj3wEEjzEVWoUtnZ3iOynEDlVt0j18o+PrZNr3aYGujvSkGvNqqxYpQSCC0q5X2qQ8qZ8bCohJ1trXbZbfMO9zOtejes3WZg7fEGRK31lIwAonSFK0EqgJSdhm+w5KyyeXtwkKd3Vs36fyZGWoyKK8wvNT6nOB0FmVEFXDPhsNAZZfnqITrV9eqaTEbbJ36LV7SbIbp+YRyTrRv/KkLqJymS2fGD1R+GYBKzDH+mj1ibMoclWxtjwdp8DBFvodpqNpFj+u+Ho+u0b45UFnnOSoNVEb1sf3JVMBAZYH3tcgObtxlGaiMUyV5mIHK5Fr5MQ1U+mqk3zZQmV4zS5FfAQOV+TQ0UJlNPwOV2XRDKgwmGKjMp5+BSleOoCIGGA1UpitPRfbjUJ8xcBUPKrfYopLnWoyxqNQhfQxMYQnGvtxOyr/uEhhU8uggrBTm2aJyaWGWLRhG26LSaefmp9zeaQuohOtXgMqX7E5yXUDltGdRCaECcQLdwv0DmiWOKClhR1Lhue9mGFReX6zSL24sBKByhmGSA5XTbFXZv296AQe++FgD3GD0MFC5wXNUtgSG7LY77F5SQeVge4EL7Jc31SndZSvYcKCyzKByZrxBJUO3VzwgLRaVAah0d1j1id5vDU+nG1KhzgNO1biuTgao5Pnb2PIZLxREVUqnzlGx2RKdo7Ah4FiBStTZPaaFG9ttesZWvPefbdA/H76jV2s7XA76kFDrlKiQrXgdFJAFg4tZgMoqg6OF2QpdWZylO1fm6NpSk87hRQxxbY2kx3v3Dl6chRSpgHs2HASVD1/hd8ius6hs7xM7Lzjw2NQ7n7XYIT3SSvvGa7GorEyz69dpujwRoDKYo3I/mGM85saphnrIaamK6lHd11huraF4NrjnAly/AlTO063lBtfZBp1lS9RKpUol9pqhbYWuB89me6bAeClgoLLA+1VkBzfusgxUxqmSPMxAZXKt/JgGKn010m8bqEyvmaXIr4CBynwaGqjMpp+Bymy6IRUGEwxU5tPPQKUrR1ARAxUGKtOVpyL7cajPPqhcDywqvxWLSgWVPEclg7hO9+AclQqMMEDlLxquYXo8Go7j7hLGC1SKbjysuc8D6NAF+jx57eaofPgKVlp7tL6rFpXTHNMJFHVMp/pkXYcDhPwN6voVoPL/Y1D5/nKTFucdqCyXeHAwdP/q6l3W78yTzg1GHw0qYaEKUMmlQ545eb5zWFq0PfhgTq1xB5VwN/yawZEPKpFvrZZaTlQLF94P1S2Nr/E0XPexFt14JB8D0rBwG3+LygBUelN7anvl59ttJ1HItWnRtEg5xfPIVks8R2VtKnD9OpoWla59w/WSQMoOw++1rV16xtaU3zKoxAsZz9faDHd1bk/UpWiOi9ifEiswWGADVJ6ZLdGVc3W6c6lB711s0qVzDXEV7uoyvu9YLqKIjNg5jlDAPRuGgMo3HqjkOVLlN8Mx3G20d1JP+Z2eOlvqztVLkwEq2V3zLv9OQT2K+w2GW5O85vSfCvq7BulxAvy+cc9Ttag0UCna2J+JVsBAZYG3t8gObtxlGaiMUyV5mIHK5Fr5MQ1U+mqk3zZQmV4zS5FfAQOV+TQ0UJlNPwOV2XRDKgwmGKjMp5+BSleOoCIGGA1UpitPRfbjZECa67RaVPqg8gHDD7gOe7OxS7AYbPP8d24A25ujMhizig5SRwfD9Hg0HDlHGM7rLCpLbFFZG3mLSlyv+zCo7HSoxdZGcP2KAXwBlZt7tBFYVMJSaz8QIPlgYLIy4QZW3XChgspr5yr0Lzfm2aUkg0q2TG2yRSWsGKZHbK7FLg+cDrp+ZYtK1hDuJXciFpXJ1EgXC7fEgUq4fh1vi8o+qOxQz3P9quXNVdP+nm6pYkE1PjBYreEaD2sHhTxQeW48Xb9+y+3bN3D9ug5QyZZ7Xmaj+rj8x4dy6+XL09/TYC8ZNmFROU6gEuUJfbV3mztcP9nt6/NN+oznRn32jp8JeyW2duxnUNt5FUTb+2i4Hh9c98+j4SIh/8ELIdBtYYYt3M5Wec7AWbpxsUHXLszTAs9XWWK3sCiXQCW2jKcC+jzF74zt7V16+W6L7uOFKVhUCqiE69dBi8rhd1srX3ItwrLKSeAlvQ6LSgGVKG/jNUelc/26Ib/fxPUrQCXX4T2xqIQ2w5XzFdMXqxRI6r7G0XDdlxcx5MUfA5Wqia0nXwEDlQXe4yI7uHGXZaAyTpXkYQYqk2vlxzRQ6auRfttAZXrNLEV+BQxU5tPQQGU2/QxUZtMNqTCYYKAyn34GKl05gooGKtOXpSL7cajPIIWHgkp2/brNFm4hqESayMhzZBenHFj0eDQckRCG6xhLULnXE11aPLDqQOWagMrXm3D9SjyQj7nv+qByQJSCdtzwfGBRya5fr56r0r+sAFQ2AlBZozIPHiqodAP6BX15ytO4wehDLCoDULkNUMlzf3LJkPKh5Sfl1x0aHeeEFm6OyvEElQBGXz3dYHgEi8ptntvTgUpkPOrIVKvkwBB1uBN71NNPjzvNMN9prYI5KqtiUQlXnLcuNcXd8Dm24p0uVfhTEn1Pu7yhAKHcAYzv8pyor9dahJcwHKhkN80KKo+cozIUy9MFm31t9ICEaHCQDCuUOVgGzo2LRSXnzf1e7tJbfoEALxJAuy+fbrJFJQOlXok6IajUDKsKaddRDBKkx2kZsAiorE/RpYUKvX+hLqDyvYvz7FKyzqDSzcF7mmUtbW4t/qAC7tlwiEUlzyXrQCXX58GkB/b0eYHfFv362bf6HQzvJ0c40sK19QCovDQJoBIWla4tVH36OXdbCh6H1MRo9HBf9GTd0MaJRWXDQGUojm1MvAIGKgu8xUV2cOMuy0BlnCrJwwxUJtfKj2mg0lcj/baByvSaWYr8ChiozKehgcps+hmozKYbUmEwwUBlPv0MVLpyBBUxsGgWlenKU5H9ONRnGchn4AYrtkGLyk1nUclz323vwqIScwYGcxxhRMp7K98NfLnhQxnscpthxnRgTAa0wlC34QYoFVRijsoxsqgEqGTdtgJQ+dl3DlTC9esGu35t89x3Xbao9LWKZD//rgwQKqjco2sMKn/+3py4fl1iaNRkPcuVCg/mO3B0rNdyRG70XsMF3WEWlQDj/hyVWn6OOH3qw2h/atXxtaj0QeUrBpUoi9BNIaVUU1YlUh2DmisFZ+jRQTH7Z4BmohvDb52j8s4VdvE3zqCyux9aVIaa9bM8KEXWvUDumoLK+QpdW3SuX2+zfpiX19VTN4cbND6tRZ4L8uUAvD15UWV1fVtA5cNXLbZEbdHzdXZ53e2DymLkOohIJITbizpbVM4z4L04X6Yb52u0cmGWVi4tyDyVZW7bMAev0+z0dDut+zUJ3+ueDcNB5SuAyl32YADXr8eUYfw+QbXDHJUOVJbZ9StbVBqoPFRx/V0H7QxUHiqVHZxABQxUFnhTi+zgxl2Wgco4VZKHGahMrpUf00Clr0b6bQOV6TWzFPkVMFCZT0MDldn0M1CZTTekwmCCgcp8+hmoNFCZvQQRFdmPkwFprtPxFpUAlS16A1C5wxaVbJHkBhN1mFBxCOdGxob74Ynzx0mQCud1FpXjBSr3GAq12fUrQOVTnqPys8cKKnsMKsm5lIRFJfLYx0MD8qhqGihS6k6SNc97x57qeI7KfZqpTCCo5LKhzEY1VFngag7LvsBgDUVxDMIDzftH3JaeD2ADFpVz6vp1ZYFuX27SlaUGLTTrzjJwxFzm7vW61Ou06fnblrjgdBaVm/SK56hst9tiAYcydHQ58mLoZlAYtUyqTk41DuX/0AzznWKOyokAlTzX4i6DSv4/oBkXu0IXaIkPQGWzNk3nGVS+x6ASLkxHFlSyCDoHr4LKRwCVz7fpxQYDzO40dXieSiwiV1SzSLnKJiiflC3BauV9AZUX5kp9ULkMUDlD5XKJSlpPBwtttq+0VCeugPttEQ8qv4Pr1w1+zjKobHNFjRazwYvFixQIgfXg4JFhe34xxTbS19lifH5mMkAl+myYvxgulKFz8pcgVEBVSBUcDMc5sbjnKc9dbBaVKpStfwAKGKgs8CYX2cGNuywDlXGqJA8zUJlcKz+mgUpfjfTbBirTa2Yp8itgoDKfhgYqs+lnoDKbbkiFDqmBynz6Gah05QgqYmDDLCrTlaci+3EywMR1+nBQya5fASrVotK73HD4KtgYHL7SoxqqCTWc9/kQrgEfWMRUMDjIg1ywLnrvfIPuXGnS9fOz7MK0QXMcDqjUtwx05UfPepJrveY+qGS3iAwq77IrzkeveN47HsRf5zkq2+y9FBaVooDK4GVfrlnDh2UgiH9YMgGVpX2arexPjkVlAMZFFs488u+kUiX62DJZeF9gZRlY90Flk3421qDSWVT2PIvKfo6P2FJBuB5iUT01WENwQECluH4NQCXPUTnOFpUvAlDZYxh25BKNokXxyIQuAvSsVeD6FaCyOuKg0rXLHQYcrZ1dAqh88rrF7VuL3eZu00uASnb92m/f9PWAvhhafoJi1T+QZosTT7Elf7UEi0qiC/Ml1q3KFpUNWlmeF/fWFZ6DN3wu6Jem+Q6Le+oKuGdqHKjcJAcq2aJyh4/znKkoT1oVD97uoyqlpuxnGSn6T5PAorI8zd4dACprY25RuSN9Noy3pAOVvk6+pn6401DrN+6FuFI3UNkvXLY18QoYqCzwFhfZwY27LAOVcaokDzNQmVwrP6aBSl+N9NsGKtNrZinyK2CgMp+GBiqz6WegMptuSIXBBAOV+fQzUOnKEVQ0UJm+LBXZj0N9xqhfMlDJcxwhLn90cFBtKjk0khF/YAuH3HGN70d2l+BbVDKoPDOGoJLnb7v73TrPUdnyQCW7fuW577Agn7KwNKqOBGl4cBirgeNefC+KbCIp4uJTVVC5WJsM168hqPQFUmWQfT8c+3HH/DCXQkL4D8owyqOAygbmqJw8UCn1G8oEFVb3oVa48DFVSRTVguqFh3F5w4FKdpkrFpU8R+UkgMoOuyLGHJWBEE6Cfvka0McTYzB+/4CGa4hKyu9i9EHlHIPKJVhUNkfTopIvGq6+MTcx5uAVUMkW9t8BVL7cCUDlNOsGQOkgpeqk+R667kvromjCSLjbBajcl/YNoPI8W1Q6UMmuX9micpFfaqnwCyxw/yrlPCr+0IuwA6OkANomLW/bXN5evtui+0/xPPVBJXswYNev7ndIv74OFh/d6+dOQ8Ly1D/kbfXLcOj6dbYirl8/uDTDL07N0sWzDTozN0MztRqXOeeiGY2Gtq/eyU5sE7qpK/UOW9S/eMtzyLJnh/vPN8R1/yueLHsTFpVtzFGJHyNQQRU57DL9yujH98MPpq+yK/U5fp5eXZqlj66xS/DlBl1ZbPBcsjOsWZVK/FKB6qXrg2exEFNgfBQwUFngvSqygxt3WQYq41RJHmagMrlWfkwDlb4a6bcNVKbXzFLkV8BAZT4NDVRm089AZTbdkAqdYgOV+fQzUOnKEVTEQIVZVKYrT0X24wRcYIBw6ByVcP2qFpX5QSUGyPwhL+RcBtr4GmBRifmN5nhOxfNnGuNnUXkAVO6xS8k+qAy52gEBjrj/A/F1kHAgUDABLI4aVeJ578YcVLLl1jbmWuS58aR8hsJBJ8236hDVLnpc9108TaUsA+1PDa5fJwRU7vJAdY8tjpBrfLhayaL51X0Xmuxv3GAy6urkgMpd2mFQ2WMAAujgFt7WwjJMJo2qxzV+NJyPQ3ecGgCkVpnuW1SOCajcajlQ+ZTbuEevt+megkpu3zoAlep6OSbvKo+/lmiBXpoUxwfCwwSIyK5fuX1jtksX5qbpulhU9kGlWNoHACSuvIanso2RVQBtfSyofKmgUl2/7jlQGeTk4C+KZFnsF1VXEKWU8R+UH7xQMMPeHebg+vVsjcYVVH73Zptes0twAZX8wgFAJXQOm7lkUh0ZS58rOK/8hgOoZI8YBiqPlM4iTIACBioLvIlFdnDjLstAZZwqycMMVCbXyo9poNJXI/22gcr0mlmK/AoYqMynoYHKbPoZqMymG1Khk2ugMp9+BipdOYKKGBQyUJmuPBXZjxMQhAHCI0FlO3D9isFEDHTpMJ8b5AuGmMOMyHnDPXefJSa+ywvXTcSf5nPCSmFutk4X2HJh7Fy/Kqh8CYvKLq2xm7rdLrt+FYtK1kszLtJ5O7qpYshaAznyQPyBSOEOXL9WywCVUxMOKsMsZ9pQVZEYZRif8QaVG/QVWx3de445Klu0u9sHlZkESpAIYEBAJRe4xkxtzC0qt+jFOwcquwCVWKS+8dovLHIg3x80mfjUBVSWnOvX8+NhUQlQ+YZfIACo/I7dv957sRPMUcmgsjdFgdG4CIQ8hotq6IfxQdkNjg0DlTiHJscWQGVoUXmOrVEvAFQ616/ValXmqZxil8T9Z1N4FbYxBgrgN8BhoPIV5qjcYYtKfqA6L81aOvqoUkOi2dXip8d138Vzofz1Ut5Qfkv8RoHMUQnXryGobIydRSXq6mueY3xzm+fhZYvKXmhRGVWomH3oGlpUGqgsRlQ7y8grYKCywFtUZAc37rIMVMapkjzMQGVyrfyYBip9NdJvG6hMr5mlyK+Agcp8GhqozKafgcpsuiEVBhMMVObTz0ClK0dQEYOKBirTlaci+3ECFDFAeAiofC0WlW3CXGV73jxuAwPSkgU3/CcoUkcENWsSGQORGtBfuwFK36JyQkDlds+BSp6n8tBlcNTUjZbGJhgUL9zj9A5UTlGjNsXWRvUxd/3KFpW7sKjsyvMmVooMgaFeQVrILqCSIcf4WlQqqNygV++2eTC6Td1gjsposTpMMq2X0Tqt4X5aaFaCReVEgMpNBpVttuDtsW5cQli0o3RTTQbiDeywWjGFDVGkvGGOynqJ51qs8csY4wEqN8WissWgkueoZPhxX0Bll7Y7U+yKMwIqw8wPF8U/4kslGgWFTd3JqpgsGVuiwqKyzK5fK+KKc+WSc/0KOFIWi0oDlX5dHaftI0Hlepu2dvmZytbPqINabqJtVphnjRAGDNtwEfEX58X5ACpn2EXB/Ay7fgWovAzXr2MKKgOLyp0TsqisqetXA5XDCpyFT5gCBioLvKFFdnDjLstAZZwqycMMVCbXyo9poNJXI/22gcr0mlmK/AoYqMynoYHKbPoZqMymG1IZqMyunepnoNKVI+hhoDJ9eSqyH3ckqJQ38uH6dVcsKtHHC0fz5NLdIN/B4f2jwjXfcBsIgDlZoPLleofWGVTuwKLymEAlFITKGFjFAH+tDFDJbhGXJgtUSklCuZO8Iqf99kO2JeToPy5lEE80CywqZWB1dkznqAxA5TMGlWsBqOQCN5DXo6U5NEYgvcSRssZ/ptlyDQPSTXbTPM5zVH79DKByV0CluH49VIng4LCm7ZC0uB941kE/cf06IaCyJaDSPRKkLRINhgkU1OEYnfpp/YNuzkCEYKvGFuPz/CLGhWCOSswZaKDS12u8t93vADcn6sAclYHr11ceqMTvEG2XUKdiFy2GsQfdMxOH9qf6EXFO1FO4aAaoXMAclWfrYw8qNwKLyj2ecxY6I4+S9yDrwzREXCz9+H2tBsMlmmvfDFQ6MezvD0YBA5UF3uoiO7hxl2WgMk6V5GEGKpNr5cc0UOmrkX7bQGV6zSxFfgUMVObT0EBlNv0MVGbTDanQcTWLynz6GajsgwYMgJhFZbryVGQ/TgaiuE4PtagcAJU6R2V/4IrvZHDx0dHCo8I1z31QCSutyXD9ukUAlWsttagMtIhKpBJE1sHYoAz6+Yc03A+TbT4vLCpr5Wlq1CcFVLIFbzhH5WB7ARl1AFVL2QFNhgSEt4A3sI32B24j52cZVF5o0s9WFuj25SZdWWrQQrNO06UKuzkthQO1OmA75PTHFoz84oM5xvZ67MKv06bnb1v02eMIqGTXr86iMt2laNmKDlhruJ4Nx6GBA5XlAFTO0p0r83TrUpNuLs/RufkZp1vJ6XZamuGaw/aNMwIL3d3dXZ6zrUUPXmzRt8+3SEDlmgOV+F0Ylg/NsL/mwhYtbxL/0ET9E8BBJfNdcSnZZFB5cYwsKp3r175FJVy/vlzv0laH2PUrBa44kVfYQfqL7kWV8+MM39azAVTWGVTOCaiERWWVVuD61Swqh4s3ZkdcG3cUqOQ6zBaV8Ozgl6ij2q2oFFoqo+E4J47JHJUCKqsTASrXA1AJ17rQWdtkbd+j+qku0n5CkyCC7uvxfrgLQTR1/XrNLCpVJltPuAIGKgu8wUV2cOMuy0BlnCrJwwxUJtfKj2mg0lcj/baByvSaWYr8ChiozKehgcps+hmozKYbUqGjaqAyn34GKoMBZJYRAx0GKtOVpyL7cTLwxHX6KFDZEotK54oTg1vDBrbS5cTFxvlwHQoq53mOyvNjPUclz3u35iwqMagKi0oZVB02OhoRDXpgiWqs4e6o/nWRMb8nLLUAQMbSovLRGn3zdIOevt6k1Q24flVQ2W8rRJMg24FEKoK31iNRsV04YBEW1RbtDwZW52cbEwIqdwNQGc2/J1GOTZRBaAbXiGJRiTkqF2fpAwaVt8cVVIpFZZd168+0qOVES5NK5hlfSZA/v6LGwVrLVxiGE/EtQT3FHJXNGQWVs3TjYpNus35LC7NUrlSoVCqLxgoBwnOc4Eb4XGCw0WaXkQCVq+stehK4fpU5KvlljK32Ps8ZiAtz4Eh1O/pSDygbJImGu2CUZliMi0XlPEBlzUDl0SKPVQyUueFzVO4QLCo32UWBgEqJG1PPojmOFidtFv1wDfPSljhshl1bLzTGHFS+whyVO+zdwc1RiRddWGRpi7zshpvabiFK1kVcvzbhoaBBH13jF1iWG3RlsUFn52b4RbQqlcRFsxP9NNu4rPmzdKZAVAEDlVFFcuwX2cGNuwwDlXGqJA8zUJlcKz+mgUpfjfTbBirTa2Yp8itgoDKfhgYqs+lnoDKbbkiFwQQDlfn0M1DZhw8YqDBQma48FdmPCwekh81RGVhUGqgcvEduUJUt3Hg+wHaHB+y3d3j+ti26+906PXzpQOVaC4OqbMkVtf5AOxqcLjpGquH6bXo8Gi7HMVjLG4gTgkoGINeXZsZvjkofVK4rqAQY72uFPB+qByIcoayCFB2QRftjoFKES/QH9wOaDYDKcw5U3mJLVFhULo66RSUDN7GoZLevXz/juhoLKlGUAtejYaFT+76+VFIvw+NBeLCvZc3V0qCeAvAyqJybKQcWleMLKp/zyxhbbQaZXaigFVXF4CBvU5QRsQKNwnqq+xp5IFLYvuFoPZjb86KBShVtotbJQWUvtKiUUqNFJ6rGYFGKHh3cD87h2jc8TxVU1ujKOLt+jQGV0Fmff4MioG13IdAh62KgMqtylm5cFTBQWeCdK7KDG3dZBirjVEkeZqAyuVZ+TAOVvhrptw1UptfMUuRXwEBlPg0NVGbTz0Dl/8/enf7IclzpwS+Sd9+5ryIvKZISJY6W0cjyLPKMxzYMwzD8UTDs/8ufDMP+JANeYMA2YHiT7XcW27JnKIkiKe77vt19ld7zi6zTnV1dWZ1ZlX15qxmJrs4tMjLjyXNOnDhPRORyuLlKI7cSlavhV4nKRo6gWInK4bI0ZjuOPmODho2o3Jw6bPjTb7+ieQQjKm8pU782IyqPTR6JHvlfe/DY5OH4FtmdJ49Ojh89NDlQRhzdRFNxdhKVV2Lq1ylReb2hKsT+umKq21HpcyTynWaaRCUCZL2JyrMxciuJyuulvlkhZtoJIhJJUHZPEpW+UbkRcZ5CMBOAHiqH7XfQSVQaUXn/GhGV7wVR+Q6iMr7Be/laTGHajKjcxGZzq0FxikKCkZjGydmUTfrm6CxReehAm6g8upYjKl96/2IZNX7+cnTUCKKy1COl0C0kymYLrNws6XKnlX7L8UJ9ln9SkDlE5YkyEnV/HVFZsNpb/8hQ14jK1z9uRlSev3g1Ov9cn1yPzj8bSwhISlHr6FSAmlSz53O/nG3tuN6u+vRwfBT15NE1Jyp1NPu8PaKyqVM36ocGnpX/p/7LtxKVK8NZM1gzBCpROeILG7OBO++xKlE5D5X+xypR2R+rdspKVLbRGL5dicrhmNUrVkegEpWrYViJyuXwq0Tlcri5SoO0EpWr4VeJykaOoCiwUUdUDpOnMdtxJcAkQNh3RGVMHZbBvHh5wx68I3UToJyd+nVvEJWXytSvArCbhd8GW54bCGfBLbLNwOrB/beVkVoP373OIyrPTj4OovJSTP16pfWNyk30xtnK0W4lsHpwj0z9+tnF+AbjpWbq1yJkIRkzspVyOFDUNrKBvqxN03xwv29UHprcX0ZUHo9vVB5fS6LyvU8RlVe3TP1ayrmDqCW0mSwx3TjeOuAY3G6NzhiHp3p678mDpTPGOk79+tJ7FyfvfX5l0hCVvnsHhVLKDdKowSXRSDBSJLcf35qej9AcoatGuJkytxCVJ5OoPFq/UdlAtCf+N37A/G9UFqIyviV77lJDVJZvVIaAFKkKxUrpSqmaBWT2fO6307nWzzmdppqpX4OovOPw5MkHDhddvTempD8VU5gePniwdKoqhJ/7U+4vaIGbKV19g/fqlSuT9z89P3n+zc8nr7x3dvLGlKg8G1O/XrpytaSTvjztWM8c+SXu8j148MDkuKlf4zvPZerX++vUr1+QaNTb3iAEKlE5ItBjNnDnPVYlKueh0v9YJSr7Y9VOWYnKNhrDtytRORyzesXqCFSicjUMK1G5HH6VqFwON1dp5FaicjX8KlHZyBEUK1E5XJbGbMfRZxHhHYnKi5fLFKfSl0DXDkGuDFxl6TKMt+145JN5bv1GpZGURydfX9sRlZfLiMo2UQnqJkCYqEzXCUqCNHO6e7chQOmQ13GoECD7A7e9RVQWGZ2CkAHh9rE2PrPnc7+dJreFt50/uEeIyg+mROV1H0UlEJZZ2cr95uzw/5EtzMoI3vi257HpNyq/Ft9YXF+iMkbwBgFiROUQFdwMzzcwkifLxvHMbIp5wS0IkEMHbgvCbf9k/YlKxJGpr3Ve2RSsLHaDyvz/m6mb83nN7PFiM+MkeWuISlPm7p+cvss3KitROR/d9Tza+AEdROVHF5tvVMaIykvxzVREJb/FMmvjZ2UoZasPKq6V/tboiGFE5SkjKteaqDxfRlQiKi/Hx2SvxwwQcJ7FrA8289Ik1iVP2KlPo144fiy++ZzfqKxE5Tzo6rE9hEAlKkd8mWM2cOc9ViUq56HS/1glKvtj1U5Zico2GsO3K1E5HLN6xeoIVKJyNQwrUbkcfpWoXA43V2mQVqJyNfwqUdnIERQFNuqIymHyNGY7jj4L+O1MVF6KgLRvBkba+O0U6MoAVpYsg4Xbjsf7zzxvu+XWyf4D+ybHY5TW3bc3Iyq//uDRQliu09Svvn/3QYz++KxM/frrQoCAGc6WKPLWJUGZHp/Z3Ug7HQcR+5liuhkZGnF0MAiQ40GAPBJT5X7nkeOTx+47Nrkrvhd47MjByb4tU+bOPsDGLXZ9I991jgC5cvly+bbnM9NvVL79UWvq15S31lOl3Mln3jJ7Pvfnpo2DzhsBcuJIBFbvOTb59umTkyfiW4sPxmiQk8cOTW69bX8ErW+iqYZj5Myvr0fA+eqVyXufXpg8++bZyfPvnJm88u7ZyQefXYgRlZdjZI2pX5U4/iVM01eesDW7eXIeOt3HYCaQfzB09djhGFF555GYonm9icoLl64EbkFUzuCUKHQd3wZwXgD4aV426a48jNRKovKeU4fKKK3H7j02eSLwu+vkkame7ityuUh2N26zSxtFv0JYcirO8xcux5TMFyZvf3xh8nqM0nrpvQtlROW5i9eiXgjyI57DL4uca8csud/sbaKW+3k+0+fxzHMLUXnyQCUqE6A9tG7qhgVEZdSp2h7iBuoP6VNHuuQn4ZnV32Y/r2pSIdvZx0ZPk6g8tCeJSoXcWvpEqv96U1enuLk0Mj10wIjKSlT2R7KmXHcEKlE54hscs4E777EqUTkPlf7HKlHZH6t2ykpUttEYvl2JyuGY1StWR6ASlathWInK5fCrROVyuLlKcKASlavhV4nKRo6gKNBUicph8jRmO24jIL3T1K8XhxGVfUuUwS7PcVvIwn698QtRiaA8NnlqLYnKcxtE5aUrMSVbjNRqcJ6iMhshTBBmj28DsUkgoFqWWNkSWC2BfETlkQNBfhhReWI9icoPk6i8PLk6nfq1lHGKxbTk3UHWxLArYR4vuOWIyr1GVF4LfFJWGuBmYWmOboIx/3yJO5ekmykdu6WZ+tWIyvhu7P13+Jbsek79+sLb8Y3KTy9OClEZo42GLogNCx1sL83h6cnfNCdvCZIyiUodCu7dA0RlfjMQDko7A0MbkqUz+RRDAABAAElEQVS2pwhOp8w19eu+yX1BVD5y16E6onIpRG/ei9SRSYxfjBkcPvjs/OSV6ITx2gfnJq9/aEQlotIUpkGMT6egV5pZmUuZyZLO6mYen3dlynAzonL/5PborLIXR1QmBkn0Fv9kenAWv0w7u96Ke15lRGXUp8crUTmLV93fuwhUonLEdztmA3feY1Wich4q/Y9VorI/Vu2UlahsozF8uxKVwzGrV6yOQCUqV8OwEpXL4VeJyuVwc5UGbSUqV8OvEpWVqFxegiaTMdtxDYEWAcJFROXnAvmmfjWisvkeWRP82xqq6l+mzaBW2SoByt+UUVoH4rt3x48ejhGV601Uvh+jPz4/H9PURVD16nUB2CzzdpQSxe4UW6/ZSBcbtl3v23dlpNZRRKURlV9SonIrVDvuwU1g9fjRkLd7jq/niMq3Y0RlfI/sg/g22aUYUWl6v/lLSpqzG1I0P2n7aCupTaQ4ws0Uf8dCV9edqDQy9UIQINdipCptaqPUhmF2u4Elwdm8buvxvCrOB24benrE1K/TEZX3HV+vEZUfnp+89H58o/KzmPp1OsJtgXlLAJZa65QBWSSvb3uePFKJyqWAXIOL1JHzicqzhag0S0EzojL8kJj6tdSpU2VNnW10L/c2C51HUjfLfh6cJiPDzvNtTEN/OKYEv/3Y4bUlKl+OOuHNj6ZTv17Y/Eblr8OHa5ZNm9Xs07YhSwNg8z+vbOqFhqg8Ft+o1IElZii48+jk9vi25/79Bya37WtGjLtTEqVD7lrTVgRuNgQqUTniGxmzgTvvsSpROQ+V/scqUdkfq3bKSlS20Ri+XYnK4ZjVK1ZHoBKVq2FYicrl8KtE5XK4uUpwoBKVq+FXicpGjqBYR1QOl6Ux23El2CdA2EFUvvHhhclHZxqi8urVq0X/XZMBJsE9S0NcNtv+5/E8kueb45tBrTwvT0H8AzFF6fEYpWXq13UeUfl+BPE/O9+M/vDtu1k8stzWGS9NVNrntm1vySimzZ0miLjq5HBMxXkivqn1yD17jajMUg7EKsHbvDyPlHUhjkIwC1EZU9WtN1F5JojKC0FUXipTvzYFTcnaUuyOnQRp9po4PnOqISpN/RpEZYx+vj+C0Os69esLb5+bvPdJ2LcYMW4EL2WcRaADsA1Y8nxel3Dl8VwXeUOA+EZljHzeGFG5tkTlpcnZC81UnOy3cicGWebNI12obKac3cor5HkL3IKoPBVE5b2nTP16eHL63vhG5X0nJ3fGlLkHQhb3FQLk1o26aTa/un9zI1BkKEg0HaK2jqhsE5W+tTid+jUkbru8KeP8o5uln0rWbLKU4agTNojKINcevOPI5MkHDpdpmu+NDlSn4tjhgweDdJsSbpE+/aHNe9y4LbjlVOpXr1yJEeLnJ8+/+fmkEJXRqeDD6Gh2NojKy1euTq6XKXODqCxl3wbAtodOd2Or/5bJlHu63VJW9enJMqKyEpWJVF3vbQT2LFHJuOj5diUMi21DzW+77bbys70bhm/MBu48satE5TxU+h/rJip/XaZF2qgU+me5Y8qsiFRcd0cPv2985fbJY/cfLz1gTkXv3APhHOo9eTMvW4nKs5OX3z0z+dU7n08+O3dlsu8232/ZnedXN5P5Q4HR7cfio9vxrY6v3n8iPiJ9bHLv7fFtmEP7b2bYNp5tk6g8M/mTX7wfvdfOTgNS4c/sDnSlUUP2fBT96KHoJXl7OIMPnZx8/4m7Jw8EjnXZ+whUonK1d7yNqIwprPazd7tor+ksm8Cm3nniYPSWjG8TPRDfdXrw5OSO4wdXK9ANunqDqPz4/OTnr34yefX9s5OPz16OUUPx7ScFjN9u+F/pHx1UXwRWj0WA6ncevzMCpMcieBWBltv4fTcIhCVvw1fVi/69kLXXA7cXYmqmdwJHsnglAn37QvbGxk49677yhdGdU+wejW87PXLvibK/Dn4KyJWjISo/nTz/1meTlwM/hMaBKNdu6+3+uMehg7dNHojAyzcfuX3y6L3Hi59y9Ab6KcqfOFh7p3Xq1wJJ739jtuPK+4h3siNRGVOxXZkSlclceHf5Pmd1Po9PI2Ibdm3zeBa39Y3KaPea+vWEqV/vWFei8vOoT2I6yc8uhV5fnVy8ci0IkK1Tv7LxjRYkOrBIw791VMPm0UiSF01TF7tYttX5vg21L76riKg8ukdGVE6/iRplzGULHnmwvU6MdkzYIE5uD0XQ2Te11p+oPD+5WL5RaepXS4LQBmje8QBtC255Xet4ZhGn9h5ReWFyHlEZBAnd7L/MJt4C4rZsyNqt6mB6akTlqYb84Aeu1TcqP2hGVL4bNq4QIGWkfSNCs4hsA2HAgWIJA9KCWxg4BG8zovJgTPsaRGXYuUpUDgD0Jk/KN9h5RCXC7dr0G5VkY5lCpZ5uvdZRz1DsW+jp4fh2sVGAa0tURhzUiMoP6Wn4b3C7Fh3SlLHA5l9C0YFjJC1L4pz7zdFN/JvjDXGpA0tDVB6vIyoTqLre0wjsSaKSoUBSXrx4cfLZZ5+Fg3R1cujQocnhw9FTI34HokeCytlvzGXMBu6858pA3Osxp/ifPfd+BGE+n5y9FAGsq5s9SkcuUnkMeF4LwkMQ60gM1xc0/evfun/yaDiAvnuyG/ecV/5Vj80jKj+PoKCG7m6VQwWjLoIRovKbDwdRGbg9YKj+sQOT/fvWh6j86PNLZQqchqiMAOC5y5Wo7CGUXzxRuT+IysOVqOzxrvZSkvlE5cXJ+agzdquDQfrljJ7vxAjaPxJE0VcfODG5L7YdQyStw/JFEZVaOeqjQlQGSfl4YPdErPcOUbnZABtTDtI/2kJUPnFXGTl0+OC+4r+M6/GN+fRNXvwFROX7MXrjtUpUDgaYr7pJVEav5w2icvc7GGwjKsPPEyytROXg1/iFXjBmO448RuRqAFHZkG5jtk2bR4hvVO4FovJ1RGWM0moRldei/aZDnpZWtEajclksPlJaZpPBafZ4pi1TSkYdcjK+qSWAv55Tv56ZvP3hucknMYIX4Xb12pSojEJmG34Dg1lwGmhKELZgNL2gyPf0XLPavNAWOT4UAenjx4IYX+upX42oDKLykhGVm9+o3JSilJSCzhZEmp08v4nP1uPTSwKvPUlUXjCi8uocXBYd6sJq9niTR/ONyoaobEZUNmTbuhCVH5+5EJ3SLkxeL0Tlhcm7n24SlcW8LYJqqXOIq0ZHyzcDg6g8dTRiBScrUbkUnDf5RWz1YqIS4baVqMwipcalFcvj3ev5KR1t7FtMNXwIUXlkOvVrxAnuPhod+9ZoROU8ojJ4Bxinc1H8kRZI8QZaexvJNo4mzploNrXzB6M+rURlIlTXXwYE9hRRaXg2glJD8+zZsxs/juX+mPLmyJEjkxMnTkyOhdOMsHRszAah+yJG33///cmbb74ZBuXg5Omnn57ce++95T6r3ouzcj3+ISr/9JfvTV6KIMy5CDoLSLct3ayxW1WQ876C25tE5QNBuB0rlU42cla9z25f3xCV8eHoD85GEPBc+cg7otJIBYHhsQfKqGSy4afiuufk4SAqT5VRgUa1nYpphPbvi5G+Y994ZCCvxzdgyNiHhag8M3kpKugXBQA3RlSOfMNpdrAzAkcPyTuC1DUP+1djNKopq4yovJEBwFVKeDk6EpjOj77+f794r+ivoHpR2pCLsfVVzpaCXyivkaeVqGww+TL9L0RljMQyIuuFtz4vxMe7MVLLyLaGqBwXjZS7kmvsbBKV8R2FQlQeLseMzlqHpSEqYffZ5KcvflTqCyMqd8teF4sQ/9SnGnN3nThUcCsjKgO/tSIqo75456MLk5+9+nEJLH989lIQ5NFTN4wSu6SMY9s9fsq1qKsO7r91c0QlorKMqFwfolJHgvc+EbA6N3n+7c9bIyqjQ1X4CmO7C+TOe4nxpoXMRZAL7umIdjqCy3eGHK6Dn8KmCMbwU55947MyolKdy+/bbb3lf+vId9iIyvBTno4RlaeNqCxE5b4bZu6SOMi1NkcdUTkM/i+GqLzU+kbluCPOmwBlm6iMb1Su64jKNlEZ7Y+NEZWmXIvXXOqUmYold50ftMQFzTVNJwedXU7GCJA9Q1TGSK2mHdLUx7BRN+eijp5d2nbFudzfTLd5ka29QlTqnPvhZ22iUok3y7pZ/hW3IstbbhETQLjtjalf3w1f5vyFi0uMqGxj3BLMAnEb+6mWhsCaNU28YK8QlWcuNCO1NvS0DcmKokbX/eh5mYozcCtE5akgKu829euxOqJyRYxvpssbP2D71K+vRjz09Q8vlg5+51rfWpR+1Zh1u/xZV8iTvB0JotI0rw9FLPTJ+3VoXm+i8pKpXwtRSamakm8lKmdpyo1k7fB9G7KN43lQtniFSlQmInX9ZUBgTxGVRk5euHBh8uGHH05ef/31QlRyXBCYpoA1kvKOO+6Y3H333YU8PH48RgTG+bGMcRKVH3zwweSNN94oozjbROWqAsWpiDhcCWD9ybOIymZEJSJkw6KFJZvayFVvt3E9JwlZVUZUxjSSTz54avLH325GVAqajX2/jRuPvCFg9UbMKY6kNFrB9GpnomJGVOoxKzg85hKvqniCZR0o3XPq0OTpGFFp+tIH74qeQzH1625PJThGeZDjRu0KAL78XhCVEfx7MWTv0w2iclzc8pk5NuTdtxME6RGVjz9wvJn69QYHAPOZlllfiR7fZy9em7z4dkNUvhaOYTY85FfQGxHCRu6a4LP7ICrv9x2AGAn9/Qjc16lfl3mL63eNegHZ9vYnLaIyeuyev5xE5YhCF/Ck3CVSiErTNSM7mhGVU6IyOmesw3ImOhe8HXiZQvKnv0JUXih14G4SlU3kYJOoNJLy8QdjRGXUGWtDVEZ9gSR/O4nK985OPkqiMs6REz7XuNLX2LskKmH1WGD2O2HvHomOLYeit7gRb+uwNCMqL05eQ1SG7JHBM+G7qEdu24Wp1r0RJG9DVBrJe2jy1SApkZWn72umfl0HPyXfrRGVz74+JSqD6DX1664RrYFbsXtx86Yj375Svz79yB2F6L0nZtEw9fqNWjIYlGt6VonKYejfWKLyfHyj8lL5hltO/erdbTZFplYyjm1ZNhNsOTxvR35+myMq9wBRGW03HSUbotI3teDjF3jNVCzNbgYJZ05uaO/s8SY7V6mptA8Px2xCiEozRHz39InSUfeuE/EJiiMHJ/ui0/NmLGFOXvNezC4cy3ed39S6EiMndVR7JghenUvLiMrPL5QRldq90sMtYyDN9ern5uHax9uP23U808hWGm3qZkSlqV9PTL59Oqaxf+BYaf8anXrrbfsD280YTOab+dyodRu3X1+PKeqvXon4wIXJs2+ejc5CZ8rnToyovHTpYhlRmUKWOG2IUT7wFhHYshMpYG7ZPL4pnSFtkSldNcXf8aOH1/wblWcn72pzlKlf2yMqN8vehuKWhKYc7P8PfnT0tpClQlRGJ3CdhHQqWJcRlZ/EiEq+3usRo3rxPSMqIz51HlHZfLu4QSNwm4HO8dlDszDm+S3HY4fcp7zpiNEQlTFqvBKV/YVvTVI2Nq6LqLww+WA61TDCTf3RyMY86dq5wClvmbKRu9DS2CBvpVNfjqiMGMHX1oyofC6+UfnKzIjKTaIy4vHbNHIrEnk+649SDbtqFrjpZd6FLPki6tMTx49FHLRO/Zqo1vXeRmBPEJWNAf5NISk/+eSTQhI+99xzZepXpKQGhOPSnTp1anLfffdNTp8+PbnrrrvKyEofiR6yyEfPiTTmrnXscjQKPv/880KUvvPOO4UYfeqppwoxuqoDzn4xf9fj31sxL/afP//+5NUIABpRKYA1x/f1WKMsbGRDVGqs7SujPH74zftKgw1RuS5LGSET2L0ZjuAbMQXOB5+FIxjBaFO/cnK7KomVyhfgNZX0LZO7IwD49a+cim8XHQvi6OjkZEyzocKO2OPG61vpXrtwsddbiMrA6KMzlwvB65tjepgKAO6LBlV0AN2Vhdwh2so3KqPhgWwzihfJaxrdEgCMNA2+u/IIK2VaVCP+0c9zQVS+8u7ZmLL5gzKvfUNUtp68JF7pdlsvLtg1dulI6KwRqEa1/fZX7yyjK4HWuvvWa+venkCA3J0JudOjWacWNu99IyoRlbulty25QlTef+pIONRHJ48GaXRvdNQ4dnjf5ACispXuZgO70dtbyijodwI7Ab6/fPnjUl+Ukai7WekxetEYEeBLss0ocsEW3+kFXElys4E2fR51KLLwcsgeuXv2tU8nb0THjI8jqEzuBJT5SsUfGtnmmXHnWjSwD8Z06qdiBP7pwOy7j905eSjqC3WId7cOi5GnfBN+ig5BJWAVHQ7KFPVRhtH9lBA59VETQAi5CznzfcVHwk8xe4HvfSJ58bw3s+zlu+WnPPfmZ0VvX42OVWVEZdgcPt7oC3WdLiX4EoS470E/FX4e/Pgp6t8x9dZ7MlVb/tptC7plybVz605UKku2tZSn/SuFHfnfjScqo05ufaOy2EdlKuI6ldlZxRtgBOTnV4jKINROBPmx1iMq34upX8OP+TQ+PZFE5W/KiMqm7hwAzULJAbnWG3krUyMiKo9Fx6vQ67UkKoNwQ1SaYrJM/Vq+faeEIWozYpb7RQxjh/y0F5hYZo9nGsklaYjK9jcqT30JicpEpXud6EI1R1RuIyqjk64ORHcGOV4I3mkn+3wX3bnv3pny/uNlW5u5TAzso5Av35F96V0zQgRRGR0lz5epX/PbnoufZyqKm4kSnM0jW7bydE79qkNBGVEZ9fBaE5WfICovTi7Ft+/aerYNny1o9N8puMV7S/tmtjSd5++7vRKV/VFcn5RkaN7Ur+J5r394YfL+5xfLN1EvXZ4lKmfLuFUC23tkKvdznfqppolHKPLWDHo5GCMqj0T7bH2JyjfKNyoDN9+oDNyuT79R2SCWCLTxSzSac1nPwsWS+81e87851ySgq5WobKNTt78MCOwJolIj1mhK066+/fbbk5dffnny7LPPlvf3jW98owyVfuuttybnzp0r5CHy8vHHH5889NBDkzvvvLOQlUNedjpkRmm2CUtOmilnP/3004lRlUZwPvroo2UUp4bOKguzptMo0ujd6On3s1c+LoTlxSvXG6KyNDfcIRryq9xozrUqOFNwGklyOKaHEHj+3uNGZh2NBtxu3HHOQ4xwCKn7Xjh/Aqh+H5+9HKOLroaDPSUqR0auqVriv42oYHyTshBtgdvdMQ3ssQjkC9przDVpRyjkyFmQpWZE5fUygvLt6J37VvwEUU1n2kxFN7bENYWAiQDqwcDoxJH9Jej30J3NtK93RQDQx99LR+qRyzxWdlARG70SvQsQlTB7JggPgWdO224tTc6NU0h34WSUjJ7g34gp6RDmq+KWDacvspG8W/jtlXz1mCd3RkK//mGMIA/bZ/TGpagzdkNvU+4SPwF6snZ/BAwejDrDVKaISqTHqvKX95hdjyGXjd7eUkaevheEkdH3z73x6eSTqC80sNjr3VgSv7z/yQgaPHSXbzsdLbor+FKIvhVuPgY+i24PGvXF5ZAxI9sQ5Oraz00pdDk6d4U9ir9SH46NorzhY/Tc8agvjMB/6isnS8cM0w0Xcn6X3t0iTPqeK+8/yoDQJWs5/euHobPIy6tRjzTf0h4XOfclF6BhF8idkdAPRGcqnYPs54jE8u76FmhAujHksnm23xTsfMfOFP869RmheiP0FhHu+6hs3qMxEpX83RXfe+Izj6m32hI+W6F9YRooHTFzSRxzrX5eZ6JSObSxtLW085TVT+dSZdsN/2N3icpLJXDPLpJRM7x8FN8MTKJSGRnIjXJNVX273qUNaGqNfP8b100PpBxYk5sDa0ZU/ibe/eVo2xuRZWTgL0z9Gh10+dCfISrDVl4NP+fX0UslyzqLQWIzdC2/pqpKotJUdciPICofvTlHVCqj56Yz4hRbRlS2icpLrW9URikTs8Qwscrjub/Tun29a7cRlY9Oicqwjes4ovLixWZE5SYuW/VvJ3y2n9+ux0lUlpGoGyMqfascUXls/YjK0FtTv14p3/bcjsDskURk4/gOEOdpHZFuDf/8cIxEPZkjKqNTQY6ovPvkkcltZeTz7tUdG8+8w0bRE/Yl7NaV6DBwPqZ43Tqi8nzUExcnn5+7WEZU8h/KEuBsjfBl6WdR2+l4qWaaLENPb4Nbi6h89O6mQ8bp+05O7gzc+Bq7WefuAFc9PQICZI68XQ154298EFNZvxKdcBGVr4UfYkSlqYZXJSo9KmlMiUxJVJvGI5S6JonK208kUamN25r69ZDPYjV6Ghds1E8jwDA4i4Jb+GWlPg0/9P0YWZ8jKjeISrhtTP3665WfN3UcZpYGtwa75huVdUTl4BdZL1hbBPYEUYmkPH8+DG2Qg6Z89X1IhKVvUn77298ua8c02DmajiMpH3744ckjjzxSRlkOeYMaskZOmmYWOen+jJnGtPwRokhTwQTfpzTFLMd207kdcrcmLaPPWF0Lh0UDTRAG0XYlplgTGMzKICuH4XfovkLeyqfRIWAl4FxGeERP+zIScaNK6s5j0Rl551IqhdhPrHLtfHs70w9Z65mmF64pS60FsExRBz957wp206JF9mUEoFFFRsocP3ygjE7NANomAkNK1KRNzGavTLxyPXu+zz5MOMmCpPCCG7lLwoNMKNtuLI3cTcr0uEbynghi1/ezfNvzRIxGPRAjZ+LRll7myZ3MiiyUcq1eMEQl+RJQgdlr0RvcSNRNjV368RdeWGAJPaJayHCBe6M7vhIjZArhMRJuHqItf4md46vInev36jJP7hKrXCt7e3sZLK6FzhpxcOb81SJ7dFfngjKCfJf0ti1WvhV4Iuyc0W13Hj9UOhscCj3eF0oxht4mjrmGVxuz9vZQ/Ng1dQPMPoiepm9PCQ+jUuLUri30Vf6eHdHL3iE+7o5prOyvQngkTh4+t63HxM2ze8YymjfsnE5V/JULQVI65n5ZxrFBlC+yEmGEHFLPItxOhvztR3AwxrEsKxeJmTza27N5rpI//eHTqWvhppOBWR9uhJ/i3fHnjsR3Fo2qNIL3VPyOxNSlOhc4B+Nll8Qs1+182pi1t9tpdtqWr+fTIc2I1I8CO99GbTpm7LLexsMFRAUn9es9oa93hM07GfUuknwMvVU+RJaAoTaMdsXJkyfLJyYSm8Q217BcR6LS8yNbtK20t7S17CdBe+jQoRI8RVqu2gk0scv1F01UKrv3VqxVY7Lm6N30xEbLr3n6Wd0pchA6YS2Ij6g0neTdtx8rnV+eejCm5IxORHeePBrHA9MSyL95puJEVJoS99wcovLTaIdcjI6mV4MAQVRuQJHQ5Atd0ma5zE92TSDfCJCjzYjKLUTloZmpX/PGX8w6defmISoPTI4fI2/HJ9969OTkySDcdHJeb6Jyp3ebQjcrjB3XtZLTYVOY5ojKB6Kz2tfisx2FqLy/RVROp8yd1fmOO+zK4ca+qHfnjag0ba6pX6dEZegxbeJnbFlaZZ97fMvBrh02M3yUKeGGqLwvZnMx+tkoVJ9PWDei0mwuZ4KovHRZJ52m3AW6Wfy6INnheIE9/hV5C9zKiMrw94yorETlDuCt4+nQ0STGN4jKmBlNxx9E5fvhM5+NqYbbU782xaSzjZL+5jdbhS912VEp4hZlcbydshwu92/krRCVh6ONceLodETlHKJy/5SoLDajnVtzjxv1n20zY8NiolI7Y/Mblava5Cxtg1vLtYkTh8o3KoOojPr0m18J2xZ1gk6Rt8e09Pv3H5jclgRvALTqc9wojOt9KgKLENgTRKVGrMa46VZfffXVQhRqyN5zzz2TJ554ooyYNPXre++9V6aFlf7w4fiIb5CV3/rWtwqZuAik9jlGCxH57rvvlulkEaQathrRG70Yg7BEWmpAHz16tDSuGYyVjUZYrTCZJeDX9CT9dQnMZeXQfs6xt91X01kFJPAiaFqm8It9x1dZirMbGeTIWPsZgIBh4pbrZe91PSobQfryiyC+KeqaER5N2ZbNt+91AqWC93ArPcNjv1To/q2wwC1/sEtZg10bv2VvIU8yhviA3RXBg1gLvq367nd6JhV1wFSCzJwbRLlfErw7Xb/ofFvuZvETEIPdGIv7lOB9fDPQaBmk741ZmvcmwAyvAyF7Avi2x1gauWgCimyfpch1BA9TBse4z17Loy13cCN78EpdzfWq9s59kORGjNNXckeH2bzd1dviXkd5GrkrsjfVWTZwjHI1OvXrUucmnil7Y+ktnIq9C+II4Vs6tBRhXM1e95Vn3yNUV+iQYaQW7Bpk++YwP12Ri6nckT/vo23vxng/IXZF1hBsMIQdG7jbS/FTojywQq6Vb1MGhsqUb23Z8sFtVu5SV3Nd7hP3WmZJOSZ3ptqH2+UYLXTD/ZSp3MGvGUnZkHzL4pZYJHbtuhZufnR3jPzdi43zfV4jymHY2Dtnlnsvruy1RPYhdqV+pa/FVwkMdXoYQ/LpqnYFotInLHSC/MpXvjI5ceLExuOlDOUaputGVHp2Zc0Zas6cic8MRMdPgSIjO7SpELTHgvzQjmO7xlxuFqJSmVJil5Uf1wecxW4J4sPqRBCSd98eUzvHyMD1JiovFaLSSC02RTktzG8btzzenF3ufxKVAqunY1rsrSMq15CoLB2smykl1Zm5bFRd00ObZ5oUbVwdyf28fmNd3kHj6x0KnT0e39Rab6Ly84lvVM6OqGzb2Y2yx0Yez2OJU+KZ+3l+Yx0vgM2+NUi3HFG5/kTlucm5mMJUh4NS7g0hm5Z6EShd5/J4yaLZgdtt4SuXb8mGjbs3iEqf2Vk7ovID36g8P0FUfn7uQhnhVuRpS5mj4LNClOf7Ho8s2MYyZW74X4jK2xG8iMoY3YbkLd8oryMqp4K65qt42RtEZYyoZ898Z3ELURkjAzemMNVem+pqquxsXdpb1CSM7MgxPRUTOBKjJm+PDlI+zfG1+/cYUZmi0gVcnu+5LrgHhrJDVJ6IToqIyqcrUdkTwZpsnRHYE0Sl6VYRh0ZNvvLKK6XRYrSkRryfxixC0ShL36408lJD+MEHH5z84Ac/KIRl35fI0OboTVO82hY8kF/2uNDARoYKvmSPZ8bZb9aB7XvfdjrGStjN+otYGE2Nm2I8V3yAxERDUyPASFVYemfIZlNbtaecWBW/xKy8jxWffZnLPX/A11Tay2Qw5xpyJ7hCDm0rm4AE7BK/ct8VX1gRtyJ7KswbL3xwy/e/YlE2UJQf3MgevYUf7AQCYZf327hgyY1N7NZfb4vuxPunp0Y8wI4NdtwoD3pLZwWg67IVARilLpI79QcMHYcZmctpdhK/VWTwi5a7dj0xps7S06xnbcOK3Alewy8x3or+sL1N7L44excFGa2+SL2FF51VZ1iTO2QH/FLmhiE1P3WI9J7wU+BmIVMpd/SW/VNXkLesZ5NwW0Vn3Wt6y2IXmrs7emOWxj8ZT+7yqbO+oLfkDkZwI3dZZ4yht+7XyN4Xp7f5/seweam3cOMfy1s9e//995dPWNxxxx0JcTlnJ+/v2nUjKtWH2k86l2qzISqT3KZ/ZEWZfbbDZzzY/NS7DSBW2CCbiNH333+/tCnd75vf/GYhhtnHtAc73SLfgeCgHvlGxpw5fymmdI7vt5lyLUYyvPHRhTL168XpNyqbkYE0UAtvukw3il7mMes8sJFwerJ13Cky6Fk8O3tl5OQ9U6Ly6w8cLZ/yuOtUjKg8YkTlvoIlBcpy5nqa+66vNnALzH4d33y6EiMVzkdg1awGz76RU79emHwao6UvBqZGoBeMs9zxhGk/V9U/1yu/mRQOH4rZIU5E8D4C+N89fTJmFTK185HJsSPaqNGx8QvuoLeBWzw0fbkWenTlyuUyZe4zr5+ZvGjq14/OxhSTFycXZ6Z+9VJnxShfdMI6HVSzPWEmyAumGVnB7uCBmEEoRqI+HIHVb5829evxyYMRnD4ZcnjrbTFqZmZk4Bcmb1PcroeNuX7NFH8XJr9488zkhcDt5QjoC+xfCttw7Xp+a7Fo17TU29Gb1qCduCZcW9eN3hV/FsFbpn6NEZUPndg29SvchtijrfdZfa8tbzni6HLI20cxA8lrH5wr36h8oYyoRFReKHrc6GVglXB1yE6vp2tdC+uip6GDhw8eKLJ1b3xyAlFZpn594MTkrpjlYN++A0VPbybcELhGuH0ceml669eCqHwpicqzvlEZMzBNO/klbBv45IEWFuXc9Hjav0y2gXskcsmGfUuisoyoPBxEpdGox1tE5f6oF5o2/Y3Wz42y1o2VEKCv9PTKtYjZXLoyJSqb+vS1+EalWUjOIipbIwOLoobwpPwUMWvJWh7PB8tTW463djxD0dOoTzeIyhgN+LXwQx4OmbsvZno4FSMDD0Vd20z9uulvfVFy55lLfRr1gpjN++G/Pf/WZ0Hynp28Wb5ReaHgliNR+XCbRc6tRCaRGrCOS10tJ/ZTm0nHn4aoPFGmA0f2njoWnfaizkhfOPHK9YA71qQVgZsKgT1BVGrI+gbla6+9Vr5PyQn52te+VqZ1ve+++0rDVlDOiMuf//znZc3gICp///d/v0wBO+StZKBPHrYZMcbMPTyLBrZGrgDWV7/61dKgTuMh3VhLmsCx8uubz3glYHhjGsDAT3BCQMWIWAEZAYjbb7+9BFCRlvDzXtcdvzGxy/clsELmyB4ZhKngjeAzDFVscKvYJWLNGh6Cp4LOH3744cZ0zrB77LHHClk5JmZ593XXW/LlR2dh99FHH5WAnmM6hgggCu4JijlWl00E4JFyh9xVT8DQAjMEuSC0qf3gxzaOtXwRb2I37F2SRYL28GP/YAU7dTr9hfGYursXsCN7fuoIda2fzlbqh9OnT298S1s9uxvLjcZwTNmDiboCdkli8FPaU3Dm7Bnr7qeMiVvKEV2EF1sHP7LnmI58fBQ/9q/qbSK2uU69VV8g7qzJmHrWjDHWuaTNy7VrYf2rX/1q8pOf/GTy4x//uLSBpEe+/ehHP5r84R/+YWkv3XXXXSWbvJ8d2zdy8dzpj7388suTv/iLvyg65xMdOlSYFYcepq1Xfm08bQTnhy6JU/s6Os4uqlu0K9nHJCqHtOMybyQaws1IhSQqX55+G+pNRGVMUXwxdIM/Va4JDLYs01eQb2LjbG7kibyoddwpu0azk5n9Qajl1K+mfE2i8s4kKmPGjaaMm3XAFyEDiiKouklUXilB/A2iMoKFn55riErf3IIb2EYV18gPdsoPu8MxAiSnfv02ovLeICpj1Naxw9OOeTclURm4BVbPvPZ5Icff+ehcISovhbzxoxoZVcqdlw2x7JK3zKJ1PjyOoj/HYhS0wCrcTGEqsLoOROWzb52dvPBWQ1R++Om56GxwMUjMpjNwU9xWYbP803WeadBNjPNoJs7jmRsfLYLzMTT/YMSPjh85PLk/AvlPxNSlTwTZhhy/88ThINuibXUTEZWbgfwg3ApRGWTbu+cmv0JUfnKu2PStIyoTh63lD20rf4nOwnXrUnJc9LQQlc2o8fuCqDSKt4yoDJm72YhKBgtucDkfRKUOBO98fGHyekzD+dK7vlFp6tcYURnEEVtYloRtITD9TzY207c9mxGVPqtz3+2IymZE5SPxre3mG5WVqOyP6s2Zko6UWCui8mIQlfGNyleNqIxvVJI536g8N/3Wonq36NQ8fWzpXa+SpszGdVFLFz1tOv7EiMrowKLTypMPhK6GP3LP7UG4BVGpUxCiUr2bBuFG+yFZtsRNfSnmr8PK829+PnnlvTMNURn2Dm4IXvg2uOXVi9fwsKgnLZvQ5tYUPLuxae9AdMQwlfpDd8eIyoeb7xab+rUSlRD8YpcvSka/2FLv/t33BFEpYGk0JZLrpZdeKg1WDctHH320EF4amhqBiMpnnnmmNPYFNwU1f/jDHxZCcwjUDJFfe7GfhJHGtG9lCrz4RqaG9JAGbjvfvbxNqf0EHlQAcHv22WcLjt6NqXv1nBZ4FoSQdhb3vYxPn7LBxFTEgirISkEelXsGAGEoiDNPZvvkv1fTwI1TQfbYDwFAhJsgoCAY+wG7itt2CYCdH51FjhvN/uKLL5aETz75ZBnpIYCvo0amrXrb4AgPWJA7Afs33nijYOgszASLBe2Rlci3Kn8Nbu3/GgzqWgF4+LF5ZE3Avt3BoMpcG7Vo5Ez1Fl7qC/Wt2SXUD9/4xjeKvcsOQVuv/HLvJW4pd+oJckcG6Sl9pbe2yaH6t8peIzNp7+CBAFLXwk+dof7VIS1/7F+1d9t1LeVPfaGNw+6RRXK3l4hKdaJ2mnJqz2nL/eIXvyiA+ESH9pR2FX9X3Zif9vAJD/pHfoYsMESeu6d7Z5DJMX4gf5p9dC/1irZIux3XV8dzROWVINQEpI04eit64gtKvxfBwc/jW7SCXEZqNXlubVtmmcJ8l2Wm6ZmnF65dgwDZF6NijsR3o28/bvTC4cnpuw9P7o2p/k5GcPDINDhomlNpl13Ia3vpi1P7GtsNUelzEzFjTJC8HwZur8Q33t8O3GB4RnAw3t21mOJ58x4Z8pvNbfi+t1ACqxEeZNM3vhl4x5EgjeK7ULG+Pb5De/jQOEH80XCLl51tm2tXr5TvHP8qSKM3Ixj94WdBfsR3yEon65D5AK6UsqCz9bVtAjYVx+lq8/iOWzI0uw+ZOzy5744g3AK3h+8+Mrnn5OHJ0ZDDMqISMTf1TXbMspVgFq88tSkLeaTf2nVw+zVbcD06TseI3Vfev1BGuL314bkYwRvTl8aIQbaCiHfdv7lbymGGohc9QyLb4OVKeRvdbNSRUbsPB3H0lcDtofjm9okgkxK3JpC/KO/t57qeexXcXMuesqWfh3y9Gzr65kcXCwFCVy9eipGocd6yeX/lzbKXU87mRu91ySHuL99bip7ui84DB8o3oh8ImfOdcp0yfG/7ViOfI83NgBvdM/pKRwv2DW6wevfTSwU72+oLdUORIgUdDs9CHOMRCm6Nfds3OR643XXi4OSBOw7H70ghyX1Ldp+R9lNifGGGC05uvvcm0bLytuAWe/LUeLixb6Gn4WfoNGVGgrc/Plf8EDL3SexfiO+781PUvY3lSju2OrTySxFuf4PX99wfuetQ1A8xU0Z0wjgeMxSoa/kqPh+zrNDP4pYlWEbusj5l3z6JeuD198+FD3chvusZMzuE/3YhRqj61ETpUECpPPUOutqkyqdasJ5J6DvifN17oiPGV+9r6gQYHo9vftLTW0NPiy3c6QE6bjkmbh232JOH2VDtA1yFn3ZDjaeM86r3DFEpcNQmKp9++unJo482RKUGLgODqPzZz35WeskKMj3wwANLEZWgn2fskqgUhEmi8jvf+U4J3C/jGI3zim/uXBhFjj8nFm5GvApoISp9g0fw2UiFDBDc3KX5Yp5O4FnQWS9wQRyyhqgUWMkAzjx5/WKe9ua5K0w4IAJjiEqjKgWoYMd+JFF58zzxzfMk9FawA1709oUXXijOkZHspqSjs4L2dZmPALmjr+otwXsL5zNHkid+VW+346euUEckYcTmJVGpzhfAr7htx80ReptEZZKV5C7tXa1nu3Fry91rMXsHGUROZj2bnQuqrzcfQ3ipa9Wzi4jK+Vd/uY/S2zZRqT2z14jKJAiRg9pP7JOOUGYGeeqpp0qHCn4uoladSc/Ul4hKM9fQw76L+sH0suoQeWm78WeyLWKbvLKVbKLOazp0eA9+lr51jHR+1+PbqVcjmCUYeOZCdPK6cHVy9mKMbrhyvdyXTzATk+pbnK3pMpONQJn7N7YfZr55fCi+R3b8sG+S7Y+g/r74lu/+CKz4DnKSRrLcyGBr/jvszWLUF6fZbBM3uCAjz1+6NvkkgoJwOx8Y+v7xNaRSBF+bEOhsDmPtC9Y239UqZOWR/ZO7jh+YnDhyIKaZjIBU+QZy4rYcZp405SqfelncXF9kyUjekLnzEZT+6OyVIELiUwMhb5fje9vNt8pjlFbIRYpL3nfsNeIbWVlIkMDt5NEDk2NB7h4IeQtmaaPcs+Xf6Tm60i+LW8pbWceIoouXyduVyafnkUjxeZDA8XoE+Xf7m8fMC3mji3A7GlidCsxOhq6eCNk7FKOeA7T428RuJ6za53cTN/bzctizMyFnMPsssKOrfKfrocdlmVWRFMDZ4+2H3mlbHlPb7Pt3vu1+JHA7ETbuZOjpicAObuzfGLglhikzOz3evPNFTtUL7NsUN1idvXit4HYh5A9ppN7Yat9WAar9JE2ucms6sURnjNDJI2HTThyJX2B3LLBL3Brslr93YpZPUMqfO3XdicBYuDV4J1kZs8OEfBU/JEZXnr1wbVLkLerZ69ezPk3F7Hy0pU408qY+va10xlAXnDrGH4mOGeGXqBecS5Jytvx9b+q6vLYta+3tvnm5Rp3qpx74NOoF2J0LXb0Y9SmSckNPdwe2jUdVnx6IDhfHQj/vKPXp/qgj4rNBUVfQY3pqybJvXNhzo+u6ZXDrecu1TwYb5KRBcWJ32iziAbYRlnVZDYE9QVRqzOqBK3ikFy5FFTA/ffr0xrRAGsFJhFlriCIqf+/3fq9MV7gajM3VbaLSsyBIv/vd75bAfdtojnGvvZRH0xCtROWy7zRHVM4jKhG9Q3uaL/sc63ZdOh8CgKb4mkdUrluZbuTzsqFkTweQJCq//vWvV6Jyh5fQlrt5RKWgaBKVO2T1pTzdJozgN0tUwq8u3QjMEpVkDVGpY9CqwYjuu67/GXLHj0Ru8O8qUTnsnXYRlTpTIZyqn7IYz71OVCZxqNMY/WKnkIPs0uOPP162+bjIyrT72lg6NebMNYsR3DyrDk6/OWciodsC7X6I4CQr2URBB0GIriDOZs6bW+6Ri227SDWBaYTllWu+FxWzesQx5EdJkBfMW+8UI9683barncoA4a0RwCvkUQS8Du43HWwT3HI8/sqvSd1k06fMTfmamSKS7HVdu1d5n3zccR5ujiHXkJNXA7OrsQ23ct/ymAsK3xRjxf8xqjIQJAvwOnSgIUOQSU1AdXtgsE95Z3GDneuQ4/nrk0+7cJmnY23cyNq1lrw1NEX7yh22ZyFOeew6Ps0O6QajglsEoZFIyKRbI6hahDLStcvY3l70RBk3gJlyus77GYqba3PZ2E55C7wEoQt2AvhB/kpeip7lz4tHXae8NQT5gdBTuMHwthnc+uKVjzcPN5gldn3z28AqMs5ta3pZ7NvV0Fcyh2ijq0mJ7zJubNhtIW9JWOqY0dg4ktjcXBn7ljNxUzb1AnmDoesFpxO7vvklVvK1TfoKboETrK6GnF0J7EpnAnVD4pYPMura3RtMSr3Qwm1/6OgGbkCVckncsk7gQ1vUpXBbJr+SwZfgH5kgZ7Dji1h0yCVzQ3CblTf5NPIW8izv0FF1KnkrdaoEuypz8g/7FrJU6oX0Q2LNL6G7qUu5LldMZdB216Jc+SNrfvZhlrrazrMrn/bxzM8x24jcrBOKH3JDcYNcU8/RT/6buoGtg1vqcruM7e12uWa3U9Zy7XzWCSlzs9fU/aZTmHoBzuyaGSDF3XUk9dNGqctqCOwJotKoHsFyjVdEJUV7+OGHy8+3TQRANHqlef755wshATYjf77//e8XwnI1GJurEZV6+yJC542o7GswxniWdcrD+6LoAhDzpn4VSE1ncJ3KdaOeNQMugjjkXOWSU79WorL7LTRORzP1K6JSADpHVObUr91X1zOc57S9voHFvukgogMIm8uprjZvu5yQOzZP4FmdpaONBWaC9pWo3I5Z+4jGR9a1Oii1icrTp08X/Nrp6/ZWBJKoVN8atUTukqjM4MHWK+oeBMgdso2Px79LolJjhN5qoGRgviK2FQE2D17zpn6FnR85rEs3AnudqMzZGdSJr7zySqkjffPaiEk/pCUyUzvuueeeK+049Sii8gc/+EFJ043e1jPkUR3CZ3ZfsqkNkm0R59hJ8ipIgyyl3/wZP9f7LbXEZcLTLkdQLpvNMveOR99YlCPJyY2DAzbSt0ss2Edkr3eUI16Ncs1OV9olmXbAbTaSwqlcH0cKZrDbOLv7GwldkQEcG1lY4raJm0uVJwlxMkcOnYcZeyjIRf7IpbTLLg3uDW6J2grZDXuMAAlOZC8xa2PQN7PZa7L9ATPBfPIFLz+2Am5NuVfDreiohyzy1/dpV08HLwspa2O3lNDJJzLxg5s2B3mjs3CCl18SSO67qrwFx9bkYS3D3V428GputIGZcq9wb5iRrZQ39QKZ4ytrq9FTGNpvL0PxKxgVrHRcaeG/2+Btw029oCSN3LXLNGQ7cUu/WTuNDwMn/l7G9aRrL0Nxa1+7ztuzOKQvAjedqSziednOWLU+lV9jH6dr++Wgf7u4zMpb275tFYVeD9HGTXnUBXw5ekreHNPRDG6pq+PVp01tekNwSzQCIzCVH9tmf0aHMumi9ew1MGPX/NQNcBPDg1nK3Kq4LXqedTsHPz/1AlmDH8zUo/xeulo74I7zVvcEUUlIjIZCNmjgaigxTL4N+dhjjxVF03AypZDAJiXkzAqoIyQI0xiLfFUoiMrXpiMqTf3a/kblGPfZa3lkhSxo+stf/rIYSiSyqTcFAdOhUTHXZTsC5B92KmXbcDL0nBPNIWQ467IdgXRq2Ib8RiWnEMnrm21j2YXtd94bR7Lhxt61iUq6y7mpROX899yWO3VWm6jk3JC/DO7Nz+HLfVTDN+ta+CVRyTnUMamOqFwsH1lfqDOQlXS1EpWLMXM2Ay58vPQj+ZnkrR1AmA1Y7Zzz3k/B5mkAq2sRvWSP3wc3Poo1OaxLNwJINZ+3IH8axXzjvfSNyvbMOC+//HIJaJrylU1HFGqz8TkQlc8880zx2RBjiMo/+IM/KB1Tu9HbeoY8ZjCLXNoW4HI8/Rr+tDYjP+bRR5spxTM4Id0ySxNcEmQSXSp/07UzN2ZpgmrTAFvstPeXfQK6nAEbckq/2UEEM91OAmRZ3DxXCchBruDWwm/Zh17iOs8Pr831Epm0LpEP+SP7fkhz7Te+TE4fpvMLfJddEq9mwEWRvGWzWvo6qFGZsXDzIHxAnUv5f/QXWSSgmkFpeut+yy4NblO5i51GQ2+cnnruTdwaPV2lPPJjv8THcvYgOgsnbQ64qYNXnaquINSoZ7lfyp/736ilaCl5ixsSgTIqcXlRKDqZ8padW+Cmrcv/s03+VlngpDJo8GrkruyvkunAa2dxS/kbmM1GcraMbrJr/BbtDTidPn262Dh1BJmsy3YE+CP8G7jpvGXhC6kbxq9PI/OiszfuXRR1DOVMHW3vl8Iu8Y99zHYaPRWbgqNYHtzYObo6Rn2aenqjcQNL6mXWp0As+C2BWV5C1tSldFU7Td7qA34I7Pjfq+CW99lLa7aL/5axd/Up+5axd+0Wsby6rIbAniAqKRjF0jhCEGpcUjhOq2+XUDDGXkOAs8ERQ4IhKvXYZbzGWDJ4qjLOEZXf+ta3ijOjwq4V8nyUGT+Vi/dmxCvFR+4KYAnGMJYcGhjWZTsCSdTTgSQqM4CqgqlE5XbMHFERc2LYBcEvAVR2Q8UsQEb+6tKNgEoZXpxBI9nZtyeffLLobjZ4q83bjh+5Y/PoK4KcY2iBGX0lf5ybVQMG2++8N45kQ0SdTm/ZPFipK8ykMFZ9vjfQ2l6KNlGpziV3OmZwqmvgYDteeYTc8TU1gBHk/BQ+Jnmjt6mzMKzLVgTYvAzI01tyxwYiMWBnXf2UrZjN7pG7vUxUqg8F5JQRUanBrwPFo48+Wnwx7ThBzyQq6aA2l3bcX/trf60E8WYxW7Sf7Q5rPzJqIaeehYzqkCAo6Dm0GcfyZ7aEA7fsLHriEc9No1qrBrfyiWDHNvKl4ab96/3xB+Hm3Y1hF7dAtWUnn2SX17uAW8YN2EU6Did1MbuoHSewuuqyAdXGxqo5LnF9YDeWvLm7todYC10le+kDqo/50PR2jKVAtodw097QZiNr2nBwEsgXXB27g+QXit3Iupp+M5KXrrJpBkKIE8CQvRtr2Uu4kTEyl/FZepozVtX2RrfEZHsDbuKifA+f1lE3kLcx6lN33zBtGxvdzzT6mZF1lB8Ct+yMwY8U49NhKge+jFovbAFwdHQWZzhyfZp+r3pBLJRPrB4VV2HjxAnqsh2B9N+QlXwSi7iANi1dtV2X1RDYE0Qlw5TCovE6O3UQw0T5pKNslE5AExlGCccKkHgG9+E8ewaOzG/91m+V+1SisltQM2DAATQyS6NDpeI9cZ69s+rQdOOnUtazVIPNNlnTwFXJaOySw7psR4BTw4nJhpvGh4oGdqYwJX916UYgiUqOtJHsHGnfkqK7dFaDZKzAXvdTrN8ZcsfmCeoJGCRRqR7K4FTit36l2/0nVo9zqjmGgtZsXgapNEjYvbp0IyDgIpjsR3fJGqKS3tZ6ths3dUUSleSODGqEZGMuicraoWo7hmwevNg6PjJfjw1k7/zG9MO3331vHIHbl42oFNA8ffp0sU382DZRqZOPNpcRlT/84Q8HjahcJBHklC+oHQdv9/3ud79bOpxWf2Y+cqnf2iB8GrgJ3OvwZ9QRDMcM5M9/ivU7CrdsvwkO+qmDYab9oW4ZK7C6fugsfmJ1iXqYXaSzCN2sS5CVtd07Hz/tDnEyNg5ucBqbGJ9/5/U+mnECPjM91V4zo0ESR9W+zX+/2YElOzTTU9+UZuNqe2M+Zo5mO5eN80ksiw5TOmapE6q8FUi2/ROXUqfy38STtduMRB073r/txmt+QF2gLmXbtM/gJv6uTmXjtG/rsh2B9N9gJybFp4OVNi2Zq0TldsyGHtkTRCXBoFRIBsE3o3t++tOfFkeMsKgMCRPHQiBTw1dAXUNApen8GEsqOkem3ROXkleishvhJCqRbXrAwFHQlIHU4KhEZTd2zpBtlYsGiMANWcsAqoqmNtjm45d2QwOEzqpokqicnVJtfg5f7qMcwiQ9XouR7IJ4epjqncvWVqJyvnyQOzaP3HGmsxcWPVVfIcoTv/k5fLmPasBp/HIK4cf+kTW2TtAafnXpRoDOaojwlazJmoAyva2Bg27ckqhMuSODfBPB5OxQRQ4rUbkdQzYPXmwe/Ph6bCB7l7/qp2zHrX2Ef7KXiUr+q3YTX0I7gB4Zkae9psHPTpEhgbtf/OIXhRBDXArc/e7v/m6ZHaeN17Lb2elVYNWzkEuf8BBYre247aimP6PdhqhUJ8NNIFWHP7hVf2Y+brBTH2u/5U8dDDM98tUrlajcjp0j6hKdFdhFsqfuTaJSnVLrk/m4pZ1FVLKn5CvjLZUYn49Zypu6p01UiiOK75E19q52ZNmOX9YL6lP1uphrzjQHs+ovb8fMEb4NX4TM+SSWRcct/o76tBKVBZIt/9SnGZfih+ACtNvMnMiHrDO3bIFryw5ZU5cmUal9pj6oIwO3wLRtR/xJLAV26lYyiKhMgrcSldsgG3xgTxCVWeoMhBjN+LOf/awonEqR8yC4SWAQlYyWHyUcc1EhE1ZCm0SliqUSlYtRTqKSgeTIeI+CphobGmoq5RpA7cZQBSPwp+Fmm+OXRGXtWdqNmwolOzgkUSloADtEpYqmLt0IcKSRbMgOU31ZTNOm0ZuOdG24bceP3CVRSe7InGMau2we+auBve245ZFZwqhNVGrEVaIykZq/TqKS3iZRaUqhSlTOxyuPptxlMJ6fokFC3rJDVSUqE62ta/Yt/XNEJV/PMf6dXw0sb8Vr3p62xV4mKvmvgnI5Iw59014z+40e8epE/oaAp57yfF4Lm/+9732vrOfhNvQYH5p8uk9+wqOOAOlGkR57VxmQ5tO0p34VIKwzRGzHD278wCQqEUd+2roZVK3tt+245ZE2Ualuyc5q6hI/9qIu2xFIolJ9AjdEJd9Pe7cSldvxyiNZP4nvxteH0gAAQABJREFU0VPtNZ+Wqu3dRGj+Wr3A51OfmnlJTNZMczpjVH95PmaOiq9o2/KJnnvuuZLQzDdJVMKuLlsRyHaGOpUfIp6cfmTWqbVe2IpZ7rWJSr413PgflXBLhOavM56iPlVHkME2UWm7LqshsKeISk5/GnfBJBVke1FBajD57UYguBKVbbT7b1eisj9W81LOEpXIoSQqBQFrz9J5qG1+o1LwqxKV8zFadLRNVOpBT+4qUbkIseZcBqg4NeSuPaKSvtYRlYsx5EALsOTItiQqOdV1ROVi7JxNx7oSlTtj1U6RcpfTG5LBJCrpbQbjaw/xNmrNdgYQ2LwkKvl9AsqVqNyO17wje52oTLtklJSAJruuLmTTjcyjXwhEPeWRmc7zbQU8dQgVbB9jqUTlMBTpNtuo/YsEEcj3foz4yKkRd6O9Pewpb77U6QeSY3JNv/0QlcgPIxn4NHVE5fx3py5BfsCsTVTW+mQ+XnmUjuZI1CQqzS6mLq5EZaK0fZ2dZHL0jLonZxCyXUdUbsfMkTZRqaMVgq0SlfOxah/NWHYlKtuoLN7OdgZfkp6SN74JH9KAIaRbJSrnY5j8hU4YOeMN/zuJykq4zceN/yaWol3bJirVp/y4itt83IYc3VNE5WzBswHleE7Zs5sjfFLRGcg6onL2bXTvJ1HJOOoBY+h+HVHZjdfsmTZRyWi2R1RqtFWichaxZj/tg8qFzqqg4VdHVM7Ha/ZoJSpnEem3nwEqcleJyn6YtVMlYVSJyjYq/beTEGDz1LkabnVE5c74pdxVonJnrGZTZACBzatE5Sw6/fb3OlEpaE631IlG5LFPbJVgiSlg2Sn2ivzw0wQ8tRMQlWbIQeqMsfCn+YKzIyr1yK8zu2xHOP1o7V/6LWjjG3iwym+We3d1qrqt2KUfSJaTpCTbcKsj3LZiNW8PcaTTAsySqGQD/Gq7dx5izTE6ivxI3BDhOc0wW1uJ8fnYpbxlHQSn0zEtOZK3EpXzMXM0iUq6mkSlby3WEZXdmDmTRCU/pD2iEm7q0zqicjt+6tSc+pUfwo/UbjMKNevUGg/djpsjqad8ER2nxOXVBwg32FXCbT5u/Dd4qU/VEWQQVnwQuOlgWZfVENjTRCVoCE0uu0lSukclKhPpYetKVA7DazZ1JSpnEem3zzZwYjTcKlHZD7N2qkpUttHov03u2DxyV4nK/rhlyiSMOIYavxm0FqCqIyoTpe51JSq7sVl0JuWuEpWLUJp/js0TTGbz6G02hOuIyvl4zTu614lK+iXIpJw6evq+kG9R8m8F58yIQ/e04wTWTQtrxB4CUUDF+TEW9/MM6pbscGoEiPtkh9cx7rNX8kh/JolKum3EFmLSDBuCNRnI3ytlHqMciVvKG9n2Q1QiPwS6jHAbS67HeOabKQ9+DP8ZZuwGeeMDwsy6Em7z31aODEzcyJdRMzAb047Ov/v6HiVv4gQZlCZfpiVXF9muIyrnv1v1Almjq6/FzEsINlOYqk9t1xlI5uMmvqJu4Ic8//zzJZEOpXCrROV8zNSp6oIkj/hv/EqYZZ1aicr52GmfaZv58X/FqAza4IfArhJu83Gjozo2snHqCDIIK/WpuqHiNh+3IUf3PFE5BIxV01aicjkEK1G5HG55FUOpl58goAq6jqhMZBavVSicmEpULsap62wlKruQWXw8A1SVqFyMU9fZJIwqUdmF0OLjlahcjE/X2ZQ7DRK9nDXs9JwU3NOY0yCpgZf56LF5laicj03fo3udqCQjfhkQfvHFFyc//elPSxBAox+Bw7+lZ0ZQIsEQlYgw58bqiJrtOIHVJCqNADF1WCUqt0tr+jOp3wL57KN3MjviaPvVX94jiZv2G19GvWJaTri1R7hVonK+jKSdgFkSlUjK/FWicj5uiMo2wUu+dBhiYwWmq7zNxy395gxKky+dZZC8laicj5mjSYCQOSPckqhUn1Z/uRu3a9eubRCVL7zwQkloCvwkKusMBduxU6eqC9Sp/BAzO4gvkzV1KhtXicrtuDlCT/kgfvwR7d0kKmGHHK/LdgTImrYZP0QdQQa1UfghFbfteC1zpBKVy6DWcU02cPW6ygaub6fUBm4HYNPDlahcjM9OZytRuRNC88+rUCpROR+bPkcrUdkHpe1pMkBVicrt2PQ5koRRJSr7oLU9TQZc+Cl16tft+HQdSbmrRGUXQt3H2bwkMuitIAK/r46o7MZs9sxeJyqVl5wINLFRApumihMAENBERAreCZjo4e37L36CKWORlJ4h23FJVAps1XYcZOYv6c+kfue0uQi3Rx55pLyrOqJyO3aJm/ZbBgjJOjK8EpXb8Zo9kn7MPKJS56FKVM4i1uwnUZm4JVEpsFqJyvmYOaqTjOkkk6iEWyUqu/HKM7NEJdyeeuqpEhetRGWitH1dicrtmOx0JP3HJCrN7KCd0f7mcyUq56M4S1TCTX2QIwMrUTkftzZRqY4gg7CqROV8vJY5WonKZVDruCYbuJWo7ACo43AlKjuA6Xm4EpU9gZpJpkKpROUMKAN2K1E5AKxW0gxQVaKyBcqAzSSMKlE5ALRW0gzwVaKyBUqPzZS7SlT2AGsmCZuXREYlKmfA6bn7ZSAqQaE9kL6FoDD/NolIciTQKYDiZ0SzYOeYS23HDUMz/ZnU70pU9sMvcSPfs0QlIl6AsE7F2Y1l+jEIN/bCCCN4CRBWonIxbjmiEm7spw5DlajsxsyZWaISEe5TE3VE5WLcKlG5GJ+us5Wo7EKm+7g6tT2iMolKg4XoqZlvageW+fi1iUr+CD+cf12Jyvl45VHtBT6veB6fhAxWojLRGWddicpxcCy51AbucmBWonI53PKqSlQmEsPWKpRKVA7DrJ06g4l6mfr2hGCi6dj0XlNR1292tNHa3M4AVSUqNzEZspWEUSUqh6C2mTYDfJWo3MSkz1bKXSUq+6C1NQ2bl0RGJSq3YtN378tCVJIVP4E6QSd6116MOONb+Bm1Z3/MpbbjhqGZ/kzqdyUq++GXuFWish9es6nSj5klKpOsrAHpWcSafbhVonI+NouOVqJyETrd52aJSsS4b1TWqV+7MXOmEpWL8Zl3Vp1aicp5yOx8rE1UaqOJy9cRlTvjVonKnTFaNUUlKldFsHV9beC2wBiwWYnKAWDNSVqJyjmg9DjEqalEZQ+gOpJUorIDmB0OZ4CqEpU7ANVxOgmjSlR2ALTD4QzwVaJyB6BmTqfcVaJyBpgeu2xeEhmVqOwB2JwkXxaick7Rb+ih2o4bBnf6M6nflajsh1/iVonKfnjNpko/phKVs8gs3q9E5WJ8us4iKvnM6mE+YB1R2YXU1uOVqNyKR9+9SlT2RWoznTq1EpWbeAzZqkTlELQ207aJStOqW+qIyk18xtiqROUYKE7zqA3c5cCsROVyuOVVlahMJIatOTWVqByGWTt1JSrbaPTfzgBVJSr7Y9ZOmYRRJSrbqPTfzgBfJSr7YyZlyl0lKofhJjWbl0RGJSqH4+eKSlQuh9vQq2o7bhhi6c+kfleish9+iVslKvvhNZsq/ZhKVM4is3i/EpWL8ek6W4nKLmQWH69E5WJ8us5WorILme7j6tRKVHbjs+hMJSoXodN9rhKV3diMdaYSlWMhGfnUBu5yYFaicjnc8qpKVCYSw9acmkpUDsOsnboSlW00+m9ngKoSlf0xa6dMwqgSlW1U+m9ngK8Slf0xkzLlrhKVw3CTms1LIqMSlcPxc0UlKpfDbehVtR03DLH0Z1K/K1HZD7/ErRKV/fCaTZV+TCUqZ5FZvF+JysX4dJ2tRGUXMouPV6JyMT5dZytR2YVM93F1aiUqu/FZdKYSlYvQ6T5XicpubMY6U4nKsZCMfGoDdzkwK1G5HG55VSUqE4lha05NJSqHYdZOXYnKNhr9tzNAVYnK/pi1UyZhVInKNir9tzPAV4nK/phJmXJXicphuEnN5iWRUYnK4fi5ohKVy+E29KrajhuGWPozqd+VqOyHX+JWicp+eM2mSj+mEpWzyCzer0TlYny6zlaisguZxccrUbkYn66zlajsQqb7uDq1EpXd+Cw6U4nKReh0n6tEZTc2Y52pROVYSEY+tYG7HJiVqFwOt7yqEpWJxLA1p6YSlcMwa6euRGUbjf7bGaCqRGV/zNopkzCqRGUblf7bGeCrRGV/zKRMuatE5TDcpGbzksioROVw/FxRicrlcBt6VW3HDUMs/ZnU70pU9sMvcatEZT+8ZlOlH1OJyllkFu9XonIxPl1nK1HZhczi45WoXIxP19lKVHYh031cnVqJym58Fp2pROUidLrPVaKyG5uxzlSiciwkI5/awF0OzEpULodbXlWJykRi2JpTU4nKYZi1U1eiso1G/+0MUFWisj9m7ZRJGFWiso1K/+0M8FWisj9mUqbcVaJyGG5Ss3lJZFSicjh+rqhE5XK4Db2qtuOGIZb+TOp3JSr74Ze4VaKyH16zqdKPqUTlLDKL9ytRuRifrrOVqOxCZvHxSlQuxqfrbCUqu5DpPq5OrURlNz6LzlSichE63ecqUdmNzVhnKlE5FpKRT23gLgdmJSqXwy2vqkRlIjFszampROUwzNqpK1HZRqP/dgaoKlHZH7N2yiSMKlHZRqX/dgb4KlHZHzMpU+4qUTkMN6nZvCQyKlE5HD9XVKJyOdyGXlXbccMQS38m9bsSlf3wS9wqUdkPr9lU6cdUonIWmcX7lahcjE/X2UpUdiGz+HglKhfj03W2EpVdyHQfV6dWorIbn0VnKlG5CJ3uc5Wo7MZmrDOVqBwLycinNnCXA7MSlcvhlldVojKRGLbm1FSichhm7dSVqGyj0X87A1SVqOyPWTtlEkaVqGyj0n87A3yVqOyPmZQpd5WoHIab1GxeEhmVqByOnysqUbkcbkOvqu24YYilP5P6XYnKfvglbpWo7IfXbKr0YypROYvM4v1KVC7Gp+tsJSq7kFl8vBKVi/HpOluJyi5kuo+rUytR2Y3PojOVqFyETve5SlR2YzPWmUpUjoVk5FMbuMuBWYnK5XDLqypRmUgMW3NqKlE5DLN26kpUttHov50BqkpU9sesnTIJo0pUtlHpv50BvkpU9sdMypS7SlQOw01qNi+JjEpUDsfPFZWoXA63oVfVdtwwxNKfSf2uRGU//BK3SlT2w2s2VfoxlaicRWbxfiUqF+PTdbYSlV3ILD5eicrF+HSdrURlFzLdx9WplajsxmfRmUpULkKn+1wlKruxGetMJSrHQjLyqQ3c5cCsROVyuOVVlahMJIatOTWVqByGWTt1JSrbaPTfzgBVJSr7Y9ZOmYRRJSrbqPTfzgBfJSr7YyZlyl0lKofhJjWbl0RGJSqH4+eKSlQuh9vQq2o7bhhi6c+kfleish9+iVslKvvhNZsq/ZhKVM4is3i/EpWL8ek6W4nKLmQWH69E5WJ8us5WorILme7j6tRKVHbjs+hMJSoXodN9rhKV3diMdaYSlWMhGfnUBu5yYFaicjnc8qpKVCYSw9acmkpUDsOsnboSlW00+m9ngKoSlf0xa6dMwqgSlW1U+m9ngK8Slf0xkzLlrhKVw3CTms1LIqMSlcPxc0UlKpfDbehVtR03DLH0Z1K/K1HZD7/ErRKV/fCaTZV+TCUqZ5FZvF+JysX4dJ2tRGUXMouPV6JyMT5dZytR2YVM93F1aiUqu/FZdKYSlYvQ6T5XicpubMY6U4nKsZCMfGoDdzkwK1G5HG55VSUqE4lha05NJSqHYdZOXYnKNhr9tzNAVYnK/pi1UyZhVInKNir9tzPAV4nK/phJmXJXicphuEnN5iWRUYnK4fi5ohKVy+E29KrajhuGWPozqd+VqOyHX+JWicp+eM2mSj+mEpWzyCzer0TlYny6zlaisguZxccrUbkYn66zlajsQqb7uDq1EpXd+Cw6U4nKReh0n6tEZTc2Y52pROVYSEY+tYG7HJiVqFwOt7yqEpWJxLA1p6YSlcMwa6euRGUbjf7bGaCqRGV/zNopkzCqRGUblf7bGeCrRGV/zKRMuatE5TDcpGbzksioROVw/FxRicrlcBt6VW3HDUMs/ZnU70pU9sMvcatEZT+8ZlOlH1OJyllkFu9XonIxPl1nK1HZhczi45WoXIxP19lKVHYh031cnVqJym58Fp2pROUidLrPVaKyG5uxzlSiciwkI5/awF0OzEpULodbXlWJykRi2JpTU4nKYZi1U1eiso1G/+0MUFWisj9m7ZRJGFWiso1K/+0M8FWisj9mUqbcVaJyGG5Ss3lJZFSicjh+rqhE5XK4Db2qtuOGIZb+TOp3JSr74Ze4VaKyH16zqdKPqUTlLDKL9ytRuRifrrOVqOxCZvHxSlQuxqfrbCUqu5DpPq5OrURlNz6LzlSichE63ecqUdmNzVhnKlE5FpKRT23gLgdmJSqXwy2vqkRlIjFszampROUwzNqpK1HZRqP/dgaoKlHZH7N2yiSMKlHZRqX/dgb4KlHZHzMpU+4qUTkMN6nZvCQyKlE5HD9XVKJyOdyGXlXbccMQS38m9bsSlf3wS9wqUdkPr9lU6cdUonIWmcX7lahcjE/X2UpUdiGz+HglKhfj03W2EpVdyHQfV6dWorIbn0VnKlG5CJ3uc5Wo7MZmrDOVqBwLycinNnCXA7MSlcvhlldVojKRGLbm1FSichhm7dSVqGyj0X87A1SVqOyPWTtlEkaVqGyj0n87A3yVqOyPmZQpd5WoHIab1GxeEhmVqByOnysqUbkcbkOvqu24YYilP5P6XYnKfvglbpWo7IfXbKr0YypROYvM4v1KVC7Gp+tsJSq7kFl8vBKVi/HpOluJyi5kuo+rUytR2Y3PojOVqFyETve5SlR2YzPWmUpUjoVk5FMbuMuBWYnK5XDLqypRmUgMW3NqKlE5DLN26kpUttHov50BqkpU9sesnTIJo0pUtlHpv50BvkpU9sdMypS7SlQOw01qNi+JjEpUDsfPFZWoXA63oVfVdtwwxNKfSf2uRGU//BK3SlT2w2s2VfoxlaicRWbxfiUqF+PTdbYSlV3ILD5eicrF+HSdrURlFzLdx9WplajsxmfRmUpULkKn+1wlKruxGetMJSrHQjLyqQ3c5cCsROVyuOVVlahMJIatOTWVqByGWTt1JSrbaPTfzgBVJSr7Y9ZOmYRRJSrbqPTfzgBfJSr7YyZlyl0lKofhJjWbl0RGJSqH4+eKSlQuh9vQq2o7bhhi6c+kfleish9+iVslKvvhNZsq/ZhKVM4is3i/EpWL8ek6W4nKLmQWH69E5WJ8us5WorILme7j6tRKVHbjs+hMJSoXodN9rhKV3diMdaYSlWMhGfnUBu5yYFaicjnc8qpKVCYSw9acmkpUDsOsnboSlW00+m9ngKoSlf0xa6dMwqgSlW1U+m9ngK8Slf0xkzLlrhKVw3CTms1LIqMSlcPxc0UlKpfDbehVtR03DLH0Z1K/K1HZD7/ErRKV/fCaTZV+TCUqZ5FZvF+JysX4dJ2tRGUXMouPV6JyMT5dZytR2YVM93F1aiUqu/FZdKYSlYvQ6T5XicpubMY6U4nKsZCMfGoDdzkwK1G5HG55VSUqE4lha05NJSqHYdZOXYnKNhr9tzNAVYnK/pi1UyZhVInKNir9tzPAV4nK/phJmXJXicphuEnN5iWRUYnK4fi5ohKVy+E29KrajhuGWPozqd+VqOyHX+JWicp+eM2mSj/mZiIqvVPLLbfcMvu4N81+JSqXexWVqFwOt0pULodbJSqH48b+VqJyOG6uuBmJynWoTytRuZy8DbmqEpVD0NohbW3g7gBQx+lKVHYA0/NwJSp7AjWTTCVYicoZUAbsVqJyAFitpOSOzatEZQuUAZtJGFWicgBoraQZ4KtEZQuUHpspd5Wo7AHWTBI2L4mMSlTOgNNztxKVPYFaMdmNasdlu0dA0s8+guPWW2+d7N+/f7Jv376y7bjgmyWPS5f+q/N5rfMW+bFXec5aennL98CBA5PbbrttFEIl/ZnU790iKpVBmdzPYj/L6RhMlM0PDspqcc51fnzWxGRe+ixL+5q8Vn5wy/eS+ZebLPEv73WjiMrED2aJg8cmBylXyuS5nLe275eykvg7Z0kM7PslVplOmpQ3a5j7jbGkH7NbRGW7PFk+65Q7ZYCbH3wSC8elS5ytXeM8DPJnP/NNvOzbhr+FvM3Lv5xc8t+NIirbWLFfcLDAKu1PYgCfXMiHNBbXyKctN5mvtcW1iZfr4AszebevK4lX+HcjiEpl8v5TZpQtfx49y2adGGWRXJM4yyNxbMuotImXtaWNJ8zkmzJXEqz470YRlcrhlzjAQBmVKXXO2pJypazO+1lck9fBDw7W0ufxfB/ulXmnPEs71uKe6ob33ntv8sILL5Rsv/a1r03uu+++yeHDh0uZVrlX4qU8trPsWU55Z/kOHjy4Rd4yvWfMn2Opf4lH+x7uI621e8hbOu/Ett+qi/vdKKLSvbJMypW4zeqdMmV9m+dS7uThOtfbdjxlKM8572ffAi/vwzrTlhMr/tttojLfu7Wy5C8x9PgpD4lPFimvhWPKUMpm2ir7mWe+C9fbzvohcZvNP++zzLoSlcugNuyaSlQOw2th6hvVwF34EGt4Mg3VRx99NHn55ZdLRXP33XdPbr/99smpU6dKpdw24GtYxF195EpULgevSk0lhjAStBdg0Rg5duzY5Iknnpjccccdy2X8JblK5X/u3LnJhx9+OHnttdeK0/Too49O7rnnng1HekxHaq/ASu7YvEpULvdG6Ww2fjXi6Cxn9eTJk5MHH3xwcvz48eUy/pJclQG+SlQOe+Epd5WoHIab1GxeEhmVqByOnysqUbkcbkOvulHtOAE1tpgfwI+yL9hy6NChyYkTJyZHjx4t245//PHHRYe0iY4cOVICWuyRZ6VXfDGBK3Ufn0uezrnWTxo6KI1877zzzpKP+63qo6U/k/q9G0Sleyijto5AlWdWLvg5luVXNhjAT1DKIr1nS6zhIr02pfT8fb6DIBlMpXdeOr6F/F3Px9AmkD4DXqtgl7jJX53ih3TzTu66667yTMriucZYlAkGZI18KJPnVxbtbFgkBs7Dwb7zAuPSukY+cLKfgXz+rOPKkj9plIW8Kg/cxgwQKou2x24Rlcqf7z7LrJwwgKGykRvvSBlh5VjKKtlhsz2n6+keuYQDrGEnbd5HGvspD/Alb66Rv/RjLJ7n/fffL/LmnvJlV/LZxpI371/+ZIlNcF/4kSX3gwNMlJkuO2fxPNI4Dmf4SAdbmMjXO0j5sk0GXJ/67J2wo2PKm/fJZ/ZO6Sq90N7wjsayB8rE7sDKWtlhY1v5Yda2b/CwwMrzqSfaug0P75WcwsMiv0zjOvd0zDacyRp7kOnLRSv88374fGTu9ddfL+/3G9/4xuTee+8t28o1xuL9kzfYpd7BjzwlbtYW8qLc5My7I/PKD+f8wQIOMCaH9FL+sFIm6eUNYzIAt5TRMcqTdmE3iErPDi/38FMu5U7bnfIAG/KW/oJyuTbllBzBJnW7nZ785D0yfzLqPvCDedantuGd8rwsfp7NsysHXXj77bfLM5A19/I+PeMYCwyUQ5ngYK2cZCDtkPtZyL9zyggX520njmm/yJDjFunzfVhLm3m369NVMSs3i3/Kkn6I5/Xu6Avb4f3TlVWWLI/7kDnlcQ/7WZ/Cq12fKlvKEBnzXNaJJdsmfeLp/TtHxqxda9t1sFMO1ygL3R1jkb/6LXVBnvJ3nzFwG+MZ1z2PSlSO+AYJrAqSM/Pmm28Wg/jNb36zVMhjVmAjPvJNkVUaokpULvc6VGKwy8qSrKlgGEqGfyyHc7mnu3mvSocLbpWoHP6eOAIcjEpUDsOO3LF55E7jDYYWepqO2piO1LCnu/lTZwOB81mJyuHvi6NPZytROQy7lDuNuXfffbc0sDSQNJTobQYT1b912YoAm6dByubRW0EENjA7o1lXP2UrZrN72havvvpqaRSre2GmQ5XGcC5wtuRaQx/Wv/rVryY/+clPJj/+8Y8nP//5z0sabZMf/ehHkz/8wz+c6Kkv+GFxTQY/cl1OfEn+7XY7Luv/9J28HzpBP+DNjngXAmt8eM8jwMv+PPDAAyVwJJgl2EOfnCcP9EcnMeneeuutkmc7CEnfXCfwdP/995f82a9VAzZZntTvsYlK+fsJBMrb2qLcbLH6zHaWTSdX5YMdPD0XfOkPrOEumCm9dhL9gSssYCcvuAoeZsBbeue/8pWvlHdjW3BV/svqSOKm/ZYBQvdUf3j/SYK5zypL3gdO8PODAxw9O18TBhnIJTP8UjiQqazfPIPnhKdnhF8+m7Ty8/zuY5tMyl85Hn744VKm9GtXwS2xSD/GPd3L83hWP+9+1YC09wIn+duGi0VZyYUFZt6Vn/vSJXrm2aThJyRmzsHCNWSUDFnkTT5h5h7kU3teergJso/Zjvdsu0lUkrf0lZQFhvx0ODrOvmWZyANZgCkdU34yBx/5uM7x9K3gRf5g6p2TP/vwci19hhU72PbJXLfq4v3sJlHp+dNWkRvYWZTTNp1L+VE+Mt7WPzL0zjvvbNhI6aUhb/B2rfTePwLH+7B4J+5hgT2/gj0kz2MsWabdJCrJChlit9m37MBAdugRDPzSVyKTykz+yBa5sbge1jCCBSJa3lmfJlbuRc9Tp+EFt7acroodeWAbdouoVH7lINfK3S674xYYwI0dsmZjldtzkR96R+5StzO9epLswc77h6k0WZ+6N8zb9SksV60X3M8793ze8W4RlVkudkiZ3At+yqpzRPoV5EeZMk5B/9h9smIbDrAhs/Kko3Bw3HuBmfPKQx7kJU3Wp/AeAzfv2rMrj5936j0rh/vRG7qyyqI8cFIm8qU8FmVzP2UjY+pS9t+9yRudy/oUjp7Psyo3G+8a6ekeDDO/xAx+WZ8+8sgjG3XPqv5BYqEs3l/qj+OwIv9j4Jb3+TKvK1E54tsnsIxWJSqHgcogMlqMSR1ROQw7qRlk2DGUKgMOKiPPUI7ZwBn+ZDf3FSo1lSDcKlE5/F2lk1WJymHYkTs2j9xVonIYdlLTWY4q5zYbAJxWDnUdUbkznpz+SlTujNNsipQ7DaVKVM6is3ifzaOzbB691WBlAzXY+SjWGt116UZA26ISld34jHVmt9tx7IhgGp/d+0zSwH3piYCLwHL++PS//OUvS/Dq9OnTJfghgMP/okv8f9ep/wRi2Pef/vSnpW6Uzjlp6ZtgkHbBQw89VOpKhJ62guPLLvKXd+q3oA37KGDneQSe6LZnWWaRP8yUFWFLD7QX4SXopbzZ7kkf4Otf/3ohKzyD8wL4nomf7xp4OAdrU+k9+eSTJeDlPs7zK7wXP3nDRzkQ+nwM91Emba1lsUvcvL8MELq3PN3LPZAFSUYsg51rst6CGxyUTf2vXBblcC/ydjrkS/qXXnqp2OoMqsLI8wr8es9kBnau9S688/zBG76O+7HtOlSQOYE7ecJeOVdZ3CcJCfcjX/DyI+OrBiLJ22sxUwy8UmbkqfzqL2VzL9g8/vjjRd5g4jjddp0O63Bpy5ugKpwFWOEgkIoI8X7YBXWk62Fs5Jm0rklicxXMXAu3tDmei+/sHcHMPVeVN+WHEZn2ftwLFsrlHJlRHrKg7vfe6LLngrNnoI/kTQcbckofYOuYNOTYPeCvDO4jD+8HruwO8mhM3DzHbhKVygMjMqMzEfmDhTKTEYvywgLBQ1/hx/aSFzjDhWymzXI9nNks5CZsvJef/exnpSypg3SeHJAB8vzVr361yES56Yr/lEtZPN9ujKiEj+d3D/bJfeBBTsgEeVZu5Vc2ZYYRu2sbRuTENh2Ej2th/dRTTxWdzPoURu5HT61dw3aybeQt69MVISuX0xXPSHfGnPrVc/t5L8oJK3UjTOige2b5YAc3+qh85APWrqELno19I5+eV3rySX6s2Tf5Seu9+JFNx2GuPpVv1qfq0mXrU6Dlu3FP5dkNojLxo6vpVygXLOFGJhBVZE352Dd+nvOpY3SXrfKcjsNQuWECG8fIs+PeCRsHX9iz1d4HnSbXY9Wn5CH9EPdmV+iGdzMG4Ua30t9VvrQ5MCNP9skXHcr6VF2h7CmjWU86Bidlh5mZ3OACw3wv8+rTp59+enI66tOsT8bQUzbGe3LftNOVqBwD2c08KlG5icXKWwSWcjDKdURlfzgZREaYMapEZX/cMqXKDnYMJScgnS9GvxKVidL2NYdD5Qg3OquygZ/KuU79uh2v2SOcBRWzRuJrderXWXg698kdm0fuOLjp3HDK6KsGEkeHU1uX7QjQ2Wz8aijRWVhxqCtRuR2v2SMaPnSWzVNvkDWB3WwkrdJQnL3XXtpPudOYq0TlsDfL5tFZNk8jWBCBDdS4ZPOs2b+6dCOQDX1+iroXZnVEZTdey57ZrXZcBrjkTw/YkBdffLHog6AL++Jc+u+CKeoz7xpx5JyAUQbs0x65ls8qvaCfOvHf/bt/V4JogozqRTbe/dWVFromuPqd73ynBNRWsfnpz6R+k09l81xjEJXshLKqs55//vlSPm0ebUZYOS/YZd+2egzBI+AlcAprAXL1HV8Lnq5L/OiRgLTrHGef4C3IqX6EDRzhJSAmyMhHg6Fzy2KXuClLBggF8DyDd+m9uY8yrLJ45+yt8vu5V+LnGeBm3z2/+93vlsCqwDgc3Fs70jm4wdECB7IoCAtf/r+8Pb/FNXCBN98MfmQTwZJBz1X92/Rj3NOzjU1UKs8vfvGLUmYyrTxw8NwwVTb3Vn9961vfKvJBN+kB+Sc7GSylC2RUeroq0CyQTS/lQ97cj457H3kvgW6YCUhLO8YCN+0OcgA35aED3plnW1XeyBK83EMczH0sqcf0ThkFmWHgnuRQOvJFr7797W8X+/Hss88W3NgRz+c6uCIw4UzHpbcNRzKLADDi6HQEpBEmsFtWR9t4e+fekXfqWd0nyQLb5G+V+8hf2Z577rnJf//v/73olYA9+VJu8qOcyssGsUfK6By76Nls+8E4cfZ+2RL6x1+A0//4H/+jkOPyIld+bA2s5E3mXDPG4pm9F/KwG0Rl2nE6R1/VE/SNvJFvC5kmb2yPMsOIHpBT9pZ9SuLI8/rRCfWINP/+3//7Up+SV+9DWjLrnXnn8PM+yK00YyyekS7tFlHp2ZWNjpIfdQQZzp8ykDk2jeyQdWWE6yuvvFKeix5IA0N42IYFPMkSmVIG7z1JdHil/LbrU5g6t4oO5TO4524RlSlvdPW1qPfINbuu7LBz3v3JCfkhb8oujecji3Aho+TMe3YcbmSOzKbvoW61SA8Xuuv6efWpe6+yeBbv04++es9sMzuQdfYq+cNJB4k33nij3EN55Dtbn8KAHpE1MqFOpAPw9lyeE3Zwgils1JP0TnrH8AjswGx9ys9j29zDexljcQ965F15PxbvUn01Bm5jPOO651GJyhHfIIGtROVwQBlExpohqkTlcPxUirBjKDkfnAYVDEOpshzLIA9/spv7Cs4BpwJunHyVDfxgV4nKnd8dR0HFzCHgsHE8OJ6cKBU1x8mxumxFgNyxeeSO85bODT2lrxwv+HHg6rIdgWwocForUbkdn52OZICPzVNvkLVKVO6E2ubIFA05QRENJsEP+kpvNejprPq3LlsRYPPgxebR20pUbsWnz14lKvugtHqa3WrHqfMzuKLe1ztcD3P2QkCaHeaD0g8+gaCL4KC1fbpjnT/XsTuC9QLygn9skGDtP/kn/6TUjb/zO78zMcUvv0x69xQ05K9J//f+3t8rQR5+2rK+Wvozqd/KMDZRqX2orhfo8vww4i+djkC9oJNnZ5eVy/Zjjz1WSCVp1HeCY9pJfHv2WvDNcYSc95J+K3tOz9wHVq4RAPvd3/3dkqfz8hQsXNW/TdzcIwOEgmze05hEpToe+QE/74g8ZcDeMwiKGjHk+N/4G3+jBJjJJRsNd3gqL7wEteFtumhkkPOu/3//7/+Vdy4/ciXQCCsBWvdXLu1RAUXXjRHASz9G3vTK+/Bu/dxLeVZZlOt//+//XUhEMk23+El0khyR8Z/EdNru/f3vf78EQDPo6pzAdbbBBe3h8Od//ueljal9KWDqPSsHosA1ZIFsug9yTltKHvAXnB1jcT/2h8x5ds/snbqPcrnXKkvWU2I5CA35IXLJBflQzv/7f/9vKSu9c1yZ4fO//tf/KvqIMKdn8lBuNgyGbDMZ/Mu//MsiewLPcJKH+7reuyJfbOL3vve9QrLQqWXtW2IhNrCbRCX7njL3b//tvy2xib/7d/9uKTuZZvPYdjjBlPyQOzr9n//zfy46hhghn9J7z/Cj9/JGNv3xH/9xeec6sqQuw45OqkfS7qQvm2VfZe35PDuZ2w2ikp6pE5HXCF469Fu/9VvFbnvn+d6s7aszldc19JudI1vkn83yU35YSAdD9al3/4Mf/KC8DzJnUa/A0Q+O6lOdEFaVNXmzrcri/Y09olKdl/YTbn/2Z39WZOR01KcIInUeXWW/vTsYw0PHJtey9+I+5NBx19FT6emf54ahvFzr+bP+yfqU7kvD75GP61fFzTO7n/vvFlEpb2VUVvaNHLFV5AcW5Iw9Z1vhwrZ5l2wXPUw/znN6XrKXI1DVWfT7mWeeKfLGbpEr8gwnttP5rE/ZPzLKfju/ykJP0w/xzr1n9YHnH6O+5k+p/3TKcR82ij/A7rgP2/eTqE9hxd8iO8qf9RU8XUN/lRcO5Nb7SALSOe8j5c05x7Ke0BnNe1LnjVWfeq/kId+rd+BduM8YuK3yTvfKtZWoHPFN7lYDd8RHvCmzYhAZJ4aHU8CAMyiMESND6ceoxG7Kwo/wUIwx7BhKlSTjzvAzlPBTidZlOwKchEpUbsel7xGOA6egEpV9EWvSkTs2j75WonIYdlJXonI4Zu0rsoFaico2Kjtvp9xpZGkwatgJamTjKYM76t+6bEWAzYMXm6cRXInKrfj02csAsEaxupd/XEdU9kFuWJrdasdp49ABvjoiSNBWgFNwSiCez04vvF8/wRRBKMEOabx/hJOADxukXSQQJjAqkCWgJdD3f/7P/5n843/8j8t9EE9/5a/8lRKoYZeM4ETCCdDK9x/9o39USATnlg0Spj+T+u3Z2UdtNgFdwTttkGV7/Gf7UCDrL/7iL0pwmN+pbQM3gU95w04g3znBP3YZbtpErk2dcU7Z2SIjtjw3nOVDpwQBTb3IN3NvAdW/+Tf/ZlmnJCVey2Imn8RN+y0DhO4tb5gJECqDIO4qC3kR0CNr7qldKAgob3UaOfyf//N/Fqz+zt/5O4UAIWvkFAbeJ0zh5Hp4kCtECWxfC3KYzEnrWcnk7//+75f3A3dtelMXKxcCk80SnHT/VZb0Y3aLqERC/Mmf/EnRGfcgM4ghnQrIiXL/m3/zb0p9hlijf/D0LpXbghxy3Pvkb/23//bfinwJ0NINMsfeCKxqR7neNToYwJF/wQ6MuWTg13PSibGJSuWkh3TLj1/0R3/0Rxv6Q94EpD0HQlanAvJI5oz0cz2s2Tf6h6xFdJJbaZADiE7y9Ff/6l8to5bIInn9T//pPxU5d0/4JYGcRMgqOJJ1z+YZYMe2IAXZE9urdlyQJx0VzP+X//JflvfCPiPHyJt7w0f5vTdl1mGAHfsX/+JfFLn6vd/7vVIfkLe0b/BmG9m4v//3/37B9F/9q39ViBZ50GfyqCy7ES9iN3aLqGSPvBc2is3+j//xPxY7RS7oF9lmu7LOJE907rd/+7cLwfFf/+t/LXjKR/2bIwfhQm/hQZbVp+7xt//23y71qTrXgpByX8QdzLM+XaVeSBnlL6gbdouohAuZUoY//dM/LbipT9lnugMTNo6v4gcfBJKy6RDA3rNPMEME0THy63nJG/2Vl3z4HY65lrwhzK2ds4xRn8pHfrtNVKoLlJNe0UXyl2Qe2WLXyIR3R47YL3LkffKL1A2ulQ5+cOCjwZH9d559Ux+wY86TZ/avXZ/yr8ix+sh9vZ9VFnrKBvntBlGpzOwXohJmYvzqOfYfPmTsX//rf13uz27Tv3wmuJEdGMGTrpEz+kuOTelK39UbcEe+qw/Ig2vcx/lso6+C0+y1laicRWT8/UpUjojpbjVwR3zEmzKrbIgyLJWoHP6KGGbYcUxVACp9lVYlKhdjqRLjGMCNw8YRhR/sOFgaIHXpRkBjiZNQicpujOadIXdsHrnj7MPQIpjHYRPA4eSPHaCY9yzreIzOZuOXs0pnYaWxkL2J17FcN+qZNZDoLJun3iBrdUTlzuin3GnIVaJyZ7zaKdg8OsvmVaKyjUz/bQFSjXl+SpIu/BSN81zgbMm1xr0AkYa74PCPf/zjEkSRRlDqRz/6UQl2ClRq/Ftc45fbZeNL9G+32nHembpKT3zEjSCXbe9KoEmwm20WaBPU4gsI4gh2Oc9eC8wgNfj8znuHRtH4kQM2ymijf/bP/lkJ+AgIIlEyKKh9pbe+wKTgt8Aq8igDhcu8Zs/Pn0n9Jp+7SVTymZACymskgrXnZ1cEwdRpnsciHcz5CTDjJ6jv+PjeB0yTXEOe8fmVQ1AtSUM+hUCidXtJHWkfG7KduHmuDBDmPeniWEQlm/Ff/st/KdgoGzmTP3+TnLEP0pAnBCRbIIDKRxAwRTY5L73AqFE1iHHpPS8SAIHOPsEW6SQN+fJOBMIFKN3bCDe+BplWvlWW9GM8g3fpXfOd/bR9lXOVBVGJwFV2+Xv/CNjTEcR3LzJiJJtyk0NBUM9CplwLW8FYOMHFO6a75IbeCpx6D/RdXuTUswusIkIEZMcg2GYxgBsd8jzKRSe8K5jRC/dcZaFrOhSQHYSGReA59YdckRc6y/awXWSOfBjZy34J9HsOxAhZQoQk5kgP5z2rDgTkCW5wR8gL5gviw689wnDVcnk/7rFbRCXfyLMjvf7Df/gPRX7/wT/4B0Xn6KO6IolKdRQ8/+AP/qD4A//0n/7TYnPpJUKSbsEToeI9szV8BSM0ySWikt5LDz/1A1kkC7Abc/Ec7ACZI+fu4b0gImyTg2UWZVLf0Tn1jQ447Jx70Ue6550rrzTSJoGN/HU9UhgRScfppfqWbrJz9NCzqS//+T//56Wd/rf+1t8q9SmM5cevanf8+Yf/8B8WUl1eq9YPnlvdQJ/YDQs94QuwJ/RhmSXrHXEH70TZ2XjlISNkgQ33/Gw7+60jCizYP/el2+TRNXSPPVc/wDl9GDLIjrHD3g/9IVvyRhzBt72sipe8lG23iUp6433wNWBHv8gx2VJm7807U1Y2/nTUF+TKs7kGnvCGBwzoAly9V9ilfyhfNg62OmrIm+1xbyOH2W52lW1kC9jvVRZ66p5+/z97d/qsy1XVD/z4c0CZh4QhYbgJQyCJioITMSRRCIgFhWhJRC31j/KFhVBBC4gUahCBENRcY1QEASWOYcgNg6iILyjKV774nU/j994+z3DO83Tv/dzznLt2VZ/up3v3Hr57rbXXXt/uPnSID6V+9fKxyNycRPe1+8Ih+Q0LcsC+wwhWznvQRP3kEJ76y+aSQ7JGp7WHrMkHS/fCB86uGX/zL92BSeZT9/awb0VUzpGKze4tonIznDbK1WuBu1Hle5yJQWTci6icNogMchGV22NnsjT5FVG5PXbu4CgVUbk9duSOzSuicnvs3EFns/i1IODwc0A5qRx2C6dK6xFIgM8CoIjK9TgtXoncWSAVUbmIzvG/2Tw6W0Tl8Tgdd9WiXWCkiMrjUJp/rdc6TgCF7RU8FmgXmBH0khIICsEm+ILgEKQSaBHUFcDyyb7xW4MhjTxRLtDMJxNYffe73z34ZgL5yABBIfaL/CACBLyV/6u/+qvDdfVNDRTGn4l+9yYqzVkC94gIwSkki7YLriF/zWtwhoXzCSRpn8C1IBcfwTVrJwE4GBsD5coH6/gVCMy8sTpfui6VENy0wZxiE+gVEDaW/BntnEuwCI7ed999w1stZEAAUNDO3u/UD0dEokC1AB8cH3rooSEoLSAIm1tvvXUIjpI3eWCOXEEeabs8fDDBfuVZHyClvOkET+XnM2zkb06KH6NeYy2ADi+btrUiKgVYjRVy4tWvfvWw95suCbyyy67xQekzn5SOywML/XSNXCHpyJ+3YOi0Y/jLbyyMC1lUHvzo5VQiZx22cOtJVNIdRAZyA3FIz7zpxwbBhEzASN/JAlJJf+GA4ERusk8we8Mb3jC8TYRMgwNZFqgnU3B13TV5lalO8ijRZ0SJMaBLc+WB/NIJ401XlafsVm9U8o34lewzfaGfb3vb2wailg0QrIcLDGDFLtFH9vDtb3/7YLMQbOcObSL5hzVbbN7RVucRA+pBVNIbD2AgSpAFMOqRyD070ZqoTNzQWBhzcyp99FtfY+PIjTbYsw3mVPaczSCfZM6DQ8pDBHqwB3FEXuUxHu95z3sGzO+8885hPGDvmrEYP/iT+ZTM2+akXRGVSB32m7ywS2SBDdJ+8sjfIHeu02OyiJx1TRuDKdtP1pyTnKcfbBi7ZwzcCzvzh/m7ddJGbaAfbDG5MK5sLT1lM+baATYA0Wjs6SR7xv7wI+gQOWNj2Td9JW/mVLi4D3YeRpCHjPHPPFAGczrJfiLAlRsM6aiynSOr7IM+ItxdUwedn5OMD92x9SAq+bv0k7yRiXOHOupBC203RubT84cPU7Kv8IIprNn12HQYZT7l15EruNPbPNgD14y7c+5RR+bTuXq5iHH8S3bV+EjaZDzi1y/eU7+3Q6CIyu3wOjZ3rwXusZWegYtxODhc9Ubl9gNqwoIdQ8mZNlF6EoahZNRNmJWWEeDUWBzCzYTIqYcf7DzRU29ULmM2PsNRNzFzFjghHAAOBufARC1w0NopGNe/r8fkjs0jdxZvcW7oKX3lnMKPo1ZpGQE6m8UvJ5bOwoojb2EEv0rrEeDI01k2z7xB1gR7LOznBKzX13g2rkTuLOSKqNxuTNk8OsvmWQQLIrCBFo9snn35KcdjWkTl8fi0utprHSeAZq4XRBGwGtsQ8xYdEMjLmxICauYyAWvHCJP7779/CMiy3a4JcgmsIoA8kc8nE3j93d/93SEQ5lN1riuf/RIMct2bmebL3/iN3xiuWzNM9dXiz0S/d0FUwoSfmc9wart6BVxhoy36q20J+JrbYmsQgewNvEJw2buHnglAmicTWEW8qa9lCm7WbwkQClQaix5EJfkRTDbPCxKSN3Wxw/DiBwiswhYu5FQQHimEAIaxN0OQ4wgSwV/thhUiU9uVL/gqEA1r8u6aNyX0E5HC12hBjMSPUe94HOmFte/cgLSAqjcqF4lKbTd2YhX6RV6QbeSJDPKt+KVwFEQWKE1b2Ba+qvG1z9pdHWSPDssPQ/pJZo1RywS3XRCVgvFsjbHwv/usqSNr5IJu6a/x0kf2ENHINlpPwoyseVuGPPl94fA8HZeHfHlDUJAalgL83pAjj8qjr/CHpTW9+uakXRCV+gCzj3zkI0Obf+3Xfm2wz+QAmS3Qr//GkJzcfvhJXTL327/924MuItJgJXbBnslHrrI+ot/K8YlFvphPSrJtLYiOddiyLb2ISn2LP47AQTyaZ82L9JSN03f2QSJD5JHtogPeiIY3/YMX20TmELrwVT6i0nxKV8mb+ZTcqocNcD87oa5f//VfH65PnUvHGGbeYkt6vVFprtR3NlqiS+YGcsLG0Um65gsMfiMqYegcuWPzYAE3MgcTmEkwiAySa/ojv7LJnHmoddJGbTBWvYlKuGUdw1YjpdgcfdQGfTcfkjfXzVNIX3Mq20gnyAxZZR/JK52ElfFw7D72K0SlMlwjt+aSvFEpj3rmJHoaP2SXRKWHVDKfImDVba6Ao7k0W+ZT8kMGJRiw67Cn5+ydmAYbZ66Rlx7zfzOfttDNMc7aQBeMV2J5fCnjUUTlGKnpx0VUTsdu6U4Cy2ll/CkKZWKQTYocp9YKstSAPT3BoJiUi6icNoBZ7DCUnAGyVkTlyViaHDlVcKOzJhv4FVF5MnZycP5NzByJC0VUbgbaYS5yx+aRuyIqN4btYkY6m8VvEZUXYdn4gDNPZ9m8Iio3hm2YK7KYs4B3bCEq0CZoZiFlsWT+rXQUATYPXmyehWgW+BbqIQ8sTCutR6CIyvXYtLzSax3HX+JfmrMEm+3ZYnZDsEZQw7HACxvCllhDxke1pvQ0PtKI3yAf4shbSYKrgjGCYwJhd99996Bntx8GsgWuE3gMqeQTiwKL3thBBMxZn8afiX7zo9lHwXXBt5CC2jYlZX2IOBMU1nftZTuUz35I9IMfyidlS1KfoKFz7klAUXAV1spOQFEZmRsFIR3DWKBrn4lKmJw/fFPB2AugkTPEhD35ggsZc0wmrMXJJrzh4Ji9hiM8kHJ33HHHEEAlzwKrSAJ5lCHeQSYF6gQ+EX5kUiJrgpAwNW/OSRmrnkSl/1FJ38i2fnmDhZ6J5cAGcQSDBJLNb+QfZrDwRl/IOHIGZxiSQ8lcKK8HCOCewOo+E5X003h7ww2hYZxf//rXDw8V6DOdCw4JrtPPC4dyKoBP7uSBk+tsI7nha/FZjYeyyfLth/aNPMqrXnaNb0u3jZM35+zH9kAbpiTjrH52hlwbX2XTGcfGd06Mj+zou7dRvTmvTz7Nrg9sKZuqf/SYrJARbyRpizfo3U8v6R7ddr982uQYXjD0Jpg3KuVHPO0rUZn4DdkxLsg8RLV5Fvlz7v8exjD28gYH4xTiF8Ho7TUYwgim5lT6bGzhRYbNp+yMT2MbD4QUbNk29SIzzXPmU9fnyEFkU/lie+xDT6KS7eFXwI0uwUD/YEan2DkYkXHEGPxgQg/IFPsWmdNmSR73s5vsIZzoj/xs3D4TlfrDf+OPWT+zZey58dc3MuMcDByTNdjKS/eQ2/Zkjo+X+ZQv4z6YezjI3BB8Ecjiku7JfErGkObmU2Pg+pxkrJRvUzcbrMz4TNoyJ5Ejb+aSN/2ElU8A0zN9ufB/fgqZp0PqJWPwZtPp8biv8FUOuZSXvGXuVZdr/IwiKueM2um4t4jKhuPQa4HbsImnsigG0QRXROW04SmichpuJjaTGYedo2uS40wVUbkZnpwvi4QiKjfDK7nIHZtH7jilMJQ4YoJuFvacQk5spWUE6CynOkEeOgsrziqnd24AbLnGs3UmAT42r4jKzcc2cmchV0Tl5rjJmaAFm0dvi6jcDj+5Ldot8vkp5l5EjYW7oGQSnKXsBQBgLdB0/pCsqP9RGaTW73ut42I/jKNgs0CXoJPzginGkw+QjQ8g4E5nBLYSFDPf0SFrJnkEFRNg9JaCoKY3bAS9BXxckwchpYxs6vSGCBkiJ7YpKf6MOVlbyWcPojJvG8GMzyRQpV8w0obMawJX+oxA0Gdtgrd7+AnID6RTiEp7uAlUpwx6Jr9zfAo4uqdlCm7WbwkQCoZrs6Anf4Yvow1zEh8TcQQD/RM4VTYclA9HsmTdo6/kH/FoDB3zVeUhd4LmfFRvYiE7yQw/AnGUOVHZ8CfHZB3+ZIN8IwBCks4NfGasehGV9NObRGwnG0pOBIzJijGy7vGGDGy80SzwCiu66V7jClM4j4kj5ZAlcqoPMKXfiOAEVWEEZ2OlrpZJnWSCzJlH6ISx0U7tnStv9F8gH24C7hIiQzBdXTb90n/1BTPyKT8ckJNkC47kKG8JkkHnPLDgPoQH3OXRH3WSWfiSQQSn6+qbat+CvbrJOplWl/bTl9ZE5Sc+8YnhjUc6gxjz1jgZI+dsX+QKseENZfr1oQ99aLDr+qy/dBAmZEc7yRxddx5R8r73vW+4D1HJtsHbtR4pazUyh0Aw/uyAMXI8R77ZUHjQOTLnwQJ4wEb5+qzv6iHXMCE3bLv2hDCCr01b5IeFsYX9hUMC5Z3vfOfwQAZiCMbss8QGsA1kHsHnU8Tm0xaJTda3nkSl9tMZMgdDmLFVeQhzJMMAAEAASURBVPjHOW2Q2D36Bj/2Xrscw4r85B4402s6x77QF+PO7rD5cKXP5u7WiTywH9psXPLQg37RU22kD3MS2dF39opssAX6StboHnISLvHN+HfawXbA0/10Qhtt8vsSGfvIBpNf8y/5JAPOIc/JLjnL/fqThwwcuz4naZO+2LRTH4yj8SUTc+drtsuDAfRUHfpNV+gS/OBjPqWrPkkND/KjLbBm17UHHvpLzuBDX+HuNxtpbNg418yn9NKW+XTuPLCIsXYZF+NmbCRYaWcL3BbruxJ/F1HZcNQJLKNM4ThTDCJDwkiaAFsrSMOmX9aiGERGxQKagTHRmBQ5z4wLpW/haF7WTnas3GQHO4aSM03WYtDhN3cC69j0y1o0p4YTATc6a7KBH+xMoCbDSusR4ESYmIuoXI/RqisJUJE7zmicG3pKXzm8bB6HrdIyAgn4cmA5pXQWVllcwq/SegQS4GPzzBtkrT79uh6vXIncWWQlKGuxRd7obQIicwIvqeus7dk8C2E2j94KIvD74uPZl59y/KhbWxRReTxGLa72WseRdzaE/SX/gncCM3SCr8mWCLawI3xQdkWww7gjmtwnyOd6glzmPgFWwR5BmXOHb5F448j/LLMX4HJewEQ+92mHegQXvSEmiDNnbRp/JvrNj+5FVHpzRb/MW7CEUQKP/FH9g4UAvn6zxfATjOWnyqPv8Ii9gTFiCP76oP0hQ+UVMLOOtyZtmYKbNidA2IOoVLZ4hGCfceFvGm/95zPxnay/HQvCW38jgmAMWxjDlKwhwd0b4sl1sixwrQ51KReesGPrrd0FpZ3Lv4aIrM/BM35MT6LSpySRZ3AjSyEQHauf/05+vBmSPtNnuk3eYA1f8gYr2MITkWI/jnuQTbEiQVd6bjxg19qfSLuNlTq1ryVRqf/wgoGgNAyMt43tIh9kw56dk9xD3hI3Q2TAxtuFMEMQa6P2kmPySW/gyHbClu4aD/1xniyT01Z6S/75zOwJ7NRpnFI/+zrHjsIAoeF/dL7//e8fdEfAnn0mA+ydPquXjLDfiDTt8iYhzIPrGGfHZFQ76TGM77333gFz5cOI7MrTIxkXdsDYkAnj04qo1F4yQab5RrAjR/pCtsiY9Q0MspErNt4Yug+2ZEvSTnjCmi6aR+DqwZ8Lh6SUuJB5Rf7Mp+y4MSFz8CQTLZJ2Gm9r3F5vVLJRPiXqgQyyzSZlg4v64adPZEQf6aM3Kskq3Waj4r84pufwcZ9ErumssoyH+bSlXo6xNhbap65eRGX8HA9OGRdyzWeke/SLnPGxyCAdNY7ykUuJrLDt8pBD5dEJ+WGj7cYEvrCDGRuQ+ZTckU3n+HfsG8yVMSdpB52w0QPto0Pa2oJw41N5kMB8StbIij5YO5M1ukjW1Re/VB4YsBvwk0f/x/Mp3Pi95FYfyDRbwC7Ak77aMp/OsdGr8C2ichUqbc8VUdkQz14L3IZNPJVFMYiMOUNUROX2Q2Rigx2Dzsli9E0wDHgRlevx5NRwSOFm4uQ0wA92RVSuxy1XOAIcVY4BJ54DkEAEJ3Xuwi31nLV9AlTkjpMLQ4lDWkTlyaNNZzmknOkiKk/GazEHZ5/OsnnmDbpaROUiSsu/I3cWcoIXZNAC0QKpiMplvMZn2Dx4sXn0tojKMTqbHRdRuRlOc3P1WsfRARu/iZ8pcONTh+ywIJRAE989fgCbIiCDiEFUujdv9vG12HH+g737BMl8alJQR2DV2yICjAJZbBQ7L/AjYCNwI/AowKOeOcEb7bKGi37zo9lHdQlsapc+8QenpKwP80YlotL8pT59gpu6YGAT/PKJOnu/zXGCY4KlbLfy2G2YS4KsAn6Cq/oCT36FdZWyjQHSI8HsKX1YdU9wU08ChD2ISoFbdteYkAl981siF8bF3KafAsiSQLQ8ZIX8kBflCLhqr7yuwZ9dkp/cGXtjIUAoIAtL+RAT1gYCrNalkfWhsol/lE0OehOV3m4he8YLQQQzyW8bEhaxRtcEqek2WaPfMGFPyI4+w1D/EUxwhb1+II/YBbjSF2Vah8Jyjm6uglZ97AaZU6d2aZ92qTN6sereTc7RS2PCrsGAvJERMkbvyAzZcBxZcC16AF/+qHIQT9ZH7Ij88sAKUQlnOk6+gpFy/CZr7mlJwKlPn3oSleQsRCUfyRuhdIacGSfYkBt9I4tkBU5sIpnTPu0kV8aR3TWmxpdMwYuOPvDAA4PckdvopXw9kvbx+cgcfdCPlkQle06OYecTpuyc3zBj/8ka3PTPXv7Io3PmP5iKQZItGGovXfSmtHa/4x3vGDBGPMlLFmGrL+RtTOQpr0XaBVGpr2y6BzLgpk/0kx3TNyn2Td/JER2MvJlb/WanInORUbqtLDZPPnJAJsksmVNW62TM1adNvYjKyFuwIzN0iu0hT8FN/9h5bcr/nHSd/LBL8sHFZh6DoXkWTh6sQlSSU+fZAHJMt8mXB6f4bsYGzrAn63OSeuOHkH/91F7yTReM5ZwUojJviyqLn6COJFjpK7tHp9JnGLvfpp0wgJ+x1r7xm5n8FraQ/pC18XwKo8wVqXPuvojKuQiefH8RlSdjtHGOXgvcjRuwpxkZxCIqpw+eSdlkx0CbyBhjxt8ExqBzPiotI2BS5FzAjdPB2YAf7IqoXMZr8YzFQBGVi6ic/JvcsXnkziKoiMqTMRvnoLOcVc50EZVjZDY7ToCPzTNvWIAUUXkydpE7izmLejIoYGFhbp7N4n7ugvHkluxfDjYPXmwevS2icvsxFLSoNyq3x23bO3qv47LeYX8FsNjgcfBEMI9Nif/OJ/UAp4AVIklwSmCK36+MEDX8VsFEMnL33XcPQR0EnGCQstgqgR/EnCCQcgR5nBvXvy1e8Wei3/xo9lF7exCVglV8JvWyueYvQSu42fQ3b2zoF3t94fBBOr5CsJJf+1xPoNme7Rb8co+9PMZC8Ax+LVNwM44JEGqfNgiuaY+69WlOIm/mLvYDUcDnZIOdg50+ymMOIy8wERikB+RGkFL/5Yc9/4HMBEPjTY4RBcZFewVc4QVD5SLQyZzy5spbsIgfkzGlE/CK7qhnToKBN43yNox+nTsMDhsXY6c+dRgremcPT30WeIYHvOgvOYWzOAc9Faimh+6nN+ZD+Gbdrg7rdmNha5l6E5X6AQNzPdvGRtE98gQffYal/umvY30kg+4LPnBBgGsvbOUhwxcOddkbvLD0piX5NB70RlmwQ+IhBJAF6myRjKO+aAN91Q8ynXEkD3PGCl7s5ic/+cmDD3zgA4MueeCCLukfvPRHv5xL3+CgTfQauUEftIO8aSPZo3fayTbqB1zhSw4zD/SKE6mHvdG+HkQlbGyxb3SPzISsZKdgwB7Zy8tOwQeG2eAIP/drr/zsFtLjXe9617AXFyJbZFRZMMt8alzYOpi3SNpjbqA7bJBE3o0hmSZvU5L+0zUYkGcPr3hzmQ3S59ggfYMR+VBvyDLjaSzh5B5Y03kyB2tlwIEdDt5kW39io5UpX+ukb2wI3LTNeOqrthunluPDBrDvxofsqZf9gZlE58wL2jSeT9kyDwzIS87YSGWQV7aLfhoTGBsj8kTfYSmPPliv56EZ1+fYnYyBcY0fol1wIwva1IqofOihhwZZVr52w0f5EtzIHHziN8qjXeP51NjCQn4ypY3KMb7kT/4Qx/EFjIWy4dQCq2Bmb15THxk3XhL9VGcL3IYCr/A/RVQ2FIDeC9yGTT1VRTGIDA6DXW9Ubj80DDfsGEqTHIc9zoZJjYGutIwAB4KDBTcOm8kGfrAronIZr8UzRVQuIrLZ7ywUyF2cUXfS0zj4HB1OV6VlBOgsZ5Szy8Gns7Di8AoccE4rrUfAwpLjz+aZN8haEZXr8cqVyJ3FXBGVQWWzPZtHZ9k8eiuIwO8TJGLz7MtPOR5LwZAiKo/HqMXV3us4uhB9EFQW4Bon/ruAnjnNxu4IgAiwCH4I3sjjvLZaO9ElgRvXvD343ve+dwie+V+Cgq2CP/zaBPPZfHmVn8DauA3bHMefiX7zo3sSleYubTfPCw6b9/VH4E//2BHX5IEZP9WcByvHsJIvQSsYjP0t15NPHteVNTUwvA7L4Gb9lgBhD6Iy8raIg/PBwB5+gseO+VTkS9+dh6n8zpM3MqM8v/mw3rBh1+EqYOhNEjYdlsowHjCG4Vx5C57xY3oRlYLQPlV34ZAYo6OC6z4FiSgIdtE5gVMy4rc+0wXjGrwib9oeeYPrOL8yx3o/vid9brGHmzEjc8ZQe4wV28JGaNecpB8wICdwsNE98kS29Dmb/upnkvucIyuOja1yYMt3QHQJ4iPGtfW22247OHdIHmc8lKU/5NU9+tIKR2PZk6hk49lNb89/9KMfHTB64xvfONhvuMFFf/RL/yI/+k4+YUzm4CW5R9/dl3vdZxzgah+5hZkx6ZGMP9tA5noQldoMg9g39ekb+ZGCnf7BQV4YOQ9Dm/47Dz/3K8t1+JA1/9ubvtxyyy3DeLBxbCV83U9eYasc51okbdSe3kQlO+cNPv1GfiH1+AuZC8hb/A4YwlbebH7DTtJ3eWAC68iUvhiPyKMyXW+dtIMuwK03UZn5j96pM/0jNxJZID8S2wEn5/SdvMBCGXBUhs2xNTk/n7zBCNmVLz6oQxnGg7yNMR4qmvFH3fFD6Ku62NhWRCXd9z8q7ZVNzl7+8pcPe82GW2SH7JGhsbzBZ918Cmf5laFs40EW4AMvG7wzNjNgWrpVu4qoXIKl6YkiKhvCSWAFEzgznqBgkOp/VJ4MMMNiIiui8mSsVuUwKRdRuQqZ48+ZyDgPRVQej9O6q5wsiyuBowuHi3lOwHWHn/SwoE9QoodjsK49+3Ke3LF5RVROG7EslDjTRVRuj2ECfEVUbodd5M5irojK7bBj8yyEi6jcDrdx7iIqx2j0O97XdVzWUT6t5X+QWRfceuutFz+zJ/jTI8WfiX73IirN9T7tZ95P8ExgFcmib4JRmyZtli6nfxrcjFMChD2IynWYqH+Mw7ZYRE9CQAgi8vsFIJHjAqw9U/yYXkSlN6n87z9v5cDGWzD+d5aH4ca4ndTHjLN8vYKlJ7VhfL03UTmua3w8V97YFSSlzQPtyKLXve51w2eZ18nuNuM0buuq495EZcaF/X7wwQcHkuMtb3nLYL8F7Nf1cVVbT9M580JvorJHf/n7YhzGwxuu7N0dd9wxjAciLyRUj7qV2ZuojP20lkEeSd5M8/YeG46k2rdE33dFVK7CZo69IV9s3Hg+5dd4MGYXL070JirzhQJfGuCreZs7b4yvwnLdORjzdaXTMJ9m3Kxt643KdaM273wRlfPwO3J3HPciKo/AcuKPLLCLqDwRqpUZiqhcCcuJJ014RVSeCNPaDEVUroXm2AtxtIqoPBamtRdDGBVRuRaiYy9kgVpE5bEwLV2M3BVRuQTNiSfYvBAZ9LbeqDwRsqUMRVQuQdLlxL6u46yj2CiB/Pvvv38IrL7yla+8+P/wegVW489Ev3sQlfrlgThvt7C/gqgIynxuT9+2ISq7CM6WhQa3y0VUaq42SFNIkAQ2jQsyTz88nG1M8gbIUHinP/FjehGViHH/C1HgWBIw9gaI/bZpDs7b1nVS/hBi9Mgajt60fKPyuPrn4MBvQKhk87CCBzF8JpD8TpHh49q6eK03Ual8ftHnPve54fOv3ph67WtfO5AUiEpB+X1M7MQ+EpXmU/phPv3Yxz42+K8eVPC2+C4+6diTqKSH5C3EmIcyJCQl+8aG9/IXesqwfl1OonJO36In4/nUGBgLLwD48kzPlPncvEBfyX/LNyoR4j4xbD4153jILP+Dd9t+zZlHtq3rpPxFVJ6E0PzrRVTOx/BiCfu6wL3Ygct0UETlPOCLqJyGn8lOAARhJGjPaeO8mZx38QTTtFafnruKqJw2FuSOzSuichp+IYw40/VG5fYYJsBXROV22EXuiqjcDje52TwLYTaP3hZRuT2GRVRuj9mUO/Z1HRd/1hPrAkICdvxYb4AJ6gt890jxZ6LfrYnK9Ivd9VS++Utf+OmCd/a9PiXXA6+UGdwuJ1GZtkzZ8/+NhXHxkDG98fk1spb/nzWl3E3viR/Ti6gkx97cY3clZN4LX/jC4U2+Tdt4GvPBTbDYuO2aqJyDR9qt7XxXQfybb755ePvrLBCV7CdZRvr7n4R0yaeGkUdFVM6RnGn3WqPz+c2nn/70p4f51L/IMB75FOq0kje7qzdRmfmafUOOSQhYbyr39Bc26/20XObUfSUqE0/LfGp8+DV8HOQxP6dnUp+6bT2ISvOphzDIG3tGznbxQFNPzJRdRGVvhA8fpDtU7G8/Ute/rjNfw74ucC/3wBRROW8Eiqichh/TV0TlNOzcFceKk1ufft0cR3JXROXmeC3mDGFUROUiMpv9ToCviMrN8EquyJ2FXH36NahstmfzEhgponIzzBZzFVG5iEif3/u6jqNjNp+fEsxnr5ArvYm8+DPR7x5E5dh+8DsFugTyBfLs9zGQH9z2lajkwxoL4+4BT/LmrS9jQuaMS88UP6YXURk7YC8hx+lTL8K/J1bjskP48WOM3y7fqBy3Y9tjbdV2G3nT7vH/0tu2vG3zq5PPbB6GHTn3EIiAu2P/D23OW53xL9lvNpQuhaDYBRG7LR6b5mcf+HzmJG/DGrcbb7xxIJgdn9Y3Rdlnm/HwQCx7Z6z9v93MOZtiMCVfL6JSW/SLvCH12DeyLeV/bUaeh5N79Ee/9pWozHxqPPgExodfk/mUrvRM9JRds/UgKsfzKXtG1vaVEB+Pg36x1x7CrU+/jpFpd1xEZTssB4PPieHM1P+o3BxYBtqkXJ9+3Ryzcc4iKsdobH4cZ63eqNwcs3FOC0cTcxGVY1ROPiZ3bF69UXkyVqtyZEFfROUqdE4+lwBfEZUnYzXOEbmzkCuicozMycdsXoiMIipPxmtVjiIqV6HS/lwCKvu4jotPqw+OBVQF0AWD5wTRj0M5/kz0uzVRqe7UYZ3oOIF7gbwc9+rfcX2fcy192leiUvvTB3Mjn9YYkLXI3Bx8Tro3fkwvolKfrHH0S9IvwWIyt88Jbvv4RqVxMCY2dsB4hDDahe73JiqjS/qGbNEnpHhs3L7K3L4SlfA2JuRtl/NpxpkcmBuQpN6wlW644YaBvEbysLFzUuQtOqUssmbr6S/MafNJ9+rTvhKV2j4eE8eZTzMmJ/V/zvXeRCU5G8+n+nSaH1TYFMsiKjdFanq+IiqnY7d05z4vcJc6s8MTHFCTchGV00AvonIabhwBk2cRldPwK6JyGm5xRouonIYfnc3itz79uj2GCfAVUbkddpG7Iiq3w01uNi9ERhGV2+PnjiIqp+G27V37vo6LX6vfCTj2DOTHn4l+9yAq9UU92dKfxb18+5KC274SlXDWh+xzbEyyDRc7/Ykf04uozPiMmx99Gp/bt+N9JSqNx3gjY7scj95EJTlK/0KOC+bHxu2bnKW9Wavt2xuVab8x4ftLkbddjElvolJ/Im/2Uuz2Lvo3VNj4j37sK1EJioxDxiXjkHFpDNeR4uhpzzcq9Sl2TcXpU/p4pDF79KOIyv6DVURlQ4z3fYHbEIqtiiqiciu4ljIXUbkEyUYnTJxFVG4E1cpMRVSuhOXEk3HYiqg8EaqVGUIY1RuVK+E58WQCfEVUngjVkQyRuyIqj8Cy0Q82L0RGEZUbQbaUqYjKJUi6nKh13Hawxp+JfvciKrdr1enPHdz2mai8nCjHj+lFVF7OvvWse1+Jyp6YbFL2LojKTdqxb3n2nai8XHjvgqi8XH3rVa85dZ+Jyl64bFJub6JykzbsY54iKvuPWhGVDTGuBe40MIuonIZb7iqiMkhst+fUFFG5HWbj3EVUjtHY/DgBqtNIVGqbdJqfcgthVETl5jI3zpkAXxGVY1ROPo7cFVF5MlaLOdiVEBlFVC6is9nvIio3w2lurlrHbYdg/JnodxGVm+EX3Iqo3AyvxVzxY4qoXETm+N9FVB6Pz7qrRVSuQ+b480VUHo/PuqtFVK5DZv15c2oRlevxOe5KEZXHobP+WhGV67FpdaWIylZIHpazywVuFjlIvvHr1D5NkE3X5LMluZY0vpbA9PicfCnbeXnyKQrHuSflTd3viqhM37LX3hzbS/kW+Lh/4zxjvOUJ1mMskl95OQ6O+d8d4/zyzUm7JCrH/XGcfi1ikXz2UuTGce6zDw7Zj6/lXtdyv+PkVdacpPxdEZXqWrfpAznSr8jTuF+ROfsxJvIGF/ldS56Mi/PBLHW0wm/XRKX+Ga/0MX0LZulXcBhfd835bK6tSilbPillB7tV92x7Lu3rTVSO+5r+5FzkAy7pY/BLf9LOYOL8Yv5FXBfLj3y2xC+E0a6IynGfFrFI/8aYBSfYBJ/cl3OL592TehxnTFriplwpAb6eRGX6kv23a/62P+FccEg//U5yHV7ZnF+VP/nspeyHH4d/3LM4Prk2ZR+52xVRmf5FdtK/YBbZcF7bpOCUvWspx3X3JOV88uS8PHAb5821qXt1hMjoSVSqR0qfsodhUuQi+OX8OG8wd02+bO6Vxnlz7HxwT9nJ79rcVETlXAQ3u3+X67jNWnS6c5F/+hL9LqJys/EKbkVUbobXYq74MUVULiJz/O8iKo/HZ93VIirXIXP8+SIqj8dn3dUiKtchs/68ObWIyvX4HHeliMrj0Fl/rYjK9di0ulJEZSskD8vZ1QI3CxxEAeNib6EoOOKf09oe97jHDUETk12CLoImIcqcs7nuvMCUlPzO5Xrul0e5+Yfy6muRUk/v/1GpHlgJ6sHQ5jibPuuff9AenIKDPO41CcLIvfofPORPXvWMN/ndJ//jH//44R7H8rdIuyIqg5n+wMIWLPSfXMBO34KpfXCSJ5i7F0YwcN2Waynfdefc55/Kt5a71IcwErQXYLEYeeITn3jw4he/+ODpT396i+EZ+pC69BsmkQ99hcEYP8eRDffJw7YEb40K1uQvwfjIaPK5V3JdmakD1i2Ser71rW8dfP3rXz+4cOHC0Obrrrvu4JnPfOZB/tl7+tGivuAQHVRm+kQ+YsPgwC7qP5zkcS1YRq7GbdJO59VhU0bGhUwH5/E9U4+1Q129iUp1RNZSp9/po/YHP/1znBQsjHHkaZw/NhJGylZuttzj2tg+Rk5Tx9S9erL47f0/Kse4kTv1qj+yQe6CW/DWr9gq+Zx3nz0MbHTQb1gpz7HN2LgG3+i438pplRLg60VUwiwyoX/6FRzGv+EWuRvLRsY3cqQsGCzmD34wk4Kh44xPyndubkq7dkVUqg8G5G4VbpGxtEv/YQRL+2DuujIk9zif8Ymtsw/OZC/zrbwtkrLpAJvXi6hUhy1yYK9f2fyOXJifYBHdyn3ywnqVngdveMifvBkb5yOn8hqDlO/a3FRE5VwEN7t/V+u4zVpz+nNFd6LfRVRuNmbBrYjKzfBazBU/pojKRWSO/11E5fH4rLtaROU6ZI4/b17g8+3r/6g8vnf9rvIvzQ3WuP/6r/86VHTDDTccPPvZz74YX+lX+36WbE611oHbN77xjYOvfvWrw3rgWc961hDPe+pTnzqsOfezd31bTU+tbW301XpJLPQpT3nKwTOe8YxB5vq2YD9LL6Ky/7gVUdkQ410scBMgUReSQPCCAyVYIjDyhCc84eDJT37yAYMsWCIfA8R4C2C5Ll+CV+4VTHFeitMvbyZK9zoW9FOu8hkwAa0WKQGl3kQlLBhhe3XCQL/gY2LT5/TvSU960hCsE9iSh3MvyOZ+mLlf/2Ehrw2uxgFewd29juEKv2uuuWaow73qa5G0HXbal/E0PpEDQccWCV76pw44kD91k0l9UydyT99gqt+uk0MTnXbAzTV4wCYBPVgoX9nuscFRfkFF5eoPOXVPiwCqdqsTbj2JSn3QV33jAOhbzsFIQmCTIfLnmCxJkT3BH5i5V9/lgcfTnva0i/nhKp/+KFcdcDU2cDM+9q30lizsgqjUDxsdDA6wlMgU3IIDW6bv8hlb18iPPhtv92l3knKTnI/swRLO8IIz+YV5K7lTr3GyeIOhpC/GP20m53NSdFVfort0Sh9hCQ/Y6B/90lfy4rx7YEXP5XWPRJbGeMuvL9F198hvc025HN3IXYvAvb5oH13qSVTGPsBMf4yXOiMb5GEsF7H1cIKr8YSX+y2ajIfzzpFH52MP1OV+OMPNmMDZ3j3OtZA9bdMXDxf0IirJgzEiC/qnn2RZ/2AIB/3RL7Ixlnc4wIBfw97RDWWxh/CWnx1zrDz55FenzTmJnMmX8RlOzvwTuaMTX/va1wY5INfaH7utn+qem+BAp+AXHFbhpn7njSecYQobG4ydg09sjPzxU6LjrjuGn2tw87CJfvndqj/qiA7RB/Wx27Czpy9zUsZfn9UVm5Q92cjcefXVVw+6RRfhlLGlG/A2xsrRd1jG/4CvMmAub/zvlK08+WFnT8+V3yKp69FHHx3mNvXDzANVZDyJ3EjZayusH3nkkYPz588f3HPPPQcPP/zwkOemm246eOtb33pw2223HQiAXXXVVcN599ik7IcfV8gf8gJr9vHLX/7yYK9hJdhFHq5ETI4berJG96LffC/2kdy/4AUvGOSKbrMllS4hENzYX/bGxvaQMbqYubGVv36p5rNxFD8GZuwh+WJ3bfwmtrjSMgJws+4gb3Djs5hLYGbuL3lbxswZ/qw5wdwAO/J17bXXDr6BY/JXc8MyduYFfmwRlcvYHHeGT2luKKLyOJSOXjOnZs1TROVRbE76RU/jh9BXPp35gB9ijWHtU2kZAeuFxF2zzoaV+bRwW8ZrypkiKqegtuae3gtcRthm8mKETWBf+tKXBpLKOQ6mxTQy7Ny5c4NhERjiyDM8HFLXKRFj7hoFs6B8znOeMximr3zlK0NQS1nyuI9jq2+CXM973vMOnvvc5w51UMQWjlkCTL2JSn1JoEdgipPOODMunE/46JtNPwXQYANbTta//du/DXjLy4kQiIIbvN0DV85scLWHm3qNFbxe+cpXDmMDy1YLAu3bBVEZmREUFcCBib5lQvOk14te9KJhcovMwQpOL3zhCwc84WYyTKDXNUEM2BsLZZNJeWApv4nS/TAmvyZPcjdX9ugSOTBOPYlKfSAH6vj85z8/9I8JiWOgHxaK8Dt3qLeCFORDggGcv/CFLwyYwVsgQ36y5w1GQVc4wtyTd+TUsb7BFn5wE3y2T9lDBTP+0B+6Y8wudHqj0hgFP/V88YtfHPpHRlxL3+AAN/2FERzIK4w4C2RGOeymsYBhyia/Uuyq+8mEPO4ldy95yUsGnNmDFnKnTnUY2zg32t6SqGS3yRxZIGv6py79Yy/IB/wstvWPfGiD8+5xLz23p4/wEgQip/QR3hbo5ICew9YYKdumLPaALSWj7m1BfGif/uhHT6JSPWRI32HAZpkfYUMGyIb+wYGsZL6Ek3HMBnNEAV0OcUtfnafXsHN/5gq2MPNKAiGwbBXoTYBPn4yTeeulL31pM/nO+ERf9TM+B3mHAz2Cz/Of//xh/owNcy85Mk/DWvuMNTlj8+gibMiq8+yp/O6j38ZAii+EgDFOLVL6NZ6/2FJyrS/G1NgZyzmJ/LAP5IJd9VQw3Oiv8tVFLvgdsHD+n//5nwd74pr+wkde2LOVxlpi/2FDBvQDvvbywS+43XjjjUP56VMLm2e82Dx6a8z00Zhqsz0Zn5O0f2zj4Kfvsefqhy19Je9kSb1wcg0O7Ak9hzlMMgfE/4AtPYSfvOQPhmyg8+yp8o2PvC39PP2J/6o+bS+ico7ErL6XHYY1nSmicjVG47OxV9FvOldE5Rih1cfBjc1ie2x8CzaHDWFLzC2t1omrW7G/Z+PHwCz2F142a20+Q6VlBODGnyBvcDP/mUtgVkTlMl45w281J5gbYEe+4p87Nv/P9ZNS11namxf4YGTuscceG+SNf8kXJXtz/eWzhNW4L/Fn+Zn1RuUYmfXH5lRrQHOqNUa9Ubkeq8UrWQOxbfTV+qyIykWUln9bL/B5rW2tGSXxDvOpNaDjSvMQKKJyHn5H7u69wGU4TF4MsACnCYyTzoFSt+AfJ13QTxDGQkeiRAL8jLjAoDyMOcUSTKRIAtXK/uxnPzvkF6BSHkdW2ZxbThjlE3j+/u///iFgLc9cRyP9EvDRL23TB86zIJb2qWeuEwivz3zmM8MiWp3K41zqI8MsEClYxvnUP44UzOBk4c3R4qjCwnn9dr98HC9tNRbwRhZxaE2Y2QQP95GohBVs9JvswYLDGcxck0cQnhyREXnJpjHljJI7k57JEEauSWTRdWUoL0Q5XJ2TN2NiXMg1458A6lDIxD/GUB3G17gaN/VpZ8tPvyaAI6j513/910M95FsAgmzATn/Jjz7aBKG1Dx7kNgHl5LcwgjPZk5dOav/5w7cmBDMleiPgQe7kszneF6JS/23Ghz7Bgb3iSBknuJEd46WfCDTywVn41Kc+NexhSS5hYRzosDLJEJuSstgc19k6GKd8YwTfm2++eRgXcucc2zHVHqlf2/VLe+LckHMyQCe0V9/mJH2jp2Q784R+6qP+sXsSmfDGCKxgaR4gS/AWqI3TOtZz+oEkIUvK1A+LAg/OqFeCsXz7SlSSB33RNzjSQbLhPOz0PfJFH2NLXJfoJ9mBD6LSXn7zMn1X7t/+7d8OY0M3jTeZIBvRXdgpo6Xess30iFzok7paEpWwIV9k4dOf/vQgP+Zy+kbWYUgGYaZf5gbzBtmLLSZ32uY33Oia/LGRCEjn6TnbIGU+lk9Z5FO55LBFMr6RCXOgYzJAX7Urc9Icf4ht0C/6aozgYO935Ir90T+yxO9wj7fk+BvaoD3B2zgYb5v7zDvya7+5PPg6Z9NHfUHAs6XGR3lz/Txla7/62BN1k/P4ePZziUq4kS9yY96kX+oha9qf8YOdB1voFhlRL5zcd+GQGHZP9JwtNMZ0EN4wIV/6waf++Mc/PuTXfvMMWYNZ/BT3wr1FMr5FVLZA8vgyyAusyQ/9Ix/1RuV6zOg2XY5+8x3YFzpXb1SejBtbw8+wsfPmD3YkvmAr+7G+Jft5JX4MzPgc5M1cxe+ysdOVlhEwR7JtcDNnki/zF8zMjSVvy5g5w0fgM5sb6Cr5Gvv/fPqpa8LVNZ6Ns+YFPh+ZK6Jy8zHle5obrMOLqNwMN74ImwY3fryYBL/f+jpzas0Lq7Gkp/FDEvMxH4hXwE6coNIyAtYLfF5rwsTyYGU+LdyW8ZpypojKKaituScLXM6MBS6D2HKBa+JSh2AFwoNiCLhY0HCQGBpOlHwW14L35w7f0GJ0/vzP/3zY+y2/xJjLL8CCQHP/n/7pnw6LTOcEtQRyOGBxNPTLtZ/+6Z8egoDqcX1OssjVZoGinkQlJwkO9vBigAWrGBSTGYdAwE9w6Y477hicUIaH8db/4KrPNhPh3/3d3w0LpB/90R8dsLJgcl4/7N2jfAv2vD0Cf0FpC6sWyTgmqMuZttA1wahXXdo6J+mTOhhjE7/AKbkh3/oFOyQc+eEQJLDJYYCd8xIstM9CKES0MXBeYEPZxkZ7X/7ylw/jo1/GQFnGxcJA0NCmnjkpgVl6RGf1T/tg15KoVKZ+CNp/4AMfGOr4yZ/8yeGtNH1yXVBfP8mFICd9hMtf/dVfDbKnv/ROfnjCK5MjHVWeMXnXu941YEjPvQUioO9esqBf7p9LgAVzcqHN5OHCYYCXTrFHArUm6rkLt4wPmyMgrH9sGhz0Bz7qzkamfMZOvX/zN38z6J8FODklo9pKJ8ncK17xiiGor92wpN/ar3w4kVH5BbzhhfCgv5FvdUwlJBLYI3cWb+qRyD19JdfwmztO5ONzn/vcIA/qoUP6pv8wNJfQW+0hK4heuLLHcIJ3FuTsJLxDuJE54wxfckB3zUsWVNr9wz/8w0OZxkSfQvCyeXCek4wlHTBmbDb9Uad+sQ9z7ULaJpBD9sgInLQdMQRD+Bg/7SAL6s4CyDW4S+RG32FpfM8d6qXfyobVAw88MOBPbukOmdIv84ey4ec8opzstUgJ8PUiKsmZ/uVzk/oeUoh+kSO4wokMkqVXvepVw7jBmtwa44wp+VSG/OSY88/eSQ8++OCgo/TVGCgLTrET5Hzu/BfMI3d0gs0wPuwpeYuMa/Mcu6Cv+m/82TVYkjt6SW/IuvPmSXWSGzqYL2u47h4Y02ObNtJt+LArcPz7v//7oR6/YWTOUY+yzfX6QPbMHzCda4/0C150hnxrg7Zpu37Yzx2nlE+2+HFsEjz0+2Uve9nQB/JD/rVHvewefNhgtgT5SJ8R9/Q0fhU5pfd00V4Z//RP/3TwoQ99aOjXq1/96qGOyJ3xylwL1xaJDhRR2QLJ48sw1rAmK2SJXY/9pRdz56/ja9+/q3SJLkf/2C/2kdwXUbl+PIMbe2tOyQa3BFXNLUUcrcYwfkwIN3YbXmyvLf7Y6ruv3LPmOraNvPHdyZf5F2bmtpK31bLBlxgTlXDiN/GfyBr5q7lhGTvzAp+PzPHv+ciJi87xl5drOltn+PDmhiIqNx9Xc6r1D9ysMcRv+CbWhJlTa15YjSc9jQ/C/4Wb+cA6qQi31Zg5a73A57W2TSzPetl8Writx22bK0VUboPWCXmzwO1BVDLAMSQ+8/Unf/InQxBGQJizxGniuOezj5oqwDQmIC26BV8Yavk5CTZExg/+4A8ORgqRYpGpTEEZgSr3MPqCZ94A8fvOO+8cgrZZEMxx0BhEk7KgkDpMNIJkCWJReou3OXXAQ7D5ox/96OAsCYrpdwIQrqv7Yx/72BDEes1rXjNMbnFMGSGOqAmP4RaEMgkKNAtg3nrrrUMwVl8YLQEvzpl2q+eHfuiHhuvug72+zO2PNksm5QTUONOCKSYYY6OtcwOA5FpfyMWFQ0dTMFM9yha412eBT5OcMcumHWRS2wQNXTfOjDfcyZiFpTywVzZMBfvgL7jq3tTrXucEP8ilcuYkOmXsjK1xNm76BbuWRKX+cdKRbf5HlDp/7ud+biDL9AG23vQVbFY/XH7qp35q0Pd77713uP4DP/ADB9dff/2AjfFAAsgPG+TMm970poEc+M3f/M0BQ0Qcstd95I/O03UybFxapN5EZewCe/fBD35wsEHGHdmjT3AkM3SNbtPNX/iFXxjkwpvhZI7NhKlNeyWE4y233DLoxT/+4z8OOBoDegJj5ZAB5/7hH/5h0C9yh2xRN/mkw3R7SkqAityRizg36qdTdGIuMaBd9BR2MLLYUa++kRcYwMQbafQyhKK64creax/bok1wp38ezBAYgoF7kJXGKTr6L//yL4P+v/71rx9sQ+abyF0Lu6d9Wfz2ICozPmwPPbPpF9uNkNB39iKbPpEPdk8eY+pBIrjCmo6TJ1iRL3ponmYz77///kHufvEXf3HQVzLg2l/8xV8MY8aGs0Xklfy1SOwR2dB+fSRrLd+ojJ9Ct8y3cDBPIG313/ghaSOXAj3mT30fvxkITz4MfGHKfzFPOP+6171usGMerIKXc2wCrNhPmJNlWLeyd5E7/SHv+mm81UNHyPqc+iJ37NYnP/nJYfyVRwb0zbG62X32Tb1veMMbBl2mq8YSRnSC/tJXbYKH+Za+6oM8ymffnDOfIvLMEeZfmzYo/0d+5EeGMYufN1X+9A1e2qle7WM3jBvs7I3/nBS5I1ceCiIXMKCDiHD1OKdvMIQnmdRP7TKmHjAwpohwcknusmmf/NrKRzU3/MEf/MEgX/SXr+0eug7Lsc2b06/cW0RlkOi7j7/LPtIFY8l+sd9sSSu/vW8vdld67Bb948vwo9kg/hHbwhZHH3bXqtNfU3CLvPGrbGQMZuwVW8LXrLSMAP+VH8N2s8fsLVuejcxVWkYg69EQvOSLrJnjS96W8coZ8sbHMg/zw+DGt+APxL7V3BC0Lu3jl5lP43dZb5hP5/jLl2o4m0d8V+tv/mcefLW+sYa3ZmPvKh1FwJwaopIfwte3zoAZ33z8QPHRO+sXPTUnsG02a0XzAcxgR+YqLSPAfzMnwM4cQQZhxQ/hxxVuy5hte6aIym0ROyZ/BNaELBjEecmTQ3MWuFnQWARyzAXzEGqMiQAyIyyIwkBzpBgcDoCgugCndghsCVoL6CuHAUoAK2/TaPMf/uEfDmUIXHEmBGaUrV73Iir15cd//MeHaxRRYGdO/0JIaHtPotJk/+EPf3gIPjC8gqaIXsE62Ln+Z3/2Z8OxNyT1i2PFUYA1x0BeBoiTapwFxBwLYAv+GysGS0DMOHD+ObMCHcbJ4n0OVqvErzdRSV4EHmABI3hY7MCHjOmzcdNf8iCoKkBBxrTN2x8Co/ASOHUPfO3JFrzIFvkjAxzY8RutFw4DjwgVAU7lGzfEOnJgTtJuk7GFrrb1JCqVD4Pf//3fHwJdb33rWweikhMgaOsNQBia8OCHqIX77/zO7wwBVHYkQR+yGrmEHTx+/ud/fpgk3/72tw/j8xM/8RMDOU6PE2CL3LVaTO2CqFQHEvfd7373IB9sFT0z9saPnmUjT3fdddfwgEXslXvJnzEmj+wZTHzamd1BvLkfpnQVlvAi2+SNzSQXdN95NlHdcxYK2q1ubaJLxlmiCwIG7Ivy2fA5icx564eskBPlsUU2MkY3zSXykQl9ZMtgTg61j57TWZjTFaQwrNgyONjYNPnZCFjT0de+9rUDaaTM2Dx9aSF72sHWGB91cg71jS7pG/zmJOXDAHEBP4Sbvb54AMDcAU/124ybh3rUbX5gwx566KHh7SdYOeeBAbJDRiVvRpG984efaiZ3b3vb2waiEv7snfnduJFDn+H05lzuHQqY8ac3UckPMl8icj7ykY8MY8UemQP5HDAxj8DAQpJ8wYdOsPPw02/jiRSW2Eg6TV/IngelyJL5mn6yjWwDf4hMKhOW8rSQOW2I3OlbT6LSPKBf5IBck2c4kL/YDNec80CAPvPxYMTPy0Ms8ET0uo5wlN/95ApJTg5gSa7IJjmGr/uNg7oRyO6fu1Bl8+is+ulMD6IyAX9+BJ0lL8aezPE32CWyAl9zMftHt+DrOPoOZw+WCUAaa+1l3+QL4Use4OQBGtj/8i//8mAbyK3fbF5L2VOfNtQblZDom8ipcWeHyAQ7QgfoT+sx7duT3ZROt6PfbAp94VPQAXMi2xGd2E2L9qOW4Mb28Cds7CPc2B7zH3tC/iotIwA38wjM+Gtw4+Pb2OrCbRkzZ+L/wY0vFt+ZH1rythozZyNv5gbYwY1/RU9j31r5mutbsX9X4veZT/n7/Ct+l3WRY3GJSssIICr5tPxY625JfAJufHXYVTqKgDnVXEBXE5OwDoKZOdW8QFcrLSNgLmDXbGwc3KyjzQuwI3OVlhHIuhNui0Ql3IqoXMZs2zNFVG6L2DH5CSzjaJHWkqhkMATJLGQsnAWXBbIsCAXnBJs4mIw0ZZGXURFcFqChKAKDglPe9NA+zpXF9+233z7sGXABHESKchF4yBHlc/gT+EdUao+3tVwTvHdvAjPHwLP2kvJMyhwZbUhwl5IL3Gv/nPJTscCST3XBkIOJtAhRaWKDkTckGRtEGEfUOcFwuMLBffrLSWDMBY4YczgbB203TvCSTJACZbDSnx5J22EXQ8nxs1gzwcBv7gSjP+RZX+FBfjif+i0QoT79NY6CEkgQDpU2WBR5e8abL5xUbSJ3Au8w4TTAUbkII8dwFCCFm3o4aT7ZK4gKQ/d5e0E9cxJ9oStw06eeRCV8BH+9sUx2EBNkD0bqppdwko+MCcTDAvHoqX7yKKAsWAZnWLjOKXNewBrhdffddw/98AYJHfUEngBtj6RudWrzhcPAuUUawoZNoLP6OWfhZmzUIaj8jne8YyDB9Ed/YUTuyL1xgwWy4o1vfOMge3TYW3AIHyST63D4sR/7sQFLgXm66kED+q2dZJds0VntZweNievk0HU2kcyTffZhSiJ3xpDcGceeRCWyiF6xEdpsTqA3xghGiFj20DEbDNM4rHTXnAaL4E1WJRjZyKN74CtoBCvnbj+cV8gCXOfIwCp8yYXxMD7q0/YEW4wN+zwnZYEIN/ghPeBkDqKH+gcX9cKIDrN3cCVjMDp/SECSO7oBa29I00cyauw9mIGoRGiyab/yK78yXFeHORDBx96yj2Te/fBskRKo0k76Y3zpg/FsMc/GcQ9Rabx8Kt7DAeowdvqvf+YVY2dOcF/+5595i77BRpsSxNV2eLJvbIMvS9Cj8YNV6Qe5bJkid2SdzugHv4u8aW9swtR6Mx+xOfwUNtXYhLCFE5mDBV0jh978Ix/qN5bwQ1aaT7WXvCGBzadwYSsRlWSTTaDX8vCF1KVc8s6/lMgdm0eu9W9q0jd4GSt6qx56YHy13X6un0J++N/8DBg6hhmb4AEDfWBrXYv/TG+NH5vHltBz40fPyR+MbOTOONAT5cCWDHtjGC70F7mZgOVUnI67T3/ojDEk+zBbnN/hLGXP9sKa78sm+aKD9YNkXD0wxRcLEef82Ga3tt3KP+2JnNIx+kQm+P1wpm9jbE57P3bVPrJmI5N0hY6TUXrEl+fz00N2vNIlBIIbu2VOgRv7BDe6Dbciji7htXgU3MyJZI98scXZyFylZQTgxS8lb+Y9OJnbzIMlb8t45Yx5IXrKH4CbOQFujuntlThfBp91+6wn+SEhKq2zYSdGALdKywhkvRGfVg6+rLmhp5+53JL9OTP2Q6wzxFesM8iadQZdhV2lZQTMoYlp8ENgaU2WNW7htoyZM/FDzKfwg5u1tLgQXXVcaR4CRVTOw+/I3QmUCAC2JioFTy2eBcgEknwajrMkwGDy4mhyljhTEqMiMMUhoDgCFYhKgSwLcPlDGAlSWRQJkPqUlXoQb96AEMRSrnsEZhCVJlBBV0EeQUPGf06A00SyS6LS5OUzeoIPCTzDTXAPUQlXwT19QnbI77p+njskSQSwXOOAwcrCCM4MEqfCxinjhAkEGh8bjHskwQHjE0PJ8WMk1WdynhsA5Jx7U0rgVKCMAyApX3Bev7Uh9QrkeatF/+Ulrz5xKECqLeTOp/vs/VY+2SLb+iC44Q0cmJE1RKVxcV1/yJ03RPaFqMzCECn2R3/0R4NuegMDUUlPBcPoZWSNzCAeYfdbv/Vbg/zBA15x7mFG/iyM4IUEYHfe8573DPcJ5DuH3IBZj8Sx4dD0Jio/8YlPDIStRQ59Za+Q2LCjf2RPW+AgWAwjmAtII8jJn8AZpyGfxPVmDNkyJvQ7i06yxW4qG/7KIHfKg6P7yB3nzT1TEnvM5pF1dcNQogvqYGfo1NTy0ybygCzSfviwCew5+WLDglH0jo3TNnnNZfZ+u0+ftQ/WbJ/5w3nn2G42D8Y2+k9e2dgeC/cs4IxPL6JSPxGVZIcMsE9kgvywbZmzYEQW4GPsjBlbiQiBPd2AtzdM6Tv8tV95CCUPKJBLRIc5B16uISrZPXWapxB9+0hU3nfffYM4svd0R19hy9e4cDif8JPME/pIHvM/eek3fSZLMGDr4GZTBj0la8qnP7BDVpqHyTYcW8te5I7tpTP0JYs4Y88PMP5TAy90TR3IMm/qwUb/2SN+lvLpGplTNx2EG3mki/TdPOItckQkzOggm+gNfXLIT1CuuUYZ5NbDC/CM/0LmPeBBxkNUsqnqn5r0TX1sHr3tSVTSWzpknIwHP8K8AUuyR5/1TxvOHfpzcIz/GT2HGfkzHsGcXYYnnPRDOfwS2P/SL/3SRZ9x6vifhC0bW0TlSSjNv05OyQ5doeeISj4HHTC2re3K/BZf/hJiu9gcOmYNBSe6RV/4DL304vL3floLYGZLoIsvay6DkznM+sr8Qv4qLSNgHuQz0Fd2Gm6w4i/YyFylZQRgxi8lb/TVHElPS96WsRqfIW9sm43MkS+YWQOVfRsjdfSYjPGXzKceuo6/n1hGzQtH8cqvrDHNpdY6knnBnADDwi1IXdqbT6On/GXxFTiSNetGPry5odIyAlnfsm18ElhmPuXDkblKywjAypqWjTO3SuYEskbmYFdpHgJFVM7D78jdBJZx7EFUMiIUgeEVZBLQ4zAJzgmemMAoRxx2xwIogjPuEzQVAENY+u2aYKAAjuCeoIwAlk+/UjpBVcSIsjmynAxBEm+BWIB6o819CWBx1KYu4DMhq6PnG5WIyLxRKUAnOIeMhRFnSmD4wQcfHI71n5GGF6fedU6pYLE+u6a/JkWGSLBQEjg3/hwLhh3G6hIkhHmPRA5gF0PJgVGX9gqgkoU5yYKGbCAabX6rQ9n6xyAbQ/3lRNkENWEGOwE9AVROqsnv3GFwUOBd8Edei3PYCwqRPePhzTeYwVedPo0KU3JuDBDlwXxq37QlepUxM9HATgBYXS1SFoZIMf9zEk7eZNAHTpO6vTVI9vWRvHijEs7vfOc7h7FFPOatJ/dwJrSfbhoHWCFIvDEhyCY/YgAx4nqPZGyMnTG+cEg60AdjI6gex2aqTdBeMqUOb6LeffimKN2ilyEqyU6CFOqx2Gav4KtN8GSvEtzVpui8csiwhzfYVPaLUwEv406/4chmJKhO740BuVMW7Kck46ZvsecwlOipsaJPc8pPm8gV0kHg3txEJwXt2Xp9JZfsG9shGK9NkmM2xR5G7Ag9pxf0Rb8dxwZaSMEbTjCL/rqnR9IG485W9CIq4aVscy0ZIN/GJA9IkLfIdsYNLu4jdx4kQnrAGNHkwR9vMNEP99FVbzZ5oxK+PllMX10jrwh2NledbNHth2+onju0my1Sxot8GHt1xLbMmcfTNhgYGz6HN86MF8KLr0K+yRZ7r39sPvzMw+4zT5AhssOm0Tl6Hry1z2/nPbjwx3/8x4MesaUhKsl2jxS5076eRCWdRFTTW/1kT9kcdoE+JsGBLtvTXfLKniEpySw83U/ufErdvMLmmIfJtXFQNqzJlnEw/5Bb46Aun7QnG2wiOZma2Dw6q36yEZtqrMiEvfrnpMgduYAhX5y8wM5DO+og+2TPAwLsrn7TP/OMvrvGTnogAV7anRS585vewNmXHowLolIdMItdyH2t9kVUtkLy+HLIER03p5El/lbsY8/xPb5Vp/sqPbGxkXSJ7y/RafNi4bY8fsGMHSZv/H32EVZ8KHYpftby3XUmssZPjZ3m+7P59nCstIxA/D/yRk/ppzmYvJnj+PWVlhEYy1vWRuaGyFqveX+5Jft1Jn6f+ZTPDy+xReuiYLdfPdpNa9k0fj0dtY6U+Kb0s+bT1WMAM3iZU/nL1mkwJGvWMGwcna20jACc2Di2LXNq5tL4cMt31ZmsO82n9JQMWitbF85dNxe630agiMqGkkBgexCVmbAYXwEeQSiBFvVZQAvOW9gkkGTyN5nZHAt4IXo4CYyRxABFmdwvUChQ440vDoVvyAuqetqekRKYcT/SiTPrk2MIA+QAw59A4hQ4tYlhVEdvolIAEPFlwhK0hx8n3eTmPMIINv63p745B3PYw8s9gn8WkLBl2C3GnbNAT/BVEFg58mdjuHokbYOdRS5DyYnRPn3Ut7kBQAsb/SFHgpwCjcYMPogfdfhNTtSr/hhvgR732eDImAsYemOBfAV7uMnLsdBuhADnQp/ULbBPRgQdkUkcXTI/J9Er45c6yD/89KElUZmFIdl6//vfP/TjTW9600BMqIvNQFok+E0ffY5NW973vvcN1xFr+b8O5FDb4UzHo8d00xuVgh7kd9/fqIxdQHj4JDUZZ6c8ICGITJ/kYXvoIhzIDhkTRCZPxtbmHP2Uj03zxh+98PAGotJiyiIAOUImybL7kEaukTV1klm6zubR7ykp9lz56u5FVLLjiEq6RcbIC53SFw4U+TKX2Oszu67f9Ezb6HD0XECfvo/1HJYwZQ9grT7H8PE/jMlxj8U7nTUm6upBVOqjfpE3+CAvyAH5YpfYc/0y/vCywQJe7pEfYcS7doK4AABAAElEQVTeRSZhjhAyZ5I7406uEZJk6c1vfvMg28rMfK0s48IW5Q3VKfK2eE/sUU+ikg3SP/Mt+UL8sF9kw9jBk70zhuYD1yUPJUSG5CWvdDoYk2GYwkUZHqwi2x64YhvUQbZ7pMidvsVW0xl2SHvYD+3UxikpduHCISmOwCbbWewgJONrxe7AAXbalfk5/od7+QXkktyFeNMusudtXjZRu82p9NtcQofda55XN3ln81L3lH65R9+MO7tifOkGPdN+bbCf66fQP2MDA7Knn2wZOTIXkiNzvP7x64wT/0U/40PBfkxUaju84Rhd1g/2zpwdotKn3PMVjh42TzvIOZnXB/YJZmzD2A+Cs5S9tsAacXu+Pv06YHPSH3IEazpFD9hnZD9Sn8z0Gt+T2nXar5O5bHRbgldhtnrkYAUn8sYmssfsF3tDp9lF6wN2udIyAsHPPolujrecr/0lBPh/5kayZh7hs5hLzI8lb5dwWjway1tkjq5G3hbz1+9vI8BfImuJ45A3DxWaT+f4y1cCvpE5Pr5E3mo+XT/y8ApRyU/mp8PO+sm60JzKn6u0jADsFje2LfLmuNIyAlkv8N/MrTDMWpEf57jSPASKqJyH35G7I7AmZMGSLHAZySj7kRu2+EH4TfiC2gKhefMgxJlgSwJlAj45thAS+PTJVs4C58D1GHPlCQYKSAkofvjDHx7yy6fdglicCfdm8S7/7YdveAgKUkJBnDlGTBt3QVR6UyCf1DMenHOkBezUL6ADWw47oggGJjvnBY059forvwnPAtJ9nHvBQAnWgphkQB3IDZtxcl+PlCCbIKAxVK826Yd2zg0AkjtGOHJtb2FNvpGG6jGG6hEUVD/c5IOF9slL5uAokSF5bWQ7ZcOfvAn6KVc9+qR8v5EfsG6Bp3o5MXBTvzarC3atiUrle1jg937v94b6vKkS/WE34GRPb8mkwLLfiAz35u0iAWN56BxMyaP2GmekSt6oRGwIzp47DEC71iPRB/ZDcPvCYYCXDRDUpjct7ELGh0z4xJ7FtYnfxukMsUZeYOE3fLQH1mTNb9fTVrrsHKJcGxHgWbTDCFYpl9zCnlx7oIGss4fkEP7kfErSL/JM7tTdk6hExIYUosdjYsVvAXt9Ie/sPUxgBUP2nt7rf+wkffE7Dph+yGNBACv6Sqe9Eb2vRGXGR//JB/zoFvlGNMbGsf90j3y5xzjCwGZcYSOv+5QFe3JkHpAXmW4+gqk3DumOvPDkP7ifvCHm6TK9apF2TVTSObbOPGjONWcii8i/PjrP/6DDHrYge/LAl67CmMzB2UKd/rGHSIQQlfCB7b4TleSI7ORNaDpKP/UZDvHryAk5Mg+aJ+SHqfz0F36OzWewgzFZVA4d9aY5nGHM5ilHPeZA87TzZE9ACdapd6r86Zf2GHP1kwljTx7Ub6+dc5Ly+al0Vv/YNvXBid7qgzkBXuTIHIKodJ7Owc+DctFzNs4xLORRjnbC13yNDPUFDud9IaGIyjmjd3ruJR/sNT3MOs7b7i3Wcaenl9WSy41A/Ax+JruVjW1ie9hFtoX9qVQItEKA78m2kTdzGfkyr8XHKHlrhXSVAwF+GZ+PzIkT8OM9+GM+dTx1HV3oFgKLCJhTE9u2xrBGtM4ga+ZU68m564zFOuv3lY2A9YJ1s7VmYnniaObTxMmubITm976IyvkYXiwhC1yByixwMyGbjAU95iQBTUZYAEYgnhFmmJVrwmeAE6ynII7Va9Et2C8JSAu6cFLzZLkAmDdglO1/9wj0KIuyuSaApW6JE3vukPzwaUmB+zzhM1yc+MdEIrAmiNTzjUpEpU/R+eyZAJ62jwNkaQcizBtp9mmXQLUxFdyDOXxhYcwFAH16zsIyQTgyAHvBPoE+Gyx7pN5EJVwsaBhjsmGMPKGvXoQjWXGdEyCoLuBsLLNZbAv02HMebK6RW/eTLwaePAv+wRQxohxyqnwyJ5CvfE4HLOcuqIwjuTbBGK+eRCW58XDBe9/73iHIKaiu7zAgh/pKpuiUTf/powA0zOHF4afn8kY3yRwZE3Clz/7HLMwE/r1hpBzj0iMZc+Ombxc6EZVkT/kINzg4JnfBjI1jz+zhCTv9F0A2rogLBA8Z8ls76bTAPbkji8q0iGITnFO2fZxeQXxvkCuL3MHfuNmmJOXql/aoN86NNiY4ZXyN9ZxEZuDG7iHbzAPqVS68HMMKbuy5/tFR+MLaPeaN2Djtgx0HjC66D/bklP7Akv1DAPg0ONymYnRcv+lsFr9k3rjpEzlXpz7MTcbIuCgfDvQQfjBgd9h2ssD2Gyv5tck9jl0zljByn/+zCkeECT2X2FBfMHAd0aYsOkX+jA+5pdd0uFW/1NubqIQDuWLLfWodiU0mYAI/iczAMWSR+dO40UWyZ44mT3A2tsHdPbDwBiVyCdFL5tg62JJLc3qPFLnTN0SVfhon7TbWdEFbtXlqIjvGhz7BIcQtXWWT1AEDesWGedOezHgDE3ZkixyRHWU5Z16DFTk9dziPkkNEHn/GsXK1357uGxPymDfXc21uv+DF5sVH0idjpfyxHzYVO+XrJ331mXX6xb5K/C9j47rxCqltvoAnWeNXe3hA+yJvdBHm2meepYvy01nl/+Vf/uUg1295y1suvhHdw+bpgzr1zXgac22qNyoh0zbRCViTHTpCFlqu49q2tkrbVwTYZzaQzTWn2NgndpYdZxfZKvamUiHQCgE+KttG1uKHmUuKqGyFcJUzRoBfxqcic3xafhh/vYjKMUp13AIBcyqbZk7l04/fqDSnFlHZAuUqY4yA9YI1mbVtYnnW2kVUjlGad1xE5Tz8jtydBa4gUw+iMgsbwScBaEaYYjDMrlnQCAYyxhxPQTSLnhhtAXaBPAtviyKOg+CM+zgOHNf7779/IIzitCrDdcEX9ynbmzIW7oKwLZLFWgjB3kTlfffdNwSYBHrgISjHqEj6KSglEJrAhEAVJ0u7OFnINHjDR2DPsQlQYE9ZZCDBOJg5ly31tMBsXIbxRUqoV9DemJMDbbTYNW4tElkToIUD4pa8kA84jAkM9cGMjGmT/gss27uHfpA77bYQh5N85FmAGqZkzFgI2sojCH3uMMjqvD4ZlzmBU3jQmV0SlcgKn3LVf/qWN6TgRV8FU/WRA6+PMEWUwOXCIcEmeKbN8CZL0XUBVHoNW/8bjwzASxBWHcaoR6JDZII90j7jqB3q1D7tdG5q0ldbAvfeVlYPpwA2yicjNn0UoBdI1n+Bagsk5Bs8XXc+cucecqbtdIdTK78yLaTYBsdkzAMLiF+yN5eIgIU+sXlkng2Oc0MOEpyCn7rmpMwTyEbzEfmQ2Dl40CFjCB/EInkhU2wYmaPn7J72yQ+P6Lm87L/8+hJdlzf6ajzmjP+6vocwYmN6EZXqhoN+sXlwICdkRn9DYOijsTKmsIGncaQDCTa6z4NFxpsswUcfkFCIPOfZR2W5X3nGBBFFnuHsHjLZIu2KqPRpTF9oMEZsm/mITEev9Enf6BVSjCzRUbKH5HQfTCUymw0uHnyBlc9pkzl207zd84GgyF1kgr2gF/pBb9mUufYhckT2yB3ijN0zV7pGttRB/vSV3GhXvpiRB6NgJI+5ho1RhuS8soyNc/qgPPMP+VIW2fXGOR3n7zlPj+fosrari6zTW/aWHqkXdvb6Nicp39iwW4hYdk//2XBYKV/f1eXtRw8B0DM4mVuj59pGtrTZvcZVG+Wlv/STnMKPnCrXgyzwkn8OTsf1vzVRSYe8CerrIeNP6Y/Huldfjuvn5b5G92BNdnqs4y53/6r+04EA+8IGsknslo2PxobwBdhec4t5r1Ih0AoB/h+fgLyZ5zL/J+ZT8tYK6SoHAvwyPh+Z45uRtyIqSzZ6IGBOtUY3p4aoNMfy3cUf+fGt1tE92l9l7h8C1gtFVPYdtyIqG+K7iwUuQ6weTqZgCaMsmJegSoJ5CWjpnuvyCcgIBtr7rRxlCEYIuF04JAE++MEPDkrnk18CLyE6GHsBRuXKaxHluEVStjYK6PYkKpFrAqeCDyYtgU1EhslLgM4CEX7655z+wQZWgqE2Tj7nXt4EpQSuxoE+190jKUOZJke490gmZdgJAhpP7TLOrYlKY6QuOFhQ6+M4qKWPFtbkxDUbLPRf0N7e78gdB9ZvZXFiswkMCqIJYpMN92URZWw4uuN6p2JKZ4w73ASlTDbwg13LT78qE2n0mc985uDee+8d+nznnXcOZCLM9E+fyJG67Z3Td1iROZtj5ySyJA/5gpf7YAlD4wQvWCnLePRI6iMLvYhKbTZG+qPvkT3Hzkn6Bjs4wkKfXTOWxpY8BgO/jYV20xGOLLKI7qhHPnrMtsFTmfIpk/zat5I740jujBcMJe1nd9Jm/ZqTyDSiw8MV6tIX/VNH7Jvy9Vv/9BmewYnMRc/dm76TO3m1VxthB9NszpE/13sk7cvitydRqR59Infwsydbxg4W+hkM9DP2hG7CNLqtrXREWXBjF5Ei3tL0YIFyfuZnfmaYi5Sd+6PXqcMYtEjGVXvIB9nX1rzhrW59m5PoJ/8EGebBILj5FLCHGPRJf9VBjtQNk0XZE9igq8Fb32Pz6CH5hZV+yON+ZStP+T1S5E7fjJ9x1Rb6qj3p29xx0i91wW3s5zmvbJs+qo8fQ+6MI7nSf+dtMI7dhKX2wkpebwMqWz5BcYQ4nVUHeWMnYGqM1DdXJrRR/fSoF1Gpr8pm7/SPfMNEXxBx9vpPLx0bN33VN/nGeg5/WLjGJsoHK+NNDpUDU/X5LRCS+aGH7CmzN1FJDoyTPme8s+/Vp9NYLjkqovI0jszZahNdY2Ni59njIirP1hifxt7w/6w7yBuflC9hPswa25xfqRBohQC/j59E5oqobIVqlbMKAXNqfPkiKlchVOdaI2C9UERla1SPlldE5VE8Zv3a1QKXMc4ix0LH5reUQFaCSznv2jgAMT7vfoEZb2kiKgXOvUEkuIPME7CSx/2CMim7VRBD2Qmg9SQqfQJR4FQAy5Px3uLw5qQgvf5L+mdbxApetgSwkl8Qa4xJ8g2FHf6BldQKq6GwhT8WuoKPgoCCZ+o0ZhYeY0Ji4bZJP9O/yIy9c+pcxEEFrul7ZMbvJA4sWdN2wUVBZ78FX73R4e2P3Jv7x+OScqbutcU4wo1M9CIqMz4J3JOZN7/5zQMZK/jp9yJOkRfnbXAeb8HFvcEm/YFHcMp+KkbH3WeRa7yM24UOb1SO6z4Jh0XZg5WU/tsnBUdkw2c/+9mBsBSIpiveSGIPQrQF2+xTxpx9xpPc9SQqjQuiUh3sK1nTPw8A0DG/06/sg1PaGKyCp3zZgi0sMj7BJeXld8s9nc3itydRqc36JQUHdeec/q+aK4JL9vJnvpDfoj2fk/UmHGLcpyM9HCTlPvvg6LhV2hVRSfZ8Sl56zWteM5Ch5iVEkaRPY711LtgGM7hF9uTPfBs8gqvf401ZrZO6yJ0AX0+iUrvHOOi/um3Okwm4BTv5cw0GY5mR36YMvoHFu7aTP/4qO2Ce5Quxf/IulqH8uUm5sNsFUal/5qME/ZGIefBp3L/gpG/O26Ln9n5L8AjWuWecXx7X5euZiqjsie6lsne1jrtUYx1diQjE3hRReSWO/uXrcxGVlw/7K7HmrNWKqLwSR3+3fTanFlG5W8yv9NqKqOwvAUVUNsR4lwvcBFESMEk3EizJPuezz/nc77xjgS5vHPofjgJaiCJvlQnyCC7K497cn33KnbMXFNoFUSk498ADDwzBOsG56w7f8PCJzLyVoA/p42L/gpd9NvkTuMq9yee3tFjOt8+2/RsibBdEpZanj2McnA92jhfTKhw4FBZNCEKf+wtRaDx8mo3s5b7F/WL5U35rP7nvTVRmIvNZ2/Pnzw/Bdm9QISYE7cdEpX6kr+nTSXiP8ydv7h1fy7lW+10Sldqcvtnn2Hl9TD/tx9dcl3LdsetsDnLap/sEgENUkjny520bTxbnvuzdPzel/t5EZd4YRVjqb4hKb84gJbx9JaVv2ad/2hkss0+exX2u595cz++WezqbxW9vojLtDhar+rmur+Pzuc85RJdPEHvrC1GOFP/Zn/3ZYS5Kffa5P/vxtTnHvYlKY4Mk0j//K1b7fV6SP8GXGD+x79qq/p2Ed+4JrsEj5/O75T5ytwuiMu1O/8Z4BLPs5c319D/7XLPn17F1dMaT7eZfD2V4eMEblQhz5eTe7N07Nym3N1GpfGPDrvtqBvtqbvVVkPGnTfVF3xb7p41SsBx+/N+f5B/fk/yyjM+P72t5XERlSzTXl7XLddz6VtSVs44A+8EvK6LyrI/06epfEZWnazzOemuyViui8qyP9OXvnzm1iMrLPw5XUgsS37Xe9OKGJLYmrmN9nTjblYRJ674WUdkQ0X1d4DLugnCCOx//+MeHJ+19Cs7/jtqFou2KqPQ/hT71qU8Nb7QIXiErE6BrKAY7L2rXRGWrDiKntV1QW3DRng4JmApqk72eKXLfm6hMgNZbHv53ouDpq171qoNz584Nn93xNsY+pl0TlS0xMvaIcW9wkzuEMXKSzJE/zkWvT0iqm83rTVQK2iMkBLjVqU/6h6TkRI0Jo5bY9i4rhJE3E3dFVLbsE2dWu8233vJH3t1yyy3DXNSynnVl9SYq2QV1IGN97lp6xSteMfSvp16t62+r85G7XRKVrdqunBB5eavS/MvWeXAhD6S1rG9cFvuTeZDeagMb6JNz7JG9h0XmpPGDT4JifAtzqwdP8v8255R/ue/tRVTefvvtF4lc4xRSVn93QcBeblwX69/XddxiP+r36UYgfmARlad7nM5a64qoPGsjerr7U0Tl6R6fs9Q6c2oRlWdpRE9/X4qo7D9GRVQ2xHhfF7iMu03g3luHAo15s8j/3ekd0N4VUZnAvTcLkBKCdLsgYhuK2Mqi9pWoFPgla9oveG/vt2A2+SN7PROZ14beRKWAMNsgOCt47y1cn9oje/mkXM9+9irbWO3q06+t+2DsQ9gYG+SxQDmZI39sXi8CWd27ICrpE5lj79SJeEWKebNSX/V5H1MIo30lKi3ckePmWw9oIMn9r2SEyi5S5F7dPrtN3lv+j8qMj/4hYyUPBOlfT73qjV36ta9EZR4MYrP1gQ0y9myCBxfySd4eOLI/vYnKjA/5pl8CFmw4e3cW/LweROVdd901vO2cN06NUxGV9T8qe9iAKvMoAvED+WnssY3dsj7w8Ij1ab7scfTO+lUITEegiMrp2NWd2yNQROX2mNUd0xAwpxZROQ27umsaAkVUTsNtm7uKqNwGrRPy7itRqVsMvAWT4K+Azy4C9oFzV0Sl8bEYRLAImArWC9T1enMq/eu9N267+h+VLftC5siaACpn1t5v47ELgjz19yYqI98II/InEChoT/bGnw5uie0uytp3olL7LdrJnXFA3LEL5E+Au9fbJORuF0Sl/rEN9urUp/RPX/V5H1MIiX0lKmPvjA35MxaxB7sYD3X6HHAvopJsGyP9Y++kfGq4p171xi5yp0+9/0dlj74YF7bAQt586zdbxybkE+Q96lUm+9ObqIzc6aO6jBcbro9nwc8rorKXdB4td5/XcUd7Ur9OMwLxAzNPmleKqDzNI3Y22lZE5dkYx33pBV/MWq0+/bovI7a/7TSnFlG5v+O3jy0vorL/qBVR2RDjfV/gCuww8oy94Kmg4i6IFAEmwVtkm08xasPVV1998bNggkwtApz6l6C9ftnSx4ZisPOiLHT3kagEVBbrZMCxTXBxF0SKushEb6Iy/Yr86bfgcAuZVtblSnRpX9+ohFkC2/Zkzha74LhXIg/q7P3p1/Qv8rfYv5597IWdculRFr/79unXjEXGRl+MQ+xBT9xSdm+iMn2MvdO/3uR/+tZzH7nbV6Iy4xLZg1VsQu+5SN29icr0z95Y2euf7Sz4eUVU9tTuS2Xv+zruUk/q6DQjwD6xxUVUnuZROnttK6Ly7I3pae5R1mpFVJ7mUTobbTOnFlF5NsZyX3pRRGX/kSqisiHGtcCdBqbF2i6IymmtO/137TNReTnRTUCzN1F5OfvYs+59Jyp7YnNc2QlQ9SYqj2vDPl8LYbSPb1Qaewl5crlSb6LycvWrd72Ru30lKnvjc1z55L43UXlc/WfhWhGVuxnFWsftBucrvZb4gUVUXumSsNv+F1G5W7yv9NqKqLzSJWB3/TenFlG5O7yrpoPh33r5NzfieV7ckLxc5d+pnIV/OXIaxriIyoajUAvcaWAWUTkNt9xVRGWQ2G7PqRF8LqJyO9ySu4jKILHdPgGqIiq3wy25QxjtI1GZPlzOfRGV09CP3BVRuT1+bF4RldvjNr6jiMoxGv2Oax3XD9sq+RIC8QOLqLyESR31R6CIyv4YVw2XECii8hIWddQXAXNqEZV9Ma7SjyJgvVBE5VFMWv8qorIhorXAnQZmEZXTcMtdRVQGie32nJoiKrfDbJy7iMoxGpsfJ0BVROXmmI1zhjAqonKMyubHRVRujtU4Z+SuiMoxKpsds3lFVG6G1bpcRVSuQ6bt+VrHtcWzSluNQPzAIipX41Nn+yBQRGUfXKvU1QgUUbkalzrbHgFzahGV7XGtEtcjUETlemxaXSmishWSh+XUAncamEVUTsMtdxVRGSS223NqiqjcDrNx7iIqx2hsfpwAVRGVm2M2zhnCqIjKMSqbHxdRuTlW45yRuyIqx6hsdszmFVG5GVbrchVRuQ6ZtudrHdcWzyptNQLxA4uoXI1Pne2DQBGVfXCtUlcjUETlalzqbHsEzKlFVLbHtUpcj0ARleuxaXWliMpWSB6WUwvcaWAWUTkNt9xVRGWQ2G7PqSmicjvMxrmLqByjsflxAlRFVG6O2ThnCKMiKseobH5cROXmWI1zRu6KqByjstkxm1dE5WZYrctVROU6ZNqer3VcWzyrtNUIxA8sonI1PnW2DwJFVPbBtUpdjUARlatxqbPtETCnFlHZHtcqcT0CRVSux6bVlSIqWyF5WE4tcKeBWUTlNNxyVxGVQWK7PaemiMrtMBvnLqJyjMbmxwlQFVG5OWbjnCGMiqgco7L5cRGVm2M1zhm5K6JyjMpmx2xeEZWbYbUuVxGV65Bpe77WcW3xrNJWIxA/sIjK1fjU2T4IFFHZB9cqdTUCRVSuxqXOtkfAnFpEZXtcq8T1CBRRuR6bVleKqGyF5GE5WeD+x3/8x8GXv/zlg+/93u89uOmmmw6e9axnHfy///f/Dr7jO76jYW1np6hFopJjc/XVVx88/elPP3jKU55y8PjHP/7gO7/zOwu/NUNeROUaYE44zakJUUlnBQIt4p70pCcdvPjFLx7k74QirujLRVROG/4EqIqonIZfCKMxUfld3/VdB0996lMPrr322kF/p5V8Zdw1Jiq//vWvD/PrS1/60mHOrXl2vQxE7oqoXI/RuitsXhGV69DZ7HyIym984xsH//u//3vwtKc9bfBTnvGMZ1wsAM5S9tYc8j/yyCMH58+fP7jnnnsOHn744SHPzTfffHDXXXcd3HbbbQc33HDDgXLc556sVbIfbrhC/mQd95//+Z/DOu5xj3tcreOukLHfZTfpmrVvEZW7RL3qKqKyZGCXCBRRuUu0r+y6zKlFVF7ZMrDr3hdR2R/xIiobYpwFbohKC1zBgCIqjwd5TFR+/vOfHwJaz3zmM4uoPB62i1eLqLwIxVYHnJpFovJb3/rWRaJyHADcquArJPOYqHz00UeH4Ob1119/QHe/7/u+7wB5dCUGOk8a/gSoEJX//u//fkDmJA+2CD4jyuH33d/93ScVdUVeD2GEqPza17528D//8z8DVkVUbiYOY6JSMN6DQC972cuKqDwBvshdEZUnALXiMps3Jir/67/+awjSs3fZ2L9K6xFAVH7xi18ciMcQlS95yUsGgjF3wVnK3vxbRGXQ2WyfdVwRlZvhVbmmIRA/sIjKafjVXdMQKKJyGm511zQEiqichlvdtT0C5tQiKrfHre6YjkARldOx2/TOIio3RWqDfOMF7le+8pWDPIkrcF9vVK4HMESlgMoXvvCFIaA1fqNS0L7e9FiPn4Uu7L75zW8OT+eStSc+8YkD4SF4XwHA1dhxagSfEUXjNyphV29UrsZsfBZRGdLjwoULAyl53XXXDYRHEZVjpI4eJ0C1+EYlzLxBXkTlUbwWf4UwyhuV7F/eqLzmmmsG/Bbvqd+XEEDsepPSJhhP7uqNykv4rDuK3IWotCBG8tJX86xjDxeYfysdRYDNE7Ay19JbRKVzcLMhK8tPOYrZ4i+4eSDov//7v4c3KuG26KfAVMq+iMpFFE/+PV7H+TJO1nH1wOnJ2FWOzRGgo/VG5eZ4Vc42CBRR2QbHKmUzBIqo3AynyjUfAXNqEZXzcawSNkegiMrNsZqas4jKqcituI/ACmIJAH71q18dFrgJABZRuQKw/ztlsSYIiGxDeHBsrrrqqiF49eQnP3kIYBVRuR4/gXrBK0QlGRScClGJ+KgA4GrsEigQPKWzAoEWcbDzZqDgaaX1CCAqkR6Czl/60pcGuXv+858/6G4RletxG8sdogiGkoAofSV/wW99KVfulTFhBL8QlbB7znOeM+B35aJzcs/Jm7mW3trMD97MMueWn7IeP3JnEczH82ALPwU5SV9DVCLMi6hcxjABhBCV/BXn6Czsyk9ZxmzxDLkzz8KOLMJs0U+BqZR9EZWLKJ78u4jKkzGqHPMRoKNFVM7HsUrYDoEiKrfDq3LPQ6CIynn41d2bI2BOLaJyc7wq53wEiqicj+FJJRRReRJCW1w3IQsmCP75JN33fM/3VABwA/xCVMLOm6gmGiQRktLbCgL4FUBdD2SISm9orSIq4VdpGYEECgTuQ/TCUvD5ec973hAIXL6rzgSBEJVID3orKAo3/1s2RJtzlY4iMJY72JE5iZ4iPZ7whCcMx0iPSssIjIlKDxiwebASuPfWCwwrrUcggXifkmT3EJXnzp0b5tyaZ9fjFqLSA0GLRCXZM28UUbkaPzYvb+DzUzwU5Fx8vPh5q++usxAgd97wg51Pv8LOFwwQvUkwlbIvojLIbL6PfaxPv26OWeXcHgE6WkTl9rjVHfMQKKJyHn5193YIFFG5HV6VezoC5tQiKqfjV3duj4D1gjiKda0HcSXxT+sz/z7McaV5CBRROQ+/I3ebkAUTEJX+9xii0qeZvKkgYFBB+yNwXfxhchEE5EALPAvCJGBPyfM5tcLvImRHDhAdgs5johLZIfgngFpE5RG4Lv4gd7boLflzLHDvc80wrLQeAYHnkOQhKp/73OcORCUMBe1LZ5fxi9xxcMwXZE5i55AdsHPsLfJKywiEqIRdiEp4cQwRlaW3y5iNz1jImSuy8VOe/exnD/NF+SljpI4ej4lKJAYc6SpfpYjKo1gt/mLz+HVsngeDsqBj77LR4UrrEWDvfKkFUUkW+XcveMELiqhcD9mkK0VUToKtbtoSATaxiMotQavssxEoonI2hFXAFghY3/JZPNz32GOPDWvbG2+8cVir8fnqCyRbgFlZj0XAnFpE5bEQ1cXGCBRR2RjQFcUVUbkClKmnTMiLb1TecMMNRVSeAOg4cC94avEmACiAastnX4v0WA3kmKh0DCfB0yIqV+M1Pkv2EG4mG/rrOKRHEbxjpJaPQ1R6K9CbHuTOG5WeIiqichmv8ZnIHX2Fo4TYZe/IH5tXC7gxYpeOQ1Saa1cRlfVG5SWsVh2FMGLzbGTNW1l0tubYVYh9+9yYqBR0sSD2IFURlesxyxX2jl/H1sGN3Enx8cy19WBG0Fq9R1SO36hEjp87fBO63qhcjdfUs2TTg3/1RuVUBOu+TRCITeQD8mVsdJzf5+Fm+m0Nx0ZWKgRaIVBEZSskq5xNECiichOUKk8LBMypRVS2QLLK2BQB64V6o3JTtKblK6JyGm4r7yKwnhyywPWGkeCLJ4e8nVWfVFsJ2cWTWbQlaA+vbIKnFUC9CNXSgYUusgjJOyYqvWFksSsAXWk9AgKogtD25JCs1ZN+6/HKFbqat6A9KQm3c4eB06uvvro+/RqQjtlH7sicBL/ME45tlZYRCFGZp3TZPPrK1vkflYJ7ldYjQN5gGJtHzkKOr7+rrkTuBJTHn37NA0HeDKx5Y72cxMdj92xSfLzYvfV31xX27sLh/3C3KPawAYLyRS960fAFg6CTuSR7us03fOSRRw7Onz9/cM899xw8/PDDQ/abb7754K677jq47bbbDjxQ6QEj943nnitxDiqiMtJU+54IxB4WUdkT5Sp7EYEiKhcRqd89ESiisie6VfYYAXNqEZVjROq4NwJFVPZG+DA2eqjY346S9q/rzNeQBa4gliefEZWCAT5HV4GYMz/8l62DFro+N4yo9Fk1suYtD0Rl3pS5bI2ris8sAohKn/DzVtujjz46BDivv/764cEMbxrVp1/P7NBf1o6FMBK497+gx0TltddeW0TlZR2ds1t55A5RSe4EYPKJdfNsEZVnd+xPQ8+85WeeRTwiKv0fd/9aAsGYlOVc9kVUBpnN91nH1RuVm2NWObdHgI56YKOIyu2xqzumI1BE5XTs6s7tESiicnvM6o5pCJhTi6ichl3dNQ2BIiqn4bbNXUVUboPWCXmzwA1R6U22m266qYjKE3Cry/MQKKJyHn519zQExkSlNz0ERa+77roiKqfBWXdtiEAII0Sl/wXt4Yy8UVlE5YYgVratEYjcFVG5NXR1QwMEQlR6o9LcW0RlA1BXFJF1XBGVK8CpU80QKKKyGZRV0BYIFFG5BViVdTYCRVTOhrAK2BCBIio3BKqyNUOgiMpmUK4tqIjKtdBsf6EWuNtjVnfMR6CIyvkYVgnbI1BE5faY1R3zEQhhVETlfCyrhM0RiNwVUbk5ZpWzHQJFVLbD8riSah13HDp1rRUCZ5WoTL8QFB4ik3x5wMNkuWYv5X+x1xenBjh28udKJCojd9as5BIG/Dlf/fHlMy8V+F+w5PFK/Nx5T8E7K0Ql2RnLDXtW/1u9p+RsXzY9rzcqt8dtzh2+CkE3fOWFTWU/2VVb5vc55Z/2e4uo7D9CRVQ2xLgWuA3BrKI2RqCIyo2hqowNESiisiGYVdTGCIQwKqJyY8gqYwMEIndFVDYAs4rYGoEiKreGbNINtY6bBFvdtCUCIU/O2qdfM0/6RPX4Syv+h7hr1g025KRAP4KoAv5bCs+M7FcaUUnPbAizb37zm8P/F/cJdXrn3+P4dPo111wzfKEAYSnAXqkdAmeFqLTefOyxx4aHLzx44d89XHXVVcO/fGiHVpU0B4EiKuegN+3e6Ld5he9sXmdX/fsx/wIKoX+WUxGV/Ue3iMqGGNcCtyGYVdTGCBRRuTFUlbEhAkVUNgSzivr/7N3rkyRF1Qbw/gd8NTRQUGFXEMHF5X5XLhogKjeRm4ihX/RfEwUFFbksgiy3RQVFrgu7qCvXCNRPGn5+55frA7Xj7GzPdPVsdfXJiJruqcrMyjr51DmZz1NZPbUFQoSVUDm1ySpjDxYI7kqo7MGYVcWGLVBC5YZNtqkCNY/blNmq0AYtMFah0soKc1Kk/h/+8Ie2wuLcc8+dHHvssW3VhRUvyE2Ept91RmYuEqFpBYnNeMBnVpAsymq8ZRQq9ZNx25tvvjl566232iecEprg8oQTTmjfFwmHG3Q3Ry17hAw/icUnEC527NjRfhLLd35gEZKfGXnuuecajrx2/7jjjpts27Zt4gGMSsOwQAmVW98PiSd+kuJf//pXe9DjmGOOmXz0ox9t94aHkMacSqicf++WUNmjjWuC26Mxq6qpLVBC5dSmqow9WqCEyh6NWVVNbYEIRiVUTm2yytiDBYK7Eip7MGZVsWELlFC5YZNtqkDN4zZltiq0QQuMVag0L/DK19dee23y8MMPN6tcccUVk+3btx8iVBL4rLogVubVsBs04ZZn12cELj4iG4HSChKr8SJWyicN8TWiIZaNY/QVsYjwklUwVriOKekLY7d33nmnCU0Es2Dv4x//+CSb68/rX8d0/Uf7WsYiVBJZH3/88YmV4p/4xCeauH3KKac0gfto27jOf9AC7vV69evWosEq9bfffrutVHdviCef/vSn2z1CrPTwx5iTcUBE2n//+9/tUl2zeGK1/tivfyv6toTKHq1cE9wejVlVTW2BEiqnNlVl7NECJVT2aMyqamoLRDAqoXJqk1XGHiwQ3JVQ2YMxq4oNW6CEyg2bbFMFxjaPQ96FrOfDbFYY2YewJ64glyKyMJpj3VVjqaO7giy/QRQxJmUIOTbnSZLHyhnnyOaYPMnbbU/yZLVN6vaZMr4nXz7Tlpx3yJ+5JvM3McWG9HPNVnpZqfOhD32oiSdDvg6YMBfQL64JUUwM27t37+SBBx5ofXTttddOTj755JYnRLLrdH3ESkKlPgx2gs9gBtZ817/d8yUf+zimDvmSN3YL/oKP2N6nlPypv9sOx1Ov7/oLMamvEJPuHWKXV0EiJYld6pNyvvbPQP4sm1AJm3z666+/Pnnsscda33nV66c+9am2Wf2DVLbyBx5s8V36Ud93fV2wE4wE96vzBgMwGpx2y4ID7HTrHghEem3GWITKP//5z5OHHnqoCTJW4X7mM5+ZfP7zn2++Ggbc6+nP9D1D6vvEOPn8Ly9cJX/8RPAXvCivjPKOrcZYyiW/T1vOkzLBmPL2OS51/epa51a/Mj5zrlZwoH9cQ+IL0YyAxh6EZaIZHz32FX66Jn2cuMwG+i848On/YCX4VM5+2My2uv9hJ/XKb9xipboHQMRFD+xsX3koiY/lWz2INOZUQuX8e7eEyh5tPLYJbo+mqarmaIESKudo3Kr6sBYwWDFR//vf/z45cOBAG+AYvJu0m7BnMHTYCupAWWATFjBQzuTX63g8vW/Ch9hDPiC/KpUF+rZAcFdCZd+WrfqmsUAJldNYafY8Y5vHIaNC1psr2MRP/gwZZawWkcWYTZIf4ccWPlOH78og9olL+U23EGOOi8dEHOcI8YX8IuDkVZ8hr4gmNmW0x/nVqW7tEteV1R5kWq7DuFObkI7yye+7tsm/CIltXIP+WGShUj975ZvrSP/4vm/fvskjjzzS+vPGG2+cWH2kD2HE8eAoQiV7qAcW4M7/+r/bv0hT5eFLPt/ZUIIdGMgmr2M+nSuigLzBEsxJsKm8vK5B/XDp2uBJG7LaTl/5jUPErHzOd/zxxzdi1tzHveFczjnEtExCJQzBGgL9xRdfbMK5Pjv99NObyHTSSSc1Ml0fyss28sOfFJ8Fo/LoU5iCC/iDV3nju/ghvs0GAxKsye/TOXzPyhtzlvi6+N5WaER/XK+HShf91a+E7gcffLCtzPUgidcF82n6EAbiz/Qn3MT/JCbCiw0OHONT0v9drDhug6lgDV7sU69y8OgTZmCKz7LJw94wHN+obnNicVV59cK5NgTXfFzqyblB0DU5hzrkH3pyDa7b9S+rUMkGcAOTYhXMwYa+7Pon/+trtuIT5fe//bASPwYziY3qls/9rJzzOAdBmL2dC6b4VasqCZXOOebEHrWicr49XEJlj/YFWGTCe++9154wMLA57bTT2tMcnPyiTKB6NElVtQUWEDD+8Y9/vD/JywDEhAkBIehUKgv0bQGDnxIq+7Zq1XckC5hoZfJbQuWRrFXH+7JAcGdi9u677zYMmoQhAcTZkFOLMKHvyyZVz9ZZoITKrbH12OZxxmkho2CI/0Kwh5RClnr63YoDPkwyrkNeIZ+UNXdVD4ITaYmA8lorr4w0v0Bgicnqdg7zEXn5TMeQX+pOGedCcpkrZ3Wa9iDEEF3dFYXOrQ1IMO3WLudQrzmONsjPDy8KocrG2s8Gro3dbK5N/OheP6JwiEnbYUKb9aP2u3fgwHZg5eHF3/72t62PbrnlliYMIZEd05cwIW7qN9hQVlyFB9iRYAE+4UWsVcYxr/GEAWW0QVtCyMOEMrDkfPADo4j9kKbOz+7KO24/vkYZ9RtXaod8+gPGQuq7J1544YXJG2+80a5d27evrCDJ5t6QV3uGmFwf0Uh/sZ12jvHVr+4v/gc+4cXvpd53333t/7PPPruJleHn2AAu+S0kPBspDzewRID2CSOwxj/CBzvGlwZ/bMnPwRTsqJet4U171A1DsJZVR+oeK0/j+sciVFohbvWY/rJKzz3v/ocB/clvJ8Z1RUFYsbEDe8AFv2dlJt/mu338UWId3wUvfJCy/lcn/6ZcHi6CU3ngCi592uIb4SrxhA90z6uPL3UN6oNd8Vd553cu7XFtyqjD9Q09sYW2uwb32LKtqIzPYwNY4/eCOb4LNuGNj9LnySfu8lH+hwu+C8blhw84sB92xF12lR8+YYYf9Mk3qv9zn/tcE/JLqKxXv/bhM0qo7MOK/61DYHATl1DZo1GrqiNaQFAuofKIZqoMPVvAoMXgpFZU9mzYqm5dCxgcZ/JbQuW6pqqDPVoguDNBK6GyR8NWVVNZwNzCSh4kk9iLbPA6RWRCEqJCyicSHmFjddPu3bsnP/7xj9vKEnm+8IUvTL797W9PLrvssrYyQD3KKWOT8tn+WZI/Y5vHuR4+yzgNwWR+CktiqP5FYsJRnoDXzfLI+5e//KWN8RBV/J9yCC55syGyJIKAMnxjV7hSDsmFWLUKxVsPEPSIUatU5EemhSRDkFmhdtxxxzVBFEGq/eY4qRum1Ymw1Q71wm/Eptaggf9xryH2zN9cn22RhEr40WYkJdEOvhDkfJNrs/+ll15qIs9tt902OfXUU98n4eXTr0hN+EGGw8DLL7/cyNXucWQ6LMinz/m/V199tdUP2zZEPRIfMSo/jPmfgCApC3/xlepwPvZWp/3IWd9hHC6NLR1Xj/O7TwgKrpMAm3tDuW3btk0++9nPNj8Ku67LNQ0xsa2+0Xf6yv08RqGS33F9fIW4+eyzz7YVlfoXkU6kJFjyH/wGuxChgmPl7dfvJ554YsMA3+SeVScRAAZ8dx7YIVDDnzcLwRTsqPett95qeJMPpmALPrw6dPuK2KWMsmNMmavBnN95hLcdO3a0a/adILYIiU/YtWvX5MDKAxj6lU8hYGs/X6L/9SM86X9Y0d+O6X+xS7/DgzL8EV/BV/EvsIZPkV+s8x0Gw7GIE84Bj3yNGKoOvhamlRF3lecP5Wd7sds54r/EWdhle3Ffm+WRV/uUV9Zx1+ccsOmah57YwvVpv/vSeMT96hr5ZNfCzmNMrt0mHop7+nj//v0NE/pWH67GG5zoc9iGHWWNCeEBXuQ3xhJXxWu+S71+f1p++GRrscR3grb87m9+TTl1jTmxmfEEm7tXJb6dfxADhjoOWKQ+KaGyx94CWAHDDWzAwyHmiS2BaRkn/T2at6o6jAUEihIqD2Oc2j03CxiEZxBt8M6/GaAb3ArOBjbl8+Zm/qWt2IA4k1+DbINtkyoTQxMyZFKlskDfFgjuTMqQDjCIpIQ3E2ATskUiXvq2T9U3XwuYW5RQOV8bq31s87hcjzmCeIlUMW4TNx3jt8RNpBTBT0KsIrmef/75RvwZ1xnT8X2IP37POA+Bj4xBoCIGX3nllfZp7Mc/OpdzmP+Kz+pHsCpr3kJAl8fxkF4IUcQiwku7/I8gQzqmfvmJK9qMGJYvJKRzL0JCKi6yUMkfEWr0C2zpT/0SshiGnnvuudaX3/ve95pQicwLoaefkKEIPQQngeiPf/xjE9HEUXEV0Qlf+hoOiNvO5bywqA44Fo/ZUn6YQOTLr21wpQ6YIwhI7gPngz3YhTX3gbyIeoKK63M+BC/MaoM8yj3zzDMTcx7XYp/7w+vuEP/BomsaYloWoRIezFH1MSHdisr777+/9S8inUiIn4MNeeEJXvhG+IMF2OLr9K9+hVeY0+9woF54UZ6/sx9u1S2/7+rj5yIEyAdXhBPiPZFbvXA0xuR62WjRhUq/UUmo5Hv4OX0rTvEz+th1wgxfct555zWf4X+YyAM2sAhT8MLvwJk64IUfdG/CFf44sVbeLr66eOTvcCwRzrVRHeqyn092noiNYrh7Qn7+SfwOrsXpPKChja4vQqZzauvQU2KP6142oRJG9C288TV8WVb9BwPGSvDG3+hP/SyeimWJd3AAz7BivGbbviI6qoNIKa6rVx7Yl18dPvlGsfLMM89s2CqhsoTKPnxGCZV9WPG/dbhR3ewlVPZo1KrqiBYQlAUKAUrgMYgyuTJYMUAZ6oTpiBdWGQZtgRIqB909o22cyV8mvyVUjrabB3dhwR0CoYTKwXXP6BtkblFC5fy7eWzzOLHS3ACxZH7AjyGUEJqIJyKgfYhzv7mFkELG+k233/3ud+2B269+9auNrEewwyEi1TwDgYWMQmSKxU8//XTroIsuuqgRnSHAEIiILwSZuUnIfoKQ7wh7nwgw/hXJav6CREXguwbYf/LJJ9v3M8444/3Xi6lT3UgzbdeuRUiLLlTq28cee6wRovpfPxBdEKA4EK9HReybg/7whz9sZDyinmgBc/qbeIgQ13fK/OlPf2pkK6ERqQqTjgXDyGf4gF19Dh++E0TVTQwnVPoNQskKTZhHziJQHZOQrRErYYaA6Typ2/lch/PbiFnm0fCvDQQB945zwidC37U7jzYoq94hJvZahhWV7i+b/oc5GPn5z3/exJtLL7204ZHvQvDDgn6FS1jWn3yiByP4TRiI0M0vwa7juBfl/Q9TTzzxRDsf4Ym4CXPs7V5wXFsiAMELUSsPWAwVL7Ni2L00JqHSvS8+8XfipU9J/+7Zs6f5pauvvrrFNH6Az4ABK/0k/8MNIUmM5XfES1iRhx/sPsCT88Dy3r17WyznY+znb5yD3+KPPVgEi+qDLXiDX8edE2YlWOO31OO8BEv95EERMdgYSPkLL7ywCVXyL0JcZaNlXVGJj9PfhG5Y4M9gTZzku8Qwvsv/+jK20u/K8WOwxBd6EMc4j380xvP2Ffm9IUH9yqsLRpT3hgPx0DhTGfizah1Gx/oARruRVv64V4wDMsa2n43Zh819rzSbBUqonM1+h5Qe2wT3kIurfwZrgRIqB9s1o25YCZWj7t7BXpzJVia/BtUG2QbYJl15inmwja+GLawFgjuT/hIqF7YbF7bhJVRuTdeNbR6HuEOUipOujR9DtCLlCY4eckRmWjFG4EGcEgWRqIhXRLpXdyKrlEOwevWlMl2ynU985JFHmrB5/fXXN5LKK8KQOIgthI0YjfTUHvuRW4SBs846q5UjkGoPgo2gSaxC7CPJiJqPP/54a8OXv/zlyc6dO9+P99ol+cz3rUHL5s/impCD5m9iik2fsBWCj60i4m3+LPMriZz8xS9+0fqQAEjc0Sf6Wb/+/ve/n9xzzz2NqPzBD37QSH0kvH6PUInodJ3ePkU883pOeCUAbF8hSBHxBELYVQ72+EF54DKvk4VHdbMXTMIT2yJcEYiESFhyDD6Q8dqoDHs7pg3aFcLRfeD8NkQvnLp35FE2QiciVnvV4R4a+oPB7r1lECqD/BD4xMKf/vSnTQj/xje+0fqMqENEs3qcTfhHoiQcww9xE96QzjBgdZF7ExbgKOKP+YfVaL/61a/afax8MKxOAjzM8GvwdO655zYhE17H+irK2D9zNfYdy6tf9be5ppVjfAssEBHhS7r55psnHqbhG/h59xwcSnwXfwYvhG3x8IorrphsX/F3MMcn8a3iAV/KZ8ELP0VIFGfVCYNEIfiBMUIpnMEUfBPKnYuwlJWS2iE/DPrUDngX953bqmP1KMef+VkA7VqUxC7a7r4k1IkXrsv1us48FLAo17ORdsbPwY9xlH5378Gg+Mx/iU8RzthFGXnYi+3gWmw0jnO/KuNNAXDOd4mz4iNMqpM/hD0Cec7JxhdccEEbD8BnCZUlVG4Ex2vlLaFyLatscp8b1qCGo6hXv27SiFVswxYQZGpF5YbNVgVmtIBBDrIqT3AZrBscG9wYDJkE2lepLNCnBSIYIRhKqOzTslXXehYI7kqoXM9KdWxeFiihcl6WPbTesc3jQmARWMwTEHg2wo/N9RqnIVzPOeecRoAi+JCliFEEn98yJVRKRMyHH3641YGoUg4hJRb/+te/bnmuvPLKJkBZGYLYQvRn40fNkRFhyiBq1YMkC7lGJHJe+xFfxpLKEBvks9KDMOaJ9S4RtkjjTcQgstD8bRGFSsT83Xff3frQihyiIcIbUQ5nSG8r2PTld77znbZqR3/DHizabzWPPkS6I+AJ0fLAClzpYySzPmYnvEoERvucE57gQp2I2O0rxDqRwP4jCZUwGPwRTPUJnFmJaXxJLCemaoe2yoPY1Q7thVftiFDpeAmVzQUM5g//Bzsw8rOf/awR7oQcq3YdgwFYhitCjrkrkUbSz/IQJ+1TxvxWH8uL1IcT97HvTz31VPt+/vnnt9fKmg8rHzzBOVzDC1wRi/i2MacxCZUPPPBAE7/4GX3rN05hwzV6sIc/hAUP6vBB/EEwxq/wizgTYho/BxfyfP3rX2+CEDFITCb6qJNvJAhZnSYRHOFUvOCr4m9gyPiQ4M53RmAkbsI3AfPAyio7PpJ/9spj5a2Q44cJSzDtwSI+zTUQYq2Mc/5FSa5pWYVK8Q7W+CMYgSErwvk+MVQ/itP8DoyIe8Z+4nH8GPsRqsVN5WA8cd3/4jOMwktWTBK7jRU9hAA7fFz3Nyr9P+bEhu7ZPODkWsUQNq4Vlf30fAmV/dix1TK2CW6Ppqmq5mgBAaSEyjkauKpe0wIGRSVUrmma2jlHCxiQZ/JrsmegjLw0aasVlXM0/JJXHdyVULnkQDhKl9+HUHnHHXc0csolEJ4IUEgtxKlJNaKC2BPBJ59H6ZKPymnHNo8zP0CkIJHESzgSM43d+DKxVIpQiWCRzyvuPCmPiL3lllsauSkfMhRhiwBFfhFwkF/+f+ihhxqRdckllzSBsUv0i88IHOdGjMmvLUjW7goPmCP+ECjtJxZJrgFRi9xHECPR1On/RUzutUUWKpHsd911V+tH/gM5GXIeWWo1GuIeWX7rrbe2VRhwhRiFO/1mvEb40df2W4UJM2ItgpM4BH8waZ95LsJfPcZ8VnsgXJGrCFQ+TJ0IWbgnliIQYRtWs6ISqaqMeuAPlrQD7pwjK+D0D4wRP9VLkJdgN8IDTCNztVU7h45HApv26wNzOHYkvGi7e41wMaYEF7DAlxEq4cirOfW5/QRnQiUsEDkINvwOXLAR/MKVfTDAThKMwxHxBwbV41XZkt8oJAbBjfMj8vk9WOYv7fcghnOof8wpczWYW+QVlYl7cCLeEa2t3IYNPoYQ7sEMPuOaa65p/tC9pN8JR/F7xhcwyB4RKuFRffaLieykHueBOX5Lsl9dfI97l4/kf2ziuePw+aUvfalhDL7gkl8lXMGz9sInrP/mN79p8d+KTX5Ove4J5eDUSjptWJS0zEJlxhPwlrmCB83EM/gQu/g8/cr38Pv62vED/xWx+TG4FLvhVHwlSPJl8PXoo4+2uYEVwOI9zIgn7g1x2xiT/4RlD5k5ZwmVtaJyVv9RQuWsFuyUH9sEt3Np9XXAFkiwMVgSpAx8TTgMWEwABY5KZYG+LVBCZd8WrfqmsYDJXCa/Jo18XgmV01iu8sxigeDOZN/kHwZNwjyhLM4iOeFw7MTTLDasspu3QMgHgo3Yi1zOqrLUiqyQ8kn0QZB5Ut7vyZVQGUsd/nNs87gIfCGS+CeiCjISNuCKT+PDEJP8GcISsWmVCGLzpptuasQUqyGl7r///kZoqQfxZVPGSkt+0Yoiwo+5CbLWd74SLrVH3QgyJL7zEZEiAGifMgSfzF3EePUj1OzTTuStskMXhg6HtBCLbMT+NqKH6ydoIAFd31CFI9jgTxCbiEwPPhAq4ch1IEnvvPPORmx6IAJJarwWwt516XdEuOvl04JRpKj7ED7YiR+z+Y5gN9cVayMuB4/QawAAONxJREFUis1wAavOD2vOg3BVV4RKbZCsCiaWWzmsHmSstvCp6o+IGV+rbn3huPza5hph2Ln8JiFBIf2lnZI2Dy2VUHlQqNy+IhbGr3gog/DIJ+pjfg2HAgvIe33Mz7g37YMpG1s6DnfuAzFWnV79KjbDFFyLv/wirAeLcKqsOsecMldjn7EIleKd/uXviD6wQKj0KmwC43XXXdeEGpgSX31K+hyO+AexDF74EysqI1TKyzepB1b4FThSJkIlvwNXcGneoU5tUKdzePiMr+N/CJuESj5PHoK7NxLAqVcV85PO7TqcQ11iD6wSquxflKT97k8x1XUt06tf9ZHrFzPZwPXDi/vO2ELKmEKc4ntgiA/rjjvEPA9cwCH8GWd5EEnMJVTyV36znFBpzNYVKp0LfsTTEiprRWUDXQ9/SqjswYipYmwT3FxXfQ7bAiVUDrt/xto6gxwDX09RGSAbFBscIw88wWXSN8SJ+lj7Y1muK4KRgXQJlcvS60f/OoM7k74SKo9+fyxbC0qo3JoeH9s8Towk2CDjEUlIKitBkEzGbtnsR7z6tG89ofK+++5rsRehj0i1KWNFJeLKK0ARnUgt9fmOVEW+wrHxIiINoeY40YBAYMyIuCVi+W5DdCvD5yLejC27QuVQhbwjoRWpyB6LKlRmRaV+sYIRORnyEqlpBZvfqNSft99+e8sDf0h2MVS/wQ9sECrNFULqG9v5DiPmGGJvBCN2sw+2QrzCFoLe3IPAJC88erWheuyzuiMrKiNSujcQ/dqNWIVB7dA+5Qmn2uL8CFiClPzyuEaCpuuzopKYgNR3XNvksQ0tuT9DXpvDab/r5g/YaVHvp8PZ2TW6xwhJ+Y1KK96IMI7BGBFHn8MNW2xf8Uew5f7kg+BLYjt4YD/9Dxf8Fxyrxwo12IQn9wS/6Bxegc2PIf6JXPKr3/lgZcxpTEKluKfv9etaQqUVlfyTV7/ya/yMB2xgRdzif/S7JL56Vbr+/9rXvtbwqCx/BY/sxt+oB15gUfzjl+AZdtSVh3nyQJJ72IpK/gg+nd+KYZ/qh0GveIdT16NOZfhP5yNkwqh8ecBoUfDp+pZVqHTtNkIln8NXwRK8wqH/+XY45M/gB8Zs8CRuwSesPv300+2BMnFZXBTfCNtPPvlkqz+/sQszysJyVg2rW34+tFZUfqzZe1Hun6G2s4TKHntmbBPcHk1TVc3RAgKFyZ8AI5gY+Bh4mHgIJBnIzLEJVfUSWsBgyGDXwBnxZFJeQuUSAmGLL9nkK5PfEiq32PhLfLrgroTKJQbBUbz0EFhZ5YNQrRWV/XfI2OZxxBRPyCMrrTAwL/AbQ/Bj3oBgtyE9CZiId6QmYRO5j6zqvvoV6X7vvfc2oRIZikS1IcS8EhZOiVZITiKIepGtmYuwr7zO4VMepJb2IM8IQkgzhJrviEdjTEQYAtd+r35VRlvlWcSEVFxkoTKvQkSC6gcEt/G//jEX9bpLwo3+//73v9+IeKJmV6hEiCMzYUPi2whBVnj4hEtzWrZCsjoPIlYd5h9IUfucA07VB0fmvOrQRriJMEpgMD82bnQu8xfnJqwrC0vmMVmRcmBlXiOfc8Ik8l5+bdEG94e8ViS5dkQvsl89BK4hpmUVKr3O1wpfuCFUWgXL98AJX8cfsQ1M8S0w5f6EpfgmxyNgwyZfBn9ESRjzG274GKuLYQLe4MO9YM7CD9rHb8IRjJRQuRhCrbhnBSIfpg/FuO6rXz2Y4VXX0g033ND8IVzxP3wInwBXsKTPCYTwAoOESnjh5+DKClz+zzH5nQ8W+SP4gjn7+Vx5+DEPVVjlrm7xkV+FOb5YnVLqygq53bt3t3EBX8ZvG0+6LkITrPJ56l+UxH7uNzZyzcu0ohI+xCnXzqfxVfqfXyMk2qeP+TdjQDiRFz74Ln3t4Q37nn322bbPGI5vEx/Nf+2HZb4LlsQ6/tQYk73Fag/ryK9cCZUlVPbhO0qo7MOK/61jbBPcHk1TVc3RAgKLgUYJlXM0clX9PxYoofJ/TFI7tsACEYxM5Eqo3AKD1ymaBYK7EioLEEfDAiVUbo3VxzaPQ5QimF555ZX2ZD3CFJGEiBdDzRuQWkin7opKoqAVafYjXhH7kt+JfPDBB5vIiEAl3iA1EbiESnMRhJdySE7EWIgt5BaRJwQa8hexSCRCvhKJkGnaZrMPAYcEUz9yl3CgnZ70d9z1LGJadKESOWnlxYEV8lx/6lckpT43J0USE/KQlbfddlsT8gjT+hJpKh+iHSaQpMEEP4d4R3qqR/8TsIk6zmEfLMANbCkLA+qBI3U6p3itDTADs+5rYqa6xHD/q1tewpJj/jevUbd2uEZtReqqnxDpGrVD3Xv27GnHCU+IWSS/+wEu5Rlicq/rBzZwrfrBtY11RWXIe1i0wtc1X3XVVW3Vowe62YOvI/SwCx/kwQvHYAi+ItooCxfEeRjsClDw6nXHyvCvfCA8wlJWVMImfKiffyuhcnF+KqEb9/QhYVEccu/wV0RCQia/fu211zY/xPfwE3BlPx/DN+h3AhJM8mF+88+DDrAjr1jt0//iIdwoL5bzX2Kf2Atf8GoerH1+kxdGCZHwBZPqUEY7lYM9eBb34ZWAL87zcd5U4JrgVx7l1L8oiY3cb2LEsgmV+lkMFa88TOGTLdgBDsU7mIADMUzfypOxlf+3rwjp8AN/bAhfcA4XMOuBi7xlAC7Uw+bqgDUJVr36Gp7hnf8cc2JXccD9xA9I7lnx1BjY90qzWaCEytnsd0hpgDWIEYDczAbPfrvBjSsILJLDP+TC6p9BW0BAKaFy0F00ysYZ0AjMtaJylN072IvKgLyEysF20SgbFtyVUDnK7h38RZlbIFRNisVehMOsKyoJCH7PCMFuUo10ME/JXCWfgzdOjw0c2zzO3ADBhMgkKiGvEEiEFN9DZJqnmq8iUhFP5rBWpBEZr7zyyka8MzPS64knnmhzDk/cww1SBiG7a9euNgdGbsEn4Ucy/5UHuaqMY3BMQNU+x7QH/hA7xCP5tAlBZpwpH6JWPis+kGhI3qEKQu3C1/njWtnH/E1MsUUU0z8Raod6ffyRFUPwoF+0vdvfiEtku366+uqrW9+bKxi3EYf0K6ITkQkf9sOnfnYP2kcschwm2AsOnJdIRICCTQSrc7ETvCDjEfVwLL86n3nmmTZPUZf8wbz/lYFX50I22lyL9rgPXJPzhLTVZkm9u1dWJOF79BcREy6JlvIMlaBcRqESgW+VmldTi52XXHJJi53uMcf4OrGVv4Ml/Y2/gzE4gil9zN/AJvwpY6WS+vgqed0LymVFEWw5DivOo474NfXCIpyPOblu9xJfwD6Eca/GZRvfF+X6u3FPfMr9DkPuKdixgly6/PLLm9AnxhERYYUN+BJ9zlexC38ofnpVK/8CW3wp4dCn2CC/eGksxi+JlexHCCImBl9exU2o5PP4IL5NeXiEOe01zovozs8ZExAqrexUz3nnnddWiRKa5Fu08Z8YsaxCJf8DPxm7iaP6OHHMvdZ9sEz8FUs9jMM38ntwDaOw576EI5ghXsOTMZ6YD2vGK/whfMIOrMgDN+IgQVwcHPsb/di8hMr5RrASKnu0L8C68Uuo7NGoVdURLVBC5RFNVBnmYAGDE4PiEirnYNyq8rAWiGBkAF4rKg9rpjrQswWCuxIqezZsVTeVBcwt5iVUIvYjVGpMCKp8TtXAkWQa2zwOWYVYz4aQCkGM2ENIITP1PzKTwKIMwgthhYxCVCGgJOM9T9wjTZHtyqrHKqOnnnqqkWVZocZnInGRZs6jDqQ/8guxZhWKY9325FWL2oPYVz9SV5sQQgg2JBjyVl7/L2JiM6TgogqV2q0/8B0ITN/tc0360/W5l5DkXkWItIQZeLAfHuBHXmXEVaQpPwcbyHx9DI/ymms4j3EfXMGFOuU1DkRQExthA/4IIfwXvFq55BNWEKv2Kw/bcAaXzqd9cObT+dwryhCu1Ie4JRSEzCUMuE+cN2KAfMSLoRK0yyZUwoq5Kl/mt3r9b3U40l6/wQ8fBFsHVgRF/a/v4EReGLGCji+CV30PozBHvILl7IdheLW6Vhm4UQeRznlgBzbsD67HHmPHIlTyHwQa+NCHRB0YIs64RvgiMPJ7/A/fxB/yV2IvzPAn8MPXSHyWfPAIQ/ADh4QjmIQdfsr5+En5YceDFYRN+9VJPPIwkniqLWKs+mAOzvg3fpG4qTzMuQ7+Vpv99qC6+Wlt1x54X7TE9q6D3flltmRvPpmdXTt7jjHBSu41vsn1i2H8PXvwaXwSW+hb/k2cg00PXMjL10mwAW+JeVaHwzksw4zxn3jPlomn8ieewppz+D9YH6PNXRP/zxZsxoaSe46vN7bwvdJsFiihcjb7HVIaYAWjEioPMUv9M2cLCEIGNRylQYtJnQE4RykwD3XCNGezVPVztkAJlXM2cFW/pgVWD8j5PINkkzYTRxO0SmWBvi0Q3JVQ2bdlq75pLGBuUULlNJaaLc/Y5nHISnME12Xzf8hxnzZzBqQTMgrxJA/SS37/d+cR9iO4EILKIsLMeZH2fgcTMXPxxRc3slS9SDMCgTmKY8TQ888/v5E4fKn6uu1xPuSWTZvUgWzUppxTPY6HJJutx49O6UUXKsVD/aH/4Mt3+1yXPsv16UOCo/4yZ9CP+lMybkvfK596HA8m7UeQWpWEIHXMSvKI1cpYGQRf6obVvHbTmFDq4sz5bOonQmqXObL/tc/mnGmn/cGi+0ObXZvz8snyqif3j7rkgc0hpmUTKoPD+HX/40b4EP3m/2CYbfR/MOJTnvib9L0+V5+5B8xnv7L6HUGvTPbL6zz+z7bIvmsjuGZb9++ir6h0HeKePtaH7vdgCAb0MT8jwRefYb/8jsWnwEF8A18mn/yOE40InsQg5fjNPLDD90jy4/e0QXJOvtGDQmIw/0NsJE7Gd9mnrXCpnPO6FucxpiSMSh4g4jsJnfGd7cCC/GFbduSbl02odO36Nb4JXruY48fgwAa7fFuwSWAT7+SRfHdcXriDG8fk5/My/oOv1b4SPpXxqYw8Y07u7RIq59vDJVT2aF+ALaGyR4NWVVNZQFAuoXIqU1WmHi1g0GKA40nDAytPohqweGLPk8UZwNtXqSzQpwVM/jL5RYKWUNmndauuw1kguEMMIBRg0ASOMI4cDYk59onZ4exT++drgRIq52vf1D72eRxCyyaFZMq1b+QTKYbQQkIjSY0BrexAUJ111lnt6X2+EGFIwJTPuNCqNCs3PN0v9dWejbR9CHlDLJq/iSk2BCCbWY2AKBZb2HNZUnCZ64VP4lHmGFb/mHdYsYaIZx/3a16tqDy7bV/5rS3Hs+o29U3zmX6RV19sZA6T9m+kzDRt6jPPsgmVG7XdLP2/0XMtQ/7M1RZdqOyjr9bzD3w/P2ZOy9/xPWIkP4ZTIU52/ZHYm7GK+Qj7emCITxR/PcghdkQUTfvFbXOZjCedT2wmSnmIyDmtFo0wmnKL8Mm+yypUru6f8mOrLTKf/92DJVTOx7aptYTKWKKHzwSNWlHZgzGriqktUELl1KaqjD1aoITKHo1ZVU1tgQhGJlclVE5ttso4owWCuxIqZzRkFd+UBUIsmRSLvUj4vn6jsl79+kGXLMM8bj3C9ANLrP9NHTYPqyFJrcyw4o1/RJZ6al9CrlrFYdUI0tWGDJUnqY/2pK5F+XTNSOMSKtfvMcQzjOFVvKIO1pD6iPqsDCGG+I5g90pED0xaiQR3q4n69c928OiY8VhC5ZERMOb+P/LV95ujhMrp7OmNaARHfs7DPeImgdLGr3kosvsARHc+wicSNy1YIGj6jUkxlv9TTzeJOcaPyniAyLjSA0RejaoMv+lci/iAjPu2hMoPerv82Ae2mNe3EirnZdkP6i2h8gNbzPxtGSa4MxupKujdAiVU9m7SqnAKC5RQOYWRKkvvFsgErYTK3k1bFa5jgeCuhMp1jFSH5maBEirnZtpDKq553CHmOOI/7MUn+j2o/fv3N8LU2BAhiihDevqtIwIlEYnAntXnR6x8xBnYpoTKI3ewuAtPxEnkOpx5xWFWn3q9HELeSkr4QrbXTwAc3q4lVB7eNnWkfwuUUDmdTfF4BErjPL6NKOlNLbasDF8tVPKL7mdlzIfFYaKjFecERw9prCVUesiDsLlv3772RiIipfw+81rZzTzgMd2Vzi9XCZXzs23VvLYFSqhc2y597i2hskdr1gS3R2NWVVNbYBmEynoyaGo4bFnGEiq3zNR1oo4FIhiNUagsP9fp6IF9De5KqBxYxyxJc0qo3JqOrnncxuyM9ERGI1et6MjvHUWotNLNakpCkk9E6loE6sbOuvi5S6icrg9jJxhDyou/CH3zXsQ9Ip5YSfyGLyt1uyt6pzvL8uQqoXJ5+noIV1pC5XS9gE/h02xsxrflNwXFTD6um/hFcxIrCI1Zsnlog7CpDN/YFTeVjz/lB8Rr5+U7s1lJuVa57rmH+t211YrKofbOONvlvqtXv863b0uo7NG+NcHt0ZhV1dQWWBahEvGBFDE4sxlMIUF8+t8gxaAs26IOtqbu+KOcsYTKDzoA9mDTINnGNploGPiv9QqWD0rXt41YwL2eye/YXv0KR64Phvg7Cam71m+NbMRmlXd2CwR3JVTObsuqYeMWKKFy4zbbTImax23MaolZIVp98pXd+IVwNVZPHFtNnm7sjOPIzW5sZP4mptiySnBZf6PycD0bW0UUZzM4k8zzbMbY8FXj7cNZ8eD+EirXt08d7dcCmatZDW0ltPt0x44dbZV9zYs/sHWX3+rGTvO/tR7s4RPjF8XbbHwhH0jYPFycVY7/NNZxLv1gy3kOV+6D1g7zm+sqoXKYfTPWVpVQOf+eLaGyRxvXBLdHY1ZVU1tgGYRKxjDg9R5/Ey3XjPjw/n6Dsv/85z9toGaQZrBl0GWg5vuiDrqmBsBRymig6+l5rxA5cOBAs7PfhtEneQJwGWxvcGySAJeerEIoI51gjy28TsVqAnitNLsF2DqT37EJlazDl8ERH8efuZfyhOzs1qsaNmuB4K6Eys1asMrNYoESKmex3vRlax43va2Ss0uYIj79n2QMmHG5z2UYE+ba1/uMzcT5EirXs9TBY+xlCyG/FsbgKxgrnK1t0xIq17ZL7Z2PBTJXK6FyffvGv+VTbj6su62uIT4wZfJ//ODq/N3/xWm+VHKOMfhN119CZbeX6/u8LVBC5bwtvOKfVm7sD2YU8z/fqM9QE9xRd+9gL25ZhEqimMGu11WY2Pvtm1NOOaW9U99rIA1QiEOEyzzBTaw0ABt64oaz5Wk6g8chC60lVB5ElcE+W8DlX//614ZRgrpXqWzbtq39bo4n5P1faXYLRDAa46tfWcd1wREfx3cRKU844YQmds9uvaphsxYI7kqo3KwFq9wsFiihchbrTV+25nHT26pybt4CxvvG+iVUbt6GVXLjFiihcuM2qxKbt0AJlZu3XZXcmAXE1BIqN2azyj2bBUqonM1+05QuoXIaK02Zpya4UxqqsvVqgWURKhF1f/nLXyZvvvnm5N13320E/he/+MW2Ys2qPnawktLKNT8ITqwkWhL7hpxCWCDCvdrIhsAgsroGn0NMJVQe7BX9BXtvvPHG5Nlnn23YhDni5Mknnzz59Kc/3VZWllDZD4ojGI1VqLRK9E9/+lMTvN37xx577GTnzp0NQ/1YsGrZjAWCuxIqN2O9KjOrBUqonNWC05Wvedx0dqpcs1kg4/4SKmezY5XemAVKqNyYvSr3bBYooXI2+1Xp6S1QQuX0tqqc/VighMp+7LheLSVUrmedDR5blgmuYLB6Y6ruKwry3f5uXvvzv2NJa712IPm6n8nvU10pl/3dvL6nHUfKl/LJn8/sH/LnWIVK/Uew86n/rFh7+eWXJ3/+858nb731ViPur7rqqvZbB4TLCJVelfihD32orWAjVCqberqYWN3HOZ++9j0p+XyuTtmXT+VSNvtSJsfy6Xja5ikw/sOqUQN7pDiR0mtDrRxdjd/UeTQ/S6g8aH395beFXnvttcmjjz7aXttpBZzVlJ9ZeRXucccd11bD6c8uDpWGgeAgeAk+fMqf1M3Xzeu4vN1P5dbK3zIt+J8IRmMVKj2IsWfPnsnbb7/dHlIgdJ933nltZW76Wd92t/R9Fzur83Z9SI4lfyARvKVuZST/r86b/1OXPN1zKJf6Vu9PmXwqK+Wz/TOwP8FdCZUD65glaU4JlVvT0csyj9saa9ZZDmcBsU98LKHycBaq/fOwQAmV87Bq1Xk4C5RQeTjL1P6+LSCm1orKvq1a9a1ngRIq17NOP8dKqOzHjq2WZZjgCgQ2q4gQdz6zDxlpJZFVILaQnCZjWSUWwlPZbrm1fshZOWKIvLaQnoytHudKudSbdqV++7VldT51yZN2BQbqzLaaXE2eoX2OUaiEDf1j0OG7/nvvvfcmzz///GTfvn2NxLfS6LrrrmsE/mqh8v/+7//ab7spJwVH+j2YCFYdX42HYE3e4Dp4CNZyzKdNCk6zL/eAa1Bn2uF/xwipyhAo/S6da/TaULh0DSeeeGJbmZeVoTlPO9lR/lNC5cEOMPHXby+88MJk165drT+/8pWvTL7whS80Md3q3uBQv3Z9TnwNH+W71MVi8up3x9XT9a1wZIOhfFdeOWXgJvmHhJ2DltvcX9eaye8Yf6Pyb3/72+SJJ55oK3T1+fHHHz+54IILmp/TrxIcBDvp1+CGfYIbefmZ4CYYg5Xk96mO4EaZ1N3FThdn8iS/T0mZ4Ng55ecjpOx3ni5OfZfstynnc4gpuCuhcoi9M/42lVC5NX28DPO4rbFknWU9C4h9YmcJletZqY71bYESKvu2aNW3ngUyV6vfqFzPSnWsDwuIqSVU9mHJqmNaC5RQOa2lNp+vhMrN2+5/So59ghuCUSAgrNgMeg1ETLiQoVZ/Wc324Q9/uL2Ck5EyUFEOESmvff5H/iHT5c/rOpGazsWeSEHn+c9//tNIT/tDaDrXxz72sbZyzr60S5vkT5vUm5VpyFQkrsmhfDbl1OuYVU9e0egavEJUvUMlTgPAMQmV+kG/6SP9/s9//rP1D4wQ8qyoROTbb9Xa9ddfP/nUpz7VhEt9Hgx+5CMfaX3JRsGRvlYvfMGELasVYcBxK+PUA5faAa/dV8nCiBS8aJfNedWdc8CM8yirDvWpF9mY+h2DS8l+YqvfpnOd8sP25z//+SZUyAeXzj8UPC67UAkfbACLBw4cmPzxj3+c/OY3v2n9/o1vfGNy9tlnN3GJL5EXDoMv/k/ib2CQ//Nd36qTWB0fC5v2O+73CtVn1TBcwZzj6vaZ8ygPf14/S/BW1v9jSO6NxJSxCpWPPfZYw5T+9EDGWWed1fwB/wIL+jT+i++R4sPgRv/7X9L38AU3fAjcwJi4wRfBEL8S3DmnMnAZ7Dgun/zySerPebSJP3MO5eBTPqte+Urntw8G/a9sNnXFj/qMj7V/SCm4K6FySL2yPG0poXJr+losZWsPH1ndzq+ddtpp7c0dfCdfV6ksMKsFMtcpoXJWS1b5jVjAGJJoZBxjjGb8aF6RMaUxWKWyQF8WyFythMq+LFr1HM4CmVuKqXgZbyUyn/3EJz7RuDa8oPFcpbJAXxYwX8DZhrNTL65DPMXh+l5pNguUUDmb/Q4pPfYJriCArDPAfeedd9pGXMmAFwnqNYdWgHxm5ZWHgoJJvZvYbwvKF0EHgYlQNYhBuG7fvr0JTsccc0wjSZGiyhGlnMsgR35BR53qQeBm1ZJ9RACBCXntNaHqQJp+8pOfnJx66qktWAlSzqk++bSfg3FtyFmCkKBG/NJ+pClyYshpLEKlPshAQ1/rm7179zbSSH8jyfVv8Ob3/wiVcAAnyoQsFyBMuNyTfr/S62IRT2wFp/oXLpRVN+zAA6EQdmAE1iU4gGnYkBce4FD9mdzBmTLabCIoX4IVct4x5/d6UJ8miOp1DfJp44EVsYsQ67hzEJlOOeWUyec+97mW7+Mf/3i7vqEQ+a6BzdN296D7Xjtdk+u2b6xJn8KN36a0mtLvCv7hD39o1+xVnYTKM888s/kTuObz9DHfo5xEwOEzTzrppDaoYS82hfNs/BM8wJ98sMhPwp97wnGDcuWIP0hWr0bWB6effnp7BS0iwv9jSBGMxvrq16yo5LP4K/HRymo+BXbcVx7S8EpY2HEcvuCA/xEvEezBmL5XHm7ENvUEM3yec4h9/BbcuK9hTd3O4zusycc3wRmcqgNGncf/sCw/n8r/ym/1O1/Z9Qv+V8amDokfJXLa+PAh+o3gTvxhZ/c/v6/NfLm4kvgwhvusrmFYFiihcmv6Y+zzuK2xYp3lSBYQs8VCcVJMsYmJGfvn4SKxt1JZoC8LlFDZlyWrnmksYJxsrmbuYm5jjLxjx4735yL8XaWyQB8WCH8oppZQ2YdFq44jWaCEyiNZaPbjJVTObsP3axj7BNekClmHMEHOI+uIFCZYCHMEKoKSCIQYjWAhz7PPPttITmQeEhIZKpgYNKccktMABvGaJ5oRocr7X15EvLLa4TwXXXRROw/ylLBpMCSv9shD1NEm7UHsEn9CoCKCDZwMpJCxNgIAglZeBG8Jle/De+5f4Iu4rP8Q5hFq9CUi3af9Br36lvh84403tgHv/v372yRfPqS3ftR3iHDE/auvvtrqNUhGKhMp85SVC4MdpD1cq9+Ax/m0BRG9c+fO90VN7XSvIxCcB25gRX7tcD8g25HYyAbtcAw2YU6dISMIldoM40TSl156qd0n7hF1W1EZsbKEyrlDcEMniFDJh0SoJFbqOyLlGWec0YTCiDYGzvLCGpzDUfpZH8NjBCFYtPGxEXMIjfwX7BJ+4FIb1Ae3PuGSn4RBmLQST17fS6jcUPcetcz60qtf+RJ9zz8QGWGD7+g+gMM/wI0YGoGbv4MbpCeM6Xc+Cm7EQX7JMXlef/31Fg/5RD7PQxriI/8Et4Rx8ZAfs5+P0gb1+l89fJu4LLY7h/w2x1955ZXmqz/72c+2eK1OvpsvJHo6H58s9vNvJVQeNdjViQduAfdd3rhgDMynGz+4p5Lcw1I+xRdxxwNSVmnfcccdbYwhj4f8brvttslll13Wxhjq6ZaTR/llS2Ofxy1bfw71et1riaPGa7YSKofaW+NpVwmV4+nLRbiSEioXoZfG0UYx1VzU3LOEynH06dCvwnwB94arwcFIOJdaUdlfz5VQ2Z8tG0mMTEDAjfGVQYIAktENiUQ34CXCZCDiJvUdoYr0QKASE4kwDz/8cBOM3LxIU6QkctXEDFmJNJX/qquuaqQoYUk553QON75zy88pKOMcF198cSNekbr2y48wReiHpBG0EK/OSQjVxt/+9rdt9RrhCzl6/vnnNxIXYZvNOZUbOlnj+thDvyCNtZnYy9auBdG9CAn5pv1Wnf3ud79rgqE+g5eI136jEpFvUm+l4e23395w8+KLLzYCPeIgbOg3+WCVQARvBCErftjEcVhgN+Ik/GoDvMEEmz7zzDPNngRxYgF8ZRCkPoS8+oiK6tEOOIQdONQeZIRj7hfnQ8xrn+vyqU5YJ8wSMpWHdXUSSLevrDaGZ3Vp21DwyFZspu36TLuIYvrM9bPPUNo6D/zrI4NiOLFiHDafeuqp1kd+o9Lr4vSb/ifM6Ff20v98HfshntUDW/K6Zx0nRtnyYIZP5WGfCI5khjv3ugE5f8mHGjSxPb/rYQv1wlj6Yx522Oo62ct95J51b/MZbEZMc93u3UVO+vjJJ59sqxGNJ9z3fB1/4J6CNyuv4UD8I1a4djiTn5+BHfiyBTfisvjH77AffwOzvvMx7lvnEmMJGzDHnjBESFSX/WwenBEm0yZ+kX+CYX5RP7kWnxkLqEc73S+OaYO2+w3Obdu2tWtS3xBTcCemuNfYjb3gTZzl7+HQPVmpLNC3BUqo7Nuia9dXQuXadqm9/VrAuF8sFDfFFFsJlf3auGr7XwuUUPm/Nqk987NA5mrmwHgYY+RaUTk/ey9zzWKqeWs4OvNLMdb807zXPM28uVJZoC8LlFDZlyUPX08JlYe3zYaPjH2Cm4mVgS6y1AAE+enT/whzZIrAgKxEYiJYHbv33nsbKSlgICuRq4g9ZYgzVlwKJLfeemt7Utz/BBAEOyITiY8ARE5H1LGfwGi/Vy4SiAlQCFcrQRCehEjnNwG0n+CExN29e3d7shzpqp1+U87gSZvUYVN+EYQWQdk1LrpQCUeuwSqcBx98sJHhCGyrcfS1ayQwI8sNevXxd7/73dav+h+WCJpWGgQv8IhUPrAipMGSFWbw5zscIPHVa3P/GkTDLhHIOXbt2tWI9i9/+cuNSFcGltQJd0h8+IFrbfc7heqKSInIdj/ArX5C9msjEQkeDZzcV447nwcctEk+woRVefIOEY/auMxCpX6z6Xer2Kyq9BuVMHTNNdc03yeIhGCGT2KHPreyzYBaGTaEN3jQ5xEr4SuCE2wSdgjnfBK/F2EHXqzE5cvUr5688tV9A4tDErg3HFhXFYhgNGahMisq+Rp40Z98DVGMLxNPYYMg7mEGfS5eyQ8rcOk4jMGNeGrfOeec07DmmEmc/fyT+vlZOIQnQik/RuTm3/hadud7xVn18rHKwRd/CN/IVliLqOre4Cdg1j4PisC1lZyuQ3xWz5VXXtnOMWScBnclVK66IevfLbFA4kjGB7Wicj5mzzxOPM0Dp3ysMR7ftQhzgvlYpmrt0wLisdhrXmAss1qoFPfFe3PsSmWBvixgvmGuCW/GZuYrYom5grlp4a0vS1c9LCCewhrMGfNHqDTX8F1MrVQW6MMCYmpXqDS/NG+DNXNZPg6XVqks0JcFjN/MzXAvtBEYjG6RRQJ9nWtZ6ymhsseezwR3zCsqM7FClrgxDXpNsoiHPhGTCFM3qFUaxD9577nnnjZQQXgSL72yTsBAcFodgphF0HsVlbIIeQMbg2aDZyKmervnM4lDIBhs//73v29igUGPvFZ2KZt2KYdw9ZoreRCxSFdtI1z5rUNtQpTa5FmUAdRYhMoMaIk3v/zlL9sA4+tf/3pbPWbSjji30pKQiWRH3H/7299+X9h2HCb0vxU98KJ/EfUwBrsEciKRsp6sMoCGEQMa2EVCGcwQgRDvDz30UAs8V1xxRdsHa3Auv7zyWXVkIOR+IJiqD3mvLSaAkmP2azd8uTfgjggprz50vgiVRFvEvt84hNsh4pEt3FeuywSEPZZpRaV+NSixoo8NrKb99a9/3fzODTfc0GwBX0RMvkYeg2a40O/EIivBDazhB45gAtZhBAZggo3ZGnaee+65httLL720CUsGRAZJ7gm49D+8WM3pE8bdB/pmLCmCketFJo9tRaWnjsVDqw5hADYI07ChP62e/clPftKum1DpYQZ9bdLP38ED2xg0ww38eSWx41Zg8n18ITx5sEKc/NKXvtRERL4TTq0MFps9aCEew6syVgAnxvOx4q88YrVNf8CzffyaTf/wa+K9/DDq3NplI7JGqBwyToM79zTb6BvX6X52z7rWIl7G4mWGdx0lVG5Nn4R4cI8bO/JbHsjgh43DhuyjtsZCdZY+LNAVKhOzzRNgzMM75g5iS60A6cPaVUcsYD7BtxnHGKsZ/xn3mffCm/8rlQX6sgBeJ3M18wdjZG8EMn+o8XJfVq56WCBCpTln+LQIlWKqeVoJlYWVPi1gvoB3EU8jVOICxFPcie+VZrNACZWz2e+Q0ssgVHL6JlMIRjenG9P/vvv0v8GHoEC0MMF37Gc/+1n7NEAhXlpZYUBM8CFM7V5Z4UiQJDwZNBMqDaYR9m50kzb5kYPsLBAhCQk+Bt5W2iE/kQjyOjdCVJscVwaZi5CVnn766baqw3F1XHvtta1Nh3TogvwzJqHSgBZ5fvfdd7e+vOmmm9oqSE6fEGkFENERmU/Y8RuVBh/w4jgMwR5i3QQfqY4kJyLBJvFPOQIj/ISUgmeBBhlPMIcJuCGMEnoIQ/CTAVCESvm6QqV2eFAB9uDQU/hw69xIfIIVDNuvjXDq2pwXFmEeaSGP67ACFEE2xFRC5cFegSGDYhgjbOtvuCQsEW9gdd++fQ2fBtJwB0u+62+f8JDNcXiAtfhY/8MooRK2CFRZlQ63cCU/zMEV/wo/YyRWIxhl8uu6xRz3kXsR2bLICV78npxP2CAsEhjhSX9aPfujH/2o+TYrvYl/xD6xj8gIi3wIXMINjBEq4ZIf4/uCPXiCt69+9avtN399h1nxEd7YNSvM1cXv8U1szNYe7hGf+V7n4Wv5TefSH3DOr/GL/IX4rz6Y1X/wDfceINKuIafgTttdKzuwF1uUUDnknhtH20qo3Jp+DLHKl/J3xpHmLMaOY4ynW2PVOstqC4jBYiq8iSnipPkKjEU4qhVuq61W/89qAXNRc1R4MyYzxjN+MS4znjF2q1QW6MsCxsmwBnMe/IG3U089tcVT3/m7SmWBPiwgpvJp5r7mluawYqz5sZjKx9WDP31YuuqIBYzfzM2M3XBRMIijwwvAnO+VZrNACZWz2e+Q0gHsWFdUIuoEASSdFTwCAacvELhBDYDdrAhTA18iDqHSTUx4Qkyee+65jdwk0BigIAIQr1Y4mpR1hacIj2509eVJGOUMcBCkAg+S1IpK7SIqeYrBqjnH9Ik2J1ghSg2cELFWrGgzAthvY1rtuYhJUGaDOEr2YUvX37Xb0K8t949ViT/96U+bSP2d73xncvbZZ7cJFAIe4Q57nswj4H3rW99qhHheFey6EUr6VKCwag3hRChSHjbhiKATPOW8sAArJmtIeAkG/W/VEnvCvPvbAIidkeuEIWQ7soGwieiHWUKR48rBn3ZYDaUdyuobZX3CM1w6ZlDvewmVQ0fswfbxffpttVCpb+GEeMPXwIcHL+ASXvhOmOOzDGj4M7iA2fgtPjdJXfwcIoFAFaHSfe9Vmp6O9pQq7BK34ArebWNKEYzGKlQeWFmdTKiEG3jhQwiV8MSPiJd33nlnEwQvv/zy9oYCPgpmxED+w2BZv8MT3BAK4eySSy5pvtFxfspvVPJPV199dRO3+SF15A0FysSnOre62F+MhTOvi+VH+UXlEBHwqBz/5bj47xXa8oj7fK+2aQOsuxc8kAG7Q07BXQmVQ+6l8bathMqt6VtxVFwWX4yr+UTjybHG062xap1ltQXEP1vwZhxpE7ONEcV08wixtFJZoC8LZL4La8Y08AVr5h+Ft76sXPXEAjiVxFPcCbxlfmp+w99VKgv0YYHMKcVU81DjN/uM3RJTjecqlQX6sgCseThbPMXbwhuOTjyFuRLGZ7d0CZWz2/D9GjIAHKtQiVQkivlNSa+mQ9iZwFs5YcBrMIKMlM/A12DEqysRLFZUIlC9ws7TycohKwmVVsh51ZwyhErEKcHHMSlCpeNZYSnwcASCjpVqiFjEKwfh6RkkvjzaxXEYDPkf2avde/bsaeKBPtNOr/Ykbi5iGptQaUUlvEi33HJLwxCHTwwiVO7du7etNkJswwv8rRYq9SlCHF4MWGDJoCWra+EC/gQR+BBsBBmf9hE74Q45DzdEbNhybyPkEfbKaQPB1MohuIJbeZD7CHiYQuobnGsHEUJ59wRMIvODZfW5R+TTTuUQ+OpWfmgDehMQ7STAui72JES4/zLhtW/syQAFtrpCJQGdj+ObiN0RcGBIf8MNfwaH9sWHqgc2YEB/wzA/B8eEK36XTS9fEai6QiUhlN8lHsGLjR+Vd2x9wGbu1TELlbtX3jCgv2Fj+8oDP4RKfeva4eyuu+5qA2M4IGTyHfwPP+e+JHzDl/L2eRBIni9+8YsNl+5J2CRU8lUe1PFKdn6mG0/5Qth0Tzt36g+G+Td5+GZ+D9bFVJjl/9Tp2H333dfuAXVlg09+nf/kX7V5yCm4Y2c+HAbZ2DXEhxuPDM1PD9mm1bbpLVBC5fS2miWnOOreNq4Wh8VeYzE+bWyxdBY7VdnZLSAmB2/mHjYY8/CPuCqewF+lskBfFjA+NGeBNWMa+II1Y8XCW19WrnpigYybYS7z2u6DuRVTY6n6nNUC4inOTkw1D8VP2Wfslpha8XRWK1f5rgW64zexVcKjiKcw53ul2SxQQuVs9juk9NiFSgNbYqRXtfrNSd+9ShWRaoDrf0SlTzcnYrUrVCL4IlQiVxF6iM/uikqv+kT8ES4RtQYxRA8DG4Sqmx/RiQT1v6CDNIxQJSARAbxaQhmDJHVon3qUQcQi/Amu+kxb8htZh3TogvwzJqESRpDn8MXpeyWhVbAEG0SdfvaaS0IhIejmm29uxwjVBDP9S/yDPWX0OQwQNWzqCKHObvBikgZHBjgwDrtIdOQzDBEt4Q2OIlTCLewgqp2PMGoQTqgk1js3sQgBr7yApR3ywKsVcL5rX4hu59UmNnAeOD/nnHPadWon7A9pUK9/SqictEk/7K0WKom2MKcvCZX8oj6Eqe0rPhPGYIJvggEitdXC/B6cwrJXuBKJ5LGK2Mo0drcyzmo1uNAH8GTAFJHSAxnq52OHhJk+XCqbLYNQmdckwQqBUZ/q465Q6RXA+hx24CzipvhHAOR3vELWK4nZ7IILLmi+Ci74IavX+R1xnK/iz2CZPyUw8oHwx586t5jpXHwmv0eohGll5IdbvpS/VIaPtt9vt/Lb/Js2abNj7hH5IgT0gY951RHclVA5LwtXvetZwL0lBhhfiAHGGB5Wce8kIWWkfPL97j/3nlXad9xxRxtvy2Nc5TfhvXZZnFFPt5w8Y4sdrulIiQ3c6zY+jw34OH6yUlmgbwuYd3Q39YuhYvQYx29926/q25gFYI1v88nX8W/BWuFtY7as3Ee2AIwFc8Ytiad83DKOL45sscoxiwXgLWM44zcJ1hJTC3OzWLfKrrZAsJZ46nhiavm41dba3P8lVG7ObmuWWgahEiluVZtXzyEsrfQg9JnIu35kqU9EJgIyQqVXvyL4vPrVisquUJkVlYh5K+R8ejXrgZVVWgY2EY4i1iDgkaSIFaS/dmgTAZKDQK6qn9AjaRsCVr0EAfmsMEG6Im8RpoTKWlG5Jqy3bCeRkHjn9aiIbSKPFTlwpE8dgxUioT5HrOdVwQRMq9H0MUwg1+EEdtSrLvhTB+IOTu2Xnwgk2PifmCRFPEQGqotIgGD3VCBSnojg0//ERsdhSfuQ8cRy+2BL200ATQ7dG1b+7t+/vwlM2qi9iH1tdX/BJ2y6h6yoJEhpgzYNKfC5N0uoPChUZkUl3PI33/zmNxuJDBPwxpf5DObgQn/CBFzxZ7CU1cKwpf/5JMd8h3sPWEiESvhXFqbhRX9EqHTP8H9jJB7YjF3d0+4VdnMPu17Xza6LnGCFqBChUizrCpUe7PFqbNftFcB8DX/IFoQMMRA2+Ay+yUMd4p2B9IUXXtj8kjzw5MEKmCRasJ04qS4+2H77bHwZ30VI57+CY/v5LdjWHueIgOIY7BJW/Ia0dsMpX0ugJKbybx7y0E71DDkFdyVUDrmXxtu2EirH27d1ZWWBskBZoCxQFigLlAXKAmWBskBZoCxw0AIlVPaIBEQeMiErG5DLp512WhMhxkAYI8KJMMQYr3JDgFq1QeyJEIPAdN0Iyu0rK0F27tzZbPLzn/+8EaB+bxBBGaHSCkzkJ2FSPddff30TmKzosEoo5D7yHxmNzPQdQY/gJHz6H7Fq5QiCXxsQn/Iog7hG2iJ0nRexjzhF6roe9ViZgjxdxOQaCCWuC1msL9iSCEYkY4dFSJ5+cg9ZFfTcc8+1/iSEIbZdg351nfLoVzi65pprWv9aGYQQd90EC+IfUY89lEH6uzeRzTAknw0x77xwBjvuXXmQ7XDsuxWRfms14hIx06s2/e4l3LG/+4DdnUs5AhOiHu60A/mP4PbpXMh++QgK7hMYVN4qJ6sf/EacdjkvYd+KB2KmMuobQiqh8mAvwJg+5RcfeeSRhs1rr7224RNO4VKfwgwc6lf+EaZ9d4/mNa6JH0Qk9zNMwKu8cMfHyX/ppZc2f8X3wVRe/QqHESvdBwQp25hSBKOxCpX6mCANA/qe37nooouaYOja4cyKc7iDA74DjvgvOINFfQ574qB8cCceRPBUz4EVQZSACZ/8VB7sUJcy/k+s5nvgkV+GNX6OKAmLzsVP80twzaeJ89oFn/yeBzOslPemBPXwj373V3vEZGWHjtPgroTKMXmTxbmWEioXp6+qpWWBskBZoCxQFigLlAXKAmWBskBZoCywOQuUULk5u61ZauxCZYg6pCchhaiCSEWmS4hG34k/CEsrMYh/iEmEKKLdCjnEK+JTXuQqYhb5ilT1CjpJfoQoEUg+YiPSEzmKREWuIuS/9rWvNZHIahLtIZ46n3zIT2WUJTbldXMEL+KoMvoMSYtYJWQuYhqLUKnPiF8IOZggWNr0J5wRLK2kCsZgy6sM4YZoSLCx6hb+bPLbR3xE+vsOo/BA0JEXFpIHAc2WzoWA1xYYkY9gaGUjTKsD1uATbpUnbCqnPgIojMOVc7ku12RzDtfjXnBMnfIGe86H1CcIuFaEPuKfkKW+EiqHd4fqJ/3qwQe/r8rvWGkOn/qLiOMYYcgn/wWf8MKnwpo+9kkkgic44OOUhR9CpbxwQ4S3Uh1mnMs+fs9xq8z5XhvcwqptTClxaKxCpYcYvF6d39K/YqUHnsQw1w5HhEyxi1/KQxLsAV/Kw428cAM/wQ1xkK+CMasbPSAEu2IjYVLiX50Xhvgd9fNpBFD4dg5l1MuHwTFfyQ/Ly2dZoclf8cH8HXx6Ze3uld/eVNa94SEmAixBcxFScKf9/L/73n3MBu5d9xtxmP0qlQX6tkAJlX1btOorC5QFygJlgbJAWaAsUBYoC5QFygJlgaFZoITKHntk7EIl0hFZhwRFoiI/7ZOQcwhxn8g6BGlIPCQmYg8Jb5UPYg+pKT9hSH1IGOQo8t0qSyswfSI+CTkIU/WFYPXKOsSgFXVWmzkvkUhdCEQkqqROYqW8WUXnGBGAYOC7tiKB5VnENBahEpb0h/7TjwhuG/wEX/AHNzAGR0h8fYyk1//2I859qs8+/awe3yX9jZR3DisiiUJsqP9hjdgDnzBCMHAMkW+FJFIdhiX1WlmkvTCmjc6Ztqkj7eAbbOpyPa6TiJX7IdhzTyHCCfjuF+2UJ0S4ax2K8OQ62FBbiSfaRYAgULh2/TCUtrYOm9MfmIQtGGML18xvwaf+0o+O8V02edOP8sKjPoaHYBYOYEVe++BLYnP5ukKkfbAon7qyjdX+EYzGKlTqd6KgT/hwLxGn9as+hiXxlB34Kr5DPljhO2CBr5G6uFFenJPPw0Ye7rDKkY/ySlgPYaifH4Md9RI1HVeG2Oj1rfwiDPJNzh/R3T5l7FdOu9XHR2ivNyQ4n/bxpR7SIFbyF4uQgrsSKheht8bXxhIqx9endUVlgbJAWaAsUBYoC5QFygJlgbJAWaAscKgF/h8AAP//uyTzYQAAQABJREFU7L1ZkxzHebads+8z2PcdJECCACVKpEhqeUlrsU3LYTvCBxJlhewI+8QO65f4zPaBjxzUG7YZ/j4voQN5CTssQpSohTI3ASTBFcRC7CCxAwNg8D5X1tyDwnAAVAM1Mz3TdwUK1V2dlZl1VXVWT171ZLVdiyl5qoXAxYsX00cffZSOHj2a9u/fn3p6etIDDzyQli9fntrb21NbW1st5cxmJpwuY2Nj6cqVK+nq1at5pj7sm/aR18wdHR15Jv3ly5cT23Z2duZ0pGXiM2byYpuurq709ttvp+9///vpww8/TKtXr04bNmxIW7ZsSQMDA+n06dPp3XffTT//+c8z39/4jd9I999/fxoaGsp5q17kSXmqF+WqPqwnHWmYqIvqlVfMsf8uXLiQjh8/ns6cOZPOnz+f92dwcDANDw+nBQsWpN7e3jmzRxwbZp0TOk46t7QjOq6cL0ycPzrefMas9TpPdbw5D/huwuyVV17J31UYwmzz5s1pyZIlqbu7O504cSLt3r07nTt3Li1btiytXLkyrV27Ni1atCj19/dnzqOjoxPnLuWpXtS3XGfKZl+YqSefUQ+dl7xXfUmj7wvrSaN02q+ceJb/o45nz55Nx44dS3v37s37vnHjxsyqr68v17mZ6jtduMrnLEyYOC85vuz/5PNZ5yrpdB6Qltek17mv85Z15bx4X26vlL/yUz6km48TXC5dupQ+/vjjdPjw4dzmwXtkZCRfL7gWzOVJbQXHlal8vHWs1T7oPCANn6mNgRETn+szzgs4wY1r7J49e9Jbb72VuX3ta19LmzZtmmhD1TaRnrJOnTqVDh48mN58883MmzZy6dKl+bcN1xnaU53zbFs+X9mW30MHDhzIM/WkLV2zZk2iveAaNRcmnXfsz6FDh/I5yG8Szjf2gWsCDODsyQTqJsDfFu+//346efJk/k4uXLgw3XvvvWnx4sUTRanN0JLvPr9j+K7v3LkzPfvss2nXrl05/fbt29PTTz+dnnjiibR169acT3k7ErG9JxMwARMwARMwARMwARMwARMwAROYKQJt8YepRWVNtFtBVIKKU6Y8C1+5U4PXmkkrSaR1SlvOR5/RqfmTn/wk7du3L2eN9KAjEHlEZyEyjk4bOmgeeeSRtCFEpsQI5ZAnE0uVU+68L5dJOpWrtKybS9N8EpVwLx8/nTesn3x8dNzK20xOVz7Wypft6Einw5mOPzrgubkAjpxjzHS800FPJx+vOceQlNx0QMe8BEH5fKNspnK9VGfVQ/ujNOUl25JOefJan+v8JU2zTPCxqCyOhs4tHd/Jx0vHv7xkSx1fLVlXTsPr8md8zjR5Hem0vrzMK+fZfxJG81VU6viXD1v5ePO5zrPyetLrM50Pk6UZ7+HGzT7M7733Xr62fuUrX8nSUGUqX9JzMwZtJVL4nXfeSfzOQVRy84baQ0lKbcdS9eGGEAQnbSxtBkIPUcm25MP7uTDpvLOonAtHa/7V0aJy/h1T75EJmIAJmIAJmIAJmIAJmIAJmMCNBCwqb+RxV+9aRVTeFaQKG6tjkwgMOgXpKKXjU5EaEkl0lHJHOZEdrOPzVpzmm6icqWOI8EZEHjlyJEfv8proSSQAcpJOejqnkeREG61atSoRxYAUV4f8TNW1GcuxqGzGozL/6yRhNF9F5XQfQUYl4GYgbtBg5saLRx99NEc4TlU2119uSOC6zDWZ97SD3CiEaCQSXRGUk7dHmOp6zvGiXSXylWs37SrbIjnnwqTzzqJyLhyt+VdHi8r5d0y9RyZgAiZgAiZgAiZgAiZgAiZgAjcSsKi8kcddvbOovCt8ExsjkIi+QBwx00nKUH/IITo6GW6NTk6G4ERS8v5mHaUTmc7jFxaVd3ZwGSYRdnQ8M3wp5xqd6XTE03mO/GapISU53xS5q4ihOyt5fmxlUTk/juNc2wsJI4vKOztytHkMH8mM/GBocIa8pn2bql2jneT6y9DibMN7rrnMGvaVmzqm2hZRSfuK4OS6TvvJUKlsx/a0r2w7FyaddxaVc+Fozb86WlTOv2PqPTIBEzABEzABEzABEzABEzABE7iRgEXljTzu6p1F5V3hm9hYAglBSacgUW6wpdMTUclQcYrGoOOTzs6pOkknMpznLywq7+wAcz4RPQk/oow0IyoZ2lWykqgfIog47zTk652VOL+2sqicX8dzruyNhJFF5Z0dMb63SEPaPZa0c3ru7lTXUdpIyUrS856bODSygZ6DOdW2tLGUg2ShXKQoc/m6PdV2d7Zn07uVzjuLyunl7NynJmBROTUXrzUBEzABEzABEzABEzABEzABE5g/BCwqazyWFpX1wKRzk05BOkfp3GTJeyaiLySRWBJJOVciMuqh88lcLCo/yaTqGp1rOs9Y0hFP5znnlc43DS3M+rnSsV6VwZ2mg5WfUXmn9LzdnRKQMLKobJwg7Z3aPDgy057RvnE9nWrSNhKWpFHbyPJ2bSJlcPMH23O91ny77aaqy2yu03lnUTmbR6F1y7aobN1j7z03ARMwARMwARMwARMwARMwgVYhYFFZ45G2qKwRprOqTMCisjIqJ6yRgEVljTCdVWUCEkYWlZWRTSREOjL5ZosJJJVf6LyzqKyMzAlrJGBRWSNMZ2UCJmACJmACJmACJmACJmACJtCUBCwqazwsFpU1wnRWlQlYVFZG5YQ1ErCorBGms6pMQMLIorIyMiesgYDOO4vKGmA6i4YJTIeo/OY3v5mefPLJtHXr1rR48eIcbU3FdCODlg1X1huYgAmYgAmYgAmYgAmYgAmYgAmYwB0QsKi8A2g328Si8mZkvH46CVhUTidd530zAhaVNyPj9dNJQMLIonI6KTvvyQR03llUTibj9zNBYLpE5RNPPJFF5ZIlSywqZ+JAugwTMAETMAETMAETMAETMAETMIGbErCovCmaxj+wqGycmbe4ewIWlXfP0Dk0TsCisnFm3uLuCUgYWVTePUvnUJ2AzjuLyurMnLI+AhaV9bF0TiZgAiZgAiZgAiZgAiZgAiZgAs1JwKKyxuNiUVkjTGdVmYBFZWVUTlgjAYvKGmE6q8oEJIwsKisjc8IaCOi8s6isAWYDWcB9dHQ0XblyJY2NjaX29vbU1dWVOjs7U0dHx8QwpQ1kOSeTTpeo9NCvc/J0cKVNwARMwARMwARMwARMwARMYF4SsKis8bBaVNYI01lVJmBRWRmVE9ZIwKKyRpjOqjIBCSOLysrInLAGAjrvLCprgNlAFvy+OXHiRDp79myWlUjK4eHhNDg4mPr6+rKwbCC7OZt0OkTl008/nTT0q59ROWdPDVfcBEzABEzABEzABEzABEzABOYNAYvKGg+lRWWNMJ1VZQIWlZVROWGNBCwqa4TprCoTkDCyqKyMzAlrIKDzzqKyBpgNZHH69Ol04MCBdPLkyXTp0qXU09OTli9fnhYuXJiGhoby+waym7NJLSrn7KFzxU3ABEzABEzABEzABEzABEzABCoSsKisCKpKMovKKpScpm4CFpV1E3V+VQhYVFah5DR1E5Awsqism6zzuxUBnXcWlbeidGefXbt2LW/Y1tb2iQwk6I4ePZrOnTuXoyhXr16dli1blmVlb2/vJ7aZjyvEAWHLtRdRe++99yYiITWJo5bwJBp1z549aefOnenZZ59Nu3btysm3b9+eHFEpcl6agAmYgAmYgAmYgAmYgAmYgAk0AwGLyhqPgkVljTCdVWUCFpWVUTlhjQQsKmuE6awqE5AwsqisjMwJayCg867VRSUSjBkePDuS5VTPjgQ56/mcawXPmGQ7njHJ8yX1jEnelwWl8iU9E2LuvffeS8eOHUv81mHI1w0bNqSVK1emRYsWZXGZE87z/ywq5/kB9u6ZgAmYgAmYgAmYgAmYgAmYgAkki8oaTwKLyhphOqvKBCwqK6NywhoJWFTWCNNZVSaAyGAISIvKysicsAYCOu9aXVQiH5GI58+fT8gzlryf/OxIBCQikyhImJEOhghKZOPAwECeGcq1o6Mjy0ryJh1RgPyu4T3f83379uU8EJ1EEm7atCmtWrUqRxPynMpWmCwqW+Eoex9NwARMwARMwARMwARMwARMoLUJWFTWePwtKmuE6awqE7CorIzKCWskYFFZI0xnVZmAhJFFZWVkTlgDAZ13rSwqEYW0+/zmINLx4MGDWSCyrr+/Pz87kqFIFyxYkCMnkY6kO3ToUE6H0ERUDg8P5zRLlixJIyMjE1GR/IYm/f79+yfE5JkzZ9Lhw4dzmd3d3XnIV4Y8XbNmjUWlh36t4ZvtLEzABEzABEzABEzABEzABEzABJqFgEVljUfCorJGmM6qMgGLysqonLBGAhaVNcJ0VpUJSBhZVFZG5oQ1ENB516qiUsO9Ih+J7vvwww/Tu+++m6MfiXxEOG7cuDFHOiIgiZI8ffp0loxKxzWDSEueK0kaDeFKlCQT+R44cCC99dZbOV/WSV5SBoITQXn//fentWvXeuhXi0pOEU8mYAImYAImYAImYAImYAImYALzhIBFZY0H0qKyRpjOqjIBi8rKqJywRgIWlTXCdFaVCUgYWVRWRuaENRDQedeqopJoSIZcZv+JcERUElHJ7w+Gb+V5kcjD5cuXZ2mJkCQakmdLliMkSY+QJAJzy5Ytaf369WnFihV5mFfSkSd5k45hXfldzXvKZshYynjwwQez5ERweujXxRNnNzKZSUue/ckwunv27Ek7d+5Mzz77bNq1a1dOs3379vT000+nJ554Im3dujVHp5a3I1H52aF5I/9nAiZgAiZgAiZgAiZgAiZgAiZgAtNIwKKyRrgWlTXCdFaVCVhUVkblhDUSsKisEaazqkxAwsiisjIyJ6yBgM67VhWViEIiJI8ePZqfGcn3DybIw5UrV2bRxWuiJRFcRECyDTO/UZBgDN2KjHz++efT2bNnE0O4EoW5bt26/DzL119/PUtMxCfDx5Iv6V566aU8fCyHcfXq1emRRx5Jmzdvzs+rtKi0qKzh6+0sTMAETMAETMAETMAETMAETMAEmoCARWWNB8GiskaYzqoyAYvKyqicsEYCFpU1wnRWlQlIGFlUVkbmhDUQ0HnXyqKSCMkjR46kvXv35kg9RTkyhKsiKRGVTPAaHR3NEZH8NkZcIioZBvYHP/hB4vu7bdu2dM899+QoSYaUffnll7PYREISackwr8jRF154IZdJfsuWLUuf+cxnLCojmhTRyzNBNSkiUktHVIqMlyZgAiZgAiZgAiZgAiZgAiZgAnOBgEVljUfJorJGmM6qMgGLysqonLBGAhaVNcJ0VpUJSBhZVFZG5oQ1ENB516qiUvuv50giHN98880sFhGKiMVNmzZlYUlkJRPRkMePH89RlHxfkZVEVP7sZz/LIvPhhx/Ow44SOYkEffHFF/OzLT//+c/nYWGJqmS7V155JT+7EpnJuvvuuy/LTSSdIyotKmv4ejsLEzABEzABEzABEzABEzABEzCBJiBgUVnjQbCorBGms6pMwKKyMionrJGARWWNMJ1VZQISJhaVlZE5YQ0EdN61qqgkSg/RKPn43nvvZYF48uTJPPwrEZVEQiIdh4eHE8+oJK2eUUlkZEdHRx7C9Ze//GUeChZRSVQgUZIIUEQlEZm//uu/nu6///78HEvW7969e0JUIkGJwmQIWItKR1TW8NV2FiZgAiZgAiZgAiZgAiZgAiZgAk1CwKKyxgNhUVkjTGdVmYBFZWVUTlgjAYvKGmE6q8oEJIwsKisjc8IaCOi8a1VRCUJkJe0+v3URlERH8sxKmDAR7Tg0NJSjHBl2VM+nJBKyq6srLVmyJIvLnTt3pnPnzuWhX4nEXLRoUc4PgUmEJKKSqMn+/v4sMHft2pVFJeKT/JGbRHFaVFpU5hPP/5mACZiACZiACZiACZiACZiACcwLAhaVNR5Gi8oaYTqrygQsKiujcsIaCVhU1gjTWVUmIGFkUVkZmRPWQEDnXauKSkVUXrlyJT97EtHId5BnVn7wwQdZPCIWEY16TiW/iXmuJNsgGNetW5fF4w9/+MP87MmtW7fmdUuXLs2y86WXXkoIzs9+9rOJ516yDZGYb7zxRjp8+HAWpAvj2Ywe+vVygoOfUVnDF9tZmIAJmIAJmIAJmIAJmIAJmIAJNA0Bi8oaD4VFZY0wnVVlAhaVlVE5YY0ELCprhOmsKhOQMLKorIzMCWsgoPOuVUWl9p/oSOQh0Y389iCikudV6vmRDPvK8KyITZ47STrSIzF5hiXvf/zjH+dtGd6VYVyJquT382uvvZblJ4JyZGQkR0yynmFm+b4znOyKFSvS9u3bs8h0RKUjKmv4ajsLEzABEzABEzABEzABEzABEzCBJiFgUVnjgbCorBGms6pMwKKyMionrJGARWWNMJ1VZQISJhaVlZE5YQ0EdN61qqikvee3BvvPcydZ8pv3xIkTOaISPkhE5CGSEVHJ8yWPHz+eoyE7OzsnhOSrr76aSL9t27a0ZcuW/GxL0u/ZsycP8cqwskRWkhfPxSRqk2FkkZ08m/KBBx7IkZhEFRLB2QoTLN9///08RC7HwhGVrXDUvY8mYAImYAImYAImYAImYAIm0FoELCprPN4WlTXCdFaVCVhUVkblhDUSsKisEaazqkxAwsiisjIyJ6yBgM67VhWVDOHKcK9IxIMHD2ZBScQkPJBoDPfKcK2rVq3Kz5xEPCI0SUtEJKKRZ1TC8cCBA6m7uzsP4crwpURVdnR05PWkffPNN3M0JsKT9fzGYcl78meblStXJqI3e3p6aji6zZ+FRWXzHyPX0ARMwARMwARMwARMwARMwARM4O4IWFTeHb8btraovAGH38wQAYvKGQLtYm4gYFF5Aw6/mSECEkYWlTME3MVkAjrvWlVU8pxJfmvwvWO4V5YM48qQr0jIgYGBHO3I8ybLEZVEQ+7fvz9vSxoEJkPBIjbXrl2b1qxZk7cj4hIZh8R86623sgAlWpL1bIPYREwSZYmkVDRlV1dXS5yhFpUtcZi9kyZgAiZgAiZgAiZgAiZgAibQ0gQsKms8/BaVNcJ0VpUJWFRWRuWENRKwqKwRprOqTEDCyKKyMjInrIGAzrtWFZXIQhgQWYmYZMk1AIHJhFBELBLhiDwkPZ/zuxiZybakYWI9EZKkZzhXlgz1Sp78niFSU2l4LiV5sSRfhCXpeU0erG+FyaKyFY6y99EETMAETMAETMAETMAETMAEWpuARWWNx9+iskaYzqoyAYvKyqicsEYCdCQTUcPwfnv37s0dzRs3bkzLli3LHcl0StP57MkE6iQgYWRRWSdV53U7AjrvWlVUTuaDPGRmop13Wz+ZUL3vLSrr5encTMAETMAETMAETMAETMAETMAEmo+ARWWNx8SiskaYzqoyAYvKyqicsEYCFpU1wnRWlQlIGFlUVkbmhDUQ0HlnUXkdZllUXl/rV9NBwKJyOqg6TxMwARMwARMwARMwARMwARMwgWYiYFFZ49GwqKwRprOqTMCisjIqJ6yRgEVljTCdVWUCEkYWlZWROWENBHTeWVTWANNZNEzAorJhZN7ABEzABEzABEzABEzABEzABExgjhGwqKzxgFlU1gjTWVUmYFFZGZUT1kjAorJGmM6qMgEJI4vKysicsAYCOu8sKmuA6SwaJmBR2TAyb2ACJmACJmACJmACJmACJmACJjDHCFhU1njALCprhOmsKhOwqKyMyglrJGBRWSNMZ1WZgISRRWVlZE5YAwGddxaVNcB0Fg0TsKhsGJk3MAETMAETMAETMAETMAETMAETmGMELCprPGAWlTXCdFaVCVhUVkblhDUSsKisEaazqkxAwsiisjIyJ6yBgM47i8oaYDqLhglYVDaMzBuYgAmYgAmYgAmYgAmYgAmYgAnMMQIWlTUeMIvKGmE6q8oELCoro3LCGglYVNYI01lVJiBhZFFZGZkT1kBA551FZQ0wnUXDBCwqG0bmDUzABEzABEzABEzABEzABEzABOYYAYvKGg+YRWWNMJ1VZQIWlZVROWGNBCwqa4TprCoTkDCyqKyMzAlrIKDzzqKyBpjOomECFpUNI/MGJmACJmACJmACJmACJmACJmACc4yARWWNB8yiskaYzqoyAYvKyqicsEYCFpU1wnRWlQlIGFlUVkbmhDUQ0HlnUVkDTGfRMAGLyoaReQMTMAETMAETMAETMAETMAETMIE5RsCissYDZlFZI0xnVZmARWVlVE5YIwGLyhphOqvKBCSMLCorI3PCGgjovLOorAGms2iYgEVlw8i8gQmYgAmYgAmYgAmYgAmYgAmYwBwjYFFZ4wGzqKwRprOqTMCisjIqJ6yRgEVljTCdVWUCEkYWlZWROWENBHTeWVTWANNZNEzAorJhZN7ABEzABEzABEzABEzABEzABExgjhGwqKzxgFlU1gjTWVUmYFFZGZUT1kjAorJGmM6qMgEJI4vKysicsAYCOu8sKmuA6SwaJmBR2TAyb2ACJmACJmACJmACJmACJmACJjDHCFhU1njALCprhOmsKhOwqKyMyglrJGBRWSNMZ1WZgISRRWVlZE5YAwGddxaVNcB0Fg0TsKhsGJk3MAETMAETMAETMAETMAETMAETmGMELCprPGAWlTXCdFaVCUwlKoeGhtLw8HAaGRlJvb29lfNyQhOoSsCisiopp6uTgISRRWWdVJ3X7QjovLOovB0pfz4dBKqISsq9du1annnd1taWTpw4kfbs2ZN27tyZnn322bRr1y4+Stu3b09PP/10euKJJ9LWrVvT4sWLb9iONGzvyQRMwARMwARMwARMwARMwARMwARmioBFZY2kL126lOjEOnr0aNq/f3/q7u5ODzzwQFq2bFn+g99/9NcI21lNEEBU0ol15syZxOv29vY0ODiYJCt7enom0vqFCdRF4MqVK/l8O3bsWHr//fdzG7dp06a0ZMmSLMc7Ojrc0VkXbOczQQBhhCTnWnv48OF8DnZ2duabMlauXJnbvonEfmECNREYGxu74bzj915/f3++zi5YsCD19fUl2jyuv55MoG4C3JjxwQcfpJMnTyauvdyEds8996RFixbdUNRkUUn6t99+Oz333HPpH/7hH9KvfvWrnH7Hjh3pW9/6VhaVW7ZsyfmwLZP+VtEyr/R/JmACJmACJmACJmACJmACJmACJjDNBCwqawRM5+m5c+cSHff79u1LXV1d6f77788d9/6Dv0bQzuoGAnSYIik5986fPz8hKpGVdKRyHnoygboJqOP++PHjWVSS/+bNm3NkBuLIHfZ1E3d+EKAznXOPNg9RSZtHG8eNGUuXLk0DAwMGZQK1E5AA4rw7dOhQ4rrLNZaZ0Qu4IYjfef6tVzt6ZxgETp8+nQ4cOJBvSkNUcs5t3LgxIcknT2XhyE1s7777bhaVf/d3f5deffXVnPzBBx9M3/72t9OTTz6Zr9sIz/J2k/P0exMwARMwARMwARMwARMwARMwAROYbgINi0ru6PU0NQFFGHEH88GDBxOd9eU7nunAUkfA1Dl4rQk0TkCCnA57hh/mPKOzHknJsK904vu8a5yrt7g5Ac4xZBFtHh2hRJAzrVu3Li1cuDBHFrnD/ub8/MndEaA948YMRi+gzeNaS5vH8IVEtrm9uzu+3vpGAvrtVj7vuO5yrnHeISs1coHS3piD35nAnRPgnDp79mwW5AhLrrvcmLFmzZosLG/V3hF5zo2TP/vZz9K//Mu/pDfeeCNXZNu2ben3f//30+OPP542bNiQIzRVw/K1+1Z5K72XJmACJmACJmACJmACJmACJmAC00tg1apV01tAk+TesKjkjlxPUxOQqKQjgefC0Hm6evXq3JHAH/7uwJqam9feHQE6TCUpR0dHc2YISjpRmTkP3dl0d4y99Y0EaMsQlZxvRBgRRc7EMNd02iPHHVF5IzO/q4cAbZmutXTCcw4y5CY3ZhBdhDBye1cPa+dSENBvN513DMPJa841rrWcewz1T5untGZnAnUR4LziNx7XWW7QYPhrzrkVK1ZkUc61uDyp/eNcRHASef7mm2+mF154YeKmIuQkz6fkWZX8wYv41MR2nkzABEzABEzABEzABEzABEzABJqHwG//9m83T2WmsSYNi8o/+ZM/mcbqzL2sy3/Q03FFpynPCaRzgM5ToouQReq0VwfC3NtT17iZCEw+7xiGjnOPc5DPEEXMdKRyHvq8a6ajN7frwvnFLFFJRBuykonOTjrukePqtGe9zz8oeLpbAjrv6KjXtZY2j3MNUSRJzvnmc+5uaXt7EeC843wq/8bjHKSd03W23Ob53BM5L++WgK63tHfcBMn1lmsvv+3KN2aUzzm9ZlvSc0MHwxW/9957eQQE6sSzpO+77760du3a/HxK/k7RxHaeTMAETMAETMAETMAETMAETMAEmoMAf+P9xV/8RXNUZppr0bCo3LRp0zRXae5mz4lDBwIdWBJGdJ4iijyZwHQR4LzjnNP5p44tOu/Lsmi6yne+rUeAc0znm9o7KNBZT3unc7D1yHiPZ4KA2jyut8ycb7R1kkV87skE6iZQbvN4rfNO11neezKBuglwXtHOMXoG11smzjkk+e1uRCM92yEsicZEeDIhOkdGRiYeEaC/U3wOZzz+zwRMwARMwARMwARMwARMwASahgD9D7t27Wqa+kxnRRoWlfxh7MkETMAETMAETMAETMAETMAETKA5Cdzqpo2yYC8LyvLr5twr18oETMAETMAETMAETMAETMAEWocAf9cxemcrTA2LSj3H5FZ//LYCOO+jCZiACZiACZiACZiACZiACTQjgan+Vpu8zmKyGY+c62QCJmACJmACJmACJmACJtDKBMp/p/Faj9ya70waFpVf/vKX85B//KGreb5D8v6ZgAmYgAmYgAmYgAmYgAmYQLMSKEtI/Y2mpYbK1qMC+GNXM/tT/kO4WffP9TIBEzABEzABEzABEzABEzCB+U5Af6exZCQcphdeeGG+73bev4ZF5d///d/nZ6XwDEb+2OUP3/Ifxi1BzTtpAiZgAiZgAiZgAiZgAiZgAk1CQH+PsSyLSf5e4+82nlHJzGc8l5I/erWUqNSySXbJ1TABEzABEzABEzABEzABEzCBliEgSam/1fT32ne/+92WYNCwqHz77bezoOQPXslK3a3bEsS8kyZgAiZgAiZgAiZgAiZgAibQZAT0N1lZVPL32uXLl7OkvHTp0oSo5I9ezfqD2KKyyQ6oq2MCJmACJmACJmACJmACJtAyBPR3GX+ndXZ25hlp+dhjj7UEg4ZF5YEDB/IfuJKUGkJId/G2BDXvpAmYgAmYgAmYgAmYgAmYgAk0EQGJSpaTZaVuMuUz/tjVXbos9QexRWUTHUxXxQRMwARMwARMwARMwARMoCUI8Dda+W8y3VCKrOTvta1bt7YEh4ZF5dGjR/NQrxKV/BGsP4pbgph30gRMwARMwARMwARMwARMwASajAB/kzGx1N9oLPW4Dl4z6Y9gCUu9zx/6PxMwARMwARMwARMwARMwARMwgRknoL/LdFOpROXq1atnvC6zUWDDovLEiRP5j1/9wStJWf7DeDZ2xGWagAmYgAmYgAmYgAmYgAmYQKsTKP99NtVr/QE8ednq3Lz/JmACJmACJmACJmACJmACJjCbBPgbTTeUElnJ66VLl85mlWas7IZF5enTpydEZfkPX9VYwlLvvTQBEzABEzABEzABEzABEzABE5gZAvp7rLzUa9VAkpL3vPZkAiZgAiZgAiZgAiZgAiZgAiYwuwT0d5pkJcuRkZHZrdQMld6wqLxw4cLEUK/6g3fycobq7mJMwARMwARMwARMwARMwARMwASmIDD5bzS9l5jUUuunyMKrTMAETMAETMAETMAETMAETMAEZoCA/j4ry0pe9/f3z0Dps19Ew6Ly8uXLWVRS9fIfteXXs79broEJmIAJmIAJmIAJmIAJmIAJtC6Bqn+fVU3XuiS95ybQOAF1NDW+pbcwgflFwNeY+XU8vTcmYAImYALTQ0C/Hadadnd3T0+hTZZrw6JybGxsQlD6B0eTHU1XxwRMwARMwARMwARMwARMwARuQsB/v90EjFebQM0E6GRSR1PNWTs7E2h6AuVrTfl101fcFTQBEzABEzCBWSSg345aqio8q7IVpoZFpX9ktMJp4X00ARMwARMwARMwARMwAROYbwT8t9x8O6Len2YlYFHZrEfG9ZoJAuVrTfn1TJTtMkzABEzABExgLhOYLCnZl6nWzeV9vFndGxaVN8vI603ABEzABEzABEzABEzABEzABJqXgDuMm/fYuGYmYAImMF8IlDtUfd2ZL0fV+2ECJmACJjATBMrX0Jkor5nKsKhspqPhupiACZiACZiACZiACZiACZiACZiACcxZAufOnUvHjh1Lp06dShcvXkxXrlxJ7e3tTbI/bXFbPnXpjDle52ks/td8LcWzfsbXe2EC1QggIznHOzs708DAQFqwYEEaHBxMvb29eV0rd7pWI+hUJmACJmACJmACFpU+B0zABEzABEzABEzABEzABEzABEzABEygBgIffPBB2rlzZ9q1a1f68MMPE+Kyq6trlmUl8hExGc84auuOeSDmrngfgvLalVhejOVoLK/GEmnpyQSqEUBSXr16NZ/jw8PDaf369enTn/50uueee9Ly5cvT0NBQHrLOsrIaT6cyARMwARMwgVYlYFHZqkfe+20CJmACJmACJmACJmACJmACJmACJlArgd27d6d//Md/TM8//3x6//330+nTp5tIVEY0ZVtfzMMxh7DMojIE5bVz8frSuLREVCraslY0zmweEpCo7O7uTgsXLkzbtm1LX/nKV9JDDz2UNm7cmBYtWpQlvUXlPDz43iUTMAETMAETqJGARWWNMJ2VCZiACZiACZiACZiACZiACZiACZhA6xJ49dVX0zPPPJN+9KMfpX379mVRiaRpDlGDgOwdF5U98ZoIyhCUKURlXhJ56aFfA4KnBgggKxGVRE9u3749PfXUU+lzn/tc2rx5c1q8eHHq6OhokvO/gZ1yUhMwARMwARMwgRklYFE5o7hdmAmYgAmYgAmYgAmYgAmYgAmYgAmYwHwl8PLLL6e//du/Tc8991zav39/flZl8+yrROVIVGlcVBJJmc5aVDbPQZqTNenp6ZkQlV//+tfTY489lu69916Lyjl5NF1pEzABEzABE5h5AhaVM8/cJZqACZiACZiACZiACZiACZiACZiACcxDAq+88kqOqERU8rzKjz/+uEn2EkmpoV8XxGtEJcO8MuRriMr8nEqeV+lnVAYETw0S6O3tTSMjI2nHjh05ovLRRx/Nz6l0RGWDIJ3cBEzABEzABFqUgEVlix5477YJmIAJmIAJmIAJmIAJmIAJmIAJmEC9BDT0686dO/MzKptLVHbGsK88ozJEZdu4qGTI1wlRGc+rnAVRqWFxy0Pk8pohRTXptZasL79WuvJS+ZbXTX59uzwmp/f7qQkgKnlGpYZ+RVR66NepWXmtCZiACZiACZjAJwlYVH6SideYgAmYgAmYgAmYgAmYgAmYgAmYgAmYQMMELCobQyY52d7enp9lyFLrkIjleWxs7Ib3+kwlThaTykdL0pW30WuWnu6OgEXl3fHz1iZgAiZgAibQ6gQsKlv9DPD+m4AJmIAJmIAJmIAJmIAJmIAJmIAJ1ELAovLmGCeLRFJ2dHSkrq6u1N3dnXjOIa9ZR1rEJPPVq1fzfOXKlRuWrJdklIycvER8Sn5SnvJku/JrSUvSKE8tWefp1gQsKm/Nx5+agAmYgAmYgAncmoBF5a35+FMTMAETMAETMAETMAETMAETMAETMAETqETAovKTmCQoJy9JiaAcHBzM89DQUOrv78/rSIuIRE5evnw5jY6OposXL6ZLly5NzKxHNjIhIzs7OyekJO8RnprJT3KSPDVTxmRhWX5P3haWULj1ZFF5az7+1ARMwARMwARM4NYELCpvzcefmoAJmIAJmIAJmIAJmIAJmIAJmIAJmEAlAq0sKiUiEXt6XRaGiETmsjxEcI2MjKQFCxbkeWBgIItKtpOkRE4iKc+fP5/nc+fOpQsXLmR5iVSkLCIxFZHJe5XLkvfUqSw+kZzkz7qyrCQ/rddS4rJ8AlhelmmkZFF5Iw+/MwETMAETMAETaIyARWVjvJzaBEzABEzABEzABEzABEzABEzABEzABKYk0IqiUlJSS4lKJKEEIiILCdnX15dnpCLSkgjK4eHhLCsRluWISkQhkZSISsQkgvLs2bPp9OnT+TXrKYsyyJ/ITJYaOlaCknwUlck25Mn7sqCUsFSZpFHZbMN6ypKgLL+e8kRosZUWlS12wL27JmACJmACJlAzAYvKmoE6OxMwARMwARMwARMwAROYTQJ0ntIBS6cuS3WqUqfy69mso8s2AROYOwQkn5A/iljTEJtzZy9mrqatICo5J5gVtci5oUhJRTCyZB1DuyKxEJAM7YqsRCgiLDmftJ7PJBrZjklyEWHINY2IyjNnzqSPP/44C0uiLJnIh22JyiQ/ti9LStIxKxIT8ajro4SjoibLUpNtSMuSOpCmnF5yU8KTpT6nXq10zbWo5Ih7MgETMAETMAETuFMCFpV3Ss7bmYAJmIAJmIAJmIAJmEATEFBHqDpl6Ugl2uTgwYPp1KlTExEjTVBVV8EETGCOEqB9QQAtXrw4yyCkEALK0ycJzEdRyfFn4nrD68kSEumIqEIYcl4Q4Vhesp7PScd5hKxUdCXvmSUu2ZYyuJYhEzVLGCIqP/rooywsEYikZVuiMjk/EZ7KA3HIdghKtmMmIhPpiZBkYnvNvGcf2a4sLJGU1IN1kpK8Zj11YCZPymI9eTCzD8ozv5jH/1lUzuOD610zARMwARMwgRkgYFE5A5BdhAmYgAmYgAmYgAmYgAlMFwE6Q5noEKXTlE7YDz/8ML3xxhvp5MmTubOYaBc6ljVpG7330gRMwAREAGlTnhAviBhExKpVqyZmhukkcq3ctpS3a9XXc0tUXg0zdynms3G4Ijrx2uVYFnJNx4/jq2uIXitSUuIRcS3hyHmieSpxyWdsV5aVvEdsUg7nH9czhKDkIOcgrxGBXOO4tiEdkYOkp2yiKZctW5aFJXlRV/Lg3CUtN+4QicmNPERmkh/bap8oW+Wz79SBmbInz7reUh/qQJ30/ExFXqrO2pa6SGDOx2swx3XhwoVp+/bt6amnnkqPPvpo2rx5c5bHZa46r7w0ARMwARMwARMwgTIBi8oyDb82ARMwARMwARMwARMwgTlIQB26dN4eOHAgS8oXX3wxd84iFujAJcKEDlk6WOdjJ+kcPGyusgk0JQGJSkkcxA7tCu0G7QnyYceOHWnt2rU5eg0p5Ok6gTkhKtsXRoU5bhFVKFF5jWFUiTK8Lio5B5CSSChJR5aKkJSclHRUZKSWZQHJ9YeZ/NieNAhO0vCe9UxlOYhM5PrGjPBTdCTXOglH6kheixYtSitWrMjXO+qr651EJZKS7RCWiEuEovavHAFKPdiWzzRRJ9VLdZE4JX9JSqQl76krSyI5mfmc99ofXYO1VDlzeWlROZePnutuAiZgAiZgArNPwKJy9o+Ba2ACJmACJmACJmACJmACd0WAzk86QxVJSUc5opL1DzzwQJYLdCIS1WBReVeovbEJtAwBiaXjx4+n3bt356gxhta899570+c+97m0ZcuWtHz58hwZ1zJQKuxoc4vKjhjrtD+l9gWxJz2pLUVEZboUy3MxIyp5X4hKRB3nANcOhKKkYllCll8jHDWzXrOiG8kPMUeeyEDSKhJTaSQBkYhcvxB+ZVGpa52EIwKQibIQlatXr85RfeRNGZSnkQYQlGyHsOQ14pCJ66LkK0vqgrhkvSIBqbtkZbk+5M1MXpo1BCxRliqL8rhGk4Z9ol7KT9dk1pXnXLk59J9F5Rw6WK6qCZiACZiACTQhAYvKJjworpIJmIAJmIAJmIAJmIAJNEKAzk+e2bV379708ssvp9deey29/vrrufP18ccfT5s2bcoduXTc0slKZ6gnEzABE7gVAYQSoubQoUPpF7/4RTp8+HCWOOvWrcuikpsgaFsY7tHTdQLNLyr7QlYuiIjBEJXtIQ7bRmO+kJdtbURUFs+h5NhzzSBakqh8hvrl+Y/l6EnklCSfohJ5L2FJWt6Tl+QcpMoClLSKykXiIf6InGQpUYnM4zUzwk+iEhlIvhKVa9asyUON8l7yk22Qh0RRcp1EHrJEcpIvdaGO7Mvk/WH/qTtpmCQVWTJLrFIGElVDwZI3chLJT11PnDiRy5eoZFu2YR/ZTvmwVN5z7TptUZlPEf9nAiZgAiZgAiZwhwQsKu8QnDczARMwARMwARMwARMwgWYhQKcoHaLvvfdeFpVvv/12jq5EIHzxi1/MQzXSGUyHK52gc60DtFk4ux4m0EoEkDPMEpX79+/PYoWoSp5Dt23btnTfffelJUuWtBKW2+7rXBCVbSEq2zv7InKwI3V1jqXuztG4PlyJ4301ZkRlEfWI7JssKomCZB3XFAQfgrIcgchr1iMLSYvA4trDhIhjIkKRNPq8HP2I7EPolSWepCDbc72TqEQ+so66cF4iKjkfJUiVLzKQ7RguVqISyUkZTOwn9SGfsrBkX8iD74EiQvMG8R/XUWaJRYnHshTluoykpEyVRzq2Y6mhYtlnydnJ+z1XrtcWlTozvDQBEzABEzABE7gTAhaVd0LN25iACZiACZiACZiACZhAExGQqPzggw9yJOXRo0dzJ+jKlSvTww8/nDtv6XCls3WudHo2EV5XxQRalgByBtnyxhtvpH379mVBhLhBCBFNaVH5yVOjqUVlW0cIt/7U1rEw5NxgyMS+NNDXkfr7roWoGwspdy2kYjHka0dHZxZ4yDsiKSUoEZASegg+zgdFHXK+8JrrjbZjOfn6Qzq2JS8EoW6iUUSihk9VtKFkINcviUrkH6IS4YckY+hXiUrqSrmUoeseApBoTGQlEZVsSzlsX64z2zGTp/aP+mrmiEtc6uhTP+paLoP6ISkpT5KS+usaTHrqM9UsgUka8tY2Kq8ZlxaVzXhUXCcTMAETMAETmDsELCrnzrFyTU3ABEzABEzABEzABExgSgJ03B47diwR8fTWW2/l6A06f+m0/fSnP52fUamO5Ckz8EoTMAETuAkBotfeeeedLCoZ/hVxQvQaQ8Def//9jqicxK0ZRWUbdYwoyfb2ePZi51Dq6l6S+gcWpgUjwzH3puHhjjTQ3x5iDtFYiMr29mLoVyQj15PJM+uZFXEoeceS9ci+4eHhCRlJOoSbhJ/SsGQbhCGikusZs6IqNRyqZB0Sj3MSEYgE5HOEIiMIcM3j3CSiUhJU+XPelkUiohKBSJnkTf3Yx7Ko5D3XTtUZjNRV11NeM0lUUjfyZNhXzYhIyqWM8nbsL/spUUl92Ce2k4Tlc7ZlH7X/ucAm/M+isgkPiqtkAiZgAiZgAnOIgEXlHDpYrqoJmIAJmIAJmIAJmIAJTEWAzkyinoh42rNnT44WoeN29erV6aGHHsqiks5aOkk9mYAJmEAjBIgKo12hfTly5EgWJkuXLk3r1693ROUUIJtNVCIpIyAw2v/O1NkVz5TsXRSSclVasHB5Wr50cYjm4bR4UU9ETXZFFCFDviLfstrMxxpBxoysQ8xJ1CmSkuuKZj5jPdcbRCHPtdQwsaxTHix5r4hFMCLuEH1cz5B9vEbSsZ5Jog6BqaFfEZV8Tj6UtWrVqhxZWY76RDZSFnUjD/KUIFRUJZGLTKRDuJUjKtk3la36k077zHaIRIQn9SZPZCN1k6QkDempp/aZPCmX/WFfSc9+cS1nVASWvCfPuSArLSo5yp5MwARMwARMwATulIBF5Z2S83YmYAImYAImYAImYAIm0CQEJotKOjfpDKXTFlGJsOQ9HaWeTMAETKARAggTRCVDSyMqmZYtW2ZReROIzSYqOzqIcOwMARfDvA4Mp4HBZWloeF1auGhFWr5sSVoaonLJYqIquyMNUZSFpBwbuxYC7krINObLWQiWI/sk7criEoHHjBxEUCIPGYYVccg1iLTMbMv1iLS8Z6IMrmUSlSyRfxKVwo3YY+hWRVQi8ciH6M0VK1akBQsW5PKQjdSDGYmma6AEIXkjEpGEvCYqUjJR2yFdqR+fSVaq3pTJ56yHi2SjIiklW9mWdOSpOokF20lWIji5diMpeS4s0ct839hXCc8yf/FolqVFZbMcCdfDBEzABEzABOYmAYvKuXncXGsTMAETMAETMAETMAETmCBgUTmBwi9MwARqJmBR2RjQZhKVCMGenu40NDgQEZQLY1jU5bFcnQaH1qaRBcvSooULYsjU/hj+tSvSdITQIyIyx2COD2daSEpEHrOEZVnclaUjAg5hhZhEUCINJz8vUqJS27GUPET2IeUk+ZCSiEpJQtKSBkmpmXqRJ+Ug0CmzHBEpQUi9mJGG5MM+IDnZXhGLqpMkpCQqddA+S1Syr+TFJMmqYVupG/vBeibJUkWXlqUpeVM+6dmOYdwRlZp5r/yoK7KyGSeLymY8Kq6TCZiACZiACcwdAhaVc+dYuaYmYAImYAImYAImYAImMCUBi8opsXilCZhADQQsKhuD2AyisizcBkNSLlmyKCTe8og4XJNFJUO/DgwuTsNDgzHka3caHmwPudgeIq8ti8q2eJ4lEZWSaAhDJJmkHusV3UdZCD1JSiQh0nBoaChHOfIaiaXoSck/tmNmKotKrmdIu3JEJZ9rpi6IOyINmXnPhARcsmRJjuJU5CJlShIiT5mpC+uoB/JRUY0SgKxnVv1Yz/4yk5717Ct5s2Qqi0rJSurPtuRFmdRJopLyEZ7sE2kQlaRn+FeGWiaSkohKZCURlshK8oULacUiF94k/1lUNsmBcDVMwARMwARMYI4SsKicowfO1TYBEzABEzABEzABEzABEaCDE5mgZ1R66FeR8dIETOBuCVhUNkZwtkUlIk1yjOFQFy9elFauXB7zqiwqh0dWpK7uZSHrhkOe9YS860wD/ddSX0jKnp5CVLI909WrY1nCSVJKWCLLJO5IpyFTJeOQlJKVkoZEH0oAsk15QgCSH/krqlJlqRzSMCuiUqKS6x/iDlG2eHHI19jncpllgYoo1Mx6yULlXa4Tr8mX8tlfZKQiJCUpyQPeyEbqi2hkRirynvWUURaVvJa0nSwqGf4VCct3DjmJpGRGXHJdJ1/2V0zYvlkmi8pmORKuhwmYgAmYgAnMTQIWlXPzuLnWJmACJmACJmACJmACJjBBwKJyAoVfmIAJ1EzAorIxoLMtKhFjROwh7IgwXLlyRVqzZlUWlUuXrQxRtzSltoWps2Mgdffw7Mq21N+HqEwRIYiovB5RWBZ1yErJQ4k7RByTRCURi4hARCVLRTAiKUkjAVomShmIQiSgykBGUlZZiJIGQcf1DmGHqETq8Z56sM8LY3hbyi6LSklFSdSyQEU0qk6SflpKXqpekrV8rv2V7GR/qCsRjwxbi3AkvdJSN2bqwDbl4WfZJ9Kyz2zLvrFfiElmCUu+h7wnb9hQL+qo+paZzsZri8rZoO4yTcAETMAETGD+ELConD/H0ntiAiZgAiZgAiZgAibQogQsKlv0wHu3TWAGCFhUNgZ5tkUlQmxkZCRLylWrVoWkXJ1FJUO/Lli4JITZghB+wyEre0OaEe13a1EpiThZ2PFeohLxhoCTqERSIuUUPViWlEQgMiHYmCUEJewkBFlSButJR1m8l6iU0JOopHzkLKKSesCBchGV1I+6sF6ikjqShs9VJx1p1YsyEZCIQWZkIuuQm+Sp7XnPfkg4UifqSj58JlnKssxCbMmbbRCVEp2KzkROMhws30NmBC1pqAtsyINyZnuyqJztI+DyTcAETMAETGBuE7ConNvHz7U3ARMwARMwARMwARMwgdzBSQemh371yWACJlA3AYvKxojOlqhEtjEj6pYtW5aQlOvWrUurV6+K1yvysKiDgwtSe8dQujTaH3KrJ+Qiz09sj6FfU4jFGPo1R1SSTzH0K3teloRIO2aEHJIMaUeZyDekHSIQQcksicdnqhtLTWUZSD7kp7wlKSVDVQfSIPQk8VhKHiIB2XdEpEQlMpHyJVKpF5+RDlGJXENwIhOZNZXrJlFJuRKQpNM+s71kJ/tBncWHdMpbSzHQPpGefSBvIjL1fE69Zh8VYUl0JdJSshLBCRPyYp7NyaJyNum7bBMwARMwAROY+wQsKuf+MfQemIAJmIAJmIAJmIAJtDgBOjgtKlv8JPDum8A0EbCobAzsbIhK5BciDGHG8KcbNmzIkhJRydCvy5YtiWjDkRBzIShTf0QH9qexa905orK/rz0NDbanvv72CVEZ4ZZ5pyXAiNqThEPcIdcQZKynbMpF2ElQSt5RJz6XnCuTZNtyvuQpUTm5DOqhtEg9RR2yRNbxGXVAUk415CxSkc+Rpwg1RXxSX+rK5+W6ar+1z5ShYV2RkJRHerYlT2ZkKOvYlu1YMikvLdm2PLOvkqBaUh77yZJ17CeyUpGVPLeS96xn+3J5ZcYz+dqiciZpuywTMAETMAETmH8ELCrn3zH1HpmACZiACZiACZiACbQYAYvKFjvg3l0TmEECFpWNwZ5pUSlJiTRDwC1fvjxt3rw5y8o1a9bG+6Vp0aIF+bPOzt50dawnnb/Ql66NIRc7I8KwIyRmR0RVhqjsaQ9pdz3qEaGGYJNYQ04yIxX1GeUrYhFZhRCUtLsZOeWJYCM/CUokoGbK4HOVo21IW4465D2fSVQSMamIStYhIZmpk2QlgpKZ+kpUSlZSZ/aJPFU/CUOeD4lApM5M5Mf2ykflsT0zE/UnvfLiNeu0b9RfgpIlZYkBn0laUjaRlHwfDx8+nJcMf8s2yjMXOEv/WVTOEngXawImYAImYALzhIBF5Tw5kN4NEzABEzABEzABEzCB1iVgUdm6x957bgLTTcCisjHCMy0qFdlHNOGSJUtiqNfVWVSuW7c+oilXxrpFISIHQ6b1RcRfd7pytTvkVk/IsuuicmQEUcnzKhGVxRCoEoMShZJrCDZJNshQPpJP0q4s/PicfJjLU1kCSsZJyEnUsV5CVNKPPJByijaUNORzhKGGfUWaaehZBCV1KtdRUZASjEojwchS9aY8yUQiGLneKqqSPNlW5ZEv9YAFXJhgpX1hKakonqxDvLIvzOTNuvIME8plGFiiKhGVRFXy/EoEJttQzmxOFpWzSd9lm4AJmIAJmMDcJ2BROfePoffABEzABEzABEzABEygxQlYVLb4CeDdN4FpJGBR2RjcmRKVyDRJSiIpFy9enJ9LuXbt2rQhhn5dtWp1Wrp0SRoZGQmBxxCnPRHl15lGL3eFGOsOsYWoDEE50JkWjHTGElGJeCMS8LqoQ6hNNUs+UgfJSgk/EZPs07K8XgIQycY1TLMkJOslKtlO+0tdJPNY8p7ykYQ8e5JoSglIhKHqJKmISGSWrOS1RCb5SFZSJvVGAFIPZKFEJa9Zx+cSoORHuZopl7zYnnqqzmVRSd3LorIsaVlPGSzZFj6UT1TlkSNHEs+r5Lup53SqPtR7NiaLytmg7jJNwARMwARMYP4QsKicP8fSe2ICJmACJmACJmACJtCiBCwqW/TAe7dNYAYIWFQ2BnmmRKUEmSIpiZ5cv359WrNmTRaWRFcWknIg5BlRft2xI53p0mhnCK/OEGgxFGoM/TrQH6JyQWc8p7IQlV1dHVmwSVZKUiLleF1eQkYCsbzkNemYWGouv0esIeAQkwg4Zg3pirCTDFSkIHmyz+TFtpp5jxRkKFf2V8O+IiBZL1Gp15NFpT4nb8ooz9SXfZZUlUylntSdOjDpWCDrNKws4pL1bF8WkORFncWSPJRveb8lKimH9ZQJI55NSSQlkZUsNfwr6WBF3tNWXsoAAEAASURBVLMxWVTOBnWXaQImYAImYALzh4BF5fw5lt4TEzABEzCBOUqADgU6FpjprKDjgonODd3lrU4T1qvDRNvwmZ6JozvBSefJBEygdQhYVLbOsfaemsBME7CobIz4TIlKBBtSbNGiRXm4VyIpEZWrVq3KQ8Ai7Yi07O3tiehBIgu7YkciojJE5ZksKouhX3NEZRaV5MdvT4Qdw5YWz1gsSzVel9+LDL9Fy1P5PemZykt+y/KbVwKOqEDJSq5nZbEn+cZvXH7vMrEtwo8lE+vLolIyUnJSS9IxIxGVhvfkLVFZ/i3NflBv/UZHBlI/CVVkIvVgYntFVcKd1+TN9oqKVHqxYDu2J08N/cq+k04zZfIZZTIz1Cu8EJbMiErYkQfbiFe5DMqZ7smicroJO38TMAETMAETmN8ELCrn9/H13pmACZiACcwBAnRQ0Amhzgde0zFCh8vChQvzneF0ftBxwkQnhDokSMtnw8PDOR0dIrz3ZAIm0FoE6KBEJuzbty/t2bMnR1nQCUuH9UMPPZQ7sXXjQ2uR8d6agAncLQGLysYIzpSopE1nqNNly5aljRs3pnXr1uVoSt5LUiLLenoY6pTnNDIUaWfIvRj69TxDv8b6ns40ONCVFiIqhxCVneOiUlGVxb4r+g/5NXkmxeR1/I6VrNRSFEkrUcm1i9+0iDcEHK819Cu/dxGRiDemsqjktzOzRCUiElEGD34/w4bfxFryOe+1jvWa+Uy/symjLCrLdaY86oRIpN7M1FVykLTkBfNCEBfPyWT/tS8syUcTZbF/5ClRyZLf98yTy1OZpBE32CEr4cfnbEcZOiYqa7qXFpXTTdj5m4AJmIAJmMD8JmBROb+Pr/fOBEzABEygiQnQiUBHA50K5buk6XShw0SdT3Q20ZlCpw7p6YjQME90YJCWDhFk5YIFCz4hNpsYgatmAiZQEwHaEYvKmmBOUzZ0GjNpqc57LaepWGdrAndNwKKyMYTTKSppL5gRXAxxym9EhnxFVDLkK6+5yU2irBBzReQkEq29vTtdCUF5/nxvtEXxnMYYEnZwcFxUxrK/vyNkHpLy+jCo7L2k182WEplaql1TffVeJEnHb1gJN4k2iUpkG7+Hy8KNfWZ/mFjPTBrqxGf8VtZ+81pS8mZLpeFzfktrJq/J9aVMhCJlUreyrFQEJPvEduTHsdGzMsmPbakr+8xr6kxajgnbkaf+JmBJnqxTWazTevKgHqTh74fJ0ZWsYzvS6HhQ/+meLCqnm7DzNwETMAETMIH5TcCicn4fX++dCZiACZhAExNAOB48eDDLBToV1HlBhwZ3YyMely9fPiEf6aDYv39/OnbsWJaVEprsIp0RdFZt2rQpLV26NHeO0FEyVUdLEyNx1UzABO6QgEXlHYKboc3olFaHMa+ZaJ/VIe62eoYOhIu5IwIWlY1hmy5RSTvBrKg9bmhbvHhxWrFiRZaULHnP70d+RyLiSNvdzVCnRBPyviddHesLydUXjRDPU+xOw0Pd8Vuzq4io7JWoLGSd2ibarVvNtG+aoaXtVOfye+VTFpWKqOT3ML93JePUbpIn7SUyke0lDfktzGsmPisiSK8P6wqDyTO/j5nLopL3sGKmHObyRJnURcJRAlH1RRpKmpIHUZ3MyEresy2/1UnDkvxgQrm8Zn/Jg2s5M681S1iyLO8v+ZCGvyd4ViUz31WGg50NWWlRWT5j/NoETMAETMAETKBRAhaVjRJzehMwARMwARO4SwJ0SDAfPXo07d69Ox06dCh3PNApwp3gdGxo+Co6nAYHB3NnykcffZTefPPNdOTIkdyhQYcM6emooHOCzpD77rsv8YwinlfEe9Koc6hKtVU3lkxa8nqqzibWezIBE5h9AhaVs38MblUDOpfLne+0p3SSq1OdtrrKpDZanfd6X26fuZbofZU8lUZ5aan15SWflSeVU16WP5/qtfIv74PSSRAoP633cnYJWFQ2xn86RCXfCdoJxBa/EfltyCga/E5kqFdmSUp+/5XFW1cX4gxh1xtzX7qW+tPFSwOpvQ2Rxo1xhagcHOxIfVlUIulujCrU9xYSeq0l32V9n1kylb/Dk1/zOeloFxFtSDVkG9GUvGadbt5TGcqTNoJ1EoYSf7ynHKTgZDGptnaymISR1mkbcSvXWXVQufzupn6IQ669tO1q39kv6sgxkqgkTybVme1JRxn6TNJRolJ5SlKWxS15cS6QB+thJknJ3whTyUrqPt2TReV0E3b+JmACJmACJjC/CVhUzu/j670zARMwARNoQgJ0LNAh8cEHH6Sf/vSn6cMPP8yd1XQ48Tw5lnReq4ODzgjSHz58OO3atSudOHEid2zQSUWnFJ0Y77//ft5TIirXr18/ISvpeGmkA5y6MdOZwrLcsaEOIHckN+FJ5Sq1PAGLyuY9BWhH6UgmGl6RLrSjtPXM3FhCB2+VSdcPOueZaavJX+0znfRTdbTfLm/yUNvP9YZ8p5rK1wQ+Zz8ok+sMc7lz/2bbkwdlaB90rVFeXLfIk/fk52n2CVhUNnYM6haVfA/4PvDbkBvUiJik3WCIV5ZqS/hdiKTUd4jvZCHuuuI3ZU+0M32Rx0B8rwbSpcshKtv700B/73hEZWcaCFHZ2xvf6Y7r0YTxdc1T+bvPa836/mopUtRXU/l7rNdsTxvAb1ikHJKS6xhtJevKolL5aMm2kn6kYyYv6qB2BFaSjyxpF/We15q1jjZY26hNo67aT5bkz6z2i3IV9cg+UAc+07HidzwzZTBpn9XGkj9lsWQd+y1RyZI8xYLPKZuJ/MlT28GMmxm5xvC3ArKSmyHLj4lgW8qfzsmicjrpOm8TMAETMAETmP8ELCrn/zH2HpqACZiACTQRAXVS0Knw3nvvZVFJByCdTTxXaMOGDVk+0nGhThU6KUiD0ERI0imizu0lS5bk92+//Xbu5KGTCtm5devWPBQYHVrkU2Uqd75QpjpF1LFBh5fqxJL3dJbQUeLJBExgdglYVM4u/1uVThvKDSZvvfVWOnDgQH5NG8/z5IiAZ2bo7qkmtqVtplOeNpmOcUXacC2gY5zP1HFNRzGigiUd2Wqraadv1lZTRjlvhAHlqO2fXC+tJz/KoDw64ylTne7lskjPXBYL7APnLPtA2XzONQVRQH7ahyr1n1w/v6+fgEVlY0zrFpX6/aWhXrlJjd9//Hak7eC3n76D3d098b2/fvNAR0dnfK8QlUWEX2+vROVgtBtEVPamocHuyIdROpChDK8a+xtOa2zsupCMr2heyXdb32mWEmAsmficearfh/pM29MmSPapTVC7IOlHWk3aTm0iafi9qpm2hIl2SNKRNuRmM+0X6ZjFj3as3O6Uy1S5lEPZzNSfmTrwvtweK1/ygwfbsy0z6eChNlMs2H/NZVHJ52xPPto/5Uk6boLhe8ooLfy9wJL3RKrSzmp7sZyOJew4J7dv356eeuqp9Oijj6bNmzfnv2s4h9lfTyZgAiZgAiZgAiZwMwIWlTcj4/UmYAImYAImUDMBdXbQCcxdz3v37k1vvPFG7txAUtJpTTQknU6kVccUwzkx5Ct3SdMZQmcKMpJnUSIs6RzZt29f7gAnDR1Wn/70pyfyouOgykQnBnnR4UEdyx0b1IeOEfKifDqR6YBR50uV/J3GBExg+gjQsUmnJG3Bnj178jBwdMLSVjz00ENp9erVWSrRrniaWQK0nzxf+IUXXsjDffOa9nPbtm25Q3fHjh152MapakW7TOe32mWe40bkDEvaadps0tBhTftM+6/oKgkM1tNWSxKUy6FubE97T4c2QpUObvLWVO5cJj2z8qI8hpwkqosoL64PnGPUh0np2QfKIF861DVz0w7rSUcdubmG+tPZjYxBzFB/deaX66L6eTn9BCwqG2Ncp6jknNd3AznJb0WeRclrvuN85/R7rLOTqMHiWYvt7UQ5F89cLIvKnp7++P5GROWl/gjNi990vURpdsX3tz1+23HzARF78d3NN0jQPhCJV0Tj4Zn0HeQ7izjTLEL6/rNUO6HPtC3v2Z62B2lHO8ZMW8C1TO/5TG2IliqPz9SusB0z70lH2bpRA3a0H+WZdXzOrN+1tD2w5D3rtR/lcilT5VKW3mvJ/pCe/VT+OjblfSYdM+nEif2StIUBbT4z+8V6ymCibaX+5Kt2ljTl9vvgwYP5mkNUJdcL8qO+lEH9pmuCnUXldNF1viZgAiZgAiYw/wlYVM7/Y+w9NAETMAETaBICdA7QSUCnATKB6EiiKumAoKN3+fLleabjl4k/+On8pUP3f//3f7OEQBAiKDdE5CXbkJbtuXv6nXfeSa+99lrujPnc5z6X7rnnnrw9HS9VJjpB6Oyg45hOcDpIJnds0ClCfuoAbyRis0odnMYETODOCFhU3hm3mdiKtp+2/n/+53/SSy+9lF/TviOQP/OZz6SHH344C+VyXXS9oD1GEtHhTDvPTS5cQ1ivznGuK0y0z+TLdQHJxzWFm2CY6Tym45zO+vJEZznXEK4zCMq9cQMNkZ/cIEN+zGXZQL2YWcdnlEPEzLp167I84dpAJzqd76Qjf64r1Jn6a1hC3uuGmHIHPJ3v1J9rH5KdWfXnM/L1NPMELCobY16XqNR3j99+fNf4PvD7j+8EopLvir5vhfTS0M/IuWIupFZ3/HbryXIrP6PyWm86f6E3giYjojDEZn9/R+TVFtISKcr3W1HchUhEVhJiGV/78TahEI20PXzH1S7QJpSFoCSc9mMyRbbn+89vTdohyUraDF7zWTl/lcc6PmMb0jLzm5X3pKEetHfsu+pDXTSzjs9oL/lNy29ZboqAM+/VVlJv9k3lqp4smakDn2kmLVOZA3mJg/ISr+KYFUKXPMhPklaSkn1jv/iM7ZQ3+fKaiW24JtBu08YiKvk7gzadm0+QmOKpOuYNa/4PnhaVNUN1diZgAiZgAibQQgQsKlvoYHtXTcAETMAEZpcAnQN0rtDhR0cwM3KRZ8jQAavOEv7QpwOFO+YfeOCBvA2d23Qk03lLB9XGjRuzsKRThQ4MOrHJ7xe/+EXuuHj88cfTli1bcscWaapM6iApi0rWqZOITg46lungocOMTjKWVfOvUgenMQETuDMCFpV3xm0mtqLt58aUH/7wh+nll1/Or2nnP/WpT2VRiaxEQJQn2l3aXOQezyZm3r17d+6Apo2mgxtJwbWD11wHaJ/pPGeis5gIfaI2Nfwe67jOkF4T6dmOcl5//fX0yiuv5OsInd3kzbVInexsw77Qoa7OciJ1Ea0M9cfNMdxIw76xDWnpbKfznOsTz2RmFAE60SmTPEhHGUxcbzQTnYn85Br42GOPpXvvvTfvE/X3NPMELCobY16XqNR3hChjhojmO83M70PW9ff35e8R3+n4uhXfyy6ezcjzFmOOJUPB9vSEkOyNCML8fe6KdqIrnT3Xma5c5RmWcYNDT1vIurhBrodoRKQkkdwSiIWQi29+ao+mg/RFG0JbUPyu5bvORH0lCGk/+L3Id5z1zOW2h22YJR1pi2jHaPc0SwaSRjKQ19qGdJKUEnp8RjmSjdRB5asOZVFJO4qkpM3hNW0MdVe7pzpSF9WLejJTlvad/adcZpVHHprL+67XpBMftmcfKadcFmVSFuv4XPmX68fnXBf4O0E3tSAqdXMLf2eURW65zpRb12RRWRdJ52MCJmACJmACrUnAorI1j7v32gRMwARMYBYI0MFApwadCHQ4Iyk/+OCD3GFLpwidKuU7pJGAyEY6FBjKkU4YImQY9kuikg4VOigkPxlakOnzn/98fk4lMrGqSKRu5EU5dHjQOcI6dQ5xR/aRI0dyfciXutBZRseOJxMwgdklYFE5u/xvVTptOKLyueeeyyKQ1xKVDNP92c9+Nt+AUs6DqHaeZ0nbT0Q9w39zvaBt5nqhZ9XRvtPZTXtNJzUd0sxMdLxv2rQpi8oHH3wwXxOIxFfHPWnYjrL2RiQl5fzsZz9LP/rRj3JeiBGuQ9SVbbgWsC/MdJazjhtn2If77rsvX5e4NlA/JjrGuWYo2v/nP/953gfKpJOd+mm4RfLj2sN5zLWGMtjH+++/P331q19N1J+yfL3JaGf8P4vKxpDXISr1HUM0cgPAPfdszt8xvpd8j/n+8N0MPRZzcQMB23R18ZxJoiTjBrj4vDu+j4jK7u5CGqbUEd+1jnTqdFe6NBoCMTbv6b4W38WrkW4svtd8x8dFZfwmvDTKkKrctIaMRKx1xve/iGzme1oMDVuIStoEfsfSBkj4sY5Z8k4k2VZtCr81uUmB36DMtBHMvEbQ6aY50mlm3aVL/GYthkcti0rK4gYIypXQo1z4MLGOz6nj8PBIjBIynEcKoc1hHfug7ahnUdZ1Kar6URc+Z6JM8tdcfq/X5SX5877MRkzgQpmSluJQLo9yKJq0ly8Xzy6mLee7yo0miEpuCuF1EcHO7/riWZWqc654jf9ZVNYI01mZgAmYgAmYQAsSsKhswYPuXTYBEzABE5gdAnQm0MlApwHRMXQi0KFMRwWdUOp0onOCTgV15tLZS8cDnRpISqJMuKOebehQoSPjZqKykYhHOi6onzpHeK1OJJbUiY5y6kVkJ+W743h2ziWXagKTCVhUTibSPO9pW28mKjX8K22pJqUnAhN5SDQi8o5OdCIvuYGFJdIOIUCHNe021wqGhn333Xfz9QXRyUTeRG0+9dRTOcISuakoRtpzOrcZmvaXv/xlFpXPP/987jz/2te+liMaae/pgC5fE8iXaxd14IYVrjVFhFd/vlZxPiIpuSkH8Um0JjfpUF+iIxUVxlCx2gc644m0JAKTbelkp+wnnngii0qiN7lOepp5AhaVjTG/W1HJd5rvF1GRfMf4DvO92bhxQ37eMDcEsB6JiahEWF2J4Vnb2y5H1OSl1N93NT5nqP6O+O4WkrIzBCb5Xr3SFtGU3enEyb5oMzqzaOvuvpKGBi+n3tguRoKNLOP34BVkJaISwYVE7Ih2pivSI0dJ1B6vi6FR402eOjuLZ0Py21Q3IfD9pr2RmCsEW3HDA20dM78xacOYy6ISETk6WgjLK1EfPqcdunqVtEX9Ll4sBCKi8sqV4hmVsCuXSeVUFq/1OW3hggULYx7J7RdMqXtXl0RlIQIpl/xpY2mjWNJ2sp66M7FfzJMnyi0+Q/IWEZQSqCw1UyfSsdTEtsX+sq9EWhaMyr/Nec1nSEjqxm912s/9+w/km12IquT7e+ZM8UgHteMqo86lRWWdNJ2XCZiACZiACbQeAYvK1jvm3mMTMAETMIFZIkBnAh0E6rylE5bODjpK6LSl44mOHTpDdBc0nQt0ItNxw2d0Tmv4L0Qh6/iMPIlaoaOZTg6G+qNTi85jOl2qTurIUSeI3rPkOTdvv/12jnihQ9qisipVpzOB6SdgUTn9jO+0BNrPKqKSdHQ4c11AdPzTP/1THiqWSEkEHcOgMsQq0YVcCxAAdHIzsS2d5ohKrgUIQoYMR1ZyjSDi8Rvf+Ebi+cW6dnCt4DNumKF+L774Yh6e9cc//nEeZvU73/lOjs4nPdcRrl+Uw8xEpzrl0zlNXTQiAOm4dnG9YDjy//zP/8zXNKQkz7Pk+rR169YsOFmnfaD+nMfUZ//+/Xkf+IyIU65nvjEmY5+V/ywqG8P+6quvpGee+V7aufO5+G7tzRHKjeTAdxPpx+9DhmxevXpN3KBQCP5Vq3jm7KL8+w+hVohKnpN4OUTY+dTbfToN9F8IUXk1vrdt8d3keYzkV0jF0Rgd+syZrnTs+GBEaPeka2MMkzoWN0KMRnnx1MpI3x5hlvE1jjaF5yOeDgkW7dLF7jR6qTfWDURZPN+ykJVEczIV7cF1UYn0o/69vYjKQvyxX6RjG5oRtSf6fUwbQBtIu8Rv4UJCFs+eZF0R2amoyiItwlAzbQ8T5UhUUh7llH/X6nN+Q3MzBL9pmakz7Zm2JS+2K0TgpYmI7wsXzue6IUaLfIv9Jz1TuTw+Z6IelIuwZeb4Ep2q9/qMNo+5LHbJg30TmzKHsTHWI3iL+tF+ch04cOBgbv8PHNgfN4kci78lTkX9L9wgV3PFavyvEJUL4jq1I98Yo2HH+VuE/SmOfY0FOisTMAETMAETMIF5RcCicl4dTu+MCZiACZhAMxNQRwMdCAznx13ORMnQMaJOWCJm6Jwh2nJvDMVH5zGikk4L0vHHPp21GzZsyEN/kZ4OGjqjiYgh8oYOFjp2N8ZzLOngouOgkUmd0FqyLa/pqCR/hh6k09yishGqTmsC00vAonJ6+d5N7rSftxOVRCVyjaC959rAMKmIStp1OtCJovzCF76QduzYkW9WYd3kjl+2p30uP7MY4cmwsdwI87u/+7tZPOpZkohFxIAiKhGVDP1KRCVDe//pn/5p+tKXvjRxwwv7Ub4uwISOZ3Wo09FOfohWou81lCz50clOpzXXJoaK5eYcriMITrZjov5szzWN6w15UJ6GukRqNno9yxn7v7smYFHZGMJCVD4TonLnHYlKIu+6ujrjOzIYv7WWpTUx3OuWkPV8F/hu8v3v6ysio+MrGN/D0dTWfj6iIU+lgb4TEVF5JmTlpXjm5FjqiiFfea5ke0ch60ZHr6QzMezrsWND0V70hTLkJoP2KGssxGJnfCfjuZcdXfF9bI/vLVGKx7M4PH++N36fDkSE5UisG4jvZkjSGAIW7dgWYpO2APnGb9C+vt4sUrnBoRxR2daGqCxYxlc7f7/VrtBGMCP/iiFdiwhG2hOGdy2iKy/nNERUFmkLQTca9nU0oj9pQ9rH26TOziKKk9Iog880w5f2j9/ViEp+K8MUcVncAMK219slRCDtEtfZc+fO5iXvWU+eUULMxY6Vy2JYXERikYaaICsLVlrCjbkQlTF0bvArhustIlHhyefkcSVHToaYDSmJtKS9VKQp72HF3xUnThzP1xH+Nti3D1F5JGR5ISovXy7XhzrVN10XldtDVP5WtPmfi5tT7snXkMnXq/pKdU4mYAImYAImYALzhYBF5Xw5kt4PEzABEzCBpidA5wUdKwzLRKc1HQh0KNMJsTGkIgKSjlg6P4gmIaqSDmvEZXFXevGcMNKQlo7ncnqGlGXIPOQlkTdr1qzJr+l0uduJuqujkqGlKJfyHeFyt2S9vQnUQ8Cish6O05EL7eftRCXygeuDhgb/yU9+kiMRiabkOY0IPoZAJTKSDvWbCTs6q5GVXEOIqEQ8MoQsdWD7xx9/POfFEOJ0ytP5LVFJRP5Pf/rTLCpp27/73e+mJ598MnfkV43M13nIzTjsA3UguhMZ8Hu/93tZlBJVyfWDaxOd1+WJelIn9oFrDkwQmtSVfeamHU8zT0DXf+QxIzgw8ZxEhDPnJMfT03UCxdCviqh8P0uiQmSNW7rrSad8hSQjEpLfWtzEsHbd2rR50+b8u27p0iXxnUDa9+fvQ3s7z5W8GILrdAzd+lEa7D8eojJe98S6iJTkO9PeESKsLaLyiL4LqXfmdGd8vyKi8jzPGB/IYmx4uC1+a4a07BlMHe29aexaR3wXz6exqyEqLzO8am9EVcbNcRcXpMtXBuNzflsSJUcEYyHg+D7zmxYJ2I+kjO8sr3Md2nkm4437H1/3cVlZSES+70hIoicvXChEJW0BEYy8R0gWacpRlUWkJdKO9oMyiFRE9LZHmXBnPdvRtly7NpbbHZ7jOTg4NC4qF2SmtDOqL3KQaWysiFZXlOf589SHulwf+pX9YGJBWdeiHCSl9mcMYRnl5s9y2pwy14X82b7gGBGpwY8hf3tD9vb2wrCInKc25Hcp/ibg74QsK4NTHgI39p22n78XiuGzT6RDhw6nD+PviP0H9sd39mhJVCJXcyWoco3TtahvT5a+23cgKp9Kj0YEP+29IyprxOysTMAETMAETGAeE7ConMcH17tmAiZgAibQXASKDopruROBjj46/N544418ZzYdfnTk0ilFZwtRl3TOsA2dJshHOigQkSy5C5zOWzqs6Zw4dOhQ7rhAaNLhvWnTphzxSOfy3Xbsqt50VBJRaVHZXOeVa2MCEJAgIhobSURbQdvBEKE8B5Hn+/F+shgyveknQBt6M1FJdCESkmsAbTkRlIhFIipfe+21fMyQi0QjPvLIIzmiChFws+NIRzzXCNrrd+NZlYjH73//+/mawnCrDz/88ITw5HpDZ/xUohI58ud//udZVNLJzLWlyqQbcRA1iEquGawjIvQP/uAP8tCzRTRYRG1FJKVkQDlveHEdRAzwmmsYM/s8Vfrytn49PQQsKhvj+uqrr6Vnnvm78aFf3y2Gfm3TUKm3yYtzPovK7vwbb0UM9bo2bjzbsGFDtOcr47fi4vhNOBxtA89SZAjRqyG2zocY/CiE/sdpMOa+3nMhvC7FdwZxh7CL6MB47uTYWAitkH1nz7ankycGQriFqGwfCTnZn4aHIhKynzyHIn0M7ToWovLahfgOHo/vI8+K7A05FhGVl0ZCVMZNDuOikghGIiqLSMZiyNquiGbs7olnY8aQr53jwnCq73vsKoPA5u85co/oQ9ovhCTPXGSo0rNnz+TrG9c4oir5nBn5h9S8QnRlPK8SCUteyMnOaCvUZnxSVNKmdOQbJYaGBvMwurSFw8NDeYjrYpjaom3K9Yv/kI5Fm1Q8D7OIpiSikeGwEZCFpMx7w+u8L+OiMtqyos7xfjy6Uu10rj91Z3/iM9o32jmePcrv9zzHMemJ9/m3fBR0KdrFLCqJMo3XHE+uHaMxE1FJxGfxnMqjcePLoXzT47EY+vXjU6czT/aD8qOw25yIjXzMTl/NN5MwLHEhKn8jROXD46JykdvvRnA6rQmYgAmYgAm0KAGLyhY98N5tEzABEzCB2SFApysdDAzNRMc1ESxEVdIZQUcwd3MzIQPpjKazGIGJmGQdndg8K5J8kA7ISjoc6JQgDwQlkZRsx2fkkTuo7mJ3KYsy6KjkmWMWlXcB05uawDQRsKicJrA1ZEsbWkVUcm3YtWtX+td//dc8bCptPQLz137t17KkJFKe91UmrjHcwMIzIv/5n/85XzvojOcZl7/1W7+Vn3NJFByd4lOJSm54+bM/+7MsKvVMS/ZjqqksD7kJ51e/+lW+tiEqqQPXps985jPpW9/6Vh72tY7r0lT18LrpI2BR2RjbV1/dlZ753rNp53Mx9Oved8YjKnme5I0RxJ/Mle/YtYhoRKR1x81rC9KKlSvyjSbrI6py+YrlOWJtYGAoJGB//A5Eal2J79jZNDQYI2oMnoo5nlMZ0ZQdHVfj9x/DinbGHOWGlyKSjuFKz53rSB+dJEJyMCTjwpCKA7Fdd+TD8xn7I313fnbl2DVuFvg4BBtCsCe27Yt5MARdX4jKEK+RN0GSkpUdIUTbQ7J2RkRjvsEg2pd25ixKoy5UIu9hXuRXRbOCrCyiKhGQl0Om0R5eCDmJqDwbN+6dPxfPhYx1V8fFXxaV42IzC07kW0yURbvWlSNJGZp2PKIyPr+WIwmLmx/YVyIqFyyMaMoQv4ODMezreNR2lqq5dsV/WSzG9gy9yhCzLC+HYBxDMEa945DxL0+0k/rdzOcSq0jNidesj/1gmFuOx+VY6nmXZET9iTjPfxsM9Odlb7xn6FxEI3Ly4iWGxL2Ub+gYDS5IS34H8Bv9o4jEPx43PNIeH4nRVriWnDp9JkeCZlFJnfNUl6wkvyvjonJxiMoH0lO/+dUQlZ8NUbkxIiotKseBe2ECJmACJmACJnALAhaVt4Djj0zABEzABExgOgjQUUFHAZFPRLsQWUnkCBMdOxKLdBQ/+OCDWTzSWUFHAxE2pGc4QPJhPR0a3E1NeiJzNmzYkMUmnRx0IJc7ke9kf9ThYlF5J/S8jQnMDAGLypnhfCel0IbeTFQS7YrEQwYybB83r3zve9/Lw6XShjNs3v/5P/8nC76NG4vnDlepg84HIht/8IMf5Oh9om3IjyFYiazkmoE0nEpUIkR5RuWTMfQr6biRRh3w5fLL1xheM6Q5w80SEcpMpznXpM/FEIC/8zu/k4cJLW9Tzsuvm5eARWVjx+bVV3enZ/7v/x+i8vn47r9XiMqQf7cXlZEk2gtEZRejaUSU37J4RiVDMa9duzq+i8vyyBv9iMruEGshKnt7rsRNbmfTguETEWl5NiIqL8f6iCyM4V4RlIhKhGLE6oWo7Ajh2F6Iyo+7o82Jm+PaIqKyuz+26wzRVAyZ2hayD+93LQQlshLBOTbWFcKyO3579uTXofsi7yLyMIvK+P4jRvkNW8zFUK83ft9vFGNEQOYJ0RfyLLcxURZSj6FNabPORoTguWhHzp49l4c9RWLy+xd5KFlZfg4kdcqiMiI5WVI+MpQ0FMd72j2e8TkYEZXcwMHNgP0RucgNgFmssk1Rs/H6RU2jXlejTD0fU3WY3C5SVrGO/SH9eGTl+PIKknL87wD+FsjyM/4G4O8AJOjlWLK9ZGVxE2N/boOpN/lnPsjJuGYgb7l2FNGU52JY39MhKj+KmwtP5BsMTxyPaMr4m+FMMLwQYhNBCrsAUd7Du3zNyTIuKuPmyu077g9R+Wvp0UceGheVCyeOxV0W5M1NwARMwARMwATmMQGLynl8cL1rJmACJmACzU2ASEqiZ3gWJZ3KyEY6UOicQFgSFUn0C0s6J+hoYEhHOoKJoKRTg/XqiKEzeceOHblDi/USnndLgQ4TOjUsKu+WpLc3gekjIDHloV+nj/Gd5kwbejtRSXQjx5CbV/7mb/4mt/UM18s14Itf/GJe8p5hwKtMdFxznXj99dfTf//3f6dXXnklXzvWrl2bvvGNb6THHnssDwtMxzyikvq9+OKLE8+oZLjXP/7jP05f+tKXJkRlFgN0cI9Pul6Rh26y4UaanTt3phdeeCHnxzbI2M9//vPpK1/5So761/Zezh0CFpWNHatXX319XFT+uCQq45mORDbeasKORXvB77fOGD51YGAwj6rB78D169emZSEqF4RYyxGV8SzJ3niOZV/vlYiGDFE5cjwiA8+lgX6Ggo3IwohsRNrFqygXUdkV0rE3JFd/PJuyL2RWbzp/gRsQBuO3ZE9s1xby7lpIzojE7CikIV/3q1fbQtAhOWP7sc5Iz4wAZEZKMuxrlBVL2gTJSknMsg9DsjGNL4o3eUXe8cgXIRgiLkQe0o52jMcgFKIyRFuIy8tENBLNGJVjRvohAxGRiM+oQfwubg/RW/w+hiVRlaRlIuqzOwD19w9kQVk8A7c/RwPyXEskLfuhOsaIuROTfg+rbJa5zuwYdY+U+SWvEK/jcrOcPovLqCuStRi2thjmOkeQZuEYfw/E73vkMO1qX0R59kdUJTKV5/QytC1l8jcDQ+EWkvJ85kQkPX8rMPQ731mWH310Mp0+czo+j4jUyJ9hcjMLDkxdrpKDlkVlPKNyQlQ+GaLy0xaVE2ePX5iACZiACZiACdyOgEXl7Qj5cxMwARMwAROYJgJ0Spw6dWq84+XyRCeKOn+JlqQDpeiYaM+dEnRCSGrSOUNazaTjznC207o6qq6OGYvKOmg6DxOYHgIWldPDtY5caUNvJyoRg3TIP//88+mv/uqv0jvvvJOf68gzLBGV27Zty8KQzuoqEx3SXF+4ueW5557L0pBnIhOZ9e1vfzvnibQkanMqUcm155vf/GZ+NibRlVxXyhFEusawnmdOEvVDp/revXvTf/3Xf6Uf//jH6aWXXso303zhC1/I5SEr161bl69PVfbBaZqHgEVlY8ciR1R+7/8rIio19Gtb2MPbDv0aScJx8f3iJrTe3uJ34IoVK9PGjRvzMyoXxlClg4P8NuSZihEZKFE5fCyE5Zk0MIioHIvhV8dCuiHn+K2IfOuN35nDMczo0ohSXJROfjSczp4fjPe9+ZmYA/2jISsvxXf5fAwrGzfOxbZXr3bGTXEM94rg7I7tQ1KGCMyCMkQlEhApiDzjtSRlloPj4vK6DQuRl30e/+V/JaixgikSjMVMW8OQqPxOPn8hJFxEUxKdzXVudPRi/j1cRCoWopLIRCIdaWuZiIrsDlGp4V+J+OSj/BzNaKd6exj2dTBHVDLsK8/mLG64KOTr1AIvrB71jow0TC0ispCViFLWF/WnMEWIsp59YuNi25xJXqeIS6IqiSBl/3LkYyyRkKTnxkPaWZ6nSTuboz6DNfufGSFz2S7+Pjh16rqk5GbIkycZ8vV0sDuTRSU8r0dUZlQ1/SdR2Zuf+bl9x7b01FNfdkRlTXSdjQmYgAmYgAm0CgGLylY50t5PEzABEzCBpiUw0XERNeS1OoBZ3moqb6dtbpX+Tj+jHDpiLCrvlKC3M4HpJ2BROf2M77QE2tDbiUqeRYww/NGPfpT+8i//MqcnmpIhWhF89913Xx4eVs8xvl1d6JAmv7feeitLQ4ZjRRwiHf/wD/8wPfHEE1kacoOLRCXDzhLRiSzlmsKQs1u3bs3PxKNzfLKoRKQwZO2WLVty5D91I6L3P/7jP3KZDDtLx/qXv/zlhKx85JFH8rP2bndtu92++fOZJ2BR2Rjz/IzKZ+IZlTt/FN/l8WdUVh36NYri1x8ysLs7Ih3j2YkrQ1Ruie/i2ngG+aJFC0NajaTePiLsulJ/71hEWF5Iw/kZlRFROTgaYutiCK4LIREvRT4hHRGkbf3xHV4cQmxNyLBl6djxhen02f54H5GH7VfTUOTB8y2Hh89Fvpdi2xiC9WpXPCdyKIYXDVk5Gm1ARFZSuywqQ1DmEUDGn0fJ62IOYZnlZSE0Icf+ZFUXbeHEb1d8HR+O/08MY3ycP2eIVeTjKM9ivICIK0QlN3NcuniheE5kRAYSWVk8L7IYQrWIqiwkL8/41KgjRVRlaOKoY1cXz+IsxB8R6jynkvdEU/J57Fwxj9crVzH+IyqTVVHLHCmZBSSRkVFXylVU51WEJZGWrNf+jucFB7LJeeXX5Ckxezmk7IUQjojFkLIRATkaApPU7MtgtK+0sUhV9ov1MEJong8uRE1+dPKjOK7HE5LyyOFD6USIytPxbEqGz82SF1EZQpe61zuVReX4Myqf+oqfUVkvZOdmAiZgAiZgAvOegEXlvD/E3kETMAETMIG5QICOm/JUtSNX21VNXy6j6mvKsKisSsvpTGB2CFhUzg73KqXShk4lKomWZFhU5oULF+Zh+hCVf/3Xf52F36c+9aks9xCVyECGh0X8VZkQlUTYEJn5k5/8JAvIX/ziFzmPP/qjP0pPxrMn169fH0LiRlGJ0KQOPBMZCUlkJVGXdOAXkUNFpzod/8hLROZv/uZv5mdoMvz4hx9+mP793/89D/26e/fuPFTtV7/61Swq2V8iOqfzelWFjdM0TsCisjFmr776Wnrmmb/LwyC///67xTMqczQlQ7FWm/iedBJNF1GVq1evyUP7b9iwPhF9PRRRgD09fTF3xdweaa6GyLoQ8/n47Hy0Ex/Hd/tYREaejShHIioZrnUooieXhQxbH8O+Lk+Hjgynj0/1hgyMIUY7LoekvBDDx15IixZeCCF2JSKkeRZiV0jN/pBcPSEru+I99UdUxgCr0SYQRd0ddaQtQJ4xd3aOC8v4rIjkHN9fSb5oD4lEpF3Mc7Z/+g3MOp4nGZGSMV8OUYmIuxBRlVm2MQws0YbxrMUrVy7nyEpkZn6+YwyXys0UkUPsT2dut3jOZ67PeMRnZ8jInpC/tKO0bUQpIippBxGVRIZeb59yhYvKyzCS+3iEZK5jHna2eO480hBxSsSibuoYY/hXcoidwn/yXwSajnNB6MIqhG58xDa020jKM2eYT2dxSb60t/0hU/tDVA5FfWmTiV5FhDJM7PnzF/INJwz1euTI0XT48OFoiw/mGwxPnT6V82TY3NGI2syMaheV7OXVzHHhQp5RuSMiKn89ROXD+bnIixcvyteQ62wjuScTMAETMAETMAETmETAonISEL81ARMwARMwARO4kYBF5Y08/M4EmpGARWUzHpWiTlVEJcOn8kzJm4lKhCCC4k5FJQKSiEnk43e+850cLXkrUXns2LH8bDwijjTUoMQCHfR0OLOeSM+vf/3rWbbyHL1Dhw6lf/u3f5sQlciAr33ta1lUIl4tKpv3PL1VzSwqb0Xnk58RTfzMM/83ROVz+SYFnhuYnxWZldQn09+4JqutWBWiMmQfUgpR+alPPZg2btyY2wGGK+2J4Ut51mJPdwxl2hsiq5/IyssRWYloPBlRggcjzcepqwPRxZCtQxGhx7Cv6yLSbmk68OFgtDmdEVE5FkOkXg5JeTEk5cXIP4aAHbgaYulaGr3UGZK1J7bpiucbdoTozMYuP8eRmxe6IzqR+vX2FjNtAs/WpN7MiLgs6NhviclY5kjDaEf0+5I4xfJEG8OMpCtEJc+qPJuHxyaqkqGteUbjaIg95F6ex0UlbRNlF/JxXFSGPEVeUj/WE5lI28TwrwPx/EdYIjSzqKSu7KaqVOzyRPVUt2Lo2Su5HgjVQpYiT3mGZkQtIiljX8sTdYMH4pGyOnKdeJZmIbDZDqF4Noa6JdKdfb4QUpboTCQw7T9tMvtAfZkY/rXY5my+hhw9eixE5aF08ODBHFl5MqIskZ7IXuqYRWWu16QdK1e04dfs51iuVxaV2xGVvxlDhz8yLioXW1Q2zNQbmIAJmIAJmEDrEbCobL1j7j02ARMwARMwgYYIqCPJQ782hM2JTWBGCVhUziju/8fenf3odZz5HT/d3JqrKGrfqc2ybMnjseNtJkE8yEUgBEiCIEGWPyBAEiD/Sy5yGVi+zEUQ5CZAgEzGwiBeRp7RjJexrJXaLTmWRC3cm0x96vBHll43pe4mKdPqp4RSnaXqqad+530PT9f3fepsqDP30LUiKoE70ZRf+cpXOhT0Tkmg8j/9p/80HWnveny0RaVY+tWyqVn6dSOgkr1nn322L8MKUopwFPX4r//1v+42F99ROS79asLcUq333Xdff/exCf5McAdUAhXgJD+9e9I7ki39KqLSOyp/+tOfdiDwD/7BP+jvqPzqV7/a3rF3e4ecGxKwKv/OFShQubFLMIPK7y6Ayo2DId8xIBDgtxT04cOHpxvaMtEiKldahB1gBbLtbkvA7t6zvUVULjX41oDlnqPtGFD5dssnG9RqYGxpXwOV1zfoeGuLtDs4vfzqrgYstzVQea7BztXpxkOnG6RsuZUru73v8WxbUnRp+n+/2dZA13Krt9wA3KwD0Aac7WxLkor4tHTqDPx29eNzZOWOeQnY/n7Mzik7uMv7HQP8lO6Ribajkv2Ly7+KqDx+HlS291W2JVGByhlQnuhLw86gEtA7N21rAM/yruCpkoYiKUV+0tI9dO/efT2aErCk4QxYG6hsviKJ/uuwsvkSvxzI87B7oSVUOyxt8O9Eg4l8mv060ZajnfWD7+b2H732eZdn17BFeHqfJriKjs7g8USDlW3J1hZdCcyKGmWHn6JAZ1ApYlXUq0hMUZXeU/lej8x/8803O6gUWWkZWKDc+Tmi8rcBauv4CqRz3S+rAzzSQeVjDVR+vUDlFVC2TJQCpUApUAqUAltFgQKVW+VK1zhLgVKgFCgFSoFNKpCJmQKVmxSwmpUCn4ICBSo/BZE32YV76CeBStGSJqW9HxKotGSr5V5BzL/39/7e9IUvfKFDRhFA60kmzE1O/+IXv5j+7M/+bAJOQMQ72zvu/vk//+fTN7/5zQ4NTXznHZVPPvnkhXdUmsz/Z//sn3WQ6v2ZJsYDFozHpLnMH/ATpAQBjjTA+r/+1//qoPKpp57qE+vedfl3/+7f7e/aBEfnifv1jKLqXCsKFKjc2JWYQeXj55d+fbF/FzdmYa4t0g7M8m5Z9wPfnxtvuLF/3ywD6nu6srKnlW1Z0D0tUnDPzgbhtrVlQj9s38c3G4B8t8HE4w3SNYC3baXBv30N+h1sgHLP9PIr2xvUahGVp7dNK7uWpptvOtd+MHGmRVYCm6datOKp9t7Ds+2dh0sNgHkfYltq9OwcESiaz73DfWH3buBvfn+iff7KogX7sqbn7xVGlOfJ+V4C9s3veLxwT4EIW33Lt6oD2oGQfenXFmUI2vm37nh7b+WJ9q5K97m+pGnz9XRbClbi1xzlGQC5vfvjOKA6g8rz73ts+97/2KM/e5TjxaVt53tcQCPLAZXn343ZqC1QOQPK490vS7CesMRqi6rsy9v2sMwZUhqj3C21MQb20gxU5TPNJHaNU7aENw0ASdDVteazus1MPx4/jrV3ebqf+76+/vobfSluS8BaEnYGnqearhf96J1dwf8ZywwqH2kRlUDlNwpUXkF9y1QpUAqUAqVAKfBZV6BA5Wf9Ctf4SoFSoBQoBUqBy1QgE0sFKi9TyGpeClxFBQpUXkVxL9O0e+gngUrvn3QNvU/yP//n/9wB4x133NGjFf/+3//705e+9KW2/OMdfbnC9bhj8t57Jn/yk5/0pVj/9m//tk9Ui5D8p//0n/ZITYARUFgLVPLn3/7bf9sjLy1La4J/nGjng4l8E/wm2Gcwsa2Dyj/90z/toNI7MSVRoUCld1Xqv9LvnwIFKjd2za4UqBR5BwoePHh9j1oWkWz5ZstrzkuX7m8/FgAsZ/AGYoGVu3adacu5Hm35gxZNeLx9P880gAcCtvdMru6ajr63vUXcLTeAurNBtRZVuHvXdMtNrZ/rzrTv+ofT2dUPGyD7sL3D8nSLqJzavSmQcrn74/s+Q0qRlHPfiU4MpJyXN23vX2z3CUBNwukSUdnf5dhg5BypPd9P1F1uOaBSJCEQ6N4owlA0pW3gUgkMHm/AEqjr0ZQd5O3p9yv+uTeJVgxUXWnRn0Cl+5nsPH0BwNlP/l70ZT42w9PZ/xmggoaizi2lOsNS/n0w+9R8PHn+XZCWh80z9DzuGRL2cTYwyjc+zQCVP5Z0nWFlbAOM7udgJVvaBALTkx/Gr/7Jk7NWlhFPVOVLL73cwOWvu2/0ZONqpQKVV0vZslsKlAKlQClQCmwNBQpUbo3rXKMsBUqBUqAUKAU2rUAmWQpUblrCalgKXHUFTNr6joqa++Uvf9kjKEzOmti2vCjIZd+EbKVPVwH30E8CleCDyWhw77/8l//Sl001ef3ggw9Of/Inf9KvIcgnWmU9yaS590X+5V/+5fQ//sf/mJ5//vkOFERminQBPsFInweg8oUXXujvsPzBD37QozotNfkf/+N/nL797W/3PvmynuTzZ9nX73//+z060+fSGL71rW9N//gf/+O+hC2AYaK+0u+PAgUqN3atrhSo9D0RlWi5z5tvvqVHL99yy639u+sHBGBlQCFY2SFcA1+NI07blk+0qL3jLZ9o8Kst57qy2o6fa8uxNvB4fKktCSpScncDlwfbub3TTTfubEvHnmntPmjA6+j09jvvNZDZIrOPLrV9sK4toXp+WVWAzz3Bcq8zJFV61yPw195NqZOW8j2fv+6+8zOoEy252mBfX0L17Op5eDYDQfcH9dQB38BIsE5k4QztLP/64QUwCBQCgtrpf37vZKJNZ1g5v0NT5KI8L5nr38MdbUnYi/ej9D+XS20ZWKC4jaLDS3zPvTygFaz0PsqLIPW336EJIKo3R46KHr0IKl1X7/MM8KWfbN992fjzzkr/NujH/kUt21shz9eZ+5nBqeVd3dO9q9J7Ko8ceXHyzmH34g50rx6n7L5XRGX7yFQqBUqBUqAUKAVKgU0pUKByU7JVo1KgFCgFSoFSYOsoUKBy61zrGunvrwIFKq/da7deUCliBuD4r//1v05/9Vd/1SfmgeZvN1j49a9/vS//agnI9SQRNeAoYPjf//t/n7yr7P777+9LybLnnZdAB5CwFqj07sn/8B/+Q+8b0DSBvp4EjvL9Rz/6UQeW3o+mH1GV//Jf/svJeznBhALm61Hz2qlToHJj1+JKgkogTaSkqEo/aHBPcB+wJPP+/fv7dxM0lEGuXbt2NwDWfgwwrTaydqbBrFPt+3aqnTvd6rSIyT3A2bn2vV9u0Yh7prPnbmxtDjT7rW17V+XqmfcaCHx3evs377bIyxPThy2acnUVpGzveWxwbxGsuTekb9GUAX8gmntfUqCl/XMAW3/P4wzxwL8ZCM5LogKV4OMMKudoRaAyUZW23bf8u6eOxDfQ1jLUdLE9+jYvrzovB8vPGaiCooBq/3+/H8b/lL1C+98MKefx5LkYOPR+ShGe4Gkyv+alaU92/860ZWkTXdl7ah2yzwe+uCfGV37nR0X60ce8/G1bUraN1X4HvE0/24GUtuf9Mx3ivv32b/rSr/4d+PWv3+rAV92rmXw2ClReTYXLdilQCpQCpUAp8NlWoEDlZ/v61uhKgVKgFCgFSoHLViATMhVRedlSloFS4KopUKDyqkl72YbdQz8pohJ4MIksGtY7HkVWPvvss32yXTSid3199atf7e+YHCfYR+eAABPYJrMt+2e5V6Dyf/7P/9knuv/oj/6o2wEN77nnnm6bb2uBSsvCApWWnQVHTJ6vJ1lu1hjASpGVIjVFAgGj/+pf/asOLE1kAxuLICD2+WQcJt0lUFO+VP20q/LqKVCgcmPaXglQ6fscmOX7AsD50YDo+Hn51+v79xLUshwo4LVjh+2LPwQ422Cf79Hy8pkOKvftXZ32H1idWmDkdOLktgYL2w8Qlq5vbdoSsi06sgX5tXdTfjB98OH7LZryaANvJ9t9qR1cmiEloMYXOWDNtuP6zw8Q8h0GK2XJeJJzPsBtrGPM0kVQOUdPgpMzrPyg37O8g9e/e+6b+uUPQAngzsvi7uv6xM/46P6pPl8kvsQvfSeP59WRpfn4HB3KxzGqkj+i2QFL23IiIQNu2R/7s80n/gF9/FWOsJJOYCVbSvd44052jdmf75urvd93332n/0DlpZde6hGVfFI/fvTBXOH/Fai8woKWuVKgFCgFSoFSYIspUKByi13wGm4pUAqUAqVAKbBRBUzOmNgoULlR5ap+KfDpKWBC1He0ln799DRfb0/uoZcClV/+8pc7gAQGTTZbqs9yrZZgfeKJJ/pysI8++mgHfH/8x388PfDAA30y3iT2YjLhbSI70ZRg4Y9//OMOPS2H+E/+yT+ZwEo2gA429HkpUPnv//2/n7797W9vCFQCCaIqQVLv2/SOTLASiNX/N7/5zQ5Jb7jhhj4xH7BhLIEBJtuNw4S/Y5m4N5E/1l8cf+1fPQUKVG5M28sFlb7LgBb4B/74/opMTkSl7w8o5zvsOzKntqxra2M50Q7DxFS2Z7fTp1enpeVz08quc83O0nT9ddumXSuWZt3eju9s0HJPe4/lnmZL1PS2BipPt0jLk+choKjM9kOBAaTxB5xU+k7ygZ/6lC4CsznyL2DM+e7XeUAYSKlMnYybHeMC5PzbBrIlitL96ujRow2kzqBSe6CPHjQCKpWJqvQjC77yUT19JOWeY9+9JTljcZ5vybN/6s1jcV7/87s02/sy2xKtfHXvCrB0LwMIpYxvtO8ezI5zud7Rl7Z8Sj+JrMw7K+kTHdnnp6VmRXPSyZKvr776an824A9f9KcNm1c60bkiKq+0qmWvFCgFSoFSoBTYOgoUqNw617pGWgqUAqVAKVAKbEoBkxkmPwpUbkq+alQKfCoKFKj8VGTeVCfuoYug0gS/ZVC9P1SkpKVW3Wfffvvt6ciRIx0uerfkK6+8MoESn/vc53o05MMPP9yjKkVXmXTPhLeJZxPQIKU2ohrBEu+mtPzq4cOHp3/xL/7F9LWvfa2/584kvrYmuq8kqDSRzp7xBpL+8Ic/7LrpG5j94he/ON19990dJpiQzxiMX3ufZZGZoK3Je3WB3ERibuoiVKPLUqBA5cbk2yyo9HmXfbdBqkQJBsCJqAQrff/dQ9T1HU72HXLMuxW9YxGLsszr8ralBusazNu7o33vdjbQCPo3CNagpvc07thu23tod7T7iOVGAc5ENGeaxQYUAABAAElEQVR50p0Xov5GQAmk+Q7rV//uRYuRfu6B6iTnuVJ99y1lUu4H6jgHrrknAG2BlSCle53j+gP43NPcI9wvlfYB3kR8xk9203+2o7k6ua/mXPyLts4na6ceH1yD+BpYyT9QcQSV+tBOcjwRkvqRXHf6xu9AYP2oz6Z+tNPGcSk2+ckXPvj3xLLf7qfgLg3jz9i2G7gC/ytQeQVELBOlQClQCpQCpcAWVqBA5Ra++DX0UqAUKAVKgVJgPQqYBDHxUaByPWpVnVLgd6OAyUvf0Yqo/N3o/3G9uod+Eqi87bbb+oSzyWcTyn/91389/bf/9t96dKXJZlFBDz300PTII4902AfemYg3qe3+bOJaO5Dy5z//+fT0009Pzz33XJ/YBkG/8pWvTP/oH/2jDgnzDjQT2/q7kqCSLybTLT37i1/8YgIpLWXrcwm6WHLW0rOWgjUG0U8mt/lict0YtLXsrYhQ47P8LMDpHZsARKVPX4EClRvTfDOg0ncApAPBfCcC3oBJgFIGLC1rmu+M7xogFTDm++w7KM1gDhRrkZk7t097du9sy7vubu33tIjKPe14e5flthZluEMk37YO+5aXRRxmaVK+OD4vLeu7KCcyMUAvgCzPinySfZ8D+ZxTL2PkHz9lkE851onNQM8RALpfuU+4L/p3T198oRd9Rp3c60A/PrMZH9M3P6ToHlA51s0YtEk99mih39QdYWUiK5V856P26UvJFxoZQ7J67OUzwHc5cJMvrrF6AY3xabweqQfsiqoEKmVwl3Y+L+zw+UqmApVXUs2yVQqUAqVAKVAKbD0FClRuvWteIy4FSoFSoBQoBTakQCZ2ClRuSLaqXAp8qgoUqPxU5d5QZ+6hQOWf/dmfTU899VTfNqErohJAlG+//fZu02S2SWaQ8X//7//dIytFR5pwBii8n05Upfom5k2YaxPgCPLpS6mNiXv9iNr0nktwMBPaOkw7y7N6Lyaw6N2SIhgt/QoSWrbVhP9GEphgyUHA5v/8n//T4anITpPulp7lBzgLPI7QRcQPAKHtM88808//w3/4D6evf/3r04MPPtjB5kb8qLpXRoEClRvTcaOgEpwCo3yfRdL5roP4IKUfGijt+877voBTIBMI5nsOPmUpVPBLYmvHebC4srJr2rtnpX2P97S8r53b2+4be5odoPLctLPDSkvN8kEWRTmDSW0DKPko65/PSXlODKwD0gLTHHNe0iZ5bOMetlhHfce0Z8tYgT/3CPcX0M228fLHPSrv8XTfo5UfR/DdebZiT3+yFO3dF8exqauO/uMfO+rRNgDUvqS+ezd/3Ff5m8z/UYf0r55/u4FD19G+c/rRR3SP/2zoQx2Jv+rwRZl7e3xhm07uqe6/vsfApc+KPvNZ6cauwP8KVF4BEctEKVAKlAKlQCmwhRUoULmFL34NvRQoBUqBUqAUWI8CmawpULketapOKfC7UaBA5e9G9/X06h4KBP7pn/5pjxIEEk3oWvYVpBRhGFCprmxiWRuQUoSkNiaaTdR3ANEmsU1Sm2TPPZov9k1qW/JQZNHhtuTrl770pR6NCAwCIAEF6psYBzn09aMf/ai/G/P73/9+hyP/7t/9u+nb7R2VmwGVJsBBhNdff71HVoquNA7LEJpo5zP/M4b4NE68Azb8/9a3vtUjMMFT46r06StQoHJjmm8UVOZzD7Zl+VKA0ncvkDKRlGCU74/vmPsBaJelUMEucMx595iV9h3aIzdgt2/fXK7satGUy7tb+z3T2faeyu3bgMqpwa5zLS+3dttbuXNuz0bLIBgf3VuS9JEc8Aak8Sug0v4I6LT1XZe01c55pf2k1Ek9ddhk2/iMM6CSBpL7BTjpvgdYulfwnV7sLfbXG7X/GZOc8Y1jzLjiGzvsBVQGDmorZTwZv/urzG/3vUBG47HtuH+75UBXNiR+6E9OfSVfnOOD8bm2xm7bMf5J7OibVgGV7r/eIRxYmc9Kb3AF/scHn1+R/4899lj/cYxIeMvx0shYKpUCpUApUAqUAqVAKXApBQpUXkqZOl4KlAKlQClQCpQCXYFM7hSorA9EKXDtKlCg8tq9Nu6hlmT9wQ9+0GGdbREwX/jCF/pSrI8++mgHEuMITGCbYDapLLLQUqiiLO07bnLepHUmtU0Cs5mJepGX995773Tffff1CEbAI5P2i/2YILc0K7hiyVlLrprs/zf/5t/0SEbbJsI3kjJhz7Z/O4DWn/zkJ/2dmSbLQQZjME51AwACakAay8Sa5LZMrPEYGzBQ6dNXoEDlxjTfKKgEmHz2RQL6rvr8+/ECSAnygJTAWz7/vvsglPt+ogvBKD86cEzynQWxfG/27d837d8HaLUIwx0r0+rZlen4iVau+rEAUNnA5u5zDWoutzo7WrmrQ84RgAXg5ZnQ93Yx8ytQLlGEgWvajcm+nPtYzqccoZZj+grcEw0IzroXGq9ztKERvYBKejo2QsT4O/ZhXHJA2lr9xu/UAwPZTg4MVS99BEzSI5kmOT6CV/fC8X4Y/8Yxg4raSj4vro0xGrPStU6UpzGwoQ923W9F2fv3Q7R63ltJx0tdn97RBv9XoHKDglX1UqAUKAVKgVKgFPiIAgUqPyJH7ZQCpUApUAqUAqXAogKZICpQuahM7ZcC144CBSqvnWux6Il7KIgANJoktm1iG3yT77rrrg4ixnaZoDYRL/olE8yiKrNsn4lrk8wmzzNxDWiYqBd9CHSAHpaMNJGdifixHxPf7PAJrDxy5EiPrgQ3/viP/7hDzhGQjG0/btuYZfZN0ns/GtsiLI0h0VCZfDexHlgjIof/tElUmXE5H+jwcX3XuSuvQIHKjWm6UVAJ8IBrwKTPve+uCOgxOhCI8h3I9wqoBJoSUeka5f2D6gBZvrvsHrjuwHSgwcrdrZ/l5Z3TmdWd0wcf7pxOn2mgcrmByp0g5bl2n9jWcgOVbZlYUJNf4FdAHLvuTYGLKRfhXIDceF7bpGwrU8d2cuopAw6dS9/gm/ugsfsxBBt8NF73O/cL4wcSx/aL9gMpleol63esazvn1NUX265HyvH+Gj345R4YKDkCS8fkRFwq1c042XDePdJ4s1Sr8/o0VvdpY1W6xxsznzKe3N99LvLviB/K5D5MPz6l34xbuZlUoHIzqlWbUqAUKAVKgVKgFIgCBSqjRJWlQClQCpQCpUApsKYCmTQpULmmPHWwFLgmFChQeU1chjWdcA810QzWJeLJRLIJZhBBNsG7VjJR7domamqMIjKxbTI7E+dsmKyOXcDPZHZAw1r2tTdJrQ+2AUv3em1EY4Keti8XEBo/2wAlDQDYTLzrHwQw+W+i3aS7yLJEkgXQBBSsNY46dnUVKFC5MX3XCyrzmfa99XkHJ/1wAagE6UE33+FAKN8R95MAqPHe4P7iOwzc+V67H2h7sN0Hrmug8roGKnftWmnfte0NgO2Y3vtwRyuByqkdn6Z9e8+1vK21ae+z3GvZWKByVwNfc1RingXZBuACGLOf0j2Lf7JjsrZjjpqjzZwf69NHSmnb+RFUupfoyz0qOroHGj+91krRXen+GbA39hN/0n5soy9AcASV+oqttM34+QcIBkraptNiVl9S0ld990nX1DhzzwcqjTH3eqVrHTibcceGtr7DIin9WAao9OOXMbJd3fidMW+0pHkt/bpR1ap+KVAKlAKlQClQCkSBApVRospSoBQoBUqBUqAUWFMBExcmTQpUrilPHSwFrgkFClReE5fhkk5kwtjEtG2T3iaTM9mdifJFA5mwzkR3ol/YkJMyQT7aZNv+GOmT+ikzMc0W28nsmfQHKccJ+rTbaDnaXwQZfJD0ydeMIYDVfnzYaL9V/8ooUKByYzquB1T6TPvM+3yD84mCBioBS9GVoI/vIVgPQqnr++J+INIuoBKgDKgEpXzffP+1ZQOo3L/f97mByvZeylOntndQeQao3LbUIi2X2vmW9/EFqGzvPDwfkTh//5b7c2DuR+zbzn5Kx2X+yY7nHjOWUdOxxTr2k1Nv/P47twgq3VMCKkWhBlQ6JulHih26R/9s51yv2P43+jues537FG0SUUlvx2NvbE+T3FtTBlJGK3W0Yd8Yc42BStfUDzyM27mAyvzQJaDS58Q5fkls6A/oBCVFs7/22msdVIKVPjNss6tufOiNN/G/ApWbEK2alAKlQClQCpQCpcAFBQpUXpCiNkqBUqAUKAVKgVJgLQVMnJgYKVC5ljp1rBS4NhQoUHltXIfNeOEeK5mgvhbTte7ftajZZ82nApUbu6KfBCoDvsAtcEcEMTgp33nnnR1airAEosBGkDIRgr6PoJLoOvf9QCwRy2BWlvMEzbQDsQ4c2N9B5c6dLXRy2tYjKj/4cFcDU34s4b2Uy9N1B7a15WF3tLois9uSrytZRnSGfSCWZ0HluJ1jKZ0L9HJM4nPuIynH4zmv/pjHugCgxH5AZaKzR1AJ8BqzsdNgtKE9O2vl8f4bf8a2uWaxEVjpGiYDhIGV6iXxOWByBJV0ctx5WdIPDZwDo0FG19RY7fPJjzjm6zpH5QdUGrNzxqce2/oDO9kAJkVSgpSy5WAThevzpM9xzPF/vWWByvUqVfVKgVKgFCgFSoFSYC0FClSupUodKwVKgVKgFCgFSoELCpi0MGlSoPKCJLVRClxzChSovOYuSTlUCnxmFChQubFLuR5QCSYBO2Bkln213KucaMoRVIqUC3gDtYAlwC7LggJOYFaAFo8BNKBz7949/b2TbCwt7WhAaud0/OTuBqV2Tbt2WjJ1+3Twuh3NF0uKNlDZ3lG5q/m2bduODs48BwamLZaeDx1LOW6vBb0Wj2Vf+9hImXPGEvgGphn3uIx0QKXlT72Xl26gHXDIxqKdwMQRWAZEpj4fxnY5zxfb2o6wMpGMNHdOnaSMJ7ByrZJu+tNOfeN0jXN9AWnQ0Xmfm4DKQEpjd3y+xku9vX7Gz4nPyJtvvtmXgBVZaRu8BDFpyv445vi/3rJA5XqVqnqlQClQCpQCpUApsJYCBSrXUqWOlQKlQClQCpQCpcAFBUxamDQpUHlBktooBa45BQpUXnOXpBwqBT4zChSo3NilXA+oBLlARFASXBNNGUjp3ZSiLC0Jm4jKADDPZCAWqJTlX8EsOaASdFIH1PKOyV27RG7uahCrQbRtOxuM2j2dOr2vnd/dln3d0frZOV1/cGcHldddt7tBTXW9mxbou/jORDCN3eRASc+IgXHZ5qfMB/njUuqyF5vKHNeWDfuO+/duLVBJL3rSbgSV2seXwMlFWBk/06dx2L5UUj+wMhGVIKEc2xk3O+xFNwDR9RuzY+kzdUFGY5VFRTquL2MDKI0XoJQt+5rlsvkc2wGVPhM+H77LAKWISu+stBwsLdnXJuO/1Lg/7niByo9Tp86VAqVAKVAKlAKlwCcpUKDykxSq86VAKVAKlAKlwBZXIBMsBSq3+Aehhn9NK1Cg8pq+POVcKfB7rUCByo1dvvWCSrDp9ttv7/mOO+64sOQr0BYABVQCQAAV8AXUAV6LIMq/AYGVoBOIqR7Wtry81ADatg4td+xo76lc3judPXewRRx6/2WL6jywazp0/a4G+HY1+LXSQeUMRuelU1dX56hJ9sZowEBFAG2EbNTy7BiYF2D3cSrGBpuxuwjN7DsHuiWCVDQgn8BBmgGViag0hjHxIz6tBSzVTZ9jGRuOZZy22dJv3lUJUoKF+l2EldrxPRq6fiCi6yTbNo6M37Y6zint8xmQ9JkYISVwmc+IOvpatE8zUZmgpCVfLQGbLKrSZ4cPGV/GvJGyQOVG1Kq6pUApUAqUAqVAKbCoQIHKRUVqvxQoBUqBUqAUKAU+okAmZgpUfkSW2ikFrikFClReU5ejnCkFPlMKFKjc2OVcL6gEJO++++7prrvu6u+mFFkpmhJoC5ACpQK/eAF0yYFXI5Dy7wAYBVQGVgJcnuO2bz8fXbmyp0G0A83SwWnHThGbu6eDbbnXQ4cCKnc16LWzw7elJdDrYjRg4Fn65selwNYIBbPNj0ulPGsG1MWu4zmXEnRLRCVQyZ8RVCaiMnBX/xKIZzuQMmWgYnxLn8qk9B3/lDmvfSAlaGhb3wDmOHZjGkElCOmaGY8MFBpLMr1z/djxOQigVCbadoSUfOIbG+zJgaE+E6Iqvc9UNKVsCVjg0nE+RPeMeyNlgcqNqFV1S4FSoBQoBUqBUmBRgQKVi4rUfilQCpQCpUApUAp8RIFMzhSo/IgstVMKXFMKFKi8pi5HOVMKfKYUKFC5scv5SaASIAO3rr/++umee+7pGay85ZZbOqgEoQAvsFIOiAKrAikDs8Yy4AuQEiHn3wXHzp5dbf0tTzt37WxLve6btu0AKg81mNaWDW2g8rrrdk03NFDpHZV7985LxS4vb+uQLaAy8GsEagGV/Arssz0CQftyUuBe9lM6HsgGlo1Z3/ZT8gGoFFUZUKlPEZVgL1BJN8AwMHL0aTyW7fgdfxbLPAuP+oOIfNIWmHSdEgEbuDzqkvFpByAaR6BygGVK101f2rDh88A2iO3zkc+IfgJE1dUmOZ+NwEr9+VzQzXfaErBAJWApqpIvxsPOZlKBys2oVm1KgVKgFCgFSoFSIAoUqIwSVZYCpUApUAqUAqXAmgqYsDBBVKByTXnqYClwTShQoPKauAzlRCnwmVSgQOXGLut6QaXoycOHD/eoSpGVAZWAG+gDTsm2ASlQDUgK6AqIShkg5d8DkZWBXqurZxrssvxri5bcva+BrQPTuen6aeeOFpW3b176Fag8cACo3NH6Egm4rQ8aqNQn+MU+m0p9ejaUgDSwLFAuUHARUqq7CMFGiHkpOJn+c54PQGWiKvmjT/BuBJWBeM6NvmU7cHIsR39sj/v6p72xB9jaNyY29TfCZfuJrOSDxMaoJXAYWLkImI1b/4CrsVkqGKhU5jNC99Eu3wIq4yt95IBsnw2wUiRlQCVoCfrqk4+bSQUqN6NatSkFSoFSoBQoBUqBKFCgMkpUWQqUAqVAKVAKlAJrKlCgck1Z6mApcE0pYOIWTHj55ZenX/7yl31pN5Ob3n/2h3/4h5P3n9k3mVqpFCgFSoGNKFCgciNqTdNGQKWIyiz/ClSKBgSlQJ/FHCgFJgFSAVBj6bh/D0AvJTgFVOJkO3a0aMmVve3fgf39HZU9onLf+YjK9o7KgMqdO7d38CewDqicwZr3JR6/ACoDs/gUGBcAuJZalwKUIwjM8ybbxijrO9tKdYxLBGBgpTHqO6BSpGoiUQNQnR+zfgNUU46+2B6zMembDwGV+qV9/NKXaxZYGchMH32zN9owDtAw0a8BliAo23TgG+BpbCBlIirHiFH9j1DStrbKfDbYi7/61ScNAUrvqnz99dc7vNSGvc2kApWbUa3alAKlQClQCpQCpUAUKFAZJaosBUqBUqAUKAVKgTUVyMRRRVSuKU8dLAWuCQVMPI6g0gSuyVGAMqAyk6XXhMPlRClQCvzeKABo+AHESy+9NL311lsdttx00019ydLPf/7z04033vh7M5ZPw9GNgMp77723g0r3atGAQFTeSwlQJYNeAYIj7AKiAqECopT+TQjwOnPmdAeVQNrOnXva9r5p9Wx7R6WlX/eLqFxpS7+u/BaopNXq6gy8jh8/0e2xyZ4EvvEpfiVq0Dk+JttPWoR/9qVAPIBNDqBMdGCOK/Pvnc+l9y0aL1/AvFtvvbUvnzuCSuMegSU/k0d/4sNavuZcfIvW9A/cU4cWI6zkh2uo/+gTsGgcohgDKu3LbIOh9DMu9owtsFI0pc8Dm9IMki9GTQZU5jh7o7+29SOq0vc576r03OCcdvreaCpQuVHFqn4pUAqUAqVAKVAKjAoUqBzVqO1SoBQoBUqBUqAU+C0FTFaYmClQ+VvS1IFS4JpRwKSjZdxeeeWVDhQCKu+8887pK1/5SgeW40TpNeN4OVIKlALXvAJg0LPPPtsjtkENzwTgpEjAhx9+uEDlwhVcL6i84YYbpvvvv78DX9HvNAWjAiUDAUXDg0Cybfdyz2aAV0ClfwMCJlMGUImobLGR58HiSoOCQOV1zQ5QKaJyBpUHz7+jMhGVhiWiEjQT7TfDLku+euflvNzrIqTkV7LPiWRfAvICCJX2czx1tJEDKJXG6Vjs8sW/d55LZX6xB/LSka6BvbSS+Tvm9M+HZL6kj+5Y+9/oY84HNAKCtAmo5KP6gZWuY5ZodUz/zqunjWsWUGlMrpvsnHGry3fX3XgCK207xh5/1Z2vzQyttU8fY0RltpXpm36iKkVU+p7zw5iid3RYT8kn0ayPPPLI9Nhjj03f+MY3+ufb9cjY12On6pQCpUApUAqUAqXA1lSgQOXWvO416lKgFCgFSoFSYN0KmAQxYVGgct2SVcVS4FNXIKDS0q/PPPNMn3A0MZqlX5UmPE3OVioFSoFSYL0KgCWJqHR/ATUky5RatrRA5W8ruR5Q6X4sKvVzn/vcdLi9p9I92jsrs6QnsON+rQysAoJkEXqOS4FUAJN/B+SAyoApEZDnzq2et7ertdpzEVRa+vXgynTjDbung9ftOv+OyhmqsR9QKeqPPf1J8Wv8dyXPi4FcgX4+Q7LxZEzZ7sbO/y/1gUD9JNvPOSVfgMrASuPlD1DpxzmA7wgqnRtz+lbGt/iRfozhUsk5vsVPpey4zGZgZaIfA5idUwdM5DdQafnXgMpozA/+xY7xyOwlStOY1OMLuJjrbZs/+sjxgEx1bKdvcDJRlbb543xg56U0WOt4gcq1VKljpUApUAqUAqVAKbBeBQpUrlepqlcKlAKlQClQCmxRBUyCmFQpULlFPwA17N8LBUZQ+fTTT3dQaRLTMniiG2677bYL76j0ffa9rlQKlAKlwMcpEKADCP30pz/tS7+CGeAJICQasJZ+/W0FPwlUujcDV+7PQK/lX4FK76fMMqHu0blPj8AKpBKpBwqxI4FRQFeyfw/ApsCp06ctI+q9g2DhjrYNVB5oAHRfg18rrd/d0003NlB5cFcDYZabTfTfcveBfXBQySewTd/5fPAhoC5lfFdXPfUXs3NjypjZCKQEzOxLzvv3yzjBtYBK42SbfougMn0u+hDf+RA/0n/6Gcv0nzr8kJMcj9+2Jdc4YPFSoPLo0aNdW/oaB43Z5RPffR5ca9fctQcrbedzoh/1Ryhpe8xs5rOgBClpCEpa/tUPEejpu21/hNIZi34+KRWo/CSF6nwpUAqUAqVAKVAKfJwCBSo/Tp06VwqUAqVAKVAKlAIXJoYKVNaHoRS4dhUwMe07KuIJqDSBK1mG7YEHHuiROyZKTc5movXaHU15VgqUAteCAoE7lnt96qmn+tLS7jUi1x588MHpoYce6hGBlnasdFGBjwOVAJR7MdgETn7xi1+c7rvvvg4t6QoCqwN6yYFhrkXaBVglslI9AMq1AZlAKDBKdhyoPHv2TMsg4/bp7LndzfZvg8rrr1/poHJlZUeDmPzw78XU27M5gkr+BPDxMRGGtuNzYBvgJvIyOfAw7SmXf5eUscdmQGWOO8eXQEqQzRjZTkSlzyON6JW+Fkv+ZwyLfmQMa5WjH/yOnYxh1MG1BBYBvIBFfbFrXK7XIqjU3nl2tQ+oZMOYZNtyoln55DPApohJ10m2Hw0TSZnPCQ1FcuofnAQpkx1zXht22V9P4lMt/boepapOKVAKlAKlQClQCqylQIHKtVSpY6VAKVAKlAKlQClwQYFMyhSovCBJbZQC15wCJqRN2HpHJVAJLJiQNHEoaufAAZPS2/vE8nonHa+5QZZDpUAp8KkqAJYAPN5f9+STT/b7ivvIHXfcMX31q1/tkE1UJThR6aIClwKVAXfuy6DaXXfdNX3hC1/oEZWW0s19mqURMgUWuRbAFViVSD377AJS7vkBlSCY7N+GU6cs2Wo5UNH0yw2ErUynO6hsS4nuFUk5R1QClfv3i97zHswZVMaX2c68pOj4b0hgHh9l+85nrD4vgFuyfeMIJBxt2ZbZYMuYALfRtm0Qzb93wJr3MavDJk2tHuDzSONFUKnvsf9ARr4mjT6MfsSHlM5pl++IUnI8mU+JfuSLvtVjg8+uz7j0KzBozHxQbwSV7CQb2zg+frCp7XpApWtJQ9nnJcCSlp71leDlCCujz8eVfCpQ+XEK1blSoBQoBUqBUqAU+DgFClR+nDp1rhQoBUqBUqAUKAUuTBoVqKwPQylw7SpgctJkI0D5wgsvTK+++mp/l5yJS5O3JjhNfI4TstfuaMqzUqAUuBYUCIgRvfazn/2s32OACHDya1/7Wo+oBIZAs0oXFVgLVNISqAKsAEnvpwQqRaUqvVeRjuCW5N4tj5FxjoNXgFBAJWjpmJQovUAoIMz2iRPH2zkQDEhsUHN113T6zP7mzwwqr79+T1/6Fag8cAAE29XO7ej/ZoBmsRtoCMSNQG+Ed45L/r0xloyZj4F1juffo9SPPW0D+ow/wDZ9KI0LSJOBvoDK/fv3TzfffHPXF8DVZ3xIeSlQmX8bR3/S5+gHfxzno8RuxmNMGYfz9vkhZ+yOOeff7IDKwGXH2GZDPf5r63qPkNL2CCr5wSbf2JBpkuvluG3HwWwlWGlb5ocsktKzfvIIK9mPNvpbKxWoXEuVOlYKlAKlQClQCpQC61WgQOV6lap6pUApUAqUAqXAFlXAxIQJigKVW/QDUMP+vVAgk5Rg5ZtvvtlhJbAg4iQTmiY+kz5pwjH1qiwFSoGtqwB4I4MY3mEH8oBqln199NFHe2Sl+4vjlS4qsAgqASAagU6WAgUl6Sjfc889k2jK/KBEPZq7p4NWAU4BZM4BWGAVSJmlRYEw93XgKRFzAWDHjoGVxxrIOjOtnmng8czOi6By354WBddA5U27p0PX7x5ApWs6R+rxwWdACdoFWuX5ML7m35V8bgIH+TtmY/TvkXqSdouZTVl/yQGHxghQZnzOs0cPIB3ETX8Bifbjj7pjjr98WfRJn+yP18G1iAbsjKCSDcl4nAukVGbc2kZT/2YbB30BxBFUqu+6BlKm9J2TjUkf0YlNNpLt8/VS2bj057y6/PDM4IcJfvSktO+48+p+XOJTRVR+nEJ1rhQoBUqBUqAUKAU+ToEClR+nTp0rBUqBUqAUKAVKgT7ZYhKkQGV9GEqBa18BE5QmPi0B+9Of/rR/b012mkjNZGwmk6/90ZSHpUAp8LtWwH0D0ACHgCBw7e67754OHz7clgw9+Lt275rs/6//+q+n7373u9MTTzwxvfjii/2eDCqBikCOKFT63XnnnX1p7kOHDvVzI8wyMM9eI2RyHRxzP1cXuALllCARaKW+fweyrOcMwbyzcn7n4OlTDZKd3tHyvgbO9ralXvk0RlSyBYBt689/p07NEMv1D6ziQ8CibQArgI3fPjMBeP79kY0fdAswzL9J+fdotDnadty4A9X0k/EZIyjrnP5oIFo1Uab5ty8+ZF/d/Hs4bueYUuKH/gPyjF/f9jNe7eXAyrTV3vGMm2/pPzb5Hkj5caBS2xFSBn7qk4984ZvsOvFRts/XUT99J/ORv7HBH1AdoHzjjTd69sMnS8EGVubaaLuY+FmgclGV2i8FSoFSoBQoBUqB9SpQoHK9SlW9UqAUKAVKgVJgiyqQiZoClVv0A1DD/r1SIBOWok1MMJrIzUTs79VAytlSoBS4ZhQIyACaRP6BQSAlYFLptxUIqPze9743HTlypEMeoIl2oie94xPsBSxFV9IT5KEv+BSIx7J7uhxQBzIFgGkDygVUBlwBVMCXDDDNQO+DBvVArPYew1PbG8TaO21roPIAUHlwT/MDZLKkrKVK52hKS8UuAq+AqsCu0T/HJP7JGQtYF2CnBOycl2LvUiWb6UNpn0+BlCOo9HkMuKVlwKAy2/oNTFQmx2f78Y1/0T7wT7kWqMyYYt94bNPAmF0rpX3J9WTLNTIG2b7+JG3ZNA7j0l62nbGoxw5/RkAZXx0ffVVfyhhzfezz1+eGrsBkQOVrr73WnyW8E9S5XAP1FxP/ClQuqlL7pUApUAqUAqVAKbBeBQpUrlepqlcKlAKlQClQCmxRBUxGmBgqULlFPwA17N8rBXxfM+Fo4tMkpUnISqVAKVAKXI4C7ivABnAigy4j0Lkc25+1tougEowSTXnDDTd0SCmSUvY+RWDHuWhKY7pG2zyDeQ7L/d09PRArkBIkckwCk4Ar/wYcOyZ/2HIDle1dlSeOt+jYk8sNijVQuW1Pi6jc06Dz7unGG1YaSAX6gMQ5Us9SseyMsCs+BCA6F1jmmMS/QDA++axkfLYdy/jGccX2Ypm+lPIIYvmnf/b0QUua6Efmh/7W0jW+Ru+xDLzTX6DiCAAd42fGqo+xn5wbr1V8cix2XSNjGHXmF1/4bEwyQKnUh/ZSdI9fbNAm2fnRz9iMJvHH8dhjQ1SlpZ5ff/31vjoDWGkpWO+s1Fds9kbD/wpUDmLUZilQCpQCpUApUApsWIEClRuWrBqUAqVAKVAKlAJbSwGTLSZUClRureteo/39VSDfWZPVtiuVAqVAKXAlFABIAnNsB5hcCdufJRuLoBKM2r9/fweTIilByttvv71HUzouKjIQir6jtov38FyDQKwArETaaa8NmHTypKVAgTBZ5B54eaoBy+UGK3dPy0vecSnSc2W64dCuFtm5o/kC6s3vpgQqT54EAj/6b4lnQv++6CNQDMByTOLDCMMC2wLGnMtnhy3+LmZ2cix1UgZU0lVOv/oBKgEz2wGV+hv7jG1ltI7PymT963NxnPp3zDnt2c7Y0ufYB3sBmTmvnfZs0U4J+maMaRO7gaDsjj6lbcr4xj+6qCvxkY3YG6+Jc8aqPjuWj/duSnDy5Zdf7rBSaUlYUZX6UH8xFahcVKT2S4FSoBQoBUqBUmAjChSo3IhaVbcUKAVKgVKgFNiCCmSipkDlFrz4NeRSoBQoBUqBUqAU2JACi6BSlJplX2+99dbp3nvv7VGVloD1bkpLlQI8AVgA1Zg8g8nAVgAasLQIx4CnwCx1tQGewK/Tp8EwS4sCl6cb3Fuajh1faZSuLR27e6VB1J3Toet3Nl+2T7tXGqjbBnRqO4NIdka/YhewMjZwS3ZcGqFYgGEAGR/Z4qOU8aXMsZSLx4E3YxojEUE5SR9AZeBt9OCP/hb77I3a/xxf1DZ19WdcAYkBgnxwnH+5FvqV+aH96Dv7AYXRQP98l9lTBiyqHx3jP3upGz9SLgJKvo22on8AZTSyr6/oyo4IYNGToiq97/pIW77Yu1aBS8vKu+Z8WUwFKhcVqf1SoBQoBUqBUqAU2IgCBSo3olbVLQVKgVKgFCgFtqACJiNMYBSo3IIXv4ZcCpQCpUApUAqUAhtSYBFUgkmgpCjK++67r4NKy8CClwFrQFKAlM48d3n+CgwLTFMnAAvwWsw5/1Fb3nM5v6/w5MnVDio/OLaz9bGzQdIGKPfumA5et735sm3atbOBu+UZiIGVwBg/wCw+BGgZE2AVYGifrwFyAWIBZPEzfgUEsi2vlXJ8LG3zKf0r7Uv6suxrYGG00Nda/cVuzgcOKmXJeOUAQkBRn7Jt59TVN1CX6NiM03n96MMxOqR0zDm6BSyqv+gPP9QLGKb56EMg52hjHJs++eea0Iaf0cg5yfjYD6gUVem5/9VXX51eeuml6fnnn+9LwXp/pf5jvzc+/78ClaMatV0KlAKlQClQCpQCG1WgQOVGFav6pUApUAqUAqXAFVTAH/pr/bGvi0xUrNXdYjt102at+pdzTF8mTgpUXo6K1bYUKAVKgVKgFCgFtoICi6AS/Lnxxhv7kq8PPvhgB5YHDx7sy8Emsi0AK89znruAJwDJtuRcIJdyzNqPGZQCpxwD0paWGnBbbUuNnj7bln5dast77pjOrG5vYHJ7g2vbpgP7tzXItzTt3AGqNTDX4dkM6fQdcBegBVbJlgIFLI2Rn3zSN2gVWJmxsRE7xrLWs2zGn3LxGdk+XQLVlPYl/ayl52grfS7aVUfm/6zXxXc7G5ccWAkSGru+HZMCKselZzPW9GWf/VGP3rj9L31kPz7wyTkwks4ffPBBj3hM/8aerB91JdsZj/4CKQNy6eS4OtEz8JNt/fzmN7/pcNKyrwGVloQVcZkxxV9lgcpRjdouBUqBUqAUKAVKgY0qUKByo4pV/VKgFCgFSoFS4Aop4I98kwPjpEJMmzjIhIbtMWUiIpMaqTtOaoz1L3dbfzJQ+cwzz/TJC1EAJt1uu+22vmzZ5fZR7UuBUqAUKAVKgVKgFPgsKABUPv7449P3vve9Ho0GMlnq1fspH3rooQ4qvZsSNAK4Ahw9x8mSZzzPiECYbN+zWEBayjwrBoAF1oFG7F8Ahju2IZ0NQJ5rkG15OvretunMae9WXGp1ltuzXCt3gX0i+mYwd/bsxUhHz5p84AtgBlAm2zdGKZAyfWd8GVev1P5nLFJK9pPVtT2m7KsfP8ZIQnVpoP8RBI7t0laZfhf70Pda/as3Aj2wzrjBPbb0Df4BlaIqjZ8vo63YVub6xT/2R5/SzjGa64fell4NHDZ+PqmTzI7EbvpxDcbrEpirDi0B13E8AdCg5BtvvNGjKi39+vrrr08iKgtUzhrX/0uBUqAUKAVKgVLgyipQoPLK6lnWSoFSoBQoBUqBdSlgQsEEg8mATDSYLJAyuWBSwWSHCQbHMgmhvgkLpckL5zIRlQmhdTmxzkrpt0DlOgWraqVAKVAKlAKlQCmwZRVYBJWe1fyw6/Dhw9PDDz/cQSWg5TnPM9xa2bNXgJz2AVJj3cCsACmAznOgZ0LPj/rYu3df2wYsd3Y4dvbsUouoXJ7ePbrcANXStL3xy8bXWp1zLbry3AAqAykvAsOAuoCsEVTykR8gWPrOs2n8zPOkcWVbKRmXeskZZz5E9pO0iSbKPD+PsDZ9pp026VeZNqPN0Yf0n/bKjD+gNnDPceddz1lzuu/t1yHQ1PmMbSwdT44v4z6/Pe+nz6NHj14AlQBjxpS2KfURPfjlurgeuSbOsc0G2xmLvy9su7ZA5ZtvvtlBpahK76gsUBmFqywFSoFSoBQoBUqBK61AgcorrWjZKwVKgVKgFCgFPkGBTJaYBLCskvfAmCgw0SGZXDDR5Nf2N998c5/sMGmRyQoTCCYqLMtkcsEkyIEDB3ret29fn4xQ/0ql+FtLv14pRctOKVAKlAKlQClQCnxWFVgElZ7v7rjjjv5+yi9+8YsdVIq4A4sCzQKc8vyWMufZSB26eTaT1JNHKMW250HZ8yFotmfPSnu23N7aLc+g8t3l6eSp9sy5dK4BtrPT7pW5nCMqL0Ycsq0rfQNmnlcDKgO0wC3+8UHf+kvEKEgWKJbnyYwpZcahXiBexpVz9pPSTp9AZbTQNmDQdmzMY5jHFB2VaTf2MfYfGzmvjbEav2d3z/G2+SB5djd+oDbjBwjjU8aXMv6ln4wxx9nkY36gSO+xX9diBLXqS+zpQ798cg0ASr7IzrGrLf/9LZFyHF9ApUhK76p86623OqhUd9Ru7rWWfo0OVZYCpUApUAqUAqXA5hQoULk53apVKVAKlAKlQCmwaQVMdJh0ACktpWQiwL6UCQWTCocOHerLhFlm1aSFCQpt1AcqTSyw5Zz63nd0++239zITQ5n0WK+zJh7GrF0mhPT93HPPdT/0VUu/rlfVqlcKlAKlQClQCpQCW0WBRVDpOQqo9H7KRx55pG8DRp7RwCLPgCNwcjw5mgWsLT6jOa8uOJVnSKAMpATLgMp9+4DKeZnZhrEalFqe3jm61KCbtmenHW2515Vdq639arPjuXKOeEzfloDVPx8TgQdWJQeY8QEQC6hT2gfM+Cjxnx7sJTuWMQTisSWlXfYz/rRVOiapEygYe0p5bJf+c2zsY+w/fbKtrvF79gYo/VgwoDLP8Poexw9aus6uS/xic+xj3I6vKcd+FwEx7aM7DZLYjw75PPibQLavPynX0t8WxgRQsmcsbAOi/t4QRZnlXwtURuUqS4FSoBQoBUqBUuBqKFCg8mqoWjZLgVKgFCgFSoFLKGCiw0SAyY1XXnllMpllWSUTByY3QEkTOyYTbrrppun++++frr/++j5Bot5PfvKT/o4YEwomIkxEmWwQ7WhC6tFHH+1wUxsTJOpkAuYSLl04zLdM3mQCKMccN2Fx5MiRPqHBPv/qHZUX5KuNUqAUKAVKgVKgFCgF+rPd48M7Kj1D3XnnnR1UfvnLX+6gEjiSPBMme55L8lzo+c1znOR5LGkEWY6lHhjGruc/z4SBlSIcd+8GzLa35zwRlUvTO+8GVLblXrettnanW+l1AqdbX/NyqvyWRlDJRzDLc2hyQCsf+Q3O6XuO5NxzAdZlTOp5zowt/ThmrLJ6eX7NWJWSeurnOVXpmKRNgGDaOZaUftJeu7Ht2N522qa+8Xp+D6gM5OODpG9jDyj2XO96OM5WxqWUHQ/ETH/K+K7kH/s09jkJHB5BJf8k9WM3tgMo7cderh9AKbPrWkRTx4DYgErRlJZ+9XdILf3apa7/lQKlQClQCpQCpcBVUKBA5VUQtUyWAqVAKVAKlAKXUsCEgwmA9957b3rhhRemJ598cvr1r3/dJ3PAP0u9mlwy0WD/7rvv7hMeJkOAzb/6q7/qv2523gTQDTfc0CeKAETHPv/5z/elxRJZOf56+lI+5biJDhMVJmL4aCLDpIXjMp9fe+21Xkc0JV9vvfXWPhkVG1WWAqVAKVAKlAKlQCmwlRVIROUTTzwxvfTSS12Ke+65Z3rooYemP/iDP+irXwBIngkD+5SewRbBWcBV9Bz3R6DlOBgFlI2gcoaUF99R2Zd+Pb40vd1BpUjGBheXz7SoylMNprW1YCfvPczz3wwAPQPmeTAwK5AR5ApEc0xdz6N8kC/2P0M75wLM2MqzpnE7bhyyehlrxkmDPJPGD+XYVrv0EXvRTr20z7ZzqZe26ds59fNsDA4CeHIgJf/Z4qtn7oDKRJPmOXwcS8aX+krXLv1nvEq2ZT7Ej/E53THnJXbZSWY3/TtvLK6XZ3z+j7DT+fRlTGAsUCma0t8f/s6w7Zi2qatdEjDrbxdRw4899tj0jW98o//g0t8quSapW2UpUAqUAqVAKVAKlAKLChSoXFSk9kuBUqAUKAVKgauogD/sTQy8++67/Y/+v/mbv+kAMNDPL+4t06VeIixNGHg/TCYKTDCIvDQZ4I9/+6CniRO/YAcPLS+mTHTmeoZksoMtkxOWluWnY5kEMjFh+VcTIbfcckvPBSrXo2zVKQVKgVKgFCgFSoGtosAIKj27AUef+9zn+o/JvKPSs9OlQCWY5BkwYCtltAvEsp9tdWQ2PTt69vOjNyVYuLKyq50T1eedmEvt+e48qDwBDp5tx89M25dPtO2T09lzLXdQeXH5V/7EL9vZ94zoGTXwyzOkrC4wNUK7gDvH6MF3yTNmbC+OJ3DL2JLUlfN8OrZVb4R99tNP/I7v2WdXPT6NbR3nm7EBg56BPR/nHZHGyQd2tAMEoz/NM97YZU9SX0qfrhdNogtb8Xv03Tj5I0dz/S+OP/1lLEp9ZizGkbEYA1vO6zOZXX8DBFSKpvSqCj9W9HcAHdjLWPqA2v8KVEaJKkuBUqAUKAVKgVJgMwoUqNyMatWmFCgFSoFSoBTYpAL+qF8ElQAjUCkKUgQlCGmywKSHX6KLZHz66ac7rLTtuEkuUY1gpQmUI+2Xzt4jAzBq/6UvfanbAj1NfqwnZWLCJAyQaiLCBEYmI0xoWPLJJIj+ZcASHK1UCpQCpUApUAqUAqVAKTBdWPr1z//8z3sUGoDzhS98oYNKwNIzH6gleYYDw+TxmSvQLqW64/a475nRs5nnvUAyz2a2HbPk6/btIhmXG9iapmMNVFr69QRQObX3UrblXpeXjzeTIONHQeUIy9KnUgo0MwbPtp4bPdMaiwSS6T/wdIamH10OdRyT7cCysUwdz9DyJ4HKxbZ8SbvYUCZFP6W+AgVdD8++xha4Z3z2jZENY/RcnnEaa66BY64Lu+mXZuxLzqmb7DPBXvzgS1Laxzd28nzuXDTSfrShved7/vKb/66T7Lqxoa0x8Ed2LM/8figJVD7//PMdVFoFRttA0vinLFA5qlHbpUApUAqUAqVAKbBRBQpUblSxql8KlAKlQClQClyGAiYT/PGfiMq//Mu/7L9OBhS98/GOO+7oUZL+2PdreCDSL5rV824YEwiOWUJMfRNRJhosx+TXzoCmiY6vfe1rF95vaQJkPcmkA1smYwBRExqOZSLEce/CNKFhkg2kBCsLVK5H3apTCpQCpUApUAqUAltBARGV3/nOd6Yf/OAH/RnPc5J3iIOUhw8f7s95IJbnKTBMBpLyzBWQRasAqJSjfo6BWiMsSxSlUh8A1LZt6oBeDUu21ykeO9ZA5VGgsgGzcw1UTe2950vHp/ZTug4qV1dPd5iG5QWaeX5NX/qV+Mn3PDuCYH7sludHbeKb59rAPM+p/Bqz59sANv0kZ4y9w/Y/NkdYZ9ux1IuPab/YTl05SbuMxzHPvK6D62FcAZXGJHsWdk6/+jCGjMvz9ggqjdO42NdnbCu1d04b18oPE7WNBqNfo3/xP+POPt/Thl+Sc+oZB98DkpX2XTvn9cnXXBfttPHjRH9fWL4YqBQdDFS6zmmrbhL//Y1SS79GkSpLgVKgFCgFSoFSYCMKFKjciFpVtxQoBUqBUqAUuEwF8se/iRxg8cc//nGPhDRZYSIrUZK2LesKXIqS/OEPf9gh4cGDB6fbbrttuvfeezssNMFhwuStt96annnmmekv/uIv+kTFt771rT4hxoYJkPUkkxWZcDKBwW4mU/jNZ8s+OVbvqFyPolWnFCgFSoFSoBQoBbaaAgGVP/rRj/oznGc3K11Ylt8S/2AOuAUoefbyXOX5CyCTHUsKfErpeMCVkg3PkAFmgWWgkWMz+ALjADogbo6ofLdFVB4/DvKdbkTrZIOV3jvYQOW5FlW52n6kdt4HUZgBX+mLTYmf/B1BmGfFwDznpLSLn0oZGBv9DShbhHUZbzfW/hfNxmfUaKGvxfbaaeNZNtkx21Ls8dd1CKQ0Ltl+sn396k8/rqM8jiOw0niilb5iX6lP57TzzO/HidrRxRgy5oxLKcf/+D7u204d4+KnvsBWcDKg0jO+cTgn6dMY+CuzY7x+VBlQ+dxzz10AlX7MSCf24wc7BSqpUKkUKAVKgVKgFCgFNqtAgcrNKlftSoFSoBQoBUqBTSjgD3p/3JvE8QtlkZJ+oWwSQfJHPrBo0sJSsA8//HCfLDDZ5ZfNibq87777+rYJDpMJfuH8y1/+sv96n50/+qM/mh566KEOFNcLKvkmm3iQM6nDnuP694tqExwm3cBK0JSvlUqBUqAUKAVKgVKgFCgF5qVfRVT68Zjos0OHDk1/8Ad/MD3wwAN9JQrPUCOU8rwFGgUs2U8KtAqoWiwDAUFJkCnAKZASaGx8q2WgzjKgXkGw1CBUQ5MNVK6uWsK0vZ+yg8pj7dlvBpX9GbA5EVCpX2BtBGmeDfntORT4ChBTyoFZGY/67EhK/gbUeZYUVRj/jUteK7G3+KwaXfiYtjkWG/ofMzv852d8VgZK5proS7s8G7Onj8Dh6B7fXVvZefX4EX/ZZlffAZXG7bUNntdHffWj7Tgex8Yx2GYrOfvKXBvP7f7uSA6k1IZtfhhDsj74GVBp6deASj+MBCoDbPWTZMwVURk1qiwFSoFSoBQoBUqBjSpQoHKjilX9UqAUKAVKgVLgMhTwB73JChMAiYL0bkmTB5k4yAQGEOidRiYSfvKTn/TJLsut+jX+vS2iErQ0wWOCxZKsQOX3v//9PqkRULmRiMqPGxa/f/Ob3/Q+TLpZqrZA5ccpVudKgVKgFCgFSoFSYCsqkIjKJ598skexeRb78pe/3Jfk9xwXUAkMgUQjyAoU89wV0BbYlf0RXgU0gWIy0CXPdmdI6RrMoBK8mhqonNrSr8vt2bNF3J22jGk7cO6DBsDWApUi+WZoyKYcf9gNgPMs6tnWs6ycfc+0Y3bcvnZsgXsgJVDn2RK082wbYKePJJrInovTb/RSh19pFx+jmfNpr9SOLyCe7NlWmWdxfUjROnayz3d9uYaj9tk2rlwHdtgzbn3q2/44fp8JoFJ7x/Ujjf3Z5ruUsSxqkf30Zzzgq7Epc22cl+gU3eI7266lFV1EVOYdlX5Y6W8Xx6NT/GGrQCUVKpUCpUApUAqUAqXAZhUoULlZ5apdKVAKlAKlQCmwSQX8UW+CwISBiRGlyQuTCH697F2Uoi2lu+++u5f21fFuSKDy8OHDHVSa1DDpYeLg2WefnUyKmcj45je/eWHpVxM+l5MyGQKGWl6WzyaUClRejqrVthQoBUqBUqAUKAU+iwosgkoRlUDlGFHp2SyQKwAqz4dKxyTlmAPglIDWmAOdUme2y4Y8R1SuNj4lotI7Ko996IdzbSnTM8ca+Xq/PZt+2LKIygbTzs5QcO77o8upftT+RXgYODZCxMA5YEv2rJusHlu0AOpkEXnAJehlbEl5FtVHICPbgX78VD9wMD6mdF5iRxsgzo8EQTcRgpas9TzunBQICTgG4LHPnhzdx/7Sl3MBjral9Buf7asfUGvsIK3PBJvxd9Z/HluOxR4bsafMdjSxT/MAZGXOdafa/2JfmTHRWDva+DElQGlFlVdfffUCqMzfLnyQpQKVXYb6XylQCpQCpUApUApsUoEClZsUrpqVAqVAKVAKlAJXWgETJpZwPXLkyPTzn/+8/wofmDSRARKaCDCRceutt0733HNPh5beaWMywS+eX3jhhd7O5Mjf+Tt/Z7I8rAmfKwEqTVrwAQwtUHmlr3zZKwVKgVKgFCgFSoHPigKLoFJE5R/+4R9+BFSCOoFZIFGAT8pAqYAk2gSEKQOVQK3AMsdSP+1nhgSKAUoi+xqgPNGWfm0RlR8ClSePN1AJUDZQufpBA1ltH6hsRHMGUBeXHh37t5399JnrN7e7+O5HP6jzrOo5N+9JVAYM8j9RlaCuyEr7oF1ssxlICcCBbjLbjqtn/AF9tuNjtvmnrnaBlFYLASr5wy47rotnZz8GlF2rteyO42c71852+lRHGv3ng311gEqA0vO9MScSM+PWNmNTjil6gI98l20HRtqnT3KOjzZsx7cc1871yo8nAcoXX3yxv6fe3ykApmsX7bWXClRGwSpLgVKgFCgFSoFSYDMKFKjcjGrVphQoBUqBUqAUuAoKmDQxYWKJpV/84hd90sTklokMcNAEg8kckzgiLS0fBkSaLDCB4BfPgKVJlUcffbTXEflo0uNyUiYwClRejorVthQoBUqBUqAUKAW2ggIjqBQ9aAUKoPLBBx/sPzbzbAZ8gVh5xgpECvSJToFTAVeBVgGU7ABryQFkaTczJBGaM0w6e3Z5Oj6CylPH2/KvHzSA914Dlh802NX2z5xs2fKkF5efZW8Ef/qR40f6TZ34bzwBX55zPc/KwKBMH8CNnURW0kcG8IyLzRHKAY2efcE0mXYSGyNUtD/6GF/44b3rb7/9dv8RXp6x9eOZ2XO0HwICh3yIzfiiXtJ4/fiR6xj9U09pDHKuMd/0px+gUp/29TOmXPOxX+fZkdNvyvRjn/bRLv2mzPEAzbSnr+sCVFqx5bXXXut/m7z++utdL2DXeaBSm9grUDletdouBUqBUqAUKAVKgY0qUKByo4pV/VKgFCgFSoFS4DIUyISCiZVMsmQSwX4mBUBHdW+//fY+SZJff5scMFEBYAKWMlvgpnMmMxwzGQZkZpLnMlzufpjMKFB5OSpW21KgFCgFSoFSoBTYCgo89dRT0+OPP96X4/ds553iX/3qVy9EVAJSIJrkGTA50CfgJ1oFeillz4Hag5TAVspALucCtdqjZEuLoHJ5OvreHFF5GqhsgHL1zHutfL/Bp2MtA5UAYgedRwAAQABJREFU12+DyhH8BY6mzDl9x1e9s+VZFdwCJwMqUwY28ttzq4jKgDvw0vE8P9MoII22MvuSemBZNLHPNyXfPMvqy/OyyEDPtWAlG4GGro3MByV/+DDayfXgE5u5folcnLWbgaTzuZ7KbLMRfwMqlfx3PEl9dfm3qKs6sakf2igvldXNOXXl+ExT2T6NEnFKJ6+kACuVflCZZXLVN1Z22Ka7H1A+8sgj02OPPTZ94xvf6O9lzY8uo1vGVmUpUAqUAqVAKVAKlAKjAgUqRzVquxQoBUqBUqAUuIoKZDLBJI0//P1KWekP/nFiwESO7Bf43mkEOJpYUFekpXb2TUz5xTm7fhnuF+DefyTa8rbbbuvnTKyY3LiclImNApWXo2K1LQVKgVKgFCgFSoGtoMAIKkEfz3EBlZb0B6RAJ89ygUOeAwOaPHdJi6VjYI+2YFYAJbgFpinlQDX15jSDSlZ7RGV7R+XR97ZNx481UNWiJ1dbROWZBipPn36v+fNByyfb9pnef/pjS45tfY85xz1zapOs/wAxQNAzMAimzHaee+nBjrF4vg0sBMDYowcwRjNt2KOvY5K+jT+gkq34pT2N9QlOBrqBlvzTp+uSaE6l/Sz9yg77cnTN87H+2eYXf3JNHTemZD5qwxd26Me+foBZUNQx55LG+rkG0TaaqKsPdRdz+ubL6Gf8VWY7/tPVNaKNvy/ASX97KO0DzGNd2zTke4HKXLkqS4FSoBQoBUqBUmCjChSo3KhiVb8UKAVKgVKgFNikAiYPTBgAk5ZotYSSLIoykxomC9QzAXXHHXf0d02KqpR+9atfTZYTS/SkeiY1TFyYGPGLfUuLHT58uE+0mKxxzkTG5aT4XaDyclSstqVAKVAKlAKlQCmwFRQIqPzxj3/cn8+ASu8O92Myz2rglAQcef6TA5ICnJz3/JVsP8mznedEQM6zHsg2ZsAoYG1uM4NK26vtHZUnTrSIyqMNVJ5oUXhn9N3ez3jas+h7zV8QSpTivJxqwJxSDvzTR3JgYOrmuTOlMRmfZ1UALDmAEjwM/OIje+CdiEbAkF7Gyx4gRq9ASjYdo5P+aZIcHejFB/15v6LnWaASdHOM3fQFGAaS8oGu7K01xjwf6z/XMn7lujqXnGs5Xr9xnPrSj/OS+hL/HFvM0Ved1E2ZY/b1H38CePnpWEBlYCU9nVPPNQEr/Z0SSEkz10t9ddVR3z7fC1RSvlIpUAqUAqVAKVAKbEaBApWbUa3alAKlQClQCpQCm1DAZIHsD3x/8IuQNFliEsAf+CY5JBMtJp5MbFnC1ZJJksmB5557rgNLbdQ3KaC+pN5DDz3U25lUyaROP3kZ/+OzCZ4ClZchYjUtBUqBUqAUKAVKgS2hwAgqwaAxotJqGYCU56rAoTwDAkqOe+6SxjLbgVae/zzrjYAy2wF0eT5slhrsmm2ebaDyeAOV74moPOH5rkVytuVeT51+Zzp18mgHlZZ+5cfS0sUISrY8V+p3MTvHrxGc8T/7fA+oDAQbS+AS8ALHaKGdsfkxHnAYYKh/frGlPW0DKvXHD+2S6aCN/tX1Q0HP355nRQjap7nxiN4EKPVnW59Z9pU9ddjSx+J441OuJ59k++O1zfUNqHS9ANJk+2w7z+dcc2MLpMx5+2tpru6Y4pvx09ffIAHF8ZGefIv/6ub6BFb6GySZDfXZ8/eIfbboU6ByVL+2S4FSoBQoBUqBUmAjChSo3IhaVbcUKAVKgVKgFLgMBTLpkMkSEyT+wPeHvkkC5006+EPfpIjJEpNbJkokEwEmVvyy2bY244SFiRXRl8pMYFyGuxea8stER4HKC5LURilQCpQCpUApUAqUAmsqMIJKAMdyr5Z+vf/++/uy/n6MBvR4jvNMGFDkWSvPigzbTsq25zvPicBZoinH0nYAnbpzWhtUNtbX+hDx10DlqbcbqJyXfj3Toix17RkzkM62rO/kHPPsGr8zBv0GpDkXEAaALWbPwYAYgOYcG2wDd55pgTzPwsYlsUU3GVyzL2kTSBm/+eq8PhJN6XkWsNQXH2mWfpQjpORDbNE8Y6ZtnrWNj8+uY+BkfHMsx/kh51lfpGj6NT79sC/leiuj41p9q5vzthdT/PI5jM7+hojW42ePb6P/9FHP3ysgpSVzaedaaceev0nsq0vrApWLV6D2S4FSoBQoBUqBUmC9ChSoXK9SVa8UKAVKgVKgFLgCCoyTGePEhePjZITJDxMyJi384S+pP/76OfWdM0mRSSul/SuV4vOnASr19btNs25XUL7f7XCq91KgFCgFSoFSoBT4VBUIqHzyySf7c1uW5gcqrX4BjAUgBbR5/kmOs4vPcvbBKs95ng9HQGnfc6PsfEDa/Fx1EVSurs5Lv4qoPHHS8yOIeGw6feqdBqms8HGs7c9Rjdu3z/2wp9/YTMkf2VgC4ZT2JefUlTJez7EysAV0pUyUn2OedyVjAfACDo2RTWOim3py4B8f8+ysbvx2Hpzz40DPsqAbaMmGOmBk+gAP7Sezkxx7nsuNK5rEp+gw+pVjo7/GxoY+Akb1y370mq+bmnOK5mOf+k1OvcWSHf4Ap7QNpLTtOjjHx+ToqXRePaASpLQSjFdX0I49tmwrC1QuKl/7pUApUAqUAqVAKbBRBQpUblSxql8KlAKlQClQCmwxBUxymMC42qBSP6tn24TKapvwauU8SdMmYa6S3kGibZ5nnkxrG9uWTaq13Cd/rlLHZbYUKAVKgVKgFCgFPrMKeJ/4d77znQmoBHrAyS9/+cvTfffdNx06dKgDqjxbBezNzzyzJCN8GrdBKjALjAMpk4E0x8AvOTCLNXbn55z5uQqoPH5iqS/9evIk0OX4ifauyqMNWomUO9b2gcI5cjN2Rz/Ytc+2bAwBccq5zxlS8iV11QuopAu4BVBmWymz4bnTWAJjE1EJ2LHvPHuAmn3JuegTUMmGeuwCbiIAs1ypdupFR2Wg5FgGfrI9amxsi1oH+Cnj1+grXyTt9Jdo0fhLKyltbTsWUKm0v5jVWyuxo0+aBjzSwjbY6Fx8jZ+OyYGb9BKBClK+/PLL/e8BNkRS0tQ1VJc+FVG51lWoY6VAKVAKlAKlQCmwHgUKVK5HpapTCpQCpUApUApsYQVMcpi8uNqgEqA8caq9d+hUm1BqJVjZp2uuFqk8f01N9mw3ubW9TRrtaL/G39G2t1nS6yp3vIU/UzX0UqAUKAVKgVLgs6pAQOVf/MVf9GgzcPLRRx+dDh8+3KGl6DnPHlKAlHIRPo1QaoRwI1izHYA2wqzRduul2Q5UnBq0W56OtojKk6f8MKtBvqVT09K5D9qz3rGWLb06RzR6RyUwx26eBRdLQAssBMLkgENtwDi+2ZYCvxJFCXaNoNJxAC2gUjvtAwrjSzfW/ufZNJAt+qUN8BfIqj6Qxr7oP/3alwIIUzcgUqnv2HE+OqfOCCpzPbvR4X8ZO/+S1XWcbZ8F15B99hbt5DOhvpz9sRy6+8hm+guAzDUydtsBkupFS9cvx9WjFVApmvKVV16Znnvuuen111/vkZRZrtc108YYClR+5BLUTilQCpQCpUApUApsQIEClRsQq6qWAqVAKVAKlAJbUYFMYFxtUAlOvvP+ient909Ov2kZtBThuDhpcyWvgd/gb2997GyQcv/u9kvwfbum/Xt2Tnt2tQmq7Xm305XssWyVAqVAKVAKlAKlwGdZgYDKH/3oRz3izPKejzzyyHTPPff0d1SKDgyUWgROI5DKtlJ9IAjcyrKkAVwBeAFZtPXsdjEBlSL8vEYAqFya3gUqW0Tl8vK5lk9P27cdbz8OO9HqgE4gFgg428hzIBgVoJVtYGsElY5L/AX25IzVuQBDACw5gBLwkkfYGWCYMS7qFd9GYKk/Osn6p4vz4NwI6fjJrvrRmv1sp282aG9/zNqNbdmTRh/H87HrWGwvXkNtk0Y72o77i/2kDT3GPF4v+ue65Xjqau+Y87meroVrIwoVqHzppZemp59+ukdVirAEMNWhKzv0LlCZK1FlKVAKlAKlQClQCmxUgQKVG1Ws6pcCpUApUAqUAltMAZMPJi+uBqhkWzL58u6HJ6cjb77f84u/en9675iJM7/2vzhpc6Wl1/vOBiT3reyYbr5uZbrnln3Tbdfvma7fvzLtbrBy9O9K9132SoFSoBQoBUqBUuCzp0BA5Q9/+MP+PkTvP3z44Yc7qPS+Sst9BuAFXo0QyrERcAVsgWWBlMqAuNRlY+10MaISqGwrdU7vHl2+8I7KHdtXmy3PXIATUAhQnepQ79SpOUpyEXJ5PsqxgC3Pio7zIyAu4+SX+qAW+AVSBnKBh7aT1ZPYMEZlQJ3jo1b2A9jYlp0HALWV6SPxTV1Zir+Oy0mxn2szjsU2e8mpkzZr2Uh7vgQ226aNc/Joh43YW6tcPJ/9jCNjpKNrI9vOeFOf7ehqW/u0cS1cJxGv3kMJVB45cmT627/92+nFF1+c3nrrrQ4qXTtttKV5gUrqVioFSoFSoBQoBUqBzShQoHIzqlWbUqAUKAVKgVJgCylg8sHkxtUFldP0/947Mf3ylXenp1892vM7LapSVKOoyquRMie1qy31eqBFU95+w+7poTuum+6+ef90c4OVe1eAyrlnEziVSoFSoBQoBUqBUqAU+CQFAiq///3v92cny3t+/vOfnw4fPjzdcsst03XXXXcBUo2QcYRGAVdKdcAscIut5MAu5wO0Rt88v0keYSz9KkIyoPKdo1MDUY6fbb6cm/bsXm3PXJZ8BShnQHXsmKVZ52VSA7oCu/JsaD8wzPbc3wwqA+gyLjaAxEAw22lvO8CSPUl78CtAb3wWG8fLF21iQ9too236T5uU+tZnQN5FvebVPNJOyZfAyXE7dWJT30mOpS1/wGXXLrA5EDc2UsbWWuVoO+dzzHjkwMZoopSdk/STsYwle+rkGoGUsohKfwMAlT/72c+mF154YfrVr351AVTGboHKXIkqS4FSoBQoBUqBUmAzChSo3Ixq1aYUKAVKgVKgFNhCCmQy6qqCyvb+JKDyF0DlK0enXzZY+c4Hp9qk2dJVB5XeSXlg9/bp9kO7p881UHnPLfunWxqo3Ld7Z4HKLfQ5r6GWAqVAKVAKlAJXQoGnnnpq+s53vjP93//7f3vkGTAFVN57773TbbfdNh08ePDCUqJA0SKg4gNolFKdQDvLxiYDQ1mSdBFaaev5TZ5B5byPAX547Nz09jtAZTs3ebdgA5V7zrV3dIuMAyqPTx988GHLH/RSZB0YFSDVHTtv3zFgLCBz7m8GdPHb+PIsGZio1EbKOTANxMw57QA+45RpEF20SUr7ADrHo6tzqRtAl1J9kZ1gpT7VSztlNFVqkzLjUi7WUyf9xT91+O9zILrW9QMsXTt2R9vj/tg/W/bHlPP6y3UInDSe5IDK+KUPWspgaUCw45L6iXr1GRBRuQgq33jjjQKV48Wo7VKgFCgFSoFSoBS4bAUKVF62hGWgFCgFSoFSoBT4bCuQCZCrBir7ZNPZ6ddHTzRIKaLyvemZ1977VEHl/pVt020dVB6Y7mkRlbcd2juDyjaFJy1ODn22r3iNrhQoBUqBUqAUKAU2q0BA5Z//+Z9Pb775ZodtDz300HTfffdNd95553To0KELEX+LoCt95rlDqQ6YBHYFUirtJypPu7FN7HiGw7fCuIDKYw1U/qaByuPHz3ZQuWPH2Wnf3qnZAg5bZOPJ49P7DVC9/z5Q+UGHVqBeQFf6se/4mMc66gXCpS4IlvqOBcwpwTagEjiUnQ+gBfZAS1qom37GUnvZMTnwThlfopd9vnz44Yd9fLYl/elDGd8z3thwPPXiT+p0I+1/8Ss2+W4MI6h0LH0ox5y+Fu2yNx7LOPkvRzuQMsdGvbWP/8YAltIkujivvusgmpI+lwKVjuuHvpLPYy392qWo/5UCpUApUAqUAqXAJhQoULkJ0apJKVAKlAKlQCmwlRTIJMiVBJXjBM45k0pnVxuoPD49/VqLpnzt/enZ19+f3u0RlVd/6dedLWpz38rydOv1K9ODtx+YDgOVNwCVu87P7M1LgG2la15jLQVKgVKgFCgFSoHNKTCCytdff73DoAcffHC6//77p7vvvnu68cYbe3QdSAR0AU+BT9lOGWgGJAVUAl62x/Z5rkq7j5bglEg/72VsEZUfTi2i8lwHlQ01NkjVQGWLqFQuNVB5+vSJ6YNWKVGVouvAqwC/qGI/MAywSp34klK9ZHUc5x9gZnwBZY6zs7g0rPMAn4hEcA9g015KH7bHPvTDN/akUUf9aqefEcSyybb+AiLVja8pA/rYtC0njf6kPlv85r9rN0ZUpk7sKHNMmRz7Y6kv4zRGGaQ0JtvGHj1Spi2box7RP32zydb4jsoxovL555/vS78WqIyiVZYCpUApUAqUAqXAlVCgQOWVULFslAKlQClQCpQCn2EFTISY5LgaoJJtoPLs6pkLoPKZBimfe6P9gvvD09OObSayPrrU1ZWUunU/AZV7dzVQeXBXA5X7e0Tl7Q1U7t8DVG7rsNKkTqVSoBQoBUqBUqAUKAU+SYG8o/KJJ56YXn311Q69QEr53nvvnW6++eYOq8CrwK4AqRFYLcKkwC4luBSwlOc0JTuOg2PK2T4w5TnGOwiX2tKv0/TO2+emYycatTznHZVnG0A725d+XV5qcO/Mqf5uStF0Y8ShZ8H+3ObZrWX7gWRKgGusYzvHlLJ2UnwzDsDVmBxTB2yzJCtQxq4xgXxyokjVdTz22IxPbIz+qEePZHWBvIBKJV/ZVCeg0nauj3Ny9lOOfuhf1p+sjsye8fEdqMxYjFu92Ei77Kd0XMr59BMQG0BpHLYDKdVLynZssB3/ootjUq5rrsNiRCVQKVL4vffe6zqrLxlfRVR2Kep/pUApUAqUAqVAKbAJBQpUbkK0alIKlAKlQClQCmwlBUxumIS40qAyEy1nWzRlB5XvHuvRlM+88f70/K+AyjOfCqgEQ/fuWuqg8oHbGqi8Zd90+w37pgMNVC4tA5XzZNhWuuY11lKgFCgFSoFSoBTYnAIjqHz55Zc7YDp8+HBf+hWsvPXWW6frrruuA6vAMAAp8CuwDOAac46rp77nqABAcMp+ABQIlrbbt4Nl2lhe9TyoPL/067TUQOX2Bip3nweVy5ZmPT2daNDr2LHjfflPwHAEYJ4J0y8g6JzS8TzbKe3zK3WVjksZbyAlyGV8ElsBlUrtMvZFSMuOlH71mex49NA+ujnP50QM6m+xrvrJI9DLNrsBe9qmT35kbNob3zjGQErjcJ6NjKE70f5nX04fi9v6oKsxRKdco1yH0Ua2xzL9jmX6H+2zG1D54osvTj/72c8mJVD5/vvvX4Ci2haojIJVlgKlQClQCpQCpcBmFChQuRnVqk0pUAqUAqVAKbCFFDBhYQLmaoHK1RZNefbM6R5R+cv2bspnWzTl8282UHnsPKg8Pwl1NSQ3X9bm7qa9O5emW67bNT3QIyobqLyxgcq9Kx1UmtgzuVOpFCgFSoFSoBQoBUqBT1JgBJUvvfRSf4a66667psMNVgKVd9xxR39PpeVMwUTACgALGAvcuggaPxrZl+eywColQOU48KQdaCQDYjt3iloExbY1X5amY8eX+tKvJ06Aam3J1wYqd+9ebRGVZ5sfoiJnCDaDvDmycYzW01/61K9s37Pi+LxkP5Ay5wMrR1/5COAZN/+d019AIiDnGNuBewFsY3/q6FNynqbR0LakDn/ZHMfETnJgZK5JrpFyPJe+9TlmffMzgDLjcz2Mc74mO3ud0Qbf5CR2Mo5x3PoKaBXx6l2SgKVx0Vdaq13sKTNWdaNb+mafnfQBVP7617+ejhw5Mv385z/vpb8JLJub68pOgUoqVCoFSoFSoBQoBUqBzSpQoHKzylW7UqAUKAVKgVJgiyiQCYyrASr7xE6bDFlty4x5R6X3Uz73qw8aqDzeQOX5pV/b5NHVSuaDgMo9OxqoPLhz6hGVN18ElcvL7T1I5yd0rpYPZbcUKAVKgVKgFCgFPjsKAJWPP/749L3vfW8CKsGcW265pb+f8oEHHphAy5tuuqlHVQZaBaiNcCtgDFiS8jzGHog05gBAII2NvAcRGFtZ2dXgmKVitzcbM6h8p72j8sRJQMu7Kc9Nu1dWp107gUpRkcCnpVHniD0QTA7YG/sNpNQ/+BUYNgI45zzv8du2NrLxBOhZEpUWxsxG6oGVsr5HKEaPEbZFH2U0YA8804e6salvY4i9ADplttUfQSVN+cZWYOU4xtEW/9XVJhGUY5lxssNGwCD/ZPsSO/qT03e0oQdAKaoRrAyo5P/YbvQ343FeltSPLuk7JY3Yfffdd3sEpc/y008/Pb3yyivT22+/3ftNXbYKVFKhUilQCpQCpUApUApsVoEClZtVrtqVAqVAKVAKlAJbRAGTGCZNrhaoFFG52iaNfn302OT9lCIqXwAqj3/KoPK6ndP955d+vcPSryIq2ySSCIRMRm2RS17DLAVKgVKgFCgFSoFNKrAIKgGfG264oUdSiqi8++67p9tvv72/z28GiXPkI4Alj1DM80eew8AwOaAw4BB485ymLqAFGInWDKycQeXODtjOnl1u8Gma3n53avAPDGvv6t55btqz0pZ+baBye4uubAuZnu+nLQHbICFYBYqN0DDwkC9g1QjIADEgLL7Hf/XU5zebxqIOmJZ3NwYsam9MY33tjDX9sTsmtqLBqGtsBVBGr7RXxsdF4GgsrkfgIn0dk/Ul8XP0KT5oww9ju3gt5msdgJj2+o1/sRU7+kz/+tUf/V0T74kU2Whbeynt9CHHZ2WOsSMZdz5T2ttOpjfboOQbb7wxWcbY+yltg5euYXxlq0AlFSqVAqVAKVAKlAKlwGYVKFC5WeWqXSlQCpQCpUApsEUUyOTNpwEq54jK3yWo3Dfd0yIq78jSr9vmpdIyGbVFLnkNsxQoBUqBUqAUKAU2qQBQ+d3vfnf6XouoPNKWywSVDh482N9Nebgt/yqDlaIqA7AArUBKMCmgLxAMsAOSkrMPKqkjaRc4tn///m57Bnbg58WlX4+3pV/feXfpfETl1CIpG6jcc25a2SXCEfAE3kDC0xegIhAGTBnLCC75AVZJ4FdgmO2MwTNUgBj4xRYAxhb/1Q3QS2QlOKi9dsasnfojrDRu55PUl7VlR8m2OtGLLe34FP/iG19SL5pmTCAcexlf+krfbCSzrV2uhWsMHPMpfrlW6kj60m/GyEeasqO/2NG/dvqJjiIqaSmqko3Rb3VlNpIXx5Cxa5uxZ5tNQPKtt96aXn311Qs5y766HvpjQypQ2WWo/5UCpUApUAqUAqXAJhUoULlJ4apZKVAKlAKlQCmwVRQwAWEi4mqDyrdEVLalX5/91YfTi28eaxGVn947KvvSrz2icgaV3lF5XY+oLFC5VT7nNc5SoBQoBUqBUuBKKLAIKoE5oOrGG2/sgPKee+6Z7rvvvum2226bDhw40M8FVAJLIJhnr0CkALSAJCArgFKdgDEQKsAvYGwGbEDVDA5XV88v/doiKhtnan3NEZX7gMqVedt7K8+ebYDwPLg72daIDahUJgOWfApUC9RTysaRkq58BreANe89ZAdw4z8/A/JmuDqDwWgRDbSPDvoNmIsGAXPgnr7po140i6/RWTvHAudGEBqbdGUvoDCQ0vlk47OdpE6uR0Cla7J4nbXJ2Ogp88Ex59iINkrA0ZiMR91ASlCRNo47L/HBOJPZSmaHPvqgoXYyHfTPtmhNz/6vv/56j6Z87bXXOrTMtVO/QGWueJWlQClQCpQCpUApcLkKFKi8XAWrfSlQCpQCpUAp8BlXwIRHgcqLk0+f8ctdwysFSoFSoBQoBUqBy1AgoPKJJ56YXnzxxQ6TgK7rr7++R1UClZ/73OemO++8czp06NAk+hHAAo8C5jx3AWiAlRyQZNtx50EmsAmIYh/ISoQm6HcRrqnjnYSg3DR9eKwt/foOUAlyWvL13LRv79R8EF3ZbC43eDUBfOeXgG3vqgTBAihBsYCxgEpyBYwFgCllxyW+j6ASZAPEPGcCaPxdC1ayIY1AUb/Rwrn0zU76pBHd1LOtn/hEs0C6+MW3QMLUdU3iW9roz3lJv3L6dIxdSf2AY0DatZGN07lFXWgRXfnkvL5dV5BTqS37zvPVNdEm18axRYDIjv7kcTy2ZfaMh01t6cBmlnwVTfnCCy/0JV8dc92iv3bRgn8+44888sj02GOPTd/4xjcmSx1b9pg+0aWLU/8rBUqBUqAUKAVKgVJgQYEClQuC1G4pUAqUAqVAKVAKfFQBExAmeCqi8qO61F4pUAqUAqVAKVAKlAKLCvzN3/zN9Pjjj08BlSLTACGwCbQBKB988MHprrvumm6++ea+LCygBUoFGoFyAZXZVga4BY6x+//Ze9PmuI7sXHeh5gkojATBmaKGVjd7stuS7XNPHDscx8fqG74RDt+2I/rP2V/8K/zFDk8R1+6WrG6pJVISBwkkQBAkxsJQVcBdTxYWuQVxAEjMeLc7sav2zp3DU+VgKp9amcgrJFFIMc5c415PzCHTWGIUGbVpS8ubLqE2rbXS24+SpV/7+/usXst5Ob58K1IzyaueHAx5FTIsxBhCK0QlY0XaFAkm0UbOIddCghGVF1KOPtHWkHLRH87RDyQbBwxoDwmxxnvqpp/kIT/1USb3s0KNNpAnpCLPRXkhKcnPs5E3JCVnnuPgPs9xkI/6KDf6mW74H/LzWSAnkdF8/ttFJW0IvvBAAtIW2k7ZwSKiMfmcKZc2xHMITj4bziRkZbbf1MHBc7QzvjPBN9pOmTxHWXxnZ2ZmjChK9qa87UsYT01NpevUkWWfCvc/EpVBQmcREAEREAEREIFXISBR+SrU9IwIiIAIiIAInCICTHAweSFReYo+dHVVBERABERABETglQhsF5VIOSQWIgdhdfbsWbt69WoSlbwmqhIRhTjKyjDGXpFoCOIqK8aycg8hRvmkEJQhzvwxP2Lfya7LMB/TPdpwIYX47Hq9Zs2BnLfNZWoN8emRnS61vDbrPkeIIbNCiIUspBbqjHEjbQ0JxmvyRRQgeysivJBt3KPN5CXBgHI4Rx9hwz0OmFBWJK6Rn/sIRV7HPcrndRzUxZEtgzwRiRh5qTsrTkNU0jfy8AxlUB6JuuM15ZOPayEqY4nfEJXRR/KFHERUwgVRGVyoN8qIaFOYcNAOnqXtISnjHJ8N5WSlIvXSrpCUfF+og2vRFtpA5CRi8ll7U1I2ddP/7EFZiqjMEtFrERABERABERCB3RCQqNwNLeUVAREQAREQgVNIgIkLJiMkKk/hh68ui4AIiIAIiIAI7IrAdlE5Nzf3RMYhcxCT58+ft3PnzqV9KsfGxqzZbD6JtkMmIb0Yf5F4HeIuRBOyihTCKc4IJ/JwxPObmyx7yjKyiMKOLS51bXZ2w5eA7V2vVvpsaLDobSi5rCQy06MxC74kqAs/xn8h8pBg2yP3uIe0oq44eIZrHLQl+hNijTKQcggv8tK3kGX0lYPrHNyjnyHUoizuR4p83CMvZVAXgo5EvmgfZ97H/TjTj8hHnXCkTdRLgi/XeZ4yEYQ8y3vqi3alRm/9oS1IRiR0iMpYkjfyUydlRTRlVuBSTLSB55CcnLlGWzhoQ3w+CM4QlZzhGyk+J56hbsog0S/OISopA7FONOX9+/eTqOQ8PT1tjx8/TuXTXvpNyh5wkqjMEtFrERABERABERCB3RCQqNwNLeUVAREQAREQgVNIgIkIJlIkKk/hh68ui4AIiIAIiIAI7IrAs0QlBSCIEEKIq9HRUUNQjo+Pp+VfeT04OJgiLpFb5ENGhQSLZ7lOQoJtTyG/YtyGxCIRNbmxwf6DPVG5hKh8vGmrK0i2TatV874kLXtoVl1WslSpR9kVt5ZQ9TFgiLCsqERoIaxCgGXFFXUi87hGm+gHKaQc5SDmeJ6D/iDMQjKSj+d7be+JzBBqSDXK5Ig6YUSiDu5xPdoQopIyuc6ZxP1IcY/7wZu2UFdISs6UTd4QlfSd9xw8t/3M84jFEJVZ0UhZPMPzyMQQlSy5Cp+QgcGGciLBguv0lyPaRDlZWcnrSNwLFjxD/fE94sx77iOREZKISSIq7927l6QlEZZI1CgHVtsPicrtRPReBERABERABERgNwQkKndDS3lFQAREQARE4BQSiIkdicpT+OGryyIgAiIgAiIgArsi8DxRGTIN0cQSsIhJ9qxEVk5MTCRhyXvuIX2QZSGU4oxUyiauk7LSKivpeJ0VlevrnbT06+M5c+lEJGCfNepFl6ZVGxmu2eBQ3UWl73FZclHp5TIGfJmoRJSRL1LUz3vaRQopR1khKnnNdfqJnOVMXsrjHuKMM0eIw4j+47k4eJ1NIe54ntRj0BOU0UbyZNtNWVFGsOZzIvFZkOAeZSPsoo2UGQdl0AfKoK3ISRKfKedsP3mG8pCSiEpEYIhKyqfcKIfnaAOyknNwoK4oh/bwXMjK7Jnr1AOPaC/PxvOUQZ7ssq/IygcPHqRlYLORnsEtVZz5Q7sUUZkBopciIAIiIAIiIAK7IiBRuStcyiwCIiACIiACp48AExpMSkhUnr7PXj0WAREQAREQARHYHYHniUpKQWQhvEJiseQr0ZQXL15My8GyZyWyEqmF+EHQkR9hFSkEE2fK42CcFoIwe97YQCCyRGlv6df19a4v+Wq+vGefizaWVS34sqQlOzNW9yjPuoumhkcAVr19RFT2RCVyC4mVjaiMqL8QgbSB8WKMGUOIhbjjTBujHMQcYo0+wAIRhxSkjxw8TyIPz5EvuHHmiL5HHVwLDjxHinKibfFcPMv17BFlRV18BiTaxzXKp8ys+IN3HPEZkZfnEIt8lkRV8po+co/+BCvKClEZQpBr3Ccf34FoR5wpB27UF32hHfQZIcnnE9GUnKO93Ccf/YhEf0KWskzx7OxsEpQs/xqRlPF582yWZfSbM22TqMwS0WsREAEREAEREIHdEJCo3A0t5RUBERABERCBU0ggJlIkKk/hh68ui4AIiIAIiIAI7IrAi0QlBSGWEEwhK1kGFlFJunDhQoqsjD0ryYPYIj/SimdDTIUwCjkXUi5EFPd7B2dEJpGKG768p4vKRZb6pA0Faw6UvU4XlSN13z8TUck+iD1pSBEh5hBeLA0a+1Qiv7hHfRzRHs4htLJtjnKQXgg5pBn9Qrptl3g8T+IZyqPPlJWVfNQZPLhHvhCJlB2Ja9zjIF+kLMfteaJdyDck5bNEJf0P8UfZlEf7IvqTZ0NUcuZ9yEXyBye4xtKvS0tLSTJSLvdpK+XBiEQZ8Tq+G+ShbvLDi2dpW4hKXpOQkcEk8nGNfNTL3pRISsb7JKQlEZ583uTj84BTtD29yPyhbRKVGSB6KQIiIAIiIAIisCsCEpW7wqXMIiACIiACInD6CDDxwcSEROXp++zVYxEQAREQAREQgd0ReJmopLSQWsgm5M758+eTpERWRlRldgnYkFE8G+OyrMxDPMX7EFw80xOcyM1etGO7vWmtlT4XhXlrIyqLeetHVI7VPJLTReVWRGW53NuTEb9HuYgqZBdCLWQlgisr6qJt1E/iyLabNiIpeR4BxrOIvRCViK4QkenhbX+eJxa5Tj0cjFejvdE2rpEnmGelL8+Qn7y0L/KGHAxJGaKSfpGPvsMj6qAcyuXzDJHIMyEoQ1Jm+0ddPE9ZiNtgG1IwygzxGeVSB4nrlEe99I22USbti88rJGW0lfq4F/f5LKibyMmIpOSMpOR6SMpgQ5vis+V19pCozNLQaxEQAREQAREQgd0SkKjcLTHlFwEREAEREIFTRiAmPiQqT9kHr+6KgAiIgAiIgAjsmsBORCWFhjhjWVCWf2WfSkQl0pJ9KxGYyC4EVci1GJMh15BHIdiyIglph8BCZJHyeaIIEUzm+bciKhdytu5Lvxb8Xr1R9GjKiovK2hNRWan0ljr1Vib5FUINcYVQIyHYslItC4p20r+sRKMMRCXRe0gwnqWdSLeIqOy1txc9Sj9CdFIWEo5+R995H/VwPysrqYt85AnW3A+OnDmyYi+eoSzaAXeWbaVtWVEZ/UD+0QfK4Bn6gqwjbzZRDn3MSkXqDqEYspBzCEXuRzuiLVlBGZyiP+TloC30mzaGjORM4lr2Op8hwhhJyV6UsSfl48ePn+yVyXPZ7xa8n3dIVD6PjK6LgAiIgAiIgAjshIBE5U4oKY8IiIAIiIAInGICMSkmUXmKvwTqugiIgAiIgAiIwI4I7FRUUhiCCZGFrGQJWCRlLAOLvCSqEukVYopnQq5lRWXIMsQVQizkGEu4Fgrsb8lzm7a23nVRuGGP58xWV1w69W1YtZJ3QVnyZV9ZurPhe1bWvU6Wfy15+3pLqiK4kGjILUQj5+y+hbQrDvpECjHIa9qH9ArRGaKSe/QNyUWbeR19pR+kEJAIuKx0ywq0qIv8HNQXEpP30absmftZqUcfg2O0CVEZspKyuU++ELUw4RrcaT9Sk8QzfG70KyspaWfwoP3IXsqCBzyjDeSjPtrx9LPsRVJyfbugpEz6k+0TZZGykpL3tJm6qBNJiaCcnJy0qampJ0u+8jnxXMheyn3ZIVH5MkK6LwIiIAIiIAIi8CICEpUvoqN7IiACIiACIiACadKDSRiJSn0ZREAEREAEREAERODFBHYjKikJ6YTMYl9Kln1ln8orV66kCMvh4eEkMZFVCCoEVsgoRFfIOq5RDmKLsrJyq/dcT96trXVs0UXl7GzXllsecdjteP5N36cy7/WXXVTWXVSyT2UjlZHPF1J91JMVlbGXIjKLe9kDaRbikDaFRCMvAozE8yH5yEubo+2cSbR7e595JsQb9SLSOKJO8kd9XKfsuMc5EvcY25IoJ/oBR/JQP+ItKyq5xn1kX4ja6AP9hDnc4pkQldGXaEvUTTkhbolsRFrSHg76wXOUmf0soyzaGAdt4oj+wIQ+kagjzrymn0hK2k/k5MzMjN2/fz+JSoQly74iMOkXz1F2lB/1Pe8sUfk8MrouAiIgAiIgAiKwEwISlTuhpDwiIAIiIAIicIoJMEHB5IdE5Sn+EqjrIiACIiACIiACOyKwW1GJdEJiIbiIqmQJ2MuXLydhyevtS8CSP6RUiC2uIbcQfpFCalG2/8+f6bqA6rqI6tpDF5WLSy6x2muWz3W8bvPozYLLyponROVAigikLA6kFSItxFqIyhBatCOEFm1B3JGyohFRRn5EGeXE0rE8G8+EsIy2h+gkT0Q/0hYS72OMShuj/uy1aEecs7KQfJGX8qIP5KHdiDdkY4hH2kY7yUsfSCEXeYb7kZdziEquU3+2btoe4haWIQdpD/mykjIrKmkXZUV/o83Bh3KfxSnqgz/1zc/Pp3E9UZSRGOezNyUSk88qyk6V7eCPROUOICmLCIiACIiACIjAcwlIVD4XjW6IgAiIgAiIgAhAICZxJCr1fRABERABERABERCBFxPYraikNAQYQmpgYMDOnDmTJCVLwF66dCntX8kSsCwpGtIr2wKeDbnGfSQXCaHF0q1+2xPRg0QD9pZ+fTi74cKKCLtVv9e2ShlZmfP6Ky4sG54GUn3IJ8pGzoVYC1mJpEN8IbVCkCG3OHiG+hFrtI+De5TBMzwbiWvxXDwTQi77LHmyKcancS3kJW3hCOFHWcGjx+RpNCL5suXEczzD55EVlbzneeqj7SEqqZeDOmLp1zjzfDxHe+gP9dFGOCAFEYck3nOfOvgcYc+zkWgTiXIoI8qhrOg7Zz4PztlrUR+fHZKSJV+Jpox9KUNSRmQnz9LP3RwSlbuhpbwiIAIiIAIiIALbCUhUbiei9yIgAiIgAiIgAt8iEBM4EpXfwqI3IiACIiACIiACIvAdAq8iKikE0RWyJ7sE7Llz53z/yOEkMbmPxEJWZVMIPspAZiG8SMimzU2EUy/isdvts9aK2dzjPltqudRaX/N7a1YurXvd5rKy5NGU1RRRGVGBlEc5yKuQixFRiGQkhRjLRuJRf7SR/sV4kjw8k5V9vEemkSfazjMcXCNxUB7346BdPEeZJGQf1+ABi5B8IXC5nn2ecqJdUQ91xLNZUQl7rnNkZSttp06eC8HIErAhK+O5aDv1BMuQlAhLyqFt8I5282y85nq0nzLod/Sd8oIB5/g8stf5zBCRRE0ypichK1kCFnlJVCftoG+US592c9BWon+vX79uH3zwgb3//vt27do1GxkZedLu3ZSnvCIgAiIgAiIgAqeLgETl6fq81VsREAEREAER2DWBmMCRqNw1Oj0gAiIgAiIgAiJwygi8qqgMQYbkQkyy7OuVK1fs/PnzNj4+nq5xD3kWQjLkFedIlBNjN6QVS75ubvYk4OZm3kVU3qVUwVZWXZh129Zn617eqsvKDZeVeRdsvb0Za7V6Wso05BziCpmGyMomrpGoK84IMtoQ0jHkYLZdIfsiMpFnQ75RF4n3PEOiX9HfkH7kCRnH85RJXtqMOIMV56yo5FkO8sWZcuI997PPx3KulBMs6Ct10XbOvOd5RCJicftelVE/HKLNPIscRFYS6UgZ3KcOyqC+aDvP0/fgGGyyfQ/2ceYeiffURT1ISsQkY3qiKnkdgpI8JPIH9wRoh39oq0TlDmEpmwiIgAiIgAiIwHcISFR+B4kuiIAIiIAIiIAIZAkw8cKEiERllopei4AIiIAIiIAIiMB3CbyqqERCIcmQVCz1SiTahQsXkqhEVrIkLCKIe0gh8iG1SCEpEWUciKaQaZ0OshJxyBKwBZdXJY+qrPh9z+tLwuZzbSsW1rycrgs983KRfNUUEYikox7axXhwuxhDaiHqOMdr3hMtSV6eoV/ZFJIt2heRlVEO16OeeE05tCGEX0g7rtNX6o7yqAtBGdGQIft4hjK4z8GzcaZN8R6G1MNzERUZHCiDI+pC7EWUKe2gbJ4l/7NkJWVTF+1FTmYlIX3m+ayohH30OdrO8yTyk7azj/e0kRSSkqhJxGQISt6HJIV9ln+WR+rwDv5IVO4AkrKIgAiIgAiIgAg8l4BE5XPR6IYIiIAIiIAIiAAEmLhhwkKiUt8HERABERABERABEXgxgVcVlZSKqEJmIX3Yr3JsbMxiGVgiLHmPrESCIdHIF8ISicazjNsQVAirXoQcS3m2Kd1FXdG6GxWXUnXPR5QeS41uuBxzUVnobL1G1D3dn5HyKZcDGZeViNQRgotzSLuIzMvKP/oWwjNEWwg/pB3Pbi+PMslDftoQfUXmURYH98gTiXwhGDnHM1ynDaRoF2cSZXDmHhx5BtHJ8yE9EYbRfuoKviEr4UIZtI3n+IwiZUUneegXy6yyFCtn+g5byuf5+Fw58z748yz5tn8OWfa8zibKR0oSTRmSMlsveYMdfaBs6iHt5qCtiqjcDTHlFQEREAEREAERyBKQqMzS0GsREAEREAEREIHvEGCiggkcicrvoNEFERABERABERABEfgWgdcRlRSErEJMIX6InhwdHU3LwCIqSbyPPSsRYIg0xFqIyhB3CCgk2toaEqy3NGlfX8kFVNXa3X7L9flzFV9mlCjKUtufb3vdXT8j63qyLERfRBKGIMtG8oVcpL6sqER+MYZE/tEf+kXioI2kKCdkH89HOdF+yiEvz9LPrHiMskLecaat2wVjSMpUuf8JCRdj3HhPHYjBrKgMaRiykDqyojLaTF84qD+iORGVfIacuUbZ9IW+Ec2IQIx9IblOHRHNSb28pryQq9RBov54DX8S7YAjCfEbr4naDEFJRCWSkvu0IcqJMxwipc7s4o9E5S5gKasIiIAIiIAIiMB3CEhUfgeJLoiACIiACIiACGQJxCSORGWWil6LgAiIgAiIgAiIwHcJvK6opMRsZB+RlchJ9qkkupIzy8CyNOzg4GCSYCG1EF0IL8TTU+EXopJyi7ZpVReXTcsXalarshdin4tKoik7LgORlebCrBdVuF1UhiSj/BCUcX5a39MoSPqB/EO2cSZxMLYMucjzIdWyso3XJOriCAEYonJ7WfSbciNfMEEOZvOGiNt+jraSn2dJIYERhtwPOZjtP6+5Tv3RTp6FHSI5KyppG/noF6IScRiikvZkJWlIWT5T7sEr5CLMot5oC2VSFin2vuQ1MhRRSURlVozyXJRL+4NH6sQr/KHPiqh8BXB6RAREQAREQAREIBGQqNQXQQREQAREQARE4IUEmLhgUkWi8oWYdFMEREAEREAEREAEbC9EJRiRa0gqhBUReUhJBCWi8ty5c0laxr6VSDFEEfk5kFCknuhb93Fcb49Kln4lorLTHXBRWXeZ5kucVohU7InKvj5E5WZPVPryr/V6LS0DG7IMQUaiXKRZpLjGdaQXUo9nEHzINwQd77nOkRVklEGEHwmxFq+jDvLyPBxiCdWQj4xPSUi8EG3kpd6oO1sveWNcG695zwFvEmXzLDxJvKZM8kX/aTP9jHpDcvJ81M+ztDeEJe3nHs8gZolsRCAiLCmXg7qQo/F58p4yo26eC+Z8vrSBM89zD36Ui5QMMYmcjEhK7sdnlO0/5QeH1JBX+EN/JSpfAZweEQEREAEREAERSAQkKvVFEAEREAEREAEReCEBJi6YzJCofCEm3RQBERABERABERCBPROVoMwKP+QVIoh9KlkCFlkZS8E2m80kt5BsPIMMC1nZ6RA51/XriE9EZc06Gx5RmfeIPxeVFReVpbT0K4KxJyoLTyIq60kQIvsYC4aoQ3aReI8448x7ZBnyLCRlRCTSLq5xxLgy2sjzIdkQaYg7ZCXl0QfyhzhE/CHEEH70kzaRKIszebkecpF8ISq5F3Vnz7QpJCX5Q1QiFqkr2k4d0W/azPt4ljzx7PbnKSPKoR740F+EImPrrKgkH8zoJ2f6QftpLyzgwrPBPMuQe5SJlKTc2dnZlBCWRFiGpKScLCv6QPmve9B2icrXpajnRUAEREAEROD0EpCoPL2fvXouAiIgAiIgAjsiEJM5EpU7wqVMIiACIiACIiACp5jAXkVUBsKQaAgw5BVSElkZkZWcWRqWJWKRayG2kFFIsY0NJGUvSrJQKPvrhnURlb70a9WXfvVHrFRcd9G2br64qUu73jKr5XIlRXJmy6S8rJzMvkbiIcPIQ5uRbM+KDKRdyDLykZBuJEQbz5NCyJGPAwlIOygvJCRCMsaolIO0C3lI/hCHtIW8HCHk4sx1EnkifwhKziSuk4c6kIQhCukH1/lctj9DG6OdITFD1EY5RDoytkYiwpGyKIc+RiRmCE7aznMhg0NUci0+A4QnUvLBgwd27949m56etpmZmW8t90r+kJSUuZeHROVe0lRZIiACIiACInD6CEhUnr7PXD0WAREQARE4ggSYMInJljgzacKkBufsERNPTMbEJAmTKJGXiY69PKI9EpV7SVVliYAIiIAIiIAInEQCey0qYcTYLmRayEr2qIw9KzkTzYbgQnbF2JExnOs5HyMi1HwZ1lLV7/VEZS7HcrEsc+p7IxZWPQ+isp3yFgrs01h9IioZZ1JWRPCFHOMcrxF4WVFJOyIyMNoUY1ieQUJG4j0SDkEZwpL3XKdexrgRqciZ/sV4lzJpV0g78od0DFkY+eOZ7JnXpHgmoho5026uc5+2hkBFFFIn5VIHnwmCMcRsPMf97XWHcIyISkQl5dHuKAtuz2JHPlKwoU28h32UNzU1ZXfv3rX79+8nUcl18pCXOkj7cUhU7gdVlSkCIiACIiACp4eAROXp+azVUxEQAREQgSNKICYNmPCIiRsmMZgYYZKCCZnsQR4mgpiUYKKCyRt+Rc8kCXl5v5dHtA9ReePGjbREFb/m59f7LDnGvkm7PaLM3uSS/wreJ08ezLfsxuSi3ZxatlvTLZtf6VjRJ9ZyPjm0XwdzNUXHVSv22XizZNcmGnb5TMPOjTasWa9Yny99lss93VNpv9qhckVABERABERABE4Ggf0QlZAJocZYDynE2G94eDjtW8l4jChLxmeMHbOCLp9n30XfL9KlW7lc93FNv0dUDnh5Pm4scK9rhfyKX1/zayzb2pOaISoZXzIm5WB8ivBiLBrSLMQZ41JEHnkYi9LG/v7+JO8Qd1yL53mGMkIwRpmMa6MczuQjDwfCjzKCA9d4zZiSPCHteE97qZM2wCLGxyEOOXPEeJQz14JtiMcYW3OP8kOk0jb6wnXqCbFIf3mGOp83Ho/xPhGQLM+KSKRcrvNMtjzKpQ+0i4M2kEJUwoe2UBZLvhJFOTk5aXfu3EmiMvbA5BnG3PRzvw7aqaVf94uuyhUBERABERCBk09AovLkf8bqoQiIgAiIwBEnwEQHEw1MMjBZEZMfCEAmnpj04IgJksjHpAaTPExqMJFBvsHBwTRBwrWYhHnd7sckDpMpn3/+uUTl6wLV8yIgAiIgAiIgAieWwH6JygDG+A4RhxhCVvLDMfarJKoSWclYkMi+kIOlElGTiDuPEKwgKhvW7vjYchN52OepnRGV7FH5bVFJPdSHFMwKQWRZNoXEIw/5EXYhTimDsWn2eca1vCdlBSZlUlaUHfliPBpneNAmDiQc+RhP8xqxR/9pA3XzOuRtjJF5NuQdZw7aTT6eg2FIR/LSxuyytLynLMpmzM44PEQl9T+rfOqg/fSZPrJUK2N/xvaM6Tl4lrpDfmbbkO1n8OFZyuEHhURRIiq//vrrJC1ZXjb+u4J69/OAs0TlfhJW2SIgAiIgAiJwsglIVJ7sz1e9EwEREAEROAYEmKhgcoGlmphYYMKCyQ0mnH7yk5+kM91gsoH77DfDMlFMyISMZMKDyaCrV6+mSSomNWKSZC8QMLmRFZUxMaaIyr2gqzJEQAREQAREQAROCoH9FpWMEUmM8xjvEVWJqDx//rxduHAhjQMZpyG7yIOoLJdLSaiVyzXHXLe19ZpLOmSaL6uaY7lXj6jsI6LSx5a+0kQ+zzOVVAYCKsaUjDcZfyLJEGBx5jXjWUQeeZB91I80ReLFuDQkHVIO0Yd4I/92AUd5UT7ykTpJPBPP8X0JFlFuiEqEIwKRNpDoA4l20ZeQlTwXbeB1VgLHM9F36o8VTWgbz1EO+eAdspL3XKc82hr9i/JpMwcMGM+TGPvDkry0gTJoN7IyRCXXs5xoA5KS/z5gjM7elIhK9qfkzDXaG0yofz8P2ixRuZ+EVbYIiIAIiIAInGwCEpUn+/NV70RABERABI44ASYN+BU0e8l8+eWXKWKRZZqY4Lh27Zr92Z/9mV25ciVNzrCc06effpqEJpMbTFgwicGkBss9MZnxzjvvpEkqJq24RzkxIfIyFLSFxCQIKd7zHK9p1xdffJEmoph40tKvLyOq+yIgAiIgAiIgAqeNwH6LyuDJGA/xxpiMH45dunQp/WANack4EHGGrOtJSoRl2SVd1cd0FVtZrbhEY6sA5BwRlb6VAEu/5ljOlGvsCVlyYVZNZSD+sqIsKygRZkhKEuKMcSlyj3EpP6KjHYxJaStjUsaUIe9ivBkCjvEtYo2yKJdEXVyLeyErKYc2xTiXMpGJXA821BsJ4YdMo20hH2EZ7eFMWfSV+7SXRB3co37aRXuiHu5TLpGU9JM+c42D9tBu8vKag3ZRPmeuURbMEJWUTV4OyghZGdGgtC36mH2O8Tk/YuS/E/jRI2d+AInAjPppP2k/D4nK/aSrskVABERABETg5BOQqDz5n7F6KAIiIAIicEQJMGHAxAyTCyyp+sknn9hvfvOb9J4JjOvXr9tf//VfJ2HJr62RmUx+8YtpJlCYEBkZGUmTELdu3UoTKcjNy5cv28WLF9OvmpnooKydHLSFyR8mSZjYiF+sx+QGQpWITu4zAcbyYoqo3AlZ5REBERABERABETgtBA5KVCLQGA8SzccqHIz/+MEaY0B+TIYkRB5VKsjKnnjL58s+vivZ0nLRx3Ps06KaLekAAEAASURBVLjh48eOlYprPl5kpY6eqOzrYz9IhF0vAjFEJWNCxoeME5FlWUlJ9B7SjXEi+RF4tA15F6KSMSnt3n4wBkXChYzMlstr6otE+TFGpRwEHinK4FqwCUnJOUQl4i/6Q14O+hXl0Ma4H5Iy+hwykfzkC5kYkpI6eCb6EhGNPE/5fF4k2kA+rtM/xvkhQekH9ZOP8hnL8z7Yw528yE0kJVKS/zaIxLWs+KQtPLvfh0TlfhNW+SIgAiIgAiJwsglIVJ7sz1e9EwEREAEROMIEmIxhAoNfPiMab968mWQlEw1MKvzgBz+wX/7yl3bFIypZwunOnTspD5MT/HoeWchEFBMWPMvkEJNS/JKeiSomrZiYYaJjJ0d2EiaWoOJaTHDQViZDmGg5c+aMjY+PpzqYnNntwYQJqTep5HX4pNOD+ZbdmFy0m1PLdmu6ZfMrHSv63kk5r2+/DuZtiu5xa8U+G2+W7NpEwy6fadi50YY16xXr80k6ogrosw4REAEREAEREAEReBmBgxSVyDLGYfx4DFH57rvv2hUfNzJGYxnOWo2lQ5GVCDKiA0u23i64yMq5+KMnLF/adXHWdlnZ9deIS6RdLok0ZGVW3PEE48L4URuSjRSSkrEo41vGTQi2kJQIvBBulMd9RF2kbLk8T5mMdymXcS6SkuuRqD/Gp4wnY5wWrymX8S/1krKiMn7EF89wjvbQtki0ifIYC0cbODN2JQ+yMUQo/aSeGHPTTtqPgOQZ3nNQdzxHXurlHv0MltTHdeqIttIf+ktZ5KVcVlNheVfG5vzokddZSUm5WUapAfv4R6JyH+GqaBEQAREQARE4BQQkKk/Bh6wuioAIiIAIHE0CyEAiFJlgYMKBM+8Rl0w+EB35t3/7t0kKsuQrspJJDCYCkJFMSiEsmbxBYk5OTqZnmbBib0smrLjPhMhODiY0mGCibmQpS0ZxjQkTDiaFqJ+JGPZBIpqSiTCJyp3QVR4REAEREAEREIHTQOCgRGXINWQZUpL9KRGV7FfOOJEfs7Ekab1ONCHLvvrSq7mijxvzvu2AufTiR2O+TGthwypVT+VNF2MsT4q4eyoRQybGOSIXs7IyRGWIOfKQnzFopJBuCDjucSZSMN7z3UCsMfYMaReikroYj5Ki3rhGXRzBI8oOKRiykvEzbaFO8nLwbEhBrvNMtCkkJeNspCOSkDopn7LgjqAMScmz3KONtB8WjPWjD9RHHtrD2JkzfacN1EH5PEcd0S7K46AtMU5nfJ6VlAhK3nOdcTxtjfE7z5EO4pCoPAjKqkMEREAEREAETi4BicqT+9mqZyIgAiIgAkeYAJMGyECWemWCgf+4Z2KDiQb2l7l9+3aacPrFL36RJp/+8z//M0lI8jHxdGXr1/JMdDC5ce/evbTH5ccff5wmWN577z178803U9QlEyE7OZgYYYKDiY6Qp0x0xK+xec1kC5M8SEqJyp1QVR4REAEREAEREIHTROAgRSWSjXEZ40HGZW+99VYSlSz/GlGVAwP9LtXYm7HkO1LmXYb1+Xhz08/sR971ceOG1WubLs76fDyac5kW8rAXXYgsCwkYgo+xIeNGxqDItYh+DCnHmJa8iDkS0YMkyoqEpOMaYjCEXJQb0i7EHdcpk8R4lPvUTeIeB+VFXZRJ2VH/dllKfYzFs89uF5XIQvpIXSH/eIZykZMwD+HI+Jz6OWgf+RGVWXlIedQRohLRSbvgRL/oS/SL97SNZ3hNOyiT8oiajOhJXrM1A3XBnjKon/w8e5CHROVB0lZdIiACIiACInDyCEhUnrzPVD0SAREQARE44gSY5CARBfkv//IvaYKBCEUmPZhcQDoiHBGSRFSyv8+//du/JbHJ0q4s6frGG2+kiEomOXgG6Xnjxg1DaDLh8Ud/9Ef29ttvpz0sybOTIyZDKI+Jjpjk4DrtZXLkm2++SdfZG1N7VO6EqvKIgAiIgAiIgAicJgIHJSphypgP+YUkYmyGoGRFDcaJjC1Zqp/VNeq+nD2istvtc+G1abOPNvy84TLLl9nfEpW1es7zsS9ib3lS8iPlEHDUQ+JgTJgVhiEV48w4krFjVhw+qxxkIW0nX1Ycxjg0xqIIO+rkzD3qQcoh7njNddqGlGTMCwteU3bUwWvaECmkIv0IoRcss+1hLEyiHtrAPcoPScnYnfeUS12URX4Ea/z4D4lIW7nOEe1EWNJe2kbZUQ91kUI6Uhb9JTozBCU/bAxBST3kgQ1lhOCkzIM84EBk7/Xr1+2DDz6w999/P60Ow/cSbvH9Ocg2qS4REAEREAEREIHjQ0Ci8vh8VmqpCIiACIjACSDApAETD0xa3Paoyf/6r/9KkxH8Ah4JyT2Wf0U48h/7iEqW7frXf/3XtP8M/7HPcl4hKpnkYGKCCMjPP//c/uM//iNR+uM//uO0TyWyc6eikgdjUiPOcY2JFyZH2AuTyRLaStn8ep/Jmt0elE+i3G5Xe1Tulp/yi4AIiIAIiIAIHE0CBykqIYAgQ5QxXuRHZCwByzjx0qVLacw4MjLs9+qeB1Fpttwyj8pzUbniIoulX4ub1qhvuqTMJ1FZq5VdvvWEHxF/iLSQTIzdEGEIMcasCDKkGmfehygLQcjziDnaF3IwxoCUSb4omzFhRA9SDq+pK/LznjoiWpHxKHWTJysQkYfZdlN+1EVfQlpyjTojUU/2iPaE+KOtyDjG1bDmzDicukLExTPZdiISaSvXKCs+L8riedjQFvoXkpI+knguIjOJnmS8v32ZV8rlvwUomxS8sn05iNcSlQdBWXWIgAiIgAiIwMklIFF5cj9b9UwEREAEROCIEWDigAkEhN9XX32V9pRk0oEJDvaj5D/wmXz48ssv7de//nUSlX/zN3+TIir//d//PU1OICr5hXxWVDKZw8QFEZXk4whRSf7diMpA9qzJGtpNHUy4SFQGKZ1FQAREQAREQARE4CmBgxaVIeEYR/LjMaIor/gWAYhK0pkz7Gk+4OPNim24qGz50q9z8zkXZ0hOlo41F5XmkrIXUVmtlnxMWnWJ1hNxES1IDxnHZiUlMg1JScpKSuQk49sQebzPisqQg3Gm3BCTvOZ6CLc4I+NC3hFdyGskHfeRj9TFKiSISl6HYOU+ZXKGVYhKztGmqJ8+RIp20O+sCIVx1BFCFPnIQR20nTJob7AJqUp76Wd8ZsGF56kvnskKSiInY7lXxvu8ZyyO1CQ/5QVH6icdxiFReRjUVacIiIAIiIAInBwCEpUn57NUT0RABERABI44ASYRmISIpV3Zi5JrTHjw63cmKbiGqGTvyuHhYfurv/qrJCw/+uijNDHB8l1EVF69ejVNRDFRwiQFS78S7UiEJuWw3BJRmohKJmte54hJj5ChEpWvQ1PPioAIiIAIiIAInGQCBy0qYRkCDvEVY0V+2IaonJg466tgDPs+lHXryxWtvV60xeWiC668SzuWMu2z/kZfEpW1GsvIIiorW7KymiIiGVsyHkSKZWUaQi1kJfc4EJs8z/iTxOts1CHlZMUgz5FCDG4XbYyVuY/ki+jE7cupUic/zOOHdIyrqTciFXkecUgZvKYv3IMVbUNCRp6IaOTMM5GfvNnyqYNnKYfy4M8RY2b6QqIM2h3CknPISu5zxLPUBVt40k9kLD9gjEjKkJX0PSQlz5CCWZxTwQf8Bx5a+vWAoas6ERABERABEThBBCQqT9CHqa6IgAiIgAgcbQJMJDBJgqj88MMP7f79+2lygckbJpWYzODe3bt37YsvvkiSkT1e+GU8+1kycYGY5D2icnx8PC09xYQHe0feunXLPvvsszRp8vu///sp6pIJAyYOXudg0oO2K6LydSjqWREQAREQAREQgdNA4DBEJVwRZiSEGuNKxov8EO7cuQnf33w8SaRqxZfr76u7LKvZpvmyrOWC1aq5J6KyWi24VOxJvFKJJWCfyjjGg4g0xp0Iyoj6y4pK6kfqhaQMUUk5RDByn3IYD1MWifEv0i4kW0Qb0qfIi5gLgRd1h/Ajf4hEIiqRiDDgGhKSsskbdVEmgpE2kZd8tIt81INMpI5s+YzVGYMjQln2leey0ZTR9mhzyEPO9I+6o2zK5zWJOrgf/41APvoZy70iKhl/IyljyVfub2eWrZ82HMYBT4nKwyCvOkVABERABETgZBCQqDwZn6N6IQIiIAIicAwIMAnBJMj09LR98sknKXqSiQYmWPiPeyYeEI6Iytu+fyXRkH/xF3+RlnplmScmNiiDyRF+Jc8+REyYMMnBvpaITyYzuH/9+nW7ePFimkxhIuV1DiY/qJeISqI2FVH5OjT1rAiIgAiIgAiIwEkmcFiiEqaMKRGCCEJk5dmzZ1NE5fnz55K4HBwcdcE2ZN2Npks89qH0SMR6oScqa4xH8y7uiLQsuszr7S0ZgpGxICItRB4yjzEh41fGoowXyYv4Y1zL+DPOvI7oQ8oJeUd5SEueDUGJNOQ110IyUke2PiRfCL6sHA2JiFQMkUh9tC/kIGXGUq7khxVt5gihSH3k5z3toO3IT0Ro1BHlR1vJF4k6ec1BffQx2pDll42OzNadFZUIShIRlvCmHMqM8lMlR+CPROUR+BDUBBEQAREQARE4xgQkKo/xh6emi4AIiIAIHC8CTCgwsYB0RCwi/pgIYfKCiR0mIG67oCQykj0sR0dH7ec//3nav5LJGCYtZmZmUn4mSpCU2UhMJj6YbOEX9Ox5icjkPZMrr3PQbiZcJCpfh6KeFQEREAEREAEROA0EDlNUwhdxh4iLyMrxcSIrz3tk5XkXl+etf+Cs3x91OdfveVjStCcqq1X2q0R09p7P5fKpHMojMYZFpjHeDHHI6xBn5GHMSUL8cWZ8G+9pU7YcxrYkxplc537koR/Z+hB0EUkZkpL7HDxDHYgyfqyHpCRlx8C0kbaSqJODNm7PGxI1pGL0DRmJmKT8SCEqaTvPkWgTKd7Ttxj/RxQpbaAvjOv5bwLOXAs5Gvf47wLucyZxPdof5aaOHJE/EpVH5INQM0RABERABETgmBKQqDymH5yaLQIiIAIicPwIxKQCkwzxy+iYcGAChMkIRCV7VBK5SEQlovLtt99Ovyxn2Seuc+ZgggRhyWQI15gguHLlSoqk5Bf03GPihgmc1zloN3VIVL4ORT0rAiIgAiIgAiJwGggctqgMxowBEXEjI8O+EseEp4s+Rrzie6BfsHLlrIu8Ib/PkqZFl29mVd+rksDCfL7Px529qMZcjr0Xed+XxoLZqL+IOGSMiKyLaErq5X02MRaNyENEHgIQYRjP8gzPRz7ycJ9xMikkJa95lucYn0a9PM84mP4iaEkRKUm9lEV7kYEhH6mLZ8iLfAzxSLkhFXmG11yjfVE29UT+YEM+6onzdmHJe+5RP+1APjJ+n5+fT7Iy+hiiEoFJiuvRd8qhPenobY3JR8T/nhzp9laWJxf3+YVE5T4DVvEiIAIiIAIicMIJSFSe8A9Y3RMBERABEThaBJhYiMkXJjNiEoNJDiYqWPYVUfnpp5+maEn2qHzzzTfT5M7U1JR99NFHSWaSl2eZMGGShrLYs/JnP/uZXbly5cmEC/co+3UO2ixR+XoMX4e/nhUBERABERABETg+BI6KqGQMiMAbGOj3MeKYi8oLdunSVRsdu+xC74KPFUet0V/zZV8RlX0uKs3zb7oshHWMe3rnkHGIypCVjD0ZI4bwQ9yReM/17YlxK6KO55F1vA7RyHMhOHmOskMsIvWQdLzPikPaFBGbPI8oQ05G4j3XqYO6qTfKCdkZbUdUhtykTI54JsbrlBPL2lIHZUdexsnRL8om8Z4+hlTlHPloBxISUckPFUNWEjkaCUEZUZb0mzLb61v7efoOo0lO5ryhJGeRhvvISU+bG85/o/fa/x7IIVF5IJhViQiIgAiIgAicWAISlSf2o1XHREAEREAEjhsBJiRY2nVycjIt/8qvtd977720HyV9IaLxs88+S8vGEpHJhAUTJkyyMHmCqPzRj37kexFN7EkkZfBjwoiJFUVUBhGdRUAEREAEREAERODZBI6KqETkkWq1qg0NNdM48cKFy741wGVf/vWKDTTP2GCz30VmxZoDOc9HRGVfRlQ+lZT0NARiyEbeh7xD8oUYpE4kXyTyk0I+hnBkbMkYNuQf0i+eZYyLzAtRF+KPM+VykJ9nEYaIwxCVvI8U42TaynM8H/WHgKQcxtwkhCXlhICMtnOOOiN6kzP5KJv7iMmQqtHuqINnQ1RyLURl7D8ZS7wiLxGVUQ5tpVyeIVEP5VjO2Rf6LOefV67YS+nzdku50fG+rntq+2tPm+A6gOhKiUo+ZR0iIAIiIAIiIAKvSkCi8lXJ6TkREAEREAER2GMCTD7ExAzSksmP4eHhNPlCVdxjAoPJCyYtYqKGSSISkzTsW8lEAe+ZsNiLgwkYicq9YbkXn4fKEAEREAEREAEROLoEjoqohBBjwWIREYesHHRJOWHDIxd9vHjV30/4srCDNjJct6HhkkdWFnwsyX6PLNPKOLI3lqQMxpUh+xgTMgbtlV1MUhBRiRxEPHIwpiWRjzFrCELGsCHxKIexLsIvng3pFzIxRB1lhazL1s3Yl7ojIpIxcMjJKDt+0Bftp5won3Zxnfq3l8PzHLST+skHB8oLScl77keZjN8jIhIZyXXaSx5S8HiWqIyoSsrgPm2DHXXHszy/mXPB64Ky4HuKlvpd9Nbz/tr393RxSSRld23D1hZdyi778rnLXueae0qXl/stKyUq09dFf0RABERABERABF6RgETlK4LTYyIgAiIgAiJwmASYLIlEO/ZSTG7vF/UwQaKIyu1k9F4EREAEREAEREAEvk3gKIlKWpbPs2Rpb3/FZnPYJeU5j6a84nJywsZGh11eDtiZMURmxYUfy6eWXCCS2Oe892O4+PEbY0IOziHsIiISecfBmDEbBchrxCDyLRLXogyEIIk6QkiSH1FH2i7qKD8kKaKSSMj+/v4kGnlf9Hb3+d6aIRWjnbwnIftCLCJNeU/9yEqEJ2Wkcrw/1EM7kyD0M6+5Rv4QoLSHNtLmkJTsP0lfgwPPkydYICP58SERlbH0K89wLSQly7x2urF0bC+Kss8Rl+r+48QR/zyHi1Ybcck74JGl9UKKsERUdlY3bHWhY63Zti1O+3K3j9q2vuQRrn6d6Ervwr4cEpX7glWFioAIiIAIiMCpISBReWo+anVUBERABETgpBFgsiQOJk3266Aeicr947tfn5vKFQEREAEREAEROHgCR01UMkbMubgrFku+6kbdanXfm7Jx0QaHxpOoHB9v2sRZl5Vn+j3CcsCXgmW/xnpaoQP5iJCLcSbnSFwnIe22S8oQjYg5UkQHhnhE3HFEWbzmWuSN/CkiccOjCH0smsa9W0Nfngu5GLKSiMgQlZ47lY2YjHwIVRLXGNeGrEQgst8jghPZhrCMCE/qoV7axjPR7hCV9D/aTpuzEZURORp9CVkbkpIISkRlCEokJ/fIl43ExCxu0j5fmrc6nLfmxYqdeaffhq/4/qLjLpYHfQnaske/5omo9GjWde9bq2Pz91dt+neLNvvlss3ddUns4rKz6mP63iq2qd17+Ueici9pqiwREAEREAEROH0EJCpP32euHouACIiACIjArghIVD6doNsVOGUWAREQAREQARE4dQSOmqjkA3Df5gmpWHZZN2iV2oTvUzlqI753JRGVE2f7fQ/LpsvKId92YDBtJUCkIvIJCYncCzGZfZ0VdowXI2owogJDVCLeuEdC+KU2YQf9fxtdIjDbSWbyXJKcq2tJ1pF3A/u2deQyy9EmSerL2pbZp7Ja8baWrVL2/SX9GoIxJCrSEYFJXyqeDyHJgXRc9Xo63jbqIAqTe/SXc86jSTlSlKLn7dJ+j3BkCdXEotCTtLSJ5+kjchLZiHwM8ci1uB4Rl0hK8nDmGvfpOwlmIUbTcq3ejJw3uTZasJFrVRt/t9/O/WjQhi67mG165GsJ8eoys+sS0j8DoPrHZa3H6/bgxqI9+MyTC8u5rz2q9ZFHdbZ6kZV7vRSsRGX6uuiPCIiACIiACIjAKxKQqHxFcHpMBERABERABE4LAYlKicrT8l1XP0VABERABETgdQkcRVFJn3p7TvqSpYV+l1tnXN4N2kB/zYYGazY6Wvboyn6XloP+Glk5kmQlS6GGrEwCzyUecjJeE6EYspI6EGwRFcl5u6BkTBlHL5oyZ+21ti0vtmxpccnTcpJ2EXmZrKA/EPtlIkkRg0SIIhbTEq8sT1twkVr06EmXlCVEY8mjI11aEmWJcK03PJLUX6e+uIT0J3ttXfOIT/aRdAmZojARsltSthdN2etTp+17ZHZctnpe8uVzW5GkXg/SEuGKhEWyIh4RkCzpml3WNSInWd41EnmzUjcbSRmsckUXqPU+G75atUvvD9n5H7tQfmfAaoMl67Z9mdfFti099P0sfU9KRGWpVrCBs5W0Z+XqQtse3WnZvf+ec2G5ZI9vr6QlYTtr3os9jqyUqIxvts4iIAIiIAIiIAKvQkCi8lWo6RkREAEREAEROEUEJColKk/R111dFQEREAEREIHXInBURWWyaebRhrm6i70Rl3m+xGulZA3f37A5kLfBwYov/epLig4P+XnE3387shIRFftREqXIeyIVuYa4ROwRARlRlSEpkZeMJUnk4YixZafjUY0eRTg/7/szLizawqLv7ejRhWtbUYW9SFAiBPNJUCIIiXRMonKrLMrzGMFUds6XP6U9dV+6NglKl5RJUHokZblUTiKTMlhLdSNJ1XaSqW0iPf19Orxcyu/x2mqrR31GH7gX0Zr0m308k6j0aEukI/KRJV1nZ2dTevToURKWiMqQlURdRqQlQhdmcMqyoky6WKiyJ2Xexr/fsLf/7EyKpmyeqyZJuzi1anPftGz2tpfnEZR44NpQycbebFi/y8qyf7Zcv/fxnN3/7YJNf7Jo89/4/p8Lvoyty8q9PCQq95KmyhIBERABERCB00dAovL0febqsQiIgAiIgAjsikBMJj18+NBu3ryZJmCazab/4n7UJiYmfJ+jxq7KI3NM9vT2+/HJGf+F+oP5lt2YXLSbU8t2a7pl8ysdK/qEUy4zEbXril7yABM6RZ+vqvmv1cd9+axrEw27fKZh50Yb1qxXfL+fQpoci4m1lxSn2yIgAiIgAiIgAqecwFEXldZXc8k15BF3NY+GzLnA27RquetiL+f7U1Y8krLfJeXQE1HZ39+fxnqxFCziL3stIhVTZKF/9si27ZKScVQkxoBPow9dTi77EqjLc7a47MugtpY8KrHl0YvrSXryTIqidLmY96Vr+zizN6Yvu8r4sG8z50ueuqgkOnCD932+v6T3YaCZ2lyrVZNE5SvZG3N61GGSjt7OTm+5ViQlEZO0myP0HVGXRGwW0j6cxbTELEvHRhRpgaVfiab0PJveiPX2epKPRFMiKRk3z8zMGKIylnolkjIkZexdCYsYF8eZdnjgaNp3stzM28D5kp3/adPe/T9n0/6UxUreVuZ87HxjwR58vmjTvsTrskdVMq6tj5Rs/HsDNu77WBJ5yTB65uai3f9kwSY/fGwPb/rytA98Cdilp0vqUt/rHhKVr0tQz4uACIiACIjA6SYgUXm6P3/1XgREQAREQAReSoBJEyZ3JCpfikoZREAEREAEREAETjmBoy8qq27AhlyElXFsvoxp24Xbqgu4DY+QLLiwrCYROTAw8OTMa36YFpKSH6xFQlpyHYGHWGTciPRj7EjiWpKLLvVYwrXrkYerKx4JOD9nUzP37fHSQ1vemLPWxqKtdVq23l3zPL7EqpeTRGVa6hUxSTRlT1QW/FdmiEvr+P6M63nbaLnV83Nuo+DStWbDQ0MpopI20ZaIXlxeXkpCkeVbfXTr/8dyqV3fe7LXXkQfto9TarcvA1vy6NF6vSdnm80BZ1Rz+elL6CJOXZRu+N6Q7XUiQ3vLvrLca4hKxs68j0jK2IuSJWJjmdesnMz+vw7dy5X6rD5atGHfm/Lizwbt3f991oYu1Wx9pet7TrZs0pd0RUDOfL5ky7NEVG6miMrRt+p2/ieDdu3/Gkvv5++vuNBcsK//85FNf+oRn3fWbHXOxWxY2WzFr/haovIVwekxERABERABERCBRECiUl8EERABERABERCBFxJg0kOi0mfydIiACIiACIiACIjASwgcD1E56Cau7DGDLhJt3aXcsgvLdReKvsdhyWWfL+lKIooSQRnSkve8HnIRODw8nBLCElmJqOpFP7o03Dp4j6TsLZHaE5Xra2tJ3N2fumc3b92whyv3rNM/bxu1FbOyC8ocS8WymGvShUluEt2Y9qb0spCUxZJLSV/GdnO1YBtLnua8L62KFbve5nJv+VrazxiWCMYHDx6kH9w9evzQWuvLvp5q13LlDSvWvNyij3OTnPT6ev9LsjK13aMmicpsDjbTkrijY6OJR9FX3LCNvHVbTm9501YXOra8sGIL84tJTCIqWf4VSZmNokRQxv6bCNTnSUrw5byKfKXP+ifKdubdul362ZC99b/GPGKybPP3V+2BR0l+/etH9gDxeHvVIyw9rNTbX+rP+zNFO/97g3b9/z5nw1fqtrbcsUe+POzd/5y1+x8jNn2vyhlksFdE2oNDonIPIKoIERABERABETjFBCQqT/GHr66LgAiIgAiIwE4ISFQysSZRuZPvivKIgAiIgAiIwGkncJxEZVo31dZcVi35x7bqOrDjYtGXOy0UUoQke1Ei/JCVWWmJqCTFXpbIS/I9XRa1kOQkgpJrpXLJozbdvLkUW2mt+nKos3bn69v2m8/+2x6uf22FCy0rjrWt1PToyzLZevasJ9JcU/o4jL0pC8VckpTFipfl0ZTtxT5rPyxYe8pF5ULNKh2PAi0P2vDIsC8BW05Ri3Nz83ZvctLuT923mUdT1uosWr7RsfKQS8Az3j4Xe0QveiXpq8u4lyP2oqzUKjbgy+EODSErh9P+l0jMjZU+W501az3YsOXpDVucXU2ict7rezz3OAlK9qtk30pSdqnXkJSpouf8yRXZn7LPBi5UbOJ6v4vKQXvjf4xZpb9gM18t29Qn83b3V49s5rNlW/im7ftO9pau5ZnKSM4u/P6g/eSvL9jZdweSkJybXLG7/zXry7/O29THS7Y45RGYPCJR+ZxPQJdFQAREQAREQAQOkoBE5UHSVl0iIAIiIAIicAwJSFRKVB7Dr62aLAIiIAIiIAKHQuBYiUqPqLTNEJUe0eiikijLiIzkjGwkWo4US78iJomkHBwcTGfeIzPJg7AkX/bMdSIrNzc2bWlx2WYezNiXd76w//70V/bYJq3/e11rXPZlThGHdayhN8vzsqwqHg1RWfAoynLdBWjVIyrLHs246tGS021b/sbL/Mqfma1bvTtiA+WRtPQrgpR9I+ceIyrv27QvM/twftra+UUrn9mwgUsFG3unYY0zZcuXcmk/yFRx+uO1IkfZo9KXeS2Vi96fklVdWibh6tjWHrucvNOxhdubtnCrz8Vf2xYXlrx/vgzr0rK1VnwZ2/X1J4IylnrdiaSkCYhKIj4Hzruo/GG/L/06ZG/88ajzcVHp0ZT3fjtv33hE5cyNli3ea9v6Ym/PyYJHYZaHXFT+XtN+8v9e8GebqX+LU6suKh/Z17+as8lfL9r8pO9p2fF+7tFWlXzGyOvr16/bBx98YO+//75du3YtyWw+e/3o7+m3S69EQAREQAREQAS+S0Ci8rtMdEUEREAEREAERCBDQKJSkyuZr4NeioAIiIAIiIAIvIDA8RWVq24H171nT81VimR0WUmEZQhLJCQJMUli2VcSEZfZFFGY5K1Ut0Sli0ck3sOZh3Zn8o599sVvbTE3Y823zRoXfZnVkWJa7hSZ1+2wd2QnLYFaahStNli0xmjZSo2CR1f2eQRhx5c8XbH5L9s2f8P3xpz1fSQ3x6xZGU4ClYjK7kbX62vZo1lfhnXBoxxXHlm30rKKi8r+ywUbeceXinU5yl6QvhVmOhj3prHvk3VRXeb5PceQok19nVjrtLu2Mtu1ha+87q82/Jxzadqx1pJHT3rE6OraapKU9KPT6aTEa7ZSoOydHCz9SnRk/7myjX+/kUTlW//TIyqbRXt0xyMqf+d7TnpE5QOPqJy/u+7Lz3ZdMvvSr/Wc1c8Wk6i8/v+cs7M/aFqhnLPF6S1R+V9zvlflvM1946KyLVG5k89CeURABERABERABPafgETl/jNWDSIgAiIgAiJwrAlIVEpUHusvsBovAiIgAiIgAgdI4CSJSrCFrCQqLpaETcu5esRiNoIyBCbnrLAkshJpGBGV7NO4sLBgs48e2vTDKV9wdsmqY31WHHAZWN50udj2pVJbtrrusq+7ZqWBvrTP4vDVho2+UbfqQNEl5qaLtzVf9nTRZj9fs/mbLtzmqtZfGrKB2qAN9PeWoiUa0ntgG50NL7djG31t8/VhLT/YseJw14ouLPMN3ysy5wKxzwWli8SNrudFMLokjbThsrHr7zdcfCIp22sdW3nkEZSTbVu617XWPV/S1iMs11c9stGjKCN6EjEZcjLOO/0qshxt3gUqInX0rZpd/IMhe/d/j9vARNWWH63bwy8XU4TkPfac/Mz3nJxte0/7rDZa9EjRqovNQXvrT87YyBuNJHYXplj69bHvU/lYonKnH4LyiYAIiIAIiIAIHBgBicoDQ62KREAEREAEROB4EpColKg8nt/cZ7eaOAa+012P6mj7ZOSaTziurvuko7/e74MgCl9FzpeN86X0CjmrFH35Oj8X/T3Ly+kQAREQARE4/gROmqjkE0FWZoUlS8JGlCXSEhFJQlyGvERQknhPnrRkqpeFxGNZ1FZr2ZZaS9bdbFvBlzjtK3ikokvKNReUrZVla2+u2EZp3Rrninbuh4N25p1+G3FZWax49KKLukd3Wnb/Nwv26MaqLfryq7Zcsv5q0xq1fmvUG75Ma0+Qloq9uvOFvC+n6su5ljxvreOpbZv1Ndsotj2G1CWkb9iYxGKKguxFQrY77dTetsvHji8j22l3/L2ntbZHMHo/HrqwnPWxxGMfUyz3Ii2JBCWSk7I4shGU2dfp5gv+6WfLzL5Cn1WH8ta8WE57Tn7//5y10TcbLlTNFh+s2n3fp3L6d4spqnL5IaLSI1PHSjb+g4ZN/GjALvx4yPrHK2kZ3bnJVk9UpojKOZv7WhGV6TPQHxEQAREQAREQgSNBQKLySHwMaoQIiIAIiIAIHF0CEpUSlUf327n7lm1sScoVj4aYX/ZojIVVu/doxZeD8wnLreXYiEjY64Ndtny1OJeSHh1RKdhQvWxnBn0/p0bZ+mtFK/sEqs8C73W1Kk8EREAEROCACZxEUQnCkJVxRlYSJUliWVjEZSTeh8CM6Evu5Xx91Q23bL2ow/W0h6THHFq+yPVuikhEVK5vrFjOZWJtImejb9fs0h8M2+i1htWGfN/JlQ2b8WjCKY+mvP8bX8L0qzVbe+DtW/flYSssS+tL0HpiudloV97DE/v8B0EkcyFqRY+gLLpMRFLmn0rKFFHpgjEtPZtZtjVJyljClTOycrVj68sbPUHZ8gVz1/3fef+HPsbN8bWLsUW8T+etf+6/88/+1vWt4UgaFhR9KdfaSN73mhyw77moZM/J+nAp7d/JvpNz91ZcOq7Y6pyPY7x+GI1cq9vw5Zo1z1V9n8uCdda69vguovKRR1TOpSRR+a1PRG9EQAREQAREQAQOmYBE5SF/AKpeBERABERABI46gZhwefjwod28edOWlpas2Wza6OioTUxMpP2JdtsHyoxyWUqr67+ufzDfshuTi3ZzatluTbdsfsWX5XKpk/vOLM5ua3t+fiaCPKjNav4L+/Fmya5NNOzymYadG21Ys16xvrxPquUkKp9P8Pjd6fokXtujHRZbbZvx79zdmWW7ec+XoFv0yAL/PrhP9NnO9L897RySEkla9snYwXrJzg5W/btWt4nhmo0MVKxWLm5VujVLuae1qzAREAEREIGDInBSRSX8kJTZA1n5vBTSkjOyMqQmzycRuBV1yA95GOt1fWUDlk7t+vKsfZW21cZzaQ9JogMjMnDDV0NYcDk3+dt5jyZkydNFW7znkY6LPl7s9uqJiM5Ybjbv4ziO1HbfZ7LP/7nNVzxVPdV5zz/8Li75P8anablWlqB9Kiy7Lic3GK+6vEzJxxHrPk5d830h1xZ9VYYlj8l0UZnGEF4a5Ww/QNdbztWjUT0qNCXfOzLnKyskqvwhUYxHTLJcbWfd28UAom/DBi9X7er/GPHo0qYNXfJ9P33fzq7nWVv2pWcfr1m75eLVl8Qt1fIpirIxVnah6Uvu+hh3fbWbIlARlV/70q932aNSEZXbPyK9FwEREAEREAEROEQCEpWHCF9Vi4AIiIAIiMBxIBBCUaLyOHxaauPLCDDxyHKv80trdv9Ry6X4on16d95F+WpPVHoB2+ZhX1bkju7z/0dIUpZ7He4v2TkXlIjxi6N1OzNUs0a1lJkATlOWOypXmURABERABI4WgZMsKreTjujKOMd95CXXIuoSWYmo5D1HGlti47Ze++aRfs3FoEdVFuqbVj+bs+FrFTv/06ad+V6/RwdiFPuM5UtnvliyyY/nbObGks3fXfO9IX2PyXW/vfl0OVokJXUmOUkdVOV7UPb5lpXFui+nOpxPez8OXChbtek/SvPr/NuPrOR//LCI6EjOiMIkL70QXrOHJftUrngE47xHMy7cW7OlKV+y1oVl8pP+/LMO6ihUve7BgjXOln2JVheJIyUr1b2dKdqTNtIIr8OF4/pyx1q+xO3SzJrL2TXfr9KjS32vylGPlhzz6NL+s760bb9zJRrVBS7PdNsbqYiC5602i2k/y2I170vVekTl172Iyq89ovJrROU3Wvr1WZ+TromACIiACIiACBwOAYnKw+GuWkVABERABETg2BCQqFRE5bH5su6goR2Phlhb79hjj6C892jZvry/aJ/cnbPp+a2ISi8DTchc4V4eseRspZS3kUbJzo9U7c2JfrvkEbxnh+u+/GtPVPYmVfe48r3siMoSAREQARF4IYHTJCoDxBMhmEwd/4b2/h3jTApJGe8ZW5KyEYQEPuZrZvXxgo28WbHx7zfs/E+GbOhizZBtrcfrds8jKac+XfB9GRds7q7vZfnQ95JsuUBMRX1bjGalaC8q0avz/SnLTY84PFu0oStV3/fShd+4S02XfVuBl6lLlJcS6hLx2PuDw0xSsLO24QJx1R5+uWSzX7Xs8S2EaacnRMn0jCNf6rNSf581xks2/EYtRUU2z/s+noNFr7u3LC3CkrZ225u+lOu6zd9fsUe3Wjb92bKtt3wp3JGiNScqNnylt6wrsrM84Mvuevs5EJVdF5abHmnJErBjvp9lZbCUIjMRlV8TUfmrOfvmVws2j6j0vB5MuicHkaxDQ0N2/fp1++CDD+z999+3a9eu2cjISPr84zuxJ5WpEBEQAREQAREQgRNHQKLyxH2k6pAIiIAIiIAI7C0BiUqJyr39Rh1iaT7RiKhcWWsnUXnfReVXU0v2u8kFe+iiMubq0nTfHrtCn3fsRVT6Em9DvlzbuaGqvXG2bpfGXFSOuKislnsRFb5/lybzDvE7oqpFQARE4DUJnEZR+Txk8e9ZCMp4nyRl5iE8YKmR80hDJGXVJn7UtLPf73fR1u9LmRZs+dGaPfxq2SXbI5eUizZ3B0npe0Wu9iIJk0H08qKeiOiMejj7P68uKj2icShvzQslj06s+zKqg4YsLLAEK7Iw/u339vj/vnNwv+vRi+z5yBK007/zFRk+X7aHN1asNcMekf7Isx70y/lyn0vFPhs4X7YxF6QjHhVJpGjN95uk7pCVSVT6kq+IWeQikaP3Pl60pYdrqZ2VAY/IHCtZY7Rk9dGy8b5QIVrVl891QYnkRFgOnK3YZd/bc8AjL3kfovKbX8/b5H+zZK6Lyq43LAY/3+nt7i5IVO6Ol3KLgAiIgAiIgAh8m4BE5bd56J0IiIAIiIAIiMA2AhKVEpXbvhLH9C2zjr39KRGVjxZW7N7sst1+sGyf3/OIiMX1XsCE9+7JROUe9pRJ2I7byrRHpU+6TgxV7I0zNbsYorLm+0h5OEda/i01IGZL97ARKkoEREAERGDfCUhUPh/xs0Rl2rfR91Gsnyna6Ns1G/9Bv0dSDtro1YbvsVjySMKuTd9YsPu/XbDJD+fskUcwtmY6vjQqS7F6Xc8Qg8+sB1Hp9VSIqJwo2vBVr+v7AzYwXnGJiKjM7BW5rQvpX2T/w5klYdsponLNZm4u2uyXHlH51aqtPHp5RGXRIyr7iai8VvOoyLoNXvCISt8jPUnKLVHKuBvZyNKyCx5ROXvbZeXnS7Yy73u3+96WRd+Dslzn7MkjTfO+UgOSkv+jbTSS6NCRN+r21v86Y4Pna76fpi/9emfZ2KPy3n8v2NQny7b8wMc9iMpn8NvW/R29lajcESZlEgEREAEREAEReA4BicrngNFlERABERABERCBHgGJSonKk/H/C71l5tpt3/Npdb0nKh/2ROXNqWWbXfJIiK3Jun0TlV5B2SMqmy4qz/rE5FVEpe9ROTHabwN1F5Vb+3f1JljTtOjJQK9eiIAIiMApIiBRuYsP2/+pK1RcHg76agNXq3bhp4M28cOmjb87kJYu7Xpk4eO7Lbvz/83avd8sJCm4NN22jktKZN53jmdcijwpotIFXqGa8/ryVhv1PRzPeUSi7+WYR1L2Vk+N7E/P3sbsv8jI0a7vU7m60LHF6bUk/FozXWt7m14k/dgfM0VVuiitjxU9GrKY+lhy2diXJGWvojTu9qjNtsvFlfl2EpYrs23fRzPne05WrNzwTnjWznrXZS31+rnlkaVtb3Ju0/e8zFvDpS8cv/dn42mfSva6ZJlaROXUb12u3vQIUBerqb0vYPYUwstfSVS+nJFyiIAIiIAIiIAIPJ+AROXz2eiOCIiACIiACIiAE5ColKg8Cf+PwPeY0Iv1JCrXbHZ+xSZdVN6ZWbYvpn0PqAMSlSVEpU9KIiqvjPVE5bnRhg00PKLDRWWKqsSU7octPQkfpPogAiIgAkecgETlzj6gEIfV4YLvF+l7Ur7bsIs/G7Yzb/uPd85Vk0Sbv7eS9qO87aLygUcVLk2v29qCR1K6yEvRgF5V+pGRRxIiEJ8XYZlaxD+tLiNzHlWJHC16VGJ5IJ+iFFnN4EVH9m5vONGLqlxbdFG4tGFtROH6i41f6m/B9+ssmxV9mduSR0QiKfO+v2SSpFuVbA1X0nKt7IXJcq4Ixf4zZTt7fcAjMiv0OonSuW9aNu9LuC7eX0/SMucytO7LwrL3JtL32v8cTRGbLPvKvp7sUfnA97uc/xqOHk754ia/CMl37klUfgeJLoiACIiACIiACOyCgETlLmApqwiIgAiIgAicRgISlRKVx/17nySlz8ZtbvRE5fLKWoqonHy45KKyZV8iKpd9ybatCbv9cISU3fE/ISrHBxCVlRRReW50wJqIyoJE5XH/rqn9IiACIiBRuYPvgEs5pFreheHQ5apdem/Qzv+0aeeuD/ZEnN9f9j0Z73+6kATblEs2pCXLvRJliWDriUn2qPTlWF0SElG44VGWITCf2QovtydIfU/Iotfv0jLn8nDLET7zkWddpO4Nl6VEdVL/ZqcnSp+V98k16iZ5QCR1Ikx7e1Nu5YhGPOlbb5lZ2sn+nWe+17C3PULyzDvs25lPkZZwIU3/bskjJNet4GUOXarZ5feH0/K5Zz0ylfHH/d+yL+Wc7/H5OEWlrhAB2vJO7OEhUbmHMFWUCIiACIiACJxCAhKVp/BDV5dFQAREQAREYDcEJColKnfzfTmKeXvRlD6J6DOL6+ttW3oiKomodFH5YMUeH4Co7PpsYdGXdyOictwjKi+PVl1U1uzc2IANNqq9iEqPqkxLv+6HLT2KH47aJAIiIAInjIBE5Q4+UJdyhaov+TqUTwLuzT8Zs4u/N2hn3upPS7Gyp+LC/dUk4WZvLdnc5EpaBhURmcyb/xuZRKFHHLL86epcx1Z9D8fVua51Vnoi84WtQAoiDbfOL8y7/aY3IR1bTYnXW1dffKI+P9Ip6t661rvjfyk/6vCXRH+WfWnciev99u7PJ5KAJKqys9a1h18s2YObSzb9+aK1HruodAHKvpcXf2/YRq7WfYnbYhKYd11QTn40Z/d9+dz5r1etveiS9SURoE/as8MXEpU7BKVsIiACIiACIiACzyQgUflMLLooAiIgAiIgAiIQBCQqJSrju3Bcz88UlfOt3tKvDxGVqwcmKksuKgcQlQNFuzzm+3H5HpXnx/o9orJqhXwhyUqJyuP6TVO7RUAERMBMovIl3wIXcwjCsu/VOHChZOd+NGBv/q8xO//jQRu6WLNyfzEte7q60LaF6dUk2laXOi7mfMlXX+KVI+dLtXbb7BPZTvtEzt5etkdftezRrVVb8b0XiXDMyr5ntmi7IHxmppdczAjFl+T87u0d1k9EZbHWZ6Nv1u3NPxm1C78/aGMIXR9HrLc6KbJyaWbNkLtwqfqem01fOpdo0bQ35VfsTfnYpjw6dfZL5/mwnSJQXxh5+t3WvvSKROVLESmDCIiACIiACIjACwhIVL4Ajm6JgAiIgAiIgAjww3Vf1sqXzHz48KHdvHnTlpaWrNls2ujoqE1MTFij0dg1JsqMcrvdjk82te2Bi6Mbk4t2c2rZbk37njsrnRR9ltvHyDKWwyr6Elw1n8whwu3aRMMun2kYewY26xVfnqvgkz4Slbv+gI/YAy8UlURUzhyyqBx1UdkvUXnEvjZqjgiIgAi8EgGJypdgQ1T6XpGVwbwNXirb+A8aduUPR23clzQlUrBUK/i4czOJyTVf7YDIwY2tpVX595wf8/jQzDq+BOzKXNvmPdrywc1Fm/E9LGc+X7HlmXbaw9L2dmXTl3Rq/26zVCzSceBC2ZfHHbCJHw7Y+PcHbOBsxYplv+kHLGBEqCbLynIdiUs06vRnix5JOW+zX62kPT7XFxG+/tDrSNZU67f/SFR+m4feiYAIiIAIiIAI7I6AROXueCm3CIiACIiACJw6AiEUJSpP3Ud/Yjr8fFG5tUelRyIc1NKv34qo9KVfL4x5RCWikojKgiIqT8yXTh0RARE4tQQkKl/y0SMqPZX6c9YYL9jQlWoSb4MXalYbLPlSp7kk0tLSrr4P5CaJX5Zx4OJccvblcimicm3RIyof+L7Td5Zt7vaKzd1Zs5XHHlHZ7eVNzxzzP7BCVlYGC9a8WEq8Rt70H9R51CS82K+SvS6RlAjIbsf38fTo0yXf43P2yyV7dLtlj++4pHSBi6TsrjnEPZaUIJaoPOZfNDVfBERABERABA6ZgETlIX8Aql4EREAEREAEjjoBiUpFVB717+jL2nekRWV26VeJypd9lLovAiIgAkeegETlSz4inJon9l5kj8r6mC9Vet6l24hLt7r/YMejB5OXzPg08seRxJ0vcUoE4Xqr61GV60lWLk2vW2vaZdySRwwi4vZBxkUbDvS8xStf9r0qmzmrj5as/1zFo0/L1nBmlYGCs/SxKkxc6hKBurrQ8cjSNZv7eiUtjdua7SQu7PG510u+BguJyiChswiIgAiIgAiIwKsQkKh8FWp6RgREQAREQAROEQGJSonK4/51l6g87p+g2i8CIiACx4eAROUOPiuXbzkXksjKYj1nlWbB92HMuaTMpWVdwzE+8ZNPXvTKRsoRadl18db2vRnXlropWrDtkrK7njGcO2jKsciCrPRIUiRuwTmVXU6ydG7VuZXr+RSF+lRUbjiPjq3OdywJysWuy0vfxgFJuQ9LvgY/icogobMIiIAIiIAIiMCrEJCofBVqekYEREAEREAEThEBiUqJyuP+dZeoPO6foNovAiIgAseHwEcffWR///d/b//8z/9st2/ftrm5uSPSeGwf66bWPA3667InzNWaRx8u+nnVz52ta37azwPxRvIlS3NFF3Cl3hnZxhKmHOm09bp3ZevvVhaiJjd9L8sNbzJystv29/sYMfitNhziG5aBzXl0ZbHa58u+5qxQdsFbyiWeaenX9oatr25Yp7XhEtf5OJv9FJSBAlE5ODho169ft5///Of23nvv2ZtvvmkjIyOWz2ssHZx0FgEREAEREAEReDYBicpnc9FVERABERABERCBLQISlZpcOe7/zyBRedw/QbVfBERABI4PgQ8//ND+7u/+zv7pn/7J7t69a/Pz80eo8Zi/qlvAbaLSltzyuai0A9zcEeFIczi7oCRiMCSlv8q+5O3Tg2c4tgInkXAIy/R+HyMGe5Uegb+Jl/NxyZsvZNhtEUvy1qNNN/yjTPt7HhCTcrlsAwMD9sMf/jCJyj/8wz+UqDwCXxc1QQREQAREQASOCwGJyuPySamdIiACIiACInBIBCQqJSoP6au3Z9VKVO4ZShUkAiIgAiLwEgIRUYmovHPnThKVuRwRb2HYXlLAvtx2kZdEFm2o+MusqCSSctmve2RlEpWYLY4Dbi9NfJXjgJv5Kk3c12eex+2AuDDGIiEqm82mIir39cNW4SIgAiIgAiJwcglIVJ7cz1Y9EwEREAEREIE9ISBRKVG5J1+kQyxEovIQ4atqERABEThlBD7++GP7h3/4hxRR+cUXXyRRWSwW0/KXh4ciRKWvG9q3FVHZV/LmEHbH0q8uKjkbS78SVclxQKarV5nXHy92eT7gZu6ydfuf/XncDogLY6xOp2Ms/To8PJxE5Z//+Z/bH/zBH9gbb7yRrmnp1/3/GqgGERABERABETjuBCQqj/snqPaLgAiIgAiIwD4TkKiUqNznr9i+Fy9Rue+IVYEIiIAIiMAWga+++sr+8R//0VgC9ptvvrGlpSU7WqLS96bsa3jyzSGTqGRzR6Iq1/19do/KAzJd8c15nnCL+887H3Azn9eMQ7v+PG4HxCVEZalUShGVyEkk5TvvvGMTExNpOdjDjyg+tE9HFYuACIiACIiACOyQgETlDkEpmwiIgAiIgAicVgISlRKVx/27L1F53D9BtV8EREAEjg+BxcVFm5yctNnZWWu1WtZut+3wRQ02C3NFIqoSSclrrrPJI1GULPkay776Sx0isAMCjLFIfMeRlf39/TY2NpakZa1WS5L+cJc93kEnlEUEREAEREAERODQCUhUHvpHoAaIgAiIgAiIwNEmIFEpUXm0v6Evb51E5csZKYcIiIAIiMDeENjY2EhLYXa73SRw0r9Be1P0HpWyJSw5ISqfROQ9ebFH9aiY00YAIYmwZKnXWO5VkvK0fQvUXxEQAREQARF4NQISla/GTU+JgAiIgAiIwKkhIFEpUXncv+wSlcf9E1T7RUAEROD4EODfnEi0Ov0bdKSanwxlL6AyNTAaJ1EZJHR+NQIhJTlHerWS9JQIiIAIiIAIiMBpIyBRedo+cfVXBERABERABHZJgAk2ogMePnxoN2/eTHstNZtNGx0dTXvPNBq+z9Euj5jAo9xut2NdXxbtwXzLbkwu2s2pZbs13bL5lY4V8/7LbJ/s2K/Du+Z1mNWKfTbeLNm1iYZdPtOwc6MNa9Yr1pcv+C/DJSr3i/9BlZsmidPE8Yatr7dtaWXNHvn3bfLhkt2ZadmXM2v2eLnjk8m9Fu3HV46yu/6n5N/pgWrexgeKdnm0ahdG63Z+rN+ajaoVCoUnEQg+w3dQeFSPCIiACIiACIiACIiACIiACIiACIiACBwaAYnKQ0OvikVABERABETgeBCQqJSoPB7f1Oe3UqLy+Wx0RwREQAREQAREQAREQAREQAREQAREQAQOk4BE5WHSV90iIAIiIAKnmkBEFYYI5MwySeztQorlk7ZDiueIRsw+s19LLEX7FFG5/ZPQ++NCgO8w4ZKbm4qzfL9ZAABAAElEQVSoPC6fmdopAiIgAiIgAiIgAiIgAiIgAiIgAiJwOghIVJ6Oz1m9FAEREAEROIIEEI2dTielti99ynsEZbFYtHK5nJaAfFazyUf+eJZnSqVSWjbyRYLzWWXt5JpEpSIqd/I9Ocp5JCqP8qejtomACIiACIiACIiACIiACIiACIiACJxmAhKVp/nTV99FQAREQAQOjQDiBNm4srLyJCEe8/l8kpT1et0qlUqSllzjQFB2u11bW1tLz6yurqYykJO1Ws2q1Wp6FtH5vGjMV+mwRKVE5at8b47SMxKVR+nTUFtEQAREQAREQAREQAREQAREQAREQARE4CkBicqnLPRKBERABERABA6EANKE1Gq17NGjRzY/P28LCwu2vr6eBCPCsdls2uDgYDojLBGPiM3l5WVbXFxMz/AaaYnIbDQaNjAwYMPDw4bk5NpeycpoL0u/3rhxw5aWllK7RkdHbWJiItW9W3BRZk++dqzrfXsw37Ibk4t2c2rZbk23bH6lY8W8L4Xrfd+vwz8Gr8OsVuyz8WbJrk007PKZhp0bbViz7tzzBY9y3TuW+9UPlftiAnzftPTrixnprgiIgAiIgAiIgAiIgAiIgAiIgAiIgAgcBgGJysOgrjpFQAREQAROLQGECVGRSEkE5fT0tD1+/DhJSyIkuc7BUq5nzpyxd955x0ZGRtI1BOXk5KQhDHkdedNN/4PcvHz5siEQkZ2UsReyMqSiRGWQ1vm4EZCoPG6fmNorAiIgAiIgAiIgAiIgAiIgAiIgAiJwWghIVJ6WT1r9FAEREAEROHQCIfwQkkRQzszM2P3795N0RCiyDCwRlpHOnz9vP//5z+3NN99MEZj37t2zX//610lWskwsIhI5SaQl91j+9cc//rFdvXo1yUqiLPcisjLaLVF56F8hNeAVCUhUviI4PSYCIiACIiACIiACIiACIiACIiACIiAC+0xAonKfAat4ERABERABEQgCIfwQlURTIiSRf7xHOrKU6zfffGO3b9+2L774ws6dO2e//OUv7Xvf+16KuPz666/tww8/TIKzXC6npV7HxsbS8q83b95MMvPatWv2xhtv2KVLl1IkJvlij8tox/POtI9oTyQo8pPXXGN5Vs5zc3OpbSw3OzQ0ZNStpV+fR1PXjxIBvr/+Jfb/bXgkctuWVtbskS81PPlwye7MtOzLmTV7vNwhSzr2Y7Vhyu76n5IvZzxQzdv4QNEuj1btwmjdzo/1W7NRtUKh8PTHBfvRiKP0oagtIiACIiACIiACIiACIiACIiACIiACIuAEJCr1NRABERABERCBAySAMEEEIvtISEpEIKISeYmg/N3vfmcfffRR2m/yF7/4RVrOdWpqykgPHjxI+VkOlv0oOVPGrVu3kvikfK59//vfT6KTqErK3slBuygLYUpbiPBEWIaojD01c7lcWpZ2fHzczp49qz0qdwJXeQ6VgETloeJX5SIgAiIgAiIgAiIgAiIgAiIgAiIgAiLwXAISlc9FoxsiIAIiIAIisPcEECYk5B8SEDGIIOQ9EZZfffVVkpWff/55Eo5/+Zd/mc43btxI0ZdEOSIfWRaWiEaWfkV43r1714i4JCKTJWB/+tOf2pUrV2xwcNAqlcqOOkJ7kJMsS0tblpaWvhVZST3cKxaLSVAiKZGVtGe3R5ZDt9uxrtf9wCPcbkwu2s2pZbs13bL5lY4VPfost4+RZf5ReB1mtWKfjTdLdm2iYZfPNOzcaMOa9Yr15QuWy+X3ZK/P3TJS/r0jIFG5dyxVkgiIgAiIgAiIgAiIgAiIgAiIgAiIgAjsJQGJyr2kqbJEQAREQAREYAcEkjTxfEQosvQrUpAIRl5PT0+nJVa5d+HCBfvTP/1TY/nWX/3qV+l+tVpNgvLy5cvpjCREIBJt+eWXX9rHH3+clo9877337K233kpRlzyzkyMbUYmQjIjKWAKW97Ozsy7uFFG5E57Kc3QISFQenc9CLREBERABERCB/5+9O31uI7vu/3+wEcRCkBI3URK1znhsT2zHScYTu+qXh6n65klS+YNTlVRiu+JkJrZnbI84kkYLJe7iAgIkAVC/c06zJYBDit0gQGxveCBsF43uVzcI+H5w7kUAAQQQQAABBBBAAIFmAYLKZg2uI4AAAgggcIUCe3t78urVKw8ZLWi0oNKCwKOjIx+u1Soif/WrX/ka/fa3v/XHbbhXmxfS5qG0ikoLIa39+vq6WNWltbOTPe+TTz7xakyrsIxyskAyrPK0oNQCULsvHPrV1tfW027PzMz48K8M/RpFlja9FiCo7PUe4PURQAABBBBAAAEEEEAAAQQQQACBswUIKs924V4EEEAAAQS6LmDB3/Ly8ruw0uafDINKq6K0qslf/vKXvh4WQFrlpYWTNuzr6aDSQk4bLvbXv/61tw+DSgsUowaVFuZYMGmVlRZYNoeUFk5ub2/Ls2fPPMC0wNSWbaEpQ792/VDhBS4pQFB5SUCejgACCCCAAAIIIIAAAggggAACCHRJgKCyS7AsFgEEEEAAgYsEwqFfw0pKCyqtYtGGgbUqSQsBP//8c0mlUvLll1/6/WcFlRYqhhWVv/nNb3w+RQs441ZUWpjTfLZw0k52n123oPTbb7/1IWtt7kuCyov2MI/3iwBBZb/sCdYDAQQQQAABBBBAAAEEEEAAAQQQaBUgqGz14BYCCCCAAAJXJmAViza8qp0tmLTA0uaZfPr0qVcuFgoFsbkmbXjXpaUlDwitkvHmzZtiw8LOzc2JtbHn2tyWFiJ+8cUXPoekBZxx56i0DfdA50QgvG6XdrZqTxtedn9/XyYnJwkqr+xI4YUuK+DHsh/Hx/p+qUm5eihbOxVZ3ijLs/WKPF4/lDf7dT3Og1dKJC77it9/vi27of+MpRJSyqVkvpSRuzM5uT1TkFuzEzJZzPn8svbDhIStQDdW4vurxT0IIIAAAggggAACCCCAAAIIIIBATwUIKnvKz4sjgAACCIyqQHMIaKGEna1i0YJKG8L166+/9vt+8pOfeBhplZYWaFowaZWMNiysBZWlUsnvtyFkv/vuOw8rx8bG5G/+5m/k/v37YpWP4+Pjl2IOg0oLUi2oLJfLBJWXEuXJVy1AUHnV4rweAggggAACCCCAAAIIIIAAAgggEE2AoDKaE60QQAABBBDomICFJjaUahgAWkhpVVThnJVPnjzxsNIqJW/fvu1zTNowsXbbnjcxMeHzVFpQee3aNTk4OPCQ8vXr1/LmzRt//NNPP/U2FmRacHmZU7i+YUUlQeVlNHluLwQIKnuhzmsigAACCCCAAAIIIIAAAggggAACFwsQVF5sRAsEEEAAAQQ6JmCBic0pWa1WfShXG0a1Xq979aQFjhYG2jCuL1++9CFc7+kQr1YVaQGltbW5KG3IWAsgbfhVCyotwLSKSnt+Pp/3SkurprT5LG3Y2HQ6fan1D4NKq6i0IWgJKi/FyZN7IEBQ2QN0XhIBBBBAAAEEEEAAAQQQQAABBBCIIEBQGQGJJggggAACCHRCIAz8LOizMNLCxRcvXsj29raHjxY4WoBpgaMN87qwsCB///d/L4uLix5arq2tye9//3sPMa1dMpn06klbN1umhZY//elP5cGDBz48bLFY9EpNa3eZU7jeBJWXUeS5vRQgqOylPq+NAAIIIIAAAggggAACCCCAAAIInC9AUHm+DY8ggAACCCDQUYEw8LMhXm2YVquatHklrYrSqiytstKqJW0oWKuCvHPnzrug0oaGtXDzyy+/lKdPn/p8lvYcq5i0x+x5Fmx+9tlnYlWYFlJmMhlf1mU3IlxvgsrLSvL8XgkQVPZKntdFAAEEEEAAAQQQQAABBBBAAAEEPixAUPlhHx5FAAEEEECgowIWmFjF5O7urs8naSGlVUNaSBnOW2kVkDav5PXr1+Xu3bteKWnhZTiHpQWG4XMspLSzPW5DxFp7e54FnXZ/J04ElYFvJyxZRm8ECCp7486rIoAAAggggAACCCCAAAIIIIAAAhcJEFReJMTjCCCAAAIIdFDAAhOrfrShXS2wtCFcbchXu8+CSgscLai0akirlrS5KLPZrN9v7SygrFQq/vyw+tLa2zlsPz4+7u1tWZ04EVQSVHbiOOrlMggqe6nPayOAAAIIIIAAAggggAACCCCAAALnCxBUnm/DIwgggAACCHRFIAwrLZgMA0q7z8MUfcUwrLSKyObKSGsfDg9rz7P2YRhpl9beAk4LLcP7O7EB9jr22gz92glNltELAX9v+XvsWH8YUJNy9VC2diqyvFGWZ+sVebx+KG/26/qeCtauQxl/y6bashv6z1gqIaVcSuZLGbk7k5PbMwW5NTshk8Xcu/e7v3+7sRIta8QNBBBAAAEEEEAAAQQQQAABBBBAoPcCBJW93wesAQIIIIAAAn0tQFBJRWVfH6ARVo6gMgISTRBAAAEEEEAAAQQQQAABBBBAAIEeCBBU9gCdl0QAAQQQQGCQBAgqCSoH6Xg9a10JKs9S4T4EEEAAAQQQQAABBBBAAAEEEECg9wIElb3fB6wBAggggAACfS1AUElQ2dcHaISVI6iMgEQTBBBAAAEEEEAAAQQQQAABBBBAoAcCBJU9QOclEUAAAQQQGCQBgkqCykE6Xs9aV4LKs1S4DwEEEEAAAQQQQAABBBBAAAEEEOi9AEFl7/cBa4AAAggggEBfCxBUElT29QEaYeUIKiMg0QQBBBBAAAEEEEAAAQQQQAABBBDogQBBZQ/QeUkEEEAAAQQGSYCgkqBykI7Xs9aVoPIsFe5DAAEEEEAAAQQQQAABBBBAAAEEei9AUNn7fcAaIIAAAggg0NcCBJUElX19gEZYOYLKCEg0QQABBBBAAAEEEEAAAQQQQAABBHogQFDZA3ReEgEEEEAAgUESIKgkqByk4/WsdSWoPEuF+xBAAAEEEEAAAQQQQAABBBBAAIHeCxBU9n4fsAYIIIAAAgj0tQBBJUFlXx+gEVaOoDICEk0QQAABBBBAAAEEEEAAAQQQQACBHggQVPYAnZdEAAEEEEBgkAQIKgkqB+l4PWtdCSrPUuE+BBBAAAEEEEAAAQQQQAABBBBAoPcCBJW93wesAQIIIIAAAn0tQFBJUNnXB2iElSOojIBEEwQQQAABBBBAAAEEEEAAAQQQQKAHAgSVPUDnJRFAAAEEEBgkAYJKgspBOl7PWleCyrNUuA8BBBBAAAEEEEAAAQQQQAABBBDovQBBZe/3AWuAAAIIIIBAXwsQVBJU9vUBGmHlCCojINEEAQQQQAABBK5M4K29kv7z1v5nl/bPFZ4SiYQk9PX0wv+xC04IIIAAAggggECvBAgqeyXP6yKAAAIIIDAgAgSVBJUDcqieu5oElefS8AACCCCAAAIIXLGAB5P6msd65fj4rdQbx355JWGlJpIWUSaTCUmdnO16GFxeMQUvhwACCCCAAAIIuABBJQcCAggggAACCHxQgKByMILK4If4V/tr/ODACX6D77/I/+CR1LsHm4PKw6Oa7FcPZXOnIssbZXm+XpHH64fyZr/uFQ22lt3YFts/Df1nLJWQUi4l86WM3J3Jye2ZgtyanZDJYk7S6bSkUifHWzdWone7gFdGAAEEEEAAgRMB+15Sb7yVmgaUB0d1KVdrclhraHD5vrKy0xWO4TdECyQtoBzLJCU/lpZxPWfHUpJOJrvy/YedjgACCCCAAAIIRBEgqIyiRBsEEEAAAQRGWICgsv+DSttHFoTZL/Ov9HTyq3zrTLNczTq/+vH0oaDymQaVTwgq+3G3sU4IIIAAAggMnYB/r9bva0e1Y9k/qMnW3qG83tqXnUrNg8u3x+EPpjr9ncq+K771SsqxTEomchmZKWXlejErpUJWchpYelVlp1926PYgG4QAAggggAAC3RAgqOyGKstEAAEEEEBgiAS8Q+X4WDY2NmRpaUnK5bJMTk7KzMyMLCwsSLFYjL21tsxwuY1GXRq1mqxphduj5T1ZWtmXp6sV2anWJaPVZ8kuhl+W62lfjeQzCZmfHJOHC0W5O1eUmzNFmSyMSyKV1g6d/g0qzdB+kW+/wq/qL/L3D+pSqx9fyS/irTMrk0r6L/LHFTGrZ+v4sl/p99vJnCzJfau9f6crKgkq+21vsT4IIIAAAggMr4B9J2lo6WT1sCY75SN5/UZHdni9K+u7h/4dzh6z776d/vprX4XsB21p/Z42nk15QHl7Oi83ruVlZionxfGMh5j9+qOz4T0i2DIEEEAAAQQQMAGCSo4DBBBAAAEEEPigQBgoElR+kOnKH/T9oh1OFQ0nN3cPZOVNVV6s78uuDh+m+W7Xqhvtda1u00LKYi4t1/SX+LOT4zJdGtdwN6tDiGny22cnW2eCyj7bKawOAggggAACIyhgYWFDh3zd1+9rm7v23a0sf365K6vbVf3hmc5VqY8HlY2d/eFX8H1eg0r9kpjPpvW7W1bu64/zbAj6G9cLWlU5pj82C4d/7exrj+BuZpMRQAABBBBAIKYAQWVMMJojgAACCCAwagJBxwYVlf22322/WEfXTuVIXm3uy3cr1tG140OIWVVjtwobLfOzTjSroLxWHJO5qXG5M1uUBf1V/txUXgr6i/x+OxFU9tseYX0QQAABBBAYTYFjrZisNxqyp9/fNrYrOorInnz9YlcrK6s+HKzNZ921ikp97TConJvKysP5YCSRWz6SSFbnyU56VaX+3G00dw5bjQACCCCAAAI9EyCo7Bk9L4wAAggggMBgCBBU9ufQr9bR1dCOrjflQ3m5UZZvX2lH1/Nt2dChw2xYr04PGRYerVZNqSMBa+Vk0oPKhWs5uacdXbe1k2thuqBVlhkrXvRTvwwf9qGg8rnOUfmYOSrD3cslAggggAACCHRLQL8g2fe3Wr0hu5VDWXujPzRbLQdB5fZBU1DZ+ZEx/Pu8fj+z74h5Hfp1TisqH84XfMqDW7MTMqUjZKRTKQ0qk++2vl++x71bIa4ggAACCCCAwNAKEFQO7a5lwxBAAAEEEOiMAEFlvwaVx9rRVZc3e4fyXIcNW3q1K1892/Gg0isq3/czdeZAOFmKhZDWyWbzUl7XisqF6zm5r0Hlog4fdkuHD5vIjenQsMEv8fulg6s5qDw6qkm5eihbOifqsga8NkclQWVHDxEWhgACCCCAAAJnCeiXKBsN46hWl939A1m1oHJtX/70ck9Wtg/lSANMm6PSvsd1uqZRf2Pm39/SGkR6UFka0+9vebk7W5DbFlRO5CSTtrnG9Qvkya/d+uV73FmU3IcAAggggAACwyVAUDlc+5OtQQABBBBAoOMCBJX9F1QG+6ThHV1bewc+N6VXVOrQYRsaXNqv5ZN67sYpeG3RoV+1ojKfkYVr415ReceDyqJM6BxHwZBhVtXZnXWIu10ElXHFaI8AAggggAACnRXQOb4tqNQw8lCDyp1yEFQ+06DyL6/2ZXXHgsrjIKjUr0+d/g5lw/ZrBurfEXM6n/hsKSP3ZnM6fH9BFjWovGZBZSbtVZUWVHb69TtrydIQQAABBBBAYNgECCqHbY+yPQgggAACCHRYIAimmKOyw6xtL85DN61ZtPmNrDpwa9eCSh361eeoLMvm3pHPP9StjNArKvWfbDopU/m03NA5Ku/NFcSDSp2rspTPaudW8Gv8funkIqhs+3DjiQgggAACCCDQCQH97mTfR+qNuhweBUHlypaN7LAv37yqyNrukQeVFij6HJWdeM2mZehifY5xq9a0oHJmQoPKGZtnPC+350pyXYPKMQsq0+kgpOzWF8mmdeIqAggggAACCCAQChBUhhJcIoAAAggggMCZAgSV/VNRGYSUoh1dx1K3X+QfHWlQWfWKyser+ov85bJslWs+ZFiXCip9/knrRBvToHIyl5J5nePIgspFO+sv8ksFCyrVTIcOI6h8/5ayDsKGuaUSUjI3rWS4O5PTuT0LYnNDTRZz3jmY0vmh3I0Owvd4XEMAAQQQQGCABcLvb3Ly/e1Af2hmFZUeVGpF5TcrVVnbOZKaDgt7NUFlUmaKGblzElQungSV2bGMfhex7yHvh38dYHZWHQEEEEAAAQQGSICgcoB2FquKAAIIIIBALwQIKvstqAx+kV/TYcMODsOgsixPNKj85nXFg0ob+rVbOZfmbd6JZoGbBZVzpazcncvrL/KLOsdRUQO3cQ0pg04uC9z6Iaz0DkINCS3gZY7KXvwV4TURQAABBBAYXYEgqDypqNT5xQ8Pa7LdFFQ+sqByt9bdoFL5LQT1ikodvn9ag8q701n9kZn92GxCrpdyEgSVVlHZPz82G92jhi1HAAEEEEBgtAQIKkdrf7O1CCCAAAIIxBYgqOzjoPLgUDZ3qvJ8oyxPVyvyyILK/ZrPP3QVQWVpPClzWlF5dybvHV23taNryoPKtM6RGXRyEVQGbzkqKmP/6eEJCCCAAAIIDIXA6aDywILKvWpQUblekaWmoNLa+g+9Orzl4Q/NvhdU2nc4Dyrzks1aRSVBZYfpWRwCCCCAAAIIRBAgqIyARBMEEEAAAQRGWYCgss+CSq8MfCu1ek2q1TCo3JenOnTY0uvq1QeVpTEdOsw6uYKhX20I06QOX5r0qkoqKsO/HQSVoQSXCCCAAAIIjJbAB4PKNQ0qV6uy3lRReSVBZSGtPzTTikodgp6gcrSOR7YWAQQQQACBfhQgqOzHvcI6IYAAAggg0EcCBJX9GFTqEKY69KsHlbsVn6PyqXV06S/yr6aiUnyuxYlsQod+1aByNudDv1pH15QGlQmCyu+9gwkqv0fCHQgggAACCIyEwNlBpc1RuSfPTioqexFU3tGgMvixWUmHfqWiciQORjYSAQQQQACBPhUgqOzTHcNqIYAAAggg0C8CBJWDE1Q+WqnIm/36FQz9SlAZ9/1JUBlXjPYIIIAAAggMh0BrUNnQOcZtjkod+nWzx0Glz1Fpo2KUZJqgcjgONrYCAQQQQACBARUgqBzQHcdqI4AAAgggcFUCBJUElc3HWjDHEUFls0mU6wSVUZRogwACCCCAwPAJ9HVQacP3zxNUDt9RxxYhgAACCCAwWAIElYO1v1hbBBBAAAEErlyAoJKgsvmgI6hs1oh+naAyuhUtEUAAAQQQGCYBgsph2ptsCwIIIIAAAgh0Q4CgshuqLBMBBBBAAIEhEiCoJKhsPpwJKps1ol8nqIxuRUsEEEAAAQSGSYCgcpj2JtuCAAIIIIAAAt0QIKjshirLRAABBBBAYIgECCoJKpsPZ4LKZo3o1wkqo1vREgEEEEAAgWESIKgcpr3JtiCAAAIIIIBANwQIKruhyjIRQAABBBAYIgGCSoLK5sOZoLJZI/p1gsroVrREAAEEEEBgmAQIKodpb7ItCCCAAAIIINANAYLKbqiyTAQQQAABBIZIgKCSoLL5cCaobNaIfp2gMroVLRFAAAEEEBgmAYLKYdqbbAsCCCCAAAIIdEOAoLIbqiwTAQQQQACBIRIgqCSobD6cCSqbNaJfJ6iMbkVLBBBAAAEEhkmAoHKY9ibbggACCCCAAALdECCo7IYqy0QAAQQQQGCIBAgqCSqbD2eCymaN6NcJKqNb0RIBBBBAAIFhEiCoHKa9ybYggAACCCCAQDcECCq7ocoyEUAAAQQQiCgQdFx8v3Eikfj+nU33nH7eRe2bnhr7KkElQWXzQUNQ2awR/TpBZXQrWiKAAAIIIDBMAgSVw7Q32RYEEEAAAQQQ6IYAQWU3VFkmAggggAACEQSOj4+lXq+/O9ttCxzT6bSMjY355ekAMgwNa7WaP6/RaPhzwvapVOdDtfA1NzY2ZGlpScrlskxOTsrMzIwsLCxIsViMsLWtTWyZ4XIbjbo0dHvWdiryaHlPllb25elqRXaqdcmkEpK8ILRtXXK8WxYeZVIi+UxC5ifH5OFCUe7OFeXmTFEmC+OSSKUlmey8aby1fN/azBRO/zuWo1pdqtVD2dytyIt1NVtTv5WKvNmvSzqZ0OPi/fM6eY2gsj1N23UN/WdMj+lSLiXzpYzcncnJ7ZmC3JqdkMlizt/z797D3dqB7a0+z0IAAQQQQACBNgX8+5sE333r9YYcHNZku1yVlc09ebZe0e++VVnfrUmtcSzH+l3Bvv93+mtc8P3traT0O2Iuk5TpQlruTGdlcSYvi/MlmS7lJZvNnPz/j6SvQ5uby9MQQAABBBBAAIHYAgSVscl4AgIIIIAAAp0RsLBxf3/fg7+9vT2x2xZSFgoFuX79uuTzeQ3J3ncUhOHewcGB7O7u+nPtuj2nVCp5YJjLBWFHZ9YwWEoYKBJUdlK1vWURVHbA7agmZQ14tzQYX94oewfh4/VDD3gtTLRTNzJCgsrAln8RQAABBBAYNQGCylHb42wvAggggAACCMQVIKiMK0Z7BBBAAAEELikQVlJaSLm1tSXb29sePB4eHvqvl7PZrFcsXrt2zasWLbi0kz1uAeXOzo4/p1KpeFWlhZkWUFqV4+zsrExMTHh42RxyXmaVw4DUgspHjx5RUXkZzEs+l6CyPcAWN4LK9hB5FgIIIIAAAgi0JUBQ2RYbT0IAAQQQQACBERIgqByhnc2mIoAAAgj0XsA6Ko6Ojjzs29zclNevX3voaCGkVUdWq1V/3MJMG1b1F7/4hdy+fdsDTAs1v/nmG1lZWRELKa2NhZp2aaGnVVX+6Ec/kps3b3p1pT3mQ0d1oDzM1tvW117fhn6112Lo16s/nloCN4Z+jbwDWtwIKiO70RABBBBAAAEELi/g30MY+vXykCwBAQQQQAABBIZWgKByaHctG4YAAggg0I8C1lFhgaRVRVqF4qtXr7xK0tbVwkcLAy2QtMtbt27JP/3TP8lHH33kYaQFlL///e89qAxDSquitCFjbTnj4+MeVN67d+97lZVRLWz9bNnhpT0v6FwRXy+bo9LWkzkqo4p2tp3vC91HzFEZz7XFjaAyHh6tEUAAAQQQQOBSAsF3aeaovBQiT0YAAQQQQACBoRYgqBzq3cvGIYAAAgj0m4B1VFj1pM1JaUO+Wlhpt22eSatUfPnypTx79kyePHkic3Nz8i//8i8eVFo4aEGlVTTa8K821GtY1WjBp7W3wPLGjRuyuLgo9+/f94pHCy9t2VFOtm6NRsOHk7Vl1ev1d6GlPWbra+tm62vD0tows1b1WSwWoyy+pY0tz84WijYadWno663pnIGPlvdkaWVfnq5WZKdal0wqIckOVIS2vHjTDV0FfQ2RfCYh85Nj8nChKHfninJzpiiThXFJpNI6T2jKK1Obntazq2amcASVMfdAixtBZUw9miOAAAIIIIDAZQT8ewgVlZch5LkIIIAAAgggMOQCBJVDvoPZPAQQQACB/hKwjgoLAS1ctOFaLZy0QDCTyXgA+fTpUw8jv/rqK7l+/br88z//s9y5c8crLNfW1mR1ddXDvenpaQnPtqzHjx97xaNtrQWIn376qVdkWog4NjYWCcFCSlu3cBhaG6LW7rMw0c4Wrlrlpt1nr21BKkFlJNqONWoJ3Bj6NbJrixtBZWQ3GiKAAAIIIIDA5QUIKi9vyBIQQAABBBBAYLgFCCqHe/+ydQgggAACfSgQVBE23lUv2u1kMukVixZU/vnPf5YvvvjCKxX/3//7f14Zafe/efPGK/usktKGhbVA0q5bsGiVmM+fP5fvvvvOqy3/9m//1qsqp6amfEjYKAwWmNqyrHrTqjYtALX7LJi0DhYLVq0C1E7z8/N+tgpOKiqj6HamTUvgRlAZGbXFjaAyshsNEUAAAQQQQODyAgSVlzdkCQgggAACCCAw3AIElcO9f9k6BBBAAIEBEbAODJuX0uaA/NOf/iT/93//JxMTEz5HpYWRf/jDH3xeS7vPwkGbh9IqGguFgoeLNiysPdcCzlQqJZ9//rl8/PHHXvlow8RGOVkoGVZ62hya1Wq1JagM59BM6FCsFlTaehBURpHtXJuWwI2gMjJsixtBZWQ3GiKAAAIIIIDA5QUIKi9vyBIQQAABBBBAYLgFCCqHe/+ydQgggAACAyBgnRdWtWjDulpIaee//OUvXkn5r//6r16x+Nvf/tarGW042Js3b8qDBw+8otJCSBuidX193YeMtXYWJP7qV7+STz75xIPKfD4fScHWIayotGFew4pKq/i0dbSKSht+1k5WzWlhJUO/RqLtWKOWwI2gMrJrixtBZWQ3GiKAAAIIIIDA5QX8ewhzVF4ekiUggAACCCCAwNAKEFQO7a5lwxBAAAEEBkXAAkGrYFxeXvaw0YZxtds2vOs//uM/eoXkf/zHf3gYOTMz8y6otIrK8fFxn1fShmT95ptv5Ne//rVvdhhUWvuoQaUFkuEclVZNaQGo3ReebThY5qjs7VHVErgRVEbeGS1uBJWR3WiIAAIIIIAAApcXIKi8vCFLQAABBBBAAIHhFiCoHO79y9YhgAACCAyAgA35aiGjzUNpYaVVRN6+fVvu6fCuP/jBD6RcLksYVJ6uqLQQ0gJFCyofPXokYUXlL3/5S3/u9PR05KDSOlHCUNKqK8Pr4f02R+aTJ0+80tLmvrSqSioqr/YAawncCCoj47e4EVRGdqMhAggggAACCFxewL+HUFF5eUiWgAACCCCAAAJDK0BQObS7lg1DAAEEEOh3AQsYbThVq1K0oV5fv37tIaCFkX/1V38ld+7cEQsEbVhXq5S0oWFtjkobctWGfrWKymKx6EGlzVH57bffyu9//3uvwPzss8/ko48+EltW1DkqzSvoSGmVs/sstLQw1ObBtHWenJz0oWkJKlutun2rJXAjqIzM3eJGUBnZjYYIIIAAAgggcHmB4Pv1W/+eXa835OCwJtvlqqxs7smz9YosrVRlfbcmtYaOZKLfu+1Hi4nLv2zLEt7qLVt2KpmQXCYp04W03JnOyuJMXhbnSzJdyks2m5F0Oq2vn/R1aFkANxBAAAEEEEAAgS4KEFR2EZdFI4AAAggg8CGBsELRqigtBLQw0ILJGzduyP37931+yUwm4wHl//zP/4iFkalUysNHCyqtnQWGNq/ks2fP5LvvvvOqzGw2Kz//+c+9ItMet+FhL3M6HVRahSdB5WVE239uS+BGUBkZssWNoDKyGw0RQAABBBBA4PICBJWXN2QJCCCAAAIIIDDcAgSVw71/2ToEEEAAgT4TCEM/q6a0uSh/97vfyYsXL7wq0qojFxcXfQ5Kq5q0MHBsbMznq7RqSQsqLSS0+yyktKFXLdi0ZT1//tzDzkql4vf98Ic/9OXYMq39ZU62znYOh5clqLyM5uWe6x1dvj+O5YigMjJmixtBZWQ3GiKAAAIIIIDA5QX8ewhDv14ekiUggAACCCCAwNAKEFQO7a5lwxBAAAEE+k3AOinsfHBwINvb2/LnP/9Z/u3f/s0rIZPJpAeMd+/e9aFdLaS8du2aV09aFaUFkDYErM1jube35+FjoVDw59TrdX/MttcCzFu3bnngafNTWnWlPf8yJ1vn5qFfCSovo3m553pHF0FlbMQWN4LK2H48AQEEEEAAAQTaF/DvIQSV7QPyTAQQQAABBBAYegGCyqHfxWwgAggggEC/CFgnhZ0t6FtbW5Ovv/7ag0qrhrT5YKw68ubNmx5Q5vN5v7Q5IC2wtOFbbajYP/3pT+/msrQA0uastJOFlxZufvrpp2Jhp4WUFmRaGwtBL3OydSao7PRMQe3tEe/oIqiMjdfiRlAZ248nIIAAAggggED7Av49hKCyfUCeiQACCCCAAAJDL0BQOfS7mA1EAAEEEOgXgTDws6DShnG14Vz/93//169bmJjL5TystJDS5qa04PL27dteJWnB4/7+vldh2pCxVpFZq9U8wLQw0oLEubk5+dnPfubPsWXZMhKJhJ8vYxCutw39urS05EErc1ReRrT957YEbgz9GhmyxY2gMrIbDRFAAAEEEEDg8gL+PYSg8vKQLAEBBBBAAAEEhlaAoHJody0bhgACCCDQbwLWSWFnG8Z1a2vLKyO/++47Dx0tULSqSquctIDRgkubX9LCx5mZGR8C1uaitPksV1dX3wWV9pywatLCzI8//tjbh/d1wsDWmYpKKirDY+mtXjnWf8ZSCZnIJmSuNCZ3ZnN61jlW5yZkqpiThFfypjoSlIeve5lL7yAMK1EJKi9DyXMRQAABBBBAIKaAfw8hqIypRnMEEEAAAQQQGCUBgspR2ttsKwIIIIBAzwWso6LRaIiFjjZXpVVJWmWknSysbA4Yw+BybGzM56S051rIac+z51h4GFZM2qW1s6Fg7TK8vxMbbK9LUElQGR5LBJWhRLxLfRtJQ/+xgLeUS8l8KSN3Z3Jye6Ygt2YnZFID3vCHB/b+1TdxvBegNQIIIIAAAgj0pQBBZV/uFlYKAQQQQAABBPpIgKCyj3YGq4IAAggggEA/ChBUBpWB/bBvvKNLw663b4/liKFfI++SFjcqKiO70RABBBBAAAEELi9AUHl5Q5aAAAIIIIAAAsMtQFA53PuXrUMAAQQQQODSAgSVBJXNBxEVlc0a0a9TURndipYIIIAAAggMkwBB5TDtTbYFAQQQQAABBLohQFDZDVWWiQACCCCAwBAJEFQSVDYfzgSVzRrRrxNURreiJQIIIIAAAsMkQFA5THuTbUEAAQQQQACBbggQVHZDlWUigAACCCAwRAIElQSVzYczQWWzRvTrBJXRrWiJAAIIIIDAMAkQVA7T3mRbEEAAAQQQQKAbAgSV3VBlmQgggAACCAyRAEElQWXz4UxQ2awR/TpBZXQrWiKAAAIIIDBMAgSVw7Q32RYEEEAAAQQQ6IYAQWU3VFkmAggggAACQyRAUElQ2Xw4E1Q2a0S/TlAZ3YqWCCCAAAIIDJMAQeUw7U22BQEEEEAAAQS6IUBQ2Q1VlokAAggggMAQCRBUElQ2H84Elc0a0a8TVEa3oiUCCCCAAALDJEBQOUx7k21BAAEEEEAAgW4IEFR2Q5VlIoAAAgggMEQCBJUElc2HM0Fls0b06wSV0a1oiQACCCCAwDAJEFQO095kWxBAAAEEEECgGwIEld1QZZkIIIAAAggMkQBBJUFl8+FMUNmsEf06QWV0K1oigAACCCAwTAIElcO0N9kWBBBAAAEEEOiGAEFlN1RZJgIIIIAAAkMkQFBJUNl8OBNUNmtEv05QGd2KlggggAACCAyTAEHlMO1NtgUBBBBAAAEEuiFAUNkNVZaJAAIIIIDAEAkQVBJUNh/OBJXNGtGvE1RGtxrmlvb+8dO7K+EdXb5MiOh/nBBAAAEEeiBAUNkDdF4SAQQQQAABBAZKgKByoHYXK4sAAggggMDVCxBUElQ2H3UElc0a0a8TVEa3GtaW9rf0XT6pV95d7/IGe0DpQWUQVSZILLsszuIRQACBVgGCylYPbiGAAAIIIIAAAqcFCCpPi3AbAQQQQAABBFoECCoJKpsPiEEPKg+ParJfPZTNnYosb5Tl+XpFHq8fypv9uliYaKduBDkElYHtqP57fPxWao1jqeu5odfttv53JSc7ntPJpIxlkpJOJSWpd3TjGL+SjeFFEEAAgQEUIKgcwJ3GKiOAAAIIIIDAlQoQVF4pNy+GAAIIIIDA4AkQVBJUNh+1wxZUPtOg8glBZfMu5nqHBexvaK1+LPsHNakc1qWq56OahpZWYenpeLdKHN/qcK8JDSZFctm0XCtmpTCekUw6CCsZC7bDO5rFIYAAAucIEFSeA8PdCCCAAAIIIIDAiQBBJYcCAggggAACCHxQgKCSoLL5ACGobNaIfp2KyuhWw9bS/oZaQLm5eyBv9g5lZ//Ibx9pdeXxcXcqeM3Q3qsWgaY0qZwqjMnibFGmJ7KSHUt7ZSVVlabECQEEEOi+AEFl9415BQQQQAABBBAYbAGCysHef6w9AggggAACXRcgqCSobD7Ihi2oZOjX5r3L9U4L2N9PO+9qOPlyY19eb1VkbacqO5WaHGpVpQ0Dm7ChWDv8wvY+tde1MNKGe52fysmP70zJrem8V1WOpYO/a4SVHYZncQgggMAZAvb32H4+Ypf1ekMODmuyXa7Kyuae2MgOSytVWd+t+RDhx/63uzufC7Zs+/FKTocCny6k5c50VhZn8rI4X5LpUl6y2Yyk02n97Ej6Z9MZm8JdCCCAAAIIIIBAVwQIKrvCykIRQAABBBAYHgHrVDnWsp+NjQ1ZWlqScrksk5OTMjMzIwsLC1IsFmNvrC0zXG6jUZdGraad9xV5tLynnTX78nS1IjvVumRSNmxhp7vw36+uroa+hkg+k5D5yTF5uFCUu3NFuTlTlMnCuCRSaUkmCSrfiwVVWja33pjum4lsQuZKY3JnNqfnoizOTchUMaduqXduFsL0+mTHmh5w+t+xHOkclWWdo3LrZI5K6yBkjspe76Hhff3w79zW3oE8fb2nc6KWZXmrKm/KR3Jw1JC6vpl0+kgNKjv7PtGj3efAtL+f9nfUAsq/eTgt9+aLMpEbk3GtqkxqZ3U/vD+Hd++zZQgggEAg4N9DCCo5HBBAAAEEEEAAgXMFCCrPpeEBBBBAAAEEEDCBsKOdoLL3x0NL4FbTue40cNvcrciLdQ131zToXanIm/26pD2A6M76Wk0AQWV8W8tKG/qPBbylXErmSxm5O5OT2zMFuTU7IZMa8FoVQ0pDXg+P+iDgjb+VPKNZwN6vdm7oEK+bu1X5dnlHvlsry8vNqmxpUFn1oPLYf4zR6d1tx5tVznhQqXNSWsXMZx9Py4N5PdYKWZ2zMuNVNRxrzXuM6wgggEB3BPz7G0Fld3BZKgIIIIAAAggMhQBB5VDsRjYCAQQQQACB7glY5woVlZ2tdmp3b3lHl+4PrwwkqIzM2OJGRWVkNxpeRiAIKY811W80GrKxXZUlDSqfrmpQqRWVm80VlfrnpTtBpWhQqVXjYVD50XV5eMOqnsclP65BpVU+2wt3+sUvw8ZzEUAAgSEUIKgcwp3KJiGAAAIIIIBARwUIKjvKycIQQAABBBAYPgGCSoZ+bT6qqahs1oh+3SrcqKiM7jX4Ld9XU9brddnQeSmDisp9H/p1a78mBydzVFqYqP919BS+Tz2o1Dkqb03n5G8fXJMHN4oeVBZ0+FcPKnXcWYZ/7Sg9C0MAAQS+J0BQ+T0S7kAAAQQQQAABBFoECCpbOLiBAAIIIIAAAqcFCCoJKpuPiTAAYY7KZpWLrxNUXmw0VC288jmopqzpHLzrJ0HlMx2m+dWbAx+i2YNKbWdVjd0IKu1vt4WQGQ0qb14bl5/fn5T78wW5XspLXoPKTPr9XLJDZc/GIIAAAn0mQFDZZzuE1UEAAQQQQACBvhMgqOy7XcIKIYAAAggg0F8CBJUElc1HJEFls0b06wSV0a2GoqUHlcdSrzfkyILK7YoP/fpcg8rX24caVDbksH6sVbbB8KzdDSoTHlT+7G7pXVBZyGc1qGyaE3Uo0NkIBBBAoD8FCCr7c7+wVggggAACCCDQPwIElf2zL1gTBBBAAAEE+lKAoJKgsvnAJKhs1oh+naAyutUwtLS/mzaXbEOHfT3UeVFbg8oj2a7o/XWtuLyioHJhKis/uzuhQWXRKyqLFlRmgnkqGfp1GI44tgEBBPpZgKCyn/cO64YAAggggAAC/SBAUNkPe4F1QAABBBBAoI8FCCoJKpsPT4LKZo3o1wkqo1sNest3HdLHb7Wisu4VlWtv9n2OShv6dWWn9i6o1CY6PGt35qi09bAQMp3SikoNKn96pxgElZN5KebHZYygctAPNdYfAQQGRODd54L+XbZK+4ND/RwoV2Vlc0+erWvF/UpV1ndrUmscy/HJ3+5uVNrbslM6eXEuk5TpQlruTGdlcSYvi/MlmdZhwbPZjKS12j6RYP7iATm0WE0EEEAAAQSGRoCgcmh2JRuCAAIIIIBAdwQIKgkqm48sgspmjejXtW9Qq+feis3tWcqlZL6UkbszObk9U5BbsxMyWcx552AqdXK8WXrFaSAF7G9mePaKShv6VYPKpZc7YkO/elBZDSoqryqotIpKDyrnCjI9WfCg0ioq0zpPJRWVA3mYsdIIIDBAAgSVA7SzWFUEEEAAAQQQ6IkAQWVP2HlRBBBAAAEEBkeAoLJ/OvK9o8tDkGOt0qpLtXoom7sVeaHhx9O1ijxaqejcd3VJ66/lu5VzEVS2994lqGzPbRCfFYaUNvSrz1F5dBQElcs7XjljQeVOL4LKxYLc06FfLaic0IpKgspBPLpYZwQQGEQBgspB3GusMwIIIIAAAghcpQBB5VVq81oIIIAAAggMoABBJUFl82FLUNmsEf06QWV0q0Fv2RpU6tCvOkelDf26pEFlWFG5U234HJVXWVH5Ew0q789rRWVJg8pCjqBy0A801h8BBAZGgKByYHYVK4oAAggggAACPRIgqOwRPC+LAAIIIIDAoAgQVBJUNh+rBJXNGtGvE1RGtxr0lqeDykMNKn3oV6+o3JdVr6gkqBz0/cz6I4AAAlEFCCqjStEOAQQQQAABBEZVgKByVPc8240AAggggEBEAYJKgsrmQ4Wgslkj+nWCyuhWg96SoHLQ9yDrjwACCHRWgKCys54sDQEEEEAAAQSGT4Cgcvj2KVuEAAIIIIBARwUIKgkqmw8ogspmjejXCSqjWw16yw8FlQz9Ouh7l/VHAAEE4gsQVMY34xkIIIAAAgggMFoCBJWjtb/ZWgQQQAABBGILEFQSVDYfNASVzRrRrxNURrca9JYElYO+B1n/URbw7zz6QXesE8ja511wen8tuJ0IH4h5eXo54dMTYktM6D8J/efdZfgwlwMvQFA58LuQDUAAAQQQQACBLgsQVHYZmMUjgAACCCAw6AIElQSVzcewdbNq/62MpRIykU3IXGlM7szm9FyUxbkJmSrmJJFKSTIZuFmna69P3kGoSeHbt8dypPMFlquHsrVTkeWNsjxbr8jj9UN5s1/Xx4M17cYqE1T2+ii4ute34y04H0u9XpfmOSqpqLy6/cArIRBXwN63Df2Aq9X1s0LPdj04hZfhEtv9XDu9nGB5QTiZkFQyIWk9p1JJSetnbLIbH0bhJnB5pQL+PUSjb7us1xtycFiT7XJVVjb3/HvI0kpV1ndrUmsc63est0Fg3eE1tKPPlm3HWS6TlOlCWu5MZ2VxJi+L8yWZLuUlm81IOp3W10/6OnR4FVgcAggggAACCCBwrgBB5bk0PIAAAggggAACJmCdKsfHx7KxsSFLS0tSLpdlcnJSZmZmZGFhQYrFYmwoW2a43EajLo1aTdY0OHq0vCdLK/vydLUiO9W6ZLrcUaeroa8hks8kZH5yTB4uFOXuXFFuzhRlsjCugVv6XeAWeyO78AQzUzj9TztRa3WpauC2uVuRF+tqtqZ+KxUP3Kyjs1v9m0FH1+AGlRYa7ZvbSVD5nKCyC0fqaC8y/Ptm71OCytE+Ftj6wRKoa0h0cNSQvaqFSIdyUGt4pWPwTye3xT5J7aSBp15NeECZlHH9QpIfT0s+G5wz6WTQjH8HXsC/vxFUDvx+ZAMQQAABBBBAoHsCBJXds2XJCCCAAAIIDIVAGCgSVPZ+dxJUtrcPmt1OB5VWUfmEisr2YHnWmQJ2vAVngsozgbgTgT4VODiq6499DuX1ZkWeruxpWHnk1Wfd++GP/a0QfY0gpJzMj8ns1LjMTo7L9GTOA8s+pWK1Ygr49xCCyphqNEcAAQQQQACBURIgqBylvc22IoAAAggg0IaAda5QUdnuMG9tgH/gKd7RRUXlB4TOfqjZjaDybCPu7ZyAHW/BmaCyc6osCYHuC+wf6OgO20FI+dV327K+c6DDsErXhmDVvxQeVKY1qLQqyumJrNyeLcjtmYLcnC7IhAaXnIZDwL+HEFQOx85kKxBAAAEEEECgKwIElV1hZaEIIIAAAggMj4B1rhBUElSGRzRDv4YS8S71bSQN/cfm9izlUjJfysjdmZx3SN+anZBJndvT5oVKaa+4z+vZrRKeeKtN6zYECCrbQOMpCPRIIAiQdPhV/Zgv65Cvr7f2ZWl5V758vCWr21WdKzKpQ8B3Z+Xsc8FeP6OvYUHlTCkr9+aLckeHoLfzZCHrQaa9ej/M99wdhdFYanCcBT9iYY7K0djnbCUCCCCAAAIIxBMgqIznRWsEEEAAAQRGTsA6VwgqCSrDA3/Qg8ojnaOyrHNUbp3MUWlDvz5m6Ndw93LZAQH7mxmcqajsACeLQKCrAkGAZLNFvpXd6pEsr5flGw0qv/h2S1Y0qLQQMalzSHbjFPydEA1DEx5UzmpQeV+DSgsr792YkKniuL5s8NoEld3YA1e3zOA4I6i8OnFeCQEEEEAAAQQGTYCgctD2GOuLAAIIIIDAFQtY5wpBZXc6KePuSu/o8hDkWI5qdalq4La5W5EX6/vydK0ij1YqOr9WXdLaqdqtgjyCyrh7LWivu42KyvboBu5ZQfhgHdIElQO381jhkREIgqNwc99qHHgsOzo/5QsLKl9qReWTbVnVoV8z+nnavaBS9FXf6me2VlSOJb2i8r5WUt67UZQHGlRem9CgMmHlnPaZ3h/fQ0IxLuMJBMcbQWU8NVojgAACCCCAwCgJEFSO0t5mWxFAAAEEEGhDwDpXCCr7o4PQO7oIKmMfxS1uVFTG9uMJ8QTseAvOBJXx5GiNwNUJ+OdC+HL6owJ52/Cg8vlJReX/Pd2RtZ1D/+FPsoshYRBUJmQ8kxSrqLyrc1TePwkqr1tQmbThwJMEleG+GtDL4HgjqBzQ3cdqI4AAAggggMAVCBBUXgEyL4EAAggggMAgC1jnCkElQWV4DFNRGUrEu9S3ERWV8cgGtnUQUlJRObA7kBUfCYEwqPRLDSrfHtdle//ARyh4tLwnv3+2K+sWVOqwrN0KKu3z1D4bdHRZDypnJsY0qMz78K9eUVnKaTWnBpUeVvbH95CRODi6sJHB8UZQ2QVaFokAAggggAACQyJAUDkkO5LNQAABBBAYbIGWDjPdFBvi67xhvqytnS08tEtrl9Rhwz70nMvohK+1sbEhS0tLUi6XZXJyUmZmZmRhYUGKxWLsxTdvQ6NRl0atppULOnSpdg4uregwpqsV2anWdW6o7nUQ2kpbB2EmJZLPJGR+ckweLhTlrg67dnOmKJOFcUmk0kEnYRerKeLgmZuttA0pydCv0eVa3KiojA5Hy7YEwr9vDP3aFh9PQuBKBPxzQV/J36/HDf1OVZed8oFYReXSq7L84dmerO92N6i0DbWPdQsqs+mETHtQmfM5Kh/cKMl1DyrTGlR27ztep7DN8di/oth3lE4t9eLlhF/Pgu/AJ9+fL37albcIjjeCyiuH5wURQAABBBBAYGAECCoHZlexoggggAACwyrgnWTWwaPBo53tlEql3oWPp7fb2tQ02KvXNeBrNDygHBsbk3TaQrXODw8Wrp8FlY8ePSKoPL1DrvC2d3TpsUJQGQ+9xY2gMh4erWMLhH8zCSpj0/EEBK5MwD8X9NXs8tiCynpNtjWofLERBJV/fG5B5VFXKyptYy3UC4PK68WM3J05FVT6dzsb/vX8H7BdGdo5L2SGDT3XG2+lVm9IwxLLKzhZjalVu9o5pT9sS+l3YLvsVgXsZTYpON4IKi9jyHMRQAABBBBAYLgFCCqHe/+ydQgggAACfSxgnRZ2tsDx8PDQw0cLHi2kzOVyYuFjc/BoAaU9fnR0JNVq1Z9j161NPp+X8fFxP1tgaR1alz0FnSrBUuy1qai8rOjln+/7xI8bKirjaDa7HWpQuV89lE2t4F3WDunn6xV5vH4ob/br3mFsy+3A2+d7q6e7jaFfv6cynHfY8RacmaNyOPcwWzUMAv65oBtil8f63arRsKCy6kO/Lr0uy1dXGFTq1zjJasA2XUzLnVkNKnVkB6+onMzpd8KMTlPZ30GlBZNHGlBWDuuyV6npiA+Nrh8iFoUm9atuWv/JpFMyPhae0xouK2ife08iQQAAQABJREFUnYLjLfze35CDw+B4W9nck2f6PWRpparBeE1qDf3Roh6THkx3eBvMzJadUrOczok6XdDjbTorizN5WZwvyXQpL9lsxn/4yLyoHcZncQgggAACCCBwoQBB5YVENEAAAQQQQKA7AmHwuL+/L1tbW1KpVDyItMDRhlW1IVUzGe2gsh4sPVkVpbXZ29uTnZ0dseeFQaW1teFYr1275qGlhZ3thJVhx529XnjdLu0cBpX2ugz9akJXf/J94vuDoDKOfrPb6aDSOgifEFTG4aTtBQLh30wqKi+A4mEEeijgnwv6+nbZH0GliFVU3nlXUTnhwVHypKKy+YdrPWT73kub35GGa5WDurzZO5BXWxUpa1gZ1lQmTq6Ft7+3gJh3aF2pL1G/mXrgltWQMj+elqn8mA7Zr+di1kNL/clRzCV3t3lwvBFUdleZpSOAAAIIIIDAIAsQVA7y3mPdEUAAAQQGVsA6LA4ODuTNmzeyuroqL1688CFVLWCcnp6Whw8fyuzsrFdI2n3Wfnd3V169eiWbm5seVloVZniyUNLCwzt37njIaRWZFnLGCSvtNexsAWp4br7PwtQnT554WDo1NeXrxxyV4R64mkvbH7qT9D+CyjjizW4ElXHkaNuOQPh3k6CyHT2eg8DVCPjngr6UXfZdUGkVlQsTPkdlWFHZj0GlhY/md3Ck83vuH8nrrX359tWebO2F3087FU9+/5iw0WXTWoWaG9OQspCR2clxmZ/KydxUXgq5jD6BoPK0mu0NKipPq3AbAQQQQAABBPpFgKCyX/YE64EAAgggMDICYSf22tqa/PnPf/bzN9984xWSFjZ+9NFH8vnnn8v9+/elUCj4EExWTfny5Uv54osvPKy04WJtaNhSqeSVlq9fv/ZKyp/97Gf+PAs7rcoyTmVlWOFpr2WVmjbMbBhY2qVVcT5//twfu379OkFlD45Y71glqIwt3+x2Oqhk6NfYnDzhAoHwbzxB5QVQPIxADwX8c0Ff3y77I6hMaEVl89CvAxBUmp0mhpWDmmxqNeV3q3vyh+/0B3hvqu/3bBfyQn1ZDdxEMjrEa1GrKWdKWbk9nZfbMwVZ1JC3lM++r+jsxlju77cu8rXgeKOiMjIYDRFAAAEEEEBg5AQIKkdul7PBCCCAAAK9FrDOCgsBLaj8+uuv5auvvvKzBYEWVP7gBz+Qf/iHf5CPP/5YJiYmfHXtsadPn8qXX34p6+vrOodM1kNKq7q0ysylpSVvZ8+xgNMqKy1MtHYWVkY5hfNf2vJsiFmr2GwOK23I2ZWVFe/Us6Fp5+fn5caNGx6IRll+cxsz8M5Bn3ezLg0NR9d0zsBHy3s6T8++PF2tyE61rp1QCZ2DqAu9XCcrY51dGeXJZxIyPzkmDxeKclc7uW7O6FC6hXFJpNI69G57w+g2b2+nrntHl9tRURnHtMVN56gs6xyVWydzVNrQr8xRGUeTthcJhH/fCCovkuJxBHon4J8L+vJ22V9BZV7nqCzoHJUDEFRqWmjfE8sHR7K+U5VvX+/JF99uyauToDL89tbpr3EWUlpAmkknZUKDSqumvDub9+9v92+UZEqHfw0mm9aBYjv94m0essHxRlDZJh9PQwABBBBAAIERECCoHIGdzCYigAACCPSXQFi5WC6XPXS0APKPf/yjB5c2tNft27fls88+kwcPHngYWa1W5dmzZ15RaZWT1ilkFZN2tjDSAsUnOiSrDSNrJwsRf/zjH8vNmze9ItMqL6OcrErTXssCye3tbQ8rrbrSXs86WOwxu9+CTwsoCSqjqHa2jXd0EVTGRm1xI6iM7ccT4gnY8Racj8X+rloV7/qbfVla1qr09X1Z2anpDzEacljXgEQ73K0fPezQj/dK57fWxfo6WCe9DY+4MJWVnywW5P58Qee9K8hEIRgePK3zu/VLR/75W8MjCHRewD8XdLF2SVDZhq+56R+wun5H3KscalBZ8aDyyyfb8rqpolKjwiAzbOMlznuK/d0MgsqEV1TOakXlnZn3QeW1iawk9Pt0ImHnTv91PW+tPnx/cLwRVH5YiUcRQAABBBBAYJQFCCpHee+z7QgggAACPRGwzgoL/2x4VQsZbd5JCyrt0m5b0PjXf/3XcvfuXa+wtGpKe9xCSgsObTjYW7du+dCrVoFpz7E5Lu28vLzsj//85z+Xe/fu+fPHx8cjbedFQWU4pyZBZSTOrjTyji4PQaiojAPc4kZQGYeOtm0I2PEWnAkq2+DjKQhciYB/Lugr2SVBZRvk9l1WR8Wo1+qyq0Hl2raOTrCyJ//3dEdWtg/eLdCCyk6f7IcYFlTajzCK4ykd+nVMFnXoVxsR475Vok7kJKnDwr6f17Pz6xB3m4LjjaAyrhvtEUAAAQQQQGB0BAgqR2dfs6UIIIAAAn0iEHRg2y/Rj/28urrqQ7/aHJRWZTk1NSU/+clPfPhWCyI3Njbkv//7v8Xa5XI5DzIthLRhXy20tMDTQszHjx97oJnJZOQXv/iFz3VpFZf2nCgnC08tCLXKyXDoVwsvbT1tnW3dbOhXu83Qr1FEO9/GO7o8BCGojKPb4kZQGYeOtm0IhH/jGfq1DTyegsAVCfjngr6WXRJURkcP3ewZ5nak3xutonL1TUWerJTl9892ZXXnUGFPlqkZYadjQt1lWo0eBJWFbEqmJzIaVObkzqxWjS9M6u1xHf1Dh+7XsDKoqOz0GkT3ClsGbgSVoQeXCCCAAAIIIIDAaQGCytMi3EYAAQQQQOAKBMKOHutACeeqfP78uVj1ZKlU8qBycXHRKyItoPzP//xPb2fB48LCgg8La0GlhZAWVNq8lY8ePZLf/va33inzy1/+Uj755BMfHjafz0faIlunMKy0ZTaHlPaYrZsNQWuPXbt2zYNSW5disRhp+c2NbHl2DobBZY7KZpsPXTczhdP/CCo/5HT6sRY3gsrTPNzusED4942gssOwLA6BDgr454Iuzy4JKqPDhm72XcRHB9Ggcnf/4KSicl/++NyCyqN3QaV+ze34ySsq9fXTyYTkx5IeVN6+rvNUekVlSa6XcpJOv59jvB+Gfw3cCCo7fjCwQAQQQAABBBAYGgGCyqHZlWwIAggggMCgClhQ+dVXX0kYVFoVZVhRaaGlVTH++7//u4eRVslow77a/JXNQaVVXX7zzTfy61//2hl+9atfeVBp7aMGlfZE77A7qfS062GHlF1ubW3Jt99+69WWto62bILKqz3qfH/4fiGojCPf4kZQGYeOtm0I2PEWnBn6tQ0+noLAlQj454K+kl0SVMYjdzt1s/kp7cdrOxZU6jy8T1Y1qHxRlrUrDiqvF9Nyy4JKrah8cEODysm8BpUZn1PdQkqCymD/hgFvSgPeXEYD3kJa7kxnZVHn91ycL+n8xXnJZjMe8vbT/J7xjk5aI4AAAggggMCgChBUDuqeY70RQAABBIZGIE5QOT09LTdv3vSgcm5uzisqbbjWMKj8zW9+03ZF5YdArVNqc3PTw1AbAtYCVILKD4l157Gwc5CKyni+LW4ElfHwaB1bwI634ExQGRuPJyBwRQL+uaCvZZcElXHRw8rAuhzqZ+puuapDv+7LUw8q92Vt9+oqKnOZhFwvZjSozJ4ElTr066RWVOo0CDb8K0Hl+31LUPnegmsIIIAAAggg0H8CBJX9t09YIwQQQACBERM4a+jXn/70pz5H5cTEhM9N+V//9V9+aZWMN27ceBdU2hyVFlTa8LBLS0vyxRdfSDKZlM8//9znqLRgM+ocleexh53uFoba8LIWVFJReZ5Wd+/3jlUPQaiojCPd4kZQGYeOtm0IhH8zGfq1DTyegsAVCfjngr6WXRJUxkVvCioPj3zoVwsqraLyqxc9CiqvZX2OygcLWhnYVFFp34mpqAz2L0Fl3OOc9ggggAACCCBwlQIElVepzWshgAACCCBwhkBYUfnixQvZ3t72ELB56FcLCH/3u9/5ELBjY2M+7+S9e/dkfn5eLMi0YbeWl5flu+++87DS2vz85z+X+/fv+1yS4+PjZ7xq9Lu8E0+Hg7X1sDCUoDK6XadbescqQWVs1hY3gsrYfmc9wTo8/fTuSnhHly91vrMuTHnW0ZW24y04U1HZUVgWhkAHBfxzQZdnlwSVcWFbg8qdsKJyraJBpQ79ultT2GCZ3Z6j8l1FZXNQqUOYBhWVKf/xHkFlsC8IKuMe57RHAAEEEEAAgasUIKi8Sm1eCwEEEEAAgTMErBryj3/8o89ReTqotMrF3d1dn8Py1atXHkranJOLi4seVF67dk0ODw/l6dOn8vr1a59Hslgsyqeffiq3b9/2IDObzZ7xqtHv8k48gsroYF1s6R2rBJWxhVvcCCpj+51+gnm+yyf1Sng9vOxUkHh6eb5cDyqDV+hGB/jpbW3ntvucvE/r9WBoxHWtNlpa3pHn6/uyslOTnWpDDusakOhG2nYEW9TOq539HLOz9bAO+nQqIQtTWfnJYkHuzxd0HrKCTBRyktGhEdPpVF9UG529FdyLQPcE/HNBF2+XBJVxnc8OKp+sVuTrlwSVZ2kGx1vo1pCDw5psa8C7srknz9YrsrRSlXUNeGuNY/1cCP52d+NzwZbNHJVn7SHuQwABBBBAAIFeCxBU9noP8PoIIIAAAiMpYB0WFjBaCGmVlBZU2uXe3p7P//ijH/1I7t27JzYPpbW1MNPOKysrPtSrBZhTU1Ny/fr1dxWV1WrVh3mdnZ31oWHt0oZ9TafTlzL2TjyCyksZdurJ3tGlxwNzVMYTbXaz+bT2q4eyuVOR5Y2yBkcVebx+KG/26+oaLNeCo06fbNkN/WdMQ6NSLiXzpYzcncnJ7ZmC3JqdkMli8F5NpU6Co26sxCU3yhwbmqwd1Y+lVtegraYVg42g4/WEzl+hU3zBMoN/ExrlWedqJp2U8bG0jOml3U7qud9O5hScqajst33D+iAQCvjngt6wS4LKUCXqZRi46Q8xdOhXC9zCOSq/fmlDv1JReVoyON5CN4LK0z7cRgABBBBAAAEECCo5BhBAAAEEELhiAeusaDQasr6+Ln/5y1/kT3/6k1dMWkWkhY02nOuDBw/8/NFHH3lYacO3bm1teTsb5tXa2bw7pVLJ194CTgsubchYG/LVAk6rrLTQw9pd5uSdeASVlyHs2HO9o8tDEOaojIPa7HY6qLRKhicElRdyBn8H3kr1qCG7Fe2Y3j+Urb1D2T+o69+zoPPVstVOx4YWU+oh74FkVsPJUmFM5qdyMqmXFlhacNkPw/o1A5pVcCaobHbhOgL9JOCfC7pC/rdNv5M1GkGF2wutel56XZavnu9phduRVyQnu/jDkeDvm0hWf8RyvZjWeRbzcm+uIA9uTMj1Uk6/x2UkefJdrn/+1oWB20lQudcUVC4TVJ51nAfHW+hGUHmWEfchgAACCCCAwGgLEFSO9v5n6xFAAAEEeiBgnRUWVFp1pFVSfvPNN/L48WPZ3Nz0akkb2vXGjRty584defjwody6dUusOtLmhvzDH/7gc1FaaFmr1cTaWhhpwwsuLCzI3/3d38m9e/c8pLS5KjvRqeWdeASVPThSvv+S3tHlIQhB5fd1zr+n2Y2g8nynDz1iw8U1dEi6vWpNNnZ0uLo3FXm1WZWd/SMdqu4kqNQFdLo/3zrxw6Hq8tm0zE6Oyz3twLewspjLSDYTVKB24m/dh7Y/zmN2vAVngso4bme1tQrew1pDjuyslbx2244HOy46faydfn17DQvCi+NpP86sgrefjrPT68vteAL+uaBPsUsqKuPZqZq7+dDWVlF5ElQ+Wd2XPxFUnokZHG+hG0HlmUjciQACCCCAAAIjLUBQOdK7n41HAAEEEOiFQNCB/VZ2dnbEqiPX1tY8pKxUKnKsgaDNG2ZVlVYhOTMzIzYPpVVOHh0dic1TubGx4UPEWlBpw7qGQ0Va+7t378r09LQvw+7vxMk78QgqO0F56WV4R5d30hNUxsFsdiOojCP3vu2xBkQ23OuOVlK+2tqXlxv78nxtXzb3LKjUObX08W5UHWm3ri87nUpKQQOjhes5+eHtSbk1XdDhcrNeVWmv208BUvg33oZoZo7K98dQ1Gv+ftXGtk8rh3XZ2D7Q4+xAq3iP/LYFlhZWdnqfh9XAmk/qKQhDp7Ry974G47OlcclpUG7HIafhEAiPM7skqIy7T8PArbWikqDyfMfgeAvdCCrPl+IRBBBAAAEEEBhVAYLKUd3zbDcCCCCAQE8FrMMinKPSAsqDgwOvkLSVsqFaLazMZrNeMWnDvtrZQkyrqgzbWwe4ddRaezvbfJQWVtql3d+pTlzvxCOo7OnxEr64d3QRVIYckS+b3QgqI7O1NLRqyqNaXd6UD+Xl+p4805DyqZ43dg81wNRKNz0uPTBsedblb1ggZSFoWivbJjSovDWdlx/fmZQ7c0W5PmHhUUbnquyv4V/teAvOBJXtHAH+ftUn2meYVew+W92T5c19reINKngPdG7Uhn4mBZ9z7bzCxc/RXehh6I1rOfnrB9M6FGdRJvJjXll58bNpMQgC4XFmlwSVcfdYGLi1BpVPtaLyayoqz8QMjrfQjaDyTCTuRAABBBBAAIGRFiCoHOndz8YjgAACCPRSwIZ/tbDRzhZC2tlOYchoFZHhOZxn8nT7MIy0S2trAaddhvd3Yvu8E4+gshOUl16Gd3R5CEJFZRzMZrfTQeVznaPyMXNUnskZdKzq3yR9tK5/rw41qNzaPZAXa0FQ+cSCSquoPBmSM6hsPHNRbd+pGaUHlTYEp1VU3p7OyaeLJblrQeVkXu8Lgkqv5tS/g/1wMrfgTFDZzv54d9zp7tzUIHxpeVueruxpFW9FKysP5UCHgbUhYIPdffl9/u6w0WOtuZrSQvLbMwX51Q/n5Ae3JuXaRFDB+379Lv/a7fjwnM4IhPvRLgkq45qGgdtJUFlumqPyJXNUnqUZHG+hG0HlWUbchwACCCCAAAKjLUBQOdr7n61HAAEEEEDgQgHvxCOovNDpKhp4RxdBZWzqZjeCyuh8QceqtX+rP6jQjtWjmgZHVXmhFW7faUj53Xr1fVCpx2VXg8qToV9vXR+XH3tQWZCZyYLOUzmmw3GmtKpcQ6N3iVP0bexGS3MLzgSV8X0DO08MdZfafKh/ebEt377e1aGGm4JKnRfVd/cl9/npqLElqNQw9M5sQf6/T+flEx1ueGYy58O/6u71Uyd/EBTfiWdcViD8+2aXBJVxNcPA7XRQWZGvX5ZlbbcWpv5d+bNsb0H7IUFa/+7nMgm5XszIrWtZf78+WCjJdCkv6ZMf7tkP/frhvRocb6EbQWXcI472CCCAAAIIIDD8AgSVw7+P2UIEEEAAAQQuJeCdeASVlzLs1JO9o8tDECoq45g2uxFURpcLO1Y1dfP5KauHGlTuVDQwKsvzdQ0qNw5kq1zzikrrNLassNO90rYOOuqsZFIJKegcgQvaGf2jW0WtqNSgckqDyvy4PqZBpQaZ/dAZbbq2zsGZoDL60Ra2DO00ilBHC8YtqHy8UvaKyq3y0buKym4MNXySQdpLexByW4ca/uUPZ94FlXkdalgjUl/ZfjneQjku4wkEf9+C9ytBZTw7SyHNz0b5ODw8ku13FZUEledJBsdb6EZQeZ4T9yOAAAIIIIDA6AoQVI7uvmfLEUAAAQQQiCRgnSs2LO3GxoYsLS35PJmTk1pdMjMjCwsLUiwWIy2nuZEtM1xuo1GXRq0maxqAPFrek6UVnftutSI71bqHEz6kY/OTO3jdOqMzKZG8/iJ/fnJMHi5YAFKUmzNFmSyMSyKV1kqtzg6le5nV944utyOojOPY4qZVgeXqoWzp8ba8UZZnDP16LqW7nXRI13TY1+rBkVe4PdehXy2ofL6pjvt1qWmSaG0tuDldoXbuwiM+YO9Rm//SKmcK2ZQHlZ8sFLxyZvaazhtYyEomnX4/T+UlK+wirtYHm5lFcCao/CDUGQ+GdvLWjin93NGKym9e7sgTDSqXNyt6vNV0CGIdKr1Lx5utkh1z+p+/xq3rOfn8B9M69OuEzGpFZX58TMP4pD7auXmg7TU5Xb2AHWt2skuCyrj+YeBGUBlVLjjeQjeCyqhutEMAAQQQQACB0REgqBydfc2WIoAAAggg0JaAd+IRVLZl1+kneUeXdqpaB/6RBUcauG3uVuSFhkZPdVjERysVeaPBkYU63cprgg58kTGtcJvIJmSuNKahUU7PRVmcm5CpYk4DXhuKMwh4+6HqqMWNoDLWYWl2drxZUFmxoHK7Is9WdRhOnS/whQaVbyqNk6AyKKbsZlCZt6Byakw+Wcg3BZXjMmZBpR5z4UHf62MuMAvcvOJIj7n1N/s61+KOB7wrOzX9IYbO+VnXgETfUPZe7bib7mVbD7NI63t1YSorP1ksyP35gg6LWNCAN+dzGqfT/fNDDDsww+PtrX7mHB839HjToFLdnq5aUHmgf980qNQ5Ubvl5uvg6xGsy81rOfns42vyg5saVE7lpJDLqun7Y83a9/p4s3U46xTEcPqIHge9OPWrS2hhx5qd7JKgMlSJehkGbgSVkcX8eAvdCCqjutEOAQQQQAABBEZHgKBydPY1W4oAAggggEBbAt6JR1DZll2nn+Qdq9rZRVAZT7bZjaFf49uFQeW+BuMtQeWWDvlXsYpKOya7FLjpcsOKSgsqb2jl8w8WLBgvyNy1CSlp5XMmEwSVYTASXsbb0s61tuMtOFNRGVc1dAuCo3pTULkvr94cyLYHld0LeG19Lb6y49nWZeHauHz2cEo+1qBybiovhfyYFlSmNZy0qsrg1OvjLVyP5stgG8xJj0VLda/4ZLXVCf3BjA0H3Y8+xmH7N7wkqHSKGP8Ef+O+P/Trvs5Ruc8clWdIBsdb6EZQeQYRdyGAAAIIIIDAiAsQVI74AcDmI4AAAgggcJGAda4w9Gun650uUj/7ce/oso5nKirPBjrn3ma300GlDf36ZF0rA7US9aTfWjvWz1nQJe62ZVvgZpWopVxK5ksZuTuTk9szBbk1OyGTWomaPqkM9I79bqxEG+tvdm+1ss0qeCtVG/p1P6ioVLcXb2q9CSpv5GTRg8qilIpaUZnJeEWlufVDKOJm5qbvUyoq4x10ZmefN2FwtK4VvDb063drFlQe+vF21MVKVFvbIOQ7CSqnxuXvHk62BJVJDSpt+NfwWAsv421pd1tbQFnXIZkbeq5rUBmElWFg2YU/cKc2J6kJZVrnjbVzyiv8u/+ap1bhwpv+uaCt/JhrNNRK/57pXIs2QsHS67J89XxP1nePvCK520PQJzX3zupnw/ViWn+EkZd7OgfvgxsTcr2U079tGZ2D10YpeH/MXbhxXW9gf9+a5qjcq8qqVo0/XdWgcpmg8iz+4HgL3QgqzzLiPgQQQAABBBAYbQGCytHe/2w9AggggAACFwp4Jx4VlRc6XUUD7+g6CUAY+jW6eLMbQWV0N2tpdu+DykNZ17k9n63sygsb+rVHQeXHGlQGFZXvg8q0duR7YNQHAa+bnbxPCSrjH28WVPrcxXUdMvdUUGlD5h5pBa828R8UdCP+ag0qs/K3FlQuaEXltbwUbehXnbvYKirDgDK8jLel3W1d0+Fxywc12T85H+m8nvpODlLYLr5HbH/Y4rM6+fOEVp8WxtOSG0t7YNndLY6/dP9c0KfZZRiME1RGdQwDt5OhXwkqL4QLjrfQjaDyQjAaIIAAAggggMDICRBUjtwuZ4MRQAABBBCIJ+CdeASV8dC61No7uggqY+s2u50OKp9rZeBjKirPNTW7MKj0oV+tonJlj6DyXLGTcJeg8gNC5z8UfN5YdZueaydBpc5R+a6ismpDDUsPgsqizlGpQWU+q9Vtmb4MKv3vnNJacGoB5eqbqrx+U5HXWxXZq9SCYWDt8fP5L/WIBbxWeaiFgTpXcdarnm9ouHttIuthZfP6XeqFOvTkcH38mKOiMqZqGLidnqOSoV/PgwyOt9CNoPI8J+5HAAEEEEAAgdEVIKgc3X3PliOAAAIIIBBJwDvxCCojWXW7kXd0EVTGZm5xO6pJWeda3NLKwOWNstjQrwSV55OaHUHl+T5nPeJmBJVn0Vx4n9kd61DDYVC5tr0vj5Z35elaWUM3HfpVKyoJKs9m9L9z+pBVNO7sH+kQzXs6FGdZnuoPC7b2Dn3oaQsTux1UpnWo17nJcfnk9qTcmy/K/LWCVqJmvHLR1rxfKlBDLz/mCCpt18Q4hYHb6aCyonNUlpmj8gzJ4HgL3QgqzyDiLgQQQAABBBAYcQGCyhE/ANh8BBBAAAEELhLwTjyCyouYruRx7+giqIxt3eJGUBnLz+zCoLKiAS9Dv17M52YElRdDndHC7N4N/aoVlX0RVD6YlI8WisHQrwNRUSkaTB7I49e78vjVrixp0LuhQaXNW6n/6albUaVVVIpYUHnj2rh8eueaPLxZkls6D68NAxu8NkHl6cPeXAZ+jkqd2zOYo5Kg8vT+DW/79xAbgFl3eL1OUBm6cIkAAggggAACCIQCBJWhBJcIIIAAAgggcKZA2HG8sbEhS0tLUi6XZXJyUmZmZmRhYUGKxeKZz/vQnbbMcLk+F5l1SGuF26PlPVla2dcqkIrs6BB/GR1DzoaS69bJOgh1Ki3JZxIyPzkmD7Uz+u5cUW7OFGWyMO5zkSWTJ3PfdWslYizXzKy39+3bY2GOyuhwLW4EldHhtKW/V7XCzY43Cyo3TuaofM4clec6utnJ+5Q5Ks9lOvMBs+unoPLGVFb+bqCCyrcaQ76VTQ0ml3TIXAspH+llEFT6x4e7d+tT1Yd+TYoGlTn5q7tT8rEGlYtzE1KyoPIkIKWisvXQt491gspWk4tuKZkH7xaK5/T72/ViRm5dy/rcxQ8WSjJdyks6k5GUzl2cVNx+OOb8ewhB5UW7lscRQAABBBBAYIQFCCpHeOez6QgggAACCEQRCDuOCSqjaHW3jXd0EVTGRm5xI6iM5Wd2zRWVBJUX87nZyfuUoPJir+YWZkdQ2SwS/Xrwd+5Yn/BWtnYPZMmqKfX87es9Dy6PLd2xc7dSSl10OEfl/NS4/PiOVaJOeFA5WcieBJWJvgiNdFX9Rxjh5TFDvxpFjFNYGcjQr1HR/P1JUBmVi3YIIIAAAgggMIICBJUjuNPZZAQQQAABBOIIhB3HBJVx1LrTNuiIpqIyrm6LG0FlLD6zI6iMRRZUoRJUxkM7aR1+3ryrtO/xHJWDVlFp1fbythEElVpNaSHltytlHQr2yDNKPSx9Dsu2dk6EJ9kACCn9Z24yKz9anAiCytkJHaEgqy+swxfoY/1Q3Wab4p8LJ5cElRF2bksTgsoWjgg3guMtdGPo1whkNEEAAQQQQACBERMgqByxHc7mIoAAAgggEFcg7DgmqIwr1/n23tF1EoAw9Gt03xY3gsrocNrS7AgqY5ERVMbjamkdft4QVLawRLoRvlft/bq5W9VqSg0pNah8YkFl2YJKGxS2qwWVHoLqiO0yW9Kg8vaEDqc+ocNx6lDqxXEPKhN9MgyngfrnwsklQaWJxDmFgRsVlVHVguMtdCOojOpGOwQQQAABBBAYHQGCytHZ12wpAggggAACbQmEHccElW3xdfRJ3tFFUBnbtMWNoDKWn9kRVMYiI6iMx9XSOvy8IahsYbnwhr9P/b16rEPn1mRz50C+tWFfNaR8srovb64qqNQ1DYLKMfnEgsobRbmj8z5PaVCZSKb13GdzPuv6+jHH0K8XHmOtDcLAjaCy1eX8W3ac6dHmx1u9TlB5vhSPIIAAAggggMCoChBUjuqeZ7sRQAABBBCIKBB2HBNURgTrYjPv6CKojC3c4kZQGcvP7AgqY5ERVMbjamkdft4QVLawXHjD36f6Xj3WasrjugaVWlH5rVZUPg6Dyv3a1VRU6pomE2+1olKDylvNQWVOR35NS5Kg8nv70vIrLTSVrCa814tprUDNy725gjy4MSHXSzlJpTKSTKW0TbJvhs19H7gRVH5vh55zh38PIag8R4e7EUAAAQQQQAABHYBFvzDZT7s4IYAAAggggAACZwrYV4Xj42MhqDyT50rv9K9tuj9sHjKGfo1O3+JGUBkdTluaHUFlLDKCynhcLa3DzxuCyhaWC2/4+9Q+qxt1PVtFpQ79qsO+Pl7Zl6dWUXnFQeXMhAWVRR36VSsqdY5Kq6hMngRuzFHZujt1txFUtpJceMs6sI4VLp1MSC5jAW9Gbl3L6rGmAe9CSaZLeUlnMhry9k/A699DCCov3Lc0QAABBBBAAIHRFSCoHN19z5YjgAACCCAQSSDsOCaojMTV1Ube0WXBEUFlLOcWN4LK2HYElbHICCrjcbW0Dj9vCCpbWC68YW5up0Fl4ySotPkpraLy6WrlioNKkSCoLARDv+oclVMTOYLKc/YiQeU5MB+4m6DyAzgfeCh0S3nAm5TpglbwTmdlcSYvi/NBwJvNZiSd1mGaE/1UwfuBjeIhBBBAAAEEEBgaAYLKodmVbAgCCCCAAALdEQg7jgkqu+MbZ6m2L7Q3mqAyDpq2bXEjqIylZ3YElbHI/HhzN/1BQb2uQyPqMbf+Zl+Wlnfk+fq+rOzUZKfakMO6VsDpWzqR0HO8l7iwtXVI2zpY9Vpah5RcmMrKTxYLcn++oNVGBZko5CSjFUfpdP/MGWgbZetsFfwElRfu4pYGwfEWVFQGQWVFvn1dJqhsUWq9YWZ28mOOOSpbcS68FQTj/vft8Ei2y1VZ1b9xFop//bIsa7s1hQ0WYn/fOn2yRVNRGV81dCOojG/HMxBAAAEEEECg+wIEld035hUQQAABBBAYaIGw45igsve70TtWLTiiojLWzmhxI6iMbUdQGYvMgw875ux9SlAZ346gMp6ZtQ6ON4LKOHL+uXBid0xQGYfO1PyYI6iMzhYcb6FbQw4Oax7wrmzuybP1iiytVGVdA95a49hDWPuRSaczXoLK6PuLlggggAACCCBw9QIElVdvzisigAACCCAwUALWucIclZ3uLmrvEPCOLoLK2HgtbgSVsfzMjqAyFtm70IigMp6btQ4/b6iojGfn71P7rH439CsVlRcJ+udCeMwRVF7EderxMHDTinEqKk/ZnH0zON5CN4LKs5W4FwEEEEAAAQRGWYCgcpT3PtuOAAIIIIBABIGw45iKyghYXW7iHV0ElbGVW9wIKmP5mR1BZSwygsp4XC2tw88bgsoWlgtv+PuUoPJCp+YG/rmgd/gxR1DZTBPhehi4EVRGwPImwfEWuhFURnWjHQIIIIAAAgiMjgBB5ejsa7YUAQQQQACBtgS8E0/nDCOobIuvo0/yji6CytimLW4ElbH8zI6gMhYZQWU8rpbW4ecNQWULy4U3/H1KUHmhU3MD/1zQO/yYI6hspolwPQzcCCojYHmT4HgL3Qgqo7rRDgEEEEAAAQRGR4CgcnT2NVuKAAIIIIBAWwLeiUdQ2ZZdp5/kHV0ElbFZW9wIKmP5mR1BZSwygsp4XC2tw88bgsoWlgtv+PuUoPJCp+YG/rmgd/gxR1DZTBPhehi4EVRGwPImwfEWuhFURnWjHQIIIIAAAgiMjgBB5ejsa7YUAQQQQACBtgS8E4+gsi27Tj/JO7oIKmOztrgRVMbyMzuCylhkBJXxuFpah583BJUtLBfe8PcpQeWFTs0N/HNB7/BjjqCymSbC9TBwI6iMgOVNguMtdCOojOpGOwQQQAABBBAYHQGCytHZ12wpAggggAACbQl4Jx5BZVt2nX6Sd3QRVMZmbXEjqIzlZ3YElbHICCrjcbW0Dj9vCCpbWC684e9TgsoLnZob+OeC3uHHHEFlM02E62HgRlAZAcubBMdb6EZQGdWNdggggAACCCAwOgIElaOzr9lSBBBAAAEE2hLwTjyCyrbsOv0k7+giqIzN2uJGUBnLz+wIKmOREVTG42ppHX7eEFS2sFx4w9+nBJUXOjU38M8FvcOPOYLKZpoI18PAjaAyApY3CY630I2gMqob7RBAAAEEEEBgdAQIKkdnX7OlCCCAAAIItCXgnXgElW3ZdfpJ3tFFUBmbtcWNoDKWn9kRVMYiI6iMx9XSOvy8IahsYbnwhr9PCSovdGpu4J8LeocfcwSVzTQRroeBG0FlBCxvEhxvoRtBZVQ32iGAAAIIIIDA6AgQVI7OvmZLEUAAAQQQaEvAO/EIKtuy6/STvKOLoDI2a4sbQWUsP7MjqIxFRlAZj6uldfh5Q1DZwnLhDX+fElRe6NTcwD8X9A4/5ggqm2kiXA8DN4LKCFjeJDjeQjeCyqhutEMAAQQQQACB0REgqBydfc2WIoAAAggg0JaAd+IRVLZl1+kneUcXQWVs1hY3gspYfmZHUBmLjKAyHldL6/DzhqCyheXCG/4+Jai80Km5gX8u6B1+zBFUNtNEuB4GbgSVEbC8SXC8hW4ElVHdaIcAAggggAACoyNAUDk6+5otRQABBBBAoC0B78QjqGzLrtNP8o4ugsrYrC1uBJWx/MyOoDIWGUFlPK6W1uHnDUFlC8uFN/x9SlB5oVNzA/9c0Dv8mCOobKaJcD0M3AgqI2B5k+B4C90IKqO60Q4BBBBAAAEERkeAoHJ09jVbigACCCCAQFsC3olHUNmWXaef5B1dBJWxWVvcCCpj+ZkdQWUsMoLKeFwtrcPPG4LKFpYLb/j7lKDyQqfmBv65oHf4MUdQ2UwT4XoYuBFURsDyJsHxFroRVEZ1ox0CCCCAAAIIjI4AQeXo7Gu2FAEEEEAAgbYEvBOPoLItu04/yTu6CCpjs7a4EVTG8jM7gspYZASV8bhaWoefNwSVLSwX3vD3KUHlhU7NDfxzQe/wY46gspkmwvUwcCOojIDlTYLjLXQjqIzqRjsEEEAAAQQQGB0BgsrR2ddsKQIIIIAAAm0JeCceQWVbdp1+knd0EVTGZm1xI6iM5Wd2BJWxyAgq43G1tA4/bwgqW1guvOHvU4LKC52aG/jngt7hxxxBZTNNhOth4EZQGQHLmwTHW+hGUBnVjXYIIIAAAgggMDoCBJWjs6/ZUgQQQAABBNoS8E48gsq27Dr9JO/oIqiMzdriRlAZy8/sCCpjkRFUxuNqaR1+3hBUtrBceMPfpwSVFzo1N/DPBb3DjzmCymaaCNfDwI2gMgKWNwmOt9CNoDKqG+0QQAABBBBAYHQECCpHZ1+zpQgggAACCLQl4J14BJVt2XX6Sd7RRVAZm7XFjaAylp/ZEVTGIiOojMfV0jr8vCGobGG58Ia/TwkqL3RqbuCfC3qHH3MElc00Ea6HgRtBZQQsbxIcb6EbQWVUN9ohgAACCCCAwOgIEFSOzr5mSxFAAAEEEGhLwDvxCCrbsuv0k7yji6AyNmuLG0FlLD+zI6iMRUZQGY+rpXX4eUNQ2cJy4Q1/nxJUXujU3MA/F/QOP+YIKptpIlwPAzeCyghY3iQ43kI3gsqobrRDAAEEEEAAgdERIKgcnX3NliKAAAIIINCWgHfiEVS2ZdfpJ3lHF0FlbNYWN4LKWH5mR1AZi4ygMh5XS+vw84agsoXlwhv+PiWovNCpuYF/LugdfswRVDbTRLgeBm4ElRGwvElwvIVuBJVR3WiHAAIIIIAAAqMjQFA5OvuaLUUAAQQQGAIB71DT0LBer/u5oZ1riURCMpmMpNNpSaVSfruTmxq+5sbGhiwtLUm5XJbJyUmZmZmRhYUFKRaLsV/Olhku912H9E5FHi3vydLKvjxdrcpOtSaZVEKSun3dOulq6GuI5DMJmZ8ck4cLRbk7V5RbM0UpFcYlkUpLMtl503a3x8wUTv87lqNaXarVQ9ncrciLdTVbU7+VqrzZr0k6mdDjoN1X+fDzdA3kWP8Z030zkU3IXGlM7szm9FyUxbkJmSrm1C31zs2Oz16fWtw0qNw3Nz3eljfK8v+zd+fPklzpWcdLLfWubm2tfZdmH49t7MELxjBgwDZbQLBE8LcR8AMRbMFiHAYMRAD2GA8GG49nPDNaR2ppRvtob7W6tQ35OXXf7uy6i6puZd5b99aToVLWzTyZec73vOec7OfJk/Xcaxcn33/tcsftI2jbMkaWnfvj7n+4nT15/eTOs0cnD547Obnv9tNdvJ2Z3NRxu6YNj5GJXVREa6uffNzi7WLH7Ucdt+defmfy/I+6uHvzw8lbFz+afPixmJy0mBu6toubmD51/PrJXV07/exd4u305I5bunZ644nJMf1f9X0rwK3i7ZOunX70Ucfugw8mr7753uSpF96ePN+11Zff/rDr3z6eXP7ox60tyfLg3Lq6lg/t74Yu5u6++fjkK/efnjx85+nJbTednpw5dXJj3Fid/k14yvMn3Rh3ZVx4671uXHin69/em7z05uXJW++Lt64P+mSceGt5aPmY5gW3n33kpslnurHhjltOTW48dXxy5PqjHdcjja30q9LHNXYff9Sx+7Dr396fPP3Su5Pvv3yhG08vtnGhi4aJLm7oWMOgFufumurk3Jljk8/fe3ry6F03trHh5jMnG7cj1U7rgH1ct3baXX/K7ePJJx23ty6838bTJ1+6MPnO8+9OXnvng9Z+xroPUR/6uOuPTCbHu3Z66403dLxOTR664/TkkbvOTG4723G74WgbT48cuRpz+4ht49JluE2Nyrc7bq90fdwzXax994cXJq++82FXsGnSMbpkp/6kA2dcONndv91649HJvbccb+PCI3ef7fq4U914erTdE68Kt2m8FbetjUrcPvz4kyt999Bttbhd37gdmdx2uou3245P7j93anL/nR23s6cmx49P/z3R7+P2P96SgxAIgRAIgRAIgXUgEKNyHWo5ZQyBEAiBEDgUBIgcPpcvX5688847k4sXL3ZG1fvN3GAcnj59enLixIn295AFrusyKp988slBjUrn/pjp+mEn5K+EUXlm8uCdnVF52+lmVDZBeqXEwamo+uNOpf/gww9jVM4Z6OKMGmyN23sXL+2bUcl8v6mMys7gve/chlHZCflNWK14G0PdnZNXP9m0/X8y+XDD4F0Jo/LuzqjsuK2qUYlf47bxUMnljt3UqHxrk1HZebzN2BlDkBb2wmg7o5LBe/0NB82o/LgZ43tqVD560+SznVF5+82nN4zKG64YlatgUl6Jt67CP9lkVHrw5709MSqZIOK46+KuNSq7h388wGI8XSWjss/txzMzKlfBqHz07jOTWzuj8vrGTcx5AGnongKF3SxluG1vVLb+pzv1GFkWax6YuqEzeGeNykfvvqkzKrvxVP/WjPHVMXhrPPUAy6XLU2P85dc9MNU9yNI9aFZGJROWOT50bRe3qVF53VWjsjPH778jRuVuWkKOCYEQCIEQCIEQGI5AjMrhWOZMIRACIRACITAagWawdLNyGJQ+b731VjMqP+xMF0+Lnzp16sosRzMczcwa6inyqbDy40kZle+9997k7NmzS8+oBMvMmU86gbAZlW9dnDzxYjej8qXpU/nvdDO1COxmGoyxEGw88c84OnV0Mrmjmzlj1sxDnah6Tzej8qaNGZWr9lR5CV0fthmVDLf3u9ltxGgzKqczZ4hQZrWMsRAfmSvHbuhmBl6ZUXmqzWR4oHsi/6ZuhtuRbiZqcVsFYRWztmzMRL1QRuVrNaPy0uSNmRmVQ+MjqtaMSkalmagP3X5icn83M/Cec2cnZhxN2+1qGUdTg3c6g9eMyte6GW7PvfJuZ7iZUfnB5M2LXfvtAuKTroBibmhRGreaOVMzKj93t3g71RmVZ9oDBc1w6zqKFmtDZ2CXjaj6TbPfr8yo/OFbbQavGZVv1YzKkbgJeeyI3frRu245NvmpB/RvpyfnOsPtzKkTbUblGLPwd4ls2kS7jGPXn1H5xA/fbjMqX+zi7e1uXPhAvLWyTY2xZa43e2x32rZM+f24m4l6YvLVR892RuWZ6YzKkxv9Wz1QMHuCffq74s1rmmtG5VPdeDqdUfle17992A13V3u1q9+GyXBxc179wO3djMov3Hvj5BFvKehm2998BrduJuoKvaGglXwj3j7puE1nVF7q+rYLk6devDD59vMXJj+6MqNyGE6zZ8FNLNeMytu6mYHeUGDm8yN3MY6uzqhcTaNyOmPcjMqX33Dv9t7kOz98rzPcPmj3V8qrSx483jpm7kP0bd6IYSbqvbec6N6I0XHrZlSeM6OyMyrrjRircB/S6n4j3j7qHihoRuW7lyYvvfHu5LnujRhTo/KDrn/r7ou7oLi+Azf0cKZfa9y6Rnqi43buTBdvt3Vvxege/PFGjFs7bieOHWv3Iqs0njZ2+V8IhEAIhEAIhMChJxCj8tBXcQoYAiEQAiFw0AmUAPn666+3V6++/PLLzaRk8h3rBAX7mYdmVX7+85+f3HPPPe11rPYNJWy5RhmV/Ve/3nXXXbt69as6cU4LQfqKUfnDdydmMnh9GEG6GZVDKzXtqlc0tPbqV0/k31VGZTej8r7eq18Zbqu1bMxk6IzKi5c2jMpXO2Ze/frSxWa4ddpdMyjGyLdaK8Pt7IkjzXB7sDONCITNqCRIH7lqVI6Rh92ds8t5958ZlWVU/rATpM979eurM0Zld4GhhdXuZW7tdZXN4D15pHv1K6PyZDMq77u9M3h7RuXuyjfWUbiZidrF24ZRef7lzqj06tc3GJXTV7+a4daMyoGzUdz0BaePH2mvfmVUirdmVHYztY4d7V573RlHg6u6S5ZFH2fG+OWNV78+yaj0ClOvfu0M3sudYtxp0qNw005rVg52d3evRWRUMkDO3XzjhlE5fV34ksUc/HDc2gMsH3ll7sXJ4x03swJf9OrXZlRumLDdlYdup7hZul62GUj3dOPCV82ovOfM5M726teN/k28rdiCm9eCf9Jx+1H3AMtTL77Tvf51ah55JThDrIiNxc1wfeS6H7dxgVFpVuDUqDSjshsXOqNy5Rbx1rhNZ7g9342nTzajcuPVr13HNuqrXzsgxuzjN0wmjErjaTMqvcK0exXn9d0rTK9bMWN8Woc/vvJq67cvXOqMyguT7zMqf3Bh8krXx9Ui1saIN/ch7ZXgx7qZgV79euvUqDSj8tzNXv26Wq/uLx7G04+6B/QudTPt33r3/clLP+qMym5G5ZMvmVHZGZXdK8GVTUwMPadSF1DcThw90hmVN0we7F796lXqD5RRebwzKru2umrj6RV++RICIRACIRACIXBoCcSoPLRVm4KFQAiEQAgcFgJm5Hjd6wsvvDD59re/PXnppZfaTMTjx4+3mY1mVdrmta9f/OIXJw899NDk9ttvn5w5c+bK7/PMw6KJw2Y4dh/XtJ4Kn1Nj7M0335w8++yzzRR17ltvvbVdxytnpdvNMr2mGZXdb2q98/6V2R/P/ejS5N3ut8iIUGPpwZXj+o0jM0AeurN7/VX3ZPldt05f8Ted/TGMIF1P9O+W1VW+Xc473h92dXTpUmcUdU/kv/D6xWYcPfPapWbwMox8xlhUNcHbb3ve2BlHfovs3ttOtNflTn/bs/sNN0blQBU3HDc0Om6d4eY3Kt/sBMKXX39v8sM33p+c/9HUAKmYgG5ofLiZyeA3Km/sDN7bu5kM93XC6j23neribfqbqNdvzIQeot6G49ZlusXbx5P3u3jzm6gM3hc6bi+9Nf2txelvVE7naw39XEFxmwrSjPGjk0fuONm4mRl4Y5sZuGFULgluOGYbGekyT5BmjvtN1Gdeeqf7XdSLk1ff/XDyzvvdLNUuIJiJLd4GDjjcuv+a1szE9Ru8X7p32r95neTpk8fbjEoz75ddipvzLN+/Tc9hhtvHH139rcUfdNxefrt7q8DGb1S6zhjcWhlaOVrYT+7ouP3kg2cmj3QPsJzrXid5+kT3ANDGjHFpl12K3RDc5KVmBhoXnn2lexCje4DF7GcP/nSeeFsGDrWNs27EW/eXsYdx9Jm7TrcHCu7txtT2hoKVHReK20eTd967NHmxG0/NDHz8hQuT17u22t5QsHwzucKp/6XaqWt0P8E7ueV0Z7h146nfDGQc3eINBRuGW/+43X6veHP8EDHnd9I/6Ay3d7u3FLzWvRXjua6dPvHCxcmPLnzQ+h/Xae3UlwEX3NyHmIl68tiRyS2nbmgPm93XjacP3Dl9ZW69WWSI0by4DcEMhjYudNze6Qxev+3pHu6ZV9+fvH7BjHH34RsPsAzcWPvcjndG5a3db1Te2822v7fjds9tN05u6R6YOnbMb1R6oGC5i2PmY9a+j7FmqDe9DBhKOVUIhEAIhEAIhMAKEYhRuUKVkayEQAiEQAiEwFYE/Bal2ZTPP//85LHHHmuvffWq13r96qVuVt0zzzzTGTAfttmU999//+Thhx9ur2ZlZhJr5lkITs7hdy9dkzlqWxmW77777uSVV15p20+ePNmM0FtuuWXiGsSbRQScftqpsPrx5O33LnevwOrMo7c64aYTpN//4OMmQpVANE8ZFklDxLfc0AkpxzpEN3Xvf72re83fubPHJ7d2swJPEqTNptwQW6apF/t/P+/1fVFWm684ZX1VIOzE/E6UfvXty03Iv3Dp4yYUd9keZyEQdmee/jbUkcnZTiBkVt7WMbvtphNTIb8TpOrVr7vJRLFybH1fnttUmPXbUH4z8N2LlydvdNx+9G43a6ubxYBbxcQY6JogTVjt9L9TnUB4U+N2tHG7pTOOTnXxxlDqz5zpl30ejpV+llsd2293tW2rdZ1nmn4ab41bZ/LiZrbW6+9cnrzevS73vcufdLMCpdmQNQeGV9z4aWaA3EyQ7swj7fTs6ZOtnfqdxfZ7XrsM+ipvrWc5zf69FTPbrj2+2mn34Ec3a/ydrn97+Y1OwO+4vdXNprzY9W9Tg3ccboK5xXNXH0yQW7r+7YFzJzqj93j7ncUTx6di9BDttMqNAVbz8pLeUsfXsW3dHpr5uHF7sRsXXuu4mRX43uXOwOw6IGlaqA0cb/LTuG3wE2+PdA+wGBvOnj7evRZx+jrJLtNX8u2Y3SzK3S97nWNRfv3j2ozKzuS9cPGDyctvvj95pRtPjQ24KZfPCMhaFurcPL2zJ2+Y3NM9iHFHG1NPtP7NazjH4LZbXi3T3f9avJlR2XG72B7G6Nrqm90rYF/vHpi66PX66qlSD7zeiDPX6Lq37iGW6Xh6+03HO3YnJzeePNZdv3sdeBtTp5nYKmZ2ylWll6b/fVputba7xfH1YBtu7uHE2g86bh4oqDOPga6NC122O2zdTNQjHbfrm+l27mwXc7d096enPDA1NceqzLWeN14qPTr1vX9s//siBIvbhx9+PHkPt+6hKf1bPYjxUTeeMmG1o6Eba5/bsY7bmY7bbd1rc8+d6e57uwcxbvQASzee9l8JvlXZP628jvHx7w//XvAgpc/R9jre1fnN0E8rR/aHQAiEQAiEQAjsLYEYlXvLO1cLgRAIgRAIgYUJ+D1KMxl/8IMfTF588cU22/HcuXPNiLztttuasfj9739/Ysajp5XvuOOOyZe+9KXJvffeOzHb0Stg51nMomRSerWra3qdLOOSIUZYYYjabptzMkv9Hibhwf5FRJtK29bdsV67dunyR92sD69E7EykS93rJD/6pImDY4hceGzI3O13gDw8frJTCc90Yv6NJ49OTnUi/rFuYxNoemLyPBxn0yhjmb32OecyT5VP2ZVA+MnkshmCHS8zUImDl3FrmSiZcDZHy//tzIwhrzE90U2tJBKePnG0fY51f09FqiazTRkueEllrE//0GW4dSdswuknncvxYRfTlz7oXmV6uTM+OnbvtN8L5H642vjcjnbcWrx1ovTp7n1/THHcmtlGkJaNju90dd3cbauYTWOkHb5RF1cNkenWnf+v/uoc07VXlJpp3cVbx+1CJ67ixqQUb0RV6cZrq9N2c7RTpU8eu37iNz7F28munR7FbUPEb+1156Jtu1f+q51O43fKbJFzbsXNb4057/sdt8tLUmAAAEAASURBVHffY4h/2JmU09mUU4N3ZG5drTBBTnXcbukE6TMdt+PdkxnE6KkBQzSGZXe1h5sxop2hO1Gxq7/bjk/53yw352yfjtulrn97uzMoL3R923uduftBizf7K8fjtNeqFdxu6x7EONuNDSc7bjd04n6Zu4vExiyCrbhVHNu36FLHMCp9107f6cbSd97vxtOO3QedKbL4WRfNxTS9eDIu3NwxM6Zqq9NxYWpQLMut2qmrOVdx201uZ7nh9F7XRrHzu56XurbKDBtvXOj62laO6QMFJ5rpdkN3H9K11e5e5Hj3WmtlrDcUFDvryvs85Za2/1mWmwYo3z/u2qix4cPuAaD3u/FUrL3ZPcDiQbPpMl7UTbl1ZlhXQbj5DePG7dSxKbdu+5GNV/evDLeOmuatnX7czahvr1Tv4u1Cuw/puH3IMMe2eqB5anexNH1uZlXe2HE77V5kY2zwwJQ2ZdkNt4pLa/9W8O8ED1b6+HdDza5cLNdJHQIhEAIhEAIhsA4EYlSuQy2njCEQAiEQAgeagN+k/Na3vjWxJtB57ep9993XXrvqdykZiD/84Q/bjMvnnnuuCQE/8zM/M3n44YcnN998c3uKeR4ADEjmJDPS71GaQflB99tqZVT6bj9DkzlpJqUnpD0xXcLEp11n23SdoOEViB91ws1HTfia/r076fzTcjHdXzKQaxAimUTXd6/lnIo0ncAlWbdtulo8J8qqvvAqjkQfvIbgViynRggjCbspt5bpJiNOvw39/yZ0MUBw6/5nxtb1XdmmxsfUrOhfs8Su/rbtviuXmPMRkxhaiFuLzBDun79Y2eZ791+LN+xw8xF/06XWG38OuFqE2yLMpMVplpvtuBEL+zMkFilSsWvcugN/3HhNZ1Ga2VbcxhRWp/mdxpXf7Wrx1sXcsvGGj3JhJ9bMIvddG63Pstw6Ylfi7eMOWIu3LhBwFA9jcqt403thdbR7TyJ21/l022ZjbPbvKfft/48dbmbg42ZcwK0/a2Y352xX3GiPbVwQc93YMG2n9k7ZKYPv4yzTsxsXjnYmCDNEWaafa6+4aBmlL27abMVatdNrz/7pf1UbrZStrXZY2sMFjZuHCcbiVFe9do3bDV283dBMj2u5LcqrzqxceLnnsfY3dmZsWc9yqONm19ulsx2mFnNdW3U/cpXbWPw2jMouk1olf6j6tzbDvgvDPq/+99lybfX3tExTbmLO/Qh2+jWmEW6LLrP86hpYaae4+R3E6VLrRa/y6emdGTNMuubZ7kHcw+nr2tjQ7atlN9z0aTU2YOY7A899iD5uN+es/FhXvGmn+jb3bzUujNevTc98DbeOl5hr3DqWXcGukNtNGXHCq/6t4N8pfirCQ5TMyt32c312+R4CIRACIRACIXA4CcSoPJz1mlKFQAiEQAgcIgJmUv7BH/zB5NVXX20mpX/sP/zww+0f/WZMMsGYmE899dTkj//4j5sA9fM///OTz372sxMzLol48yxEBTMqGZRmZxKfCVslCNpnu22EGuf1pDTRYVa42u5626UjhpCUrAlOtJK2oVOixpO5Nuey04muiEf9vC4q1jhziTW4vfPOO80EcR7cvDLXun+Nzbm5umW7dFty2zhsL7mVuGu9VV4X4SfexBjTSAyKb+dk7i5ivF+ltzlPfW5irX0OMDflwQ0rIn49UFCiqocbtNlWboWdc5mty624zXmqXSebze001jbaqbN2G2ZjXT7nWaTTTvV9Ys1DGr5rm+LNWv+26IytLbm1OKt+bp7cDZOmSGA07d+utoet8rnIVR1vBr6xSZs1HvkwQHAbJN72m1sH7go3BmkLtmsjbt54K7bSi7fXXnut9XNiDTPtFLdFl9l6NHhqAm2GduO36BmHSX9NW+1xW5RX5UY5680OxlULdkyQIcbT4iZ/dR+Cow7m2hpvlx7tf9txqwsuwg8zY4Px1LiAm+/GA79lLu4WXWbjTX5gauuNePO3Za+5TdltjAn+6C2LcKv7N6yKmfGVQen+DbdpeaukvQtt83UrbpIWtxZzM8deW4KZnQP9CdP0M+W2XT7nuVyfm3izGBP8W+See+5p93BibzcG+TzXT5oQCIEQCIEQCIGDTSBG5cGuv+Q+BEIgBELgkBMgGPhtyt/7vd9roiZBzj/2H354alQS54gnBM8nn3yyGZpEj1/8xV+cfP7zn2/iwLxClGuVSUR0JtjbRniwZlL6LUwCK0FVXghdRIihlib5ELp6J9wLoaZ3uQ0hur9ld9+L5dtvvz156aWXmqCvbjxd/sADD7Qny3d35s1HHSZuWxnmWIo5v7/qifyhluLmfBVzBzXeihtT3G/aEvTNmsHrzjvvbA8VLCqsbse5z227NENtr3qp87X6IazWhiXX1U71bx4KIUxroz71QAGO9Sq8ZS7XTI9lTrCLY4tf8eq68sEW44LZ948//ngT872SnCBtbDDuLGrwbpexfec2YLxVGcXb008/3cwjD/xgpp2OMZ7WNfdjPWS8yb/x1BskrN2b1HhqPeRSMTfbfoa8xk7nGopbGUceKHCfKO58F3MeZhN3Qy39cWG/uA1lKte4wGzDzLjq3le/9uCDDzbDbaj+Df/ZeKs6GbC7rlNuvR6ojxNv/v1QD7Awees+5O67777ysFmMyq2rIVtDIARCIARCYN0JxKhc9whI+UMgBEIgBFaaACHY61x/93d/t4lMhGC/PfnII4+0GZVlVBKLn3jiick3vvGNVp5f+qVfmnzuc59rv2M5r1G5HQh58DFr5nvf+16bcUnAN7NTXhhIWTYTIHQxPd54441Wh4QuRhEh/9FHHx1UINx89YO7hchFHDS7jbBK8MKSEB1u29drcSOqmmFNICQGaqvaKX5DCqvb5+Rg7al2Ktb81i+Dl3ivnfoQ9HEcwqg8WGQ+PbfGBQ9h/Mmf/Elrp4RoZpuxATcCtT4vy2YCNZ4aF7TNGk+HfBBj81UP/hbjqXZqLf70b0Mbbgef0tUSMI48xMLY9Rvn7hV9F3Nf+cpXWtxdTZ1vRaDGBe0Ts77B697a2JD+rWhdXePmXsT9m3jDTzv1AIbxQXs1CzpG5VVm+RYCIRACIRACIXCVQIzKqyzyLQRCIARCIARWjoB/4DMqv/71rzfThjhiRmXfqCQKbDWjssSUIYxKYhdh9bHHHotROWeUEAdnjUpmBxPkM5/5TIzKbTiaIVxGJYGQUYml177GqNwGWre5uBHwtdUyKnErozLC6mZ+1U53MirrNxc3H72+W4xNPoxKv6Gsnd51112bjMoYvFvHyKxR6e0Efns6RuXWvGpr36h0X2I8jVFZdDava4Zb36gsczxG5WZetaXGhVmj0oN57t88NGg8Tf9WxKZrRqV7kTIq/ZREjMprGeWvEAiBEAiBEAiB7QnEqNyeTfaEQAiEQAiEwL4T8A98ryM0U/KVV1658hpHRmW9Jo4oYAaVWQbf/OY3m3jycz/3c1fMMLMul1nkgfhAyDejkiDN/MgMkJ2pltDlNZzqkOCVGZU7M7O3DDdCVxmV4i8zKndmh5u2aeZHjMqdWfX3aqdmUerfnn322SszKmtWpZmBMSr7xKbfjQtMEGMPo9LDBWVUMt1qRmWE/M3sbNFGv/vd77ZxoWZUxqjcmlV/axmVxlUxWDMqPcSVZTOBMiqNpx4qwK2Myp/4iZ/IjMrNyNqWun8roxI/46t+LUblNtC6ze7VPKBXxngZlbjl1a/bc8ueEAiBEAiBEAiBKYEYlYmEEAiBEAiBEFhxAl6f9Ed/9EdNZPK6JAI6o9I/+gmcRHa/Y3n+/Pn2G5LHjx+f/PRP/3T7HUv7vWZpmaUE6ZoBEqNyPpoldBFWGZUEG0al+jMzkMCaZTOBMioJXURV8VZGpbgPt83MbCluBNW+UakPMAvbOjMqN7Mro5Iprg/Vn4ox7dQnRuVmZrbUuMCo/NM//dP2+2312tf67eLMONqana2zRiVmQ/8G7/ZXP7h7YlQuVndlVBpPGZX4lVH55S9/uf3O+GJnXI/Udf+GFWZlVHqFqfu3zKjcOg7cq7kXwcvYUEZlvfrVQ47+jZJXv27NL1tDIARCIARCYN0JxKhc9whI+UMgBEIgBFaegJk+Xrn6wgsvNNPm2LFjzaQkbPpHv6eXGWHSXbx4sRkSX/jCF9rrHons0i+zlCBNWO2/+tX1vVIyr6rbmm4JXWa4qbsyKpkgDz/8cAy3rbG13zcyO4uwSiD0nfglznAT81k2E6jfqCQQ6gsYbowi3OqhhhiVm7mVUckU98AHbmJMO/WJUbmZmS01Lpjpb2agsYdRaVyo3/ZMvG3NztZ68Ec/5yECbyjIjMrtedWeMiqtxaA2mle/Fp3N6zIqGW6MI9w8/GNc+OIXv9gMt81HZUvdv7lvw0w7dS/CcHMfUr9RmRnj18ZKGZV4GRv6RqUZ9/q6GJXXMstfIRACIRACIRACVwnEqLzKIt9CIARCIARCYCUJ+Ic+o4uI/swzz7R/+JslSUD3j37CAEHAQuw0e+qBBx5oQsoQgkAJ0syPxx9/vIlcrltGpd/sybKZQAldjCMzGYiDZlQyQR588MEYbpuRtS0MN6ZHzWTwvYxK3MRels0EihuBkOlWRqX2yUAiTMc42syt2ikx2gMfHvwQY9qpD2E6r37dzK0/LniAxW+iGhPMNDITFbfE22ZutcV46lXqNbutxlPtNMv2BDz44z5Ie2XCMSrzm8/b8yqj0n0kcxw/9yLGhc9//vPtPnH7o9d3j3sOYwFWYk07dS/iN9/dh+jj9G8xKq+NEdzci+Dl3yXMXWMFbv59on8b4t8l1141f4VACIRACIRACBwWAjEqD0tNphwhEAIhEAKHlgCxxD/6GZXf+c53munl1UpencSstBChCABf+tKXmklJLCYUS7OskNIXpJ944okmPBDyXYMpGqNy69AroategUWwYVRix0iO4bY1tzLcxDRRlQFSRmVejbg1M1uLm76CsNo3KgmE2mmMo838qp1i5oEQ/a2+tMxKAmuMys3calzwytwnn3yytVOzjHyYRzEqNzPrb2FU1hsKxFsZlRlP+5Q2fzcm+C1Zayachwm8ipNxlGUzgTIqGW5izv1IGZWZibqZV22pccF9m1hzP1JGpfsQfVyMyqJ1dY2bexG8xJtYK6NSH1dGJXZZQiAEQiAEQiAEQmCWQIzKWSL5OwRCIARCIARWjEAJTV7bxahkWJo1ZSaQmZX+wU8cYEb4bUoiSonrjDGfZZZZQZrwUEalV0pGWN2abgld9QqsvlEZw21rZrYSuZiThC6iahmV4iyvRpyP26xRSSD0UEOMys38qp0So/0ecBmVBFUmiL50iAc+Nl/5YG8xLvgwKp966qkrRiXDiIiPW+Jt+zrGbdaozIM/2/OqPcaErYxKMZdlMwH3j+4V3beJOfzcixgPYvBu5lVbjAv1u8/GBvzKqHQfYmzQvy37IGBd77CscXMPhxejUqyVUenhRvdxZlRilyUEQiAEQiAEQiAEZgnEqJwlkr9DIARCIARCYEUJlEDHsGR+EQMI6CWWEAE+97nPtZmOQwrrfaOSID1rVNaszhXFtm/ZKgNEXXnlWt+ojOG2fbX0jUrsiIPEVgJXfhN1Pm6E1f6MSkZlZrhtza7aqf61jEqx5mEMnxiVW3PrG5VPP/30FaOSYRSjcmtm/a1MI69S90AGUzxvKOjT2f67dnr+/PlrZlQ+8sgjLea2P2p99/SNSq8EnzUqY/BuHRt9oxIz7dRDU8aDvlG57IOAW1/94G6tByvx0sf1jUqz7WNUHty6Tc5DIARCIARCYC8IxKjcC8q5RgiEQAiEQAgMQICB4x/9DAjfCQI1Y9LaU8pMQ+vaPsBl29PQrkV0iFE5P9EyQGaNSqJ0ZlRuz7FvVBIIM6Nye1b9PcXNq18ZlWYGeoiBMBijsk/q2u/VTsWaV7+aRROj8lpGW/1VD7AwP4wLxqV67WuMyq2IXbstRuW1POb9SzutGZVi0My2GJXb0+sblWLO/Yj7yJpRGaNya3Y1LmCFWRmVJ0+ebPdvNaMyRuW1/PpGpRmVHjTTThm8MSqvZZW/QiAEQiAEQiAENhOIUbmZSbaEQAiEQAiEQAj0CJQgHaOyB2WOryV0lVFpJipRyyytGJXbAyzDjTAYo3J7TrN7iluMylkyO/9d7TRG5c6cZvfWuNA3Kr32Na9+nSW19d8xKrfm8mlbY1R+GqFr988alfgxj8ywz6tfr2XV/6vGhRiVfSqf/j1G5aczSooQCIEQCIEQCIHtCcSo3J5N9oRACIRACIRACHQESpCOUblYOJTQFaNyMW5luMWo3B23GJWLcat2GqNyMW41LsSoXIxbpY5RWSQWW8eoXIxXjMrFeFXqGhdiVBaR+dYxKufjlFQhEAIhEAIhEAJbE4hRuTWXbA2BEAiBEAiBENggUIJ0jMrFQqKErhiVi3GLUbkYr0pd3GJUFpH51tVOY1TOx6tS1bgQo7KILLaOUbkYr0odo7JIzLeOUTkfp9lUNS7EqJwls/PfMSp35pO9IRACIRACIRACOxOIUbkzn+wNgRAIgRAIgbUnUIJ0jMrFQqGErhiVi3Erwy0zKnfHLUblYtyqncaoXIxbjQsxKhfjVqljVBaJxdYxKhfjFaNyMV6VusaFGJVFZL51jMr5OCVVCIRACIRACITA1gRiVG7NJVtDIARCIARCIAQ2CJQgHaNysZAooStG5WLcYlQuxqtSF7cYlUVkvnW10xiV8/GqVDUuxKgsIoutY1QuxqtSx6gsEvOtY1TOx2k2VY0LMSpnyez8d4zKnflkbwiEQAiEQAiEwM4EYlTuzCd7QyAEQiAEQmDtCZQgHaNysVAooStG5WLcynDLjMrdcYtRuRi3aqcxKhfjVuNCjMrFuFXqGJVFYrF1jMrFeMWoXIxXpa5xIUZlEZlvHaNyPk5JFQIhEAIhEAIhsDWBGJVbc8nWEAiBEAiBEAiBDQIlSMeoXCwkSuiKUbkYtxiVi/Gq1MUtRmURmW9d7TRG5Xy8KlWNCzEqi8hi6xiVi/Gq1DEqi8R86xiV83GaTVXjQozKWTI7/x2jcmc+2RsCIRACIRACIbAzgRiVO/PJ3hAIgRAIgRBYewIlSMeoXCwUSuiKUbkYtzLcMqNyd9xiVC7GrdppjMrFuNW4EKNyMW6VOkZlkVhsHaNyMV4xKhfjValrXIhRWUTmW8eonI9TUoVACIRACIRACGxNIEbl1lyyNQRCIARCIARCYINACdIxKhcLiRK6YlQuxi1G5WK8KnVxi1FZROZbVzuNUTkfr0pV40KMyiKy2DpG5WK8KnWMyiIx3zpG5XycZlPVuBCjcpbMzn/HqNyZT/aGQAiEQAiEQAjsTCBG5c58sjcEQiAEQiAE1p5ACdIxKhcLhRK6YlQuxq0Mt8yo3B23GJWLcat2GqNyMW41LsSoXIxbpY5RWSQWW8eoXIxXjMrFeFXqGhdiVBaR+dYxKufjlFQhEAIhEAIhEAJbE4hRuTWXbA2BEAiBEAiBENggUIJ0jMrFQqKErhiVi3GLUbkYr0pd3GJUFpH51tVOY1TOx6tS1bgQo7KILLaOUbkYr0odo7JIzLeOUTkfp9lUNS7EqJwls/PfMSp35pO9IRACIRACIRACOxOIUbkzn+wNgRAIgRAIgbUnUIJ0jMrFQqGErhiVi3Erwy0zKnfHLUblYtyqncaoXIxbjQsxKhfjVqljVBaJxdYxKhfjFaNyMV6VusaFGJVFZL51jMr5OCVVCIRACIRACITA1gRiVG7NJVtDIARCIARCIAQ2CJQgHaNysZAooStG5WLcYlQuxqtSF7cYlUVkvnW10xiV8/GqVDUuxKgsIoutY1QuxqtSx6gsEvOtY1TOx2k2VY0LMSpnyez8d4zKnflkbwiEQAiEQAiEwM4EYlTuzCd7QyAEQiAEQmDtCZQgHaNysVAooStG5WLcynDLjMrdcYtRuRi3aqcxKhfjVuNCjMrFuFXqGJVFYrF1jMrFeMWoXIxXpa5xIUZlEZlvHaNyPk5JFQIhEAIhEAIhsDWBGJVbc8nWEAiBEAiBEAiBDQIlSMeoXCwkSuiKUbkYtxiVi/Gq1MUtRmURmW9d7TRG5Xy8KlWNCzEqi8hi6xiVi/Gq1DEqi8R86xiV83GaTVXjQozKWTI7/x2jcmc+2RsCIRACIRACIbAzgRiVO/PJ3hAIgRAIgRBYewIlSMeoXCwUSuiKUbkYtzLcMqNyd9xiVC7GrdppjMrFuNW4EKNyMW6VOkZlkVhsHaNyMV4xKhfjValrXIhRWUTmW8eonI9TUoVACIRACIRACGxNIEbl1lyyNQRCIARCIARCYINACdIxKhcLiRK6YlQuxi1G5WK8KnVxi1FZROZbVzuNUTkfr0pV40KMyiKy2DpG5WK8KnWMyiIx3zpG5XycZlPVuBCjcpbMzn/HqNyZT/aGQAiEQAiEQAjsTCBG5c58sjcEQiAEQiAE1p5ACdIxKhcLhRK6YlQuxq0Mt8yo3B23GJWLcat2GqNyMW41LsSoXIxbpY5RWSQWW8eoXIxXjMrFeFXqGhdiVBaR+dYxKufjlFQhEAIhEAIhEAJbE4hRuTWXbA2BEAiBEAiBENggUIJ0jMrFQqKErhiVi3GLUbkYr0pd3GJUFpH51tVOY1TOx6tS1bgQo7KILLaOUbkYr0odo7JIzLeOUTkfp9lUNS7EqJwls/PfMSp35pO9IRACIRACIRACOxOIUbkzn+wNgRAIgRAIgbUnUIJ0jMrFQqGErhiVi3Erwy0zKnfHLUblYtyqncaoXIxbjQsxKhfjVqljVBaJxdYxKhfjFaNyMV6VusaFGJVFZL51jMr5OCVVCIRACIRACITA1gRiVG7NJVtDIARCIARCIAQ2CJQgvRdGpWtZrOvj7+uuu+7Kp/Zb11L7/T17XKWZ3Vd/Wx85cqSd33fnGmIpoWtMo7LPq/LcL79txabWs+nmSd+/Tj99nbPWde5l1nttVFZ5ai3vVR7r/lIcan/93U/T/177++e2v46vdf+Y3X4vbmMblbNlkd/+tipTras8labWtb3S1Xr2fLN/S9dvr3We3a6rne6VUVnlr3Xlu8pVf1tLY7Gvv8xur78rTf1tXVxrXWmWXTs3UXovjErX6pdJ3uvvKlet++Wq4+SzlkpXa9srXX2vtLUWbxbHDLXslVFZZat15V9Zqly22V/Ldmxmyz97zjpHv33OHlPX2O16bKOyyiB/Vb7+tmJT6345+unrmEpX6zpvresYf1eaWtu27LLXRmWVp9aV/yqTdS3yttX2rY51jO21ru/+7p+jf/6WeJf/q3FhbKNytqyy29+2XdkqTa2rmFuln03T/1v6fnut8+x2rU7di3jQ7LXXXptcvHixlefUqVOT2267bXLmzJnJ8ePHJ9dff/1uL5HjQiAEQiAEQiAEDjGBGJWHuHJTtBAIgRAIgRAYggBRg/gwtlFZ4olrEYl8fLcQNepDWLGv0hNZ7LOubR999FE7zrb+9v45q1zOd8MNN1w5Rwk97QRL/M+1Ll++PBnLqKyyYuS7pcpUfEqEqvJV2epY6epjWz99seufV5p++uLu/NI7ftmlDLe9mFFZvMSLcmFZDKpss/GjfEePHm3piqN1f3GO2lfxXLFsX8VyXaN/7G6/F7cxjcoqE16+Vzln2c3GmzIVh378FOuKH6wrbfGyro99mOFvPcQiP9rpXhmVyoKfj+84KrfyFIfibG3pty3b5NlSDPxte9VHnbe2VX30z9NOsMT/nNt1xjYqq7xVplrb7qNMFQ/K72Op/PVZV7z100vrnPVxXC22OUb6YlfnrzS7Xe+FUVmMqs1VnMhztSPl6jMtRlVe+/QtdYzt/fPabynejp+Nt6GYuc6YRmWVq9bK5tPnpvyz5euXX9pq27ZLj3V9+mldp+pmNm3xt32ZRf7l58KFC+0eDj/m0enTpyePPvro5NZbb13m9NccW9yqTMWtYqr6N39XvipeqrzFxP5aqp+zzcd5a7/ji6218wyxuIZxYUyjUlmrPL4rS7/8/laeijd/+1gcp16LRR1f6a2LhbTFq77X35hVfzgEN+eNUTkEyZwjBEIgBEIgBNaTQIzK9az3lDoEQiAEQiAE5iZQYsrYRiXB5YMPPmgiGlHt/fffb3/L6IkTJ5qwdvPNNzfxhdAmLVHk2LFj7Slt6xKXHC/fxDgiTG0nOjnOPiKP70Sgs2fPTm688cbJyZMnrxGl54a0RcK65lhGZfEiCimH8igXcU35/a1snmRXPmtPshOvcJOOGYgJns4nPdaeesdOesKY80vvI61jXAsv6aR3HNGrhLQtkMy1qQy3sY1K+Vdm5SHeVmzgoyx4VfzIePFSRuKueJNXnK2dT9lLLC3Gly5duhLLrme/84o39VKM54KzQ6LiNpZRqXzKpDyu4XrKokzY4YCd8lS84SimLPg5TmxWvDleGizwFk+uU23f9cSev13P39jdcccdbb0Djrl3yb+8jW1U9vnpE6qPs10sKddNN93UvsuT8vpgigtWFtvwE2uYWfxtu23FDDfb9H/aZzH297Jt1DWrPGMblcohviomlEmsYaSs4s24oHy+z3LCWdw5jwXLfnrb1L/z+1QcO7ft6uaWW25prLFTH0MsYxuV6qdiW3xg6IMfRmJnth+TvtqkcvuOwRtvvNFYa9e2Fxvnw9XfjvVdexfH6kPbLmZDxBzuYxmVxav6c2VRdv2dj+0VbzhoU2JJ+RyLq3SYiDfH2o5XjaeYix9p6/xVN9LaX2MDdvgvy02e5V07EHNjGZXyX3HhWjgoWz+mipkYUX55UT7tC0vf5dVx9jsfBtq1BVNtFGf7ndu5cHNuH2mXZeZazu16yqG/dj/i2vJ5//33tz5kmfrByzWUxbnFhLIov2taixVxUPGjbNI4Vt7kq8/ZvhpPxZHv0sp38XIdf1vbh92Q46k6c25lyoxKkZQlBEIgBEIgBEJgEQIxKhehlbQhEAIhEAIhsIYEiBnEh7GNyhJeiBs//OEPJ6+++moT/IhOhOV777138pnPfKYJfy+//HITaYg5RJyHHnqorZ3jzTffbMerqnvuuacJMbYTxV588cV2zhKICIqEp4cffridn2DjfASfZQXpErrGMiqViRikDCWi4eF6+BGjiFC333775JFHHpncddddTUAmrBKScPrBD37QmGDufIQtr+d64IEHWnpCNg7OiR/BWlocxQReGBPupO0bBbttKiWkKZNrKgeW6uW+++5rJthuz90/DitlFmdPP/305KWXXmoslffcuXNXykZwt8jLc8891+LvS1/6UmNZgmzlEdv6EAYxxoyZozxESYzuvPPOVhZxh5sYX1ZcLW7iwXWVjZCKmxggXC4jrKoD11CW8+fPN+FbvCjTK6+80sRl5yc6i5+77767xZIY1IfIk+OqbcsfEV/51av0uEtbbV8a3PQ9Jd6K469+9ast/bLM1Gu1U/X7wgsvNEEXM2aLj/wP1R8Uv2eeeaZdS7lcn/GhXOLBd7EjppTdtbXJ4ijm8FF2zCxiFx/8HSsGKjb1bw8++GDr31xD2YaItxoXxjYqlUW96HdKmBcXyumjjj73uc+18hknxKRF+cWleHv++efbscot3j772c+29LhaxKa6wBV3zNUV/tr/5z//+cZaHWjfQyyu9/jjj7drqHOxry9VP0Ms1b+Ja+Olj36bCVR90Be+8IXGT+woN67Kh5E8iR3Hf+9732s8apzFRl1sNZ5iZLwxXiuT8gzRfoqJ/Dz77LOtzsSgOnc9/c4yS/Vv+Kj3GvOqD7cdU/GmnRrz9OPK61gM9fXaon7OcdW29Yd4GC/xFZviWnoxh6OyaJ9iQH+oPEOMp8Zp+XZNMYefshgPhpxRWddR7rqvULbq5+u+QkyIK4yNu9qk9mWM0n/Jm+PwkW9jRNWtvqY/njq39o4bZupFPAzRv6k751evYkF+tRF5H8KodH79lzK5r3CdGk/FUPVD2mLdj2EojVjBucaR/v1bjaeYbDeeSu96zmMM+Zmf+ZkrY8kybcix4qD6TtdRn66jnci//mCIuF42nzk+BEIgBEIgBEJgNQnEqFzNekmuQiAEQiAEQmBlCBAZiA9ErqeeeqoJXsQ6IgiRo4yc3WbYuYk2hCACFZGGME1QI3i4PgGLyMcgIhQRoO1nkhDciFTWxB3nIf4QdAhXRBF5J1gTxggnJWYRwxhTBBTCDkHfdYi0jltmKaGLyKVchEICGnaELtdYZlEO5SIG+Si3a9rumlhgS+Rj8LomMVC5cHPM97///ZY3abGWXr6kJZhigj3WJcAqC5FLPeBORJMeQ+cmSi+zyEfVo+v6rlwELtdblpsyEkCLEwH/fCcsixnCpHgjJuNGXCb8KSuB9Lvf/W4r38/+7M+2fGBR3Amq6hYD15B3AmQJ1kRJZXFu8YdvGcjK5vhlluImpl1TWdSdcw9hVGIm/9rRn/zJn7TyKa/FNfFURqzEjbgghLt+xSoe2q74lD9tT3qMsWYgyTNjSXw6r4/0ro+t8/7ZP/tnGz/tadml2qn6GtOo1GbEi/JrS+KtYke5xLW+R9xhVqzlTz9rGzZ4qAPxwmgSr4wk8Sm2MKm6cn6MxZs2+sUvfrHVjXS2L7PUuKAcxgV9inr0UQbCtPwuW0fap3gTE/pQbavEeoywYxgpn/4eJ7GlDahPzOVRDMqzfBm7tD/pcdSPSssAk65Yi2flwU16Ywx2QyyuOaZRqRzKLd4YYb7jV32cOtI+tWHxYrsYteBruz7dMX/6p3/a2Bl/jffYOiejxHWkVc9iU507Tsxqq2IaT/U0xKKdjmFUFgPnx6zaqTZkqf5VfyXWfIxHVfbKl3jVX2HZ7w/FqA+2Nf4qR42n4rLGUucWd5gtO57WeKcexZx8qrOhjEptykdZnRc39xXW2qft8iAGlMtY5Lv8PPHEE+3YvmHqGHms8QED6W0/v2EAO5/z2qY9YlXjKcZidNnxtMYF9ag+xYHrVf3rU5fp3yqemLrf+ta32jWc01J1JCbFRY2n7heUTZ6Mj8ZT8SN/eOjTpcfDeOq+Tx4rnTLUmKp8YlHMefAHvyEWdaNsriWPYqL6Xf1C9QXylSUEQiAEQiAEQiAEZgnEqJwlkr9DIARCIARCIASuIUBkID4QucYwKokaxAxCKtGmRGUCFFGLAENosTzUzZwk1BCMfAg2xBxinjwSAH0nHhJ3CNGO/853vjN57LHHmjBM+Pq5n/u5tt+xBH4iuOMIO8QghqVzLLOU0EVEIiYR5oi5zkuwW9Zwcz6iKNMCH+KWusKHSOz6tisjsZiAh0kJperTfscQXOWNgUIgLPPIeaR3LSLh//2//7fVFa4YlRCtTI4ZwgApAY/QpUzqWVkIXEMYleJB3Cg/c0L9OD9hU524HgFZLBEOsSP6YfDHf/zHLe1XvvKVFi+Mb+ci+mMrPp1HTDkHg0V5zPhyLnWBb8VpCYpDzAgqboTIMYzK4qbMv/u7v9vKKD60UeVXNvFYIrN9v/iLv9jiXbvGRLlLTBV3mMorQZOI+Uu/9Eutnhkg4rrEbm0WK+1GXfiIh2VNMO272ql8j2VUKitj5//9v//X4k4brY99YhAfHLAkHNuuvcmX2BI/2pi+Ehtt7hd+4Rdav/f1r3+9nZ9IXTErDqUV5/o35/2Lf/EvNjN4CMNN/vS5+usxjUrx9tu//dvtIRPitnhj0Gqr4sf11ZvyiR9xJG6U2z7xpg1jp43Ypm3q1zxw4DiMxagxQvsUX47Bsh7IwVv/NpTALn9jGpVVL2JFbMm79mNtn35O+8FFOX2vvsN+fPRLxkXjgnQ/8RM/0crPYHryySfbRzoPDohRfYT2XOOpmXLGCtfFb4hFXak/1xGD8j5E/6n/wsRYzWhzDeXAi0Grv6k+zHVrPNIvYaPtfvOb32x9HONN+xafzomLuHGMtM6pr/zDP/zD1kbl39iBk/gecjzVRpVNXsQcfvqFoYxK59euqi2KFfcVtmtH2p/2qF3px3HRRvW7GBgTbLdgZRFTWKlb+5xf/8nQUxbjqXbrnDWe6u+ctx5AsH+ZpcYF47v+eWijUkw4p77z937v91o8y7v6V27lVGbX9t2+n//5n29cajxVdnzVJd7ar/rVfvVbf+7P/bnGWzw7puoB3xpP3VM5N95DLBUPMSqHoJlzhEAIhEAIhMD6EYhRuX51nhKHQAiEQAiEwEIESpAmco1hVBKnCIBEmfOdGea7axLrmGHEGDM6pCN8EVgILYQkIg7BUt6Ib0QSYhUhm0AqPfHm//yf/9NMI8Kw/X/rb/2tJnYBQVQj9BPaCKtmz/gQjJZZSuiSxzGMSuUi0Ms/Y8ffBFCCMUHZYgYgnngR9olXBD3ioO2OJwR++ctfbsc6D6GL+CU9EYv5QVQj3P7+7/9+Y8yAIt4SuIhqzkmI9VnWPCrDbSyjUhwR8gj4BHf1LtbEBXPCdQnO+BBVxZB4UJ8Mb3ykx8Q5lJnITGxmdBP2bWd6/NEf/VHb/+f//J9vpq46cX77rbFy/iFmNBQ3caBNaDcljotlYuYy9VPCqtl7//W//td2DYIxUV471WbFD3GfWI3nX/gLf6HFDxaYixNxxTRXdtvEoH6FsPyrv/qrrX33hVXnlX+xaMalY8WkWF821tRHtVP1Ki/KSbRVxz6upU61id0s8u+jjP/5P//n1hcw+Esgdk68tC/xqI3qn1xXe5MnZdVO1aF8qFvtnHCt33NeMaVvdF71gjOR3Xn1f47/K3/lr7R2q80yTJbhp0z6W3WobPoU1/dRl/K/TLwVa33Yv/k3/6YZldqjeCPAiwNtiJGp/9bH/Y2/8TdarIgf44k2rC71h+Kx+rH//b//d+PIuGWoazOEfO1bubR37Rlj5XEO/LBfhlmVyXoso1L+xbSY0o+pf7Gtzv/Mn/kzLYbsY6zZLg70X+pLX2ifONSfaNfqUj3qG42rGOrXsMIeV9zFnGuf78bw3/md32lxaVzB0rgqrodY5FmZhjYqlVdbUn73CWKoDP6/9Jf+Uos3sY6b2FJW/ZLyY2uM/1//63+17f34lF/HOD/WYrifHhPpjadiUZ8z5HiqjY5pVDq3to+NPsinf1+hLO5RcJAXfZh2pb06ru6P1Cf+4lAfZlzEQ99rn/rwoIe/f/mXf7mZus6n7bqm2LXPcR5AcOwyS40LYxmV+nBlN57+t//239p37Uv/Znxwfe1LvyR+tB/3EWKoZtBjK160Tf2SdNgbh8XlX/trf63Fmv7ZecSoRXqmuPuV/ni6DK86Vp2IdX2z/KhTbUW9ylP1pfqULCEQAiEQAiEQAiEwSyBG5SyR/B0CIRACIRACIXANASID8WEso5LQRFwhZhFmiMuuR9AgOhE9CC0EUn8zx6yJUsQxxxFuCGFEQAalGR4EHyKzNH/wB3/QzE5iFwHr7//9v9+EQeIOUfI3f/M3m6hCUCWu/tRP/VQTcK4BseAfJXSVEEdIcz3CEpGIkbDM4rzEJ2KqDwPOubEhelbZ6il6fxOLrIlkeBFciV0MIDxL8GOMEKj7M2GkJVCXUEhUI54RHp2z/1mmXOpbWQhd8ug7luKBgLksN/VgJoeYMkMSxxKcxYv9RHjXJyiLJ7/hpJz4EAEJ+cRTnMSTGZZEP+zFrpg044jBTpBl2DE+mB2OE3PO5drimRDuOsssxW0so1LMYIINY4yAq52JNXGi3NovE0E8ijWCsbjAWf9BZBWjeDN9tE3xKY5t+7Vf+7UWUxjb7nqYYeQafeNkt8bhLONqp2JNPzSGUYnNt7/97cm//Jf/srEhRCuPeFYO8YiZcouhf/SP/lETk/Vr4pT5Iy6xIDKLNf2bmMJJ/8VgcV77GHM4iy/HMzz14x4wKNNIX7mMkVjjgjp0jbGMSgwYleJK+9TvMCq0O0xw/Q//4T+0OPt7f+/vNTFcu8MSE+32p3/6pxs3dYsz41YsMsbVQY1BDHXt1UxVfaLr4aSdD2lSisGxjErGkbaqnGbkqhvX0n8a16wZCMZZ8SFOxIu+XB+iX2IISVNtz35xVayMpwxk/Zy2iTtD0oLhb/zGb7S27TjjqbW4HWLRTscyKvtjg7Jpl/L9K7/yK61d2q+vEnP6cSyNk/ouvMw0x1C/yACqcQxr/YxYkp5Rh73xVGwZH8Q1I6fG6KHGU33PmEalNqWf1tbEmrjz3X2FmLH2tzQW5X+oM8bFm7bGPBOnuMqn7WLGsfpCHBzv3k27xscDBvo+9yLYijl9t/pxzNe+9rV2jWXircaFsYzKMmmNp//lv/yXFive9GE81c6wcA8h1t176cfch+h3azy1zT0KZhbt3HggvfjrP/gjlpUFc4yMPz7qwzgw1Hgq3rQB9R2jcpkIzLEhEAIhEAIhsJ4EYlSuZ72n1CEQAiEQAiEwN4ESpIkghChiEHGTOEIsJr4tsxD8iMr18bfFeZ3f9QkvBBDiCxGHSEXgI2AxfQinBHPCMsGUUEVodg5CGGG6hFXH/4N/8A+aGEYEIxT9q3/1r9q5iPhEVTNPXHuZpYQugqQ84OZ62BGIljXcnJcoSOAj0hG2CKbOrewW4h5GmBKviVE4lrhISCIkMoCsMbSPmEUAw4AYK9/KgKF9ZoC4hrL4Wx1ZpFt2KcON0DWGUVkzMBgg4sbfYkI8qxPiMmOojJeHOlGV0WEfUZRYagZEidhmEhGm8SBqy7/jsWdoEhMJ0Ywl5gjGhG6mnrohWv/6r//6lTrbLb/iNqZR6dzay3/8j/+xlZNgzKQVJ+JPmbVj4qr2qS2JJ5z1H+KlhH2x6HzqGBPscJLGeWxzTvGnfoirrrNsfzPLt9rpmEalaxCX/8k/+SeNjZjS5hiLOGmH2peY0n7/7t/9u62sWGpzXhFpnzbK0GbUEbS1d33Av//3/76dQyzq/wj94lWZiN1meokP/RoDQD+KK6F/twJ1jQvqaUyjkgnx7/7dv2vlFCPKV0Yl4R3Xf/7P/3kzgv7hP/yHbQywTdtjuimnGcvKjAGO2r44YkBpf8ogrXKIPw8OGAu0XcyH6Ndm4057MHbp59SV/ofpJ7/LLMqIizZo3FMm11Je7VG/rt25rjHJmPmTP/mTLZYcq+/6xje+0dbarjj9q3/1r7Z2Lt6wMiMVQ+OOeK0Hf+Rb//gv/sW/aIaJ6+kfnB//IRYxPYZRqay4iQ/tTtmUVb7Fg3IaQ10bH/ckjFw8xZCxgVHpPAwlx+m/jCNYW8RT9X+upS81fopn44e6d84hx9OxjUrlUyfuQ7RV/RFudV+hPHX/wRATb/o/fLQ1xzHqxKl+Uh9fD8DoHy1MY3WiL8OojErnFttesW7M0I863gMv+rlllhoX1JN7LXXo3sAYLhbU+zIPehQ3MeDBH9dz36q9iCexhol4wwhPfZ/t2rVyiyU8Gdz6cfUgr3jjazzFy72GOnENfY38G0+lGXo8jVG5TNTl2BAIgRAIgRAIgRiViYEQCIEQCIEQCIEdCZQgTRghnAxtVBJQSpAhchGECEAEFkIKgcY1S+QiVjMuLAQcwqhX/xH6CTfEUU+Sm6FAVLKd0F9P3RODGQFEH0vfqCT0O57QSPBaZimha0yjkoBHUCWuWuQZG0KUxX5mGMYYyhPxtC+gEvuIVmYqlJlJPMWfGIc/AVv9E83sI/wTsKWxf8jFtWomCuHNd/l2LWUjtC2zqA9xo94J7oTIMm8wcC1pmBMlQhNWldt2oqgZXLiLJXFkBpbYIeQTqnHHyke+GbvOgbW6ELNEV/kQ4/XqxGXKVdz2wqgkrBIkGRjaC6FU7JzvTA5lttZemT1e/UbYJ55quxhJL26wqg/BlfnmOP2B9Iw0YrZ6x0kabXrIpdqpWFOn8qPOXNdHTCxr6LmG2VP/+B//48aHKeFhC21IefR54hArInX9Vqc4Eiv9V8A6FndmpP4O7/4MNv2eWCRAiwUiNf7qR125rmtoR8uWSwxU/13GvjqSL9zU5bImX814ZyTq38SUV96WmG/23z/7Z/+s9WmMStfW3zMqmUnasJmk2qq6xdqYoG5rprRxxwdvsemViTgvw+fTYlR/OoZRWWUUF2KOwcNEE9PqX/1Io7/wwaviBRvx4jfzGD/GDPHyd/7O32kPa0gr34wSeTd7y9hhv3qx9I1KMcp4GeINBcVTO1U2/YN7E/Urj/K2zIKJ8ooN/ZeyuZY+S3s0pmrH9jEkxUv1T2LL2KCdYerBFrHKPJJH4612oK0bR7QN11I3tjHmjA+MTA8uDLmMbVQan8UXHsxrbU4/ZqzDRZkwsejztTts1Jd9GJgR7v5MOsdor8ztessA5vURx+rDPqzEozFZ3Lq2fuGv//W/vvSDPzUu7IVRyagVJwxWbUYbxVUb1Ce5T9CX6pOwNR6ITWUVn1hiq48Xx/LuHO5NxJ17Denx0l7Ech039Hgao3LI1ptzhUAIhEAIhMD6EYhRuX51nhKHQAiEQAiEwEIExjYqiVxEJoIMoYkwROQqQavMI6Igcb1MBEJOiWNeHUZgJLoQTAn5RBrpCVllVBJ8iGR/+2//7ZZO2WrGjqfxy6gkNBKBlllK6CJgyhthkmikXISiIQw35SfgMyOVRZ6Vj7Hjb0xdWx6IWESkEi79jSG+xGb5kT9iVgmqRFXp5R0f/OzDx2wHQuNBMyqZN8qh3sUNg0XM4FbmCmEZB7GGC3FPfWJJjDVDjTnMDHqoE5i94rTOgRfuhFVCI0aMzJo9J95d237nUGeMdTMul1mIvOpzL4zK3/7t327i+1/+y3+5mYtEUfGkTCWsigtt0HaGMKEUY4aj2WO41CLusCZSqw98cNIPEGHFM4NT2xGTQy7VTuVvLKNSTJjl90//6T9tsYcLc1zsiCGM1J82q7xiwXcCMxZm4Gp/YlIceUVgxZv2T+jXzzmv7Y7HVyyIQXEuphlG9hGp8SZ+r/qMyjIqtVmxo49Wft/FO5PCjHi8zOzDD2tmt/bKeGRsiCEMCPliDHcGl7rBmfnlGtgMMcP502JUfY1lVOqvtUNstEn9vz6Maah8Yh6vanf6IN/FGIPXDGhjC1bMdDNPmZyYayfiSd6NO9rs3/ybf7PVi3PqH//tv/237VyO8XFdxw6xuP5YRqV7DzHj/PpmZqi+Sv9tjBQ/uOBjf994wxgX8cQAl97+Muu1M/cwODuPcog3Y46Zztq179r4kIv8uJ6YEHOuy+zSP+gL9N3LLNogTvohscPg1b7qvkKfXWXXn9tum+/yxqj8H//jf7Rj8dfPa9/iRvvEDyfxLK12y8jEyz4PSog5cS5+jTHu/5RtmaXGhb0wKv3mMxb93xDGVXnFm/tivPTtthtPxVs9wKOPK8NRfOGtf8dCenzUkbFTfffHU+cdclEOY5mHHtSHWNMvqHf5lC/XVHdZQiAEQiAEQiAEQmCWQIzKWSL5OwRCIARCIARC4BoCRAbiA5FrjBmVBBdCFJOA8EnkImQQCImkjCKiEZGP2EGEIb4wMkrAOt+JVPJH8CecEqbNziLUEOvNADGDjiFKqGVUEryVS5kIRfJhNpLjStC9BsSCf5TQ5bzEzzGMSuWvj7ITAAnCuCkbkwInC36EKkJWifU4lmBPRCIeEUoJrCWqEpoITuqGWKgOCKuu4Vhph1zKcCN0EVXlEUsCF4FtWYNXPSiLWGAAuZ6ZCuKGmF9GmPIT/zDRBsRXzdJyDvVKJCX4MojEE3HUcURb5ycQ4kN4ZU6LXceJV0zFesUr4XWZpbiNaVSqk3pVnbgws025xZzrizfxSKxWTwxciwcFlJshIq21/SXkS6PN20ZQLWMOyzIqtVvfq34cM8RS7VSsjWlU4vav//W/bmzMmhFzZgVps/IgxrRNHLQr8YYbsRp3ZoN94k2cihf9lHz/1m/9VjuvNkncZ4JKZx/ThXGHpZm96otYXX2pa+5mqXFh7BmVjFqvflUO9Y+bcjA0xLpX4+q/lc9MeWXDWl+l/NLVjDh9onz7YImjc9TYoN2KzzIqd8tmHp7qdwyjUpzoV8SydqQ9MluMpww0bU+8KZu+vvp7MYYx0xYP7MQhfuJK34a9c4tLfRvG2iWj0j7nNZ6aIYYrs0ms12uK5+HyaWkqpvUT6lEdDjGjUv+FEwNSP8Yg8l3MiTdxYb+xQxl911/Zj618md2rj9cvYibG8LUNb/k1nknr3K6hHeonpddmpR9yEfNjGpXuO9S18oifGtf69xUVZ9baad3DSSs+xZy2p01g4D5MPIk5DKUTa2LO3/i6F8C04sGx7mtqPDXeLrPUuKCejV3ah7pzT+Dc4s69wW77CO3IebU1D/74myHOjNSmcBVnuOhjxZr7LvWp/cmPsmrP2qj9+FaejJPumbQTDyzgY7wVr9i5hjIMPZ7Kn7Ykf+otRuUyUZhjQyAEQiAEQmD9CMSoXL86T4lDIARCIARCYCECxDXiA6GDcMKkIa4T5xiBBKllFkIGIYYASLQiPBHWiCzEeGuiEUGFmSYvRDFmCCGEkGS/8ziWGOM44iWxi3hFqCfWEIUIYWaCEQZdh4lIMLb0DadlZxqU0EWMGsOoJA7iRcBjZrgOkYr4RIjC6XxniBG8cPNRJkJhscLR3w91M7sIWAREQiBGeBO+CGjy76l+1yLUEbzxU/d4D7mU4UboUp9DG5XOx7wlABJIXUc5xDQ+ykd8VC7bMVCXhP9vfvObLd7El20EbfFFxLaNsIqbuLSPcC3GCPaEQXWCp7rTrsp8Jebbv8xS3JybOKnexYNrEDLVaYmYu7mO82GlDXn1K4baF9FWX4CHGLEdYwIqQVk8EZi1cfGFL87yVTFmu/zJp2toq2JObKoX8YyxGHXMkEu1U7E2llGprsWD3zbVb6lr5fEhMKsXH+UVdz7SeXVnifc4SqOe5RUrBop4YgzJO67OqU5w1lfjyEDA0W+7MSqxd77diuz417gwtlEpdszQ85CAtigWCPb6M7Ei5oxL+j0zqcSI/lD71g5xUGa88PXBkUGEl3MwO9WPY9TNQTYq9TFiRPwweZlh+gPtiwmCm7qr/q1iShrlNyaIGWnwwct4oY0z1MTMt771rcbMuKmPrN9w1Ne5ru3atH4Ne4a8ehliEfviWX7lUb0PZVQqp/sQsSYeznfjp/5GvMl/37Cy3f0PRmJJO9XfY8+Y1f9hrJ35lImkH5V3nIynB92orHFHv68Pwq/uK9wj4CaG8FJWHCzSe5gHCzGFvb5E/KpT7ZUR5xjM3Ovga78HNOzXd+v/1ItFX6p/EHf2L7PUuLBXRiUO2meNp7hqj+LdPYZ48opgMaVPFG+4atcYa4f9OKvx1P1AjafYS4uRfg7nocfT6n/0qzEql4nAHBsCIRACIRAC60kgRuV61ntKHQIhEAIhEAJzEyhBeiyjsgShEjiJkAQa24lNhBQCKIGF6ElYJUTLDwGHAG8GEZGGMELsIvpIT2AkzhBOiTVe2Ub0ImTZ73iCKjGM6EgoIq65LqFnmaXKNaZRSRjES/kI9owJfJRNvWGkHMqLkbIRuohfGDJwfSdCE/SJTFgwOwhZPs4nTQmr0tWrX4ljzjfkUsInoct1hzYqxRJxU5wQP/EjSrsO0VkcqTvccLDNd+K9mbnEwa997WvN7GGQEFwtzB8xhI8Yc16viLW/tos3PHHzakAzR9QJzsvGW3HbK6OSASLOlEWZLbjio00yJwirvhOuifKMYdy1OXHjOLwxIUprr/b5nTtt2XbHjymsVjsVa2MYlbhoi9oPk/f8xuzvYqV/U/dY+I4nruJRXMof8dp2sSsOtXfHMB612f/5P/9nOy/BH8+aXeNY+30Y6b/8y7/cTB1p1NMyS40L6lM7UB5txUf+9SPqbxkzVP6YbYxK5pj+DSdl0d60J3EkRh7qHrbQLym7fk8/M5IrAABAAElEQVRd4id/uCmvfY73XZvT/sSbdOJTf6Av9Cpm/eWyed+JrzyqR/2cfLmuNqBcyyzqxUe5GUH6+fNdzDGCxEzVvetg4G+xVh88xJr98qUdGjfVpfixHS+xzEjX73jAR53XeOo6yoKvulI28TDEop3qW43xyjmGUckIqo8y6aNxcm1r/ZH+zf2CcmtfYsdMN+1cjIlLzHwXn/Ipre2Yyb8+1H79Hk64lZE3BCvnkDfxr+2IOWWoWPBgjfa6zOL8+hmGnvOLOXGhfM6tfOpJ2YynWOjnxKc2rY1ptwz0MgWNmbg71nFiDle/nWqfsdM+fU6NETWe4jjk/VvlSTvVRlxXf6w+l+nf3CO4N6wHf9zzig9lxcdSY4TtzHgPCthvPD3ftWnjKc7yIa4cZy2G8PCqb3zNrFY3tsu3fRj5XtdaJgb6x4oH8R2jsk8l30MgBEIgBEIgBOYlEKNyXlJJFwIhEAIhEAJrSoDIRHwgdIwxo7KwElwIoERIojExjWBKXCGEEWvM4CIyEwWJPEQYYisjjoBDxPJEPxGHYGMmGxGHIEwU8xtSBLAyKglBhDDCk0+JqrY5fplFnkuMYi4QCuWXUOdaROBlFoZUGZVmgBCvCGnK69zWGBKjlJcoSWBVLuxwNkNQvRLfcJXnMiqJij7SO4/6IRYSatWDfXXcMuWYPbYMt7GMyornElZxYIwpXz/eigMBVZ0ReKVTZgaReBSHBFfcCfy44yjGxLDfknLeYkUwdjzhnvhdrycegmNx2wuj8j/9p//UYk85tDsxIt7EN4ZEUG2JUYEJ1oRYgqn2iY+0YsmxJawSrG1jrojRMk1KVK1rzcbMMn9XO1W/YxqV2r82isP5rn8SNziIF8zEm7Iro9gSp9qm/foLafQn2iDzDjPCtT7sv//3/97izTaxJK1zWfD1t3MQrpnmVVfLcKt2pBxjG5W/8Ru/0WJHHCmfsuBkXNI2mUbak5jDUToGG84e4MBR23OM/TgZNxhNhHr1gquP83lVs75SjI61yNMYRmXlV/+prYk3fZF+SKwrk5jSLjEQH2IIn2q/ths3sNImvv71r7f9HjzAixllFqoHXRxXRqX43G48xXyIRTsd26hUL/XRRsWCuNOGxZDxz3jqg6NyY42Jtb+LtePElDZtLMEUZzEq7rRTD2AZI7BVN0Mu2og6GsuorLziJOa0OUal9qc8+hp5EBfiCgfb9e8Y+27mPWPbGKZ+xay148QN5u6hvvGNb7TtZhfiKr2182r/4lD/NgTHGhfUk7wq2xhGpXsKr37VTxnnlFWZq4/Wd4sNfZv+yn516d6jxtOKNW25PsxI46m/3VcrQ401NZ46t/1DLupMveAlBtw7GivEvfiXfzGvfFlCIARCIARCIARCYJZAjMpZIvk7BEIgBEIgBELgGgIlSBNWxzQqiXtMFh+iCrGDUeFD2CM+Ebn8TQQps4MA4sl9ggvBlbDkHIQQRorvTEpPrnsCnVBSr6oro4DYaLuP6xCKXGeZpYQuZRnDqHReYpVZGdYEIWIgUVS5CXjqzhof2+3HRblxKSEJXx+Lskvng6306kL9uIa/ncs+wteynGYZuxZBUB0TK33HUt0QjNXVsgsuriNecOibjcWBmIYBfphgJp0yEwHtJxg6h/w5p8Xf8q1OGEr2EbaJqWKrPsqjXorxshyLm/IwJdSXunIdwmTVfZVvUYbVPrUjRqWy1yuAnRsj5674UU/MDu1S3rRL/Bznb4KmMuPpmMqnbcrgenU+dYC3dMo05FLtVJ2NZVTKrzJrP8qvfOLaNmXEAKf6KKt4EkvKS1BWdnl1DvWLk3Yuz+qD6cHA00aI/o4pxmJOfRCK1dVuY6DPXf6cf2yj0kMYv/mbv9nMMXFQM4XEtDaJizhTPmXGkjFTfQjemGGHpfQ+OBhPbLO/PtgzBpxrzGVso7LanPKXyYKBReyINWVUfryq/7Jdn4QDlo4XY/YziLRjD7jo2zxQYCzw4IbYUx/Opy60Zx915jyuOcSinY5hVMq7sjGrlUv/zTySfw+UaDviHRfxptw+/XbpQSm8qn9TXmWvPr9YaH/VH+BVprDvQ3Eq1vK8F0al2FImseZ+R3uyVF8jrrS5iiv9u35MmZmL9smr7epB+1Un+kDfGcBiThwydo2nzul8uIo146nzOGddtzgsulaeyot7rWpDriXW1dky11Eu5y2jUpm/+tWvNsPVNcSVMhQ35XNNf+Mszmo8rfYrduRJGumND7a5jrL4bp/z4+YaY8RbxUHdX6ozfUqMykWjMOlDIARCIARCYP0IxKhcvzpPiUMgBEIgBEJgIQJEBgLS2Eal6xCHiC4+vrsusaYEmBJvSphREMJMbZe+zlEiD9GMsErwJj4Sof0GmdkQzmNxvPOUoLqsyOWcJXQRicY0KonIxFX8iHcEfOsSoZWpPiVKFWuCEga42abcxcFaetvsk8bH0uc0BKt20o3/yVOZDGMZlS5VZap4qZizveJNOXFQRmWXxncinzSOld7Hd8IqE0p9E7nPd7NLpDWzl4FUQm3//MW4z2A334vbmEYlsdasIb+JqLxf616BW7M8lNOiPBVvFUPFB7/iXDFX6bVBH4s0jrHgXXXgu8+Qi3IQccc2KpVHmV3PpzjYrkzFoR8Pta/KX+dwbHExu+23fuu32mxyM4q8svSh7jWoTBTXka7qA1/nH2KpvIxtVHo4xm+immXFiFA+M/v042Leolz9tlp567OueOvzcIy/q16s8XE+zMdcxjYq+wwq1oqBcimf8iuvtD5Y+NhXcYih9mHRxvVtZsx56Ic5zrz8tV/7tfZqyn599M/vnEMtYxuVzBX3CcZURhpzyKtZmWnFptpTccKu36Z9x9pSx0hb6W23v9L0Y9e+IRfXUP+MLTGHHxPRWOQeSJsaYqkYUnZxYO1TcTUbD7ZLhw/zzH5LnyUzz3imToynHspi+vqNSuMp86v6tKqTittlyyTve2FUGk/9drF6+pVf+ZVmwrp3Uy5slKfKVvHTZ6RufWyzSC+d430s9ju/81U81nfrIRfXUa/uFWJUDkk25wqBEAiBEAiB9SAQo3I96jmlDIEQCIEQCIFdEyCAEB/GNiplsK5lXcJLZZygQoSpdP3tJbrUcXWs9IRVT62bJUFYZeIRhLwusJ+uzjGUcFNC11hGJQGPSalMBCFlJd4rnw8hsspkX30vfsVK3da2StNfF+dK7++qB+mGXohce2FUyneVyRoHa0u//P0ySuPv4lnHO0Z9y3sZlWaAEFcJq4xKM8H6MyX613D8sktxG8uoZMISHxn+fhMRi691RqXZomZvEJst/XLVd9v7jPvcKk0xrbTWtUhjqXVtH2Jd7XRso1Jeq9yz6yrXbLuSrvhUWetYf9vntX6MY/kvo9KMH0altJWu+Na12o4l/ufcYmBso9LsOa+21c+ZkfNw1297ha0+zvUt/bIpX5W7WFU6fxdP6+Jd6QpHna/+HmM9tlEpz1Wu/rrK0udgfy213d++26eNWBggDDxvKFAvymDM8YYC9VKci1//XO0EA/xvLKNS/2lWm3g2rlrr78y6FW8M2SrP7FqxcFL+Yt0v6nbpi3vx6h8z1Hd52gujUn6r7LPrrcov/Wy82GZxfI2ndf+k/RtPmZOMSrPGja1MPEsxdK0hlhoXxIQ8iAX3JUPNqGSCGquNp7/zO7/TsqwdmS2qjEOPp5gWm9n1ELzqHOpUW4pRWUSyDoEQCIEQCIEQWIRAjMpFaCVtCIRACIRACKwhAQIH8WEvjMox8Hodm1k5JT4Su/0mEuNozKWErhLazGggEDEQGAlej7fMQkAjGhNU61WQZlMqnw9B7SAuZbgRuojSxEEsmWFDvfp1DC59QVidaC8MZGJqvarOjKSaOTJ0HorbWEal85uJc76bJfqHf/iHLftmtvltMKZ4zd4Yulxjn6/a6V4YlWOUxSyj3//9328zpsyQ0reY/TX2q0trXBDrY/5GpX7bbwu7DtNIv80c18cd5GUvjMox+KgHsynVi7FNPegHxh5PqyxjGZXMPGNNjdfW+jvjtd/a9RrNg7j0xyUxh98YMyqHZjObb3n3cV9jFr8HFYw5Y42nNS6MZVTWq4aNp2Youzd0X2o8rQeahma6F+dTbzEq94J0rhECIRACIRACh5NAjMrDWa8pVQiEQAiEQAgMRqAEaSLRmL9ROViGZ07EIGTo1e9HEfCHfOXZzOWu/FlCVwmfQxuVnsh3boYeMY3QxcyrTz2RfyVDB+RLGW4HzajUTtS519Wpa/lnGhJTzcYhePtu5scYS3Eby6iseGa+mp1seeSRR5qAz4CtmS1jlG3Mc1a5DqpRyTgyK0fcmd3GOPK6Sgb5mEuNC2Mblfpts0a1KYY4s3IvjNgx2Tm38fTxxx9v/YSHVhhhZonpv1d5UQ9mi6sXxl6Np+plL5axjEr9QD2MoQ81pjKTzG47CPWyHftZw++gGJX98dRYWh/3Ndq/NmPMGWs8rXFhLKOSMe4eznhqdrLFfal+YMwHmraLk6G2x6gcimTOEwIhEAIhEALrSSBG5XrWe0odAiEQAiEQAnMTKEH6oBqVxCDiqlkEvhOBiPkEyDGXErrGMioJXcwJZSKoWpStPgfVOCrD7SAaldpKCZDqRf0QUomqjCOzP+q1a0PHXnEby6gswZtwy5yyaEfMozEF46E5zZ6v2ulBNSr1a15vLe6YRurDjJyx23+NC2MblcrnGvo4Rr9+m5mnnzvIy0E1KvVr+mb1os9hHO3FeFp1PZZRWcaYONNvW+sbxJyHTA5qvFW/7R5IzB0ko7LyLuZqPNWv1Xhar3ut2BhyXePCWEZlle2wjafKpV/QRzBh9RPaln5bP1F991gzYYeMgZwrBEIgBEIgBEJg7wnEqNx75rliCIRACIRACBwoAiVIH0SjsvJOxCc8EVGIW0THsYWSErrGMipLWFUmH4uy+YxpiI0dvGW4HTSjEhd1UjFX8caYJK6OXSfFbSyjssqmLZUxzqRQNmUcy4AdO96qnR5Uo7Lyrw9QF3sRaxXrrjm2UdkvX/Vtyuj7QY05/A6iUVl9236Mp9UPjGVU9mO6+m7lFWdjzoSvco211kbV10EzKqs+1EHVh7Ls1Xha/c5YRuVhHU/VUYzKsVpzzhsCIRACIRACh59AjMrDX8cpYQiEQAiEQAgsRaDEyYNqVCr8fgjaJXSNZVQuVakrfHAZbgfRqNxPrMVtLKNyP8s25rWrnR5Uo3JMNjudu8aFsY3KrfLg2pb96Ne3ys9uth1Uo3K/uY9pVO6mHlf9mINsVO4n2xoXxjIq97NsY147RuWYdHPuEAiBEAiBEDj8BGJUHv46TglDIARCIARCYCkCJUgfRKNyqYIveXAJXTEqFwNZhluMyt1xi1G5GLdqpzEqF+NW48J+GJWL5XQ1Ux9Eo3IVSMaoXKwWYlQuxqtS17gQo7KIzLeOUTkfp6QKgRAIgRAIgRDYmkCMyq25ZGsIhEAIhEAIhMAGgRKkY1QuFhIldMWoXIxbjMrFeFXq4hajsojMt652GqNyPl6VqsaFGJVFZLF1jMrFeFXqGJVFYr51jMr5OM2mqnEhRuUsmZ3/jlG5M5/sDYEQCIEQCIEQ2JlAjMqd+WRvCIRACIRACKw9gRKkY1QuFgoldMWoXIxbGW6ZUbk7bjEqF+NW7TRG5WLcalyIUbkYt0odo7JILLaOUbkYrxiVi/Gq1DUuxKgsIvOtY1TOxympQiAEQiAEQiAEtiYQo3JrLtkaAiEQAiEQAiGwQaAE6RiVi4VECV0xKhfjFqNyMV6VurjFqCwi862rncaonI9XpapxIUZlEVlsHaNyMV6VOkZlkZhvHaNyPk6zqWpciFE5S2bnv2NU7swne0MgBEIgBEIgBHYmEKNyZz7ZGwIhEAIhEAJrT6AE6RiVi4VCCV0xKhfjVoZbZlTujluMysW4VTuNUbkYtxoXYlQuxq1Sx6gsEoutY1QuxitG5WK8KnWNCzEqi8h86xiV83FKqhAIgRAIgRAIga0JxKjcmku2hkAIhEAIhEAIbBAoQbpvVJ49e3Zy7ty5yd133z05c+ZMWG1BoIQuRuUrr7wyIXhdd911k5tuumly//33TzDMsplAjMrNTObZUtzKqLx06dLkhhtumNx4442T22+/va2vv/76FoPznG9d0lQ7jVG5WI3XuFBG5fvvvz+57bbbJrfeeuvklltumZw6dWqSeNueaYzK7dnstKdvVDJFxNojjzzS1jsdt677YlTuruZrXOgblRcvXmz9mvu3m2++Of3bFmhjVG4BJZtCIARCIARCIATmJhCjcm5USRgCIRACIRAC60mgBGnC6pNPPtkMtxiVnx4LJXQxKl9++eXG7ciRIzEqPwVdGW6ZUfkpoGZ2FzdG5RtvvDG5fPlyE1I9SBCjcgZW789qpzEqe1Dm+FrjQozKOWBtkcR4+thjj030czWe3nvvvXnwZwtW/U1lVOrjyqh89NFHY1T2IfW+l3F04cKFibaKH8Pt9OnTE9w8WJBlM4EaFxiVmGmnHsbwAEaMys28akvFG16vvfZaizVjxcmTJ9vDje5Hjh8/3u5N6pisQyAEQiAEQiAEQqAIxKgsElmHQAiEQAiEQAhsSaAE6ZpRSbgpYdWMSjO2smwmUEIX42jWqLzvvvsyo3IzsralDDdCF5OXOIglgYuQn5moW4Prc9vKqCRMZ4bbZnbVTonRL7744uSDDz5osSbOzH4mTJuZ6iGDLFcJ1LjA/Hj66adbOzWj0gy3zKi8ymm7b5lRuR2Znbdrp+fPn5+8+eabzag0sy0zKrdnVsaR+zYxZ0xlVLpvC7ftuRkXjAVbGZXu3zKjcmt2FW/u38QbfsYK46jxIUbl1tyyNQRCIARCIARCYEogRmUiIQRCIARCIARCYEcCfUH6+9///pUZlUSHO++8M0blNvRK6GJUvvrqq1dmVDJAMnNmG2jd5jLczAApo5L4RVgNt+25ffTRR80sIhAS8WtGJW5e0xyjcmt21U4ZIC+99FLjRkz1iVG5NTNbjQs+jMpnnnmmxV699pWIbwZNjPHt+dVMVO1VrBlP8+DP9rxqjzHhueeea7PcjAva6MMPP9yMo0qT9VUCGBkbjKceYOkbleF2ldPstxoXmLqY4eehKf2a+xBxl/5tltqkPTzgHk6/po/DzziBWxmVx44dy4zKzeiyJQRCIARCIARCoCMQozJhEAIhEAIhEAIhsCOBMiqJXM8++2wTHgirZs0wKhkgWTYTIHT1BRsil9+oxC4G72ZetaUMtzIq/dZiGZX5TdSitHmNG1ZmMBBWzQYhpJrJwECyNitQDGa5SqAEacwYlbgxd8uoJLBmRuVVXvWtjEoGL+OIMc6grJmoJ06cSLwVrC3WxlMP/ujnxJs2mnFhC1Azmzz488ILL0ysxaB2mt98noHU+7OMSuOCmMPNvYj7tgcffDAGb49V/ytu7t8YbUw3/Iyv+rW77rqrxV2Myj6x6ffipl/byqjU18Wo3MwtW0IgBEIgBEIgBKYEYlQmEkIgBEIgBEIgBHYkUEalWVoEacINcZAo7bfvYlRuja8EQryI+YR8JlG9AosBkmUzgTIqy3AjDopBcZYZR5t51ZYy3IjQREIiK2OSsMo8inFUpK5d1wMFjMpXXnmltVOxpo/DLUbltbzqrzIqxRqDV7wRoX3wI0bHGC9am9fGUw/+6Ofw8uDPHXfckfF0M6prtuDlt++sxaDxNA9MXYPomj/qPgQv9yFMN2MEbg888ECbGXjNAfmjEShu7tuwcx/iu37NzEBtNv3b5mDpG5WMcfe/FvcfuBkfjh49mhmVm9FlSwiEQAiEQAiEQEcgRmXCIARCIARCIARCYEcCfaOSsErkIjaYARKjcnt0uJV5RKwh5DMqCV34WWfZTKCMSgaI2R8EQuIXZjEqN/OqLSWsmhGImdireGO2EQf97ZPlKoFqo2LNb8kSo8Waj9f7xai8ymr2mz5OrDFAtFtitM/x48fbLNTE2yyxq38zKutV6mItbyi4ymanb9qntmot/sSadmqdZTOBGheYbWKub1SaUYldls0E6v7NfZsx1cd3s+s9xCLe0r9t5tY3KmtGpVTGUf9m0Ne59zUbNUsIhEAIhEAIhEAIzBKIUTlLJH+HQAiEQAiEQAhcQ4BgQ3wgOjz99NPt6XIzjTwd7RVYhIcsWxModkR83y2ews+rJLfmZSsxsGYFMkB8F3/EwfxG5fbcxFfFG14+hFTxVq+oi0m5mV8ZlWLtxRdfvPLq13qFqZlHaa+budUW/LRZ8SbOfGqmUeKtKG1eG0+feOKJNvtZ3+Z3ZPMgxmZOs1vEWcWb/k68eQhDzGXZTAAv9x/1Kk4mL9PSfZvfqGSQZ9mawOx46m99Wo0H6d82c6v2yRD/0Y9+1GZU4sao1Mfp62JUbuaWLSEQAiEQAiEQAlMCMSoTCSEQAiEQAiEQAjsSKLGG6PDkk082kctT+BFWd8SWnbskUEYloauMSmYIgeu+++5rr+Pc5alzWAhsIiC2zM4Sa377zswZsaaP84lRuQlZNgxAwHj62GOPtRluNZ7ec889LfYGOH1OEQKNQN+oFHP6OW948OrSRx99tM1yC6oQGIpA36j0imaxNmtUmo3qAYMsIRACIRACIRACITBLIEblLJH8HQIhEAIhEAIhcA2BvlH51FNPtSfzS1jNDJBrUOWPAQjEqBwAYk4xN4EYlXOjSsIBCcSoHBBmTrUtgRiV26LJjhEIbGdUeuDHW1g8BBSjcgTwOWUIhEAIhEAIHBICMSoPSUWmGCEQAiEQAiEwFoEYlWORzXm3IhCjcisq2TYWgRiVY5HNeXciEKNyJzrZNxSBGJVDkcx55iEQo3IeSkkTAiEQAiEQAiGwHYEYlduRyfYQCIEQCIEQCIFGIEZlAmEvCcSo3EvauVaMysTAfhBgVD7++OPt1a9+D9Wr1PPq1/2oicN9zRiVh7t+V610MSpXrUaSnxAIgRAIgRA4WARiVB6s+kpuQyAEQiAEQmDPCcSo3HPka33BGJVrXf17XvgYlXuOPBfsCMSoTBjsBYEYlXtBOdcoAjEqi0TWIRACIRACIRACuyEQo3I31HJMCIRACIRACKwRgRiVa1TZK1DUGJUrUAlrlIUYlWtU2StU1BiVK1QZhzgrMSoPceWuYNFiVK5gpSRLIRACIRACIXCACMSoPECVlayGQAiEQAiEwH4QiFG5H9TX95oxKte37vej5DEq94N6rhmjMjGwFwRiVO4F5VyjCMSoLBJZh0AIhEAIhEAI7IZAjMrdUMsxIRACIRACIbBGBGJUrlFlr0BRY1SuQCWsURZiVK5RZa9QUWNUrlBlHOKsxKg8xJW7gkWLUbmClZIshUAIhEAIhMABIhCj8gBVVrIaAiEQAiEQAvtBIEblflBf32vGqFzfut+Pkseo3A/quWaMysTAXhCIUbkXlHONIhCjskhkHQIhEAIhEAIhsBsCMSp3Qy3HhEAIhEAIhMAaEYhRuUaVvQJFjVG5ApWwRlmIUblGlb1CRY1RuUKVcYizEqPyEFfuChYtRuUKVkqyFAIhEAIhEAIHiECMygNUWclqCIRACIRACOwHgRiV+0F9fa8Zo3J9634/Sh6jcj+o55oxKhMDe0EgRuVeUM41ikCMyiKRdQiEQAiEQAiEwG4IxKjcDbUcEwIhEAIhEAJrRCBG5RpV9goUNUblClTCGmUhRuUaVfYKFTVG5QpVxiHOSozKQ1y5K1i0GJUrWCnJUgiEQAiEQAgcIAIxKg9QZSWrIRACIRACIbAfBGJU7gf19b1mjMr1rfv9KHmMyv2gnmvGqEwM7AWBGJV7QTnXKAIxKotE1iEQAiEQAiEQArshEKNyN9RyTAiEQAiEQAisEYEYlWtU2StQ1BiVK1AJa5SFGJVrVNkrVNQYlStUGYc4KzEqD3HlrmDRYlSuYKUkSyEQAiEQAiFwgAjEqDxAlZWshkAIhEAIhMB+EIhRuR/U1/eaMSrXt+73o+QxKveDeq4ZozIxsBcEYlTuBeVcowjEqCwSWYdACIRACIRACOyGQIzK3VDLMSEQAiEQAiGwRgRiVK5RZa9AUWNUrkAlrFEWYlSuUWWvUFFjVK5QZRzirMSoPMSVu4JFi1G5gpWSLIVACIRACITAASIQo/IAVVayGgIhEAIhEAL7QSBG5X5QX99rxqhc37rfj5LHqNwP6rlmjMrEwF4QiFG5F5RzjSIQo7JIZB0CIRACIRACIbAbAjEqd0Mtx4RACIRACITAGhGIUblGlb0CRY1RuQKVsEZZiFG5RpW9QkWNUblClXGIsxKj8hBX7goWLUblClZKshQCIRACIRACB4hAjMoDVFnJagiEQAiEQAjsB4EYlftBfX2vGaNyfet+P0oeo3I/qOeaMSoTA3tBIEblXlDONYpAjMoikXUIhEAIhEAIhMBuCMSo3A21HBMCIRACIRACa0QgRuUaVfYKFDVG5QpUwhplIUblGlX2ChU1RuUKVcYhzkqMykNcuStYtBiVK1gpyVIIhEAIhEAIHCACMSoPUGUlqyEQAiEQAiGwHwRiVO4H9fW9ZozK9a37/Sh5jMr9oJ5rxqhMDOwFgRiVe0E51ygCMSqLRNYhEAIhEAIhEAK7IRCjcjfUckwIhEAIhEAIrBGBGJVrVNkrUNQYlStQCWuUhRiVa1TZK1TUGJUrVBmHOCsxKg9x5a5g0WJUrmClJEshEAIhEAIhcIAIxKg8QJWVrIZACIRACITAfhCIUbkf1Nf3mjEq17fu96PkMSr3g3quGaMyMbAXBGJU7gXlXKMIxKgsElmHQAiEQAiEQAjshkCMyt1QyzEhEAIhEAIhsEYEYlSuUWWvQFFjVK5AJaxRFmJUrlFlr1BRY1SuUGUc4qzEqDzElbuCRYtRuYKVkiyFQAiEQAiEwAEiEKPyAFVWshoCIRACIRAC+0EgRuV+UF/fa8aoXN+634+Sx6jcD+q5ZozKxMBeEIhRuReUc40iEKOySGQdAiEQAiEQAiGwGwIxKndDLceEQAiEQAiEwBoRiFG5RpW9AkU9jEalNmS57rrrVoBwstAnEKOyTyPf94rAQTYq05/tVZQsf50YlcszzBnmJxCjcn5WSRkCIRACIRACIbCZQIzKzUyyJQRCIARCIARCoEcgRmUPRr6OTuCwGpUl4Flbrr/++skNN9zQzEvbtDNGps+RI0damhibDcOo/1tno1LMKb8299FHH7WPmDt27Njk6NGjLUYrJkethDU8+UE3KsWOz2zc6NfSb61OQBtb1NGFCxcmYu6tt96aXLx4cXL69OnJo48+Orn11ltXJ7OHJCfaRXH3XXswptfnMLcP5TaevPvuu5PXXnutxRoGp06dmtx2222TM2fOTI4fP97GlkNS3SlGCIRACIRACITAgARiVA4IM6cKgRAIgRAIgcNIoEQXItdTTz3VBK+bbrppcu7cucndd989ufHGGw9jsVOmfSJwGI1KKC9dujR5++23J5cvX27CJbHu7NmzTbCzj8BHyGReMol8j0k0fhCuq1GpX1f2999/f/LOO+80YZmZIf6YF2Lz5MmTLRYTh8PH4UE2KtGodsP0Ej8MSnHDkKi+a3hqOeOiBMowi1G5KLndpdev+hjnmXXaifHcwx/1AMhhbh8xKncXNzkqBEIgBEIgBEJgSiBGZSIhBEIgBEIgBEJgRwIxKnfEk50DEzisRiUx/wc/+EET9QmVjKB77723iZcMTLNeStA8ceLENWblwIgHP12Js9a1lLm16rNHynAx0+iFF16YfPDBB23Wh4cxfBgvzDt1dpgW8cakVO5XXnll8sYbbzRhXezdf//9kzvuuKPFqL+rLg9T+fe7LAfdqNROyvx68cUXW39lhp5ZU0zLw9Ze9jtednv9GJW7Jbe74+p+2Xj/8ssvN8PSGGIGq4f6PPxxGMeTohWjskhkHQIhEAIhEAIhsBsCMSp3Qy3HhEAIhEAIhMAaESjhJTMq16jS97Goh9WoZAZ9+9vfnrz66qtNyL/zzjsnX/7yl5sRZpsZGGZZlqjJIDoIgn/1DyWIl1kp7zUzdB/D6VMvva5GJZPy9ddfn7z00kuT5557rn0Xg8zZz3zmM5P77ruvmU7iMUblp4bRwgkOulFpJqUyPP/885Pvfe97E/3VL/zCL0z+f3v39izLWZdxfJFgBEvlJAVEMYFAAAXkoATLRAUTq+RURThZAuU/RsEd6IWlV3iICARFoiaewSSguQC8oDiDQIoL96f1h52Vnb3Xck82Mz3frpo9s7rffrv7+z799uzn6bfnhhtuOIjz/twNdqArTL88oXKPfn18G9L1zzXFY0//7d/+7eTb3/72yVOf+tSTpz3tactTSLb+6NOCysdXX9UegQhEIAIR2DqBgsqtt3DHF4EIRCACEbhCAhNEbDGonFBl3gfVjAKb95m/Lrf+bPmUnffT65wuP8un/LxPuXk/S93KnKX8bHOf37cSVGqPaRNt+/nPf/7kk5/85DJqz4gKQdBrXvOaZdSaEUke/2q0hZEXRlsy/pWz7tQz7TZamfdZPu9Tbt6n3Pw976fnn15/ls/8eV+vz5gUcNl/4ZeRetaz/45D+Dr1zHr79H6sQaXA4qGHHlpCSiN9tR8TXYAubPIutNSOp9tv/p537TnamPd1G0+5eT9dZv23MlNuXcfWPh96UCn4EnI/8MADJ/fee+/Sd91xxx0nN9988xJUulHBNG057+ZNe8+7eTNNuXk3/3S5039P2Xmfutbl1p9nufJep5ddqh7rnl4+9e3je0Hl/7XKtPO8z5LRwfw9y+d95k+7z/vMX5cb3q7pf//3f7+MUvczCUaoP/vZz14Cy7m2z/pbei+o3FJrdiwRiEAEIhCBq0+goPLqM2+LEYhABCIQgYMiwIRhPmwtqHRcc2zz2TsTan5DaN412Lrs/G2eaYyuKT9G1tSL33xer2Nd68x6/rZcea+ZZvm8Tx3rOtflmcSzT7MvU9e+v28hqDzdhtrDyKOPfvSjy7sA8nnPe97JrbfeuhiXp4NKgdHazBSmTZtPu44WtOdsb60H86fsvJtnGk2s39frrucrP8vWOrZ9x+URkB5d+9WvfnUZlSfwMt8Ikp/92Z9dwq7T21fnvkzHGlR6LCEj/d///d+XdhNKvvrVr15CSo8oHP1pu2l/bTZtOfpbL1/rwzoXK6sOy+Z9XbfPoyvrbnk69KDS7+95VPJnPvOZk0996lNLUPmGN7zh5EUvetFyc4I+wKQdL6YVbb3Wi7JTbt7X2noszVxsvYuVNW9eo0v76PP62jnLvJtmHe8z7/T7UnBP/3Fsbh459hGV0454jO6mrenNa6ZZPu9Tzvtam8pPvd5Ncz1xY5IA33nipg8hpUe9+x3XefzrssLG/sHMdzjHbVSpkdfYGJnvsdBbH1G6sebscCIQgQhEIAJXnUBB5VVH3gYjEIEIRCACh0WAycB82FJQ6ZgmEGOkGAkmcGEyCZGY9Ea2MVWuu+66xaCc3+Qyekw55p8XNswr6zD7mVAMUNuwXHmP/7KNWXcML+UYOB4NZjv2SdCjvLLWn7qVEyAoZ5v2wT4pZx3v1ve4TfuhvLJjxh6K6qZdGF1GfeHmWLWFUYhG6e3rNOeKdqEr+68NjSoURv7VX/3V8rtVtOXxmr/2a7+2BHoMf8c5baz9tN3oBwvtq37ta31lac5kXdvzGh2bTzu2rbz1ZjKfLszz7mVbw3qWORdM6rSM2T37Ydt0q72MrHJ8joN21WsEidDiOc95zqLbfR1ZSVuOSVsdw29UOl5tZjQlPT744INLyMxM//Vf//WlzbSrtteveNGVdejPfH0c/dGVv/VHGNKpsj6bR0d0fFqvlqtv9mX+tg5dGYGk/i1Phx5U+g0+58u//uu/Ljpyzt9+++3LDRja0TXOPO2p/b2PVqZ/1J/Ql76HVta60l/op2hE36OMybt11OGzMrOea4RtWMcyGlOOvrx7jS6tQ+e2S7PWsa7tzr7SuzrW/aIy02/atuPc58nxYnHsQaV2dG3S1t7X2nG91ea0sC437a6Npx+jGy8aMH/0od7pL+nNo97vv//+RXMCSt9d5nHa1qejLU70hklB5RZbt2OKQAQiEIEIPP4ECioff8ZtIQIRiEAEInDQBCZ82UpQOcfDSDGqyEvQIqhgZDKRmPbMpeddGPVmZBhDijHrt9xwYEQxprzG8HrmM5+5mPzWZXrZjuV+B86jFRlXPjOzxuxUTpDzcz/3c0u4OGaiu/GVZZQp607066+/ftkfQZ1tqts+feUrX1n2yYg2xyTMFBApP48QPQRDdU6SMf4cyyEFldrbizbGxGdU+syoN+rQb1ZpUyGeNr/tttuWdqcr7akc09SoC23m+OnGaEztq92173Of+9ylfWnONpWZsFA5DM1nrtKv8rRgHi0zZGlPXQIh5eyXOuz/hFCWW8e+O0/+4z/+Y9kP82iSzuynsOtzn/vcMjpPuzmHjCB58YtfvASyz3/+839wTLa/T5OAwjEfS1Cp/6ER2vzEJz6x/L6g30ilO48ifsUrXnHykpe8ZNGivs4yba9dmdA0qo/Tv+gjaYTe9Ff0IbD2WV9KY7SsHD3Qvfa3XH24e9eHebeOMq961auW+vdNK7vU7aEHldprgko6ch790i/90tLfODZ/05Q+SkAz4bP59KeveehCWG7Ulb5HQKjtaUV/RTfCIHrVvwmXTPpJ6zhffdZ3zXqu1/SoPuX1vfou+vLub32oeulRH0yjjsN2aFt4qT59oH11nPMbwsqabxsTVun/9nkqqPyfUY+jV23tRTt0oK1dx7zTEt34vqYfo7spR8u06fua/o8GtL3yc+1VL53RjfVolE7o8sYbbyyobETlPncV7VsEIhCBCERgLwgUVO5FM7QTEYhABCIQgf0lIJRgdjEfBRLCNKYN43FGS+3v3j96zxwPI4mhJABiMjEimadjSo25ftNNNy2mFHNSmXvuuWcxsMZAVw82XgIhgQyTlellO8yuCUOZpYxSJipjX1jEGPV4zF/4hV9YmDLwbcc+MbmUY4YxSLEW/OBue4w39SvLUNMu9osxzIBlDjNdGWXqmH1+NJH9muOYMXJ8GPiM876PqGSArwMvpqXAmQHO1KQ3QaUyDM+XvvSly4hKyz772c8u7SdM1GbTxrSjDoa+9mXm06a2pTGaw2t0oLztmGc72AkUhU/qNA9LWqBp82zTZ9p0fjNe7QPe9k35CRYEqtqFwcuspUfaEnpNWGk5zU0A7zfrjB61HzS/bzp0fOt2E5Y5dn2cFzZuFnCcW5gmqKTFj3/848uIOP2HYxU0vfzlL1+Me6Y97a37RqxokD5oWP9Ig/o6/ZvHyNKiz7ajrWlhwnIBqHoFAcKmCZJGs/RB2wJT7/umlV22/xaCSm39L//yLyd333330t76NDrSbzuPnDfa3jVuQh7nEU1Zd278cd0137oTbOpf9BnOzXk88fraoD9cr0cvrnu2N+ezbeBMX8pa37WTXvV9yo/OraMvtP31jR10qn/Tl+pzracPdu3WD+57v+B4XZvwwkLbYGH/nb+OZcuTY9cX+W5Fb67L+jv6xMB1SlBJb9rT9cv3QtqhU9dDk37PdYGO6YY21e3aqD9Tr+9iyuu36MU2bWNu2HEe0I/vc86NLU705jzD0blDa84311HMMMRSX98UgQhEIAIRiEAEThMoqDxNpL8jEIEIRCACEXgEASYD82FLQaXjYZKvRwoxs5grY2gyU5iW89tCTPs/+IM/WEwp5taYlgwnJiBzhvmivOARN78BxxRjZjLBGFQMUdtlcjFGGWQ33HDDYlwpa78sUxdzZ8ozVAU+9sm+MRwZqEwywYCyQiFmmH1g2JpnPcbZoZj+azP6kILKMSaZnH63jSa0AWNOu2vzT37yk0tb0c7LXvay5VGJjExmv3ZX3ot5jAPzk1nK8NTmRmZof/VpzzEELadbuqY7y5iE9sP2X//61y+aUG60artGxTEPBZIM13/8x39czgFmqnm0bRv2jQFLi3Rs/7yUs5wBPC/121fniOCCtpWjxX3U4bEFlXO8RsB+7GMfWzSi/6ArI3yFF8JF7f3QhYCcrmlotKCfXAcdo0d91gRCtqEMTambFvVzr3vd65YwyradJ84J21G3kIoeBU76MGHoofRZj7hgnvGPLQSVgpl//ud/XoJK7SjA0Yb6FjpxzRQIacsbL4woc110zZvwUBl9Cu1Yn170J9N/qM/k8bKuc67PJlpxXaMzddAdjQqE6NH11jY+/elPL9qjI/2Zftd11Xb0lfbFpH8zqcP69KcfpkvbFcTaT/vvZiT9mWPcx/5sOZDVP44TX/0yJscWVDpu36tcR/VDdGbSx7n2uY5pa3+b6GkY0a6/9WXYucYpqx/zHYuOzHce0KZrL325Ppvvemoe3dDmaMdy2triRG/zvaSgcost3DFFIAIRiEAEHl8CBZWPL99qj0AEIhCBCBw8AUYL84HxspURlY6HYcVIYcQzjRiRTHUmFQPKcTMyGa/uuGdYvv/971/MVyODhIJMfaYqNswwxjujk5FlEgSYxwxjilnG/DJCxPYYnowvxqv5gin7w8hirFrPfj7wwAOLKcvsZfjbL+YsA9c6zDjzf/mXf3kxw5iojDdmLuPskAx/JhcGjL9DCirtr7YQHv/d3/3d0v5GqBmdox2YnHfdddcyUkOb//zP//wSVArwBNrac4xT7/QgzKEry7S54E8AqD7LBY9edMuMXre7ffEbhHTwlre8ZdGxUSLK4qssQ555r24a/tu//dtlW0xY+mP42w4jn2HrvGHmC5No13rai1GrbsEELdo/++kYnT/2wTm2jzqc4I7WMHO8jtE56aV97Pu+j5xaBHeGf/RrXvRoRKW+hb60k35Lu2Gh3/IoV21uFBG9aGtlaVJgzXi3nn4KI7qiQ+0854M6nBN0dueddy7v/p7HxCrvfFCPGzFsn670q/uolzMgPlMR55tRrTg5XucTBrR3CJP9ds6vR1S6+cI1URvqB/RrAqLRiiCcDlxfhT/a3vlHY/ol10Xv5uk3f+VXfmXRlXqE5vohfYmbH2jGOUmPtGTSn7nGOmf1i4Ii9enL7JNrNl3pr/BXn32wf/osL+e8/k8frF30wR/+8IcX7ftNYdd+bWUbhzBh6xi1h2N2bq9vNHDubnnSxr676c983/O9iwaE58437awtaYDu9P++cymnn5zg0s0Vro808da3vnXpE2mDvjGlxbkm+v7mWqh/tb5R6hNU0mdBZSMqt3zOdWwRiEAEIhCBKyFQUHkl9Fo3AhGIQAQicAQEmDXMLibXVoJKx8RUZ2IyXJlJDC3HyERlfpqYWcxPd8QLMd73vvctJuerX/3qZZ4RH0wvyzy+0+gS6wiGBIR/8id/spRnzjI/vRhhODIP1c2Ytj/MLuYWE5FxZj4z1n6qWxsIKhmuzEXz1cMoEyAIj2699daljG17Mc8OLWA51KBS+wl/GPd//dd/vbThG9/4xsXYZoAz4RneD10w3BnnTHPBEP3ce++9i+aYpoxMoTMODHgmq2CIQc8kZ9IrQzPmezFhlWei0o36hQiMUlp629vethirtk3jjFjBEz1N8G2+Rxs7Djpl2Ns3emX6m+8csZ6Ait6sS1+W2w/6VTfT1zLnB+2PFvexuzy2oFIb0A5dCbIZ8NpMGGnkrf7LPHqlF/3OBJX6LP0lw5+m6YCGaIFW9JvqUk4/yshXj2BSKPCe97xneRfQWab/pQ19mmDKiGHngNDT/C1PzretBJV/+Zd/uejkta997RLIaE9tS1/6RHqjk9/4jd9YtERD+hJ6mXAbD7rTlyhPW3fccceiKzd+0Ku+iD5f+MIXLqEQndgOfalT32c5DQnm3Mjj3XVUv+nGCf2h/pIuLRdI0d4E8XTnszroWln9tvPA/qhDfcKmQ5js9zEHldrftcl3NFp0HaMvOpkbLbS166v59Kjvoifa8E6TvoO5PiqrH6NxI3Z9/8KXJlzTXZvp0rY+8pGPLOub7yYM50BBZY9+PYR+o32MQAQiEIEI/LAIFFT+sMi33QhEIAIRiMCBEGDeMLsYiVsJKh0PA4sJybgSDjJOmZ7MU+aU0TzCGua5YFLZD3zgA4tJf8sttyy/+8e0ZNAzUQVUHu3JLH3nO9+5hDUMTgbZhE/MesGIAED985hF+8D8F/QwyiaoZKYxwdQhEJpwiCkmXGWGMdVMlhlRyXTVZqZDHJF0qEGldmB+G8XDoBca/s7v/M4SaDMumfD0IHykGW3/q7/6q4tOGPE0YESGgE8wRKPCHC/6Mmlb7Wy5OhmojFIjhJiqAkrbpQ/LjAAx781vfvOiQfvHtJ0RNYzTCSXpj37th3kMWXXZD3VZz3boUMBuX7wY9s4lZSwXVNk3xyFYpf19no4xqNQe+hqBOk3QA10JKvU9NKyvV0Zfoi3pyGc600dq99GBdQU8rhH6UYGS88H6+jp10crv/u7vLtvxSGJBuu0KCNa6prljmLYQVLouCfv0d855+nFN1Hfolz524YkC9OUaJii8/fbbl4CHNlxrvWhg+hh9pHmuW4Kd3/qt31r0+Dd/8zeLXvQ9+khPONCv0Jx+R+CLp0nIKESiU+GS66R19HVunLBcX0eDn/jEJ5ZyRry51s53AX2CPlQg6ruAGz5sS9BqZJxrvD78ECZsjzmo1P7a1bWJvlzHtKl+jib1XTOSme7okXb1cT7rz+jVuq7TbrzRj+kT77vvvqU+N22ox29B055Jv/cXf/EXy7Zpz7a8JhR1vmxxojff4TB2nmHouoG3Y59zx/nUFIEIRCACEYhABE4TKKg8TaS/IxCBCEQgAhF4BAEmA/NhK0GlY2FEMlE89tBxmcdcF7J4CV6YVuugkiH6wQ9+cDFSX/Oa1yy/MchMNXqM6W40JUOToS+oZMD/6Z/+6RIymcekEQCNuWq59RlWDF91MMgYa0IA6zBPlbdPzDDrm2dd5axnGQOXQWbkpzKHPB1yUMnMFPIwwBnd733vexfzkpYElB79ysCkGeblPNrQiMoJKrWj4EYZGtXG1qVJuqBJJqnPNMtQHd3QCN0YcYujgFN5I20Z9AKjdVAphKcX+2q+kVGCAvO87Iv9sG/0yfinN9uwD0akCJbsB2NSHYJ261heULm/ZyJtrIPKGVFJV/SorfV5+n99F/1oV1rWf+qDaITWaIDe3MzB2J9ghC7pl+aFmoJ7oYBgibbVzbimd3qxHYb2MUxbCirpiG6MIBcGakP9hX5Q29OD9ndjhr6G9rS/Po1uXOP0Mfo57zQmJJyg0g0XtKReQZGbPNSjnBBJv2a5vsd+rG/0UL+gUpCkTjq2DaPh/uzP/mzZN/2j+uYarA9UTv/l2mrEJr27EUifaT+cC4cw+W4z5yPNOXbhkePDceuPftVX0YBjF5rTntASE23uGnfjhWuxz9ra9c11DCffsbCiU+u5tuunjKjUVwnQ6c71UB1uaFOffs1NZK6n9KnPo0n9nPVoqaCyoPIQ+o/2MQIRiEAEInC1CRRUXm3ibS8CEYhABCJwYAS2FlSOccVIMoKMGc/IZB4x9ZidTCpGlnnugv/FX/zFJcD5/d///WWZ3x70e4FMS4YT40tQ+dGPfnQxAN/1rnctZuaf//mfL4YVU1PAyKxicgp2GPTWNzFzGfr2hanGRGT+M7UYr/bZCA5mmr9NgiHGKmPNPggNjGZh5B7y5Pi1gePTDj47fryYzfs64oohyQj9h3/4h8WgFBC++93vXsJjBqg2pgfvAkXG5jqoFDhqb0YnU59OmKQ0YWQa85xxSqP0NC+8rEtTtmM95rNtWEYzY7gz9IVJE3raDuNVGYbr3XffvRirdIq15bRou2P0Chj8bTuMV8cpaDDPvmov+0GHzhHvAgWvfZxoy+grWnMOzrnpuLwcp/PLMW1pmqBSuE5n2tyIMf3L/CYgXWlbuhwWdIWFcs5FOsRMQEkbWOqraM85MQE3XesX6U1QSSvq1i/SGp3YxvRvW2J9sWM59KBSeztfBH6CSv2ER54LKmnEMkGN/k6fpY0FfZZ5+oD2pxPtbV316Z9oyLVXiPaGN7xhWTZBpeuxkZT6TroxTb+rPppTxrXQeW00Om3SL32r0/nsXKdBNxaZPIJbH6i87wVetOlGIhrXt1muPxM62W/LD2E69qBSG/kO6/uE72n6I9dTfRs22neuY65Rrl9zHfO3MnRlHSMoaUk/5hpLl/Q618JXvvKVi/Zsk5bmCQU0Q/9e1qfRgsrDOH+0ZVMEIhCBCEQgAlePQEHl1WPdliIQgQhEIAIHSWBrQSUTlPHILP3DP/zDxcDy21rMT6Ypo4qZxchy7EwmQSVD9EMf+tDybkQl05LxyXBS/mJBpUd/GSXC8GKszqO/GJ3+HkOVMSpAEkbZJlOUIcvcd7c+05UxyuCyXDgpWLIeA05YoOzLXvaygsof0lnG/NbWM6KS+e63IbWJ9n7ogmH/x3/8x0ubaUcm+Dz6dUZUMjFpgqkutKGtCdHojz6M9phATRl108dMNExbXnTEUKUl9QilaNW+0pNwSEjvxXAVVAoMnAu0ylhVB31Z3/at71icJ8JjRi+N0qV9FHo5x9RthC9d7nPQd8xBJSNduC64ZtYLKrWp8Ikxbz590SMN0dQEicJKoRO90IP+aMprc8v0TQx74T0d3XnnnYvW5lGd6qBhOpvAfOofPW/1fStBpUe/CiqFNRNU0ox+wohK16j1dVRw+LELj4SlFY9w1c/QFq380z/901Je/0SPb3rTm34QVKrPdZSO9E90Y9Ln6HddE/W5+iNl9EOCT/P9TcO2R1/6UJq17/42opK2hff6SPq3vj6MlunT/nipa5/7s9Pny7EHlbSnjxc2uobpr/RVNEBT5mtTOphyvh+65rl20rVrpX5MsO3a7bpuGb2qiy5p2GOBaUa/pv/0SGS6plcvN/4UVPYblafP0f6OQAQiEIEIROD/CBRU/h+LPkUgAhGIQAQicBECjB5mF2OV8cdgZBIKN5h4QopDmiaoNGro937v95bQZh7pxuhkZgkNGZwmBqcRlEKaywWVDFh1ePQrg+sjH/nI8ghFZhhOc0e9ZYxR5jxDDFN38bvjXwDENGOe4as+/Jm/1mGKKc8gY7Yy3tZBpToPecIKAwYfw9pnBuK+j6jUDjPC6FOf+tSyzwJuRjxzU9sy7rUbk5Npftttty1tN0Glc4rhqQ21u7amR6G5F1NeoKguOlaPQEgZ3MyjhTHrmaKMU+Gnuuh6RpXQN8OdLm3T/jsnBAkTIFgXd0at+rWJ9T0WVLuof140qn46tp/0a4SJUcP6C5q2D/s2HXtQKZihBX3THXfcsbSncInRrq21Kx3Q2gSV9EB3tEMT9ED7+kiaojftzfBXh5tC6OStb33rojW/D+h6wtCn1XVQ6Vw5hmlLQaXA2zXNb+Ea1a/tLxZUejQmTRhZrj9zs4a21/8IBgXY1qNHoc48+tVvA7rWKUN39EV7vpsoK3SiuxsvhEH6UNdK/ZCbh7z723x9kfX1obavD7PtV73qVUs/R6u+4+gH1Umf+m/9tD5Rn0af5u9jX3ax88b567rgOwPNuabi6PpyDI9+dT2jEbobDvop1yrBtP5f/0a/PtOEa6E21t/hRM+0IXikO/2Y72b6STrBVHka058pj7UbNGzz5ptvXh6TTp/6QRqi1y1O9DbfFZxntOY8xdex+z7hfHQNaYpABCIQgQhEIAKnCRRUnibS3xGIQAQiEIEIPILA1oLKCSaMpvijP/qjxZAXDDGYmEeMFqaWz4wqv8PHYGV4evSrEGYe/WqZckwspujH/jeo9GgwZpXfqGTKMwmVY9QwVPrqrAAAF8JJREFUvphawhshKMPL9hlkyjLPmKiMUOGAeuyTz35ji1nqb+aYsoICdTNvjd4rqHyEfK/aH9qPATqjKmmC1rSNNqcBo9e0HfOboW8kL8NOUKnNmebamTacd+tgknlq3amPYcr0o1UhEWOUFuhmgkF1GxFMv/ShTkGpEUP2xT6q03YtY7raVzoSIPhsvnrtn/1xjLZlmWNgvtLcBA1+91X4Rbe261GQAgna30eDf/oD59OMVMUVQy8GK+b2fUuTvsNIOG2lfY0W+83f/M2lT6I1+vAbbDQi3GEua1Mmu880o+3Nowv16b98Vp4OaWtuqqD5d7zjHUsoMCMqR6v6QRqaIGhLnB/rWJxDOGDv2iPkcK7S3iFMNDI3ZqyDSuc7TQgcPfp1PaLylltuWfoTN3IYoUYf+ijnmHADE+80JtwxwpfWBJX6FXXq79TvnNQ3qcM5TEMepY2hOtR/zz33LNvXN88IOevRJObCR32TwG507Hyge4GlvlK4+du//dvL6HD7Yn26PZQJnwno8D22oNL5pQ/Tlo7f9cvL9UyAqc95+ctfvvRLWPme58YLIRtu+n16VA9t+L729re/fdGna57A03nsXHBO0Ibvjda1jLY9fcN3txsv9JfO89HvoWjoPPuJYUHleYhVNgIRiEAEIhCBNYGCyjWNPkcgAhGIQAQi8CgCYwYyebYwonKMlIcujNBwh7yRQ0zMCVHGhJw7wBn4DE0m1F133bUYVn7zj7nJHGW0YsPEEjgxM/3mle0YOcL0Yloxu5j7ljNbcWXoC4SM6GBeqYPhLzAyEsA+2DeToId5y+iyr0wy5pt3+2D5FkZIMLkYy4xBpqrPjGgGPlYM5n2c7DfjU5sIK7Ujg1Rba0cacDza3GghBjlTn94YndZlcDpO5bU/s5QWGJ7amUnu+Ncj1hiutoOXbYzxj5nQiC4EBLRhXduZMNE5YDQms1bd9lU4R++0ap79YOzady8mv3IC1QkqfbZ/999//9JH0DGNehQezTJp7Yd9m/NrX9rwWINKunJzhf5P2wkK6WSCHvoV2HjHyEQP+i99mVFIbtSgFxqhUQE4/SuvfzPRJL3oR2+//fZFB3RHt7Qg8FYXDdKn+o9hcs045KBSP6J/0td5BKbrl0eo0oQ+zjKPwaYLWtCfCGz0AY6broRC+h+6UmY+6weFOkZkmy+o9N2DZl0P1G8ddXl3jRSEezKC7egLBe2CSlqzjnI0piy90rvrrn7ROuqynvKu4/ZR2Gk/3vve9y5BJb3uW/91uXPF9xDnoH6b5vThznfn3Ra+L1zu+LWp6yMd6psmpKQ12qKD9SPzadJ1zDqY4Ufbru/WFVTqx+jcNQ5TN2g4D6yDNf3SE93p01zr6dO6ltGv5Vuc5vs17voAWsN5vk/7fqOP3+rxb7FNO6YIRCACEYjA1SRQUHk1abetCEQgAhGIwAESYDIwHxgyWwgq53gY5e6CZ6iYZ2JCMp+8GJoMKsaKkIWxxcRkcvp7QiVlGVJCIWYYVoxPhpcRJbgxSAUAjEEmDcPLSBPGPiOL8cXIYoYxduyb7TBzvGyD0SMQ8u7vKevdfgsPbMc+H/LkePDULkxVnwUf+x5UjkE37acNhduMS8YkjU2bOhZtRUfajlYct7ZTlvYcM6N0XpbTgnamGfV7pCZ9mW/ePBJR+6uT2W4SFtKX5epXp32jQ/s3xqFt2gf7NTqz3DFpB+/2w2Q5nduuz47NNtXpXDCph2a922/H6rVPk2N2btPaMY2onKDJeaaNtaE+ah6pSR+MdzqznH71O7RGQ8rpA+mF9mmKMe1d2Snv3fp0cuOF0Ed5dWJOC7Ru2/ShXvUfw+S8PeSgUj8wfZ12125uJJmQxjLBEJ3RwPQXNKSPoAFaoQ3r0gId+Uwj9KLfcF7ed999y7mpLI3YBv3RjL5LCCkwp1+f5/o6/SON6W+F5dZzzgvqJjhSj207F/QBgifXZ/vvZqQ3vvGNSx+6b33XWc4Tx4Ub1jR3bEGl65JjnxddunECF1qkBzdK0IPJctcxWtBHmWiKhq2nvH6MBunBdXG42sZcQ9XtM33SHj1bd661h6ilBcZl/sFV34BfQeVlYLU4AhGIQAQiEIFHESiofBSSZkQgAhGIQAQisCbAoGE+MGO2EFQ6tjHPGU+MLMdnHvOIwbR+MamYo8owsbz7m/lpmXUYUswZppV3ZYxUMiJJaGMUJBPXXfiMKkaYER9+C4uBK6g0im3CHPulHtPsk22tTa45Bu8mBq998n7I03BkdB1SUKkdvEYLjoPRad60ic/aczRFR6bRoHKjPRpSx7ysa9noQCjuEYreJ1xkxDNE7QOTXRDCpPb4QssY80JS9SizNmztl5d9sF/e/W0/1LHeD8tmP6asco7Xy/GY6JFm17pdFuzRPzjY52MLKrWpdvKu7bQnHXk3mT/9o8/0ZxoNKjdtbz59KL8uO5qyLi3o66yvrG2apj7vXtY5hunQg0ptOn2d80e7uX5pZ5Nloy9/6zP0A8qZTwPWo4Npe+V8VnZe+je/9YcXfQok3SAh/BEumWcd2/VZvcq6/vq+IijV5xk55/orYBq9218atr59mZuNHrpwQ5KQhZ7d3GGkpv7zECfH6pw81qByjh+D0SNtmmhsrlHT71mm3eloytEH3Y5u9GPWM08Z2lHea67Tlq23obxtTF3Lwg3+gxEOBZUbbNwOKQIRiEAEInAVCBRUXgXIbSICEYhABCJwyAQYL8yHLQWV2sMxzcsxzjQGk/d5MZfGmB0j6rThpC6mFUPMi1HqkXWMVgaVu+mNEGHWMsIYh0aVMFz9lh8jlNHKPFOP+kyzP7Z3epv2ZfZdudPL55gO6Z3JJfA9tKAS42kP72ttjY6mraYtvZuUtWzKLTP/d/4sWy+nEUa631n1u20CdHUx7sd4N885y4w3opK+BOYTFs0+Tr2zzbWOfLZ8XXbKW2Y/RnNTZq3dOU7llN/HaYzmYwoqT7epdpn2nHZSZtpyyk85ZaZtpzydKq/sxSblRwej6XV96pm6Lrb+1uY5Nw95RKX20Nba0ss07evzLBs9aFvLTbPO6OB0248OLJ9HFAsRXUON0BY6um6uA6PRo9DIjUBGAwscXWuFkwJHo8oFnabZH+v57HrsWj0v9ajfevpP1+5DnDA85qCS/uY1ultrcnTj3WTZut8zb/Q4n+l4XX5dr/WVX68z2zg9X31bm7AoqNxaq3Y8EYhABCIQgatHoKDy6rFuSxGIQAQiEIGDJMB4YT5sLajcRWOsDa+pb4wahucESYI3k9BIaMmwcoe9v40Oed6FEW+MUPPGzJ36ju39kIPKq9lWQm6/80ZnQksjh2iPtmZ0CIOaSc9on9GUM6ppbaRezf3et20da1CpHdLAD0+NWwgqH4vexa6Lj1X2sebry5ybOPlNXTeuCCr1Z66ZAkc39kxfN/UIJvWN+kQ3cbiezChMj4Y1uvJik/qFm9YTijo33NjhsaDrR4NebN19nofjMQeV+9w2W9y3+f7biMottm7HFIEIRCACEXj8CRRUPv6M20IEIhCBCETgoAkUVJ6v+fDyMkLL71wZEcI4ZaCa7+56YRHzVHjEDPVZaGnZsYcHBZVn05sRP8zAGT3kd9nmN7KMqvSiMwH4jRd+U8tIpBlteewaWxM+xqByffx9/uEQ2HJQuQui873Db6UKHN3soz8zatz10mhH/dnpG3tcP9y04ZqrT3R+Kzu/eek6e7HJOkJK6/nspiGBqGDUNv19iFNB5SG22uHuc0Hl4bZdex6BCEQgAhHYBwIFlfvQCu1DBCIQgQhEYI8JjGHYiMrzNRJjVYjE+GS2+tvIBiERg3UeR8d0ZYIWUv4P34LKs+lsbUAz2AXjgkoaoyemPlN+RiAx22mvkPKRfAsqH8mjv64OgYLKS3P2vcNrRki6Lggl9WtGUrqGGk3purme5ny23owyV1ZfeKnA0Y0frtMemW1b+lB9p/Uutp31Nvf58/o6QXOuE44Ri5tuumm5gWWf9799OywCBZWH1V7tbQQiEIEIRGDfCBRU7luLtD8RiEAEIhCBPSNQUPn/axCG6cMPP7yYnoxPfzNxBEXMVUYow7WQ8pF8CyofyeOx/hojXzDJZMfNZxqjLy/GPpOeue9zIeWjaU6wcUy/UfloCs252gQKKi9PfL576NvW1059mf7tYjdezDrOay9/T184653e8qyj/5zrtHVcm6ffPNS+EzfH5SaWgsrTLd/fuyZAb87XHv26a7LVF4EIRCACETgOAgWVx9HOHWUEIhCBCETg/01gTDwm14MPPrgYXkYBemSpR5f63aimCOyKQEHlrkhWz1kIFFSehVJldk2goHLXRP//9fmOYzrUMPJSR15QeSk6Lds1gYLKXROtvghEIAIRiMBxESioPK727mgjEIEIRCAC5yZQUHluZK1wBQQKKq8AXquem0BB5bmRtcIOCBRU7gBiVVyWQEHlZRFVYIcECip3CLOqIhCBCEQgAkdIoKDyCBu9Q45ABCIQgQich0BB5XloVfZKCRRUXinB1j8PgYLK89Cq7K4IFFTuimT1XIpAQeWl6LRs1wQKKndNtPoiEIEIRCACx0WgoPK42rujjUAEIhCBCJybQEHluZG1whUQKKi8Anitem4CBZXnRtYKOyBQULkDiFVxWQIFlZdFVIEdEiio3CHMqopABCIQgQgcIYGCyiNs9A45AhGIQAQicB4CBZXnoVXZKyVQUHmlBFv/PAQKKs9Dq7K7IlBQuSuS1XMpAgWVl6LTsl0TKKjcNdHqi0AEIhCBCBwXgYLK42rvjjYCEYhABCJwbgIFledG1gpXQKCg8grgteq5CRRUnhtZK+yAQEHlDiBWxWUJFFReFlEFdkigoHKHMKsqAhGIQAQicIQECiqPsNE75AhEIAIRiMB5CBRUnodWZa+UQEHllRJs/fMQKKg8D63K7opAQeWuSFbPpQgUVF6KTst2TaCgctdEqy8CEYhABCJwXAQKKo+rvTvaCEQgAhGIwLkJFFSeG1krXAGBgsorgNeq5yZQUHluZK2wAwIFlTuAWBWXJVBQeVlEFdghgYLKHcKsqghEIAIRiMAREiioPMJG75AjEIEIRCAC5yGwDiofeOCBk29/+9snP/mTP3nyUz/1UyfPec5zTn7iJ37iPNVVNgKXJHA6qPyv//qvE+YXnf3Mz/zMor1LVtDCCJyDwOmg8nvf+96iMX3cU5/61JMf+7EfO3niE594cs0115yj1opG4NIEBJWf+cxnTr75zW8uenvmM595cv3113c9vTS2lp6TwMWCSt/hfvzHf/zkpptuOnn6059+zhorHoHHJnA6qKQ105Of/OTl/wy+x/3oj/7oybXXXvvYlbQkAhGIQAQiEIGjJVBQebRN34FHIAIRiEAEzkZgHVTef//9J9/61rdOnvKUpyymQ8bq2RhW6uwEBJXCSQb+1772teXzBJXPfe5zCyrPjrKSZyAwQeVXv/rVky984QsnE1Tq47wElT/yIz9SUHkGlhU5O4EvfelLjwoqf/qnf7qg8uwIK3kGAhMc+d4mHHdNnaDyBS94QUHlGRhW5OwERm/f+MY3TvRxE1S6jrq5saDy7CwrGYEIRCACEThGAgWVx9jqHXMEIhCBCETgHATWQeWDDz64BJWNqDwHwIqei8DpEZXf+c53ToRJjag8F8YKn5EAbT388MMngsovfvGLPwgq9XETVDai8owwK3ZmAj369cyoKngFBCY4ElR++ctf/sHNPzOi8mlPe9oV1N6qEXgkgdGbG80ElW4683+IdVB53XXXNaLykdj6KwIRiEAEIhCB/yVQUJkUIhCBCEQgAhG4JAEmgxeT63Of+9xyh7TQ6BnPeMbJs571rOURYpesoIUROAeB73//+yfCScbq17/+9ZPvfve7S1DJWG0E7zlAVvRMBCaopLX//M//XEJL/ZuXsNIj6woqz4SyQucg4Ho6N/7QmkdwepS6fq4pArsiIDhyTTWyjeaMdBMe0dmNN964PN56V9uqnghMUDnBuO9y/v/gOqqPo7uCynQSgQhEIAIRiMBjESiofCwyzY9ABCIQgQhEYCEwQaU7pI04Ehy5O3rM1Sc96UmRisDOCMyjOOmMuWq0G/OL0SUc994UgV0RWBv5X/nKV06M6KUxfZzX/J7WE57whF1tsnoisARGHjXMyF9fT+vfEscuCfj+5ppKZ77DCSk93tr3tm402yXp6kLA9XStN1qjQeFkN/6kkQhEIAIRiEAELkegoPJyhFoegQhEIAIRiMBiNMxINybEtddeu/xuW3dGJ45dE2BqjdlFcz6bR3MTGu16m9V3vARoy4vWJhSnNS8jKb0XUh6vPh6vI59HXHc9fbwIV+8Q0L/RGc3NNVW/JqzUxzVFYFcE5no6evP9zXTNNdcs/2eY62nX1F0Rr54IRCACEYjAtggUVG6rPTuaCEQgAhGIwONKgAmxnjIb1jT6vGsC6W3XRKvvUgTS26XotGzXBNLbrolW36UIpLdL0WnZrgmkt10Trb4IRCACEYjA9gkUVG6/jTvCCEQgAhGIQAQiEIEIRCACEYhABCIQgQhEIAIRiEAEIhCBCOwdgYLKvWuSdigCEYhABCIQgQhEIAIRiEAEIhCBCEQgAhGIQAQiEIEIRCAC2ydQULn9Nu4IIxCBCEQgAhGIQAQiEIEIRCACEYhABCIQgQhEIAIRiEAEIrB3BAoq965J2qEIRCACEYhABCIQgQhEIAIRiEAEIhCBCEQgAhGIQAQiEIEIbJ9AQeX227gjjEAEIhCBCEQgAhGIQAQiEIEIRCACEYhABCIQgQhEIAIRiMDeESio3LsmaYciEIEIRCACEYhABCIQgQhEIAIRiEAEIhCBCEQgAhGIQAQisH0CBZXbb+OOMAIRiEAEIhCBCEQgAhGIQAQiEIEIRCACEYhABCIQgQhEIAJ7R6Cgcu+apB2KQAQiEIEIRCACEYhABCIQgQhEIAIRiEAEIhCBCEQgAhGIwPYJFFRuv407wghEIAIRiEAEIhCBCEQgAhGIQAQiEIEIRCACEYhABCIQgQjsHYGCyr1rknYoAhGIQAQiEIEIRCACEYhABCIQgQhEIAIRiEAEIhCBCEQgAtsnUFC5/TbuCCMQgQhEIAIRiEAEIhCBCEQgAhGIQAQiEIEIRCACEYhABCKwdwQKKveuSdqhCEQgAhGIQAQiEIEIRCACEYhABCIQgQhEIAIRiEAEIhCBCGyfQEHl9tu4I4xABCIQgQhEIAIRiEAEIhCBCEQgAhGIQAQiEIEIRCACEYjA3hEoqNy7JmmHIhCBCEQgAhGIQAQiEIEIRCACEYhABCIQgQhEIAIRiEAEIrB9AgWV22/jjjACEYhABCIQgQhEIAIRiEAEIhCBCEQgAhGIQAQiEIEIRCACe0egoHLvmqQdikAEIhCBCEQgAhGIQAQiEIEIRCACEYhABCIQgQhEIAIRiMD2CRRUbr+NO8IIRCACEYhABCIQgQhEIAIRiEAEIhCBCEQgAhGIQAQiEIEI7B2Bgsq9a5J2KAIRiEAEIhCBCEQgAhGIQAQiEIEIRCACEYhABCIQgQhEIALbJ1BQuf027ggjEIEIRCACEYhABCIQgQhEIAIRiEAEIhCBCEQgAhGIQAQisHcECir3rknaoQhEIAIRiEAEIhCBCEQgAhGIQAQiEIEIRCACEYhABCIQgQhsn0BB5fbbuCOMQAQiEIEIRCACEYhABCIQgQhEIAIRiEAEIhCBCEQgAhGIwN4RKKjcuyZphyIQgQhEIAIRiEAEIhCBCEQgAhGIQAQiEIEIRCACEYhABCKwfQIFldtv444wAhGIQAQiEIEIRCACEYhABCIQgQhEIAIRiEAEIhCBCEQgAntHoKBy75qkHYpABCIQgQhEIAIRiEAEIhCBCEQgAhGIQAQiEIEIRCACEYjA9gkUVG6/jTvCCEQgAhGIQAQiEIEIRCACEYhABCIQgQhEIAIRiEAEIhCBCOwdgYLKvWuSdigCEYhABCIQgQhEIAIRiEAEIhCBCEQgAhGIQAQiEIEIRCAC2ydQULn9Nu4IIxCBCEQgAhGIQAQiEIEIRCACEYhABCIQgQhEIAIRiEAEIrB3BP4bsC9+k3lY/EEAAAAASUVORK5CYII=)
![[[@Lindquist2012-jr]](http://dx.doi.org/10.1017/S0140525X11000446)](data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAA5AAAAKcCAYAAACe+VXrAAAK1GlDQ1BJQ0MgUHJvZmlsZQAASImVlwdUE9kagO9MeqMlRDqhN0E6AaSEHkBBOohKSAIJkBgSQrMjiyu4FlREsKzYQBRcXQFZC4KKhUWx9w2yqAjPxYINlTfAI+zuO++98/5z7txv/vnvX+6ZO+cfACgBHIkkC1YDQCTOkUYF+zESEpMYuH5AAkSAA9rAkcOVSViRkeEAkan5r/L+DoDG55u2477+/fl/FQ0eX8YFAEpGOJUn44oQbkPGM65EmgMAqhbRm+TlSMa5C2GaFEkQYcU4p0/yu3FOnWA0fsImJsofYT0A8GQOR5oOANkS0TNyuemIH3IIwvZinlCMcD7C3lwBh4dwC8IzRaLF4/w7wpaIvQQAChlhZuqffKb/xX+q0j+Hk67kybomBB8glEmyOAX/59b8bxFlyadimCODLJCGRCGzCrJ/9zIXhylZnDo3YoqFvAn7CRbIQ2KnmCvzT5piHicgTLk2a274FKcJg9hKPznsmCnmywKjp1i6OEoZK03qz5pijnQ6rjwzVqkX8NlK/4WCmPgpzhXGzZ1iWWZ02LSNv1IvlUcp8+eLg/2m4wYpaxfJ/lSvkK1cmyOICVHWzpnOny9mTfuUJShz4/EDAqdtYpX2khw/ZSxJVqTSnp8VrNTLcqOVa3OQl3N6baRyDzM4oZFTDKKBI3ADgcAJuAN7AHL4+TnjRfgvlhRIhemCHAYLOWl8BlvMtZvJcLR3dABg/NxOvgpv6RPnEaJfmdbxUgBwOo4oj03rREYAnE5FQi6Z1pnFIfeDAHQArlyaO6lDj18wyPdAFdCQL4IBMAGWwBbJ0BV4Al8ky1AQAWJAIlgIuEAAREAK8sBSsAqUgDKwEWwFVWA32AtqwRFwDDSDU+AcuAiuguvgNngIFKAfDIJh8B6MQhCEgygQFdKGDCEzyAZyhJiQNxQIhUNRUCKUAqVDYkgOLYVWQ2VQOVQF7YHqoJ+gk9A56DLUA92HeqEB6A30GUbBZJgG68Pm8CyYCbPgMDgGXgCnw9lwIVwMr4cr4Rr4MNwEn4OvwrdhBTwIj6AAioSio4xQtigmyh8VgUpCpaGkqOWoUlQFqgbVgGpFdaJuohSoIdQnNBZNRTPQtmhPdAg6Fs1FZ6OXo9ehq9C16Cb0efRNdC96GP0NQ8HoYWwwHhg2JgGTjsnDlGAqMAcwJzAXMLcx/Zj3WCyWjrXAumFDsInYDOwS7DrsTmwjtg3bg+3DjuBwOG2cDc4LF4Hj4HJwJbjtuMO4s7gbuH7cRzwJb4h3xAfhk/BifBG+An8IfwZ/A/8cP0pQI5gRPAgRBB6hgLCBsI/QSrhG6CeMEtWJFkQvYgwxg7iKWElsIF4gPiK+JZFIxiR30jySkLSSVEk6SrpE6iV9ImuQrcn+5GSynLyefJDcRr5PfkuhUMwpvpQkSg5lPaWO0kF5QvmoQlWxU2Gr8FRWqFSrNKncUHmlSlA1U2WpLlQtVK1QPa56TXVIjaBmruavxlFbrlatdlLtrtqIOlXdQT1CXaS+Tv2Q+mX1Fxo4DXONQA2eRrHGXo0OjT4qimpC9adyqaup+6gXqP00LM2CxqZl0MpoR2jdtGFNDU1nzTjNfM1qzdOaCjqKbk5n07PoG+jH6Hfon2foz2DN4M9YO6Nhxo0ZH7R0tXy1+FqlWo1at7U+azO0A7UztTdpN2s/1kHrWOvM08nT2aVzQWdIl6brqcvVLdU9pvtAD9az1ovSW6K3V69Lb0TfQD9YX6K/Xb9Df8iAbuBrkGGwxeCMwYAh1dDbUGi4xfCs4UuGJoPFyGJUMs4zho30jEKM5EZ7jLqNRo0tjGONi4wbjR+bEE2YJmkmW0zaTYZNDU3nmC41rTd9YEYwY5oJzLaZdZp9MLcwjzdfY95s/sJCy4JtUWhRb/HIkmLpY5ltWWN5ywprxbTKtNppdd0atnaxFlhXW1+zgW1cbYQ2O216ZmJmus8Uz6yZedeWbMuyzbWtt+21o9uF2xXZNdu9mmU6K2nWplmds77Zu9hn2e+zf+ig4RDqUOTQ6vDG0dqR61jteMuJ4hTktMKpxem1s40z33mX8z0XqssclzUu7S5fXd1cpa4NrgNupm4pbjvc7jJpzEjmOuYld4y7n/sK91PunzxcPXI8jnn84Wnrmel5yPPFbIvZ/Nn7Zvd5GXtxvPZ4KbwZ3ineP3orfIx8OD41Pk99TXx5vgd8n7OsWBmsw6xXfvZ+Ur8Tfh/8PfyX+bcFoAKCA0oDugM1AmMDqwKfBBkHpQfVBw0HuwQvCW4LwYSEhWwKucvWZ3PZdezhULfQZaHnw8hh0WFVYU/DrcOl4a1z4DmhczbPeTTXbK54bnMEiGBHbI54HGkRmR35yzzsvMh51fOeRTlELY3qjKZGL4o+FP0+xi9mQ8zDWMtYeWx7nGpcclxd3If4gPjyeEXCrIRlCVcTdRKFiS1JuKS4pANJI/MD52+d35/sklySfGeBxYL8BZcX6izMWnh6keoizqLjKZiU+JRDKV84EZwazkgqO3VH6jDXn7uNO8jz5W3hDfC9+OX852leaeVpL9K90jenDwh8BBWCIaG/sEr4OiMkY3fGh8yIzIOZY1nxWY0ivChFdFKsIc4Un19ssDh/cY/ERlIiUWR7ZG/NHpaGSQ/IINkCWUsODWmQuuSW8u/kvbneudW5H/Pi8o7nq+eL87sKrAvWFjwvDCrcvwS9hLukfanR0lVLe5exlu1ZDi1PXd6+wmRF8Yr+lcEra1cRV2Wu+rXIvqi86N3q+NWtxfrFK4v7vgv+rr5EpURacneN55rd36O/F37fvdZp7fa130p5pVfK7Msqyr6s46678oPDD5U/jK1PW9+9wXXDro3YjeKNdzb5bKotVy8vLO/bPGdz0xbGltIt77Yu2nq5wrli9zbiNvk2RWV4Zct20+0bt3+pElTdrvarbtyht2Ptjg87eTtv7PLd1bBbf3fZ7s8/Cn+8tyd4T1ONeU3FXuze3L3P9sXt69zP3F93QOdA2YGvB8UHFbVRtefr3OrqDukd2lAP18vrBw4nH75+JOBIS4Ntw55GemPZUXBUfvTlTyk/3TkWdqz9OPN4w89mP+84QT1R2gQ1FTQNNwuaFS2JLT0nQ0+2t3q2nvjF7peDp4xOVZ/WPL3hDPFM8Zmxs4VnR9okbUPn0s/1tS9qf9iR0HHr/Lzz3RfCLly6GHSxo5PVefaS16VTlz0un7zCvNJ81fVqU5dL14lfXX490e3a3XTN7VrLdffrrT2ze87c8Llx7mbAzYu32Leu3p57u+dO7J17d5PvKu7x7r24n3X/9YPcB6MPVz7CPCp9rPa44onek5rfrH5rVLgqTvcG9HY9jX76sI/bN/i77Pcv/cXPKM8qnhs+r3vh+OLUQNDA9ZfzX/YPSgZHh0r+of6PHa8sX/38h+8fXcMJw/2vpa/H3qx7q/324Dvnd+0jkSNP3ovej34o/aj9sfYT81Pn5/jPz0fzvuC+VH61+tr6LezbozHR2JiEI+VMtAIoZMBpaQC8OYj0xYkAUK8DQJw/2VdPCDT5LzBB4D/xZO89Ia4A7F8JQJwvADHI2NmG9CDIvQrC4+0QooOdnJTjXyJLc3Kc9EVGukrMx7Gxt/oA4FoB+CodGxvdOTb2dR+S7H0A2rIn+/lxCbdFWmRd+/AYx56ODPB3mez1/1Tj32cwnoEz+Pv8Tx0kFWc+ZYWOAAAAVmVYSWZNTQAqAAAACAABh2kABAAAAAEAAAAaAAAAAAADkoYABwAAABIAAABEoAIABAAAAAEAAAOQoAMABAAAAAEAAAKcAAAAAEFTQ0lJAAAAU2NyZWVuc2hvdJPiH14AAAHWaVRYdFhNTDpjb20uYWRvYmUueG1wAAAAAAA8eDp4bXBtZXRhIHhtbG5zOng9ImFkb2JlOm5zOm1ldGEvIiB4OnhtcHRrPSJYTVAgQ29yZSA2LjAuMCI+CiAgIDxyZGY6UkRGIHhtbG5zOnJkZj0iaHR0cDovL3d3dy53My5vcmcvMTk5OS8wMi8yMi1yZGYtc3ludGF4LW5zIyI+CiAgICAgIDxyZGY6RGVzY3JpcHRpb24gcmRmOmFib3V0PSIiCiAgICAgICAgICAgIHhtbG5zOmV4aWY9Imh0dHA6Ly9ucy5hZG9iZS5jb20vZXhpZi8xLjAvIj4KICAgICAgICAgPGV4aWY6UGl4ZWxZRGltZW5zaW9uPjY2ODwvZXhpZjpQaXhlbFlEaW1lbnNpb24+CiAgICAgICAgIDxleGlmOlBpeGVsWERpbWVuc2lvbj45MTI8L2V4aWY6UGl4ZWxYRGltZW5zaW9uPgogICAgICAgICA8ZXhpZjpVc2VyQ29tbWVudD5TY3JlZW5zaG90PC9leGlmOlVzZXJDb21tZW50PgogICAgICA8L3JkZjpEZXNjcmlwdGlvbj4KICAgPC9yZGY6UkRGPgo8L3g6eG1wbWV0YT4KJjM13wAAQABJREFUeAHs3Qm8rlO9B/CH6jbduqXxUjkyhcyERKeUhIpQQnUk0UCluRC5ZShDiQpFZW40FFdkKJmnSMqQWdEtKqVB7vtd/Ld1Hu+797v3fvc+e5/z/5/Pe573fZ71rPVfv/+w/v+11vPs+R7oUJOUCCQCiUAikAgkAolAIpAIJAKJQCKQCIyAwPwjXM/LiUAikAgkAolAIpAIJAKJQCKQCCQCiUBBIBPIVIREIBFIBBKBRCARSAQSgUQgEUgEEoG+EMgEsi+YslAikAgkAolAIpAIJAKJQCKQCCQCiUAmkKkDiUAikAgkAolAIpAIJAKJQCKQCCQCfSGQCWRfMGWhRCARSAQSgUQgEUgEEoFEIBFIBBKBTCBTBxKBRCARSAQSgUQgEUgEEoFEIBFIBPpCIBPIvmDKQolAIpAIJAKJQCKQCCQCiUAikAgkAplApg4kAolAIpAIJAKJQCKQCCQCiUAikAj0hUAmkH3BlIUSgUQgEUgEEoFEIBFIBBKBRCARSAQygUwdSAQSgUQgEUgEEoFEIBFIBBKBRCAR6AuBTCD7gikLJQKJQCKQCCQCiUAikAgkAolAIpAITEoC+cADDzTxmY6Q98N7P2WmY98nmud///vfRTcmup2x1k+u//rXv5r77ruv+dvf/tbcf//9w1YVeuA4mfT3v/99RN7Gws9k92MsPE73e0JnRuoHW/nnP/9ZPiOVzeujR4Cds6PR6Pxoyo6Go351YjR1jlR2Tvhi/dTudKM5gdWgMZob+jBoTEZT31TSXbL8xz/+UWKV0fShn7LhixxHInzkGDUSSnPP9YEkkBRLcP3Xv/516CjgNiCjv/zlL82NN97Y3HrrrdMSudtvv725+eabSxLRrQMM5pZbbin9Y0DjocARfnMz0Rn6csUVVxT9mKp9/cMf/tAcfvjhzdprr92sscYazY9+9KOi4734/eMf/9j85je/af785z8PFenH8Q4VHsMX9vWWt7yl+f73v18GkTFU0fUWAxLdJ6ekiUGA7G666abyGamFq6++ujnggAOa/fbbb6Sij7hOB++9996SID3iYp4oCBx99NHNu971ruauu+7qK4nk6+++++6BT9wYN3/72982N9xwQ5HZZIjHGHbNNdc0d9xxx2Q0N9QGrK+66qqhWGHowhi/6EfEIRF/jLGqnreZRLz++usLVoMYp9kmXvHtM1F81x0yUXL55Zc3f/rTn+rT+b1PBODHRq+77rpJkddIbF155ZXN7rvv3hxzzDEjFR319d/97nfFF91zzz0j3nvttdc2Bx98cPOZz3xmxLJZYPojMO4EUmK15557NjNnzmxWXXXVZuWVV25WWGGF5hWveEXzpS99qQyu5513XvORj3yk+fznPz9tEJPInXrqqc3LXvay5g1veEPzute9rvTx05/+9NAKgMF2t912a17+8pc3m2222VCZE044YdgkoxsIjHTfffdt1llnnWaVVVYpGL7oRS9q3vnOdzY///nP+wpoutU7Fc4ZFH/961/PFrwaeH/yk580m2yySTlOBT678SCoPPbYY5u11lqr2WmnnZrnPve53YqVPkjiXvrSl5Zk7sUvfnHReQPMfPPNV+xAMmqiYRBBR82ERO+ss84qietIK6T1fb2+q2vTTTcttky3V1pppeatb31r8+Mf/7jXLeM+L/CD1WQHNBJ+E1sSuTlBF154YbPLLrvMNuDiReL++9//fjaWYMOOfEZDAtRf/epXRYaHHHLIaG6dp8oKfn7605+WpG2kSR+J45FHHtmsu+66JaHnv0dLEoX/+7//Kz6BDQfRSUHYxz72sZLUxflBHLUp+cF/kETY5MQ222zTfOUrX4nTE340Lnzzm99sttpqq8LTeHwXeUm4jZfiEGPoiiuuWL5/6lOfKhPYg+qQiZj3vOc9JZ4xWTheOvnkkxvjhQlKvC+77LLNBhts0Bx66KETMnlHBy677LLi4//3f/93tnF5vH2ZV+63IHLggQeWCafRjll33nnnwCdmxRaXXHJJsaNBy+CII45o3ve+95XJ86ibDYh/9aUmY5dx/Je//GV9Or/PpQg8erz9MhCZyTIQCjaf8pSnFIf0X//1X80SSyxRgucFF1ywrOA87WlPG29zj7jfwCFAHzRJbiS8j3/840sSqQ0rBY5mnx7zmMeUgF37Auz//u//bszQfOc732m+9rWvNU9+8pPLff3yZcUKjoJGScjTn/704mTMCu+6667NXnvt1bzgBS/ot7opVc4ga3Zs7733bhZZZJHCG1wXXnjhZuONN27ox1QlTpluz5o1q3nhC184JPvgl/w5UoOJ2fRXvepVzTOe8YxyTkAUySKdOf/885tLL7201PWc5zwnqhj3EQ+DWl0S0JnFlNDRwyc96Ukl2BQgn3HGGcXGx81wlwoMwgK9rbfeelR206WqUZ266KKLGp9Xv/rVxY5HdfMACvMbL3nJS4o/ieokMuecc06z0EILlSAvztMnchntLgf6wTeZGHjWs54V1U3KUdsT4Z8ngnlJHDvqB1/JyimnnFISL0mXicbRYiuBkrBagdt+++2bGB+NHSZi1RfnBtVfAZ5JUBNE66+/fql2/vnnL235vfTSSw+qqRHrefSjH13ae81rXtOIF8ajJ/TM2PnDH/6wWWCBBcqELp/CJ3/rW99q/uM//qP43Wc/+9kj8jVSAWO/id4ZM2aUeGek8iNdN7nDB5lMVScbF4AfddRRxW5NXJLRoAjOxihjr/Ye9ahHDarqeaYe+iruc6RboyF+Q9y83nrrNUsuueRobu1Z1tjAn4gzBk3LLLNM89jHPnYodlO/BNp4wle9+c1vHmqS76S/9YTY0MX8MtchMO4EksI84QlPKCuOtlb53iaJD0fVJsouOBdkP+5xj2sMKAxB0Pqf//mfRRHNxkpKw0gNFMozFuWcp7AGRr/V6R5O8ZnPfOaQc3TdLI37Gb06hyMJpM9JJ500FNAyegNSOHNJ0Bvf+MZm8cUXHwoA9f+ggw5qfvaznw3dN1w7cQ2OjHRmZyX3s5/9bOFRUikZNftjkBVQ6qO+6McTn/jEkqgYEJ761KeWZFd9gYFVVLgKQgI/1yNQYvxIn2AKF/V0I9gZkPUddnFvlNWWevFENmTgu/KSMIP4m970piEe3SdJ/sQnPtFVFtqCN1y0JSioSXuCPXU44o/M9RWO/ZCgWhtIn/QfwZZO6YPEScBArnRV/XWgQ1/N5p599tnNu9/97pLsl0o6/9meGwGLlUdlbIEVHEZb5BmOXxs17/jTFj3DQ5AZZP0NmbnGdvQf3/hXJ2xqgqUgS5v0ou6Hcvg9ojPbeNttt5WE+JWvfOXQ7aeffnqZWVR33Oc7/PBJzmTEboPiOv7pX8hJ22xTPcrQ51/84hdlNYIt8RfKBEb6imflbRvST8G1PtN1faIPsFNvjSFetKEMncKH4En9cCQTOw3IVZ3adE3d7jM5AEuyqW0s+ugIc2XwF3zGedg4p93ALewTHwKIelUbRiYZ+B1YWOFiV2QJ4/io3+xv6EBg5Xw3ghnZ6BvSN/finZ3Cgh9AcKj1rZzs/OceNgFH+NTl+Cp1wMjqqe+1PtA9/JKT9lxTR03BAyxdgw/c24QHOhFl6FdgqyybDGz0WVv63yZY0wv6wk7JCL51Xe174rddIQJ/iZ9xz0SfVa9uBC88q5d+hgxMrJ3VCcJMFpisMUEFH+VMKuiX/sOWrNr9gBesyCXkry3YwEDfazvTX3x+/etfL9gKgLVFNrC0ekdX24QnddJD/ESdUQ4frjtPn2DqyKbIuhfBW+Jt1S18lTbYB3lpi//1u9uY064XDsg4Y1xB8LCax86tSJKzcvxW6BYbhaOJTDxFf/il8Gv1eEd+2223XdGb8DX4VicM8GusIAMYRDuFoS7/wZ+uf/SjHx3SIUmGbYBf/OIXi47VfpUusFX1sze8tXUWD2RCduHPQifI+PnPf35pz/1tO4SHD5zUrUxNsPIhMxjRbXXQz8AjysOSz8Zz2CKMhyN9o/P6RO7t9t1Ll9k4HmGs7trGw987FxjVbdY67V51kHWbYAZH/dQ3Pk+d5EyvwibiPuXxBR+YwChsAK/OG2/0UV3aJR86Eu3Dky7Byf1tTLWlXToWdoYnPrutB8EXfSBTfkK9yuHHeTZC/+JefYCPupW3m3BmJy4NPSZLix0/+MEPCv8bbrhh0UW86oNP6BQeyYrOdZNj8JfH6YnA7CP4OPrAUVCUULK6KsGz4JQSmblDlNCsG0Wk2IyEEVA82wU5e4ZoC6HZ0li5otS24FjNodgzOokpJbU6YuuHVUJtmdl3HwdoELr44ovLgM/wrP5YLTWYaLNNHLp2OJYIrvDFmfkE4bEm91lV0xf3j4bCeMPxM1x42Tpri5ttawwd6Z8+6+NZnQBEOQOxwJuju+CCC0qgAFfXrATbHhODIDwkx8svv3yRmYAV1nj3rN/znve8IVzgZZuybci2+nEOBh99J5PAz6oJHrVl9tSWKLPZnAZZIUm1tvUVVj70xvbfWIWMAUd7Aiy/yUuwo+4YfARvggLbn8jWMwBoueWWa2Z2nF2UKydb/3GQ6rZ9UJ84Usm5NpZaaqlSmoOnU7Hl9MQTTyx9oVP0Isi9gbOAryb4IgOotvCsLJkZJCQIgk5Y2fZhyzJsER7JiN7Ra/whesVuJK0GWeXxrDxZ0B/JmDJvf/vbZ7NHMrbCuNFGGxXZhZMvFXf+E1jSn3e84x1NnTy6ztZ8gsgF3/jQP3pBfwSDbDJkDEN6Sg9hjjc+Qv30VT36bnssmdEdq/iSKqu58P3ud79bZA93gZX7BXD0h+6SPXuAP730MSAiuLiunMSYDeBx9dVXL4G71R92ZZJDfbbf0yE2rH9kIICgx4L6ts1rg0zhAF+z+uED6Zbt7GGb6iQfegAX8iFDvBn89Vc97JudwFTf+QKDdNgaP2sF7MwzzywJLrzZ96KLLoqdvggu7JW/pFv44jf45dVWW63YQu3ryEnShD84kgV7obP6ywbJ0az6t7/97YIpnk2W0CkygLXATr1sg+6HnZIB2atf3+Fh5tv9sVpPF2BHVjd2ZsDd6zp+I0iDJz61RScEaGSK1zopZtf8Er9FJyRw5BgYDwcirPTH8UMf+lDRVzJsy6Dml+5rn26tueaaBRM2SgbapQ/6xr7pIfmwJ7rDZoyTVgjZRYwVJjf4SYEoDCTusKEbeKOzoXvqdN5EED0kK89Mqwufrgsa4VAnwuSlb3igd4JENsDnxHgCZ37l9a9/fbEVuJIn/8ZnDBc4khXfOGvWrKJTfAkc8KT/7M85fo7Nhn/sJR82ox9BeKSj8BeUI32FtzEn9No1jyDon/b1GZ58pPFQTOKIjIm2fsKdb5Y44fG4444ryavxkj1IFkwQwYD/60XhK+lu+HH6SqZ8gfMx5mgHZiYCyINt8GfaCb1gw/SALekXedNrfaF7bIZcv/GNbzSvfe1ri5zwBovQKXjBSf3KL7bYYkO2yk7PPffcMvbyGWwN7mya/oSd4c81Okof+GTxn3G8G+GbjrIL9ql9vo386v7xq+SDDzzzE3RDPAc/142x5CA5Y3PajfFO3eyX7fNN5Od+Oq18EDlrh37CkR+mBzHRQzZ8ofHU2KOP5OIePkx7YiD2xG7xZoUcX7ARD9ER/ilW+dgl+9EHOGoLrvoWRL/xRMf0H9/qUz85dCNyjcnraIuPIB9x1Nve9rYhO9VXckXiZLZvnIAjm9Zv/kid8Pve975XfIGEOvwnWYrT6Yn+0iP+kRyT5h4EBpZADgcJReJcBSqRQJpZMxPDsBiZGXcGIbDkPJEgwUwiI4oEklFzoF4mYYDhYDhV24gYGQNm6JSZk+SEDE4MjSNy9FIUA94XvvCFrttCGSGHaTC33ceziX6PRJyBQVlgzOmOlWIgcD8e/NYnTioSGwMgJ2MAgx8MYLHPPvuUgUFfXTMAe/70k5/8ZNkeaObMfZ5PVZ4jhbc2JOuHHXZYkVUkdGbHP/jBDxYHwXkKdjz7w1GQKVx854jUqX4Ox3mOWaLE+SCDEqelLXIwQMOLQ4v2OFDOjNw4dU7UTBddOP7448s57RlkbO2hC+QsSTGg7L///mUF2JZEA2c3EkTZnixQ2GKLLQrGtqAKwHfbbbcSdHHI9FZ/OWZ6pT/kURNeOHj8CpzpqrZrUhcnqr8crnoFv+qDA2dN/wVaESCpT/JA5mTPdug+3iVPdN/AY3A2iKlXnQZAQemHP/zhMnAJLmBo0DGofvzjHy+DQgymNZ+uC4oEnSMRWVhp1zdJE5z+53/+pyRB0TbcPD9K3oJMfaJrknF9U96EhWsGc+UNSnQSLn7jm2zgDmfBlAQE7rZ1S6IEWgYoOmjW3oq9LdOCUPXaGsYe+R524vkydZEJ7NTpO32hh/Dim2ypFSy7NxJM12FUE14FDWxPkkk2SIDwgQ98oMz0v/e97y22IbAga4GErT/alzCTmwSSjrBPg7g+siVtasNvR9set9xyyzKgw/3LX/5y0QX2GIlUzV+37+oRBBj89Z3vxDc722OPPUqdAmqkLD+w7bbbFlkKKhCsyZKv4Y/snKALdJGsBQxkoIz+m+wRoNMJZeElgKW3gtkjOqvfZMl38APkbjz46le/WmQCG0k33d18881LwMbnGBt8YAFL+seHSB7UyQfRFbs4YMj++HTXlIMxfSRDejQSCd7oDDuhC3ihe3xRncQL4OmiiQg+gW8kN/Lis8mZvqtLf/EPN78FaWyeDxdUm9whB7ZPP5FA0/hIT2FjBUudMKVPxjh2qozE3kSIdtgVPcQX29QmfVPWWBEJpIDYrgp+UvDIN3leTzvkRz/YDB02yWms0S7+1MuXwtmKILl3I8GrukyUSpLookkXGJMVOyU/Nks3jFF8Yr/EZ8IJ74JfpP9sxVikbv1kt8ZWPlAfEf2FmVXmWZ0Ygg7TTffRHb6BbfLjxjbPrIph1M8OxQNkQ49NPPLjIxHcEL3wYZeRgMMZnsYZk3x8I9nSEfgbLxGb4F/wRm78DTzZLTyRuo0FbD5wMcaxLzzouziI7dIpbRpLnDNexdhr0oye8tnGVGMCXaMDdJZd0yft8lsmm+gCzNrEXrzEyrjGPowLxnMy5OfZD9vgN+hIbO31yIX2xHx4ZWPsmL6IDST7IX/+jX/gm40LsOVf6JcVeO0HiYW0z+7Ijx3wXfyemIXPkyDx23QSH54xDx/GbtUrbmKL7MzYTM/0ic6wHxMTjhI8soOp9ujS5z73uTIx4/GfmEQQt9rebFJCW9rlE9l0xNfRhzjCMrbcs19+iw7oD/2kQzMfmnjnH7VrAtDkB7vm/9kAXTGGSsLZBlvgP+mpPuGD/hsvxVb0jx3YSUdP2Ba/HHoe/OVxeiIw7gSSQ2FgAjXOnXMw+8BZCrwM7IIIZXyQ65I427MMsAyB8xMsuEcQjtwncHSsiSFSVscgSsqxCUoNBhyDexmXgYhz4Ng4XY6GcXAAlLrb7KDrnBXDNavG2Xg+q70CwbEZBDkDCQzDZKAC49FQ9FEwFTO7nB9HYYDTLqdrwONgDAheXqRP2mSQnBnnIul13uy7QYez49w5iXjuBQ4cnGDRwIA4GIOlPnlxEJly1PCR8EvSOAhOgHM1WLg/BkaORRAUQZ3zZO7+CJgFm+RBXvoggYykTPAQA7tBwqCjrMBOn7TJUQnSOCnyJ2cvNHCOHAxanCJnyCm3idPj5GEqETawaAMG+iOYoT8zOsm1QZpOucdgpT8hm6iX/pswkKDD3pZmAzV9o9/kwgYMCDAXoAjE6TtZ67u+ONb6rP6wmdANPBu86NasjjOHJcwM7jBiEwYZM7wcvbYczfwaWA3Q7qUvNalfACnwN5vZvt4uK1g1gQMjQZNJILzSLxgYkCMI1SeDJn0ScBro8QAfg7jvsCAryYzBWDmBqA/MDFLwIRc4+u4aDOAtsPAbjgY/tqhNwZEEG3+SFwkEm6J7BnyBxo2dgEJSrl5BPn2iDwZvA6yExDn34r1elQtc4MVn0RNBhnbZKB70X2DAd+CTPUo09BsPIWOyQ+wTf37TaQGiMsqqi/3jD+4CLbYleISlAC10Lngb7qg++g1zNoRvfdBXAYAXewiY6d3OO+9c2iMbcqArEie6Rn/w68Pn7NZJHATQgjg2KWHiE+kADAQ57NSz3ezUmCGBsDrCvvUV1nwJnwIvEw9kqb98Mz8lKFKnQBkJdNSLV+fxon7BC560637b6dmLcnSIDQuQBE3aDHvrhZ0+wYlOsW/4sUE6I4EOMhbRa74Mz+ToSK76aUwha/3SH7ZHh/XZC1QEafSfbrlHG3wHcl7SCgPy0FeTJoifJls2KEEmY+OB3+yEz5X8Cpbxjy82ww7wgxyNnexDm/CEk/bI0qQefaFv2mKjZCYB0Q7ejRXGFPbdK4HUNr0JzLVhXIIvHyMA5dO0acLDGCeo7UX0zpgFH3XEZJRdLjEx7V48m4SU3MJeG+ycr8QvHeTv8YZHds1/sgcUfjv4Vh/ZwRfmxi73BN50O8bJUkH1H5z4X3qpj+yNLbNz26ONE4hfNSYb99ko2emT8db4jm/8HtFJbiUtMfHLjum+GAKeKGwW38g4on19laSKGbQPI5Mvxkw6S+bK6Kt4ZIcddijjmx0kkiQ6Y/yjC/QOTsZGOPCvEsj2GFoY6PwnaYx2yENZti32wAffyBfAhz6b3OCHTJhEQkp32NNpp51WkmB6TDdD/8Rz7ucz2Z/+82N0wCQSv8ze1MMGfedrZ3TGulghNc6JDWEAi9ABcaU4gv2FDzPByob4G8nejjvuWGRoHDKByA71k12Rm/5IuumhMcMERMRhxiuT8XCmXyYA9IsPIyN+I3gJTONojHKPeJLd+2184+fpDP7oh3a1AW8+k+6FDegrYvMxLvM7+IAj/eZH4Evu9A8exky+1VjKz9IDOp80/RGYf7xd4MQoDwOQQDFwH87TYIyUMTD4IAopkKJwHBLjZ1yIczHIB7m3Tc5xbnGN0RioGePMTsAgYKe0DNygqD5lBdGMjPL6jUezNt2IszSDwuFwiIIGM5CxGhL3MP4YeBiz2RjOiyGNhvSHgUrevA1P2z4GTA4sBj99YtQzOg7NliFBB/ycN2ut3wZYAYfvBgKDv37qL9J3Dh3mEbTpr1lHCYeZaY7P0T0CJAMP56KvEmqOwUCtHNK+/nMuBi/8BS4cKjIY+C6IdN1v/Q4SdOgvGUqO1KcO3w0YcCU/ROfI3HmOj944GoTIgAPrRgIKQThc9Jcj1Q4sBQfuNZjC2OSCgARW+IVRzW/U75wAmMMVYBscOE/OX130PtqhLwJrum+QobthG6HPUW/7PBkKUuEPY3UYzAyE6iUD9bEpMjN7jBcksaLD+KQXbSJH9xv4fXoR3CNZFSjCHH6wEYzhCYba1R/80NHQCZgKZvSNLA3KggVJnnPqohd0OrBmF3RA0ER/zNiqm7z0H7Z8Cpm7z3ezvGToGls8q5NM0y86DB9t6affsMM/uZA3/0CfDJIGPX4MT8p2wwYvJiJszxHUG0DJ3X3wZpfaRoIJgzOfol144iVkDwv2wWfpC1nBBJEP/gQ19B4O7CNkoJ/w7pe0KQAQYJKl/umDxJQ+wBCRk0CDzc/s2KZ7YKgPeMM/GcGarzGhQp7q5z8ExfAzRghOnNdP40X4X+X1g3z1kwzJQN3adw8Z+S4IsVqCjxkduyRnJGFn33QM79qiQ+yMDCT1yPikDZMP+FKvxE772h2O3CcwpiMCIWTcITM2pk1E/8kZf2TluraUpWehm/pAD/SfvEM/ndcGnearyER99ArpP+zITn/Vwc58+CuBM1sUPPN3KOwMJtqCdei99tURRPclf/wp28UXnkwYSJLJVUIVRE6Cb+MJnVROYksnJUe9yH1h58rQ3+hL6DhejX/qHWlcZU/0Bsb6ZDyDgQlMOhykHXWatMYr2dATNuW3e8kajmQHw/Cl6sA3nQwKu2MTEjl2Sxa+00sJRy9SD775C0G95Fw/2SS91Jb6+V36z874EzGUdtgqnSYz+muCj6zpHuyM89rgU+rYCu7RB3or8aKf2mXX8HYvjOzICFvFKxnRf0kJvGHJ7vm/GHvdzw5NDJIBmeCHXrdJ/+grHwN7ZYwNdBnPcFEvWajLqrKP/vNF0S9HvJnkhrt+sA+xAhlIzOm98d419eGbHeEhdJof4a/EPcYe5egGW2AH/B251Hqgf3Bo+zB9wT/CH57Jh86xKz6dPZEtv0DvjBPaV9aRz0HwxSec+AW6gEfjPVsO7Evh6j/8k6VkVN2Iv+KDJdB0y4SLfrmuXfUH1f1kJ+ojL+XgoR8I767DlH7QQWX4Vv2VmCuTNHcg8EhLHmW/KDNHxGgoKEVi+AwllKpdJQUzMAmGrRYJBBms8upRn3qDwsnF7/aRQxPYMXSGGYQPA60BgWMw+KqXk2WAFJph9CLG6c8XGBTNTlkBs3LKaXIk+FI3R2Sg4SzN8HCYnIIkggPsl/DGORpwOQLOVp/MdDNYxPg4Q06Twwhi/AYUwSXDrklZmHNEBht8c2oGuNqZK+OcurRvIHKP4AXGgZUBg2OAZwRc+NKuwaeWl/uiTO3cnPOp5WyQNnBKwqIt/cAjh4SfGIjdR+b1nnrlyF+Zut4aC06TbpJbTXiGMQfKkZuEwHt8yAXuvYiTlOAYiAQrdMW2aoOOmUT4qLvGI+rCa41ZnI9zgYUABh8G2Fqv8Krv6oGxfphEsBqov2Rq4ICt5I2c20RftcdmtNGL8G8bHDulVzXhyWBCLwQbMFEef6G/ypORgRKvESSEjsAoKGQIO3XM6OhcTfCwim3wDbsnOwO5uvXHgC4oMnCZDKJHBluBpevK4dGxloMkXT/NrDvyFwJkiWI3YquCR0G+4ER79HVWZ3ZYYHZjZ1Cm3yax+Ao2pX1ttyn0RL9rUpacAr+45rdgTb2jIf3l42o/og3nyCVkQpa+m5Ri+23SD7yyK4FC4Cig5Uv4HLongYs+q4+vCN1mswJ8flof6bN71Es2SKBEf01sWZ2iE5IMkwjKaMsMfmAfbemTQF5bfKuPNthmkKCTjmrPfb1IIkqWdJvuwYk9aYONkTW70FfyELzCs6boMwxC1oG1cr6H7B2Nh4I5+NIvumvspOuev4r6BJeSdT6bLWqfDFyHD3+pbvKp7QzvzjsGKa8+votNBynDn8A5EnLX1GkchyNSDkZ4CPmVCyP85z79NKbSgSA6HvWFfsW1+sh30QdjSOielTWrrjCxwwnBFaa1DjgPBzKkY0EmxEwKaXckIpt6nDAm0v+QZ7f79ZkO8VV4YssmkvHPZ/hNpmyE7uINT8i99JZtwhn+5ENP7QgzzilPhuKdGtOaF76UXvNXtf7HGMuX6QeCQ5yPOtRLRvQicDIe4lkyJ1ERy/Cr+GiTNslOvGYnAZ2HicQGdvrmqJ/8uVhLLCaxjZ1r2p/RGSOs8tFd20/5WX6bX+LH4CfObMem5MbPi6EQLPDZHuPw1aboLx1hf/wY/wIj4y8Z1famH87VdhETQ/TPzjh+RVvqNN4bb5FE3/f4HbwYl9hHLz0z3uqjfkus4cF+6eesjsytSkaSS9ZiXvWh6F+05RjyaLcXugw78ggiSzo42jEq7s/j1ETgYQ89Rv4oDOfBUQmc2smLatsKaEBT3iDIuQsEGLxtZAK1uCccWX1/nCuFqv+UYXy1Qsc5PBo8GDXF5+ANMBLB9uBeVTn01cBo3zwjsMXCDKHZVbxw7j5BDFNyKfA0uEvo+iE8cjjuteJoEOxG0X9945RisHK/vjn61KSc8nCP+12Hl/NBgRfHpZz7lGnXF21E2+5X3nk81HVG3f0cyc4g0m5PfXghwwiW1Bf8Oka/HIdrXz3aabehvjgXdTk3WuIkbU0RsAqQzZLaelNj1a5Tn/BU8+1c/ds9eEft8zXm6tGWgRMv7MpAYMbaQGTipt0/v+mw8gZmg6ckrBdpr81vlMWb+kJOvisfvEc5R2Xbfamv19/d71MHQCZqTNg4L4jQz8Ai6pUc2TXgOSABh1VzwYzthZK+NhbRJtkJVmypdh+btzNAcNLNxxksBSq2RsEvkhgTClblDfx2QwgIBYoRwASf0a7jcLjQ0W73xH11Pf18J8daNupuYwJTpI/ta9GG83hjvxIA5LePa+TGf/quDcngzM5qJp1E/KWteAJZSSG/HLIsBTr/SV7JTYBFJpICKy+2dSuvLXoXbcV9ZKlOuiCAUQ61cYRDr/5FXQJW9wmm8cA/atNHgCr4NaMfvtWY0y3ojPpGOgaPAkSBMx0yUSFBkFT4jny3VdhKhUkSiUiNf7tf7d9tPrRLljAJHpRxn3N8jPGqpuhz1O1Y31uXHe47vmudVDbqHKk++sy2+DIfJNaws0XyLTh2XX3d/BL8JB78pUk4degXCh7Kjx7/9cJgOL7xLPGTYBn/6arEn+8RT9hCDBN10CW40/GoU5zBV4Xd6a9xR2wVk/NiMxP8vUj9MfbW/dQG/si7fb7ua1wLnrRjskcya1urbZd8sEl14yO7aJPExiSf5BgO+qEN7SM8sjN+2ESA7eq29ZKZZ3fJy8QDH2H3jWsmmuJxn5Bju12/teEDX32QwBkTu/HZvj/6bmeEZx75AZOUkjw6pt4o0743fiujrPa1qZ8IBuRm3GYTysChTf20Y7zjQ+B8VmdHjnv85h9NVkieTXBKJGN3Rbud+nct6zivn8637Teud7snruVx+iEw7gRSl0NhGF234CpgCSMSTJlZMtAaAM3S2OrFUMyUIAYUAYdZLArJaVJ6szKMKOqL+ttHdVmhMvPCgTr2Q/oTih6G7D4OgaOujSOcWtTL+PGNopy6otxIPBscYiCIOvs5Cu44T07aLGCQduHtiH8Dle8+tszAM8h9ZgA5FMmamU11mlnn4AILzsZAbHCIvgZecYw6HeM+bbneCwMJgBk/7dGlcN6CNQE57GNWrK6//b1X/cpZEdBvqxs1aU/wCkdBLZ5h1A9FOe1G2/qhLtfqAcQg7XeQ8vpJxwUNQaHndD5IMOM3R09W6oen/mgn2lZesG1LKSzNbrrmOaC6TNQbRwmQANnHBEnIylH9MHE/fAwyBpua2CVe+AAzkO7zGY6CnziymWh3uPvMJJvMkehKImPlQ3/rWXB1sSfbwW31FcgIyDxPYpY18K2DoeBZAOHZGB8rMYIyk06euWlTBIBmdT3zCAe8saGZnUTJ9mGJpUF8pMEZFnAYLuBpt+93YNjt2ljOBQ6xGsTm6WjYfOhEr7rpNXuFjSRa0hi+oL5HnXaiWFGTHEbgzybM5msniF/yrJEATTIFS/Kg2+TFJgTg3SZK1GFMUIat099YUWaT+BgOQ2UkrYJy2zjZATm5h855zkeSoh8wwyt9YBexcht26p5oKwL36GO3Iz0SIHtuVGCuDTYW/LNZgbctxN4Mi+hgJLlRZ+DPvwwnP+OQRJSd8/cxoQQDNqZ/fOlw1I8dD3d/fS10sT7X7TtMa31Rxm/9wXuMyd3udc7zdvRVEskfIruP4F2PleVCn/+FnIcrrn/qxx+fRIcF+pIku6DIXxJH5nSv3jUQ9eonX89mxDsmLtVhJVBs5f5exHasxpItfQwbp9fGXjo/XGzXrV59wq8XiXnG2gLBWZ3EhV7xiTWRjckXjwyIDU2AIHotma8xpMMSSB8rZ5IgE4MeSYmYQTzpY1LFM9BWQbVvLDBJyv7pcJA+sv9FOosZiL6TuU+Q/sC4PSGEN9e8hIqPl0hG3bA0eeF6EP7VU/t3sRnMxTgS7G5EP8idH5akBqmbj1FfjVNcjyP58yEmFkykiVFipZF+qJf/gne8RCnubR+1Q2b6Nxoajr/R1JNlpwYCD05zTDIvlEhwIKgSlHFSlNY21tqoBBIMntOJ5XWOwAqdwdGgMBy5lxPllLRVk+0KBvdug4IkyuyfYIFTQVYOOKmYHeMEBD1mzmNrB4MyYHPCHJEEi3Fryyx5m4ean9F+rx2Sezkgq14cGN7NSCMDisGQ0zabifDO4XgxjcGRE/Bb/ww4kgdBloCPk7Ey4HwQp271x8Cm3HCkrShj5YfcEQfaphmd7SccmofXDSJBnJqZPQlAe8tRlKmPbWzqa+o3yJAbvQry4LqtPoKGWBUZrp64D3a2isIunHqsThikzCAaROk8HOpnsdRBh+kKnYntOpIjLzLwDJLvMETqkcCYJGAryPZNLzJgQ+oKngXKnp+xyk9+kk5B93DkBRjaUF777mGPbI8OGazZi21IAh1bdeOZEfUKvuiR2WPBRvAyXJtRRoBNJ9hK4NhNR6KuCLZgLflH5MfGDIT6737+he0iAyifEN8dyURfyAXO+OEXyKNOkA3u6uzlc8hX0EE+Xnbifjggdmkl2JYyvLGv4UjShQf2gtpBb2DWrqPXeeXiWhzb99a/lfEJ/PkA/eJH41kc/tcMP9wEI6GjdT2+84F8j1VggVuQe+iONnzI07nwpcYDshTYwR1ZSYhxQJAdk0m+a99uD7bEHtlNEJ9ja5l2BH/6wm5NJgRJ/mIiohtGZBDP9pOnCQnyZQs+gnq+gx3imz82EeflRsYsZHzw4i7PYRlX6JRPbDsNXhzbPAjwrWZL5qzS4EdSHsRO+Zvwr85rC161zkYiC1e6iODSbk8SJTG38s7fx3XjC99AriPpsbrjPt/bNNy1dtl+fkd9YS/8Al/Cd7Elk8nGD+WibLteOmjsjHGfnhvnjaOhh+17hvs9XFvt+2qe6KhdJGyE3iH6hj/jY038tPGZDfBDdFAyJnFhT/yY8SKC/bqdqMdEhCTCYxe1T+c/nTMxMdyEQV0nHtifsYkfQ3yvDwxj0i7adgwfQI9hjvSVLbEPdbpPbMa3BokRw675EHptDAni391nMscuBeM/X8zWg8QXxm++wU4WbdFt2BmDtYnYnpcauZ/d1kTntE/n4I/Y11mdGAM/ddLJbxmTol595ivYnLiEzwhSL/n6sGOJHd00ecYWEXl5UZFjm69S4KH/4M+HWLCxKkwv7IbRX/1WpzfWSn7FLc73IuO7PsAChc31Kh/naz2Jc3mcvgg8vLwxxj4wXgrNQHopB+XiDHwQQxaUGFAFFYI6hm9FgYIbnAUDFFmyaAbJrD4nJ8C1/VSSFIGjehg7B1nzwHnY3sG5qpvjpfjuY8S2PQgG2sQpcRaMzItQOB+8u8fqhVkcxPAFzd4QxzFyIM5Z+cEjg/WbwXrDJwcwszXzFm0HjurwvRfpK/71t+4r/CWIHIlBX4KHJ2Xg7XzM6qlDnwxIkmSBreBDHTCROCJBAlkYhK0WR7DNSQmYvIQiHBa+8eTYJoG15MWbwgQ/EhFvUMObvobzUU478LLVBf94MjBycFaN9AXBtY2B+ugAB15jU/NDhrYhShjNigqotE82ZjLJriYBWR2U1dd852StKpk48EZa/CI8v//97y+zv35LWg3QdEvfBTNWTWzHITd4Sjwk9HCwgkI++A09990WScGuN/1plz0IXNkHuQYZNOm29hC9F5AMR3iiywImwfURRxxRBl9yMPNJ78nbx9tB2aDZ0sCQTtEJM71BZIT/mjeYCRL4jZCTvsHD1iOJilUmb69EyrK/ekCzDU1Qgwc48Q14VI8j/aBbAkdv24x71QN/iQYycAu0BAZWv8gDnoJ8fkfgHn3QngG8F9EhGNlCBXeDM5JAhX9gH7Ucwje6HsT++D9BDn+oPNuhWzBr+wd1Oq/PvQjOknvlkN/uYyttm2VDZBZ2Sb677bZbedMj7MhfX2MG22+4RuBX80C3vcFYsmlCg3yC/5d3dp/Ahm24ZqeEpIhsBKt0xvfQf0E8bLUj2BLAxKMI2rQFkG2zRfWwD5jg1aod/8cuyZFNH9HRbwEi/iWB5CUoC/7qftABuqnP6lFnm9ib4I/eaMPY5e2YxhBJjLatJMGDrRgf+AT91w/2Z6WbryQDbdZ2E7buZWL0g54G8V3OSxrP6gSt7N0KjfvJMfpEluyTrksM4UL32/oBE6tYgnU2ZozAJ10RXOI3HrNQt492ws7U5zf9Cj0KXuOoLNnGBI/z+KVLtT04r46RdBymMNYfzxC6Rz3GLXYtFgjSRtsvuQZ/Y3X4aH7UmAOP2nbCJwSuISfHbhjE9Wi/PuKDTqsLbgjW8KUvsRUV5uqmhyYR+UCJGv74TnLxnY6Sk9iKvO22EfvQNe9ToDvawlPYvsTa2KesN+PTTW3xOfRRHKA9RDf5kuDVOd9j7PXbvcYzsQibh5e2/Bma2v8pi9gT++UTyICu44md4lfd6qQrfImk2iSNCRJ2Z7JDHGnClI+gCxEjSMb1wf0mW93PRxjj9JOe8Ov6z26QtsUg9N4KrlgU/x6xcg/8yI1++45/Y5bYxepx+DB8G5uUDRLLmCw21kpcve3daqmXNB7R8UniEvLn4+gpXMieTzS5bUXXGG2sESvASv/0t544i/biSJZ0hE6Jt+kcX0Z2Emt6r4/tMU4fQn5Rl/gEru6BPxlZJIEDO8F3TbX91npTl8nv0w+BR3UCg93Gyzajo4gSjrbjVzfDZYAch60Qtlj4GPRsQWKsnIEkxsDGCTJUymhJX52MSbmZnQTMwKZMBOTawINgUBn3Ie1yoByqcz4RYArStIGvNnE+2vThLASR+GDkBqEIyl0zYBlktMOYDeoSHX11DXEw6mKYnEk3wquBjmH6GLC6kXIcpe24grNIWJTldARk+om3kIvgzEosDJFAzcweR6YtZeHgN+ctAEfq9l2/kPo5MU7egAVX/CBH1wRh7onzrsFTWTjBX2ADf7/xxBHCFF4GKU5S2zAgZw7KAEbHol5HjhMvgZVz2prx0EomzLsRHcGr9tyrnpkdvSJbbQSpTxmOkq7UWNdl4Oya9shGX9UnuaI3UY++6yce6TsdIUf46zMs8COQZBecOn7ojHu1YWAiB/fQSzpPFuozaxrY4Y8eCKQECuprDwzRh/qofu3RRTpNL/BDRvQDH4gdu4Yn/MBU0A5D/Q8iY/00cKkzCCYSP/qqjA89U5cPX4JfmPooqw73BSkfuITuSQQMuPoAH30X7MAWv3SPjrtXXWRHJnD3na6RCbkbCPEiEdF3NqSd4Ugb9J9ekp961aV99ZIDmwvCg/rDfzpPP5zzcZ9r7Cp4DB9R10H+Ak5yqDGKMnjQPzqCD2XITn9gIqhAzitLLmQAw9A7+kDv8ARfOuFefCF6os+13bmGJ3ahHkd4s4vA2nmY4UFw4TubI3MJNF3jF/hRAaX61YE/q+b6ow58RVv6oC3ycL+2tOm8c/DCm7roPL3VFzpGf1yrcdS2oEg9xh1YtMk59eJfHXghQ/KEG52jF+pQVv30V9/iOp8OW/3BL5lGW2xE/WQmWGX7yiH16Bd9ZfcwYY/6Q+Yw0HcffLBFeLkOZzy6l/xCP7TLlpVTJ8wFinyxPoSc9cN3NhJJhnP45b/VibduhJ8ZHX89s+MvYeA+Olfbg/uc56dDx7vVhUd4qw9OeNdvmJugoy9BcOvmlwIbeOCDTksg3Kt8+A39ZZNkBB/80RkYuA85pxxeYMz+upFydBAG7vUbkZH2ZnT6QyawhQE5wU1f4WospW/akZhIorRHR+ige6yqxvZw+hu8sWF257ej8urWF3rM/iRW+h98ORpnxG/KIed81w/3kGVMHuBX3XyfRKeXLui7etmavvFB6uLTyYHe440d8uv0UXnJFX3UpkSFjyA/9m/cZtv0BtFH/Qo7gSmbo9PGG/Uj/WGH+HCOLZCBWIpukiu50y/30z11+/BhjuRCDvRfOecQvtUHL1joI9xhR4YI/9E/eoZ//Qm+2Dv7givZmxTSBvt07EX0Xtv6xj+ETdBNfZrZ0UF6o96gaJP+h33DGn8+7jMm8Z901ofsAnP1qMO9MGGXgXO0kcfpicB8HWV/eHP2KPvgVooxGuJUvKHS6p5nNWK1gkMwY2UWgzFbCeuHxsJDP/UOV6bfNsdTrt97h+Oz1zXbW6wgWe0zIxhBSK/yI52fSF5HajuuTwUegpfRHPvlu99yddsGYpMtBhsz6gbTOUlj6UMvfgdRVz91wHDQg10/7fbqd/v8WOrqdk+3c9rqdb7Nx3h+D6KNQdQxmj6022v/Hk1dI5WdyLq7td1ve/2W08ZoyrZ5Gs+97bq6/R5N/aMp226r273dzrmv1/m6zrZvkliJrayaWYkXRyHl7OoQWxkP7GIZJI3E60jX8dJPmTbPYsbxxi7tOsf6eyz8j7Wtke4bDS8jlR3p+ki8uD6IOvppJ8tMPgKPHk+To00etWXWxKyR/eG20JiVMCNiuwXHZ7aln5WS4HssPMS9Yz322+Z4yvV771j6oG5yYNhm6yQY46GJ5LVfvqYCD/3yWpfrl+9+y0XdggZbt81ImygwezinabR9GI7fQdTVTx2DTh71qZ92h+t7fW0sdXW7p9u5QfNa811/79V2XWak74OoY6Q26uvt9tq/67Lj/T6RdXfjrd/2+i2njdGUbfM0nnvbdXX7PZr6R1O23Va3e7udc1+v83Wdbd8kmbKS5cUvnmWz8mhlyeqj2Mrqj9XoQdNIvI50HT/9lGnzPVWSx7Hy3+7PoH6PBsuRyo50vR+eB1FHP+1kmclHYFwJ5FjYtTRvK6gA15YKz4fYdmDpnnNzzVaBpIlDwPYNW29sn5hKTnjiejzv1exZBsmjLSm2Gtlik5QIJAKJQCIwdyJgW6HttlbmvBDFs4CeixNbmZT3nKCtoEmJQCKQCAwCgXFtYR0PA7ayelFNPITM+VkJs+c8k5rxIDvyvZ7tipcXmaFsz2SOXEOWmOoIWF0282xbE5vyXEJSIpAIJAKJwNyNQDu28oya2Mrzahlbzd2yz94lApOJwBxLICezk9nWwwjkfvSHsZiXvqXc5yVpZ18TgUQgEUgEEoFEIBGYOAQygZw4bLPmRCARSAQSgUQgEUgEEoFEIBFIBOYqBB58Z/Fc1aXsTCKQCCQCiUAikAgkAolAIpAIJAKJwEQgkAnkRKCadSYCiUAikAgkAolAIpAIJAKJQCIwFyKQCeRcKNTsUiIw1RDwDGZSIpAIJAKJQCKQCCQCicD0R2BCEkh/QsCbPmvKALJGY/jv/sTJ17/+9eZLX/pSA8ugO++8s7xZM35P5NEbPL0l159YSUoERovA3//+9+byyy9vvvjFLzY33nhj17/zNTf6hHvvvXfo7bejxWwQ5fmOsdKckgdf88c//jF9zVgFl/clAolAIpAIJAKTjMDA/g6khPHUU09tfv7znzcSHcHIs571rOalL31ps8oqq8w1f0ZAPwWJXok9UX8g1d9xOuWUU0oC99a3vrX8Db977rmn+fznP9889alPbbbffvuB4fmnP/2p9OPxj3988+hHP6wOX/7yl8vf6nzjG9/YLLHEEpOslg83J/m45JJLGq8if9WrXlWOD1/Nb1MVAX9/7Fe/+lVz1FFHNSuuuGIzY8aMR7DKfn7xi180xx9/fJlwiskKkybPfOYzm1e84hXNi170oin36nm+7b777mskyezGnyBCzvsD3vrkb9r6+5uTRb///e+bH/3oR80VV1zRvOlNb2qWWWaZ2ewZb3fffXfxJfWfdGH/N910U+Pv807039/lw/wZAW3VvsYkg99veMMbJpyHyZJHtpMIJAKJQCKQCMzNCDycMYyjlwKT7373u2XV7NZbby1BvuBQEHnttdc2Cy644BxNQsbRtUfces011zTnnHNO8453vKMEj48oMIATgj1/v0+iGisKElcBucAaroOiM888s3nCE57QLL/88qXuqPfXv/51WRH485//HKfmyPGGG25ojjzyyPJ3rATkEsmk6YEAnZXYDKev559/fvOpT32qefazn93E3ySVmC2yyCLlj16vuuqqU66zbJJ9+Ky88srN85///MIjn3f77beX8y94wQsmle9f/vKXzS677NJcf/31zROf+MRiy/6AeJCk/IQTTmgWWmih5pWvfGWcLvxK8p2f6ATyxz/+cfOkJz2p+JpnPOMZQzzwaxJIf78uKRFIBBKBRCARSASmPgLjTiDNxJ911lnNNtts02y77bbNQQcd1LzwhS8sPZds7bvvviUZqqGw0hCJkRnp9h+3dU0S5Q/cO1qRc+xWVr2uRZ3uEYzUq4NRh3qdV49yQXE9+FCXc1FXlBPYmuXfeeedm9e85jXNc5/73KG21I3PdtvO+6i75knZ6Jd22jw5V/MowD788MPLOQEiCj6DP8eoy/fol3aQNmo+8HXooYc2gjmrEpLT2DIbqwJWWGqKOt2Los66DL7wob/Kad/3xzzmMXWxvr7DU3/xUePXvrnGM9qqy+MbX3gInpzrxr+6oz7f63rwg+r+lxMPnXO+Lbtoz1Fd6qhlG/dHm/iK68rX9UU/HF3Df5SNevo5tnlq46B+/KhbO74H/73kGPy7Hveq1/29iEye/OQnN0cffXTXFTt80El8qCsILz5jwabue/Qbj4GtczWm7bZM7px99tll0uw973lPSSDDbvjBt73tbUO6rn/qDhyD/25tBV+uRX+Hwy7qcpSESR75JSv2M2fObCKBJAvbRD/96U83a6yxRtkVEljaLXLeeec1iy22WKlOP9ry1Qe8ocCr/Hjov+ija1G2zb/zX/nKV5rnPOc5JYnkcwIzux20Oad9Td2n/J4IJAKJQCKQCCQCvREYdwJp5vsb3/hGs+aaazYf/vCHhwIRTdr6aNtlbPFyTsL505/+tKxMWk2SbNriVq8sWXX63e9+V+5X3oqf5G2FFVZoVlppJdXMRsoqc8cdd5RgbrXVVpttNc0K6YUXXtio1yqHbXERMKlIEKUfeLFacu655zZ33XVXs/TSSzcvfvGLC2+2X3mmyxZdwZGtak95ylPK9txFF120tC0QM7uvDYTnm2++ubnuuutKPXFe4HTZZZeV+gSjz3ve88pKxsILL1zu6/afIPCiiy4qQZYg0G98WuHxXaApCJMEwsg2Mc8Vaefqq68u15daaqmC9QILLFC2DOrDb3/725LgmwTwXaAngEVWVqyk1Fve4KJOWGqDTOAUMtY3fOmL1QYr0HATzNqSWNfVrZ9jPXfllVc2l156admmBwOrldokK0QHzjjjjGaDDTYowfTPfvazhkxhRf+inLL6T5ZXXXVVwY2cI6heb731ykqJ/tPZWh9tt7UCJdmv9ct5K23wpQO2N9KZSEajTTpqCyJ5qgMfZEUGVvER/PEe7dgeTn9GS3YKwAtvVqDJ0Sp0yJHdRWIhsKdDPiZNyJHs2yRxYR/w0n+Y9ZMA6Se765a8sA+TNlbI8BhysO3yN7/5TcElVvskSXi+5ZZbyqSI1csZra2zEiF9Vs4zvmxCOXKxYnr66acXvxN14o3c9F1d+n/BBRc09I3f8B0W7g2if7aQOg8TdqA+OCPyjXutYKpTO7fddlvxjeyW7dEtdY1EfJc+2eL99re/vdluu+0Kf2uttVZpi6wlvLDUxve///0hfTfJhxf6/4Mf/KA0tc466wzJlw/jd/RXf+gbzEJ39YXvffrTn178AD9iZZb8laW/dlLwNfw0/6IM/a19DXuAUUyOYQRffA0508HwNTFWhK+2Ws2vSKLpH1vhhwPvkfDL64lAIpAIJAKJQCIwOgQetVuHRnfL7KUFs1YdBS0zO7PeMbgrFUlNBOcXX3xx88EPfrAEHAIQQfixxx7b2Nq09tprlwHfPQJ9Samgzey04EzS6RlLQVC9rc3Kxfvf//6StHg+UBCkXs9f+qhDUCWR06ZA/uCDDy4z4WbDBaSeWfrCF75QAp9DDjmkJGUCsu985zslyJaMaPcnP/lJ4U2i6hlIdQpeBIDq/cAHPtBsuOGGJXjVf9s/bRH1Mhx1uEe9VhJtyxSsCzT7G1gAAEAASURBVMAEdCeddNJQoiCY/Na3vlX6vdVWWxVMBXIf//jHSzAlUHTfD3/4wxJgCaYlHqeddlrpuwBfv/VTQCw4liyRk/YFt/pt65oAWN1IQO264O6b3/zmUBItOEQCedsNycuqkcBu7733LgmVANA5stpzzz0LpvoguBRAkpNg1TY5OPRL+iYIh9W66647m35FHe973/sKFgJSuEjEDjzwwJKk4QtZnaGj6sOTgNh3ExySEzLUhmSBzh1zzDElKHXft7/97RL0SgRMKAiQ9Q1WdD4IPieeeGLRs2WXXbac3n///cvKC/2hfwJc+iCRCT2WOFnJEljTfzzQEe3gCWYCcbZmdYu89JU9wFWwLmiOVaXgp9fRVkarQRIPfZJwkSPbpc8Cbwk3OeKBnrBdiY32TIjQoZgQkWzaacCGQoeOO+64Uj9b2WSTTcqEQjd+yJa9e9ZX223CE7yUwZdJKX2Hg+f3JFn0CX9bb711Ocd+lD/iiCOKDsAG9rZIfu973yu7JWBMBmyP7bJPfX73u99d+hWyUZdEm+1o32QJO+ML4KZe+k0XfOgAf0KfTAKonyxNGpAhwged4quWW265IjuPAOywww5F5/QLzmd19EF/1TMc4cekjckRn8MOO6wk8fogIaPDnqmGER3hj/HOf5ngIWOY0jVJmwk439mrnQjkz5eaCPJyL3YvOVUXXdx9993LaqzzEkW6TmYwNilHp/k7iaikD6ahJxJavt5v2IZvoNvqZat0lE+no/yL5JqvgSMdpcfGERjwuWxXMrnkkksOYT4cfnktEUgEEoFEIBFIBEaHwLhXIAUJAh6BkGCqFwnWBUyCkve+970lWRK0CJzMjgsAbP8SpKlPQmR1TTIkEIpkUwAj2FROQKNOAYvgy8y4wB0frgtKBLWCDcmHGWzBnfIC+xmdFQUBjqBEwKEvL3/5y8s2L/2Q1EmkvNzB6szqq69egknJhxluCWskKBI0gbW6gvRP0C1IE2Qjs/gCG1u4zLjjTSAmCNQXPEbCHfU4CnZhIACTJOmjlUh1+H7yySeXhPfVr351SQ4FXVZaYac9ZCUCL/B+85vfXIJNibl+SOAlBZIZs/2SRDKL/ljdgZngb9NNNy2rduTkIyERHAu+8Sk4hZEVCFvqXBPICiIFnoN+KQ/MrCJKpCLYlGBIJK3waF8/BMqSlPXXX7/oij7BQ5BPdwSw9FMA7n7JoeRAObrwsY99rOiVIFXgGwlUAbfznwRE38k8SPJH5uQkOKaT73znO8sK2Ote97qSPNBjwTWd2nzzzUsCb7VG4mIVlY7RT0mflV0yturow34E1pKU+rmyaL/b0aoWPbdSyAb0j11IitixZIfukj986D39iK3EAnSTGJIkFJMVbEwCo7+SMvpMVyUQvYjush12LQEwsUHH6YjEE0nu9FG7dF45tmY1WDmJmwRPougcPZBkSPL32Wefsk3bZJFVRziZMGDT+k5P2Ru91i75W+UNcl6iwvYkV2yDXsMF35ItPop9w4wfowPqoB8wYtvkhy+kPi8OYpdkYWLAb77IBJTVNPe674ADDmi+9rWvdfUJwSPdYdvwJ1Oyop/0X336S97slM6wyfAx5K4vymy88cZFXvqo3+xCIm4Cjh9h63DWnokpdqQc/NVBNnClh3wpvVbuLW95y5Cv0V/8sTW+hq7op2Q2fA3sTEhIYjfbbLPie9kWvE3qsSPPoNMt99E//Hlhm/7TU0kv+w3/HFjlMRFIBBKBRCARSATGj0DvjK+PugVcgiEDvaAngpJutwo6fAzyEsUgAUUEkN4eKPGTUDkKuD7ykY+UogIgQZEkRnAtEZDkCJxe//rXl+cvFRT8SmJ8bEE0k7/XXnuVLbbaEphJACQIZuYFbfgWNAkyd9xxx6FZa4GQIFQwLIGMREWQKICO2XKBTMzs1xgInJ0XJMV5M/kCuJoEyPphuxa+3dON4GxFRhn1CWSRgEugJ0nZYostCnYSFx+kvEBP4oF3uCABloBSP32XQAYJzKNf7hUM+lhB9hbYIImXANpqiaAOX5IAgZzgz3Y4pG4rogLxblsVo76xHGfNmjV0m75KsgTnAn66IoHEF1wllFtuuWWRndUT2HkOVDlBrRUziaZAOFbE6IkVEfeqg1zJQT9rcs35Wn4C+KDoN10SLEvQ6IPkAn/0wLW4bhJFAKw9MrO6KYmkh5J+gbI+fuITnygBe78JpIRQsI/IVmIgGVA/PUTaDLu2Cm4yAuHH6i7eEVuiFyZ7yF8gj5THmxUpMulF+o1ggA/YkYv2gzbaaKOSNEocTQixR4miD3KvRFpyQ0bskj+SrO+0004lIWfbVtsl5ZLIkIsJDRiQDV7YaluucT54kjRKNCWxcLR9P4iumKCKPrtOVuQHEzLix/ThXe96V0kwY3WObNkdueKBHlrJ5fPU0Y2s+knWrFKGvprgkXiaHNJPkxCxtZ5tSDSD7C6wk0HCbXU9SNtWLPkAibz+8J2x2mfCis7ABD4zOsk4vxZ6IqHGA8wlkPwAGUio6Tm/EaTPYTfa4Qet1O+6665Dfl1Z/s29JsCMIdp1H1/jbdHGlihn5RQ2YXPlQv6XCCQCiUAiMGkIGD/EWMb2GBMnrfExNDRf03kXQucfqr+PoaoB39KJNefDVyeWeqB3PNVPoxHHxMJSP/f0KjOuBNIA7kMxJJMCsV4kcTCLLBFULjohoBK4mzFWBzLoC2xmdpIadWsDCRTcRxmRWW9BroC7pigvGYjVIltJkfoEcFb+oj38SCAFJwKSIGUEi1GPRERwq7wgMRJI5Yfre9TniH/JgzoF674LRiUwvtfY1PcN911ALCC1ChurQuqBk75qS0Bnpt7qGHyDrJr4re2anMerj+tWagR9kZRGWUGhJMgkgK2v5AZjwajkNEgADGPt+LSD9Cg3lqO+6WPIiZ7hP/qmTnj44Cvk5rrAm46QK8KjyQtbKx0F8xxgrBSXQqP4D3YSKXII7B3Vp00Jk8DcOYmQQJteWFmBZeCkDv1yXlJGLpyzJIldWCntV3eU0yc8SfzITrBNX2CCAi+rRfUWSjYgydKevsEOv1aA6wSWHUnO2B196EXag7kkQ2LLnvULPjVJYvFqCzVMTApJmNTNdvCOD1tIA5uQG30z0eV+9fM37gs/oXzgXLcZ312vidx88E73gtTJpzjCD5m0krBJiPhAspZw48NEAMI/OUo+9QFfMMB3TLCF7ys3VP/BFy8maPTRPeyOPkmW4QJL5/GF51pPnNdWyD2qpg/OsQEJpn5ZLaZ/eCf/IOXIQvIeFJM22g/SDp2pMXPN/fRFv12XbOtPrNjG/foFD33ja/RLXyTw/FCQtsPXaGs42cY9eUwEEoFEIBEYLAL8r51Idv8YQ9pj6WBbm4trm89jX16G2Rl3H3g4fh9Lj8nAR94wXhpXAimgEIwb9OOlCHUCVjMn0BRMGNhrEjTEAB9BjM4JDNQbgZh7QvninIDQ/WbY20RZI8ixRQ9/yL3usbIp6I1z0Z4gK0h7EdBGm3GtfQze2ufbv+Fg9t4LhyS+Zt8F8IJIAVG/9US9DFMQKdmYVa3Eqc/zfVY3rMpKLAWF2qpXyKKeXkf8wMR9sAhZ1eXVJziEOXIP2TkXBHMEx5GwjHv6PXqezOqTQNokAH70X/ttPEOnnPfBl/6F3AXCVqWttL72ta8t2zvhZjXV6g0aDf9ndVZmrdjZKmgFWCIgSbCSpx48WLWj+5/73OdKko8nWxp32223oh9wFegrK5HUh+BDYm4F0gqS6/2QiRf65yhpY8O+w6+tG8PJUVuuszOJdk3wDKzr8+3vyvEj9JMt0LFuZItl2LmZM0kCMkDBBsEmZOOIJ88NW7nSN3xKftUT+lhu7PwX98XvQR3ZCz0iSxMEknEJJH2CO57wBgcrifwnOeLHhI0EXmLeS7Z2YZg8ou/0VD36RseQbcm2Ffe6v1c/TZD5SEpNNNBPPOHPjoPYrRC4RXIY+hNH/IyGlOdryIletEm94WuiT3gblK/Rn6i3/t7mI38nAolAIpAIDI+Asc2jFOIZY12MC8PflVdnR6AzJs2/eGddtDNZ/W+P15i07i/Wm72eh3+JEeZ4AokdgZ9g2FbP2FL2MJsPfxPweT7NKlEM0K4KUgQM6hE0DEf1fcqZjRY8mr1vk7oE68j2LdubBCSRVAiEIpmNIMixTpAEJrapCVh7BbbqZxTuE/xEcO98twDIli4vjfDCD1spBcaSCwZmlSJ4cX8vChyUtSJjlcBzpXHefZ4ps93sox/9aNmChn9b2rRd89irjTivDcom2PVsV72ioAw8JcUCY0ms8v30Ierv96hv7aDfvYJuL9IgY1trrYZKuGzBI9+RAtjALOoW5Fvh2G+//UrwTAfot9WiKBuyjsSl7oO2o5zrttKpw7NbEhc2QF9hppxrXnBiS7QVF8/xkY9t2bYoW7FCtmQq77lJ+kjn9A3ftS7XvHT7TjaeJ1TvHnvsUVY88WmL7hGdl87gp19SF72if5G8xL10fzR6oD/DDS62XrMPdmNG0/N8ViXDr2jXdk3PcNbYkBXsJJr0wZG/CdsPfgNHvyPxhXfwVPfFd5+RdEtd2rfrQsJOJ9XpxTBht/AjPzzaNk1/lVG/tnvJ1nWJnWRUomlLJ1t03j0mFqyiw2qkBNJ97b7QD3I1qeUlVfRV3XjTp/CHzg2SyIGvsdKpzZropoQWRrHTob4+iO98sWeUteNlWnagJCUCiUAikAiMHQHjS3zGXsu8e+d885mItbPJ4kH/MVovxIzjg6AHl4XGUZOZaM+feH7KipeECwksrGrs1pl5t3XLap+BX2JjtjxIMGVLnuds2kFdlIljO1jxvI2B3gqUIAjZtualFNq2miTYNgtvJkRCKTgRdAoiI1gW7EqMvDnQ8zVBvgtarRDFM0gCJ20KcGpynVCsCEaCpl9Wn7QTCYrkAW8CNLwJHK2keu4ngtW63m7f1aVOL/gxq2OLXDzTFuU9owZnSYvkDn+2sAr068RW+8rhAQWOvsOb0QtIbf20wmOLYJRV5ohO0iEoJ19ttANRZXoRnLwVk3ysEnej4EF/teO3JA2fPmRh658JAytNeCBL24JdGwlT9fmEQQlard4IzumMvvvU9dBj18hMG8h2Qc/hkW2UlZjd2HnBCH4lNnGfRAKGysHL1lTl6K6y5Gv1TLtBdNkqI/zZiX7SZUcJrz4gdZzVWfWUYIV+Rx1xlCTjTcLKJtiDiQ02EbxH2eGO8HcvH6DfnlEMYgf8Af2s9S2u18cYWKIP9TXf6a0+4fVDH/pQ2e7KVukNgo2+eA5YWzU2sZ2ZfigDW9vlQ0/pjrrhpn64a488kO2UVoaVi6RJHZIr52pbKDe0/qNXVhDZj2dazfrRHdsuo77QW7IN/+TIR5Bt6GZdNezxzX5N3PGfjla5Hb1obMaMGcWfqSN8a23r6tMPk3hsuyYvI5M82roOC3IO3mDcTu7qe3t9x6sJn8BMH4LI3m+Y8GUmGuFRTw7Cjs8wmQObXvoSdY7mGHVpz0RbPNc7mjqybCKQCCQCicDDCBi7jJc+SeNAoIPjA/M9qlPBYHDsFlOMhbtH7hEaZS0SFNuxBNMGfIGkVTWBqEBeouUlGJ6PsUrkRRG7dZJK2wEFUgIU5a0mRJAjSJAstANg5wXsEfzZrijBs41LYGlFUiAkYbKiI0GzKiWRs9rit+AHX4IZia+yQfjx0govjxBICB7xbCUoVkcFqz6CSm8p1HeJrOdzrA55U6SAS72SOCtOEpLgWVDnTw9EgC9ghZnEIcrgR/8FdkEE7rfzsJUA+JMj2hBIChJhI+mw2mHbqgDXGxxt31W/wBiGvgcJbK1K6o/kUl/IKspEkCdJsCom+bAtUDCOHy9GIod4KYlADI/aqfujPfxFvX6Trz5oj+wEqG2y2qE+7XipBh3x20qddq0OWuGxTcJWTs9C0QF1wyz4x4tP/NZOBK2wxC/CgyCWPkvYtA83z4F5aYjJhFiRFGTSO7yoQ1AteYm6yF6igDd/4iUwkyxbcVdO/fTSSiXZSTacNxEjIYW5baa21UoKTGrQX/aDf5hajfFWXbok8Laiye7YQLfkzXOW6vFyFnXTHQkUXMPmajn6HuR74AVL8qDPVvhtl2bPEmWJggTEqnXUGXXUR/xJMumBBBAe+k8OdNPEBN2ELVuz2uZlPVZ0TT747ZlGK3D8wCc/+ckiN23ARlLIt0g4PJPJpu2WIIPAi87Qb8mUREwCQcf1ix8w+Ol36C6etYlXfVYGH/RDX2sco69eMPPZz3626JKX3ND3INtU+Qxv+iUHvkMd/BE5479NMDqiM3njOlvnn8JHKUtP8WRlWX8kllbT2JHt2fpGZ/gteu68twzz57CkhyamTKTwnfrLZ9I9WJINGdV+yfcgthZ9iHNswaQXHEwq8jXkC1c+LWyTrtN7k0DkgD86gkf+GM4o2nB/raOutX2Nc/1QyFjZ0Ft1133rp54skwgkAolAIpAIDAKBBwyt5b+Hx9hB1DveOsb9dyANrIIhwZmg0SBv9UGiISjwcgwJmGBSsCP4EvhZrTAj76UsXv4i4IlZCgmP5ERwIeiOwVswagZdACYwFKwLeASDkjmrKAIcAZVgRYBoJt11wWWsKEo6Z3QSOXwJ8iVOVi0lQe4XyEoiJQq2muExeBBk6oNk2T1WlgRjgiu8asMHFgIkQb+gXgAXs/gCL6+314Ztpp79gp/+Wz1QvwRGHwWefguWlMe3wE5w41lKQaYEFe76JdmBDzyjL1bHJBQSBzKAWaxY+i7R9BylIBbmEhqJALnGM3J4gJcAV9/1UYCn794mGS8EEQQK0mEmKNcHhH+rzeSiDcmSxMWzqAJGASl5tkkQCj/65AMv9Vstg7d+Cnbh5c8B0BEJlfNWdpSBtwTEs2L+VAEMEZ7wIEiWbEkkoh3JqYkNOkQ/JGWwFcDSZdgIaq28SyT1SSKgzxIoATf9V079kgMTBfQEVvSGfkkW4KjuOE8PJCXeYilBIw8Jp/6wDUmBvtiiKBCnP2QDUzhIdn03cUNuNdFjssGLFTzlYSlxco9+uE6vYC0J8WEniN6RfTwHJ4mCBx4lKnTDpAZeTd6wV1hKsruRFUK80E/bB8mWX3Cfe8hKkiZBZUvaI0/8WcnyHW7wpid0gG/RLyu9+gIbyRVdlzDRbXzil4zYHj2Hjb6yA/Xgh3z8jU5JhD55bpl86By/QgbaklTpJ/1jA3Sa7gW5Hi/R8WcxyCyIjrBL1+mJ+vgy/aNPbKxNEki48Cv0jp62CUbwpccwghXZmFBzZAv6pz9kcNZDK7H0hp+DHV1Wnn/QV5jAEl6uwSX8Ep2AJ2Jv/JK24YK0Fe1on67Bgd8iGwkuH0nGZMaHkhO/wS6d86eC4m+s0kX80D3yhzEKXwMX8lLfcKQPkSRqxwo14kfxF74/jsPVldcSgUQgEUgEHkRA/CaeNubWk3OJzygRmN/fYu9sY33AYx2D2cJqsn28NF9n4Hx4eWG8tT10v+BGtQLSXmSGWJAieBkEaS9mx3vVZ+WEEkegEeUER/5otRXJWZ0X0Qi6BYH1jH6UjaPgTEAhOGkHFtoQSEUwFfe0j4KsCMzb1wb1Gy6wHqkdMsN3t0C0zYs6YUl+YyWORaBvdeWrX/3q0KrCWOtzH/5DJmOpR+BuNVYSIIiMLYaSZuc9OyhRF+wH0RNtjqTH8BXYRp1xv0TJyrWJglgRck1wLDF3n0mYCMLjPjJFEtKa/P08K5ef+cxnykTKcDoIL+2MR451276rDx7Dtdu+Z9C/JfawHk7n6R89ZuNt+8WP+2HfllebV36ADgwSQwk93xk7Mtptjue3Pod/qicXnDdp18v+Y9XZ9fHKVvuw7dVW3b9B+Jq6vm7ftUHe+uWZ0dhNYTdKTMLQkW560q2+PJcIJAKJQCLQlAl6u0jEU+LdpDEi8KgXdJLHzi65f9/eqWD2v5gwlhrFq+KW8dLsyxPjre2h+0ea8VWsHfiOt2mDezsxbNcpIOsWlEUAIYgQACszUh+GCyyHS5xrnoYLcOty4/kOl37a0d+R+hx8qHO8AbMVCM807bzzzmVlIeoez7Ff3Hu1oU8SCyt8Vj6s2giyrUZZ1fMyknbQ228w3Qtb91vBs8pjFcYKNH20CmelfrvttptttSp472Y/7rFCaXUo/iZelO92hNd4MWvXWycl7WuT9bsto27tDjc5pDwHO5yNR539lImy/R5H8mP91tOtHNvtxrPzw+HWy3d2a2Okc6PRu0H4mpH4cV07bWKHSYlAIpAIJAKJQCLwSAQmJIF8ZDNT+4xg0nYsiVa3QGJqcz89ubNd1cqbRMd2wKlAnrWyCmgiwVtxPa8l2KYXtq7iN7bkDopfyaMk2hZsSauPJEwS4e2XVkB6bf9s8yCp9OyYLbxJiUAikAgkAolAIpAIJAKJQI3AoCZHM4HsoCpx2HbbbZsZneetus3O18Dn98Eg4Fknz3JO5GrLaDm1Sui5VUmdZ/E8HymZk8B5xsrLkwZNVmM8q6VNz+xp0+qX5+LwYlWyX1LWs5KDXlXst/0slwgkAolAIpAIJAKJwHRDIHYdib3suBGLec+BBQUfi0vx0bfYuTjd+hm8D4LveT6BpARe7uCTNHkIxCrb5LXYX0v48gIUn8kk22V9xkPj3VY8nrbz3kQgEUgEEoFEIBFIBKYyApEEenzId4mjhNFLDe3eMgkvlvJSPi+2804C7weImNWjHp4f9Eynj/dReEeAumJlL5JLx/g+lTEZK2/zfAJJ6EmJQCKQCCQCiUAikAgkAolAIjB3IhCrjB5Lsgsu3ndi1dH7J2LHlxfx+XgDureySyY9IuSN397kLqF03tvjvS1cImnnV6xWelFcvHjOC+O8Dd0L9yKZVG5O0qDynnk+gZyTQsy2E4FEIBFIBKYmAoMaZKdm75KrRCARSATmHQTihYWSRX+Sy59S811ShySG3sdhxdFfdIi3q1uV9HJNjxlZnfTnuSSX6pNUerzJn8hyHUke1WELrCRS4igB9dvHd585SYMa2zKBnJNSzLYTgUQgEUgEEoFEIBFIBBKBRGCgCHjHhqTQu02sNkoGJY3+5rctq16K6M37sUVVGcmhVUPnJJcSRb+tMroW76iQYErEJJfa8CJOL+RU1rOTksVY8ZRUSiT92TZ/N9zf/nafRNXfe8bDdKRMIKej1JLnRCARSAQSgUQgEZgrERC4DmqVYK4EKDvVFYHUm6b8OTrvNLHKKFGUMCKJnRclWi2UWPpLABJGmEngItmzkiixVF7S59lHZDVRwieJdN3fA/espERRvb6r230SRs9GSiglrba3asOKpXasYmrX3xiXTF599dXNH/7wh9LOdPovE8jpJK3kNRFIBBKBRGAgCGSwNRAYs5IJQEDyKAj1nJWViqREYDgErJJJaEb6+8bD1THdr+m7bag+SyyxRLPiiiuW5E1iJxG0giiJU85Hoig5ZF9WKH33cV4iaAXSKmM81ygxlDz6qEeSqqztqur3ch0fdqse9/pONngw3jh6m3/8zWUJpLacv/LKK5s777yz3DddZJEJ5HSR1FzIJ4NiMBnIzYXCHXCXOHFOdiL+7Av9M7vogXhOf+LIC7se1fmr9Z3PA4JCD9LnH6ufOLy710zGXqBgdjopEZiKCPB3F110UXPUUUeV565yNXIqSmnq8GTs2nTTTZuZM2eWFa6pw9nkcGIF0J/j03+Jo2cVbSuVyKFYTZTo+S6BtBrIzpyzEmllUCzgHvWhSAIlmT4SR4moLapexCP5i9VK9UoW1eN+35UXW0g0I2l1n7a1q06rkRJK584999yyKomv6UCZQE4HKc2lPHoz1QEHHFCMNP/+5lwq5AF1yyyfgWHrrbceUI0PV8OJX3zxxc1GG21UtqMI1gYfsEkU5+8kj09v5pv/6c0D9/+28/uezkfCmm+C7oAwaWRw3mKLLUpwPmmNZkOJwCgQuOKKK5oDDzywOeGEE5p99tmnBKQTO7k1Cuay6JRBIJIeEw233XZbSURWW221KcPfZDBie6jVV383WxJpa6qETaImIWQ34kvn/Ob/JYkSPnGFVUIUyZ/rrvlEWQmmeqxKeo5RPT7ujZVI3yWFYgft4cuWV3GuiW8fiaJ6yE1d7pFYWpV0j+TUxJF7pgNlAjkdpDQX8mjp/jvf+U4xUrNFjJ2xJiUCbQQ4W4Pj9ddfX3Rmk002aRcZ92+6J5FEMTiMu9JHVCBZ7Aww9/+rs+5o0IqtabkK+QioJvhE+poJBjirHxcCAlHPcK2yyirNrFmzSnCbOjsuSOfKmyOBNHZ5GYvVrnmJJIX+Zvfyyy8/9Cc4Yqsqe2FHMJG42XXiaHyXoNn9BjMJneTOiqXrkjoJo5g0tp9KCpVXpy2s7pdgWmFUnxVHCaPyklO7pdzvT3yIdSWHPlYs1WUFU3JJbtryQVYjb7/99lI3vqe6zWcCOS9Z2xTqK+NjQIx6ZmfbwUILLVSMZgqxmKxMEQQ4Zls7Lr300jJjOBFsceo+E07zWYW0hbXTlk9uYZ1wyLs1MCmy7tZwnksE+kBAUGplwgtABKJJicBwCNATic28Rp53fNnLXtYss8wyQ3+HUbwgWZPISSAlhBI7yRq/L2H0ghsvrRGHSuoWXHDBkkRKBJURlyovRpUkqlOybsXSURn1SlYlk5LPKKMtCWGsOnrJjkRVUokv17TvXuS8c9ohR3+LMhJUbUxlygRyKktnLuaNATJwH8brTVVJiUAvBDh5Dppznt7USRolkSV5zARyessyuU8EJg4Bqw+xjU7QmpQI9EIg9KTX9bnxvEWHVVddtaxASrrEkuxEkiZ580xh4OIocWRTEjXJtt8SN8miBFBs4ZzYVKxRbzc1oRP3uKaMc5JMdTv6HcklXqyO+tMekkCriba+uldbzuEDr/FxTZwjKbbaOR3eyppeaW60rGnUpxgkpxHLyeocQCAc9BxoOptMBBKBRCARSAQSgSmCgARuhRVWKM89SrqQ5E3iZuU+VvskZc5HYidxs4po9U8dkkwr/LHyKIGU2FkhlDB6TtFvSaI6om7tuN85iadjJJC+uy4BlUDG34dUp5VIq5ru9UHuj3otpCy++OLlmU68Bc03RZ9yyRXIkFAeE4FEIBFIBBKBRCARSAQSgURgyiKw5JJLlpVHq5BWESVvkkMJn8TPd8lYJH0SNAmb5NEzhlYD3RfbR93vugRSGdetAEo0Y7VQEqkeL+mRINoe67cVRL+ROiSEfkdZ5UyAe+5R3dqUtLquXYsoEl2rptp3r2TTs5P33HN3597Oy3ym6GalTCCnrIkkY4lAIpAIJAKJQCKQCCQCiUAiEAh406y3stsOKoGzWidZlDxa4ZOARVJmBVBSJ4mTwLkm0bPF1J/gkNC5LpmTJEoaJY8+yjqnTvV746rkT5vqk4T6rS33KxNJZOyuc79zElHfJYq2zUogJbzOuQ8vyqjXiyWtsEoeb7rpug7fD741dqotRGYCGRqZx0QgEUgEEoFEIBFIBBKBRCARmHIIWJ1bYoklyp/rkIhJwiRwvkvSfDxvKBGT0LkuObQNVUJoFVCCJnn0whrHSPp01vfYxmrl0G9JoiRPfepC6pU0Sj59XJMwurfmw0qmN7GqK8rj1/ZWvyW0kkg8qUdi6n5bav2d4sc85tHNcxZ6VnP55f/qrJze2Pzjn1PrLxVkAlnUIf9LBBKBRCARSAQSgUQgEUgEEoGphoDkUVK13HLLle2pkjmJltVHyZg3l0rIJI/KSv5sA5WQORdbUp2XcMZKot+R9KnTx/2SREmh70gdfksGfY/nJiWPsfVUOfUpZzVUwipBVNazmhJHK434jRVH39WhHQmk+x2tnFqpfNoC/hzIHc2dd93aSSDjT39pac7THEkg4+/VEKIZAARw2TqACZZAkxKBRCARSAQSgUQgEUgEEoFEYN5FwLOH/jaqJNKKYDyfCBFJ2h133FE+cgd5heTR34+WkFndk+RZnZSwuV8SaOXP78g3JHpyEPdHYhiIK+Mef5YjEkn3SzitUko4ox712/YqgZRwqjMSS/dbEZUwShDd46NO5/AQK5H+zNf99z+t+c8n/6V5QqeOv/+jk9Tef2+Ht+Bqzh4nNYEEPoH++te/LjMFSy+9dONhWGD/8pe/bG699dYiBOdmzJgxZ5HJ1hOBRCARSAQKAjEwGlSTEoFEIBFIBBKByUJAcuZPdXg2UIJlpVFSZhuqpNCzil6O4yOZkyxKMG+44YaSPHp2URIp2ZNvWI302+ql8sY3Y5u6rQyqU0InQVSX+xy1KzGVy8TLeiR9Fr3UFeOj+ySJkkgJJHLNR93a8/ym+ySwEmDXJJl+48P99933t04//tLh5V+dl/c8pXPf05o//uG+Dl/3jwv64HNclXRuntQE8vLLL2/233//5lWvelVRBsxbdr722mubvfbaq9l2222bk046qbnsssuarbbaqsw0jLeDeX8ikAgkAonA2BEweBpsDXoGNYNnUiKQCCQCiUAiMBkISLS8VVWyJpEzHlkllPxJhuQR/nai5NB5Y5YkzN8Y99vqpXuUjcRPPZJE5yR8xjf3SUyVd07yp5wET92S0nhBj/KSPWXqeiSB2sazbauSTuXwqn4fCex1111X2sKX8q5bkVRWMvrXv/6t9OvGG29s/nT335vHP7az+tip8567/fXFeSyBtJRriXn55ZdvtthiiwISxbvmmmuaq666qtlyyy2bmTNnNl7Le+655zZnn312OddLOa1WEiahEgBFcUxKBBKBRCARGB4Bg5gBznMWBmWDpUHQeQOgGV/baGJLDd9qoOR3zcAaVN1jMHZfUiKQCMzbCAjErbiI6Uwy+c1/CMaXWmqpgYGjXrGkXWv+6PrCCy9c4kn+SJxpQUIZAX58/PZBylm5sgMOXXTRRSV4V5dkI2lqImD8kbxZTRTv28pKXnRNgoesUsZ54xL5S85iVS+SQeXj2Un30Q3XfNzjmvtjBdH5qCOSQWWMi34bM/Hh3hgzJYN0yqqocdVv/NI/9xpHfbciKQHWPwnnjM7uS/UusMBTO+3f/VD/7mvuuvN3zb1//W3n9z+xPC7C5yBo0lYgPfdo6+rVV1/dHHLIIUXgG264YcnOvaXIa3kFNIsttlhz/vnnN7fcckvP/hE2sAUvsRx90003lQSUoJISgfEiwBnYbk3HECdkBiz+aG2v+uklx+A+A5tAfCSKyZB4aFp5Oq4ejiUck7qU4XzaxA4MqiZVaufACdqSgfduvNx8883FwWmPszOBg/yu62m3l7+nDgLkZACLAdOxG/GNPgYn22foksHYOXrrPkljDJAxMKpLG3RMMBg2EYOfZzrYilnVpEQgEZg3ERDcn3baac1BBx1UAnHjjUCdvxDYb7fdds0666xT4rzxIMTXHHPMMc3hhx9e3sh5wAEHNDM6QbdA/IILLii72YxffGIkBX7zZwhP6667brP33nsXv7bHHnuUt3pus802zbLLLjse1vLeCUKAbH3IUJ7gGGOSXTHGImMaPfMRI8WYaNxCfiOxXIxzviNjn9iJbkQCaCXSebqjbeOka+pxpN/G0oir6Lkyfrse46w63CMOwztyFJNpT90SVTmSOnx/kNcHk9FyQ+dZyL/9TdkH49EHz439/0HFdpOWQHIuAmEBi5fnCDouvPDCAiCAKYQj4BGF6EU6L5snIPdwKBJTgk5KBMaLAN274oormqOPPrpsi+Bsaidg5nKttdYqz++22zL7+vnPf75s9VtllVWazTbbrOzbb5fz2+BmUmXfffctieLGG2/crL/++kPbJ9QjwWMTdB5fnNrqq6/evOIVr5gt2aP/xx9/fNkWgd8gicFLXvKSxmTNIossEqfLKv/pp59eZmvVixf3+dtDr3zlK8tOgaHC+WVKIkAv+EGTGgat8JkmHWLmlE+MgU8ZA6t7JI/xDAbd4pvpuAFVOR/3OVfrBx2hK+7xcZ0vV15b2nU9KRFIBOYtBPgQ2/LsHjPurbTSSmWB4De/+U1zxhlnFN/AT6y33nrjAsaOtZNPPrnshrAKKTGd0UkgxZDiy7XXXrv4IvHhT3/60+acc84pL1+Z2dnhpoyA/YUvfGHxXxg577zzysoW35U09RCQTJGlccsEtzGKrkm+jFHGJ3J1vZ4oNw4pR96STOOTcmIo9Rk/1e286zFhT0eNqbaUKqOeGAONeRGPQQoveNAGfoyPEkX1K6stq90xDhsv3e88XtSLR7rnt7HUllfl/MafBNjn3/++v5NGTq0cZ9ISSEAQyjLLLNNstNFGnT+OeVPz3e9+tzzkyviBpgxBEIgkczgyEx5ECLYtJCUCg0CAw/C87qGHHloc0qqrrloSPBMV9JSDueSSS5pZs2aVlfO6zV/96ldlZpRj+cEPflBWIT343Y3MOJ144onNkUceWVaAzEq96EUvKgkkHg488MDivFZeeeViO8objA2evr/tbW8rZTmnX/ziF83Xvva14gjXXHPN4nC0yTFKKGIWTlkDqqTVBA7nZtXfdQNzTOrYap40NRHgHw1UBkuy4z8lhvwg+drRQT9M2jlnIHOe3hrc+M6YDTWYGaxiINZjZd1HB31cR3FUH+Kv6bn6Fl100TIw2jliEOTHo3wpPA3/08/ALvo8DbuRLCcCk4IAG+EP0A477FAmWX03BnnHxe67796YtFxjjTVGjO/c14suvfTSsmiw/fbbl3dq2IJqvLKt0fjpE3TssceWnTmbbLJJs+uuu8bp2Y6Cfb6RD0yaeggYoySHdkiZ9DS2WKlzNA66LnYyBhq7rCQav8Q0dC9W9ZyTV9jiasyiq3RWAqeMMdMW6Eju1KsOiaEETxljmna0K8Hz0aa40L34oEuuO0+nJKLGYh/jaSSGvmsDH8ZjdfsYP/HtPu3PP/+Dz3PO36lrvvkkvFNHRpOWQEoeAX/99dcXwdvuBGiBh0BIACy5tIIjMPHHQvul6R6o9NvPLDd5CDBsDmDHHXdsdtppp9Kw3xK4PffcsznssMPKVutTTz21JGHtAHPTTTdtjjvuuJJozuzMfNL/NrGFU045pTwTLInkbOp6OJYNNtig2WWXXYae15DYmt3VvlUkL5tC7mUHW2+9dbPffvu1mxr6zUmaseWk9MsnBn0OUr0catLURMDgY/AzKWEgtE2absVAZlDiTw08PjEDa4Cjv8oZIA10PvSN3igX91i1NoC5xhfHx3Vl1eMTeqJNQdjiiy9ebMEqhETSQK5c0uwIwNAHvnXQCkcE79oPzH53/koEpj4CxpIg44tFg4MPPris8ljpGWmBIO5tH02OXXnllSXglqSeeeaZZQXRqqMEsk2Cf3bFn/mIOZOmFwISP/EY2RmD5A7GMd+R7/woWbvmfCSQxijjkATQWOQa3TMG0kt+mF647hMLWJGcmnw3dtJZ9eAhHiOS1BpD1SGe8sGHtmOMpHtsgd667jd+EX75evz4SDod8eGFQMo+7nGP7ay6/nfzf7/3oqC7mr/c++DkbalgCvw3aQmk2QMBj0DZSo7X626++ebNa17zmsaqjSDZtkF/C9I2PnvlkxKBOY0AZxD0/+zdCYymx3kf+Op7uqd77iGHMzyGp0iREkWtjli3GDmxFrZlGxsbieyNldhYObCNIIs4iLJJFjKQxFYQwA5gOIYN2/JmAzuEJVtaH5Isy5JCHZFEkRRJkeIxPOcg5+pj+u7e51ffPMOPw7k4nOm5qsia9/3qeKrqed+u//uveqrKR911111Xfv3Xf712HNZ6uPfuJjDpTHQQzExzg6hvfOMbdefhlOOqg0EGjZ7+83/+z+tsZXdZmZa8/FAXxsT0x37sx8of/MEf1PUeSSAzvU7neE7HZO2ID/x/+2//bfkn/+SfHCGP8uhUjeq2j/7jafDchxuptE6HGTXwAjgG5oCh54b8eY+8B95XYcDQwAEA9D4BLYDsPXWVVn7gCtSk987JLw1gS5nd7xdZfieQkwFUzYqSuyN2jwO+zb1UA6xl4J8NFVgX6DvonAWAjxnHWLEMaK5p4ELVQPcAiHsfz/om77w+63SdDRbhmFlG35MGan/1V3+17tz/nhiobe7i00BilkFKmOc9EgareA4Oecf0n4lJ4twjdXDMjB6shH/CYZp3U558P8mFffLKZ1YRicRLcobRAK4430tkyYu4Jl6qD9ni4G+WpRz9vDJ42Kgt2qW/N/ALN+EtWXB1aGgwBka2BXFcVXY+91R5Ye9g1K/TZuWca7diBFJDgaU/dkrljaJ7CHboYuIgjLMIliKbaxo41xrId7K7HjoiR9HcfffdlZD90i/90hECmel0AsxmrKNktiN9tzOKasDErDtS6CNSp3K0O1b5RlrTvOHo9Dqe4zkE4k/+5E/q3xszIp3x0U69mzv/NACMrP94/etfX98X9/msPTPvA6ffzNlG4cAJaErLiwOQPIASBgyRP+m9g2n6mu8jmQmG8ngnM70ygSUHpBNYvc+AFvgqC4iSIz+w5ck51vtdhV2E/9Dnxz72sfq3bq2WZ0MPPqqF5wcEqwQDOT6Q6b25poELSQP6FJMEZlG837DO+709TO3vvPPOaoZ4uu1h8WNG573vfW/tu2yGYzmGMpTXBl5OV7PnZz7vkgG1t73tbXUdq28WYWYE7Q+BvOEQiWcwDA5Jk3iWxFE/y8MmuAO3Eu9gHpdp4Zsw77HldjYoRAA54dIpk0uSSqYwJFEZZHAwUXqeDN47nMQSMSXTwIjBFZipbqtWDVfrotHRwYgfK7fd/vo4vOO58vB3vxnEszP7Wgs4h/+sKEvz8H0AHe0otu1+dbRW2u/zWQM2pLEQ//d///fryFT3qKqORMfE/PQTn/hEMQNpK+duExvrEBFIs4nZKR6rvT66dYjdzvpL4d2b4ojXaRrR4nRQ+XGuI9WZGVWzfgSIG6Rp7vzUAMBJkANI7llw3HHHHbX/RB49U8/ToABSpg/1ngAn4cK8I+nNXArzfng3gBQAQ2LId813zdW7BMRcvTu83+QnOCtTXTlEx7047536+ZtQVo4Yq5845jxmwV3V4VJw2umDx4eITUQMptKRZ2LnSHq7+eab68eQj6IvfOELdXbFczmWo1fPPp8RM2byyGmuaeBcasDfuf0DnOntXdYfeY//6T/9p5X4nW7dfGjDLxMPTFY535MmIMzos+aBuc1dHBrQl7FosRmTby0DbDAG1uj7ktz57Z1DDg0g6AelE+aKYMIiVzimL07yp//UB8PAxEr3ZCJ1sCu9dJk3iaA80itbnPLIJF/9fQeSlSRXnDBOXv04MmzwA/H0nShM3bdssZvsWGB7f1kT1xtiud/i4q1xDuSzgSXPHPm+O5dPe0UJ5LlsaCu7aeBMagAgWtCtcwBs1qLpPLodYPPR7wPxL//yL+v6RPE6Hzu/GXGy82oSzu687nVARnCZvCmPGaK1wna0Y8LznqNMdnRkQNRorzI4nR/CyOuYEAc2/D7+mzv/NOA5+0DyjLxbQMUMolFYg2zII3ABUADOM5UOMAIk4WYZgRkgBVjiyAXG3jXp5QXC5HsXgCOQ886RT5Y48siRJ8FWGk4aaf0Wz3NZhr8HZZPhPkE6wdS76ONS+ovd0aW/XTrzAYE8e64+imxcZcdmZu9041ncdddddXmH53Ys5/l1j4r7ALGfgA+Z5poGzhcNeE+9mwgAqxcDWafj9BF2VNVfeM/1j/6O9Ctmp2xG96UvfakRyNNR7nmcB/HzrA2y6zuT3LFu8V75PoJj0omDReml15/qE129K7CIh4/62cQe8cJ4LskimcpKTE2cU4ZvK/hGBozklOG99P3lKp0rnFQPacnIdoiHg+rkO1JbpDHZYP3j4CDLIJv1DJSNmzaWa+e2l6efuirwY08tvxZ6Dv9pBPIcKr8VfeFqIDup7IC6W5JhOhcmNrYaZzpqgxvuCzG7YJMR5quIASKZebrl6IysDzaLiXDK8/nPf752qD/90y/fARYRMDtpo5+Up0PSkTpnVYfHJSHoLqvdn1sNeJ8QCpuHeVYGJLw/gMzz8+GFAAKedAgdYuJZe6bS+cDy0Qa0yANMwgGY/N4RYAl4vXcIqPcigZN8pIVcspRBPtBTR/cJvsrgOfWURlmunDjp5ROW8UmcDMAYcVXHi93Rj/NetdfuzNpsoMDxOp5FflDQg2fpudDd8Zzn6TnmO+Lv3gdKI5DH01gLXykN6E+csWiW0Azkf/kv/6X8y3/5L+umcnYOPx0zU30J0259if7HWsgcPPO3Y3DV4Km+T//S3IWvAc+a87w9f99D8Eufp/+EZ4il90CaxEBp5ZVeH5mDcDANsYNp8utD4VVimnjvj9+wSjrlKcNVfGKZOnjPyZFeWeRxylCflJ/1UT9EV3ph5CtfHXllK0N9vcPCxI8MB7YPxiBuwMGGDRvDemxr1OmRml8559I1Ankutd/KvmA1gJgZAdNxGC1yPdrpXHJ01NlYNtVhpvbJT36ydi5mH0/kdFY6KSCs45H3/e9/fz2n0ajc0U66n/u5n6u73emIOJ1VdmQ+LslU75yhPFpG+31uNAAwPF/ni9ogx7Py/hj59Ky8B4DS8/M8AVASNXHAxzvouSMrnjtwkR4IATzpgViSSCOfOQsoL68e8pGhXF5eZabcjHOVR3refTrgyAHDrGfGkWUNoLqSz4RH/ovZeV70wxLAxnGeix0kHTPQrcP8uPBRLM3xnFFxnqNHfcuJ0h9PTgtvGjgbGtBXcd5RA6fWLtoPwHr/0zkH0mAXK54dsTGXAVlnJB/t9GVmIWFkcxe+BvSLyBtMghl+u8IjGAbXeH2l3/rYfO+S9MEffSp8FAdTuzGRTPkSY2FpYpFwfSsvn3BhPBnyZBg56pVePdXLVfqsu3qqD3xUJ78NqCCLBhKVI5/fZEk7FGshh0dizeVsZ1D4iisur2sj85uA/HPlGoE8V5pv5V7QGmCC9sgjj9TZQB3csZxOgskhM9bPfvazdU2ITsHoKXPEk+00LD+zn5//+Z8vzqLUWSEAxytPZ6PD4nRMRzsdk5ktAIxsNneuNdCZYUKwDBDY4Xf79u1HjuUAUoAHgAEXz8xMpOcvXLx7RA4YcfmOACLvj99JIKX3W17vUYJUmpEirepBnvzK9b5K571KMuk946TxniUoCiM7iaU8yuaUqz5ZL+EGXtQpd7hDlrMdNdNF9A89Mle1VkvbfeD4e+T0EQZ1fBBI556ZXuruZGqgw+aaBs5XDehTPvjBD9bdxs0S2gBH/3OqTr+TFjgf+chH6rFX+qDsh/wtOTPZmkuz+6dDIP0N6buaO1saOL41xbFK1KXBo5tvvikGGy+LgcjhGIhkAeNoi+H6e2xsNGbk1tdwGKrflGbdurV14AJW+j0zMx3WNgcrhiKRwrx/nnl/f4f0uZ+fD8xcDIIYWOWdm5iYjAHW/TG7/Xy9dghbf+Dw3oqTTEvzHYxdAKq8eCsD6+Cc99MsY2f5xvx8p4/W569axXrEwGpPlKFNLHXs/joaWGotpvewE6Z+TFnXr7NfAFPXpajPlfU7YO/eF8JK6MXjco6lx+OHvbLncTw5jUAeTzMtvGkgNHD07Aml+Bi2DhGBdKZifggerbDsXCwCt9j/937v96qNO5nCjjWL2C1DfgTBB+aprh3JMrvl5D2gtcmAejB5s+aqfXymdlboGkBRAgQDMaLATieeI5QJamriY8Z75eMryRcSyXsnhHueRliRLqPviBvQNeqf5qfCETkkz7NG4AAhMqc84eSJ8xtpQRyVKV8SyKyjOiUxJINPgikN+d5vnsxMk3HqpXxOed5tbUEid8TswsVKIOnYYedM2X/zN38zHn/HjMn5eJ7Xb/3Wbx1Z62y2xQe3Z9Nc08CFrgF/52Yd/82/+Tf17OT77ruvDojqE+AQ0meQ9QMf+MAxm8rah/mqjcSQT0s/jnaOfiPrC7E8RJ93qnhJjr9F/Q7zcqawyAOnD9Y36WObO10NwDjkKXeTzuuJ5Q0NmZm7olyx9arApzWBJza/YR5qSUcssxgeK1su3xZYYrfSjkWVfnNkBDZuqH51PLeZwJrpaZvf+JaLXcAX4lkHmVyIAKRtKDBudfTNw8MxUxn/LcW7sBg4OjtrrSQrH8dqhCnpQCz1mDMDaZOcOMNxejbi+ioBhZlwsr9/MMI6azAXFpYi/UzphauBmQORprd3Id4pA7pIoTWZg1XW+Phk4N8LZWdMSszMzMUA8qZoM3zfGO9xtGXds2X/gYMxC6mckSDDoxG3IeTZ5RXZTB2fWKcvxnY/jxdDT+euEcjT0VrLc9FrAKj4APYxzZkd0UFZ6AzwLNq3fvEf/sN/WDuPEymESeJ73vOe8q/+1b+qZhTWuPGn4nzI+yg/Ew65cPaqDTrsHgvYAS+Q5ICnNZY6Q+dLan9zZ1YDPQE6gzFy2VNibcYw0OkN0re2zgxb0+GjChFMooYk+pDJ0XHvn+ckHeImnQ8c76d3VZjn6dkKYx4qXDrPkwd4yIl8nrUw7zuy5x1HUhE96ZJE+sDyLspHNoIoff6dSCceOcywLE9azm/1lk57yPKhp77yy+tj0QfgxebowOy/dWHa6W/aMR36AXEsDDxberk2dnjWZ9BXc00DF4oG/N3ruzh/4+m8x9b7vuMd76gb4SCD3nfp77nnnvKf/tN/qnhzPAKJ2MEsFjvkHMux6EEsf+M3fqMeryWtfkofpf/Leh0rr34IqXWuM1xXd/2UvhdeWlbQ3KvRAEuLjeFjQKznZLO81h/G2sUgSJdddlv0h1sDi9YGFlmDOBDfT33heyoB6+m9IjAslmXEjOT0DMwIM9bFzdG/ri9Th2IpxsJgEL2lwBTY4qiPWO+41B/Ezi6uk/WdiO62DB2YD/yZqQSyLwhjXx+yZ3O4NYHVC/EezMWso9nGAzVMnaYPxYY4S53dXmEo/NJ3J9bNx3tX37ngdrF8sQyvMiDc8UNDA/F+8ZaK2CjvUOysureMHzQ4uza+B64MAnlNlHVZDGwMRPhomZwYLbMxk9rTAx+XQh9Xlr7+50OfKFz8zZ1Ur5EsHVh5Jekz3zGujUAeQyktqGkgOwM7qH784x+vH7Y+bnfELIlzHR2D8Qu/8AvV7LBbW4AHaLmmY+Nuto9MZwrJB/C6nc6mG3TFkSMM0J7MSZPpj5dWJwfEkcM/+IM/qOtIHn/88fphK79NPgAoIP6pn/qp44lp4aehgb6+3jqSunr11jI4sDUAYKBsu3I0QHJTzERvqesgzEQhepzn4cPLO+O5OaYBofMb0fBeiJceAeuY3EzUMB9O0vpwQgg7ZjsdEudjyeY15CGQAI8spEZaM4EInHBkVBnIJDLq/VJWxzSot+bJdw4hcq8e8qonYHXltUec8tVLGgMa6qBsaf3NSGfXYYSVvIvJ0YNNRI7l/sE/+AfHCm5hTQMXjAb8DZslZF1jMOxoZ7CVGb6//3TMuh1ps3379gx62RU2kmfDKWunj+Vg7Lvf/e66kY4BNn2HflB6gzSWB+hzjuWc0WyHV9huAzpOP6SfM+PZ3KvVgEHKwJISa/pihu5Eric+m/oHBgMrt5T1a7eVhblVgT9m+qy1tybQGY42vuHH4hkvBTkLktgfG7XFf/NzcdYiojkZO64OhgXMvDWI1kHG7GCfQVP5Y1C2f7zMTMUa/7nZcmgqsO9g4FaUMbq6s8ncYLA+/TVMGuiPmcqByWiDGcsNkWZL6e91bndvkE6Dr4GhCx2M64t6BDuLmUx1DSIaKeYiPnYuiPBgkqU/2hHYGLOVy5HUrGd/XyxLGdtatvWvD0xcF9+WWyo+mw3dvftgWVq4qowfGC3jk/cH9j8X32mHyuqRy+K74YbA9mj7+EQUGSWdWLUvVXuQ9DMEIwIAAABAAElEQVThGoE8E1psMi4qDQAagGSG4M/+7M/qBgA+pm1yAoiswwA6zO+Odj72AY8PZR/f6eT74R/+4Xqkh9FXadLlx7QyfPin87HO9K07LOO6r91EQrknc//xP/7Hurbyd3/3d6s5HeIAMM2o/v2///frqOvJZLT4U9eA57t167Zy62tvKes33BijjcNlcemZOL/sinLDDdfF2rjLgiR2CBTA8g55Hj6ckC7kzMeZ98GHkTjeeyHcuwTskC75E/jIQdKULx8i6F0RnuRRGfLlDLt7YWT40OPVAQkli+82eRUunk8iKo330Aec+iWhpTFp1KUCc8SJV17OhkqLFNt9OInyqWu6pWwaaBo4Vxrwd8/0mj+WQwD5dPoY5yDzJ3KOuHEEzsncj//4jxe+28Fc/kTu13/9108U3eJelQZ8Ax0MvNoRVCnW6S+++E10LLEIVRxyUUaGYn396IYgeguBDUsxgHB54J81+iVmBA2qzgeW9cag51Jg0nydJXxhbCrSDZYrr1obBCyWXwQ+9gT2BSRWsmcd4vRMbEwYFspTk7Fj+XRv4J6N6pynu1g2xADuli2jZSDI42wksnYSGfQZd/Dg3rKwtDuO0lgKs9qtkWa64hqs6o/32FrHTno7strcJ6yLBmLwlMVR4O3AAFNblkUGVINKB5/sjfofOBCb1PVPRppYmxlytmwZju/KqMPAUsW/srwndmDdW1aFae78/K5gnHuiTgfK1deMRl1Wx4zrXGDlU3UQVl1P2b0itnl8qS9+rR4/TYtpGrikNOCjlgmMUUkft5wPbx/sQM9HLn8sZ40hsOueTZKO6c3v/M7v1D/0o0dnEccvfvGL9cMZCHPkm/30Ie6D/URO3awxMct5KgSSLDtBvu9976vEAHlQHpMddTle205UhxZ3bA0gb47meO973xOE8eYgg2vK44/FOsAAs8suWx+At74+X+YsCB6CJY8rcmbEHuFCEj0fz5dHKj13ab2T3hFAZTYPmUtS5n0STlaaSHqPpRPmqlxEkTwzk8gdj6QqW37yyfJ+KJNTrnty5CdXvHp6h8hLWdL6u8q88ovnss3ieGUyozXjT25zTQNNA00DTQMXqgb080luTs2aqj+wYn2s/0MS9+x5PrBI2zub3gwNrQp88S3m/GPr+EcDn2Zq2oFgZstBjswM9oQ5qjWJ1iZ2NrYxIBtrIg/NxVpIOGsH1yBhS526bQjss8QEntmcZmlpObCsg5/WYa5ZMx0Edl39ToO/+6cOVJzbtGlzDAIznZ0JQmgpyaHA35gBX7IhTsckd3h4ddSzs+Skp8fgcFiW9cLYw7qJdlr3uW7d+orvhw45RstaXKR3tux5Ps6H3jYQ5HhNefDhUp58ojdmILfGrsZ31LIefviRirW0dGoun8eppT5eqkYgj6eZFn5Ja8BH89FE71QUkh/4R6f1AY1UHsv5kD7aNEcYM9lTdUbCjpZxorzax5vVshtmc2deA57JjTfeWO68884w67ojTEc3BiHsC5OZIGwBNoGRAWxMnju7njIdRf6QKM8fscqZxgxzRdSQLA5BE+YqrxlE4CaNWT3vo3iyPG/5pPFbmiyHLGmRxJSX5E9a95meDD7rkgMO8nvHkUh51CPTaAenvCSeGea3cL8rMAdpRkIRzkYgq9raP00DTQNNA5eEBnx33XjjTdXaC+YY5OT37z9QMQiBtBOrdZGusAJuwBGzfzatGQwiCX/hEFzkpwL3YK97WAObclDTICpLLAO1o6OODhmLvDa96RwTct1118f31RUV25A8g+5mF80awkYDtghn4pgZzZ4eR3+I75DIxNWBAXsLdCYgtAVJ9R3QGXx11Brz0s6mdoxcrXHsCZK75/nYNG/15bF3wmjUxVEipVrJ0Ys1nTt37qztPZWXJLH3VNKeKE0jkCfSTotrGmgaaBo4DQ0gU2YerTm1xufyyy+ro6YzM3Fkx/rBMhaL/UvZX4Gxv78zy5YEDVgCGwBIjnDkLwkWsASMnDBkDgAioOKklS/zSgMozSKSnXLIVU4CaeYRzwFDLstQDvBF6twjqGSQC/zkB+bKVw+O7CyHvPQ18vA/0siDiKonb2DDbCg5ymyuaaBpoGmgaeDi1QAcsCzIbtW33XZbHYxECmEAj9Al0UtskwcOIXHC/E48giewClZWfAzSx6qFLDgEl8TBN1hr8NOaXPfCOjuxdix6pCObk5/stO5xD7/UAb5OTnaO1lAfWCacvMRRades6av1Jg9hTqyHn+oGX+WvG+1EeQjq1DSLnrASC7wdXd3RS6ft6tqxKPINoJyTuVNJczIZ4huBPBUttTRNA00DTQOnoAGdPwBDHpkUI4+IEDc9PRMdfezYFiY1I2PrYgE/U9POeYsIGNIEjIAdsAIqCUwABXAZJUWoAJOyAJtweYCIcLIyn7TS8cLJT7PsTKMsPn8nwJIFwBP85FMGn3lcyZTO1W/Ob065CZx+A1Nt4YRLB8jztzgyrEFWnjQ2d8o8NWH7p2mgaaBpoGngotIA8mQDpre85S21/9c4+AYfEof8hmmIIAezxMFOhA7eCEsyCcvkRyQtB2GemngmX+KlmUfkFYYl1popRFot/bD5oVlH2G45EnxyL73ylJFYCOOUy8EwdVJWYph7aVyznmSpi9/yiIez6j07GxZLIa6vdzmO8lgVhDP2FxgcrvHanATaso8HH3ywfgvUwk/wjzqdCdcI5JnQYpPRNNA0cMlrQKcMyIygIo+OYkAehTMbZUIzHluKL85vChCIs6GGRgPU9lVAzBFIIAKQEkQASK5VFAaEgBXgAECASrw4QEIOUPNbXulTpvQJHEAU4AEqYQnSQAtAi5NGOFnCXaVXPzLF+S0dYO+OF56/yZJe+SnXy+K3uqozGcolRzuAdI7Gap/jSM6VS52dq/JbuZe2Btr7d2k//1Np/cXwjiBwdoC34WASRRgGS2AEn5hlpg2JhBUw1yweXKIHaVzJgFGctPIIg0XwRhoyncdtZ3KyEv+UCZ/IJA953BG79MIk6YWTIb20iB8SqA1+79mzp8apP0dW4qpvArgnf+Jo1kc6eIcMdmTM1I2E+vri7MfVq8rGDf3hWf4MVXnkKJvTPmas6qSdJ3Jn6n1pBPJEWm5xTQNNA00Dp6gBYGAn2/e85z2VPJrxSwKmc9+794XY+W0kwGMsQAZR7MzcWVCP6AER4IMUSg9IAAEiBsSAFpkAg5c2Zx7FCwNA0gtP8EqipxnykKmsLA+YyMeLI8s9py4JSMKBLHnyyC8+zYKyfvKJz7LITLmZz5UsMsWps6uPAQBLFh2QYQSYLrIe5DfXNHCpaMDfSnNNAyfSgH70Qn5PYKejzRwDA+MSDxLH/OYQPngDF+BP4iDzU+3nE9dgqoHbXbt2VQIoP8xRFuLFIYRkyAfLlONeGjqFdfaIQCDdS6+O8ohXlnKSHJJvdlIY/CJLOnKlgWUwjpdGPKyThsu08G4q1ms6J3osNvBZv87xWbH28Yrhctlm2N9b26Z98qs7YkuuvMLJP9uuEcizreEm/7ga8MfjD84fY3NNAyfSgPckR+1OlO5cxXmXgZuNjBz/kmarOnHEB5jY2a03zsOy7fdcAMrCYpiXxn+IJHCSFmACGQCAUAnXdqAKKPwGMtIiiIDJb3FJAMkAVhwglEf9ODJ5v6UHxDwnjI6l54STn/VQjjzSiQNaWZbf6iC/+kp7tMu8WbZyyKYfQJzPOOugzeTy7p1Z6ow3dTpbTj2OdsLUubmmgZXSgL8Lf1vMt//8z/+8/j0d691cqfq0cs5PDehnvRf/+T//53LttdeWd77znednRU9SK+0wi8iyBmbp84XBVHgAO3ht9bcBe9wnVkmrj+aTrCFScBeeCvOdycsLH80YKg8plB+m8u6VxblKc/3119dZSnVRT+Wqozq4kk8up87yKEO4PN04jIyaXVQXaeGeq7IS62ygKB7srFsbO5/3bIi9E0bK5ZcNRH3NZs5XIq2+9MHB32uuuabW/6mnnqrLXWrEWfynEcizqNwm+vga8IduPdcf//Ef1z9Gpgv50Xv8XC3mUtSADtaZlY888kj56Z/+6fNSBd5nIJCL8P1OsEMGmaQsxXqK1aMBhAE+M7NhdjobM2zLiwEUI7VNAM86DX8HAAeAJDAmeCJbCXSAA5ABDunE8T48E4ClQTQTEBNw5EvQkycBTroEUTJ4LkHVbzKzbfIBYuHqm3UmRxmudMF3h8kPcNWNPM+YHG3JfGRz0qoTPQJf9V4Jp87NNQ2cCw3ccsstcYj47vKZz3ymHueU/cm5qEsr8/zVQPZR+l3k0bKJC9XBz/T6fJjDwQ3h8CDxEN64R/4M1sIMTjgcRRrhIAdTrHHkYA5S6dvTQK/ZRbqjx27c9JsnVzm5u7h7dXFVFlxSV2nzqjy/4ZVyuvOL0w5lwTKymM9mHmkRTrOY69dviLwlBk8HY9f2wNZVloKwHIoNdaKN4+MTVQ789M3Aw1NevVbCNQK5ElpuZbxMAz4W3/CGN5S77rqr3HvvvfWPeqU+DF9WmRZwXmtA5/rEE0/UTvz1r3/9eVlX4KHjz1kzQAUMdOrIYwWLoc4sYRz5FGQyOn2b6ES65eBoAAcQ6PjJASzAixxABHASQKXjkmgBMiQQcCiPDGWLJ7fbSZsuQUdeaTlgrS10LszVby7r50o+n7OGwtRPHn/HylE2edK58tII1y7lZnvpjSwu82WdmOYwG+rM4nZmZbT1bLts99kup8lvGjhaAz6K3/72t9e/Z31Bc00DJ9IA4uh9ScuXE6U9X+P0+/p8hM4VNgjjhfHwAybx7jMctkgHh44M2AYG+duBK67wBubBJ/hhgAbukGHgF+4qs9uRCQe68dTvrFde1UUZZCnfQDDTWUQv24TEKk9a+Cgdr1yzmok3iZMdjGVOG5vNxTfDdNR9//5DUfahsMTZX8uAo/Ihqsq0V4DZTe1cCdcI5EpouZXxMg0gkB/4wAfqR6WP6+aaBk6kgZ+OmUcgeccdd5wo2TmJAyJGQhEdVw5IAQAAgvgAjHo+VQBfQGIFFcAyNTkRoBUmpXHgMSCRH4DICwzJAQ7Az98MMFSeeEAEPKQBGDnyKB7AJrlTjrScevDyJREUn/Kk8xsY8tJpA68O2a4EbNeUJ617IJyyMx35nDhefclTT21WV2nFkaEevHuga3MC7cp2GFCQtrmmgYtVA8zhP/jBD16szWvtahp4iQZgC0Kkn2cCiozBCdiRGAYvcoAVFkjLwR7pYEIOToqTPzHIb/mRRZgClxNHnNdsllE87CKPgz+8ML7bScOTLx+Z0iJwTz/9dMVtbUjLmszbLQsGwlZtz3pqB7nCFhbsRD4bg9DLQSDHo877o7GI5/SRGVNpc1aVPG1Wj5VwjUCuhJZbGcfVwA/+4A8eN65FNA2c7xoASkZ9mZyZUbcGARniAJkRRp25NQ2bNzO1WVMOTcbsYRxQ3N+HJJmVjMOPgzzK1w2CZh2RR4AAnBA6LoHMVfnyADDOb2mNdgJh4EU2AObUhZc3ATDlJEi7kslLC+AAImD0WzgnfwIg4AaWfMZJm069pKcTRNfVB4A6qm8SQ2kAp/YoR93F+5h2JRNYGnRSn+aaBpoGmgaaBi5sDejXYV1uMgNHYCbc4BLnuklWYkoOOsKLTE9ehsM3cfLaZIZ5qHvYDNcc0aHcG264oZYJexIbYRFZ8CrlZH2k4WAXbJbO2sOHHnqozgYyTTXwCbeUL94GQFlH9SPDFcbD/4xTnhlF1kvz84dC7lDUYSrK2Rt1PxR16a11yjbKD7fhJZ+yawXP4j+NQJ5F5TbRTQNNAxe3BoCE9buO7rg2NjEwm6YDBwRAAGggcxs32iBgLEYNh8rB8cUyFeTQ6OLwsJm+jolNggxSBmiQJCQSCUS0yPIbOACsBDTAAXCNrAIhI6nSq0eSwXwK8vBkpUtwdeXTaQOAA5DaAqQAKp8AiwwKz3zkkp95pXPPC1d/eQC4upKd8a6csshLMAT6SZDpUju1EeiT2VzTQNNA00DTwIWrAX0/80ubpDHnRLpgGDyFBXCEc4UxiRXCuu8zLYyCDdInBsIaWEqmq3j3jz76aCWRX/rSl+rxUTfffHNdl6gc+aXjuzEziVuWh+whjza9EmbTHeQxrXgSJzNfyoRxsFsdu9sG+5HbF154PtrnLEqWTXaInQ3c7K31lw9WwtTETLiqTN8QdHC23YoSSCMMFJzKs67Fy8Jp8AMPPFAfnJEHCj1V1/0CnWqelq5poGmgaeB0NaDPMZJp1tH249lnGdXUv4lHrDhAODo6VknQxESQx8mZcqj3QIDkclkdJGpoqLNZThJHM2wpB0ggTUBB/ykcMBwNhoBQOdLxQEw66YGXex7QZP2ADTnSK8c1SWfqRTuEIWxATR0AlroCPDI5sqQVLp5cXj144CsugQ6AI5AAVRiCyGedEWjplZvtyY8BBBJZl/fZZ589Upesc7s2DTQNNA00DVxYGugmaIlRwtxzcATGuHY7YVzGwSuYkQOV7nkYB3cSe1K2dDxz1scee+wIdsFUeJQ4B2+yDsqCeWQ4e/H++++vV3zmpptuqmdZskySRn5WPEhm4jfMIwMG5gAp+Ul2tVk+O7SPjASZHhuNshfLps2jwZkGoz0jR7BSPtipfcpXn9RZt57Oxv2ps7RXWboHYWrXrmIUrNE+eDTYR8Cf/Mmf1A+yr3zlK8UIwDve8Y764XK8YslLJeWH2vHStvCmgaaBpoEzqQFAAiCYrDr03ogpp19DtPRPPAABEgkkc3NxRqPznQZm4mwn5M3oY2eGD2lisoKkARj5kjySSx4g1GfaPQ65Aj7SARBlSJMENE09c41F9pdACsjKk0TPPU8+mUBWGjKzDQBVGnUDuMBTXI6eKluc8uTlu/UiHmgCOrKUnTOtCab0Kj/yqAzy6IDTz5Np97ysN70ATOmaaxpoGmgaaBq48DSALMEcnABmwAi4AjPgTBK2JFh+p5eXc4Uf8sPmxCH4CDcSM6QlV3oyknjBIrN+JrlgLIzKtLAUHpEBj1KeMhBPG0GqG2yCn3CUyzopQzs45UjLa2/GJT4rA84KZ6W0Zk0Qw7BgWr16IGY1e0JPdnidrmQUTsonj+8B7aaD1Ekt8Cz+s2IEEvh7OBi+7YYp0wvjAXz7298uiOOHP/zhOgvp3igzs7DjOdPdHgTluW+uaaBpoGlgpTSAOCXQASEddjeoAKjs2DsdurOinAnpsGJHd4TZaZDHwcGBAInlIEcW0x+qwJYgmfL1lQgSIEQcr7hiawy2dbb+llY5Zv2AnFFOZkC586v+Vd0ADEDS5wIndZKPXL57dtBmQExwzKrKp3zlJFGVj1z5yNJuaRYXAeSL53Tp83m6QvTynkxh8ijXqKz4ycmpKK9z9Ify1Fu7pJNeeeoP2Hk6ZsaqzeKaaxpoGmgaaBq48DQAQ5Ep5IzXv8MAuJAuCSPsyvtuogSXOLhpaQkZcCPTS4sEdpNH9zCIQ/rIxSeQSPiXRBBGwVe/eZinbnZa3bFjR+UiTFbhE5nwF3aRp3xt4bNd0sBBbRaf2Ji4B1e1A84qZyh2cN+wYV20yyZ3s4F5ExXzEhulTWsd+wVwBqK192y6FSOQHh7lWS/05je/uT4AysLcne/2Mz/zM3UbYqZJn/zkJ8uXv/zl4xJIsnwkedAeBCV7UM01DTQNNA2cbQ3oa3TwRilXr+6YkuiHgMfIyHAFBWRrOc7n6OvrECIEZ37eOY12GO0ro2GSMrSqs1h/fl5cEKCe5Tp6uRjnRXJDQy8eLmz9JGK3ZcsVIX8sZMe23gEOL+wNsrh3X2wos7uCGUKFiHYIFeLY2eUUoFqHyZQWSTMTChSnpjpbju/bt//wzOlcDN5trmAHyLWD0+ciu/LqaoHa9HTHlHV21szgi2awiCpQlKZDYDsHLatXEkfAR4d0CfiWlhZDN/QwX3FCXeGDMO0EsAA3Abg38tG/EVd11I4Oga3Vbf80DTQNNA00DVwgGkhMhR08ggZzOHH6fngBV9KJhyOueS9eXlfpkSgersAjPgdLYREvDsYgiPAK1iCQMBMf4dQBQRXHI2wsjUyIIYsmvJBB8sggTx3USxgsS5InrfYIk0YbeL+Vk/eIan9/Xy1vdi5mGiNN1KT6Vas6y0q0R70SI5WHY8FGFp/Cz6ZbMQLpoWrMX/zFX9RZRuweafTwjCD7EKA4HwiU4GEez1HybbfdVtO5f/zxx+thu8dL38KbBpoGmgZOroEOYJ0sXXQ5QVqGYte2a8uNN15fSSQyNjExFaRsPPow6yzGAgxK/D4UANDZNCawovT2xCzgarNojvnojH729ARp29gTI6/ro4+0s6lNZDojsvrF0dEwZV27LkhnZyOd55/fUx5/4vHyxOM74iyrXdFXxmY7Qc4WFpwJZT3iQFkdeTZt2lhnLLdtY1azLuqzFGC3PyxBnquL8ycnJyrYTUwgksxGZ2p+6cbH95eF+ZmAK5sHdMyIFhetn3TMhvUZzFUR1Y5ZKrBbs2ZtAN76AEegiEBa+7EUbQLKDk5mmmuH1Y4Z0JYtl1eyOja2qfb98IHs9Rs2Rn3XVECdjZlZ+RBE5r7r1tpUoS8e0WK5LHa1veKKy6tXFxYpgacVF072DM2UNtc00DTQNNA0cO414JsfGcIB3BsAdY8c+Y1c5cyfb/4kZ3mPpElLhjCOjDyLUVzKcw/vUqa0SSLxFOHymtiCKWQjnYioWUUTVwZX03qme4dw5UunDkkIkUVOXuRTewwGa4979eGkky89AowPLS0NxqZ7sa9BHCANf22qI87Aq7qpuzK1CTn97ne/W9utrLPtVoxAavC73/3uur5Ro//wD/+wfPWrX63K8zudF8PvfAky/OirdOk8qOaaBpoGmgZOXwPRn/Ssiez6FffHkRS8I1ZOBFCtC2I0HJ32qujMgzgNWB8YJGzOVuSL5YXnYx1kgMlCdPgb1g/HOsl1Nc1TT/UGAAGwAKW5junqxohfu3akjE+UsnvPZEgfisXym8pll28JwrQ2RlRjpq53IEBhoq63eOihB8qOHU/WgTezgExjl5YcNhwjnTHjOBqgMzxyeQzKXRUjkVuiLx0J4rhQJqKAF/bGgcMHYn3hVJiIFoTPGYzOjZoqSwux4U4A5dTk6pjV7IlZzcXS1x/AvhjtnV0sO3dPRx3mgtD1lYnJ2E324GCQTIStlKkBu84aEY4zMUejPw8CvBRtX14KMhz9s42CRlevL4urglAGoQ7OGzOwsa36GiY/iLFR5Fj3En3/4sJwAGKMNocmlpYCZEvkX2XmdChGYWP32RiIDXgOkrxY1q2fCL8vCLFR2L1RV6asnbUmx3mCUWHnzo4eN7pFNA00DTQNNA2snAaQKHuhWM9uJg9xS+KFICF/ZiZzpi/jEDAuyRo5GZZE0cBkcgq8QdokbmTjGuTZFA8xFI9IyoOEZTzCZ6AYKZU/6yLeb9Y/0kunrjhPklZ1VB9ET1namGUo52jeQ6b4ShIDg6dn1tR6dDbi2R14frDWk9WTdMp3Ncu5I0xqHU2CzJ5ttyLMS+MohDJ4zloeyvSwKNNDMato5lFaDP1UHfnNNQ00DTQNnL4GEEeL5q2vCNOT6IOO5QIqIj4GuIJ8TUz0BuhNBPHrqbNjQ4M689j9bXG6jE8dDLDpANeGdWvL9KE4wzBkT0TamRlmLs6CjJ1SF4bKePSBCOKhQ2Hy2hNHfoQJKfOZ1SNRn+X+IHSx1mL37vLww98tj8To4tMBtNYLLgVALEbX11s6u50uhmlsX/9wGRm+PIDw6pi53BoEbiQIa9RnojN6Oj8fu6JGGSOj68pyMLG5ANCgsWVkaTraFesVg9z1BJnbuXOyPPjQs0E2Y2OCALipIJ9PP/dseX4PU1dnQ/YHqMU6kJAXlrfRjiB+wdvMUs7Nr6ogGfwxuFwA4fBA6AVgxm6yAayADdAuzPcFkYxR1chXPwh6Yr3lcuhoHCAHsB8eTOzviw0JYuZ2JgZqZ2bR94hfAtCbYyOia4Kg74n7g5EnPh5KmOwc+9G9+DjhRchrrmmgaaBpoGng3GsA0Xruuecq8UkzVkQMsUpSiCwJMyuXpC25RfeEEll+wxRpETk8w2/pxcMfxLJjeTNb5VkKQT5SmTN8ZiLlkRduCU9TVJvlJPdQHtJrVlL9yXJVz24iR444V06+JJDZJuHu1ZmMvv7YRCjCzGDOze0P3rQ3Bnf3SRYY39kTAHnULvm0Sz1Wwq0IgdQoZNEIA5ZOYWyMX/Oa14QJ2I31pfnrv/7ryqBNv0r/pje9aSXa38poGmgaaBoIDQSpWNbpTsddkMn4eSwX82SlP8je2nVDMdM3EMRpMma/OqOKa2MgjPnq2Ji1DNZrxBqHHiAwFJ1/mNAsT5aYTIwd1eyAOh/EJ4jYIQCDkHXOdrzyyk2xznFrBQ7gND09EwTwudho7Fvl/u/cV3bt3BUzl7YA7xC36CqPgFD/oNm//hh8W1O2XrEu6hjHYsThw1NT+wMUZ6JMO7h1Rkblm5txhtR0OTQds4JDsSYx/MChOFYjTFkPjoeZ7OOxGcD+nUF0+8vBGNh7PtZbzkfZgGppiXkrXXVGOc08zsUaz8k4oqQ3FvmPhLmp9Z8A17lVq4ZihrTfjnIxOzq8poLqoUOTVSezMbtp1rU/1lfGstEyF/pYihu/+wdiHWjsUrsULHM6zFmZ8ADfVaHIsbG+MF9dV67YsrY8EmUsLh0Kwhpe407orAtZ+U136C136HXvY8WIdzpraXwsGWT18XCqDl421zTQNNA0cKFqANblhmhIHYcEJXlDupAkeMLpJ7Pfk4ZH/FyTlCFfzEsROP0p3uEeEZNGH2xG0W+YQr5wBAwZRD71xcJ4ZSJsJreUrSzy3ItXRhJOv/Xv6u0+SWISyAwnU705+TMtmekXYn+EyUkb3cX3w5BjvDqkVtmIMJnSao86uV8pd+oo9Spr5OWwu6odV70EFP2DP/iD5Y1vfGN9UH/6p39aPvWpT1WAFX6iHVhfZVVa9qaBpoGmgaM0YB3C3vDjxyWPmYE55jXX3FhuvGEsRiMHw2pif4DQC9HJ74nOe01dv7dmDROUGBGMdY5ja4JUBXgNBLEaG1sIAjUTYDBRtm4bif7OsRW9Yco5GL83lG1Xxs5vAWQ9Ma1nLeTc3J7y3M5vB5n7H2FW+kwAxGyQULOlHWetZG/MZA4EyXJe1OVBpq6+OmRtBVox8zh+MIDsYFkbZHcw5A4NxUY4g7p98nuj3gCzL9IthklsbESzEGvPo4zFmNo8cCB2up7srC2ZPtTZdRUptLC/L9rV3x+zoEthpnoYtHqWkeHYLXV6X8ykjgTYOrB5bbTZaDBTodhlNcxx1q21CVGsiZ8ZiLLtshpkNEB2OWYfEffgxmUg2iXNYJit9vVbTzkX5qlTUT+bDcSW7INjFaD7I27blQNhrjsXetsXBDfsgI9D/lNn9bocz3kFHR35WPmbv/mbqi8fCj5u3vrWt1bgf/TRR6t5Mly0m9/rXve6+mFwvCqSl96HRHNNA00DTQMXsgYQR4QKuULU3OvbzPjZC4DJKFIoHZKE8PH6QaQviaA0yBgCJ086fas0yFw68hFV/bFykDGEzoSXAT2/hUunHHL5rKt7dSGXI1t+MhG6zgBqh/yKk16dySUPUVUW+cLVWdvdJymEG/DZOv/FResaOzOiZEivjGwTmco9mZPuTLgVI5DXXntt2b59e1WuBqaCNMIs5O/+7u9WpVOKh9lc00DTQNPA+aYBnfrq1WvioOAbqhk+83s7hgIB3oYv42HumWBRwSeAcFUAx+LCYJie2PQrNotZfj4AwQinnVxXxzq+9TGbti1IUGfXU2sCbTizf//BsNZ4OkZn90X/2NkkB4CoB1dxIIibjWWAiY14zGqZ/QSqNvCxhrKSMbuohnfuZCWekacv0oW9bqQNkhv3y3E/O2uDnA4A6qsBonWW+mX9c18QVudTAT3xqtIBJP12ENOo98LiZK2bsvjZ2c7Or0ju6iCW/RX8YjY0gFJZyueZoQJEu8cC5f37D9Ty7WjbwYU0CTLiSgNhTLx2fd2d1s5zgP1UAFTOlXQ+hhxj9cd//Md1cBRJ9HzozaYM/+E//Ie6hMMz/MQnPlF+8Rd/sf72wXEsJ68Pi/wAcs134ljpW1jTQNNA08D5rAF9pP4b+cpNZpBJ+IgU6it3x1IO/R4ssMxDny+fPhQZY1YKP5JUJVFCHslmAaKv7GDaizuziodtytMHu+7Zs6fsiPWECJ3y9K+uypM+10vCfUvv1FMa+NPB3pmKYfpw9ei+uidHna37JIPTThuMWo85PNzZ4d1GfFNTZioR56moQ8fklgx1lUe9tCkx4WTPOfVysnQniz82Op0s12nGU64HezyHVDbXNNA00DRwvmpApw04dPDIC6KHnNmB1PpHxKdDJDtABUxsiCMt09jFxSBf8wFEMXM2PT0fZIxZSgckkVFAIA9wURbwApSIpmMwABMnXTr9KsLK5AagDg93RkwRsJkwU02wWliQx/mUHTKmnIUoxxbhzqFEWJWbZTOT7ZBEpjqdzQnEcYhmZD983zl7S9naAKQRV3mANsKJOMoLaBMDgF8FwZjRHRpcVRZWdwipo0K0T1uriW20A6DbSVZ6cdqrbmQCfh8c2w8PUPrIyHp2anju/03Avummm6rlzWti+QanjZ/97GfrEVbvete76kcIsym7lTuL0zM9lnOMld3HPWPP0QeN96C5poGmgaaBC1EDCJBZP/iZXAE5cg9TePihr4MzcFgeGIPsIZ7wIdc96nP1jfACVoiXPp3fwuWRlpWk/lh+8uVVH3GJO3DFb2nwFf2vPMijPH4jhbnGEjapv/jEbPl5GMaTzSF/2ihOviwTDs8Gnk7Fko+enun4Juisj4Sj6iCd8ugAJtDPyVzW5WTpTha/ogTyZJVp8U0DTQNNA+ezBnTaSB0g01mnJQWwARR+Aza/7ZimY7d+z8xcbgoDNObDRNSRGsxfzSwiDcCJfHIS+BCjN7zhjgjrmNg89dRTFaDEAxpAAEAAFDMfJE4dAB8PMJVHbgJSggcZQBRoAW3AA7AznXoIE0eO+snDuScHCVQe8DRyyiwTEAN1gAhczSIik9LSCRnkqa98wFIdyeGU23GAt0OMALb02sHlB4H6y2vkWdsRzRfzd6ScD/+qk3raGv63f/u36/p/yzdsHPe9732vfN/3fV/Vn/pfddVV5Qtf+MJLPnaObgNd0Bc9eEb06n1ormmgaaBp4ELUgL7cPikw7uqrr659fWIFYgZT9Hf6PXiQfT0s4vSx0iWZhMPCsu/N/jJlyuM+cRSewULlwCUeFvotnXj4k5inLmTDHmEpSzvgvvT65voNEPnhHKe+wtRHG2B/OvWHjfJxyhsdXR2bzy2U2Dev5uvpWYy8qyueqDt5MJreLIWAuyvlGoFcKU23cpoGmgYueA0ACZ08kxmdN+KmkwcOQCTXL+jUAYlwQML11XV9NpUJ89TYqXRm5kDIcgajHVA75jFIGEACLMALiF0b5v/ABtAgrXasBhjqoT5In1lKZBOJUg8AlKO15CRJUw95+KwjOeqqntoDnJEdaYCREdXMl4ClvuqEyEivDq581okMZahLd3lA2m8y1ItXR7+7nTB6IMc12+C3e3UBxNJpMy/ufHTq5Rm9973vrfqw4yC9e3dcs/2pEx8RJ3KeE8/5kEl9nChPi2saaBpoGjhfNQAT9Iv33XdfxZFbbrml9nEwRBx8QgrhIKyDNTBA3wkzET19qSs8g0k8p39MU1R4xGefqa/tELXRikHyw05yleFqIJSDMfpd9YE9+mt9u3rAyu6+WBm+E+QlP+srjTLlEa495GSYe2m0Wx3XxS7uGzdaAmPpy0SU3Tm3WXr14Bzbcf/999dvA+WulGsEcqU03cppGmgauOA1AJjM9ui8de4IHmDID3pXnT8QAjZJDCoABjD09w+W4Ugz1rsxTFI6a/6s/XvssceqXMcbIRods9X1RwBQOcwfETbmi4DJDKP6KAewyqMu6gVsxSGaQCrrAbQAE++eV1+ghRAqxxUoAkjglwDu4WmHtORpa56dpTzlileeq3qRpwz3mde1MzO5/0hZZCuTbI48zm/h2Z4cufVhoJ7ZLmmVQ/b56LTDs/vQhz5Uq3f33XeXz33uc/VjiR59+ORHgzb6qEgdnKw98ml7c00DTQNNAxeyBpCwBx988Ehfbn+UxFcEEFHUN8ITuIQs6fNhgX6T1x8acBSv39U3whB59Kmw2ZWXV/qMT0yBT+rit/JcpSMD9uiv5VeGMtUjvwnUV7niuq13uvFJHvHSkqm+cFM9hBu09Z1hw7qRkc53Rk/P5dEW5x0/HwR3sparLdI5veKBBx6obVvJ598I5Epqu5XVNNA0cFFoADEy24e4uUcCEChgAGgAjt/AANgsLVnTGDuUxvrH/jir0ZEf6zc4G3dNgMF4BStgY3buoYceqvIQjjTLTHABWkkSlQu4gA5Q81v5QBGxUj7QUh9OXg64qZ+00kkjDIgBS2XJA8QSRJUhHRnkKg+Ya7ereE6c/NJK4145PLAjT/3EA+mcyVVe6lAZ6sNJx3N0iRT7Ta66K1deOiabTPLPN6fO6q+udOG5eV68gQHHWtlMwUeSzXbswqqNzTUNNA00DVxKGoCDMFBfyCKHh0vwgdeHGjzV18MD/ar+VT79KhzTr8I4cYklcAVeuCaWkAVH9MkIoD43y0qsgWkwVxlkCZcP3shDHhnK5JLEyqcO6cWRAZ/UE2ZJo77kuidLOFy0L8Hk5ER8A4zF0VobAmfXhU764kzpocDWzprQHbHJD10hj5aOaNdKukYgV1LbraymgaaBC14DQEinn0TAjCCTF+GABvDxthDP9YWTk0GI4miOQ1MjZebQmjK8qgTgABi7sI69ZBQTAAEphAKxUA4yRiaQUnaCGeBJT7EABLgCIWCoPpx7QJZgifTxwsgCiPJqA3mALIFUvPoATHVJYkuW/MJcyZBWXbXBlby8CkuSpw3K9xGQO+sh47yPBPkAbbfPMG1TR3I5dTMKa+2L6/lIILX785//fPlCrG308YCc33rrreX9739//fD5yEc+Uo+5okvP+V//6399pH21ke2fpoGmgaaBS0QD+vgdQY4MptmxGo4ZgNRvwgxk0W8O8UqcgCuJI8KT7MEqPtMhfvphOKfPhSfwSTwZcFYYJ1xacTCIHNfEKffkpWw46l46deDEJ7mDk2S6kiu/tgjLMqWdmmIS60iRvWXP7kMx4GypSG+cPx0b/MSRV5kW7lk7StZKu0YgV1rjrbymgaaBC1oDOm7AkAQsyZdOH8ABP6AAnMwgijcDubgYo45TQSIPzQbZsf6RqQ1TzI4JCyBLsgYkkVJksBvolAGMOGn5BI4EMiRKGHnqKRwpAXjCeIDst3hgp47kAtwETLKlswZTHKIn75VXXllNV+XJ9iKogJcs+gGeWR5yiUhrCy+fcsnXnhxJpjce8Qbg2TZ10gZlcPRihBaBJCvNeXNEuCY6z/6hGxvl2FmVLrUHEXdF1P/ZP/tn9SOC7jwr701zTQNNA00Dl6IG9OvWQ1rXZ2mGPjMJIPIoHh7AGPgink98FA+L/IaFrvpWPuPcwzNXGAOH4E6WI4+04vXTrspI8gefhEknr7T6brgvjpOPbARRGvUVl2RRHdVPPPwiKzHb1Y7ky8thYeOIq32sV5ajjqxuOjucW/to8xw6yTJX8n1pBHIltd3KahpoGrigNaDD19HbzIbpoV1HE1iACzAAFAiT3zlDBwhWrRoMgBgqs9O2756K+JnDI6ozFQwBIvABYO6BG8DxGwFJ0gWogEX6JH3KkxYJE4dgySMvIuiefHXym/wEOzLFAzzOb20Rj+Bs3769ttNvI7wIDrBLcJRXnXkyAKk6SK9edAYk6Up+JFHbACkvHd2mF5bgK4zcjNM+JJl85RulBqLC1Ol8dNpivWjn+JeX19Ban+aaBpoGmgaaBjoagKN2FWXCCh9gBnyDIbADLsEZfat+H8bADYOaBiphEqwTL3+3yzBXjkxpYUxiN9wiQ1ni4Y1yhMMd+AiX5CPffTppOflhqXzq5l466YX5rQ4pO3/DZm0bDlOlvj7HiVgWYp2l9sG/8XLvvfdW01UzkOp9LlwjkOdC663MpoGmgQtWAwDq4YcfLm9961vL9iBWiBowARCAxj2AQWgAhZnKVauGAogc+DtcRkat5XAMhtFJ6zg6C/DlA5AJKIAG2SJfGMBRtjLSA540f0mwSxmADfiRIU5eaclL4JM/wc4DUY58CXrAEjgJA2ocWfJLa8QW4KobOcrOtAma2kWO9EkeESnpfSRknLx0CPyFS58ylceTJUwe6exIizwy9T2fCWRVXPunaaBpoGmgaeCUNcC6xBo/A56wxjXvYRsHq2ATXDITBzssK4G9yCdcSQIIO4TDliRx8E1+OE0WDMqBUfkMWCbZk18YLBJGDqds5QjjM0w68vObwFW9DeqSm8Qv8yuXfHKlWxPtLT3zUT/fFQZ14d6e2DTn4fLlL3+5HgFFzrlyjUCeK823cpsGmgYuSA3o9AEVcNN5AwPAgqAxrwRACUrCO+AxE2knI12sj1jsLxvW98dahnUBWKsD8Dq7nQIv6XnghojxCJ84I6w5OgqseE4dcnYPMcuRWiCUwCQe0bLQ3shmgiD5XAKkemccksaTL582S5+gR75R4AQ8ZXGu6k8PnHtASk/q3CHUnY1kpAXe2qyNWSYgJZ9XZoJzFRj/SI9Amgm2/gN5lKe5poGmgaaBpoGLQwOw84knnqjkkdWLAUge7sAseAufhMEVeAEbkEiYJRwRkzbJmXiegzniEtdgI4IKq5BQ+xBYupEO1omzjMMVFiZukUkWB+/gmvrAPw62aY/f6grTpEuX6fzOuJ44+kv7JibsXD4ebXomLG4eKl/72jfq7KPvgXPpGoE8l9pvZTcNNA1ckBrQqSMvN998c13LBlgQJus23AM7R3IACKAiPRJ34IADgQ+W/sH50tPLNLWzmynQAkYJbvIAG+AkjEzxwpKYAT1xiCtyqSwglMAlHS+OZ+oprbpkGuvwEsS6iaEwspWPnKk77zdZABr5A6LqChzlT9LnHmBLqzz1IFM7Abp6apMPAV68cnwQyIs089pIjrooUz71oGezjq6IJL001zTQNNA00DRw8WhAv48kORsSLvCvf/3rjww6Jq7CCTOUMAO+wAyYAptgBuwwAOo3meLd5wBmYidMgmFwLa1raBOhI1s6ZSlDuiSEwqVJHHTPK4NPuWRpQ8c8dbjWRRsSI6UjG06SPz4+Ufbu2132vNBT9j6/M/ZOuCdmHe+Jcx+frVhO3rl0jUCeS+23spsGmgYuOA0AIOBkDYL7HbFbnBFJgAMYgEmOlgIRaYTPzU0H+ERzd8V6v4V9QZYOBhly1lRnrSPQSgdkAAiylAQK2CnDNUczAZh0THXMPjpgWXlGT62NkF884udeHId4CdMOM5YIJaCVlnNNoMx2IXoJoEBRHaTh/SYvZSgHGPLu+QR36aVzFaYsRBO5JUebEnATiNVJnDRGhc2mmgGWT7nNNQ00DTQNNA1cnBpIEqm/N9D4pje9qWIhDIIV3TgBW4QlFkljYBIeuodF7sUbQIWnSTAzXH5pkbk0g5Weg81JGF39JlN6ctzDNHjFJ2E060ge2XzieJLdlOtbIeu0f/++8sLze8MvlUceebjseOKesLh5OAhnZ1fYc/20G4E810+gld800DRwwWkAMDBt+cpXvlJn9gDa93//9x/ZVAch4xLEAM3IiBnD3rIwH6YrQST7emM3ucXOzF0Cm/QAKEFIPoTK7ySn0iCDHABEHI1YkmE2Tr2Y7yBbwKnbAVpO/QAWWTxA45STTjiSp1zkFtnzGwi6Zh73nLyAT3iSUyOrQFW4/OQkWaVD9fNxAOB9HGiD9rqKc5+mSe61zUwqs1UkGQHWjuaaBpoGmgaaBi5ODcAWM4j2HoAVBkDtaH3VVVdVSx+YklgJVzi4BMNgj0FQMmAKPMr0TFzlk044jEnSRwZcFZ+YBs/gFNzNwU/pYJD8rsojDwZmXaSFo8pVD/gGi6WXJgdB5c36JBZq967nJsqTTzxerW4mJgwEK/Xcu0Ygz/0zaDVoGmgauAA1oNMHZkAJEL35zW+uoAIgEBseIPD9sQvr0CCzltiMZsQOa7GxTvx2ztP8gtnGzu6rRi7lB0DuXf1WVoKY38BHmWYcARNQQ6yYdXYTK2k56eV3ReLsHnv11VdX8il/lit9N4AKB8RATTsR15ST+YBiEkryOTLIUv+Md00QlYYsHoEkWxlpuqO98gN0wKxMQKqNti43UgzME3jJa65poGmgaaBp4OLVACzYERY/BkhhgyM+cnATziamwBOkDQmDIzn7Jz/Mhh0wRRq4hMhJy0mTeAv/Ep+EiTMwq6xMQw6f+d0ndqsHDBQGG5McKkeYuIxPIuma9/LbKO7JJzvrMScC56M5541rBPK8eRStIk0DTQMXogYAhxHRb37zm0dmIIUBAa5DwgZjrZ8zprYEWdoUoBcjlTENuWrY7F2HLCboyCc/8EgwQUaBI/MdZBFhYyYLxKzFtNGAOsgHMHkuQSqJHbA1anvbbbfVuvoNDIEYECUPMLt2119+5ZoBZDoKRM18AmYe8QPGZMknvZlKv9XFtaOHoXpPvnBlaou06qBssrtHZqUF+kxXkUezj+rSXNNA00DTQNPApaeBtK4xIAl7kEAO/vFJCOEGl0s/YAkcgovuebgDf+RJogeH4BNZBmThTWJpYhN8dp/4luQQ9vHKSZnKyDByyHXlc4BVWhhvoFT5yjRg+sADD5QdT86Ug7EecjnSnE+uEcjz6Wm0ujQNNA1ccBoAQmb+Pve5z1XTlgSOBBrxjvG44oo4B3Dza8vaNbcEyQpyVyzIN0PZMZEBMoDFKGeadSKMAMxVWAKZ2UdhyBrQyVFTZSNyScYAUQIVALOO0XrNbdu21fCsI6UnyLnPPO5TpvxAM+uGyCGP6oJMWitiRjRnI5FG5asjkOUSlIVJZyZUmN8J6OQp328fCggrn5vmNPJYVdn+aRpoGmgauGQ1AD+QthychGV+W9NvwBV+wEjYCLeQSDjjXhyi5jfcS/IH67jEI2nIkAfRywFXZSbOKRc2wkLYS4b8ZjlhG3IKd12VC79gtjzIr6u6KScxXT74es893yr3339PeWHfZUFGz7+1/o1AXrJ/fq3hTQNNA2dKA4iSDp+5ydEOGAQfKk/seCpAZl/ZvOGFct11Q2Xrtthprd8ajc6W34AoZxmNnCJqfJqkIpdkARxAZgaSCQ/ClbORwBCRA2JmC9UJmPmdQAasAKZw8lw55SNuZABI9xygzJlG+QAgcNRmxI5sa0Kuu+66CpC56UDNHP8kKQaeZEpPPlnaog5kieeESyOMoxMmq8CVrOaaBpoGmgaaBi5tDcBGpqxJFuEGnIJvsAzpg1XieQQQwRPfPUgrLYyDP7x0cAnuCIe1MAom8e5hUZrPys/BNvmkUR78UpYBW6QWzsoHM9VbWgOusFPdzabCObOOrIl2hKnuk08y132+LC6sL8vg+AyZr2rDmXCNQJ4JLTYZTQNNA5e8BgBEkrFjKWN+fl+A0mDZt7+37Nrj7ComLAhhh0wBIgQJeQReSapcUy6Q4oEncAQ+N9xwQx3JfPTRR+ssHZDKkVKgJB3ZCXA5Ykum8PSATxoEMstxFU4eMMxRVwAEbBNY3ZOnruoOEMWRTR6wlAawCiOHUxcu6yBP1kG4j4Ak0fRCP+Kla65poGmgaaBp4NLUgOUM999/f91Ex+AojIFBfGIOfIRJXA5MJr7lensYCVM4cXAKZsIacl3Jliaxs7ss2AjPYJjBXvnFw0vEUTnqBPs4JFYY6xpYnRjpN69djzzySF2yceiQXdiDoMZ3Apuljifl1bkzhZ+NQL6659ByNw00DTQNnJIGcJ75OeYzUwFQO2Mk8lDkQ+w6C+lTCNIEcE7kkrAZrQSAQMk6SDOO8gM+o5tmJoFbpgdWSdCAZTdxc88lQUuQQSDJYIYD+Iygqh+QNFqrDGdeGpHllM+RT4YrkCWHy9FZcQA5yxPnXl3VBRFFGu0sC1iNGis36yV9c00DTQNNA00Dl54G4JCjtOASLEIWE9vgCmwShshxcAOZRA4NSsoDu1jwcIgibHKVFuGEQ7DUb7jmN6yFuTARtuUAK3lmFuGWcPmkg2dpRis/DBUH2+AaqyWy1cvMIwJpuYY6clF0bVeHPHaIbo04D/45JwSSEinUA+oegfah4OH42PAiNNc00DTQNHBRaSDIooHE5cWFMruIaJ3+TBpgsoEOANJfJkgBO2AGDIUjdoiX9ADUNdMkIfNbv8yJ54TJn2kBNcBVXs4ykn3NNdfUtYz6bjL47L9Thn4eqAp3VTdenTnx6ixe+UAcHvhIMEqboE5ec00DTQNNA00Dl7YG4IzzgO++++6Kb5ZQmPFDGGERHMElEEm4ArOQMsQN9sCcxBpcxD2s6bbYgTfwylU8sginEEPmspkfjiKrykpZ8ghXT/jlXn5p1AuRVH+bwxn4Vb/HHnus/rb2kux0L5qvnhn8S3xO+ad7XXECSYk+ekzRvu51r6tbyXuYwiiPUm+66aY6on26jWr5mgaaBpoGzl8NBIMMQDsTixoQwjTR6W4v8MkNAIAV0BNm7SKAlAeQATdAxyeouE+XIOg30EQYkUiyyHUPsIGxtEkeyapNPBwmHBAbteXECzMCS5b86qOeMEK49SDIqnvpxal3d/2qsPZP00DTQNNA08AlpwHcwdINV3j32te+thI4BBLewJYcWDU7KA0cMRgKS4QxO4U9ZgERSA4JhDPkyp8YC7+S/CUOpoUMHISPZhfJJQ/emuEUxiU+qgNMg3uI445Y72jPAmQSyZWu2/Uc/hko3R182vfKPhNuxQmkj4GPfexj5etf/3r5d//u31UCec8995Rf+ZVfKR/60IfKXXfdVUe0f/Inf7JuBnEmGtlkNA00DTQNXGoaAExMYYx6OroDICKPAA+QAkJAwgMsXlpO2NFEDcgCSJ5soAigyRRn8O+lViUdQokQmmjt6e2UBTwBvFFZ3ogwR17OYiKPRmeZ9ABU5TTyWNXU/mkaaBpoGmgaOKwBJM/yjcQImOKMY7ORSCQCCO9gFsyztAPOwC0zfzAyiSX8goGwBm6laanfMA5ptOM4UkgGjFQuEooAChMnLdySX32Qy5yZTLIr3kTagw8+WK9p0qq+RxPIshyWS2eIPJ7JF2dFCSSFfetb3yp33HFHPdvEg/Kh4JyTH/3RHy3vfOc768NBLr/4xS+Wn/iJnzhmWz20r3zlK/Xhe7AekIfTXNNA00DTQNNARwP6SQCGMAIwBM/vBDtgiMxJxwMtoJkk0m/gyIs3WgocAbMRVk4cQNQPA2v9sLQLc2HusxjmPouxxjH+W1iOHVgPE0jx+mz5YALgVZYZTl6cEVlWKeLUWbxweZtrGmgaaBpoGmgaSA3ALUQQVlj28JrXvKbcfPPNdWdw2GdwMq1lkDxkDVGDafDMNbmEK1wjM70BVbOPwyPD9SoeHpELE2EZmWYc4axwJFIamJiWN+QhrvDt4YcfrmdHu8JT9ZFfmpe4YGnLvzBeysH50vtXccblk12xlwexvHNtKW9bH7vzxODv7tnSc9fOsnx/7Fb+ogVsV4Yze7uiBNKI8l/91V+VD3/4w+Uzn/lM/eCgONO2P/RDP1Qf8K233hpnn9xTFXyiptoalw0yZaeME6VvcU0DTQNNA5eaBgCd0VXgtHHDxkrAAKyF+gDVaCwSmQRNf3r0rGSS9a+wHQAAQABJREFUNukQvNxMJ3eQA5IAlHWJsO898L2y7vLYfe61cb7lljiHcra3bB6/ugwvWovSWxaXOmdXIrL6fgQXaVU2Qum3EWWDi+qt/KzDSj6/1MNKltnKahpoGmgaaBp45RqAdQYcYZHZRkQQebMkzvpExA6RM9DJI3jIJAyUHi7ufG5nOTQ1XRbmg32FlSdcGl5ll9XBjoVNf+cYK/nNZvIIKgJa9wjYuKFiZMXU5dhtPM5/7u/rr+XBSeTRxndf+9rXyje+8Y26bA9mqisMfhl5HItK/KPLSvmZraV8bX9ZvjdMbBHICO7dFpvgffjKUv5ubF7XHwEvxG6zf2dzWb5ltJTf2FHK52zSd3bdihFID9ZULYUjfx4ApflA8MA9WAqkeE7c8RxgN0WdzsgDU6fmmgaaBpoGmgZe1EAlhrNB8KZjlnBhrvQP9JfpQ9OVuAFVAAkA9cfuXZcC+KrJafSzff1x9lV4s4j+H6xAOlpHWA3cATxlAODxg+MBjs+Ub/7Pb5Zt7728jL17dVm6Pna6e2Z1uWXnYFk3uy6IYCnTM4cqSbS+EVkE/Pp0RBeBZFYEzI0mN9c00DTQNNA00DRwKhqARWYCYQoO4WpW7/rrr69EMc8zzlnCxDz4hYc8u7SzzM8Epi2Exc1gfxkaiDX7gU29iz1ldoYlznjp7euthHRufq6MT4yXsLMp85PzZSCsbeZjlnDfwX1lII7mGhtbE9g6VuYDdyenD5W9+16oA6OsML/0pS9Vs1WYp84vc1Fsb2Dn0vuDHP7cNaVcP1LKdydLTxBFNjhRpbL0oxH3fwQPmogZy997uvQ+HaavP7KllPduKuWZ2EDvkR1l+aljyH5ZYacfsGIE8jvf+U41SXrjG99Yz27x8eEDwog2Umn02seLh5gfNafaLMy+uaaBpoGmgaaBozUQgBNgaJTTKOnG9Zsq+Xt+z/O1P0YAEUn9sEE8RHEuCKfwvt6+OoJrUC+65vDxu3ewrBrqjLwKB9AGAfXfiN9Djz1Y9q47UJ65NcxUX3N5KesGyzsfmy2Dzzxe1h5cU0d2xyc7RBGwm1lEHvX7ZMAF/lyRx3Mx03n0E2u/mwaaBpoGmgZOTwP6cFjyve99rw5Q7ghz0WuvvbZs3769EslbbrmlLpWDh5z0c7Nz5emnngne9USZWhskcSg2bRscKCODYYa6sFgOTU+V+f0xozg1Wrasu7zuOv7o9x6N5SB7gwDGOZGrYofxukTDxj3zZf2GdeU1t7ymXL/p5rL/+f1l11O7yvceebTcd9+91cJS3eDf8VzvVX1l6X/fFgQxyOPcYRIYOB6ju50sQSDLG8N0Ncos/z0mzz76TBwIFu6+iVL+a8xAviGOLnltkM6nJjvpz9K/K0Ygr7jiivqh8u1vfzuUeF/92HAIqI8XZ4h997vfLbfffnud0kUkHY7dXNNA00DTQNPAK9QAcLEXjnX3AY7zi3Nlz/N7wjTnUNk/vr+MrotBu+GRALDOtua56B95NFO5MBvrF8PUdPXI6jK2OjbGCWK5aniobNu6rYyu6eRl+oNAAsFqivrcrvL0zqfKxKapsvSL15byt2MU9KrhUvbOl70HD5SexxfL4K7BshhgPBf1MerKvChnHs1EMu9BSJFX4Rczmcu2aWe6Y4VlXLs2DTQNNA00DZy6BmAJy0cYZcmGtY7MWc36OXoqTVhdd+/aXT5916fLg29/sOz4u0HMrgnyhazha66r+8vmz+wrW/7bQHn92K0171/+xV+Wb371W2VxexxJ+L9dXuaHAG6cg7y4VDYtrS43D19Tbh+9oyw8OVumPz9ednx1R3nk6UfKnt17TmhhSUbPtatKuTmI4KNTYYr6fCk/e02cddWFFdgi4vididL39f2lrppU/HjczUfkQBw9MtipD3lny60YgbzxxhvrQ0uQ/NznPlfe/OY3lx/5kR+p9sC//Mu/XA8FtSbm+7//+8v73ve+s9XmJrdpoGmgaeDi1UDgXd91sabwfw0Tl9eNleWvxNqJ/xYHHF8RR3j8yFgZviVGUu+NLcP/dGfMQu6IjQECrCLPzNx0ObgcM5U3h9nqxsHSvxDGOQfDjLVvoKxeP1ZuuebWcsOGG8vGvjh8ee26Mrp6tI70siQxozmydqRc9Z6t5TtvjC3LgdiumUDTOAsrSOzsbBDTMAFaWojwwDVWJgCeuZE+34hxmvLAiMSJi/EhmW3VZm10dhnH3MpmcuJsAGF9anNNA00DTQNNA69OAwYmmajm+nqbs23btq3uoWJHcUs4DGA+uyfOYrwnzERjDeRyzCiWmcCqa2IQ9Mdj/eFIX3l+96Ey/dD+MrR+oEzOTtXlHG97+zvK/Fv2lc//RBC34cgTWWLHuLIr+vZd/QfLl5/8mzL2wMGy+Suxyc79U2X/bOdM45O1aPm+OB/5/36kI29D0LQPxlrHvhcJJLzu/euo6/+IAdu5+BHOWOTyT0ZdtwWe/397Snns+DOcJyv/VONXjECaLs4pY5X79//+3x9Zx2jjnI985CNHRp2NDuSuRafakJauaaBpoGmgaaCjgcVpCBP3RjEBysGY2bOO4u9sKlPzy2UqyGP/rpiRXNpfevtjtHIxiF5fbFt+bSDg2wOs3jBWypoX4aFvcLE8uOG75ft6ny1vW769XBWjsIB377691fx1VQDt6g0jpTcI48bf+V5Ze8Vo6f2+NeXAtjgfEu7V6nTMfJbiUCsmqkijjXJckclLxX3qU58qn/70p+uZZf/iX/yLagL8a7/2a1WfiPVnP/vZ8o//8T+ux1ixxjmZg6vdM5knS9/imwaaBpoGLhUN5ICkpRYG6iyRMICHY5h9dIU/e8McdWLPeBn4dux0GqRsaUusQfzlm0rZEKaun9xV/czOxbJzLs5qnDxYLXBWj4yV/m8tltufiEHaUOhcyLGkbjZ2IV+cjTOWD82XiQPT5elnI24qMPYUt0Zd2hs4/EJ42HkHZhjS3Xe5peCYBc5zsWZy+cdiJ9afDLNXm+l8/oVSHj77mPriF0KnGiv275ve9KZalodr/c3b3va2FSu7FdQ00DTQNHBRa2BPgM+3Y+vv18daiDvDnPQfXdUBwjBtKX/9Qun5WpiLTscxHQAt96qJqJ79fWXzw7MxyzhfNl++qqxfM1LGwnyVuelY72jZ3Le5XDl/ZVk9urqaBNlEAPgyO0VEDzxzoMx/Jw5C/l8Gy8D1sRnBFUFuYp1Gf2wqYE3l9Ox0mZqZqqTRiLDRYXkvFWe2lsnvjliX4+OF++Y3v1lHx2+77ba6PtXHjWOsWOcwFT6WQ7xtHIeI07+Po0Yij6WpFtY00DTQNNDRAKzh9cHd/aWBurB7KfNB/Ix29q6Ls5HvDCuQ7wtS9tU4p/j/CQb49ZmyON9b9u3fV/Yf2F/z9/UOlNErh8rIhlVlqTcIYk/Ij+tCmJsuxYzk8urAv8WFMMSJDW4qxex6Eqtjc547hspyDPD2xqwlS9me8dhQ7ptxlMf+DjHsiaUoy3ZY7Vrq0CWh3vbETq3LP7SulJ/fHnsOBNmNHVj7/9pGQkenPPO/zxmBzKZ0P8QMa9emgaaBpoGmgVehAaTw3hii/KsYibw1ZhPfFWAYhLH8ZZi2fHp3Wd4Z90c7A557AzzvjjMdH18o/Zv6y+D6NWVobF1ZMzpWbnztzWXb5VtLzDWW2ZH5cvmWy8tCgKNBwDqTOD5RxvdOlLndAZgTAZhzq8vU4mDZGPELAdqzh+aqqdCBif11PSbyI++l4ny4mH1kovr2t7+9rs3R/rvvvrsOoL7rXe+qBBImfuITnyg/8AM/cFwCiXgjkLkRERLZsPRSeZNaO5sGmgZejQZOOGgZY3ZLPxp4+fPXlgCwUn5zR+n9m8kSe+UEBVyqg3XdZY+PDpbyY9dXM9cjs4RmC21wY+Obrx8Ia5/nytKho7BuTZDM98cRHe/cUBano5zA3+Udh8rAkzuCQB4eVCUnyeNR2dWhEl11/dmrS7klcP4Pnys9v/dcWdixMoOy55xAUkJzTQNNA00DTQNnWAMHAnHuiV3ZvrSvlBtjXeJkDEkGmBlJPaaL0c6l2Dh11zt6y67ts+WRwefi7MbnYj1+b2ymM1iu2fJguXnzDeUNh24v10++pmzauCl2Zu2tgGqXV+v3ZmKGkcmqjXuWgtTMxprHiZm5svj8wTL13HQZ338w1lrOVOJ4KREeRNHRJF//+tfLnXfeWXcDNPOI+DGpYkbFfJVO3DPrPRG5tnYy10+Scddddx1ZQ3rMZ9sCmwaaBpoGmgZOqoHeH1hTlj50VWdN4689UXr/IixqJo7B3g5L6j20VJYejt1ObaKD8GVS93jcU4GJh9cpHs7SuQh7PNYpjgYNm42ExnR3x6BqmqW+JPGxfyz9VBBQx3xcFjasn9hZyv/1WFl+IStw7DxnMrQRyDOpzSaraaBpoGngfNFADIyWzdHFXxdrH+3ItjoYop1Rr4j7R48xQgnwRiLOxgGxdnI+Ng6Yly/WgCzGxgHLQ/NleGFnuW726jBJ7YtlF2G6E0cwJfmxCQ5f4SvOkuxxE793T8+XoRd2lfl9QSTrluQdgDsRQTpfVHim6mGG9uMf/3jdHI5Zqt/MTu0IiDjSm1Hx1AmdnirBRiAz35mqb5PTNNA00DRwqWmg5z3DZcmOp9sDM/97ELKP7y5LBmJP4JZjrWHfx545dorI6kzJGE99udsX1j7/b5jS/tHeI3HLAcuLMycuryaG1X839jf4qStLCWwuv/90Kb/6VKx/PIW8R0p79TeNQL56HTYJTQNNA00D550G+rbGTqxvC/OW28K0JUxj7CRX/las6Xgi7h8NU9ajXYBXz1NBYv7rc7GBTsyGreor2zdtKG9+3evKG+64Lc7O2lKGe9aU1TPrqimP7GYgHeeBBLl3FEhlkLEpz7JFHcxy5sN8NWYhl4MgXaqOjqz1t528Hf+++tWv1nWPro64Mjtp8wVk0P21115b9Xqp6qu1u2mgaaBpYCU10BNsaPkfhSno+2LPAO7dGzsDroFlxa6s946Xnk/sKsv3vPTc+eUw7Fk8wQxlR9gx/gWPNsI5MmX58jR1AzrHd9jhNfC7roeMZL1rw/z1/7yulDfHkSOxWU95T9R1a6ypZzZrt9b7xstg7MQ6981jMdeXl3O6IY1Anq7mWr6mgaaBpoHzVAO9W3vL4vsDVD6wpYNP/y1IoZnFt8Zi+x+JsO/F+VKfD9+90D4AbTl2iivfCYDsmS/LwTcnNwzHQcgLZWo81kPevLlcvm1rGRkdKcvDsQFPkB1rIM2o2RCmezfQpaXYby5wrIJZgFpd6hiiL1U3NjZWj6dybAldOQvZjoB2IL/qqqvq7OSzzz5bZ3SZtNpAJzfZuVR11trdNNA00DSwUhqoZO2xwMQ/j8FVpBExQ8jg2GG3nOsRM+BsX6MK5fnA40/FXgY7YqO02KMAjAqu5q+O63DciGOzOHUFtnGt+FsDz94/jUCePd02yU0DTQNNA+dGA9uia3eEh5FTu67+UYycbo91EtZp3BTrId8WM5Ffi5nIg8dgdYIOo9TBfRPlge8+WCYmx8vOPbvK7W94Q9l25baYbeypJpd5vpbNXKyBnJ6ZjmUfi7G5SxBM50A+G3a0+wMA4+iQS9nZxdZZyOn+3t/7e3U2cvv27dX81MY6dgZEKoW9IfR8qiasKbNdmwaaBpoGmgZOUwNhINMLJz8XZM0sXgygHiGMfu+bLz1PxsDqaYo/rWx44TNRsd86VHon4niR56IeXGzI0/vbT8VAblSSU1dkV+Xi0rM/ljU8/dKZ0pruDP/TCOQZVmgT1zTQNHDhacAashPuzHammlQXOZj24w+PGp4p2V1ylizEv2e8M0r55TijyplQgKgntiO/PY722BPbtB7Goq5sL7udm5+N3T6fqf6JJ58oO57aUa688spqskpfdlJlkrl79+66rs/RHM66WgR0X4y1HU8FSd0ZZVlHcvaa+7J6nyzAmsNz5bxrd8ZGOukQxZ/92Z/Nn+3aNNA00DRwUWhAX6evZa1y3ruAhKWHEpuPXdsVJY+qoMDx5dLz1eG4jcHfGhCXsExd+p/HN089WT3PFP41AhnPormmgaaBS1sDPuKt4TvrrifKUM5SdL3LyjtLRObh+dL7RBA4RcSi/FpKmKf2fCF2i/tKmOkEl1w+elvxkzTexi/IovV8qavc+MX1JaBkZvOLU6XnfwSBZA4Ug6EnA7WTFH9Go7P+Z1ToKQprM4unqKiWrGmgaeCC1kDiKsxo7vQ1sGwG1JqSpcMzjqcvquY8U/jXnuqrfBAte9NA08CFrQHg9pa3vKXce++9LyVBZ6VZAACrCwZ3tsjjWan3xSMU0V2zJmZhm2saaBpoGmgaOGsaWLt2bfnoRz9afumXfmkFsPWsNeM8EDxwuA5mSM+fodhGIM+DV6NVoWmgaeDcacAoqY1grr322nNXiVbyimuAeVWbDVxxtbcCmwaaBi4RDZjp2rx5c/WXSJMvqWY2AnlJPe7W2KaBpoGmgaYBGmjksb0HTQNNA00DF6YGYvyv7J2aK8+Pz8VmqaewoL+rmQYPN4wOlPUjA2Wgn0VQc6ejgUYgT0drLU/TQNNA00DTQNNA00DTQNNA00DTwIprYDGOivrOsxPlLx7YX4b646iouisc884XyaS7l4Z0qrkQZxS/47o15S3Xrikbx2J38uZOSwONQJ6W2lqmpoGmgZXQwGJ09EYaT8cZlOyztXVzTQNNA00DTQNNA00DF4cG4qPAzq6P750un3psvKwZOPbmMscjkLPxXbF5tK/ctnU4CORg5+jEVziLeXEo8tW1ohHIV6e/lrtpoGngLGlgbmGpfPmRfeXgdOzwGYBxqlQQ32TSsnlsoLz9xvXNVPEsPZ+LXWwzcb3Yn3BrX9NA08CFpgHmpxjf/NxCWVpYLAOxs3mfXUqPzDXGoHP8Ot73Qk9E9oZHQOfmbQ++FOJ62nfCabwIjUCehtJalqaBpoGzr4HZAIg/f3B/eTLWOCxEh98ZIOwGis5+pkfXBL70x8zjrZetKn8rzFT6+41OHg9Ojs7dfjcNNA00DTQNNA00DZyvGlgO0jcf5A8JrCdjHaGMAf5dWB/UMPY874n9zoV3XP2OiM+BxcWFshAyluIIqtLbqFDq55Vcm9ZeibZa2qaBpoGV0UAAxOzcXHkuyONTB+froRc6/oSHhIOkhRmucmGdUmx6vWm4t8wHwPT3xSL5Dvtcmbq3UpoGmgaaBpoGmgaaBs6KBgwSL8UaSEcy9cQhx36D+M6pyh3SqODcHievwmraIJWLi0vhF+N3pG/fB1Tzil0jkK9YZS1D00DTwNnWwHKAA/OS3rA3sUB+MTp8IHCsEcWj6wIg+oNqGnecn5svQwNx8D0S2VzTQNNA00DTQNNA08AFrQFGqh1D1QD7w863AfNUbunwzYvfDJ3w/JcJaySvZPJFCRnbrqeqgfZVdaqaaumaBpoGVkQDRgSNLDJPce+/HqwwrsxR8uo+O7BO+Eur15ExX2W8NKb9ahpoGmgaaBpoGmgauGA1EOsWDRMzYTWB6BuhXvO+fjN0wrvbWGcqk2l2R7T7V6yBNgP5ilXWMjQNNA2cbQ3o+3kQUf9nmxJAsRwdv77flUMg+c7MpJBY2xC+ExtENIRUEhrXtilKVU/7p2mgaaBpoGmgaeCi0kA3vuc97Hfvms4QdP2GiGUy+aWQce36yjSwogRyZmamPProo2XXrl3l9ttvL5s2bTryUTc5OVnuueeesmXLlnL11VeXoaF2Nssre5QtddPAxaQBLBH567TJKGMMOB4eZYybwxFLAQ4IZJ2hNLR42NW7KuJF4Mi4dm0aaBpoGmgaaBpoGriQNdD5RqibHoB5oJ9w/+KnQFfg4bZGXF0r2fW9cDimXV6hBnx7rYibmJgo999/f/nGN75R9u3bVz760Y+WL3/5y7Xsxx57rPzKr/xK2b17d/mjP/qj8ulPf7pMTU2dcr36+1eUB59yvVrCpoGmgTOggejwkcdKFA93+kYV+c5ahsOzi0hl+N6KIpAk0eQM1KGJaBpoGmgaaBpoGmga+P/ZexNwS46y/r/uMvuSWbLNZJuEhCSE7IRAEpOQCFFA8I+KgIggGHlAcX0E9fEBHvz5BFBB2VxAQREBWRQJJEQgZIFsZN+XyT6ZJJPJLJnl7v/381a/3X3OPefcc+/tPrfvOW/NnFvV1dVdb33rrbfet6q6qkIISB+vigEkiZIg//WXBKK6gK4g0ckPtSDGV6gY85SUjhmQTCEvW7YsPP/5zw8vetGLwjXXXBPuu+++sH379nDrrbeqf+SRR4aVK1eGe++9N9xyyy0tIX3qqafCgw8+GDA+H3vsMVUmWz7gNx0BR2B+IoDAb2oL1t7gKu7E5ubj/Kxsp9oRcAQcAUfAEWgDAQzGGTgGnmf46Axy695HOjZ1h2F47LHHhh07dgSWqx533HHhgAMOCBs3blQj8I1vfGM46aSTwsEHH6yzkNdff30444wzGiKPMfrEE0/ojCVnwezZs6dhOo90BByB+YwA5qD9kiBSnyiT/uIz8ci3kkS7cwQcAUfAEXAEHIHuR8DUAC1pVARioQmnLh+WyJqH0kQemAECHTMgoe3xxx9X4/CnP/1pWLduXTjmmGPCs88+G7Zt26Yzj+yauHjxYlEIJwJLXps5lq4dccQR4ZBDDtG0zEDecccdzZJ7vCPgCMxzBPT7RytDXQdgy1HM19FFmYasS2ZPu+8IOAKOgCPgCDgCXYAA/Tw/zETr81OTsf4G5U1v1oW5525aCHTUgNx///3DG97whnDOOefoN49XX311WLt2beAbRg70NDcwMKBxdt3IX758uUZjTLIMNr/LUqP0HucIOAJdgID1EvlOIO06LFLmIzu2OL8LMPUiOAKOgCPgCDgC8wwBJhonxuNme9gC0Q6I3zxSFK5jvF7F0ulIc6IrmMUZ7/jfaSLQUTVr4cKFYf369eHUU08Np512ms4yMgO5Zs0a/Y6xv78/bNmyRSv8oIMOalkUmIIfzvyWD/hNR8ARmHcIIOZp5XlBNUnmJ3JABAHCQFLzVNJBSMidI+AIOAKOgCPgCHQfAtbTZ5NI2dFdlDbGW6qoT2RX3YdHJ0vUsRnIZ555Jtx1113h4Ycf1s10HnnkkXD66aerMclmOBdffLF+G8kxHxzv0ez7x06C43k5Ao7A3CKQGovJSGJcq5Kd6WgdAXajhb2LmNs689ybI8AO5E8//XRYvXp1YEVO3m3atCnwTT+fdzDY6s4RcAQcAUegOQLoB6mO0CCZ6gWJYqBjy5JGZyUbpPWo6SPQMQNyaGhIz3+888471YBcsWJFeMELXqAb67Ac9YorrtDNdHbv3h2OOuqo8LznPW/6pfEnHAFHoKsQiJ2DjCiOS0g+btTlKFLCtNNgDQsu31PIZX+WQm/7H0dgLhFAaeFTi5tvvllX2YyOjobDDjssnHnmmfr5BruSs7M48atWrdL4pUuXtkUyK3d8FU5bUHkiR8AR6CYERBHgjGjr/jMjcepC6s7u2ajz1A94ikkIdMyAZOnqL//yL+uPzXLo9MyxGc5FF11kl+47Ao6AI6AIpPI9ERe2TMV8g6n+Oj8faWncdwTmCoHh4eHAqhs2fGOn8S996Ut6HvJLX/pSjfvc5z4XTj755LBkyZJwySWXBAZVOe5qwYIFDUlmzwB+8P3evXsDfao7R8ARcAQcgQwBG18mxsID2e3cSHQ+0sPtItAxAzJPUN54zMd72BFwBByBWgQm0u8fsyGn2hT1V6OiS49L4nbT1z/v145A0QgsWrQoHH/88fpjtvDuu+8O7EbOktZLL700vPzlLw9nn322Go4sbcWIZIXOPvvs05CURx99VM9RZsUORiS+96sNofJIR8AR6FYEZIS5X/p7dl5XZ35yydacGIwMr9ktFjPh2Nnd3ewQmBMDcnYk+9OOgCPQKwgg9FX4i7BXXyJsuQoY0AdgKFpHkY/Lugxi3TkCc4sAht53vvOdwBnHGI7nn39+4FMOZibPOussNRZZtsrO5HwPmd+ZvJ5yvqHkUw++mWTZK8deTZ6Fr3/Krx0BR8AR6E4ETAfIG4sYj8SjJ6Qzj6JDoDOkRqeE3c0MATcgZ4abP+UIOAIdQMAMRNmpW51eS5hOAmezjMSbIy5/bfHuOwJzjQDLV5kt5JvHjRs3hieffDKwvJWjq5iZ5MexVlMtScXwxNi0Jax8W+kG5FzXrufvCDgCc4WA9fn0/6YfQIvqDHVEWdp0WrLuvl+2h4AbkO3h5KkcAUdgDhBIBb3kbcYiZOTDja6Jc+cIzAaBojemYYnpSSedpD82lbvuuut0KSvLVJlBZCYRw/G5554L++23nxqVzejnXbZk1Q3HZih5vCPgCPQKAnmdoFkYLDAux9VyZDlTr6BTTjndgCwHV3+rI+AIFIKASHiR+Crn871CIe/2lzgCEYFmRlhRRiSG4Y4dO8KuXbt0o5yVK1fqmcgsUz3uuON0NvKOO+7QbyCZmWQDnXaP8mhGu9etI+AIOAK9gwDLlNAUkuVKFBxrEb0hNyWpqWRJUy6KlO5mgIAbkDMAzR9xBByBziDQJ50B//T7hsSQtG8dUgqsg6gxMHOdSJrQA45AcwTKNMRYtnrllVeGb33rW7pMFWPy2GOPDRdccIHuovq+970v3HjjjYHNdtasWRM++MEPqqHZnFq/4wg4Ao6AI6AGY2ooJv1+jS5Qi1H8LrJv0iqm2lR+1Q4CbkC2g5KncQQcgTlBgKUmjCnaB/BycIGE+8SgzBmINpSY+Lpd95iMMNq+3XNCuWfqCGQILFu2TDfNefGLX6wb37D8lKM68PmWEYOR4zhwHOXR7hmQWQ4ecgQcAUegxxCQPn6c2URRByYYYMZHYUh0AdUV5CbnQusqJrkVdYc+S9JjgBVbXDcgi8XT3+YIOAIFIkCngKmIIcmgIj9mJPFtsQp2Yn5nVtLj9KDgGPS/jsCcIsBSWIzCZoYhu6q6cwQcAUfAEZgGAqIE9IkyQP8vu4jp6o4YjFoAxmQfRqb46Aw4/GHVKkxT0Gj/MwMEDNMZPOqPOAKOgCNQLgLWGbC8MBlUVGOSXFX8N+0Dmt4ol2B/uyPgCDgCjoAj4Ah0BgGmH80xmswvcaYzqBEpcXadaA+WzP0ZIuAzkDMEzh9zBByBMhHAdMQlHYIMMdIJ0DWoURnNR02hqbI+IzfzmItMU3rAEXAEHAFHwBFwBOY7AtiKLEm179fre3x0BnMWJob02R1L4f50EXADcrqIeXpHwBHoGAJsomOmJJnyXWO/GJP57xvtGt+dI+AIOAKOgCPgCPQCAvFzlmYzikxO9otakJ+k5DodmHYzclZM4gbkrODzhx0BR6BMBOIooXQSEkDuI/wZSYydQMw5GpXRuCSGQUfdvTU+HBP5X0fAEXAEHAFHwBHoGQRMTzCfgpsx6ePNs2cDNyBnj6G/wRFwBEpCoE+kPctNOLpDP9gWo1DtwpxxiGEZjcZIRLxFwuyLh5LI89c6Ao6AI+AIOAKOwFwgQDcv/8wonIoEdAjb0R2dwd3sEPBNdGaHnz/tCDgCpSKgPUT6XaNu0Z3k1ywcb3vvUGq1+MsdAUfAEXAEHIE5RAADRlcmJTRw3cqoYUg5Go6mH/C0u5ki0Arrmb7Tn3MEHAFHoBAETMybsZg/mqNZWDsIe7AQKvwljoAj4Ag4Ao6AI1AtBKSjn+ZCI1SDqB64kjDbuvQlrLNF0J93BByB8hAQGZ/fL02XqsqgYevlJ3FUsXWa8kj2NzsCjoAj4Ag4Ao5AuQhgO8ZdWGWXdgmbSWjzivH8aNl0L73DjKXcbXfNa7nkz/u3uwE576vQC+AI9C4C1lHQcVjYNtCx695Fx0vuCDgCjoAj4Ah0LwJxGSV7JYgOkHT6NnjM5jnsoZDfRKd7keh8ydyA7DzmnqMj4Ai0iQDGoM5BimdGonYUuefT+Fxcv6S2s6Fy0R50BBwBR8ARcAQcga5BIFEOKA/KQN7ZtflyjxVN8fMXQrkb+ec83BYCbkC2BZMncgQcgblAQMW7/OlLPng0cW++0VR77XOPhov7joAj4Ag4Ao5AtyJA3x/7fzUNk2JaTLycpB/IKLTNUrq2MHPO6OgmOmNjY2HLli1h48aNYXh4uIZq7j388MPh2Wef9ZmDGmT8whHoXQR0tpE/+W8WTPI3gyU5+qPZbY93BBwBR8ARcAQcgXmOgOgCE3ndQE3JnLmY1xVy4TjzWGtkznMk5oT8js1Ajo+Ph02bNoXbbrstbN++Pdx+++3h1FNPDQcddFDYsWNHuOmmm9R47JNFzEcffXR4/vOfH/r727NvecadI+AIdCECIvT5AH5C2nhfrgNIhw/zRU7kAOlr0ubTeNgRcAQcAUfAEXAEugQBDMGc0dioVKY74Ku5MEX6Ru/wuEkItGehTXps+hFbt24Nd911V7j//vvDwoULw3vf+95wySWX6Ituvvnm8Dd/8zca//3vfz984xvfCI888kjLTDBI+ekh4zJ76c4RcAS6EYFaQW/fNeJP+iEP5IfxiGCbmJjm/t7dCJ+XyRFwBBwBR8AR6FoEREdAH2AmEgMxCdvMpPlp8UljLhe0KPfbR6BjM5Br1qwJL3vZy8K5554bBgYGwsUXXxwGBwfDtddeq4bl29/+dr2/YcOG8IMf/CBcfvnl4a1vfWvDkqA4YmAykzk6OhowTt05Ao5AtyKAsSj9QlI8MyJbl1ZS+8KE1hD5XUfAEXAEHAFHYJ4iIJqB6AbxR4ef6QatO//83Xx4nsIwZ2R3bAaS5agLFizQWUYMyCeeeCIsXbpUjcnNmzeHww8/PCxZsiQcfPDBsmFGn95vhQqzj3w3yY+wO0fAEegBBGz0UC1KTMrkp/HZNctYM5OzB3DxIhaOgH8aUTik/kJHwBFwBApDAOPPfvT3mTEY+/9oUGZGphqbrFQSfaFfkpCelO5mhkDHZiCNPGYMP/e5z4VTTjklnHjiifr9IxvqMBuJIYiPsYlh2MzRse+7775h5cqVyggsiWVzHne9i8CO3SPhkWeHwlM7h5ue+YOgyAsMEzasfDhi3yVh/apFYeFgx8ZUereypltyFfQ5MW9G5HTf4+kdAUfAEXAEHAFHoCsQyLQCQvHQr9qCxfgYl6VmyonZS3ezQ6CjBuTevXvDDTfcEO68885w4YUX6kY5N954o848svsqhuOuXbvUX716dcuSYTya2717twXd71EEntoxFC6/Z2u49pFdYcGAmYa1YJjxWH93eGwi/NKJa8PqJYNuQNZCNudXUciLoJ+QWquvOKjL9wFJGPtSk+bvzXlJnABHwBFwBBwBR8ARKBIBdIRs7tE6/XplweIlZ1cOCoO/owYk3zt+5jOfCW94wxsCBiKzjCtWrNCdWG+55ZZw1FFH6S6tGJLHHnts24X0JaxtQ9V1CVmKwBmBu/YOh8e27Q23bdkblgwiIeoFSIwxMWJ3ud4zOhF+ZsfesFdmwvdZNiiz2pI22dGz6wCbbwWSyqA+zBGk7tIoC+QiST+u50baTXvafUdgbhFgBQ4/PuPgk468YyUO8ox4+kB3joAj4Ag4Ai0QkC6evfLo85l/tB6/di7SYuN7uEI9SI6WbvFyvzUVAh0zIB966KHwve99L/zoRz/SzvE///M/w1lnnRVe9apXhdNOOy28733vC3fccUd4+umnw2te85pw3nnnTUW733cEVOGaGB+Tc0VHZAn0mM4gMgPJN3D9iREZv4eLAkMnsmrlSWDVKkrdyIgocOOLJaErb9ViraRjQOgLYdoBJARaVdIZpOGUeItJIzzgCMwZAhiH7EL+wAMPhP33318HSZcvX670cJQV/R+rdDjGav369XNGp2fsCDgCjkCVEZBDvaS/p3+Pc4+qF8iljftjUOK4VuOSBInLBS3K/Rki0DEDkg7xPe95T3jLW96im+cwa7hs2bKwzz77qBHwqU99SmckGXllx1Z3jkC7CMBLI6Mjue9m4zEOZlKk5qBIDhUedRIEITM2JgakGJG8q38gfaJdEjxdiQjkz3Q0k9D8ZIxAOwkLQ0r+mRJJ81c7Am0hwOzi3XffHa666qqA0fitb31Lv/f/5Cc/GXbu3Bk+9KEP6UocZh+597u/+7vhsMMO05nKqTJgNtNXTEyFkt93BByBbkGA/r9GjZMINR4TxSAzJFmhlsxHmlUpzzKYl+oQ3QLKHJSjYwYkG90ccMAB+mtUTjpLd47ATBDg7D8MP877UcGAMJEXISBqhEyDSBudGh+T5/khWOTnCtlMaqKEZ6ignEPsa4eQxPcxpSxO45Nwmrz+Or3hAUegswhg5K1bty6cffbZugHcww8/HO65557w+OOPh1tvvTWsXbs2nHrqqWpcYmxyHvKb3vQmHWRtRCkzltu2bdOVE6Tn5zKrEVIe5wg4At2GQF6vi2qd6Al5VcHC6HNWeNMl8pam3XN/Rgj4VMuMYPOHqoSAGX1IEBjahIv5U9OKiEnFzNTJPcUcITC5jnLdg9KUX8rqVTpH1eTZTkIAA3K//fYLL3zhC8OBBx6oA6mLF8tyeXFsJHfCCSeEk08+OZx00klqSN58881qFE56URKxZ8+e8Mwzz4Qnn3xSP/tgCb47R8ARcAR6BQEWr6IR2CdKrcs9WXdwna81Yu3c7dgMZDvEeBpHYEYIyIgSM1EYjP0iUuIMVbKkQeJs4KnRu22pQ7zXSMg0esrjOoUABiIDBOqkgnVQQK5tcIBbpKEedSBBEnIsiz4nf905AlVDYNOmTXrOsRmSzCaydNVm1lmtw87iKd83KADGJxvRMfM4MjLS1lLXBq/xKEfAEXAE5icCdO+qAGhALtAKrM9PNARTFCihhU2fIC5Nrxf+Z5oIuAE5TcA8eQUREJlhqxWREf0iIMbFosDHEcahkJmSphHJH/1erkao5O96eE4QkCqLxiMjjLHuWtGRrz4OCOZpd45AFRH49Kc/rUtV3/Wud6nxt2jRIjUEmUVkKT67k7M/QKslqewdwA83NDSky2BbGZxVxMFpcgQcAUdgxgikBqG8AR1QNIU4jRDD2XtrdQHdgCevMGQJPTRNBNyAnCZgnryCCIggwQhEgRoT8vgxMqV+EsbD1StZAzHazY0Eh8p5yH6Efe1U8RRkWodh/hTJ/bYj0AEEMA4vuuiicOihh4ZXvOIVuqSVGUSuN2/eHFiWylJXwhxpVX/MRzMS62Vas3Qe7wg4Ao5A9yGgSkIywRj7fLMtG5VVVywlN1xDaIRQ+3FuQLaPlaesOAIIDftpwKSDSROuCSfx9gFw+kzFy9eL5CHs1WFEiqurQo2r/5MkjYZn/c0euB4ZHQ93bXoubNo+HAb6jflbFTzBWJIw63XgyoXhiP2WhqWLbHil1bN+rx0EOJ7jiiuuCF/4whf0+A6O8sBwfOlLX6pHVn384x8Pd911V1iyZInOKF544YUabufdnsYRcAQcgd5FIE4e6OoyASHf4zG4Zis5bKBNr1ESuNe7oBVScjcgC4HRX1IVBDAK+ZmhAV0sgTQjMR/PPZydFxmv/O98RoB6To3O+VyQGdJOJzk0Mhqu2rgtXP3QrrBkQeTuRnyfZRG7UXDrFwPypYcsDWuXDYoBuSRL4qFZIYDSwtmPGIarVq0KLFvlOA++Y9ywYUO44IILwtatW3UGkrgjjzxyVvn5w46AI+AIdDsCuvIMbU/6Pfo4HD46IHofzgzHeBX/Wtp8nIenj4AbkNPHzJ+oMAJRjLCpTkZk/CYuXufjidHv61LRkz3joaogIKOE2jmIkRPtnBrCLIrqtjAJctVfk77bL9SAHBoOG7cOhRue3BNWLrJ59lzJBRzMyvFky1ozMVnyPSjxBy3rC3uGRuRqkXS+0pbE+HE3OwQwGNlhlV8j9+pXv7pRdClxo3Jc0RMyO71j79ikleHUdKu2MyC8sG6fhWG5zE73tzW7XUoR/KWOgCPgCCgCqU4neoI5Mx5VwFl80o9Fg1I0RUnOz76btGfdbx8BNyDbx8pTVhoBhIcJEPGRIPlhqCa080Qf23a6qyQC0XSJwh4CMWboAMyosdFFi88KwTO9Va+Ul+/s+K5uQBrAsoH+sGRADEUBpb4pqO0oWKadr6SZkIT9smp1fHw0jIyKASnvCn2+jDXjqfkfgke27x4JX7r+yXDtI8+FpYMNBhiaFJODQhaJ0fjbZ+wfTj50ZViyyNWHJlB5tCPgCFQBgbwOkIRVK0j+YFP2lpZQbKV4D1Asnv62OUQA+TA+xl5coggjGJhSET8aIXJP7mu8+CY0RL8OQypYLGYOC+BZ1yCgRo5UC/VplWhGofn2QM21PGN1bvd7xZ8QK3A42c1zIrENzEQwX7EQcHXkNW9BSuPgZK3x0bEwJr8JDEjZ1MVddyBAG+G3V2aon9o1HB7cORxWyBLnyS6OyUvqyCNJglFpV4sH+8LOvSOBWUykKKLTBnMmv8djHAFHwBEoGwERQuKQbSaLUn1ALUQVUikR6AbINnezR8ANyNlj6G+oCAKcAdkv5gY6cTqpKGHsSNQkBIfFRwPDZmcQJi5QBITKOQwiNSSnaRFSm/GY4coVqVSCUOgnRLnXzhRmb8bW2ovKzRpc4/W4vGRccNd3lEqtv7zTCHBEyJDMUI/KAEGf2IDRDIxskLIKTIQzHolXOjg31t8vm/zsledlhnpCFjz3NTJAkwfccwQcAUegRASQX/RXajCKvMoMxyRTZFmdHBOh1bxfLJHWbny1G5DdWKs9WiYEiUkGBAtzJ6YU2TV+pvKgKKt0yRLKfXcVREDqlpqy+sNv6OgbtNLjTHTDNF0dSWcq6IBXyuh0otZpAk4ubAeoGiYJdnbpfvcggHLFrPKIGJA6MCMzzimL5ItJQ1PJKYHcDDWswqDMiGzShAEalbVs1D//Cg87Ao6AI1A+AtphxWxywVTx404+nsuoIEy+QVp300LADchpweWJq4qAGRQybyL/VAOqlxs115lMQalKrrLIqhaz9+iiTlB8VXXFFyNSjKGsE5AIqpsbNUaSpE07CrnXYw5I9CcGADBYiwCmBCwN1YaTWzUtJUnmXhcgENsPs5CpzGtYqsgl9bfU2BR+GpcZbr61pX3R5Nw5Ao6AIzA/EWgs6+ZnWTpPdU8akMxUbXluJIyJ4ZAtc6MnNGayXpFrC8fKYee5xbLxwMolPQld5zm0zRxRbqgtfhiTDJyPS9XhWw1yjzC+Oa5Z4trDtoZBUVlfl6gIddQnFaiGIWEclZlWbFazFqVpevKPLOcW+dYv00bg1i8Mru0hwcKU/8j3CW40HGlI7Lxa00h6Er/uLLS2HapbK14CSdVPKi3tizTWznIJ1Ay153PxHnQEHAFHoJMIqH6HmEKUiazCxxF2Vz4CPWkFPSfbl/+/7z4smwGMhwWT9oiAA/Pcl4VHRSHbR7bFP+WQ5eGNpx/Y5iHd5Vei5wAC1Jv8GBRIqsyEieGj+pDcU90nidSPrl0ZMogq6WctUMjTuqqrxGhVys2YEiVZj6ioZGk6T5QdY2O+zdZzzeAJDuRsECYbVNNb/qcbERAhiZzkFwcT6mb1VQMTBslpYropk0TFVtaNoHiZHAFHYL4iwEDzgEgn+jcdKGtREFlDEVUJkWfuZo5A7xmQ8n3Q3uGhcN+2vWGbGJALdV0OANItRm6KexFGUPOHzA8Lg+4rBuTB+wzqJgIDCxckz8W0/ncOEaDqpH7kfyI8iDBVJ9Yr1HE/7+I1Y+rcqLuZT+jhOUIg1gkbJDEgkJ9NywiyejM/u9O7IeP9yQikIk9u5cfPQI/r5k9OfpfHzE8EmJEekB+DCCwJjzPUWc03MirZdEdnqF1Ozs9Kd6odga5EgJ4L4xHHRoqZY4CUeDUqk+h8n5el9NBMEOgpA5JOcUK+/+CcNJavMqOYdZmiOMk1bkJ6Ve0sNaxRej0swSHZy3xoRLYxl40EFg7KeAcHp7mrDAJsDkGt6uHzUrn4qYvD7RKZr3XSxBQoUr3mhkbGw+PP7g27h8d0N7OaBjEFGJzysGrpYNiw79IpUs7sduwW4l90VoxHq07zG71Zn6CtN7rZK3EKkFTQOKa38Lj82sFDsSN17zWFXuEMLScKFe0XX41HYoVnUL6IwxGfn6HuBEvct3lX2Lx9KM6MKhXGvVwYBxs3ZxRxPNOBKxeHg9csDosaHk2iL/M/joAj0CUIWOtXvV77uwYFk0Tcyi2kiNea1ORJg+c8qi0Ees6AZAMBdpGD+TD9OAcwdfmhiyScsphcD2jPKp3qGAdtjwojLnQ9KwVvbgPUU1y2IIaDXFCtKljET1VoEzLmyz2qVE0NeQaFqdfctt3D4f/u3hoeenYofhOcGNFgEk+Da47IHjE+Tz54eXjrvkskUb4hNX9mZneEGqErr9xablZjWt/Jy7UaNYHdnVmurZ5ioOLbt28NT+0cCYM1QmTyU0YrdxijWiDa7sErF4TTn7cqLC5N2VWuVt5WI4G85RcHVvIUQRU4Meginso4osrDTt7uroIIWPfXqJ1BLlzDvXru4V5R7icP7gg/lt8AFqG6yJu1Ye4pN4uhKUvWhFdpU2dtWBFWLRsQA3JRTO5/HQFHoOsRQBLEbquRrEju2S3xVXJo/0aku9kg0FMGJEBxvtmoGIDqMk1TLoWZ8n0WCeTaZqc0TJw8wy50upOdhDFS7PBSbrubSwTiCDpKDvWiPmElqbGw0CrXmWfuGwPoA139J/LtRNi5Zzjc9OjO8NOnh8KI4JAt2c7wMFSymAjN9qHxqOhNyGBK4Ew4S1ksdFKV8n6+a8yMSOqWWDklUv5GxdZmS4jXdsuNMpwQhAz44g1Ph2uf2BVWyofUdEoMVEAP//PfG0ZaIyFjQtPyBaLsHrQ0HHfQMjEgF6qtViR21AJ4scoC7EAoGw4AG4kEm5gwEiaAQZvUojwTu+N4w/92IwLwpP0oX55H89f5eNpX2h+SqEAX5dFYeGDL7vDjx3aFhTKwAns2djWMqwbkIgZlVvSHPUPLpYEV36Ya0+GxjoAjMPcI0JnlnV2Lb99yc7tmGjKf3sMzRaDnDEiA0i3MVUmCwSZDZ3qwKl9JL0aYzrRPelA6O3uwSMVvMiUe0z4CUTHmnDNmq3ANqrbx60yhbny3e2MFqzGZSR+QtrB0sE8MDjEeEn5HfTMzwpRG7lkYUEYk/aCs9R6TJeF9YkSV0hakbvjgHcNMZ88k36z1QQVXQnviE2MpkqLEqAL/Mgg1MjoclokhuHbxgPj9ihXyYSrMxO4MS+XT6UGxMO0w9snq++yJRcZNjHNcg5i1Wm9ynQOEelTk+INLfGSb7sIaY/1vFyNgVa/T4sz46UBaqwLDQMngTKtkM7infDcyLH3seFgismiB/Kw9mRGbb1vIeJNFtKmFg6wMYnXRiJRDTN0+/7RkBtXgjzgC8wwBk2L1fixGHJgSqaW3Yxokg17bI/OsxFUitycNSCoAZdcMxXhtTCZXOcZKZxJQwkjIPQ1w4a5KCKBgRAUYqqKyE+mLFUZ9Izjy9U76zOXDWWy3hjC2mY0fReFKnClrXKbhRFOTFiPgZRjpzBbGlChtgwOLkjYVsbb3zdbXDkBewlv5kbv5EtQwsTE+o417MXUMFfVX+UsU7eGhYZ35UEzk5YaV+YbTJMySBLoMXhTmiXFZbtefPlUUmfIew0J8GgagmQZOLnaNn7g+LEwTeBbpfpciwGqDyBsMvvB5Bq0IFoEbYQukgplhcBNLRXF8ZZ40PL2e7R/aFOdKjsmnJQzO5N+dbxlpWGRQvl31yQ2uaVOjMiDGu/oGjPLZUufPOwKOQFURQCIhlpAhtHhdJSG+yQqu6dKQa6TNXO1VFu+h6SAwJwbkQw89FNasWROWL18uulOs6j179oRbbrkl7L///uGQQw4JCxaww2l7biYzH3zvxhlpaTeT5yfrRck+GZWFTI3mOp+2PRI9VdkISJ1Qpyg+/FCGok/G1BxL8wgjbKLPX0vba5WqhpAoa2MyfD+uPA1G4tJpKkACncxT4JIoAxGlknewo2MZjuwsy3rf8jNK8xQQNoXX0hXjR2VXv4FWw1tw0/MNeLtRIkEjFlwsnAoOUXZHx0TZlY2LRNktxX4UEtRAMFDMh0ycXZsvURgGaj9aXEq3PuF/ugUBqVeWKTNwNC7tnWpOWr+EdVgo5WSNhx/0GUmnY00SYTxSECZmRGp/K2up++KGAw3eLoRAcE27kiiuRQ7pzLuESeLOEXAEuhsB2jk6vOnx5lup7doMSuSZDphpAht4ttTuTxeBjhqQjz76aPjud78bbrrppvC2t70tnHzyyWpAYlB++ctfDkceeWS46qqrwjHHHBPOP//8sGQJm3M0dhic9h3irl27GidqEWsK9ARbOzZy1qPSMeHkmqBFx0j/WzUEokiIVNXoOKIwoVagHqGB5O+p0lETU7VSlUSPMDSzkGYM1qBmShpoJWApcgac3Ec467JSZg1KctQnr8eX1ZjqciSktaa0JWRET2Ks7RZIG69UWsBNXCQpoagNzEgfv0GU5+UdyKFSnNYtlEaZle9ACefjqV/GDTLyS6KplIL6S2eCgPZ/MnCkbYu6T3ggfsebXVvbhzc0jaSzuJnk2+qZ2CdH3lOzVjOsf0KkOLQm9HBX+dmWl9Qn92tHwBHoXgSQBeh26SDuFEVNhBfaYHTmT/Gc326IgOkVDW8WHYnR99hjj6kB+dRTT6nytHPnTp153LhxYzjssMM0jpnI22+/vWn2dDQ8/+CDD4b7778/bN68WZfPNX2gwQ1UK94DI9X/rCMjHvbih7qIrhdjGrzQo+YcAa03lCKtKCor94M6ETSqfKjWJDWqgof0hOec/I4TkBWZ5WyiGDb5cc/u14fV6CiTcqqQyiHjOmf025E7dhuaSneSOTrrdDBjdo/0Za8SBRf95flfwjojm/OtfSABNawz0enTpUPoGcwVAtQxLvqJTjXJQLR4S6oDH3pR1p+EHmnrsW3Fllz/uUk+d5VLdM7iomdli3H+1xFwBLoUAWnqUYen7U/W4xvF0SUi9tApoqSIMqZLESq9WB2bgWS2kBnGd7/73WFoaEiXrw7IdwoYgffdd194y1veEk477bRw6KGHhq9//evhmmuu0etmCOzevTts375dv78iPG2XKFf5TjJdvZe8LH9Pw8pxyn3Tzs4fKBcBlAcVDgiGKBkaZohQqXXMS0pcvrJrE3T9lSphUsp+AZHvidTVw8T95BZehCsK4Y6IYCo4T0COFht8TP2oSQqRuURFB3k3GEleKVlTYKYkSPqJAWZRyiQO2qgb4XYBhQ2RuIp0RsIh1eKVLi1M0oZaNaCY2P/OYwQimxqzml9fIGPwyKe6VF2SqMLQ7JH6V8zgmtz6ZSDDjlRigVA+zCthT5twZEUtzX1QebZEwsjYnSPgCFQQAdp9+21fJZo+0v4zFSx0JUjqmAGJsYhbtGiRfujOyAHfP7L8FENw1apV+j3QsmXLpIOYCDt27GgKECOSxx57bHqf2cuf/OQn6XU7AZbu8c+WsOoSrkTxbMRW3Nc+Sr7P0BmrdjLxNB1DIFXiVZCoiGgvb63U9pJ2ayodxROmRxmzT49oCvb9gLUHUNVPoJJ7OnFbMn5x1iNSwEwjAwBxh1ijSohp4nS35Sb3iogGN76nAAfDJo+ZYWiYwaMY3lxj0JXryIE85C91ZPLL8tXrGG90aDmkAPoUf9x1LwJav/In/lf+ra1yuzI/QqG8VCIq1qYiF2I80rIiDTEs7QcDU3aNpX3TxmhTyAR3joAj0GsI0L9FCZFJitgfgwSSIx9PXP01ce5mhkDHDMipyLOOCeMQY9M215nqOe7PZBMd2Ei/s0gysBmVZvnF+/RCIc4AAEAASURBVM3uevzcI4CoYEBABEr9msZWxIk0gfeiMsI7es1RZvkJbuohXcWpoZML24SZtQMUN0FN/5K+fEde5vJhlMc4C2GDCJxnOCGEJuTbQ8X7wjccdqJnP8rb85iRmV0bZlCtZy1Cn1JNqhKcyFCVp3xjSUWBBL1sDhFo0hj+JE6DWrH6kEW734UI0HZVVubrPldOZhz7hY9sIyrCKla1URnTFN3CeF+yIiRlQcsL4mJYB2lEXpGaMEl199Yy25Tk4c4RcASqikDURoy6yVLD7uBHOZOP8fDMEOi4AckMIzusskEOht8+++wTVq5cGTZt2hSOP/74sHXrVo1ft27dzErUzlMYDBOyAx27tg1YJwjL5cO8KM9occRT9TDtstrJyNNUHgGVNHlxU3mKCydQDWhtE9mrI5/H60bhONMmqlv+ZvZ4YaE4ARE7h3GRF9FYjLOQlgn0o+Cq0aSR0m6p0pKrVZeI6kY40XSth8KuzTcDF2OdJazMppbiLMMUA8vH/AyaLCZWJcuAI475O6VQ6S+dSwSoXviEJizByT0ffSTx3I08oWNMwrfExFi9VfwfyQjD1WiaKgMMyAGhyAZqpkrv9x0BR6CbEIjSiAEu+4zF5hC4zsdTagbO1JUuyGI23fy3owbkk08+Gb7//e+Hm2++Od1hdb/99gtHH310uPTSS3VX1XvuuSdgZL7kJS8pEXfpHIWJMCI5aDs6uCnOSSbsJdcZh2Hsan8rUZmimjzqXkUQwNDgX1aPeSUk1nAkVetY/lD7PNHTDsbWXy0KfYmg5dDuCBE4RUQ1ubahzmCHcqiqq2QfFUWNULIj1RkdHatPy1LA0Bk9aEObhaPATBwYWhgMdddauaXJNEVJf6xOteIgNNKTVKTGEmNFECLlQmhVvySa/LWKAN/sc3zL0qVLa1basDEc5xjSJ5Z9jJV0fNqeYv1H/siHY1XBpcY3xMR05fKuDsvo4ErsmWOekZ5IYaQpxit18K3upm73Y2r/6wg4ArNHgCXjIzLZMjoq7SsvDtp5tTyyaEF/WDBoenY7D00vTX4/ATMckVH5HM2ItP45pzhMLzNPXYNAxw3I66+/Xo1HjvS49957ddMcjvO47LLLdOOc5557LlxwwQV6lEcNpYVeoCzFHTlzKpTkIB1Rs3xQyNRNtwU1e6HHF40AVTQm9apKsLzcagxBgkCxa/KNNR3r0mdcBB0BJxppoFPrandfjChiW8aRvDyqtc8VcZV9A0ldYeDwVkxJCLAcGNzhItanGpp2q0yfPAWIaNBG/Cy7PGYWBjO+mVwIqbEglrxwHzxUFReDUO3/JD/CauySY4Q0yVuJSstSOEH+Qh0gfeKJJ7Sfw0hk0ziMSIxJdh5nh/KRkRE9C5l77BfQzPEMP+p5eHg44f9mqevilTfgDtq9mWnUvzkJp4NGEqfNirYHr9vi63x6e272fvpWXXttA4ExNrZulQJJS5d4/Z/4MdnsifA3OAKOQIrAzr2j4f6ndocHn9kbBuXb4+m4Uen0XnToirBh37jicDrPTpk2JUXkkogyVvTkjUZ0PjYGxMcRNmd9sl27PzMEOmpAnnDCCeFjH/tYQ0o/8YlPaAdrm+00TFRYpPU0sePR18KMFl0f5hb39H7sRPUZ/1MZBBAe/FMhIX+sKvFtExOItfhIOJugiLmhgqX2Trzf3X+j/AU1+FswAweVwNYAYooMNYtPcOEBrKJEnUtiC/XUEELhlayNmowey8rqLvrRoNNSWYJSfN1MS05Wj2dQGTZGpdFk8RkJKkvSbs3SZ/eLCQlm4EZm8b/il1yqgaADAMlMKXlqWrnOOteyaCumhPPtLcw8fvaznw3f+MY3wqte9Sr9ZAMDEqPy7//+78OJJ56og6vf/va31Xg86aSTms5E2uZzGJwYkMxczmwvAOpd2UH9FFMi1SXMI0xEu2JpaSZd09TFBchWjWMEkeQp/Kh8KVdWPq7z8VF8R4NYpVkPsS3id1z+jFFfaZ21Ux1xYGlwQHZp7iG82kHG0+QRmAhbtu8N373jmfBfd20LS4VfYk9MQ8VF5uFvbYzeDDtHx8Nfvnx9OGzNwtA3UIK5Ac+LvICOuJFWpMI21UJW2aZ7KickZbyO9Pnf2SFQQo3OnKDOGI+RvriEFQVLrvmjI66R+VJNlctUuCYdmS+VmXkFl/gkSq+Njsdz9vKVF+tVv6GTdPjmen5H3WTJIjggYPOjdIZRI18VOBHOqrA1SlBSXL3y2CgbLYfcKJs2+A2Fmh2d23GqAMOKyqDxK9J2nptpmrSrp23kFPH0fRJvjhbBfE+pxoFl1oM+fMuyVI6xwl+9erXOIHKk1fe+971w3nnnhbPPPluPt2J28pJLLtFPO5otZd22bVvYuHFj4Gxl3o0hqfzVLrZS9Yz/8CwuxwrtvqGkdNBjv4w+MjNa68NcY9yWfb4q+VTNPbJld7ji/m3hrif3hIVqCYJdXp2H4vx17PsQ+7IqMbz9jAPDEfstrVqxnJ4KIEB7o0dAtsiGIWGhGI+DgzbAGPkottXGxNIrLpRd48ZkcGt0VJ7vlz4vp3s1fmrmsdFYjM9bP0YfmI/nrl7TTNzNGoFKGZCzLs20XhANjrTnVIaKXEVnmjaPhNG41pkNbiZx08rOE3cIgVg5KvySSmR0No5nRyVEFZFUkJFejIAerVOMLDWABIA4c9deNSlqGE6lA2f1A6XRxSztSuLoqWJPoQkwhNAmp1Oe+Obp/qWDFTpypLR6A3RrKcS33S1bpZ/5vUy2aTuQjCExm1lMIJN4ZjBsdlfTygCZtg8ecFcYAhh3Cxcu1O8b2TiOwVLimDnEEDzzzDPD2rVrdUnrAQccoAYk95o5jMzly5frqh1mIJ955hk1SJulr4+HIxg0YvYcMan8oYmyUP0z8Gw/6WHkshxWrVCj7arNPHT5muimzSlv80XzKJm2VRFw23cNhbuf3B2ufHRXWIpyr+jFgiTdX8NSgdWILM153a7hEPZdLMIBAerOEcgQgMfGxXAcFgMyrnCgm5W2KYwVe5RaPVnjc6KB/SnhSJ7FCB1cMChXIj9IWJAjOwxCfjikR60jL+5leSqnZ5e1yf1qWgj0rAEZZw/oQI2TcpwvENZexQYTR2wTBWtaMHviTiCAwMNY5KgErcG0EhEwEoNATAjRDpiwVD9h+ZPc6UUPZdIgiDgg+Cd1EhEuBUgxo5V0HDbLUHytzKT95pfSSmFsA6AyazPtuBIY6BgjLll3BbWZhIkJ1WCDRm6W5KCDb+Rkuj1WkVzH7KCG+o7X+jeSpfdj6hIJK6m88+G1djSVfbtoNDMLaQYlPDQ4OKhKl91v5C9evDjww6lyJs9My8F/UuM65yzVrcofL6ipei4S7lUv8s208plmYnJkdVDMjvwibbyGME5pzccTxzU3NU1CM9dd6rT9inK/Z2g4PDc0Ij9p75Q9lYMKRNPSg+WI/NkrzzND1D+4oFDFvmnGfmNeIUBbHB8bVUNSGyBsRfMy9rKmVu9TSknTJwO5+vyYjFbYs9wrwSEDTJ23vpX+LZJrfpJxRwa/SyhkxV45zV6nYtTPiJzI6XH0NXZQvCYyGSEYLUmTxBOrDqmrmw7QEtxVDgGpH1OM44RUrCcbW+WKmk2NSK6pUuq1J521BeF5nY2I+AAFgtdcFkbltHgJixCmHZXtdLl5roWycXK21BZ6YjmMjtSwS2m1OwX7KGscBdQfOSzPRzU4CR3aoSWab9zp1HAsmKb0ddJxS778Ij75/GI4nl+Zi7ek+kwuPn1nbwUw5vKDAkWVHgPRfhiOzEiyeRwj9RiXfCu577771uzQ2irvMVHO8rzXKm28Rx8HT/IjlMjB9MF83Sdh8UgPR5XpYksWrtVPRWJO9eJZRY4kTEWPFkDKgQwrk7iKvXtMZI8uXRZ/kczALhAhg3rShz8FEFGmihE5KrNLYiAsSL5Pg9/dOQKGAC1edSphKGYf6UvyvGXhdFCHFLRN8fnRM9IuOac1Pi0RBTvtW3PvrL22XClJ5uIVMfnY7L6H2kOgBw3ICIyOcqIAJvJS2SgN55mqXqByL3+/PaA9VXkI5GsIgaVCS+q2n29CxE9rS66pd02QVLzdtTT5d5VHcZXeTMlFuKONyX+dHRMQ6BhMmTDlNHYSESmUlNgO4nU5JYp5UEfQo7SRK3VY4+za/Jqb5V2AGR2jZGuGRjPMoKxfCyEB9UuklVfLj2/C+I1LXUUfKLhpLhdOaeu9FmBo1PtWl/XxM73GONy7d2/g+0W+bdyyZYvuuHrssceGRx55RJeycoTVQw89FNhAhyWvZbnIgmIOEpCGnS6pNk2QjPNhucxkZ0lUwXranoQuWUZC9nkSCOOU5MS366jkctU7jgE8vi1jmSEzyf0iILG7ta0DQx4vwlLVtHiCrFjt4/s0GXzgtyDhA7nlzhHIEIBn6G+FP5Rv5E+eVQjj4DnbrBDVgLQ6BsR4Ex13EkfaYp28GNoSQmqNR8tJ0kxyUOhutgj0rAGpHK2KM+aGNAD5xaWPepn7kzCfMCjH4IyzJEwEt7vqIUBN6Si0Sq58PZmwyAuSZuHqlat8igSLCJ55sQMwoZwQgJKZIskjakDlcSyeUsYtydO2nKGN8m1F47Ya89c+S+krlzYFiz+Ci2FjirjhZPFQFunCGGYUN47IpoBG0gv7i9HNkSGiXmrGYxA0hdgCLRttLowQf1GKwI4dO8JFF12kZx4z87h58+bwute9Lpx//vnhz//8z8N1112ny1KZkfzABz6g30OmDxcZUP6DQxj8EMZI2rlm0Sxs+UcmsatSfG07iSaoymtiOaYDR3Kdj4eIOMhVcnsvpbQzfyk1SKNGkmBCosAPSBQ+GKLUpy4XRgzwU81HjYM0lQccgckICDNJi0t4KmGkHD/xABsysUErfEUYR58Nj0Y+1aji/wgdmkNebuVyqR/Y5ZbJilLpytHQzcGeNSDhN5Q7nRWQGrbRk1aVTSPiv7vqIaDyo0aIIMVUFRE/SjTsHWLimUFZGfR7uZpns3tdH4KlRYlgFBvBb6P8+XLTEUQEwQ8EYyPok4GUKIzzqYsPUzWmHJnxiI/jHrRZvEYSb4ES/aiAJ0p4XZ75/GvCctF4lLRIQpM6Ahzj6zwRRWY1zXeNiuW/fc9I2LlHjp2Y5rM8sFzW6q1YsiAsLPFg6umS1W76FStWhPe85z3hHe94hz7CER4Yi/gYkCxdReHheuXKle2+tqvS6QCGyBVdpi4lU04WHq7hFeVleDspuvo1KboKk1aFic17QpR1fpMVeYxEkFH5mbxIDUf5E+EzEJOb7jkCdQhEA036WVEQJn32kKRFL2DVAM4+AYs6g/wlupFioaln/kdzE+bm1fA4dJJ3pCL5q57ciZeaGWlNn5l57v4kCPSkAYkCxwwATNRIAa7lddgNl3FgFop3/O/cIxBrCZWef3SR+VqKYepdJyflrinxcQCe+/n0c1+ezlEgwlUNwUTQ5zI2zrfBFa4tTLJRe84S5p4tLJhWjQQkH2ZMGPRRlVKuGX3H6UxKEqZuYzcS7xVGS/2LeD3KrfyQJXDdVB0THRkbPTWSO/Wvn9W15EMeVI2SKX/Zb5MwceBGZ2th3YvTCjGrjFs/vG33cPjhvdvC5Rt3hMV6ptjk9HkcCeMgjQGOszYsD2c+b2VYt3qJxs+nP8w6rl+/viHJ69ataxhfTmTkCJhhurxrq8fhmzKczsxL38y3mTgoxZlfexVjo9Ep6RkhVL6OqXrlry7fQxaL4qK6jcAAMhoGBIFGw/lKE0HO+ZG1uPYKYl7OaSEgTBJXzIhupQwDIxnnGFPR9vLhmENcNUBaSx/ji/pLP6btX99P/0Y+0GG02DU5Rho0xgQZ0e5mjEBPGpCgpcqVCN30FF3hN5S76GA+uZD/9h0MjMn9dMlPmtaeKcYnjxFGeoTBrQm0/WbpQAbkOz9+tUZw22+Y1wmpEgTWhOywQr2xMQyOjjXv6EyjS+o0ESwW23N+ZGxFIcNG2V+hgPejEwFN2gTOqOzZvXJQQzmiW2A5VswpGj42AGQ1y3LNaA4JeUqjUG03SyFNXi75kFXswEQhi1FJboYLRAhtpLP73OLB0hx0yftFjsTNSEAmq0Vy5hpnsfiMLg/QsSbtRhMU+Ufeu3vvsBw78Fy49IEdYcVCMVtjE20rlyGhjRMHTlwvf1azAynYupsuAlrzwh8oX8DPtfnN3qX8IgmtuvQdzRLPIt7aU2zD1mBqXwivRo4mXq6EGP1pSWrTdvsVWNDW2VBnTPo5Blxseb+1Dq7z8YYJA1kKnEW47wjUI5AwkfZx8q1s7GOJNAlgXMZ1Pj5eqj4Gn5XiYr+akpLmYbREeRUHIaO8g0auo+GZPuCBGSLQswYkLKRMlACXV5y185J47SzzzI9xkmqBM0S8xWM00r3D4+Hmx57TZV6NB+jrG2x8IWRyyOshqxaFw9YumZdLvFpA094twYBllX12tllSd4YYL8nXc7xG2MiPtAR70GnR4x+BIIeWBRNcJgw8w8n8EjGLbZSMICafoYTlfyQxmkexo4idAyQZuYTLcOwsp7/k5Qm7xSuhTTsqIcK+RdL70Cy/2rIkLyjKE1C0wxeAwC/LSzPO5ZJdx1SkFiw1OruXe2DGQaVH2ia7Rg5MjIVVC2Wp5rR6nwk5lFowneBQajkfUZZcT/TFsxRnTFSvPgh/8E/4N87cZxzSDBK4Qb/nFdnKs2W52J6hjRyiNCIY4+tzzegww7Y+RS9c6woSBsPlvw6WKnjSjg00uY7xoJFhpstJItC9AJOXcTYICJ+gF8dmKYxmg/I22Mh1PkxekjjuF5LjudnQMOnZhMHhYQY+1ZlvV/EaygllO8lyVZs2PuF/p4PAtLrw6by46mnhOX6Z0pkPR8ZiWVdUwGJpkuaTsGLxJWSXvq279oZPXf1EuO2pPXow8ORc4hgQtMRQTDEiJK9a1B/+v2NWhl87fZ0YkAulfJLGGvrkF3VZDHjITyoUsYJylCJFRavTO7lwkjYVPsmtnvNQJBMFJLF2mBnSE2sEC13eKL5+M6zoyj3BjBleG2wpCzI9uDhpcbG7iHVI3epH+lK3/BsQPoc+HCPualKm9V40dZES7TAFA6Br5BivRciorwmiBBkXza7MjbgiCvyNPwxWlMm8QW3XqZIpqfVbYGk/auzFlzQq1ozieCfyDQNybES+f5TdI5FNRgcvNVoa0ct9do0cGRnWd/CuPlkS6m76CGgLQuYxQ82KFYnQNiO+DnrIK2k6Fm85KPfC6wXzhr3ffHiF2TH4QMhT3RCa811ZbEnJE9BDeSC6x5wNTgkAUnJpTwoBfwQha1AaBpgMnyiz5Bpg3TkCLRGI/KT8I0HVK5P0hNWZz0UuHAepiNNU5fxJeTifSQzHHgLdMG7qEwmI3wunj5VDVU+8tXcNSBQQNg7J9Ur577uo/XhtTIlwFqGs69SK5w3eHRWs4TAo0n2RtLhFDbLBoIUi/WA+1yqJEwp1dH5ElhpYh9LgFV0YhSigw5SfaEITybq4iFRWXJY5qtEhUeBogwcq5OS651yELRoOSflNscBAi98vgGwUteBpYbDiPjiW6eiLVNeVfNimPvZE9XkKHRqVxRvFZdKmRZcsySvyU37AaTKtym8Y5mrJZbSWRSPGvY4a41OfeYwkXHMpclApFrmoLgJYKGkq4zjrUM87FH9MhgckHzNsbak5gwYM/8TR4kgCz4I0GzmMMfuoZSqUvN56mYEO1soIUnzxk9pXLNL4BJnIEpa4HLh0IEteTd3TXjBskdnQZfTEvi+2O21TkkaPbFIOloQ95aTwIrS1D7MP/NsqP/IAYdRWYk/U4wggfsekbxhP+gldjZDDhPumSquolntcqypmEbn0RQaREfwiM1vHpRGTskFe0NMhS2KKxukmPegRDRHoWQMS9qk3MBoilETqTIvwWnm6n4y6SgMdHhoWQ1KWaGmXaY0hEqEzQiLziSWc/36IkUgU7FEZ4ecXxuUMMWvRSRm62UMMUJ8ItmzkNStxVIxqZ4NiHB1pVLQ1dS3k2Qu6NpQgJxIVI8iKb3hR7LyRmIURwsxAktKeKgMk5XbJR/xU6SUf6M67yddlUkXOKkFQ3kRxEy5SivCjA08U4Qy/iLQ8lwTKm72VDLQd0BZiZuqltClRRpySy8x9SjAxPFaCM8xYaq5AqFVgNUWmgmCs8ujnaNBHkI0SiOXK3fRg+whEltD0QG18SoSF6+O5pwNKTAmW5uK7mQnvk84t2kP1gzIM7CZ8Ir4OlcplZCeo70FH49Zf+2VXaZXIhvaf8pQ9iQDNTdqaylxYLQHBfMMk9jF2FVmSD8WiK09u0HXVd1+Wa0ZNDEX5pr2QRNSXoD61X0+FQO8akCi+0uv09cN67Tk60Amz4tp7pP1UwsvMQGL8caTCBLOj2oPS8GB08ZNWoY1FwkY5Sj3FQZFVA3JU3iERfaoVtk9CN6QEogSmFsVpIDgaRLV4QdfcotiIU1CDhzJbMJuhtaWhDEhkRg/82AFFPlWMoDHXFpRmicoI1no3G1PLxZ8SnY76C2jMlkUXAYydWcyce4ZZSpMkMMOuFPKs3FSoyAQuzcC1/BRJuW2UA15MJynqNQF7qABfu26ha0K+g5zgrAbJKw5bRKL5mxqQ5JcQiMc28RMTsjRfKeWmuxkjIEDDg9p36EuioQb+cDHOwvAzrBTrTmtI75fyR+mSWWbxdXkmg6DwiA6GJjUPf9rgqPIPSWI60vaOS7CRMtsAt+KiGMg9+a+O6wRHrkGIny5ZJ8KdIzAVAsJD6YoQ5a/4gLFWnr+sbcZkxm34JTjokl9+4Dafi/J5EqF9rrQD5JtSUxJJZDcyOh72jIhuEEFIKJjaQ38ZlG8HFi3oFx/JW23XswakKn90n7ZulbqynrNRncn9eF4gzFc85/FGGJzlWcxA6k9O/2bmhSVvtq05NqE2APmD/mVOjdtROUx4HONRKNRGZXd7xNdq4Y+o7eKpIipXZlREFDSRBuN9EsY7vfqXwQb4R0f+8zyVAGJ6SK1yRisAu3LBi22NfKDC8jKfmBjmrtFZW98l1irtTJZU9snmVZmLFGUxGa2WhpUD2plZRME+efN+qobcmeSzb8nISqtYEhHPd27RJYn1gqdKcmQj/DYu30CON6yoxnkTqzNgIh9TNiiJxF54LVyqfQSDomCr/2LJuYZH8JWXYrRE8AzMaxHF+5FvI030c5pn4tfkBnOnLh9OI3sikGLEFCzKsfz0mCOpuLgSBwVb2rrEo2Rrk9M2L2mR+70LXU/wRxGFVNmgOoL0dfAYTCO+OpUJ9C9ZPPxnfBkHKUhfBCWN3xHpS2iCHpLl6CPMAAvxsW1o40heVg5hjz27N1z/8M4wLCNh08mBPnndysFw3PplYd2q6h9V1bMGpHKTalDKbtgcrZ3cjywo6ZNHWj8ws7vC52GCXQZlJpIOgDyZSYwjGXJTjMrYIUjHIAYjDt2aojDbMTEWq5TuoqccuCl4dIqi/nAtANBh4jd0cgPFWpWihgl6IzLOkCmDT90OcpDYMSm5qBKCtAFzsQVGVSjGGpdzlYblIv+UPV24L/wWR/4zClvlQSo1gnQZYHvPtHpfq3umiEe5kXTu+oCExec7MhxUKG4S0PbDqodySRMeo42OChZGhZIy5R/9vhlBVzZ9U1Iy3xNoZWs90GdgLMITOrCQFM2u8TMH8NbKstiiQtmbyWc6lUx5oGI6zxRF9Vy/R2SiKvcyICOrqegCqTN8c/lrg0l9jE7BzHRtS+++I1CLQOQxVsaNiS6qOpX80cGIJOFYei2TIFlDFlEfv1evfV85V9p/Ja+2sH4vTT8t8dbb0GdbuGiZQb4Y2Xdv3hU+8+Mnws74cabknwMllVNZnG3YyfYlZxy0JKyQDTExIO195SA2+7f2rAFJxagukghaUWmEqWKFRjWVrlJm9HKVTfryDwEXunQWMi7xgiJZ2Kq0xOqWBpow6RijwYmTKN1RTxXUpEx2rzd8hMSEbEAkSwCoJw1HZdnqsxEO1ClCJEuTNepG6bsuTsoP73DuKLJuQPGQa4FBl5BpgZNIQdgELiPaOtOtSkjJqAiB1iHozp3C9/jmTMhaGlqFtgwKVoqT9yY0MdY6LmCRn3ZWLfLTWR15VFa3iCuLtkgAbQGq+C4aHo8DJVmutvBCSUloNriYmchSJjcL8xSxRMbxUuOprD5j3nad4aTGsBJJnN0vjLAee5HhmrQtaU/WfgDCBmnyio+2M2Eme7IMwOA93WEXAdSmgx52k83T3+aj8z6ZKvPIIv4lDbi+fuzafApN2Pq+eQ+CF6B0BOAv9E14Bj0ZXqOHsUEn9AfdCV0oUR1UfK7jWaNlkgcn269RPsgR7mfyJA6qkpb4MpzsND42GhaIQrBYMFCMLCslJ6Mlyz0mkGkj6bNHw5is0FEtVvrvvK6Tpa9GqCcNSKpKG4Aov1k/hWITK5amYV2oMRlT9Go8KjsYN5RQibwaumQoYkI0eMsp9ZOAkCNlyPKHtlH9piivEmb3eyEEHKOCwyDACD4qvHJtlTq3xmidbcSRJ3Ng9gJYVkYdRozfEfCNA50CTme5E0js/EddBmJ4CpZqQNp7SvNr68XqzXzLtv4aA6pMBwzkSYdqHZIZZa3y1Y2vpGPRZ1slnPG9WEE0AfjfaIvKYrsvLRc7FIwx+WEotONSlpNndLal5Lpth6b5ngb+izNXiWoFw5hDKIIxcQa+xCScFe+VVQfQpe+O9MAhqZIqYRzX+XgllZh8GTRlj/xRqERyS/lp86kin+KR1KfBIfXLI9ICLaan/Gs3bg8/kR8YKDLgpDwPKAqmMXuKi6V9/v5Lw2kbVob9VzbaIz9N3mWBnBAQfLJJldj/UVhtkwl21l6RMf2lDkbWwqyrgZJ61LDcZmIocn+UKlax1i/WvmH2V5S5T3T3UTluSqzA0J8ZGC1fbrO5LLEdkz1MeH5C3tPXv6Dlc3N9sycNyAg6IoFfJkSNxex+FCWx8WhnSwOBIeKNmKzgvzD+uDDe+JiMYkxzgx/Zy1xH9kslsODyFvk6VcpFYFFN6sy3TOQ6rWPumVzkgfq09ky3+xRdyq+ji9IU+hJJpnAkmICZdgrJkmogYWaLNoGALtVpZQoFUldaXWRHwLLNd/wWlpspD5RIHLhxniPfJ0MSBqQujZELyx+SlC+VjoTwkmUIWVE3Qpz41p0rAYqjzSwblKo8URYiANqwjY8U+1fzENpklQXfx+hlkgPo4BKU9F4N9Zo+kdeWOD7if6eFQAKeVDimhFW31YUpM8oIdpP3yxIFlkkqo+Tjp5X3VImjIZSXK0mNy4ORM5RPJUg8tOr1VK+d4X3aLoOONz28I9z0+K6wZc9oqPnkWd4LFM3YkYHntUsGw88dtybst0J2Ri/ckbvIIOpGR6dYcWCIGV6xDFAKVtF8RH5zzfO94WJdhnD/lt3hsvu2xXqLAjqrwAQO3eU+CVsSUB0aHg3HHLC4xwxIAUL+s3O2bmYFLgnDG/vQRpSXxLe2gC4bIYx/y+Ay3qwD2QkhytP5jHK0RmIkQgksj6YxJn/kp8dVCSPRh6n+kmRJuN4RBX/p4KqsAR7leXQLEXA26VH/TBWuK2NAAtbmzZvD8uXLw4oVK0oDjYrip6wNg0/JRzEB6UnLsrDYUKi+BpxA9Ayc0QUTqQEpoxATfPso1/l7jV4Ng4o+pganKo4kKo60RllWK46yUk1amRpIwnJDq48/CSB6nZAv4Vj/RGY3qtxgE8pn7anATzGBt5PR6wyGFDLd9IUcEWaixOPoSPgGkk6iDFazbii2O/KIuZA7oZRMrcBaCmIUT1oR5Gm0gAIc70lpkfLrRhTWSCVD7Rzkj8oI8pNsE8h07EnEnCjh8T7t1t5VAGmTXwEACkZ2K3bqQhRwpPQSSK7xDd2CMOONmZMMBATtXOsGyDRrSWg+z3CgETjhmLXEYMcVU5v6qt77Q5tVvsgjHWFQXJVh5R7rni0siMPTevxKiYil+Us9T26zGb2pkWv8nd9RrnD6JsKdm54L37x1S7h323BYBC71ztoK9CRhgoPCvEfuszCcftgyNSAVQ0tb/46ZXEsm9Pn6OYsMTEW5HHGynWu1ndNiSEse8geJam1pJtnOx2cUexnm2713JDyze0Rmd5pLX9WpkkJaGOye2zMchobkmDTm4cQ4mMyj8xGZKWiWgqvxyMCEDUgqI2XPYcRpqxA/Suh4T/vHLFkpIUgxcsyflFH9jfrrSQ/MLAIe46dnFYsRiM6kWcmfNMs0kOUh0k4uJC36QWI80l6r7iphQD733HPhrrvuCps2bQqLFi0Kz3/+88Phhx8+7cY5VWPW+ybcpYNSAZoK81iBVJhVWxYjkXKhzME0A7VMBAmKcgkd6qk1KB8ss4RViGGnRPrxfMPMZ4vSqoxHopTpRDimZcun7q5wszKCFYKfuuTHX7oLmjN+EpXExMte+5uxB3wjiAnvJN2AImSsBFqKofxJkNO2oMtASJS9qBAIYzvN8iJzzd+qLWl+lhmzFVrXquxmBoc+QyIj2h4owldQwIyRwvhCohAP9c6yJxlLzPpE2S11oMcyNF/R4yIiQi0jTKg2rWO5JKwprNIlNddFOerU2ipyl05ydJTldpEGZBh1iNNvceUavIwcpVXSI4PdFYFAgmXSZhq+seYeXAP2RXLF5FwxDLWfJW9jiLpkUFBLCVf6YHKn7oFZXEJLn/TH/TKUsVxWk+0nm1ssyLYuzr1ZlEa5GpCUyCOlSBh4UIyUJQNi4MmqoigdmhQq96bpBhnUYwCe79JxtJu84/vQPtktWp2EI23JQHgCXcnVmidnzsLU5bgMzI/J8sAF8p0ZxpDA0dgJLlrN4CPOkvHsCOdsg2fdAFhM2Y1/ExDEA4fY3xKqNRaTVCkAOiFO51h/I01RVCBmoG1VO4rY/ni7UpnQneamyfkTn0vjCwjQx+mbhdc4qspGjy2nSI9cEUi5Kss4yj+hTPQxntFk2e3KhebcgByVHUdvuumm8NGPfjRceOGF4eKLLw4333xzePOb3xwOPvjgpoAxTWzf0GiDlpRDQ0Nh4cKFaXz+Ye2UpFJZWzwsv6XSDEb7x0XAm0C3Ks7YisozYYyg4QPXRWLUDUrLGOE9kl//gCzFSgR3Pr/phinL0PCwnuO4ULqiJX1jYTFNVfJVwycdcm385lG5v1g6rz45xmNU3gMWRdHWOMdqxIIbdTomS34XCF7LpX4WCRZRyFGXmTDJajapa8GW3a8YXeM3Kp3LsGDXLx1+EXVaDYQaUwFuEyyTljIPiHK0VNpCNH4iNjW8L68glp+1FtrFmLSDQUEazMIAdSCKVtqeJMEMHW0VumjXCySPJVKnqpxpW4gyWTt+uZakKV2ZvhsNE6VN3kE7HeO4jYLaKTJreHhE2ttYWC70LU6UMrAxeQF+hqFhBn4YkEuRIYL5sNIm/Ca7PhVBG50OmI2MyAf8QtsyoWapaUgAlRClmMklt6CJaxzHavD9VJ90fJwniwwJ/cXWKdihjC/tG5GfLvid3EtCp7kkjAwcEB4dmIjfhwwNDQdppi1xg48GBgbC4OCcd3NWmkr4VDdGurYzq/wcZfArePOtU7axXEwfkyUMk3umqKCdgaxMkTXomtdb7qkvAdEGtDwWV/PALC9GhGe1j5E2wWYhtHHtl5PMCOPootmBEp8obqNDsnvwCDJS5GPRRgd5UE8oq7JllvZnse9LiIKuHI41Ya173tD9Dl7Hqd4oxjzHL1FJDdg/gkFyq8QYo3/REfgxw9Qn8qsIuZ17fUWDYhQJHmrURNjgOKU147ImpMdkTW7OMlqJStq99H3ILBuYJWwuC1lM+T78ho5F08vnn8IhASSJ0Wl4yiMyli9PpAnLp3U2Ocx5z/rAAw+EO++8Uw3Gc889V43GK6+8Mlx++eUa16hwVM4111wTHn/8cVVyEAoosd/85jcbJdc4EyDUzKhsg/iK/fdoZzCVTmn1CBMQ5puGRcM7wvcu3azKclECBPr4jcjuSy9aMhxOPAgB1bQ4k24YbUt3PxuuuPxxPYS0KNomZVahiFivE2Hv0Gh4Xv9IOOiQiBt4GHxWh43I1nqVBEt37AnXXPlwuGEwKrXdjp3hNjwyFk5YMhSOXYdaVOsMtzyO+bCwa1gysTd8+7tPCq9GM6kd3EgT86/NL39FRzAqnfxLVgyF0WWoSLX1aXTUPJO74P7CwV3h/y67rPB2iiRAkTxl6d5w/GG1/NYKM8izciwc3BmuuerZcIMeFizdyHQae66c+SCY8kMeHi6G1oEbxFhrBFSODqPX3kPyJRM7ws3XPhXuvDF2D0XRRh7Doozvt2ck/OrBsrN0E9qMlnofWpcNbQ93/PTxcN+ttNPWuIHDfvvtF4455phw+OGH17+u4XU9X7bDqw1fNA8ircVT5nw5I09ElcYUG6us8pc9RqYw1rBNYaCp2cYXsRzCCwikSHxh6Ft7wgAcHR4K4yPSvkTUZUtFRZYZsXW5YkiC38iwDJkI34+Jdlj8N00U2Aod2z8dH0Yk+UdHwIiMkWCGYZs+mqTsZo+61NUPKPZy1nb/qMwWCyxZXcYwGGjdJZAZjjzPjtv4veWk3AxQCA+Paz8PMIZBBMliEsj0PinUUGcUpURHnpq/1Iv1KVPVkfKCFaEM2uAR6X80nxbvrydBv+dUpqy/0+Ilc3hrzg3IrVu36rePL3nJS/T7xyOOOCL8+Mc/Do899lhTWOjsjjrqKDU2URKeeOIJNUJf/OIXhwULZB4KM76F4/lBWZ+QMXuLxHW3qFblDVEgWzEpMzHMBjz11FPhtttuC5RvzZo12pHUvbLmUmkTrc8aQs3NKS6mSxsYs2z4ec97Xli/fv2UtE2RfSG3KT/u7rvvDrt37w7r1q0LBx10kCrFrTLoF8ww7pspzK2e5R59KUZBqzolHbMZ1Ce0HXrooeGAAw6Ykt94rhOO2fc77rhDB1ZOOeWUsHLlSuGj1h2eYqbYzYxCRtj027QpOlXaJQM+rC4455xzwuLFizXDVnjPpi3wckganaJOjd+2b98err766nDWWWeF1atXT9kWZkNbu+2UPJBlzz77bLjxxhvDscceGzZs2KBypVVt8dxAyXVKHtQdsvfRRx9VQ41PD2w1SDP6iqBNZ4Gm4DdmHhmc3LVrV9i5c2czclrGQ2u3Oqk9ma1il0SRefIvKslwZlQNgRdZmsIs19S3qoJpZMHoKNxRButRAHId6YtEQS8OGhg+4Ep7egJy/lDrXl/SzNBRbp1NkLZIWMa2xClqyRshYDKvKH0SzWSXzoyUhRvvZQYm+T7NDJ6EOKXZWBn6cfylBOomkx7ju/SvHpMms4giJKUNRCyAIM8/OiiQwEMY2DSNTDEbhl0Kz6RiKc8LAHruM7wsSCA/gIc5tDh0HHEkLrJTBC8OOMXwpBcXEgElSf1YPYmf5/fYV5GZ0REp5NpiCiElfQnvl3czUAFuml2Mi0my/NNHkoDKCuRMSZTV5zfb6zk3IONysOF06SkKOhVOfCu3//77p7dJj8GBUVk1h+IMbYcffnjYZ599KkUexveOHTvCIYccokZalYh75plnwrZt25Suww47rEqkqSEEn2JAYuBWyTFgAXZV5DcGVTBwjzzySB3oqRJuGJDXXnut4obhXSX39NNP66AFgzy01So56hQDl4GUqtG2Z8+e8Mgjj1RmgGfu680UF1GwRHPSDZWwhqT/jEu/5L7qOcmScblAkVHlWR7At1+5ZUGtQ1ONymHc8CfmnWinql6Zwq/KFiNZJSpdE7JkcUJ3R8++NazHQFam64wWZKBUQ5/adKMJ7lGTrH9sVtfUhzC4jJQJfVQqVr9iIfGExanBK8RFw1ejYt2XjFnMqWJ/qQMBghlIhiGiccjgQ1JHDXjIjEwMJ/igTD6rGFpCTsQlyopYcswbMxqJgc85P5qJM9pA3kXjMx9TcNgEEr62L5NXVp+WX44wbTS5a0tSlC9ZU24dqBABMK1mL8/pzuni15egKPKKfM+cG5BsmsNsBMYCyggzO/jTMbYwNqkw/WZKDLaqOOjBSMPAnWpkvtM065IaoQ03lbHeadrIz2aRq0gbvIarIm3gZgMw0Em4Ko42UMV2Cj7WPs2vCmbUJ3yGTESWVM1ZG6gibSZ7wc5dLQIYjCwPjaJM5JmKtESpQm5Icq5M1sV0SWTtqwq+ghboEj8xWi0DpY4/ItNUtklQdxolPUZULIQlL9QHL47W4vOXZjIVklGiFTvCQgGsN6azVpmJUihhlDkpPwYiP6s7IDHOJ77G6TOGGE/0kKMuWVZZj0kOAgwhnYWUOOxskqqxCahyHVHG7x0Xl1dmsoGSR86JRiTjFwqNxEefdsqsfckYaQaJLEsyy2dpMiyjIn83iy0qFNs/5ZZBHWGe6bQuaJ0Yt/UVRVFU3nvm3IBcu3atzuIwM3HcccfpbB0d/tFHH912qRHozYR62y8pKSFlwUCuohLDLBoGPMu9quaYueVXZdqqXKdVpA0eq2o7rSpdYEZdVrWd0j5ZOo0sqZqDpqrKkEZYYfCiQIBpZ/gRRUp+eX0qCavSI7SY8iNqjdLWr4tHG1FfRBxnGIoyKkrnmFhijH2Rv+n5RgvnrhrRegarPKOmkvpF0DH5HWpAMgspCmGyUnRSIrkrZMVZU+hjMwylmSNG8hhPenIWEVJmaNMp5aSOlHckPoeSZGAERBT1njyH0t9rTo+WkLamxr5Z2HUgWF3GaOqSUBy4yLCse6hLL+O3xZHPmjcxGClyXMZS0YAsC6/IyWQrIRtUIizOjMZIkUbFtkhQ09hAQLxXxl8GtaBD+SwDJc0KMWbtT0Wa3FH5ArMp0LEs6QMVDMx5r8/3dyyBev/73x9uvfXW8OSTT4ZXvvKV4fzzz28bLjpcjLTOdLptk6X0oMQsW7assgYkuFXRSEMphbYqKqZGW1VxW7p0aeXaAq0GQwjsqtZOoQ2aqihDDDdkCMZQ1Rztc8mSJZWkDbyo0yriVl+PrLxhEHXv3r06eJr/RKM+bRHXOtItisqEaS7tvFSUGp1RSI2Rdh6aXhpdKqfvF40r/k9fkOlgWYhEzBDV0lW84qXKoHw3NyGbjsXzFVOymgYErTAiy0jHx2xGoZyBbnTN+I1mJAUjPO+gI1Ok4z2SRCU7S1tFuZwvRzFhQQLjUTZE6h9oX/0FQ13/MSYyOMG3N/AS1DG4pMz6DSTskjFTDFMxGg9TcWGOi5oIu1GMnzcWoY+3Wt0kOZC7kZtSImmMMn1G/yQPFOTpbKwMOLFoiNfrcVXiI6vsFCD23UhWmWs8WTNOocvly8SNjApy7beggjJs9BpmHj/ykY8kAi2EAw88sJJGTSPa24mrsqCpOm1VpA+aqkgXvFhl2oy+dtqMp6lFoKr8ZnVaVfqq3h7Aj3OQP/zhD+tGRBi73/nOd8I73/lO/aa0nZUEpKGcU6XlvqZVrYXZgUTfQpsyLQuCGjiS6Oo9CWhebECiK1dmr30Z7dDGQk8dgNcZH8nU6MpnU09vco0Kae+SQINSTD8KmhgoBCtmiMfkiJx+0wCneB3PQM+oaIqGWZF0aV3K+3WWQzRTNpdq6CQ6HScgiUCjAwikl/AAddlfvVVIDcsyw0jjiz4pKxBwrNOEnAXZLptQl/DmGMwp7UcHj9t9eIY0z/Vj8NeE/HBqQAsITXmsCbGWnnf1C/axQTdJ3Ga0tSXeyfuQS7r5jPBy/YyeDkhJPcX4yPy0BZ7BpXxRUF0qZiIveP2ofJdMQPmNPDXH7I/ylMTjCOO0CUthhGKVr30J/vFu9f5WwoBkBBsj0p0jUI9AHCWtj/XrZghUHa+q09cMV49vjkCV67TKtIEox0+xeRMzuS94wQt0tQobOv3whz8Mr3/96wOrCRo5dr9lgyA2CsKwwb/99tt193L7fjz/HDjw4972nbvCQRNbw3lr9+pB9/l0rcLMbHFu8nObHwg/vX5LWLp4ERqYKmHNnlOFKsm7URqjS2mTTWpW730ynLN2l+4i3Ch9ozh5vRhAfWHF3qFw121D4YkHZQWG0Ily2MxF5RNFsl6ty54w2saFrrEtz4TjFu8Kh+u5z1maqUIogksW9ocH7ro17HziAcErGvHNnoNmflZXjcqQ0iUzHJue3h4O7tsRzlkzNC3MyB/lfvODd4Urdz4uBlHcvLBRfkar1SWYtUpn6TvlG13g0sgZXnqfzXNk5/8zVm2fURn2H9sZNt49FHZt3jitumxGWyN6OxFn/G/fiTfKk3pm5v3pZ58Lq4e3hXNX75k2j7EL+l6RFz+55pl09VEr3rG6bMVjYMl9zinu2/ZsOG/fnWEBgzo0d1gg3+zz18Yech+Dc/nu8XDPHbvDU4/eL7zQvF2Sn602I99WTu9Lu3xu07Zw2oqtk4zGVs9yDxZe279A2uVw+NGux6fkMWhjv5iDDz447LvvvlO9vvD7lTAgCy+Vv9ARcAQcAUfAEag4AhiQ1113XTj11FPD6aefrgYkcV/72tfCa1/72qYGJJsXsdyVH0ogmz+hUGFINlNyUDZQ3pYvWRTOO369LKUSQ2UG+AzLjBozOHskb1wjhZA46Lj//vv1+CqOxjGltT5L6LL3nHH0AeFnjs1rgPWpm19znNCI0LZ3aFgSxbI2Sk1+fCqDY8dllocbDfXpDbNjD1kjZ76ulbLWp5jqGsO9LwwLXnuH5eiIpA4aPUVeHDvDLtrQxf4QU9XlujUrwiH7rUyXwjV6b6u44VHOxYZ/4q73jeqS56HtwQcfVLrYcRmeq4KDXujiG3HOfG32yYvVL+lPPHRNOPXwtTMgXwwXYdXIY1KXLXiMl2/ZskUHiFasWKG4GQ0zyLjQR1guD20s72e131Q8tnrF4nDmqvXh7Cbfi7YijuaCvOBMZ2QVrhmPIR8efvhhrUMMomY8ZjjynhMO2zecesR+05Zj0IVxy+qAoSnaJXRxVBX5wmPwWjNHGug64oCV4ej1curCDOQFPAZdU8kL8qEe2aUdvncDslmteLwj4Ag4Ao6AI9BlCKC8sQcASokZWIQxJExRalRkjkyxY1MwHr/4xS+Gk08+OXCOcpXc5z//eZ1ZPemkk6pEVvjRj36kih5nq6IUVsVxNvP1118fjjnmGD33tSp0QcdXvvKVwFmv8FmV3Fe/+lU9RuilL32pznJVhbYbbrghsJqAfT42bNhQFbJ0gIIzjzE4zjjjjMrQBSEs32fVxbnnnlspui677DIdpOOc6Cod88URgaxEsd3QOw3aDMYUOk2i5+cIOAKOgCPgCHQfAowis7EURiAj7hiNGJXT2dCJZ3mu2Yj9XKFGOYwuZlWr4qDLfnOleDXCAqygxzBrlGau4vJ1OVc0tMoX+mgHVXJV5DHwMR6Dvqo5eL+qdFn7rBJmJvPpR+bC+RLWuUDd83QEHAFHwBHoeQT4toblWiypZIkXS5EIH3nkkU2X4zUDDQWnao7yMLM6VwpOMzxsqWPV6IIeNlKy2ehm9M9FfBX5CxzAq4qY0bahy76fm4s6a5Yn/F9FuqrKY1WVF8iJuaxLNyCbtTCPdwQcAUfAEXAESkSA5aove9nLwqc//emwceNGPRKFXVnf/OY3a7idrDE6mMWsmkIIXSxHq+LRPfYdU9Uwg54qHotDXaKoVtGwBS94rGoOmuCzqvKYtYEq4VZF/gIfsKoi/zNAAW34c+HcgJwL1D1PR8ARcAQcgZ5HAOWS78r4fovNU1BSuGZH1nadKff4VXNVnYEBd/CqGmY2o1BFRRqaqkhXlXkMPqsaZiYvkDVVc1Vsk2BkWLm8qOWY6nFQLX1+5Qg4Ao6AI+AIdC0CKCVveMMbZlW+qi79gq4q0lZFmowBnDZDoj2/qnhVlS5DtYr0VZEm8KoqXXNNm2+iY63JfUfAEXAEHAFHwBFwBBwBR8ARcAQcgZYIuAHZEh6/6Qg4Ao6AI+AIOAKOgCPgCDgCjoAjYAi4AWlIuN+VCExn6cF00nYlWDMslOM2Q+D8sRQB56EUCg84Ao6AI+AIOAKVR6Bnv4G85557wl133aU7GL3whS8M69evD1dccUU4/fTTdec4DvTlgOfjjz9e0xFme3UOEeUw0TJ2sOL8Gw4F5aPr++67T/NnQ4UTTjhBGemJJ54Id955Z9i2bZseTsvhzGz9zuYLDz74YNi8ebPStv/++5f24fYDDzwQbr/9dt184JRTTgnkdccdd4Q1a9aEww47TGnYvXt3WLVqVeDsr0cffVR/7EjGboPLli0rpVGAG4f2kid1C20cqk2+xF933XWKJ3V91FFH6VlIjz/+uJ4d9dOf/jSceeaZ4fDDDy+FNl5KHV111VVhaGhI81qxYoXGUdfgBs3E8T3U008/rR9t33LLLbqdP4ddL1++vBTaqKt777037Ny5M2zatCkcffTRyvN8/M8ZQxy4bccKvOhFL9JvAahveHXr1q1KM/FlOdoddbdjxw7dWIQDtm+77TY9RH3t2rWBdgrfn3rqqdpOaRtcr169Wg9JLqOdYuxAD3nR7qgvDiM/99xzFQbaJG2EtkL7OOecc7TewYudNp966iltpzxT1gYL5EE9wXe0A9rDTTfdpPXFERHQCI6HHnqo1inth7bK+YPnnXde2zuATrfeH374YeU18ofv4B3aHRthgCeHqFPnyDwO4OZsN/iSdn3jjTeGs88+W2mebr4zSe9G7UxQ82ccAUfAEXAEegGBnjMgUQq2bNkSrr766sB26bt27Qoo0S9+8YvDDTfcoAoUygxKC8YOh67+0z/9UzjooIN0O2YUH557zWteU/gObuSFkoQihUKPwolit27dOjUgUABRuszQRDmlPD/4wQ/CzTffrEbwySefrPfLUEwxuFA6UZpxKJxs/nDNNdeEAw44QGm57LLLVFmFLhT/+++/Xw1ylEKMIBR9DKWiHYoneVFPDz30kCrvv/Zrv6Z0ocwzWABuKLCvfvWrdSAAgw66MADArSwHbtCHsYMB+b3vfS9gyMJ71Pdv/dZvaRyDFxht3MeQg1bo3rNnjyrOZdAHn3/qU59SRR5+g1Z4nPZw+eWXq8GDscQ92gmDJ//7v/+rdYpxhpJflqPcYEHeDAKQvxkU1Cm8RPtgYOXEE08M//iP/6jGOPTQTminv/ALv1A4efARRvUll1yidUOdwXv77LOP0kFdY7xBL3UOvzPAAr8xKMC5f9T9vvvuWzhtvBAeQ1bAO7RD+OjAAw9Uo5YBFXCF5/mxBf6tt96aHiFBe4BeDM4yBi2uvfZabZ/INOjCoObICrBAtlg7pV6RsdAHbvAAcrvMwYr6yqjajnv19Pm1I+AIOAKOgCMwVwj03BJWDDMzNl772tfqbAvK3t13361K83e/+93wsY99TBU/tlZn5PvLX/6yziT8/u//fmA26OMf/7gaI0VXGoopyjD0MJvx67/+62rgYOyieKHoMXv2S7/0SzrLQjmYYcBHWf2jP/ojVaCZPSrDffOb31TD+ud//ufDy1/+8vD1r39d6WMmCEXw3/7t33TGilkijPCf/OQnaoy85z3v0VmY//mf/9GZ1TJoY8aWGR+2v3/f+96neWNEoHxiCDGj8Zu/+ZuB2UZ+GCQ260K9HnfccWWQpe+Ehv/6r//SeqNO4TEUZWagmOH77//+bzXGmd1GucfIwOCgHCjOV155pRoFZRAIv1Gv8Pqf/MmfKD995StfUeProx/9qOL2zne+U9sBAyko1fAhRub5558fXvGKV5RBlr4T2uApaHvd616ngyoYbaeddlr4zne+Ez7zmc9oO8UYt3bKSoI/+IM/UMP27/7u79SAKppAjDLaGwM3zOb9zu/8jub37//+72pYUp8YuvDbIYccEr48Jv/SAABAAElEQVT2ta+pYc4AFfHQx8xfWe0UQ4yBCfL41V/9VZ3xZJAHYx++R35BI8YYgwA//vGPdaCMdsoM3ze+8Q1tz0Xjxvtok8gKVna8973vVd6m3bL6g3bCqgpwg16MYAbTMDQZ6PjDP/zDgGxx5wg4Ao6AI+AIOAJzi0DPzUCi+DF7gVLyyU9+UhV2Zho554WlZp/97GdVeWYmi0OQUZQxPpgRQnlmdB7FD2OvLAWQ5ZTkT37MjEErMxn4zP6hbKFYYWygOKN0MVKPs/NqymArjDQMGpQ58maZIHS8/vWvV2PnS1/6Uvjc5z6ntDCbijGHcYkxyWzRv/zLv+jSvzJooz42bNigMyfkDYbgh6GG4s7Awfe//32tTzDiGvpRmJk5KtOBEYMA//AP/6DZMDNGfWG0gt1b3vKWcPHFF6tyzEwWdMGLOOqW8oB5GUtsmWWCx1jmCD/TFuB56peBDAw4ZotYYsuMNw4D6md+5mf0vDqNKOEP+T8kM8lgRZukjpi1Y7kvhjfLLDFomfl++9vfrulopyz5pt4pV5ntFAwwbKEHwx/58B//8R9qHMFbGJfMmCFvWA3ALCU42xLustsp2GGoMbjErCLY0QaJox1j/EM7gyi0U2YoaafQCN6UoQxHO4XX4GuMaWa0wZJ2Cq0YuKxigOepP+hAhjCgBs7uHAFHwBFwBBwBR2DuEeg5A9KMQAyHn/3Zn1UFFOUOhY5laSiqzEYyKo5DuUGp4TkcPkqtXWtkgX94L8qzGacoUBit0MioPbMtzBywTI3ZAwzLMo3ZfNFQhDHMWFpmy3sxdjDI8ZkZYpaR2UnwRGmFdhyKNM+whK4MB24opDjyZvYKejHUNohhiZHBMjmwoo75lpS6Beey6tLKyfuZqWJGERqpXwwfeIyZFpbaYqCxhBV8oIuBAeqXGWa+SzMc7Z1F+SjqLFHFeMBxjQHJNXleeOGFOutOGmiDfugrY4l0vky8n/wxct7xjncoDfAQdUv+DApQt2BEGyE97dTowqed2nX+3UWEqVMwsfeDGQYONGFoswT4rW99q9IAvcgWZt+sXRdBQ7N3UHcYhcgLeApeou7gfeoX2pnlJh4HrtZ24E2Mb+OHZnnMJh4acORBOwUTcMOwpK4xGK2dMptKmHovu53Opkz+rCPgCDgCjoAj0EsI9JwByeYuKFXMrKDYoSyx7A1FlRkiRuYZHWcTim9961vh537u51TB+sIXvqCzSHw3xHdVpjgWzSy8l9kqZhpRtMiPJXIoyiyFY2aDTUIwKJmlwseY5Fe2o9x818hM4wYxylA8icNoxCh717vepTSy5BJjDeMHRR+fWQ9mbMqYRaPc4INSzLJLwsz6oYiSHwroF7/4RTW+MTaoYxRqlH4ziMrEDkUeY5GZHYwLDGlmmPmGFCWapaN/+7d/q8YktDLL+4lPfEIxxbCE3rK+lzPj5q//+q9VSccIY3aR2XeWSl966aWKK8YcSyIpCwMDZnCUhRt1yPJPBiZY5ssgBRiAHwM8LMekbYIrWP3e7/2e0sUsN7zJ7B+8WZbRwXuZzWPZKt9p0k5/8Rd/UZeYMzhA+2XpO+Vg2TkOg60T7fQlL3lJYBnyD3/4Q8UBOcKM7Ve/+lWlgyXb8BVYIQtpnywXRa6AJ/WPPCzDMRjC99rwG7yPEU47ZTDv//7v/7SdMjNPO2UgijRgBnbwqjtHwBFwBBwBR8ARmHsEes6ARKFjdJ5vGXEowygoKCcsjWNJFcozhhL3cCg9KDIoYhgkKGNlKaYYEOSPwyhDiWZ5HIYls48scUS5YtQeejCI2UCEcpXtmH1E4YMGZnfIH0MChZ3ZA2ZduGa2gHvgZnShVIMbfhnOZimYycCoxrhACUY5pR75Zg6fGRYMR+JZ0ge+Zc8KoRBDE9+EQgP5QQPLBtnQh3qF7zCOWFKIw8hEaQbzMjcOga+oJ6uzM844Qw0IDOw3velNOmBh/EY7IR0z0BjAZTvaI0YEy3ehgcEe+AvawI12wWAFM3u4fDvFwGTX37Ic8oI6Aj/qkzYAf2NoY8Axc0ZbgWZwg+8ZmILGsh2b9LCcl1l2cINGcIMW5Ne5shyUNsxAFeVo1E7Bvgxn7ZQ8aRN8Tw29fPOLTGHAwtopuEEHG/p0op2WUV5/pyMwFwjQrsvSUeaiPJ5n9RBwHqtenXSaop4zIAEY5eW3f/u3J2HNTJA5Uz5R9JmhZAkksy9lO5Q9lGaMm/pZp1e+8pUNs+cbtU44jEFmWXB54cESTHMYbjhmMlD6+KaKmaCyHYopmDFbW+8wsPmRBnzNMbPXCcfSRgxEfs0cG4fgWNKHccaMWt7l8c7HzzbMwAiG2B//8R9PMqQZZLGBlnw+jdpO/n5RYQwzDMV6x6yZOWbh+WF0YIiwo20nNlpBOcNII294K+9Yys2v3mFkdsohQ/jlnfEYccyK8qOdghv0NpMv+XfMNkwbJK9G7RRDkR+GZX5Qp6xBp9mWZT4/z8ACAxzwMbP8yHZWbdA3MoDJQBt1hdxBZrLahB+DJhj7ZRkn5EHeDHgwCMmgGgNGOO6xOgPHwAI0M8hA24dGnmNg0NJrwoL+wJOszmCQA4zI35bIgxc4seqBPttWtvAMacDXBpsKIid9DXmRD4ON1Cm0kB/tB/yoY/BhYJXBZgYAeQbcKA9xDCzl+8X05bMMUF/wFNhAE7jZgBYDbUY7NJGG+oNWeAt8y6hHikS+0AaPgRt1Bj44cKEe4X14CTxJRz1CFzQysE980Y48rV3SL8P75Ee88Q8rXagz6pxy4PL1mJebRdEHJmBgbYB6pD6hEdqsXdAeoY164xnjMeqaXxm0gQHf+MPr0EQ9UrfQCi3QwD3ytvqDx+A36p2ylOHgK3iGvKERmUkdQgNxrHA0msEGek0ek560xFfd9aQBOZ1KQbBiBJXFaHlayAtlrqzGls9rtmEaQitHQ0bx69TGF+0sjS2jk2yFwUzu0XFiACFc6FjNTYW3pZuuD6/B33QQZQj46dIz0/TQTjnKUjrydFEXKF0M3JShSOTzKjtMO2UmnM62E46VClPV0Xzmw05gONs8UPoul52p2YwNJZCZc1aPsKs2sgcjng2NqCeULRyz/CwjZ0DprfJtb1lyneXqzJyjOKNkMThJu8axKojPSqD/V37lV/STCAZA2BwN/mVJ+W/8xm+EDSUM1LAyhCXWfCbBAB8DVex0jGMgCVn9r//6rzowwo7p7E2AUsiqE/C94IILVMHVBwr8w0Zn4MTAI/jQvtiUjTbNkUysvGGDKj4/ADP6ZPBlJ3LqmDhWnaCwFu2ghxUF9C0MyLOahmtkDisPuE+fDE0YbSxtpz6pXwa7WClWRp9N2akX8uFTAzBg1QgGECtH2DEdI4SBbwYIwZddqzFQoI/d+1lNUrTjEwLaJbSRFwO18DSGBauU0A0+//nP6+781C140f/wWRN1yOB9GZMJrFZhA0fooH0yofLGN75R93Wg3r797W9ru6AewYYBaT5Zon2wyoWd2vlEAcO7aIdMQiZAG59eUF/UL3sOsOM9n1DRDjHGqVNogjbkC0fQMRhdRn/DZzZ82gL/IjsZnIU25Cb58xkYn44wiYFsoI0QB0YPyQZ4TDacKyuFqu4yDbXqlM4RfXSwHI/RCQOSvJg1gOnKYOpOQkgHRuMoy/CpL0snZjnr8yzjGuWNme688VhGPvZOlvky+0jnOZ8d9FMOlJOyHW0TJRVFslP1VFaZ6PTp3DvVTm0FQ1nl8fdOjQDf5lLfKKgoXn/1V3+lO0Iz8o3ig8KIoshxLyhBKGEso+cYlX/+53/W41YwBspQCNmbgPb1l3/5l6pgfeADHwgcx2Pnmv7Zn/2ZKoJsSEY6Zhg4hgaD5G1ve5sqt1MjML0UGBrQhRH2oQ99SL8t5vvhV73qVXokE8cgoQTajAZKKgo3hgnGI8cgodijQBbtMDo4/gZ59Bd/8Rf6PTgGCEv4MXZwF110kZ71y4ZtfE6BAcmO0Xw7znNl6Dbsi8DMFJ9BMEgJ77CRHIoynySg9POdNt/+U4/sUYAx8qd/+qdqQHGftDZ4UCRu0AY/ffrTn9Y83v/+92u/QT1jKEIr7QPeYxYJQ5ZPJdDNKE9Z34dzfBJGDu0RQ5I6wvCmHqlbjrKizYEndQjfw2O0lXe/+906qMl10Q5DB57C+PngBz+ouDFIAjYY18wAQjNthEEBBr+pY85l/vCHP6wDLGUMtDJwhLGF7gcfcwwV7QyDkQEfBivYd4J6xPgGT8rxkY98RA1JZBwzhWUYarRJ8gMTPvkBH/JitRKnP7DakUFo9gJg8IfjtqAHw5E+krYyH9zgfCByLmlEkFDRnXDkNd8VecMJI7iTZSmjE7SydNJnEIFfpxydd6f4u8wydbKdUg5w4zffXafL0S3tdD7XOzNPKHksB2WJIT+Ud5RPFBoU0/+fvTsB0+wq6wR+e9+XdPa9QxISEkggrIGBhB0VRFBAI6swjIgoOgoKPI+AOAiKZBRwHhiGTUWBcUBAdlmjiCAQtoAQEghZu9PpvTu9zfs7n2/nS6WqU1Vdy1dV7+m+de9371n/95zzvv/znnPuC17wgqasImgUZko0AkUBotAa5JoMAkk5Zi2gELLCUF4RCaSWNY2CKE8sR+5TylhKETUDOvqBiXSmutnsTHoOyilySGk2LY1liFJqgypEV3tCypWBdYMc9GkhZMox0ZjBwmCt96L8lnGkUs9KlRu1GRgw2MVPzjoxhX2y2iNLrbThBEN1Tb5YyZBvRBzpodiblmm6KAJuBlZaKpGQySCQrK2W1yg/0uh9sUjJh+8MU97VIxuNIf3aCn+IBgvvZPT7BgJYGxEiWMCOFQ050vZ82ghu6pj8y7P3aGNCdUodMwCkHcNxIp06pi2ytKvX6ph3w9LnvSKz7373u9v3ob0/bVebZKVljZys2QreDyKrjskLDL1Dlkhnhh+7envX+gp4wdJvGCJsMPdeJ9ppV9qaARt9gHeDcMuXQTkzOQx2s3jDVv2CMwJpFshMGZguAjnRNafiKwQKgUKgECgEBhQB66yQQgo0R9FDQijNFBckCan0m/JDQaXkGBA02ITUUXgmw1E8k2RJi+KHgCCKlDIKGSvNb/3WbzXyxjJEEaO4TpaTNgsKDCigSARskFkWF8RQXuU9nd85gJpn5ZhoJ05pIR0cJZlSb+qx/CBDlnfAx2wDWHn/60PJn0wllbLsPSIQ0oEZqyjylelS/F2rY95tzh5JwjFZdcwAbaYh7SSI6pr8qWOu7SpvaitiIowBlMkgj95b1m91CoHwzrxbmBnwQRjznfLPqZPZVtQxZZmMOpZpJTFVx+CgHWoH8ggbxMygCmxZ1dSxrPstwxP8Bx7qDDKm/tg3AsmVNwc81DFnv+Ull2rIo/v6uslw6rV2KQ1OXZaWwzvyjrVPsybkH8GUR+0028dk5Gui4ywCOdGIVnyFQCFQCBQChcCAIoAwUj4phEgFpQYBeXusr6IEmk6FGFnPRPExHYyVzdonU/9MZ03FdaKLSCm1Zo/FjOJOoaIkUqiRWvmlaCFyCB3F20Hhn4yZGxRkVrJUjinF8KKIwoUFg/UHGTc12GZ7lFkWJFNG5ZUynYr1RONF2RS/94MYssiwRkqPBRKRS0sb6xE/LDcsbpNFNpQRCaPge0feofdjkMJ0X+kiZ9busWZT7JFwa8NYZFi04MyCMxlO2Vm0rR+Fh3aQhMIGX3DyDin28uw9y796NlmESPuDk3ql7Egt7FgUva8klvB7ZqxB1iZYs6xjFkZ9066994l26rB8qFsIkRkKBgQMDsgDa6SBCv6kL+8wQ4q0lcly0lT/9UUw8460U5ZFeJgmaio0/PRd2oJ3fvnll7f3L6/imAzHmggD/aX6nv2t92xmgnqlzmmb6lv/jIrJyM9kxVkEcrKQrXgPG4HJFHCHnbmKoBAoBAqBGYiADTDe+MY3NkWLwkXZsX4IcTP9yzQq3zdl+fDMlE2KPcsD5duavhzJn+jiUwiR11e/+tVNYbcRBgXr4phmhmS87GUvaxYPCpnNYuSDoo3oTZaTBpJoQxCbdkjPpkNICMX+F37hF5qyzALiGvnw7K1vfWtTuBEhCm5aIyYyn/DKDVUo9hR9m4NQ7n0H1hqwq2JaJOXZdEgKs+l7lNfJdE94whM63862jhZ2LEGmRdtQhBKNBCEYlHl5Us+8d4MEyvO7v/u7zcI8GXk00ICsmq6NQBookUfEB/Gwzs/7UsesO1Tv5Xky65h3Ig+m0FqzynrF+gkL+YAHomSAwqY06phn2ql8eQZHJG6inTpm1oG1vaYmG7CwNtP0Wb+t3ZQXAwbu28dBnpOUT3R+Mj7rjqVtWq8BC3UfPuq7d6nvUO+9aw6Z1Y/p+/Rpz33ucxvGGd9EnmGGWFtnrO7Lqz5DXdI2rA3Vd8DIXiFmhBjImIz3N5HlGhrXrCGQg0w2Km9Dq93M/13vdOa/w6ElGOR3OjSvg/Z7kLEb5LxNx3s08m4zGmSQokV5pVTbjIMC47A+jbO5CVLJusHqQDHqn6o50fmneFqL+dKXvrQp9ywt0kTCKHymfLEEuc8ahFwivPxMprOxhZ0UWTkQQelTSCnv0lbH3ve+9zXFGQmCGULJoqpMk2EdVV4kzJo07wdp9G7y/Vi/h4TAS/rwkneDBg95yEMmLU/yRVF+3vOe13ZTRQ7VMfmyLo1z7bn8UOYp00996lOb0s/qLa+TpUyLGwmj3COQyBh8vEO7/j75yU9uhNZ9fuUHSXE9mc46XnXZ+4KL9KWddUza1tqqe9bRWX/7yle+stUB9/K9T3QevT+WMu0SUYNDpmUttEEKfrQDz7w31lQzFdLfROdJfNqVwTAkXzvwW3pwgRtnt2RYInMGfgzwWBcpr975ZPUb4tb23vzmN7e8yZd73qW+xAZcMHPfe3bfpk6uZ5Kb3F53ipDwYiZ7FHI8RVEpVBAjmJPVGY4nX8LImzypsIOWN/nLztO7HTTnnTpgOGgObka1BvGdwmwQ26l3SJAYZZ4sgXI49YQANCI9qG2BcjjZytV48NMG1LlBxG085ZmoMPot9ZyS3+/632G2A+3VNdLpPU+2o3gihVx//uTZe3RQFNPJW+Y1703GWV2ipDvS9fex8pf5kn8yP+VE+p+MM4z0D95PP17Scn9ont2X7/68uzfRLuuYd9Nfb2CSLtuleif//E6FAm0adNaZfsz68zz0PU82XjDxvpT/UBhkvsgqJOnO/CfWh3P2/ljHvKOhU9dHyvNU1DFlko52l23PPXlKl+9ZHcu897/z9DfRZ1hZGyp9srvfuefId5nPhv7O+4N8nhUEUkPC5gdtpFlFVmmt1xjEygEvo6iDhpsGk2tz+pWaQWlIMMsRrkHJU+aD8oJAqm8E4iA5uBkpHEQHLyP5g9pOjUrDb9Cc/m0qFOXxlFsbHeS2Op4yTXUYVgT9CEI0Fe6xj33sQA5GjKXspqslORpLuPH4tWkIK/JUvZ/x5PHOwlC2rT2cKnnFMqs/ncnOTrVTJUtNrc31mTMVMzoRK/1UvXe7B8NstrtZQSB1oKaKDJrTIU7FCNF4yo00UrBMyRlEMjR01GY8ZZysMOqbYxCJN9wGFTvKvHY6iLhR+Cy0H0QHLwNkg0ggB7V/8x7hBrNBxG0Q69lweZqsjUyGS8v7sjsh53qqCMVweRnvPfk23W+qnM04ZrozaDdVyrb3Y0MmbibXMbueTpVjfZzpjiHCNOmpcOqVtKYqvako00hp3GbrHcnHDLhP0MxEYTMI0BZuY38LVd/GjlmGqPqWSIztXHVubHj1+64614/G4F73v6f+68HN8R1zNlPzfceSzM47/e+n/3omlXam5nsmYXw4eZ1L72dWEMjDedkVthAoBAqBQqAQKAQKgUKgECgECoFCYHQIFIEcHU7lqxAoBAqBQqAQKAQKgUKgECgECoE5j0ARyDlfBQqAQqAQKAQKgZmMgHU3jkFz1tcPYr4GDafMT77HQcRsEPMEt8QsMRyk8yDmTZ4GtV0OIl5ZxwZxr5Cs63CbDjcrNtGZDuAqzUKgECgECoFCYBAQsI19blk/CPnJPMjXVO1ImmmO5pxYDdp6JZ8lsOFH5m80ZZkqPza7GcR3CS87QQ/au7QLv/fY/1mJqXpXh0on69ggvkt17FCfMTlUuSbzmfoFt0F7l1nH5G06XBHI6UC90iwECoFCoBAoBCYAAYpzKjgTEN2ERmG3SJ+YGETFa0ILOkGRIdx2b/TZgUFzg1rH7J6NeAwa6Vbntc1BI7YImjo2iJ9IU8cG7T1qhz6PxgI5aKRb3cp6Nh39RRHI6UC90iwECoFCoBAoBCYAAdOXbrnllin7xtlos0y5udvd7jZa71Pqz/fg5G+6pn4NV1j5QRwHkTzK7+bNm9sH5YfL+3TeO+OMM6Yz+RHT9qkvn0kbtM8IIdvr168fMd/T+WDLli0DaYE8+eSTpxOWEdNWt/Rl6tp0uFoDOR2oV5qFQCFQCBQChcAEIYB8lCsEJhOBqmNjRxdmhdvYcCu8xoYX39OFWRHIsb+rClEIFAKFQCFQCBQChUAhUAgUAoXAnESgCOScfO1V6EKgECgECoFCoBAoBAqBQqAQKATGjkARyLFjViEKgUKgECgECoFCoBAoBAqBQqAQmJMIFIGck6+9Cl0IFAKFQCFQCBQChUAhUAgUAoXA2BEoAjl2zCpEIVAIFAKFQCFQCBQChUAhUAgUAnMSgSKQc/K1T22hb7755u7SSy/tbNF83XXXdd/73vc698qNDQFbNX/zm9/sfv/3f797+9vf3t1www1ji2AO+P7rv/7r7p//+Z/nQEkHv4jf+ta3uk996lPdT37yk8HPbOWwECgECoFCoBAoBEaNQBHIUUNVHseLgG/VfOYzn+l27tzZvleza9euSfs2krSuvfba7qtf/WonHW6QvvU1XgyFQyD/7u/+rrvvfe/bPfjBD24f6D6c+GYLLv0YfO1rX+t+/OMf99+q62lCwCDR97///TZwNE1ZqGQLgUKgECgECoFCYBIQWDgJcc7aKH1I94c//GF3/fXXdytWrOjuda97dStXrmwKklF2hOWmm27q7n73u3cnnHBCt3///m7jxo3dN77xjUZiFi5c2D7geuqppzbF6uqrr27hzzzzzG7dunXtQ72sdCx0xxxzTOcDuT76OlMdCwQlEm7O8EDwfFw3yct3vvOdRviOPPLIg+VFAvhfsGBBd+yxx7aPFwsD6zVr1rT4tm/f3ggUfOB74403dieddFLzg2QhEQ996EMb0RL3THfIo3J++MMf7nzU9ogjjmhFgjHCfPTRR3dnnXVWqy+w+Pa3v93q413ucpdOfePchz/M73GPezS82oNZ9Ecb1D5ZallozzvvvFZ/du/e3dquNrd48eLuggsuaPVn69atrf2qX9/97ncbrj5+vmrVqvZxdnX3Rz/6UftYL3zhrg/gR/30fMeOHd3y5ctb3ZxFUB4sig8VG5BR1tNPP73Vp23btrX66HzKKae0e9qnNv6Vr3ylg7d2LKx75QqBQqAQKAQKgUJg9iBQBHIM73LDhg3d5Zdf3v3gBz9o1qBFixZ1D3jAA5qyjrScffbZ3X/8x3800vjIRz6yKZgf//jHmwKKYPL/iEc8oimn//RP/9SJDzFAhu53v/t1lNuPfexjjRD8l//yX5qCP1MJ5Ne//vXus5/9bLdp06b2kVPEmLvllltauZE6CumHPvShjhKK0Kxevbr5Ne2NtfLKK6/s7nnPezbMkB5EGyG46qqr2jPX8+fPb9ZN5Mi7QK7+/d//vZGlo446qr2T2UAgESOk2IDEv/7rvzYcDGKw7MIRDgjNXe9610ag+IGh9/BzP/dzHQw+//nPN/wRJQMUCPdsc8ghQm0aa9aTSy65pGFxxRVXNCIEJ/VRGzWl+stf/nIjPOqb9mhwA0GE0xe+8IU24KGtGtQ48cQTG/7qKuKEWKl72qvBjdnmEEADWp/85CcbRvA1qKMefulLX+oQcBg87GEPa+2P349+9KONNObAhrpZrhAoBAqBQqAQKARmDwJFIMfwLinsrFpPfvKTO8Twsssua0okaw/L0K/92q81pfQv//IvG0mkYP7t3/5t9773va97//vf35RTiue//Mu/NCX3RS96Ufd//+//bUr+0qVL22j9e9/73u7P/uzPugc96EEzeuT+9a9/fVMof+d3fqcp4X/+53/eykPZvuaaa5rFlfLJSvSrv/qrzfpI+fy3f/u3Rmye8IQndH/0R3/UiBEr2kc+8pFGgij3SClCTplHCljffvZnf7Y77bTTmmL7rGc9q1nZHv/4xx+0vo3hNQ+kV9adiy++uFkZX/ziFzeyDRPWnd/7vd/r/uRP/qRhN2/evIbfc5/73GYte+ITn9hItXpr4AP+6h2iPVsdTB74wAd2j370ozv16FGPelTD4sILL+x+5md+prW93/7t325kG2HUflkQX/GKV3T/83/+zzYAARsECMbveMc7Wr0y6IOQs8J97nOf657ylKc0Mqke6xNmo8sBCwMT6p+ZEga8WGif8YxnNOvu8573vIaHAR79ncEg5Pwtb3lLs1Kqk+UKgUKgECgECoFCYPYgUARyDO+SBdG0LFYJU1VNZ6OAsu6whlGgONYiFg5TVj1n4aB4smqYKueadeOP//iPm7XIdFfT4BDUn/qpn2qkh9LFUjJTnSlsyuOgfJrWCz8E0MGyygIGq9e+9rVNAefHfZZLmLCgUewRAnikIuosDqT7oosuakQTKf/FX/zF7j73uc/t/M5U/EbKN2uOeoWEIz6mayLaplUuWbKkYcJq9J73vKfVKZZbGHsflP773//+TekfKf6Zfl/7YrVWl1gXWaHVH2VnKUP8tEnEiDOll0XNlFbYmglgqrBpq8IiTSyLBiuE1U4f8pCHdH/wB3/QnX/++S1ecc9WZ3DGmts//MM/bH2fNoYgqmMGu8weYInUlg3sGND5lV/5lTbAcc4557S2CONyhUAhUAgUAoVAITB7ECgCOYZ3+cEPfrARv+c///mNBJoqSWGleJralQ65oaQiOkiOaZssQEim6YZIESXUKD3ChFhRclmI/J7JxDExUOZ0iA0rTRJB+HBr167tXvrSl7apu9ZNmXaJOCLicKKsWneGAMFS+CTl4s81pSxHpixSXqXjPv/S5aTn92xysDjuuONanTJ90G9WSsQnFXpWOGRIfaPEq6fql/Nsdd619sR55+qCesAyhjT+8i//csPKIJA2ymV9dI0YwUcYmKp7cOXEp87C3eDGP/zDPzRiee6557bns/EP/PRbr3nNa9rAmcEvU+3hwhpperSp1Uh01rH+tpZtdzZiU2UqBAqBQmA6EJiNOs104FhpHh4CRSDHgB8F1KYlNuhg8TGVi6OIskykozRRpihSRuVNy6SEUkzXr1/fFDLrhMSFLJrqevzxxx+Mh+I60x1LF+sXxd20SZYKDlawgReyw4pmqiBskW0KK3xZ1kyzpMjDjBXzAx/4QHdVrH+0syOMWdUQT8o88gg35JMyiyAg9aa2Uvhng1M+OCo3Ym3AAVbWMyI+LGLqGOs4JR9xdA1vAkc4/voJ02zApb8MWS/cg5d6przuW+P4xS9+sdUzz2DlsL7RFHP1Ub0xfZz1TN004+Ctb31rw1GYi8MiqU7y8/d///fdM5/5zLaZVn8eZtO1ARk4wNGZJZZl1gAPMqlf01bdZ600aPHmN7+5rT22XtwAG9zKFQJzDQF1X99CB7AMQ5vR/87m5QOT8Y71L5/+9KfbLAeDWbNxrflYcaNbwsWeG5Zm0JvKTQ8CZjrRLegEc83NXlPEJLxJ09sQI9NPCQJKvDWNlKf+ymPKFyFBYCAxFCsNHmlkETLN0tROlQ6JNB2Rko8IsGawEs1096QnPakRN0okgmc9IkKDzNldlhCgnCKZ1jOaFgc3G5Ugkgg1BfUTn/hEU/5h5n4SI9MUYZ+kHjH1PtxHyKUj/ty8Z6bjKf+IC2sjpR1+Nm1CgNRHOOrITMckZNU3wgUmphci1a49nw0W7pHe573vfe+2FtZz7YjlH26ELFKIdLOYwVG7zAENdcUzdZQ/mGmTyM9VMWhhkMPghHg4Zxhrx7NZeMPHAJB+Sl9mGr5+UB2DqzoGKwoyi+RjHvOYRjZhqX3yx2pbrhCYawjoOww0W16hf7Hen/yaLKe/0h6dZ4sjv+kBBooNhtZg1G1vllyyh0Zhchsm03FFjzeYPJsH5kfCtYYtRkJmyH2Vw7f3HK4pj+koopSqdP/tv/23RnrsJupTHzaboGy9+tWvboq+aV/WUA2NB+mhoM0GhwgPN7XPWrN+1+/HlGCK56//+q+3jUr+3//7f01oGMWl1NsoZqhDRnWgiFS/u/TSS/t/zoprJPCVr3xlK4u6o245hpbfQATiNNSxxs52Z0OmdNYoa2dc7sibz/JM4fKZEwMeyE7WI1YDRN00cxsSIYrpDB6xLvz8z/98q5d5fzaeYfi4xz2uKaX9Aw/IogGfoU57fN3rXjf0dv2e4QggJWYwOLQRAy2miud9/ZE2YZDPwfmdMwDIS/6FMyjhvn6LXMwBU7MEhOHc718W0m7OkD/ZP+hDLLUwkAynfp2BHwOr7im/Qaj+2RN+w0ubExYWMIZdYglD8XjGP3L6p3/6p50lNvqy2eD0z5Zk2L3+4pj9oW4pt3qozPpoePgNP3UKHuqZ+/DK+/wLn338TMdHuVghDU5kfcnyZdsb2sbUF7gkJtn+1DGH5zBT72Crjoo765/n8HNPPLPJKb+6pfxw1C7h5ND2/Hafgy9s4EI/NcA8F10RyFG+9f7G0n89UnAVjcVH5/eyl72sdWzIp04w3WjiSb9z4czCg1zbsVXDZGWDF/J4KDdbBMKhyjj0WX/dmYvlH4rHeH+zkiHcBGk/pgSk+ufZUHwJGFOmfTbGjqxzwfWTx7lQ3irj7RFgPfMpF+vUtZWnPvWpbeMpCqylBdqO6fQs+z/90z/dyA/LkUHAnFVikPWXfumXWttxn2Xb52/sYCy8eMyk0O7M9DEdeiY6GMFKuQ1OKRvMEEr3KKM2irOrs7LCi+yDieUuyBCLvtkUrPssHHQHiqydysVn8MYmVqbYm73DvzTtDi8du5/3D2rPRBwtEbIBoV3s1S0KvJlJliHQq8zyyt3G/bZjNoslLC0tgK1+2n3EwOC8AUGDYrPB6ZMRGfXIBm/qgbpkHwkz3dQPVlsbDT72sY9thNCXA9QfZ8+0P4On2re6pc2ymtsY8gUveEFb50+P1R5hKJx3oL71D6rOdDwRbUtY8hviBizMmtOmPhOfSlN2dcc9zoZ8rL+5kab2PRddEchJfOuEh9FAozaEhg6NFanc8AgY8aKUW+/IGYEtvIbHqu5ODALqms+/GLAgWNMRjgQy4ZmjjvnMb9ZHfuaq4EgsZttZP13ujghoH2Y7mKJsSiYFihXa8oO/+Iu/6N74xje2nY8p975lTDF1/yUveUlTSk2vt7QAufGZq2c/+9ltYMbUxL/5m79pm1uZoUPRpQSbaj8THYuZDd0MLPlk0Lvf/e6mcJNlyDHSjGTDRnlz7wPriy338HmhN73pTU3useYjj0gQBR6houQimLC3FwBCboCVgkt+Ilw+q+NdzXSnfzVDyeeYDCbYWft//+//3eqGT1fB+V3velf7nBK8fb5LH/62t72trde2zMDayb/6q79qmK5fv35W6RPKympmYMGsLe1KPXjhC1/Y6gIC6Ln1/TDwmTl1Rp2CqXDasnap7tivgjy0i7aBDNjSXdVdpNQZtq961atmFXnUTlgTtUkEUZuCB+wM4PhsmvYJN7MKLacyywZOlmf4lvlclRtFICexl9XgWDDKjQ4BHaKpJ45yhcBUIIAMDiWI0mV1NOLtGOo8M0JbbvYh0D+IMPtKN/4SaSNG6Vk6KFMULoqUtuCZwRbTxCnsFC0KvymVyI516qwlrP2esY58/vOfb8q89dsGVjkkFRkyaDNTnbIhPjY0YwlzVn4OAWQhU04KuV2cfaMWSYKTdcYwZqU0qMW64X5OoaOkUugptgZWkXF4w9+UVfdhyTpE95gNTnks7WFthB8rmTpER3BtYAJm6iFyrc4h3Dm9Xr3LJUazBZN8r8ptAFNZtRmzZnL3e7jlBoOsitqmvk1dQhhZY82kgZc6a+8IgxLqjrrkmenQT3/60xvOLOb2+WDxnI2D+uoJ8sziqPystuqLtoxM6/O0S5hruzAwwGPgwqARq+1cdPPnYqGrzIVAIVAIFAKFQCEwOgSsTadccZQnCleuFTJKT/mkoDqQHmfKF4uHQRgbfrEqIgGIJOUsFXvWSg5JQBZmsqPAU94NMCF2sEFukD+YObtv6iV/pld+JqbIeYYUUVSRcUqse8ImURCWAg87OPlmNGxZkUzhhLnnzrPFKbtDXbPWD2GCk7plsAEB55AkijxyzioEK2Fcrw/L42wjj8oMF+XKZRRwYTljhVSnrL1V1/jhl1M3tDNtkH+DFEil54wdOTCa93NPjvxMGhIp3Gxz6pVZRQZuzARQXoMTPt1n53u48mMAB776rhxcdq2+zUVXBHIuvvUqcyFQCBQChUAhMEoEKOimpl4ca9JZPHKKF8UzrWKi8rtfWbW5BMWUEk9pRSJdW6ogLgQgN6ZDPBGgmewsW7GpBryQZwciQ3GHi2uOFcN3jlnNrFv0jNJ+VeysyRqUm8PBGYbiQZDEzdIIK8T74Q9/eLNEsugiTyzDLHSZzkzGUt4p5srLITfID6uuTeIcpjsrKwJtcMLaNYQqyXpi1SKYZX+SJGt/nLIqtzpiPS2sfG+cxdEzddChHhmosK5U/VPX1B31jFWXVdIUdQRdnTQIYr2k9q+9IqWzzSUBvOSSS9qgg/KzKr7zne9s1lo4mm4OP/XQmlFtEmbwcn8uuprCOhffepW5ECgECoFCoBAYJQI+VcWa89rXvraNyLNCUiQRn36LBEU0LR6mWFJSKWOULusnKWKsZ9ayUboouQhkWjZMw5vJztQ3m5p8JixApgNS3Ps30oGVKa0vfelLG54IIkz4s0GJ8psuaAMcBJNFBCF8/etf3yweSDj84Pqe97ynralEAKxpgzv8raEUT1p2ZzKeLLDKhRgpNzyQZYRRfTSYYSCCNQjmqdiznAkjbFqKZjIOw+Vd+bTBJC8sZO4hfdZ/vuMd72htU32z+z2SxK91yna8N1hjQMf6SSQIdgY+zDQwQPGKV7yiDfrYzAm2Noeajbvbw5YVVruBCVKt/RmIsO72//yf/9MGKbQpU/MRcrMHkEt1kEXSFOG56GZ2bz0X31iVuRAoBAqBQqAQmEIEkDyWMdYeJIViTzGnkL7lLW9pirrs/MZv/MbBtWosGC9/+cubkmpnbd9Ks4GMTT5YypAmSq/piJxNQBCqmewomTZ4Ub4kPzBCHK0nU2bk+zWveU1TVl0jiJTzF73oRd0Tn/jE9l1fZBJRespTntIUVbiLO8mUc1qWkAi4IQ+UXco/xXY2OEQQaUyrl13a7YZJ0Vde5fbMp9Oe9rSnNczhrk45+3QVcj0bnY2VEJwkyNpVlv05z3lOW2+rfXH8ZJtTx4RDyD13qF/aK7y0UfU065D6a1r1bN7PAz42sDJdFYY5QPaGN7yhtSdtL9ufM3KNdLrmn4PTXHOzgkCq7DrNcmNDAG464pxyNLbQc9e3zrZwG/v7V89Mt8npImOPYW6GgBfcqp2O7f3r37RV53KHhwDSmOujhsbUP/pOcXdYf+W+nR0pWZR8a4w4Sv9wRHGk+IemN+i/KZ+Ooa6/zP2YsVSy/tigiNWIRYPVMj/DMRIu1l4NdSP5HepvpvxGlB3p1KMkk3nPeSTMEfd+C3l/mJl+neQvy8E6nS7bYf52NsWX5QxxNC01CSL5on2zRpoRwILZ71gnPber8Gx1yN9wdSgxGlpuWPfjPfT5XPk9KwikTpOZPUcC5srLO5xywooQ0zlXQxgbktavUOgLt7HhRjBpp/0KwdhimHu+CTbKgJ0aRxJmcw+V0ZWYXKBszlYFcnQoTI8vCiqF02crDLbZJZLVo9wdEUAsWXdZNGCFcMMKiSxXCEwUAggnq6X6llbLjJt10bOh5NFza//IHrsKlysE+hGYFQRSpR+u4vcXtK5vjwACaec8R7mxIWAEr9zYETD6SVEqN3oEkkDO5E8bjL60E+uz+reJxXMssVFQZ8MavLGU+XD8siY+7nGPO5woKmwhcEgEDEzYDXmoI2MQxOEGKFknrbEtVwgMh0Bv8u5wT+peIVAIFAKFQCFQCBQChUAhUAjMOQSQy3KFwEgIFIEcCZm6XwgUAoVAIVAIFAKFQCFQCBQChUAhcDsEikDeDo76UQgUAoVAIVAIFAKFQCFQCBQChUAhMBICRSBHQqbuFwKFQCFQCBQChUAhUAgUAoVAIVAI3A6BWbGJzu1KVD8KgUKgECgECoE5goCNLnyupD7zMvoXDivru2BXbnQIVB0bHU7pSx3zCaFql4nInZ/hVZ9dunOc0oe6lUfem8pzEcipRLvSKgQKgUKgECgEJhABO2rbhdw2/eXuHAHE0addnO1MWW50CNilsz6JMzqs+LITcX5jcfSh5rZPn0Yb7jufcxuVkUuvz/eZr+nq++dFBa8huJHfTz0pBAqBQqAQmMUIEIFGcRGxD37wg93jH//4VtoPfOAD7dMK+ax2JJzFlaCKVggUAoVAITAmBGoN5JjgKs+FQCFQCBQChUAhUAgUAoVAIVAIzF0EikDO3XdfJS8ECoFCYM4jMBrL4mj8zHkgC4BCoBAoBAqBOYNALQCYM6+6CloIFAKFQCEwHAILFixot/vX37hGHPPZcOHqXiFQCBQChUAhMBcRGBeB/MIXvjAXsaoyFwKFQCFQCMwyBHINJLL4la985WDpvvWtb3VHHnlk2wjD+shyhUAhUAgUAoXAbEDgggsuOOxijGsTnZrOc9i4VwSFQCFQCBQCA4zAunXr2o6A5F3JvAF+UZW1QqAQKAQKgTEh8OMf/3hM/ofzXARyOFTqXiFQCBQChUAhUAgUAoVAIVAIFAKzDIGJ+ADHuKawvve97+127drV7dmzZ5ZBWsUpBAqBQqAQmEsIEKQpTL/2ta91b3jDG1rxn/nMZ3bnnHPOwSms6WcuYVNlLQQKgUKgEJgdCJhJs2jRom7x4sUTUqBxEciHP/zh3c6dO7tbb731oOCdkNxUJIVAIVAIFAKFwBQj4FuP3N69ew+mfK973au76KKLDhLIgw/qohAoBAqBQqAQmGEIIJDI47JlyyYk5+MikClkbSxQo7IT8h4qkkKgECgECoFpRqB/sxxCdtWqVQcJZK2DnOaXU8kXAoVAIVAIjBsBMsyu4kuWLBl3HP0Bx0UgV65c2Rmx7Z/60x9pXRcChUAhUAgUAjMBAXKMPCNcybZ0yKONdDxDLItAJjJ1LgQKgUKgEJiJCJBl/QOlh1OGcRFI7LUsj4cDe4UtBAqBQqAQGAQE+glk/8is6xUrVhSBHISXVHkoBAqBQqAQOGwEDIRO1GDouAjkRLHXw0aiIigECoFCYBQIIAkT1WmOIrnyMsAIDK0L+Vv96JdtpvosXLiw27dvX5v2U/VngF9qZa0QKAQmBIHsDycksopkWhGY7Hc5LgI5rYhU4oVAIVAIjBEByv8111zTfexjH+s++9nPDtiUxANRmnlxLO26+Sd13f5r4np7HAviKHe4CJiCauOA+9znPt2TnvSkNi21P85+YjjctXv99/vD1nUhUAgUArMJAX2dQbMrr7yye9Ob3tTk5kTt2nlHnGLzsnkh9xaeFTuYhdw7sCW82MiMPCw3XgS2b9/ePfKRj+ye//znjzeKUYUrAjkqmMpTIVAIzHQENm7c2H3qU5/q3v3udw9sUebNu3csD7g88lefSJrol3TzzTd3j3nMY+5AII3S3pmb7JHcO0u/nhcChUAhMFUIGHS77rrrune9610duTnZbv68C7v9Te4ZOC03EQh4h5NNIOdPREYrjkKgECgEBh0B0xOXLo3RzoF1sTPaouUx+LpqYHM4UzNmBN2axrIkztQ3WPkuBAqBqUJAP2n6/po1ayY/yXnLugNLQubNXxtplU1rogDv3xBuouIcGk8RyKGI1O9CoBAoBKYFgZi2Y+ZOCO9yhUAhUAgUAoVAIVAIDCoCRSAH9c1UvgqBQqAQKAQKgUKgECgECoFCoBAYMASKQA7YC5mO7IxmDdB05KvSLAQKgUKgECgEBhWBkp2D+mYGO19Vbwb7/VTuRodATTgeHU6z2pf57lu2bOmuuOKKbu/evbVOaFa/7fEXLjcSsZ7sHve4R9vZMu+NP9YKWQgUAoXAzESA7LTj4be//e0mO6s/nJnvcbJznfVi0aJF3bnnntstWxbr/urTUpMNe8U/yQgUgZxkgGdC9Dt27Og+9KEPdR/84AcbMZBnnVu5QqAfAcqS7b2/+c1vds9+9rO7iy66qG1M0u+nrguBQqAQmCsI7Ny5s/voRz/avfe97+3OO++8VuySnXPl7Y++nGSnenH55Zd3T3va07pHPOIRA76h2+jLVj7nLgJFIOfuuz9Y8i9/+cvdO9/5zu4b3/hG2+be9r8lBA/CUxf/iYBd2Yy2f+QjH2nWx2OOOaa75z3vWfgUAoVAITAnEfja177WPnVw2WWXdY9+9KPb7B3ys1wh0I/AggULuj179nSf+MQnmm517LHHdve97337vdR1ITDjECgCOeNe2cRn+NZbb+0e9KAHdZdcckn39Kc/vSsCOfEYz4YYCcHdu3e3ovimHoFYrhAoBAqBuYqAPvA+97lP97jHPa57xjOeUQRyrlaEOym3T0iZvWNg/oYbbijZeSd41eOZgUARyJnxniY9l8iB76RxOrtyhcBwCFjDoZ5UHRkOnbo30xGomRcz/Q1Obf7VF31hf59IlpYrBIYiYAZPfz0Z+rx+FwIzDYFiCjPtjU1SfglCI2TlCoFDIaCO5EjqofzVs0JgpiGgDywCOdPe2vTnt2Tn9L+DmZADM7tKds6EN3XnebSm1WD6XB8sKgvkndeV8lEIFAKFQCFQCBQChUAhUAgUArMMAYSw/xg6mLh8+fK26RHSaMYBvzbQ8vWCuWx4KQI5yxpCFacQKAQKgUKgECgECoFCoBAoBEZGIJfkJDFkUUQOEUgW49wQ68gjj+yOOOKIDpE0DdnzH/7wh92uXbvm9HrWIpAj1616UggUAoVAIVAIFAKFQCFQCBQCMxiBtBzmJpHIoJ3kHStWrGjEUfGQQ44/30X323c7Hcijtazu20nXpoIskIgk4sll+PZjlv8pAjnLX3AVrxAoBAqBQqAQKAQKgUKgEJitCAwliFlORHH16tWNACbJY2lEHE877bTuuOOOa58lQw45BNCBGNplGUlM0pmkMi2Pni9durT5EcYXDUxrFcZv/mezKwI5m99ula0QKAQKgUKgECgECoFCoBCYpQgghosXL25WQNZBB4fEsRgiiSeccEIje4jmqlWr2r0TTzyxW7duXQsrvGfccETRmsdt27Z1mzdv7m655ZYWr2mt0kIkkcqtW7d2N910U/OHRPLnvnzMRlcEcja+1SpTIVAIFAKFwLQh8JOf/KQpE0cffXRn/QxnRPvqq6/u1qxZ05SWHA2ftkxWwoVAIVAIzHAEkD7kDynMqaj6WiQQocv+Folcu3ZtW8eoXzYFFQEURnhWySSQ/aQwyWSSQ8RQWiybwuaUVuQSYdy4cWMjmb6Vfd111zWLJBLJOine2UQmi0DO8MZT2S8ECoFCoBAYDAQoEVdccUV37bXXtl367na3u3UrV65so9Bf/OIXux07djQiefbZZ3fnnHPOQYVlMHJfuSgECoFCYOYgYBDOBjimqJ5yyinNqmgqKqKGsOmPkTzkkaUR8UMoWSORSL/T8phTVlkTk+y55pBIz8Wb01+POuqoli4C6Zm+Hcl0bNiwobvhhhtamvLgHkKJYGbczjOdTBaBnDltZU7kVIPSSI0EOcY7Sp/xCJ87a40EoE6i31+OXukYRkpf/Pz1dwAj5Xk4v5kX8UtnOJcjX8KnvxwhG85/3SsERoNA1ml1SR3rr8PC99c1z7SPoX5Gk85c9HP55Zd3l156afdzP/dz3fHHH9+UE0rDd77zne5Nb3pT97znPa/72Mc+1n33u99tI9h3uctdRoTJu3Fw3snh9IcjJlIPZgUC6gl51O/6LSr998dyrd2PVs71x0uGcykT+5/ltbjTX97jX76HcymXh3s2UlkTF2dxH0qmDxdv3ZteBIb2efpBhM85ZZLNbRDE8847rzv11FMboUTmEDfhzQBBGjmWQOGtW2RBFFbdSbKJ+CF7qRNmOvlcnAjrySef3OJFVvnh1OX8tIf4DRyeccYZrf2QAWalGFhEJDdt2tTIZK6VbBHMwD/Da64zsCCV5dmBgAb4uc99rjVQC5w18vE4HcBll13Wds26+93v3kaKhotHx/CpT32qKXtnnXVWEzBXXXVVU/Ae9KAHtSkOw4XT0fzHf/xHm57guY7KiJQ8mxbR7/j93ve+1zq0vJ+dn47tAQ94wMFOqP/5D37wg5YPHY7RM+VYv359eqlzITAmBAhKBwFIwFGmCEuKWQpK1+6b3qNuah9GUtNf1tsxJTxHPN94443d9ddf393//vfvfvEXf7H1CYr+rW99qx3/9b/+1+6hD31ohzR+4hOf6D7zmc+06+HggfOPf/zjNhVKX2YU3XodSkkqLMOFq3tzDwH145prrmmW72zHlOSTTjqpO/PMMw/Ww/EgQxH/5je/2eqhvkG8ZJG4WXBGcv/2b//WBj/uete7DusPobOW7Gtf+1pT6sWj36Hsn3vuuU1u98etPXz729/utLFU7j13n+wVRvvod8jCj370oyZ7rUtDMu5973u3zVP6/dX1YCKgrqkPdEDv2IGw6QPJMO+efOLIK6TOO/bMRjaeI4h0JhvmIHjqgTotnPosTo7eqW6Zcuo5OZkyUjh1yX31NvMgH+lHWsig5+pxPpN3ccmPsmgzSCQ5YTkD3VB9nqmuCORMfXOzNN/mj7/gBS/onva0p3XPetazxk0gdT5G/I38/PZv/3b3Uz/1U3dATCfyla98pfvN3/zNdhBC27dv7z7+8Y93f/qnf9q9973v7e5zn/vcIZwb//iP/9i99KUvbcSUcqdzIRBZHZ7ylKd0v/Ebv9GELb+Uxd/5nd9pnY0Oh5DX0eh0TGOTjk4mHWvFq171qu4LX/jCwa2jjXzx/7M/+7Pdi1/84pZO+q9zITASAv2CN6ftEGqEHEGoHvLjrO4TdOongawuE8DqNaFnYCUFbApedTKd66G/89lcOOu7TF/96le/2v3hH/5hI936AooFReHBD35w6xPgyiHmh3IUF8qFM/y///3vt36sv684VPh6NjcQQJLe/va3d+985zuboqz9qouUYoOOv//7v9895CEPuZ2MGS0yBj4vueSSFpZSLM607FDKyVYyiQzU9qXNve51r2v1ltwbjmjqaz784Q93r33ta1vfo08SL3nNvzz/zM/8TJtiKD79EzktP/wiCVymqb098YlPbPf8+ed//ucmw1OG6s8MyAhnFoDBHAO+5QYLAfVH/7Zs2fI2UGEJgHWL5JV3p54ZwCCXOPWR7kUe8WcqKx2L/qduZBhTVcWtP1X3MlwSPH00OUcnJBfFI4x4kUdh5Et48buWJ/mQVvbV6mkOwnomv/wJIy1xOsQvrtQbB+stjD43RSBHj1X5nAIENGCKkhEa14fjjEZZd/SRj3xkWAKp8X4mrADS0vnobKRJ2aMsez6SMx3BQTCzKuhQrrzyyu6zn/1s98Y3vrHNsX/84x/fOhDKH8X753/+57uLGFUk9wAAQABJREFUL774oOKugyPEUhmX1r/8y790r3nNa5oS+tM//dPdPe5xj9bhsEJSTo3YErKphI6Uv7o/txEgrNQRbYAg89s15cxv9ZVQIyzVQ8RRfVZPCVqj+aeffnojkoQjsmPE1GEUlwDmtBlOmCShFEEKLKEr7rnilFdbPv/887v73ve+rT/4+te/3torfCgS8IA9fxSNkRz/FHT9kjCUX8dcwnMkbOr+7REgp8xWQSR/7/d+r1k/zBggS8i3V7ziFd2b3/zmjjVwrE6/QK49+tGP7h7zmMe0uMkifcWXvvSl7n/8j//R+oSnPvWpzSqZ8Quj39AXDOc809d84xvf6H71V3+1yUVyjdx93/ve12Sg9vILv/ALLbh6b2CGvHzCE57QWUPcrx8YiOXEa1D4ZS97WZsm+LjHPa4NAlPaWZhgom+iYxSBbJANzp+Yrb9g4fyY+bIyZNeJbTqqdY1kjP6QDDMQkDJNH6oOkEUORI4fcs0zxE8fSy4JnyROveJfG+GfDETmkvi5p9916Kv5zXizPktXXfNcfH4jodqia/cNuJC10pYf95RFejkYg7TKp7Az0RWBnIlvbRbnORtbdhqHU9THPvaxjUCaTqNh6wT6HYHFknjhhRe2KTCeEVQ6BE6jP5QzisSyyVqa7m1ve1v3ohe9qHvrW9/aXXTRRY1AKpMOgzB7xjOekV7vcNaZEJ7/8A//0P3Wb/1WG4XtF3KI7Xve856D8/nvEEHdmPMIqLOmnlKwrAlZH9N31PsUrgSaa+3LNWFG4BlEIRAJS/XVKC8CY1oQPwil6dmmyl0VSh7/2grFzCgr4ew35dIz0zb5JRznitNvmL6OpFO4ld/sAoo97GDj/VBCnGF8KAfbdMhp9kt5r86FAARSZqp7rGvpDEAieG95y1saUdOWtfmxOHFzBkkRPe2c00/80z/9U/eSl7ykI/PIt1//9V9vz/zRZ1CYHSO5fGZg9RGPeETzRolHEF74whd2n/zkJ5sMTQumPssU1Oc85zltau5w8SKJf/EXf9EZuMmZRbn+jX/E0SCYPqvcoCAQg4whOxZE/7l06bLWhyKOpl+rRzkw552ZBupQD9UfMof8Uh9d86+fRBrVF/fpfghaPlOn/c5n5J40hBGWvySb2otDHMKl1VBY6QrrPjJKn0yCKYw2IQ/8iVuf32+RFCYHWul24lKGmeSmhUBSKoBFkXFwgPNSvECgZ8c1k8CsvA4WAoSNTsh0FiTSFDKNOJ0RfVNcTKMxvY8bbb3jL+usDiLrMcuDNVDSVJ/T6UD8zo4s7/efPxOjxZ///OfbtKM/+IM/aNMc+p8jk7/2a7/Wf6uu5xAC6lwe6jEhlYqQukW4IYv3vOc9W103/YfypE/tD+e3+kpIikdYApJfgyLqtWfpxz1CVb+s70aEDHbwR2nNDQWkQYiyuKvvHL/6euHkT5jZ6hBuWFmr9cAHPrApq95RziL493//9zaqbk0Z3L2f0brEc7T+y9/cQ0Dboghnn2AGwsUx4wWBRKy0QfVxPE5Y1kj9AiceU0y1acTxAx/4QNs4ytrI0Tr9BSfP6bQLU1FNgWWhZOlMAskPWUvxHsmZ4vrud7+7DdQa2O0nj8LY1MRRbnAQCKnWLZq/IN7Vum7t6pg5c8JRbRCBvkNfImvUN32r90kukXPkWMoqvw0+qFOpazmTa0kkxeGeg0xSl9Q9B7mlLuegh7jJPCRQnXeff22AE4e0hZEGIigOeZCOfMqTdJKsikPePefUawOzmba4xTfUDbLMnHICCWxTDEyDMu2Ags8ZtbaDnUqjgQ/diGQoqPW7ELgzBNQl6xo/E8TMRjk2xSGgOIothU6H436/kLqzeA/1XMeSHYgOo9/pCA7VGZhuK192cNRZlpurCNyRZKmnBJJD/WKdWh/WRbvOEWxG1SmJpvg88pGPbPXeSC2BJWwe6j/h6FA/U5DyR/HU73qm/rmXftVbipuR0hSihKs0cm2lt8WqJqxw7hOsDvlDLCmis9Upr5Hzv/7rv27y7aqw1D796U9v7RmpdN/uqzC0bsz093KFwEQhMFS+aKc5JZ1VL8nfRKUnHrN8/vIv/7K1b1NMx0IgR8pHymj9j2OoG0mG6mcs8SCDkVtlLjfYCMyPMYRFSxYHUVsbev/K7tRTlobcODqmsJ7Q5IjdStXjtAqqw3n0G5rUCeSNLGQFdEbcnJEyck694Mgm8hAXEcZ91+oa2epwTT4m6ePfQKtnwpjeSv6JF8ElG+WBDCX/yGf5kxfPyD35zjxnHRYf3VM8XMbTfsQf/oYjlfl8us9TSiDNd9fZ2JgkRxdUDlN8rPsy3/1v/uZvGoG0ix3lpFwhcDgIsAha+2EqmfUhOg9OnUMgPTe9TAcxFpcdgPj6hRwF8dOf/nT3qEc9qin74hS3ToeAM60tR6aE1eHkKKk86aSsPSs3FxEI4rg/1sWF0FiyxE6pvR3cclBCvbBTMCUN0XMQrAYdXFMWCbUkcISV385ZXylnqaClUM2zeqxfFhciqP8Vfwpl6yD11+JHlJzFL4w6LI0UyIQlMsuP+ORBfWdR6Lc4zLa3TLax+MCU8pHraEwp7r8Pl3KFwEQhoE1rewYqtEWy5v3vf3/3d3/3d23phFkxlNWJdto0opZTQ8cTv74hHVn50Y9+tBkU9DEGyNJ5pu9gbGCAyMEofQ0lHAkwSMXpIyejvJmXOk8MAstXLA0rnO833i2WXCyM923znNWhI61ocgp5oi+RQ+ST+qZfVRcceU1Gqv9klXpAj9IH06+QOXWEX3VNfOJVl8gz91OWaTv88+O+eIUR3jNpOJOP6hfrIZLoPj/Ceu5ZP4n1PGUw5JBbeci0peNa3ZYnz5SvX1b3h58Y9A8/likjkF6WKYOUCaNDXpgK8cMf/rBZHiny97vf/VpFsVmIKYDWjI3kjLarKCqJl2X6UH9HNFK4uj+3EGCBpNS94Q1vaOuy7nWve7VOCNEzfdWaRI1+rE5jV//+9V//9eBIlGmydnDVgViLoq5z6qUO43/9r//ViKxOQQdB6Fmn8t//+39vHYXRp1Tix5qf8j/zEVgwf0n0ZWEFXHtyd5fTfOz4pFYv9J0EJ2XK7AxT01LIESo5IAEBgohilX2juqZOue9Il2RP/+kg4PgTd07F9NszAo1iRilV7ymMFDTC2W8kkl/5NBrLHwFOALpPsIqXEiCvrJFGlmejU0ZlHer0AWURGYpK/Z4oBNQviqzdv7V/skT7tEbfvcmsezlARJkeq9Ov2XhOX5Gz0P72b/+2uzim3tITlSGdvoRfOqM+J/u0Cy64oO1yrk8UB2fgptxgI+Adnn76qTFb5t7d8mVndKefsaA74XjWyCUh9xY0nSnlhzNZk6Qqr8kt9cCZzNEGyCvkjR6mXTirG+QQeYj8CU8G8itOeVFnPEt5xi+9jT/X+nbPpScuaUiTLuc52eY+/ymD5YcMlX/hpSucQ97IWvmTDzzG8ySf0uFPWx5UN2UEUsPW8AFqgTNgkEaKBGJphy9KiRdNwUAiRyKQ+eKF9fL8Fj9y4EWWKwQSAXPM1QuN1fpCFhyDFzb5MAih3mn0Y3Uauk7LtGsdBEFIybauidC2BoofTv3krBVjDZUX9VaHpc7rwDhncYqv3NxBwOtfuXJ1CM8zuqOOPi8E6d7urmceGcLluCasKGaECUsfckJhI4zUW/fVJdbCJHCEl0M98tyZn6xX2UdmXXNfHSW4xJMKoThYN/WtLId+E6DalHxku1HPCVrkkX/xEabidJ9T103xJDAJaN9QTb/ZPubOG6+SFgITi4B2rh0ahOess9Vm6Vtm2GSbn9hUe7Hpd6Q/Hqs6ufnlL3+55dOgF2XZkpJf+ZVfaXsJ9OdXGvof1nyzG/Rf+g7LoPQprlPmZl/XH76upx8B9ZCes3z5spjFcmpYHc+P93l+t33HifFu54f+H5/eWNzTg/bt29nesXdKZ3KoL87uqQ8InPqn7iBhZCU5xB8ZRe7gFOoHmemQvrP6Ig6O7OMPmSO7+CG7xI+r9Lcf155pW9IVR8btWYZJeeye9KXBpbzVXsha+ZBnZ0RTnsUrniTDyXNaBAP0Z8oIJCCs/QCUtV6UbS8AaF4GcL0IlYOjrIzkvFwvxFknQsExPdb26f0veqTwdX9uIYA0Ejof+9jH2jcaTX8xmEGwqjM6m7E69ZZjRVy/fn07GzEVX/+0G340fiOpvj1FMI7kdBzqvQGVcnMHgZUrV8VI7F1ioOP+3bq1F3bn3J2Vj6DsbS+ur1OHUgipr/q5PBA5wo8w4k+/yDnrUwlAwizJXApiddgzYcSV8XjOL0uiabGmruq31WFpqKcEZipr+uAUluovIew5Aeh+CnDCMTfccc8gDpnAX7anufPWq6SFwMQhoA1TSP/8z/+8RWoQ/uUvf3mbxmpn7yc/+cmtTU5cirfFRJfTfk05HavTP7GO5o6bZDLDQi5fEm/2Z8p4cVgmfZZkuA2oLAFJ67/BYX70a+UGAwHygowhx7zzs866W7yjs0MmHBdLe5Z0K5az7NlEZnfTg7Zu29oIVPKCtOKRT+oEGUUuGYh0ZlByJk+kYbCBP+mqB87icHYfSSOvMn55c5BdZJqDnyR8KaOchRE2OYx4HZww7pN9DnIz864OC4vv9KdFpgsj3wZREGHyV1rK4xBu0NzYNedxlsDLMIJOwbZJiCmEvs/ne0GsMEDnKDResFGAQ7n+KRleTindh0JrZj5LwXG4uWf5YGl89atf3QYb7NSm03FPZzEep1OhBPvW47Of/ew7jUJHoMNzjJSmNW6eWZtZbnYioE4TZlm3CZfTTlvfrOTnnHv3bu+tx4bydCBIGoG3sAkvgoMA0c+pR/pIzm8HQSQeCmT2n+oRoSQd4d03iKcf9szhmfuEGr/6XPGp2wQxAUao6Y8RR0JPuKzLnqnPfuu/XcsLAqmM0hK3eA20OLsnrkzLwB8FlDAuVwgUAuNHQDtMJdQgpl1ItS/LN2ys1a8zjT+V24fMTaHMLBgPgdT/XHLJJW266u1j7v3KfjKf6Zv0hcM58jhn9BicomsiEcM5WA2Nezh/dW9iECAPyA9ywCDiXe5yRhx3iYHTtd0xx94SROv6bt0RG7s1a2M94d7t3c4d+2LgclGsdd14UFaQTWQRWUWmkTnIlvpArqQMQzD5I8OSbPrtkI+8l3VAOPc597iUm9IgN8kxdc9ZWvIiD3nNv3g9d8iTQxjPpEF2ygM5SUaSlRySKT4Dsw5tgpFDOHFIX9ziyvy1gAPwZ8oIJGAoOUa0OUACFak02uRDuD67YJMRwBmJGq0bNFBHm+/yNzIC6kA26pF9je6JTss3GV/1qle1HYANXLB8+Fbbodxo0teB6DA08MN1F8fo6uc+97k21dbaShsfDHUUBB2PtlNuZiGgz9MPGiWn2CTZ0geqowuizu+OTXQWLLT5jY0DljWhoZTqF4FFiKTgIpAQMYLZyCVhpy+UDjKZ9STvCWvghIDyTFjtjPDSN1PAhCWUCV/+CDoDMIQeYSYu9zlCjj9OOxB/5kmc2k/mL8urrXDikbbfyCOLgfDlCoFCYGIQMPPGrBjfgkQk6VnjlVPDyWNt+I1vfGMbAPrlX/7lzlrEfjdcmP7neT3Wdi/d4ZyBqQc84AGtz3rHO97R9EnfeB7q9Df6MH1kuclHIGUM2XfaaevD8nhut379yUH2V8aa/p90Jx1/ZZCoI7pTT7umO2rdrSEP9nTbtt/a3XjDPcMStytkSO87w2QLuUL3IW+cyT5yhpySjvtkFVnoPZNPKZtS5vDPDznHaikeso9/ZI181U7E55rc45+scohPPjznyEzXzo58Jqy0PHNOeZxxu6ceqv/iTdkpTnmSH5ZI9/kdRDdlWijrCpKowwGKNTCUetuZsx7+2Z/9WVv3iECax/+whz1sEPGqPE0BAhoe6wfrtIaj8XIaptErU1X+/u//vvvOd77T/cmf/EnrPA6VLQ3WdJbcCdG6RbukWqc41OlkUkCZBrQ+pqfqjNLlBiN+p788p5/xnq39IPCt1bS5z4tf/OKOANTxwuOyyy7r3vSmN3VvfvObD651GW9aFW7qESBc1F/9HlJGoKjT6rg6tHv3ghh53d5t27owBIvd4nrrNoRTh41UqotJ4FJoOXsuPs8InxRSwnLuSUN4gtNzQjMJHkHMj/CEKH/yxQ9Fy8BFCm1xuhYPEul3fzrar7w4XCsjgip9wtJvhNIItDKJm5AmzMsVAoXA+BAYKoesVfapmFe+8pVtZ1PyT//jcxv0LWv1yRvK9505bZNuRqnV5s3isbO5ZSGmxz71qU+9QxTaNVlNjlKQ5U8fQ4Zmn3eHQId5g+HhhS98YSPNv/u7v9tmB+lv9T8GqegN+reHP/zhbQO7w0yugo8SgbQ+nnDCiaHPHBN1YHt31l2/1Z15+qZufpNfMYV1RSyz2HOg++739nff+PpZEfPikFM9Mke+kHP79vU2g1N/vNMFC1gje99/9Jx8UUfJE7Jl8+YtB2ULuZNkjHxTPx3ixkmEEyfZZDBCXSX/pLl7d29PAXIv25n0XItTOId8STflYfpxFt+KFb1d0RFbaab80z7kr0eIe5/X0i7FmXkeJdRT6m3KCCQAzzvvvM5IFaBMsTDX3bQHo+c6Ifcp+ne/+92b0jylSFRiA4GABqjB/OM//mMTUhqRTsF9C+f/+I//uI0qEVysdS9/+ctHJQCRMFNWL7300haXnVk11qFOw1cPuT/6oz/q3v72t7f0czTpN3/zN9ugh45E56Ph35mjlOuwUoiO5F/aT3jCE5pi/p73vKcNtvzVX/1VU/Z1VDDQbmrkdCQEB/e+OkV581F5/RslSj0nVA4cIKB2dxs27owZGnaGI9gQvNUH6yiyliOV+lJhETREMNuH+sGpqw7+OHXHb/7cU+8dwiOQ+l/xqdMEGwXLNT8pFMWRJFVZPOek6bfwma66Lqx4peFa3XY/67F45cWmUoS9NmJAKC2aLfL6UwgUAneKgHanDWm3Q51dmw1Mvutd72ozbvRBV111VUeuUJQf+tCHHlJ+6p848sj3u8ljbV17pgDbbfwpT3lK+6Z3f9ravr0uDPD6NFv2De77NirDQcpO/dJonD5QOcU1kpM/g68Uc99+ft3rXtfKqp/T/+i3kGo4lBsNAr1+fjQ+h/r5T/ETBNFsE3oSy/G+7sSTv9ade87W7swz9ncrVy/r9uy+X7do4YndgvmXh7zYGHre0u78807rzjn3mLCcXxf1xNK22FRm19awTBq83BVys0coFy82NdTg56KQRQikT3ksDXm6NOqKGToLoy5si/QN0lp/uDv8sh7ui2eLgmesiC9nkb9bQ++6OeoIErgv5J7dYG0Yd0zMDlrb5LP6qv6Q1/v3K5MS9wZvV61a2WTlrl12V3U/LIt7wvAQRV6wgCw2/XR3+DYNN6a+7rVjbO8bl71B3wUR3mwi028t/+jtRrtsmW820/0ijohseNu79IZz4393w8U23L0pI5AUB1O3dDZDnY5suPtD/dXv2Y8AcvTc5z63bXJDWGiwFM1Uhgkb9YUQoIRTTkfjKLlPetKT2uip6YLI5HBOfKZSG9Bg9Uuluz8P6jIiYO2GaUKpuA8Xn3s+JfJLv/RLzQJK8B7KrQ+L54te9KL2uYZPfvKTTdhTqpUZ6VUGSkG5mYUABcZ7M0BmyqqRTXWtp4zNb6Ofe/bcEsLCKGtP0SGg1DsCRlswQuo38mWE1JmCR6FSrzxTX9VPZ/ddO+dv6WljwichTDJHQVPXpKe9JLH0WzsUNskgYSq8uJIoukdJk1fxyJNyI5Gc/CXhdBZe/sTtkE+7b8uH++UKgULgzhHQlslDbVOb6nf6mhe84AWNQHrOmf1g7b4pp3mvP0z/NZ3tOc95TpummuRNmzd74JxzzmlWvGzf/eHsc6GPM8g6VIZnX0IuGjAdDZnTd/g2uDRzg53+9PJav7I+ZOhLXvKS1teSoWa4mbqPXOasN/1wuTtBYF7Qg/nHhKeb4zyWnepDX4t/CxfGRm/zenIp9vgOIrcmlg4t6nYEcdq5w3KKmMK55/iQGQ+O67UxeHpSbCJ3bWyms6E7/a4ru3177t396MdXdFf+4PqoR3tjp9ad3epVZrvs6uaHbFkcdX3lSksvFjcyqO4jdvv22XwOiVwWcsRnYGyEY8Dfun/r/+OTU9tWxbP4tNTiNU3ebd26s8mdbdvs7goXMnRhyDf+QoYeIE+DQO4NYrjPZ69MTyXPrKtcHLI5vpe8J5Z/bNkXOubCKM/S7pYtQUaj3nbzlje/O3fujnz21mFu3rKgu3nj4kjTLJ2YPdQGZVZHvKbOzg+i7FuR6+L3MfF8U6AZZHR+DBAFeR2V2x9LS+ZN/qdspoxA6gDKFQKHQkBDRZRyJ7lD+SUgxuJ0Lqas9k9bld7QekmZNr3FcShHADvuzEnDaKtjtI5ARk4d5WYuAuoWgqfuUfIobml5RNCQKwdBFL19+NveLVl0oFsV6x8JRn6QKsSRhVJ8lDIKFIu6eN1LMsY/0pYjpUnM1EGOvySQ2hnl0T3PKXqmqSFv8iwdh+ski+qlNBDKnKZDmZQXcbkvPnE45E8cnDQyfb/lU/7cN7MAuVZO90yPk5/MN//lCoFCYHgEEDAzY4Zz5JnB+Ryg16asrX//+98/nPfb3eP3tNNO697ylrfc7v6d/RDu+c9/fjsO5fexj31s5xiN01+87W1vG43X5kef9KxnPasdbhhA05eVGwsCMRixYH3ImJAfsSP4aF1IpG5JkKrFQYoWBcFauCimogZpW7hwZbdz+7ruu1eYnjqv23DL3pCHa7rdO7fGJ2d8eiNk0BH2dzi5mzf/yCBrdwkr947uumt3BHmM3byvjd1VVwQhXWANfszCCevjypX7YuOdhfHtZLNlemR1376QgXtNA43ZPcjkvpUt67t2xu7iQfTUg127Fsf1mpBtsbbx1pCvu6yZ3NLFKSyHy+P3wiCD84NAWgdpVs/8uG+gd163a3esswxZh9KQcUuX+najvQdis7rdyOOqkH/HR7xrIlys8d8xv7t5g8+TxMyc5b1PmMyfH+WOvOzc3luSsiPI9q7dy7uFUc8PHLB5zqKYUWBtZGxStz9mDnXKvTXOkZnRUKlb49uRC+74PeLRvsPR+qsWNVqkyt+kIzCUzE12glOR3lSkMdk4VfzjQwCpsvbb2iOCBlHK+oBMIWNI2b42oskCR8jEbmwhRFn1jG5yBJ64HBRCB4dwiYcTvzCIKksBophpee5aPBQxTjhhnFkNTX8zSi9O5FIayF9//BRDJFH8CCSyxx8SiQgLi2zykxZPYZBg6Suv+Fw7+MvnnpkZwLIhfuumpFGuECgEJg4B7W60bix+++Mcb7j+OCb6usjjOBA9EERt77ejj/5OmMNGM9PLQGVQnSCMS5aFnAqZsDD69dUxGHrkkdYVLojzjphSvai7x/n7YxOd2MhoVUxL3XZjI0xnn70pCOUp3eKF9wqydE7IkGUhl2KDtRuu7fbfcGMjoktXrOmWLV0eRC0IXMiypcvXdEcedVx3ykldty4GVufH9M9du2ITHcQsyNeCkKGL5CcI7OLFZJ7lH8jl0pBVsV4xCOPm2Khn48Yd3U0bbgmSGJ9ci29UHh2G1+OOX9oduc4Mn7BGxv1dMYi7LUyDW7du6bbEIQ+soGvXrumOCJm5KqyV88N6uH374iC+SyPvli7t7m6J2Ti7du6ItFk0Dbge0azh5OemTT3LJ5K+cmWsg4x8zgsCuWvXztADtoa83BxW26vDEvutbv/eTUEmTUvt4XzoNxqyc+/tN7U6tP/xPS0COT7cKlQhUAgUAiMigBia3vyIRzyiW79+fSNWBAZLoumdCB4/PaLXI3jI1YLFsaZx354QQKaS+n5ob9E9goZwOaw94pA0YZJcIpBIYb/FkOLEXxI1ZC6PVPQ8TxIpf0ihZ+JC7Dh5ZxVEEpMoIqPSdPAnXXHz65k8K6M8SF98rsXNn7IIwyGrLCmZf8983kO+yhUChUAhUAhMNQJh7Trg80qmTY6mHzbLJAhbrNmzuc28uF4TaxzXrVsWFsKlQbJWdCeedHR32uldd+pJN8S9n3TzF+3qDuyNCa7LTo+wK4P0HRey4rj4eLZN4zaELNwR8mheEMsVbabLEUesi7hj6mvIKYRu0aI9QRJj47hFdgBHrmLGzgGflNrUZEdPhgXpXGaw1reT94cMijW3sSZz167YvG3HppBr1j5uijh7cnXefLOG9oV/A6S9AdoQad2SsDxaazlvXkzD3RGbze26OYivWWw7u6XBr1etiiUdMU16eZDnBfN3h4zbHkeE2xnyNy6sZ1y3bnkcdpVdFGF9s3lnyMBeugsWhHU0xnfJxi7uz5u3O/IbJDgsrvKyZHFvcLY9H1VV6G0+OSqv4/RUBHKcwFWwQqAQKASGQwApZEmzZtWGOSx6iJJpnQgRAkgIOPhFwJC2latiBDKE444dt8SGOru6lbsQs95zJBG5E4dzkkmEzHQt1jtrfMTFJTlE5qTNP8uec2/aLGHam74qH/wlkeTHPXEggtJ2T7riIpQd4mHxRBIzrDSEQwjlR9h07kvTkSQT8VR28cm7uBBUGCHa4kM+yxUChUAhUAhMJQL63ex78zxy+tG9Rz++MGRW0IoDNrPZGXLgpJgGvb7Nvlm1al2s/2eJvCGmcobMORDrHHfFzuF7DD6uD//HhuUQkTTDxiYzMWUz4uxZL30jcU3E59MrdhPv7dptQxu/ncmgPXtujXBbQk5ubn5Y/BYsMBjqs4HWH7LAR/iY3bJx44a2bMMmO/tjmuiyZaa3Loopsavb2sdlIYt68fa+Abl3r52EycVeert39z7vYRfYJUvtLbAq8rC8DQDvvnV3EOBtYYX0GZE9EbfvI6+KMqyOMqxssnRHTMsVBydvvXLAzRIRG+nZDX1b/N4dmJK5C+K6V94WaAD+FIEcgJdQWSgECoHZgYD1iaeddlqzPiKRCBYyhYghStYZsuQRdo6eZS6mlgYRXLlib4xcxjfKTM/ptrcNCJA2JAz54he5Mt3U9E7h04qJ2HkunXRJDMUhHNKGvCFtCBv/mQ/xI7N+O3vOr/ickTtnZeDHtbT95idJp7jlt3+dZj6ThnidheGXgHeWf/lEiJFGnwq4/vrrD27qk2WqcyFQCBQChcDgIWDzl57ciHV8kT3ywSZMp59+Ruxm6rNVNo6LqaSLtsXnOtbFrqpnBnE7IdYrxo6pO44MsrQ8ZJIBR5vF9T6nhjSSNRy5Qj6QJzbFSRmG9JFl5AbZumHDxiYjhSPLli3zTeXejuXi4Tdlo9+eLQ0Tovj4J69zjwHxkbdkHkeGkqU9GUg29j65YWDYfYd85ACo3+Il5+gC8iKsgWADpHv29OStspGDZGPmLbIZcrK3h0JPJlsCsquR3ZaZAfhTBHIAXsKgZEHlLVcIHAoBir+j6srtUYIJS6MdfO3Q6xrpIgx0/q4JJla7XNuXm88QEiGZgjTOiwX/exuRikUnISx2NCEjDvE7c84OwlR8DkLKOyEc80irImJGaMlH5kVYpI2wI+QIWxZDebJzI2KYAk2aBJywnPg56QknL/wKLy8UB+VNYpiE2W/+pa084nQWRh7hIi5rRX2uxjrI6667rikMLcH6UwgMIALaQfWHA/hiBixL+j793WyrK1ke/boykg/KSY7ox32P1OZxS2PtYrO0HTgxSNlRsUnOKfH7tPAf6xFvXRDrEWPZxp6NsfvqzrbeEek66qijQyYdcVDmkFnkGjkhLXKPXHGPnLGO3+75SKb0c1ZOv+ySXzKKrHNfWPfIITLJM/FKS3wGM8knjj/lE4aM5N9v+SFjpUuOIZ1JHuXBgKqz8AaRxcuPcNLNtMWrPH57Jg/CCuO+fLnvGARXBHIQ3sIA5CEbRTaGAchSZWHAENBh6yB1jIPSgU03RISmjp2w8l1b31wjLHX8hI6RSUIBdvwSAEYiOcKnJ4D2N6GzefMtsTjftJhbYr2DDyBbR7G/CUbCyrX2KR6/CT/CJclqCpdsy/LgfQknj5mesPkuEUj+xJWCjGBFACkBHKHHiT+vMy/S4o9/4bOsGb9RVqOxwlEIOM8cnLAEr3j4kU9EG54p4JvH+lMIDCgC6q72oO6rw36XKwT6EdDf6edmm+xU3/XhykeuuE55k1a3lFVLltixNKas3npmYBFkOr7HuHixnbjjExkBlo1mNt9yU8i9LSGPejuEkwP9ZErc2cbIQYffKW9zmQg5Qn6QJWSbNgl/4ckx+Ui5RUZ6Rj66zz9/CKlDnEgpWSk9+ckBYWkLmzJNOvyR++KQPnlPTotbfJ5l3CkvkUtxy5t8uVYG8dMnbrzxxkY6hRskVwRykN7GNOVFRVfpr7zyyra9dwnAaXoRA56sjk09ufTSS9t3x3Ryc9Xp2JXfxi++Tea7YmeeeWZ3yimnNCGANDkINp0+/9qZ39oXLAksAsa10ciNN2/sdm2PDQViW/AVQciCasXI4y1N4BDC0iOsCJ0M7z6B5pn7Q9suwUYgSotwcs0vf/KTQjOfee5afofGxb/7/S4Fp3vpn5LkyGk6rikT0nWIAw5GanNKL7It/w7XqWgNTa8/7bouBKYbAXVen3jVVVd1H/jAB4ZtN9Odx0p/+hHQX+vzXv/617eBxkc96lHTn6kJygEZgFAhQUnMyBGySV/uUHaEqLn4NuT8mHVDFtgQZ++esOjF2kXtyCDqpk03hkzr7QJOboiTIwvgSD5J028ySfzu52CptOVHWPJN2kib+PlDLMXh2qENp+yRjjjJH/JJWDJXWHJcGcRJ5oqHX+mKjyzPMgrj8FxepaO8/PHjWrril39xuBYvv8rmeZYl77s3SDJx7mqAakq5hsCFF17YFNiPf/zjrRIXLIXAcAjouHSKdhf1oWifp5iLTqd+3HHHNQFljcdJJ53UNrEhWHT+KTwJIQci5UyI5TRNQsVi+qNjv3AbA9jVbceOwHefEU4bDfSmxbFCGqGFOwFG0PTC9kZWpUkguUe4pPPb/cwPgeWeIwWQc4b1PP2IJ+NyJgT5zTQyfPqRN2VTTuVzELgEIz+En0NexQELlkmjqjfccEMLJw4u4+YPrqb6EO4bNmzIotW5EBgYBGyUZfDnQx/6UKunA5OxysjAIaAv9L1b34+2udpMd/poMgPBQ9YQSKQKcUSUHGSWPl7/jmA5PO9Z3MhL8mBft2NXzJTpNjdL3+bNm6It9Yihvl9YcsyRBMyZS+IHW7IsZ8LIh8N9sob8IJOkTT6TaZ6JG3FzcOQrf+SONfgGOPkRLuNmUbRpnTKTV5k/fsQrfrJOnPKXBFH+OOVw3a8vwFE45RKHfAgrDvnk10wj9w1Mk7fu59EinoY/RSCnAfRBS1KjNvVOA1GJyxUCh0LgkY98ZFvrp1Oba856RYKJAnDiiSe1a0RQx++zGylMYGMHVaSKMOJ27rT4fk8jUT1BsDmEy4HuhBP2xTektncnnfzDEA5ruzPO3N4dHwR1/rxlsdHA1m7TzWtjauu88Esgme5phPK29R8EUlCvuCcVf2ynnlNEexa/3u5znvdIGsEnnDO/SdwyfO/Mb2/KaS9k7y+hxTkTcoSZMt5886bIY2wCFEIODuJYsaK3UythCyPkkVC2rsTZb+l7lvHKl6lLPYHam8bzzW9+MxQBa0Zs1V6uEBgMBMjMBzzgAU0hLNk5GO9kkHPxsIc9rLvXve7VBtQGOZ+jzRvik8RQW0AM/c4+XZsgI8hBRAqhXLfuiGgvdvHufS5qz16Dp9u7bVtv6rZt3xpy4LblEuSIuMiGlFeukSzOPbIDIctrv+XDPemSNeSTMJ6RV+QOJy5pkFmc/HkujMNvsgtpRBBd5yH+JHnCisvvJNVJ8tzLNKTrcC9JJNzEmZZWOAkrbfmXN8+tJRU3Wa0s/IlnOl0RyOlEf4DSZklxlCsEZjcCt1npxlpOPO2oo9bElNWzugsuOD+mq54cI4N2jhOn6SZL47ePFpsWY3QWAestrtfpL46PGK8IobkohKfvPN588+aYrhoE7pSbuxOO3xDfgIoNY3af3Z191q5Y9yB3dpzb3t10w97uR9esDiGEZC1tpM43pXppsRzyS6DmZh5IoeseiUQCPWslj7wmaWx342GSR1uU3+ZaoBau3YuPG3seY57tnqf7Iy5bjG/fviWEnem6O0MosqCmAhE7y4ZwtAkCQk2Ibtq0IUZ2r4tzb13k2rWrmlAkED0X/cLYCp7iccwxR0Y4xNynT6xB2R7Cd7wCs/eOWlnqTyEwgQhYo/SYxzxmAmOsqAqBmYEAEoTcGFTV1yNBaU2z7s9MHcQL0UnyhBCuXMkauTruL4jNdGLjuF07Y5B0Y8itPfEZjVWxZnBdI1gscOQCeeBIwic+hNHMFoTVmRO360zTICXyaFZMEjSkTJzyKY4kfsKLl8XSrIJMV/mSGEtL3MpNZiXREy7JHWLHiVt++u9LGzFULuVJ0k0+JkZJHPl1yB8spSsuhFIaiU1LbJr+FIGcJuAr2UKgEJhqBNCenrXutvNo83CgCaD160/vLrzwITEV6cwQmjHKGaSRoFgYU3EWLeptRINo7dhhHR8htSTChYUyOv4lIXR8L2peCJX581knb4nvYV3dHX3k9u7oo/Z1O7f7juMZ3coQgEsWrw4BRQAFmTzu8u7mLbtDEK6N+OxuGtNkIs4lS3wvKwTwQU6lfEkI44xExr8e58sprkGk4v++IISIZSOX8afHgYUf6nr+Pd+PQMaFOAnG/XFPGbdvJ8xs2249Zm9NphHmJUt8vLm3eyvhuDWE8vXX3xRTmmKnvfi9Lj4KTZEgiE0bwh8XNIFrW/XlYYH1DbC9MfJ6Sgxundhd/aPr4uPR8hOHco3aAcj0oRJ3o4asPBYChUAhcAgEyD3EB7kyiOLaYKQD2UEe7RGAuCE8pl4iZoiP/t6MHAOE+0OQ7InPcmzfviPI6PxubZsKe0SLhz/+kzyRO+JnSUSmkDuH3wgfYofkyVtu4JaEUT7IHaQyCRwSJz4yyDNxIIVImjgRY+RNGsrnHhmYJM7mOo4kpJ6Ly3PxppVRXhFoRNYz5UAeWTblwT3YZHn9lh9xSFf68uxektC0ZLo3Xa4k6nQhX+kWAoXA1CIwL9ZNzDsmqMfdYqgyiOSoOQj2FBRkybqwGJ4XxOac7uYNR8WIaaxjCKG0ND4S7Lwovkd14437ui1hXdx4C4EY0y8PxDegQqAtC1J4K0EaH00W1+ZbQiis+GE8uzFGJJd03/v++fEtrAd3P/zBCREmhPJaax98ZDi+u7jvyO7qH27vrrkm1kTcGoJnVYy2zlsbz9Z2mzZGfCFkIlD8VqDewfg4P/7Mb8961sd5IaiR2/3hxbOehVK4dEEyXYafeSEk2SMPiCPi9HtvEFrcLeOcH9fbdy8Jgby827alJ3znz4/vcy3eFZjM69asWhbftlzSLQiL4u5de7qbNuzpfnJNrEnZEjvdBRb79x3dbd66LATilsBjR7c7PpK8ND7mfMTq2OUulAhCd9vWJYFLTHOdt6lbtDQ2JNh5c3cA01TM0bpYTzMviHw37+gIUSJvtLCVv0KgECgEDoUA8oMonnbaaY20JUFEpJAnB4ugAxFCBJEtJCsJEZlC8CBVi0JWIH+eI53ic+1ZTktFppC4JLDkBBLlLD/O0kiihZRx4nPPb8+F519exY9gyhN/4pdf6w5zOYV8IG38cP1pSF9ZheX4SfKnzNLItPlNUuiMsJouK1/SFU9ac13LG/w8E39aVbMczvI2Ha6k6XSgXmkWAoXA1CMQxKgLUnagizWJsY6wx5YOnQ38y4eKjz/p+Ji2em4c58V3CleH8LH43prG+HbVbp28DQF8EyoWze8JYbBjQ5C93TFyGOsb1i6PRffL/tNiuSTu7417u2I664GwrN0UI6YxankgPkAcU19Xr97RHXHk4hBcQfr2B4nsNnS37r2pO25bfANqYXxUeUHEt6YnkBeHoGQPDGMiihf/eucemXQd5Q1iaYR3XxyIH6skd2C/tZQgiG9q7WsXPYtlCKv9IeBANT+sqoTxwhC0TcbHtFwEVMDGSyOu5bf2LKubFs8LYhjflIxR5W3brCdZFQL/uG75yrBEhjVy84EdgVFM2V22M4R27IR3xMruqKNj2/cg8kuXRJ6irFu2bO8WztvdLV+xLKyyLJqLuu0rQyjvWhR+gy6vvLXbFusgdwdWLUOtJKP5E5bT/UEgGyFXknKFQCFQCBQCh4MA2YDY2FBm/fr1jYwlaUN6ECzkKAlQTgNlXUuyZ6nF/BiJRLB6RMvOp7c2soWAiQOxE4ffSRoRK3Fwaa1zj1+OpY/FM62QyBfihtghpYhgkjXpImxJHqXpHvLIQihez0wtdSBx0kbapOMs/8hglltehROHPPEjnLy6Fp5fv2Hm8Fu6PRx6FlVlkVd5yucItPwqHzwc0lO+qXZFIKca8UqvECgEpgkBHez2OGJHTwRyFE6nfcLxJ3cPfOBdY/v1c7pT158QHfzKJgR2hPAgUG69NTaD2RkCYZ7vK8b0msXbgjxFhz9vYVgSl4QQWRSjtM69HVNvDQK5bt3qIJALQ0D5mPExQfDO6HYs3dStWhNTO9dt746MtZYH9i+NgyDZFBNVj4tprjGtZ/7aEEi2BUf0rEgMQnfQhZCMDQn2NgIYT0Jo7olNe/buNaIaAs1oa5A5gmZHjFruIcyCEO4N4UiYGo1VNrN8EcUUiosXhZUT8bwd94ofbu2f361euTTCxTe0NmzvNmyM3Wa3bY+wPhGyOsjxnjjbtp2Fcn5c90ZjbaSwJqYARxRBmmM9yOLtMUU3iGEIwmOPnd8de4zvYIXVN74TtnHjnvCzr1sT/jZuMAXKqHQEvF1+DoIwzEV43m/DJ+++H687eoXD4TgKxXXXXdcwhvMJJ5zQpkGJkxLy3e9+t02JOvbYY5tCcDhpVdhCoBAoBKYDgSRzSJIpo4gSAoUksoghPYgSa5k+kR/TQflBkHpkLwZdEamQDUtjaceSpesizJ62WdqWLduaPEJQk1ApZz8BE4c+Vp+dhM1Z2kiiw/pHYaSd00XJdHlCyjJO/uQX2eOX1ZF/+UX8xIWM9uT9rQdJm2fSR+hgkJZC+UISxYfgwSQPYfiXj/QjTfI3p8qKUx4d5LCz8jrgIR1xi4dTZmlOtSsCOdWIV3qFQCEwTQggB6afEBw94XFnGZkX36w65phV8cmS9d2ppx7fHbE2pmWuNNK4oNu9enEIllhrsSV2U91lHeC2njCLNNauXdqEKaHSE7AW+sdOcRFuSVjcbDSzaNFZjWgtDMvioiCG+/ctDwFh/WOsnQwSF6siu/kLr+8WL4yF9GtPaWH2H4i1gTFNlICx1XljcUGK9sYaEkLEd7SM+rofRscQfrvaBgWLY30mwrYkLHG7wzK6OabZEmi7d/d2wCN45XX+/GMiT6YEwcdW4whxTCsKYTeUWkWQcMhskNx9iyPd+OZkfOfYd70QyIUL90TYGEFe1lvzuWjREZHP1VHG3jfClobARIFDDjb/qwJP5TryyDUhSO3etzcUA6PV+wKzsEhGXhYv2d8t2BGEN4T90PzIzaGddz/2UIeO87anFBIE8dOf/nT7HqjfFBDKgU+WfOITn2hCnyLiu6E+AUE5GMnBIh3lplwhUAgUAoOAAOKDYPnUUk7xRIAcyI3BMvIF4UJ+kDi/PSMPESVx7NtHiMxr/eLSZUeEf5/c2HTQL//60CR9SBJZ5b5z9pFpifNcekn2/Ea4TLM1aCdNBI6MlEfPyU35dE/+yUHrHhE0fpBg1j7xCiuM9ORJ+vpwaQinXBx/DjJAvAgoK6NrWDg8kx4SqIzScyiXsOLW72cc4kUgE2dn4cTF/3S4IpDTgXqlWQgUAtOEQGM9o07bOkFkcf58XaVpIj2ytnhxbwc4O6/aCIYA0okTkoQUIUlgEUTZ4YtrXwgEAmLRothwoLswpqceF1NQfxxhQjDtP687EERsz56gjjtizeG+qyPstUHRYgpMZ6SxN23Gp0AIMcIlomr5IkR8RuOaa65pwi5HNwkfQpCQsnnPkiU94WkX1F2x8922sBYStvyLz66y8iu+JUsIOCPG1lBKqDcdlmBThvw0SNsEYY+t0IOgBWv1qROYsZDuC2LLeGkzHQfXC4fg+txIb5OAo48+6qDwlT63q5FypG9e5GNpI54LFtj0wEeYe9uuN4+j+iP/DaxR+R6PJ+/+Bz/4QffVr341duo9p+GpLOqFez40/7znPa/78pe/3F177bVtgIG/kRzFRZzei40ayhUChUAhMN0IkBWIDPKo/zLLwj2yBrFKKxlrZE5p1Y+lVZC88eyI2ERt3bqQa7Hmf+FCcoaMWxH+DEbuaDKOLNWHkmHC6UsRNoNyPTm66D9lUW/Hcf4QPemSYfw5TLNF8DIu/uSXHM0wwpHbCDF56TniqB8m35M4ypMyKrMDmUWm01Iqj/Ivv0k6kVTXPZk9r+XNc7IUaZQvhzLJD//yL80MKy349mTvbdfTWR+KQE4n+pV2IVAIDDwCdovT2Tt06ISJjpwgI2gchA5hQFDq4AkFgoU/goAw4t9vgqAngFZFmDPDQnhCtzNIws6YBrtjx4KIP2aRhvVu//6Y2rrvohCwQTxDyNqBdOeOW8JfbySUoJGWuJzlK/MpHQJJWpx8E2qEWwonVkvh+BXX1q290WJCLoWlMnH9pNHz/vvKJl7WTL6znPztCWK5L4jkokW97dKlk3nwnF/5d1616rb8pgKB9PWe9UatWUcH1cGEYnTve9+7861Uigb3jW98o7viiiu65z73ud1FF13UnX766e3D85///OebAjZcecT1ve99rxFN9Qdu3lO5QqAQKASmGwF9EcKU0zz15fr1JHwscfp1Z8SJbMrDb30aubp7t+8fHhEDjbF+MZZXzJ/fmw4rfnGRo2QR/2Qb2cXq5hm50G+JTLkiHfKDkz7iSEaLyyF8vwwTf6YhTnKTQ0KTPPKfJE+5XGce4IB4+i2MwT7EUz78Thmb8lhfLj3+5FNYh3vicFZOR8rLxJY895wjI1IOtxvT8GdwpfE0gFFJFgKFQCHQjwDBoZPPjxETbNmBEwwEmzMBZaSTf891+ClACAHPCVvnJHXWJd4au6pu22a9w462LnHzliOCKIYFM2YsHtjPGhlrFiNOO5GyGt4Sx9ZtW5twkZa4xEmQygsnj4QVYek5f4itPPntOSEqvwSQfBJ4OeIqv8Lz5+CUMc+EXAovcQsnbfELKwz/0nJuO7+GP7hQHqw3oQxw4pLPzG8KWX6F9ZzAll9CVln5kc50C89WgL4/8qV8H/7wh7vvfOc7bYT+mc98ZsOGssAirUzqgbwfyqrI3wUXXNCdf/75LYWrr766+8pXvtKXWl0WAoVAITB1CPT3xwZMza7RL2d/j3w59Nn8Jgny28Aav8gWSySZui3k2IEDu+PegZB5a4yadstiZs7Klb2dWvWR4tbX8y8cOeO+uMgtz8gZh+u0dJJFZJN83CaHeoOh7pEv5JaDP3lM2cK/fryXx57lkYwif5RDGcgkYbK84kAWyTa6gryJO/MqbnFwcJHP1B2ElR9n+c7nypNlz/wKlwTcM/FPpysCOZ3oV9qFQCEw8AjosAkF69gIDsIghWV24ISJg1BIQYKUEQg5Uotg8EOA9YRGb32FeHt+F8Z5Xgiu5SFETAlKC17v21UbN25oxCuFqHjykA9xOsujIwkkoUMoEUIELwKDPCKd/PMnHvlOoecZgZlCj79My5lzFqeDMBOWI/RTWVDm9O8ZPwSzw335TCHpWrh+nFL48ytP8m1UGK5wGCQn/w984APbehvvGZH84he/2MqsHA44wievD5V/8aWDS7lCoBAoBKYLAX2Wfo08Qx6RKXLQ/SRS+in9NJmQ6/70d+SIsM7kFDlA3mzfjsCZURNT9WOmypJYby8uz8ktfvTz4tqwYUPr96XLkQH8pnzxm3+ymjx1X1j546TrkEcyp5f+9pZXfS15p2zyKS5++OdSpvGT1lX39Of8Zv4QSPqCcOLkl7wVr3TlKYm1s/DKwK9rYZFPccIg5YW0lCXLSH767Uh/LaNT/GdKCaQKYdRV5QKwecmUMc49o6wqB5NzVorR4DEWv6OJr/wUAoVAIQAB/ZLOm1DUyeu33NPnpNB0dnD6OILu+uuvb517ChBChABJoSMeAixHLG1+s3dvWLC2rQ1hZO2hHecI0d4W3r6VyC+hI44kZoQHYSJdeRIvIU1g8ZN59SyFJBKmn5Uf/jmCKMuk/3UQllkuzzgCLZ1r9zM/+nLxEMCuMw8EOH/ODv79dsirfOVzzxz8yZs0lFG8HP/yne+CnyxD5mu6zsrAyujgvv71r7e6oDzqAcXm3HPPbcqG8pF/o3WDUsbR5rf8FQKFwOxBIPtr/Zb+PeWCfhnRImv0ydmf67NTNnneHyYJF7K0eLFBNQOmlkBY/tBbDkE2GigUhzT9dnBkgvgyXs85eUEcDcgidCkrEDJ5y/zJq3iToJKp4pAfsoWsTzkkTD5X5iwn/xmH2TRks7TFAasc7NTHk7Xy675+XFi/peWe38qkfMI7Um5LkxzN+2Q9XJTPdRLJ6ZIPU0YgAXLllVd2l112WexqeEx7+fe9732bogH4L33pSweBO/vss9vaEMCO5MQHdE5FKlcIFAKFwEQhoIMngBAha9buete7dieffHK7p1MnPHTa+iDCJgWEvkhnr2Pnz1Sf448/vgkBwkK/5Ugh2hMa1ocYtYy1DyFFN28m0PjpTcmRFiFGEAqL2MmXtKVDmHguvf/P3psHWXJV576r5up5HiR1t7olNRKSQBMgMwghBGKQEVz88MXYJh7X2MbhsM0NRxjihf95fzgcLxzhZ78wJuzr6/fAF8tcLmAuF5CQERKyJoTG1tzqee7qoaq7xlN1znnrt3auc7Kqq6qruqtOnaqzdneezNzjyi+z9pffntJbUT0cknSiJQwhy55rgwzJg/zdD7KC8Fz8uZjDdnek4Zw9jnJJQ1yuizDKpwxwwYEVduTrbfwhUQQteeBIS1zywXauj2sjHWSOnYgxMKZ8wurFYSuNB9wPuIuXCp6ZLVu22FxGuA9h/vLLL1u4D0+tF/vDjkAgEAgEzoeA189wF+LJR7OQjjA26nH4hfqfepG48Az1OEKPup3jzk7m/HVIr9bnPWdKyn+swnrCOpqoP0mHIy51KsKMvD0fwpx7CYdH4C6O2WMbXATPYI/zDLbAR9gC1/g1wWFwDJtfA3FJB/+wx5+08DHXj37BTvKiDHiPcvO9tM5n7ElLnpTl+XIOTvAa2GA/+VCm8x3hhMEvvuFHWudi8Kilq5mA5MIhzgcffFA+9alP2TX6g8AKdV/5yldshbr77rvPlkLnQbniiismxIKbRp7ceB60cIFAIBAIzBQCCCJ6i/jcAiJg69at9vJPhU69Rd2DeOHYNypxKnPqJEiNOozKHyJzgoA8CGcPWZAHeSVBhH9Bj8kXAVWwcPKFnCApyoLcyNsFFGTKBnnhD2FSPgRHPckeMoKw8Mc54eAHueLvpEU++HPum6ehfD9mzzkkiE3k49ePH/kQzvW6/XmydSyJB0Z+PVwnxIzdECRYcU2s9oednh9lkS/nfj1m3Bz90OvMpzqeeeYZw4R7y6I5t956q82J/Na3vmWt47Qg33PPPfKOd7xjjiyNYgOBQCAQmD4C8IxzF+/dbDSSIpioi6nvqYtdGMF/nob6kDhsOLiFMOepo0cQiyxAk6Z0UPcTBq/AxzR+kp+LQfgD7mAPX+EIxx4cXOhcCf9xzh7ewBYXc8R1XsQ2uJj8PM3Sg+0AAEAASURBVG/C4ScvlzDiwdvOT9hA/sRjcz6Em1wII/pc8HHdYESe2IQjnOsFX/wcQ/LyssiLcOz39wdLPEc/NROQAMbNhUw/+clP2oPGNSMq2X77t39b7rjjDhONkPBDDz00oYDkAT1w4ICRsd9If7GZIxyj2EAgEFhACECIb3nLW+S2224zcoRMqNS9YqcSp+7hHGKjsoeQqNTZQ0hU/ITjh2hgCCMVP2QDeXIMobB6KaTW1KQCUldGpeeRj96fPt1jpEIe1G+UB3lR/5HenRMpIos6FlspE7KhTPb4kxY7cdTH5AnZQXw48iQO/vnNArMfynbB5jYQl+tlI9zTEs4xWEGAiELswg97uC4I1NMRB3uPHTtm8bERe4jLCwQY8XLA9XJMvpSH4OR4rt22bdvkc5/7nPzar/2a2ZV/Abn66qvl61//ut1DrpewcIFAIBAIzAcEqGepj6nT2cNdCB64hUZB+BLn7+PU0dRz8A08yB6ehDvc8dmngQHySI2xh48c1aAuXZWc+jxNhSAdUwJoPCQ9+cIJnrfXsezhFfJn4xh+gd/Iw7kKnkCkwUNwL9dCnnAgecLdpGUjDbZzDAexEZdz8iRvHGnxg8s4plzwIn/wYdoe5cFZbh97ymIjL+K7KOSY6/HrJE94D34kL+wkTj24mglIAAHEH//4x3LkyBFboe5Xf/VXDTReGnhR4+Yw3AvHS8REjpu1fft2gbABEjH5wgsvTBQ9/AOBQCAQmBYCiJ09e/YIL/4MQUScOdFQl1HvsDmRcQx5slE/ER/Cg1QgBhdzkADEgINEIJWWlrSSalG/AXnyZBJRHR2stJpWnCMuZAYxeXnux7mTFiKL8vK2EA4Jsnfxho0MpXQCJG827MJ2jj1/jv1afW+B+sM5jjTuKAfn+XCNkDTXTbmQJfa4zb4nPvk5yVIuOLOBO+ekxzGvhA3e4Jr8utweizQHP1wDtrKNddjP/WMLFwgEAoHAfESAOpa6nHoe4ejDTRFX1N3uqAPhQPhobL2HiHPuW7qUOr4gXce1QXFxjxSGVDwWaGSk/k9TQBCQcCm8Qb6UDS+QL/lz7Jzq5XMO11AnO/9gN1zs4hfxhh3EhYdc4BLf+YfynIPID3/yxAbsoXzywN/Lg6fABrFHwzGClTwph/zYE590YOFCFvxIS/74O1+SBq52EYutpK8HVzMBCQg33nij9TQC9OOPPy5PPvmkAcVDyQ1h7+AC+GTOX36Iw8teuEAgEAgEZgoBCOC1116zRqqtOnyVyh7Coo6igqcSp05zcnIhQ8VPHebklq/XICJIAsKg9ZZzCGjx4iUaX+d/DGovo/ZGQsqrVhXVPw1FpRwnOupOzimD9AhBbMCfc/xdaDnJQXTYjW2QJ3Gd7LkWyGgiQiIdZU9UH5OezXEB/7xfPj15EIajPOxjw2Yc14WtTqTEIYzysZsXADYc18rmdlNOuEAgEAgEAoHZQYA6ljqcDVFE3Q0P0vvI+zjh1OFeJ8M31NHwDY5w6nb8qdNxibcQi9q41sGK3GmlbsI9b4So5+P1PmFwJzYQlzw5pnzyxAa4Bi6Eb9EIPoSUuPh5GuwgHdfB3rkE7iEf8vVrGhvm8fF3XBCNiEfKRKTisBt7wYlj8gML+Axb4DfK41rxZwNjroEyiM+ea8Me8uAdw+NaIXPwUzMBCQDMafR5jYcPH5aDBw9WQKOFnjiAxp4Xo6k6fxinGj/iBQKBQCAwGQJUzJAA89oOHTpk5EMFDwEwnISNyh6Cgih8Ej2E4ZU+lT1p2NN6SD1F3UYcCANCYIMM+Fai9klquAoyPcKPHkXCcaQhH0jDCZUwJ1AnNuJSPumJB0lSJuQGUWEDNpHW7SOcjTzYiEMenqfv8cv7UxbO06Wz9Otp2JPG8wcvr6/xxwauAQfm+PFygD/nxEecY7uTJfG5NsSmp02lxm8gEAgEAoHAbCHgdTd8BO9RT7uYg1dYOAaOxN+5gjoc/oHzqMc5hg+ou6n6tZo3P/Is6kmbeiIo0QBwIFxGfBx5wl3wIH7esAg34EfZ2EEY3IeeoDHYewLx4xqca8gL++ES5x3CsZGyyIf8/FqwwTkNP5xfG2WhY9h4H8Cfa0Q4wldslEGZOGzgmr1hlGPHyIUleZDOrwf78cMmcCE+5863lnENf2omIP1FAJAAHiBQ/HRPMxSJhQe2akv/iy++aGCxEmu4QCAQCATmCgHqKSpnKmnqLeowKm2f7O6ikHDIgLhOZBCFEx/EBhE6oUEG+EEkEAxkiysU+E6jis0Vq7VuZFgrRJx6H7EF0oDoKJd0XiaE56TmZEfZbjvhbJRDOOnJBxvIh7zHEhB+vpEGm/2cuONtXANxxu5J69cKVk7i+GMnNuDnQpy8sRc/79klTyd60nA/mObAsZdpBcdPIBAIBAKBwKwiQB2NyGG0DLxCHc6WF3EYAGeyIa6cNz1+Sws9hggw7Xks6AJsJXrhWm16wrLlK4wzEU/OW5SJg4/Y4AfyZIPrEI/sieccAacg6IgDXzr3er6kQfQiVglD4MI55M31eDzKJV/nGsr3Y+cpRCobeeDAAs6Cb72XlnTYSBqOCSce5VAuOJEvcThHiMKF2OLXij/8hz82OS5WaI1/aiYgmad4//33y9NPP20vE4DxhS98weY+Mn/xT//0T23IGA8kK9Sxel24QCAQCATmCgEIhdZDyIU9FTdkiAhiHjeVOyRA5e7DUxBKEIITBmRAOCRIhe9iE8Jw8UQ4ZbW360eOFy9ScccCMyOaLrXuUgbxydsFE+QNiUAehJO/Ew9E6YIXomKjbNJDZvSe+nAdsM0TI3mQH/aw53wsSbkfJIfjnPiexjz1x4mNcH+xAD9scxJlz4bNYAOBur0eRloELzZTBi3K8AdYhAsEAoFAIBCYGwSosxmlQ/1NPZ8Xe9Tb8A4ijk4i9pxTbxOfTWlLtxVyppeRPSy0s0Y5Kq0kTnrnTt/DYZRDPvAIfAEfkBe8wR5/4sMfiDls5Bju8N5AGnTZnIdBD/vQH+RH/nAO6Tj2vOFEzwNbcJTpHOr8jI2EIxAph7xIiz0IWuwmjcdhzwZ+bBxjt3MicbHNh8by7sH7CHbNpauZgGQhClan+9jHPmbXC0hMWsWxVP7f/d3fVW4ygIcLBAKBQGAuEYBwEFybNm0yEkD8UGlT+bOntRGioBUT4nCyYM85JEY4+eCciMiHMEiCMI0irRBTJ+JTh56SX3ua6+ciDDKlzuQcgmQjv5Q+iTwnT0gHgelCDBuwCQGGGGYPCeJPGvaQm2/5PPP2+73ABsr2a2LP9bCR1sM9X7ebMiFQtzufHixcIDo5YhvXDR9gN9cAoYMf+GIvzjGyk/gJBAKBQCAQqAkC1L1wzfHjxyt8xzm9etTR8BTCjA1ewlFvOz8tXky9f6mMDG+TzkXU9TSWpsXGSA8vsJEGHvF6H27wxkg4Bc6gUdFH15AG24gDD8JLpIdDsI0NXiE/8sI+roGGYfgdLoNrnG+wnXzgNHgM7YL4JJ7zHGXDV6TBUR7nlMkx5ZAPZfHuwDl2ko6NY+x0R974gRWilqk0TP3zxePIy3nY09R6XzMBCYCA7qIxf6GE+eqref84DgQCgUBgrhCAtCADKmpIg8ocsUgPHgRAyyvEBeFQhxEOIUEYpIUMnBA8LS2REB7kwZbipWGf5XLq9WvWVVlLOieypIQFAUISkJQTmp/nBZsTnBMm5IlNbNhMuWwudt0uz8NtpRzPn2Oc712osccW7MeRljicYwd4EI4/eLDhsAVsPD/8uX7OsYNjNtJCmhxDwNgMfvh7i6vH97ysgPgJBAKBQCAQqCkC8AH1PlMK4A440XvcEJDU5eyJxwZPuBhbsWKJptHFZRbTM9muPIvoSpxB3U+973W9cxZlwHNs5MXeG1Sxg/j4wxf44zwPOMcFG/k7b9EgjABFqJEHYfAPdnJOGc6L8DfXBJc5vxGfdwSuG+fXyjFxveHU7STcr4F82eA5yvMN+3HYhnhkoxcSOz0/izCHPzUTkHN4jVF0IBAIBALTQgAyQChCDAw5oYKnxRI/9hAFpESrJUQAETkRcMwGAUA+Tp7kuX79eiMX8oRYCIckCdMkUiouVvGoAg1SKaYeRvJhg3QgUcolDY4ynUDZE8eFF7ZDShAm+ZOGMGzDsXfxxzl5+d6vIR/upOh74np+kJ73ytLCCoGTFhvAKl1f+rwJfuSRvw7K45yNY9L7tZAP5+DINYIb524v8cMFAoFAIBAIzB0CiBr4kAZEuMbrcecuzqnT83X7yAiNs2tVQOpwTp3v364ikrQ0Njq3cEWkc67j2IWdCzLSwHXe20cayqUsnHML+cI/xHdegYOxGV4hX8ohrQs59thOfMqmTE9DPnAb7wSEUw5l4IgHJ4ILeZAWR14IT/zgM+JQpnMadrCRng0+pdeRRuux3GcZzuFPCMg5BD+KDgQCgfpEgOEpmzdvtkW+8mTihIQogzAQR1TqHENgPiwGsoA4EIoQgAspRmDkiSiRC8KP1VF1cbHCMhOO6qGkklopQYhySQdhQawcYxcExMYxxAR5QWjYwjVAqJwTn3AcZMXmhEZajgn3vInPRjz8Pa4fs4csiQ8RQsBOcpyTjnxpLaVF2kW3kzd2kCdx2EhDXlwn/h6GPxvnYMVGKzEvKi5UyStcIBAIBAKBwNwhQB3tDabwg3OFW+R+nBNXpZRySBJaMBNz/tt0YyEd4sIhLqo8Lenwh/PgFsQXDh5mWCq8DC95HOyBXxBs8CHxOHbegbvhE/JCrMF5cCwbXEq5xIebKJs4pPF8KYfysBObSEcaHH7wlQs/4pInG+Ww9/yIR77E4ZrIH97zcOwlX9JgD2URd65dCMi5vgNRfiAQCNQdApCCC0IXP1TcVPKIF/cjjgscJx4qdhc6zKlA6EBekBBz+dg8DWGDg6lHrVAYkuERlulG3NFSmWCBPCAvynfRRgjlsDnJEYZAoyzsQjg6WRGPfCA1Nsp3kiLMiZM0PsyUMtm8LPZcAxvx2cgL8egT/CE8ygQLwjjnGiE8NuziOkiLPYQTj3PKJoy0xINISeMt2tgLGe/du1deeeUV2yNOsT9cIBAIBAKBwNwikK+LXUg5h3Du3OGcVizSQ6ijSVRMKtvoMY2KaXQJcXHsPS/OyS8vzOA8+HjdunXW4AuP4OAV+MP5BW4hDH4hvU8/QTyywS+EEc8bX+EyeBV7SUPZxMMRRjzKZk9a4sFbxOMYnmXv/Ic/10JajrENO0gDBxIPf46dI8kXcYy/Y+H25PE2o2r8EwKyxoBHcYFAIFD/CCBUECmXXnqpbVTUVPQIHiryjRs3WgUPAXjrJRU9lT5+EAdiE/FEGkQZlT75QHgMZYVEILRTp06aSEpkkEQbJIOAHBlJog8SY6MM9jiOIRQnFs7JD/FFGU7A5AvpIcTYsAtbfMMOH/pDmnw6J24nND8nT64RTBCPbJAg4RCwi1BIFwzYO+lRHvE4hzCxB0frMbiQlgV1uC7SgS/lE2/fvn12X7g3HHOfsGMmHOWxhQsEAoFAIBC4OATG1qWcw43u4IDkpzwwrD17fS3KJ7qCqo4ALRWrI2LgEzjP41PfwwvkhT+cB3cgIGmchQe9LOLAM/AHcdnDUz4yiGN4xUUh+fhaLYhC51d4023HDvgVniIOvZocUyZ5kSc2wpFwv4tC9p4P8bGFcuFQ7CF/8iCMayIcrvRF7/y6yJvNy0vvDY5qbfchIGuLd5QWCAQC8wABKnYWBTh48KANzcyTF0QEGUBiEARLayNyqNDZiAvxULG7cPIwyAwHQXBM+pQPnwfROZN6PjzCB5WVJEaqhAQ5kickAqngvCzKwyZIjQ3C9fI9f0gNG53ASMPm4hGSQnhyDnF5uB9TFo5rYuMcEiNPyI89BOjlQ6zkxcZ1Ui7XCcF6nuRHPviTF7b69ZEPLbvYwRAjBCo9vzt37pSXXnpJ9movJHmSt9tGfjPtKD9cIBAIBAKBwMwiQL09ovX3oHLp0GBBhVRR2toRezSaVnkSToDfcHAM3AsHwXNwFjyB6OOYMLiOPXHhB7jFuZP6nHC4A3/O4Sgc4pGePkQoeVEm4Z6H2wF/ueMasIU47MkXLuT9AT8XhZQF97FhGxuOeHlbuCY2L4vyKQ9bsAlexF7CeYdwPGaTA/1ax9uHgBwPlfALBAKBhkcAotm/f78JFkQSLYNU7hANBOACBuJggyTYcMTDOUlBAGxU9AglKn7IxEkOIjl9WoeCdiHUjmlZfO4ikRcCizKdSDwf8sfPCcXJB9KhDIiMjWPypzxIDTKFiCAgrgmxxzn5OmE6cXkZfo2kZ8NxLZRPueSFIy8InXPCcKSlTOzwtOTveRLHMSEvt49wygBrH7r6xhtvmKhHVNbKYWu4QCAQCAQCgZlFAH4c7EdU6Tcd+3QhnQ56FWmETaIJfnQegDtcfMEvcI2LPh+xAk+wOdeRljzgFNIgvAijXOcZehA5hgvZOPb0HLPBQy7aCGODU3GEYZvHgy/czzmO8rCDjWPnO66HtM7HcDk8TF7On6QhHuWxkZ58CSfeXLoQkHOJfpQdCAQCdYsAFTUCEkJgaCXCiG3Dhg1GABxT4UMWxOHYK3TOqfghB1oOIR/iITohDydCzhGnfBLk6FG+RTUkZ3v3KKH0al7VyfxeBqRB3r5BNi4cCSNfH1YKkUI4TmyEExd7fNgNdpEXdmKXEx7lEZ9wyI1j4hDutpAOwsUPcuac/MHBCdKJjnjY6nmwx1YwxkYImXzZSEt8vx7HDCHNcFjihgsEAoFAIBCY3wgUtadxcFAbVFVADg1pQ6x+ygoO6exMwzjhFDgBvoAr4AT4DH5wEYl4REjCCzQsspgbHAj/5vkYHmODX3B+7o2elAsXkQc8Q3nOZ+SDDXAb6SiLzXmOMOdAeA7exFbCCXMR6JznPEu+5Ek58DJp/JqJgw3wHlzOiCj2PmLH77zH9/Na7kNA1hLtKCsQCATmDQJU+gybhIyo3J2wGE66bds2G+4C+RDme4gAEnAyg5AgE4gDYvLhM1T6OIjKv/HU3a2tsL36IeIS8y36lFSKJrogR8+XNKSlDCcqF4GUib17dXgnJIr92Ex69i4cEY8ckw8EhW0QHbaykc7JmutiWA+kCZHnHQSHH7ZhC+eQJdeUF3qkJZx8faNMiBBswYTycPhzThrIlfzxI2+Ilnwcu7wtcRwIBAKBQCAwvxBI4lB7CYeVG3TeY1tru/IVDbUrR3EAV5XiJiEJByAi4Sf4DH6DN/I8BqcQh7ikhR/hN+I4X8FdhLGHuxBohw8ftmkp8A7+cA975yTOceTPsZ+zh6PgOBz5cuy8SrkeRl7k6XzGnnN4nQ1bSOc8SQMzApJ3CBeQ9cCDISDtVsdPIBAIBALjI+AtjYRSae/Zs0f2qkiDvCACWkDplURYQmQQAMIIMUdlj9jcvn27kRkEgjCCLMgXYjh8+JAJrlKJb0i1a36rNB9aMFuNHMkXEeitmpCNt1qSF+VRzoEDB2y4LYvLQF6kIR5xsJF5ItiMHw5bvHUUssJmJy3I0fMmDfHYyJcNsnQ/jskLgqS3kA0yx4/rJD0OMnShyDnx2bguNrfF8yYOjuvzOODN9YcLBAKBQCAQmP8IlEo6RLOs3JJbTJs6H55i45gN7nWO4BhxiHiEE1ysOUeRzht9ietcA08yrYMw0nm+hCPOfDQQ/M68e/IlT+LDoXAZ/IM/aXDwFfkQjzD2cB3hLlqJA4eRj18XcfMb/p4f6QlzkcreNziavN029nPlgonnCvkoNxAIBOYNAhCAOwiCVkonH4jMycX9IA564ajod+3aZfP2rrnmGlu9FVEFkREOKdDaumLFKs1+ibaAbpFLLqFXk2Gaaa4ik/opw8nGxRR7J0d6HF9//XUTkAg47CGc/dq1a23Li0euAQf5IPQgJWzmmA0HoTmJeRjpsNlJjHPywI9rYuOYsiFo8nCCxc+Jljg4XgCclCFNhKeLWK6N8kmDQ9DixxYuEAgEAoFAYH4jAK329+v8fxVrp0+1yPp12tC4cqkKsNRLSF0Pb1D3s8EpfuzcQRx4gw0OgY/gDReYxIdT4EX4Cv4lH6agwK2cE4awRDTSEPvyyy/bKt/kx0bDMPFplHVeg9uwAY6F57ETWygjz6PY4nZTlnM5cdmc3+BJeJH0zpmkgx9JQ1y3h/yde+fyCQgBOZfoR9mBQCAwLxFwkYXxVOqQF0SFcxKBDCACRBUExjlxGRKKgzg4hlSS0OqUQwfXyyWXKmmogGQRHcjHCQQSIy5ERXwvjzwRo/QgOjHymRE+QcLcTcqg5ZP4kCubO+wjvYs2wrCfMsiLY8iVDZJ0gnOS83zAgzDKIC02syd/wtg7sbLHcS3YBTmCBdfq8SiLdPixYaP7ESdcIBAIBAKBwPxGgKqcOZCnT+u0hzNtcuasjro5syjjqtTAyRXCQ3ALm/MgHAgnwHveYOs8Sxriwi/Ex5884A78EX4+qoc8GC3EyKLdu3fbZ6Jee+014RvOcBB8BT+RD1wFr8FdLh4Rls5j5MlGXMpxu52v4TIXk9iIc86jHOdm0rIRHxsQtpSb52+Pm3KZm98QkHODe5QaCAQCCwQBKnIEzkQOgqLHEiKA7K644grriUTk0XoJ2bS1MVylTbpPL1YyYu5GsxKLGIEQjiCDpFxsOjlRppPbJZdcYi2qxCVfJzbSQ2SU76RDGhzkhfBjI5xz4hIO6bqwZJ8Ph9ggNIjRidkJkLRsxCdfysyfEx8SdXL3MNJzXdiLc8EItohwRKz3hFqE+AkEAoFAIBCY1wgUi9rIqvxy4kSPTukoKAcwooWVRxMvwR/OSXCFN1TCD/ABjbfeSAtPwWEu8uAY503ygLdID4/i+AQX4ozVvRnBg4BkCohPP8kDS1n0Urq4Q0Ay7BWehW+ZagKvkTfncCMOmynbN87zzv3JlzDnYM65dq6TayDvPN/m85ir4xCQc4V8lBsIBAINgwAkwNAYyA5S8A8eQ3TNzWmSf1kngRCmP4ZLS0tzRWhBVpAIBAjh4IjLBmExBxPhiGiDhNiIyx5h5hvxSY8/cbELQvVwiNiFHPEgMz+nHG89dQGJH3kSh7hs5Ec+5I0jDuVBiIRzzVyP91J67yJhni/XSnpEI/NS2CO+yRe7wwUCgUAgEAgsAASUzorlYRVtR3URnVPKH/0VUQgHwC849mzwBGKOMPiADeFIGDxDAyT8grDjGNGFPw5+gW/Y4BUWsEM80uMIP/tKp3DYeA7ucf4hDra4aKTnkXIp07nXxSF5ue1wJcfO4x7HbWSPH3vielr4k3wJo9x6cCEg6+EuhA2BQCCw4BFwUoA8IDBIL7WYpl684UKbFJSgSqWyia3OTuZdrLTWTBdbTjIu2AANP0Sak5uLVIiNdJTr5AXxcEx6iJe4TsaINF+9FZHnREh88qJVlSE8OPJhg0S5FvZOdvhjE3EhPReOhGMLeTvBssdhO2ny8SF+8qKVF/HICnnYip9fkyWuwx8wY8NONpz7cez3keNwgUAgEAg0MgLKejp8VRtXD52Q/oH0/WL4CZEIb8AP7Nmcb+CuvMCEV+AcRBxcBfdRzzovwlEcw7lM+dirC+Uw1/HVV1818QjPwIGUMRVHXnAUI4lYJI+9cx5leL1PXs5XcAD85S7PefgRDifCg36tXCf2ggdlEqdeXAjIerkTYUcgEAgsaAQgBuYjbt682QRZVVxki8Ko0OhobdPeyVW6kA5kmIatIt7yQ1cgFhdp7CFUyI/hOBAgfghKyuPYCZFjCAgR5nMmff4I5ERahug4EZMegqN8VnClVxBig4jJgzTkzXUQz4mPOBx7+ZApaYjDdSBMIVry83SUiYMcicuGo2zyg0QRkFxrvTuuARz5hijzUH3OK/fo2Weftd5iVuYFg3CBQCAQCAQC2gtZKmo936Mc1WsiD36hznf+QCzCB9SvHMMF8IaPZkEwUqey4Q/fkQdcQlzy4dzrZoas0vPIkNV84+RU7wW8RNmsM8AIIBarg/Oc19nDjy742LPBeTjCcZxjKzayce72Yyu9pPSMchwC0iCLn0AgEAgEGgsBxBNCbNOmTUY6tJIioiCLjg5WcFsihaE1SkRNKiLpuUurz0GWCD3IBTKEeCBSNggF8vNvRHHu4s0Jy4mMtAg/iIjhO8zzoMfReyEhWNJCiNgEEVMGJMkx+ZEWoUk+xCUcscrmIpfyiEt5bDjSE8fFI+kgSo9DfJyLXMjZ7acsrt/Fbp6ULVGd/YDjX//1X8tzzz0nf/RHfyS33367Lczw1a9+Vd773vfKgw8+KFdffbV8/OMftwUXpmI+WIcLBAKBQGChIlAs6tQEFZGlEkJqxLiGz1zBEdSpcCWcAL/AT3AJU0EYqso5dSScAgfCaTjiw5+k95488ty5c6fNd0Sc0ZgKx8A903FwlC+ag03wM37Y4fbgj2N6CiOLcE06ZQVHedgHnw3pQkJcJ7ZgrwtKGh1ZzOd413GzvzCkq8zq9zLrxQUr1cudCDsCgUBgwSMAwSGenPgQVosXLzIyLJU65PhRndxfZsVRliOnty0RE+RH6ynkgoO4EKOQFuIRUiQMkUYZEBMkxAahEsaGCKOnEuJkwQD2+EGwEBn2EA/RyKI8kDP5UTYkSzyIDj9v8XWyxM9JkT3nkLwLx3w87MMu8qI84uPc348JQ7SyednEqVfH/aBlmz1za1z0vvDCC4bX9ddfbwKcRZV+8YtfyPvf//5xL4VrPHjwoKXnvnDP/GVk3AThGQgEAoHAPEaAOo9eSPsupApI+IbGSngEIUUDJTwBf1522WW2eZ1IWrgEh59zBHtvgPSGVniPT2vRgIpgg1ecf6YKH2XAa86BlOP54M85cbAZQVka0d5H3ZqU5pr0ehCUOrnBrhdRWBooWXr4Apuc8yrnXWelt0fneyomhdL48zOnavtMxgsBOZNoRl6BQCAQCEyAAIIJIqTXj5VYIcK0LdIWyw4lEMTikC5nflZKZYQi8+dSLx0k6CISQQFZ0tqJwEJcQI44/JxoPR5ixoen+sIBEChCB+FJvtgG6SHyiMPeW3YhRsogjpMmrcGQJ+eUCVk6cTt54k8+XKP3oJEHNpMndrHn2vAnPS8L7P3aTp0+ZXiBG/HJu54dCzE8/PDDcs8999hwVfDBjyXiP/GJTwjfAqWH91/+5V+sh3IiAck1ck9ZIZB7jYAPFwgEAoHAQkaA+j1fx8MLCD3qQrgBB/fAEfATdSMc4ryHf97BgXAf6eFdGuVo2KPhFH84hTIuxMGzLhDhN2xAiFImApCeQxfATS1qV6tO92hGQSrPtehUlTXrNI2udj6ijbvDqQGX/BCNiFyuuziiXN+v8z+X6mJCb2UOqCbfrRiduBCLZz7NnAhIHhBA5oHwhwI/biY3gW3sgzDzlx45BgKBQCBQOwSo3+h5YuI+k+7phUwEOGD1IaNu+vpadetVwYWgSst6U1f6BtlRVzpRkZ6NvPFDsCFaOMYfcYhAg4zYIE7m5iFovIeLuHlHCyjlIfSuvPJKq6MhRAQjvZL0TrJR1lgCpe7GeR3udmCf20mLq23q1z/QL2d70/BcuKC9LRuKpMK5W+04sPeAHDxwSE6dPHXBRJ+/ttk8BjNeTtj+8A//0Hpz8UP88SIBfuDB/eEeTiYK4T+ekW3btpnJ3LPnn39+Ns2PvAOBQCAQqDsE4A42d15veuMl/IaohIucG9mzIS7hHepfRur4JzoQaYSN5T4v43x78oaLyZs9fMdoG4QkdsGzjz/+uNXZw0Mj0n61zo28Rnl7DQ2g+jmRRYvlulXX6PeedcTPcEGWnl0k13W9RVa2rpSe584Yh9B4uKh9kTSXtMfyAx3S/7kt+m0rVZBf3SdN9/eKdmLOuau5gOSG0cLNC8zll19urbH4ARZzRviWCi8tDNEKFwgEAoHAQkIAQeetoIgwFljxHrpCQReoKS7VYS00sDE/ojpHAqJig7gQbogRCBNi5ZwNwYdgcWKDJCFX7wGjFxEh6PMfCR+PQMmDNOSPY7hQfu4iZI3dDHeFOL311e8TeULOXCvEzd7tHxnWhsMR7a1UHiyWdYGB1rLlQ/7kZQKy1KLDdfrk7LGzcnzfcTlxsEt6z059dTy3o9b7Rx55RBiq+rGPfazS+gzezHUFIzDAgY9dq96zyRyYuOOYex8uEAgEAoFGRgB+ojeRupZGURawoY6ljqRuhf/gH4QmceEvOBb+Y9QNnESci61Pqc/hX+c6GkDhZezAj4ZEdI6aJPIbl4h8+lKRS3SKivK76DzIfe3HbT+oba63d7fKLY+9TV5/bKe8ofMzT+vIG1yxpCN23q1Dd393s8h7Vos8e0ZkWc1lm9ky3k/NLQHYv/iLv7DhPV/60pfkzjvvlBdffFH+6q/+Su6++2554IEHrOX1U5/6lM3xGc/osX6QcbhAIBAIBOodAUiHYaOPPfaYCaurrrrKlv9esZK5hktkZLhDh63wrSm+MZU+bwHZIcLYI+ogKTZaOqlP2SPmIE8I04UkApGeK4iMnk+GgRIf0kNYEn8iRxiilAa9rVu3Wrmc+/Ah8qCXzEVsPh9swFauFdKmYZBe100bdPGg69dL4cpBOb28WxYN6mqse5bIyR0n5eiBo5bflm2Xy7obN8jpq3Shg0teljdW75NjbSdleFAVZ50M28lfa/6YewIXPfPMM3a9vDwwzxE/cKT1+4YbbrCWcF42WI13qi5G5EwVqYgXCAQCCx0BOIaGUEQiIhKB2NbcJi3aULdkqS74pv8QmfAUjaEdOkUE/uvp7lH9NjHvTQc3ONI3ysEW6mlsYRQPw2ZLKha3XLpVCm9fKkevXqLisUnecbJTtvattLgaXRtRdT7l0Rb5xePPyiM/fFj279tvDcii7YeFu3Sl7j/aKnLTCh0Cq5EZxau7enE1FZC8tLz00kv28kOPIy86vOTs2LFDeJG6+eabrVUA4n3yySflwx/+8Lg40XKA6PT5IbQoBMGOC1V4BgKBQJ0hANEg6qgPEVhX6DDFdTovrq19hZw6sUVFGt+3QuhVh8hQvyHeEIouHrksCKxZ51FAWi4MEZsIRYbseCso9Sx+EC/1J+kmcwgc8qRllxZeRCotqwhGBBF5UH+zcewbeVIG9rJB4gzl2fHCDrnxHTfJ4Dv0cxxX9chLy7pl5S5dUXbXkJx4+JSc3tMj7etbZN/GnbJ20wY5c3VZdnTo0KMbVoncrD111+uqrfefkPJLOg8yrSM0mflzEgaHcW+4v7h/+qd/siG/73nPe6y1/J//+Z+t8YD5LYyyue222+bEzig0EAgEAoH5jgAcRl0LBznfNJVbZMmi9HmoM33dJha7T/ao7mqSYQSf/jMBNgODOQqDOj/zUJcc3nDYuBIbnAdpJD59Sudt6tzHNdetk97NTXK0s1ne3t0m9xx5l2w6tUlt02GwMix7Du6TXzz2tLz+s1fl4L6Dxp+tb26VkU+sF/mIbu/SnkfvI8PuGbB9pu59TQUkLQEPPfSQLV/+85//3F5GaBmnZfauu+6y+R6sLAjRIhAnEpBcvN8o9uECgUAgEJgvCFBn0ei1Vz9kjEBjUZUWbU3t6z+lQzeV6lp6VSwOKDGyQMCQzUEkDnERci7gmGCvo0Cltdwm7S36GQ39huSZkTPW+nni+EnZt2uvvLHrDek63VURjlPFqFnnIHqP4/IVy21oqYtH8oC8vQ7m3Oth30OmkDt1O+KRxXAOte2XwvohOazfuNzdovX2cK9oF6lc2t8u6y/ZKG03iOy7eUSe3qr+fRq+c1C0WVnk6qUim3R4kg77aTl1VIp79aLr0HF/2Nx9/vOfl3e/+90mwFl99c1vfrP1ytIb+Za3vMU+5+JxYx8IBAKBQCAwfQRosEQUNr9D5/5v7ZSmNj3Zpd9/fEKnDCiNFJfoiqc36bzz7culuU8588kzUj6kQnKCtXOalmrj53Jd6EZncEyqLkZKcrp00hpqN122SYeWJl6kY8sW0LG1DPTTIpc0ycjitABQywmdD/mKNgS3LNeVWfW7lz3aiPzoKXnx/uflyJmj2ZowujrrO5VH/mBbGvK6UxdH6FQFyfDXOup95E7VTEDS+s23V1Dmb33rW02xM8yJlwxazH0IFi8pOFrnJ3K8nJCHO4Tpz372Mz+NfSAQCAQCdY8APYY46jPqx1Mne+VMzwptIz2mvX192qNYtF5FegLp/WPOJMfEp9V1WIe5DgwiNPWzH7p6a6Gg530DNlTnxKkuOapi6+QZXT21rEw5KROOA5UmaSo2S5sK0w7dKIO6mrIRsLbgjQ5pZW4ffu7yx5DoSy++ZOKxZVGz7L78jLx2ic5tX6R1/IAapMuatw2X5Yarb5TrbrpeBt7TIw9c+bqcaFLi/3GXLP6erkCqw3iGP6PzR+5YK3LrSik+qavNHtEevlnqhXQB7NdzMfsvfOELlpw8WXzoT/7kTy4mu0gbCAQCgUAgMA4Cut6clH5JRdfvXC5DCK3/dVyaut6Q8hsqLm/T70f+ZxVj710jped1DuF/fkk5ZBICub5dSrdr3OWTyKOM8oYf6ZXeQ73S39cvq1braBl1DK1FROJUikp3R7ccatYw/X7j7v4BeWLgYTl6+RYpDelaAd39sqP7Rek6eyLN/29rVZ7VUT0ITr73+FSPyDcPi3BtdyoHZt+QtMzr4GcShGbWOub8MHz1ox/9qI1JZnEBxiT7UvG8EOEQlbyg8MI0VUeacIFAIBAIzCcEEBYMMT1+/Jj2OC7VhWk2ay/ilVY/FgqHtVev2epCegJpZGNeBUKEYZK92rq59+heGVo/KM2r2mSxKq2WkzpMZvdx2Xdkt3R1dsmxrWeksKQszfuapNStZDTeqFUlQtN/VQ2o5Wo93Kyfjhjq1ZVQT8qpbhbfOWsi1VeZy+8RjT4klrobkYsgZi7gjhdelNZ2JcObO2VYBaBs6tReRS2M7zxrOZd1LJX1SzbK8s2Lpffyw/L6CjXymV7p/DcVvo/3SZPar+N9RLbqXJC1OpT1mqXS9MpgakGeJzc7L6rniclhZiAQCAQC8wYBRuLI91W0IbKu1REruuBM+Vc3StP/e0TKv71FV6lZo8pOGyYf1wbIl7XhNrXdjn99b1sh8oXLRdYo30zmlrTI2eIu6fqvXdo5tksXwBu2ldXpJEPf4IraGLprcbcOQVXuU548vr1DvrNFbWhTP8TgSV2BtemUdBzSRfH26zcrCzrqRom66fFuKX/5FZGXdDTODrX7/1Y9VGfikeurmYBkaCrDr5jbyIsF8xyfeuopG8bKHBvmhdx0003W9QvhskJruEAgEAgEFjICTLTv6emW1rYOHZ3RJx2tS6Vv8BVtSGM4S4uNzFi7dq0REkuD0+C2ZPESGdQ5knu7d8nA3Tqf8ZZVsqRdewV/2C3dT5yRgWu1N/M/bRC5Tsn0sVNS+r/2iHSPGfaJYFymP7ctlfJ7dY7FZSrstEfQnO6GNWjP8iH5yfqfyq7WV+SyV7bJpWcvleX6r1l7ExGKDB3yOZXec0fdDXmyQt5TTz4lw9oruu2WK2Xn3crY79RWWFp1Bxk+lNRsa3Or9pzqdyaHtce0+SxNtiLHtDd1T5+UdLiRid7dqja7tIGRoawqJFtWNMsIQ5DCBQKBQCAQCAQCUNdu5Yt/OiSiDaryDuWaz1wmZXjtZhWEHdqI+YQKt/9yQORMxnMToNb0s9NS7n1dPzjpEw8niqia7oluOXGoLMP0Ip48Zgt/0jHWrZze0dwhnUvaped6HXUD76kJtgLrPh1Bs09HWL5PRa02qPYy11GnhLT+n4ekaCt1q30vFqT5VV1Ib1CPoW54ka3OXM0EpH/3zIdtffvb37aFBD70oQ/ZSnV///d/b6KSOTMMT33f+95XZ1CFOYFAIBAIzCwC9NzpfxWM/SqkTmiDmg7bLOhKcaU+FZBpqCq9eQg0Nnr+GObfVG6Ss8M90qfzCendM5K8vkXa3qFzP96r5HmjTsjQITKi8yeajup8xfH0FrU/vXrbdXW4K7SHrzCaWE/qpP+THWU503dMFpdXyfriBuUw/XavLtpDryijRBjKaj2QWgCrrjK383jXcXn8iceFeZhbrr5MVt25VHb+kra4ru6QW4d0/ocOjT2k8x9fV0LUy5Ce3h4l7BHpGlZSZdjOmRFpVsGrVttQ1aYu/XzJWWXRRUroK1qlrHaFCwQCgUAgEAgERiHwgA75vFFHrNyoonGb9tqt0IZUROQL2jj5Q/1sxqvnn85ReoXF3U6lkTmjMh99AluWhrQBVzsIT585rQ2/ffb9R18foEWHo5bhWOiKufzdWva/dYl8/aC0HdIpKAja39wk8mZtGL1ttYxcc0RH32iumh8LxZU173p3NROQDLtic/fFL37RVl1lXg8LDPA5D4Y+8XHtW265xZY997ixDwQCgUBgoSNQKuoHk1v41mNSeywBztB+Gt28h88xYJEbhprKL5RprjohcqnO+3j7ShmGrLapGGTY6IMnpPkHXdqTNwER6TzEFp0TUqQ3ECHpPZAUosLu0mWLZduGNXLlqk2ypneNNOuQ2mH9LpVOULfvMrKyK3Zof6ROa1wky5dq76Sev/7cTjmmQ2k3rd8kV79vm3TdflJF6rB8sH+JXH3mCil0jsjZjj2UIgMjQ3Lo2BHpPqI9sH1q8xoVicM6hEdJ1Jya1qRCuIyw1BXtpBXBmoXFLhAIBAKBQCAQcASYqnG/ijSd6iB3a8/eJSoeu5VMfnhMmh/ksxoeceJ9sy6i07xGeaZ9EqLRIPIqd2lD58m0qjkNvWw4WydAJ2YO92uj73ePyshB9ddPUbXoENriIwM63FXjHDgiRcQt0zPY/9IKaXlFv+Ws6wLMF1czATkWkE9/+tPmxYsRq9KxYl24QCAQCAQaFwFlJdQRW45DxopH8Kl8y+qgnvxUBdrlSkK/cVmaAzKgzMZiM/92Qkovq+Abz5E/HX46TEYOae9gB4xYjahnIivXyZKr9DMeb9kuq5avtp7H4ZJ+Q7KnXw4ePChPPPGELUXe3tEq6y5ZK9uv2S4bN2yUFw48I206KvaKm7fKap33+NybDmOwrHhOhWafzoXUuZlnN2m5Wvf3qEAeOnBIOre0y6E+JXzR3lCcGZAOK7+mmCtncRAIBAKBQCAQCIxGYIeKtUe0F/LD67T3T4lOp0SINpSWGOI6Bdd08yIZ+WXtuVytQ2FzPDwqKfwET39Hew1/oPMdlc7yzqZ3MPYU+v1fGni/9oDqMe2g7opdevK09pi+X4e0btQGYOb3t2sv6TxycyYgHaNYYMCRiH0gEAgEAtNEANGn31OUf1fCvF6Hrf6SDl/t0XMVj02PnZ2Q/yilSTsdy9coSTKEhnkjOXZDqx3WFtj2tQekTclte/9VsnZgvRR1ZdjegV45sP+AzWEvtelKqlc2y/Ir18p+/R7WhjXr5am3H5Kt7YvlzIZu6V11Wp5eqvMfnx+QPY/tk7PNA1JYXJQ31qufLk3ep/MZC9ItZwd0uNGQzgkxw0arRzpVTduauNaT0cGWJH4CgUAgEAgEAoHmjS1SYvgqw0ZxLIZzqfbwrVDi6MkpuBR6zm+RKR18f3EdBHlOcNWDOZJ8YuP+cwVkJZKa0KTUak6PbbGfSqAe0Dt6WjcEZB0ukpM3dbzjOReQ4xkVfoFAIBAIBAJTRACeZGVTNlyr7hnuOZmDzFbpDwvo/NplacgPcybzToP3qtepwyek+7snZP3rl0p7oVMKTUP6uZGzsv3K7dK6QRfN+cCg7PvQSum6WklalExv3yZvnCrI4Rd2SdtZpZgVupjPdUvl6bUqGssarvMYbVulgvcPtsrwpqNJ9PZriy0kqnMcy/SI4jRqcakSNYsgMNS2oFtO6KZI8RsIBAKBQCDQ8AioWCvdo+LvNzYlLoFGmJ7xv12SPt1xr47MOY9rfkAXnmORm8XKOxMJyIyeml/Uxd406rhOKatpg3LZJ5T/lmleT+pInyc0snaIumvSef1l5vbjdJTOfHMhIOfbHQt7A4FAIBBwBOCe7UqQ71YhyLwP3Go91xbU8h4dGvPflbTGc2jFHp1r+KiuOEcrKK20+TmQpIEkVayd0RbSp14clmVHd+ncxCZp62yT5cuX23cp165aI9cWl8jtu9qkeLwsg/qdq7MDgzrvsk0Gelvk5Br9/AhLp/fo+B26NSFLJVbRlVytPF08x1phWWWVbUBF5LJWGVmt8fbosfaClterbepnefD9rl6MDxcIBAKBQCAQCOQQeL/2HvIpj6XKF7u0d1BH4sgnNqYVwFlQ7iFdsO3I5PxR2l/Uhec0LTx1HmerpE4WZ7lm8p826xQT7RH974d1OO3+tJAPaZTWym9bmUYOndDG1WfP2OI5k2VXb2EhIOvtjoQ9gUAgEAhMEYGmjdrCeYcSpn73SujB+4kSJkNwtiphfXCtNL3cJ2UVf+M6nSpS1m9iNe1Rgad6bewUQ5uKSaOoaryCLgZwstBr2bDyKgvosCgaiwYMF9bLst0rpFkXuBlWgVka1G1EFyLQz5C0txZkxY+7pOfskHYm6oI7+pHnwruVNG/Voba6qAD2tuh8Ff0iiNqs8zgZfnuV2n+D9k7u1JXw9FMjZV9Rr0tJdne/jJye/AVg3GsNz0AgEAgEAoEFi0DTOuWK/3CJyLuUW2gU1cVrWr5xWIpw4SXaK6mfkSr/R93/l2P6HeJJevs0aXkmFrJRmmo6XbQVxm047N06r1IbUlu/dSSt3voh5e0P6MbIob3a2Pvzbiny2Y7xXNbjOV7QXPqFgJxL9KPsQCAQCAQuBAEIZaUS5vtVjH1IFwugl07nQTb9/X4p0wL7H7TV9bY1Ut4/KC1H9PtSp5SYxuMmHVFT1tVYx3Pj++pIG11+js91sPEdy+NH9TMfi5ZIS6uOH2rSz41oryWL/JR1wsfg0IAMnlHhWRjSEajN0nGFqtYNautb9NtYCN6Xe6W8UxUqvZ9bdaGBAxrOd7vuUfsZ3sMwIkh2laZ5ShcceF3zYqW9cIFAIBAIBAKBgCLQpBqx/B+VB1k4h7mJjHr5zlEp7lA1+D0VjFs0AmsEsNDcsz3SpENJ+VTGbLuyrtAq/3xIbVKpdZNynn62Y4TeSLjvLrV1kx6/qpz2fbXxDeVBNXdcp42zrBlgUznONz1l3AxmxzME5OzgGrkGAoFAIDB7CDC8ZpsKNuYwbtAJ+C8pCf2oS8qP9KkY06bPN2kv3i0qxG5eLsW3nBR5VJXiROR0EVbymZHjQ12aA9tEzptP+VakkiTDVM+oMZCofrLDhgopmbe80CdFiF97KY1sEZJcJx2OL6u4fOSkNO8qaO/mROWEfyAQCAQCgUCjIdC8uEmKTOOgbREO+Z8qyJ7LFKLO37fvHK9QvmRhnRv0c1PPDUqxBt9ZtC9yfVP5FwE5pHy3XaeZ0OCLw9bXlLf/xxFp+vpR7amcpGGU6R1MSTmgPK7TPiaJaVnX6icEZK2QjnICgUAgEJghBGx46VklJHrl9iqpvK7zO36mcyi0A0+e0Z+v7tOFa7TFlQVnCnNNN9XyS6yCp0N1pE9VIHa91i+lzL7iIVWK/+2ItsSqCNbhRjYMl6S7lDgf1VVmdVXZ0olqXjMEZWQTCAQCgUAgMI8RKOoUC/nL3an38ZRyyyH4Jbsg/VxG81cOSOl7ulgbxNk9Iha/FtcLXZ3Vn/9Hh9Ped0KKtygn0+tImypTMl5RAfn6kJTPw2tN/99hKWsDsQwqRx7Qxtc6ocEQkLV4iKKMQCAQmHMEGHrZ369ipG6dCj/mGbJS6XlcmV64nSog9yqpUIszFNXTINIeUBH2U93owXMi9fC53J9R2x7T63xSN3oWK0brMde0Xz33q8D8NxXG2qpsjpbZixy2WigU7N7zDIQLBAKBQCAQmBgBvj3MtwyZ6z7rrjwgTUM6V994DxK4AKf8J09NTHSlAwivGoxZHc90OE634qt6ba9q72iLbjil76m68i6NzDYNV4t3nRCQ07ghETUQCATmLwJbt26VP/7jP5ZPfvKT2hCZiZO6uhyd49CsPW9lJe2pTNBQUoKbcJWr8QPlS8LsMt3PYtbBD4blDc/bp/62mA/C1/3Vrwndh98FOF6GWlpaZNOmTbJ+/foLyCGSBAKBQCDQOAi0trbKtddeK1/72tekp6fH6s/ZuXqIQGVI81qt+LXh1IbQODlMs0SSsTlv+J5sPIxj/PNh+NXCqQ02pDVXVoWfz2eP2+/xfJ/LK39Ig+lVV12V95qV4xCQswJrZFprBM70D8veU0Ny5MyQfgKPv658TcIxbqx/8uXzO1etWySbVnXoqpEX+JaasorfOkUAEbFixQp5+9vfbludmhlm1QABnoX6bECowcVHEYFAIBAInAcB6sc1a9bIRz7ykfPEjOB6RmC2uS4EZD3f/bBtyggc7h6UH+44KQ/uOSsdfEh9GgJySFeA/N9vWSsfvm61rF6qC3iEW3AIhGBYcLf0gi8onoULhi4SBgKBQCAQCMwTBGab60JAzpMHIcwcH4HUwlKWgcKw9AwW5ISudLVIv0GXeht9vhPnjIDTzwvYkU6qtn36GdBVK/sGh9MKkfp1V4bQzfYfXq74OAwEAoFAIBAIBAKBQCAQCATmDQIhIOfNrQpDx0MAAVkujghjvosjRRu+yhBW/QqdikT2uDSEld8kJd1HzzPP4WEVkJqHlDrVMy8vLYP4CQQCgUAgEAgEAoFAIBAIBAIBRSDelOMxmPcIsLLiiH5frlgqZgIxiUdkIg84QpIN53s70R+mS7KNqAgdHhmxj6R7WOwDgUCgcRCgMSpcIBAIBAKBQCAQCJwfgRCQ58coYtQ5AiVe/FRElnU1HA79PZDeR3oi3fnDnkRlaj3xuJaWPNQjXiQdsdgHAo2FQP5vP4axN9a9j6sNBAKBQCAQmDoC/k499RQRMxCoNwRU9CWhWBWLnDM8lY3jFD5aUPplNFnvJGlVPLpn7AOBQCAQCAQCgUAgEAgEAoFA4BwEYg7kOZCEx7xDQMeg2lxGFYJNesCQVLohKz0I3s1o8VCVaTgr15kOXTZW/ecdBmFwIBAIzDkC9GAyn5qPcLe3t4/6fhphQ0P6mSH9JiXfWavUT3NudRgQCAQCgUAgEAhMD4EQkNPDK2LXIwKq/8pjtF/+5axy7KLSBWV2LU2cu4asx+sLmwKBQGBeINDb2ysvv/yyHD16VK677jrZvHmzdHR0mKDs6uqS5557TtavXy9XXnmlfZd0XlxUGBkIBAKBQCAQCIxBIIawjgEkTuchAioerQcSgWhiEDWIKMw2U4c+PDUpxSyGJmRY67kCdB6iECYHAoHAHCJw+vRpeeaZZ0wk0gP5xS9+Ue6//36z6MUXX5QvfelL0t/fL9/4xjfk3nvvlVOnTk1q7bFjx2T37t3yxhtvyOHDh6PHclK0IjAQCAQCgUCglgjUVEAytGfPnj3y9NNPCy21ecdnGF544QUjSsg3XCAwPQRSF6T1RHpvJHs/5tCPdc+xnZue9IDplRixA4FAIBBwBDo7O+VNb3qTfOQjH5EPfvCDxnGIRMQfApIeyVtuuUVuu+02C3viiSc86Tl7hrueOXNGTpw4IcePH7fjcyKFRyAQCAQCgUAgMEcI1GwIK3M/EI87duyw4TyPPvqofOADH5Brr71WaGm97777ZOnSpfLzn/9crrnmGrn11lulra1tSrAwpyRcoyOgSnBaOhDlmBI0aw/ltJI2OtRx/YFAIHAOAosWLbJewl27dlmv4dVXXy1XXHGFDWfdv3+/3HXXXXL55Zfb0FVEJUNdP/rRj56TDx4Mu1+xYoVxIGKS8/P1WI6bUXgGAoFAIBAIBAKzgEDNBGRfX594hbNDAABAAElEQVQwxAci3Lp1q/z5n/+5LFu2zFpsn332WfnRj34kv/d7v2dDgJg/smrVKmuxneiaGQpEjybk2tPTM1G08G8oBKw7MV1xVR/m5jfmPYmm5/o8hgsEAoFAYCYQQOQ98sgjAqfRI7lmzRo5e/as9TgiMPlmLYvr4AYGBiYtkrmS7pqbm204q5/HPhAIBAKBQCAQmEsEaiYgV69eLe985zvtWhF99DzS4/jSSy/J66+/Lr/zO79jQ3tosf3+979vJMyQn/Ec6V955RVr2WXoK72blYVSxksQfg2BAFKwGU2o+7xU5NgcYrESkAlHfZbSAjwEVGJmCWIXCAQCgcDUEdi4caP8+q//unz4wx+WL3/5y8JImw0bNpiYHBwctIxo+GTUDIJyqg7hGS4QCAQCgUAgEKgXBGomILngQ4cOyXe+8x15/vnn5bLLLpObb77Zeg+Z57Fu3TrDZOXKldarONlwHcTitm3bBLJGTB45ckR27txZL5iGHbVGQHVfWZ8JBqLaFx/1nGeEZwM5iW7EcZ787cz8OLeFd0x2ZhEtJH4CgUAgEJgeAvQUMrKG7fbbb7fpGsz3pzeRBXHgPHiQemfr1q3TyzxiBwKBQCAQCAQCdYJATQUk4pAFBFja/Fvf+pbNh1y8eLFBAfHiaJn1Y/OY4If5IWw4WnaTWJggcng3AAKoSARjctXnQXsYq94TPCe5CJ5B7AOBQCAQmAYCzGukx5HF4JYvX26LxX3yk5+UO++80xbR+cpXvmJzI5kPiZC84447ppF7RA0EAoFAIBAIBOoHgZoKyCVLlsiNN95o21NPPSUHDx4UhrYiBFlpjiGrzGdEQHqP5ERQ5RfO4aPM4RoYAe045DMe9B/6NlU0rFfSIiMiQ0hOFbeIFwgEAqMRYM7jpZdeap/qYJ4jQ1ff9ra32TzI66+/Xn75l3/ZvgnJ6JubbrrJ/EfnEGeBQCAQCAQCgcD8QKBmyotFdFhtFYHIh5UhWwTlpk2bbFGBf//3fzchycp0CEgIdqqu2ts01RQRb6EiQD/22G/TMHtorJ9fPx+MQXSGdnREYh8IBAIXggCNoe9+97ttG5uexXQ++9nPjvWO80AgEAgEAoFAYF4iUDMByTzHn/zkJ/LYY4+ZUOzq6rLhrO9973vltddek29/+9u2TDkrtd5zzz3WcjsvEQ2j5w4BVYKIxZLuGbaq04zMcYxQ5GMvLhhdUOI3bLHiJxAIBAKBQCAQCAQCgUAgEAgEzodAzQTkli1b5HOf+5ytUDcyMmK9kL6cOR9f/vrXv26rqfLtx6l+//F8FxfhDYaACkXrgfSRqL7PwRBfDM2BEYeBQCAQCAQCgUAgEAgEAoHANBGomYBk1TnmKo43X5GwEI7pzg0MjchzB3qlu39Yh/JmXWhTuKkM421vaZZNqzpl67pF0t7qfWxTSBxRAoFAIBAIBAKBQCAQCAQCgUAgEJgCAjUTkFOwpeGj8K2vs4PD8sNXTsmuk0PSoYIwObrSkph0SVn1STGK6rGkvUned8UyuXRle+MKSMar2qRHRWqyyY8Zsuz0Qx65szgMBAKBQCAQCAQCgUAgEAgEAoGJEAgBOREyNfcvS1EFZP9gQXaeHJQdXYOyWHsRXSiyx7nUcf/kq3P7dHLf8s5muXp1mwwOj8jSRS06D5BvIHoKj7mQ946Sa0c9VxFZmrQnV78cWU22kMGJawsEAoFAIBAIBAKBQCAQCAQuGoEQkBcN4cxkwBDUoqrAQqEg7Sr6Fre2yKLWJtU/ZZ3Xl/YTlcQnLEZUJ7brBL/iyLAMDw9LudSmarORZvwlFahffdSJkE16/ex1MR0VkHY8EXgan07LWIZ1QoAiIBAIBAKBQCAQCAQCgUAgEKggEAKyAsUcH6iIKWsP5AjiD9VjfY/++QlEZNX5yEz2ODoZm4iACNUFitgQpNOZQ2kZLYAfxDRdii301eqe/lc71n1RMeUYkenYJYltCnIBXH1cQiAQCAQC9Y/AxbTZNdSgmvq/lWFhIBAINCgCISDr5MZDqIg+5kGyT5v6KVs2cZ6xJjqRzUUk5hM3KSLtqeQ42/BvlCGsOlrXcCjqNVeHpIIqEpK9OtOUYJlO+TW8qqdxFAgEAoFAIDCLCBRGSrK3q1/6CiVpydXF5ysSzmvV0SWrlrbJhmXt0lpZI+B8KSM8EAgEAoFAYKYRCAE504heRH7IHBODuk/CL4lHO0YUqiupkERAIiqt65FjxCWe6vC2fPR4Gtxsaefzj/U85i4giWc8EOMpADwMtnRa/c3Cqx5xFAgEAoFAIDDTCNBA2tNfkG880yW7Tw1J5wSrhVtdrYXnOayoCnJJe7O8d9tS+fD1a2VpCMiZvj2RXyAQCAQCU0YgBOSUoapVxGoPWVNOFaERET/N4yogtc3GZJK2cdVQ6p1l2Kq/figuCocjgr8ilHDMenSBM/l5rFrd5ygnEAgEAoFGQiBbKG6gIK8eH5TnuwZkyQQCcjxURnRoycqOZrliRYsMsVBcJwvFZQ2o4yUIv0AgEAgEAoFZQyAE5KxBe+EZW++iyR5vf03ixjQPh0aa7JN/paQxpxX/BjlIQ1cVBHDIaUhfhJXhqhwTb9TQVTxIwxYuEAgEAoFAYMYRYFQIPZDDw/qJKm3tW9bGQnGp7fN8C8VhTLHYJB26sFy5OKJrBRQ0IQvFZUNvZtzayDAQCAQCgUBgMgRCQE6GzlyEZZrROhInFDQInpxCMrWEsRMmmIsrqXmZ9CSyUI5+0WQ0FBksYEpHbb6TNs2HbGzcan6josBAIBBoSAQQkIVRC8WlKRlU2HkpSD3NuQ2s8WM8VISmheK0ptfF5pqaG2mlcb3+cIFAIBAI1AkCISDr5EZUzFAtYwvCoA91c5nIHpekjg/SzHy9JzIblpliNvYvrxUISd45HDttwNZVWBO+/mJiYjIfScPDBQKBQCAQCMwsAtAUK42X9HNV9Eamjfo4zfU/30Jxtji51ubD5MGmeTClo1EWipvZuxG5BQKBQCBwcQiEgLw4/GY+tYocRI0NY4VwsxJ87wWOOlcCRgPROmt7j9SIe1WG6GgEIsLRcWLPeUU46nHF4ekRK55xEAgEAoFAIDCTCFDNIhxxCD8Xj1NZKM5Hq8KPUV0bhPETCAQCgcCcIRACcs6gn6BgZUaEDiLIHccZ51oY/hBoJUoW2NjiEUTyqFSPGdp6rgO98fzPjRk+gUAgEAgEAjOHgH9KqZk62IiMnsSUP0udmbdV0dTRWV2dtf418kJxM3cHIqdAIBAIBC4OgcbWHBeH3ayndkKd9YIWVAG8cLDh8sfJp/rrcTSWCfDqeTVOHAUCgUAgEAjMNALVheLGyfmcqtjHjWhcb0kdJ1l4BQKBQCAQCNQOgYbsgWTuxInegq3GmTV6ThnxZlV1i3T1OJYQnxWnBpnssZ/UEwlnVsSkHhPEuXMp18CWo9lZMa3uM1VAWFihrM3bNjwqA8jnyCAU8/5cTwoD0XCBQCAQCAQCs44AZKXO5p9PWPVCdEScMILlET+BQCAQCAQCc4NAQwrIMwMj8n/8z31ydmhEqp+hypNVxnBJqlXuTFEFyMqOFrl1yzL5zXdulNZZ+pAx3cK+VfhzLI/mzkuZubMkaSvXX9cHOTyw0+fZTHZs1zMmnfnFTyAQCAQCgcDsIKB1biwUNzvQRq6BQCAQCNQKgcYTkNpDVdBvSB3qHZLugaK0VxQkSsKFYxX+vM+wdvH1F0pyqm9Iv0ml4rNFv0M1Tppq6tocNavp1d7HxlREXDVzHa1Vu6K6z49/SsOv/svf7PMnjRiBQCAQCAQC00VA69npLhTHSBGbb6ONuDHvZrqAR/xAIBAIBGYegYYSkPRKlXUJ8cJQGr5aVPE3kr4+r8iiHiYXX5rUlhDnO1bDwyMqPltm/TtUZbWvqVmXFdA9IhHydEsregfPyU3XCA3gchg4blw1x7bPXlzyQhHodNCrhcdPIBAIBAKBwCwjoNUt3MU0jAqHcTBBNWx1tHI3rqFH2RgC8RMIBAKBQH0gQN3cMA4BWVQVOKwCEL6CjFr0wDcjND3P7wHIN/xVjeiHjIsyMjI8apikhsyQy3rDELuZ8HGB4zfLeZY9sd1lHOunDbpXme1AsM8wTGAkrMb2UnJbcb5PZ/EbCAQCgcD0EIBfTpw4Ibt375ZCoTAqMWH79u2T06dPzxJ3jCqu7k981IzzmvNs/pyL8HgcV9mOs3CBQCAQCAQCc4VATXsgEXADAwMyNDQky5cvl5aWansiHwY+c+aMdHR0SGdnZ7a4yczDUkIAalkmvUxouGyYYGiMB2sK/w4V6YtFFmtR+aabL9IyU9Z6vuRH8ZjLXpeGMcGI9dr3Kfo5Zj1SgasiiSO7nAZmWFb2Y4EkcMgvllO5P+YPTrmhrtoqwHkC0OCMn0AgEAgEpo0AHHb48GHZsWOH9PT02P5tb3ubXHbZZcZtzzzzjHR3d1u+11xzjbzpTW+S5maXS9Mubv4myDg1NeSly6AKdjeeYEz8N1pMevzYBwKBQCAQCNQegZqyF+QJiT744IPy0ksvSV9fn10xLbV79uyRRx991MK7urpmFQmI3kRDRTUkIXiuiqiymsXPerOqaZNQmVFjq0VayyuCCEeZiN9UNh1rVZrlyM+SlLQkDfWTvZNUrtlxwoNjP/e9Rxx77v6xDwQCgUBgOgicOnVKXnnlFXnjjTesIfTLX/6y3HfffZbFs88+K3/5l38p7e3t8tOf/lS+853vyP79+yfNfnBwUHp7e+Xs2bPGlQuprqK+5uXDX0CqTcnn+ltYVsHn400KXgQGAoFAIBAIzCoCXn/PaiFkfvLkSfn5z38uTz75pPVAfv7zn5fvfe97Vu7zzz8vX/rSl2xo6Te/+U35xje+IcePH5+yTZUepimnUJLSniedWljRjPnjiqdpN37oZdSdoZUdT6OsaUc10UgvWRKPviefshkyOkfC83FGhzbGWRLOGQ4ZfqoaFbB0/9I9BYuEaWOgElcZCAQCtUJg9erVcscdd8gXvvAF+fjHPy7vfOc7pa2tzTjv1Vdfld/6rd+y8N/93d+1ETgPPfTQhKYhFp9++mn5wQ9+IP/6r/8qTz31VGP2VmYIsVBc1Y06qXrHUSAQCAQCgUDNEKjZENZly5YJw3muu+46Wbx4sbXA0sL64osvyssvvywf/OAH5V3vepesWLFCnnvuOXnkkUfkV37lVyYEgqFCtM4yr+RCeixdVrge47xybKKDbwmiP3JkRTcf53m/CS2cqQCzjIItQ4RiyQezqhfDgJgjWTGpcjBT5c/TfBJcyfjKsR/4PqFq4ttwq/rP06sOswOBQOAiELiQxkgvjuGo+SGpR44ckUWLFtlUjaNHj5qg5HzTpk02xJ7wiRx2wJVXXnmlcRB8hwhdME7r2yblU5/n7225cJmyWnaZ1eOs7TaxYFTTC+YxiAsJBAKB+YtAzQQkQ3fWrFlTQYphpAhJRODBgwflox/9qKxfv96G/jAMaNeuXZW4Yw8Qdf39/TavZGRkpDIUdmy8Sc81DzRDXiBW6YqUOVE2aUazEUh/WmJJZj3q4q82+xESxWZsM7HLNXCaOQtLEdwr9lNBIMN0KlEjTiAQCCxcBPJ8cKFiEk76x3/8R7n55pvlhhtuMJ5imkZra6vOZy/ZHqFJ4+dkbuXKlZVgFn5bOAIy8ZuzF+2y9ikq143GalTK2ojrUzWy1t0S00iC4yrPRRwEAoFAIDBXCNRsCKtfIAT6k5/8RBjuQ+sqi+YgBlk4hzCG/OBYaGciB7FDrhs2bJBLLrnE8poo7sT+KsJy8su5K9FWtQ0Ufw8jr/zxxHlfTEi2qEtGlC4KbcEBEzop73M41MNm38CLubjZTwsODo7vJymVZ8ChmyRaBAUCgcACRuBCxeJYSBhV88QTT9ioms985jOVhXLoeWT1VYQjc//Zr1q1amzyCc/PJzYnTFinAQj1/MiZfB2cpmSMMVzjk8bordE5bgw0cRoIBAKBwFwgUHMByQIDX/va1+Szn/2svP3tb9fPYej3FLV3kp5ISBzhyOqsS5cunRSPtWvXyubNm+Xyyy83ITlp5HECmYzPRsMmG0Dkj0ni/ham5zUDK2NT9A8b4pG9N776OXvfsNci2EED/oCRAmSvJQoWLxu2aaNE5dj9xuw1ZQMCFpccCAQCjgB1xEw45vj/zd/8jbzvfe+zRk74jekbDFtlrv+xY8dsj4C89tprZ6LI+ZdHBWoOVBQa9mmfjtMl5Y9TzORfST7/rjwsDgQCgUBgwSBQsyGsIAaB/sM//IN8+tOftvmQ+PE5j3Xr1snOnTttyA/fyYJc6Z2cqrso8s+xkb9DVAYWaZgfIzY5zkWfqnnTj5drYeXaENZ+jYhaHLb6sR4lz9pYl5VVhzuDQX8AJg+Q31jCuYEWz+3nxNewdb/YBwKBQCAwPQTgrgceeEAefvhh47B7771X3vOe98jdd99tjaUsFMfq43wn8mMf+5gtqDO9EhZObJr6aPAraassC9jROGoL2dklegV9TmW9cACIKwkEAoFAYJ4jUDMBeejQIfnud78rrLLKsuSsLMcqdbfddpuJyT/7sz+z72YR7/bbb5c777xzVqEtqcgoKU8hDHF2DF9N4qA1p7ZJol10kPUqGqMmLYQYqopFtNFoUYkqwnT1Nd100QbM6wwUCb+PLhyz6zHcuIMWJUWyIawaLyE4ry88jA8EAoE5RIDpFH/wB39go2uY3+/z/FkYDve3f/u3Nu9xusNX5/CSZq1oRvPAvez5VJV917hSX3sFTvHVY4JZQE6VJwHhAoFAIBCoOwSsGst9cm+6BjbpB+ebqq1p001e0/g1E5AM46Ellg8oM3eRRQE2btxY2X7/93/fhq4y12P79u2yZMmSWQXChscoIUFg5qo85T5pj5DDcUOJ7+fJd1Z+ETM2P4Tc3S7d8wkPH9aTeiQ9UG2DUysEPCtm1X+mYGSQOS5jTFZ8ygqVOcfKeyvHRI3TQCAQCASmgwBTMeA2tvHcli1bxvNuQD+a7airq/W08ZnXzRpm0zT0nH3F6fGo80pAHAQCgUAgUB8InO4vyKO7euSxvb3S0Zqr1HLm4UvVlg/lvKj64tYtS+UdW5fJ+uWduRT1eVgzAclQVeY8so3n7rrrrvG8Z80PInLxiPayltCsNBeJJRWNHicFIexq4XiUxj5ekKf7Y8PYRy+zy4VRdtooO9PPYGbXn8emURCI6wwE6h+BoZGSHOkekiM9BR3mOT17+dPesKxdNq5ol0XtPnZkenlE7PpAgOGrvCyVadHT/7TLUmvbKTQ3kSPiZOETpQv/QCAQCARmHYGy9A0MyXMHz8o3X+yWZe2psWxssVbXZZ4c4+C3Ia3f2vT7Rlev61AB2aG+HmpR6u6nZgKy7q48ExuMhvFb5LxU2UNWFZeOp/nOU0k93YNkU778sTmMDeNcU431HptsAZ+DGX+uCTt+HYzkM2occDYmOPXkVp+BBQxPXFogMOcI9PQV5MHXTsmPXu2WRS3Uplm9ZZbl/17z/snsgjLsXVetkA9eu0o2r1k859cSBlw8AmlaQa6m9kdAa/HUYJrqbq+nUw1/8eVGDoFAIBAIzCQCVkepoGDhNETgqs4WWdKmU+WU43RQqhY1eQcUnVoISP3YU/rEE8NgSZmfvzaTBs9AXo0tIO3lZWoolnQFHevcIk2F5KaWdrqxmhCutlEUgshm6k2YjT1gapx9lDnTShNGXqAB9irKrcm20TfpPDcM7NLNXaDoxGUFAnOLQBIKZRkqDEvX2SHZebogS9rHb46DbKFOXCLeZDvk2tU7KINDw+qhMbS7qp7JNVkdvxMhwHtRmkEwdk5/atBL3JdS2zsUj4TW03X8PjXRpYZ/IBAILHAE4Liyrvo/rByHiGzKxtsnlnM+mwQE6kP9NzIyrHkUpKTT+ZqskXWSNHMc1LAC0jVaEeGgW7OykosIX3mVezOS3SCbAqlR03l6GGb33uXLyB+fW6o9uHhzDfYvEfC5McPnHAS4/7gGFd7p4uM3EJh9BMraojqkxDhSHJEWrVBb9G9u3CkB+jeJSEj863Wf+unhsBJzYbggZSVXaW5Y+pr9mzXrJaRGO6pfqt7UwFAt1MUje3O6Y7RQWR8ar7KrseMoEAgEAoG5R4B6jHVcSkUaOKnjtH7TCo49w/OrTaZW66nB7JMjPuekJY+xdaLHq6d9wzIwLQW80DRzz9SlleDSMQLSb7S/4DRx9yEzWr3tRqe4s/FL67s9fFnm2XNVKYpzfyjx5Nh41q6FHzsgqOFc9Y8WMZ3/81RUFDjvseAYl6DT48aFzHCIn2kioM/PIzu75XTfsImhqabmMUM8rV3aLm/dvFQn2XtNM9Uc5mc8/t4gRlpm2VudpZcy7tWrUqQl1v44K3+Y+Cg5jxRlZFjz0Pp7unMo5ydyC9NqhGEa0NVsd3hs9YtY5OGwvR7qwoTqqKf1ucjqbnzCBQKBQCBQLwgYz5m20DpLSc5HB9pxvt6iNZRKL7WSmvlwIrUhefhWL9c1kR0NKyC5e/zL3lIMH7+/SVSkEI5xFpfo47eZEzAzTo2oCFQrL8tWj/nUiD9v8KvbZpdBNC4gnyZL2jg7rj9h4DCw9/tJmPtbPH+LrXg2DlJxpReBgD5HCJgHXjstr58YlM5xhpn436Y/f14aIx86VTRet6FD3rS+UwVkuwfN2H5EBdr+U4Nyqk97+tyQSu6Vv4aKz6gD/ZtYvbhFNqzolM62ceXdqOjTOUkiUltnFTv7Q+RvtVqLpay8gqv+pSZ/jcqfa0nnEpCevMLNZwSy+5fdx7FPglXNfs+5TAjP77nu4+7P53sftgcCjYMAdZnVWLp33ZjqMgtQIKjoMjyyim2+1G8NLCBz7zB677h/Y29a/pxjNp0by+wbPaqNszK1KNvrjz9neFSONdzeu0ZdRD60NrbObSkg5Jteu79sZL4V23L+FsffVGp4Tyu2xMG8QyC1DKaetN2nh2SHCsglKrR48vJ/fumvLw0pt940u1KEp8hiXdp7eXtZBvVTRit0yvwE/XAXjM3QcFEefr1bfnGgVxZN0MPptiY7U1FcA0P5b75skdzx5tXSqSJypp0JP+tWGotYVtJY74qBBPhfeDrOUsRuviGgt4/bml8k59ymiuqDQBhnftdtMJCeVR6N+Xb9YW8gEAgscASsxevca9RKi3qL11ATlv4+6pXbuSnq2qdhBaTdwKLetQwBv6Hpbtktzm5cbWnqnOcIj5w5durnmWnWsm/WqkelBT8zv6F2DtbUL9p7Mrj/tWwYmLqFEbPeEGCxqqHCkK60JraSaKd280EXvOiyx/mfYfp2a/Ljl+HxdOyVtSeNoZgoSotjDRnVeBd6RO9cQSfxv9LVLw+qgFw+bi9i+jwRz3t+kRr+Bpjrjbi9ddsykRUdGdFlFc2FGjVOOi7XCXTssHIbpmhyQSPxJ63O4NHjZMnM25NKid9aIcDfEM+b17+jyiUAx22uHPMskIZEFho/gUAgEAjULwJZXZWvrkYde91Wv1dwXssaVkDCTYmf0i21GTaVGzrqNldAxDcRWD68EjxzB5a9/lT2ZmmWvy8iAJniX3UV86teDXZUxYOhgrbwkSLguPDlOF7gieUt3kDoL/vqHS4QOC8CzJcusEoaD1b2yPnz5Ht/qKz3sfKA6ZlGYHU2hmKy2hqCr6llZr5pSN3EvO7C0JA0a76LVHV1jCNuuUBMGituealv1vgscjOstulKNRpzZmzLg2p/f2or889LamMzOKrjGMd1ICqtrjWf6t9wdhq7BYFAuu/eVnCOMMyeC7vU/PE5ERcEGHERgUAgsEAQgMlgTjamnhnXcW2J4jLuS/54u/MGaD+v931jCkglI6grr7+Sj98uQrM7XSErPUeVzPz7lBda2ZfoC6NnAgWkbqxtbp3785Jq76h2UZVsGu6AF06wK5Wb9TZpr5C9dOhqjxkSuq5V8tdzjkEX7DgOFwhMDQH9q1ORlubx6XPDMzbe41OpPjQwf6yFQBKkL+pW0jplhvSjmU9+Qyr+iir+vP5wUet7i6gVBs++C13z0/qGQbclFqrRxW6og1RRWtBM/gAX0tTkqf3NptxdSOLPMTg5tBUIU9T4necI2J8NNze7wZz7PebS7Fw92ONoW4DjbDG7zC+FxG8gEAgEAvWLgC/Uafyb1V2J+9K7AF7UfYTjP59cYwpIvUMm0OwFKdGW38TqzRvNUibW8FJGc+FWjTt7R+e0xuda6dO4LjVJ/yGWkEcVxp09k+o2Z+6kb9Z7YbdWcfFbqef4e8OBeRtudXtJYVidIVB5lrJnqirKxvSa+UOmf5WVB06fP8jEG3vsGSV8Bh152hLiKiTte7LnZK9GmAFaKBdjfyNuQDp3YUteo4I92gzswS2fdxXHqj9+3iLLcTXNORc1AxZFFrVCAKbSh0/vf65uNh/94aHIbm/lb0293M+eB39+Rz1BRAoXCAQCgUD9I+Dcl+c4KrnqlJL5wXENKyCTqtCbBCM5adld5cb57eUwO6b3sUYOfjShM16R9tKXsw+bMqal1b7y3lojW+ulGOZzGVwZZja/q3KcrKR/BX/ubx7BdJxFrpcLCjvqGwF9aHgB5tlhSOqol12zfJznyb0scpqHOOMXSR1Aw5ju8894tRw1Qv9TrWGOvYtnESvnFpLiVdPN3JFhptlBnjjfp7PR5/kw8E5We8zYz1sE7G/AnrjRl4DXOM68LY0GThBnnGThFQgEAoFAzRGgimKz91Bl4rTHjES21rjLCEMaet3XyM5TErf+XcMKSBsSo/fHZIfes9FzbriJyXljp+lII7D0AHj4bOyTyMGGVFbqqaiW5MO7fMiXaWAn5Krp1QQL/MjvCPcyCe8EgrVW566dv08fHucLd/AEpBfTXMQ4DASmgIA9d0oAzdpqk+Y5aO9f9WE0AjGh5i++mqctQMqo0JzfFIqaepSsfHZpviVF6TOuhng9Qhg9oT5cxudn0KyS6pQaViI0zGVEOulFZsP5Zw23SQuPwFlDwP8OeCjzj52fs8dpmPEiLaT5eCk0fgOBQCAQqB8EtF5jmko5eyGwd1O1zqs142JeUDUe1Rlbs8a1kZFeJ6pfvbsGFZB602ilZzNlyH2sspIfcbOTmNNo6kn0osXjzs+mw4LMCspLb6GVAr1031ts/bFrsJNK1IY44JLPdfjm3j70LN077qm67H7z8mwv9Y43YeECgakikHWR8RzhfO9/m5y7UONp9B4131uiWflRYWviVp93LdganfzPQctjAWqvVtw+FtEpqmH2p5Fdz8ybphlTAMJRQbJ5ybqHRh0z9LU19OjeTS5q/OxPduZNihxrjoA1zHLDed70H3Mb2VecH2Z7fw7SqQdWYsdBIBAIBAJ1hQA1mukG9moZtVa+jstrDgx3Hva6Dr96dw0qIPVG6tsI4rC6kim312+d05Tfdm58OuaBMPKbtTubHrr0tCU7zvfmhGUMozP7Z9e4WbvqmcjY7p79ZLgZiJPnzMu1x548ZoQGAlNHIC8Q88vQmDCyZ3TqeV1sTBe1vnehNlbcejkQ3+y5lDt1L/O2gaLSSJcVin34Y4VbYud+ksWL3fxFgMXOWIyqrAuepYYMXY3YG3O5LJ4NPa+8ZOmxfTeSVoQa//3MX5TD8kAgEJgTBIyrEmHxSu5Ty/z1nPO8PzbSgDvfXMMKSG5UupkuDeGldAP9NnLux7zK+M33F7BZu9lKkvYPsqy8SiXWzDgWfjXirdjghvq+EtA4B7xs2BAAxu6N43iR93vnMVJPh/o2MG7jQBVeU0DAXmhtJEOzPT4mciZJ5++99pxW+iInSXBRQV7auZn4s0+I/03gV9If6peJU56b14X4UHfxd1ryYannyQTbsMlwm6W/UxcrFcFyHpsieCYQ4GZqnZ3d07HY58/TvU/x7Y9tlp6DmbiqyCMQCAQaHQHVCyoIeRW1d3WFw2qvrN7iPZQGXH8fdbSMG+dR3dbAAtLvElINZ68ofh9tj7/Hqr5UzdLiF16yFpjK9F8XuJWAZJueVlps9amz2PPt6fNrnoG9vcwrCtynyhzRLF//IyUUpHghdVTT8QwYEFk0HAL0nPF3x68Rgh6zT8+i+me9JuyJxVxJi+9/r+o7ey5ZRv552yYrr0m745uU1exvo1rhTZbkwsIYM66bfZLhnBzOLdjqNkNOwyztOYnCY14hoE8YfwPZZqanP5F0mLWSEu4cV1GZGkO9wwUCgUAgUNcIWP1m75xqZlZnVaouKFDrPOuFVE+qPM6tFxKOmyeuYQWk9z76jeV+pddB9snBaVWXreCpgbNPYBSSKwdDkpftUUl6akPAKvYZGY+2uBLWEAdpZVX6guylA7wUDu81BgIfKpf+PEFQe2A0TvoESjo3z/gJBKaAAE8MdQb1AX95DIfOP29NRgQMvas+cRaXlkk9mFWaoD7gnxaS7Ez7iS7L4mCcG4VHrR1lGoCg6QakY0b3VIf4eFitDYzyZgoBr4vJz+6w3lL25ngO1dk5x5mg5A/NG2csQvwEAoFAIFCHCKTGMeWsrC4bxVh+ovvKoR6kN9jMxwPq8NryJjWsgLRbpzdXKcnwyA9XdSLjHlaONW51DtEs313NngevMjBOj9PDlUSQnurQs6bKojBcAAvEeI+IXVAD/VTvETgkLLhzqQUIICoxsuPq/SN+ehbSjrNwgcBUELDh0sWiDsVs1oVE9XlDGGYvuzRK4NICo9VjnjXXaPR+z5ajRIZnU5aJVbVrspdv4mMyfymJ9JLNejrDLsuXwtL/VKaXYsFZHPOrHifRUT33JLGffwjw7POs+fPG342/bJ1zNdnfEuH2d5WdnxNvBjwKwyU52V+Q/kLJ/nanm+WKRW2yvLNVWmmZDBcIBAINiEDiKGvAnYSuxgZVeZeQsaH1CWPDCkh7f9Gfqii0BeHsLuVf8PyYAG4wi+7M5otftZxMCNEiz7OU79rQ86L7kyB7abUXxIblLe4NWwaXggZsOBoHcPxy7P74pRcYjtQ1LHbp8uP3AhDQ+sD+FPXBS89Serr8UeKZ9GOeQKtvLAo/+SfxAsqeIInZoXMMyyMqIbVusJ5RrcjYuzOb9dyrFRs+o+ZYWk5m1fE3WP07HIsCIsEEudqQSBUhnglg/sDDzXsEuI3W5Jk9a36fJ70wfQZmk3ux4cCpfvmHJ4/Lz/adlSVtLnP9mUt/F/Y3oobm/0r8mf3NG1fLh65dLeuWd056KREYCAQCCxkB6gwlXeo33s+p8NhPxc0jjmtYAQl9GYU5N+hZXixyn0edT/Xmk3AGXM6s8+eWf+Dyx+dPuWBiOF5QPhvn/rKRD+OYP2P/U+Yee++uHoYLBKaOgA1NhSQqYwXsuSMDf778ecOPP0160RCR2r8xun4hwow6SmZTlx7yxGHqZZyW8+cwfUZBD2pRf6gN9iFloKNsDFDnkpJzFxSVMLUL01zwWoI6/tm7d6+sXr1ali5dKs3aQ43r7++XF154QdavXy+bN2+Wtra2Or6CWTSNZ1BvJsOSvac+Xxr1MYjZJ16yp4NH2J4JfXj9Ocmnudhjs6dUlKHCsPQNjsjZwaKMpOEpZgvl4yZ6/ujtZ0Gqfk07TMPNqCdbT8MFAoFAwyAAr7ljpFKaVgWHpRDj4CzSqGOvaDxxne/nREB2dXUZsXZ0dFTIdWRkRHbu3Gmku27duor/VPDzYWNTiVt5XeHm2c30V5TzpLa4moj/2UNwnhQXFZyeLe29SEVaubrgub2AkjH+EDDXwyc80kuXnqaEBDSk4+8v/Q3ymuH3NgNFA/RdPzk9btLxhem0wUFryCfl4i+av8H0OpsGV9rfqj9yWfb+ZHmYzUm0P9b0lF68FRPnYLLWDNACtc5KptGTl0vDsRmXVR1mWxaeEuQiz8whPU8Ih1SNjvk7tTLdiLwBWR2ntuZ9Z8aimcvlwIED8sMf/lCee+45+dznPic33XSTcRmC8t5775Xt27fLI488Im9+85vlzjvvlEWLFk2p8JaW/MdgppSkriNxD/nbsTvNs6kNtM6rvFDZX4f6p15zi2UPKKJyNtpyTUAWSzI8XFCbStLe0iwdEw5DtT+ahC8PsZqH1iR6caQgIyPDaqs1FaUXx7q+E2FcIBAIzCQC1A62+IC+oJe02rbaK5FdpRhrQM0qMjsmRCNadaI/pJkPrqYCsvv/Z+9OADU5qnqB90wm+wIhOyQhYSeEsO8gEVBANhdUcEHcHg/R58JzwQVxee4+8OFDQVHc8KECKigii0EhsmOAhBCWJCzZ95lJZp9Xv1Nd39f3zv3u3Dvz9b3fUjXTt7urq7tO/eucU+ecqu7vlltiYP3Yxz7WPO1pT2vuc5/7xOB6ww03NBdccEEcb926tbnf/e4XA++mTaPJ27lzZ7PHJ/RT2r59+wFgzXDJHeZmfVn6mFNRzLuBv9G5nkq6pccUlKXnZ2PPgJqX2g4ZC4MNnJ9BQ1LuAuuwRxIn9tHAgE5JC/tqQ+nYdDneWUtOJONh7mErcNX9ihFgIvqB+xzIyVqh6BBcRz4LN4a2SYLsPPNbcQi6vLriqvdfMBGCv7PGwPTq4bqlGdDO3UFf+lNKpoMo1yky1sN4fHpiRqI8uuSm885hPsl0Kwnb2BYWKg+ZiP0dd9zRfPWrX41xTqBUX992223NRRdd1Fx++eXNU57ylNg7NxP5yEc+ciTdN910U4xtxrnrrrtuZLmpvZD602sXC7pcY+S1YpGv4uIhHwQTjL3RSTYSzrvYFem95sx7Kin1tgQtoFYDMiFlDYJA+O60mXVIBk2+WP9WBCoCc4PAQFOkINIGv4010CEgKFfTYSi+jvZrz43WS2hFN09cGu2h9UDql770peaNb3xjc+GFF4bzKBq7Y8eO5hOf+EREZ1/84hc3H/zgB5srr7yyOe6445r73ve+I6ngdFoStDspewP1alNEHFOH2ZfUNQTludLpboUjsLBgDCk3j22f6QkDa0AbKkr+sKJCW/aJDLKpTC42LDRnRwZym5keP0kgErAcJOXLjuX9sILpnMFWm3sACMTXVQ0Sfgw9pA/vZQ4yAOA75/mILlmOEw+AgGVuUVPWbRzcRFciqywXpC8Ma2U/oIqDG5GUuHuZpx/4pcXyRcV1g3f5yQOK0mk5bmW6P9IOvFHtncaie93rXs1LXvKScPyOPvroxsyh2Uera17wghc0j3jEI5ozzzyzefOb39x86EMfGulA6juzmcY2Y6SxbnUrbQ66Ob0+QPv0PR6UuuNw9Hl0e+IWhVxvpchpmxX5Y/sTz00BoTQLmTzJqHHAenGAjpZ77RAxYOZ0nG7ZkGJCnE+O6ILLYyOyPqgisLYI3HZHWpKdDOPM8mRgBSkpdOPNpjSLf/Rhh8TxCu6aqSKho5JOGOq1hdjlMS/nOZaibMoaDME5e6L/rpkDaXA1s/iyl72sefWrXz14N+Syyy5rPvOZzzQvetGLmvPPPz8GYEuA3ve+9410IAHNgRSVNRN5IIOrpTBpIVXqnBwl5DyWOQG5knOxSOkQxlUqE11ewqP50tj/7rX8xSCUSNuNu9Jx7NUUnJn2mQPbuhNtqQHuy9eDyvbafO2ijyISkDHJrc8SmmdfhsuAXdP7jJO2Z2XVVBFYBQKZd0L5JzYbDBjdQWFwnB6reGI65fqa9Q7pT8+P+cakEsKJTbqhOLca52rXuZUnBf1Fx+Sssf8tMlqqKfv9VjThI2tZZurVDDNRsPT+o1U1t956a3P88ceHc8GxdM3M5KjEWbz73e/enHrqqXHPVVdd1Vx66aWjik9hfsIGT6axVPAuYn0DOQkOboe4cpwuJsz6kpniDOoXw7vhNQ+xqc5cdcI4G9JojXceM2mB/e4k035/WFbO7lycwt6pJFcEyMLrP3BVc8WN25rDN2VLKUnFgMPzMZzwuvycmF+HbdrQ3OekI5vnPuzk5pj0VeK5SvRU8i0gEmglPVeCf2Gjp/zQJKkc5WInZQSdZD0TmRP+Z8161kBqgL3zne8cgypc5G3evLm58cYbm9NOOy2un3DCCTG4chBHJZ1h+ator2TG0rLY1aXca4QkUuq99p350pP5PPdqOmaOScoXtzIy+v1T6Cv7UtuC87YN5doc7rNtmQbwBf1UgBjiw4ncJ8Fyiex9ytWMikAHgWAZSqHlHfEbxmakLj91j11M52b6eo1DeX4QlmXC6DTQdWgo54VeZCU52MC5jWOF+kgaL3SXJDEBoHr1dchozxbmRlsCMPnTlYrxUF65cG4sLB/XGdUaq3BKEiSdncRAWtyPw/6GD2eMaA2CunR0SplPFt8bl8b2h4NYtnjoEtUVukqlC/m35NZ9RWA6EeDomPT5zPXbmk9ec3tz1OE5/EgU9sfrJvCPSg7koUnGt+3YlRxI0rK/u6YTp1FUU1dljNP27DgOMXAeo18qmMfp7rhbckY9fXLy18yBLIOoQbMYMvIwqWitAVV+GVhLmVFQifKWtNIPEZTysU8dvOBnL7qDxMCyS5mD4wV393YyIKPlNTt5hfXK9ZI/JKSUKiWGV+bnqNv2gofW53wqMDuQ5eM5hNxMTE0VgdUjUNR8V1e1du7Ih7keZTibXXYdeccBXEh6FU1ezg/aSj3tfqBL0nk5VkumK+XEwQHUu5JbBjTkKKuqErmRFlbbFnQlXQ+HIxXgVCwg2vmEJTOMvrBqXDLGcQRtZhEf+MAHNt5tlC9oulzqOpjd4+XumZZrZiniTcNkSOlpfV/kCTuU88IThuFSLguOs34SOhJZsaonaEnV2Jek5pKPH40fzvdYtxdUpl2v6UDa3m1Br8TVh085AsaMPUlAfVtkW/qi8LY0s3LIruTwJGbH4xsJ74hEHsrlrdt3xkepmr3pa9PWd89LakUNjkV/7asXirYDyhBP5Yf3TD5ga+ZAFig4fj6OY28Q9ZlzA653Pc4555yYkTRYnnjiieWW/e5LZHe/BRcUyAZMN6t0aVG1ulUkVBoOYPvGTqPAQf4pdcZjUpWMywV5LiTpNEOwOKHQRnjt5zOVltsX5JbKg063zHyiVVt98AiEgxZSN0IoVYEVCxs6T/rEzFuvo0SMQqpIFZfRvENDHLbnod5aGuO2NPUSgbUFRCN8TKkVTXWUnw6h6+iuYSrEpn1LYJAoahtEDktO2tG1117bvPvd7473+o84Iv8WoK+Ke5//ne98ZwRLvbZhzHv0ox89aeSvIT3koA0IlP5vu90uDks+qtprQWD3ODLG+ye+EJw40hwBycabXQkv54Vn7fFwfp/eQT8EXr95e3PJ1VubK2/e3hwSzmoruNH8LFj75sTFJtn/zYPvdlRz75OOao48fI6M+dz8+neVCHiFamd699r3JDYldokPEichCLbrCsOi57qE13DYnvSzODvTF4nLB6XY+/OQiH8e3+iBrkQu1gud83RTnLm5J/3RB/Zr6kD6Qt2nP/3p5uqrr24Moqeffno4kGeddVZ8UOCUU05pLr744ljac+655/bR3sEzs9Gis9oszN0eF0fSlTJIuJaNl7ZcuW/wxPEdqD/4qDX+/NxE+dgLgrwbKRo0+EmKtuqgb3xkTNmTCKou1DHZCN6b+nSISb5S3P9B96Xb8sdFBjnxnPqnIrA/BHKEkSBm3sNrmecSl1Ew0gK2Kie4MPNjLtTD31AgXQLQWOrPR4Mc2e3mtvKxnR6oinoCp1RR1BPy19YUOnhI46B+ZdNJyPISlwflJuCAA/nRj340HERfY/XxHO8y+jmPd73rXc2HP/zhCJI+9alPjW8CTADJ60iCzsyysyIiYjxM5XvlAQ/PG37zfmY4h3LbYxSX/EI30sKuJkC9pL3NV2+6o3nbJ29o3vL5zc0xaYlgxk59jnNyVCgY5qbfp9y5p3npY05q7nrMpuRACmx0r7Y3111FICEQgZ0UrNu123vcxreWWxLLGLWo6UgdRhuwfbqWJzmS7CQHcnfyJk3wpDmhuUp5XUW2DUJ/tJgVs6ALRtYr2R6IMS5dHODZLTiBx2vqQHIeX/WqVzWXXHJJfARny5YtzbOe9azm4Q9/eHyVzlfnLO95znOe0zzmMY/pDS6dGOuTaf32t54IyiGtUs2db0BIS2wH6hg5RWLKvicS0+Nj9qClZ+A8RnVJgFlT6ZoZEGlvhIXcFKfz+ye1nwAWISz7AkhZrhrwtWB51yWUIuzmED+KalfiI/vMc6sBIb3LlXhv08jfSyvIz+Y+D64GyPwuH77Cc4GgP8UhakfcPDCn6wZTA7NUBuN8Nra/6uJ27U2VZTI6A79agr5236l1qQGuc/mgD1UbtKX255nYziMx4aiUcKLulikx6s41zT/vvPOaV77ylUvW6eNxXtnwmsa8p6xvFrJhEYXSx87LcbE/M4uU3PGjmOlKspOM3liSmqoa1DbiGG3Zphg/PZ5IXrx8sScZ9IemAeuENIN4JAcSsWVJUgZmSQLQf3iy4DemZ+zc4zcqD8/iP/AElrytZs45AsFSwfyZ8fG5ZedFFgfwRJl8RkdvTENbsGV6wJ7Q850Cg5tm+4A+yOOcg/gfOmJJJNrrbIEyNvpt92lIa+pAisK+9rWvjUHUMlXviRx22GGB0xve8Ib4oqrB9YDeaVwl2oSDiRVGYLpXdw2dxTyUpQ9yp9x8HCWcKriMsk5Xx5C8lxcVxV8UOJOG1BhUcgpHcnGB9tr87TIqMeh2DHc4bEzn8u1LinLpBC/MY7phy/bm7Rfd2Fxx87b4TcMBMjAqfC6zwNPBbluKaj/o9GOa73rUqfMIXcgfaAKehJUlOvgpUsGvnLfllB3gGgX7+pN1W3HSgqrSh6pEq9G+05+hD9P5MPAS1PZF4OqeW2gPx7ucrO4Rk1C6Oo9FlejDvJXeLPvSTwvPCy8uzC1lx7kvtlveqy8kfGQVSvgAn7BAR9xHll/tBTqF87hrx840o7M7iWzGIFRMq5iLGKs/8nORqCrOUxN8sX5n+qhJfOE9WfjzsqRwtXjX8kME2KGHtEztF27SL3MkOyFfj8BJOowvEre3mGnEeljUto+z2Zab7R0E0hYYpJ8GSmeHpOPiPmg7XSGfZikYGY4zevbTkdbUgfTuo3cel0rHHnvsUtm95RXmjz5LtXSHiOy85Q4vx6VcbwSVB6eKYsAQcWzzunXnuQ5sxoFEdTel827h7qWZP84uIKO4DOIDY760fXih5OyD4ODCXBzsbbZu29lces3W5qIbtsXvPe0zmYifFrNZi83mbXvSciiSlEIvMdM1ouCMYhkOV+KpwmdlH80tvOYkHRtMB68+xrX+BXWg41JV2djMtAwszCAh06HngsYu3Wgfe1Jf2tJ/jiodh87u4Lq4SrQpkyV88dV6Pi0IDLRDsEDmg7KKZn9tCFlrC7Urxvd3y4FdDyFIs+NpBjKv7PEYtEpaUI4jY/DHovQQ8BHXBwUP8MBybx8bNIONRDqlS0k5HjiPidSCN5GWv9uSwnS/ZzmvqSKwPwSMaTHJkn6nBsuI3wX/OW63yC8PSl6RfDw2z58nTNojMPDrDRJnsZvKuat54ioLZNj8BdiBBHfvnKzjNXUgJ6rplGh0bu64rhDo/EGKHk1CkSyYGMTS6NW5Oig23gNCu+8TY2xra8/0dgqlG+KepW7c91GzmZO024K+W9TKglbZl8srNWJK+ZnZJ7x2pKj25uREbtmZXnhPWm2wuK5Y9fs0dojeljQDebuI9s4dabnXYfHu8jxFtYlaHmBhUjRI2sdhwSnP6JUSIb7LeUv74H1gGaHbEoGDfSIqaEh59rb8UwmIzefaY/lRyJACPaXc/Ow8qtP7ZFFdqTPwS5Vn0tK1rNsy5Sk/yrUXe6KxPrZ/BLw+kJOD3OntcFsuLNjjlTKjvuDCOE+ijsRplrC2nmqMux126547LsGZ4E9E9pHSc3ez3tNGkgWPnQ5WsAIu6WyYMk7RRYVLLplFiuBqateSxkWUrH8qAgsRsATVF1n3GBikwt5ZXHPeor8xk5auE4Xl7LFFt83WaeBE1vzvfvU/yyUZli85NgY6yj5JAVneZKe5dSDzTFXqwjxvHN2ZPzGVurHk+XhN+ifFe4eUcJyuTQcbBCTjWDkOjoxcHDekI5ajoa+TF8Xm7E+eMc6CuU/T6cAlHKMFM0f73DSbGdFmX0lLy5qaPendmgSZ5ScD5BwUC2TAU4OrAcqhyqdnbN++o9l0RPpU95wlGFq6lqWwyGKbkRVFAFpkMt63TZexYbncD2T0FNpao1OvsjZjqMqVoylM0cjPVGiLHs6DWM7r5W9LVzgDqYLFWMQHwhIh+V3vTMFuCtD/Dr190DaPuqAPHPf/TPyXN2XpbRw6KoXaxiixjSo1rnz1+OLvQAF2xt9cx3A8HpKU7upNdjS7GJf2fkohHMUWNHUDMPbocN6maIXCITtZG3Wvl3J1XxHYB4GQN0w2lIUoE/yYSy+0T3NeNqELN+a8ufkb2KQ/g+YXC6DFJu3In1z7kEU4u6H4HulsGtL8OpC6LzqNcOTfByxBlhI1GUTjUwkT+AN+6Ltn0RWMlGvsDlbDqteMmmGVU3DUjqcrp7TVi70bzSunaM1KRpQ9OYB70xbOTQKvRLSXZvaFPBezlYlX43dcQ5bWjPQJqCjmAPKwqu15RGhHgy55Q8zw5vBseNQtPa5jOoyzVZy0bs25jjKoDemIGaE8mo2LjH2fk6oja4J0+M+SaY4rMSzL6rrvk3XlOQfJ8sC774NrzjQhYOzN4y/+G/4+b25D4ckiVHKDMdO+XMslx/83PR9/ejC5XmlSNjFwlir3FXpX+oD9lGsfBzNftdyT6pJVKCy1lZrtyzVPHtLlrKaKwAoRwNdp29c+GnLf0D5tuS/Kp+d3AjArrG1miuWxntTBL/0fwhVtLOcQ290RXstZp0lW59aB1Gel3/Rw9iN0ZzfWUvIF93CB83RXLhZl+/jj8cMBdmENhWZluscDpuuZtoXUTMoZJLLgWW6xmwW6RApjNUVifbS2YJdeCUmpU75cWOL+g8/q9tpKn3Yg9+z/2WGIJH6GVz7OS58OaS12uwXLWT1SZutwe5E+F0VfBz/l5iXFQJlQSM1Ph/nF+MVQdM7z8l54A6hzYdx4pUdzBsu2J43wju0XpoXn+TdviwAsvLbwvoM56z4/8V9bDV7qkid/AQVxkv50Cx0MGfXeiUFgg0AHhyhmyJBVeCS4IugM7U5wkuXluK8U7JXqyQGYVIs6RTay0C5bLT26r6G97C2rvhh2QWp//ATOEnejf/GMkPPQ8+DsD7olqKlZU4+AYEX6gNMey5NWkCKsgQfTvw17yj1FnlfwgJko0raXzkgbkSvDVhG/GI9TfiDUZuZy6e8KdM2kwDS3DmR0ks4tPRo9suBkYR8ZRIjFYJBbeHmcZ2rCWG2NaT8cNNvYaLoaIhrVRnl8Nxh4I3uu/ug5OHES86xGMppj4Ofo5H7d3YZ9yrstMFZ2R2DXwpWLjhW7L914R/O+y25pPnnN7VFfS06nwkSETix1O27THTv3Nk++z52ap51zl+awQwcuXbk8hn1e5riRYx04ZCIEStLbjfF8+ZHs08YO8VWxZJVENDyuzeGfTjclXBIgXUOzXIRTexzGn/OU4bjPRDuIZsaXpFOHRZRTlaOqTTTqV6sueiUtjaQ5GJdhiTpbIBxL5LKbL8892bh3VtO0IxCykPRHdLTGdJluIDuJWctxt8HFGuvmjfE4aEuRxb3JaFa9me8Y/ts6Foq6a6lUS+oo8RoLeSrmbEfUczEwQWkWb8W6FTpP94bkLb6tW64eVwT2QaC1KrvyuU+ZYQYuCzZNPFfsruHVOTlqB6osc3nSCRa0SRE/OmVjmskwzpVX5vzeO/AWyO6EQza3DmRm9JV31YLIf6p61gAAQABJREFU4gqF6cD7Hl1BYfuILp2d4zZ8LyeMr3BuO9cPnICpvBNicCC4kdI+cBm0pmCT966VSBC8IxUJH9xzsAd7m82372guS186/eCVW/J7hkvWUTLRUY6bZvP2Pc09Tjg0LRM9LjmQJaxwsDQtvF/kP2Z8Cjxx2cCREE2kmG1Us9SyXLqSrqaTAdb58pz8zf0TS8kYwQk3OfF1w0AgnRUejPwMS2CouAGmcz1fHedf1LR1lHoW9O0SdS24vuBkicIHl1WaH/yTKY0Hllrty9a2JHgx11pK5bP6dzoRwAN0b+5f2mQYJI3Oj2alAvl/LkffyC883UvTVUhI05b+R/0tEwbfpmM0lPd0kRCGIppKACmIdGW8yWNzXfQ1olaWIgCIPiT2RNvKKKmlpg2B0NFpjPOV9QWp8HpktgLSFgjxDHbzHvGCu+bmJNqdgIhXNVKrO9qtoBQqJktxBinUTmiX6QFtbh3IPAwElw+ZuoQZQwL0utGi7UzHYWUr3m8HGyR8AAND7VtbqbsVWqfITLtMarket87XnxYEHweRWviWxKCFLWMGuB5g048bkqWxI31+fXv6WM2udG6JaNHFVPLSNA6J2ZkI25l++8vHbvYefmh6RzG3bclGHWAmOuNra6na8vQBBW1eOS/X0b0neZbundcUuBlc0yfU4MK8jaNksRXZ9U5p/hhMvgqtMrj0ipt+w39BV+7Xbk918wd0pE7dENbmIGf8B4kIdOSVHNkoWVoGsmx0P4oV73TG3eMnqz5xjRFoLUtSsW/q5mUezooS89j2vWNcOR5Nrr0XLjYUqVtlOjbjaHwmKpbe5jK5JXFvzunlb8h0skMG76qvoBZtCtslS94K7qhFKgIZgdC5bZB0OIoknieHA0HMo4msMhsfx8F4c4hkajdtUGwjs48tmi1m+aus8he65d1y04Hb3DqQWdHnLizRzxItyUNBEoaWEXRlHCepKGNKf92L8bJ8jnrPIdddmC2dtXQaaju5/ZE4iU/W8ASc97jyu1z7Eul1R4tAo2jaU3t5+O8PNcuNdiXnb/func2mtO6Twii1DetPmYNcx8O0KeXvTg7ojvQMM16W5eZ36YZlDvQo2t4yW/zuGebOgKzokSkOnshe5U0revK0FNKfmZtKn4bu6CiJ4kjmFpVSfbcvD15707vAIsiMzUKHAESZ+Rvmo6ssp0mHpXBfZOK5hFsZQAtcZTB17tg+X2vlJt+WclNaBZ/mG+rfSUIAC2TDKndkyE0hsMsQ3WMKHA/0mdLzwzZo98Fnpc4wEBK9iXhZSMs/hRNZA13QH3lqRViq2Xf/QTegra3VeSc/LgfJ7iuF27J1VxHYLwJ4Jo8nQ2ZbzEfD8yI7wZfzzG8tJFnHLQZ5iFfokAWXi8JLmeR4wtPcOpDB4K0SLoNX2Yf1kjqOoTU4zhKRctfOSWNE4aHi+KTDTHbac4Tk55ToLHw35M1ycWx7j4YJWApWhceXqzbKJMeH89PDBNqwfYWYYc6Co/IGoWLlWIFiuC4oPIaTUKbJ6eMA7k6ziHt3pU5SedpKsCLOZcG0pX9wTX6630vsnrEcxgdFLoOEsl9lBbF2f54HiQR6loMcWCpOj74oPEUsu8cFLrwx/GiIO8aY8FF6Pgc/ZuOLblDFqOO287FApnGVzODZK0pRQyqZ93BIEZGERdEo6TT9c2ZfUlDTKVPy6346EdC/8WPjAhypCd0xbtCiDq+GigqeyZwzKDP2g8KfKk/jlfHOuIVP038GoStl/C0kxpiYGhL83NLZB2nly9nxFdZEV66PCGVZCb3SyUeDa3tjtUihduyU1QfOKgJZ8WLsFbVQKTxoyWteMZIyhmp8Rc+Y/kKBAiRiW8q2G45vjoYpgr7djOGliTyaXwey7dwle6WrZ9vjHPxk9BGOvns4M15E6NW1aEBgkCoxMExj8DAQ90nX3ua627Y377n0luaz6WMwh25i9CU6knKwD1oTBXxyS3vsS4IX5/Hx9zyueeiZxzXHHdnnbwbCgHGS6Eg0xJk/SyQkgq7X/kxttzx0715LonaHLo06U91Yq/Rhl8RyrOtjtogTGRZKubJEYw4oKxvqZjbNlOZ3+Dodt+QzMw1hlLgvbfOaqH5o4H16IYw6nZZSNouNncq0mLa7OO9GCeKO8f1BQaYsH3XltNRS5LYELeSHHLS6JLeilB7zPmHEoB1EcVrMBrXASd5iVlxcbnBDPZg2BHRtCeLFz7akjPLxbHrb9ZKvbc5DZNpr8vpJSWLxZ9K3dFt8fCodDz61r9LEmoOPUrVEoE9+3ylkNLzpja2+zjWW1UroyHp8SEkma3719BCJerRaBPIS1rycG+8dksYHy7dHpnSdqZIkJ8lRkfCRpWf7QgKCXSDtM9zLD6WRLhY827E3bpiSP3PrQOo7WxEFx4MBbInOM6iJ5ud+L3ctUXAcWamS7lLMUUsyS1WuFx4ctqhcPfh9DFoJKR+DufjqLc3708dgjooPuizEMGoqoNq3iUKB31nHH9acc+pRPTmQq++TuCMBZ2hNCLbUdggvDTjYvUcnY8RsIo8xZraTsgiohh23by2pTI4cF9r2LXLQOS1t4eTqqFBi+39qNrLSrChjZk5TGJnJEdqQgikSTIbck2USnFl+2vEiQZyDAf2DZtDKAa+27qAx16vbsXzs02EEMuK8q0ty2fH/JW1qzn89v9ARdcUJYhy0iAabpT+Rt6B03FL/TBMCeExgLW8oj24tTdD17XE3P3SOYFxo7FJ43HuVJ/qSLgzuHBCS68GNthjTOlWT6fIVxSC+ZdtOkTEcIgZ2SMyEFfLKw7MGCspDpmMFQntPBLmCuFK67isCyyGQmJjNYmsjjSVQUe7ChnmMk4P70j34Mwos5s5y1xzsAZNwG/4O8+I2F5A6GMWSQvnyOvmLb52g87l1IPNgpLNyb5TuihfjOx0Udks6F0vJgwpTq5TuFBzzYdCXnmkcclyWqKhG7ZHfnmxowxvhmPRE2+5kKG/btr3ZfMeO5tZtu5pdu8vcGSJyQldBpxzDz5C/KVF8+x3b4mMwTXN4CFa3Te0jDm6XcBoaJSJhpZ+gleloP8weMhqKL+WXKFE//ZrnoDhae9Is5IbhuuOgqVA46FS5+lsPp/8xO8hJy02Ie8b/J1FB2aVqNra8pN9K/xRjBXb6Uop+TnSWa5E5d39y+0M+U9v1dOmoBf3aYmYwIaPxHmKb1x9kaCsDmOPCQJmy4MPUidHdqbP3ivBIhfCyz7nj++u5aRsZFGOZD4hQ7ZCQ0G9kepilQE1TiIAupJ+L7rU2w/vEsbJBH6eEIxPntsdWvKQz/JGzIn/8f8iKChItSSfSg2WkC52owiQr6EglBu8Xk/2QoJZ2xcad6N6QaVo4vsQ2BEK1g/EsVewKqOQ5iY+kRe64qarPm2UEst2S+S7aibEWDRUDtkrXSEUWnyQRDIp5Ta0OCbuqhQx0Cz+gmN/tD0DbcQ9+AaDCU5Dm1oGMwQiDlx5NnVUGiqX6DT+UgSV4Y6lCY8kjeGKsBDFVmoQ181IMV5EX5/Lzhag1yrclyx1jISc9BB270xLHHTt2pEF1V3Noom5Tt/JlKirvitjvTPfH10QtX9s4/uUNSIqv16YDTnUhsTjYYSx38l1nsMAuY9ZqxmXac8CXGCPpXUb1qaV0XT7LGMdg71q66Ho4aem+mLkc3HHAFIy8MYwS9KVKBwggoCUolFo6jeEg5duHc5nuiUFicNPIKmbuQm4ykBI6cGh7VY6uCuhSB6ZLYWRmANKFwpRRMOeO/W9hoKgLfWjIFcZxW2H3WOcrEV93LDSOnTAVZN7mPIQzsKiOUazUJ1yLSKina4FA6n9Ol6TPw3l00vKsQ7pRCfrGdfs8XPfHDZn1yUKuNVUZyZkUuWHsOcuyVfLL77k5H3eCUbZZEmVp/EThYhSGtCudr8qLMS509eI7xk1lfd6sIYDfQ1dn5spctYCNyKiMVnO35SJrICezhsp+2pPggAjsyr74FsbcsOvStSKlBcF46pRhNrcOZHEaOEc6OZaoOmiPHZIFPzQv6VcdnZV4zuvjr+fnFBQM7c102h0yBsUULrcMD9pnjGkHozQA7drZfgwmvcuXZzSGIhA1FTpKdls9upkLu3f5Gql7s7k9Juo6jykE6DvHmZCCVRgkg7rbjk3nRbg7DxrrYfAMT8Igjqb0f0hprirO28xyLRTQXstEU85gBmmspKWHMUhS/6TZUUQt0F+FkIJjVJ1xM5G6W5sKuOMma6Kfpz/IRMIuYTD8TbYBYPEesJ/vgKefbomUjkO+03kOCvTXyMwzLY0tDboKKfidgxu05e7MhKTriw5Kxpj3KSCVnjhcIbDw8YckfktaItMZvIduujeVm0t+W4jPtJ/pR2Npdhp1anEVMzN2WdJxrP5JxfC0j9rQU+0fB2NP6rFlGyHLzLAS19JZEqCQsfaCPCo6XdWcnpIKVGQMDSlO9SBGKpU6L3lxIZ+xXBfl56v1b0VgKQRafkp8EwHm4C95eKvw2jCwQ5YHKcloBFATrw7t2cHV2T9o4TGZEWNsarHxjsSWaRPjm3GOvnCtpFBvZHxK0lw6kLpHx0VkJe0lHVfSBkvN4oQADIUlGzClVI/7GCTS81fISIaGeK8KezJOO20ZC5UtBLEMJs1AmknzwZDQEmgMbbFMTamM8rvD8UTt+FNpsp7LYknNlV4s9ZVSzttraReDftr3pewYIswRTBazoeoqJLX77nk5pmTykq1M66JbxnIa3ce4QJNd603blZ4qg4PzwXG6EV59YTaWxq3hQzYmDPG4fUmwhaO9ZOc9a0WUGpZ0dXwJ/+A4/JOdtPLhg8JZI+pq6QxCRxQ5+OxSSX4SIzxms9NpMcjRrlQu2ZZP2NI/Nc0GAmSCLGT+bPt4H8Yr+dqcebd/Dkh14jOrZIq1twjy4M6wEYb0bUhfjdvbzvIvKj7G01Qz4BgrSVkbV/IrGXRJpqW8ZlDyS+Wx5LZVRPvRAuWWuq8IJH5rHcgkDuxK5nDXvsx8V1ZxZcCwp3dv9+7lXmS+nC8oAZV+ci3JG3sAAq1ZNUBD3sJxLquboT6cDtzm0oEcMnOrSilWTlCrYBleJZVjOTHgudaWK2X63BcDqxhXS9WF9HCaOChRoG3XUoUPIs/PDuxNzuPeNJO4J35EbiFuueoWR0S1qbQhTT+Smh6TAXVQae5Tp21/larLPpek6BbmtE8Y4y6pivhh6sxNeUltwWd03XqTs06Jl9JjJArTpP+pfhHtRJ9IWFmGlb8Xm2nLx2rmlAwpybPJTLphnlLzk/RQHiS0OduPGbP4m/4QAwOqVEQiEGvz8pXx/zXIG7w2phHfDKl3W4PalB/167J0vCvtQ2bSMe7clDJiWbKLfaUkb5m2toJF8gcvKz9QULAjw+TUv5pmAQH92G52LT+Oaln0fHR9/FlgxI6658Dy02xo4rP4iR2CMjJlOgaXCw/bl+PBxTEdqDLJtHfpzUBmyFo82iril8lS/XvCylcip2hPX3SVSup+9hBICnhjCqZsaIMpXedRY3GfMTBzYW5+YtFmTxpUunn5ypz8bRueg+1Lo1BiUyTUsVIx7rWoLX3X5OE3vw5kKPrE6WXKpaNcS+d1FXTpOvGEoVouuePdE8eI6Qj3pLSvg4PCTF0UGFBUKM+54/7r6dkRMgs5cNUWVEPBtGSHQW1AA20sBaZZekxBn/rbviz7QZXyixUfZRJxieDsRMXdg6JjO8hVBAh+xmMjxaqqFSQzgrHsNeFWMF3BbasqEq1Of8yQ6p7yLRV8vtSHTgrp8S5nGDL99umqGrPGhS1DjfcfB53TopN4bB/eQxuco/PJTkFy3ETTC56e/qW6cn32Oa/UVngw5LXNLLLr3t6SRwcxhbZOTUhHZ9Aah3GsRHbEE97pYq/0dciph/0hMODNltWcW8g6SPLTaebffOyamTR82l+KtSLDd+nV2VbWoW5B9a4Lvm2KtWil9IIiYzpJz07/Y9VHjKVDitRKRuijrJsZpEPdHMdFsMZEzbQ85vbtu5vPXru1ufWOXcPOHBC/fH/tTBCectzhzXmnHzO4Yz4Osl0kmLgrxrnEXGyXhIe5A5xXkIt9y2qOiyzHKx7zAda+rUyyVr7Hse/F0Tm9B29HV31AV+bWgYyBiUIdiMHy+GVVXcqHyCx/w0FdDXN+P7QNadCWztl+7jsIwlI9eRlrioIOx6YFDzQLw2mMfXtF0VA4QeeQ0gU3HuRJ7p9UdyBRzkY8NPrdNbSUsgnD4lyOuO2As7WbEo6ZvjzjU+ArS43Qza2QyjXk+X3GbET1g1uuL/VrDBILpaHUiLuGKAWJ4Wl6BzIvuWjz5maX0dBn8W/ATxmAPICmY46ksGLLV5EPVPzQrxUcPKNvbKrvkNH2kjYgZtiz+K73VXhRa6YrYCtkpPxhajkuCgxz81HhysX59XzSEVjQc3GS+rnlz8yKuUTp9sKZ2uU4An1RsOTYjz9lBy3ThiLxoQiuOOkS1akazcplXd25MMbDoCVZ8PHTTsmKb6Ukaohr7VFeryJnmHx4h96JNKINw9KzdXRL+vmx937mxuYLN25PztBwnF3QSpgUyAo+6Xxr8iAfdfaxyYE8OuAry+0X3DvDJ/g5b4l/UoAEb220FC+l+JuwyoHHDJpxBl9G7CL+zDA4yzQtMIBdAJXxysUdFwZb9ICUHXqkyOmiy5N4OjEOJAP29tvTD9Qfemhz2GGHRRStD8CGXRddHMIxqp7c7fmOYIWUEe+JDTp4+LRRzzig/JbfymBEaZVjzyvnC5RZS1N760gePSB6BjclFNJAtCc5NXmJzOBCe5AMwzhauCYeTZAKY7qctHeMdReN9yfRmXYM50FXpdygIWUs6LVBgWHuAlzHRKD+27PLV1izEi6PDZLbk4xduh5WvM/aJ53d43ujuVpYJaOEM5i0fweFGAgW0zlwstuX+QbwlYIzvl+KN4psFrkEgX5l8MJniGl7RV7b8QuvuT6mpOJ2i6qcdh6t3kRd0BgUprJBE6aL+zqFezlEzSKiWgIDEzSklPEpOJbybcEoUf9MCwILeD2YTT/mvtWGtssHzVlwvk+X75MxuO/gD8gChyu7rCErpbq0F/zJ30ZItKeL8c2EROzuReP0wdPRfUJQkTDa2+xKtOWvoOe8XMpxSYXY4XnoqAWAlmuzvk925fYdzeXXb24uuib9jFga4w4pkdoFTS/4wW54fOv2Pc2px6VFhvExu/TZEwP4vKRgI3/IqLEiJX/Scc51xXv/xe5TIKX2ej6Z578JpYQFlBYm5wFkm52Po6iyUXzxPQufMClnE+FA7kqG9TXXXNN86lOfak444YTmXve6V3OXu9ylF4xytww7Zynjr+gIHVp+AmLY4a0gBXXD5xwssd0nEc9wtpLeyvLaqVNGuh67zGlt1Z7QeUrnsC1w0LugaXd6/3Hn9vSsJbXwPnUgw1R+Nq7TEpIujfuUPoiMtr120W/t3hMzVo4WHuec4d/yQZhQloUJhpcP+Cj6k5OWtojgJQKDzkRYmfEps7YqQT+crIjKvwPZNk52D6k4kBEcGYA1rLNAga7yER1kMLK6H43pgbSJe2TWFxkbx3nJCdAEeYQACoCuIb/kO0796R4o5os9SEOmTZ+VrfBzqRMdmQPt8xmq81fhShgjLvTzp8Vt+NuUK6sm04/igvHK7pukUqUP5soQXdQBB8JhgZvpjY7s9MEGOEtsjJPm3WHnql3AcSG7ER4KAQqO9EfD7PtK6dlw8BpJvGQWVJUKM4Xla/Ih+y0ddE5+HYHUK5/aVZR6X7ROwHNhJZS9c0eaedx2e3PIrh0JO3qxi1k+3jcnN+CQXSlYvuOIZpefMNt0xAS0qn8SuqwR/NYZ10IOCwlAIwtFSNr8uCe94jSAuZSfpz05Tbgt/LUCABRNQlk4DhBdiJTxxbelXLkymfuJcCA/+clPNr/927/dfNu3fVvzxje+MRzI5z3vec2JJ564YtQoxP0pRdc3KpeeujH1H+Od0RRJP7bHFK7kGucnJwyRjjGGCCQps40pBf3pWV7J9FKtzdPVHpMC7XE7xZcv4sGS0nHIedyRMrVzTPRtKIveE0FhJKcPu6zkdxwNViEKGlHuDXrHSVvug8Ap9c2h6fnwMujDUH4XpnS6KCUa2wLaGTziRcUxpMwj6VmJCLO26XdQol+69Iw6HlSfsN6QDIaIeKemjq9P42FRzQYvyYdRkgjNcA6qdwDDbkKzwSE+8IO2MfFZt45JPdbW0CAJp1YdBEAcaw7/IS1aaeiIJuAkH6eR4s50mGfi81Kq8WOnX1NlqZN8ECk+iiTLccrOMyZoSnKSMqIN8m3phF4MAyCdj5O2jUmm9pIrRKQEF4f4qNAkf1QKeoPIVCLt28eMKj6R+bfeemvziU98ojnllFOas88+uzniiJUbpKUv4DitaUOifWPSsRsT46UFD7Htwqf6M3VoOhzwgzYW3ijt3Z0YZXfx5jwr+Gk8nLAx/S5xeZ4xLj7Br66RaeE15bULY+qr8qyRt6/igmdl2RGMsvw7O7mO4RaUpGMpzlvSHJdk5k32IS1thZ/K9VncRxsTU5mg2L5tW7Nj+85mp35aorEtZHGlHINvx/Z07/bt6SfIdqfVcR1Al3jGrGSxg8gDefWayq40xu+kgNs0OGx5rjAdeXVMFpLbnfZ7Er9tTDO+2a4q98/qvsgp3Iy25NSqLnoNz9m30prOh7O2jikO42AOMCdLYUrkdN0dyCuvvLL59Kc/3TzqUY9qHvnIRzaHH354c+mllzYXXnhh8+xnPzsgX/yH4rzooouaa6+9Nn6Y/o477mhs73jHO0Jxu75Ukm+p3u3bdjSPu/ONzc5jkzJZQieUu10qx57nsZuSV3L8tq3N+963pdm0aVMeKJaqrM0rjLC/mSS0+arl1ju2N+cefmtz9im70rNFObHWQjqWqi5oS+WPvn1L88ELb22OPDy5UokJlxsoXLMFLh4wIrmO/lu33NHcfe+W5riTdialsARwI+4v2Yfesrn5+Ievbz77ycP2S1sMwOnGldCmT2+67fbm9GZr87STd0SQoOCm7tEty5QR8kNv3dF85MPXNZd88vCx0sY7vfG2hFuzuTnuxOQM5ipX/Nc7G8ds3tFc+IHrm8MOz0u7l+tTuLmOl5ZLcEXbbVu3N/c+9Jbm1BO8a5l5bbn7utfQdtKu7c2737slxRM2Rb3L0eaabX992q1jLY9jAEggBDYjKs60p6h2+k3Ucw+7qbn7KVkWioyW/i08V/I9rhwTnTtt2Nx89EPbmouPODRhsvwACzO0rUSHKLMtLdm62+7bmmeeun2knKKv0LO4qSfu3tJc/InNzZc/mxycts8WlynnK+3T0udbbt+W9OeW5hmnbAtjtjxnf3v04s8Tdtye9MfNzdVf/PR+cVOnlSxnn312c9ppp+2vil758rLLLmte97rXNY9+9KObCy64oLnvfe/bPOtZz2qOOWb0xzluueWWZkea+dCOm2++Ofr/6quvjtc89PMkpNL/9vvnz8Sbaey9+eYbmxM2Jp142LbmMAPqSlJi1qRumiM3bUgzSVubG6+/rjlk97Zk5GbHD0aLU6GtyM5ymLm2c+fO5rZbbmyO3Xt7c/Zh25vDDy2m3cIxpMiNfaRUdczyJcHeuGNLc/ONNzRXHZICa4zwhMuodMghue1o3x9te1LwceutNzVH7d3anHHYHc1h6at0Sz0ZDFFl2ncR2Zms+43bNzc33Xhjc3iKEm1MdY+DtlFt6zO/jHHL8Zu2xRiYfnbslhsSbjtvbk7euyPhnJ3uxfTl8BqdyNHOyDq+UzL7j0wLrq66+prm0COOavTZcri5VvhtKZ5cXO9anq+UNnRzurdu2dycsIEsbGsOX1JOC4fBKx/7iwfx59F7D2luTLJwWJNssk2HBi6jMCl9uj9ZWEu8Sl0rpQ0/mii4JbX58F1bmjMP394csSm5jkApwtoeDxFbeEmAzL23Jh159dVJtx3CPig3F4oW7o8//vjwmxbmrs3ZujuQlq5++ctfbp7+9Kc3d7vb3WJAveKKK5rPfvazyyJw9NFHN4Cj9AFsgHVOwJdi0mHehuakpOjvc48zln3+UhepbP0vwrIzhU7LM0d1MAXGCLj88svDYDj22GOXHCi6z/Gs+92TEsqG9lJ0LJWXaUtOaCJwV6KtzKIuR9sNN9zQ3HTTTWFcWTq81CDWpe2UUzY059w7DeCJn+Gw2uQ9fl/0osSlUbS59pWvfCUif/rUTPRytBHBkxNt5ybMRFhzrNVT9peyYJLPnQm4+CjMfmijTPCnqOTJJ5/c3OlOdxpZScHupJM2NufeJ/Xp8npgyefAaFeibWcCrzxPwaWww/tf+tKXktK+sbnnPe/ZHHXUUUuWc3951omJtvve88wDpo3C29GRheVow29f+MIXmvPOOy+MYGUnJW3durW55JJLmgc84AGhg5bnt4z/WWfcbRVyqvOL1IhO5j7dnyygA22f//znmzPOOCNmr5YKDpT+hCfdcc+zzgjnsZu/UqyJAHkoGCzFa57l2fSHfiUHd73rXQf3dOvq0kAW7n2PDc2mVcnp8Gn02+5WVuWOok0+Z8v4Qu+uxIEc1jLeo+uuuy4Cnmi6//3vH5gJnH7sYx9rnvjEJy5ZGczIsfGDMWd8M9YZK0eNcUs+qOdMPEIX2uAsqNrt71J95KU27Upj4rYURHjwKZuae97pqOBV+jvLRlGQQzkpMlNKCFyefExyqJORtm3rran/Rwdf1Om7CmhjL5jxHUWbfG3Zdcftzf1P2NiceviRWbYTZV2qljrWRvdb3XTykbvD+Nu9fcuytLnntttuCycHXXT1UrQphy6zonu2397c5/gNzQmHHpGevS9qykKuS6M8Kd3enHzErqBtV6JNKWPZUgmfbt68OfgObauZKV/qeePMQ5vJAn165JFHxvcyRj0/9Fdq+O0pSHr/u+xtzjhaQKDw1uK7CmoL83enpZgnHbOrueqaayNIWpyJhaXyGdoEfOhrskBOJynRI1u2bNmvnOJD286dO5oHnXxIc9YxyXFehfECY3bYcUduTPx2Q7PzjtsSr412vNUFN/1KDkwijZKF9cATXdvSDDa66JFRtGU53Zv6f1tz5nF7myffI+m3hMNSSe5AVtuT4M2UefKdNjTbt96WdH3ybdoJgSWfkZ5Nh/hmzN3vfvfmzDPPXKpYr3nr7kBiHEqeMtABPqIj6bBRiaDe+973HlzmoDEUHve4xw3yJuWAoWCm9BGPeMSyzsZ60Guml6POoOe8T1L66Ec/2ljypZ/XQzCWw4ICQduDHvSg5tRTT12u6JpfY8CRKbP5xx133JrXv1yFX/3qV8Oof+xjHxuG5nJl1/qa/uTcws3gP0mJgwY7zu097nGPSSItgikXX3xxDGDnnnvuRNFGt9G/Sznca0kop++LX/xi89znPjf6kM5405ve1Hz84x8f6UCijywzDhitNgag80lxII3DnFpBBGMwfWhGdRTe9Obh6Z6jjz6qOeXE8o2DUQb90j0U5lhyGvd3F9rgRa6Nv+RGMFLeUglt7mHA3vWUE7J1t99alnhSoq2k8sxyXvbqcY1cczYEN8yUL4ebe04/8qjmjFNPGGnAlucvvU/IpWeUNIo21/EXfkUbu0BwSPlJSGTiqquuij41i89ehM2ohO6TjziyOeWEO6ciq2uD0vnJeeKg1DGqPrRxvAX7HvrQhy7r3JZnreWe020lxEMe8pAICizHb+g6ItnkTziv2BCrwy63a4jbKMzko4NjCzffP7nzne88Uk7XEi+04Z/rr78+eO6kk06KyYz96RC+zMknJn4LyFaPm6BYuWs5OTUm4Dd6hM23HnbyujuQhI4S4ERKgKDAePorTToU0Aaz4oCu9N4+y6HHJpV9n/Wt5tkwK7iNEojVPG/cZSkVAYVJw0070Vb4bdztPtjnlUGBHC2nfA62ngO5v/Ql2sj9JKUubZNEV5EBg9kkymnRIYXvJgm7gtco42WtaBXBNtALTOjPMhPGQB+V0CyqXBLjyuqCBz/4wSNnjkrZtdzrd0662d5zzjmnt4/fHUibYM2YgyVnYz0MrOXopnM4uFaLnH322csVXfNr+NXshqDQpAWXTS6QnQc+8IETxW86iYwKppDTSZq5RZtAj1U25FQwZZKSIA/syEF3cmgSaBSIZ0vRH5MWJGVH8ZU4t+uRhuGy9ag91SnaYImiAZJHLVpL4U+aQj1QeDjDpr61adIS5isR7kmjTSAAfXhi0hK69hf5XC+a8VuZpVgvGparlyKuaXUI0B2TKqf4jSzYT1qaFNr0Hz1WghQcm0LbSjErcuPeSUrosaFvUmkr9E0Sbmgp2E0abmjTn5PYp4tpK3IhfxJS6cuynwSaCg2FprIv+eu9R08JQE4abbBBU5GH9cZqcf1FRtdLDtZ9GkD0zRTx7/3e78XHcziQPjTwpCc9aTFWI88NxpPopDEc0CZCYD9piSMkQjuJtDGY0YbGSUt4zezGJOKGtuXefVxPLBnR+nUSgymTTFvRIZO0uqLwEdpE2ieRNjSRh/WmzcyjQKmxzTJP4x1+O/300wuMK9qTm/UyFEYRiCY62tJVbZq0pO/pw0nV1ZM8xk2ybcCmmkR+w/9kdBLHuEmkqeiLSR7jip283uNIwaq7RxM5ZVetR1p361zDLUV4/vOfHyCYIrYt93GSxUARjElVJmWAXUzzJJwX3CZRsRTaJrFf0WSbRNzQNYnGUuH3SezPSacNnxnEJpHf0ITfJrFfJ4W2s846q7nf/e7XvPWtb40P43jXlvP4+Mc/vrDefvcwZixMIg+gTRBhEnkAbWwM+0lL+tM2ybRNYp/ShZM4YYC/yCf6JjGhbVJ1CD6jQyZRFooOmcR+Rdt66pCJ4HTrsX1gYFbTpEWNpwnnit3qemvS8Zp0+laH9tqUhtkk41ZpW54PvENj5tGHwbyHxFDyHlL3Hcfln5AN00k0rgrdk+hooI3RPKm0oQ9tkxgUmGTcYDbJuE2io4HXJDpkEvlt0mmbZH5bT9omwoEMzq5/KgIVgYpARaAiMIMI+BDJy1/+8hlsWW3SwSIwyQGYg21bH/dPOl6VvgPr9UnGrdK2dJ9O3ksLS9NZcysCFYGKQEWgIlARqAhUBCoCFYGKQEVgnRGoDuQ6d0CtviJQEagIVAQqAhWBikBFoCJQEagITAsC1YGclp6qdFYEKgIVgYpARaAiUBGoCFQEKgIVgXVGoDqQ69wBtfqKQEWgIlARqAhUBCoCFYGKQEWgIjAtCFQHclp6qtJZEagIVAQqAhWBikBFoCJQEagIVATWGYHqQK5zB9TqKwIVgYpARaAiUBGoCFQEKgIVgYrAtCBQHchp6alKZ0WgIlARqAhUBCoCFYGKQEWgIlARWGcEqgO5zh1Qq68IVAQqAhWBikBFoCJQEagIVAQqAtOCQHUgp6WnKp0VgYpARaAiUBGoCFQEKgIVgYpARWCdEdi0zvWvW/XXXXddc8011zSbNm1qTjvttOZOd7pT8/nPf745++yzm0MPPbS55ZZbmttvv725613v2lx77bVxfNtttzVHHnlkc4973CPuGzfxe/fubW6++eZmw4YNDfruuOOO5uSTTw4a1KX+q6++OvJPOumk5m53u1uzc+fOoO3GG2+M62g79thj4xnjps/zbrjhhqDB8RlnnBF1oemoo45q7nKXuwQNaDriiCOa3bt3R3tgecghhzT3uc99msMOO8ytY09wg5c69RfaTjzxxKh327ZtzZVXXtls3bo1sITdnj17oo/tv/SlL0WfnnDCCWOnqzxQ3V/4wheCPn10+OGHBz36Gm5ohpnzzZs3B91f/epXG7SecsopUb48a5z7HTt2NNdff30Do1tvvXXAbxs3bgyMyAR60AFTSX/jVfKhHWeeeeY4SVrwLH16xRVXBH2nnnpqYHHVVVc1xx9/fHP00UdHH5ILNJBn5Z3jR7JMvsedtB1e6iJ3W7ZsaY455pjm3ve+d1SF/2FEVkr+rl27Ai958LznPe8Z1/R3H0kd8MB3+FrffeUrX4n+0peFRjgWvUOG6D5yat9Huummm5rt27dH/XScfkMf/QBPcmpP75Jf8owv7b/85S8397rXvaLv+6CtPrMiUBGoCFQEKgIVgZUhMH7ramX1rmspRtV73vOe5uKLLw46Hv/4xzePfOQjmze/+c3NN33TN4Uzeckll4Qhw2j9y7/8yyjH+GHkP+MZz2ge8pCHjN1JY9R98IMfbBj1l156aRih55xzTvOCF7wgHNdPfOITzQc+8IEwqBjH3/Ed3xE0fuxjH4v7GGbf+73fG0ZWH4Yzww59//mf/xlth8GTn/zk5l/+5V/CmX3MYx7TvP/97w+HiOP7xS9+sfnUpz4VDgoDUDvud7/79WKcfuhDH2ouv/zycBw+8pGPNA984AOb5zznOY3+u+yyy5q3v/3t4cwyjr/hG74hHIv/+I//CGdDe/7bf/tvYcj2wZiMeX33rne9K/rriU98YjgQHDfO0HOf+9zmne98Z6OvGfMXXnhhGNCwPP3005unP/3pzbnnntsHaeG4/smf/EkY5fCDz1Of+tRwqD/72c82//zP/xyGe8k/66yzmre+9a2BH8eJA9CXA0kOPvzhDwfPc7we9rCHNV/zNV8TeXe+851DBj/5yU8Gn33P93xPyCnHV1nOpX7Go+NOgg4cRHyGx/UjJwgN+utzn/tc8+///u+xl/+d3/mdgdd//dd/hfxwJl/4whcGD/Qhp9oLF/xNZ9397ncP2Xvve9/bHHfccQ19J2BBb8BHW5SHm7ZpR19OJFnjCNKjH/3oR5vzzjuv+cZv/MYIXOC3f/qnf4qgwH3ve9/Qs9pCDgQF6J4f+qEfqg4kUFaQBDlsghQCn4ICZJZskBPyRS+7Rrc7xpuc+VJmBdWsqgj9pl40Ga/UhxfQIBkD0YIPBRsFghwr5z7XBWX6CnCgAV1o6OLmHC4CZmhAk/PSHnnKo01+H6n0lzr1K2zQo159x66xh6dNOXTZC/ahra8ArvbqI/ylPvwDB3kSOtFcsCl4wdU19JZrccMY/+grddurV334DW5oRXPJh6l8dLkGT3TJ7yvBSP90+079+sr4UPgepvKVcw+6+pJTbS31qF99aDE5IcESLa7Jx1vwUo78uBdtfcqpPtOv6tSn+giOcHFeaCw8X3DUv9pBB/aR1KP98FhOTvEgOpXTlkJv33I6zjbPnQNJyTJcGHk//MM/HIYTw44C43T84z/+YzgiDPbnPe95IQy/93u/17z85S9vfuAHfqD513/91+YXf/EXm7//+78fu8LDQGbDOI/qxmD/7//9vzAEzRSgk6P7gAc8oPmDP/iDcDTMLHDgCM6rXvWqEOa+FPHf/M3fBO99//d/fwjIj/7ojzYPf/jDQ+GaqUI7I/W1r31tzLIwBuH6Mz/zM80FF1zQuP/bv/3be3GGBAMYoJxUBin8zPKY9fv4xz/enH/++c1DH/rQ5r//9/8eAYIHPehBgbNZmpe97GXNWckx6iv927/9WziIv/zLvxxK6yd+4iear/u6rwtnWnCCk8vBZEybrRUkEBz4zd/8zebHfuzHwqi+//3vP3Z+01599prXvCb4/ru/+7ub//2//3fzx3/8x83P/dzPNb/yK7/S/PRP/3Sjbk7mr//6rzeve93rwpA32/asZz0rZKYv3DhmaPulX/qlULTvfve7m7/7u78LhxtdnBx9LJhCUb/yla9sfu3Xfq3Bn3jvFa94RcipAWOcicIXFPjrv/7r5tu+7dsauOF7uP2P//E/Ih8/wY+++LM/+7PmKU95SvCAweV3f/d3oy/7klPBr+Kc0Rf/5//8n9AhZvUECX7jN34j4HhhcmINrm9729vCsYQpXtUuTq9+H3cSULoizSiTUwEe+oCzSOdxYp/0pCeFU0vXcr4FnD7zmc+Ec4sn+wpWlHbq25LGzTfluWuxZ1ThPbpEHwsKCRb84R/+YfP85z8/HHYBBEGDZz7zmc1b3vKW0D1mgK2IoH+MLePGAF2CCHAWHLLK4tGPfnTUB5eLLrqoIedmp9FLj5M1POA+Abcf/MEfjNn+voIvAi9o0Ha4CVz9wz/8Q+DxuMc9LmQL/fQ2HWVcFqRkLL/4xS+OgE0ffSxIKvii7ve9733N13/910cAl21ApoyvgkECRMYX9oQgF5uHPnzRi17UPOpRj+qDtDDi6Z03vvGNUR+9WFY9cDTQw06hFyX9jO53vOMdzYMf/ODmaU97Wm+46SP2GiOdnqET8Tw+d+3P//zPw05gy3zzN39z6ELyIAm0WaFGZvpIHA00oQ/PCy4LypvgsFKJTcB+wV/f9V3fFeOfYJr+tpKEnApajjuhi2yyqfQVvlMP+4nzhc/oFzqEThaAhy866Ry4kVMrRvqSUzafoDydwJ6DDxyNG8Zbtp1xmfwKEsCR3c9pI6f4s49U9BRdJ/hJh+hTqxwFnMkpfATD0Qk343WRU34Jmqch9eOCT3DLdRJjHaMzhikwwqATdSYliPEoDYwoMVAZORjAzIdnUM59JIJLkVG0lqhyajF9oZmxx1C1HM3yOcoZbQZhkQvCMe5Bv7ST8Wdm5U1velMoXbRSwAZ5GHK0HVMw8KQA0W+gQB/hMSPSRxL1YiCbBYUDAUSfwZ0Qm+H7/d///ehbzoZlccoZhN3XVzRKWykLBgknTN8x0vCPweHZz352YEaZGEjxlkiagaQENUTyKPM+kjo4Yjb9hH/ggw4zPn/1V38VRqdZ0cJXnCD9SWn3NTigAb/gqze84Q1hmHz605+OgE7pZ4MsXL72a782jCr81pVTQZW+5NRztV+dBlbOFvkgq/rbwMb5NZiQU/zJGYKb/uxTThnbjBK6TTDH4I+mRzziEVGvgRTvCwJwYrVBwEz/MzAFXSxn7SPhHXjBgZ6g6+QVOWVkvvrVrw69UuTUrCk5pZP7lNM+2rsez4Qbow8PcMT1uz7noDOaOBMCLfiVDsKbglXkW3nyJWjTh+x4Jv7SzxxEwT6GIDnBt8Y5csXZEeyQb4wR2NUeM9DGxb74QD3q4zB867d+a9gIxgpOLGPadXYDmaH7GKz0FGOZDhDgEkztI+lTepjj8NKXvjR0Iuz0qzGO7AqGCtYy4gUj3cMh1q99rWLRVnaBtus3AVLHeA5G8MJraGJUc4Dobnz3Uz/1U0GjFUJ9Jc7W7/zO74T+fclLXhL99frXvz5sJ8E0Y68JAvZCkQMOOj4VdKZ7+krsIeOEVW0Cn3jNSjiTBII99HcJ6sALvQJC+A0fkNNu0GtcdHomWoy9HG2rUuhh52TBxAZ9gd/Yx5xwdJIPuJFTeqSvICndwOlit3C4BMLJBhrZBALhf/qnfxrX6QoBAWXg5h5OHLntI9Fh9JYAkwmAv/iLvxjoNjQ+9rGPDdzYCGwsYy05JR/kp4/AbR/t9My5m4Esg6KBkxHDaMJg3reh3EQRRTQYXCW5h9Enuc4wLcZ0KTPOPeOy1KceQog2S9EIK+XC6OLgiqyYqSQ4Up90UWCMTo42HMwgEEYCwOmh0AwUlgGjGS3aIhlISl5k9PAHTUVhoVX9+o7jj2Z7gwUc9bGy5T3JHsgZPNLAJPDAWeRYc3ItV+VYUHacHhjqUzTZSv/rd+3Qtj6SgcJW+McxA7QYaJwzfV4cH/QoA8tSpg+6PNPzvf+pz0QyBUs4PvAxGMCr8Ls9ulyX1kJO0Uf2pMLfjmHDwGQco7nwIVlwTSp0x8mY/8CFo4AGes45PYenSv8yctFvQ8tayikDqNRdeI2cotFMBV4UQDkrBdH0s/6m6+xr2j8CxgTGFYeLYU/HcCboGTO8P//zPx9OknO6EI/iXzJGnzPO8Ic+GTfm+E99Zi0YSvhOXYJDZEkAV9CIftb3gh/4Fm94LQKNntGX/Jjlgx8abbDjUFoOz+kww2bW3LghiEvf4E3BIRhadcCRQ+e4E8dLfZ5PjzDQGfOC4RxIso5WBrx9CbhwyAUn+1yGiWcEJvCUvmEHCBAYd9Fl5oqD5JjzaEw0E4j/bMZrgXtG97gTerSdA+35zvWrIKkgGxzpSg6vvpfIC10k+FLG4nHTRS/DDe9zWAXUBCns2XkCJRwydHCS2A7GNeMxOVVWX/clD8Z6Np1VPhxpeHF2jcd4TiKzjs2emizgbMLTPX3RpV56Ao+RA7YT55VDyNk3WYDH8B8dQxbIjb4np8YecmqmXr+PO3G89SE5VS85JLto5DCSAf4Feq0AgRk9W+RUUH9a0tw5kDpUR/H2GS+EmMFCqYii6HSDA+OeQuGs6XzT9ZSKaJ+oOSbsI3muaDy6DOCispYKYjCDGyF2jQI2qBrktINR1nfi+IiYUfRmEdDHABA9hKEoJ8El3BQdDAkJIdcOuFMufSTtp1BFhNGERvUaAChCg72BAM2UHiE1aNnk9ZkMBniGYUSxUBjqNXBQbr/wC78QS6QMvpQufhOFZDCZ0aK8HfeRtJ0yU58+NIBybmFk2QV+o6Dhhd/Qh3Y82GfSh3iM0lcXQ4kRgA5GMf5iyEmWAQtaUNxmNrSHnJqF7ktO6QsDgplle/10flomfVZyegwYkn6UnJMbcgq7vhNDycCkL2GH/+k0Bp68b/mWbwkDyvut6EUbQ51uJLsG3tKGcdNKThk9lgTqQ4YHOWVM4i1yyjjCl6W/tWEt5HTcbV2v5zE08R6dwalhRJEbeoizA0+6WbmSyDZjUSJ7xpe+EtlhHOt3iT6kG41leJCORKelXwxRfEIPyJfc31cyxhsn6EIbpwcuhVa4wdQmFcfWMRmDaZ/YoaPQUgxPek+fohNWVgHRffLocf0vv89UZFSglL6hm9lS9K9+47DZk39yjq6yhBDO5L3Lj+OkVV/hJ/0jsaX0ETroQzxH17BvYOoaHjsr6ca+nEd0qAfPaz+ZEyyFn350jm72Ad3nHK74rbSDTPfJa2hEkzolfGcMYxOou/St8Qat+re0Qfm+5VSfoQlmdIX+Mm6hhRzb9C2ZLbiiCy/gNXj2kbQbZvpVcgwrcqpe/UsOLD821uLBIqdon6Y0XdSOAVkMxZgXpbC0QxLREaHjuInOUhoMQw4k4aBwrPXmDBmUv+/7vq834cB8BlNGHWbC+Og1ADCKTXWLzGJIdGJEQkOY+k5m0LzPxfCk/Ck/dRMMTodIGaEUVXPdMcNaee2x7IaQ95H0EaO59KlIEweEoYJOTgZDHx0wI8ScWQoIln0mgzklYekvmigWhhGan/CEJwRunCJlKGj0ej9IonwKD/ZBo7brJ+++qZezajmNAcIHmXwwx2yGQQumPuhjX5RjHzR5JjnQP9qufnjgHdFqEU/vsMBVhNaSEQ6ktigrkIAv0d9nghe9gMcNAJZjGUDJgkjj3/7t3wZ/GSTIg/bQP30nkWC6DBZ0CUNJf8GJMQkreoSzTQ4YITCzFyz4yZ/8yYFxN25a8TyjCL9LsCqBAis7vH95RVpyRk6LgbBWcjrutq7X88iuMc34YOUA58tYQqYsm4I5PSNowIHHG/AW5MOnjH339KUXPdesI1nBm8ZVPCuRI44u54OxRf8JOKCXEd13gpsxwSwB+sgEp8KSTIand4MvSEtEjYWcHsap8Q+Wxmd6Xbk+kj4kzwKP+lid8BJwgQ+9TEdKaBAMgiEHBM59JnqEnSJgi072CvkV9Eav70bQlfQPHUDW6XT8Rt7R7Rl9JONbCSoaF2BlPIMh3Yxu/S6RGeVtnOI+EzuEnGm7ABoayCjs8Dw5tNxRH7NrjM1FTt1nlt7ePX0kzxXQs5JBX1ouTS5gyK5iC5uNhJNxTT8aE9fCFrWCgb3EHvBectEVxjU06FNBNOf6GW54ju0sD9bkuo+E/01gwEsfl8kLckqfFDklk+TUTCp+m8Yg6dw5kBhGpMRackxHSHS4ZK1+SWV2g7IxkDAOKUed3pfAqtugZOkbZcGw6iYzfOpHN8YsqY9p+PLs7p6S8A4BGjE7xSb5+EZJxWhnIBBujrl3SfpO+onR4YM9FFoXH8swbRRIV7mJAK1FMliZ9bGhk0JbnCypkDhEFPRv//ZvB87ddiy+ZxzneN/Mp3dEDJ7dZPCywZPRV+TEeyJrkQyk3gnAbxQvwwMdoo0l4S2bqCjc9D+e61tOPZ8DKwCA77rJByFsBgWp4MZwX6vkwz4+KtDVFa94xSsG1dNlNs6vwVhbfECi74T/DfiCSYvl1HI322I5pUdqWjkCnC58KRDAKTReMdz1tzHuV3/1V+PjTwII3nG2PA4/Cw6WlQ8+MtGnAym4IUCrXg4sGujm3/qt34rgggAvun1sifzQmfRB34mD6N0z78QZVxmp3unzMRrBLIGrP/qjPwqDnhHqumARI5vRKABdnJFx08qBFuihq+lCdgxD3lI8cu59OI64AI0gNwPf+KFs30mggpH+Iz/yIxEA0KdlOSung11Al6MfPWaH4KyP0ci4XmpcHAfd6uBM4C3jBKeRrmO/eCfOskx1c2D1sQCba33j5vl0LyfQ9xHQiedsnH+BXLOiApEC4GxCY6GyAm3GO0uD3ddH8lwBC+/weWcQRgIBAjzotmxaEMAkBwdYHtzKeNcHTeWZ9Bv9gTYfymEf4HnvPRpn6TgyCzf2tH4XJO+u+hLs6SORU3qXnNKh8BEELUEx4zCZELzz8SGyQgbg1ldf9tFOz5xLB7KAuZIBSYeKGJSZgz472LMpMQxVnLNCa9kr07dTUeoatScUo+gr9xAijpNo7lokyrT0zSh8us7jWtC0VB37GyQpGwp6rfoZJvh7ucFyFJ5Lta+PPPy2P2PWde0oTnDhhT7o8Wy4MdDI6qi0FgPpqLrlr4SHyKkoKKdjLZKBVJ3SKL6aBDldCyz6rEN03YckBPoKv8KbA0nHkBdBBvsyy8apZ7C6B1/3JUOcHTN4jHX8YBwu44kApQACQxm9xl10mKnqi55uPxizfvzHfzwMTjSgi6P2sz/7swM60Qg3M4Do4jAKDgoYGe9G8XW3ngM5ZgSbWeHQwoiuK/hYGcKRFJQhX2hGo4+foGclts6B0FTu0Y8CVhwz/UuG1StAUJKgPT3jNRN2jkAkjNFb7KtSdpx7YxunTH3qoZft5XtnDm54vuTb+3J1nzSV9rEHfGjFrFnBQp6Atz0MBTV81AmW+FNgSDBIXy83/pQ6DnSPHs4255Dz47zYc5wy2OlrOOJFuAka4Lu1SJx/kwDogpP+EkTB6+jxHj3azdSajGEfoJuc0oF9yYSgErx8kb3MyKINLgJQ6CpyWvSsQJk2wHCa0nRRuw7IYjLRUQ5K30ld56f3qIoA9F1fn88vyxuWc0zGWb8BdBaS90TPSkta1koJM4BEjSmvaU6MEO1gyPSd9A3cDEIMtWlOVlcY0PoaTBdjI6q+Fo7A4nrn7RzG+HNxYoCWVIItjGfReEYWfu6Tp9HFCDXror7Fy8jQVOgqdNqvlWGFPsbeYuehGM5oKfTROfDUhrUIwKiH7tGvi/sWLbbFaXE7Fl8f1zm6OI3LBX8KRviL/qR71iIJAnDyS7CiW+co3BbzZfeecR6P4rduoLnwm6ABOYVdnzJa2gcbdi85Xcxv6OvSWO5Zq3FEfUvxW5dONo3NbLcgKZu08GCht4+9utA2iXI67vZWB3I/iFKMZapbBIjA95XU1dd7AH3RPOq5oxTzqPIHk69f1mowOhg693evdjBUusbK/u452OsG18LfB/us9by/K6d900EHLDV49V1vH88np2ulc/A3x6GmyUKAc2b2aC36Rl2i8/TOcs7GZCG0NDWcDEu/ydBaJO+bzUIyE8QBMnNEb/ed8DX+7tN267sNns8Z0o5uEKivegX+OY9eYatXXccAAEAASURBVFkLvdBXOzyXnJq5p3PWIp2fJoHWauJkLdqzXB3VgVwOnUXXpl0BLWrOzJzOSr/MSjtmhrFqQ8aKQOXvscI5tocxdsxCS30HSdXlfbdZSJygtZgJKv2ylu9Q99U/+Mt7m7a1SOoz68Son/Yk+FLa0bec0tVwW4sZu777Za3l1NL8eUn9h3/mBcnazopARaAiUBGoCEwxAtXJn8zOm5V+Wet2rHV9a8U9s9qutcKvr3rmrV+qA9kXJ9XnVgQqAhWBikBFoCJQEagIVAQqAhWBGUNgJhxI0/m2SU3W+k9qmlTcJrlPJ522ym8HJm0Vt9XjVmRhUvWIFk0ybatHvN5REagIVAQqAhWB9UdgJt6B9OWntXoXYbVdZt16+YTvau/ts7ypdu+j+ADAJL7w64Vxn1ueRNrwm20SlyvAzccp1uLjBKvlT7JQPlu92nv7Lg8vtE0iv6FNn04ibeTAxwnsJy3BDc/Zpj1xgic1wIE2unpSHfVJpUt/+gmVSaQPTZNIFzmeZNzQN6m4oYucTmqaVJ5DF56bVP27njw3/SNrKw2TaMwzYDi2foNorb7UNqnKYbV0+RQyg36tPkO+GvomkdcK/XDjROK3SaMTPZNGU8HNJ8l9pGKS5XQSsaPffBBjUr+oOYmYFZ5bzZ4TXD7nv5r7+i4LXzLjy5CT5qijzSa4YSyetKQ/0TWJOqcEXiZRfuDmy5qTGLQS5KMLJxE3ePmi6qTJKbkscjCJQVLy6cv4a/UV19XoKbjBbL1wmwkH0o+Z3nbbbavBfU3K6lwO0CQ6QaIqop9+A8x+0hJHaFKTH4HFczCctGRgtU1igtmtt946kbhxIO9///tPImwR+fQbYJMYPZ5U/aYjRYzx3CTitlpGYyAwnCfNMDXGMa7IzqQaWJPqQPrpKbxZHcjVSYOfHRK4orMnTR44Z3TipNEFYY7tQx7ykImUU3iRg/VyhJbjQPqtBOWXK7ce1+hfPGe/HmkmHEjMN4kCux4duto6K26rRawJXqu4rR43d1TcKm4HhsCB3zULPCdodcMNN0xc8AW2DHnbpKUSJL3jjjsmMogwiTPKpQ8Flrdt2zaRy/YmOWgFt+uvv34iceNo3PWudy1dPFF7kxibN2+OgN9EEZaImVT9BicBKDy3XkHS9XFbJ41DKj0VgYpARaAiUBGoCFQEKgIVgYpARaAisF8EqgO5X4hqgYpARaAiUBGoCFQEKgLzg8AszNzPT2/VllYE1h6B6kCuPea1xopARaAiUBGYYASq8TzBnVNJqwhUBCoCFYF1R6A6kOveBZWAikBFoCJQEagIVAQqAhWBikBFoCIwHQhUB3I6+qlSWRGoCFQEKgIVgYpARaAiUBGoCFQE1h2B6kCuexdUAioCFYGKQEWgIlARqAhUBCoCFYGKwHQgUB3I6einSmVFoCJQEagIVAQqAhWBikBFoCJQEVh3BKoDue5dUAmoCFQEKgIVgYpARaAiUBGoCFQEKgLTgUB1IKejnyqVFYGKQEWgIlARqAhUBCoCFYGKQEVg3RGoDuS6d0EloCJQEagIVAQqAhWBikBFoCJQEagITAcC1YGcjn6qVFYEKgIVgYpARaAiUBGoCFQEKgIVgXVHoDqQ694FlYCKQEWgIlARqAhUBCoCFYGKQEWgIjAdCFQHcjr6aaqp3Lx5c/N3f/d3ze23397ceOONzZe//OXmtttum+o2rQfxe/bsaa688srmNa95TfP2t789sFwPOia5zne9613Npz/96UkmcW5ou/zyy5uPfexjzQ033DA3ba4NrQhUBCoCFYGKwDwgUB3IeejldW7jtm3bmje/+c3N1q1bY7vpppsaeX0kTtYtt9zSXHHFFc3OnTv7qGLdnqk9//iP/xjtOvbYY5tDDz103WiZ1IovuOCC5tJLL51U8uaKrq9+9avNf/3Xf9VAx1z1+nw29o477mg+/OEPN9u3b28ETAVK5dW0OgSM37ATCPz4xz/ebNmyZXUPmIPSn/rUpyIIPwdNnfgmXn/99Y1AKZmfx7RpHht9oG3m9Nx6662h1Bjvp556anPYYYfFoGFGbffu3THLdtJJJzUM/L1798Y5JnO8cePG5s53vnNzpzvdqeFEeZb773KXuzRHHnlkPEcdrh199NGRv2nT9HaRdu/atav5yle+0jAmHUuw2LBhQxybnSB8Rx11VHP88cc32nvttdfG4KvMMcccE9dgC6sjjjginE/OlGPlr7vuunBM4Sq9+93vDgfySU96UvOgBz0o7osLU/4HjmZyf/Znf7Z53OMeF/wEY/jhlxNOOCHw4KjDFd6F32Ap3wDt2sknn9wcd9xxU47IvuSTKe1jhJBJ7cRb+EdgwfVDDjkkZBf/MPh27NgR191DDk888cTgGbLounvcD1/Xb7755ihDnpWBM31w+OGH70vQDORo+zXXXBNYkFFyBjNyZ+/cRj7J9tVXXx3y/aUvfSmwm7VAzgx0aW3CmBGgg1/3utc1v/EbvxF6lo64613vGvpizFWFjNFLdA87gz6blUSXvuc974mVCw95yEOaM844I2yAWWnfONrx93//980DH/jAwGYcz6vPOHAEPve5zzVf+MIXmkc/+tEhiwf+pOm8c3q9k3XAmwHPOfnkJz8ZBvsLX/jC5gEPeECDiSwpZMRffPHFzbOe9ayG88JY/8AHPhDRNMYsA/NpT3taXPunf/qniM4bAL7hG76hedjDHhaRjI9+9KPNhRdeGAz5zGc+M5zIdWjqQVfJ4HzLW94SwsWAZ2hyYgy0BldOtgHwTW96U+BHIT7lKU8JDP/sz/4sjFBl4fKYxzwmzs8666zmfve7XzyTcXrOOefE4PLXf/3XzWWXXdY8/vGPb+573/s2f/VXfxVOBEcc5uedd95Bt2e9H8CJ+eAHP9h8/vOfb1772tcGTgyUd77znc1nP/vZ5m53u1vz3Oc+tzn77LMDC7jiObjiR33wH//xH2HciO7+wA/8QDih692ucdevvy2R1v7PfOYzIW/4Ci9p97//+7+HUfct3/ItzaMe9agIblxyySXBixek2UtO4vOe97zmXve6V+B60UUXBX/ipac//enNPe5xj+a9731vlOFoupcjhRfPPPPMcTdn3Z9Hh1111VXNn/zJn0Q7n/GMZzTnn39+5P35n/95I4Bxn/vcp3nqU58aGMBCWc46/oPJLBm4694hPROgvwUM7OlrgRF75zYBAnuBu9Kvzhn+JTAo3ybPNfc7F3Apz1CHc/muT2vSRljQLwxJwRLtspV2FRxKe7W14AWzgk3BT7mCU3mO58oTpPEqiKXhX/ziF5uv/dqvbe5+97tPK3z70G2c+8u//MvmF3/xF2PsL1gVPoJ14ceCIfxsknJS6ZeSH5kz8keA/fTTT4/WsKtK+/EPubLBCFb4R34XF3ldHMt9HuhZrnuG44K1MoUXZwTGBc3QviKnsNF2mC3mMXiUsnQjW1bgWUBnHlN1IFfR6xxEhtJ3fud3Nv/2b/8WxihHyFILSs9mZuitb31rzI6ZofzTP/3TeGftHe94RzCZGbUPfehDYbhSkpYkGgwwrBmiP/iDP2h+/ud/vnnCE54w1TMaIrEMyx//8R9vPvKRj0S7CCPHkiNu9ovwMfZf8pKXhMHOEVfWgPiDP/iDze/8zu/EgHzPe96zed/73hdOEAfRDJNlA4x9DsFpp53WcLY5TwzaFybHnkPJceLgz0Iyw3N+Mtxh+jM/8zMxgLz//e8PZQbrV7/61c1//ud/Bo+J2v7ET/xEzD5+//d/f8zC4UvBDc6APrn//e8/C7As2QZLxx760IeGDOEts9Dk7mu+5muCJ/Dgr/7qr0YQg+IXtDEY/Mqv/ErMIsCRweZ90ze+8Y3Nq171qnCeBI4sHzKrK8jz7Gc/O/iPsyQwNIuJQyhoJhAhkGO1hMAPPH/oh34oeOt//s//GWUYNfAiv9/93d/d/MVf/EUEPAy6NU0HAnhegMTSY7PN3/u93xt6lb6lg8mFoIkADJ1CpwuyGPPwhVn9Jz/5yREkfdvb3hY6SaBFkPTcc89trkivFgh44SkzTPLx1DQm47UVIZbMC9DBBq9z8MrqB4a4ACfdQeeyH+hyQRZYuw6HBz/4wTEekiFBKs6hVTv0vfJsC7ib6TAGvvKVrwzD1Tj3/Oc/P+RzGjHs0gwzfPGJT3wiZnJf+tKXRgBKHh4T1Pumb/qmWD3iPfe/+Zu/iRUO+FCQkOHPFqP/8d73fM/3hM7q1jELx4IIeOf1r3998MS3fuu3Ng9/+MOD58zesonw4wte8III6rGx8BNc/uVf/iVsB8FmeprNZYMb550dxd7yHM+l511nc7ErZjFIiic4gWx1MkdG2VrsBGOYMZA9Qa/BjNxbbcDWLEFSzvU8pvls9QH2tOVrDPA3vOENMZgSTIMmYSVcnBXGKoMSc4lUmLngMJk54iQacBmsBgMRfM6kJV/KuY5RDRAEl1E7rYnwcbgNiGZnKCXt0X5RM8ecb07kGxKeBg0DJcwMtmYYYaucgcHg7JrUjQCZXeRIGaQN5IwRBj78Zm2JJj4zw2ZZJoXOyKP44We2DW8WRWbQNROrjIEB5gYegQkD8TTz1nIygb/wmtlF/MWg03Y8Qc44OFYLMGTxlPL4zgCMd+w5lQZceOMvs7sMNxFJg4ljASFl4G0Vwawm2Nz73vcOg+xv//Zvg8fk4UMOBR4zM47HDML48ZGPfGQMtBwG9zKSa5oOBOgFqz5++Id/OFZ7WEFjPNOvf/zHfxxj09d93dfFyhrjGD1tHBMwMG4J5pEJeseMnJUO5EpZOsq48Pu///vhMNFFZGxak6Cdmcdv/MZvDMNae/G6GSK2gb08DvM3f/M3R8CJ8Wm8IkMCnfQTuRFsFuCjo+gZ9zLcnVtlAqdv//ZvD0dJ4JSRSw/pC/puFpLxmiNN337Hd3xHc8opp4RdADOBQFizl+juslJEoJQewl/0NjtCAOQ5z3lO8O8s4LK4DXiGvmVzClwKtLMH5Av0/ciP/EjwJIeIfMJNUJRMCggJcMCRTWoMI7/nJ4dJgFBAwvjoHhMbEofU6iX9MYsJdnjGii4y+YhHPGJgLwlY/NRP/VTgogwnXLCMPAogsyvgWOyuWcRnuTbVGcjl0Fl0zWDKuXnsYx8by+AMEgxQUVcKrRjlHJyytMA15RhSlKPyBhlLMW2MLAYo5cnppBQ4qtOevPzOIZYMlkXAKDnJOYVEYA0IlBVs3aM8zER8YMSRLPdxJg2wng9vzqlouIikwcNg47ryykr6o9wfGVP6pzjQ2mcQwCei2gIXItUcS3xnMOUcwU7EkLHvnAOJvwouUwrDsmTDBv+U/hbE0G4rBsyikEM4/sM//EPwiYfBp8yCwBQPeo492RSM8DznnsdgY9j967/+a1w7Ky2tntUED041J4IxTMYYvCKveBD/mfmHCwOOrMJfkgfbmqYHAX0mmMkoMsvBYGJM0h9kgHNJf+B91/EHo5OcKGu2kW73QRn5nCDlyI5gIn5QlpEmml902vQgNKSULHDkBKuM83SIdsLB+ESHaC8D1fik/XSJoCn9AQfl4ekYzvS3ZF/GMkYqB97zYEbP0+3O6f1pdsKHaOYjRryZb3xGlxR9Q+dyhmBVZmXLzBjbQdDKOIhP6SOvs8B0FhP7kZPN4aOD2Ur2AqYCNOStBH3wIUzIGVzYoJxGwWYyTqbhJF8ZwWj3c47MVuIzup7dOqu6nB1J73GUySdsjWHkC0acdXyoDFlW7kUvelGMi2wK/aHsPKY6A7mKXrd8jSI3EBgoJUKHgQwaJRG2wlCMKoaXQbUIq0itgVfU0bPKoOA+z/G8aU/aZDmAqLMZCgoNVnCxwYVgGgwNGpxnM4iUnYEDZgz/4gwowzESveVoMnLgpA54UoJmes0QUQKiviJuBmG4z0KCH9zwCb45KzkujAfYODfIugZz+MAWFowU97oG28Kbs4DJ4jbgn2KEabNj7RfdZ3AI1Aj2yJPsGXSWpDJEDA5k0zIygzIsRV8tiSKbVgi4LnAhD29xsGY14TeyaWWFRN7IJueArDGai4zaC4gxPOhJcuremqYHAQaTQCkHh+zQsfQGuWGA0rP4v+hwBj09Tb7IEr1PJwky2OMXBpmZfLyiPOeAUyRNs26GTzGq6driPJd2wYVeFnA2oyiIZYzTZkY5e0JAyxJWcgY/12AkeT7H1EwjHU/m6CmY6hN9QMfNUsJv2mSPh+hgBr4207kCVjCiyznl8uEFO7ofz+Gv0i+zhE1pC2zwA1mElfEMBmTXUl8OJf4qWCpD3jjeEnmEKeeSLONBuNHfnoNvyatVcsa+otdL/bO21z4yJnDBUbThK440HhO4kPAUe5K9IBAvkXt2/KzJYTRuBX/qDOQKQCpFKC/RG+8vUPgGRApeZIJAl4SpDLjKEFLMWBwaSw5ELL37yLhShgEqYkaIyyBbnjWte0tQLIX4zd/8zcACdhSTNhJUDrVBwM97cCQpLFFpTiDDkzPEoDfgvjDNUsLsZS97WeBosGDMeAbn1HP0CyOXIjCQEH5LDorxMq04dunWbu3RboaJ2UW8KJpIsfnAiUGCEWepJifSQGKQoPwMIvCfZqOti8dSx0X2XIMXnCR8Ydmp90YMGDAio/CxlSV3IrgMPsaK6KLB2PtHDMCv//qvjxkH+Jt58CEsWHrOrCbYXJA+LsRpJFM+JERXMS68B0J26TeOo+WLItf/63/9r1giBZMyAz5t+MyrQWDpu6CJvhWwszyQo2KjV7sJRvifPIjM0+Fmgeiis1Jwi67CD4JYDFbOgMCCMY9xO+3J2I//4WDcggGcYCKPkydZesoI9fVM7/jDib3AQSq6hq6ml8gS590mT/KsF7/4xaG/jIfOGa0MWc8oOi4KT/kfbYMh/mAncG7oV+/muQZzBj5HGgaWGNJPxjjXbfjLM2Y1aWPhrdJmtpX3cc1iezcdD3pH1HXyyQ5lJ7GbyLfy9DVZNc7hN/LpuTAns3gUz1ombfyc1YRX2EU/+ZM/GR/f44iTQ8F2OFiizpFUBv+RNxjiOeXIILznMVUHchW9bjBkQBlIMRLhZKQSxCc+8YmDJ73iFa8Io1T0xtIKH+cwgL785S+PKKSyvqSJQd1PmO1FNSwlcDztycDIMYaRthsQ7L3wLlFqEoODAHPCrdO39MmafjNAFCLn6Iq0xNWg630rzyOsMPIM91N8XRyV8RVOSZ2zkhgYHMbSJh+FgTM+wkPyYULxC1SUSLY9zLxUD6dZVnZkrLTPkp4/+qM/ChzIqPdDDJBkF8/By4w2Z9I7M64bJODFOHNM3st7XIXvyD+8vdckMDLLCYaCQWUmQLvxGOMN7xUeKzqL/hK8kOClrG1aEt1RUve45M363hjEySFHggd0TtEf3X6EjXNyJOhi1pIx6v1qgSwG50//9E/HLBv9JABIX3WfMe1Y+pjeP//zP4d80BUcHXIAG3vt5oQz5DmDVsoIPMHV+MSYtxTYl1Q9C2Y+SCXQBVfBF86Q5faMf7O8xkFLF42n7hXY8sG5skJg2jEt9NOxJWBPh+NJzhFjnm0g0G4sFJAQ/PPV+y5vlTGgPG/W9kU3aafNuGZ1jKA9uSO3dLWxzky21SJW15RXOX70R380ZmrxmllLH27kDHltCC/CEtY/9mM/FrYtfT+rSeAHboKkAjT0F11lhcAf/uEfxoQGu1Q+mRS0+PVf//Wws8g4h3LW+W1U31cHchQyS+RjEkJp6yb5XeXl2GzPWSkKq+yv/dqvRXGKr0RyDMqiG93kOcUg6+ZP47G2dJeRFAOz7EubRHFKEvmiwAwYHEpK0nIUTqK0GHd56lkKR4pz1pK2djFwzgkqDmVpL4wZNIvTrPDW4nZ1z7ttLPiU6/ix8GThQ/xnCRB57c4kum6JkIFTfve5jBvGnGAIg3nWU8Gi2046bikeW8yj3Xsm+ZiuYYRZTmjGWfsY/CU4M8m0j5M2QVJ8DQ+6Rt+TD7NBXSflF37hFwIvq0Vg9su//MshI7/1W78VS5h/7ud+rvnd3/3dKIMnGLTkyQoBMsMBmPbEaeHQlaWs9nASWKEj6BaOoDbjLe/8w4vjxwDFX1bgCFaZBeIwveY1rxkEXtwPt2I3cBrpIv3huT5qpI88f1YSPSy4XlZ0MeThor14Ed/AxV5Qgg4S4IOBY86P691xclawKe2wEqvoXsum/+///b/BW/iCbHnVwnU8eFayQc3WwtPKEVgay5yXJdImNAQJ2Vz4zTPxq76w6mbWxzi8JIDsNSvJOV0leAov/CSQAw/jgkkhH9CCI93G1iT385iqA9ljr1P8PrFdlrk4t6ynpqURILjnn3/+YEDkGIqIGzhqqgj0gYCl0oyPrvOoHoOGDwiQ18UOOuOEkc25NODWNP0IMARsjIaymgQPSOXa9Ldy+RZwGsnBYllwF57v5pcZCbMWZMBMHD1NXzO6JDP7i5PnFON38bVpO9cOMxKLE+Ozm8w0lsSgtzzejIalcmYm4WQWg55ZyljHh4sNVFhzQGctaVfBCz+yARYHiLUZHosxkV++TeF4VlOxIeHDiSl44Z+lljPDlF4ToO/yDKcItuxSMtt1ujmf+NPsePeeWcQUhpxvWzeN0oXkEZY1NU11IHvkAhGdbtS2x6pm4tGiOwYAUceaKgJrgYCoom1xMhgvNagoZ8ApRvLi++r59CHAQZT0qxnqcu6Y8VXOp69lq6P4QNrJkKKvLZFjdAqslBUjq6t9PkpzvL1e4P18s9scRkvnayBq3/4/EH7c9ymzm7NSfAR1OD2LnWt5nE9OJNurm+g9s5Tke5ZmuLttrMcHj0B1IA8ew/qEikBFoCJQEZhyBBhNliOVGWfntppGI2DViA+c1LQyBAQpzEJ+3/d938puqKUqAgeJAEfQtjgJkJlBX2oW3Wyk2ceaKgLLIVAdyOXQqdcqAhWBikBFYOYREM3nLIrEF6dR3kqj/DMPUG1gRaAiUBGoCFQEOghUB7IDRj2sCFQEKgIVgflFoDqN89v3teUVgYpARaAisHIEFi58Xvl9tWRFoCJQEagIVAQqAhWBikBFoCJQEagIzBkCM+FA+sSzTxLXtDoE4OZzzb7mVdPKEfA59YrbyvEqJfGbD21UfiuIrGwPL7jBr6aVIwAvsmqb9oQHav+vvhcrbqvHzB0VtwPDzV1VTg8MOzxnq2nlCBTM1gu3mVjC6munXgSu76usnPFgBTefd57F30xcORKrL+mLeV5At9W0cgTKZ+/LO2Yrv3N+S5JTfHbaaafNzM8frFVv0m/lN73Wqs6+6iEzvppY08oRIDs+WuODIIu/Mrnyp8xnyYrbgfV7kdNqi64OP/JJTqttsDrc4OWjb+uF24bkuVaXf3V9VktXBCoCFYGKQEVgzRC48cYb48fmH/rQh1ZnaIWomwnyO4s333xz/NB693csV/iIuS3m9z2t6vJTRhW3lbMBXvP7iQ9+8IMHX3Ne+d3zWZILsnXr1tBvfuNS0K+mlSFwww03NFu2bBlMBq3srvGVqg7k+LCsT6oIVAQqAhWBisDYEWBk2epM2uqgLbiZEaqzQivHDm4lVdwKEvvfF36rcrp/rLolCm5VTruo7P94veW0OpD776NaoiJQEagIVAQqAhWBikBFoCJQEagIVAQSAjPxEZ3akxWBikBFoCJQEagIVAQqAhWBikBFoCLQPwLVgewf41pDRaAiUBGoCFQEKgIVgYpARaAiUBGYCQSqAzkT3VgbURGoCFQEKgIVgYpARaAiUBGoCFQE+kegOpD9Y1xrqAhUBCoCFYGKQEWgIlARqAhUBCoCM4FAdSBnohtrIyoCFYGKQEWgIlARqAhUBCoCFYGKQP8IrJsD2f38bP/NnI0aKmaz0Y+1FRWBikD/CFR92T/GtYaKQEWgIlARmE8Exu5ArnTQ9nsvfqz2y1/+cuMHf2vaPwIw84PSX/ziF5tdu3bt/4YxlPAjpZdddlmzffv2MTxtdh+xUr5fawQ+97nPNR/+8IfjR3rXuu5SHx669tpr40e9S17drx6BSeWx1bekCX1y5ZVXxo8g96X/6Us/tHz55Zevmb48ECzcc/311zdXX3114HGgz1jpfV/4whdiHBk3P23evLm55pprmltuuWWlpMxNOX173XXXNTt37pybNk9KQ0fx+e7du0Pm9Mt62Dej6JoU3A6EDrr8K1/5Stgc/7+9O4++far/B76/yywic4Z8hCISmcpQt8gUFamoRKWUlKalQfMspTlDgytRaVZExI0UK4RSlFY3raxGolCt/vidx+Z1f9vbOZ9zPvfzOe70eq117jnn/d7vvV/7+Xq+hr3f7/O5N998c4HxMIHDXXfdVeO0GDJOUTeHL/73v/8d51BLRN9LvaMnMzlTSfvWW28tN9xwQyUSY0VytFiUYP7973+XhzzkIeXrX/96uf7668ujH/3osswyy8ykGottX5deemn53ve+Vx772MeW5ZZbbuT/HJmTss0w6bazePzc5z5Xttlmm7LSSiuN1MewMRaH8xH8A1PvAufcuXMrRg960IMW6DQF7uuuu66cf/755ZprrinLL798ecQjHjElnbpcmNLFTWOL2KuuuqrccccdZaONNmrOLFwfZ2q+w2Y1v+NEbLWBdPfdd5cHP/jBi6w/yglnnXVWzQNywdJLLz0MtjI/uF100UXlggsuqPFy2WWXXSjw6jePH/3oR4WfiOlrrrnmUCwGNdA3ibjUr91HP/rRstRSS1VfnMn/8PyXv/xlufrqq+siaYMNNrjP0Io1+f9Xv/pVWXfddctMjnufgRbSL9/61rfqJtraa69dVlxxxRnXsp/d5aJrr722PPShDx3Jv4Yp1Y+3bH7bbbfV+g13Fxa56aabat2J5yussEJVq6u/GHrhhRdWu6y++uq1vunqb2Fpo4tvrrfeepP6Vffaft8tqm655ZbaT7dGsHhiM2Opmy3GcMUcFhW5/PLLy7nnnjtvw/phD3vY/Wr7LlfVKmKD9cAaa6xR+Mj8StfG3X5sZv/gBz+oG4vi7Ez6orHV5xapq6yyyhIR42b8DiSDKRbf8IY3lFe84hXlyCOPLM973vPKC1/4wnLUUUeV17zmNeXUU0+tIEvsZ599dsmdgC7NB3//6U9/Wr7whS+MvFMtCCnc7cILmIME6d3d1K7dNfL95JNPHsuO9SBdFoXjdi1vvPHG+6hqYf++972vXHLJJfc5viC+2NF717veVTdoFOirrbbayGqwv6KA7Wdix1xShIliZmEV87Tghts45c4776zJEr7zc+fNZsCHPvSh8rWvfW2B7JrPFDZizVe/+tXibtgo8V9y/sUvfjHlu1sKmjPOOKPAfWERxa2nb1rBvSuuuOJ+x9s2wz4rdm3QdvvuXif/yiPzw79uX+138/rhD39YN4/b4z6z91e+8pVaE4xi7+71i/p3tY7ichx3Z8Uud9otdKI4h5cNmuc+97l1Q789Pr9Y2ky2sGnFZsRJJ510v1zYtnmgP5vr5z//+fKpT32qbp4aX077+c9/Xmuh0Ie/iA82PW6//fY4fJ93G12nn356Ofroo6flLzgvn3784x8vr3rVq+oi6z4D9b7Ike7nvPjFLy5HHHFE+djHPlb41KIicDe/2bNnl8033/x+C0fz0MZGO1zjrq84JG5985vfrAvo6cy3X2xt+5Pff/KTn5Sf/exnZabvdqqhDznkkDr/cdcR7ZwW5Ofh277zod2OO+5YPvnJT1aCSByHHXZY2WOPPcpb3/rWajSrczvOyOR9st3S+Rh+sb7EbpS7taNi5m6vxHXaaaeVt7/97WWLLbboi4+C9pxzzql3rE455ZSy8sor13Z2imO8JW3XuC9Q9x6UoL785S/XRzVi53X//fcvT3rSk4rdzAUtfMtdDQGdXqHjKHrZcPj+979fdwQ/8YlPTGtH0Hh4g7cLM388YrvrrrtWH/A+LnEHRgH/hCc8oTzjGc+Ykl3otO2225YNN9yw2L2eik3HNZ/57RcXxP5ROaHYkENOOOGEcvDBB488LN5Fjhk1Zo7c+Xw2fMlLXlK22mqr6pvRBRziFcem+m4BceaZZ9aNwuOOO27g5eI5XGYaD/oP8nO7/YceemjZZ599FmneDgR1yAm4jANzw9rMVHzbuJeXQg7r1V277bZbWWuttWbE1s9//vOrD773ve+NIcqb3vSmOq+FIeeFUnj9spe9rNaXsXHqzpPcPLu3uNlvv/2i6Ty+DopD7oa99KUvLQceeODIsWpe5/d+sHnlTuc73/nO8qhHPapuZnU3tNxIseD3hI5FuXrMQtKTQ94X5id3Yr7ijzt64vTTnva0umjH+VZsdlgYr7/++uXpT396nRd7eQ2KHe31wz73i63tNe04g2zetp/KZ3HVkwY442m9JUHGsoC0+PCy6yO4IYa7IB5rcSwMp8j1WBFnmjNnTt0pe/zjH1823XTTeW0Ywe6QnSIE9RjBrFmzagHVGsguhp0NO2QKrHXWWWfeaTtPxnTtqquuOu94fLATYRfK4xh03WGHHepCCyHslFx22WVl5513nvdoEeemr4CkP/rZ1XCnxeM5T3ziE+st7OjfItqjVIrUTTbZpJijRXQrdi88+uvxBjjZkfYow8Ybb1ye/OQn900A8OMQdh/tKHs8QsCkl0cB7HqZu8WAd0nGDih8OHCI4HrllVfWW/va2Sn2WLFiNZyaneyQ2UVzLTwEtxCBAX708MiIArn7GFO0VQzCFS50gQ29FFXbbbddNKsJUZFsLt/97nfLIx/5yHmP0pozfWBKn1122aVyzMW4oE921M4Cmi3h2PJCW9ezr3YetzDneIQML+zmsw3Rj+DI/uyNb7AKHNhD8Gg5Zq4w9cJB87M75zPMYGCMhz/84fUOHQ7hiPlMJmwLb3ekiEeabQ6wl7l84xvfqHzy+0f6bLbZZmViYqK2jX/MC8fMi7AZ/d0ptGPujo+7RDDx+CsMtbXA8vgkMZbH1Z1XRBjfpgX+ejnuzrfEgqv09ohL609iAj64G0XPfsWIGKE/OBpbYYQX+mRvd2/gYbGw/fbb10RdFez9Y9EmoMPcGHbSt9566/KYxzymzknMcEePvT3ya05sxA765NN2Nr08DmZsvAkO0Bf++BmiD74Q1+y+++4VF3doxBpzNid2hhFc2Moc6fq4xz2uHo/+4l078QbXFIb6wEPfxVi+zmf0S++uRMxw3C4sPPFwyy23rLjBiMBJ38YInxGb2Amf+C3uevRIrPdYLUye+tSn1vPs0hUxnC3giif4RKKt/n/84x/XGA5TPBNDxMXvfOc7tX9x1/yMj5c4zA76xQM2EP9awQntJHc6myv7xwIcJmwCe1yd6PkJe4l5RL9yj5gCf/zia/FonDY2a+iAm87z567wC/bHCWPiHA7P6uUzGPARO9d4w6/Fw66ejvNPMRtvdtppp8oXdhEPxTJz/fa3v10xgkerZ+jkmHxh3ub0lKc8pca9sAVf0J94RC9Y06cVOtDHkxjGwWmx0Ny6AkN8gn+ch7fPsPT0Bp8Rs8UA/Ahhf5jQJfzPBgqbmCs76cv14rM27IhLsBbPcSniun61jd+G4y//xYsQm2jOi21s5Hr1Q4hz4qOxXWth0IqYYnwY7bnnnvUUHg4S3Ah/hCO7e7W205c2eMYH2Yy9xL45Pb/gg1/60pcqr/XhXCwetTFPOa4tbsV5x/ik3E5nc8dfucr1OIn78McXvGW3sCPbtzFbfjSe2AA7/bS1gH7ELDHKwko7NtcuYkLghJviAt3kDHizuevNl68HZ9la3II3rmlnzvShMx55fNFxOkcNhEvugBnDWG3+CN66NgQGYgfMzjvvvNovP+zWrdHeeOb/wQ9+sOpiA1HMbYXd1ITPetazag7GVRu/4rPxLCDV0uatL7w0X2PC0XzNjZ7ymp8cEZxRX7CV/uUH9hdz2Izv4o4+cRxeXXFtYIz3fnZGjIGjBG58WG7FTbrIoeqJ8GU6qh9sdPBdtjWmdoTtYRU1ifyF43E9zHAFR9l0ohencYa/94ut5qcWj5xWB+n9Yy4RPyJu6EfNS8QOucwY5oKj4Y9yrznA1Dm1vBo9RFyBrzH0Yy70c514wZfwi90WB5nx30ACBXheDAdEDiM4K7wcc45YFEj+2iAXcigutRFQOK9A7C4IggoeCjrXMWgUsfpCcs7jsT3OHcnO8de+9rX1mEQVRYNriIRGB6TUP4ezG6QvSdMCwI4JsjO685ydroo8SdjCDDmQkrNzMM7Keej/nve8p+rHQfTDIdpkRA/jmJu7f4IvwtGNwwoU5kM/hYoCwm6u+RvPIsb4Ej29fBe4OaAga34c1neBiWPFzpyxzcnYkrRCZKLnmLHgh7u7lxxUUeE8/SQyzsmWrrdgUeTpl3MpYMy3n6OwtT4++9nP1rnom37GFyQj+Dn/xS9+sc6JMytO8IID+12mRGd8AUkgcF5SEGg8Qq0/gUA7SYvdJA52Eahc9/73vx8EtR9Y00vyNA9zZVvJR9JmI+fMT3FFV0HHfAQG/cGbDuxrUXziiSfWohE32NSCTCEhYNOd3h4RZkfn8cli3hyM1a/o0A42FjuCFbtEUldYw/Piiy8uc3qJQXBjc3ww71bccXbHQpFgbooONjNvnMFd/cGCvpLsG9/4xroQd4z+Ehl+TPQ4I1DCwRzZDT9xV9LSFw6znx1swRdG/JH+insJ3Jyda4UfmC9+aW/++G5M/oTvsFBs4Ymkzg78kzgnubEtPdjL3WPj6IOd4KX4Vgjqx6KJ/nDVHzu5ll/AVEzAKz4pCeOkQgyW2sGWHWGPJ7AxLr+hO7uZiyIEjsZ2Dd80B77MXvpvBZ7syl4WEHSHXfs4KNvhUcSg9npjuQaWrvNd/BVX6KAQwgEFo8UBPcyL4Lc2eGLs+H007Ob2/Iv/KXTgpt8Q/cNR3MYPPIIpHBQIMNcXnsAK58VQ3GILmNAHxhM9e7EB2+Ek/9SOTnATG/knPzcPWMgpfAJXxHVj01fypyfcXUdPL/MWj8yF38oNn/70p6sPiRt8U6HFB/TrsXX96ct1Fons7XwUP7AwPn3MBa7m4ry8BFsLbAUHf8AVvDZ/PiEX0hFG+jFf/IK9OIyL+sBT5+nO/njc5ZBHDuUJcdtYOCXnsWnw13dt5GJzYys68V8CR4/2ifO4zAf4GRsoYKMYq417/xjL3HGAzdlY7BHH8UHhpThmG/qrF+RuxzyyzSau0U4sFksU1+zPH+goLnj6CRZ+TwUfumkHF/6GM2zGnsZ1zDwtpORHPhccwTf+Kx7inPPw4jtwpw/94G7sWESKM+KLcbWRK2Khxr/01Qp95SrzwAf6KcD5hMUBoYNcoh/25MNyNh6ah9zlGK6wkxc+wXzvvfeuOsAD72CAT2KEJ8XYnJ3Zwvh4jMPmhqP8mt/jbYsDvR3TlxiBb7gkvovZ/A8vxF7n+bH+9W0xpbbg7+IljMR/PtkKvdj72GOPrZsD/D7y8mc+85m6YWVOeAEb2NtEMKYcwAfpT08xTj2CE3Tho3SBvZghNpkzTmkHF8foxu8sTGHjLq/8Ru/gbVu3tvr77Bq48R12EU/gGvlJm7e97W3Vb9zl1d6Lj+OEOcMFn9QK9GVr9jMvnORXcLGAdh5v8RWXIz/Ag2/hljilP3MT0yKuwAQ+rdCFneBiHPwL24sLFkT8Ht/EKCJHij10Z0MitvJD89cnu8AYH+GohoQ/jHzHOeM4z8bi/OzeHWT94RF7mtNED0vn9QsPnx3TLjbrqwK9f9iTX+OJOYnlcBR3+L75a8O/1TXmA2vx1BzxBW+Nj9t8kDhP3vzmN9fxxUAxwZzwEk7mIr7jppjfYlMvXgT/uceyY1JcAhBciM9dYWDGQXIJwW6EgCcRaa9IkggEUsnNbXGOjewCpqQUoi9Ow1ACOrISgQEZBGWFTVc4lGv1K9Duu+++NRkxOsd0jSIaETgP5+SQCMABJHmksoPheotObRUlZG4vsVlAcyi37M1TYO0KnDiNQMSJ6GN3jTN/+MMfrsHVNRFcfBZgFAOw4CjusFl8cWQYWjxZzChSBTA6S3LdBKZI4qSSj4BrR9X4HJEdBBgBR190Mq7iQeIi5u+l2IGBNuwgUMG/K66HC6flVMZz9wsPFOY4QQQ4fcDFeTpxesUNG7vOnBUr5itpE1gKEgIQR8cruAvGjgtQgoaiVgCKzQ0LGgW+RBqFoqCkH4l6Vm/XDsdgHYsfeLO5wkAxIoDTkeAAXNhbkScJCE5+k4VHAjEbCSiCEo7bgcddiZ2e/QRuAiWBi7Hx2PwFVYEbNnjN9uZH71b4gz7ot9dee9WkK4DigrnhK54IhIpxvBBA4ccvQtgKZmwHa3ixBRvTzfiuY0PnvYjEhVNEseF3vfwJ57rCJ/gj7rMBX9MPG9kdlFTobjwvY+GIZEVwEC7GwyF4RVxQtMDLPAkbwA7X2UFBBG+2Yj+2N67CkB58Gr74TBfncB9niF1kNjU3i3g+JmlKOK7nd6ELf2ILc9QGhl2Bt3joGr7J7jYf2AU+dIS5GAm3fiIpekwK1rDgQ7gCM7wkdMFldg2BKzzC7yVIHNfOmMYWA+jRCl0VjGymIIYxbPEi8oN50EdRJQZbCOCnBQoe8jM6wo2/icnwkSPYVHHnBWdzE1eIa4xFZ3HCXI0Lm9hJ1wc7uOvoEUv89/spcZgoNujhWvHN2OwPfzEKBuzl0TjcUViKj8ZpRRyw8Smm44J5xIIMZgpAmE70ih8xS2zgn2KUeSh2FIX0wCv9KOosgMQn3KK7Igi3FO142xV90RH2uAlbj8/xYxI2oStbzOrxke/YxAkRl9knYhtMxD36678r7EtPi5joX0GoH7GYD4mRuAxPtjVfGyPyGbzMmw0dJ+KGesE18hr+ydcWeF5iPnvilHHk6hA68nk1BTuIv/xXfI47IOIv/9avYo/eCkaLTxjjm01xdpO3oh4Ry/gj/PiXsXABD7u+QR/zwSV216cx8Ve9EbnEZ3izp/yKZ67hb+wMf4six9mezs6LxfoX42AgDhJxio4w5Jf0gt2snq2Nz+fgCntj4Ih3PIeXMbyblxgKK7ZnL/HPXPiT3OG8jXFjEfGALhaG8gy74joOim/0DqEXjuGmuEfieraPY2IQ2/BLIh47Rj+5i/764iP05u8RX/GSbfEBv/BSzWg+juO+eiP0EjfYAme6dWtwsypx7z/qgliUha3b82wtp2rX8oMNHI96CH5yEX3FW7Y2PzdYYIJv/Iheahs+x87syO7sjI94ItaLaRb9/E7cxBUbNf1EPlNrwlJswhM214drcEdcl4fFIHjTv40F+CNWawN/eIt/5k3M1XzUKvQU04w5t2dLAn84yvE4iotyGd30YcxubG3xrJ3c+w8MI4fjA/6p0/mya3AJ1uwvZ8k7+IKLFsxiPR3V3ngboq4Rrwn85cs5vbqTHeLOI/9wTKxZ1GXwMxUPwMwEVIHI7hJCEcRnKMkbGez2+AM8jCyJaC/pSUScNxzTtQjHsAK584IocjGcwKBfwQCpkRGRJQIvju06JOeYgofAZ4xjjjmmJlK7PxZhHCOCvATPCSUq10i4xx9/fCWggsuYgjvn5WQKAJ+7grT0QlB3z8yXcEY7j4Ka4BAOidicQKB7wQteUDz7TTglvQU5BTQ8OBaiI3wULLXxvf9wPo4NI84aC1enOQGM3XniSPTk5AonAdp8LKjZkg7aRjGg+BRczKEr+oHjBz7wgYqL8xxK4SspsLG5st0rX/nKWrhrw1mNddBBB9W5OWZ+Eh2M6E8nweE5z3lOOfzww+uiCJ6CjV0jwda7JPaRj3ykBkT9SC4CqTsKfq/LTopLnHMXuRUB3JizeoE0AqDxcSjsayHqsRt/SArHiESu+BPQXWuOOGfnCrcIbOxEGrufzO4tmLR99atfXRc62uj/2c9+di2CYhGib3yAYVcsIC0WQy8LpuAG+7oOhyQXfhJibsFBx3w3f+/0ttDjS/F7D5zTXsJlX3P1Q3MFFxtEwpPk/F5TUuiKO8eKpNe//vU1STlvlxkP+J/EYwc3xDg2caKINL5Cxm8OPWJJ/K5FUYIvzklK9McTOhOJ2NwkZtzmIyFilnH4IdzwVeEZBQJsFZ38WX9R2FiAmot5x2OSdHA+FvnGc10/gbNz0R+dxTI2DgzELHhLVF3BKX6Aq2wVC3a814/CwuLNvI1jvBCfFbAxNlz1x8f8obR+InFKqLjkjpP5E/5DP/rjIr55KUp8Zw+FkQRMFJnGxfuwj+P8BOa4AD+xwFMF4hbRl+Tv99/sTPymyCKF/4t5z3zmM2ucZk/zEefEu1iEmrP8BB/+GzywKIWhPujOl/BhVs+vcRnHWt+Bp9yCF/zNOCH0xSML1AMOOKAe9oc0xG0YWCBZCOGZGMM+7KwAj40R320AmAcbDhKYiBX+oJ3cRWcbVvKVBYt4LefCFB58wiaXgj0ETxSicKID3eQ6MdE1XWFn9qN3CDzhJo6JvYTP8meLSrHAXQCx/kUvetF9sNQWZuyBUzhIYCO2imFytO/iodwtZynC6S2nwwF3jM0WeOoYvfgD38Bxc6M7LutD3LCYZke5nY3EG/lYgS2WsQX7EQU3/rM/rLtCn9iYNXcxEo/pNnfu3IqDTUfc98dcAivzImol/MBXC9oQ9mIn2OtfTI47JrCziUI3+PEROUgNoz0fU1vgHgzwTn0ldrkmRO533NzkVXfm5M/YzOFz6jZ/zMfvEtkLvvjP7uZJxAK2V1M4RocQ+Vwdow1e4wbfk5vwHT/Ny0JLPCDsRafgnfmbB75F/IjNMTnaH3yEoWPivnnrj230hQ8h9FcT0B8eJOpWvii29RPXdcUx/kraWOs7/XEgYlnEfk/UqW1xG1/Ffb9FNS5+80M3XvDVnHGOvm95y1t0W+0l97IN/yX6tsmHb12hn3oaLu9+97vnndYvXtrcwC12k3fYGM5dEUflNrEUbmI8LpsjERvETHUbEU/V/7HxIGd64Tm78kv5Qt41T+P3i621s3v/gbdr+Ym6zPjiFZ/DT7Wm+A478cfTD2F7vo5/8IA7HtEvNjEMoR92CzEneuIXzsNcbBIP9GNNsCjLAl1AMibjtCBaVHAATsOIyKHYVERyZEnNLgGSIk1XFIh2EtwJQSgJHlkYT1LnbMZDGndsFHEcw06KnWbGdS0HDVEMSgacBJntMCKhgGkOxlDQ0ptOCOV6RFIwKAj85S1J5eUvf3ltay6t6Me1SMvRQhBQ4uAoRBtjc17BSuA3t1Ycs3CDk37hqH04ats2PtO9X7vQi118Njb9FGuciBjH7pV5R9Cmm8DeDYra60c7fbQ25Px2sSJg0NmxdgEqeeCMObaiiKG/nXUJh7BzYGl+AoCdRI5rDH05FvNSMEgYAhS89ccedOiK/lwH0yiKtHeM0F2QVvhKoCF0c84Y+oAPHEJP7cwP/7XrJ3DGJbqFxEJQAUDC1nTqJ+Zks4OOAqGC0UsSMm7MJfrp10e/Y4ooRVDYQBtFqiIjdJnVK7D9RWaLGNyhg2TS2rntW2HHvt3zxoKbpNIKPPGVXxM2YQM6EHrY6PAe84v30FE71+En/wsbx/USgJhhFx2P2FMhoh/Fk0LIxg+/EDvEJTiwOXy9a0t/yVbcUcBZzCi2FPhRnBhzkOjLXIJ32kXCi8KjvdYiQQwVG9sYpA/4WlzSjZ+PIhO9Ow18ZpDwI0UZDCXXEGOzCf7DmP4Kb3fUFG/wUJjHog9W2nTto53iXVtFpbjTxhzXiDWw99m8jOs77hE6WlDaPVaMW0RFW+flD4sGRajfKFmssBmcbNaJ/wos/KKn/lzf6qGfkLB/fPfumMVH5ELXyzuOhx3FW0W2O0dyD/9hy+AUbHz2mkz0Tdewh/5joRv4KiTdsVOY44y42cYx5xVfcZ3x6E9n/Y8i2uFqG199NmdzhaX8H/7c7dM8nYsFvfPiv/mwh8W4NuwgH8rR+iT8Fpb4zubmaBPGefEQjxTUNhnYm18SeYOv81Ecxl1xiB6u1RdMur5rXpNhw0fEE7VH4N3mdT5BP3VPV8LmwZXIJcGDyNkWfsaBjb4V/op1uRRnLf7pgFvGww98D2EvY7QSY8DBgkZuEvtDBxjRWU5wPsRxcUN/rpV7xWx87orYxPdsDISP46nFhoUbW7AvO8TGSZeDYffgd4yhndwpRhDfjYd/5tYvDmoDGzEkJOrWqIni+LB3/Zs7joafxzW+iwcRE7TFs8hFsKMrfEN/1/oMR3p68XXvIc7rJ/xOP/wQNoFTtPXu7pzr1RytuF4NwldaDgYn2rbx2RjG0ybGCt3ED1iEwBgG0R+O4q5NC7aWY83FxnBI9B3fB73TG++IPnzHLfbTB8z4ReRS7egu5uIcnvMlsbjdtNEuxLyMIR4Ej7yLA+xjnEVd7ruKWQhmA2Av4HsXaF73utfV4C8oIZNznKp14FBdEecRE7tv7goqLvxeUEB3p04SYVR9KJYlSG0UqYpaRZTEIdi1wUZxgVycdU7v9nPcJUAGQdg18aiB6+iHhMayY2QRKWB7tMlOmh3QfiKQIHQIDPQbQYPeJLCxGGr1dI7TwYoDau+lX9cME+36Sffa9rtAoGh2ly6CFTsZP4JUt8/Qv52raySaNoho146FDySCSvQLB3qwBzFnGLT92zQQEGAZwQN22sYYvnNs7QQsEufql84/0U97WHvHvesrAoVx9O8F58Bau+4Y2g6Sbr/awSP6HXRd97hdSY/7WQx59MhvV9xpw20Sc2iv6+oJ37CJdsEzuoTAW5uYk6Tnd1B4GoswO8yt3eNa766jS+AV54zN5mzfSoxtnEESdgudvMc43WvaOWsjZhx22GF1x9LdEQlWoal4DVuLObFDbuNK0enOK5310fZJF8WdO3Rs4W6QZGbh2RbHXb3a721/7fHuZxgaX3xr7UZvHIJZ21fMRz/8Ka6PfrVt28fxeA8OGS8wd67lDbtLygoD8VtByP/cTejatu0DpmKrgtWC22JTQnedOYa4BgdDzy4/LNydw0kLw7m9xZGiINrpl2+wKT+xK01Hd2Xobmzx3WZU3H0zZruJErrEe+gS3+O9PR5zhZXcgx+e0IixPXLpjk9X4rru8cm+u8bYYrYnV8zXBqAdeQWQp2rkr5DgTmDkeD9+RPvJ3ts5+xzfg2vGGjSntr0xXMMmFo/uEuIIoafcpEi16LdZxmbuymjDfy2aYj6KUpu/FibuhNuYwwl53Z1iedw44g8foSOcFJY4K7a1woaD5kAf3FccuwtiE9XTKxa3rf+ZW+De9h1cdz6wa8/HZ4swd5LFXHMR523MEZsF7tK6w+SYDRM1T/BZm27ccozEceMTcSR08j0wbWuYrp5dO7ouRExyF0d94U6xfmzM8lUx0sLX2PQWN9qxow/HQs84Nug9dOvXz2TXxHWD2gw6jlP0V/vAO3CykeVcu0HXHWMy3Nrx2uvimvaYtoPmSzc8jHoo+g0786upSNihO74+JjvmbrrYi6NisjjlCaSWo4P66OoH4/Alc8NRsS843K8fj7S64+rmkJyBl+4sdnNUd6zunHwfhHX32oX9+/1v4S0kGgNY0LXrZtfPDo9FoF0BO592Q4MArcqOKYyR3fPT2kvsgjdHtBPukQ2PK0h4djglDsHHdXajkEnCiP6R1E73i3uPPlmUeqzGrou29NTWQmmitxvvZWFqRwMxBQXnBGu7efS2k9YVpOKQ9HEnNERCstsSd1/0aUzzoTO9FJ2tcDQOItFpzyn0MSiBuVYb42vHkVoxntcggSlnMldYw4BuCqgIhu219LADoxB35yDEnSh3BdiMxFzbsc1Jwc4mIYori3LJXPLVnu0UBXaaCZvacWYXNpDs6egOUeBiR8sjSrN6d8i43OZ0AAAWiklEQVRwI+bdjh9jelckdIOX49FegYkn5knY2GNDHuEwDzJsjNqo8w+eWmAoMEIUBubGFiR0iPPddwU9ffDKporfApm/F5u53p01OIT4DDcccS2xY45/MZ4FD+wVQyHu0Aj2bV/ufuIuH8V3OnR5F9crFOz2tTz3WbGgiGPjuJvkGpxyDP4kdKtfBvwTiaNNBq6LV1yGtzBiV/3jG57HXXO4aGMjgy3cucBn7RUEYhox99jsMCbfE9cspMUKMUVh2ZVBc+ke736Pfuw8K6LFFa8QMXBOb2PMDj472ZxTWIcO+nOXzg5sa0fHB42lb3FBnLRBF305zkY4wS9h5lE03xW2YjNswv+156Patvbx2cJTzPEkSTxm2uoDbwt7m4Qh7nSau0dPtXXXE/dsgk30YhdxPDhuXBuH7pJbdLA134MRP6Y3TPGArb3oFLauHTb/mJeY1U9a3eMzbuILnxK73CGjD2zdrQoxnhiMZ1OVGEsfuKpYUrSLNXw+isXoVw6NuOuY68U2+avlR7Sf7D3Gjn7iXRyxiBM/2TDE4obghGvb691Zcx28ujnZPLTFRS/2j8c3w3/1KY+IgYpUOdu7u3LmK8aKNc6LnfjCb9Umjvkutra+Nbe3IYHv7BJxJubi3V1L+Qt/bSTRUzsxL0RMgIEcGYJ3fEAclIfo2HKuxcU1eAMbT1DJFzZM+KZYo1/+Lg7Li9r5Hj7gerwV2/oJfljssJdHX8NPbX77bux2M7mrW78+4xgs6KQGtPiVl2w0iWVsYPFLr7j7GNe17/rgx63+w3Ro5z6sbTvWoM/6s9hiI3mrFb6GV2JTiMdQzdmdP9iTrh6+d4/F9e172yau6R5r27efcUQ8FvtxNQT/xCS4wzd0jPOD3rXFucA33rVvdYrrtVdr8Sl+HRzF+34cHRRb9WcsY9gMk/OIvtXkuCumxzy6uphv/GzJU0/43I2LtcPmH310+2lOL/If77mdM+ZpWMx1d+QMydBdYwuEjKI90vrtg2RmB1SwJwL0rF6Rb6EmaHfFThtjCzYec+CEg0QiRgSOazeXsTmKRC3gWKy4u2g3li6SgODrsVq7lxaUdoJ9Vkwin6ApEQj6kp+7oRKPvszNorifBLk9R84xzNP1Hn8TQAl8BGdtJRoFJ8cy/kQveQlCigpOprjhfBKTvjxSR1+JU5HTCscRKCQp/4eYgK9gMo7xWicwB4EYPubrN32CnR/KW4AJjpKfQlAB0iaOGFOiZns2YisLIkW/363Qg7TBPq7THye2cPH7SXaSoBWAdiUFFf3q30LHjpE27GYegp3Eg0t2mXFL0YUjkqjPno0X5M0P3v24GwnNb6s8DuiuAKy0dx05rHeXSnEKd/zRDy7YHccP3wPLLr5wbY/F/L27k6VfO8aKKePpl08owIlrFUL67yf4ZWEnGeMmnln84QrOSMyKKn/ASb84q0DGt9mzZ9eCB65hb+PQWRGOf3RTiNLDGLChTwhu8SPj6l8Rpk0/8fs683UnT6IVT9jZ/83lWn3468uCOttbkLKPMUjYsMWCLo7HMfzBLXcW8JCfmB8beYUtFFYWC/CxIYEz+GfOYhHsFIPuXPjsZQz+psByPYwVPWygQHXeAorufIUPW4AYoyswpk9gSa9WP+3hyHf6+Q99xCX+7fe/YgFf4YPiidipjaLF78dtRvEFWIkNxgt+04XOgWFX19DF3PHSHSEFB1/DW3FCH4T9FVfwF7f0q1iBA4GbopG/KiLiD3TwPfGADvqNDamwl+OKFTaxe01sPllo4BLRh2KCn8oHPrseLuatAMYx9mBvfiD30MkdEQsnT73gHw7QfaIXi+kbmwR1oHv/4Us2QPyOVjyK/zONHQPLwDm4qx+LFIWOmCZGmgdbuwaOkctsnJmLjRevrg7iYPAn9KKz8dkW783PXTGbJeYpJ4atXCN2+j26u+X8xbV4G3pHv/Eex40TYm7td8f14zj9LEQ9PmrjQhySX+kuV8Fan6Fz9Okad/7FLncJYqMOB/DeOTyBJbs67lF3+Ud/uG8RLJ6aL+yMyR/4LY6yOX3kEOflVsfd+faO6zjpbxnQmZ+bJ26EfUNf7za2+Z5Y6jdVYhE/idihjaenxEBPjIhTYqB2Yrd85t2iWJ3kPO6Zj3Yh4g9/Mld9GUu8oRtO04+Piq02SOgdvq4PXGVrfx9A/JDH2MmLzfR9WC/nuUvO1+DFh3HIT3foSLQXm+gX4hg9+uGjDZzFczUa+7tTy9bilZ8sqUHoF4IXXtGfGAYT86aT+ki+i3bBbTqFLnEs5hd909OrFfMPP2yP+4w/4gd/xS21Js7A2eaEjUh3xPmTDVWx2DXypidccI8O+u9i5BgsQ1fjsVnEfvMxxxZrn8PHtCf9jt1z5p6nRWDLJ/y+nO8Zw2Pt/EgOFivpol+vQYLTYpK54qH54Sv9u3Ojk3l7sbWYZ9MMb/GBv6i1cTekja1wtVFtzJB2np4mEYfVhuaizhBDA5uujfGZr/k5Hf+AgRon6lVjsBt9ibFg73vYJ+ZkjDhWGy+i/yz1jp6MU3cgKxYU0N2Fk6QuWQm4CEI4mAINSQVWDsaRBGRGFowEMeRAHMmtKwIlcnFYv1tE8pZEbXtBjbNZBM7tFYKKJP0qLhQ1CGD32t1HOiEwkrlzZQdPQkIaxSTdzEk/EofkZXFnYeWcIKJPj13QsRXjmKfgoviQsBUCiiyPEznGSfWN2IoO5yRFCyWFFVyMwSmdF2gVGMay+w9DCyTFAUxagaMEonhSOPpuAUgccydFstGf5GyByJ4SXxR57OMRspgHnBSoAmUr8GZnxRhdXCM4Gc9CWIInbBhFrQRAzFm/go9Fq3lJCJK3edMb5yzwPWoAK3cpFAnaWACZJ2wUhMaY07vzYtHBuRWm+oki3K6uABHFZlWi949CQd8WwApaQVSgEBjMQ7DBXQFO38aAo/l6FM4Ch80VMAovXKITgS0eWSgKlF3BLYWBRYciCMcU6dqHzeiC/3jgXFdwFr9gEzvbihQLW74IZ5y1UJcY6O0Fe3NmaxjRBQfZmS9N9HhqbDbl9+ZmYa/Y4utxh1TxQ3fnBe42+Xd1NQZMcAQ3+YZ4YkwvuOGC4xYQEq6EHPPmM/xWP/RnZ+PCn158gW/px6JaUjEP/biW/fmyPsQR/HQ9feAAL0mRfygu4WZxbCOIb7Ih7hnHvOFmBxT34AFr81Io8wuJxQJWwdMVfFHY6YtOsMZR2LbtYY+D+NwVdmJLxRQdYCbmikvmQfTHT8UjcQOPFYx4Kz7Cki58z7jt77W645k/XzBnnIU7fO34whi+fNF4+Ahb48NBG5wkcLeAUlzTl88ZV+yjJ7vKGd4PPvjgai/20w8cPDmCl3yED8KPLcQvPuoc/4UP//BoqnEcV7jyE9zgUzb1xDXxyfzYml58XByGvf7Fmq6Yh3b8z/g2XdhQf+wSGwfBUYUu/FynOMcRhYu+xWB5CYa4BVvjsynd6IBfrfBpsRH2eA8f8+IL7Os6XPY3AuBnfnQzhjuwRB5jf/bAteAHuwY/2jHFZHmDjfFInOav+jb/wMkiSGzFQxygP6zC98V9fKMrfMQmuniFwIu++O1asVSeoJdYxaZ0YS92D57Jk/o2Bk6JBebPv21W0YUN2MLiGmZe4i/BMfZ0nm7a4Aw+0VehLCZFnA999QkPNUXkMzqqcYzpeu82oAMHHHBe/IChuOSYOAI/HJfr6cEXIn9qhyPihj8CgzPmB38cjHwivsAUF2FG6Ilbc3q5TH8WYcbEl8gN3uV1PkMXHME18Th4KGY4btFHd5jJz3QS1/XnWCt82pjwseFjo5mYDx6JBXjku2vZjT3FqegPp+lucw+X4YdvzrMZu9ABNwh70Q8f8Bdv9T1Z3cqn4NSKeA0Lm6ryARz4i7Fx3DVws6gUa2HnnBhjo4bextcPXeRT1+G+fmxemju7aydX6ksOgRu8tY/8ILY5Ju7Bm2jP/ngaObqdgziLC+KzGpPfiBXqYtgRNQmO8rNurR99wY8uwQ/2UlvxQTr5jHtEfYXv4q14iqt0ZEM8pA9fZ0t5gsA+YqvvOGqMENzXh3HkafEUV/iLGwewNq5FJV3hGmLxKF7a/GUHY2njWvUbidgqfsklxqIf/8Ulx9iHTvjn2KIs/9cj2P/fBlrIZ8Kw1EWkyQSRTzjhhLqbLcBH8JzsGuc4VgS5YW37neeEgrFXVwRzeksU/YTOCv539NbzHjFBOGTv11e/6x0bpj/8EHcYHvqRgDnLVEUAM45ie5AI0AKRoszCPTBpHX3Qtd3jCpIuH4wvOHpWXuKK/jl7PxGIyfyMLyCw0aC+Yzy46H9Yu2g/6vt0+xW4cbNb1MT48DW/LjYw6x6La+IdNlEYxrH23V+OU4D4w1aS6CjSz95x3TD+R7vJ3hUQeD9sbvowP740yE+ch2+XnzF+93pje00nBkXfo77zFfYdNAf9DLPjqGNpF+lmkB/Ayyt8tts3vpP2vD6Dj4P6dc0wP9c3HAZhoegd5Cf6hxP7TRb7tCN01nYq8f2eK+8prIbZTD6ZbnEyzBahz0zyI/rsvrMd+0wWT7rX+C4n89F+18mvbN1yqe3DeWMO8ke+M6weCV62/U72eZQYZlz5uZ9M1+6TxVfj4YQ5DashRuVOvzmM81i/+DHO8eanb3FhGL7z0+9MXTOMI6OOwxb8Z6pzHTb+VGIrHfC5X3wYNI+FlduD9B3n8f6rmXGOOI2+BwXNbpd2Re1m2C2dCjkHJYpu/4O+T1Y42B0aJogpaXmRqRYXw/QfFb9h/Uw2D8l4Mhzi2pgrB56K88b18T6oOIehQEwmKyqdH2WxoF0/GVX3QUVKvz6ncmy6/SqgunfD2/EH4TsKZpNhY2fOnQt+MerikV6D9HFuOrx1PZlKvJhsfvqa6nljT2V8Y0xXRokJw+YxFR2G+eJkCzjj9OO7Pvsd7+o1jLPD+pjMT4wFp1GxovNU43vMZxSbTXfxaKxhtgh9Rp1ztJ+fd7YbZr9+/U6Wi4bh7/xkbUaxw1R1HiWGTTbudO0+WXyF76icGLVdP5uN89gwHx/n2KP2/UDngFH1inbDOBLthr3Pry2GjT+V2EqHqeqxsHJ7GN7jOL9ILSBHBUCAdXs4bs+Pet2CbIfEE73Hfzw+6TGsxVnYx2NXfv81VecdBRd9ehzBYwScPWVyBIYV9ZNfPX9n3fX0WLTHO1ISgUUBgQXhJ4sCLqljIpAIJAKJwJKHwCL1COuo5vEctsdqFKcL+25OzMkdOY+v+M2CZ8gfiF3dGPuBfvfIgOf5PRLmefeZXuR5hMFvCCzEPXKWhd8DbeHh43mMOR73m+wuwfCeskUikAgkAolAIpAIJAKJwAOJwGK5gHwgAcyxEoFEIBFIBBKBRCARSAQSgUQgEVhSEMjn+5YUS+c8E4FEIBFIBBKBRCARSAQSgUQgEZgmArmAnCaAeXkikAgkAolAIpAIJAKJQCKQCCQCSwoCuYBcUiyd80wEEoFEIBFIBBKBRCARSAQSgURgmgjkAnKaAObliUAikAgkAolAIpAIJAKJQCKQCCwpCOQCckmxdM4zEUgEEoFEIBFIBBKBRCARSAQSgWkikAvIaQKYlycCiUAikAgkAolAIpAIJAKJQCKwpCCw9Dgn6v/j8/8bhvj//tr/k8954t2rPe+6ON8ej77a8/qc6f9LMMbJ90QgEUgEEoFEIBFIBBKBRCARSAQSgXsQGNsC0n8S/rvf/a7MmTOn/PnPfy7rrbde2Wmnncpmm21WR77tttvqf/buP5XX9pJLLikHHnhg2XTTTctdd91Vrr766nLFFVeUFVdcsey2225l4403LksttVS99l//+le58MILyw033FAXjltvvXVtE+fTuIlAIpAIJAKJQCKQCCQCiUAikAgkAjOPwFLv6MnMd1vq4vH4448vq6++ell77bXrYvCyyy4rG264YVlnnXXKn/70p3L66aeXM844o/z617+u7SwS//e//9Xj55xzTtlyyy3LX/7yl3LyySeX5ZZbrmyxxRblH//4RznttNPKX//617LGGmuU22+/vfb9m9/8pmy//fbzFpnjmFP2mQgkAolAIpAIJAKJQCKQCCQCicCSjMDYfgO50korlZ133rnsueeeZf/99y877rhjufvuu8ull15aH0111/Gqq66qC8Rdd9217LXXXmWDDTYo5513Xl1Q7rDDDmW//far13qU9corryxz584tyy+/fNlkk03KrFmzyr777lsOOOCAsuaaa5ZTTz213slcko2Zc08EEoFEIBFIBBKBRCARSAQSgURgnAiM7RFWdxkPPfTQ8vvf/77ccsstdeG36qqrlptvvnne7x0tDC0sDz/88HlzvOCCC8p//vOfss8++9QFo982rr/++vUx2Ouvv75MTEyUvffeu/ztb38rt956a71jucwyy5Tf/va3xeOwKYlAIpAIJAKJQCKQCCQCiUAikAgkAuNBYGwLSI+WemTVbyD/+Mc/1kda3XXcfffd60z8EZy11lqrPt7aTu3vf/97ufzyy+viMH7T6DeU2223XVl55ZVrU4+8nn322eXGG2+s7fwW0iOuKYlAIpAIJAKJQCKQCCQCiUAikAgkAuNDYCwLSHcCLRyPOuqocsopp9Q/nmORd+655xZ/ACek/UuqccxfVPVY6nHHHVdWW221+ldc3am0mFx22WXrHU397rLLLuXoo4+uf3TnzDPPLEceeWR0ke+JQCKQCCQCiUAikAgkAolAIpAIJAJjQGAsv4F09/EPf/hDVddCb5VVVil33nlnvQs57L/bcKfRAvTaa6+tC0a/eVxhhRXqZx3ecccd5aKLLirbbLNN/SM7+vOIbPvfg9x0003195X+0mtKIpAIJAKJQCKQCCQCiUAikAgkAonAzCAwljuQHjXdaKON6sLx2GOPre8WlX7b6EXcVfRHdeJ7TOeggw4qZ511VjnxxBPL+eefX6/1l1jjj+a4K+kP7px00knl4osvrr+PvO6662o/+iTudF5zzTXliCOOqL+xjL7zPRFIBBKBRCARSAQSgUQgEUgEEoFEYP4RGMsC0h+12XbbbcshhxxS/OEb/6/j5ptvPu+/2XC30B/U2WOPPervIFv1t9pqq/LPf/6zLgjdWbTAjMdeLRD9bvKYY46pj8b6Qzrx/0v670I84kosMtddd91657LtOz8nAolAIpAIJAKJQCKQCCQCiUAikAjMPwL/11uU3XPbbv77mPaVVGgfQW079P9CLr306Ovcyfpq+83PiUAikAgkAolAIpAIJAKJQCKQCCQCU0NgoVhATk3lbJ0IJAKJQCKQCCQCiUAikAgkAolAIrAgEBjLH9FZEBPJMROBRCARSAQSgUQgEUgEEoFEIBFIBMaLQC4gx4tv9p4IJAKJQCKQCCQCiUAikAgkAonAYoNALiAXG1PmRBKBRCARSAQSgUQgEUgEEoFEIBEYLwK5gBwvvtl7IpAIJAKJQCKQCCQCiUAikAgkAosNArmAXGxMmRNJBBKBRCARSAQSgUQgEUgEEoFEYLwI5AJyvPhm74lAIpAIJAKJQCKQCCQCiUAikAgsNgjkAnKxMWVOJBFIBBKBRCARSAQSgUQgEUgEEoHxIpALyPHim70nAolAIpAIJAKJQCKQCCQCiUAisNggkAvIxcaUOZFEIBFIBBKBRCARSAQSgUQgEUgExovA/wMMte/d9NXl8wAAAABJRU5ErkJggg==)
Amygdala as a ‘hub’ for fear
Our meta-analytic findings were inconsistent with a locationist hypothesis of amygdala function but were more consistent with the psychological constructionist hypothesis. Our density analyses revealed that, as compared to other brain regions, voxels within both amygdalae had more consistent increases in activation during instances of fear perception than during the perception of any other emotion category (Table 1). These voxels were not functionally specific for instances of perceiving fear, however.
Anterior insula as ‘hub’ for disgust
Our meta-analytic findings were inconsistent with the locationist account that the anterior insula is the brain seat of disgust but were more consistent with the psychological constructionist account that insula activity is correlated with interoception and the awareness of affective feelings.
Orbitofrontal cortex (OFC) as ‘hub’ for anger
Our meta-analytic findings were inconsistent with the locationist hypothesis that the OFC is the brain seat of anger. As compared to voxels within other brain regions, voxels within the OFC did not have more consistent increases during instances of anger experience or perception than during any other emotion category. Rather, as compared to voxels within other brain regions, voxels within the left lOFC had more consistent increases in activation during instances of disgust experience than during the experience of other emotion categories.
Anterior cingulate cortex as ‘hub’ for sadness
Our meta-analytic evidence is inconsistent with the locationist account that the ACC is the brain basis of sadness, but more consistent with a psychological constructionist hypothesis of ACC function. As compared to voxels within other brain regions, voxels within the sACC, pACC and aMCC did not have more consistent increases when participants were experiencing or perceiving instances of sadness than during any other emotion category (Fig. 6).
Fear
Animal models

http://www.cns.nyu.edu/labs/ledouxlab/images/image_research/fear_conditioning.jpg
![Adapted from [[@davis1992role]](http://dx.doi.org/10.1016/0165-6147(92)90014-W)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQEASABIAAD/4QB0RXhpZgAATU0AKgAAAAgABAEaAAUAAAABAAAAPgEbAAUAAAABAAAARgEoAAMAAAABAAIAAIdpAAQAAAABAAAATgAAAAAAAABIAAAAAQAAAEgAAAABAAKgAgAEAAAAAQAAAo+gAwAEAAAAAQAAAaUAAAAA/+0AOFBob3Rvc2hvcCAzLjAAOEJJTQQEAAAAAAAAOEJJTQQlAAAAAAAQ1B2M2Y8AsgTpgAmY7PhCfv/iGyhJQ0NfUFJPRklMRQABAQAAGxhhcHBsAhAAAG1udHJSR0IgWFlaIAffAAYAFwAUAAIAFGFjc3BBUFBMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD21gABAAAAANMtYXBwbAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEWRlc2MAAAFQAAAAYmRzY20AAAG0AAAEGmNwcnQAAAXQAAAAI3d0cHQAAAX0AAAAFHJYWVoAAAYIAAAAFGdYWVoAAAYcAAAAFGJYWVoAAAYwAAAAFHJUUkMAAAZEAAAIDGFhcmcAAA5QAAAAIHZjZ3QAAA5wAAAGEm5kaW4AABSEAAAGPmNoYWQAABrEAAAALG1tb2QAABrwAAAAKGJUUkMAAAZEAAAIDGdUUkMAAAZEAAAIDGFhYmcAAA5QAAAAIGFhZ2cAAA5QAAAAIGRlc2MAAAAAAAAACERpc3BsYXkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABtbHVjAAAAAAAAACIAAAAMaHJIUgAAABQAAAGoa29LUgAAAAwAAAG8bmJOTwAAABIAAAHIaWQAAAAAABIAAAHaaHVIVQAAABQAAAHsY3NDWgAAABYAAAIAZGFESwAAABwAAAIWdWtVQQAAABwAAAIyYXIAAAAAABQAAAJOaXRJVAAAABQAAAJicm9STwAAABIAAAJ2ZXNFUwAAABIAAAJ2aGVJTAAAABYAAAKIbmxOTAAAABYAAAKeZmlGSQAAABAAAAK0emhUVwAAAAwAAALEdmlWTgAAAA4AAALQc2tTSwAAABYAAALeemhDTgAAAAwAAALEcnVSVQAAACQAAAL0ZnJGUgAAABYAAAMYbXMAAAAAABIAAAMuY2FFUwAAABgAAANAdGhUSAAAAAwAAANYZXNYTAAAABIAAAJ2ZGVERQAAABAAAANkZW5VUwAAABIAAAN0cHRCUgAAABgAAAOGcGxQTAAAABIAAAOeZWxHUgAAACIAAAOwc3ZTRQAAABAAAAPSdHJUUgAAABQAAAPiamFKUAAAAA4AAAP2cHRQVAAAABYAAAQEAEwAQwBEACAAdQAgAGIAbwBqAGnO7LfsACAATABDAEQARgBhAHIAZwBlAC0ATABDAEQATABDAEQAIABXAGEAcgBuAGEAUwB6AO0AbgBlAHMAIABMAEMARABCAGEAcgBlAHYAbgD9ACAATABDAEQATABDAEQALQBmAGEAcgB2AGUAcwBrAOYAcgBtBBoEPgQ7BEwEPgRABD4EMgQ4BDkAIABMAEMARCAPAEwAQwBEACAGRQZEBkgGRgYpAEwAQwBEACAAYwBvAGwAbwByAGkATABDAEQAIABjAG8AbABvAHIgDwBMAEMARAAgBeYF0QXiBdUF4AXZAEsAbABlAHUAcgBlAG4ALQBMAEMARABWAOQAcgBpAC0ATABDAERfaYJyACAATABDAEQATABDAEQAIABNAOAAdQBGAGEAcgBlAGIAbgDpACAATABDAEQEJgQyBDUEQgQ9BD4EOQAgBBYEGgAtBDQEOARBBD8EOwQ1BDkATABDAEQAIABjAG8AdQBsAGUAdQByAFcAYQByAG4AYQAgAEwAQwBEAEwAQwBEACAAZQBuACAAYwBvAGwAbwByAEwAQwBEACAOKg41AEYAYQByAGIALQBMAEMARABDAG8AbABvAHIAIABMAEMARABMAEMARAAgAEMAbwBsAG8AcgBpAGQAbwBLAG8AbABvAHIAIABMAEMARAOIA7MDxwPBA8kDvAO3ACADvwO4A8wDvQO3ACAATABDAEQARgDkAHIAZwAtAEwAQwBEAFIAZQBuAGsAbABpACAATABDAEQwqzDpMPwAIABMAEMARABMAEMARAAgAGEAIABDAG8AcgBlAHMAAHRleHQAAAAAQ29weXJpZ2h0IEFwcGxlIEluYy4sIDIwMTUAAFhZWiAAAAAAAADzUgABAAAAARbPWFlaIAAAAAAAAGFsAAA4MgAACkJYWVogAAAAAAAAbxwAAK6VAAAWmVhZWiAAAAAAAAAmTgAAGTkAALJSY3VydgAAAAAAAAQAAAAABQAKAA8AFAAZAB4AIwAoAC0AMgA2ADsAQABFAEoATwBUAFkAXgBjAGgAbQByAHcAfACBAIYAiwCQAJUAmgCfAKMAqACtALIAtwC8AMEAxgDLANAA1QDbAOAA5QDrAPAA9gD7AQEBBwENARMBGQEfASUBKwEyATgBPgFFAUwBUgFZAWABZwFuAXUBfAGDAYsBkgGaAaEBqQGxAbkBwQHJAdEB2QHhAekB8gH6AgMCDAIUAh0CJgIvAjgCQQJLAlQCXQJnAnECegKEAo4CmAKiAqwCtgLBAssC1QLgAusC9QMAAwsDFgMhAy0DOANDA08DWgNmA3IDfgOKA5YDogOuA7oDxwPTA+AD7AP5BAYEEwQgBC0EOwRIBFUEYwRxBH4EjASaBKgEtgTEBNME4QTwBP4FDQUcBSsFOgVJBVgFZwV3BYYFlgWmBbUFxQXVBeUF9gYGBhYGJwY3BkgGWQZqBnsGjAadBq8GwAbRBuMG9QcHBxkHKwc9B08HYQd0B4YHmQesB78H0gflB/gICwgfCDIIRghaCG4IggiWCKoIvgjSCOcI+wkQCSUJOglPCWQJeQmPCaQJugnPCeUJ+woRCicKPQpUCmoKgQqYCq4KxQrcCvMLCwsiCzkLUQtpC4ALmAuwC8gL4Qv5DBIMKgxDDFwMdQyODKcMwAzZDPMNDQ0mDUANWg10DY4NqQ3DDd4N+A4TDi4OSQ5kDn8Omw62DtIO7g8JDyUPQQ9eD3oPlg+zD88P7BAJECYQQxBhEH4QmxC5ENcQ9RETETERTxFtEYwRqhHJEegSBxImEkUSZBKEEqMSwxLjEwMTIxNDE2MTgxOkE8UT5RQGFCcUSRRqFIsUrRTOFPAVEhU0FVYVeBWbFb0V4BYDFiYWSRZsFo8WshbWFvoXHRdBF2UXiReuF9IX9xgbGEAYZRiKGK8Y1Rj6GSAZRRlrGZEZtxndGgQaKhpRGncanhrFGuwbFBs7G2MbihuyG9ocAhwqHFIcexyjHMwc9R0eHUcdcB2ZHcMd7B4WHkAeah6UHr4e6R8THz4faR+UH78f6iAVIEEgbCCYIMQg8CEcIUghdSGhIc4h+yInIlUigiKvIt0jCiM4I2YjlCPCI/AkHyRNJHwkqyTaJQklOCVoJZclxyX3JicmVyaHJrcm6CcYJ0kneierJ9woDSg/KHEooijUKQYpOClrKZ0p0CoCKjUqaCqbKs8rAis2K2krnSvRLAUsOSxuLKIs1y0MLUEtdi2rLeEuFi5MLoIuty7uLyQvWi+RL8cv/jA1MGwwpDDbMRIxSjGCMbox8jIqMmMymzLUMw0zRjN/M7gz8TQrNGU0njTYNRM1TTWHNcI1/TY3NnI2rjbpNyQ3YDecN9c4FDhQOIw4yDkFOUI5fzm8Ofk6Njp0OrI67zstO2s7qjvoPCc8ZTykPOM9Ij1hPaE94D4gPmA+oD7gPyE/YT+iP+JAI0BkQKZA50EpQWpBrEHuQjBCckK1QvdDOkN9Q8BEA0RHRIpEzkUSRVVFmkXeRiJGZ0arRvBHNUd7R8BIBUhLSJFI10kdSWNJqUnwSjdKfUrESwxLU0uaS+JMKkxyTLpNAk1KTZNN3E4lTm5Ot08AT0lPk0/dUCdQcVC7UQZRUFGbUeZSMVJ8UsdTE1NfU6pT9lRCVI9U21UoVXVVwlYPVlxWqVb3V0RXklfgWC9YfVjLWRpZaVm4WgdaVlqmWvVbRVuVW+VcNVyGXNZdJ114XcleGl5sXr1fD19hX7NgBWBXYKpg/GFPYaJh9WJJYpxi8GNDY5dj62RAZJRk6WU9ZZJl52Y9ZpJm6Gc9Z5Nn6Wg/aJZo7GlDaZpp8WpIap9q92tPa6dr/2xXbK9tCG1gbbluEm5rbsRvHm94b9FwK3CGcOBxOnGVcfByS3KmcwFzXXO4dBR0cHTMdSh1hXXhdj52m3b4d1Z3s3gReG54zHkqeYl553pGeqV7BHtje8J8IXyBfOF9QX2hfgF+Yn7CfyN/hH/lgEeAqIEKgWuBzYIwgpKC9INXg7qEHYSAhOOFR4Wrhg6GcobXhzuHn4gEiGmIzokziZmJ/opkisqLMIuWi/yMY4zKjTGNmI3/jmaOzo82j56QBpBukNaRP5GokhGSepLjk02TtpQglIqU9JVflcmWNJaflwqXdZfgmEyYuJkkmZCZ/JpomtWbQpuvnByciZz3nWSd0p5Anq6fHZ+Ln/qgaaDYoUehtqImopajBqN2o+akVqTHpTilqaYapoum/adup+CoUqjEqTepqaocqo+rAqt1q+msXKzQrUStuK4trqGvFq+LsACwdbDqsWCx1rJLssKzOLOutCW0nLUTtYq2AbZ5tvC3aLfguFm40blKucK6O7q1uy67p7whvJu9Fb2Pvgq+hL7/v3q/9cBwwOzBZ8Hjwl/C28NYw9TEUcTOxUvFyMZGxsPHQce/yD3IvMk6ybnKOMq3yzbLtsw1zLXNNc21zjbOts83z7jQOdC60TzRvtI/0sHTRNPG1EnUy9VO1dHWVdbY11zX4Nhk2OjZbNnx2nba+9uA3AXcit0Q3ZbeHN6i3ynfr+A24L3hROHM4lPi2+Nj4+vkc+T85YTmDeaW5x/nqegy6LzpRunQ6lvq5etw6/vshu0R7ZzuKO6070DvzPBY8OXxcvH/8ozzGfOn9DT0wvVQ9d72bfb794r4Gfio+Tj5x/pX+uf7d/wH/Jj9Kf26/kv+3P9t//9wYXJhAAAAAAADAAAAAmZmAADypwAADVkAABPQAAAKDnZjZ3QAAAAAAAAAAAADAQAAAgAAAFYBLgHrAsQDgwRTBUAGJwcaCBgJHwo9C04Maw2QDrkP7BEtEmUToxTjFioXeRi/Gg8bVxykHfMfPCCEIc0jESRSJY0myygAKTMqZiuXLMot9y8oMFExgDKrM9U1AjYxN104ijm2OuE8DT04PmA/iUCxQdxDCEQ2RWNGkEe4SN5KAUskTEdNak6PT7RQ2FH4UxZUM1VPVm1XjViuWcta4VvwXPdd+18AYAphGWItY0FkUmVfZmVnYWhTaTxqIGsBa+Bsv22gboNvaHBQcThyIHMIc+901nW+dqh3k3h+eWx6WXtJfD19N342fzmAQYFLgleDZYR0hYWGlIekiLOJwIrMi9aM343nju2P8pD1kfaS9ZP1lPqWBJcVmCyZR5pjm3+cmJ2unr+fy6DUodqi4KPppPSmA6cVqCqpQqpcq3eslK2yrtGv8bERsjOzU7RztZC2qbe6uMG5w7rAu7u8tb2vvqm/o8CdwZXCisN8xGrFU8Y2xxjH+cjgyc/KxsvDzMHNvM6vz5jQc9FD0g3S09Ob1GbVNdYF1tfXqNh22UDaBtrH24DcNdzn3ZHeO97j343gO+Dr4Z/iUuMD46/kVeT55ZTmLOa/51Ln5OiF6S/p5Oqo627sM+z17a/uY+8R77XwUPDm8XzyF/LG84/0bvVh9mb3ivjk+sX9Lv//AAAAVgEjAbACagMyA+gEsQWABlgHPQgtCScKJwsiDDANOg5RD2MQfhGfEr4T6RUQFjgXYRiIGbQa3hwLHTMeWx+AIKEhvyLbI/UlCyYfJzUoSSlgKnkrkyyxLcwu6DAEMSEyPDNaNHQ1izakN7k4zTniOvQ8Bj0bPjE/TUBsQY9CsEPQRO1GCEcdSDJJRUpVS2NMbk14Tn5Pg1CFUYZSiFOKVIxVilaEV3lYaFlUWkRbP1xGXVped1+VYLBhwmLKY8VktWWcZntnV2gwaQxp7GrQa7psqW2bbo9vgnBycVxyQHMfc/t01XWtdoZ3YnhFeTB6JHsdfBl9FX4QfwZ/9oDigcqCsYOVhHeFXYZKh0SITIlhioGLpYzGjd6O7o/1kPeR9ZLzk/CU7ZXslu6X+JkQmjmbbpymnc6e45/poOWh36LZo9GkyKW+prynyqjtqiGrXaycrdmvFLBOsYiywrP5tS62ZLebuNK6Bbs0vF+9hb6mv7zAvsGqwpDDfcR3xYDGj8edyKPJnsqOy3XMU80tzgTO2s+y0IvRatJR00XURdVR1mLXddiE2Yfafdtm3EPdGt3s3rnfh+BY4S3iCOLm48PkneVx5kDnDOfU6KPpf+pp62TsaO1t7nDvcPBw8XLydvN59Hf1afZH9w/3s/hF+Mv5Wvnt+tD8DP3a//8AAAArAMQBSwHYAmUDBgOPBCsEzwV6Bi8G6QelCGgJLwn5CsoLnwx3DVIOMg8OD/YQ2RHAEqkTkxR9FWcWUhc9GCkZFRn8Gugb0hy+Ha4eox+bIJYhlCKPI4YkdiVhJkUnICf4KMwpnypzK0gsIS0CLeku1y/IMLoxqjKYM4A0YDU5NgY2zzeVOFg5GTnZOp07ZDwyPQY93j62P49AaEFEQiZDEEQARPNF50bWR7tIlkllSiVK4EuYTE9NB03DToNPSVAYUOpRvlKTU2hUO1UNVd5WrVd8WEpZFVnfWqhbb1w1XPldvV6AX0NgCGDRYZxia2M8ZAxk2mWnZnFnOGf8aL9pgGpAawBrwGyBbUNuBG7Fb4VwQXD4catyWXMBc6l0T3T0dZh2PHbed4B4HXi9eWB6Fnrae6t8g31Yfih+9n/DgJOBZ4JEgyqEGIULhgCG9IfjiM2Jr4qMi2GMNI0HjdqOsI+KkGuRR5ISksaTW5PslHqVQZZCl4iZCJqKm+SdGZ4ynzmgOaE1oi+jKKQhpRmmEacKqAmpD6ohqz2sYK2Hrq2v07D8siizWrSStdG3FLhZuZy617wFvSm+R79iwH/BnsK+w9/E/8Yhx0PIZ8mMyrHL1sz9zinPW9CX0d7TMtSN1e7XU9i52iHbld0v3w7hd+SL6HntTvL2+VH//wAAbmRpbgAAAAAAAAY2AACTlQAAVo0AAFbpAACRtwAAJrsAABcKAABQDQAAVDkAAoeuAAJHrgABa4UAAwEAAAIAAAABAAQACAAPABYAHwApADQAQABMAFoAaQB4AIkAmgCsAL4A0gDmAPwBEgEpAUABWQFzAY0BqQHFAeMCAQIhAkICZAKHAqwC0gL6AyMDTQN6A6gD1wQIBDoEbQSjBNkFEgVLBYYFwwYBBkAGgAbCBwUHSgeQB9gIIQhsCLkJBwlXCagJ+gpNCqEK9wtOC6gMBAxiDMMNJQ2JDe4OVA68DyUPkRAAEHEQ5BFZEc8SRhK9EzgTthQ6FMMVUhXjFnUXBheWGCYYuBlOGegaiBsxG+Mcnx1iHise+B/GIJUhZCI0IwYj2ySzJY8mbCdLKCspDSnwKtUruyyeLX4uXC85MBQw8DHNMqoziTRoNUo2LjcVN/847TneOtI7yjzFPcQ+xz/OQNpB6EL2RABFBkYIRwhICEkJSgxLFEwhTTROTk9uUJRRvFLlVA1VNFZZV31YoFnEWudcC10wXlZffGCkYc5i+GQkZVNmhWe8aPpqQ2uZbPluY2/ScUZyvHQ0da93LXixejx70H1vfxyA1oKUhEyF9oeUiSyKxYxljhWP35HNk92V/5ggmjmcTJ5foHmioKTapy2poKwwruCxobRetwu5rrxdvyvCH8VGyKLMDM8o0fbUkdco2dHcnt+X4tLmYOn57RLvrPIA9DD2Off/+Yr6uPvU/Lv9o/5w/zf//wAAAAEABQAKABIAGwAmADIAPwBNAFwAbQB+AJEApAC5AM8A5QD9ARYBLwFKAWYBgwGiAcEB4gIEAicCTAJyApoCwwLvAxwDSwN8A68D5AQaBFIEiwTFBQEFPQV7BbsF/AY/BoMGyQcQB1kHpQfyCEEIkwjmCTsJkwnsCkYKoQr8C1gLtAwSDHIM1A05DaAOCw54DucPWQ/OEEYQwBE+Eb8SQxLKE1MT3RRqFPoVjxYqFssXcRgWGLYZURnmGnkbDRukHEAc5B2RHkofDh/dILMhjiJpI0IkGCTrJb0mkCdmKEIpJSoSKwgsBS0ILg4vEjARMQgx+TLoM9c0yjXBNsE3yjjbOfI7DjwwPVE+aT90QHJBaEJYQ0dEOUU0RjhHR0hfSX5KoUvHTPBOG09FUGtRhlKUU5dUlVWUVqBXv1jxWjVbgFzNXh5fdGDPYiNjZmSVZbVmzmfkaPtqFmszbFRtdm6ab8Bw6nIXc0Z0dnWndtp4EHlNeo972H0mfn1/44FngxCEyoZ0iAaJh4sEjIiOG4/DkYKTWJVDlzuZO5s7nTmfKaEEosakdqYbp72pYqsVrN2uxbDPsvW1Mbd8ucu8D75EwHLCpsTpx0LJsswxzpzQ3dL01PHW59jj2ujc9N8B4QrjFOUk50jpluw374rzpvds+c77hPzD/fT++f//AAAAAgAIABEAHgAtAD4AUQBmAH0AlgCwAMwA6gEJASsBTQFyAZgBwAHqAhYCRAJ0AqYC2gMRA0kDhAPAA/0EOwR6BLoE/AVBBYoF1wYpBoEG3Qc+B6IICAhvCNQJOQmdCgIKagrVC0QLuww5DMENUA3mDoIPIQ/AEFwQ9hGQEi0SzRNuFA0UqBVCFdwWeRceF84YjhliGkIbKhwUHPsd3R64H5IgbSFMIjAjGCQGJPkl8SbvJ/Mo/SoOKyQsQC1gLoAvnTC4MdMy8zQaNUk2gjfDOQw6XDuwPQc+YD++QSJCk0QWRa5HWkkRStFMmk5vUFVSMlPaVVpWzFhIWdNbZlz4XoNgAWFwYtVkN2WcZwlogWoKa6NtTW8AcLdyb3QedcN3dXlke+5+pIB5gdaC/IQChQiGEIcziHOJ1ItVjOmOiJAukdmTiJU7lvGYqZpenAqdp583oL2iP6PApUSmyqhQqdSrVKzQrkavubEospe0CLV9tvy4ibomu9C9gr82wOnCnMRPxgTHu8lzyyvM5M6e0FjSFNPS1Y/XStj/2qzcT93o33Xg/eKC5APlgucC6IPqBOt/7O7uRu958J3xjfJ180Tz9/Sq9U712/Zo9vb3dvfp+Fz4z/lD+av6Dfpw+tL7NfuY+/H8Sfyi/Pv9VP2t/gP+WP6s/wH/Vf+q//8AAHNmMzIAAAAAAAEMQgAABd7///MmAAAHkgAA/ZH///ui///9owAAA9wAAMBsbW1vZAAAAAAAAAYQAACc3wAAAADKKtWAAAAAAAAAAAAAAAAAAAAAAP/AABEIAaUCjwMBIgACEQEDEQH/xAAfAAABBQEBAQEBAQAAAAAAAAAAAQIDBAUGBwgJCgv/xAC1EAACAQMDAgQDBQUEBAAAAX0BAgMABBEFEiExQQYTUWEHInEUMoGRoQgjQrHBFVLR8CQzYnKCCQoWFxgZGiUmJygpKjQ1Njc4OTpDREVGR0hJSlNUVVZXWFlaY2RlZmdoaWpzdHV2d3h5eoOEhYaHiImKkpOUlZaXmJmaoqOkpaanqKmqsrO0tba3uLm6wsPExcbHyMnK0tPU1dbX2Nna4eLj5OXm5+jp6vHy8/T19vf4+fr/xAAfAQADAQEBAQEBAQEBAAAAAAAAAQIDBAUGBwgJCgv/xAC1EQACAQIEBAMEBwUEBAABAncAAQIDEQQFITEGEkFRB2FxEyIygQgUQpGhscEJIzNS8BVictEKFiQ04SXxFxgZGiYnKCkqNTY3ODk6Q0RFRkdISUpTVFVWV1hZWmNkZWZnaGlqc3R1dnd4eXqCg4SFhoeIiYqSk5SVlpeYmZqio6Slpqeoqaqys7S1tre4ubrCw8TFxsfIycrS09TV1tfY2dri4+Tl5ufo6ery8/T19vf4+fr/2wBDAAICAgICAgMCAgMFAwMDBQYFBQUFBggGBgYGBggKCAgICAgICgoKCgoKCgoMDAwMDAwODg4ODg8PDw8PDw8PDw//2wBDAQICAgQEBAcEBAcQCwkLEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBD/3QAEACn/2gAMAwEAAhEDEQA/AP3M8a/ELwD8NdKi134jeJdM8K6bPMttHdareQ2MDzsrOsSyTuilyqMwUHJCk4wDXmH/AA1j+yz/ANFk8Gf+FDp3/wAfr5v/AOCiGk6Vr+lfs/aFrtnDqOm6j8X/AApbXVrcxrNBPBMt2kkUsbgq6OpKsrAggkEYr6Q/4ZO/ZZ/6I34M/wDCe07/AOMUAH/DWP7LP/RZPBn/AIUOnf8Ax+j/AIax/ZZ/6LJ4M/8ACh07/wCP0f8ADJ37LP8A0RvwZ/4T2nf/ABij/hk79ln/AKI34M/8J7Tv/jFAB/w1j+yz/wBFk8Gf+FDp3/x+j/hrH9ln/osngz/wodO/+P0f8Mnfss/9Eb8Gf+E9p3/xij/hk79ln/ojfgz/AMJ7Tv8A4xQAf8NY/ss/9Fk8Gf8AhQ6d/wDH6P8AhrH9ln/osngz/wAKHTv/AI/R/wAMnfss/wDRG/Bn/hPad/8AGKP+GTv2Wf8Aojfgz/wntO/+MUAH/DWP7LP/AEWTwZ/4UOnf/H6P+Gsf2Wf+iyeDP/Ch07/4/R/wyd+yz/0RvwZ/4T2nf/GKP+GTv2Wf+iN+DP8AwntO/wDjFAB/w1j+yz/0WTwZ/wCFDp3/AMfo/wCGsf2Wf+iyeDP/AAodO/8Aj9H/AAyd+yz/ANEb8Gf+E9p3/wAYo/4ZO/ZZ/wCiN+DP/Ce07/4xQAf8NY/ss/8ARZPBn/hQ6d/8fo/4ax/ZZ/6LJ4M/8KHTv/j9H/DJ37LP/RG/Bn/hPad/8Yo/4ZO/ZZ/6I34M/wDCe07/AOMUAH/DWP7LP/RZPBn/AIUOnf8Ax+j/AIax/ZZ/6LJ4M/8ACh07/wCP0f8ADJ37LP8A0RvwZ/4T2nf/ABij/hk79ln/AKI34M/8J7Tv/jFAB/w1j+yz/wBFk8Gf+FDp3/x+j/hrH9ln/osngz/wodO/+P0f8Mnfss/9Eb8Gf+E9p3/xij/hk79ln/ojfgz/AMJ7Tv8A4xQAf8NY/ss/9Fk8Gf8AhQ6d/wDH6P8AhrH9ln/osngz/wAKHTv/AI/R/wAMnfss/wDRG/Bn/hPad/8AGKP+GTv2Wf8Aojfgz/wntO/+MUAH/DWP7LP/AEWTwZ/4UOnf/H6P+Gsf2Wf+iyeDP/Ch07/4/R/wyd+yz/0RvwZ/4T2nf/GKP+GTv2Wf+iN+DP8AwntO/wDjFAB/w1j+yz/0WTwZ/wCFDp3/AMfo/wCGsf2Wf+iyeDP/AAodO/8Aj9H/AAyd+yz/ANEb8Gf+E9p3/wAYo/4ZO/ZZ/wCiN+DP/Ce07/4xQAf8NY/ss/8ARZPBn/hQ6d/8fo/4ax/ZZ/6LJ4M/8KHTv/j9H/DJ37LP/RG/Bn/hPad/8Yo/4ZO/ZZ/6I34M/wDCe07/AOMUAH/DWP7LP/RZPBn/AIUOnf8Ax+j/AIax/ZZ/6LJ4M/8ACh07/wCP0f8ADJ37LP8A0RvwZ/4T2nf/ABij/hk79ln/AKI34M/8J7Tv/jFAB/w1j+yz/wBFk8Gf+FDp3/x+j/hrH9ln/osngz/wodO/+P0f8Mnfss/9Eb8Gf+E9p3/xij/hk79ln/ojfgz/AMJ7Tv8A4xQAf8NY/ss/9Fk8Gf8AhQ6d/wDH6P8AhrH9ln/osngz/wAKHTv/AI/R/wAMnfss/wDRG/Bn/hPad/8AGKP+GTv2Wf8Aojfgz/wntO/+MUAH/DWP7LP/AEWTwZ/4UOnf/H6P+Gsf2Wf+iyeDP/Ch07/4/R/wyd+yz/0RvwZ/4T2nf/GKP+GTv2Wf+iN+DP8AwntO/wDjFAB/w1j+yz/0WTwZ/wCFDp3/AMfo/wCGsf2Wf+iyeDP/AAodO/8Aj9H/AAyd+yz/ANEb8Gf+E9p3/wAYo/4ZO/ZZ/wCiN+DP/Ce07/4xQAf8NY/ss/8ARZPBn/hQ6d/8fo/4ax/ZZ/6LJ4M/8KHTv/j9H/DJ37LP/RG/Bn/hPad/8Yo/4ZO/ZZ/6I34M/wDCe07/AOMUAH/DWP7LP/RZPBn/AIUOnf8Ax+j/AIax/ZZ/6LJ4M/8ACh07/wCP0f8ADJ37LP8A0RvwZ/4T2nf/ABij/hk79ln/AKI34M/8J7Tv/jFAB/w1j+yz/wBFk8Gf+FDp3/x+j/hrH9ln/osngz/wodO/+P18P/8ABR79nv4BeBv2MfiH4p8FfDTwz4f1qy/snyL7T9GsrS6h8zVbSN/LmhiV13IzK2CMqSDwTX3B/wAMnfss/wDRG/Bn/hPad/8AGKAD/hrH9ln/AKLJ4M/8KHTv/j9H/DWP7LP/AEWTwZ/4UOnf/H6P+GTv2Wf+iN+DP/Ce07/4xR/wyd+yz/0RvwZ/4T2nf/GKAD/hrH9ln/osngz/AMKHTv8A4/R/w1j+yz/0WTwZ/wCFDp3/AMfo/wCGTv2Wf+iN+DP/AAntO/8AjFH/AAyd+yz/ANEb8Gf+E9p3/wAYoAP+Gsf2Wf8Aosngz/wodO/+P0f8NY/ss/8ARZPBn/hQ6d/8fo/4ZO/ZZ/6I34M/8J7Tv/jFH/DJ37LP/RG/Bn/hPad/8YoAP+Gsf2Wf+iyeDP8AwodO/wDj9H/DWP7LP/RZPBn/AIUOnf8Ax+j/AIZO/ZZ/6I34M/8ACe07/wCMUf8ADJ37LP8A0RvwZ/4T2nf/ABigA/4ax/ZZ/wCiyeDP/Ch07/4/R/w1j+yz/wBFk8Gf+FDp3/x+j/hk79ln/ojfgz/wntO/+MUf8Mnfss/9Eb8Gf+E9p3/xigA/4ax/ZZ/6LJ4M/wDCh07/AOP0f8NY/ss/9Fk8Gf8AhQ6d/wDH6P8Ahk79ln/ojfgz/wAJ7Tv/AIxR/wAMnfss/wDRG/Bn/hPad/8AGKAD/hrH9ln/AKLJ4M/8KHTv/j9H/DWP7LP/AEWTwZ/4UOnf/H6P+GTv2Wf+iN+DP/Ce07/4xR/wyd+yz/0RvwZ/4T2nf/GKAD/hrH9ln/osngz/AMKHTv8A4/R/w1j+yz/0WTwZ/wCFDp3/AMfo/wCGTv2Wf+iN+DP/AAntO/8AjFH/AAyd+yz/ANEb8Gf+E9p3/wAYoAP+Gsf2Wf8Aosngz/wodO/+P0f8NY/ss/8ARZPBn/hQ6d/8fo/4ZO/ZZ/6I34M/8J7Tv/jFH/DJ37LP/RG/Bn/hPad/8YoAP+Gsf2Wf+iyeDP8AwodO/wDj9H/DWP7LP/RZPBn/AIUOnf8Ax+j/AIZO/ZZ/6I34M/8ACe07/wCMUf8ADJ37LP8A0RvwZ/4T2nf/ABigA/4ax/ZZ/wCiyeDP/Ch07/4/R/w1j+yz/wBFk8Gf+FDp3/x+j/hk79ln/ojfgz/wntO/+MUf8Mnfss/9Eb8Gf+E9p3/xigA/4ax/ZZ/6LJ4M/wDCh07/AOP0f8NY/ss/9Fk8Gf8AhQ6d/wDH6P8Ahk79ln/ojfgz/wAJ7Tv/AIxR/wAMnfss/wDRG/Bn/hPad/8AGKAD/hrH9ln/AKLJ4M/8KHTv/j9H/DWP7LP/AEWTwZ/4UOnf/H6P+GTv2Wf+iN+DP/Ce07/4xR/wyd+yz/0RvwZ/4T2nf/GKAD/hrH9ln/osngz/AMKHTv8A4/R/w1j+yz/0WTwZ/wCFDp3/AMfo/wCGTv2Wf+iN+DP/AAntO/8AjFH/AAyd+yz/ANEb8Gf+E9p3/wAYoAP+Gsf2Wf8Aosngz/wodO/+P0f8NY/ss/8ARZPBn/hQ6d/8fo/4ZO/ZZ/6I34M/8J7Tv/jFH/DJ37LP/RG/Bn/hPad/8YoAP+Gsf2Wf+iyeDP8AwodO/wDj9H/DWP7LP/RZPBn/AIUOnf8Ax+j/AIZO/ZZ/6I34M/8ACe07/wCMUf8ADJ37LP8A0RvwZ/4T2nf/ABigA/4ax/ZZ/wCiyeDP/Ch07/4/R/w1j+yz/wBFk8Gf+FDp3/x+j/hk79ln/ojfgz/wntO/+MUf8Mnfss/9Eb8Gf+E9p3/xigA/4ax/ZZ/6LJ4M/wDCh07/AOP0f8NY/ss/9Fk8Gf8AhQ6d/wDH6P8Ahk79ln/ojfgz/wAJ7Tv/AIxR/wAMnfss/wDRG/Bn/hPad/8AGKAD/hrH9ln/AKLJ4M/8KHTv/j9H/DWP7LP/AEWTwZ/4UOnf/H6P+GTv2Wf+iN+DP/Ce07/4xR/wyd+yz/0RvwZ/4T2nf/GKAD/hrH9ln/osngz/AMKHTv8A4/Xt+k6tpWv6VZ67oV5DqOm6jDHc2t1bSLNBPBMoeOWKRCVdHUhlZSQQQQcV8QftN/syfs26B+zb8V9d0L4UeEtO1LTvCWu3NrdW2hWEM8E8NhM8csUiQhkdGAZWUgggEHNe4fsnf8ms/Bv/ALEzw9/6boKAP//Q+/f2+v8Am3L/ALLN4Q/9uq+/6+AP2+v+bcv+yzeEP/bqvv8AoAKKKKACiiigD5l/aT+Kvjf4XWXh248EW9vdTX89ybiKdC+6C1gM7hMEYbarY616b41+ItpoPwrvPiLo2LtZbGOfT1xkTzXYVbVMDrvkdBx61578aYIbr4kfCW2uEEkUurXqOrDIZWs5AQR6EV5L4AF5qfiLw9+z5e75E+HOsXt9ds+cyWFiVk0nJPBDtcxkD0h/GvraGBozw1GbjrG8pecby/KyS/xH5ti82xNLH4mkpu02oQ7Rnywat680pPyge4/Bz4oXfiD4O2vxB+JV5Z6dMkl2l3NkQW6fZ7mSEcsxA4UDryenXFeieEPiN4F8fJO3g3XLXVja481YJAXj3dCyHDAHsSMHtX5+Rx6g3wU+GMv2u3sdJi8W3zXU97E01nFKLu5+zNcorpmLzODuYKCRk17rp0eq/wDC8tI17X/Ful6rq+maRfPcWuiadMjzWDKNouJfPmUbZdrRBsEnIHUY1zDJKPNVmnZ3nZK9lyt6bP8ANWujmyTizFcmHpyjzLlpKTbV25xTcruS2vtyy5mmk0z3C++N3wj0zXW8NX/i3ToNRSTymiadfkkzjYzfdVs8EEg1t+MPiV4C+H/2ceM9dtdJa7z5KTSYdwvUqgyxUdzjA7mvgXxN4gutW/Zs1PUrCfw14X8I6hFO9npMSS32pSSvKSEeaSZQJi/zMwRio56Dj2PVYzN428N6z4L8XWeh+Nh4VtEktdYtjLZXtiW3gpMSrKwkLb/LYkjGQMEmanD9GLTbdryT9YpdVF231+JLuXQ41xVRSUYxbahKPTSbeylNOWiuruDf8vR/UaeOfB0ul6XrcOs2sun61PFa2U6Sq0dxPMSEjRhkFmIIx6gitS517RrPWLLw/dXkcWpaikslvAxxJKkGDIVHcLuGfrXwhc+KrDUPhv4Z1a40qw0C38N/Eezj1CXTnLabIYZiZryF26Qsz9TwCD9K9k8Q+JtC1v8Aaa8AadpF9Dey2Ol6u83kyLIE89I9gJUkAkITjrjB6EVx1ci5ZNa2Sm+n2Vdbaa6HqYbjDngn7t26SW6vzys7J66a29Op7HpPxU+HOvazb+HtG8RWd5qV1HJLFBHKGdkiZlcgf7JRsjrgE9Oar6R8X/hfr+vnwto3iiwvNV3MggjnVmdk+8qHo5HcKSRXyr4D8OvF+yR4m1PwtZqNdv01xzNEg+0ORczRNtYDdnyk2jBz6Vz+o2N7q3wl8HaWPHXh630pptMGjLp+lzyahHdo6GMRBLp380NkSnZnlt2DXZ/YOHc5xUnpJx76rrZJ6eWmz1PM/wBcccqVKo6cW5QVTsrO1o3lJWfeWqV0uXqfZvin4t/DPwTqcei+K/Ellpl9IocQzSgOqt0ZwPuA9i2Aa722uba9toryzlSe3nVXjkjYMjowyrKw4II5BHWvjnxHBNa/Efx7qPw58V6PDfXC2w17RfEluUt5NluER47jKyeU0WOgZAc84IFe7/A7X7HxR8J/DWuabpA0G1uLbEdkhLJCsbtHhCeSjbdyf7JFeVj8shToRqwv0vfu1fayfo9U1rdbH0WT59Vr4yphqtlbm5ba3UZct7ptPdXTUXF6We5ZsfGtvp174su/FGvad/Zmj3kEKCIsr2azIgEd0W43u7grjjBFXND+K/w28TeIJvC3h/xJY3+rQ7t1vDMrOdn3tuOG2452k4718Z/E9BJ4J/aHjbo+saSD+P2SvdPi7oukaJ4l+EA0izisxZeIY7WERIE8uCS2l3RrgcK20ZHfFd9TKKNo3bvK9rWsrU4y173bt07nj0eJcUnPlS5YNXvdt81apTsnfSyjfr2tbb6ZJAGTwBXy18L/AI36/wCL/iHJpmtQwReGvEqX0vhqdFKvMumTmGYOSTuLriVcAYUGu3/aD8U3/hz4a3lhoR/4nniaWLRdNUHBNzfnywQexRNzg+or528d+A/jT4G+H/hzW1Hh+W0+FjQX9uunpdpdyQWy7LhGaVmQrJEWaXgE4yKjJ8vpTpP2rSc3yxv5Ldf9vOK9LmvE+dYiliV9XUnGklOfKrp3ez/7cU3bvys+wfE3xI8B+DJpbfxVrtppcsMCXDJPKEbypHaNGAPJ3MrAAZJwa53xH8Q9K1P4aX/jTwD4l0uOGPYI9RumMllE3morCXZ8wODgDggkZryzS5PD3jP9p3S/FdvHHe28ngW3vbKR1DbfPv3KyLnOG2MRkcjJFeNfEK3gsfAX7Q9jZosNuus6dKI0G1Q832VnYAcZY8n1rXB5NSdSEHfm9xu9re9JRatbz/Sxz5nxRiY0a1WKi4fvIxte/uwlNSvfra2ivs79D7T8S/FH4eeDpbm38U+ILPTJrRYmkjnlCuBPu8vC9Tu2NgAHoazL/wCNfwk0xdNe+8XabCuroJLVjcIVkjZiofIJCruBG5sDIIzkGvONE0nTrz9qbxZqN3bpNcWnh/TkiZ1DFBNJJvxnpkKAcduK8j8LeGdAtP2cPjCbfT4ULXniTJCLnFsX8kZxnEe0FR2PI5rOjlOGajzOV/cvt9tenT8dtNzfE8R49Sn7NQsva2unf92+uv2tult9dj7E8XeP/BXgO1hvfGOtW2kxXBKxefIFaQjkhF+82O+Aa84+Ifxg063+DHiH4lfDTVLPV20yIGKVCJohLvRSrqCCCA2cHB5BryNPEt7P4y8GaH4XtNHtfE1r4RtLp9X1x5mVLWbarRW0EboGfcu533DjjOMivF7m8W7+HH7RG3U7XV8XGns9zYwi3tZJSFWRoow8gA3KRncd2N2ea68BkFO8JTvdON+zTmo7W036u77HmZxxlX5akKVknGok1upRpud7t6tNW0jZae9fQ/TS3dpII5G6uoJ+pFfLPwC+Nfirx/4u8TeFvGcdvG9s0txpbQIU821guZLWbfknLI6p0/vZ9K+pLP8A49IP9xf5V+e3hKeLwX4U8OfGFz5dt4d8V6xZak4BwNN1O6aF3fHJEUoicD6/j52UYWnVo1oSjeTso+T95q3q0l8z3OJcxrYfE4arCbUIqUpro4pwTb/wqTl8j3HxP8ZfEtp+0P4e+F+iRQHQpcQ6lK6Ey/apLeW5SONsgDbGqMeD9/6V614i+L3ww8Jayvh7xL4nsNO1E7SYZZlVkDfdMnZAe27FfKmi2F2fEnwg8X6pGYtS8Z67q+tzq3VEu7Nvs8fr+7t1iXHbBrufgzqngfTfB/jy0+IFxZwaoms6q2vpesod0aRjGXD/ADNEYSoj6qeQvOa78bllBQhJRb5Uk+Xdvmkm+umn4pHj5Vn+LdWpCU0ueTknO9ox9nCSja61tLvpaTs9j6V8Q+MPC3hPRv8AhIfEmq22naadoW4mlVY3LjKhCT8xYDIC5J7V5b4O+LNv43+K2o+HfDmoWmp+HINFtr6Ga3O9jcSXEkUis4PYKPlIBB6182fDL7RpafAm98fhrfSI7bWo7JrzKpFcySZsPMZ/lDNajEGcH+7zXR+IJrDUfir8YG+GjpNqr+DQsjWZDM2ofvR8pTrLs8scc7uvOaUckpQc6b1dn732VafL+l7362t1KnxZiKypV4pRjzRvDXnd6XtPu1ta3S9+h9O2Hxg+F2p+Iv8AhEtP8UafcauXMQt0nUs0i9UU52s47qCT7Vn+G/Ges6r8XPGngq68v+ztBtdLmttq4k3XiymTc2eRlBjgYr40ksrnVfgD4c0d/HHh+z0KZdPSxSy0yeXU4r0OhQRCO5ZzciXiQqmfvZGCa+mvAQkH7QnxKExDSDTtA3EDAJ8ufPFZ4vKaNGnVcXeye99GpQV9lvdq2tu+ptl3EmKxNbDxqJRTlF6NaqVOq7NKUtE4ppu3N2Vj6Jooor5M/RwooooA+AP+Co//ACYp8TP+4L/6eLKvv+vgD/gqP/yYp8TP+4L/AOniyr7/AKACiiigAr5n17xT8Xdd+MWtfD/wLqmmaVZaPp1peFr2ze4d2uCykApImMbfSvpivljT/EOgeH/2nPGUuvalbaak2h6YEa5mSEMQ7kgFyMke1exlEL+1lyptRurq+t10+Z8xxNUt9Xg5uMZTSbT5dOWT3VuqR0/hjx/8QNB+IVl8NPixb2Mk2uQTTaRqemrJHDctbKHmgkikZmSREO4HO0jpzWJpvi/4s/E7xH4pg8CarpXhzS/Cupy6Xsu7V726uZrfG+SQCWMRRMT8mASQCc1ma/4k0b4ofHrwFpvga9i1e18HC/1HVLu0cSwQ+fD5MEXmoSpd2zlM5289jjDm074G/FTxHrmo+InfwH450e7ms7qS31E6ffFYTiOcHKJKjoAyuUbjAzjFezHDQjapOnaTim7Rvyvmavyt7NJel7pao+WqY6rO9GnX5oRqNRvNxc0oRbSqJO7jJvtzWs37rOs8R/FX4leHvgb4q8X69o0Wj+KvDMzW3zRu1ndbJY0FzAGIYwyK52jcSCDk16N8HPHl945+H8Gt+JFjtdZsJbmy1SNPlSG6s5Gjk4JOAQA/XgGvkvxL4j13Wf2cvixpF7rb+KtH0G6itNM1mTBe8hEkLOGdeJTEx2+YPvV03j671LwV4u8WfDbRS0T/ABagsZNMZQdsV3cMtjqLZ/vCHbOT7fltUymE6bpKKU3Nu6vslC611slJyae1mctDiWtSrxxDnKVNU4pp21k5VUnpo23BQTWkuZM9J/Z9+MPin4reIfG8euQQ2+m6ZJZTaWqRlJDZ3omkiaUkncWiWNug6133x58ca18OfhZq/jDw6Ijf2L2ax+cpePE91FC2VBGflc45615J4K0nXNH+J3xi0P4cfYrS/sbfwtb2Iv1ke1SOKzK4dYirnEYIGD1xnjNZ/wAe4/iVH+zt4tHxLn0qe6NzpnkHSY544xH9ut87xOzEtnpjjFYSwVCeYU3BJQbp+71s1Fv89fmdtPNcXSyWuqjlKpGNf3+icZVEtejXKradj2vWPHGt2Pxt8PeAIPK/svU9KvLyXKEy+bA6qm1s8DBORisz4d/FCbVtM8f6141ubeysPCniPUtNSbHlolnaCPY0hJOW+c5PH0rC8R/8nR+Dv+xf1L/0alZPwQn0K20n4uT+JpLaLSk8ba2blrwotuI8Q5Mhk+QL654rmlhKX1ZS5deWO2+s5J/Ox3QzHEfXpQ9ppz1FrtZU4NX8k22fTtrdW99aw3tnIJYLhFkjdTkMjjKkexBzXz7qXjr4jeO/G+teC/hQ9hpen+GXSDUtXv4nuc3bru8i2gVkBMY++ztjJwAOCfoGzezks4JNPaNrVo1MJiIMZjI+UoV424xjHGOlfKHww8TaL8K/iR8QvAHjq8i0e417W7jX9NuLtlhhu7e+C5SOVyFLRMu0rnOc4zg44csopqrOMeaUVonr1V3brZf59D1s/wAVKMsPSnU5ITlaUk7fZbS5unM+qd9LJ6npfhTWvi1onjSHwb8Qba31vT7+3lmtNa022lgjjkg27obuMmRY2cHKMHAONoBOcdPrvxe+GHhnXF8Na/4nsLDUyVBglmVWQv8Ad3nome24ivnKy8Ra3pXxp8G+FdA+Jtz42gv5bx9UtFW1ljtoI4GMbSPbR/JmTA+Zgenrz5z8Pl8Rw/D3xta634q8O6Uj6hqn/CQW2qWMs14JWkcMZG+0xlwyYMWE9lywNezLJYTftKrS0Xwpq93JXtyu22yVnpqrny8OK61JOhh021KWs3GVlGMZW5lNJ/Fu5NpJ3Tsfd3iHx14N8JeV/wAJNrNrpYniknjNxKsYaKLaHYE8EAuo+pAHWk8JeO/B3jywk1TwdrFtq1tC2yR4HDeW2M4cdVOORkDivknQ/DVm/jH4A6XrE/8AbsVjo+ryQzXNu0RkVIYfIdoZcspVCuA3IIBp3j+z1SDxh8c7fwnG0d9eeF9PmKwDDySbZ1dgF5LmMEAjmuX+xKLtTUnzWvfp/E5Nt/Pfyseg+LMUr1pU1yJ8vKtZX9j7XdO2/u6LXe/Q+mdI+L/wv1/Xz4W0bxRYXmq7mQQRzqzOyfeVD0cjuFJIqzrXxT+HXhzVTomu+IbOx1ATRW5gllCyCWZQ8alevzKQc9ORnqK+LNRsb3VvhL4O0seOvD1vpTTaYNGXT9Lnk1CO7R0MYiCXTv5obIlOzPLbsGvdfAehaPf/ALRfxb1S/s4rm5tk0GOJ5EVzGrWhZtpI4yUUnH90elViMmw9NTnzO0U9Ot1KMd3FfzbWdrbkYLijHVpU6ShFSnKKvurOE57KbenJo20pJ3srHp2v/Gj4UeFtYbw/4h8VWFjqCEK8Mkw3Rk8gSYyEOP7xFUNf8c6nY/FbwP4R014ZdJ8R2ep3Ez43MxtUiaIxuDjB3nPByMV8q3Him4134Y+O9b0OXw54P8KXs+qfaLe6jlvdVu7jLIxlDzRok07D5Fw5XIxnv1Pw+lM3iD9nh2bcf+EY1EZ/3bW2H6Yrd5HTpxcne6U00+6g32smmtrv1ORcW1681TTVm6ck1p7rqxi1q7tNPdxjfXQ910j4teGNA8I6frHxF8W6T5l/cXkUV1bsY7aXyJmXYm/ndGu1X/2ga6rwp8U/h144up7Hwj4hs9WntY/OlSCQMUjBA3H0GSBXy78Ovihp/wANfgHo4iijv/EGsanq1tpVk7hBNOb+bLOzEBIY8hpHJAA4zkitDxJ4Xj0L4FePpdE1yDxH491ywkutWvLWVJJZQMCWOJUO5YIoiyRqAAB2ycVnXyalzyjK6bm4p6W+KzdraJeu+i622wnFOI9jTnBxklTU5Kz5l7l1Hm5tZSfTl0Wr3XN9F6H8Xvhh4m1xvDWgeJ7C/wBTBYCCKZWZyn3hGej477SaueKPid8PfBV/b6X4s8Q2WlXl0A0cVxMqOVJIDkHlUJBG44XPGa+ONWsb3VvBHgTTj468PJpy3mmnQ10zS52vVnjKmNYwl07KcArKSvHO7Br2D4f33hvSviN8WofHM9rbapPfRysb1lQSaObZBDgy8NEpDhsfKGyDWdfJqEE5xbaSenXRpbuK766OzVvM3wfFOLqONKajFya9525dYylaym3f3dG2rp3tpY9H+CvjbVviB4KbxDrJiaf7ff26mFdqGK3uHijPU87VGTnnrXOfHX4ieL/CMGj+H/hxBBdeJ9ZN3PFHcIZEFrp9u88xIUg7mISNMkDLVkfsnHTT8HoDooxp51LU/sw5GIftcnl9efu461xGmN8TPH/xl8VfEf4evpH2Dw/nw1atqq3DqTAVluniEDKOZTtLEnIAA701gqccdXbSUIN6PRXvZJ/Pp2TJlmteeUYSMZSdSqo6x1la3NKS+Ssn3aPpTw94/wDD2vfD+y+I7XKWukXNkt9JJI3ywps3SBz28sghvcGs+w+L/wAL9V8Rp4S07xPYXGsSHC2yTqXZsZ2jsWA6qDuHpXx/p0uueBvhx8ZPgj4sNul5pul32s6eLXeLY2OoRu0iQCT5xHDMSvzZOWPJ613/AMQNB0fRvgr8LxplnFbGw1bw3JCyIFZHd4w7ggZ3Nk7j1PeqnklBVOVt2lK0bWtZpNPz31X46EUuLcXKjzqKvCF5pp35lJxlFWemzabv00d9Pqey8WeGtQutWsrPUoJJ9CYLfpvAa2JUsPNBxtBUEgnggZzXIr42tta8ReEZPDOvafJo2uRX8nlNuNxeiBU2tasONsZJMhPUEYr5S/aItl134hXz+B7S7u10bTY/+E2FhKIvtOlmaORLTod1x5au/BU+VlckHA9W1i/0HVPi38FL/wAIPC2kT6ZrrWRiGIhD9lt/LAUdABgbeCMY6is45PCMIVLv3oydnbS0G9fXeO2mu+28+J6s61Sg0vcnBXTfvc1WMbryivdnv711ot/X9U+L/wAL9F8Qf8Irq3iiwtdV3rGYJJ1DI7fdVz0Rj2DEE1ynjL43+HfCHxS8N/Dm+u7WEavHO91LLIQ8DAKLZAvTMzMQCT/DgDuPkTwDFrg+A+vWHiHxb4e0uyd9RTXrbULCWbUUu2kkEvmkXKF5jwYiEz93aCRXtFsY/DvxC+Blvrep+cY9B1O3F1dK1s07eRbBNySncrsMZVvmz15rrnkeHpVJRbcrKa66tRbuvd/DXo7tHmUuLcbiaMJpKHM6Tvo7KVRRcWue+z+K0dmrJ6n1PpPijw9rukPr+kahDdadGZQ86MNimAlZAx7bCpBz0xV3SNX03X9Mtta0a4S7sbxBLDNGcpIjdGU9wexr4d8cvqvg/wAR+M/gZopaH/haF7aXWlOoyIU1JjFqx9hGsTPgdA+fr9yaXplloumWmj6bGIbSxhjghQdEjiUKqj6AAV4eY5fCjCM0782q/wANk9fm7eqZ9dkmdVcVUnSlFJ09Jf47tWXlZc2vSUS/RRRXkH0p4B+1j/yaz8ZP+xM8Q/8Apuno/ZO/5NZ+Df8A2Jnh7/03QUftY/8AJrPxk/7EzxD/AOm6ej9k7/k1n4N/9iZ4e/8ATdBQB//R+/f2+v8Am3L/ALLN4Q/9uq+/6/OD/gpNqOu6R4V+BereFtI/4SDWrL4s+GJ7HTftCWn266jjvGhtvtEgKRea4CeYwKpnceAa6D/hof8Abs/6NH/8v7R//jVAH3/RXwB/w0P+3Z/0aP8A+X9o/wD8ao/4aH/bs/6NH/8AL+0f/wCNUAff9FfAH/DQ/wC3Z/0aP/5f2j//ABqj/hof9uz/AKNH/wDL+0f/AONUAfd15o+kajdWd9qFjBdXOnuZLaWWJXeB2G0tGzAlCVOCVwccU2DRNGtdUutctrC3h1K9VEuLlIkWeZYxhFkkA3MFH3QScdq+E/8Ahof9uz/o0f8A8v7R/wD41R/w0P8At2f9Gj/+X9o//wAaq/aSta5m6ML81lff52tf7tPQ+4ovC3hmDRZPDcGkWcekzb99mtvGLZvMYs+6ILsO5iS2RyTk81U8NeCPBvgxJk8JaHZaMLkgy/Y7eODeR03bAM47Z6dq+KP+Gh/27P8Ao0f/AMv7R/8A41R/w0P+3Z/0aP8A+X9o/wD8aqvbzs48zs99dzNYOlzRnyK8dE7LRdl2PsS3+F3w1tb671O28KaVHd36SR3Eq2UIeVJgRIrnbyHBIcHhs85q3rXw98B+I7Cz0rXvDun6hZ6coS1hntYpI4EUBQsSspCKAAMLgYGOlfGH/DQ/7dn/AEaP/wCX9o//AMao/wCGh/27P+jR/wDy/tH/APjVX9bq3Uud3XmzJZbh+Vw9nGz3VlZn3H/wjHhs6F/wi/8AZNp/Y2zyvsXkR/ZvLznb5ONm3POMYrP0XwF4H8OfZT4f8P6fprWRlMDW9rFE0RnCiUoVUFTIFUOR94AZzgV8Wf8ADQ/7dn/Ro/8A5f2j/wDxqj/hof8Abs/6NH/8v7R//jVQq87Nczs/M0eDotqTgrrRaLReR93aVpGk6FZJpuiWUGn2kZdlht41ijVpGLuQiAAFmJY8ckknmucsPhv8PdL1xvE2m+GdNtNWcsxu4rSJJ9zfebeqhstnk5ye9fGn/DQ/7dn/AEaP/wCX9o//AMao/wCGh/27P+jR/wDy/tH/APjVEa81e0nrvrv6jlg6UuXmgny7aLT07fI+zvEPw78A+LbyHUfFPhzTtXuoBtSW7tYp3VRzt3OpOM9uldbDDFbxJb26LFFEoVEUBVVVGAABwAB0FfAn/DQ/7dn/AEaP/wCX9o//AMao/wCGh/27P+jR/wDy/tH/APjVKVWckot6LYqnhqcJSnGKTe7tq/Xufb1z4R8KXsOo215otlPFq7pJepJbRMt08eNjTAqRIy7VwWyRgY6CtC+0jSdUltJ9TsoLuSwlE9u00ayNDMAVEkZYEo4BI3DBwTzXwj/w0P8At2f9Gj/+X9o//wAao/4aH/bs/wCjR/8Ay/tH/wDjVHtZdw+r09fdWvl53/N39T7sv9F0bVLizu9TsLe8n06TzraSaJJHgkxjfGzAlGxxlcGrtxbwXcElrdRrNDMpR0cBldWGCrA8EEcEHrXwN/w0P+3Z/wBGj/8Al/aP/wDGqP8Ahof9uz/o0f8A8v7R/wD41U87012LVOOrtvv5n3BpfhTwtok8NzoujWVhNb24s43t7eOJktg5kEKlFBEe8ltg+Xcc4zTbrwj4UvYdRtr3RbK4h1h0kvUktonW6ePGxpgVxIV2jBbOMDHSviH/AIaH/bs/6NH/APL+0f8A+NUf8ND/ALdn/Ro//l/aP/8AGqv28783M7mX1Sly8nIrdrL0/LQ+7otI0mHU59ahsoI9RuY0iluVjUTSRxklEaQDcyqScAnAycVVj8NeHIdNvNGi0q0TT9QMrXNusEYhna4z5pkjA2uZMnfuB3d818N/8ND/ALdn/Ro//l/aP/8AGqP+Gh/27P8Ao0f/AMv7R/8A41U+1l3/AKRfsIfyrr077/f1Ps7XPh54C8TQ2dt4i8OadqUWnKEtkuLWKVYUAACxhlIVcADaOOOlWW8EeC3t720fQNPaDUYoYbqM2sRSeK3GIklXbh1jHCK2QvbFfFH/AA0P+3Z/0aP/AOX9o/8A8ao/4aH/AG7P+jR//L+0f/41Wn1mpZLmdvUxeAoczl7NXe+i7W/LT0Pv4AKAqjAHAArnz4S8KHR7jw8dFsv7Ku2d5rT7NH9nlaRt7s8W3YxZvmJIOTyea+If+Gh/27P+jR//AC/tH/8AjVH/AA0P+3Z/0aP/AOX9o/8A8arONSS2ZvOjCXxJPp8nufdU2haHcTWFzcadbyy6USbN2hRmtiy7CYSRmPK/KduOOOlYms/D3wF4j1SLW/EHhzTtS1CEAJcXNpFLKAvQb3UnA7c8dq+MP+Gh/wBuz/o0f/y/tH/+NUf8ND/t2f8ARo//AJf2j/8AxqqhXnF3jJr5mdXCUprlnBNb6pbn3VrGhaJ4h06TSNf0+31KxlxvguYllibHTKOCOO3FUNE8HeEfDLK/h3RLLS3SEW4a1to4WEIYuI8ooO3ezNjpuJPUmviT/hof9uz/AKNH/wDL+0f/AONUf8ND/t2f9Gj/APl/aP8A/GqSrTUeRN27DlhqbmqjiubvbX7z7Lt/hv8AD2018+KrXwzpsOsli5vEtIVn3nq/mBd2455bOT610cGkaTa6jdaxbWUEV/erGlxcJGqzTLFkRiRwNzBMnaCTjJxXwj/w0P8At2f9Gj/+X9o//wAao/4aH/bs/wCjR/8Ay/tH/wDjVOdecvik30+QqWDpQ+CCWt9Et+/r5n3/AEV8Af8ADQ/7dn/Ro/8A5f2j/wDxqj/hof8Abs/6NH/8v7R//jVZHQff9FfAH/DQ/wC3Z/0aP/5f2j//ABqj/hof9uz/AKNH/wDL+0f/AONUAH/BUf8A5MU+Jn/cF/8ATxZV9/1+IP7evxl/ax8V/sneOdA+Jn7Ov/CCeG7r+zPtetf8JdpuqfZdmpWrx/6JbxrLJ5kipH8p+XfuPCmvr/8A4aH/AG7P+jR//L+0f/41QB9/0V8Af8ND/t2f9Gj/APl/aP8A/GqP+Gh/27P+jR//AC/tH/8AjVAH3/XGa58OPh54nvzqniXwvpWrXpUIZ7uygnlKr0XfIjNgdhmvjP8A4aH/AG7P+jR//L+0f/41R/w0P+3Z/wBGj/8Al/aP/wDGq0pVZwd4Oz8jGvhqdWPLVipLzVz7o0Xw/oPhy0+weHtNttLtc7vKtYUgjz67UAGaxvEXw98BeLrhLvxV4c07V54wAsl3axTuFU5ADOpOPbOK+MP+Gh/27P8Ao0f/AMv7R/8A41R/w0P+3Z/0aP8A+X9o/wD8apxrzUudSd+99SJ4OjKn7KUE49rK33H3HN4Y8NXGhnwvPpNpJoxQRmyaCM22wHcF8krswCM4x15qxdaHol9f2Oq3un29xe6Xv+yTyRI8tv5q7X8pyCyblGG2kZHBr4U/4aH/AG7P+jR//L+0f/41R/w0P+3Z/wBGj/8Al/aP/wDGqXtZd/6e/wB5f1en/KunTtqvue3Y+7rbR9IstQvNWs7GCC+1Hy/tVxHEqyz+Su2PzXA3PsU4XcTgcCk1fRtI1/T5NJ16xg1KxmKl4LmJZonKMHXcjgqdrAEZHBAPWvhL/hof9uz/AKNH/wDL+0f/AONUf8ND/t2f9Gj/APl/aP8A/GqXtJXUr6lexhyuFlZ3uvXf7+p92yaNpE2qQ65NYwPqVtG0MV00SmeONzlkWQjcFYjkA4NUv+ET8LfYdR0v+xrL7Hq8sk97B9nj8q6llx5kkybdsjPgbmYEnAzXw/8A8ND/ALdn/Ro//l/aP/8AGqP+Gh/27P8Ao0f/AMv7R/8A41TVWS2ZLw9N3vFfd30f4H31b28FpBHa2sawwwqEREAVUVRgKoHAAHAArF8Q+FPDHi6zGn+KdJtNXtlO5Y7uBJ1VvUBwcH3FfD//AA0P+3Z/0aP/AOX9o/8A8ao/4aH/AG7P+jR//L+0f/41SjUlGXNF2ZVSjCcXCaTXbofbnh3wf4T8IW72nhTRrPR4ZMb1tII4A5HQtsA3H3NZ2s/Dj4feI9Vj13X/AA1pupajFt23FxaRSy/L9352Uk7e2Tx2r40/4aH/AG7P+jR//L+0f/41R/w0P+3Z/wBGj/8Al/aP/wDGqtYmopOak7976mMsDQdNUnBcq2Vlb7j7um0fSLjULXVrixglvrBZEt7holaWFZQBIsbkblDgAMARnAzRFpGkwalPrMFlBHqF0iRzXKxqJpI487FeQDcyrk4BOBk4r4R/4aH/AG7P+jR//L+0f/41R/w0P+3Z/wBGj/8Al/aP/wDGqz55bXNvYwvey7/O1r/dp6H2XYfDf4e6XrjeJtN8M6baas5ZjdxWkST7m+829VDZbPJzk966O20jSbO/vNVs7KCC91Hy/tM8carLP5K7Y/NcAM+xeF3E4HAr4R/4aH/bs/6NH/8AL+0f/wCNUf8ADQ/7dn/Ro/8A5f2j/wDxqrnXnL4pN9N+hnSwdKnpCCWt9Elr39fM+yx8OPh8uvSeKR4Z03+2JWLPefZIfPZm4LGTbu3EHk5ye9atn4T8Lac2nvp+jWVq2kpJFZGK3jQ2scuPMSHao8tXwNwXAPevh/8A4aH/AG7P+jR//L+0f/41R/w0P+3Z/wBGj/8Al/aP/wDGqcsTUejk/vJhgaEXeMEuuy33v96T+R9j33wy+G2p2ltYal4T0m7trIytBFLYW8kcRnbfKUVkIUu3zNgDceTk1Y0D4e+AfCl4+o+FvDWmaPdyRmJprOzht5GjYhihaNFJUlQSM4yB6V8Yf8ND/t2f9Gj/APl/aP8A/GqP+Gh/27P+jR//AC/tH/8AjVU8XVceVzdvVkLLsOpqoqceZdbK/wB59l6X8N/h7omtP4j0bwzptjqj7ibqC0ijmy/3jvVQQWzyc896ua/4I8GeKri3u/E+g2OrT2ZzC93bRTtHzn5S6kjnnA7818Uf8ND/ALdn/Ro//l/aP/8AGqP+Gh/27P8Ao0f/AMv7R/8A41SeKqc3PzO/e5SwFBQdNU1yvW1la/ex936bpOlaLbGz0ezhsbdneUxwRrEhkkYs7bVAG5mJLHqScmmaToujaBZ/2foVhb6da72k8q2iSGPe53M21ABljyTjk9a+E/8Ahof9uz/o0f8A8v7R/wD41R/w0P8At2f9Gj/+X9o//wAarN1JO93ubxowVrJabeR9wal4U8Lazdvf6xo1lfXMls9m0s9vHLI1tJy8JZlJMbZOUztPcVZutA0K+srbTb7Tra4tLNonghkhR44ngwYmRCCFKYG0gfL2xXwt/wAND/t2f9Gj/wDl/aP/APGqP+Gh/wBuz/o0f/y/tH/+NU/bT01ehP1anq+Va76b+p91adoWiaQ95JpOn29k2oTNcXJgiSMzzP8AekkKgb3PdmyT61Q0/wAG+ENINgdK0OxsjpfnfY/Jtoo/s32j/XeVtUbPM/j243d818Sf8ND/ALdn/Ro//l/aP/8AGqP+Gh/27P8Ao0f/AMv7R/8A41R7aeur1F9Vpae6tNtPO/5pP1PsrUvhv8PdZ1pPEer+GdNvdVQqRdTWkUk2V+6S7KWJXHBJ47Vsa54W8M+JxbL4k0m01UWcglg+1QJN5Ug6Om8Ha3HUYr4d/wCGh/27P+jR/wDy/tH/APjVH/DQ/wC3Z/0aP/5f2j//ABqq+sVNHzPTbXb0I+o0bSXIve30Wvr3PpPTPAvi/WfiqPiD48bT1tfD8N1aaHBZGR3CXTjfcXDSKu2QxKqbFyoy3Pr7ZXwB/wAND/t2f9Gj/wDl/aP/APGqP+Gh/wBuz/o0f/y/tH/+NVWIxMqrXN0Vl5L/AIfX1IwOAhQUlC7cndt7t7fkkl5JH3/RXwB/w0P+3Z/0aP8A+X9o/wD8ao/4aH/bs/6NH/8AL+0f/wCNVznafQH7WP8Ayaz8ZP8AsTPEP/puno/ZO/5NZ+Df/YmeHv8A03QV8P8A7Qvx2/bO1f4BfEvSfFP7L3/CP6Le+GdZgvtS/wCE20q7+w2sllKs1z9njiDy+UhL+WpDPjaOSK+4P2Tv+TWfg3/2Jnh7/wBN0FAH/9L79/b6/wCbcv8Ass3hD/26r7/r4A/b6/5ty/7LN4Q/9uq+/wCgAooooAKKKKACiiigAooooAKKK/K/xB+0f8LfgL+3v8SZPjH4tHh3SdQ8K6FHZLOtxLE0yvI0mxIUkCnbjJwM+tAH6oUV+T/iT9qT4O/Hn9s39mvTPgr4zXxBFp1x4qOpxW63MKBZdJJg8xZUjD5MbleuCO3FfR3iT9uj4WeHNe1qA+H/ABJqXhPwvqX9k6z4ss9OWXQNOvQ6xyRzT+aJWWF2CyvHC6oSOTkGgD7Sor5C+J37ZvgD4TeI9S07xN4W8US+G9CmtbfVPFFtpfmaFYy3gjaNZLkyK7gCVN7RRuqlgudxwOg+KH7VXgv4X/Ez/hT83h7X/Eni2bQRr9pZaLZpdveQG5e2MMQMqMJVMbyMXCxhB/rNxCEA+naK+C/Fv7cf9gfEDwT4J0f4S+M9afxXocmuyQQ6ckeowR4TZCLaeaJWeMuPtB8zEZ2qN7Ftn25qetW+kaBd+IryKRYLK1kupIwB5oSNC7LgnG7AxjOM96ANiivirwd+3N8PPHOj6d4s0HwZ4wl8LahpV1qn9tDR/M0+I2cLTS2jSwyyFrkbDGFjDoZf3QcuGVer+HH7WnhT4geI9Z8EX3g7xV4Q8VaPpDa8uj63pfkX15piyeV59rHDJMJP3hEezIYOduODgA+qqK+GfgV+2/ofxV+Hms/E7xl4J13wJ4Z0W21TUJda1CCI6S9rp949uI4p45XlkudijfGIQPMWRELYUt3Hwu/a68G/ErxnpHga/wDCfibwRf8Aii0mvtBfxFp6WcWsW9sokmNo0c0vzpGRIY5Aj+WQ2MGgD6uor55/ag8aaP4B+FEniHXdX1zQ7RdU0iA3Ph77P9v33F/DGiD7TiPypGYJN38stgE8HhPHP7ZfhHwd498cfDfT/A3i3xXrPw9jsbjVv7F06G5gjtr62+1LMsr3EYwkfDI22Rmz5aOquygH2DRXyx4r/bA+E3h3wF4H8c6NHqfi2T4lKD4c0nRbQ3OqajhBJLsgdowggU5maR1CdCc8Vn65+2H4J8G+AvDvj74ieEvE/hGLxD4ki8LrY6lp6R31veTQyTpLJEszb7crGdrwGVnJAVCcgAH1vRXx7fftm+EdH8Mpq/iDwH4x0nW77WG0TS/DtzpKprOq3KwrcF7OATFHhWJg7ytIqpyrENgG7pv7YfgbVPBet+IoPCvij/hIvDupQaRfeExpm/xDFe3KebAn2ZJGjKSRZlSXzfKKAncCCKAPrWivkrSf2zfhRd/C3x18UdfsdZ8Mx/DaVbfX9K1Oy8rVLOeQIYkMKO6N5vmKI2VypzkkDmug0v8Aah8Ir8OvE/xO+IXhzxB8OtH8JQRXV3/wkNisDyQz58prcwSTpOXYBAqOWDsqkAkCgD6Vor5g+FX7VHhf4meNY/h5qnhLxL4B8QX1i+p6da+JrBLJtRs4mVZJLZo5plLR71MkbFZFBztwCR9P0AFFFFABRRRQAUUUUAFFFFAHwB/wVH/5MU+Jn/cF/wDTxZV9/wBfAH/BUf8A5MU+Jn/cF/8ATxZV9/0AFFFFABRRRQAUUUUAFFFFABRRRQAUUV+V/iD9o/4W/AX9vf4kyfGPxaPDuk6h4V0KOyWdbiWJpleRpNiQpIFO3GTgZ9aAP1Qor8n/ABJ+1J8Hfjz+2b+zXpnwV8Zr4gi0648VHU4rdbmFAsukkweYsqRh8mNyvXBHbivo7xJ+3R8LPDmva1AfD/iTUvCfhfUv7J1nxZZ6csugadeh1jkjmn80SssLsFleOF1QkcnINAH2lRXyF8Tv2zfAHwm8R6lp3ibwt4ol8N6FNa2+qeKLbS/M0KxlvBG0ayXJkV3AEqb2ijdVLBc7jgdB8UP2qvBfwv8AiZ/wp+bw9r/iTxbNoI1+0stFs0u3vIDcvbGGIGVGEqmN5GLhYwg/1m4hCAfTtFfBfi39uP8AsD4geCfBOj/CXxnrT+K9Dk12SCHTkj1GCPCbIRbTzRKzxlx9oPmYjO1RvYts+0fEfiay8L+EdU8ZanDN9k0ixnv54kVTN5dvEZXUKWC78KQAWAz370AdHRXxL8PP27/hh46vfCsGseFPFngaw8cvDFoWp+IdKW30zUZrld0EUN3bzXEQeYcxhyu7oOSAeq8d/tf+BPB/jDWfBGgeFvFPj/UPDAT+2n8MaS2owaW8ieYsdxKXjUybPmMcW9wOoBBAAPrCivnW0/aj+FGrr8L7rwzc3Gu6f8XLm4tdGvLSNfJSS1geeX7SJXjki2iNkKhGdXG1lGCRmeNv2r/h14DuPi1baxp+qzN8GrHS9Q1jyIYGE8WrxvJALTdOu9lEZ3iTywDjBbsAfTtFfKngL9rjwT44+I2h/DW78MeJPCl74tsp7/w/c65p62ltrENqglm+zESvIHSNhIUmjjbZzjkZ+Uf2xP2uND8XfAT4veFfhh4X8Ua5YaTb3OlXPirTrSNdFtb+F1EqC5aeOZxEfleSKJlBIwSDmgD9WaK+J7H40eFfA/jfxTPrWqeIryTwb8NrHxHfaYotn0sWcAnkaW1DMsrXsnlsreYyxlQvzf3dLwL+2v8AD3xv4l8FaFN4V8UeG7H4iwq/h/VtY05LbT7+Yw+ebdHWZ3WTZnaWQJJgmN3UqzAH2NRXwz40/b4+GvhTX59G0nwX4v8AFlnBrf8Awjn9raTpsH9lPrG/yjZpd3l1bI0gk/d5HyluFY8V9wW8pnt452jaEyKrFHwHXIztbBIyOhwTQBNRRRQAUUUUAFFFFABRRRQAUUUUAeAftY/8ms/GT/sTPEP/AKbp6P2Tv+TWfg3/ANiZ4e/9N0FH7WP/ACaz8ZP+xM8Q/wDpuno/ZO/5NZ+Df/YmeHv/AE3QUAf/0/v39vr/AJty/wCyzeEP/bqvv+vgD9vr/m3L/ss3hD/26r7/AKACiiigAooooAKKKKACiiigAr5B8H/Cvxrp37Z/xG+LGqaUE8Ka74a0awsbwywv5t1au5mTyg5lXaCPmZAp7E19fUUAfIHxn+FXjTxT+09+z18RPDWkrceH/A8/iZ9ZuhLDH9lXUNNWC2Plu6ySb5AV/dq23q2BzXxbB+y78RvhvqHjT4d3fwOT4veHNe1jUL/S9WXxa2kWy2uoy+d9m1GykmBJhZmDSRxuZB0XPJ/ZGigD8i/2q/gv+1F8U9G+JHw6tPCV7rugz6VpUHhBNO123sNHs/Iiia9W7tpJ4Z7q4MyMsJnWSILtIMbDJ+pvC3w++Imqftg6d8ddf8LNouiSfDSLRHM1zaTy2uqHVjdPasIZXYsIcM0iBoj90OTX2dRQB8SfHvwh8YtE/aE8AfHz4W+DR49t9E0bVtEvtNj1G3064Q3zwyQzI90VjZA0ZD87gOQpr6k12313xF8N9QtbiwFprOp6TKj2azLKIrqaAgwib5FYK527/lB68Cu3ooA+Qfgl8P8A4t/C/wDYp0L4eaFYW2j/ABK0TwvLb2tvcPDLBDq/lOY/MeJniYecQWIZlOSSSM186fAn4Z/H7Qv2m/CvxZ8aeAtcjt77wZcaBrmpav4gsdSuE1N7uC7kuPKjuXWK2YxFIYbVAoyCYo/mNfqTRQB+UHg39nv49ap+zN4t/Yn8X+DotG0lodaNh4vXVbea0uZbnUn1GyAs483Khnk2y71Xaq9GLYrrfgL8LfiBB8Q/Bd749/Z8j8Maj4Y843Hiafxe2qQRyNavC0mn2fnSybrgsAyyKgRGYEsQM/plRQB8q/tl/Dbxp8V/ghL4Q8Aad/amrtrOh3Yg86KD9zZ6jBPO2+Z0T5I0ZsbsnGACcCs34a/C7xxoP7R37RPjvWtLEOgeO4vDKaPcGaFxdnT9Mkt7keWrl49kjBf3iru6rkc19eUUAfkn4C/Z3/aG+Evw5/Z0+Img+ELbX/Gnwn03WNK1jwxcahbQSy22rucyWl5ve1WaIKCcvh0YruBGK9w+Kfhv45fH/Q/g/rGu/DNvCl54T+J+j63e6dPqtjfSRaLY28+69keN1j3CWUL5EZkkGAwzk7fvyigD4Y/bM+BHjT4m6p8N/id4A0WLxZqnw5vb95dAm1F9J/tKy1SBYZ1ivUZfJmQxoyFiFI3AkjKt5RY+APj1ovw58ba/8Hvg+/wu8S6/e6ElzCPEkGqa7qmmWkkgvfLubiSW0tp0gk2WxeQkbnJ2sI8fp9RQB+V/wy+Ffxk8HX/x/F/8Fp9c0Tx+ukXVhpmu+IrS9bUYoYIbS7s7q7luLmT7S0Zll3yFog6hBIVKtXNaB+yl8c/HX7O3xh+CmoWVz4F8KavLpdz4E0PW9Vj1i50qWwkW6nglubdpAtpLNFGsSGSRo1LNjd979daKAPzp+AXw48Zr8UfD3iPxj+z4vgS90CC7WTXrrxc2tBHnhMTJp9sJpW2z/LvMoTaox8zAEfotRRQAUUUUAFFFFABRRRQAUUUUAfAH/BUf/kxT4mf9wX/08WVff9fAH/BUf/kxT4mf9wX/ANPFlX3/AEAFFFFABRRRQAUUUUAFFFFABRRRQAV8g+D/AIV+NdO/bP8AiN8WNU0oJ4U13w1o1hY3hlhfzbq1dzMnlBzKu0EfMyBT2Jr6+ooA+QPjP8KvGnin9p79nr4ieGtJW48P+B5/Ez6zdCWGP7KuoaasFsfLd1kk3yAr+7VtvVsDmvi2D9l34jfDfUPGnw7u/gcnxe8Oa9rGoX+l6svi1tItltdRl877NqNlJMCTCzMGkjjcyDoueT+yNFAH5F/tV/Bf9qL4p6N8SPh1aeEr3XdBn0rSoPCCadrtvYaPZ+RFE16t3bSTwz3VwZkZYTOskQXaQY2GT9TeFvh98RNU/bB07466/wCFm0XRJPhpFojma5tJ5bXVDqxuntWEMrsWEOGaRA0R+6HJr7OooA+JPj34Q+MWiftCeAPj58LfBo8e2+iaNq2iX2mx6jb6dcIb54ZIZke6KxsgaMh+dwHIU17z8WLjUbv4AeMrvWLMaffzeGNRe4tllEwgmaycvGJAFDhGyu4ABsZwK9irF8SaDYeKvDuqeF9VDGy1i1ns5wjbX8q4jMb7T2O1jg9qAPyG+FXhf48/tFfs5fAT4RnwB/wiPgnQ18N6vfeJrvU7O4F5Y6UiTRJY2sDNcJNcEKd0gTyxkHOa+hND0T9pL9mrxr8TLD4d/DGP4peG/iB4ivfFNhewazaaXPZXepInn2t6l4QTGjx/u5It+E4IyQq/b3w98DaF8MvAugfDvwwJF0jw3ZQafaCZ/MkEFsgjTe2BubAGTjmuxoA/KyH9mH43fCz4c/CLxXoOmWnjXxr4G8War4o1nRrK6SyhmGvLOLmCwmutsY+z+au0OVD4YjBIByPGXwP/AGj/AIl6V+1brmq+AV0PUfitonhm08P6eNVsrl5n02KeOaOWVZFjjkTcpbcRHlsI8gBY/rVRQB8b+O/hT481n40/s1eL9N0rztL+H/8AbY1ufzoVNoLzR/ssPys4eTfNhf3QfHVsLzXyF4n+Cv7T/gD9mnx/+yJ4O+GC+NtI1ibVf7G8R2Ws6daZttRujdoL22vZIJfOjLsjFWcHAAOACf2FooA/N7xr8BPizq/jH4uarp+heba+J/g9H4W05/tVsvn6ysd0ptsNKCnMifvHCx8/f4OOh8RfBP4mXnw4/ZS0W30YNf8Awz1Xw7c+IENxbj7DBY6VJbXLbjIFlCSkLiIuW6qCOa/QCigD8IPD/iP4ezePvEvxL8eWnjaL9nxfGZ8R6JNpNq0vg+W4WZGOoXzTKL5YzfF5CkaC23j5dy/e/d1HSRFkjYMrAEEHIIPQivkV/wBh34ASQHQzaawPCLTG4Phca7qQ8PGYzefu/s77R5O3zfn8rHlZ/gr67VQoCqMAcADtQAtFFFABRRRQAUUUUAFFFFABRRRQB4B+1j/yaz8ZP+xM8Q/+m6ej9k7/AJNZ+Df/AGJnh7/03QUftY/8ms/GT/sTPEP/AKbp6P2Tv+TWfg3/ANiZ4e/9N0FAH//U+3f+Ck2gf8JX4V+Bfhb+0r3R/wC2Piz4Ys/t2mzfZ761+0R3kfn202G8uaPdujfB2sAcHFdB/wAMC/8AVxvxm/8ACv8A/uWj9vr/AJty/wCyzeEP/bqvv+gD4A/4YF/6uN+M3/hX/wD3LR/wwL/1cb8Zv/Cv/wDuWvv+igD4A/4YF/6uN+M3/hX/AP3LR/wwL/1cb8Zv/Cv/APuWvv8AooA+AP8AhgX/AKuN+M3/AIV//wBy0f8ADAv/AFcb8Zv/AAr/AP7lr7/ooA+AP+GBf+rjfjN/4V//ANy0f8MC/wDVxvxm/wDCv/8AuWvv+igD4A/4YF/6uN+M3/hX/wD3LR/wwL/1cb8Zv/Cv/wDuWvv+vK/jF8ZvAfwK8HHxt8QLqWGzluYbK1gtoXubu9vbgkQ21tBGC0kshB2qOwJJABIAPlT/AIYF/wCrjfjN/wCFf/8ActH/AAwL/wBXG/Gb/wAK/wD+5a+iPgr+0J4S+OMuuafouja94b1fw79ma+03xDpc2l3kUd55v2eQJKNrJIYZACrHG05AyM+m+O/GOj/DvwR4h+IHiHzP7K8M6dd6nd+Su+T7PZQtPLsXI3NsQ4GRk96APir/AIYF/wCrjfjN/wCFf/8ActH/AAwL/wBXG/Gb/wAK/wD+5a+sfDHxh+H3iy+8OaPpmqKmr+KtDj8R2NhKCly+mSeWPOK8qMGVVI3ZznGQCa9OoA+AP+GBf+rjfjN/4V//ANy0f8MC/wDVxvxm/wDCv/8AuWvv+oLm5t7O2lvLqQRQQI0kjscBUUZJJ9AKAPgf/hgX/q434zf+Ff8A/ctH/DAv/Vxvxm/8K/8A+5a+2fBnjLwx8QvC2m+NvBeoR6toesQie0uos7JY24DDcARyMYIBFdNQB8Af8MC/9XG/Gb/wr/8A7lo/4YF/6uN+M3/hX/8A3LX3/RQB8Af8MC/9XG/Gb/wr/wD7lo/4YF/6uN+M3/hX/wD3LX1p47+K/hT4d+IPBvhnxD5/2zx1qR0rTvJjDp9pWF5z5pyNq7EPODz2r0ugD4A/4YF/6uN+M3/hX/8A3LR/wwL/ANXG/Gb/AMK//wC5a+/6KAPgD/hgX/q434zf+Ff/APctH/DAv/Vxvxm/8K//AO5a+/6KAPgD/hgX/q434zf+Ff8A/ctH/DAv/Vxvxm/8K/8A+5a+/wCigD4A/wCGBf8Aq434zf8AhX//AHLR/wAMC/8AVxvxm/8ACv8A/uWvv+igD4A/4YF/6uN+M3/hX/8A3LR/wwL/ANXG/Gb/AMK//wC5a+/6KAPgD/hgX/q434zf+Ff/APctH/DAv/Vxvxm/8K//AO5a+/6KAPgD/hgX/q434zf+Ff8A/ctH/DAv/Vxvxm/8K/8A+5a+/wCigD4A/wCGBf8Aq434zf8AhX//AHLR/wAMC/8AVxvxm/8ACv8A/uWvv+igD8Qf29f2Q/8AhV/7J3jnx1/wur4meLf7M/sz/iV+IfEf2/S7jztStYf39v8AZ037N+9PmG11Vu2K+v8A/hgX/q434zf+Ff8A/ctH/BUf/kxT4mf9wX/08WVff9AHwB/wwL/1cb8Zv/Cv/wDuWj/hgX/q434zf+Ff/wDctff9FAHwB/wwL/1cb8Zv/Cv/APuWj/hgX/q434zf+Ff/APctff8ARQB8Af8ADAv/AFcb8Zv/AAr/AP7lo/4YF/6uN+M3/hX/AP3LX3/RQB8Af8MC/wDVxvxm/wDCv/8AuWj/AIYF/wCrjfjN/wCFf/8Actff9FAHwB/wwL/1cb8Zv/Cv/wDuWj/hgX/q434zf+Ff/wDctff9FAHwB/wwL/1cb8Zv/Cv/APuWj/hgX/q434zf+Ff/APctff8ARQB8Af8ADAv/AFcb8Zv/AAr/AP7lo/4YF/6uN+M3/hX/AP3LX3/Xj3xx+N3g79n7wMfiB44hvbjTvtdtZLFp9v8Aabl57p9kSpEGUsS3HBz7UAfMP/DAv/Vxvxm/8K//AO5aP+GBf+rjfjN/4V//ANy11en/ALeXwWTVLHTPHeleKPh2mpzpbW934o0C80uyeaQ4RGuXQxJnP3nZVHcivtNWV1DKQQRkEdCKAPgH/hgX/q434zf+Ff8A/ctH/DAv/Vxvxm/8K/8A+5a+/wCigD4A/wCGBf8Aq434zf8AhX//AHLR/wAMC/8AVxvxm/8ACv8A/uWvv+igD4A/4YF/6uN+M3/hX/8A3LR/wwL/ANXG/Gb/AMK//wC5a+/6KAPgD/hgX/q434zf+Ff/APctH/DAv/Vxvxm/8K//AO5a+/680+MPxX8K/A/4caz8UvG/n/2JoSwtcfZoxLNieZIF2oSufnkXPPTJoA+S/wDhgX/q434zf+Ff/wDctH/DAv8A1cb8Zv8Awr//ALlr7/ooA+AP+GBf+rjfjN/4V/8A9y0f8MC/9XG/Gb/wr/8A7lr7/ooA+AP+GBf+rjfjN/4V/wD9y0f8MC/9XG/Gb/wr/wD7lr7/AKKAPgD/AIYF/wCrjfjN/wCFf/8ActH/AAwL/wBXG/Gb/wAK/wD+5a+/6KAPgD/hgX/q434zf+Ff/wDctH/DAv8A1cb8Zv8Awr//ALlr7/ooA+AP+GBf+rjfjN/4V/8A9y0f8MC/9XG/Gb/wr/8A7lr7/ooA+AP+GBf+rjfjN/4V/wD9y0f8MC/9XG/Gb/wr/wD7lr7/AKKAPyg/aF/Yk/4RT4BfEvxT/wAL8+LOsf2P4Z1m8+w6l4p+0WN19nspZPIuYfsy+ZDJt2yJkblJGRmvuD9k7/k1n4N/9iZ4e/8ATdBR+1j/AMms/GT/ALEzxD/6bp6P2Tv+TWfg3/2Jnh7/ANN0FAH/1fv39vr/AJty/wCyzeEP/bqvv+vgD9vr/m3L/ss3hD/26r7/AKACiiigAooooAKKKKACiiigAr5z/aZ+B9x8dPBWlaVoHiFvCnizwxq9rr/h/UxGs62+p2IfyzJC3EkTK7K6+hyQwG0/RleO/Gj4KeHvjfoGnaNrWr6x4eu9GvV1HT9T0K+aw1Czuljkh3xSqGU7o5XRldWUhjx3oA+TdB/aD+LuveEviN8Kvifqei/CD4rfDyLTptR8Quq3miyaTeS8alZRzsuWZFdVimOFlZA3JaNfHNN+MXxF8Z/Br9rj4W+OfEtx4zsfBXgqW60nWb7RjoN/dW+r6RfOwntDHCNqND+6kEa+Yp38gjH1DbfsJ/Ct/BXjTwx4k8QeJPEus+PZLCXU/Eep6gs+s7tLlWayEMvlCKNIJFBRBER2bcAANCz/AGMfCseo/EPVNX8d+KtZuPijoDeH9ea7urI/aIjBLbJcKI7NFSaKKZxGFAiGcmJjQB4d4C8ZfFK/8T/Cv4MeCPEY0C31f4JLqNpL9ktZzb6un2SC3uS00MjMqbvmiJMbDqpOCKPwn/aq+Kfx18R/B/4TeG70aL400oahe/E0fZ4He1XQnNi9oyOjpF/aF5hsxhXjj+4w5r7L8L/s7+CvCXjnwl8QNNvdQk1Hwb4UXwfZxyyQmGTT1eJxJMqxKxnzCvzKypgn5OmNTwR8Bvh18P8A4j+Pvin4csjDrnxHktJNUYkbM2kRjAiAAKCQlpJOTuc7j2oA+FPh/wDHL4xaB+0Z8OPAmrfFe3+Kmi+PLjWbDVvsvh9bHSdNv9PspLpY9L1GJQt0I3Qxyq0srKBl9rtxpfs1x/HzVPHXxy8aeIvi3eajpnhHxdrenSaM2mWf2a5a202H7O8crq81skRdCsMb7CY8tuLyFvWvDv7B3gXwrd+B5tF8e+Lobb4cX1xd6Bam8s2gsYrsOtxaqDZ72ikWRlLOzSheFkWvQE/ZP8IWXxN134k6B4r8TaHH4ounv9V0Sy1BY9Ivb2SD7O08sDRM5Ypyy+ZsZgpK/KBQB8pfD79oD47/ABL+HX7Pfwz8JeILPRPGXxN0K+1nV/Elzp8M32W00wqrLaWKCK2aeVnVcFQkaAttJIx0uu/Gb4+fBu/+Lnwa8d+K7bxdrei/DjVfHPhvxJDp0FlcxfYxJbmG8tED2zMk+x42CBGUMGBztX3S8/Yw+Fs/wv8AAfw2sdU1zSrr4Zoy+H/ENjeJba5ZeYCsu24jiETCVflkRoSjADK5ANWfDP7Hvw80Pw/490/WNc17xTr/AMSNJn0TV/EWsXqXWrfYJ4Xh8i3fylhhRBIzIqw7d2C4faAAD5p8K/E39pTwXL+zn8RPHXxCi8XaL8ZJNN0zU9HOk2llFaTalpzXcFxbTQgTb1ZD5u9ijEnaka7VX7Y/aS+LF18DPgT41+LNhZLqN54b06S4t4Hz5bzsRHF5m3B2B2BfBB2g4I61nah+zl4I1Lw58KfDE99qK2vwfu9OvNIZZYRJPJplo1nCLsmEh1aNiXEYjJbkEDivYPFXhbw/438Nap4P8V2Mep6NrVtLaXltLnZNBMpR0OMEZB6ggjqCDzQB+VPjLwd8b9J+KP7L/iv4vfE5/Gk2ueJ1mk05NMsbOysbmXTZ5D9lmtkErxopMY8x23/f4JrnD+0n+1f8RD4w+KfwvutZkXw/r97p+keEbPwqt7pV7aabceS8d9qhX7RHczhSx8tlERwMYY4+xvD37EfhDQPFngvxJJ8QvGmsWPw+uxeaLo+palbXen2rLC0CIoe0+0FFjYqoM2QMDNT67+xR4JvPF+v+KfB/jnxl4CtvFl099q+k+HdY+w6deXc2POnMRid4pJsfvHhdGb1HFAHAah4z/aA/aA+P3jX4W/DXxq3wk8P/AA107RZb2VNLtdR1G+1HW7Y3iI63gZIoYIxtYKoLOGG4g4RvjbxP+0pcftIeDv2dfDPj+DTVv/h7JqOsauNKtZGGo213HBJqFvbyAgPK3yLC0jRIshbazIM+wfEP9kfwZ418ZQfEXw14q8S/DzxOLCHS7vUPDWoLay6hZ2/EUd2s0U6StGMhJCoccZYhVA7fw38APDHhz4jeH/ikdZ1jVde8PeFz4Ujl1C6S5NzaNPHctcXLtEJZLpnjG6TeFIJymTkAHwr41+K/7SX/AAtxv2eLLx7q9rdeBPD1heatr2geErfVb3WNRvy7I7Wjh4La1jVQCEALvuXcAcL2F58Sv2iPEnwk+H2qePPHjfBTxXeC/tLvT7bw5/a2ta3qFrNsga006VJZBA8KieVY4yy7wpZEGa+lvi7+y34Q+K3jSw+Jlj4j8QeA/GNjaHTzq/hm+Wxubmy3+YLe5EkcscsatkqGTIJzngYxLv8AZC8Kyv4M1Kx8ceLbLxB4LXUYotaGpR3OpXsOrMr3cd3LdQTKwdlG0xpGY8DyyuBgA+V7j9of9pjx1+xn8L/jZ4ZNxZXeoX9xF4xvtA0uHUNStdOs7i5tHurOxut0bZeFXlG1igJKqFBxr/Ej9pT4g2Hwz+Cvhv4X+P8A/hLtT+LF/qSHxlpmhJc3AsNODzssGkjMX2zaUhZXGFaOVjGp4T3bw3+xboXgv4daR8OPBPxN8baBa+H729u9PurPUraO4hjv3Mktqw+ymKaAyEuBLG7hyTv6Aalx+xP8GJfhDovwgt5dXsovDuoyaxp+t21+0WuW2pzSNJLdx3gXiWQu275NmMfLwpABV/ZT8Z/HDWb7xt4R+L0Oq6lp2hT2cmheIdX0caJdanbXUbGaKa1QLF5lrIm0ugAdWViAc19i14n8HvgpF8I/7Wup/GviXxxqOsi3Wa78SagL1447XzPLjgSOOKKJcyuW2oCxI3E4GPbKACiiigAooooAKKKKACiiigD4A/4Kj/8AJinxM/7gv/p4sq+/6+AP+Co//JinxM/7gv8A6eLKvv8AoAKKKKACiiigAooooAKKKKACiiigAooooAK+B/8Agoz/AMkM8P8A/Y5+Gf8A0uWvvivFPj78CvC/7RPgH/hXvi3UtS0izW9tdQjudJmigu457N/MiKPLFMow3P3M+hFAHQ/GS18AXvwo8XW/xUWBvCLaXdHVPtOPLFqsbGRjnoVAypHIYAjnFflZ8G/iT+0f4f8AgP8Asf8AgXw5ra6Vqvj5tYsLx761juQdNghnksJisqhz9ntRHLGqOnmbVRm2sa+ul/YN8A6zfWz/ABT+IPjv4naTbSJMNH8Ta6brS5JIzuRpbaCG3WTacHD5U9CCMivojxf8GPCXjLxr8PfHN7Lc2d38NLi7uNLt7Roo7ZjeWptHSZDGxKLG3yBGTBxkkcUAfKvxQ8SftBfsweHPA3xH8dfERvHnhPSNfe08XyPpNlYt/Y2qMsFpdsIE3K1hKVLmNlEiv8y/LmvTf2UPiV8QPjda+NfjHrmoE+Bte1mW28Haf5EUYj0nTibc3hkVFmdryZXfbI7BAo2YU1iftwX/AIk8Q/DbS/gB4Itnm1/4xX6aB54hMsdjpjfvNSvJeCoWK3BUZIO5wV5FfV3g3wloPgHwlo3gjwvbCz0jQLOCxtIR/BBboI0BPc4HJ7nk0AfGXxJ8R/HTxh+1/P8AAf4f/EI+B/Do+H8GvySRaXZ39wt62qT2u+FrlDtLIqBg+9Nqnaiu3mL8r3fx1/a3tv2Zdb/aZvPiJZNcfC7WptKvdFg0W2jtddg07UFsrma5mcvLFLMrFgLfy0XaAFydy/qKnwg8NJ8b5fj4Lm7/AOEgm8Ox+GTBvj+x/Y47x70OE8vzPO8xyC3mbduBszzXlt3+yL8Nrz4F+L/2fpdS1YeHfGuoXupXk4mt/tsc19di8kWF/s/lqgkGFDRsQvBJPNAHnniLxr8cvjT8f/G3wc+EfjO3+GuifDOy0mXUNR/syDVb/UL3WYXuYUjjuj5MdvHEvzNtLl8gH+74prP7T/xy0bwBqnhfXNVs4/HHgP4k6B4T1LVrC0jFrqmm6jNC3m+ROJVgllhk2yqh/dsMoy5GPrj4n/sr+EviH4zh+JOh+J/EXw+8WizTT7nU/DN8lnNfWcZzHFdJLFNFLsJOxigdePmwFAor+xv8Gl+C2p/BEJqTWGs3o1a81Zr131qfV1kWYai94wJN0JEUhiu3jbt25FAHNePviH8UH/atuPg34X8RLo2j3Xw01DW4Q1nBc+RqsepxW0d0N4V2KROwEZk8snBZGxX52ata/FvWP+CVfib4kfEL4j3ni+28T6XY3EGn3lnbIbKdtciaST7XGouJy5znzWOM8V+oXw3/AGVvDPw9+KC/GO98X+JfGHiptCk8PS3Gu3sN0klnJcR3P+rjgiCMrRADy9qkFiys7F681h/YB+Gdv8OvEPwhi8aeMB4I1y2+yQaM2pxSWelxfbYr8/Y45LdsP5sWA8vmsEZ1UjcTQA3SPFHx0+Ff7UPgT4Z/ETx5H450L4n6VrVwITpdtYDTNQ0dYZm+ytAPMNu6S7VSZ5XGMlyck/dteVeKPhB4a8WfFTwP8XtRubuPWfAMOqwWEUTxi2lXV4o4pzOrRs7FViXZsdMEndu4A9VoAKKKKACiiigAooooAKKKKACiiigAooooA8A/ax/5NZ+Mn/YmeIf/AE3T0fsnf8ms/Bv/ALEzw9/6boKP2sf+TWfjJ/2JniH/ANN09H7J3/JrPwb/AOxM8Pf+m6CgD//W+3f+Ck3inQvA3hX4F+NfFN19i0Xw/wDFnwxqF9PseTybW0jvJppNkas7bUUnaqljjABOBXQf8PRv2E/+imf+UbWP/kKj9vr/AJty/wCyzeEP/bqvv+gD4A/4ejfsJ/8ARTP/ACjax/8AIVH/AA9G/YT/AOimf+UbWP8A5Cr7/ooA+AP+Ho37Cf8A0Uz/AMo2sf8AyFR/w9G/YT/6KZ/5RtY/+Qq+/wCigD4A/wCHo37Cf/RTP/KNrH/yFR/w9G/YT/6KZ/5RtY/+Qq+/6KAPgD/h6N+wn/0Uz/yjax/8hUf8PRv2E/8Aopn/AJRtY/8AkKvv+igD4A/4ejfsJ/8ARTP/ACjax/8AIVH/AA9G/YT/AOimf+UbWP8A5Cr7/ooA+AP+Ho37Cf8A0Uz/AMo2sf8AyFR/w9G/YT/6KZ/5RtY/+Qq+/wCigD4A/wCHo37Cf/RTP/KNrH/yFR/w9G/YT/6KZ/5RtY/+Qq+/6KAPgD/h6N+wn/0Uz/yjax/8hUf8PRv2E/8Aopn/AJRtY/8AkKvv+igD4A/4ejfsJ/8ARTP/ACjax/8AIVH/AA9G/YT/AOimf+UbWP8A5Cr7/ooA+AP+Ho37Cf8A0Uz/AMo2sf8AyFR/w9G/YT/6KZ/5RtY/+Qq+/wCigD4A/wCHo37Cf/RTP/KNrH/yFR/w9G/YT/6KZ/5RtY/+Qq+/6KAPgD/h6N+wn/0Uz/yjax/8hUf8PRv2E/8Aopn/AJRtY/8AkKvv+igD4A/4ejfsJ/8ARTP/ACjax/8AIVH/AA9G/YT/AOimf+UbWP8A5Cr7/ooA+AP+Ho37Cf8A0Uz/AMo2sf8AyFR/w9G/YT/6KZ/5RtY/+Qq+/wCigD4A/wCHo37Cf/RTP/KNrH/yFR/w9G/YT/6KZ/5RtY/+Qq+/6KAPgD/h6N+wn/0Uz/yjax/8hUf8PRv2E/8Aopn/AJRtY/8AkKvv+igD4A/4ejfsJ/8ARTP/ACjax/8AIVH/AA9G/YT/AOimf+UbWP8A5Cr7/ooA+AP+Ho37Cf8A0Uz/AMo2sf8AyFR/w9G/YT/6KZ/5RtY/+Qq+/wCigD4A/wCHo37Cf/RTP/KNrH/yFR/w9G/YT/6KZ/5RtY/+Qq+/6KAPxB/b1/b1/ZO+NP7J3jn4afDPxz/bHiTWP7M+yWn9malb+Z9n1K1uJP3lxaxxLtjjdvmcZxgZJAP1/wD8PRv2E/8Aopn/AJRtY/8AkKj/AIKj/wDJinxM/wC4L/6eLKvv+gD4A/4ejfsJ/wDRTP8Ayjax/wDIVH/D0b9hP/opn/lG1j/5Cr7/AKKAPgD/AIejfsJ/9FM/8o2sf/IVH/D0b9hP/opn/lG1j/5Cr7/ooA+AP+Ho37Cf/RTP/KNrH/yFR/w9G/YT/wCimf8AlG1j/wCQq+/6KAPgD/h6N+wn/wBFM/8AKNrH/wAhUf8AD0b9hP8A6KZ/5RtY/wDkKvv+igD4A/4ejfsJ/wDRTP8Ayjax/wDIVH/D0b9hP/opn/lG1j/5Cr7/AKKAPgD/AIejfsJ/9FM/8o2sf/IVH/D0b9hP/opn/lG1j/5Cr7/ooA+AP+Ho37Cf/RTP/KNrH/yFR/w9G/YT/wCimf8AlG1j/wCQq+/6KAPgD/h6N+wn/wBFM/8AKNrH/wAhUf8AD0b9hP8A6KZ/5RtY/wDkKvv+igD4A/4ejfsJ/wDRTP8Ayjax/wDIVH/D0b9hP/opn/lG1j/5Cr7/AKKAPgD/AIejfsJ/9FM/8o2sf/IVH/D0b9hP/opn/lG1j/5Cr7/ooA+AP+Ho37Cf/RTP/KNrH/yFR/w9G/YT/wCimf8AlG1j/wCQq+/6KAPgD/h6N+wn/wBFM/8AKNrH/wAhUf8AD0b9hP8A6KZ/5RtY/wDkKvv+igD4A/4ejfsJ/wDRTP8Ayjax/wDIVH/D0b9hP/opn/lG1j/5Cr7/AKKAPgD/AIejfsJ/9FM/8o2sf/IVH/D0b9hP/opn/lG1j/5Cr7/ooA+AP+Ho37Cf/RTP/KNrH/yFR/w9G/YT/wCimf8AlG1j/wCQq+/6KAPgD/h6N+wn/wBFM/8AKNrH/wAhUf8AD0b9hP8A6KZ/5RtY/wDkKvv+igD4A/4ejfsJ/wDRTP8Ayjax/wDIVH/D0b9hP/opn/lG1j/5Cr7/AKKAPgD/AIejfsJ/9FM/8o2sf/IVH/D0b9hP/opn/lG1j/5Cr7/ooA+AP+Ho37Cf/RTP/KNrH/yFR/w9G/YT/wCimf8AlG1j/wCQq+/6KAPyg/aF/wCCj37GPjn4BfEvwV4W+If23WvEHhnWdPsYP7J1WPzrq7spYYY98loqLudgNzMFGckgZNfcH7J3/JrPwb/7Ezw9/wCm6Cj9rH/k1n4yf9iZ4h/9N09H7J3/ACaz8G/+xM8Pf+m6CgD/1/v39vr/AJty/wCyzeEP/bqvv+vgD9vr/m3L/ss3hD/26r7/AKACiiigAooooAKKKKACiiigAooooAK+CPjt8VvjFqn7S3h39mj4YeMtK+GX9peHX15dY1LT11OfUbhbloBYWcEskURZEQyyfMZNpyoAU5+96/Pj9srxx+yDf3sfwm/auiudCC2SanouvmzuQIp3aRXFhfW0cjR3UJiV3iPDBo8rJ90AHtx+NVh8B/AvhKy/av8AGug6f4s1q8m09bu3b7FZ3ZWZ/KnCTt+5UW/lvcMzCNJCVU/MgPreo/FX4X6P4Us/Hmr+MNGsfDOohWttUn1C3isZ1cEqYrhnEThgCQVY5HSvx01HU9W1P4C/s1fGT9ozTpNS0/w/40kg1DWtY0/fdyeHCt5HY3WopsZljdfLLF8hjtkYlnyfVPFvjv8AZ3g/aK8M/GP4sWdrefBHUPCIsPBepXWlyvoNjqMV7Kt7GbdoSsE8yKpileJQ8a4RiMUAfqJcfET4f2nhEeP7rxPpcPhchCNVe9gWwIlkESH7SX8r5pGCL83LEKOTiudi+OXwVnt9eu4PH+gSQeFiF1eRdUtSmnktsAumEmIct8vz4+bjrxX4u/Eez0LVP2bf2p/Ffwr0qXSvgp4k1nwe/huFrd7OzubtNQs49WurK3kCFLeWTywCFCsVO3G0qPszx98IPhVY/t4fBzTrPwhpMFlJ4U17dbpYwLAxsmgFsWjCBCYg7bCR8ueMYFAH3NL8XfhTB4Hj+Js/jLRo/CEwBTWG1C3XT2yxUYuS/lE7gVxuznjrXQaF4w8JeKPD6eLPDWt2OraJIjSLf2lzHPasicswmjZkwMcnPHevxy0jxB8P/hL4H+KVjrHhHSNU0PTPjpfWOkjWYs6D4dkvbKIfb7iJVKpbW4llCoAqhpFAaPh18u0VH8Tfs0/tM/DT4c3S6+8XjqHWLzTfDlpJYNd+HpjYPePYWe92ihnjSQxoHbeisFLjqAfqf8W/2l/Az/An4oeM/gb450PxF4h8GaFfX6f2fe2uo/ZpoYmaNpY43cbdw/iGD0r0X4RfGrwH8QtJ0bQ4PF2kap4zGk2d5qem2t5bveQSSwo0rSW0bl4wHbkFRjIHcV8I/ET4q/sc+P8A9mj4w6f+zimjrrNn4B1MzrpukvYywWCxbRDNIYIgpWQr+6ZtwOSFxk1Z1jwV4P8AA3ij9i3WfB2iWWi3r3DWTy2kCQvJb3ehyNMjlAC4dvmO7J3Et1JJAP048ReJPDvhDRbrxJ4s1S10TSLFQ9xeX08dtbQqSFDSSylUUZIGSRyQKjvPFPhjTtX0nw/qGr2dtqmvCY6daS3EaXF6LZBJMbeJmDy+WhDPsB2qcnAr5m/bw8N654u/ZC+KOg+G7KXUNQm0oyRwQIXlkEEsczhFXJY7EYgDk9q+Y7n48/C344/ta/stXfwv1j+27fT7bxabmWOCZIoZJtIjxA0kiKvnLsO+MEsgKlgAy5APpv8AZf8Ajlf+Nf2fG+K/xh1ux0/7LqmtW9zfXDRWNrFb2Woz20JdmKxrhEUFiRk9eTXu/hr4s/C3xn4ZvPGnhHxhpGtaBp2/7VqFnfwT2lv5a7n82ZHKR7V5O4jA5PFfipotlYxfs3/BXxd8QtNuNX+FHhn4j+Jr3xXbw27XcUcf2+9Wzubq3RXMltDOcygoww2MEkA+7/tL658HfjH+zXq+qfsqwWeqeHNF8U6HqHjCLQ9IcR3enWzb5t9uEgW88tRG8iqW+WPaxXGQAfpn4Y+Lnwq8baBqPirwd4y0bXNF0gOb2+sdQt7i2tREpdzNLG7JGFUFiWIwvPTmo9O+MHwl1fVF0TSfGuiXuovp6aqtvBqNtJMdOkjEqXYRZCxgaMhxLjYVIbOCDX5P+D7r4L+KX+MHjv4VfEa28U39z8MNastQsNF8MSaFpqQwJvt5bo8R/bE3tHGjDzPL3ggKlfQPwu+DHwA039g7wxr/AI10GK0t9U+HlsmrazZ2Ym1eO21ewia7eOWON5225BVQGwiKu0qNtAH3b4L+J3w2+JEd1N8PPFeleJ0sX8u4bS76C9EL5IxJ5LvtJwcZxnFdxX5h/sA+NdIvPEni34beHzovjTSPCunabHYeOtE0v+zf7Qs/nWGw1ACNVa6thk5RmBBJID7s/p5QAUUUUAFFFFABRRRQAUUUUAFFFFAHwB/wVH/5MU+Jn/cF/wDTxZV9/wBfAH/BUf8A5MU+Jn/cF/8ATxZV9/0AFFFFABRRRQAUUUUAFFFFABRRRQAV8n/tb/Fnxp8I/D/w61DwTcR203iPxzoGhXhliWUNY6hI6zooYHaxCjDDkdq+sK+Jf29fB/i/xH8HtC8S+CdHuPEN/wDD/wAVaJ4ok06zTzLq6ttNmJmSBP45Ajlgo5IUgZOAQD6A+Pvi3WvAHwK+I3jvw1IsOr+HPDesalZu6CRUubOzlmiZkbhgHQEg8HpXnnw5/aO+Hdt8IPhl4g+MvjzQPDvibxf4Z0jVpItR1C006S4lvLWOSWSKGWRDsMhYDaMA8DpXy18dv24fgT8X/gV4v+GHwZvr7xl488d6PfaHYeH7PTbwaglxqMDW7G4SSJViWASFpGZtuFOCeteUeJYvgx8Dv2kPg94R/aXk0gaHoHwch0V21a3S8sjf2t3BFwHjkXJWKTaxAyOM5YAgH626r488D6F4W/4TjW/EOnWHhzy1l/tOe7hisvLf7r+ezCPa3Y7sHtVey+JPw71LwjJ8QNO8U6VdeF4UaSTVYr2B7BET7zNchzEAO5LcV+MmmeF/hdqP7O3jPUvFWr3Xwt+F+p/E3+1/hxfXOnSXNlZPBCZILmSyljKpptzKkpSKVUj+YA4LDLfHPiPxL8W/2SPC3jJfCtjpfh3wp8SoZPFE2g6PJLpOtabZAoddh01lie5tGkaJ5IWB3lCTwpAAP1+g+OPwcvPAWqfFHTvG2j3/AIS0VHe81S0vYbm0g8sAsryQs438jCfeJIABJANj4XfGL4Y/Gfw/H4k+GPiXT/ENqYoJJls7qGeW1NwnmJFcpE7GGXHVHwwIIxwa/KPw/wCGfhD8UvCP7QesfCHxfbfEfUfEfgC8hn07SPC76Vpi6hbW839nzAEGM6hmQqqqPMA5IBHP2t+xB8Q/gh4u+EOh+HvhQ1outeH9E0WPxDDa2L2jw3xtvLZLlzEivOrxSBxuZlxk8MpIB9J+LPit8L/AWpWOjeOfGGj+HdQ1P/j0t9Rv7e0mn5IzGkrqzDIIyAeeOtcd4i/aS+BPhH4jx/CfxT440nR/E0lql39mvLyG2wsrqkUe6V1UzSl9yRAlyoLY2kE/nL+274g+GPw8+Nt58SdK1DS9Z8eR6NZ6ZqfgbxFpMl7a+JtLeUvGunTeTIY7oF2X93uXIHmIACX6v4g+Ivgj8KP23I/iN8bNDtfD3h/xN8PLFrKa+0o3CtrMWpSySQgRQyk3qRFMhQXIAAPQUAfpF4p+Knww8DalaaP428X6P4fv78A21vqOoW9pNMCSB5aTOrPyCOAelXvEnj/wH4Nks4vGHiTTdCfUUuJLZb+8htTOlpH51w0Qldd6wxDfIVyEX5mwOa/J3Q/FP7MXwy8b/G7Rv21NHtX8ceIPEupXdtPrOlSX76r4elCrpkemSiKXMccQ8sRRMGRxg/MOOc+GngHW5NS/Yl8HfGPRJJUF544mttL1qMTz2+nx2MtxpcVyku795BEsJVWGY9qggMvAB+qtx+0F8B7Tw5p/jC6+I3hyHQtWma3s799WtFtbiZCA0cUpk2O65G5QSV74re8X/Fj4W/D59Oj8eeMNH8ONq5xZDUr+3tDcnj/UiZ139R93PUetfml4U+DnwqvviL+2jo914S0uS0tY7JbeM2kRFv8Aa9D+0TGD5f3RebEhKYJcK3VQR4+3jfwJc/Bj4QeCfGOiaBBrvib4U2+PFHiWwfU2lt7VTDFpGmQqUL3rtKZMiQFRtbZKQqgA/afxH408HeD/AA8/i3xbrthouhxKjNf3tzFb2qrJ9wmaRlTDZGOee1fKvxu/aTsNL0r4QeKvhB4r0nVvD/i7x9pmganf2s1ve2psJre6luE85WZI3UxIS2QyjrjNfnloPibwRffB/wDY9+JHxQDeIvhD4Is7+x8RsLd7+x0/WRZxQ6fJqEKrJujhbegbaRE5wTztPuvx01n9nD4weE/govwas9K1TwbffGjRrXUorTT/ALNY3V3/AGfcGbdG8UaTBo2jDOFZW+6SSGAAP018E/Ef4e/Eqwn1T4deJ9M8UWdrKYZptLvIb2OOUdUdoWcK2OcE5xz0rs6+CvhroujeEv2/viZo3hixg0qx1bwToV/cwW0axRyXUd3cwLKUQAbvL+XOK+9aACiiigAooooAKKKKACiiigAooooA8A/ax/5NZ+Mn/YmeIf8A03T0fsnf8ms/Bv8A7Ezw9/6boKP2sf8Ak1n4yf8AYmeIf/TdPR+yd/yaz8G/+xM8Pf8ApugoA//Q+/f2+v8Am3L/ALLN4Q/9uq+/6/OD/gpN/wAJV/wivwL/AOEF+xf8JJ/wtnwx/Zf9peZ9h+3eXefZ/tPk/vPJ8zb5mz5tudvOK6D/AI2m/wDVGf8Ay4qAPv8Aor4A/wCNpv8A1Rn/AMuKj/jab/1Rn/y4qAPv+ivgD/jab/1Rn/y4qP8Ajab/ANUZ/wDLioA+/wCivgD/AI2m/wDVGf8Ay4qP+Npv/VGf/LioA+/6K+AP+Npv/VGf/Lio/wCNpv8A1Rn/AMuKgD7/AKK+AP8Ajab/ANUZ/wDLio/42m/9UZ/8uKgD7/prokg2yKGGQcEZ5HI/I18Bf8bTf+qM/wDlxUf8bTf+qM/+XFQB9/MqspVhkHgg9CKR0SRSkihlbggjIIr4C/42m/8AVGf/AC4qP+Npv/VGf/LioA+/6K+AP+Npv/VGf/Lio/42m/8AVGf/AC4qAPv5lVgVYZB7GkCqpLAAFup9frXwF/xtN/6oz/5cVH/G03/qjP8A5cVAH36qImSihdxycDGT6mnV8Af8bTf+qM/+XFR/xtN/6oz/AOXFQB9/0iqqjCgAZJ49Tya+Af8Ajab/ANUZ/wDLio/42m/9UZ/8uKgD7/pqIkahI1CqOgAwBXwF/wAbTf8AqjP/AJcVH/G03/qjP/lxUAffqqqAhQACSePU9adXwB/xtN/6oz/5cVH/ABtN/wCqM/8AlxUAffqIkY2ooUcnAGOTyf1p1fAH/G03/qjP/lxUf8bTf+qM/wDlxUAff9FfAH/G03/qjP8A5cVH/G03/qjP/lxUAff9FfAH/G03/qjP/lxUf8bTf+qM/wDlxUAff9FfAH/G03/qjP8A5cVH/G03/qjP/lxUAff9FfAH/G03/qjP/lxUf8bTf+qM/wDlxUAff9FfAH/G03/qjP8A5cVH/G03/qjP/lxUAH/BUf8A5MU+Jn/cF/8ATxZV9/1+IP7ev/DfX/DJ3jn/AIXX/wAKz/4Qz/iWfb/+Ee/tr+1P+Qla+T5H2v8Ac/67y9+/+Ddj5sV9f/8AG03/AKoz/wCXFQB9/wBFfAH/ABtN/wCqM/8AlxUf8bTf+qM/+XFQB9/0V8Af8bTf+qM/+XFR/wAbTf8AqjP/AJcVAH3/AEV8Af8AG03/AKoz/wCXFR/xtN/6oz/5cVAH3/RXwB/xtN/6oz/5cVH/ABtN/wCqM/8AlxUAff8ARXwB/wAbTf8AqjP/AJcVH/G03/qjP/lxUAff9FfAH/G03/qjP/lxUf8AG03/AKoz/wCXFQB9+CNFdnVQGbGSBycdMmvny/8Agxqd7+1Lpvx8N9B/Zdl4RufDjWTKxmaae+juxKDjbsCptI65rwb/AI2m/wDVGf8Ay4qP+Npv/VGf/LioA+/WVXUo4DKwwQeQQacAAMDgCvgD/jab/wBUZ/8ALio/42m/9UZ/8uKgD79VVQbUAUcnA468mlCqv3RjPPHqa+Af+Npv/VGf/Lio/wCNpv8A1Rn/AMuKgD79KIzKzKCV5BI5GeOKUqpwSM45Hsa+Af8Ajab/ANUZ/wDLio/42m/9UZ/8uKgD79ZEYqzKCVOQSOh6ZH4U6vgD/jab/wBUZ/8ALio/42m/9UZ/8uKgD7/pCqkgkZK8j2r4B/42m/8AVGf/AC4qP+Npv/VGf/LioA+/VRFXYqgL6AcUKqooRAFVRgAcAAV8Bf8AG03/AKoz/wCXFR/xtN/6oz/5cVAH3/RXwB/xtN/6oz/5cVH/ABtN/wCqM/8AlxUAff8ARXwB/wAbTf8AqjP/AJcVH/G03/qjP/lxUAff9FfAH/G03/qjP/lxUf8AG03/AKoz/wCXFQB9/wBFfAH/ABtN/wCqM/8AlxUf8bTf+qM/+XFQB9/0V8Af8bTf+qM/+XFR/wAbTf8AqjP/AJcVAH3/AEV8Af8AG03/AKoz/wCXFR/xtN/6oz/5cVAH0B+1j/yaz8ZP+xM8Q/8Apuno/ZO/5NZ+Df8A2Jnh7/03QV8P/tC/8PI/+FBfEv8A4Tr/AIVN/wAI3/wjOs/2p/Zv9vfbvsP2KX7R9m879353l7vL3/Luxu4zX3B+yd/yaz8G/wDsTPD3/pugoA//0fv39vr/AJty/wCyzeEP/bqvv+vgD9vr/m3L/ss3hD/26r7/AKACiiigAooooAKKKKACiiigArnPEnjHwj4Ot4brxdrljocFwxSOS+uYrZHcDJVTKygnHOBXR1+Yn/BRm78MWPiP4AXnjPw3N4v0OLxbK13pMFgNTlvIhZvmNbMgiYnrtx2oA/SrSdY0jXrCLVdDvoNRsp8mOe2lWaJ8HB2uhKnn0NaNflv/AME+k8O+I/iV8afiX8I9FbwV8LdUvbCwsPDsrRwzQapYxMt9PJYRO4sS5KgRnBcYOAFArfT/AIKIW8qj4hReBkPwgOrDS/8AhIjrdqNRMZufsf28aPs877J53GfM8zZ+82Y+WgD9KaK+MYf2qPFuu/tC+JPgf4J+HqavB4NutPt9WuptatrK/WG+SOQ3ltp0se6e2hSTLuJVLdFUkgHjdT/bF+LS+MPiRYeEfgjP4q8LfDDU5NP1TUrLXrZL1hDClxJJDp88EZkIjbIRZiWPAOegB+gFFfFWlfGn4cfEb4+fB7VvDOmXOo/8Jn4R1fVdM1X7fNbx29pvtjJBLp4BikkYkAu7ZjKkDPUc38Gf2w/ih8bfCvh/4i+FfgpeS+Eb8alHqN7Fq9vJNbXFgZ8Ja2zxRSXayeUqbh5eJZNgDBC5APvmivjHwB+1B8RNQ+K9j8H/AIv/AAqm8Da54g0m81jRlg1i11X7VFZFRJBMY1iWCbDZGWZO2/vXjXw8/ac+NHxV+F/xf8R/E74XWtt4L8PQ+K7aW4stf8m4/wCJTE8baeEjh81XYK4N2knDHciABQAD9M6K+FdM/aYGgeBfhD4C+D3w/vfF3jDxf4R0/XLPQRqaxx6bpC28IEt9ql2pJVWcRLIY2eVwSQCQCl/+2rLoHw2+KWueMfAN14e8e/Ca0hvdT8NXV9EyT290f9HntdQhjkjlhkAI3iPKsCrKOCQD7ror4y8GftS+Nb34seCfhv8AE/4Zy+C7X4m2V5d+G70apDqDyvYQLdTQXkMUai2kEB3jEkgJ+UEndt9y+Ofxk8MfAL4W638VfF0c9zYaOkYW3tVD3FzPPIsMEESkgFpJHVRngZyeAaAPT9Q1HT9IsbjVNWuorKytEaWaed1jiijUZZndiFVQOSScCpre4t7u3iu7SVZ4JlV45EYMjowyrKw4II5BHWvzp+NXxE+PHjL9l74023xd+FCfD+yPgzVLm0uItct9W3uYGBt5o4oomjlUHdkbkPIByOdz4L/tHeONCuvg38M/iN8OJPDWg+PdIitdB1capDdyyXFjYLPsvLSOMC3M0SFo9s0pzhWwd2wA/QCiviTTf2rPiHpvxQ8IeC/ip8JbrwXoPxCv59P0LUpNVtrq6aaKN5o/tljGga281F4AkkKE4bBzjj/EX7afxKtB8VtV8J/B19d8PfB3Vr2x1m/Ouw2xktbGFJ5ZraFrdneVI2Z3iOFChdsrsxVQD9C6K+LvHP7W2tJqegeG/gV8O7r4k63rnhWHxmYH1CDR4YNHuW2QM00yy7ppHyoiVSRjJOOa87s/279W1X4e/DbWdI8BWtx4y+Kkd1eaVpJ16OGxt9Os1Uy3V7qU9tEIirNs8sQsWcFVJ4JAP0Vor55/Z2+P1r8eNB12S70Y+HPEfhPUpNK1jTftUV/HBcKiyo8F3CAk8MkbqySBVzyMcZP0NQAUUUUAFFFFABRRRQAUUUUAfAH/AAVH/wCTFPiZ/wBwX/08WVff9fAH/BUf/kxT4mf9wX/08WVff9ABRRRQAUUUUAFFFFABRRRQAVxel/En4d63qzaBovinSr/VEZka0t76CWcMmdwMaOWyuDkY4xXaV/MWdQ+FesfCbxB8M9P+HVxH8Y/E/wAQNStfDnjOSzi0y0trv+098X/E8dkYtEisPJUsckADJBoA/p0or5a+Ov7QU3wD03wT4Tt9KHjLx34zkey060ku4tLt52soVkvLq4u5gyQQxqQT8rMWZVVTkkeOXP7etnonwj8f+NfE/gt7XxX8NtR07TdV0a21OC6swdWZBZ3S6oiCIWsivlpGjBQqysmcEgH6EUV8QeNv2qviT8PvgpbfE7xD8MLd9a1HXNN0bTtOtfEVvcWOoJqZRYLuDUYrdx5Rd9uHhU/KT93DHT8L/tN67qviXWPhH8bPhxd/D3xc2h3ms2Nr/acOoWWqWNsNs4t7+1CFJUJG5CiuqkOKAPsyivzb8EftXWXgb4Zfs6eG/APw5vNRX4saVqC6Xp39sNc3FnJptuksMMlzdRZmWRnxJM7R+UgL7XwEr1DXP2gP2mtH8M2epj9n2R9Ut7e7vNYhk8SWcVpaQW0rogguvKb7TLMkZlVAibFKb2BYUAfalFfnV8SP2zPiBP4I+B3jb4F+BU1y3+Lt/bwiPUr6KyZGaN5WslfEu2RvKcNOUKKq/KGZwU7vT/itpdh+0lCnxN8Kt4U8TWfwwm1/VLwa3Le2VhZQ6mFmtPsyRpBKyMPN+1hQ5UbNu2gD7aor8+2/bc8Z2XhK0+NWu/BfV9O+Dd55Uy+ITqFrJfxafcMFiv5dIQGVbdtyuSJWcRnftIxnsfFX7UHxHk+MPif4N/B74Xp44vvDukadrIvZNch061lgvw5CEyQSFXO0CLbvV8ksYwoLAH2nRXj3wC+Mmi/tAfCHw58XdAs5tNtdfilLWtxgy289vM9vPExGAdk0bqGwNwAOBnA+WvFv7Q3hz4P+Mf2oPGum+D573VfhxZ+F7q/aTWJvJ1X7XZsYViheKSKx8lSVYxq/mn5mANAH6DUV8beDf2ofGt58XfB/wx+KPw0l8FQfEixvbzw3eDVIdQeZtPhW5ngvIYo1FtKIW3/LJKp+6CTkjJ+Hv7VHxQ+MWvxa18JvhHJrXwtbWjpH/CSz61b2k80UNwLa5vrfT3iZpLeFgx5lV3VThQwK0Afb9FfDOl/tY/E3xn428a+C/hh8IJvEr+APE50LVLg6zBZxC1xHtuo/OhG+U7nZrdc7EjyZcuingvEP/BQhdLuPEHjHSfAY1L4W+FNVk0u/1w61aw6i/kTCC4urTSGQyz28UhIz5is65YKNrAAH6R0V8P8Axr/a08efBefVPGWp/Ci5vfhToU9lBd+JP7WtobiVbwxL59ppxQyTQxtKFLGRCxyQNqk1qfFH9qbxZ4T+Oq/AX4e/D5PFmuRaPHrcgutattHkuoJJJI/L06O4jcXUimMl8vGq4IJ70AfZlFfFnxc/ay1vwl8ULv4QfCzwVb+MfEGh6dBqWsNqGuW2g2tml1uNvbpLcJJ51xKqlgoUKoKkv1A98+B3xf8ADnx5+FegfFfwrDNbWGuxO32e4AE1vNBK8E8MmMjdHLG6Ejg4yOCKAPV6KKKACiiigAooooAKKKKAPAP2sf8Ak1n4yf8AYmeIf/TdPR+yd/yaz8G/+xM8Pf8Apugo/ax/5NZ+Mn/YmeIf/TdPR+yd/wAms/Bv/sTPD3/pugoA/9L79/b6/wCbcv8Ass3hD/26r7/r4A/b6/5ty/7LN4Q/9uq+/wCgAooooAKKKKACiiigAooooAK+ZPjx8F/FPxQ+IXwc8WaBdWdvZ/D3xE2r6gl08iSSwG3eLbAEjcM+5gcOUGP4u1fTdFAHyTqvwE8W+Gv2nbD49/CO50+y0zxPa/2d440q7eWFdQjh/wCPS/t/KikU3kOSpEm1Xj+XcpZmr5l8HfsbfGr4Sx3Hw48A6R8MvEHglby4l07VvEelTT69YWlzM0phkSOIw3TRbyImaVBgfNhcKP1QooA/PL4y/s4/Hn4s/Gfw/wCLEbwjoln4W1u11DTPFNn9uh8TwabC4eXT5I1XyJ1lBZDvl8sqTmPcST4t8PdL/aX8Q/Ez9pzwv8EJPDNhpms+Lp7S61LWJr1b6wlmsIEaa2ht4mjlKxkFA7piQc5Wv12rkfDXgPwh4P1HXtW8M6XFp954nvDqGpSR7s3N0UWPzXyT821VHGBx0oA+UPh9+ydffC74j/BvU/DGo29z4X+GPhPUvD85nZ0vbm5vXhcTpGqNHtZkdnBkBXcAA3Wu2/Zq+CXjj4L/ALLekfBbVdWtIPFGm2uqxLqFgXnt4Zr27ubiCVPOjiZvLEykhkHzAjkcn6nooA/Ln4Sfsm/H7wJ8V/hZ8SNYtPB7XXhO11HT/EOoQX2oXGqa2b9YVl1Keee0DTXLCMlYpH2xk4EhU7U7LQv2cf2jPCmgfFj4Q6NqPha7+H/j+bxXfWt1cNfR6tDN4hgm8m3dEjaBUhndS8oLsybtse7GP0UooA/Psfs0fGv4fD4V/Ef4O6voTePvBXgmx8E61Yav9pOkanZWqRyZhuIY/PieO4RmjYxfOrAMFwQ3O+J/2RvjP8SvAnxn1/4ja7ocvxO+Kmj2uiWkFj9pi0TSrGycvFCs0kb3MpdmZ5JDF97AVQK/SaigD5Z8Y/BDxX4h+KnwD8cWV3ZJYfCz+1v7TSSSUTTfbtLNjH9mAiKtiTlt7R4XkZPy12P7SPwX/wCF+fCPVfh1b6q2halLLa3unagsYl+y31hOlzbyGM4DLvQBx3UnHNe60UAfDOveAf2xfiv8JfiF8MPi03gazHiHw1faVYT6PNqRaXUrlBGk1wZ4sQ2+0tuVEkcNjBwMHqvFPwB8Y65c/s9TWl5YIvwmvIrjVd8koMyR6a1mRa4iO8+YwI8zy/l5znivryigD8oF/ZA/aRg8X+EvGd0/hDXfEfhTxhNrs3iG+vL86rrFlN9oWKGc/ZXFulrHMqR20TtEdoKmPBL/AEToH7OHjjSvAn7SPhe4vtOa6+MOqa9e6QyyzGOCPVNOS0hF2TCCjK6kuIxIAvILHivtaigD8TviT8M/FPjLx/8ADr4G+GfCMfjXVfgb4J07S/GEVrrR8NG8W+tIkhgt9TUrdT2x8mV2iaJIssNzBiQPXI/hhp37TvgD4V/HH9nLQdC8O6p8N11Pw+PDHim2W90WS1jb7LPaSyWvm7vJlhElvPEWB3bm5JA+8/iP+z58D/i9f2+q/E3wNpHiO/tVEcd1eWkb3CxKSRH5uA5jySShYqcnI5Nej+HPDXhzwdolp4a8JaXa6LpFguy3s7KFLe3hQknbHFGFRRkk8AckmgDxn9n7wN8QfBmjas3xF0fwjoF7qFwjxWXg+0kt7SKJEwPOlmSN55SSTu8tQowADyT9A0UUAFFFFABRRRQAUUUUAFFFFAHwB/wVH/5MU+Jn/cF/9PFlX3/XwB/wVH/5MU+Jn/cF/wDTxZV9/wBABRRRQAUUUUAFFFFABRRRQAV8LfDX9j2FP2c/G3wC+MrWWpW3i3W9a1JJbBnk+zLf3BntZUaaOMrcQNtbhSoYYywzX3TRQB+bHiv9lP49+O/ht8L9Q8Y6x4X1r4rfCO5vIrO51OGfUNE1zTrqNYGGoxPCkqTSRpGXKCTEiF1YlgU9B8PfCr9o7Svh/wCJ4I/DXwt0nW9aks400ax0+6XRrqxjEouYr+fyklleQSgJiEpHtPyuHavuaigD8Tfix8CfiZ8Bf2Zl0e+udIt7zXPid4e1PRdCsJbubRNFM11GFtopZwJzC0w3yBUVVySijJr7J0X4DfHHx98VLj4zfHm+0Gz1LR/D2oaD4f0jQJLqezhfUhi4vLm4uoonMjgKgRI8BMEksOfrvxl4D8IfELTrXSfGmlxatZ2N5b6hDHLuwl1aOJIZRtI+ZGGR29q66gD8/wD4afsp/EPwbc/svzapqGlSj4KabrtnrPkzTsbiTVLFbaE2e6Bd6q4y5k8ohegJ4rt/2iPgh8Ufif8AEXwxr2gHQvEHhHT9PvLS60DxFLcpYx387q0OqLBBFKl1LAilEim2hcko6Mdw+yaKAPzI0P8AZP8Aj7o37O3wf8F21/4bi8d/BfX4tU08NNdzaXqFtDHNH5U8vkRzRPJ57HKxMFAA5JJHqcX7OfxM+IHxavPiH8bbjRPsfiL4a6h4I1e20SW6GJ77Umn3W4uYuYhasFMjuGMuSIwpGPuSigD8z7v9nL9sLX/hJb/ss+IfFfhJfhxHaQaNP4htobz/AISCfRbcKiwizdTapcPEgjeTzWG0lgC3NfR/w5+BWs+BP2iPGvxPimtB4Y1zw/oOjafbRySNdxHSRKrearRhApV12lZGJwcgd/qGigD5x/ZO+EHiX4E/AvRPhj4uubS71TTbrVZ5JbF5JLcrfajcXcYVpY4nJEcyhsoMMCBkYJ8C+LP7JnxG8eP+042kajpUI+NNh4ctdH8+adfs76RbGGc3m2BtgZjlPL8wkfeCniv0LooA+XfGnwT8VeI/jH8BviFZXVkmnfC4a2NTjkeQTzf2lpf2KL7MojKttk5feyYXkZPFeS/B74M/tWfs9R2fwj+HWo+D9a+FWn6nLPY3Orfb4tas9Murpria08q3UwTSJ5jiKQyKM8soGEH35RQB84/AH4QeJfhVr3xZ1TxDc2lxF488YXniCxFq8jtHaXFvbxKk++NAsoaJiQpdcEYY8gfI9h+x78Z/hlrfiTw/8LdH+GviXwfrmqXepWN34s0yebV9KF8/myQAQxNHcxROzeUGkjP95sHA/UaigD8rf2j/ANjX46fGK5+JFha3PhbW7HxPZ6bDoF9rEt3Hc+H1so4vPtbK0jt5YIUuZYyzTRur4YqyuMY7r9pb9nz9pD9oGystEns/A1oEitLiz1rztSj1rw3qSbWnl064iiUzoWUGPcYCT99SAAP0YooA/PD4qfsqfExfjHP8avhbB4O8YX3iHSrHTde07xvZO0Es+nr5cN9azW8UrRSMh2yIUKkDucbfsD4OeF/E3g/4eaZoXjG30S11mNp5LiHw5aNZaVEZpnkVLeJ/mwiMql2wXYFyBnA9PooAKKKKACiiigAooooAKKKKAPAP2sf+TWfjJ/2JniH/ANN09H7J3/JrPwb/AOxM8Pf+m6Cj9rH/AJNZ+Mn/AGJniH/03T0fsnf8ms/Bv/sTPD3/AKboKAP/0/t3/gpN4W0Lxz4V+BfgrxTa/bdF8QfFnwxp99BvePzrW7jvIZo98bK67kYjcrBhnIIODXQf8Ouf2E/+iZ/+VnWP/k2j9vr/AJty/wCyzeEP/bqvv+gD4A/4dc/sJ/8ARM//ACs6x/8AJtH/AA65/YT/AOiZ/wDlZ1j/AOTa+/6KAPgD/h1z+wn/ANEz/wDKzrH/AMm0f8Ouf2E/+iZ/+VnWP/k2vv8AooA+AP8Ah1z+wn/0TP8A8rOsf/JtH/Drn9hP/omf/lZ1j/5Nr7/ooA+AP+HXP7Cf/RM//KzrH/ybR/w65/YT/wCiZ/8AlZ1j/wCTa+/6KAPgD/h1z+wn/wBEz/8AKzrH/wAm0f8ADrn9hP8A6Jn/AOVnWP8A5Nr7/r4V1j9srxVc+JvFUPwr+DPiD4g+EvA2pXGkaxrWn3FpEwvbPH2qOyspnE935RO0lMbm4HGCQDK/4dc/sJ/9Ez/8rOsf/JtH/Drn9hP/AKJn/wCVnWP/AJNr7/ooA+AP+HXP7Cf/AETP/wArOsf/ACbR/wAOuf2E/wDomf8A5WdY/wDk2vv+igD4A/4dc/sJ/wDRM/8Ays6x/wDJtH/Drn9hP/omf/lZ1j/5Nr75uJltreS4ZWcRKzlUUu5CjOFUZJPoBya5rwN4utPHvhHSvGNhYX2mW+rQidLbUrZ7O8iDfwzQSfMjcdD25HBoA+K/+HXP7Cf/AETP/wArOsf/ACbR/wAOuf2E/wDomf8A5WdY/wDk2vv+igD4A/4dc/sJ/wDRM/8Ays6x/wDJtH/Drn9hP/omf/lZ1j/5Nr6b+Onxu8K/AHwMPG/im1vNSNzeW2m2Fhp0QmvL6/vH2QW8CMyKXc5PzMBgHqcA4PwL/aE0X42z+KNBbw/qng/xV4KuLe21jRdYjjS6tjdx+dbyBoXkjeOZASjBsnB4xgkA8C/4dc/sJ/8ARM//ACs6x/8AJtH/AA65/YT/AOiZ/wDlZ1j/AOTa+/6xPEuuweF/Dmq+Jbq2ubyHSbWe7eCzha4uZVgQyFIYU+aSRgMIg5ZsAdaAPhv/AIdc/sJ/9Ez/APKzrH/ybR/w65/YT/6Jn/5WdY/+Ta+6dA1iHxDoWm6/b29xaRanbQ3SQ3cTQXESzIHCTRPho5FBwyNypyDyKxNX8a2WjeMfD/gubTtQuLnxHHeSRXVvaSS2VuLJEdhdXCjZCZA+Igx+cggcigD4x/4dc/sJ/wDRM/8Ays6x/wDJtH/Drn9hP/omf/lZ1j/5Nr6Y+APxksvj18MrL4l6fpkmkQXl3qFqLeWQSup0+8ltGbcoUYcxFgMcA4r2egD4A/4dc/sJ/wDRM/8Ays6x/wDJtH/Drn9hP/omf/lZ1j/5Nr7/AKKAPgD/AIdc/sJ/9Ez/APKzrH/ybR/w65/YT/6Jn/5WdY/+Ta+/6KAPgD/h1z+wn/0TP/ys6x/8m0f8Ouf2E/8Aomf/AJWdY/8Ak2vv+igD4A/4dc/sJ/8ARM//ACs6x/8AJtH/AA65/YT/AOiZ/wDlZ1j/AOTa+/6KAPgD/h1z+wn/ANEz/wDKzrH/AMm0f8Ouf2E/+iZ/+VnWP/k2vv8AooA+AP8Ah1z+wn/0TP8A8rOsf/JtH/Drn9hP/omf/lZ1j/5Nr7/ooA/EH9vX9gr9k74LfsneOfiX8M/A39j+JNH/ALM+yXf9p6lceX9o1K1t5P3dxdSRNujkdfmQ4zkYIBH1/wD8Ouf2E/8Aomf/AJWdY/8Ak2j/AIKj/wDJinxM/wC4L/6eLKvv+gD4A/4dc/sJ/wDRM/8Ays6x/wDJtH/Drn9hP/omf/lZ1j/5Nr7/AKKAPgD/AIdc/sJ/9Ez/APKzrH/ybR/w65/YT/6Jn/5WdY/+Ta+/6KAPgD/h1z+wn/0TP/ys6x/8m0f8Ouf2E/8Aomf/AJWdY/8Ak2vv+igD4A/4dc/sJ/8ARM//ACs6x/8AJtH/AA65/YT/AOiZ/wDlZ1j/AOTa+/6KAPgD/h1z+wn/ANEz/wDKzrH/AMm0f8Ouf2E/+iZ/+VnWP/k2vv8Ar42+IP7VXinS/id4g+Ffwb+FOrfE7UfBcVpL4gmtLy00+CxN9H50EMbXTA3EzRfOUQcAgZJyAAcT/wAOuf2E/wDomf8A5WdY/wDk2j/h1z+wn/0TP/ys6x/8m19veEfEB8WeFNF8UnTrvSDrNlbXv2K/j8m8tftEayeTcR5OyWPdtdcnDAjNdDQB8Af8Ouf2E/8Aomf/AJWdY/8Ak2j/AIdc/sJ/9Ez/APKzrH/ybX3/AEUAfAH/AA65/YT/AOiZ/wDlZ1j/AOTaP+HXP7Cf/RM//KzrH/ybX3/XHeCfGtl460++1Gx07UNNSxv7vT2TUrSSzkkezkMTSxpIAXhcjMcg4Yc+ooA+Mf8Ah1z+wn/0TP8A8rOsf/JtH/Drn9hP/omf/lZ1j/5Nr7/ooA+AP+HXP7Cf/RM//KzrH/ybR/w65/YT/wCiZ/8AlZ1j/wCTa+vvit8TvCnwZ+HWvfFDxxM8GieHbY3NyYk8yVhkKiRrkZd3ZUUEgZIyQOa8b+CX7Uuj/F/xlf8Aw41fwfrvgLxXaaZDrkWn67DFG91pU8nlJcwtDJIuBJhHVsFWOOSGwAeS/wDDrn9hP/omf/lZ1j/5No/4dc/sJ/8ARM//ACs6x/8AJtff9B4GaAPgD/h1z+wn/wBEz/8AKzrH/wAm0f8ADrn9hP8A6Jn/AOVnWP8A5Nr7O8AeNLL4h+ENP8Zafp2oaVb6krsltqtpJY3sflyNGfNt5QHTJXK56qQw4Io8XeNbLwdNoMF5p2oagfEGpw6XEbC0kuhbyTq7ia5MYPk267CHlb5VJGetAHxj/wAOuf2E/wDomf8A5WdY/wDk2j/h1z+wn/0TP/ys6x/8m19MfCP4yWXxZ1T4gaZZ6ZJpzeAfElz4dlaSQSC5ktoYZTMoAG1WEwG05PHWvZ6APgD/AIdc/sJ/9Ez/APKzrH/ybR/w65/YT/6Jn/5WdY/+Ta+/6KAPgD/h1z+wn/0TP/ys6x/8m0f8Ouf2E/8Aomf/AJWdY/8Ak2vv+igD4A/4dc/sJ/8ARM//ACs6x/8AJtH/AA65/YT/AOiZ/wDlZ1j/AOTa+/6KAPgD/h1z+wn/ANEz/wDKzrH/AMm0f8Ouf2E/+iZ/+VnWP/k2vv8AooA+AP8Ah1z+wn/0TP8A8rOsf/JtH/Drn9hP/omf/lZ1j/5Nr7/ooA+AP+HXP7Cf/RM//KzrH/ybR/w65/YT/wCiZ/8AlZ1j/wCTa+/6KAPyg/aF/wCCcP7GPgb4BfEvxr4W+Hn2LWvD/hnWdQsZ/wC1tVk8m6tLKWaGTZJdsjbXUHaylTjBBGRX3B+yd/yaz8G/+xM8Pf8Apugo/ax/5NZ+Mn/YmeIf/TdPR+yd/wAms/Bv/sTPD3/pugoA/9T79/b6/wCbcv8Ass3hD/26r7/r4A/b6/5ty/7LN4Q/9uq+/wCgAooooAKKKKACiiigAooooAK/Lb4yaPr/AOzAfGf7X37Nniy21LwhPqUt94z8IXc6yafeXazi1vZ7G4GWtb4SjEiHO5xg52rEf1Jr5N1z9hn9lTxH46uPiNrPgG2n1i8vP7RuV+03SWVxebt3nzWKzC1kcsSWLRHcSS2Sc0AeAmLXv2qv2n/iH4G1n4geI/BfhLwBpGgT6Rpfh7UX0a6vn1q1+1y31y8Y8yQQsViVclFYDIDZ3cn+1bZ6h4asdc03wl8T/iDq/jr4feCl1K3ttG1CK0s7CLT7eXGqa27GGK5ku3icumXkdQwjhAO6vtv4tfst/An44a1Y+JfiR4YF9rOnQm2hvrW7utPu/s5JYwvNZSwu8WScI5KjccAEmsXW/wBjn9nLxCLFNS8JkR2GjReHljg1C/to5tKgBEVtcpDcItwseTsMwdgeQcgEAHyhrWs+PfjL8XP2b/Dmo+N9d8Oad438AXmsa1HoV8+n/bLj7PZy5Jj4Q75Dh0AkVcqjKCa8c8VePPi34Y/Z6+Kvw/sfiBrjXvgP4qad4d0nXJLx31ZNLubmzdYprnhpygmZWMmQ6nawKYWv0/8AB37Onwf8BXvhLU/DOiyw3ngawutM0eaa+vLl7azvCrTRfvpnEgYqoBk3FAAqFVGKj1T9mz4K6zYeIdM1Lw751t4r1uDxHqSfa7tfP1W1MZiuMrMCm0wp8iFYzjlTk5APlLSbPxd8E/2rbz4YaZ458Q+JPDPizwFqOtyQa9qMmoy22q2F0kQntpZMGFXjkOY48JnkAAKF+fPAXiH4mfE3Rf2LPC2o/ETxLpieOdI8Wvrt3Y6nNHeaiLO1ilj8+ZixdhyFkOZI9xaNkfDD9YtV+FngPW/HVv8AErU9M87xJaaXcaNFdedMu2wu3WSaHy1cRncyg7iu8YwGAzXKeHf2dvg54Uk8AS6B4f8AsrfC6C+tvDh+13T/AGGLUoxFdL88zed5iADM28r1Ug0AfnXfeMvjV8OPhh+0v8OPC3jTVdTj+Fut6QbLVtQuo7rWbPQtQit7q/SK6uyBJNBA0vkmUls8KS2wV6l+xfoHibVPix458e2fiH4h618OVstPg0GXxvf34a4uplZ74w2VyUV4k2xbJjCpDM6ozDOO8/au/Z81LW/A/iL/AIVF4cl1Cb4ja7pc3jy2tLmEahqui2cJhkSxOpObWG4VVi2bfKJCkhhJgnJ/ZZ8LfGXw98U9RNva+M9K+EH9iCKKz8e6ja3+pLrK3C7PsSwzXE0VstvuDCWTDMRtXAyACr+3fc3nxQk8Ffsk+DrKE+MviDctqlnq11O9vFocWiMk730ZjIkkuBysUadfmLEDr85fsWfF7xnpHxv1P9nF9OtdW+Ja6rqepfEnxTf6gt6uoW9kEisn03ynTnEiRCEoFtlU7k3FlX9MPjT+zl8GP2hbLTLH4u+G49cGjStNZSiee1uLdnwHEc9tJFKFfaNy7tpwCRkAjn9F/ZG/Zy8M3HhC88L+CLXQ7vwJNJcaRcadLcWdxFJMAsplmglSS5EigCRbhpFccMCCaAJP2q/DPxF8R/BHXp/hFq97o/jPQPL1nSjZTyQm6uNOYT/Y5VjZfOiuUVomifKMWBYcV8H6d8efGX7SnhH43ftJeB9b1bw74I8FfDq903RrW3up7VH8RTaY+oX12yoyZnsC0dvHJg4OXQg4Nfr1Xlel/BP4X6L8ONb+EmlaDFaeE/EQ1Nb6xieSNZl1hpGvBvRhIu/zXA2sCi4VNoVQAD8vfil498b3Gl/BfTPHfjLxzpnhbxN8N7S6sn8ESyS67qXikQxNMbowiS7kTyXRlJAjMrEyOO/oHwn/AOFyeAPiB+yV4K+IWv68ur+JNG8XXniSy1HVru+Nxdx2Vo8QufPlfcYSxZY2JELs4TFJ8afgX4rtfjNY3934V8Z3/gLwhoOn6R4Hm+H2o2dpqGlkJ5d5Fey3NxBeSBjFFsZpJIQo+cbixr6b/Z5+E3jO+8JeA/HP7SdnJffE3wS2sxaVd3N4Jry1sNTk2CO7azZbS4uGtkiWRwjDK5UhixIB8LfBeLRtV/Zf+F3hXUPGHi7Sb7Wdf8Yx6foPgq5Wx1TW7mPVbmQlrotGIoLSMO8m+WKLLqXbIVWSH4nfGfxR+yn8HLfUfHGtaV4kvPi9ZeE7vV7e5VNSksl1K6tdlw8eYZyEChw6vHIUBdXGc/ovc/sk/AC58MeHvCK+GpLbT/Cd5e3+lNbalqFvdWlxqUjyXZju4rhbjbOztvQyFCDtxgACPQP2RP2e/DGmWmi6J4Xe3sLDxDbeKraA6hfPHBq9m7yQTxK05CKjuzeSP3RJO5DQB8hal4t+IP7MPxX+NHgrwz4m1rxppGlfDCTx1plv4hvZdVmtdTt57i3ZUmmPmeS5jEjx7sf3cV61+zd8JNbsvA3hP9oW++Kfizx54l1jQxql7ZzawZ9Ev7i9tTJ5MNltEUSRSPiHywpUrz3FfXL/AAw8Cy/EO4+K0ulrJ4ou9HXQJbppJWV9MWdrgQGAv5OPNdmLbN5zgtt4ryHwD+xz+zj8L/HNt8RfAXhEaPrNjJcS2oivr02lrJdxvFM0Fk87W0W9JGUhIgAD8oFAH5u/Aq//AGqfif4J8G/tKeFNX8Rarr2qaqt5qsl34ksk8LS6cLtoruwXR2kH2cRRAojYEokBbPzDH7e18p3H7EX7Llz45l+Ij+BYE1i4vF1GVY7q7ispbxX8wTyWKTLau4f5stEeeetfVlABRRRQAUUUUAFFFFABRRRQB8Af8FR/+TFPiZ/3Bf8A08WVff8AXwB/wVH/AOTFPiZ/3Bf/AE8WVff9ABRRRQAUUUUAFFFFABRRRQAV+evx4+F2v6F4z8ZftQ/sxeOLbRfHfh+1jTxZot26z6RrEWm2wnjhvkyGtrlbVl8qYEEIV+4GZz+hVfLnxN/Yt/Zn+MHjC48e/EDwXHf65fLEt5PDd3lmLxYQBGLqO1miSfaFUDzFY4UDoMUAfJ8nxQ8Q/tffF34V+Ak8Uax8NvBXiP4fr41uLbRb5tP1HUb+e5+zGxF4iiRobZQzkR7S6ncRjBX0D4wfDyy8PReBPg2vxR+JGta79l1J9K03Q9VSHWb8rP5rXupajJ5KNDZrJHEnnyIpGOJXyK+nviX+zR8Dvi5oWheHPHHhS3uLTwsoTSGtZJrCfT0CKgS2ntHhliTaqgorBTtXIOBXNn9jz9nf+yPDOiQ+F5La38IfbP7OaDU9QhuI01B/Mu45biO4WaeOdzmRJndW5BGCQQD8/wDSfib8XPir8AP2SLnU/HWsaLrHjjxLPpms6jpdx9luru3t1vYSJCoKMzLCpyyth/nA3AGtfxn4g+Inwd0j9rX4VaL461/U9N8I+FtN13QL3UdSnutV02fULefz1jvmbz9u+IMmW+Tsclifvjwl+yf8BPA2meG9G8NeG3trLwhq8uuaRE2oX0q2d/KjRs8Yknb5NrsBEcxgksE3Emun8SfAD4SeLr3xpqPiHQftdx8QtOt9J11vtVzH9ssrUOsUWElURbRI3zRBGOeScDAB8O6JY+O/hD8X/wBnDW7b4i+JfEsPxVgu7HxFZ63qDXlpPIul/bIZ7e3IEds8cq/8sgMrwc5Yt45fePfirr3wm8I2dt4+17Sr7Wv2g7jw7JqFtfSG7TS3ubyMWqtIXVoUCrthdWiG1coQMV+sGo/CL4eatqHgvVNQ0nzbr4eMz6E/nzr9jZ7c2rHCyAS5hO396H9fvc1ykX7NnwVg07TdJi8O7bTSPE58Z2qfa7v93rxd5Dd5M2W+aRz5TExc42YAwAfF9vJ8YvhF8V/jf8Ffhj4w1XxM8fgCPxR4eHiW/OozWWrSSXFuVS5uzxG7oH2yt5YbGcLurg/2Q7fxX8R/jd4b8e+HPF/xM8ReB7Tw0bvVrjxRf3tvpcmvyuiJFaQApb3UZjaVmVEkiTCMr52g/Z37Svwd1/WfBvjvx58ENLgf4s69oUGgJcTzDbcaWl0JZ7VY7ovZq7RPMI3kjxvYb22jj5p/Z0+G3xX8BfFbwXpPwt0Xx94W+GGk217a+IrHxzqlheWYVbfbZJpUEFxcyK4nwWeMpDsBAHOKAPc/26fGVnZfCO2+D8WgweJNc+MV8nhXS7W7mNtZpcXalvtNxMvzKkG0OAmXZ9oUdSPz5+BfjX4l/s+ftHv+zh4iFr48+OGv3WlaZH4o1HUPP0+Hwda27TGFIBJHLDcQiN2+zn55pG8xmI2l/wBifi78F/hj8ePCTeB/ivoUWv6P5yXCRu8kTxTxghZYpYWSSNwGI3IwOCQeCRXk1h+xH+y1pXhiy8J6X4BtrK20/UYdWgube4u4dSW/gJMc/wDaMcy3hdcnGZiOelAHsvxc8G6v8Qfhl4m8F+Htbu/DeratYzQ2WpWNxLa3FpdbcwyrLCyuAsgUsFI3LlTwTX54fs4fG/4mftR/FfwJot7d6l4fh+Dej3DePLeGWW2S98UvJJp8VlcKhUSRILeW72ZZMsqsCRx+qYAUBR29ea4vwp8O/BfgjVPEmteFdKi0+98XX/8AaeqSR5zc3flJCZDknGVjGQuBuLNjczEgH46aB8VPiTefsofs3+IPHnjjxVa+EvEl3rdr4n1XQrqSfxLdXQnuBpcUcp827eMsjiTyVZwqqOF5HU+EV+M3w7+G/wAE9Q8S+JPGVveeMfjFZ2uPEWq3r6lc+HWW/wDskV9BLKRD50So01uAEYqrMm6vUPj1+zXc+H9S+G3g3wd4P8RXvwS8C2d7c2tj4Nv4Y9f0/XpZ2aO4FxfzpdvAYppQFgn3BzlgVCivXP2fPg/458V+GmX9pCw1a/svC3i1Nf8ABEfiLUY7nXrKK3t9kEl/Pp0nlSurySlY5GkIDYk3YXAB84+HrrQoNH+Ptr4i8Y694QtdR+M1zaIvhhjHrOrXE9jbJBptpIoLxvPLtYspU7UYF0Qsw88174n/ABi8Mfsb/tOaRP4m8SWOq/DrxBaWejXmpaismv2FpcPYSCCa/s5G3uokYF1lYkMV3EcV+n+ufsv/AAN8RaPr+h6r4bL23iXXh4nvGS9vIp/7aVVRb2CeOZZbeRVRQvksgHOByc8vH+xd+zcmjeL/AA8fC00mm+PEgTWoH1XUnW7Nu8ciOS1yWWUvEjPKhV5CPnZsnIB4c2l+NPgJ+1r8M/DWneOvEfjDSPibo/iNtVstbvjexte6NDBcR3FrGQsdq0hk2FIVSMLwFAAxgfsfeCvFHx78DeF/2pvHXxX8UT+J9Y1Ge/m0nTtWaDQbWK3u3jGmNpwXyzGI0Cy7h5hYk7s4Y/fmt/DbwX4j8b+GfiPrOnfaPEXg6O+i0q686VPsyakiR3Q8tHEb+Ysaj94rbcZXBJz4x/wxn+zYnxGX4q23g5bTxCuoxauWtr29t7RtRhkEqXTWUU62rSh1DbjEcnlsnmgD86NPv/2o/j1qHxS+IXgXVPE8HjDwr4r1PSdHhsvEVnp+gaUmlShYrW+0qVwLjzFG6eSRTvDDb905/aDw/d6hqGg6bf6tFHBfXNtDJPHC4kjSV0DOqOCQyhiQCCQRzmvnTx/+xf8Asz/E/wAZXfj/AMZ+Co7nXNSCi9mgu7yzW9CABRdRWs8Uc/CjPmK2cYORX0xZWVpptnb6dYQrb2trGsUUaDaiRoAqqoHQAAACgC1RRRQAUUUUAFFFFABRRRQB4B+1j/yaz8ZP+xM8Q/8Apuno/ZO/5NZ+Df8A2Jnh7/03QUftY/8AJrPxk/7EzxD/AOm6ej9k7/k1n4N/9iZ4e/8ATdBQB//V+3f+Ck2v/wDCKeFfgX4p/s291j+x/iz4YvPsOmw/aL66+zx3knkW0OV8yaTbtjTI3MQMjNdB/wAN9f8AVuXxm/8ACQ/+6qP2+v8Am3L/ALLN4Q/9uq+/6APgD/hvr/q3L4zf+Eh/91Uf8N9f9W5fGb/wkP8A7qr7/ooA+AP+G+v+rcvjN/4SH/3VR/w31/1bl8Zv/CQ/+6q+/wCigD4A/wCG+v8Aq3L4zf8AhIf/AHVR/wAN9f8AVuXxm/8ACQ/+6q+/6KAPgD/hvr/q3L4zf+Eh/wDdVH/DfX/VuXxm/wDCQ/8Auqvv+igD4A/4b6/6ty+M3/hIf/dVH/DfX/VuXxm/8JD/AO6q+/6KAPgD/hvr/q3L4zf+Eh/91Uf8N9f9W5fGb/wkP/uqvv8AooA+AP8Ahvr/AKty+M3/AISH/wB1Uf8ADfX/AFbl8Zv/AAkP/uqvv+igD4A/4b6/6ty+M3/hIf8A3VR/w31/1bl8Zv8AwkP/ALqr7/ooA+AP+G+v+rcvjN/4SH/3VR/w31/1bl8Zv/CQ/wDuqvv+igD4A/4b6/6ty+M3/hIf/dVH/DfX/VuXxm/8JD/7qr7/AKKAPgD/AIb6/wCrcvjN/wCEh/8AdVH/AA31/wBW5fGb/wAJD/7qr7/ooA+AP+G+v+rcvjN/4SH/AN1Uf8N9f9W5fGb/AMJD/wC6q+/6KAPgD/hvr/q3L4zf+Eh/91Uf8N9f9W5fGb/wkP8A7qr7/ooA+AP+G+v+rcvjN/4SH/3VR/w31/1bl8Zv/CQ/+6q+/wCigD4A/wCG+v8Aq3L4zf8AhIf/AHVR/wAN9f8AVuXxm/8ACQ/+6q+/6KAPgD/hvr/q3L4zf+Eh/wDdVH/DfX/VuXxm/wDCQ/8Auqvv+igD4A/4b6/6ty+M3/hIf/dVH/DfX/VuXxm/8JD/AO6q+/6KAPgD/hvr/q3L4zf+Eh/91Uf8N9f9W5fGb/wkP/uqvv8AooA+AP8Ahvr/AKty+M3/AISH/wB1Uf8ADfX/AFbl8Zv/AAkP/uqvv+igD8Qf29f2vP8AhaH7J3jnwL/wpX4meEv7T/sz/iaeIfDn2DS7fydStZv39x9ofZv2bE+U7nZV75r6/wD+G+v+rcvjN/4SH/3VR/wVH/5MU+Jn/cF/9PFlX3/QB8Af8N9f9W5fGb/wkP8A7qo/4b6/6ty+M3/hIf8A3VX3/RQB8Af8N9f9W5fGb/wkP/uqj/hvr/q3L4zf+Eh/91V9/wBFAHwB/wAN9f8AVuXxm/8ACQ/+6qP+G+v+rcvjN/4SH/3VX3/RQB8Af8N9f9W5fGb/AMJD/wC6qP8Ahvr/AKty+M3/AISH/wB1V9/0UAfAH/DfX/VuXxm/8JD/AO6qP+G+v+rcvjN/4SH/AN1V9/0UAfAH/DfX/VuXxm/8JD/7qo/4b6/6ty+M3/hIf/dVff8ARQB8Af8ADfX/AFbl8Zv/AAkP/uqj/hvr/q3L4zf+Eh/91V9/0UAfAH/DfX/VuXxm/wDCQ/8Auqj/AIb6/wCrcvjN/wCEh/8AdVff9FAHwB/w31/1bl8Zv/CQ/wDuqj/hvr/q3L4zf+Eh/wDdVff9FAHwB/w31/1bl8Zv/CQ/+6qP+G+v+rcvjN/4SH/3VX3/AEUAfAH/AA31/wBW5fGb/wAJD/7qo/4b6/6ty+M3/hIf/dVff9FAHwB/w31/1bl8Zv8AwkP/ALqo/wCG+v8Aq3L4zf8AhIf/AHVX3/RQB8Af8N9f9W5fGb/wkP8A7qo/4b6/6ty+M3/hIf8A3VX3/RQB8Af8N9f9W5fGb/wkP/uqj/hvr/q3L4zf+Eh/91V9/wBFAHwB/wAN9f8AVuXxm/8ACQ/+6qP+G+v+rcvjN/4SH/3VX3/RQB8Af8N9f9W5fGb/AMJD/wC6qP8Ahvr/AKty+M3/AISH/wB1V9/0UAfAH/DfX/VuXxm/8JD/AO6qP+G+v+rcvjN/4SH/AN1V9/0UAfAH/DfX/VuXxm/8JD/7qo/4b6/6ty+M3/hIf/dVff8ARQB8Af8ADfX/AFbl8Zv/AAkP/uqj/hvr/q3L4zf+Eh/91V9/0UAflB+0L+23/wAJX8AviX4W/wCFB/FnR/7Y8M6zZ/btS8LfZ7G1+0WUsfn3M32lvLhj3bpHwdqgnBxX3B+yd/yaz8G/+xM8Pf8Apugo/ax/5NZ+Mn/YmeIf/TdPR+yd/wAms/Bv/sTPD3/pugoA/9b79/b6/wCbcv8Ass3hD/26r7/r4A/b6/5ty/7LN4Q/9uq+/wCgAooooAKKKKACiiigD80v2pvG+uSftM+DvhF43+J+r/B34b65oMl1Y6vo8kVk+o+IFuihs5tQmjkSFY4NrrGQA5bBbcUFet+K/wBoLwb+x54R8DeBvjn401Dxnr2t3k1rFqQsGku5LQzSSRz3MVmj58mApESiF5nUsE++UT9p34iaJZXN58L/AIsfBDxF8S/AOtafHLFeaFpg1lDeb5Ukt5YEZZbaVAI2hnBGWc4ZCma+QtI8A/HLwH+zx+zx4n8YeFdc1qT4c+NJdUudJgRtS1uw8PSreR2iSxqd80lvFJGpRBuVdq7RtIAB9xeI/wBt79mDwlJpEXiLxn9iOs2cGoRb9Pv/ANzaXJIhlu8W/wDogcjAFx5Z74xzXonxA/aK+C/wuvdM0/x14nh02bWrGfUrHEU863NrbmMO0TwRurMTLGI0B3yFgI1Y8V+f/wC0h8Qvi18WdB8ceENF8AeMPDHhzxV4Lu5tOhtPDXnajrWqulxb/ZtVmaOdbKKJUiZYz5c7o3yyBisR6rwB4S1/xZ8Yf2U/Gd94Q1e2sfCngfVra6m1DTbi2/s/UEtra0VZ/NQeU8m2Tyg+C6/MoI5oA9p8Uf8ABQL9mrwyvgK7bXLq/wBP8f8A2uS1uLXTr2Vre3s1mDyzW6wG4GZ4vJCCIvuJYgIrMGfD34tX3iP9rHx1b/2/M3gWHwPoGuWUNyXt7eAXkk7vcGOYI0TNGFL7wpUDDAEGqH7VNn4k8K/GH4EfGrSPCuq+J9A8B6jrw1aDQrNr69iTV9Na1ilW2i/eOokOXKg46nkivG/FXw6+IXx2+I3x0u9F8O6v4Wh+Inwu03T9Ml1e0ezAvnN0fsk0nzxrINyrMiszIrcigD69+Gv7XH7PPxe8Wr4G+H/i5NR1iaOaW1jktbu1S9jtziV7Oa4hjiuVTBJMLPwC33RmvWPiT8SPBfwi8Eap8RviHqP9k+HdFRJLu68qWfylkkWJT5cCSSNl3UfKp656ZNfl38C9FHiDxF8I/D/jjwL8WLTxV8PpbfYmp2ttD4b0mWC1+zzPFerGiSWrJlI0jdnddoKjGR9bf8FAPDHiXxn+x/8AEjwz4P0m713WL61tFt7KwgkurmZlvYHIjiiVnYhQScA4AJ6CgDubX9rf9nW8k8YrH41to4vAMK3GsTyxTxW0MLyGJHjneNYrgNKDGvkM+5/lXJIBs/D79qn4CfE/SvEOs+E/FcZt/Cdqb7VFvre502a0swhk+0vDexQyeQUUsJApTHfPFeSftsfBfxH44/Z5svD3wq0t7m68Ialo2pxaVaMkUt5ZaRJk2kJkVk3LGS0SsCCygYJOD4V4FsvCXjfxP4q+IGr/AA3+LPiO+j8F6lo2oR+LbKCzF3YTyI8mk26Yha4nkbeUKNsUg/OCwoA+wPDP7Xv7PXi/w14k8X6F4oeTSvCWntquoyy6ff27R6eAxFzHHNbpJNEwUlWhVw3bNO+Ff7Xv7OXxp8Qp4S+HXjSG/wBbmiM8Vjc211p9zPEo3FoYr2GBpQFBY7A2FBPQZr83be8+Jmlfs6/tAeALbT/FUnwe0fwLeN4fuvGumtp2r2FyYnSTS4zLtkubaKNQVkZfk2iMEgAn0jSdU8SftITfs0eHPBHw/wDEOhr8Nb7Sdd1fxFrelyaZbwWdjZeW9paTy4Nx9tLKMRblKhWbgZUA+1fHv7Yn7NXwx8Wz+BvG3jq0sNZszGLqJIri4SzMv3BdTQRSRW5PXEzpwQehFdJ8RP2lPgh8KpvDkHjjxTFZv4vt57rR/IguL0X8VssbubdrWOUOSJUMag7pNw8sMa+H/hv4p8Tfsz+H/iF8GfHnwZ8UeOdX13Xta1K2v9G0r+0tN8SQatK8kRu7ofJA+xhFOs4wiKCNw4ql8HvgP8Ufhn42/ZA0DxfpVzfzeCtB8Yx6vdwRPc2mmS38UUltbS3Kho0KK3kRkth9hCFhzQB9u6b+1T8ANV+FV/8AGq18XwL4Q0q4NneXUsM8Utvdh1T7NJavGLhZizqBGY953AgEEGr2h/tLfBHX/h1rfxXtfE8dp4Y8NOYtTub+C4sHs5QqP5csF1HHMrsJE2rsy5ZQoJIFfDWtfDm0Rf2ndP8AiV8PPFGteE/FHi/S7q2XQLKX+0CBZwP/AGlp33TMbe5QMzRbzkEFW5Q8Jrvg79pz43fspa9o2s6dreuTeCfF2nan4fl1e0TSPEHiLRNOZJWS5tpQwFwu5vLeRAZiikpuyGAP0m+EP7Rvwb+Os2pWnwy1/wDtG80lY5Lq0uLW5sLqOKbmKXyLyKGQxOPuyKpU9M54qb4t+Nf+ES1z4d2P/CWweGP+Eh8RQ6eYJtOlv21XfBM/2KN4zi2diu8TP8o2YPXB+U/2cLfQ/F/xzHxJHhj4oW2vWfh+fS59T8c2kNhax273MUoskARGnk80FwU3Im1vm+YA+n/tV+GPEviLxn8Abrw/pN3qcOjeP7O8vntYJJltLVbO6Vp5yikRxhmUF2woJAzkigDW139uH9l3w1rGqaFrXjZbe70LVJtG1L/QL9o7C8gcRuLqVbcxwx7ztWZ2ETkNtdtrbe/+LX7SHwX+B8Wkt8R/EaWM+vbjp9rbQT393dqg3M8VvZxzSsgBGX27BkZPNfEWj/DPxk37Nf7ZOhXnhXUP7T8UeK/HF1pNq9jMJ9RintIvsctrGU3zpJIv7lowwZh8pJqGys/G/wACPjD8Pfjz4o+H/iDxfoN/8M9J8LzNo1jJf6noeoWzC4mSaxXEoimBwzgFldSrAA8gH2VfftY/s+aZofhTxJqPi6O203xxbahd6PNJa3Si6i0sKboYMO6ORCwURyBZHchEVn+WsW9/bQ/Zs0/wNoXxDuPFxOleJp7m202OOwvpL66ms2KXCpYpAbr9yw2yMYgqnGSMjPjHjG01T4y/tF/st/FGHwJrumaJpf8Awmlzdxatpr28umTPbRRWr3qLvS3aeSMvAJGDNwQNwIHLftVeBvGnhf8AaU8IfH7TdL8V6p4TTw5deH70+CYorjVtOuXuzdi5+zPG7PDMGKymMbhsBPZXAPqXWv2uP2d9C+HWh/FW78Ywz+HPEsr2+my2tvc3c91PHnzIktYInuA8e0iRWjBjPD7TWrN+098BoPhRb/G4+MLWTwXdzC2hvokmkaW5ZzGLdLdEM7TbgR5Qj38H5cV8O6De+Jvhf8P9P1b4ReB/HGh6b4z8a6hca3rGtaImq+J7CG9tYnnvrTS4YnaJbu4jWMmWFwpDSPE25BXnPw/8LWX/AAzt8RPAPxX+HXxDuIU+JF/qtrc6fpUsOuWS3UslxY6va+WiLLJC0QMptldUMg+QqcUAfpZa/tMfA65+F+o/GR/FMVp4R0iY215d3cFxavBcLs/cPbzRpOJiXQLH5e9iwCgkitL4RfH/AOEvx1g1OT4Za5/aU2ivHHfW01tcWN3bGYFozJbXccUyrIASj7NrYOCcHH5peKvCf7UHxk/Zf0HWPFuj61qur/Df4hWusWKyWsWleItf8O6chjiums5A6RX4ExZFcDcYgShYjd9Hfsw2WgeI/jHrPxNtPDfxKs9Xm0FNMutT8dWkVhE8aXKyx2sUQSN5ZY2MhMgBRRldxyMAH6B0UUUAFFFFABRRRQB8Af8ABUf/AJMU+Jn/AHBf/TxZV9/18Af8FR/+TFPiZ/3Bf/TxZV9/0AFFFFABRRRQAUUUUAFFFFAH58fsjftARH4XW0/xi8T3F3q/ifx5rnh3SJLlZrh5po7iU29sGjRgirFG20uVQAY3Zxn6o8SfHr4T+EdS8X6R4h137Jd+A9Jj1zWk+zXMn2TTpQ5SbKRMJM+W3yRl3GOV5GfyR+GuieKz+z/4I+JPhnw/f+KLb4ffGfVta1Ky0qE3N82nCe6gllt4FO6ZkaVTsXJIyegJHo/j6y+IPxZ179qLxlo3w88T6Zp/if4b2enaMmo6VPBc6jPEt0CkMGGfzCzYEJAlAwSgDLkA+1rP9uP9le+vZbCLx7BG62f26F5rW8givIchf9CkkhVLt9zBRHbmSQt8oUkEDe039rr9nvVPhX4g+NFr4qx4T8KXAtNWnksryO4sbgyJEI5rNoBdKxaRAP3XOc9ASPBPGPgDxPdeM/2Pru38OXk0PhR7gak6WkjJpqtonlj7SQuIAZVVBv2/OAOvFfLX7XXhTxDoPg79uHXb3Rruw0TXW8BzafcyW8kVrevElsly1vIyhJSknyyFCcNw3NAH6afCb9qT4CfHDV7rw98M/F0Op6xZRefLYTQXFjeCLODILe8ihlZASAWVSBkZIyM+d+Ef2pvhX4T+E3hnxn8UviXaazD4m1PU9PsdYi0m70+G7ntLm4H2cW2yR42hWIxBnwJWTK7i658jl1TxJ+0f+1L8IvHXhL4f+IfCOifDFNYudY1jxFpcmkTS/b7U20OnWyTYecFyXlIBjUAENk4PhPwq+FnxHsvh5+y1Yar4Q1aCfw98QvEV9qUU2nzq9jbS3OovDcXKsmYY33oyO+FO5SDyKAP0q+HP7RPwa+K2ga/4m8GeJIpbHwo7pq/2yGbTpdP8tDIWuYbyOGWJNgLB3UKQGwflOOf+Fn7WPwA+NHiN/CXw68VLqGrCBrqKGa0u7I3Vsh2tNatdQxLcRqeC0RYDr05r47+LfwH+IvxL+If7Wnh7wvplzpx8eeFfDEOmXssTwWeoXllFPvhS4ZRG5wqwyYY7A43YFZPwTtbfxb8Q/hne+JfAnxYh8V+A4roRjX7S3ttC0lpLFoLhI7kRRrNDMAsUSRklgUZlUKcAH278Pv2rfgD8UvEcvhXwL4sTUr2OO4lR/st1DazxWhxPJb3U0SW86RnhmikYD6VZ+F37UPwM+M/iCXwx8NvEv9r6hHBLdIPsd3bxXFvA6xyTW808McU8aO6qWiZgCa+F/wBnGfxx4M+KMvg74VeBfGdr8HbzTdVvNT8OeLtLNrFoGo7TJFa6RdTNieO6d2QwqzxqG8wOdxxc/Ze1b4g/Dnx9q3hf4Z+DvHB+DNhod9qK6N4r0trS90XVYmDx6dpNzOwa6jnBYCPc6IdrCU5YkA/VkkAZPAFfNHg79sP9m/x/46h+HPhLxpDf6zdyzW9qRb3MdpeTW+fNjtb2SJbW4dQM4ilYkcjPNbfh7xVH+0v+zcfEvhmK58Pf8LB8P3K2q3Y2T2j3kLxKzbcj5WO4MM5GCBzivzL+C3gm+j0b4V/Bn4qeBPisPEvw+1jSZIoYba2/4RW1u9NmCrqMWoCMJ9mC73YbzI4LIA2QxAP0e0D9r39nPxP8R/8AhU+ieMoZ/Ej3U1lFE1vcx29xd25Ilggu5IltppEKkFY5GOeBk8VoXn7U/wABrD4mH4Qz+KFPidLyDTnhjtLuW3jvrkqIbaS7SFrZJnLACNpQ2eMZ4r83fij4h+Mnizxf4H1XxR4J8V2qfDv4pWlyNC0fwzI2j2WhWklxHHexXUUDy3s9wrozNDL5S7mBiT5Wb0rxBceO/Cf7VC658AfBvjPS9Q8SeJbaDxhpmoaYz+EtU05WEUms298W8q3uFiVWDI292AVozht4B91R/tJfBST4a+JPi6fEyReFPCF1c2OrXUttcxPZ3VnIIpoJLeSJZxIrsqhBGWYsu0HIz63DrulS6EniVp/s+mvbC7MtwrW+yAp5m+RZQrR4Xlg4BXncAQa/Jv4hfBJvHH7etx8J/DGohvh7r0ek+P8AxzpKpmIajpbSwWkbH7gF8/kyTRfecRmQ8AV+rHi8xL4T1pp9KfXYxY3JbT4wpe8XymzbqGIUtKPkAJAyeaAPBfC/7Y/7OXjOx1vUvDvi37RbaBYTarcSSWN7bq9jA6xvPbmaBBcp5jqgMO/czADJIzxfgL9vn9m3xz8Or34lPrlzomnaYsD3Ud9p16kkf2uVoreOMrAUuZZCv3LZpSPoM14B+yra/EXTvF3iP4UeBdD8X2nwXbw9cmwi8b6a9heaDqkkgRdNs7iY+Zc2pjYtjDrHtUCQ87+X+F3xQ+MXw2/ZL8O/C7QvhD4mh8dfDyCytr65v9Aku7S0X7WIJ7rTlVwb+eK3laWNIcgjO5sAqwB98+HP2ofgV4s+GniH4u6F4oS48MeE1lbV5jbXKXFj5KB3We0aIXKsFOdvlZPYGuTn/bX/AGaY08Stb+Lxdt4T0w6veiGzuyPsqssZMLmIRzOJHWNkRiyudrAEMB+fMPh3x/qnhv8AbF0ODwz4z1e68d6JY32j3ur6DLaXGqtFpq28oWOC2hjE7TMdlukSyBB8yBlav0gl+Hl3P+yY/wAM/D2mCz1GTwO+jWloy+U0Usmm+QsJ34KfPgMGxgjnpQB8q/FT9sXw/wDFj9n3RPHvwN1rUtI1H/hIfCEWoxta3VlLbx6tdx+ZbebNFHHOCodHaBpEOMhsMpP1f8U/2sv2f/gv4kXwh8RPFa6fq4gW6mt4bS7vTa27nCzXRtIZVt4ycYaUqCOenNfAeqan46+IH7IXw5+D1n8MfGGmeIfh5d+BLPUVvtGmiikfTbiCG5e2YbmmjjEJkeRV2KjKS3UC18QfBfir4WftF/F7VvGOh/EPVvCXxSawurK/8C2kWpJKkVn9lmsdRhMMkiFCCsP8BRjkgk0AfrhpupadrOnWusaRcx3tjfRJPbzwuJIpoZVDJIjqSGVlIIIOCDkVdrxv9nvw1p/g74KeDvDGkaTqeg2Gm6fHDb2GsvHJqNtCCfLjuTESnmKuAQCdv3c8V7JQAUUUUAFFFFABRRRQB4B+1j/yaz8ZP+xM8Q/+m6ej9k7/AJNZ+Df/AGJnh7/03QUftY/8ms/GT/sTPEP/AKbp6P2Tv+TWfg3/ANiZ4e/9N0FAH//X+/f2+v8Am3L/ALLN4Q/9uq+/6/OD/gpNp2u6v4V+Bek+FtX/AOEf1q9+LPhiCx1L7Ol39hupI7xYbn7PIQkvlOQ/lsQr42ngmug/4Z4/bs/6O4/8sHR//jtAH3/RXwB/wzx+3Z/0dx/5YOj/APx2j/hnj9uz/o7j/wAsHR//AI7QB9/0V8Af8M8ft2f9Hcf+WDo//wAdo/4Z4/bs/wCjuP8AywdH/wDjtAH3/RXwB/wzx+3Z/wBHcf8Alg6P/wDHaP8Ahnj9uz/o7j/ywdH/APjtAH3/AEV8Af8ADPH7dn/R3H/lg6P/APHaP+GeP27P+juP/LB0f/47QB9/0V8Af8M8ft2f9Hcf+WDo/wD8do/4Z4/bs/6O4/8ALB0f/wCO0Aff9FfAH/DPH7dn/R3H/lg6P/8AHaP+GeP27P8Ao7j/AMsHR/8A47QB9/0V8Af8M8ft2f8AR3H/AJYOj/8Ax2j/AIZ4/bs/6O4/8sHR/wD47QB9/wBFfAH/AAzx+3Z/0dx/5YOj/wDx2j/hnj9uz/o7j/ywdH/+O0AfXXxd+Hdp8W/hd4r+GF9ePp9v4q02502S5jUO8K3MZjLqpwCRnIBrovB/h2Lwh4S0TwnBM1xFoljbWKysArSLbRLEGIHAJC5Ir4j/AOGeP27P+juP/LB0f/47R/wzx+3Z/wBHcf8Alg6P/wDHaAPv+ivgD/hnj9uz/o7j/wAsHR//AI7R/wAM8ft2f9Hcf+WDo/8A8doA+/6K+AP+GeP27P8Ao7j/AMsHR/8A47R/wzx+3Z/0dx/5YOj/APx2gD7/AKK+AP8Ahnj9uz/o7j/ywdH/APjtH/DPH7dn/R3H/lg6P/8AHaAPv+ivgD/hnj9uz/o7j/ywdH/+O0f8M8ft2f8AR3H/AJYOj/8Ax2gD7/or4A/4Z4/bs/6O4/8ALB0f/wCO0f8ADPH7dn/R3H/lg6P/APHaAPv+ivgD/hnj9uz/AKO4/wDLB0f/AOO0f8M8ft2f9Hcf+WDo/wD8doA+/wCivgD/AIZ4/bs/6O4/8sHR/wD47R/wzx+3Z/0dx/5YOj//AB2gD7/or4A/4Z4/bs/6O4/8sHR//jtH/DPH7dn/AEdx/wCWDo//AMdoA+/6K+AP+GeP27P+juP/ACwdH/8AjtH/AAzx+3Z/0dx/5YOj/wDx2gD7/or4A/4Z4/bs/wCjuP8AywdH/wDjtH/DPH7dn/R3H/lg6P8A/HaAD/gqP/yYp8TP+4L/AOniyr7/AK/EH9vX4NftY+FP2TvHOv8AxM/aK/4Tvw3a/wBmfa9F/wCER03S/tW/UrVI/wDS7eRpY/LkZJPlHzbNp4Y19f8A/DPH7dn/AEdx/wCWDo//AMdoA+/6K+AP+GeP27P+juP/ACwdH/8AjtH/AAzx+3Z/0dx/5YOj/wDx2gD7/or4A/4Z4/bs/wCjuP8AywdH/wDjtH/DPH7dn/R3H/lg6P8A/HaAPv8Aor4A/wCGeP27P+juP/LB0f8A+O0f8M8ft2f9Hcf+WDo//wAdoA+/64H4n+ENd8eeB9R8K+GvFN74L1G+8oR6tpyxtdW4SVXcRiUMmZEVkyRkBsjkCvj7/hnj9uz/AKO4/wDLB0f/AOO0f8M8ft2f9Hcf+WDo/wD8doA+pvgh8GvCPwC+Gmk/C/wU1xNp+mea7XF3J5t1c3E8jSzTzvgBpJJGLHAAHQAAAV6zXwB/wzx+3Z/0dx/5YOj/APx2j/hnj9uz/o7j/wAsHR//AI7QB9/14v8AtDfBnT/2g/g54j+D2q6lLpFr4iS3R7qFFkki8i4juAVViAcmMDk9DXzR/wAM8ft2f9Hcf+WDo/8A8do/4Z4/bs/6O4/8sHR//jtAH3/RXwB/wzx+3Z/0dx/5YOj/APx2j/hnj9uz/o7j/wAsHR//AI7QB9/0V8Af8M8ft2f9Hcf+WDo//wAdo/4Z4/bs/wCjuP8AywdH/wDjtAH3/UNxbwXdvLaXKCWGZWR0YZDKwwQfYivgb/hnj9uz/o7j/wAsHR//AI7R/wAM8ft2f9Hcf+WDo/8A8doA+9bKystMs4NO063jtbS1jWKGGJAkccaDaqIq4CqoGAAMAdKtV8Af8M8ft2f9Hcf+WDo//wAdo/4Z4/bs/wCjuP8AywdH/wDjtAH3/RXwB/wzx+3Z/wBHcf8Alg6P/wDHaP8Ahnj9uz/o7j/ywdH/APjtAH0d8JPghp3wv8TfEDxxPqkuu+IviJq/9pX15PGsbRwQxiKzsowpP7m1jyseSSdxJNe418Af8M8ft2f9Hcf+WDo//wAdo/4Z4/bs/wCjuP8AywdH/wDjtAH3/RXwB/wzx+3Z/wBHcf8Alg6P/wDHaP8Ahnj9uz/o7j/ywdH/APjtAH3/AEV8Af8ADPH7dn/R3H/lg6P/APHaP+GeP27P+juP/LB0f/47QB9/0V8Af8M8ft2f9Hcf+WDo/wD8do/4Z4/bs/6O4/8ALB0f/wCO0Aff9FfAH/DPH7dn/R3H/lg6P/8AHaP+GeP27P8Ao7j/AMsHR/8A47QB9/0V8Af8M8ft2f8AR3H/AJYOj/8Ax2j/AIZ4/bs/6O4/8sHR/wD47QB9/wBFfAH/AAzx+3Z/0dx/5YOj/wDx2j/hnj9uz/o7j/ywdH/+O0Aff9FfAH/DPH7dn/R3H/lg6P8A/HaP+GeP27P+juP/ACwdH/8AjtAH0B+1j/yaz8ZP+xM8Q/8Apuno/ZO/5NZ+Df8A2Jnh7/03QV8P/tC/An9s7SPgF8S9W8U/tQ/8JBotl4Z1me+03/hCdKtPt1rHZStNbfaI5S8XmoCnmKCyZ3DkCvuD9k7/AJNZ+Df/AGJnh7/03QUAf//Q+/f2+v8Am3L/ALLN4Q/9uq+/6+AP2+v+bcv+yzeEP/bqvv8AoAKKKKACiiigAooooAKKKKACiiigAooooAKKKKAOMl+IfguDx/B8LJdVjXxXdac+rR2GG81rFJfJaYHG3aJDt65z2o8ZfEPwX8PjoQ8ZarHpf/CTapbaJp3mBj9p1G83eRbrtBwz7GwTgccmvjfXCE/4KaeGC3Ak+F98q+5GrKSB+HNO/bxBkm/ZyiQbnPxi8KNgddqLdlj9AOtAH3zWfY6tpepyXcWm3kN29hMbe4WGRZDDMFDGOQKTscKynacHBBxyK+DdG+OH7VXxO+JPxR8D/CnR/CltZ/DfxHDp/wBs1kXyi7s5IY5TCgt5G/0hcuzSnaigxqI2LMy+b69+0t4u+Gngj44eMvh/4P8ADlvrWhfE+38PpGttLbx6kLs2UTXN68cu5rphLjzhhRhSY2wQQD9SqK+G/Dvxr/aK8P8Axr1j4H/EzRvDmtaxqPhO78TeHJdDa6s4JJ7SUQNp90128pyzuuJ1CgLyU52rzPwn/aW+M178e7j9n/4kXHgnxFrF1od9qdtceFbm68mwvrKSNGsL8TNM6n94GLgBgP8AlnzwAfoDe3tnptnPqOozx2tpaxtLNNKwSOONBuZ3ZiAqqASSTgDk0+3uLe8t4ru0lWeCdVeORGDI6MMqysOCCOQR1r8oPhj8Xv2m/Ff7NXxj+K3xcfwh4o8L6Vp/jIJpslhdtLLdaYZkS2mUzLC+nlYmQoFErIRukLFmPs9t8ffinrr/AA3+C/wA8N+H7fxTqXgrTfFOpXGp+fDoej6fOiQwQQW1sfOkZ5AyxxiRRHGoJJByAD7+or81fGn7Y3xc+HXwp+NNv408OaNZfFX4QW2nXm23e4udD1Oz1SVVguYVLxXCjG9XjZ8qwB3EHA9L8NfGz4/eGPj74G+Fvxr0zw62lfFHT9TutJk0L7X5+nXOlQx3E1vdSXDFbhTE42yJHFls/IBxQB9v0UUUAFFFFABRRRQAUUUUAFFFFABRRRQAUUUUAfAH/BUf/kxT4mf9wX/08WVff9fAH/BUf/kxT4mf9wX/ANPFlX3/AEAFFFFABRRRQAUUUUAFFFFABXMeM/GXhj4e+FtS8a+M9QTStE0eIz3d1IGKQxA4LEKCcc9ga6evkP8Ab4/5M4+LP/YEl/8AQ1oA6T4cftjfszfF3xbaeBPhv49std1++WV4LSFJ1d1hQySEF41X5UUk89q+mK+FPgT8Qv2kX8OeHYPGHwd0vw34atdCWRdZi8Qw3suIbLdATaJbxv8AvWVQw8z5QxOTiuB+E/7UH7SHin4I6P8AtS+O9E8Mab8N7fQtT1PV7O3+2rrMrWEUzLLZh3eBI5pYgiRyO7bCJC+W2KAfpTRX5Y/CD9tz4peIvHHw+g8cjwrqehfEi4S1Gn+Hlv21Xw/NdRGW1+2yzkwXCZAimeNYwrtlcrgM3xn+1z+09rvwk8cftIfBzQvCWn/DTwnNqEdl/bwv7rVNVi02Y28tzGlrJBDFG8isEVnYjacnoaAP1Qor5X+Hnxx8V+Lf2g9W+E+o2llHpFh4O0bxCksSSi5N1qMsiSozNIyeUoQbAEDA5yxr5hv/ANu3xtZ/A/wl4new0O28a+OvFWseH7GS7+0w6NZW2lXU8b3l0qSSTvsiiXckbAu7ZXaBtIB+o1FflRc/tz/E7RPgx8Z9bvLbw9rviv4X22m3thqmmwXo0HVbbUZli/1M0izpLAdyyoJsbipU7evuGj/G39orwt8cvB/wt+LWk+GZ7X4naXq13oZ0h7yNrC90qFLiS1vJpy4mjKOB50cUZzkiPHFAH3PRXwB+xR8Qv2n/AIpW2veLfivq3hy+8MJrOt2Kx2drdRajFdWV35CJG7SmD7KiowUGPzCMFmLbifcvjH8X/Enw8+Kfwd8D6LbWk9j8QtZvdOv3uEkaWKK3spLlWgKSIqsXQAl1cbc4APNAH0bVHU9T03RdPuNW1i7hsLG1QyTTzyLFFGi9Wd3IVQO5JxXwr8S/2pviB4N1X9pex0zT9Llj+DOgaJqukGaKdjcT6la3E8q3e2dd6K0ShBH5ZAJyScEcX8Uvil+0Wv7Nvjj4ufE/wn4LvvBN54Si1Ox0O5hvLq5FzM0LCDUo3dYZI2jdmZYmBjcBAz48xgD9KlZXUOhDKwyCOQQadXwNrXxo/aI1346v8CPgzZeFNOt7bwVpviL7brEN46wTXM8kBhWG2mQPGdqhB8nljLEvwlZ3h79pH47/ABL/AGf9C+JXhW38H+CdUtrzVLHxRqHie6nXR9Om0q4NqRAsbxyN9ocbkLyARgFWLkqSAfoVWfcatpVpf2ml3d5DDe6h5n2aB5FWWfyhuk8tCdz7AQW2g4HJr8q/G37XXxz8a/sfab8V/h6mi+HvE0niqPw1qdwjTXNrkaitmJLEEBvLm3KWZ23IhYJlyrr7zqvi/wCJHh/48fs9eD/jDo/hLWPE/iL/AITAy6rp1jcq2nR2dlHLH/Zz3MzyQmZGCXO7eHAwNooA+66K/OXSP2jv2qvib4D1L9oL4O+EPC8/w1tWvZtO03VLq8TXtXsNPkeOSeOWIfZrZ5vLcxRyI/bc2ME3rv8Aam+K/wATfiL8N/CH7PdnoMWmfEXwXL4pW78QR3TyWLJPHHtZLWVRLt3+W0QK/Od3m7V2sAfoZRXzN+y18ZfF/wAYvB/iZfiHptnpvivwN4l1Pwvqo04yGxnutNKEz2wlLSLHIkqkK7E9eew+maACiiigAooooAKKKKACiiigDwD9rH/k1n4yf9iZ4h/9N09H7J3/ACaz8G/+xM8Pf+m6Cj9rH/k1n4yf9iZ4h/8ATdPR+yd/yaz8G/8AsTPD3/pugoA//9H79/b6/wCbcv8Ass3hD/26r7/r4A/b6/5ty/7LN4Q/9uq+/wCgAooooAKKKKACiiigAooooAKKKKACiiigAooooA+ZPjz+zh/wtzX/AA18RfBvi29+H3xD8Heemma3ZQxXS+Rc4E1vdWs2I7iFsZCMRhuQeSDw/hf9ljx7rHxI8N/E79ov4ozfEi78FyPcaHp1tpUGi6Za3bqUN1LDFJM08yr/AKtmcBDnAr7TooA8N+EnwY/4Vb4t+Jfin+2P7T/4WJrv9t+T9n8n7H/o8Vv5O7zH83/V7t+E6428ZPhniP8AYw/4SDwx8QvDf/CY/Z/+E88dWnjXzf7P3fZfsr2r/Y9v2geZu+zY83KY3fcOOfuWigD5r+Iv7O4+IPxaX4oN4km0of8ACIat4UNvbQ4nX+1ZEf7XFc+YNjxbflXyzk4O4YwfFfhd+xx8Qfhz44+Gfi2T4k2F5a/DjSJ9Ajsrfw4lmlzpsxiJyy3j7bmRogZZiGDEkrGrZY/f1FAHwTY/sceOdG8M/EX4YaJ8VXg+HfjuHxEI9Il0WCWazuPEMcquxvPOV5YoJJfMSNVjLH5WfFdL4h/ZP8RW03gXxj8JfiDL4L8feC/Dlt4Wm1M6dHf2Wq6ZbKpEV1YyyrjEqmSNkl3IWYEvxj7RooA+BtY/YgufFHwp+Knhjxj8QbjXPHvxdSyj1bxNPp8aRxQ6e6tawW2nxSokcMahgFEu4sxZmPAH0B4x+CX/AAlvxh+FHxY/tr7L/wAKwj1qP7H9m3/bv7YtI7XPm+Yvk+Vs3fcfdnHy4yfeaKACiiigAooooAKKKKACiiigAooooAKKKKACiiigD4A/4Kj/APJinxM/7gv/AKeLKvv+vgD/AIKj/wDJinxM/wC4L/6eLKvv+gAooooAKKKKACiiigAooooAK8h+Pnwq/wCF4fBvxZ8Jf7U/sX/hKLJ7P7b5H2nyNxB3+Vvj34x03r9a9eooA5LSvC39meBLPwT9q8z7Jpsen/aNmN3lwiHzNm44zjO3d7Z715T8K/2fdE8Bfs36Z+zd4lvf+Em0i20efRbyfyTafa7e5EiS/uxJIY8pIRw5I6g19B0UAfF3wo/Zw+O3wxuPD3hl/jrd6v8AD3wu0KWmkyaHZx38lnbALDZz6kHLPEgVQSsSuw+XcBxXy/8AtU/s5fED4N/s0fGK1+GnxGWz+GV5b3+qv4ZvdJS7mtmu5fOuLezvxPG0UEkjMwV4pCmTgkktX641S1LTdO1ixn0vV7WK+s7lSksE6LLFIp6qyMCrD2IoA+NLn9njxrrPjDwv8cPhP8QT4H1m88LWGhatFLpcWqw3dnD+/heJZJIvJnjZ2Ac71IxlCAQ3Oab+wvFpfwQ8NfDO08f3tv4s8FeILzxJonimGziW4tr27uJZm821d3imRkmZJFJCyddqj5a++I444Y1hhUJGgCqqjAAHAAA6AU+gD4f8Xfsu/GP4p/Bvx58Lfi/8ZT4iufGNpa2lvcw6Bb2Nppy286ztItrFPvmklKgMzTgAAbVGDn2jxl8E/wDhLfjH8Kfi1/bP2T/hWMWtx/Yvs3mfbv7YtY7bPm+YvleV5e77j7s4+XGT7xRQB8l/Bn9nTx18FvGurSaF8R3uvh3qWqanrC+HZdKg85LnVHaR0OoeYXMUcjb0VY1YnhmI69b+0F8C9Q+Mlv4U1jwr4nfwd4u8DaoNW0jUvsiX0SSmJ4ZYp7Z3jEkckbkHDqQcEHqD9D0UAfBDfsX+KNW0z45Dxn8TG1zWfjdpGm6ZcXp0iO3TT206GeEPFBFcAPGRMNsZZWUL80kjEtXvfxX+B3/Cz/2ddW+AX9tf2b/amjw6T/aP2bztnlKi+b5HmJnOz7vmDGete+UUAfPXhf4D/wDCN/He6+Nn9ufaftPhSw8MfYPs2zb9huHn+0+d5pzv37fL8vjGd56V4Lb/ALEut6LoHg+18M+P4YdW8GeK9a8T20l7owvNPmfWnlJjmsjdJuktllPkzCUFW+YKCePv+igD8/8ATv2IdZPwS8a/BjxF8RpL2PxD4iPiXTNSt9LjtrjTr43ovy0iGZ47jMoAIAiXbkKFJyPSNJ/Z1+It/wDEX4YfFH4ofEhPFWt/DqbX5D5WjRafFdRa3Zw2qwosU7eUtuYy+5vNaQsQSoAr64ooA/Pz/hjL4meHdD1f4V/Cz41XnhL4U6zLdMdEGkW13fWFvfuz3NrYak8gaGJy77C0TtHnIJOTXsnhf9mHw94I+LHgj4geEtRNjo3gXwhL4RtNJMBkZoXmilWdrkyA7lEWGUxksSWLDofp+igDw74JfBj/AIU5N8Q5f7Y/tf8A4T3xbqXinH2f7P8AZP7QjgT7N/rJPM2eTnzPk3bvuDHPuNFFABRRRQAUUUUAFFFFABRRRQB4B+1j/wAms/GT/sTPEP8A6bp6P2Tv+TWfg3/2Jnh7/wBN0FH7WP8Ayaz8ZP8AsTPEP/puno/ZO/5NZ+Df/YmeHv8A03QUAf/S+3f+Ck3hbQvHPhX4F+CvFNr9t0XxB8WfDGn30G94/OtbuO8hmj3xsrruRiNysGGcgg4NdB/w65/YT/6Jn/5WdY/+TaP2+v8Am3L/ALLN4Q/9uq+/6APgD/h1z+wn/wBEz/8AKzrH/wAm0f8ADrn9hP8A6Jn/AOVnWP8A5Nr7/ooA+AP+HXP7Cf8A0TP/AMrOsf8AybR/w65/YT/6Jn/5WdY/+Ta+/wCigD4A/wCHXP7Cf/RM/wDys6x/8m0f8Ouf2E/+iZ/+VnWP/k2vv+igD4A/4dc/sJ/9Ez/8rOsf/JtH/Drn9hP/AKJn/wCVnWP/AJNr7/ooA+AP+HXP7Cf/AETP/wArOsf/ACbR/wAOuf2E/wDomf8A5WdY/wDk2vv+igD4A/4dc/sJ/wDRM/8Ays6x/wDJtH/Drn9hP/omf/lZ1j/5Nr7/AKKAPgD/AIdc/sJ/9Ez/APKzrH/ybR/w65/YT/6Jn/5WdY/+Ta+/6+f/ANpT47p+zv8ADhPH50CbxNLPqNlpkNjBOls8k19J5UeJJAVHzEdePcUAeAf8Ouf2E/8Aomf/AJWdY/8Ak2j/AIdc/sJ/9Ez/APKzrH/ybWnqH7Yvjv4fRR618efgX4l8C+GBIqXOswXNjrVrYq7BfOu1spWkjhXPzOFbHoa+3NJ1bS9e0u01vRLuK/0/UIknt7iB1kimhlUMkiOpIZWUggg4IoA+EP8Ah1z+wn/0TP8A8rOsf/JtH/Drn9hP/omf/lZ1j/5Nr6z+MfxV8NfBD4YeIvit4uWV9K8OWxuJY4FDTSszCOKKMEgb5JGVFyQMkZIGTR8JPGPjbx34Lt/Enj/wVceANVuJH/4ld1dw3kyRD/VyNJB8qlwclDhlPDDNAHyZ/wAOuf2E/wDomf8A5WdY/wDk2j/h1z+wn/0TP/ys6x/8m19/0UAfAH/Drn9hP/omf/lZ1j/5No/4dc/sJ/8ARM//ACs6x/8AJtffNxcQWkEl1dSLDDEpd3chVVVGSSTwAB1JrlNc8a2Wg+KvDfhK407ULmfxO90kNzbWkk1nbG0h85jdzqNkAcfLGXPzv8ooA+Mf+HXP7Cf/AETP/wArOsf/ACbR/wAOuf2E/wDomf8A5WdY/wDk2vv+igD4A/4dc/sJ/wDRM/8Ays6x/wDJtH/Drn9hP/omf/lZ1j/5Nr7/AKKAPgD/AIdc/sJ/9Ez/APKzrH/ybR/w65/YT/6Jn/5WdY/+Ta+/6KAPgD/h1z+wn/0TP/ys6x/8m0f8Ouf2E/8Aomf/AJWdY/8Ak2vv+igD4A/4dc/sJ/8ARM//ACs6x/8AJtH/AA65/YT/AOiZ/wDlZ1j/AOTa+/6KAPgD/h1z+wn/ANEz/wDKzrH/AMm0f8Ouf2E/+iZ/+VnWP/k2vv8AooA+AP8Ah1z+wn/0TP8A8rOsf/JtH/Drn9hP/omf/lZ1j/5Nr7/ooA+AP+HXP7Cf/RM//KzrH/ybR/w65/YT/wCiZ/8AlZ1j/wCTa+/6KAPxB/b1/YK/ZO+C37J3jn4l/DPwN/Y/iTR/7M+yXf8AaepXHl/aNStbeT93cXUkTbo5HX5kOM5GCAR9f/8ADrn9hP8A6Jn/AOVnWP8A5No/4Kj/APJinxM/7gv/AKeLKvv+gD4A/wCHXP7Cf/RM/wDys6x/8m0f8Ouf2E/+iZ/+VnWP/k2vv+igD4A/4dc/sJ/9Ez/8rOsf/JtH/Drn9hP/AKJn/wCVnWP/AJNr7/ooA+AP+HXP7Cf/AETP/wArOsf/ACbR/wAOuf2E/wDomf8A5WdY/wDk2vv+igD4A/4dc/sJ/wDRM/8Ays6x/wDJtH/Drn9hP/omf/lZ1j/5Nr7/AKKAPgD/AIdc/sJ/9Ez/APKzrH/ybR/w65/YT/6Jn/5WdY/+Ta+/6KAPgD/h1z+wn/0TP/ys6x/8m0f8Ouf2E/8Aomf/AJWdY/8Ak2vv+igD4A/4dc/sJ/8ARM//ACs6x/8AJtH/AA65/YT/AOiZ/wDlZ1j/AOTa+/6KAPgD/h1z+wn/ANEz/wDKzrH/AMm0f8Ouf2E/+iZ/+VnWP/k2vqn44/FO0+CXwk8UfFi+099Vt/DFm149rG4jeUKQNodgQDz1Ir5dj/bD+L2k+H4fG/jb9nLxRYeFXgS7e90y/wBN1eWO2dd4mNrBMspUKdzcZAySOMUAQf8ADrn9hP8A6Jn/AOVnWP8A5No/4dc/sJ/9Ez/8rOsf/JtfUFt8cfAer+CPC/xF8ISXXivQPF95Z2dlc6PayXoU3j+WJZ1QboYomBE7OB5RBDANxXr9AHwB/wAOuf2E/wDomf8A5WdY/wDk2j/h1z+wn/0TP/ys6x/8m19/0UAfAH/Drn9hP/omf/lZ1j/5No/4dc/sJ/8ARM//ACs6x/8AJtfTHw6+Mll8Q/iH8Sfh9baZJZTfDi/s7Cad5A63TXlql0HRQAUCh9pBJyRmvZ6APgD/AIdc/sJ/9Ez/APKzrH/ybR/w65/YT/6Jn/5WdY/+Ta+/6KAPgD/h1z+wn/0TP/ys6x/8m0f8Ouf2E/8Aomf/AJWdY/8Ak2vv+igD4A/4dc/sJ/8ARM//ACs6x/8AJtH/AA65/YT/AOiZ/wDlZ1j/AOTa+/6KAPgD/h1z+wn/ANEz/wDKzrH/AMm0f8Ouf2E/+iZ/+VnWP/k2vv8AooA+AP8Ah1z+wn/0TP8A8rOsf/JtH/Drn9hP/omf/lZ1j/5Nr7/ooA+AP+HXP7Cf/RM//KzrH/ybR/w65/YT/wCiZ/8AlZ1j/wCTa+/6KAPgD/h1z+wn/wBEz/8AKzrH/wAm0f8ADrn9hP8A6Jn/AOVnWP8A5Nr7/ooA+AP+HXP7Cf8A0TP/AMrOsf8AybR/w65/YT/6Jn/5WdY/+Ta+/wCigD8oP2hf+CcP7GPgb4BfEvxr4W+Hn2LWvD/hnWdQsZ/7W1WTybq0spZoZNkl2yNtdQdrKVOMEEZFfcH7J3/JrPwb/wCxM8Pf+m6Cj9rH/k1n4yf9iZ4h/wDTdPR+yd/yaz8G/wDsTPD3/pugoA//0/v39vr/AJty/wCyzeEP/bqvv+vgD9vr/m3L/ss3hD/26r7/AKACiiigAooooAKKKKACiiigAooooAKKKKACvgf/AIKM/wDJDPD/AP2Ofhn/ANLlr74rzP4tfB74c/HPwe/gL4paT/bWhSTxXLW4uLi1PnQHdGwktpIpBtPPDY9aAKXxy+IPw6+Gfwq8S+KvipdW0HhyGxuEuYrllAuleNl+zIjf6x5s7FQAlicYr8idAufjn4D+AH7Jv7Ol1PrOkN46j1ufVBpd/FpGrSxWxN5p+nR307AWxaKdd4ysjCPylwSVr9FPB37B37JfgXxLaeMNC+H0EurWDB7ebUby+1QROOQ6JfXE6BlPKsFyDggg817f8V/g38Mvjj4X/wCEN+K2gQeIdIEqzpFMXR4pkBCyRSxMkkbgEjcjKcEjOCaAPyD/AGjvh58adP8A2RPixoHxa1XXrfRPDms+H7/w0l9r0WoanJZ397BbT2upzWzN9phhlLSWxmywba2SYwF9O8eWfxS8T/tH6n+zjpGt+Nr/AMPfD3wzptxp8ek+KItJ1XUJb0v5uo3t9cOs12YmCxKpJRWALLyd33FpX7Hv7OujfD7X/hfZeE8+H/FU1pcaqkt/fS3N5LYyrPbtLdvO1yfLkQFQJAo5GMEg9B8YP2Zvgh8eL7TNW+KHhldT1LR1aO0vYbm5sbyGNzloxcWcsMuwnPyliBk4AyaAPhDU9c/aL14/s8/s1/GDxjdeDdT8XSeIh4j1jRr6GHUr9dCANnaw3luXWKeZHQ3OwhiQwB6qfZPit8M9A+Gngzwv8NNQ+KvxFvV1rWLn+zNO0zVTN4j1a4e3Vlsk1B/LdLW2SOSZvMlRAG/eSgBQfftX/ZT/AGfNc+F2mfBnU/BlrL4R0WY3NjarJOkttcF2kaeK6SQXKSszsWkEm9tx3E5NYdv+xp+zraeFLLwba+GriGy07VH1u3nTVdSS/j1GSJIJJ1vluRdBniRUYCUKQBxQB+Z/jzXPiF4+/YE8SQ+PvFeuC+8F/EP+wHmlvI1vJ7SDWoLVEv7iEETPCsp+ZX2l1V8ttU19meJNH134OftC/szfDHQfGniTV9G1i+8YvqH9r6vcX018ItIaaFLp3b98kMnzRK4IQ8rg17von7JX7Pfh/wAH+Lfh9p/hJG8N+OJxc6tp9xd3dzbzSq/mB0SaZ/JIk+cGHZ82G6hcW/BX7LHwK+H194a1Xwr4dkt77wjd39/ps8uoX1zLFdaparZXcrtPO5mMluix4l3BQAUCnmgD4d+HenfEH9oT4I+JP2oPEPxs8R+CPEcFxrcttp2nX0Vtonh9NJnmjhtb2xKMlwQkSvOZzuZX4wPmah4e8cfEf9pX4k/s+23iLxXr3gyx8b/D7UdX1mz0G/l05bu4ilgVXGwkxhyd6OmJFQ7VcBmz9l+L/wBib9mDx34wvfHPibwPDcanqk63N8kV1d29nezqdwlurOCZLadyeWMkbbiSWzXs7/Cr4fv440T4jjR0j8QeHNOm0nT545JI47exnKGSFYEcQ7SUXBKEqBhSBxQB82fsU694vm0b4o/D7xZ4gvfFCfDvxxqug6bf6nKbi+fTYooJ4EuJ2+aaRPOYF25IwOAAB9q1w3gz4beC/h9d+I77whp32Cfxbqkus6m3nSy/aL+dEjkmxK7hMrGo2ptQY4UEmu5oAKKKKACiiigAooooAKKKKACiiigAooooA+AP+Co//JinxM/7gv8A6eLKvv8Ar4A/4Kj/APJinxM/7gv/AKeLKvv+gAooooAKKKKACiiigAooooAKKKKACiiigAooooA+Q/2+P+TOPiz/ANgSX/0Na9k8H+JvD3g74J+HPE/ivUrfSNJ07QrGa5urqRYYYo1tkJZnYgAV1Pj3wH4U+J/g7VvAHjmx/tPQNcgNveW3myQ+bExBK+ZCySL06qwPvXy3pH/BO/8AY40bU7PVYfh1FeTWDK8KahqOo6jApXpmC7upYWHsyEe1AHwd8MdW8X+E/wBnzwp4x8IXWoeF/DnxE+PlpfaJbwSSWbHwxql+ES2ZEKn7PNscmNhtdTnBBBPd/Gbxjr/hD4oRfEX4W/EHxv4iutP+ImnaHrskt3EnhG1ivL1YpNEisXZTJLDDIiNLDG+GDM8qyHaP1L8Z/C/wJ8QLDQtL8WaUt5aeGtTstY06JJJbdbe+05t9rIogdMiM9EbKHoykcV4x4q/Yu/Zv8Z6nrOr694Xma41/UF1a7EGq6lawnUlZX+2RwwXKRRXDFBumjVXYcFiCcgHztc+EfHnxv/a2+Ovw1u/iV4m8LeEtC0zwtPb2+h6lJZzQXl1bTlXgfDiKMlXeZIwvmuIy5IXBytB+Hvj/AOMn7Sv7QPww1P4r+LtB8L+Dh4RTTl0nVXt7pLubR0dpfMKsEQsrSSRxqqTSOWkBKrj9APC3wp8A+C/FeueN/DWmG01vxJbafaahcGeeUzwaVG0VopSSRkXy0dhlVBbOXLHBq14f+G3grwt4y8VfEHQtO+y6/wCNmsn1e586V/tTadB9mtj5bu0ceyL5f3aru6tk80Afl5YfCa/8bfFb9rPULPx34l8Mz+HbrTpbI6NqL2GbyHRIpEuLloQr3BBRR5bt5eCx27iGGH4h+NHxk+MWmfsy+A2v9Zih+IXhSfWtWOganBoWo6zqFnDErQpeuY/KQZaeWOIguG2jAAx+qWmfCX4faPqfjPWNO0rybz4gukmuP587fa2jtxaqcNIRFiEBf3QT1+9zXBeJf2VfgD4v+GXh/wCD/iPwjDe+FfCixrpNu09yJ7Lyl2qYbsSi5VgOp83J4yTgUAfGKWHxN8NfCeXwB+0L428W6DA3i5LXw5Do2qQXni7WrO7t2e10qe+tiWDrKHdpmaPdEg3uqZNe2fsH+K/Huu+DPiP4Z8f6jqmpXHgjxvqmh2Ta3PFd6nFYQQWs0MF3cwM6TzRmZg0iuwPYkAV6Rb/sb/s5WfgpPAFp4TaDS49Uj1tZY9Qv1v11OFPLS6F+Lj7WJVQlQwm4XgccV2/wr/Z++E/wV1DXdT+GujyaTN4klW4vg17d3McswUKZBHcTSIrvtG91AZyAXJwMAHs1FFFABRRRQAUUUUAFFFFABRRRQAUUUUAFFFFAHgH7WP8Ayaz8ZP8AsTPEP/puno/ZO/5NZ+Df/YmeHv8A03QUftY/8ms/GT/sTPEP/puno/ZO/wCTWfg3/wBiZ4e/9N0FAH//1Pt3/gpNr/8AwinhX4F+Kf7NvdY/sf4s+GLz7DpsP2i+uvs8d5J5FtDlfMmk27Y0yNzEDIzXQf8ADfX/AFbl8Zv/AAkP/uqj9vr/AJty/wCyzeEP/bqvv+gD4A/4b6/6ty+M3/hIf/dVH/DfX/VuXxm/8JD/AO6q+/6KAPgD/hvr/q3L4zf+Eh/91Uf8N9f9W5fGb/wkP/uqvv8AooA+AP8Ahvr/AKty+M3/AISH/wB1Uf8ADfX/AFbl8Zv/AAkP/uqvv+igD4A/4b6/6ty+M3/hIf8A3VR/w31/1bl8Zv8AwkP/ALqr7/ooA+AP+G+v+rcvjN/4SH/3VR/w31/1bl8Zv/CQ/wDuqvv+igD4A/4b6/6ty+M3/hIf/dVH/DfX/VuXxm/8JD/7qr7/AKKAPgD/AIb6/wCrcvjN/wCEh/8AdVH/AA31/wBW5fGb/wAJD/7qr7/ooA+AP+G+v+rcvjN/4SH/AN1Uf8N9f9W5fGb/AMJD/wC6q+/6KAPgD/hvr/q3L4zf+Eh/91Uf8N9f9W5fGb/wkP8A7qr7/ooA+AP+G+v+rcvjN/4SH/3VR/w31/1bl8Zv/CQ/+6q+/wCigD4A/wCG+v8Aq3L4zf8AhIf/AHVR/wAN9f8AVuXxm/8ACQ/+6q+/6KAPgD/hvr/q3L4zf+Eh/wDdVH/DfX/VuXxm/wDCQ/8Auqvv+igD4A/4b6/6ty+M3/hIf/dVH/DfX/VuXxm/8JD/AO6q+/6KAPgD/hvr/q3L4zf+Eh/91Uf8N9f9W5fGb/wkP/uqvv8AooA+AP8Ahvr/AKty+M3/AISH/wB1Uf8ADfX/AFbl8Zv/AAkP/uqvv+igD4A/4b6/6ty+M3/hIf8A3VR/w31/1bl8Zv8AwkP/ALqr7/ooA+AP+G+v+rcvjN/4SH/3VR/w31/1bl8Zv/CQ/wDuqvv+igD4A/4b6/6ty+M3/hIf/dVH/DfX/VuXxm/8JD/7qr7/AKKAPgD/AIb6/wCrcvjN/wCEh/8AdVH/AA31/wBW5fGb/wAJD/7qr7/ooA/EH9vX9rz/AIWh+yd458C/8KV+JnhL+0/7M/4mniHw59g0u38nUrWb9/cfaH2b9mxPlO52Ve+a+v8A/hvr/q3L4zf+Eh/91Uf8FR/+TFPiZ/3Bf/TxZV9/0AfAH/DfX/VuXxm/8JD/AO6qP+G+v+rcvjN/4SH/AN1V9/0UAfAH/DfX/VuXxm/8JD/7qo/4b6/6ty+M3/hIf/dVff8ARQB8Af8ADfX/AFbl8Zv/AAkP/uqj/hvr/q3L4zf+Eh/91V9/0UAfAH/DfX/VuXxm/wDCQ/8Auqj/AIb6/wCrcvjN/wCEh/8AdVff9FAHwB/w31/1bl8Zv/CQ/wDuqj/hvr/q3L4zf+Eh/wDdVff9FAHwB/w31/1bl8Zv/CQ/+6qP+G+v+rcvjN/4SH/3VX3/AEUAfAH/AA31/wBW5fGb/wAJD/7qo/4b6/6ty+M3/hIf/dVff9FAHwB/w31/1bl8Zv8AwkP/ALqo/wCG+v8Aq3L4zf8AhIf/AHVX3/RQB8Af8N9f9W5fGb/wkP8A7qo/4b6/6ty+M3/hIf8A3VX3/RQB8Af8N9f9W5fGb/wkP/uqj/hvr/q3L4zf+Eh/91V9/wBFAHwB/wAN9f8AVuXxm/8ACQ/+6qP+G+v+rcvjN/4SH/3VX3/RQB8Af8N9f9W5fGb/AMJD/wC6qP8Ahvr/AKty+M3/AISH/wB1V9/0UAfAH/DfX/VuXxm/8JD/AO6qP+G+v+rcvjN/4SH/AN1V9/0UAfAH/DfX/VuXxm/8JD/7qo/4b6/6ty+M3/hIf/dVff8ARQB8Af8ADfX/AFbl8Zv/AAkP/uqj/hvr/q3L4zf+Eh/91V9/0UAfAH/DfX/VuXxm/wDCQ/8Auqj/AIb6/wCrcvjN/wCEh/8AdVff9FAHwB/w31/1bl8Zv/CQ/wDuqj/hvr/q3L4zf+Eh/wDdVff9FAHwB/w31/1bl8Zv/CQ/+6qP+G+v+rcvjN/4SH/3VX3/AEUAfAH/AA31/wBW5fGb/wAJD/7qo/4b6/6ty+M3/hIf/dVff9FAH5QftC/tt/8ACV/AL4l+Fv8AhQfxZ0f+2PDOs2f27UvC32extftFlLH59zN9pby4Y926R8HaoJwcV9wfsnf8ms/Bv/sTPD3/AKboKP2sf+TWfjJ/2JniH/03T0fsnf8AJrPwb/7Ezw9/6boKAP/V+/f2+v8Am3L/ALLN4Q/9uq+/6+AP2+v+bcv+yzeEP/bqvv8AoAKKKKACiiigAooooAKKKKACiiigAooooAKKKKAPzU+H03xj/bbu9e+I1v8AEXV/hl8J7TUrzS/D9l4aMFvqmppZSeTLf3V9KkxRXkVlSKNRgDnkbn+lvh54J8Wfs/aR4s174j/FjVfHHgzTrL7dEdbtYZb/AE2O0SSW7ke6tY1kuVZApCGPcuzC53Yr5W+FmofEb9iGDXPgr4q+HXiDxv8ADL+0L6+8Na34WszqctvZ30pnezvrSJhNE0Lu2JVBV88AYOPFvhB8KNThb9ozXfhf8MfF3gvwHr/gOfTdF07xGl2dQutTFvcCUQ211cXNyfMLLsHfgAAnFAH6M+Ev2t/2dvHPiCPw14Y8ZQXV5LpH9uh3guYLYacIkneV7maJIUMcciNKjOHjzh1UggV/AH7YX7NfxP8AFsHgXwR46tL/AFq88z7LC8VxbLeeV9/7LLcRRxXGBk/uXfgE9Aa8C134DeMPF/8AwTX0b4LeDdLOj+LJvBmip9guVNpKbuNLa7vLObzNrRPcOkkUgfbhnO8jBI4L4k+KvE/7T2m/Dj4T+A/g14o8Ear4e8Q6Nqt1qOt6V/Zmn+HbfSpFeb7Jct8k8hQeVCkAw6Nk7RxQB9jfE/8Aaz/Z4+DfiM+EfiL4zt9N1mOJZ5rWOC5vJLaJxlXuRaxS+QpBBBl2Agg9Dmtnxl+0n8DfAGi+GPEfinxda2+leNC66NdwrLdwXpSIzYikt0kUlk+4CRvYhEy5Cn498JeKPEn7LPxH+Mmm+OPhb4p8aL8QPE114h0rWfDelHV476zu4o1i064ZCGge12tGqy4QqSwwPmbzf4afAr4p+CV/ZltfEPha4s/sPjLxNrl5p9pG11a+HbLU7e5ltbaWWJTHGsZdVBJC+YdoOcUAfXk/7ef7Kdt4dl8Sy+Nv3FrPLb3NsNO1Br+1aDb5rXFiLc3UMab1zLJGqc4DE8V3+rfFPRr74lfCax8O+ObOLSvHdlq17aaclhJdnXoLe2hnSaC+Q+XbLbrIJDv/ANcHCryOPEfAPgLXIf2k/wBqvWNT8PXMek+KrDwvFY3Mtq62+o+VpM8NwkEjLtm2MQkgQnDEBua+fvgL8OfiRpeofsTy634X1azHhTQPGEGsNcWU8f8AZslxawJbx3ZdB5DSFSsaybS2MLnFAH13pf7c37LGs6lpel6d45R31e9OnQzPY30Vsl6JWhFvPcSW6wwSMynaszoWXDjKMrHovi7+17+zn8CdaPhz4o+MotK1RIknktobW7vpYYpTtjaZLOGYxBz93zNu7tmvz5n+E/xDb/gltr/gMeDtV/4S59auLqHSxp8/9osf+EoE6Srb7POP7geYGC/6v5vu81zn7S/ijwjaftLfFr4cJ8Q4vBHhD4jaLpUXjPUX0pteVbiGJ7T7NFJagjTpRaKNxuyQC3mIo4JAP2Z8J+KtF8b+HLDxZ4deaTTdTjEsDT281pKUJI+aC4SOVDx0dFPtXRVzPgr/AIR7/hDtC/4RK9XUdDFhaiwuklE6T2oiXyZFlGQ4dMEMPvZzXTUAFFFFABRRRQAUUUUAFFFFABRRRQAUUUUAfAH/AAVH/wCTFPiZ/wBwX/08WVff9fAH/BUf/kxT4mf9wX/08WVff9ABRRRQAUUUUAFFFFABRRRQAUUUUAFFFFABRRRQB8x/tneKvEfgf9lr4leLfCGozaTrOl6TJNa3du2yWGQMoDIw6Hmvmf8AZ11L4YvqWh+ItO/ar1bx14jfSZbqXw/e69p+pW/mmzZ5meyt4kuH+zfNJtD5Gzk8GvpD9tnw9r/iv9lH4neHPC+m3Osatf6RLFbWdnC9xcTyFlwscUYZ3b2UE1q+EvhF4I8O/CSxu9I8FadpniNPDoiMkGmww3wney2SISsYkDsSVYdSSQaAMXQv2k/hn4J+AXgz4o/FD4g22s2viGCGO21W3025tX1e5kzj7LpiJJc7mx/q1QkYyQB08V+A37UPhvxZ8Vf2j/Het+L5ofhp4Ufw09i+qxz2MWnJJpxF2ogukiliL3KkFSgLvjaDkZ8H8B/Dr4l/DjwR+yb8ZNW8B6zr9p8M9G1bT9a0O2s3bWdOl1SERxXcVhIY5JGQrslXG9UYMqnHHH/Ee+tfFXhj9r34q+MvDXiDwhoniZ/B114eXXbFtEu77UtGtF8qOGLUECTH7VAoaJQ5dDtUbmWgD9Hvhz+2R+zj8WNZ1PQvAnit7240W1kvb6SbTNRsra1t408xpJrm6tooEGz5hukGVBIyAasfDv8Aa/8A2c/iprdx4e8FeMY7m9t7Wa+Vbi1urFLi0t/9bPbSXUMSXEaYO5omYAAn7vNfn34M8PRftPfDr43Cz+J1r4m+NXjvwpa6fLpUelz+HFsbWx3tFE1td/vZRLNIY5bk5QBwo2rjPZfArTrfxZ4/+FSeLfA/xXs/FXgPeqHXLS3t/D+jubI29x5d2sUazW8igRxJGSzDaSq4OAD6C8Mfty/BX4v+E/Hw8AeI7rw9qXhrTtaulvr7Rb6aC3t9LjIbUCoiEUqI37wW4l851GCisSo7+7/av+Cnw/0XwhpvxC8area74h8PWut2r22l32/Vbd1RWuLa1hilcNIzbxbjMqLkldqMw+MfCL+PfCnwD+PP7NOqfDfxU/iPVF8f3VjfQaVJNpV1BqEV1NZmG7QlZJLgsI44ow0hcgFR29M+DngHxnp37Q3wR1zVvDmoWtho3wZg0y6up7OVIbTURNaZtZZGQLHcBQ+Y2IfAbjANAH1jD+058BpvhMnxyHjKzTwS7mEX7iRD54cx+R5DIJ/P3DHleX5n+zTPg3+038Fvj/d6pZfCbW7jWpNGJW7Z9M1CyjicEAp5l3bwozgkZRWLAEEjHNflt8StNvPhZ8FfiX4g1uzuvDviTTvjvfeIvBsVzb7DdSmWFY7qCzmXffW3lSzPtt1ZmALIRtJH21+xNfeBdej+IPjTSviHB498aeKNUt7/AMR+Rp8ujLYyLbLb20I024/fwqI4iPMkyZSCSSQaAPumiiigAooooAKKKKACiiigAooooAKKKKACiiigDwD9rH/k1n4yf9iZ4h/9N09H7J3/ACaz8G/+xM8Pf+m6Cj9rH/k1n4yf9iZ4h/8ATdPR+yd/yaz8G/8AsTPD3/pugoA//9b79/b6/wCbcv8Ass3hD/26r7/r84P+Ck2na7q/hX4F6T4W1f8A4R/Wr34s+GILHUvs6Xf2G6kjvFhufs8hCS+U5D+WxCvjaeCa6D/hnj9uz/o7j/ywdH/+O0Aff9FfAH/DPH7dn/R3H/lg6P8A/HaP+GeP27P+juP/ACwdH/8AjtAH3/RXwB/wzx+3Z/0dx/5YOj//AB2j/hnj9uz/AKO4/wDLB0f/AOO0Aff9FfAH/DPH7dn/AEdx/wCWDo//AMdo/wCGeP27P+juP/LB0f8A+O0Aff8ARXwB/wAM8ft2f9Hcf+WDo/8A8do/4Z4/bs/6O4/8sHR//jtAH3/RXwB/wzx+3Z/0dx/5YOj/APx2j/hnj9uz/o7j/wAsHR//AI7QB9/0V8Af8M8ft2f9Hcf+WDo//wAdo/4Z4/bs/wCjuP8AywdH/wDjtAH3/RXwB/wzx+3Z/wBHcf8Alg6P/wDHaP8Ahnj9uz/o7j/ywdH/APjtAH3/AEV8Af8ADPH7dn/R3H/lg6P/APHaP+GeP27P+juP/LB0f/47QB9/0V8Af8M8ft2f9Hcf+WDo/wD8do/4Z4/bs/6O4/8ALB0f/wCO0Aff9FfAH/DPH7dn/R3H/lg6P/8AHaP+GeP27P8Ao7j/AMsHR/8A47QB9/0V8Af8M8ft2f8AR3H/AJYOj/8Ax2j/AIZ4/bs/6O4/8sHR/wD47QB9/wBfH2sfsmXKax4wn+HnxO8QeCdE+Id9cajr2l2cOm3MU1zeRLFcS2015aTTWzSqo3bXYA8qF4xwP/DPH7dn/R3H/lg6P/8AHaP+GeP27P8Ao7j/AMsHR/8A47QB9lfDjwB4d+FfgLQPhx4Rjki0bw5Zw2NqJXMknlQqFBdzjLHqTgDJ4AHFdrXwB/wzx+3Z/wBHcf8Alg6P/wDHaP8Ahnj9uz/o7j/ywdH/APjtAH3/AEV8Af8ADPH7dn/R3H/lg6P/APHaP+GeP27P+juP/LB0f/47QB9/0V8Af8M8ft2f9Hcf+WDo/wD8do/4Z4/bs/6O4/8ALB0f/wCO0Aff9FfAH/DPH7dn/R3H/lg6P/8AHaP+GeP27P8Ao7j/AMsHR/8A47QB9/0V8Af8M8ft2f8AR3H/AJYOj/8Ax2j/AIZ4/bs/6O4/8sHR/wD47QB9/wBFfAH/AAzx+3Z/0dx/5YOj/wDx2j/hnj9uz/o7j/ywdH/+O0Aff9FfAH/DPH7dn/R3H/lg6P8A/HaP+GeP27P+juP/ACwdH/8AjtAB/wAFR/8AkxT4mf8AcF/9PFlX3/X4g/t6/Br9rHwp+yd451/4mftFf8J34btf7M+16L/wiOm6X9q36lapH/pdvI0sflyMknyj5tm08Ma+v/8Ahnj9uz/o7j/ywdH/APjtAH3/AEV8Af8ADPH7dn/R3H/lg6P/APHaP+GeP27P+juP/LB0f/47QB9/0V8Af8M8ft2f9Hcf+WDo/wD8do/4Z4/bs/6O4/8ALB0f/wCO0Aff9FfAH/DPH7dn/R3H/lg6P/8AHaP+GeP27P8Ao7j/AMsHR/8A47QB9/0V8Af8M8ft2f8AR3H/AJYOj/8Ax2j/AIZ4/bs/6O4/8sHR/wD47QB9/wBFfAH/AAzx+3Z/0dx/5YOj/wDx2j/hnj9uz/o7j/ywdH/+O0Aff9FfAH/DPH7dn/R3H/lg6P8A/HaP+GeP27P+juP/ACwdH/8AjtAH3/RXwB/wzx+3Z/0dx/5YOj//AB2j/hnj9uz/AKO4/wDLB0f/AOO0Aff9FfAH/DPH7dn/AEdx/wCWDo//AMdo/wCGeP27P+juP/LB0f8A+O0Aff8AXj3xq+C/h/43+HdK0TWtRvtGu/D+q2muaXqOmvGl1ZajYlvJmQTRyxOAHYMkkbKQTxnBHzD/AMM8ft2f9Hcf+WDo/wD8do/4Z4/bs/6O4/8ALB0f/wCO0Ae1fDz9nWfwz8TP+Fy/EDxzq3j/AMYw6XJo1rcX0NlZW1rYyzLO6RW1jBCm93UbncsTjA2jivpWvgD/AIZ4/bs/6O4/8sHR/wD47R/wzx+3Z/0dx/5YOj//AB2gD7/or4A/4Z4/bs/6O4/8sHR//jtH/DPH7dn/AEdx/wCWDo//AMdoA+jPjH8DNN+LeoeFfE9vr+o+E/FXgi4ubrRtW0z7O8lu93CYJ0khuop4JY5EOGVkzwMMOc5/wk+AMHw38YeIfiZ4j8V6n438a+Kba0s73UtQS1t0W2st5iht7azhhiiQFyTkMxPJbrnwP/hnj9uz/o7j/wAsHR//AI7R/wAM8ft2f9Hcf+WDo/8A8doA+/6K+AP+GeP27P8Ao7j/AMsHR/8A47R/wzx+3Z/0dx/5YOj/APx2gD7/AKK+AP8Ahnj9uz/o7j/ywdH/APjtH/DPH7dn/R3H/lg6P/8AHaAPv+ivgD/hnj9uz/o7j/ywdH/+O0f8M8ft2f8AR3H/AJYOj/8Ax2gD7/or4A/4Z4/bs/6O4/8ALB0f/wCO0f8ADPH7dn/R3H/lg6P/APHaAPv+ivgD/hnj9uz/AKO4/wDLB0f/AOO0f8M8ft2f9Hcf+WDo/wD8doA+/wCivgD/AIZ4/bs/6O4/8sHR/wD47R/wzx+3Z/0dx/5YOj//AB2gD7/or4A/4Z4/bs/6O4/8sHR//jtH/DPH7dn/AEdx/wCWDo//AMdoA+gP2sf+TWfjJ/2JniH/ANN09H7J3/JrPwb/AOxM8Pf+m6Cvh/8AaF+BP7Z2kfAL4l6t4p/ah/4SDRbLwzrM99pv/CE6VafbrWOylaa2+0Ryl4vNQFPMUFkzuHIFfcH7J3/JrPwb/wCxM8Pf+m6CgD//1/v39vr/AJty/wCyzeEP/bqvv+vgD9vr/m3L/ss3hD/26r7/AKACiiigAooooAKKKKACiiigAooooAKKKKACiiigCnqOoWWk6fdarqMogtLKJ5ppDnCRxqWZjjnAAJr5Z8K/t0fsmeN/Eul+D/CnxIsNR1nWbiO0s7aOO4DzTzMFRFLRAZYkDkgV7t8VP+SYeL/+wPqH/pO9fnJ+wh41/aMT4H/CXQbf4PabN4L+yW0R8QHxDCtyLMyEPc/YfsxfcoyRH5mTjrzQB+q9Ffnv8H/jv+1J8ZrrV9f0XSfCeleEvCHjLVND1WW9F8t3d6bp9zseWzVJHRZIoSCWkYrLLuUJGq5fwXw7/wAFEfiHrkek/FRbfwxL4E1fVo7QeG4BfSeKoNMlufsy3rTAm0aRf9c0CpgR8bwwJAB+wVFflp8WP23fiNZ/Er4g+GvhdP4S0zTPhpKLSWDxEL6S/wBdvo4hNPBZm0Kx26rnykeUPukwcBc43fjb+05+0d4a+EOi/tB/DWPwjaeF/ElhpZ03QtbstTufEFxq2oqoWwjFtcQxPJ5hKgBQQqsx4FAH6PS6tpcGpW+jT3kMeoXaSSw27SKJpY4iokdEJ3MqFl3EDAyM9RWhXwvpfjbx/b/tB/BPwj8WvDnhqXxlrvhTXL3UtQsrOUT6fPE1uWtbGaSeUrCd4SUkt5hXcuwHbXgPgv8AbH/ahu/gZ8Pv2n/GWh+Ej4G8QapbabqdhYLfJqSxXWoNYC7hklmeKPZIB+5YSZHPmDdtjAP1mor8uf2if2yPjf4H1b4rS/DK28H6bo/wiksIL6LxJLdSarqT3sMU/mWVtBNbqECyhYw7nzSDgjoP0l8JSeIZvC+kzeLZLeXWpLWJrxrSGS3tzOVBcxRTPJJGueiu7MOhJNAHQ0UUUAFFFFABRRRQAUUUUAFFFFABRRRQAUUUUAfAH/BUf/kxT4mf9wX/ANPFlX3/AF8Af8FR/wDkxT4mf9wX/wBPFlX3/QAUUUUAFFFFABRRRQBwvxD+JvgL4T6FF4n+I2t2+gaXNdQWSXFySEa4uW2xJwCcse/QAEkgAkd1X4r/ALdfxQ+A3xR/aKsP2dvjh4vTw14J8FaFfajeuI7iYy+IdUtzBYJst0c7rOCX7SpOFJba2QcV6t8Hv2ydevf+CeeufGPR57PWvG/wvspNLvftQlkt7m6sGSOK4kVWhlZbiBklJyhLlh2IoA/VKivgvwx8e/2ifAfjv4e+G/2jNG8M3WhfFCb7DpereGHvITZ6g1ubiKC7tr0yEiZVKq8chwwORg1414e/az/a3f4Va9+0hqfhvwfrnw78NajqcN9ptib+y1xbHTLt7aWeN5pJ7Z3VUMhU7cgEDBIAAP1Zor4F8QftDfHjxl8dtU+DXwE03w9LbP4Q0zxPY6rriXYhi+2yupWcW0m6TzF2CJFVNvzuzttCN5V4/wD2zfjJYeP9f+Gnh268E+HdX+H+n6d/bb63/aE8Oqazd2q3MtppotmRooIs7POnySxX5cBjQB+kN9498Iab400v4d32pxw+I9Ztri8s7IhvMmt7UqJpFIG3CF1zkg88V11fkjqfxQ+IPxn/AGgv2ePiV8KtLsdE1/xR4I8QStBrwne305vOtlud8cPlyzmN0KIoaLfkMWUZFeiXf7bfjbwR8CfHXiL4haHpc3xE8FeMD4IWGzlmh0m8v5jE1td5kLyxW5ilLuhZn+QjK7htAP0qor8yvAv7ZHxMN3478LeLpfDHinUdF8H6j4o0rV/DcF/Fpnn2CEyWF5FduZN4Yq6skih48/dbOLfg/wDae/aSsrP4FfEP4o6P4XPgv403Gl6WLfSRerqFheatZtcW05lnkaJ45CjFotmYwQnmSEF2AP0qrP0/VtL1YXDaVeQ3gtJntpjDIsnlTxHDxPtJ2up4ZTyO4r4P+Hn7SPxk8cftL+K/hFqA8LeHbXwzq89sNB1Fb6HxBfaLGMxatZzlvs06Sg7vKWP5FB3OON3nGk/tLeJvhv4Y8YfEfT/Cnh7SfBnh/wCMOteHfFbWNnLBMNNe5S2XWHdZyrXQmeM3LsjCRSNqpjNAH6j0V+bPxJ/bg8T+EB8WfFOg6Pp2peE/B2s6Z4O0CWUyxtqHia6x9tM84cotpZ7gGCRb2Kthx2639nH9p/x746+Ld38IPiJc+HfEbz6QdYsda8KxXsNkjRSrFPY3Ed6XcSrvDxurYZAcgNkAA++aK+Sv2tv2jNQ+AHh3wxa+GYNPk8S+NtVTSrCbV3lTTbNQjSz3d35H7144kX7keGYsMH18Z+Gf7Sn7QHxN0j4i+B/CcfhTXviD4Ngs9Q0rUbWC+TQNbtbjdvttk0yTW90hQoWMroGZWxtyaAP0bor4I+CH7WvjX9pr4h2GkfCzwwug+F/DFuf+E2udcglNzaaqxKDSLFUkiDTRsjNLM6sgTHyBiFbxbwr+13+1mPgtP+1F4l8N+D9a+HGm3l8l/p+mG/s9chsrG+kspZ4zPLPbSMmwyFCUyowCCeAD9YKK/NL4v/tn+L4/i7q/wu+EGqeE9Bg8L6Zp9/e6j4rW+lW9uNUh+1W1taxWTIUUQlGklcnG8AJxltGD9sX4nfE/w58H/D/wW8LWOn/ED4p2eo310uvmdtO0S30dzBeSSpBsmm3zApb4KhsqWwDQB+jlFfm1+0F8Rf2zfh54M+FmnG/8IWXijxT4607Qbm8s4r57KeK5ZpbZTBKfNjifymW6CylyPljIBJro/jn8X/2rvg14Ph8U6re/Diwi0fS3ur6TUJr63/trUkaZzY6VCZg8ZEKJgyvIXkfAUKCaAP0CrPj1bSpdTm0WK8hfUbeJJpLYSKZkikLKjtHncFYqwViMEg46Gvzb+JP7QH7RXjDxD+zLcfBCbQvD9n8XtPuNVeDWEuLtGmTSGvTb3PkGJvIjWTKGNwzyqpbCKVbr7z46r8K/jL8TtU+JugaJ9r8D/DvR9a1XVdJtHjv7243ziW1SWaVibfzVP2dHG5Sw3uewB+gtFfnNrf7Rv7VXwu8EaX8fvjH4Q8Lw/DW7eyl1LT9Lurx9d0ay1GRI455ZZR9muWhMqeckaJznacZYaXiP45ftSeJPjH8bPhx8HrHwlFp/wpj0aeCfWIr6S4u/7Q01b1rcrbzIm5n3BZcqEUAFHLF1AP0Horx/4AfFVPjh8F/B/wAWUsv7NbxPp8V3Jbbt4hlOVkRWwNyh1IUkAkYyK9goAKKKKACiiigDwD9rH/k1n4yf9iZ4h/8ATdPR+yd/yaz8G/8AsTPD3/pugo/ax/5NZ+Mn/YmeIf8A03T0fsnf8ms/Bv8A7Ezw9/6boKAP/9D79/b6/wCbcv8Ass3hD/26r7/r88P+CiGraVoGlfs/a7rt5Dp2m6d8X/ClzdXVzIsMEEEK3bySyyOQqIigszMQAASTivpD/hrH9ln/AKLJ4M/8KHTv/j9AHv8ARXgH/DWP7LP/AEWTwZ/4UOnf/H6P+Gsf2Wf+iyeDP/Ch07/4/QB7/RXgH/DWP7LP/RZPBn/hQ6d/8fo/4ax/ZZ/6LJ4M/wDCh07/AOP0Ae/0V4B/w1j+yz/0WTwZ/wCFDp3/AMfo/wCGsf2Wf+iyeDP/AAodO/8Aj9AHv9FeAf8ADWP7LP8A0WTwZ/4UOnf/AB+j/hrH9ln/AKLJ4M/8KHTv/j9AHv8ARXgH/DWP7LP/AEWTwZ/4UOnf/H6P+Gsf2Wf+iyeDP/Ch07/4/QB7/RXgH/DWP7LP/RZPBn/hQ6d/8fo/4ax/ZZ/6LJ4M/wDCh07/AOP0Ae/0V4B/w1j+yz/0WTwZ/wCFDp3/AMfo/wCGsf2Wf+iyeDP/AAodO/8Aj9AHsPirRP8AhJvDGseG/O+zf2rZ3Fp5u3f5fnxtHu25Xdt3ZxkZ9RXn/wAA/hV/wo/4N+E/hL/an9tf8IvZJZ/bfI+zeftJO/yt8mzOem9vrXPf8NY/ss/9Fk8Gf+FDp3/x+j/hrH9ln/osngz/AMKHTv8A4/QBc+AvwXHwR8NeIPDp1f8Atsa94h1bXvM+z/Z/K/tScz+Rt8yTd5edu/I3ddq9K8B8Gfsk/Fz4Vg+CvhL8brvw38MUvJbm30J9EtLy8sYZ5mmltbXUZX3JGzO20vE7IPunOTXuX/DWP7LP/RZPBn/hQ6d/8fo/4ax/ZZ/6LJ4M/wDCh07/AOP0AeXeKf2YPiTpvxR8TfE34AfFaT4dN44aCbXNPuNGt9atJ7uCPyluoFmliaCUoAGAJViMsDwBzHxT/ZK+NHxB8afDvxnp3xqSzuPhvZ7bJdR8OQ6n5+pyoUn1KZftdvCZmBAjAhCw4ynJJr3j/hrH9ln/AKLJ4M/8KHTv/j9H/DWP7LP/AEWTwZ/4UOnf/H6AOU8Ofs++OR8TvAHxb+I/j+LxTr/gvTNZ02d4dHTTkvhqsqPG4RLiRYfISMJgB/M+9lelcJY/sX/Yv2RfDn7K/wDwmO//AIR+8tLv+2P7Px532XVP7S2/ZftB27v9Xnzjj72D92vZv+Gsf2Wf+iyeDP8AwodO/wDj9H/DWP7LP/RZPBn/AIUOnf8Ax+gD8ofil8RbG9/aC+KH7QWt3Xw8ttR+HVy2jWPhbx7K0msMuh75EvNKRFjaFr1nD25EdxubBDjgD9q/h14ruPHfw/8ADXje60ybRJvEGm2eoPYXPE9q11CsphkyAd0ZbacgHI6DpXjl3+0n+x7f6hbatffFPwJcX1lnyJ5Nc0t5Ys9djmYsufYitb/hrH9ln/osngz/AMKHTv8A4/QB7/RXgH/DWP7LP/RZPBn/AIUOnf8Ax+j/AIax/ZZ/6LJ4M/8ACh07/wCP0Ae/0V4B/wANY/ss/wDRZPBn/hQ6d/8AH6P+Gsf2Wf8Aosngz/wodO/+P0Ae/wBFeAf8NY/ss/8ARZPBn/hQ6d/8fo/4ax/ZZ/6LJ4M/8KHTv/j9AHv9FeAf8NY/ss/9Fk8Gf+FDp3/x+j/hrH9ln/osngz/AMKHTv8A4/QB7/RXgH/DWP7LP/RZPBn/AIUOnf8Ax+j/AIax/ZZ/6LJ4M/8ACh07/wCP0Ae/0V4B/wANY/ss/wDRZPBn/hQ6d/8AH6P+Gsf2Wf8Aosngz/wodO/+P0Ae/wBFeAf8NY/ss/8ARZPBn/hQ6d/8fo/4ax/ZZ/6LJ4M/8KHTv/j9AHz/AP8ABUf/AJMU+Jn/AHBf/TxZV9/1+UH/AAUe/aE+AXjn9jH4h+FvBXxL8M+INavf7J8ix0/WbK7upvL1W0kfy4YZWdtqKzNgHCgk8A19wf8ADWP7LP8A0WTwZ/4UOnf/AB+gD3+ivAP+Gsf2Wf8Aosngz/wodO/+P0f8NY/ss/8ARZPBn/hQ6d/8foA9/orwD/hrH9ln/osngz/wodO/+P0f8NY/ss/9Fk8Gf+FDp3/x+gD3+ivAP+Gsf2Wf+iyeDP8AwodO/wDj9H/DWP7LP/RZPBn/AIUOnf8Ax+gA/Z/+Bn/ClNO8VXGra3/wk/iXxtrt7ruraobb7J501ywEUKRGWYpFBEqoi+YQOSMbsD48/ba+Bf8Awh/w6/aF+N/h7W/s2neOfCdrbavov2bKzalYTIkF+twJRsYQExNH5R3feL54r7D/AOGsf2Wf+iyeDP8AwodO/wDj9UNS/ad/ZG1qwm0vWPiz4Gv7K5XbLBPr2mSxSL6MjTFSPYigDyf4bfs1ePNf8UfD74ofHL4j/wDCbxeBbXz/AA/plppEWk21vc3VusZubopNM1xKkfyp9xVPzAZJFfJH7M/7PPxI+PPwB1TwjrPxNOkfC7WfE+ujUdCs9JiF/PFDqsrSQLqbTFkjmddzYhLAZUHaa/RuP9q39lWJFii+MXgtEQAKo8Q6aAAOgA8/pVHS/wBpn9kPRLX7DovxX8C2FsXeTyrfXdLiTfIxZ22pMBlmJJOMk8mgDT8MfAmz8K/HjWfjNp2pqlpqfhzTvDsOlLb7Vto9OlkkWQT+YdwIcKE8sYxncc4rzbx3+zL4/Pxa1z4yfAX4nSfDnV/GEFpDr9rPpMGs2V+9jGYbe4EU0kTRTRxELlWwwAyOufR/+Gsf2Wf+iyeDP/Ch07/4/R/w1j+yz/0WTwZ/4UOnf/H6AMHR/wBnvXLf4pfD34reJfG8/iHVfBWiappNy9zZRRy6lLqckcrXBaF0jgEZTasSxMNuBuGOeJ1j9jHw/wCJfC/xY8M6/wCI7hv+FkeJl8U2d3aQ/ZrnRr6FIBbtExkkErRPAGLEJuDFdo+9Xqn/AA1j+yz/ANFk8Gf+FDp3/wAfo/4ax/ZZ/wCiyeDP/Ch07/4/QB57o/wD+PepaB4n8OfFv42v4vsdb0LUNFt4YfD9ppscT30XlfbLjyZWaeWNeiK0Sctxkgi7qP7Ln2/4Y/Ar4cf8JN5f/CldV8Pan9q+xZ/tP+wbSS18vy/OHkedv3bt0mzGMNnI7X/hrH9ln/osngz/AMKHTv8A4/R/w1j+yz/0WTwZ/wCFDp3/AMfoA8i1v9lLx941+L3hfx/8Q/ieNb0PwRrsmu6LZR6JbWmpQFi5Syk1NJC8lqoYKyeUpkUDeSVDV3fg79mDQNE+H/xZ+G/inUv+Eg0j4r+IPEGtXK/Zxbm2j14gm3XMku9ocZWX5cthgikV0X/DWP7LP/RZPBn/AIUOnf8Ax+j/AIax/ZZ/6LJ4M/8ACh07/wCP0AeXaV+xT4Fsf2UdM/Zel1O4jXTViul1u1UQ3a6zHN9q/tBAS2G8/kIzN8nybuAa9A+Enwv+PfhTxD/a3xa+Mb+PrG3tXt7exh0K00iMu7KftFw8LyNLKFXAC7EG5jtJxjR/4ax/ZZ/6LJ4M/wDCh07/AOP0f8NY/ss/9Fk8Gf8AhQ6d/wDH6AH/ALQXwItPjr4d0a1tdeuvCfiXwrqcGs6HrVmiSy2N9AGUMYZPkmjdGZXjf5WB56V53H8Ef2mb7wV4m0LxF8e2utc162isrbULbw7b2cemwbj58kEEVyGN1Kp2iZpSIuqR5Ga9B/4ax/ZZ/wCiyeDP/Ch07/4/R/w1j+yz/wBFk8Gf+FDp3/x+gDzDwH+xl4W+Cvjrwt4y+AesP4Nt9Psk0zxDp8lub2DxFax5ZJbn97D5d8jszrdAMfmKlCmVPxX+yv8As5/EP49/sw6f4T8TfEs2Hwo1bW9Ye+8PWekxJfXEdtrNw7W51QzFlillj3tiHfglA22v0j/4ax/ZZ/6LJ4M/8KHTv/j9Z2lftM/sh6HZLp2ifFfwLp9ojOyw2+u6XFGGdizEKkwALMSScckknmgDivH/AOzD4wk+KFx8X/gD8RX+Geu6vp9rpmsQPpUOr6fqEFiCtq7QSyRGOWFGKqyPyoAwBu3ReNP2W/G3ifTvAniiw+Kt9Y/FjwH9uEHip9OtpY7yHUnL3Ntc6apjhaAjasSKwMYVSGLcn0v/AIax/ZZ/6LJ4M/8ACh07/wCP0f8ADWP7LP8A0WTwZ/4UOnf/AB+gDzfxp+zV8TPiX8LtI8NeP/io1/438N+JLTxRpOv2+i29tBaXdiCLeE2CyFZYV3PndLubdy2BiuN8R/sh/FfxX4jXxnrvxehvtf1Dwze+FtUupvDkDKbS8uJZjJp8S3KrZyBJRCzHzvMRFL5bmvev+Gsf2Wf+iyeDP/Ch07/4/R/w1j+yz/0WTwZ/4UOnf/H6APDn/Y/8bxfDf4P6JpPxLTTvG/wWDRaPraaMj2r2r2n2FoZrF7glt0ACs/nglssAMgDsLL9k9dY8U+MPEfxW8VN4yXx14Qs/Cmqx/YUsGmFu0zSXStDIVQyed8qKg8sqCGY16D/w1j+yz/0WTwZ/4UOnf/H6P+Gsf2Wf+iyeDP8AwodO/wDj9AHz5/wxn8TPEmiaR8Lviv8AGm88XfCvRZrVxoh0i2tL6/t7B1ktba/1FJGaaNCibysSNJjJIbBr6B8I/Ar/AIRX4rfGD4nf239q/wCFrjSB9k+zbPsH9lWH2H/W+a3neZ9/7ke37vzdad/w1j+yz/0WTwZ/4UOnf/H6P+Gsf2Wf+iyeDP8AwodO/wDj9AG3+z58JP8AhRHwY8KfCL+1f7c/4Ri0+y/bfI+zef8AOz7vK8yXZ97GN7dOteyV4B/w1j+yz/0WTwZ/4UOnf/H6P+Gsf2Wf+iyeDP8AwodO/wDj9AHv9FeAf8NY/ss/9Fk8Gf8AhQ6d/wDH6P8AhrH9ln/osngz/wAKHTv/AI/QB7/RXgH/AA1j+yz/ANFk8Gf+FDp3/wAfo/4ax/ZZ/wCiyeDP/Ch07/4/QAftY/8AJrPxk/7EzxD/AOm6ej9k7/k1n4N/9iZ4e/8ATdBXh/7Tf7Tf7Nuv/s2/FfQtC+K/hLUdS1Hwlrtta2ttrthNPPPNYTJHFFGkxZ3diFVVBJJAAzXuH7J3/JrPwb/7Ezw9/wCm6CgD/9H9zPGvw98A/ErSotC+I3hrTPFWmwTLcx2uq2cN9Ak6qyLKsc6OocK7KGAyAxGcE15h/wAMnfss/wDRG/Bn/hPad/8AGK9/ooA8A/4ZO/ZZ/wCiN+DP/Ce07/4xR/wyd+yz/wBEb8Gf+E9p3/xivf6KAPAP+GTv2Wf+iN+DP/Ce07/4xR/wyd+yz/0RvwZ/4T2nf/GK9/ooA8A/4ZO/ZZ/6I34M/wDCe07/AOMUf8Mnfss/9Eb8Gf8AhPad/wDGK9/ooA8A/wCGTv2Wf+iN+DP/AAntO/8AjFH/AAyd+yz/ANEb8Gf+E9p3/wAYr3+igDwD/hk79ln/AKI34M/8J7Tv/jFH/DJ37LP/AERvwZ/4T2nf/GK9/ooA8A/4ZO/ZZ/6I34M/8J7Tv/jFH/DJ37LP/RG/Bn/hPad/8Yr3+igDwD/hk79ln/ojfgz/AMJ7Tv8A4xR/wyd+yz/0RvwZ/wCE9p3/AMYr3+igDwD/AIZO/ZZ/6I34M/8ACe07/wCMUf8ADJ37LP8A0RvwZ/4T2nf/ABivf6KAPAP+GTv2Wf8Aojfgz/wntO/+MUf8Mnfss/8ARG/Bn/hPad/8Yr3+igDwD/hk79ln/ojfgz/wntO/+MUf8Mnfss/9Eb8Gf+E9p3/xivf6KAPAP+GTv2Wf+iN+DP8AwntO/wDjFH/DJ37LP/RG/Bn/AIT2nf8Axivf6KAPAP8Ahk79ln/ojfgz/wAJ7Tv/AIxR/wAMnfss/wDRG/Bn/hPad/8AGK9/ooA8A/4ZO/ZZ/wCiN+DP/Ce07/4xR/wyd+yz/wBEb8Gf+E9p3/xivf6KAPAP+GTv2Wf+iN+DP/Ce07/4xR/wyd+yz/0RvwZ/4T2nf/GK9/ooA8A/4ZO/ZZ/6I34M/wDCe07/AOMUf8Mnfss/9Eb8Gf8AhPad/wDGK9/ooA8A/wCGTv2Wf+iN+DP/AAntO/8AjFH/AAyd+yz/ANEb8Gf+E9p3/wAYr3+igDwD/hk79ln/AKI34M/8J7Tv/jFH/DJ37LP/AERvwZ/4T2nf/GK9/ooA8A/4ZO/ZZ/6I34M/8J7Tv/jFH/DJ37LP/RG/Bn/hPad/8Yr3+igDwD/hk79ln/ojfgz/AMJ7Tv8A4xR/wyd+yz/0RvwZ/wCE9p3/AMYr3+igDwD/AIZO/ZZ/6I34M/8ACe07/wCMUf8ADJ37LP8A0RvwZ/4T2nf/ABivf6KAPAP+GTv2Wf8Aojfgz/wntO/+MUf8Mnfss/8ARG/Bn/hPad/8Yr3+igDwD/hk79ln/ojfgz/wntO/+MUf8Mnfss/9Eb8Gf+E9p3/xivf6KAPAP+GTv2Wf+iN+DP8AwntO/wDjFH/DJ37LP/RG/Bn/AIT2nf8Axivf6KAPAP8Ahk79ln/ojfgz/wAJ7Tv/AIxR/wAMnfss/wDRG/Bn/hPad/8AGK9/ooA8A/4ZO/ZZ/wCiN+DP/Ce07/4xR/wyd+yz/wBEb8Gf+E9p3/xivf6KAPAP+GTv2Wf+iN+DP/Ce07/4xR/wyd+yz/0RvwZ/4T2nf/GK9/ooA8A/4ZO/ZZ/6I34M/wDCe07/AOMUf8Mnfss/9Eb8Gf8AhPad/wDGK9/ooA8A/wCGTv2Wf+iN+DP/AAntO/8AjFH/AAyd+yz/ANEb8Gf+E9p3/wAYr3+igDwD/hk79ln/AKI34M/8J7Tv/jFH/DJ37LP/AERvwZ/4T2nf/GK9/ooA8A/4ZO/ZZ/6I34M/8J7Tv/jFH/DJ37LP/RG/Bn/hPad/8Yr3+igDwD/hk79ln/ojfgz/AMJ7Tv8A4xR/wyd+yz/0RvwZ/wCE9p3/AMYr3+igDwD/AIZO/ZZ/6I34M/8ACe07/wCMUf8ADJ37LP8A0RvwZ/4T2nf/ABivf6KAPAP+GTv2Wf8Aojfgz/wntO/+MUf8Mnfss/8ARG/Bn/hPad/8Yr3+igDwD/hk79ln/ojfgz/wntO/+MUf8Mnfss/9Eb8Gf+E9p3/xivf6KAPAP+GTv2Wf+iN+DP8AwntO/wDjFH/DJ37LP/RG/Bn/AIT2nf8Axivf6KAPAP8Ahk79ln/ojfgz/wAJ7Tv/AIxR/wAMnfss/wDRG/Bn/hPad/8AGK9/ooA8A/4ZO/ZZ/wCiN+DP/Ce07/4xR/wyd+yz/wBEb8Gf+E9p3/xivf6KAPAP+GTv2Wf+iN+DP/Ce07/4xR/wyd+yz/0RvwZ/4T2nf/GK9/ooA8A/4ZO/ZZ/6I34M/wDCe07/AOMUf8Mnfss/9Eb8Gf8AhPad/wDGK9/ooA8A/wCGTv2Wf+iN+DP/AAntO/8AjFe36TpOlaBpVnoWhWcOnabp0Mdta2ttGsMEEEKhI4oo0AVERQFVVAAAAAxWhRQB/9k=)
Adapted from (Davis, 1992)
Amygdala circuits
![[[@medina2002parallels]](http://dx.doi.org/10.1038/nrn728)](data:image/gif;base64,R0lGODlhWAJNAsQAAMzisP///46Srv/xxWFpaewLCvOvjqiechITE6mrxHGyTMvM2AceZGFrjP/44NrsyHyCf7LBmtXW4/bRugltK+jq6kJJU+j121tVTb6/vOXl9DIzNPro29/h5b291vP29CH5BAAAAAAALAAAAABYAk0CAAX/YCCOZGmeaKqubOu+cCzPdG3feK7vfO//wKBwSCwaj8ikcslsOp/QqHSa7AggEEHmQ+16v+CweEwu6yQECyZt2VgS5rh8Tq/b7/PE+goh+NsZeIKDhIWGh4YdCQQCjVgNfmyBiJSVlpeYmTeLAgkRjXx9fxuTmqanqKmqZB9YGQsZCQmOj34YbRWruru8vb44CwQJHRKwEZ60ogQYGwS/z9DR0qkSEBIfxLCxs7SQy20C09ALcOLm5z0fGa4l6xAJXDPqBB0BHxXZGbGfod63pOh4ESgVo4K+EQZLuUuQK6BDVBA2IJiIAGAHCxQnhouR4N01Efc6LNAmK1mkWxbq/z2ECKFGhorxXiK4hyEjgo0rc1JaMNHCq5obAkBAQAAf0Bh9MnToUIHLh5Aj93HDooxZS52ZEmxYUCMBTBFeEQQQgABDhQpHsaolRHZDvA4SOxAgCjILDFcjJTB1iq+YvpLdTgJcu3ONA5cTY04UStSpXcKQ6wy9CpJLWKIC4rWo4CeBvlcSzvLNtw9UrT8WNEcWxBlDORpe3YJdLLOsgIarc5cZirME2Yy9U6AVJkvfAr1NRfSNCjhUJEC68XwgcADDYcRiRcjMdXki5ejgu0we8eHKUpECCEicuYIzo0bFQe+1h0+kMWTO/zgLP6fVgQMEzRDbCNvptUB666nG3/+CTYTFVQBhSSBRKYmtAIEFVMHnmXzJheTXNqZhoR+DcWRwgAEQXIfdg7F9INFrEuFG4oxIVEARAUPRNdcGjaSVwgcW4IiFAAd04lle83lIEj/OrUHjGNUYgIFKNnQw0QZYeMcYlulV+OSXRWSw3kRmBYCWTcGZsEAzQoJyjHHIOXVPMSTNQlUfToLpxXQG/KeiDb9RZEEu09n0nZ6I+lBBeoyUsAiOD6qQQZB+ZGgknEku9xeTVFmQZqJOQHAiBhLssIAowT0KQaSgttqFmJFYKsCbSHaoKYi0WFCqq09c0ecBvAbLy5prVDrkpa8ssJdTS0V1DJM4CttEBCf2KaP/tNhqgpIfDVga30hJYmPfNrNEm20S6Rmg7gF/nusuJVqpwa23Gx6XKWme+KHgu0F0gIG6UgbI78CEMINBsRkWSau9tmZDDoYEF1Gtugq0G/HFdcAlr7FZaAhaaHLWJ8DBVHnGFMY7JKAAwBiwivLLcoi5cbcdy1JvnPbck14GEsRyRSQ4agHyvjCnsMC/AFdc9NJxxFuskDXfPN+pw9D5V0nKAA21Fh08xbR2EAAsJZXifm22eBtvrfDNTS2ygLgfesJNP7ZY0MYGEm31tQRIUxyBme5s8NrZhDuhMaVbI3tcNRCMJkHc+NFd990Co/xB3+picAAEmv93aOGgKxHR/8yW0pqBa17fs1ydRIZSFTNFmU2k2AYoMPEBBFgc+u5GGIywtxnsLNpo+B4TIgTeqPc5zB1MTDvLRPMuvRBouRHrkB1dmMDj89HXbJ1zU5UGBp++3EHYz4utwADTt3/EdNZz7EcwyeLs/bgZPAtKep5GjzJa1QJA+jBQOfcZ8AdaccMa1BAOucBiceFa3dVAkYYCwkxU66oO5hTwtwN6cAhW6EMWVLIO4zAsdfUphgfyJ7c+XItpnDuAAkQFAOqoCwCa+6AOlXCFz9QqHkqqkx9euDS0QCACEzhMBghww//s8IlHwMK3lCUa5UhwEY1rn78Ahjv/QfGLOmDEFJFjq//l9NB9FcAgxTgAxjb+AC0akloVVVcNC4LuPwDLohv3aCryGSkCJrRfAOh3QA1MDAMA4KMic8CJItkMU/NxmwfViIEOLvKSNBChm+rFsDTqsQT76hooXRaALWRLDxQDFiZXKQMRdkxxyqIaCgSwK/J8rhqg/GSwttin5bHylyi4HM1A8UhYjBAFp1JQLEzgmRKcD1toSCUwp8mCi9wpC44Mnmt+FLsSQIBKI/hmOwbnqstlkJroTMGkCDBMN/WhlieAgMDUYYI0mkAL0pqdlHSXTnQKgFI06xiOvPgYZwoseCaAABERxcuW9fOhJLgQx66wzR8V1JvwFIE4SfBMYVX/q5L8hCgw0xCJbnUmBR0IXgFnYQJcOoqcoDJRnywp0n66aGO3gEcJKgALPrwNBeuIJzjtoUtQHU1KAAhpTVkpIbuNL6NgS8ACFiqCYOxLAmlCKA4+8Cg7SsOcAFqqSOPVBgJ4kQVoIKJSEjrUGcgFR+qBqjkwmCKxQjQieFOoDYLRVjN97lQ4OF8GelnRgJxubHa9a0VgGoNq9FUoAlvVDQA4gc1xzpfRQAtSE3tXUu01eI9laTvK94JWDKSyohgUOvwTVs4+lKotWIcAHisUxmamtFiQ0ekOMAETiWgw5jARBibg2uKqABsdoaUKLjoCWNxFDWXSKPkM4Ns/kNYX/63wrHGLW54sJKBIJaOtCK5wgunANpdYwABlIgKB3m7OD1g6B+e8ul2Irg5ksS2fPGEwnU49KCK87eWFBBfcHNb3wPztCNFE+wLOIA9PubhQe92hHr1NAy1TQrCG29OIAr5DBtOJ1TLU44cDfPdCZaVvDbABC25I9bxOqEAzYLzhlz3lxl6TwhJNGU/GsiDEyEvDBtbwL1EdoA0HIykpV2yFb9hNDU8OUhY+MzeGNMFuSiUBjs/K3xrnJoF4C/N1jXCq2wJ1zCpIAJSH3CkT4a4ZG6BZGt5xMhAHz8lP09rkosxnedKYBwLYwJ/xKubSLtlMqm0lZr2MCp6eCgHJEv8vCLN3aLAsmgViasadlmGA7wZJ05u+xRrCK5w7b6ukek61qtkQpNv0Tq4oWMNIwPWCo6HARlw+gR8YDQ0x7VQfDeFCBbZHAnJIGkICoOKN+xI8LETgvFfI9QkWNepr+sEdB8MQVQIqoudIWaot7oPdELbqcpv7G1j6c1d8HOtPSUCqWtZHPNQsSnVMwkYp5THgrDyCd+9q1yIQJa91oRXNjOwPXBmx8ohK5EsbhJg2u4JnYBw8VcrgQMu4prW1pnGaBVTO2R43udl57pKXWw0q7soFYKDenY5YvVw4nS0YUhMC9IzIpPoAUZaht4OnIRytmXMAdo0G7Q5cFQXXzgb/VGLrW3CBJ2MJigh4JIRW6FQGrdG41rd+TY+7EkfcfjAkskZyk5s9ElSXgnqejIHptH3qjUNAOV4iEgyIIE+NgrQIliGhXaHF7cLeAOPgguajWyLpY/mOWVpuppkczGZqCMI6Kr2Ct3L98nfqVkChJkKTproPJk1e2c9+bmaknAiPL46LSvFPrZDAU3wbLwJWJWwENKT1+xmvmErxFM41w/C7iIBsAlAkEpCqUWYSdFkjwW4byPbYKOCbkMLMecxr3evsFCHYRXQaoJGd9CZP+xPIp2ULQ4h8FjD+ASSQfu0wQ/A6b8iRO0ICE5tfo0RBAOWBf/jhL8L4l3MVuMZ4/6VEU4yUBbDWAgsgZRGBAAzAAAjAgNa3bZm3aQ8GVxgIV6AHfhwIXwQ0fsFBYNL1EpohOLYGISCRBjJ2exYQAXYXTmvyGtbAOUM3fPx3Cl4RDxKgdxDiDIyHa16RC1byKcnRYAeSBep2ApOSBVgAgRMBgdo2gdbGfXJ2gaEnCqLXgWb3NGW1f0JAgBq1dI0HB/HFGB+wJnCnUYNie2FoJXOXfhGRCy/hHvbwezd4CqdDAv+UXlzxYfbwdn1wcCgQH/XzOIYYFfrQYUmohFHYBw5IEVJIgQ/2CFSofdxXKRqYgVmoheZmN8olOuTkdulxFUeDBT4RANaUUtpGQJczb//OoGZZ4Cn24AcjEw6NUFUW4IV3eAlnwQL44EWqw1PkYmKeERpFCAzVhxFXUn2RSImTuH0YOHabiIncwomqRmRA0waXhgTYcC29SAJF+I0q8ItaJnAmIG27+C5GpHHX04xbJyRVaFLdZomjZ43llmfYuHDpuI/pQIvuKIWaR4nQeIHZN3pYaI8lVywKqY/82JA3MFH/uHVh13X0SJDaR40IuWpPs5BrYIcO+ZExMDJMGJGYN5DyOJAYuIH16A3TiJAbeRIkBZIy+WOu1AjgMCQk+QgSKD4UmIlBlokk930ZaQswOWJB0pGFN5MNyTlXwChsQFI46Y7qoV4fJ5CdZ4n/noeRQ6mR+qFwRCaGShmWIoAGoVBWQ6FeKLGTE7iTVQk1mZg8obeV96hnxXKUDCmWM7kOc1ETB7NzGMIHbBCJavCOmyaPY3eR2gcJLCmXMJmPHCloeKmUctETQzZkoraTUymRQaaNkvhxbnmRcJWVjJlq+KgfSRYkEBOZM7l2F9ITGGGKHINiHMc5UNaIzjiJF5ibJ1kpY5eBo+mYRPlypqeaMrmEV2A3WDI+vsdmP+NU6aFeEygkJkmPvSmUo0mXCmeXC5RoxOmQaoYjbCZlGKFtbSBlXFEk/tKMYeeTvFl2QQaXv6lwXblARnl63clrp/KUQOEGFfGX5Ecgc6aZ/2K3bdMJdphondcJNPmonYF5nzJ5D9qQUt80DxuxObhRPZH4mSdJjweqlWaHOOYGnAupnanhoHgpF5NQfKVkcyMQL8xIgVVZkW95kKnWknrGJme3kdh4Czyniya6i1bHBZ/AFdXwIP6SBmE3ka7Ulu+ZkiS3mGeHoyHqlVRKKai5jT+6j0skbFgwXhvxoh33jJcYZBeZlQgaolJqcjr6ckdpN4uYpcBHf2DBohbqSuzIjIaZm06KkWeqak6VkPKJOAyKlHCKl8nWbwTAFetQbV2HJbrZkxVZndWoNTYKNGmqpkSJjaj5DblXqDN5D1omWez3ogFKkRq6gTSaNZWqNf9/CqgvmalGyXNvugJbVqs35qk1hQ9gAZ3XdAuUKI+1MKahCZp9mmpDxoEjGqubClw6IAAP+Kx89mQNMK3UOq37Iwse4AH2smW4Ch5PcQEX8AAAMK7jGgEAEAHomq7quq7oykHo2AJ2eidqaZWgx6GqKmLXCZyN+XKc2gPO+qwAG7ACO7AEywBRVq3dAh/aao7dygsfEK7iqq4KMLEUW7EWe7EYW7HvSqu8ek0MmKfBGqmY2JuUmqCaqh+bepRg2awF27Iu+7IFazfWKgAegF8Niwgf8AAR264Z27M++7MUu7FpZpudUn0DWYkp6Q3KsKouSZr8Sp+pqQMegLBUO63/fAazWJu1FmCtUsWwN0sG4XquEQC0ZFu2P7sDl0OqjnqBjXiYPimp9XhurZqjRDmoy0Cf/wB9NqAgtjonIpGtzUGtT5a1WLu1DUCzH/G1UpCzYmu2ZTu260qu46qzOguulguuOZYyRGuKRAtq0rmh0cincatnc6uFJ4uaC7QGEahvY3BjGrAAHoA1Vku4BXu4UqUBaPujwWZmPxa2jeu4Ewu56kquDwCuIEEF6al1viqJvup11MmevjmpN9qBO6qg/EpS5WkwnUBFt3I1j/QKQlt1EOoBoCC4FkC7Bmu72jqr5KAc97lRkvUjv+u46Dq5FxC+UqAHWleqjcqT22dS/yMnvSV3qdT7tOOznQoacmzHkQejkP0jB2c4C7NLuFubbNJmDX51n/hUSmnCuGNrtvVbvIgQr4/gBhIYUFLGpLbwnio5usZawCJKUgt5rAipBgnICh0QuwLQABTcANvjP60Qc0nZkAeiHDhxAecKwgAgwpfAfuyYi6XEox4LnaCLJ8SqieZGwOBXmlC7duAno7Tpo2UgAeTbAOcLs1vLb8W2ERtMnEEsAuYKAI87rvjbuiPRITJnbSlBHvxDKbTJfbqZcVDJcUDTkmlDt9VrpbGKEmRXkVMIyNq3x5SQwzt8xi57uB4wbx1UVJH5iSrzs+b6AHVcBjKTZN+gcQfTVv/3sA5OhSHxKEJr0ABu0Hku7Kcgao91ya92w452Gr0jBsmBmAkfgFVmDLM0q1cdRZzhekTagbEhbAoi0RGba4FntShQ6chslzXFqjVaPKUnS5/KqkCmnMDtiMrV94GnoAESbDcFux/tG5kfkMQT684YELwAMMp1sCbTPIUNxjF5apZA2ZtMu3zWyIUK5wYZ8HCigDBaMFW3OkqxeSF6i7NWwMMAK3dgwQA+PNGG9wAfXLFlwhmivAvYMK+N2nwmsESPCl+hWXabqJiji2SkV70LaZRYsi+Z+wJ806tYigkV4AEX8oC3B7ANIMaM5tEZOxAahc+CEBFK+hzo/AJYJJ3/D7bLmyesVRFl/4AhcnHLdBvOSg1omiMq6cXUhMBVuUcAAvsGuLuL8nyx5hpUY2Gf8PKcy0BRntJhQjuKZGqKJqzV0cp2jABI5IAguThVEYHIRckMbwAEUPMfDvUMWAUSG1C7HsB/D9CzEfAA4fR0Q4wJrdAMD2wD9UqJGFIBcaMFyfJAU3WMATCqmoY3CV2eyBplrgYEeuA5uIPSqLDBwTO1tftTXobUcL3ECoJPb/wLZ3EcOACLX/cG+Js9qiY+2XsSgf1kszxxQ9BfrUMd0GBmGzXMCWDRAZvGZo1JxG2x5mo0bGzU00AMmnE+J3whdF1s2nacbdKUTIghQubH/4DpSsJtBFfwH5vz2bx4b8uDVZYMsBZw2dslx3DN2cd1FSZCGHwDXXA1mM7Y2DUwPncbK/rtCPvBU64NBbhTJLv9DAahHaTFVeRN1LyNTulNsZvtAhs1q79gBWn6xD2dAq5wOpSiXj/zHDHeBAMO2TecCvHQxingAQv+rA3g4DX1AR9dsffcYOe9CkuUbXlVgd0ii5M1FtoWNFYsN0kOBVhgYrgjDuoG3ALLu/0E4RW73okyMguJJ5yXJ8DA39c0W3GABmoe1ZEhAcXM4J9ITRdQ5Ry0cq2CFvz9BxiQPL66AxIiDOTRH5qTACjS4w/BVU+u0WfeRnJO41mOFedDUv/xw4B+zgNThQdYYADUYlYLUh6fLuWXROXqLeHSsihlIWvosESdVkIzogEv/oAGbkAfcLFXfi56sQ44bgpLhK59El0zkgAC2wChfkC4XrG67qkjcyLCle2rkUYCW+Ttk9lz3u389xT1sRRV5AI98+r6sDn1HRluDuXPbjaKru4Dp9CX6H13ciR6cR491QcAQl16me+QYe0MbusfpOiMfnQhVGL/EQHUdfETcPHU4jmNbGS9RV2oJSzsF7DmTjiKXurUZFoYNFjz/nU4YmIG8PGDNQE0T/MxnwG9RfMRYC7C8gHFfuxLg+4Uy+81Fgwn0lstjyPDcBb4QGGikgA5z/L/SG/zuBPWpxSwQP8yF2CxiTRw5VFJ7oUijMC69dRsL3/wg9VLOFLvT8Lwz1ryMCP0i35063AiuKPaAe4CqI2IPpS4BOPkAJv1BLP1Gkv3THT3HN0+Ph/4vLPtE4vy0yQqjKDwu0PsAAv3BFPlET9w6yDuO1QBL+7wXyP3Bqi4S/MBAev5/KL5pk84bm+wkA8mo775rf81HsD4ZmPltQ86//qAiY8to777oLP4D9gAX8Ptwg86EnD5SyPnpZ/8X9P77WdjH0300H/65C34ieL81x86FQCwv98q1d/9oZP9KEP4E0v75B/9ABv74BH86184y/+sov8uH/388b805M3p/60CAoqoPIF5oqm6sq37wrE807V947m+873/A4MmD6PYECKTSt9lpPgso9IpNfjpdBaLDLe7kHQq0Cq5bD6rPkWGZYx+w2XNUbxuv786mQSk3z8ABgb69QlkSLjhKS7aNawlMkZSPYxESF5iCukJ/B1MTAyEio6SfkYc+AlIZLK29gisrbrO9lCKWNLm6pp0cEIcRICSDhMTT6BCGELuMrfCFnk0S8PYKgBMY+PpZUT8CheDh49OdB9wyWan2xEVJai/V1+/z1NJ9AV/i+vvD5D3JSyjJzAJOwbuBkqLh3Dhjwz3+EGMGOoYgYMML/YoaBHjrGq4OIKM4RCCAYkmJf8eIJAhJMsZGlu2UghzpokKnPKdzKlvAgSVNH+eeAk0ksyhHD8kIHBAJ9OIPCF0MApTqFQ8RasOrJABUNOuEU9liIoVI9WxcTyaFbgAQjCvbiF2ExAwbbqydM9cvTtNAAGcb/+GS7lA7zy7hKvkPZxLglLAjvnxHKwYm+HJURJbZpWBQITHnndWzNyssmgkaEu3cuj3M2tSfSqgpkU69g/MtO/YW9p6N7hTAm6zmg18h+3hbzZ35q28GCrjkoQ7v1E8Opmkq5djP7CReh3o3Gec/n4mKfbyxfqI775me3ob09sjWWt+/rDm8M94v8/ivX4fqukDKAoq6PQnRX4FosD/H4I5/BKggxNBtWAUB0oYnoQ/CKDbgw7ydKESFN6XyFVzeSgDKhuiaF+JOyRSFon6RRBBCQHI9EAE8qxYAx8o8phMjjp4YMFBQtnDHoK2yAjACADYKMIFP9JQAQQ86uPAchAYCSUMH1jAQAMeaOSBIwxEs2IETqB5i5YydADBdSdFkMFn5BQjp2NsranjGnvu2UaOc6Q5wot5BlABZ38hsIE4VobjAKOjOHqAosM44OZjB/xGqAxd8rlnlgueGagCH2maxqFuOSApAhE82s8Ejk4AwACthsKBlZ88eusAESgKKyiOWmplBm+adABspb6QQKdr+PljNWk+iawKGQK2/8EBGGBwKwQYbIDAL91iMEAfVlbq5gbnYuBmquduQEC4A3B7LgCVygoBuxo21Y20LqixLAOZQikqqfsGsONbDkSAwK4IvDoBAktlkOgED2zAlgWzDrAqBgQM8AABCie8VMIbOEDABg/Am62bCXfG61tYEszCmJ2KBaWSaeJIsKHEmuQAtlY+7MAE2VpZsaMEEODzq5IOPauVF2/saMkkAxOKvZWSo1vEPEdEgGQxo/AMn0es+UGg0cbsyV8ORLwBtt0OAADHoVhwwNFJA+JA3RxMGQrWdZN7wMUZbHytBanG6m4fDL/F00pgn9DBsjVreXMlkCeAr1smA8BNyBMk/f+03SUnHYEFLjftKLwTWAABufZmPOW6WGNggTAIoNw4AcdCHoDYRmhqthM5S9tBX4A9ELTTJoMuCuCkF92uo0aLqzDs0yvKsKMWUA2A0Qgz/pe+vQcgAZ9f5xmqCINqyRZgqSo8SsSoqP480sBqD74FFmzMwQDnbmsDUKvY/h7mpgN4y10ImNtfIPC43s2MbJpS0I8MBhguDCMYsgrFsG4lq3VhDBQxGkBfrJQST9jphLsa1qxSwpljOKZD5AMRgoSnALRJSz6eaRW5MOa3XCHMWuRK1ye29UOpkUtqGGOUElXnmDiRLwBdalapzkS8Uu1sN7TymyiSKK7RzUputcP/wKt6yEQukmKLniEA5WJGBAmWqgk4RBa1WHNGMy7Rh04zIxPvmEc88rA14+udkAg2o319QHPv62Ia/7hHQCLRaY4EJBfVeKcHgs0DGohiqbihxUpO0od8pKTqSJlHSOkxhgRgHydbWaAPHO8xPQSlKUsZSj7u8ZanRKNjtOPKX+Yoc7JEZS0f+cgkIhGXyNQlLx/DkzYCM5r30eEOG0lKZeZylLYs5hlB+UQIsFKa4rwNSRwzyyVes5TqbGIytblOSeIxlZ45APrGqY4KJEAAmcvnPvWpHX/y85/9HKhACxrQgwI0oQRF6EIValCHMvShDR0o71hCzWqKEp3bXCZH/zW6TXh6MZ6OBMwEMAmTDAggpfnUp0pbytKXrjSmLeUnS2lq05ri9KY6zSlPdwpTl8r0p0JdqSGCsADOACOpp1iqUpvK1KXGKKpSnSpVq2rVq2I1q1F1Klef2lWvzcR95tSjHzfK0XaiNa3ZvOZujhHOgfSkAXKdK13rale6QqABhNhrIZLh15QCNrCCHSxhC/tXvhJCr3m9K2MZizSjQgAAkp0sZStrWco+QLKZfQBnO+vZB1wAtBcYLWlLa9rTova0oP3sZ5k02cxeNrayhUA9OXJRVDUSniB9Z1p7G8nffnSWlASMA3/CicIil7D8TABzm9tcMCUATNKdrnQX4P8BLWA3u9q9LnW7C13ngpe5yR2vYNEDhLXINr3phS1r29vZ1MIXvu6d72bVa1/K0rYlH7DUWNOp23X6NsBoPSY2c+sWnryVHskgL4MDG97nMte7ErYudyssYepGN8IPFmqDkatXyN73vvWlr2fja2LUina0JOasZkMc4vyyRJhrs+YkkSngG6s1uE4cLmB8ORMBNKDDyX1weC883e0iObtGli6RwSvk5Jr3B+h1sXpHTOLQYjnFJ47vakO7YhbDlsqxhXFI+DvjWvIWx2oWMDp3eU6vgC7B71jwkwUrUyJ7IMMT5q51tetnLVwX0EvO84ZpWmfBfvi8kRVzbDf75Sz/bznSKs4yiV1raUZXlsxHSc7BiKnRNq851O3UpR8vZVKQAPnQg13ug/O85D/DOskXznCTX6pqwEbZB1PG9GUdPV8sSzrYpF1tpVnM60zXliES4PSZ/2hjUUMbx6BmpCVPIsOW0PnWNW11hC8c628jedbfJbKtVZ1oKS/62K79cpe1LOwTt7vSl1a3pjFi5q68GZIejTa/IRnPajel3hxJtbZZ3WpXdxfQ4F44di08XQg7Wbza9gWI1d1iJtEX2O/eOLHlHWZeC3wht8WtWQnc75P/1tkjhTMEYJLtQ9uU23sGtwS+YPOag9vI5Da0uVuuaIuz98qidffGTTx0L893/97HDjlCBMDspgiXlrtFOcqNGXXPMH0hBL+1wZ2L8IQH+s8SGPvNyw5rPndXz4WeeK57sGt1+/rX7/7AaOku7KGvWOmMzjo9pITRdFI98B797y6bgilsc128eI6ud/usXZzj/Atjn3zNJx9rCdN657c+t67TjWkrZ/wCfZD0fuN0ALoTnctId+/F6Z3sgWyF5KJ0Z+D5ZqvAFzOGB2jJ1hncAHZtQKVd1/CE/XzdylM++WS3ufEpnPbMN7fch247D97++XVnPGIIAMCJP+CAD6QrAgQIAK8uYHfvG13F9G09pvk+j3IOM5v7jnYAJDW6qqfclG6JgJyx8fLxNgACWP+AXJ3L8DEedTWcFlieBCTABkje2GEB5UWg8kWekmHeuIVXz1Uc3GGce5GWu1hA0oTWApCWBIDWB9hIAlxAulzAAjwABKzKA6BgBCRABjhAaHEfF6SYZiXdx4Hc69FDm3QamuEesBiASuDep72Z+BAIRvTeeBUgS21AA4jXP9DaceVZSgWaBwDWF8CgAHjA2PETGNScSlmeSn0BdKVUn+UTlriaL+gTUelVYP0fclHfDlgflYHer0UMACTMvGTAyVwA6QRAIPbEuTgdZ1jAAj1AxKSLtQQABOzPuRyAimFABqwea63b0gHhPFQNvnka4J3cUh3h7RUh4YkUUxiAz4H/RB0i1yIKn0rBYAFBgJgI4Mdwj8lYgBbwjy4uAAYgAAYIgAaYjL044AK4jTFKgAcEYzF2ALeEYDAuQANiCwKAoQC4zSJWhAAlyhQCFgIEme+xYudZXAd64GgRwOFcQMWQnyAGwGO1iwkgkHZgQDuOlreYQMJcgAAsUMlYwGjJjfq11yZyIkbsF9dAhCgWIQdgQAQYAAD8QhJiE8DpxCqFxBNCWbcIEBXm0wKBycckhQVcFwxCAAMiQD5tgARoADPmmQNqAD8ugAZIQLskgAVogAYgY01iwE0GyQIsogf0AgL8YkpKALd4wBSCyRd2C5akYz4FYIfdoQ7koYs5mjmy/1ZoWctoSSLbbEAFDCLSJM8DfN8HBJ/4OUADfkD5kZa1IJD5AcD2JRIEZKIm+uDedaI6jFxOBFJIzR+/eUMwGAAGSEAR5t/KGd6pLYQrJlcDcE+iJEAAOpe3yBWY8CN3bUANMh4MMpcDdkAIyqQGJMBJeksYbsFJUthlhmAWsOQXquFJEprbCJB4WUBKCVCHcZ7beR6jBd05jswkbsAFBCJovaPXdGXdhV89Jsx+/SPdgV8y/OZotY7PRIBAsl5dUpn7pYPTNRvtBR5jkMN30o9EoiJgDJIThiODLdhKNeNlRtgU9gFlboBlqoLJcONmfkHFyGQWCCU2JgqWdCO7JP9AutQcSy7A7wlgRz7XJFYESiaDNULlOOIm0F2Z6J0OIJhO5/wmFGxMWH5Ah1pLUpzlBqTlb36fAwSfpNSdpAQi93VcdRYkQxwkblFbjVHdLwDAJ8TKMZiiROYeUyBYKwrZFMaUyfyec1XMZHJhfFrXZZoMaW7AHqRkZxKABnQAaCJAWOAkNsLgIWhBAvhkAwwoAiAcNgafa0aXANGkPu2PyQjZbVZfbuoh9rWXl2EAADBnImFA8iQA+IDTxtDdF0YAcuJOBbSLDKKovZQWMMoldWqi3l3nXWaDBCjSSeSKsxFmpLhJrNxorABCAPAo4fUYYg4ERr4iOH5jRRxoTV7/pl5RppguaU1aQBZ8jD5daQeEZgJogCE2YEzOpADUzgLY6i6m5jKKqVEW6EmGYJ7NKk0yFwya6oNu4PVdmemY1gLgzixuTMs9wCTWjnasUgeciwtyD/dcgySSVlpuH6Wtn3W+GKRigyfJaB9ZKsp5Z45+wqZ6zSmyE48xRXkyhGIW1iIiwMAKCUpqJBiy6gJgo2Vu5rm4iwMu5QdgozZS6SKOa82NqzVqgIBuQXweJbuAYQOCbAMuqD5FoW1C6JseW9zNqZep1iA2YorRXQYsgN3V3Q2eYAtGQAfYXYpF4j9OWg9aHHa+K0JWCbX10aV6w6Zu6sSYJWHuEcosYVP8/6h51lkhOJk+QVfC9dkyMhfZLV+V7kFMYsFK1iAWdAAD1uDyNVygaSGTCZ/ixWG7PFlU5sBUxumExtf3md/NjmWHdqj5Be4gDm5pAcA8Bu26Oup9Ee00tIVXAJIfflARrsUD3OvlWi5PWILgSU2BNsDtzWjANSFCACxyxRzEHSACZhcFRmDZUinaVimVTt4EWp6SCdrzORksPpmb4iGcUqVVXqWwMWffDm/h2uxppWXrlFaLDiS7Mq67SkMiyWibLYnJQRv54YPlctbEcC8AEA3uvWARCECpeYWPXQSpMliTeZufUSAEwu77pm3aKt/Z4e6D2ct5QuvPceCKdd+5Ev+v4BLv4B4vvCnuiyKEEEKd1M2K+ljD5PJbm3yC9k4ABzwABU8MB4Rnvw3ANM6MBYilLclTTlzbvxZcHB6c6mIX8rEu/Mav/LZvuGEYoUWc9IljtIoZywYv6dXd/wauzfowAc9lZxFk+0FvM8iYXqLRKA1AoNwIv3EGBXNAFEOxFHPABOMJv2UAp+zJ+AJY4Z0EBFRU05VuYRlg6qLw6rIuGLAw7c6v8aWdDDsXTl1tyvYu0AFviWlc9/HtnQbw+QVbsdFbBgDuIBNyIRvyISNyIivyIh/yJ34GwgjMB6sZ+UFAFUNxBUdxBWtyFRPAIIYaBwiAFvdJBwyAFJdyKVP/sSlTcD90DCuDwie4Ciw7ECPTci3b8i0n8hgPmQkX2Rkr4APO7uuqMdoOc/uyrQU+HBxHH1HtLh1Lpe+GGA5fJWgdbnwxZwSQYPEO7w5LGvO2Vn19HgSIyjiTczmb8zmjczqr8zoLzLzgGAf0xRSn8jxfQKeqWZv4Sz7r8z7zcz/78z8DdEDn82zCHC97XePp0+MlXwdwYTG/rzFX4O16V62pmt3iAN7+7q+hAnAKojW3TgDUnR8D7v9umTd71uKGmAIQADuzdEu79EvDtDm7s2/V31Js8gXMcyZH8QRwsYDZg0ADdVAL9VATtT+XsPrmmZD4JAFENDD3QpQ+dAu//3BTO9/W7hwzo6wNx+kdd1alHI5aapndBUCHposJMCdIQ8ENjnQAk9YNDhujNqrz2pc4x3Rd2/Vd4zWaTCdNG88EdEBO4zROU3EFB4OnBlgGzExRK/ZiM3Zjr0HwqVoZU9dJSsAAMvTNkV3ZYqNKyiRnx27a3mQW0K4CRh6YVLXmVbQz3y00R7OcflagXiavoAIBQIAMQqQfyGC6xGVP8B9KzfZW9MTpbQVpOd0FdANvBzGYyfVcj8qZOPeoQPdzS3d0U/d0W3d1Y/d1a3d2Qzd3b/d3czc5x4gk+5b7pHJg20oUC7YUO4SaIbYod8oA7g9jyvf+2Pd943d+67d9M/9mf8u3fwP4fwt4gBP4gBt4gSP4gRs4QUe2QXtdUixQKLcO0pSkwiJNRTy1rV44AYShXxFASvVERdhDGDIgbT2m4jCZMi+zBuqvtHI1Z6VjIzLiBdC23hAAZ4VgAAijvUhAC/4jDEaAlHhLaOUpio5WuryljCAQi7Le0NZs/7nCvQ2h0zDw8EwyIOi0enOA93G5o3DAeqdKy6nZB9gDfBdBB0jN7e2ol7N5ep+y/8D5KY/CAXgAFNj5WOP5net5nvP5nvt5nwP6nwt6oPP5cRU0nrVmLcJiS64FZcOgBIRyr46du0iANCJjfNqDmFI2pIsp/+SZR2JgHGO1kFn0DWD/tIi9OGhtC/hwn3FjQD3L4ArWY7pkwDUQueiJKPkhgPlF4h4A7QqW5MOMlgC8+BDbJULsF8ntq5WkSROHmgBMsBRXQBQ7wJaX6Ads+ZdTMVitWQD0Qqf0NF/uH8A0Hf4+meId3K0udeVxOHOlbWgqbEomgPymo7XiamdCgEwyYyAmgBjGJ/dc19tCn6izOLrZsTSL3qsH52gFKgqm4yLOuutsC/f85rZ4XwPWnQMJDmmlC/y0C5PTJUo/7wH3DVNs0TYxsIyIWvkcwBSbZYeOpQMUzAakNxV3Q7SBshZbAOh2E0VChmqrgy6TcdyCl6uFpgCsexhSYZmfy35ugK3W/w66WCuwPqMApC1LBl8ZxmcD9qdVZ+CokzrQmzpr2xd7cXXCB5FoJQXFNCSNR7zJJACTdLLFJ1JxVgo3/DoLhpbprEpyi/zIw96k9kyb5VIlkHeoOZAUp8r3wryGXim2r7e4rAS/PWY7FFgIS8QxnK+5n7uDd9vRf6kCcngxZgAzKuwu0qTCYsDUj50wXv1JXubYLex1PSao15rE1RnvPvPBe+C2GHdHi1/EPAlZYkBzbgAuzGTCXzzumB9bNGAJJI/FDOI6ymXzsp+YNe4uHLFOVGo2KYnK059DdAwFO4CvxnyHBgAy9oSXazsHbAbK1eTnKpFh6gT/kTDXeb7aWf9jZTM1CHhSk2yQpgnIImwZ8aGYtaySZEGd5CGeuYMgSoIbYeNJJBPMpiAhiEqn1KoAAglot9yu9xtYQADksvmMLj/Ij7b7cYkgIJEN/HIgPGaEvouAtdFHgHEAgeHwUQfnQPdgYdGXsxc5A3Dx9gawltaZBrEAJjpKWmp6yiVgMMDa6voKC+sw4FBLa3tbq7vL2+vwQDAxwUHscID4kZgYgEHQgZDhcEH8wDEQAeGrve2wYD1rO8sqHlseG/GBqr7Ovi7QYBVvxfTUZJ+0YWLRsOAhlgBhA4aBggTQsNCs2YYSIiRgKMLDR8BmFpBMrAjBwxJ7TqDI+/guSztRYjz/mTSzJpPKCAceZLgAcw0mQy8zsFEEoeWDBQ8kxHwZE84DQxFkxskpVCWbk0wBgBoJNSo7CBPMWXUFbty4rNy66gpAZ0C1ag4EYEiGNgyCDgEiBSAGtxoor3R30dJ6NW+rCBWk+v1LCgJIkB45NkmSRDATjQsSLGgMqF+CDBl4QCAhIUOSBTcYP3YsQQCgxZBJbDQcxePgeFgAbynZ1GTKTSrvwMR0G2ay2x94f+idO7jwC9Ju266tiVPsT6FcO/f7gare6eCq1/X6QtiEahyMIUuU7shjIR0cUONQNUOEANe53a2uldx0WOie2/f7bvVHeoaVKNEIoGeP3TAggTfs/3BDggM+tmCD/TAGoH/9qaZfFQ2I5Bpsy3mCnEoxDQdiiCIah0mHs23oyVP3rahOBRDMZw5X773Xni8fEBDBdsNw8EBZiCjjQAX5DInAAeadJwwE3dXYSy53PQmjOXyxSKU6glU4T2oTBsilgAx+CWaYYj7WZYQT0pMallS05pyGKJ4xm4lujEgnnUKVKOeJb5qhYpV+ftHBi1HGSCOT2gRgyADacWcMAew5oMUBaxVzhDRxbXfAAY8aalc44mQ1qCsRsPVnqV/kpyYV/J35H5cPjgkrrP6UGeFpHVGYakjPubmnGrSZ+KFtdQ6LW1ImbtIrc6YuG8YBoWL1pIzWGf8aRjATAKAjWQdsEEEC3daRRS0fZDAENcI8IAwBC3Cqyy3uQvmsqM0xu+yVuaq6Zatl9hNrvwzOWqaEZ967JoaA8ZrsUnnixjCxdRor51IJk9EnvVVm4Gy8r0jL7i50XAsyutVkMFDJzZT3QTFYxIUuthHA0LF17soYrwEJWLwsqgQ/oeWZtAbo75gA/2yrPTzjmuqFu44xscJ5ttGwwyHe+TSyyjVdMc4reqtxfDN6yi4HB4C8KFxL9gLXNEeiqx3ImW7K7tddi6qp1n/au3M9Pvv3M7/+BD00rQL3x3PeUbCZIdNNW/0rsFRLHdzjVes5cdZ2P3fAKnN7DR+7iB7/gO21oW/Ho9lmT2M6NS0Lgy0GD8Bd48xyyzfoBAJc7qfOhkORL9EA+i008L4jxmrhOyvdpuKLW/10sZCTWOzkEi9uOe6ACRBB16BOm0tXbfASQAcY5Jhj6CIPw13qcfHItnahA2CIL5l1cLZ74WzeygQGW28f3jvzziq+DW9WwiPg8AKItLwh7mAtmVhKnias5xELYse62p4eUD3+SUU6+OOYL8QCgAiIUAEKAEAvcjKB8rFOR6QrnQvPY7Y2kI0MBpgAATIAuw4wgAH7EEwGvLENGs0oXvrT4IquYDgpAJAjxPMdv/xlwAMGMInIyxCORIjFLGpxi1zsIhbVk0X1/1AmApQpoxltgsYzjlGMYfSiG9/4RnUZ0TkcfJa03NWGMmCRhHzkYwTA9wIDRMAAACDkCrOFvvVx53xtC50gJxC/XmRgh5Sk5D609MMewWtuRZzjcy7TgFCKcpSkLOWFBINKJKqJd0fTm9FYCUssoRIL8LiQKW+Jy+dcIAEH4F0vn/BLXvpymMBcIgI50ssmdMtbzATIICDAhG8dk1XJDKY1iSnMYhazL578Sx3nJqMI9HGc4/wjL/YgyHQWkgyHPF8bXChDtmFrniks5CAPIAHYdaOS/OSnBaLRuU0OCgLp6CZgMuDKaXIkTaxs5TCZkEyISlSYAKFkAyJ6TYpO1P+hDK3H0RSKGhwadAuQes4D+AgAMJTUFCsdqRGjA85o0QgA5Kxpj3YBlgMMcpDrPGTIWMhIFr5PGDlKJ0uy4QsB9LOfFpDA/QQaKv0V1KVUjcoCKHmzqmrVpCjdqldLEZ2qdLB7tDhpTf0oSQwYQJ2ENOQK0TUAa8i1FXJ9gKLk6b62GuCG+nRAA5ZqSacGFC/xggA3v4rYUXgAq4ltLDvMqoD1ONaxMNXYHbl3VhIShxeEGIRnIdAH0GJhtKP9LGlJCwjPenYg2tAhYBvQrjvOzbCTra0WFrvDrNp2t16AbEp569Ww4u9runiAOM96U17wCKiIdOcEJLAd8SFgugf/6ICOoOvc7NaPFxAALAMacNMhEvZZtAVuYnHLAN2al7e+Xa9WXRTTT+UihJlVQHK5wZ78Qmq/JNVCIxCQD5Gmg78E1q9XKvBX7+ogtlAd6GHdq1X0qhfCk20vhUcK3w5WB7KZNWHMehEAAgDYBX3tWAUs4N0dCuBd4w1VeS9MVQnDuLaQleyM55jheIHqFjQtp3HR2rFk6De/RwjwkMUVs0lSMjRMBWiDo/TiG3dTxlJOrIWrzD/hbq4WPy5nLY6rgM3WCC2H+q+RcbqLZHBKqTykxQISXEkB4IJ2DsbylBlrZ69eOc92q6xl7wLmPnrYAT0eNLuEXOAik5jAylAz/7v+agFHs7mSN8SFxqLMZ9xROdNU3TOn6aVlHQ+gx+M09C8ie2gg+SLEI8ZhVxxdow7sgxcZgDMlIZCBJ+sF05/G2aZ73c0aA9tifg7VLEgt6A9GQMxBrsWRFe1q/Toa1oZCqC84MGmLajKqBB223X7tbQ16Otx++iaMCB1oEhYltrq4QPfaM20kg0/EZ+4FtTlVDE/dYgEorqQFEqBrc3Sb3PQCN8EvN+6Dr8jc80k3CT1cqA/rgtpHFsKZ2UNmiWuDRg+w9Q4rHaVOKtxUBh+5xRJucjqKFUZd7qM5p6VxXmQcp/RetI1U/WHZ3SID/a4kBOw6H5GnvEolH7qphP9t9Psw/CrI5uO2ZxdzexM4IBdvNM6j7qRcZHuHNAj6/pJun6KDvUooHztUdDqfpqs7WlyJOpBm/pWau9rtG7+fpbvhcQbgWi8GuJ3Z7yP2v9+n7IJfBy/1ovYS3j0+MKe7moUM7Ylfne5DpB0Gm3wVlhT+OYHfvGsI73lTYOwqLfejTOXbObdnHNZyDwC1H491u9OMFjzv54qlJNLQ+6XzupcK0nvPjgVkrBwOL2E52t54uiM5xEOaO9xjP/tYzGLr3wVAOQyQe+CPhPfabwfou+8FCQgKFonPnvQ5927V45zV9VY+L8hq91gkoOe3Pkc+wd8O7uMfFd/fvxZy7Ar/ahcB1icLrcAx6ed4uhB5SBZvlLdjeUF9//YKU+J/6qB/FUgK/ed/oUZf5USAhAJ16id5cddqrud+nTI7AfcA88dPOLIXU4WBpXCBMQgGv0eDX8BBxfeBeYF8Ead8iLaAMud27yZeiJd3N3QND3aDYDCDS8gFGuh/ELAKDmd+eiEfyJeAINZ6I+h+p+c1elF7/EQHMOiEXtCEZRgAULh/vFRTO3gVqCdEJ+gLQSiHRPiAMBKBGoCGTIhne3gKagh+pbd2UQKHMuN+sMZ+i3ZvQ3hZARcLYVhJE+aHZ1iGNuiEcUBOAyhq6CeHnNV8ncg9d8ht9PddCUAqk9iHflgK/4Coe4qQiX+Wgp3oX5CyhSV1gnJThKECialYhpTohKzoeeW3ZYw3M+4XDY8SeVoAAOvigLKFPweQdw1wik7oi0sIjIKHiaWGdsN4eskncQRgAQfQFxZHYi9gc4zYiLPVASzITwJAhjFYjTdoif7HYXx0AQFweLBIjN4ocQFALvkAWkNyBRuAAASgfMXoiCHXbR8QgfNCg/FIg9cIdtnocgUlfmPFdlgYdRYAYCNGJAQ5d83ohXSmkFzwZv0kif4HkTEokUNHkX70OltQbJsYUHKIj0OCk/lQkAdZk8PFCm/TBQbBTySghPi3khjYkiZHhe8IFlUIi6inkTGHAR6Zk/8hyZPOeGn31wUfwI5xxpTAd5QVOI+6V48kFJNfkI/DmHoniI9UqZM/woiV55OsMHBfwJD91AB6uH9hSY9dpX0vqW73KAoXGVNyGZUSxwEcmZMboCm3CB+5+CxCBwZX5U8pqXt8CXxwsAVXhgnBaFNgNXzxJTs2SXUfWQEOSFYJCSNASQoIhpcOWXj9sAWbpgFaqYoBQFNFkYZd9WO/9XeuWGqokCNz+SmyR3cSQJA4aZCOuRW4qDEHAJuKlXfu6HkSwEOhQJnpFQBMZpll+AEuB56RJXiCWEKCeQoZsHKi1nY+qHylSZByFpf6pprTIVXrcJL+JgBFCXZstg+W1HP/+rmHZTlOZ5l0gBlZ5okKC+CU3HhZdNcBObld0CdeJDkoEeB37JAATNWdKacBKaZ3t7mV9aUAZiegBKoOhFmYIymHU0mQSBV7Oidfz5h96+CaQxmdRpd3lWSbIIqbHQZ2wFmRUBFqhSl77Clx5MiMXThYFFpnVpV3HvB32YmXPNoFF5BZX0lwieebUBEWDLqPJ5gBBLmcL0ozMTo3BvB17fAB0bijQ0eKvEilxWdjJkeeAICl6jB6GNmI/MguUwlQGgd/WDk3BzCjUeEB03mjCpejkUalIUpOJkpwdfocweClHmSMY0qmMjWX9ekcGtBdQ3mn3iallASljdoFDheq/5xmoMtmHwIQmvpIXMQVczyCjk4Sf0wKI9iwIuhlSaVqcmvqT6bqBVbqciOnpSsiBj7ZoAgIikxiqzGKq88yKizSAWyachkaZ8LqBYGGoN4GpH5EJUOqp7HYrIYShxM6XBTIIh7wphd6cBpAf3qprVtAanPqbcdaJWmpPZpqiOVqru9XpnN5AKkKHdQnjSM3aWkqrMRqX+F2AQL4AATrFwDIjZVnqdchfFiQKRtrCIQaodcRi5q6OQfgrlWSAnjpq+EGrzskr/O6BeJkr5/GsILGLK46l+SKby9DABwrSDq1saH1p14Rcc45n7s2jX9SrXjZpr2mVA3gsl5AU5DKZ//fqm4S6xwoWrG4eLHXdgw5sVaDlE5hC7Z58HOxs6dziaa+RooWkLLApkMKq62KYLUUpqVzq3LECXXpx6wOEAEYIIVGJUg7JbiBu1bHYCRBtJ7Rd7OEqjXAOpQt22sN0LZP63q9tqp2+xzqcbPQUojFyAt5MLgixFZ6BbZYtFfq0iQfFLCiSEQwcDldCaeZhrmUC2MC2q3EtqD7uj2jyQ0YRABfC7wpJLwDSEjkA7bpZLiqm4JFGyW9ZD0n60+JSrvTex+KkG4DmGVjs7lfuKe1kAeZI7rEG0Luc0hFFUKAmxPGmY6sqzGc+rwGO7nUK7+AIaB2akQZMH7DtWM12Qv/eQC8bSW815IjIENPNHS+O4WmLpq4crm5zjtHI4CXRzu/EzwSAji7fkKp22uYbde3hDu+A+hTBTxDRJVC6VQIqbe8zJsXNnTBfgK7KkbB4ZoZakTDNWzDN4zDOazDO3zDkyEBLWwK9TtS+jpWUNmNA5ATpVtDAzxUo9NIJFzCCIwBThawBtjAG4o70FtJDQDEplABC8DDYSzGY0zGYrwAAPon+AsIOcGxbezGbwzHcCxMvUTHc2zHdUzHrhrHe8zHbowFgFCofrGqt/tSS5e152oMmePBTBzCBDzCwxvFx7Akscq+I4vGGjSqPITFI1EBotEMBBBKoCzKDTDKuGRL/5dxWqmcyleQSq3Myqysyqp8yqYcyqRsy6XcWfm5LGJAByjyQLURNRIkzMGCHMiyIdiQQezwARCrVXmKtwwMDgswPqL7yObTXOQ7wghsAIkSfxrsCim0VSlAiiTQxTL5AoCQKsbERPrSJU/kzkHzID+DQLlyGQSQAJfsGlewOL/cPBEEOXcAQXlEPSWrzIlHyCPVpc9sxfdTCGtVvG5lPiJTOouEV2RzLYS7s2cbreS1tCPFlWsrvajgEPDwPwnVHwMUOCjdJYsxIa10L6RczqSQAATQNPy8MMH8z84zOZSzJ/YMFUKMWBJAABp8h977uyJUSI7cXKlTDduBV2RAPv/A+yKVfLO6mliOG2eQ2yIYQNL3sionPUBP9Dey4juDQzgJhCUwzSILQNMO5DTA8iE4ndMUVMzGPDFytA5Ue6COlSnejH47i8Dja83pMxbwRAxO3TbaEb4HMMlEO5cE0NFVxZUaOhKgnEQBhBh9Q0CxMtYpvdJFcys7AwgsgiNunUf9PMwiQtd1bUFv8jLrQJ5S61UaMNTb+4AOEEiPNMDku0jqsz7osTofjLx/1GLbawi2VaNbHNlgwHN5g0CZTStkAs//4jeCA9pGYzhNdR/4u89ssDB3ktrQQzURI9CVE8h2uZS7tY1+DQ5JvFOsgy3u5NtMvVxkUz49i1QLrcH/BIDPW8Vv/RS/YHAj/rNKSmQYS0A07zzdYCJFC8UfxwO3UdHLCcMJzSPX4R05eNIh05MsVj0KdSrbV/2qyhotNuTQ67TbLXQ608DiaWMu7eMyYYsBYrXRs3XejcWVHmcBukwKL7A7rJLgCr7g1O3ZCN7SaI0lGLDc7RAoy2PTHgI1/ozhJLLaSmHXvcLfoyCABz1ZNuvXtPAyhLvb8Z1IpoMW3YE6ZqMjoxPVdFDjgxrhlIWt/obFl1HS6szS0A00Yw0rEsAZf94vRGPWr3Q8BC0VCOPLzAPXUz4secI4lZOokgphN5Kem+u9OvVISl3YHEAcJJUWcMM+5BvVLGEk/1/OwhfmIraHpVjg3CZ9GOwMIX3OGX5O63/u52LyNwFz3Q1FMFV0MMrTK08O5ZADHMDR6AxD3le+J5aj16x6Yfj75Y0AvoFLNhINF6C1s2iBv6cFAMDtyPa0zdnw5WR0Y7zKdQF+566e59ENJrXu7gny7sGj67tuPKmyQH+B6MtR4Y0DzHiwbMQSAQsQQsY+QRpezK39JlkD1DOWE+ydxJnuyNxxATVgAgHhulNJCCXjDUGFwEf15QMwsFKWtEPZpqyOJRUBCR510rDuJWFC6/AO8wQC6GEizy1d70kT58EH7BfE78AMANMl5cHxGx/gtwmAAf5IALnxGw8TMcqOIv99ssyZGNNVVQEZrMEPv1NOXDoO0AclxQEmwAyOsguXIsJgO9Rv/iwka2fsipJBmdYABgkb8G+urue5/kQGYvSzfiAxjyAxj+uCM+geRc85b587vyFxAteEEAnFISxw0BsiRPQQMPF4sAIX0BsY41QfQgbEXEFYEwrkqZt2Jn6V/tjA28TpEwEWsCTpkAE30wyzaB4yVL7Im78NTJ129gGH6k+3b/KrIfcetQEkwARYICEecAWOYfwC8CXHfwMq4A8SsI4CkAF9zzvU/wR+PitP4DdXoBH+QfyrAuFLY9py8rAIEEII0CNh6vh9kIZxPwPYIxjTtQE8kQ8YQGKHIAT/CPDve/ASIPCIIwk8JpCqK9seWaTIc3QFN57rO9/7PzAozEAGxiMyqVwqIYeIIRKZACZW68MBwTgcH6/jhrFEEgdBhKMWXaeAqIHAnNOXE8JHqN/z+/yPAIPgoIVHgECDgOIiY+PGhkBCJESkwOOjgAfEJUKDxQYCwcICwYZF6ALoBoTG5imGhMTnKUFHqilkx9gjAsRCwiWkBzAvBDAGaO9iZ2PzIgSEH9ACRIv19YkJyfbDhffmxQOCgEPGRncAAUGABdcHBMJBAtfB+cUGxsVH/QW8hQSBfBcAEOjGbQSKbNcWqoAw4yEAaRInUgxyYEKdjHScwKHi0YQaLe44/3zJEwADglkbCKyxUqUKFCgHimisqURUxZw6KXZoMGhQA0rOhjZAkDJRAkkpk37yUCrBgqKiBCDwYGmBhARVgXnooDWBBg+9gEnQoDUDMqwCYJ1KIAEeqQ0LPJjy0G4YgXGgLKxtIKnoUGdBd+KgxvBwQm0Hu907cCPghwgbKlxwoO4CggwfPmBGgyEAsA8ZEDzYjDnCAdIOIiAAsA9CuMUiUCBmCIHAQwU2CPPu/eOtzeBGnMSE8hILB5EOSHrpcOADhnXL1aiZ8MDlxygzHQi3acw3+PA57P5kACnwUGRGLSSAlzTpuAYN5lL1MBfBewm/8ANboKGdBh1oYMlXZf8lIFYCHcTCngW13OcUfr8Yg98wwJTCniR8WaIhes9Ew5thtVlzAomydTOaOgFtcIE56KiTijc2YCDPZ6y9YwFn+mBAyYrerJLOAd6YSKKI1zg0Q0TiKembE91p5MABB3R0nHXWcSBZF8kFsAl00k3HwQMcVEkFHDLR5GQdMy25Jm8a+DTIeR02IgkinTz13koQNOCBVRvYR5cAb30CSlJySbCKBhJ0IBYpCOCTwGgWSNoWBg3oJxafp2ywJ1Xv2fUJAUlZoggzcioyGIjVFNlCibJ9YwE00FgAgDnenFQQaabBGgEBDoTGT44WRNnjPhuYA4CQQ9K2akMyRGASm9H/UvTOAWjWwWtxE0zhkZUcEABBcg50AOutAVAHppjXfaQtFBBEwJ21TEwAAbTS2itNAm8yYOoiFvgVSQKfNLDBe8DoCQGflvxJ8CaZiJUBWeMSkKhZmcUiQClUAdyeXRBcmtmB+ZrS6TCfriRJwA3GaSo0vYXIbAolKrZNjBgA0EUYtwGQWQAL9BLASjekRmMAqemzShis9dOjN7Q4EJtsMcO8ghP3Wj1Rk/EygQEcBlBpgpUTPIIBShB0gZKkuKShLkxdR4ABRlovQe/VdffR08pygqKIJKZohQGfeXkQVMIEz0UwgB3kBZ9/VC3wwaEECGBBB5vxBWAFC1igOQGx/2Aq6cX4bconYI8mBU+p/KJK2MtTt7rYBRFYwJg3LealYjSpBPQJGutIkBKtppQSwZYWxLhPZuEkG7VCMPtiN/RCfEBABHI3IaUUUXBbZZhXQvNuchlEOf5FbLehrQEzWd9EAtG7HwQ0/AqwiaOXTJLS3k4hvIDCh/c9xqDMAQkNlKIUG4jFIwjwiQUIsBSsgNXHhpGSU8xnQxQEhl9Qtjf5ze9DrFOV62YGuxTEKEYZ8EZ7TtgNBzyAEhfowAMqIAF9LOAAHQjAA2aimXAsIEYOsESMlNW8qT3vfUbUQQUIELf1rYYAcIDJuq7TvSxRpwtfmI75oBiTKMFrfQNQ0/8Rw5gDRHAQEQpEynsI4Bc+JYVPJRvFgbIiOf24xSoZEND8BCAgBeVxAR2wBcYCJQE+jeIXoxhcqNyIMckNg28A8xsHV7eT1oXQVbQrYQmtqA9v5AEMm/GCPgLAmU7mCJMAQJryoHaQxBBxAWIUYw29eAQMxER7VKoSus6lyzBJETttk8kS1wfGV4pRKGUsGDIrdCA3/sk+hVRULKD5RwUJSAMBmqY1AxSLP2ozmoU8ZCGZ6UZkigowZWxZqqYmtSFdEpPu5OQmRxnPUr7zeJKZXRDZuSznuZKYR8yaF2fyRO15RIq8RNdBwzQmbk2hlqiR5Rfp5s8w5rGMi0BZMpf/Kc5vjiKaHo3mNEMqUgV9VD8cPaQ4Q0bO04XCopLUCSVh9jqaiaCemMyRPDczyp3m9J01DeLyVjlEZhVxou77QHsgSgDsaS8Ct7yCQtHFPexkp5YEKIAsz1Avo7qPjBatxEpDJk5CnrSk21TUSKdpVm9+s5nMVCkyK/FVdH5QnQkZkk3ridOemgaeeXWnEPdJ1H5y1X3q8yJaHOrLgmJhqmOiknHgUI8CGGB9qClsGI15zI1ldKzO/OZat5lWaob2pChN6UrB+tWX5iSmzJopTf96PL/mlKfxlG0qTcRKfmL2fYcV5hPKpC2qcktdC4Us+mKygQIwN5hoelsFemtE/69+FaOdHatpS0pSao42tCYN51w8m1o6WZSukwShTEXIjdz+Faf6sK0ob4rboArVrikoqnTtNr+Ack2xw/0aVY8Dkzc49KrMZa7WeHXD/LpPsxy07nU3ylHvchet1iTpWk0rXmWS86un8uB57XtX2LUzr/KE52Ze0NP5Miaw6sQvg68WJesZgLlOdOi2uFUFHa/LCpFtFwQmcOACVAtNcVhwjKFHXYtC2FMalTBotStNbmYgUKT9KFY46ta3hhVgcwUxTNH7WvWuN5Ul3GsoOblTL3zgR++11ZvraRDdDnWwSYYeQNEkZOb2QrhNXewtG+pQ7chhADU+cGW7o+A7R//PwQ/mbDKxG2WsbJek2szmgDbwx7Kcla2xCOdY4Vowua4WzK0V85hhSzNMPiAgASFeq6NTkACgZi15WUVkorOjftArHBAo8ZyZZ1cYM1paonGXnoecvqUWp6kN/a+23vCG4dYyz4dmbqJtIo+tFvteS97sSkvm2Y4GbAyi6MDkzB0LY5DtFRkgYDsABxAPrHs+oEbteD1s3jDblUgmeqcWNgCADsDjAWnphwW25KgXrKSHq6jAaF4Qj5MYr4TBrm+di0TsbkcLINXrzpCxOoAJzKhMHXl2TCZQpkHvKAl7PrBzN+IYjtvN0fJrcsE8+1m6+EIrE/KFOfQIilA9RQL/k7MPMmSxgYoJ4LQp5fB7SO1SU1fEtUVS9XovKYqnXUByrfGCOKqMAFe2ORpEwIHs3tGL1gBWldzYLW9pbjVvFVnbykbCt0yevVoSuKEGKNMBaLmEayM4I3don9zr9u0H47xCOhfUNjHWIIoloHOhsKYl/sieD1hTGKFYyaXC+1aoj5q856Q6Rax+9ZiRmNUYoB2UNoBTDAgAWPeIRsBuow7jOQAlz7G429+e8VVtPPFrqr1NXl54JASeqYOu5crTt6OYHyHk1EfC2xYQBuNbzeb8olNYdd5Rspl0QXr8NAZSAZZMI1DWtxuQo0yqc9KLyvRTdxmqV0/mrAdRIJwE/wdnOICwABFnkEvgxYq7+EgvcB1Q6dPw1UbxcZ94vMOZ1AHhUV8EbEFwPV8U1NJMUE9GhBwdzER0SaDVLN6jhd+TuZEEjEH5aU6CoB9dJAi6/YPPfEtQGAMBOQq9bdk40R9G6RvqTYTqiUg24JWZUc8XPICwbADx4FBm2N6PZMA63EAGXgDZsEaQCAl9vZ1gxZ0JSosZXAQdEF62LQEAQAP5RAlqrOFM1J0F3p0S1FoYXo33fR+k5Zz4WcI2BQT5YV7nEMwfWUKA8AXntaBSTI6fiN7odZn9lRFrVV3+pRc7lRBrfAs+IA8mRgM46MgqYOEq5MUBSIo+HIvFsRORDP8bYdUhm4gGAQAAE5hhTfhYlOieE1yEFQSHA8jiEcwLBCAZK0oLCt4cznnKuDlQW9BFKOLH78Tg33iAOaiRKZiDW2jO5uhcWNWfh3UQ/okY1sVWjPDKjHASFc4IZ8BAEEGDr0XHCf1aOJ5Q8K2SiDXEKgbjmhAcHBqB8okcyCFaF3UHLx4A9dhj922jXKmg+DlFg7jFIEFAO1SjAGRANN1GgqzFjtCH/CFMqIla1End/aXTPI7YQSgP7G2SD/WVSXIS15WENzAg8MWj8IlYBBKkb7wDCFbfkFnLASCa1lgfCdKktyWCQRZjG60gIbEVVpxVUn4aWpVUR2nZ0wGhqBj/ZCTyxCSu3jeOgJnNF1fKmYvNo8cApbRQQ5AZmhw6yVkaWchlgFjei54MJViRk7iN22eVVZbdJV6elA9yWZd5JCQKQG9MT0iKpCV1pWE2ICp+IczgQVtKSwbuZE6iiQOkJZpYX2MKo1BuI1EWpfiZlmd+JnhhY8mM1yOWEQGwZW+YgTey3v7VlEEcJouRZGA9oIicwWUam/XFC2RSVrxMwBOEHLfdpm/ogkE6EkIapRvNBWguJzjNn1ROZXEyZm8kUUgeISqSJGzKGRcmJm0WiXQK55IQ3lLFC2XaRAae5g0M2cyBp3gMYwpmo3P+CXNqmHKKJkdqY2lyEHuER3tU/2dWuuZWZmfbcafU2Nd3sKeSKJ8BcEDlkWE/Yptw+CY0rKKCImh4VACHeNhmOp5zKud8glN9zl82aiNVfmdgOsFg/qdWYqeAYicqzkZ3IsYwWSh4AOcNVEB7LNX1McFuniEdaMtMJEAJ5sAHXCCN+kYGZKhm5mFRImdyElJd0icjhtpoOmJ+ys8/KMn0HMBqvuhrBihXfunFCVuMHoZADumR8gbhccAOeIUa5mIdlKfLzQQ0aIYPcEDIpWlvTM5QbuYyOelGyaeg7qV9OmJxTg5qaimKVtJ1vmaJtWiLwaQ8Tk0GBqeeUoQZ+oADZEWs5BicGsFk8qQ+XgGQqqEEWP+qDlTope5EwGRmdY3oXCZkfWpZc07pho0odEKiBSTqkohGB0kBsAarsAYrADiVsQ4msqZABhQrs0pBsQ4rtAKrGtrpqu6EGaIqkdpCBsAADNSiEwzZUo1PBGxrBowCMOpBplZrTuCoJ8iHfOhJULzru1aUnKBMXOIqvo6oanWIUFBCvP4rvMKrJxiDBFhNB0RAByGgwi4swzasw3YQxFKCxOZR/FSsw14sxl5sRJ6rulLEPrKpRGxGBVRAB1SAsgkItv6BjXZsRXwA/2RsDmaszM4szdaszUKDW6Qsy+4sV+0mZfFGj/oGniKazvKs0R4t0jKaqhLGkE0AeBhp0kb/rdRObX6FHMgSxrUZQHhYLdV2rdd+LfQUadP6xthu7ZBdLdimrdqurbWWbW8ErXjkKdvOLd3WLRCkK9nCnJLgrd32rd+q7dAemHhkrZZa5t8eLuJKreHWqN4mqNwmLuRGbrXuo5LA7d6ereRmruay5wc8rtk215pwANRuLumWbjB6bngQ7pp07pAVrem+Luxi1ujGbeOurnrGLu7mbn4pqOtShOWuCd/qrvAOrxGt7JK47er6LNoSL/M2r7SsabSobrRQrvNWr/WmrrJJC/KySfBer/d+bx+wruBKy+9GL+aCL/qm7w+I7YEt7/HWbrRsBuqqL/2mb/eGJ6JdzdLW/y//em/gEpnVbO/zCnD/FrDw/m8BuO+alK+9+KzTGjAEC+/8RgsBa+/5RjAGm+79cm/+2s21ZjAIa+4+9m5vVLAFt28IpzDkLq7VMPC9iC+RkbAKz3DU7m8Aw2/d2DAN7/DXKqgCDzC2vc8G8zARHy0MF4AM5y3oHtXsFrETG63PJrFvuPDVHPETXzHPDrGSiK7WpmfjqlwX53D2YjEZX+r/GoAU6wRkLui1WQFk/nD8hlwYlzEdt+VWsfC97GPIBXH0ILAOqEEdB3IYqhy0aDEH7/EFQ8/+/qwgN3Li1VgX6/ALI7AcH1G64ul6OrImM9r4AmcaqykiL7ERQe/4bv+yKccYokGvEIdyJr8PAr8cHJ+yLIsRJfNx8SLyHBuRHtvyLPcyMRFeBx/RLvsTMPOyLx+zEfmssqHxEe1xLh+VyoUyEiMzNR+VNJuwvezjM9tNLe9xLFczOG/xNWPbJ/OGFR8R+17zNoczOw/uNV9EGI1xGImuOrezPQOvNDNzGF3bN9fNB+xyKd+zQJcwLvvT0K6zJYdyPw80Q+uBHi+oUdVYOQOxPDe0RUsDME/ARK+JykmXb1b0RYc0ugYz26ZzAYg0SusBSc/tP6NwSr80D+ApRB8uJMO0TRPpQq8tIN80T/e0T/80UAe1UA81URe1Ue9B5pCrUm+rBKCp3Mn/0FIrdVNLIOQwUFSXa+Uctfci1RZwhJR8dfp0dQJs9PtQw46ANVpvgeQ49Z29BdmgdfqkT+DNkVY3bwXYmpT8nV7vtV4HHu0VW+WVHF8P9t9tgQCQdd0QgWATNl8fIFvX9evOAwQwNmX/nUDWI2YtANlUNmUHHq/2llvnNWc3Nu0hNmSD7VqMdmUHHmBK1zyodmXvCMdOlGaLNmzzNdlg9mlHruJwzW1TtjrMthi9g2//NmF7Nma9tnEzNgYg3m5Lrhoud2cfqD8RgW1LN1+HilG9xXVjd18P4XPX7RZ4d2V3jj8pN3kT9l8TUwt2d3obwL6Ft92ixWirnBUYwH2D/3FsP3b00Ddn2zeA4/doY8Bnv08FLDZjB3h+Dzh4yzfYvkNl27eAT7iE6/dgC6Rp74QTUXaF/x0Ydzhlw4IYLRWHC/iHmzh+W/hgN7eD2y00lDiAk+oVTHiIt3J/u7eHm7iMz7iEM3bgZXhFEMF/6zipUnhs63aLe61/D3aF53eRO/l9qzeSW82Bl3iKx7iM6zhjU8I/RTiW7ziPq3hfm2iSe20LJjiRg3mWczYGCLe9UAthf7mar3mIF/jV1F6Cy/mcn/hx23iZTy1HxPmV7/mcH3eDs8mS83WTEzqYb7n7VLmgDzqj8/iWA/mfN6YuoPmkN/p+R8+GM7mkb7qYN/82fwvjZIO6qO84jTd2wV461QoUqqe6mh/3lLPJmee5rHP6Xh9Ara9ucTN5rj+5mNumq0/tpwP7nIvJBCi7rl84QrMJniN7qFuBsjO7qvv4oSuJZse5nlO7txc6c1t6sdvjre/1ol/BuXg7dYB7Y9sNdEQ6mIuJvFf7nqu3uOtBoCP7jlfHsve7tV/7XhP4uCdttCv6vovPReiSbyY8nQ82mY/lr5t7vPs7tdP7mou5wFfxsev1jOs4vyt8s194tg/8keY7x3s4qaLNIxwAdWwCL8BNyBuAiMuYe3/5TMw7BzjkunNAdPz7cbe2wUa8ueeFURiFEq1Cus9ILzB7QET/OV+PPMlbKI7vODJcATwcgBbEw7KPBswv+GBDgJ2zicnnuIznPLicyyZUhxq0A7Oj+K5DfW8IucFbQdX7WC9swbkETGogwILS/QaMuszfe9Qb3z4IOqmmRgZUEdlMQDwsx7IbAAJ46qpbNtC7Zc1L+s0ve3Kk/bmQjbVPfmHXDawD++JT+yaYQ+c39yVcwRh0/F4TQKkPPo0WvqJLOttrvpgQwQboEgdIScP3tZ+LveGTqtnrEudT++0XOWE/PLR3N4B7fkhsAslpPuM/wd+HgpgsPnPntOzfJu3HutiwPM6fBAZQB52CvbA3NtzzhtCnuRuffUikBtnMP/bHvETd/0uU5HlexAoygMDhQAQ3btyBTBlycNg2zUZtE0ug73zv/8CgcEgsGo/IpHLJbDqf0KhU+TnYbLTZbAPhTDhgLwETHmMQJe3kajtApkkIG6v+QrphDkQ2WcwsZGpzNx9wRwJWbGoEaBgYBBlgGSgcXA4HMilkjmuDGBOGoaKjpKWmp6ippVWDBnWOYF8cERh7eWCcWq5zBwKqPB8QiVeddW5eYCe3j151g2+/O26tM7C3Xi4tE1WZExsHBHydVxgV0efo6err7O1Gw8SuWiqRYZwuDsgTaM27c8LphA2qYwdPGFuxOABqRmOQm0LnMshRJC/XDGQwHpFJwUcPgg2fWv9hgOiupMmTKFOq5CFwzho1MSJNgIAgwQgXXiahETTuRg50B+DVKKbGjRo9lGTBIsgL2rkJGKhBtVAnTAucKVZoOfOppw0IJFeKHUu2rFkkLeMRjGFhw7eDHy0guENAl9caZNIliOqyqgoEgNFA+JTwkSCRGdJV4Etxwhgt+ry4lYVJEIKQTc9q3sy5s8kMjNU6O5BBlukM8gjeNXBAHYxWQ1VXle2MDYYO6tISe0lQ35ffqofOwZDAs/HjyJNL+RBa9FHgzRjWdmhgnW7htCNfDN7Kzbq9UmmbFr/aQF7l6NOrXx/gOpbU5Gn7u0J6HWhq8OPLZ3OguOvmwuWnn13/87XhFHsIJqjgSve5JOCADa2GgQTrLDbQgxDCxhoH7LiXGobxFdhGYguWaOKJ6AAYIIR2lWcAAe1Mgx+LvIlYA1jsNBgejTbWcIA5KAYp5JBT6KgIbyxqiAGJ62ig4i40NiRSa+14mEWSPbKGI5FcduklEQc8CSWIWbw3HJXtGBkPlOKNWR6M7rw2I5ktwnbbl3jmiSdzGtYYXJmrhXmSh9ghydNLSv7kTgZCNfagPy6ah6aelFZ64gJiQuqmpnYqGicBfWK3aZZ4+WJSB5luimiNsIFj6auwKhiUhqqOqaR/J4VJ66qjwnZHSuDtKiqiGobJYazIJoscDI3CFikb/wCl9AEBE9EaoLWsERCWSboKS6onnior7rhlVdAttuhiMClKVaSK7hXCHKtSBgQ0+25ToJCr775inXuvSLiKBZq9/5q3pVgZFdwKCdvy6/DD9qmrMDlLNqxSB/VO3Ia6mvk7cS3yQizyyNFwMIbGtVgsVjASK/xIuGS1SzCtjjBJ8s04m9Kuu/Q5AvNZ9LacbsWeLTDGzLyoq3LOTDcNhQMJHG1Fo1M/ksDSQA82K9WsPfJjcjPVMjV1jkCAm9Nopz0FB6glYAAEBWBQgEAJRLIe2wa4HWbcBVhRd77p5cSaG3LLbUUGC4Ss9uKMN/GBAQVEXl2QkUse5AeVFwB44/+cd64E5JFvbiLomguZ+eSep666EKSjfuLpplcu+uq0r0767AseIHvsltfuu+2Vu24i7JTv/vvxnd8upPLF947882q3znsBwpeYOe7QZ08y8yjqHvr01WsvPsTSN0/99NiPrz653J/Y/vDBry+/w+WjSLz9xs+vv7jvl+h96eYL3/4GSKn6vS5+5ksfARfYpf4tyIEKuh8DJ/glA8LPeQf8HgU32MD8neh/CmSPBDlIQvchEH8YvCAAS8jCEkEwQS9E0AhbSEP1WNB6J8zgCmvIQ/TEkD0gBF8Ph6icGy5ohkf0IBGXSJYZ7IB5j1NcEeXFvGawxwDyIp4VmchFlHD/QHKPk10YCyBF5IzRAJiTHQdAV8bjnDEA11tD5NrYxTqiI3N4DB7WPDOBPOYRjezpox9Pt0c7GvIUpBtkCI0zyOslqJFKPKQkVZFIPypIkI28JCQLMMlOngOSAkQPJBd5nFF68pSpqKQjNTlIF2YSlbA0xSuTmEdSIkeRsczlKFR5Plfm0YS/1KUw4ZBGPBYyOcXMHDCROMxmLqGSoVwPNE+UzDk685pOwKQGTaRNMlLzf5HDpjiZ0LpjilKPKNJmNMfJTh9gko4IIh081zNGb7bznmCinjnRI0hABqmf+8QnNsNoS/Vss3gFFag4+8glyDWUkwqNqA8+ME8FVTRB/xeVqEY3ytGOevSjIA2pSEdK0pKa9KQoTalKV8rSlm4mAXe4gwACCgTEceYDAogpBEgkgZ8ZIQNAQkcGKLSDClzNpbH8wAaoFVOa/qAWnFmAYAaDhgDktAkWsFk0uMADqQYVqZ5kTig+4FR2SJUHLZCAd5iAAZ+iYgOm0oFXwYpKsfKgAwLoAARmKoCyQaSvGPCFXnVaAcR9IAOAxRHLlhTXdpyVBwjIAEx1ELXA7kACY4CEDhYrAAu49RTf4IEEEGCOoBGgOBXI6x0yQC8LQGABELAAAc7WWs1KoG5jwFVfNRsAjL2Mrqr7gAUEkIDiLkCqgTUaDmBrgcUslwACMP8qazdgAeaABQECOK5rK8AF7SKgrKKQqkxj+4arjgFxBKguphKwgAS4VrivXYAAEPBZU1jAtTkVACMOawH2ZsC1o00uSBAXEwnsIQDuHWqCW/DaFhTnZf8FywYSIAEBbICowOWcUh8xBgGMNjHMIYlrNwARCZBYriMJQGCntYOoWXgH8zUJch0hFxjltAOk3cGESbCDSbiXBxuobynaYgYMyKU9uMKUVEmUVcpiYLPffW2LBdACiFALxxBZwAYqEFm5fjXDiwtxV09cAUAAIgaMcISRvxsA7pIosBVwCkwH0+MTu0OqJOFyf3a6AR7Uglo7+MBcDtRWdcBVtFE+WwD/ptUCIN0JwXDicgUIEFS1/ncH1GqBmuVyWLfwFsyNE/MOtFyISR8XcfINMqqZ/GQdrBhOkI5ti+3s2BzvYDA51bKfSeCUDnAB1gHwrDpc29UoY3jSjXY1kPaiA+7GOajyvbQOMq3qUxfisIzQKqjRJmq5nli4QbWACs4mAQu0p887qEWcW4wD+k6bzXeG96K5AFNBkwjH8jW3DvZAaigLmRSHHjUCrOqUvo7W0WeLWrOjjKu9SjsABKA0Aog6iYfLdtuM63YA+t2ewFY2GMONWt3mUtwDYGymcoYRrvtKa7PORQAwT28HvNNZmD+izdCF6QYS02Gd/3sUwy32B8oN/4EEWHioti4HZSNN4gWEPLYdePivYgvz4cLX6E3G+OIUvVmu3/YAisIr2Ht76uNW4OxtDnQHKrCAD3SgAwlAtzuGvgDWLgBIaG9zBBKg6A9EgMokUSvfKwDeKKwdGEQ9LGmujeG+K5qoFfh7Bkqt6LfrYOZ8b/Hitc5DHPvn4pwP/c3i3hali/70IvvAcVHP+ta7/vWwj73sZ0/72tv+9riXpASiTgISAVW+bW6qDugl5UXr1z91b/HbO1Bhc8y58LmP/hJIf4edBzsGbQVJbJ9s4eq3XftVjTiUWVv6YHChLV+WvvpDgQkgYVa4G8ANoPeNqVazdrQ5WLf4501+Cv9NAkj7t34CaAhXlW4dEHTBJluPMGFatlQ5MC2X4R/7p1SsBQ00kVluAX0DuIE8UICLVg7EFmzjlV2LBlPYpQMGloE8xn9Gt29wJVMBw4EyuAQHoG+Lpi0BRxw9Ri9nU3PQQIEriGMVqAMvNnwHMoNIeAQJMBdnB2hcZVX0VVhbdl9r9wgOZlQTt4S7dwaS9YNMKF5JGIaHQC0RR2ntwSSD0WG9VYazZVUaURwsw1SnNmoRB1VieIdDcFU01TAaiId+2ANr9YeC6A55N4iGeIiImIiKuIiM2IiO+IiQGImSOImUWImWeImYmImauImc2Ime+ImgGIqiOIqkWIqmeIr/qJiKqriKrNiKrviKsBiLsjiLtFiLtniLuJiLuriLvNiLvviLwBiMwjiMxFiMxniMyJiMyriMzNiMzviM0BiN0jiN1FiN1ng8EnGNtWM0vTZ/QEBWUhA1bHiERHB8oTBfYREBCoAr6hgBiqEAB6ONybGENSFXHxEEDCYFNBEYgNFqRnAG5AgFFhYWBEABFEAiEEABCpAOHWCQ6SePnnEVbCZVcrdoJJGPREAS4OgDNLEkJthlm4U1t4VhG2kEZNUw6MgDCmCQFFAIK3kg+6SRfMiHgbYAAKCRP1CSgTZREKkO9FhVFEmEG/AR2vKTEwIIhRBbxeFaNDFTZ4AGXNce/3ORbvUYDIBhho4gV/ilg1J5GTkgcwjWZP81IeHwEfGICQTJkguZkAvpdwaJAwEAAASwkAEQATjQAQogACspAW6pkERllwapAGcDmAQAAAFwARCgAIUAmBRAIR8AjyupkNcWmbyFlwbpjj0ZDRIZWTjWZ/u4B3OhEw4IGObACG8QGHB1BmPwEVx3gaylE3zJFWewZdkGcWhgmrb5CICBKSc4lDCSZoxwfvUIY/IWACtZkBRQAS9ZlwoZmQ8QAQapAysZAQDAkoX5kpFpmIEpAJLJnQp5mQ2ZnNCpkAX5AB/wlpHpjscZmRcQnvCokJn5C0sYWIAxX33WXt5lbvkoVf9sBpABsJtQCAk6cSD7yI+JYZ/TNRc4pi27aZoBxlpcMV8WgGMfwWX0NZtFB1Qd2HIFGQELoJDcuZAGuQAVRgERkAHRGQwmKgEGeQF9SVbQqS0GKQAnqgEPYJC4AQA0Gp4oepALAJ1gYZDmuZKKSQFwgpcSQKQ/iqPxmQotYG5PeY9Ox4/6OZX8WQi4iQD6VqCBAWw0MVz6dY9cepX/WRP3aJr8yY9BBhg0MZT7uHFDCRhB1oHFSaTGqZYawJIsGQHjKZ0rCp9WBagfSgHMuaeD2gM3aqJ6ypLnmZwIRgEQUAGAugOLOqJNigpLaG6jJaeChgZGo6UBsJ+kGWxzIWj/cNKUEtBTGdCaU6kDQ6lXH5GqdUchUfoGAClVs6Wq3xcY80WmHUCiFtaqOoCWPFCQxcGiMxoAClkBD9ABANABfdoeB1mdMOKWcBKjzPqr2WmQhbAAChAB4TmeEtCezdqo5kCkkgoNeAkACRldFSABN3mpp9AC6LaPW1afKgaqT4p0JMem/9lqo3VhUxowNOEU2bap7CUXiVGgX1mqH7EAGDOV9kqhXSYXxbGEB5KSO2CndwqfKzl50JkB1fmnf6qYAZCoAICsCiCpCympLRmYlgkBF+CQgRl5Jmquj/oG4LmSS0oBD9Cj8noKmbpZQwml/Aiqm0pacsGP1wVrXFpd1ywhrDRRHFyaYleBbrj5kxUaqmQap0l5tDYzkMVKAf6RAIGpeotqDpUqska6A+P5lliqpws5qIwankOXtsrqqHb6tpDaZpFpkNoWtIZQWMdmU4dlXDYVABLAWiXIXlGHG0OFVsSlbY+7A5W7ccVlM6x1NpfbXoA3aqvaW5ErVwnQH+GisTrwAAvwAJcFABpweRngoRewWT/6rDepeqxrubHLlzpwAT9qWM32oz/xAQ8AecILEcUbvJwbuwswu4v2u84ruLWDutLbUmJbvS6lk9i7vdzbTCEAADs=)
- Direct (fast) pathways via thalamus
- Indirect (slower) pathways via cortex
- Input and output (behavior, physiology) specificity
Specificity of learning stimulus/response mappings
![[[@Pellman2016-xu]](https://doi.org/10.1016/j.tins.2016.04.001)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQEAcQBxAAD/2wBDAAMCAgICAgMCAgIDAwMDBAYEBAQEBAgGBgUGCQgKCgkICQkKDA8MCgsOCwkJDRENDg8QEBEQCgwSExIQEw8QEBD/2wBDAQMDAwQDBAgEBAgQCwkLEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBD/wAARCAEEAjADASIAAhEBAxEB/8QAHQABAAICAwEBAAAAAAAAAAAAAAcIBQYDBAkBAv/EAFsQAAEDAwIDAgYKCw0HAwMFAAECAwQABQYHEQgSIRMxCRQiQVFxFRkyOFdhdpW01BcjM0JScoGRwdHTFhg3VVhigpahprK14SQlQ0RTkrFUg6M0NqJ0dZOz8f/EABwBAQACAwEBAQAAAAAAAAAAAAAEBgIDBQEHCP/EADsRAAIBAwIDBQcDBAECBwAAAAABAgMEEQUhEjFBBhNRYXEigZGhsdHwFDLBByNC8RWisiQlM3KCksL/2gAMAwEAAhEDEQA/AJ6vXEzm9l0NynO0ag2yy3/OdWZuHYlNva2UwLDDZm+Kdo4HNkcjceHJfVzb7rX8YFZbEuJjKs1044b9XmL+2p3IsvTg+Y2+IttUaXLejyWHHBy7gckqM283ynbkc84VUp4LwrYXjmQzbzkrbF/Zh5Ze8nxmO8hYatfsqhBltLa5i28e17coWpO6UukDY7kp3CvhrOaYxecU7OzWG1ZpL1AutpQHFonXlcNUZlxvdXKwhJWpxSEJ2UoJ6DckgThSlKAUpSgFKUoBSlKAUr4VJT3kCtduGo+ntqUpFzzrH4ikHZSXrkygpPoIKulM4CWeRsdKxlnyfG8hbLtgv9uuSEgEqhykPAevlJ2rJ0ApSlAKUrjffZisOSZLqGmmkla1rUEpSkDckk9wA89AcFzutrssNdwvFyiwYrXVb8l5LTafWpRAFRhlHE/pPj7yoVsuzuTTminto1kDb5ZCgditxS0NJ326Dn5iOoBHWoK1dy+26lJk5FMyh5uy3WLJbsLsKWeXxdLLhD0VQ8nt1tBbwKfLIG3UN9IRhYNeomYRLzgt0QrEVx2Apm7LkOSiv7YHVsLcKlhpaCyS2vlBcR2g2689G1LtZOjKpC3ilw5SlLOG14Y9+Pjtydv03s5SrqEq8m84bUcZSfjn5ly0cXGDq91iOUo+ItRPrFbFb+JPSOXEZkS8hftzrygjxeXCeCws9yN0JUhSviSo1VT2O+IVxv2ovMONIWptS0KSFp23SSNtx8Y76rVv271OEv70IyXo1/P8Fkr9jNNmv7U5Rfqn/H8lrLRxMaQXucYcS/TWmwstplSrXJjMqUDsdlOISeUbHyyAjbqFbVKSVJWkLQoKSRuCDuCK87MYxZuLlkKFfrg4trxUGFKM5TG62uXtkPNDlQ4FhRKQSpKAghKEbbm9WlUpqdpnis2O6HWJFmhusLG/lMqaSWz1678pTV47Pa3X1dz72KSWGms9c7PLe/w9OpS9c0qhpjjGlJ56p48t1hLb4m1UpSrMcAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFUw4htfdVMQufELf8MyV63p0wteH2m0xUxW5LfjdxkpdlSi0sELc7F9ptIPTyT03O9XPqE8r4X8WzjO9QrrlUhU3FdS7NZod7sza3Y7xnW19a2JTchpaVI8hTaSkbHdpJ32JFAV+h8SWrzeA5Lfciyq4C5aY6o47CkxbhYxZ7pPsM9yOypE6HsUIDnjbq2lo25uxQrYEEG9tQHfeEHA12WDjuIvSoEWbl9oyfK5VzmyrrPvrduV2jEdyTJdW5t2jbAG6iEoSsJSCrep8oBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpUL6sa/s449IxjA249xvLfM1JmOEqiwFjcEED7s6COrYI5SPKUk7Axru8oWFJ17iSjFfnvfkSLW1rXlRUaEeKTN01Y1Rs2k2JyMmubBnSE+TDtjL7bcme7tv2TAcUAtZAJCd/NVV8i101mynI5V3hZhNxvH5LTXi1hiMRVuMkJ8suSlM9qST12QU8vUcyq1+a3cLxd3cjyK5SrveH0lDk6WQpwIJBLaAAEtN7gENoATuN9t+tPFT+DXyzXO2lxePutPzTh4/5P7e7c+k6N2Tt7Rd7fYnPw/xX39+x08hmXXLmTGyq4S7yyoj7VcHlSUd+48lwkdD3VoOmeJy7VdcpyhlsQLfkc5l6Bb2k9mhphlkMh7lGwSp7l7QjYHYp3677SBc44ENSXGedpa0NvAjoGlLCXCfiCCon4hWV8S9AqofqK86c1Uk5cWzy2+TT6+eN/LzLV3dGE4uEVHh3WElzTXTyya8q1sKlsz1R2zKjnmZfKQXGz6Uq70n4xW+4trDqbiUhlUPJJE6Kjo5DuRMltxPm2Uo9ogjuHKoJ9KTsNsH4l8Rp4l8RrOzvbywkp21Rxa8Ht71yfvMLu2tL6LjcU1L1W/x5os5p/xBYxloEO+siw3HcAIeeC2He4eQ5sPOe5QSfRvUppWlaQtCgpJ6gg7g1Q19pmIw5KkvIZZYQp1xxxQSlCUjcqJPQAAEk+avmOa555DZLGnc+8uQkjkbmPJbEHlPUKaS/uXUn8JtGx8yutfQNJ7bVWuHUIZS/wAlt8U9vg/RFF1PshST4rGe7/xe/wAGt/j8S7WXZnjeDWlV5ye6NQ4/NyNhXVbzmxIbbSOq1kA7JHXpVRNXdXcg1YK7ZIbVbMYA2NqK0nxo7ghcpQ6LA26NjyAdyechJTp6p+oWRXORedR8obv89RUI8hKCylhokfakMjdtoeSnco25ykFW5G9fZdtZnRH4MpHOzJaWy6kKKSUKBBAI6joT1HUVy+0Xa+rfSdtYvhpdZLnL7L5vr4HT0DsvSs0rm8XFU6Lovu/kunifnTmzXq54nHkZPdF3KC9cZN2s0RxpIRDjPOOLZ83MtRQ5zeWTycwSkJ5dzufsePwa6mkcaSMJi2a5XCVOn2V123SZEuQp994oVzNuLWoklS2VNLO56c+3mrZ7vjz90t7sKLOkwH1bKZkxz5bTiSClW3csbgboUClQ3BBBIrh16fe1Xl7dPToTKVdUIYSw+vr1+Zg/Y8fg09jh+DUTaaXjiUtWRWuJrm7iLNtnXAxGXok1kPzklEgNvMMtpG7a3HoDfKVFYICiNlnawCreeU8qRzbdN+7evbnTnazUXJSz1i8r4nlvqiuIuWGsdHzNPuOO2+7wX7ZdITUqJJQW3mXUBSHEnvSoHvFWh0OvK73phZ3FtJb8R8YtaQkbAoivrjpO3xpaB6dOtUD0HdvzWoeQRtQNeGcquoaflLsjLbzKbSQ5yupeaClMshktrQBv9/uT7mvQHRi0PWbTm1svsrZVLVIuAbcQUrQmS+4+lKknqlQS4NweoO4q9dk7OpZXdWkpZhwp8mll8uaXnuVLtFe0763p1MYll+Gce43elKVfSoClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApVQ9dvCM6c6eXi64FpTYZWoWYWp9yFOSysxbXbZCCUqRIlKB5lJIO6GkrPQglJqmOW8TfFblGVQNSpeqz0a6WKUifbcctKTEsZ5TuqO81uVyUrTzIKnVEgLJHLUG51K1tJKFWe76ffwLHpPZLWNbpSr2dFuCTeXsnjpHP7n02zvzwexVKj/QjWfFtftLrLqfialNx7m0USobit3oExB5X4zo8y0LBHcNxsodFCpAqdzK6008MVjMnedj41dn2HFNutQX1oWg7KSoNqIIPmO9ZOuKVFYmxXoclHOy+2ppxP4SVDYj8xoeFG+FrP87vngtL/nd6zS+3DJWMWy99q8yri87OQ4yZnZLD6lFwKRyp5TvunlG22wqDc31L1Xb0A4Ysr1HzHVJOkM6yyXM8vmJz3vZVcvmUmOqVJ3K+yB5D5R8oBzbdQRUvWXg74z9N9Ksi4WtMtT9MnNKr8udGYu12izPZ2Bb5hV27KUNjsFKIW51JPVaiCjcBMw3PR3id0gwjC9OeGHIdN7njGPY41Yp9tzmHKBfkIKuaYhyLuTzhXlNK8kbdN96AkDhWm4HcNGLTJ001evmpOOrekKh3u+TTKnhJcKvF3lqSlfM1vy7LSFAAb+apcqBeC/hpmcLmkkrCbxf4l2vN7vcrIbq7AYLMJqS+htBajIOxS0lDKANwNzudgNgJ6oBSlKAVjbJkmO5K1Kfxy/W66twZbsCUuFKQ+liS0dnGVlBPK4k9FIPUHvAqF+MvXC7aO6VC14InxjUTPJaMYw6Ik+UZ8jyTIJ7kpZQS4VHyeYIB2Ct6rhoHY5/AXrJjOAXvIZtx051gYjwZVymLUtMLNEo259z7hEtIIAO5Kkp3IS3XFvO0GnWGpW+k3FRKtXUnCPjwrL9PLxw8cjZGlOUHUS2R6E0pSu0axSlKAUpSgFKUoBSlatqblxwjCrjfGXGUzeVMaAHgShUt1QbZCgnc8vOpJUQOiQo9wNYznGnFzk8JbmUIuclGPNkc636ry47r2DYjNWw+ABc5zKtlMg/8BtQ6hwjbmUOqQocp5juiBWLczFZRHjNJaaaSEIQhOyUgdwArMIiubFT8h6Q8tSnHXnju464olS1qI2HMpRJOwHUmv14r8VfEdb1WtrFw6kn7C/avBfd9T6zo9jS0qgoR/c/3Pxf2XQxPipp4qay3ivxU8V+KuN3Z1u/MR4p6a+tOhqU1CkIID+4ZcA8nmA35Feg7bkHuOxHftvlvFfirrXGzRbpDcgy0KLbmx3SrlUhQIUlaVd6VJUApKh1BAI6isoQSftcjCpWco7czl8S+KniXxV3rCmRNtEaRM2VIAU08pIAC3G1FtagB3AqQogeYV3XIobbU4UKIQkqISkqJ29AHUn4hW50cPBGVzlZI0yO2s5Hf02KY0ty2WttuVJZP3KVIWT2Ta/wg2Ec5QehLjKvvay3i/wDNNdfGHXbm5fLk7HfjiVdS6iPIaU08yjxSMkJcQoDlV5O+w3Gyh133Azni382sLmLUu76L8ZItaqcOPq/xGL8X/mmni/8ANNZTxb+bTxb+bUbgJPfEX5rdslwTJ7HlmLwX5KjOSqbFLqURZzKW1Nlla1AJjv8AI66tp5aktHkU2sgrb3lxrINJ9dbNKwVN1efVcWPt0JK3ocpPLsoqacTylRQdvLaUpO46KI7+mI5B3A2NZ/Sqy23JdQ5llvuCMXq3+xCg/KloadjsJU4khK21c3MpakJCAQCAh89w623R6i1OpSsnHhnFPE0+i3WV1xy2aKlrFv8Ao41LyMsqT3i11ezw+njyIPv/AIOPS64Xux3az5fmNoVYBGRFSm5KfdZS06Vkodd3UgkciUhOyEcpISSomsJxC8V+sOh2WjE29FrtNsdsSx7I5g7b3Xo8ltSUlTrCWuzZSvY7FKlgBYUNgAKtrK4VrPCnvyMA1VzzCIEh5ySq02mXFfgtuuKKllpqbHf7JJJJ5EFKB3JAHSvzprw5yLheLxd831LzHL8eL0Y2Zi8vRUsvFsczjxjsMNsuN9pydmVoJ3bK0nYoUbpT0S4qVU7qUasY8s5T39F/LKfLUIQg+4Tg3zwaJwrfuj1RjW+551p+/aS9fZt2kRXoaoZchIbKYb0qOVqLbjqi052S1K3KFKG6QDVzaxOL4pjmFWVjHcUs0W122NzdlGjNhCElR3UfjJJJJPUmstVgsbKFlTcIdW3+ehza9eVxJSl0WBSlKmGgUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgK28SHA1phrvJfzKyOrwnPyjycgtjKSmWR3JmsdESU/GdnBsNlgDavMaYu4Y9nl90zvk6yXW546vs37pj00TbZJ7vcOjqhwb7KaXspJBHmr3KICgQQCD0INUM4h/By26CLjqFwz3CBjb47SZcMTuL3Z2eVsOZa47p6wlkb9OrXd0QBvXI1bTY31JuEV3nR8vz3l57EdrKvZ69jCvVkrZ/uilxL1xnbza38maX4LrL7TjWo2oOCZVmbtuv2UPN3C0492Abt85plGy5Ud0qJXJ5dg63sk8qEq8sDdHpRXgTdcuTcMenZDZGZjF9xSZ2jTsCWkPW+ayv7sxJZKkrCCCrnbKgpI8+9e3Og12yu+6M4bes4yixZJfJ1njyJd3sYUIM4rTul5rmAOyklJPkpHMTslI2SM9JuJ1qPd1I8Moey17vz+DR220uhYagrq0q95Srp1Iv1bym8Ye+fPxSfPfaUpXUKaKUpQClKUArjffZjMuSZDqGmmklbji1BKUJA3JJPQADz1yVU7jlz2/ZInHOETTe5OxMq1VKxdpzKtlWjHGjvMkH43UpUykHordwbg7bxr28oafbVLu5lw04Jyk3ySSy38DKMXNqMebIHxTiJ0v1f4voWrue5O0xFmvycP0jtLp32YTzolXdY35WhKdSqO0o+Uvco2PISJi40rnoynReRiutGSIsUPJ5jVutNw5VKXCufVyPJHJ5SUtKRzLUOgTuD0V1rlqH4L++5bq4xm1o1YhY/jNudgM22BEiOIk2+BFQ2hKGnQrlS7shS+fb7ooqO5JrXcl0D1c8INbmsgybVHCbTLwi5TLVGj2i4Iusd2C4GlsPuBh5YbfVssKKiFKCUboQU9fyhrlXs/2t7Q23av8A5RwpU1F1cJqVF7ulGHs7+1tJpSXFmXKWF3KSq0KUqHBlvl5+OS+fBTrzc9adK12nOJDf2QcGkewOVNJUk9o+gHspaCnopuQ2A4lY8lR5+XoKsHXmtZMAyjgHm6b60y76LzY4kRvDtUHobK0tvW4vFFuuPZEk80YKaaUeqilISBso16SRZUabGamw5Db8eQhLrTragpDiFDdKkkdCCCCDX6V7K9pbLtVpyv7GopxTlFtbe1F4ez3Se0op78LWTj16MqE+GSOWlKVYzSKUpQClKUAqF+IdbsufilpS+pttpc26qSk/dFNNoYCT6RtMUr1pTU0VVnV/VBF/1vs2J2qMzKtdvZm2l2WgnmFwWgSHB6C2hEQNnbr2iyOnId+J2irKlp1RZw5LC88818MnU0alKpeQaWVHd+WOvxwY3xX4h+anivxD81ZfxU+j+ynip9H9lfIO7PovfmI8V+Ifmp4r8Q/NWX8VPo/sp4qfR/ZTux35iPFfiH5qxeTTnrJajIhxUSZ0h5qHBYUSEuyXlhtsKI6hAUoKWQCUoStW3k1tfip9H9lQ9hd8ves+Xuar4rb0O6faeXKRaIjzrrpXc7qpRjPzWWo7bqnWmWHHENpVyc5kKVuOUVP0/TKt7UfBHKisv7e/kQr3UoWtPMnhvZff3Ep4xiS8cgORHbzPujr7pfdfl8m/OUpCuRKEpCUkp5uXrsVHrtsB181vicUsyZTTCJFwmyG4FuiqUE+MSnD5KfxUpC3F7dQhtZ81ZODmLIvHsBk+PXPGpxZQ4hNyQkMuLPumUvoKmi4kKaVy825S6nbchxKNM1czDF4uUYPjci7QzJdyB3xmKpQDzIagyVpdWD1Q2HOyHMdgStA3IOxkysK1Oq1Vg8pN4x4LP8EKF9TnFKnLm0vizswrcuLGQ0672zu3M69y8pdcPVSyPNuok7ebfaufxc1lPF9/TTxY+g1wnHieWWKNRRWEYvxc1wyHYkRbDcuU0yqU6GGA4sJLrnKVBCd+9WyVHYddgazXi3rrTpthcybIIs1ySs3GI/IjY3biwpYRNAWyqWtvdJeICiAdwhDalKKgVFSd1vauvPgXy336fngaq12qMeJmSfRc3LlBttstkqcZSyHhBbS/KaR02WlhSkBae8FRWkIJSdlDfaxOnNkhWLG0xYmMzrKpxwuPN3BxhyXIc5UgvPLYWtBUQkDorYBKUgJSAkcmDYHZcFthjW9Dj0yTyrnTpCw5IlOgbcy1hKRsOuyEpShO55UpB2ruXmRDn3S24a7IWhy9JfcdQ2opWYjKU9sQobFO6nGUEghQ7XcbEbj6joej09Lhw7Ob5vHy9PhnwKDq2pz1CfE8qK5IxNvRP1RnKLCvF8KiuqadWpKg7fFp7w2enLEB6FfUvEKA2aAU9JqEJbSEISEpSNgANgB6K/MdhiKw3FjMoaZZQG220JCUoSBsAAO4AVyVZ0lFYRwm8ilKV6eClKUApSlAKV8NUpkaiX/WfNLpMsOoF8tEa3P9vzWu4qR4sguuIjx0N9WeflY7R5S0rJU5yA8vROqrVVGPEydp+n1NRq91SaT8y69KrjZ9T9W7lAuWOOXqAyYEtLCL8YyFzn21NocI7DkTHbWntAkObLSeU7tg1xw3MztchEq2anZQ2sJ2cRJfZmNvH0qS+2vl69ftfJ6OgrOMlNKS6kavRnb1JUp84vD9xZKlQ3ZNab7Y1MR9QrYzLhFAS7ebQwsBlflbqeiFS1pR0QOdtTh3USpCEAqEt265W+8QI90tU5iZDlNpdYkR3A426gjcKSpO4II84rI1HZpSlAKUpQClKUApSlAKUpQClKUApSlAKUrWdSdScJ0hwm56i6jX5qzY9Z0IcmzXG1uBsLWltICG0qWola0pASCSSKA2alVV9tE4Gvht/u9dfq1PbROBr4bf7vXX6tQFqq6l2tNsv1qmWO92+PPt1xjuRJcWQ2HGn2XElK21pPRSVJJBB6EE1WD20Tga+G3+711+rU9tE4Gvht/u9dfq1AQ3rt4Ny74oJWU8LUpD8DdTz+DXWTs2N+qhAlL6tk/9J0lHU7LT0Fap4P8A4m3dH9R3eFPU9ufYrddZixYIV5aVHk2O6LJUu3OIX3MvElbKh5POSkE842sf7aJwNfDb/d66/VqiPiB4nvBa8Stlbt+pGpINyhj/AHbfYFguke525e+4Uy+Iu+wPXkUFIJ6lO+xqNG0pwrOvT2b545P18/M69TW7q4sVp9y+OEd4Z5w/9r54a5xe3LGGj0GpVRMc8JfwRWLH7ZZJnELLu79vhsxXbhMx65mRLWhASXneWKE86yCpWwA3J2ArI+2icDXw2/3euv1apJyC1VKqr7aJwNfDb/d66/Vqe2icDXw2/wB3rr9WoC1VKqr7aJwNfDb/AHeuv1antonA18Nv93rr9WoCzWQXmPjlhuWQSo8qQxbIj0xxqKyXnnEtoKyltCeq1kDYJHUnYCvNXQbiJxaLm+d69ay4rn7OdZ1PLTEJvDrlITZbKwrliwkOJZ5SdgFrKeijyk9Qash7aJwNfDb/AHeuv1antonA18Nv93rr9WqvdqOzdt2t02elXlScKc2uLgaTaTzhtqW2cN+nhk20KzoT44rciTXDifwTU7SbJ9P8Smai41c7/AVBbuStPbo+GkLIDqeXsgfLb52+YHdPPzDcgCqy8E0WJwuZ5dsov2aZfOtN0hGHItFv09vSkSlBQU06pbjKQhaDzbbJV0Wsbjfer6+2icDXw2/3euv1antonA18Nv8Ad66/VqqGm/0m0fStIuNDtq1VUK7zNPu23yX7nSyuSx4PdYe5Inf1J1FVkllev3NIyziw0CzfGbph+T4fqHOtN5iOwpkdzBbps404kpUPuO4Ox6EdQdiK2Dwb2qGSX/Ty+6QZLb8lda06nCFj96u1okQvZKyLKvFAe2Qk9q0EFCk+ZPZ7b9TWX9tE4Gvht/u9dfq1PbROBr4bf7vXX6tXV7Ff090zsGqsdLqVHGpjMZyTWV1SUVh42fiseCMLm7ndY40ti1VKqr7aJwNfDb/d66/Vq5I/hPeByTIbjI1vbSp1YQFOWG5oQCTtuVKjgJHxk7Cr2RS01KUoBXwkAbk7AV9qG9bs+kNc+F2SQUFSP94OJ6KAUAQ0D5twd1H0ED01B1G/pabbyuK3JfFvwRKsrOpfVlQpc38l4mD1l1pkyg/iWCzlst7qZm3Jk7KUO4tsKHd5wXB1G3k9fKFeChrEJdvyO3Wd6W/bj2DDSHVbJC0qQPJJ2O5UE7gFZ5gBzHYGQrJid6yW4ItVgthlyVDnO6+RppP4Ti9jyp36dxJPcDsdpZtXCpp7NgxjqM05k09iUxOaJecjR4j7SkrQWENqB6KSDzLUpR6jcJPKPndG01TtVcq6qvgpJ7c8eiXV+L/0Xmrdad2etna0lxVGt/H1b6eS/wBmuQ2nXorS5AaD/KA8GVc6EuDotIPnAVuPT0rm8V9f5qkW9aV9pe1zsaeh2+NPHPNQtpSwh5LaENrbQFJGxS2lKk7juCh15t9Wk4Jqbb3VREWi0Xcg8yZjEpUNtaN+oLSw4pCxuABzqCtieZPRNb7nsxeUZPu1xLO2PscujrdGaXG8Mwfivr/NQRCTsAT+SuW/27KoEhuBKwPKktK2Uqdb248hpJ/B5UOLdI/9utTvGHYPa7i7lGTpyhQmNpguN3GPdPE0hSgTzoWgNJBKUgqX5IAAG253570i6p/+pTkv/i2b/wDk6T/ZJP34NR1Tx3W3WvDrjhnDS5aGFzu0g3DLbhO7KFDT1S6zHU2hxbr+26SpKOVvf3XOCE6vw+cOHHzwm4QnDMKkaQZnjzc164u26TMnRpq1uBPMlp8tBsb8oPljp/ZUyY9h2mdsd2xBi1wwpXahm0zOyZ5idyoNsqCASfPt1rdbDqVcbdqHjuBRFTLmbi6tueylh11MCN4tIcbkOO8pS2C5GDQClDm7Q9CQKteh3tpQ4bOlTknLm34+fl9DiajSrVc3E5ppcl9iB7Rxf66z7ldMK1V4C86WqFsiULI41dozg90Oi0IbUOnmWr8/StWw7iSwC9okW3P+CvVWy5RObXbFMxMfVMjulAW4ltkuLSEDdJWE9nypPnO259Dq+FKSQSBuO6rY6UHucbjZ59aM5s/Fv8/TS7YJnOK21sNyMYVl0BTEmQypvd6Kp0lSXnm3EurBCypba99h2aqmfsKmvVzSrHtYsMfw/IH5sT7a3MgzYT6mn4M1o8zEhspIBUheyglW6TtsQRVe8XmZZZbu/pnqgw2xllrZ7VExpHJFvkQK5RNjDfp96HWu9tagPcqQT8/7SaF3E3eUF7L5rwfj6P6ls0fVeOKt6r3XJ+PkY3VTI1YVp9ecjbgXOa6y23HZYtqUqlLefdQw12QUQkqC3UnqQOlSxojp1Kxpufl19sibZdL43HSITjqZD0NltJASpzchKlbgrbbUWgUAjmUVrVGuYaeP6g5PiuPXq93GPh0y4ITe4sBSWXlutntYa+3250I8YQ2lYQQoktbFIC+abPsKTrlFu9pyfVHLZtpuEh1UaHFmeJrjR1JSkNeNNgSV7cqjv2o6uKG2wAE3svpsHQ/Up+1lrrsttmvEi61eydXun+3C+O5+811b0209tV7u+WZlaYSMeguXG4MKmNmQ0wjbc9lzc+5KkpA26qUkDqRVJeCDjHzvio4zMhm3hhqDh9sxu4x8ct6Y6AqKHZURQ7R0DmW643GKiCdhyHlAAO9WvCPcFEzhmyqPnGA3G+XPB8rStD65shcl+DJStKiy+7t5baiWy2pe53SQSSATq/g0c/ueF8Q0aOysotktlMu5OJHlNMRlha3OnUgNF5JSO8L9VWuov0tN1FvjHPwys/BZZxIf358D258vHG3zPfSlY+DkFiuikott6gy1KBKUsSEOEgd5GxrIVLTT5EbGBSlK9ApSlAKUrXdRMonYTgeQZfa8dm3+ZZrbInMWuEnmfmONtlSWkDqSVEbdAT6AT0oCFuI/WrFsbYucbJsp9hMOx0NIyKakr5ZMl/ZLcI8iSsoCXEKcCfdFxtJPKHUnRH7dKiXFvKsBs8O6QbzaYcJhMeQhhlhLReWw96CypMgglAKwEo5ULBPLhIt7xfUDAWYea4zCvNuuykyroi/NMqQ9JW6HHFuMtpcbSrt1bhJ22PKB5qkXH3mHoY8Qm21+C0EsxkQW+VDKUjbk3CiOg2GwCdtu6sJwjUjwy5Ei1uqtnVVai8SX+j8Yvj6MbtQhqlLly3nFSJstY2VJkK92sj70dAEpHRKUpSOgFZelKzSxsjTKTnJyk8tiufBr/I0/yqKywVKx7IpqIsuLzeTDmvL5WpLQJ8kOOKDbiADzKWhwcpS4XOCtcz27221Yxc3Zl0aiPsQX58fdSe0Co6e07RCSpJVyKCVdCNunUd9DFLJaulQfC4pbILbFk3fEJqZTzCXXG4F0t77SVFIJSlxx9oqHXoeUAjr0rsxuK7TMMrVe4V+sz5QpbEZ+IiS5JIG5S2YjjySfWod4qPG7oT5TRJnZ3EP3QfwJnpUe6cXzUzNpjmXZTYlYjYFtqbtdhkJSu4vpKv8A6marbZhWwATHRuU7qK1qJCG5CqQRuQpSlAKUpQClKxGJ5bjWdY7By3D71Fu1nuTfaxJsVfM28ncgkH4iCCO8EEGgMvSlKAUpSgFVT8KP7xzUX8a0/wCZxatZVU/Cj+8c1F/GtP8AmcWgO9w08MnDbfuHPSy+Xzh700uNxuGF2SVMmS8TgPPyHnITSluOLU0VLWpRJKiSSSSakn96dwsfyadKv6m279jX64VPev6Q/ISw/QGalOgIq/encLH8mnSr+ptu/Y0/encLH8mnSr+ptu/Y1KtKAir96dwsfyadKv6m279jT96dwsfyadKv6m279jUq0oCKv3p3Cx/Jp0q/qbbv2NP3p3Cx/Jp0q/qbbv2NSrSgIq/encLH8mnSr+ptu/Y1Trwq2hOh+nfCwch0/wBG8Gxm6/ukt7Hj1nx2HCkdmpLvMjtGm0q5TsNxvsdhXo1VHvDD+9APyptv+F6gJV0M4X+Gi76J6fXW68O+mU2bNxa0yJMmRiVvcdfdXEaUta1qaJUpSiSSTuSSTW7/AL07hY/k06Vf1Nt37Gstw+fwCaa/JCzfQmq3+gIq/encLH8mnSr+ptu/Y0/encLH8mnSr+ptu/Y1KtKAir96dwsfyadKv6m279jT96dwsfyadKv6m279jUq0oCKv3p3Cx/Jp0q/qbbv2NP3p3Cx/Jp0q/qbbv2NSrSgIq/encLH8mnSr+ptu/Y1Snwteh2imm/DNaL9p3o/hOLXN3MYUVc2y4/EgvqZVEmKU2XGW0qKCUJJTvsSkHzCvSuqFeGe96hZPlxA+hzaAvrSlKA6N9ujNjslwvcgEtW+K7KWB50oQVH+wVWNiLOvchMh5lSps9ztHEJ7y84rmUB5uqlHv6dasLqXEk3DTjKoENBVIk2Scy0kd5WphYA/ORUUaetpuTtqyRI2Z5GZpbPQqCk8yfN5jsaqXaa1nfVra2X7ZN5+X0WTv6Jcxs4Vq3VJY+f8AOCXcKxKDh9lbt0ZtsyHNnJbyRt2zuwBV167dNgPMAPjNZ+tccyxfczCSPjUvf9FdVzJ7kv3PZI/FT+urVSpwowVOmsJbJHCnOVSTnN5bNtpWlLvd0c91MWPxdh/4rrrmzHPdy3letZrMxN87u+vyXGh7pafymtAUpSvdKJ9Zr5QG13vFMRyeOqFkWOWi6sL6qamxG30K9aVgg18xjDMQwqG5b8NxWz2KK852rjNsgtRW1r225iltIBOwA3PXpWq1+kuuI9w4pPqO1MAkGlaKi5XBv3E14fFzmudF+uqO6WT+MkH9FAbnWqaiaeWfUG1MR5o7C42x7xy1T0D7bDkgEBQPnSoEpWg9FJUQfi/Kcnuae/sVetH6jXIMqnedhg/kP66xlGM4uMllM9TcXlEPXKxP3q2rts96Tbn2n0JkeKrRztvMuBRSFFKhtzIHXbcpIPTepAs+ok63utov7wfircS2p4oAcb5lbBR5QAUgkb9Bsnc79NjhL3gGL3bOGdRolrYtV+7FyLOlQmkIVcmF9n9rk7g9pyllooX0UkJKQeVSgeu9p7DlXidc5eQXp6NPjsxXLaX20xEIbUpXkhKAsFfOoLPPupOyT0AAqtvo99YXH/hKi7vOcP6denU61W+oXNL+9H2+WTftXdM7DrJpjkul+SoBt+SW56C4vl5iypSfIdSPOpCwlY+NIrx4wPgh4peFPX6z5ZdNObzkmNQpC2XbxiLqJSzGdBbLiWfdlSQrmLTjfKrYjr317CqyG7HukAepCf1Vxqvd1V3zF/kAFWirSjXpypz5NYfvOVCbpyU480QyzjN9fesLmItKXOamMuMpdaWw60NwFuFXL/sxSgrUQ42QsAtFPl7VZaKmSiKyiY6hx9KEh1aEFKVL26kAk7Anzbnb01pvsvcv/WOfnr6LzdB3TXP7KgaXpdPS6bp05N5ed/sSry8nezU5rkbtStMTf7sn/myfWhP6q5U5LdE9621etH6q6ZDNupWqpyqePdMsH8hH6a5U5Y8Pdw0H1LIoDZaVr6ctR99BUPU5v+iuROVxPvozw9WxoDqu6YaavX4ZU7p7jS72HhJFyVaWDKDw7nO25OfmH4W+9QLqRw6616i3K5XB/K8XtT893fxy1OSoL6mkqHZtuFlIU6ORCUKLi1q5eYIU2SCLDfuqgf8AQkfmT+uvhyuF97HePr2H6a8ayZwqOm8rHvSf1IWsmnmtSkMQrhjeOMiO0lt6U/kDiy8sDbnQlEQ8wO2/lch691ZRzRnVV5a3Gs7xOIj7xpVhkyT6ivxtv8/LUnryxP8Aw4RPxqX/AKV1nMpnK+5tMo/ISf8AzXpgRbO0b1sTBkG35vhSpaWllgO2KXyLcCTyJVtL3SCdgTudgd9j3VGGOcNHEBk+f41e9Vp2NOWuyXBifIQ8Wp6Hm0c/aR22UMM8hcS7y87i3QnkUAD0UuzDl+urnfKKfxUgforhN1uR75z35FkVi4qXM2060qWeHrtyN4AA6AbUrRxdLkP+ef8A+81t1pcddtzDjyytak7knvPWsjUdulKUApSlAKhnX7iVtXDhcMeumf4beHMFvDpiT8qt48YasskqSGhLYA50srBOzqSrYp5eXcjeUMryKPiOM3TKJVvuM9i1RHZjka3RlSJTqG0lRS00nq4sgdEjqT0FaPphrboTxN4tP/cFk9myy2qbMa62yQz9taSsbFuTEeSFpB6jZaeU7HbegJBtd1tORWiLebLcY1wttyjokRZUZ0ONPsrSClaFpOyklJBBB6g1D/B1pHP0E0KtOkF4yq332fj825do9CdKm223przzSNleUk8i0kg9yioDcDcwVgeK2bW+Te8bsYlY7oPg95nWDH8VtE16MzfJLLyxLlyXUL7RUUPLdQ1GSoN7J3UkjZI3m68KuhsiKk4tg0LCbswk+J3vEkiz3GIvbotD8flKtu/lXzIP3ySOlfGO039cOz/ZnV5aRUhOo4PE5RSxF9Vu05NdcYxy3ex0aOm1a1PvE0vA2riw13zXTWFjmmOitkZvWquospcDHozyCtiAygDxi4yAO5plKknr0JIJCglQM4WCHcrfYrdAvV1VdLhGiMsy5ymktqlPJQAt0oQAlJUoFWyQAN9gAKrrw/66qTE1CsXEFcbQzmOjATFvGUKjNx03Cyvt9vGm8qdy2XEN/bGkeT2jfkjchKcho1xV5Vr9nKY+nOguSRtOGS4H82yJ72MRI5Uq5fE4im1OSApYA5ipASNyrYgJP16zu6N/b07u2lxU5xUotdU1lP3pnPlFxbi+aLE0rT87zxOOKasVmbalX+cgrYZc37KM3vsZD+3UIB7kggrV5I26qTrOkb09GZZdCm5Lc7wExbZJU5Me5gHnDJDhbQNkNJIbR5KQB5IrpxtJujKu9kvnul/JAnf043EbaO7ez8tm/jt9yVqqn4Uf3jmov41p/wAzi1veea0XRyBKyfTuQst4TdjGyW0TIgQ9IYUrsw4knyko5grY9D98rogoVG3hK7zAyHgDze/Wt7tYdwZsslhfpQu5RFD1HY91eVrSrQgqk1s9vR89/c015GVvf0bmpKlTe6w/JrdZXo00/BomfhU96/pD8hLD9AZqU6izhU96/pD8hLD9AZqU6jEwUpSgFK6aLxaXLo5Y0XSIq4tMiQuGH0l9LROwWUb8wST03223ruV4mnyApSlegVR7ww/vQD8qbb/heq8NUe8MP70A/Km2/wCF6gLRcPn8AmmvyQs30Jqt/rQOHz+ATTX5IWb6E1W/0ApSlAKUpQClKUAqhXhnveoWT5cQPoc2r61Qrwz3vULJ8uIH0ObQF9aUpQHwgEEHz1FuM6O3TCrtclWrLpdysUz7ZDts5psuW9xTrq1pbeQEksgLQhCFhRSlsAK795TpWE6UKjTkstbryMlJxTSfM1dvFZqvur7SPVuTXZbxNkfdZi1fipA/XWfpWZiYlvGbYj3QdX+Mv9Vc6LHakd0NJ9ZJ/wDNd+lAdZNtt6fcwmP/AOMV+xEiDuitD+gK5qUBwmHEV3xWT60CuNVrtq/dQWfyIArtUoDHLsFpX/yvL+Koj9NcC8XtyvcreR6lD9IrMUoDAqxNk+4mLHrSDXGcSPmn/wDxf61sVKA10YkfPP8A/i/1r9pxNke6mLPqQBWfpQGFTisAe6efP5QP0VyDGrYO9Lp9a6y1KAxf7nLV/wBJf/ea+HGrWe5Dg9S6ytKAw6sXtx7lvj1KH6q41YpFPuJTo9YBrOUoDXlYl+BO/O3/AK1xKxSSPcS2j6wRWzUoDVFYtcR3OMK/pH9VcZxu6DubbPqWK2+lAad+527f+nH/AHp/XX0Y5dT3soHrWK3ClAaojFrgr3bjKf6RP6K7LeJnvem/kSj9O9bFSgMO3i9uT7tbyz8agP8AwK5hjtpHfHJ9a1frrJUoDGnHrSf+VI/pq/XXfZZbjtIZaGyEDYD4q/dKAUpSgFKUoBUcnh20WRqnH1sh4BBt+bRw6k3e3OOw3JIcBDnjCWVJRJ3B/wCMlfcD3gbSNSgKP8BkFWG6SXrRq6yVryHTjK7vZbuh0EOFapTjzT/X3SHW3ApKu49du6rJVGHEjhR0ozePxb4i0WmobDVr1GhNjybhYgoBM/l878LftAe9TIcRvsAKyOqmoqMc00ym/YbOh3K/QYao1sjMOpdK7o+QzDZUBvsVyHWU7H8KvxJ/V3+nt1Z9roVLNcUL+eY+VSUkpJ+XFJNeTx0LJYXcZW7Uucfoapwn25eX8Suv2ssBp0Y+7LteGQX1jZEyTbmVJmrT6UodWGwobg7K694Eha557k+M5vbbPacsm2iG9aVylIjNR1Fboe5dyXW1+bzDat50L0otGh+kmMaW2dYdbsUBDMiTy7KlyleXIkK/nOOqcWfjVUBcZ1szaTmmOzMUxi73RAtbrTrkKC6+lB7XcAlCTsfXX7h7HaTbabC30yGO7pQUFxeEY4Wc9dig9prmrTsp16eeLK5Zzu/Lcxqs+bivSpj1yemTZrnaSpchQLrygNhvsAAAOgSkBIHcBXYxuBK1Gx7US7Wi7TIlzsEa13WA7GdUCX44mLCCAdlbjcDfuUUnzbGrF1v9+iTHrfdhKhyWFlDsd5tTTjah96pJAIPxGpo0C1MhYNpXqbOe7d643cQLRamGWytx6Y83LSgAAHu6qPpCdhuSAfo19Yu3tnOhvLMceH7l8vlg+b6fqaurtU7jaOJ58f2y+effkkzH8hj5nqTg+TKQhLOqWNzrJfkM7pC32EONvOJ9BKmmAPQkfH1iLiRvMy5+C9z+BNB7Sy3KHA5z3r/3rEd7vNt23L/RqTMQsRwvUzSnCJrqWnMBxu5ZLkaisKEPxkOOLSfSUqLSTt5lA1Euvcab7VvqJe5ieUXy8RbgyCNjyeysNrr/AEmlfk2quar3f6V8HL2cf/aoo/8ARj3YLXojqfro8f7sT4vXhpOX/Xn35LncKnvX9IfkJYfoDNSnUWcKnvX9IfkJYfoDNSnVWLuK1DU7U2x6U2BnI7/bbxNjvykQ0t2uEqU6FqSpQJSCNk7IPX0kDz1t9KxmpOLUHh/E8llrYpBZ+JXCYvFRftR3Mcy5VumYmzbER02dRlpcS+2oqU1zbhGyT5W/ftVudO8/tGpeMM5XZIF0hxX3HGktXKIY7wKFbElB83oNaTaNNsnh8UF81VeZj+wE/FGbSy4HgXTIS82sgo7wNkHr6qlyudp9GvTc3VltxS2xjrz58iPbwqRzxPq+gpSldMkiqPeGH96AflTbf8L1Xhqj3hh/egH5U23/AAvUBaLh8/gE01+SFm+hNVv9aBw+fwCaa/JCzfQmq3+gFa/n+bWbTfCr1nuQtzF2ywwnJ0pMOOp98toG5CG09VK+KtgpQFPvbTOGr+JNSP6qPfrrZtM/CFaD6r53Z9O8ZtWdNXS+PmPFXNxt5lgL5Sry3NyEjZJ6npVm6UApSlAKoV4Z73qFk+XED6HNq+tUK8M971CyfLiB9Dm0BfWlKUApSlAKUpQClKUApSlAKUpQClKUApSlAKh3UHjC4Y9Lb25jWc61YxbrswvkfhCWHnmFehxDfMWz8StjUIcYvFmifCvfDrw83SVc8/lBEa83a3rCImOxisB4OSt9kSVIC0oSgLUk7kgEAGAsN0nwnHceYtb+C2FD4B8YUUJmreXv1W4842lbild5JA6k1yNR1elpzUWuKXhlbHUsNLqXyck8LxwegeCcTHD5qc+iHgWs2H3mW50TEj3Znxg/+0VBf9lSXXk7lvDdoxmLCkTMHgQJBO6ZdsbER5B8xBQAD+UGshpXqtxC8H17hBzKrpqXpEuQ2i5wrkXJN0sscq2W9HI3K0oB5ilPQ7EcqfdVrs9etbuSg/Zk/H7md1otzbR417S8vseqVK0fSXW3SrXTHP3VaUZtbsit6VBDxjLKXY6yN+R5pYDjStuuy0g1vFds5ApSlAKUpQClKUApSsHmmcYdpzjkvLs8ya22CzQU8z824SEstI9A5lHqo9wSOpPQAmgM5Sq8tcaOL3weNac6I6z5zbVfcbnZsOcZhvj8Jt2atgLHxgVJelOqFw1OhT5dw0ozjBlwHUNBjKYceO5J5gTzNdi+6FJG2xJI6kd9Ab3SlKAUpSgFKUoBSlKAUpSgMDn2Px8swTI8VlobWxebTMt7qXGy4kodZUggpAJUNldwBJrzJ4NLdpTl3E8zEjydAHrhEkrkGJZbLf7JeVuxB2yOwizEpjEodZQ6pKfL2aUo7AGvVSuuIEFMsz0wmBKKeQvBsdpy+jm232+KgOxSlKArzxT8MjGrtsXlmHtNR8xhNbJBIQi4tp7mnD3BYHRCz+KemxTHvg+7PMiDUO05JZ3Y023zralyNMYKXGHkB8jdKhulQOx9PdVyK/CWm0KUtDaUqWd1EDqr1+murHV636GVjPeLxh9Vhp49PocOeg271KGpU/Zks5WNpZTWfJ7+/wCZW7VPRnKrNb73ZtMY1yu1/wBVruWL5kc5xBFqt2xUWvJKSG+UFAAT1Hkk79mK0/wi+L2vCfB5ZhiVlb5IVpjWWKzv3qCblFHMr0qJ3JPnJNXEqqfhR/eOai/jWn/M4tRK15Vr01TnyW/r039EsLwJ1tp9G1qyq0+bWPRZy8erbbb3b9CWeFT3r+kPyEsP0BmpTqLOFT3r+kPyEsP0BmpTqKThSlKAUpSgFKUoBVHvDD+9APyptv8Aheq8NUe8MP70A/Km2/4XqAtFw+fwCaa/JCzfQmq3+tA4fP4BNNfkhZvoTVb/AEApSlAKUpQClKUAqhXhnveoWT5cQPoc2r61Qrwz3vULJ8uIH0ObQF9aUpQClKUAryX11mcVejmvWTW7OeJfPMag3q7PXDC7ibkRZpsVSyoReRYLDbrQUkFtewOwAGxBr1orC5fheIZ/Y38ZzjGbZfrTJGzsK4xUSGV+g8qwRuPMe8VrqwdSDjF4fibKU1CSlJZXgUBxvij42sRip9nZ+A5vCbbC/GpVtXDlrT5yrxZfZ77bnyUbdPjrabVxtcTN8hGZA0u05QgLKApd6lqCiPddEtnuO47/ADVKF28Hbw/uLccwubm+C9oSeyxzJZDTCT/NZe7VtPqCQPiqJ8j8F3fXZzkzFuKHJX0L7mMqtTN3SPUQtoD8iRXMdHU0mlVi/cdBVdPe7pyXvPtz4z+KizxVXK4YPpeiOlSU8omT991HYAr5dh185+L01w434THOoU5DeoPD8mVAV7uTi17TLfbT51mO6hCiNu7lUSe70Vpdw8GrxDY+1Kdx/NtMspLzxfLU6JcbO53AcjbkZxQQOnQd25NV+jXDJ+HnL28S1PsWRW7JbPGLlytE8IltTogQVPXG3y0gcyG0NuOKQ4VdN0ghdap1dRtouU8S932ZthTsLiSjHMff9z1z0a100v18xb912l2Ts3aG2vsZTRSpqTDd2+5vsrAW2r4iOveNx1rfq8srrk83QnKbdxIYAtUdcB+OMpiR+jV7sy1pDodT3KcbSorbcPVJB7xXqPElR50VmbEdDjEhtLrax3KSobg/mNTtPvoahR72Kw+TXgyFfWcrGr3cnnqn4o5qUpU4hiqo8e/EXf8AS/F7No7pbL7PUbUtbkG3PoPlWuCkbSZx27ilJIQenXmUN+TarXV5VNZgNfeJjUvXZ4qetdomHC8WKlbpRDik9s6jzbOuHnB/nqFQtRu/0VtKt1XL1fImWFt+ruI0unX0NL05w3HcZzm5YZcJJYjYrDjXNpiQvZV3kOhReushR+7FKk8idyQ2Uk95BE4tOtPtoeZcS424kKQpJ3SpJ7iD5xUQcSU7DbPjsK63eb7G5ElbiLJcWZfi0iKQnmdXz96mwkDmb2IWShO26gRjNKeHzjc1ExpvJrHppAbs0lAetsy63mTj8qcwrql0xI0jkRzDr1QnfffrvvVNp6fW1aPfxbz5/Nr1+pa6l7R02Xcy/PL3HczzR/Mpt8XNxHJ7q7Fkw1MR2Zl9lJFqlEq2kNeUSR1QeXqR2ew8lZ22K3Xq+aZ2pkat6oWCe3yhKX3Yfikl0ge5SlKyHTv5koBPxmu9D4KuOrIgI8q24HYmVdFOXHM7zKIHxJbcUD+UVu+EeCvzmRc27lqfrvBgNEjt2MOsLUWUtPnSJ7wLo9fKa6MdEuq0VCs1heSz8d8fmSBLV7alJzpJ5fm8fDb85EUcGeW3e++ERj/YysEm0WF6xTU5cygFtMhtLThZfkNbANudsqMAD5Y367EqA9bqjTQ/h00h4d7C7YtLMUatxlqDk+e6svzZ7g+/ffXutZ3JO24SNzsBvUl1aaFJUKUaS6LBXK1V1qkqj6sUpStpqFKUoBSlKAx2R3kY7j9zv5tk+4i2w3pfidvYL0qR2aCrs2Wx1W4rbZKfOSBVSbrh2o97tjfE1rjpPcs6zJUhpnAtL42zkDGi+rlZdllX2syeoU/KcBDI6IA5RVx61rOc9suB21uXcUPypcpRbg2+KkKkzHQNylAJAAA6laiEpHVRFAQfZ+ENzUZo5XxWZxfM0yeb9t9jLTe51psdk37mYTEV1tSuXuLzpUtewJ27qsHjVggYpj1sxi1KlKh2mI1CjqlSXJDxbbQEpK3XCVuK2A3Uokk9SSag+4XTULM30zMlySVZIaXCtq0WKS5HSlO2w7WUnlfdV1J8ktoPTdB23PRfxRSwpyFmOZwZRIUmSxk05SwodxKXHVNr9S0qHxUNndsshSq9xtccq0qkMq1emMXjD3XAyrKGI3YyLUT0Sq4Mo+1qZJ2BktBAQSOdpKOZxNgGH2JTDcmM8h5l5AcbcbUFJWkjcEEdCCPPQwaa2ZyVomK6l+y+oGV6e3qCLfPsUlkweboJ0N1hDiHU7nr5Xap6bAlpYHVC9t7qEeJDCUuQmNTMVlsxs0tDBiw4rhPZXxntEueJOhPlAhY52nx1YWVL35C4FAibqVGejGoMnNnsltyX1XCDj01qEzcj1Drpb5nWOb78tbo3X5+0AO6kqJkygaw8ClKrprNxm2TTnVBGh+n+mGWan583CTcp9px5pHLbYygClch5Z2QSFIIGx6LTuRzJBHhYulVfyfjptFgxrBRG0Tz+VnuoMydAteDPxG4lybdhqKZCnlOK5ENjvSvrzJO+wAVyzTpBqJeNTsPGRZBptkeC3JqU7Dk2e+tIS+hbZHloUglLjSgRyrHQ9fRQG70pSgFKUoBSlKAVVPwo/vHNRfxrT/mcWrWVVPwo/vHNRfxrT/mcWgJZ4VPev6Q/ISw/QGalOvN3Q/wsfC/p3ovgWAX6150u541jNrtE1Ue0sLaL8eK204UKMgEp5kHYkDceYVu3tyvCT/FGoXzNH+s0BeylUT9uV4Sf4o1C+Zo/1mntyvCT/FGoXzNH+s0BeylUT9uV4Sf4o1C+Zo/1mntyvCT/ABRqF8zR/rNAXspVE/bleEn+KNQvmaP9Zp7crwk/xRqF8zR/rNAXsqj3hh/egH5U23/C9XU9uV4Sf4o1C+Zo/wBZqtnH/wCEP0F4m9AvsZ6cwMsZu/s3DuHNc7e0yz2TSXArykvLO/ljYbemgPUDh8/gE01+SFm+hNVv9edWlXhb+FrC9LsPw672vPFT7FYLfbJRZtLCmy8xHQ2vlJkAlPMk7HYdPNW0+3K8JP8AFGoXzNH+s0BeylUT9uV4Sf4o1C+Zo/1mntyvCT/FGoXzNH+s0BeylUT9uV4Sf4o1C+Zo/wBZp7crwk/xRqF8zR/rNAXspVE/bleEn+KNQvmaP9Zp7crwk/xRqF8zR/rNAXsqhXhnveoWT5cQPoc2uz7crwk/xRqF8zR/rNVf8If4QPQrik0Mt2nWmsHKmbrEySLdnFXS3tMM9g3HktqAUh1Z5uZ5Gw27t+tAexlKUoBSlKAUpUAa1ccGgui093GJF+k5Xl6d0oxrGWDcJ3OPvXOTyGf/AHFJPoBrxtJZZ6k28In+lebGZcfnGXf1Ov4NoNjOD2cglMq/yXLjLSjzKKGlNpSrb71SSAem/pjp/XXi7zWW0zmnEhc7XCfdCHUYza4cAIRuObleKCvpudz5tt+oNQqmp2tP/NP03+hMhp9zN/sa9dvqeqeU5liOD2ty+ZnlFqsVvaG65VxmNxmh/SWQK8t+NzXbDuKnUXEbbozAXPsuJtXWFcsrlMqjxpbU5lMdyPEKgFuAJ5/tmwSD5Q3SCThr/pxoXizyM41Sv71+mqIWi45VeHJ7rqvNyNqPKs+gJQa1HUDO7zqBcrTiGnWATm3JclluEpEPnvExokBRiW4bKCAnr2kjkRy+auZLWJXn9q2pvD2y/wCF1/NjoR0pWv8AcuJrK3wv5fT83MjqFkly1UsTuiOJwyi53l6Jah2B3LZfWG47Kkq6guLG59DSHVHYJBV7EWC1IsVittkacK0W+IzFSo/fBtASD/ZXmToi3w68GGQDUTXO/C46tvR1M23C7As3q420L5+dclbP2tcxwOEFayhCEkobG25O737j64sM4cWjSPhysmJW9ZKW7jmdxW48R+F4s0UKQfiPOKl2dOhpVHhqSUc77si3U62pVc04t422PQ6leZkvPeNrKx2mTcTkawpX7qJjWMxkJR8SXnQV/lIrV8kwTVW+xXXrxxKar3l4NLPi67/4u24vbdIAbSkJHeNvjHUbdcJ6/YxeFLPomZx0S8ksuOPej0h13zJWnmimd5y252btix24Tmleh1uOtSP/AMgK8meHeBlVs0qtePWSUnxqUwi4OKWoIDReV2q+YnytzzBPkpJHXuHKVRrrVp7boOAXO4KuuVz7q2mOw2Lnd33i4868hvZTaln8Ikd3cfdA+TYmz2WLpgqwWq3MSXm1sR4ThXylKSQEEjuIJKUdANvcjuAFc3V7+ndUIqn1b5+SX3OhpdlO2rSc/BcvP/RjsFxCdq9xIYbgeSIh3Cxt3WNarkpb6i4Vw2XLpJbS2UkKbcKYSFKKgftQGx3O3rilKUJCEJCUpGwAGwArxUw+53xevlnvmlGYz8WyxByG8zLiu3GUylCnm4rSUsyB2KwptKklSOvd13HS0tr4yeMDTx5IzHTnENULQ1tzybI8u0XMp86i04VtLPoSgDep2nXlra0o0KklGT3328vToQ7+1uLio60I8UeW2/n69T0HpVfdF+ObQHWe5NYoxf5OKZgrZK8ayaOYE0L/AAW+f7W959uRRO3eBVgq7iaayjjNNPDFKUr08FKUoBSlKAUpSgKi2jXTIsazbCM8zXLpxs+f3+42SXb3VJTCtMbs31wyhsDdKmzGbQ66pXUvOqOyQhKNkmZG0pN11Wz6UiFzNrU0HiSm228K3bYSAPdq8lS+UErcVsNwlAGi5jov4lqWsXC7h5nE5l3uNtguRQtmVFvDwkIKt1eStl1uSwOigWwD0KiBukyxt5Bb1229tB6I8n/6dYB5yNlIUfRsoAjz7gGsW9zfCK5kZ4XkGdWsacZzfcqm3eHqcytqXBUhPi8GTIjLnQ3GBygobbaYdjqSSedTjZJJBJnFhLiUDtFbn46jWNjb0Z7T3A4CyLfiT8ifIfO26whL6GI6U+ZI8ZG6t+gjpTseclMkvvdk2VDqdwPy1lkzRjcuEdWMXQSW0uNmI6FIUNwrdJGxHnB7j8VRnpDxGQNGcGiYfcsjxKRaYgcXbo94vLtulWyOVbiLuGXkvNIJUG1boKW+RBB5eY7Tq3cp/sHCw/HEtP5Nl8v2GskVxXKHX1NrW4tZ8zbTKHXVq7wlshO6ilJmzSXRDA9IcbgWiw2K3uXNlkCfeVQ20zLhIPVx51wDmJUrc7b7JGyRsEgDzfJhUa5Mhtniry3UBzxTS3C7/eF8hKlWmwSFsLUR5PZ3KUluIP6Y69O6u9bdFNZtU5SZ+qt+GIWR5A7a2W+YZd5lNnYlp+WPtUUbjqiP2gIO6VtkVZelemri8DGY3jVhw+xQ8axi1R7ba7e0Go0ZhPKhCe/8pJJJJ3JJJJJJNZOlKGIrzzwDU3CeGDwguvCdfL01jMfUWNbbpjV8uSVIiyI7SCFtJd22HVYT12G8cjfflB9DKwuT4ThmbR2omZYjZb8wwrnaaucBqUhtXpSHEkA/GKApnxM5fwWcTVrwzI5/EcvT/IrYu4TcJzKKt6DyqZeDMjkcdShDqA62noFpWeXdCtid978HNrDqprHotebjqheRkZsOSzLJZsnEXxf2dgspRyyeXYc3lFQ59gTtsrdaVE2JvGnOnuRWiJYMgwTHbna4G3isKZa2Ho8fbu7NtaSlH5AKzUC3wLVCZttrhR4cSMgNsx47aW220DuSlKQAAPQKA7FKUoBSlKAUpUPI4lLKtce9fuEycYVKvacfay4iJ4gqUqV4ohfZdv4yGFSdmg8WuUkhX3MhdATDVU/Cj+8c1F/GtP8AmkWrWVVPwo/vHNRfxrT/AJnFoDa+GHSDSa5cNmlFxuOl+IypcrCbG8++/ZIy3HXFQWSpalFG6lEkkk9STUm/YT0Z+CPC/mCJ+zrAcKnvX9IfkJYfoDNSnQGl/YT0Z+CPC/mCJ+zp9hPRn4I8L+YIn7Ot0pQGl/YT0Z+CPC/mCJ+zp9hPRn4I8L+YIn7Ot0pQGl/YT0Z+CPC/mCJ+zp9hPRn4I8L+YIn7Ot0pQGl/YT0Z+CPC/mCJ+zql/hadN9O8V4TjdMYwLHLRN/dNbm/GYFqYju8hS9unnQkHY7Dpv5q9BKo94Yf3oB+VNt/wvUBOWg2jukc7Q3TubN0sxCRIkYnaHXnnbHFWtxaobRUpSijckkkknvre/sJ6M/BHhfzBE/Z11OHz+ATTX5IWb6E1W/0Bpf2E9Gfgjwv5gifs6fYT0Z+CPC/mCJ+zrdKUBpf2E9Gfgjwv5gifs6fYT0Z+CPC/mCJ+zrdKUBpf2E9Gfgjwv5gifs6fYT0Z+CPC/mCJ+zrdKUBpf2E9Gfgjwv5gifs6o74X/TrT7E+F2zXPFsEx6zTF5nBZVIt9rYjuFsxJhKCpCQSklIO3d0Hor0WqhXhnveoWT5cQPoc2gL60pSgFde4XCDaoEi6XOWzFhxGlvyH3lhDbTaQSpalHoAACSTXYrz28JjrNf8jvln4SsFvC7cLrD9ncwmtvFsi2gqCIe4O/2zlUpSfOkI8xUDhUnGlFzlyRnCEqklCPNmm8QfG3nPEVfZ+mmgl+dw/TmNIMO4Zg2SideiFcq0Qv+kz6XPdEDfce4PzBdN8O05t3sfi1nbjqX1kSl/bJMlZ6qW66fKWSep3O2/cBUSwLJDs8CK1aEQ4kNpsIjtNI2CEDlSny0kFXXywB17wdwNk4rUbVXKbtMt+mNuXLUu4pJnG1JPsg812iG2o7Ck+SHH3FpaCgPJ8o+qmX1W41aoqcHiPh+c3+IttnSoaZTc5rMvH85Eg6la34pYrTerXZWZeQ3CLGebkotyQpiIrk/wCO+ohtvv7uYq8229RXieLcRmtltWnSfHblK8ZQhUdyy28rhtK7kl25SuzaHKQNw2hwjbvFXq4aPB0YZhtqhZDrlGYyW5h1ybCxVSy7YrCXHFOdklpXSW4kq2LroI6DlAABNzokOJb4zcKBFZjR2UhLbTSAhCEjuASOgFdqx0SlbR9rd/nu/OZyLzWKleXs7L89/wCcjzj018FrlN+bRL1lzODj7LhBeYsS13K8OpCgtIVcpQ7NkhYBIYjgHqNxUban6tYXZMivOgvCPBTiOLwXHIOYaisLMq8Xl1CCXY0WU4SvbdJSp3mPXYpCUgBdwvCQ64X7Rrh0lQMJkvM5ZnU5rGrQ4wvldZLoKnnUEdQQ0lSQR7lTiT5q8+8GxeDhWNW+xMeMlEFBD/IPKfd2JJIHUbr5jv37EjfyTW6/rxsafDSXtM1WVGV7U4qr9lEm6R2LSfHY64eEQkIuBUoypUw9pNkuffKW6dyokncgHbr3CpKqvcZbimkusyJEVTYKiVbpIQnvCdvSFJ2Pd0A22G46E7XrMmXZ7GKJiuQbRKYiXW83pe0C3qdeQ2lPMNlOuJ5wVJ3GyUqUdyfJptWyq3NRyi8vz+5aqd1ToQUWsLy+xZOsTkGTWvHGOec+A4pKlIaGxUQATuR5k9Ntz061XtEDVXVO7Qm8MvOrWo8ZUlYnqw2zG1WtLfZq5UIk7bH7Z2e6lOEcvN59q1DVTTBzCM6gYhetF4tzyq7w1x27FJzt65XCK8ogplzUsEhlCU7DkUtJIJPm3EqloU2uOo3jnyx/3OP0I9TWIJ8EMZ9c/wDan9Ta1NR9UdW8axFtpL7NrfTk98cSk7MdmN2WR0A+2OqSog9dk7jbrvYrI2bYLa9dLmyhabY2uWlSvvORPMfybDqK0Th/0Wh6MYd7GuvNS73cFB+5zEA7LWBsltJPXkQCQN+/cnz7VlNbb5AsundwYuEvxZF5Ui0dqAd0JfPK6vp18hntXCfQg1DuKka9xGjReYrZPx8X+dCVQhKjRlVq7Se78vBfnUinh5uEm8an3qXfUx2pFkx63WWJ2IUUK7VS5bwKlf8AECnE7j4/z2RqjWnOfexUmRGt0ASTk16kz4sSSp+LcfF+QpZUzJkKLcpPI0nlQVJVzq2BINW6xXUfE8ix5V3iT3IwhK8WmRZyS3LiPDp2TzZ8pK+nx794JHWt+r2tSFXjxtsvz1eTVplxCdPhzvv+e5Gnas2zBs5kv2LKsdRJRBQlSJ7Cw3OjuDmPMw4NyFII35VbenYgjffuF3jay7Q7MIGiHEHkErIMKnvoi4/mk4csmApQHJHmnchTe5ADhO6eu+46IjTILoLhcbjNtqUtxZICkIWyoEK6cw6EBKioFR2/CG5O/k65lFjh5bYp1gvLLbrUxvkWjsT5J2BCkbDcKHknfzH73cVO0y8nY4j/AI9V/KIeoWsLzL/y6M9oELQ4hLjagpKgClQO4IPnFfqqY+DL11u+c6aXjRHNZypeT6USUWsSVrKlS7WoHxR1RJPVISpv8VCOpJNXOq6RkpJSXJlRknF4YpSlengpSlAKUpQFb9Zsrn23Xu3YhDwy4SWb3jTUqTfGz/s8MR35JQ24OUjyisjcqSd1J5QvyyjVp2od5teqWK6fwdPLvdId+jypE+/NIcES1BptamwtQbUjdaklIC1oPlJ5ecnlr7xTaiXjAtYrPkF+wyTHwi2WJyDcMlbK1tNuTXkKZD3khCEpdh9mCVkgygVBCSCetH1Dn5q1CxvR5pnJMlnRUPtI7T/Ybchfc/Peb3DSB1PID2jm3K2n3Sk49TdB+ybflWU4Vgsb90WXXu3WhtX2ht6S8EKeUSNmm0+6cWTy7IQConYAGowz7LLvk+n8u+2nLnsPlOP9hjVpLSFXW8zAeplRllLsWIlJKlA8j3L5SuQJDbsvYtwvpwCx3TO0T0Z1rM/b3kRcov23LEkKSeVuEyoLbgsJUeiG0kn79S6hHTjhr1cu8hrHLlZGMbhJswkG4zIrq3Eqecd7WH2qXw4HFLS04FJ9wlIKgFFKE+tHneZNu4RsIvOcaq5Hrrk0m4zrRYGF4ziDs1YUl11S+a5S2htsQVIZjpWDseyfA6HrcOsHg+PHEsLsOLKEQKtFtjQleKMBlgqbbSkltsbBCSQSEjuFZyvVsa28vIpSlDwUpSgFKUoBSlKAUrD5JluP4nHaevc/slyV9lFjttqekSnPwGWUAuOq267JSSB1PTrWqq1IyeHc2kXbBAxEkNKfbhM3Rp+9IYSQFPrhJGykAnqGnXFjpslRVygCQqVjbBkdjyi3puuP3NidGKi2VtK6oWPdIWk9ULSehQoBQPQgGslQHxSgkFSiAANyT5qq9hum+a5Pg9p0YhX3Gl4BDn2/I03AqkovarSqf7IRIbkRTQaacJaDSnu2J5ElXZJUobS1rtmON4/g8yyXu/QoKr+WrU4HZojutxJDiWpMhB35gWmFPOAjzoArIaSdhcsfl5nHVzM5VOcuUQhPKnxEJSzDKB3hKozLLmx++cV6aA3iqp+FH945qL+Naf8AM4tWsqqfhR/eOai/jWn/ADOLQEs8KnvX9IfkJYfoDNSnUWcKnvX9IfkJYfoDNSnQClKUApSlAKUpQCqPeGH96AflTbf8L1Xhqj3hh/egH5U23/C9QFouHz+ATTX5IWb6E1W/1oHD5/AJpr8kLN9Carf6AUpSgFKw1rzDGL1f7vi9qvUaTdbD2IuURtW643ap5m+YfzgCRWZrxSUt0zxNPkKUpXp6KoV4Z73qFk+XED6HNq+tUK8M971CyfLiB9Dm0BfWlKUAryKyyM5qRxjax5NIHO7DyU2VL6juWI0NhtsBKT03JHnBHUnzEH11ryritKxri04gsKks8jy8kbvzRPepqW0Fjb4uqfz1ydblKFjOUfL6nT0eMZ3kVLz+huTFot0e3i1ojJMYNlspX5RUCOvMT1JO53J6kk1GmgenzSvCTYbHurfb2tmxyLzAbc6jtWGHktn4yhaioeoGpXqPM5zV3RjU7T3X63QH7g9hs19q6wYzXO/ItEpvs5JSO7mQPKTvt98fMaqWhXHc3seLlLKLPrNDvbR8PNbnqzStW001OwTWDDrfnunOSRL3ZLk3zsyY6t9j50LSfKQtJ6KQoAg94raa+hlFPObwpEucvWXQW2Ps9rbUvXaUELBKFPhtvv269AE/nqLcFxZi+ylTrlblogs+UyObZL/mSSU+SpPRShynbmIPdy1bvwjGhmY6r6SWjNNMLabhmmm13Rf7bDSjnXNY25ZEdKfviU8iuX77sykdVCq1aYalYnqdjTN7xeQ2kpSEzIJ2S9Ce++bcR3gg79dtjtuKqnaRVYcNWK2xjPh/ss2gSpzUqcnvnOPE5G9M8WSXA7HeebXtyoW6RyEb9QRsTvzHffffz1GOEcP+T6Z39q72iTheZRbatx+0wMwtUh9iI+talqeLbLyG3XTzAc7qFkBKeXl2qeaVWrfUbm1fFTl8d/qd6vY0LhYnH+CL9TNc+OfJIpxyXqpYcIsr20d1OMWZMXkjbALWJDilONbb7DlWnp13HdUfYHhMbAVKnW2K8/dlSSufdJbhVKccKEFbrqnNykeVzHnHnOwG5Jm7OZ8Buzu2t3kcmTU9nGa25lJWSEpc2BBASpSTuCDvsAdyK0rF8Mevz7U1+Kpm1pIKUl7dDqeUe5AA5zv99skDvHNsOXpz1Svd0v772+pAhptG2qf2Vv8AQ3S15vAltMInMvxpCj2b3M0UobWB1KgTzITv0HMAeo+MiK89gZJrtrXiuiWETWy3dJj1llqQ2pammSlKrlKCgdkdjH+0gkHdT7yRsRXZ1FymCLvE0k0NxdWU6lzFdlbrbACnUQFd5kSOvICgEqBX7k7KOwG9XZ4GuC1nhosMnLc5mRrxqNfmA1NlMqUti3Rubn8UjqV1O6yVuL6c6+vm3M3RdL45q6ksR6J9fP0ImrajwQdvF5fXHQm7LtCdKM904Y0ozDC7fdcbiQm4EaM+3uuO22gIQppweU2tISNlpIO4768htXNNLvofrFlWN3bJLhcZuE3G0wocp/kAuWOzSRGDqkpClyGlqCS4on3Gw6b17b145cTuUY3r5xqZVbMPIujltulltfjbBJajRLYFOTXlHflO8hxLSe8nkVt0PWxaio9w5y6fTr8jhWDl3yjHr9enzM7YcGyidZHHzIYYVIZPZ9tzlw+UroT96dgNlDfpy7e5Brtw8EyGS+uDN5YqE8qXJSPK507A7o6956b7jvCieiusn0r507ubbL2raCwR3wm3RenHhIIuJQ5vPHzXEH48pA3AW602t9ClDcjmAikDbuCvXXq1XjzwlzGc28IHiuXRo8gqm3O9vxJCkgtLtMO1vxG3G1A9y5Bd3H4tew1fRtO4lawU+aRQr/hdzNx5NilKVNIgpSlAKUpQHXuEC33aDItd1hR5kOW2pmRHkNpcadbUNlIWlW4Ukg7EEbHeuhi+IYlg9qTYsLxi0WC2oUVph2uE3FYCj3kIbATufOdqxupeWzcCxGXmMazG5x7StqRcmkKIdbt4Wnxl5tIBK1NNc7gR3q5CkdSKzy5hk2pVwsxZmF2OXonK4Oze3TujZY38k9Oo36HegO1uD3Gla9p1lrOf6f41nTEbxdvIrRDuqWebm7Lt2Uucm/Tcjm27h3VitN89VkukFj1Kyow4Jm2Vu6z1NApYYT2fO4RzEkJABPUnoKA3elavpjfr/lWn9hyjJ7e1AuN5hIuDkNtCk+LIe+2NsqCiT2iG1ISs77FaVEAAgDoafZpc89u2R3aJHipxOFLFrs0kJV2099hS0zJAVvy9h2uzLeydyWHF7qStGwG70pSgFKUoBSlKAVqmpl+u+PYw1JsbzLEybdrVakPOtdoGUy5zEZbgTuAVJS8pSd+nMBuCNwdrrRdY/wD7Xtfyrxr/ADiJQHewXGccgsryKD43cLpN52pV2uSg7Nf5F8qklWwS23zI3DTYQ0D7lCayuS4nj+XwRb8gtyZKG1Fxh1K1NPxnOUp7Rl5BDjLgBIC0KSob9CK6uBf/AGwx/wDqJX0hythoCLHdMcoi5M3cYV+UtbxUkX5kojXSOgJUUtymwgx7i10S2ntEJW2k8wKlkuD7gluybVDCLNlmaZrcGmb7a2ZDtqsW9sjoLgC9w8hSpYUNwN0vpSfwetSlWiaEfwL4R/8AsUL/APqTQGxWHDcVxcPHH8fgwXJKUpkPNMjtpHKNkl1w+W6R16rJPXvrD4ZaLjhk+RhqIy3McbR4xZJAUNobe4CoKh38qCQWiOnZnk6dkCvcagLi3wh7UVrSzEot0k2uVKzkvQ7jH354MxmyXV2NJG3f2b7bS9j0PLsehoCe+dG+3MN99u/z1VTwoxB4HNRgCDsq0g/F/vOLWi4xkOoGdznb4xa42O5cvXiHGmMXKE8/Eizo+Gsx5WyErbU6z2rTvZrCwFJKFb7HatU4uo+TR+DPikRmEu3yrqc/tJfft8RyNGc/2fH+UttuOOKSNtgd1q3IJ6dwAt/wqe9f0h+Qlh+gM1KdRZwqe9f0h+Qlh+gM1KdAKw2ZZA/ieI3rKI1guV8etFvkTm7ZbWu1lzVNtlYZZR984sjlSPOSKzNKAp9+/wDM7/kH8Qn9WlVn8A41MxzbNbLiM7g21wx2Pd5jcRy7XSwFqHCSs7dq8s7cqE95PmHp7qtFSgFKUoBVHvDD+9APyptv+F6rw1R7ww/vQD8qbb/heoC0XD5/AJpr8kLN9Carf60Dh8/gE01+SFm+hNVv9AK0XVPUe+adxbfIsmmWR5iua44hxuzNBZjBIBCl79wO+w9RreqVhUjKUWovD8f9nkk2sJ4KQaY625ZZ9eNV8li6EZrcZV7VajItcZgGTbezYUlPbjzc4O6fiFXFwzIJmVYxb8guGOXCwyJrZW5brgkJkRzzEcqwO49N/URWlYBpVeMR1i1I1Gm3GG9CzM24xGGubtWfF2ShfabgDqT02J6d+1SfUHT7etRi+9lneW2F1k3nbx5+8j29OcE+J9X9RSlK6JJFUK8M971CyfLiB9Dm1fWqFeGe96hZPlxA+hzaAvrSlKAV5m+EJsMjRPilwziEaT2OOZ3bRit9kDfZiU2ftbyvU32RHpDKxXplUX8SmguLcSej190qyltKRPaL1vl7EqgzkA9jITsR1So9Rv5SSpJ6E1qrUY16cqU+TWDZRqyo1FUjzR55TsNyhfZAz3ZqQ05yvJlFWxJHKrZRB7iegKubolSu9Stfu8K7tNpcyJ6WG2XlhJlOLQ2SW+7mKQAgk9FJO4JICU78iY+wHU/VfQq+XfQLWqwrFyxEqYblqStxxqKnYNuciUkvR9vKS6nfZOwVsNiJbx/J2tQo71lukyFNhXdt4sOR0haUABJACgOU7b77Hc7cu569aHXt7iyqONVLC6rwLrQr0buClTe76Ea4mvU3SjIE5lwz549Zr5KW29MtSRzWy8ggb9pHCS2FDmHMrZPLzjYkjtFW8058KTYLG+zi/FhpxdtN70rcC6RI65tpk9ehSW+ZxP5O0HT3Q7qr3bzPxC/sGZ2TfYOMsKbSkoR2J3TzJSeZQHMXSkJPKDufOQJVuFutd9t7kC6wI06FJTs4xIaDja0n0pUNjU2nrtWzajJcUPmQ6mjUrrMovhl8i9mnmtOkmrUQzdNNRsfyRoDdQt89t1aPxkA8yfygVB2uvg9tHtW8gf1Aw+4XTTfN5C1OvXvHHOyEpw+d9jfkXuepI5VK67k1SvIeErRO9yDcLfj0nHp/MFIlWWY5GU2f5qNy2n8iRXdgad8QuKNtMYFxjak26NH6MRZ0pUxpA8w5VLCdv6NdWn2hsayxUyvVZ+mTmz0O8pPMMP0f3wSNdeDfj4wVShi+o2nuokFvo2m5tOW+WoebcJRyb+tw1hHNJfCONHkHD5hD2336MlYAP5DIBrqIzfwhdsY7K28WltmJQPJEzEoBWfiKiwo/nNai7xKeEPbdWy5xG4+hTaSpe2M24bbKSlQ3VG23BUd+u2yFHcAb1gv+FuHlKPwa+xn/AOb0Vht/FM2eNwz+EVyG5Ldf0y04sDjx28buF3RIS2Ntttm1uHbb+aak3GPBza85isHX3iccYgKA7W1YZDEULG/VJkrSlRSRuNuT8tV/c1t468nns2y88W4gMPJWp12DYIUctJSvl3JQ02R3pPugQDsdj0rIztJtbctjGLn/ABf6nXqI790jMXFxhpYP83tFJ/sr2VzpFo08LPTZv+DFW+qXKay8eqRfvDsM4ReCrHFsW6ZiODNvpCZVwulxbE6Ye/7Y86ouubnry77A9wFRDqh4WThvxVarPpfGvupd+UstNRrPDWzG5+4cz7qQSCe4toXVWLHwgaG2l5My42CbfpgVzqkXWe66pavSpKSlCvypNSZbcFwuyWtdns2LWu3QlnmUzEioZTzeZXkgdR6e+tNbtPQjtSg367fc3UuztaW9SSXpuQ3qnxWcafEo85ZZVwGlOIPJUl6Dau0bkyGwCSFugF5Xk9FbFCBv1HXasjobaML0jtS8Xat5ZmOBDsy5uN8z8palKG7pTvsgHyQN+hCtx0UquteEKRPMSG9zRoRebbBZWEujZCtyrdRUByjbboFISVDYJFbVbsStsVuNKya9M29K2kvGMlwoW4AT0cKienKrl6ddiRv1Vzc7UL+d5DhqvCfRE+ysoWs+Kmt11ZJbMhiS2l2O8hxCwFJUhQIIPnqK+IPPbjYrBHwPD1BzLsxKrfbWwdiw2ro7JUfvUoST5R8/XzGtd1WvlnwHxFzFb4zKuNzmpft9rivFcyVIUoIACuY8rZUAkqO3UqG5KjWvaYaGarcSGsPsBYspceu8hhUTPMjhIS5b8ftjm3LbYqzuFP8AKlYJSdyVK338o1H03TO+qxnzXRNeHj5ePjyN9/qCo05Q5Pq15+Hn4fEn/wAFNpk9kmaZDry9aHItgxuyxsGxjtFhXMtAS5PdSR0Vu715x0JdWPSK9Mq1rTbTrEtJcEsunGC2tFvsdgiJiQ2E9TyjqVqP3y1KKlKUeqlKJPU1stX2nBU4qK6FJnNzk5MUpSszEUpSgFKUoD4pKVpKFpCkqGxBG4IqPNI7JesFVetMpUB/2AsLzb2MzSN2zbH+dSIe/wCHFWhxoJ/6Pix3JKqkSlAaVo9jN3wzAo2K3lhDSrZOuUeGlCwseICa/wCJncdx8WLO48x3HmrUZOlWUscPdg0VSqLKdcj22xXx9l3la9je0QJ5QVAKVzxw62npvu4DtsDUx0oDS9VYeZXbGmcUwbtYci/ykW2ZdmXEoVaIKkqU/JRudy7yILbXKDs662ojlSqtlsNitGMWSBjdggNQbZa4zUOHGaGyGWW0hKEJHoCQBXfpQClKUApSlAKUpQCtezvFn8vx8WqHcUQZTE+Dcoz7jBebS9FlNSGwtAUkqQVNAKAUk7E7EHrWw0oDR7Tm7Ngej47nFkbxiU852caQhztLZLdWrolmRskJWpSujbqULUSeULA5qy2UZi1jq24EO0T7xdZLanGIUNsbco33W68shplHT3S1DfYhIUryazkyFDuMR633CIzKiyW1NPMPNhbbiFDZSVJPQgg7EGtORo/iRUmHKcucyxtHnZsEqat22tK6f8FXu0DYcrSyppGwKEII3oDU7Pfs+za7t3exT2bghlR7B6K441j0PcLSVB0cr12cCVnonkj7pH3NxIVWcxjHtSdNcbt2NWsWTKrZaITcVhJ7S2zeVsAAbkutOqIHn7Iek+epGACQEpAAHQAeavtARVqtrXO04xKzZC7ijdvevF8Zsji8kuCIEG2FbbixIlyWQ+lDRLaW0qG4LjzSSU7kjc8Vg3WQo5VfpafHrnDipVBiTTJgwwlBKksLKEdrzLWs9qUJUpPINgEgVpWV5NlOZXAYZiMy3Wl2XcJcVtd0tAukebBjslEt5+MXmFpZEhaGEqSslSyk8pbVzV0bNpPk2m+GYjgOI3OfLtWPRkxW5sGSmJOjPKWeZ7sHCYsiMA4f9nWB2SWx2faK5QkCZNh6Kqp4Uf3jmov41p/zOLW33jiGzi1Y3kupkPA7VPwDCrlMtl2nruzjNzkogPqj3CZHihhTfI062/yoU8FLSyogp3SDqHhR/eOai/jWn/M4tASzwqe9f0h+Qlh+gM1KdRZwqe9f0h+Qlh+gM1KdAKUpQClKUApSlAKo94Yf3oB+VNt/wvVeGqPeGH96AflTbf8AC9QFouHz+ATTX5IWb6E1W/1oHD5/AJpr8kLN9Carf6AUpSgFKUoBSlKAVQrwz3vULJ8uIH0ObV9aoV4Z73qFk+XED6HNoC+tKUoBSlKAgrig4RtP+Jq1Q5V0lSsezCx7qsWT27YS4St9+RX/AFGie9B9JIIJJry1z/TPUfhKyOS3rJZrzj8l6Y7KgZ1jzZmWe7ulS1JEmMryG1nmKQlPZq2J6co3Pt/XSvNls+RWuTZMgtUS5W6a2WZMSWyl5l5B70rQoEKB9BFaK9tTuI8M/wA/PB7G+hcToSzA8jce1mytjFo2WZhiduv1oTGQ/JveJy0S2oo5QpYdjrV2qCj77lK9tiem1b5jGrOnOYOiLYstgrmFXIYT6jGlhXoLDoS4P+2pi1c8FDorlcu43zR/Jb5plc7g24h6PbHC7bnQtOxQphRBSg9d0pVt16AVAeqXBRxsMRYsLIsR0+1htVvkNyA5FeRCnvIRuShXbBATv03CCd9tuu9Vq57O5eafy2+Tyn8UWC313CxU+e/zX2ZJHd318qpN5jZNppeIse5aZ656UwW0vCWhlh+bCQ75HZhoOBTZT90JP4u1Zqxav5dc7o5bce1dZdhMxmnG5ORYi+0p11SlhTZU0EBPKEpPNtsebp3GuPV0SvDln3p/xxL5nUp6vRnt9Gv5w/kSTlWT3a9TTAiKkR4aX+UNtJU3IWgHYL5T5Q3UlWxIAHQkb9/z9whFsSvKb0xbWHxyBp0c4DqiNio8wTsAnuI69N9uUbanbNSNR27rdA2dKHHIpaQqe9PfhJmcyO03QSk82xUQf529a9Pz3UzJ7tc0OO6ahyA6iPucoAaO7SXPI5kbrR5ad+7qlQ9NbI2taPsrEUvP7mLuaUvaeW35EnXTTO8NwHmbdcEz4zrSk8nNs45s3ypJPQK327t9upB7+ZPWx+43vGr01AbadU2pxLL8c9ezRuTz7HbkTuVBJ71lB22AKRH2Oa46kwMWevi5GmniqW3n0RJmTqEhJbKgpKUBG/lFO6U7+cbbAgDgyDiIyVti0XCTnGkkJUt1CSph2RcH4KVNqUVrQNiNtgkgdd1D469VpcyzCaTX54ZMXdW8UpxbX55lpK45IWqM6EAFXZq23PTfbz1Tu9cSRlzodv8As8z5zTodEn9zGK9k6lQA5Ajxjcq3PMCfN0rmsmGakat39X7mNAdYdS4jjLCIZyaQ/Fgof5nO1WsjlZCCC1sOYbcqt++vKWg157v6P/8AXCe1NZoQ5fVfxxG/nUHDMZvsNvJb7AmPNdohu0WpPj0t9zlRskNMFZ90kdFBHlN79wTWpaiZ5k2UMuZ5LuVr09x2TDC7a/d3UybpNSEnYsRG1K5CVEg79w2OwPNvPukvg9OK6c5NRdL9h2jOP3aSiTJg2FInXFGzaUFKFjyEbhIPRw9So7datjoH4Pfh20HnMZLHsD2WZW0Q57O5CoSnkOjrztII5Gjv13A5h+FXft9GxLjns/j67cl/1HEr6snHghv8vnzfyKPcN3BlqdxDWdEqbjVx05wK8oYcvmQXrZ7I8hKdllMdKx9oZ5wOVZSE7bKTzdRXqNpHo/p5obhELT3TLHmbRZYW6g2klbjrivdOurVupa1HvUT6B0AArc6V3KVGFFYgjjVas6zzJilKVtNYpSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQCog1Wn3u+at4BpQxlNzx+y3+2Xy8T5FsfEeVMchGEhmIh7YqbSoTHXVchSsiOADy84Mv1r2aafYVqJAj23NsahXdiI+JMbxhHlx3gkp7RpY2U2rlUpPMkg7KI7iaA0Th+n3OYvPLReLu7fl4llcjHLfe5aUKly4LcePIS286lI7Qsuy32ObbclolW6iomXKxWMYtjeF2SNjeJWOFaLXDCgxEhspaaRzKKlHYedSiVE95JJO5JrK0BB924cLpcYmRYRH1Icjad5Zd5V3uth9ikLlKMt8yJkZqZ2gDcd91TqlpLSlgPOBLid08uh+FH945qL+Naf8zi1ayqteE5t8+58EWo0a3Qn5TyU2x0tstlaghFyjKWrYddkpBJPmAJoCUuFT3r+kPyEsP0BmpTrzO0W8LPodpto7gund50v1MkXDF8btlmlPRbdDLLj0aM20tTZVJCikqQSNwDttuB3VuXtzfD58E2qvzbB+tUBf8ApVAPbm+Hz4JtVfm2D9ap7c3w+fBNqr82wfrVAX/pVAPbm+Hz4JtVfm2D9ap7c3w+fBNqr82wfrVAX/pVAPbm+Hz4JtVfm2D9ap7c3w+fBNqr82wfrVAX/qj3hh/egH5U23/C9WB9ub4fPgm1V+bYP1qq7cd/hDdLuKXQr7F2DafZ5bLn7MxLj292gx0Mdm0HAobtPrVzHnG3k7d/WgPUnh8/gE01+SFm+hNVv9ebWl/hc9CsI0zxLC7npbqc9MsFigWuQ4xboZaW6xHQ2pSCqSCUkpJG4B28wrZvbm+Hz4JtVfm2D9aoC/8ASqAe3N8PnwTaq/NsH61T25vh8+CbVX5tg/WqAv8A0qgHtzfD58E2qvzbB+tU9ub4fPgm1V+bYP1qgL/0qgHtzfD58E2qvzbB+tU9ub4fPgm1V+bYP1qgL/1Qrwz3vULJ8uIH0ObXV9ub4fPgm1V+bYP1qq3cevHxp1xd6N2rSrTjTnPoV4ayWJdAq529gNuIQxIa7NPYvOKKyp9Gw5eux6924HshSlKAVj7nd2rYUJcZWsuAkbbbdKyFYm/W2RcUM+LhJUgnfc7dD/8A5QHRcyx0/cYaR+Mvf9VdVzJrmv3Kmkfio/XvXM3iktX3WS0n1bn9VdpvE2B91luK/FSB+ugMOu83Rz3U1wfi7J/8VwLmS3PdynletZNbQjGrWj3SHF/jL/VtXOiy2tHdDQfXuf8AzQGllSld6ifWa+VvabdAT7mEwP8A2xX68UijujNf9goDQiARsRuDWIuuH4jfUlN7xa0XAHoRKgtOg/8Ack1KhhxFd8Vk+tAriXarav3UFn8iAP8AxQFd7vwucNt9WXLpoNgLziupcGPxULPrUlAJ/PXPZuGfhxsCkrtmgOnSFp7lrxiE4sf0ltk/21Pa8ftK/wDleX8VZH6a4F4vble5ceT6lA/ooDT7DbccxdAaxzDsetaB0CYVtaYH/wCAFZ8ZVNA2Mdj8gP667SsTZPuJix60A1+DiXon/wDxf60BwHKp/mYYH5D+uuNWTXM93ZJ9SP8AWu2MS9M//wCL/WuROJsD3cxw+pIFAYtWQ3ZX/MgepCf1Vxm9XRXfNX+TYVnU4tbx7p18/wBIfqrkGNWsd6XD610Brvsvc/8A1rv56+i9XQd01z+yti/c5av+iv8A7zXw41az3IcHqXQGBTf7sn/m9/WhP6q5U5LdE9621etH6qyysXtx7nH0/wBIfqriVikU+4lOj1gGgOmnKp490ywfyEfprlTljw93CQfUsj9FfpWJfgTvzt/61xKxSUPcSmj6wRQHOnLG/voKh6nN/wBFcgyuJ99GeHq2NdBWL3EdzjCvUo/qrjON3QdzbZ9SxQGV/dVA/wCg/wDmH66/Ksrh/exnj69h+msV+527f+nT/wB6f11+hjd1Pe0getYoDvLywf8ADg/lU5/pXVcyi4L6Ntso/okn+019Ri09Xu3WU/lJ/RXZbxMd700+pKP9aAxi77dXO+WofipA/RXEbpcj3zn/AMiyK2JvGLan3ZeX61bf+BXKMetI/wCWJ9a1froDWBdLiO6c/wDlWa3KApaoTC3FFSlNpJJ85IrqHHrSf+VI/pq/XWQbQltCW0DZKAEj1CgP1SlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlQJlnGHiOD6rOaUZVprndteaQJbt8lMW9m0t28ym4xnqfXMCkxw66gElHP16JJ6UBPdKjmPxH8Pstb7cXW7BXlRYJubwRkEVXZxBsS+rZfRsAg83dsQe4iujkHFJw+Y9hsnOndX8RmWxlcthkw73FcVMlR2u0cisfbAFvcpT5G/36N9goGgJUpUeYZr9pJnzLbeM5xZ5V2VaG709ZE3CObjGYU0h3Z1kOHkUlLiArc7JKhudiCevgvEforqA7Y7XZdRscRf7/EalxrA5eoTtxAXHTI5C2y84FKDS0r8hSklJ5gSOtASXSoGxzjR0ayibJtlsN5E6Dnx07mRXWGUOR7h9v5ZCx2u3iq/FXuVwEqJQRyAhQEhY9rhozlymUYrqtiV4VIffitCDeY75cdZaLzqE8izuUtAuEDuQObu60Bu9KjR7iZ4dY7Lkh7XTAkNtMIlLUchi+SytLSkLPl+5UJDBSfOHUbb7isozrhozJvNvx6PqviDtzu0RM+BFReo6nZUdSFOJdbSF7qSpCFqBHQpSojoCaA3elRa1xN6ITpltbsmo+N3e3z/AB/trrBvsBcOB4mwHni+ovBQHIoHdCV7DylcqSFV2neJTh5Yg265v644I1Eu2/iLy8gipRJ2c7M8hK/K2X5J27iCDtQEkUrRpGuuisO4X60y9W8PZm4s0Xr3HcvcZLluQFBJU+kr3bAUpKTzbbKUkHqQK/MXXjRKdNsVuhauYe/KydCXLMy3eo6l3BKlqbSWQF7r3WhaBt3qSpI6gigN7pSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKotxx2iHc9TsgbmJK23tIzHcQdilSDktuJBBHn229VKUBW/iVx2yw+InW7StmG77E26x3LOIrhmP8AOzOmQWmXW0Nc/i6WuVxW2zQc7klwpATWS4s7zJ0l1Y1AzHEmYol5Pkk3BJjMphLsdu33DHbSX1tN7DkeBYRyr6jqoKSoHYKUBmuEq6SXeJ29YLG3g26x4JJuShEccbTcHZNvhgmQzzeLlSN+jrbSHVf8Rbld3hZsNvjYppk+2gl1GrOJkOEJ5tk4S2kJ32322J6ekmlKAZbovh2TtYLfJK7hEnX3WTMMZuL0OQGjJhPTrmtIX5JBW0pTpaV3p7Zzv5qgnEc/vGMaZWjXmxMtMZTaM7Y0/aW8/IlMuW42mTE7RaX3VqDobfWU8qktJITythI5aUoDf8UlDSThLn68YvEiu5Pp/luK3O2Gayl1h5ZxeDF7N9I2UtsIkuqACklKyFAjrvjYGMQJOrWmuiLMmbBtef4U1eLxcIEhUaa4ZUGc47H2b2juNcz6+TtWXHGwEpbWhI2pSgOhbchu3EVptCyXPXY8aRZ42SWyKizQmIIDVntEZyMVuIR2rilKbTz86yAN0thoHastp41BzfR3O9RrhASxc8w0kyq5z0IfekIS+bpCZKkLkrdeJ5Y7Z3W4s77nfu2UoDtZfdJ70nXDSNT6E2HQ3xvL8dWhhtE6TOkXmPOUmTMSkP8AZh0qSAytlRSU8ylKQ2pOOwa9uNaocOLlrii1ytUnol8u8yHKkpkNOi9yXVtJWp0+MNLVvzJl+MEbqUlSVqUsqUB//9k=)
Figure 1. Evolutionary Influences on Innate and Learned Fear. (A) Predatory history shapes prey’s innate fear responses as illustrated by Peromyscus maniculatus austerus deer mouse’s freezing to weasels and Peromyscus maniculatus gambeli deer mouse’s jump (Jan Gillbank, ‘Drawing of a grey mouse’ October 27, 2012 via Wikimedia, Creative Commons Attribution 3.0 License) to gopher snakes [2]. P. m. austerus deer mice live in the coniferous forests of western Washington State and P. m. gambeli deer mice dwell in the arid grassland of eastern Washington State. (B) Ecological history predisposes fear learning. A classic study by John Garcia [3] found that rats easily acquired conditioned fear to bright/noisy conditioned stimulus (CS) paired to footshock unconditioned stimulus (US) and conditioned taste aversion to saccharin taste CS paired to X-rays (or LiCl) US. However, rats showed lack of conditioning to bright/noisy–X-ray (or LiCl) and saccharin–footshock pairings.
- Specific stimulus/response, \(S \rightarrow R\), patterns
- Visual OR Auditory \(\rightarrow\) pain
- Taste \(\rightarrow\) nausea
![[[@Pellman2016-xu]](https://doi.org/10.1016/j.tins.2016.04.001)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQEB9AH0AAD/2wBDAAMCAgICAgMCAgIDAwMDBAYEBAQEBAgGBgUGCQgKCgkICQkKDA8MCgsOCwkJDRENDg8QEBEQCgwSExIQEw8QEBD/2wBDAQMDAwQDBAgEBAgQCwkLEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBD/wAARCAYhBiUDASIAAhEBAxEB/8QAHgABAAMAAwEBAQEAAAAAAAAAAAECAwQICQcGBQr/xAB3EAACAQIEBAMCBwgLCQgMBBcAAQIDEQQFBiEHCBIxQVFhCRMUIjdxdpa0FRgZMliBkdMWFzhCV5Shs7XR1CMkM1JVVnR1sSg2Q0ZHVJOyJzlTYmRzd5KVwdLwNDVERWNmcoKXpiUmKYSFhqXkScLDxNXhZYOi/8QAHQEBAQEBAQEBAQEBAAAAAAAAAAIBAwQFBgcICf/EAEgRAQEAAgECAwMGCQkGBgMBAAABAhEDBCEFEjEGQVETYXGhsdEUFRYiMjNScpEHJCU0QkOB0vAjJlNzsuEINURiosEXY/FU/9oADAMBAAIRAxEAPwD1TAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeWntkONXFLQ+q+HWi9Ea7zvTmW4nLsXmeJjlOPq4SeJre9jTh7yVOSclCMXZN2TnJ2va3UvIeG3tEdUZFl2pcizfidi8tzbCUcdg68dXVEq1CrBTpzSeJTScZJ2avuffvbc/K1w2+juJ+0nbrl1i/vfeGO/fRuS/YqR5+o5bwyWe99jwfw7j8R5MseS6knueaf7TPtKP8AnnFD64T/ALSP2mvaUf8APOKH1wn/AGk9brPyFn5Hl/Dc36H8mOl/ay+r7nkl+0z7Sj/nnFD64S/tI/aZ9pR/zzih9cJf2k9buhjpY/Dc2fkx037V+r7nkj+0z7Sj/nnFD64S/tJH7TPtKP8AnnFD64T/ALSet1mLMfhubZ7MdL+1l9X3PJH9pv2k/wDzzigv/wA8J/2kftN+0m/55xQ+uE/7Setk4tqxCi0h+G5t/Jjpf2svq+55J/tOe0mXfG8UPrhP+0j9pz2k3/PeKH1wn/aT1scWyFCV+6H4bmfkv0v7d+r7nkp+037Sf/nnFD64T/tI/ab9pN/z7if9cJ/2k9belkODfiPw3M/Jfpf2svq+55KftN+0mfbG8T/rhP8AtI/ab9pP/wA84ofXCf8AaT1rUGvIOLH4dmfkv0v7WX1fc8k/2nPaTLvjeJ/1wn/aR+057SX/AJ9xP+uE/wC0nrTKLVt0Vs/Qz8O5Pgqey3S3+3l9X3PJj9p32kv/AD7ih9cJ/wBpH7TvtJf+fcUPrhP+0nrQot+QcHbujPw7k+EPyW6X9u/V9zyX/ad9pN/z7if9cJf2kftOe0k8cdxP+uE/7Ses/SxJOz7D8O5PhCey3S/tZfV9zyX/AGnvaSf8+4ofXCf9pH7T3tJP+e8T/rhP+0nrN0shxZn4fyfA/JXpf279X3PJr9p72kn/AD/if9cJ/wBpH7T3tJP+fcUPrhP+0nrJ0P0JUWPw/k+DfyV6X9vL6vueTX7T/tI/HH8T/rhP+0j9p/2kf/P+J/1wn/aT1klEjpY/D+T4N/JXpf28vq+55OftP+0j/wCf8T/rhP8AtJH7T/tI13x/E/64T/tJ6ydLKyTash+H8nwbPZTpb/by+r7nk7+1D7SL/KHE/wCuE/7SP2ofaRf5Q4n/AFwn/aT1g6H6DofoZ+H8vwjfyT6T9vL6vueT/wC1D7SD/n/FD64T/tI/ai9o/wD5Q4n/AFwn/aT1g6H6GfSx+H8nwjZ7J9Jf7eX1fc8o/wBqL2kH+UOJ/wBcJ/2kPhH7SD/KHE764T/tJ6udLIlF2H4w5PhG/kl0n7eX1fc8o/2pPaP/AOUOJ/1wn/aR+1J7R/8AyhxP+uE/7SerXSx0sfjDk+EV+SPSft5fV9zyl/al9o8nb7o8T/rhP+0kftS+0d/ylxP+t8/7SerElZtEPdGfjDl+EbPZDpP28vq+55UftTe0d/yjxO+uE/7SP2p/aOf5S4n/AFwn/aT1W6WV6H5oz8Y8vwjZ7H9H+3l9X3PKr9qf2jn+UuJ/1vn/AGkPhP7Rz/KfE/63z/tJ6q9D9CsoteKH4x5fg2ex/R/t5fV9zys/ao9o3/lPif8AW+f9pIfCn2ja/wDnLif9b6n9pPVPpZEovYfjHl+DfyO6P9vL6vueVv7VXtGv8p8T/rfU/tA/aq9o3/lPif8AW+p/aD1QsxZmfjHl+EV+RnR/t5fV9zyu/ar9oze33U4nfW+p/aCf2q/aNf5T4nfW+f8AaT1McH1N7dyelj8Zcvwb+RnR/t5fV9zyw/as9owv/nTid9b6n9pJ/as9ox/lXid9b6n9oPUpxdyOlj8ZcvwV+RXRft5fV9zy1/au9ov/AJV4nfW+p/aCP2rvaL/5W4m/W+p/aD1IcW7ojofoZ+M+X4Rs9iui/by+r7nlx+1f7Rf/ACrxO+t9T+0j9q/2i3+VuJ31vqf2g9RWiHFvdGfjPl+Eb+RPRf8AEy+r7nl1+1j7RT/K/E363VP7SP2sfaKf5Y4m/W+p/aT1CcHZ7op0sz8Z8vwip7D9Ff7zL6vueX/7WPtFP8s8TvrdU/tAfDP2ia75zxN+t1T+0HqB0spODb7ofjPl+Eb+Q3Q3+3l9X3PML9rT2iX+WOJv1vqf2gPhp7RJf/PHE3631P7QenfQ/NFZRaY/GfL8IqewvQ3+3l9X3PMX9rX2iX+WeJv1vqf2gj9rb2iSe+c8TfrfU/tB6c2aKtXZn405fhFT2E6G/wB5l9X3PMj9rf2iK751xN+t9T+0Eftce0PXfOuJv1uqf2g9N3ErJbMy+Kcvwip7BdBf7zL6vueZX7XHtD/8ucTPrfU/tA/a59of/lviZ9b6n9oPTDpY6HZ9h+NeX4RX5A9B/wATL6vueZv7XftDV3zziZ9bqn9oH7XntDP8ucTPrdV/tB6XOLfaxDi0ZfFeX4RU9gOg/wCJl9X3PNL9rz2hv+XOJn1uq/2gj9r32hi/+fuJn1uq/wBoPS1pruUcW22Z+Nub4RU/k/8AD7/eZfV9zzV/a+9oUv8A5+4mfW6p/aCP2v8A2hSV3n/Ez63Vf7QelEotWvYq4tpq6H425vhFT+T3w+/3mf1fc82lQ5ltP3wHETjBxNyXNf8ACRw37KMRO9F7RleNWXdqS7+HYPS3O5nf/wB09Ea34m5rktb/AODYz9llaPvbfFns66atNTjuvA70a04KaL17nCzzUFHFzxUaMaCdHEOC6IttbJPe7e5+h0tpjK9GZDhtNZLGrHBYPr92qs+uS65ym7vx3k/zHt5vGOP8Hx+Tx/P9/wAHg4PYDHLqcsebLXHPTV7/AOPZ56vQXtB131BxLX/53Vf7QR+wL2g3+cPEv63Vf7Qej8ouT2aKuLSuzw/jfl/Zj6U/k68P/wCJn9X3POH9gntBl/xh4mfW2r/aB+wT2gq76h4mfW6r/aD0caIlF2Y/HHL8Iqfyc+HX+9y+r7nnH+wX2gn+cXEz621f7QP2D+0D/wA4uJn1uq/2g9Gekq4ton8cc37MVP5OPDv+Jn9X3POd6I9oGu+o+Jf1tq/2gj9hPtAv84+Jf1tq/wBoPRaUH5oo4uw/HHN+zFz+Tbw6/wB7n9X3POz9hPtAfHUnEr63Vf7QR+wz2gC/4ycS/rbV/tB6IuDfkUa9SfxzzfsxU/k08Nv97n9X3PPD9hvP+v8AjLxK+ttb+0D9h3P946m4lfW2r/aD0NcG/EpKDSvcfjnm/Zip/Jn4b/xc/q+557fsO5/f85eJX1urf2gr+xDn8/zm4lfW2t/aD0J6WZOLSu2jPx1zfsxc/kx8Mv8Ae5/V9zz7/Yjz+f50cSfrbV/XkfsS5+131RxK+ttb9eegj7Gdn5mfjvm/Zip/Jf4bf73P6vuef37E+fr/ADp4lfW2t+vD0pz8r/jTxJ+ttb9ed/3F3M6kX5j8ec/7MVP5LvDL2+Vz+r7nQN6W5+F/xp4lfW2t+vH7F+ff/OriV9ba36878uLI6H6Gfjzm/Zi//wAWeGf8XP6vudBv2Mc+3+dXEr62Vv14/Yzz6r/jXxJX/wCdtb+0HfaUXdlJxbtYy+Oc/wCzFT+Svwu/3uf/AMfudC/2Nc+n+dnEn621v15D05z5LvqziT9bK36875NNdzOpG9rGXx3n/ZjpP5KPCr/fZ/8Ax+50R/Y7z5f52cSfrZW/Xkfse58f87eJX1sr/rzvd0O3dFOkfj7n/Zip/JP4Vf77P/4/c6J/sf58F31dxJ+tlb9ePuDz3rvq7iT9bK/6870Si1dtmbRn4/5/2YqfyS+FX++z/wDj9zo08i57v87+JP1sr/ryPuHz2rvrDiR9bK/687yOL33RScW7WsZ+P+f9mKn8kfhXv5s//j9zo88k57Vv+zDiR9bK/wCvI+43PWv+OXEf62V/153ea8DOVNt3Rl9oOef2Z9bpP5IfCffzcn/x+50l+4/PT/nlxH+tlf8AXj7kc9P+ePEj62V/153XcWldkE/lB1H7M+tX/wCH/Cf+Nyf/AB+50n+5XPQv+OfEf62V/wBePuXzz/558SPrZX/XndOcGl3RXpZn5RdR+zFz+R7wi/33J/HH/K6W/cznm/z04kfWuv8AryHlvPKv+OvEj611/wBedzyGh+UXUfsxc/kc8I/43J/HH/K6Yfc/nk/z24j/AFrr/rx8A54/HW3Eb614j9edyuhlJR79uxP5RdR+zj/r/Ff/AOGvB/8Ajcn8cf8AK6b/AAHniXfW3Eb614j9ePgfPAv+PHEf614j9edxellJQbbd0L7R9R+zj/r/ABXP5GPBv+Nyfxx/yunvwTnf/wA+OI31rxH68h4XneX/AB54jfWvEfrzuB0uWyM5RavujPyj6n9mfX96p/It4Nf77k/jj/ldQnh+d5f8eeI/1rxH64j3HO54664jfWvEfrjt30sxndSexl9pOpn9mfX965/Ip4Nb+v5P44/5XUn3PO3/AJ9cR/rXiP1xHuudtf8AHziN9a8R+uO2rTfZFGncn8pep/Zn1/eufyJeC3+/5P44/wCV1M6Odpf8fOI31rxH64i3Ouu+vuIv1rxH647ZTi3bsYulK7YvtN1M/sz6/vdMf5EPBL68/J/HH/K6pX511319xF+tWI/XC/Ot/n9xF+tWI/XHaqcGrFHsZfafqZ/Zn1/eqfyHeCX+/wCT+OP+V1Y6+dRPfX/EX61Yn9cR7znT/hA4i/WrE/rjtNKLbvsZyXdGflP1P7OP1/eufyGeCX+/5P44/wCV1cdXnSXfiDxF+tWJ/XD3/Oil8oXET61Yn9cdn5QbfgZyT3VjL7UdTP7M+v71z+QnwO/3/J/HH/K6xfCedD+ELiJ9asT+uHwrnP8A4Q+In1qxP647MtNdzLol6E/lR1X7M+v73SfyD+B3/wBRyfxx/wArrV8M5zl34h8RPrVif1w+G85n8IvET61Yn9cdlHCXoZdLH5U9V+zj9f3qn8g3gV/9Ryfxx/yut/w/nLXfiJxE+tOJ/XD7o85P8IvET61Yn9cdj5R3MZR6fEy+1XVT+zPr+90n8gngV/v+X+OP+V11eZc5C78RuIf1qxP64r91Ocf+EfiH9asT+uOxMotu9zGzJvtX1U/sT6/vXP5AfAr/AOo5f44/5XXz7rc46/5R+If1pxP60j7r84q78SeIf1pxX607BNXM5RY/Kvqv2J9f3rn/AIf/AAG/+o5f44/5XwD7sc4i78SeIf1pxX64fdnnDX/KTxD+tOK/Wn31xdn2M5xbRn5WdV+xPr+9c/8AD74Bf/Ucv8cf8r4L92+cL+EriH9acV+tI+7nOAv+UviH9acV+tPu7VtjOUSfyt6v9jH6/vXP/D14B/8A6OX+OP8AlfC/u/zf/wAJfEP604r9aHn/ADfL/lM4hfWnFfrT7jJbvcrKJn5XdV+xj9f3qn/h59n/AP8A08v8cf8AK+Ifsh5vfHiZxC+tGK/Wkfsi5vPHibxDX/50Yr9afa5K0nfwOLiKvX8WHZd35m4+1nWZXXkx+v73PqP/AA/+znT4ebLqeX+OP+V8qwOpOaFKX3T4oa+qxlFr3T1PiZxaa36l73e6drfpOHqXXnMfkWWyzT9tbXuDwuHUYyjHU2JUYq6iulKr82yX/rPq/RKTcUtz8dxjp9HD/HpL99T/AD/GR6Ol9ouo5+rw48pPzrI+J41/I54T4d4F1XW8Wed+Swyym9d7Jvv2ekHsgeJuv+I/AHU0de6uzTUNXJdUVMLgsRmWJnia9KhPDUKjp+8m3JxU5SaTbt1NLayXfA86vYmfILrv6Xv7Fhz0VP3L/LgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPH723XytcNvo9iftJ295dFfl74YfQzJPsVI6he26+Vrht9HcV9pZ3D5cv3PPC76GZJ9honj6z9GP03szfLzZ/Q/e2QsjYHzdP2fnZEWTNeh+hKi0NHmY2QsjezRA0edj0ryKuKTOT0spKDbe6FjZmxUU2S4xs9jTofoOh+hmjbHp9R0+pt0P0HQ/Q3SvOxshZG3Q/QdD9DNM8zCUEyrgjk9D9DNxab7CxUyZ9K8hZGnSx0sab5mXRHyIlCNnsbdLI6H5jR5mHRHyDjGz2N5RexCi2Y2ZOPZFlGNu38pt0P0Di0ZouW3HnFXK2RtODb7oKDXiLFzKaY2RVxS8Dk9LKSpu3dDRMmFkLI16WR0szSvNWdkU6I+Rv0sr0sWNmTLoj5FZxSWyN+lkSi7GaVMu7jWQsjfpY6WZpXncVwjd7BQj5G8oNt7ojofoZpUzZe7h5GRyuh+hn0sWKmTJLYiUV4m6ViJq5mlTNx+iPkVnFK2xv0srOLdrWM0uZMLIWRtawGleZxmkLI2cG3fbcjofoTpUycdxjfsOiPkauLTfYdLGlTJxpRV2RZG7i990LMzS/M4rhG72HSvI2cG290VasYuZOMOlPwN1FsicGnYzSvO41kRJK5yOllZKz3FmlzJx2tirSfc5Ddikld3Ri5kxlCPkZuKTORKLsVs0ZYuZMeleRWUI2exvJNqyKODs+xmlTJx+iPkQ47Oxt0shwb8SdOkycayKtK5yXFryKyi9xYuZuNJIr0m8oPYq4Oz7EukyYSivEiUUk9jXpZDXcWLmTqxzHa91npriDTy7INT5jgMM8uo1Pc4evKEHNymm7J92kv0H2HgZmmY5/wsyTNs6xtfG42v8J97XrTcpztiasVdvvZJJeiR/f1Dm3DfAY9UNV5rprD433aahmNehCr0NuztUadrp28O5/TyWtk2MyyjidOYnAV8um5e5qYGcZUHaTUulwvF2kmnbxTvvc+p1PU48nR4cU49Wf2vi+B0HRZ8PiXL1F5/NLv83fp3nztqkVF7Iq9zktWKyV0fIsfppk4/RHyKSW7OR0szcG272MsXL8WNkRKKSexq4O/dFZQdnujLF+aMLJmRyellelk2OsycZ7MpNWRypRKTjt4GWLmTihpNWZv0sy6H6GWOsyYzVnsVcYtbo3asVkrqxi5kwlTjZ7GfRHyOT0NK90VMs2uZOLOK6nYpKK8TkTi229jNq5LrKxcY27FWjZwlfuisotW3Rli5kwaTTfjYokn3OQ1s90U6WS6TJxpRjd7FJxSOQ4NX38Sri32sZp1xycdr4v5jNo5Di032KSjYmzbpjk48kmuxRwXgcmW6M2rGWadZk48opJ/OU7nInFtPdGbi15GWOkyYOEbvYzmrPY5DVrlWibHWZOO4p+BlUiovZHI6H6ENWJsdJk4rSasyklaTRypxco2VjJrpdvImzbpjk49SCitl4+ZmcqSbVkZzg07GWOuOfxYSSM5RW7sciUXYo14Gajrjk4zWxVq5yJr4q7Gb2JvZ1mTDpS8DOcVd7HIlFlXF2ZljpMnFsjOUF1bo5DVu5Dg2m7ol2mTizSVrFGtm7HIlEq4tp9idOkyca1yrS32NpLp7lXHvbxMsdZXHkk7XMJJb2OY00ZThJ3s+7JsdccnGKySs3bc2lFxdm0UafcmzTtKwsmZzildrvc5N0jOUb3a8WZ6uky7uM0nu0ZtKxyJwbd9isoNJu6J1XWZMLJmDSsciSsyHuibNu2NceyZlUS6UcmcG3fYze6JsdMctd3FasZzikuxynFopJXRljtM3EdiskrdjkuLuYWZNjrKydrspJK3Y2kilmS6ysHFeKKOKfgciUb3MZxb8ibHWVjNK7sZtHIe14vvY4GKrdX9zptW8WvH0+YTC5Xsjm6nHp8PNkyxNZSbhD87Xj6HGUHKSUVuzWMJVJdEVdv8AkOXHDqnFKNn5t+J2tnHNT1fL4ePl8Q5PPydsXGjShCLSV2+7Pw3GhJcPse/++p/9dH0B7d/A/BcbN+H2Of8A39P/AK6O3hV313Fb+1PteP28xnH7Kdfjj6fJZ/8ATXfP2JnyC67+l7+xYc9FTzq9iZ8guu/pe/sWHPRU/rz/AJ0gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/Kax4qcMeHSjPiFxG0vphTSlF5znGHwSabsmvezjdN7J+Z+V++x5WPyluFX1yy79cB9VB8qXNfytSajHmU4VtvZJaxy5tv/pj6qAAAHj97br5WuG30dxX2lncPlwSfLzwv+hmSfYaJ089t18rXDb6O4r7SzuLy4Rvy8cLnf8A4mZJ9honj6z9GP0ns5dc2f0PodkOn0LJWZJ85+w2qCen1JSsDarV/Ajp9C4N1abVHT42J6fUkw2zasuzFnfsXl2IDZUWQsi3T6jp9QbVshZFun1HT6g2rZFHBeXia9PqLeoJky6F4odEfIu1cW9QrzVT3a8mVcVd7M2It6g81YuN/wB6yOl/4rNun1HT6jUb5mNn5MdL/wAVm3T6jp9TNHmceULPdMjp9DeULu9yvu/UeVUyZdPoRKN/Bm3u/UONt73M03zOM4q+6HRE2auR0+o0vzsvdryZT3a8mcjp9SjVjLCZMvdryZE4JLs+5v0+pVrwM0qVh0v/ABf5CLLyN+n1I936jS5k40k7vZkKL8UzkOnv3KuFle5mm+f3Mun0Mun0OV0+pn7v1FiscmagvJlZwtayZv0+pEo9tzNNmfdx+jfsyJwStszfp9Ss4X8TNdlTPu4/R6EOHozf3fqRKNrb3uNVcz7sOhq+z/QVat3RuUnC/iTpUy3WLi23swoxtujdQ27mdSNmtxYuZMHFXexDVuyZso3fciVP1M0uZMOn0Kygr7o36bO1yso79ydLmbHoV+zKzhvsmb9PqVlGzGlTJh0v/F/kKyp7/is5DVirV9zLFzJxpQVt4vuV92vJnJnG6tcr7v1M0qZuM4bdmVlDwSZyZR2M3DxuZY6TNh0v/F/kIcNndPsb9PqUnteP8o1YqZbcfoj5FZR3dk7G3u/UONn3J06TLTjyT8mV6bvdM3nC/iVcLK9/5DNKmTGcVtZFHHbszaXxreFiOm/iZqOkycZQ9GUcd3ZM5ThbxM3C7e5ljpM3V/mJ4Wa/1jr6nm+mtNV8dhFl9Gj72FSnFKalNtWlJPZNeHifXeCOn830vwvyXI8/wM8Hj8N8J99Rk03HqxFSSu4treMk9n4nF4i8etHcNNQR05nuXZzXxMsPDEqWEoUpw6ZNpK8qkXe8Xfa3bc/VaL1bluvdL4PVeT0cTRweO957uGJjGNVdFSVN3UZSS3g2rN7Ndux9bqubqcujw4+TCTD3V+e6Dpuh4/E+Xm4eS3ku94/DvH9dxT7ohwVnsaqFla5D3dv5T41j9RMmPu15Mq4Ru9jZqxSUbXdzFysJxV9kVlFWexs1crKOzVzLHSVh0R8jPpf+L/IbNW9Q1ZXM1Vy6ceULPdMrKKa3TN2up37GbVzLNrmTH3a8mZ9FvA5L2KSjsTZp1mTjygm90yriktkchwu73M3CyvcyzbpMmDXgykopL4q3NpQvJ7lXCyvcyzTpMnHcbp3TKdC9TkyjfxM5Rtb1Js26zJx3FpvZ2uJRTt8Vm77FGrmWaXMq47iu1mZzTXZP9ByJdyso3t4WMs26Y5a9XGs/L+Qjp9DkShZPczasTp0mW/Rx5R3ezKSgnbY5Els9zNQv4madMcve43S7tWdirgvFHIlGzZSUb73sTY6zPbGUV0uyMpQta6ZuRKN7b2sZY6y6cacNm0ncz6UvA5Ml3RnKF3e5LrMnHcduxVxvu0ciVPbuZyVna9ybNOky249n4pmcoJu9rnInG6tcr7v1M06zKOL0y/xX+ghwX76Lv6m7dirj1u97eBNjpMnF6X5MzlF9T2ZySkoXu7+HkZY6zLTjtJ9yjgr7I2936lWrOxOo6zJjKKa+Kr/MZSj4WZyFCzvcpOO7JsdMcnHlHzRRp3slt8xv0+pWUd2ZZt1mTjuC8Ymc4yu0k7HJlG9vCxRxtfcyzTrMtOLKF7XT27Gbi07KLOW438TJx7+hNjrjm48oW7plGt3szkSje3hYylGze5Fm3THLbjzhdr4rM3Hd7HKavuZThe7uZZp1xy9zjTir9itl2sbyp7/jGc4WT38SbHaZRjOKvsjOUW+5t0+pWUNnv/IY645ONKKfgZuO2yZyZK2xnKFl3MsdscmDXmjFxsuzORJ9O3cpJdW3YizbrLpg15mco+mxyJU9/wAYrOFle/j5GWOky047jHxMJ07L8V9zkyhd9/5CsldWJ07Y5acNprazKuK8jkTp2ff+Qo438SbHeZOPJbtWKSj2sjeUbNq5/NxeJvelSe3aTXj6L0GOFyuojm6nHgx82THF4hSbp0nt4yXj6L0ONCm6slGPf/Yawozqz6IK7a/QvNnNhh40Y9MX87fds6Za45qPmcPHyeIcnnz9IwjRhSXTFNvz9SX6m7j33MpRvbc813bt+i45MJMce0YzgrNqLPwHG1NcPMfs18el/wBdH0KXa3kfgOOHydZh/wDV0v8Aro93hX9e4v3p9r8v7fX/AHW6+f8A6s/+mu93sTPkF139L39iw56KnnV7Ez5Bdd/S9/YsOeip/XX/ADrAAAB/NxeotP5fXeFx+e5fhq8UnKnWxMISSe6dm0+25l+y/SX+dGU/x6n/AO0B/XB/I/ZfpL/OjKf49T/9ofsv0l/nRlP8ep/+0B/XB/PwOeZLmlSVHLM3wWMnGPVKNDEQqNLtdqLbSu1uf0AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfFecnjRmXL/AMtWuOKeROks4yzBU8PlbqJSUcXia0MPSn0vaXRKqqjjvdQd9rn0rO9faF01jVl2o9aZDlWKcFUVDG5lRoVHFuyl0zknZtOz7bM+RczGG4OcwnArWHB/E8WtIYSeoMAoYTESzrDdNDF0qka2HnJKd3FVqVNyS3cbpdwPNPla9nDxD5ydN1OYTjBxaxuV4HUuLr1cNWqUJY/MszcKjhUrznUnFU4OcZxi31N9LfSo9Lf3z8CDwx/h01R/6Mw//tH9H2aXNppbSOmJcoPGPMsDprVOh6+MpZXi8ViqccHmOG97OrOEa9+hzg5zcWnadPplFuzO937b3Cb+FHSX/pvDf+2B0Bj7EPhgpJy45aoa8V9zcP8A1npJg8JRwOEoYLDpqlh6caUE221GKSV2+7skfmVxd4USajHidpJt7JLOsM235fjn64AAAPH723XytcNvo9iftJ3I5brfe78Lr/5l5J9honTf23XytcNvo9iftJ3H5cP3PHC76F5J9honk6v9GP0ns5+tz+h9GaXgRZ+QXcsfP0/XbVs/IK3iT1Id90NGxpeBFn5E2tux1IEu0WfkSkvEdSIbuNFuhpW2VyOn0JWzJbuhol2qATZsyRqATZoG6YWfkQ+xbqQt4jTVEvMmyJasCdG0WRWzv2L9LHSzbDaln5Cz8i7ViDNN2rZ+Qs/IsAbUa8yGvIu0RZoNlUs/ION1ZplwCVn7teTHu15M0AV5qz92vJmfS/8AF/kOT0szDZky6X/i/wAhWUbd0zkWbKzi2trGabMmHT6Dp9DXofoOh+hml+Zg479irimbSi02iOljTZky6IlPdryZyOllGmhYqZMnDyRWULW2N7MrKLfYzSpkw6fQiUbWumbKEr90ROL27GabL3YdPoRKKdvis16WRJMlcrHpj5FZQW1kzZxZVxa8hpUyY9L8mRKCb3W5v0spODb7oyxczZdEfBblZRW2zNuh+hWUHsZpUy7uP0N3+KyHT84s5NrFZRcns0ZpczYdMfIpKO+yfY5Dg7PdFOlixcyceUHZ7Mqo+aOQ1dNIp0smxcyYuPoyOleRrKLsR0szS5k43S/JkOGz2Zv0siS2aM06TJx+n0KSgrt2dzkdD9CsoO77GKmTj9L/AMX+QhwV90zezRVq7M0vzOPOFrWTKuGz2ZyJRexWUW4vsNVUzcXoj5EOEbOy3Nvcy81+kq4tOzsTp18zHpfkyrivI3aaKODb2tuxYqZPhvGHl4xvFDVkNS4bU9HL4RwlPCujPCuo24uTvdSXfq7W8D6Bwz0VU4faGy3SNfHRxs8B769eFNwU+utOorJt2sppd/C58o49cd9ccNtcQ07p+llksLLA0sTfEYeU59cpTT3UkrWitrH1Hg9q3Ntd8Nso1VncaCxuO9/71UIOEPiYipTVk22toK+/e59bqcernR4Xlu8Pc+B4fyeH3xLlnBjfle+7/jH6z3a8mZuC6ns+5yehtbWKODUm/VnxrH6eZOPKG+6ZWUduxvUW/gVlBqPgS6TL0cfoiVcduzN+llGmjLHSZbceUN90VlC6ezN5Rbd0Q4tK7MdJk47jbsmU6I+RyGjP3b8zLFzJx5RXU9iHHbdM2lF3YqJ9K3Xcl0mXo47ik+xRwv4M2asVs0NLmTCUN38V2KSj6HIabuU6WTY6TJx3HfdGcoLxOROPxvAznFu1rE2OsyYuCtsijg/J/oN3BpXuipljpMnHcFd7Mq4peDN5J3KNXMsdJk47Tbs07fMVlBbWTN5Qdm7oo4teRNjpMvewcduzMrNeDOU4vfdGcotW3RljpMnHlDZ7Mp0v/F/kOS4uz7FGrGWOkycZwV22vEpKPkjeabuU6WTY6zKsOmPexlUjurJ2sciUWrtso0Tp2mXvcbftZlZRv4G3Q/Qhw8yXWXTjuKt2KNWZv0P0KSi7syx0lYTgkuz7lLW2sbvdGc4Nu+xOo6TL4uPKFlsnco0+zRyDOVNyk2rbmWadZl8WEopL4q3KOKvutzkShKKu2vzGTg2200TY6Y5MLPxTKuKfdG9Sm7LdGTVnYl2xy2xnG1rJlXHbs+xtNN23WxRySTj4mWOkyceStbYrJXT8zWUXLtbYo4tMl2lY2fkZzWzsn4nInFu3Yo4vfdGWadJfe4k+pWsn+go4t90zlTg9t0ZuLTJsdpk48lZ27FWlZmtWPxluuxm9iXWXfdlKKv2ZSUYtPbf5zdq5lKLTvdE31dMa481Z2SfYo1fubyi5O90ZW2Jsdsb2ZSgr7JlHFWaNzNwaV7mOuOTjzh/3pjZ+Kf6DltXMJwdu6MsdscmVr72M5LY2e2zKSV1YizbrKxaT7IxmrLZM5PSyk4O27RLrjZHHcbq77mUlZbJnIkrN+h/Lx2M606FB/F/fS8/Ren+3/amNt1E8vU48GPmyYYzFdd6VJ7dpPz9F6f7f9vEp0p1ZKEYt38/BeZpCjOrUUIK77+iXmz+lToRowUYbv9833bOts45qerwcPHyddyefPtGFOhChHoirt93buTKN/A2ku7MzzW7719/jxmGOsWUo+aM3FeKORKN7mM4t+RFjvjdsKkbXtF99j59xvT/a6x+37+l/10fRX2a8T59xyTjw4zD/AOrpf9dHu8Ln8+4v3p9r8t7e3/dfr9/8LP8A6a70+xM+QXXf0vf2LDnoqedXsTPkF139L39iw56Kn9cf87wAAedXNH7KXUHMNx41TxkwHG3C5RQ1JUw1RYHF5PPE1KDpYWlRcfeKtFSjeleKsrRajva7+U/gOtWflEZT9XKv9oPWsAf598/5Hc2yLnJy7k/nxEwdbG5h7m2erLpRpQ95g3id6HvG3ZLp/H3bv6HZ38B1qz8ojKfq5V/tB/X4h/8AbstM/wD4n/Qkz1UA6P8AI57OfOOULilnHEjNOKuE1KszyCrkkMHh8qlhlB1MTh6zqOUqsr2+DpJJL8Zu+1n3gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOknNf7MnT3NNxdxPFrH8W8x07XxOAw2BlgqWU08TBe5i4qam6sXumtrbWe++3x38B3pL8ofNvq7T/tB6enxjnI1vqrhxywcRdcaJziplWeZRk86+CxlOEJzoVOuMepKaavZvunbut0B0p/Ad6S/KHzb6u0/7QPwHekvyh82+rtP+0H8PhR7YrKdB8F9H5HxE03qPiDr6nQxP3ezFVKGBop/DK/uIKSi/eTWGVFtqCTurycuq3cLlS5+eCnNjiMRp3S/w/INWYSjLE1cizVQVSrSTSlUoVINwqxV1dbTV23G27DqyvYeaRTXVzDZu433X7HaV/wBPvz02wuHjg8NRwsJ1JxowjTjKpJyk0lZNt7tu27Z0y9o1zrau5Zsr03w/4RZfRxev9Zuc8NVq4Z4n4DhlJU4yhRtarWqVH0wTTXxJXi7pPq5kHPd7QDln1hkea832jM1zPRWdV40K3w7IMJgasIWvJ4athadODrRT6vd1L9SUl8VvqiHruD+fkOeZVqfI8u1JkOOp43LM2wtLHYLFU79FfD1YKdOpG+9nGSav4M/oAeP3tu/la4bfR3E/aTuRy3pPl34W/QvJPsNE6b+27+Vrht9HcT9pO5PLf+534W/QvJPsNE8nV/oz6X6L2d/W5/Q+i2QAPnv1x0+hVprtcv1egtfcCu97MWRbp9R0+oZtSz8iUvMt0+pD2DUNbFTRK4cdnuDemZZdh0+pIbsauQ0vANXFvUMRZ+RPZdySLeobse/YizW5KViQbVuybq3cdPqOn1Bse/Yiz8ie3rcdXoNG0WfkLPyJ6vQdXoG7Vd15kb3sy9r7jp9TNNVshZFun1HT6iwVshZLt3LdPqOn1GhTf1/QV6fQ16fUqZrRLpVK3gLeZYiXYN2rZCyJJ6fUG/ixlFOT2Cgn4M0cd+4StuFSqe7XkzPpf+L/ACHIK9PqFTJj02u7P9BRxv4HIa7op7v1M0qZMlHfsRUgnbY2936lZwt4maVMu7DoRWUFtZM36fUiUe24sXM+7jqD8UyJwStszkdPqVnC/iTpXm24/R/3rKyhZ7pnJ6bK1ylSO63GlTLbDp9Cs1a2xuo3fcrOn23M0qZd2HQvJkSi07JM2tZdyGr7maVMmEk0uzKWv4HJlDZq5n7v1M1VzNi4Lwiyko7/AIr/AEHJcLK9yjV9zLFzNx+leRHRHyNnTuu491/338hmlzLTi9PoQ4Ld23Nen1Dhd9zNWOkyYdPoVlFXexyPd+pSUd+5mlzJg4LyKOFvB2ORKOxSUdrGWLmTBxv4NlXDZ7M3ULeJEtrr+UlUt9zj9PoZzheT+KzkdPqVcd+406TKuPKL2+Kyso2TdmjkSj23KzjeLVzNVczfgtX8G+G+vM2jneq9N/DsaqMaCq/DK9K0IttK1OcVs297X37n9zTemMk0fkmH05pzAPCZdhHP3FH3k6nT1zlOXxptyd5Sb3btey2sjq5zV6n1Nk3E+lg8p1HmeCoPKqE/dYfF1KUOpyqXdotK7st/Q+68uuNx2a8G9PY7MsdXxeJq/C+utXqOpOVsXVSvKTbdkkld7JJeB9Tqul5cOkw5cs9y+7b4nRdf0/L4jycGHF5c5veWvXvH0G1trFWt3sbShd3uVlCye/8AIfI1X6OZMJQT8PArKOxt0+pVx77k6dJkxUVbdGfRb96zkSjv3Kyjs1czVXMnHkvQo47HI936mfT6mOkyY9MfIzkml2ZyJR37lJRvtcyzTpMnHcE3dpmbUn3TOV7v1M+n1Js26zJx3D0ZE4JLs+5yJR37lZR2M0uZOK4bPZleleRyGu6M5Qsu5jpMmE4Ldpb2KdF+6ZyHHZ7lOn1MsdJk4zTu1Z+RRwStszkuG7dysoX8SbNukzcaUNm7FFBeKORKPgZyja3jcyzTrMmM4veydjO1/A5DV1YzlG1vUmzbpMmLjbezM5pO1lf5jkSV00ZqFvEyzTpjfe47i7v4rKyhZ7pm8nYo1d37E2bdZawcI27FJRtayZu499yso9tzNOkycaUNnsynQl4HJkr7GU42drk2OmOTj9PoQ4J+Bu47dykn0u3cyx2mW/RxLPxTIcL+DN5R2M27OxNmnWZOP0+hEl6G0o9W3Yr7v1J06zJxumPkUlHfZOxyJR2M5LwMsdZk47V1ZpspKLu7RdvmOQ423vcqTY6TJxmk1Zmc4K7smbuNt73K+DRNm3WXXo4/T6Gc6cW+25yJRtbe9ykoXbdybHTHJxnFeCKyj32ZyGrFZK+5mnaZONa/gUce9kzZqxVruS6TJx+lvunsZzTV7JnJasZzjs3cyx2xycWUb907mThu7JnKlHfuVcO+5Nm3WZacZxt4FJRvfY2qKzRR7k2Osu+7HoS8DKVNWfxX/KciUd+5V7k2adMcnEnHpfZoo1s0cmpC7W5k43Xcmx3xy7MHF32T/QUlFPwN5Lp27mTVvEyx1xyYTpq/4rMnF+Ry2r7mModO97mWOuOTBxv4FJR27G7Vz+PmGO674fDy+L2lLz9F6f7fm75MfNezOXqMeHHdY5hjFPqo0H8TtKS8fRen+35u/BpUKlaap04tt934JebNqVGpXmqdNXb7vwS82f1qWEp4WkoRd2/xpPu3/wC/gXbMO09Xg4sc+sz8+fo49LDU8PT6Iq8n+M/F/wDv5EThbtF/oOTKO7d/5Cko9Vjz3v3fe49YSSejjOK6XfuZSja1kcmdOze/8hlNWJsenHJhZ+RWS7bGzje5RqxLpMnHcV3sfPuOsUuG2Yu2/XR/66Po7je58748w6eG2Yb3+PS/66Pd4XNdbxfvT7X5b28y37MdfP8A9Wf/AE13j9iZ8guu/pe/sWHPRU86vYmfILrv6Xv7Fhz0VP60/wCegAAPPzmb9rDQ5duN+pODeH4D1dRfscnhqc8xqal+A++nVw9Os7UfglSyXvelPrd7Xsro+Xfhzl+S7/8Abt/+oHpJn/CDhLqvNa2e6o4W6RzjMsT0qtjMwyTDYivU6YqMeqpODk7RSSu9kkl2OB977wF/gR0B9WsF+rA8QNQ8833e52cs5xP2rvcfc73P/wB7v3b6vee7wTwv/wAK+Dq179f+CdrW37nar8OcvyXf/t2//UD0V+994C/wI6A+rWC/Vj733gL/AAI6A+rWC/VgdbOSn2jtDnB4mZvw2lwfqaRrZZkVTPIYpZ+sfGrGniKFGVNx+D0nF3xEWmm7pNWWx3SPzGl+GfDjRGLq4/RfD7TWQYqvT9zWr5XlVDC1KlO6fRKVOKbV0nZu10j9OAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADrD7S/NsXkvI/xQxmCcFUqYXLsI+qN17uvmWFoz28+ipKz8HZnZ4/E8ZuFuR8bOFeqOFOo6s6OA1Pl1XAzr00pToTavTrRT2coTUJpPZuKTA6y+zt5YOB2QcsGjdZ1tA6ez3UGrsuWZ5nmmY4Cli60pVJStQhKpFunCCSg4Rsm4tu7bb6783nC7RHLLz58ANfcE8jw+nMRq3O8NDMsqyyCoYaT+F0cPVdOlFWpqtRxE4SjFKLabSu5N/jtKz9qDyJ08Vwh0Tw6zDXGk6OIq1MrqYLIK+e4GEZycpVKEsParQU3eTpVOlKTk1FOTk/zGf6c9odmHEjTfPHxc4GZnqirpfHUpYPIsZg50pYOlTjKVOSy+jJYihThNuopuN1UipTUk3cPv3NFSWpva58C8ndCM1lmT5bV6arvBypYjMMT1pW2atG3rFfm7I+0t03kuouSziK84o0pSyvD4XMsHVnG7o4mniqXS4vwbUpU7+VRp7Nnm5nOf8AO9x845Ztzv6N4N5jleL4e08JVwuDlga0qUKFJdDw1GFRRni24zq1Kqhuo1JtdN4I/tcxXtH+JHOVw1y/lx0JwWq5VnGqsZhaGZwwePljquPnSqxqQoYemqUHTi6kITbk5NKLi21eTD0f9nnmWMzbkv4VYrH1pVasMnnhoyfdU6OJq0qa+ZQhFL0R2LPmvLfwsqcE+BGhuFmJqU6mL07k1DC4ydK3RLFNddeUfNOrObT8V3PpQHj97br5WuG30dxP2k7l8t1vvd+Fvb/eXkn2GidNPbdfK1w2+j2J+0ncrlvV+Xfhb9C8k+w0Tx9X+jH6H2e/W5fR/wDb6Nt6Cy8glZkngfrEWXkSCG7bASRdeZF77WHT6gWAAAh9iQBSz8hZ+RcBW1Ut90S+zJIDN91QT0+pIbtWz8gXIBtUE9PqOn1BtFriy8iyViPEG9osvIrYv0+pAN7V39Rv6lgGq7+o39SwArv6i78yWr7kW9QF35lrLyKFur0CpUPuRa5L3Ee4ZtFvQmz8ixHV6CTTVbehDXoXtfe5DVtzLGyqWfkLPyLAwt2rb0It6FyJBsqtl5ESS8kWEo9tw3fdTpX+L/IUnHyj/IbqOy3IlHtuGzJx7ehFl4o36bq1ysqfqFzJg16FZR33Rv7v1KzjZ2uTpcyY29Cs4+htb1KtXMsVMmDjs/iv9BTp9P5DkyjsUcLu9xYuZMJLbZFLPyZyZU9u5X3fqTqrmcYyWz2K9K/xV+g1I6fUxUrBp22RWz8mbNW3IFi5k4/T/wB7/IUnFqT2Zyun1M5R+M9zLNLmXdx5JrwaK233RvON1a5Rwsr3/kJsdJkzmttkZtbPY2bvsVcb+JmlzJjb0KuKv2/kN/d+pSUN+5Nm3SZMuleRnKO90tvmN5R9Skl4GaXKwkvQrJPp2Rt7v1KyXdGOkyflc+4h6G0rjllmo9WZXluKlTVVUcTiYwm4NtKVm72bT39D+lk+cZVqHLqOc5HmNDH4HEdXusRh5KcJ9MnF2a2dmmn6pnXnmL4GcRuIfECGf6VymhicFHLqOHc6mLp0n1xlNtWk0+0lufX+B2k860RwuyTTGosPChmGC+E++pwqRmo9eJqzjaUW07xmns/Gx9Dm6bgw6bHkwz3lfd8Hyul63qeTrc+Hk4tYT0y+L9nKLv8Aiv8AQVlHb8U3lsyrV1Y+bez7cyYdK/xV+gycd38X+Q5Xu/Uo42b3Ms2uZ93GlGz/ABSrW3Y3lC7vcq4WV7marpMmHS/8X+Qp0ryOQZ29SbNukyYTjv8AilJR27HIauZv42xlmnWZMXH0KdK8l+g2lG0nuVlHYyx0mTKUV/ir9Bilvujk9Jn7v1Ms0vGsJR3do/oRRxfjF/oN5Rs3v2Kyaat2J1t0l7dmLjs/i/yGVrd0bt22KSje29rGV0lYSXfYrb0N3Cye5Tp9SbNukyceUd38UpKK8jkyj33M3C/iZZp1mXZxpR7/ABSjXmjkONm9yk4X8SdOsyYOPeyKWt3Rv02drlJRv4mOkyceUe/xf5COj/vf5Ddwsr3KN2M06S/Bxppb2W/zFOl+TORKO7dyrVidOsy7acZqzexWUb/vbm013KJXJs06S+9x7PyZSUd/xTkW9Sso33vYzW3WZONNK2y/QZOKv+Kcnp9SkobvczVdccvcwlFW2SM5bG3T6lZU+p3uTY7S6YTjtsjJrd7HIatuUlHxv+YizbpjWEo/96ZST6uzOQ1cpKO7Ms0645OPKO34plKLu7Rf6DlyV1YylG0u5Njrjk47i13RVpX2RyJQ6rb2sZSXS7XMdMcmE428DNp7qxyZRvbe1jKcbN+hNjrjkwlFO1kv0GUlu9jcq4d3cmx2l0wlH0M2vBnJcL+JlONr+hmtOuOTCcfJGbT32Nyk47N3/kJsdccvcwlFN36UzHpfW9trs5LVikod3cmzbrLpx6kW2rJ2+Yza27HJasYvcmx1xrJpX3X8hjKL6XaLv6I5Eo79yr3JrrK4rT8Uyjj5x/kOTUjutzGT6tuxlm3bHLbCcd9kUkttkmbtWdj+JmWYKV8Ph5fE7Sl/jei9PXx+bvklt0cnPjw47rHMsf7xuhh38W1pyX770Xp6+Pzd/wCfSoVK9RUqUbt9/JLzfobUcPVxNRU6Su33fgl5v0P7OHwcMHTUIbt/jStu3/V6F2zCdni4sM+sz8+fowoYanhabhBXb3lK27f/AL+BazfdbfMbyjdt3/kKSjt3OF7932+OY4Ty49mE473tsZNW8DlNd0ZSha25Dvjlpx5K/gZSj2+L/IchxsUlG9vQyx2mTizj8Z2W3zFGu2xyKkbX3/kMWrEWO2N97GV1fufPOPHya5j/APV0f+uj6PKN/E+c8elbhpmL/wC/o/8AXR7/AAv+u8X70fl/bu/7sdd/ys/+mu8HsTPkF139L39iw56KnnV7Ez5Bdd/S9/YsOeip/WH/AD4ADz75uPapYbghxUxfBzhNw7oawznJaqw+b4zFYqcKFPE2TeGo06cXKpKN0pybSjJOKTabQfz+abgR7SrV/HjVGouBnFjG5bofGTwryjCYfU7wMKMI4WlGpH3N/iv3sajb/fNuW17L5R97F7YT+GzN/ry/6z7Tob2xfAzNuEmZ6z1zpjNMl1flVSlQWmcJUWJlmMp36amGryUIqmulufWk4KyXvG49X9vhV7Xvlj1zRzJa5o53oLEYDDzxNNZhQ+F0sXGNv7nSnQ6pOq7u0ZQSdtm3sB17+9i9sJ/DZm/15f8AWdw+RDhtzZ8Ocj1dR5q9a4nUGLx2LwksleIzl5hKhThCoq1pP8VNyhtffpbsvH51lftk+UvMc7hlWLyfiHleFnVdN5nisnw8sNCN7e8caWJnW6bb2VNyt4X2O6ekNYaW1/pnLtZ6Kz7B51kmbUFiMFjsHVVSlWg3a6a8U0007NNNNJppB0I5U+X7nn0Xzmag4h8X86zmegcVVzZ1auJ1TTxuGzCnOUvgkaeGjVlOCi3CUVKnT6IxaVvxX6IgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHzbjZzFcGuXTJ8Bn3GXW9HTuDzXESwuCcsJiMVUr1Ix6pKNPD06k2krXlayurtNq/9FcaeFj4V0uN89a4CnoatgaeYwzqq5woLDzajGTUkpRbk1HpaUlLZpPY+b83fJzoXnA09kOSaw1HnOSV9OYurisFisudOTtVjGNWE4VItSTUI2aaaa8U2n/dnyt8OK/LTQ5V8Zic5q6QpZXRymdf4TCONnCnUjVU3UUOlTc4pu0Et2kktgPz/AOEB5NPyg9L/APn1f/YH4QHk0/KD0v8A+fV/9g+M/gb+Ub/n+vv/AEzR/UD8Dfyjf8/19/6Zo/qAPs65/wDk1k0lzB6Xu/OpVS/T0H2rB6U0rgM3rajy7TeU4fM8Wm6+Po4OnDEVlK13KqkpSvZXu3eyOl8fY48oyabxuvWu9nnNG38wd4sNh6GDw9LCYamqdGhCNOnCPaMUkkl6JIDYAAeP3tuvla4bfR7E/aTuZy3fuduFv0KyP7DROmftuvla4bfR7E/aTuby3K/Ltwt+heSfYaJ5Orm8Y/Qez/63L6P/ALfRAT0+o6fU8GtP1cukWfkLX8CwGjavT6MWfkWA1o2rZ+QLENXY1YbQCyVtwYbVBPT6kPYN2h7Ijf1/QWSuTa29w3aLPyBPV6C197hm1XfwITfiWasQG72XXmSV6fUkA7+BG99/MsQAuvMWRHT6i9trATZEW3tbYlO5IbtFkLIkA2q15BLfdFiA3aLR9Cln5F+n1HT6g2ql5jZbkvYhq4am68ytn5E9PqSBW7RK32Yauxa24bssipbq9Co03aSLX8Cy3Q7etxY1Vxt4Mruy9/QgyzRtF5Ebsvba5BhtUjuXI6fUa02VRrYr0p91c0tfYWttcKlZSgul7Mp0v/F/kNyOn1Cplph0ryKuKT7HIcL33Ke79TNKmTFxVnsyvT6GpHT6mWaXK43S/wDF/kHS/wDF/kN+n1HT6k6X5nF6L7dLKuFtrM5ChZ3uVqR3W40uZe5h0ryM5RTk9jkSjsUcLvuZYuZaYSjfwZVwdnszkOFle5Djs9zNLmbi+7X+Kx0LyZyOn1KSVpMyzS5kwcH4JlHFX3W5yG7FHDqle9idLmTjzitrIq47dmcipC1t/PwM+m6tczTpMmHT6FHBXfxTkOFvEq499ybNukzfFeLXMdkvCjVMNL47TONx1WWFp4r3tGtCEbTcklZq910v9J+74da0wvEbReX6zwWAq4OjmPvemhVkpSj0Vp03drZ3cG/mZ+E4vctOA4s6sjqnE6txGXTjhKeF9zDBxqpqDk73cl36u1vA/e8NtCUuHGh8t0Xh8xnjqeXe+tiJ0lBz95WnUd4ptKznbv4XPoc86SdNj8l+n73zOlz678Mz+W/Ve70//r9B0Xd2ijjv2ZyVCytczlH4z3Pm6j7Uy248k09rkSirN2dzaULu9yrh8W1zLFzKdnH6W/BlWtt0chQsrXKe79TLHSZRxpRs/irYq4uz+K/0HIcOl2vcq47dzLHSZOP027pmfRbtFnKcLu9zNuxNjpM3HlBXexRx27fyHIau7lGrmWOkycfpS8LFHG3gzkShd3uUlCy7mWOsyceUb3utynS/8X+Q5DjsyvT6k2OkycZ092+llXC1tmjkyX8hlL41vCxjpMqxaVnsYuNvBnIcb33Kyja3jcyx0lYON+6KyivI2cb+JScLeJLpMnHlBWbUXe5TpfkzkOO3co1YmzTrMnHcbN7MpNdrJnJktnuZKN/EzTpMmDjtZozlHfZPsciUe+5Xp9SbHWZOO0rPYo4ryN3C7buVnC3iZY6TJx3BW3iZyjZ7HJlB2e/8hjONna5NjrjkxcLLsynSvI3burFJR37madJfi48o2V0mUa37HJlHYylC8nuZY645OM4JLsyri/BM5Eo7FGrE2adZk404q3Yo47PZnIlT/wC+/kKOPqTY6zL4OLKFldJlHHfdHKlCy7mbXgTY6zLbjNWXZlJRW7tuciULK9zNomzTtjkwat3RnOCd3Ztm8o9XiUas7E6dZXHs13TRWUU03a7ZvKHVbe1jKS6Xa5ljpMtuNODXZP8AQQ4q3Y5Eo3tvaxm6e73/AJCdOsyYNW7JlJxTXY3nG1t7mUtk2ZZ8XXHJg4JP8VlJRTVrGsn1b9irXfch2lcecN9kyrivI5DVzJxt8buZY6zJhKFn2M+iPe1vznIl8bfsZVI/FtcmuuNYTUb7GfT6G3u/UiULK9/5CbHWZT0YSim+xhKnZbROU1Y/hZpmXvb4bDT+J2nJfvvRenr4/N3TG29mZ884ZuuPmeYqo3h8O/idpyX770Xp6+Pzd/51LDzxM1SpRu34+CXm/Q2pYWriaqpUldvx8EvN+h/dw2DpYOiqcN295Sa3k/6vQq/mzU9Xl48c+rz8+Xoww2Dp4Sn7umm295Stu3/V6F5RTXY3aszOULLucb3vd9rDWMkno48ou7snYpNJI5DVzKcLLv8AyEWO8ycdrxsVav3RtJWM2rmWO0rGcVvs7GMk12TOTPa6Mmrk2OuNYONzOUErXizkSVrvyM5u9ibHbHKuLKO+6dj51x8VuGWY/wDjKP8A10fSpxumz5vx9VuGWY7/APCUf+uj2eFz+e8X0x+Y9ur/ALs9d/ys/sru37Ez5Bdd/S9/YsOeip51exM+QXXf0vf2LDnoqf1h/wA/A8yfZH6Uy3VPEnjnxr1Bg6GM1HPPFgsPjKkVKpQVeriK2JcG7uLqS91d33ULXd2emx5DcIONmG9mpzccWuGXFzIc1eiNX41Zll2MwVBVakKKqVamDrwi2lUg6dadOp0tuNSnazcWgP6/tPeEPDWpzd8EXgNNYShjuIOY0MLqSGFj0SzCDx2HpQqTjHZ1JRq1YuduqSik2+lW5XOzwu0Lxf8AaF8HeXBaay3T2namT0amPqZRgqOEr4inOWIqTp9cIp2VLCQpwTuoOU2kru/57WPHHJOez2i3BSrweyrM6mntDY3BY6eMxVD3U60MJifhuIrODf8Ac6aUIU11Wbl6yijsB7Sfli4w6u1Pozmh5c6OMxOutB044avhMDFTxc8PTqyrUK1Cm01VlCdSqpU7NzjUSSaTTD7Vqf2eXJ7qbSGJ0cuCGQ5TTq4f3NHMMspujj8PJJqNSOIu5ymm0/juSk18ZSV0+uvses1z3JMNxs4IY/M3j8t0FqTDPBVVO8XOvLFUargrtRg3gYTSTavOT7tt/I5e0o59OIOXLhZpDl/w+D1rmCWBePweR474RRnO8feQo1ZOFGa7uVRuEbNtJLb6L7EihSraG4rZ5WTq5hjc7wEMRiZycqlVRo1ZR6m93aVWo9/GT8wPTAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHwnnD5qMq5Q+GGX8Ss20fi9SQzHPKGR08Hh8XHDOM6lCvW945yjLZLDyVkm25Lwuzpx+HE0l+Txm31ip/2c9H9Y6E0RxEyf9j3EHRuR6myv3sa6wOcZdRxuH97G6jP3dWMo9STdna6u/M/EfencrH5NPCr6m5d+pA6L/hxNJfk8Zt9Yqf8AZx+HE0l+Txm31ip/2c+e+1h4T8LOHPE3gzgOH3DTSul8Lmnwz4dRybJsNgqeKtiMKl72NKEVUspSS6k9pPzZ6VfencrH5NPCr6m5d+pA6Lr24ekW11cvObqN93+yKlf9HuD01weKp47CUMZSU1DEU41YKcXGSUkmk0+z33R8zhyp8r1GpGtR5buFsJwkpRlHR2XJxaezT9zdNM+qAAAB4/e26+Vrht9HcT9pO5vLb+524W/QvJPsNE6Ze27+Vrht9HcT9pO5vLb+524W/QvJPsNE8vVfox9/wC65cvofRgStyen1PC/VbLxIe7IJSvuDzVAJt6kBsqbMdier0Fr73BtUEtW3IDNrXiQ7X7EAN2W8ibMJ2F/QzTd7RZeRNmQTf0NZtDXmiGvIs3cCxsqlmWsrdi3T6kGaFZEWfkW6b+NrC3gZZoVLWXkiOn1JGtm9oa8kV3T9PnLkNeJhLpBXf1/SWJ6fUN2qtluLomUd+5CVmGpAK9XoBLa8SO+yQtfe5KVtwIsybLyJAbvarXkOl+RYA2p0+gsvIuV6fUN2iztsitm+5du2xAarZizLACLrsQ7eAfcJXDZagmzJt6khs2pYhrcs+5BOmyq2a79ibxJI6fUCGv0FWtzS21irVg2VTpX+L/IOlf4v8hcBe2Hu35Ihxs7NGpEodXjYzSplti4rwSKyjfukbe79Ss42drixUyYyp7fimbhu9lY5Et0VcdnuSuZOPKLt2IcHbsbdPqQ47PczTp5q4/T6FXC77I3936lJQs3uZqqmUYThe1kijg/I3mrFfBozTpMqws33Vyrj3sjb3fqVlG0nuZqx0mTDofiivSr26Tfp9SkoWvK5Nm1zJ1c5j+PnEPhpxCp6c0vicDSwTy+jibVsLGpLrlKabu32tFbH17gdq3Ode8K8l1ZqCdKeYY74T76VKmoQfRiatONktl8WC/Puf19UcKOHetMzWcap0ngsxxipqiq1ZSclBNtLZrZNv9J/YyHTuS6VyjD5Bp7LqOAy/C9fuaFK/RDqk5ytdt7yk387Pfz83Bl0+PHhhrL4vm9LwdTx9Xny8me8L6Ry+n0KuCu9kbSVna9yjXifNs2+1MmLivJFJQduyN2r7lJK+xlmlysLW7oz6X5HIdPfuVlCye/8hlm3WZRx3BvwKuHojfp9SnT6mWLmTCUWn2KSirfio3lC7vcrKFl3MsdJkw6V/i/yGfRbwRyOn1KS+Kr9ybNOkyceUd+yKOP5zdq/oUULu1zNbdJk48ovq2RWUWl28TaUN73/AJCJRvsTZp1mTjuN12KdK8kbyVtjOUe25lm3WZMXB3bsrFXBeMUzZor0+pNi5XHlGzdkisoX7o2lH4z3Kyj23DpjXHklaySv8xnKLVvio26fj/pE4X8SHWXTBx77Io428DVrexSUe24sdJkwnB77LuUcbOzRyHHbuZVI7rcmx1xy2wlF3e2xWSfkbyh8W9zOUe25NmnWZMHF+WxRwv4HIlHZ7lPd+pNm3WZOO4pd0jOUb7pK1jeUdmrlFCytcnWnWZe9x+l+RSUX1PY2atuVauZZt1lYdN+6Kzg2/wAVdjaXxVfuVtfclcy97jdL8jOUZJ3schxsr3uUkrpmWO2OXdx2roynTk22onIcLLvchK5Nm3aZe+ONa+1jOcHfsjfo6d73KyjdMmx0mXdxul+RSce+yubyja3qUavcyx2mTj9EvIpOG72VzkNWKSj3d/DsTZt0mTiuLj38SrV07G8odVt7FHSsm+rwJdZkwcdvjL/1mU497pWORKN7b2sZuN/ik2OuOTHpj/ir9BnOOzsl+g2lHodr3/MVaurGWO0vvcZxdysrO6srms1Zmbju3ch2lZTi77Ip0+htLzKNWVzLHSZMZQv2ijF7qxyW7n5zOMzU08NhJ3h+LOa/fei9PXx+bvmtty5pxzdZZtmaq3wuGfxO05r996L09fH5u/8AKp0qlWapU4uUpOyXma0qVStUVKlFynLZLzP7+Ay6ngqd9pVJL40rfyL0/wBv6Ett8ry4Y59Tn5svRGFwVLB01TppSb3lNreT/wDUvJEzg7bR8TkvYpKN1a5yvfu+vx6wkk9HF6WvAo4+aORKNn3MnG/iTZt6JkwlF3ukUcU1ayN5K2xm4WV73JdpXHnB3eyMXFructq6ZhOn23Msdscvcxcb32TKSi9rJG9rbFJRIs26zJx2rXTW5nKF/wB6jkTju3f+QzMsdMa40otN3R835gElwwzKyX+Eo/8AXR9MnG9/VnzXmCjbhhmW/wDwlH/ro9nhk/nnF9Mfmvbm79meu/5Wf/TXdX2JnyC67+l7+xYc9FTzq9iZ8guu/pe/sWHPRU/qz/AIfN+MnLtwU5g8rw2UcYuHmW6kpYFylhalaVSjiMP1W6lTr0ZRqwTsrqM0nZXTsrfSAB8r4K8r/Abl2hjVwb4b4DT1bMko4rEqtWxWJqwTuoOviJzq9F0n0dXTdXtc+qHnXzTezT4zceOPGqOK+mOPGDybLM+nhZYfL8V8K68OqWFpUXH+5tRt1U21Zdmr3d2/lH4HPmF/KVyj9GO/rA9az5TwD5ZeEPLPlec5NwhyHE5XhM9xixuMhXx1bE3nGLjBRdSTcYpNpJfnbZ5Fczns8+Zblu0B+2THiFU1lkuDm/utLKa2JjVy6lZdNecJu8qV21KS/F2bVm2v0nA/2b3EzmC4Z5PxT4e80GWVsqzaDvSrRxka+FrxdqlCrFSajUg7ppNpqzTaabD2nB5WaN9kdx805q/ItQ4vmPy2pQyvMsNjasKaxrnKFKrGbUU5JXaTtdrfxR6pgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHXHnr4PcdeNvB7LtIcv2s/wBjWoKGocPj8XifutXy732ChQxEZ0ve0U5O9SpRl0tWfRd7pHQz8Hj7TL+H3/7fsz/9gD9b7Zb5WeBX/wCPfacIeqh4v6m9ljz6a0xGExesOIuQZ7XwF/gtXMtV4zFTw92m+h1KTcbtJu1t0vJH9/8AB4+0y/h9/wDt+zP/ANgD2BB4/wAPZ5e0zU4tcf8Aos/xv2fZnt+iF/0Hrxg6WIo4ShRxeJ+E14U4xq1uhQ95NJJy6Vsru7su1wOQAAPH723fytcNvo7iftJ3P5blfl24Wu3/ABKyP7DROmHtuvla4bfR3E/aTujy2K/Lpws+hWSfYaJ5eq/Rj73gN1y5fQ+iJbkktWRB4X6natmStkSSkDaBZeRLVkQDatmStkSSldBqBZeRLRAPUsvIhryJJSugzaqW4fZkgNUJsyelEgVsxZlgBW78yU14jpQ6UGy6G/JlS3Sh0oHmqoLdKIB5qlNIqCUrhsqLLyI39f0l+lFRZst0rZizNEg1ZGaazsyLLyLkdKMs0S6RZ+Qs0WIl2MN90AAN2q0/AJNFgG77BXf1/SG7MsCVHz9xZeRIAh9mVLkdKDZdKhLyRbpRD27Bu0NPcrd+Zdu/cr0oNE14kPfsT0olKwbLpWzILXb2Y6UDaLMi3mXKy7mabvsiy8ir2LES7izTZdI6fQWt3RYiXcxsqrXkVcb+Fy63ZPSg3emLg/Jdyjg7fim12x03TuFTLTjdPoVad+xyOhepSUUpOzZljrMmThbukUcbu9kbSV1uUasSrGspwvb4qM3Tav8AFX8hyGkyHFWZml7scbp9Crhd9kcjoXqZyirvuZZpcy3WEoPayRSUJWexvNWKNXVjLHSZV0l5vcFqutxXpVMlwebVMP8AcjDpvCwqOHV11Lq8Va/b17H3/lnoZjS4I6bp5rTxEMUvhnXHEJqov77rNXUt+1rX8LH6bV3GHhloPNlkmrdWUMux0qMa6o1KVSTcJNpO8YtWbT8b7H9zTepMi1hkuH1HprMIY7LcZ1+5xEIySn0TlCVlJJq0otbrwPo9R1GfJ0uHHlhqT3vl9J0/Hx9bnzY8m7f7Pw9HNcfNIrKHxXZI3lFXKyirM+Xp9yZMPdvyRm47vY5BVwTd7vcyzTpMnHcX5FZR23RvOKTKtK3dmadJkw6V/i/yGVreByXszOUEk92Q6SsWrvsikldbJM26UU6EntcWOkunHcJX7fylZwdt0by2ZE4JqxmlzL0cbpX+L/IZ9LXdWOTJJOyM5wTVibNukyceS77Fel+Ru4JJ7sz6UZZp2mTCUHd7IpKL2+KjkSS3KSj1d7k2OkyrjtK1rFHFo2cVfxInFLsT6usyYSj6bmbi/FHJcVvuzOUfUyzTpjkwlDZtRVynQ/FHIlFWZmYuVx5Rs3dLYrKN7WS2N5RTv33M7W7PuTZp1xrDpd90VnFX7LsbSileW9zOyZNm3WZMZK6aSVzOUGrfFRyGt3uysop23Zmq6TLTjSWz23M2mnZm7irvuUnBN92ZY645MZxSj2Ri014HIburMq4J77kWbdscnHcV/ioynHfsclq5nKCu92ZZp1xyYzimtkv0GfTbaxsQ4p+ZjpMterjyjdbpFXFbrpVzVq5SSW5NmnSVx5wdt0VsltY5DSaszKUVdk2OsrCcNtkjJxbvZHJaTVmZy2bJs27Y1x3Ta7pGcovqe2xyHv3M5bMyzTtjkwlF7fFRSUXd7bG80mZyit92TY6Y5MJRbtZIo13TN0jOUFu/nJs27SsKke1or1MpR2dkrnIkZyileW9ybHXGsOhvdx/SUlBq91axuUqRTi++5mo6zKuPaLd2rmc4W8FY2kknZFWrqxFm3aX3sGkvBGco3XY5EoK/ifnc+zR9TwOHmrdqsovdv/F/r/R4NNJa3LlmE3XHzfNVVvhcLL4nac1++9F6evj83f8AjxozrzjSo03OcnZJeP8A7+ZrTo1MRVjSoxcpydkku5+ky7K6WXUupvrrS/Hl4L0Xp/t/Qlt/N9HmwmXU5bt7McBlcMvp7pSrSXxp/wDqXp/7+SWso9Ku0jlSs+5lOCa8Tle76vHJhNSMFFvwM5J2OT0qOyv+cznBJbNk16McmDXmkYzhK2y8fM5Mlv3ZTuRZp1xycaSsndK9jM5E4q779jGUUltcmx2xrGUXe6RSSXkjdoznFK3cx2xrjTi97JehnuvxjkteBlOC8bmWO2NYys/BGcovayRtKKV7NlWrkWbdZXFlGz3R825hUv2rcy7f4Sh/10fTpwW73PmXMKrcLcyf/wBFof8AXR7fDP65x/TP/p+a9t7/ALt9d/ys/sruh7Ez5Bdd/S9/YsOeip51exM+QXXf0vf2LDnoqf1R/gUAAAHmzzYez65vONPH/VfEzhxxtyDK9OZ1PCywGCxuf5lhKuHjTwlKlKLpUMPOnFdcJtNSd003ZtpfJPwU/Pp+UJpL62Zz/YwPVjiZq3QOiNCZ1qPidm2W5fpjDYOqsxqZhOKo1KLg1Km1L8dzV4qCu5NpJNtI86vYl5tnGJwHGLKcL76Gl8PmGVYvAUKs21RxFWOKjPpVmnJ06VBTd7rohs73X4DA+x25ntXZjRp8VePelngaUlarQxmYZrWpxf43TTr0qKvbsutXu7tePpHy0ctnD/lb4Z4bhtoGFevB1njMxzHFWeIzDFyjGM607JKKtFRjFbRikt3dsPrQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8fvbdfKzw2+juJ+0ndPlrf+504Wb/8Ssk+w0TpZ7bv5WuG30dxP2k7pct37nThX9Csj+w0Ty9V+jH3fAu3Jl9D6Ts9itmCepnhfp9oAJSDZdoBPSh0oCCrW5YlK6ArGLuTZk9t0E7sCLMPyLFX3AqluWAAFbO5YlK6ArFENMs1YBsulAW6UOlBu1bNkuLsSkkH2B5qzsywAJdoa3Isy6RDXgGqAt0odKAJ7BvYdKDVkG7VAAJdALJXQaF7t2qTZkE9TMk0S7OxBJA01DW5N0COlGWaN7Q1fdLuQk13L9lt4FW2zCXQ9ytmWAV6gIbsE7g3tL3K2ZYBsulBZstZMJWDdqvsVsy/iT0oG2dmStkSA2VD3RFmS3YjqZt7tl7oJsyelDtshe7fpRZkFu+zHSidHqzBPSiGrGK2h7ohxdiy3YfkGysXFtFXCVnsb9KIklZ7sLmWnHsysk7myin3bKyirvuZY6TJhOLexTplbsciSXmyrimmSuZOO032KyTs14m6il5mfSupq78TLF7dVOZXl64k8T+ItPUmlcJgauCjl1HDOVfFRpy64zqNqzXa0lufZeBGjM84fcKMj0jqOlSp5hgPhPvo0qinBdeJq1I2a2fxZr890fi+N/NEuDms4aRWhXm/XgqWM+EfdP3FuuUl09HuZdui97737Kx9I4Ua9XFDh/lWuXlX3M+6fv8A+9fhHvvd+7r1KX4/TG9+i/ZWvbe139PqMupvS4TkxkwnpXyukx6OdZnlxZbz98fp5xbexVp9jZ7Mr0K97s+TY+5K48otPsUcZXexyJrddyjimZY6TJhKLuU6JeRyJQjfxKNWRljpMmEotPsVaduxu4pu7bKNK3dk2OkyYWsZpNbvY3ezKSimrGWadJdsZXcm7GdmcjoXqZ9KJdZWUou5nUTstjeUVczkrrcyz3rlYPa5nNXW3mchwT8ysqaSvuZe7rMpHHcJWexRxaOQ4rfdmU1YmzTpMtsXGXexVq5s0V6F6mXu6yuPKMrvYpKLRyJRSezZlNJk2adMcmMkZzTfY2cU77sq4pdmzLNusyY9EvIzmntsblZRTtdsl0mTjtO3YobygmmnczlBJ+JljpLGE4vdlLNG7V7oznFK3cl1lYzTs7GfRLyOQ0rd2ZvZk2adca404vpdiqi0jkSgrPuZySTsjLNuuOTjpbkSi2zVwS8zNuzsS6ysbMrJO5q0Ul3J1p1l249mikk97G84JK+5RxW/cx2mW3G6WvAWZrKJm3ZtImzTp5mEoyt2KSi7M5DSaszOSSbSMs26yuO013KNN3sjeUE7bso42fcl2mW2EotGckcmUE/FmU4pPxJs0645ONJMq0bOKZm0t93sTY7SsZJvsZyvZrxN2rmUopK5Nm3XGsJQk3dIo15nIM6kIpOSve5LpMvdWE4tvYzlF77G5/BzzOFS6sHg6n90d1Umv3vml6+b8Pn7ZraryTj71hnOcqj1YPCT/unac0/xfRevm/D5+356nh6leao0aTlObtFLx/q+c0hh6lerGjSi5zntFJdz9TlmWUsvpbvrrT/Gn5ei9P8Ab+hJezjjMufLd9GGWZXTy6lvaVaa+PLy9F6f7f0Jclxa3ZyZxV/EyOdfTw1jNRhKLb2Rm+xyZRV/EycETY745MZRb7IxUWu5yX3MppJbEuuNYzi29jF9jkmHSiXbGs2u5nNPpNpJXZRpNWZmo6yuLKMrt22KnJlCNn37HHmlHz3Isdscmc4t3Zi0cloznFK3cx2xyYTjJ3dtrGMos5MkYtXJsdscmUtk7nzHmI34V5n6VKN/+kifT5RVnuz5jzEJLhVmlv8AutD+cR7fDZ/PeP8Aej817b3/AHb67/l5/wDTXcv2JnyC67+l7+xYc9FTzq9iZ8guu/pe/sWHPRU/qT/BQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPH723fytcNvo7iftJ3U5a0vvdOFd1/wASsj+w0TpX7bv5WuG30dxP2k7q8tn7nThX9Csj+wUTzdT+jH3fAv1uX0Po1l5Cy8iUrk9KPFq1+mZ2YtLwLAzQjf1/SN/X9JIFmhUJPwJ6USYK2l4izLAAQ1uSDboQtnuLxEuxBhLpay8ivZk9TFk9/M2Tbdo3YsyXt2HUzDzVFmSrDqZANjt4AAN2iy8ien0J6ULtbeQEWsVs7lm7gCtmLMsA2XStmLMsQ3YN81V6fQWXkT1MgEu0NPw7BX8SyV0GgbRZeRWzLANRtazDaYsmGg3apa8QldFQeapt5Dp9BdodTGmy7Q47ditmXu2QLNtRa67BK3exIaTJpvsqCWrIgxsqrT8Bv6/pLpXRD2YNqpO+5Yqndlg3e1H3BayY6UFSxXvsLW7otZIWTBtTf1/SQ1Y06UVlFXDUXQuiFuyelBuy8SHa/YnpRDVg2XaA1clbslqyM1Ws7MejLCyZlmiXStr+BVwV38VF3t2Ie/cKlrGUb2skVcHb8VG/SiHFWYXMnG6fQj3e7aib9C9SjVm/QyxcyfFOLnLBpPi5qqGq87z/ADfBYiOEp4RUsL7rocYOTT+NFu763422R+64c6By/hlorLtEZVi8Ri8Llvvfd1sR0+8l7ytOq79KS2c2lZdkjrnzU8d+K3DjidS09ozVX3Oy+WV0MQ6PwHDVb1JSqJvqqU5PdRW17bdj7jy86s1Brvg5p/VWqsweNzTHfC/f1/dU6XX0YqtTj8WnGMVaEIrZK9ru7bZ9DqOLqMelwyzylx+D5vTc3TZdXlhx46z99fvum/dFXHvsbSSTsirS3PmWPtysHH/GSuRKPxW0kauKfe5DXgSvdcfpfkVqR+K7JXN5RVyskulmai5k4vRLyKdEvI5PSinSibNusyceUWnukVlHbsjeaV13KdK9TLNOkyYOJnKO1kl3N5RV2VlEmzbrMmKg7bxRm4eiOT0oymklsZZp0mXdg42b2RSVmjaSTTKdC9TLFyz3sJRd3ZbFZw7XijkOKTKTXbuTp0mTjuKs7JX+Yzaa7m7VrlJR6u9zLNu0rGUb7pGdreBtLa8fApJerJdJkwlHbsjPpfkcmUU4u5nKKja19ybNOsyYOPfZFGjkOCs3v2M3FPzM1t0mTjuLu9tisot+BvKK37lelGadZnpxnTd3su5WcO14o3ezaIlHq73J1t0mVcRwd3tsUlFp7pG8tm7eBR7u7JdpWLV12Mpwd9kjdqyK2TFjpLpxmr9kZSg7tpI5EoqK2b7lbJkWbdplpx3HzRScLvZI3krK6KN33JdZkwlFNdjKUXftsciUVFd2Uavcyx1xrjyg2uxR03d/FRvJWV0UJvd1mVcfp9Ck499lexu0Vavcm9q6SuM4td0UlF7tI3lFO27KuK6XuzLNusycZppfGKNJ32W5tKKdt2ZyVnYmx2xu2LjbukUlDvZI5Eo37mTW9ibNOsycaUfQpKJyZRTtuzGSSuTY7Y5bYTi77IzktmmjkbNn53P87jQc8Fgqj97uqlRP8XzS9fN+Hz9s1t0vJMZusc9zn3DlgsFJe87TqJ/i+aXr5vw+ft+co0K1erGjRg5zm7JLx/8AfzNaNGriKsaNGDnObskvH/38z9ZlmT0csoOTanXkl1yXZei9P9vfySyzTjj5ufLd9GGW5RTy+ndpTrTXx5+Xor+H+39CXI6GtzlFJQXS92RZvu+hx3yTUcczcXbsbSSTsipFm3olceUXczdn2scia3XcxcEZe1dcctsZRbd0jOSXikchqxnOCS2uTY7Y1x5Rd9uxj0S8jl9KMpRSW1ybNu0ycaUXfsVcXbsbyineW9zPuTY6y7YTi7vy+czkr22TORKK9TGaUfPcmzTrjXHmmm9rFEvM5UqcZJve9jCaUfPcmx2xyZTi7vZWsYyi9rJHJfZmTVzPV2xunGmrJ7HzHmK+SnM35VKH85E+pTgmm7s+X8xatwozTvvUofzkT2eGzXWcf0x+c9tr/u31v/Lz/wCmu43sTPkF139L39iw56KnnV7Ez5Bdd/S9/YsOeip/UX+DQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeP3tuvlZ4bP8A+l3E/aWd1eWtX5dOFf0KyP7BROlXtuvlZ4bL/wCl3E/aWd1+Wv8Ac58K/oVkf2Giebqf0Y+54H+ty+h9GtbdjqRJHSjxv0t0kABiGrkdLJbsR1MNl0nqQtfdFSy7Bu9+gkSAGbV6WSlYkhuzBdj3RBKYsmPUl0qTZsnpRJmjarTRBZpMNWQ02XapDV2SBY0SsH2AM0Kll2I6USYS6Q0LW3ZJF29mDZ1IdSHSh0oN2h7sB7MJXDNhPSx0odTCvT1QCUr7sNAQAAKvuQWsmOlBUsVBLViAS7Q1dkNNFiGkw1C7livZ7DqYbLofcgtZPfzIasDaAAZZtu0NXZK2QIbGtepLtD7gd3uT0ozTdoABipdoHUgOlAGrhbIkA2olcNWLWSFkwrardiOpEtXI6UFS6O+yIs0TZIl9gxQhq7JLJXQ02VVKwLdKDVkNKU6WVlF3Zcq+5LZVelkST6X2LkS7NBW9PxOq+H/CzUuZrMdZaa0/j8eqUaaq46lTlV92m2leW9k27fOz+3p3JdP5DlFDKdL4HB4PK6HX7ihhIpUo3m5S6VHZXk5N28Wzp1zj8PNfan4t0sy01ofUGb4RZRhqTxGBy2tXpKanVbj1Qi1dJptXurrzOw3K/kmb5BwL01lGfZVjMtx9D4b73C4uhOjVp3xleS6oSSavFpq63TT7NH0Ofg8vTY8nn38zwdP1Pn6rLj8mvn+L6fKJDi7M1lFXKtK3dnzbH1Zky6WZSW7N0kUlBXbuzLHSVjKJVxdn2NZRVyhNml7rJqxnKDs90az2e3kJRTi7mWbdZdMFsjJQa8jk9C9TPpRlmnSZMJx+M+xm90cl003e5j0omx1xrKUSsouxrKKuVlEmx0mTJrZ7oo9ldmr8ik4JqxlmnSX4sJpu7XijNRa7tHJcI2fczlFRta+5Lrjl7mD2vcpL41vQ2cFu9zPpRlmnWVi4u5WUXsayS3KtJmOkrjuL6n2KSi/Q3lFK8k3e5m9ybNOkrJ+RnOLdrG7gr92VnFK3cmzbrMnHlB2e6MpROTJXTRlKKuS645MmnZmc4t2tY2a8CskZY6yuPKnJp2sZyg4uxyJPdoznZu7Js2642sWrppGbVjbpRWcE33ZmrHSX3OPKLsVNWrlHHd9ybNusvurGVN+aM2mnZm7Vyk1uu5LrK481ZblWro1aTVmZy2bXkZZp2xrKUGl3Rk4u5u91ZlHHd9ybNusrGasrsykm22chpNWZnKEd+5LrjdMGrlGrM2klHtfco1e5ljpKxnFu3bYynB77rsbtWKSindtsmzbtjdOK4tENXVjaSW27Mns2S743bGUWrbopKLszdxUu5+a1BqD3DngMDP8AuibVSpH956J+fm/D5+031Xc/LO7HUGeOg5YHAzvV7VKkX+L5pPz834fP2/M0qFXE1Y0KNNyqTdlFLu//AH8TWjh6uKqwoUIOc5u0UvH+r5/A/XZVk9LK6V21OvPac/Bei8l/t7+SSzSMd813fRjleTU8sp3vGdea+PPy9F6f7e/klzWje3mZzgld79yHuw7TUYzhKUrqxkcgycEk2myb2rtjfcylFyezRk4uz7G5SUEk92TY6y6YNWMXTaRvPZ7eRVpNWZFm3fG6cdxdyj3RvKKT7sylsr+pLrKxlFt3Rn2Nik4JLZsyx1xvuYzi2m9t14nH6H6HKk7JpGK7k2bd8axaa2ZRq5rNfGZRqxLrLvuxqQk79t0YuPR3tucpq9zGrBO3fxJs7u2GXuYSV22Zyi5djVxV7eBWcVG3fcmx2mWmEla9/A+Xcx6/7E2Zv/6JQ/nIn1SUVZvc+WcyKtwlzP8A8bQ/nEe3w3+ucf0z/wCn5z21u/Zzrf8Al5/9NdwfYmfILrv6Xv7Fhz0VPOr2JnyC67+l7+xYc9FT+nv8JAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8fvbd/K1w2+juJ+0ndflqTfLnwrsv+JOR/YKJ0o9t18rXDb6O4r7Szuzy0/uc+Ff0KyP7BRPN1P6Mfb8E7cmX0PotmCw6UeN+jt2qLNgtHsG+aq2YLDpQPNVQS0EroG1XuiLMu0QDYASldA2gXXmS0VsmCVJNmQTdoKuSLNB7kttkBm9q2ZN/UN2Fk9/MG9p7h7kJJEhs0rZizLEN2YNi2RFncdTJTuwS1FmQXKGabLtDW5FmWJSuNNUswadKKdKMs0IsxZlg9kYK2YJ6mGgb2izFmhdoXbDdgBPSg1RrcmKJasAKvuLMmyZIVtm4u7EUXsmOlA2zs7izLEN2DVSyewsnv5kPZhsuk3QbKgXubTZ+RVrcut9mQ9mZpSlmLMsDLNEumdmStkSDGy7QLomyezIcUg3aSH2JKtvcCCLfGvYst2GrIK2gBq4DUSKlmkyH3BvSboh3e9ielDstgqVFmiHuiy37kGdq18z4g8wvC7hhn0dNaxzrE4XHyoQxShTwdWqvdyckneMWr3i9r32P1uitZZBxB0zgtX6XxU8RlmP957ipOlKm5dFSVOV4ySatKElut7X8Tr3zI8r2v8AjBxCp6s01m+n8NhI5dRwjhj8RWhV64Sm27QpSVrTVne+z2PsfAvQOb8MOFWSaIz7E4PEY/LPhPvamEnOdGXvMTVqrpcoxbtGaTuluna6s37ebi6fHgxywy3lfc8PBy9Rl1GWHJjrD3V+7l3M3B3e3ia2THSjwvpS6ceUWn2KtOxvOCb7srKCSe7M0uZMOlvwKOLaN0rGbVjLNOku2EotPsVe6N3FPvcy6SbHSX4snF3KTi7G0oq5WUTLHSVh0S8jOzRyWrGcoR9SbHSZMJRk3dK+xQ5FktkZdC9SbHWZMJJ3bKvdG8oqz3ZToXqZZp0lcdqzKTTdrG8oLqfcq4peLJsdZk47XgZzi3bY5DgrvuVlBbbsy93WZacfpfSUkmbySTaRSSW27Mq5fe48oys9ilmjkSinF3M5xSta5NmnXHLbjzi2nt5lEmchxTT7mUoqNrX3Js06zJjNPcpaxs4p97lJJJ2RNm3XGuPKLu34XKtXN5RTTvczklHtfcyzTrjWDTTZScW3sbtXRm+5NjrjWHS/Iq4u/Y3lFWZlLuY6421x0mt3sVldybsbTS6Vu+5n0om9q6y77sbNFWtzWUSj2Zl7ukrjSjK3YrKLszkTgmrGcorch2xycZpruQbTgmrGTVm0jLNO2OW2Mou3Yo09/M5DSaszKcUm9ybHXHJx5QltsQ9l0+Js0mZSXxn3JdZd+rFq5SSdzeUVtuzOUe+5ljrMmE03axm4u72N2rH5nUefrD9eAwFR+93jVqL955pPz834fP2mzbpM9TdZah1B7hywGAn/AHXeNSov3nmk/Pzfh8/b8pRw9TE1Y0KFNzqTdkkt3/UvXwNKOHq4itChQg51Ju0Uu9//AFL18D9rlGR0cqo3k+vETS95Pw+ZeS/2934JSzHed7+jDKMmpZVSu1Gdea+PO3ZeS8l/t7vwS5007OyNpxV7XKNeBNmnsw1PRxpRdyrWxvNbruYtJXJsd5ltjKLb2KG7VzKUEk92TZt2xrKSbd1uYyV0zkpWM5U4xTavsTY6S6cZpp2Zl0teByXFN3bZk1cmzTvKylF3M5K6sjaez28jOWy2JrrLtjKLuZHIe5k4Il2xyYzi23YyqRl0rY5DXgUqLZdzLHXHL3OM4ys9jOUJbbHIlBJX3ujJtsmu2OVYOLRSRtPuZyiTe7tL72M4tttLuYTjJWujlNGdSKdu/iTY7Y5acY+W8yq/7EmZv/6LQ/nIn1WcUrvfufK+ZXbhJmj/APouH/nInr8Nn884/wB6Pzvtpd+zvW/8vP8A6a7eexM+QXXf0vf2LDnoqedXsTPkF139L39iw56Kn9Pf4WAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4/e27+Vrht9HcT9pO7PLT+5z4V/QnI/sFE6Te26+Vrht9HcV9pZ3Z5af3OXCv6E5H9gonm6n9GPs+DXXJl9D6QT0sjsLs8b9HtPUiHuwAbE7E9SIW7LWQNqPdEqLsuwF2DaelkWaF2LsNl2EdDe+xIuwS7QotbtokXbAAq1dlgBVbdxdPbzJ7iyAjpZBco+4bLoBKJt6A7IaaCdkRdsBsukvdkEpbE7LcG0dLKdLL3ZAbahbINXJJW7DZVOlkGtkUsg2XYuwl2Id1tclb7MM3VQWsg1sZrXo3aoLJbBrYzVaqR0skD6TaE7bMdSJsiGjDZdPbzHSyF3LBUu1SGrl7IhoCnSx0ssA2XShDReyIezDdqdLHSywBLtXpYasS9kFvuw1A6WOwuw3Z0sNWF2A2XaHuiOlkgmtVABjZQBbsPZhu+4VauywDZdK9LIa8CzdivcNl2hbdxa+6J7kPZbBotu5BK37hrYKlddOPXNZmPBnXMNIYXRmGzWE8BSxnv6mNlSac5TXTZQey6O9/E+q8G+INfipw2yjXmIyyGXVMz+EXw0KrqKn7rEVKStJpN3VNPsrXt4XPzfE7lo4c8WtSR1Vqqtm8cbHDQwiWFxUacOiDk1s4N3vJ738j9xw+0JknDXSOA0Vp14mWXZd733LxNRTqP3lWVSV2kk/jTdtltZep7ea9PeDGYT8/314eHHqZz5Xkv5nuf32iLNF3syH2PBp79qlWrssAqVlUjuuxRxdn2NpJXK2T8AuZMellLM2mrPYo0rGWOkyYyi29mjPofociyK2RNm3SVx5RabRn0NLexyXBXexkTZp0mTJqxm4tK7aN5JXKyimuxlm1zLTB7xaMehpb2N5L4zXgVkkS6zJx5Re5VRb8jkOK32M5pRtZWuZY6zJhKL3VynSzZpeRWyIs26zJx5Qd29ijVzkSit9inRHyMs06zJxeh3e67iUXsbTiknZeJTuZp0mW3HcG290UlFq26N5LZsptLur2Ms06Y2+rCSbTSMpwadjkSSV7FGk+5Nm3XHJhLtbxKSi9jaUVdlJJEu0rCW94+JnKLTNpxSu0t7lHv3Js0641i1cylFt3Ru1sVsmZZt2lcd7opKLubSiktilkS6ysJO6sijg2bVIxilZWMya6y/Bh0srOLd912N5RSWxm0mmTZt0mTjuLSu2ivQ3vsbSimuxRpLsZezrLtxrOO7KSTbZyZwjZbGMkl2Jsdsaxkunx7lJRb7eJtOPmUezM9XXGuPODVuxRrZ7nImk7XPy+pdRLDdeX5dU/uv4tSpF/ieaT8/N+Hz9ovZ1mep3Yak1C8N1ZfgJ/3XdVKkX+J5pPz834fP2/JUcPWxNaNChTc6k3ZJd3/UvXwNqOGq4utChQg51Ju0VHu3/wCpeN/A/b5NkVDKKN3aeImvj1PL0Xkv5W934JTJsxvmvdjk+R0cqo3dqmImvj1LbJf4q8l/K+78EudJNPc5DRSUY9LfiZZt6cbqajjTTb2M5q8WkbySuZSW7RNj041jZrZlJxbT3Rv0p+BnNWuTZp0lcaUXco1c5DVzFrYmx2lYyi27ozSszeXcpKKSexNm3aZe5jOEpO6sYvdHJOOTezrjWclZ7mXfZHIaT7mEkoq623JrtjVJRMXGyvc3KSirdibNu0umElcynFtG8lu0VklbsZY6zLTjyi1fczcGzeSW+xmTZp2lYyi0/Ayn4G813ZhPsibHbCs2rmU4t27Gz7FLXJdpWEotI+U8zHyRZp6VcP8AzkT6zOK6WfKOZhW4RZpb/uuH/nYns8O7dZx/TH532yu/Z7rf+Xn/ANNdt/YmfILrv6Xv7Fhz0VPOr2JnyC67+l7+xYc9FT+mP8NgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8fvbdfK1w2+juK+0s7tctH7nLhV9Ccj+wUTpL7br5WuG30dxX2lndzlp/c48KfoRkf2CiefqPR9jwf9Zl9D6O1cjpZIPJZt+jR1Id90OlEpWAhKzJAEmmeoVfc06UQ4LzFmzaiVxZotZIGSNVFmyelEiQR23YbuiWrkdKFggBqxKV0NEukJ2JbuiGrAxuxOwJ6UQDYnYm6e3mQAbT0sdLHUx1MN2hqxD7Fu+7DSt3YGZcjpRbpQFJdyC8oq5Vq26AdLIJ6mQFS/FDV2O27LJXQlEEsV6kWs0V6UTKTsFWoauyLNEp3JauCaULkdKJBv4KvuEmybJh7djNKl32Va8COlliBZ8RHSwtu5IaTM7URdPbzHSxZLfyHUxY3Z0shqxPUyG7mNl2hq5HSyyVyelBsumYLNbvdjpQNyKkNEgKVs0LNliO2yAjpY6WOpjqYbKdLA6mB6tl2h7orZosCa1UE9KIasY2UBDdiE7sNWKvuWKvuFY+qASlcNK3dg26Vc3vF/iZofivSybSWs8xyrAvKMPXdDDzSi5udROVmnu0l+g7Bcsupc91bwP03qHUuZ18xzLF/DPfYms05z6MXWhG7Vu0YpL0SP1OpcBwvxGYqpq/CaXq4/wB0kpZlTw7q+7u7K9RXte9vDuf2MioZBQyqhT0xTwEMsXV7iOAjBUFeTculQ+L+N1Xt43vvc9/Nz4Z9PjxzDVnveHh4M8Ooy5LnuX3fB/Q77ohp2ZZKwPA+jLtn0snst/At0oq14GaajvuiCyViGrIxsqjREouzLlbthW9MmmnZmbi2buKbu2ysoqK2b7mWbdJWPQ/QzeyuzkGdSCsu5lml43v3ceW7uinSzfoXqUcEvMmx1l9zBqzKyTasjaaSuZGWOsYyi7sq4t9mjaUU222yjVibNOkrGStdGco38tjkSgnd7mbil2bJs26zJi4Oz7GUom8vFFJJbbsmx1xyYyi2mkZyXT49zWT3aKyj1d7k2adcaxavczlF7HIcVv3M5xSta5jpMmMouzMZfFdn/IciXYzlCMndt/mMs06434sZJtOzW5nJdPj3ORKCUXa5nKKdrtk2bdMa48k9yjVjVrdorKKuS7Y1hKDsUasbmc1uTZp2lYSi7FTVq5Rx38TLNusrCUGl3RnKJyGk1Zmc4pPa5mrHTHL3MO+yM5Qd29jf3aW5ScU7rcmx1lnucd7oq07s1lFRWzfcpZMn1dZWUk2rIxnB79jkNFJJNMh2xunFkunx7lWtjaUU7bs/K6m1J8H68ty6p/dd1Vqx/wCD84p+fm/D5+2V2mXvY6l1J8Hc8uy+p/dd1Vqr955xT8/N+Hz9vyFHDV8VWjh8NSc6k3aMY93/AFLxb7JG1DC1sVXhhsNTlUqVHaMV3b/9S8W+yW5+7yTIqOT0ndqpiZq1SpbZLv0ryX8re78Eo71cu2GSZDSyij1S6Z4ia+PNLZL/ABV5L+Vvd+CX9GTW6/MbyivMxkl1PuY742MZRexm9m0ciSW27MpQV27syx2xrGUXJ7NGU4tX7HIezKSgnfd7k2bdca4z2M5R7+pvOmrrdmco99ybHfGsJRKSXkbSirmRNmnbG7Yzg07XM7M5Eo33MmlbuybHXG+5jKLb2aMpxco2VjkPZmcoqK7smzbrjXGlFxdnb8xV7o2nFN+JkS7S7Y1YPqXYo2chxTd22YOCXmTXXG/FlJbP5jCSbRyZK+3gzOVNJXVya743TjuLTKSi32N5JXZnIl2xrGUH5mU1e1jkySaZhKJlmnXG7YTpyd5XVrGXSzkS8vAp0L1Isdsbr1YSR8o5nF/2IM19KuH/AJ2J9ZnFK6v4nyjmdVuD+a2/7th/52J6/Dp/O+P6Y/Pe2V/3e6z/AJef/TXbH2JnyC67+l7+xYc9FTzq9iZ8guu/pe/sWHPRU/pj/D4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPH723XytcNvo7ivtLO7vLOrcuPCr6E5H9gonSL23XytcNvo7ivtLO7/ACzpfe4cKvoRkX2CiefqPR9jwf8AWZPo4LWQtE8j9AqBZ+QAhq5HSyws/ICelkODfiTv6/oF2GbU6WOllibO/YKl0p0sdLNLIh7MEulOlk2a38iRZ+QYq3cgv0+hFkGy6Qk2LNFuwYNqNXI6WWs/JkDTZdiVgSEt+xrVZEF5JEWRNm2eqtmybW3ZIfYa7NR1IgWfkLPyMLdlmxZoboNvxDd9gAm0Q2XaAHa+xLSQEFelliHsg1UFkr72IasuzBtBXpZaz8iWtg3aqdtmE0yen0ZDVu1w2WJIauE34khv0KkFrIWQNqgtZCyM9PRu2bT3ZMexLXdW2FvIzWmz4j7FSwsjK3atmw1YX9SVvuw2XasYu6JasW2W5UCtmhZsu1GxUNlUe6IasXS33IaVw3agLWQsg1UFrINLwDZdKkNXZIMsUq00C1r+AsjNEulSLpFmiOnxsYre0dSF0x0ryG3gB0r5v+D/ABM1xxYpZ3pLReY5pgVlOHoOvh4JwU1Oo3G7a3Sa/SdgeWXTWe6S4H6b07qXLK2XZlhPhnv8NXSU6fXjK043Sv3jJNejRjxQ5meG/CTUsdK6rpZxLGywsMUvgmFjUh0Tckt3NO94u6t5H7Xh/rnJeJekcDrbTkMSsvzH3vuViaahUXu6sqcrpNpfGg7bvaz9D6PPzc2XTY4ZY6xnveDp+Lhx6nLPHLeV9z9CtkGrsmz8gfOfSVasSnZE2TK2d+wbLoe7IJVvElpeAqlSOlliHe3YnQpKJWzeyNFutyOm3gFSsXB3fYrODS7o3sijSaswuZMGrGc4tqxyJQ32T7FHFW7E2OkyYOD33RRxaV20bteFmUlG62Rlm3WZOPKLbb2KtNHI6FZ7b2MpRta6ZNmnSZb7MJRd29vMqbOK3VjOUe1kybHSZMJwbk2rFJRexyHH4rbi7mcorbZmWOuOTjyi0m20Vaubyj8V7Myat2TJsdsbti1ZlWrmzjs7ozaaJsdMcmEk02UaN5QW+zKuEfK35yXWZOO1a5SUb27G0497J9zOzXmjLNOmN2wkmmykouT2aORKMWm7bmTVuyZjtjWBSSu7o3lBdL2Zk4tPsybNOsy24807FOlnIlFNbpmbikRY7Y5OPKLsVasciUFbZeJlKO9mmZZt1xy2wcWldtFXumjaUU12M5RSTsmTZp1l2wlF2MpQbb3RyLeZRxV+xlm3WXTjPdFGrM5Cgr7xZ+S1XqaOGdTLcsqf3bdVasX/AIPzin/jeb8O3ftN7ertje+mGqdTLDuWW5bU/u28atVf8H5xT8/N+Hbv2/GUMLXxdeGHw1J1Ks30xjHu3/6l4tvZJXNsPhq+Lrww2GpyqVajtGK7t/8AqXi29klc+gZFp6hk1BuSVTFVF/dKngvHpXkv5W934Jc9bdZXHyPIaOS0bytUxNRWqVLbJf4qv2X8rau/BL+jJNtvzN5w7XizOUbOyTFmnbGsJRatuirizecU7GbT32ZOnWZbYSi/QpLe8fE2av3MpRabaT7kuuNYVKcrrsZyg9/nN5Xv4lJJNGWO2OVYuDfkZTg990chq3ZMyknd3Js2641x+llHFtHIkvQys/Jk2O+N2wlFplJdmchxTe6MpQ+K/ismzTrjk4zRlOLasciUbPZMpJImx3xycdqxScXKNlY3lBPujJ3S7Mmx1l2wlFxdnb8xSfb85vKKk7vuZSjt8aOxDrjXHlFtt7FJRdjkOK322KTjFIyzTtMmD2MZROTKPfYxnHtZMmx3xrCUe5lKPT38TkuK3ujKUV3sTZt2xyYyje5lKJvKO7stirivFE2Osriza7b3ufJuZ75Hc1f/ANGw/wDOxPrjhFtu3iz5NzQxUeDma7f8Nh/52J6/Dprq+P6Y/P8Atjf93+s/5ef/AE12s9iZ8guu/pe/sWHPRU86vYmfILrv6Xv7Fhz0VP6U/wARgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8fvbdfK1w2+juK+0s7v8ALO197hwpv/mRkf2CidIPbdfK1w2+juK+0s7vctKb5cOFNk3/APeRkX2CiefqPR9fwj9PJ9Jbvsh0sJNeDLHkfe3tHUiHuybIdK8g3aE7E9SHT6CyBLo6kRa+6JsiLSWyTt8wLdnbdk9SIfV4pkWfkCXS3UiG7kWfkSl5g2LZk9SIa2Is/IG1upEWb38yLPyJuw3Z0sWa38iYsPsDZ1Io47trxJAJdoSsSSibKwaz6WCw6fQEqtmxZota3ZMNeYbtQFrIWQNqh9i1kLRtuv5QbZg0tH0K9PoZY2XatmxZot22A03fZUFrIrZ+Qs0JTsg2El49w0kY3aABZ+QbLsKyLEdwKkp2JsiGrdkwraAACUI6kSRZBvqd1sFshfe19iQ2b0r3Y6WWsvIh38DNNl2o4vfdErZFrehD2Y1psqH2KlhZGa36N2jpZDVi115jZ7mE+ZR7ojpZdrYgNV6WQXIsn4Bu1HuiLNF3bwI9GGy7VSuOz3LWSI6b32Dd6OpEd3sOxAVKSi9h2W5N2xs16/ONbaq9+xXp33L7Iq++xBLp1r5hOVXUXGbXsNX5XqrLsuowwFLBujXo1JTbhKbbvHaz61+g+t8E+H2N4WcMcm0HmOYUMdiMs+E9eIoxcYT95iKlVWT3VlUSd/FM+PcxXNPq/g1xAhpHI9OZRjcPPL6OMdXFe961Kcppr4skrLoVtr9z6/wO4gZjxS4XZJrvN8FhsJi8z+E+8o4fq93H3eIqUlbqbe6gm7vu2fR551H4Nj5/0Pc8HBen/CcvJ+n737sq1dliHe2x859KI6WQTv6/oFtvUN7qtC1t2SRvezDUPdkt3QsiobsIl2LEWv3TMsUqV6WaWRWz8jFb2o1Yz6GlvY3tfwKWv3TCpdMGrMq90chwTT2fYycbeFjLHSVi4u5ScG7WN5R72RS1/Ayx0l13caUXd7lHFryN3Hu7eZRrzRFmnWVk1s0ZSi1bdHIlHvsyjjfujLNumNcaSbTSM5Qe26OQ4pSfxXYiUVtsTp1mTjtbNehlKLVt0chxtdtMq4r5ydOsycd+RlODb8DdxSb2Kyjv2Mdpk47i990Zzg/Q5Ela9u5Tpb/GT27E2Okuu7jSi7PsUcG/I3lHd7PuV6fQmx2mTjN3RSUXJ7NG8oRSbUSjSXcx1lnucdq5SUXdmrTW6TKtb7kOsumFmUlFyd0cicVbsYzTT2T/AEE2Osy2wa23M2tmjeUb+DM5KSbVnb5jHbHJhKDS7oochxuj8fqzVCw3vMsyup/dldVqsX+J5xT/AMbzfh2W/ab29XbHLfZjqvVPwbryzLKn923VWrH/AIPzin/jeb8O3ft+Lw2Gr4yvDDYWm6lWo7Riu7f/AKl4tvZLdl8Pha+Mrww2GpyqVajtGMe9/wD1LxbeySuz6JkGnaGSYduVqmKmv7pUS2Xj0x8kn+dtXfglz1cnaXTHI9O0cloJtxqYmov7pUtsl36Vfsv5W1d+CX9JqzN2m+9ykkt0b7tOkvxYTi3btsZTg9+xyHG3gysl3uibNOuN04kl0+Pcq1t85yKkE7fFZk1a68ETZp2xrjyi1bdFJRdmcicU7GUk0uzJsdscmDg790ZSTTZybJmc4xs7LcizbrjlXHaMpRe6uu5yGrdkykord23Msdsbpx2mnZlJQ2e6N5QT8PAykpWez/QZrbrjWElZlZu0WbOKbu0ZSi2uzJdsbtg92Yyg0tzkOFtrMrKKa7EWO0y16OM1YznFtWORKO/Yykn5E2adsawlFplKv4q+c3cU92ZyjdWafcmx1lcZq6fqZzg0u6N5xabsnZFGk1Zk2bd5XHltsVaubTprdqLMrNeaJdsbL6MZQe/YylFnKaTT2MpxXgZY645MZRvcxnFm8la+zM2k+6Js27Y3Xdx3FxV3ax8k5omnwbzW3/dsN/OxPr8opuzPkfNJBR4NZrZf8Nhv52J6/Dv63x/TH5/2wu/AOs/5ef8A012n9iZ8guu/pe/sWHPRU86vYmfILrv6Xv7Fhz0VP6Q/xQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADx99t18rPDb6O4n7Sd4uWZ25b+FC89EZF9gonR323Xys8Nvo7iftJ3i5ZrPlv4UfQjIvsFE4dR+i+t4R+sy+h9Jvfaw6fUmyQuvM8b7u/gqTHsTZANt2EdPqSLrzDN6VLLsVs/Istl3Ns023Y1crbexYjxMJTp9SHsWKvuCWgC3ZayDdqkW9S9kVs79gS7QlYE2fkLO/YCvT6jp9TSyKvYCvb1uL+ha1/Ajp9A3apZdiGiV2BLobtsRe+1g07jdBu+50+pBO/r+gmyBLtUEteRAajp9SQLPyAq+5Bfp9GQovxTDdqktW3JshswbUauyGrbl/m7EW8xY3apPV6E2QsjNN3pV7gNb9iUvMwiA3ctZWKhUu0EdPqWCt4glVt6kGjSs7f7TOz8g3e0W9QlYvZeQsg3fZUjq9CWu6tsRZBu9pIavuFe/oSG/Qrb1ILPsRZ+QbKr0+ovbYtZ+RDXmTpsqL32HT6k2SA03Y1cq14E7+v6ArPv3MbLtVK24auyzSRAJVen1DdtixV9w3fxVauRb1LWfkGvMK3tQFrIhrfsBHTfxtYdFt7/yE7oXYs2qV8s4l8tnDLixqKOqNXYbMqmOhhoYRPD4t0odEHJrZJ73k9z9joPQ2RcONKYHRmmYV4Zbl3vfcKvUdSa95UlUleTSv8abt5KyOnXOdxF4g6W4v0ss0zrrUOUYN5PhqnwfAZpXw9Jzc6qcuiEkrtJJu13ZeR2P5V86znUPAXTGc5/m2NzPH4j4b77FYzETrVqnTjK8V1Tk23aKSV3skktkj3c/Dy49NjnllvH3R8/h5uLLqcsMcdZT3vqrVgS07kHzn0pQi/oSRaIai19xb1JSsH2Ct91SOn1Js/IBqErBuxJXdhssHuQTZ+Qs/IyxSCrVty5BmtG9KX3sVnG6tc0aj5b/OVe4VKy6O+/b0M5Gz6t9nb5ikoWtdMyx1l7sZR/lMpRtb1OQ16MpKF7bMl0mTBxum7+HkZtWOS4pXVvzGU4WtZMyx0mTGUbpmUo2t6nIaVnsUlFO3xWTZt1xyYyjeL9UYtdPje5ynFWdkYzh2vFmWadccnHcb39SslZ2vc3cFZ2W5m4L99F39SbHbHJhKOzKtWNXHvZMrKFrXTJdZWE47N38TNqxvJJ7NGco77J9ia641hKns9zGpT37nJa8ykoJ+HgZp2mWnG6fUpKF23c5DiktkUcN/xWTrbpMnGauVlHfuayjZXSZRrzRNmnWVg10q5nJXuzeUU12Z+O1bqpYXryvK6n923jWqp/4Pzin/AI3m/Dst+0XU7u+NtvZjqzVXwXryvK6v923jVqx/4Pzin/jeb8Oy37ficPhMRjcRDC4Wk6lWq7RjHu3/AOpeLb2SV2aYfCV8biKeFwtGVWrUdoxj3b/9SS3beySuz6Np/TlDIqLcrVMXUS95UXZLv0xv2S/S2rvwS5/p+r0yzGa97DIdN0Mjw/VJxqYqokqlS2yXfpjfsr/nbV34Jf1HC67nIlFNdjKUbbJbWN1rs3HJg4W8TOcbN7nIa80Zygnd2fbuZY6y/FgUkt27m0o9rJmbj323J1t2lYzdjKUbp7+ZvKKdviso4pX2Js0641xpRtbe9ykuzOROMXay/QZSja+xNmnbGsJRMXDu7nK6U/AzlH0dibHaZOPKO5ScNm7ms077JlWrqzRLrKwasZyj8V7m8oq+yMpLbsybNOsrCUd+5R9jkdK8jFwsuzJsd5WLhfxsZShZdzeaaeyf6DOSuibNu2N97FqxlKFvE3nF32TMpq62VybHbGsZKxlPt+c5DXmjKUNuzJs07Y1g1dWM5Q6Ve5tKLTdlsVmlJWauTY6yuPL8VmMo3tvaxyJRs2rO3zGco9rJk2O+N16MJR8CnTbxvc0mnd7Mr3JdpWM+5nKJvOC3aW9jKSe2xNjrjXHlH98fI+aX5Gc2X/0bDfzsT7BNKzuj5DzURS4MZtZf8Nhv52J6/D5/OuP6Y+D7X3+gOs/5eX2V2j9iZ8guu/pe/sWHPRU86vYmfILrv6Xv7Fhz0VP6O/xWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADx+9t38rPDb6PYn7SzvDyyr/c38KfoRkX2CidHvbdfKxw2+juJ+0s7x8sy/wBzfwot/mPkX2Ciceb0fV8K7Z5fQ+kEdPqWs/IWfkeV93aATZ+QVvEG0EdPqXaXh3Is/IM3v0QQ1dlrPyIDdiVtwSGvMzRtXq9Ba+9ybIGmxK24AM02dwAht+BmhICv4h7DQEW9SLssLNG9CVgHfwITfiNWiJR7bkFpEWfkY3fYSvuGrK5K2W4lZruDfdTq9CxFkXsg23SpD7Eyunsnb5itpPZp2+YG+yCer0JshZBsukdXoL32DW/YgN2np9SvT6lrsmyBLtUiXYvaJDStsrhrMlq25bp9AG7UBaysQg2XaH2KmjSs9itkLNku1VuGrFrIiROm70gnp9SLNbi7DdgBKSYbLtUhqxe0SJJAVttcgs1tZEK3iG7VTuyRZeQC5e4Vl3LDpv4AiqVyen1JskDNFqhDV2WBlmlK9PqLepL2RVt9mYS6ALPyFn5BW0N2Iv6FmvNEWQJVQWsiGt+wVtWRFtrlmvNANfk9Tay4XZDmKwOsdV6Wy7Hukqio5ljsNSrOm20mo1JJ9Ladna10z+1kOaZBnOU0Mx0vmGX47K63V7ivgKsKtCdptS6JU24u0lJOz2aae6Z1C5ueB3FXiFxVpZ9ozR+JzPL45Th8O69OtSilUjOo3G0pJ3SkvC252A5aNKZ/onglpzTGqstqZfmmC+GfCMNOcZOHXi6043cW07xnF7N9/M93Nw8eHBjyTLd+DxcPPyZc+XHcdSe99MFt7kvZkHhr6A1fco499y4su5BtRKxIa32KpvxCpdrFen1LFd/X9AbNl7bEFlutyGtg3aCE77Eld0FbJdyC3f8AG7kO3gK2XarV2On1JD2ROm7Ve1yrXV42sXttdrciKQVL72Lha+/8hU3kluU6I+QXL2ceS3buVaubSit7RdinT6E2adca40lu9yrVzedPZ/Ffco4R8rfnJsdccnHa7lZRvbe1jaUe7t5lFHzTMsdZe23Ha6W33sUkup37G84KzfSzJxafZk2adcbti9r+hVrq8bWNZQ2ezM2muyf6CbHXGsJQ3bv/ACFbM2cb32ZRq3ZEuuNYShZPf+QzasciUVYynDfZPsTZp2l2wcLK9yr2NWn4oo4q/Ymx1lYSjsZShd3ucnpXkfitZatWFc8pymp/dt41q0X/AIPzjF/43m/Dst+02yerthvK6jDV+rfg3XlWVVf7tvGtWi/8H5xi/wDG834dlv2/DYbC4jGYiGFwtJ1KtR9MYx7t/wDqSW7b2STbJhSlVqRpUoSnObSjGKu227JJLu2/A+k6U04slwkquKhB4ytvNrdwjZWgnez3V21s3tukmcdXOvXucc0ad05QyPD9T6amKqL+6VUtkv8AFjfsr/nbV34Jf1ZQ3e5q1ZdmZy6up7P9BdknoyZWsGrblWr3NZRv4MzkmnbwM1t2l2xnHtuVavFxv3NpRTtsZyVnsnYi9nWXs48o9Nt73M5Rvd38O1jkOKf43gUnGKbSMsdccnGauUlHur/yG8o2tZMzaW90TY7Y1x2unxvczlG/iciUE7bGUk1fZk2O2N2wcLeJSfaxu15oylG97p2uTZp2xrBq5nOPxWbyVnsmUlFNPYmx2lcZqxEoWTd/5DZwXkyj3JdZlv0ceUd+5nL8VnInFXMnBNdmTZp1xycdq+5k4WV73ORKLTskzOUU12Jsd8cmDdjJwsr3OS4K/YynHbZE12xvwcepHdbmcuxyJRv3W5lOCtsiXbGsJK5m42V73NpL0ZRpPZk2O+NYTezRicicVdmU49ulP12Js0641jOF7u5i4W8TktPs0UnFK2xNjtjlpx2vApKN/E1knduzsZzv4J/oJdsawnHur+J8g5qVbgvm/wD4/DfzsT7HKOz2Pj3NWv8AsLZv/wCPw389E9Xh/wDWuP6Y+D7XX+gOrn/68/sdn/YmfILrv6Xv7Fhz0VPOr2JnyC67+l7+xYc9FT+jP8XgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8fvbdfKxw2+juJ+0s7ycsv7m7hR9B8i+wUTo37br5WOG30dxP2lneXllV+W7hRb/MfIvsFE483o+p4V+sy+h9JW5PT6izRF35nlfb2ENX3LWZANoUd+5bp9SBd+YNhDV2SSmvEEu1Urbk99iW0yA1DhZXuOn1JAJUdPqRb1LAN2oC9l5Cy8gSqAvZeRFlbsDanT6kk2ZA1tsoCbNizDNoBNmhZmabLtDV9yOn1JkpEJPxFadPqOr0JFl5CQtE7h9hsthdMy+oqT0+oa3G/r+kxW0PYr0+pdWXfuG0wS7U6fUsAG70q+4TsWIsvIG0dXoQWsvIXiGyqkJWL7ESDUEdPqSWvEEqnT6kNWJd7u3b5wr+IbtUjp9S7t4EWaGttl2r0+pDVi9mRKLuZY1UCzXfsTeJhEENX3LXiQ92Fb0hKzDV9ye4swSqEp22JsvIhrcK2qnfYkdNvAhpsNlnoNXDdtiQGxVK5DW5chtbrxMs22W7VbsR1ehIsvIzXdsqO/pYW9SXfwI38e3zmEQCbxF4hSrVyLepae9un85W0g2XT4lxh5qtH8GtWQ0hnmnc5x2Jng6eMVXCe66FGbkkvjSTuuh32tuj6Dwx4gZbxS0Pluu8owWJwmEzP33u6OJ6feR93WnSd+ltbuDas3s0fJuO3KU+Nmt4ax/Z/9xujA0sF8H+5Xwm/RKT6uv30O/Xa1trd3c+pcH+HP7U/DnKNAPN/ur9yvf8A99/B/ce897iKlX8Tqla3vLfjO9r7Xsvbyzp/kMbhfz/e8nFeo+Wymf6PufrwWa3IXfc8T2oD7FrxKhW0R7C3qSCdNlVBay8iG14GNlQCLN9iEmg0fcNW3LFbS8QqVVq+4tbctZlVfxDS99rENW3J27LuRaXiGy1BVqxezHS/IyxUrMjp9TSyRF4mN2za7ozlG1t73NZLdtLYi1/C4XMmEt00ZSjfxN3Gzey7lWr9kidOsy16MWu6v6GU4W8TkOPfZFXG9tkybNuky04zXgZyhd3ub1ISs2kZNNPcyx2xrFruiko9tzeUXZ2SuZuLXdE2adZWEo9yjhd3ubyjs9kUtbuibHXHJxrepRo5E4Wi/ioycXfsTY7TJhON1a5m1Z2ORJbdj8PrPV/wSVTKcpqf3feNatH/AIPzjF/43m/Dst+0XtN1345c7qMdY6t+CueU5TWvX3jWrRf+D84xf+N5vw7Lft+AjCdSoqNOEpzm1GMYpttvZJJbtt+BpCnOrOFKlCUpzajGMU2227JJLu2/A+j6U0fHJ4rH46EZ42a2V01RTW6Xg3bZtfMtrt8NXOvdLjxY9/VjpbSUMnprHY6MZ42S9GqKa3Sfi2tm/wAy2u3+gnDdu/gcmy8UjKau3ZbHSTXZzmdt3XHauVcLvubuFlskZvuzLNOkyceSsr9zOSvdnIlC63SMpwd3ZbE2O2OTjyj23KNeByJQb7JGcovfYmzbtjkwlT9TGcLSe/Y5TXmjOUW29kTY645OO1czlDu7+ZvKDVtijtutjLHWZfBxmrFJR2ZyJRvayRlKNm7oizbtjk49SO63KNeByJJN9kZTi92ltcyx2xycepHdbmcoWTdzeS37FJKO6ZNmnaVx5dzNwsr3OTKF3skZdN/Amx3xyceUb73sZvscmaUXZpL8xi47PZE2OmOW2LV9zGULK9zkOLv2M2vNE2O2NYNWKTVkciULu6SMZxdt0TZp2xyYSj1b3sZNWRyXH0MaiukopXuTY745MJxvvczlHY3lF2asZNeaIs27Y1x5w3bv4FDkSXfZGUovayRljtjWE47t3/NYylG9t7WORKLs1bexi4uPfxIsdsaylDZq5l0+pyGrpmM4S22t+cmzTtjfcxnHwv4nx3mtjbgrm/j/AHfDfz0T7I15nx7mwX/YVzd//RsN/OxPV0E/nXH9MfC9rb/QPVz/APXl9ldmvYmfILrv6Xv7Fhz0VPOr2JnyC67+l7+xYc9FT+iv8ZgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8fvbdfKxw2+juJ+0s7y8srty3cKPoPkX2CidGvbdfKxw2+juJ+0s7z8sivy28J7L/iPkX2Ciceb0fU8Lus8vofSL+hBe3oRZeR5X2d7R1egtfcmy8iQ3atvUgs+xWz8gxPT6kPYvZ+Qt6AUBZrbZEWfkwIJt6k2XkQ732uGy6GrbkEr1J29AbVAs/Islt2BFQS7eCIsw3ek9PqVce+5N35gG0JWJHYA2hq4vbaxIDd7Ra+5FvUsAKFun1JsvIrv6g3tEo79yErMtv6iz8g3aAQ738STLFIaux0+od77XCvfe4s2SnT6jp9SQZZo2i3qQWFl5GN2qV6fU0svIWXkDai2RHf0sWa77bFXfwQbKdPqOn1G/qN/UKlOn1DVhv6hW8WCXavT43Ei+3oQ7MNV8CG7l9rdkRZeQbLpQjp9TTYWXkDbPp9SGrF7EW8xpSq2J6vQlrbsVs/IyzRLoBbb0KvvsYqK3vsOn1LWt4ACgFn5AKlS1bcq1dkgKlR0+pD2LFX3BL3QCbPyFn5GaUr0+pD2L2fkVfcy6EJXJt6kb+A39TGyuqHM7zO8SOEPEmnpTSlHJpYKeW0MW3i8LKpPrnKonuppWtBWVvM+18ANeZ5xM4Q5DrfUcMNHMcy+Fe+WGpuFNe7xNWlGybbXxYK+73u/Qtrzl+4R8TM7jqTW+kXmWYxoQwyrfD8TRtTi21HppVIx2cnva+/fsfqtH6O03oDTWE0lpLLlgMqwHvPg+H97Uq9HXUlUn8apKUnec5Pdu17KySS9vLy8GXBjjhjrKe95OLj5sefLLLLeN9I/qhw8b/yFrLyId9+54nul2r0+pBYBqgL7EO1trBstVI6fUks7W2sZYpS9tirdi5DSt2JJdKdXoWKtb9iwUhu2xUuRZeQVKpbe46vQu0rPZFbLyDZUdXoOr0JsvIbBsqnn6kdPqX29A7eBlilOm6tcjpt43uWZXfxMbFJQ7792Zyja297m5WSXkmFy6ceUbp7lGrG0k7v4rKyi/Iyx1l9zCcbp7mMo7nJa9Ckor/FX6CLNuuOWnHcdu5nOF/E3a3e2xWS7bGWLmWnGa7opKO/c3lFb7b/MUaXiidO2OTjN3Virhd3ubOO3Y/K6w1dTyWDwGBlGePnHfs1RTWza8W1uk/ne1k4ysnevRhvK6jga41P9zaDyzLsT04ypb3korelBp+N9pPa3dpXezaZ82jSqVqipUoOc5tRjGKbcm9kkl3bfgbWr4uva1StWrT9ZTnNv9Lbb+dtn0rSWkIZLBY7HQU8dJbeKoprdJ+La2b/Mtrt+fvyV9CZY8OPzsNJ6QjktNY/HRjPHTWy2aop90n4trZv8y2u3+jl3ORJJ+CZlOO/4p1kkmnLz3K7rjyhZXuUcbq1ze3mijS3sibHSZMKkelLe+/kZOF3e5yWk1Zq5nJK7skZrbpjlpxmrlZQ77m042W0f0Iza7k2adsawasZzV2zeUX4L+Qzku6sRZp3xrjyj23KPZM2a80Ukk77GWbdpXHl8a3hYylCzbucia8kUaXaxNjtjdOOUnHZu/ibTjbtEo1s9ibNOkrjyjv3KSV9jeS9DOS77E2O0rjzjZ2uZOFm3fxORNO/ZlGlurE2bd8a472djO3qbTi7/AIpSa+K9ibNO2N2484XfexRx27m9vMzmvi9ibHaVi4Xd7mUo7HIs/JmNvMmzbpKyasZS+MrdjkNX8P5DKa22Xj4GWO2NYSjv3Men1ORJPyZnNJLZEV3xycebs2rGcl1bdjaUb3dikl/3rRNjvK48o2bKSORJKzdlcxkr22Jsdsbv1Yyje7v4GUodVt7WN5rurGdmvNE2bdca48l0u1zOZyJru7GVl4q5NjtjXHlG12fHOa/5E83flXwv89E+zTXfZ2PjfNj8ieb7W/u2G/nYnp6D+tcf0x8P2tu/Aer/AOXl9jsv7Ez5Bdd/S9/YsOeip51exM+QXXf0vf2LDnoqf0R/jcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHj97bv5WuG30dxP2k70csaty28KPoPkX2CidF/bd/K1w2+juJ+0nerljsuWzhPf/ADHyL+j6Jy5puPp+Gfp19JI6fUu2vAqeax9iVUGjStskR0+hmqbUW5PT6k9PjYDRtHV6C197k2XkTZ+Q1TelbW3F/Qs15kWXkNG+6oL9PoQ1bwGq3ajVxb1LNeQSd+xjNoINOn0KtO/YK2p29bi99rFul+Q6fRBm1en1Frb3NLLyKyTs9g2XardyCUvMlrYCoJ6JeQswCV9xb1FpeAafiFbQATZg2gLcAGyUd+5QuLLyDVbepBcWXkGy6UBey8itmCWotvcEtOxVJoyzbZdpAAsahvwILDZEm+yttrgsmuyFl5Bu1LepHT6lrPyIkmGo6fUdPqN/X9I39f0hs7HT6hqw39f0jt+N3DZYrbe5JO1rIWYagrLuWIa3AqCbMWYVtXp9Re2xIuhpu9Kt32BPfZIizJ0bT0+pVx37k3fmApXp9Rb1Je6ItICALNAKl2EX3sSHFtdgqaQ3Yq97k2b7kpbDW2yqpWBLt4EGa010U50dRcQst4wUsNpfPNRYTB/cfDSdPAYqvCl1udW7tBpXsld99kdkOVXG5zmHAXS+L1Bi8bicfU+G++q4yc51pWxtdR6nJtu0Ukr+CVtrH6jV3GThdoPNVkesNaZflWPlSjXVCu5KTpybSlsmrNp/oP72mtTZDrHJcNqTTGaUcxyzF9fuMTRu4VOicoStdJ7SjJdu6Z7ubmyy6fHC4ak97xcPFMefLOZbt9z+mlfcOO3clbbMhu58971WrEFyLLyDZdKNX3HT6lnsQFI6fUqXukVswb0gADSpdqvuCwsvIhsulQS1uVe6CpdjV2R0+oSfiWCpVHsRIs1uQ15oG1Sen1DW3Yb+v6QqXSLepDVybtbBGWNl2oC7j3dkVa8jFSqNWuyjXV42sateBS1vALlYTja6uUszkyS6Xsr/ADGLi79jLHXHJhOGzdzNqxyZW3VijivJE2OkycZx8bmcoXd7m7jbw2PyWtNY08kg8vy+UZ5hOO7smqKa2bXZya3SfzvaycXtN13w3lZIw1jrCnklKWX5fKM8fOO72aoJrZtdm2t0n872sn8vtiMXX/f1q9afrKc5t/pbbfztsu1XxVftUrVq8/WU5zb/ADttt/O2z6bo/R0MjprH5hCM8fOO3Zqgn3SfZtrZv8y2u35rLyX5n0Zcenx+djpDSFPJ4Rx+OjGeOnHzTVFNbpPs21s2vmW12/0ko/Ge5vOFlsrfMZtb7o6zGTtHHz3K7rjtW3KtG0ou3YrKLT7GWOkycaULLuZtd0clxTXYznB3b6UTZp2xycdwsr3MpR+M9zkNeZVxu79KM06zJxmrmc4bt38DkSjdfFW5nJWdmtydbdca45Sau2byg9rJGUotN3RNmnbHJhOF/EzkrOxyJJbbGco3u7Imx2xyceUe25lKHd3ORKLRnJehNm3bHJg1cpKGzdzaUe1kjNxlv5fOS6zJxqkd1uUa8DkTjv2M5R2dkrk2ad5kwlHfuZSja7ubyi0+xWSVmrImx1lcaXczlH4r3ORKN3dJGUouzRNjtjk47hd3uZvsciSS2Zm4eiJd8axauYSjscpxs7NGDi13RNjtjWLVjPp9TeSV+xlOLtsibHWVlKF3e5jKOxyWmnuZzSt2RNm3XG6caS7vyM3urdjeUbp2SMpRa8CbHoxu2E43urmTVjkSi9zNx80TXbGuPLuZyjfxORKO72VjKSZFjtjWE42vG/5zGUem297nKa2e25lKN7bJk2O2Ncdq6sfG+bSPTwSzh3/4fC/z0T7PKLV3bY+Nc2q/7CGcelfDfz0T1dD/AFnj+mPh+1f/AJF1f/Ly+x2R9iZ8guu/pe/sWHPRU86vYmfILrv6Xv7Fhz0VP6E/x2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADx+9t18rPDb6O4n7Sd6OWT9zbwn+g+Q/YKJ0X9t18rPDf6O4n7Sd6+WP9zZwn+g+Q/YKJx5vR9Hw79OvpALpX8B0+hwfX3pXqY6mWsvIWXkG7Vu2QWa27EWYbKnpQvbZDf1/SRZhm099mLJb+RFmLSBs6mSlfdkWZK2QbKdKFkt/IPdEWkNQ2dTJaT3Kk2kNbZtALLbuG14GaaqH2BNn5GaaoF3L9PoOn0GjaCr7l7MhrzGqKkF7LyDW3Yxu1OlEizFmg2Xasu4SuGtwluAezILlbMCAOwDdgJ7bNEBsCGiya8Q7PyDVUroqXFl5A3pQiRpZeREktthqN2zLJ3Fl5Ayzap3A0mQ0/AlX8TLNCGrIgsLLyMbKqRZMl9wGqtW3Q6mWKyDdlk9/MPZk9kQ92G77IIbLK19xKz8EGq9KKyirmr7FQ2XSlkgSQ4y8F/KG7R0ohqxZp2HdbrczXdsulQBZmNlJRKtWRYBsu1CbtE2XkH2YVKo3Yi7ZbuLLyDZVUrk9KJS8kHfsu4bNunvNZy5cVuKnE+lqbRmSYbF5fHK6GFdSpjaVJ+8hOo2rSkna0lva25925dNFag4dcGdP6O1ThYYbNMv+F/CKVOrGqo9eLq1I2lFtO8Jxez2vbuj8lxq5r8h4Laxho/MtJZhmVaeDpYxVqGIhCKU5SSjaSvddD39T6Twp4h4PitoHK9e4DLa2AoZp7/ow9aanOHuq1Sk7tbO7pt/M0e/ny5702OOeOsfdXi4ceGdRlcbvL3v1nfdjpRId7bHztPftHSiCd/X9JBjdosmQ1ZFrNlUn49g2VWyZJay8ioVKjpQaDTZFpeIagE9ibxCtqkdKLPd7EPdGWNQ1ZEC7WwJVAhpMXQ7gVIbsXaXdkOz7WCpdqpX3Ye3YkiQai7ZAAVLssmVkltuy9n5ENrxJbKya8COhN73NGr3sivS/IOkrCUUmyjSZyJRunZK5+O1trKGRQeXZfKM8wmt+zVBNbNrs5NbpP53tZOcr5e9deOXOyT1Y611hSySnLL8vmp5hNbuyaoJ9m14ya3SfzvayfymSxGKxNr1K9etP1lOc2/zttt/O2zZxxGLr3Sq1q9efrKc5t/nbbb+dtn0/R2iYZHBZjmEIzzCpHZbNUE1uk+zk1s2vmW12/N35cvmfUlx6bHv3tY6O0bDJKax+YRjPHzWy2aoprdJ9nJrZtfMtrt/ppwTfdm7XmVav4I6yamnmvJcr5q4rVyrppu9zeUNtku5m013Ms06SsOlFJrfxN5r4qst7mTTT3JsdsawaKSVzkSjdWSRlKLuyXXHJhOCS7vuZtWN2vNFJQbk2krEO0ycZozlBN9T7m84u2y8TNrwZl9XaVhIpKKd22zapFu3SkZuPmjHXGuPJLbdlH5G80vJFGlZ7K5F7OuNYTXbuYtWuciSfkZuPfZE13xrCRSSSTZtKLVviozcfTYmx1ljjySbM2t3uzkTSv2RnKDabUTHeZMZwTfdmM4pXN5xlcpKLS3RN9XbG6cd7MrJLpZs4+hnJNpkWOsrjTgm+7M2rHIkrPdIykrxdu5NjvjWTin3uYTgmrHJaa7oycWldomx1xrjzirmUjltR8UjFwst0jLNO+OXxYWTMpwTVjkSjv8VbGTXmRZp1l+DCatsZtJqzOROK32V7GUotLt4mWO2NceUUm+5lUVrWucia77GLV/xlf5yLNvRjWL3TRnJerOQ0t9kZTj22JdZWE4rfuZSSj2vubyi9zNpPuibHfGuPUXxWfGObX5D85/8AH4X+difapK7d0rHxjm5jbgdnDSStXwv89E9PQTXU8f0x8T2rv9B9X/y8vsdjfYmfILrv6Xv7Fhz0VPOr2JnyC67+l7+xYc9FT+gv8fAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8fvbdfKzw2+juJ+0nevlkX+5s4TfQfIfsFE6Ke27+Vnhv8AR3E/aTvZyyK/LZwm+g+Q/YKJx5vR9Dw+6zv0Ppa7liqW5Y4Pq72oC1l5Cy8g3aoLWXkLLyDdqgmzFmGiVxb1FmEn4gQC1l5EW37bBm0AlpWIDUdPqSCbMG9KyILNeaAN7R0+ovbaxIBvSOr0HV6BryIsw3aer0Fr73Isyb+obKhqxBL3Y7maN90ES7FrMOz8hrs1QLcta/gT0+hmm7UasC1vMWXkNM2o1dhK25ZryIs0YraGr7lSWmybLyAi3qGrbliHug3sqCbMgN3sIkTZhrzQJdqAukvIl9PhYK3pmCz7EIG0EdXoXaVnsitl5GabtFr73IasWuhszLNN2qQ1cvZeQsvIxu1enbuVasXaf5iLeYbFbepBciy8gb0i/oEr7izFpeAVKqlbckWaAaiXYjp27l+l+RRp37BsukJW3D7E2YcXYabLtQCzLJbdidNl0qQWkQG7V6fUh7F3bwKtbhu9Cdgt2LMhqSWwVK+EcbOU3IONOsoaxzLV2YZbWhgqWC9zQoQnFqEpNSvJ3u+t7eh9J4U8OsHwp0DlegcBmVbH0Mr9/wBGIrQUJz97WqVXdLZWdRr5kjrXzWcxnFfhXxPpaZ0ZneGwmAlldDFOFTBUqr95OVRN3lFu1ora9tj7xy5a21BxE4M6e1lqrFQxOaZh8L+EVadKNNS6MXVpxtGKSVowitlva/dnv58OedPjc8t4+6PHw58N58phNZfF9IasQXIfY8D37VbtsQ143JBP0tVTtsC2yGxjZVSvT6l2tyrvbYN3pDVirdi6W25Elt2CpVHuQXsvIWXkGqp22CVw9hHuFSolHdlTRpWKNeRljdqtXYSsW2Ss+5BKpdoIasTdeY7hsukW2uVauWafh2Is0Gy7Vt6kFt7+hNl5BUVvtYhq5Nn5CzFm1KrZkS3L28bH5HW2tqeQwll2XSjPMJrvs1QTWza7OTW6T+d7WTm2YzdVhjlnlJjGetda08ipyy7LpxnmE1u+6oJrZtdnJrdJ/O9rJ/KWsRjMRe9SvXrT9ZTnNv8AO22387Ze1fGYjtUrV60/WU5zb/O22387bPqejNE08ipxzHMIRnmE1stmqCa3SfZya2bXzLa7fm/O5cvmfTlw6TH42sdG6Kp5HTjmGYwjPMJx2WzVBNbpPs5NbNr5ltdv9S1c5Eopp2irmUotPsd5jMZqPJeS8l3b3ceULJ7/AMhm1Y5LSatZGU4XeyRlm3SZfFhKOxnKF5Pc3cWldozlF9TstibHTHJx2rlJQu73OQ4WWyRnJWe6RNmnaZfBx2rblJRvd3N5xdtl4mUlJNpomx2xy2wlHYobyjZdkZSi3J2WxFm3bHJjKFle5jKG7dzkyV0UkkrppXMsdZk4zVjOau2ciUe2yM5R73RNjtK404X8TOSs7HJml5IzlG93ZE2bdscnHcL+JnKNmzaUZKxV2s+1ybHbGuPMylHZnJlC/gYyW722Jsdsa48oXd7lX4ryN5JeRlKO2yVybHaXfqxkrvuYzV7q5yWrdyklGzslcmzbrMtOLJWdr3Mzkyin4GLg7P4pNjtjkwlC7vcyt6nJlF3M2lZ7ImzTvjXGk7PsZzjscicU3tFGUlt2JsdsawlHfuZ1FZHJcUvAxkr+pNjtjkwMpROS4+SRhKLsRZt2xrCWzKT7fnN3FeW5SSXkjNadpXHlG6ZjOFrbnJnF72RjOMtrf7SbHbGsZQ832Mpq5yH237mco3tZLYmx3xrjyj3MpR7bnJnFq6sYSiybHfHJhKNru/ifF+bp/wDYNzledfC/z0T7ZJKzVj4tzeJLgbnFkv8AD4X+eiejof6zh9MfD9qrvwPqv3MvsdiPYmfILrv6Xv7Fhz0VPOr2JnyC67+l7+xYc9FT+gP8hgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAP5eo9Sad0fkmK1Jq3P8uyTJ8BBVMVmGY4qnhsNQi2kpTq1GoxTbSu2t2l4kab1PpvWOSYXUukNQ5bnmT46Lnhcwy3F08Thq8VJxbhVptwkk002m9014Af1QD+bqDUWQaTybF6k1VnmX5NlOAh73F4/MMVDD4bDwul1VKlRqMVdpXbSu0B/SB/F0prLSGvMnp6i0PqrJ9RZVWk4U8dlWOpYvDzku6VSlKUW14pPY/tAAAAAAAAAAAAAAAAAAAB4++26+Vrhv9HsT9pO93LD+5s4T/QfIf6PonRH23fys8Nvo7iftJ3w5Ynblr4TfQbIfsFE483o9/h9/Pv0PpLdiOpmmz2K2ZwfWt0r1MlbouQ1uC9kAWYDNq9TJTuSA23QALMEuwq3ZlrMdLswbV77MnpQUWnuSGo6USCtncMJEFookNl0jpQ6USQ90DaGrEE7R7u1w5ReyfcN2giyZazFmGq9KFki1mLMCCOlFrMq90A7bIJ3ZFmLMN2myZUkWYNoDVyewWzJ03avSh0os92LMaao1YF+xA03fZm3Zhos+4MbtC7ESLENXDdqgm3oTdBs+lUhJIl7koEqrbew6UW8Q2Gs2t3uwlYmzuStkG7VBZ9iplhtDdmO+7JAvoqVDVkQWe6K2ZLZVepjqZpdEPfdBsqnfZk9KJlF2K2YbKsVfcWYsw3aA+ws0Ra7vYNlVBeUXYja1mFbUW/ch9y9myrTTvYyxp0ohqxLZBjdhBI7hsr8Vq7gxwu17mqzzWGi8vzXHxpRoKvXUnJU4ttR2aVk23+c/v6a0xkGjckw+m9MZXRy7LMH1+4w1FNQp9c5Tla7b3lKT+ds6Pc7uA1liOM1GpkWDzmrhvuLhl1YOnVcOr3lW6vFWv2v49jsxymUczo8v2laWcUsTTxa+He8jiVJVVfG12rqW/a1r+FvA93NwZYdPjyXPfzPJxc0y5ssJjqz3vrbYvfZkkb3uzwvd32dKIJug90G70rZMhq26LWZBNikdTILC6MJ3VIl2L3RD32QVKzJaLWZSzQbtFkx23RNmyFtuwqVF2yCb73Jug2K2TI6UWe7FmjLFKOC73YLPsUauS2VJDSYWyJDVCG7F3uiLMKl2ghuxJ+R1trWnkUJZfl04zzCa3dk1QTWza7OTW6T+d7WTXKYzddMMMuSzHFGttbU8ipyy7LpRnmE47vZqgmtm12cmt0n872sn8n/AL4xmI/4SvXrz9ZTqTb/ADttt/O2y0vhGMr7e8rV69T1nOc2/wA7bbfzts+raJ0VSyGnHMcxhGeYzWy2aoJrdJ9nJrZtfMtrt+X87my+Z9OXDpMP/dVdFaJp5HTjmOYxjPMJrZXuqCa3SfZya2bXzLa7f6xqxZptPYrZneYzHtHzss8uTLdqj7kOKbu2zSzKy7mqlYdKKT2e3kciVmnuZSi34GWOmN+LCcE1Yyas2kcizRSUW2RZt2lcZq5ScE33ZrOLsVSstyXWX3uO1crKnHeW9zecXYxkmn2Ms0641jOCasZNWbSOTNXW3mYyTuybHbHJjKJnOKu3dm7TXczn3Is2741xpFWr3OROLZjJNNku2N2xlFO27Mpq2xyJq5SUXZmX1dMa40o9Xe5k4q77nImmrXKS/FZNm3aVx5qxlOK9TkNXMpJ2ZFd8a48oq5SS/kOQZSu2/nJdsb3YOKfe5SUI2fc2afkZNNNt9rmWadpWMkk7IzaurHIna5nKLSuTY6yuPOKT2uYyinF3OROLb2M32Is2741g4pGLVzlNbmNROyMsdscmE9nt5GUoqK7s3nFt7IpZomzTtjXHl3MWrnLnFt7GE4uxNjvjkxcE77sylFdzWUXdoq4tEV2l0wlsmZPc5Mk9zKcW7bEu2NcecE79zO1jeSafYzmrmV2xrCfd/MZSjc5D7NGcotEWO+NcWUV6nxXm9X/YLzn/AMfhf56B9tnF77HxTm+T/aKzn/SML/PQPR0M/nWH0x8X2pv9B9V+5l9jsL7Ez5Bdd/S9/YsOeip51exM+QXXf0vf2LDnoqfv3+RgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAP5+d55kumcoxmoNSZxgsqyvL6MsRi8djsRChh8PSiryqVKk2owilu22kl3P6B8+4+cKafHPg3qzhJWzyWTLVGXzwKx8cP794dtqSn7pyj1pOKvHqjdX3XcDqr7RzmE4B645M+Iel9E8cNAagzrHfcn4Nl2V6lwWLxVfozXCTn0UqdWU5dMITk7J2UW3smz+X7Mzj9wI0FyhaY01rrjXoPTub4fH5pOrl+bakweExNOM8ZUlBypVakZpNNNNrdNNbHVXmj9lN97ZwK1Nxr/b5/ZF+x34F/8Acz9i/wAE9/8ACMZRw3+F+GVOnp9/1fiO/Tba91xOU/2W330HBPKOMX7en7GvupiMXQ+537GPhnu/cV5Ur+9+F0736L26Fa9rvuB6v/fY8rH5S3Cr65Zd+uPjHN9zecqD5c+IOnZ8YdGarxmfacx+W4DKskzfD5jXr4qrRlCjtQlP3aVSUJdcrKKi5K7SR1n/AAGK/Ki/+0n/APXzm5N7DjJaGZUa2oOZLHY3ARd61DBaVhha015RqzxVVRfq4S+YD9b7E2tnlTgdryhiZVXlFPVUfgd7dCxDwlL36Xj+KqF123Xmz0aPwnBjgzoDgDw8yzhjw1yh4HJstUpXqT95XxFae9SvWnZOdST3bskkkklFJL92AAAAAAAAAAAAAAAAAAAHj77bv5WeG30dxP2k748savy18JvoNkP9H0Dod7bv5WeG30dxP2k748sKb5auE30HyH+j6Bz5fR7uh/Tr6XZIE2ZBw0+qAteJD3Y1sQR0okCRiOlDpRJNmZpqvbdDqZZrzIsvIaEdTJTuLLyA1Tehq5DVkS90Qk/EzRKglK6JsvIhp+AJdIl8W3qVTuy6V/xtybLyQVtUCzJdreoZtWUervcjoS3u9i1mxZhqvUyxFl5C6AkhuxHfdMJPx7AOphomy8g90FbVJSuSl5h7LsDfdWUVcABqr7kFxZeQFCeplrLyK2aCpQgsmvENpg2rZMjpRJK2ZOhRqwSuXa3vZWId/AzWm7Q4rfdlelFrSIsw3ekdKIasWsw15oNUBZ9mVDYAsmvEh7sN2rdvZjpRIDVSCzXkO2zQbvsq3YjqZPfYWt3QbEdKHbZEgnTd7Vu2CWvIizMbEPZBO5PS/IWsGyol2IttcdvxuxN0FS90XbK2TL3iLxDVHt2KvfuaOz7WK2VuwJVOlBqyJkmEn0jStqgWYsydNnZ8719zB8IeGWeR03rjV33MzGVCGJVH7n4qtelJtKXVSpyju4va99t12P1OjtZ6c1/pvCat0jmPw/Kcf7z4PiPc1KXX0VJU5/EqRjJWnCS3Sva6umm+s/NByv8AEri/xKp6s0nWyaGChllDCNYvFSpz64TqN7KDVrTVnfzPuHL5oLOuGPCDINDaklhZZjlvwr3zw1Rzpv3mKq1Y2bSb+LNX2W916ns5eLhx4Mcsct5X1jz8XJy5ctxymsX0Hvuw0rd2WdvArvf0PG9e1elC7W3kWexAV6oW6HSiQGzar2ZXpRdrcqZpU7qvZhOxYhryMsPQ6mRKI7C0vExUu1e2yEolul+QDZdM2rIqaPuRZeQbtUNtk7EO3gFS6QR0okdjLJFKtWIbsWd3vYhrzRLZVepjqZNl5H5HW+tqWQ03l2XSjPMZrd7NUE1tJrs5NbpP53tZNbMZuuvHheSzGTdRrbW1PIqcsuy6cZ5hUW72aoJ9m12cmt0n872sn8o/vnGYj/hK1etP1lOpNv8AO22387bH98YzEf8ACV69efrOdSbf5222/nbZ9X0VoinkNOOY5jCM8xmtls1QTW6T7OTWza+ZbXb8v53Nl8z6f5nR4fGo0VoqnkVOOY5jCM8wmtls1QTW6T7OTWza+ZbXb/VlrO/oTZeR6pJjNR87PO8l3b3Vu2QTZ+Qs0LNsl2h9ilky4svIyxsumPSh0Jve5ey8isotv4q2+cx0lYSjsU7G8o2XZFLLyRljrMmEoRt4mU4pPa5vNO2xTp80RZt1lceXYzlFO7bN5QaW6XczlF3e2xljrjXHaM5Lc5MopqyS7mTg0+xNmnbHJhKN1ZmU4pXs3sjdppXbKSje+xFmnfG93GKuCd22+xs42/eoo1Zk2O0rjzila1zN7XOTJLa6RlKO7aStYl1xrjygpd77GUopXW/iciUZeBSSST23Ms07SuLKKja19yjV0chpPsk7GU4veyIsdsawlFXMpwSu9+5yGrbNGc43T2Xcyx2xycaXcpJJ3TNpRae6RnKPd2Isd8awnFXKSd00btJ90ZWXkiXaVj0oxaVu7OVKLb2Wxi4q26MsdcawezMp9vznImlfZIzlC/dImx2xvxceyZlNJLY5E4u+yMmvNEWO+NYmMuxyJrfZWMpRduxNnudsbGDindtspNfFW7N5xsmrbmNmvxlt6k2adpWL2uZyORPp3sl+gxkr2sjLHbGsZRTu2zKS9WbyW1mZSTIsd8awlBXbuzKq7WOTJWTujGaT8CLHbGuNJ3uvU+Kc4EbcCc5f/hGF/non29we76drnxPnDVuBGc/6Rhf56J6eh/rOH0x8f2pv9CdV+5l9jsB7Ez5Bdd/S9/YsOeip51exM+QXXf0vf2LDnoqfvX+SQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH8rUmmNN6xyTFaa1fp7Lc8yfHRUMVl+ZYSnicNXipKSU6VROEkmk0mnuk/A+aag45crPLpg6Gg834h6A0LRyyDVDIMPicPhpYWEpdXxcJS3gm5uW0EndvzZ+Y59+N2dcAOVzV+utLYieH1BiIUcoymvFb4fE4moqbrJ2dpU6bqVI3VnKEU9mea/KR7L3VXM7oKjxs4kcTK+m8r1BiKtbA0qeCeNx+YRVVxqYipOpOKpqU1Ozam5fjNJNdQeln4QHk0/KD0v/AOfV/wDYH4QHk0/KD0v/AOfV/wDYPKXAcielsXz+ZlydS17mscpwOFjXjnSwtP4TJvKqWNs6d+hLqqOHfsk+5+65v/ZT5pwB4Y1+K3DLWeP1fl2RqVbP8HisLCjicPhbpfCKXQ2pwhduaaTjH426UrB648L+L3DLjTkFbVHCvWuW6myvDYqWDrYnA1HKNOvGMZOnJNJxfTODs0rqSfifsjqP7MbSvBLJOWfC6g4I4vP62E1LmFTHZzRzzF0sRisFmUKdOlVw3VSpUodEfdqUH0JyjUUn3SXbgAAAAAAAAAAAAAAAAAAAPH323fys8N/o7iftJ3x5YW/vauE30HyH7BROh3tu/lZ4b/R3E/aTvnywfuauE30HyH+j6Jz5fR7uh/Tr6Xe+zHSg1cjpZwfT2gskSTZsMVasippZogChN2iXuiOlgLtkE2aC7gQC5V9w3aAADYWSuipNmw2XY1YglpoA2gWTLdSDd0YbUe3YXb28yQZpqOlFWt3uy4N0KxiJbEtC1t2Y3anUyXsi3Ug2G7U6mLtlgDahbpRJDdgbs950ohohxbd01v5hQa8g3YGrk9LJe6DdqdKDRbpYs0DbMt0okh7oNHtsipazW/kHv2DdqgtZrfyHUjNN2qRIvZvfzKyi9jLNNlVI6UW6WOlmK2r0odKJfkV7bMCASu5LaQbtUiyZaze/mQ1YN32RZIWTJDg35BqgJ6WGrBu1UyQBpQVfcsQ1dk6Iq1cdKLJE3V7GKU6UQ1YtKLsR2W4IqCW7hOyCtqtJjsi/UiG7hW2YfYs1cgG3XTjzzc/tI64hoz9r77tdeBpY34T91fg1uuU109HuZ9ui97737K2/1Tg3xG/ba4cZRxB+4/3K+6vwj+9PhHv/AHfuq9Sl+P0xvf3d/wAVWvbe13+F4y8qOjuNGroawz3Umc4HEwwdPBqlhHS6HGEpNP40W7tzd97bI+i8L+HuWcKdB5ZoHJ8dicXhMr997utien3sve1p1XfpSWzqNKyWyXievl/B/kcZhPznn4vlvlbcv0X6l7MlqyIs2LNHiexFkypcgEqpDdi/UioVKr1MnpQukOpBUukPZkF07kPdGWNlZvuT1MsQ3YlsFuiGh33RFmgpFkyGiwlF2AzsmQ1Yv0sNNBW2YaTJ8SepBUVIkXtfdH4/W+tqeQU3luXThPMJx3fdUE1tJrs5NbpP53tZPMtYzddMMLyWTH1RrbW1PIacsuy6cZ5jNbvZqgmtm12cmt0n872sn8naxOMxH/CVq9afrOc5t/nbbb+dtl17/G4j/hK9evP1lOpNv87bbfzts+raK0TSyGnHMcxhGeYzWy2aoJrdJ9nJrZtfMtrt+X87mvzPqbw6PDt6q6I0TTyCmsxzGEZ5jNdrpqgmt0n2cmtm18y2u3+uKuO7a8SUrHokmM1Hzs88uS3LK91X3Bcq0GSqt2YW/ckhq4VtBBa1t2OpGaUo1ZEFmrkdLMbKyaTVmZtWbSORZoq1dhcycfpXqUnFXNnFpXbRSXcyx1xrjyV1uZyj33OROLa2MpQe7uiLHbGsGjOS3N5K6KNWdiXXGuPUpxsu5lKKSe7N5waXdFGtmiXbGuNIo0nc3lB7bopKD8ybNO+OTjygna7Mpq2xyZGcltYmx2xycZpMzklun43RyJRatuij7O/gibNu0riyj0Wt4lH8buciS67W8PMznBpMmx1mTjzW67mcopJvc2nBt90ZteBNmnbGsJq73M5QVn3ORKJlODd+xNjtjXHlFXMnBLxOS1YykTY741hJ2dkZSW1jkSi5O6MpRbRLtjWHQm97mUuxyOlmc+35yHaVhLuZTgkvE5DRlOLa2sZY7Y1x5wTfdmRyWmnZmU+35yLHbGuPPdszlG6szkNPpZlKLsTZt3xrjzik/EyasciUHd9jOaaJd8awnvdGfSjeSv28TNxa8ia7Y3swnFNvv2MJxSscqcG2+3YylHp7smx1wycaTdmj4lzifIPnX+kYT+eifcWrXPiPOOv+wPnP+kYT+egejop/OcPpj5HtPf6F6r9zL7H3v2JnyC67+l7+xYc9FTzq9iZ8guu/pe/sWHPRU/eP8mgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOnHtY9IY7VPJpn+NwFB1ZaczXLs3qxXdUlV9xOSXjZV236JvwP1Xs29WZLqzkx4dPJ8SqssnwlbKcbT7So4mjXn1Ra9YuE15xmn4nYPWWk8h15pTONE6owMcbk+fYGvl2Ow8u1ShVg4TV/B2bs+6dmt0fB+Tjk0wnJ7hNV5PkfE/ONS5PqTFUcXRwONwlOjDBTpqcXNOLfVOcHCMpJRT91H4uysHT/AJgNVZXy2e1w0zxc1tKGA01qvLcI8Tj6iap0qVXBTy51ZSvZKnOjCU/KCu1um+93Mpxa4YaM5dtY6t1VqzKfuJmensfhsHNV6daOYzq0J04UqCTarOcpqKSut220k2vynOpyYaQ5wdEYLK8dmjyHVOQTqVckzmNH3qp+8S95QrQuuulPpg3ZqUXFSTa6oy88Mh9ivzE4rUSwWo+IegsvyWFSKqY/C18Viasqbe7p0JUad5Jd1KcFfs2twOwPsSYZwuCfECddVvuU9VU1hb/4P4QsJS9/b16Xh7+nSejp835fuBWjOXHhXk/CfQsK0sBlcZzrYqvZ18biaj6qteq0rdUpPZLaMVGK2ij6QAAAAAAAAAAAAAAAAAAAHj77bv5WeG/0dxP2k76csCvy1cJr/wCY2Q/YKJ0L9t38rPDf6O4n7Sd9+WBr72nhL9Bch+wUDny+j29FdZ36H0rpQ6UWbumkU6WcH0doJu0aC6QNs732ZPSi90wDanSiGrF+pC190DbOyezJ6Et1fYs4tkKEr90G7VIsma9LKyi7sEu1OlDpRbpYcWgbV6ULtbeRbpY6WDaq37hqyLNNEA2oCelkvsGqglpoLuG7QC5DV2DaoJasQNG0dKDRJKVzNK3tVIdKLNWIauNCOlFZdi3SyTNHrVF2JJauyLNBuwqmWFmg3ewhq5DV2EgJ6UOlEgN2q4rfdlUkizT3CTYbFW3uVLkNXDd7SuxDSZFmgk2G7S1ZFS1mt/IdSM03bN9yLJmtm9/MrKJmmy6UskGky3Sx0sxW0FZdywAoT1Ml9ioVKEWTJJs2GqtFS8ouxXpYbtAJ6WLNGWbbLtD2RXuyz3QSsNNl0iXYgs90VJbFWrEFwGo6UQ1ZDpZAVsIsmWiJJ7hrpLzg8cuK/DrizS0/ovWWJyvL3lGHxDoU6NGadSU6icryi3dqK8bbHYXlj1ZqHXHA7TWqNV5nPMM0xvwz4RiZxjF1OjGVqcbqKSVoQitku3mfp9T6J4V5/mSx+s9J6TzHMFTVNVs0wGGrVlTTbSUqkW+lNuyva7Z/c0/lenslyjD5ZpXLsuwGV0ev3GHy+jClQheblLojTSirzcm7Ldtt7tnt5ebDLgxwmOr8Xm4+PPHluVy3Pg5yVirb3ReXcp3Z4bHr9UpENeBPSyDFbR0odKJKpWYbLuIlFXIUU2WaICpSyWyAAVFX3IauXKGabLsSsGrgjpZlmm7LJb+RF2ybNb+Q6kYrajdmJF27opINirXdlS5+R1xrankEJZbl04TzGa3ezVBNbSa7OTW6T+d7WTzLKYzddePC8lmOPqjW2t6eQ0nluXTjPMJrd7NUE1s2uzk1uk/ne1k/kzWIxuJvepXr15+s5zm3+dttv522T/fGMxH/AAlevXn6znUm3+dttv522fV9E6Jp5FTjmOYwjPMJrZbNUE1uk+zk1s2vmW12/N+dzZfM+n+Z0eHz1GidEU8hpxzHMYxnmM1t2aoJrdJ9nJrZtfMtrt/rZF7p7Ihpo9MxmM1Hzs+S8l81vdQEtNbkN3JZKhuxHUywDVCG7F2rkdLCpVb32Y6US9u5HUgqXSoLdSHfdDWms7tlW7M0KvuSqKTgmrGU4pPa5yCsu4dMbXFaTKSindG84NLujNqzJsdcbtx5RSXdmcldnIn2/OZ9De+xNjtjdMJwTVjKUUm+5yJRdjOWyZNm3XG6cabsUaTi2zdq5lOD37diXbGsJJbbszkjdpopLuQ741hKPV3uZSit1v4nIlF7GTg++xNmnbGuO0o2t4+ZSW6dzeRnNOzJsdpXHaMpwSu9+5yHBvyM5JpsmzbtK48u5m97m8+5SW6Jsdca48oJvdmTSt3ZvODb7ozfYmx3xrB7GJyZGck2rImx3xyYuCe+5hNbeJyJRaZSq1YmuuNcZ7MpKJs0UnBpd0S7yxhOCb7swnBNWOVKJj0NLexNjtjWElZWRlNJLY5E/Exe6Js0741i1e5lNdu5yHF7u6Mqkb9mibHbGsJRW7MpHIcWrmc3cizbtMmEu5nNdu5vJdzKcW7WsZY7Y1xppJP5z4hzjr/sC50//CMJ/PRPuclZs+H85PyCZ3/pGE/noHfov6xh9MfI9pr/AEN1M/8AZl9j7v7Ez5Bdd/S9/YsOeip51exM+QXXf0vf2LDnoqfu3+UQAAAAAAAAAAAAAAAAAAdC+Y32ruk+X3jNqLg9U4O5pn9bTlShRrY+Gb08NCrOpQp1WowdKTSXvFG7e7TdrWPm34cTSX5PGbfWKn/Zzu1r7lB5Z+KWq8brniDwbyDO8/zFU1i8fiKc/e1/d0404dTjJJtQhCKdu0V5H5/8H9yafk+aX/8AMq/+2B1F/DiaS/J4zb6xU/7OPw4mkvyeM2+sVP8As52g1byacgmgclq6i1xwm0Dp7KqH+ExuZ4l4WhF+Cc6lRK78Fe50i448wnstOH/v8q4T8s+U8Rc1h1QVelCvgMthJLxrVW6lSz/xKTi0naaumw/e/hxNJfk8Zt9Yqf8AZz0I4NcUMm41cLdM8VtPYPFYTAany+njqWHxSXvaN7qUJdLabUk1dOztfxPA3AaC4v8AOBqj4LwP5bsnyjLqdbpVLTOVyoYLDXdk8RjsROW6T3UqsU2m1FWsvdnlp4W5lwV4CaG4WZ1jaOLzHTmUUcLjK1C/unX3lUUG0m4qUmk2k2km0r2A+nAAAAAOPjMXhcuwlfMMfiqOGwuGpyrV61aahTpU4puU5SbSikk223ZLc80OYH2zeR6Z1DjdMcv+gMLqWjgqsqP7IM5r1KeExEls3Rw9O05079pynBtLaNmmffPan8Qc10Dycano5RiXh8RqjF4PT0qsZNSVCtNzrxVl+/pUqlNp7dM5eNj+H7M7lY4Y8OuX3SfFTFaXy7Mtaawwcc5r5vi8PCrWw1Go26FCg5X91FU+ly6bOUm3LZRjEPm/If7SjjHzNcdqfCfiNo3RmDwGLyrF4yhickw2LoVadWioy+N76vVU4uPUrJRabTvtZ9xOaTjnj+W/grnXGHBaEr6ujkdTD/CMvo41YRqlVqxpOq6jhUajFzi3aDaTu7JNrovwp/7dJr3/AELFf0bhz0B5g8gy/VPAjiLp3NaSqYXMdLZrh6ia7J4WpaS8mnZp+DSYH87lq4+6d5meDmR8XtNZfWy6lmvvqOJy+vVjVqYLE0qkoVKUpRSUt4qSdk3GUW0m7L6mecvsTc+xuK4Ia903WnKWGy3VEMVQTf4sq+Fpxml5L+4Rdr2u27Jt39GgAAAAAAAAAAAAAAAAAAA8ffbd/Kzw2+juJ+0nfblhV+WnhJZf8Rsh+wUDoT7bv5WeG/0dxP2k778r/wC5o4S/QXIPsFA58vo9nR/pX6H0tLfsWsgDg+jLsuvMq9yeljpYZtC7liLW3Y6kDaCy7EdLJSsDaN/X9A39f0Et2F0w1G/r+ggsVfcAnYlWffuI9w09wFokAWbBZtErvtdkWfky627i6e3mBWyIs/Jl+li+1g2X4s7X8B0+hdJsnpYN7Z2fkC7TRR9w2VDTuTaPT6/OTZshK4bKrZ+RL23RbpZV7oCN2Pn7ErZB9g3ZaJUE2aBtAknbsWTsg2G7Z2fkWBPSzLGyqS8bELq8UyzVmI9zGyos/Ii68y5n0sN2O/h2JQSsG0gFkLIXTJDVH5DsS1dhpoNl2giyXckhq4ai/qT33bI6WGrBsuktLw7kWfkE7EppmWbbLtm07vZiz8i77kGWNl0r32I6V5FrW3ZD3Zit91Cbslq4WyAi7ILS7FQqVD2Q7Ld7lullZRd2GxUE9LHSw3aBZE2aIM02VDXkRZ+RYEqlqtn5FZJ3fcv1Ihu4arug2/EsQ+wbLp0M51+G/ETVXGOlmmltBajzjBLJsNSeJy/K6+IpKanVbj104tXSabV7q68zstyn5JnWnOAGlsm1Dk+NyvH4f4d77C43DyoVqfVja8o9UJpNXi01dbpprZo5vEbmP4T8KtQR0xrTPMThMwlh4YpU6eCrVV7ubkk+qMWr3i9r32P2Whtbae4jaVwWstKYueJyrMfe/B6tSlKk5e7qSpyvGSTVpwkt1va/Znv5uXky4McMsdY/F5uLjwx5ssplu/B/badyvT6F2rENXPA9kVDXfbcnpZDdu5uttVCtfcPdgi9my6TaJWyLdLKtXCkNb7EFlshLsGy6VIsiQFSqteQLEPdGWbVLtUi0SWvAjpZlb2Q7X2I7luln4/XGtqeQQlluXThPMJx77NUE1tJrs5NbpP53tZOcr5ZuunHhlyXyw1xralkFKWXZbKM8xmu+zVBNbSa7OTW6T+d7WT+Sv4TjcT/wlevXn6znUm3+dttv522X/vnGYj/ha9evU9ZzqTb/ADttt/O2z6xonQ9PIKUcxzKMZ5jUWy2aoJreKfZya2bXzLa7fm/O5svmfVlw6PD45VTROiKeQ01mOZQjPMJrZXTVBNbpPs5NbNr5ltdv9a0Wfcq1c9Mkxmo+bnyZcmXmyQN2Ta27HUjWRX0YsiSDKpWV1eyZX43qaFXuyWyhDdiSGrhqrV+6KtW7JmnZb+BD3YVLtSz8iVstySsu4VLpLSRWyJSuDLFKWfkQ15lyJdzGysmm/wAZbGc4q7VjeSbVkZSg1fdbB0xrCUL+DKONtrM3bsUau20Te1dZfi484pLZGcorfY3qJ9KMmRfV2xvvceUbWsmUaVndHIauZTg9+3Ymx2xrjuC22KSglfZm7TRnPe6Js27Y2uLPq2tf9BVo3cW+zRnKLu9yXfGsHBbbGNROzsn3OU1YxkrtpEO2Ncez8mUmlZ+fznInBp2MZwbu9jLNO0srCUE/DwMpxavs7XOQ1Yzmr7E2bdsa47XmZyhGz2N5wadjNxdn2Jsdsa484q+yM5LbszkSTuYtXId8aycU3ujGUE1ujkyiYzi2tjLHbGsJQV9ltYznfpXc5HSzKe628ybHXGuPJb9ikkjdwb8jGUXYmzbvMmM4xd9tzFwSXZm8oO7exnLsR6u+NceSabW9vmM3HzRvJO5SUW7Eu+NYTS3+YxlFbbM5E4u73RlJWJrtjWEk7OyZk7+KZyZfisyauTY7Y1jKKs9j4Xzlr/sB51/pGE/n4n3WcXax8N5zItcA86bt/wDCcJ/PwO/Rf1jD6Y+R7S/+TdT+5l9j7n7Ez5Bdd/S9/YsOeip51exM+QXXf0vf2LDnoqfun+VQAAAAAAAAAAAAAAAAAAdf+YPnn5cOWnF18k4ha0eJ1FQhCpLT+UUHi8faUVKPXFNQpNxakvezhdNNXTTPPfi97YjjbxBxv7FuXjh/htLLFz9zhsVXpLNc1rSey93S6fcwbv8AiuFV3Ssz6Jz5+zG4u8W+LepeO3B3O8pzqef+4rYjT+MrPC4qnUpYenRao1Z3pVFL3XVacqduqyvZHXHgBzQcafZ459V0XxH5b8phTxFSTr1Myyf7mZxVp9SbVLMIwfv6S3t1KpG9umSV7h+h0d7Pznp5tc4oa047ajzPIMHW+MsdrHG1a+MhTbu1QwN3Oml4Qn7mNux3f4Geyh5YeE6w+Z6yyzE8R88o2lKvnqSwKmrfiYKD6HF/4tZ1e737W/VcCfaT8rPHJYbLoa1jo7P67UFlWpnHBuU34U693QqXd0kpqb2+Km0jtLCcKsI1Kc1KMkpKSd014WfkBx8tyzLcmwNDK8ny/DYHBYaCp0cPhqUaVKlFdoxhFJRXokcsAAAAAAA68c+/BPN+PvK3rDQ+mMNUxOfYenSzjKcPTb6sRicNNVPcpfvpVKaqU4p7dU4t9jqJ7PL2ifDDRHDLLOXzj/mdXSmaaWlVwWW5rjKM/gtbDqcpRoV2k3Qq07ygupKLjGN5KV0/UI6l8y/s1OXvmPz3F63xFHMdI6txz68VmuSyioYupaynXw804Tl4uUOicnvKTA6e8M+NHCLBe1i1rxXxvEzTOG0bisDiZUM9rZpRhgqt8uoRShVlJRlJyjKKim22mkrqx9b57/aX8HMNwn1Dwp4F6opas1JqjBVsoxOYYKMvgWXYWtF0601VlFKrUcHKMFTbScupyTilL57ifYc5h90enB8yWH+At3c6mlJe+gr9ulYu0mlbe6v5I+68BvZHcvvCbO8JqnXWa5lxFzTBVFVoUcxoww+WxmneMnhYuTqNeVSpKD8YsD+n7JzgrnHCflhhqHUuXzweaa9zKeexpVY9NSngfdwpYZSX/fRhOqv+9rK9ndHdUrFRilGKSS7K1kiwAAAAAAAAAAAAAAAAAAAePvtu/lZ4b/R3E/aTvvyvv/c08JfoNkP9H0DoR7bv5WeG/wBHcT9pO/XK7+5n4S/QXIf6PoHPk7yPZ0fbK/Q+lAs1cjpZx1Xvt2ghvct0sdLGqeZS7BfpY6Wbpm0AnpZKVjNVtu1FvsybItPdbeZRRe26GiXSSr7l+lkODfiNCI9w272uOhpb2JXYaIWQI6WLpbeRjdkgkrXsSmmLpAl7q3ZLStexJFmt2G7IkvYJpgCt2yVCNu38pLVyOlgT0ryKOyvZF1siHvsgKXYa2L9LIcWw3ahD7Gii0HF2fYN33Ylpdi3SxZoG1Utg1sGrsizQagm7IAbLoFkgSlcWbbLtAsielkGWfBqr2ZHcs1dkpWM0RSyJLPdNGfSwraxHcjpYW3cB4hom6ZIbtQWTJSsxLuGy7UtvYt2ACpdKvuSlsQ092SmGyqlXszS6YaM12azBYhozTZe6osiZRdivSzGl2Ts9yLNCzYbLoa2K3+Na5IDZdol2KvY06WUl3aCorFkkpNgN33RZCyAukTpu0NFG9+5ommH2MbK6r8yXKfrLjTxDp6xyLUmTYDDQy6jgnSxfvetyhKo2/ixas1NW3vsz7PwH4eZnwp4U5HoHOMdhsZi8r+E+8r4Xq91L3uJq1VbqSeyqJO6W6fgfIeY3m11JwT4g09G5VpLLMxoTy+jjffYitUjNSnKaatHay6F+ln2HgbxExvFjhZkmv8xy6hgMRmvwnrw9GTlCHusTVpKze7uqabv4tnv5vl/wfHz/AKLz8XyXy2Xl/S9791LuQTLuQeB699wo0rvYuAqXSlkNluTLuQ1cWKVuy1kR0sgkl0PZkS7EhK4UoC5WzQVtBD2RZMSkFS6UW73Ja2F0z8jrjXFPIKby3LZxnmM1u9msOmtpNdnJrdJ/O9rJ5llMZvJfHheTKY4zuprfXFPIKcsty2cZ5jNd9mqCa2bXZya3SfzvayfyW+JxuJu/eV69efrOdSbf5222/nbZe9fG4n/hK+Irz8nOc5t/nbbb+dtn1fRWiKWQU45lmMIzzGotls1QTW6T7OTTs2vmW12/Ju81+Z9b/Z9Fh85ojRFPIaazLMoRnmE47K6aoJrdJ9nJrZtfMtrt/rbtgN3PRJMZqPmZ8l5Lcsr3VfcguDWS6UIsizT3ZAV6q+JLRD7kBsugWQBljfVWXjYiLJfcEqlQLIkBqgsmT0srKIWPbdEdyUhdIWbVKqLJk2aIFaqQ0mnsTZoEqlYziktkZNLsch7K7KOLbumt/MzW3THJx2k1ZmU4pNr0ORKLsYyi7tku+NYT27FHubyKNN3JvZ1lYTilbYyklvsbzi3a1jNqzJvq7Y1g0UlFWvY2qLrtbwM3F2fYmx2xrjySMpRSV0tzeUWrbopJE2O+NcaW7KTSt2ORKLbumjKcXutiHaWOPJK5nJJXN+llJ7Jku0riz7lGlY5DRlKDs90ZY7Y1hJK5g1scmUSkvxWTY741xpdzM3l3M5RdiLNu2NYztcxkkbyi27oya23Jsd8axm2nsYy7HIl3M5O6sib6u0rFpNO6M5Qj5G0ovcynFtbWJsdsawnFLw8DGe3Y5Ek1dMymRY741hLuykop22N2n0sylF7E12xrGcVZ7GLVjecG7vbsZdL8yXfGsJN3Z8M5zfkCzr/SMJ/PwPu0la6PhnOcv9z/AJ2//CMH/PxO/Rduow+mPke0t/obqf3Mvsfb/YmfILrv6Xv7Fhz0VPOr2JnyC67+l7+xYc9FT9y/ywAAAAAAAAAAAAAAAAAAAfw9XaJ0dr/Ja2m9c6WynUGVYj/C4LM8HTxNGT7XcKiauvB2uvA8yOc3mX9oroTmR1fpTg1hdV4XRmAnhFlKy7RWHzKhOEsJRlOaxE8LVc26kp3XW+l3jZWsfEvvxfazf9119/8AY0wf9gA7fcdfY78DNefCM44N57j+HubT6pxwcurH5ZOfe3u5yVWld3V41HGKtaDtZ9YY6H9pt7P6oq2mKuZal0Xg3d08v687yZ01s1LDySr4VWUbyUaV9kpPdH5/78X2s3/ddff/AGNMH/YB9+L7Wb/uuvv/ALGmD/sAHaLgR7Znhfqn4Nk3HrR2M0dmE5RhLNsrUsblru1ec6f+HorfslV2T38D0OyTO8n1Nk2C1Dp/M8NmOWZnh6eLweLw1VVKWIozipQqQktpRaaaa8Gf54+K2Xc33G3NPu7xI4O6jx+Zym6lXHYThvSy7E15O6vVq4TB0p1Xu/x3Lw8ke3vJRpjUmjOVLhlpnV2T4vKs3wOQ0o4rBYuDp16DlKUlGcHvGSjJXi0mns0mmgPt4AAAADp/7VXF60wPJ1qDFaMxGNoqGaZc81q4SpKE4YH3y6m3Hfp977lPdKzd3a6f832Smc5DmvKLgYYHUVXMs5pZ3mEs8pVsQ6tTC4iVS1KLTbai8PGhJXsm3K17Nn2HnQ4xYLgRy46r4g5noTC6xwdOGHy7E5NjKvu8NiqOLrQw841X0y+J01XddLvstr3Xhlw75n9W8BuLmO4ncuuFr6KwOOn/AHXT2Jx0szwdWhe/weq5xg6tNNvpb/ukE9p3vJh/o+B42fhtOPX8E2gf0Y39cPw2nHr+CbQP6Mb+uA9kwdYOQLms1fzc8Ks913rLSuWZLi8p1BUyinHLVV+D1aaw9CqnepKT606rTs+zi7K+/Z8AAAAAAAAAAAAAAAAAAAPH323fys8N/o7iftJ355X/ANzPwk+guQf0fROg3tu/lZ4b/R3E/aTv1yu/uZ+Ev0FyH+j6Bzz9I9XTfpV9NAW5PT6nN7doAAN7AAGl15i68yOn1HT6gNmLIJW3JAAAN2EWXkSAS6Vs/IdMfzliLeo0eaijHwKSW7LpWDV2LDam/r+gl3t2Zbp9Q9kZpW9qLxJurdw1cjpu7XM0EWG34EtdPje4MNie24su5Eu5K7BsvcK3ZYqG70Xl4B3t4/oCdtiwN7Z7+v6CWn5Mt0+o6vQCqj5orbzNSso7Bu9M7Iiz8i/T6jp9Q2Xattu25G6LPYWvtcNVuyCzjZ9yGrbhu0pLxIdvAgA2CyAGlb2i0SHFP1Jt6jt63JIq428GFfxLXvtYdPqFbUbfgSt1uSASq23tbYNeRYBu1CrXkaW9SGrBsu1N1uLssR0+oVLotEh7MglK+4btD3K2fkXatuQTY1SSduwtsaSjsUexjZVUvMNbliH2DUXZFru9rgm/oFSqtW7XIs3uXbuQGyqkWXkWSsGvEN2rsg90GrgzUU+P8VeVvhpxh1PHVuraucxx0MLDCJYTFRpw6IOTWzg3e83d38j91w60DkXDDR2X6G01LEyy7LPe+5eJqKdV+8qzqyu0kn8abtstrL1OoXOZxl4paD4vUsj0drbM8pwDyfDV3Qw80oOpKdVOVmnu0kvzHYvla1Nn+seBGmNR6nzWvmWZ4v4b7/E15JzqdGMrwjdpLtGMUvRI9vNxcuPT45ZZbx+Dz8XJx5cuWMnee99VsiGlbYlq+4ttY8D1ISTIJasLeoVLUdN/AhryLXtsQGyqlbRLtX3KdPqFS7Q1vsCb22IMsbs7kEgxUu0WRUufjtc62p6fpSy7LpxqZjOO72aw6a2k12cmt0n872snmWUxm66ceGXJl5cYjXGt6WQ05ZblsozzKa3ezWHTW0muzk1uk/ne1k/kdSpUrVJ1q1SU6k25SlJtuTbu2292297ipUqVqk61apKdSbcpSk23Jt3bbe7be9z6doLQPwNU88zyj/fG0sPh5r/BeKlJP995L973e/4vlvm5rr3PsYzj6LDd9UaA0PVy2cM+zaE4Ypp+4o7p0k005TX+M02rPsnvvtH9215ou9rkd/Sx6McJjNR8vk5cuXK5ZM7b9iWvItb1Ibsb6JRbbtuR2LESMVFSGixDVw2KWQsl3JIauFb2WiLRIt6kBUo0rkNeRa3qGrGaaoCW/Aq3bYlsqN/X9A3ZYhu2wXKqQ0vEs3fYq1dhqN2LPyJ6vQdXoNRssVt5iyJbvsQZr4KVkotWKNJdi7Vtyr7mKxrFp+KM5pbnIkurbsZSjZszTtje7CUFtsZTjZ7JnJkZyjfe5Nm3WZONLw2ZSUU7u25vOFvEzlHvuTY7Y5OM1bsmZvqu9nY5Eo9tzOUe6v8AyE2adsawaT7mU4q7SNpwt4mcoWTdyL6u+NYyVvAxmnvs+5yZdyjjsybHbG6cZrzRlJPdW2ORUjutzKasmTY7Y1x5K3gZTTcdjktXMen1Jd5WDjvumZySszlSjv3OO47dybNO2NYSjv2MpJpdmcpqxjJ3XYix3xrCS3MpxSWyOQ1cznG67k2O2NceUE/DwM3TtvZnIkrO17mVR7E12lcacWm9naxnbzOQ97mcoWV7ku0rjzit3YxlFO2xyZxvdXMZwt4k2O+FYSTu0k7fMZyXayOQ9rmMvi28bk2ad8awknd7MpKK8jeSvdmUo9tybHaXbjzUfjbHwvnPS+9/zv8A0jB/z8T7rNW6vnPhfOfty/52/wDwjB/z8Dv0f9Yw+mPle0n/AJN1P7mX2PtnsTPkF139L39iw56KnnV7Ez5Bdd/S9/YsOeip+3f5bAAAAAAAAAAAAAAAAAAAONjcdgsuw7xWPxdHDUYtKVStUUIJt7Xb2W50B9pfz+an4BYzC8EeDGJpYbWOZYNYvNc3lSVWWV4ao2qVOhF3j7+aTk5STUIOLScppw6b6Z9nVz4cyVLD6917Xngp5jH31LGa6zyu8XKErbypdNWvTvZfFnCL2W1rAe42Dx+BzKisTl+NoYqi24qpRqKcbrurptXOSeIebey856uCinqzh1nGVZjjsJBV+vR+oq9DFrod/iKtChKUl3Si232Sb2OyHs8PaE691lruPLTzLVKr1O5VMJk2b43D/BsXUxNFPrwOMg0v7taMumbSk5RcZ9U5JsPSwAAAAAAAGGKwmGxtCWFxmHp16FTadOrBSjJLfdPZ72P5/wCxDSX+a+U/xGn/AOyfw+MPFXSnBDhnqHirrWvOllGncI8TWVOzqVZNqNOjBNpOc5yhCN2leSu0tzyA1R7Sfnq5gtY4vKeAOU47J8FKXVh8m0xkEc1xlOin8V1q06VSbe28oKnF7qyV0B7N/sQ0l/mvlP8AEaf/ALI/YhpL/NfKf4jT/wDZPG39sb2yP+TuKv1Wof2cftje2R/ydxV+q1D+zgez+Cy7L8tovD5dgcPhKXU5e7o04wjd+NkrX2W5yjq17PbOeaXO+Emd4rmsw2c0tQLUFWOWPOMDTwmKeCWHobOEIxXQqnvOltXbc92krdpQAAAAAAAAAAAAAAAAAAA8fvbd/K1w2+juJ+0nfnleVuWfhJbx0LkH9H0DoN7bv5WuG30dxP2k798rv7mfhJ9Bcg/o+ic8/SPT036T6Yu5YA5vYiyIa8iwBvSqW/YmyJAFLPyFn5FwG7Us/IFwGKAnpZKVg3atn5As3Yhu6BLpAADbdId/AlXtuCbNg2gAA2iyFkG0hdMK2iSuSkrLYNXJGjasl6EPtsXKtPdixsqu/r+ggsOh+hOm72qWI6WSYS6BZEdSJDZTZbB7kNXZN0w3e1bPyBYqBDS8QreBDV2WgrPcNVl3ILuDbbTRRK4btVrfsSl5kgEu0NL85Fn5E23uOpBUulRa/gSE7Cw2jp9AWunt5kdLHb3qRaJV7Mlxe+6HSydG9IIuvMlxdn2K9LCtrEWTJIukBDWxBPUpbJPfzIasFS7Q15EbovZogNlV3ews/Intux1IKlRuyLLyLdSD33Rlm2yqNbENeZYXV7GVstUshZeRNmiLpGNV7Al79hZoKlTaJWXfbsAGyq9iCzVyOlhsfktUY/hThsyVPWmN0nSzD3aajmdTDKt7u7s0qjv03vbwvc/t6fq6dr5RQq6Vq5dUyuXX8Hll8oPDu0mpdDp/F/H6r28b33udLuc3gzxS15xeo55o7RGZZtl6yfDUHiMNTTgqkZ1W43bW6TT/ADo7FcrWmM/0bwI0xpzU+VV8tzPCfDff4WvFKdPrxtecbpN94yi16NHs5eHHHhxzmW7fc4cfJllyXG46nxfVnsydrepFmyelnhvZ6orZMh38C/SyjfgY2U77thpeBACpQhrYt0sOLaDWdkxZEtNOzICtqvZglq7PzmsNYYXTGFUYKNbH1k3Rovsl26p23UU/Du2rK1m1GV8s3V4YXPKY4964OuNcU8gpvLctnGeYzW72aw6a2k12cmt0n872sn8jq1KlepOtWqSqTqNylOTbcm3dtt7tt73FSpUrVJ1q1SU6k25SlJtuTbu2292297n03QWg/gXu88zuh/fG0sPh5L/BeKlJP995L973e/4vmtvLl8z7MnH0WG76o0JoL4E6eeZ3Q/vjaWHw8l/gvFSkn+/8l+97vf8AF/fXZYr0s9GOMxmo+XycuXLl5sqhvzCS8CJRd2ErFJlS+xS1/AuR1Iyzaor22IkWbuQZY2K/P2Dt4Fn2KmKiGluV7FyGrhsulH2Is/IvZordIKE/MPfsQE7BUqGl38Str+Bci6ROmqkWTJMYYvC1K8sNTxNKdWCvKnGacorbdpO67r9Jsxt9FStbIh7Msfg+NHE2lwl0NX1W8CsbiJVoYXCUJS6IzrTTa6mk2klGTdt3a217rr0/BydVy48PFN5ZXsb13r9xZCyPjPLrxyz7jPHPIZ1kGDwDyj4O41cLObhU977z4rUm2mui90333ttf7PZrZnTrei5fD+e9Pz/pRWOUym4i0RZEWaIPIvYVcVfsWBl02XTOWyujNx2d0avdFWtmjHSVg4p22KSi0nZM2kunx7lJPZoyx1lriyT2umVa80Wr4jDUakKNXE0oTqbQhKaTk27Kye7322E4tN7onLGzu7Y5SsXFO2xk099nYzzfMsNkmV4zN8a2sPgcPUxNXpV2oQi5O3rZM6/8MeZ3PuIXETB6SqaTwdDA5jOsoTp1purRhCnKacm9pbRSdku+3gn7+k8J6nr+Hk5+HH83DvaXnx47McvWuwE4raybM5JWZu00Y1Ivd3Xc+XY9mNYzir7IymjfpZnPa6Id8awkrvsyk4Rs7o2a8jOcfivsTY6yuO1FGUo7Oy38DacG3dNdijVlcmx6JWDUvFMx6fNOxyZ7vbyM5RdmTZt1xrjySMZQVtl4nJlB+aMnFpXbRNjvjXHafgmZyT8jky7mEuxNnud8aycU92ZVEmkbtGMoNLuibHbGuPLZuyKSu14m0qbu3tYzlF2Isd5WEl32Zm4p90byVm0Zyi9ibNu2N7MJxirmM4rY5E4Pft2MJxbtaxNd8Kxku+xnJPyZu1Z2KTJsd8a4so7vZ9z4VzppLl+zv/SMH/PxPvM1fY+Ec6kbcvuebrfE4P8An4Hfo/6xh9L5PtJf6H6n9zL7H2j2JnyC67+l7+xYc9FTzq9iZ8guu/pe/sWHPRU/bP8AL4AAAAAAAAAAAAAAAAAAPIDTWQZZxM9s5m2G1vRhj6OW57i8XQoYhKUZTwOXN4RWlfaDpUppL/ua8D1/PJX2k3DHify48z2Rc73CrDS+BYuvhKmOxNOi5UsFmVGnHDuniEntRxFBKLat1N1U2pSi5ffOFXthOWHVuTYWfEqGe6EzfpSxdGtgKuPwsZ7XdKrhoynKO+zlSg9nt2bDvceRPtOclynhrzxcJuJmkaKwmeZosszPGxwsF7yricNj+mnWaTu5yhGEPC/ukr9ztHrj2uHJ1pjL6uJ01qTP9YYpU3KlhssyTEYfqn4Rc8XGioq/dq9leybsn0i4RYriR7SnnuynidqHIZYPSmlcRhMdi6FOTnhssyvCVXVoYN1Gl11K1W6dknJ1Ks1GMYtRD2wAAAAAADzX1Z7abSGmdU5zpynwEzjFQyrMMRgY15Z9SpuqqVSUFNwVGXS303td2va77gfXfa0ZNn2bcmmeVcmpValHLs4y3GZjGnFy/vVVeltpPsqk6Tbs7JXaSV1y/ZVaf0JlfJxpbNtIYXCrMs3xOOq5/iYRXv62NhiqsFGq0k/iUlSUU9lFpq/U2+t2pvbPcNdZaezPSeqOWXMMyyfOMJVwOPwlfUFN069CpBxnCS9x2abWzv4p3OpvKxzxZ7yk8SM5xWgMozHNOGed4qdXEaVzPHxdWKatTqwrxh0xxEElFzUEqkYpSimouAf6AweYX4cTSX5PGbfWKn/Zx+HE0l+Txm31ip/2cD09B8A5N+bXJeb/AIf5xrnKtHYrTUsnzieU1cHiMXHEuVqNKqqinGMVZ+9ata6cb33R9/AAAAAAAAAAAAAAAAAAADx99t18q/Df6O4n7Szv5yu/uZuEn0FyH+j6B0D9t18q/Df6O4n7Szv7yuq/LPwj+gmQf0fQIzm49HT+tfTRZ+RKVmSRI9culQAZoCLrzJtfYe79Rq03pF15i68yfd+pDjZ9xqwlLrzJIULu1yXtte9jG7AV6vQsldXGtsl0gWRLVtwDfdFkLIkBW/iiyJAAiyIs79iwBvSjXmgkttizVxb1DdoaIJkLeobKq20T4bktWIBtFokkdPqSGlkyGlYN22JGttl0pZEb+v6C/T6lTLG7Fe3YiPcuuxXo6d73Ms01De5Fn5Ey7kmE2hLzD2WxICto39f0EJK5YqAcY9yHHyTJttcmIN6Us+zRHSvI0a7u5Ru4VtRqwVvEu1sV6fUN3tHjt5i7Xcm3qJBsulSSCU7bBsu0CyJIM1W72q9h038CbeovbYwl0r0pPZCyZICpUENeRYhu2wbvStvMiyLN32IClbPyG/YsQ3ZhsukJb7kSSu9iW77Ayxu1N2On0LtW3IfYzTZdKdiN7+hYgxUqHbwIJ6fUlbINiEvNEEt3IDdvj3Fbml4Z8HdTw0jq2jnM8dPCwxaeDwsKkOibklu5p3vB3VvI/dcOte5FxQ0bl+utNQxUctzP3vuViaahVXu6s6UrpNpfGg7bvaz9D4PzG8o+pONnEGnrLKdXZZltCGX0cE6OJo1JzcoSm27x2s+tfoPsHAzh1jeE3CzJOH+ZZjQx+Jyr4T14ihBxhP3uJq1VZPdWVRJ38Uz2cuPBOHG4X85y48uS8lmU/NfvAAeKx6Qq1u9iwJJdKWQsl27ktWAUrv6/oF2WKBux79yGtiyV9z85rDWGF0vhVCKjWx9ZN0aPgl26523UU/Du2rK1m1mWUxm664YXksmM3arrDWGF0vhVGCjWx9ZN0aPgl26523UU/Du2rK1m18XxuNxWY4urjcbXlWr1n1TnLu3/ALEkrJJbJJJWSL43G4rMcVVx2OryrV60uqc5Pdv/AGJJWSS2SSSskfSNBaB+Be7z3PKP987Sw+Hkv8F4qck/33kv3vd7/i+W3Lny1PR9fCYdFhvL1V0FoL4E6eeZ5R/vjaWHw8l/gvFSkn++8l+97vf8X9/aJLV9yp6JjMJqPmcnLlzZebJay8iovtYGpl2hrbsVJb8CAqUIsiSWrBUulGt+xlWxOHw7gsRXp0nUfTHrml1PyV+79Dc6K83+e4TOuOmDyPMcQ4YDJ8LhMJiLXfQqknVnO3n0VI9vCKPreDeFXxfqLwS+XU38S13lqSjCDnOSjGKbbeySXdt+B/Ly/VGmc2xLwWV6iyzG4iN70sPi6dSasru6i29l6HUWVDjRzcZpisRl+LenNDYaq6VKlUnJUPi9k4xs69Ts23aMb2TWyf4LjVwB1DwInlWdUNSxzHC4us4UcZQpSw1WhXglJJpSlZtJtNSv8V7Kyv8AZ6f2Z6fPknTc3USc19MZN/xvxN31ehtkfzM01Jp3JqsKGb59l2Bq1GlGGJxVOlKTfayk03f0Oo9Tmh4g6s0Lpjh/oKhicXrjM6ToZhjqcE5xcZyjHouklOUIxnOo7KKbad7uNMTyT8Qc0ybFZ/nOusJidQ1YOs8JOFSt72dm+meJlJPqdkr9LV/GyucMPZzj6bLXiPNOPd1J62/P80V5t/O7j0MRRxNKFfD1oVaNSKlCpTkpRkn2aa2a9UWaVzo9yba9zzJ+JMOH9bGV5ZXnVKvbCTk3CliaVN1OuKe0W405xdrXur3aVu8TVj5fjPhWXhHU/IXLzSyWX47VjlubVa8j+VmeqNNZJU9znOosswE3b4uJxdOk91dbSae63Oq3HXmP1ZqnVM+GPB6tiYQ9+8FVxeC3xGNr3s40ZLeME011Jpys2mo9+Np/ke1XnGCWY6w13hsrx1f+6ToUsI8bKLe7U5upBOXnbqV+zfc+lw+z/DwcOPN4lzTj83pNbp59+k27eYDM8tzWi8RlmYYbGUk7e8w9WNSKfleLaOTZM8+uIXDXiPyzaly/Ocr1JNUsU5fA8ywLlTU3GzlTq022k7NNxblGSfd2aXc7gxxFXFHh5lmrKlGFHF1FLD42nD8WOIg7Tsruye0km20pJNtq55/FfAp0XBh1fT8nynFlfX4fSrHPd1Zqv0Gr87p6Z0pnOopyUVlmAr4ttq/+DpuSVvHdWt49jpryY5RXznizj9QYqrOby7La1ac5O7lWqzjFXb73TqNvzXqW5juA2odNTz7ivnercFjIZlmrlSw8ac1VtVm3CF27JQgrWV7KNlsfmuD/AC2ah4v6axOpsvz7B5bQoY2eDUcTSnJ1HGEJOScfD46Xzp+R+n8M8P6LpPCOXO881yaly16fN8/qi23KdnoC9kfwtY6J0xxAyZ6f1flEMxwDqxre6lUnBqaTSkpQakmk2rprZs6qfeMat/z4yf8Ai9U+N8W+GOK4S6qhpTHZ1hcyrvCU8VOeHhKKp9cpJQalve0U/mkj5XQez3R9TzSdJ1m8537Y33f4ul5LPXF6FaL0BpDh3l1TKtGZFRyzDVp+9qKEpTnUlaycpzblKy2V27b27s/twxeFq1ZUKWJpzqwv1QjNOUbO26Tutz+Nw9yarp3Qenchr9fvcBleFw9Xrd31xpRUr/nT28Ox0l4R1K+juaHDZfWm+qjnmOyyrd36nP3tLd33+M016pPc+Z0nhWXiuXUcmXLblxze/Xza3867n5dT4u+/f8bsVnOnThKpUnGMYJylKTskkrtt+CLNXOsnO3rV4DT2TaCwtdKrmdZ47Fxj3VGk7QT9JTba9af6fm+F+H5+J9Xj02HbfrfgvLKYy2uy1CvQxNP3uHrQrQbaUqclJX8roufGuVDRU9I8J8Jj8VSdPGahqvMqid7qk0o0l8zglL/69n4Lmb5h82ynM6nDPh5jKlDGwahmWPw7fvYzklahSa3UrNdUlum0k00z18fgmfV+IZdF0uW5j65e6Se8vJJj5q7G4/U+mspxKwebaiyzB4iVlGliMZTpzd7WspNN3urfOc+lWoYmnDEYerCrSqJShOElKMk+zTWzXqjpxpbky1nqTKY53qzVdHJ8bi4++WFqYaWJrJve1WTnHpk73aXU1473S/P5vhuMfK9isXk1XGrE5JnmFxGHpThOU8JUnODSqRTs6daDalaybta7W59TH2b6LqcvkOk6mZcs92tb+ip+Wyn52WPZ3khXwtdyjRrU6jpu01Cabi/J2ez28SJR8Yxdjq7yPZC44PVOpZp2qVcPgaXkulSnP579dP5rep+m5jOBWoOJObLV0NV4PA5Zk2Vyi8NWpzbi4OdSpNWdrtdKvt+Kk+1z53L4N0/B4neg5ebUmvzte/Xpp2x5bcPNI+Ia0qVOI/NFPAKs6lGeoKGXp91GlQlGnOy8koTe2zbb8bneOUd79J52cJOFea8XNRYjT2V4+jgpYbBzxlSvXg5QSUoxUdt7tzVvmfkfW3yR6s3/APv2yjb/AOgVD9X7Q+H+HcmXF03L1M47x4ya8u9/O5cHLyY7ymO912tzDAYTMMJWy/G0I1sPiacqNanLtOEk04v0abX5z8hpLg9w30JmNTN9K6Wo4LGTg6brOtVqyjF91F1JS6b23tbbbsdROLvADNeEOS4LOM01LgMc8bivg1OjQpyjP8WUnO8trKyT9ZI7AcoeUVcv4UTx9TrtmmZ168Lvboio0tl/9VTlv/Uj8/13hGPh3ht6rpequWGV1qSyX6//AKerj5rycswyx7x9nmmrWRhia2Hw1CeIxVaFKlDec6klGMVfu23ZH5Pi7xPyrhXpeWeY2n8JxdeTo4HCKVnXq2vu+6glu34bLu0n1eyLSfGHmVzCrneb5w6WU0arj7/ENwwtJ/4lClHaTSaTfzdUr2v83wzwC9bw3q+o5Jx8M999/wBEenk6qYXyYzeTtvQ1VpTF1/g2E1LlNetfp93TxtOUr3tayk3f0OfJJt7HVzPuTXPMFllXE6f1nh8zxlOLlHC1cE8MqllfpU/eSSb7K6Sva7S3XB5b+LGocp1ZhuHWo8ZXr5fjZSw2GjiJNzwddJtRTbuotpxcfBtNW3v7Ob2b6bqOlz6nwznnJ5O9mtVvH1ueOcw5sdbdrmvJM4861GVV4dVoOql1On1LqS87d7HKls7HRfJ+I2GyjixnPEvEUFjK0KuNxOBouTUKlSq5U6ab79MYTb87RsnezPm+CeA8njWPLeO6uE/jfdHq6jq501x373dbGYrBYCg8Tj8TRw1GLSdStNQir9rttI42CzXKM2pSnlOZYTGwjbqlh68aiW/i4t27P9B1ty3gxxV40dGrdf6meWUMSveYWlXpupONN7pwoJxjTg1a12m+7Tvd/O9Uaf1VwI1/Ro4PNl8Lw8IYvDYmjeMa9Ftq0437NwlGUW2nZ7tWZ9fp/ZTpOpuXTcfVS88m7jJ2+jbnl4lyces7x6xtd3ehPwMpJpdmZZRmKzbJ8BmsabgsdhqWJUb/AIqnFSt+a5ypfis/D8mF48rhfWXX8H28MvNJfi4s+q+1zOW6ORLuZShZXuc7HolYSSMHH0ZypR37mMlZEWO+NYNW8DKe62VzkS7mLVtya741n0/Fd0ZTjFLw/SbyWzMZxurXJdcXHnHduxnJejN5K2xnNWIejGsJp2e3gZWX75fNc5EvEylG9t7WJsdcb2YTjG97GMkn6nJnDZ7+BjKNrb3uTY9GFceSSufCOddW5fM7/wBJwf8APwPvcod3/IfBudf9z3nn+k4L+fid+jn84w+mPle0d/ofqf3MvsfZPYmfILrv6Xv7Fhz0VPOr2JnyC67+l7+xYc9FT9q/zEAAAAAAAAAAAAAAAAAADgZ3keTalyrFZBqLKMHmmWY+k6GKweMoRrUK9N94zhJNST8mmdM+InsiOUjW+OrZlkOE1ToutVbn7nJMzjLDKbX/AHPEwqtRvv0wcV4Kysl8V5wfaG82XBbmM1hwz4faKyKenskqYSGBqYvI8RiatWFTCUaspurGpGMk5zk0klZWTu02/jf4WHnl/wAyNL/VjF/rgO2GlPYy8r2SY2OM1FqfXmooxe2Fr5hh8PQmttpe5oxqX2e6mtn2vudy+GPCjh1wa0tQ0Vwv0fl2nclw7clhsHTac5vvOpN3nVm7JOc25Oyu3Y8fMf7XHnTyujGvmemNH4SnKSgp19PYmnFyabSTlXteybt6M+h8BvaPc9XFrX2ksBhuF2U5xpnM9QYLLcyxuWaWxsqVKhOvTjWbrxqyhTcYTbcpbR2bVgPW4AAAAAPjWY8nHKrm+YYrNsz5fNCYjGY2tPEYitPJaLlUqzk5Sk/i9222/nPsoA+J/eTco35OWgf/AEJR/qH3k3KN+TloH/0JR/qPwftP69fD8jHE2ph686UnDKKblCTTcZZtg4yi2vBxbTXZptPZn8v2VGMxeM5KNHfCsVWre4xua0aXvKjl7umsbVtFX7JXdktkB9Q+8m5RvyctA/8AoSj/AFD7yblG/Jy0D/6Eo/1H2w+D87+fcO8p5WeJmB4i5/g8vwuZaYx+HwdOriIwrYjGOi/g0KMG05zdb3VorZu17K7A+ncPOGHDvhLktXTfDLRWT6YyqvipY2rhMrwkMPSqV5QjGVRxiknJxpwTb3tFLwR+qPO72K+p9Q5twI1lp7Mq9evlmS6ki8vlUbap+9w1N1acG9ulOEZdK7Oo3++PREAAAAAAAAAAAAAAAAAAAPH323fys8Nvo7iftJ6A8riX3svCP6CZD/R9A8/vbd/Kzw2+juJ+0noByuy/3M3CNeWhMh/o+gRn6O/B619Pt6C3oQp2fYn3noS9XdS3oLehZR37kt2du4Japb0Js/Inq9B1egNos/Ii3oW6vQkG1Lega2exolcOOz3BLpjb0LLsi3T6jp9TJNG1XuVd7PuadPqHHZ7izbdsd/Ub+pfp9STJC3SiT9SVt/8A3LFXG8hZpsq3xfNFR7v1D+L+YzWjewFer0HV6Bu09ySE7kglCGtuxIDdqWfkC5Xp9QIBPT6kLcKlBZeRLViAb+ICG7MJ32DSy8kVLkdPqG7Ud77XCvfe5fp9R0+pOlSqlTTp9TMa0W7SmrE7FQlcxso3+gJL0Jt6kBu9rNK3ZGcl22L39B39LAZjuXcPG5Xp9QqVSwLENX3DVSbPyJtbe46vQNlVBISvuK3e0PsVLFWrGaaEbFyso79zFbRZeQsvIkAUs/IW9B7z0JW66u3oFSosvIjb0JItvcKl0jf1I9GW6vQWvvcNl2rZeQsS1YW2uTprN3XmPAtIgxUVVvFE7egauRbewbK6n8z3NhxF4LcSaejtLZLpzFYKeW0MY54/D151eucqiavTrQVrQVla+73Z9s4AcQs64rcIsh19qHC4LD5jmnwr31LBU5Qox91iatKPSpylJXjTTd2927WVkuHxI5beEvFbUMdUa1yPE4vMY4eGFVSnjq1Je7g5NLpjJK95Pe19z9joXRGneHGlcFozSmEnhsqy73vwelUrSquPvKkqkryk23ec5Pd7Xt2R6+Xk4bw444TWU97jhjyTktt7fB/cs/IguV8TyPTEFX3Zp0+pVx77hu0LdbkFkrAmtl0qUs/I06fU/N6w1hhdL4VQio1sfWTdGj4Jduudt1FPw7tqytZtZbrvXTDC8lmOM3aav1jhNL4ToioVcfWTdGi+yXbrnbdRTvt3bVlaza+L47G4nMcVVx2NryrV60uqc5d2/wA2ySVkktkkkrJDHYzFZli6uOx1eVavWfVOcu7f+xJKySWySSVkj6VoHQHwL3eeZ5R/vnaWHw81/gvFSkn++8l+97vf8XyW5c2Wp6Ps444dDx7y9ajQOgngnTzzPKH987Sw+Hmv8F4qUk/33kv3vd7/AIv76XYt0+ocLq1z1Y4zCaj5nJy5c2XmyZkNehdws7XKm+qJdqgsVatuZZpqbRtulco15IsDFS7V39Ssr+pchu4bFV5Puecud5XiuNfMXmOU5dVkoZ3ntajGsld08JTk06lvG1Gm5W7bWv4nf/XGerS+jc91J1qLyzLsTi433TlCnKSXrdpK3jc6b8kGQvM+KGZ5/Xi5xyrK59Mt3atVnGKbfrBVf/dH7P2ZyvRdH1XXe+Y6l+e/6ir31HdPIMgyrS2R4LTuR4SGGwGX0Y0KFKP72KVt34t7tt7ttt7s6z8+GcUqWQaU0/s6mIxmIxnqlShGK+ZN1X+j0O1J0S51c7nnPF7CZFQTksqyyhQUErt1aspVHb54zpq3oeT2Vwy6nxXHlzu/LvK/6/xLez7Byb8LcFp3RH7YWYYeMs21B1KhKS3oYSMmlFeTnKLk2u66F4O/37Nsxo5PlWNzbE39zgsPUxFTe3xYRcnv4bJnE0lkVPTOlsn05SSUcswNDCKy2fu6ai3+dq9/G5+I5k8+jpzglqnEqbU8XhFl8EnZydeUaTS/+tnJv0TPn9RzcnjHim8rvzZa/wAN9lTs6r8nuWV8944QzmdNN5bgcXjqkkrJOaVLb1brPbyv5M7T8yGtsToThFnWaZfVdLHYyMMuws07OE6r6XJPwah1yTXikfGuRDIEqOrNUVItuUsNl9KXgklKpUXr3pfNb1P0HPRUmuHOQ0VNqE86UpLwbVCqk/zJv9J+k8SmHW+0nHwZd8cdT+HcnbHb8jyP8P8AC4zF5zxIx+HjUlgZrLcA2k1Cq4qVaS8moShFPynJeJ28fpc+G8mSpftLw6HHqeaYrrs97/F7+trfmsfdGrH5/wBpuoz6jxPk83uup82l4ekdc+eKVL9q7JoSt715/ScL9+lYfEXt6Xcb/mOXyU0MRR4QYqpVm3CvneJqUk77QVKjFpf/AF0ZP87PlXOpr3D6g1fleg8qre+jkUJ1MWobp4mr02ht3cYJbLs5tPdbdmeCOiqnD7hdkOmcTT6MXTw/v8XG1mq9VupOL9YuXT80UfX6yXo/Z3i4OTtlnluT5v8AWv4snfK2PivPXn0aWR6W0vCbbxOKr4+cU9kqUFTg38/vZ2+Zn1DlhyFZBwS05TlFqrjqVTH1Ha1/e1JSg/8AzHBetr+J1r5ys5q53xhoZFh5dayrLsPhY00/+FquVRvfs2pwXzJHdfT2UUcgyDLMhw/SqWW4OjhIKKslGnBRVl4KyRz8W/mngXTdP7895X/X+KsO+VrnHQniE3xJ5qq2VNqdCrn+Gytq91GlQcKVS1/SE3bzbO92a4/D5TlmLzXFu1DBUKmIqu/aEIuTf6Ezo1yp4DEau49LUWMj1zwdHG5rWbba653p3u+76q9181/An2Vn4Pw9V1v7GOp/j/8AxvJ31Pnd7ToNxkjLRHNDic2VoQoZzgM2jNtJNNUqsm9u3V1J3Xg+/j35OknO3k3wPiXlecQhaGY5RBSfnUp1Kib/APNcER7G8kvXZ8WX9vGxvJ3x3Pc7rznCnCVSpNRjFOUm3ZJLdtt9kdCc9rYrmP5iVhMFUnPLMRi1haM1dqll1Btymu1upKckm/xqiV90faeYDjXHJeDeSZZlmLbznWmU0Krkm+qlhKlKLq1G73TndxV93eT7xN+UDhTLSulauvs4wrhmeoKaWEjNWlSwV00/T3jSl80YPxaPR4Zx/iLo+bxHlms8t44T3/PTK/KZTGej7Jq/PcFoLQ+aZ9ToQhh8ky+pUo0UrL4kGoU0trJtRivnOnXKlpSWvuL2I1XqBPGLJ4zzSrUq7+8xk52pyfr1OdRPzgjsXzWYqrheBmfqjdOvUwlKUldNReJpt/ptb5mfOORbDYdZPq7GRa9/PE4SnPzUIwqNfpcpfoM8KyvS+BdT1mP6eV8u/f7vvrcu/JMfg7RTStsdbOd3PIYXRGQadUoqpmGZSxVnZtwo02n829aO/pbxZ2Sk4wi5zkoxim3J7JJd22effMtxNw/EniNWnlVdVcoyan8AwdSLvGq026lVekpNpPxjGL8TxeyHRZ9T4jjzf2cN237Ivmy1hr3u0HKdkDyXgvlmInSUKmbYnEY+Stu7z93FvzvClBr0aP7HMZnsdPcF9T4nqtPFYVYCCTs5OvJU2l/9bOTfomfrdCZCtL6JyHTqg4vLstw+HmmrNzjTSk36t3b9Wz4fzt548HoTI8ghUUZZnmTryV95Qo03dfN1VYP50jj0n9J+PTL13nv/AAl39i7fJxP4PJBkPThNU6mnFv3lTD4Gk7dulSnNX8b9VP5rep2hkmrrc+R8puSRyjgtlmJ6VGea4rE42du9/eOmm/NuNKL+ax9fn3seX2l5/wAJ8U5st+l1/Ds69P8Am4SOofO1njrZ9prTUZ2WFwlbGzj5urNQi2vT3UrfOz7/AMIMiWnOF2mMqcbThllGrUXlUqL3k1+aU2dUOOlWpxA5jK2n8PJzjLHYPJaXxns/ixn8yU5z+bud4lShSpqnTiowgumKSskkrJI+v49PwXwno+j+M81/x/8A7WdPfNy55uj/ADEZ5mXETjYtI4CfVTwOIo5Lgad9vfTlFTbXg3Ul0t+KivI7gaY01l2kNOYDTWT0VDCZfRjRgrWcmu83bu5O7b8W2zpdpCarczlKriZKTnqrESlKT7zdebT+e9vznemXiPa23pun6XosO2Mwl+m/6+106P8AOyyzvrtxpKV090dGMtpSxXMylgZWj+zKpUi4qydNYuUmlbwcU18zO5XELV+A0HpDMtU5hUilg6LdGnJ71az2hBK+7cml6K77JnVLlZ0ri9UcS6urMZGVShktOeJqVJRup4mqnGCe1r2c5ejijPZfG9J0HV9ZydsfLqfPf9a/i6dXflOTj4567doOJWdQ05oHUOctpTw2XV5U97f3RwcYK/heTS/OdTOWzh5h9ba2lmeaUFVy3IYwxVWEleNSs21Sg/C11KTXj0Was2fdea7OHlfCupgY1EpZtjqGFt4uMW6r/Nekk/nt4n83lIyRYHh1jc4nFqeZ5jPpdnvTpwjFf/8ATmb4Zy5eGez3P1OHbLky8s/1/FfLJzdXhh7pH2qXc6c80mOWacVYZfh4uVXBZfhsJJLducpTqJfPaqjuQ+9zpZBLiBzJdvfUa+oG7d1Khh5X/Q6dL9B5vYrDydTzdXl6YYW/4/627+J3zcePFPfY7d5NlqynJsvyptSeCwtLD3XZ9EFG/wDIbtXW6OTUjutzKUdj8XyZXlzuXxu32eP83HTj1Ivq2Tt8xlNO3ZnKk7O3cwl2ONj0Y1g15orNK3ZfoNJ9zOUdiL2dpWUkr9kYNK3Y5Mo79zBq25N9XfGsJrukjKUX5M5E/EzlsibNO+N7MHDZ7bmTS8UciTuYzhfxJvd2xvxYztd9jJryRvOna7uZNWJsdsaxmtnszCS7bHKnvcxmrE2PRhXHa7nwfnaS+96zy1v/AITg/wCfgfe5x2bufBOdpW5es9d//lOC/n4Hbo5rqMPpj5XtFf6I6j9zL7H2H2JnyC67+l7+xYc9FTzq9iZ8guu/pe/sWHPRU/aP8zAAAAAAAAAAAAAAAAAAAAAD+FrPROkeImmsdo7XOnMBnmS5lTdLFYHHUFVpVI+Ds+zT3UlZxaTTTSZ5AZ9PPPZlc+GXaV4a6jx2P0Bqz4DjMXkles6vvMtxVepRdKok/jVqM4VJUqjSnZQTupzUvZw88uaDJOSl89+hcv4u4DiNmvEjUuKyL4BTwFSg8joTeIVDCU8QpyjVUZVIJzVNSVpNu12gPQ0AAAAAPmPMvwz1Dxl4Ca44W6UzPCZfm2pcpq4DC4jGVKkKEZya2qSpxlJQaTi2oydm9n2PpwA8F+PXswOPnLvwnz3jFrXV3D/G5Lp/4N8JoZVmGNq4qfv8TSw8OiNTCU4O060G7zVkm1d2T43L37M/jtzJ8Lsu4taG1ZoPA5RmdbEYelRzbH42liYyo1ZU5OUaWFqQSbi2rSe1r2ex7j8TeGeh+Meh804bcSMihnOnM5jSjjcFOvVoqqqdWFWHx6Uozi1UpwknGSd4opww4WaC4MaJy/h1wy07SyPT2V+8eFwdOrUq9LnOVSbc6spTm3KUm3KTfrZJAeQf4FTmm/z94Vf+lcx/sJzsl9ihzF18yo0tQ8TeHGCy+Uv7tXwWJx2KrQXnGlPDUlJ+jnH5z2bAHyTli5cNF8rPCjBcLtFV8RjIxrTx2ZZjiIqNXMMbUUVOtKMdoq0IQjFN9MIRTcmnJ/WwAAAAAAAAAAAAAAAAAAAA8fvbd/Kxw3+juJ+0s9AOV1L72bhH9BMg/o+gef8A7bv5WOG/0dxP2lnoByu/uZuEf0EyD+j6BGfo7cPavptkLIkEvT5qtsUnu9ty3T6jp9QbUs/Jiz8mXXcsGys7IWktknb5i3T6i9trBm1fjrz/AEC8/G9vmLp32DV0w3e1brzJXT4tFen1JUL+IJZSVktmr/OVv6lvd+o6O+/b0Brv2RaIsipddgd/elRTvdFJRtJ7Fuq3he5Dld/OGy6RZ+RDiu7RcSjePcEumfTF9g4x8F/KWULeJPT6maP8WTTXZMku1Yrb1FikASjv3IUd+5JtIDdnbuAQKruWAUiyZWz8i4ApZeQskWauyOn1DdqttEk9PqQG7ARbe5IaEdEfIkhuwbtSS3aSdiY332aLkNXMsbLtV9ipdqyZQzWm+noEoldgYqbH2djPf1/QaPsVArZ+TI7dzS/oQ11O/YN2oRZLuWcbeJVq4bKNK2xF2TbawvbYNVFkyb+hAV71ml4FGncv0+pDVjLq+jd9lATe+xEo79zGy6V6I+RNl2JatuRbe4ahrYhrzLlW/AKlqtkSCenbuG70pIXfYNWC7hUqGvNCyLNXDjs9zNd2s3bwK+JZqwJbNOhHO7xJ4i6U4zUcr0tr7UeTYJ5Lhqrw2X5pXw9JzdSqnPopySu0km7Xdl5HZflLzvO9Scv2lc51Fm+NzTMMT8O99i8biJ161TpxteMeqc25O0Ukrt2SSWyR+q1jxF4QaVzZZXrjVWnMuzJ0Y1VRzCtTjVdJtpO0t7Np28Nmf39L53pvUOR4bOdI5hgsdlOI6/g9fBSjKjPpnKM+lx2dpqSdvFM9vLzebgxw8mvnccMPLy3Lzb+Z/UIt5dyzdyFseJ6do+N6kb+pdu5W/oGoIJbufmtY6wwmlsKox6a2PrRbo0X2S7dc7bqKfh3bVlazazKzGbrrx4ZclmOM3anWWscLpfCqEFGtj60W6NFvZLddc7bqKfh3bVlaza+LY3G4rMcXVx2OryrV60uqc5d2/wDYklZJLZJJKyQxuNxeZYurjsdXlWr1pdU5ye7f+xJKySWySSVkj6XoHQXwL3eeZ5Q/vnaWHw81/gvFSkn++8l+97vf8Xx25c2Wp6Ps448fQ8e8vU0DoH4F7vPM8o/3ztLD4ea/wXipST/feS/e93v+L+939f0FgenHGYzUfL5eXLmy82VV39f0Et+RJQ1zl0ndlLPyZdO2wbvsFSswWa8SrdgraGvIgm/oQnYy6bCz8irVuyZfq9CG7mKfGebbPJZJwOzmlTTVTNKuGwMWvBSqKUr+jjCS/Oj8LyK6eng9G6i1ROi4vM8wp4Wm2rdUKEL3Xmuqs1deMWvA7OVqNKvCVKtShUhL8aEopp/OnsyKdOnRpxpUqcYQiklGKSSXkkux9nj8W+S8Ny8Pxx75Xdv8PuVPit3W50BxdOfErm7qU5wnVpT1Qqc7ptvD4Sai7q10nTo237Jq53+Tu7GSwmGhWliadClGrPac1BKUlts33fZfoM8J8VnhXymUx3llNb+DfVo0vA6088+ezwegMiyCnNxeZ5o607fvoUabun6dVSD+dL1OytvUyr4bD4qHusTQp1Y3v01IqSv52Z5fDesx6Dq8eqyx83lu9fOq95p8a5QtOyyPgnl2Jq0/d1M4xWJzCSd02nP3cG/nhSi16NH6Pj7wwq8V+HGM07gZQjmeHqQx2XOo7RdeCaUW32UoSnG/ZOSb2R9FatuOr0N5PEuXLrr12HbLe2z01Xnzw24vcSOW3M8fpnNtMzlhsRVVXEZZj1KhKNVJL3lOaTteKSbtKLSTXZM/aam51Nd6owryXQ2jaWU43FJ041o1pY6um096UVCKUvJtSt5eK7j4/LMtzWmqOaZdhsXTT6owr0o1En5pSTRnl+RZHlNSVXK8mwODnNJSlQw0Kbkl4NxSuff5faHw/qM/wjn6WXk+O+1v0El91dXOXnlmz157S4m8WMNVhiYVni8Jl2KfXWqV23L3+IvdppvqUX8Zy3la1n2tsi77FT4HififN4rzfK83u7Se6ReM12dEObvSeocg4v4vWcsJVWX5xDDVsJilC9NVKVGnTcG90pp0+qz7qS77n7nS/OBxC1rneT6UyDQWWPMsfiqNCrV66tWCg5pTmoKzikrttyaild3SO2OJw2HxlGeGxdCnWpVFaUKkVKMl5NPZnGwOTZRlPX9y8rweD95bq+D0I0+q3a/Slc+1faHp+fo+Pp+q4PPlxzWN2yYWXcr8FzGZ69PcFNV4yEnGeIwawUbd37+caT/km3+Y+L8iunZKGqtVVaUkpPD5fQm00nbqnVSfZ96XzfnO184QqQlTqRU4TTjKLV009mmn3RFKlSoU1So0oU4R2jGKSS+ZLZHzODxb5Dw7k6HHDvnd7/h21/gvXeXfoh7I6uc9GTurkeldQRpv+9sXicJOaStarCMop+P/AAUrfO/M7SFKtKlXpulWpQqQltKMkmn86ezPL4T198L6vDqpN+X3Ks80sdB+AfCnOeNercNmOpalerp3IKdCjiKlRNKpCmkqWFg1ZJNJXtuldt3km++9OjSoUoUaFKFOnTiowhCKUYpKySS2SSSSSJpUqVCmqVGlCnCO0YxSSXzJbIs+x6fG/GuTxnmmdnlxnpi3DCYPyXFTRj4hcPc80fCpGnVzHCtUJTXxVWhJTpt+S64Ru/BbnRvh1xH15y6aszDCYvIZQnXjGlmGWY1SpdfS24TTXZq8rSSaak9ndNeh5w8fk2T5t0PNMqweMdNPoeIoRqdN+9upO35j0eD+OY+H8OfSdRx+fiz9zc8PNZZe7qBmXF/jjzGp6O0Fpz7j5ZXap4+vhpycVBpXVbEtJRha7cYpSkrq0k7P5/DhthMu4+ZPwywFSWLhgsxwWGxleUWnWmlCpiZqPhFN1FFf4sVd3bZ6C4bCYXA0IYbBYelQowVo06UFCMV6JJJEqjQVZ4lUKaquPS6nSuprybte2y29D38PtVj0ky4+m4ZhhZZJvvu++33p+S82rburdzrXzr6PzzO9O6f1LleCq4nC5HVxUMZ7uDk6UKypNVGluop0mm+yclex2UB+c8M8Qz8M6vHq8Zuy+n1O2WPnnlroxw25rdR6A0XgtE0NJ4HMXgOunha8604O05uSUoRT6mnJpWabSXjdvuDojM8+zXROU57q7BUsDmmLwUMVi6FOEoRpOS6rdMm3FpNXTbad090f1aOnshwmKWOw2SYClibuXvoYaEZ3fd9SV/5Tmt+Fj3eL+KdJ1+Uz4ODyZW7yu97bxY5Y+90O4C0Kuv8AmIo6hr0pzh8MxmdV3Zvpb65Qba7f3ScO/wAx3mauXWHoUZTlRoU4Sqy6puMUnJ+ba7vd7sOO3c4+N+Lfjfmw5McfLMZJJ6+jrwYfJS/O6Wcw/CTVmi9e4niRpXB4mrlmLxSzOOJw0HOWBxXUpSc0k3Fda6lJq26V7rf+rlfOnnWHy2FDONC4TG46MUpYmjjpUITaXd03CVm3u0pJeVjt1JXtZn8avpbTNarOtW09llSpNtynLB022/Ntq7Pp4+0fTdT0+HB4lwfKXCdrvROnyxy3x5a26aZhjeMvM7nVDD0cudDKMPU6oKEJU8Dhe6c5zd3Una67t97JJs7U8N+HWT8MtL0NNZQnUabq4rFTilPEVna83bstkkt7JJXbu3+vjGFGMaVOEYxikkkkkl5JLsVn2ufM8V8dz6/inTcGE4+LH+zPf9L08HBOPLz27rqtznZu3jdM6fhdKnSr4yp5NycYw/R0z/Sj7NwXyOWnuFem8sqU3TqPBRxNSMlZqdVuq013TTnb0tY/dVsNQxEoSrUadR031Q6oJuL81fsys4bNt+Jx6rxj8I8N4vDscNTC7t36+v3u3FweXmy5bfV/I1HmUcjyDM86kk1gMHWxVn2+JBy/9R1N5U8nq5rxKxOd1oymsswFWr1vf+61GoJN+bi6j/MzuK4fG7mFOhRw8XChRhTUn1NQikm/W3iT4d4x+L+i5+lxw3eWa3v0duTp/luTDO3tiznZLqk7KKvd7JI6VZbqTWGveM9GeU59mL+G5y5UfdYifRSw0ajeyTsoRpp3XZpO/dn2bmV4nVsny+HDnTc6k83zmKjifc3c6dCbsqaS366j2st+m+3xkz+rwJ4NUuHeVPO86pQnqHH00qvZrC0nZ+6TTabbScmu7SS2V39rwn5LwLw3k63qpLnyzWGN9fp+hx5/N1XNjxYXtj6vqU1G+/8AtMZxSWyORKF3e5lJdW3Y/DZXfd93HswlFX7GU42WyZvONna5nLZEWO+OTjyT8jFp+KOTLuZzjdWuRZt2xyceSW+xlKPmcidO13cylHYmx3xrGUVvZGNVNWsn4nIfYym7ku+NceV7O9zJrzRyJdn8xl038bWJs0741jJLfYxnHtscicbN7mco9tybHbDJxprZ7M+Cc7aa5ec92f8A8JwX8/A+/T2ufBOd1f7njPH/AOE4L+fgdukn+3w+mPme0N/onqP3MvsfXPYmfILrv6Xv7Fhz0VPOr2JnyC67+l7+xYc9FT9m/wA0AAAAAAAAAAAAAAAAAAA81uf/ANpPxH4R8U8RwC5f8vwVLO8sjQhmucYjCLF1Viq9ONSnh8LRknBtQnTblJTvKXSopxbl17w/M97YjGUKeLwmScUK9GtFTp1KfCrDyhOL3TTWX2aa8UfacNwE4maH9rZDiZm3CTUmf6KzjNq+Pw2f4XKK1fAYSeJy6UYVKlaMXTg6Vabi+tprpUluonqIB4rffLe2S/zd4q//AGJ6H/8ATjrxxU4i83eoOYPS2s+K+X6opcXMvrZZLT9HHaWhgsfKrSxLngvdYJYeCrN17qKdKXW/ivq7H+jA8y+cDgjxg1T7S3g1xD01wz1Jm2l8FjNMVsXnOCy6rXwWEhhszlUrutWinCl0QXU1NptNNJ3Vw/O8rvHf2o+p+YDROQ8YNP8AECGi8bmXu86lmnDqjl2FjhvdycnPErB03Ts0mmpptpLe9n6rAAAAAAAAHwfnbz/jhpflw1LnvLzRzOrrXB1cFUwv3MwMMbilRWKp+/dOhKM1Ufu+q6UJPpcmltdefvCf2yfFjReLjprmG4YYXPZYSTo4nGZdB5bmNOS7+9oTTpSkndWSpLt5O4evQOunBf2gPKrxydDB6b4nYTJ83r7LKdQpZfiuq1+mLqP3VV99qc59mdiYuMkpRaafZ3umBYAAAAAAAAAAAAAAAAAAAAB4++26+Vfhv9HcT9pZ6B8ri/3MvCP6CZB/R9A8/PbdfKvw3+juJ+0s9A+Vz9zLwi+gmQf0fQIz9HXi9X09K/gOn0C2J6vQl37oswWDV9wbZ2lfZfyk9My1rb3HV6BUsqt0Q7vexb3fqLW2DN7VS3LPcACtmN1sS3Yi197gE/Mm6fj3I6fUWtvcNl0jp9AT1egtfe4bvQreKX6CrXxtlsWasLeoJUEPq33dvnLdPqQ14At0hEvsQlYkNQvEnp9CErE9dvD+UCHH0RHT6Fr33sA3bOS37EWZpa+46fUyzbZVO2zQlutu5Mo79yErMaar22kSTKHV42IatuZZomWgEN2YTvsY3aSGvIkA2rslZ9yCzV2ErbhW+yjUrkt2LNXZRq4bsuuxJFrb3Cdw2VJForwRJDVw2XSuw7k9PqR028b3J0re0eP5ydkRa29x39LGNlR3ewtLwJWzJDYj4tt7EPpVuxLXiVauBDa8CC/u/Uhws7XCpVGtuxFmX6fUNWDZVLvzAt6hK+4bLpAAGlb2ENbtpbEgy6IoLxJlHYra29zFb0lryI3Q6vQnwDZdK9yGnfYslch7BSN/X9JFpFiG7MNl0q15oNbdiW7kGWKldKub7l24x8UeLFLUuhdH/dPLY5Th8K6/3QwtC1WM6jcemrVjLZSW9rb7PudgeWbRepuHvBHTekNYZb8AzfAfDPhOG99Tq9HXi61SHx6cpQd4Ti9m7Xs7NNL+Bxo5t9F8EtYQ0bn+ms6x+Jng6WN97hPc9ChOUkl8aad04O+1t0fRuFfETK+LOgss4g5NgsVg8Hmvv/d0MUo+9j7qtUpO/S2t3TbVm9mvE9nNnzXgxxyxkxcOOcfylsvd+p7bNEd3sXlHfuVtbe54XpLMq16blur0PzusdY4XS+F6YKNbH1ot0aLeyXbrnbdRT8O7asrWbWZZTGbrrhheTKY4zdqmsNYYTS2E6IqFbH1k3Ro+CXbrnbdRTvt3bVlaza+LY7GYvMsXVx2OryrV60uqc5Pdv/YklZJLZJJKyRbH4zF5ji6uOx1eVavWl1TnLu3/ALEkrJJbJJJWSPpOgNA/AnSzzPaH987Sw+Hmv8F4qc0/33il+97vf8XyW5c+Wp6Ps4Y8fQce73qugtAPA+6zzPKH987Tw+Hmv8F4qUk/3/il+97vf8X6DZeRMu5B6scJjNR8vl5subLzZGyKlmr7kdPqLES6UafgTeJL2K9PqY1N4kd9kh0+otbcKlqCrs/Is97kdPqGq2XkRIv0+pWUe24qt7VFmTa29x1ehOtNmkWZVryL39CApS29w35stb1Kyj23DZdUIa8he21gncL2r6MbItb1Kyjv3M9Gy7ReIvEdPqQ1YlW7oKtblluGrBsqra8CrW5a3qQFTShZNeJDVtyAqUsvIhrcs1bcgzSlQErh7EqmkWXkQ+5Yq+4VEENb3RZK5DXgFRQFun1Iew1ts7oauVa8yxDXiZe1VKo43tsmZSg99ttzZuxSW6Zi8b3ceUX5GMovd22ORIzkr3RN9XfGsJJX7GUk91ba5yHT37mUo2bIs7u0yYSg77LwM5rZq25yJdzCXeXzk2O2NYySWxlKNlukciUOrxsZS7Mmx3xr8pHh3ounqeeso6dw0s6qS65YyblOalbpuk20nZJJpJpdj9BKL6WrG0o7mTbL5ebk59fKZW69PmdMJjj+jGMotPsYzi7bKxyZ9zGSvt2PPXqxrG11vuYteaOTJWdr3MpQsr3Jdca480r9kZyizaULu9zOSsu5N9XfGuPOMruy2MpRlbscie1zKTuuxFjvjezCS8DKaXkjkNXuZSjfxJsd8awkl0vZXMul+RvKG7VzOUbW8bk2bdpdMJp3exlJM5E13foYyJsd8awkkr3R8E537fe757b/AJzgvtED77JXuj4Hzvxty7Z67/8AyrBfaIHfpP1+H0x8z2gv9E9R+5l9j6x7Ez5Bdd/S9/YsOeip51exM+QXXf0vf2LDnoqfsX+bAAAAAAAAAAAAAAAAAAAdKee72jOV8qWa4XhzojTeE1LrrF4WGNrwxtWUcFllCbfu3VUGp1Kk0m1TUotRtJuzin1R4V+2n4s4LVNCHGXh5pnNNOYiv/fE8io1sJjcLBtJun7yrUhUUe6hJRct05q6a/pac0DkHEz2x2rMk4t5TSzajhMZiMxwuBx6jKlXdDL6UsGpQe1SMaSpzUd01CLd0mn3n55+BGh+L/LNrfB5rp/LnmenMgxWa5Hjnh4xq4KvhKMq0I05pXhCSg6corZxm9uwH3TSeqcg1xpnKtY6VzSjmOT51g6WPwOLotuFahVipQkr2aumtmk07ppNWOtnDjn40trLmm1Byqap4fZrpHUGV4rF4TLsXjcXTqU8xnRTmkopJ0/eUV72nvJSj4ptX/J+yK1njdV8nuCyvG4qVf8AYrqDMcmpOTbcKf8Ac8VGLb8F8KaS3SVltay65+0ynhuFHPPwX4waccsHm1eGXYrHTg9qzwuO6U2u7cqUvdyXZxilbvcPWIAAAAAAAA+e8VOX3gnxuwbwfFbhjkGpLQ93DEYvCJYqlHyp4iNqtNb/ALyaP7vELiLorhRpDMdfcRNRYTItP5VGM8XjsS30U+qShFWinKUpSlGKjFNttJJt2Og3GX20XCnTrxGWcEtA5rq/FxvGGZZo3l+Bv4TjC0q9Reko0n6gZcZfYt8L8/8AfZnwR4hZppPEyvKOWZtD7oYJu/4sKqca1NW8ZOq9u2+3UbUuqedr2cmpcPpGlxoyutgYyUaWU4XPqGb4Jwitk8BXfv8ADRcUvje6ptqyUrpJf3avHv2lfPRiq2WcPqOocPp/FSdOpS0xh3lOVUk5WcKmOnJOS2s4zrybSaSe9/sHBn2K+cY+rSzvmH4qRw3vbVKuVaaXva8m221PF149MZLa6jSmm27S2TYf1uC3trctxHuMs5gOFtXCTdozzfS8/eU234ywleXVFLZtxqyfe0dkn304Nc0PAPj/AEVLhNxPybO8Uqbqzy73joY+nBd5SwtVRqqKbs5dPTfs2fJc39l/yaZpoOjoWnwxqZe8N1SpZ1g8xrRzRVJd5yrSclU9IVIyprwitj5BwF9lpn3LlzTaT4xaK4s4fNdJZJLGyxOEx+FlRzJQrYOtQVNOClSrJyqJuT920r2TaVw9DgAAAAAAAAAAAAAAAAAB4/e27+Vjhv8AR3E/aWegnK0r8svCLb/iJkH9H0Dz79t38rHDf6O4n7Sz0F5WXblk4R/QTIP6PoE5OnF6vpzVl2RF4l/xvi9rlfd+pOvi7l0LOW8exHT6l47I3WhVKSe/b5xZeRZ9irdtjO9Eb+v6Sfn7kdXoOr0M0bTZeQ2I6vQJ32sNB8UW8iXCyvcG/QS6QtnuH02tsS1ci1t7mN3Pejp9CV02tsOr0Fr73DZUOz7WIJasAb2rv6/pG3j3+Yv0+pVx77glQ9+wsyUrEhu1bMhrzLN2Itfe4N9kAlqxC3DZUpxS3sUu/Mu4Xd7len1DRbrcbBKxNr7AVe72DXmXULK1yrVw2XSjjv2I6X5GnT6lTNN32VsyLouV936iwlQSQ/iu3ewTvsSvaSNkSQ1cG9oabvYqk13Lhq4bvspvf0JJt6kBso722IV/EkBW+yvjYhxl4FunxuSZpsrNJpht3L29SktmZpqFu+5ay8iq7lm7GEukO638PnC3W4e6EewUhqXgirTT3Lp3ZLV9w2XTIh38CQG7+KtmiCzVyGrBqAALNqlA0rPZGeKxOGwWHqYvGYilQoUouVSrUmoQil3bbaSXqz+fp/U2Q6sy95rpzM6OY4F1J0Y4mg3KlUlB2l0TtaaTuuqLaumr3TtvyeXl8+rr4rm7Nv6Nl5C6Wxbp9SklaTIJdnfsQTENd3cNiA1chO5IbtC27hteAauOn1Ctut/H/k+/b013DWv7Yn3E6MBSwXwb7k/Cb9Epvr6/fQ79drW2t3d9vrHBfht+1Dw0yfh392vut9yfhH9+fBvce997XqVfxOqXTb3lvxne19r2XXvmo5p+JvBvibS0jpGhks8DPK6GMbxeFlUn1zlUT3U0rWgrK3mfc+XfX+fcUeDun9d6mhhY5lmfwv3ywtNwpL3eKq0o2Tba+LCN93vd+h7ObHn+Qx89/NccLx/KWSd30aXcpvf0NGr7n5rWescJpXCdMemvj6ybo0fBLt1ztuop327tqytZtfPyymM3Xs48LyZTHGbtNY6xwmlsIoxUK2PrRboUX2S7dc7bqKfh3bVlaza+LY7HYvMsVVx2OryrV6z6pzk92/zbJJWSS2SSSskVx2PxeZYurjsdXlWr1pdU5y7t/wCxJKySWySSVkj6boDQDwXus9zyj/fO08Ph5r/BeKlNP995L973e+0fJblz5ano+1hjx9Dx7vrVdAaAeDVPPM9oXxO0sPh5r/BeKnJP994pfve73/F+gNNeBN7bC/oerHGYTUfK5ebLmy82VUasS1t2DV9ySnNUh7H8vOdVab0/isFg8+zvCZdUzGUqeF+E1FThVnGzcVOVo9W6tG93vZOzt/U2aTTTvv8AOVePLGeey6+PuWrZstsgRe+xzptWV77diLS8SzViG7GKRZizHV6Dq9AraGnYrZvuXv6FW7BW1fRkNeRYgerUbWsyHbwJa8SDLNLl2FZFrbXIauYKgAKl2O9tirVixVu4VLpDvbYK/iTba5DdidK2fN3KtWJ6vQhu5ipdq2kLMsVqVIUacqtSajCCcpNuySSu2/zGyW3UbLpXt+N2Idr7HwzhTzSYTivr39heX6JxGDoSp1a8MbPHKbUIK9501BdN20rKTs2j7p0+p6+s6Dn8P5Jx9Tj5crJWy77xFpeIJ6vQWvueNc0pLZbbEFmrkW9Sa2XSACG7MxSLMh3/ADlk77EPuFSqbolNePc4mbZpluSZdiM3zjH0cFgsHB1a1etJRhTiu7bf/u20lufwtIcTNB6+q4mho/U2EzOrg0nWp0upSim7J2kk2r7XV1fxO2PT8ueF5ccbcZ72zLXZ+ndvAq7lkrh7HCrlUa80Q0t9l2LSKvsYqVlKN7WSMpRe+xyDGXeXzmWOuNZSi7mcrWeyv8xq3czlDvv3Jvo7T52Ekr9jKcU07JHF1PnWF0vp/M9RY5SnQyzCVcXUjGyc1CLk4q+13ay9Wj5nwb48x4vZrmeXUtJVcrhl9FV3WeMVZSTkoxTXRHpbV33a2e56+Lw/qOfp8+qwx3hh61XyuOOUxt9X1KUWn2MXCSV2tjkz7mU3eLVj59j141x5xbd0vAzaVnsjkdPqYyhs1cmzTvjWLSfdIxa8jkOFna5k4WV7kX1d8awnFt7IxqJtK3mcmWzMZR2JsdscmDTT3KVLdK2RtLZmUl1bdia7SsZdNnsjCSS7pWORONm9zKcbq1ybHfGsJp72/MZOL8TkSja+5lImx6Maxku+3gZW/wAb+U3l2fzGLVybp2x+djU7vbwMZLfscicLt7mUqfqRqu+NkceS32R8D54U/vdc9v8A85wX2iB9+krM+B88X7nTPf8ASsF9ogd+kn+3wvzx8z2gv9E9R+5l9j6p7Ez5Bdd/S9/YsOeip51exM+QXXf0vf2LDnoqfsH+bwAAAAAAAAAAAAAAAAAAeN/tLONvDzRvNzlevODGI1FkvF/h7XwuHzjGSwVFZXjYqhGpSan733kpxp1fcVIypdFSm+m9orr/AGmrvbJaR1zwkzzRWccF83w2d5/p7FZViMRhsxpPC08TXw0qUqkFJdfu1KbaTbdtm293351/ygcs3FHVWM1vr/gzp7Oc+zHoWLx9elJVa/RBQg5uMkm1CMY3avZJX2R+f/B/cmn5Pml//Mq/+2B5h8jftF9McofBvOuHGacMs01Lj8x1FXzylXw+YU8NRjCphsLR93JyhJpp4eTuk18ZLzZOjdW8V/aZc7ukdTZtpz7m6c0ziMJiMXhsHKdTDZRlGGrOvKE6zSvWrTTgp2Tcpq0VGFl6fUOQbk4w9aFeny96UcqbUkqlCc4trzjKTTXo00fX9F6B0Nw3yaOnuH+j8l03lcZdaweU4GlhKPVazk4U4pOTsrt3b8WB+hAAAAAAAB1957NE8OeIPLTqPTfFTiXS0Dp2picvr4nPamFeJVB08XSlGCpJpzc2uhJO6ck7NJp/HOT3kn5CMZpalxD4aYajxYWHxU8HLOtQ05V6UcTBKU4xwlSnCkrKcGm6cmla0m7s/fe0r4Q6y4zcqGoch0Hg62OzbJ8Xhs8jgKMeqpjKWHcve04K15TUJymoreTpqKu2k/j3sheP3DbOeCGG5f1jMNlmtdNYrHYueCrPoqZnh61aVVYild/HcFL3c4reKpwbVncD0BwuEw2Bw9LB4LDU8PQoQVOlSpQUIQiltFJWSSWyS2NwAAAAAAAAAAAAAAAAAAAAAADx99t38rPDb6O4n7Seg/Kyl97Hwi2/4h5B/R9A8+Pbd/Kzw2+juJ+0noRysJfex8IvoHkH9H0DKvD1fTvm7jf1/SWasiCXVQmzJ6USlYN2hJ+PYPp8UiSLJgl0fF8l+gfF8l+gdKHSgeaofTbZL9BCsnexbpQ6UbOx5qXRVxbe3b5yzRF2jPT0bLtCTX43Ytt6foHfZhqyM1v1N6Q+lW7CzIaTLLZJG3s1V7dwrd0WlHq73IcUovuZ2oi6Fk9you0LBMiF3JW/cWS38jDZJLbYizDbYu0G7sRKLuQrrexfvuw0rd2G7Ub8hZk9KD2QEdtmg7W27+Fh33YatugqVW0/X9JG6L9THSvUG9ojKKSTe5GyDikyLthXb3J2ZFmTHsR1MHoiy8tys1ttsaqKau29/IicEl3fczWyaY7+v6S1mT0okaVvSvYB9wS3YReJJHSgbVBbpRDVg2VABDdgqXaSJRuSAqXSjTXgR27l32KtJmWNl2XT2BXs9iy3RLZdABDYV6ofT4JEdLfZFmrIpUqwo051as4whCLlKcmkopK7bb2SS8TZLb2aizQPjvEPmw4Q6D97hKGby1BmMG08NlVqsYyW1p1W1BK907NtWe3gdYuIXOXxT1c6uE03KhpfATbjGODfvMS4vwdeSTT9YRiz9B4f7Ndf1+rMPLj8b2/7vTxdPycnf3O7GteJeguHeEeL1lqfA5YnFyhRqVOqvUS/xKUbzl+ZM62cQueylH3uA4ZaX63vGOYZrsk+140IO7805TXZXi+x1GxuOx2Z4urj8xxlfF4mtLqqVq9RznN9ruTbbfq2cc/b+H+xvR9NrLqL579T3cfR4Yd8vV9UyzN+KnMnr/LNK55qfG434bXTqQv0YbC0Y7zqKlFKCtFOztduybbZ6J6d0/lWlchwGm8kwyoYHLaEMNQprdqEUkm34t92+7bbe7PhXJ3wi/YVox66znDOGc6lpxlSjNWlQwN04R9HNpTfmuhWTTOwx+R9putw6jnnS9NNceHwnrfe8vUckyy8uPpEb+v6RbbdK5IPy3yeXwcJ2VtbwBK37hxW+7M8mXwar8UizfbsT0oDyZfBs3FbNB3tsWaTKjyZfBU7vwGt+A/CbiPnS1DrfReGzXMY0IYdV6tatFqnFtqNozSsnJvtfc/S6S0lp3Qun8JpXSeV08uyrA9fuMNCUpRp9c5VJWcm27znJ7t9/I6Lc9GP1ph+NdGnkGMzulhfuJhW44OpWVPr95Vu7Rdr2tfx7H33l11tidO8tukp5vUxGKzvELHuNPFTk6lvh2ISnUcn1KKSSSbu7JKyTa9nNwcnH0+PJldz4I4N8vL8njj3fYNZaywulsIowUK2PrJujRb2S7dc7bqKd9u7asrWbXxTH47GZni6uOx1edavWl1TnLu3/sSSsklskklZItjcbi8xxdXHY7ETrYivLqnOT3b7eGySVkktkkkkkj6XoDQHwF089z2j/fO08Nh5r/BeKnNP994pfve73/F+Lcc+fLUnZ+iwx4+g493vapw/4f8AwH3We59Q/vnaeGw01/gvFTmn+/8AFL973e/4v0ItZMjpR6ceK4TUj5PLzZ82XmzUa3I7l3syrVt0V5MvggsyGnYnqYu2PJl8G7fh+MPDbBcVNB4/SmKcIYmUVXwFeX/AYmCfRL5ndxfj0ydt7M6KaU4zcXuEOZVckwee4qnDL60sPXyrME69CE4NqUOiTvDdNPocX6npDZM6ec6HCT7n5hQ4rZJhX8Hxso4XN4wirU6ySVKs7eE0ult7XjHe8j9p7J9XxZZ3w/rMfNhn6bnpf+7vxZf2b6V+r0BzsaTzb3eC1/k9fJMQ7ReLw16+Gbv3cUuuHfslLs3fwOwWndUac1bl8M10xnWCzPCTX+FwtVTSfk7O8Xs7p2atujyrP6ORaiz7S+YQzXTmc43LMXT/ABa2FrypTte9m01dPxTun2aZ9/xH2I6XqN5dLfJfh6z73W8Uvp2eq90Vdn5HSnQXOrrTJ3TwevMqw+f4ZJReJopYfEpebSXRL5rRb8WdktBcfuFnEX3dHJNS0sNj52SwGPth6934JN9M3/8AUOR+F8Q9nPEPDt3kwtx+M7z/AF9LncLj6voewvEWTHSj4SRteBXuS1YgxUu1WnuQXI6UFS6VIe/Yl+R+F4scYtH8HckjmmpMROricSpRwWAo2dfEzSV7JtKMU2ryeyuu7aT7cPT8vVck4uHHeVVO79xv4g6ZS5m+Y7iNiK9fhnoR0cBSk0ng8snjXDdNKdWScHK3gox2vt4meWc2/GXQGfU8q4taQVWk0pVKVbBSwWLUG7dcG7Qktm942bVk13Pv/kp11x/NuNyn9nfdTuheJDt4H83TOosn1fkGA1NkOKdfAZlRVehNppuL8GnumndNPs00fw+J3FDSfCfTs9Q6pxUkpN08LhaVnXxVS1+mEW1e3dttJLu90n+fw6bm5Ob5DDG+betNj9c7WK7M6XYzm34364zOpQ4aaIhChSd1RwuAq5hXs7262lbsvCC7Pdlsk5xOK+j86hl3FHR9OrQck61KWDngsXTj2bipfFdu9nFXatdd19++yfXzG/o+aTfl33VL8Xc9tdivc4+U5lhc6yrBZzgXN4bH4eniqPUrNwnFSjdeDs1sfGeNvNHpfhXi6um8nwazzUMIr3lGNVQoYVtbKrNXblaz6Er27uN1f4vS9B1HW834Pw47y+xu5H25rbsVszpRS5heazU9JZ1p3SeLlgJtyhLL9OVK9BrvZTcZtpLxv49z+zw951dRYPOoZLxXyPDLDOqqNXGYShKjXwsr2bqUm2pJPukotJOyb2f2eT2T67HC5YXHKz1kvc80dv214H4jjXn60zwm1XnCdp08rrUqcr9qlVe7g/zSmmfssNiKGLoUsVhq0KtGtBVKdSEk4zi1dNNbNNNNNd7nwXnVz9ZXwkpZPTq9NTOc0oUZQ2vKlTUqrfzKUKf52j5XhHTXqOv4uGz+1N/4Lvpa+Z8iuQe/1LqfU8qe2CwVHAwk/OtNzdvVKir/AD+p3HPgPJTkCy3hNiM4nTtPOc0rVYyt3pU4xppfmnGp+k+0as1Xp/RGQYrU2p8xhgsvwcLzqTd229lGKW8pN7JLds+j7ScmXW+LZ48c3ZZJPo7fa3DUj+pZoHT/AFLzj6/1TnDyfhNo1Rptv3Lq4aWLxlZJ91Tpvpje6urSttufxMVzJ8zGipUsZrDTs6OHqT6YxzbIqmGhUduyaVNt7N7M6YeyXX5Y/nXHHK+mNvc+Ukd2yGmnv2PmPA7jrkfGXKsQ6OEeXZzl6i8ZgZT610vZVKcrLqg2mnteLsn3Tf0579z8/wBV0nN0fNeDmx1lPc6SyzcQ2mQda+M3N5hNLZpX0rw3wGGzXH4eTpV8wrtzw1OotnCnGLTqNO6bukmrJSR87++S5ltNxp59qLTk3lk5KSeOyKpQoSi3ayqJRe99nd7277p/a6f2V6/qOKct8uPm9Jbq1nykl07sFWtzqnrLnQq1NG5PmGh8uwmGz3E16tPMsJjoTrLDRhGLTg4uKkpuWze9oyTSaudjOHuc5zqHQmQ6g1DRoUsxzLL6GLr06MHCEXUipJJNtrZq6bdnc8HXeC9X4dxY8vUY6lup8ey8c5bqP4HHHh7nHE/h5jdI5HmeHwWKr1aNWMsT1KlUUJKXTNxTaTaTuk90tvFfjeXPl/zPg9VzXNtSZrgcZmeY04YenDBOUqVKkm5P404xbcn07WSXT3d9qczHHfUXCN5Hl2lcLl1XGZmq9avLGU5zVOnBxUelRlHeTct3eyj23uvpXC/UGdat4fZDqbUVChQx+Z4KGKqww8XGnad3BpNtq8HF93u38x7bl4h0fhEm5OHkv+P+uxJjc/nj9Q9+xG5+U4lcTNL8K9Pzz/UuKaUm4YbDU7Otialr9EE2r27ttpJd3uk+seM5r+NWtcyqUOHGi4QoUndUMNgKuPr2d7dbSt2XhFdnuzzeH+BdX4jh8rxyTD45dou8mOPau43cq+508yXm74p6SziGX8TdI06tByTq05YSeCxdOPZuKls7d7OKu1a67rtzleZYXOcrwmb4JyeHx1CniaLkrNwnFSV14OzWxw8T8G6nwq43m1Zl6WXcVhyTL0chryM59Nnsr/MfEON3NBlPDTG1NL6YwVDOM+pr+7upNrDYRvdRn0tOc7d4pq192nsfHVzE8y9LDfslrZDVllL+P7yeQzWEcdn/AIRJO1vHr7Pv4nu6P2X67q+Kc3bGX0811s+Xxxrua1d7JGc00mjrFjecj4Tw6q5hluVYTB6tp4mlh3haynVw86clJyrQs07Lpt0t7OS3kmfZOCesNR694c5bqzVFDC0sXj51nFYanKEHThUlCLabbTfS33s1ZruePrvAus8P4bz9RNTevp/7PRx8+Od8uL8zzS539xeDua0lK1TM6tDA03t41FKS/PCE1+c/E8mGRfB9J5/qGcLPHY+nhIu27jRh1XXpes1869Djc7eedGXaY01Cpf31evjqsPLoioQb+frqJfMz6Zy55Esi4N6epTh01cZSnjp7W6ve1JSi/wDzHBfmPt5/zP2Yk9/Ln9U//icb5+p38I+itLxRi1s9jkyirmMoJJ7s/DWPp41x5RbfxVt85m4Ss9jk9KMZdmibHfG1hKLuZNeZvPZ7eRjdsizbvjWdS1127GE0rbI5Lin5mNSnGy7mWO2NjB28UjKUWl2ORKEb+JjN3VibNO+N+DCcZO7ttYycW0ciS7oxeyIsd8axktndbmMkttjkySd7mUoLzJsdsa40luzOaW1rHInFJ92YyjfuRY9GFZPszCafgclxSutzKa7dya7Y1xpxlZs+Ac8af3umff6TgvtEDsFPszr/AM8n7nPPv9JwX2iB26T9fh9M+183x+78K6j9zL7H1L2JnyC67+l7+xYc9FTzq9iZ8guu/pe/sWHPRU/YP85AAAAAAAAAAAAAAAAAAA8uucH2hvNlwW5jNYcM+H2isinp7JKmEhgamLyPEYmrVhUwlGrKbqxqRjJOc5NJJWVk7tNv43+Fh55f8yNL/VjF/rj2pAHit+Fh55f8yNL/AFYxf64fhYeeX/MjS/1Yxf649qQB4rfhYeeX/MjS/wBWMX+uO3vs6ucTmE5mtXaxyXjFpfKsDl+SZdhsVhMRgcpr4RqtOrKLhKU5yi7xTaVk/iu21zvYAAAAAAD4vzgft9ftBag+9p+Efs+97g/gHwf4P773Xwmn7/o+Ef3O/uuu997XtvY8etQclvtEdT65q8S8z4R53S1RWxKxs80wGKy7A1vhKd/fJ4erBRqN7uaSk3u3fc97T5/xV498GuCODpYzixxJyLTKxEXKhSxuKSxFeKbTdOjG9Sok1ZuMWl4geS37XPtkf8o8VfrTQ/tA/a59sj/lHir9aaH9oO+P4VTke+HfA/22cb7q9vhX7G8y91a3e3uOvvt+L39Nz7pwq5g+CfHDD1K/CfibkOpZUYKpWw2ExSWJowdrSqUJWqwTuleUUr3XdNAfHfZ65HzSZDwkz3Dc1eJzqrqGpqGrUy1Zzj4YvFLBfBqCXx4SklTdRVLRbumpuyTTfaYAAAAAAAAAAAAAAAAAAAAPH323fys8Nvo7iftJ6EcrLtyx8IrP/iHkH9H0Dz39t38rPDb6O4n7SehHKwl97Hwi+geQf0fQMqsPV9Q+fsTeJDVkQS6S6AW6UOlBm9KglqyIDZQABuwAA81BZeQAJdoa22CUv/dkgNLW8CGn4Fm2yASq7olNW3LJdXe+xVxSYs23Z8XyX6Ctr+BbpQatugS0St3sG1Z9gt+4cVvuydN3tS8Sen0I6UXWySNs01Xt32Gz2LOKbu2yrikrpvbzHYVsw4tonqZIsb5qolZbklrJkdKMNq2XkVu/M06UV6UGy7RZsJJbtEiXYNl0h28ClmiwauG7TGSSW4updiOhW7sj8XdBs+ZLVvArZote+4auGy6ZtMrZvsauK33ZVJIyxsu2aUr2f+0tZlrJhuws1Gz51SO46t9ySVRWzvf1Ds+1ixHSg2XVVIabJAbLsFl5EN2Cdw1Dje+yIad9lYuRZMa2qVU/g6t1zpDQ2A+6er9R4DKsO0+h4isoyqNd1CH4036RTfof3n2PyeueF+gOJGGWG1ppfBZk4w6KdeUHCvSV27QqxanFXbdk7X7pnTp5w3kny9vl9+vVeFx3+d6Ov/EPntyHAOrgOGunKma1VeMcfmN6NC/g40lacl87g/Q6x6+418TuJc5w1ZqvF18JKV1gqLVHDR8l7qFlJrwcrv13Z2G4h8h9SHvsw4Y6o6+8o5dmtk+97RrxVn5JSguyvLuzrTrbhlr3h1ivgms9L47LG5OMKtSn1UKjV9oVYtwl2vZNs/qngXH4H5Zek1c//d+l9f8A9Pr9PODX5nr878uAD9a9gAAOas4zeKUY5rjEkrJKvNJLy7k/drOf8r4z/p5/1nBBy8nF8IzUc77tZz/lfGf9PP8ArH3azn/K+M/6ef8AWcED5Ph+EZqOd92s5/yvjP8Ap5/1j7tZz/lfGf8ATz/rOCB8nw/CN1HO+7WcP/52xv8A08v6yPuznH+Vsb/08v6zhAfJ8PwhqOb92c4/ytjf+nl/WPuznH+Vsb/08v6zhAz5Ph+ENRriNU4vDT91Xz7FQlbqs8RO9v0k084xmLisRDM8RUU/3zqyd7bd2/Sx+czvKMZjsYq+HjBxUEt5JO6b/rP6WVYarhMvpYeskpw6rpbreTf+xnzuDmz5Opz4uTCTCelebi5OS82WFx1j8X9RZjmEZKSx2ITW6tVkmn+k0+7Ocf5Wxv8A08v6zhA+j8nwz3R6tub92c4/ytjf+nl/WPuznH+Vsb/08v6zhAfJ8PwjOzm/dnOP8rY3/p5f1j7s5x/lbG/9PL+s4QN+T4fhDs5v3Zzj/K2N/wCnl/WPuznH+Vsb/wBPL+s4QHyfD8IdnN+7Ocf5Wxv/AE8v6ylbM8yxFJ0cRmGJq05WvCdaTTs7q6bs91c4oGOHFLuaOwADq0AAuvePp2guY7ivw+6MPgNQyzLAQslgszTxFJJdlFtqcFa+0ZJb9mdk9A852gNQulgtZ4HEacxcrRdZ3r4Vv/6uK643fnGy8ZeJ1Z0FwS4l8SJQqaZ0ziHg5P8A+HYn+4YZLzU5WUvDaF3utjsjoHkj03lvusbxDz6rm9dNOWDwLdDDL0c3/dJr1XQ9/Tf8N7QcfgEl/CLJn/7fX7v4uWXknr6uxmVZzlGoMFDM8izTCZjhKn4tfC1o1actk7KUW1fdbepzEfytOaV01o/L1lOlsiwWV4RNN08NSUFJ2t1Sa3k7eLbb8z+qm0fyzl8kzvye/L7nJL7Mo7+Ba7ZBzVvbi5nmGDyfLcXm2Y1lSwuCoVMRXqPtCnCLlJv0STf5joFpzDZtzVcfniM7qV4ZZOUsTWpqVnhcupNKNKLWybbjFtfvqjlu7na/mnzqtkfAnU9XDuSqYulRwSa8I1a0Izv6ODkvzo6kcBeW+rxuynNc3erXktLLsTDDRX3PeJ99Jxcnv7yCVk47b9/Da/7r2a4uHpeg5uu5c/Jb+bLrev8AD/FePabd9KMNLaJyfD4CjLLckyzCU+ijTlOFClTgrdrtJd1d+bu92dOecfitoXXuJyPT2kMdRzavlNSvUxOOofGpLrUUqcJ/v7uN243jtGzbvb9XhOQXA06jeYcUMRXp+EaOUKk/0utJfyH1Th5yucJ+HmLoZrhsrr5vmeHanSxeZ1FVdOa36oQiowTT3TabVlZ33PL0nP4X4Rzfhc5suXOenbXf59k7P6PLdpbNtH8GNOZLntKrRxnuquKqUKmzoqrWnUjBp7xajNNp7pt33OpGvM3zXmW5gqOQZXjJrLp4t5dl7TbhRwdJt1KyXa8lGdT1uk3ZK3dXi3nVTT3C/Vec0ZqFbDZRipUpN2SqunJQfdfvmtu77I6nci2S4fGcQ87zutTc55dlXu6TcbqM6tSKbv4PphJJeKb8jv4Ny+Xi6vxnOfn99fNb/qKl27haO0Zp3QWQYbTWl8tpYPBYaKVoxXVUlZJ1JtJOU3a7k9381kcfXHD3R/EbKo5NrLI6GY4aE1UpdbcZ0pJreE4tSi3azs1dbO6P0ad2Jdz8d+E83yvy3mvm+O+6o+b8dOItLhFwwxud5dGlSxrUMvyul0pRjXmmotLs1CEZSt2ahbxOtXKVwcwnEXO8x4j61wyzHAZdiXChSxN5xxeNdpznUv8AjKCabTum5q97NP8AT8+mbV4UdH5FBtUak8Zi6nrOKpxhb5lKf6UfYOWDKcNk/A3TEMPFdWLo1cXVl4ynUqzd352TUV6RR+u4874X4D8vx3/ac11v367/AHfWb3X1KNOMIqEIqMYpJRSSSS7JLwOmvPZprK8v1FpjU2Fw1Oljc3w+KoYqUYpe99w6XRJ27ytVau97RS7JW7mJ3Z0m559T0My13kelqM+t5LgJ16tv3tWvJPpfr0UoP5pL83i9kPlL4pjcbdSXf8Pv0q+jsPyz5jic04GaSxWKc3OGFq4ZOT36KVepSh+bpgrelj4Pz25+6+odL6XjPbCYOvj6iT7urNQi2vRUZW+dnZTgzpuvpPhVpfIcXBwxGHy6lKtBqzhVmveTi/VSm1+Y6a8wVWfEHmVxGncNOU4yx2CyOj3um1CM0vK1Sc/9p7fAcMOXxvl6ifo4+a/X/wBy+mncTghkH7GeEek8olDonDLKVerHt01Kq97NP1Ups6n8z+vM44p8V6HDXTs3WwWU4yOW4ahCW1fHzkoVJv1Un7tX7KMmrdTO8FaVHLcBOpTpJUsLRclTjtaMY7JfmVjoNyt0FqjmAy7Ns2tVqw+GZlLa6lWdOVnvftKfUvVIz2euOXL1XinJN3CWz6btuXujuTwp4Vab4UaXw+R5PhKMsW6cXj8d0JVcVVtvJt7qKd+mN7Jerbf6HVGm8q1fp7MNNZzh4V8FmVCdCrGUU7XW0lftJOzT7ppNWaP63Sj87xE1NQ0ZofPdUVpqCy7AVa9O/wC+qqLUIr1c3FL1aPyuPP1HV9XOTzW55Wfx26TXo6O8omY4nBcccow1By6MwwuMw9a3ZwVCVVX9OqlH89js7zT8ScRw94aVcPlWIdHNc/qPL8NOMrTp02m6tRNbpqHxU1unNNdjr7yV6er5lxTxOfKD+D5Nl1WUp22VWq1ThH53F1H/APWs/o88mdVcVr/Ish670sBlLxFk/wAWdarJSVvB2pQf6D+gdf03H1/tFxceU7Y4y5fTN37kY3WD9FybcHstxWCq8VtQYKGIqqvLD5PCpG8afRtOuk9m+puMX4OMn3aa7WYnD0cXRqYbFUKdajVi4TpzipRnFqzTT2aa2afc/LcIMkpad4WaUyilTUHSynDTqJf91nBTqP8APOUn+c/TZlmOCyfLsVm2ZYiOHwmCozxFerN7U6cIuUpP0STf5j8V4z13N4h4hnnLfXWM+E3207YSY4vPDjNorIcu455lojRlF0MLWx2Gw9Kkt4Uq1aNNyjBLfpU5tJeFrdkmeh+CwlDL8FQwGGi40cNShRpp72jFJJfoSOhXCOrX4o8zeC1DXpNxxOb4jOZRkrqlCn1Vaa/M1Tiu+9vnO/Z9z2v5c8Z03Scl3ljjN/Te3/0zhnrXRPm+zSvqDjYsiwyc55bgsJgKcI+NSper+l+9S/Mju5kmV0ciybAZJht6WX4WlhKb7fFhFRX8iR0WyL/sl811PEK9WhX1LUxUb3alh8NOU4p38HTpJW/MdzOLGbz0/wAM9VZzRn0VcNlOKlSk2klUdNqD/wDOa27vsjfaHjuPH0Xh2Pa+Wb+m6n3nHe+WTpnrnNs05j+PdHIsrxcll08U8vwD3cKWEp3dSsl2vJRnU9bpN2St3a0jo/Tuhcjw+nNM5dSweCwsVG0YrqqSsk5zaV5Tdrtvd/NZHUjkiyahi+IGdZ3WpuU8vyv3dJ9N1GVWpFN38H0wkkvFN+R3Rfc4e1nUXg5MPDuG6wwxn+NVwzcuV9a/P610FpDiHlccn1hktHMMPTmqlPqbjOlJNbwnFqUb2s7NXWzuj+Fxm1zT4W8Mszz/AC+nTp4ihSjhMupqK6VXnaMLLs1FXk14qLR+9tc6uc9GcVKWU6T0/CfxMTiMTjasb9nThCEG181Wf6GfJ8D4cvEuu4el5LbhLvV+bvf46dOSzDG5R+M5UOFWG4g6kzDiBq2i8fgsqrr3UK7c1icbL47lO9+pQTUmn3c4t3SafdFxj0uPTt2tbax8p5Wcmp5LwTyKcaajVzGVfG1Wv3zlVkov/wAyMF+Y+ryas3JpJXbd7JLzOntH1/J1niOeMv5uN8sn0fBvDjMcXQ3mx0xpbS3FBUNM4OOEeOwFPG4yhTSVKFaU5q8ErdN1BSaW13dd2l3F4b5EtM8PdO5E42ng8sw9OrdWvU6E5u3heTbt6nSzV+a0eMnMWng2sRgcyzrDZfQezjLCwlGm5rzTjGU/mb+Y79yirN3Z9j2oz5eLoek6Tku8tbv+v4s6bXnyyjo7za5pWz/jHDIsKuuWW4LDYKFNb3q1G6v6WqsV+ZHcnJcppZHkeAyXDpe6y/C0sJTtsumEFFfyJHSjJLcR+aWGIV6tGvqOpiY3u+qhhpSnFO/g4UkrfmO889kzz+1V/Bul6TovS447v+P+q69L+dllm48k7mUldM3l3MmrJtH4ix9PGsWmu5nL8V7G9lLdmcoLpe7Jsdsa47t4pGLivBI3nFJ7XM5R6Vtci+rvjWLi0zGcZW3/ANpyZdzGfb85Nd8a484yb/8A7lHHbdG0u5nNJLYix2lYTSu9l28jGaVuyN5RV2ZzgmrE2bd8bpx2u9kZzi9rL+U3mrbGUibHfGuPJMycWjkzXdmMibHoxrKSsncxavY3lu2ZySj2vuTdO2Ncea2eyOvvPMrcuWff6VgvtEDsLUikm97nXznnt97jn3pisF9ogdel/X4fTHzfHr/RXUfu5fY+n+xM+QXXf0vf2LDnoqedXsTPkF139L39iw56Kn69/nUAAAAAAAAAAAAAAAAAAHl1zg619pzlXMXrDAcDcv109DUquEWTPJshpYnCum8JRc3GpKlJybqOp1XbtLqSskkvjf7Y3tkf8ncVfqtQ/s57UgDxW/bG9sj/AJO4q/Vah/Zz9fwg1/7WzFcWNF4bWWXcRv2P1dQZdTzZY/TdClhngniIKv72fuY9MPdud5JppXaaaTPXo6o8b+fHL+DfNVoPli/a0xOb1dZ1sqoVc6+6scPHBSx2LeHg40PdSdVRaUnecO7STtdh+b9qPm3MnlHB7Ttbl2ravoVKmduGd1tKxr/D4UXRl7r42HXvYU3O6bi0uroTe6T/AH/s+M044ZxyyZDjOYBahepni8XGnPP6U6ePqYNVP7jKqqiVRtrqtKa6nFRd2mm+yYAAAAAAPk/NLxwwvLnwG1bxcq4ejiMVk2DUMuw1a/RXx1WcaWHhJJpuPvJxcrNPpUndWueSHLTyT8bfaB51m/HPinxIxGW5Ji8fOlXzvGUni8ZmNeO86eGpdUYwpwuoXuoQuowjLplGPe/2veCx+K5OcdXwfV7rB6iyyti7RuvdOU6au/Be8nT387LxP3ns2szyDMuSvhrHTtSk6eDweKwuKhB708XHF1nWUlfZucnLfupJ9mgPh9D2KHLlHLlTxXEziRUzDps69PE4GFFy81SeFbt6df5zq/zM+zy4vclFDD8wnBDiXjs4yrTuJp162MoUXhMzyhuSjGpJRlKFai5NRm1aymlKDj1SXtYfK+ajMMjyzlp4qYvUlSjHLlo7N6dZVbWn14SpGMFfu5SlGKXdtpLcD8vyScyD5ouX7JuJGPw9KhnuGrVMnz6lRSVOOYUYxc5QV24xnCdKooveKqW3STf3w82/YjQxy4Q8Rp1FU+BPUmHVG/4vvVhY+8t62dK/pY9JAAAAAAAAAAAAAAAAAAAA8ffbd/Kzw2+juJ+0noXysNLli4QL/wCkPIP6PoHnp7bv5WeG30dxP2k9C+VhJ8sXCB//AEh5B/R9Aytnq+oN3TSKdLLtWRBK5U9SHUh0odKDJo6kOpBqyIBdK9LLKLSLWSIbsw3aHFsjoa3uti3UyG79wS1HUiG7k9KIasGoSuTZoRV3Yu4pg3pmC7il2bI6UG72qCWrEAl0Et3RADdgABsKvuadKKNbvdhqpK7k9KDVt0G7SUbsT1MnpXqZr4tliFuroh9iySSsg0rd2Pe1mT0sdKJFmiXSoLWTIaMVtRol7okAUCVy1kyO26DZdIcG/EXS3ZZO5Vq4bs77+ZPSyOxHW/QNJRd2VlF7Fupt72DSYbLVX2KtqPfxLlJpME2daeyvuCLJEk2aVsKy7liLJmKm1SWmibJESDZdK2a3Y77okjtsg2bLW3ZKdwErBUulH2OPjMDgcywlXA5jg6GKw1aPTUo1qanCa72cWmmvRo5TimRZLZCXLG7ipfg+BcQuTLhXrB1cZp2FbS2Pm21LBJTwzk/OhJ2S9IOKXkdYuIfKbxe0EqmMoZRHUOXQbbxOVXqzjFK950mlNbXbaTSs7vtf0ZIbP0Xh/tT1/Q6xt8+M91+/1eri6vk4+19HkHUp1KNSdGrCUJwk4zjJNOLTs00900/AoepGv+CnDLiXCU9WaUwlfFyVljaKdHEx8n72FnJLwUrr03Z1k4h8imeYH3uP4aajp5nSV5QwGYtUq6XhGNVJQk/nUF6n7fw/2v6Hq9Yc35mXz+n8fve/j6zDPte1fSeUXiNlHEPQn7GM6wmEqZ7puMKE51KUHPEYV3VKpdq7aS6JPfdRbd5H3n7i5P8A5JwX8Xj/AFHm7pHG8QOXHiVlufag03mOW1KE3TxOHr0nGOLw0rKpCE38Se26abSkk77HpHk+bZbqDKMHnuT4qOJwOPoQxOHqwd1OnNJp+mzW3ddj8d7TdHek6j8I4Mt8efeavpfe8vU4+TLzY3tUfcXJ/wDJOC/i8f6issmye7/+5OC/i8f6jnEWTPzPy3J+1f4vNMq4P3Fyf/JOC/i8f6gslydv/wCKcF/F4/1HNasDPleT9q/xbMq4X3Fydd8pwf8A0Ef6issmye7/APuTgv4vH+o57bZ+a1prLB6VwnTHprY+vFujQvsl26523UU09u7asrWbU5c/JjN3K/xdcMc+TKY497XB1lnOnNLYRRhlGArZhWi3RouhGyXbrnZXUU77d21ZWs2vi2MpUsdiauMxlCjVrVZOUpOnFXfokrJLskrJJJJJI5WOx2LzLF1cdj6861etLqnOT3b7eGySVkktkkkkkj6doDh98CVPPs+of3ztPDYea/wXipzT/feKX73u99o+P5fm5stTK6+l9vDHj6Di3ld2ut2uuNunOAWtcPk2quGcs3xlTAUswoJYinSjRU5TS64OErzXRdJva+6utuyPB3VuQcXeHGUcQ8NpTDZZSzb3/ThZxhVlT91iKlJ3kopO7pt9la9vC5+A448oWneOGtIa0zPWOY5ZWhgaWCVGhh6c4tQlJqV5NO762reh9Q4RcOMHwj4eZTw9y/M6+YYfKvf9GIrQUJ1Pe16lV3S2VnUaVvBI+rlycePBjMLfN9NfDz5ubl5blnez9D9xMo/yTg/+gh/UHkmUW/8AinBf9BD+o55Vt7o83y3J+1f4qmVrgfcXJ1/81YP/AKCH9RP3Gyfb/wC5WD/6CH9RzLJi7W3kPluT9q/xVMq4byTJ3/8ANOC/6CH9RX7i5P8A5KwX/QR/qOb1Mgy8vJ+1f4t3tw/uLk/+SsF/0Ef6iv3Hye3/AMU4P/oIf1HOKGfLcn7V/i2WuE8nyfv9ycH/ANBH+o6k85vEbLadfD8K9PUMNTlTcMXm1WlTine16VFtL5ptf/UeTR2W4qcQMt4YaGzPV+YOE54an0YWjJ29/iZXVOmrb2b3dt1FSfgefWQ8P+KnGfPcXnWUZBjs0r5hiZ1sVj5x93h1OUm5N1ZNQVrv4qbdlZJ2SP2Xsl0ky5L1/VZaww9N31v/AGenhn9q+kfhDk5fl2YZti6eX5VgMTjcVVfTTo4elKpUm/JRim2/mR220ByO4GgqWO4kalniZ2TlgMsvCCflKrJXkvBqMY+jOxmkeH+itBYT4FpDTOAyuHSoynRp/wB1qLb8eo7zn2W8m3sfo/EPbXo+m3j0089+PpHW80np3dMNAcnHErVCp4zVFShpjBTs7Yhe9xMlfuqUWkvHacotbbM7J6B5YeE2g3SxSyR51mFNqXwvNLVmpLe8adlCNnunZteZ9al3C3Z+G8R9pvEPENzLPy433Tt/3cby5VCSjFRjFJJWS7JLyI7bss1Yhq5+dytvesQ990QWSsVasS2VD7FSW+6IDXy7mb0xj9WcEdSZfldKdXFUKVLHQpxV3NUakak0kldvojOyW7aSOtvJ5xo0pw9r5xpPWWYwy7C5rVp4nC4yq37qNWKcZRm1dRTXS1J2XxWm90d5V2Pg2vuTfhXrHMa2cZVPG6cxeIm51IYDpeGbfdqlJNRv3tBpbvY/UeEeJ9Jj0fJ4d125hldyz3f606yzWq+iYzjPwjwVD4TX4naW6HHqj0ZtQm5K7V0oybe6a2T3TP5+ieOfDniPqnE6T0XmtfMsThMHPG1a8cNOFBQjOEGlKaTbvNWsmmk3fY+RYXkM0RTrKWO1znlaimuqFGjRpye67SakltfwZ9k4Y8F9A8JKGIho/LKsMRjFGOIxeJqurXqpNtJt2UVd3tFJN7tOyPN1XD4Pw8WXyHJlnnfTtqf4t7e5y+LWncTq3hlqfTeDg54rG5ZXhh47vqqqDlCO2+8kl49+z7HR/ld4r5Nwm17ipar66GWZvhvgeIxCg5PDVIyUoSlFJtxummkm1dPwafodZM+I8SuUnhnxBzavqChPG5FmWKk6leeBcPdVZvdylSkmlJvduLjd3bTbbPR4F4p0vT9Py9D1svkz989zZfi/SZjzF8E8swUsfX4i5VVgo9ShhpuvVe2yUIJyv86VvG1mfyuDXHd8aNSaho5Lp6eDyHJqdBUMViJf3evUnKXeKvGKag2km2rK73svw+Tci3D3CYuNbOdU55mFKLTVGmqdBS81JqMm0/Rp+p940horSugMnp5BpHJcPluCg7uFNNyqStZynNtynJpLdtvZK+x5+tng/T8OWHS+bPO+lvaRsvwdcOe3S2YYvJ9Naww1Oc8Ll1WvgsVZXVN1eiVOTt2TcJJt7XcV3e/9Plh4/wCgKXDzLtE6s1DgskzPJozw8ZY6oqNGvSc5ShKNSTUU0motNp3V1dPbsZm+TZVn+WYnJ86wFDG4HFwdKvh68FOFSL8Gn62afdNJrdHXzUfI7w2zLHTxWRZ9nGT0pycnhlKFenDftBySml87b9T2dF4l0PVeHY+HeIW4+W7xs/187ZdP1vEbmk4VaHyqrVyvUGD1Hmbi1h8HllZVoSlbZzqxvCMU7X3bt2TOt3Avhvqbj3xPxHEjWVOdXKKGN+HY/ESi1DFV004Yamr9l8W6V1GCSum1f7Ppnkj4ZZTjFis+zfN87jGSaw85xoUpJWdpdC6328JLZ9vE+/ZVk+V5Fl9DKcmy/D4HBYaChRw+HpqEKcV4JJJL/wBb3NvinQeE8GfD4ZvLPPtcr8PmVJvvXIqVIUacqtSajCCcpNuySSu2/wAx0D4B06nETmZw+osTFyhPMMbnlVtdtpzg7eFpygvQ755tltHOcqxuUYipVp0sdh6mGqSptKcYzi4txbTSaTbTaav4M+ZcIeXDRnB3O8XqHJMzzTH43FYZ4TqxkqbjTpOUZNJRit24Ru23srJK7v4/B/EuDw/pepmd/wBpnNT/AB23vbH1LF0IYrC1sLOUlGtCVNuPdJpptfpPOrhfqKrwE43U6uq8LVVPKsRXy7MYwheapyTj7yCfdJuM1bult3TPRi7Z834ocAeHXFmaxuoMBVwuZxioRzHBTVOu4pWSndOM0trdSbSVk0ifAPFuDoZy8HVS3j5Jq6/186rN+jnQ458HauXLM48S9PKi11dMsdCNW3/im1O/p03OrXMTx9fF/E4Thxw4w2LxOVzxUHOpGlJVMxr3SpxhC3UoJu6TSbdnZdKv9BjyJaRVRSlrvN3T6ruKw1JNq/a/ZO3jb83gfWeGvAfhxwqk8VpzKZ18xcel5jjZKriLWs1F2UYJ736Er33ukre/g6jwTwnP8J6e5cuc9Je0h3s16P53LxwkfCXQkMDmKg87zSaxeZSi7qE7WhRTTaagm02tnJyabTR1654MgxeE4gZNqP3c3hMxytYdTa2VWlUm5JO3+LUg7d92d2Grn5rX3D3S3EvT89Oasy94jDOaq0pwl0VaFRJpThJdmk2vFNNppp2Pn+GeOZ8Hif4d1G75t+b6L9yrjLNR834T8xPDHMOHmTvPtXYDKsywGCpYTG4bF1Pdz95SgoucE/xoyt1K13vZ7po+R8eeYDEcVcJjNB8L6WLlkmGo1MXm+YuDgq9Ckup2TScad0rt2cm4xSV7S/YU+RjREcYqlXWmeTwim37pQoqo4+C6+lq/r0/mRTmE01oTgrwLxOktH5dSwOI1Hi6GFlNzc8TiIwkqs5zm3dpKCTStFe8SSV7H3Oj/ABP+MMc+jlzzyvaX0x+f59M3lrV9H4nkbyB4vWmodSTheOXZdDCxb7KdeommvW1GS+Zs7Z62zxaX0fnmonJReWZdiMXFvdOUKcpJW8btJW8bnxfkp05UyrhljtQV6ThPO8ynKk7W6qFKKgn6/H96vzH2bW+ksDrvSmZaRzPE4nD4XM6XualXDSSqRV07pyTXdJO6d02vU+R7Q9Vx8/jWWXJfzcbJ/D1/+3Tj/QdPeSrIpZnxQzDPq0XKGVZZUcZvd+9qzjFb+sPenbPitp7Eas4bal09goOeJxuW14YeKTblVUXKC233kkvz9n2P43CDgjpfg1h8xp5BjcfjK2aypPEVsZKDdqfV0RioxSSXXJu922++yS+iHDxvxbDrPEp1XB3xx1r/AA/7tww8uOnn9yy8U8m4V66xMtUddHLM1w/wSvXUHJ4eakpRlKKu3G6aaSbV0/Bp9usx5hODOWYJ4+txCyqrBR6lTw03WqvyShBN3+dK3jazPz3EXlS4b6+zWvn1GeNyPMcTJ1K88C4e6qze7lKnJNKTe7cWru7d22z8vlHJDoDCYpVs41PnePpRaaow93QUvNSajJtP0afqfY8Q6vwLxjOdXz55Y56m5J66ZjOTDtPR+14Pcc/24tR6go5PkE8JkeT06Ko4qtL+7Vqk3LvFXjFNQbSTbVld72XzDnmyHE1ct0tqalTm6GGq4nBV5WuouooShd+F/dz+eyOxektF6X0Lk8Mi0nk9DLsHTfV0003KpKyTlOTu5SaS3bb2W+xyNTaXyLWOR4nTmpcup43L8XFRq0Ztrs00000000mmmmmk0fE6TxPp+h8Ux6vpsNcePbXv1rV/x97pcbljqvgvLfx34f4Xhnl2l9UajwmUZlkqnQaxlRU4VqTm5QnCT2dlJRavdOLdrNH8njPzGR1pFcKuDMMRmePzqawdbMKUXCLjPZ06SaTd02pVHZRSbTd7r+jjeR/RFTGzrYLWGc0MLKd40ZQpzlFeKU7K/o2vnufUNCcDOHvDrLMTgtOZdVji8Zh54fEZnWmp4uUJKztNq0PB2ikm0m02j63UdX4Fw9Teu4vNnnbvy30l+dmOPJZ5b6OpXKjp2GacbsFNyjXpZLh8VjXKG8JWj7qMt97dVWLXqkd1ta5z+xrSGeai61H7m5fiMVG/jKFOUkreLbSVvG5+P4TcAdJcHsfmGZ5FmOZ43E5hTVByxkqbVOkpdVoqMY7tpNtt3srJWd/2GttJ4HXOlMy0lmeJxOHw2ZUfdVKuHklUirp3Tkmu6Sd07ptep8/x3xbp/FPEsOaX/ZzU+904sLhhY6gcmmRyzLibj88qpyhleW1JKT3aq1Zxit/WHvDujJppn4LhNwV0xwdoZhTyDG4/GVs0lSeIrYuUG7Q6uiMVGKSS623e7bffZJfvpLY8ftJ4lx+Kddebiv5skk/w/wC7t0+HyeOr6sZpt7GL7HIa3MXFW7s/P2PbjWLRlKDs90bvZmcvxWTY641x5U23dWM3srs2bMpK6syK9GNZT3e3kYtm8oq5jKKitm+5LvjYymm3sYvdHIsmYtEO+NYTi7vsZyg0u6N5q7bM59vzmX1dpXGlB3b2KNpG0u0vmMWkyLHfGs5tXb9DCbucicU7rcxnFImu+FYT8TOUW7G0orfdmbViK741jJbNHXvnqTXLjn3+lYL7RA7CyW519561/ucM+/0rBfaIHbpf12H0x87x2/0Xz/uZfY+lexM+QXXf0vf2LDnoqedXsTPkF139L39iw56Kn65/ngAAAAAAAAAAAAAAAAAAHlDz/c7XMNnHMJiOVzlqzLNMseWVqGXV6mQwvmea5hVpxqTpwqxvOlCmpqNodL6o1HKTVkvmGG5cvbGYqhTxNLUXFlQqRUkqnFSlTmk14xlmCcX6NJnZ7D8o3HXQ/tOocx2ndDYfPeH2bY6ticTmcsywlOrgFisvlQrP3U6iqtwqzk10Ql1Qdrpt29DwPFb72n2yX+cXFX/7LFD/APqJ144qcOubvT/MHpbRnFfMNUVeLmYVssjp+tjtUwxuPjVq4lwwXusasRNUWq93FurHofxn09z/AEYHnvzSco/HbiR7QHhRxz0bpXDY/ReQYrT2IzXMXmeGpSwccFmMq9ZSpVJxqzfu7Ne7jO90rp3sHzHld4De1G03zAaJz3jDqLiBLReCzH3udRzTiLRzHDSwypyvGeHjjKjqXbSS6HZtPa116rAAAAAAAH4vjJws03xu4X6k4U6tjN5XqXAzwdWpTt10Z3UqdaF9uunUjCavteKumtjxjynV/OJ7LDiHmemauXQxulcyxLnGnjaFSvkmcJbRxFCpFp0q3QkmlKM1ZKcZKMT2n4l8TNEcHtEZpxI4kZ9DJdOZLGnPG46dGpVVJVKkaUF0UoynJuc4RSjFtuS2OBoTXXCvmF4dYXWGjsfl2rdI52qkac6+Ek6Vb3dRwnGdGvBSi1ODTjOCeydrNNh5rYf24+cxy1UcVy24KpmHTZ16eq5wouXn7p4RtL06/wA58C4u82vNn7Q3OcDwZ0lpNQyvFV4VVpvTtGfRXnGSca2Nr1JO8IOzvJwpRaUmrxUl6+4zk85U8fj3mWL5c+HUq7kpycdOYWEZNO93CMFF3fe6d/G5+lzStwh5c+HWbameTZNovSORUPheOeU5R7qjQpK0er3GFptytdL4sG7egH4nk05caHK3wGyXhjPF0sZm8qlTNM8xVFWp1cwrKPvOjs3CEYQpxbSbjTTaTdl9yPw3CDjTww49aQWu+Emq6Oocj+EzwbxNOhWoOFeCi5U5060ITjJKcHZxW0k+zR+5AAAAAAAAAAAAAAAAAAADx99t38rPDb6O4n7SehnKx+5i4QfQPT/9H0Dzz9t38rPDb6O4n7SehnKx+5i4QfQPIP6PoBs7PqPcWYHUzJNN2WYHUw3c09fQ7izA6mD09UAFkroywl2qCzRUSaaENbkgyzRLpWzQLNXFkjC90RRN0G7EWT38wTSboq2rk9KIt8YNhZkOL7vsXDV1YG2VmybMuopeYcVvuwbZgt0oqFTLaU9hKzT3IANq2ZL3RIClbMnZ7EkWSGtt2izLAr1Myxu1ZfjMR7lulS3be/kRJKO6G+2mhWzJ6mSNEujZbFZtNbFrJkNGK3uaUsxZlgBGy2IleVrK9ibJhJIKlU6ZJp+CZbrj5kvdNFOlAt0dSuw9+wUU2Wa6e19wSsys1c0smRJLbdmVe9s1styVv2JcUyG3B2Xz7masbKWYI629tiTFb2r3YsybJEhsulAAFeqtmLMsBe7fNWfS14ErZF2rkdKM031VDfhfchMP8YlU17nCzfJcnz/AVMrz7KsJmOCq7Tw+Koxq05bW3jJNPu/A4ultKZBorJ45BpnBSweXU5zqUcN7yc4UnNuUlDqbcYuTb6U7Jt2STsf2GrkWS38jp8rn5Pk93XwV5r6b7KdLXgHF2LSk7FbtnNvmqsUS+wPzetNZ4TSuD6IdFbH1ot0aL7JbrrnbdRTvt3bVlazazLKYzddOPDLlymOE3VdZ6zwmlcGow6K2PrRboUH2S7dc7bqKd9u7asrWbXxLHY3GZni6uPx1edevXl1TnLu32+ZJKySWySSSSQx2OxeZ4urjsfXnWr1pdU5ye7fbw2SSsklskkkkkfTuH2gPgHus9z6h/fO08Phpr/BeKlNP9/4pfve73/F8duXPlqej7uGPH4fx7y72nD7h68D7vPs+of3ztPDYaa/wXipTX+P4pfve73/F+hO1reJLbZFkj1Y4TCaj4/Nz5c2VyzrpfzdcyvFzhNxVpaV0TnmGwmXSynD4p06uBo1X7yc6ib6pRbtaK2vbY+/8tGudR8SeCWnNaasxVPE5rmPwz4RVhRjSjL3eLrU4WjFJK0YRWy3tfuz+1rHglwp4g5us+1nofLs2zCNGNBV68ZOapxbajs0rJyb/ADn6HS+ldPaKyHDaZ0rlVHLcrwXX8HwtFNQp9c5Tla7b3nOTe/ds9nLy8eXDjjjjrKe958ccplu3t8H9JrcWZYHkdp2UIe6LS7kBSFtsw90LJkdTCpSzKWZfqZBjZdPz+p9B6U1niMvr6qyWjmkcsnKrhqGIbnQjOSSc5Um+ibSVk5J2TdrXd/7NHDUMLQhhsLQp0aVJKMKdOKjGKXZJLZJeSNypWXLyXGYW3U93uVLvsp2JsybJkdTObZpBD3RLdyHsg3aLMgm7ZAvdcLodxZMJJGWSNG9u5Rq5ZruyrdiVQWyIkOphb9wqVV9ipo0rd2V6UGos/IrIs5d0vAqt+4qojuLMlq26HUyb2bLpFmVkWu2Q0mFSobVrEEtWQSC5VFsw92T0odKF7q9OykuxFmWauRe2yIaN7EE9KHSgpV7o6vcweluYXB8R/wBn/DSvmdXLqeDpUKVPLq/XOn0puUZ4dt+8Tk29oyTur2aR2iKy7n0PDuvvh3N8rMZl7rK3W+zpdT4/819Kn9z6mh8XVxN1DrnpquqvVsrWSSvfw6e77HHyzgXx4466moag4sYnFZZgoWjKtjYxp1Y0r3dOhh4pKLd+8lFXbbu1Z91+lC9tkfavtNOGW9HwY8eV7b97Jj8a/nafyDLNL5JgdPZLhlQwOXUIYehTTvaEVZXfi33be7bbe7Oe9rk9TIe/c/LcmeXJlcsru27tdYqVa3LNWBzXLtEUQ+5YiyYbLpUi6LNWK2TChsqW6UGrIVU2qVl4liGluZZpsZmc4vd22NZdysvxWY6yuO4u/YxlCVnsciXcylsiLHfGsWmnZmb7Gsnd7mcoqzJvausYz7mMtkchxTd22Y1IKy7k2O+NYy3MZRdjeSSdkZzSS2IrvjWLi79jCUZW7HJkYz7fnJr0Y1i9rmcuxrLuUlEl1l0wqd2YzVzkVIrd3fYwaTIsejCsntdeJjJM3nH4z7mc1Ymx3xrjzTu9jKSZyJru/QxkQ741g9rs6+89rvy3Z8v/AArA/aIHYOf4p17561/ub8/9MVgftEDr0v6/D6Y+d47P6L5/3cvsfSfYmfILrv6Xv7Fhz0VPOr2JnyC67+l7+xYc9FT9c/z2AAAAAAAAAAAAAAAAA/CcddE5zxJ4Ka94e6exqwma6k03mWVYGtKp0RjXr4adOn1ys2ouUkpNK9m7b2PGvLuJ/tIOQatTwGpcFqWhprCVFCOFz2i83yOa2tTp4mMpRpJ2T6aVWm93dJtge55+R4icWOGXCXKXnvEzXmR6ZwVm4VMyxsKDqtd1TjJqVR+kU36HjRxi9rtzOcRcFDKtDRyjh5hJUI08TVyqn8IxdabVptV66fuot7pQipxt+O+5/R4D+zi4z82zhxT4k8ecgngcdKM8VjIZ4tR5rJNuVpunUlTg3eW06zlF3vC6aA7Q8avbNcGtK/CMr4K6OzXXGOjeMMwxt8ty9d0pxUouvUStdxdOndNWkvDrzwp5rvaC83XG/R2I0zUz7B6RwepcBVzOhpjL54PKsNhYYmE60cTiXdzh7pNOnVqyUkrKLbafeLgv7L7lN4Qqhjsfo2rrnOKNpfDdT1Fiaaknd9OFio0LXtbqhJpJLqe7fa3AZfgMpwVHLcrwOHweEw0FTo0MPSjTp04r97GMUkl6JAcoAAAAAPz+vNc6V4Z6PzfX2uc3hleQ5Fhp4zH4yVOdRUaMe76KcZTk+yUYxbbaSTbR+gPx3Fzhfp7jPw11Dwr1ZiMdQynUuClgcVVwNWNPEQg2n1U5SjKKkmk1eLW26a2A6Dc+nPpyn8aOU/XPDThpxW+7OpM5+5nwLBfcLMsP733WZYWtU/ulbDwpxtTpTlvJXtZXbSf872fHPTyr8DuV3TvDrijxS+4mocDjcxq4jB/cTMcR0Qq4qpUg/eUMPODvGSdlJtXs7PY/J843svOBHL9y36x4v6D1TxDzHPNPRwM8NhsyxuDrYaUauOoUKrnClhITajTqzkmpqzim7pNPiclnsw+CfMRy8ae4tcRNRcQsqzfOa2Mi6OW4vCUMNKjSxFSnTnCNXC1JtNQTb62m72stkHcf8KNyKfw5/wD2s5x/ZD5TzQ+085S864F630fw/wBV4zWWdamyDH5NhcHTyXGYajGeJoyoqdaeJpUl0RU3JqLbai0rNpk/gVOVn/P3ir/6Vy7+wnOyb2MvKdleZUcdjtQ8R83oUpXng8bm+EjRqryk6GFp1Ev/AKmafqB+a9ijk+e4TgZrnOcZCrDK8x1PGGBUrpTqUsLTVacU1Zp9VOPUu7g094noufn9D6G0jw00nluhtB6ewmSZDlFH3GDwOEp9NOlC7be+8pNtylJtylJttttt/oAAAAAAAAAAAAAAAAAAAA8ffbd/Kzw2+juJ+0nodysO3LDwgT/zD0//AEfQPPH23fys8Nvo7iftJ6HcrCvyw8INv+IWn/6PoAfUG77IjpZayAEdSHfdCyJAhKzJAAr0slO2zJKvuGy6T32RFmgL+oYgmzZNkQ7ra4bLos0CVvsxb0BtHSybpbeRF2Tb0DdobuQWshZGWbap0snst/AtZFX5DQh79glZk9g9jNGwh77Ii7JtZXXcwOllG/A0W6I6Y37Bu9erMt1It0R8itkGyoe7IJezIDZdIaEuxIDZdoXYdSJIsg06kV6H6EvZlh6eipWd+nby8h32RLSv2Jilcy6brsr0sdSLPZkWRhNIe+6KtXJd728CUFRSzRBd9yLIEqjVwot+RL2F2gqXRZ2t+YKLXdoXI65eYb6ji990VlEv3Fkw2Vl0Nb3RDaRd+RWSRljYAskrLYq9mZrs2XSGrkdLLAxSgACpQhoPZBboNR0shuzsXKS7t23MsVLtEuxK7Fbti7JbLqrPdEdvi+IT3PzetNZ4PSmEtBQr5hWi3Qot7Jduudt1FNdu7asrWbWZZTGbrrx4ZcuXlwneq621lhNKYRQioV8fXi3QoX2S7dc7bqKd9u7asrWbXxDHY7F5ni6uOx9edavWl1TnJ7t9vDZJKySWySSSSRbG47GZpi6uPx9edevWl1TnLu328Nkkkkktkkkkkj93w+4e/DnTz7PqH967Tw+Hmv8ADeKlNP8AeeKX77u9rKXjyuXPlqej7/Hx8fh/H5svVzuHWhKWHp0NS5vCNSrUiquEo7NU00mqj8HJpppeGze9un6MncmSRB68MJhNR8bm58ufLz5VVu5BayDWxTlNV57c9eiNaah420cwyDSGd5nhlkmFputg8BWrU1NVKrceqMWrpNXV77o7RcoGU5rkvLrpLLM5y7FYDGUfh/vcPiqMqVWF8diJK8JJNXTTV1umn2Z/d4g8xfBvhZn0dMa81j9zMzlQhilQ+5+Lr3pTbUZdVKlKO7i9r323W6P1mi9a6Z4h6YwesdHZn90MozD3nwfE+4qUvedFSVOfxKkYzVpwkt0r2urppv2c3LnlwY4XHUnvc8McfPvff4P7TVgSt92Q9jwvQFH3Jb3Jt6BsukWbK2tuyz22RAUjqQbuiHsyAqUI7bsulsVJrVW7kdSJezIsjFHUiGybIqG7A3YEySCmctwlYl7MhuwbLANXK3ZYmzSlWvAhKxL7gxUqH2ITsfz881Hp7TWFWN1HnuXZVh2+lVcbioUIN7bJzaV914+J/M07xG0Bq6v8E0xrTJM0xG79xhcdTqVUkm2+hPqtZN3tbZnedPy5YXkxxuvjpT9E/Eq00WexW7ZwVtDVyrViwsmLNqQnZB79iHsLtEql2q4vfdFZRLXu7XJsmGy6Z9LHSy63ZWbaewXLpHSwLsdzPTuqK9LIasLsEtlmizRVp7lrtkPsGyqkNXZICpdqpXFmgL+oVFZEWbLySKu62uTVS6RKL2I8CbtgxUqgLWQsrBW1Ft3Dd9kT3KruFynSyri990aFJeJljVWrEPsSQYplPuYvscmUVfdGDSsZY7Y1jNNvYzlB2e6N5JXM5fismx1xyvo48omE4NLujky7mM+35yK742sJRM5SNpdzKSRD0RjO19vIxascrpT8DGSRNnud8awm+69DF7o5E4rd2MWtiHfGsKi3fYxnFu1rHImu7MpGWO+NcecJK/Yotu5vLtL5jCRFmnfG7Uqb3t4owkrG8u5nKKb3RNdsbpxpdjr3z3fubc+/0rBfaIHYea9PE69c+CS5bc//ANKwP2iB26Wf7bH6Y8Hjl/ozn/dy+x9E9iZ8guu/pe/sWHPRU86vYmfILrv6Xv7Fhz0VP1b/AD6AAAAAAAAAAAAAAAAGOIoUcXQqYbE0YVaNaDp1KdSKlGcWmnFp7NNOzT2NgB1Y40ezU5TeM7xGOq8P46Qzmvd/dLS81gH1Xu3KgovDybe7bp9T3+Mm7nSPX/smOZngznE9Z8svFVZ3UwzlOgsNjZ5Jm8ErSjGE1P3U3dWb97C7SairtL2DAHjDp/2jPPbyu5rR0jzCaHrZ7RpPoVLVGWVMDjakF/3HF01GNVf9/KNW9+53J4Ae1b5deNGb5To/UGFzjQ2pc4xNLBYbDZhS+E4OtiaslGFOGKpJ2vJpdVWFNXfc7f6l0rpjWmUVsg1hp3LM8yzEK1XBZjhKeJoVNrbwqJxezfdeLOred+y95V8VxF0/xM0lkmcaPzDIc2wmbfA8nx395YqpQqxqKE6VaNT3cW4JNUnBWvZX3A7dAAAAAAAAHWPih7SDlB4UZ1itN51xShm2a4Kbp4jDZHgq2PVOadnB1qcfc9Sd049d0000nsYe0o4s51wh5RdXZppvG1MHmufTw+nsLiac+idFYmdq0ou6fV7iNZJxd4tqS/FOmfI77LXh1xj4PZRxm416izyX7JOvEZZk+U4inh4UsLGo4Kdeo4SlKU+htRg4qMWrtttRDsx+F85OP8q6t/8AQUv/AGh+F85OP8q6t/8AQUv/AGjpNlvJtwVxPtNM15V62Ezh6EwmEjWpUlj38KUnk1LFu9a12vezk+3ayPrXOj7Kfh/oHhBmPE7l7q5xHHaToVcwzbKsxxbxKxuBguqrOlKycalKKlPp3UoqSVpJJh6BcvPMpwt5n9I43WvCnMMbicBl2PlluKjjMJLD1addU4VLOL7pxqRaabXdd00fVDqT7MfNeC+oOWPCZ5wd0LhtJVK+Y1aWp8uoYvEYpRzinSpRqTjUxFSpU6J0lRqQj1NRVS27Um+2wAAAAAAAAAAAAAAAAAAAePvtu/lZ4bfR3E/aT0P5V435YOEH0D0//R9A88Pbd/Kzw2+juJ+0nohysNrlh4QfQLT/APR9AD6hZogXYAEpk2QsgIbvsh0snZbkXYEE2bIF2BNmiBdsAACUtgIBLWxAESIs2W7gCvSxZosAKFk7IWQsg2XR1IdSDRUNlW6kQTZFXsK1DRFmiwMsFSwsgZIW7Un3Kl5JXCSuNd1e5XpY7bs0sis1ZCwlirdyCUtiDGyglF2Au2Gy7QnbZhsmyIsgqXSvQ3vsOh+hLutri7DZdii0vAhq5ZborPbsZ61qOljpZF2WFjdqlZF2kV7mNl2qS3cmysVDZdKtPdloOy3JSTW6Ky2e2wUmW5VpomN3JK5aSQbLpjJPcRi7F5q0dkVj2M031QCZWSexEEmtzLNKgVaLAxsu1ErhqxayREu4VFbpkiyQDUT3W3mUs1v5Gh+Z1rrPCaVwnRDorY+tFuhQb2S7dc7bqKd9u7asrWbU52YzddeLjy5cphj3tRrPWmE0rhFCCjWx9aLdGi3sl26523UU77d21ZWs2viGPx2LzPGVcfjq861evLqnOXdvt4bJJWSS2SSSSSGOx2LzPF1cdj6861etLqnOT3b7eGySVkktkkkkkj93w+4ffD/dZ7n1D+9dp4fDzX+G8VKaf73xS/fd3ttLwZZZc+Wp6P0HFx8Xh/H5su9OH3Dz7oe6z7PqH967Tw+Hmv8ADeKlNP8AeeKX77u9rKX1a3gS9uxB7MMJhNR8bn6jLny82X+Cri15CzRdb9ykm7teBTjtWRUv3Ia2CnUDmq5VOJ3GbidS1fpCvkkMBHK6GDaxmLnTqe8hOo3soSVrTVnfzPuvLpw+z7hZwb09oPU8sLLM8s+F+/eFqOdJ+9xdWrG0mk38WpG+y3uvU+XcxXODmXAzX9PRWE0Lhs3hUy+jjXiKmPlRac5zXT0qnJWXRe9/E+v8D+JNfi9wuyTiHicpp5ZVzb4T1YWFV1VT91iKlJWk0m7qmn2Vr28Lnt5rzfIYzP8ARc8PLM7r1fuWh3+L4kh2tfxPDdO6rVisot3s0WK3dzFS7TFNKzIs0St0H2DZdKgAKR33RFmiXtsgt9mFS7VK2aLvZlLtk3TZdIukR32RNkyHtujFFmiCbsgK2i6RDdy1kRZBsqoD2BliptDV2fw9caqwOhdIZvq7Mk5YfKsLPEygnZ1Gl8WCfg5SaivVo/sYzF4bL8JXx+Nrwo4fDU5VatSbsoQim3JvwSSbfzHUvmg5iuGeueFtfSehNUvMsbjsdh1iKSweIopUIN1HJupCKaU4U1ZNu7TtZNr6fhPh3J4h1OGGONuO5u/BeM3dPleg9F685teIuZ5tqLUNTDYTCJVsXipQdSGFhOT93h6FO6SulKybSSi223s/vmhOTXKtA6+yPWmX64xeLpZTXlXqYWtg4qVWXTJRtOMrRSbTacXdJq6ufguU/jDwg4W6Ex+E1fqxYDN8xzKdepSWAxNVqjGnGNNOVOnKL3U3s7/G3O2mktW6f11p7Caq0tmCx2V47rdDEKlOn19E5Ql8WaUlaUJLdLtftZn6X2h8Q8Q6Tly4OHG4cEnln5va9vjp0ts+h/WauyHt3Py2veKWhOGWDjjNaajw2X+9TdGhvOvWt/iUopyavs3aybV2j5tl3OZwQzHGxwdfMs1wUZSUVXxOAl7rfxbg5NLtu0rX8r2/K8HhnWdTheTi47cfjpmt+j7i3dFGj8ZrDjNw20Pp7L9UZ7qWi8tzeXTga+FhPErENK7cXTTVku7dknte9kfotOahynVmRYHUmRYh18vzGiq+HqOnKDlB9m4ySa/Ojz8nS83HhOTPGzHet1Uf0LNBJsVJwpU5VKk1CEE5Sk2kklu22+yPkmqOangppXHSy6rqp5jXhJqostoSxEINNreokoPdPs2/Mzp+i6jq7rgwuX0Nj63Zoq0fLtHcznBzW2PhlOXapWDxtaahSo5hRlh/et9lGcl0Nt7JXTbdkmfs9a660pw7yV6h1jnEMuy9VYUPeypzqOVSV7RUKcZSbaTeydkm3ZJsrk6DquHknDnx5TK+k0p/O4icV9C8KsLg8VrbOJYJZhOcMNCFCpVnUcEnNpQTskpK7dlul3Z/a0xqXJ9Y5BgtTZBiJV8vzGkquHqTpSpuUbtbxkk1umt1va6urM6Lc2PFjS/FHVuTT0bmrzDK8sy9xdb3FSl/d51G5xUakYuyjCnvazu14HYHh9zH8ANJaF0/pmWulCplmW4bC1UsrxjvUjTSm7qk025Xe22+2x9/q/Zvl4PD+Lmw48ryZfpTXpPo0qV98bsR1I42V5ngM8yzB5zleIWIwWPoU8Vhq0U0qlKcVKEkmk0mmnZpPc/lax13o3QGXLNNY6hwmVYeV1Tdabc6jXdQgk5zavdqKbSPzWHDycmc48cbcvhFdvV/cIl3PhdbnO4LU8R7iFfOqtO/+GhgGoePhKSl/J4n0PQPF/h1xNhNaP1JQxeIpR6quFqRlSrwXi3Tmk2lezkk1d2uenn8J67psPlOXisx+hu37K6ZDdrh7bo/IcQ+K+h+FuFwmK1rm7wax85ww0YUKlWdRwSc2lBOySkrt2W68WePh4eTqM/k+LG5ZK3r1frepEN3PxuouMHDrSmlcv1nnmoaeHy3N6EMRl96c/fYqE4qUXClbrd003dJK6u1dH8bhvzC8N+KWdVNPaZxGPhj4UpV1SxWFcFOEWk2mm0rXWzae+y7nonhvV3jy5vk75Z79NlkfSOljs9z+Jq/XWkdBZcs21hn+FyvDSbjTdaTc6jSu1CCTlNpbtRTaW58onzk8F44r3EcVnMqd7e/WAfR3tezkpevbx/MV03hnWdXj5+Hjtn0K3J619ybuQfntE8QtF8RsuqZnozP8PmVGk1GrGF41KTabSnCSUo3s7XSTs7Xsz9E+54+Xhz4c/k+SWWe50llnZRu5Vq7LtEHK9iBR9y5DW3YxcVK92WIdrX8QpHSxfawuyWla9g2aUaIJb3DStexKoo0Y2aOQZTVo7IOmNYzW9zKS2NpGb3W5N7V3xrCXcybOT0p+Bg4peBFdsbGU3ZmEuxyZRTfYwkkTXfGspdzCUXY5E7XMZdiK9GNYyW7RnUT6Vuu5tJK7Mp9vzkWO+N240oO7exVpo1l4mcidR3lZT7v5jGauciUU7t97GMkiXXGsJNJWMZq5yKkVdmUkibHoxrjST3R1658F/ubM/f/AIVgftMDsPNK7svE688+H7mrP/8ASsD9pgdel/XY/THg8cv9Gc/7t+x9C9iZ8guu/pe/sWHPRU86vYmfILrv6Xv7Fhz0VP1b+AAAAAAAAAAAAAAAAAAAAA+W6z5oOXXhzqTF6O13xp0hkOeYD3bxWXY7NKVKvQ95TjUh1wbvG8Jwkr91JPs0fxfv2eUb8o3QP/puj/WB9sB8T+/Z5RvyjdA/+m6P9ZzMl5veVzUecYHT+Q8ftD47MsyxFPCYPC0c5oyqV69SSjCnBdW8pSaSS3basB9gAAAAAAAB1H9qZoPHa55NdWVcto+9r6axOCz7oUbt0qNVRrNeXTSq1JN+UWcr2YnEXItfcnejMHlVWPwzSqr5DmVHqTdKvSqSnFu26U6VSlNXS/Ga3tc7RZtlWXZ9lWMyPOcFTxeAzHD1MLisPVjeFajUi4zhJeKcW015Nnxvlu5QOEfKpV1JPhTPP6dHVFWhUxWHzDMXiaNL3TqdCpR6VayqyV5OUmkrt23Dovzd6r+9R9p7ovmI1BhK70vqPLaEsdXo0upuksPPL8SkkrynSh7mrZbtOCT3su1HMHz38rGScBtT5xkvGHTWpMbm+S4vB5blOUZhTxGNr161KVOEZ0YtzoK805SqRjZJuzaSf07mc5XeGXNZoOOiOIuGxFGrhKssTlWa4Nxji8urtWcoSkmnGSSU6bTUkl2cYyj0ZyL2H2VUNRqrqXmExWMyGFSMvcYHTscPiq0P30XUlXnCm12Uuid+9l2A/YexPyLOsHwO1zqDF0qkMtzPU0aODcrpVJ0cND3s4p91/dIR6ls3FrvFnowfk+F3DHRfBvQWT8NeHuTQyzIcjoe4wtCLcm7tynUnJ7ynOTlKUnu5Sb8T9YAAAAAAAAAAAAAAAAAAAHj77bv5WeG30dxP2k9EOVZW5YOEH0D0/wD0fQPO/wBt38rPDb6O4n7SeiHKvf72HhBb/MLT/wDR9AD6i1cjpYTfiWAr0sdLLFW2gFrbsnqRG97MmyAqWXYWQ2WwBuxF09vMnZiyAjpZBchpeIELZi6e3mTaItECGmgnZE9yr2Alu5AJ39QHSyCd/X9BAENXI7P5iwcfQCOpCze/mLIO4EdLFmiVuhu3Z9g2XQmVLWj4lTLNmxxb3TQ6Gt9iyasH2Gmy7VABl00KvZXZLe5HfaRhFXuyDS0fQr0+gVLtUlbMNbkBUsS3cp0stZ+QDVSYk2RD27BsuklWmheREmydKQ1dkpWLW2W25HYG+yj7kNXLtbdioVLtQlMmyIa8g1Vq7drbslR6fHuTutxdsKl2ghoPquTG7BLqs0rMiUXJ7NG0oxUW0tyhmm7U616i6e6IcduzI3RK9pbsQ92SLINioLWR/H1RqfLtK5e8ZjH11Z3VChGVpVZLwXkldXfZJru2k8tk71048cs8pjjN2qao1Pl+l8veMxj66tS6oUFK0qskuy8krq77JNd20n8KzrO8fn+Y1MxzKr11Z7JLaNOK7Rir7JXfq222222753nWPz/MKmZZlW66s9kltGnFXtGKvsld+rbbbbbb/Z8PeHnw90s/z6h/e208Ph5L/DeKlNP954pfvu722l48ssufLWPo+/w8XH4fx/KZ+qNAcPXj1Sz7PqP97XU8Nh5r/DeKlNP954pfvu72t1fVOz3NSjir9j04ccwmo+Rz9Tl1GXmy/gh79glZk2t4A1ygLq9h2Kt73QaSi9ihot+5WSW4bHxDjFyk8OONmrYay1XnepcJjYYOnglTy/E0IUuiEpNO1SjN3vN3d7bLZeP0Lhhw5yXhNoXLNAacxWNxOXZV773NXG1Izry97WnVl1OEYxdpVGlaK2Svd3b6jc5nHvi5w24vUtO6I1ricqy6WT4bEPD0qFGadWU6qcrzg3dqK8bbHY3lZ1fqPXnAbTGq9W5pUzHNsd8N+EYmcIRlU6MbXpwuopJWhCK2S7eZ7eXj5ceDHLLLeN9ycbjc7JO76oCRtb1PE7TaLpFWm/FblrJgyzTYp+LsxdMmSVyEl4GKLpB9hZMjftZhsQQ2SRZBSpNmibRI3ZiorLuVbsXa8yrStsrmWabtXu9hZolq3huRuzFCdkJNbbMgWT7hUqpDVy1kGgqafzc/yXA6kyHMtOZopvB5rg62CxKhLpk6VWDhNJ+DtJ2fgzoHzU8LdA8JtQ5HkGi6OLhVxWDqYzFrEYl1X0up007JpWV4VPn/ADHoZZHn1zG1anETmbraawspyj8MwGRULbtNqCmkvC1SpP8A2+J+v9j8+XHqstZWYTG2u3He77Zw/wCTvhXm2hcgzXUeFzV5rjstw+Kxfu8a4RVWdNSaSttZu35j6jqfN9KcuvCGpXy/CdOX5FhlQwGFlUblXrzk+iDb3blOTlJpbLqdtrH0SlRp4elChRpqFOnFQhGK2SSskvRJHUDn21XW+E6Y0NSnanGFXNq8L9226VJ29Eqy/P4W38vR8vU+P+IY9P1GduG7dX01P9aJbldV884T8M9W80mv8z1brTOMRHLaFSMsxxkFaUm7uGGoJpqKSXk1GNrptpPsZnfJhwVzDKHgcry7MMqxig1DHUcbUqzcvByhUbg1fuko3XZp7n6Plj0nS0hwU05h4U1GvmeHWbYiVrOc69pxb9VTdOPzRR9T79y/FvHuqnWXj6TLyYYXUk+b4quXfs8uuJeldXcPtRV+FmeZjUxdLJ8VOrhKVKblSl7+MGqkIXfS5xUG13TVu92/SzR2RQ0vpLJNN0oKMcqy/D4Oy33p04xbv4ttN38b3Oh+KrU+LHNup0puthcXqaEYyW/vMNhpJXXknTobPwTXkd8dXZ9DS2lc51LVinDKcBiMa002mqVOUrbbu9rWW+59D2p5eTm4+l6fLXns3fpv/fbcvc6j82nGnPNR6qnwX0VWqvCUqtPDZh8Hb68bi5tJYdNbuEW0ml3k2n+Kr/t+GXJZofK8moYriXGvnObV4KdbDUsTOjhsO2r9CdNqc2m2nJySdtklu/jvKJp2trnjfU1RnLliZZTRr5tWqT395iZyUYt+vVUlNesPzPvu15o5+N9Zn4Lx4eG9FfLqS5Wettb6dnV3idySabzLCLGcLMbLKcbGS6sHjq86uGqQbSdptSnBpXe7kna1l3PsmruDeldf6NyXResKuOxeGyZUXCrSrulOpUp0nT65NXu2m27+L7n71peHc4eb5jRyfKsbm+J/wOBw9TE1N7fFhFye/hsmfnc/GOu6m8eGfJbcb2vv7/O2V5z1+H+nc15hv2tdO06yyZagjljU6rnP3NOajXfX3btCo0/mO1v3mXBNNXwecP8A/KD3/kOv/KFltfVnHh6kxtNVKmAwuMzSrOzUVVqWp3+e9ZtL0v4HfWyZ+m9qPFer6Pm4+n4uSzWM39Na/D8R9b5Fwa4d4jP6tCEaGXUIYTL8HF2VWrbppUV4pbXb3ajFvex074acN9bc1Ot8x1ZrLPK9HLcPNLGYuML2b3jhsPF3jGy38VFNNptpP9nz3aprVs+03oqlWao4bCzzOtTXaU6knTg36pU528lJ+aOwXL7o/D6K4QabyynRUK+KwcMwxbsk5V66VRptd3FNRv5RRw6fP8SeEzrMP13Le1vrJ/r7VTve789S5ROA9PALCS0riqtVQ6XipZniVVb/AMaymoX/APrbeh1J4oaQzfl04vUqemc2xF8J7rM8rxM7KcqMm10VErKSvCcJKyUkndJOy9G7I6Ic7eYYfG8YsNhqEk5YDI8Nh6vpN1K1RL/zakX+c6eyviPV9b1eXB1GdzxuN3vu2zXo7t6ezalqDT+WZ/Qio0szwdDGQV72VSCkldd9mdNOeLP/AIdxDybT9OfVDKsr97JX/Fq1qkm1/wCZCm/znb7QWWTyXQem8mqQlCeAynB4WUZXunCjGLTv4qx0X4rYetxW5nMy0/hqygsbnVLJozW6pwpKNGcu3h0Tk++9+55/ZXh48fEuXmv6PHLf9f4Nvo/Y8LeCefcw+YviFxBxuKwGmqEIYLLcPQaU6tGkumFKldNQpwSs5WblLqtu212C4Z8uug+FOqMXqnTFXMp18ThHhI0sXWjUhRi5xlJwaind9EVdttJNJ2bR9IyfKMtyHKcHkeU4WGHwWAoww+HpQW0IQSSXrsu/d9zlNb9j5XiPj/U9Znlhhl5eK9vLPgqYyPPrDzzHmb4/QwecZvVoYDG4isqNmm8PgaUZSVOmnspOMe9rOUnJp7p9qXys8DPuesveiYP4ih7/AOG11VbX77qVTvfftbwtbY6zcVuCXEXghrKet9EUsXPKKGJlisBmGCj1zwSbf9zqxSfSkm43acZJ2fdxX0vhZzp4PMK9DJeKOWU8FOdqazXBpuld7Xq0ndxXi5RbV3+Kluv1XivH1nVdPxc/g+f+yxxn5uN1d/OzHU7X1fXOEPA3TnBrEZ5VyDMcZi4ZzUpNLFKLlQp01K0E0l1bzk27K6SVtrv6UnZFKVSjXpQr0akKlKpFThOMk4yi1dNNbNNO6aLO19j+e9T1HN1XLeXntuXv36u0mu0Q1cq4ssHseW+q2bi15FhdsGKQ1cqXKu35wqVBXuyW9yA0asVauy633ZWSe5lXKpKIfYndgxUYz7mUlsbyUW97fpMnutzLHXGsJpt7GL3RyWlc47sRXoxvwZyiYuLSu2jeXcynutlcmx1xtYVItyVrGdmbST8mYy6vJkWPRjWc+7+YxZrNPd2f6DLd907fMS743symndsykciUVv8AMYySId8awm+69DGRvOK6ntsZVIpWsia741jODd3t2MnFvs0byTszKV0TY741x5U3vuu51358YuPLVn/+lYH7TA7FTTs2kzrvz53+9qz+9/8A4VgftMDr001zY/THh8bt/FnP+7fsfv8A2JnyC67+l7+xYc9FTzq9iZ8guu/pe/sWHPRU/VP4EAAAAAAAAAAAAAAAAAADoTzG+yi0tzA8Z9S8YavGPM8hq6kq0K1TL6eT08TCjOnh6dJ2m6sW03T6uyt1W3tc+cfgO9JflD5t9Xaf9oPT0AeT3EH2JGZ5fpXG47hnxsWcZ/Qj7zD5fmuUrCUMVZO9P30Ks3CT2s3Fxvs2k+pfAeSDgrwdxXMplnC7mExesdEcR9O6goYjK8JKph4YHGYvDVI1Y4Ksp03OnObguiSm41VJKLUnDr93zym56uXzjrxa589L6n4ZcI9RQyvARyHC1tS4bCSWFlWp13VnipV38SPuoThBt9vcLZ+IerIAAAAAePPEj2o3OppziHqfT+VcPtOYbBZXnONwWGo1dP4urOnSp15QhGU1VSnJKKTkkk3dpLsvYYAeK34WHnl/zI0v9WMX+uOBL2v3ORHG/c55HolYvrVP4P8AcHEe86n2j0+/vf0tc9Oeebj3mvLhy16n4iabjH7vzVLKsnnOKlCji8RNQVVp7P3cOuok005QSas2db/ZV8reQ4bQlHmy4h//AHxa61piMXictxeOl7+pl+HVapSnWUpXbxFacakpVG2+iUYqzc+oOsv4WHnl/wAyNL/VjF/rjjUva5c6VfNqOQ0NM6OqZpiKsKFLBR09iXXqVJ26IRpqt1OTurJK7uu9z2yOlntKeUHS/GvhNnPF7T+CpZdxB0Ll1XNaGYUYKFTMMHh4OpUwtWS3k1CMpU294zSSaUmB++5D+M/HzjhwqzjU/MJoVaZzrB59VwOBp/cutl3wjCrD0JqToVm57TnNdafTLsleLb7KnU72a/MlnfMdy70MZrPHfC9VaRxryLNMRJrrxcI04ToYmSStecJdLfjOlN+J2xAAAAAAAAAAAAAAAAAAADx99t38rPDb6O4n7SeiPKv+5g4P/QLT/wDR9A87vbd/Kzw2+juJ+0nojyrq/LBwg2/4haf/AKPoAfUQLPyFn5AALPyAABWvuTaIEFWrssAKpFgABVq7LACvSwlZlgAKtXY39f0FgISsSCAHUiLN7+ZNkQ7gSlYXXYjf1/QN/UB0sgnf1/QN/UCACVa+4FJdx0ss0rgCrViC9kyGvICpPSwTv6/oMs23asou7I6WWAvoqVXpY6kWKWfkSW7HFt3TW/mQotbtosr7Kwnstg3at0ipZK+9iHbwDUEtNEC7YVLsK9LLCz8g2VF0tvIiTW2zJs79vEOMfEyyRUu1fD5yOlk7fmJfT4WMb6KWaBLu9vD5iLPyCohq4SsTZ+RDuu1wSqve4i1HZgmyfcKl2hzT8CO+6DjZdiLtBu9Di7PsUasabPYiS9DLNql2z6kE7kyikvirc/i6o1RgdK5c8ZjLzqzvGhQi7SqyXl5JXV32Sa7tpObfLN1048bnlMcZu1bVGp8v0tl7xmMfXVneNChGVpVZLwXkldXfZJru2k/hWd51mGf4+pmWZVuurPZJbRpxV7RivBK79W222222zrO8w1BmFTMsyrddWeyS2jTir2jFeCV36ttttttv9rw94e/D/dZ9n1D+9dp4fDTX+G8VOaf73xS/fd3tZS8WWWXNlqej7/Dxcfh/H8pn3qvD3h78P91n+f0P712nh8PJf4bxUpp/vPFL993e1lL6uWsvIq+56sMJhNR8jqOpz6nPzZf4IumSLJAtxQ1co/xjQhxW7Ms2qXarVyrVizdiLX38SWy6OlkOLs+xZX8RdPYK9X43VWhuE2oMzWYa20hpHM8xVKNNVs1y/DV6/u021FSqRculNuyva7fqf3NPZVp3JMnw+WaTy3LsBlVDr9xhsuo06WHheTlLohTSirzcm7Ldtt7tnR3nd4Q8T9ccZqWdaP0FnecYBZLhqDxODwkqtP3inVbjdK10mm16o7Mcpmms90ly+6V09qfJ8VlmZ4T4d7/CYqm6dWn142vKN4vdXjKLXmmn4ns5eKY8OOXm38ycMt5WafWmiLNFpXTskVbfieN1lQASrX3FVLtWXcolZmk1vsivz9ib2bLoKuSRZ7MiyfgFI/G3RBZJJWRDSQVKo+5PUibIqFS6T33RRIumrBpIyzalGnuQS739CCVS7Vauw00WKyDZZpBDVyQFKVJRpU5Vak4wjBNyk3ZJJXbbfZHnxwDhPiRzR0NSYhOpTnmWOz2q2t0l7ycHbwSnKmreH5junxt1E9K8JdW54pKM6OVV6dKTdkqtWPu4P1+POO3j2OsHIPp74RqfVOqZ09sFgaOAg2u7rVHN29UqCv8AOvM/X+B/zXwvquqvvnln+v8AGOuF/Ntd07pnQvnopYqPGHAVK6fup5Dh/cvw6VWr3S/+uu7evqjvpt4Hw7mf4A4zjHk+AzTTVWhS1Bk/XGlGvNxhiqEt3Tb3UZKSTi3tvJO1018/2a6zi6Hr8eTmusbLN/SYWS930vhpjMFmHDvTGNy2cZ4ark+EdPpd0l7mKt86tZrummnufOOZvjfgeGGj8RkmUY6EtUZxRlQwdGnNOphoSTTxEl3SSuoXW8krXSduuWk9Jc5Gi8E9JaVyzUeAwTqSUaKnQdGnJt3cJzbjBNtu8Wk273u7n77JOU3WGX6e1BrrWWN/ZJrrF4HEfc7BfCFONPE1Kbiq1StVaVSrHqvHdRjKKacnZr6k8J6Ho+r/AAjqefHLG5bknv8Ap+E+Lp5ZLu1+A5JNPPNeMU84nSTp5JldeuptX6alRxpJLybjOf5kztzx/p4qrwW1nDBX94sorye1/iKN5+D/AHilv4d9u58x5P8Ag9q7hpgdRZtrbJZZbjs0qUKOHpVKtOclRpqbcviSdrymlZtP4qdrNN9hcbg8LmODr5fjqKq4fE0p0a1N3tOEk1JO3g02vznk8f8AEePl8XnNhd44a1r5u5buulPIhmmX4bXWocrxFWnDFY3LISw6lJpzUKic0lezdpJtd7JtbJndpu50R1zyo8XeHuqZZ3wyhic0wVCs62BxeBxKpYzDpt2jKLkm5JbXhdNbu12l/QoZNzu6vpvJMTidQ4TDzShUq169HBJRezbnFxnJbbpXbv23PqeL+H9L4xzzruHqMZMpNy3vFal77dxsFqzTWZagxumMvzvCYnNcupxq4vCUqilOhGTaXXbZO67N3V02t1f8XzJ5zPIeBur8bS6r1cCsF8V72xFSFB/mtUd/S5/I5deA9bgzlmY4zOs3hmOe526Txc6SbpUlDqahBys5Nubbk0r2Wytd/tOK+hFxL4e53ol4pYaeZ0IqlWkm1CrCcalNtLdrrhG9t7XPy2OPS9J4jhOPPzceNx7/AMNno65chGVUZPWOdSd6sfgWEh6RfvZS+e7UP0ep25ezszoDp7hXzU8J82xlHRuR5tg54pRp1quAqUa9DERi30N3bi7XbTaTXU+12jtVy96Y4o5FprMMfxdznF4zO80xirQw+Ixar/BaMYJKK6W4RbfU3GGyVvFtL7PtR0vFzc2XX4c2OUutYy9/RTrFzuYetS4v4WtUpNQrZLh5U5PtJKpVTs/Rr/3ujuXw5zTBZxoDTmZ5dXVXD4jK8LKEk/8A6HFNPyaaaa7ppp9j8RzDcB8NxnyLCzwGNpYDPcqc3g8RVi3TqQlbqpVGk2k2k00m009ndnW7T2jeb/hL7zI9JYDNqWE63NQw3wfG4Zt95QjPqUW+72T81c9Gun8d8L4eDHlmHJx9tZdmy6d1dVapyPRWn8bqfUeOhhMBgKbqVZyau7LaEVf4027JJbttJHQ/h/k+bcxnMBUz7McG/gNbHLNMx7uFHCU2lCi3Z3bUYU14vd7JO36Z8EeZ7jZmWHqcR8ficFgqU7+9zOtCMKXdN08NSf41tr9MU7pXS3XarhVwl0vwi04sh07RlUq1WqmNxtVL3uKqJNJya7JXajFbJN9223z4+TpfZzps8eLknJz5zW56Yz6Wybfq8yx+HyrLsVmmLk40MHRnXqNLtCEXJv8AQmdEeVDBV9Xcf4ahxkXUnhKONzas3dpTmnTu2+76q6a9VfwO5PF/LtQ5vwx1LlOk8E8XmuOy6rhMPRVSMHJVF0zScmkmoOTV2t0vE+M8ovBnWXDrE6hz7XGRvLMRjadDC4OE6tOc3BSlKq2oSdk2qaV7N2btazfl8I6ni6TwvquTLKefL82T33/W1X1jskkZ1MXhaWIpYSriKUK9ZSdKm5pTqKKTk4pu7Sur27XV+5qfAeZrgvxA4h5jkureH2cKGPyOlOnTwfwh4eopOSl7yjVukpuyTTcVaKafgfnug6bi6rnnFzcnkl99+Kt/B97aTTTSaat850h50dG6L0tqvJMbpnL8PgMfmuHr1sww2HioQajKKp1VBbRcm6ibSSbhdq92/wCnhtQc8WW4d5THAZtVcGoRrVcBhK047/8AdHFqXztv5zm6H5VeImutTR1hx1zaoqTmqlbCzxSr4vEpbqm5xbhSh6JtpXSUbpr9h4T0mHgHPer5+pxuMl/Nxu7l/gy25dtPvXL/ABzFcF9IfdOU3WeWwcXNu/um26Xfw924W9LH0ArSoUsLQp4bDUY0qNKKpwhBJRjFJJJJbJJJJIsfieq5p1HPnyyfpW3Ttj2gk2Q14E3aJ2au+5wqpdqdLKWd/wA5o7+BUhUQ2kR3exNkxZLt3CopKJHSy+7Kt2QUdtmG7ojdkCqlLpFGrl7JkWiSqXTKUSkoOz3RtNb7IpL8Vh0xrjyXiYyg0u6OQ07mU7tbIix3xrjTg2/AobtPyMZqyukybNO+NZz7mMuxs1fd3KdK8iK643TCfj8xhLsciol1NeBlOMUibHoxvZhJXvbxMpRatujkSS3MZrsRY741hNOz3XYxlF7HJmu9zJxTtsTY741x5Re5jOL2OTOLTeztYxmn5Mmu+FYS7HXfn1/c1Z+/LF4H7TA7EyWz2Z1259l/uadQf6XgftMDr0v67H6Xh8b/APLef92/Y/eexM+QXXf0vf2LDnoqedXsTPkF139L39iw56Kn6l/BQAAAAAAAAAAAAAAAAAAdKee72jOV8qWa4XhzojTeE1LrrF4WGNrwxtWUcFllCbfu3VUGp1Kk0m1TUotRtJuzin1R4V+2n4s4LVNCHGXh5pnNNOYiv/fE8io1sJjcLBtJun7yrUhUUe6hJRct05q6a/pac0DkHEz2x2rMk4t5TSzajhMZiMxwuBx6jKlXdDL6UsGpQe1SMaSpzUd01CLd0mn3n55+BGh+L/LNrfB5rp/LnmenMgxWa5Hjnh4xq4KvhKMq0I05pXhCSg6corZxm9uwH3TSeqcg1xpnKtY6VzSjmOT51g6WPwOLotuFahVipQkr2aumtmk07ppNWOtnDjn40trLmm1Byqap4fZrpHUGV4rF4TLsXjcXTqU8xnRTmkopJ0/eUV72nvJSj4ptX/J+yK1njdV8nuCyvG4qVf8AYrqDMcmpOTbcKf8Ac8VGLb8F8KaS3SVltay65+0ynhuFHPPwX4waccsHm1eGXYrHTg9qzwuO6U2u7cqUvdyXZxilbvcPWIAAAAAAAHWL2jnBvP8AjZyn6p0/pPA1MdneT1MPn2BwdOHVPEvDTvVpwW7c3RlW6Ut5SSivxjpj7O/2k3DbhLw1y/gJx5li8mwmR1ayybP6OFnXoKhVqyqOhiYU06kZRqTn0zjGScWlLp6by9azqDzD+zB5b+Pue4zWVHDZlonUmPqOti8ZkMqcaGLqvvUrYacXBybbblD3cpNtybbbYfSafPLyg1MF8PhzE6JVJQc+mWZRjUsr3/ubtO+2ytd7WW6Op/O57UTgvV4V6l4VcB83r6rz7U2X18oxGa0sPUoYHL8PWhKnWnGdSMZVavRJqHQnBOXU53j0v8piPYa4aVecsJzO1KdFv4kKmjVOSXrJY6Kb/Mj7DwZ9kDy58Os5w+ode5tnXELEYWSnSwWYxhhsu6lunOhTvKpZr8WVRwa2cZIDD2OvCrUOheXPNtbahwlbCLXOc/Dsuo1I9LngaNNUoVrNJpTn71rwcIwktpJvvsYYbDYfBYelhMJQp0MPQhGlSpUoKMIQSSUYpbJJWSS2sjcAAAAAAAAAAAAAAAAAAAPH323fys8Nvo7iftJ6Jcqv7mDhB9AtP/0fQPO323fys8Nvo7iftJ6J8qtvvYOD9/8AMLT/APR9AD6iR1InZ7CyAjqQ77oWiO34vYB0sdLCb8exN15gVJTshaJHzdgJbIJS8ybKwFSU7IgAAFuyWl4AQAABVq7LAARdIkiyAJpkkdiQI6kOpCyFogR33Q6WO34vYXkA6WQTdk2iBUEvZkAQ0OpE2fkRZAQ3cLZk2Qa2Dd6Q92QTZ+RNomabLtUPdE/N2IldeDHZsqE7bMhQa8huyd/X9BJtSXdog06U92tyHBeCDdqFupENbkdgqWaG/Eh79iR2CvVVxZHQ/Qu9iE/MVUqt7beQTuWtEhpLsZ201AauA7+BhGbVrkF2m/Mq1bsmFD+MreZXpZZdydmFS7ZruWIcdtk7n8jU+qcu0rlzxmOfvKs7xoUIu06skuy8krq7tZJru2k8tk71eGF5LMcJ3qmqdT5fpTLnjMa+urO6oUIytKrJeC8krq77JNd20n8IzvO8w1BmFTMsyrddWfZLaNOKvaMV4JXfq222222753nWYagzGpmWZ1uurPZRV1GEU3aEV4JXfq222222/wBrw94d/D3Sz/PsP/eu08Ph5r/DeKnNP954pfvu72speHPLLny1PR+h4eHj8O4/lOS7tZ8POH3w50s+z+h/eu08NhpL/DeKnNP954pfvu72t1fWCXGK7IJRS7r9J6sOOYTUfG6jqcupz82Xp7kN2I7vYtLpt4P85FkU5eiLNFWrsuVl42DZoIbW6Cv4j4oXLpVq5C22ZeyKtJMyxvqENXZN15kJtv0JVK+Q8U+aPhVwd1NHSWs8TmcMwnhYYtRw2CdWHu5uSW6a3vB7fMfuuHmvtP8AE/R2X660rUrzyvM/e+4lXpOnN+7qypSvFt2+PCVvNWfidduZzlC1pxx4kU9aZBqfJMBhYZbQwXucZ773jnCVRt/Fg1ZqatvfZn2zgBw4zPhHwjyHh5nWPwuNxmVfCve18K5e6l73E1aqt1JPZVEndLdPwPXyYcM4sbhd5X1TLl5r27P30u5V9i81vtuVa8zyOvooSGvIgNS1Yhq5JAWq1YFrIhryJJdIK2aJd15h28QuXSpD3Re0WtiobLtXpYs0S20RdhUukES7Eh7mXalSrTRaz8g15olSh+P4mcVtH8JcpwmdaxxVelh8biVhKSoUnVm5uEpNtJ3slF3fg2l4n7KysfkuIfC3RHFTLcPletsoeOo4Sq62Hca9SlKnNqzacJJu62ad16XSZ6Ok+QnLj+E78nv16qx9Zv0dSOZvmeyLifp6hoHh/Qxs8BXxEK+OxVek6br9DvClCF22uq0m2k7xiku5995V+GGN4Z8LqFLOsLLD5vndZ5jjKU42nRTio06UvFNQSbTs1KUk90f3tFcuvB3QGPhnGndG0FmFF9VLE4qrUxM6b8HD3kmoteaSfqfRrJdz7niXivTZdJj0HQY3HDe7b611uU1qeiiVmSAfmsvVhdIq3ctZdyGvJGNmlHvsh0slra9ncjf1/QYqXSX2KNpFt+1mRa/gapUgs0rFTFS7R1Ih7sWfkCVbQ90R0ssVbfgGi2Ye7IAXKN2Kt3JbVioVJoKvuWeyK7sm963ehK4JWz3Dtu7mKikij7mkijXpuFRUkWfkN/UK2NNFH4ovuyjTu9mTZpSErEjsDFBR73LlXb84bFe2zDd9kLIOy7dwuI6WUs0ab+v6CplkilTNq6aRo9mVa8jFy6ZNNOzMpqyuzaad+zM3vs0TXXGspdzjy7HKaVzCcYpE12xrGXcwbscmSVzGcUlsiK741x5xbbkntYycWzkyS6X8xg9kTXoxtYzi7vdGckzae9zOS9GRXoxrjzW77GT27m807vZ/oMZp+TJrtjZ6M5tWbMG7nIknZ7P9Bg42ts0S746Yza3OuvPv+5o1B/peB+0wOxck23szrtz8JLln1Btv8LwH2mB16b9dj9MeHxq/0dz/ALt+x+49iZ8guu/pe/sWHPRU86vYmfILrv6Xv7Fhz0VP1D+EAAAAAAAAAAAAAAAAAAA8b/aWcbeHmjebnK9ecGMRqLJeL/D2vhcPnGMlgqKyvGxVCNSk1P3vvJTjTq+4qRlS6KlN9N7RXX+01d7ZLSOueEmeaKzjgvm+GzvP9PYrKsRiMNmNJ4Wnia+GlSlUgpLr92pTbSbbts23u+/Ov+UDlm4o6qxmt9f8GdPZzn2Y9CxePr0pKrX6IKEHNxkk2oRjG7V7JK+yPz/4P7k0/J80v/5lX/2wPMPkb9ovpjlD4N51w4zThlmmpcfmOoq+eUq+HzCnhqMYVMNhaPu5OUJNNPDyd0mvjJebJ0bq3iv7TLnd0jqbNtOfc3TmmcRhMRi8Ng5TqYbKMow1Z15QnWaV61aacFOyblNWiowsvT6hyDcnGHrQr0+XvSjlTaklUoTnFtecZSaa9Gmj93k2O5fuCWOyrhRkWYaA0Ljs4lGWWadw9fB5dXx0py6FKlhk4zrSlKLjdRbbTV27gfRQAAAAAAAcfGYvC5dhK+YY/FUcNhcNTlWr1q01CnSpxTcpyk2lFJJttuyW55ocwPtm8j0zqHG6Y5f9AYXUtHBVZUf2QZzXqU8JiJLZujh6dpzp37TlODaW0bNM++e1P4g5roHk41PRyjEvD4jVGLwenpVYyakqFabnXirL9/SpVKbT26Zy8bH8P2Z3Kxwx4dcvuk+KmK0vl2Za01hg45zXzfF4eFWthqNRt0KFByv7qKp9Ll02cpNuWyjGIfN+Q/2lHGPma47U+E/EbRujMHgMXlWLxlDE5JhsXQq06tFRl8b31eqpxcepWSi02nfaz7ic0nHPH8t/BXOuMOC0JX1dHI6mH+EZfRxqwjVKrVjSdV1HCo1GLnFu0G0nd2SbXRfhT/26TXv+hYr+jcOegPMHkGX6p4EcRdO5rSVTC5jpbNcPUTXZPC1LSXk07NPwaTA/nctXH3TvMzwcyPi9prL62XUs199RxOX16satTBYmlUlCpSlKKSlvFSTsm4yi2k3ZfUzzl9ibn2NxXBDXum605Sw2W6ohiqCb/FlXwtOM0vJf3CLte123ZNu/o0AAAAAAAAAAAAAAAAAAAHj77bv5WeG30dxP2k9EuVdX5YOD7v8A8QtP/wBH0Dzt9t38rPDb6O4n7SeifKqm+WDhBt/xC0//AEfQA+oJWZIAEdPqSlYAAR0+pLvbYjf1/QA6fUkENvwAluxF/Qd9mibICpK3RNkAI6fUW9SXsiFcCCbepNkQ7gQ1YErfuTb0Ajp9SBdruAAAAEW9SQBCVgSQ722Ajp9R0+o39f0FgK9PqLW3uG2hv2swHV6ECz8i1ogVBPzdiAIbtsVL2TIsgIv6Bu+wa37AK2gN2AtfumZYbA3YWt4Ed+5mmquN7yv+Yhu5pt22sR0xfYxu2YLSja+xUKlCOn1JAUjp9R0+pLvbYjf1/QGy6Q1ZXK9XoX3ez7Doj5Gdvere1Or0IbuWaithZGEumfT6jp9S3z9j+PqrVOW6Vy94zGP3lWpdUKCaUqs14LySurvsk13bSeWyd668eGXJlMcZu1XVOq8v0rlzxuMfXVndYehGSUqsl4LySurvsk13bSfwbPM7zHUWY1M0zKt11Z7JK6jTim7Rit7JXfq2222225zzOsw1BmNTMszrddWe0Yq6jCKbtCK8Erv1bbbbbbf7fh3w7WP91n+oKH967Tw2Gmv8N4qc0/3nil++7vayl4ssrz3U9H6Hh4uPw/j8/Jd5VXh1w7+6DpZ9n9D+9VaeHw01/hvKc0/3nil++7vbaX1tqxDSS+L3ITb7vc9XHxzjmo+N1HUZ9Tl5sv8ACEoWV7mUo7s2b6u7uUklfsXZtxl2zjuy9vUhqyvFbkdUuz/2GWabLEkNXZIMVFJR7bkW9S3fuH32Cpaghq6ZPYBUumfT6klmvJFN7k31VLv0dWeZfnA1NwN4i09FZRpDLMzoTy2jjvfYmtUhNSnOonG0drLoX6WfZuA3EjH8XeE2RcRMyy2hgMRm3wnrw9CTlCHusTVpKzlu7qmm7+LZ/D4p8rXCnjFqeOrtZ4XM55hDCwwieGxjpQ93Bya2Se95vf5j9zw80Dp7hfo3AaF0pCvDK8s977iNeq6k17yrKrK8mlf405W8lZeB6uTPhvFjMZ+d8WYzLzW29n6OPYiS2buWWxE03HY8i1Er7lWrXLfGXgyAqXagJa8grX3CpdIavuC1olXe2w9fRSJdysl1bdi3f8buQSqXaErKxDd9ixVrYNmkES7EgKVttcgtslYWiZraoqVbuWdt7FTLNKmkENWLPYpdsxoRIkmyff8AMFSswWaW9iLPyCptBHb1uSO5lipUX9CrdtizXkUkt+xKodXoSncrZ+RO6CpUPe5VqxYWTCpdKFW7lhaPiLKpF77EPYDuSqXaJdijV9y5DXkGy6V6fUdPqE2yQuXSrVtyC0uxUy9rtU7ol2IJl2IJbKFWvEtZ+RV3u1YKirdh1ehPcq15BW9J6vQq/MFrKw1tUu1O/pYr079yz27FW9myGxJVrxF5eBPhuFxRq+4tbe5e0SsuzCkdXoRb1Fn5Bt+IVLtDV9yLeobaIbfiSqbVl3MGrm7TuZziktvMy93TGsZbM48uxypJXMZQj5E2e53xumLV9zKcbq1zeSs9jFtWJsdpd+jCcLXV/AxlCyvc5E/xmZzimtlcix3xrjuPd3/kM5G81a6sYTUttmTXpxu2cuz+YwkbyTaezMZJ+TIrvizl4mMzdpb3RlNLa3+0muuFjjzdrnXTn5d+WfUD/wDC8B9pgdjJR3e2x115+0lyzagt/wA7wH2mB06af7bH6Xj8Zv8AR3P+7fsftfYmfILrv6Xv7Fhz0VPOr2JnyC67+l7+xYc9FT9Q/hQAAAAAAAAAAAAAAAAAAOqfN57Q7hLym5rQ0djspx+q9Y4jDrFPJ8BVjRhhaUr9EsRXkmqblZtRjGcrK7STi31X/DnL8l3/AO3b/wDUD5zqfhjpfip7X7ONBcc6Mq+T5pnc63wTETdOOOp08rVXA0HOLVoSUKCsneSXR3lden+H5R+VfC0KeGpctnC+UKcVCLqaSwFSbS23lKk5Sfq22B0G/DnL8l3/AO3b/wDUDqVxv51v25ea/RHM/wDtafcf9huIybEfcP7s/CPhfwDGPEW+Ee4j7vrv0393Lpte0ux7afencrH5NPCr6m5d+pPMLnD4W8MdMe0y4P6C01w50vlOmczx+k4Y3JcDlGHoYDExrZrKFVVcPCCpzU4JRkmn1LZ3WwH37gD7Xanxx4yaT4S1eX6WRrVGPWA+6MdVfCnh24yal7r4HT61dJNdasm3vaz9FD51pvl15ftHZ5hNTaQ4FcPcjzfAyc8LmGW6YwOGxNCTi4twq06SnFtSabTWza7Nn0UAAAAAA68c+/BPN+PvK3rDQ+mMNUxOfYenSzjKcPTb6sRicNNVPcpfvpVKaqU4p7dU4t9jqJ7PL2ifDDRHDLLOXzj/AJnV0pmmlpVcFlua4yjP4LWw6nKUaFdpN0KtO8oLqSi4xjeSldP0yzrPMl01llfPNRZxgsqy7CRUsRjMbiIUKFJNpJznNqMU20rtrdpHUbjlyA8qPOHjMZxO0vn9PLc/xtVwxef6Tx9DE4bF14pX+EUk5UpzSabcXCbunKT2A6pcM+NHCLBe1i1rxXxvEzTOG0bisDiZUM9rZpRhgqt8uoRShVlJRlJyjKKim22mkrqx9b57/aX8HMNwn1Dwp4F6opas1JqjBVsoxOYYKMvgWXYWtF0601VlFKrUcHKMFTbScupyTilL57ifYc5h90enB8yWH+At3c6mlJe+gr9ulYu0mlbe6v5I+68BvZHcvvCbO8JqnXWa5lxFzTBVFVoUcxoww+WxmneMnhYuTqNeVSpKD8YsD+n7JzgrnHCflhhqHUuXzweaa9zKeexpVY9NSngfdwpYZSX/AH0YTqr/AL2sr2d0d1SsVGKUYpJLsrWSLAAAAAAAAAAAAAAAAAAAB4++27+Vnht9HcT9pPRTlVduWDhArf8AELT/APR9A86/bd/Kzw2+juJ+0nopyrJfev8AB/6Baf8A6PoAfUb+hBZpeBUASlfcWfkN0At6kFld7PsLRAqAWSXiBUFmkioAAmwEJ32A6beAAN2A7kO4Et2I6vQLfuTZAVbuC1kLICqdib32sTZLuRt4d/nAhqwG7Fn5AEr7k29SN0G34gAABDdtiOr0JsmLICOr0HT6k2iSBW9trBu+wfcJb7gQCXsw0kBBF77EiyQEXttYdXoTZFbPyDdptfe5WUe25O/azDb8Q2XavT6kpWJ2t6kO/gTY1D3uuxHTbxvctt3fcbMw38GbXiQS920nsGrWCp86CG7bEiyYVKjq9B1egsiHbwCpdDhd3v8AyEWttcsuyP4mq9U4DSuAeMxb66s01QoRaUqs14LySurvsk13bSc2ybt9HTjwvJZjjN2mqdUZdpXLnjMY+urO8cPQUrSqyXgvJK6u+yTXdtJ/Bc8zrMdQZjUzPM63XVnsktowim7Rir7JXfq222222753neY6gzGpmeZ1uupPaKV1GnFXtGK8Erv1bbbbbbf7Xh3w6+6Hus/1BQ/vXaeGw01/hvFTmn+88Uv33d/FspeLLLLmy1PR+h4eHj8O4vPyd7VeHnDxY50s/wA/of3rtPDYaa/w3ipzT/eeKX77u9tpfWbX3JsgerDCYTUfG6jqcupz82SpWXcu0kRZMtxlIx37kSjux1SW4TT7hWtKt2KON7yv+Y1nGNtt9/Ara21hrZKonfYkWS7AhU7oauQ9mWIaW4VKhu5As/Iht37MK2JWIfclX8RZCql06Dc73F7ihofjNSyXR+vc7yfAPJcNXeGweLlSpuo6lVOVk7XaSTfojsxylakz/V3L/pTUOp83xWaZnivh3v8AFYqo51anRja8I3k93aMYpeSSR+x1Zj+EmGzONLW2M0jSzH3SajmtXDKv7u7s7VH1dN728L39T+5p2rpuvk+HraTq5bUyqXX8Hll0qbw7tNqfQ6fxfx+q9vG997np5eWZcOOEx1fiyS+a3b+kL72Ib3Jt423PG6y7Gr7mbVmzQrKMd3tcNmveoVt6l3sym/azDZdoBIVr7hUukNX3Kl3syLIm91KgO19gFTaGr7kNW3LFbsKij3ZPT6iy723IvLxDUNeASsTu9x2Mq9qPe5Xp9S7S38yrv4Eql2h7MN3G7I7Bu9ItvckEO67XCkPuQB2MqoES7kiyZLZdKEd/Swd7tW2+YboLl0W9SCVd7PsTaIbFOn1Kyjv3JvIBUUSuT0+o7fi9yLsmzSpdIbsR1ehItHxCkJXIe1yd0QFxVu+xBZrYqNbVOw1ci1t7kh9iGxHV6EPe4AUq1Yq34F2l4lZJXewVLtQm+1iUvMgLlQVlHZl+xV33VibNNl0qlYB3XmSYpVqxBayZV+QVLtHV6EX9CbR8RaIVFSL+he0StkZZpSjdtikl1bdjVqN9/wDaUaSMXKxnGztcylE3n3MZE12xrGpHdbmDjZXvc5E+5lJO26Jr0Y1hKPd38DNuxtJOz2fYxlfyIsdsbtlUlu9jGbuazTu9mZST8iK9GDOXiZTdjSSld7O3zFHFPuia74sZu9/UxlH1OROKTa9DKSirXJs2743TjT2udc+fv9zNqD/S8B9pgdj5xjZv1OuPP4l97JqB/wDheA+0wOnTfrcfpeHxn/y7m/dv2P2nsTPkF139L39iw56KnnV7Ez5Bdd/S9/YsOeip+nfw0AAAAAAAAAAAAAAAAAAH5HN+GPCvMdU4fiRqDh7pTE6jypKph8+xmU4aeNwqgnaUMTKLqU1FXs1JWVz49rz2iHJtw6x9fKs8445Ti8bh24yo5Ph8Rma6l3j7zDU5001ZppzVmrPfY6Y+0747cU+LnHTKuSLgzVxU6dV4Slm2EwlVU5ZnmGJiqtOhVndJUKdKVOcrtRvKTmv7mmuZgfYz6L0jwwzjVXE7i1nmYZ9lmT4rMJ4bIqFHDYSnWpUJzVPrrQqTqRUlG8rQbSaSTaaDtxov2kXJdrnGU8uy7jdl+X4qrLpUM4wWKy6CbdlerXpRpK9r/j+O9j7Xi9CcKtc51kvEnG6O0pqDNsvpwqZNn9XLsNi8Th6d3OEsNiXGUoK8nJOEkrttd7njPyNez70hzgcIdVawzDiBnGm89yfOZZXg3Rw9LEYRx+DUqkZVKT6ZtqU2naorq1kmrv6Tyqa841ez85q8v5S+MOY08XovWGNo0sLKNWU8LTqYqTp4bH4Rys6cZ1kqdWLSStNtNwTYevoAAAAAAAOoXtUdH6v1hydakpaSwtfFfcjHYLN8zoUYuU54ChNurKy7qDcKsrppRpyfhdfkfZA684dZvyv0tBaex+HhqnIMzxuIz/BOaVaTr1nKhiVHZum6KpU+pXXVSabT2OwHOHxE4p8KuX7UeteDeko6k1ThJ4Slh8BLLquPUqdXE06dWToUmp1EqcpOydl3d0mjxEweA5stN8VJ8YeGnBTWXD/PpVnXhDTGl8wwuFpOW84xozjOPupPvSd6bW3SlsB/omB4rffwe1T/AM1dUf8A2OV/Zh9/B7VP/NXVH/2OV/ZgPakHVn2enFbmK4vcJM91BzIZLjsBneF1DVweAljMm+5lSrhFhqEtqPRC8VOc0p2s22rtxdu0wAAAAAAAAAAAAAAAAAAAePvtu/lZ4bfR3E/aT0U5Vlflg4Pv/wCkLT/9H0Dzr9t38rPDb6O4n7SeivKq0uV/g/v/AMQtP/0fQA+oW9SC+3mLLyAjq9B1ehNl5DYCL32sOn1Hhta5G/qBBNvUWfkSuwDp9SLepL2I7vuBBN/QtZeRVrfsBN77EW9R+kd33Agm3qS15Ii7ANWIJ7kAG7Al2ZABq5Frb3JAEdXoOr0JsvIWXkBHV6C99rE7Dw2tcCrVgN/UWfkASvuQT+kAV6fUluxIAgNXJIfYCOn1IJ39SbLyAi3qGrbh3vtcb+oEAEu1vACCrdyxDXkgIJ6fUjf1G/qNbVKhx77lWrGm1vAr3JsaolvctKN7b2sS1t2EfG9zDuzcbeJBeTVna1ygVKlxuvnKuFvEm7P42qdVZfpTLvhmNfXVndYehGVpVZLwXkldXdrJNd20mtmM3fR148MuSzHGbtV1TqfLtKZc8bjX11Z3jh6CdpVZLwXkldXdrJNd20n8IznO8x1DmNXM8zrddWpsoraNOKvaMV4JXfq2222225zvO8w1DmNTMszrddWp8VJXUacU3aMV4JXfq222222/23Dzh38O91n+f0P712nh8NNf4bxU5p/vPFL993e1lLwZ55c+Wp6P0XDxcXhvF8pyd8qnh5w6+6Hus/1BQ/vXaeGw01/hvFTmn+88Un+N3fxbKX1hqw39SHe/ievDinHNR8bqOpz6nPzZKhK+5a3oQ077FPPv4KdPqOn1LFXe/iFDhdWuV6N7XJu/Mb9/EKl2dHTve5El3I+N6kpN+dgS6rNK46d73NGttkUlffuZpc7oBXf1JV7bkqlSVa8SwDZWbdhfa5Z2ZVd7W2Cpduh3OxwU4rcQOMlLPtF6FzPN8vWS4bDvEYamnBVIzqtxu2t0mn+dHZPlQ0tqHRXAHS2m9VZTXyzNMF8O+EYWvFKdPrxtecbpN94zi16NH8/jHzc8NuCGroaM1ZkepcXjZ4OnjVUy/DUJ0uicpJK9StB3vB3Vrbrd+H0PhdxGyPi3oXLOIOm8JjsNl2a+/wDc0sdCEK8fdVp0pdShKUVeVNtWk9mr2d0vZzZ8mXBjjljrGe8xkmV1e79Ra+5L2REr37NC916nhvZ0l0hu4tfYlLbdE7ehivVnJWdr3ILvfsrlA2W1DV9ypcjb0ClQ1cl9yBe6to6fUgsQ7W8CVT5lW7bEJ9Tt2LEWXgkGqvYiXYs0yAtW/oT39LDb0Dt4MNm1XHvuVasWFvMmzSpdKlZFn3I2MUqtiW7kMBUu0W9Sso9tyXdX7j9IVKra29x1ehPoxZeRl9FKgNO7BKlW77Dp9SxF15hUVKtWLENO4VPgpa25D3LW8xt6CzalSGr7kglUu0NXKvYnf1Hj/WFRBDV2WdreBAVLpVq25VvwLb+pBl7d1KlvAbehV+S7EtisiC3zh2s7WCpdKN2I8bkyIs/IKlG7kFkisl3sjLpUVbuCUlbco73fclUqxR9yy7bjb0CpdM5dyen1LStfsiHe2wUq1YgkgVUuxq+5n0+poUfYlUUlC7vcxlHY2le5lJNLszK7Y1jKF33M5O67G8luYST8mRXbHuynvcxkrLubSTu9mZzTt2ZNd8awntcxlv6G7T6t1+kpUUdrW8SL3ejG6ceas2ZSOQ7Wd0r2MpJbbE2O+FYTXd+hhNXOTK1nsYyS22Jsd8a481eLOuPP3G3LLqF/+GYD7TA7ITXfbx8jrjz/AH7mTUHh/feA+0wOnTfrcfpeLxi/0dzfu37H7H2JnyC67+l7+xYc9FTzq9iZ8guu/pe/sWHPRU/TP4eAAAAAAAAAAAAAAAAAADyh4MYeivbPaz+7UL4qGIzepgnVd5KTwC6XH/8A0yml/wB62emHF75Jta/R3Mvs1Q6WcxPKdx7w3PxoXmv4Dacwmb4GVXAfskU8zoYSpQ91B4XE3VWUXKFTBOME4Kck1K8bWv3d4j5Vj894e6oyPK6HvsZmOTY3CYandLrq1KE4wjdtJXbSu2luB0O9iZ8guu/pe/sWHPmftUM1y3WfN5wT4daQqwxGq8BPC0cTGglUqUqmLx1J4WlJPbqXTKai3e1SLaSkm+u3BfKPaZcHMjzPhpwc4b8UtMZbneMlisZCOjnSU6/RGm5rGV8Pen8WEUnGpFbXW92dx+Q72c/EPRPEuHMjzQ494nV2HqzxeV5TVxyx1eOLqRali8ZXTlGdRKTcIxlK0mpOScUgPSMAAAAAAOqvtD+bXH8qXBuhjtJqhLWmrMRUy3I3WgpwwyjHqr4twe0/dqUEk7rrqQbTimmHZPPtWaW0vClPU+pcqyiFduNJ4/GU8OqjXdRc5K7XocvK82yrPMFTzHJczwmPwlVXp4jC141ac16Si2n+ZnhJwq5C+cLnIwEuM2ZZphKeGztyqUM81nmtdVcwSduqmo06tV073UZOKg0vitpDWPAHnZ9nLnWD4o5bmksFlccTTpVc409j54rK8RO6aoYujKMJOErJJVqai20k+pKwe84PivKFzIZZzT8D8o4p4TA08vzGVSpl2dYCnNyhhMwpKPvIxb36JRnTqRTbahUim20z7UAAAAAAAAAAAAAAAAAAAAAAePvtu/lZ4bfR3E/aT0U5Vlflf4P7/wDELT/9H0Dzr9t38rPDb6O4n7Sei3Kr+5f4P/QLT/8AR1AD6glZkgACGr7kgCErMkAAQ3ZkgCL32FvUkACG7MkAVbvsF3LWXkRZLcCSr7k3QsnuBCVyen1JAFWrEEtPxJTXiBCVxb1Jb8mVAAAA1fci1t7kgCOr0JFl5ACJdyFuWD2XYCOn1IF35izAAdgAAIAN2ZF77E7PckCvT6jp9SxXf1/SBD2JSuLq25AB7ESJK2b7gLeoasPjDdhu0Am3oTeI1tsu2Tjdv5yVCytcs13sj+LqnVOX6Ty543Gv3lWd1h6EZWnVkl2XeyV1d9kmu7aTi2TvXTjwz5LMcJu1nqrVOXaUy943GS66s21h8PF2lVkvBeSV1d9kmu7aT+D55neY6hzGpmeZ1uurU+KoraNOKvaEV4JXfq2222227Z7neYaizCrmeZVeurPaKW0acU9oxXgld+rbbbbbb/bcO+HXw73WoNQYf+9XaeGw01/hvFTmn+88Un+N3fxbKXgzzy58vLPR+j4eHi8N4vlOTvlUcO+HXw/3Wf5/Q/vXaeGw01/hvFTmn+88Uv33d/FspfWJdyxKt4pfoPZx8c45qPi9R1OfU5+bL/CK9XoQ3ctZeRDW5bjLpJRuzt3L3RWTSe5kVO6se5LV9xdENNmaIo1bcgm0vEm21rbhSErhuz6f5Rutyr8X4hUsWatuUl3JUn4tk2urhU7M+n1Kt2djSzRV9O90v0Gajd90EEJ+ZJKpUNWILNrxKhUunWDmN5O8z478QYa3wmu8Nk8IZfRwPwergJVm3CVSXV1KpFWfXa1vA+x8DOGlfg/wsyXh1ic2p5nVyj4TfFwoulGp73E1aytBttWVRLu72v42PgXNLzdcSOB/E2novSeR6axeClllDGupmGGrzq9c5VE1enWgrWgrK193u/D7ly78R874tcHtP8QdSYXA4bMc1+F++pYKnOFCPusVVpR6VOUpK8aabvJ7t2srJezlnN8jj5/0fcya8116vosu5VfjF2tyrX6TxWOs7jdtiLX3uSltuHsiVTaL22KvsTu9yfi+S/QGqJX3ILtrwKO9tgqKtWCVye343cgKl0hx37lSwsvIzVbLtUEtblLSRi5dpI6fUlXtuHug2XSr2uVSuWIe3YKLW3uQ3cd33Ja8jKqXbNrf5yHC3iWadyGn4kqlUJTsLWd/AO3gFb0h73Kp2LCy8gpUhu2xZ9ioVLtF/QgtZeRDXkTWy6VfYqTZ39CdkYpF/QglLzD2C1b32KuO/cl28CDdbVFQnbYlryIJs03egq14liH2MUo1ckAKlkQnfYiXcsQ0xra5ranT6kPYmRCTZDUNXIt6l2vIhxdgqdlCL+hNmiGtnsFbOr0Iv6CzQ7hUVl3ILSi7lbMmql0hq+5FvUsDFKAtZeRUKl2i19yLeob32JuguVUjp9Sz3ZTdGXTYSjv3Mp9vzmpnOLsYuX4sZdzKWyNpRafYyktuxNejGxhN3bM5djWad3sZyi7EV2jCfd/MYTOVKO26V7GTUfFImx3xrjSXcykcmfSm7JdjGfTta36CLHowrCXZ/MYSORJbszmk7WSIu3fGuNJ9zrlz/r/cx6hd+2LwH2mB2Qkkr7I64+0At97DqG3/ADvAfaYHXp5/tcfpePxi/wBH837t+x+t9iZ8guu/pe/sWHPRU86vYmfILrv6Xv7Fhz0VP0r+IgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeSXtoliM44ycItNV8VOODq5ZiVCKSfu51sVCE5LzbUId9vir1PW08mfbO5Dq18VOF2qck0/mGKw+FyrEwhiaOEnVpLEQxMZ9DcVa6Ti7PdpgerOT5RluQZTgciybB08Jl+W4elhMLh6StCjRpxUYQivBKKSXoj8nxw0blfELg5rbRGc4anXwmdZBjsJONTsnKjLpmna6cZWkmt04prdHjx+FG5+P+b5d9VF/UY432nPPfmGDxGAxWFy+VDE0p0akf2K2bhJNNXSutmwOwnsO88xWI05xe03Ny+DYDG5Ljqa6rr3leni4TsrbO2Ghd3327W39Qj/Ovy48xfMjyrfsh/akyieF/ZP8ABPuh8NyOWJ6vg3vfddPUvi29/UvbvdeR9o/Cjc/H/N8u+qi/qA9wAdU/Z0cfONHMRwdzzWHGzLaNDMsFqKtl+Cr0sveDjXw8cPQn/g33cZzmupbO9u8WdrAAAAAAAAAAAAAAAAAAAA8ffbd/Kzw2+juJ+0notyqJPlf4P3/zC0//AEdQPOn23fys8Nvo7iftJ6L8qn7l7g/9A9P/ANHUAPqXSh0oO9tiN/X9IE9KIezG/r+kdvxu4EAteIvECpKV0LMbrYA0QTuxZgQAAABNmBXpRIAEN2Cd2FfxFktwDSZBN0Rb0AJXJ6UIokCOlEDdCzAgFu2zQvECoLXiVAB9gHugKk9TIsy1l5AQlfdhoh7PuN2AIauTZktpgVD2RIAr1MsRZeQugIfcgl3e9hZgQS1YWZHcCH2ESSHfwAl9ihNpH8bVWqcv0nlzxuNfXVndYegpJTqzXgvJK6u7WSa7tpPLZjN30dOPDLkymOM71GqdVZdpTLnjca+urO8aFCMrTqzXgvJK6u7WSa7tpP4Jnme5jqDMamZ5nW66s9opbQpwV7QivBK79W222223OeZ3mOocxqZnmlbrqz2UVtCnFN2hFeCV36ttttttv9xw64c/dD3WoNQYf+9dp4bDTX+G8VOaf7zxS/fd38Wyl8/LLLny8s9H6Ph4eLw3i+U5O+VRw64dvMPdagz/AA/967Tw2Gmv8N4qc0/3nik/xu7+LZS+tNKXfwLNbdiIo9eGEwmo+H1PU59Tn586q4pXKmrVzOSs7FuMuxqyIAC1RZS3Zay8iJbPbYNl0rJKO6I6mWs36jp9DN67VSrRBYO3iY2Vndsi1y8rNbJX+YrZhXorKKitm+5MXsT477ja/oFS7VaTKyirvuXco+BV3vfwDYo4pdmyC7a8Sr7trsTYpDSZDVkWW/YrZ39DFS7fKuJfLLwg4uaijqvXGRYrGZjDDQwqqU8fWor3UHJpdMJJXTk97X3P2egdB6a4aaTwGitI4Sphspy73vwelUrSqyj7ypKpO8pNt3nOT3e17dkdE+fLXOtdO8cKOX6f1hneWYV5HhanuMFmFahTc3Uqpy6YySu0ld2u7I7T8neaZrnnLjpHNM5zLF5hja/w/wB7iMVWlVq1LY/ERV5ybbskkrvZJLsj2cvFnjw45XLc+DJZ5tafZiLJkpNdw722PG6xSTs7Id1v4hrzD7bEKglYhqyG/r+km6CtqWTHSi14huNg2XVZyirlDR7spZoKQWaKkiqlQRLsGmx2/G7GXbYgBtX2BilX3KtJlrbiQVLtTxLEWd/zi6C4iXco22XbuyLLyMsbLpQdKLvsUd/AlUu1QAFRVt7oguRsgqVRO7JIad215kq/iZVKvuRZMvZLcq2rkql2r1Md92LMWl4BUqHBLzKtWL2ZAVLpR7Iq3cWl4izMu1C3YfkAYqXarRBZX8SH3CogPsQ9gn5hUulWkyL22RdryKtdzLFSo6mRdslEtbPYlsUaTKvxJ39f0iz8gqKrfuGrbokizvuFxDdyjb3RpeJVpXfYyqZthO7LtXfZFWvQlUSUJ39f0kWYVEdKDVkGmwrrd9g1UiXYveJWUXYyxapWey28y1mQ1dGKYyd3uZSNpxbey8DKUWTXbGsZru/Qxl2ORJPe5nNK3ZE12jjy7S+YwkcqSW+yMZpO1kibHfCuNPdsymrHJmleW25k/WxFj04Vx5eJlN2ORNK7tYykl5Iiu+NcZ7tnXH2gSS5YdQ2/55gPtMDsjOyvZK51v9oH+5g1D/pmA+0wOvT/AK3H6Xj8Xu/D+b92/Y/V+xM+QXXf0vf2LDnoqedXsTPkF139L39iw56Kn6R/EwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAePvtu/lZ4bfR3E/aT0W5VW1yv8H0vHQOn/6OoHnT7bv5WeG30dxP2k9F+VWz5X+D+3/EHT/9HUAPqKd2WAAr1Md92TZeRIFWrIguReIEdTCV92TZeQukAskH2D3WxFpAQSldCzJXYCqdibtk7Ih2AgE2Yult4gQBZsm3oBXpRJa8SoAi7uSAIaTF2tvIO/gEtuwDvuw1ZDZbC6AqCXuybxAqCXuyABHUyRZeQEJX3Y7bokN2AjqZBN4kWYAE7Wsx32SAgjpRNmS2mBUN2JI2YEXb28yelBrbsRv6/pAgE2Z/G1TqrLdJ5c8bjn11al44fDqSU6s14LvZK6u7WSa7tpPLZjN30Xx4ZcmUxxm7Uap1Vl2lMueNxr66s7xoUIytOrNeC8krq7tZJru2k/gme55mWoMxqZnmdbrqz+KkrqFOC7Qir7JXfq2222227Z5nmP1FmFTM8zrddWeyirqNOKvaEV4JXfq222222/2/Drhysw91qDUGH/vXaeGw01/hvFTmn+88Uv33d/FspeHPPLqMtT0fouDh4vDeP5Tk75VTh1w5+6HutQagof3rtPDYaa/w3ipzT/e+KT/G7v4tlL66krEWfkSttmezDjnHNR8Tqeqz6rk82V7e5BDdi2z7FWvNFa7OCE7sOKbu2yXsuxCew9xFGrIg0lZIo92SqVBFkyQFqp2HUye+yRFmNKlS0VaW7LNpLcrt3Rndqj2RF29vMu1fsiGvTcxUu1Wh07bizIDZdKtWJXYNN9iUvAKVkltuyrimaNeaKNq7M0qUSSvu9yGluRK7t0t+u4u/Eyqlj8fqvi9wt0PmiyXWGvskybHulGusNjMZClUdNtpSs3ezaaT9Gf3NPamyHVuTYfUOl83wmaZZi+v3GLwtVVKVTom4StJOztKMk/JprwOmfOTy4cZ+KvFylqfQOjfuplscnw2FlX+6OEoWqwnVco9NWrGWykt7Wd9nszsVyuaI1Nw54E6Z0brPK1l+cZd8N+E4b31Or7vrxtepD49OUoO8Jxezdr2dmml6eTiwx4pnMt34Eu7qx9S6mSGlfZEJNP0PKuXZZMrLYuRt2Ms2qKrdEdKLSsvQrv6/pMvZUQ9mS4pkpbbld+92YpDikUl2NCgVFC3SibLyK3fmFS6HsyGrk7vcWaFUo4Ld73RVM0dt0V6beBKtqt2YbbLEWXkGzsq+xQu+5GyChK6IasWIkFKXb2ZDVi3zdyrT8SFRR9wWvEh28A0asitkyRdILVIbsS1422I7hUu0XvsyJRVy2y3Kvdk1UV6mOplnF27Fe2zRioXbIJaViApQhsu2mVCpdqgltMgls2lorZMSu/Fkb+v6QpDVyLJb+RNmgHSVXqZD37lmvIjsG+qr27EXbLNXK29CGwKt90XvEh+NkFM27EXbLNeaItZ+gWqC14kWfe2wV7lWyH5lrorLxsQ2IW6D7BbIkLUIfYs92VV1u+wUJCXYXRFpeIvdU0q2GhLZkWZN7Kisu5hLsbyTb2/2mMoyt2MrrjWMtkzGbutzkST3uZySS7L9BljvK40m02jObsbzSu9kZTV/A5u2NjCaunLxsYvfucia73RjNLaySJsejGsJqzZlI5Ele7t4GUkttiLHowriy7y+c64e0CS+9g1E/LF5f9pgdlJKKvsjrd7QS33sOobf88y/7TA6dP8ArcfpePxfL+j+b92/Y/UexM+QXXf0vf2LDnoqedXsTPkF139L39iw56Kn6R/FQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAePvtu/lZ4bfR3E/aT0Y5VFble4P8A0C0//R1A85/bd/Kzw2+juJ+0noxyqJ/ev8HvoDp/+jqAH1ICzAAAlPYCo6UX2exUARZMkWYEWSJFmgAAKtbgS1cWSIS3LPcAVa7sWZN/UCIkvsQ9+wswIBNmLMAlcgmzFmBAAAiyY6USAI6UOlBrcPdAOlBqyIsxZgQAXAoGrkvdkAR0okFbMA+5Me5Oy2Ie+yAN2ZAsxZgCEkibMWaAFeplj+LqrVeW6Ty74bjZe8rTvGhQi7TqzXgvJK6u7WSa7tpPLZjN30Xx8eXJlMcZu1GqtWZfpPLnjca1OtO8cPQUrTqzXgvJK6u7WSa7tpP4LnmeZjqPMamZ5nW66s/ipLaFOCvaEV4JXfq2222224zzO8x1FmVTM8zrddWeyS2hTgm7Rir7JXe3dtttttt/s+GOg6GcW1BnVPqwlKo40KEou1aateUrqzgntZXTaaeyafhyyy58vLPR+i4eHj8N4vleTvWvDrh18PdLUGoKH967Tw2Gmv8ADeKnNP8AeeKX77u/i2UvrZa/qRZns4+Occ1Hwup6nPqs/Nl/BVuxDdywLedVdyWkyLO4sw2XSGvAjpRazIlF3MsVKNXVjOSSdkWsyy2RLZWQJ6X5EBsqLJEkt7EBatk9mVfxXbyLWYswqXaqZNkybMq07k3u2IlEq1YtZk7JBU+lRNoq5NMs2E1awVKq5N+RDipbtu5dNLuQ2rhqiSRRv41jWRV9jLNql974Hxt5wNE8DdZw0XqDTGd4/FTwVLG++wfufdqE5SSXxpp3Tg77W3R9L4TcSMq4u8P8q4h5LgcVgsFm3v8A3VDFdPvY+6r1KTv0trd021ZvZrx2PkfH3k5yvjtrmGtsbrvFZPOGApYFYengI1k1CU5dXU6kXd9drW8D6twZ4aYfg/w1yfh1hc2qZnSyj4R04qpRVKVT3tepWd4JtKzqNd3e1/Gx6OT5GcU8n6TZbb8z9m9mQXIPKvapFkiZNXAUhxTd22UTuy7W5UyrlACFdb2JbLUPZlDW6Ie+yDZ2ZlDRp3ICordoXbD3YWzCoiyvcrdstLxsVsyVAD8iIoKiGu7KtJl32KhURe2yIbbJaYiFyqhpMs+xSRljVWrXISuXurFJEqncasitkyQFSjVyrViX2KhUCr2ZYXRlipdK3bKy7ml0Ve7JVFe+zDViLNCzCpUdKHSibNFZdwqK9KIasWe6K2ZlVAAGKit2yC0uxCatYKirdhZPdkgK96jdit2zRsqTrupUq3Zml0Ve5ioq22VfYtINqwVKzJu2TdFXuFQsmR0okPsTZpSr2ZBDW5N0Yo6UUfYuVadgqXaEiSoCpdEkmyl2y9mVl2Jqogyn2/OXa3Kzi7BeNYS8TOSutzeSaumZWaJrvjXHmrNmUjkz7v5jGTsRY741x57tmU127nJlvexk9u5NjvhXHku6MJHIqfjMykmiLHowrjz/ABWdbfaCfuYNRf6Zl/2mB2Vl4nW72g/7l/UP+l5f9pgXwfrsfpeTxa/0fzfu37H6X2JnyC67+l7+xYc9FTzq9iZ8guu/pe/sWHPRU/SP4uAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADx99t38rPDb6O4n7SejPKq/9y9wf+gOnv6OoHnN7bv5WeG30dxP2k9GuVO/3r3B76Baf/o6gB9Qu2Cz3RWzAnpQ6URZizAlq26IbuSrrexN0BUXaBZdgK3bBYX9QKkpXRFmOwABbMtdWAqRZMkm6sBVJIkB7gAVsxZgS3Yi7ZMUS9wKEpXQsyAJezIAAFeplgBC3QfYkAUJu2TdEWYEpXQaI7BbMCAS92QAsmLJAACOpklbMCy7FW2x2P42qNV5bpPLnjca+urO6w9CMkp1ZrwXkldXdrJNd20nlsxm76L4+PLkymOM3ajVWqsu0plzxmNfXVneOHoRaU6skuy8krq7tZJru2k/gmeZ1mGosxqZnmdZzqz2SW0KcFe0YrwSu/VttttttznmeZjqLMqmZ5nW66s9kltCnBXtGK8Erv1bbbbbbf6fh/wAPqupasc0zSE6eVU5bLdPEST3in3UU9nJeqW93HwZ55c+Xlno/R9P0/H4fx/Kcnqjh9w+q6kqxzTNITp5XSlsrtSxDTs4p91FNWcl6pb3cftVKjSo0oUKNKFOlSioQhBJRjFKySS2SSVkkTTo06FGGHoUoU6dOKhCEUlGKSskktkklZJE2Z7OLjnHNT1fD6vqs+qz3fQslv5DqYt6E3R0eVUB7kp7AQQ3YmXjYrZgSndkS7kFk9gqVUhskiXcy9+7ZUlZRVyUtw1uS2dqzBLexAVAhuzDV1sRZ+QXLpMm0rop1NvexaPcs9zO0bLtQhq9yZRdiri7GNiOlENWJig3t3ClJBJOxJDjLulsFS7WkltuzNrwLLbuH2Cpfc6W83XNHxY4O8VaektFY7LqWXTynD4txxGCjVn7yc6ibu3e1oLY7ActGvdQ8T+COnNdarq0Kua5n8M9/OjSVOD93i61KNorZfEhFPzd34nP13wB4QcTM7Wo9c6IwubZjGhDDLEVa1aDVKLbUbQmlZOTfa+5+m0hpDTegtO4TSWkMrp5blOB958Hw1Oc5Rp9dSVSdnJtu85ye7ffyPRycnHlxTHHHWU97ZuP7JBJWzueRcu0SirgmzICpUNlX2LkPdGWbVLpndos+xNmQ+xlmlRCRNkiE9gluYqKyk7sq1c1M29g2VR7Mgs1uVC4sldFXsgDKpVvxITuS+4MbKFWrFgFS6UfYiJdvbuUkFSj7FWkybMWaMtVLtVqyKtJl7oh79iVSqLuT0ohppsgLg14FWrFgGqEdKLS8W+xR7sKielEPZlir3ZCptW7Yu0S90RZhU2XbKS7l29iAqVQE2ZD2uFRHSiGrAEqiGrlZbE2ZHYKiE7kizZDTuFy6Q1Yq29y8URLxMu27UJsrEFvAlUrORBbsH2CtqdKHSiJdyboKQ9mQ+xZ77oq+xl0tUjpRNmVS3JbKlKxF2yxGz2CopZMhOxMou4ugpHUyGrlnvuiLMyxavSisuxaXcpO9jFRlPdszmklsaSi93bw8zNxbJrtixmu79DCZyKkZXe3h5mTTXcmu+FnqweybM3uciXiYzJrvjXGqbSdjKTucqSdn8xic674VxpeJ1u9oP+5f1F/peX/aaZ2Um+7OtntB7Pld1FZ//LMv+0wOvB+tx+l5fFr/ADDl/dv2P0nsTPkF139L39iw56KnnV7Ez5Bdd/S9/YsOeip+ifxkAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHj77bv5WeG30dxP2k9GeVV/wC5e4P/AEB09/R1A85vbd/Kzw2+juJ+0no1yp/uXuD/ANAdPf0dQA+op3ZIAAhskARe+zHSiQBHSiQQ1dgS1cr4kpE3SACyYIauwDRBPSyOzAnpRBYq+4AExJbW6AqAAIbsQndlgAIsmR0snst/ACHsyCydyHvsgILdKI6WT1ICHsyCXuyAI6USABFkw0GrsizQEFulFS4FWrEEvuQBDdmSDgZ5nuW6dy6pmeaVuilT2UUk51Ju9oRXi3Z7dkk22km0tkm6rHG52Y4zdrHU+f4TTGT1s3xUevotGnSUlF1Zt2UU3+duybSTdnax1+zvPMx1DmNTM8zrddWeyS2hTgr2jFeCV36ttttttvl6q1XmWrMyeNxr6KULxoYeMm4UoN9l5t2V3a7aXZJJf3OHvD6rqSrHNc1hOnlVOWy3UsTJPeKfdRTVnJeqW93HwcmWXNl5cfR+j6bg4/D+L5Xl9b/rSOH/AA/q6kqxzTNITp5XTlsrtPEtPeKa3UU9nJeqW93H7ZSp0sPShQoUoU6VOKhCEUlGMUrJJLZJJWSRalSpUKUaFClGnSpxUIQikoxilZJJbJJbJInpZ6+LjnHNT1fE6rq8+qz3e0QQ3YkHR5Vbt7eZPSiSvSwJ6UQ9mT2W/gR33QEAm1t2T1IClkyHsyxWXcAlciUVcE2bF7qlV6mOpl27oo0TY2KyiorZvuVLy+MrIWa2ZipVE7B79wAqVHbdEOTL2aK3V7CzaoqpN7NIEuLZW/Tt5eRlnwUdKKyS3LdSIbuY2bUJ/e39GS2kQ3cKl0o22St9mG0iykpbJO4V87z057lxFXG2gtLfsj+BfcPC/wDxf7/3XX7yrf8Awe17Wv49jtBygLOly66Seofhv3R/v/33w3r99/8ADsR09XX8b8W1r+FrbWP1GvOP3B/hnncdOa51vhMpzKVCGJWHqUK037qTaUrwg1ZuLXe+x+l0jq/TevtO4TVukc1p5llGP958HxVOMoxqdFSVOdlJJq04SW6XbyPVy8mWXDjj5dfP8Wyd9v663QbsLpEniVOyFuir3uXKPuFCVgAFobKXvsy7RUyxUu0dKJeyI6kOlkqlOpkOCXmT0so2G9x7Mq0SStmFswWk1uVCpZCyZVqxYJpk1sulSG7EvxZVyS8wpBDSZN77gLRdrbyIbbLEdSDYrZMh7dizj3a8SjTRNmlSKt3uQWIe7MVBqyIACpdol2aKdKLtrcrdILlCr2ZPUiU7k1sulSrdmWk7qyKGKlW6UQ9mOlkrZBUZ9TIe9yz3RXpd73QVFQTLsQTeyghpbskh9gpCbRAAVL3Q3Yq9+5ZtIN3Rlm1+nZTpRIbSIs3v5mVUqJJbbsr3+L4F1t3Ky7GNVlFXI6USQ2tws7bIggs3dBUu1GyCWiCFQIskQ0OlhUJdyrRLVigUtHsJuyIs2OlhUV77sq1cvKJR7om9lRSd1sZPZG7i7PsZOLSu2jK641jNvcwm2zepG7ZlKD23Ivo74+jGXiYzN5xd3ujKcW7WsTXfGxlLszBqxtOD3V0ZSg4Wu1uRXowsYT8TrZ7QhJcruorf88y/7TA7Kzi990da/aEx/wBy7qL/AEzL/tMDp0/63H6Xl8Vv8w5f3b9j9H7Ez5Bdd/S9/YsOeip51exM+QXXf0vf2LDnoqfon8aAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4++27+Vnht9HcT9pPRrlT/AHL3B/6A6e/o6gecvtu/lZ4bfR3E/aT0a5U03yvcHn/9IWn/AOjqAH1IE9nuT1ICoLdSHfdAQlcnpRHSx0sCACydkBUWTLNlQBDdmSAITJsr3AAjqZBYXSAiJLS3ZD37BKzAgFir7gAAAKtvdFm7gCI9hZIkhq4ElCelk9SAqCydw+wFQAADVwAI6UR1Mm6RIFQt2TdI/nZ7n2W6ey6pmeZ1uilDaMUk51JtO0Yq+7dnt2STbaSbS2SbqscbnZjjN2pzzO8t07l1TM80ruFKG0YqznUm07QivFuz27JJttJNr4FqrVeZaszJ43GvopQvGhh4ybhSg32Xm3ZXdrtpdkkk1VqvMtWZk8bjX0UoXjQw8ZNwpQb7Lzbsru120uySS/ucPeH1XUlWOa5rCdPKqctlupYmSe8U+6imrOS9Ut7uPh5M8ua+XH0foen6fj6Dj+V5fVHD/h/V1JVjmmaQnTyunLZXaeJae8U1uop7OS9Ut7uP22hSpUKUKFCnCnSpRUIQgkoxilZJJbJJKySJpUaWHpQoUKcKdKnFQhCMUoxilZJJbJJKySLHq4uKcc1PV8bqurz6rPd9EN2Y6mSR1I6PKgE9SD37AQCelkOLfZoCH2EexPYhO4ENvdEFn2KgS1ZFbJkgCOlEpWISsxdINl0qC3UiHuxe6le26KykXe6I7bMhsUaILKLW7aIfcNlLtlbK9yQFS7V6mQ0m7su3YhySC5dqdKIfcu31dr7FWrsmtl0iSW27KtWNDOX4zCoq0mQnZ7FiW7oKldLubrlb4tcZOK1LV2isDltXL4ZTh8G5YjHRpT95CdRtWava01uff+WjQGouF/BLTmhNWUqNLNcs+GfCIUaqqQXvMXWqxtJbP4k438ndeB+E4+85OV8BtdQ0PjNBYrOJzwFLHLE0sfGikpynHp6XCTuui97+J9W4NcTcPxi4a5RxHwuUVMspZv8ACOnC1Kyqyp+6xFSi7zSSd3Tb7K17eFz18mXL8jjMp+b8Wzs/ZdCb3uRaxYq4tNy2seOxYVaW5Yq1dkql2gE9LICpQhxTJIbugqXSkkk7Il7Ihom6ZNVLoTuUUUy5WzRilZJJ2RBcq2GxR9yelEvsUC5ofkEkgApV+Jm0mbPZNlOpEqiElbuyelDutvEhpoKiJdihZq5HSw1PgVaTJIMs2qIaVu7KPZl7NbsiUlclUVSuGrEEp2C5YrZMrKKuXluV6WFS6V6USlYENBSFuw47vuLNAhU7q9THUxZoJhsukENvcPdENbPdBcVl2IJ6WOlmWKGijb3LLZkt3RiozJSuiWrlQqXuiRUuAuVRpMi7W3kWbuVb7onUahtsglJsN7b+Bip3R0oo1u92We7ICoq9mEriXcJ2CoPZlC73ZQmzSgAGKRZMp0os0Vl2ColKxD2RFmyzdgqKt3KtFnuyrZlVFG3uUn2/OaPZNmU2mtjHTFlLxMpuxtJd2YzVya74sptttmTbZtKDs3dGUovYiu2NYz2bMp72ubSg7t7GVSDla1tiK9GFYTbs0da/aFfuXNQ/6Zl/2mB2WnFpO9tjrT7QqNuV3UN/+eZf9pgXwfrcfpebxW/zDl/dv2P0PsTPkF139L39iw56KnnV7Ez5Bdd/S9/YsOeip+ifxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHj77bv5WeG30dxP2k9HOVL9y9we+gWn/AOjqB5x+27+Vnht9HcT9pPRvlUV+V7g99AdPf0dQA+ptLcqWSsyQI6UO2yI6WStkATuySGrkdLAnpRDVh0sdLALdktWQ7bsN3QFQCbNgFuw1YdLFmgIFkwACSQAAjqZBYAVSuS1ZEtXIs1v5AQCepEAALNizQAjpRIAhKxF77MsQA6UVLdSKgATZsdLArZMkNWP5+fZ5l2nstqZnmdbopQ2jFbzqTadoxXi3Z7dkk22km0tkm6rHG52YybtRnud5dp3LauaZnW6KUNopWc6k2naEV4t2e3ZJNtpJtfA9VapzHVuZPG419FKF40KEXeFKDfZebdld2u2l2SSU6q1VmOrMxeNxr6KULxoUIyvClBvsvNuyu7XbS7JJL+7w+4f1dSVY5rmsJ08qpy2V2pYmSe8U+6imrOS9Ut7uPh5M8ua+XH0foen6fj6Dj+V5fVHD3h5V1LVjmmaRnTyqnLZbqWJknvFPuopqzkvVLe7j9rpUaWHpQoUKcKdKnFQhCMUoxilZJJbJJKySFKlRoUoUaFOFOlTioQhBJKMUrJJLZJJWSRfpZ6uLinHNT1fG6rq8+qz3fRALeBU6PKEdKJAEdKD27Bq7JSsBHUx1Ml9ioB79yErE2aAEDpQHUgKgE2bAJXIlFXJ6WQ1YCqVw9mLNEphs+Cr2RVu5o2VJ1VI77MpLZl+pB77oxsrMlK6Jl2KhcGrlbb2LtlWnuFS6VaUbW8fMhO7LxDTdzN/FSrdijSbuy3SyGtnujGyxVqxBLTQXcKjrxx45OtO8ddbw1tmms8yyuvDA0sCqGHw1OcGoSnJSvJp3fW1b0PqXB7hrguEHDnKOHOXZnXzHDZT7/oxNaChOp72vUrO6Wys6jSt4JHVHnM5kONHCni9S0voHWf3LyyWT4bFOh9zsJXvVnOqpS6qtKUt1FbXsrbLdnY3lZ1vqjiPwH0xrLWeafdHOcw+G/CcT7mnS957vGV6cPiU4xgrQhFbJXtd3bbfr5MeWcONyu8fg33vqb2ZWWyNGrFHvc8ip3UW6JF1HZi9yL2VLoKtWQaICpUNjpRLVgFo6UZp2Ly7kTVluZYqXYncNXC7Ed9kSuXZ0oo0WasHsrsN9FCGiZblbNBUoGrCzYaaCvd3Vu3s/HYhxS7NktNbkN3J1pUqCJE3SIe/YKiCG7FrNEBSpBa6RDdzL3VEPsUsmXauVasZZpsulAS092QYpLVkQAFoasioAVNjVyrViz3RCdtmQqKtXIezLSi7FGrBsqWiAlcPbuFS7Q0VLNlGrsLiAS2QSqUKvuWaaIbW6CpdKN2Iu2WAVO6gsmT0sh+RlixJIhq6IaaC7ktlVezDVkXl3KNPdhUUl3ILlWnuwpKRTpRIC1Xsyqd2Xl3K3TIVElZRLFAqCViJdg0RHuFS6QVW7NSj3RlUq14FJQjbxLteBR7K7MVLWU1bYykjWW7ZnJWJrvjWM7rYykbTfdehjImu+LOXaXzGLdjaXaXzGEiK74MZtu/znWr2hf7lzUP8ApmX/AGmmdlpbNtnWr2hdnyuaht4Y3L/tMC+H9bj9LzeKf1Dm/dv2P7/sTPkF139L39iw56KnnV7Ez5Bdd/S9/YsOeip+hfx4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHj77bv5WeG30dxP2k9HOVNN8r3B76A6e/o6gecftu/lZ4bfR3E/aT0e5Unbld4PfQHT39HUAPqIJbuiAABNmwId7bEb+v6C1miABDb8CQBHfZoWRIAiyJBF0gD2RF32JumLb3ANeRUt1IWb38wKglpogAQ2/AkAFfxAAEWiLRHSyAJ7fi9iG34kx7EvsBUAACu97MsAIshZEgCu97IXl4k3SP52e59lunsuqZnmdbopQ2jFJOdSbTtGKvu3Z7dkk22km0tkm6rHG52YybtM9z3LtPZdUzTNK3RShtFJJzqTd7RivFuz9Ek22km18C1VqvMtWZk8bjX0UoXjQw8ZNwpQb7Lzbsru120uySSnVWq8x1ZmLx2NfRSheNDDxleFKDfZebdld2u2l2SSX93h9w/q6lqxzXNYSp5VTlsrtSxMk94p91FNWcl6pb3cfDyZ5c18uPo/Q9P0/H0HH8ry+qOH3D6rqWrHNM0hOnlVOTsldSxMk94p91FNWcl6pb3cftdGjSo0oYejShTpU4qMIRilGKSskktkktkkWpUqNClCjQpwp0qcVCEIJJRilZJJbJJKySLHq4uKcc1PV8bqurz6rPd9EWQ39f0EkdSOjyou+wJe+6ITsBALN3RUAHfwAAi8iOxYq3cBd2BLd0QBXduz7E2RIAi0SO34vYNXY7bMBeRBLd0QAKteRYi6QEWfkLPyJ6kOpBW1GtiPQs3Yjv8bwJbCSRR7Mv32RSXdoxst2gABcQ9uxW87+NvmLJpkvsFS+5Qh3/MS2kO6Mqp2U7jZbkiS2MU/G6r4QcLtdZos61loHI85x6pRoLE4zBwq1FTTbUbtN2TbaXqz+5pvTeQ6QyfD6d0vk+EyrLMJ1+4wmFpKnSp9c3OVopWV5Sk35tt+J0A59dC631FxxoZhp/R2eZnhfuFhafv8Fl1avTU1Uqtx6oxauk1dXuro7U8nGU5rkfLdpDK86yzFZfjaHw/3uHxVCVKrC+PxEleEkmrppq63TT7M9XJxXHixz82/mVH2m9yrT32JgrrYS7M8qp2VtF99zPs7eBotkVcWm5bWMsJe4RstySGrkuku1QT0sgKlpZMq99mixD3Rlm1S6U3vZE7LcNpOzD3RKp2RLd7bkd++5ZbIqtmFeqrVir3Lya3KhUVvIXbLPsVTsFS6R6MWRJDaRlVFWopkJLwJlFtuW1iErGKQ2914EFn2KhUVad2FbxLFWmgqXSBZE2aKtGWKlGlbwKNeSLPyKrbuSqIBNm9/MhqwVEWRD2ZLd0VCp3Qm2TZAE3SlbtlWnctZoGKiu6IavuyWwnZBU0q1sUbd+zNCGrsLlUaSIFmgS1EmyLN7liG1ugqVCt4h2vsQAqVDv4FXf8AOW6kRZ9zLNrxqq37k2S7dw2kR1IlUqG7lXfdWL990Q9u4azd15kNvxLvdlX2C1Q727As3dBUZt3IskWaKtXIVKXXmQ1sOlkt2Coo7rzI3Rd7shuwUhboqW6kVFqhpblJK63TL3SIbJVNsnFK7tuYyVzeXaXzGEjK7YMpRTu7bmMkvI3k+7MZu5Nd8aylFb7GU4rayNpPexlNpEO+LCpFW27nWj2hkbcreovP4Zl/2mB2Xm0rs61e0N35W9RemMy/7TTL4P1uP0x5vFN/gPL+7fsf3PYmfILrv6Xv7Fhz0VPOr2JnyC67+l7+xYc9FT9A/kAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPH323fys8Nvo7iftJ6PcqdvvXeD30B094//AOOoHnD7bv5WeG30dxP2k9HuVNX5XOD30B09/R1AD6k0vDuRZ+RNrb3HV6ARZ+RO6HV6Dq9ACu9n2JtEi99rDp9QIJSXiQTb1ANJEE9PqLeoECyAAWSAAEWS7jfw7fMGrkgQt+5G1yxDXiAtEWRUm+1gDt4EEpXFvUCHfwFvQAAA3bYi99rALRDSQ6fUgCUkw0vAhK+5PT6gQA1Y/n55nuW6dy6eZ5pW6KUNlFJOdSb7RivFuz27JJttJNpbJN1WONzsxk3arn2eZdp3L6uZ5pW6KUPipLedSbTtGK8W7Pbskm20k2vgeqdU5jqzMXjca+ilC8aFCMm4UoN9l5t2V3a7aXZJJW1VqrMtWZk8bjX0UoXjQoRk3ClBvsvNuyu7XbS7JJL+5w+4fVdSVY5rmsJ08qpy2W6liZJ7xT7qKas5L1S3u4+Hkzy5r5cfR+h6fp+PoOP5Xl9UcPuHtTUtWOaZpTnTyunLZXaliZJ7xT7qKas5L1S3u4/baNGjQpU6FCnCnSpxUIQgkoxilZJJbJJKySJpUaWHpQoUKcKdKnFQhCMUoxilZJJbJJKySB6uLjnHNT1fG6rqs+qz3fRPzdiCy3RDVtzo8qCLIkhuwEkWS7i+1x39LAGl4FSbepAEkE32sQAFku46vCwauBDS8AkmLepCV9wJaXgQTb1IAFWncsAK/P2D2ZJHT6gQLJglK+4FbR8BaPiGulX7kN3Agh38OxIMsVLtC2e5EldtpEy7BP4tyWy6Zglu+xAXDsRvf0JIvvYLhaLKu3ZWsW6fUq479zLpU7+haPgVlezSLJWIa8TGy93ybiZzO8HuEOpI6U11n2KweZTw0MWqVPAVqy91NySfVCLV24Pa99j9poHXemuJuksDrfSGMqYrKMy978HqzpTpSl7urKlO8ZJNWnCS3W9r9mdZearlA4lcceJ1PWmk880zhMDDK6GBdPMcVXp1euEqjbtTozVrTVne+z2Xj905deGuecI+Den+HupMXgMTmOVfC/f1cDUnOhL3uLq1Y9LnGMnaNRJ3it00rqzfp5MOKcUuN3k19HT6ezsS0mvX5yJR37kX3seZaHsykm97GjV9yrXgZZtUqie24d7bES2ZYlsqu/r+gh3t2LkPsFxRPbcPZBq+4bsFKtXe5Ce5Z7lY9yatJDSRLdtipitlotXdr/OVaSJtd9w4WV7hsUbRCt4kuO/cdPqFDS8Ctr+Ba3qG7D0Wrsroo7+BaTvJkEtlUvJuzvb5h2LkNXCpdK72uR37lr22sVkFSjasVJttcgyqg1323KteZYrL41vCxKptW/qQTb1ICtnT6EWj4lr+hSXcLm0ANXBl0pW7BN77ENWJVFXbwIJtbcXvtYKmkC68yXC6tcq42fcKmlXuVLB9jLFKkNLxJSuQ479zFRDt4DZr1+cdPqQ9gqWIsl3Ib8E9iWrjp9Qua2q0vEqWlHtuRa29ya33o3Qb82T1ehDh3d+/oYqXar2ZW7vuW6fUhrwCpdotEqS1YX9AqIKysk7dyxFvUmqiq3QDdtiOr0MUNeRWXYt1ehVq4VNqlmth0+oauLNrm1WtuxR7Iu9rlUrmVUqkrtPuYyT8mciTtdGU3cy93TGsJp2ZhJPyZyZeJlN2JsejG1x5R3ezMpp+TORN3bMpEWO2Ncaav4M60+0Ojblb1Ftb+/Mv+00zszPuzrT7Q/fla1E//DMv+1UyuD9bj9Lz+J3+Y8v7t+x/Z9iZ8guu/pe/sWHPRU86vYmfILrv6Xv7Fhz0VP0D+QgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8ffbd/Kzw2+juJ+0no9ypW+9c4Pb/8AEHT39HUDzh9t38rPDb6O4n7Sej3Kmr8rvB36A6e/o6gB9T2ewsglZkgRZC0fEkhq+4EOy7dxv6/oJSsyQKWfkWXYkhuzAPZEX9Re+xNvUCGtiC0uxF/QCAS3fYgAAAHYi6ewauLeoCyDXkSAIXiTsQ1ci3qBNokNemxBN9rARZMWRKV9xb1AgrZ+RYAFsHsg3bY/nZ7n2Xady6pmeZ1vd0obRit51Ju9oxW127P0STbaSbS2SbqscbnZjJu1Ge57l2nsuqZpmlbopQ2ikk51Ju9oxXi3Z+iSbbSTa+B6q1XmWrMyeNxr6KULxoYeMm4UoN9l5t2V3a7aXZJJNVaqzLVmZPG419FKF40KEZNwpQb7Lzbsru120uySS/ucPuH1XUlWOa5rCdPKqctlupYmSe8U+6imrOS9Ut7uPh5M8ua+XH0foen6fj6Dj+V5fVPD3h7V1LVjmmaU508qpy2SuniZJ2cU1uopqzkvVLe7j9rpUqWHowoUKcKdOnFQhCCSjGKVkklskkkkkWo06WHpQw9ClCnSpRUIQjFKMYpWSSWySWySLHq4uOcc1PV8bquqz6rPd9Fd/X9BNkSDo8qu67EXbJb8AlcCCZKI6fUgARsiSGrgTsRaJFvUgCWt+w7E+A7+lgK2XkCbepAEO+6sFstySJdwJfYqFuT0+oEAEN22Ajf1/QN/X9BKd2G7bAQ15DdbFisu4EPfuRZEkN22AglJWuQ3fYX2sGy6Q1ch26WS1cq9mTqqlVtZ77D5uxZ/G2DVk0Y33qEWXe25JNtr3C4pdruQ3Jvs/wBBLdyV2F7Kiqv4p/oBaRUzVvdTq5zJc5WZcBuIdPQ+D0Fhc5pzy6jjvhNTHyotOcqkXHpUJKy6E738ex9j4FcTq/GLhVknEfE5RTyurm/wm+EhWdWNP3WJq0VabSbuqSfZWvbwufjOM3J/w0446vhrXVue6mwmOhg6eBVPLsTh6dLohKTTtUozd7zd3e2y2Xj9E4XcNsj4RaDyzh7pvF4/E5dlPv8A3NXHVITry97WnVl1OEYxdpVGlaK2Svd3b9HJeH5KTGfnKl2/V7sqrdW5ePYq493f+Q8yxtX2ZWV92kSoXV7hrb5g2M3v3QLNX3KJWZNmlIbaJfYkgxStn5Anq9CAqbVa37EWsXKy+MrdjKqI2YsirVixKpo2RE9lsHsyG77BSjvuVuy7jfxKyja297hUu0XYu2Cen1FVNq2RWSt2TLPa4TuZYqXTNdXiWD2uV6vQxQ07sjuW8CoVLtD7EWfkS1bcdXoFy7VIaJb8SO/pYmxqje/cglq1yDFBFkWv6FJdwuUla23chXtuS1bcgXsqUskRLuSRLuQrH1VFvQjq9Bf0CoXZHcAKQ0kijv8AmLS+MrdirdtgqbRdodwCVzaG7Fe5MiA2HYhvZ7lm7lWvELiLtlHcsQ1bcmqRZ+Q38exZO4fYxUZt7h9tu4l3ICpFW7hLfsJdyer0Coh7MjYlu5QyqGlciyJBKkWRUuZy7BUuxvci8vEglu+wXKgh7bokiXYlsUe7exnNLY0btczm7h0xZS8TOSuaSZlN2JrviynZN/MZSNZu7ZlIiu+LKdt/M6z+0P8A3LOov9My/wC1UzstLvL5zrT7Q/8Acs6i/wBMy/7VTK4f1uP0vN4p/UeX92/Y/texM+QXXf0vf2LDnoqedXsTPkF139L39iw56Kn6B/IwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAePvtu/lZ4bfR3E/aT0f5Uf3LvB76A6e/o6gecHtu/lZ4bfR3E/aT0g5UVflc4Pb/8AEHT39HUAPqQJ6fUdPqBAJt6kAAAAIsiSbeoFXstiE34lmrbgCBaJJDV2BDt4BLfsSlbcALRKk9PqOn1AgE9vW4vfawEAlqxAAEJWJAiyKvYuRb1Ai7Qu2GrBbALPyILdXofzc9z3LdOZdUzTNK/RShtGKSc6k2naMV4t2e3ZJNtpJtLZJuqxxudkk3aZ7nmW6dy2pmmaVuilDaMVvOpN3tCKurt2e3ZJNtpJtfAdVaqzHVmZPG419FKF40MPGTcKUH4LzbsrytdtLskkmq9V5jq3MnjcbLopQ+Lh8PGTcKUG+y827K8rXbS7JJL+7w94e1tS1Y5rmkJ08qpS2V2pYmSdnFPuop7OS9Ut7uPh5M8ua+XH0foen6fj6Dj+V5vVHD7h9V1LVjmmaQnTyqnLZbqWJknvFPuopqzkvVLe7j9upUKNClChQpQp06cVCEIJKMYpWSSWySSSSRFKjSw9KFChThTpU4qEIRilGMUrJJLZJJWSRqeri4pxzU9Xxuq6rPqs930QkvHuQS1dkNW3OjyhF15klen1AlpMhJ3JDdgJKE9XoOn1AgB7AARZEgCrfhfYXaJt6kNWAXZAAAWTAAbLcrdln2KgCLJk32sEr7gRZCyZIAi68yHuOn1HT6gQLX8CbW3HV6AVa2Klm7Bq+4FQ0tyUrk9OzZlipds3ddrkNy8bkt32JS+LYlUrMm77BqxAVDpXkVd1tcu3cq1dhcFv3Ky8bFkrESjsyaqd3Q/nV5guMXDPjHS05oXXGKynLZZNhsS8PSoUZp1ZTqpyvODd2opd7bHZLlT1jqXX/AHS+rdX5rUzLN8f8N+EYmcYRlU6MbXpwuopJWhCK2S7ee5+s1Xl3CLE5oqmuMHo+rmXuopSzWlhZV/dXdknVXV03vbwvf1P7unaOnMPk2Ho6RpZbTymPX8HhlyprDK8m59Cp/FXx3Ju3je+9z08nJjlxTGY6vxVNP6G62IfVYs1fchvwseZu1eqSDTa7dyJdyynslbw7hW9KO68yrulcvLuUburGVaFuiSErAls7oa32KvZF0rFG7BcSr23Ia2JAVGcluS9kJK77gm+qpotfuiPik33sVlHYxU7krK9mij37luja9yGrBUulXbwIuw+5AVLoavuxsib7WIauZdqVl4lUvMvb1IMVLtHgV7FiG7hUUbe68AWISsFY+qpD27Fn3IbsTdK2o+xWz8jRrxuVbtsYqd1bPyDXmT1egbuFRBV7MlO+xD7hWPqpdkrdbkpW3BNqlLIWViZfFV+5W/oYqVABN/QKm1bIrJK72LNXKva4VFexKWxDdyb+hKpdIkkVezLN3KteIXEAhuxIVO6GvIq+xZuxVvxMulRXdC7fiT1ega8bktitkyHYsUfcLiLIqWavuVClkkyjXkSR1ehlVNoAbuCVwK2iyWr7kNW3CppElH0K2fkJdyU7hUVIl2LteJVq5l0pSyfgVlGLLva5QxcrKcUm/IynGLsb1O7MGZXbBlOMVcxkkbzXd+hjIiu+LKcY7tr+U6ze0QSXKzqJ/8AhmX/AGqmdmZ/iv5zrN7RD9yxqP8A0zLvtVMrg/WY/S83if8AUeX92/Y/sexM+QXXf0vf2LDnoqedXsTPkF139L39iw56Kn338lAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB4++27+Vnht9HcT9pPSHlQ/ct8HvoDp7+jqB5ve27+Vnht9HcT9pPSDlSduVzg766A09/R1AD6o722K7+pKd2SBXf1JSVtx1egtfcCdvQbehFvUgBZ+Quyer0Ie4ABK5NvUCAABD7Eb+pYACu/qS3Ynw+cCv6R2fYlKwAiRBPT6jp9QIBPb1uL+gEFXe7tcsRf0AL1G3oLX3P5+e55lum8uqZpmdfopU9oxSTnUm07Rir7t77dkk22km0tkm6rHG52STdqc9z3LNOZdUzPNKyhShtGKSc6k3e0Yrxbs/RJNtpJtdf9V6qzLVmZPG419FKF44ehFtwpQb7Lzbsm5Wu2l2SSTVerMy1bmLx2NfRSheOHoRd4UoN9l2u3ZXla7aXZJJf3eHvD+rqWrHNM1pzp5VSlsrtSxMk7OMX3UU1ZyXqlvdx8PJnlzXy4+j9D0/T8fQcfyvN6o4e8Pa2pa0c0zWEqWV05bLdSxMk7OMX3UU1ZyXqlvdx+30aVLD0oYehShTpU4qEIRilGMUrJJLZJJJJIUqVKhShQoUoUqVKKhCEIpRjFKySS2SS2SRY9XFxzjmp6vjdV1WfVZ7vohr0JBHV6HR5UPuP0k2vvcWtuBFn5Anq9CADKpO+5YARZeQuvMkr0+oB9yCen1Hb1uBAJv6EAVd7u1yV6i/oO/pYBt6Db0It6kASyAAD7FbPyLACP3pBLfgQAKu9/EsABVt3HV6C19wH6SLPyJtbcdXoBV7kb+pYhuzAhbPcN/oDd9iA2Fl5EPva2xLdiOr0JqpdErbdijtfYs11eNrFXG19zFSoDACohX8WJPZpd/JCRWK+Ne4171x5/c8vB3iprrjTRzvR2gM8znALJMNQeJweDlVp+8VSq3G6VrpNNr1R2e5RdM6g0fy8aU07qjJ8XlWaYT4f7/CYqm6dWn146vON4vdXjKLXmmn4mnFjmr4TcGdUQ0hrXE5pTzGeFhjEsNgnVh7ubklumt7we3zH7vhzxB07xT0Zl+u9KVMRPKsz978HliKTpzfuq06Urxbdvj05W81Z+J6eXk5MuHHHLHUnvVux+gd7+IW7LNWK+J5GjV/D+Qrt6Fur0K9N337hadvRlGtuxe1trlA2M5Xv4osVluyxF7LmwrJbdixVu+wagABaGncz39TUoZVRG1iN/Ul7MlO+xKpVd0trlN/E1vvYrKPbcKnZm+zI7FvP0I7+lgqVUiRZ7ENXFm1RUE29SCdaVLAjYkrIKiJFd/UsVl3CizFvQX9Cer0MqozlffuV39TSXYqSuKALYlu4VNRV9tiN/Uluw6vQNVADV9zNRavfuRYlq25BKlbPyFn5E9XoOr0Coq+xV+Ny5DXiFys7PyBZuxV7syxqH6XI39SzViDFK28wS3cX2sFSo+cpLxLNXIMq58FLPyLfO0Cr7ktl0PuQ1t2JIClHe/iRZ+Rdu5AWqRZeRLdxb1FipdosvIqWKS7ENl0kh7iPYkKUt5lS8u5Rq24Wldisr27Mm/oG77BanzlZeGxpbe5WXxreFiVSsandmEjkThu3fwMZRMrtgwn3ZjP0OROHd38DGUfUivRjZ6sJ/inWf2iH7ljUf8ApmX/AGqmdm5xunv4nWb2iMbcq+o33/vzL/tVMvhn+1x+l5fFLL0XL9F+x/W9iZ8guu/pe/sWHPRU86vYmfILrv6Xv7Fhz0VPvP5MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADx99t38rPDb6O4n7SekPKik+Vvg7t/xB09/R1A83vbd/Kzw2+juJ+0npDyor/ct8HX/wDSBp7+jqAH1MAALLyAAAbegI6fUCbLyG3oCGrsA/Qjf1LJW3AFbPyJVh1egtfe4E7ehV2vsGrbgCO5JPT6kACrv1eNiwAAFH3Al79iCYkvsBUA/nZ9n2W6cy6pmeaV+ilDaMVZzqSadoxXi3Z7dkk22km0tkm6rHG52STdqc8z3LdOZdUzTNK3RShtGKSc6k2naMV4t2fokm20k2vgOq9V5lqzMXjca+ilC8aGHUrwpQfgvNuyvK120uySSaq1XmOrcyeNxsuilC6w+HjJuFKDfZebdleVrtpdkkl/d4e8Paupasc0zSE6WVU5Oyu1LEyTs4xfdRT2cl6pb3cfDyZ5c18uPo/Q9P0/H0HH8rzeqOHvD2pqarHNM0hKnldOWy3UsTJPeMX3UU1ZyXqlvdx+30aNHD0qdChShSpU4qEIRSUYxSskktkklZJEUcPSw9KFChThTpU4qEIQilGMUrJJLZJJWSRdq56uPjnHNT1fG6rqs+qz3fRD7kFun1It6nR5UC3oCW77AQAGrgNvQqGrFmrgVAewAAAAVe/YPuIgQCz7FQA29ECGrgNvQO3gOn1IasBO1vAqTba5AB3tsV39SwArv6kpK24v6C19wJ29Cr7k29SAIsvInb0QIavuBEt1tuRZ+RZK25IEbehWS77IACq77h9yZdiA2It5h237Et32KtXZN9WxV9Xhf8w3tu9/UulYrKF23cxUqhO1vANWIC5o+ch9n0/yBq4SswqOqHM1ycao47cR6et8o1hlmWUIZbQwPuMTQqTm5QnUbleO1mpr9DPtXALhnmHB7hJkXDnM8yoZhico+Fe8xNCMown73E1ayspbqyqJO/imfD+aLnF1vwK4l09E6e0vkeYYWeWUMc62M997xTnOomviTSslBW2vuz7dy+cTM14v8IMg4iZ1l+EwWNzf4V72hher3UPdYqrSVupt7qmm7t7t+B6eSc14cfP+j7l930MhrbsS3cg8jRLbdFXte2xPV6CS2buFS/BCu/UhrZ7BStta4fYK96rS8bGavfxLy7kKd32MqgjYlu5EuxKkO19iAAqXYQ7W8A1fciUdjKuK2u/QnYW2sR0+pKptEt27Ebr8Ytb1ImGqPxK/pLENXCoqT2D2YbuFSoIdvAl9ipKpdKu93a4V/FFgFKtO7I2Jb8CrV9wqG3oHbwICVwvavj6epD7lmvAq1YhUVdrbWCtbew6fUNWCoO1vAo+5dq25R9wuaQCWrbkN22MvdW9oXqRLxDd9gSqfFSz8gXKvuGzsghtFkrlZR3YVO6vzlX3LNXI6fUyxaO5DaJasRbe5ipdq9gTIrb1Cp27pKtMnt63D3QVL3UbdydvQNX3I6fUhUQ+5V337l+n1Ie1wpQrv6l27kBao39Sb22Iv6BUmlW3cgmXchq25NVAAGNRsZrvuXl3KNW3Cx9wL+gbvsKqIbRR+lyzXiQSqM5dmYyN5dpfMYtXMrrj6M5eJjNdjae1zCbvYiu+PdlLxOs3tE/3K+ov9My/7VTOzEl3Os/tE/wByvqP/AEzL/tVMvg/WY/S83if9S5f3b9j+r7Ez5Bdd/S9/YsOeip51exM+QXXf0vf2LDnoqfefygAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHj77bv5WeG30dxP2k9IuVFP71vg7t/xB09/R1A83fbd/Kzw2+juJ+0npFyou3K5wc2/4gae/o6gB9TsyCb+hAAFun1HT6gVBNvUgATZkE39AKpNEgALLyKvZ9yxDV2BG7FmWStuAI39f0jbx7/MOr0Fr73Ah28AGrACHfwCW3YkAQ7+BG/j2+csfzs9z7LdO5dUzPNK3RSh8WMVZznN3tGK8W7Pbskm20k2lsnetxxudmMm7TPc+yzTmXVMzzOsoUofFjFJOdSbTtGK2u3Z7dkk22km11/1XqvMdWZlLG419FKF1h8PF3hSg32Xm3ZXla7aXZJJTqvVOY6tzF43Gy6KULxw+Hi24UoN9l5t2V5Wu2l2SSX93h7w7q6mqxzXNYzp5VTlst1LEyT3jF91FNWcl6pb3cfDyZ5c18uPo/RdP0/H0HH8tzeqOHvD2rqarHNM1hOnldOWy3UsTJPeMWt1FNWcl6pb3cft1KlRw9KFDD04U6VOKhCEElGMUrJJLZJJJJLsWo4elh6UKFCnCnSpxUIQhFKMYpWSSWySSski1vU9XHxzjmp6vjdV1WfVZ7vogh7okjq9Do8pv6/pIafiSncPsBUAACEmi1vUgARv6/pJI6vQArLv3DaYtfe5DVtwBD3RIAj5+4d/AkhuwEb+Pb5w7eAv6EAATb1DVgIK2b7liG7ARv4izYv6BO2wCzIJv6EACJbPbYkNX3AqCbepAAi6RJDV9wD3WxG/r+klK24btsBFmLMdXoOr0Ah7d0Q2vAtL4yt2K9PqBC2Ydr7E9PqQ9hVS7VkVfV33sWkE/CxNbOylm+5HYvL4tvG5R7mKQ7+BG/e7LEPsFx8d4r8qvCbjNqiGr9a4XNKmYwwsMGnhsa6UPdwcmlZJ73m9/mP3nDnh7p3hZozLtCaUp14ZVlnvfg8a9X3k172rOrK8mlf49SVvJWXgdI+ebjDxT0Lxqo5Jo7X+eZNgHkmFrvDYPGSpU3UdSqnOydrtJJv0R2f5RNT5/q/l40nqDVGb4vNczxfw/3+LxVR1KtTox1eEbye7tGMUvJJLwPXy8fJOHHK5bnwW+w7IrLxsWavuRb1PERmSvXsJdxb1C5tWSu/i9ibPyDdnbuOu+1v5QpWSV+xWSSXZItLuQ1dWCpVCRa21wQubRsUe6LtXZDVtw2KrZB7okhuwXFXtsQ91sS1d3ItbcmqSr2KtPxLX2uR39LGK9zN9wS13Kt2CoNIqTf0CVwqIIa8iXtcjq9DLVKNSuyVfxDl32CdzFDSKln2KhUReI79iH3CdgqbQLx8Q97kdPqTdqmlX2Kk39CDFJ3ZDXmSnYq5b9gudkEABU2bIq7X2LNXIt6kKmlXuiLMluxHV6BUJbLbYq73buXl2KN+AUqQ739CSG/AVYvEl9mVTsTf0JVNKSIJkQGxLs+1ir7BO5IX8FCGn4FmrFb+hNULZbh9Nha+5BilWtyCzdtioVNovHxDa8CGrBbhUQRLsWasUbvsZVKtNhJosCVKy7kEy7kBaGvIizJbsxe+xt7qlVe5WzRZ7XKt3JqopJOzt5GMkzkPdMymrGXu6Y1jNPe5jOL2sjeXiZTdibHfG9nHmmr3OsvtE/3K+o/wDTMv8AtVM7OTezZ1k9on+5X1J6YzL/ALVTN4P1mP0vP4n/AFLl/dv2P6nsTPkF139L39iw56KnnV7Ez5Bdd/S9/YsOeip95/KAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAePvtu/lZ4bfR3E/aT0j5UF/uW+Dt1/xA09/R1A83Pbd/Kzw2+juJ+0npFyou3K5wc2/wCIGnv6OoAfVGtuxFmSndkgRv6/pG/r+kkhu2wBXW77C8Re+1h0+oFSy7Dp9SQIbTKgAACbeoEC67EtW3ItvcCGvIm/qCLeoE7MWVuxHb1uOr0AizBPV6H87Pc+y3TuXVMzzOt0UobKKSc6k3e0Yrxbs/RJNtpJtLZO9bjjcrJJu1Ge57lunMtqZpmlfopQ+LGKSc6k2naMV4t2e3ZJNtpJtfAdV6qzHVuZPHY19FKF1h6EZXhSg32Xm3ZNytdtLskkmqtV5jq3MnjcbLopQusPh4ybhSg32Xm3ZXla7aXZJJc7Q+h8bq/GuUnOhl1GSVfEW3b2fRC+zk01vuknd3uk/Bycl5r5cfR+j6XpePoeP5bm9fsacPtF1tU5pGti8PP7l4aTeImpOHvJWuqaaV227N2tZN7ptX+80aNPD0oUKFONOlTioQhFJRjFKySS2SSVkkZZfl+CyrBUsvy/DwoYehHphCPZLv47ttttt3bbbbbbZyT1cXFOPHXvfH6zq8uqz36T3RXf1/SGn4kt22Iv6HV40Cy8gS1bcCAABF4kWYasWArutiCzV2Q1bcCLpENeRLV9wA2Ww2exDV2LW3AnYrZoPcN32AEK/iSQ3YBZW7EWY6vQdXoAu1sR3Ja8bkARaVye4AEO1vUJKwt6i9tgDtb1KkkAHe2wWyAAXRF4jp9SGrACU0hb1IAAhuxHV6ALMjsT1ehD3AEXtIkhq7AN7EWb3J6fUN22DcfVVq5WzLC+9ibtSs2trlXa+xeUb23tYo42vuYqCt4kO35gAqV+E1rqbgXlGcLC8RdQaEwWaujGSpZ1isHTxHum30tKq1LobUrPtdO3ifodK4zSuZ5DhsdobGZTi8kq9fwWtlNWnPCytOSn0Ok3B2mpp2e0k097nSjnX5feMPEzjHS1HoXRGKzfLY5NhsM8RTr0YJVYzquUbTmndKSfa252W5T9Hal0BwB0tpLWGVVMtzbAfDvhOFnKMpU+vG16kLuLad4Ti9m+/nsenk48ceKZzLd+C31jttIjdsmSu+5Frb3PLWxFvMr2ZZu5VrxMUq4yfZXGy3LJ22Kr4zt2uF72PfdFXe2xe1trlA2d1Xt3YIn3JJvargVb2LFWrbmNQG7EN22DVwtD812KvfZFnsmrFY9ybpSFsyzv4C29wncxcUalftt84+KWl+KzMNiGt20tiLNFirdwpA2RNtrkNXMsl9Fqtd2kVW3c08CjVzFQuiHux0+oasGxWyW4vEPsVC5dLNrwK3SAavuZVKvp8EiNkWt6lJdyVTSLNAlu+xAVNK2YLFZdwrH1Q90Vae/csH2JulS6Z2l4izLAxStmQWl2KhSrt4FWtyzViBZtc2hbdw2vASIMvaqlR3It6bFmrFW/AxUuh28Ev0FW+++5JFvUKndBRrfsadPqVl2JvqtCewdiAYraLoq722LNX3Khsukdvxu4bXh3DV9yFuFy6Q35lbNFpR37kS7E1St0hdMNX3CVtzFF0UexaXcq1cKk0izY3W5YiXYLkVIkSVbuZdVsVd/zGczV9jKZjpizk1ZrxMpeBpLxMpuxNd8Yxnbf5zrL7RW33qupP9Ny77VTOzMnZN2Os3tFHflV1Jt/8ty/7VTN4P1mP0vP4n/UuX92v6fsTPkF139L39iw56KnnV7Ez5Bdd/S9/YsOeip95/KQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAePvtu/lZ4bfR3E/aT0j5UV/uW+Dn0A09/R1A83Pbd/Kzw2+juJ+0npJyo/uW+Dv0A09/R1AD6mAAAAAj5u439f0kgAQ0/AkAVWz3JdiWrlfEBZi0ixDdmBFmQTdsALMq1K7LdTAEK/iNgcfMcwweVYKtmGY4mFDDUY9U5y7JdvDdttpJK7baSTbSFsk3WyW2SetcfPc9yzTmXVM0zSt0UobRit51JtO0Yra7dnt2STbaSbXX/Veqsx1bmUsdjX0UoXjh8PGV4UoN9l2u3ZXla7aXZJJW1hqrGarzepjK1WawtOUo4Si1ZU6d9rpNrqaSbd3d7J2SS5eh9D43V+NcpddHLqMksRiLbt7PohfZyaa33STu73Sfg5OS818uPo/R9L0vH0PH8tzepofQ+M1fjXKTnQy6hJKviLbt7PohfZyaa33UU7u90n95y/L8DlWCpZdl2HhRw9CPTCEfDx7vdttttu7bbbbbbGAwGDyrBUcuy/DRoYehHphCPh4vvu23dtu7bbbbbbOQeri4pxz53yOr6zLqsvhIAFW7M6vEsVdvDuSt0LJARZhvYsUAEJPxJAEXQe6FkyQI39f0kWZLdmRdsCAABDT8Ar+JIAWXkQ2mSVAEdyQBFlbsRZlgBVtWtcK3iQ+4As2vAqAAIa3Ddgt0BFmu/Ym8Q+xUAQ02SAK797sE3vsx0oCXuipPUyG7gCLLyJAEXiRZvt2J6URe2yALZ7huNmLtkAQ3sE149x0ohqwbLodvAq077FkrkS2MqpRX8SsmrsnqZDV/jNu9iWy91AAFyvhnGfnB4Z8DdXw0Vq3I9TYvHTwdPHKpl2FoVKShOUklepWg73g7q1t1u/D6Nws4k5Jxe0FlnEPTOFx+Gy3Nvf+5pY6EIV4+6rVKUupQlKKvKm2rSezV7O6XwjmS5Msz488Q6euMHr3C5NThl1HA/BquXyrNuE6knLqU4qz67Wt4dz7NwH4Y1+DnCnI+G+KzinmlXKPhN8XToulGp73E1aytBttWVVLu72v42PRyTinFPJfzlzUfu9137h9hNtPYi99mebvWqNNi0iz2YMUr22aI7Ey7lX2C53OpeZVp2LJB9g2XTNrzIumXsmUskTZpUqSJdiSGrmKlUa3JIk7OyJC1ZJu5VLc0fYoZYqD7FUn4Fn2IiSqXuhqVmUs0alJqwbIq+xQ06dtyjVgudizsVkWu2Q0mO+lS7PAqWKy+Lb1JXEEPdi99mT0oNl0o90Ft3JIaTCpRteBW6RbpRWUVcyxapVrcsCVbUHYJ2AVj6Ib2IJaKhUR23bDfkS1chqyJulRV7ojf1/SWKt2Zii0vEWY6mLthc7KyKlmkyjdm0gqXZIgNtgmtiWyra38yWrFH3CkO/gEn4lkrkS2C53LpFHvclu5DdiaqTSGtyLMst0G7GLk0qVbXgTKRUNk2hpsizRYi99mFIIe6LdKKhatmQWbKd9mQqXabx8Stmi3ShLsGy6VuiG9iH3IC5dAsvIAyxSsvExmbS7lJRTtdsxeNYSas14mM1c5E4pPxMpImzu74VxpLY6y+0Ut96rqTb/AOWZf9qpnZ2UEk+51k9osrcquo3/AOG5f9qplcM/2mP0vP4nf5ly/u1/R9iZ8guu/pe/sWHPRU86vYmfILrv6Xv7Fhz0VPuv5UAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADx99t38rPDb6O4n7SeknKj+5b4O/QDT39HUDzb9t38rPDb6O4n7SeknKgk+Vvg63/mBp7+jqAH1RbMm6HSh0oBdEPfdE9KI7bIBZizHUx1MCAW6UQ1YCr3RFmWAAXQIsmAb2KlulCyQEWZBc4mY5jgspwdbMcwxEKGHoRcpzn4Lt4btttJJXbdkk20hbJN1sltknrTMMxwWU4KrmOY4iFDD0I9U6knsl28N222kkrttpJNtI+Ca41xjNX43pj10MuoSboUL7t7rrnbZyab23UVdK923Ot9cY3WGNUI9dDLqEm8PQvu326522cmm9t1FOyvdttD6Gxmr8a3Jzo5dQkliK9u72fRC+zk01vuoppu90n4uXly5cvJh6P0HSdJh0eHy/P6/6+tGh9DY3WGNcpddDLqEksRXtu3s+iF9nJprzUU7u90n95wGX4LKsHSy7L8NGhh6EemEI9l4+O7bd227ttttttsnAZfgspwVHLsuw8KOHoR6YQitl4+O7bbbbd2222222ciye/meji4pxz53y+s6zLqsvhIRJCSQfY6vEXRV7gACHuiQBWzJuiSOlALohvYPZkAQ1uSAAuiG9hZMhbsCAW6UVAB7oACtmWBHUwIfcAJXAEdiWrENJgLoh79ielEPbsAsxZjqY6mBAAAh9ipa7ezHSgKgkgALoEdKAl7ohbbMkiXcCAAAKvuOpkAASt2GrICCr7liLJgQiJeJLViBVS7VJW0SGrEr8Uhsm1GyCWrBK4XHTPm25tOKfBTinS0bo6hkc8BPKqGNbxuDnVqe8nOonuqkVa0FZW89z75y2cQ9QcWOCunNf6ohhI5nmvwv36wtN06S91i61KNottr4tON93d3fjYvxG5auCvFfP46p1/ox5rmccPDCKv90cXQtSg5OMemlVjHZye9ru+72R+u0PojS/DjTGC0XozK/udk2Xe9+DYb31Sr7v3lSVSfx6kpTd5zk927XsrJJL08mfFeKY4zWS9zUf3U0tnsVbTYn3CS2Z5fnUhrcq+xcq9rmNlVsyGrdyepkSewXIlSivEq+wSD7BXvVIb2JIaMrfegh7okErUIS3LS7lUwqDe3cq90WsmRJWV0ZVxC2RD37C7e3mSkkS2K2ZWRdtvYiSW27CveoOxLVisgsb2KkpXDVhbpUQRJ2JKzMtVFXuml3KpWW5ZdybJmNipDVySG7BaLMgte+zIezCkPdFSeplZSIVELbdk3RF2wldBcQVl3LtFJdw2KpbkvcAyxXqrZoC7YJXES7FS0uxUNiJEWfkTIeAXLpSRBMiCapDVx4FulEBSji2OxZuxTq33C8btEu5BayZAVPgq1uRZkti99mRe7ZpBVvYs9mUC5pDW5Us2QlcKndAe6LdKKvZGVapV77Is3ciyRKpsWyIb2LFA2TYARLsFy6LqxVq4BlVKr2KyLS7lW2zFY3bOp3ZjI2qd2YyJrtj6MJd5fOdZfaLbcqmo/8ATcu+1Uzs3NKzZ1k9ov8AuVNR/wCm5d9qplcP63H6XDxK/wAy5Pov2P6HsTPkF139L39iw56KnnV7Ez5Bdd/S9/YsOeip91/KwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/mah1Hp7SOS4vUeq8+y7JcowEPeYvH5hi4YbDYeF0uqdWo1GKu0rtpXaP6Z+A48cIcp49cI9S8Is+zTFZdgtSYWOHqYvCqLq0XGpCpCSUtnaUI3T7q6ur3A/Y5RnOUagy6jm+Q5rg8ywGJXVRxWErxrUqi84zi2mvVM5x5H5j7LvnJ4A5jW1FywceKeOtLqdHB5hXyPGYiyuozpOUsPUW1mp1UnttZu22H59vaIcsk1hOZXgdV1DlVC0amYY7K5YJyUXZqGPwkZYWTd1duEnvF333D1qB0c4Se155XdfyoYDW/3b4f5jVajL7p4Z4nB9T7JV6HU0v8Avpwgl4tLc7xgAAB4++27+Vnht9HcT9pPSTlRT+9b4OW/zA09/R1A82/bd/Kzw2+juJ+0npNyofuWuDv0A09/R1AD6kk/EsABXf1/SSltuSAIa27EWZLdiOpgN/X9IsyXsiOpgFs9yW14EXbIAh7olXtuW6UQ1YCA+wOLmOZYLKcHWzDMMRChh6EeqpUk+3h4btttJJXbbSSbaQtkm62S2yT1pmGY4PKsFVzHMcTChhqEeqc5dku3hu220kldttJJtpHwTXGt8brDGpRU6GXUZN0KF92911zts5NN7bqKbSvdttc63xur8aox66GXUJN0KF+/ddc7bOTTe26inZXu250NojG6uxrcuujl1CSWIxHi3s+iF9nJpq73UU7u90n4uXly5cvJh6P0HSdJh0eHy/P6/wCvrRofQ2O1fjXKXXQy6hJLEV13b2+JC+zk01vuoppu90n96y7LsFlWCpZdl2HhQw9CPTCEfBd+73bbbbbu222222ycuy7B5XgqWXZfh40MPQj0whHwXd99227tt3bbbbbbZybJb+R6OLinHPnfL6zrMuqy+EiOl+RBPUyH5nV4h38CN/Ht85KdwBF4kE9KHSgIs2Hfsu5PbZEAV39f0jf1/SGywFbMWa79g2LtgTeJR7okloCFe25DW2xJD2QDf1/SVJ6mS0BUAALoWXkLJgBsQ35MNvchK4ABqwAq0/AlX8SSG7AH2ZUm7ZAAAhsCdluRdC99mOlAVe4LdKIezAgBuxHUwIu/MbvcnpQ7bICLNAXbAEWXkLLyJKt2YCWy22Ce/cd9mGrIA2mQCG7MCGn4ja3qG2yAqU7lW9++xZuxV7v5ya2Ds+1iuyZLSja3j5lTFPOnn713rfTnHShl+ndZZ7leEeQ4So6GDzCtQpubqVU5dMJJXaSu7Xdkdq+TXNs1zzlt0hmudZni8wxtf7oe9xOKryq1alsfiIq85Nt2SSV3skl2R+61Xxh4WaFzRZJrLX+RZNj3SjXWGxuMhSqOm20p2bTs2mk/Rn97Tmpsg1hk2H1DpfOMJmuV4vr9xi8LVVSlU6JOErSWztKMk/JprwPXycvm4scfLrXvdN7j+kknu9w+w7bIhydn2PHfVqrTZDtbfv8AMWW6Kve5ip3Ve72I77FkrENW3QVFWnfZWCUm/wD+4lIRk7oK170SVnYpuzSTu9zOXxVdBULMBSbRD2RCohrcq1tsW6mQFwV7bkNXRJD2RlVFGrO9tg9+xLe+/iQ9uxKk+F/Qo7vtcv4fOR0oK9zN3/OVs33Ly2ZAVKrZizfcluzI6mFxBDV/C9iSHt2MsbLpDVt2Vk/UN3dn5joTe9zFSln3KyLdlbwRVpMKiFsw92S1ZFRe6oNbdij22aLt2Kvd3ZNmlxSzQLNXIasYqXSL+ZV7ss1cjpQVOyoe62DVwZuLitmgWauVasSqXaq3e5NlbsRHuWCsfRRrzRV33RaUnsRZPdhUVV/Eq9mWbsV7vcyqLvzD3J6UQ/ExU7KNPxBLbYasgud0XSKPctZMgyqkkRdEOz7dybJlV3MrYNWI2L2TKGKRKz8EVvEs9mVasgsuir3QLNGVUu2dmQWbKtXJVKXRDaZPSiGgqKu9/QhbvcsR23QUPsyvYm7ZDSZlWq97vwKT3t0/nLvZW8CrVjFYsZd3cznbaxtU7sxkTXbH4sJX338TrJ7RZ/7lXUi/8Ny77VTOzk0rNnWP2i/7lXUb/wDDMu+1Uy+L9bPpcPEr/MuT6L9j+j7Ez5Bdd/S9/YsOeip51exM+QXXf0vf2LDnoqfcfysAAAHkzzfcy3OxV55tS8u/LrrfMoe5p4Gnk+SYHDYJOpfKaWNry668N38atJty7RSXZI/h/wD4bL/6bf8A9CAewQPH3/8ADZf/AE2//oQf/hsv/pt//QgHsEDolyGx9od+2tnc+bV5wtILT1VYJY95dZ5k8Th/d9Kw3x7+6WIvf4u6vvY72gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/mah1Hp/SOSYzUmqs8wGTZRl9P32Lx+PxMMPh8PC9uqdSbUYq7Su2u6P6Z884/8AB/L+PvB7U/B/Nc5xOU4bUuEjh5Y7D041J0JQqwqQl0SspLqgrq6urpNOzQdeuKvtW+UbhxGvhcj1PmeucxpKyoafwTnS6vBPEVnTpNeLcHPbwb2Oteae1E5sePONr5Byt8tjVGpJ0XiZ4PEZ1Xgv8aUoRp0KV13VRTSu1d7M/F6n9kNzK8K8+o6s4P6s0Prt5bU97hsNmmAo0qtWSvbqwmMhVwkl6TqNJ2a7XX0HLefPnh5YsDRyXmD5S6FXIcu+K8dluXVMsoQhduVsRh1Vwbte9oRj697gfNMn9l1zhcwOq3rvjfjtIaGnmDhLFyWHwscS432lHCZdCNBzs/384y2Sb2svZHAYOjl2Bw+XYZNUcLShRp3d2oxSSu/HZI6U8Lva78qmu6tDA6tq6h0JjKslTbzbBe/wvU7WtWw7m0rv8acIJWd7Lc7vgAAB4++27+Vnht9HcT9pPSflPduVrg7f/MDT39HUDzY9t38rPDb6O4n7Sek/Kgv9y1wcf/0gae/o6gB9TAAAAAAAAIukSRZMB1Ii6e3mT0oWSAjpZBc4mZZjg8pwdbMcwxEKGHoR6pzl4Lt4btttJJXbbSSbaQtkm62S2yT1pmOY4LKcFVzHMcRChh6Eeqc5PZLt4btttJJXbbSSbaR8E1zrjG6wxtoqdDLqEm6FB92911zts5NPtuoptK9221xrjGavxijHroZdQk3QoX3b3XXO2zk03tuoq6V7ttofQ2N1hjXKXXQy6hJLEV7bvs+iF9nJprfdRTu73Sfi5eXLly8mHo/QdJ0mHR4fL8/qaH0NjdX41yl10MuoSSxFe27ez6I32cmmt91FO7vdJ/e8uy7BZTgqWXZdh4UMPQj0whFbJd/Hdttttu7bbbbd2Rl2XYPKcFSy7LsPChh6EemEIrsu/ju22223dttttu7OQ3Zno4uKcc+d8vrOsy6rL4Se4bSDd0Fv3HSjq8SoLdKHSgKglqxAEdLIJ6mLJ7+YEWbFmie2yF77MCAHswBF0h1IWTIasgD3Ys0QTdsCCLpkkWSAkjqRJHSgFr7oizRN7bIi7YEEdSJI6UAe+6ITsWSsR0oA3dEEtWRAEXSCaYsmEkgD7FSW29ielAVAAEXSHUhZMdKAWb38yGrFisu4EAACOpDvuh0odtkBFmgLtgCG7EWvuiWrgCO27Dd0S1chqyAgAAQtu4bug0mGrINlqoAM7KQ9+xR7Nos242t4+ZV27+LJbK6Rc4vK/wAYeL/FulqzQuQ4XGZbDKMPhHUqY+jRfvYTqNrpnJOyU1va252J5YNA6m4ZcCtNaH1hg6eFzfLfhvwmlCtGrGPvMZXqwtOLad4Ti9nte3dH4vjzzk6c4D64hojNdF5lmteeBpY5V8PiKcIJTlOKjaSbuuhu/qfVeDnE3BcY+G+UcRsuyuvl2Gzb3/Rhq81OdP3VepRd2tnd0m1bwaPXyZct4cZlPzfi67upt+x7bMN3REu4SueOwlVaF9rFnsyrS3MVvuo0QXKtWQUpLuT0Nb7CyY629tgv1RLuUl8ZWRsop7tsy7boNndCi0gHNrwBC5YGcuxZshq4XELsS3YjstvAi7YVESlu0vEhbdw/xiWkyFRFn3/OSmmVu+xMQpWUXdlXt3Ly7mc22FQbuioAVAh79iQkkZde5UVe3cjqRL3uR0oxX0Is+5BZ9ioVENrcJkPuQFQI6kSVezIqppEuxUnvsyelBcmzqRSSu7okhsKiLNFWrstdsgW6bAq+5YWTIXj6s1sxdPbzDRC7hUuktNFH3LyK2TC5pVq5UtL4tvUqZYqdwAGKirTRVKzLSICpO6LpBu6Fkw1ZBajRL7ENjqZDYhqxXpZZu5DdgpVqxWzRZu5W7YWgPdAs0ZWzanbZlXui0u5BKohbIkAKVl3KWaNGluykuwWLsQ3cXaIFUNrdFGrl2luyjdiVRlOD37djKUXsby3TMZGV2xtcecWup3Xc6ye0W/cqaj/03LvtVM7PVOzOsPtF1blT1H/puXfaqZXD+sx+lw8S/qXJ+7fsf0PYmfILrv6Xv7Fhz0VPOr2JnyC67+l7+xYc9FT7j+WAAA8q/wD9uv8A+/8Amceqh1hr8k2msPzoz51q/ELG0sbSouVTJZ4SmsNFrKnl7m67ldRVP49nH8bxtsfr9Sc7HKZpLM6mUZ3zAaMhi6TcalPD5jHFKnJOzjJ0epRa7NN3XigPt4Pn3Dbj/wAEuMLdPhjxU0xqSvCPXPDYDMqc8TCNu8qN/eRXq4pbM+ggAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD+VqTUuntG5DjdUaszzA5Pk+W0nXxmOx1eNChQprvKc5NKKvZbvu0u7P6p894+cH8r4+cIdS8Ic5zjFZXg9SYeGHq4zDQjOrR6KsKqajLZ7wSafg3uu4HWzip7WvlM4fOvg9MZrnWvMwpJxjDJMC4Yf3m+0q+IdOLjsvjU1U7qye9usebe1I5wuPeY1tOcsPAiGBlKTSrYPAVs8xtFbpSlPpjQprdN9dJpNLe179suGvsx+TTg1hFnefaTeqq+Dj7yrmOr8cq9CCTu3KglTw3Te349NtJWvu79hNJ664IZdSoaQ0NrDQ2Gp4ZqlQyvKcwwcFTbV1GFGlJJXW6SXbcDyh0/7LnnD5hNTVNf8AHnUGntH18w6amJqYhUa2Nmm73WGwcVRi0m21KcGnZNd7eyGAwdHLsDh8uwyao4WlCjTu7tRikld+OyRyQAAAHj77bv5WeG30dxP2k9J+VBX5WuDnpoDT39HUDzY9t38rPDb6O4n7SelHKf8AuWuDn0A09/R1AD6lZoFhZAR1IdSJshZAQ3dEEtbEAAABD3RFmt/IscbMsxwWU4KtmGY4iFDD0I9VSc3sl28N222kkrttpJNuwtkm62S2yT1qMyzHBZVgquYZhiYUMPQj1TnLwXbw3bbaSSu22kk20j4FrnXON1fjemPXQy6hJuhh79326522cmm7LdRTaV7tudc65xur8aoxUqGXUJN0KF+73XXO2zk03tuop2V7ttofQ2N1hjXKXXQy6hJLEV7bvs+iF9nJprfdRTu73Sfi5eXLly8mHo/QdJ0mHR4fL8/r/r6zQ2hsbrDGOUuqhl1CSVevbdvZ9EL7OTTV3uoqzd7pP73l+W4PKcFSy/LsPChh6EemEIrZLu++7bbbbd2222222Muy/B5TgqWXZdh4UMNQj0whHw8e73bbbbbu222222zk3Z6OLinHPnfL6vrMuqy+EnpEE2ZBN35nV4kB9gAKAtZFXsBKTYF2iAAukBZAR1IN3RNkQ1sBBD3RIAhbINXJIeyAjpZBN2GtgCdkQCHsgJKE3ZNkAXYizQd1tcXbAgN2AAjqQe/YmyHYCtmgWFkBUhtIl7EdwIunt5jpZNvQi7AgAAQ1cjpZL2RF2AJTFvQh7MCW7oqABWzRMexJD22QEvdFWrC7AAi6RZrYrZAR32Qs0TZIkChKdkTZEPZgVkVL9yLKwbJtCTZD2uTdoh73MtViqn0977kS3TsSkn3IkrJ2JV73W7mA5Msr4967hrnGa9xWTVIYClgfg9PL41k1CU5KXU5xd31tWt4dz6zwX4Y4fg3w0ybhvhs4qZpSyj4R04upRVKVT3uIqVneCbSs6rXd3tfxsdYeb7mq4ucGuK9LSGh8fltHLp5Th8Y44jBQqz95OdRN3bva0Fsdg+WTiDqPilwQ03rzVtWhVzXM/hnwiVGkqUH7vGVqUbRWy+JCKfm7vxPXy48vyWNyv5rp31NvqF0h3WxEu5DbseSzZEtWKPuT1PzDStexlip39UFbNBvci7MUEKLZJCbuFyJs1syjdi5nLsY2aQ5bvuLNFX3LXbJVOwVs0WIl2ClGrsluxF/jWuSFqtXTaKJ2Lyb3XgVRNUnwKliGjFzuq+xRq5ra6vYo0Gzt2VfYjpZL2RF2FxD8UREkh7djKqVD7gAxUu1WrsNNEvZFbthU2izW7HUiG9+5AVBq5VqxYiXcm1UulZdiou2DFAJa2IC1bNEXSEpOxVbvcKm0tkFrIq9mQqIe6Is1v5EvZEXYVB79irVmTdoBSjaRDd0WkkUezFulgBNlYluPqo3cq1cs0QFbV6WCxDSC5qKhq4K3dyaqdhqxBJBikPdkEvZlX2CzqRUAKmwo90Wb3IJrZdIWyHUiSrWxi5pEtyO27JIl2Ci6e3mQ1YLuTIVUZvuQ2kXsiskiVSqSfdmLaRtKy6vmMJGV1x9GcjrF7Rr9ypqP/Tcu+1Uzs5PZN+p1j9ow396pqS7/APluX/aqZXD+sx+lw8S/qfJ+7XO9iZ8guu/pe/sWHPRU86vYmfILrv6Xv7Fhz0VPuP5aAADyk9oZxS4y8yfM7l/ItwVxdWhgMO6EM2pQxDpUsdi50FiZzxM0r/B6FGSbhunKMnaUlBL9pob2JXCfDZZQ/bJ4watzPMZU71vuHRw2BoxqPuo++p15SS3V3Zu17R7L8jojOcp4M+2O1dDiDVo4Ojq+NfD5VjcS1CmquNw1CrQak7pOThKgmmrzlbbdHqqB5Mcc/ZC6m4WZDW4m8s/FLOszzrTieYUsrxkI0MfJU11OWFxNBxTrKzag4R6r2Uk0lLtf7Njmnzvmb4HVf2b4r4TrDRuKhlebYnZPG0pQ6sPipJJJSnFTjK3eVKUtlKy7Q6r1Vp/Q+msz1hq3N8PleTZPhamMx2MxM1GnRpQV5Sb+ZbJbt2Su3Y82vYwwrZtnfHfWmBwlTBZLm2ZZXHC0HGPT1deOqdN12dOFWCsnb4+/gB6eAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAH4vjJxQyPgrwt1RxV1JGVTL9M5dVx86MJdMq80rU6MXZpSnNwgm1ZOSuftDqt7T7LszzPke4j0ctUpOlHK8TXpxpqTnRp5lhZz3f4qil1trwg14geeOjuD/ON7UnUeZcStV6xo5NozCYydHC1MdOrDLMLNbuhgcJC/vJQTipVHZvbqqSkrH1bHew5zuGW+9y7mRwVfMOhP3FfSs6VHqtuverFSdr+PRv5LsdwPZoZxprNuSvh1T0zOgo5fQxeDx1Kk1eljI4qrKqprwlJzVTfdqpF9mjtCB5BcGeMPMx7OXjvkXAvmNzetnHDjUNSnh8NiJ4yeKweGoSmqaxeCq1Ep04U5Ne8oNR+K2+lNxk/X08zvbd5lpuPDvhnk9eVJ6gnnWNxOFV11xwcaEY1/G6TqSw/hu4+jPRDh9SzShoLTdHPFP7pU8owccYp/je/VGCnf16rgfoQAB4++27+Vnht9HcT9pPSflRV+Vrg59ANPf0dQPNj23fys8Nvo7iftJ6UcqH7lng59ANPf0dQA+ppWZIAAhokARa27HUiRZACr7lnsji5jmOCynBVcxzDEQo4ejHqqTn2S7Ltu23ZJK7baS3sLdd62S2yT1RmGYYLKsFWzHMMTChhqEeqc5PZeHhu220kldttJJtpHwPXOuMdrDG9MVOhl1CTdChdbvddc7Ozk03tuoptK9221zrjG6vxqUVKhl1CTdDDt7t7/3SdtnJq+26irpXu22htD47WGNcpOdDLqEksRiLbt7PohfZyaa33UU7u90n4uTky5cvJh6P0HSdJh0eHy/P6/6+tGhtDY7WGNcpddDLqEksRXtu3s+iF9nJprfdRTu73Sf3zL8vwWU4Oll2X4eFDDUI9MIR8PHu9223dt3bbbbvdk5dl2CynBUsuy/DwoYahHphCPh493u227tu7bbbd22ciyPRxcU45875fV9Zl1WXwk9x1IjxJshZHV4htIqW7lXsBDVwlZkh7AQ1ci+1hdk29AISbFmi3Yi7v+cCoLNFQIaIs0WI3vZgQTLsTZFbtgCLpkkbLcCGg2LsNbAE7INhLYgAAAKvuErh9wA6WQTdkACHv2JHYCtmgWFkBUhq5L2AFbNb+RPUiSLICSsu4v6kARdMkWXkQ3YBdIdSFvQWQBu6KlmtioAXSAsmBVK5PSyBdgAABD3RFmiwAoC1kQ9mBBV9yxV9wJ6kVe9yUH3Mt0tQl/ik2REvxWSPwOtuBfCPiLnEdQa30Jlub5jGjHDrEYiM3NU4ttR2aVk5N/nP0mlNJ6c0PkGF0tpPKKGWZVguv4PhaN1Cn1zlUna7b3lOTe/ds8/ufrL9cYrjnQq6fwWe1cL9wsIurBU60qfX7ytdXgrXta/j2O1vJpRzXD8tmj6Wd0sXSxsfuh72OKU1VV8fiGrqW62atfwt4Hq5OK48WOdy3Pg667S7fabpBu6JsmQ1seWzbZdK3SD7ES7k90ZYrfdWzZW1t2Xe2yKvsYoTuZlrtE9KS7BUu1Vsipco9kL2VJtWT3JbKPd7liFT4IukVLWTIa2ClbpEiy8iHsgtD7kFml0tvvYzuzKta6RVu5a3oQ0SqJ8DORp4GcgKPuQWaRULWurFW0i1lYpIzfwVEAAxUQ1chpohN9dvDcmQXKgJ3IfYi7QVLocXZ9ijVi92RZMmqUl2KmkkitkYqXapF0iRZMKl2oCZpJbFLsVcWbsR1IkWRDZpSXYqWFlYLiG7kAhvcKJGb7l7tlH3MqoJNgXaD8zFSqyCdgt+5D2Col7sq1dkkPZBfohqxTxLkWRNUJ3KuLs+xYrdmKUasC1kyj2QVLsukGyCArYQ2SQ1sTe1VNIIbsQ3uSYpDdysuxMvGxW7YWX2sGrBdyZCqijT3KtpFm2V7krisn3ZhI2ns2vCxjIyuuDKTsdY/aMtPlU1H/puXfaqZ2cn+KzrF7Rj9yrqP/TMv+1UyuH9PH6Xn8S/qXL9Fc/2JnyC67+l7+xYc9FTzq9iZ8guu/pe/sWHPRU+4/loAAOovPjyFZPzb5VgdT6ZzfDaf4hZFQeHwOPxEZfBsbh+pyWHxHSnKKUnKUKkU3FykmpJ7dK6OX+2V4E4d6WyWvrLNcvwcfdYetRp5fqGEqUWlHonWjWqxVrWi1FpbWSVl7HH8HWGutE8PsqlnuvNX5NpzLoXvi81x1LCUrpXa66kkr28LgeQeL5Zfajc3mOoZDxtznOsq03LEKpiZ6gxtDBYGm079SwOFs6k1u42pWTsuqKdz1B5Y+XPRvK5wmy/hZo2rVxao1J4zMcxrQUKuY42aSqV5RTairRhGMU30whBNyacn1/4s+1v5UuHvvsHpHH51r7MabcYwyfBulhVNPtLEYjoTjt+NTjUW6tfe37HkW5zc45yMv1vneO4e4fSuD03jsJhsFTpY6WLlVjVpzlJTqOEE5RcE9oraaVtrsO1AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB/G1hpTItdaVzjReqMBHG5RnuBrZdjsPPtVoVYOE437q6bs1unZo/sgDx0zzlw59fZ+a1znOuWjEZxqzQ+Y1JVLZbglmMatJX6Fi8BaU41oKydWnG1k7SSbiqV/aZe0PzXDQyHLuDGV4fMZScI18JozMZ4qU99lCdWUG15KHh857HADyS5deSDmd5neNuX8f+dN5thcpy6tRxfwHO4xpY3Mvdy66eFhhIqKwmGUm3OLjC6bUY3k5R9bTof7UPm240crb4YS4Q5xl+BWpKmbzzJYvL6eJ98sN8D93BdafSv74qXcbN7Wasd8AAAA8ffbd/Kzw2+juJ+0npRynpvlZ4OW/zA09/R1A81/bd/Kzw2+juJ+0npVynfuWeDn0A09/R1AD6n0sdLJeyIuwIauFsgWSTAqLpErvucXM8wwWU4OrmOY4iFHD0I9U5zfZfm3beySV220luxbJN1sltkk7mY5hg8pwVXMcwxMKGGoR6pzk+3h4btttJJXbdkk2z4FrjXGN1fjVGPXQy+hJuhQvu3uuudtnJpvbdRV0r3bbXWucdq/GqMVOhl9CTdCg33e66522c2r2W6im0r3bbQuh8drHG9Uuuhl1CSWIrpd/Hoh4OTVr91FO7vdJ+Lk5MuXLyYej9B0nSYdHh8vz+v8Ar6zQ+hsbrDGdT66GXUJJV69u/j0Qvs5NNb7qKabvdJ/e8uy7BZTgqWXZdh4UMNQj0whHsvHu9223dt3bbbbbbZOX5dgspwVHLsuw0KOGoR6YQgtl4vvu273bd2223u2zkJb9j0cXFOOfO+X1fWZdVl8JPSAJaSIOrxAIeyITfiA6WOllit3cAtu4buhdsgAVae7LdiHd7eHzAQk2Olkol7AV6WVcXvui12QAWyAAEdSILWj4lbPyAWbIJ3WxD7AOpBuxWz8iwEWvuhZoO62uN3sBBXpZYAV6WT23Yb3I3ewE9SIs0LPyF2wKtXY6WSFfxAjpY6WS9it2AAAAjqRJFkBFn3Fmxv8Am+YbrYBZoq2kTdkSAkholdiG9wIs0QTdsWfkBXpY6WSQ20A7bsdSI3ews/IAVauywAr0sdLCbZYCvSyCby8SO4APsQ7+BDb7MCATZ+RBlm1REiHJdNt7ltmVlFWbS3JbHzrXnMDwd4Y52tOa61xhcozKVCGJWHqUK026Um0pXhBqzcWu99j9Vo7WWmdfacwmrdIZrDMsox/vPg+KpwnGM+ipKnOykk1acJLdLt5bnUPm+5VuLnGTixS1fonA5bWy6GUYfBuWIxsaU/eQnUbVmr2tNbnYPlk4f6j4W8D9N6D1ZRoUs1yz4Z8IhQqqrBe8xlarC0ls/iTi35O68D054cePFMpd34OmpNXb6hJXd0LW3ZJV3PMqJ6kQQ+q+yf6ATeykS7lS0u5ErNO3f0YVO6E7lH2LbrYh3s9mFS1EZKK3T7iXYRUbfG7+ol2CverdIzj3LPuRZE1SSqRYh7IxSH3IACoh7pootu5o+xRpeJldJ6BEiSJEqivS277WIkaeBnINijT3ZBLb3XgAqdwiXxrehayt6lW7GX0Wq00rsjqRLd+5FkYqHdbENNFiGl4hsulH2CYl2diAqUs0RdIs+xWyJq4o90R0ssVbfgYpAACojqRDdybR8CGrMKiCGrsm68wQqXTOzRC7mkkij2YWSKtPcm7YClCLb3LtFG32FVDqRFn3Fn5D43/uiVxDaRWW5ZpeJUNivSxZosRe7t4BfoqQ2tyz2ZV2/OZapAFn5AlUu0XSKNXL7Mq722CojtsyFsxuyAqJe7KWaLFbsyqRdIkiyZJKlWm7lOll97+gla23cKilmiCbvsBbpcQ+xRq5dtb7lHfwJVFJbJmMjaXZmMjK7YM5ux1i9ow0+VTUf+mZf9qpnZyZ1i9own96pqO//PMv+1UyuH9Zj9Lz+Jf1Lk+iuf7Ez5Bdd/S9/YsOeip51exM+QXXf0vf2LDnoqfcfy0AAHkNz/8APrzMaR5itV8B+H2s6GidPZPiMDhYYrD4SEMXUVXC0asqtTETUpwi5VXJOkovoUe+7f8AC4H+zxxfNRjoay4nc6OmdUYzERdfE4fI83lnuaJOzcK0684ujLdt3hNbK107r0+4ucq/Lzx1rTxnFXhLkOd4+pBU5Zi6Lw+OcErJPE0XCq0klZOdl4WOmPGv2SXLZp7CVNX6V45ZnwsjQ+PTr55jaFfA0ai3TU6kqVSPq3VbXdeTD7nwq9l/yf8ADCNGviOHtTWOY0rXxep8U8ZGVrPfDxUcO02vGk33V7N37RZHkGR6Zy6lk+m8kwGVZfQVqWFwWGhQo01/3sIJJfmR4U5tzY8yHK1qajpzhnzmYHihlVJbqn8JzPC0oq1qcnjqNo38Pg9WSS7yT2PSj2c3OFxD5uNGarzDiNpvJcvx+l8ZhcLDFZTTrUqOLVWnOTvCpOfTOPQm2pWfWrRjbcO34AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAONjMfgctovE5hjaGFopqLqVqihG77K7aVzg/sv0l/nRlP8ep/wDtHw/ng5Wcx5uuEuW8NMs1pR0xVy/UGHzt4qtg5YmFWNPD4ik6ThGcWruupXu7OHbe66N/gOtWflEZT9XKv9oA9VP2X6S/zoyn+PU//aH7L9Jf50ZT/Hqf/tHlX+A61Z+URlP1cq/2gfgOtWflEZT9XKv9oA5ntvc3ynNf2l/uXmmExnuv2R9fuK0anTf7m2v0t2vZ2v3sz1fPJnC+w91GsVR+G8w2W/B/eRdZUtO1FPov8bpvXte17X2va56zAAAB4++27+Vnht9HcT9pPSnlPduVrg4vPQGnv6OoHmt7bv5WeG30dxP2k9KuU+33rPBz6Aae/o6gB9Uvfaw6fUWS7dxv6/oAi/oEr7k2XkcXMcxwWUYOrmGYYiFDD0I9U5y8F28N229kldttJbi3U3WyW2STuZlj8FlGBrZjmGJhQw9CPVOcuyXh23bbdkldttJK7PgWudc43WGNtHqoZdQk3QoX3b3XXO2zk03tuoq6V7tudda4xur8Yox6qGXUJN0KF92911zts5NN+airpXu240NobHawxrlLroZdQklXr23b2fRC+zk01vuop3d7pPxcnJly5eTD0foOk6TDo8Pl+o9fs/7mhtDY3WGNbl10MuoSSr17bt7PohfZyaa33UU7u90n98y7LsFlOCpZdl2GhQw1CPTCEfDx7vdt3u27tttt3GXZdgspwVLLsvw8KGGoR6YQj4ePjvd3u27tttve7OSeji4pxz53yur6vLqsvhJ6QD7BvbYi8jq8aALPyJ2t6gQQSAK9XoLX3uTZC67AQnYX9A15IJb9gKtXJLWiVbV32AhuwvfawsmQ1uBPT6kC7AEAACrdyer0Foi0QIbuA9mABXq9CxSz8gJtfe4TsStl3FogQ3cr1ehZ7MrZALX3uLW3Gy2Et1tuA6vQqTZ+RNogVDdiXa+xFkwIvfaw6fUhrcXa7gAABBHV6FiLRAjq9A3cm0SHswIAAEX9Ba+5D7slbLcCFsT1egtEh7MCAncDZARa246vQS3W25Fn5AATaJDtfYCG7C/oNmLegBq5V7E3l4kbvcAVezLq3iQ0rsCOr0Kve5a0SPm7Cqm1GrFvANLxIfay7+hDXWPmL50f2g+IFPQv7W33d68uo4/4V92Pgtuuc49PR7ip26L3vvfsrb/XuB/E/wDbl4XZLxK+4f3H+7Hwn+8vhXwj3XusRUo/4Toj1X911fiq17b2u/nnHHk50Tx21pDW+otT57l+KhgqWBVHBe5924QlKSfx4N3bm7722R9O4R8NMq4P8Pcq4dZJj8XjcFlHv/d18X0+9n72vUrO/SktnUaVktkvHc9Od4vkpMf0ve6zWp8X68r4liLRPM2Ic7O1iO+5NoeNv0lXdXS7eBnZRLuUtbe5bdh3s9mYpVu4ctuwVvEStZ2Cora+4l2IV/BMmXYNVM49y7e5WyJqptJDdiSHuYuXar3AAVApNWLlJf8AfGVcF2Ddh4f7CFv3JVC/oUkW2uJJBU7M32ITsWdtyLRCkENXA7CqiLepBMns7Pcpv6/oJXEkN3J8NyHbwCoghKweyIuwqIDV9yzS8CjbRN2pBDdmSVfcxU7idg3cgBUqErbkS8SXsirbb9PmCsfVVK5N7bWJskGl4k2qird9irXiS9kPAxWtISuQ9ri7QCoq3ci29y1ku5VrujdbXKjq9Bf0I7E2jYhc2q1ci29iXfwKu/5wqRLVirXiSm33KtvdeAV3iCGrbkkO+6sZdKOr0KlktiCVTavT6kEttEO9tgqBQm7Fn5BUQRe+xYr23TJqkNWBEnIjf1/QYqdxvwIJs3uQFRDXiQnYnuiLPyC5UNXdyrViz2uVu2ZY2M5vuvQyaubzirPzMJNxJrthd+jKe1/Q6xe0Zd+VTUm3/wAty77VTOzk22mdYvaMXXKpqO6/+WZd9qplcP6zH6XDxL+pcn7tc/2JnyC67+l7+xYc9FTzq9iZ8guu/pe/sWHPRU+4/loAAPIP2gPPpzK6O5jtVcDdB65p6H03klbBYWOJw2BSxU1VwtGrOtVruE6qjerJr3Ki+hRspPd/z+B3s/8AJuanMKWrOKHPJkutsyq/HxGByLMqmaZlZ3clOrjJRq0nZN70JJ2bV0rv004vcrHL1x4c6/FXhTkWdY6pFQeZe5eHx9kulL4TRcatkkkl12VlsdLuKPsWtA4yvLOOB3FnO9M42lJVsPg84pRxtCNRO6UK9P3dWklZNNqo0167B2G4V+zV5P8AhS6WJw/DClqjMKTi/huqKzzGUmls3Rklh0773VJbvySS7L5XlOVZHgKOV5LlmFy/BYePTSw2FoxpUqa8owikkvRI8k8Rof2ufKVvprP854gacwfxYxwdeOoqM4LtCOHrxeLppWT/ALnCKVmk7Np9yOQLmy4lc0OndX0uK+g8FprP9G43C4KusJRr0I1nWpzlaVCu5Tpyj0K6cndSWy8Q7YAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADx99t38rPDb6O4n7SelXKd+5Z4OfQDT39HUDzV9t38rPDb6O4n7SelXKf+5Z4OfQDT39G0APqoBxcyzHB5Tg6uY5hiYUMPRj1TnJ7Jdvnbd7JK7baS3Fuputktsk9TMcxwWUYKrmOYYiFDD0F1TnLwXbw3bd7JK7baS3PgWutcY3WGOtFSoZdQk3QoN7vuuudtnJrt3UVdK923OuNcY3V+NUV1UMuoSboUL7t7rrnbZyab23UVdK+7bQ2hsbq/G9Uuuhl1CSVevbdvZ9EPByaa33UU7u90n4uTky5cvJh6Pv9J0mHR8fy/P6/wCvrV0PoXHavxvVJzoZdQklXrru+z6IXVnJpq73UU7u90n98y7LsFlOCpZdl2GhQw1CPTCEfDx7vdt3u27tttt3LZdl2DynBUsuy7DwoYahFRhCK2Svv6tvu2922292ciz8j0cXFOOfO+X1fV5dVl8JPSFmiCbtkHV40JEkkAR1IPfdE2RV3vbwAAAAVa7sl38Be6AiJLa3RHbsQAIauybPyAEJWJfYEO/j2AqWbuhaJUAGrgO9tgI6WOljf1/QReXiBPSyBeXgABDdiSAItfdCzQd1tci7YAAAQ1dhIkhvbYCShN2LPyAgJWAAFW7liLRAN3RAAAr0ssQ7+AEdLHSxv6/oG/r+gCAS0+7IAEd90SQ9tkBFmiCbsgAQ0SAISJDe2xW7AAACqRYACrYTsibRIdr7AQRdXsSQ1423AjpZN0tmRdk7Wuw2KSIXcs0vEqu5NU6Mc6PMpxr4TcYaWluH+tPuVlcsnw2KdD7nYSverOdVSl1VaUpbqK2vZW2W7OxvKvrjVHEjgNpjWmtM0+6Oc5j8N+E4n3FOl7z3eNr04fEpxjBWhCK2Sva7u22/7+uOGXBjVucxzXXukdMZnmaoRpKtmNClOqqSbcY3lvZNu3huz9FpTItMaayHC5Jo7LsBgMnw/X8Gw+BjGNCHVOUp9Kjsrzcm7eLZ6eTkwy4pjMdWOkssk0/rFWnuy72ZRvfueVqsu5K7EqKa3KtNN2va/kL3Wm6RDat2YW63KvsSqVD3ZV9iSWlbwCoQ7fnKy7E3a/F7ETul2YVFGrsokafP3M02yapYhuxJWW623MVNjdyAAqIukVl8a3oXsikla1jKuF12BG1r+IiyVS0fchpoO++xVuXjcNQ97lWmixEgqIs2RKL2Ju+wbfiFxQEtIo20SqJfYhJsnutyHt2C4rLsVLvsUDZ3TZoglt+JBNWrZoi6RN2yr7mKQlcNWACppDZUs0kVCpsIauySG34E3Sogh9iSrbva5ikAtZENemwVJpRu5BayXcrK24VESIDb8QTVRDaRUu15opJPcLgQ1cLdEu9tgtVqxF0iW7lXb85NUdSKlkkw1GxilepFS1o+JUKir3ZPUiHsxZ+QVLoe7KWaLlLtmVSLpEkWTJJUhtboo90WfciV0FRXsyxX1F2u4VCXcqtu5LfiV3ZlVPgrLdsynF7djZ23v3MpvsY6Y9nHkrK7OsXtGWnyqaj/ANNy77VTOz89/wBJ1h9o1FLlU1HZf/LMv+1UzeH9PH6XHxG76Ll+iub7Ez5Bdd/S9/YsOeip51exM+QXXf0vf2LDnoqfcfy4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHj97brbizw3t/m7ivtJ6V8p7f3rPBvb/iBp7+jqB5qe26+Vnhv9HcT9pPSDlhzHB5Rym8IcxzHEQoYejw+09Kc5PZL7nUF+dt7JLduyW4t13pJbZI+qZjmOCyjBVsxzDEwoYajHqnOXgu3hu23ZJK7baS3PgeudcY3V+NtFSoZdQk3QoX3fddc7bOTTe26irpXu22udcY3V+NUY9dDLqEm8Ph34+HXPwcmuy3UV2vdttDaGxur8a3JyoZdQkliK6W77PohfZya7vdRVm77J+Lk5MuXLyYej9B0nSYdHh8v1Hr/AK+s0NobG6wxt25UMuoSXwjEW7+PRDwc2u73UVZu90n98y7LcHlOCo5dl+GhQw9CPTCEfBd/nbb3bd2223uMty7B5TgqWXZdh4UMPQj0whDsl3fq2+7bu22292cl38Lno4uKccfL6vq8uqz+EnuSQ3bYK997k7HV41Cb+hBZWtvYCG77EX3sWdreBV29ABDV2Rv6lr7bvcCrVtwF33LO1nawFSLepIAr29bi+9y23oyNrWsrgR1egv6EWfkABF77WJFgKtWFvUb+ouwDViCSqv1eNgJbtsVbvsWIdreAEJ22Dlt2JSVg7W8AKdXoS3YWXkHuBFr73DVtyVsu5C77gQCX3IAhq7Frbkjb0YFXuT1eg29CLPyAPcgABfexVqxb1tuV39QJt6kNWF2O4AhuxG9/HuT3Ajq9B1egdrbWCSsAv6FW7Fna21iO4Ahq+4fYLtuBFvUgtt6Db0AqGr7gARa25Ddy23oxsBVu+wFn5AARfexJHrbcCGrbi3qN/Ui7AlqxVvwJ7kO2/a4FSb7WILbW8A2KtXIWzJfpchJ3M96nnjz58L+JesOONHNtJcO9T53gVkeFovFZdlOIxNJTVSq3Drpwauk02r3V15naTk7yDPNM8uWkcj1JkuPynMcN8P8Af4PHYadCvS6sdiJR6oTSlG8ZJq6V001s0f0+KHNBwb4QalhpLXef4rB5lLDQxap0svrVl7qbkovqhFq7cHte+x+y0Br3TXE7SOA1xo/GVMVlGZe9+DValGdKUvd1ZUp3hJJq04SW63tfsz08nJnlxY43HU+Lpu6m4/RJX3KtWuSDyNVTtsQ5+Fi23oUad27Ow7bWlO5R9iy2W4dreBNJdMyH2LPYSSs7pB0iidti0nddiV0+hWXYNnqo1dlU77EvuSunwsTdLQVtbcs+5Euxioq97kJW3JZCvfe4VElJS7bFyrszKuK2vvchOxO/ra4dvBEqkL+hExv6h+obtQiRL8RsFTaobuGQFQfYqWKvYlc3UX9CG7liNgqKkdPqTLxK3fmFF/Qhq+4Bl0tUhuzJKvuSpABKStuFRXp9SHsS9irfqFQBXf1LLtuTYpWUtit97liHbfsYoTuQ3a5ACojv6WKtb38iz27FHfq8bBWPqhu5N/QSIJq8fUbuVb8CXdeZUKiG7bC+1ySrvv3C+9G7lWvEkbE1SpNvUnb0K7+piohq+5UuZvsFQtfcdXoQQFRLdyjVtyw2JqlG7bEiVr9kV39TFTuluzIl8ZW7C2133KvsFQtbe5Ddx87Zay8he6ooR1ehMl5FexlmlTSJK92ZTVjSV9zOfh3MdMWMv/WdY/aNfuU9SLzxuXfaqZ2ckdY/aN/uU9R/6bl32qmbw/p4/S4eI/1Pl+iuX7Ez5Bdd/S9/YsOeip51exM+QXXf0vf2LDnoqfcfy8AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHj/7beEp8WuG0Yq7ensSlbvf4SdtuGGe4/EcDOGWmJZjh8TgMi0hk2Cozwkp+4xLpYGlT9/FzjGTU7Nx6oxai0rJuV+r3toYSwfFfhnmWLwc1ha+Q4yhSxDi+n3kMQnKKfa6U4N+KUkfH9J+0U1Dw70rp3Sc+HWVZjhMky/C5dHox1SlVr0qNKNNScumSi2op36Wr9lY8vPc8r5MY+z4bhwceF5+S959T1C0NobG6vxrlLqoZdQklXr23fj0Qvs5NNb7qKs3e6T++5bl2DynBUcuy7DQoYehHphCPZL/ANbfdt7tttu54/ZT7aTibkuW4fK8DwR0lGjhqapxvjcQm7d5O1k23dt2V22/E5X4bniz/AlpL+PYr+s68XFOOPF1nV5dVn/7fdHsDb1IPID8N1xZ8eCWkv49if6x+G64sfwJaS/j2JOrxvX8hq+55AfhueLH8CWkv49iv6x+G54sfwJ6T/j2JA9f+n1Iex5A/hueLH8CWkv49iv6yH7bjiw/+RPSX8fxP9YHr8Lb3PIH8NxxY/gS0l/H8T/WPw3HFj+BLSX8fxP9YHr8Q1dnkF+G44sfwJaS/j+J/rH4bfix/AppL+PYn+sD18atuDyD/DbcWH/yKaS/j2JI/DbcV/4E9Jfx7E/1gevpB5B/htuK/wDAnpL+PYn+sn8NtxX/AIFNJfx7EgevXb1uL73PIR+214rv/kU0l/HsT/WR+G14rfwJaS/j+J/rA9fOr0IPIR+214rv/kT0l/H8T/WPw2nFf+BTSX8fxIHr03YdXoeQv4bPis/+RTSf8exJH4bPit/AnpL+P4n+sD176vQhrxueQv4bLiv/AAKaS/j2JJ/DZ8V7W/aV0l/HsT/WB68A8hvw2fFb+BPSf8exP9Y/DZcV/wCBXSf8exP9YHryR0+p5D/hsuK/8Cuk/wCPYn+sfhsuK/8AAppL+PYkD14vbYg8iH7bHis/+RXSf8exP9ZH4bDit/ArpP8Aj2JA9eAeQ/4bDit/ArpP+PYkfhsOK/8AArpP+PYn+sD13auyGrbnkR+Gw4r/AMC2k/49iSX7a/iu/wDkW0n/AB7E/wBYHrqDyJ/DX8Vv4FtJ/wAexP8AWPw1/Fb+BbSf8exP9YHrq1di1tzyK/DX8Vv4FtJ/x7EkfhruKz78FdJ/x7E/1geur3J6vQ8ifw1vFX+BXSf8exP9Y/DW8Vf4FdJ/x7E/1geur3IbseRf4a3it/AtpP8Aj2J/rD9tZxVf/IrpP+PYn+sD1z6vQdXoeRf4azit/ArpP+PYn+sj8NXxW/gW0n/HsSB66teNyDyM/DWcVrW/aW0n/HcT/WR+Gr4rfwLaT/j2JA9dAeRf4avir/AtpT+PYn+sfhq+Kv8AAvpT+PYkD1zt6kXtseRv4avir/AtpT+PYn+sfhquKr/5FtJ/x7E/1geuIPI78NTxV/gX0p/HsSPw1PFX+BfSn8exIHriQ1fc8j/w1PFX+BfSn8exJH4arir4cF9Kfx3E/wBYHrhb1IPJB+2p4q/wL6U/juJ/rI/DUcVP4F9Kfx3EgeuAavueR/4ajip/AvpT+O4kfhqOKn8C+lP47iQPW+1tyG7nkh+Gn4qvvwX0p/HcT/WPw03FT+BbSf8AHsSB64dXoQ9zyQ/DTcVf4F9J/wAexP8AWPw03FX+BfSf8exP9YHrc3Yjq9DySftpuKj/AORfSn8exJH4aXip/AvpP+PYn+sD1u6vQNX3PJD8NLxV/gY0p/HcST+Gl4q2t+0xpT+O4n+sD1tIauzyS/DS8Vf4GNKfx3Ek/hpeKn8DGlP47iQPWzp9SHseSn4aTip/AzpT+PYkq/bR8VL3/aZ0p/HcT/WGzT1sF97Hkn+Gj4qfwMaU/j2JI/DQ8VL3XBnSn8dxP9Zlm27dqua/k+4mcc+KFPW2ks80xhMDDK6GBdPMcViKdXrhKo27U6M1a01Z3vs9l4/deXPhtnnCHg1p7h3qXF4DE5jlPwv31XA1JzoS97i6tWPS5xjJ2jUSd4rdNK6s35+5N7YPWuZ4N4jM9F6PyyspuKoTWLqtpJWl1KaW92rW8D+bmvtk+I2Bx9XC4DhZpPMKFO3TiY4jE0lO8Ve0W21Ztrvva56M8+S8cxvpF3L82berpDfgeTH4Z/in/AzpX+PYkh+2e4pP/ka0r/HsSebR549Z0r7kvZHkuvbP8Ul24NaV/j2JD9s/xSt8jOlf49if6zLLVfKYvWZq+5U8mvwzvFL+BrSv8dxP9ZH4Zzij/AzpX+PYn+szy1U5MY9ZWr7kSlszyc/DN8Uv4GdLfx3E/wBZV+2Z4pNfI1pX+O4n+seWqnLi9YS0ux5OL2zHFH+BnS38dxP9YftmeKTXyNaW/juJ/rHlrflsXrA1dkKFne55P/hl+KX8DWlv47if6x+GY4pfwN6X/juIFx22c+D1gasQ1c8n37Zbik/+RvS38dxH9Y/DK8Uv4HNLfx3EE+Sq+Xwerz2B5QfhlOKV/kc0t/HcR/WPwynFH+BvS38dxA8lVOo43q++xRux5RP2ynFB/wDI5pb+O4gh+2S4oP8A5HdLfx3Ef1i4WtnU8cer3gRE8ovwyXFD+BzS38dxH9Y/DJcUPDg7pb+O4j+sz5OqnU8fxerxWZ5R/hk+KP8AA7pb+O4j+siXtkeKEv8Akd0t/HcR/WPk638K4/i9WirVjym/DIcT/wCB3S/8dxAftjuJ7/5HdL/x3ED5OqnV8Xvr1ZDVjym/DH8Tv4HdL/x3ED8MfxP/AIHtL/x3EGfJ5NnWcU971YtvciXc8p/wxvE/+B7S/wDHcQR+GM4nt/I9pb+O4j+sy8WXwVOt4p73qvfexEjyp/DF8Tr3/ae0t/HcR/WQ/bF8T3/yP6W/juI/rHyeTZ1vD8XqsR0+p5U/hiuJ38D+l/45iB+GK4n/AMD+l/45iP6x8ll8FTruH4vVW/oQ1fc8q/wxPE3+B7S/8dxH9ZH4Yjid/A/pf+OYj+sy8OV9yvw7h+L1SauVex5Xfhh+J38D+l/45iP6x+GG4mt/JBpf+OYgz5HP4NnX8HxeqSdg3c8rPwwvE3+CDS/8cxH9Y/DC8Tf4INL/AMcxH9Y+RzbPEOCe96otXKtbnlf+GE4m/wAEOmP45iP6w/bCcTX/AMkOmP45iP6x8jmqeIcE971QIauzyw/DBcTv4INMfxzEf1j8MHxO/gg0x/HMR/WT8hn8G/jLp/i9TmrblXuzyzftguJvjwh0x/HMQQ/bAcS3/wAkOmP45iB8hn8FTxPp/fXqb0+ot6nll+GB4mfwRaY/jmIH4YDib/BFpj+OYgfIZ/BU8U6ae96lyKnlr+F+4lv/AJItMfxzEEfhfeJn8EOmP45iP6x8hn8GzxTpvi9SW7k39Dy0/C+cS/4IdM/xzEf1j8L5xL/gi0z/ABzEE/g+bZ4r00971KbuUfc8uPwvnEr+CPTP8cxBD9r3xKv8kWmf45XH4Pmr8a9N8a9SUr7kS7Hlwva+cSv4ItM/xyuQ/a9cSfHhHpn+OYgfg+ap4t0s99/g9RSvieXX4XniV/BHpn+OYj+sj8LvxLvdcJNM/wAcr/1mXp86r8b9L+1XqP0+pDlt2PLr8LxxL/gj0z/HMR/WQ/a78SX/AMkWmf45iP6zPwbk+Dfxx0v7V/g9ROr0IPLz8LrxJ/gj0z/HMR/WR+F24k/wSaa/jdcfg3J8FTxnpJ/av8HqH0+pCVzy+/C7cSf4JdNfxuuR+F04k/wSaa/jdcfg/J8G/jnpP2r/AAeoTVijVtzy/wDwunEp/wDJLpn+N1/6w/a58SWvkl0z/HK/9Zn4NyX3N/HXSfH6nqADy+/C5cSf4JtNfxuuPwuPEn+CbTP8br/1mfg3J8Ffjvo/j9T0/b8CDzA/C4cSf4JdM/xzEf1j8LfxJ/gm0z/HK/8AWPwbk+Cp430c/tfU9PXsx1HmD+Fv4j/wTab/AI3XH4W7iR/BLpn+OYj+sfg3J8G/jzov2r/B6et+JVu55iP2t3Ej+CbTX8cxH9ZH4W3iP/BNpv8Ajdcy9LyNnjvRftfU9O32MpnmR+Fs4j/wT6b/AI3XIl7WniNK1+E+m9v/AAuuZ+C8nwXj490U/tX+D0yl27nWP2jX7lLUn+m5d9qpnWZ+1l4jNfJRpv8Ajdf+s+e8eef7WPHrhpmHDXOdA5NleFzCth60sThsRVnUi6VWM0kpbNNxsyuPpuTHOWztHLrPG+j5umz48Mu9lno7zexM+QXXf0vf2LDnoqednsT6FeHADW+InRnGlV1fJU5uLUZNYPD3s/G11f5z0TPpvwwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPz2s+HugeI2WwyfiHofINUYClUVWGFznLKONoxmv3yhVjJJ+qVz8X96fysvd8tXCt//AJm5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrP5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA+VfencrH5NPCr6m5d+pH3p3Kx+TTwq+puXfqT6qAPlX3p3Kx+TTwq+puXfqR96dysfk08Kvqbl36k+qgD5V96dysfk08Kvqbl36kfencrH5NPCr6m5d+pPqoA/lac0vprRuUUNPaQ09lmR5XhrqhgctwlPDYeknu1GnTSjH8yP6oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB0J5ifa18MuFOtMbw44VaGxnEfO8vrvCYnFUMcsNgI4hOzp0pxhUnXlGV0+mCi2rKT7r5zk/tms1yLPMNguMXLFnGQZfirS9/hMyl8JjBO0pQoYijTVXuv+Ejbzd9g9PAfmeG/EHTPFbQeRcR9GYueJyXUWCp4/BVKlNwn7uavacXvGSd014NPv3PPPgdzJccc/wDaias4O53xHzXG6Lo5rn+Eo5LVcHh6VKhSqSpKC6bxcXTjunfum3d3D0yAAAAAAAAAAAA/EaI41cIeJecZlp/h5xM0zqTM8mu8fhMrzOjiauHSl0tyjCTaSkrX7Xsr7gftwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA+P813MFDlg4J5xxjnpOWpPuViMJQWXLHfA/euvXjSu6vu6nSl13/Ed7W2vcD7AD5Byp8wNLme4J5NxipaVnpz7q1sVQllssasX7qVCvOk2qvRT6k+i/4ite29rv6+AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPl/M9nGodPcuPFDPNKKos3wGkM3xGEnTdp06kcLUaqR85RScku7cUle9j6gcbHYLCZlgsRluYYalicLiqU6NejVgpwq05JqUZJ7NNNpp7WbA8pvYl6e4aY7MuImf47DYHEa6y9YOGBlXUZV8Nl01UVSdBNXSlUUYzkrtJU07KXxvULWug9F8SNPYnSev9LZXqDJ8ZFxrYPMcNGvSls0mlJPpkr3UlZp2aadmeUHMR7OLmB5cuIlXjVybZhnGOyyjiJYnC4HKMRJZzlMZO8qKh3xdDfpSXVNxdpwkk5v9Jy/+2H1PkGbUNC812iXfD1FhcVn+WYWWHxeGmnaUsVgmrSaf43ulBpJ2pybsB6e6I0TpbhvpHKdB6IyinlWQ5HhoYPAYOnOU1RpR7Lqm3KT8XKTbbbbbbbPJfgbm2V5D7X3XedZ3mOGwGX4HONVYjFYrFVY06VGlHD13Kc5yaUYpJtttJJHrhpjU+n9aafy7VelM4wua5PmuHhisFjcLUVSlXpTV4yjJd01/LdO1meHeo+A9XmV9pdr3hAtQvJMJm+r84q47Fxg5TjhaEp1asYR7OclC0er4qbTd0rMO+uuPbB8qGkdTVNP5ThdZ6rw2HqOnWzTJ8to/BW1dN0niK1KdRKSSuoqLTbTkrX7KcAeZLhFzMaVq6s4S6mWY0cJONLH4OtTdHF4GpJNxhWpS3jdKVpJuEul9MnZ2/gZHyU8qmQ6CXDmhwK0jicpeH+D1quLy2nWxtd2SdSeKkvfOo+/Wppp26WrK3m1yN5bjeXT2nefcCMizPEVclxuIzzIJJyuquGoUKuMw0qie3XFYeC6krpymk7Sdw9lTrZzB+0G5aeXDN6umNY6pxecajoK9fJcgw6xeJoelWUpQpUpf95OopWs7WaZ+l5z+NeO5fuWnW3E3JqkYZxgsHDB5S3HqUcbiasaFKpZ7PodT3jT2aptb9n0Z9lbye6C4m6VzbmX415BQ1dmGZZtiMJk+FziksThviNOvjakKiarVJ1ZTinJNRdOT3ck4h9i0t7ZTlUz3NIZdnmRa905h6lVxWPxuV0K1CnC6SlNYevUqp2bbUYStZ7s7raM1ppPiJpnL9Z6G1Dgs7yPNKXvsJjsFVVSlVjdp2a7NNNNOzTTTSaaPk3HDkv5e+OmiMdpHO+G+n8pxlXDzp5fnOV5XRw+Ny6rb4lSnOnGLlFSs3Tb6JWs14roP7I3iVqvhvx217yq6oxcqmCksZiaNFScoYbNcDWVKv0XtaNSn1Nu27oQ2V2B6A8y/Nvwo5UcDp/MOKkM7dHUmIr4fBvLMHHEOLpKDm5pzjZJVI2tdvfY/AcxPtIuXTlyzrDaWz3EZ1qXP69Cliq2XZDh6VWeCpVIqcHXnVqU4Qk4NNU03Ozi3FKSZ1s9uL/vT4S/6xzf+awx9J5GOQDhdlnCvA8V+POlsq4ga21/haec4mWfUIZhQwVDEx95CEY1VKMq0ozUqlV3l1O0WkryD6Pm3tLuWjIOF2ieLeeVtUYLKNfSzGnldCeVKeJhPA1KdPERqxhOUItSqwtack09nszzT9ntzT8KOW/jjrrX3EvFZpRynP8AK6+EwcsFgnXqOpPGU6q6opqy6YPfzsexGE5X+XTC6VyvRFXgjovH5DklbF4jLMBmeTUMfSwVTE1PeV3RWIjN01OSTai0vixXaKS8sPZWcMeGvEPmR4lZFr/h7prU2W4HJcRVwuCzjKcPjaGHmsfSipQp1YSjFqLcU0k7NrsB6e8uXNbwi5qMtzvNOFGNzOvS0/Xo0Masdgnh5RlVjKUGk27pqEt790fZD8vofhjw34ZYXE4Hhvw901pPDY2oquJo5HlOHwEK80rKU40YxUmlsm7ux+oAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAdPvaw/uJtW/6xyj7bSO4J0+9rD+4m1b/rHKPttIB7J79xNpL/AFjm/wBtqncE6feye/cTaS/1jm/22qdwQP52f6hyLSmTYzUWp84weVZVl9J18XjcZXjRoUKa7ynOTSil5tnSjXXth+VDSmZVsr0/g9Z6u9zLpWMyvLKVLCzaaTtLE1aVR7XaaptOy3s7nxH2wnF3Vup9f6F5UdHV6nucyhhs1zDD0m18NxmIryo4OjNrdqDhKfTazdWD3cVbuxwB5JuAPATQuXaXy7h7kGd5vSoRWZZ9mmWUcRjMdXa+PNzqRk6cG79NOLUYqy3d2w/K8BvaUcrvH/UOG0dkeoM101qDH1VRwOW6kwcMLPFzbsoU6tOpUoubdkoOopSbSSbul2oPLX2rPJpwz0bw6w/MZwm0xgNKZjlWZUMLnmEyqgsNhcTRry6aeIVOmlCnWhWcE3FLqVRttuKv3D5B+NWb8euVnR2tdTYieIz7DUauT5rXl3r4jC1HTVZvxlUpqnUl2XVOVlYDk6W52uBeqeM+seBlPMsyyzPdD0cZiM3xeZ4eGHy+FPCzjCvJV3NpKLmneSirJu58W1p7YblQ0pqSpkWU4HWuqMNQqyp1czynLaCwztdXp/CK9KdRXS36UmndN7J9EM+4H47mO9pZxC4N0NVVsgwWf6rzd5piaV3OWCoVJYipTUVtKTdGPSpfFUlGTT6Un6zZHyU8qmQ6CXDmhwK0jicpeH+D1quLy2nWxtd2SdSeKkvfOo+/Wppp26WrKwf3+APMlwi5mNK1dWcJdTLMaOEnGlj8HWpuji8DUkm4wrUpbxulK0k3CXS+mTs7fUTxq5G8txvLp7TvPuBGRZniKuS43EZ5kEk5XVXDUKFXGYaVRPbrisPBdSV05TSdpO/oR7Q7W2oeHvJnxO1LpfG1MJmHwDC5dCvTk4zp08ZjaGFquL7xl7uvOzW6dmmmkwPw/Fz2q/Kdwl1Ni9JfdLUWr8dl9eWGxktNYClXoUasbqUffV6tKFSzVm6bmr7X72/gZV7YnlDzHBxxOMp65yuo5NPD4vJacqiS7NulWqQs/C0m/NI/N+yQ4C8KqXLpR4t47SmV5rqrUWZ4ynWx+OwsK9XCUKFV04UKTkn7uL6HUdknJzV21GFvjftiOC/DLhvj+HnFjQumsnyLPs3xmKwePw+FwVGOHxyoRp1KdapQ6eicouTjJuLUozipXSSYeiHLpzQcKeaTTuaam4U43Ma+EybGrA4uOOwcsPUjUdNTTSd000+6fdO6W1/rh864B6d0bknCnTmZaN4e6e0bT1FlmCzrG5dkeVUcBQWKr4anKbdOlGKcltG7V7RSfY+igAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5JzW8P+I3FLl+1fofhJqetp/V+Ow+Hq5Tj6OLqYWcKtDE0q7pxrU2pU3UjSlT6rpL3l3tcD62dLfag8vPC7X/AC56r4s5pkOEwesNH4SnjsDnVClGGIqwVSEHhq0lZ1KcoyaSk24ys423T61ZXzse0r5dKK0lxn4A4zVkcH/caeaY7JMQ51WuyWNwbeHr2TV2k5PZuTbu/wAxxL1/7Qz2iODwnDDLeC9fSGi6+Ko1cW3gcRgMDVcX1RqYrF4l3qwg49ap0k3dRfTKSiB2F9i3qzUGccvWqdL5nOpVy3T2p5xy2c5XVONehTqVaMV4JTvU+etI+Gcu/wD25LWP+vtUfzFc9GOU3lxyHlZ4K5TwrybGLH4unUnj83zFQcfhuPqqKq1VFv4sUoQpxXdQpwvd3b6K8BeEPFbLPay6x15mXDXVGE0191tQ4qOc4jKa9PAyo16NWNGca8oqElNySi03fe17Owepx4/cL/8At2WN+kWf/wBCYs9gTyq4d8JOKmB9sLjdeY3hrqmhpn7s51jPu1UynELAe4q5RiadOp8Icfd9Mqk4wT6rOTUVvsB2I9rblWOzLkxz3FYOg6lPLM5yvFYlr95Sdf3Sk/TrqwX5zf2TWocuzrkq0vluCdH32Q5pm2X4zo7++li6mJXXbx93iaXfw6Ts5xP4dac4t8PNQ8M9W0Z1co1Jl9bL8V7tpThGcbKcG00pwdpRbTs4p2Z5J6S0dz1+zM1/n1LRHD7Fa/0HmtTrq1MJl1fG5fjKcG1TrzVBupgq6i2mpNJ9n7xKLA9ljxx5DVDiN7UTX+v9OylXymnmOrM+Vam7weGxGLnTpNvxTeJg1+nsmfqddc8PPhzP6exfC3gvyyZxpZ53SlgsdmeGwuKq16dOatOMcXWjSoYVSi+lznuk7xlFtNdrvZ7clH3pOg8wzDV+KwmO17qr3U81q4b49LA0IJunhKU2rys5SlOSspSsldQjJh149uL/AL0+Ev8ArHN/5rDHf/l9+QXht9EMn+xUjpJ7ZHhrxE4iaV4Xx0BoLUWppYHMsyWJjk+WV8a6Dq08OqfWqUZOPU4tK9rtNK53j4JZXmOR8GdBZNm+CrYPH5fpjKsLisNWi41KNaGEpxnCSe6akmmn4pgftzyD9j1+6m4q/wCocT/SNE9fDxbxujOaL2cnNRqniNoLhTmmrNIZxPG4fDYmGCr4jA43LsRWVWlCpUoJ+4r05xpq0kneMrJxldh7SA62clnHjj3zA5DqbVvGfg/+1/l9DF4ejp3DzwmJoVcVSdOTr1JOu06iUuhKcYRi7tK7TZ2TAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAdPvaw/uJtW/6xyj7bSO4J1a9pZoTWfEXlD1XpjQWl8z1DnE8Xltenl+W4aeIxNWFPF05TcKUE5Taim2opuybtsB/D9k9+4m0l/rHN/ttU7gnjHy98wHtHOWzhdl3CXQ3J7nGOyjLK+Ir0q2bcPs9q4mUq1WVSSlKlUpwaTk0rQW1r37n1jT/AD3e0+x+fZbgcz5MvdYPEYyjSxE/2v8AUFHppymlN+8niHGFk2+qSaVrtNID5l7QjGQ0F7TLQOuNRUadPKqFfS2ddddf3KeGoYy1RyvtZOhUT9Ez2ROm3tG+STMeazRmV6k4f1cNR17pKNWOBpYiap0szwtRpzw0ptpU5qS6qcn8VNzi7KfVHqzw257+eXlr03g+F3F/lnzzUzySisFgcdjsBjMJipUoJRhGVeFOpSxEYxVlOKu0k3KTu2HbH2rmostyXkk1lluOrxp18+x2UZfg4uVnUrRx9HEOKXj/AHPDVXbyTfgfy/ZG6dx2Scm+VY/G0qkIZ7nuZ5jh+vxpKpGhdLwTlQn/ALezOqGoeHPPR7TLXeQy4n6Ir8NOHOT1nODxWBrYPD0Iu3XWp0q799i8RKPxYySVNWavTvK/q/w/0LprhjojI+Huj8CsJkunsDSy7BUtm1TpxUU5NJdU205Sl3cm292B5S8u/wD25LWP+vtUfzFc9fDyx4C8IeK2We1l1jrzMuGuqMJpr7rahxUc5xGU16eBlRr0asaM415RUJKbklFpu+9r2dvU4Dx+4X/9uyxv0iz/APoTFnfj2gGvNHcP+UrX+P1vkSzrBZpgFk1DL/eOn7/FYmShSfWk3H3cmqt1uvdbb2OmfDvhJxUwPthcbrzG8NdU0NM/dnOsZ92qmU4hYD3FXKMTTp1PhDj7vplUnGCfVZyait9jth7SqPCWryk6pw3GDOcbluCrVaCyeeBjGeKq5tFueGp04SajO/TPrTaSpqpK6cU0HRHlO0f7S3gbwgynWnL7prKNT6L1zTWb0snxWIoVnhJtuCqulVqUp05VIwjJ+7lKLi4OVmrL6LlvJJzkc5PF7JeJHOzi8DpvS+T9MYZJhcVSdWWH6lOVDD0aEpwoqo0lOpUn7y1tpdMVH5ryi8Kvaeah4L5dnXBHil+xjRFeVT7jYTO8fD+6UlJpzw1OpRqunRc+qyvBSd5JNNSf0HiFxP8Aa98s+nJcTuJOZ5DqnS+Vyp/D4rAZZiKVCDkkpVlhadKuoNtJzTaV7tq6bD1Rw9CjhKFPDYajCjRowVOnThFRjCKVlFJbJJKyS8DY+H8nvM1lPNfwWwfE7BZS8qzChi6mU5zgerrhh8fShTnNU5veUJQq0pxb3Smk7tNv7gAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAOjftaeBXEbjJwJyTNuHGUYvOsRo7N5ZjjsrwkJVK9bDToypyq0qcbupKm7NxSb6ZTa7NPvIAPMflo9rTwU0Vwn0vw04t6K1LkGc6TyvDZJOrleBp18HVhhqcaMZqLqRqU5OME5Q6Gk7pN9l/C5r/aX6d5iuHeacvPLRw51dn2c60gssrYqvgEpug5pzhhsPSlUqVJzS6byUelSbs2tvSnUHCbhZq3HPMtVcNNK5zjH3xGYZNhsRV3t++nBvwXj4I5umNBaG0TGpDRujMiyGNXaossy6jhVPx3VOKvuB189nTy2ai5ZeXfDaa1vQp0NUahzGrn2bYaE4z+BzqU6dOnh3OLak4U6UHKzaU5TSbSTfaIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB54e0u54+N/LDxG0nojhPVyTCYfM8knmuMr4zALE1ak3XnSUF1PpjFKlfZXbk7uySA9DweRuB5qva7ZpgcNmWXcHczxOFxlKGIoVqeiHKFSnNKUZRaVmmmmvRnLw3M37YKWJpJcEMyn8eK6KuiXGEt+0ndWT8XdWW913A9ZwAAAAAAAAAAAAAAAAAAAPJ3V/G3n8pe0DqaKymvrWGlY66o4PDZZRypyy2pkXwmMVUk3SUXTlhm5zqt/FbcupNJoPWIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPGz22ny9aE+iC+24g9kzxs9tp8vWhPogvtuIA9ZeEPyTaK+juW/ZqZ+uPKnRvtpNLaV0jkel6nAHNcTLJ8tw2AdaOoKcVUdKlGDkl7h2v03td2v3Z+54d+2X0fr3X2m9D1OBOcZetQ5thMqWKjnlKt7h16saan0e6j1JOSbXUtr7gejp1V5j/aQ8uXLfnmI0dmuY5jqnVOEvHE5VkFKnWeDmu0MRWnONOnK97xTlOPjFXV/0HPxx6zTl25ZNTa405X9xqDHulkmTVvGjisS2ver/AL6nTVWpHuuqCumrnRT2WPJXojjFlWbcxXG3KKepsLDM54HJcszD+60MRXgozr4zERd1W+NNQjGd1eNVyUrxaD6Xl3tueFlXMvd5vwQ1VhsB1R/u+HzHDV63S18Z+6l0Runsl1797rsd1OAHM5wa5mtO1tQ8JdVwzB4Jxjj8vrwdDG4GUr9PvaMt0nZ2mm4OzSk2mltrDlj5edeaeqaX1RwW0disvqU3SjGnlFChUoq1r0qtOMZ0mkklKDTSVrnjhmuEzn2bnPxSwmQ51i6mnMtx+HnKdSacsdp/F9LqUqvxUpShByj1dNveUVNJWVg91cxxtHLMBicyxHU6WFozrz6Vd9MYtuy8XZM6q8DfaUcA+NOnNdavxGGzvRmScP8AC4fG5njc+hRUJ0q85xpqmqFSpKVRyh0qmk3Jyio9Tdjs7rD/AHpZ3/q7E/zcj/PRyc8vWseaXilS4OZNnOLyvTdd0841NiqTvCjhMM3GM+ntKq3XdOmmmlKq21ZSaD0dwHtoeC+Za8w2nMNwq1f9xMVi6eFp5rOth1WtOXT7x4ZSaSTadveNtN7Jqz72cQeI2huFGk8brniNqjA5BkWXJPEY3GT6YRbdoxSSblJtpKMU5NuyTZ8lynkL5Q8o03lumKHAjTNWnlfu5UsdVw18fOpDpanUxaarTbcU2nK177JNo/T8yXLjonmi4fUeGvEDH5thMqo5rhs1c8srQpVpToqS6G5xklGUZzT2vvdNNAdTNf8AtouAOn8fVwOg9Bar1ZTpbLGTVLL8PVfg4e8cqtvWVOL9D+lwo9sby663zmhkevtOai0HPFSjCGPxShjcBCTsrVKlJqpBXdlL3Tikm5OKOz/DnlQ5ceFWSQyDRfBnSuGoJRjUrYnLqeKxNdpWTq16ylUqPv3k0m3ZK7Okntd+WXhXk3B/LON2jNH5Vp7UGW5zh8tx88twlPDU8dha0Jpe9hBJSnCcKfTKzfS5J3VukPS3B4vC5jhKGYYDFUcThcTTjWoVqM1OnVpyScZxkm1JNNNNOzW5+J40cceGPL7oqvr/AIrano5NlVKfuaV4yqV8VXabjRo0opyqTaTdkrJJttRTa+Fey51xmmtuTLRqzjEVa+IyCtjMjjVqbt0aFeToRTu7qFKdOmu1lBK2130b9pznGccZOezR/ArMM1q4XJcA8myTDR61GnRq5jVpyr4hN3Sk41aUW2rWox22bYfds29trwcw2YVaOScG9Y43BxbVOviMThcPOa83BSml/wCcz+9R9tTyvyowliOH3FKFVxXXGGW5dKMZW3SbxqbSd92lfyR2+0Xy+cEuH+jKHD/S/C7TeHyOhQjh54apl1Kt8Iilu68pxbqybbblNtttts8gebHhhw75avaGafwGjNE5HjNL5rispzavprMMBSxeXqniqrpYnDKhVjKKhLpnKKs/dua6LKMUg9beW7mO0JzScOP2z+HmAzrBZYswr5bOhm+HpUcRCtSUXK6pVKkGmpxaam++9mrHy3PPaF8J8h5paPKnidJarqagq5phcmeZQo4f4GsTiKcJwe9XrcEqkE5dKad7RaSb7G6S0Zo/QOS09M6E0nk+m8ooznUpYDKcDSweGhKbbnJUqUYwTbu20rtu7PHHiN/25HCf+UDJP5jCgez+Y42jlmAxOZYjqdLC0Z159Ku+mMW3ZeLsmdXuXv2jHAvj9lGt9Qww+baLynQmGw+NzLH6klh6ND3FaU4wcZU6s/jdVO3Ta7coqPU3Y7Kaw/3pZ3/q7E/zcj/P1yQctGoObDijW4W0dU18k0rhqNPPNSzpVX1VcPQqKnCNOn+LKs3XlGEpJqClOTvZRkHoPrP21PA/Js9qZdozhnqvUmX0Z9DzGrVo4GNVb3lTpy6ptdrdag93dK2/ZTlc53OCfNjhcVhtAY7G5fqHLqKxGNyHNKcaeLpUrqLqw6ZShVpqTScottOUepRckn/YyPky5V9P6L/YDgeAujKuUuh7io8XlVLEYmsrW654monWlPZNTc+pNJpqyPI6vpOpyU+0xyfS+jsdWpZPlurMtpYZ1KzbllGYql10ZzbXU40cTOm5S7yp9Tv3A92jq7zN+0R5feWHOZ6P1Bi8y1JqynGM6+TZJThUnhFJJw+EVZyjCk2ndRTlNJpuKTTf37iPqeronh9qjWdCiqtTIMmxuZwptXU3QoTqJNXXdwt3XznkH7Kfg5pPmJ46a84qcZsFQ1ZjtPU6OZRoZp01oYnMsbWqyliqtKStVcfdVGrpxUqidrqLQdgsm9ttwWr41Q1Dwd1tgcH0turgq+ExVVS8F0TnSVn4vr28mfqsn9svyu5zm2CyilonifQnjsRTw0atXK8B7um5yUVKXTjXKybu7Juydk3sdhOZDll4M8YuEWodNak0HkdKrRyqvLLcxoYGlRxOW1oU5SpVKNWMVKCjJJuKfTJXi002n50+xuyfQeudVa40lrnhho/UNTJ6OCz/ACnM8zyLC4nG5fXjVcJKlXqU3OKb91JJSXTKm3FJybA9hgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPGz22ny9aE+iC+24g9kzxs9tp8vWhPogvtuIA9KuFXAbgdjOF+j8Xi+DOha9evkOX1KtWrp3BynUm8PTblJundttttvufssr4JcGcjzHDZvk3CPReAx2DqqthsVhcgwlKtRqJ7ThONNOLXdNNNHJ4Q/JNor6O5b9mpn64Dzy9tdDHvlx0ZOF/gS1tRVb4yt714HGe7uu7dlV7bLe/dH1X2VWIy2vyQaFp4Hp99h8VnFPGdPf3zzLESV/X3cqX5rH7zni4A47mT5b9T8OcijTeoIe6zXJPeSUYyxuHl1RptuyXvIOpSTbsnUTeyZ5oezw558Byj43P+CHHjK82wGmcVmU8VGv8FnLE5HmCSp14VqFveOnNU4XUU5QnBvpl1ycQ9qzxN9sxiMFjObHJMLl0YTxNDROX0cVGnD4zrvGY2STtu26c6VvG1l4Hf3WntR+TPSmnMRneV8TqmpcZCm5YfK8ryvF/CMRO11FOrThTp32u5yilv3ex5/8uWhuIftEudnFceNZZFUw2kMqzehm2bT+M8Nh6GGUfgeWQnZKc5Rp0ozt0tx97UaTaTD2Ox2FxOB4a4nBYxWr4fI50qu97Tjh2nv47pnkn7Ev5etd/RB/bcOevmsP96Wd/wCrsT/NyPIP2Jfy9a7+iD+24cD2TOu3O3zd5NyhcLaWqZZVTzjU2eV54HIMsqVHCnVqxh1TrVWt/c004uSVnJyhFOPU5R7EnmN7bbh7qTNtI8NuJeAoYjEZNp7FZjlmY9KbhhZ4tYeVGo0l8VSeHnBye1/dru1cPx/DHLvam86+S0+JuX8ZqfD7SOZSqfc+pSxUsqp1YJuD+D08LTlXnFSTSnVlu02pOyPnPOVyn84nB/grX1lxr5o8frvTNPMsLhp5RW1FmmNjKvNy6KnusT/c30tPd7q+x265LvaD8sWG5cdFaO15xAy3RuoNIZRhcixmAx1GpTjV+D01ThiKU4xcZqpGClKzupOSaSs31y9ozz1aa5lNKYngrwEyTGag0zktWGeag1JLCVYQUaMlCHuYys4UVOrFSq1EuqUoRirNOQdq/ZB/uOMD9Isz/wCtA6m+1myzIc15tNC4bhFWzPHcVK+BwtHH5fgKDm411WUsulBrd4iSlK8UrKEaL8Xftl7IP9xxgfpFmf8A1oHT3n2wfEnlP5+MBzR4DC5fmmEzzE0M3yZYyqpRquhhKeFxOGqQUlOPSltJJJKpCzbTSD99k/tmOIejMhWkeKnLlWr65y+nDDV6jzKeWxr118Vzq4WeHlOlJtXcItpttLoVrfl+W/l35hOdrmooc03MBpfFae0tl+ZYbNnHF4OphqWNWGa+DYHB0qvxpUU6cVOo7ppTvJzkz9rlftx8JLphnXLXWp2prqqYXVin1VNr2hLCR6U9/wB82tlv3PovDT2zvATVOcYbKOIGhdS6Np4mpGn90OunmGFo3lbqquHTVUUrNuNOT77bXYehJ4rcRv8AtyOE/wDKBkn8xhT2ayTOco1Hk+B1DkOY4fMMszPD08XgsXhqiqUsRQqRUoVISV1KMotNNbNNHjLxG/7cjhP/ACgZJ/MYUD2S1h/vSzv/AFdif5uR5B+xL+XrXf0Qf23Dnr5rD/elnf8Aq7E/zcjyD9iX8vWu/og/tuHA9kzw/wDaH/8AbJ4/6bpn+aw57gHh/wC0P/7ZPH/TdM/zWHA9qtVTyCnpjN6urKlCnkkcBiJZlOvLppRwipy965vuo9HVd+VzwV5Z+NPEnlh4p614ucu+kM61nw0wONlleZ/DMBUjCtl06lSWDliJ01L4NW6abkp7pO6aal0v3C49aCzLipwS13w2yfHRweP1Np7H5Vha021CNWtQlCHW1v0NtKVvBs8X+VHne15yDYzWXCjUHC7LtSUZ5xOWPwf3VWFxGFx1KPuakViKcK0Jx+JFNdLs4uzV2B9l4s+1Y4tcxGi8w4RcBOAeZ5ZnGpcHLL8XisLjKubYyFKsnCpDDUqNCDjNp9KqNtrqdoqSUl2f9mJycam5aNA5xrPiVh44XWWtlh3Uy9NSllmCpKTp0Ztbe9nKo5TSbS6YLupHxjAe3E01Up0nmnLpmeHm5WqrD6lp1lFX7xcsNDqdt7NLfa/idn+WD2h3APmizmGjtPV8z05q2dN1KWS51ThCeLUYylN4apTlKFXpjFyabjKybUWk2g7RAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAfFOO3J1y98ymdZVqLi/omrm+Y5PQ+C4bEUszxWFfwdzc3SkqNSKlFyk3drqV3Zq59rAHDyrLMDkmWYPJcrw6oYLAUKeGw9JNtU6UIqMIpu7aSSW7b2OYAAPifG7k05buYbFyzbihwxwONzhxUPuvhKlTB41pK0eurRlF1UlslU6kl2R9sAHTjIPZOcmOSZlSzHFaLzvN40WpRw2YZ5XdFtO6clTcHLtum7Ps0ztXo7RWkuHunsHpLQ2mstyHJsDHow+By/DQoUaa8WoxSTb7tvdu7bb3P7gAwxeGoY3C1sFiqaqUa9OVOpB9pQkmmtvNNnxrgVyccvfLbn+a6n4P6KrZPmWcYf4HiKtTNMXiksP1qapRVapJJKUYu9nLZXb3v9sAA/m6h07kOrMkxum9UZLgs3ynMqMsPjMDjaEa1DEUmrOE4STUk/Jpn9IAdPNQeyh5Mc8zuecUdD5xlUKklKeBy/O68MM3e7tGblKKflGSSXZI+u5Xyf8uORcJM74IZBwwwGWaT1HSjSzWhhataGIxnTJShKriuv385RlFNOU3a1uzaf2YAfheDfBfh1wC0NhuHHC3IpZVkeFrVcRGjPE1K85Vakuqc5TqSlJtv1skkkklY8iNGYnJuaH2pWNy7mEzSnicpwepc1y/L8qxtT+96kcBOtHB5eoz+KoOVNOULJVJOat1VN/bE6G83HssdKcedcY3i5ww1tPROr8zrrFZjRrYd18BjMQkv7tHocZ4eo2lKUl1qT36VJuTDvHiclyfGYajg8ZlODxFDDpRo0qtCM400lZKKasrKy28DzV9slo3gPp/hlpbNMFkGQ5XxHxudpYSWAoU6GLxOXe6qvESrKCXvKSqe5tKabU5Wi0pTv/LwPIp7UjT9COR5Dzf4NZanONJy1pnP9ygklFWlhnKF0laMG0nft3f9vhl7IHOM+1fT11zY8bcZrTFdUZ4nAZfXxFWeMa3Sq4/ENVXC904xhGTT2nFgdgvZdU9T0+SnQv7JXXtKpmEsuVa/WsE8ZV933d+n8Zx7fF6bK1m/32Z8l3LpnHHGlzF4/Q1eprqljKGYrHLNcXGm8VRjGNOs6KqKndRhBWt0vpTcW22/smTZPlWnsowOQZFl1DAZbluHp4XB4XD01ClQowiowpwitlFRSSXgkc4DDF4ahjcLWwWKpqpRr05U6kH2lCSaa2802fGuBXJxy98tuf5rqfg/oqtk+ZZxh/geIq1M0xeKSw/WpqlFVqkkkpRi72ctldve/wBsAA+F8ReSnlt4r8VcJxo13w/lmOqsJPD1PhSzPF0qdWWHS9w6lKFVQl0dMbbJOyUupbH3QAdavaKcStX8KOULXWqdD42tgc2qQwmW08bRk41MLTxOJp0alSEo7xn0TkoyTTjKSad0kdc/YxcOeGcOFGpeKmGeCzDXeJzirlmNqT6ZYjLsFGnCVKnFNdUFVbnNyW0+lLd03bv5xE4faU4q6GzvhzrjK45jkWf4SeDxuHk3FyhLs4yW8ZxaUoyW8ZRTW6R5p6h9kPxr4c6oxWoOVvmTlkkK0v73+H4vGZVjaNO+1OeJwSl72yu+pU4Xbt0ruB6b5po3SGdxrwzrSuT4+OJ/wyxWBpVVV/8AqlKL6uy737HiBxNyfRWVe08yvJuWCjhcPgcPrfJY4WnlDvhKONUqDxkaSi0lRjU98pxTUElNK0EkuxeY8gvtMddYWeQ6+5u8FLJa0pUsTRWq82rKtT7XdJYeEakWv3s5L1SZ2T5PPZxcLeVTNVrnF51iNY659zOhTzbE4aNChgYTVprDUFKThJpuLqSnKTi2l0qUkw7dgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAPkPNfxywfLpwD1bxVrSpvHZdhHQyqjOzVfMKz93h42s7pTkpSVvxYyfZM+Aey01vzD8WuGOpuLfHHiJmOocBnWaLCZBh8VQowUI0E1iK8HThFqEpyVNRTUU6M3a7udf/bW8WMVi854f8A8rqudOnSnqfMKMLylOrOU8PhFZbXSjitt2/eLsu/ovy78L8LwV4HaJ4XYWlGnLT2TYfDYnpSSninHrxFTba8q06knbxl4gfRgD+DqLXWiNHypQ1brLI8jlXXVSWZZhRwzmm1FWVSSvu0tvF2A4XFTUuY6L4Y6v1jlEKMsdkWQ5hmeFjXi5U3Wo4epUgppNNx6oq6TTtfddzpr7MvnO4z81Gb6+ynixUyStDT1DA4rBVcBgXh5xdaVWM4StJqUV7tNXV7t3bVku1nG/M8tznl44iZnlGYYbHYPEaPziVLEYatGpSqL4HVV4zi2mrp7p+B5u+w6/32cWv9XZR/O4kD0G5q+ZrSXKhwt/bP1blGPzWlWzKhlODwOClCNSviKkZzS6ptKKUKVWTdm/i2tvc4HKPzX6S5u+H2Ya70tpzM8jeVZnPKsXgsdKE5KoqcKkZwnB2lFxqLuk001ayTf63j3wP4YcwXDzEaA4t5fPE5GsRTx6nTxUsNPDVqd1GrGon8VpSmne6tJpo4nLxy/8IuXXQs9H8G8tdDKcdi55hiMRUxksVVxVdxjBzlVbd7RpxilG0VZ2SbbYfUgD8nPixwsp5t9wanEvSscyTs8E85wyr36um3u+vq/G27d9gP1gB1g9pdnOcZByScSc1yHNcZluNpLKY08Tg686NWEZ5tg4TSnFppOMpRdnum09m0B2fB0R9jnqXUWp+WHUWL1LqDMs2r4bXWNw9GpjsVUryp0lgMBNU4ubbjHqnOVlteTfds73AAZ1atOjSnWq1IwhBOUpSkkopbttvZJdz8nT4v8ACWtmP3HpcUdJTx90vgsc7wzrNuXSvidd+7S7d9gP2AAAA4uDzLL8w958Ax+GxPuZdFT3NWM+iXk7N2foz+Pn3EPQOlcZSy/VGudP5Piq1nSoZhmVHD1Jpp2ajUkm9k+y8H5MD9EDDDYnD4yhDFYTEU69CrFTp1aclKM4vdNNbNPzRuABWUoxi5Skkkt23Y/OYbiRw8xuZRyfBa905Xx86jpRwtLNaE6zmu8ehSbbVndWurMD9KAAAPzWf8SuHWlMdDLNU6/05k+Mm4KOHx+a0MPUk5fipRnJN3tttv4H9rLM1yvOsFSzLJsywuOwdddVPEYatGrSmvNSi2mvmYHLAAAAAADx05dNfa6xvteM509jNaZ9XyueuNa4WWBqZlWnh3QpUcxdKm6bk49EHTpuMbWi4RslZWD2LOsfOLz3aD5PMTpzKtRaUzbUeZ6jp18RTw2Bq06Sw9Ck4xc5zm+8pStFJO/TNtqyT7OHxPmN5QOCXNPHJZ8Wclx1fE6f99HA4rAY2eGqwhV6feQbV1KLcINJp2admru4fseB/FzIuO/CfTfFzTOBxmCy7UmEeJpYfGKKrUXGcqc4S6W07ThJJp2aSe17H7s/M8OeHmkOE+iMn4daCylZZkORYdYXA4ZVJ1HThdybc5tyk3JuTbbbbbZ/crZjl+HxNLBYjH4aliK/+BpTqxjOp/8AUpu7/MgOUD+fnWe5JpvATzXUOcYHK8FTajPE43EwoUotuyTnNpJvstzj6d1ZpbV2ElmGk9S5VnWFi0pVsuxtPE002k0nKnJpXTTW/Z3A/sAzq1adGlOtVqRhCCcpSlJJRS3bbeyS7n8DIuIegNU42plumNc6ezjFUr+8oYDM6GIqRsk3eMJNqyabuvFAfowAAB/H1Fq7SmkMLDG6r1PlOS4abajWzHG08NCTSu7SqSSdlu/QjTusdJavoSxOk9U5RnVGFnKpl2OpYmMU1dNunJpXW68wP7J52+1A408zXLdrfh7xS4V8Q8ZgtG42MsBjMpeFoTwrx9Kbq2quUHKSrUW4pN3j7ibi02eiR1o9ovwqhxa5Q9eZbDCutmGn8ItS5f0w65Rq4NupPpXe8qHv4bb/AB3a/Zh9u4YcQsh4scO9OcS9M1HLK9TZbQzLDqTTlTjUgpOnO2ynFtxkvBxa8D9SeefsZOLtbVnAzUfCXMcUqmI0Jmqr4KDbvDA43rqKKT7pV6eJba/7ok0tm/QwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8ReZHErjV7V7Caary68vpa0yDT/Q5t9NDD/B44hJq3eSryS2s5Wvtc9ujw20dPHT9rriHmEOiquLOZqKslemsTXVN7ecFF/n33PckDqT7R7m2zPlZ4O4aGiqtGGttZV6uX5NVqKMvgVKEU8Ri1B3U3BTpximrKdWDd1Fxl0H5XfZtcTOcHS9Xjzxb4rY7I8HqGtUq4HE4rDTzHMs0cZuE8RUdSpHopuUWotuTl0tpKPS5f1vbYZ3ja/MLorTdSb+CYHRlPG0o37VK+OxUJv86w1P8AQetnCfT2X6S4XaQ0tlVGFLBZRkWAwNCEY9KUKWHhFWV3bZLxfzsDyO4xct3NN7ObKM+1Fw51tPWXCjU+BxGT59FUJUqVGOJpSoJ4vCdclTmveWp4mEmupKMulSUJ/svYdf77OLX+rso/ncSek3MVhsPi+X/iXh8VQp1qU9IZx1QqRUou2DqtXT2dmjzZ9h1/vs4tf6uyj+dxIHar2r8muSbVyTsnmGUp+q+HUtn+dI/MexxlJ8o2JTldR1dmKS8l7nDP/a2fp/aw/uJtW/6xyj7bSPy/sb/3I2K+l+Y/zOGA+G+0+4/cWtdcwGnuTPhdnuJybA5k8uwWYQoV5UHmmYZhUjGjSrVIu7w8IzpfFWzlObkpdMOn9VgvYicOv2NxoZjxx1H+yB0l1YqjltD4HGrZXaoN9binfb3qbVt0Pac8kvFfW3EDL+Z/gLgcVm2b4HC4eGcZZgHbH06uFd6GMw0VvVaioxcI3mnTg4qScuj8vwQ9sjn+nK1DRnNBwzxMsXgprCYzOsnp+4xUJxfTKWIwVWy607OXROFmnan2QHbvkT5dOMXLJo/U/DvibxCw+qsojmsKumatHEVpqhhPdJTj7uqv7gnOz93GUopqTT3bfRn2gvs7M20JgeK3Nr+2zRzLA4nPPuz9xquVyjXj8PzCnT90q/vWrU5YmyfTvGmlZX29UuFPF3hvxu0dhdfcLdWYPP8AJcU3BV8PJqdKokm6VWnJKdKok03CaUkmnazTfwb2o/7hTiZ/+Rv6YwQHmTyc+zlzvm84Y5nxKyzirgdM0stz6tkbwlfKp4mU5U8Ph63vFONWKSaxCVrP8Vu+9j1h5J+VzF8o/CDFcMsfrj9lFbG55iM5lio4R4anR97So0lRhBzm7JUFJu6vKpLbxfX/ANip+5Z1T/5QMd/R2XHoAB5qcdOTbnV5uuO+sMJr7ig9HcJsDmk6GRYeWK9/RxGDXT7udPA0JpVJuO8p15Rl1NpbbL+XqD2IWlHkmKWleO+bLN1Sbwv3Qymm8NKolsp+7mpxi3tdXave0rWfafmZ9oHy+csGYz0vqrNsdnuqoU41JZFklGNevQUleDrznKNOjdNPpcuvpaai01fq9X9tLicW1iNL8p+d5pgpXUcRU1E6TbTaslDB1F2Sv8bZ3W9rsPmvs0OPPF3g7zN1+ULidnGYYnKMRWx+TU8txdeVeGUZng4zmnQcr9NKao1YdMLRk5wkrWbfrZq5uOlM6lF2ay/EtPyfu5bnhZwL4kVuMPtLdLcT8RpienKupdaxx88rnWdaWFlODvBzcIOTvu24Rvfse6esP96Wd/6uxP8ANyA/z68o/HDjpw4q6n4X8ueT18TrXih8AyvCYvDxU8RhIUXWlN0oyXRGbVXetNpUoxnLZ2lHuLQ9izrbUuncRqLXvMVCprrHweIrx+5dTG4b4Q0m41MVUrRq1bu6dToTV79MrWf4T2KOFw1XmB1tiqmGpTrUNISVKrKCc6aljMOpdLe6ukr272PZcDxP5GONvF7lF5saHLFxIzPEPT+aZ5+xjMsprYiVXDYPHVZqOGxeFb2gpVJU7tJRnTq3auouPtbOcKUJVKk1GMU5OTdkl43fkeI3P5VeU+01p5ngIwpYmGaaWxXWop9VSFHCqMmvFpQit/BJHrLzW6ixekuWbipqLL244rBaPzaWHmnZwqvC1Iwl+aTTt42A8rePXMDx29otzEfe/cD82r4DQ6xVfC4PC0cTKjhsXhaTaq5ljpx/Hg0uqMGmknCMYucm5fV8Z7D2mtPt4DmHlPPFRTjGtpzpwsqtruLaxDnGLe3VZtd7PsfxfYeabwWK1dxa1fUjH4XlmXZTltKVt1TxNXEVJpPwTeEp3XovI9bAOkPs7dFc3XCDPte8F+YSeOzLTGn6eDq6czbEYieKw9SU3NThhMRNKU6Lgot05WdNpLpi5NP5r7TLnV4i6a1ng+U7gBisZhNUZrHCwzjMcuqSjj41cU18HwOGcbOnUnGdOUpxfVapCMWryv6UH+fzVfGfO9M+0Q1VxmwGja2scdkmu83r4HJ05xniIYepWpUFeEZyTpwhCeyaXu/Bdg7fcPPYsZfmunYZvxr4zZx+yrHwdfE0Mmo050cNWlu4yrVlKVdpveSUE3e11u/iPE/QHMF7J7i5kGqdAcQK+odD6hrTqQpTU6OEzH3fSquExuGUpQjWUJLoqxd2m5RcWpQX3D8Ljx6/IyzH+N4z+ynwbnH5wuMfN9w3yrh7mnLBm+m/uVnlLOaeOo08ViqkujD16To2lQjZS98pNp96aVn3QeyfDDiFkHFnh1pviXpio55XqXLaGZYdS/GpqpBN05eUotuLXg4tH6k6xezZybWGn+THh9kutsjx2U5hho5g6eHx1KVKv8Gnj8RUoylCaTinCa6U1vDpa2aZ2dAAADqjz48kmY85WX6Mw+W8SIaVq6SrY6bp1sBLFUcVHExoq7UakHGUHQVnvtUl28fH3hzyr5hxD5usXymUNZYfBYzC59neRvOp4KU6UpZdDFSlU9ypppT+CtJdW3Ut3bf/AEYHity0/wDbks4/8oGu/wCYzQD99+A61Z+URlP1cq/2g62c6XIfm/JvlWlc0zTiRg9ULVGIxdCEKGWywroe4jTk23KpPqv7xLa1reNz3+PMH24v+9PhL/rHN/5rDAd1eS6vXxPKZwjrYivUq1HpDLU51JOUnahFJNvySS/MeZPtTdWz0Lz66P1wsK8Y9OZPkWaxw/vOj3vuMbXq9ClZ9N3G17O172Z6Z8k/7kbhD9EMt/mYnmB7WWjSxHO9p7D16MKtKpp/JoThOKcZReKrppp9002rMD9xoflB5nPaP1/2/eYriXX0bpnMHUqaayuOCnXaw0neLw2GlOEKFBq1qsnKpVUbtNOM38H45cFOOvsyONentUaL1/PE4bMYzxGT5zhaU6FLH06Uo+/weLw7k00uqDlTcpRcZwaad1H3npUqdGlCjSpxhCCUYxjFJRS2SSWyS7HnJ7bbAUKnBjh7mk+r32H1RVw8O1umphKkpX2ve9KNrO3e99rB8qhnfNZ7WTU+NwGnsyfDTg1ks44bF9NSdWjUruCbjVjFweNrO6kqbcadOLjdqTUp/N+bn2b+uOTzSGE448O+KeKz3LMpxtCnjMTSw0sux+VVak1GlXhOnUkpQdRwh1JxlGU4WTTbXpH7NzDYbDck/DD4Ph6dL3uBxdSp0RS6pvG17ydu7e133P6ftBaVKvyZ8VoVYRnFZH12l26o1qbT+dNJr1QH5/2cnMjqDmU5c8Ln2tcSsTqjTWYVcgzXFWSeMlTp06lLENLZSlSqwUmtnOE2kk0lb2gnN5V5S+EmGzDTeHw2J1nqqvVwGQ08Qr06HRFOti5xtaapKdNKLaTlUhe6un139iD8mPE3/X2C+zyPhHtpdT4jMeZDSulveSeEybSFGtGDvtXr4vEObW9mnCnRXZPZ99gP0XLr7N7iVze6fw/MJzL8YM/wsNUwWLy6k/76zLFYaTbhWnUrNwoU5LenCMZJwcWlFWT4XM17O7iHyXaflzFcuvF/P8Rh9O1ISzBRj8EzHA0Z1FFVlUotRr0ruCqQcIpLdqUerp/d6d9qrxt0zp/LNN5byYZhDCZVg6OBw8VisYkqdKChFW+C7bRR/F4r+0y43cVeGGruGeP5P8xwdDVeSY3Jp4lVcXVeH+EUJUvfKDwyUnBzU0rreK3XcDuxyD809bmt4HUNVagp4ehqzIsU8pz+lQXTCpXjGMoYiEP3sasJJ2WymppbJHYbO8pwef5Nj8izGLlhcyw1XCV4ru6dSLjJK91ezfgeaXsVtJcQ9LUOK9XU+kc4ynJ8fLJ1ha+PwlTDwqYmksX7yNJTSUmo1IOVr2+Jfuj08A8VvZB5/jtEc3OoeHuOqS6c40/j8vq0vxV8KwtenUjJp2d1GFdW7/Gd+x7Unh7yDzlQ9pvCjlMZYjBzzjVNOdWoumUaCw+McJtX2bkoJre3U/nPcIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA8O+Ld+EXtbI5nmFsPhv2xcozOrUcnGKw2OeHq1J3bd0o1533tdNbdl7iHj57Z7hlmGl+MeieN+Uwq0qOf5b9za1eHaljsFU64Sb8HKnVhZePuZPwZ6ncGOIeD4tcJdH8TMDOEqepslwmZSULWp1alKMqlN2bScJuUWruzi14AeX3tt+H2Pw3ELh3xUp4ScsFmGTVtP1cQm3GFXD1514Qa7JtYmo159MvI9DuTzjFkfHLly0PrfKMwpYjExynD5fm1JVeueGzChTjTr053bkm5R6l1buE4S3Uk3/d5huAeheZThdmfCzX+HqPB4xxr4TF0WlXwGLgn7rEUm9uqPU009pRlKL2kzzFwvs0ufbgPqXHfe+8XcLHLcZJr4ZlGoa+U1MRTX4vwmhJKPUruyUqiXdNMDvbz6cyvDfgLwJ1PlOqczo19RauybGZTkuS0qqWJxU69KVJ1nHdxow63KU2rbdKvJpPpZ7Dr/fZxa/1dlH87iT6dwM9ldn2K1LiuK/N7xDXEDVtSnN4XL5YyvjcMq1mqdTF4iulOuo7WpKKgmrNzWx+19m/yR8WuUjPNfZlxMzfTOLp6ipYLCYFZRi61aTVCdWUqk1UpU+mLVSNldvZ3S2uH6f2sP7ibVv+sco+20j8v7G/9yNivpfmP8zhj7bzwcCtYcyHLlqHhToTGZXhs6zDEYLE4eWZ1p0qElQxEKsouUITabjBpbWu1dpXa/i+z/5c9d8r/AWXDfiJjcnxOb1s8xWaN5XXqVqMKdWFKMYuU6cG5p0m3ZNbqzYHYDK9U6ZzvM8zyXJdR5Zj8wyStDD5nhMLi6dWtgasoKcYV4RblSk4yUkpJNpprZnynmZ5ZeBHHzQ2cR4r6ayqhXwuArVaWpfcwpY3K1CDl75YhJS6IdKk4SbhJRs00dUOZ32bvG3POOOecw/LDxm/Y9nmfV/hmKweIx2Iy6vSrNRU1RxOHT6oScU+iailunKStb5jnXJd7U7jFgZ6I4pcb8PS05jLLGwx2pqk8PVgrNxnSw9Nur2uoyXS2ldrug/PexM1Hqahxp17pLD1qz09jNLrMcXT3dNY2ji6FOg/JN06+IXql6HeH2ocJz5FuJkYQcmlk8nZX2WcYJv9CR+i5NuTXQ/KBonF5PkuYTzvUmeSpVc7zutRVOVeUItQpUoJt06MW5tRbbbk229kvr3E7h3pvi3w91Bw01hh6lXJ9R4Crl+LVKSjUjGaaU4NppTg7Si2mlKKdnYDo37FHOMur8ues8gp4qEsdg9bVsZWop/GhRrYHCRpya8nKhVS/wDqX5HoLjKtahhK1fDYd4irTpynToqSi6kkm1FN7JtpK78zyAh7MTnj4Gayx2N5d+K+EhgsapUVmOVZ/XynEVcOpfEhiaVkm1e9lKpFNXTTsegHI/wN4t8BOEOP05xt19+y3VedZ/iM8xOM+H4jG+5jVoUKaouvXSnUadBybsleo0r2uw8tPZwaa0nzA86+LzjjxHDZ7mFfBZnqOnhMzpqrTzHN/e0241KcrqXTCrXrKLTV6K22PcWrUyrIMqnXrVMLl2W5fQlUqTnKNGjh6MI3cm9owhFJtt2SSZ5qcy3sqtf1eKuN438p+vcHkGY4nHSzdZTiMTVwNbBY2UnKc8FiqSainJtqE1BRu0ptWS/kYXkU9orx1hDSXMdzGyy3Rkq0ZY7DRzipjauJimntQpRjSqWa297NJOzSdgPgeh9baY4j+1hy7W+jM0hmWSZtxAdbBYuEJRhXpKLipxUknZtNptbqz8T201h/vSzv/V2J/m5HnppH2XuouFfOHojiZwwx2S4fhppV4LEzWPx9WpmletSoONaU4KkoSqVKt5XUowSnskkoL0TzzA1M0yXMMsozjCpi8LVw8ZS7Jzi4pu3huB49+xL+XrXf0Qf23Dnsmefns5+QvjNyo8TdV6x4mZvpXE4LNMmeU4WGUY2vXqTn8Ip1PeNVKMEoWpu131XavFeHoGB4f+0P/wC2Tx/03TP81hz2J426GnxO4Oa54dUVH3+ptO5jlNBytaNWvh504S32TUpRab7NJnRvmr9nbxp43c4eD456T1BpLD6arV8prYhY7F14YqgsJGnGovdxoyUnJU30Wlu3aTitz0bA8VvZD8Xso4T8w2o+FmtsSsonrfBQy/DPEy92lmmEqydPDzvZRlKNSvFXd3NRik3JHtSeeXOh7KrBcb9ZY/i3wQ1Ll2mtT5tUeIzXLMxU44DG4h/jYiFSnGU6NST3kuiUZSfV8VuTfw+nyKe1I1Fl0eH+fcc8XQ03NOjUWM15jKuFdJbdMoQUpzhZK0HFrtsgPTzSHHrhLr/iHqHhZozWeCznUWlcPSxGbYfB9VSnhlObiouql0SmmrSim3FtJpN2PH/iZjnyje1RxWvtWwr4fJqmr6mo6mJSk+rLs1VR16sEt5KCxNeLS7ulKKv4+i/I5yJac5O8rzfM6uqq+pNW6jo0aOYY1UfcYWhSptyVGhTu211Sbc5u8umFows0/wC7zi8k3Drm+01hKGeYupkOq8nhOOU5/hqKqzpQldujWptr3tFyfV03Ti7uMleSkHYHK80y3PMtwmdZNmGHx+Ax1GGJwuKw9RVKVelNdUJwnFtSi00007NO6NMZi8Ll+FrY/H4qlhsNh4OrVrVpqEKcFu5Sk3ZJLdtux5MZDyI+0z4F056W4L8bsNLT3XOdGjl2pa1DDQu7uXwbEQUac33fQnd3vJn9X8G5z18ealGjzK8zMaWSqopywNTNsXm8oNbuUcL/AHPDJ37NTvddtlcPVTD16OLoU8Thq0K1GtBVKdSElKM4tXUk1s007prwPk/FXm05dOCWqKGjOKXFfKcgzrEUIYqGDrRq1JxpTk4xlN04SVNNp26mtlftufu+HejMBw40Bpnh3leJr4jBaXyfBZLhq1dp1alLDUIUoSm0rdTUE3bxbPFvnqymtzQ+0QxnDThO5ZnmNatgdMzqRhenSxNCnbFTbW/u6P8AdOtu1vdT7pJsPcOlVp1qUK1KpGcJpSjKMk1JPdNNbNPuaHEyzAU8ry3CZZRnKcMJQhQjKVrtQiopv1sjlgDxL4B5pgdP+2HzPF5vXWGpVeJGscLCVR2Tq4mOY0qMVfxnOrBLz6la9z20PNznf9mDrHizxTxvHbl+1NluX53mk6eMzLKcfXqYZvGwUUsRha8ItRnLpjJxn02mpSU91FB6RnmD7cX/AHp8Jf8AWOb/AM1hj9dyj8pfPRpzjtpnipzKcacZmmQ6Uo4ynSyjFaoxWZ1MVOthKuHi+h3pJJ1VNzlJybhHZ919D9pLyg8UubPTeiMFwvx+n6GI0zjMbWxVPNsVUoe8hXhSjF03CnNNp03dO3dWv2A+w8k/7kbhD9EMt/mYnmF7WH93Lpr/AFDkv2uuesnL1w7znhLwM0Lwx1FisFicz0xkODyzGVsFOcqE61KmozdNzjGTjdOzcU2u6XY6ac8Xs/uNXMdzLZBxa0HnWk8PkeFy3AYLExzPG16VelPD16lSUuiFGalFxqK1m3dO6Ss2HogedXts/kF0J9L19ixB6KnUv2jPKzxH5reFWntJcM8dkeHzHJc+WaVoZtialCFWl7irSahKFOfxk6idmkrJ73STD9B7OD9xNws/1divtuIP6PtAf3GnFf8A1DL+dpn6blO4R6h4E8vGiuE+q8bgcVm+nsFUo4yrgZznh3OdepVahKcYyaSqJXaV2m7HL5nuFueca+AOuOFemcbgsJmupMqnhMJWxspRoQq9UZLrcIykotxs2otq97PsB0l9iD8mPE3/AF9gvs8j5r7bXhvmuH4g8PuL1PD1J5bj8mqabrVkrxpV8PXqYinGXgnOOJqteapS8jtp7N/lK4m8p2htXZJxOx+Q4jGahzShjMPDKcVUrqnTp0nBqbnTgk23dJXVu7T2Ow3GvgvoDj/w6zPhjxJyn4blGZxTUoNRrYWvG/u69Gdn0VINtp2aaumnFtMP4fK9xp0zx94H6V4iabzWljJ4jL6GHzOkqnVUwmYQpxWIoVVe6kp3av8AjRcZK6km/rB5KYr2YHOTwC1PjM85WOOtB4LFSUX7rM6+UYyrTUm4Qr0kpUKqjfu52bTair2OfPku9qbxWpTyDiXzHfcnJai6MTCtqnEyjXg7XTpYWFqvn01Glt3ukB6oYHM8uzSFWeWZhhsXGhWnh6roVY1FTqwdpU5OL2kns0914lsxx+DyrL8TmmYV40MLg6M69erLtTpwi5Sk/RJN/mPiPJryt4HlF4Q1OGOF1bX1JiMbm1fOcbjp4VYaEq9WlSpONOn1ScYKFCHeTbd3teyx58OJ0OEvKZxH1PDE+4xmKyeeTYCUXafwnGtYaLhut4qrKp6KDdnawHmN7J7L8Xr3nczHW86LSy/Js3zutJ/vZV6kKKV1ZXbxL2t2Tstrr23PMz2KHCivlmh9ecZsww8ofd3HUMiy6Uk1ejhoupXmvNSnWpxv50pLzPTMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA678+XL5U5kOWzUmi8qwccRqPLFHPNPrpvJ43DptUo+tWm6tFeCdVN7I+IeyG11ravwTzzg7rrTGd5XX0LmPvctqZhgKtCE8Hi5TqOlFziryhWjXbXgqsFbZnfcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB+Q4s47X+W8NNTY3hXkNPOdYQyyusjwVTEU6EKmMcemk5TqyjBRjJqTTauotXTdzqb7OPke1Dy7YPO+KvGjC0KvEvUs50On4TDFPLcG5dU06sW4yrVprqnJNrpjBJpuafeEAAAAAAAAAAAAAAAAAAAAAAA8y/a8ZvxN4k6h4c8tvDjRef5rDGV453jKmFwNadCtias5YbCU/eKPQuj+7yldtLrg3azv6aAD5zy9cHsr4B8F9I8JMpnTqR09l0KGJr049KxOLk3UxFZK10p1p1JJPdJpb2PowAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAD5Bzb8QtVcJ+W7X3EbRGOp4TPMhyp4rA16lCNaNOoqkFdwmnGWzezVj6+Y4jDYfF0ZYbF4enXo1F0zhUgpRkvJp7MDwe/Cxc7P8IeUfV3Bfqx+Fi52f4Q8o+ruC/VntHxa0ppejwr1nWo6byunOGn8xnGccHTTi1hqjTTts097nhX7OvC4XHc6HDDC4zC0q9CpmGJU6dWCnCVsFXe6d13SA+vcIvaf84ereLGitK53r7KquXZzqHLcvxdOOQ4ODnQrYmnCaUlTTTcZNXTTV7o9ujpJ7R/mG1Vyl6R0VqThTpXR7xud5rXwuIlmWTqsoxp0lOLh0Sg4u773f5jqTn3tkeM8+EmW5TkuS6dhxAxtavPMs3jgZrC4DDqbVGnRw85NTrNLqc5OUEpJdLbfSHsiDyu0X7VjV/DnlhpZ9xVVPWXFvN8+zHDZbgKuFpYClhcFSVFwrYqFGMWoKU5xglFSqODTkulyXySHPf7T/VOR1+K+nMizdaOoe8q1MTl2hIVsrhTV237+dGc3CFneXvWl++fYD2sB0S5BvaSUOZjOP2qOKmUZfkevVQnXy+vgOqODzinTi5VIxhOUnSrRgnJx6nGSU5Lpt0nwzi17VDjhwo5qdQcOM0wGmauhNM6xll2MVHK6ksfLK6WJUaqhN1lF1nSUrNpLqttYD1gB4s8TfaUc/moKEuKOkNK43QvD+ddSweIo6VWLwUqTaUYVMdiqMoVG20nKDhdvZLZHan2cvtDdT8y2f5hwi4v4LLaWrMHgZ5jl2Z4Gl7iGY0ISiqtOpSu4xrR61JOFoyipfFTi3IO/p/N1FjK+X6fzPH4WXTXw2DrVqbaulKMG1s9nukeanPJ7VPVHDfiFmfB/l1oZU8Tp+tLCZzqLG0VibYyLaqYfDUm+i1NrplUmpXl1RUUoqcvmGnuf3n+4Y5RS1TzE8MM7zrh/ndOeFq47MdJvKnCFZOMZUa9OlTp9VpJwjUTU0rJq/Wg/v8AIv7Qjmi44c0+ieF3EXWeXY/T2d/dL4Zh6OTYWhOfucuxNeFpwgpRtUpQezV0mnsz1tPAH2XH7uvhn/8Aln+h8adneYb2pfHXVXGbF8G+UvT2DVKhmc8mwOO+56x+YZviYTcHOjTnenTpNp9KcJScV1Nxu4xD1hB416u5y/amctNTL9Q8c8pdTKcfWjToxzzTmBjhKs7OTpOtgY03Gbim+lzUrJtLZnpTyjczOnua7g5guJuS5ZLK8bSxE8tzfLZVVU+B46nGMpxjJbyg4zhOLaT6ZK6TTA/A+0X4y8buB3BLJNWcBVX/AGQYvVWGy7E+6yqOYP4HLCYupP8AucoySXvKVL41lbZX334Ps4eN3Hjjnwv1NqDj8q6zfAZ/8DwXvcojl7+DfB6U9oRhFS+PKfxrPyvsc72i/MtxE5WeCWS8QeGVLKKmaZhqrDZNWWZ4WVel8HqYTF1ZNRjOLUuqhCzv2urb3Ph/Bzn445665IOLvMLneF0ytU6IzSjg8sjQy+pDCOnN4W/vKbqtyf8Ad57qS8PLcPRkHjfor2wfMlmuUZ1kNXQOQag1jmksNhdNUcsyuv0UZNVHWqTpRqynXmrUlCnGybcm21FRl+MzT2mPtBOFmslR4mVFg6sujE/cDUOkKeATo3a2SpUq/RJprqc3unZ7NAe4Z1j9ofxw4icvnLliuIvC/NqGXZ5SzjA4ONevhaeIiqVRyU10VE43dlva6Po/LHx7yLmX4Laf4u5HgpYB5pCpSxuAnUU5YPF0punVpOSS6kpLqi2k3CUG0r2XX32vn7jjHfSLLP8ArTA63ckntR+K2ruOGWcP+YzUuWYzINUJZdgcdDL6GD+AY+TXuXOVOMU6dR/3N3TtKUHdJSb9aD/L9guH+qcdw+zHijl+BlWyLJs0w2U46vTbbw1evTqVKLkl2jJUaiUuykknZyjf3G9m3zZPmW4K08n1VmKra70SqWX5y5zvUx1Bp/B8bbxc4xcZvv7ynNuylFAdLOHntH+bDUHNdpnhXmmt8tq6dzPiHgtP4nDrJMJGcsFUzKFCUFNQ6k3TbXUndPe9z2OP86nCH93pon/yu5b/AEzTPXLnu5+dPcouWYLTOQZRh9Q8Qc6w7xWDy+vNrC4LDdTisRiXFqbUpRmoU4tOThO8opJsO3IPFP7+/wBp/m2n5cXstyLN4aKpRdaeKw2hITyr3S3c3XlRlP3as05qrZdm7tHc3kG9ovg+abHV+GfEbJsBkOv8LhpYvDPBOSwebUIW63SjNuVOrFO7puUrxTlF2UlEO74PKDP/AGnHMhlvN7mPA7DYPR707hOJFbSkJSyuq8T8Cjmjwybn763vPdpfG6bX3t4HppxV1LmOjOF+sNYZRGi8fkeQ5hmWFVaLlTdahh6lSCkk02uqKurra+6A/Vg8yeQv2ifH/mS5hcDww4h4TSlPJsTlmNxk3l2XVKNb3lKClC0pVZJK/dWPl+jvbA8bMr4qPBcUcr0zX0fgMRjo4ylluWVKeMrKlTqujTpzlWcYynVjSi5NNJSbasB7DA8fuC/tQeYzirzKaYp6pzvJNLcN8VmrjmuBw+W0Xh8NglSqTl73F1YyqpqMHJyUoX6W0ktlrxm9rfxz4hcQP2FcqWkIYTL6mKeHyyrPKXmOb5q12ccPaUIKVm1TUJTtZuSd0g9fAeLuH9pH7QLl91NgsLzBaTqY+hXXX8A1LpmOUVcTSvdyoVaFKkrq6XV0zSezi3c9HdK87HCPV/K3mvNRlssSskyLB1Z5nlknH4VhcdBRXwKT7dcp1KSjLs1VhLZOyDsKDxsyz2gvtEOaPWWY5Vy2aZw2V08FD4Q8Dk2UYbGPC4dytB18Tjoyg5PtdKmpWbUVZ25emPac84vLzxJp6I5rtHwzajSdN5hhMXlVLLsyp0JvatQnQUaNRWu1eDjJppSju0HsQDr/AMx/OZwx5e+BmWcaq0pZ/S1RQoVNL4DDVVCeayrUVWpy6mn0UVTalOpZ9KaSTlKMX5r4b2hPtIuOuY5nnHBPTmMhlmX1PeVsLpfR0czpYaCu1TnUrUq0nJppvdN2vFJXQHtQeX3PPzg86XCLmOz7QvBmOMelsFhMBUw3utLU8bHrqYaE6n91dKTfx29r7dtj+Type1s1zjOIWX8MOaHJcshhswxcctWoMLhXgq+AxLl0L4XQu4OHXaMnFU3T3bUknb+tzz+0d5hOXXmOz7hVoDC6TnkuW4TAV6LzDLalav1VsNCpO841Yprqk7bLa3cD0h0DmWaZxoXTmb55f7o43KcHicZ1U1B+/nRjKfxbLpfU3tZW7H6A6x8xPOvpflo4BaW4lauwEc21Vq3LcPUynJMNU9ysXiZUIVKs3J9Tp0KbmuqVpNdUIpNyuefGH5+/aY8YI47XXCjSWYx03gazdWnpzRKx+DpJd6cqtWlWnJpNOVpppJtpK4HtGDzf5GPakZrxe1xl/BbmByvLcv1Bm1X4Lk2e4Gm6FHF4l/i4bEUm2oVJtNQlBqMpOMelNpv8fzq+0q5iuX/ma1lwj0JhNIzyPIfud8FePyypWr/3fL8PiJ9U41op/HrTtZKyst7XYeqAPHLi/wC0t51eIjzfXnAHQ+caT4aZbOcaOb0dLLMbUYSa97isVWpVaFOTUXeMbKNmryacn9a9n37TDiJxo4rYTgjx1o5VisXntKs8lzrB4ZYWo8TSpuo6FaEf7m1KEJuMoqLUkotS610h6Zg6Rc+/tGcDys46lw04eZLgdQa/xWGjisQ8ZOXwPKKU96cqsINSq1JrdU1KKUWpN2ajLpVU57fagS07Hi9HL87jopQdb4bHQdF5U6V7+8eIdBy6N1Hq97bbve7A9sgdIeQT2iuG5qMwxPDPiHkOCyLXuDwksbQlgXJYLNKEOlVHTjOTlTqxcrum3JON5J2TS+R833tXNR6a19jeD/K5kuBzTH4DEvLsVqDE4eWLVbFqXS6WCoRdqnTL4vvJqSk7qMWkpyD06B4tY/2g3tJuBuY5fqHjTpbGRynMavVh8LqfRiy7DYhNKTpUqtKlRn1qO9nOUo3+Mn2PQ/hhz28KuI3K9nnM08PicvwulMLVefZK6kauJwuNhGLWHhLZTVSU4KlNqKl1q6i1JRDssDxsy/2hXtCuaDW+YZJy16awuWQw1N4hYDKMpw2MnhcP1JKVfE42ModTbSvaCe9op9uTkftNedHl04jYfRnNTo2jm1KPuquPweNymll2YrDSbXvsNUw6hRlezs3CUZOLjeLu0HsUD+TpbUuT6z0xlGsNOYuOLyrPcBh8zwNdKyq4evTjUpzS9YyT/OAP6wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA/I8Xvkm1r9Hcy+zVDwY9nD+7Z4Wf6xxX2LEHvPxe+SbWv0dzL7NUPBj2cP7tnhZ/rHFfYsQB3p9t98mPDL/X2N+zxJ9jHwT0XT4Xah46Zlk2Exmp8ZntXJsDi6tNVJ4LB0aNGTVJtf3OU6lWfU1u4wgr90R7b75MeGX+vsb9nifQ/Y3/ALkbFfS/Mf5nDAecntLNSZtqTnY4hfdX31SGV4jB5bhKE7Low9LCUrRjZuylKU5973qN2T2XaPKPaMc72n8nwen8l5KaGDy3LsPTwmFwtHS+cRp0aMIqMKcV1WSUUkl5I/ke1y5RtY4fiDV5ndE5JiczyHOcLQo6lWFpOc8uxVCEaUMRNR3VGpSjTi5WajOm+prrjfbh57anUGnOHGA0/rXg0tQ6oy7BQwv3WpZ38Ho42cIKMa1Wk6MnGTavNRk03drpvZB1g4TaQ4+5BzQaW4u5XwD1jp+ENaYbOJYLCadxtLC4WhUxalVw8G4JxoqnOdOze0Nm+7OBzPZVg8+57te5HmMZSwmYcQ6+ErxjLpbp1MYoySfdOze/gdxORTiZzuc2nMB+2NqjiJqHK+GGV5jPM80oYVe4yyrJNypZbhlJNzh1KKmryapxfXLqknLqNzDf9sG1h/5TJfboge6vGbTuRYrgTrbS9fKMLLKHpXMcJ8DVNKlGisJOKgoqySSSSta1la1keJfsvMwxOV83WTY/CStVw+RZ5VSf4snDL60kmla6uk7eh7jcXvkm1r9Hcy+zVDxG9k9+7Z0l/q7N/sVUD8p7PrT2VcTed3h1gdcUo5nRxeaY7NsQsQlJV8Th8HiMXTlNPaV61KDaa33R72cSdG6f4hcP9R6H1XRw9TKM8yzE4HGKuk4RpVKbi5tvs436lK6acU001deEfMXwe4o8hHNJT1TpfCV8NluX5y860dm88O5YTE4fr644eTVlKUIt0qtO6bSbSUZxb7Ia49rZxI46aElwa4R8Cq+C1zrLDvJfhOHzGWNalWg6dR4WgqUZdbTbi5yagt31WuB1z9lx+7r4Z/8A5Z/ofGn7bjzyC82PLpxhxfEjg7p3OtSZXg82qZvkWeacp/CcZhf7o5wjVw8U6kakE0m1CVOSTabTaX4b2YdH4RzxcOMP72pT97DO4ddOVpxvk2NV4vwavdPzPoeJ5ouc7kq5lqeU8ddX6v1fleSYqtCrlea5rVeDzrAzjKEMRQm+qLdpRqQdn0zioySakkFsf7TnjNjcvjw45qOBWi+ImWYatSxGJy/PMpnl2MdWK+LNqzpQlZys1QX4zttseknIfxR5cOLHC7MNQ8u/D3B6GpQx6p59kdHCU6E6GM92rTl7v4tSMoW6KitdRaaTi4rzs56/aJcI+anhXhNCaV4N4/C5xTxtLFLO86WH9/l8INuUMM6TlJ+82jJtxVr/ABW7NdpPY78Ctb8MOFOruImtMnxeU/s/xWAllmExdOVKrPB4WFboxDhKzjGpLEz6W0nKMFJXUothv7av9yzpb/ygYH+jsxOqvLV/2qzmN/19hv8A+QO1Xtq/3LOlv/KBgf6OzE6q8tX/AGqzmN/19hv/AOQA/q+xT03k+Ycdta6kxuDjVx2T6aVPBVJb+59/iIKpJLwk4wUb90pSXZs+1e2409ga/C/hvquVCl8NwWf4nL4Vej+6e6r4dzcVLyvh4u3nZ+d/lHsRPlZ4k/R3DfaT7b7bP5BdCfS9fYsQB+t9jjKT5RsSnK6jq7MUl5L3OGf+1s/o+18/ccY76RZZ/wBaZ/N9jf8AuRsV9L8x/mcMf0va+fuOMd9Iss/60wOt/sjuG+lOL/BLjxwz1xgFjMk1DVyvB4umrKUU6eIcakHv0zhNRnCXhKKfgdX9H59xL9m5zh1qGa0qmJensW8DmdKmnTpZ3klZxl1QumkpwUKkHv01IJO7i0dyfYdf70+LX+sco/msSfVfapcpf7d3CZcW9G5V77WmgaFStUhShepmGUq8q1GyV5Tpu9WCv297FJuaQHlpwLzTBZ5zvcPc6y2r73CY/irlOKw83FxcqU83pSi7NJq6admro/Q8/Opc31LzucRcXnGFqZjLA59Ty6hgqnVaph8PCnSp0oqLuoyUE7Rabc29m2fP+U791Pwb/wDKBp7+kaB3J9rByla40xxXxnM5obKMZjtNZ/ToVs6q4Km3LJ8fRpxp+9mo7xpVIwhNVOyqdabV4dQfrcL7R7niwOXUcmwXJXh6GAw9COFo4WnpXOI0oUYx6VTUFKyiopJK1kl5HVnlp0Nx20fzZ6A4j4bghrPT2CWs8JVxNOhp3G08Pg8DiMSqeIpxcoLppqhVqQXU7KPd2udmdKe211BlugMLlmquCdHONXYXCqhPM6Oc/B8Ji6qikq86Kotwbd3KEZNNp2cU0o/2vZ96952eaHjbU4pa64jajwnDDK8ZXx+Ow9P+4Zfi8RPqdLAYaLTcqcJSTkk30wgoyleSbDp1q/8A7ZDnf/lvxP8AT0j3W5gvkF4k/RDOPsVU8NuerQGueAPObq7Uk8LicDLNdS1dZ6fzCUL0q6r4h4lTptq0nTrSlBrwcGns03994k+1J4v8znC7M+A/D7gRQy/UOqsqrYDN8wwmazxUXhXSfwuVKg6UPcQdNTvKpVmoQlK7bXUB8z9kT+7Lyr/UOafzSPiXBbhtlnF/mr03w1zurKGWag1esJjumTUpYb37lVjFrtKUIyin4Np+B9t9kT+7Lyr/AFDmn80j8Dyb/u89A/TOf/WqAeq/tGqOX8LOQvW2n+H2R4PJMtcMtyqnhsvoRoUcPhquOoRqJQirJSheD7X627379S/Yh6PyHMtZ8Utc43B0qmbZFgMqy/AVJxTlSpYueJlWcW9038Fpq68G12dj0r5iOEGB4+8E9YcIMwxUMLHUmXSoUMROHVGhioSjVw9WSs7qFanTk0t7RdmnZniXwQ4ucbfZqcfs4wOsuH9So8RQeX5zkuLqSoU8ww8Z9VLEYbEKMk7NNwqJSi1Kaau7oPUH2qGi9Pap5MdY5vnNHDrGaYr5fmmWYirFXo13i6VFqLe6c6dapTsmruS72s+g3s4+GGb8xXBTmJ5dKOd/c3D6iy7I8dhMTVj1UsNj6GJqVaTkk+pRnKlTjNpN9Mb90k/5fOR7STVnNzo/A8INGcN6umskxeMpYjHUfhrx+MzOtCSdGjFRpwUYKdpdKUnKUYbpJp9jeAPJLzBcH+SDXeZ6MzTNtK8ZtX1cJnWEwmX4l4fF0cJg23SwE5prpq1IVMRJxvZSqUoys4uwdStNcOuf7kI1ljtT6X0FqPKVWh8FxuMwWWxzbKsbRUrxVSdONSCV94tuE1dpWbaP32We0s0JxP1NluP5vOVHQet54SjHASzjAYPpxuGw6nKVo0sTKpGpZzm+hTpq8m01exyuVT2n/E7gLn2o9MczNLWOt8LiKsUnjMVJ5nlOJpdUZ0vd4hxXTK9pRbi4yinvuj49zicdcg50uOWT5vwZ4M4zKMdiMJHLlQoUIVcyzrFOpKfvasKCalJJqK3nK0W3K1lEPo3tV9daQ1RrPg/lnC2tg/2vqHDnCZppyjg4OnRo0sTisRTcY03b3doYSjFwaTi4OLSasvT3kB0bkGi+T/hhhNP4ajSjmmRUM5xc6cWnWxeKXvasptpOTTn03fZQik2kjo7za+z44jYblF4SZxpnJqmca04aZDPAaky7BpVa1TC16k8VNUlFN1Xh61WrG0buUZykk7b/ADjlM9qnqHlx4TYfhDrThlLV2FyJ1IZLiaWafAq1ClKUpvD1uqlU6kpyfTJWcYtRs0lYPQfi/wCzb5X+N3EXN+KGssizqlneeTp1Ma8uzJ4ajUqQpxp9fu1GylJRTk+8pXb3bPLn2qmGWD5zdTYSNSpUjQyrJqanUl1TklgaSvJ+Ldrtn87TGleNXtLuanF6lxeUSo0M0xdCedY3C0msFkWV01GEYKb2clTjaEW+qpO7tvJrne1Ny3BZNzjaiyjLaEaOEwOT5LhsPSjsoU4YCjGMV6JJIDX2lups2zfixoPIcZVl8CyHhxp+jg6f71e9oOrOdvNymk35QS8D7Rwh59ucbhjwu0roHQfJ5Rq5BkeVYfC4CvS03m0liaSgn79yhLplKo26kpLaUpt+J+q9oryi6x4k8I+GfMNw2ySvm+L0/ozLsq1BgMLTc8RLBwpKpRxMIRV6nQ6tWM7JyUXB26YNr8Ly3+13zvg1wkyThXrzhJ+yeppbBU8syzMMJm3wKc8JSj00aVaEqVRXhFRgpxavGKvG6bkHWniXp7mP4hcZ80424Dl21bpjN8yzWnnUcNlWmceqGHxkemTqwVSEneVWLqO7a6pO1lZH6L2mOKq5hzr8QcfiMO8PVxFDIqs6TvenKWS4FuLvZ3TbW6XY7DcvfHXnj55uY54/Teuc90fw9o4+jXzyllFR0sBleAh03w1OrKLc8RVimlu5OU3O0YRaj8C9qP8Au6+Jn/5G/ofBAe4/CfReQ6W4O6U0HgcrwaynAafwmX/BY0V7mpTVCMZpwd1JSu273v1Ntttng3ys4Sjpjn00JlWUp08PgeIEMBRUn1NUliZUkm/F9O1/Pc/0CaP/AN6WSf6uw383E8AeXn/tg2j/APymR+3SAtxPprjP7Q/Ock1rUqwwuouK6yDFrEfEnRwX3TjhIwnd/FcKMYxe+3T4H+gSrkmT1clnpueVYV5VPCvBSwSoxVB4dw6HS6ErdHT8Xpta23Y8Xvabcq/EDgxx3zHmK0PlmOq6T1NmEc7eZ4Sn1PKM2clKqqrividVVe9hN2Tc3G7cXf8Ad4z21+ta/DCWTYbg/gaGup4N4Z5190m8DGv02+ErC+66r33VN1Gr921sw7KZ5yAcv3K/pPVvMDwkwOqcPq/RGls9zTKJTzmc4qsstxMVG3Td3U2l3e67nk/ym8TdecIuMuC4hcNuF9DX2o8uwmIeEy+tgcRjHQc4qM8TGFBqalGMpRUuy6342Ozfst+UbUvFriZW45cRMjxX7CMrwmOw9GvjFNfdnG4mjOhNU27OcYQq1ZSqJtKagt31dPy7O9L8bPZlc0uH1VQyGpjcFluJxEMox2Loy+A57llVSi4OpFWjUcGuqKfVTmk7NWuH1/j/AM4POLzF8Ks54Ta25O61LLs3VKUcVh9L5s6+Eq06kZwq0nNtRmnG17bqUl2bT/m8hPLdxb4h6R458BNWac1LozLdcaVwlbDYzN8oxGHwyzHCY2nUw1+uC6leclJRd+hy2dlb9Zxn9szrTXGg8RpXhDwvqaMz3NIe4qZxWzZY2rhE7J/BoRowXvHdpTl27qLbTX2HhLwy59dX8meuNaar4t62wvEnPoYXMNH5dVxzoYrD4PDyVScWo9LhWxUXOKhPdKNJuzlJIOlunuFvP5yGa4xurdK6A1HlslTeFxmOy7L1m2VY3DpppVZU1OCi20119E4u9ulppfvcv9pZo7iZqXLcVze8qOgtcywVJYCWbYLBe7x+GoKTk0qWIdSNS0nNuCnTV5Nprs+Tyue034r8vepdS6X5m8NrPW1DETpw93j8ZJZllGJpdSnBU8Q0umakuqLcWnBPe7R8l5zuP+nedTjVkebcHODeOyvMKuGWWqnToQq5nneIc24SqU6CacoxtFK85Wvd2SSD3Q4O5zw81Dwt0vnfCOlg6ejcZllCpktLCUlSpUcL02hTUF+I42cXDvFxae6B+M5O+FOf8EeWbQHDLVFlnGUZY5Y+mqrqe5xFerOvUpdV2n0SquF0+n4vxdrAD7MAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAP5GsP8Aelnf+rsT/NyPPrll+XTSf+k1v5ioAB9957f96el/9Y1v5tH6Lkp+R6v/AK5xP83SAA+74z/4JX/8XL/Yf5veMP7pzM/9fYT/APhAAf6CuAPyJaG/1Dg/5qJ0i4h/uj84+lT+0IAD0E1h/vSzv/V2J/m5HRHlD+XTJv8AR8Z/MTAA+ze0f/co6o/9/wDgqp0G9jH8teof9Xf/ALswAOxHKh8v2l//AMd+xVz997Vv9zZX/wDGVv5pgAeVfIj+6S01/wCNh/PUz/RMAB1355Pkmyn6RUPs2JPknDH9yRxL/wBY0v8AbhwAP6/Il/vs1R/q+j/OM/cc9HyeZB/rpfzFQAD+vyU/I9X/ANc4n+bpHL5yfkUxP+scJ/1mAB+G5Dv/AIp1h/pOC/6tU7S4z/4JX/8AFy/2AAeY/CH5WdFfSLLftNM9Ms4/+Kcb/o9T/qsAD/Nxrf8AdG5j/r5f9ZH+iDgv8kWi/wDUOB/mYgAdSva8fucsP/ps/wDZE67ex7/3u8Vv9VYn+bpAAfd+Tb5a8N/q7F/9VH53gj+6B07/AK6n/wDvAAejh0F9sP8AIJln+k1f5zDgAdSvY+/L3iv9Hf8ANzPa8ADxs9sh8rWA/wDF4f8AmD9R7F//AHzak/0af+2IAHrWeA3tNf3VOov/ABlb7RVAA9VPZtfuXMn/ANNrfzdI+P8AN58umc/6Pg/5iAAHe7R/+9LJP9XYb+bieAPPx+6u1J/pC/nZgAezvJB+5q0j/wCLq/zkjrPzX/L9qj/8S+xUAAPQTJ//AIpwX+j0/wDqo88OHn7o/J/pUvtDAA788R/k81R/qbG/zEz/ADnYX90nL6TVP51gAf6NtCf7x9Pf6qwn8zE+He0E/cuar/8AFv8Am5gAeSns0v3Tmlv9a4T/APiHvyAB5B+2d/39ZB/4uj/N1Dl+xh/34ag/0Kt/1qYAHriAAP/Z)
Figure 4. Foraging and Risk of Predation. Foraging distance and time away from the safety of a nest are positively correlated with the risk of meeting predators, which can result in injury or death. Motivational factors, such as hunger, reproductive and parental state, and ecological factors, such as food availability and predator density, influence foraging behavior (represented by a horizontal arrow) and thus predation risk. Fear elicits immediate species-specific defense reactions upon meeting a predator and exerts enduring influences on foraging strategy.
Circuitry
![[[@Brandao2008-hw]](http://dx.doi.org/10.1016/j.bbr.2007.10.018)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQEAcQBxAAD//gAuSGFuZG1hZGUgU29mdHdhcmUsIEluYy4gSW1hZ2UgQWxjaGVteSB2MS4xMwr/7RrIUGhvdG9zaG9wIDMuMAA4QklNBAQAAAAAAAccAgAAAgACADhCSU0EJQAAAAAAEEYM8okmuFbasJwBobCnkHc4QklNBC8AAAAAAEoAAwAAAEgAAABIAAAAAAAAAAAAAALe//8CQAAA/+7////u//8DBgAAAlIAAANnBSgAAAP8//8AAX//AAEAAQAAAAAAAAAAAAAAADhCSU0D6QAAAAAAeAADAAAASABIAAAAAALeAkD/7v/uAwYCUgNnBSgD/AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAF//wABAAEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADhCSU0D7QAAAAAAEABxAAAAAQABAHEAAAABAAE4QklNBCYAAAAAAA4AAAAAAAAAAAAAP4AAADhCSU0EDQAAAAAABAAAAB44QklNBBkAAAAAAAQAAAAeOEJJTQPzAAAAAAAJAAAAAAAAAAABADhCSU0ECgAAAAAAAQAAOEJJTScQAAAAAAAKAAEAAAAAAAAAAjhCSU0D9AAAAAAAEgA1AAAAAQAtAAAABgAAAAAAAThCSU0D9wAAAAAAHAAA/////////////////////////////wPoAAA4QklNBAgAAAAAABAAAAABAAACQAAAAkAAAAAAOEJJTQQeAAAAAAAEAAAAADhCSU0EGgAAAAADOwAAAAYAAAAAAAAAAAAABB4AAAamAAAAAwBnAHIAMQAAAAEAAAAAAAAAAAAAAAAAAAAAAAAAAQAAAAAAAAAAAAAGpgAABB4AAAAAAAAAAAAAAAAAAAAAAQAAAAAAAAAAAAAAAAAAAAAAAAAQAAAAAQAAAAAAAG51bGwAAAACAAAABmJvdW5kc09iamMAAAABAAAAAAAAUmN0MQAAAAQAAAAAVG9wIGxvbmcAAAAAAAAAAExlZnRsb25nAAAAAAAAAABCdG9tbG9uZwAABB4AAAAAUmdodGxvbmcAAAamAAAABnNsaWNlc1ZsTHMAAAABT2JqYwAAAAEAAAAAAAVzbGljZQAAABIAAAAHc2xpY2VJRGxvbmcAAAAAAAAAB2dyb3VwSURsb25nAAAAAAAAAAZvcmlnaW5lbnVtAAAADEVTbGljZU9yaWdpbgAAAA1hdXRvR2VuZXJhdGVkAAAAAFR5cGVlbnVtAAAACkVTbGljZVR5cGUAAAAASW1nIAAAAAZib3VuZHNPYmpjAAAAAQAAAAAAAFJjdDEAAAAEAAAAAFRvcCBsb25nAAAAAAAAAABMZWZ0bG9uZwAAAAAAAAAAQnRvbWxvbmcAAAQeAAAAAFJnaHRsb25nAAAGpgAAAAN1cmxURVhUAAAAAQAAAAAAAG51bGxURVhUAAAAAQAAAAAAAE1zZ2VURVhUAAAAAQAAAAAABmFsdFRhZ1RFWFQAAAABAAAAAAAOY2VsbFRleHRJc0hUTUxib29sAQAAAAhjZWxsVGV4dFRFWFQAAAABAAAAAAAJaG9yekFsaWduZW51bQAAAA9FU2xpY2VIb3J6QWxpZ24AAAAHZGVmYXVsdAAAAAl2ZXJ0QWxpZ25lbnVtAAAAD0VTbGljZVZlcnRBbGlnbgAAAAdkZWZhdWx0AAAAC2JnQ29sb3JUeXBlZW51bQAAABFFU2xpY2VCR0NvbG9yVHlwZQAAAABOb25lAAAACXRvcE91dHNldGxvbmcAAAAAAAAACmxlZnRPdXRzZXRsb25nAAAAAAAAAAxib3R0b21PdXRzZXRsb25nAAAAAAAAAAtyaWdodE91dHNldGxvbmcAAAAAADhCSU0EKAAAAAAADAAAAAE/8AAAAAAAADhCSU0EFAAAAAAABAAAAAk4QklNBAwAAAAAFLoAAAABAAAAoAAAAGMAAAHgAAC5oAAAFJ4AGAAB/9j/4AAQSkZJRgABAgAASABIAAD/7QAMQWRvYmVfQ00AAf/uAA5BZG9iZQBkgAAAAAH/2wCEAAwICAgJCAwJCQwRCwoLERUPDAwPFRgTExUTExgRDAwMDAwMEQwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwBDQsLDQ4NEA4OEBQODg4UFA4ODg4UEQwMDAwMEREMDAwMDAwRDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDP/AABEIAGMAoAMBIgACEQEDEQH/3QAEAAr/xAE/AAABBQEBAQEBAQAAAAAAAAADAAECBAUGBwgJCgsBAAEFAQEBAQEBAAAAAAAAAAEAAgMEBQYHCAkKCxAAAQQBAwIEAgUHBggFAwwzAQACEQMEIRIxBUFRYRMicYEyBhSRobFCIyQVUsFiMzRygtFDByWSU/Dh8WNzNRaisoMmRJNUZEXCo3Q2F9JV4mXys4TD03Xj80YnlKSFtJXE1OT0pbXF1eX1VmZ2hpamtsbW5vY3R1dnd4eXp7fH1+f3EQACAgECBAQDBAUGBwcGBTUBAAIRAyExEgRBUWFxIhMFMoGRFKGxQiPBUtHwMyRi4XKCkkNTFWNzNPElBhaisoMHJjXC0kSTVKMXZEVVNnRl4vKzhMPTdePzRpSkhbSVxNTk9KW1xdXl9VZmdoaWprbG1ub2JzdHV2d3h5ent8f/2gAMAwEAAhEDEQA/APUqqqqKmU0sbVVU0MrrYA1rWtG1jGMb7Wsa1TVS7qvS8e11ORmUU2tjdW+1jXCRuG5jnbvoqH7c6J/5YYv/AG9X/wCTSU3klR/bnRP/ACwxf+3q/wDyaX7c6J/5YYv/AG9X/wCTSU3klTr6x0i2xtVWdjvsedrGNtYXEn81rQ73KxkZFGNQ/IyHiumoFz3u0AASU0eq0UZGV06rIrZdWb3kssaHNkUZEe18qs0/VF2Tfi+jhsuxrGU2h9LWAW2jfTS2y2tldttjP8HU96VF3U77en29ToZi3OyrvTpY7ftq9G/0fUf+da5vuf8AQVjI+rfRcnKdmX43q5LnNd6j3PdBZu2tra55ZXV73epRW30bv8NXYkpr1O+plr2sq/Zzy/YGbRSQ42myuhlb/o2Ptfj3NZWz3/o0xs+prTcHtwGHGea8jeytvpuH5t29rfS/kb/5z/BotP1V6JVVVWaX2moMbvstsc5/pkln2iX/AKf6XvZb+jf6dFX81j47Kueysz6uOxC8dKzMd1PqWsZRFVrWYuS31/Q2XM21/aLqP6J/g8v9U/S03fZ0p6AV/VEv2BvTi/eatoFM+oB6jqY/0raxv9NQc76ltYLHHpjWODXNcfQALbA40uDv3bfSt9P9/wBN656v/m9V61NnQMu91T7oYPcw0+vk+njtZZZj1txGto+1U9P9H7NRken/ANrv066Hp/ROjZVbOoOw7WZFjQ132myyy0Guattj7Lrt+33fpN/6X+cSUp5+prK32EdPLao37W0uILmuurbsYHP321MdZVX9O1n82i4VGFV1bdhVV01XYjX/AKJgYHDeSxztrW/vfnIY+qHQCHCzHdaHO3APsfDRrtqrY17WMpa52/0/9L+n/nv0ibqD83DynnpOK3LyacSsU4rnisOaLmtsZ6z52O9Hfs3JKdtJDovqyKxZU4ObJaYIMOadllbo/wAJVY11djPzHoWT1Lp2JYKsrKposI3BlljWuLZ27tr3D27klNlJUf250X/ufjf9us/8kl+3Oi/9z8b/ALdZ/wCSSU3klR/bnRf+5+N/26z/AMkmPXuiNBJ6hjADUk3MgD/OSU//0O+w8ujGy+oi0mbcsRAnRuLhmyx38itv03qy7rXTGuDTeNxIG0BxIktbLhHtaz1K/Vc7+a9Wr1f52tBwKKL8jqTb62WtbmMc0PaHAOGNiQ4bvzkfLw62YpOJiUWX1DdRW5rQ3dPbjYkpY9b6UAC7IazcC73S0wAC7duHs2bmb9383ZZVU/8ASW1pn9c6WxrXm8em54rFoBLNxY+7+cjZ9Gl7P+P/AFf+f/RqnTj9TbuNuBiOZ9F1bGtD3slvtbuf6bfZ9Fln+i/we/8AR3MDDLsTbn4tDLnPLnsra0sMSyp3H0vS9iSkHUs7Dy8UNx7W2mrLww/brB+1UAf9Qi17+o5QvMjAxnfoG6t9W1pex9z2n+cxqv8AtJ+Zbd+t/wCDw7kLreDi/sp2NXWKasjIxmWej+jP6TIore5rqtjm2e76aL0zJtqf+y8wNZk0MmlzAG13UthgvoY32sdXuZXlY7f6PZs/wF2P6iUk6hRl2W4l2K2ux+Pa57mWvNYLXV20+17K7/dusb+Yq/2/6wf+VVf/ALFD/wBILVSSU5X2/wCsH/lVX/7FD/0ggV9V6lZlXV1dOx35dAY3Ia3LabGBwNlLbv0G9jX7nWVLcWbg4+PX1nqdtdTGW2+h6tjWgOfDCG+o9o3Wbf5aSkI6r1s3OoHTKzbW1r3t+0jRry9tbp+z7fc6m1T+3/WD/wAqq/8A2KH/AKQR6Qf23lnt9lxv/PmarySnK+3/AFg/8qq//Yof+kEXEq6hZnHMy6q6AaBW2tlhtdO4vdvmmlrdv8l1i0EklNHKrtxXuzsVpc3V+Vi1MaXXnayttrXTW77VTXUxjPf+lp/V/T/o76YYtjLOtZb2GWuxMWDx/hM/mVYzs1mHSHlpttsd6dFDI322EFzaap/ktc973fo6aWW5F3p0VWWLN6LiW4nVc4Xlpvvpx8i8VyK22WWZu9lDXf4P2fT2frFvqZNv6W5JTpZPUcLEeGZNoqLmlwLpAgHaTu+j3Qv230r1W1HJYHvgBpkauLWtaZHte7f9B3/Df6C707VlFNv85W15jb7gDoSHbdf5TVi1Y3WCXXPwMJl8i3VokvADv5xrnO3+t7vX/M/0fs32JTfPXekgP35LWGt7qntfLXB7S9m3Y4bvc6v9H/pP0ez+drT9c/5E6h/4Vu/89vQ8PEddbddn4VFZNgfj+1jngfvWObu/S/nfSROu/wDInUP/AArd/wCe3pKf/9Hvvs/XMbLy34bcaynJtbaDa57XAiqnHc2K2Pb/AIDcpb/rP/osH/ty3/0ktVJJTlb/AKz/AOiwv+3Lf/SSW/6z/wCiwv8Aty3/ANJLVSSU49tH1gy/SqyW4ldLbqbbHVusc6KbK8naxrmMb7/S2fSV/Owq8ykMc51dlbvUovZG+qwAtbdUXbvdtc5j2O/RXUvtx7/UottrVlJJTR6fn2Wvfh5jW1dQxwDY1shljDozKxt253oW/ufTx7f0Fn+ltDX1fKvY27GwXvos1qe6ythc382z0y/cxr/pM3/pP9J6f0EH6yMx3P6WX1X25Az6PQ+zODHNG4OvsyPzn4LKmfrVex/+D/m/5+mfSI/ZOHH+hYkpL+0eo/8Alef+3qv/ACSFXkZteRdkNwHl9+3eDfVA2Da3br5qv1HF65bm139Py6seiqp9ZpfuO99u7dkPG19W7D9PGfi7mWep+tV/olVtZ9bqQ5jLWZIFZfvY2lr3OBqa3HqdayutlljH5dnqWY9ldXoYf876t1SSm7Wx1XUruqM6UG5uRWym231qpLKy5zB9L+X7v+Lq/wBGrX7R6j/5Xn/t6r/ySxvsX1vtaS/NordcxjbADGyG3ep6LKqD6Vu66v8ASsyHv9XG9b1fQ/Va9TprOpsruHUra7XutLqPS4bTtZsre706dz/U9T83+2kpL+0eo/8AlcT5C6qf+qRh1TF/ZrOpv3Moexr2tIl5L4bXS2tm7ffZY9tNdVe/1bv0dakz6bfiFmdE6W2xvT+p2X2ObThsqpxCQaWWe7fmtYRu+1uqs+z+q3/tP6tf+FSU3en4mQ+49S6gIy7GllVMhwx6iQ70GOb7XXWbWPy7Wfztmyv+ZopUMvE6s3qL8vp9mO1ttNdVjb2PcZqddY1zDVZX/wByVppJKcrb9af9Jg/9t2/+lktv1p/0mD/23b/6WWqkkpytv1p/0mD/ANt2/wDpZBzMT6zZeJfivuwmsyK3VOcK7ZAeCwkfpv5S20klP//S9VSSVbJ6l07EeK8rKposI3BltjWEjjdte5vt0SU2UlR/bvRP/LDF/wC3q/8AyaX7d6J/5YYv/b1f/k0lN5JVaOq9LybRTj5lF1pBIrrsY5xA+l7WOLlX6oTmW19Ir4vG/NcPzcYHa6vT6L85/wCrV/8AA/a7av0mMkpbpn69e/rDtWWNNWAPDHkON/8AWzrGNu/8K14X+F9RVbqa8PqOH0ui7PrZlttdV6Ia+ikVBr/TsutqsfUx+/8AV27/APgf0f6JbgAAAAgDQALMsGU3rVBvex1bvW+zNYCCGbKNzbi5zt7/AFm2fQ/waSmGRh3U3Y1X2/Ld9psNU7qhtiu3I3f0f3fzGxH/AGVd/wCWOX99X/pBB61lMqzOlUBxZkZGRYMd3pvsbuGNkibPSG3279+19tP6P1P9HYtUTAnU90lND9lXf+WOX99X/pBL9lXf+WOX99X/AKQWgkkpx6sS63KyMb7dlt+z7Pduq928bv8AuP7dq1MfHpxcevHobtqpaGMbJMNaNrfc/c939Z6zun5uNb13quLW8m/GGObmbXDbvY51fvc0Vv3N/wBG5aqSlJJKrkdT6bi2elk5dFFkB2yyxjHQfou2vcHfmpKbSSo/t3on/lhi/wDb1f8A5NL9u9E/8sMX/t6v/wAmkpvJKtj9T6dlWeli5VN9gBcWV2Ne6BALtrHO9vuVlJT/AP/T9VWNk35eP1fMtxKPtLxj4m+sHaSwPz3P2E/4T/Rsd9N/+jWysezqXTsHr2UM3KpxTZi4xYLrG17oszd2z1HN3bdySmf7V6hMfs6wCYJJJEay/wBjHO/Ru9j/AKdm/wDo1eRjfrKizrHU3lrR0uxrpHqNc7VoO7b7tvovd7fzLv0f/F/Zn5NXLzeg5OUb/wBv1VNOz9FXlVho27txZ+k9nqtftf8A+lFGnL6DVdW8/WGuxlTg5tdmVU7WGtdue95e5vt/8EtSUmyuo2l+Jk5eJZisxsp7gD+ke5gw8u0uZXSHu3/8HX6n7n85+jT9M6lgVMsycq4fbMt3q3gBztgjbTite1vuZjVfo/8AhLvWyP8ADqd3Vul5vUOm1YeZRk2tyHvcym1ljg0Y+U0vLa3O9m57G7lspKaGH1zpmdm34GLdvysZjLLq9j2w2wTU7e9jWO3fyXLOxMS/I6Li5/rZF2ZbTXY79KebA117mMa6pvta5zvRrdX/AKNb656uz6y9J6U1hxcK2jAojf8AaLQ9zKW/S9P7K9rbHsZ9D1f7aSlg3PDN5oyHDRpLc4fSdvHs3nbt9RldX87/AIX/AAvvWhjdMc+lll9uVTa4S+r7Q923+Rva73LJZ0TqjWxX07ErDmNrJZnZAOwNNbR/Q/p+m+xvrfz36S307VpC/wCtffDwP/Ym3/3iSUxx8CyzpteQ2/KtvNQfs9dzd7o3beWMZvVQt6i2S6m0Ma/a956hADQdr7f5Oz2e1Wcb/nVj49dAxMBwraG7vtNomO+37G5VHdJ6u603jp2Ey10zYzMvBlzjY4n9T9/6Q7vf+5T/AKCpJToYOBkXVl+WcnFcHkMq+0ufLIb7nvY78525N07q+NV07EGbkE5DqWPcXAlzgfaHu2N+k7amZZ9aa2NY3CwdjAAB9quJgfynYTle6XjW4nTsbGuLTbVW1thZO3dHv2bvds3JKQu670prXON+jQXGGPOgG535ir9EzcXPyM3Pw3erj5Ipspsgt3NNQ2+14bY3+01bCxWdU6Zg9X6izNy6MV7zS5rbrGVkt9MN3NbY5u5u5qSkh6p1Wqms2dNfba+sPeKiAGv2se6o7i76HqbN/wDhPSu9Ov8AwSa3rWZVXbc7AsFNQJLnEg6Ocwez0zZ+Zvf6bLP0fp+l66z2W9Ca9pP1jYW1ya6xlVhrXe/a7+c3O/nPoP8A0f8A6D/q6X2jo/phv/OZu8Ajf9pp03Tuc33btzfZ6W91lf6P3121+pUkp1ch2/qvTXlpaXV3naeRLavaVpLEq6h03K6r0+nBzKsw01Xb/Ttba4Nipm+0tc93u/fettJT/9T1VQdVU8y9jXHxIBU0klI/s+P/AKJn+aEvs+P/AKJn+aERJJTBtNTDuYxrT4gAFTSSSU0T1vpYc5v2gEtc5pgOI3NJre3c1u32Pa5ip9W6piZPS8vGw76/tN1NldPqts9Pc5pa0WbA1zWfyksN/UWdIrPT62W2/aLi9tjto9Nt1z3taf8ASWtb6NX+j9T1ER2T9YmttLcOqwwPSBeGR7QXbvfdv/Sb2fTq/wAGkpnjdbwm49Qyb2HIDGi41ssDN8fpPS3hz/T3/Q3on7c6X/p/+i//AMihZF/1gOQ6vGx6m0uaA257gdji2XO2b/0zd5/dp/m/8J636uzcn6wnZuwqW7jDgLJ2aH3nje3e5n0f9H/wv6JKTftzpf8Ap/8Aov8A/Ipftzpf+n/6L/8AyKH9p685r3fY66y11Xps3h5c0z9pa526nY5n7/8A1zZkfzSg3J+scBz8OmXvaPTFn0Gx+le+yffz+j2V/mf8L+iSk/7c6X/p/wDov/8AIpftzpQ19f8A6L//ACCA7K+sW15bg1SAXNabAZ7sq3S33u3bX/4Nno/4X7R+raySmFVtd1bLanB9djQ5j2mQWuG5rmn+Uk6ml53PY1x8SASqnQ/+ROn/APhWn/z2xXklI/s+P/omf5oS+z4/+iZ/mhESSUwZVUwyxjWniQAFNJJJT//V9VSXyqkkp+qkl8qpJKfqpJfKqSSn31n7Hmz1Ptnqetdv+yftT0J9W3d6P2f9B/xnofovX9X01L/If/mz/wDhsvAEklPv/wDkP/zZ/wDw2S/yH/5s/wD4bLwBJJT7/wD5D/8ANn/8Nkv8h/8Amz/+Gy8ASSU+/wD+Q/8AzZ//AA2TO/Ye0/8AKfB/8t/BeApJKfqDpe39mYm309voVx6M+lGxv8x6nv8AR/0e/wDMVpfKqSSn6qSXyqkkp+qkl8qpJKf/2ThCSU0EIQAAAAAAVQAAAAEBAAAADwBBAGQAbwBiAGUAIABQAGgAbwB0AG8AcwBoAG8AcAAAABMAQQBkAG8AYgBlACAAUABoAG8AdABvAHMAaABvAHAAIABDAFMAMgAAAAEAOEJJTQQGAAAAAAAHAAgAAAABAQD/2wBDAAMCAgMCAgMDAwMEAwMEBQgFBQQEBQoHBwYIDAoMDAsKCwsNDhIQDQ4RDgsLEBYQERMUFRUVDA8XGBYUGBIUFRT/wAALCADuAYABAREA/8QAfQABAAICAwEBAAAAAAAAAAAAAAYHBAUCAwgBCRAAAQMDAgICCQkPEgQEBwAAAQIDBAAFBgcRCBITIRQXGCIxQZbT1BUyN1FXlJWh0iMzNDVER2F1doGSk7S14RY2OEJFRlJTVFVWcXORo7Kz0SRDcrEoYqTBCSUmY2aChP/aAAgBAQAAPwD9VaUpSlKUpSlKUqiOJ/FrTm+QaM2K+QmrnaJ2XqRJhvb8jqRa56gDsf4SUn71bQ8Hei5+t3Zz/wDov5VfO470W9zqzfi1/Kp3Hei3udWb8Wv5VO470W9zqzfi1/Kp3Hei3udWb8Wv5VO470W9zqzfi1/Kp3Hei3udWb8Wv5VO470W9zqzfi1/Kp3Hei3udWb8Wv5VO470W9zqzfi1/Kp3Hei3udWb8Wv5VO470W9zqzfi1/Kp3Hei3udWb8Wv5VO470W9zqzfi1/Kp3Hei3udWb8Wv5Vfe480XA6tO7P+Av5VceD5lEfhxw9lscrbaZSEJ9pIlvAD+4VctKUpSlKUpSlKUpSlKUpSlKUpSqd129kLQz7sXPzRcKuKoDj2vumuXZtKw6yZ5j11ymKVpdtES4tOSEqR68BAVuSnY8wG5Tsd9tqn1dUyWxAiPSZT7caMyguOPPKCENpA3KlE9QAHWSa+x5DUyO0+w6h5l1IW242oKStJG4II6iCPHWDkuTWjDbFLvV+ukOzWiGjpJE+e+llhlO4HMtaiAkbkDcnx0yHJrRiNt9UL3dIdogdK2x2TOfSy30ji0tto5lEDdS1JSB4yoAeGtlSlKUpWsybJ7PhlimXq/wB0h2Wzw0c8mfcH0ssMp3A3WtRASNyB1nx1sW3EPNIcbUlaFgKSpJ3BB8BFU/widfDvif8AXM/LHquKlKUpSlKUpSlKUpSlKUpSlKUpVO67eyFoZ92Ln5ouFXCslKCUjmUAdk+Dc1+ZWjb5xrU/S3GsOYvGQQ4WTSen06zTFC3ccLbc6YvTEXRCEpKUkjYKUsLDgAJO+3LTPU7V2RYMwnXnLb1GyWJhmSScntMyRcFqt8xttwxXGUKioYgrQtOyEtvKC0HmG5TudXNu+UaraP5harDluoeXW2ZpG7ccoRd+yR2PfEhpxllgqbTt0qenStprdC2hv178xkuU5xfLZYOxWc31ItVujYFbn9N3LI3LdF6u5S8Xm5JDSi+4HUx2wy9skNEnYA81WnxYv6y5PwsZ7bb9idm9S5GJxFrft0x125ruJEZTzXYqWygIDheAKXFdSUnxnaN6lp1CwS56x4za8jzG+2qFdMJl2uTcZDsp5CpE7lnpad236MpbSVIT3qAo7BIO1RuLqnl83WK9RrHmmaXLOYur7tohY0VSHLQbAl1HZQWOjLXIhC3CVKVzN8rfLy7jmx9I9VMxvmfYiLJmeb5Fm7moV4gXOwXEyHLOnH0PykqcJKeiCWylsJc5isL3b9akBObpBqFqrcb7hq7PkGa37VB+Le1Z5jOQR3k2q2uoYeMUNoWhLTH/ABAYQ2Gld+lR33G5rswzVmTatD8nurGUay5VmTGGx3cnt7gcZRbLouQ2h4Muvx1mK8grd7xlCkhlCl8pUlJrU4jlOp+RrGPRcuy1Nkd1RsUVi5QZ86Sv1JkW55UlLcuU0h12OXE7dItISFAKT1cpqX6YJ1Jx/OMImycvzm7x06o3zFnId2luvRzZGm5fQKdSU/NDzIQoPr3V1gBWwSBAuHTWXMrqzpddLVlmo2bZO9Dvz+X2aUl5+M3BZakdiOMdK2Wg6XG2EtKSVlalqSvfYpEIyXLc8z7SbVa2MzsryDG7xpuxcWoF0mzbs43cU3FlK0F16OykSA2shxphAbSQAB1V+rlhQUWK3JUCFCM2CCNiDyiqr4Q/2O+J/wBcv8seq4qUpSlKUpSlKUpSlKUpSlKUpSvH+qfGvktg4mMd0asWEm3y511iRpF3vLqXOkiuODmdYaaURsUcxClL3G3fIBBFW9rt7IWhn3Yufmi4VcVK1uS47bswxy62G7x+zLVdIjsGXHK1I6VlxBQ4nmSQobpURuCCN+oiueP2GBitgtlltbHYtstsZqHFY51L6NptAQhPMoknZKQNySerrNZ9RLPtXMI0qRBVmWW2XFhOWpEU3ec3G6Yp25uXnI3A3G58A3G+1dsjVPDIkW9yXstsbUexssyLo8u4NBEBp1HO048ebZtK0d8kq2Ch1jcVDsPzPRvAMxv+KWHK8dgZPd7hIv1ytPqwlyS7KcQlbzpQtwlJKEJUUJ2ASN9gOuojojr/AKD2nRl3JsayazYnhjt+nsOKu91QjaeuU4t0qLjquUukl9Kd/nbiVAJSeqysn1+0ywoQ/V/ULGLP2ZGZmxhNu7DRfjuqKWnkbr75tRSoBY6jynr6jW1w7VHDdRJl1iYtldmyOTaXAzPatU9qSqKs77BwIUeXfZQ6/GkjxGpRSo9p/p/YNLcPt2LYvAFrsVvC0xogdcd6MKWpau+cUpR3UtR6yfD7VSGleTMX1ZuehnAxas4tdnZv6rQ465Jt7rymi4wq4ONrKVgHlI5wrcgjYHqq1OGbiLh8TOCuZNb8au2PRmnuxybiEFp5wev6FaVbrSk9RUUp6+rwg7W/SlKUpSlKUpSqkyniAXZ9QLniFjwLJs0uNshx5kx6zGGlplL6nUtgl+Q2SSWV+AHwVjnXbKvcNz78bafTadvbKtvYNz78bafTadvbKj9Y3Pvxtp9Noddsq29g3Pvxtp9Ooddsq9w3Pvxtp9Noddsq29g3Pvxtp9Noddsq29g3Pvxtp9Op29sqP1jc+/G2n02h12yrb2Dc+/G2n06nb2yoj2Dc+/G2n02pZpNqnF1asNwuDFouNhkW+4v2uXbrqGunZfZVyrB6Ja0kb+MKPgqb1BNYtRpOneMxzaLYu+5Td5KbbZLWkEJkS1pUQXFD1jSEpU4tZ22Sg+PaunDtF8esaW7pc7Tb7jlkicLzOupbU4o3AtdGtxlTilLbQE7oQjm2SjZPt1HtdhvqFoZ92Ln5ouFXFSlKV5j4ueH/AC7VjIMfveFsMM3aDbZtuNyj5A9aZjQeLZCFbMPsyIxKO/Zcb3JCdlDrqmdR9IbrkvEJpLp96ss3efc8cgRdT24EJTMV2Pa3WZcZ4hKQhAed6RkJAHer2AA8FgN8Meotq4lBldgXbbHi8nKl324pTenZUWZHW1yu/wDy5+OoNTF9SS+0+hIH7XxVDIPB1qjYMd0/bhRbF2dg98yJxpmDelRDdIlycU4h/pVRHAy43uGygoXujm2Wnfati3wO5NCx68QGGLH0z2jr+DQt5jrqY9xdlSH1BKlo5uhAeQAs9fe7coFXPoroLddMtXbhf1s26NY3cKsePssw1nnTIhl/pd08oHJs4gA77nY9Qq/aUpSqH4bsVtWccKdlx++Q0XC0XJidGlRXCQHG1S3wRuCCP6wQRW6veOWrh0tN8yvC8PflwZb0Zy8WSzu9GhmO0goclRooBSp0I5OZCeUuJbHWVABVmY1klrzLH7fe7JOZudpnspkRpcdXMh1ChuCD/wC3i8BrZVA9a9Ue1BgUjIG7Wb3LTJixI9uEgMdM4/IaYSOkKVBIBdB32PgqMdsnWH3F4nlfH8zTtk6wn6y8Tyvj+Zp2ydYT9ZeJ5Xx/M07ZOsJHsLxPK+P5mnbJ1hI9heJ5Xx/M07ZOsJ+svE8r4/madsnWEj2F4nlfH8zTtk6wn6y8Tyvj+Zp2ydYSPYXieV8fzNDqTrD7i8Tyvj+ZrotuuOYwdQ8SxnL9OG8ZZyN2SxGns39qbyrZjreIKEtJPWEbb7+OtdpR18U+r32lsX+pcavulKUpSlKVSHDF6/VL7ubx/rmrTzfNrLp1i1wyLIZyLfaYKOd59SSo9ZASlKUgqUpSiEpSASSQACTUQ0uwm/s5HkGa5fOD19vCuxoduiuuGJbLc2tRZaQlQSVOK3Lji1JB5l8oACRVl1Tuu3shaGfdi5+aLhVxUpSlKUpSlKUpSqd4Q/2O+J/1y/yx6riqns6vbvDyi3XW0Y9BRpiHpDmQN22MoSra485zmelKTstkKK+lSE8yQrnG4SRVtw5ke4w2JUR9uTFfbS6080oKQ4hQ3SpJHUQQQQR7dUZxkdelcT7fWb86RavmlKUpSlKVSGt3s36FfbS5/m52sPSgf+KfV77S2L/UuNTGJqxdL1cZb1lxKRdsZhXddmfuTctKZCnm3+gfcaj8p52mnQpK1FaVfM3ClCgBzbFeuGCtm48+RxWkW9DjrzrqVobKG3UsurbWU8rqUOKShZQVBKlAK2JrAvvEFhNkxeReTdi8GkydoXYzyZIWwgLdC2ijnaCUqbKlrSlKQ4gkgKSTrVcRVkZjRi/HW0/JgvzmwCtbLCWoEeYsSHUoIZPLJQBvuCOvwnlqR3XWrCLJcLnDnZDHiu21t9yStxCw0joGi88gOcvIpxDQK1NpJWEpJI2Brra1wwh+3LmNXsOoTKRCSyiK+ZDry2i6hLbIR0jgU0lTiShJBQlSgSEkjCv+v+HWhFmTEujN2kXZ+2tRkROZSCibKRHZcU4ElCNypSkpUQVdGoJ3I6mX622rDdSIeLT2jHjm0vXibdZPSIZjtIcQ2hCNm1JcWtawnl5knwbcylJScudrrg1utLdxkXwNsK7I5m+xHy+yGOXp1OshHSNJbC2ytS0pCQ4gkjmG+9smfY/kt9uVmtVybnz7aEdloYQtSGSpCHEpLm3JzFDjauXfflWk7bEGqu4ZnENDVRa1JQhOcXgqUo7AAPncmueHuxOJK/Rcwn21f6icfuJexUuOqCLq8lJQq4OMkbFCVb9AT1+uXt3yNrupUL1P0ns+rMK0MXWXdYDtpm+qEKXZp7kKQy90TjXMlxBChuh1YI+zVYZZwzy0stiyag6jIWfXKcy+arb73PUXPDLl5Hslah+VMv5dO5ly8/XK1C8qZfy6dzLlxHslaheVMv5dO5ly7b2StQvKmX8uncy5efrlaheVMv5daLEtHbjnkSdJsGrud3RmBOftslbGVzCGpDKuVxs9/wCEH+8EEdRFb08MuXe6VqH5Uy/l1Hsz0dv2CtWdy46kakqTdbpGtDHQ5PKOzz6+VBVu4Nk7+E9Z+wakPcy5cfrlaheVMv5dDwy5dt7JWoXlTL+XQ8MuXbeyVqF5Uy/l07mXLvdK1C8qZfy6dzLlxHslaheVMv5dO5ly4/XK1C8qZfy6meOcMxXbwbtqBqOZPtozCakf3c9Wtp7gdq0xw224xZA+LZb0qSz2U8XXTzLUtRUs9aiVKJ3Pt1Iq4uNpdbUhaUrQoEFKhuCPGNqqm63rLdONU4oktSMi08yN5mGwIkUKex6XypbQlSW07qiukDdZ3Laz1nkV3se4yPYrifb6zfnSLV80qAT9Z7DY8kvlpu612826VFhMuBC3lTHn4630ttttpKirkbX1AEnlNc0a7YE7MtsVvJYrrtwaiPMdGhxSQiS4W4ylqCdmw44lSE85TusFI6+qtVlPEjg+M2S83BM+RdDbGlPKjwYby1yEJfRHWpg8oS8lDriErU2VBJUN9txUmtWqWLXzI1WGFd23boC4gMqacQlbjYBcbQtSQlTiAe/bSSpPXzAbHaMM8QVhTmGW2ac3IgMY/cGbQH1R5Dj02YuKiUpDDKGiXAlpfMSkqOyVKKQjlWrejWjCXJ9tiNZBHkruKYyo70ZC3WD2Qdo4U8lJbQXSQEBSgVEjbfcVtcKz2w6jWhV0xy4puluS4WhKbbWltSgATylQHMBzAEjcA7jwggSCqQ1u9nDQn7aXP83O1h6UdfFPq99pbF/qXGpUxpFfLROnQ7HmS7Pis+8LvL9vZt4M1DjrxfkNNSukAQ066VqUC0pY6VwJWndJTo2OHKUpm3wZmVJk2ezMvRrRGbtgbdaaclMPkPudIQ8pIjpbSUpbGylFQWdiMm+6ATJGY5Bk9kydu13e8uTUOKlWwSm2o8mHbo7qEp6VHzQKtjDiVklI5lJKFDY1hT+Gp16yS7dFydLAmRX4L63bd0m7LtpjW9QSA6nZYMRDoUdx3yk7eBVcMy4aHM1s92sTmVBjGpMm83CHGbtwVIjy7jGlsPKW8XNnG0mfJWlAQhW5QCshOx22o3D61nmQM3tF2ajT4siPIjtyoi3mAW48hhSXEtvNKUFJkkjlWkpUhPWoEg6m08Nr2PswoNmytmLbFyrTOukZdpQpUmRAkofCmSlxKWA4W0pWkpc2ABBCioqkOreg8PVqeuVKubkNQgNxWUJZ50pdanR5jTitlJKkhyMgKQCndJOyknYiFXfhPlXeyP21eWsMx7h2d6pRGLW4iI6qQ2wyh1DQk7l1puOlKFvrfHfq70DZItzTvAU4DDubImCa5PktSXFhnotlIiR4/g5jvuI4Ph6ubbxbnybhuRXG6aqXTT03OTimP3rPL1OlXpl1TLlwcYkhSLYw6nrbcc251ndJLaClG5KtvbjTSGGkNNIS22hISlCRsEgdQAHiFcqUpSlKVpsytVzvuJ3e22W7CxXWXFcYjXQsdMYi1JIDoRzJ5infcDcdYFeUv/h+6B3XRtzUB4Zku82dV7mWh22Owej5pER8tplJX0quUrTzBSNjvunvjyivY1U7xLdVt05PtZ3ZPykCripSlKUpSlCNxXhPWy8XLDZV60uTdH8tsFvudjusS4Puqek2UO3eNtAlOq359xutoklfICFDYJUfdlKr53SJl3Uc5abkorNzYuXYhYBAU3b5EMJCubxiRz77dXJt49xGsc4b42O2+5Rk3xcjs1NsSVLiABHYdwkTQQOf9sZBR9jlB699qjMvhMl3K4XGbcM27NuEi0XC1MzX7etyQrsiVFkpdkLXIV0hbVEQnkbDSOVSglKOraU4Xw7MYhn6ciNziy2W7jcLu0x6nqD6ZUxTqnj0qnVpDYVIf5QhtCgFhJWoA83Rm3DjHya/3K+tXtqNc5WQG+MpmxFux0b2uPAWytCHmlrBTFS6FBaCFdWxAPNqHeFJTkvHkJysuWmzSbNLYiPW8lTK4Ehp8pYCXkssoeU1urZkr3We/KQlIt3TfC29OsBsGMNSlTkWmG3ETJWjkLnKNubl3O2/tb1JKpDW4b636FfbS5/m52sPSjr4p9XvtLYv9S41fdKV42uOU57ktvn2+83jIH0QcgsM1Vwt8JLbUdpN5bS6hTa4Ta2VJb2WpsqfASyFFwpJ582x3nVB+2rTBu92x5iHIsLDUG32SIhpYm36VGmuKSqOo7ojdG53vKEkBatwo7yS9anZlg+OZJFuFzvdwuSbTfW7LLNmDrj8tia83GUehY5OcNFk7FISpKSvlICyNrodGu8HVbI2JNucj25b98fRIdhBKlLVeXSkB4p5ikoIUE83Kdwrbx16DpSvNek2nll1SxbV3Hr6wp2G/nN2Wh1lZbejPJkbtvMrHWhxCgFJUPAR/XUy0r1DvVhybtZahyEuZZHZU9ab2EdGzkUNHheSPAmQgbB1rxHv07pV1XFSlKUpSlU7w1fSzUQe1nd8/KSauKqc4meqz6en2s7sP5Wkf+9XHSlKUpSlKqrWHU26Wy5QcEwZDM3UO9NKcZLyedi0RQeVc+SP4CT1IR4XF7JHUFEVXrtpja9KNAoNotzj02S9kdol3C6TFc8q4y13SKXZDyv2y1H7wAAGwAFeqaUqkeJrLsqxCFZ5WNXOZDQGZjj8aDH5nZS0JbLSW3FRJKOk618rCg30pJ2cTyEGrX52bWjKc1dtT13RcVyMgkxn59paecgKWxblRQ0otetIK+9CilXRbHco6pVlM/VLFJt3Rbcnv+Qlu8TbVCYftcQhTP6n3ZrLyi3HSVLEwJbSoFKCOVBSVbkxvUXOL3qRl1tkWmLcrhYLc6iTDTKs6koS+5jt/S9uFtjnHOY7ageZIWQjwkg+rbJzmy2/pEhDnY7fMkJ5djyjcbDwf1Vm0qkNbhvrhoT9tLn+bnaw9KOvim1e+0ti/wBS41fdKUpSlKUqkOGL1+qX3c3j/XNT3VXS62asYyLZNdet86M6mXbLvDPLKtstHzt9lXiUD4R4FAlJ3BNRzSHVG53W6TcEzlpi36iWZoOPBkcse7Rd+VE6Lv4UKPUtHhbXuk9XKTatKUpSlKp3hr+gdSB7Wd3r/WB/96uKvKXHFr9i+lbmBWi/N3Jp8ZFa76h5mKVsuR4stC3wle/WtKQDy/8AmT7delcOyiNm+KWnIITEqNDucZuWw1Na6J4NrSFJKkbnlJBB2PWN+utxSlKUpSq71h1WVp5Bt9ss0EX/ADi+uKjWOxpXy9O4Bup10j1jDYPM4vxDqHWoA89H9KRpvbJ0y5zzf8zvbol3y/Oo5Vy3gNkoQP2jLY7xtsdSUj2ySYPxkDfSyJ9vbN+dItXzSlKUpSlKpDW72cNCftpc/wA3O1X6NXMf0Y4mNR5uUpu0eJdrRaG4b8GzS5qHFMuTS4nmYaWAQHW+o7euqbnjY0r2+jMj8k7p6PQ8bGle30ZkfkndPR6d2xpWfqzI/JS6ej07tjSs/VmR+Sl09HoeNjSvb6MyPyTuno9O7Y0r2+jMj8k7p6PX3u2NKz9WZH5KXT0evndsaVn6syPyUuno9O7Y0rI+jMj8lLp6PTu2NKz9WZH5KXT0evnCVcE3yyZ5eGGJbEK65dc58QzYrkZxxhx3mQvo3EpUAR7YFXzUA1e0oa1MtcGRCnLsOX2V0y7Hf2EAuwn9tiCP27Kx3rjZ6lJPtgEY2j+q7udt3Cw5DBRYM+sRS1eLMFEpG/rJMdR63I7oBKFeLrSrZSTVkUpSlKVCtL9PndPWspQ7MRM9Wcgm3pHIgp6JL6goNnrO5G3hqa1W+t+gWKcQFnstuyqKZDNpubFyZKQN1FChztK/+24jdKh9kHwgVY6UJQhKUgJSkbBIGwAr7SlKUpUN1V1QtulGMC5TGXrjcJTqYdss8Mc0m5S1/O2Gk+Mk9ZPgSkFR2ANR/R7S+5WOdcM2zV5m5ai3xtKJbrJKmLZGB5m4EXfwNIJ3UrwuL3UfEBaVURxjxpcjSXmhQZNyfjXS2yzFiI5nVoZnx3XOUbjchCFHwjwVr+7TsPufag/A7Xn6d2nYfc+1B+B2vP07tOw+59qD8Dtefp3adh9z7UH4Ha8/Tu07D7n2oPwO15+ndp2H3PtQfgdrz9Dxp2Hb2PtQfgdrz9O7TsPufag/A7Xn6HjTsPufag/A7Xn6d2nYfc+1B+B2vP1GJWriNbtb9KF2nEcotMaxzZ8iZKvUFDDSUuQnG0bFLityVEDweOvQ1006tF2mLkvtczi/DuKxe1TYtvnPxCnapsW3zn4hTtU2Lb5z8Qp2qbF/E/EKHSmxH/k/EKdqmxfxPxCnapsW3zn4hTtU2Lb5z8Qp2qbFt85+IU7VNi2+c/EKkFjsEXH4hjxE8qCd+utlSqz1h0plZg5bsnxWY1ZNRLCFKtVycSS0+g9bkOUB1rjubbEeFCtlp6x17HSTVaJqlZZRXDdsmS2p7sO9WCWoF+3SQNyk7euQod8hwd6tJBHjAj3ErkV4s+JY9a7JdH7JJyTIYVkeucTbsiMw6VKdUySCEuFLZSFEHl5tx1gVEhwu6eugKlQrxcJJ+eSpmRXFx11XjUpRf6ya+9y1prt9Jp/w7cPP0PC1pqf3Gn/Dtw8/Q8LWmv8AM0/4duHn6dy1pr/M0/4duHn6dy1pqR9Jp/w7cPP18PC1pqR9Jp/w7cPP197lrTXb6TT/AIduHn6dy1prt9Jp/wAO3Dz9O5a012+k0/4duHn6+HhZ01P7jT/h24efr6eFrTU/uNP+Hbh5+nctaakfSaf8O3Dz9fF8L+BspK7c1fbLNSPmU+3ZHcG32VeJST05G/2CCPbBqccNeWXjMNJoki/TTc7rBuNxtLs9aAhcoRJr0ZDq0p6gtSWklW2w3J22qYZ/ntl0yxKfkeQSxEtsNAKilJU44snZDbaB1rWtRCUpHWSQKr3SrAr1k+T9tDUCL2PkshlTNksKyFox6Evwo9oyXAAXXP6kJ71PXcdKwrrZol6YDUpsOIB3APirUnT6yn6lHxUOn9kI+hR8VO1/ZP5KPip2v7Jt9Cj4qdr6yfyUfFTtfWT+Sj4qHT+yEfQo+Kh0+sh+pR8VO1/Zf5KPip2v7Jt9Cj4q74OGWq3SUPsRgl1HgPtVvKUpSlKUpSlKUryVxv4XqehVuyvRewzRlnYj0C7Xm1S2m33IKknZgsK63VBR50LT37ZB5fXHbsud/u2T6B8PM6/xJsG/DJrRHuDFxaW1ITJaQ806VpWAoErQo9ft16BI3FKEb1QmivE2rVXUvJcfk2pu3WlDT03G7ihZPqrEYkuRn3evq3DiAQB+1Vv9k5kbjI0vnR2Hotxu8pEmIubELNhnK7LabOzpZ2Z+adH1lfLvygHfbY7STujdPjarrcvV4CDbLNHv77yozoCoL4PROtgp3XuUlPKnchWySATtWFf+KHTzGrrLt8+5TkOw1x2ZTzdqlOMR33+XomHHUtlCHSFg8hII699iCK0GovFVYscyG2WHHw5d7mvKYONzXXIUgQ2XHnUoebTJCQ0Xm0qB6Pm38PV1HaRP8RuIv3m4WOC/NVeWmZyofZdsksRZzsVBU+hl9aAhzk5e+5VH7G9aCx8V+Lw9PMRv2WGVbZl3ssa8TBb7bKlRYDb2yQt11CFJbRz7gFR36qvNKgtIKSCCNwR46+nwU2p4apLTfPO1dwj59liVIS9aLnlMpgLG4U8m5zOiSR9lfKPv1D+EfVCVxk35Gb5tCcju4mGkWiyMRHRbBJKNnbh0qhyuPE8yUNkksp3I3KuavZtKUpSlKUpSlKUpSlKUpSlKUpSqb4qdfZvDlpfIyqFiczKFpWGeZlaUR4ilEJQt9W/OElRAHKk7nYEp3BqlbFqhfdaeHTQnNMkVHN3u2eMuOJitdG0hKZMxtCUp3J2CEJHWSTtuSTXp4jehG4rQ55ZLlkuFXy02e5ps1znQ3Y0e4qaLnYylpKekCQoblO+46x1gVSOP8GVkwG64Dd8QvVwt12xpzo5Dk+VIlszIq2lIkNIZW7yslZIWCgbAj1p6tsnDuFh/FLZgEU5G1JOLY/dLIpYhFPZBllJDoHOeXl5fW9e+/hFV7kXDlNlZpoxhUaLdJVux21twcmvoiBm33GAw41IajklRJUZEdPeDcgOKJO1ajWDTTN5ea5vimM2y/Gx5RkdtvK0OWdtyG46lyOp54Tw98ybCWeYtqb5+ZCQDsTvZ1w4Wb87fFMQ8yiMYgM1ZzdFudtRXLEkPpddYL/SgdGSFFPecwKhuSBsdfZ+EO9xc+g5Lc8yiXZ+C5dgmW/bnVTpTU1l1tKXnlPqHzLpEhKUISnZJ6tzuNRlnBJesmw624yrOYr1ri41FsaUT7Y68IrzG5MmM2JCUIU51BRUlagE9SvBt6viMqYiMNLUlS0ISklI2BIGxIFdxG4oRvSq54aLBbMo0SuVsvFvi3W3PZTfy7Dmspdac5bzJUnmQoEHZSQeseECryjRmYcdtiO0hhltIQhptISlAHgAA6gK7KUpSlKUpSlKUpSlKUpSlKUpSlUZcx29NcmrWn5thGnklEqafC3PvhSFMs/ZTGQoOKH8Y43v1oNY/Evb7XhuI6fSI8aLZrDa82t8uUtltLMeKhxx0LdXsAlALro3UdhuvcnrqyW1pebSttQWhQ3SpJ3BHtiuRHVTahG9COqm29NqEb02oR1U2ptQjeuqVJZgxnZEl1uPHaSVuOuqCUISB1kk9QA9s1C+ERYlaLomtHniT79e5sV3bqeYduklbTifbSpCkqB8YIPjq6aUpSlKUpSlKUpSlKUpSlKUpSlV1rnqNM0+w5tuxMtzsxvklFpsEFzwOzHAdlqH8W0kLdWfEls+2K2+lOnMPSnA7ZjkR5c1xhKnZc9/57NlOKK35Dh8anHFKUfa328AFSW5WyHebfIgz4rE6FIbLT0aS2HG3UEbFKkncEEeEGqYvvCxohjtpn3afiUC02uEy5KkONSpEdhhpCSpauRDgSlIAJ2AFdsDhO0ZusCNNh4szKiSW0vMvtXGUpDiFAFKkkO9YIIIP2a7+4/0h2/Wej3/L87Wgc4ddDm88Yw84kr1aetjl2SgTJfJ0CHUNKPN0vh5nE9X9db/uP9If6Ho+EJfnadx/pD/Q9HwhL87TuP8ASHb9Z6PhCX52h4P9IT+89HwhL87TuP8ASH+h6PhCX52h4P8ASE/vPR8IS/O07j/SH+h6PhCX52ncf6Q/0PR8IS/O1oIHDrodc83vOKMYko3e0wos6SlUyWEBqQp5Leyul6zvHc3Hi6vbqRMcIejzL7bi8Ggy+jUFBqc69JaJHg3bcWpKvvg1bseM1DjtMR2kMsNJCG2m0hKUJA2AAHUAB1bV2UpSlKUpSlKUpSlKUpSlKUpSlFEJSSSAB4z4qo3SlJ1m1LuWqsn5pjttS9ZMPbUO9WyFcsu4Ae28tAbQf4trf/mGrypVCcb+C5HqHw7ZRbLDfI9jjsRnZ9zW60pa5MZhtTpYRsRy86kJ3J8QI2IJrZ8IGDZFp1oDi9jyG+sZB0UZDsCQ0yptTUVxCVoZXuTuUcykgjYcoSNuqroqnZvVxg2f7OCTviuEX/eripSlKUpVOYj1cWGpY9vFsfP/AKi51cdKUpSlKUpSlKUpSlKUpSlKUpSlU1r/AHqdlEmz6TY9KcjXrLErVcpjB2XbbOggSn9/2q3OYMNn+E4SPWGrYslkg43ZYFptkZuFboDCI0aM0NkNNISEpSB7QAArNri863HZW66tLbaElSlrOwSAOsk+IVRuda24ZqTppq9Y7Ff7dOn2uz3CN0bE1l0ykm3JeLrIQslTY6cIKthstCx4qsnSVXPpVhh9uywj/gIqV1Td0WGuL6wqJ6v1CXHfq9qfDqf4PqNYNSIUmVj8t+WxGcDTin4T8YhRG+wDqEk9XjAIqS0pSlKVTuL97xZ6hj+FiVhP90q5iripSlKUpSlKUpSlKUpSlKUpSlKVq8qye2YVjV0v95lohWq2RnJUqQ54G20JKlH7PUPB46rXh+xe5y4931HyiKuLleYqRI7DeHf2y3IBEOH9gpQorWOr5o6v2hVvUr4pIWkpICgRsQR4aqrUXTyyYrplq1cbdEQ1KvdsmzJCuVPerFvTH2RsByp5GEHb2yo+OpPo6rn0iwc+3Y4J/wDToqX146yvimw+zcb1osj8O+Ku0S0SsZXERAJWuXIlxHGSjvuttSEFXP4NiD469i0pSlKUrx1gfFRh1+41ckscSHfPVa52uBjyIjkApW1KiSJy5Bc77vUJQ6k83X1A+1XsWlKUpSlKUpSlKUpSlY9xfVGgPuo250IJG/t7V5Y4ceHvB9SdGrBkmRQbncb1cFynJMk364N85Ep1I71D4SNkpA2AHgqy+5E0s/mS5+UVz9IoeETSwj6SXPyiufpFDwiaWH9xLn5RXP0inciaWbfSS5+UVz9IoeETSw/uJc/KK5+kU7kTSzb6SXPyiufpFVrxKcOGBYJoHnmQ2KBdIF5ttpekxJSb/cFlpxI3CuVT5SfvgivVEJwvQ2Fq9cpAJ/r2qktQB27tXoGnrXzXEsXUxecoUPWSZO/PBgH2xukSHE/wUNA+vq86UqK6jan45pRZY10yWY9EiSZSITAjw3pTjrywSlCW2ULWSQk+AeKqszfiS09y7C7/AGNuVksddzt8iEl5WHXdQbLjakcxHY3WBzb7V8wfiS0+xLCsfsTkvJZLlrt8eEp5OHXdIcLbaUFQHY3Vvy77fZrdniy07/jMm8kLv6LVb3TNdD7trnaNVn4+THJ7bbXbc2r9R925FcxHK6R2L69KC6gH2nP/ACjayO6y07P/ADMm8kLv6LQ8WWnf8Zk3khd/Ra6pfF5prBivSZMrIo8dlCnHHncSuyUNpA3KlExtgABuSa7E8WmnS0hSXclUkjcEYhdtj/6avvdZadn/AJmTeSF39FoeLLTvb55k3khd/Rad1lp2f+Zk3khd/Rad1lp2R88ybyQu/otVvaM00OsmuN71UjR8mTk92tzUB1X6j7tyJ5T3zqR2L69aUtJJ9psfwlb2QeLLTv8AjMm8kLv6LVgaf6g2LVHF2MhxuW5NtTzrzKXHozsdYcacU24lTbqUrSUrQoEEDwVI6UpXnrX7H4Wea5aRYteeyXbHMN1dkRY0x6L0qkReZG6mlpV1EA7b1Ie5E0s/mS5+UVz9IoeETSw/uJc/KK5+kU7kTSz+ZLn5RXP0ih4RNLD+4lz8orn6RTuRNLP5kuflFc/SKHhE0sI+klz8orn6RTuRNLP5kuflFc/SK0OiGJ23TviG1SxqxCUxZWrRZJTcWTOflBtxZmhakqeWtQ35U7jfbqFegqUpSsO8/SqX/ZK/7V5k0xym+YXwL227Y07EZvzPStw3J7RcYS45c1tgrSCCU9/1gEGpdaOJF643J2Oq0q9Vw1AtisdLiUOxrw7IlNvMuPHqDaUsBwL2PM3stIVzpB3Fo1/l5JdnrNZ8OmT77bOlVeoAmtI7DS3IUxs0tWweUsoWtA7wFKe+KFEJPXhmus3LJYhWjHpd+MNzmukovsx1Rm3J0mM2UNk7OFIjOLUOZOyACOZZ5K0uNcW0LLLIm52/FLnJamtwX7UhpxP/ABLUqYxFa6VSglDK95LThRzLHISQolKkjCyXiXu8G/OQfUVFtjwLZdHLqUupkPMTIsqC0lLJPKlxtSJgVuQkkLTuEFCkmQZFxR2XHoL63LPNemxVuxpURK0bxpIuKIDDThG+wecUtaVbEdG0tWx70GN656hI1K4O9VLiLc/bH41tmQ3mXQooUtCQedpZSkuNqCkkK5Unr2IBBAtLVHUhvSvTNy8ojG5XVwNQrVa0Hv5850hDDCf+pZG58SQo+AV90V02XphgzMGdKTcsjnvOXO+XMDYzJ7x5nnP+kHZCB4kIQPFU9ri44hpsrWpKEJG5Uo7ACuVUTxNH/wCo9Gh/+bwf8rlXtVXagzr5cNXsRxi3ZTPxi2TMfvNzlOW9iItxx2PItjbW5kMugJCZT24SBuSNz1VDsH4ir3c8atU6dZ4NwiNSLZb51zamFhyS5MkdAzJjxwhYLKudh3rcHeuL236Pvu+x8Sd3cbxKTkOKwrFbr5brfcHJiLm9Kaidl8/RtOLRF5W1gpQN3S2hZWQhaikg8sS18u2c5njVp9Tm7Kh6fGW/0ReWmVFkW25PITtIjMrTs7CSrmSnZQCdlbFQq+agmvY30L1F+5y5fkrlS+zddng/2Df+UVodUX73BwO6zscW56sW9CJzMdpCVqlpZWl1cYBQIHTIQprfwjpNwQQCKLyDiiu1ru0u72OAjI8buUadNtClrLLBh24MIkO9IGyUqdfkOJ51nkQiOhW3fmuOfcTeWw8Yvsm12a2WwqF6j2yW9KU8427b5TbKlut9GEgLSpZAClcpSPXBR5drk3E1frBjl2uYsdoddjSb6mNCEuSp16PapK48h1fKwUtJKg336lBKAtO5USE1ZWjWW3DMrdk0qfIMgR7/ADIsYFKR0bCSnkR3oG+wPhPX9mrBqleEzr02vn3XZD+dZNXVSlcVuIQpCVKSkrPKkE7FR2J2H3gf7qo7U4f+KLRb/ovH5HV51QuGaw3y73fKZlznyE26yXW8R+wI2Lyg05HhuvITtPKuhKylsE9Y77dOwNd73FbaLfd7dbLpj9wtc5+PEmSIciRGVIjsynVtsENocJdVsgrWlvm5E+NSuqsnFuKOx5rc5sSzWuRNCI0mXEdRMigPtsOpbcLm7g6DcK6RPSbcyEqJ5VAprAPFC2mNarirFr4hu6R2VQbSW4wfldNcWITDyXS+EJQtT6FgK23QebcdQOXduJ1i1WO7zf1HXmVLsEKdcb5AYejdJBYivOMrUkqcAd51MuqQE+FKCTyq5Um7GnA80hY8CgCAapbBhvxY6rfaCwf5p1XXSlKVh3n6VS/7JX/aqS4Wcdt2V8LGK2y6xkzILqpC1sqUpIJRPdWk7gg9SkpP3qsW86NYbfr3eLxKsqBd7uiIiZPjPOx33DFWpcZYW2pJS42Vq5XE7K2O2+wAGtnaB4C9BYbkWhxDEZDwdWLjJQqWhxzpnky1BwGSha91KS+Vgkq3HfHfHtWlGnWSOw50C1PrZYQzIbU29LZiy0KeVMaKxzBuSlLrq3EhQWEKcVty7kViZDw1Ylc8eNqtiJdnbMu2upUibJdEaPEnsTBGjJU7/wAMhRYCfmPIE96QDyJA+zdLNNDdI9hftcuVcYTC+dCXpzzi2p7qlKXJdCiXUuOwioqdUrlUyg7p73eUXHSLELo9kjz9la7IyJ2M/cn23HG3XnY3L2O4FpUFNrbKEqSpBSUqAUDv11W3ExiVswvhJ1Pt9qaeQyq0S33HJMl2S864sbqW466pS1qPtqUeoAeAADaal6L5FqJleF5LZs3bxxWOMOqiwpFnROa7IdRyGRspxA5w2ShO4PKFK22Kq5nTbWEj2Z4Y/qxBjz9fDppq+fDrSx97EY3nah+sWjOsOR6TZna2dUxkD02zy46LQjG4sczVKZUkMhzpO859+Xm8W+9d/Bro1qtpBhnYepGdHIELaSmJY9uyPU7wdXZSu/V1dXIO8Tt3pNbbia/XJo1928H/ACuVe1aPJ8DxrNjFOQ47ab8YvN2P6pwWpPQ823Nyc6Ty78qd9vDyj2qpK0664bf2cey+56fiLcZOMKv1glLRDkzVxvmCUx0FKitlxapjKEpOySpwp5geoyI5ZiWQpwafYsCh3vIpVrdkW5h+PHYcs8VkobeQt5QPQ8i3UtdGjclXMAOVKlJmmneNYy3YLfcbVhULE3Vd/wBgiBHZdjOI6RsjdrdO4DjwCkkgpcVsdlneZ1BNe/YL1F+5y5fkrlS+zddng/2Df+UVmVTOruY49hj0WzScOsN4hWi3rvbbNxkxoqIyWlEDsdLqCnpBsdutAG474b7jhE1pwS9TMgiXGxJhWW32V+7vTrhEb6J1osMyJ7ZRsSFIblRi4D64uqHWUKrbY0mLl8q2Wu/6XRLTbiw5c4HZSYskRXCpJUh1oD5g+rpObvecEpc3XuBzWXBtsS2MlmHFZiNElRbYbCEk+3sPH1CsivPGgFqvt50XyGLjl+axy6qzG/lFwegpmJQkXaSVDoitIO46t9+qpKdN9YT9eaF97EGPP187WusB+vRF+9iEfz1Dpnq8fr1ND+rEo3nKpfio4ddcdRcJsNts2o36pbi1fGJLaU21i0iDytujsovtrKwE822yQSefqBIrdY/h+YYNrDoPac5zA5vf2W7x0lzMZLOw7C6kdXWvb+GrvleE16xqJWmz4q1+qLDoKE8zvS3C5wA44Vf8c4+tayoncBxaX+pJ6tjsANq0+ZafYRbnmMivJk21uOmFBcWxPkssyUofAitvttrAdCXXABzg+vIO6SRXRcuHTB7va7pbJcO5O2q4NuMuW9N5mNsNNuPpfcQ0hLoDSVuITzJTsCndHrCUnvuWkGFWyMLtcYtynm1oRJTImXKbOebSw+1LQEhTi1q5XY7awgb78u2xCiDFdZOGtnUqHMhWqcxYo90hz4NxeSqWJCky1BTqkqakNpWN+c9E6lbe6gQAApK7tbbDTaUJ8CQAN/sVS2DfssdVvtBYP806rrpSlKw7z9Kpf9kr/tVR8HH7G7EP/wCz8sfq6K885toper7rk7kMSywpUGaQ3Kl3NUeQz2KYK47iEAtiQyslXL0SVrYUlRWpIWpVRnDeGu7S5uKM5Fi9pYxyEmGmXYnHGn446G0PxVfMwORY6dbah1e0rYEVxwvh5yzT5vEblBtER2Ra7VjLt0gRZSG3LhcYzVyauDpUdkrdUJcdXSLI6To9iobCt3iOieZwLnZZlwZitOMXOHMkhuX0gCG7heH1gHYFXeTo+24G+56hsa9J1T3GF+xd1O+0cj/LVs276Xxv7NP/AGrIpSleeOLzILdib2lN6u8pEG1W/MYciVKcB5WW0pc3UdtzsK3vdoaJH641p/xPk0PGfol7o1p/xPk1UlnzXhesun+L4fHzexotlnTBTMW1GDTt3ERvZrspSGwVfNQ28SNu/QPETXSdQeH+0peOOat26zpU5OS1DkRezISYszolSoi2Fo79pbzRe6yFBTiwFciigzXS3iI4f9JcHhYvaNSLWuDFdkvpKm+iHO/Icfc5UNtJQhPO6rlQlICU7AeCpZ3aGiR+uNaf8T5NQ/WPi60dv+kOcWu359a5dwm2KdGjx0dJzOOLjrSlI73wkkD79Sa18ZeijNshtr1EtKVoZQlST0nUQkbj1tZR40NEvdGtP+J8mozceI/h6vGYt5FPzyxzpTUEQmmpLJcS2Ok6TnTu2SFE7D7wqGSc64YZsRhMnPLU/OMydKnXAtlL9xTMS8iUw+oNgqaWh8p5erl5G9tuQVnYprJoXj2bQsnnayx79coEBdtjPTIrKHi0oo3LzzcdLjqgG0gbqA8JKSo71ZZ40NEdvZGtP+J8mh40NEvdGtP+J8msbg4uMa86RT7jCdD8Kbk99kx3kggONLuchSFjfxFJBH9dXnSlKozU7r4otF/+i8fkdXnXnfUzSS+X/WK8X2BjQlvTodiYteRpmNtm0uRZkh2SspKw4N23UAdGlXSb8i9k7mq+Z4cs9uMW5QrpFlSZL+7dynrmRmmrwTdYsgO7tkvOFLLTpSp0tqZ5ujQFhRKZBqdoRkcn1TsePYvHdxd6RcnLc0w4wRb3HosFLSmmnnQ0yjpkzVFxLbjrZ2LaAHFbsn0P1HvGK5HaGnlqiycVlzoiV3DZ/wDVE/a+wSyVc2waT81eK99i7I5htyb1xzzRTOr0xlMe02osZJKeyJ9WWeqDaPVOHKhzG4EAd/0qeiW/DGy0pbR2HzpUSqvS+M45AxOww7VbIjcKHHSQhlrfYEkqUdz1klRJJPWSST4aqjBv2WOq32gsH+adV10pSlYd5+lUv+yV/wBq8q8MfEzpXg+h+OWK/wCd2a03iEqWiRClP8rjSjKeUARt1dRB+/Vp92Lon7pmP++v0U7sXRP3TMf99fop3YuifumY/wC+v0U7sXRP3TMf99fop3YuifumY/76/RTuxdE/dMx/31+iqv4n+KLSbL+HrUCy2XPrLcrrOtD7EaHHkczjrhT1JSNusmvVdu+l8b+zT/2rIpSlR3M8FtmeQmot0QpxhtRUEA9RP2ahp4bMKP1B8QqsuIvRPG8O0xNztTK4stN5s7HSNnlPRu3KM04nq8RQtQP2DVm9zZhW30B8Q/2oeGzCT9QfEP8Aah4bMJP1B8Qp3NmEkfQHxCh4bMKI+gPiFO5swoj6A+If7U7mzCT9QfEP9qHhswkj6A+If7UPDZhR+oPiH+1O5swr+QfEP9qdzZhW30B8Qp3NmFfyD4h/tU/xnG4mKWhq3QuYR29+ULO5H2K2tKUrzhxA5vYdO+ILR6+5LdY1ltDAuqHZktXK2gqi8qQT9kkD79Sw8YuifumY/wC+v0U7sXRP3TMf99fooeMXRP3TMf8AfX6Kd2LomfrmY/76/RTuxdEyPZMx/wB9fop3YuifumY/76/RTuxdEyPZMx/31+iovodn+Oal8Smqt6xa8Rb7ajZbGwJcNfM3zpVN5k7+2Nx/fXoulKUrrkx0yo7jK9+VaSk7VAHtDscfcU4tnnWo7lSkjcmuJ0Kxk/U4/AFDoTjJ+px+CKdorGT9Tj8EUOhWMn6nH4AodCsZI+hx+AKdorGf5OPwBXZH0Rx2K8h1poocQdwpKQCDU/aaSy0htPrUAJG/2K5UpSlKp3i0G+iMxX8C9WNX912iVcVKUpSlKUpSlKUpUbyzA7bmSmjPSVhsbBO24qPdorGdvocfgCh0Kxk/U4/AFDoVjJ+px+AKdorGf5OPwBTtFYz/ACcfgCh0Kxkj6HH4Ip2isZ2+hx+AKkmKYPbsOS6mAkpS54U+ACpDSlKUpSlKUpSlKUpSleWOPjhuyDXnA7RJw2S+1k9rmNoEVEhTTUqO44gK5wDtu2oJdBPgCV7dZFX7pXgEbS3TuwYrFkvTkWyKllcySsqckOeFx1RJJ3UsqVt4t9h1CpVSlKUpSlKUpSlKUpSlKUpSlKUpSlKUpSlKUrzvmmKJ1O4ppePXO+ZHAtEDD481qLZL7LtyC8qY8krUGHEcx5QB1+ICpKeFnFz++TUDy4uvpFO5Zxfb9cmoHlvdfSKHhZxcj9cmoHlxdfSKHhaxcj9cmoHlxdfSKdy1i+365NQPLi6+kU7lnF/6SageXF19IoeFnFyP1yageXF19Ip3LOL/ANJNQPLi6+kU7lnF/wCkmoHlxdfSKHhZxc/vk1A8t7r6RTuWcX/pJqB5cXX0ih4WcXP75NQPLi6+kUPCzi5H65NQPLi6+kU7lnFz++TUDy4uvpFDws4uf3yageXF19Ip3LOL/wBJNQPLi6+kUPCzi5/fJqB5cXX0incs4v8A0k1A8uLr6RWl0mxv9QHEdmuMQr1f7lZkYxarg3Hvl5lXHon3JM5C1IL7iyndLTYIG3rRV+0pSlKUpSlKUpSlKUpSlKUpSqTtw34zb39wsT8ueqZa2agdrPTa6XlqRFjXFZag29cwjoUy33EssqcG43QlawtfWO8So7jbeoHg/EVJyfFcdjW21ozDMXmriiaxbZLcdhZtzzTEp1ClkgBxT7CmkHwh9G6kjdQ1zHEpdLfer8q/WJ2LAtcu7cseKUKWYcNMNTjrhKjstCJC1lKdwdiB1gb7TLeKu049dJEGDYpt5dZcnEFhwJ6dmI4hl5TQ2JUsvl1pCTsklhZK0gp5pxlmpy7TKx+32Oyv5Bdr1DkXJiIHkxuWIwGulcJX+23kMISg7bqcG5SAVCqtL+LL9VVnxKJJs79zvMq3Whu6SIgKEouE2DHlJCGwk7M7SWuZZUOTnHUoJUU4mnfFLPatGPRMmtkq7X2bCgXW5LgpTyw25yyWG2W0J3cDaAFqCiFBBGxcXumpM1xQlUZ+S5hN2THdgT7hbSy62+5LbhzGIcgrbb5lNpSuS0sHZR6PnUUgp5TnWriLgXGLaLoqOhdvlhLb5t8xiZHjlU9MPpumQetAUtKiCEqSkkrShSVJGsncWtpZuseLEx64TmCymc860oFRhuS347D7KEhXS9ImO49y7pIbKdiVqCK3mp+rd+wDUOPBg2I36ys43Ovc5pl5tl5sR3mASgrPfq5FrCUdQJ23UmolmfFrFRaMxGN2uU+bdDurUG7FtS2xMiRHnlKda5RyM87Kmwsq3K07FKQpKlSGVxDyrLaFi9Yy3Zr0l6K2iPcrxHZjrRIZdcbWXQVHm+YOJLbaHFBW23MndY7MY4mrPldutDjFnnx5t3RbXIUJ0o53kylPBakkEg9CIspa/wDytbj1wrCwbiUayPHrBdPUaZIsrqrNbrheXFtNrZn3FiI4wnoASVJJnRgtSTsku9QUEqKce0cVAuFuwi4zMSfslvyyPFmwX7ncmGUuR5BbDXIsnolPbOBRYLiVcu3LzqIRV90qmsf/AGXubfcXZfyy41ctKUpSlKUpSlKUpSlKUpSlKUqk7d+zNvf3DRPy56rau+N22/TbRKnxUyX7TKM2EpSlAMvFpxrn2B2J5HXANwdubcdYBEYv2ieHZHcp9wlW19m4T3VPSZUC4SYbrqlMtsLBUy4k8qm2WUqT4FdEgkEpBHba9GsNst4fuUOyNtSHkvJU2XnVMJS62026lDJUW0BaGGgQlIB5N/CSTgSNAMEfsFnsrdnehW60wnLbFbgXCTFV2K4UlxhxbbiVOtrKUlSXCoKI3O5663uW6cY/m7MBu6RHgYHOIzsGY9DdaSpPKtAWytCuRSdgpG/Kdk7jqG2ns+hGEY/Ntki22dyELc3FbYiszpAjExmksx1uMdJ0brjbaEJS44lSwG0dfeJ27HtEsOeftTgtj7AtkaJDZZj3CSyy4zFVzR0PNocCXw2dykOhW259s12ytGcOmQYsVVn5G4keVFjqZkvNOMIkSGZL3ItKwpKi9HZcCwQpKmxylNcYmi2Gw7Rcram0F6Pc40iJOVJlPPPSm31czvSOrWVqUo7d8Vcw2ABAA24ztEsNuDtuWq1ORkwIkaA2xCmyIzLkaOoqYZebbcSl5tslXKhwKA51DbZSt8jNtI8W1EnNTL7AfkyG4jsAqYnyIwdjOqSp1hxLTiQ42soTzIWCk7dY2rCuGhWE3WbcpEm0uuouKZAkQ+z5KYilPsqYecTHDgaQ4tta0lxKQrZauvdRJzsi0lxfKJ8adNhSG58ZbS2ZkGfIiPtltt1tHK4y4lQHI+8kjfYhZ336tvlo0ew6wvY47AsbLDuOxn4lrXzuKMVp7bpEjdR33223O5AJAIBO+BbNBsGs0y2PwbMuMm3JiBiIibI7FKoraWozi2Ok6NxxtCG0pcWlSx0bffboTt9k6EYRKgWSCbS83Bs8GPbI0RifJaZVFjkFll5CXAH0IKRsHQrwq/hHef0qmsf/AGXubfcXZfyy41ctKUpSlKUpSlKUpSlKUpSlKUrz3rbohdsx1KbyqwZnd8QuXqY3bXXLVIDXSspcU4Aoch/bKJ8PtVDO0JqOfr25h7/Hm6HQPUf3bsw9/jzdDoJqPt7NuYe/x5uh0D1HP17cw9/jzdO0HqPt7NuYe/x5uh0E1G29m3MPf483TtCajkezbmHv8ebp2hNRj9e3MPf483TtCajH69uYe/x5uh0E1H923MPf483TtB6jkezbmHv8ebp2hNRj9e3MPf483Q6Caje7bmHv8ebodBNRtvZtzD3+PN07Qmox+vbmHv8AHm6doTUc/XtzD3+PN0Ogmo/u25h7/Hm6HQPUfb2bcw9/jzdWVoFo3NwHI7/kF6ym7ZbfLjFjwVTLrIDqkMNLdWhCe8TsAp1Z8fhq7aUpSlKUpSlKV//Z)
![[[@Pellman2016-xu]](https://doi.org/10.1016/j.tins.2016.04.001)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQEAcQBxAAD/2wBDAAMCAgICAgMCAgIDAwMDBAYEBAQEBAgGBgUGCQgKCgkICQkKDA8MCgsOCwkJDRENDg8QEBEQCgwSExIQEw8QEBD/2wBDAQMDAwQDBAgEBAgQCwkLEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBD/wAARCAGBAjADASIAAhEBAxEB/8QAHQABAAIDAQEBAQAAAAAAAAAAAAYHBAUIAwECCf/EAGkQAAEDBAECAwUDBgUKDw0ECwECAwQABQYRBxIhCBMxFCJBUWEVMnEWI0JSgZEXM2JyoQkkJUNTY3OCkqIYJjQ1Njc4VXaFlLG1wdMnRHeDhpOVsrO00dLjdYSWtkVGVGVmeMLF1OHx/8QAGwEBAAIDAQEAAAAAAAAAAAAAAAMEAgUGAQf/xABDEQACAQIDBQQIBAIJAwUAAAAAAQIDEQQhMQUSQVFhE3GBkRQiMqGxwdHwI0Ji4QZSFiQzcoKSstLxFTSiQ1OTwsP/2gAMAwEAAhEDEQA/AP6p0pSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUrXXvI8exmGbhkl+t1pip9X50pDDY/wAZZAqGnn/iV/tY8qVkhHr+TVvlXrX4+xNu6/bQFh0qvP4U8kuHfGuFM2nNq7IkzBCtrP8AjIkyESAP/En8KfaHP109xnF8Fx5CvR6ReJV0cA/lMIjx07/B4j60BYdKrz8iuXLn/r7zWII+WNY3Gif++qmf9VP4FbNN97Jc3z2+OH7ynsolwkK/nMwVMNEfQo19KAnc24QLawqVcZ0eKwnupx91KEj8STqobO534RtrxjTuYMKZfHbyVX6L5hPyCOvqJ+gFfIPBHC9vfTMa4sxd2Wnv7XKtbMiST9XnUqcP7VVMoNtt1sZEe2wI0RodghhpLaR+wCgIL/DtgcjtY4+UX0n7i7Ti1ylMrPy89DHkj9qwKfwlZxN7WfgfLyFfceuM21xGiPqBKW8n9rW6sOlAV79tc+Sv4jjvB4KFeipOWynXE/i2i3hP7nK+fZ3iBle8cv4+tgP9rGOzZxH+P7az/wCrVh0oCvfyS5mk/wCruYrYxv1+zcVQ1+7z5D3/AF0/g3zx3u/4gs1bPyjW2yIT/nwFn+mrCpQFe/wRzn/9c+YOQZvz/shGi7/5NHa/op/AdiTneVkfIEhR9Sc7vTYP+K3KSn+irCpQFe/wC8ZL/wBWW28T/n7fkVyl7/Hzn1bp/oe+Dlfx/FOMST85VubfJ/EuAk1YVKAr3/Q7+H/48G8fH0PfGYX/AGdfT4eeAVK6jwdx+TrWzjML0/8AN1YNKAr3/Q9cGo/1JxNi0FXwVBtjUVQ/BTQSR+yvh4Lwpn3rTdcztSh9wQsxuqG0H6MmQWv2FBH0qw6UBXZ495HtX5zGeb7w8U9kR8itMK4Rkj5fmERn1fiXia+flJzRjf8Asi49teUxU9jKxi4hiUr+UYU0oQkfzZTivkD8bFpQENxzl3A8luiMebu7lrvqwVJs15jOW6esD7ykMPpSp1A/XbCkfJRqZVqslxTGMyta7JluP268wHCFKjToyH2+oeiulQIBHqD6j4VCjx9nOE/n+LMzdlwm+/5O5RIdlxVD9ViaeqVGJ+azIbSBpLQoCyqVBce5ZtU27sYlmNrl4hk0glLFtuhR5c0gbJhyUEtSRoE9KFeakaK20b1U6oBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpXjKlxYEZyZOktR47KStx11YQhCR6kqPYD6mgPalV2vnTDritUfAYd4zuQCUj8m4ftMUqHqkzllENKh8UqfB+lfPaed8m/1Nb8WweIruFzFuXqcpJ+BabLDLKx8w6+n6GgLFrSZPm+GYTFE3Mcts9ijqOku3Gc1GSo/IFahs/QVFf4GY929/PeQMxyk/wBweuf2fF+qSxASwh1H0eDn1JreYzxXxlhkoz8S4+xyzy1DS5MK2MsvL/nOJSFK/aaA0v8ADphckf2DteYXsq/ilW7E7m4w7/NkFgMEfXzNfWvn5dcqXf3cb4Ukwv77lF9iwW1D5pEP2xz9ikJP4VYlKArv7B5xvfa7chY7jsdfqzYrIqTKb+iZUtxTSv2xRT+Ba2z/AH8sz3PMhc+JfyF6A2r6KZt/s7Kh9CgirEpQEMsnDPEmOTBcrLxrjUWeO5mi2MqlKPzU8pJcUfqVGpkAANAaAr7SgFKUoCF5nzDx5x9mOIYHl1+FvvGdyJEWxNLYcUiS6yG+tBcSkobJLraU9ZHUpYSnZOq3GN5pjuWyr9Dsc1T7uM3RVmuQU0pAalpZaeKAVAdQ8t9o9Q2O+t7Bqm/ENw4/zFyjhFmlwZzdnGL5RGdu8Zs/2KnrdtTkF9K/RDqXY5db+amT8jVFRsI5+vnHdzm8iYNJhvSOXVXLL7cmyLuzFzgNWRmJ7W3BbWhU2Iqc008GkknpSD0r8spIHehUkDqJAHzoSB6muMInGrFjYwE53heT55xax+Urrlj/ACMkoRbrhJkRlwSbMouvCM22me2yFJUGvOT2QOkjCx/ju82dvCk89cR5Xl+KMY3do9ks7UBy7vWWW7dnXYjT6EKV5UhNvVEZRIUQGiw4nzEb2QO3NjetjfyrW5FktixO0u3zI7mzAgMraacfdOkpU64lpsHX6y1pSPqRXIV24+yCblt4ZXxhkiuVnuSYl0suZGA4qPEx1M6O4E/aOy22wiAl2OuJ17W4V/m1dfXX5yfw4W268C8k3m78RrvOYXDPbtdUpkwTJnP25vJXH20x0r2ry1ROtaG0aCvNUUglw9QHaJIBAJHf0psfOuH+TOLLnk+fXEsWK9WHGbhj9mj4DKg8eSrjKx8NoX1ojFDjf2RJQ70uEutoCgWwpRDZQJrj2EZpE8VDmZyMPv6eM5V9uCbfBUFFEPI/Y20O3tTHTtEV5KJTCCT0h1Sntf1wFJA6spSlAKUpQClKUApSlAKUpQClKUApSlAa3Isbx/LrQ/YMoskK7W2SAHYsxhLrS9HYJSoEbBAIPqCARUEOJcjcdfnuO725k9lR3VjmQTFKkNJ+UO4K6nPnpuT5gJ0A6ykVZtKAi2F8j47m6pUCH7Vbr1bgn7Rslya8ifCKt9Jca2epBIPS6gqaXo9C1AbqU1Fs346sOcoiypa5NuvNtKl2u9W9YanQFq1stuEEFJ0OppYU2sABaFDtWkxvPb7Yr5F4/wCV240e7y1Fu0XqMgtwL50gkpSCT7PKCQVKjqUeoAqbUsJWGwLEpSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClVLc4Ny4JJyKwvzJ/HqD1Xe0OuLkO2Jv4zIalbWY6PV2MSQhAK2enoLTlrR5DEthuVFfbeZeQHG3G1BSVpI2FAjsQR33QHpSlKAUpUbzPkLF8EZjC+THVzbgpTdvtkNlUmdPcA2UsMNgrXrYKiB0oHvKKUgmgJJUUy/k/DsKls2i53FyVepaPMiWW3MLmXGSneupEdoFfRvsXFANp/SUkd6jwtfLPIn5y/3BfH1hc9LbbXW37y+j5Pyx1MxdjsURw4sdimQk9hLcQwLEMDiPRMTsUeD7UvzZb46nJMt3WvMffWS6+5r9NxSlfWgIsJHNuZ94sO2ceWxz9OZ0XS8KT/g21eyxl/IlcofNIr2i8G4M9JauWZpn5vcWlBxEnJ5PtyG3B6LaikCKwr6ssoNWFSgPyhCG0JbbQEpSNJSBoAfIV+qUoBSlKAUpSgFKUoBSlVzzBzzgnDMWIzfnZdzv92V5Vnx21M+03O5u+gSyynvrfqtWkj570CBYqlJQkrWoJSkbJJ0AKo3MPFxxjCusvC+PWL/yNk7QLblvw2CZ3syz2Bdk9o7Wj69SyU67j4VH2uIOYfEQoXXxGXl3FcPeIWxx5YJpSXm/gLnNRpTxPxaaKUenfexV84jhmJYFZGMawrG7dY7XGH5uJAjpZbB+J0kDaj8Se5+JoDjbw98heP7kHkm72a9IslvxGxXN2JNuORWxl51PQsgxmlQlMokPp+6tSNNpUlWyDpJ7gccQy2p15xKEISVLUo6CQPUkn0FaXMczxDjfGJ2XZne4VlstuQXZEqQoIQnZ9APVSlE6CQCpROgCTXPbdp5H8YjiZmUR7tgvCqlBbFoKlR7vljfqFySPejRFfBsHrWDskApIAl2Pc/5Dy3yVHsHBuPQ7thFklqbyXMJylphOlIIMW3dP+qHQoglz+LSBrv1JNXnWvsNgsmLWaHjuN2mLbLZb2ksRYkVoNtMtj0SlI7AVmuuNstreecShtCSpSlHQSB6kn4CgKs8Q3NFz4Ww8X21Yi9eJMhxMZhxbqG47TquyQsdXmLJ9QhCSSAdlPrWX4dL9l2W8S2jMc4nmTdr8p6esBAQhppTig0hCR6JDaUH59ySSTuozg7C+dOQTy1dG1Kw/GnnYeIxXE+7MfB6XrkpJ9RsFDW/QAnQPrc9rtkCy22LaLVEbiwoTKI8dlsaS22kAJSB8gABWtw6q4jEPE734drRXl63jna/DPiVaW/Uqdrf1eC+ZlVyYfGqzgHivyzw98pQyi0CRFcsV6jsE+ypeiMulqSlI/i+pxWnQPd/T933k9Z1zh4n+IItntc3xG8cWVP8ACJh9xjZQp9P8bcIkVgMyYJPr5a4oc90dyodvWtkWjo1C0OIS42oKSobSoHYI+Yr9Vp8PyqzZzilnzPHpPn2y+QWbhEc+KmnUBad/I6PcfA7FbigFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBWqyjF7DmdilY3ktubnW+YkBxpZKSFAhSVoUkhSFpUEqStJCkqSFJIIBra0oCtsUyK/YVkcXjDkG4OTxNCxjWQPAA3NCElSosjWgJraElWwAHm0qcSAUupRZNQzmPG15TxnkFviaRcmITk60v8A6UW4sDzYr6T8FIeQ2r9mvQmt/i99YyjGbRk0VBSzd4Mec2k/BLraVgfuVQG0pSlAKUpQClfNj51ospz7B8HYRIzPMLLY23Tpo3Gc1H8w/JAWoFR+g2aA31Krsc02u4/7EMFzjJB6+ZFsTkNlSf1kPTjHacH1QtW/hXjE8QfHdzaQ3Y1Xu8XP30ybRbbPJlTYDqFqbW1LQ2giKsLQtP55SASg9JI70BZVKrxXNlghd8gxHObOj+6yMXmvtJHzU5GQ6hsfVZSKkuK57hGcsOScNy6z3ttg9L3sE1t8sq/VWEklCvmFaIoDfUpSgFKUoBSlKAUpSgFKUoD4pKVApUAQexBqs+OEnj7LLjw27tNqTHVesTJ9E24uBMiEn6RXnEBI7BLMmOgb6Cas2q45vQbLYLfyfGHTIwKei8vKH6VtILVwQf1gIrjzgT6Fxlo/oigLHpXwEEAg7B7giq0yTJb/AJ/f5nHXHFxct8a3r8nJMlZAUYBIBMOJsFK5hSQVKIKWEqBUFLKUUBlZNyBebrfZHH/FTEWdfYxSi63SUhS7dYgoAjzukgvSCkhSYyFBWilTimkqSpW2wnjax4W7Ju3nyrxkNxSE3K/XJSXJsvR2EFQAS20CSUstpQ2nZ6Ugkk7bFsVsGFWONjmM21uDAihXQ2klSlKUSpbi1qJU44tRKlLUSpSlFSiSSa21AKUpQClKUApSlAKUpQClKUApXm++xFYclSnm2WWUFxxxxQSlCQNlRJ7AAd91zRds/wA88VV2lYXwheJmNcaRHlRr7nrKemRdCk6ciWkkenqlUn0Hfp3odYEg5G59yS/5XL4Z8N1riZHmUfTd4vUgk2fGUnt1SXE/xr/Y9LCNnYPV90pMl4c8PWM8WSpWW3W5Sstz28J/svld20uXIJ9W2h3EdgeiWkaAASCToVLuOONMJ4lxSLheAWFi1WuJ3Dbfdbrh+866s+844rXdSiSf3VKKAVCuWuXsK4XxVeVZpPWhC3BGgwYyPNmXGUrsiPHaHdxxR0AB2HqSACa1XNXOWPcOW2FHVb5V/wAqvzhi49jVuHVMukj5JH6DafVbqvdSPmdAxTiXgvIpGVI5x8QE+NfeQnWym3QmdqtuLx1f97w0HsXNdlvn3lHsDrZUBqMH4dzPmbJ4PMviYt7bSYS/aMWwLr82HZB+jIl/oyJhHzHS38BvsnoylKAVTHMN2uPImTRfD/iUxxj29kTMsnsHvBte/wCJB+Dr/wB0D1CdkjR3U35W5Eh8Y4dJyJ2MqZOcWiHa4CO7k6c4elllAHckq9deiQo/Ctdwvx1MwTHZE7JZCZuWZG+bnkE0d/Mkq9Gkn+5tj3Ej00CQBvVUcS3iJ+ix01l3cu+Xwv0IKr7SXZLx7uXj8Ca2i026w2uJZbRDbiQYLKI8dhsaS22kAJSPoABWZSlXUlFWROlbJCvy42282pl1CVoWkpUlQ2FA+oIr9V8JABJOgO5Jr0HP3hFLmHM8hcBvrV08a5Q+xa0qOym0TR7XDBPx0HHE/ggV0FXP/GLjLvjD5ndtywthePYsZSkEFJeLcooOx/e9a+lXleL5Zcegrud/vEK2Q0EBUiZIQy2kn02pZAFAZ1Kh38JUS4dsRxm/5H8nosP2eMR8FpkSi006n6tKWfoaeTyteP4+Zj2MsnspEZDtzkFP6yXVhlttX0LTo/GgJjSoeONYMr85kOU5TeXj2K3ru7EQR8izD8lkj8UEn41+v4I+Lj7z3HuPyV60Vybe0+s/ipYJP76Al1KiJ4n4/b/1vsH2Sr9e0ynrer/KjrQaxX4ua4MDMt06Zllkb7vQJISq5R0frR3QAJASP7W6PMV3IdUQG1ATilYlpu1uvtsi3m0S0SoU1pL7DyPurQobB/8A9HvWXQClKUApSlAKUpQClfFKShJUpQAA2ST2FVzP5khXiQ7ZeJbSvOLqhZZckRHvLtENwHR9pn6U2Ck/ebZDrw/ueu4AyeYsgnxMdbwzGngMnzJS7PaQB1GOFp/PzVD+5x2ip070FKDbe+pxIMwsdngY9ZYFgtbRbhWyK1DjIJ30tNoCEjf0AFRnB+P5NjuUvMcuvCb7l1zaDEmelnymIscHqESI0SryWArudqUtagFLUrSQmaUArSZplltwbGJ+U3ZD7rEJCeliOnqekvLUEMsNJ2OpxxxSG0J2NqWkfGt3VecqIEzJuMbQ/wC9Fm5f1vtn0cMe1z5TW/5r7DLg+rYoDFhcfZxmbKLpyjnN3gGQA5+T2Mz12+LDB7htUxnplvuJ9FOB1ttRGw0msscA8Rr7zsRFzX/dLnOkznPx633Fq39d7qwqUBXv+h/4b10nAbeU730EuFP+T1a/oreYvxjxxhD7krDsCx6ySHRpx+BbWWHXP5y0JClftJqTUoBWNDttutypCrfAjxjLeVJkFlpKC86oAFxeh7yiAASe/YVk0oBUXyzjDj/N325uT4pAlz2B0x7glHkzo31Zkt9LzR+qFpNSilAVucH5LxH87x/yG5eIiPSy5b1SkFP6jU9A9pbJ+K3hK/m1+hzGmwfmeT8JvuIqT2VOLH2ha1fNftcYKDLf8qSlg/SrGpQEMtnNPDt7V5dn5Yw6cveumNfYrit/LSVk1Lo0qNMZTJhyGn2ljaXG1hSVD6EdjWJc8fsN6R5d5skCenWumTGQ6P3KBqu7/wAD2lu8Wy88UzI3HMovrbvcmwW9hly5QVNLBaKCgtFwO+SpLy0LUgJWE66zQFqUqujwfYGUB205nyBb7gnuicnMLjKUFfrFmU67HX/NW0pP0rxVeOWuPveyS3Iz2xI+9cLPGEe7x0D9J6Hvy5Wh3UqOULPoiOqgLLpWpxfK8dzSztX/ABa7sXGC8pSA60TtC0nS21pOlNuJUClSFAKSQQQCCK21AKUpQCse42+HdrfKtVxjokRJjK477SxtLja0lKkn6EEisilAUVasoyZrg3GcTt96ejZK9d2sBeuekqebciylxZctIUCPMLEV95vqBT1KQSCOxuDFsXseGWCHjOOQUxLfBR0NNhRUokkqUta1EqWtSipSlqJUpSlKUSSTVG5KBjnKky2rH9btZvjuUt/qtoucZ61dI/GTHUs/V7fxroegFKUoBSufLNZ0cqRZObZldrzI9tnS0QYEe6yYkaDGafW02hKGFoCllKApS1bJUogaAAr9v8VYUj7rd7H/AJQ3D/t6A6ApXOD3GmII9Bex/wCUFw/7avD+DrEd63e//wAQXD/tqA6WpXOLPGmHr9Re/wD8QXD/ALas9jirCl623ez/AOUNw/7egL/pVAXC1fwWOWrLcQu95ZZTdoEG426TdJEuNMjSZLcdXuPrX0OILoWladH3dHYNX/QCsC+36y4vZ5eQ5HdYtttkBovypcp0NtMtj1UpSuwFYuYZji+AY3Py/M75FtFmtjRelTJK+lDafQfUkkgBI2SSAASQK5isWLZh42L5FzjlC0zLBwrbpCZeN4u8S1JyRQ7omzwDsMfFDXbYIPce8sDYtt5Z41ZaX5IuONcEMu7bZ2qPcM06T95XopiDsdh2U4O/bY6OmbPZ7Tj1qiWOxW2Nb7fBZSxGixmg20y2kaSlKU9gAPgK940aPDjtRIjDbDDCEttNNpCUIQBoJSB2AAGgBUZ5A5T4+4tthu2e5XAtDJSVNoec289r1DbSdrcP0SDWUISqSUIK7fBHqTk7IldVVzrz3ZuHoEK0W+3qyLN8hUY+PY1FcAfmu6O3Fn+1MIAKluq91ISfjVG8reN/PUXCyYnw1w3dX7tlbpYsbl7a8p+aPi81DCgryQNnznFJQACSOxFb/GfA1Yslvq+S/EHlNzy7Mbq0k3FtiQY8JsEd46CgBwtp+6AlSEkD7g3V30KNL/uZqPRetLyWS8WmS9ko+27e9mNx3kPDvEF5ncn848w2HKeWshbDU563LM0W1ne0W6CwyFqaZTvW9BTh95X0sf8A0RWRXz3eP/D5yJeur+LfnxGrTGcHwIckLB19emrCw/i/jrAGUsYVhFlswSOnriQ0IcV/OcA6lH6qJNSise0wtP2IOX95/KNv9R5vU1or9/7fUpYZN4tL171t4vwHG0q9BeMgfmKT+IjNAE/ga+Lx/wAWctCnrhyhx5ZkgFS/YrBIfSgfHu88P6auqqS5+yC75Xc7V4e8KlrZu2WIL15mNdzbbMk6ecPyU5otpB9dkdiQar4naawtNzjTjfgrXu3ovWvqR1cR2Ud5RX333KtwnBOdudb8jkmTzrEbt2LT5EXGZ/5JsLamKAKHpSGC70gdQKULUVKOiR01aw438Tcb84x4lbXLV/c5OER0pP7UPA1a+OY9aMTsMDGrDDTFt9tjojRmk/ooSND8T8SfUnZNMgyTHsTtbt8ym+2+z25jXmy58lDDKN+m1rIA/fTD4qtTh66g5PV7kM3/AJfBdBTnOK9a1+OS+hVH2d4v7T70bJOLr+lPqiXAmwlqH0U2tYB/ZX5Vytz/AI37+Z+HVy4xEfxkvF76zMX/AIsZ0NuH9hNSY8qXrJCWeL+PbpfEE6F1uvVZ7WPqHHkF94H1C2WHEH9cdt6rI7FcGbYu+858zi1WlPZyBZJBsUH5hK5IcMxxY9PcfbSv4tfCrMcUqj3ZUoyfRNP/AMWvgSqe87OKf30IlfvHDw/jk22wb4xfrTLemJj3OBdLU/Fm25paFlMhTZSQ4gKSEkIUVe9sA60bMhZ1keZ21q4YJhqXbZOaDke53eeiKw+yoe64020HXldjvpcQ1v5iuas1xHw9ZlO8/BPD87eelJSifNmSrLauo/edDaVB15aiNqcDW19trPY1+8WxHJMYsf5Nw86uVksvmKeRZsdkPRYzJV94JedcdkgH1911AJJPTsmtlU2bQq0ouF4T4qVn5WzXikTyoQcU1k+uf34kR/qf3CvLMDGrvyTyZn93sWOZGmMiFb0PIYlXGPHb8qO87JUPOaYS0OhpDa0EpPV2T09XXloi8KWK4puMO4445dkbSJ8y4olz/qDIeWp4/gVVzkrj7BCtLkzG4lydSAA7cuqa52/lvlSv6a9hiOEpBSnD7IAfUC3s6/8AVqNbKpcZvyX1MVQhzfl+52Ew+xJZQ/GeQ604ApC0KCkqHzBHqK9K4dvHD+C3lry48edZyCoo+zJrjDaCfUhoEtD5/c9a3uIjDcDDNt5O49tN9sKAEjIokRXtUUfrTGAT1p+brXp8UAbNQ1dlTSvRlvdNH80/O/Qxlh/5Hf4nYtKrq38O8KXu3xrrZcWtLsWShL8aZb3lI60nulaHWlA6PzB717OcMWGOkrxzK82sMgb8p2Lk8yQhre/uxpa3o2u/oWiPpWqaayZWJ/SqzgZjlnHt4h4zyxMiT7Xc3kxbTlbDSYyFvq30RZzW+ll5etIdRpp1Z6OllZbQ5NpWV4zCss7I5N/t6bXbGXH5ksSEqaYbQkqWpagdAAAk0s7XFjS8fhES4ZnZYxAi23I3PJQP0DJixpjo/a9KdV/jVMKgfDUO4Lxmdlt3t8iDOzC6yr8uNIb8t5phwhuGhxBAKHBEZihaD3SsKB9KnleAUqIZ/wAsYNxqmKzk12WblciUW60QY7ky5XBYHdMeKyFOua+KgnpT6qKR3qJIuviMz5Pm2ex2DjK0u/cevY+2LypB+JisOIjR1emtvv6/SR8KAtyorlPK/GGDu+RmPImN2R49ksz7oww6o/AJQpQUo/QDdQ5fhysN9koncl59nGcOpbU2pi43gw4KgpSVEGHASwwtIKBoLQvt6lR71it+EHw9224OXrD8CThl3cSUm5YnPk2WSfj7yoriA53Po4FJPxBoCQ/w0225How3BM3ydfzi2NcJlQ+aZFwMdhY+qFqr4q586ZJ7lsxnG8MjL7iVd5a7rMSPkYkctsg/yhKUB+qfjpnE8v8AEK0zZF5mcl4a0CZYkRmm8htrYBJdQWEIauCE9ttBtt/pBKVPrIQbAm51iMDDDyE/fopx32JNwRcGleY26wpIUhTfTsr6gU9ISCVFQABJAoCJyOG8dnsOXHljKbtmjbKS681e5KGbU2gDZ6oLKW4y0jWwp5Dihr7/AMa+QuYLLNitReK8ByLLoDCA2zJs0RiLbQgDSfJky3GGXm/gDHLg/dWPbsNvfK8hnJ+WbcuLY0rS9asOdIU2gA7Q/cgCUvv+igx3ZaOv4xxIcTaQASAlIAA7AD4UBXoyTnaR+dh8UYkw0e4RcMzeaeA+qWbe6jf4LI+tfhzlXIMaBXyRxherPER/GXW1LTeIDf1V5IEpKfiVqjhCR3UoCrGpQGHaLxacgtka92G5xLjb5rYejS4jyXWXmz6KQtJIUD8warbky5zcgzrFMSwy1OXG+41d4OQ3CQXUtxbZBc82M55qj3U67HdmBptCSSUbUUJ0TqeSLpb/AA4SpvLECG8MOuanPyktURO0tT1pJjzWEeiXHnuiO6kdlrfacOihZXOuLsQuGK487KyN1p/Jr/JVd7/IaO0LmOJSPLQT3LTLaW2G99/LZRvvskCY0pSgFKUoBSlKAUpSgFKUoBSlKAUpSgK6yrjvJmswj5vxVd7Pj90n9UfIPb4bsmJcGQ3pp1cdp1rrkNqShKXOtJ8sqSSoBIB2z+ICGgyo2fYLdHWx1exPYxKhof1+gH0znSzv06y27r16VelWLSgItgmexc0YmxJNtfs9+s7qY93s8lQU9DdUNpIUn3XWlj3m3U+6sb9FJUlMpqs+W2Bh822c1W8eW5jhEW/9PYSbE6seeV/P2YkSknuQGnkp15qt2X696A+0pSgOf/EHEks5PMuFvYU9MRhEvIIzSBtT0ixXODNYR9SVvaH841fUOZGuENifCeS9HktpeacQdpWhQ2lQ+hBBqveUfKs+YcfZdJSkQ27s/j9wcWPcRGuDCkNhX0XMagt/i4K9eB3HIvHUXEpKyZGHSpWML6jtZahOqZjrV9XIyWHf/GCgLDrGuVxg2i3ybtdJTcaHCZXIkPOK0httCSpSifgAATWTVJcvPyeV84t/h/s7ribYlDd4zOS0SPLgBW2YfUPRb6x3HqEJJ7gmgNfwtNjXDi+03CE6HY8p2a80sAjqQqW8UnR79wRUllqPeo9w3HZj8aW2PHaQ200/OQ2hA0lKRMeAAHwAFSOUgndAU14nL7dcd4NzG9WW5TIE2LA62pMNxSH2z5iQShSfeCtE613+VU2/e+QbE3MlWu55tBxKTmWHw7OcjeeFwWXbi23cEAvH2j2ZaFNJCXfU+bodJrpzNcPsucY7PxXI4qpNtuLflSGkuKbKk7B+8kgjuB6GtTm/HuP8hWT8nslZlLiCVHmIVEmOxXmn2HUutOIdaUlaFJWhKgQR6UBKYqydVuYqj2qK4fisTE7S3ZoE26y2W1qWHbncn575KjsgvPrWsj5AnQ+FSyKj0oDQ8qHeHsf/AG9Y/wDpSNVrZlnOJcf4xdMxzC+xbZZ7M0XZsl5fZoaBCdDuVHqSEpAKlFSQASRVP813KHY+Onb1cnfKiQLtZ5T69b6G0XKMpR0PXQBri7i2dyTyvzfkXInPlpiwuNrVf05XNseVXY2yNGfkt+VCWqOW1uSlpjRQG2VJCVdAOwFnrzhTlUdoK/cepOWh07heCZb4v8jh8u84Wd+18ZW2R7XheEyAUmfr+LuVxTv3+od0NH3dK+KSS70PnvJvH3FVm+2M6yaBZogBDSHV/nHdfotNJ2tw/RINVEeYeXeblG3+HzFvsHHF7QrNcijFDa0frQoqved+iljp+CgmpVx/4acJxO7ozPLZk7Ocy7KXfr8vz3G1DuPIaJKGEj9EJ2R6dVXPRKeHzxUrP+Ve148I+N30JezjD+0fhx/b7yIr+XniB51T5XFmPL42xV06OSZDGC7jJbP6UWH6JBHcKWdEHYINRvPbDw74WYEPK7jbLlyTyzkTwhY6m7vGdc7pcDoIDSVdmWkKUnqWkApSQNklIN084c14twVhLmWZA29NlyHUwrPaIo6pd1nL7NxmUgElRPqdHQ2e/YGAcA8IZUnI5PiC59cYuXJd8a6YkUe/GxiArZTBjD0C9E+Y4O5JUNnalLxnjZ7rp0VuR5LV971fw6Hjqu1o5L74mb4euBr1iVwncx8yTmb7yvlDf9kZifeYtUb1Rb4Y9ENoGgoj7yge5Gib0pSqREKVEci5b42xa4myXjMbeLvrqTaYqzKuCx/IiMhby/8AFQa1Ks+5AyQBGA8YTGGHPu3TKXvs1gJ/WTGAXLUoevQ40zv9cetASbOsys3HuI3XM7+8G4Nqjqfc76KyOyUJ/lKUQkD5qFUHwbmFotyL1nNzj3DMOScxeEu4WzH4/ta7Wx/3vAdfJTHiFLeiUvutE+h30g1hTMTynmXnFGC5jnb+T41hyW7hk0RiC3EtC5yu7ENtgdbigAOpfnPPfpJHR6Hpy1Wm12K3MWiyW2Lb4MVAbYixWUtNNIHolKEgBI+gFa+n/WsQ6v5YXS6y4vw9lf4ivH8Wpv8ACOS7+L+XmQX2bmvLyDLuFqwC2qPdqElN0uyk/A+a6kRY6/gU+VJT8l1lWzizjnEJJzK7tG5XWChTy7/kU1U2THSBtakOvkpjI9SUtBtA/VAFTK5XO22a3yLteLhGgwojanZEmS6lpppAGypa1EBIHzJrni53F7nSd+UN2e68EjPk2S1JP5q6dCtCdJH9sQVDbTR90J6VqBJHTucNh+3bcnaK1fyXVlynT383kkb++835FmIXD4hgssW0npVk90YUWVj4mHH7Kf8Ao4spb+I8wVDGsMtouicjyCXMyO+p9LndnA+638wynQbYT/JaSkfjUwf0kdKQAANAD0ArVSVetbmju0lu0lZe997+WnQsJ7qtDI11xlezsOv635aFK189DdV4zycxPseGXlFuAOXuR0eSJAJi+bFcf2Tr39eX0+g9d/Spjdp8FbciD9oRkvFCkdCnUggkfEevxqn7Nw7bLDasDdt1ix+Nd8fcjrus6LHQhyQExHWnOlxKApfU4tKve1vWz3FbPDU6bi3U1/Z/OxNCMbesZ9u5dfkyID15x5u32u6yp0SJMRO85QXF84qLrfQnoBSw4QUlXoAdbrYYxmmRZGYdyXiSYdmuLfnRpK54VIDZT1IU6x0AIChrslaiOobA760GO8Y2m02aQzeIjTtzlG4NrkpeW55bUh5xWm+vs2ShaQekDZB9a/eIjOLRHt+P3iNaFwrawmMZ7MpwuyEoR0oPklsBCjpJV76h669e16dOi0+zWn76Z930JWo/lLMZk7+NbCO/vXftUbjPE671uIrhOqoThYhaMqwxMnw5+Q5xvl72Px5iy7Ity4qJcEuk7LiGl6LSjv3vLUkK9SCe9SRGW85vDpPJFiQkjXU3jOlj8CqQR/RWpiK9K28Y9hVOraTvKKb5tJvzsYyk3m7eSMBzAoWSyhcORrtPzOUD1JTd1JVFaP8Ae4iEpYT+PQVfyjWpzrA8MxaFF5Es+I2xo4tKYuc6ExEQiPPhsrCnEutJHQ4ptO3WlKBKHG0EarZ5ryViHGttbuOU3PylSFhmJEZQXZMx0kBLbLSfeWokgDXxI7jdbnHuNuTOVrAqTnUteDWO6trbVYo7CHbo5FWCCJD6iUMKWn1QhBWkK0VBQIEbquCUqsrQ5cGuKSX0sgpNZzdo/fAszNuaONeP1RIuQZK0u53FHmW+z29pc65zhre2IbAW84PT3koKRsbIFRZd0575QKWbHaE8VY64ffuNz8idkD7ev7TET1xYm/ULeW8oeimEn0muA8V8d8Xw3YeBYhbrP7SeqU+y31SZSv1n31bdeV6DqWpR0AN9qldc2+hQIZx/xFg/GypU6w296TebiB9o3y5yFzLnPI9POkukrUkHuGwQ2n0QlI7VM6UoBSlKAVQmE4TebnytkmJ3GF5WAYJkP21Z2Cdol3KZHZlBBH9zivPSXUpO09ciOU6LAq+6gMCLds9uV3uYyS52mxxZ71tiRrcptlctcdRakPuuFBcB85DjSUpUkBLQV36x0gT6lQS8cePWu3rumE3jIBe4KkyYrU7I58mPKKFBSo7qHnVoCXUhTfX0Eo6+tI2kVK8fvUPJbDbcit3X7JdYjM1jrGleW6gLTsfA6UKA2FKUoCuvEY20rgPkR95CVex4zcZqNjenGI63UKH1CkJI+oFWLVe+IQhzhLNLcPv3e0P2hofrOyx7M2n8Sp5I/bVhUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQEP5hulosvE+Y3O/Npdt7FinF9ojfnJLKh5YHxKthIHxKgPjW5xCDcbZidltt3e82fEt0ZiU5vfW8lpIWd/Hagaz7hb4N2gSbXdIbEuHMaWxIjvthbbzSwUqQpJ7KSQSCD2INQPiCRMsyb7xddJb0l7DJiGID76yt1+0PI8yEtSjsqKB5kYrUSpaoilHuqgLEpSlAajLcXtOa4zc8Tvja1wbrGXFe8tXQ4gKGgtCh3StJ0pKh3SoAjuKqDw6ZJdpmY8gY3kjiFX2K5bpd4KUhAdnpQ7bnXkp/RQ63a48hKfgiSjfrV7VzrfSeNfGdjdzWktWrlazTLWCPuKu0VlDqlOH0CjGhsIbHx/O/KgL7v17tuNWS4ZFeJAYg2yM5LkuH9BttJUo/uBqs/DfZLirEp/JeSRy1feQpyr9JSrupiModMRjf6qGQjQ+BUa8PEyty945jnF8ZSg5neQw7VI6TpQhIV58lX4dDWj9FVb7TTTDSGGW0tttpCEISNBKQNAAfAUBzDkU3kPGMrv2P8FWuNklpYuDz01m6ISzHtsx1XmvMMSPNSXR1LKijyyGysp6/0Rp3so8Vp+/xjiI/C4//AFasbi5ZVYb0tRJUvK8gKj8SftSR61v5Tnr3oCjnMm8Um/e41xP/ANIf/Vrx/KXxP7/2tsU/9If/AFauGS+Ruq0sPOOM5FPsECFBuaHMiuF6t0UutoAQ5bHnGnyvSzpKlNKKNbJBGwn0oDXt5N4pB93jTEz/AMYf/VrMZynxWj7nGOIn/jH/AOrVnRnydd620VwnVAU+9dOZbxLt0flCzWrHGBMZNnXHHn25d16x7IZ7iXVOJbS70KShKAlbiUJUtINY/G/gYv8AYuSG+S+R+QrNl0pyUqXNhz7KuQ3IdUdlzqU8kBY79JKCB290+lWTy6QrjS8gjeksKH0IfbIP7CKvSrmGx9fCQnTou29k8le3fw8CWFadNNR4n5QhLaQhCQlKRoADQA+VRvknkbEuJsKumf5xdEQLRaWS684e6lq9EtoT+ktStJSkepIre3G4wLPb5N2us1mHChMrkSZD6whtlpAKlLUo9gkAEkn0ArkzHnmvFTmJ5+5IeRaeD8Aedk4pBuH5lq8SWdhy7ywvQDKOkhtKvkd60sLpkRJODONcu5ZzZnxS88Wwxrk40U4Ri73vN47b1naXnEkDcxwaJV6pBHp7qW7dvXLVqZvEjFcLs87Mb/EX5UuJaijybevW9S5K1JZYOiD5ZUXSDtLaq07DOZcyJEuc5dMQwd4BTENtS4t4vLZ9FvLGnILCh3DSOmQRrrUz7zRsOwY/YsVtEawY1Z4dqtsNHRHiQ2EstNJ3shKUgAbJJPzJJoCGtWbmzIh1X3L7FiMdZO4uPQzPltj4amzEhpX4GGPxr9J4UxecfMy+9ZPla1dnUXi9PqivD+XCZLcQ/wDmasCtJlma4tg1uTdMqvTEBlxflMpVtb0l0js0y0gFx5w/BttKlH4A0BkY/jGNYnAFqxbHrZZoSTsRrfEbjtA/zEAD+io3zPyTD4m44u+aSAhyRGa8qCwr+3y1+60jXqR1HZ1+iFH4VrU5Hy3nPT+SONR8MtK9EXTJWi/OdT/erc0tPl7GtKfeQtJ+8wfSqnTx4xyD4jo+O3vJb9l1rwGGm5Xx28Sw5GfuT4/rZgRG0oithCNr2hoKPcLKqqY2rKnT3aftSdl3vj4K78CGvNxjaOryX301N3wbmGE4HgrVns8m453llzecul+exqEu4oeuTx6nEOTED2VtSfuaceR9wn13VgGfzhlSim3WKxYLb1E6fuj32pcun5ezMKTHaVruFe0PD5oPxsRtttltLTLaUIQAlKUjQSB6AD4CoTzRkV0x7j6cjHZfst9vb0ew2d7XUWZ015Edp7p/SDRd81Q/VaV6AbqejSjQpqnDRZEkIKnFRWiKTPG0/m3LpUK2Zbf5VhxmcuJcc0urjb0ybcGVadj2eP5aYcMNKCm3ZrbAc60qbbPUhTidlh1q/g+ut34ffdeUjH1+1WZb7qnHJFpfUpTJK1EqWW1hxlROz+bST96r7xHFbJg2L2rD8biCNbLNEahRWh6htCQASfio62T6kkk9zXOHiGzjGrveLZmPHP25csswia/DdRCsM56Jc4pV0S4KpDbJbKgpAKVdRSlxBB1skbbZqnUm6STafufBv4eNy1h05NwS1+0TKT8a1UketbCNLRcrfGuLbD7CJTKH0tvtFt1AUkHpWg90qG9EHuDWHIR61fhdOxmiAX/jbAL5Len3bDLNKlPHqcfchoLiz81K1sn8aisrhrjhslUTGkQl/BcOQ8wofgW1irUlN+taiSzWypYmrFWUn5k8as1o2VJcMOzDHFCbhuVzJ7TZ2u13l3z0Op/VQ8R1tn5bJG/XtWyx6+QsjYW/GDjMiOvyZcR0adjOj1Qsf8x9CO4qbPxiSe1VxyDbncXuUPP7BBlSbn5rcKTbobSnHLoyrfuJbSCVOoAK0kAnSSPT0vwqqst2evP6/UnjLtMpa8/qTaKgjXat1ESe1RrDcrxrNbai643dWZjKh76UqHmNK+KVp9UkH51KfNiw0ebLkNMIHqpxYSP3mqlW97MhldOzNrET6Vk3S8Qsdsk2+3JZTFt8dcl0j16UpJIHzPbQHzqCXnmvinFVNM3TOLYuS+4lliLFdEl951R0ltLbfUSokgAfEkVZmFcYZfyLcLde85szuO4tDeanNWeVoz7k6hQW17SgEpYaSoJV5RJWopAX0gFJpVd2l61bJe99333kbju5zyX3oZvA/h8TZrsrmnlNtF1z+7I64yHffZx6Koe7FjJPZKwk6cd11ElSQdbKr7pSuarVpV5upPV/du5FKc3OW8xSlKiMRSlKAUpSgFRHicAcf2kEfnQHhIPxU/5y/NJ+pc6yfqal1RHi7tjMtPwTkF+SB8gLrKAH7qAyc8yCbZ7Ui22FKXcgvSzBtLShtKXikkvOfJppILiz8QnpG1KSk7XH7LExuw23Hbf1+y2uIzCY6ztXltICE7PxOkio9ZEC9ck5FenfeTj7EewxUq9WnHG0S5K0/RxLsMH6sCpjQClK+KUlKSpRAAGyT8BQFc8jq/KnNsN42Z95n2wZTd9eiYkBaFx0k/BS5qoqgD95LD3yNWPVbcOpVk5vfMEoEqzJ5v7JKvVFjj9SYIH8l3rel9+49s6T90VZNAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUAqvJY+yvEBa3GuwyXEJrUk/NVumR1MD91ykmrDqvLsr7T56xyGxopsOM3ObLI9UKlSIrUcH6KEeWfxaFAWHSlKAVWHiIxJ7IuPRkFqiqfvmDXGLl9nSgbW5Igr8xbCf8Ox58c/R41Z9fCARo+hoClbxcYOa+I7jlVvkok2+1YrcsljOIO0uCSWY7ax9OlaiPxNTzljkKLxhg1wyx6KqZJaCI8CEg+/MmOqCGWU/H3lkb13ABPwqjOBGXrZ4i8gwWU2tJ4/xQY7F2DoQftBb8IJPx1DdiJJ/WQofCrA5rSmZyhw1abh3trmRypbiT90ymITq42/r1k6oCL8CJvyeNynKVtKvQvd5FxU1roMr7Qf8ANKddtdfVrVTOUD3FRKwZLjnHk/IMLza9Q7FPRfrncY32i+mO3MiSpTkhp1layEuAB3oUASUqQoEDtvNkcm8ZK3rkXGD/AMbx/wD56A95aSd6rl+04FybhL+DX1rjy4XxyxZFmEuZCgToKHksXCZIXGcBffbbIUhxCiArqG9EA7FdDv8AI3GyvTkHGj/xsx/89Y38IXG+/wDbAxv/ANKsf/NQG3sUiVNt0WZMtr9vffZQ47EkKbU7HUQCW1ltSkFST2JSpSdjsSO9SKID2qHMcjcbJI3yDjQ/42Y/+atlH5N4zTrfImMD/jeP/wDPQH75bH/c1vX8xn/2yKvWudczy3GM6swwLCL9b79eb2/HYbYtslEnyGvOQpx94tkhttCApRUrWyAkbJAr28SfLGWSLzbfDhwlJT/CJmDKlyZ6e6MctXo7OdI+6vWw2OxKu479IUBFuTrtc/FryjK8PeGT34/GuJSEL5DvUZZT9oSEnaLOwsfUbdI9NEbGgHLSt9itmf5ajHIUGPH4+42eYhRreykCPPu7KUqSkoHYsQ0loJT6GR1bAVGST7WLEsT8LfBMi04VaVyWcfguPNtnvIu1xWNJK1fpOvvFCd/AqSBoAATHjrEU4JhNpxYyRKkQ2OqbK1oy5jii5JkK/lOvLccP1WaAklKVXGdXm+ZbkY4mwq5P25wsIl5Jeoyul21wlkhDDCtECU+ErCT/AGpCVOHSi0Fgfb3n+Q5NeJuF8RR4Umbb3PZ7rkE5JcttpcB0pkJQpKpUod9soUlKP7Y4glKF7XEeLcexa4rySU/Nv+TPtlp+/XdxL0xSD3LbegluO0SAfKZQ22SNlJOyZBjuO2PE7JDxzG7YxbrZAaDMeMwnpQhPr+0kkkk9ySSSSSa2NAa3JL7BxfHrnktzV0xLVDemvn4+W2grVr66FVl4XsfnQ+Nzm19bAveeTXsknK+On1bZSP5Ia6CB8Oo15eLie/G4NvNtiuKadvUmFaw4D91L0lsL39CgKH7atu3QItqt8W1wmg3HhsoYZQPRKEJCUgfgAKof2uNz0hH3yb+Cj7yD2q/91fH/AI95k1W2Y/6YOZ8Bxn7zFjj3LK5Ou4DqGkwoyVD5K9ukrH1j79QKsmq6wgKvHLXI2Sq30W9VqxZkH0IjxzNWtP4ruZQfqzo+gq+Tk+mzIluhv3CfIbjxorSnnnXFdKG20glSlE+gABJNfy98Mn9UMwPL+Qcuw3MyzYLbeMkuNxxyQvaWkMSJC3EtObJ6SerqJ9ApSj2Sfd6D/qqHPKeHPDBcsZtc3yr/AMiLVj0RKVaWmIpO5rmv1fK/NE/Avpr+RPh68HPiD8TD5k8W4W6u0sPeU/fJ7oi29lY9R5qu7ih22lsLUNjYG6sYfEdg3dXT1X3xJKc9zXNH9zvMjy2ESorzbzLqQttxtQUlaT6EEdiPrWBIbqjfDz/U/vEBxRam49/8W0xCNDdrtto9pjNK+JDshwdfx7loH/mq7Z/heuUppyVk3iPz9phsKW6uAbfAQlAT6k+zq1r1J/8A+1so4nDJX38u7P6e8m3qaz3vcYb7O99qj14u9itIJut5gQgO59okob1/lEVFpPE/h8lSFsw79y9y2pH5tZbyBbcFSh2O5iDGaV3Hfy3FEfL0FbWz8eYjZ1bsvhI458tJ6kqv9+XOf3o/FcR7R7/BZqB7dwMPZcpd0cvNNlR7VwcfZk5d0W15q6I9L5g4xTPTaYWVxbpcHd+VDtaFzX3D8koZCia+8f8ALGJ4pyM5yFzBhHI+LWWyQ1s2aVcsIuhiIdcB8+Y842wpDWmwG0lR7BbpOt9rnx7lLI8NY9lHh7t8KCNdacVusZzsPQhp1uNvXyBJ+VWHhfM/H+czvsS33R2BegkrVZ7rHXDmgD1KWnAC4kfrN9SfrUVTb9KvF0qS3XLLO9/C6XzPFtahV/Dg7N87p+F0vmReRw34Zud2WeSoGP4/f0XUeam+WKctgzPh1LfiOILih6HqJI1o+mqjWU+Evwh4ZZ7lneY4U1FtlniuTJ0u4X64usNMtjrUpSVyCk9h6aJPp33o+/JPDc7jG43DnTw621dvyGP1TshxWCkiBl0dI242Y4IQ3P6QS1IQkKK9IX1pV2/dg4jzzmSfb898Rs9ce3sSWrlaOOYLg+zYCkHrYVcnAOqfKQQhZSSGELT7qF9IWYI4qvGO5GbtyuyVVZpWTdiL+G7w4YW5k7fiEu3EllxB5xpTeF483bGmJFmt6/8AvqYQOpdwkJ0pRWVFlspaSQfMKupKUqFtt3ZgKUrzffYisOSZLzbLLSStxxxQSlCR3JJPYD614D0pVePc02G7OGHxpabjncr082ypSbc2d6JXPcKY3un7yELW6Pg2fSsdWB59nZS5yhlabba1p9/GsZfcaZXv1RJnkIkSE/RtMdJBKVpWKAzrxy7ahdZOMYLZp+Z32IotyI9q6RFhOD9GVMWQwyodiW+pT2jsNqrHGH8o5afNzjPk2CGok/ZGJp6D0nt0O3B5Jec+BCmERlA/P4zCHCxfBseTEgxrZYbHamD0ttpbjRYrSe5OhpKEjuT6CokOaLNeNDj7GMjzVJPuybRDS3BUneutE2UtmM6n/BOLP0NAfpXCWJpa6YuQ55HeTsofTnF5cUk73vpckqQrv8FJUPhrVfk45zFjg6sez+3ZTHR39kyeAliSvt90TYSUIbH1VFcP1+J9EZFzY4PPHFmMtskjTbuXOCTogeqUwVN7BOtBw+nqa8H+YHMa6XOTcEvWJwvRd4cWxMtbR+JdfYWpTCB8XH220fyqA9ovJWR255MPOOK8jta+4My1JTeIK1Ab0gx/651/Kcjtitzx1bbhbMTYF2iqizJ0qZdH4yiCqMuXKdkllRBIKked0EgkEpOu1SGLKizozUyFJakR30Bxp1pYWhaSNhSVDsQfmK9aAiOC6F5zdtQ/OIyL3z89wIak/uQpI/ZUuqI4h/W+YZ1DX9966RJ6R/e12+MyD/lRnB+ypdQCq55kmybzBgcTWV9xu6ZwpyI+40ohcO0o6fb5Ox3SQ0sMoUPR6Qz8N6n1wuEG0wJN1uctqLDhsrkSH3VBKGmkJKlLUT2AABJPyFQHie3zr69ceXshiOx7jlaWk26K+gpct9mbKjFZUk90OL61vug9wt7yzsNJoCwYsWNBiswobDbEeO2lppptISlCEjSUgDsAAAAK9aUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFVrxTIbyXLeQc6WrTrt6GNsMrHS7HiW1JQEuIPdJVIemPI3rqafaUOygTZVVrkbCcG5YsWXwvzVvzVwY9fGx2QZaGluQJZ+AV+bcjE+q/OjgnTSdAWVSlKAUpSgKDlZLa8X8aTFkmNFpeYYSy1HeSn3VymZEhYQo/MtNOEE/3MD4ipb4h8VvF/wAATfcWaLmRYfOYyO0oHq69GJK2tfHraLiNfEkVBOYcPkZN4gbRDt0xuDdbjhclVmmrSSmNcoM9iVHcUB3UgL7LSPvIUtPoo1c/HuZMZ7icLIkQ1wZK+uPPgOKCnIM1pZbkRlkdipt1K0bHY6BGwQaA/OG5PjPKOGWjMrWy1Lt11jpkspfbSotqPZSFDuApKgpJ18Umtt9gWL/eWB/yZH/wqjMXkHw98xP8e3BwtYLyBKcn448r+Lt10UdvQifRKXD7yB6bIA2So10FQGB9gWL/AHlgf8mR/wDCn2BYv95YH/Jkf/Cs+lAYH2BYv95YH/Jkf/Cn2BYv95YH/Jkf/Cs+sS7XW22G1zL3eZzMKBb2HJMqS8sJbZaQkqWtRPoAAST9KArrnXlvGfD5x9KysWhqVdprqLfY7PFbCXrrcnOzEdASNnau5IBISDoE6B0vhm4UvHG9lueccjy27nyZnTwueTXAe8GlH+LhNHvpllJ6QAdEgkdukCBcH2i5+JXlNXipzWE8zillL1v40tElJHSzvpeuziD2DjpGkb9Ej49KFHqegK/5YQLhNwLGnf8AU94y+KXuxI/rOPJuLfp/fYLXr2qwKgWeFtPIXGinkHSrtPbbUUkgOm2SiBv4HoS5+wKqe0BGuQ8zbwTFZN9TBVcJyltw7bb0L6Vzpzyw3HjpPfp63FJBVrSU9Sj2Sa8uN8LcwnHPZLjOFxvdxfXcr3cekp9tnu681wAklKAAlttBJ6Gm20DskVHrV08jcqyMiKw7j+AOO222jXuv3paCiXIB9CGGl+zJIPZbsxJG0g1ZdAKUrxkS4sRBclSWmUD1U4sJA/aaArDxP4xc8o4XvrVkjl+42ssXeM2NkrVGdS6pIA9SUJWAPidVN8FzKzcg4jasysEhLsK6xkvo0dlCj95CvkpKgUkfMGs+4X2x2q3i7XW8QYcFXTqTIkIbaPV9331EDv8ADv3qiGcOumLZJccj8Lud41cI85wzLth8mel6EtxWup5hTZKo6yfh90nXwATVOrSrUKrxUIOUWkpW1Vr2a56u610sVpvs6m+s7rNcejOhqr/hBtxeHTrrIPU/dskv05Svj0KucgMpP1SylpH+JVeR/GLiuPznLBy1iN5xG7RXPJk+WEXGIhfxAeYJJPr26divXGedeDMNwS9s4FyTBvEkO3W82623Oa3EWuTIedkiIhTyWghHnOlKOs+6kgFWk7GSxlB2TlZvg8n5OzMo4qjLJSXwOTOYeHbn/VCfHXcsZmzJDHE3CrTVnu0pklPtU4nzJMVk/B1S/wA0tQ+4iPv1KN/0cxXFcbwfHbfiOIWSJaLNamExoUKI0G2mG0jslKR/z+pOydk1S3hlh8UcJ8SW3Fn+WMQul9nvP3nI7q1eIxFxu8pZclPb6+46z0JJ79CEA+lW81yFgMhsPMZxj7iFeikXNkg/tCqlVek9JLzJVOL0Z8zvOse46xuRlGSSFojMlLbbTSC49JfWdNsMoHdbi1aASP6ACRQ1wiZNyjITfOVgGrf1B2DijTvVDij1SqUR2lPem+r82k9kpJ94+9wvKeVuQpWaOPB/HMXfetmONg7bekJ2iVP+SiVBTLZ+CULI+/W1fe9dmtfUn6U7v2OC59XzXJaceVtNVqemNuX9mtFz6vmuS048renmtsoS0yhKEIASlKRoJA+AHwrzMrv96tc/K0fWsVUw7+9RzsHOxvEyd/pVhX7HsfyyEmBkFtaltoUHGlHaXGHB6ONOJ0ptY+CkkEfOsJuYSfvVnMSer415dTW7LNHjcai3ZK6NhhnJt/45uMTF+R7s7dscmupjW3I5BHnw3VHSI84jQUFEhKJGhs6C+5CjfFc/y4UC9W+RaLrEblQ5jSmH2XBtLiFDRSR9RUl4Fym4IauXFuRzXZVyxcNLgy3lbcnWpzYjuKJ+84goWys/EtpUfv1Nh6rpzVKTunp06fTy5FjC1pUZqjJ3i9HxT5eWnlyLcpSlbA2hCs7zu5We4wcLwq1M3fLbu0t+PHfcLcWDFSQlcyWtIJS0lSglKEjrdWelOgFrb18Xhi2XiQ3deVbxIzq4IUHEM3BsN2qOsEkFi3pJZBST7q3fNeH90NTpm02uPdJN7Yt7DdwmstR5ElLYDjrTRWW0KV6kJLrhA+HWr51H845GsuEmJb1xpl3v106xa7HbkJcmzinXUUpUQlttPUnqecUhpHUnqUNjYEnUpiIwVKLbLLKNknSUoSB+4ACq4Xypds1WqFwrZGb4ySULyacpTVkZI+LSx784j5MDytpUlTzahqvjPG19z5xF15qlx5cTYcYxGC4pVpjkdx7SohKrg4P74EsghJSyFJDhspCENIS20hKEIASlKRoAD0AFAQGFw9arlKjXrk25vZvdoy0vNG4tJTb4jo7hUaCNtNlJ30uL8x4A6Lqqn4Guwr7SgFfCARojYNfaUBB0cJ8Wx5bk224fGtTrq1OOfZTrsBDi1dytSGFISpROz1Eb2SfXvWQMFvFs74vyFfobae6YtwWi5sb+JUt8GSfwD4FTClAR7GcZuFquFxvt9vLdyutzSwy64xF9mYbZZ6/LbbbK1kaLjiiVLUSVnuAEpEhpUE5Gyy/NTIXHmA+X+VV+ZcdRKdQFsWiEhSUuznkn7xSVhLTXq44QOyEuLQBp8wkHlvI3OLbQhTmNWqS2vMZ39qe6QlxFpQf01ObQp/1CWSWz7zw6bTrS4diVnwbG4WL2Jt0RYaVbceWXHn3VqK3XnVnut1xalLWs91KUon1rdUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUAqveek+XxpLugHezXK03gf/AHS4x5B/eGiPwJqwqr3xAac4gyKF+ncG2Le2Pm5IfbZQPxKnE0BYVKUoBSlKAp7lb+xnOPDuQHs05Lu9ndPzMiH1IH+UyK2GWok8T5VL5Qt0dx3F7x5f5XRWUFSojiEBDd2bSO6gltKW5CQNlpDbg7sqS5r/ABM/2Ox/DswPZvGc1s899fyYU6WF/s0/VwEAjRGwaAiHIeB4rzJgcjG7q6h+BcmkSIc6MsKUy5rqZksrHbY2CCDogkdwTUJ4Z5Pv8e8L4S5fUmPnNoZ6oko/xV/hJ2EymVH1XpJ60+uwT8FBOC/bLn4YpEi743AuN34pmSFSLhaIzZfkYmtatrkw20jqcgbKlOR07UydraBR1NolvI3HWJc5YnbbvZ762zOYCLljeS2t5Li4zhAUh1pxB0ttXu7SDpQ16EAgCx6VTnFfMd7XkB4h5mhsWfPIrZXHda7Q75HHpIjK9OrQPU36ggkAaUlFx0ArlrnK4y/Ejy3F8KuLynk4pZPIvHJNwjqIHkghce1JWPRbqgFL0dhI7fdUmrR8SXM44S41fvlsh/aWT3d9uzYza0jqXOuj56WUBPxSk7Wr091JG9kV+fDbwsnhPjlq03WZ9pZXe313nKLso9Tk+5vHqdUVepSknoT9BvW1GgLNt1ugWi3xrVa4bMSFCZRHjx2UBDbTSAEpQlI7AAAAAfAVk1qsmynHcNtD1+ym8xbZb2PvvyHAlO/gB8ST8ANk/CozyBfswlYbDv3F95xqKxMSmS/drytQjRoKmyv2hOtAnXT97t32ewNS06Mp24J5XenmYSqKN+LXBanzmCHc2rLas1ssJ6dNwm6ovyYTI6lymAw9HlNoT+k57LJkFtPxcSgdt1Ccu8RuL5FdbJxpxJlsOVkGWhKWruy37RGtLCk9ReUNaW/078to9gvXmdKeyv3wbyJjGVYNmF3xbm2FyvKtEt5mfLU43FgNSkx0rDDa0pKEMkKTtxJWjuojuFCuZeJeTvFNeOQIH5B4bx7brJAW57Xi+GWvz4rvU2pKUXC8u9KAtClJcBj9SV9AG+5rCtiMHg95VpOT/Lu8fBpyfdbyIalaUVF5K/B5vwSOmOQc+k+Hew2vGMZxmwRsdhQU/wBnckymPbozbnUrr81TxC3nVH84te9qU5skkmsPkjxBxcP8M55ZvGRMOzZ8Nl+I/hTTd0U8l99KGnIjMhaA+OlSSrZ7DrPwqucs8BEXn/kZPLfiPuVqeu3srURNtsTCiw0y2VFCet8qBV7x2fLJOh73YauXEPCZ4fsKZZat3HFullhAQlVy6pY0PTSHCWx+ASBVf02vUhFUaCjbjJ5vvXrLwy6mCVWbbV7Pm7eVlfzdzmDw2eMBi753crtyU/yNBtogFNvcvFxZkpmvqcQOgWuG2osqCdqC1Oa7ED1qDcuWbmbmLlHIMwxmAtm2T5W7a9A47uFxk+ypQlLXnJkyG2PM6Up6ilPSTs+h7/0ntVgsVia8iyWWBb2/Toix0NJ/ckCs+svSNpufaqsou1vVjbLl7XyPPRW47kmrd1/i2cfc2T+XuVuMrZxxxtxnnGGogyI6HZMrG7ZMbfjNsqQGfImKUhr3i2oLT3T0aCvWtZ4a8a8SvCrt8TkHFErNHbghhuHIMWw2ByK02V9aVLjJHndZU2SFqJBb7eprtSlQ9ni91x9IlZ6pKNv9LJPR3dS3s+6P0P5u5H4c/GhkfIErLZ1sRLgXC6qnyIEm24wFKZU71GOZAQXT7nueZvr+PrVk+JfizxOc9Wm0WvF+LLdgSoEhx+VJjXS1z3pm0BKElTrQKAn3j2PfqHyFds0rPdxd4yeJneOns5d3q5GPocc03rrlH/acHcFeHbmTi7HLvaOReALDyXPuMoOx7lc7rbo7kRkNhIaSgNrH3+tZUOknqAP3Qao29+FPxb4VAkZXmViw921xCHJa2Q005pSgnSEIfUnZKgAAnsSNDtX9Y6rHxH9+J5aT9xd4sSHP5hu0QK/zSagxEK0YVa8qrlJrNyjB3ssr3gU8bgaccNNr8qbWSXDoiBY5YIeI4xasXt6QI9riNRUEDXV0JAKj9SQSfqTXyY7rfetpK3utJM3s1XaUI7sdERuKhFRjoiGTeRccZupsi3Lj7aCr80LTLOwlQSVAhvRQFEDqB6e47969rbkMS8xTNhKWWg+9HPUNHradU2vt8upBrV3KNKPJMSaI7pjpsclpT3QegLMhkhPV6bIBOvoar63WKfaXLTeIluuAmu5DcxKHU4NxVLlqSCknpSgkNKB0Bsg+qtnWzrVIzs1df8d/MpTm0y2Lnf4ljtMy9T1LEaCwuQ8UjqIQkbOh8ToVIIj+9Hdc4sRb5OtWQCLZprCJ+LyQ5ERClJ1L2nTalOk+c6ApQ6kgb7+vw6Fg76RUmGrus3ll/wAmdKbZI4rmwKxFzDjnJ+CZY0elMmc7jk3XbrjzGyW9/PUhmPr+cfnXrD323Wrz7YtticR/Gt5Xjxa169X2rGH/ADE1bqu1Ny5Z+WZNWdqTnys/J3OnaViyrnbYICptwjRwo6BddSjZ/aayQd9xW5OgszQ5/laMGwa/5kuKZX2LbZE5McK6S+ttsqS2D8CpQCd/Dda3jzj1rD2JF4vE0XjLb0lDl8vS0aXJcTshppJJ8qM2VKDTIOkgknqWpa1brL8at+aYnesOuxcEG+26TbZJbV0rDTzam19J+B0o6NaPijLLhlGKJYyJKGslsLyrPf2E+iJ7KU9S0j18t1Km3299y28gnuTQ8JnSlKAUpSgFKUoBSlKAVXPFaRf8izjkZ0df2peV2OAs+qIFsKo3R+Htn2g4D8Q6Kntxcls2+U9AaS7JQytTLavRbgSekH8TqoZwM1Db4TwRcF9T7cjHYEpT6h7z7jrCHHHVfylrWpR+qjQE8pSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKrnlt1N5uWFcex/fk3nIYl0fSPVqFbHUTXHiP1POaisk/rSUD41Y1Vhf2l4RzNbs5nkybTl8SPi7jzvc2mWhbrkboPolmSpxTa/iXkxR36vdAs+lKUApSlAVx4jbA5k3Bma2tlJU8LS7LaA9S4xp5Gvr1NipZhGQN5ZhlhyhpQUm722NOBH98aSv/rrbSozE2M7Dkthxl9CmnEH0UlQ0R+41U3hYkvNcSs4nNcK5mH3S4Y7JJ9QY8hYQP/NqboC3qwrNZLNjtuas+P2mHbIDBWWosNhLLKCpRWopQkADalKUdDuST8azaUBDuT+KcR5ZsSbNk8VxLsZfnwLhGX5cuA+PuusuDukggdvQ6GxVZ4/yvmHC94jcf+ISSJFskOeRZM3QjpjSh+i1M/uL2v0j7p7knsVm/aovxn5mvFeCrnZrbbItyv8AmkqNilhhyWw4l2fMX0IV0kEdSEhbie33kJoCIcao/wBEh4krxzVMHn4Pxat/GsNQoBTUy6HQnXBPzCdJaQe4I0RopNXnnHKeK4DcbLZbs5Kk3S/y0RYUCEz50hYKtKdKAdhtPqVfTsCe1c/R7dmvgttmPQrZfjmeIXNxqE/YnelFxanrT1OuwelIC0rc6lFsgaKgPVRVWuzbxJ8U2PI4N+4DxxzkjmfklSYVpsbr6gu2tNJAdclFXeFGbAKla11nej0gqTYpQhBdpXT3Wna3F9/JcdeRFOUperTavx6Ikviiz7gzivMLPkmexrxnOcT2UR8SwCCsy3ZcjqIDzcUAhAJGlOr933DoKUNV98QD/GHJvFuE2HxCcfXd7I5rkS8t8d2u6refenBpQMZ8xlBLzCFqO1HSSUAgEgpMj5NzWOM8g2DjXA7FeOZHbYITt08hDqMfhOHrX50soC/L6j1Jb7dR0SnZCVTHirhSzcduycjulwfyPMLoOq53+cOp90n1Q2P7U0PQJT8AN70Nayvja+LaoUXfdy3nmo9EuL9y48iLWTjRWfF/er+HuK3wvw43TLLTCj8oQoGMYnEIXbuPMZSmJboqQdp9pU1rzl/EgaGxsHuRXQdnstox62sWew2yLb4MZPQzGjNJbbbT8glIAFZtKyoYWnQvJZyerebfj8llyRNTpRp5rXnxItnPJGM4FY7teLnK9qds8T26RAiLQuV5PUE9fllQ6UbUNrVpKRskgAmq/wCWL3cr9wze5F4uLVmutmXCvNxiWG8qXIiW9ElDp6nWwlQUqO273A6SQekqA3WnwHMONMCsOR4/ySbZEzGNOnovDMptKp18S+8tTLjKT78pDzam0pSneiPL0OnVag2SXw3gszFr2xbImFXGQ8zBtsWN51+vyZTZKbesdkIW2FqbU91LJQ2CS2ElRgqV3KLbeTTv07/vhxudtgdlwoV4whFupGcXFtf2iVm9zhbjxupJtxUXedcX8g5sh4WXObDeJFmc81VnyyTCEYS47aOrqmsA9UVet6WtKEua30oJCTlXPxJcbMvuRMZVdsvfaUUrGPQFymQofAyT0x9/TzKpye1eMrlxGeXVKFqabaTa7C0+py1RgkaCJCvWS8kBA63vdKlaQnts+/J05y04LJNpdXCLL0JDZjKLfQkyWgQnp1oaJGh8DqqjxtVQe5ouev7dL3fM38f4YwNTEReIV5SsrQdoJ31vZ736lDcimnuu1iyVeITIlnrj8HZP5Prt65Wxtev5okH/AJ6yIfiXxZsBWVYbl+ONH1kybamXHT9S5DW8Ej6q0PnquHPHzerfc1Ylx3m/Ms3jDCL1HuEu4XiNZpNzE2WwqOlmEtqOpKwkpdec7npJQNjYGq28EErFePObonHvC/iDm8o4tfLPPmXyM5jky0s2h5gteQ8lMhSupThWpBKdfDq37urMfSOz7VVPBpfKxo6y2QsasBLC2vZb0ZzTu/7zmvcf12xrK8ZzK2IvWJ3+Bd4LnZMiFIS6jfxBKSdEfEHuK2tcvy8QjpupyjELi/jGR9j9pW8BIf16JktfckI+iwT+qpJ71aPFnLz+TXBeD5zCj2nLorJeCGVH2W5sDsZEUq76Gx1tn3kEjewQoyUMZvyUKqs+D4P6Pp5NlXav8NPDU5YnBSc4RzkmrTiueWUo/qVusYq17PqEc3Y/NyfibKbTbGy5PNuckwkAfeks6eZH7XG0ipvXyrdSCqQcHxVjkKtNVqcqb0aa8zny13aHkVkt9/ty+uLcorUtlX8hxIUP6DWNMaJ32rEi2pXG2cXPi6Wny7dKW9eMYcPZLkNxfU9FSf1mHFkBPr5a2z8DW4kM77arVwbnH1tVk+/706Gmpyc4et7SyfetfqulmRmUxv4VrnIm1elSV+KdntWIqJ/JqOUCKVO5qWI3SewraxWSNdq/bcTv6VnsRta7V7GBlCFj3it6A7Vrr1ao+WZrg2CSIzcliZdVXWey4gLQYcNpTiutJ7FJeVGRo9vfrdJ8qO0uRIcS000krWtatJSkDZJJ9ABW04Csb9/n3TmO4sLbavDKbdjzbiSFJtTairz9HuDIc/Of4NDNSOPaSjSXHXuWv08SRxdScaK1bu+5O7+niVdZ/AvimE+Iy0chWa0MXjDJCpLz1rmq8w2qZ0FTTqeo/nmusEAK2UqUk+8B1JtPPeTudePsqmvt8IDMMK2gxpWPTwq5sp6E9fmRXAPNV19eg3oa1s7q5qVdjhYUk1S9W7vlb7sdlW2xWxlSM8alV3Y7vrX0ve901n148bmnfyyxW622y45DcI9j+1lNNR2bm+3HcL60FYY0VaLoCVe6kk+6rW9VFstx7IrDlrPJmAWz7TkyGW4GQWZL6GVXOKgksvMqcKW0yWStfT1qSlxtakKUClpSN5nnG+Ccn2X8ns/xeBe4AX5iGpTey0vRHW2oaU2rRI6kkHRPetRxhxDYuHbRcrPhlyvciHLd8+LCut0clMQtJ0GmOrZbb33P3js+p0BUt5qVrZe/yKW7h5Ud5SanfS2Vu+9/NeJ+BlvLtyUpdm4fhwGk70MiyZqK6r5aTCamJ7/VQ/6q+JyHnGOz7RcOLMTeCSetm2Zi888Rs66A/b2UE612UtI2fX41FMK5U5zhZXb8I5f4TW0q4OFlnI8YkiXa9hJUVOocIcjp0k91kknQA71cKbpbVXFdnTcIxntMpkLih1PnJaUSErKN7CSUqAOtEg/KkKkaiuvfkMRhamGluys7q901JW7039SM4zydZb/efyVudtumN5F5S302i8soaeeaQQFOMONqWzISnqT1FlxfR1J6ukkCphUQ5SxCXl2KrFjDDeR2d5F2sEh49KWbgz7zQUsAqS257zLuu5adcT+lXvgfImP5/blvW11UW5wulq62eSUpnWuRr3mZDW9pUCDpXdKxpSFKQpKjIViUUpSgFKUoBVccRH8nJ2VcWvHp/Jq5KmWxJ9TaZylvx9D4Ibd9qjJH6sUVY9VryUoYVl2Ocsp9yBHJx/IlegTAlOJ8mSr5+RJDWyeyGpElR9DQFlUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFV74g2lHhLNZ7I/rq0WaReYR+KJcNPtUdY+qXWW1D8KsKoB4gXixwTyKtABcVit1Q2n9ZxUVxKE/iVED9tAT+lfCpIIBUAVeg3619oBSlKAVTeB/wClDxE5/hh9yLlMGHlsFH6IcH9bS9fUrS0o/jVyVTPPX+k7KcA5ka9xmw3cWe7r9ALdP00pa/mG3fKUPqTQFzUpSgFc2Zu0eT/GxhGHOJLtn4ux2Tl0xJ/i1XGUv2eKkj9dCUqdT8O5rpOuYOLnrk9zv4n85t0B6ZcYarTaLc0hIK1qi2xS/LQN/pOLSfrsfXXsY7zSR43ZXN54i8rwfgxxzxLZlJl5HMgw2rFiONsAdUq6SFK6W4xAUS676FXSSlCVnStACE8zYja7jyXiOR8Q4zJxLnXJYDb05+KplPsVvUAp4XMpC0OJSvaQRvqUj1VpAPlOx7h7DuNOP77nUgXXOuG0JvUXGGbgnz27tc0q8gS4o2rq61FSFKAKS2op3og2RiFqVwfht45f5P8ANvGd5Q6wZyI/SpxT7qwiNbYwJ0EpUpKfXXYknpSNV9qYmar+jQndQ1lwUE3ay5yzt4vviwtGrjKsMNho/iVGsuN3wz4vTp4EZ4OzGycHOnjLmDHXcVyi6ylvu5JLfMiFkL6iT5ntZ+6rv9xetb9epRFdNpUlaQtCgpKhsEHYIqqLXnGFcwJl8X8mYT9mXSQyp02W7eW81MaHq9FeQelzp33KelxB7kDsaqzkZeaeEWxuXTEM1avOESeqOxjt6kn263qUNAwX9ErQgkHoUNAD4k9VMO6XZpUfZRfxWz62zKnYV4brtdaNNPimrpp802nzOq6VTnhh55hc5YE3LluNNZJaQiPeI6dDa9e6+kfqOaJ+hCh8ATbkSdCnoW5BmMSEtrLa1NOBYSseqTr0I+VTlezaueU5m1M/2YuLMVJgtrWJLqE7ZRraj1n7o0O9ce3zMsxz7kS05tBRb2k3+LPRjybmy44m22xnyil4NJUnbsnq617IIQW079whV8+JO4vJ46RicV1Tb2Y3ONYCpJ0oR3SVydf/AHZp8ftqiecOQce4jsVuy+Tj/t86O4q3Wtltzyko8xIKwTohKelofok9gBrZNaTaleMXaTtGNm+/9l8VyPqH8CbLq1YxlRhv1q7lCCyyilna+VpSdm8rKEk8pNGLjOe5Nkj2PDJ4FnEHJYUiQ2xGS4XIz0coOytR0sK2VDSUlBA7q9az79HYZtq7HfXXHrGtTRS+XdOR/LUFpStR7lO20e8SVFStH4VRtx8WvDmD8XwOQMhx2Vb5dpku2m22iI557y3FoStflKUUjo6dFSl66da7kp6szgXxj8d+IWfMx+zwLhZr3EZMn2Gf0Hz2QQCttaSQrRI2CAe+xsbIo01KtR7eGcXx++p1eNnR2ZtN7MrpU6sXnC+ad21Zq6vu2s7565lpcw4vgGd4DdLbyRi7F/s8RhyeqKvYWFNIUoKbWkhSF62ApJB7kehNUD4PcgwLFsrm4Xh3F9qx5F3bW8JcR19+QoNgqCHnXlrUpOt60UpB/R71YGd5DPjSxxni1xQx9oNBUyQRs2mIohCiNADahoIB2oqWSSO1SjHMS444rhSb9brZFtqY8bUmc4oqX5aQN7Uon10CQnWzrt6VQniK9SqlTnaEfaXM6jD7I2Tgtn1J4zDdpiay/Ckvaiub728lm3bgmm7hjyd671i5PYHMigMuW6cq3Xq2PCbaLige/ElJHuq+qDspWn0UhSgfWqx4veyTLLxK5Lvj0uFBms+y2W0lwpS3E6t+e6n0LiyAR8k/PdW/Ec3qtjRqLEQu1k/u5x20MJLZWJ3IyTnHW2aT4x5O2j4PNZrW2OKM+TyPhcXIH4ghXJpbkG6wgrfsk5k9Dzf83qHUk/FCkn41MKoTiKace5lvePJV0w8ttCL02j0SJkRaGHyPqpp2Nv8Awe6vut7hKrq0k5arJ+H11PlH8Q4CngMfKNFWpySlFclLh13XeN+NrkR5N43tXJePptUyS7Anw3hMtdzjgefAlJB6XUb7EdyFIPZSSpJ9apSPfrvj93awrk+E1aL8slESUjYgXgD9OK4e3WR3Uyr30/JQ0o9M1DuYbdbblxblTd0tkOc0xZ5klDUuOh5sONsrUhXSoEbCgCD8NVlPCOtUTpO0nl0ff9fjocpisLdutTdpWz5O3P5P4rIrVyN9N14GKP1TVe8DSo3KlkMbA+Q5OPZDb2wZeP3lo3SEpI7ebHUtaJIbPYdJeV0dhrWiqyXMF5+irKE2fArkn4OpvEuIT9Sgxndf5RqLF4XEYGq6GJptSXj5Wzt1sa+nVdSCnuOz4rNe7PzSPJMX5Jr5Ol26ywXrpd50eFDjJ63n33A222kfEqPYVFeU/wCGnjXDHs3yI4lbbVEkx2pqramRc5LDLriWy4hLns6CQVJ7Hsd1OeIePuIs5gwuQmsuf5GkNqC2ZNzWkswXvXSISUpbYcHbupBcHY9XxqNUa8478IO2l3kvr7l3mce1qS3KcLP9WXu193ijS4xhl25tfak3WBKtnHjag4pEhCmpOQ6O0p6DpTUT0JKtKdHYAIJKrizTkPEON4MYXuSpD0j81AtsNkvSpRSPuMso7kAa2eyUjWyBW8vt3i49Y7hfpvV7NbYrst7p9ehtBUrX10DXPeKwJ09lXIGUafyXImUSJLhPUIjCvebhs/qttggaH3ldSjsmt5snZcJKVSq7rK/NvkuSXj5u5JPewb7OlnUlm5PRJdPHJX5ttu99zdeX+Wb5s4/jtmxSIr7rl2UqfMI+ZZZUhps/TzV1FJ165PmKKrlzHf0/3u3w4UVA/D8ypf71GtxcXCAe9ULZORrq6uJOm5C3co6mpr11ZTHQBbkNdRQrrQBrfSE9Ktk72PQ12uBwNKabp04q3S/vd37zXV96/wCJOTf95r3RsvcWq3LzRKtt8u5olXzMthX9CmSP6K3dty3mO3EKgcnouYT6NXuzsOpV9CqN5Cv296o61ch5BAsV5fvzbL10iNMzI7QAQny5AAbQrX6jnWgn5J38amWMXWdDyVmzXbNXJk11tSlwl20NNKISFEsuBI7DfoVLOvkat4jARUXvQi/8KfyIadr+rKS7pSXzLutPPt0s/Sjk7DlQYo+9ebM4qZDQP1nWikPMj5kJWkfFQFZ/IHA3FHNrkPOkPyYV88lCrblOO3Ax5iGwD0FDyCUrTonWwrse2qjEBZOq00LL7nwhlUZjH8Wul+x7K1vFVmtiQt6HPbQXVux2z2KXG0uKW2Ne831DupW+VxmyKOLTjSjaWtuDtrro1rrblbjusHtTFbMqKr2jtpf8yvlnbVPRq3fdaWtk93zLijjW3OWXG73yVcrYmPGm9D7Tc2S0EnzZOj2WvtsNpGyVAfM1H8QyPgnxOsSLonFkSrzjqhFlNXa1qiXazuL6iEBzQcaUelXdpfwPfdZ9g8THEF6li1z8jXjty7BUG/RlwHUE+gJcAR+5RqxWvYLnDdl2mY1qa3oTIhQsntoKCtFKiPhvYrQVsJiMNNdomlya+D/5N3DF4bEUmrJzvfeT8045rutu266EGc40y7GgX+M+S7tHCO4teSuuXqC4fjt11ftjZPoCJBQn18tWtGA8m868tcaiLf5/FlyfYhvsRLzbIscSYSmHCsquMe7daEMtN9PStEllKtlBKWkEvVuuPbP4jePcziYllF7g8gYRM83y7/IUmLdbZ0oKkIfQB0yASAkKT72ySopGhVxR7lb5cmVCiz470iEtLclpt1KlsKUkKSlaQdpJSQQD6gg1DTqb6u1bvM8Th+wlaMlNWvePLro0+jSfg0fLXdLZe7dHu9luMWfBlth2PKivJdaeQfRSFpJSoH5g6rKqAyeKIlovDuT8a3RWKXGS4p6ZFYa8y13FavVUiJtKesnuXWi26SB1LUn3TsrTnLrdzYxvNbT9g3eSSiKfO86FPUASRGf0nqXoE+UtKHNJUQlSR11IViWVi3O2W+9W2XZ7vCZmQZzC40mO8gLbeaWkpWhST2KSCQQfgayqUBVVhyC4cQTYuCZ/PekY2+6mLjmTSVlQSFHTcCe4fuvDshp9R08OlKj538ZatY1ytluvNvk2i8QI86DMaUxJjSWkuNPNqGlIWhQIUkgkEEaNVy3ifIHGA3xzI/KfG0f/AKtXWYUyoaP1YM1e9pH6LEjae4CXmkJCaAs+lR/C83sudWx6faRJYfhyFQ7hAmNeVLgSkgFTD7f6KgFJUCCUqSpK0KUhSVGQUApSlAKV5SZMeHHdly322GGUFx11xQShCQNlSiewAHxNfwr8e3jK5QzHxCZzjvF/Ot/PHbL7EKDHst1WzCeS3GbS/wBJZIDqFPB07JUFb2NjVAf1R8QPj18NXhzTJt+XZy1dshYBH2BYumZNCx+i4AQ2wf8ACrQfkDUG8D3jD5P8YuY5rk68GtGMcbY60zBt7RcckXGRcHFde1vbDZSlpJKkBsaLrelK7mv4MqUpaipSiSTsknuTX9z/AOpB2q02/wAHUKZb+gyLnkNzkz9evnJUhpIP/immv30B2xSlKAUpSgFKUoBSlKAUpSgFVxy05+UdzxXi2Kep2/XRq6XAD+1Wu3utyHlKHxS48IsYj4iST+ialWaZpYMCsTuQZFKU2ylSWWGWkFyRLkLOm47DY9511atJShOySa0PGuLXxiTcuQc5ZQ3lOSJaDsVDgcRaYLfUY8BCh2V0da1uLHZbrjhHuBCUgbvMsGx/OoLEO9tSEPQnxKgTYchceXBfCSkOsvIIUhXSpST30pKlJUFJUUmLqt3OeJNeZbshsedwo/pEuUP7MubyPn7Wyoxlua9E+zNJUfVSAeoWRSgIxhPIWP503KZt4lQbpbVJbudnuDXkzre4oHSXWtnsdHpcSVNrA2hak96k9RHOOObbmDsW9Q50ix5Na0qFsvsEJEiMCQVNLB915hRA62XAUK0D2UlK04WFcg3CVeFYByDBj2nL47SnkJZKvY7vHToGVCUruUjY8xkkuMqUArqSpt1wCd1oM9w62cg4ZecKvCf60vENyKtQGy2SPdWPqlXSofVIrf0oCrPDnnN1y3AfsXKlkZTiEpzH72hR99T7B6Uunfr5iOlXV6ElWvSrTqgeVGX+EOT4fPVpYcONXwNWnNo7SSQ2neo9w6R8UEhKj+qQANqJq+IcyLcIjM+DJbkRpLaXWXWlBSHEKG0qSR2IIIINAe1cn8f5Xc8A5G8Vt4hsxpsmx3O23hqMpWgoLtoX7xGjrpR+9J1uusK5S5chQeK/E/EynJmk/kDzjZk4Tf3yotpi3RpKxEcWv4BxpRaSfhpR2NVJScY1Iuaurq/dxMZpuLUdSAZxwZzVnHI/JHKeM4q2uJndrwy72aY3dI7JbnwY7iXEJSpXWQA926ukEkHZKdVYjeSZtl54wxLkywzrZfrCq4XS7NSm9JlOxGm47D6VD3VhRmde0nQWhXyry8VmO5fyTxI94dPDbyLjuN3+xyrXHnuXa6vRn48ZsoVGaZWGHVLWpwMfnErQpKkpHUSrVarB184Rcxtdi8QqsekZjYMNZjvz7JLceYnodlrAkLStpvy3V+z++lIKdp6hoHoTqNp4PsqzxCbW9ZWejV95Pvt10Zv/AOEKCntJ4hSu40535XcXFeKcif5jj8DLLeiLJfeiSorqZMCfGV0SIMlP3HmlfBQ/cQSCCCRU04f5Nk5ciXhObIjs5dZW0mWhCelm4xidImMJP6Cj2Unv0L2k9ikmGvytE96i2TxLg+/ByTGZqIOSWJ0yLZKVvoJI0th0DuplxPurH4KHdIqqqzoT7SPiua+q4eR3M9m09p4b0Ku7cYS/kl/tlpJcPaSbVn7PQYvI1+yG8zG7fi1ntlxnWaVAs0dMOfLQw6Uq9umJAd8tYSlwNNlKelSSpS91F7JzTwjw5n1qveH5ZjkfHLy2m1X232txCkNdIJjzQ21v3kHbaz6lDgPfoFbyfmVpvD0Pmyyw1QrbkD6Mcza1ukKVars3pth5z4aOwypXYKSuOr0Br3n45hpiSrf+TNqQxMaWw+lqI2jrQoEKBKQD3BNVak6kmp05LeWd3d35WzyTWTXejeYPDYSjSeFxlKaoyW66cHGKi1lNSvGW9OMrSjN3bW5K9mjWcx+JPjrMsiwZeCM5FlDdsuEyStu3WWQPNcMN1tHQXUoSsgOLJ0ew2aiF+wnMOcX455KswxvFIbhkR7Oh9Lk6U90qSlx5xPutABR0hPfud1KJGZzH8Ix62ZBLU9eOMMmhMyZLh96XZpjbsNiUfnrz0IcP67KifUVM7/drRZIqpt6ukSBHT952S+lpA/aogVHOLxTlKtK6dm1ay0tnm3k01rbIvYWrHYMKVDZlFxnDejGblvzXrOScbRhFOUZxknubyUlazVzkfxDcE5RhtoxfMPD5hNhv1xxJc9L9lvsBm5NzWpaGkre8qRtC3UeQjQ9dE9PcAGo+O7nzxdOQLHmPI/E+I4pcrRHnRsftVlxuLaJNxkSUJbW5IQwAosNJSVdTmu/ZPqop64u3K8rLHnLTxBZl31/Zbcu76FNWyKfiS4QC6R+qje/nX6xXjlrGXZN6u09y8ZDcAPbbm+nSlD+5tp9G2x8Ej6b+GvPTJ9j6Nhkt3S/BLpwb9y9x7/Rugtprbm2pSda6l2bfrSktHP8ANGPO7UpaJWbkqfRyHifE12kY9fjNu19nLS/fLi2hJHmrTsJGyCUpSRpIHYH5kipDYrbceRLjb05JFlQMRZUZFttshASqcUkFvztdghI7pbOyoJJOwKqnxSZp4feN87jP5s/kEu+zWm35VrsyWlbaHupW6pwpDfUE60FbIG9Deze3E/ImC854E1k3HUxZYjueSlp5vynoUhtIIbcSN9Pukd0kgpV2PetdTwOJV3VX4d8uvf8AebO0xv8AFOxJuFPAVL4tx/Eb/I7W9RaK2it7MbWs81bUD3UpAGgANAVIIROhUax6Qm4Qm5AGl/ddQdAoWOygQCdHfw3sVKYiCNVt6WeZ86x2UnF6n5tC1Nc28eqR951F3YV/gzFCz/nNo/oro2ud8CjKvviBgBA2ximNyZTyh3CX5rzbbST9fLjvmuiK2+z/AGJvnL5JfFHz3+MGliMPT4xpK/jOcl5xkn4isG+2eJkNkuFguBc9lucV2G/5atK8txBQrR+B0TWdStgm4u6OQaTVmVJg/EMvjDk2XKwmLCZwi8wkuSIK1nrgz2wEJWwNHYcR97Z9QST2ANt1+XEJcbU2relApOiQdH6juKqfEcCzziq9NRoPIUSfx435zj0S9pJl21GlKSlmRv30dWh+cPup3rZ71bnUeMvUrTSkktb+tbrz011531hhDsGoU43TfDhfpy+BL+VMFZ5L48v2CPSRHF4iKYQ8U9QacBCkL18dKSk/sqj+I7DYvDHylO4wvECI1Z8rCpWM5G8y2h5zp0XLfIeAG1JPvI367+oA3WaeMDGYM26WLjDFLrntyt6WwHrSA7B6l9veeb6ldj27IIJ7A1CMy4x5D5k4yXyBzper68yY4k2vDsVtPS5DkOfm21uId048tPXtSSRoBWla3UKqYiFONOnByjU0zSjfg975ZvobSls6jvyqY2qqW5qmm5vjZQyemjk4x/UemS+M52/5hM4cxniJGTzbi6u1JVFyBtceSlY0opcbaUOjpKiTsdIB3rR1urE/eMI9h4z5D8uLeIrCWbdMCiY12YQkBKmnCBt1I0FoICtjqA6VCtF4assx2y55bsIzzjiHiOaW/H2LVHlPFEYy0eYpxAajdIIcdSpSnFhRBU2EnRIFSXm7hfmTlGZkLV+z2KzhkCO5OtNthRh5776EKU2hzsDsKA94qP0AJJF/YuHxOArTobSqrdlmunJxsnd55p2uuTRU/iTFbNx8aVXYmHcHBWk223NvXeTdorLK2jvqj3uLRINVvMwOD+SycRMmQqGFDrUenqcR5vmKQe2tK7pPb0JqdcY8TX7KOL8ZyvEeTZzLs62suSId6jC4x/PCelwIX1NvIHWlXYuKA+A+FZkzizm2KSDY8RuqfguPd34yj/iOMKA/yzXT0cbQoTdNVVdPjlmu/LybOVmqsoqU6cs+XrL3XfmkVIvjDGnZa3WbazFZkRHYUliM0lpD7ayk7V0gHqSU+6R3GzW9x7BpEW7wbrccmuVyNsQ43EbkIZHSFgJUVKQgKWdADZP7z3qZN8dc0KUQOPLSj5FzIUhP+ayT/RW2gcPczzyEzLniGPNK9VM+0XN1I+gIYTv94/GrFbadLd9arG3en8Lsjp05X9WnJv8Autf6rL3nxuRFt0VybPlNRo7CSt155YQhCR6kqPYCohe3uc74YvK/C+C2u8RLAzIbs7N3fWw5PdeR0rmMtHp620o6kI6loKutah1Dp3cOP+HvE4kpm6ZncbhmM9hQca+1VJERlY9FNxWwloEfBSgtQ/Wq0QAAEgAD4CuXx+0KcqcqWHbvJW3tLJ62T5rK7txy4m5wNCpGtCtiILdi09x53a03rNZJ52Teds7ZPinw/cg898sX662vm3htrK4f2q1bZ/2g5Ehox5sp6t/Z7jfmr3sq8wklXSACOk7lvLNg8P3CWQRW/t3OON3bi0JH2rj5lKtqD1FIS/oOJBJT93Q7aNdRPuwIRVMkusMFSQlTrignYGyASfgNn95rA/KHFLgFwvty1SQ4ChbXtLa+oH1BTvvWjwlTGYOl2dGq/HNeK4+J0WPxGA2hiu3q4dRjbSDs9NU3e3gl55lL4re+ePsONk/HueYhy7jkjflOOpECWoA6KEutktdQI0evuD6iq2ziXiqs6Zz15vL+B+QHnG0TLquD7VaLqgEbbf6D5T/YdlkJ0dE70NdPZLhJuGCTsOwa8qwxchs+xzrRGaSYa1OeYpaG9dJ6ldXV6E9ajsE7qJ8Vp5tW5d8H50stgvMKIygxMiglKWrshRILbsQj3HEgbVoBHcAA+tTzxVHENU8XRT/VH1Wn4ZL/AC+JDh6dXCqeIwFfdtluT9a8XzurSXNX67tiQxeZuLZWT2rC4ueWiTeL1EVMt7LT4WJTaTo9C07QVdj7m+rQJ1oGpRebLachtj9nvdvZmwpICXWXk9SVaIIP0IIBBHcEAjRFUnnng44Sz1Sbnarc7js4K85mVZ3QhtLgO0qDZ2lOiAfzfQe3rWtv/L2e+GeFZrdy+wjK8ckOpt8TIrcHEzAoD3RKbc2grUPQ+YOrR9TupPRqdRt0aityl6r+O6/NN8itvbyjFQlv8besu9WzWWqadufKzvtO88ZnyskmSLtiaezd4dJclWtPwTMPq4yP/wBp+8kaL2wFvGdNOtvNoeZcS424kKStJ2FA+hB+IqLYLyjgvJEdxzE78zKeY7SYbgLUmOfQhxlYC09+2yNfImsB3Hrzx8tVwwSIudYyorlY4lQSWfiXIBUQEH5x1ENq9UFs9XmV6lOdKW5UTT5MxjOM1vRd0Tula+w3+0ZNa2rzY5qZMV7qSFBJSpC0kpWhaFAKQtKgUqQoBSVAggEEVsKwMiruWrZfsMXK5t49szt0vdogKTd7GwelWQW9rqWGkn4SmiVqZUQd9TjR0HepEI4b8f3hV5sjsJx/lK3WW6ugBVoyFabdLQs/oDzD5bp/wS110RX8qPGt/Uo86y7lOXyR4aolpct2SyFSbpY5MtET2CWo7cdZK/dLKztRRvaVEhIKSAkD+oM/LcVtVmdyO55LaolpYR5js56Y2iOhPzLhPSB3HxqCyuTM5zDcXh7AnH46+35RZMHbdbUj9Zlgp9qlfMaQ20sej1UB4Fv6nXiHhmtLWZ8jR7XkvJMhXme2JSXYtoT8G4vWB7/r1PdIV36U6G+rs+gKfPh2hZktM3njL7jyM4FhwWmSgQ8faUDsBNtaJQ8AfT2pchQ+ChUpyDg7hbK7amz5NxJht0hNthptiXY4zqG0gaASFI93Xw1rVTelAcich/1KzwbZ426uDgc/Epjuz7Tj9zda6Sfk095jI/AIFZHhr8PuTeBK037FrfdbvyBx1ebgLo0uJA3dLG+UBDqlx0EmU0tKGtlgeYlSOzSgoqT1nSgNTjOV41mdpbvuKX2FdoDqlID8R5LiQtJ0pCtfdWk9ik6IPYgGttUKyXiTFL/dnMntyp2N5I4lIVfLG/7LKc6RpIeGi1KSn4IkIdQPgmtX9r8x4N7t+scbPrSj/v6ypRCurafm5DdX5L+h3Utp1Cj6JY+FAWTSoziHJOFZ0uRGxy+Nuz4YBmW2Q2uLPh79PPivBLzW/h1oG/hupNQClKUApStPieXYznNiYyXEb1FutskqWhuTGX1IKkLKFp+ikqSpJB7ggg0BuKUqHcjZ6rEIca1WKE3dcsvilx7Hauvp890AdTzpGy3GaBC3XNe6nQHUtaEKAj2L2ay5LznmucPWqHKkY7GtmMwpjrKVuxnkNOypPkrI2gKRPjpX0kbLej93QtKo3x7hreCYpEx8zl3CWFOy7hPcQErmzXnFOyJCgOyet1a1BI7JBCR2SKklAKUpQCo/muD2HPbSm1Xxp5CmHUyoU2M4WpUCSkHofjuju24nZGx2IKkqCkqUkyClAVvjucX7FL1EwDlh1n22Y55FkyJpoNQ70dbDS0j3Y83QO2vuOaK2j99tqyK12Q47Y8sssvHcktce422c35ciM+jqQsb2PwIIBBHcEAgggGq7bvOW8Mj2PLnbhk+EN9o9+ShUi42hv4InoSCqQykdvakArA7vJ91b5Asy52233m3SbTdYbUuFMaWxIYdT1IdbUNKSofEEEiue8Wvt08L+TxuM81fek8bXeSW8Yv7yifspxZ37BKUfRAO+hZ7a+m/L6CtN3tV+tsa82O5xLjb5jYejS4jyXmXmz6KQtJKVA/MHVY2S4zYMxscvGsntUe42yc35ciM+naVj1H4EEAgjuCARo0Bs0qStIUlQII2CD2IqI8tcXYtzNx9eeOMxjFy3XhgtlxAHmR3R3bebJ9FoUAoH5jR2CRVStJ5M8L5UwzDuWd8Wtnqb8s+bd7A18U6/t7CR6fFIH6IHe6sJz3D+RrG1keFX+JdYDoHvsL2ptWt9DiT7yFfNKgD9KA4nj4zcOQMnRwRztm0HE+S8EglUHIjDURlVnbBW1LDqnUAqbS3taTtSVJWrfZYTZvsmS27lu8Qssv0e9XG3YrZYirgy0WhJT585SVqSSdKII6tHRPf46qX+Lrwwu+JfEbdbLTlUfHbzZXnH4U5cEPKPWjpU15iSlxtCh2V0kg9tpOhqp+ObJyHiWQzMQ5TXEfyOx4xY7e/LiPl1mY0y5ObZeSogK2ptKOrqAV1dWx8TBtXGVquGpYebvGMssllk+OvvOq/grD044zE1IrN0s/8A5KZOpcrp33rUSJ2j96vee4Ruo7NkKTs7rnKkz6lg8MpI1N0nwcPvM7JrhCVMxPI4v2TmtvQkq82GQUomoA7+axvewNlG9dwCNbeLLdLmF4Zds2vUWdjik9E+0zEtC7QXUBUWX1dKgpLjYBOvRYcHwrdIkJfCmXEhaFgpUlQ2CD6gioYtUjG32bIpK3HMajvTLKRtS5lhKuuZA+anIitSGh3PleYkVSm8rPT7v9V1uvzHS4Wm1NTj7aS5cFaLz4perL9G7JtKkiOZbxmI1vdv35XZndW4yQ3c467oS7It3WFPNI0BsjpDgSdpKmwNfGrB4j4v4pvllRc7rYIt4v8AbilqdJnyHZiXiUhbUlCXlKHlvNqQ6jt6L16g15XLO8etklu2WNt3IbtIQFswLaA6opI2FOKHutp7g7UfQ71UaxXiXML/AJAvDr1JbgSV25U6yY6zKLUa6w0SFuyLY5KR0q62/N6mwepKQs9wneoqdOEaqcIKX17+fv5LMtY3aeJxGBlTq15Ut3O6e7ePG6VrxWrbsrZuSjA3PK3MU3H0ew8TJt13bt3S1PDNvceZhlSglADyFhvZUpKQgAnfx+FTbB4OaLx5mZnkhlV1lfnVx2WghEZJHZvt3KvmSfXsPTZkmIjD5lpVYrFZWrWbU4hEqzPRgy/AfSQoB1r4K2AoK7hWgpKiNGt45E38K2UKMnLtJSunwWiOMrY+FKmsNGlutPOT9p8vrlqrZvV8DeJDGOVuNeVb/neK+GnBuXbRmJhvB++Yy9eJVrdYjoYLCEtOJUhtXQF76SCVa2CKtnwhYFnFtt2TcgZ5xxjvH8vL5ERTGMWGCYUWGzHZKA4WCtZbccKipSSd9gTreh017H3+7XoiJr4VflXnOkqTWRzFHZmHw+Olj4t7zu7cE3qR62xTbr69G0oMz0ee369KXU9lpHwGx0q0O5PWTUhnXC32G1yrzdpKI0OEyp991fohCRsmtVmD0Gx2f8p50lmK1Z3EyXHnD0gNb04nfQpXdJPYAEkAbA3X4VxpfeabWm+XrIpuDYdCdamQHHGUIlXFxtQUiQ6h4aaYBAKELHUo6UQAE7rxjK/Z0leXBffA29erR3Fi8ZPcpLKUuq4RVm3Jq2STtq8k2YV84P5Qyfge/ZnheR3vE+UL1K/KS3exzFxlpabb6I1tfCSAtKmB3SsFIeWSfSoZ4J/EjyjcrzFwjxOZzPjXjIIwfxRi6WViK1c2uwV5ctvXmPJUFJU0tIUDruVEpF5o8PWWXEedN8TXJL7D6UqSYc1mPsdiCFJQRoj5VgXXwW8a5KI/5a5jn2T+yOqfj/a1+LwZdJ2VoHQAhX1Gq3+ExVShhfRXQuuDbV0+Odnq834nzvaccDtHGTxlbF5yekYSaSWSit5xyikkuiRdl6yfG8bYMnIsgttrZA35k2U2wnXz2sgVWuQeK3g2yOCHCzJGQXBZ6WoNiYXPedPyT5YKP3qFU3kmPeGrhXmO34byZwuqFZMibaFpza9TnZ8GXcST1xX/ADVKSwoAApUvQV73oBs9P2S3YHjVulScVtNlgxYaVB8WqK2kI6U9RSUtD11o9Ot9x2rCVLHWTajFPTWX+0qKWxaP/uVH/hpr/wDR/Aqz+E3xGZ77nHXDMfFoTn3Lpmcotr6T8fZGfziTrv3JFIvhmfy99u68+ch3fOZCVhxNrQswbSyr4dMdojqI9OonuPUVJMT5x/LzIYdvxLjfK5FlfUfOv0yGIkRCekkKR5hCnAToaABG91l59xjlue3wJc5WvNjxgsJQ5a7Q0hh91zv1FUruoJI0OnXzqb/pKjUUMdPrnmu7djx6S8yP+kdSnBvZdONPheF97/PJuS67rS6G0vd3wLhPDfbG7Ki12iOtDLMO027ZW6s6QhLbSfvKPbZ0N+pry44zXLM6bn3C+8d3DFrZ7n2YZ76PapSD1dSnGU92SPd0CTvdSbH7FBxmxwcftpfMW3MIjMl95TrnQkaHUtRJJ1862NTqVGFN04xz58l0Sy7736czTydWrU7Scu/jd9W9fcUJk/hOx9dvVf8ACciu8LkCJLFyg5LcJi5L6307029vsWlA9JATvWt9WiDJ+GeZnM5cm4RnFsFgz+wAIutqWdJdSNASY5/TaVsHsTrqHcgpUbUqtuXeGIXI5g5JZLo5j2aWE+ZZr5GT77R7nynR/bGVbIKT8zrsVBUrxDxGWId+T5ft04cCNUlSzpLw5/uZX8LdotvJz/GGSQTZXnY7T9llyHQGbqCNOIbOgErQrQ6Sdnvr4bn9URima2Pkm4t8QeITDLfBzi0rRJYjyEBUW5BB2JMJw+oPTtSAd62DsBQTYvIOFZRlpgO4xyTdcTdheb1GGw08iR1dGvMQ4O/T0nX841lUpUnKMPYds27td6td592T6ZLyE5pN+1y4PufDImC1pbSVrUEpSNkk6AFfUqSoBSSCD3BFRPELBmeP4zOt+Z52rKZ6nHVsTjamoym2S2kJbLTZIcIUFK36nq18K5q48438S3FeM5BmsjNU2m0WmDMuTGPvtplmQGm1uJbDIUW4wVoD82skemu1Z0MFTrRqN1opxta9/Wvyyv7vIxqYicHFbjd9dMu/h7y5uTOeTj1/d4943x0ZVmDLaHZbSpHkW+0IWNoXNkAKKSoe8llCVOKHfSUnqqq7xaOUMv6n+R+ZL6UL7m14ss2SC2D6pDjRMtY+G1P6P6o9Kz+I7HGsvGtmkpme3zr5GRe7pcVd13CdKQHXpCj8epSu3ySEpHYCo14gb9dcZ4myi/WWe7CnQoKnGZDSQpbR2B1JBBBOifga3eDwdKi91K75v7yKVatOpx14Gok8KcPed7RcOP7Vdnvi9d0G4uH8VyStR/aa/KuIuGHx5TvEuFqT8jYYvb/MqpMnzHObZimWzLTkOVSbGzPsLFpuVwtwj3Ba35rbc1tlK2Wy4joUgJUpH3lrAJAGtPKzjkJ/Hb/j9ly27266NZVZ7bbX7xDaTPjRpS2E9TyAgIW2pwvdJ0SUggnY7dDCn6upr5RdzoC18P4FbVB3E2rvij4O0u47eZdt6T/MZcS2ofRSSPpUzteU89YCjrhZAzyZZUDTtsvSGoV2DfxDE1lKWXFa9EvNgn4uj1qneKOTLvm2fIhyVqiJYxpKrjazo+yXRuY4y+kn17FGh30U9KvjXQNvJOq1eMownlNJ9/11LNGU4PJ2NR4cccTCv5vnCWbvJ42kOyWr7hd/bc9vx24hPV5LIVtTR61AqQslJSepJWFJUL6vFtw7kWw3XF7u1br5a3i5b7jFK0uoCxrqbXo+6tOwfgpJ0ex1VC27/Sn4jcJu1kUGl5zGn2O+R0ekpuNFXKjSVD4raU2toK9emT0/KrCf4RcsXKrHJ/GeQDHDdH/9Ndq8jzIV4b0fzvlgjy5IOvzo9dkkHZCuLxmEjgpKFJXi87ck8vj7vI6mhjZbRk62Jnu1IrKVtWs1drO7X5s3e1+ZFc84z4P4wxDFomT3S+Wl2NMastpypuQ57fCW51FkPS0jQaTry0+ZtCR0jt61uVZZy1xCB/CBC/LfFW/W/wBqj9FwiN/rSYo7OJHxW36AEkb7VY99tuG8iWa94TeRb7xCcSYN1heYlZbK0BQQsA7bX0qSoehHuqHwNaTF0Yrw7YMU40vGcPyn5KlWyzuXiQgyphQFLDQIACihGkjtvQSNkkbypYydNKlVSlSWieq6J6r4X1TIa2HhiI9rByVdtt8VJNXvbpxvwzTWZixRb8nbHJ/Dd9t8l+cAZbPmEQrsEgAJe0Cpl9KQEh4JK0gBK0rSlKRJcYzO05Op+EhD8C7QgkzrVNSG5cXewCpIJCkEghLiCptWj0qVqqB5n4wzPiW+yOauCpzlvacV52Q2dtsuR3E+qnwyOyk+pWB7ye6kkd9TLBuSsE5qRb7HnNjZteVtMCXBT56m1PNrHeTbpaClfSoDv0KCxohXoTV2vs78FYrDPehx/mj0a+ay45Gup4r1+xrK0vc+76F10qIfkFdIfu2LknKYLKe6I7zsecjf8pclpx9Q+nmj9lSthLqGW0Puhx1KQFrCekKVrudd9b+W61hcPSlKUApSlAKUpQClKUBHMv47wvPEMHKLCxKkQyVQ5qFKYmQ1H1VHktFLzCv5Ta0n61GDYuXMEHm4vkLec2lv1tV/Wli5IR8mJ7aQhzQ9ESGypR+8+n1qyqUBXKOfOOoSfKzOZOwqWns7HyaGuAlCvilMhQ9ne/nMurSfgTXxzmq2X5XsfFVguGcyT29qg6YtTX8pye4A0ob9Us+c4PXy9VY1faAq+4cdcjcgwZELkjkFdntsxtTTtmw8qi7bUNKQ7cHAZDnr2WwmKfpUWh+HvMeK7jIyTgTkV5kux2mH8WyVtD1nlpZQG2iHGG0Px3ghKUeft33EISpCwhATfNKAofGuYua8luwwe9YLhOC5WtCymLdr9LlrcQke8/FaREaamtgdyGpIKQQHPLUdVZOD8dxcTlTshulzdv2UXjp+0b1KZQhxxCfuR2kJGmY6O/Q0nfcqUorWpa1bXLcOxvObOuxZTa25sUrS83tSkOsPJ7oeZcQQtp1J7pcQUqSe4INQyz5Fk3G16g4byJcXLvZrm+mHYsocQlLheV2bhXAJASl5XZLb6QEOnSCEOFAdAs2lKUApSlAKUpQClKUBU904Nfx+6y8p4Nyo4NdJrypU62GL7XYbm8o7U4/C6k+W4o/eejrZWonay5oCvWz8qZzZcis2Jcu8cN2V2+y1W6BfbRdW5tpkSg0t1LavMDUhhTiWl9IU0pPUAjzCop6rTqPZ9g9l5FxSbiV+89EeX5bjUiO55b8SQ0tLjEhlf6DrTqEOIV8FIB70BIap/MvDjZJt9XnXF1+l8f5aruudakAxZnffTJinTboJ7n0JPc9WqzeNuSMhi3pPEvMSIsHNWG1rt85nSIeTxG/WXFB+66kaL0b7zROx1NqSs2nQHLnIviM5p8P9tYY5XwzHrwuS+23Au1onhtuYlK0l1K4y/wA4hfldXvpBQFlPYg1rByFivJ/K9wzDD54kwbniVoWQezjDqJM5K2nE/orTsbH1BBIIJ60UhKvvJB/EVQHK8URud7c/0BIuWJOoT9THmJJ/d7SP31Q2ivw4vk1818zrv4Nnu42pD+anL3OMvhEjE9onfao9MjFRPappLi9W+1ah+Ds+laWpC59MwmKUUQe7y2setr11ehy5KWukeTEZLrq1EgAJSPiSR9KiV6xHkrkm3+1vOMYsmEFyLXHQormKf6FJSXXUkeUCFEEI76UQat9Nv7/drPiwta7VXeH7TJvI2a2s8PacEt7m8/C2mfErzgoY3CaVZLPaW7exc0OXGChSfzyFIX5cuE6s+8tyM/tIKu5acZV8TVkZph11vFqj3LGHkRcksMlF0skhXoiU3vSFf3txJU2sehSs1BLhid0tmepi470tvZG8btYus9LTWQR2j5jC1eiG5kRLjaj+u2lXqBVs2208t58oRLPjr2BW0AJlXO8ttvTirXvJjRkKUjt6ea6rp+IQsVYw1NuDpOLbWWS+0ua5JrianbWKhSxMcfTqxjFree++Gjus5TTzjJJNycZ5brRKI+MYF4g8Rs2fPQZdovS45bTPgPeRcLc+lRS9GU4B74Q6laVIWFIJST01GrhxbzbjyiLNdsbzGIn7on9dqm6+RW2l1lZ+vQ2PoKt3BMJs/H2OM41ZHJTzTbjsh2RLd8x+S+6suOvOK0NrUtSidADvoAAAVIK3focasVKplO2bXP4PxR8v/pHVwNedLBvfw6k9yNRXtG+ST9qPXdkrvU5tVB5rQry18Hylq9OpnIIBb/epaVf5tZcXCefL2oNt41iuLsnsp+4XJy4PJ+oZYQhJ/a6K6HpWK2fHjOT8vkkyxL+MK9vUw9KL52qP3SqSi/FNFVYp4frBbrhHyDOrzMzK8xVByOuehLcOI4PRTERH5tKh8Fr61j4KqrOaPCbmGU5DHyjHOQLxeI6JTbrtru0vznGW+oFXsy3D5Z0PRCwB27qNdUUrcbNxM9kz38LZN65Xv33z8dTltr4mttxqWOm5W04JdIpWSXRK3QiWA4fk2JtzU5HyHcspMktlkzYzLPswT1bCfLA31bG9/qioy5w3l8q9Kus7nfM/I9r9pRDilhhpKArYaOkHaddu/qPWrTpXscXVjJzVrv8ASvdll4FN0INKLvl1f1ILyrwrx9zVb4dn5GtT1zt0Na3BDEpxpp0q6fvhBBVrpGu/bvr1qB5qznXh3gWtfAnB1tyTDWQ89kFrt85TV369NhD0ZLm0vdKEKBQSVrPSBrWze1K8hiqkYqnJ3gvytu3kmZOlDe30s+fEiXHHJlh5LwSDyDboV0tECYHApi9RFQ5Edbbim1pcQv0IWlQ2CQddialtaXMcOxrkDGLjhmYWtFys11Z8iZEWtSEut7B0SghQ7gehFV7xR4X+MuFcicyDAJOTRULiKhJt0q/SZUFtsqSoFLTqlAKHSADvsCfmawtSabu0+Ctf33+RlmW5Sqaz3w3P53ls/K0eITmTGxPLZ+y7BkrcWBH6G0o/NNFhRT1dPUfeO1KUfjqpXlvGE3JeNInHUHkzMbK/EZiMjIYM9Kbs75ASCtx4o0pbnT+cPSArqV2G6OFPL1tdctBd8idUqtOKOH77xpPnzbvzTnObomspaQxkUpl1uOQd9bYbbQQo+h3vtUazPw3ZTl2RXS+RfE3ytYY9yeU6i3Wu4x2o8VJ/Qa2yVJSPh72/rXqp03Kznlzsxd20J7yfxPh/LFlRasniuIkRV+db7jFX5UuA8PRxlwd0kEA67g6GwdCqztXKWdcGXGPiPPrhuePPOCPa84jtENnfZDc9A35S/h1+h9STpShYfKfD9k5dxaDiWR5Hk0CLCkolKds10XCfklLS2+h1xHdSCHCSnttQSfhWu408OHEXFFpvllxXHX3Y2TeX9sC6T5Fw9t6AoJ6xIWsei1DsBvffehUtOpTVPdqNvpbTqnfLyaMJQblvRyZrrp4ufDXab/Dxd7mLH5V0ny24LEa3PKnKL7iwhKVGOlYR7xGyogD1JArYcz5/yjiAtlp4w4Qlcgy7wl9LqjdWIEOCEdA3IcdB7K6zpIHcIUKr1zhHJ/D7f5ud8A2WFeLPNc8664rLSkSNAAFUKSR1hWh/FrJHy6vdAtji/mPCeWYLzuNzHWLjCPRcLTNb8mdBc3opdaPcdwR1DY323sEVlOFOFqlJb0eN38UrNeefM8jNv1ZZM5lwi2czcAYk3A5kw6EnEvOecgS8aeeuSccjKWVIhy0FAcLLYPSh5AWlCQErIACjJcggYjy1hUqBGu7F0sd5Z8sybdKQtK0bB91xOx8BXVtVZlfhl4byq5P35OLuWC9SD1O3THZj1qkuq/WcVGUgPH/CBVbDD7XinetHPmvo/qVamDf/AKbKMz3CIGbWJ7Hro7KZYdeYkB2M4EOtuMvIebUkkEAhbaT6H0qLReFsbbLj0yTdJ81+4wbk/Olyut91yG4HI6SdBIbSoH3UpA95XxJNXZM8KuRRyRjXiGzJhr4NXWBbbgE/Tr9nbcP7VE/WsVHhg5RJ0/4inOn5s4lESv8AepxQ/orcQ21hVH2/c/oUngq97Je9EFxvi/GLLnV25Et0R1m73yM1GmkOHynA36L6PQLICQT8QkVJb9yDi2Gux4FymOSrvM7QbNb2lSrjMV8A1Hb2tX1VoJHqogd6lVv8J0F8hWacx8hX9J+/HZnMWphQ+X9YstO6/wDGVZmCcUcccZMvNYJhtss65PeTIZa3Jkn5vPq246fqtRNUMTtig/YvL3L6+4sUsFU/Nkc12rP7Pw9yrA5C8TkWbjFxv9rdhYowlr2uDZoZcSZDb7rRUTNcUlguFKS2hHlpSs++T1PimZYnnVpRfcNyO3Xq3rPSJEGSh5AVoEpJSTpQ2Ng9xUN584LxjnzD42LZGC2YVwjz48hOwpHQsB1Gx30torR+JSfVIr8cQYDeuFMeuuN3zN41yxC0nzLE7MaDUm3wgkqcbkvbCFpR+ivQISDs+gTy1bEYitiHKpbd4W4dNdPtnTrD7PWAi6Larp5p6Nc1lrnmm+DtyPHIeGZcfla28ucb3liw3OS63GymKtpSot6gj4rQkjUhH6Dn7Dsdq3nLvGlk5hwe4YdOlezykqS/b57J/PW2e3pbL6CO6VJJSexBKVEeiqmcWVGmxmpkKQ0/HfQlxp1pYUhxBGwpJHYgjuCKqg8aZbhvNyeQOPHI68ey8lvMLVIfKEtvobPlXCP2P5w9IbWkfe2kn4qTFOEYppRupPPx4/XzPaGIqVZqUqm7OmvVbX8uaTfde178I6FkY5EvETHbdb8nuLFyurMRpqdKaZ8puQ8EALWEbPSFHZ1v41z9knG+M4dlTHH2UxHE4NlM5T+MXFhZbfxu8LPUWGnR3bQ4odTfw6gUkEbNTfk7DsztnJ2Jcv8AHMZ+fKacbsGR2oPpSiXaXXN+ckLUEhxhZ6wexIJG9DRnXIeD2zkXDrliN2JQ3Oa028n78d5J6m3U/JSVhKh+Gq2WzsY8PUcJXUck38JLqvqtGazaGFjUpxqwak3d20s75xa5PK3DS2aygFhz/KeLr3FwTmaaiXAmrDFjy0I8tqUr4MSx6NP/ACV91f4gk3FVWcbXVjlXA7ngPJ1sjy71Y3DZcihvJ2l1xI9yQn49LielxKhrvvXpWkgXrIfD1PjY9mU+Rd+PJTqY9svrxKpFmUo6RHln9Jn0CXfh2B7a1cr4ZV5uEUlVWqWkuser1to/y8ilTq9nFSbvB8eK6P68OPMu+lfltxt5tLrS0rQsBSVJOwQfQg1+q1JdFKUoBSlKAUpSgFKUoBSlKAUpSgFarKMZs2ZY9PxfIYntNvuTKmH0BRSrR9FJUNKQtJAUlSSFJUAQQQDW1pQEB4yyW9JkT+Ns3l+fk+NIbPtikhP2vb1kiPPSBoBSulSHUgAJeQvQCFNlU+qD8nYleLmzBzPCw0nLsXLr9sDiuhucysJ9ogPK+DT4QgdXfocQy5pXl9JkOI5PbM1xa0ZfZi4YN6hMz44dT0uJQ6gLCVp/RUN6I+BBFAbelKUApSlAKUpQClKUBHM84/xXkmwqx3Lbb7THDqJEd5p1TMmHIR3bkR3kEOMvIPdLiCFD51Xacx5I4S/rPlJqZmWGtdmcvtsIruEFsen2pDZG1pA9ZUdPT8XGmgCs3PSgMCw3+x5TZ4mQ41eIV1tc9oPRZsJ9LzD7Z9FIWglKh9QaqPxBxfYco47ywjTSLlLsT6v1UzGOpG/oXozSfxUK2964IiQLvLy3h7JZHH1/muF+YmFHS/aLk6fVUy3qIbWo/F1pTLx+Luu1RDkq9chTsMn4hzLxtPZjlTUiNlmDtrvEeM+y4l1mQuDoTWylxCSUNtvp0FAuaO6r4qm6tFxjrqu9O6NxsHG09n7Rp1qztDOMnyjNOMn1sm3Y9X4u/hVGSc1yGPmEpld8uPkM5U3aDHetaRbm4ikt7KpXlDpd2s9ILhJUUjp71Y+N59frvaPtePjrWY2ttamVXrCpCLgz5ifvJdilQksOD9JsoWUnsTUOlq4xVcJ675mOV22Fcbki6y7NcrS7DYVISUEbLkVLvT1NIPT5mjr4jtWiru1k3Z9cvj+59Y2TFy3mo9rFrJwtPj+nes7cHuvnYkWH3Z+XbL/cr5LT5VrvFxY8xSQkNx2XCADoD0SPU9/nWr4lyq+X6fPt2SymXX5kZm+25CEoBZhPlQEdXT6qaKQFE9/zg3Wqfc4mmtXi3Ncx3Q2y+SJEiZbI6WFtqL6tupBEcuhJ2Rrr9DUlgIwh3IbfkWAYFlEqbAbfY6bHjL0diS26E7Q666020QFIQoe+NEeuid4Rkm1aSy66+RarUZwp1FOlJb2jcWlG2au5JJXeT4WN5yTAlpwi43i2EIuVhSm929f6smIoPI/YSjpPzCiPjVp3PLb/AJDmuLY9jl+ctMDJMVn3gPIjNOutuNvQfKUPMSofckOAjWve36gVDGOMOTOS2zbswiM4Zi8gdM2G3KTJus5o/eZUtvbUZCh2UUqcWRsAp3ureVhFr/K6z5cw44w5ZLVKtEeM2EhnyX1x1k61sFPsyANHWifpW0wtKo3KSTSdumjz66HBbcx2DjClRlKM5x7R5WklvRSim84y9ZXybS45tpVFa885Js/H94y6bmP21cnMmk4la4suBHajNv8A2uYDEhwspStWkgLUnqAPcDXbVuYlYswsrkgZLnS8iadQgtebbWYzjLg316LWgUH3dJKSoaPvK321K+IcckYPdsDmSpzsK7XSZd1PpcDb8eS/NVMStpQHuqbdUCgkH7g3vvvbYljF/sC5D1/z67ZI46hDbYmMRmUMpTvuEsNI2pW/eUd+g0B33bpQnFreu8lx48b5nP47FYavTqdjuxblLLcSbj6u7utRytZ39nXjwklKUqyaIUpSgFKUoBSlKAUpSgFKUoBSlKAUpWBfr/Y8Ws0zIslu8O1Wu3sqflzZjyWWWG0+qlrUQEgfMmgM+qy5P4Jx7P5zWWWW4ScWzOCNwshtnuPggdkPJGg838ClXw7AgEg+SvFL4b2ULM3nPCIC0AlTM+9x4jw76/i3VJX8Pl8vmKm2H5zhnINqVfMFyq05Bb0PLjLk22WiQ2l1OupsqQSAobGwe/cfOs4TlTe9F2ZjKKkrMqfHecsk4/vEbA/EbbmLPMfV5NuymMD9k3Q/DqVr+t3ddylWk+p90a3eSFocQlxtQUlQBSoHYI+YrX5Fjdhy2zycfya0RbnbpaOh6NJbC0KH4H0I9QR3B7jvVDybZnPhadNxx4XPLeKhtUu2LWX7hjyfi4wo93I47koPdIG9+qjNuwr+xlLlwfdy7vLkR3lT1zXvOiqVqcVyvHc2sMTJsVuzFytk5AcZkMq2CPkR6pUPQpIBB7EA1tqrtNOzJk75oUpSvAKx7hb4F2gyLXdIbMuHLaUy+w+gLbdbUNKSpJ7EEEgg1kUpqeptO6Kx4q4pncNzL3a7Zl638CcCZNotE1JW5Z1kqLzaH1K7x/QpSoEp2e/YlViW25269W+PdbRPjzYUttL0eTHdS406hQ2FJUkkKBHxFe7zLMllyPIaQ606koWhaQpKkkaIIPYgj4VWXF3ENw4jye+RMZyFA4/uSfbINgdbUpdqmqWS6I7m9JjqHveWQdKPbQ31QqLpWjBer8P2Ls6ixinWrz/Ey4e1wea/Nxu9c7u+uDxXL5CxTkbKuLMycut8tPfIMcyCQ2pwGI+6Q5Bed10hxpwnoBOy2ewASAMnhjkXJL/fc0455Bcj/lRiF1UOtlrykTLZIJchyEp/mbQdE6KO52alXJWQZJYeOb7lGB22Jd7tb7e5NgxXlKLUkoT1lIKDskpB6dHudDfeowOZeN4uJYny+q3H2fOnbbam7hHjIU4wZBPlIkubBS2hwqSfXpUT271HlSko72mefJ/R2LXrYunKoqSblaKtqpqzvb9SvdaN5rSxgZ3/ANzfmPHuSWvzdoysIxi+/BKHySqFIV/jdTZUewSoVa90tduvVvkWm7QmZcKW2pl9h5AUhxChopIPqKj3KmGN8hceX3EVaDs+IsRlnt5chPvsr38NOJQf2Vj8O5m5n3G1iyWVtM52N5E9ChpSJbRLbwI+HvoV2+RFbmo3Vw8Ky9qL3X8Yv4ruSOegtyrKm9JZ/X5PxZAbdcLl4cboxjuQy5E3jWe8GrXc3lFblgcUfdjSFepjk9kOH7v3T20au9C0OoS42sLQoBSVJOwQfQg1jXW1W2+W2TZ7xBZmQpjSmX2HkhSHEKGikg/Cqdxi6XHgbJInG+WTXZOFXV7ycXvD6uowXD6W+Qs/D+5rPqBr4e7lJLHxc1/arX9S5r9S489db3xX9Ve6/YenTp3cuWmhdtKUrXFsUpSgFKUoBSlKAUpSgFKUoBSlKAVr7DYbTjNqYsdjiCLBjdXlMhalBHUoqOiok+qj8a2FKAUpSgFKUoBSlKAUpSgFKUoBSlKAh2VcT4blVy/KJcORachSgNovlnkKhXAJH3Uqdb15yAe/lOhbZ+KTWoCubMK91xm38jWtHotCmrXekp+qTqHJWfmDFSPkasilAV0Od8Bt/uZp9rYW6ns4cktjsGMg/L2xQMRZ/mPKqYWTKsXyaIJ+OZJa7rGUNh6FMbfbI/nIJFbWojeuIOJcklmfkXF2I3SSo7L02yRn3Cf5y0E0BBuO7BkfJuMIzDIeVstYmyJs+K7FtUiNGhRXI8t2OtEfoZ6nGgpo9K3FrUpOlb76EgVYOZsQPnY5mMPN4Se6rfkjTcOZr5NzYjYb7D0S5GUVH1dT3NT+Bb4FqhMWy1wmIcOK2lpiPHbS220hI0EpSkAJAHYAdq1ec5ZCwPC77m1xjvPxbDbpFxeaZ0FrQ02VlIKiEgnp9VEAepIAJoDSY3y1j15uzWLX2HPxXJXgot2a9tpZekdI2oxnEqUzKAHclhxfSCOrpPapvVRX/DuXOTcfNvyiTxlFts5KXvs96yzLylo/eQpEkSou1pOilxDaCCAU6IBqx8Ss0/HcWs9gul9k3qZbYLESRcpI07McbQEqeX3PvKIKj3J2fU+tAbalKUApSlAKUpQClKUApSlAKUpQClKUAqvOeQE8eJmuJ2zbsgx64yN+nkR7xDedP7ENqP7KsOq65tX9p2Wx4Gx70rLr/AghHzisuiXMJ+Q9ljPpB/WWgepFAWLUIyji+NdL05mOJ32biuTuIQ29cYKUramoQNIRMjL23ISB2CiA6lOwhxGzW5y/OcUwSA3ccqvDUJEh0MRmghTsiW8RsNMMthTj7hAOkNpUo67Cou3lXMGU/ncW4/t+OW9f8XMymYr2pQ+ChBjBWkkfBx9pY9FIB7ADDunJHIPG9qmXblDCI9xs1sYcky7/AIzJSW2mG0lS3n4UlaXWQEgnpacknQ9asm2z2LrbotzjIeSzMZQ+2l9hbLgStIIC21gKQrR7pUAQexANUrnquWJd4wzA88s1luuOX/JIvt14sqXWEMpjIcloYkxXVLKW3XozLfWl1SVdRQpKeodV50BReY4fd+Dr3I5X4qtr8iwSHC9leLRvuOt/pTYiPRD6B3UkaC0j4Ebq4MZyax5jYYOT41cWp9suLQejvtHYUk/0gg7BB7ggg9xW0qhrrG/0OGdt5JbSWeNcumhm7xB/E2O5OkBuU3+ow4r3VjsEqIPyFWU/SFuv2lp16d/Ly5ETXZO60+BfNK+AggEHYPoRX2qxKKUpQCvCfBh3ODIttwjtyIstpbD7Tg2lxtQIUkj4ggkV70oeptO6Kp4T4+zTiqRfcBlTm7lgsNxt/FX3pBXMiMudRchOAj3kNEDoUTvpUB8NJxMowHiHIsXvHhYs7rdjel2lV4ZhRmV7iNrlFSZTZV7qumSOrpCux+QIqfciY3dsuwi9Y5YcjmWC5zoi24dyiOFDsZ7W0LBHfXUBsDuUkjt61UmD2LNOU5vHfLl8iO4vmWFP3CxZNGlRFpRcWSktvBo9gtsuoQ62obSCVDZIqpOKhako3XyvmvD4G6o1Z13LG1Km6075fzqLcZNcd5pptaN3eud626O9Et8aJJmLlvMMobckLSAp1SUgFZA7Ak99D51VvFI/JblLkjj1XuMPTmMnt6T26m5aNP8ASPgEvNkf41fnhOw5Nh2ccpYtcrdNFjfyFN/ss55J8p5E1lK32W1H4NuoUNfDrr95sPye8QPH+Rp9xrIIFxx2Uv0G0pTJYB/FSFgVtdnz7aFSk1a8X5xz+Ca8TSbRorD1YuMt5Jp36SS8tVdcGi2q1GV4rYs2x+bi+SwETLdPbLTzSv6FJPqlQOiCO4IBrb0qvGUoSUouzR40pKz0Kg46yi/YBk7fC/I9wXLWttS8Xvj3b7UjJ9WHT6e0NjQP6w0fXuq36i3I/Htn5Jxpyw3Rbkd5C0yYE5g6fgykd2321eoUk/XuNj41HeJ+QbxcpU3jjkNDcXNcfQPaOkabucXekTWPmlXbqA+6rsQN6F2tFYqDxFNWkvaX/wBl0fFcH0eVem3Rl2UtHo/k/lzRZdKUqgWRSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFflaEOIU24hK0LBSpKhsEH1BFfqlAVw5wnZrItUziy93DA5O+r2a1FK7W4fXS7e4CwAT95TSWnD/AHQUGZcpYifJzrj8X+In/wDTGJHzOw9Vu299Yeb+iWVySfpVj0oCHY5y/wAbZVcE2S15ZEavBGzaJ6VwbikfNUSQEPpH1KKmNarI8UxfMbcbRluOWu9wVHZjXGI3IaJ+fSsEbqH/AMCtrtPv4DmeW4eR6M2+5mVESP1UxJqX47Sfo0hB+ooCxaVXn2fz5Zvei5LhWTtI7JYm2yTaX1D+XIadkIJ+qY6R9Kfl9ybavdyLhC4ydfeexu9Q5zKR8/65VFdP4JbJ+lAWHSq8HPfG0QhOT3C54od6UvJbRLtTAV8hIkNpYWf5rihU5ttztt4ht3G0XCNOiPDqbfjOpdbWPmFJJB/ZQGVSlKAUpSgFKUoBSlUfyF4rMOsF4ODcaWyVyLmjvUhq02QlxllST0kvyEhSW0pV7q+kLKCR1hCT1AC2spyrHsLskjIsnujUCBG6QpxeyVLUQlDaEJBU44pRCUoSCpSiEpBJAqi8ezTNOSc7m5ViWOtPXaMw7Z7cbgSbbjEVa0qfXLW2f64uDqm2yqIyo+UlpttxxlSlqX8x3gjknk69x858ReTdKmepULG7Q6WmoaVDRQXUKPl7SelflKU4ra0mQtlZZHQFptFqsNsjWWx22Lb7fCaSzGixWUtMstpGglCEgBIHyAoCMYbxhaMYuDmTXWbJyLK5TZblX64hKpBQSCWWEgBEZjYGmmglJ0FK617WZlSlAfFJSoaUkEbB7j4ivtKUArW5Hj1oyyxT8av8JEu3XJhcaSysdloUNH8D8QR3BAIrZUr1Np3QavkU/wAG3y843c7vwRmM5cu6Yk229aJzp9+5WZZIZcPzW2R5Sz8wn17mrgqnfEFbJ+N/YfOmOxlu3LBnlLuLLQ96ZZ3dJlNH5lA06nfYFCj61bFsuUG822Jd7ZJRIhzmG5Md5B2lxtaQpKh9CCDU1ZbyVVcde/j56kdP1bwfD4GVSlKgJBSlKAVTviAzPIrG5YsGgdVugZ+ifj7V9jyVsyLZdVx1Kg9JTrQWtKhve9gVcVaHM8RxXMrQiDmFtRMhQpTNyQFLWgtPsLDjbgUghQKSnfY9xsHYJFR1YynBqLsy1gqtOjXjOqrpfTJ+Dzto7ZlO8f8AO2SuWDhFnIbKUDOWpNovEqWFofj3KNHV06SfXzXGXfX4Ea9d1J/Ed/WGNY1lvoMayy03Favk2XvJWPwIe1WlY5W4a5gvF2GQeyIs3G0m1ZTbb47cC1Ge8xpzy5SSOnpShXmNqSokE9iD6VJfEdGRd+BsvMZaXEptntja0nYIaUl0KH+RurGx5f1mneV05JeHsv5mW3aajTf4Tpys21wzbnHPpFxVsrWWVyzqVg2Oem6WWBc0naZcVp8fgtAV/wBdZ1YNbrsykndXFV9y3xzMy+JCyXEpSLdmWOLMqzTj2Cla9+M782nB7qh8N7+YNg0qSjWnQmqkNV92fR8TGpCNSLjIh/F/IsLkjHftERF2+6wXVQrvbHf42DMR2W2ofLfdJ+II+OwJhVQcoY7ecEyMc44FBckvsNJZye0sjvdICf7akfF9obKT6lIKd/A2bjmRWfLLFCySwTkS7dcWUvx3keikn/mI9CD3BBB7ip8TRjZV6PsS/wDF8U/k+K6pkdGpK7p1PaXvXP69fA2VKUqmTilKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAfFJSpJSpIII0QR2NQW5cG8TXGY5dG8HgWu4vHqcuNlK7XNWfrJiKbdP8AlVO6UBXf8FN+tn+xTmXNra2nuiLMejXVkn+UuYy5II/B4H6199i5/tfvN5DgeRIH3WnrXLtLhH8p1L8lJP1DQH0qw6UBXn5Wc0Qf9dOHrVMH/wC5cqS+f3So8Yf019/hMzVPZ3w956Neqm5thUn9n9kQo/uqwqUBXp5TyYa3wNyECRvXVaDr90+tHmPNGd49ZlXVvhq4WtKnW47P5QXWMlUh5w9LbTDNuVNeecUT2R0J332UgEi3qjOe4X+WluhIi3iRaLrZ5yLnargy2lwxpSULb2ptfuuIU2662tB1tLitFKulSQKRXxJzzzmevmfN38Uxd372OWVLbL8lH6j/AEreQkEbSpK3ZCFg7CGVDtdfH3GGBcWWf7CwLGYlpjK6fNU2Ct6QpI0FPOqJcdUB2BWokAAegrRJyjmXGx5WTcbwsoZR7vt2K3Btp5z+WqHNW2Gh/JTJeNfoc9cbw/dymdc8SUOy1ZLaZVsYSfkJL7aWF/ihxQ+tAWJStPj+Y4jljAlYrlVovLJ7hy3zmpCf3oURW4oBSlKAUpSgFK0WSZ3hGGtefl+Y2Oxtn9O5XBmMn97ihUIyPnmxybFOVxTFuOZXsMLNsbttnmSbfKkAbQ0uchv2ZpKj7vmKdATvq0QNECzpsONcIb8Cawh6PJbUy62sbStChpST9CCRVSeG2ZJs1ivvEN1eWufx7dXLY2pw++7bnPzsN0/i0rpH+DrdNct3WxpSzyXxlklhdAHVLtcZy+W9avj0OxEKeSkfrPsMiqwuHMfFWP8AiNtOXRs7tES1ZPj8i1Xl2XIERuPIjLDsd1/zenoKkqcbSVa36etT0vWjKn4+K/a5HPJqXh5nS1Kj+M8g4FmqnE4bm9gvxaT1OC2XJmUUD5ny1HQqQVASClKUAr4RsaPxr7SgOX8e8KDeLYnyNZ3rswq5ZRbLzj2PQ2pQEcW5116TEDiVICvPbceWCQopCAPrq2ZGP3lHh5dxbJkNi6ow5UCeG19aPaBCKHNK+I6t96pbmRfI0jxm8b220CRcrTAYYvLcZKW0iGwtb8Sc4FHRUOlyOojZ+gq0OGc1yLkjhW6TsufafvMWXerRMcaaDaFqYkPNo0kdh+bCB+IJqpgZQp4hRgmmn8LP5nQ7Yhiq+CjiK01NTSk+aclJW8o59eZLeF56rnxDhc5fdbtggdZ+agwgH+kGpnVc+HV3zuDcKXverQyn9w1/1VY1bTGR3cTUiuEn8TmcO70ovovgKUpVYlFUgd+HrOv1OOMvmfg3Yro4f3IjvH9iVfIet31rcjx6z5ZYp2N3+CiXb7iypiQysdlJP/MR6gjuCAR6VZw1dUm41FeEsmvmuq1Xlo2Q1qbmk45SWn3yZsfXvX2qh4qyG8YVkLnBudzVyJcJkv43dHj3utuT6IUfi+0NJUPUpAV9Tb1Y4ig8PPdbutU+a4P70eWplSqKrG+j4rkxSlKgJBSlYF1v1jsXs323eYNv9tfTFje1SENee8r7raOojqUdHSR37UBn0pSgFK0+UZjimE203fL8jt1mh9QQl6bJS0laz6IT1H3lH0CRsk9gDUGfvWf8rbgYlFueF4u52kX2fGLF1mN/FMKK4OqOD6efISFjv0NHaXUgTG0ZvZr9lF3xWzpkS3bEltNwltoBisSF9xFLm+7wRpakAHpStHUQVpBkFarGMXsOGWOLjeM21uDb4YIbaQSolSlFS1rUolS1qUVKUtRKlKUpSiSSa2tAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUBFL/wATcWZW/wC1ZRxrit4fPfzZ9njyF/5S0E1qP4AeKGv9bcaftI+AtF0mW8D8BHdQB+yrCpQFe/wF4YP4u+8goPw1yDfjr9hmEUPCGMHf+mrkIb//AI5u/wD/AJNWFSgK9/gNw9Xd3IOQVn/h/fU/+rLFP4BOMHf9c7Tdbyn4ovN/uFyQr8UyX3Af2irCpQEYxvi/jTDnfPxHjzGrI78XLdaWIyj+1CAak9KUAqnfFEk2vBLVyCz2fwnIrbekkeqmvPDLqN/ItvK3+FXFUE52sf5R8M5rZwjqW9Y5i2h83ENKWj/OSKmw8t2rFvS5hUV4M2mUca4DmyGjkuK2+Y8wSuNMDflyoyz+mxIRp1lf8pCkn61Hk4hyphqt4NmzOSW5P3bRlq1l1A+CWrk0lToA+JfakLP64qRcY3w5Nxvi2QlfUq5WaHKUf5S2UqP9JNSaopRcW0zJO6uV2OSs2tPuZZwrkjQR/GS7JKiXSKP5qQ43KV+yPX0c74AntJjZhEI9RKwu8s9P4lcUD9vpVh0rw9IAzz9wu48iNK5KsVtfcV0oZukoQHFn5JRI6FE/QCpxCnQbjGRMt8xiVHdG0OsuBaFD5hQ7Gv2+wxJaXHksodaWOlSFpCkqHyIPrUHmcDcKzpLk5zivFmpbp25JjWtlh5Z/lONpSo/tNARTxQ83wfD/AIXFzZFhZnXibNatcBx2P1Np6lBbqXFhQUkeUhwjW9qSNipZjd948m4tlTnHjUJqLap9yi3NMWEY6Rcm9mT1ApT1qKjsrGwonezVe8xeFbhPJsVYVcItvxq3Wa4MXe4THVueWuIzsvNLUXU+WlaCoFYIKfWtLY/DfxzhXGOfX2949jl6RMlXjIbRJbbLyWILjRcjoCljuUgb2NjvsE1BRVV4tRt6rt3/AGzbVfQ/+lKSbVVN35NZW8lfz6ln+HFryOC8KQfjaml9/wCUSf8ArqyK4H5v5H4V8L/BPHM9zgqJmuf5laGXYFtiMlp14ojtLkSX3G0qX0pLiN6BKirXYAqGP4Mee8B8SGQzePs44YyrjPKWork6CIWR3eLDuLCCkOBspcaKXEdaSUe9tOyCNEVsMbLexNSX6n8TR4dWowXRfA/oBSq+PBeD/C652n1+7n19H/NMr4eFLCz79szPkKC5+v8AllcpX+bJedR/m1WJiwqVXn8F2WNdoXPvIDCR6IUzZnh+0uwFKP8AlV9/InlmL/rfza5I+X2pjsR79/kFj/qoDN5W45b5Dx9tmFNNtv8AanhPslzQPfhy0d0n6oV91SfQg+mwK8uJeRnM8ssiJe4QtuUWF72G+20nuxIH6afm04B1IV3BB1s6rH+xef0dk8kYA6PgV4XMSf6Lpo/0VW3JHHPN9kmv8w2XN7O9fYEMRpsbHccVDenQAoFY/rmTKbcebAKm9t+o0ersmr+HksRD0Wo7fyvk+T6P3PPS5Wqp0pdtHxXTn3r3rwOi6gt35r47ttwesduvLmRXlhXQ7a8ejOXOU0v4B1McKDG/1nihPzIqJ2HDOJ85s9iv2TZhe87YyXaYLd/ualxZSw2txTaoDIbh9SUtubBY2Ckj6VbVps9osFvZtNitcO3QY6elmNEYSy02n5JQkAAfgKpThKm92asyxGSkrxZAzN5szP3IFpt3HttX6v3FTdzu6k/EJYaUYrCviFqdkD5t1mWnhTAYipMzIbarLbrOYVGmXPJOmfIeaV95oBafLZaUQCWWUNt7G+jdTylYnpXaeCcIg9sXnZPjKB/Fx7NkU2NEaHybieYY6B9Etiv0eG48jaLpyVyFNaPq3+UTsX9nXFDS/wDOqwqUBEca4l46xK5fblnxaMq8dJQbtNWubcVJPqlUuQpb6h9CsipdSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQCtdkcX27HrpC1v2iG+1r+cgj/rqL8j81cc8S+Qc+u0+3NyAgoeas02WyOtwNoSpxhlaEKUtSUhKiCSoaB2KkeMZNZ8zsTGQWNUtUGX1hsy4L8R09KihW2n0IcT3SddSRsaI2CDXqdncEG8Mcr2zgHBXd76bQ01/kEo/wD6as+tZjeN2PEbJExzG7c1AtsFBRHjNb6W0kkkDez6kn9tbOsqslOcpLi2YwW7FJilKVgZClK1eT5Pj+GY/PyrKrtHtlptbCpEuXIV0oabHqT8/kAO5JAGyaAgPiitU29eHjkGBb1vJfNikvJDJIWoNp8xSe3chQQUkfEEj41zLd+V+SsW8GPH8ObIXPk5pCmwp1wkNA+TC2pLLKdAAFTCkgKOzpCvj3HVls5x44vwsbdvnXBZyK9uY5Eal2iVDdE5MJ2YW3GpDaHEDyGVqClJ0dp0TuphPx3H7rbPsS6WO3zLdoD2SRGQ4z29PcUCnt+FS4CVPC4+GMqx3ox1jz5PwLGIxMq2y5bOirNy3lLisrNePecCci8O88eKDhTj3l7hDKDh/JXHXt9lgOsSVwkXO3OIbbWEOjflrHlhPf3VHzASntqVeCfwz+LTHc7HK/i65TuN7lWeJIiY9Y37wbgYzr4Sl6StYJQk+WnoCUlW+ok66QD23Dhw7dFag2+IzGjMJCGmWWwhCEj0CUjsB9BWox/NLFk95yWw2l51cvE7i3a7mlbZSESFxGJSQkn7w8qS0dj4kj4VliqsK9edWEd1NtpcinRhKnTjCTu0tTe0pSoCQUpSgFYl2TdVW6QLG5ERP6D7OqUhSmev4dYSQrX4Ht9fSohYObOOMpy6Vg+P3afNusGZKt8jy7NN9makxyoPNmUWQx1JKFA/nPUaGzoVOq9Ts7h5n89MvkcrcRcz2TIc2x9Fks0fIftRpm2FxVrJdKEyVMFRPSVoSSUnRGz7oB1X9CkqStIUkgpI2CPQisK9WSz5FbX7NfrZFuEGSnoejyWg42sfUHtWREisQIjMKMgpZjtpabSVE6SkaA2e57D1NbXaW0o7RhTvDdlFNO2jXDrfXmUsJhHhZTtK6eefA9qVrMbyG15ZYoeR2VchUGe2HmDIiuxnCnf6TTqUuIPb0UkGtnWpLopSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUApSlAKUpQClKUBUniqtVzvXCs+32e3Sp0pd7x5xLEZlTrhSi8wlrUEpBOkoSpRPwCST2FUZfIuTSMzvSDBzwcvjkyKuySW2rl9moxkT2CCHEj2IQ/s4PB1JOy8V7BdKa7OrQ5bn+CYDHjzM7zWw44xLc8mO7drkzDQ85+qguqSFK+g70BzBjGEZbaL7inIjLeYnIJnM+SQJ6n5k1xpGPuSrsG2zHUotIi6TGcQegJ6ihST3FYfhdsfISs7sF2znL7nDy2Mxck5hancbvSFXB9ZP8fLfeXC6EOBK2FtISC37rekqIrqaVyTx1Cv8AbsUmZ9jjF7vDaXrdbXbowmVMbV91bLRV1uJOjopBBqR0ApSlAKg/Ntqw++cWZBaM+g3mVYJbCG5qbNGkSJrSS6jpeZRHSp0qbX0ubQklPQVaIBqcUoDiti359y9c8GxfL5WW5Vhdk5OSm15PKtUmx3G4W38m7iXTKDaGVBtMhYjl8IaQ919OiSSpGtt4swxnG+UYGcPcV2XLM3t7rTSLnJWlKJqRZRJLHVIdiBgy0tqV1N9QY2eyK7SWtDSFOOLShCAVKUo6AA9STWuRk+NOQLZdW8htioV6U0m2yUy2y1NU6nqbDK96cK0glPSTsdxugONbHFvUdOGNc927kl/j823JTYWWmrw9OadN33akzxD3KEkWwthrzveCvMB07W9sVoze0+J/IL/frNkznHM/NIaYzcZqQhbd3Vj1pbiTZutqkxAUPs730NSNKc6tBTXUFp5BwG/ZFPxCx5vYLjfrUCZ9riXJl6XEAIB81lKitvuQPeA7kVIKAUpSgFKUoDmXijCOR7ZY+WL45lt+iQJOVZk9CxxVqYbS75kl8svtveX7QrqJStJSvR7a7VXt9wDKMT4u4UbDt+bsV2gfaOeP3dm8XhTl3ctzHkKmsxn0SEt9YfT07DSHAyCkaTrtyvytaG0KccWEoSCpSlHQAHqSaA4UyrGeRG2sMseWcgXq4YmnD5bVtvcrFsiWRc1TnSkqYjPolNSW4pipYcklalBtxST1KVufWWLDneI3C4OSy819vxSwQFTb0bVeI8fKbw9DDaStASY0dlpvqdcS4UkvPNpJ2ysHoCBzFxHdLVc77bOU8QmW2yKSm5zGL5FcYglSulIecSspaJPYdRHftW7xvKsXzK2JvWIZJa75blqKEy7bMbkslQ9QFtkp2PiN0Bxrh/HGXZ1jthh56xnBTA4gdeS17dPif2ZEt7y1L8tSSuSlOikK2obB1sJIhuWX7IcqvcNvlGbe4UO24xg0q95C8q9f6XHAhMq4pU1BYWw2uQk9DplOsKQlQUrqbCK/ohUDyXgvijMMgkZPkeIMzJ03yPbgZDyI8/ydBr2qOhYak9IAA81CtAAegAoCdp6QkBPprt+FfaUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFc13vJeN8J8VmW3fnS8WGzRp+H2ljEZ+QvtMQ1R0PSzcmGHXiGw75io6nEA9SkeUdFKe3SleEqFDnIS3NiMyEoUFpS62FhKh6Eb9CPnQHIN7zDjvjTlJjK+IMwwTJoma3fF4Nx4/Ww0m6x9tRo8OZa0oIW2hqKpl8sraLYQ24tK2z1b7FrwMGEqWmeqGwZKUeWl4tjzAn9UK9dfSvegFKUoBSlKAwr1Hdl2efFYT1OvRnW0DetqKCAP31xtgGb4jlfEnhf4ux3IYM/MMeutgTerCy8lU+1/ZtueRNMpge+wGlp6CXAkdSkAbKk77WrwagwmZLsxmGw3If0HXUtgLXr06lep/bQHKvAeU47hnK9p4i49zTDuRsUuse93SDPtgbVesYBkJedYnraUpLjTjrpQlaw051oCVhZBUOsa8GIMKK68/GhsNOSFdTy0NhKnD81Eep/GvegFKUoBSlKAVTfi/i3OZ4d8rYt0SZKZKreq6sQ0KW89aEz46rkhKUe8rqhCSCkdyCR8auSlAcneIHMeDM24LvzPCmRYHdLrHXjLaXbMGJSIsZV7hBht7yCClvqGw0VJJAVrWiRLvDPb7jZeSOZLVmv2Wzmy71bptwas0UxLY/b1wkogy47KlKWkrDT6HStayXWFgHpCavpq3W+O2tliBHbbcX5i0oaSApe99RAHc7AO69g02HFPBtIcUAlS9dyBvQJ+Q2f3mgP3SlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKVx74us85x4v5Tx2ThvNl1tmO3uwZNfH7GzZbW4Gvsa2IlBpuQ/GccHnq6gpSirp6vdHbVAdhUrimV/VCMzs93et9+8P5aTBYtzs8RLzIkqY+0IhlRD5ghBjpDZbDvU6lQUpzy0uholWysHjozy+ZLYcCc4ZsFvyTLItnu1o9qy9abcm3z7dMnAyJPsfU2+lEFxAbS2sKUsaVpJJA7EpXEvhj8b2dZs/x3hvI2MRZCslYjQ3cpVLKRNuL0V2UEIajRjGaUEt9HlOPNOLG3EJUn1/WB+OjNpTMhqVxyi8Wuw363W6+Xibf22piG7pkE62RizHZhIac8sxQSCpBKAAVKVtZA7YpX8/eUfF7zvgGfcm48/cAmwL5CsuJ4fdGbews2+QHbY7NgvAtkKD8OXIW24vagWHQCD0amuCf1RF7ML9jEKXw7JtlpzK7QIlquL899pLcWTKcjlb/nRW0eclSEK6GVutnrUnzQpBBA7NpXFts/qg+TZG1PnY1wrbX7db5tvtq50nK/Ka9ouF3mWyGSRFV0tKcieYtz0ShRAC1dIVlP+P6823IWLJeuJoMaLa7s5ZcmurF8elw4klFx9i2wpmIpRbUSFockJjtqVtrq60nQHZFK4jyPx8X6HiOJ8jXHDoVssl6eZvkWJZcoiTrnJtBiXF1TUyG7G8xlZMNP8UenrPR54KFpV72nx/cgX5+LYLXwHDTfZkSZc2vbb/KiQVRGbcJwKXHoCXVLKAtBHlBBUG1JWpC+pIHatK4zT/VEFOYRceWEcYW1GI29nyPJfytlm9PXD7JauHQ3CWyA4zp5KCtDinAlK3fL6EnWJcP6oXlVmjld54ORCFqkyRfZjtzmpixYraYikvIQYHtSUqTL0XH47TSFN6K9OIVQHa9KUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBSlKAUpSgFKUoBXI3jj/2WYZ/wI5G/6ETSlAcx8/f7rbiX/wADz/8A0ZKqPeKz/aUX/wAGeJf/AHe50pQHh4XP92/wp/wJT/7pJqf8df7FeSP+FOD/AP52u1KUBYXMH+xfl3/w+47/AOxtVchZp/te+MH/AIc2n/pJ6lKAtrjH/cgeIP8A4FW7/pm+VXVp/jPBJ/O//vBpSgJPwB/toci/+GuL/wC43qvngl/2HWP/AO1uQf8AoNmlKA+4R/umh/8Ay7Rv/wAutVTjP+5x8O3/AA+vP/todKUB/9k=)
- BLA, basolateral complex of the amygdala
- CEA, central nucleus of the amygdala
- ITC, intercalated cells of the amygdala
- PL, prelimbic cortex
- IL, infralimbic cortex
- HPC, hippocampus
- Thal, thalamus
- PAG, periaqueductal gray
- PBN, parabrachial nucleus
Stress types
- Acute stress
- Short duration
- Brain detects threat
- Mobilizes physiological, behavioral responses
- HPA (Cortisol), SAM (NE/Epi) axes
- vs. Chronic or stress
- Long duration, persistent
![[[@Kim2015-dd]](http://dx.doi.org/10.1101/lm.037291.114)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQEBLAEsAAD/7R+iUGhvdG9zaG9wIDMuMAA4QklNBCUAAAAAABCloxzzoeUqjatJhLeoNI9yOEJJTQQ6AAAAAADlAAAAEAAAAAEAAAAAAAtwcmludE91dHB1dAAAAAUAAAAAUHN0U2Jvb2wBAAAAAEludGVlbnVtAAAAAEludGUAAAAAQ2xybQAAAA9wcmludFNpeHRlZW5CaXRib29sAAAAAAtwcmludGVyTmFtZVRFWFQAAAABAAAAAAAPcHJpbnRQcm9vZlNldHVwT2JqYwAAAAwAUAByAG8AbwBmACAAUwBlAHQAdQBwAAAAAAAKcHJvb2ZTZXR1cAAAAAEAAAAAQmx0bmVudW0AAAAMYnVpbHRpblByb29mAAAACXByb29mQ01ZSwA4QklNBDsAAAAAAi0AAAAQAAAAAQAAAAAAEnByaW50T3V0cHV0T3B0aW9ucwAAABcAAAAAQ3B0bmJvb2wAAAAAAENsYnJib29sAAAAAABSZ3NNYm9vbAAAAAAAQ3JuQ2Jvb2wAAAAAAENudENib29sAAAAAABMYmxzYm9vbAAAAAAATmd0dmJvb2wAAAAAAEVtbERib29sAAAAAABJbnRyYm9vbAAAAAAAQmNrZ09iamMAAAABAAAAAAAAUkdCQwAAAAMAAAAAUmQgIGRvdWJAb+AAAAAAAAAAAABHcm4gZG91YkBv4AAAAAAAAAAAAEJsICBkb3ViQG/gAAAAAAAAAAAAQnJkVFVudEYjUmx0AAAAAAAAAAAAAAAAQmxkIFVudEYjUmx0AAAAAAAAAAAAAAAAUnNsdFVudEYjUHhsQHLAAAAAAAAAAAAKdmVjdG9yRGF0YWJvb2wBAAAAAFBnUHNlbnVtAAAAAFBnUHMAAAAAUGdQQwAAAABMZWZ0VW50RiNSbHQAAAAAAAAAAAAAAABUb3AgVW50RiNSbHQAAAAAAAAAAAAAAABTY2wgVW50RiNQcmNAWQAAAAAAAAAAABBjcm9wV2hlblByaW50aW5nYm9vbAAAAAAOY3JvcFJlY3RCb3R0b21sb25nAAAAAAAAAAxjcm9wUmVjdExlZnRsb25nAAAAAAAAAA1jcm9wUmVjdFJpZ2h0bG9uZwAAAAAAAAALY3JvcFJlY3RUb3Bsb25nAAAAAAA4QklNA+0AAAAAABABLAAAAAEAAgEsAAAAAQACOEJJTQQmAAAAAAAOAAAAAAAAAAAAAD+AAAA4QklNBA0AAAAAAAQAAAAeOEJJTQQZAAAAAAAEAAAAHjhCSU0D8wAAAAAACQAAAAAAAAAAAQA4QklNJxAAAAAAAAoAAQAAAAAAAAABOEJJTQP1AAAAAABIAC9mZgABAGxmZgAGAAAAAAABAC9mZgABAKGZmgAGAAAAAAABADIAAAABAFoAAAAGAAAAAAABADUAAAABAC0AAAAGAAAAAAABOEJJTQP4AAAAAABwAAD/////////////////////////////A+gAAAAA/////////////////////////////wPoAAAAAP////////////////////////////8D6AAAAAD/////////////////////////////A+gAADhCSU0ECAAAAAAAEAAAAAEAAAJAAAACQAAAAAA4QklNBB4AAAAAAAQAAAAAOEJJTQQaAAAAAANRAAAABgAAAAAAAAAAAAAB5gAAArwAAAAOAEsAaQBtAEwATQAwADMANwAyADkAMQBfAEYAMQAAAAEAAAAAAAAAAAAAAAAAAAAAAAAAAQAAAAAAAAAAAAACvAAAAeYAAAAAAAAAAAAAAAAAAAAAAQAAAAAAAAAAAAAAAAAAAAAAAAAQAAAAAQAAAAAAAG51bGwAAAACAAAABmJvdW5kc09iamMAAAABAAAAAAAAUmN0MQAAAAQAAAAAVG9wIGxvbmcAAAAAAAAAAExlZnRsb25nAAAAAAAAAABCdG9tbG9uZwAAAeYAAAAAUmdodGxvbmcAAAK8AAAABnNsaWNlc1ZsTHMAAAABT2JqYwAAAAEAAAAAAAVzbGljZQAAABIAAAAHc2xpY2VJRGxvbmcAAAAAAAAAB2dyb3VwSURsb25nAAAAAAAAAAZvcmlnaW5lbnVtAAAADEVTbGljZU9yaWdpbgAAAA1hdXRvR2VuZXJhdGVkAAAAAFR5cGVlbnVtAAAACkVTbGljZVR5cGUAAAAASW1nIAAAAAZib3VuZHNPYmpjAAAAAQAAAAAAAFJjdDEAAAAEAAAAAFRvcCBsb25nAAAAAAAAAABMZWZ0bG9uZwAAAAAAAAAAQnRvbWxvbmcAAAHmAAAAAFJnaHRsb25nAAACvAAAAAN1cmxURVhUAAAAAQAAAAAAAG51bGxURVhUAAAAAQAAAAAAAE1zZ2VURVhUAAAAAQAAAAAABmFsdFRhZ1RFWFQAAAABAAAAAAAOY2VsbFRleHRJc0hUTUxib29sAQAAAAhjZWxsVGV4dFRFWFQAAAABAAAAAAAJaG9yekFsaWduZW51bQAAAA9FU2xpY2VIb3J6QWxpZ24AAAAHZGVmYXVsdAAAAAl2ZXJ0QWxpZ25lbnVtAAAAD0VTbGljZVZlcnRBbGlnbgAAAAdkZWZhdWx0AAAAC2JnQ29sb3JUeXBlZW51bQAAABFFU2xpY2VCR0NvbG9yVHlwZQAAAABOb25lAAAACXRvcE91dHNldGxvbmcAAAAAAAAACmxlZnRPdXRzZXRsb25nAAAAAAAAAAxib3R0b21PdXRzZXRsb25nAAAAAAAAAAtyaWdodE91dHNldGxvbmcAAAAAADhCSU0EKAAAAAAADAAAAAI/8AAAAAAAADhCSU0EFAAAAAAABAAAAAU4QklNBAwAAAAAFtgAAAABAAAAoAAAAG8AAAHgAADQIAAAFrwAGAAB/9j/7QAMQWRvYmVfQ00AAf/uAA5BZG9iZQBkgAAAAAH/2wCEAAwICAgJCAwJCQwRCwoLERUPDAwPFRgTExUTExgRDAwMDAwMEQwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwBDQsLDQ4NEA4OEBQODg4UFA4ODg4UEQwMDAwMEREMDAwMDAwRDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDAwMDP/AABEIAG8AoAMBIgACEQEDEQH/3QAEAAr/xAE/AAABBQEBAQEBAQAAAAAAAAADAAECBAUGBwgJCgsBAAEFAQEBAQEBAAAAAAAAAAEAAgMEBQYHCAkKCxAAAQQBAwIEAgUHBggFAwwzAQACEQMEIRIxBUFRYRMicYEyBhSRobFCIyQVUsFiMzRygtFDByWSU/Dh8WNzNRaisoMmRJNUZEXCo3Q2F9JV4mXys4TD03Xj80YnlKSFtJXE1OT0pbXF1eX1VmZ2hpamtsbW5vY3R1dnd4eXp7fH1+f3EQACAgECBAQDBAUGBwcGBTUBAAIRAyExEgRBUWFxIhMFMoGRFKGxQiPBUtHwMyRi4XKCkkNTFWNzNPElBhaisoMHJjXC0kSTVKMXZEVVNnRl4vKzhMPTdePzRpSkhbSVxNTk9KW1xdXl9VZmdoaWprbG1ub2JzdHV2d3h5ent8f/2gAMAwEAAhEDEQA/APVVz20v+tkO9UemQ9ge5+2DTtsfjsbjOp9Cx/pMt9TO/pGP9D9N+sPi5Lv+ct7LLLH1ve6ukmx4aHtqqtfjDDbd6XsZuu+12Yf+E/nvU9P1wsY2z61C51b2ltxa4lpB3toe1r677KG7sX0P+01eR77/AFMj0/T9b1Ep0updXyMPqODh14VuQzMe5rrmGsNbDLLNPUtrdvZ6e9/s/mv5r1Lv0Su4l7sip1jgGkWW1wNdK7LKWn+16aOqnTP6M/8A8MZP/n+5JTbVK3rfRqc0dPtz8avNJa0YrrmC2XQa2+i53qbn7m7Pah/WLqVnS+iZmbSN2RXWW4zSJDr7CKcRm2W/TybKmKPT/q/03D6S3pdlTMup3uynXtFhyLne67KyvV9T1br7P0j96SnTSXPO+0fVWtrmmzL+r9elgcXWX4TO1rXu3XZfTa/8Jv35ODX7/wBNi/0bfrsrtrbbU4PreA5j2kFpaRLXNcPpNckpkkkkkpSSSSSlJJJJKUkkkkpSSSSSlJJJJKf/0O2xepOyPrHZXU3FcxpfjvewbryKxvPqWtdvrZTf+i2W1enZZbZ/3Hr+2Cmv/nearC1suFldQc073jH2faXV/bGvbkMY19P/ACfZ+rs/67jyxnWu+szw1lxx9xtY+CWH1KQx1pyfR9J+K70am14rcy39N6dvp/ofTw541Yu636l/UK8p1V1no0Bl7S3a19ewt+1/YPXp3Wb7vsXv/rpKd9VOmf0Z/wD4Yyf/AD/craqdM/oz/wDwxk/+f7klOf8AWg2Pb0rEZ/2q6njB/wDVoL+pu/8AbFbaw+pudkfWno2Ix4jFZk59zZ1ADB06iW/y3Z923/iVuJKUsDYPqxaX1iPq/c6Xs7YVjj/OV/u9Ltd/OM/7z7P0n9C/om+mc1r2ljwHNcIc06gg9ikpQM6jhOsBjrPqy8VWkv8Aq+4xVadXYJP+Bu/e6X/obv8AvP8A5q39S/SYm8CCAQZB1BCSl0kkklKSSSSUpJJJJSkkkklKSSSSU//R6zpGA3p/1hux6ca3ZvfZ676q2iLW732faa8Gljm+oxtWyvqFmQ/fvyK/570pZTq6vrXXfYy1oa8MNha9876dtfoxQ6uvB3b632faf6bbZ+jVinOyH/WZ+Mcx7qmOcwY4YAwTWLfSt/wnqv2+tTb9Cuqizf8A8o4yC0Mb9cbHO95eWhrmskM/Qfzdl78J/pPd9P06uqU1/pa/0HqWWfaUpu9Uw+o3dTxbccONTDUWvbZsbVttbZmG6vc31vteH+rVba7v+sst9VVsbH+tH6Z2FnYX2d+VkFleRi2OfW31rG7RbRl0Nu9wc/31MXQqp0z+jP8A/DGT/wCf7klNLp/SupYVluQ/Jry8zJ2/acqxjmlwYW+lVVTXZ6ePj01vyvSpb/hLPWtsttsv9S5t6vH08ef6j+Yf/wAJ+96X/giuJJKae3q37+P/AJj/APg/+E/4/wD8CS29W/fx/wDMf/wn/Cf8R/4KriSSmk5nVnNLS/GMgggseQZDef0n73q/9BZH2TrP1f32dPYMzpRl1nT6wTZjQSfU6Yxz91+P6X0+mb/Zb/yf7P1NdIkkpysDPyupYjcvAysTIqeNHta/R21nssZ6m+qxlm/1KbP0tf8ANv8A0itbeq7vp4+2f3HzG53/AAn0vS2f9cVTP+ruNkZLs/Dts6b1JwG7MxSAbIaWMGZRY2zGzWM/M+00vsr/AMDZUgHqH1l6aD+0MFvVKGz+tdOO22AGx6vTMp/9f+i5uU//AIBJTobesbfp426P3HxO0f8ACfR9bf8A9bUtvVd306Ns/uPnbu/4z6Xo/wDgqr4X1l6HnX/ZactrMudpxLw6jInn+iZTacj/AMDWmkppbesbf5zG3R+4+N23/jPo+t/4Epbeq7vp4+2f3HzG5v8Awn0vR3/9cVtJJTS29Y2/Txt0fuPidrv+E+j6vp/9bUtvVd308fbP7j5jc3/hP9H6n9tW0klNIt6xtMWY26NPY+Jiz/hP9J6H/gqupJJKf//S7DCyGW/Wm+Mzcze+tlLXtfudXWPUr9mW9rK6/U99H7Oov+0Y/wCkv/7kTt2u+tFZLWuay+Bk7WhzbHYuvTtv8471KP177d9D/tEj9P6V1ajOZkZF4IBeLdtj3B7NvsnGextLb8nJe/Lut/7RV043Tcb1sb9IqGXf6f1sbYy6ptoezHFlp2wx9XrO6ds9Xc71P6dVlfZv579U9X/RpT1SqdM/oz//AAxk/wDn+5SyLOpNyKW4tFNmO4/rFltzq3tE/wCBpZj3su9v799Cp4vUMfFZbRe25tjb7yQKLniH22Wsc19dT2O3Vva76SRIG6qJdVJUf2zg+F//ALDX/wDpFL9s4Phf/wCw1/8A6RQ4h3CaPYt5JZ56509vPrifHHv/APSKX7e6d43f+w9//pFDjj+8PtVwy7H7G2Mmg5FmMHj1qmMtsbroyw2MrfP0fc7Hu/zEhlYxrFouYaiNweHDaWwX7t07duxu9YPUHdMzr8hz8nKrozcYYmVS3Gt9zB9o2uZYcffXZ+uWKrZgdBsZ77ckWEPL3Mxrmtda605bMgt9Av30usyKWfpf6Nk3V2+p/OJccf3h9qeCXY/Y9K3qGE5l1guZ6ePHrWEwxu5jb2u9R3s2+lax+9M3qOE52SPWaBhvbXkOd7Wse5rLmsNj9te707qne13+EWDXX0erDdj13ZDXG6vJa/7LcWttq9MtPpehtdW59W9zHf8AW7GW+naq93/NrH6c7phyr6g91drHPx7N7TXXVisezbj17Hbcbfvr9PZZ/N/of0KRyQG8h9qhjmdBEnyDvZZ6B1Kh1ed9ly6K3PDmX+m9rXVO9C722ztdTd+if/LWf/ze6ThuDem9RyekmlwaaaMndU02fzdf2LqH2zEq3/4JldFazLn/AFXt9R5z7ja57XssOPZoWvyb9rxVRV6jX25+R/4H+4oPH1WIcyvqF1TCWGsNxngsLWvqNtZ+z+zIcy57fUZ+jt/7VU5P6b1h7uP9+P8AjBd7OX9yX+LJ2vsv1xxWAU5+H1EA6jKofj2Fvnk4dltO7/2nqX7X+sND3Ny+hPsYBIswcmm4H+xm/su3/wADU/8And0D/uQ//ti7/wBIqLvrh9XmNLn5LmtHJNN0f+eUvdx/vx/xgr2cv7kv8UsR9b+m11l+bj52BtMO+0YeRtH/AF+iq/G/8HR8f61fVnKLRR1XDe9/0WevWH/9tOcLP+iq3/Pn6rf9zf8AwK7/ANJIGV9avqTms9PMtqya/wBy7HseP82yhyPHH94faj25/uy+x6RrmuAc0gg8EahOuHtt/wAVtjg9lVWO8cPxab8Z3+fhV471tU/XT6r/AKOmvNJJ21sBruJJMMZL31f9J6XHH94faj25/uy+x//T9VWDTh5dnXnXX4lzMVji2p73MexzQ0n1Ld2bZZ7sgu9Fn2L9HX6H83/gt5JJSlm2EfaXTxv1+C0lg9VyM6jJnFxjlVku9UN0eDuqazZJazb6brvU/c/nf8HYos4sDzZcJ1l5OLRT9ZG147bPWcaQ9rnOsBNnrvqv327bGuc7p7PUxKf/AEYxXelftc5Dvt4saxlDWAODdm8P0eyxrnPvtsx9j8ix/wDhd9f/AAdcx1LOLQf2dY07dz9xdAnbt27KXvtd7t2xlfqV/wDGK/W5z6w59bqnESa3Fpc3+sanWM/6ShJNagMoAvQnRhd9Jk8d/vC5xrPrCw1n9K4UvufJc15e282OY19bn/pf2e1v6pU/2fp8X/Rrb6rbk1VMfjV+rZugt2uedvfa2st930fpO9NUT1LL2epXgWWMJIbtdJO07Hu+ht9P6fo2f4T+R6ii1BNAG+7LoQLJFdmrZb9YS5rmsfLNxY3YxrXVkFjLsiHud9u/P+xNZ6PqempPv+sTaPUbXvtJAZWK2AQ2v1N1u52/9YuPov8A5n0tn/adWsfNzbrWB2E5lNuzbb74AcNXOmpr/wDPrr/4b0v0fqQHUs50Or6fZYx2jSNw4L9znF1f/F/4P9/0n3Ja/uxVp+8WGE/rTctlWQC7FHqbrHtbuIm4tfvr+i7f6FdP7+L+k9L/AAlVfr39Mr/4of8AVPVs5/UtjyMB4fE1g7tfBrob9L/SfQ/4H1bP0ay+s5V1tldzaDIGw1zJLGWWMttr+h+c1/o7lDnBMdgNQGxypAnuToS5WU3NN7nUepqwhkGGA7H+9vv9Pd6mz9FfRv8Aoelek6zqYEtbuJ3FjSxoH0nNDbnbvZ+hbXaz/SXP9NTbk5Qa31MY7tA6CYLj+bX7Hf8AkP37P5xP9oygGg4xnTcQSRxy32/+Ts/4NQ+qgOGBrT9FtVGyRKY4tf0v+9R/aMtmVVRYQd7wBDWy9h3eo98O/R+nHs9P/rqNm/0S34D8rU1GW+2w1GpzOd0mY2xLXe36W57Pof8AfE+b/RLfgP8Aqmpp+eFxEdtvP+quHyTqRkNfm6abepxXb/VBh5bDduwwAZ9/qf2dv0kNv2sNAOsNBJgEk+zc3n8z9L/XU/UuBg1EjxHhLv8AvqRst7VH8eP81XRfYFpGu8h9rGco9u8RAGmh3uIP+e1WME2HIx/Und61UyA38+ufol3t3IPqWf6I/wCo+CNhPLsqjc3aRdUD4TvYT/1SRvsPokV3J83/1PVUkkklKXO9ZcPtIY7N+yNO57wCWuc1llDXPY5v7rrK6P8A0IXRLmuvOwhksGXivymzZJZuPptgM92z2fp7nVUN9R7P9N/gVFl/R82TF18kFdeW9zmu6w2x7Pc4MDA2D++G/m6s/P8A/Piga8ywAN6xWBWCC5pbq8NNm/d9Bu2v9J6XvZXWhPu6G3cbMS82B2oJc50gOY+zf6x91X0Lv8Kz9B/hPSRun4/Rcpz2U4zq20kBosc8b9rt3trL/fVXYyi3/jn/AOkUX8tgzfy3SdSdaMXHnKZTa87W3j6Lzss9NzB7tznbftHpf+fEKp2Q3ID7cquzGFQrc7e0E2y1u7bp+k9tn/bnp+mreVg4r6G41jN9ADoYSYAI9Pa3Xd7WP/R/6NVWdJ6awgsp2xJADngAkFu6N30mte5tf+jUEiLP8GaIlQ/i1zhsvrrNuYA4MZTZbVY73msOse5jrH7PVtqfXY922z6CJe6rIruc7LqFBqbXY4WOOx7HWObaLGOZ9Nv89/xSJ+yOnGllBp/RVOL2N3Pjc6Nz/pe76DVI9LwHOLnVbnGSXOc8ulxDne/fu+k3d/6sehxDufsTwnsPtR42LfXf6rLm2VOc4loc4ja7c9v0jZ+l2W0/R/R/q1X+mWb9ZQ85Ne2wVu9NvvP9azx/le7atvHx6sar0qp27nPJPJc9xsedP5TlhfWc0i+v1mF9ZraC0CfpOsZu/wCkospsd9R0Z+XFS+h6uYx1jLWF97HVNZD5cBLjrMf1fzkMV3gmc0aySBtMEaTr+bu/9JILn4JqcW1WtbtEPZqQHe/2O3PZ+k/PexTs+w1vNXoWQYcXN3TJb/X3/wA27ao+HwP+LDp/6M2OLxH+PPr/AOisyyw6HNG6S7kEARHj+9/Y/MU8ifsNkvFhAhzgZ1DggGzBa4uNFgIJdY4ky2A4us0ed22HN9v/AFtGvrqZg2+k3Y0tb7J0EFrY7+5u3Y7+ogd43e46RH/RTHadUdDfqkf+k4/p2gyLdOYInu53/fkxZfHtsk+HARUlZ4j4fY1uEeP2o/Tu/wBNrPgOEXDZaMzHLrNw9WuRH8tkJkTG/pWP/wAdV/1bEuI+H2K4R4/aX//V9VSSSSUpYfVKerPvnAcK2tLzZuB95cPSYGu2P2elvfkbvz7a6qvob1uLF6tT1OvqDcnp5tf+r2kVuc51HrB+PXjmzHZZVv8A0NmVZ6fqM9TZ/pNibOHFXguhLhvxaja/rMdrn2VNPuDmMYdv7zHe7c/6X6P6X817/wCdTNZ9Zi5psFDhLd0NcNN/6UNnf/gfoO/9G/zc7eo/Wn1MhlGM0tqc703vqPu9MZn6NrfXb/SH42Bst3/QzP8ASfo8V7c/6xiy0txpbXY9gsDHhrajZS31BVu9XJvox/Vf+jqfXa/+jWXfpKkz2fH8F/u+H4p8iuwlsMeRr+af7kL0rf8ARv8A8139yj9s+tD31G2oU1ufR6gqqLiGh+B9oG6yx3svZk5/+C9THrxv+D/SaNWTnHq763Mf9hfUQwlkBllThL3Wf4T7ay/9H/o/sX/DqM8qCb4vwZBzRArh/Foelb/o3/5rv7kvSt/0b/8ANd/ctm92c1w+zVVWMjU2WOYZ8gym5C9Tq3/cfH/7ff8A+8qH3Qfvfgn72f3fxcv0rf8ARv8A8139ywvrBjdRdk1nFosc702gk1vLY3P3tljT+aux9Tq3/cfH/wC33/8AvKl6vVv+42P/ANvv/wDeRCXJCQri/BdDnTE3w9K3fOfsfXo0ofoIDfRf4eOxOcTrsiMewNhsxVZ9KP02jme5u76C9F9Xq3/cbH/9iH/+8iqdQyPrQw437Pw8SzdcBkizIeAKtljnO3/Z2OY71W1s9lWT/wAR/hK2fcI/vD/EZP8ASUv3T/j/APoLxFOH1YgjIxrToNG1WbZl3Et/d2KObhZxxbQMW8kgaCmwnlv8hd/1unNvxqqsPcHOvq9XY91X6IOm3dbS5lzG7f8ARLNr/wCdWPZ9iDhdWz0625Lm7iWOdR6uRL3fzlHq5dXp3XW3Prxqsi31H/0hf6PjxXx9boRT/pOXCY+30qzLV89/Z/Uv+4WT/wBsW/8ApNN+z+pf9wsn/ti3/wBJr0b1/rQxnqWNDw5o3VsraHML22FzwfWs3/ZX11fmWPf61v6HJ/RUqDcn62/Zxa+poeYrfXsEtbFLrMwOa+1z7fdkenR6P/Wv9JL91H7xYvvp/cH2vnn7P6l/3Cyf+2Lf/SaLjdP6iMmgnDyQBbWSTRbEB7f+DXfet9arGsdY30276d4qrbu2NdhW3vHqWOd+lbZn02V7PoV/o/0n89dvs6gOtY9tTbXYwc7Gupj9H6b2DIZ1Dd9H1K8mv7Js/wBFdY9L7qP3ir76f3B9r//ZOEJJTQQhAAAAAABVAAAAAQEAAAAPAEEAZABvAGIAZQAgAFAAaABvAHQAbwBzAGgAbwBwAAAAEwBBAGQAbwBiAGUAIABQAGgAbwB0AG8AcwBoAG8AcAAgAEMAUwA2AAAAAQD/4gxYSUNDX1BST0ZJTEUAAQEAAAxITGlubwIQAABtbnRyUkdCIFhZWiAHzgACAAkABgAxAABhY3NwTVNGVAAAAABJRUMgc1JHQgAAAAAAAAAAAAAAAQAA9tYAAQAAAADTLUhQICAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABFjcHJ0AAABUAAAADNkZXNjAAABhAAAAGx3dHB0AAAB8AAAABRia3B0AAACBAAAABRyWFlaAAACGAAAABRnWFlaAAACLAAAABRiWFlaAAACQAAAABRkbW5kAAACVAAAAHBkbWRkAAACxAAAAIh2dWVkAAADTAAAAIZ2aWV3AAAD1AAAACRsdW1pAAAD+AAAABRtZWFzAAAEDAAAACR0ZWNoAAAEMAAAAAxyVFJDAAAEPAAACAxnVFJDAAAEPAAACAxiVFJDAAAEPAAACAx0ZXh0AAAAAENvcHlyaWdodCAoYykgMTk5OCBIZXdsZXR0LVBhY2thcmQgQ29tcGFueQAAZGVzYwAAAAAAAAASc1JHQiBJRUM2MTk2Ni0yLjEAAAAAAAAAAAAAABJzUkdCIElFQzYxOTY2LTIuMQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAWFlaIAAAAAAAAPNRAAEAAAABFsxYWVogAAAAAAAAAAAAAAAAAAAAAFhZWiAAAAAAAABvogAAOPUAAAOQWFlaIAAAAAAAAGKZAAC3hQAAGNpYWVogAAAAAAAAJKAAAA+EAAC2z2Rlc2MAAAAAAAAAFklFQyBodHRwOi8vd3d3LmllYy5jaAAAAAAAAAAAAAAAFklFQyBodHRwOi8vd3d3LmllYy5jaAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABkZXNjAAAAAAAAAC5JRUMgNjE5NjYtMi4xIERlZmF1bHQgUkdCIGNvbG91ciBzcGFjZSAtIHNSR0IAAAAAAAAAAAAAAC5JRUMgNjE5NjYtMi4xIERlZmF1bHQgUkdCIGNvbG91ciBzcGFjZSAtIHNSR0IAAAAAAAAAAAAAAAAAAAAAAAAAAAAAZGVzYwAAAAAAAAAsUmVmZXJlbmNlIFZpZXdpbmcgQ29uZGl0aW9uIGluIElFQzYxOTY2LTIuMQAAAAAAAAAAAAAALFJlZmVyZW5jZSBWaWV3aW5nIENvbmRpdGlvbiBpbiBJRUM2MTk2Ni0yLjEAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHZpZXcAAAAAABOk/gAUXy4AEM8UAAPtzAAEEwsAA1yeAAAAAVhZWiAAAAAAAEwJVgBQAAAAVx/nbWVhcwAAAAAAAAABAAAAAAAAAAAAAAAAAAAAAAAAAo8AAAACc2lnIAAAAABDUlQgY3VydgAAAAAAAAQAAAAABQAKAA8AFAAZAB4AIwAoAC0AMgA3ADsAQABFAEoATwBUAFkAXgBjAGgAbQByAHcAfACBAIYAiwCQAJUAmgCfAKQAqQCuALIAtwC8AMEAxgDLANAA1QDbAOAA5QDrAPAA9gD7AQEBBwENARMBGQEfASUBKwEyATgBPgFFAUwBUgFZAWABZwFuAXUBfAGDAYsBkgGaAaEBqQGxAbkBwQHJAdEB2QHhAekB8gH6AgMCDAIUAh0CJgIvAjgCQQJLAlQCXQJnAnECegKEAo4CmAKiAqwCtgLBAssC1QLgAusC9QMAAwsDFgMhAy0DOANDA08DWgNmA3IDfgOKA5YDogOuA7oDxwPTA+AD7AP5BAYEEwQgBC0EOwRIBFUEYwRxBH4EjASaBKgEtgTEBNME4QTwBP4FDQUcBSsFOgVJBVgFZwV3BYYFlgWmBbUFxQXVBeUF9gYGBhYGJwY3BkgGWQZqBnsGjAadBq8GwAbRBuMG9QcHBxkHKwc9B08HYQd0B4YHmQesB78H0gflB/gICwgfCDIIRghaCG4IggiWCKoIvgjSCOcI+wkQCSUJOglPCWQJeQmPCaQJugnPCeUJ+woRCicKPQpUCmoKgQqYCq4KxQrcCvMLCwsiCzkLUQtpC4ALmAuwC8gL4Qv5DBIMKgxDDFwMdQyODKcMwAzZDPMNDQ0mDUANWg10DY4NqQ3DDd4N+A4TDi4OSQ5kDn8Omw62DtIO7g8JDyUPQQ9eD3oPlg+zD88P7BAJECYQQxBhEH4QmxC5ENcQ9RETETERTxFtEYwRqhHJEegSBxImEkUSZBKEEqMSwxLjEwMTIxNDE2MTgxOkE8UT5RQGFCcUSRRqFIsUrRTOFPAVEhU0FVYVeBWbFb0V4BYDFiYWSRZsFo8WshbWFvoXHRdBF2UXiReuF9IX9xgbGEAYZRiKGK8Y1Rj6GSAZRRlrGZEZtxndGgQaKhpRGncanhrFGuwbFBs7G2MbihuyG9ocAhwqHFIcexyjHMwc9R0eHUcdcB2ZHcMd7B4WHkAeah6UHr4e6R8THz4faR+UH78f6iAVIEEgbCCYIMQg8CEcIUghdSGhIc4h+yInIlUigiKvIt0jCiM4I2YjlCPCI/AkHyRNJHwkqyTaJQklOCVoJZclxyX3JicmVyaHJrcm6CcYJ0kneierJ9woDSg/KHEooijUKQYpOClrKZ0p0CoCKjUqaCqbKs8rAis2K2krnSvRLAUsOSxuLKIs1y0MLUEtdi2rLeEuFi5MLoIuty7uLyQvWi+RL8cv/jA1MGwwpDDbMRIxSjGCMbox8jIqMmMymzLUMw0zRjN/M7gz8TQrNGU0njTYNRM1TTWHNcI1/TY3NnI2rjbpNyQ3YDecN9c4FDhQOIw4yDkFOUI5fzm8Ofk6Njp0OrI67zstO2s7qjvoPCc8ZTykPOM9Ij1hPaE94D4gPmA+oD7gPyE/YT+iP+JAI0BkQKZA50EpQWpBrEHuQjBCckK1QvdDOkN9Q8BEA0RHRIpEzkUSRVVFmkXeRiJGZ0arRvBHNUd7R8BIBUhLSJFI10kdSWNJqUnwSjdKfUrESwxLU0uaS+JMKkxyTLpNAk1KTZNN3E4lTm5Ot08AT0lPk0/dUCdQcVC7UQZRUFGbUeZSMVJ8UsdTE1NfU6pT9lRCVI9U21UoVXVVwlYPVlxWqVb3V0RXklfgWC9YfVjLWRpZaVm4WgdaVlqmWvVbRVuVW+VcNVyGXNZdJ114XcleGl5sXr1fD19hX7NgBWBXYKpg/GFPYaJh9WJJYpxi8GNDY5dj62RAZJRk6WU9ZZJl52Y9ZpJm6Gc9Z5Nn6Wg/aJZo7GlDaZpp8WpIap9q92tPa6dr/2xXbK9tCG1gbbluEm5rbsRvHm94b9FwK3CGcOBxOnGVcfByS3KmcwFzXXO4dBR0cHTMdSh1hXXhdj52m3b4d1Z3s3gReG54zHkqeYl553pGeqV7BHtje8J8IXyBfOF9QX2hfgF+Yn7CfyN/hH/lgEeAqIEKgWuBzYIwgpKC9INXg7qEHYSAhOOFR4Wrhg6GcobXhzuHn4gEiGmIzokziZmJ/opkisqLMIuWi/yMY4zKjTGNmI3/jmaOzo82j56QBpBukNaRP5GokhGSepLjk02TtpQglIqU9JVflcmWNJaflwqXdZfgmEyYuJkkmZCZ/JpomtWbQpuvnByciZz3nWSd0p5Anq6fHZ+Ln/qgaaDYoUehtqImopajBqN2o+akVqTHpTilqaYapoum/adup+CoUqjEqTepqaocqo+rAqt1q+msXKzQrUStuK4trqGvFq+LsACwdbDqsWCx1rJLssKzOLOutCW0nLUTtYq2AbZ5tvC3aLfguFm40blKucK6O7q1uy67p7whvJu9Fb2Pvgq+hL7/v3q/9cBwwOzBZ8Hjwl/C28NYw9TEUcTOxUvFyMZGxsPHQce/yD3IvMk6ybnKOMq3yzbLtsw1zLXNNc21zjbOts83z7jQOdC60TzRvtI/0sHTRNPG1EnUy9VO1dHWVdbY11zX4Nhk2OjZbNnx2nba+9uA3AXcit0Q3ZbeHN6i3ynfr+A24L3hROHM4lPi2+Nj4+vkc+T85YTmDeaW5x/nqegy6LzpRunQ6lvq5etw6/vshu0R7ZzuKO6070DvzPBY8OXxcvH/8ozzGfOn9DT0wvVQ9d72bfb794r4Gfio+Tj5x/pX+uf7d/wH/Jj9Kf26/kv+3P9t////4Tt7aHR0cDovL25zLmFkb2JlLmNvbS94YXAvMS4wLwA8P3hwYWNrZXQgYmVnaW49Iu+7vyIgaWQ9Ilc1TTBNcENlaGlIenJlU3pOVGN6a2M5ZCI/Pgo8eDp4bXBtZXRhIHhtbG5zOng9ImFkb2JlOm5zOm1ldGEvIiB4OnhtcHRrPSJBZG9iZSBYTVAgQ29yZSA1LjMtYzAxMSA2Ni4xNDU2NjEsIDIwMTIvMDIvMDYtMTQ6NTY6MjcgICAgICAgICI+CiAgIDxyZGY6UkRGIHhtbG5zOnJkZj0iaHR0cDovL3d3dy53My5vcmcvMTk5OS8wMi8yMi1yZGYtc3ludGF4LW5zIyI+CiAgICAgIDxyZGY6RGVzY3JpcHRpb24gcmRmOmFib3V0PSIiCiAgICAgICAgICAgIHhtbG5zOnhtcD0iaHR0cDovL25zLmFkb2JlLmNvbS94YXAvMS4wLyI+CiAgICAgICAgIDx4bXA6Q3JlYXRvclRvb2w+QWRvYmUgUGhvdG9zaG9wIENTNiAoV2luZG93cyk8L3htcDpDcmVhdG9yVG9vbD4KICAgICAgICAgPHhtcDpDcmVhdGVEYXRlPjIwMTUtMDMtMTNUMDk6MTY6NDUtMDc6MDA8L3htcDpDcmVhdGVEYXRlPgogICAgICAgICA8eG1wOk1vZGlmeURhdGU+MjAxNS0wOC0xMVQxODo1NToyMCswNTozMDwveG1wOk1vZGlmeURhdGU+CiAgICAgICAgIDx4bXA6TWV0YWRhdGFEYXRlPjIwMTUtMDgtMTFUMTg6NTU6MjArMDU6MzA8L3htcDpNZXRhZGF0YURhdGU+CiAgICAgIDwvcmRmOkRlc2NyaXB0aW9uPgogICAgICA8cmRmOkRlc2NyaXB0aW9uIHJkZjphYm91dD0iIgogICAgICAgICAgICB4bWxuczpkYz0iaHR0cDovL3B1cmwub3JnL2RjL2VsZW1lbnRzLzEuMS8iPgogICAgICAgICA8ZGM6Zm9ybWF0PmltYWdlL3RpZmY8L2RjOmZvcm1hdD4KICAgICAgPC9yZGY6RGVzY3JpcHRpb24+CiAgICAgIDxyZGY6RGVzY3JpcHRpb24gcmRmOmFib3V0PSIiCiAgICAgICAgICAgIHhtbG5zOnBob3Rvc2hvcD0iaHR0cDovL25zLmFkb2JlLmNvbS9waG90b3Nob3AvMS4wLyI+CiAgICAgICAgIDxwaG90b3Nob3A6Q29sb3JNb2RlPjM8L3Bob3Rvc2hvcDpDb2xvck1vZGU+CiAgICAgICAgIDxwaG90b3Nob3A6SUNDUHJvZmlsZT5zUkdCIElFQzYxOTY2LTIuMTwvcGhvdG9zaG9wOklDQ1Byb2ZpbGU+CiAgICAgIDwvcmRmOkRlc2NyaXB0aW9uPgogICAgICA8cmRmOkRlc2NyaXB0aW9uIHJkZjphYm91dD0iIgogICAgICAgICAgICB4bWxuczp4bXBNTT0iaHR0cDovL25zLmFkb2JlLmNvbS94YXAvMS4wL21tLyIKICAgICAgICAgICAgeG1sbnM6c3RFdnQ9Imh0dHA6Ly9ucy5hZG9iZS5jb20veGFwLzEuMC9zVHlwZS9SZXNvdXJjZUV2ZW50IyI+CiAgICAgICAgIDx4bXBNTTpJbnN0YW5jZUlEPnhtcC5paWQ6NEQzQjY3NEIyQzQwRTUxMUExNTlBNjZDMzlFQTk3MkI8L3htcE1NOkluc3RhbmNlSUQ+CiAgICAgICAgIDx4bXBNTTpEb2N1bWVudElEPnhtcC5kaWQ6MDNGQ0UwODY5Q0M5RTQxMUIzNTBBNUQ0ODdGQThEMjM8L3htcE1NOkRvY3VtZW50SUQ+CiAgICAgICAgIDx4bXBNTTpPcmlnaW5hbERvY3VtZW50SUQ+eG1wLmRpZDowM0ZDRTA4NjlDQzlFNDExQjM1MEE1RDQ4N0ZBOEQyMzwveG1wTU06T3JpZ2luYWxEb2N1bWVudElEPgogICAgICAgICA8eG1wTU06SGlzdG9yeT4KICAgICAgICAgICAgPHJkZjpTZXE+CiAgICAgICAgICAgICAgIDxyZGY6bGkgcmRmOnBhcnNlVHlwZT0iUmVzb3VyY2UiPgogICAgICAgICAgICAgICAgICA8c3RFdnQ6YWN0aW9uPmNyZWF0ZWQ8L3N0RXZ0OmFjdGlvbj4KICAgICAgICAgICAgICAgICAgPHN0RXZ0Omluc3RhbmNlSUQ+eG1wLmlpZDowM0ZDRTA4NjlDQzlFNDExQjM1MEE1RDQ4N0ZBOEQyMzwvc3RFdnQ6aW5zdGFuY2VJRD4KICAgICAgICAgICAgICAgICAgPHN0RXZ0OndoZW4+MjAxNS0wMy0xM1QwOToxNjo0NS0wNzowMDwvc3RFdnQ6d2hlbj4KICAgICAgICAgICAgICAgICAgPHN0RXZ0OnNvZnR3YXJlQWdlbnQ+QWRvYmUgUGhvdG9zaG9wIENTNiAoV2luZG93cyk8L3N0RXZ0OnNvZnR3YXJlQWdlbnQ+CiAgICAgICAgICAgICAgIDwvcmRmOmxpPgogICAgICAgICAgICAgICA8cmRmOmxpIHJkZjpwYXJzZVR5cGU9IlJlc291cmNlIj4KICAgICAgICAgICAgICAgICAgPHN0RXZ0OmFjdGlvbj5jb252ZXJ0ZWQ8L3N0RXZ0OmFjdGlvbj4KICAgICAgICAgICAgICAgICAgPHN0RXZ0OnBhcmFtZXRlcnM+ZnJvbSBhcHBsaWNhdGlvbi92bmQuYWRvYmUucGhvdG9zaG9wIHRvIGltYWdlL3RpZmY8L3N0RXZ0OnBhcmFtZXRlcnM+CiAgICAgICAgICAgICAgIDwvcmRmOmxpPgogICAgICAgICAgICAgICA8cmRmOmxpIHJkZjpwYXJzZVR5cGU9IlJlc291cmNlIj4KICAgICAgICAgICAgICAgICAgPHN0RXZ0OmFjdGlvbj5zYXZlZDwvc3RFdnQ6YWN0aW9uPgogICAgICAgICAgICAgICAgICA8c3RFdnQ6aW5zdGFuY2VJRD54bXAuaWlkOjA0RkNFMDg2OUNDOUU0MTFCMzUwQTVENDg3RkE4RDIzPC9zdEV2dDppbnN0YW5jZUlEPgogICAgICAgICAgICAgICAgICA8c3RFdnQ6d2hlbj4yMDE1LTAzLTEzVDA5OjE4OjI3LTA3OjAwPC9zdEV2dDp3aGVuPgogICAgICAgICAgICAgICAgICA8c3RFdnQ6c29mdHdhcmVBZ2VudD5BZG9iZSBQaG90b3Nob3AgQ1M2IChXaW5kb3dzKTwvc3RFdnQ6c29mdHdhcmVBZ2VudD4KICAgICAgICAgICAgICAgICAgPHN0RXZ0OmNoYW5nZWQ+Lzwvc3RFdnQ6Y2hhbmdlZD4KICAgICAgICAgICAgICAgPC9yZGY6bGk+CiAgICAgICAgICAgICAgIDxyZGY6bGkgcmRmOnBhcnNlVHlwZT0iUmVzb3VyY2UiPgogICAgICAgICAgICAgICAgICA8c3RFdnQ6YWN0aW9uPnNhdmVkPC9zdEV2dDphY3Rpb24+CiAgICAgICAgICAgICAgICAgIDxzdEV2dDppbnN0YW5jZUlEPnhtcC5paWQ6NEQzQjY3NEIyQzQwRTUxMUExNTlBNjZDMzlFQTk3MkI8L3N0RXZ0Omluc3RhbmNlSUQ+CiAgICAgICAgICAgICAgICAgIDxzdEV2dDp3aGVuPjIwMTUtMDgtMTFUMTg6NTU6MjArMDU6MzA8L3N0RXZ0OndoZW4+CiAgICAgICAgICAgICAgICAgIDxzdEV2dDpzb2Z0d2FyZUFnZW50PkFkb2JlIFBob3Rvc2hvcCBDUzYgKFdpbmRvd3MpPC9zdEV2dDpzb2Z0d2FyZUFnZW50PgogICAgICAgICAgICAgICAgICA8c3RFdnQ6Y2hhbmdlZD4vPC9zdEV2dDpjaGFuZ2VkPgogICAgICAgICAgICAgICA8L3JkZjpsaT4KICAgICAgICAgICAgPC9yZGY6U2VxPgogICAgICAgICA8L3htcE1NOkhpc3Rvcnk+CiAgICAgIDwvcmRmOkRlc2NyaXB0aW9uPgogICA8L3JkZjpSREY+CjwveDp4bXBtZXRhPgogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgIAogICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgCiAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAgICAKICAgICAgICAgICAgICAgICAgICAgICAgICAgIAo8P3hwYWNrZXQgZW5kPSJ3Ij8+/9sAQwADAgICAgIDAgICAwMDAwQGBAQEBAQIBgYFBgkICgoJCAkJCgwPDAoLDgsJCQ0RDQ4PEBAREAoMEhMSEBMPEBAQ/9sAQwEDAwMEAwQIBAQIEAsJCxAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQ/8AAEQgB5gK8AwEiAAIRAQMRAf/EAB0AAQACAgMBAQAAAAAAAAAAAAAGBwUIAwQJAQL/xABpEAABAwMCAwMGAxANBwkGBAcBAgMEAAUGBxEIEiETMUEJFBUiUWEycdMWFxgjMzhCVVZzdoGRlrG0NTdScnWTlJWhsrPR0hk0NldYYoIkJjlDU3SDkqIlY3ejpMNEZNTwJ0VUZrXBwv/EABsBAQACAwEBAAAAAAAAAAAAAAABAwIEBQYH/8QAPREAAQMCAwQFCgYCAwEBAQAAAQACEQMhBBIxBUFRcRUiYYGhExQyM1KRkrHB0QYWNFNy8EJUI2LhwvGy/9oADAMBAAIRAxEAPwD1TpSlESlKURKpDi710yTQrTKHcMDsce8ZllN5iY3jsORuWlTZBOy1gEcwSlKjtuNyU7nbervqguMrR7OtVtPLHdNLPNXMzwPI4WVWWNKWENTHY5VzMKUSAOZKjsSQN0gEjfcEVT8Rms/EnguEaPaE2jIrKxrVqXKTCud1gtJVHiIQQHHWkqRskEuJ3VyHYNubDfY1IOHrPtfdNtfJfC9xD5lCzV24Y8clxrI2IwYdeaQ72bzDqQB1B3IJ3I5e8hQ5Y1oxpvxCa2cVEXia4gNM2cFgYbZF2rHrIuYiQ45JWFhTu6SdkjtXTzEDqUAA8pNZ/QLENc9WeJS58UGtmBfMJb7NY3cXxawvPdpKLa3udyQ50B2OytiQknnGw2TuSlbd0pSihKUrEzctxW3XqLjdwya1RbvNAVFgPTW0SXwd9ihonmUPVV3A9x9lEWWpSuGXLiQIrs2dKajR2EFx151YQhtI6lSlHoAPaaIualcbD7MlluTGeQ6y6kLbcQoKStJG4II6EEeNclESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREqg+MzXrK9BtMLfL07s7FzzLLL1GxywsyElTSZL/ADHtFJ3HNsEkAEgcyk79AavytYvKDWG5XjRS2TcZxHKL5lFlyWDdMfVj0Lzt6DPZDi0Putd6mOhQvbr66SO7aiKFRdI/KfTY7cudxPYDBfeSFuRkWRtwNKI6p5hF2O3d0qzdBNPeMfFs2euWveuOM5djaoDrTUC22tMd1MorQUOFQZR6oSFjbf7IdKqWw8fPEAi0xm8n4E9R3bmlsCS5AZfSwte3VSErj8yQT4Enb2mrY0C4o8/1kzd7E8n4Ys40/iNW92aLrekqEda0rQkMjdpI5lBZI69yT0opWw9KUooWBvue4Ni8tEDJs0sVolONh1DM64sx3FIJICglagSNwRv7jXn/AMRmZYTc/KT6H5LAyuxyrXBtrKZFwZnsrYYIdm9FuhXKk+sO8jvHtrZviC4HNE+JfMYecakLyEXKDb0Wxr0fPQw32KXFuDdJbVurmcV139laE6xcGejmA8Z2mPD9YlX44tmMNuRci/OSuTzKXJSezcCAEjZlH2J8fbRSF6p27U/TW8TmbXaNQ8ZnTJKuRmPGu0d111XfslKVkqPQ9AKhPF2Arhb1XCgCDiF03B/7suqx0x8mxw5aS6gWLUrFHMrN4x6V53D86uaHGu05VJ9ZIaBI2UfEVZ3F19a5qv8AghdP1ZdEUl0MAGienwA2Axa0/qjVTeoRob+0pp/+C1q/VGqm9FCUpSiJSlKIlKUoiUpVdak8ROhukDojak6pY/YpRG/mj8sKk7e3sUbube/loisWlUFZ+PPhDvktMKFrnYmnVq5UmYh+Kgn9+82lP9NXbYshsOUW1q8Y1e4F2gPdW5UGSh9lfxLQSD+WiLIUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSuvcJiLdAk3BxClojMreUlPeQlJJA9/SvPbRTTDiY4z8Xla9ZNxV5XglqvtxlIstix7mSzHjNOqbG4S4gDYpKRuFKPLzFXXYEXojStLxwCawAg/R4arHYg7do51/+orcuM0piO0wt1Tim0JQVq71EDbc/HRFy1VnEpr3ZeHLSyZqFdLY9dZan2rdabWwdnJ894kNMg7HYdConYnZJ2BOwNp1q/5QLE8tuWl2MajYZYnL7L0xy235dItTaSpUqLHKg6AkbklIUFdAfVCj4URQGFG8qzmcMZY1fNMcNRKSH2Mfkxu0daSeoQ4rs3SFbd+7m4Pft3VY/DBxJakZnnGQaBcQ+FxcY1MxmIi4gwlbw7tBUoJ84Z9ZXUKKd9lEHm+xKVJH7xfyinCHkmPMX2Rq1DszrjQW9b7lGeblMK8UFIQQog+KCoHwNV5ofnDXFTxnzOIHArPOY09wTFHcXiXiVHUz6XmPPdorkSrrypBUdj1ACSQCvailbr0pSihKqPNeGPTnPdcMV4gL3JvScmw9hEe3tx5aURFJSp1Q7RsoJUd3l9yh4VblKIlVHxdfWuar/ghdP1ZdW5VR8XP1rmq/4IXT9WXRFJdDf2lNP/wWtX6o1U3qEaG/tKaf/gtav1RqpvREpSlESlKURKUqjuMvXxPDtoPe80gvNi/zdrVYW1bHec8CEr28Q2kLcP7zbxoi1g4/ePq9YbeZmhGhF0Uxe2D5vfr7G9d2I4rp5pF23+ndRzLG5QSEp9bcpqzQzyWWpeqFvRnWuGYycRF0/wCUiD2Pnd2e5uvPIU4rlaUd99jzr/dBJ6Vk/Je8NDeoOUXDiR1EjLuESyzVtWMS/X87um/M9MXv8ItlWyT/ANopR70CvVCiy0sF565D5HjT1+EoYrrLksKYE+qq4QY8lpSvelAbUB8RrXPLeF7jR4Jrm7nenl2uEm0Rj2j12xZ1bzBQOv8AyuEob8vtKkLQP3Vey1KKJK8/uF/ypuM5i5Dw3iFjRMburpS0xkUUEW2SruHbpJJjKJ+y3LfXqUDpW/kaTHmR2pcR9t9h9CXGnW1BSFoI3CkkdCCDuCK034sPJv6e6zNTcz0qahYfmywp1aG2+S23NfiHm0j6Us/9qgd/wkq7xqBoFxX65cDecO6Oay49dZeMQneSVYpat5FuQo9JEBwnlW2eqggEtrG/KUnc0SJ0XsZSozpxqThOrWIQM70+yCNeLLcUczUhlXVKh8JtaT1QtJ6KQoAg94qTUUJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoi/LgJbUAkK3SfVPcfdXnXpNbfKH6ESb/ZtNuHPHhhtzuki5wbBcb/HfRbFuq5nEx3kvoWG1K9YIWFcu/TxJ9E3nWo7S33lhDbaStaj3JSBuTWhuO3Xi644Z12zzTjV0aQ6VRbi/b7EYcXtbjdEsrKFSFEFKtiR+7SkH1QlWxUSkKUDWbymW434UMGA3G/8A7fR3fymtyIynlx2lyEBDqkJLiQeiVbdR+WtDcqlcYnBE5A1LzfWI6waXpnMRciZnROyn29l1YQJCFEqVsCofZkb7ApG/MN8osqPOiszYjqXWJDaXWlp7loUNwR8YNFCxWaZBJxPD73lEOyS7y/aLfInNW6ICX5im2ysMtgA7rURyjoepFUldeKnL7boHZdaWuG7O5Vzu1zVb3cSaYX6SiICnR27iez3CPpQPwR9UT1rYR1TiWlqabC1hJKUlXKFHwG/h8dee+v8A5QHVO3Gy2awaF6k4NOt2Xw2LlIlwW3GbnHbcWl6AysoUla3ttkFHU7bg0QLG3vXPBMku67/ffJX5ROuLi+0ckO402VuK/dKPm3rH3netieGriUvOpeTjTn6FfMNMbRAtrkqPKuMHzaCkoWhIYQkNISFHnJAHgk9KgR8o1fQSPoONZOnttSv8Nd7h+1n4g9euJqXlUvT/ADPBdKrZjKoyrXf4vYokXIup5XUkoSVL5SroCQlKeuxUBRStw6UpRQlKUoiVUfF19a5qv+CF0/Vl1blVHxdfWuar/ghdP1ZdEUl0N/aU0/8AwWtX6o1U3qEaG/tKaf8A4LWr9Uaqb0RKUpREpSlESvJzyq2o9y1A13xnRHHSqQnG4zQ83R17W6T1J5E9PEN9iB7O0VXrC4tDSFOuLCUIBUpROwAHeTXjpw0ML4lfKKOZ3NbU/Aav1xypW45gmPGJTEHxBRjAfFRSOK9VND9MLZoxpLi2mNpSnsrBbmozq0/9dI25nnT71uKWr8dTmlKKEpSlESqf4kuF/TTibxE4/mkLza6REqNpvkZA87t7h/ck/DbJ25m1eqr3HZQuClEXi1Ysh4ivJpa1uWe6xTNslxXzvxAtQtmQxEnYPMqI+lvpB79uZB9VQUk9fWXRDXHT7iBwSJn2nl1EmI99LlRXNkyYMgDdTD6N/VWPyEbEEgg1+tbdENP9f8El4BqJaRKhv/TI0hvZMiC+Bsl9he3qLG/xEbgggkV5L3S1cQHk0Ndmp8F8z7NcVEMv7KTbsjgpO5acHXs30A93VSCdxzIV6xZar2mpVb6B6+YBxF4BFz3Ap/MhWzU+A6QJNuk7bqZeSO4jvCu5Q2I6GrIosUpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlEXBOhs3CFIgSAS1JaWyvY7HlUCD/Qa8+tHOI9/gKtkvhw4jsGyNiz2K4SnMZyi1wfOIlwguvKcTv1HrArUfVJI35VJBTufQyuGVDiTmTHnRWZDSu9t1AWk/iPSiLz61p4oV8cuOnhr4Z8FyOdGyiTHayHJbnB83hWyAh5Li1b7nqeQfC5T05UhRUNt/7NbGLJaINmilRZgRmorZV3lKEhI3/EK5YkKHAZEeBEZjNDqG2WwhI/EOlc9ESqB409R8b0y0rtt6uWnkHN8hkZDBi4jZ5bfOhd7UVebu+0FvZat07HfYAjfcX9WvPG5pRnupGmNmyDSmM3LzLT7IoWWWiEsjaauOVc7A3IBUUqJA3G5Tt40RT3S9rXaRowyjVaRYI2pL8SWVrgDmhMPrUsxwQAdwhJbCtubcpOxV3mp+HDXXWtrVW68M/E/a7QnN7fahe7RfLQOWHe4IWELWEbDlWlR8AnuUClJT60Kg+VJ0ztcNNv1I0g1LxvKGE8ku0izh3leHQpQpa0KI37uZKT7q7/AA7o1K4iuJOTxYZhp9ccIxKy44vGsSt90TyTZqXXe0dkuJ2GyfhAeHrJCSrlJJStx6UpRQlKUoiVUfFz9a5qv+CF0/Vl1blVHxdfWuar/ghdP1ZdEUl0N/aU0/8AwWtX6o1U3qEaG/tKaf8A4LWr9Uaqb0RKUpREpSlEVX8UGanTzh31EzBDnI9Ax6YGFA7EPuNltrb38601or5HPCUrvGo2oTqOsSLBscdRH7sqed2P/AzV/eVMylWP8KM+1NO8i8jvdvtu2/VSAsvqH5GK6HkpMVFj4X135bPK7keQzpnNt1U23yR0/wBLKvy0U7luXSlKKEpSlESlKURKhOsOjuB66YJP091DtCZ1tmjmQseq9EeAPI+yvvQ4knofjBBBIM2pRF4sXi0a8eTQ19auFvfVcLLPJDD5CkQMit4VuWnANw2+gHu6qQohSeZKvW9Y9CddcC4htPoWoOAz+0jvfSpkN0gSIEkAc7DyR3KG/Q9ygQobgiuzrTovgmveAT9O9QbYJUCYOdl5GwfhyADyPsr+xWknv7iCQQQSD5JoVrr5NDiC5VBVxs0/vHVEHJLaFd47w2+jf3qbUfskK9YstV7T0qEaNax4LrtgNv1F0+ugl26cnlcbVsHoj4A52HkfYOJJ2I8ehBIIJm9FilKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIvytaG0KccUEpSCVKJ2AHtNax6O8YN71RjalaoTtOlWXSHDYslyz5A9IBfuzkUr7chskbJKUgp2BAJ5SoncDZW5QkXK3yrc4tSEymVsqUnvAUkjcflry1ulw4h8X4dZXk/LboNlLuUP3R22sZDHjKVa5NqcmF/tw8ByjmB5VEkAIJ3IPq0UhWHjPGrxfw8SgcT+daZ4wvRS6XJMZ2LCKk3KBCU/wBiJQUVHnAX03I2UR3IBCq9C4smPNjMzIjqXWH20utrT3KQobgj4wa0Y4oYeVWzSbEfJ86Rac3e83a/2G1xZd7QxtbYMBp5KXX1ufYqK2FKPMEgBXQqJArd2xWpuxWO3WRpwuIt8RmKlZ71BtASD/RRF36qTie4hrLw06YOZ9crNIvU2TMZtdptUdfIubMd35Ec2xKUgJUokAnpsASQKtutTPKSLsdl0bxnUG432Hb7nhmY2292ZmY04tm4ymu0V5oooSrkK0JWQtQ5QUgEjfeihRljiC8oxc2mrjH4L8fbbdSHGkyr0hDyEnuCkqdCkn2ggH3CrR0B1P4vcxzd60668P8AZsKx1NvdebuMO6okrVKC0BDXKHVdCkrO+32NQOw+VX4Trnao0y73fIbNMcbSX4T9oddUyvbqnna5kKAPiD19g7qs/RPjV0B4g8vdwbTHIrhOu7EFy4Lbftj0dIZQpCVHmWkDfdxPT30Uq9aUpRQlKUoiVUfF19a5qv8AghdP1ZdW5VR8XP1rmq/4IXT9WXRFJdDf2lNP/wAFrV+qNVN6hGhv7Smn/wCC1q/VGqm9ESlKURKUpRF50eWMyPscX00xJDv+dXGfc3Ub+DLSG0n/AOeqtqeCbG1Ypwo6Y2lbfItdhZnLG2x5pJU+d/42tB/K+XpV01vw7FmHOY27GS7yDwckyVgflDKa9R8AsacYwPG8aSnlFptEOCB7OyZSj/8A5opOiz9KUooSlKURKUpREpSlESq4170GwPiJ0+maf55C5mnN3YM5oDzi3yQCEPtKPcRvsR3KBIPQ1Y9KIvFvEsq1w8mnxBSbDkENy4WOcUmdDQoph3+3hWyJUcnoh5HXY96VboVuk169aYanYVrFhFs1C0/vLdys10b52nE9FtrHw2nE96HEnopJ6gj8dRLiS4csF4l9PH8IzBnsJbPM/aLs0gGRbZW2wcR7UnoFoPRSenQgEeW2k2qmsvk5Ndbhgme2uRJx+U8k3i2NKJYnxidm7hCUrYc4A92+xbXsQCkstV7QUrA4LnWKal4lbM5wi9R7rZbuwJEWUyrcKSe8Ed6VA7hSTsQQQQCKz1FilKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiKheIPiE1W0byCDasQ4Zso1FtlxhqfFzsUonzZ1KiFNOthlRQduVQO+ygT4pNax8OXHFxDx9KLci4cMupOqT3nU3fJozpU3JHnC9mwUsKG7Q2bPXvRXolJeYjR3ZElYQy0hS3FK7gkDck/irU/gyzvVrVu9XfUW1Y3i+G6FqM2BiuPwISGZMh5MkbzlcidhzFLoUNwOZR2SduYlKrLWHiW4wdZ8MXp1pVwo6naeX66y4yEZC5JW0IjYcBX65ZQEBQ6FRUNkk95rfe1MS41rhxrhID8pqO2h90f8AWOBIClfjO5rVjie1J4k+HLNzrtb7laMk0YaVBh3zHDH5J1tQpQbcltu7bndax05iPggp23UNqoM2NcoUe4wnQ7HlNIeaWO5SFAFJ/GCKIueqo4k9WLLo5gUbKb7pfec8jvXNmELZaYCJjyFKQ4oPFCugSOQgn/eHtq16wOZZ7hOndqRfM9yy049bnHkxkSrnLRHaU6oEpQFLIHMQlRA9xooWliuPPTZCStfBNqMlKRuScVY2A/JVmcLHFhp5xA5nPs2CaD3/ABgW+K6qTeZFtjtx23EqQDFW40SUuHm5gg7bhBPhVp/RT8Nf+vnA/wCf43+OtdNL9RsN1B8overroRdGLnjj2C8mYzbed4Uue3IHYO8w9VbiUqSjnG+/rgE7Gilbu0pSihKUpREqo+Lr61zVf8ELp+rLq3KqPi5+tc1X/BC6fqy6IpLob+0pp/8Agtav1RqpvUI0N/aU0/8AwWtX6o1U3oiUpSiJSlKIvHvjSB1H8ovDxFs9qhN0xuxhPfsFFlax+V5VewlePWFA6m+VTcmNEvNMZ7OlJV3/AEuA25yn/wCQmvYWikpSlKKEpSlESlKURKUpREpSlESqU4qeFvC+KHAV47ewi33+3hb1jvSG+ZyE+R8FXitlewC0ePQjZQBF10oi8aeHvXvVbyf2slz0q1YtM1WMOygL1aUkrDXN0RcYRPRYKQCQNg4kbHZaRt7A4tlOO5vjluy7ErxFutnu0dMqHMjL5m3m1DoQf6CD1BBBAIIql+LvhJxDikwjzKQWbXl9pbWuxXrk3LSj1LD23VbCz3jvSfWT1BB87+F/iY1J4GNUbjozrPabg1ivnvZ3a2L3cctbyj0mxfBbahspQT0cT6yfWHUstV7G0rH4/kFkyuxwclxq6xrlarmwiTDlxnAtp5pQ3SpKh3gishRYpSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIuGXFZnRHoUlPM1IbU04n2pUNiPyGtANOdWdX+AS3TtEtTNEMpzHBbZcJL2MZRjbHbgw3XVOBp5HwQoFROxUlQJI2UNlV6CKUlCStaglKRuSTsAK8+9VeMHinz/Fc61V4b7Bj+P6WYGZLaskvDQel3hTB5XFRmlbp5d+7dPxqB3SCkJqVrVq5x5WBOhGkmhuVYpiV9lxxkuVZLH83bYgtupcUhpPcpZKB0Cio7bbAEqG/dqt0az2yHaISSmPBYbjNAnchCEhKf6AK1E1I4odc9GbZpXq9kWKW+/6T5BjdnTls9hIRNt9zlJClyUpTsEt7LR6uxSTundBKd9wWH2ZTDcmO6lxp5AcbWk7hSSNwR7iKIoRrtnN70z0azTULG4DM26Y7ZZVyix30qU2txpsqHOEkEp6bnYg7A9a1a1G4oNANXeHXS+8atWTDM1kZVfbRHu9gRexGFllPBaHpS2youpQwSsEL2BCgebuNbtyY0eZHdhzI7b7D6FNutOICkOIUNilQPQggkEGtTNWPJscPmYN2l3T7CbLiUxi/RZ91dbTJcRNt6FEvw0th0JaDgIAUkDl26URcJ0E8mKDt5lpN0//ALmb/wD1FXJofB4YMQTIxDQCVgUdx9PnUmJj86O9IdSnYdo5yLU4sJ5gN1Egc3hvUM/yd/Bn/qOtv84TflqmOlfCdw96JZK5mGlum0Sw3h2IuCuU1KkOEsLUlSkbOOKT1KEnu36URW5SlKKEpSlESqj4ufrXNV/wQun6surcqo+Lr61zVf8ABC6fqy6IpLob+0pp/wDgtav1RqpvUI0N/aU0/wDwWtX6o1U3oiUpSiJXWuU+Pa7dKuctXKxDZW+4o+CEJKifyCuzVRcXOZfMDwzak5Mlzkdax6VGZO+306QnsG9vfzOpoi83vJiwH864wrxnb4KzCtN2vC1HwclPoQPyh5dev1eafkcsPCU6lZ440QAbfZY69u/YOPOAf+ZqvSyil2qUpSihKUpREpSlESlKURKUpREpSlEStc+Mfg8xbiixAPRjHtWc2dpXoa7lHRQ7/NpG3VTKj+NBPMn7IK2MpRF488KPFVqDwV6iztEdbrZcWMTbmlm4W95JW9ZJCj/nMfb4bKtwpSU7hQPOjruFevFjvlnyazwshx65xrjbLiwiTElxnA4080obpWlQ6EEGteuMvg0xfigxb0jbvN7TntoZULTdinZLyRufNZO3VTRPcepQTuOhUlWhvCnxX6h8FmoE3RHW+03JrE2ZpZn294Fb9jeUd/OI4G/OyrfmUlO4UDzo67hRZar2FpWPsGQWTKrJByTG7rFudruTCJMSXGcDjTzShulSVDoQRWQosUpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpRF1rjDRcbfKt7i1ITKZWypSe8BSSNx+WvLK4a023QnhO1K4ItZbVOx7NLTCuUPHpCoi1RL7HffU60424kEAnnPVWydtuoUCkXfd+Md7hy4qtVMU4jL/kMfE7i3AmYWEwDIitNdiO17PkTzHmUevUgKQoHY98W088olw15vplb4PFaxb73lTUmWX2FYn5zGbaLy+wKAUqSCWuz32O++9FIWF1J4k8W4geHfDeDnQKJNy7Msns1ktF2eahuIiWdphDCpDjjiwN+VTWxUN0gcx5t9gfRuw2pFisVusjbpcTb4jMVKz3qDaAnf8AHtWm2JeUM4BMEaXGweP6AbfIDiLZiSooc9nN2aBv+Ot1I77clhuS0SUOoC07jboRuKIVw3O526y26VeLvOYhQYTK5EmTIcDbTLSAVKWtR6JSACST0AFV59E7w5HqNdsC/OCL/jqe5BYLPlViuOM5DBRNtd2iuwpsZZIS8w4kpWg7EHYpJHQ+Na255wgcBGl2Jzs3z3SfF7LY7YgKky5D8nlRuQlKQA4SpRJACUgkkgAUUK1foneHL/Xrgf5wRf8AHWbxHWbSTProqx4PqZjF/uKGVSFRbbdWZLoaSQCsoQonlBUkb+8VoCrULyO6eYjBYpSk8vMLPdNv01cXCTmXAfe9XHrbww6fvwMl9CyHpFyRAksNJhhxoKbKnl9SpRbIAT9ieo8SmFudSlKKEpSovcNUtM7Tfk4tddRMZh3lSggW6Rdo7ckqPcOyUsK3Ps2oilFVHxc/Wuar/ghdP1ZdW5VR8XX1rmq/4IXT9WXRFJdDf2lNP/wWtX6o1U3qEaG/tKaf/gtav1RqpvREpSlEStFvK2akt49ohYdNo0gJmZfeUvPICuvmkQdorcewuqYH5a3eu11tlitcu93mexCgQGFyZUl9YQ2y0hJUpalHoAACSa8Z9Ycvv/lCeMC2Y3hKJCbA46mz2lakH/k1qaWVyZzg+xKgVr2PX6knvopC378mlp6vBOFLH58pns5eWypOQOgp2VyOqCGd/jZabI/fVtRWPx+xWzF7DbcasscR7faYjMGI0O5tlpAQhP4kpFZCihKUpREpSlESlKURKUpREpSlESlKURKUpRErXLjD4NsQ4osX86YMez5zamVC0Xnk6LHf5tJ26rZUfHvQTzJ+ySrY2lEXjlw0cUOqfAvqRO0Z1msdyOKtS+S52hz13rWtR/zyGe5bah6xSk8qx6ydlb7+u2J5ZjWdY5b8vw+9RbvZrqwmRDmxXOdt5s+IPgfAg9QQQQCCKpziy4RcF4pMREW5Bu05ZbG1ehb821u4wT17F4Dq4wo96e8H1k7Hffze0Y101y8nlqxO0y1IsUuRjjj4cudjLnM24hR2E+3uH1SSB4bJXtyq5VDdJZar2bpUW001NwjV/DYGe6e36PdrNcUbtvNHZSFj4TbiT1Q4k9FJVsQalNFilKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiLQ/JeK+0aLcUuqeD8U1wnHEZTcGZhKJFnTLjMNlkB0tlLZX6yj1O5AUhYOxqveETjE4RdKtC7RhWqMplOQxZtxefCseclHs3ZbrjX0wNkH6WpPTfp3eFekN3xywZAlCL5ZIFwDe/Z+dRkO8m/fy8wO1aJ3ziG4SeD+2zNEcH07vep03EnJUi7PtW+NKFvU46pxxEiWpCUjkUsp2CSE7BJO4Iop1UxHlCOAMkAS4+5IA/5pO9/8VW5Ed1p+O0+x9TcQlaOm3qkbjpWskviN4arDm+n2DZpp9AsEjUjHoN8tU+TaI6oLbkrfkiOvBPqub7Dm25fWG5TuK2eACQEpAAA2AHhRQvtaheUkiW+fgul8LKXOzxCRqTZ2shUpXK2Iig6D2h8E9TufCtvaiWq+n+A6oafXnCtTrfGl43Ojkzg+72SWko9cOhzcdmpBSFBe4223oi1c1pxHT7TzjJ0BvELEsfi47l1uu+JyYzcBlMR09mlyMOQJ5CedaQk7e6szhlqxzQ/jylaaacQ4ttx7UjC15DdLNEQlDEK5RpBQmQ22no0HGyoFI2BI32qps+yryc0vRazcMl04gJi4+KS1ybLf4wfly7fJLrigtMltktrSA4UbD1SkJ7iARdXBnw6aPYOu461YZrDO1YvWQxxAVk02YmQWo6SklhIBUpCt0o5uclXqpGyR0JStpqUpRQqL42tX7vofw05hnOOSTGvfYN262PDvZkSHEtBwf7yEqUse9IqsNLfJ4cPdz0Ktls1Dw1N5y7IbW3Pu+SSHnFXFM59sLUtt0q9XkUrYDuPL6wVud+byqMGTL4R7pJYQpSIF7tkl/Ydzfa8m5/4lpraTC5ke4YdYrhEUlTEm2RXmlJ7ihTSSCPxGilaz+Ts1Cyu+acZXpJnV1fud60oySTjXnj6ipx6IgnsOYnqSnlcSN/sUpHhVrcXAKuF7VZIG5OI3QD+Trqh/J9EXDVPiayKL60GdqC40wsfBUUOSSrb8Tify1e/Fygr4X9UwHltbYnclFSdt9gwokdQeh22PuNE3rH8L2tekmfaUYZYsP1EsNzultsFvhS4DUxAlMvNxkIWlTKiFjZSSN9tunQ1dleHWk/ATxDapYVdM+s1hcsLtpQzJgRbwxIiPXVhxouJdhuBBC+gAHduVJ69akWGveU10/tiX8ZtusjFvjEIEeXCdmJR032DMhKzt7wNvCimF7R1gs1zrDdOMelZZneS2+xWiGkqelzn0tIHTuG/VSj4JG5PcAa8ml8R3lQ7yg2hi258l1Xq7x8GQ27/AObzbp8dRa48KnHJrnZbzqTqTasqmmxMPOtR8kfeVPlOISD2MOHsVbq3ABCUIPXr02oohTLi+418w4sL9H0L0Js91+ZWfLRHQwy0oTsifB9UKQOrbAI5gg9+3MvYDlG7nA7we27hiwpy65F5vNz7IWkG7y29lIiND1kw2VeKEnqpQ+Gsb9wTtSfCTpJl3D/wn5VrDZdCJR1raXKaiRLtbX3JkhlLqA22hncLQhSSokN8vNy7knarxtetXExJ4Q7hqzN0f7HVWO4tMfF/RUodokTUtJPm5X2p3ZJX0V4b91EPALZilazy9a+JlrhCj6ssaP8APqq44lLmLeipR5UmaWifN+ftfqOznwvf3V8yfWviatvCVYtVbLo953qlNcjpnYx6KlK7BKpC0LPYBfap2bCVdVeO/dRQtmaVrNqFrXxNWDhYxPU7EtHvSepd0chpu2N+ipTnmiXEuF49glYdTylKPhKO2/XvprFrXxNYjw44PqHp7o96c1AvXmXpyw+ipT3mPaRlre+koWHEcjqUp9YnbfY9aItmaVrNr/rXxNYHolgWZ6U6PfNJmN8bjKv1o9FSpHmClxC44OzbWFo5XvU9Ynbu76+cS+tnE1pxp1geQaM6PfNXf742FX6B6KlSfMFeboXtyNLCkfTCpPrE923fRFs1StauKXWniW0xsuCTdENIfmvmXwO+nmfRUqV5hyoZKejK0lG5W4PW3+D7jX74p9Z+JPTG84ND0P0j+a+Jey56de9FypXmAC2QnqytIRulbp9bf4HuNEWyVK1v4mNZeJHTjUfBce0Z0k+avH72tIvs/wBFyZPmAMhtBPO0sJR9LUtXrA/B37q/Wv2snEfgmtuBYZpVpL80mHX0xRf7v6MkyPR4XL7Nz6Y2sIRytev6wO3eelEWx1K1y1h1j4jcS4jsG080+0m9N6f3vzL07fvRcl7zDtJK0PfTkLDaOVtKFesDtvuelfnUbWTiQx7ikxTTLENJfSmmt0TEN2yP0XJc80LinQ6O3SsNJ5Qlv4STtzde+iLY+la4ZRrLxIWzi2s2lVk0k870umIYVNyf0XJV2ClMOLWO3C+yGziUJ6p6c23fX6l6x8RrXF+xpKxpNz6UraSteU+i5J5VmEXSPOOfsfq4CPg+O3fRFsbStcbVrHxHSuL2bpNO0l7HSplpxTGUejJI7RQhpdSPOCvsju8VI+D4bd9fnDdZeJG8cWF/0syLSTzHTCAiSbfk3ouSjzgobbU2O3UvslbqUsdE9eXpRFshStcNMdZOI/JeJ7MNNc10l9E6cWkTTZ8i9FyWvPC240ln6ctZaXzJUs+qkb8vTur9aKax8R2ZcQmd4DqRpN6BwSx+e+gb56LksekOzlIbZ+muLLa+ZoqX6oG+246URbG1UvEfwz6ccTOFKxbN4ZYnRgpdpvMdA86tzxHwkE/CQdhzNn1VD2EAiE8POsvEjn+sWeYjq1pJ8zOJ2NUgWG6+i5Mf0gEyy2j6Y6soXu0Av1QN99+6nDBrLxI6l51nNk1r0k+ZKy2RYFimei5MXz8du6jfmeWpK/UQ2r1QPhb9xoi86bXd+JDyaWsq7fOY87styXzOxypforIYqTt2jSv+qfSD37c6CdlBST19V9AOInTXiQwtvMNPrrzLa5W7jbHyEzLc8R9TeRv8fKsbpUOoJ67UTphJ1U4vcU1F0+4udA2bHZreGTYFG1SobzzpL47Zlx1aiHEcjRCkbfDIO4VtXm3jmO8VPCpdI+s0DCswwpEGSmGudPtrjUZ8K6hiQhQAW2vb7Ibb7bEK2ostV700rSPhJ49Mi18xrObblbGMQs/tUN6ZjVjgNPpVdEojqVslK3FF1XagAoQQrY77eNWHoTrXxM5xoNnec6oaPfM7m1jRLNhsvoqUx5+UREuNfSnFla+Z0lHqkb7bDrRYwtmaVrPpRrXxM5Twz5pqPnej3obUSzqnCy4/6KlNeehthtbP0lay4vncUtPqkb8uw6ivmC62cTV74VMn1QyfR70dqbbXJabXjXoqUjzpKCjsj2Cll1XMFL7lDfbp3URbM0rWew618TM7hFu+q900f821TiOOpiYv6KlJ7ZKZaW0HsCvtTu0VL6K8N+6g1r4mPoQfnsnR/wD/AIq9py/Mt6KlfB8+7Lfzfn7X6h9M+F7+7pRFsxStZr/rXxNQeEa0ar2rR7znVOW40mZi/oqUrsUqlKbWewC+1GzQSvqrx37q+ZzrZxN2ThTxjVHGdHvSGptyciJumNeipS/NUrLgdPYJWHU8oSjvUdt+vfRFs1StZtVta+JrFuGfC9R8F0e9M6iXhUIXrHvRUp3zIOMOLe+koWHEci0oT6yjtzbHvprrrZxNYRoNgec6YaPfNFm18REN+svoqU/5gVxFOO/Sm1haOV0BHrE7b7HrRFszStZuJDWzia080qwPJ9HtHvmpya+Ntqv1t9FSpHmCjFDihyNLCkbOko9Yn2d9frih1q4l9M8UwO6aJ6QfNdc74lw36N6KlSvMCGmlJ9VlaSjdS3B62/wfcaItl6VrZxUaz8SmmD2DI0O0i+bBF77X06fRcqV5hylnl+orTybhbvwt/ge41ycT+svEhppnWDWTRTST5rbLe1lN9m+i5MrzBPbtI35mVpSj1FuK9YH4O/cDRFsfStcOIbWTiPwDWHAsR0l0l+abE76pgX+6+i5Mj0eFSktrPaNLCEbNEr9YHu37q/WtesfEbhvEHgeA6b6TensEvnmXp6+ejJL/AKP7SUtt76a2sNo5WglfrA7b7npRFsbStcdTtZOI/GuJ7D9NcK0l9LacXcQjeci9FyXfMy446l76chYaRypSg+sk7c3Xvr85nrLxIWfiwx/SzHdJPPtMJ6IxuGTei5K/Nytt1Tg7dKw0nlUlsdU9Obr3iiLZCla43XWPiOi8XsLSaDpL22lTzTan8o9FyT2ajDU4oecBfZDZ4JR1T47d9IusfEc7xfvaSv6S8mlKG1KRlPoySOZQgh0Dzjn7L6vu38H3d9EWx1K1wxfWXiQufFtedKr3pJ5ppbDQ+qFk/ouSnt1JYbWgduV9kd3FLT0T15du+vmnWsvEjkHFLlWmWX6Sei9NbWmYbVkfouS352W1NBodupZaVzBTnwUjfl6dxoi2QpWuWj+sfEdlvEdnOnmoOk3oTT+yefegr96Lks+f9nJQhn6ctZbXzNqUr1QN9tx0r86A6ycR+d63Z7heqmkvzN4dYzK9AXf0XJj+kAiX2bf0xxZQvma9f1QN+8dKItj6Vrfw0ay8SOo+pGdY7rNpJ8ymP2RahYbh6LkxvPwJLiAed1akr+lpQr1QPhb91fOFjWbiS1OvWcw9cNI/mQh2RTYsT3ouTF8/BW8FHd5agvZKGj6u3w/eKItkaVrVws60cSup1mzqZrhpD8yEuyBr0E16KlRfP+ZDxV0eWor2KGx6u3wveK4+GjWviZ1H06zzINZtHvmUv9jQVWGB6KlRvPz5ute3I6sqX9MCU+qR37d9EWzNK1m4fta+JnPdE88zPVfR75msxsbclVhtHoqVH8/UiIHGx2biytfM96nqkb9w600d1r4mcu4cM31D1B0e9B6gWXz70HYPRUpnz7s4yFs/SVrLi+dxSk+qRvtsOtEWzNK1m081r4mr/wALGWanZbo96M1LtbkxNpxz0VKb87S2lssnsFLLquYqX8FQ35endTGNbOJq5cJV91VvWj3mmqMJx9MDGPRUpPbpTIQhB7Ar7VW7ZUrorw37qItmaVrPE1r4mXeEKRqy/o/yaqtuqS3i3oqUOZImhoHzfn7X6huv4Xv7ulLprXxMxuEOBqxC0f7bVR9xCZGL+ipR7NJmKaUfNwvtRsyAvqrx37qItmKVrLl2tnE3aeE7HdU7Bo757qhcHY6bjjPoqUvzdK3VpcPYJX2qeVKUnqrx99fdTNa+JnG+F/D9S8M0e9LakXZcJN4x30VKc8zDjTinj2KFh1HKpKB6yjtzde+iLZmlaza1a2cTWG8POC5/pvo96ezy9iF6dsXoqU95gXIi3HvpTaw4jldCUesTtvsetOInWvia0/0fwLLdI9Hvmnyu+NsKv1q9FSpHmClRO0WOzaWFo2d9T1ifZ30RbM0rWfic1q4mNNsGwS96LaP/ADW3q+IJv0P0VKk+YHsG1j1WlpUj11LT6xPdt4VycU+tHEpphFwV3Q7SH5sHL323p1PouVK8w5Qzy9GVp5N+dz4W/wAD3GiLZSla38UusvEjpllODWzRDST5r7be1rF9k+i5MrzAB1lIPMytIRuhbh9bf4HuNbEIdfM91hTezCWW1pXseqipYUN/cAn8tEXYpSlESlKURda5pmLtstFuUEy1MOBgnuDnKeX+navNXSm44UnyZ2sOPQlR2c7gRr182Ud0gTjN84Vs48D65TycqQT03Ch3g16Z1rPrp5PrQHXTJ5ecXCPeMayG4p2nzrDKSwJp7ip5paVIWo7DdQAJ8SaKQqj4wbtgSPJ9YVjV+VFfyy62LHG8SiAhUwz+zj/TGU/CACOcKUOnrBPeoCt4cZauTGN2pi8KKp7cFhEpRO5LwbSFn/zb1rrov5PTQXRvLIWeld/y7IbZy+j5eQzEyEQlJ+CpppKUoCk/Ykg8venY9a2dooStUPKMuyZeleG4lcL3JsuIZTnNps+WXFhZR2FscUsqC1dyUFaUbk9NwN+/atr6qfiZ0PufELpi9prA1ClYixMlNuTpDEFEsSoyUrCo621qSOUlSVb77goG1EXYxrhk4dcZx5jHbJoxhhtzbQQntrQxIW6nb4S3XEqW4T+6USTWvmluD43oZ5QG7ab6LN+Z4pk+EG+5JYoyyqLa5yHwlhxKdyGytJ6J9jh29XYDEY9wW6lYXZm8XxvyiGW2u2xk9k1DaSgIYSPsUBUolAHsTttWNxDyeOW2K73W44Nx1ZTEut7UH7pItzA85mlPcp5aZXOvbm+yJ23opW/lKqjh40czXRnHbpZc31ryDUqVPnCWzOvKVByK32aU9inmcX6u6SrvHVR6Va9FCiWrOmuP6xab5DpjlAX6MyKCuG8tvbnaJ6ocTv05kLCVj3pFan4ziflH9LcEb0NxSFp1f7dbmPRlmzaXcXGpESGBytlcc9VLbTsE+qrbYA8+253dpRFUXC3w/W7ht0nh4CzdTdrpIkO3O93NSSkzZ723aLAPUJACUp367JBPUmvvFz9a5qv+CF0/Vl1blVHxc/Wuar/ghdP1ZdEUl0N/aU0//Ba1fqjVTeoRob+0pp/+C1q/VGqm9ESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESulerLaMjtMuw3+2RbjbZ7SmJUSU0l1p5tQ2KVpVuCD7DXdpRF5ZcWfk3Mn03nvatcMXpGVbITvnzlijPL9I2paTzdpCWDzuoSeoRv2ifArHdJuEnyn6f+SadcTcjsnUERouWhrlBIOwTObA9RXgXkjb92kdVV6T1qVxY+T1054gvO8yw5UfEc8cBWqa01/wAjuS9u6U0n7I/9qn1/aF9BRTPFbWwLhAusJi52ubHmQ5TaXmJEdxLjbqFDdKkqSSFAjqCOldivGHANbOKnyeWajT/NbFJfx5ThcNhuLilwJTe/rPW+SNw2T/u7p3+Gjfu9NeHfi50a4lLYleEX0RL602FzMfuBS1Oj+0hO+zqB+7QVDu32PSiEQrppSlFCUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlYeFmGKXLIZ+I27JLZKvdrabfnW5mUhciK2skIU62DzICtjtuBvXeur8iLa5kqG12j7MdxxpG2/MsJJA/Gdq81dI7xh+kfAFqDxOsTQ/qbn3pK23S6vSFKkmc/LW02wATskoCu27ubvJO22xFu1jvFjw45ZnytL8d1fx+bkwfVGRCbeUA68kkFtt0gNuL3BHKhRJ26VbVeZepvDlpho/wCTesmX3LHrdbNR4jFsvsC9IQG7iq6yJDbiWQ78NQS0op7PfYdnzbAjevSHGJU+bjVpm3VHJNkQWHZKdttnVNpKxt4dSaKVk61O8o/ccqRpBi2L2TI5WPWfLMztliyG7R1lBjQHysHmUCNkFfJv1AOwSeiiDtjWrXlEcwyO06K2vTvFbXa5c/U/IoeIBy5sJdjx0yOYlZCgUhW6EhKiDy7lQ6pFFAWBY8lRwmtsobegZZJcSkBTzl9cCln2kJSBufcAKsfQzgj0J4d8yezvTa33pi6vwXLctUy5rkI7Fa0KUOVXTfdtPWqbxrgS4m8esUKzQuPPM4TMRlLaI0eI6tpkAbciCuRzco7hvt8Qq29BOHbWvSrNnsl1C4p8m1FtjkB2Im0XKKW2kOqWhSXgS6v1khKh3fZnrRSthaUpRQlKUoiVUfFz9a5qv+CF0/Vl1blVHxc/Wuar/ghdP1ZdEUl0N/aU0/8AwWtX6o1U3qEaG/tKaf8A4LWr9Uaqb0RKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlEUW1I0uwDV7F5GG6kYrBvtpkdSzKb3La/BbaxsptY8FJII9teZ/EF5MXUzS65nULhovdxv0OC550zb0yOxvUBQ6gsOpKQ/t4bcrnhss9a9WaUUgwvKnQTyo+pGnExOC8SOOTcgiwV+bPXJtgR7zDKehD7KuVLxHjvyL8SVGvRjSTXfSXXOyi+aX5vbr22lIU/HbXySoxPg8wrZxs+HrJ2PgTUe114UND+IiGRqJiDSrohHIxeoJ83uDHs2dSPXA/crCk+6vPHVfyafENonePm50AyiVlDEBRejuW182+9xgOvRKVBLv8A4agT+4opsV620ryY0m8qBrvpPc/mM19xFzKG4KgzIMlj0beo23T1wUhDhHsUhJP7qt9NFeNTh212DMPEs7jwLy6B/wCxbxtDm83sSlZ5Xf8Aw1KooIhXnSlKKEpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpRErTFfBnwS6g675Aq2ZDJl3mxzEXzIMOgXj/ANmMSFrPrvMhJ7PmUFczYWPYQAdq3Eujkxq2S3bejnlIYcUwnbfmcCTyj8u1eYumOZ6caQ+TtzjVW2S7enVHM3LlZbxMcWlVycnvylt9kdzzgIaUXuXu71d53opVr5PnfAdxKcUOMKyHVaddLpjSW7bZ7E52rNgmzm3lKQsKUjkcXuoJACkpXypHrjYHe2vN3WrRzTPRjyalitF+s1qhZ0uNbbla5AZQi4OXp99t1YQrbtCUtrWggdAlA9gr0Pxddycxq0uXkKFwVBYMvm7+27NPPv8A8W9EKydUHxr6Xag6w6LKwnTTGbJd7y9dY0lC7nNXENv7ILUmXHdQQUvoWEcu+4IUoEEGr8rXrja1nz7SDTGzxNJ22Pm0zbIYeMWZ19tK0R3X+Yl3lV6pICdhzAgFW5B22JQqhsLnlZbLao9slW7S27rjoDfnc58du4ANt1ltSElXtISN6trQC4ccUrOHm+IuxYDDxYW90tOWJ1SpBmc6OQEFZ9Tl7Tfp37VrJfMP468f11xPQy9cYzzVwzKyTLtCuLVrbVGD8Y7uxeUoCj6vrBfTp9jVj8J0LiCxribynT3iR1vvN9vdlsAm2a17A226QH3UJM9pQ5Ppja2+zKFN7jtD12opW7tKUooSlKURKqPi6+tc1X/BC6fqy6tyqj4uvrXNV/wQun6suiKS6G/tKaf/AILWr9Uaqb1CNDf2lNP/AMFrV+qNVN6IlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoigGq+gukGt9s9GaoYFa74EpKWZDrXJKY+9vo2cR+JQHtrRDWjyRDiS9d9BM+CgndaLNkXeCOoDcttP5OdHxqr0upRSCQvGiDrNx88E0tqz5izfRYo6g23FyJk3K1uJH2LMtKjyD2BDo/e1tDpB5W3SvI0x7drDiFyxCYvZK58HefAJ8VEJAebHu5F/HW982DCuUR2BcYbEqM+kodZfbC23EnvCknoR7jWsOrvk3uGTVMyJ9uxh7C7u9urzzHVhhsrPiqMQWSP3qUn30SQdVfOn2rOmeq1sF303zqy5FFKQpRgS0OLb9y0A8yD7lAGpZXknqB5LniN0uuZybRPMouS+aqK2HIUpVouqAOo23VyE/E6N/ZWEsfHJxw8Ndxax3Vu2Trmw0eXzTMbWtt5Y/wDdy0BKl+5RLg+OiRwXsPStFdMPK16K5IGYep2J3zDZa9kqkMp9Iwt/E8zYDqR/4Z+Ots9O9b9IdWYyZWnGo9gyAKG/ZQ5qFPpH+80SHE/8SRRREKb0pSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJXnxxEx/JoaQa3PZZqDYn7lmzcoXK4WOxh6THTJ35+1kMBSWErJ2UUFQ333Uk79d/rmuW1bZbtvbC5SGHFMJP2TgSeUfl2rzR0xx/BJ3k2NXdQ0x4k/UC8R70vLLjIQlc9E3zhWzS1H10J5ORQT0BKie80UhbBZm7wbam8SWmOUakyZBzm7YvBu2JW+6qU3AdZddW5H50DdBlBRVshatjsnYKIFba1oTxg2HT93yfuFZZfY8RnLrTYcb+ZOclITNE4tx/pTSx6xBR2iikdPV5u9INbyYy5cnsctT15SU3BcFhUsEbEPFsc//q3ooWSrX/jT0jznVHTG03XS1liRmeB5DCyuyxXlBKZbsYq5mNyQAVJUdtyASkDcb7jYCqB4zdZM50l03s9u0sajnNs7yCHitiekpCmor8jm3fIIIPKlJ23BG6gSCBsSKnNdrjrZlWJ6J8YNq0TvEPKdPbnNcyDDN1LnCC/uw9yDkCiNmuYDlJCXQrYgE1zaA5bqBxP8WjXEK9pbfcIwvD8SkY9DN5ZLT9wkvvBagAQAQnqTy7hPKnc7q2Gyel+neoOLaMM4Bnuqc3JcpXDltScm7IoeDr5WpK0AqO/Zc4CT03CE9E9wozh6yjWrRriDl8KWtGoD2fwLjj6sjxPJJbfLNWy272brEg7kqI6ndRURy95CgElK27pSlFCUpSiJVR8XP1rmq/4IXT9WXVuVUfF19a5qv+CF0/Vl0RSXQ39pTT/8FrV+qNVN6hGhv7Smn/4LWr9Uaqb0RKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURK6F7sFiyW3O2jI7LBusF4bORZsdD7Sx70LBB/JXfpRFqrqf5NPhb1EL0u1YvLwy4Obnt8fk9i1zHxMdYU1t7kpTWpuofkkdYMXkqvGkOo1oyHsDzMNTAu1zk+zlcSVtk+/mRXq5SimSvG9OqflIeFUhnJRmZtMU8m17h+mbfyjwEkc/KPicTVs6c+WGuTYbi6r6RR5QBCXJuOzeQj2nsH9xv7u1FemxAUCCAQehBqp9RuFDhz1WK3s30hx6XKc33mx43mkrc+PbM8iyfjJokhQDT3yjHCdqB2bKtRDjUxwgCNkMVcPYn/3vVn/11sLYMnxvKoKbni+QW28Q1/BkQJbchs/8SCRWjeoPkh9Irz2kjTfULIsZdVuUx5qUXGMD4Ab8jgHxrNa937yZ3F3pTON50qyW23dbRK0P2G8uWyZ0/wB1woG/uDhokBevdK8c08SnlHuHhQZzhnLHITJ2PzT2Hz1hQH/5pKeYj3h2rPwTyxN+a5WNSNHIE0DYLk2G5KZUPaexeCh+LtBRIK9PaVqPhHlQ+FTKy2zer5fMTfWBum8WtZbB9naMdonb3natgML1y0a1FbbXg2qWL3tTnwWod0ZW7+Nvm5x+MUUKcUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoi4pUliFGemSXAhlhtTrij9ilI3J/IK8+7TwxaNcbNkyDXHQTN820uYyybKtuQ2/sEmDdnUEFxa46XQFJUVAn1uUq39UHff0GeQFtLQUJXzJI5VDcHp3H3V53aaaHeUX0XuF8b0ji6ZWTG7zcXLj8zyp65MCM8vopTAcSXWgoAbpDnL07ulFIVraVeTsxzGMtsOZ6watZPqjKxVLQsUC7Et2+B2QAa5WSte4Ryp5U8wT6o3SdhW31aVPTvKqRmy/KZ0XZZR1WtbriQB49Sdq3Qil5UZkyCkultJc5Pg823Xb3b0QrlqkuLnQi969aWtWfDb23Z8vxu6xsixyc6SENz45JSlZAJCVBShvsdjynYgEVdtUlxc68XjQPStF6xCyovGXZFc42PY5BcG6HJ8gkIUsAglKQlR23G52G433BQqSicWXG7i0VOO5rwO3i932OkNLuNmnnzGQsdO0HIhxKQe/ovbr4VLeG/SjXTLtZrpxUcSttgWC+PWcWDGsYhu9qLVBUsOOLcWCRzqPhuT6yyeX1UiIQOEHjQyiEMjznjlyKzZDJT2y7fZoqvMYyyN+zHI60lQHd0QB8ffUs4ZtVNc8R1ivPCrxK3WHf79EtIv8AjWTRmw36VgBYbWlYAAK0k9+wPqrBKtkqJStsKUpRQlKUoiVUfFz9a5qv+CF0/Vl1blVHxdfWuar/AIIXT9WXRFJdDf2lNP8A8FrV+qNVN6hGhv7Smn/4LWr9Uaqb0RKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpUI1o1fxTQjTm6aoZs1cHLNaCyJCYDAefPaupbTyoKkg+ssb9e7es/h2U2zOMSsuaWVL6bffrfHuUUPoCHAy82lxHMkE7K5VDcbnrRFmKUpREpSlESlKURKUqtdE9f8F17YyWTg7F1bRil6esU/z+MlkmS18It7KVzI69D0+KiKyqUpREpStdNcuOTS3RDPk6XO4zl+X5S3FRMl27GrYJS4jShzJLm6k+sU7K2TvsCCdtxRFsXSsdjl5byPH7ZkLUKVDRdIbM1MeW3yPshxAWEOJ3PKsc2xG52INZGiJSlRiHqZgtw1Bm6WW/JYknKrZb03OdbWVFbkWOpSUoU7t0QVFQISTzEddtutEUnpSlESlKURKUpRF8UlK0lKkgpI2II6EVWOd8MPD3qXzqzXR7Fri85uVSRb0MyCfb2zXK5/6qs+lEWlmb+Sf4bsiDr2JXLKsTkL6oTGniWwn/gfSpW3xLFUBmnkfNR7e4uVp7qzj93DfrNN3SE7Be9w52y6nf39K9UqUUyV46/OA8pdoP1xN7N1w2j6osGQC4MED/8ALlZO3uLddiH5Q/jh0lkpg6kWZqYGfVU1k2MOQnDt7VtBrr79jXsFXBMhQ7hHXEnxGZLDg2W082FoUPeD0NElebWHeWNCkIbz3REqP2ciyXcKHxhp5A/r1d+I+VM4VMjLbd5umRYw6rbm9J2ha0JP7+OXBt7+lXBmPCPwzZ6pTmT6I4m86v4T0eAmI6ffzscit/x1SOXeSl4YL/2juPuZXjLq+qRCunbtJP7yQlw7f8QollfOIcUvDnngSMV1rxCY4v4LK7o0w6f/AA3Slf8ARVmRZcWcwmTCktSGVjdLjSwtKviI6GvM/L/I43EBS8G1uiyP3LN6sxT+IuNLP9SqzleTr44NL5Cpenl6jSuy6pcx3KHISzt7Eudl+TeiQF7BUrx4Gf8AlSdHd13JrUt6Ox0UqdaUXlnYe1zkd6f8VZKy+Vb4m8QdELOsNxW5rQdlpmW6Rbn/AP0r2B/4KJC9daV5xYt5YywPJbRmeh9xjn7N603duQn8SHUN/wBarexjyp/CnfShu7XDJ8eWr4XpGzLWhJ/fMFyiQVt/SqbxfjG4W8xUlFj11xErX3Ny56Ya/wDyv8hq0rNkuOZEyJGP3+23NojcOQ5TbySPjQSKKFkqUpREpSlESlKURKUpREpSlESlKURa/wDES/xnwMhgvcOETBLpYZENQnMX5K25EaQk96VhwBaFJI6bbgpPeCNtUeFmT5RW0aMWuBpTh2CKxxEyeqOciU61P7QynC7zpUtJCe0K+Xp8Har14h+JPXOdrQjhk4T8YtU/MIlvRc79ert1iWplexQkgnlB2Ugknm+qJSlJO+2V4yeI7UTRXD8KwPTiHDnaqaiS2rVbgUB1qO4AhLrwQdgr6Y4hKOYcvrFRBCSCUqoNVNJvKQcR2JnSnUyFphYsaucyO5cJkCQvtkIbcC+gClk7EA7AAnYDcAmt+bXBTa7ZEtiHVOpiMNsBa/hKCEhO5952rQy+51xp8GUrHdQtedSbPqXp7eLixbsgRHiJakWdbx6OtqDaCUjrt9iduUpSVJVW/LD7MlhuTHcS406kLQtJ3Ckkbgj8VEK5K1443tJ871K0utN+0rjol5lp9kMPLLRDXttMXG5uZgbkAqKVEgbjcp28a2HqqeJfX208OemT2dTbPIvVxlS2bVZbRHOzlwnvbhpoEAkDookgE7JOwJIBKFr/AGzyqOj8C3phakabahYzlDCezm2g2cOFDw70oUpaCRv3cyUn3V3OHJWo3EhxJTOLPK8DuOGYhZcdXjOIQLmjkmTUuu9o7JWnYer8Ib/B9ZISVcpNYpETypmZoay8I0mxXtEh5ixS4/bPNJPUIW5yuEK9v0wdfZVncNfEtn2bZzfNBuIDA4+Iam47DRcezhu88G6wSoJ84jklXcSncBSh17wQpKSlbI0pSihKUpREqo+Ln61zVf8ABC6fqy6tyqj4uvrXNV/wQun6suiKS6G/tKaf/gtav1RqpvUI0N/aU0//AAWtX6o1U3oiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiLWHylH1m2dfvrd+vMVbXDl9b9pp+CNo/VG6qXylH1m2dfvrd+vMVbXDn04ftNPwRtH6o3RFLstyqxYNi91zLJ56IVossN2dNkL7m2W0lSjt4nYdAOpOwFaj4lxGca+vFpd1I0H0Swi2YM4456IOW3F5E+7toUU86A0oJbBIIG/q777KUBvVleUBh3Sdwe6ls2hC1uotrLziUAklhElpb3d4dmlZPuBqlOHzRXiby/RDCMh0642F2zHZVki+Y29nEIjqYSEoCTH5yrdRbUFIJPUlJ3opVw6QcU+Ra06aZ4iz4K3YdWtP2341zxS5uKW2JyELLQStOylNOKbUkEdQfaNiqV8Jevv0SWilq1Kl26Pbbst+RAu0COpRRFltLIKBzetsUFtQ36+vVMcJel9qxDiX1OyC48S0DUnOXbe1b8ngMWTzFyK8hxvs3HChRbJAQUbJHfvudwarm26ixuBvW/iHw2aERrBfLG5qHiTKxs2uYrdCo6AenV1fLt+5Yoi2I024n7pqDrjq3i4tdphab6UsJZmZAtbhednJTzPJPXkDbYbf3IBPqp9tV1i3E3xc8Q6Z+ZcM+j2Hw8BiyXY9uueZTX25F4LZIUpptojkSSNuu4B3HNuCBHcD0byTA/JnZsVsvuZhnGO3LK7urb6c45Jb7TlPiSI4SCPaVVhOEzR/iPzfh5wu/6X8ZJx7H1wVNMWdnFIkgW5aXFhxguKVzKUF8x3VsTvv4iiLZDhm4nJes9wybTnUHC3ML1Lwh1LV9sanu2aU2r4EiO59k2Tt7duZJ3UFA1Uvky/2K1r/wDiVcf0CvvDxpO1i3GHk2R5XxTwtQ9Ro2OeYX20NWIQn245MctOOKbUW/UAbG22/rDfqK+eTL/YrWv/AOJVx/QKIt0H32IrDkmS8hpllBcccWoBKEgbkknuAFab2Xis4mOIi8XqZwl6U4qvCLHMXATkuXy3mkXN5HwhHaaIITsQevNsCnmKSeWtl9boVzuWjGe2+ypWq4ScZujMVKBuouqiuBIHvJI2qj/JnXSwXDg8w6NZFtdtbnZ8W4oTtzIledOLIWPaULbV18CKIu1oXxXZrnOY5PoPqxp1Fw3V3HoDk6NATKLluuzfL6rjDnUhO6kEjdXqkkHooJ1k0kv/ABXI44tUrladO9P388ftFtbyKC9dnxBhxNo+yozu3MtZSEEpOw3+Krb1MkRb15UrSuFjbiFz7HiE1d8LXe00tuUUJcI7ujiNt/8AtE+0V2dDv+kt17/Bu1/1ItEW6VaoZnxMa853rXlOiXC3p/i1wdwRLKMhv2UTHWozch0bpZaaa2WojZQ5uvVKugABO19aR5zw8YhrJrfm+ofCpxLXDBNS7S83Cy6Hb0rcjKkjdKQ+jdBBPZHm2507p35QSSShWFolxI6yXTV+58PnEBpXBx7Lo9rN2tl0sjzr9puLI6bBSgS2o9SN1deRYISQN9bdIL7xSJ49NS5sTAMFVl0q12tvJoSrq8IcS37xwHYzm3M47yBJ5VbDckVauluufFHpBxGYtw2cTUvHMtj5nFfdsuQ2hvsnwWkLVu6kJQCN2ylQKEkFQIUobiu1ox/0mOu34K2r+pFopX71U4ytaLLxEZdw2aTaS2vKMkjRYDmPKcfW2j6Yyh6S9MUVpQlptKtkgFO5Unr4GQ5RxY6m6JaHWXIdedMIY1UyW7vWWxYjYZJcE90L5W18+7nKkgpUeUr+G2B1VsIzpVHZc8qHrK+42lTjOFW7s1EdU8yYW+3x7Coh5QW1ZHK4n+Hl63Zn8x7UqTMhW+/uRESWrdcFONcqy24QhRO7Y9Y+O/hRFMsk1+479I8cOqurOhGCXDDoiRJu9ux66PG622N9k4StSm18oO6uXcdDuQNyLO1h4v8AC9O+Hi0a94pbnsnYytUSLjkFCwyZcqSDyNuKO/Z8vKvn6EgoKe81WWecP3FDbsIv8zUDj0DGMt22T6YclYZEDQhltQe5iF77FBUOnXr061H7XpjwvM8EGH6Qama6R7rid5vDqMXymNGciKM9Ul1TYYQoL2KSpxBC/VI599u8EWczHXvj40dxlWqWpuh+nl2xaIW3bnbMfuck3OEypQG+6ipCyCoA8oUPHoNyLL134tIemGk+GZvhuISshv8AqVIhwcXs0lRiqckSUBafOCRu2EcwCh38xA3A3UKMynS/jy4VsWumaYFxE27UbE8ahrmSLTlEQ+ciGynmUEqUVE8qEnol5Pd0HhUs1CyLQXjH4dNKpmseRrwG+Z7LS9irkd1RkMXhtZYWllRSQpBUdvX5d0qSdwQCCLizTiJ439BrZF1B1r0UwW94cqUyzcm8RnSXLhb0uKCUnlc5g51IHQEE7AlO+9bjWq5Rbza4d4glZjTo7clkrQUKKFpCk7pPUHYjoeorQnPLDx7cH2JXLUa267WXVLC8eSmRNg5HFUmYIwUATzklZ23G+zxPiEnurdPSTUCNqtphi2pUS3uQWsltUa5CK4rmUyXUBRRv47Ekb+IG9EUtpSlFCUpSiJSlKIlKUoiVjL1jON5IwY2RY/bboyRsW5sRt9O3xLBFZOlEVK5TwXcK2ZLW7etDMWS4vvcgxPMl7+3eOUGqiybyVHCze+0XZm8rx5xXwfMbuXUJ/wCGQlz9Nbj0okrzdyfyONrcSteH65zG/wBwzdbKh0fjW04j+rVW3jyUPEzirhl4TmmJ3JaeqDEuEm3vf0t7A/8AHXrpSimSvHk6N+VF0mATZZOojjDXcm2ZGi5NH4my6vf4uWvp4yvKMaXEM5lbr6ptn4Qv+GbA/G4htsn/AM1ewtKJK8oMf8sDrBbloj5dpbiNzKOizFkSILh/EouAH8VWrjnliNP5JQjLNGMjt+/w12+4MSwPiCw0a3kyHTHTbLgpOVafY3eOforz+1MPk/jWk1VWRcCHCPk5WufofYYzi/s7d2sIj3jsFpH9FEkKB455UfhLvhQi4ZBkNhUrv9I2R0hPxqZ7QVaeNcY/C1lqkt2XXbEStfciVPTEV/5X+Q1SuReSg4YLsFrssrMbC4r4Ii3YPIT/AML7az/TVWZJ5HG0uJWrEtdJrZ+waulkbeH41tuI/q0Sy9C7LluKZI2Hcdya03VB6hUKa2+D+NBNZavI+8+ST4iMfWqVh+d4dcVpO6OzkyYDx/8AlkA/8Vbv8B+jurWi2kFxxzWidJkX+TfX5LaXbsbgluJ2TSGwhZUeUEpWeXptv3UQhbI0pSihKUpRFpBpTk+PaUeUQ1ssOol2iWiTnNutdxsEuc6GW5TTbYCmkLVsN9+YAb9S0od4rEs3i0cSflMrVcMblsXnF9HMeWtyZGcDkcz1c49VY9UkOvpHTvMdXsraPXHhl0W4ioUSLqthzVzft4UIc5p1ceXHSTupKXWyFcpPXlO6d+u2/WshotoFpRw+449jGlWKM2iLKcD0t0uLekSnANgp11ZKlbDcAb7Dc7AbmimVrt5UXOLGxoCjSGNIal5Znd2gQ7VbGyFSFBEhK1O8g6hPMlKAfFSwB41ttilreseL2eyyV87tvgR4rit991NtpST+UVSOAcD+i2Dav3TXOUi75Jlk+4yLlFfvMoPM21bril7R2wkAcnMQlSuYpHdsetbB0UJWp/lHbhasV0kxPUx+8MQrvhOaW292ViTHcdYuEtoOK81c5ASjnQF7LI2BSAdt962wqvdeNWtO9EdNrjqLqaEuWm3qQluOGUuuypKtw2y0hXQrUd9uoAG5JABNEWvONeVY4W7tYWbjkEnJLDcS0FP2520uSFIc26pQ41uhY37junfxA7qoa0ce2hmS8XL/ABA5iq+WSxY5jK8Zx2G1bVPy5/avFx2TI5CUtpHUJRuT1B9tWoxxIcXWVMoyfBeAGIMcfSHo6bpLbamPMkbhQQoII3HUeoe/pvVy8NvEPp/r7LveH3fS5zB8+xflN5xm7QkB5lCjsHW1FCStvcgElKSOZPTZSSSlTfQLiP004lMfueTaYyLk7CtM0QJJnQ1R1B3s0r6JJ6jlUOtWjXXh2+BbkKbt8GPGQs8ykstJQCfaQBXYooUb1B1GwjSrGH8z1DyOLY7JGcbadmyebs0LcUEoB5QT1UQO7xqqfo8OEL/Xvjv/AM7/AAVOddtE8V4g9OJumOZzblEtc5+PIcdtzqG3wplwOJAUtKhtukb9O6tYv8kdw4fdZn384xv/ANPRSIW0mluuOk2tjFxlaV5xb8katLjbU1cTn2YU4CUBXMkd4Sr8lR3i6+tc1X/BC6fqy66XDVwo6ecLUG/wNP7rfZreRPx35Rusht0oUylSU8nI2jYbLO++/hXd4uvrXNV/wQun6suiKS6G/tKaf/gtav1RqpvUI0O/aU0//Ba1fqjVTeihKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpRFXev8AoxaeIHSm8aUXu9TLVDvJjlyXEQhTqOyeQ6NgsFPUoAO47jWuUPybs63RGLfb+LrWKLFitpZYYZunI202kbJQlIOyUgAAAdAK3RpRFS2g3DWNGLPk1iv2qeWaixMnDSHm8pkedJZaShaFtoB3HKsOHmHjsKq5rgAnYXKuUPQfiZ1E02xq6PLfdsEFxEmMytfwuwUshTXsB6q6Dqdq26pRFVHDxw16d8NmMzLFhInTZ13fEu8Xi5PdrNuL/XZTi9gNhzK2SAAOYnqSSYzxM8G+nfFDesSvuX3S4W6Rizq9zCS2fPoy1oWqO6Vg7I3QdiNiOdXtq/aURcPmkURfMfNmvNuz7LseQcnJtty8vdtt02rVGRwCnE77drhw+cQudaU2q+PKkzbHa1IkQQ6e9TKHCC106DqSBsAQAANtKURU5w68Lmn/AA4wLq7j0u53zI8ieEi95FeHu2nT1gkgKVsAlPMpR5R3kkkqPWufh54dLFw8RsujWPI7jdxl+QP5A+ZjbaSw67tu2jkA3SNvHrVuUoiVqtf+A+JaszvGZ6A64ZjpGvI3S/drbZezegPukklaGV7Bs7knoSBueUJHStqaURUpw+cKWBcP0u8ZPCu15ynM8jO95ye+yO3myhvzFAPchBUASBuSQNydhtkMO4dbFhvEHm3EJFyO4SLlm1vjW+TbnG2xHjpZDYCkKA5iT2Q33PiatulEStbdVOCu1ZdqdK1o0s1WyrSzMro0lm6zLCpC2LikAAKeYXsCrZKdzvseUEjfrWyVKItfNGeD6yabahOax55qPk2pefebKhRr1f3EgQmFAhSI7KfVb3BUN9z0UoDbmO+N1T4KYGda3DXrDNYMvwHIZTMaPdRZnEhu4NM8gSlXcQFJbQlQ3KTyg7b7k7K0oiqbGeHmx4xxFZdxFx8iuD1zy60R7Q/bVttiOwhkMgLQoDnKj2I33O3rGstrpoPp3xD4M7gWo9scfidqJMSVHc7OVBkJBCXmV7HlUASOoIIJBBFWHSiLT2X5PzIMpt8XD9TeLPU7K8HirQfmffcbaD7aCCht18bqWBsO8bjbpsdiLw1H4atI9TdHGdC7zjiYeLQWWWra1AV2TluUyNmnGVddlpBI3O/MFK5t+Y72lSiLUB7gFy6/WhvBs74wNUMgwZIQ07YVraaMiOkjlZcf6qUnoBsR4eFW3qlwnaO6qaQ2nRi4WZ20WbHEsmwP2xzspNqcaTyocaWQdzsTzc2/NuSeuxFyUoi1DlcA+SZhGjYzrDxY6l5vhzDiFuWCQtuMiUEEFCX3UkrcAIHf18QQetbYWaz2vHrRCsFkgswrdbY7cSJGZTyoZZbSEoQkeACQAPiruUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiVql5RNiLbtMcL1EnuW6TDwTOLZf5NonSm2RdmWu0547XOQlbvKVKSjvUEq29lbK5jlVrwbEr1ml87b0dYYD9yl9g3zudiy2Vr5U/ZHZJ2HjWueqfElwq55w4WLWXVLEZ+Raf3e9CLBiTbMH3kzUF9AWpgq9Xbs3QFb9x99EUwxnjh4U8psLOQRtbcbgNuNBxcW5yhElMnbcoW05srmHd03HsJHWtarPxP8PeYcdEjWePqRYsexXGMOVji7lPdMZV/luv8AOC0hQClNNJG3OoDqBt0IqN/RG+Sq7/nAn8zk/wCOrG0Iunk5+IrNHsC070Ct3pRiA5cV+kMZTHa7FC0JVsrmPXdxPT46KdFtzpvq5ppq/bZd40xzS15JCgv+ayX4DvaIad5QrkJ9vKoH8dS6opp7pTpvpPbpVp01wu1Y3Dmv+cyGLdHDSHXeUJ51Ad55QBv7qldFCUpSiJWEzaxWfKMPvWOZBb2Z1suUB+LLjPDdDzS0EKSoewgms3XTvH7EzPvC/wCqawqmGEjgs6Ql7QeIWm7PBZo3HZbYj3fUFpppIQ223mc9KUJA2CUgObAAdAB3V+/oMdIft5qL+etw+Uq9h3Urx/nuI9s+9eu8yw/sD3KifoMdIft5qL+etw+Up9BjpD9vNRfz1uHylXtUa1H1CxvSrCrnn+XuyW7RaEIXKVHZLrgC1pQNkDqfWUKluLxLjlDzPNQ7CYZoksEclV30GOkP281F/PW4fKU+gx0h+3mov563D5Sp5o9rTgOuuMP5bp5PkyIEWYuC8JMcsOtupSlWxQeuxCgQfGpJluU2TB8XuuY5JK82tVliOTZboTzFLSE7nYeJ8APEkCpOJxbXZC4zwUDDYVzc4aIVP/QY6Q/bzUX89bh8pT6DHSH7eai/nrcPlKlml3ELptrBhd7z7CpVxctGPrdRNVKhKZcSW2Q8rlQT63qH8vSuvE4lNLZ2iDvELHmXJWHMhRW95irzj1Xwwdmd9/hn293WszWxoMZnaxv1OgWAo4MiQ0aT3BRv6DHSH7eai/nrcPlKfQY6Q/bzUX89bh8pVzY9fbflFgtmTWlTioN2hszoxcRyLLTqAtG6fA7KHSvzkuQW3EscumVXhTiYFnhvT5RaRzrDTSCtZSnxOwPSq/O8TOXOZ5qzzTDROQRyVN/QY6Q/bzUX89bh8pT6DHSH7eai/nrcPlKtPTrUDHNU8KtWoGIuyHLPeWlPRFSGS04UpWpB5kHqOqTUjo7F4lpgvMjtRuEwzgCGCOSon6DHSH7eai/nrcPlKfQY6Q/bzUX89bh8pV7UqPPcR7Z96nzLD+wPctc7hwh6Vxpa2mr9qGEgAjfM7gfD75XW+hK0v+3+oX553D5Sr0vH+fufvU/orp1yau0sYHkCq7XiV1qWzsIWAmm3TgqX+hK0v+3+oX553D5Svn0JWl/2/wBQvzzuHylXUkFSgkd56VU+nPE7pJqnm0zTzE7lcFXqEiQ4tqVCUyhQZWEOBCidlEE77ewE1LMbtCo1zmPcQNbmyh+D2fTcGvY0E6WF1jvoStL/ALf6hfnncPlKfQlaX/b/AFC/PO4fKVdA3JFVhjfEdpZluqk7RyxXKdIyO3LktvjzNQj8zH1UB3uOx6fGDUMxu0KoJZUcYubmwUvwWApkB7GibCwuVhvoStL/ALf6hfnncPlKfQlaX/b/AFC/PO4fKVdFKr6Txn7rveVn0bg/2m+4Kl/oStL/ALf6hfnncPlKfQlaX/b/AFC/PO4fKVdFRDVHVTENHcZby3NnpbVucmMwEqjRy8vtXN+Uco8PVPWs2bQx9RwYyo4k9pWL8BgabS99NoA7AoL9CVpf9v8AUL887h8pT6ErS/7f6hfnncPlKuhCgtKVjuUAofEa+1h0njP3Xe8rLo3B/tN9wVL/AEJWl/2/1C/PO4fKU+hK0v8At/qF+edw+Uq6KU6Txn7rveU6Nwf7TfcFS30JWl/2/wBQvzzuHylffoStLvt/qF+edw+Uq6KU6Txn7rveU6Nwf7TfcFS/0JWl/wBv9QvzzuHylPoStLvt/qF+edw+Uq0YGXYxdciuWJ2y+RJV4s7TT1wiMuc64qXCQjtNuiSeU+qeu3XasvWR2jjm2NR3vKgbPwTrim33BUv9CVpf9v8AUL887h8pT6ErS/7f6hfnncPlKuilY9J4z913vKno3B/tN9wVL/QlaX/b/UL887h8pWDzbhg07sWK3G727IdQEyYzaVNleYz1AErSOoLnsJrYSovqd/oFefvKP7RNVV9qY1tJxFV2h3ngtjCbLwT8RTa6k0guG4cVqD86+0fdTm350zflKfOvtH3U5t+dM35SphX2vJ9P7U/2H/EV9B/LmyP9Znwj7KHfOvtH3U5t+dM35Snzr7R91ObfnTN+UqYV97+6nT+1P9h/xFPy5sj/AFmfCPsod86+0fdTm350zflKfOvtH3U5t+dM35Ssbkeu+nuOXR2yGXNuc5hRS81bYpf7NQ7wVdBuPHYnashheruDZ5LVbLJcnWrghJUYUxksvEDvIB6K29xrovxH4kp0fOHOqhmsy7Tjy7VzGYb8L1K/mzGUS+YiG68OfZqv186+0fdTm350zflKfOvtH3U5t+dM75SpHcr1abOqKi6XFiMua+mNGQ4rZTzqjsEJHeT+iu7WgdubWa0ONd8HTrG66I/D+xnOLRh6cjXqttzUP+dfaPupzb86ZvylPnX2j7qc2/Omd8pUwpWPT+1P9h/xFZflzZH+sz4R9lD/AJ19o+6nNvzpm/KU+dfaPupzb86ZvylTGvlOn9qf7D/iKflzZH+sz4R9lD/nX2j7qc2/Omd8pX3519o+6nNvzpm/KVL6U6f2p/sP+Iqfy5sj/WZ8I+yh/wA6+0fdTm350zvlKfOvtH3U5t+dM35SpjSnT+1P9h/xFR+XNkf6zPhH2UO+dfaPupzb86Z3ylPnX2j7qc2/Omb8pUwpTp/an+w/4in5c2R/rM+EfZRAaX2jcD5qc2/Omb8pVXO22al1aRmGX7JUQP8AnHM8D98rYEd4+OqMf+ru/v1fprs7I2ztCsX+UrOMRqT2rjbX2FsyiGeTw7BM6NHYsf6Pnfdhl/5xzPlKej533YZf+ccz5Su7TvrtdJYz913vK4vROA/Zb8IXS9Hzvuwy/wDOOZ8pT0fO+7DL/wA45nylYq6Z/jVqlqgLkvSpLfRbURkulJ9hI6f01zWPNcfyCQYUKUtuUNz5vIbLbhHuB7/xVtGrtRtPypL8vG60xR2M6p5ENp5tIga8OfYu/wCj533YZf8AnHM+Up6Pnfdhl/5xzPlK+XC9W+2S4MGY4tL1xcLMcJQSCobd58O+u9VDsfjmgONR0HS5Wy3ZuznuLW0mSNbC29dL0fO+7DL/AM45nylPR877sMv/ADjmfKV3aVh0ljP3Xe8rPonAfst+ELpej533YZf+ccz5Sno+d92GX/nHM+Uru0p0ljP3Xe8p0TgP2W/CF0vR877sMv8AzjmfKU9Hzvuwy/8AOOZ8pXdpTpLGfuu95TonAfst+ELpej533YZf+ccz5Sno+d92GX/nHM+Ur8x75bpV3l2Jl1ZmQkJceSUEJCVbbbHx7679Zvx+OpmHVHDfqd+iwZs3Z1QEspMMEjQajVdL0fO+7DL/AM45nylPR877sMv/ADjmfKV3aVh0ljP3Xe8rPonAfst+ELpej533YZf+ccz5SvTfgJbca4c7Yh2fOmKFyuH02bKckOn6eehW4SogeHXpXmrXpdwIfW72z+Ep/wDbGu1sHGYiviiyq8kQdT2hed/E2BwuGwQfRphpzC4AG4rYWlKV7FeBSlKURflaEOIU24hKkKGykqG4I9hFVpr5rJpzw76Yy8/z2IV2yK8iPFgxY6Fuy5Tm/I00k7J5jsokkgABRPdU/vt8tOM2Wfkd/ntQbZa4zkyZJdOyGWW0lS1qPsCQSfirQ3je4iuHbVHT/Frxg2ruKZFcsEyy3ZQqxed7KusdlRDrCOZOxWUr3APfsR40QKQReOXUm5R259p8nzqLKhSEhxh7sCO0QeoVt5se8e81aPD5xGZ1qtnT2M5JwnZbptEbtzssXi6tlLK1pWhIYH0hHrKCirv+wPSu3YeP3hEv1qjXQa02a3l9tK1RbilyPIZJHVC0KT0UO47Ej2EjrU5064lNB9XL8vF9NdULJkN2ajLlriQnSpxLKVJSpexA6ArSPxiiKy6UpREpSlESuneP2JmfeF/1TXcrp3j9iZn3hf8AVNV1fVu5FWUfWN5hVgO6lB3Urwy9ulUVxyHbhTz8j/8ApI362zV61RPHL9ann/8A3SN+ts1fhfXs5j5qjE+pfyPyVKcG5OkOs3ztXB2Nn1Owi0ZdaE9yPO0Rk+cIT7ye2J/eCrV4zJ0vJ7Tg/D9ZnimfqhkceFK5T1btkdSXZKz7uiPyGqw1ihysL0P4eOI+0MqMrTpmzJuBQOq7bJYbQ6k+7cAf+IasXTWTG1n4xMt1NjOCVj2mdpj4xZHgd0LmyR2sl1PvCSU7+wiui+HVPOuAM/yFh9CuewltPzbiRHI3P1UI4C342N6M6vyYkBp6NaMlui2orvVC22YoKW1f7pCQk+6vzqLqS3q55OTJM9axK14ym4M8otltG0dns7k2jdPQd/Luenea4eCz9orXP+H75+qGohYf+iiuX7x7/wDyqate0GuX787fkq6biKIbuyOWw9210x/QHhjwLK7vbpF1nTrHabfaLVGOz0+WuKgpQDsdkgDcnY+AAJIFVxqprTxP2bSXJbjrHoDarZit9ssuCuTZrr5xMtC5DKkNLlMqJ3RzKSFEbcu/X2VHtYFogy+Dm8XxSU4/HdgIkLc+pIfLMQtlW/Qd2/X2GtrdeHrVG0Vz92/qbTBTjlxD/a/B6sLAHxlRSB7yK1oZRLDlkuJ48YstiX1Q4ZoDQPlN1B+CMEcKungPhb3h/wDUu1eFUdwRbjhU085u/wBHvb/H5y7V41pYr17+Z+a3cP6lnIfJKUpVCuUevH+fr+JP6K6dd28f5+v4k/orpVxq3rHc12aXq28l+mvqqPjFeYenfaaeXJjiGiJKW8Z1QlWa8qT3qt8pIBKvcPpg+NYr08a+qJ/fCtIuHXT5rVPQPXTA1oCnbpk1wTFJHwZKEBbJ/jEprr7Jqto0qrn+iS0HkSQfArk7VpOq1aTWel1iOYAI8VtjqlnkHTbTbItQZDqFM2a2uy2Tv0cc5dmUj28yygfjrSzhnwmbg/E/grF55jeL5gsi/wB0Ur4SpMsuunm94SUD8VZqLqBK4gtKtEtD3HVquV6uvY5Ujf124doP0zn/AH4CD18RVg3BLbflErIhCAhtODrCUgbBKQHNgPxVsUKRwVGrQd6Ra8nk2w95nwWvXqjGVaVdvogsA5uufcIUr1G4isoZ1Kd0X0O0/bzHLILIkXV6XK83gWxBAIDix1KtlJ3G42KgBudwOlj3Edn+K6hWfTXiK03hYrIyRfZWW9WuaZNvkvbgBpRO5Qokgb79CpO4AO9YDhMW1G1u1/tt1IF+OSJfUlf1RUTtHuUjxKfWT/5k+6vz5QJcZ3AcItcfY3uXmEQ2xCfqhISoLKfHvU2PjKapbQoecNwRYIIHWvMlszrEDhGiudXrebuxme4J6togGI0me2dVYGtuv1z0/wApsmlunOFqy/PshbL8a3F7sWIscEjtn1+A9VXTp0SSSOm+uPFzqTrDJ04g4FrTphCsMybeoc62XSzzfOoEgNFXaMrJ9ZtwBYIG/Ub+zerTx4+ZeUKylu+qAkT8NY9Elf2aUpZ7QI/8ju+3sVXa8oG7bEaExGpqmxKdyS3+ZAn1isFZXy/8HNv8dWYMUsPiaFMMBLgDN5k8LxA5cVXizVxGHr1C8gAkRaIHG0yefBWTrbrhZND8Ut1xl2qVerzenkQLLZon1adJIHTfY8qRuNzsTupIAJNVteteOJvTe1JzvVTQSyoxJspXcBZbuXp9uaUQOdxBJSrbfrtsPaRUS4tIOSy+IfRBNkyWPjrr7T7FtusqKJDEWcVp2Km1eqondsDfxIPhUszPS3ia+ZG9pzXilxxqwGA+i6LkYq0htMUoIc5j3gcpPUdfZ1qujh8PTpUzUykvuZzTrENyiPrPYs62Ir1KtQMzAMsIyxpMmTP/AIpvrNxBQtPNFYGs2IRIl/t1ykQUx+2cU2hUeQernqgkKSPsfaCKwumet2tOeZcxdbpo0zYNMJ8eRMi36ZL5ZKYzaCpt5xvc7doACElI2B33O3WldasYt2IcANnxy05pByy3xrzG81usJCksPtKlOqASFEnZJJT394NbgTr3aML04cyK5tEWyx2MS3m20gkstMAlIHd1A2AqurSo0KMMZmLnOaCZmBEWte+8dyspVa1etLn5QGtcQIiTM34KjbFxEa76wom3/QDR6yS8TiynIsa7ZFdDHVPUg7KU22nbYfGTt3E77gSXRfiDyjUS8ZdpzmOnyMb1AxGP27luErtI0oEbIKXPsQVFHiRyrBB79oNptkHFdrjjEfNcFu2CaZ4jcnXTbIqbX55KLQcUkuEbcm5UFdfV3IJ2rC8Oltvdm4ztSbTkucOZddomOstTLquOhguu88YlAbR6qQjcI2/3etX1MPRyVW5WgsEgAuLhBA6x9E9sdyppYitnpOzOIcYJIABkE2Go7PFYfh2v+tY4ldUHW9ObGZdyusAZY16YHLaUbq3Uwrb/AJQdio7dO4e2rlz/AIh8uc1LlaN6EafsZfktqaD15lzpfm9vtgO2yFrHVS+o3G46nYbkHaOcOO30VHEGT3ekYO//AK6q7Q7FtZ77q3rRD0+1bt2HXWPk7rlzjTLMia7JbLr3ZOBSuqUDcjYdPWB8RVtanSrVn1KgaMjGa5ouG6xJgbo7JVNGpVo0WU2EnM52kTYnSYEn7wrxwziEzm1akW3SXiA09iYpeb+lRsdztswybdcFjvaCj1QvwG57yAQNxvfVafam6RakTMnwFWtfFLjBeh31qZYI7liTEekSULb5m21NkEk+oNj03I8a3CWd1qO225PSuVj6dFoY+lFwZiYkHdmE/Yrq4GpVcXsqzYiJibjfH9hfKi+p3+gV5+8o/tE1KKi+p3+gV5+8o/tE1yMR6l/I/JdnBfqaf8m/MLXGlKV4tfTErguDUl63y2ITnZyHGHEMr325XCkhJ/Edq56x+RXc4/YLjfRBdmej4y5JYaIC3AkbkAnx2Bqyi1zqjWsEkkRz3Kus5rKbnPMAAzy3qjtDs/wjT20SMMzP/m/kbEt0zXZbJHnBKvVJc2O2w6deniN96sDIMJxjUi82LNcayOIzOsspL5mQOR4voBB7NZB6dx6nwJFd6yKwbWXE4OST8dgTmZSCFNyG0uOx1g7FsrHUEe7bvFVbqngNn0cXas/05mSbTNNxajLgB9S25SVbkpAPUjpsQdxsfA7V7Rj6WP2k7ybn0cU4kEGHMmCCOIB7QQO5eGdTq7P2WwVWsr4NoaQRLHhsgh3Aka2LSeZWQ10nZanUDCkxMfhOMR7qldqWqXymU+QjmbcG30tIOw5vGpZm2q130+t2MS8kxthMm8POtT48d8umOUjcBogeuTuB19tY7WglWZ6Xkp5Sb7uU+w/S+lfNc2m3ss00adSFJVkHUHx6t0wzaGLZgKFemC3LUJiZ6uffO8iT29llGKfiMHU2jiMPVIdmpATBAzZN0bgco7NZN1l8dz/NmIl3vmp2GsY1aLfEEtp5MjtHFAn6mU7ndW23s6kDbrWKt+o2sGUW/wCaTE9MYHoZzdcZE6eUSpLfgpKR0G/h/RvXc4k0SXNJLsWAohL8ZT3L/wBn2o3/ABb7VPcbdhyMdtL1tKFRXITBYKPg8nZjbaua6rhqeDGPGHaS9xbHWytDQD7Uy6dSd1guq2liqmNOzjiXhrGB2bq5nFznDXLGVsaAb7lR/BdSbdneOzbtEhOwp9r7Rqfb3z68d5KSeUnxB2Ox28D03FQzGNY9QtQbQ1KwXT2G8+0CJz8yYWorbm55WmyditXLyknuHNXFgC2pGqerMq3EKg9mltSkfBL4Qrm/HuF1leGZCUaSQClIHPLlKV7zz7b/AJAK3MThMHs6lXrtpBxBpFocT1c7C4gwQTHb2StHCY3HbSq0MOaxaCKocWhsu8m9rQRIIE6mO2N0ZjTPUqZmky7Y7kFgNlyCxrSmXFDnOhSVdApJ+P4+8EE71iX9U8wyXIrjY9K8Rh3SNZ3CxMuVwkllgujvQ3t39x6/j222J6WJpcVxB6goZ6OKtcYI/fcre39NcnDItoaeyoR2E2Ld5SZqD8MOEjbm/EP6DWOJwmEwjauNZSB6tIhpJyg1GyTrJAiBJ3rLC43GYx1HAPqkS6sC8AZnCm6ANCATMkgbua7GOaxXy6agQNO75hvoi4rZeXPC3ysIUhJUhTRA2WhQ8fD31aVVFkEiE7xMYuwwpJksWWQJO3eNw4UA+/br8Rq3a5O2qVGn5CpQp5M9MOIubku4yYtbsXY2HWr1BiKdepnLKhaDAFg1tjAAm5ntnklKUrirur6O8VRj/wBXd/fq/TV5jvFUa/8AV3f36v0mu7sTV/d9VwtuaU+/6LjqOag3l+x4nMlxFlL7nKw2od6Ss7Ej37b1I6i2plueuWHTEx0lTkcokgDvISfW/oJP4q9XgAw4qmKmkj5ryO0jUbg6ppellMe5d/EMfh45ZY0aM0kPuNpckO7es44Rudz7BvsK48nxKPkbkSY3KMKdDeS43KbRuvlH2J7t/D4vx13seuke9WSHcoywtLrKebY/BWBspJ94NdXK8ojYrAalux1SXX3UtNR0KCVuE95Hxf8A+xVrX4p2MJZPlCT/AG9o8FU9mDbgAKkeSAB7txteeV5WA1GnItt4xae6246GJbiyhtO6lnZPQD2k1zXHL8wsjAu14xFhu2hQ7QNSud5pJOwKh3b0zQdpkWG9q3ylU5Sig9djsk7fiNZTUD/Qu8f92P8AWFbzHU8mGpVGB2aQZnQuItBXOqNq+UxdanULcsERFyGA3kH3WX6ybKW7DYmL6wymQy86yn1lFOza+vN09g8K4LLkORXGaX52OJg2ZbS3mpTr30wIA3SVJ8Nx18NhUfzBIVphZ0q6gmED/wCWpdlrbqsVuzcYHn8ycCQPZy/3VWaNGnSazKCXOc2TNgCBOut1cK9erWfUzkNYxrsoi5IJg2JiywsXLslyDtZWKY5Hdt7aihEibILZeI7+VI8KyeM5Sm+vSbbNguW+5wSBIirVzbA9yknxH949tfMDWw5htoMYp5BGCTt+6BPN+PfesS0UuavL8269lagmRt+63GwPv6pqH06NR1ai2mG5ASDeeqd94M8lNOpXpNoV3VC7yhaCLR1hNoEiOZkar7Fzi83aVOtdjx1EiZDkuNKU49yMobSdgpStvhEg+qPZXdsOV3CVe3cayG0JgXBDXbtlpznbdR7Qf/33HurpacJAkZMoDqbu4Cfy1+5+3z1rV77W7+lVW1qVDytTDtpgZWyDeZAB4xHZCooVsT5GjinVCS54aRAiC4jhM75lZO33wS8tutiFvZbMJlpzzhJ9d3mA6Hp4b+2ulIy27XC7SbRiVmammCrkkypD3ZspX+5G3UmuGw/tnZH/AN0j/oTXHpUUiyXBpfSQ3c3g+D8Lm6bb/wBNYVKFKix1bLMNZYzEuFyb/wBlWU8RWrPbQz5cz6kkRMNNgLRv1ibLJWHKZM26v47fbYLfdGG+2CEOc7bzf7pB/wD3/QakVdFy52lu9NWlbjYuTrJdQns91Fsb7+tt0Hf0JrvVzMTlLg5rMoIBjdzE7veuvhcwaWOfnIJE2nkY3jkOSV6XcCH1vFt/hKf/AG5rzRr0u4EPreLb/CU/+3Ndf8OfrD/E/MLg/i39AP5D5FbC0pSvcr5slKUoi6d3tFrv9qmWO9wGJ1vuDDkWVGfQFtvMrSUrQpJ6FJBII9hqrRwhcLo6DQPB/wCZmf7qt6lEVQ/Qh8Lv+oPB/wCZmf7qkOD6C6LaaXleQ6faX43j1zcYVGVLt1vbYdLSiCpBUkb8pKUnb3Cp7SiJSlKIlKUoiV07x+xMz7wv+qa7ldO8fsTM+8L/AKpqur6t3Iqyj6xvMKsB3UoO6leGXt0qFa0aYQ9ZtMb7plPu79rj3xptpcthpLi2gh1Dm4SogHfk26+2prSsmuLCHN1Cxc0PaWnQqAXHRyx3jQ06F3Wc9Ithx9uwmYW0h3ZtpKEPhPcFBSUrA7txXV4e9DrFw9aft4JY7pIupVNenyp8lpLbsh1ZA3KUkgBKUpSOvcKsmlZ+WeWlk2JnvWAosDg+LgR3KntH+HC1aQ4VmmFwcrm3NrM50yc9IejIbVGVIa7MpSEkhQT3jesXB4UrLC4YZHDGnM7gu3yAsG7mK2HxzShI+p78veOXv7utXrSsjiapMk3kHvGixGGpAQBuI7jqq0y/QDB9QNGrfotmSX51ttkGJFjTW9mpLL0dsIbkNnqEr2B3HUEKIO4NVW7wZ5bklpTh+pvExmWV4lFaUiJZ3Y7bCSsJIZVIcSoqfDZ5VBJIBKR1FbP0qWYqrTs09u7w4dyPw1J5lw/v1UM0b00i6O6ZWHTSFd37oxYWFsImPtJbceCnFL3KUkgdV7dPZUzpSqXOL3Fx1Kta0MAaNAlKUrFZKP3j/P1/En9FdKu7eP8AP1/En9FdKuLW9Y7muzS9W3kvqTyqCtu471Wmh2iVv0PgZDBtuQyrsMhvLt5Wt9hLZZWsAcieUncDbvPWrKpRtV7WOpg2MT3aI6kxzw8i4mO9U1ppwwYdphq5k2rdpukmTJyHt+wguMpS1b+3dDjvZqHU8xG3XbYdKzknRO3yNfImvRyCUmZEsyrMLaGE9kpBCvphXvzb+t3bbdKsmlWuxld7i9zrkZe7gqm4Si1oa1tgZ7+KpnVHhqtub5ozqlhOb3jA82aZEd262sJWmW2BsEvtKIC+gA336gDcHYbdLCuF5qBnkLVDVfUm96i5LahtbF3BpDESCfBTbCNxzA9R12B67bgEXnSshjsQGeTDrRGgmOE6x2SsTgcOX+ULbzO+J4xpPbCq3WnQDH9YX7Rf0X25YxlePKK7Tf7WoCRHBO5QoH4aN+u24I3Ox6kGt8l4L7jqLC7XVbXfJcnvEdbXo6W5EaaYgNpUFLDcdJ5StzYArJ32Hd41s1SsqW0MRQaGsdEaWEjkYmOzRRVwGHrOLntmdbm/dMd6gmr2jGGa14gjEcwafSIziZEGfFXySYT6RsHG1bHw6EHcEfECKnuHCRnGWQWcV1I4mswyLE2VoK7UIrTDkhCSClDrwUSvuHUg+3v61snSsaOOr0G5WOtusDHKRbuWVXBUK7sz235kTzjXvVXar6AYxqZpJF0cgTXMas0F2IqKYTCXOxRH35UBKj1336knfx8asGZYbbc8eexe7MJmW+VCNvktL6B5lTfIoHbu3G9ZClVOr1HNDSbAk951PgrW0KbXFwFyAO4aLXHG+E/OsCiu4vp5xMZZYcScdW43bEwGHno6VndSWn1fA+MJHXrtvUk0u4WsU0h1Lf1DxHI7qpM20m2zYU4iQqU6Vha5S31HmLilpCj0279tquqlXv2hiagIc7XWwvztfnqqGbPw9Mgtbppc25XtyVGXThhfTrTK1lwrVe/Yw7d5MaTerXGaSti4dkU7oUeYEIWE9QQdt1bd+1dvU3hot+YZynVTAs6vOn+ZloMSbla0pcbmtgAAPNKICjsAN9+oA3B2Bq6KVAx+IBDs1wI0GnA2v3ypOBoEFuXUzqdeI4d0KjMG4XkWzUCJqpqpqVetRMmtieW2OXBlDEaCf3TbKSRzDckdwB67b7EXnSlU1sRUxDg6oZi3ADkBYK2jQp4cZaYie/3k3Sovqd/oFefvKP7RNSiovqd/oFefvKP7RNaeI9S/kfkt/Bfqaf8AJvzC1xpSvteLX0xfKEJUClSQUqBBBG4I9lfa+URVRL0Dat10kXXTzObxiZlq53Y0Y9oxv7k7jYe477eFdywaJRo9+jZPm2XXTLLjBPPF89PKyyodQoI3O5B2PftuO6rLpXYdt/aL2ZHVNREwM0cM0ZvFcVn4d2Yx4e2loZiXZZ45Jy+CiGpGnETUWHAbXeJdqm2qT51DmRgCpte3XodvYD3ggiupN0vdu7OJG95bOnTMVmGYJTjSeeYoqB2X16dwG4qdUrXpbVxdCmykx8BsxYWkEG8TBk2036rYrbIwdeq+tUZLnxmub5SCJAMSCBeJ3aLr3K3wbvAk2u5xUSIkttTTzSxulaFDYg1WcbRS/WWO5ZsU1bv1qsbhPLC7JDqmknvS24SCkfFVqUrHCbSxOCaWUXWN4IDhI0MOBE9uqzxmzMLj3B9dvWFgQS0wdRLSDHZoo1iGn2P4Pjj2OWFDqUSQsyJLqud59xSditZ8T7B3CvunmERtPMWYxaJcHZrTDjjgedQEqPOrfbYdOlSSlYVcfiazXtqPJzkOdO8iYPdJWdLZ+GoOY6kwDIC1sbgYkRpeB2qL2fAotpz69Z43c3nH72w1HXGUgBDQQEgEK7zvy+PtqvbbYMLz7I7zkeCZne8NvTUlca7Rmlttl1xJ2Ky0SR1O/rDpvv0Bq6qiOS6SadZdcFXW+4xHdmr+HIaWtlbn74oI5vjNdPAbW8m9zsQ9wJa1oLQ0wG7i0wHCI1uCJXL2jsfyjGtw1NpAc5xDi5sl15a8S5pmdBBBiyq7D8bsULiBixsYukm8C02t568XJ9/t1uynOZPrr7t9lJGw7ttvA1f9YjGcQxjDYSrfjFljW5hZ5lhpPrOH2qUdyr8ZrL1r7Z2iNpVmvbJDWhoJiTEmTFhc6DQLY2Jsw7LoOY6AXuLiGzAmBAm5gAXNyZKUpSuSuwvo7xVGP/V3fviv0mrzHeKox/6u7+/V+mu7sTV/d9VwtuaU+/6L8UIBBBG4PQg+IpSvQLz6iK9PUw5TsvGMhnWXtzzLZa2W0T7knurntWCRotzRe7zdZd5ntfUnJOwQ2falPtqT0redtHFOblLtbTAmOcT4rnt2VhGuDgzQzEmJ45ZjwWKvGPsXi42q4uSXG1Wp8voSlIIcJ26Hfu7q7F8tTd8tMu0PPLaRLb7NS0AEp677jf4q7tK1xXqDJf0dOy8/NbJw1I55Hp+l22j5cFgrriUa6Y9Ex1ya621ELJS6lIKldmNhuO7rWdIB3BAII2IPXcUpUPr1KgDXGwJPedVNPD06Ti5ggkAdw090qKJwR+3PPHGconWiPIWVrjIQl1tJPeUc3wayeOYvAxtLy47r0mVKVzyJT6uZx0+8+A91ZilXVMdXrMLHusdbCTzOp71TS2fhqDxUY240uYE6wCYHcAsTYMeYsC7gtmS48bjKVKVzpA5CfAbeFfX8fZfyaNkxkuB2LGVGS0EjlUFb9Se/frWVpVZxNUvNQm5EHlEfJWDCUWsbSDeq0yNdZn5rFQ8eYhZDPyJElxTtwabaU0UjlQE7dQe/wroT8MK7o9erDe5Vnlyf847FKVtun2lB6b++pJSsm4ysx2YO3AaCIGgI0KwfgcO9uQttJdqQZOpBBka7isHYMUj2WVIukmfIuNylAJdlyNublH2KQOiR3fkrOUpVdWs+u7PUMlXUaFPDsyUhA/vvSvS7gQ+t3tn8JT/7c15o16XcCH1u9s/hKf8A25rtfhz9Yf4n5hed/Fv6AfyHyK2FpSle5XzZKUpREpSlESlKURKUpREpSlESuneP2JmfeF/1TXcrp3j9iZn3hf8AVNV1fVu5FWUfWN5hVgO6lB3VqbxA6w6qaYcSdvumNSZtywzHcQaveT2Bs8wdhKmLYelNo26utBSF77/BQfDevGUKJruyt1Xsa1YUW5nLbKlUfrXqHOEnRG74FlTybRlub29h56E7s3cLe6w4vkV7UK2SdvaKkOZcQ2F4nlU3CYVjyvKr1amESbrGxqzOT/RraxugvqSQlKinqEAlRHXap83eQCBrPgYUeXYCZOn/AOqz6VXvz+dNHcWxPM7feHp9ozS7x7Ha3o0dSj548VJDbqTspopUhSVcw3SRsRWdvOoWOWLPMb04nqlC85W1MetwQzzNFMVAU7zr39XoobdOtYeSeDEcfDX3LPyrCJnh46KS0qn8T4qNL82yVGN45FyWRyuy2JtyNoWm325yP2naIkySeRo8rSljcn1SknbmFdOLxeaTyTEuTkHLYuLz5aYUTLJVheasrzqlciSJB7kKV0DikhJ9tZ+bVhbKVh5xSicwV2Up7wQR7RSqFelKUoiUpSiKPXj/AD9f71P6K6exNd28f5+v4k/orVSRfndTdaM4wvPNdr3p8MbntQ7Fj9ruLVscmxi0lXnqnXEkv86idkpOyQO6uc3DmvUeZgC53740C6LsQKFNgiSbDdunUrZ6lU7jd4z/AEWxnNbnq9fZeU4ljnZTrLfW223rlIgqT9NS+03tzFpX2f2Sdz4VOMs1LxTDsXt2X3OS+/Au8mFEt4hs9s7KdlqSGEtpHfvzA+4Anwqp2HeHZWdYGwI36H3303K1uIYWy/qkXIO7d9FKqVUl24nNPrddL5ZLfY8vvs/GZj8S8MWexuSjC7L4TrhSeUIPXl67q5VEDYVm7nrvppbcEsmoqby/PteTLbYsrUGK4/LuL699mWWAOdTg5VbggcvKd9qk4SuIlhvpZQMXQMw8WVgUqvcI1wxTNcokYOqzZJjuRMQzcBbb/a1Q3n4oUEl1rclK0gkA7HceyovH4uNK5toXksC2ZfKsEVfZ3K8MWJ1cO2L5ikpkLB6EdCrlCuUKBO29Bg8QSQGG0eOnv3KDjKAAJeN/hr7ldVKozUDXW4Y9rhp9h1mt96uGP3qBLmy1221GSmfzNI83Uy4PhNo5itwp+CCknpWfxzUnDcatmpOU5BqBdZVrxvJJMe4OXVkJRbXEob/5JFCdy41upPJ0KipZFZHB1QwOjUSPfCxGMpF5bOhg+6ValKrXEtfcNyvJYOIyLLlGN3O8MLk2lrILQuCLk2gcyiwpRIUoJ9YoOytuu1SvPM2sOnGI3LOMncfbtVpbS7JUy32i0pK0o3Cem/VQ/FVTqFVjxTLbnTtVza9N7C9rrDXsWfp1qrbLxHafXq+Wi0Kg5JbI2RPGPY7vc7Q5Gt10d23CGHld5UBujmCeYfB36VjMM1CetmouuMrNspWzjmITLa4yZbn0i3xzBDjvKPAFW5I7yas8zqgOzCCBPO4Fu8qvzukSMpmTHKxPyCuWlVHbuJzAplxssW5Y7mdjg5HIbiWi7XewuxYMt1z6kkOE7p5/seYDfesnlmvOJ4rkdxxWPj+WZHPsrTb13Fgs7kxFuStPOkPLBACin1uQbq267VHmdcOy5TKnzuhlzZhCsmlUBqzxHsWyz6ZZFpwbtdbNmF9il6Xb7SqT5xBBX2kZG/VMhSkgBvbn2Svbbapxj+W2aTqxlMdzLb62uHjttuUqzXFhLMG2MOBxQeSo9Q6oA9oD8HkFZHB1WszuEa+BjuusRjKTn5GmdPET3qx6VUNt4pNMrk/b3/M8nh2G7zEwLdksyzOs2iU+pXKhKX1dwUobJUpIST41GtQtXH8ExDWvIcUyPJLzesfnJitx5NuD0WxyVRUqQGwO+MBstS1dApXWpZga7nZC2DbXmB9Vi/HUWtzAyOzlP0WwdRfU7/QK8/eUf2iajuC60WG+rw7GrtCv1vvmT2h2bEF0tqo3nKoyUh89e4nfnSNvWQdx0pqJnuPzjlGm0RUl28WqzxrpM5WfpDDTz4S2lTm/w1cqiE7fBG9aeLw9SnSeHDRpPdpPvsuhs+vTfiKRB1c332Me5UnX2vlVBEy7JIGt9xhzrxIdx56c3Z0RFn6VHfcipeaWn2cykrT+OvKYHZ78eKnkyBkaXc4iw7br6Dj9o09nml5QE53Btt0g3PZa6t+lVPr1luRWaHEtmK3V63yWGzc5zzJ2UI4cQ0hG/hzrc/8AQamt/wA8suPXCPZFx7jcrrIZ84TBtsVUh5LXd2igOiU79NyRv4VZ0TiDQpVmCfKZoA1AbEk8BeeSqG2MMMRWoPOXyeWSdCXTAHE2jnZSOlRVOpuIqxeXl3nb6IVueEeahxhSH4rpUE8jjatikgqBPu610laxYezBTc5rV4hxHJaYbD0i3OID6lIUtKmweq0qCTsQOpKRt1qtuysa+ctJ1jGm/hzuFY/a+AZGas0SJ1GnHlYqb19qN2DPbNfrsuwGHc7Zc0s+coiXKIqO46zvt2iN9woA7b7HceIqCYJrHb4eEx5mTKvtxciuPpuNyagreZjfTlhIdcHTonl3232G29W09jYyo1xawy0tEbzmmCOy0cyAFVU23gqb2BzxlcHGZsMpbIPb1p7ACSrer7Udv+d4/jwt6HVyZ8q7J54ES3sF9+QjYErSkfYgEesdh1rE3HPrffMLymVYX5sG6WaBIL0eSyWJUR3slKQooV8W4UNwdqppbNxNUNfkIaSBMWuYnlNp421V9XaeFpFzM4LgCYm9hmjnF44X0U3r7ULYz222HF8ZXenZs+63eAwtmLEYL8qUvsklawkeA33KjsOtZOwZ3j2QM3BbTr8F60dbhFuDJjvRU7EhS0q+xIBIUNx0qKmzsTTaX5CWgxMW1ieU2njbVTS2lhajhTzgOImJvpMc4vHC+ikFKgTmtmGx4aLpKiXyPbpBCYk162OIYlkkABpR7999xvtuASN6lNtyS2XW9XTH4xeTOs5a85bcb5d0up5kLQfskkb9faNqVtnYvDtLqtMgcuBAPuJAPAmEobTweJcGUqgJPA8QSPeASOIEhZSlYyxZHbMjTOctSnXG7fMcgOuKRypU63tz8h+yAJ239oNZOtWpTfScWPEEblt06rKzQ+mZB3pX2vlKwWa+jvFUY/8AV3f36v0mrzHeKox76u7+/V+mu7sTV/d9VwtuaU+/6L8UqLZxNv8AFfszePP8sh6Q6S0T6r4Q3zdmfj2I+M1+r5kXnmBycgsshxham0FKgdltL50hST7COoNetZgaj203g2eY5Xi/OD7l4+ptGnTdVYQZpieYgExykTzCk9Kx1yvsG2PMRHUSJEqQkrbjxmi44pI71bDuHvO1cScntCrVLu5ceS1BJTJbW0UutKG3qqQeoPUVSMNWIDg0wdPl81sHFUGuLS4SNfdPyvyWWpXVn3OJbWmHpSlhMl9uO3yp39dZ2Tv7B766MjK7Wxc3LQG5j8ppxDbiGI6l8nMAQpRHcn1h1+P2VDMNVqeg0n/xTUxVGlZ7gP8A3T5LMUrCSMvtMdySlDU2S1DUUSZEeMpxppQ7wVDv28dt9q617yhUK5WNqCh+RFuClrWuOx2odb5CUhBHjvsSB4VazBV3uDcsTOvYJ+SqqbQw9NpdmBggW7THzUkpWJm5Nb4cxy3tsTZkhhIW8iJHU72IPUcxHQEjw76+ryeypt8W7CSpUOW6llLyUHlQsnYBfijr0O/ce+sPNa0A5TfT+9u7is/O6EkZxbW/cfdv4b1laVi516gJenWoSH0PxYapD7jKN/N0kHY793P4ge6uP5oLdb7ZbXHZEqWua0jzZKWiuRI9UHm5R47dSegG9BhargCAZO6OyZ933Q4uk1xBcIGpntiPfbnbVZilR+dklrl2ec6J063LhrbRIAZ5ZDBUocvqnwV3b9RsTXxWRvpzB+yKiSzHahpWOSMSO0Kjuvm/c7DYHu33qxuBrEExBE+EfdVO2hQa4CZBiI7Z+x+XFSGlY9u/2x2x/NGl5QghlT5WU7KCR3gj27jbb213WXQ+y2+lKkhxAWAobEAjfqPA1rOpvZ6Qi8d/BbTKrKkZTMie46FfuvS7gQ+t4tv8JT/7c15o16XcCH1vFt/hKf8A25rufhz9Yf4n5hec/Fv6AfyHyK2FpSle5XzZKUpREpSlESlKURKUpREpSlESuneP2JmfeF/1TXcrp3j9iZn3hf8AVNV1fVu5FWUfWN5hVgO6qFdZZk8cD0aSyh5l7SwtuNuJCkLQbiQUqB6EEEgir6HdX47Fnte37FHa8vL2nKObl9m/ft7q8VTqeTntEL2dSn5SOwytFcvxjJNFta9LNEPNn5OBSNRImSYdMUSrzBtQcTKtqifBDjqVo/3VH8WXs7sbTzV3Vqz59xG3vSeZcsmeyGEOwhJi3eA62ns32npDKy4tASWy2lW45RsnvrdNxll0oU6y2stq5kFSASlXtG/cfeK4J1rtd0S2m6WyHMDSuZsSWEOhB9o5gdj8Vbnn0iHDdftvM3B71qHBR6J3292i0jkRrBjOhOIalWx3L5GLtawxcruFyv8ACaafMVTikOz0ssJARHWvZY3SD6xOwBFWRctU8E1M4vtH1af5BHv8K12/IESLjB3ciB1yMhSWUu7cq3AlPMpKSeUKTvtvWzi223W1MutoW2tPIpCkgpKe7Yg9CPdXDFtttgttMwbdEjNs8xaQywlCW9+/lAA238du+sTi2uklt77+I5LIYVzYANrbuHetU9JsVvOW8FmpOLYm0fS97uOXMx0NnlU++qW6kJ39qgkIHxiupnGumkuW8IzmmeNzWZuW3jG2MXh4c0yr0m1cuzQ12S423M32biSorICQE7791beNMssJ5GGW2k7lXKhISNz3nYVwotlsbnrurdshomuDlXKSwgPKHsK9uY/lqPOmlxc5u/MLp5q4NDWndBsqfRpBratNncgcRt0sESHb7dHetDNhhyW0rZYbQ8O2c9dXOpCyT4c3SrpPU7gbUpWs+o6pr8gFssptp6fMpSlKrViUpSiKP3fb0gvfu2T+itXsl1F0TybJMh0+4r8Rxey3S0T3UWeRdYqgxcLWerL7EpQ3Cj15khQ2PcO+toLx/n6/iT+isVOtttujaWbnbYk1tJ3SiSwl0A+4KB2rnNrNo1XFwOuoMEX3FdF9F1Wk0NI5ESDzWuHDdacbuWUajY7p1Kulz0YlQ48SAJzjrsYznErTLbhre9ZTPIUg945iNt6jui0LIsm1HxbRbJWXnIegjs9+W84DyzHistWlW579o61L+NNbVXeNdPQMuFir8KBcBGWi3uPx+eOy7t6hW2kjdAO24BHSohpJpvdsFj3y9Zbf2L7lmV3D0jebhHjlhklKA2ywygklLTaEgDc77knxrZ8+Dm1HnUwBvMxBcTHAme2FrDBFrqbNwknhEzAE8QI7JUT4b2203vWd1KEhbmolwC1bdVAMtbb+3bc/lqk8Ozq54foTo7bodztGMw7zfrzHk5dc7ciWix8kh8o7ML9Rtx3coC1EAAGt1UNNNlRbaQgrVzK5Ugcx9p27zXE7AgvxVQX4MZyMv4TC2UqbPj1SRt3+6q249ocXOZIJB9zSOHbPC0FWOwLi0BroIB8XA8eyO+Vqhgt4t9x4ssYVD1YuuoSE4rd46rxJZZRD7fmbUqPFLKEtq5UgKXylWxI3PhXU0Y1x0mxrhYl4neLrHZvjMe7QzYi2TLub0h54N9g0Bu8HCtI5k7gEEHbatu24UNlptliHHaQwkpZShpKQ0COoSAPV/FUG0Z0sa0uwCzYjcX4N2nWdcpTdwTECFAPSHHdkc26k7c+3Q9dqudjqNSn12m2WACBMZr2Eb9IVLcFWpv6jhfNJidcttZ3ayqJt6fnN5Lw0I1OuTdpTa8XudpmSZSiG2ZjkdkNsKV3BW5CAPaKw+S224y8O1Tu0W2yLhExzWtm+XWIw0XFuQY5YU8QgdV8oIWQPBJ9lbkSYkSYgNzIjEhCVBaUutpWAodxAI6H31+0NttFRabQgrPMopSBzH2n2msBtOCH5b7729Iu7tSFmdmyC3Nblf0Q36SqoGuulucZ3iWN4Km353PmuvTDMglDqbBHDR3lOrUk9kpXRsI3Ssk7VwcXuw4a88Km+0AgNEo/dDzhrp+OrYh263W8uG326LF7ZXM72DKW+c+1XKBufjqJ6z6fSdVdLsh09iXRq2vXuOhhEp1ouIa2cQvcpBBPwdu/xrXo1aTcRSc0ENaRMmd88PkFfVpVXYeo1xlxB0Ebo/sqlNX9S8D1lwbDdONLruzc8lut9s8iPboyD5xZ24rqHHnZCNt44aQhSTzbdTsN66uRZTcsOuHFDk1nxuLfpMObZlIhS43nDCgYTaVOONbeuhAJWU+IRWztvtcK3pC2IcZElTSG3322UpW7ypA3UoDc93ia7KWmkKWtDSEqc+GQkAr6bdfb09tXNx1OmMjWS3tP/AGaTuHsx4qp2BfUOdzod2D/qQN/bPgtH9Wssg3bHsOdVxHydRJKslsct+32qDFYtcBkSUbuvhlBU0ASEIDiwdz3HY7T7UObgdr1lzKbbNbb3ovlqPNnpqrh2C7VkLYZAblNsu7pd5QOzVyqC9x8Gtmo9ptMNlyPEtUFhp1XO421GQhK1e1QA2J95r9TLZbLl2fpG2xJfZHdvzhhDnIfaOYHb8VZ9IssA0wJ9neQdMuXdw7VX0c+5LrmOO6Rrmnfx7FqTftQLvddF9GNStRocKzRrbqHGfmzY8NUWKqE2qQhub2W27SHE7K2I+y38azGYW2dqBqVrvbcKfEyTf9MLULYtlfSR2qJBb5D484IA/fCto348eUyqPKjtPMrGym3EBSCPYQeho3HjsnmZjtNnlCN0ICTyjuHTwHgPCsRtBrbtZBvF7XcHcOyFn0e50Bz5FptezS36rXq0cQukS9JcSxq3W2NlF/dZttnbwgNJ8+blt9mhaXWHEnsgyUqWVqAA5NweoqJZJGkzMP4u40Vhbzy5h5UNpKlK2trROwHf0Bra5Futzcxdyat8VEtwcq5CWEh1Q9hWBzH8tcqWmkFakNNpLp3WQkDnPd19v46xbjadIk02G5m5n/IHh2RxusnYKpUAFR4taw7COPatdNQLtZ8s0JwrW3Tq6MXeRppIg3hD0QlXOwy2lm4R/bv2Sl7p9qBXJpvFfvmkmday3JlaJupNzXdmA4PWbtjbiWYLfuHZI59v/eVYerenmaZxj7eFYTlNqxewXRDsS/8ALbiuS5Fc5QoRVJUlLa1J50kqB+Fv3isrnNrt9j0tmWS0xkx4VugsRYzKe5tptSEoSPiAFUY7EsOBexupzcw3WDYDW9ltbNw7htGm92gLeRdMSO6y17qmblY5V+m6rs28f+0Ic+3XGCR3iQxHS4jb4+Uj8dXPX5CUglQSkFXeQOp+OvFbP2g7Z5c5okmPBzXd85Y719L2js5u0WsY8wBmntljmd0ZpnsVDZHPOYaY5jqWtlbaL05BiQULTspEVh5sHp73VOn8QqXR71asL1XyKTlktq3MX+FBXbp0k8jK0soKXGe0PRKgo83KT133qy+zb5Oz7JHJ+55Rt+Svy8wxJb7KSw08jffkcQFJ3+I10H7ap1WOoOpkMMiAbhv/AB5QDGo8mJMXk6arm09h1KT2121QagymS2xcPKZiROjvKmBPVgXOioLK3Wr/AI9qbl9saWbJdZFqjxXigpTMWy4hLrqQe9JJA5vHap1rFcYFlXhFyuTalQ4eRMrcCUFZQgMuetyjqQn4XT2VYpbbKA2W0FAAATyjlA+KsBleLuZJMx6U3NRHFjuqLkpKkFXahKFp5B16H1u/3VZT2xSrV6XlG5WMzb7x5NrAJjXq6xqdBCrq7FrUaFU0nZnvy7oEiq6oTE6dc2nQaklRWXe7Tm+qOKO4lPZuTVhamSrhMjHnaabdbCENFY6FSldeXv2G9Y3DUITw53AJQBzW+7KI27yVvdTVsssR4yS3GjtMoJ5iltASCfbsK/QQ2E8gQkJ/chI2/JWsdrMbTZRpsOVhYRJk9UvJmw1LzyA3rabsd5qvr1HgueHgwIHWFNoi50FMTe5O7RU3YblCxPJ8TyXJXhFtdyw2Lbo097oyxISUrU2tXcjnSRsTtvttX5vU+JlV7z3KseX5xaI2IOWtya2D2UqSOdeyFfZ8iem46ddquVxll1osOstuNEbFCkApI+I9KIaabaDDbSEtpGwQlICQPZt3Vd03T8p5fyZzwG62jNm0jWBGsb4VPQVTyfm/lBkku9HrZi3LrMRJnSY6swqftVwhYnlOIZNkboi2mdh7FsYnOj6THkgpWULV3I509xOwO21dTKI7upV6zWfgxMyKMVFp87a37KZL7XtOyQruXsgFO46bq2q61tMutFh1ltbRGxQpIKSPiPSutcLcmZapNqjSXbeH2VstvRNkLYJGwUjboCD1FZU9tsbWGIDIqQGzMtAzZpyxJMW17dVjV2C91E4cvmmSXQBDicuWM0wBN9J3TCpzUbUjDb9pYizWp4vT3PMW1QAwoOQSh1vmLqSPpfKRyjfvJAHfWc1UuUvT/IIeo1tjrd8+t71jktoG/M+UlcNR2/8AeAp39iq78nAc3yGNEseYZba5VojvsvSVQ7epqXP7JQUhLqyopSCpIKuUddvCszf8TvWS5LAkXO8RPmct0huei3IjHtnpKAeXtHCdigKPMABvuBvW43F4DDvpsa4Fg8oXCSZDg0BoOVt7SLWsSVouwe0MQypUe0h5FJrCA1uUsLiXEZ3WGaDeTJAEXPdwTG/mSxC12Baud+MwFSV+LkhZ53VH41qVWdr7176+V5StWfiKjqtTVxJPMr11CizDUm0aYhrQAOQsEpSlVq1fR3j46ox/6u7+/V+mrzHeKox/6u7+/V+mu7sTV/d9VwtuaU+/6KOZD/pFi/8A3x/+xNRvP4cmwxLi/CaUu23zlEhtPcxKCkkOD3LAIPv2qxOh2JG+3d7qEAjYgEewivYYfHnDupnLIaII49YuHKDpyXjMVs0YplQZoLjIPDqhp5yAZ59kqF3ZPmOZOTJt8kWmPNgNNMSUpb7NSkE8zalLSQk9QR3b11JEZm4WLK5lrm3C6LejoaVIcbQEPlsb/SuQDm2HQnb8tT5aEOJ5HEJWk94UNx+SvqQEgBIAA7gOm1WN2lla2G3GUbo6pB4TuG+N6rfsrO50u6pzHfILgQd8bzundzhd/wAis93jWRm1zUSlquUN1Ya3PZJCvs/3J3O2x671lrAAMnydY6KMmMOb3diP76ziW20b8jaE7nc7JA3Ptr9bDffYde/31U/Fs8kaVNpAI3mf8geA4QrqeCqeVFao4Egg2EaNc3ieMqHYzfrRj2PqtV6lNxpsBx5L8dzo46StRCkJ718wI2I3roRkmwW7DHryfM22JEguF3oGQ4hZQlXs6ECp+W0KUFqbQpSfgqKQSPiNFJSsbLSFD2EbirfP2Zi7KesSXX4gi1rekTedyp6NfkazOOoAG24Oab3v6IFo3nlFbPdbbYLnfIl7mNQnn5y5rTjyuVL7Kkp5SlR79tiNh1FY9mWyxjN0cdtqnxkFxfFuhOJKS+HNgkkd6R0KyfAdanS223Nu0bQvlO45kg7H8dfSATuQCR3HbqKxGOZObKZMTf2dIta/2CzOz6kZM4gZotfrazeDAmLdpUEs7K7BaL5i1z2VcTEeliTuT562WyObc9d0Ecu3s2PjXJBkM2mZjN3uSw1CcsghpfX8Bp48igFH7HmA23PsqbkA9SkH4xXwpSpJQpKSk9CkjcfkqXbRDyS5vpa37Ita3Hw0WDdlmmGhjvR9G3B0ib34TY79VC8qvEC72C9m2xw60x5shU1AHI8rtU7oSe9XL7e7rWScmRoWoRRKfS0qZbG2o/N07RYdUSke07eFSIJSEhAQkJHQJAG35KFKVEFSQSnuJG5HxVh54wM8mGmL773Dez/r4xZW+Y1DU8qXjN1TpaxcePB0d0mVBHozyby5gYbV5pMnpuYO3QRPhuI/jUgf8VTRVxh+kRay+BLUyZAa2O/Z77c2/d39K6NqtE5i4ybxd5zMqW62I7XYtFtDTIJOwBJO5J3J9wrIGG2qcmeoqLiGiykb+qkEgk/Gdh+SpxdenWcGkzA1G9283HKeSxwWHqUWlwES7Q7mAmBY7pMa6wuavS7gQ+t3tv8ACU/+3NeaNel3Ah9bxbP4Sn/2xre/Dn6w/wAT8wuZ+LP0A/kPkVsLSlK9yvmyUpSiJSlKIlKUoiUpSiJSlKIldO8fsTM+8L/qmu5XTvH7EzPvC/6pqur6t3Iqyj6xvMKsB3VCtRNZtN9K3YETNci81nXTnMKBGivS5khKfhKQwylThSPFW23vqajurXq1XWy4vxp5jIzafEt0i94dakYvJnOpaQ7Hacc88ZZWsgcwcKVKSDuRse6vG0GNeTm3Cbb17CtULAI3mF2NWOIizMYTgWoem2eW8Y9dc4gWi8T1oSENwz2hktOh1IUypIQObcJUnb31O8A4gNI9T7/IxfDMr85uzDBliHJhPxHXo++3bNJeQkut9R6ydx1rX7Vi86RZ4myfMBjbJtzut9mjXq4JZT5lfJ4bX2rrSgoh4D1UKVsASNuvfVm6roP0WOib8fkRKctmUMpcI2UQIrZSkn9yFHfburadRp5Q2CDDju3Cb8fBaza1TMXSCJA95iykuS8TuhuI5FMxi+5001LtjqWLi6zDkPxbe6e5EmQ2hTTKuvULUNvHas5m2s+l+nKrcMzzGHbhd4rs23qKVuJlNN8nMWy2lQWT2iOVI9ZXMOUGtSND38js+guQWu88QWG4lGtMq7s5dYrxjDL8xmQp5ztu3UuQlbynEkFJ5fWCkpG+1SXBMdtMDOuFK0IuMy9woGO5BIt0q6W8xJCm0sJUwtTC1KLZShSQnck7AGsnYOkwkSbT3wCeEbu1YsxdVw0F48SBx7exXSjiz0Acsbl8bzsqDUpUJyCm2yjcEPJTzqBidn2wASQoqKOUDxrtZvqpjd70hhai4Lq1AsVpuNwgtxr/AOj1TW1BcpLao5ZI5kqcO7W5AKCdzttUa0xgwE8ZOstxREZEwWPG0dsEDn5VtuFQ37/W5Eb+3lHsqi5SWmeD/I2WglDTesTiEJT0Skenm+gHgPdUMw9IuGWdW8D6QnhuR2IqBpzR/lxGnets9RdcNL9LbnFsuY5Gpi6TW1SGLbDhvzZamUnYulphC1pQCCOYgDoa+ydctJImn0bVN7PLb8y01aWo09ClLD7xJSGUNpBcU7uCOzCeYEHcdKrTArzYcT4stZWs2ukK13m8RrNLsb895DPnFpbjlK0srWQClDwVzpB6Hqag2T6p6aSnsOjaK4HisB3INQLsm2ZRkkcptEe5MtAyLjHCFjtVOk8jZ3QFFKvZWDcK1xDYOgM7rif7rvssziXAF0jeI36x/dOav/TzXfSzVG8S8dxDI3HbvBZEl+3TYMiFKSyTsHQ0+hClI3IHMAR1FQTR3X+DH4fLLqdrTlrLUm4XKfAS+I30yU4iY820y0wykqcXyNjohJJ2JPiar7FrtepXG3iUDKdVrHm15iYdd25K7PbW4keBzLbUmPulxwuKIBWQpW6QR0G/WJ4NFxCZw+aOm76iT8DyNrLr2vFsibiIfhxp3nMkKaldp9LCXEEpHMRuegPfV3mtMCNxg9ujuyd3D3qrzqoTO8Ty1b29vFbaad6wad6qpnpwfIfPJNqUhE+E/Gdiy4pUN0dow8lK0hWx2O2x276mVa46NZzlTOuuS4PqLIwPKLxExhm5yM1xqN2DvmyHilEScN1BCuqlpAVtsCduvS/ceyOwZbZYuR4veYd2tU5JXGmRHQ4y8kEpJSodCNwR+KtGvS8k6BpbxW7Qq+UbfVY+8f5+v4k/oqrLPl16zDWq9Y9Zp6mMbwSG1HuaUJSTPu0pPOloqI3CGGQFEDbdbo36J2q1Lv8Asgv4k/orXvQ5q4k67W2Ioov3zbXUoJOyuZyK2Yp+Lbl2+KuS1gJqvOo+pifd811nPIFJo3/QTHvUruXEnolab8/js7OmEPxJQhSpKYr64UaQTt2TspKCyhW52IKxse/arLBCgFJUCFDcEHcEe0VpDps/LjcLTttveveJ2HHoVulwMhsMzF2np0aQVLD7LgU+lbr6lElJ5d1EpIraHS+649iWB6e4ddss7WfPtLEe1i5t+aTbilplJJ7BRKgsN8pUnckeNW43BMoD/iJJBI37t/oj3X5qrBYypXP/ACAAEA7t+7U++3JceVcQGkOF5I7ieRZg2xcowQqYhqK8+3BSv4BkuNoUhgHff1yOnU7CpTiOYY3nliZybErq3crXIcdaaktpUErU2soXtzAHopJG/cdunSqI0LzTAcAtWp+Pao3+02m/x8tu06+s3J5Dbs2K8rmjuJSvq82pkhKQnf2Ada7/AAnZRieMcNeMXK8XaJZbbKuk6JDMxwNDndnPBlkb/ZkbAJ79xTEYJtOmSwGQWidxkEyLdlrmymhjHVKgDyIIJjeIIF79vAXWwSACtIPcTtVU8NOc5RqHpvJyDLrimbPbyC7QUuBlDYDDElSG07JAHRIA37z41a6Bs6kHoQoVrPwn6kad41pbOtWRZ7jtrmoym+LVGm3Nll0JVMWQShSgQCO47VRRpeUw9QtEkFv/ANK6tU8niKYJgEO/+VaWpOJaqXeau8YRrK5ikCNCJXATYY00Ouo5lFztHDzDcbDbuG2/jVecOY1z1Jw7ENU8k1zU7BuhMmZZE45ESl1tDq0Frt07KTvy78wG43q8lXa1X3GJF3slzi3CDJhvKZkxXkutOp5VDdK0kgjcEdPZVYcGZB4acBIO480e7v8AvTtWtqubhHS0SCB6LZgh03id3NVuptdimw4wQT6RixbG+N/JQzSs8QGsFvyHJY2vq7BHgZNdLPHhIxiHJCWo7xSg86tiTsR3jw76nWhufZ1eckzzTHUedb7resFnxmRd4EfzdudHkNFxsraBIQ6kAhQHTr7tzUvD1o21nmP5bfVao6gWH/ntfGDDsd781i+rI+Fych9Y79Tv12FX5heA6c6D4pdXra+qBBU4u6Xm73aap599zb1n5D6+p2HxAflrYxzqLS+kACbAANAi43i5tbfqtfBNquDKpJAvJLiZtwNhe+5Tuqu03zfJcg1k1ZxK7XBL1rxeVaW7WyGkJLCXopccBUBurdXX1idvCrKhTYdzhR7lbpLcmJLaQ+w80rmQ62obpUkjvBBBBrX7T7NsMxHiK1xRleW2azKkzLIWRcJzUcuhML1innUN9txvt7RWjhqWdlURJDR//TVu4ipkfSMwCf8A5crb1CxnPMljQWcE1KXhzrDi1SXU2lmf5ykgBKdnSAjY7ncd+9UfpizxE6h3XN7c7xFrgjD8kesKVpxWE550ltCFdqR05Cebbbr3d9bFY/lGM5ZFcnYtkNsvMZpzsXHoEpEhCF7A8pUgkA7EHb31UPDUQcm1rA8NRJv9i1V1Co6nQqAtEtiJa0m57Qqa7GvrUyHGHTo4gWHYVi3J2tOfa6agYLjOsZxa1YlGtK2G0WGLMLqpMfmWSpzYj1kk95+Ft4VlsYzbVPTvVmzaUauX63ZPb8ujSHceyCLAEJ4SWE8zsWQyklG5R6yVJ+L4sLh+Z4fiPFLrMcryuz2YSomPBj0hNaj9qUxlc3Lzkc224329or83/KrBrZxHadW3Tu6R73bdO1Tr5fbpBWHYrDjrPYsRg6PVU4o7kgE7D4jW05hJyOYMmQGcoEHIDOaJu7tvMLWD8oztec+ciMxMjPERMej2WiVYmHZfeoWquVaU5TPVMcQw3kePynEpSty2vKKHGFcoAUWHgUg95QtG+5BJseqVyoKe4u9P0Qt+0i4be3ZxT4R1PMpb5v8AxAdviq2MeyTHsttib3i96h3W3rcW0mTEdDjZWhRStIUOm4UCD7xXOxNOA2oBqATzuPGJXQwz7upk6EgcrHwmFkqi+p3+gV5+8o/tE1KKi+p3+gV5+8o/tE1zsR6l/I/JdPBfqaf8m/MLXGov6bn3XUE47bZJag2OImVcykAl557cMsbnuASFLO3X4I9tSmoDgIWjN9RmFnaUbrGcST/2SoqezPxdDXm8DTaadaqRJa23e5rZ7gTHAwV7vHVHCrQpNMBz78mtc6O8tE8RI3rJ3PVLA7PcX7XPyBCHoqw3JWhlxxqMo9yXXUpKEH98Rt41KULQ6hLjS0rQsBSVJO4UD3EHxFUNgLtyhad3CJP1FsloahuzEXqBNtSHHm3StXadoVOArKgdwduoIAqy9PpFnx/BsWtT99U4mVHSxb1zW/N3pI2KkpDaiSFBG3T2CujtXZFHBtPkCXOa7KdTNiZHUaBpMAusZm0nmbI2zXxrgcQA1rm5hdoi4EHruJ1iSGXBESYHbvuo2GY1cvRF4vQblpbDrrbbDj3YNnuW6UJIbT71bVlbHf7PkkE3OxT25kUOuMds3vylaDsoAnv6+I6HwqA4LfbBjV+zuFlVzh2+5LvTs14zHEtl+EpCexUkq+GgJBGw326ivmjF4sFs04eujkpi3WtV7nhhT6g2lCVyCG09e4ncAD8VRitlU6WHc6m1xc0sAOrXZ2k9URPK5kKcJtepVxDW1XMDXCoSNHMyOA6xzRvvYQVZ9Q2/6pYba13G0pyBtNxiNuNlQaWplmRyEoQt0JLaVb7eqT7qmR5uoSdlbdCfbVP4Pd8Wtmjd2tGRy4jMqGbgxeYr6kh1UlS3O9B6qUrdPKdjv02rW2ZhKVdrqtRrnQ5ohuvWm5sbCIjeSLhbW1MZVoObRpOayWvdmdMdWLCCLmZmbAGx3SzHc6hQ9NsfyzNLu0y9cIbCnHCj1nn1p35UNoG5Uf3KRWYsWa4xkkeXIs9zDvmA3ltONLaeYG2+621gKAIB26dapuMJ0djSG4em4toiiyOsMTpkYPsNTFITyhQKkhKlJBCVE94NSeEwX9QbpLm5zGvV2i43IZlNwbYGmQyo7o7V1K1DnBBKUnrsTXXxWxsL13yQTncImBDy3LAZG7XMIJFo142D23i/+OnlBaMjTMSSabXZpLwZvpkMgG86Scaw6bqdjNjJ2imV2YQ92LvYpUsAoStzl5UKII6KII361lskzXGcSVHavty7J+Xv5vHaaW886B3lLbYKiB4nbaqx81itcJ5bbYbS2bAHyABsXCoKK/j3671nbPMh2rV55++yWo7lxxmCi0vPqCUqCCovoQo9ObcpUR3kVTU2ThAajqYcRTNQESJdky3By29KTYwAr6W2MaRTZULAajabgYMNz57EZr+iADLZLhZdzFtR4dyn5rd5l8Zex6yuRVxX0N9GmVMBS99hzE8++4PUHpXYnajY1eWS1jmatQ3IdziRpD5hLdQ52jnKGUkjY85BTzgnYiq/u02yz8f1ul2F1lyI4lklxnbkW52ADikkdCCoHqO871LdRo8eHgGJxozSGmmrxZEISkAAAOJ2AraqbOwgxDDlILi1oHVgRTpuOYFpkmTOl7rTp7Sxhw1TrtcGh7ietJmrUaA0hwgDKI1tbtHftupkW56hX/DfOEtNQI7YiKEV3mLwSsvFaiNtk8o27gduhNZG2ZlZLTg1syTIsujzWJDaQm4iOWvPFqJ25GQObc7fBA36VhIM2Kzq3nsN+Y02/JtEAstLcAW6EtO78oPVW3u7qh2JvxYFu0au16cbbtLUOWyHnSA01MWj6SVE9EkgKAJ8ards3D12jK0tGWmYES7/AInPMHLMkiN4mTBsBa3amJoOOZwcc1QSZAb/AMzGCRmghodO4xAkXJt7HMxxvLBIFiuPbOxCEyGHGlsvM793M2sBQB8DtsazNRmBecRuOezYlrjNyr1HtzfnlwYCVoQyVnkYW4D8LfdQTt3VJ683jKTaNQBjS0EAw6JuOIiRwsLbl6fBVnVqZL3NcQSJbMWPAkweIk33oO8VRj/1d39+r9NXmO8fHVGv/V3f36v0munsPV/d9VzduaU+/wCiiua3q+Wdy1eg20vLffc7VgpBLzaEc5SCe47A7bVy33Iv+Zb+SWKQN1NIcZWUg8u6wCCD4jcgj219yE7ZHi3X/wDGP/2JqMZwy7jEG5xWWybTeyFNpSOkaXzpKh7krAJ+MV7zCUKVcUGQM2v8hmIIPcLd44L59jcRWw/nNQOOXT+JyAgjmTB7YPFTm43q2WhLXpCV2a3+jTaUKW45t38qUgk/kr8t5BZnbY7eUT2zDY37VzY/SyDsQpO24PXu23qPTRNbzxxSLtHt637a0mKuRHDgcAUe0QglQ2O5BI8RWPuDKHbPmMlm7puLy2G0yCxF7NoOIG5KSCQpXL8Lb2VgzZ9FwbJMnKd+8gR6MCJ1zG4iFZV2lXY58NEDMN3+LSZ9KTMaZRYzPGdS58OChp2W+G0vuoZbJBPMtfwR09tdKXk9hgzPR8m4pTJ7VLJaCFKUFEAjfYd3rDr3daw2U3a2SolhaizmXlyLlDcQG1hR5Arqo7dw6gdfE7V3bA2381WTvBKe0L8ZHN47diDt+WqW4SmykatUHQ20/wAgOB4rYdjaj6wo0SLkCddWuduI4D3rtTMsx6BJciSrklLjBCXiltakMn2LUkFKfxmu1cLxbLVGbmTpiG2nSEtEbqLhPcEBO5V+Ko9h8+0wMVej3SSwy5FdkC5IeUArnK1ElQPU7gjb21i8fJtjmJy7yewi+ZSmo6n+gacWsFsEn4JLfQb/ABVacBSlwg9UkfygE2tbTtsR30DaVXK09XrgHf1Zc0da99f+twe6Y27ILPdXjGt85LryUFxTXKpK0JB2PMCAQd/A9a4E5dji5IipuiCoudiHORXZFzu5Q5tyb7+G9R+ZJj3LK72LE6h2T6AW0pxo7hTvMdgFDvIBA/orpJU29gTbMjLILVtVFTHUwi3guoV0HIE8+5cCvdvv1rNuzqJgukTltvEidzTPYLTxVbtqVxIblMZr2g5TG9wjtMmOCmlwyCz2t8xp05Lb4QlYaCFKWoEkDlABKuoPQeyv3AvVqucJdwhTW3I7ZUHFn1ezI7woHYpI99YaCwEZzu6S46zYmUhxadldXCCfcTt1rCX5l5yJm7cZJ287iOOhKeb1ORBcOw7+gJI8aqZgaNQimCQYaZ3XIGkds6q6ptCvSBqkAiXCLz1Wl2sxujTtUsg5TYLlJREh3AKcdBLXM2tAd27+RSgAv8W9J2U2G2yFxZc7lcZALoQ0tYaBG45ykEJ6e2o/dt5rFrRKzCG+29LYXCTEgAuFaTunl5VkpG3QnboK5XQtq53p/HclixVB8rnwbiyOyK+QbqB3CglQ269RWYwNCZkx38QNcn013rA7QxMRAmRw0gn0c/Z7WmgUgenNruNsQxdW0ty0OrQyG+fzoBIIKV/YhO+/v3rgXl2OIkGMu5pBS52Jc7NfZBzfbl7Tbk336d9YO3zk3Gfhk1EFEMOsTClhA2QgBsD1f907bj3EV0DINqxyTMst7gT7M0pxSrXcmQHEnnJU0CDvzc3cFA+FZNwFMkMfM6cL5nDUA3tadeMALF20qgBeyI13m2Rh0LgYveLi3VJJVhV6XcCH1vFt/hKf/bmvM5hwvMNOlstlxtKuQ96dxvsfir0x4EPrd7Z/CU/+3NW/h0RjSD7J+YWt+KzOzwR7Q+RWwtKUr3C+bpSlKIlKUoiUpSiJSlKIlKUoiV07x+xMz7wv+qa7ldO8fsTM+8L/AKpqur6t3Iqyj6xvMKsB3Vg8swbC89gt2zN8Ss9/iNL7Rtm5Q25CW1fukhYPKfeKzg7q1v1k4jM00x4jMUwNqHa14RNhQX77IdZUZMbzqWuKhxK+bYIDhZ33Se8+2vGUKb6rop6i69jWqMptl+ivhrDcQZtltsrOK2hu32d5Ei3REwmwzDdRvyONIA2QobnZSQCNzXck2WzzbnDvUy1Q37hbg4mHLcZSp6OHBs4G1kbo5gADseu3Wq24mdV75o9phIvuJRIsvJ50tq32eNJSVNLePM44paQQSlDLbqz1HwRXbwjWGyHQ7EtWNUckstgRerREmy5Mh5MaP27rYUUoCjv7dkgk1Ip1CwVOJhR5WmHmnwEqQXzSjS/J761lGSac41dLwzylE+Za2Xnxy/B9dSSTt4b91Z2RZLNLucO9SrTDeuFuS4iHLcYSp6MlY2WG1kboCgACARuB1qLK1n0yk6f3XUux5xYrpYbSy4t6YxPR2KXEj1Wlr/6tSlFKQFDfdQ6VA8K16k6oaY4BnONZZhNnn3u8QYd+gzZKnA0p0KUu3sEHcSynk5Qr3nbrUilWcJMwLclBqUmmBqbq5o9ms8S6S75FtURm4z0NtypjbKUvPpbBDaVrA5lBIJ2BPTfpXSOF4cq1uWNWJ2c256X5+5DMJvsVyefn7Yo25S5zgK5tt9+u+9c9hyXHspYkSsavUO5sxJbsGQuK8HAzJbOzjStvgrSehSeorCztWtLrZjBzW46hY/GsHnDkQXJye2mOp9CilbaV77KWFJIKRudwfZVYFSYEqwlkSYXdy7T7A9QGGI2dYZZMhbjKK2E3OC3I7InvKecHl38du+v1d8Ewi/463iF7w6yzrEyEpatkiA0uK2E/B5GynlTt4bAbVgWtcdKLjg961Bx7P8fu1nsTKnJciPcEdm0vb1G3Ff8AVlatkjcdSem9QjCte39T9KsHzzG8swqzXG8XiBDvsOdJU6hgu8yl29og7iWU8nIFe8kVY2lWjeAD7iqzUozuJI94VpWbT7AscFvTj+E2K2eie18wMS3tNGL2oAc7MpSCnnAAVt37DfeuR/BMIlY2vDpOHWR2wuFSlWtcBoxCVKKiey5eXcqJO+2+5JrG5xq9pXprJjw8/wBQrDj8iYOZhm4TUNOLTvtzBJO/Lv8AZbbe+srLzPEINogX+ZlFqatd0dZYgzVS0dhJcd+pJbc35VFXhsetYf8ALZ17rOad22suHGsCwrCbVJs2EYjZLDEkhRcZgwG2m3FEEbrSkDn7+4+HSuHTfBrfprhFqwi1vl9i2NrHalpLXaLW4pxxQbR6qEla1bIHRI2A7q6uH6waVagXSXZMG1Fx++3CCCqRGgT23XEJB2KuUHcpB6cw3HvqX1DzUEtfPG6lgYYcxR68fsgv4k/oqt4uC3SxaxTc+sC4/onKLY3FyCMtZStMyN/mspsbbKJbUppY3B2CD12NWTdxvPX8Sf0VrhgfEjd5+v8Alekeb2+LEtabw/asYubTZQl6SwhK3IjqiSC6ULSpJ6b7EbHcVy2U6r3VTSvAM8p+mvcum+pSY2kKlpIjnH1071bk/TDTW65CjLbnp/jsu9oUFpuD1tZXI5h3K5ynckeBPWs1LstmuFwhXa4WmHJnW1S1QpLzCVuxisbKLayN0EjodttxUEf1BvzfETE0rSiJ6GfxB2+qV2R7fzlMoNABW+3Jynu27/GsvN1i0ntuUJwm4akY5GvxcDPo924NpeDh7kEb9FH9yevuqt1OuY1NptJgfRWtqUBOgvG4SVXmTYJrf81FzmwbBppmTbstcjH7zkTJan2JtfcyUoZV2yW1blBCkqPjU1020gsGD6YWHTi9NRMhTZ1edOPzIqVJdmqdU6p9KFbhJDi1FPiBt1qXuX+yMX1nGHrtFRd5EZcxmCp0B9xhCglTgR3lIUQCfaa6svM8RgPXiPNyW2sO4/HRLuyHJCQYLC0lSXHh9gkgEgnwBrJ+IrVGBgECxsO4Hx3Rc8VgzD0aby8mdRc95Hhv3BZnc7779fbUQe0d0jkPLkSNLsSdddUVrWuzRypSidySSjcknxrmtuqumd3lXeHa8/sEl6wNB+6pbntnzJojot077JT7z8XfXDiGsOlGfXNdlwvUSwXqe2kuKiw5qFu8g71BPeoD2jcVW1lemCWgiNdfFWufQqEBxB4aKR22z2iy2xqy2e1Q4NuYQW2okZlLbKEHfdKUJAAB3PQDxNfbTabVYbezabHbYtugxgUsxorKWmmwSSQlKQAOpJ6e2oNoxn99z9rNFX1EVJx/L7lYonm7ZRvGYKOQr3J3X6x3PT4qleV5niOC2v01mmTW2xwOcNiROkpZQpZ7kp5j6x9w3NRUpVG1DSNz71LKlNzBUFgu7a7LZ7Gw7Gslqh29l55clxuKylpK3lndbhCQAVKPUnvNdbLLAjKcXu2NOPNNJusN2GXHYyZCEBaSnmLa/VXtvvsem+1dbDs+wjUKC5c8Fyy136KyvsnXIElLvZr8EqA6pPxgVjYOsWlFzyk4RbtR8dk3/tCz6PauDani4O9AAPVQ/cg7+6oDKweTBkXNtOaF9IsAkQbC+vJSGwWaHjlituO27n80tUNmExzndXZtICE7nxOyRvWIu+mOm2QXB2737T7G7lOfILsmXa2HnXNgAOZakknYADr4CoJjnEJj2X51qDgVpv8AjsaRjDCRaZDk0OCY4I63JDq0j/q2VgJWE7kcqtz4CaYHmcK82mxWu75djt0yabZmro+m0vfSZTRPKqTHSr1iwVdAqrX0a9AlxkG3HeJVbK1CsA0QRfwMLM4/i2M4nFcgYtjtss0Z1ztXGYEREdC17AcxSgAE7ADf3Cuxb7NZ7S5LdtVqiQ3Lg+ZUtUdlLZkPEAFxwgessgD1j16VFdRs4j2awZBBx7NcXtOTWqA3O3vbx82htLcCUvSUpPMltXrJB8TtUdncQGMWzWu36Qz7zY2u2shmyZapoCxPU4hLURKPBSkKLgB6kFOwqG0K9YFwk6k8hBnxUur0aRDTA0HeZEeCml50004yO4OXbIcAxy5zngkOSZlsZedWANhutSSTsAAPcKy1msVjx2Em249ZYFqhg7hiFGQw2D7eVAA3riteT45e7CjKrPfYM2zONLfTPYfStgto351846bJ5Vbnw2Ndi03e13+1xb3ZLgxOt85pL8aSwsLbebV3KSodCD4GqnGpGV0wLK1opzmbElQ7DMEucLUHLdTcqVHXdL2pq2W1llZWmHaY+5bQVED13HFLdXt0BKRueXesxp7g8DT3Gk45b3kvJVLlTnVoYQwhTr7ynV8jaPVbSCrYJHcAO+pJSj6z6libW8BAUMospwRrfxuUqL6nf6BXn7yj+0TUoqL6nf6BXn7yj+0TWpiPUv5H5LewX6mn/JvzC1xqPPY7Lj5wzllrU0GpkMwLq0pWxWlBKmHU+1SSVJI8Uq91SGsPLzPEbfMRbp2S21iU5J8zSy5ISFl/YHs9u/m2Unp/vD215TCmuC4UWl0ggiJseXDUcCAV9CxYoFrTXcGwQQSQLjnx0PEEhfufieK3Sei63PGrXLmo25ZD0RC3Bt3esRv0rvSIMGW7Hflw2HnIi+0jrcbCi0vbbmQT8E7dNxWLn5vhtruqbHcsqtcW4KIT5s7JSlYJ7gQT0J9hrv3i9WfH4SrjfbpFt8VJCS9IdCE7nuAJ7z7hWTm4s5GuDr+jrp/17OSxa7BjO5pbY9bSxHtdvNQ++WHO1XyVLjWrFciircD1uXd0dm9bTsN0ApbV2iOb1h1B8KyuIYPEx/EmcbvBj3Zan3JktbrILbkhxwuKUEHfYBR6eI2FZW25Njl4tjt6tN9gy4DAUXZDLyVIbCRurmI+DsOvWuKFl+KXGVJgwMktsiRDZEmQ23JSS00QCFq69E7EdffW5VxONqUvIZMoaRMNIMiwnhruiSZMladLC4GnW848pmLwYlwIh1zHGY3kwBAgWWYrGScYxubc03qZj9tfuCByplORUKdA22+ERv3V1LbnuEXhYbtWW2qWtTyY4S1JSolxW/KkDxJ2O3t2r9XvOcMxuUmFf8ptlvkKAUGn5CUr2PcSO8D3mtRmHxlKpkYxwcRoAQY321hbdTE4KrT8pUewsB1JBE7r6Su+9ZrPItossi1Q3beEhAiLYSWQkdwCCNgBX5tdhsVkiLgWazQoMZwkrajsJbSvcbHcAdento9frJGjRpki8Qm483/NnlPpCHvVKvVVvsfVBPxA117Fl2LZOp5OO5Db7kqP9VTGfSso9hIHXb391RlxXknGHZJk6xPE7plZZsJ5VolueIGkxrA3xHgu56KtZt3of0bF8w5Oy817JPY8n7nk222921cd0sNjvkVEG9WaFPjtkFDUlhLiUkdBsCOn4qwluzS1W6wQp+YZdj6Xpa3kokx3wiO9yLI2RzHclI2CvfvWSs+YYpkD64tiyS23B5pvtVojyErUlG+3MQD0G5HWrH4fGUCXgOhpPWGaJ0Jn66qpmJwWIaKZLZcB1SWzESBHfpouyiw2JqM/CassFEeShLbzSY6Ah1CRslKk7bEAdAD4VzyIEGWy3HlwmHmmVIW2hxsKShSfgqAPcR4HwrDw9QcFuFzTZYGX2iROUooSw3KSVKUPAddifcKx0XUi0S9QbhgiZkBK4cRtba/OAVuyVFXO0E+1CUgkd/XwrIYTHPLiWulozGZFrCb93dyWJxmAYGgObDjlEQb3MW7+/tKkkiyWaXcGrvKtMN6cwkoalOMJU6hJBBAWRuAQT+U0XZbO7bPQjlphrt3LyeaKYSWQn2cm2234qxlmyJlnEY+QZPkNlUnkUp+fFd5IavXIHIpR9wHxg13LJk+OZLGcl4/fYVxZZOzq47wX2Z/3gOo/HVdSliqc+kWsMSJgEHcd3Zobq2lVwlWIyhzxMGJIIvI39uosue02WzWGKYVktUS3xyrnLcZlLaSr2kJHU13KwCtQcERKjQl5jZ0vzUpXHQZaN3Eq+CR18fD21IKpr067HZq4IJ3mb+/VXYerh3tyYdzSG7mkW92iDvFUY/8AV3f36v01eY7xVGP/AFd374r9NdbYmr+76rk7c0p9/wBFxLZZcWhxxlC1tEqbUpIJQSNiQfDpXx+OxKbLMlht5s7EocSFJO3d0NfulehDiIvovPFoMgjVcEuDCuDXYT4bElsHcIdbCwD7djX7YixorCY0WM0yykbBttASkfiHSuSlTndlyzZRkbmzxfiuoxZ7RF5vNbVDZ51BauRhKd1A7g9B3g91dlLTLa1uttISt0guKCQCsgbAk+PSv1SpdUe70iSobTYyzQAuq/abVKkJmSrZEefRtyuuMpUobd3UjeutfIl2k9iq3eZPtJ5hIhzE/S30nuPNsSkg+7brWTpWbK72ODtY43/vcsH4dj2luk6xY/8AvfrobLB2SyzI1yfvFwRDYcWwiKxFhg9my0lRUepA3JJ37gKyPoi0+d+kPRcTzrfm7fsU8+/t5tt9/fXbpU1MRUqOLpjdbhwWNLC0qTAwCYM3vfj/APncvyGmQ8ZAaR2pTyFzlHMU777b9+3uohpltbjjbSErdILigkArIGw3Pj06V+qVTmPFX5RwXUjWi0wn1SodriMPL35nG2UpUd+/qBSXaLTPdS/OtcSQ6jolbrKVKH4yK7dKz8rUzZsxnmsPI08uTKI4QvwWGVLQ6plsraBDaikbo3Gx2Ph09ldddntC5fn67VDVKB5u2LCSvf277b7126VAqPboSpdTY70gClel3Ah9bvbP4Sn/ANua80a9LuBD63i2/wAJT/7c13Pw5+sP8T8wvN/iz9AP5D5FbC0pSvcr5slKUoiUpSiJSlKIlKUoiUpSiJXTvH7EzPvC/wCqa7ldO8fsTM+8L/qmq6vq3cirKPrG8wqwHdWrWr+AtapcR+W6fuEJcvWji2Izni1JFy5mHB7Cl1KD+KtpR3ViBiONJy456LOwMhNvFq9Ide1807TtOx79uXn9bu33rxlCr5FxcNYXsa1LyoDVqVbM5kcRFo+aaehXJpnpjcnLs2ofAyaZGejOIP8AvNtR3j7u3FcekEaz3TUHhzt2bssP2tnScysdYmAKjrvAU2HVJSr1VPJY25fEDqK2qtGmWAWCBkNrsmJwIUXLJD8u9NMoKROeeTyurc696kkg7bd52ro33RjSzJsMtmn19wi3S8fsiG0WyGoKHmIbTyoLLgIcbIHTdKgdq2/O6d2gED5SD9StXzSpZxIJ+dx9AqSyGBYofFLnNsw+HDTGmaWSZOWxozaew8+S6oRFuoHqh4t82xI5inrULt7cNPDJwsORW2EqczDGlOqaSkFS9ngSrbvV08etbV4JpZp5pla5VnwXEoNqjT1lyZyAuOS1kbbvOOFS3DsSPWJ6E1h7Dw+6MYxAatdg0+t0KHHvLWQssNqd7Nq4tAht9CSohKkgkAD1fdQYpgEXtHfAIv7+1DhXkza891wbKhdVMxufDnqDqpYrA0rtNWLWzecRaT3HIXHEQZDaf94l1l8+5JNZO94va9Osq0d0MwPTqy5RnWN43Ll26ZkFxcj2yC0OVEqSW0JV277jpURsnmSNzuK2LyLBMOy662O+5NjkO5XDGZZnWiQ+glcN8gAuI69/Qd+/cPZWN1C0h021UTB+b7FI11dtilLhSC44zIjFXwuR1pSVpB2G4B2NYtxTOrmB7fdA4bvmVk7Cu60Hl75PHetbMFZvJ4jtZbdly8NeuqtOGFXONjUZaITbwW4UJcDhJW8lJSSogHZSeg2rH21uGnhT4Zlxm2AXMwxdTymgkFS+d0Eq271eHXrW0OKaOaW4NPTdMQwa12mWICrYp6M2Qp2MpztFIc3J7TdfrFS91E+NdCxcP2jGMW9NpsGn1uhQkXlnIG47anezbuLW4afQkq2SU7nYDZPurM4umTv3eAI4rAYR4G7f4kFVzw/W3Hrzq1rvMyyDBm5Y3lyojwnNIcdbsojt+aJSFg7MFJWenqk99UNcLZZLtopfMWtbaZGBHX2Jb7E0hRMc29clAeaZP/Y9op0Dbp1O1bh59oNpDqfdWr9nODQrlcmmfN/PA46w8tn/ALJa2lJLiP8AdVuKy0jTDT2TjVpw5eH2xFjsUmPMttvaa7JiK+wrmaWhKNtilXX3nqd6huLY05r7u6OH9Cl2Ee4Zbb++eP8ASql1bx6w4/xIaAzLDZYFteVNvltUqHHQzzRRAJDJ5QN0AjcDuFX1HnwJjr7EOdHfdir7OQhp1K1Mr7+VYB3SfcdjWPu+JY3fr3ZckvFnYl3PHXXn7XJc354jjqOzcUjY7bqR0O+/SsDgWmsTCcoznKGTES9ml3buTjUVpSEoDbKWwpZUSVOqIUpShsOoAHTrrPe2owSbge+5+hWwxjqbzAsT7rD6hZW7/sgv4k/orVDHNLrfq61rzismWqBPb1CXOs9yb6O2+4NRmVMvoI6jZXQ+1JIra+7/AOfr+JP6KjtkxXHMbl3adYrQxCkX2abjcnG995MkpCS4rc9/KkDpt3Vx24g4d1TLqTbuMrsOw4xDaebQC/eIWqWmuo+WZ5r9JN5ti4Ge4vpvcrLeIoRsPSbMoFDjfgUOgtuJ2/dfFWG03xrK71wnOLlP6QtYnPtsp++XC6sTFT2ZRUrt3pK0/wD4lDnUbddwnYd1bfNYNh7GZvaiMY9DbyWTDFveuaUkPORwQQ2o77EDlHUjfoOvSordeHPQ+9ZA9k9002tT8+RIEuR9USw++Dv2jjAUGlq367qSd63RtKjoGlo6psJuJsLi17HdwK0js2tqXBx62pIsYvob2uN/FVfg634muuj7N4yJi7yn9K5TDdyCVtpuC0vMnnQHAFkqQObYgK8SKwGeXOBcck4q3bbMZkJjYbbozqmlhQS8iI9zoJHiD0I8D0rYvO9K9PNTYkOFnWKxLoi3LLkNaitp2MogA9m42UrQCAAQCAdhXXtmjGldmtV0sdpwW2RIF7gItdxjsoUlMqKjn5W19ev1Re5+ESokkmq2Y6iDnIMwBFos8O15WiFY7A1SMgIiSZvPo5f7dU3mlo0ywLhvwm2PaaQbyvIDYLbFgJeEFE+e4EutGZISN1NBzmWoL5gonuO9YjM288ga/wCiCc/i6fWq4LvkpMWBjaHjKRH81WF9q64E7tb8qQkIA38fCtj7/gOGZTiYwXIsbhXCwJZaYTBfQVNoQ0AG+U77pKdhsoEEbd9R+x6CaPY2/CmWfA4LUy3zG58aY4469KbfQkpQrtlqKyAFKASVcvU9KUsfTa0l8knN2+kI427bX4pVwFRxAZAAy9mhnhfsvbgorwxf5vqf/wDEi9/pbrqav3RE3W3DMUxTALPkWcxrRNukCVf7g4xbrdEK0tuLDSArtXlEADZPMkDvAq37BiuO4qm4Jx20sQBdZztzmhrf6fKd27R1W5PrK2G+3TpWIzvSjTzUwwl5vi8e5PW0qMOR2jjL7HN8IIdaUlYB2G432NUDFUziTWcDBB+UcRPKVsOw1TzYUmkSPvPAxzhawNyM1t2o+v6WJONnLGtOWnHW8VZcbjtSkh3lGyiVKfShQ3Udj1HQVZMGz8PVv4X8MuGTQoScUjxbRKjSIKT5ybiVN8q2ltfTS+XieblPN8IHpvVr4rpVpxg1wF1w/DrdaJghej1PRUFKlx+0LnKvr655yVcyt1EnvrD23h70Vs+TN5fbNObUxdGZCpjKkhZZZkHqXW2CrskL368yUg71sVMfSqEekIg2gEwI427DeJNitangatMH0TM6yYkzwv2i08VWdjjW9GovEuUxYyHkQYSm/paUrSFWlwq26bjc9/tPfUatjacJ0g4e9d2fpbeLQIFovrgH/wDJ57aWnFK9zbvZL93WthrhpLptdMwe1AnYjCdyGTCXbn5+60uPR1tltSFhKglW6CU7kE7dxrvqwLDV4T87hzHYi8YEIW30WoEs+bAbBvv32AA8d6x8/piLH/GeQblI137ll5i8zcf5RzLswPdvWrWWRncl4ZNa9b7g0RI1BfLsArHVFoiyEMQ0j3KSlbn/ABg1Zrce3ji3xxb0aL2itNFupK207qcExvZQ3HVQHj37VbM/BMPumGnTyfj0R7G/NW4PowghnzdG3I3sDvsOVPj4V1Mi0t09y+92PI8kxaHMumNrSu1TFFaXYvKoKASpJG4BSDyq3G47qg45jgWkEA5u4ENAGu6FIwL2kOBBIy95BJPvlaw5HIueFNZzwlWRS2JOZ5LF+Zop3HZWa6qU7NKf91nspKfdzitvbXa4FjtcOyWphLEK3x24kZtI2CGm0hKR+IAVUmMYXmuZa4/Pe1FwuDjrWMWt+yY/HRObmPyVOukuy1rQAEJ5ByoQeo5171ctV46sH5WDXV0GesQAbjsA7yVZgaJZmedNBIjqid3MnuASlKVzl0EqL6nf6BXn7yj+0TUoqL6nf6BXn7yj+0TVOI9S/kfktnBfqaf8m/MLXGq0wSzW2Tnmo11etsaROau7LbDjrYWUbR0KSE7/AAfW2PT2D2VZldODZ7ZbZM2bAhNsP3J4SJbid93nAkJCj79gB+KvM4TF+bUqrBMvAAjscDfuC97i8H51Wo1DEMcSZ7WubbtkqkcBtuQ3HS+c68MKXClqmKvTtzbfMlL3OrtS+oHopPh7ABtWRxaMw7l+n0HI7rHvMZjGHl2qUtKuxkyg4kdokOAErDO2243261YNz0xwC83Jy7XPFob0l9QW+r1kpeUO4uISQlZ96gayd9xbHcmgN2y+2eNLjMqCmUKTyllQGwKCnYoIHToRXfr7foVXOgOAeXEwAC3M0ixnrRP/AFkCI4eew/4exFFrQS0lgaBJcQ7K4OuIhsxwfDjM7jWuTtw4moWWM2NDTaHsKkO3hDIAR24KgypYHQLKeb37Vj7xZrTa9CMZWzAZYhSTafTLyGwFuRFuIU92ix1KSSObc91WracLxWx2yVZ7VY40eJOChKQASqRzDY86ySpXQkdTXDkNouLOIuWXDodtDjDLbMeHNb5ozjKdgplQ8AUApB67dKrpbZp56NKnMNcy7jEhoiTExrbWABeysq7EqeTrVakS5r7NEwXGYExIMX0zEmwlQ/UyNirWS6dqhNQG7h6eYTE83CATF5Fc23L3o35NvDfurgtMqdeLjldxwzHMbh28XJ9i6XK+POPOPutpCXFFsD1WwBsElQHuFcVo08kzMjskqNptbsPt9omi4yXEy0SH5TiEKS20jk35WwVE9SPiqbTNNsEuF2dvkzGYrkx9YceUSoIeWO5S2weRZ96gauq4zCYOmyg55eQ03s6OuTBDX5TxHWdGsSbU0cFjMbVfiGsawFwtds9QNkFzMw4HqtJFgYF6fx6HCu+nOmECa21Khu5W8go5fpa2wqQQkA/YdB09nSrEuUOJD1xxp2HFZjqkWG4IeLSAjtEoW3yhW3ftudqlLGH4vFZhx49kjttQJirhFQkEJZkK35nEjfoTzK93U13XbRbH7rHvj0Jtc+I04wxIO/M22vbnSPcdh+StXFbbp16rnNDspFW1tahcRv3SJ5LawewamHotY4tLgaN76Uw0HdvgxzuqpwvKLdi+kFmcetiLpc5kqZHtduCEqckvmQ5sBv8ABSO9Su4AVlFY7asBwHJcozRAulzuURbt5XDPZc6COURmSnbs208wSCNvFRqTy9M8BnxYcKZi0Nxi3hwRUHmAa7RXMvl2P2Supr7bNNcCszj7ltxaEyZLC4zwIUsONK+EghRIIOwrOrtXBvc57C8ZnlzhAuM0hubN1RvMA9a94Cwo7Ix1NjKbwwhjAxpk2OXKXFuSHHcJI6trSZqvUJnIYWB2VV0tGJWO3In230dFhqW9LB7VBSEObJSCE7lRAO439u9TmC1HGtuVbtNBfzPwVp9Ub787u5Hv9prKRtKNOYrLkdrE4im3EhOzqluciQoKAQVKPINwDsnburKzcRxq43yJk02zsO3WCjs2JZ3DiE9em4PUdT3799K22cLUpmk3NEPEwB6RYRbMT/jBJJN57EobExdOqKzi2QWGMxPoh7TfIBo8EANAtFtVTOGtxJMLSGHfUoVaVouS0IeA7Fc5KldiFA9CduYpB8aml+ZgxNZ7MLM201MkWK4el0MgDmYSE9ipwDx59wknrUvewrE38fbxR+wRHLQz1biKSShs7k7p8QdyTuDv1r7j+GYvizchFhszEUy+khzdS3HR4BS1EqI929MRtrD1qjqwDgYqANtlIe5xk31GbSDJaLphdhYihTZQJaQDTcXXzA02tbAEXBy6yIDiI41PabBZRwuSnfRcYuv2aRMccLQK1PBSilfN37jlG3sAq4LFJbVZrSh6Qjt3oLKwlSxzr+lp3IHeffXxvGbA1j5xRu1MJs5ZVGMMb9n2St909++x3NdSVikSRkdivraGGk2KO+wykIJc2cSEBAVvsEADfbbckJ69K08XtCltDOKhIl9R4m+o6o13kAHcFuYLZ1bZopmmAYZTpmLeiTmdpuBJG8rPDvFUY/8AV3f36v0mrzHePjqjH/q7v79X6TU7E1f3fVZbc0p9/wBF+KUpXoF59KUpREpSlESlKURKd1KURKUpREpSlESlKURK9LuBD63e2fwlP/tzXmjXpdwIfW72z+Ep/wDbmu/+HP1h/ifmF5j8WfoB/IfIrYWlKV7lfNkpSlESlKURKUpREpSlESlKURK6d4/YmZ94X/VNdyuneP2JmfeF/wBU1XV9W7kVZR9Y3mFWA7qg+tOpTekmm12zdMJE6bH7KLbYK1lImTn3EtMM7jr6y1jfbrsCanA7q1i4hr9lWYa8YFpthGFLzBGDgZzfbYi4tQ0qc3LUFK3XfVHKsqc5diT0rxuGpipUAOgue776L2GIqGmy2psFbuhmqEnVnA03+7Wlq0X63z5dmvlsbcK0wp8Z0ocbBPUpI5VAnvChUvRkuNuTI9ubyK1qly+cR46ZrRceKDsvkTzbq5SCDsOhB3rWnSLJszwbipvdlz3T9zC7frDGF3t0NV0ZnNqu8JCUyClxrZKS60QopI33SO/eojo5gOM2jQ3U/XG14tEnZ/ZLzlz9lubzfavwFMrfS2mPv0QAVuL2A6qUSetbL8K3MXTAMRF9e3sNlrsxLoDYkiZm2nZ2hbhjJ8ZVeTjicktRuwG5gCa15z/Fc3P/AEV3PPYfnno/zxjzvs+2837RPa9nvtz8m+/Lv03223rUe5aKaLw+Cs55FtNvRe2MSTlDWXJ29Jm7diHhI87+qFZePLtzbdeXapJpddLtfuJzGL7kKeW7XTRK3ypfMnlUp5cxCnOn74kkeG9VnDNLS5p0ndwWbcS6Q1w1jxWyjU6E+8/GZmx3HopAkNodSVMkjcc4B3TuOvXbpXStGUYxkDr8ewZJabm7FOz7cKa0+po93rBCiU/jrVO7WiDld84s8e+bq34om5y7HAN4lPhphh0wkJ7N1YIIStQ7JWx32WQOvSvuK22Pppqrpu3qpw32jBLquaqxWLJ8HuDfoydIeZKRHlspSl0oWEkgOc2x6+BNZeaCDe8adwPG+u7msfOzItb/ANI/sram65bililtQL5lNntsp/YtMTJ7TLjm/dypWoE/iqJaiamz8Lz3TXEIdrjS2M6u0q3SH3HFBUZDUVTwW2B0USU7demxrXSHY8Z1GiagZLgfDna82tF1ulz9IZrnl5YbLjjW6HRGSGlvIjslJDYHJty/jHV0ynSrjhvBxKmS3ZTnpK5t9q4sqUUohyEoG56nZKQB7gKsbhGtue21vZJ3G2m9YOxbnWHZ8wPruW69KbUrmropSlKIo/eP2QX+9T+iulXdu/7IL/ep/RXSrjVvWO5rs0vVt5JSlKqViUpSiJT8VKURKUpREpSlESlKURKUpREpSlESlKURKi+p3+gV5+8o/tE1KKi+p3+gV5+8o/tE1TiPUv5H5LZwX6mn/JvzC1xrrSbpa4b7cWZc4cd536m06+lC1/ECdzXaql7tis/0xlV0awyw5/a7nMcMhxMpKbjBKUAKjpKwQOTbdISQRvXndm4KljHOFV+WB2CTMRLiGjjc30C9ztPHVcExppMzEntMACZhoc48LAxMlXI7IjxyBIkNNEgqHOsJ3AG5PXwA764G7hGmwXZlplxZiUpVyLbeSpsrA6AqBIHXvqoZrWJ5rM0nbQJLtlktTUtMznCpx0NsgBl07+v6yNiO5XL41lp9tt+NanyLdjcRmFGu2LTZNyiR0hDXO2Qlp3kHRKjupO+3UCt7oem2GOcfKQ50Ftoa5zSDeZOU99t8jQ6aqPl7Wg08zWyHXl7GuBFogZgOUndBsG0XGQ9aIUq+GDFmvsdq80xJDjSVAbq5Fn4SR7a7cOdBuLXnFvmx5TW/LzsOpcTv7N0k1Rlms1uv1v0WtV0jh+G5bJZdZJ9V0JZQoIUB3pJA3HcdutZu4RWcJzzLvmOgNQAvDVXER4zYQ2ZLbi0pcCB0CtvYKtrbFpGo6mx/XOYgR1YFQsiZJ3TvjS+qpobdreTbVezqDKCZl0mk2pMZQN8aidbaK1hc7aZvo0XKIZgG5j9sntf/ACb7/wBFditfY+JXOVpbHmxsKxuK45CbuDeTu3zlkokEBXnS3C1uDzd6Srbwq+LU7LctUJ6Ypp2UqM2p1TSuZC3OQElJ8QT3H2VpbT2bTwQBpvzdYtPo6iL2JieBghb+ytqVMeSKlPL1Q4eloZt1mtmOIkHcuG63qDbWZKTNi+etRnJDcVbyQ4vlQVfB3326eyupimSC/YbasruCWIQnwm5boLmzbXMN9uZXgPaarXDMRxbJ9Kbjk2RwI793uPpCVOuTv+cMPoW4AEufCb5AlIABA2+OsK0Lld4WlOOJs0S7wnLCud6OmSvN48qQ2lITznlUF8gJUEEbePhXTbsXDua+g1xzMeQ5xAFmscTEuiCRviNTZcp23MS1zMQ5gyvYC1oJN3PY0ZobMgOvEzcC4V8xpsKbH87hzY8hg7/TWnUrR07/AFgdq/HpW1ecNRPSkPt30hbTXbo53EnuKU77kfFVWWqzXmzZbeXFWWxY9EuGPSFyrTAuQdLrqQeSSGQhPL0KklQHXpUeewvGYvDezkjdoYN4atUe4IuJTvJQ+FJ5SHPhAAdAkHYAbbVS3YmHLmh1Uw5zGiADd86kOixG4mexXO27iQxxbSEsa9zpLm2ZGgLZuDvAg8dTes24262oS5crhFiIWeVKn3kthR9g5iN66t5n3KMxCesrMGT28tlt0yJIbSGFH1loP2S9u5PjVeZRYLrcc1+aO12bHcwWi0R2JVmuT6Uvwwd1do1zApHabnfcDfbvrBOyrG7hlgt1ks8+0ei86iRpNumO9qqG/wBoVKbQoEjkHMOUDoBTD7GpuFN7XZiYkQIEg6jMHDdBiDuPGcTtuow1GOZlAnKZMmCNDlLDvkTLYuNYtC25xabjk19xwOMsmxhjnfckIAdLiSo8o36BOwB9/sqQOPMtNF915ttpI3LilgJA9u56VVtgw7GrtqLqTAlWSEG3mojHMlhIU2l+OrtSk7dConc+099YS3z5mVWPGNIrmormw7o7EvYPeYlvUFAn3ObsD37mlTZFCs8eQcQGhhdI0Bp5y4X7DbiQAopbZxFFh8u0FznVAyDq5tTI1ptbUXvYEnerv3BG4pX0+3bYV8rzS9Qvo7xVGv8A1d39+r9Jq8h3j46ox/6u7+/V+mu7sTV/d9VwtuaU+/6L8UpSvQLz6UpSiJSlKIlKUoiUpSiJSlKIlKUoiUpSiJXpdwIfW8W3+Ep/9ua80a9LuBD63i2fwlP/ALY13/w5+sP8T8wvMfi39AP5D5FbC0pSvcr5slKUoiUpSiJSlKIlKUoiUpSiJXTvH7EzPvC/6pruV07x+xMz7wv+qarq+rdyKso+sbzCrAd1RnHtOMUxfLsmzq1Q3/TeXOR13SS9IU6VhhHI0hAV0bQlJ+CnYb9akw7qiWq+ott0n09vOf3OIuYi1tJLMNtYQ5LkLWltlhBO+ylrWlIOx233rxLMxORu+y9o/KBmduuuTNNNMRz+fjd1yOG+ubiV0ReLRIjyFsuR5KRtvuk+skjopJ6HxrlwfTzFdOrJLx3FoLjMCdcJdzfafeU9zvyXCt47q39UqJ9XuA6VjtGdToWsOCw8wjWp60yVyZEC4Wx9wOO2+Yw6W3WFqAAJSU777DcEV+NHNT2tXsSfyuPZXLYli8XC09gt4OlRiyFMlzmAHRXLvtt0323NWOFVrS06A/36qtppOcHDUqIt8I2ijchLXoy9qsTcz0gjGV3ySqyJf5ubmEIq7Pbm9bk+Bv4VJtR9DMB1QutqyG+Iu9uvdlaXHg3WyXN63y2mF/DZ7Rojds/uT3eG1WACCOZJBG+24O/WuOS83EjuyXyQhltbqthueVI3Ow8elR5eqSDmMqfIUgCMohVxYOHHR3G7Hk+M2/Eg7asyZZZvcWXJdkJmdmgpC1FairtDuVKXvzFZ5t966+I8Nmm2IZBaslbkZNepePhQsqL7fpNwZtZKeXmjtuqKUK5fVCiCQO41LdMs/tOquB2bUPH4spi231hUmK3KSEuhsLUgFQBIBPLvtudt6k3fvsQeU7HY9x9lS6tWaS1zjO/5f+KG0qLgHACN3zVRK4V9IjcrjIbj5AzarxMXcJ+PMX2S3Z5MhZ3WpcRKgghR6qT8E+I26Vm7HoLprjsHC7da7bMRH0+lyZuPoXNcV5s4+FhwHc+unZxQCVbgeHdVheITuN1fBHifioegKidgO8nuFQa9U2Lj/bKRQpC4aFA8A07kYjnOoWXOuJQ1mNzjS2Y6ZTj/AChlgIU4efYNlaifpaPVSEp6kk1PKbHpt493voNjvsQeU7HY77H31W5xeZKza0MEBKUpWKzUeu/7IL+JP6K6dd28fsgv4k/orpVxq3rHc12aXq28kpSlVKxKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESovqd/oFefvKP7RNSiovqd/oFefvKP7RNU4j1L+R+S2cF+pp/wAm/MLXGonc9L8Vulxm3PmukF25bGci33F2M3LO227iEEAkjoT0JqWV92PT393vryWHxNbCuLqDi0ngvouIwtDFtDa7A4DiJ/vBVxl+nkO7X7CLTEsa02C0CYh3zZZaEQdiAypKgQpKucDYjrvUmseCY9j4nuRW5UiVdEdlMmTJS35DqNiAkuKO4SATsBsKkOx9lPYfb3Vs1dqYqrRbQzkNA4m8uLpPbJ8AtWlsnCUq78QGAucQdBaGhsDgIHidxWAg4Njdu9AeaRHU/Myy5Htu7yj2aFpCVBX7roPGu78z1q+aBeUFhRuDkIW9SyslJYC+fl5e7vPfWS2Pdt79qbHr7u/3Vruxdd5lzybEa7iZI7yZ5rZZhKDAA1gABB03gAA9wAA7BChPznMF37HzWf6P7fzn0V5+75h2m++/Yb8u2/Xl7vdU1GyQAkAAdAANtq+7bbb+Pd76d3QimIxmIxUeXeXRpJn+9qjD4LD4SfN2Bs6wAOXu3KHz9KcMuE2XLdizW2ri5202EzNdbiSXPFbjIPKSe8+09+9dubp5ik/H7bjciE8ItnCPR7rchaJEYpGwUh0HmB26b79fGpLt3e/u99dCdOuMa5W6HFsrsqPLW4mVJS4lKYaUp3SpST1VzH1endWw3H42qWgVT1bjrREA8TrFuJ07Fru2fgaQc40m9axhszJGsDSb8Bqd5WLsmAY3YFT34rUuTLujXYS5s2Ut+Q43sRydoo7hI37hsK5nMLx53Dxga4rpswiph9j2yufsk7bDn79+g61neU+ym3efZ3+6qXY7EvdndUJMg67xoe7dwVzcBhWMyNpgCCNBobkd+/jvUaven2O3y4M3dw3CDcGGBFEu3zXIzy2R3IWpJ9YDw37qMad4lHtMKyM29xMaDPRdG/p6y4uUk7h1xZPMsk9+561nLk9NjW6TIt0NMuU0ypxlhS+QOqA3CebY7b9wO1dfHr5ByWyQ79bVKMea0HEhQ2Ug9ykKHgpKgUke0Grhi8b5AOFR2RpjXQwY+sd8b1ScJgfLlpptzuE6aiRPO8T3TuWOk4jZYuRyM9jRZyrt5uUutx5KkolBKCEhTe/IpW3QE+6sFgFhlSssyHUm6449ZZF5DMWLEk8vbpYbSOZxwJJCVLUB037kCp9X2obtGsKTqZMlwDZJMhoIOUXiJA7rKXbNourMqgQGuL4AEF5BGY2mYJ77lYC2445CzK85OV8qLlGix0th1SuZTfNuspPRHeE7Dv5dz31nqUrVrVn1yHP3ADuAAHgFt0aDMO0tZvJPeSSfEr6O8VRj31d39+r9NXmO8VRr/wBXc/fq/TXY2Jq/u+q4+3NKff8ARcdKUr0C8+lKUoiUpSiJSlKIlKUoiUpSiJSlKIlKUoiV6XcCH1vFs/hKf/bGvNGvS7gQ+t3tn8JT/wC2Nd/8OfrD/E/MLzH4t/QD+Q+RWwtKUr3K+bJSlKIlKUoiUpSiJSlKIlKUoiV07x+xMz7wv+qa7ldO8fsTM+8L/qmq6vq3cirKPrG8wqwHdWsXE9lsy86t6caW2rDb7lsSxyRnGRWyyNNuvqaYUUQkKDi0pCC+So7nuSNga2dHdUNxnS2wYxqDluprMydMveYCK1KXKWlSI0eOgpbYYASOVHUqIJJJ67143D1G0nF7twtz/t17Cux1VoY3jdUXoNnU+wcTeYYheMEyTDrVqcgZTZoN+ZbacNyZSlE8N9mtaSFp5XO/fdJ6VVcfI8mh6DYngmPR7u+xm+rN+tdzYtExESZMiIlvuqiNPrUlLRdKQkq5h0BAPWtxs+0ox/UK/YflFxmz4F1we6+lrXKhLSlRUU8rjLnMk8zS07BSehO3eKjZ4Z9OXdNH9LpT13dgLvcjIos5MoNToFwdfU+Ho7qEjkUhayE9D0Ox3rcbiqVnEXtPdIn5LTdhavog2vHfBj5qptPcUy7CtbMLuGnOhGQab45cFyIGUQbhkcSRDnMdnuy+hgSFrL7bgG6kjcpUd96xWgOhOAaj4HlWoOoEe5X26RsiyaFAD90kpZhxUSHh2SG0LCdipSlknc7keAAq8sQ4f7dY83t+o2Y5/lWeX+yMORrM/f3mS3bUuDZxTTTLaE9opPQuKBVtUl040vx/TLGJuJWWRMlwrhcp9zeMxaVLLkt1TjqQUgeruogeO3iaipiwAchva4nt3kzvWVPCGRmFr2Mdm4CFptYsNtkLhB0P+ZyTOsU3MM1snpSfAluIfWtbkhorSSohCuToOUAA7Hbered05xTQ7ie0wiaYRJlogZdb77FvcFM995mf5tHS6y4tLq1buhR+H3mphjnCtimO4taMKGb5ZcLJjmRw8jssSZJZWLeuMpakRmyGwexJcJIPrd2xFT/ItOLLkuf4lqNNlTG7lhonCC20tIZc87aDbnaAgk7ADbYjr371NTFNc4gGxzeIt7vBKeFcGgkXGXw1960xwC0XjWbC7nqRmOgudZpkmRzZ7kXKYOTxYgtQbeW2y1CbVIQY6WeQAgpBJBJ3Bqb5PjGtecYDpHeM+w1rUFdmt01OVYY1kTMV+5vghtieFNr7OSUpTuUcxTzOHx7rWufCzY1yL7GxHU3OMPsGTyHZV3sFlmtIhvOu/VlNc7alx+068wbUAdz3VlL/AMNmCzIOIt4ZcLvg1xwWIu32K52F5CH48RYAcYWHErQ82ogKIWDurc77k1m7F0y4EHwNrRGtu7mq24SoGkH5i9+V+9VDgjdtvelWr+m+jt2y7TzJ+ybdRjmXPmOMbceRsBFeJUUsPcqgFhZCVKBAHjitNo2Gadao4NHyfR7ONGsifkm1olR7j6RseSvuNFIjyX+dYUpSgVoJSFc32VXlbuGnA/mdy+yZlcL1mMzPWmmL/drzJSZchtofSENltKUspaPrICEjZXXrXBYuHCHEvtgvGZap5tmzGJvpl2O33uUyY8SQlJS28sNNoU+4hJISpwnbv76w85pw4TY8+Ed9+I7dVkMPU6pjT7/bh8lcFKUrmLpqP3j/AD9z4k/orpV3Lx/n6/3qf0V0641b1jua7NL1beSUpSqlYlKUoiUpSiJSlKIlKUoiUpSiJSlKIlPx0pREpSlESovqd/oFefvKP7RNSiovqd/oFefvKP7RNU4j1L+R+S2cF+pp/wAm/MLXKqSyjHpbOQZDc8swi55TBmSFLhXa0T+aRbGQgANJY5gUKQdzunv8auyobL0zjG4XGdY8rvtibvDhenxoDzYaddI2UtPOhRbUod5SR7a4mxsazBVHl7ssgD/LiDq0gjxB0Ihex25gX46mxrG5oMx1eBGjwWkX7CNQZUVdkwM6u2C4m3f58/G5lokXJ5xxwtPXJTJShKHinY9CSVJG25HWuw5bo+DZ65jWNreYtF7x2dMdgdqpbcd9nYJdbCiSjmCiCB0JFSidpljUiz2e025Uy1Lx/wDYuZCe5ZEYkbK9YghQV9kFAg1yWXT+3WuVcLrNutxu91ucYw3p85xJcQx1+ltpSAlCeu+wHU99dA7UwoYQx5yQ4ZCNSXEh5jqyAQZ1kQBC5w2Viy8F7Bnlpzg6ANAcwT1oJBAGkOzE5lUTuKxIWh9iz6POuCcmS3AebunnThdbC3Uo5Egnl5Ag7cu3hudzU1m4tasI1KwtzG/OmF3dybEuJXJcc88CWOcKc5id18w33qUPaeWV/BYmny5EwW6G2w2hwLT2xDKwtO52270jfpWTuuNwbve7NfpLryZFjddejJQoBClOI5Fcw269O7bbrWVbboqlwc8lpNa3/VzYpjkDMDRuqwo/h80Q0tY0OAoXtOZrpqHmWwCdXaFVXMgY1lN0v0hjEsizd9Ux5n0iqQmLFhFI27FhanEgBs96kpJJ9tTbRmfOuemGPzLlKckyFR1IU64vmWoJcUkbq8TsAN6/LWlkCK5Oj2/KL9CtFxkOSZNqjyEJZUtw7uBK+XtEJUe8JUO81nsTxi34bYI2N2px5cOGV9j2xBUlKllXLuANwCoge6qdp7Rw2IwfkKTiSHNImbANII9kGYs0Adp3X7K2bisNjfOKrQ0Frg6Iu4uaQbdYiAbvJN9Bvq7K8dmIybILrlGD3HLbbMdBhXC1T95FraCAOySxzApUk7q3T1O/Wsii6wrjdNJpVmvc24w3XJqBJlHlef5IxH00DbdYI2PTvFSebpswu6XG6WLKr5YfS6+1nsQHWw285tsXAFoVyLIHUp2rtRtPMdg/M4i3iRGbxftjBQhzfmLqChZcJBKidyd/aavdtXCupMDnEkNIsDAmm5twSQDJHoGCJJEwtdmyMU2s8taAC4EkkSYqtfYgAkZQ70wXAwASJVe5Tg+KZLkysFwu0dhLbUJF9uyJLyhb21Hfs0br5S+vwH2I6mv3k8dyZqE1hPzK3S/2LHrJHdYtkechpKnFqKe1eK1pLuwSEgbnruTUmtGkDFhZdj2bPsrhtvvLkOhuSzu46o7qWolvdRPtJrJ3LTuJcHLbcmsivMS9WyOYqLuw6jzh9oncoeBSUODfr1T0PdVvS9Cm9rfLF7Q0gE55DjEvJkEaZQGmWg2kzNXQ1eoxzvIBjnOBIbkgtEwwCCDrmJcIcReBAGJ0tiZDabvfbZIsc202L6RItsObObkuRVqBDqElK1FLZICkg9252rm0qUWJWa2hv/Nrfk0kMDwSHEocUkf8SlflrK2bG7Pp9Bu99L9wuc2SDLnzZB7aVKKE+qgBIA2HclKRt1r86bWCdY8bL14QEXS8SnrtPT39m88rm7P/AIE8qfxGufjMVTr069UaOyAWIkgXdck7jJJk5pMErpYLCVKFTD0TMs8o43BhrjAbYAbxAAgZSBIAKlVfK+18rzy9GvtfKV9oiDvHx1Rj/wBXd/fq/SavNPeKo1/6u798V+k13diav7vquFtzSn3/AEXHSlK9AvPpSlKIlKUoiUpSiJSlKIlKUoiUpSiJSlKIlel3Ah9bxbf4Sn/25rzRr0u4EPrd7Z/CU/8AtjXf/Dn6w/xPzC8x+Lf0A/kPkVsLSlK9yvmyUpSiJSlKIlKUoiUpSiJSlKIlcE2OZcN+KlQSXW1IBI323G29c9KggOEFS0lpkKGfMHJ+2TX8Wf76+/MHJ+2TX8Wf76i+rfFboBoVkEXFdVtRI1guk2IJ7Ed2JIdK2CtSAvdttQ+EhQ2336VCf8orwacvP8+yDy92/o2dt/Y1z+isL7PiV0OlMV7XgFb3zByftk1/Fn++nzByftk1/Fn++o7pBxRaEa9Xe4WLSXUCPkE61xky5bTUWQ0WmlK5Qol1tIO6unQ71alOisL7PiVHSmK9rwChvzByftk1/Fn++nzByftk1/Fn++plSnRWF9nxKdKYr2vAKG/MHJ+2TX8Wf76fMHJ+2TX8Wf76mVKdFYX2fEp0piva8Aob8wcn7ZNfxZ/vp8wcn7ZNfxZ/vqZUp0VhfZ8SnSmK9rwChvzByftk1/Fn++nzByftk1/Fn++vkTWHTidqnN0Vi5K25mlut6brJtfYOhTcVXLs5zlPZkfTEdArfr3VM6dFYX2fEp0piva8Aob8wcn7ZNfxZ/vp8wcn7ZNfxZ/vqZUp0VhfZ8SnSmK9rwCrubpbNlSVPpvDCQoAbFk+A+Ouv86Wd9umP4k/31ZlY68ZFYMfMJN9vMKAq5Sm4MNMl9LZkSHDshpsE7rWT3JG5qh2wcC4yWeJ+6vG3cc0QH+A+ygnzpZ326Y/iVf30+dLP+3TH8Sf76sylR0BgPY8T91PT2P9vwH2VZ/Olnfbpj+JV/fT50s77dMfxJ/vqye3Z7bzftkdqU8/Z8w5uXfbfbv238a/dOgMB7HifunT2P8Ab8B9lWnzpZ326Y/iVf318+dLO+3TH8Sr++rMpToDAex4n7p09j/b8B9lWfzpZ326Y/iVf30+dLO+3TH8Sf76sylOgMB7HifunT2P9vwH2VafOlnfbpj+JV/fT50s/wC3TH8Sr++rLpToDAex4n7p09j/AG/AfZVn86Wd9umP4k/319+dLO+3Uf8AiVf31ZdKdAYD2PE/dOnsf7fgPsq0+dLO+3TH8Sr++vnzpZ326Y/iVf31ZlKdAYD2PE/dOnsf7fgPsqz+dLO+3TH8Sr++nzpZ326Y/iVf31ZlKdAYD2PE/dOnsf7fgPsqz+dLO+3TH8Sr++nzpZ326Y/iVf31ZlKdAYD2PE/dOnsf7fgPsqz+dLO+3TH8Sr++sVlehdyyPHZ1kayKKwuWhKA4qOpQTsoHu5vdVw0rF34e2e9paWWPafus6f4h2hTeHtfcGdBu7lqh9BbkP3e27+QL/wAdPoLcg+723fyBf+Otr6Vo/k3Y37R+J33XV/Pe3f3R8LfstUPoLMg+723fyBf+On0FuQfd7bv5Av8Ax1tfSn5M2N+0fid90/Pe3f3R8LfstUPoLcg+723fyBf+On0FuQfd7bv5Av8Ax1tfSn5M2N+0fid90/Pe3f3R8LfstUPoLMg+723fyBf+On0FuQfd7bv5Av8Ax1tfSn5M2N+0fid90/Pe3f3R8LfstUPoLcg+723fyFf+On0FuQfd7b/5Cv8Ax1tfSn5M2N+0fid90/Pe3f3R8LfstUPoLcg+723fyBf+On0FuQfd7bv5Av8Ax1tfSn5M2N+0fid90/Pe3f3R8LfstUPoLchHUZ9b9/8AuK/8dPoLch+723fyBf8Ajra+lPyZsb9o/E77p+e9u/uj4W/ZaofQWZB93tu/kC/8dPoLcg+723fyBf8AjqV6+cbelnDvncLTvMMdy+53a4W5FzZTZbciSktKWtG3VxKuYFtRIA7tutV9/lSdA2Ppt2wbVC2xh8OTJxsJabHtUQ6Tt+Kn5M2N+0fid90/Pe3f3R8Lfsst9BZkH3e27+QL/wAdPoLcg+723fyBf+OthNMtUcD1jw+HnmnGRR7zZZpUlD7QKShxPRTbiFAKQtJ70qAI6eBFSun5N2N+0fid90/Pe3f3R8LfstUBwW5ADv8AN7bv5Av/AB1X7nk4ctW4tY1TtA5lE/sY74n75W99K2MP+Ftl4afJUyJ/7O+618R+MdsYmPK1AY/6t+y0O/yb+Xf607P/ADY78pT/ACb+Xf607R/Njvylb40rZ6BwPseJ+61vzNtL2x8I+y0O/wAm/l3+tOz/AM2O/KU/yb+Xf607R/Njvylb40p0DgfY8T90/M20vbHwj7LQ7/Jv5d/rTs/82O/KU/yb+Xf607R/Njvylb40p0DgfY8T90/M20vbHwj7LQ7/ACb+Xf607P8AzY78pT/Jv5d/rTtH82O/KVvjSnQOB9jxP3T8zbS9sfCPstDv8m/l3+tOz/zW78pT/Jv5d/rTs/8ANbvylb40p0DgfY8T90/M20vbHwj7LQ7/ACb+Xf607R/NjvylP8m/l3+tO0fzY78pW+NKdA4H2PE/dPzNtL2x8I+y0O/yb+Xf607R/NjvylP8m/l3+tOz/wA1u/KVuNjWqeBZjmGSYHi+RxrlesRLCL2xH3UITjvPyNLXty8/0tW6QSU9N9t6ldOgcD7Hifun5m2l7Y+EfZaHf5N/Lv8AWnaP5sd+Up/k38u/1p2f+bHflK3xqicJ4qMXzTijzThxhhjtcXtceQxLC+smWknzxkD2thxgdOu4d9lOgcD7Hifun5m2n7Y+EfZUJ/k38u/1p2j+bHflK2n4e9J52iumkXA7jeWLo9HlSJBksMlpJDrhUBykk9N/bVlUrYwuzMNg3+Uotg6alamM2zjMfT8lXdImdAPkEpSldBctKUpREpSlESlKURKUpREpSlESlKURR/INPcBy2Yi4ZVg+P3mU02GUP3C2MyHEt7k8oUtJIG5J27tya0etGn2BL8qVfMUXhGPqsjeCtvotptjJipc5GfXDXLyBXU9dt+tegNaO2X/pab/+ADf9RiiLYXVq96f8LulGV6yY1pbZe0skNDj8W1RWIDstBeQgILqW+gBXv1BHSqp0243s21dz3HLfgXDTlM7Ar3IZgv5gp5SIsd8o5n1DdrlW00oLRz8yeYp6AEgVL/KAnbg61OI+1jP60zUr4XI9sx7hg01DSW4sOPiNukOHuSneMlxxR/GVE/GaKVVd4418uy/N8hwjhl4f7vqenFJSoN2vRujNutzclJIU0244D2hBBG/TfYkAjYnPaBcYLuqOpl10P1M0qu+m+oNrh+fi2TpCZDUtgbcymnUhO+wUlQ6EFO5CjsQKh0n1r1+1eRf5vBZoLpth2nnph9oXq/FUc3GUAkOSBHjBPUpCN9wrwBVuCBGdOoGrNr8pjjcTW3MsfyLKDg8pbjljgmNFisqDpbYCVesoj1lcyupCx7BRFcuY8eTdg1ezHQrGtFsiyrMrBIjRrRAtclKjdS4yHXXVko5YrTSSnmWoq3KhsO/bl0z41cjnaxWvQvXzQy76Y5JkTanLI67PRNiTSATyBxKUgE8qgCnmHNsk8pI3h3DnEjOeUY4ipzjKVPsW63NtuEdUJWhkqA+PkT+Su3xrNoHFHwpvBIDhy2Sjm268vNF6fFRFZWtvFfPwLUuHofpNpRddSdQJED0pJt0SW3EjW+KTslx99YITv7Nu4p3O6gDidMuMe9XLV63aD68aL3TTHL77HckWUvXBqdCuQQCpSW3kADm2SrYDcbjYkEgGNa8cPuW5XxAv6wcL+vdjxbVSFZ2oV8sktxuQiTETt2anmwFqbBHZj12yk8qCCkjcxiwcR2v2lusGCYJxl6LYq6vJJ3onH8zsobcLchxSU9xKuUFSm+YJ7NQBCuVQFEVw2LUzTyXxtZJpZF0lgxsxg4ozcZOYpW35xKiqLO0Ujk5+Uc6e9ZHqDpWN1O4yZ1t1Sm6IaCaQ3bVPMrO2HL0mJMbhwLXvtsh6QsFPP1AI6AE7blW6RBMR/wClYzj/AOHUb+tFqkuDG2cV11uGr100TyjTe3THc2lpyFvKIkp2cqSFrKSC0Ng36y9gevMF0RbbaNcXa801Kc0P1g0vu2mOoRjGZBts+QiTGubKQSpUaQgBKyAlR227kq2JIIGJ1X45bZphrZedCWdKb/kuQx7bDlWVi0vBx67ypGxEdLfJ9KShHOtTqlEBKD0qrcw0V4nsr120hzTW/V3RSBccUvaZVrjW92RCnXFgutecMNJdT9OJSAAkdxX176zWOQosnyr2VyH2ELciaeMOMKI6tqPm6CR7DyqUPiJoikWMccGZ2XVPHNL+Irh4vGmbmZSBFsdzVc250V19RCUtrUhKQklSkpJBJSVJ3AB3qpOObP8AUGLxY6IsxNHLlMbxXIFyMfWm5MpGTPLTGUtpkEfSChXqFTnQ77jpU88pYhPNoO7yjnTqVAAV4gHbfr+Ifkrl41PrquFP8KpX9eLRFtlgd+v2T4dacgyjEZGL3afGS9Ls0iQh9yC4e9pTiPVUR7R0rPUqq+IzK71ZsEaxDDpRYyzPZzeMWNxPVUd18K7aXt37MR0vvb+1tI8aKFqdkPEDmOOcauE6zXeR2Wk2cuT9O7Q6F/S1IjyEoVLV4ALmeshXi0kmvQWtZOL/AIbLTmnCNL03wm3dlLwOAxcMaQ2PpiXITe3ICOpUtoOJ38VKB76mfB7rSjXrh7xTPH5AduqYwtt4G/VM9jZDpP7/AGS58TgopV0UpSihKVg84xqTmOJXXF4mTXbHnrlHUwi6Wl4NTIhP/WMrIISoeBINUN9B1l4+Dxla7A+031g/0djRFsrStafoPM1/2zdc/wCeY/yNPoPM1/2zdc/55j/I0RbLUrWn6D3Nx8HjO1yB994jn/7NPoP88/20Nb/51jfI0RbLUrWn6D/PP9tDW/8AnWN8jXz6EDPh8DjR1uHt3ucU/wD2aItl6VrR9CDqF/tp62fzhF+Rp9CDqF/tp62fzhF+Roi2XpWtB4QtRh8DjV1pA986If8A7NfPoQ9Sv9tfWj+WRPkaItmKVrP9CHqV/tr60fyuJ8jXw8Imp4+Bxtayge+RDP8AT2NEWzNK1m+hF1S/229ZP46H8jT6EXVL/bb1k/jofyNEWzNK1l+hI1ZHRPG9rAB4bqhE/l7Gn0JGrX+2/q/+WD8jRFs1StZfoSNWv9t/V/8ALB+Rr79CbrD4ccGrP8VA+Roi2ZpWs30JusX+3Dqz/FQPkafQm6xeHHDqzv8AeoHyNEWzNay8Z/Fvd+E+RgV1j4jGyG05HLnR7nHU+WJCENIaUlbK+qQr11bhSSD06p76+/Qoa1f7ceqf8kt/yVaW+Ur0mzfS+y4C9mOvOV6ht3CZcEsN31qO2mEUNslSm+yQnqrmAO+/wRRSBdegGg3GDoTxEMNsYPlrca9lHM7YrntGntnx5UE7Ogfumyoe3arrrwo4feC3iD16lQ71iOPPY/ZEuJcRkl1K4jCNj8NjYdq6oeBbG2/2Qr2V0L01y3SrAYmJZnqvfNQLgwdzc7s22laRsPpaOUc5QPAuLWr/AHvCiEQrCpSlFC0b1buEC1+VL0rnXOdHhxm8KlBbz7qW0J3RPA3UogDc9K2xyfVHSC12GdLy3PcUatCWFiX55cY6mlNcp5kqSVHmBG45djv3bGtKuJ/S7C9ZvKO6b6c6hWxdwsNzwt5UmOh9bJWWvPXEeughQ2UkHoauaB5NHg2gS25fzrHJJbO4bkXiYtB+Mdr1opVReTTu9nxXDdddSULdtOmDeSvz7O5ISpLTUVhDq3VJB/csmOkgdd0gd42qX4fxCcavETb5GougGl+CWDA+2dRZ3cvkyDNvCG1FJWlLJCW0kgjqNgdwFK2JqyuLXA4GO8FuoeE6aY7GtcK34675rb7cwGm22G1JcdCUpHihLhPidyT31Q/C9odrnn2gOE5Lp1xt32yWN+1tts2mLjcN1FtWjdLkbnKuZRQsKG6up7/GiLYXhg4mZetzuTYLnmHqw/UjBpKYuQWQvdq3srfkfYX3qbUQfbt6vVQUknXfB+NPjE14gXW36D6J4pcLti9zlt3uZMkLZhdilZTGjtdo8kqfWlC1LPNsBy9B3ma8MGl2M4nxWZ7e5vFC9qXqJHsibZklvcsQiLYSFsdmtx1slpSkhCEco69Tv1Sa63ksWWk6U6ivpbSHHNQrkFqA6qAaZ23+Lc0RdzUPjF1yw/XqHw94/pHZcjym74pAnQYzElxpDd2dBVILzylcqYjSUOq3A5jskbjfesBf+Mfid4dc/tWI8TelGOXiNlsZ4487hTrpcenJ5UtxdnlHfmcW2gkgFIcChzAEVk2mm3PKvvLWgKU3pqFIJ+xPaAbj8RI/HXY44EIc4keFNDiQpJzZ3cEf78Q0RYnUnig41OHxu06m646TYCNP7hcGYc+HZJ7z1wtaXT6oW4pXItYAPVKSgqG26dwaunib4m5OilkxO34FhzmY5nqDOTb8atgcLTLiiEkuvL29VA7RA26b83eACRX3lT/rQ7z/AAza/wC3Fcev/EJqXh900f4e9ELfZUZvqBb2XWrte2y5FtsdDYHOED4S/VcPUHYI25VFQ2IsLqRrZ5QbQvFZGq+oenelN9xa28j12gWSXLRLiMKUAVBbh2OxIBUAvbv22BNWHxC8W0vTbhgxziM0/ssGcxkL9qWiNdErIajS08y9w2pJ7RA6d+24qi+K3RziGxvh4zTM9ceMGfeIseAG0Y/arLHtsKfIcWlDbC1D13ElR+CACQPcawPEmUr8lXpftsQUY6P/AEKoite/a78fF2xKRrVguiWDwMJajKukSyXeY85fJVuCecPLCFJQham/WDY9Yb7bKPfcWnHFfgmZ8MaOJy9tOWWzRYD8m6RebtVx32VltbKDsO0KnAAju5udO+2/Sx7o2hGmUtpCEhCbC4kJA6AebnpXmRj9vu1w8j9ePRTS3Exb+5JloR4x0XJBXv7h0J9womq2RxPW3j81mxxGq2lukmnNixKakybNasjnSDcrlG39RZWgpbRzgeqVco6g9Rso2toBxQxtctM8nv7uMu45mWELkwcix6WorMKa0hRA5tgVNqKFbHod0qHhuaf0k0A4g8x0uxPJsK48shZsVws0R2AxHxiEpuO12SQGQeb/AKvbkO/XdJ361+eGPTnDMSyXXu+WPiNVqhks2IuLlSfQvmQizmxIHOpaSW3FKPaD1OnqmiLG6PcWvGLxP4LGu+iekuD296AXGb3e77Jfbt65fMVIjQ2kqLiilotla1EgKXtsPGyeHDig1T1VnajaQ6g4DaMe1a0/jlXmrchardNUtKgyvfdSkI5+z3IUrdDiVJPgML5K1CEcIVmUlASV3m5qUQO89ttufxAfkrHaJADyluvWw23xm1H/ANEWiKruC+48Rw4ptZ+xxvBSqVlUT5v95kjaEed/f0d0+md7n1T2Jr0frS/gq+up4rR4/NXF/ryq3QohUC101MGkelt8zZiL55co7SYtogj4U25PrDMRhI8St5aB08Nz4V588RWk134Ksx0Q4lbQ47OmQXhAzqW3vvcZ7qlvSnlnxLyHpSB7mmx4VuPkpOsPE9ZcMQS7jekDDeS3cDq29fpKFIt7KvaWWS9I9ynGTUq4ndHo2u+hmWabONoMyfCU9bHFAfSpzX0yOrc93rpSCf3KlUQKx7TdLffLVDvdplIkwbhHblRnkHdLrTiQpCh7ikg/jrt1p95MbWCTnugitOcgcWjIdNpZssll36oIhJMfm36+rs417uxrcGihKUpREpSlESlKURKUpREpSlESlKURKUpREqhoPDRPicYNw4oTl0dUSdjqbGLN5moOJUEtjtO259iPU7uXx76UoimXEVpPJ1z0VyrSeHe27O9kcVEZE5xgvJZ5XUOblAUkq+Bt3jvrNaa4InBtK8a01uMtu5psdji2Z94NlCJIaZS0pXLueUK2PTc9/fSlEWsOFcHnEjoNJu2NcOPEdaLPgt0nOT2bZfseE9+3qX3hpfNso7ADc8oPKCRvuTmdPeB29afcQ2O8Qi9ZLhk96aiS2cndvUTmdurz6FIC2S2oIjIQktpS2EqADYG/XopRJVg6ZcOU7T/iQ1N14eytibH1AYiMtW1MQoXD7FKRupwrIXvy+CRtvX71z4dp2sGqukeo8XKmLW1pleXbq9EciF1U4LLR5ErCx2ZHZHqQrv7qUoii2tfCXlGSauscQ2gmqatPdQDCTbrk47ATMg3VhIASH2ie8AJG+ygQhHQFINYvFuEnVfMdUMZ1W4pda42bv4S+ZdgsdotKYFujyehD7nXdxQKUnu70p3Ow5SpRJU6s3DnOtXFxfuJlWVsORbzjTVgTZxEIcaUgtHtS7z7EHsj6vKO/vqE6g8H2c2rVq7a5cMGr409yLJADf7bNt4m2u5Od5dU2T6iydyeh9YqIKSpW6lEX70w4Pcwc1hgcQHEtqwNRMwsjRasUWNb0w7bat9/Xbb8VDmJB2TsTzHmIBE0tPDnOtnFxe+JlWVsORbvjLePps4iEONqSWj2pd59iD2XweXx76Uoi+cUHDlO4hRgHmWVsWT5isoj5C52sQv8AnSWv+qGy08hP7rr8VY/ix4Yr5xAnC8kwjUJWG5fgNyXcbRcDE84b5l8m4UncEEFtBB6joQQQeilEVsaYWXN8dwGzWTUjLWcnyWJHKLjd2YojIlucxIWGh0T6pSNh7KgWpOlermQ6u2bU7CM2xWEzYLPItsCBerLImpYfkOJMiSktSGtlqQ222Nwdk8/7o0pRFy+geLQ9DqTpaR4g4lO6/wD19RPhM4X8w4bbrmxuGd2m7WPL54uzVqgWx2K1bpZUrn7Lndc+llBSnlPUdmnqaUoi2NpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREpSlESlKURKUpREqIZtpHptqReLDfM9w+3X6VjDjz1qE9vtmozjoSFrDavUUrZCdioHbbptSlEUtQhDaEttpCUpACUgbAD2Cv1SlESlKURVJkHDji+Q8RmN8Scm/XVq94zaXbRHgN9n5o62tLwKl7p5+b6eruUB0FW3SlEXG+wxKYcjSWUOsupKHG1pCkrSRsQQehBHhWqEnyflqxu8XSXoZr3qNpZab08qRNsljnBULnPeWkr6t9Og6kgdBsAAFKIrb4e+GjTbhusE61YO1OmXC8PCTd7zc3+3nXB4b7KcXsBsCpRCQAN1E9SST+uHfh4xnhwxq94xi98ul0Yvl7kX15y4dnzoeeShKkJ5EpHKOQbb7nqetKUREcPGMo4j3OJYXy6enHLAMfMD6X5p2PMFc/wefn6futvdX61a4fMa1fzzTjP71e7nCmaa3ZV3t7EXs+ykuKLZKXeZJPL9KHwSD1NKURc/EToPjvEhplK0uyi83K1wJcqPLVIt/Z9sFMr5kgc6VJ2J7+lRnXXhHwLXSz4si4Xy949keFJQLFkdnfS1Ni8oT0J22UCUJVt0IUN0kbndSiKFP8A1gzK3yYuuetmoepr3mj8e2+mJ6URrY64go85ajtjkU+nfdK18wB8KkmVcGmHZhw2WDhmu+aZAbLjrsdyNckBgTFpYWtTaFepybAL5eiR0SPHelKIr0k2diTYHcfW64GXYaoZWNuYJKOTf2b7VW2h3DfhOiGjitEIcmXkWPvLmGQLuhtapDckkuNrCEpSU7EjbbupSiKnU+T0j40mfZNJeJPVLA8SuTq3Hsdt1ySuM3z/DS0pQ5kAjpv1PtJq6dH+HDTPQzTSXpjp9AkRodyQ6Z859wOzJjziORTzrhGylbbAAAJAGwA60pRFy8Oug2O8N2mMTS3FrzcrpAiSpEpEi4dn2xU6vmUDyJSnYHu6V18T4esaxHXvMuIGFfLo9eM0t8a3S4TvZ+asoZDYSpvZIXueyG+6j3mlKIoJP4JsZa4gneILDtTcyxafcbjHud5tFulJEG6OtKSSl1JG5bXy+sgkjdSiNt62QpSiLXq38KWUWK85HecX4mNQbKvKbxIvlwaixbYpK5LuwOxcjKVypQhCEgnYJQAKyH0POqf+19qd/JLR/8Ao6UoixuhXBtYtCdU7/qzadTMovVzyll1F4YuDcVDEtxbgd7UpZaRyrC+Yjl2Hrq6da2HpSiL/9k=)
- Homeostasis vs. allostasis
![[[@Lee2015-mh]](http://dx.doi.org/10.5483/bmbrep.2015.48.4.275)](data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAA1IAAAG3CAMAAACJ7pQtAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAMAUExURf////v/////+/n6+f3////9/////f39/fv7+wAAAFxcXPr//0xMTP/7///9+//9/QQEBPPz9M/Pz/3/+/f39/77+Pf///n7/f/6/v/7+/3//f//+fv9/fz5/ezs7P779vb29f39/ycnJ93d3f349//++Pv//L9RTvv9/xMTFP/88vHy9Pj+/HV1dfv/+///9hoaGpY5OObm59bW1//19cXFxf718B8gIbCwsPX7/P/+9ZA2MwkJCW1ub4o1Mfn8+HwxLv/5+/X//5E4O8FOTPP7+hYvS4M0MDs8P0hISDQ0NKKjo1hYWLe3t5w1NPDv7qmqq52dnfv7+IM3OL1RTYo2OJ42PZ4/PYeHiNLS0pxjYpeXly4uLgkNF4kvLPj++cRRT4GBgfv7/g0iPTk5Ofbg2eXv9YkvMxIqQoMuKvX/+7e9wnF7hcrLy/P//ZAvNZA+OFFSVOnT0I+Qkffi5JYxM+76+5lZWO86EIdHRfv/+AgbMo5OS49BQvrV0I8tLLlLStvk6r+Ska1IR0NDQ+zy9+/U1GFhYXczMfnt6cOdmfn5+E9PT7hRToMvMqVwa/svAWR0hNnZ29C3ubuIiJhQUSM2TXt7e+///ZZCRKh4eN/BvePq7m8wMMHM0wwMDPXIv4yMjDVHWv7u7/j2+OfHxuPj4yc6U80AAdy1s7KHhczV3Hw1N4mUoKm1wOHh4Q4ODoJBP9KrqJg+OQcVJ/woAuJqRgMDwf3t5Yc9PMXBwHpDQXGDlqdhYSUwQNKxsEU7Pudpbet2XsNOTPb79ktccLN9ftTg6IydrsTR2syend/Hx2ZmZsmppe3e2jlPaGhoaK27xV1eX729vSc9XmRkZLJ1cs7V7aowNr+/v0dUZp2qtOo/I+BaPZSSkqaHh+LPzGpqavKvqN0BBDAx2YpYWqyy8/OOi6611wQDzFNjgJagsu++wZuc7sTE2Pg7DeQgFNmenYGD70xK3YSTrgUE7t6HiLfD0MdJRgkTkmRg1nx73s0dGWdmcZeZ5mBpeBIWbR0AACAASURBVHgB7P0LWFTXuQcOr5nZt8lkLjjMfYaZwYFnGEduIjqo4MCDGC+gwwcqGvmLF5QqoEGIqPGTR0108kFjvOClsVGMUUm8YBttYmz7TNSYaG7G1KRNc2mak6a3c9rTnNPTf/P91h5FjYCAYNXOTmTP3mvt9a619vrtd613vRdCOj14kvqxQ+g0OZIQ6YFID/SsB3gyQF5AZHzPnorkjvRApAc66QFA6jAg1Ulq5HakByI90MMe4MmikVEkwqR62G2R7JEe6KwHwKU0g0hkMdVZ/0TuR3qghz1AIRXhUj3stEj2SA903gMRSHXeN5GUSA/0ogcikOpFp0UeifRA5z0QgVTnfRNJifRAL3ogAqledFrkkUgPdN4DEUh13jeRlEgP9KIHIpDqRadFHon0QOc9EIFU530TSYn0QC96IAKpXnRa5JFID3TeAxFIdd43kZRID/SiByKQ6kWnRR6J9EDnPRCBVOd9E0mJ9EAveiACqV50WuSRSA903gMRSHXeN5GUO9oDnBkHQw9CWByE0F/33hGB1L33zu7TGkvrLZZEswQHw7JKpZJlJBFI3afvOtKsO9IDUotFKuVENkUxxUYgdUe6PULk/u0BtUoNHOFgwKfo1C8y8bt/X3akZXeiB0Q4sU6GkSgU9+aUL9xLkbXUnRgtERrd6QEnhRPwpJBIrCKmIKG4B48IpO7Bl3Z/VtmqV3i9XmtWVqmMF6zW8NzvHmxqBFL34Eu7P6vMe71ZwUBAZ0/z21w8Cwk6FaTfe0cEUvfeO7tPa8wH4jhCOJ3Lb2SJk+edEUjdp2860qw70wO8mjB7Pjm1YdPpAw3nP4kihOfptpS4NXVP7flGuNSdGTARKt/pgfD+E8TlavGQBcie3x+oOzG6sXxpVeMTRZtP/zCdVPicej1vVLLZzffQpm8EUt951ZHLO9MDFFJUU0LCq9Umncuc/nRT5oLMnJSUqlVLc1Lyly7NbNprMcqys61Go1KRHYHUnXktESr3bg+EuRT+WiXeRC23o6S1ceXKxtbWFSV1X9S93JpZvbKwdVMNCVRmO5XgU/dQQyNc6h56WfdTVa9AihCJImRZ39BaVVw8emXT0SU169enr193avuR0duKV9XttZQCU+BT9xCmIpC6n8bpPdQWCilonPNqiZXMLRldnbJq26YlaZzUbJYonE6n/5PThVXFS6uPEkOl3mn0RyB1D73bSFX/FT0g6vHxPCuYjOy6zVXFIwpPzyWcXaciVoXCaDSq2LgddQtSluZtYA3NlV4hAql/xVuK0LyHeuAKpJRKl3LJMrCjzfs9nvr6eEgAGadTqXRKeJ2lZnteSkrrBmNpZbP3HmpaZOJ3D72s+6iqIqRY1u9i1y0rmlK1eUl9qFlPtfvCSuhUGJhu8TQU5uTknbNkZVdGJH730cuPNKU/ekCElFHpZy9srto2ummux5ObC/5EoaSQaLX4ybI2F7ehsHhE9Q4tNCnunSPCpe6dd3U/1dRMTXedPFvzRXlxXslxjyeRLRNtpSiotFpqLoXpn0t7bnTbzG3r1AKsqKzYE74XTKgikLqfBuq90xZzPIsdXG/aparivLoabaKCUUHB77sH70o/PXrKqpM1dpvTQCHF3gOGVBFIffc1Rq7vRA9AhO5U8kxDa0rh5nUmk0RiVqlupmvN8hw/OTNl9Ha7UW1QCLCevwe0/SKQuvlFRu7cgR5gFHqBnGpdmfPyJyqXWy9hOoIUb9AlzV2RuXL0edYl8UoopO5A1W6TRARSt9mBkcd71wPO7NykdStzqvP2E7/Bp5eoO4KU0yp4PDuWFVVtvqCz53qxwuI6mB72rgL99lQEUv3WtZGCu+oB3lt/vKS8uHCTlrdm6/USrkNIYUJolzYUFldtT7eHIpDqqkMjaf/uPSDYMw60tpVfSufhbALuW1iI824+sOTi4tbXVU1pPVWf7oWQMMKlbu6kyJ1ID6AH+Giyv7WtaPMFJQ+FCbpC6kjliJOqiURG9h5JyVwxt14A7qSRiV9k/ER6oKMeEMwXVixNKdzBGsxYRanUHfvBtCRBbJ5dKm1YMCXvACeQCKQ66svIvX/nHsC8jeqfw7WEdHvVL6oaiEHPWVQmk0qNLd6bO8aSZFI6syvjjp9c1fbyEkbNkgiXurmXInf+nXtApWIkhLfZBOnR8uLik+leq9/I0aNj17KcFAlWa4A8XdS24FKaK9saZ7nruy8i8bvrX9H9VEHq9JwYBD83d1l18cp1zlptWkdrqBubzAuM/1J5dd4O3pAbgdSNfRO5+nfvAbMWq6bsXFPa9syc1nNKVxbn70jQd0M38TwRlG8UVueV7FFmlWpvSLsbLyJc6m58K/dtnRTxSiVTG9Lux45UyR6Xza8231ofAphyuQ7knSjcD0jF3219g9rBPxr+xyHQykUgdbe9ovu6PgqFUqnNDa3fVlx85BNVAE4lvB1IJW7sAoHwjCruwoqV5Sf32CvuOmNEXggSGS8TgtFBmSya4osM0EThb+SI9MAd6AGFVclyIc+BxuLyBlXIqzSy+ltCilaLC3ENrTnlR00hxR2oZY9I8AIwJf4hAk4RSPWo9yKZb7cH4KjFrKvf0ZpTVbfeUeGFHmz3IGUOqS+sGLH0ZJpw63ni7daxp88DUUOnBqMdl58jFFERLtXTDozkv50eUCr1ldo9dYXVT+ywBuOhYES6ZwBlzg0pN5WnzNx/902osI5KTU5ILVijGTaZyCJc6naGR+TZnveAUVlZq9xQ1Tb6QH1AZ4OpbjeZTnzIp1y3LT+zxM/ebRpJvIyM16yRfSwfKT+8iEQg1fNBEXmiFz1ATeBpgHhGqcwtvbCtuGrZBUuWysaHgwh0o0Cz1+v3n17Q1rqDWOO9Zg56FN0EYzcK7zqLVks3ojkzrPc7PHgHeV8+jSyWjx8qHxyBVId9FLnZ1z1AEQUMwKuE05blubSquPG8tJMB2hlpVsKb1q2szrukC1VUeKUmKDB1lrWP72u1UjU04LWdQUoWJG/KUw+NjC2QyedEINXHvR8pruMeuAIpiikZ2V9YXV6SrrvlDu8NRcH1iyTOdbqxrWgJyfKWcuo7Byk4vACHhe1JJ2oe2Ix6T/7AQ/JPyUD5BBKMiCdueHGRi37qAZFLYeIn8cr21JVXH3mD6HpGiVU6KwLcjhM5mZukwfpEjjK9npXQ69xKWOdLYKOvVHZcBATn0Wvk8pFDoibIx0e4VMedFLnb1z1A5304GKZUeq6quPXXGenOnpFgjU59hTa9qXzp5guqQIDGR7yDkFLA6hiY6rjKPB/kY2bNmQi5X+xzVCAZ2ertuKMid/u+B6CwI72wsjjli7n1hk5mUZ0RBaQU2SHVqcLi1vM6O4XUHTPvtckkubWVFV5DJ18BPkhGTYjBZq/jkUVQo4hAqrN3GLnflz3A0N0nnmUFC/QmGs9pm72dfPI7I8qqOEl2dtzxuqryuj1pPCClvlPCdIkvqzTkKfX5DJ1ULppslT9JogSBx7ZUBFKd9FLkdp/2ANYi2IDiWRe390ROecn6+gp3DyHFWTgiyZbx58q3Fe4nQewRq6R9WsXOC9NbAxkZnnSVtxNVKOxL8U/OgJ6fwyHjI5DqvCMjKX3YAyKkGJ63lTWtyizckRSqcPYQUuZ6KcsqvJa5K1Mym9RldxJSkiC34cBce5rQGZcSyOVJcx7H1I8IVHkispbqw5ETKaqTHuDMZmzzql3ce6On5G33eLTuXkBKyUpyA54Dq3KWLaFzSEs/m/dipgpjY/wV+LlfPLFsgyuIOV1HhxBFJkBzIvnNZxcdkkXWUh11UeRen/cAl5SkhruJtON1M1MACCfDmHu404vhDYGGEGQHFBWP3mC2KZUZSX1ezesLZBSM2mTRQtBnIOcy235wXmKPuz792m+IJ2bNSJaLx2Iii3Cpa10T+dVvPcAlWdROxiU9NzqnsYGFHhKBlk/PqVEBgL9kaV7dHqXTbcnoeQE9eAKQUposiQqnUb2+buaIL9ZLXR34bKcF8tjeFQYOnzxrzjBNRHuiB10cyXo7PUCVHawcvJtXr6gxwR0fK+0VpKjZ7LnGtrz9jC/b0gnTuJ1qXvcsA/6ksnj1jI2cX9CWuUEtWOk6qaNDgP0hvS9EDRka2ertqIci9/q8B2AWJdFrtZvypow+T1w0shTXG0hRY3SmZnPVqkuksllr6vNqXl8gw7BKtdbrNZM9JzMzt9UYrfpOJH5AkUAG/WT6oihwrIjE7/pOjPzutx6A5oFesCw50raqZI9O8CLAIWfuDTGe43nz6aLilReym613AFJmb2mAOzqzevQGo82Q3YmeLjWUHxqr0YyMnc4HqQwjoj3Rm3cbeaZHPcAay3yepEujc0bsV4e8Cl6pVPcKUiDKkB1FxeUb2EqZv0dV6GlmcCmWjS8N2NNPzoTzaKUR8U47LgOT0dRkeULy4ZGaUSQCqY47KXK3j3sA6kRBz97C6saTabxVVJfrfZwoa1rJ0qqSPZBt93ElbyyOMQNS5iyPfX/mlCcOECPr7ExIia3eyfJng4NiLsfKfxJZS93YjZGr/uoBG+9Jv7Qg58gOIkggnTYqnZ188m9dAat6w4KUwr3E6rt13tvIwZip0pMuPb0pM3PZBVLm7HQjDJCaID9EtfsWwQQxIkS/jU6PPNr9HoDD5lN5bTlNGQIrUfCm24GUgaw7Ul2+yWio7D75XuQ0U0ixLrLj5Zy8A0mlCicb18lGGPalpsrfg2SCOCImiL3o6cgjPekBhg5LejCB9JIFU4rWJRrgbYLn4XG216YZvIxryszfVqNv7klNepzXTG2yjEZd0+iiI3s5K4VUJ+oawNIAefKkGIF/RL41MvHrcU9HHuhZD4QRxcq483ltM08TX3bPHu8gt5UnTzemZB4llb1GZQel3nQLerqAFCJbVeed9tix/mMtnenpBgWoosuTY4dpNENJdETid1NfRm70ZQ9Aiwg8iifr64pSts1V+Xq9hGqvlMTK1qyA7izpbJuoPeft/eBQcaNq++i2wjekgWxF55ASJecfx2rkmuTJxBERot9et9+Jp69szcPr4q1EXKKr06tVunX263OKLh2v3ujpGXQ7qRoABeN4iYSXnm+csqBB5S9z9rTwm/JbfUblppkjVlxgOyF60xO9uyF+CZYsq17V5PEk6gGpzqxJ0Pih00jB0I8npZLoiCZ673r7Tj/V4dihN9sTwl7uv1Mv8eYN95BffOTKs1dO15XTnlvMhsy0DKSHL+kPeuvadXsdaEK4nOuTw16RJLz2+Mri4s019V5bD2062utz7Uei18ruLUTYj3BTriX07S84u5Co+AOjU0b8PslekY2Jn6oTrUR+GnlALlrIBweKH5fIVm/fvoo+Lg2vZ/pwjFdemPjxEOhnYuRSx9vwd0DfngzTeOjA8LxDcARJwfTh0TQDMst4mh2ZkExz00y4jzMceNNk/KD2ckGBHzR5fCqZOEnMgiJxExprYqH0L71CTtGLKiAjPhiuAE0TSZGC8eNjhGAwmtYrXGzYkTHmfIxE4c3SObiGBcVV50huthJ86zYPbbyC7KkrKipJA5tCFfrpAKRg1b+sKPOkJy6xUk91qDqGFE+mDkuWj09FBzuGbcUrAtQjYQb66a30RbECGSzfNQCb8sFYzXCcMGQBiqAD0BDwNyhEO4LARVCI4gselE8HCnCJTGScfBIZRDYOD8LNnMxBUcLz0chInwk6kEfM54gmAzWHp5HHNfjM4qB/BIcD0JABFkKUAwSDDpI6PEocu7wjKuiIBpCBLZQVpAErZGSofFgqcTiCvIPeiI4mMcMLwkMdEnNFVrRdN3cZ9YHONev7AFKWRL3b2dC4MmcdhXxf9HGHZbBGt92yqXBl5n6iUmDeBzdJHYtDeDIwQX5YPuzBtycughA9AqkOu/PuuYlB/oAGG4gOMu1wwnM8DbUCjWcABP+wqQhc4A/lIVgMfSoPu+QGcwryqcN2DSRkuuYRJNOHIOm98hfAomVQRKA4uj85J4pM1kwPr6xRELLD5w+wJK4LZEI0eW/X21Q4LB40mYI2PJzxV4geNHwRIIYq4cEobHwOTZhwhX2ASSnq7emqA41FraeUZdkSdc+8912hecMpzqL3kjeKcmY2sKij2CE3pPfRhdMZsDy2IqW8Kd2ODwOjpp7HOi466Bg4RzNMLtfADvGBiBC94066e+7yZFCsXB4bEySPaJILMGcbMhmLYNmiWdB8iRo/NZWQic8iXgTmgWQ8/DI6SHDAx9M38lHksuZwQeojsfI3h0cJZMj04THY1p82azgZ8GwMiR7w8UQHGFsQV4vIs/KxhAyXQz0N45PCb8jk6UOCFKTBaR9PRJykQZPnyBdPGiQuXSZPPTRoMm6iJqRg4iPTHPTX8OmXScEj02PIe7Qug8aPlT84qUDMDkQpAq60Cy8vLSrZA8NDrlNBdLe7nLPH6bNZaIePqEPdxM9Ft5/tUUarPWnDguKc82YBYGLVRgCr4+cFB5mVMGDApMfHHdZcjnCpjjvprrmLETNAI0/GhI5s1YBTDJoA29HYn4gIIBPl8gEk9XDycyJDSR2WcEhGNo7F13LkVPAn+WLs6Y8cCZzFPIiHZiwi5FnN4EkjRz6XOk4jl8/ZiJc/GAlbJ8iRfSLyUUxBA/TdXXL5rodiCDmEsuRjN5JJuJaPo+OXDEqWP5Agly8uwMOTqSHrmuewETMW9RsiHzZxjlyesAieV5E9VoQUZ6bSPlfagdZthU87DQyrvn1IkTjMw3iyoTGl8A2Sa6WBCm5/fXbjCxf30ki09viylKUn9zS7KQmlmu0o4j19EB8f4VCQA5secDnidOzGrrzbrsA04BtYkzxeM5bnH8LIFybIP534tnwOf+jwYkKepJCapZmMFRbybYUadLRjrHzwxK3yXdNwOYtMGyWfMWkjeUg+auIE+RqBvC1/cJhmetQ4zaxJD2oeB8Tk44Z/nDBSPpzwE+WxYH6YDJJZ8sXvfTwMIIkZJ986/En5u2TILGy4LAKiBDIwWTN2/MfJeIJM0mimTnxX/qBAosbJF/HD5btip06aIX+cDHx2ZMLkoSKTklqgJiGoPjkxorFpT0CBb30fQMrMweSeLHk5ZemmpNpKhCmVqG5/Ntn+6rFgCu9OM4b6DXnF5aeYWn17Ysc/6IeoYPLkQ5iDg2cLEfFEx910d9zlsTDZqhlL1sgXRR3Gamc4RiyJSpBfDiYkk41gF4uihyUXYN4XJDFUCzr43NBJg7jUw9A4Gyt/hCCgxCiMffonZhc29hM0CRODANKzmNzJE1JhkXCZQR45GNhE+UgKKSyfYmn2RUOfI4/QfNPkCRu5ZzUPUrhhrTRJnoy55qeoRsEM+ccEPDJhCKQTmo2A6+GJFOWA+mTMJMV1GOFU+MDbwgE34gRoIPQJpKB851SnNc3M33y8okKEVB+zKYopYnWvXzEis65G2ZmvsfYhAkQ51sg1CeNmjZ8ECVIEUu09cxf+oEP8SfkEbpL8zQHyhGlklPyB4ZMnH4YsLzmBbE1+XH5oMtgJVlIyoG0GllokdeIjk9+Xz4iO2bULoY7WyCcS4QH5qOHjp4+UTxQSaOboBzVbh0+aKpcPHCpfQxzcJM2MGB5MJhlOs6gxHVJmYIUFtqgZPHzSs5QTvg94OkTZ+lbgLQbVmAMcJcfwwqAEzXj1ePlDUeRxrMwLwBDHEzIYDNKBjzXVm2BhHv/JkRGrmtI9pdjb6QtIUUMmRsdvKEwp30G8FFKd+f/v9RuFNyfGwJ7PzG88x9qMtyoGvT9ZPgxLXggoIr4nbtVb/+J0IAqwGE8KEoY9II+NAgfZlZCQHDtjkTo2oSDh/aHyoTOSCxhAigdPeZwujg7LD2Mm9xAZogE7GZKMsEfBZM3hZDw07tA0efIhrJUQWCz5cHLs2OBg+YMQs8+Co9QgXXqpIb2DTDw4acZIOWSMz+2CVV1yQuzYVGEGJnqQQ0B68SYq4wB8PgW9WEgOY0aCN34MnMlE9EaPk0PJjdbYIfYcxj5C2GxvTVm6wyJul6o72y7tQUczsAlmAtIly3LKD1jgfMzJdLxf1IMir8tKfa1TfQ8rVKiqlm6uUTD+W00rMUeYAE6/JnZrMno+MvG7rjfvup+UaQw5TOdlsyCfnUP4NZiLQVKN1zYqear88gAN5R/AHSCFBROdviUML0DssOmAyDgegoKxDsKPhWwd71mg9yAoj4nVgHXR6wnU6zCZI38Tf9+mlj6Q8WErl8iGvivfdSgYS1kcTzhM/pIhywCXIrKxYJCyqDma8cBRLEKVDZXHOkB6KpmWQKu5SKM5hH0u8FO6pACXUloF6Y6Xq1ddSrKX0u1SdSfbpT3pekCKMIJ9T1NhZt16nRVXfXmAsSK8iNcQdJ4q/MXSDaRW0R1IfSofSB54iExKGIJewjIystXbl++kD8sSJ3Sa5ENY+iRrYDcA1vAghvGTl/FrZEKsbNpI+bACYIC+RMgSKPLGEvIeHd2DARGoR78NBD4rfwBCjgcXYb4G2MiwEvqUkPETpkHqPqOATN4FnhIsGIbll8ikPp6DFdQAKqwQ801aTPOhVKo3QQYelj/pQIF4buDhke+RgjXyWSRqGPagL0PQT2efaxyQW8yAR+MwpMr4uPSm0TnVv5eqvNA8VbKwQrrdg078zIkB6fnW6qL9nBWaQt2L9ts9upRLSbxWgU8rKV+6ba6z+daQwhpzuGYwmTCORKE7IpDqXkf/a3LR3abpkG5j4+NBuQb8IXWcfMY4TTL4wWS55lk6wmeRljCkHgKkooGxGbGQWmwEBxpPmVpy7FQycIZ8xhxN8jQyAUWA7QxIlo+bIR83kMo3hsGlo2YRcWIlhnkdD0WLyVhKjUumoodFV/N9Kk+IHQyuhm0seWxy8jjN4ffoHFOeMCdZ/mAQtTj8HOQWizHZe0T+Jg8Z5OHYt0GHHlZZ3I6ibYWnM0yMAkyqUw2EnvTvFZ8Q5nXLRsBaxAt91j6HlJV3OZ8uzCnfpNQ3S3SqW9UOSl0zNM/Nkv9kSGQtdau++henUxW9rdSqDVxmJCIrQ3lo1K7Dg1MxoCdqkgeSmORhh2geOoubinxRZPg4zZvTDydMc4zFpiOZtvgwFcNtHLUrYetzlKVMxIQsWhjyuCb5WbqzdHnNrjeHD8OiSxiLJKgY0fna0PexIphEQGzAuyNpvujUtxMwv4wGvsfL1wxcLH/wMhQl+OD0hzRrHoHi0XsjZ2CvE/NOxITBxFRI/XQYkB6k2jtWLywPVxXnLJHw2NVRQwOhM5coPehpEVKhkMPflEnXOtmsUtGxplAPyryWFVxKIhFsfmNTVc6ydUSvNyd1ZiZ19SG8Am7I+wVD5SN3oeF4Hf098VNzZuzFiR8tuJg2o/V9O/m92rD75Iw4y6KrYtoc3ioIgwYhyAprDUYVYEMXAoEoTKowB3MUFEArqGCQ4BB7k4dQEFpLAguGxkc5ZLKoAplMUOOnleowRUVBwTZ6UDQUleh/fAEKaeGtjCwKc75oB/SWHqdSchp7TCDOaOjzQdbo4PE4CUYD0FHRoq4teRdzRuSRQZ3PCg+rg6Aoiko5UJWogmjsyTgKHFZMfBwOcXuGEEUF93Rh8dLT9bVOul1qVBJA6voWUodHODCOw8eNabhnNsMteRgxYhqeppCSVIRc7NGZxY2izE/kUte67WqhV8rsGUGED2UYZ5rtjZeLF2zKCEgU10OK6SRMHP0YOfjBGvnYQ3cCUq5Ac7OyzGozWuO1FrgalCiNVzvwapsj56s9wAtqk8qlE3S8YPYavD5G0PJlRr+xrEypNJp0Jp0jTh3AcI83wHlrouAs413GMrfbZoOg4E3iUAUhtpN5Ga/BLFhb3C6b02mwGYN+tcrlMAVYXmuWeX1eq4wrs7n8tjKfTemSERfUPrF0CsRnGRQ+a5bQYlC7WsrKymxBl5p3uaKlVFFJlmvwWjH9nERa/GWGMptf0LkCUlcAuNaCnNdLzFYD71c6y1AZq8TgKzPwNmW2N/3kiBH42IcMTr8ftbHxJrGFKrRBbKHXmqg1lPn9Rre7zGYMt57qDaI0q88ar8oNaQPYfDLanDaTKU7nUgWQapUpcr0Cmds2Iq9kvd/mCulMKh1NMwm0hV5vfCIvEkTPGP0oNM6lEkwCT7vUKwkTtF1HMNzdXi8Iau0B3uY2tKQ1PZFyYolFkKngZFYb71N4FfFa1MjgtlLV/KvvC2eeDB8fDYUxTBUGTMTnD1+w/uZSdntzs9FUkyazciqVKjEefnEjkLruldzwEyMOwyPoEvhAIFHIig/U69wGl8tU5nMLLl5wAW8Y4IGANgvvNaDjaZrVp+BdJv+McRhVAq8LagVZlhAM8O6gS8f7FG6dnw/qLCaXIKiC2kRBFh8MaGma2uB2+10OQTdt2MifkAAlGLKCICNzudQgaAwTDIiV0QpZMmlqrHyoM90liAStqIxIMIA0StBcFiboRBVlZU6+xW8U2P0pxQsaLAFWJOjTgyAqE6dyuQR1uIVZAZ2bEjT43CYQDKD14Rai9VmB+sRQmj2xlAIDrdepdEjTgaBM0AazPE0LqkefUrYEPegSpKGiaKEgPqgKt17v1lGCKpXYel04zRImqAdBNSXoCohdSrs7S2dxycoUArOjcMTo0yabAd8MpyASRAeZDT6DwoCaBK57aZD3aA69vWbw1EXYBIdY5g5M/CwmX3PZ3B//UG/Ah9aoprtofTj1va5t98NPTPVahJYyfImDdoc9oAs4XLmGuPR6ny83aC8TWlpkNhbv3+VwiWkyn9aTIfPlRtux2zi1xcYL1oAr6LDrAtHpslzeY5flIk2wBltsLVRrm6YFdEEUarV7Ark+RyDQwiqHv7lIPcjoClwjSNNyAyJBg00tVsYVCEhTZw1eLUVlaKGoaIsgYynBaIdOFwiGCWb5aqPtdnsIXVYPXgAAIABJREFUUz/e7tLtqasq3rbO4/CJBFHRgFAma5G1OHnDdS30xoUJ6tBCAAotROvFFjpcNl2azmVrSRMrQwmWoYU0LdrBncubsqApzWEVysqC6DYilAkul8NuRwsDaL0nQHuGtl5oQWgrWqgu3HpDhkjQRQnKgle6OyB2KSDk80XrtudVH/nEH+XzWWTWgN0RtAdAMOBz+6wyp0xQhYUv4ojD5sSMggfoPu+u2MHT36MC2P7mUonGssqkvz3/M4+AkNxGcChFheR+GP3904ZoxLB1KgxlVhQP6wt88hh9KCMAPwuqAA1sybiRRj9JQZh1cIw+yx4wYykRLHPjpttgsDqxFhMCLZj0K7yYxMAgISCLpmmYSVmx3uADLTziCXpDdoFRED7a7SYEPKmFbne1E7QLRMHxQdlVgrStQZUJe1SGkCfEU4Iy6kUvTJBw1MYEBEtBELql9lA6Nl8lAFvZ0y9vK2/IsHOoTMgej+iiKlkLmuamraAEAy1ooUSPytAWigSdV9LQQqSxSr/LxUoI7wleJcjQFgaDnJoUrMjPP/EJEQxW2kIJWs9j3wutR8zRKwTRekoQhVqvEKSt1wczBPSMSka79CpBlhLkiNtX6VMvOTIir8mPHSpFlnC1u9Emsb2YH6Bi1x8ysmjys2OTD98p7QmVNdv52J8///wz1qmQOBHwPj4Cqevfxw2/y8jOhQsHP+P07Zy1cOH3IClYuPCp75mDg55d+NRwxqDAaRKT63th4cLxrIHg9AKJjh//1MIXOAMZ/+3CR7J9kuFPzXrWZyMvDl74yiAr972nFs6ax5KhTy1cuFPhXrRw4azVanJw8MKFZzhh48KFC6cRJnXhwm8HMBLQXTiUMGspQSIreGXhwomM2zeV0vVFg9Ik1uqe/tRT45loSncy9HUmg67E5560cOEjZWXs8KcWvmKQkYlPLZy6VkW+9+3ChWsTdUPH/vSnL9c4B+BqNeGngcRGQgY+Neupg8QwEFcDWNrep57hZDtnvfLUZcbtQOvfIblo78LhrMGN9n7EOPwvLHxqvNnATl/41GSFzz1+4cLpbgP70eCFL5xe9YsRX8ya6isjE78FXb3kewsXTo1xkmdGob2EX4RmP0fC7eXdtL1DnGQ1CA5gZM8tfGXhUIa2d/A3nC9mKtrLlhloexW1oV9/UdhWuENB6UqiDZPRXmcZQT9/LPEpJkEP60YZYHjDQCgYOH3WhPfvhHgC8749P/v8R5//ak9Wc22zQqnUlt7+dt8N4/A+usBu6rBjD39N3Kmzlz/8Aps979XlD7/itNXitJDI9GOPPfwtaeF2H3v4NTaX+/DYsQ9JC3nt2LGHIMD+9uGEfQofeerY8ldrfdwrOK21khceXj57J8OMf/jYsDOEfHRs+bGDhLyD0zOEfIPT9zDgEpY//CJhBi5fDrrenZQucceA4FRiyH312MMLmWjDPtB15jK7jx17jWkJ041m3n0YdHOZb48d26fPdS58ePmr+Oa/8vAx0OVAd/5qhn3h4VePjXeC7mzQ5cJ0JYseXv4wpQuCLxLzGZGuM3X2MUp37YxjlO7aLbS9udn7jh37HXHE715+7F0SzX348MMfsrks2rvbmcuB7u5Tq/JHvJwcazBIpj58bP5at/qFY8dm79Srvj42e/nqq+3lvgeC4faCrvngsdlX2zuemHeiu18gZfPmo5+Jj/bzU2xtxk9/cGLEJZeZ9jM2E946dpXuQ8THwN5M0F035ATyyOOPTN4YLd7ioylr7e+Jn9Gl/eHnP/rRn5//UhmqbW6G4Ul9BFLXvZIbfgrk0LFXH57MkJ2zZz/8AuebN3/2samMzffq8mMLlUFm7PKHvzVHEwyx17i15MPlyz/kW8i7x5bvZqPJqGPLx+p97EJgyVfLTj02e/YgMLKHZ8emKsn4h+cfG8iQFzG0pykAqfnHhjJkEa6+x5Ihx2If/ogjh5bPPjaZ6HfOnn8MQ2zQ/OXHXmEF2avDjj3FBcm+5ce+JbXu3cuXv8a1EEB5sbOFeRdjGlffUrq1oDubQgp0Y9fayAsY0zvNaaha8pls0J1/7CBHvvfw/IcXsaAb+/A7HBmyfPbDHxHFmeXzj01mWLG9ausg+gnhDALa+xSp1e+bfezb43MtlC42khcfO/akJBqfkOW7VbUs6O6rqVuVsuynM2wtZCoautZL6Q7bmc1+fWz+crR3OFo4hAXdVx8W2zv/2Dus5CAIfgRjMJD/mnXujEU/S8uiaD87fbVbZh9bqA6lr6iuPrGDI/iEPACu/OGx5R+q1pLXlh9bI/GRZ2OYGyZ+MihzwaRtxthnJy2KEXXD+h1Syozj//P5zz7/2S//8Vh6qDbbqVQnRiB1A46uuxDI6rG7X/2IsDG7d786njjXPrhvywukxfbh7lenghN9OHbLQtKi/nbfq4NJkPt2yxbwLHbhln34hpOF4/Z9KGnhX3h194OY+OG0e1CZc9Kru/fFKCXDZ+weC6bx3pgHX91IuG9e3b3ldWwEv7r7VUz1Ns7Z/SqYRuq+3a9OIs55Y3ePGU9sgx7c/erHWIcs3hemu/tV0GX+3y1bBoMt/r/7tvy/lOCr+94Fz3ply9gPnS1lL7y678OrdAXF+Pm7d6eScz+NjR07UKF+Z8buLRuJcxEIDiDMAJAfig3oLbtfnUg42t4XiTJmDW2vadBDa159ARtctL2sTP/ulnFbf/6fnncv7hsscXDfztnyLUb4wi1j30Ut0OwPpUvqqnKKnowOor1rdq91k/Gv7h67E2wRzQaX+h4IYqY5dMbuV18nzDQQpO3Fze8RltKdRNQxOKG9DnT3C6wh98N9rz5LkvbnFI/enmSXPL5PbO+3+7a8j7kBbS+41KQCCBave2U8uTxr1LDDsOqUa8atuSMKSf6kL5//8WfPf/nl83/bkx7KJkozJn7tBs5hDTX+uip256f4VHcy3mt5LDph7dqdaxmVbl5MTIHLzq+t3bmWi+Njds4bBJH4oNp58+J51dqYAqSpBq2dt5PXkUE75xUE7Wqk7VSUSQftjIlJc0mjUIxBJ42aNy/G5VLV4qrWqQruXBvjSJMGB9XudKg4A9Ki7CofJShRCTFrY6KwI0YJ8nHOeTvn4Qrk582TUIIxBR6Xai0IMjpuLQgGQBBXercqCoWm2aWDUIyg46J3gqBHWzuvdqdqbl3jy19s9NvU0fPW7gyppIad8+Y5XKogJajghHkFqEycjF6pQHDezii7zjoPBLHZgtaDvHpt7fov//yruTHz9sSsrVehMlgQqgbNoz2jilpbuzaUAVP2olN+j9qBynit6hY86EqTtqDQoEoqQ9WiXWhhLW29DASj0qUy2kKVlAdBh8tE2ztPojIViC00FdTu3Mmkn6xaunJHkkdAZWKcRoa2Hp1IW6hwS2QGng/cOGBhKjpYPuPJWFE8Eez3iR+rqvnV548BUnP/5x+fpXvilUqzN16sEWAVPtNxT39imRf+S6/oXj5kyfgnileogSu9Q/Ux8T+tNs0l/gwvD+819HRYX07KQx7FOAlm66o4e5yOZa1Wi0UF0ZMqTsrzbq/EqTbxjEplj4vjCWPVWlQ8o+BpGqOQuNUqlrA80tSQuGktcTxjtcZZ1EZGr3ezEP1i19MeZ2Ld7niLReBooRye06NQHeFomo51ukFQcIYJqt0KN9J4coWgxGq2qMxmkSBEfBJEiG5PU4CgBaVZdRl2i1lfSjaU57RuMKbbtTyj91otcQKR8JY4NU9EgpCuiZUheIISNIuVcbrRerUujlhV6Z4KQ+Jjv/z+oz/32GzuCtpCvYK2Hl5cxBa6vaGMC9u2UYcrRrcEaSxvhXGiHS1E6+Np6xVuSpDVoxWcEG69gBbS1sOCQ2y9m7ZecDIqtV1lgwjNS07lQeHDEsrA42VOJetEGu1uWCVa3QwPsSKEhtcfEHcukm+N4lMnzokdfwf2pUzky+d/RgAp/2fP/z92Tyl0JxTQ5Bw3bDq0omVkcAKsgcbOGIALihkHUAN1G7q3jk1qYIb6hxPRRK1Ksc+H/2Dx8OxhqvhGf1OYicC8vo339G98J2iDMICc9OTEEKRXsA2np6tp4Sv6YbmWBu8H9IqhJ/yhJ/y5cqIX9HnxdOXm1SsiUuLF/cJ2gmz45g0EAVexlHBlaOntBJGPXrGoKZeoMhqtFTFf5Od8UZOWrjJLrtbiCkGxFFrTcGWutDBM0EnzSD31CqM9o7nZ8/Mf/en7v/xhfchQRimZbyDIS7JLTZtaV7bukBolXl5NrRHD/YSq0Jzt7b1KUFxxXGshVTkAQGlOhRXdzeK3W1hfkpmfso5U0sevVE3s0nDlaebvHHw0LMieg7sCMoSateC5/jXuYJjf/6zG/dnnXxr3/O2zjFI9tqaoa7qRVCWNlcHAbjjM5eBOC7piUDGLoo7hKFDgfQ7OroY8PolAYQ08iyJKTEByENpU0wlkK9RiDp0n9sl3mhm5/Bf2gBkO+gXJ+R8UF56HsoU2XtTB60l9pB6zW51RWhv3w19+/09/evTnx+sN+pufB0sR2L3LTuRdIv74UgHBS3uvRCBqIEC7jxHI+da2mZvULdk3E+zsDlXLn06RVKBZfCeMO8qcnoLmzz7/mzo93RNq1rNGG3TKNOPgJoTadUPFBYZzsVRdkx5wVAdGRGuHn6SAGokC9eIltMnCP8CtKMMC0CiWgD/6YOS4i3oAY93K1axYVl633pWeVY/dX8oPenBw0F1NLG0uO/6r73//T9//06OfWbwdhOlglWXxOtf21uptF9isCt/tmY5QTMGYVxIHhY+UlXNVBroz3L0DY3dgsnzC5OHD34ZVrzh6+9cEURkIzcsGlyIeIAqQApPaKl88XDNjEKxCfyJPOITLBwGc9wYnj4XMH/WbtnVY8qfvETJVM3LkLnjEnfRkwrhnqandxAcSxm0diM3C2NhphB/6ZnLs4AHioqt7TY/kukM9oHRXetmGxm2IHu8IZFEm1UNIgeFY6psrpV8+CkQ9+qMf/ep4acXNYmK1kYc23v4jOeUNxJWrgIZ679sHBXcKKLPVsqG8uLxBW+o2drsw6udmIvXAhgPmM/TD37+QMipDzfVYS8UHzAqFlzcGWUD6vYG7YCcXducIH+2TYVOn0QxLkK+JQRSEw5rkBA10nt8fKU9IwEl+eNhI+fs83NDtSj6sGXYIJqTjHLDdGZmcQP3GUd+rkeNu6gGloTk0d1txed1xaX09FKV7DCmiVCaFKvm5lEn96dHvf/9HX3o8HUGKSBSW9JKipV/UJFFnMbcDKap8ZNUGuOMrliIegrbZrex2j2KFGiSHpj54OGHxZDF0R79DypoRKvv9838jOiVYs80mYwbL1/BwEjcAkJoFU244bIIT1F2aSdwg6tQbxqdYZo1aM0nAom8rbMTnjHsOFtgjUwsSYMy98aEJA+CS50HimAG71ZjFDw7F1K/bjY9kvBM9gPDxWZZN5cWN5zkhnn77e7zEgUv/pNL69C8f/dMvf/z9P/38T4/+aq7n+r2gK61QcZAWsucbQUkagNtyAhW9Xh7U44QkS5tOGvKKG/+TVFT2BFJYk0A2QQpSCRcVXrb0L5dSKjM8akBK6ofarFLpcB96GNCADxBIGERvc0NGagZArg/XcJgCPklTZkyKAvcUeOpQZJBodTdJPieYmiB/cCjg44AfrlGkIFY+9j0HcomCwV52ZOSxfukB3mq5sCKlqmS91Ao40aOnZJycNpH74S8f/eVnP//znz778Z8f/Xm99qYyzFoO2wj8+s05RXV7XAYww957tqCIirek6S6syCms26PUZzO38ot0rTp0+e8Y/+SMSRD6UXlZv0/82HiPx/T7z39GBDiLgikbM0qTMGfsQ8mQTzjgCWQAjOfgx2cNJn8OCE7gVHEAtsyGPbkInrwTdh0i6kUPxA4bdlgOfcTxhzWHZwyGdetYiknMXkfOeBNG3lckG9eaGPn1L+4BRSDpQF5K0SliCIMJC/+egYqRxMc7XT9/FLK+Hz/6px9C7vdLhOn4bqviS83YtnOQhtFTQMsKbhh/8+zwuw91ck0hpdWlpf26sRjxEPAlkEq7zfGAqKgJGg2chLwLb2t3AFJEH/JoKaQYBcOZvRXwAUzXcRoIRyhonoMDkYdIzGFM46LIKDhVRJikSXDiHRuD1dK4aLIoQT521lTqFRyOQx6ZIcdMEWJChE4iqdPhg5sKLSNS9E7GyXW3YdaBTSP8pWc1TM3hrY6hVujX5bndn6LrYxTC5OrWVadkluzJ8mKrh65wJNQ/Qk+O+MR4Z9qP//HLH3qAK/z5858/Ce8fXV9IfEU8y7qFuLkr8qsQbMrplvQeUpSVci7d3pdzqkocQStRWyzd7xrRMenW2AlkTkIqRX5/r6XM5noP9/vPf05EuxMHs1UTO3jUqFGL5eMGUc9y0bC+hjPvh6iIAk6BL5NDE4ew3FAaSHgw9X//KXUJnDps5EbZRriow9QwFgur5ILgwOGp1Jd+AvSCb/p8Xd/vkd/E6WTVKouKZZUqnFQ6k8tvNLL67AqvVtXBCqW3PYZPphomphInybVsmjnliaOWwFUg9XTmZ05M1KqYCxs+SXL8/PuP/lB7/D/3pzlvqhfURQhr5I3k1+UphdjuddIbvTvM2J1WenMztlcVv7yflUERQ9UDSDkgYRtK3l8Dj4fUk1S/Q0rLSz1E5FJoLYwXqI96oABOuafBrc9g6roRXnw+RpT7RWvkE+CzSj524tCpkGBExWpGDY0ZLJ8zdPga+bCoqDXyN99b9AAkFuM1DwT5BzQTJi4apXkfq6mIeKLrcQS1AM4C//6Y3FjN8FHHq3joqllFJtX9T3HXNJAKi0YlIAVgIXhgcXnJ8Qy+tyI4LhG6TTqpVJqWRiHFeZIsSTfXFHM1CM79fnJhZcroJo9foZD0duJnToTuhWB+4+WlVU3pArS+egIpuGqbBbe+j68hMZo5WL70P6ScHIUUuBS6nYdfqjWyaEc0N0AOh4qL4cxq4DBUh4+Gt3u5ZjEWSjGL4T9YHgsXW09idjgc7vPlh2fIEwaRn2DWJ8ZhGQWRYHDaHKTSWCzQqbjl2/43z8ASjE44GtJnZ/sMVixUKpprvfBXBLd6fdgzMJOF/gLc9fFxB1pzCvcnRfc6MC+n1UqlKl26XWcPQyo9vYOlDYUUIX6jTXkps/rEJ2xzNm/qbXvwueH59KbWKSd2SIO5lEt1QLCTwrG/ughzqbcfgh/DTyFzuwOQUoUhJTJlx9RRl6lqESnYOngRmT5qkXrgrK2IGcHLBjw7dZqocxScNn0r4lgGSdSkwdODzkPTp6YuGjUVpihRl2dtRRbZeOwDDyLRQ6duHYrol1gcRmZ+nbzs8G21WmrRmrFYUGQbMDdbvyc9Dst/NYVUb9lIR/QYMzZJJHq9luwtyjmBuAKJLcaO8nXjHodPgNRi0ersIqSIJz2xA7tVEVJqk9LFvlGYsuqACsZ4/m4U3lEWVgl9QbK/qChve5InMbtnkILQWYCj0MMJsZqRUD24I5DKECd+7RoeIgooDOhKTlwJQVOW6h1RNccr8ziATuQ+gIuTo36mqfc6MQu9TRUBw1cUTpHFFO2Yzg+VxWKJj7cS1qRiL5y7tGLFiqbzF4iOV0gQ8Kzzx3qcosXmjiK71B53aVV16w5tqKzXXIpQTGm1iYn14sSPhJIU2TczVCpSoEOG5f0lVfkrLmQ3G7u/P3tj61i4lbUeP1mVc2SJ1O6Fq2l4b78xRxdXvIDdqEeo1G0YfGHTQd3P2hNa5xUuFe4TqptHDz6sDUsREZ64AUvhFHGvScwT1ouAukdY3E8TaJarcLpalJg38qezHuBUFqnZrNW6/HHHN61oLK8qXlpVvm1Dmja3Ut+XPvo5LccylfqAZW9bfl6TTithlL0d4dQ9KGc2Q4KnoxM/opJio+jm9omYwm2enCovntkg8fp7yxaJsSzXcq6quPGANC4xm7qaVnd/WUYVvh0IszoVQb+hy31HIEW5VHgtdXO3RO70ew+I6yi4udP+cPPoEzk5q5ZWVf0ip7XkgjbkhUOlPjs4yMio5yXP6VUQwGGf6DaU7ijzpIAxe0RICSpJZ5CiuRh3Wt2IohXr1f7eYphV8pa5m5dWrZhLeFheUaX27gsPKY6u8AU4WbozkKJrKexL9dnLixTUox6gyjbE5LKfOjKirai1blPDprqqqm2tX6zTuToLP9uj8q9k5rBek+gd6h0n2jKbXEYeoW+6r4HQCUH1FS5ldCo6yCHCiWo8+dijC1Lyzilt3Wct3ynO7ZI2jG5r3AA30zR8vJF2WXcPqCPJBs948NnhiwaBQ905SEW4VHffUJ/nA6QYdZrl6A9W/qJ88/50bL7u+c9tM6dUbYZj5Y5Gai8rYKaQUvD+prwRJ/YSG8hIpb0sSvz80l1iNryWwjZaR/oXFFIUUYps9fHNmVV1e5j29XoPCTPe+sdoGNH1jJWCCftrsK3ubhlQ3xlPhdEazbgHp94R8YRZZSd78/9GesuUu9uySL5OesCkrdTblEcLq1e2nq5xqrKsTs4ytyRvSvnJuU6ZEubfajWcAHfycPdvc3GIzJsl3XGkrXW7Kg3hCm6XSwFRYS7FqkzOjiCFuomMitELcRtmTik/qszFKsiohrPQ7oldwJNoWAEJYw15Ts9syztPZOLqUrzZ/ZZHI/bQhFETHsTuzx0JLBqfqPb7/QUtab0VcHa/aZGcHfZAnKOilN27LaXtBw02uGegaxwhcX0JVK6b0luUZQaJqPTQ4aM9uanWmZQs72nKW/ryJ0aXWdIXzu85189/BPFEnK7DiV977RhzEl0J1R2PVzh7AinU0ajmsUVsTXqjqHhVSbq0N2wOxrLj5Ycw5RsydDqCIfe/eMJbkahCZIY0121PrNt7MPKjRz3AWwNYezeuHN1gs6WViSoHZab6uXVFK1sPMEar3sr3yaYvdIMQ4fbpJ6ozDxhtfsopbj+ehNpPxROsSgf/zl0crDvD0jAzpfU88TrVNPoSdAu7yN6exAD2SkDKii8BdG93JMmcvZDXYOPHMRYBXtul0f0tRNdXejm1v0VW6m1vSeTHHe0BBY/Ato0rW7fbjLYyA9wasXBVbE9at6zoxModROf14kYf6FHQ7VLtnpON1dVLWLuAJU5fQMoOfy635lLKsnrP3GVFsPHQ0j3t6wJWdd3RmPhhd7oCgXWOPrGt8ZLHYe3NVhoNHjnp8IThqQ4qnbgD4gkJ3iEoGSr1XTcvktpfPaCw+RtWpZSX7DEi9JNXhJTb53BZYGJeVVdjzwrBHrYHO5udVVOplNSSUznFjaelAo84ThSoneXt7n0uzKXiTF1P/JRl8D/WMDpl1VHOHq/ofnPMsA4BpBJVxzePKFq2zhNlsPWiylBTuLwLG70JsW/fmcgdon4/C59o3ZdKdrfDI/lu2QP0o2kj+0eMGLFindIG4RiGOt6IG3Mdu31T4ZTWA0nppd0fg13RY42K0PqSquKV66jtEpALpyxd5e9OGgcu1Y21lNKp90rnrhiRX7feTi3mleruyRrNVOFDXxokDTOnjG6QBrzu3sxV4ZboTfmw2CuRO4L9v5aCTQF0yfrgi9WdVxDJc2MPAFE876+py2zL24/IH5iNQUSGLFiRM7y6Bu6Pl+2tz4KNRB9wKWILEBitF22yaEU6UCrqvlLPjdVuv7rCpW65lmIVegE27vlV57isSmhtqLXd2qGC6TDcVwS5JUdSMuuOq6y9cwkDIfpkeFKJGfDI1gc+vRPiCRFOEUy1j5I7/ANrZ9Om0dvyDmS4wn60KOfg4jHjcZeR/SeqFzSlB6D+183PeleVZ3yqmrqinGUXqO4mNXeUwqDkNg8q8evGWkrFSfRe9YUVq1JK1msrEHFJ1T2/+9pEbKWVqfdsL2xbcIoVYBzSm68AvLcKY4dD9RuNddyJtZS4Ged290KScpuvI/I4eoDqQ+4oyincfDzJ57vydSOMud6ClU+lTLd9Zv4Tpzge7lNue/QjFB85B6lbg5N61KMlQv/9dt9B99ZSXFKSWWRTC1LKj7Jep9HYzYBL2kRw7RayvzUlryldh+0FaZKl51WGEH3i4NjxqfTJsIZ3P6vNSuBqrEJBJPE9r2vkidvoAY76fCVWgaTXjaA+jkPeaGMYU4jTDkixZdm50iUr8zO/SIeZBw/XKL07IDVj1dT5s9V7fPPSnBU1boSPB6YAKe1t4/TKWkoX16V4gkuymCUKQYeJbNWK9fGwBFN1YAvSQfO09WZoOa7/ohyhrk29hpQQha3ekfJhEwZPnIawHv2/lkqysBkZq//wgeW216od9EnkVqc9YK5XKZXubIHbsKq49dfUd5D5RtTwTkHgGsqrR2+Qusqy6m9M7LTYmxIk2CnGjpSCCajOlRfnbUjMurq30weOLbrHpQiUCSGz9/MbWoufOB8fEFwqXbeao4rjEeL3160pq35NEGCbTqi6tQa7sRPAmR55CJayVCVpzR1w4MxZVEpB8R//+08IniLHHewBEVJOH7mwLGXEsrmY3BHtd+dhvMBeQHSyFTVKQ1ZSt8ZgB/WnW7pqxNSwsnNX5IxYMTfpivVOB1l7fotyqT/9kI3rmkvRjV0g22YctGLE6C8uIJhOoHvzN54xBFSfvHxi1eb1REBIAWCqF4pZPDwN8akDhk9/cFwCPHz1O5fiLGqljfmPl/5psUWWUz0fU71/goMEwikJ2k83VudtIPUYKtx3vsCQBgpsQ96IzA3EkNgD7yU31knUkoB0QCCbCnMazyXVY8l2Y47buOq2xA9gsMJi/tyC6tENarjJpKKCWx8IHu9JL8nb1rpf6ldjytY7SAFF1PCQQil12h2AFElSKW3cH16KcKlbv+G+zUFtOtTSvSfa8krSPV4Iyr8LKcjXCbmworiqbr02oOotl6LbpQzit5N121I/oZCvAAAgAElEQVSK6o6nl/p6s7fTSdOv7kupulxLhbmUT2bTpa8oyt+2jsvN7t7n252NoG8zixu3WwQErRKVcDupSJe3xZBMYkgZfKSQs78VkjKS2DJCIVUmVkuMH0V/USve7n1LxOfa/9A642mx7ldu0hgeuBE2ur9y79/+xCoRP0OX3jSz+MgOS1YphAjf8cUq9n5Ii6VW4flEV68doVApCEKyBVSbVqUUHdWWVhj6ElJXtCdupeNHPx9eg9WatD+veNXpNK/e0C1MGXzaT44szd92AZL0Xg3G746ycCH9DCkuwyMV2P946Q+UI+K4WnOcRX2o71bqltcUO4CjaD55JTMfDMqEaDDu3kD0lgTv1QxKt0LF7C9Mmbndbk+kenzfgRRtl7lSVrO5OPOL4xZjb5up1WIho5dxn7Tmz4QqNzwk9eXE78pa6tZcSrScik/ynFxaXPgGkRm61Rzekd5E94fZSkXYuqVvqt7PkCJJSUl25rcv/ZML0FYKgmPjQIAL084hqeEZaLcaf10mihwHpq9hiNIE0VKZhAu+LuO/908ExOQ9e0oKq6vXkaBEgclRR4aq3pDyXHnO6KNaWW8NBRITMZqtiPPU2Fa4n8N2KZzZ9lnPX5X43YJLifSo6VRIuiMnp6gkXeieLExHNjyBze49bDa0SURt295iinKKdhbR35BSs1pPWTuXioYLdPhmxrRtlGYUGBeqEWYt9Cz+vO7GFaZzbUJHGRSqDpnSwEOi52a6CwBEkZghMiiFrBGvrhRIEyjjog9cLfPGX/RFXCn7KmVawNVHwmWLf+nde+5gzAHmaOGJ0ZuUNidUSaFyLbm5DVrBWLN5xMy6dDFW6M3pt74jQsrG7jiRM7Mp3cVLiLTXko6bialFHT827hZciorpxD03icx0IK/6iXNhVYaby7vxDi9dsqy4bdkFrsUGM0S6wYbjxizdvAqPGwwdMX9/QwoiVo/7Dy/9gRPXUgKZhOCLdIg/AC/o4aF+BQNXBj8qJaKAnmkNwws+8ZdYYSyZiGPrDBpf4MpNnowfNhxxBxZPulLgNf4lFhHORxvbXkoYLmJ59L54XMsWJnz1/r16DgoZx0vycpY9pjLybh6Wu2BVNx2qgFK5aUF+5imEow3PfW7Kcosb0EBAhNy0psacE3stfni2tWTc9g5vO8lucinUXIQDazWo163IrF5xgb3KdDuXujBqJr0kp7jqnCUU8ouyvl5Ciof6xMbxcCk5JIpEi0Osn7Un1Kw64KRcSoQUDWv6PnEEefg/f4+GOUgdEk2BwA8ZEEVSB2LQx0wTiAxTQn7gQHj3g8SfDBpQIGbBfG/AwCB4Uup0eew06tA9dRq8zfIFl4dphj8nG3iI+kfnU6fFXJV8bFwUQw4NQRkgAiDCiw0/5DnCOygYU6cFCdZfQxYhvGKQnzaAbnsPHIJoJnz0gGkOfsjGAQOiIEB5bpHoOL79Hd87P1hblmVDXvGCBi1iqBNAqkPI8FY3RHUjMkvSrN6KikQOe6Y93JkxBSXegPrp1uKZvzbBiQthoDjXZ73EpYd1/FRKxa2qJXIYPhRKOt9KTZ9KrSyURQjpSJJJ7U7MFV6H/9yqlMbtrKyCN4Vjf/eOSWHdkTosOVWYMHLc8DviIYlTq4VrEj+BzEFAAQzfjfKEgTy5/FCCfMazGOqDhsnfG7xr5FbZp4giNRAzwknjRu56aAC8ym58d5g8+XEwJeG5wcPku8ZOUkfN0MDKfzge3iVPHlwAn9CHEU1bSIBvdX7gqAR57FTRlSYPUhMn7RpLBlAi40FUmDxHfvhJWuhE3Ip9Fkh+ErzSgclociqZOHaXZs5lFCFP3jhYPmsGihOCKALQvScPnlu/IjN/21xnV8bfTokhwG6C2SA0lioqSlXqDv2mdNV+o18Syjpel5mycgljxDKql3s7HZOA2qzox8+k7B7SpaH6QFpT4crG/ZaAYCyDqpTqZpYJOyNWaa4IWXaU/2LEFxecVgl3W9EWeJl6knxcwXR4FB+5kUrh+nviB0gFyFUuBZYTK581efr0yTQUInlvl/yBUbHyZwnZmDxy7OIH5YfXjH08GbEHuEny2FFj5TMKhOdmyNcMHqt5CA4y39c8NOohuWZaDB55aHBw0a6R7749TD6LICjOjDeHDBw5rIA6WF88KhkwAU9CIJCRgzXy9w4lj3wX4X+GIgqIZtjgNfKxUdzEkZoJo4bRwCCzQDzIr4F/9vcOJ7z9oFwMWzrjfXls6gMArXMijSF3b0KK8XHnR7ctOMc6uvq+YzcY+gMrcxZcUtWHKszwY9K9sXsNAUpnRUh9tDVnwSbGZvT3MaRIGqIgwqq3u5Di+XhBueQItDhqVKFsLxYdHVh5MF4YiDllqrlfFKXknVLCAJlT9W4JFe4F+MV8V74Ien6PDNdsvQMKSUSE1NW1FE8WHabKUFCHwmAtmEPRdFk+IwayBfkj3EYa9gq85W3yXHLyIRrocDxY0EMOgtBuQ8nGkZgqFkydfoikJmimYe00ajINjjOWxk0EOIbLY6MQOucBQh55ezimfgJ5T66JHT4ATZ1IyNtwAP+ePGEIGfLuIwOjxwGIZKgGdKfTX9PhJD5mHKLH8WsQ92CURvPk0I2Iajo5HFUENmX34mGVrN+cAgUhu6xrSCm8dtP21pSiJepAaTzdY+oqewcdobTm1tdsTslZMZdBjB1ML3ul1NNBweItQOrRx4i025By+vR+Y0Nj8czTrtLaCijQdrBzwJViS0FpczVB1eKAHRIVBl+SzirQjftYirwrT03dlZzKw0MShssd4FICuFR4q5caax1OHpacDBPIZzHGh8UI/ECN/CfgFsMELOoQpieIILxgpIujBmKUbyWxFEdkHGLgxMyQJ0+FZE+g4a+jgjISPDRtEUVTDGIpQoKINVpq8shFonSdxzwRhdBoiYcSEp47FDVYPoNspUGAMckEmIYVCIGNGs1lFPUkolcBoe/Jx8UMjHoQmJ4A5oj4C6PALIciKLAoyehGx95NWcBXoWsEr8Z5DZYQ7+yiaja1Ql9qgaOw8gMqe2I8FbX3EFJsacjSgMC8GzgBI7PXSj0d15F1gUs9RizdhRRbptCX+feUlCMGY9K8Cnggg9rIdw8u0StR2oybRrflfXHcbqf+bW4XUhPloyZg/MVgvGDRcQcgFcBaKiyeEMizNEBbkC5SLiO81FiM3Wly4GAwjXQ9UTOngBQM02zEsB6J/zTyydM0moGQiM6g7Az4kcvHLVIj5shiEowZRfkdwuUAIQmI6/E+uMpweUKBzBGMdkA2Tkk9CJ6MyPPykfAMsJifA6YX5XAUgC79mIDuAFA7TAZrwKm2ivQ0qMZDdC7owI05ZLHmJ2EZznffyl1+DbmKoDsOo/UVc+tDXUZ5ghxQr69PayqH6aA9AK0l9jv66rdsKS9DYN6lS+vWYx2GzJBF3zyIb1lIpxlELiXtPqScsCUKuH6/MqdqxYWkeq3fKHTwhSi1ltmMp44Ujlj2CUcdz9wupAQSM0cj1ywS3kewzjuzlrrGpXiA5RECDrMxAVGlEGsKHOgn8jnRmHFNpgubBzEfkyM64wPyxTEDhqRCZiA/nBqUOWIRf17gZYveTtY8yIsTPf5t+bhJAxAQcSCFTlAYNA7gmk5Du2E8RYEXyUBqFnEg/HzyxmnTUlMdBQkaMbBCMExXvUg+J4onwxImjoxFjG2EhkudtjE1VXguFqwS7gSGy8cN0TzEXK+l0emLv9sSqHoJc7SxuLzBHAp16aGEk8Lpcql5f+O2xgapA4Eq1T21a+WDlu15OVX72RBzxVFlB4O4tx1EuRQmft2GFN3ulXjT2Q2jqwtL1ut0Rh4GYzcdMlm6csfKwm2j9xOXisr6eifpu1ouOpsMeTN2krhcoTaI/c6leP4Kl3I6zSQqAZMxiBqoiAR4iYV4fDGYxHMjNYvomgis6hEaxHeS/CGIzMcPIIdiwc1wb6wjZvjgIRyiJY7jB80AHAcN00yiqyCID97Gt4FKEIdgGpdcQBYNnuSAlJwPDgPIEOA3+fBAMMDhvOxB5ONnTZ0WpissBuKisWQaJp/ERdEd6AIim3SZ/ojhZWCemtgnsQqDCP4ePHjeaj+ZmbOsJi6k6DKchVYLb6u+YHpJ+YjNNSorVB96CikDwkmljCjZ43djt5RumPZ05thl9/aQS8FqinXqa0NUayRvk9/h7dDlkc2l3rssZ+UTDQRaFlT41MsN3is150k4jnxw0CyM7TvBpTgurOOHfSlsB2ERR9UdpsvfJELUHPnYqWvALMi0kbEIgDiHrn1mYaQLqTPkT84ShReDNclbH5fvwhLpIXns1sFUYgGxw/tbn1ssXzPrycNY+xQMg/xvEeLSRwsITL94ayymlvhMk5jkBLgAFfg35TOmTtA8OYgdr4nd+qQmdiDYtHzO1DWaOQ5w6cly+QQhKshHj0WY7QexfpoIVgndXnLoMJWgUM3Be+/Ad9O5Y1V+6yZ1wMx3Gc4isV6tZL1ZlnOjqWNJQIrG3+jR4fY3Lc3P26G2OWHAh0lUn0Kqp1wKro2VbEVtLgM/FL+Ab5fa7A5az9qQnJlffsmP2FQ2J+Lu3I5wAqOaTgoGBql6XThcU39zKWbtB0ryW2hPmD6Yp6RyuRY3VjhvY/AGycbFCfLkd1NFpkXjy8PPDHgJZoZk44MazbDx2JaWDU7W7Bp7GbdiPk1GwNFHBDLo3ZHyWMeAcRrNhDl0qjg1QaNZNB2/HGTgkyM1sXD1DvEEFfAV4BcfPfiwfNfgQQGBIDD9rsWplHM9mCBPeCCV7iRPlCcArjKBDJywS5P8SBBrqHexeBP4QclyDT46lJH31aGmToOkeOlqUe9ZbTLBsw/kTR1tR/aWphlsh7USu277qpRlSxjBrNZ1VZSWjsEyWdz6L0ZA6VVwGpWQUXTn4OhsSclbrQQTzJmXPBk8Ver5V3MpGnOXVVQ2W7n9J1KKT+yX+GClx0PbANGq4rWIxwuXAPpcc00JVPtO7rHZnM4y+J25XUghZNv7a6L44YufTY2iBpj9DCmn8v/+97fSf770B/aDr/7OeC6890dDLlY70yZuBLR5cmgoFP6CgY3vDRWEgqHvRQWDiyYeokAQBiC0iBjsMGYR1CiwOIJixaKhg5AmkGnvgf9EQdI94L0BWJg9d3kIP2TiACADjVkUJT7Oc6mXwyFMgZah2CkO4rnUn4BFioVRuigpmo+ZOI1q3VJtpGmXaelDJiJ0MNZSqckUrn13YD4kpfEIVSaj0Wk1eAXeBAGuAgqbfQopke1IgpYdRcWF23mBYbhbz+TgREk6eUH+zB2c0WasrOhWmyFuVxqNNmvo+OacnBXrOK2p1+YhXdBjxIkfUXVX4tdeVL12wxNtVTm/Z3N9eLcmtRQhFS0qo9HY4sutmFs3s62opMbY5ZS4vahb/sBX97L88KFFkJatwUe8/yHF/falv5ggRFf+/X//aQt5tCrBF00nnIIMM1C6dkRUUbqkQxBeepMOeAGDXEzBVE7MEnTgh6h6Dm0iJOMpeongiFS3PVwIvROkKx/YfVC0hNmwWBR+CtFiySK1K7q0DvAmmQwPUGLITx+gCkn0hiOq4FP5SKhS3LI/e5ABnr3E2H7wb2MwWAUhqz4eLufAs2496LtPJVEM7xlIupQ55cQncLPSnZkcugZR2aesarLYjEZ99wR24IWEt/EusmHBttYGpUOr6w9IkXQqnuC6LZ641k9B+wEIyVe8obTptIFEiDE5tcopcde2+MmSFQvaMjdfMAotzmv5b+MXvr9b5R+r35Y/NEE+6Q5AioQsf//qD9Ce+OB/f+Pz1ae3GGUhqPDRcHHgBXQIY9QDBmBIggNTUTrMwb4QqpHeocOcXtM7GPYyRCPFYwKk8LiDIqg4Hk878GS0+JiYE3mADJkDin2I+QjXAGJ2/ApGB+k1TQfoRIJIw4PoUEBa5oDWH62UwMM7h+ZZ2j19dtDZBdbvEkllcziiHxcfCvH40iudPVVY6KpOiOkkUZRK153IKW9KE6wwj++G7yPau6dnTilaQgwM3y0NPSi2I3i8Qee/sCKncXONOsuq9ndVrd6mhblUzyHFCLr07aNXFp7Yb7LbtaUKCZz7KSXNzbWhjB3Lqqcs2LxOFWhu9va2Wjc8h+XJA/JUfpx8WhAibHyj+3niR0oT5331m//66r/+8hKmfJ4Q5q9Z9SxGNtABzgHy4EzUoFAGZVkKJfzDWfxFuQ1+4Q9FAf0nHuJ8EMq2ApgXzQAw4BGUgJ9U4kKfxU/6/5XbYoJ4k6bRTCiUopWaMsowX8Q9ek3rEaTGjPz0sQ+8R+eRfXZwWDBRQDHu3Hiy/o395zbsX5LFufxGJSDQh2JnNbxE6rOkmxphied3IRQhjWNxi0PskzdO5K/aZK7Ud28CB0gpsisrXP7TjdsKTymjvV6l8RZUepUscqnuC9HbaTA+IT296YltmUvP7YnzBLzwI4uPQGVzafqGEfkpo0+uS/NUVPpy2/Pfzg+whI/lQ8fDO1LUndGecPts//zv//7v37z0d6evPgO+XfgkCx2qGNUYxvQX3ii9FEEjJoRviikY5jjTPPhHz+H/8Sju0L+0DLEAChLcupIhfP/Kc/RB8QGaWfwdRhB9ULwOlxPOTUsN50KVaOa+OTi4XmWsBpufSNdt2lxYXjQ6b1nT/jQ24M31+vqGhFgKzKKyvVK4RWpsSrNDCEUDkt2yeLTTZGpaRaOy5wLj3TgopCqbA7o3jsDib08Atrz9A6lec6lsvT3peNPotvyqk+vi4uIEq1UwmpiMvU2jU/Jbm+ba0xGeGzy8Lw5wgyEJml1U1QDqPhiv/c2l4n3CvN8AU199QNxxSVjV0sF11x+UxfVpJdFqBDCJSmNrGqrzioqLl+ZPqapqbFpiCeVWuPtmTi/WF0O9IkAaCnNy9tvs9druQQpPwiltY/GCDYwPmq/dOEQuVYogO+UrW98gXj21Gu7GYz3O0msuBa/vlvr0TXkjpjSubFiXToecOm3v6Zy8lKK801GurFIz9dXf4/p09ABlDo8kQz3nUAJU66hcoJ/tperLPJ7fAlL/RRDPEmiCG9Ie7nx01Ix77h5n0TJKv82494sFKdsyG3M2b2ssL972xLLzSQ6hL7kU61QkWmo2F5WfTNMFEulGbje4FHqTL0urW7V0cwzpHqQIVNgZHbaztpUf8AteibN/EEU10XuiPXFtWNBtZ5Xdc/RI4bac0cuaGvbveLqhZGVOSnHry0f9tqwKaKn3FaSwbIA60ESH4FgzmKox9DukOBvWSH956Tc2fNikltt3k32t1+6pX+ZEiPdsxnOtRW1FhSVHP5m7bsemlY3bigob0iylt56adbutrFuiI0fLty04R7JKvVpE4e0epFgDOT/zF43nEbe6W7SwKuSVcIaZv7LGVlqJxX+fqva1V6G3XAruMQlU91yqdU2j83JSqjLzWp8oKsopLi/cvoS1+fXZVqVRqVC007mdH3ThDlNZBx81kMrP+h9SSpvNpvvgpd86xRjjFouZdO+d3U4j77JnITrnuHgFfDc2tOakFNbtxzyExShcd2lB8S9WNehKr476PnB5DPs63Z6TVdWba8xCRalWpWa6J/xgFfFzN+dXlaRDy5YivLOXJN6HNp9C4bQpT49uW3XOGfD6YEXRP5DqLZeisbiJ2w3n756jdYWrlhbnt6UsrSo/cXK/X2nz29z6MqXSWZndNwMlGE2emzVj1yMEpuGOOwApQpxqpfBii02JFQN2qvumFfdKKTS8pgn7jBa10VqffroR/nt+HcOaVDzvdFoV2g15+cVVDSTLa+WNmNnf5GK55800+3xk/4K2BQ1mLTGLkQa6yQLdpQh3W1z4NFw9o9LUVYXkZurUGl20MjSXeg3KHScyC0/uISozpCBhIN78xG3e6S2XwkgLe7pUJXKuHb8uWdZYtHJFyYZPBKLGxgUqi2glPfYJ0ElbwKKg4kZt/dZMvWJaNEAThelfvx5qE/WP+294SBSs0SRNxKKmjPeAKVWdOGpkqZwffcH4EpPOQaTbeooLCDysullsKt3ukZgr6Jpm5h8JR3nqfmmsMzHpsRX55U0eEwwkGLWR7cj7SzukEku9yj0lM6tb9/bv0Oktl7rWcIXP7XTuufDJ3iWHWiSMVgxMfC21L35BiA7vXJPmPEme1Ayh3dHf4gmx0uKOT19U/14rQ/QYbtYrnHypHZupKSv3YyIcRhThnUGP5VzeyhPLlhCd1YCsqqt+fXrfTEUpgc/mBZfUhh6W4bRmJJ2eua1wh5qH4iFU5TqayYlMStyxrhCUDY1tCw5A6NRDQj3K3msu1U7FyaoFlx2reIvfbw/Eo5f7WpICSE2QT+PeX0OGwCDdcWcgBUSJe6vt7fy3+QGP4dgpgoxZSDrQ+Iv8EzvYXGj3hb/samrNw2xqrZ4J055ABWLMdm9PqMvOqwg5D+SNqN5LeggpVmkt9SxZVjSzyaXCJ4AAUh2oUYQhRTHlZZccqZ65uUbSR5s7nTTq9rmUEbMCITeUWxtKNwUQG7sfIAVrV9gmvf8QGYSt3juwLxXuKyoS6aTX7uvbUqkaBjzZjJ89P3pK/rYdbKiCbmqLfaH28/psnx/TtNGb4tLjKzFH7NY2a5cd5pWuW1GViWihYa2nLvNen8gq3bn1ltN5bUt3YPuZemHuNHyvWH1bWklRfuZRaXMfLfGvr8t1v2+fS6lNAqSe+srKbCtvVUBW0/dcygHjujmp74+FffioO7EvdV3//Bv+lFqwM6T3tTD7T4xIKdofFwpd0y5VmdSKbJ8JkdDbCndIHV4Ftkpuu4sC0obGlML90qwe6q8hKoHeS5Ysy1/VlG5PzMY+UwfuuuiKHzXE1qbfea4V08v0xH6G1O1zKbPZfMWPLNUHg2y1r+d90I9DzDaooe/SwMac6ovekbXUbY+Ue7UAEVKKoHLdiqVto8/rQs3117bs1Srsy2bLuP2ZI4rq1mdAQ+b237aaO7551YiSdF98D13TsZC26wXz6aL8oh2Ciy7iO4JU2P0RVQVbd2Rl5op1amtlB3LBPnxZt8+lzOJcT4ltKGodSb8Ifb34g6poFLk8NlmeMGco8BWBVB++/w6KEiFlDaxvKp/SeACRYhQ0CObVg4VD1IpSu//AzLYnNkh1ZvoNvc2D508tmLLgPFtr7qFdIxSmJHofWXIkBRrsAQGQ6khvTIQU7I+M/pOZ1aPPk6BB10Po9rB9t8+lrnAmmB/ShSC21fp8H0eIxjrqkDBk0jSoUVATpAiX6uFb7lF2qRSvkbcfeCJl6cn0kKBUqr3xVwtARDW/DlNBoWbzKir1g7Oi2//k20pm5k+pqWw2+K9B9yrBrs6IpS4Bm+JO5+UUnmLLAKmOJPoUUgwvGMmGxikLmtKtlb50e1el3nba7XMpzFUZMwwQE83QQAKssFt327X6TgFB+NwSvY0F4XEBy+QIpL7TQX16qbJIEHLzVF7bgi8uSAOQo/HXYMMwMJWvL/UGpafyqjO3s2V6b2LvicMDn95q4t4orJ65SeWtNPQMUYSLg68khWBctywnp26P0ioxdxTFja5KmOwQIl4gADCsq/Rq+93OpWiPwko+PhHRGulxU3zV3nf5lSepod6EWAfvcMDYKLKWuu3+vEUBalduLvz45yzN/AniLkH01NHkjotrWpCfsoP11mfcorhOk2G2DgMmr8t0aeaJE+tYeFDtNGvHCRhs0Ab0pzkRlb31PBvMlun8HeSEBEWSGzpet6qt9SgHT5qi8lIH+frqVh9wqb6qSqflwBZ84uSxT4q7vNRMKMKlOu2qvkhgFLLAnhLsicLBFQ1ZHl7ff6dk3ry3Or+xpKUs1PtPPiCh0Mv4dSsRi8Ll76Zi33X1ECFlhT5m6rKcomU1nM+rcl2XfOWnmKvecyCvbfR2OJWEgWM3bIZvLqb7d25/LdV9Wr3NKW71jpQnLN763pCYCJfqbTd29zmFr0XVAJuipnSIzLHHS20ObnqWMegurYJeEhPd631TTMhYic+ubihcemI/q0Usws70Xm+iHr4hgsVr8NuUG0ZPqdrkD2XbjDdnBW6zBfXRwuqiFRdcJgPDSpNuX4fqZjLX7twbXIp8vAaeFeix5s5t9V7rpH+vX3qf+fcjRhQduaByseIWaUeOIxk9t656ROYX6crbghQjqGq+yKkqSQtv1fasowEpIkEIaZtqfV1VcdXe+FxDB3vz2LyqUGMem3lkh9Hl0zNqSz9D6l7gUvDwMIjEDB0+/aEZCWMjkOrZuOtZbjqttrrX160akXeK+Jgr8oKOuAc2pzYtaGs9hVl4Lw/sYBJrQHq0tTjnnDKUfYOsvltFUkhBPiGxmi37VxZXlawPyTrYd2Zt1sTjgFzrBrUf8klGZUnSdqv43ma6F7gUdqUGj4PvPLjfihkYWUv19lV35znKlWwI2wktA7vPF95mpCP35kMhIB4mQhGmdyQRuDl7B3eoKI5DtOvynBU1llw9Aiv1dEOTQopKTyQZnkvATAOr6wDfvE23Zzs8xVxKt0HkDz1fSx+GPOygWb236u2osP65x5NHBo+TQzjBk4J3L0cUkvqnl8OlQjuWJ2+cWJm5eW6SQU93dKh+WQeY4qQyg3oTlvxHe8um6AqNs0t3tCKygC3UjC0Yde92X1DHLPO6FUVtJ3Yg0t5Nh9NhbJiZU/hFjdEII0TCqqGbcFOmvrxxD3ApGlxJviv5gckTC6LkYyOQ6svXf3NZPJ9WV5TTut8SihY3SakQoSNIZWRVGmq25a8qSe+ltowIKV0SGMjKJUm1ufAOyHtvFoPcXMHr7iC7WDsWDn/Oj05ZVVejvZlNJSrPVxUvXfYGHD1zEliAYG3VN8bm11Xkhp/3wloKjvVjNQi3JI8dC/HEHfDjd0MP3R0X4rgOywk6GOB9UUdqTouZngZSE/cAACAASURBVI6cW7WtcXuG3RHC2h+cRBy0NxHgPNpsH9swom3BKSmVTGOPv6d4kEiggbTk5RNLL/kRIBSQcvZ0qF/loWyZO+S5lDdl9PZ6KY9YZpioQu8UeFMTBF0+VZiSX3TK4nLDP6CZGlVd27m+qVl9ceMe4FLwUUwe0Eyf9WYsLHun//tBCgPdCV5BvY90OsJvfyjA5QG2RLO95iU5vyhaUUN4BurQnRfLabUMT2pWZC49uUdXmQ3P3U59F9k7KIjRV8a7tJtGV6/aS+AHHVoavd+DVSi00povCqtXNcQFXEZ4DGHiqbGHscWlI/uX5uRANJEuhK5Womf1vPpUt8/3ApeCKWDqeLjYjxk6dXC0499Nxw+eWOkBJ290adPJPKzb77vTjOZ6eCaXZAtx21flF553o5fhCaLzg0vEhinPNDwxZeYOc0WlE06dYdfTk4NRZGdlzK3LK6pL69mDHRDxSng/t/fIieKZ56UumUGvYDgz+k1hS/OfWplf3drgT+OFHpqOdECme7fuBS4lRA8i3KAoeBuD2Yv4t5/9+HWv6+5QLqo+mZhYWlrqpUwqvHroD9KYvTkl0ZgmtUG5lFpIdQkpEg+3UfD0vyylqimttNINQ9QeAsNp0IfIucZto49yNy+AethAnrHyLv+pwqK2wnOeUG6zHr6c4Fq2ot6+oTBz2w82uVx+p9BNb389JH1z9nuDSzn4Zyc4yICtw+GiH/3/b6U2K1XRA34ILOL6gGLq5tfYB3fAARleVbN5RPWRvRlBrKG4LgVwZvBLYjcdKM8p3MvmZlPu2cNK8N7S9JNVxZuPJ/UQjDfTwUxPYbX7NzTmFC/YdNySCOdNkHbEs3O358H1zCbBocguU94pSN0LXAoWmdPkCQMRK1f+uOik+N8GUnQTTjBi5Y55lZpj6GW/HU631eCSbsorfqJBm14K7aCuIUXlFk6THVrgVaftjmz4xO8xpLIsOwohQa8P3XablGpJZWXIldaQOaItr26vOJNUkz376/La8hsbXC5fc7akjyIz3bqu9wKXgsLEs4hAuFUeO04+8d9Ke4JiSJG7Fv7zDIbsXOzD4gaYya1fay9yOA2+gGnHkeoRdevr6ysQQPYWkAJbYhJdaduLIASXlkKK1lNMCXbP9sal29aVVvSitjc+ooY7l2y9Li3tXGFm24iqS/uXPHdhybmT5UvbyqvPOf2CPjsbYbF6ivkbaXT76p7gUtRD0nMIajs0SrP432pfikKoubY5V0Gde/gCahsg1qFieLffd+cZrYay9D1NedV5+81BfQWYTkcevK49Tmd6kvg45RsnihHYXRevQIixnh3BuHVHcspPS3P1PXuug9xSqRniG6XfL92/Iq+4uKp12csvv9yaU5xSVLdDm2YqM2AZep1pcgcl9OGte4FLQYi+VXPoJ/KxfJR83L8Vl8KqUQgkWTzHl6yrSVdBs4HnO7a16HBEOHs0T1QoXNy5xpzCS/Yya3YFeE7XkKLbVYoKsKmT5StX1JhU8T2FFCNwm8qrW9+wCLfPdbVabKqBS2KJULP9SN6InJXLVq6sLi9ctimd05kFmw1xGy09DZLdYZ924+Y9waXo2ik5Wf4sWQQ/fv9WQnTE91u/Y1Pd5m3bttWdfqMFDiqpTtvtj8EORoa+VHoBMWyrl0jLFJUKDs7+u9x5pZu7kgqvUXl0Zk7rOQIvFF1KM24maJXUrCxe0OSJg2/i2z0SEzmE4cW2mt5mrt+xfUXmqlWrFoyo2zTXklhZ6RUQ4VapstT3lI32slb3BJcCm3pfLh9XkBqLtdR9rT1BtXTo95ZlTSarvtJK0naUZC5Ymr8UOjUzf1CyX0dyKw3YyhSPrl0SqOwf/NbnjNNiVdTF2DCHCcILfLYgPV1U3LghqZ5lrAwHx2O3QC5VA4c2wp66zBGbj2uzu2nmTj0tw08gU2ajziBan5aG7LdvviR6UqfOZt0Q4XBJNTuOnj+3f8lxqaVeoB8gI/Cv7gPn7V3043VJ9wSXQmxn4b1ZqSQqdjF1kHQfC9EBKRFThDVJrT4DWdeUt6otc8GqETmrFlS3ZY4+fcGcazAaw7DrGlIWy29emhfnqVd0uSy/QhBh0qzkkxMjVtWlh3q2JepkEXdg5n62UoH4Gd04aPBBxANQ8M6ausyZdXvMuR5LNx7rSRbEHAFDQuyR+DvEl26s3L3ApSA4p9tRAnFcjkEw9/sZUlSeRw+JRO8VON35Za0rlxYta/r1f/7tdNOyvJTq0St2EBdV37n19hTP/eV/5zlNt4DUNYKmtKbGtqL99lDPbAqx3bti1aomu8Jn6NbMT4JtWDXvlRjI0dbqBUeVPm9W3I1D8raveA7Rk0wYNT1aS9422asF3BNcikaXDg6S0dWn2E/3875U2ABIomj2SmtOFxZtK9+2aUmF9Pj/fBa3ZNOyxikjVh6NC2DdAthRVnX1NXZw5hF0bh4LSDm7ykWdsWJ1JpFQ9dIpCILhanF2UFgXt4ymAzOrV+4lvopbTBTDZTCYiLJWeFl3nazKhC6hXsFfc2bbBZkeJFEkUVnpvwhS9wKXouHYqVduAIoGbr+vuRRdoMBQ1Ww2kAsnZ7blNDatMyqb2b89/4+5RuXeksbi/BP7k+xSrRZKrV1DykkAKaXxVpACwXi4t9JKqeXryiWct6UDy9iuxrPAIexG3iZ4pe0qV3saghiwxFsR5HaUp8zcRGTZClvXkG9/sic/qGz0X8OjCLmOS4ngxtgVqy5GregidMW1+oq/aP3po9du01Lo8zfeuaFbrtK64WYHF2BSQayhKKbC/XQfcykKKW+pLs7lXLe5ccqqlUfTTLq1xsf+/OfPf5YWZI5fmpk/4sh+iysuDkE/bwWp33w1j6Fcqkv3eBRSiaGA3byhsbhqE6nQ99TJeWKSp6lxxMq5XG63LPvMNBavPtcegHrusnXEmy3pcq3XwXC4229d41IY/yKyqQsiyghE3tkhIjBTDWcSGyejDwajRd07pLQ3WCwMf65gtP1++w+RRPtVpz94sih2qOgSs/27cz9DCkwqMc6V5t+77URO0eYdcXbYEu352ec/+vzPj2WF4DsrM6XoyF5tehxCU2Pm12mvQYRHACmWcqmuIcVI4ksD6dILR6qLNtew2T2HVIA8XZgzs4HtngGhVAxvHW355MiImaeVTq+Xud8gdY1LYfTLyLTLQ6cBRjw59N5GGRnwk9T2YXzdy6OuKq9dwmM5LgUZP2RoKtY8VxOgMy4MuTyQTtmu3vrOOXroItGhxHduf/dSRgbLU7fC90QQs75wWfc5pAL2dP8nWEQVljwmdZVWVKT9/h8/+vxH//h/7OmhkG5TUXXmFxfSPHYaQbgrSLHkL1+tJt1YS0ni6z3p9gN5OXAvWaboWub+3ZeDa2lASP9ixNLN6yWdvegbHpJaOEgXW1ynR1cfeYOUxddz9xukruNSYCipyWG3XgI8Jr9PyFj5eMjZwHqudBb9hZ8yBykYSrEjpvAETpUXAU2x8uEk+lpuWTR5Vz6LOPDI1eevJVJ29pxm5IAOueANrwAV2CofNU5+SMwaxtT9DSkmkM4vWTaibXRTDWvzen3O9F8+/7PPf/arz3+fFqqtTLtU3vZE0550QAps6sauuuGKrqU+uDWkoMNttq+XvvEyXLNEuUTp/A3F3PLCHvARyNEzT3VPWUOEFI8QNjmjm2xGpzbD0uWH4Zbk774M7VzKKxP1fnbJ54iiklHQVSiYIV8EFNBKi5gK/6HS7InDtobnhXR6KCMDH1wD3DzybCpFSntuPvgQ1XIV8SRC4SoexOLgUUI+g84Wb3XwZNpIuUYzbtb44dOiwkzw/oUUazRn64W0JSuOVC9o2qMsc+sNfv9nn//4h8//7LHn/6fG3lxrrKmbubK1IS5dm2i+hdoaxBO3hBRsxxlzVnpa0xMpRxAwCsuzrrVlb35b6havuWZFfmZJGou4SFT40OUesZZCyG9uGN3W+rTR5tYmIZrVzYXey3fauZQi6CCpyZrHNbGwnyX8HDCoRbvAG2TEsWjoIAodQG3AxFSga9B7Y+Vbp0UT+NZ7bxASpo2SrxlakDp0AOaABNM5PC4i7dAw+QASzcumDS3ANe2l5y4PddDlFSl4L5VMD4c07EbvTVx8mLqe0BweN2cwRff9CymTq7S52bhu8yrKiVx2xJpRM5BNPPb7539m+tnzX3oEd4tySduIlMIdcaHSrA5dgF/tT5FLMcr4roXoUouUZSBAbyxuPa1Ng9sG4OJqCd07w3BRR+CmLG8HF/CWalVqtksPRCqV001NFzNHlLT4EbD6FtPX7tXhrsrVzqUkmM7Nko99bpf8PYTDTR2ZsEg9ST7WIZBHYjXyYc+SoIwMH6fRJKzZSMbLNbvgV8XxaYJcM2wygR3TLvmu1EfkDwEs02fINeOgNSSAeQ2Vx6byZPK4kfLkWVgJkdTFu8KJZHqyfOTkN+VbaeDdWx8g/YB8/Kw38fidDCx664r1fQ61KSvXuaekqG1Byfo0Fzag4Chl/y9+Ztz7/M+Yx/7x/02v9NlM3NHMbXl1NZZQSO3qsgZ/+eoDYrwFpDgpjPfUe74oylm2JM7F9wZSrERQLTmxcuYlVSCxNBHaRl3qIKpMbr1V2jC6uvFpRNUO60N12Yp7LrGdS2GvL3WYZjxZDPYUJAPksQWIPDiKwFIpYdbWZPlkQoZo8GuGfAK5/K48eet7yLnm2fflh4ccQvKnk7mt8o8JJAnDpr4vHwZeBov24fLFBLHgRw6eNUM+lQgFc+SLn31SPiMG94bNejwhAWsvavZ+68MhPAffE0Q4NG04Xdzdx1yK5Frt6ZcKT4yum8unueqpTQKp+XIuh4kfm/7733sq3WVOTwZc5pdvigtVOLvSOAWX+mqe+1ZcSvQYrtxQntPYYPEEsnoBKaVS4bWkX2pMObGOC9YngtF1CSmTSVHJ1GyuzqvbY/OLGnj351pKZVIqHEDPmiBA8QjnwKRsnBNGSlvJgMPgWmS8ZrGDfCp/iJBp0wcMAnoWEz51ErTuBiUAge/JkwtI9BrAbujIXdNIzODJh6DlECSj5I8DpshALsvHRZGN05+Fea5Gs9ExQz6JAJHyoRQftz4gR6QLOByYUNKl2f078XNnq9IaRuesXLGEszuCIUCKVfo9x93rnv+ZNOTXlVaoHa4sBHapKs7ZIc212brsPHAp5624FKvMbkbkm6XVK2rsGVDvg01Ht5QgrhFWKt36LOf+Ezlw9G8P0GAzXUPKr2gmR1dNyTvPtvA+Lxp4v0EqzKVUKqWXjv5nhwwcBSu/aEy0tpKCZM1wcKrFJIpMlB9+DkxHPnY8ROwygXKyaG7QwKGXn5UPew74e5xEp47ctQigfJNEBQkHmQQEFeM008lwzThM+abJ5XgweshPJg6WjyWXNTOiojB9BDPrHqQoPgdOHjVpI4SNmFDep5Ciy01FkJzKTIEVkdIW0GZRBy6sxODxJAJSqhDvlGh1Lp21NGl/9S+W1q2vsBqvjeybfjnJ3yHxuyWXatHnSk8vLW48ytlD3njwmJ5CSg2biuz69SU5VStqdA7trSHlrUyvy6+qqzFjU5uG67jfICWupbigyejFSgrxpeVY+GBzaTH4yHMJCYfAqQASoCkWUWgmz9DINe86SEys5ifgXMOQWyMfx0NWvhXsS4Op4hpIzaMFhwOAEqA3DhyNQvjCGNg5JQwksyChh4fLBxAX/kGg9mP5Q4DeTSOhgxtQmiXjR0I+MXJwFO+4b7kUZKd8kF1yom1E4XklVhn0wPhWeJMyCNZSRKh0w+rH5Fcqsjynq4qPbKB2Lp0fYUjdikspbQayI6e4qilda/Z6E1WwkOghl4JhlT7bqz7fCMMQzsXDK3OXxoh+v8CeKvpF+QaCWR9tX5c7AZ037+5NucKldH4GTCp8JA8kh5I1Q8kk+bBogGQUKaB8CNyBGzRx8UgwKIgdBtIZ4axpjgnAUNQwzURIISA5IDM0U0WJg7htPHTkyEOwc1qM54djzjhcnjB5SME4YHWC/EnAbA12vrojRAdTEsjGkbvmjB2XLH8y+v6d+FF1K1NNXWFO6ya7qywMKKqVrq/XKcGlLLZsp1Fthu8fiYFdt2zEiBVz2bIuBhZPudQt11JWtyutqTyl8A0771QAUrcQLnRATyrlGL1Xe3wzlD3W+2Gi23VINHVAl1ZStXRbDaKm9xS9HVC/C2+FuZRO1UJmaeYsWrRowMSEkYvITyCrww7rA4RM12B9lRqLKeB7o7aqeT4WK6aPIaLAjBD8aeAM7DwtGpmM/aL34XAFCzHwq61vDwUK4HVZvkbAxG9GAYmao3lEBBfk8pohKHNONLm8S/4IiQLLufUBSD0LwQlUOtbcz+5c6GyZvVS4rbEp3RGIxu4OZkQYderEkACJ38+h6u03upsrlUazu4VsGJ1SdbrLebPIpW65llLkkqPlKY2n1UElXElYVCZW0i1VvWtvTYuNJYk+i2tYgNmjmkKqoyDU7fkVucpTRSkLGiQ+uoqik74ecsX2ku7WHyKXYlU69U4q7gMLkA2D5G64/EEeczaI8MC7HnozVvMmRAwj8WuNZs4gzApjJ0weoNEsXow4aosArsMTRkVNoCKHy/KET9fIhw1BQWGvRkLUGvm4T2PlTwa58fKEBx5MlifEkAEj5Q8uHiYfCeR1q1vAICdgh8yBBZXm/fvSnQs1lXNSzZTzjTk5m2u0pYk8lWdTTElM9uYKJfal1LWldkCqWaHUusvijyMK+4m9XXUgT/6rG1xKn3u8bulS6K8asEckQqrLXaUO3ldiPfyMQ+g3d/PSopI0RNzlLB2Y6bYzJG+Fq6QqZdtcZ6VTbCGa2EGh9/ItkUsZgy7wpDlU9zUYPUwzSmRQjrHySZD9TZuQIJ8zNRoQGYpfCe8PJGToWDnQ9vEwzZzBGvll8tyEXfLFBbGQ3zmwdSVPfncgEIX8EyA5H0Q2vpssn7HVQQQHZO0PjpIfThX48cPkYz9O0Dx3ZQP4Vv2HsqZjvokR58B2GJZf953ET6qCUzyrLgO6pDlH3mCs1znCZ6TSeC8DITrJ0qrVCnjPYjnekFi/vzAl82S6jbdaqZuIDoRsTuffv/qt01jv7UiHjgrauCyvlQ3oNlQV5zVwAVhVMaI5bPvgv9VruZIO1/0IvWYNMJtWtR15mmTFs66b1SEYhUSNg3GyPump8uKZ8KnkDn8zuknlHsoWXksFTM6NQ6hiBI6B+JGKf8LAIVEYxjyJGUhn+VQeFbMReq44B4ccwjK0YKBaNmQj1CfIoY1B5A7StOiBuIEsyP/ctAKKLVKwUUyB2P0QGTRko4PwzoKBxLFxY7dkE6gRIHVIM3LWtIGHRtH9rfsRUlJ4Y7CqjsOVY+sGpU0BR5PtY0hrZq2EQsrebvbNB+MD9u3l2xYcVdl8Ep56Xrn5U4+1FLZ6lfWlHUIK2rF8fEUpa7+wIqdq83qLrKNc7XW49Y9ogVyoXprXZLd7JTDk/+7B6BWsERNEq41bX1eesmJuhnCthd/NfI9fh9dSEKJ31g6Kj/CcXfwVhgvNLOJG/EF/0R846FnMJ16Jd+gtCp5rT157tD1Xlz/owmw6Ffhp5AlDICvE8wM0gHuXD91TiVjhSxiX/XRe9ehLabYyBZSI2kcchZTkRkjZBEmIXXcCXiz3KINWTBE7gpTT/V9f/Za1aTuGFLgUX+G1urjTT2xrPK8L0MAZt3PIBIPqdF7OiTe0IXgkuqkkcClEOcRfzG2f2LZgU0ZAuE2CN5G4a26EuZQFW73WK0OU8iORJ4l/UNGr52u/wnfoX/EX/tBfYpvEG+HWXf159UyzXPlNT1ce6E5P0GnkxCeT5YfXXA4LFO83SHHw7Kijzhgyv5hrSsPIw27N1Y7pgEv51RXeQBxiEGaeUgboqqujiR/vBKSIsmNIUdkHk1sRJHthlVWyxx8N6cRVgr0544XyZO+RnKpL9lBuVgdF0ZkmUejN6uObM4tWzPXIym6PYG8qeYeeEbkUAaQ6mI3foSrckgw0JoKwGkl9b4CMGpLch1yKSLwh7ZJlmW2ZO+JcKlghXrdZ0wGXMqqtlaWWC8uqcr5Yr9MJcG8Uf/PEz1lGIWXseOJHvcFIKhLT05tmbivcwcmyIZ245XvoIgNeEa/1bK8qLnqjvkN/MFRFnfF67epzM7eNRhQo7x3zp9xFrfsnqZ1L3cWQElsuMrUrnO2+m/jxsqyM9JK86pkNFruKxw7odS+7Ay4FOOj1WVIEqig8x7k4QKqD6GpXuFRihxM/IMop8QaliBDYeCktTg9b3tuCFK1vvPSNouLy7RmhrOtqf/WnVopK5wbjLmxeOmLF+nosrW6f4NWy77LzvcClrnQZllRX56b328RPKPUkbcjbVngynePU1PvRdWK3DrgUdYyuyEqqWVGEeIUqFVhaBxYZYS5l65hL0SCiEpmlpqQ85cQSS5b++sVbb0eoxGfaXl5d/YlFcN9UBEchZRWiYQUyZfT5OK8eS6vbxvBNVO6OG/cOl7quv+47LlXmMn9yJCdl5TpIQ53UqSM1erhydMCl6MKE8WZZzhUuzWsw84hbg5CE3z26XEtx1FpdUDcUprQ2ZDhKoclw+wOc8ZE3WqtnXuJkN28W070qlpGplyzbNrPuuFqvN6vulIvy7/ZLv1/fK1yK8idxDUx75L6DlM7hgD+xqnOk1u1G0OabIPUdiZ9CgXmatzSjpm7piWXUf0dHkHJ3sZai7lgZN3thRUrK5gseMCnG3AEoezj4WKc2qWlBfs5eorjpyTCkbMrTjTl5+zlBLzEnJd38FbjpsXvyxr3BpWB9NYRucVFY0T/3x8QPszs1ArEpnE67peGJleWXjC3ZYE5qHmzqVlwK6kOJdhgjVmOb1tqsiLvZC9IVLiUkduQaMy6OtdnYX48uLj/Pp+eiIh2BsqcD2pmV9MYTbVWX0nk3vKrTCPXXWF9cnApi2x1HisthrSzTKyR3LJBGT1tx2/nDXEp7l0v8HNFPjhvETx47+BC8j90/kKIe9xFrTGIjO2ButHmdUua8+YXSiZ/5xn2pK5nUxFEyIn/K3ObmSrvqpgedRNyXqi/twFc5lxGArdWOJ0YsPQk9ZLpr2AcHlxjKyNjeuLJxP1tmsHrjOfV1LgHj4nQqU1RJ3i9WvqF08VYazLsPSN6VRYS5FCB13YL4bqsodPzekyc89x5sQ8ZRxxj3DZei209Mhd7K7oH+edFRU7qb8uDvHB2spa7m4MmpvPxVDWxtbtzNkIKOH1VI4juEVH28zZYGEWPe77H5ekWOerXU3p55od5j2bsyp6huD8sbfFieXQcpXtDauQ2tK2duMtqMtitmHb2ldHc/186l7mJIwVxqMHT83pdP+BR68PePCSJiolFxuFfGbcpb2Xg6PU0ouzZTah82HUr8rqQy6+uW5m+uIblxNy9MOAopJSR+HXApRl8pY4/mtY0+TTeU+opNsV6vJ/F0YcrMcwiFAOtCurt79XC6s+p/uHLEiRVwpMZboc52/x73BJeKhgpuqjBDPkSQj6V2d/fJWopQD12K0gD3xomUwhVzXboy27Ux2D7kuuBSJMCdKspp3MDWl97s1kjkUm6jtWNI+RLnbs7PX3GhrF3m006w1z+UboVgQTzspStqXDoBioo0Js2Vw+nWHm9aVdx6ygy/9lDUuX6ddTXPfXJO//GPHn2M3N0TP3Cp6fLhH8M+OEoz576DVJZnfUnRlNb9Urv5Os2+a8OrKy6ldaTX5eSsmBtXevNn30n+2SmXIvpcy6YFv1jV4KzEgx3MNq+R78Evp5Pq8J1rLH7igMulo8ESREhRXSXitHHnZuav2m5RWK28lSKqg69HD2jdxVnvCS4lkIHJMLEfD38Vs+6jtRShrMUaiGt4Ylv5aa29k7gBXXEpcxZ3qjWnvEEVvBlSIpcq65hLEQPcvcLp8p7s2j4cmxDMS3jVnpIRK584RVxQq7oOUi3sGwhWv3KuMxsTQpFH3VzjPqzKv7KoewFSAlwMTo5FmN5DCbB4vH/WUpwWmOKln5xIyVlxQRuCBoP62kypfVBQSHUs8YM/8hCEDJkjts3tgNM4nf/86g+drKWIMb1pQVvR00pfRTud2/7Bwds5K5FxbxRWV2++YDQxEi5siwg2pfJf2DyiOu8Ua3OLayzoblzbJrhtyndVAUz6r77/6Ny7fOKHcQcng0MRXvRJmEvdPxI/c7wZvozTSzLzq04pZc0KVmWx3Dw6uuJSqhB8o4zOn9nQAaR48oeX/sDYOpT4QVS4ID9zO/E5OiB4cxW6d0ebEYcI1LVB0jB6yuimNCfPcFfkkIifm95UPqV8e5K5xeYWI45ASfF+ZVMipDj48btrJX5wnj78UIEDuOKjDgnB+yiwqFnLO22K80/kL4WRlK9S0jmkvqM90T7EpVoru/4kPLvUEB6Bn7E+ufYanQwgxdoSrxdPiNuvDMPDifmI/G0XFM1R9vaybvuHtj7JqGRqc8mek6tW/uAA3hfsjdWYDDJqXnt6QfHSkvUVhhYjakgVDBGz97Yp3p0FRIlcKvEuhpSMvC3XJDywdfqQAnQh77h/uJQ0I2hTrlt2InPbOqX4SRPDNn93nHTFpQhnhT/zvJzMTWqDwiqwSqLXX31e5FJOm9N7zY2/uVSlVDp92QJ7elX16A3Epq+8WXfo6vM9PlNnY8A0ryaPbSta2brB7/IiRoI1u9kb9DfQUHPrEEW0XVn2GvZ7TOgufwBc6pdzSSJ/93Ip6pVd9IeWkDxu8PjLEP/dL0J0bYbJaNw+ui3vKOKrdXpQSHXGpVi1OV69p6QxpfoCEQyAFKtoxwhD/km5lEShaldWYhRQaLDmyti91fmFJetVvELfnr1T+j1PsClPtVbnFJ3zW6NNvNUX0qVveiI/P3OHmvp66WC12HMKd/MTdC312oRdjgAAIABJREFUp7mk9C6GFLpPeG7i9FkzDtOYOHPuI4kfn2VznmpNyStJM3Yx0LriUtBn8gbI0cac8k2sUOrF3irbPp0SuZTb5rwOUoRuvnpl9XtKyhFow6zrLxFBAI4yU4rLN61XKxRWhlsHZdrqvPNpEAIyXDu+72ZY3Fbdwmsp7V0NKSqRwMFvfHbU4vfvI0gxa21Y07SdWEe6cnFJIdWZxI9VMtle456moqoV69ShCniBaZ9YEYa7iUuJixivQ3uudduC0x6HV3+9fsNtDaMbH87NTWsoLy4eXXf+kD9t3bkpC/JXvXxelUaDCsC66z4/WIfIpbx3NaTgwFmGiFTDYzjCR90n3max/almfDB2yF9wztKBgdG1cdcVl2KVTn12SL3/yLbG08ZgLiByDVIdcClkV2RnWZYsg+niXHt9RTZznRbeNYq3/Ss71+Q/eqK8rTxv2Rcrtq3KbMurPmUJ8FYJJDD3PaTIHrqW4u7mtRTYEzxPTEKUq8f5iXBWdn+IJ6hmnY08jRjQJ4+rs7ta0XS5loK5u95r8pcU5az8xJMFT2LXIOUk//HSf924lmKVimyFznUpr7r1nM7uzdYz6k49Y90OrpxOXse98UXj0vyU/Pz84hNPlCzhEFQATIq7trC7HQJ387P3xFoKEolp8MoJf9KxD4nG8veBjh91jejcU5dXXb1fbdNbuxgjXXEp6pWFUYTYp18ubtye7gegrqn58OS3L/2z7Ia1FC8oshEapDUnE0ZLOsgmmP4Z4Urov4eSjjdsXrpq1aqZhSXno2AABkDBLOua+LGLJt/bSffCWooGEYZPzAfWINiOGOXtPoAUNpGilYgFmHmA2Pi4Djwetw+rrrgUAxcOSmuFzr99AQ2gY4RhYrsZEi/97Xe5lDnRWyrUbC6qXrmEccEZJ8Nou6LcXoWe/jAa1RUVIYupZn/D6e0N+9cjDhLshrFLdf/L+/CRuyfWUkHEuxpI3l9DYjRr/nWuMftUViWA9X6SkzJzxXG1wezRdTFou+JSEj1rNNpyQ8wbmQhA77cZrxOiUy71X5YbuBQXKg2pthduKz8Xl+W0gaUx8douKPc6SaVTqRT6XAdUrCCADNgDwWhYKqtU4In373ZUe2/dE2spAYF5JpM3HyIbYdxxJ9ZSVBoNhTuMAp0Jm6OGxCy/Tekt1dmyy6jDQzVGh1TNJmklKrXR7qGmd0o1rto5RHv3dvRDCzcP8KXiWt9U1VZ4Sm2VmDswILz2IIVUZxI/kUux1ngt14T10X6jT2GBd3VOarFomTILJn5c0KnIqKfWw1IcfEVU3Kmi4qqSPRYDtN4xSezAs9I1yr3+xcHHO3FKFNleq1XQaqUqHn3JqdFl/waQuifWUkDRoGGaqWtmjB8nn8zdAbVZ6osVkFJzOrtUMPgMZWySaU/NcY9dXWlwwv14GFLKjERFvcloD5XWH/dYVR7ttW3WLgdjYj2W6kwaRNlTGrereIbp2u9DV1zqKh2erDvSll+3x5Dt0bEsECU1M2WUS8W5nJK4REj2TBatRcX73HNXLC0uXufsavF2tdDIuXc9cG+spYLcAMTdwTEBU4g7oj2BLzj9pOqxJshmjevOlWzevLluw9w4nYzKreiygFVmxCsyVEqZzBfy1CvVGd3lUvAoppDY1BdWZI5YCaWHWwVk6mot1f7OtdqGBfmN54hCFocQalRqwfLgUv9FbIxC6aT+yM2IncEKru2NxeUNzkD7hnB7CZEffdQD98ZaShAQzXTyhOQJ0+F64g5wKU7KcdQ9pUTfXOHxeN7Yvq0qM2/EL/JnLmuYa5HiPnVdySotkkRPnLFs7dqy+ixT9yd+mBLpFa607QvaWs8RhxVMj4Kg06M7XAozOGwaQ3tWrSjNsMBYGIcgff2l/1MHEAxHycFHmQQic5cKkd4aS9JcXeg/dVqPSEI3ewBc6q7fl4L2xKzJ4E3Y5XUE74SHJKlFqoXbL8TJ9dUn7W16uXXpL36xasGqpW0jZtZ9QnSEgYMf7KqqzfUek437+1/Wah02pYVjbrZW7+gtsE6FXuBOFbYVlaR7QlSLqEtBWLe4FFMpPZqZv2ATMXjrE2EyAqV2j+SPL/2f2eMJKZUqiAKxI8Vze0+kjFjxicVRFsFUR2+mT+45Aam7XsePp74n3kdoRRn+uxPaE7Bb0iYmqkxqF7N+U9vM6hH5v3j+fzZdqmtN+UXesk+0aXwYUqwkyWOxkd/8bwwbULJStbl7DlSVNr3DXlNXlHNiryVQWnErpaBucSkm25d+cmZ+5hucAZvIvMWe7vHUf/DS/znTQwVJWkDKmZ1t8IOTpVQd5ULZHbn265PxFCmEkHthLSWTkUVr5Ie3IhhckIaq73dNdKyUtFqLzuUy7oUHouKZOT/+x+e/Op6U/vTm4raXly1RuRCuhlFDBl2fpLQl/ealQYp6CDNYdTchVRaKkm4anVO4iUuvz42n7sm7WtpQSHUm8WsfwQhALX1jdFtm3R4bL6jqPUn1oSzpb7/6P3Mo1OzxIKi1Ux+0pUFtony7LlAZgVR7x/X9jytc6q7X8eMd4xGfdKJAoO3X/5CCeNmsjUu3p51fuaC4qrDp91/+40eff+lJd9Zcggr3irl+HkbgliSO0aqUAQ6QsnogpmbV3fNJXFYm0+44sa28rgbhzUKl5BZB3LvFpQgCjKoOLGjLO8Da/HEeLaSUBvLBV//H2HJrQx4EqGasaaZzT4zIq1tvRGSQyLyv76HUXuK9sJYCb4L3iVQEBn5oGpHdAS4FviHh7HHHDzyxakrjF08ba375+Y8+/8djgVx9zaVV+TOb0kwMJ8Wkz8kojSHVX15aW+GRqtRKVfcg5XRq00uKioue1rq0pRVasJAOwoK2vyDSrbUUZ9c2Z9dsXrq08A02zeORGf/4lw8IuJRy7d//r6Ie+7mMkLj/RFv+kTfiZUrl3RxM7FrD79FfDLZ67/q1FFwMizg6NFa+6+P+3eoN29WyZRKJTje35Acn2kZfqkn0/O35P//oR5//zJ5bWzn3/7eqbXQDI1NYPICUWanMkv7mK3CpenCCrid+4mYXhgmryOXOj06pusTZTXxiKeTxpEvPJt3iUpzdXllL9sNFS10Nb6+3Of/jf/9CuZT7Dy/9vT4XIkAb+WRZ5tKio+pohJKKQKof4Ern7xC0ShTiWop4E+/mXobS9qCBAyZuffshubw//fhh09ViAdOAB36fw7TuZGbbEy9vSAt5fvj5L7/8/Mv/+fNnIZ+RXMDIzHmDqRU8FlZrYZVlBBO/+FIOkNKCG3R6MHoaUx1bXWxt9twpxSNWXIiDfXI3jm5xKYJoBQped2B024JLfpvB5G/5y0u/hXii9jdffcC5DTa/cu6KvLbCA0Y/TCzgj7yrxVs36hTJcnMPUP1KaFnqm1t+9aM/1UiaS7uefNxcwB29EySPixu9mrGLn+1HLkU3XS1SjiVl3ijLks3lKZkrduhb7J6fP//lZ/T/X6Ub/V6yP69tZpPN4LWr2EStWgmJ31dR0EawqNXUh1inByCFmOqIU83GB05XFTeeT3KB9Xbj6BaXwt6uRMEbazbntS3YwAZDIfcHL/0GXOq/Xvo/YmPdpdKaupy2zKY0ZYsSVksRRHWj33uahcYEgxIzL0n/1Y9+WWMK1V+zs+lpUf2fH8HzJhwe99DHQ4cE4c4F9PpJEx1SCSjCIbh7RVTS3mVwklW3Ln6P3/D7f/zq+H8+/6X/Z8//JwmUllouzVzaep4YZGFIhblUNyCFmOrUAzrDc0+3bis/edzDd2XLe61XKaRuKfETIcX4dXuPZBav3JFRUFur+7+X/vLfv/nNb9ZqA8qW+uMlmb8orluPrygPDahrhUd+9VUPmDEysPeiMvnXi2spj8fYL7ZofVNfbEVtfE4sinfIqE/8/oOUWavVmpkADF/xuS9Zx7r8Bv3f/vFZGrgU+/t//M0WwMJk7rb8EctqFLmMyKUAqa+i2G5Ainrdx+xMRWo2V+Uc+UQair5m29RVP3WXS0Gwb/O7zrcW54zY4Wmu9cb85r//+7+/+kN2uqfFOLfpiSlLEWzHxkIySLWVuiIYSetND5grEi2W+gqv0ghIwTVmRobxbparwjM9GfjI1mkAUz/68aMjzRyfaImzq5Zg4THzZI3SR5xu357f7/H85+dfMqEf1jjtnkqf+T9nti04wPr4MKTYv2AtdWtIEepDT+HLsqs2rdrW2sDqcg3dG9jdW0tRIaXEaXQxB1rz86fsSMxtcf4BkPpLbUWBn11XN7ptJhBlgD9yxJ+PQKo3mLnFM4zCrFZ7m5sNRtG4g8nSuam/+bv04KGFNHUXdGbJ9AH9aCjPUtsDqyouPe6xuhFTFpw8Hl+bzeitTrfTwH32+ZeKWo9NaY/Lzk7cA2eUR5YQIQwpJ8QTyltDCurmmPf5AtIdsK74oobjsarqVn93j0shEi4jcbJGk3r76Pz8baeUxujmv/z3f/+2Porjnl5RNGXVsiVShZuuoiKA6la39zgTTKoZRXNtdtkerKWOq6NC2c1d+T/ocfl9+gDWUkM1mnG7YCg/I4qGgeiniR8EEwyJdnlq6qqmZJYcD9U2V7KVcXxzc60NE7/42tpmVie1GnzSHZltC7azpqsTv25xKUSrBaSsrj0lM1MK9xN4C4L8rztH97gU1fJl4POBr09vyiwuLtq0XlL2wUt/T/IYCza9fKJ46cm59bVePuyPnDqA7Q7lSJ4e9AAkExJFhiuN94lcinjSW+5qLkVGIWzHm2sQuWNyPwbDgWUgo05zrS9pbSsvqRFqm71GZaIno7m5jAWkmNzmZqNX4Y/TZ7uaRiw9sZfheyLxowGgnUwL29AKObcWTiKh2dqVAnr72+wel5LEa6Uw9WMk1vj0A61Lixu/OF9j+791yr0b6p4oOlF0soY0VwRcMJLE7Ja5f50nt/faHf/BKt3NIc/xmjTPcaqJ7j9+fE9at97vHa8pJQjfE2/CUP7xNWRQf8aX4qQQg6pdab/+wcrGunVG8Ch/Gmvx1JcZbezR578kLfW8DffSs5uZvZn55U2E1dbTcLTY6tVa1XRfqishOoUUHJcvWZaTueKCSoAJU1wHoQs76F8KqVtK/LhELI6xp+ZkbDalbcPMVfnlrStOXjrdtKwxJ39EUYPf6fPaXC5z2MW/IlvSAaHIrdvpAVZZ0exJ//J/PhMh9Zjns//P3453bxpyO1R7+yyW1O/Jp3JvP0SGyh/qv30ptU7b3GxTNmRWty5bp/S7syWwiVAlJcK7g5JyKaNJpczONvkZhRsLlpTivaxsvcflyoBCks4R5/FkKLuye+LiTLzM6G9qzc87z7h4SIO6aaTePS5lNkPxg34VGbfVKt2/eebS4hy4J6qqyilaUPeG1ahE5EGTOjzfowZfvX0Xkec66QFzaX19fc2v/vzoD0Uu9dgvH/2fdVxfBcPrhGbvbwNSg8bKJ8wYNmpYf3pIUutKa33sqcyUES+/wdrU4kqeUSWalUYvAaSI0ZKh1sfDtwO8oLzxcv6q02woVGBLo5roiR5ZwOM3dqXjx6n5Mhd/6gfVq5palEY6/equqwpwqc58onfQpyiWtyTVnNt8YsECQKr8xIpz681BGHx0kDdyq+96wFyRpGIUP//+9388lwrRv8SPNPaunflB/5zEPIl48nL5YNKPhvIyX4DsXZZTnXdU4bJBfkBlOBRSSp8UkHKJkFKIkJJmbF+QP2KdsyIR8QR/81KBz6BUGm1xHksXr8hpDdjnLjsBh1+s3+jHsq0HkCI/fP5nxN6tF0TBqrVnxB1/etP2kqbtDTv2GM0CZe2Ro397wGmptxIwp0e//PGjj372p0d/+Vn/0rut0mHOGy1Yf7J17Nb3ongZNOP6R+LHVHrtczcXrXxig1nngnDsGqRu4lJa6Y4T+QsusY7/+6fe+5eX5jk/+PsHpKze08WgdxoqPZ6mqmJsSfn9rKlnkOoBlxJluRWhEGz94/akGSnUreBRNCxX5OjHHmCV0hC8KvznP/4kHo/+4+cef/d2SfqxUp0WzcuoLro4JoL9ZSiP0iWVxvRLq1Iaf+13yXifKG92hrlUKd2XCnOpeGjVsoTPQGDO/MxPuP996QPzb76al/2X//1AWVbqqe+0DYQYcrXnF+RXlbQIRhOmYf3Gpej2iF6v0BuCvLNMWebLrvRRQ98IpLp4N32QxBqdoZDWM/fH3//+nx7904/+9KvHkgJ3L6QoW8I/OHKZNkToH98TGHKC1UA2zZzSeLKGGulRbW1ObQ5DquK6tRSFFGu2p+04MmXmAZgk/R0Tv7W/fekvSrc7sb4LLkXKVBdWrMrftlfiBSYhRsDRrTfZLYnftZIgpKA7VNgm4bQqFs6YsiPivWu903+/lDZzc2likuezR7//ffr/l55Sb/9R66uSaTzExf0T/pqHfNtFThXmZK5cwiF8HQAFK0N4wxPXUuBSV9dSIpdizSpdWknVtmU1xr989du/vLQaJhRE6ewyHpnaZTo9um1Vg9KtF+HUI0j1YC1lFgXlmO8ZVRapVKuNj4/vSrbfV+/m374ccCl9fX1l1nHKpr7//V/NTdJnd++b+a/sOhn5FN5m+2UthXlRtHPJkaKcwv1SnZV6WIHbIil8toqQun4tpaYBZxiZTfl0FVT1lL/96i+/+eq/vvov4hFnhJ13j8Dsz5uSeXKPvwy+yClicXSe+7oUrdlpZbovnqACcgTwMGIVRX2Qw38a4n1Gjn7vAU7FO6VJpT7LD38JHvXoZxkuva97L7jfq9YBAYgnxKWADEoUvd6XurKc6OQkRAvM+rqibY0N6lCWg6f+wnmVrj4RkBJstlLy2ed/M/lVEKJ7URcla/UJtj11VZlf1JC/fwXt1N/Ma+E9FqNNrGbHJLi5m1OKC99ICqUpJRIrDkDqSk4OZzT7uivMDKFbFL4raKHnTiFFhR+4R/cPr6ZdyXLTCd5o6gUeonoWSdSGpKsH1Ncep7rTuAKhq3XDVXcI3oLEjeRvbC8liMdpeyn2b8zaN1dXWnhdQ0HIjLKv9DNl47dLSeWxGo0eT2VL3M8f/dOjP/eE0pzZN5ZJCV4jdNsEbyjtRko3XqHZIHvzwcMBhUPkUr0w7sAUjm5uMgqwHRStSrLQEw9lA9xMhFsWIpE4hAOty1p/DccNpTTNRNOIIl7cHS01Y+LH8eqMeloMa1JbGadSeXR09RPDyQfUhuK3SoPWIwYj46X0OUTEuEIQ0zAcAJvk1605VQ2cV8+aEkVPzwqtVoIk9fWVIRKLmd50x4s15OKSdLiyGtQUUro4sxtXEmtcPE4knEZMKhO9UkkVIMxYLaJlMaz4qYtmozqcZlHQB63aK63PEN+sykSXdGaLhCp3WrVxdFxxSUm0FWqTSdz4t4gE3fFx4jtpJyimabXUYwYIir6gVSrxvZlgE02fD1+hg3GB0lT0eZzoJb7mYfIq2lE3phGVipbGozSc2PBzOInF4CZ9gg+nXSOIe50QVMEz+/UEwznxPIP3K6apVWqxMmESYmXgqfQKQZOYhg4WyaMyTjT4WmXog+1X1A83a/Rn0F2Uub/8/i8fK61Id4kTPzj77pDgdZW5kWB7ey3iq9CJDvOdN/QFFxcn9p7OYqG1UJvoX9omepKGPezzcaK+GxdHX7rEqrte+00g46dOHDAImenEb02vJn5xDsXXrzx1tjLEffTKKy+UpSe9+NQrr2S5yPdwyk2XfoNT7f7C1p/ufmptWtKAp15ZuNpChix85ZUzFeTMK688dZD57M+/2PrKEGMoFVfPcKoCnP7YsuyJn3648Jt/fvXfL/39leESv+GFV576iLiYFxY+9RHj0J8FQUWI+3rhU2cNLcqPfrf7By/XpVO69R7mHUowxFHygzwZz+CUaiEDQHB1EnMQZZ8xJ60++/VfD2V4Us9+/cIZs+f3n/8/0w9yobVnz/71dSYj9+uzf/1j0vHQR2dfeEbtMk46+8I3Kof+xb/+9R2Fx/zi2b++KD2e9M7Zs6gMufzC2Y+sIfM3fz37UXOW9Jm/nv26xeMZgFOUjgx54ezZggzJmb+ePTuv1LL6ha//murxPHf27Aur1aqYF0A3LmseCB5kMgahMgcz0kNfgyBxCZQgF6r86Oxfv/F6vN+8884il8v2zDsvvm50+Re98843QRdz8J0XnzHYTa+/88730tKYjbjZ4jJtfOebFwelkzPvvPNOlEq3+sVvXoxJU+/EVYFRl4qr1WncTpS2U22fh5wDXaZB37zz4mqTK+p737x4UKfLxdVGib0FWQa4XAIIHjS6XEPfeecZl8v9Ogi6rxLkD4KgzWUaQimlUYLfRKns814/OAS1wOngIJNu0MGDB9e61GsPHnx9EEjgqsCkizr4+usxRntwyMHX5+l0LUjbaXQJSEs12W1nXn99td/lH3jwIE5lZ14/eAYkVr9+cKDRpV79+utnWvym1IMHN1o++8ffNr5+MD0UvxNXLS7VztdfPxgVJhgFgiJ5OyU/SGcaRAkq7aB0cJ49TFDtsuFqZ1ycsHP16rU6u2vn6jOojBNX89DBuNppz1DNW716J59hpyeji925eufqMjRt9erVWo80BldBu2UtrgweC26u1adnXOfFAUIJjUY+Z/Gnk6aJkOoFlyIBmfmp2fNf08sYnN7KDXGvzB6ze63V+sLs+Rd3ZpCzs8fsm7Zs5Q9+um/LB4R7cf6Y+QcJ873ZY+Z/w6j/OH/M7Hecp54f/eryF4l345Yxsz8iZOerY2afjW/4wU8vLn9B9ZuX/vfi7IVEVrl7Pk4G8uH82U8RmfZ3s+f/ThIg34Kuz6ZcOHvf2Jd3gO783Wtl7hdmj7k4z8qA7sXVGeaPZo959QxhXlw+ZszrhLwDus8Q8szsMbO/kXoOjhkz+0VS+vt/tB77iOhX7wNdopuH0wtmz9rdY2a/QmwGkW505WugKwmS1+bP/53ZE//U7NmvlQWdC2fPfssXJCLdANoLuh4t6G5ZrWQ/Wk7pml+c3U73dWnGM2PGzP8eUR9EC18kijPh9q7G6WuLZyelS0xrQfAV0tL81pjZTzEyw2vzl39LbNzv5s9/jRHIt/Nnf6gQCO3nWiFMt4y2d988lqC7t6wmHNo75gxxvjh7/qsH0V508B+J+plXxyz/hpDX549BP6vP4OojwlK6Zwkbs2/+8heINUzX6ntr/vKFrAC6s78lRrR39muMTIJ+/jCb0p39oY/SxfvleRDcN88cpstQuq+eMRK0F++X4P3OfoZVPwPy3yPk4PwtoEvC7VWC7vKzLL923xjQta3d/eryV0jQ8BZ9sS2SD2l7XSJd0sLQ9hpa0M/zdxtaxPdbMPfLEnRwTAV3Fi1cbSMiXVaN9zv/IBumS5hnQJDSFd+vMdxe506Rrn/exTHHXiCGqNcubnmB8ObX9m2h5J/at28hOvipfVueUkSThfsufpstSKZu2fctCXKv7Nv3msLGv7Bl94c+g/rslotvDTJLv3519+55HDN8y75vSNb10mhwqSfXwFoKR3KsfE2vrHptBvbbMWM+dMvQ5WPeqo3mXpk/Znetj7ywHK/aQM7OH7Pvi6JtP311zJgP9OjyfWNeZ8g36PLX9ewiDLFvmHWXfjp//n8Y9We2XJz9NZHsHIMuZy58Mebi7GfN//H3t/53/iuJody3xsx/ivFJPhwz+3eMgX1tzJjXwFK/nT//rewW57evbnl1vJK+6otrbcwLs7fsm6ekUN6yWs9+NB9DTCG+6g8k5Jv5++b/kSHPjLk45j8Id3DMvtkvSvRLGn86+0Ul2blv3+yvGeXO3Rdn/5WU1u4eM/8V4qv9ECe3wf0ayDtbJL8bM+Z37pAZ7X1N72OfGjPmLZ+NvIJTrc35wnxAysDR9u7Uk4/mb8EnhLw4f98YOrRB8I8S8sz8i2gv8/qWLbNfZDDE9tFPyOqL+2afZePngeBfrb61u7cAUmX6t7bMf0XZokBDnyK5Tpx+R3Ipwd9V1jJPjdn3VnYt6G55C1peL8y/eHGtj3w9f8vFedlo78UtZ7LJi2NAF/08/yLt7mdw9Q2jODjm4vx3JBLQnf8RUazed3H+12b9vN375v+VaV6LE9pbSek6c/WULpNL0NDfMZTgltdEgmM+9PnYV0BepLtl97xsvN+L++Y1k4/G7Nt3xg26F7ccVJBvtmzZ8rqbeWbM7jHfKMjBffvGvMMyeL9jPmKYnbv3jfmaoXTHvOD21r61ZcwLvM/31sUxIK//3ZYxTyl8im/HbPmdIpeI7fWRhWMuvpXtI69sGfPWvOb66WMu7p5XSs6O2bJ7tQJ0L+6j73fLvn10XM3fPeZ1N3lmC9pLmIMg+CJhzoD8Rwxt75iznGLeW/vG/FWrr6XtZXyVH4rNzv5wCzrYx4D87/Qi3df0uQxe7GtO9Pr8LbuzfU5xWOeSV9DetbTZ+/btdALR+94hN9m7Chsvj39yXCxUkub0Sm2WbyGvfPjWUwab+a9vvfWUL1p79q23frfWYP5aPLEf7X7tpy/nFL/82lu/O6Mn77z12lv4eD+D00Gn8iBOryuN65587eIfWf7Mh6/tfgeQeu213S+yxsk/vfjqr5l58556662z2oIQTvi26HHCN5x95a0PX5HYlC+AoJ7Z8NOfvvrhHj9D6fpa2K/feu21tWXMR2+99do8meTFt177cDUTpqtU/ZHSJezruy/ue4YhB3e/teU/mOyBP3hozH+wtp1vvbXla3w237q472vSUvvaW/teYH3Nv7u47xWnw/3t7n0L2Rby1MWLT0mi3K9cvPg7H97xxX2/80WxL+zb/draFufZixc/XBvFfX3xrbd2GswvXry4e3UL+719u3cDWc9cvHjx9TLu9Ytv7aNDG+S/IcbVIP8iAd2LFz8ipWtB/izrq33t4r6/khbb73bj69kioUPb6ZNgaH8r8TnzT7gQAAAgAElEQVSfmg8o15pFDIP8/DFvrfUpXhgzZvc8N6C85eJqveSjMVu2nNEzL1JEM+R7wBLlymO2YIhlHwSWXmTAHbfMx6Rg58V9888StwhlUkuHNoZYNv2EsD4RUsRn/h2+JJJaPYb2a5W5koUgj28lyO9ea5DgE/LWPB+lu281htjy+bPPSAiY8+yDhHsHV98wkm9mz1+ON/o6TiB4BkySzgbmz17+V1K5cwvljr61+2Yvf4Vpad49f/lToLt79vLfkVr9a5gGAFKvzV7+Vna0BNwRY5osXD774rxa9oXl87fM4xmcxqx2k6+Xz59/hgHd+bNfl0jemT1/9jMM881ySpd5fTblyoqN8+cv/5rJXr1l/rGzRL9zCyXoq91HZwOG5t2zMf3I1b81G5/qXAWlm51LMAl6KzvXTek6c81PLQf5XGbh8jFban1qSnetHuTHzN/pJuOPzX+HYChcf9D4Ajj4guGTRw3uFaRULONbW2twErWvtrZWWUZPlZyKza6t9el97hblGy+vrNq20bZ2bbabOJEFAfv0OCmsrLK2dl6z177HN6/ZZ/N78WCZSi1prq3N9inXffHyyyXptbioba731OPkM6sIClU4nSC41uDkMADnNcOko/XlD+H6wYk0PTawUBmfkqgVKKXMJtaJUREJnnfyxFC7tlbpJNFndp5pcbKy1WfORCn1LTgNUnKG1asHriVqnM7MY3kbbs5zKvX0ivDunWfO7HSydMq9E3J+OuWWsAyuVkvUzLzVZzADITG4KlOSQWfOnPE5mXlnVp/Rqwmm3ANbCDto9c4zBiUbjUJrWVKLtLWEtKDstSyRiSelDSTmcU490ubBLDn1zJkYhCc9+MwzZzAhOfPMMwexu0dPTJl4gvxiNa58Sgan1w1mshMn0KWnWobMw2ktR9Y+8/ozoLT2mWeemUcYejWPIbmvP/PMTjcboieO9eK0mkh8r7/+zGrsQFCC8Epz8HXxFCYooScQpCe9kjz3zOvgBJQ8CILu65Tg6hffeWetmuzEqYARTyC483vvvLgT5N/Bqo3hB+F0CORxcyDjpqeDiIiENdxBc5kVpwEML6GLRs5AT8/AQcLBF995Jl5gXn/xnW+sakLXcM3Nko0gEVKSMzjlOkW6g7BaoORB8MXvvbgTrccybydxUoKrr5An7loQ3MgwPpzOcHz0i389+0deafjo7NlvWLfvHaxgFTzz4tmz7zjL3N+c/es7GEPfnD37Ilum+OaFsy/WlhGslT/C+Poj1tGDlOQglsy1CsXBs5PPuL3omxsOKhXEGgoHbKfwhwzQRIkqFeK9W//BDpNT4XYzFguNua42+SHEUds90LRzcvGOehOsDlc2HkXIGAiDqF4s0Xl0agSYikuyOJ367ArgBUo+Rr8nEXlMHjvLOp1ss814oDVn6X5FyBOvYLkMKVxcqux2WgYNQ+iEcy+tBTbB2a7jJ8u3tZ53muwq6j1RjXg6brfbTCvDOmllsEkcJgg/glKzW+I0S2kayyj91MuO2p7EsvpsViU1wxMztstQexZyHsizGD4uzgxxPB6k8TpYjkZBRIBDkxF+NJi4DLh7orWRwr00y1o4hBdgYUxPBWGWDBUknTSN0Ki9Uinai7rRniEqSK6wawYTLwbeA9SoDG6iULghhEApLpxmgYgQ4eGoQYt4F9UgqIxYqEq0NFGrRGsXyKfoTRCk4k+ViiNm/EfPiN8bl4HgQpw0wyK+cSqWgpkKBHF0O9ySQaOhcBlxNA2RFQmHfWw1nhMJoopmtF5LbWTg6xdJSKOFEhCkm35xGehu9J4Fj8HxBydWCmmwW+MgD8NfdCWtDPKgGTCNsWTgZDZnoEspCTMtCyJKMU2FVqBwtJCa0aikqBA9Xy2UNpi2Bf6AEs1Gp1kCA4Z65KCVQV4OXYr9QVBEibRStN84VNGC59B6pKC7ofiCxqGitG46u4DKEJ0JgkYMSIj4cFsN+S7jJEad+C7UOrpPwpowVhFYHFnpC1K6dHDJQJR2PxpG4ku9DByoKBAnkF5+5wCqoqOh7dcbSEGPoD6R2r2aLPUWld9vsvAmuyeOd0JoUurxHBidM/pSGsKF6iwqlSWO5+0eO3ZK4jKSkuqt6Kb6+opSJav0Z5QKSqXOA2iwcXHZNtvewvylJ42mDK2Vx+YqzzpN8OnvZNQZCJyGh0BQW5Er829ald/YBGGNyqLm1Up0p+VKZVTaOFqZOGc7QTGtngYBVSVmWYxGU5zVClssi0LvjoM2RCArgL9qlTbLYoL9lgXPeVQKK0aIVFovUIJqk660XkUfVNHn3F6MAlQjkIW/EBl7E2maTqXzeOKcXokJdy1ZWSCo4lX1IkHE2YXtl1mhQLzTdoKmxCwpCOoowQwVTRMJBkAwTmU10CrazWZaGVyo4nQqqDFiYy8uzupFfEiPnfaoh17gVjgNQ5T3etUZngwdg8pk8F4FbqmkeMsm/IB6GI9CdWYBaZxXIcTF6ei+Ek3Tua8QFFt/hSD96nZJMA6Feg0g6BH4ACrDew0o1KKChxupBWmGCh5pAZqGytC0OBXdEaUErT4zrYwVlfGoDF6BtuJaGm1hhkArY0EL6+1+pS3dU2rk6zNAUHUDwSS0MFHgMcZA0EcJojIZVwmqVAKfqFLFZdi9Bh7DLGgQUCicAJWhMiaZoA2Y8APPYZjZZV7U1MSXCaakOFNAJohpgo8OQU8WTdMZDQZdRn19lkyIT6TRbDs6oOYHDdpeQApQxtcH4FUqtaVep9KohiMIbL4o9NgIijbvz8tfuqLG4g1kYJMIrEVNgS/RS6Rx2sR4Mw0kVQGW5JSo4psVYHI6FaNXaC0GpT9p++jinP3ECOszpxJGvqzSBKMZDDgwhTBBdSCg3FG0dMSKubo0K32KsgsxDTHflOZSL75mSil/hSBlXPQ7SRmGtrQ0HqQt8XoE4lA7ldoMsVAOrWDhNC4Rj0u1+DLGmWH3TosNE2RJaWkpDFIQIcuSkSFFCxE49wpBlk0U0zitWZqRoaItvC4NBGlbLVouA4xBrzBTTnmlMubSClRGiRdzE0GVFBHoVBngM5QgRwnGUZe9iGWiirPQcugjYDkZyCimiYkYVag4TeMYVIYS5PDc1QdpOfQRRiSo1zMY/FfTvkuQsXSTIKO/RtCCWoMgJ/J4ThVHCcZdI/j/b+/bfpu60r6X48PeC5NkO3Gck2McJ4SYxNTYBIdiAyEgchwnnBITqxCUqgIpNCkzLwIVDWqiktYJlVJGgaIoisogDYqUUG4G0GguQFxMASGifPq4oeWif8B3N5ff71nbCQzQFofY2PPuXRrvwzo861nPbz2HtfbaYtCg1ZIleEaEqy0kconda7HdIz3LomfmErR+qYU56+VL1evXVxXZ7y22XrTw1QoxaMUrjLMbo1S8wg/OmM3GgjU55bp8wE227azGKI5n5SBmDeyloxiDSquOf/Dh+oLywkL0pMkg2aE8135Yu6UaquLDDYfX4plk23LmQ9pFSIJQvTwv9RK2MILp6S2FhA0/WHuwXQhR+HTZzmr80HZC+ELMTlu+VP3HX7Yd/PRvbPfOqtLSDTDwNtAzZt5pK8mil5og80bbpaPYm9ew8xIsVUgLrUXNwt7iVT/fOfjJP9YXHb53D/uOn7lHWNDhGYm9OLLLC9iXn2Od07emTVSiDtYhwLdEjG1LIUoXxBhATEkurjChKQg9ftyGuqpsO0/c+wB3cu4ZaP9S+1EJdRw/fkKHnypdVimQYUOFWAkg2SU01FArnpVU4eMiwP5OMz6BWgamkcmLD6WeMKPdVTqYnSX4iltJriCGKiw3xys0Q2JM+GginokNUykjKtwJm1VXZRBCRBWqGaUyWSJOURYiRlSIARECB/vGZCLikMWwVCENlkvPwCn1GQQVhajPyMo0lRDhazHxLSoE42AZLRYKwlFh6WKFgMaLZxB4G2WhT6tAotHCVypcJCZeIToYBwy8lyuk1qPCF89eqfDFMyzrxLPFCkXrS9Yev3Tiw3vHs49eAtwgJ/HWv7lCbM5QkiXYrYNFGCemdAMagQlgnc0O4xfE2GEBY3srvB1hyD+DbQ5y136Ybzux4YMPTNU289ocg11XbdaZDTmAmzl37fq1tcfzAbRqiHK+EeIGofr1bTpJvS8DUjBG1QOODroaIo2NJGkctW/YsKHy4kf4fqBJrgZjaZgwkFegJqReRGpciJen4BobjIvPMOTpzuRfHLn5p7+Z7l06DqHccJyeiRqwKB2IKyqvLTy66R+f3Bz5gpXl5lLZeLxEDAxZPVbixLDfJyRe5GPiNRbs/4nZeIiTKn4YDUED6S40XWLl+OgCCKGiDHjnRaalTQwr9MnLlI/iJomIKA2iRs+MhCj1UAulm/FnS8Qgx1KF6jNkXGSbJMPaUQt9+RmGOKx2wG6bdkncXip0sTqVDHH7zc/idCwS8yLfC/oXK3zTM3IvBFNfPBSXdPvtKkRykUMtR9T1osKlh5Rk8Vm8wlefvagQw4g5qxQeqk5I0yutfznfi0LpLtoQf0iqkxaZ4hoeG92Nt5PIILVKjhwERH2GQYNu4R/cZzjr9AyVZpG80YAienGxJ18w6j/PEtZS/5ldXIEC+jD82jVZV3Zs/POpv3+4ARGxRA6CVI7p0Y2BP59af2b3bjttOkVsEQf8KCxdLbcjFnTxQtexQ5vL3lC6pGdOf39Ylixq6AU5sewKlm0MJny8oFd+JFaR57MvJccH7OLneuwe6oiExYjzSh7ANK/f9+rNhK4J42/IgDdDa/IiTvGBojc81m5lDgdWAlLAbgkc8E1GvMjbeOvKB9ulNwj9b7IE6qBk7QcXj3XteWi6txv2lEG3lD4Xloe5UBeLsYczjfs+/7j06ptEUt/TN8ejnrB45ZbcXSbpLcza7xTfJSYxXhJl8RSQYy1KK0Q4/kSyxpi/gmKf2OtwOsCnnjKh4YgONbs407t4Ox6o1C2+zblU/Mt1/GcSFKLWi+WVzn4Ra42/CorSiVwna+ct+CQ5Xgx9U/vU0rS/GcCBlYAUHAtz1fbiTQ0X8WX3L9jh3VUifvz2rYctprNtWPvx59sasYuY0/ZCRaEMlL5z5/+YK9mVgVu3blwpt6zZ/lrJMpsWa0J4HyAkRJKAwuqizyH+QkLpr/qAdJE4k3smKqgk9YkkWUKBcbqIsdm5oKetjl4nU1OqicRfn3eOcEdlqnMRdEYXohT6E79Qz4TiE3/UyiUUWucKIbnIJxKr2fFSaKhDhoO7lJ/ua0fmcWBFIJVtMNeeqSzHJhIHTn1ffemSlCikKNRxfE3+/5s5u+9+cWWtTSdeg1C5mV2k23nphB1zvFsPzDzMbti7/bV5AYmFXdwTiXROYdsnCHC4DsaZXNO/Wgn5w2TQVcz72Ww/Cb6Mae66WaSBcZfnt7LI02vMiuQym32meG/PwrRjvhAP5NXA/PPhASme8fkKVlGHrLRlm1dmeX6CkMws/X7clJhjPk/SR5CW9dMTukNF0okc6a+hegGe8aeUwtK/mnv8TsYseXX4YjJK0dMJVGMEJeiRoQ66SgAu86RJoxgcWDFIbS9+dGfy2L4rWNVjE3OICbAXX4vS7TxxZs3fD+1rPH3FuNd8XKZpUvUoKio8sdNa8PEve7buub6pLLv89ak2uEVB2GP4dpbSyfTh827uDoVZPVemeJ+FIPWcr1qlBJsrmFV2nPcqU8PYEJ718JYYa+YdnW7u7WDOPh5UeAecr1gTnbWaIqu93P3EgU0F2nhP+1wUvlkM2nCww6VMTTCL3hkKKEofVdvBB9uj3FvXH+V4gjpWEQWWmJV1tgS5q5vBap0fDnIvvuYqqGqJSRNezl2dzBI76eI84LHIMRft/9vaonDv43dz1xY5p/2+Fw6sBKTI9dmwl/3c8/XFH4qPUhyaJq0TObAuyWYr3MS+vdGIDSty5ON28XaSKKLIWF1dYPrjoR2Xj32zqawMsfDDr81eS2wVj05XSLEwQn/DfLDXxSfY01WKO1QPCY9JbXxuVSjKF5hsaVZauge5a1Zi3byXOVp4dNgzzKM1jmkXH+31YQLc0R3lq3ot4wE+2j1ML2rqm3jIDXm3IgjiUVx9nkEezGOWVYqr9zx3Q/md5IFhTwt39S20cSXCnM28bcKlTDDWyadCngDvYdLs1JTHE+Wj7OkToK2eLfA+ZA72YyPtaO9jNz/JrnmnrpESnMAWixMY6rQjQzmwEpBC002mysoGQMvuLJUQ4n4RXHgrtoitKapprv/+scsXLhbtrZJfoAZb51UWf//Nvpt7Tn1cqcf7ioba1wqV2Gwb53MhqCF2UullrF1xOSDpw8wZg/fviPJpxuqVqAOqqc/B2ChvzZVGcfM256st0DLuChb28tssLCP0EHbjEl+LPA8TMqD0szyokfprKAgGXBMfDqs6sYN7xxm2w0ZtgOQs6+XBduzhi/TPBAXcFXZ4ofagl5otbII3MRaZrgtDzzUz5uvwVDB9AGS0cI/E6nsi7KnidoCwNsbmT+ZpiHqtjzPmxkpACtv05eZU5pxxbsGmd2vWFBcbbEsh8LdihGlDFUL+ZZK+5GN8g23ft0x/T7j0IvOWqoKir77b1/iHy1+W3cNyDUyWvl46cNM+HFQC/aQiOmYrOrgyDk3Qgw/+YJIpjweuMYg7H4eh18kc7An2snG4lXZDPZ8CLjr5cAyPow58cgvJIwi9SdcCHIUxF582AHdw0YA1iV0L8nkm5XG3nz1QJkw1gEof00cJVyECTT1vqQHC6sdnWzmf7eDDeifBbBzhPD7cCkcrJgmqmPNapK5HcY8zDw8OtpMT1c1HJYwCynB9haaj3kps0jTRSkAK81JYP4s9Gmpttdv35mC/cF1ircWqHkBKksq3l/5t38HGs1/mlMWwYwOmqnMl+4l7VX/EWtnGgb/i7YdasQPMK6VTFMCpZzH/eail2SCn/7jLEWtR492EFqgrMqrCPi9/ymrgHD1gFUrAB6XVxixQHDDSepAIEQV4Xk+VYeg57vY5rXIAgfZWIAPhAwKnn7tqJOi2PtkZxRdQrEDEKJv1wtpDmc8YeVWIF+I9T/zzxkKidNR7TZI6o9jj90mNZAFVUEu4ApUuvRT2uBU+dZKxXmWa6Z3dAdzX7L5XejijLlcCUivYYH31FxcOfvr5I+NVe6HcUEQo+x/p43/86eCnN36k5UVvOgCCPJcXYTJTP+cVpHwcFeGaMJRNYJwmg/TsGQS9hiILrGKK1+EUeiQXTgyDC+aBgiATEJDoxSmFAnvJ+oIxGLbIDje0WAgmoJWsvhjZjcBRLyk5MurCMA+fEfrCzEETVmxBCTH/HG93zDos9PAJkrTyQYTGDeH20aDSwyKK18/GvTzUH15A5SjW19nC58ZBUyuuYBIOK6DxhZZ+U5O1e2nMgTSDVFn2pm9GGvGxbKOsr9bVmgv11VlXPr9wc9vpH3P2/gobJSvzuxVPf3/dE95Uox+Facf6nzkAitUmml6NMYQjHCZflCJqq2EMwjxz+RAFPM8QZetAtN0LNIRbIMhwYaDyRJprXqWOwa4bZKxFhCaATj1syRD0XTPFD56QapsPzkVgsiFRRJmDXedSUN4o1dE/7YBB2WRl4T4ouroJD7OjoJMA2GqyNF165uhD5fXPQW0Fn4rUKMF+qT0EdVXjBjkapH6lt9P/dppBKvtqge+XPQc/+fxvWLqK8HeJ8asvDhwYGPn8RyzF/RVukq3mUWd6lXpm6uBTC+enXH5ooMD5HlrhY2rhgWioBcrCComeWngSpKhBiLyqgOJnptkgTEDEIAafY6oXkGqGv0QmXSC0mnsjzBLleViLQUYhVFk9SuyD9Yhoh7LquZusvV4KR8zzJieiGUGYgB18buF8EBT4XLwZYb42TDrPKatDgzzqgxvnetLpn+JtT6IKipnmU6tCfbBDIwhngDplMDQMp0oLT/xKZ2fA7TSDVPnRLR9+fOpY16f7vvvx7wVrN3/29S+f/OHmjkOffXDv0q9BCrJuZc+G57hrtINhjV9HC8QU4Yj5PgW2HBb5kU02yN0LDgQHWGsfn2uuhy6IAh39ypwDjg13WWXnhJsHHKSkfCIwITk9Lu4d9EP/TCk0A0zqDiXdNrBxN756p2f1TcHgcCeerCZcdZP2qUN5eNIaVaaezAKWkUE3VknV4FZd2xyfawPe+gVVPQHcV0gbdTZhvmrCATRFoZrqsTWI+zlNCWtHpnIgzSC184RuzQePvhk5ePCTxr98882ps580btx24/rHH36I92x+hceqtcbGI4iO0zlz0kIGYMMZGderKx7mfKxiVtxDJGN8lhLF/HkWU02eH+kcefC5EM3LC4sarJFIjCCEhbgCYpZIhK4IbVZkYpKTEiCFfraCooDSOGX09aPYmogfCXOZNU4BbvtjVC/uOsQ9hmfjsOrCkZg+QhSz3NlIGKX48tRAnw83Kbl2ZCoH0gxSJy5hV8T1f//iwJ//sPHTP2/duvXmn2ZO/Q0vV+bjfaff4rEQQlUU6a/4X02PbaH4KNyguKBSwjeJ7Gv3xI3X7r5EAz2DLfiGY6kOOlHdoqXS1BJFCnoYvxQJ6fLFPTrTjgzkQJpBymzDi3yHN5Re+T+3duw5cOHCvgunHn51ZvOZKgqe/yZ7SRUsCmZ8iZx6R0KMwUP2Xzz7YjpavKdmoTMc8Z9FMadfNc9SuYtpFm/EE4iM6p/FHIu1LVZGxS8+Eyf0R71DQckXtceviB7tyEwOpBmksKcItoYojDHnlw+/+wZfHvzx78W5+qrjeIP6V974/122S2y6BS9x/G46LYHGgRXhQJpBqmBNFb19X7j7EvbUKSsrx95mMXxuymw2YxuNd2jwotZ4hyK0rBoH3ooDaQYp7HlCbyljNyhYReXl2ABnaQEuXnZe5rFk0S0zv5ZN40ACHEgzSCVAuZZU40BackCDVFp2i0ZU5nJAg1Tm9p1GeVpyQINUWnaLRlTmckCDVOb2nUZ5WnJAg1RadotGVOZyQINU5vadRnlackCDVFp2i0ZU5nJAg1Tm9p1GeVpyQINUWnaLRlTmckCDVOb2nUZ5WnJAg1RadotGVOZyQINU5vadRnlackCDVFp2i0ZU5nJAg1Tm9p1GeVpyQINUWnaLRlTmckCDVOb2nUZ5WnJAg1RadotGVOZyQINU5vadRnlackCDVFp2i0ZU5nJAg1Tm9p1GeVpyQINUWnaLRlTmckCDVOb2nUZ5WnJAg1RadotGVOZyQINU5vadRnlackCDVFp2i0ZU5nJAg1Tm9p1GeVpyQINUWnaLRlTmckCDVOb2nUZ5WnJAg1RadotGVOZyQINU5vadRnlackCDVFp2i0ZU5nJAg1Tm9p1GeVpyQINUWnaLRlTmckCDVOb2nUZ5WnJAg1RadotGVOZyQINU5vadRnlackCDVFp2i0ZU5nJAg1Tm9p1GeVpyQINUWnaLRlTmckCDVOb2nUZ5WnJAg1RadotGVOZyQINU5vadRnlackCDVFp2i0ZU5nJAg1Tm9p1GeVpyQINUWnaLRlTmckCDVOb2nUZ5WnJAg1RadotGVOZyQINU5vadRnlackCDVFp2i0ZU5nJAg1Tm9p1GeVpyQINUWnaLRlTmckCDVOb2nUZ5WnJAg1RadotGVOZyQINU5vadRnlackCDVFp2i0ZU5nJAg1Tm9p1GeVpyQINUWnaLRlTmckCDVOb2nUZ5WnJAg1RadotGVOZyQINU5vadRnlackCDVFp2i0ZU5nJAg1Tm9p1GeVpyQINUWnaLRlTmckCDVOb2nUZ5WnJAg1RadotGVOZyQINU5vadRnlackCDVFp2i0ZU5nJAg1Tm9p1GeVpyQINUWnaLRlTmckCDVOb2nUZ5WnJAg1RadotGVOZyQINU5vadRnlacoAgFZb02qFxQOPA23NA/g0wS6w/6GTSb6TQHmkc0DjwCgd+CzDQUrx5dHS1dmgceBMHRt9083/7vdHh3lcg9vKlxByeJ6uerFrZA+U9oWNFCkYpK1bW8pq5os1JkASqm/5b6S76PTKoPrD9/O+l+9/4/Elb98sYeu38t3TYa4nf/obRmMuM+G8FDhRDZb3Xw2ikxrwHItRq30PFJvDbaMw20q92JMQBvSUWi8lv75m9TcqyWINI1nBVfteS5auVslWvj1VeLXubmpOSRr7aEEPBsYarK82o3yW3MlYp49ALNhIRqTqowjJ0n6xvUCtPVcUpqkdeYmbi0h9zxhKC2IokLq+uqtq+fbv96LvrwKOF1sOHD2/fUmhfEcqWVcjRLfe2Hz58b0tZ+bKyv0smg85sO2Gx68uMMendmZkAJbrjcm5RsW2nrTr1jU6AzOUllSRJH5MoLCdJGDyWV0hKcxnM9vwPLWsKam3vjgPzFn3B+s1rZJs5pU34j8oKt+xdu3l9gd2m+4/bKbgwmG02s/New/eVxXbIQQpqXKyi5N6aTd/nHL+0s3rxzn/Rr2yx6J1CT8UkOeZMJV+XyUWD2cqu/PJoTeWWd5fBWn1B5Xc/rSnWvzs6l9kaxrboi/946GFJwW7DsotYZkaDTqcr2Lvmp/ubDLKkT2XX56wpuP/TmjO7bdnvwZFbJrfeOtvLg1MqmfrWBL6WUNIbrpye/OVRcezdcSDlVt6/M3Kx+GUuvFZhkm9I7I+HJu/8k0kphxQallOQdX1m5L7eJLPcJLfzpeKl/E3XJ29cX3NY/i+ElMTqH3s8nhD9m42cf5a+qDLl5BoN1QadXWZX7sycvfCXRw05dskoLSNSZsrKMjHjUemoJFVeH2k8MPlFQYm9UJKNxhx6kpJD0ABryy6xrw6hOTPAFNHAcpNOAgKMJlSfZTLbiitzLk7eatzxU8Ne2WbT5RYlV2mYsnKNRoMk51Re3wG2X89qKDtqNrBsEJQqvie/cyXWxhUujuBsJ29jklPP9IjFSJJTlugHrnStqBoAACAASURBVFaMftDb7/XIKc416qq3wOj/8c6tPwwd3HHqM0kPGcQgYEhwfDeVQp7Ky66WlW26PvLpUFfj5PUGe2Hh0fLcnNKcFEEqCzQYJdBQ/sdDI38Y6to38k9WuKVMMiadBiAKElxaWpr1P5f2Vl4caezqahy53+DcfWl3dkNyIVVampNttDsLYvcvfHS5a2DmesNV205zeVF2NrrkvUrXylUusdhCS1OL4m7qaxmVLONhWEASBfFk+iPFrLKVkATBxX8rV+1ySsqFkjKbpYYc0lFj64YGYPsZK2U7/IEEEcVYrqmkxBBraKj8aWRf15FzXWdHrmc3NOw1lJhyU2b+gAaDPdZghNV3a+juua4bdx6WNxTLRENy5VpAqqSkxGSy7836YnJr19g6wpQs22sNlckNwJXko217K9dcn7zVNTaGYeSLMlk2lJcTxv9bICXifKyDt5CQWycWbhvyBntj/aFOmeU97r3GYjKraD3fc40Rpt6vUYiJQbPNXllw5fS+s0NHzo0NDez45VFl5fbaWrMhURk0InpcVVm56frMga5169btv3xr8otNlXurbLBCUnTAADLXbqhscByauYXmHBm6fOHOt8VEQ+IjRIIkY2zSmc06g/Fq8UUMKWNHxsYO3hq53iDJttrktp9Gv5y166/PDABR+9d1nZ68WNxQWQ5iWE7KxrIEubWM5DEne8DPs1iM+aaCeayTrw7BEuzuhT3YYpFYnRcn0QgpqmUUvoJZjNkGs66y9G+n9x2EEKzbPza0cR/ifvcOA1KJVgNxtp35YPP9ycYhQhQ6t3HyYsHaM3AnEi4r0brj6QWkzqyF1Qca9qNBYwcPzPyQ9WGtDfKV3CPLhOkosyFXaiBE7QcHjuzvatxxf1NR4c7k1m02s5zSzT+N/Llr//79d+/u7zowc7Ghsti808xSZx4kl7lUuiyzQd7BMCUVCbpqmIfPeWafcD7o71GUfuZ3e+vHz8PHgluVfFp+q4bscklas/bK6U8Onjt3BDhYt27o05lfvtx87zDcq9/K+IZnZfYt29dvvj5y9uCu/Udw7Lo7dGDk4vq1h3cXFr4heTJuGUHD3g8+JkTdXQcS7u7adXDPzA+lew9vKTyajAqXyjSV5utstsKiq8aLM41d647cvXt315Ej/7dx5v4mabeO3OalpCt9Um0vy1n/0+S2oSNA1L//fWRd14HJi5uKMS1Yklu00pW9t/IwHxFu4XUspmfTsP9MUT7I2AQP+FgdD/pNIV7PcOaehemXPFa/TevLj5YVmK7c2bdxTMjf/l3rjgw17vjlsw/X703UzTNmX21Ys/n65LabKOrIrl30p2vPyPX1lXJh9dvQsgJpjGVXK6GjJge6BA2QbPh0O+48LK1EmGQFyv/1IghSZntZQwMQdXls3S5A6u7dI0fgT/206WqxXZ/Et3HsVxsKfpoc6br77/277v77yJF1Y123Zi5WFuSU26X/IsNPz/q5d5yU1QPezcIu3srYc95mYvW8WQbaVnk8bZyPMyyxeK9HeXllzpUbO84OjZ0DCnbt2r9/DP7UyKlHBZUIRCd2xMrgR00OQKAwQANSu+BO3IRhv6myLGXrR642SLD6bg2t27ULaAKoQcPAvjs/FGPdXWKtSTA1gmsGQ1lZ2ReTZ8EAWH1gAA5I9+T9SszQScljgZy96buRG5fH9qP37mIQQRdevjFzvbKoAbPOCTYjjZPHCDsiRt4G+2/cPedj0ijOWA88LD9Xok19TatDFiwCS1RwV7bRJTIQtW+g8eyBIVh+u87tH9u4devWgZlfPi6SCxOsSs5FZOLsQFdX17n9MHyOnBvqutm17ezMF2VScYJFLTt5AfvjNyNnb6LiMVJRu84N3QQNtzDnW5zcAdtUamK52UUXZ241Xr7chSjBunW7oCG7ugZOz1wswqxR0vrZKDUgMjGwdeAPQ/CldsF03zpwc9tZxP2K3uuSsGX34a9kdLJeZRXpoGtuPo/oRMDCKub4UyY18wnmi/J+RoMmRk4Kq7/HQ8769s7MrQM4EC2CzYD+2Lpt255biKUnDKms8HeTIxeOffTRRigJlLV/aOtHxy7smJz8aXNOotHDZbJEypn9ZXLm2LGP/nwTkCIaurbuOXZsZnLyYUHOMst8u2yAlDG7DPbXsR0XPtk4JDC1buhPHx2b3LFjsrdGFv39dkUllkoybp6e3Ldn27aPthGkKMKEDtw3c2fyp4ZCmy2xwtI5dYw18WciOsGjPvhTIUnKU+BKyS7giiEyEbZMRP0iiv5+mxHr//bb299e+ce2LsQmdqFDbm67eGX+22/b/cVFiVEmGX/s/QnHN1tvIuIH73zd0MaB/0N3en/EcoyUHMaGh4IGNEf4MzREXL5PNPw0m2QaSnSSMdJNNX0303iOAp6ofOO2b+hOd17yFmdJzPct+uvKwxtbKT6xH87wtrP/pE69HS4qShHfU9C5kmRxQRPBwyf7D17USYazYcYq3EGfxPxRPhXkDyzwWd9zm+25uQZmzC6/f4wGVni2YzeP/ZCF1weNuVJ2YoySyorw/tvR7L8eaIQzsUtYIJcfVZcdzc7OPZqiKLpUhinX8qPGfx2D7QXXcN3Y0Ef/2HQUR3V2ciN+5LZkZ2eXV2dnj5/+RIXU/nPbTn9WZS/PzjbKSezoYrxrWW549Pm2c9DK+/cDUp9b8C5idnZRQ5LXbSQmIO+a2tfcdI0g1RPwYC2Ft53l9gQmGOuPNtfg7my3a7D9PaNJtFBXXVzZUFhW9t2OrnUwlaClBrb9YLKUbSo4U5ggpOy716zZrLcar9yA4YcIBQbpxtM/F13du3nN9t3vvhb3rTqkdkvBpk36q8avZ8idEZDac+qrMqLh8O63KmHZiQwGiWU3lJVdbfjrjcZzEO39R/af23rj5w8qKyuvwspPXm/nFJQVlRkfAcikpWAcbPvLVwbcaijKllJkcC+bawlkxAKJ+IEABAUhZPhVmIXCaAVHVX2Ii/d+mHdilZ/5qAmQEnGqsbGNe35gJ7Ib9h42JygEup16moNiP85AS5GGWDd04PSXbPfhtWsOJ3myc4mN5tq969ce38K+HoEvpWqpHYe+N28580Fl7YnkakoYHFghDD1d/vONA+d2Qbjv3h3aduNn0+GqnFxa17lE5Eqf5KNiSXp054Aw/KCltn4eZljWCKl7z8GvFW0pXjxUo7ZWp94aq8GL7TEs7LPSbbAXqyokGQtoV7TKZRVmtpXkSLZCBkhhrYHQUhd+MFwqLy49bktwIkdny1+ztsrGfoRRrxp+Y58AUraqtWuqdiZXnJeabrblrFlrtrGHIwcRbRGwvnDo+3Jb/to1tcmlgTw1yVCi09nZj6cPCC2FKQmCVK1NR5KdvM7eUFJukNmjO7eE4Ue+1OdfgRYDVlwmuvR5iZFpeEJ6STCR5s2x5hwaShbaikYOuof/0wVSBlsttNSiLwUtZQKk8o/bEoWBMbs4p8Rc/tdbjcKPOTI21Hj2S2YvySk2pqxrjbnFOTp7+b8uUHhCGH6fHPoeNKzNKU+0OQlKFRwaRm8f2tnPN7YJLbVrFxl+DJBKbtVVgI5kWNRSgNRHf4GWApzoSLAVWvJ35oDZbMoBpEhLLfpSe1QtVZswpOAOFxvs7K/7aI0fyTNBylhtyGlI2StxRtCQC6n+1wVoKdX4vPCP74vMJcXFyaaBEEUHIBXXUnfXnTtAkEp8tWRi3ZpfQjoQWmqIwhPkS0FLqcRoiEqMlSuR2mzOzTGaqwCpoZd8qd0w/GrNibq2pKXMtdk/QkvFfalPoaV2mqGlErQhl98yY1Fxic0GLUWLglQtdep7405d6miQjD/f2EpaCsOKgJRdlygjE2x+vskIpQQtJcITwvAjX0ocCRalJX93Duh0ucVGDOvfIeqMVTQ0L3UB4QljQ2ltdYKSYNAZc/J3XjIuaikR8fsy+9LOqixDsgfqRUYYDMacqks72b9mDi5G/OBLVV+y5WOd+GKi5PySQiApLoLhp/pS0FKNN342SniBJjlVxkull3pJSx04R+v/d6m+lIooska1I7UcEJAyl5i+24Op3v00LwVfyngiu6i0NtHBFf5wfk71ifIf96m+FCD1KcITO6vz85PsTLxgGdEgmXdWP9yxFPGDL1UNGqqSTYOwsSDIkjD8EJrAVC9BitkNSRbsLBNqNTw6jYgf+nAxPKFi6gVrtLMUccBgyIUvZWf3jy36UhtJSy0HUkZj1YbcbLvhx300LwWji+alvjQfzc7ZUJVkoVpilrG8Nr8ou9b4NflS8Yjfqc02CTQkOkIslZngCSBF+gJW511h+BntyV66Cl9KaKnGpXkp8qW04z1xIA4pEfEjww9aat+7QSr/x1t/WvSl/nTjkSG1kGIEKZvhXzTNpvpSHx36ypxiSJ3edu4utBQMv08pPJESSJU/Ov3p0oIkmpd6TwKlVbuopeBLxSN+wpdaMS31WYohVV5bCi3Fvt6DVcDxiN/70VL78fbjXTXilxJICV9KRPzIl/pek+z3xoEXkFqK+F1Ypi8VN/zyYfip6+uwEPt0qiEltFRtycMdB19oqc0p11IHzuF9deh8dV4qJZDStNR7w9ArFS9CCr7UUsTvn8sLT7zRl3pUklrDb1FLYapXrPFbN/TRqbAuxYYf+VKkpdQgemoMP2gp+FLrMJhhqlfzpV4R81ReLvlSMPziEb/M11Jm9vX71VLwpaCl4EuJ1ROp0lJqxE+sRMeCpFRKkVbXSxxY1FKv+VI5CQfR36ilPmPVKY74CV/qIZSuiPiNDV04tbk25VpqbNd+hDzvnsMavxSFJ6ClPlUjfuLljhRtRfqSJGmncQ4QpMpt9hJ6X+qViF+iqycIUsXZ9qyXfKmtt95LxA/hiZe11FcUyD+cwiC6iPhhR5f4gqQUaimsf9e01HsGNyCVi3kperlD7JaAvSewEn1LeXaOLlEZBKSqMC/FrtxaWom+9fRnJnN2blVJyualcqvys+MRP/hSdzE3duHQVzqDoCE1rI7PS0G21x0ZI8Mv6QuSsG823tUiX4pWTxwRqye0xX2p6e031GLCzLvOnF8KSCHqjNmUsQFsI77dpqPJ9zek/41b9MWCcl1V1pUbB8S+D1iJvuf0Z1l2XTl2JE+wrN+o5jcfoSJs0Lkh6+sRrDOkvSfWde049PcsM5qTql0iCVLbxu4CUdgPOyWrJ9Z+sMFmwyuIfxoS8yAYRWgl+m/ySXuYPA4ISOkKBKQQLMKGVX+YeajbC0jh7e7EqjVBnMt1dkBq2xjJM6aNt934rLREQCqxopadGjufgwa59F94BRG704CGoT2n/l4KRQFYL7vUhDKqC5LIAlu3buzA6Z+N2NM5oQISTlyKt9Rs7NEtepcY24fgzU/MS2mQSpiPK5ShBItZDIYcghRCVLswV7gRWqoAYpAoohiDwsM3WQCprUAUtoWEBXL6yywJS9xS9kFmGiGMhqzSr6F0sd0tMDVEWopoyE2Rx05aaus5mJw4VC2V7NWFubk6m7n8S6yeIBjDOPhE01IrBI/lFGPQYfk0a8i5PwLDb93YubGxrTP/NFfSmmqG13ASOwiGxaa/4q3eMSoMu18RpLCxSJKH6ZepBOHGYpPQUmPnQEPXnkNf5YCGRM3Yl8tM6FzCa80DeJ+T3pXBgqTPsu3JhhSZ7nb26PQ2YjuOv3x6WQuiJ9RnK5pYQEqqlH6aOXu28fLBjY0DjXcesgIJ3/uCAkvwAHaMxfj0262BgY03N24cuHkD4YliglTqvGXCr8y+vjMgaBgYuDxz6KtivF+RMhrI8LvVOLARzNx488Dpz5g92V84AKTwKv6j0/sGBm5uvNl4thE72GiGX4Kyu3LJCVJM2tvw3aQ4RiZHJic7ssskbBSWX5VoNRBnbFFzBSVROShp8s7PhqP0vlBKIYUvOn5NrYkT8UvYKKeQBjL8qO7JEeyJOTn5c3l1sl8Wg6mrs5s+Q62oluqcvKOFJxKV3ZVLj/cGjUxv3xT59sXxJb6JJBlzSjck6H2QzWWvbXDcpqL+if+wP2M4t9aeOqNLfQmwsKShYj5OAojI+z6/lt5xTRGsTTlfCQbEWeAwJltJwag16PI/ELUSz+l4z3sYr5x8ZmBJeA3WaKglkTOYxIGtQYqtOrsxG5BKsD3YcMtYe/wwLK/8rHx8EdGET/dmHz6OslLmyNCGkKBBb8zGZ2pLGL5KWFJiPHpc0JBga5adHC/ZMuxOhOoxJCEAaS5J8hdz4fVu2LB9rSkfL06B6aYUjR3LZtB/eUZsNWssqT1hPtqwd7t67K0sqDpeS5A6k2DbCVKHL9kKG2KFhbV0FMbK7LZLVSmFFKKLx0+Yy/ABHpUE+9Uy84njJSmkoWpDZWXh8d2X7h3eoi8ry9bp8vEFgmQe+Hxo/pkN9/bKZUfLCgu3FNZW1SazOq3sN3KANo7GrmfY+cwOR8NehRPa9E0c+AY7YlTG3KzfDk+IjdOoBPyL78BIUUKz2Y5Beqkofe0WpEnJ9mq0NyURZdfXFuqz5cJyu53IQHPMZizfeCXqSESL49WP1OCzNYuPlrexJH1ESLLX7tbZ8W1WaH9S/6ITaANHFP1KhbhOuB7axI66T7Bez1BBSX4+1RHne3W8PmoI0ogr7U+SOUDdLlv1MafAFfbCfaWjf7d66ihsTIhuxR6gvwoZmfYE1Ttj+JZq8g+nNYYvi8ck2oYUG/z+KlEqJUQ6ZPxV2glQMfyPdqGByyYbRetfyUyVEVmv0EVUvJLyd1klgIIG6t9Q3ovM1EJJr3+VjhcJtLMV5ACN6Gpx6GX90ta4CdRAu1CjR4XUOX9tJ2oMkvjiD76umoojXo3k1GNXX4bB/7clVZbhwwN+kPSXD2xgCnKBKTy0vvLs5XS/c/46dCDgTqowvgvxUn5A7/fQv5R28YS6D4SD8tfLW0xDjaAvwyz19IsH2lkSOIBeZI7WVX2hOup7vcwghAkd1FXoUAzl3d6T8T3eXy1AQqm+4WE/qws8iSV9okRiNe0TTQ/qnQyq93y0HZX/J1heIc8aY9OBadpK+z8eQLwFzfOBkCVx9bFU1OuiDBCwXlfPaxUy2SlGnqWsb3FCfJctMusOvKG8eH6JOYaHI+ypa5XzLUrUkrwrB9DjFX2ccyXogakkOzyt2JReWPpQO/FRk0wUtbPpxqINRE/FFX7zzvczfOW7E9JLSUR+qAZVdYmCpHHuvYYv1q1KOqJkFqEGcaWvAkLqwnfynBBVoF4ciz/gG12LhlnZY94Dw29RS8Rvy0zQ3AOaoe3Ug9itui/UNsqu/pCXSIikH7XhxBv8g/HY+txB9+kCiUlfxFgbvi9LOMV/lA6Ql5nP08lIzeOOoI2Si8ciI/lMdCIOlCIukU3qXxC8R3nCxlWTxduLxLimdvjA+ydJ571K3f/yvxjlPNz1IDSo4GOn1lgzb2fwkmk2wymsNHS3avRAEETXw4wQ4iHMJSQjAfEH3WEIAnlLuMZNCAU9cQrZoAs9PqrVwmriZeFhkg7y+SMBHngSWnDz84z5vVP4kCuMP5JfYR8RSWQKUXPILIQ5CnEmK0+oMmxbL4hHG+iTYH2MhfC1Sigw8lfwEXNqJo0YQKv6Aw8FF2gOnBV4btR+8Cn+vRZqb6cyCEVipQZb9XLMiZSspg9fRCfOOlEoHeSHDivt9ClMYp2aPRaDRS0eA0uq9UC044DvKn6J91NuKFFQR4/orkgCUp1IJVqsl6gd6JxEDRBRhfYnUQ5AQgL4jFyMreIPWKd3SvF65OHo+HiLhzl6hltOXmMW2dHa5HoOqMWkugeB1T0YcyGSeivL8wSGT84y9sAdnArMz7qgFqbdzxwTLdPytd7oc5/M+gNtNTJrd4csLATJso4G6oX4JUrlW6eHxOuHeRM+Nw48RCtYB3c5OluG60BwW+D2teHzLNyxyuV5ShJ63vvU53F5HPh8XrQlAgGsb45OjDMri7U/CQy3WmiwARpaoMFC3l4WZvXuEI0wUshbbx0clWP1KKkC6VcFOjr6mucxIslyRxuVgaGlLuRq6XYwX5+bu2Hy+p41tfTUQBE6O/r62iM86AdJVNrsyajLU8dYh3eOBzzOpui4PwrePxuO9jgwJoH3gQewXWMyeD8seC8QlRdy9U3PMgm8DwbaK4j3J909Dk/TNHNMuEI+fMDM21YjsXZvKIZRYZTJqwOthFjtSDYHMCQP8sFIjEV6+vGNRu7mnQ7F2xpUOsLDytyUstrCLH08OKco3Yx1c2UuyFuwyAWjJmtVkIAHrln6lODc3Pg8H7ayB7ytKcj5BLLgU4+5rXw1RO4kXyCzsJX53FN+MZImrVUYttt5sJ/VILY473fgQ67Nw/jMZGCWOVy8M4BPvI7yoFcBLZZYlHuiUwrpsjreV8N8w3zOrXgh6s950K1ANyGtoHkcLVjFnGxY6YD+0EvDSg/ysvNK0M1dflbjUgLeIFeeQn8v8Kk57hqnj5sr7iC+e/l0TgkG+/SzUY7LJ1a95EGKuUHeDErJda0I0GcwYSEsIAPvqZly9wSVVmgxJB+0xMB7FMjB+2lRYFQsMIrldoD3Qe6dDffhtnu2nTc5Yb229RHvm5Sggg9u1iuj6KQU8j5pnZphBcPG6w9wd1s76NZjOIOA1XN39Mmzkid82up38XrpJMwGudfVyyq8/DaraxmMiAHWEuUdJn9Tcx1z4IyxZxA7R1Rx1ZnQzZ2slwcc7Al/DEgtQDwhJPOsf8prSS5/YHZNk/jDzEFNMsTM28ryCEJPg3PRwW5IdJMPugu05c1x7zxGCa+D9SirTUi62hlrhhrtj0K9tXJ3jdwEMxg0O5BhFMoWCJERyxifC7oGe2PP+AOHZRBNmw3wkOwYBcjAg/g93yA+dxkJos1PlSA+gjkKVZfnRXHtHKCd4FB/ZD/qMYotsJoHfT1W9lwB7zv4XPTJSbhaJyV/QKmXunkTY73RXmnWzftz+1tGaUiC4Yh+YePEe19U6TABwKtYOMoDdQy8b0WjAmHBeyfGuF7iPVoV9IaTy3utdJUDEBLn7AM356M+GN4thI1eHqxnLAIg6aFb2oAzb6eT3CO/whcq4Feo/rPDDe0GU0J48bMyG4Z0+YMIUeB7322k0ZokKYq+lKwB9Ho/D0Zwry+5Soq8wGaM6bDArDG4GJYADfC+ILyUTg4VBT2p3GaSE8ItdXAMEBDGFita+RhAcfuZ7HNa4ZU4ZUCqieXxKUGzxJ7yYQopwPgFCto5ocfpUmAyTvMoAdQpS3RmjQZha53kLbLeVAOnjrsj7JkyKEMNNoNTw8jXDCqkcS94hbiB3slW82h7TCLPh+QeYAPvTePk+4Cq8zA9vZ1hKEbB+3EKQEjoNxPxPo+8Qebn3gqUrHjYuOD9MPG+F7xHX9bjq+sB9Gge8X6a91G/aUfSOQD7A0dkuoWvjjFfgMNmCaFXLBhw3X0tfW5lFfM1ce4afAosdcPW6DtJ3+omldYDL6Gl10HDdx9ErYXfNtRx7ziD+uphEIZu06yXj5sgxnOzSDPsxL1eIDCph5W5gCK97IR3LmNsn4sgRsFdPohUE2y3dh6okWVIZCtuNCMy0QthlPoAllalBZ8qx4y0NB7qi7rc/DnRTP4UQIkBYdYPSxYCLQFCrrDMbvOplpY+RELAqRD41c1d0IRBcc/FfL1NLa4ApBj5JyDjihcP3Dxk9fI8ADfKMWcBLWqRIy4ejJ73I2Af4BWUetDkBCjdIvkqNt6nKMR7i9yr8l6gQpZ65gTvCd99GD2i/Clg64VjF0XLAMxuNutVKmgImaugdsCfguWeZN4ntWMzp3BJ3/O8AkNvBYwURIZcsNVXo1ecsEhcvR7PdHeE5codg5xDNiVTv8fLVVhApbFIr4vzBxKMi/MQk4AyLsb2eGcipE7lYc6kToHJ0UOjZxt0WJK7VYIDNIGwuTO3ox7zYNwFf0UE7hYgZhaM3y4EqmenoJE80BkxtqA8Yz4Xj7DHShOzAIdw63nfcw98SmFOsUEYUpLPy/0h4I5ggDIWwJ9WPueZ8HR3t6OgkyjZA2DRPQ/uzYdbuGthIoqIjyrjq3gUzOzu9tVxxS+xeQXRCZSFGKnk6KQRywciYazJq5WeJd53g/fMWt8GBxe8Z5EJwXtkQ0Y23u3iClzU5xQav+Zd5H2Fm4DUhjGN+lLO7efemng70KtadCIFwJTYNS/4L6OXoWS6ucfAZqd4nZGCD824jdi4Lw/zHtdcfJqNP4U1HoKBQ9JguVaHsBJcEQsMGnhiMH9q4LGjl1s5IOSbgx9xkodMMlypYZiFygS7NoXBNLmQAtJ7uAsBamvPVLAOw3UzJHSB1K4LFpATPrvLKcF36rPCKO1RTUCM4nCXegEpNts/i9RPGAvT6E6m7LU5hLut8Kp6pobj0twHCDmZf26uxoQgB5OaSA/Bk2llkTm3xVRusYCGKMR3GCyoCGJKCNejUOvQbXDr/KS0AkLVS3pfHi5vB8HAabImwaE6CH4raCHeM0ekn+XCVzvJKp46l3jPYr6nMDrB+/AS78l3Au87qCkoZR7lPUA/nefDJtYE3lfMgfcapFIBKRnjnLupqc/LRw2xPv7Y7/PzqAMdCsOhd7xnaoHVB10d4/NTU+POJt6bN96G0R6wkGKDPJRXEYIXAi+81a/vhKUEz6CD5Oc8w+xi1Gfq5cP9daunKAfZViiZJomSeQBSsDa9TU3RIInoY1RNDmEnIO7FzC8Lu7hntnMKxPi8ZBNeUwIUimiDQ6i46/NWAx0LfHR8fpi7rMIejCjQIHo0bA55qN1M7wJWEdRezVf55wN9FliR7o6KNt5sofh9m7/d1Sdg3b8QVCJAb6AuYhp385P+Z0GP09nHPeOwmM8LPugx6PTN+8mShAP6fNwBDkFV0TDXPd459djUoQTax9unpvyI7PVGxtsUMpzheY2qvH/A28HYAgAABddJREFUIlO8NRJrJd6PEu8pRsQqSOsJ3g/OYSSU++BK+hUX9ap2JJ8DWI7UpnCF8z5MI43ipBUBWUzEIxjt5YrSF2HhB4jM8mAP1E+AViWsFnFcWPFRusJ8yDjMvzk/+UkIVNeRHPeSqmrKZe1TWMQQaoZE+91kmmDwFe51EpuF8p+CHg63HrO5wxAzGVTdNsyTEsXDeTeeTSGGApPIEYPWipKLF0JcslcBFwbDLA8pplzwkSrc8Cw7yKuH+uXwpMiPQaBgDiE8QDcSBbcCCAkGA89RZp8fSgARUnHvWgvugD8Rhmi3Ao+sdQ6caIaz1IMHrmjcp9RLvmZcc+9TmmSI856W/tW7cRezWeHH9Bi8lzq8dLY6TPxDK0RP9M1SDJ7PYWAj3ivg/QLxvoM3CbND8L6HAc9+6g9TfP45idzXiiYOyCwWmX7QE6nBmaX1wYSzfdVJmv+3ShXT59uxMABW/LOFkxW0aMBRH/LMw/4X+XJr2j2hdgtuV0wvdLJniMP7PA98TO7BmTS/iszJ/lD3rDzdlscqnofCrL3tWbJtD5CmZz4izIdwSk3vk3E4Qp4Hs6z/CWSNpNHX+rj1GiZs8s5P6GPs6apuSe4cxLIFJ2aue/Mwz8sqJkKz820TNXGaMQ8ehruIkZ5WQABSjz0UeJNYuGehE/ZXKyzh26Ee8A/3HJ3nWy1ouGP6cb///AM/g/u5gAkr5u9dmJcJiXXPeyydbQg7orCYLFn7uxdaHYiT1LQu9Frm206SfwWd2b3QjlUdMSlycqH7GqtB6KgjFLqN8CyyoaZwu+dxux4TBbPTC63sZNu8yedZANJ72uYZa181zTA0gPfSdFuEjT8POcD7afSg6DkqQTuSxwGxVJaKR2+r0SQ6J5ERYzLWCuABDlzGV8Go17D8RP9gqY9Y+I2LeA66TadIQEMq5IvWA0FDxLC3hRpgpAKTc1CNKr1oEEQTIXxahiOraECd4iHQRgTSKIE7eI5GCurFyiW6qac5I7pPCWCJRZVhQl38ABbxXDTOCufyOfBhouulMggwovZ4AYJntJ5dLPUCkeWipDgFqA6YogPZsNxLZTktJxLZBfRUuimKTulkyUB/CXz0S5VRjbR0nrqOiKcqRWK1fUiGNCIzZdGO5HGAegBL0uhNA4g9ZkixMg9goTVnuKK7WOzstDqxag0JY05nTH11grLRFbpVb8WJ3ollbHonrf/DUjeacnGiDOREClzH8CthAlYVxCQ2B0UTYXqx3JBagIkmEBfDajohfngI+rAWCFSj4Xp60Qh3iAmyhV7wwIFHlEM0RI+5o85mzKJGEKRQxRnJYwREBK/BDdiEnWi4YA9KwfoIrANEViznwx9iHhVK03/gCr3FhdtOi1MUpTIR82dUuxMKHxPUSAXZjyfHiRN58FCSwEp0Bw0QBA4rmIkrvd6ix12qglYYUumos4Z6jwBGBRLboXzRXBViyWO+VrLgAFhP4xktu8QvupB6V082AnUtTumEBES9Qf24KA2QTrqiHPiDHJRdjJL4QT68U0FloDDcFPnpT/L5Dp0oSEFl+EdwIY0VrxlNIoqpaXRPJBHJ1CsiFNgjiEFwcQ9SiRBDsBlTQ0LDgFXITIMOiqbnNVHELQg6VIrIQoXjWtSEM6TCEzyH7qARS9RMPCctggciE3ABdlFOkRpLcOkR2L6URL0fV8HxVKK/FpuHklA6ZaOkoiNQGsYSoJC6UZSVfO5rNdDIBTGnPzijQ/zET+mR+F/8Xbwpkqn3KbeaRs2tFqaek+Uhsi/dRH/H8yb3B7WIf6hOpQDXL1etXog7Km3xNsZ/BJGLfyhBGItD1JGE6F4sUj2XZ7F6YbHw+KOXfl5OjaLUJ/RXHCp9cWpxB88Xb9FNSh4vms7imRazLl7GE4mMIo2aUTwWBeJMzR4vazGj9qtx4P1xQBPG98d7reb/Qg78h4p7tX0a2l7liHatcUDjgMYBjQMaBzQOaBzQOKBxIC048P8B/uRUBUv5a4sAAAAASUVORK5CYII=)
Glucocorticoids
- Released by
- Adrenal cortex
- Other areas in small amounts
- Cortisol (hydrocortisone)
- Increases blood glucose levels
- Aids in fat, protein, carbohydrate metabolism
- Suppresses immune system
- Reduces inflammation
- Receptors in body and brain
![[[@Kadmiel2013-zz]](10.1016/j.tips.2013.07.003)](data:image/jpeg; charset=UTF-8;base64,/9j/4AAQSkZJRgABAQEAcQBxAAD//gAuSGFuZG1hZGUgU29mdHdhcmUsIEluYy4gSW1hZ2UgQWxjaGVteSB2MS4xMwr/2wCEAAMCAgMCAgMDAwMEAwMEBQgFBQQEBQoHBwYIDAoMDAsKCwsNDhIQDQ4RDgsLEBYQERMUFRUVDA8XGBYUGBIUFRQBAwQEBQQFCQUFCRQNCw0UFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFBQUFP/AABEIAeEDLQMBIQACEQEDEQH/xADWAAEAAgMBAQEAAAAAAAAAAAAABQYEBwgDAgEBAQABBQEAAAAAAAAAAAAAAAABAgMEBQYHEAABAwMCAwMECREDBwsCAgsBAgMEAAURBhIHEyEUIjFBUWFxFRYXMlWBkpXTCAkjN0JSU1Z2hZGhsbTBxNJXlNEkJTNicnOCNDVDk6KywtTh4vBEVBhFRmPxJ0eEZIOjs8MRAAIBAgMFBAgFBAEDBAMBAAABAgMRBBIhBRMxQVFhcbHwFCIygZGhwdEVIzNS4QY0QvFiJHKSJYKy0lOi4sL/2gAMAwEAAhEDEQA/AO5eDXB7S2oOD+hbpcIs6XcJtigyZEhy6y9zrq46FLUfsviSSfjq4e4Por4Nl/Osz6WgY9wfRXwbL+dZn0tPcH0V8Gy/nWZ9LQD3B9FfBsv51mfS09wfRXwbL+dZn0tAPcH0V8Gy/nWZ9LT3B9FfBsv51mfS0A9wfRXwbL+dZn0tPcH0V8Gy/nWZ9LQD3B9FfBsv51mfS09wfRXwbL+dZn0tAPcH0V8Gy/nWZ9LT3B9FfBsv51mfS0A9wfRXwbL+dZn0tPcH0V8Gy/nWZ9LQD3B9FfBsv51mfS09wfRXwbL+dZn0tAPcH0V8Gy/nWZ9LT3B9FfBsv51mfS0A9wfRXwbL+dZn0tPcH0V8Gy/nWZ9LQD3B9FfBsv51mfS09wfRXwbL+dZn0tAPcH0V8Gy/nWZ9LT3B9FfBsv51mfS0A9wfRXwbL+dZn0tPcH0V8Gy/nWZ9LQD3B9FfBsv51mfS09wfRXwbL+dZn0tAPcH0V8Gy/nWZ9LT3B9FfBsv51mfS0A9wfRXwbL+dZn0tPcH0V8Gy/nWZ9LQD3B9FfBsv51mfS09wfRXwbL+dZn0tAPcH0V8Gy/nWZ9LT3B9FfBsv51mfS0A9wfRXwbL+dZn0tPcH0V8Gy/nWZ9LQD3B9FfBsv51mfS09wfRXwbL+dZn0tAPcH0V8Gy/nWZ9LT3B9FfBsv51mfS0A9wfRXwbL+dZn0tPcH0V8Gy/nWZ9LQD3B9FfBsv51mfS09wfRXwbL+dZn0tAPcH0V8Gy/nWZ9LT3B9FfBsv51mfS0APAfRWP+bZfzrM+lqCvPDfh5aVFsW6ZIfHTYi7S8A+k82rNWrGlHNIyaFCeInkgQD+ktGNnAsTwPmN6mZ/8A9lZls0jw8lOpal2mZDUroFm7y1I/TzelYUcdCUsrVjb1NkVI088Hd9C3NcDNDvICkW+UoEeIusv6Wvr3B9FfBsv51mfS1szn2PcH0V8Gy/nWZ9LT3B9FfBsv51mfS0A9wfRXwbL+dZn0tPcH0V8Gy/nWZ9LQD3B9FfBsv51mfS09wfRXwbL+dZn0tAPcH0V8Gy/nWZ9LT3B9FfBsv51mfS0A9wfRXwbL+dZn0tPcH0Uf/wAtl/Osz6WgHuD6K+DZfzrM+lp7g+ivg2X86zPpaAe4Por4Nl/Osz6WnuD6K+DZfzrM+loB7g+ivg2X86zPpae4Por4Nl/Osz6WgHuD6K+DZfzrM+lp7g+ivg2X86zPpaAe4Por4Nl/Osz6WnuD6K+DZfzrM+loB7g+ivg2X86zPpae4Por4Nl/Osz6WgHuD6K+DZfzrM+lp7g+ivg2X86zPpaAe4Por4Nl/Osz6WnuD6K+DZfzrM+loB7g+ivg2X86zPpae4Por4Nl/Osz6WgHuD6K+DZfzrM+lp7g+ivg2X86zPpaAe4Por4Nl/Osz6WnuD6K+DZfzrM+loB7g+ivg2X86zPpae4Por4Nl/Osz6WgHuD6K+DZfzrM+lp7g+ivg2X86zPpaAe4Por4Nl/Osz6WnuD6K+DZfzrM+loB7g+ivg2X86zPpae4Por4Nl/Osz6WgHuD6K+DZfzrM+lp7g+ivg2X86zPpaAe4Por4Nl/Osz6WnuD6K+DZfzrM+loB7g+ivg2X86zPpa4z+uJ2kcJvc+9qE+7WH2Q9kO1dlu0r7Ly+zbM5cPhvV8o0JR2hwC+0Tw4/Ju2/urdXyhDFKAUoBSgFKAUoBSgFKA1ld7lqJF1mJY1A4yyl5YQ0IjJ2JycDJTk9PPWJ7Kan/GV3+5sf01oJY+opNJI3scBTlFO7Hspqf8AGV3+5sf009lNT/jK7/c2P6ap/EKvRFX4fS6seymp/wAZXf7mx/TUbqXWd+0rp253mZqOSqJbozkp1LUFgrKEJKjtBSMnA84p+IVXyRVHZ1OTSTZW9M8cZGrhp42/U00i9MSX2ebAjp5PZygOod6d1QKwMdR0PXwzNx+Jy5cB2cxxFt70JpwMuSUdkLaHD4JKgMA+iqnjqy5Iuz2Sou2vl2+h7Q+IMu4xGZMXX0STGeeTGaeZTFUhbpGQ2kgYKiCCE+PWvJniW5J7fyeIkB32PBMvZ2Q9mA6EudO4PXio9PrdEUfhcejPW48QZVobkrna+iQ0RtgfVITFQGisZRuyO7uAOM+OOlely1xcLNa0XGfrmPCt69u2XJajNtK3DKcKIwcjw89PT6vREfhkXbjqfjeu5zzXMb13GW2H0Rt6W4pAeVja3nHvjuThPicjz1BXnjlEscdp6TxIhKaXMRAKmURXA28o4wvAO0DBJJwABk09PrPkiqOylN2imTaOIExy6+xaNexFXIN83sYRFL2zGd2zGcY65x4Vgni2yG1L90217EFAUrfDwCsZR8odR56en1uiC2UnwT8+4sXsrqYjpqV0/wD8mx/TT2U1P+Mrv9zY/pqPxCr0Ra/D6XVj2U1P+Mrv9zY/pp7Kan/GV3+5sf00/EKvRD8PpdWPZTU/4yu/3Nj+mpPTlwv717iIlX1yVHKjvZVFaRuGD5UpBFV08dUnNRaWrKKmBpxg5JvRGxaVvjRilAKUApQClAKUBTOIGsEWSMuO26G17cuLzgpHmHpNaOl3y43pbwaKorYOApGd2PX/AIZ9NcvtKveWVcjvNi4VQpb2S4kc2yp0EPyFL2AHClkk4Pf8vjipLe7b2yWVBbYzltSyoH9JrRpuLudLnU/VZsbhPrcPy2rY8tQbdBDAc8UKHUoPxZxW3a7fB1d7RTfHgeabUw/o+KlFcHr8f5FKzTUilAKUApQCtecS9PxLxcIa5BkbkNEDkynWRjPmQoA/HWFjJShRbi7MzMJCM6qjJXKd7SLX553zjJ+kp7SLX553zjJ+krQekVv3M3vo1L9qHtItfnnfOMn6SntItfnnfOMn6SnpFb9zHo1L9qMW6aMis2yW5BRLfmoZWphpy5yQlbgSdoJ5nQE4FapTr2BdNPafm22K8l+bZJt4ndouUtSIIjt7SCEublDnnZjocJV5aqVes/8ANmVSwNGoruK8p/YsNn1XpO4R5KVG7yHbdb0TbhJivzOzsksB8pBLmQSg5APnxnPSs9V80itRbjov06R7HRrmmPEemOuLZfUUtkJC/HKTnzDqSBTfV/3MPZ8E/ZREQ9SWbUGqdHRLOJyrdd1XFqWmXMltyGHYyU5RgujaQokHIPkxUhdtT6OtE2/x1ovz6bC0XLg/HelraYwgLKSoOeOxQV5vTnpUutX4ZmVPZ1O6ioq9tfjY+Lrq/Q1rluxQ9eJslEiPFQ1DlSnC8680XkIQQ5gnljcfNkeUio1vX2lmdQaqgXG33+2w7Aww85OekzNrnMAwnbuylRJASnqVdceFN9X/AHsLZsWvZV7fb7mU5rrQ7URpakai7WuYu3+xyVzVSkyEtc3YWwsnqjBBHTr4+OMnRmrtD69ubEG1OXvmyIqpcdyU7NZbfQlQS5sUpYCihSglWPL58GjrV7e0xLZsVFyyrQvPtItfnnfOMn6SntItfnnfOMn6SqPSK37mYfo1L9qHtItfnnfOMn6SntItfnnfOMn6SnpFb9zHo1L9qHtItfnnfOMn6SrxwxsUWzLuJjF8lwN7ufJce8N3hvUcePkrLwtarOtGMpXX8GLiqFOFGTjFX/kvlK6E0ApQClAKUApQClAKUArgH66v48L/AM6fylCUdhcAvtE8OPybtv7q3V8oQxSgFKAUoBSgFKAUoBSgNa3ZIN0l/wC9X+01ilIrlJxvJnWQl6qG0U2gireUrzDaKr/ELTsjVmgtRWSGtpuXcbfIiMrfJCErW2pIKiASBk9cA1KiVwnlkm+RqeZ9TrczqmfJgXWNDs8+yy4rjCd+9idIZQ048gYwUq5SFHJBznp1rxtvAG7PWdmPcI1sTJE20qklVwdlNyY8RwladimUhOUkhKevjgqAqu5s/TYtLroTr/BGavik9dGJcWLpFRM9uAzuS61cOzmMFpSBtCQg7gc53AdPLVYjfU86hm6eNrmOWS3GFpyRYosiApxSpq3FIIdfBQnaBsBKRvO5SjmlyI4yCSve6S971uZly4O6xvZvlynNWZN5muwXofYrk+z2JyOw61zA4WVbj3x3SnBClDpgE3HXvD++al0BYbc0uJK1HbuU4LgJS4QakJZUgvN7G1gjKj9jKcFKiOnSjsUSxNNyjbl2crW6lRZ4aXVXGuzIkrMq2It0a73RaIxbjPXJhtcZtSSBtGQoL2DqOWD5qx2eBep2tNqgpFoDMG6wbha7RIluSGmkMqUXWi+WQsIXu7qSlW3HicmmhdeKhG1+i8/AkrbwSvEbXMy4SxDlwXrm/dmH03F1tcdxxko2ckNYVjO3O8DbjpkYqKY+pslt6dbhqZshlo0a9Y+YEnAnKXuD2eXnb49/33Xwpcj02K4X5fXtN8WeEuBaIMV1QW6wwhtakkkFSUgEj9FZm0VbymqcrsbRTaKZSMw2ipHTqf8APUT/AGj+w1dpR/Mj3otVpXpy7mbDpXVHKilAKUApQClAK+XnA00tZPRIJPxVDCV9DmbXUqRqm/8AL3FLCHOcVAnO/qQPHwCf1msaans8B1yK0l5aUZSk4AWfPXBYiTnVbZ6vQgqdGMFyRQrjPnBxxT7ZUV5ALashOfEHwxmrJpWe5c7c+uWgMuNAp75HeT6PPiqZQurlEKsVKx5wpr1tccuURW5uLIbUFIBACgckY/8AnjXXUGSJkRp4eC0g10+y3ei12nK/1Ck68JLoe9K3JyopQClAKUAqn63TmZG/3Z/bWDjVeizOwWlZe8rW3I8abfTXO5Tosw2+mm300yjMNufLWtbTwJtNoc1opmdJUNSMPRghYGILTpcUtDQ8xW6pWD6KqSsX6dd000lx+h4p4C2928sTZN0febYta7UltqO0y4tpTAZIccSnc4AAVJCs7SfQKjB9Ti09AdZmapuE95USHCQt+OzsSzGcK221NhIC0HOFJVnd1JNVXMlY1r/Hp1JzSHBC36Pm2KSxcHnlWmRPkNoLLbaVqlbd42oACQnb0CQBWVN4Rw7lr53U82WqSVRnovYkRmmuY24gIUh1xICnUgA7Qo9CfHpUXLbxTcnK3Jr4u5TdH/U/vN8HGdL3aWmNNlPrkTS4y1M8U8ttIUtJwpDaWwFpxgp6dOlTl0+p/t91avUVy9Tzb7tb4cOTHcS24pTkXbyX96kklQ2jIPRRJzU3LssZ6zcVz+qt4GdY+DES0P2N9U1ov2q4OT0iHbmIaHFKYUztKGkgdAonJyc+XGAMjSXCCFpGXpx9ifIkKskOXDbS4lI5gfdS4pRx5QU4HrqG7lmWKcla3nX7l92+mm301RlMTMNvppt9NMozDb6atWhhhU31I/8AFWZhI/nR9/gYeLd6EvPMtdK6Q5sUoBSgFKAUoBSgFKAVwD9dX8eF/wCdP5ShKOwuAX2ieHH5N2391bq+UIYpQClAKUApQClAKUApQHOf1S8qTC4dX9yIqUl9U6I0EwXyw8tK5rKFIQ4FJKSpKinO4e+8RWrY+ntbWnWOlYrE42Vc25XN62xb6+5dlwIghsjluKDwLii6lxYHNIQHAATtxWjSWt+rOgu3a3RElA486gmW/SSFwLe3dNYWm3SrM2lCyjtC3EpnJV3sqQ024h4AEHbuBzjNej/Em9yLsi135EVq5tagt3IgsMuNITDeluNtvpfbfUl8KSgdFBGFJVuRggCndJFW8bK3q/i1qK/8LrlIlP2tyDqbS99kswYTC25drLEdZTzHC4oLxnYs7EbXCMeara/xX1BFucp2O7aharZqC26dXZ3WFmdJ7QmMC+lzmAJI7TvCeWoFDSiVd7u1OkuHnoQqjKkxxp1NrTQ+r7uv2OkWWJZlySw1DkRVsyS8QhoPh8KdTy05K0JTknxHvRe2uKd0Np1ZqGberLarTbZdwt8e3OW91+W05HfLKXF7X08zeU7+WEp7rrffHUmJUktEFUbK9pjXl61jr7TEW8oQzMs+p59sWWmgxzUi0c8FbaXXUpUC9tICz70HunIGRqXU+otI8T9aXa3SICrTGfsDEqDKjuOOPJfd5KuWsOJDRSHN2SleSADgVDgr27PqM7av2/Q92eMeo1SbfcSu0O2253a7WhuzojrEyGYaJRS845zSFZMUb07E45ycK6d6Cc4xcRINil3WRI0261E0vC1W4y1bn0qUh5TgXFSS+cEBokOkeJ95ip3UQ6kraFkPFrWcnW18TB08iXpyz3R23SWwllLuxEbmc0OGSFlalFOGwwcoUCFeWo+38WdcTIFvEM2C/wBwvenUX6G1AZLYhkvx0FpRW/teKkSFcslTW9bKgdoV3Y3USd5J8D2s3Em73TWtouJ1RCTpxOmp82Yw9aHWSXo8lLTpKC8SlSFZBxu96oDO4EYNr4ya3uzF+iw4kSbKgi2zESBbktOmHJS+pSkRTLVvUkMBQSXUK2rPdJSAqd3Hz3kbx8jKXxNvc1V5umlkxbo26xaJSpqGHVqcivRXXVvNRXJCBnCUkNpUlW0qJ3qSAd0aZuzGotN2q6xpLc2POiNSWpLbSmkupWgKCwhRJSCCDtJJGcE1anCyLkJ3JLb6BTb6BVqxcuNvoFfKGLw7cIYs0iFGkBZ3KnMLdTjB8AlaT+urtJLeRv1LVZt05W6Fg7FxA+FtN/Nj/wBPTsXED4W0382P/T10RzQ7FxA+FtN/Nj/09OxcQPhbTfzY/wDT0A7FxA+FtN/Nj/09OxcQPhbTfzY/9PQDsXED4W0382P/AE9OxcQPhbTfzY/9PQDsXED4W0382P8A09OxcQPhbTfzY/8AT0A7FxA+FtN/Nj/09YF/Y16xZJq3Ltp4oDKshFufCsY8h55qmbSiy5TTc0l1NC3mJfrbNU6/JgYfwrmIiO4SAkg/9J09R84qWalOx7S65cEtylJycRYygMY8ylevy1w9Z0nL1E796+x6ZTWIcVnlG1uSafiym2SLG1JMXGRBdXHSglZbSQU4IAzgn+Plq5W60uIcZc7I6IsTcrb13q6EYx4+BPm8BVp6tFdKMcrdynzeKU6dc73YJmi58KGxlli5IUHEvEpJC9uAcdACQVYJFdG6U4msnT8LNg1E6eWMqatbiknp5CK6bZ0FBSSknw4dxym2006d0+fFdv8ABLe6Yx+Lmpvmh2numMfi5qb5odrc2OXuPdMY/FzU3zQ7T3TGPxc1N80O0sLj3TGPxc1N80O090xj8XNTfNDtLC490xj8XNTfNDtPdMY/FzU3zQ7SwuPdMY/FzU3zQ7VT1vxKiN7ZL9mv0VhhlS3HH7a4gJSOpUc+QCsbExzU2jKw0stRM5s0FxXvNslakvExV2PtisczUVtjXqK+3HjPMFxSI7W8AFJjLjEhBxllxX3VSup+IuvPYS4Ql3i0QbiyNNXNudDt7iUNszp6mHGFJU+dwTyeq8jclShtSSFDVOlC+q88DcKcsuhddB6ovUS76vhzba7qKWxqFENyZZkBlpKVQIbgdW3IlK2AB3GGiQduduSSaEOLOrLbpm7vWX2IjR7LZ7vfXW5zMiSuQY8+SjkpUXwUBaWuqiVbSeicYApVOL49hU5tLTtMLW9skXXiHqK+T3YcpcC/aVYtx7KoSIaHZ0YqSh0uEBKgtYUAkbt3XoMVLXL6oK/QTql+GYN0gMaaul9tUlyAqMhSojzbQSR2ha3EEuYKilrJQSkEHu15FNJPzoinO4t2JeTxG15YNQ3KNcplgmwrTerRb3xGtzzTklue6y13SX1BstF7IJCt4GCE+JxrbxH4h6gulrYiztORGLw9emY/Ntj7ioogyS0hSsSAHC4B1Hc25yM+Bo3dO17ebXKs8+Gnl2PFn6oG73L2kT4QiORbqLGi5wBCP+SruGzAEhT6ckBwKCUtL6J7xGcp9NL691ixa4Fvk3+1SbleNYXaztXGVAWG4Tcd6aoJKOf3yoR0obTuTtB6le3vS6UUuBCqSb0ZmaY4ma31zdrZardIsMN4Rbo5JmuQnXmpS4k7siFMpDydiHPf5Kl7fAbvGrLwe4oTuKbs2R2VmJBt8WLFlthJ5jV1KVKlsZzja0C0nwzlSuvSqJU4pO3IrjOTaNm7RTaKx7F4bRTaKWA2im0UsBtFWfRAwqZ6kfxrLwq/Oj7/AAMXFfoy88y1UrfnPClAKUApQClAKUApQCuAfrq/jwv/ADp/KUJR2FwC+0Tw4/Ju2/urdXyhDFKAUoBSgFKAUoBSgFKA0Rx21Q1orRt8vLsKDPTHlMAM3JeyOFLlNoC1qwcBBUF5x02+SqjpfjJa75a/ZO7wI0ifHur1qtr9iC7i3cFBlLi3IiwgKKAnelZxgKaWCo4GdK6bd2upvVO1l2HtAvWhdTaw0a/aRLkvWy2rk2+NBgLEWGzJy0lx0Bv7ErEd5tKSU7fsgUnwxhab4l8KTb5Vyt9uYtUV+Mi/B9dicjduQFpSmQ0C2C8sOOISCkFW5xOM7hmlwm9Cc8TJRrXhtb2rtONhTDuFwmJtNxiGwLTOlPOtlxDbrQb3uBxGSCcpOT18cZcDWPD/AFDqKw3OHaBO1DOadbjy27MtUyI2092d0Or2b2UodJbUFEYIV5ASDhPjcnPHgUPScXhZoSbJlO3K5T4lutUyFHcuttT2Z2I062JKGltx0GUoLbaGVlxRwSknKib/AKwuWlNP6A1TxCf0cmS83bJDk1mRaQ3NltNpILTu5G4oO0dVZTt73vRmqpRm2QnFLQxGtfcO9GNRW2bKbMiM03PeEexONotiXQWUOvlLeGNyUFOVYOxOThIzVo1VfNN2K7wrfcLcqbcrz9lTHjW9Ulx1EdTf2VYSk91tTjfU+BUMVQ4SuVZ42KPA4m6EYuepLzJ0q7bLm1djYHJqbItcm4ulPdQlSW9zm7bjb1x3c9CKn3NdaENzNnehJbS4hu0uuP2paYqCGi8iG44UbAQhRPLJwCrb0UcGXTl1IU4mGnXGgJzto1gmwOyLrcl9nts32AcM+Sjkl3c0eXzC3ysnI6Y6ePSvx678MbbeLjpr2Agtt3Ca1BuK27H/AJC7McCVtsvuhvllw70YCj75aQcKUAWSfC4zRZjwuIvDi4MWR2FYnZSYrzsOzhjT7pKdqFB/so5ediUoUlRR06AdSQDdBwv0am2uW8aSsQgOKStcT2NZ5SlJKiklG3BIK1kdOm4+c1DU48wnGXBGZdNEadvkZUa42C1z46y2VMyobbiCUAhBIUCO6CQPMCcVMNsoZbS22lKEIASlCRgJA8ABVtpvQrvbgfW0U2ioyk5htFSFgSPZiL/tH9hq5Tj68e8t1JXg+5l8pW+NAKUApQClAKUAqI1arbpuf/uiP4VbqexLuZeoq9WHevE05MZZmRZSH0hbKklopPlHh+2qxO0o63Z3+zzZTigkhKAhKlHPTHhXCSVz0tSai0TumNOo05aG2koCXF990gdVK9PqqRKFJiL5aXc7cYT1FVcCwmmV+5uoEF0lICSk90+Q1ubhU8ZGgbOsncSwjqfL3RW92UtZ+40u3/Zprv8AoW2ldAceKUApQClAKpuvYbE9SY0lluRHeZU24y6kKQtJyClQPQgjoQaxcT+kzLwv6qNM2HiLpXXFz1PbpVsaYjaXKyJNwabLLjH2aO68144QFMyWVeHRJ8ihnG1BxS4XI0zcbi/Ls95tanIdpmJjtNvhaHXNrCFp8FNArWoeKcBZTk5FarJO9kbjPC12Wbh7f7DqJm8K08zAbgxJiYpdgKRh1SY7JBUlIBQoJKEBKuu1CT4EVgJ1bw4W3emROsKWocZ1NwStLaUJjqcId35GFNlxSt3inco56mqcsr2ROaNj9OvOHkyzv3j2Usr8FUxqO4+AhW6S0A40kjGStKQlxPTISAodOtRcO9cNkOXO4sQ9NM2aZbEz5F3QlhKZbUh1TZCxtypK1NJ6nIWcDqRVSjNEOUGSszXnD72E9nJN0s6oEuSEqkuBJLkhjvDcMbt7XL3dRlGzPTFesbWWgWp9njsXGzNypiOdASjYkkSVEhSTju85QOPDeQfE1GSZOaHUwLRceFuoNQoYt3tZm3q0x9rfKZZLsVqOsApQrGUpaXt6A9wlPhkV4O6g4UXmy3eU4vTc63S5DRnhcdpwSnlDLSlo2kulQBKVYOQCQSAanLUIvTJBnWekrA/IU4bVa4dvZjRokllbeVtvI5iGkISNyUkIyEjxCcgYFTWiNLWvS1mdRanVyWbhLkXRyU4pKlPuSHFOqXlIAI7+B06JCR5KpaaWpKcW9CwbRTaKt2K7jaKbRSwuNoptFLC42irPooYVL9SP41k4ZfmrzyMbEu9JlopW8NCKUApQClAKUApQClAK4B+ur+PC/wDOn8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUIyKA03xa07B1Np27xLlPTbLe1Ibmvy142toYfQ+SrJwE4awSfAEmtV+5DpLiOZc3T99tVyYj3pd1YhvQ2p8OI87HDb7S2SoZS4TzsZSoLUVA4JB16p1Msqii8qer6G2unZX1LI9wTQ5d9LTG5sGC3YUJ5aLbam4rqiNxU2hxChy2FlQ3NbSDgdQcmqFprgS7D0Foy2a31K81e2rNFsVpZjx2wYElJZlKG5O5LykrgtkKICdjODkqKjTBOTtFa+bkvvLzE4WMK1RHlXfUXshqhy4s3t1TUYMIW3HZVHQ2hvcragc4kkqUSpR64wBjTuB0ZJsUdV9Zjoi3uVdWXlQ0CbzX5zk1bcd/cFNghSm1gBW9tJ6Dqahpq2hOnUxLVwKs+hFymY0mxMNXFUiLBXK0+w5K3vqU4WnXs5fRt3p2kJJT75RIzVmtfB+ND4SXbQj9xddiXONNiuPMthtMdEneChhslQbbQF4QjJwABk1Er2TfMKy0Kjq/htHD1xg3rW4io1DaA3fo6LeEiZGjENrcaO48glMhDas78ggpAIJq68TuGB4kx7dHNwatzUV3m84Q0uSmlZThyO9uBYcABG4buijkGqpQlFRm1o+HgE73VzDTweIvsiWbytUFeoG9QsxOzAKafDRbWgr3d5KuhHQFOD1OekQ9wislg4lm8ok2pqXfrguUyiXZWpExMoRzu5Mk9UI2tFeFA4OQFAKAqIpzdorl4ah2Rhv/AFOPP0pdLL7MQWm7jMMtaGbOhEeIrlBsORGuZ/k72QV8xKvfqJ29cVMngeRdXUe2KSvTr92jXx+1usJU65LY5SkHn5yEFxhtxSdpJUDhQScVTmuLHlfOBCLtw5t2kGrs21GiKeV2x6Al19tS1KUHWFbgWXkbztcGceUGtpNMcppCNxVtAG5ZyTjymqZesipOx9cv0inL9IqjKVZhy/SKcv0imUZhy/SKzrGjF2jf7R/YarhH1kUTl6r7i8UrbmmFKAUoBSgFKAVEavaL2mrglPVXKURj0dat1FeDXYXaTtUi+1Gl3XBJtbxT75SioD0g5/hWTAWChJzkVxElyPR+MWSCXOelQB2kdAa8HmmgnCpTgV5wo4FQWY3TslcquqCEwUgqCyV9VjyjBP8ACt3cLmkNcPrCls5R2VGP0Vvtl/5Gm25rCn7/AKFppW+OSFKAUoBSgFUviK3MdiuJt62m56ozgjrfBKEuYO0qA64zjNY2IV6Zk4f9RHOzf1OTmnrXBbtmpJK1CwTLDOVdHCtDqH28hxIAG1QfCV9c9Fr8p6yl/wCClxnt3JcK4Q25DtrsEWMl1tWwPW2Y7JyvHUIXvSnp1GCcHoK17kmbPJbRFgsnC7bdr9cbxKeU/cbu1dm27ZOkxUNqTCixyhfLWnmJ3RlEBYIwoZHjWvpn1PWqL5Luz941GxPflWmZa0vvOyXS4XpTDyXC2tRbZASzsLbSQPA5PglGSQlFs8+MHDrUMbVCL3aJUphubfkXBUy3QnZT0JKLWuL1baIcVvUcZT0AV18le9r4M3m+2LT10ZYiaXn262WlmJZ5AW820uDKcdQlw7t21aFJHiVIV1O4p6zmSSIyttkHxF4Y6ph3OHcjIeRLu1wn3C5SbHBkSWoxXEjRW46ENKDpC22ernQbgokJJRif07wTuM+4W29TbZGgRbgxapE2wPXOYlNteittgNoQ04GnwnlIxvHdUknKgcCXJW0IytslZHAF6fp6BaJF0baZbi36I+9HbIXtuK1KCkelAV1z4kVhscDLmmwyEPxbW/fFdkbbnqu9yLoSwHtjjbynFOMKSp5RSlGQAtxJJCsinMirIZNx4KX2VBmrfucO93iQ3bv84SFvQnkSI8ZxpclC2DubcUpZVgdNpUk9FVtjT8CZbbDbYdwnG6T48ZtqROU2GzIcSgBbm0dE7iCcDwzVE7SVkVRTRIFJoU9KtZSu58oWl0EoUlYBKSUnOCOhFfW2mUXG2m2mUXG2rJo0YVL9SP41kYeNqi88jHxD/KZZqVuTTClAKUApQClAKUApQCuAfrq/jwv/ADp/KUJR2FwC+0Tw4/Ju2/urdXyhDFKAUoBSgFKAUoBSgFKA0jxzs8/UOjb1Zra2tyRdJDUBZS3vDbLr6EPOKGRlKWlLUeo6Ctea20LdbVPYlsy7rf75cpjTsxiJHfixZTSUhpLXNYWOzhtKlOJU4pQKs5ydu3KwlWEIqm7XbldvktOT0burrutzM6Sb17jBuTus7ncNesWJ6c6zp2VMfhPNPbzKkvxmyhlAJwoMc18lCum/lD7k4zblotm9yrAYMHVHsJAYn3R0TpUtEl+ShlDDTeVr5qC4l147Rt3bTkYUc5eanSy5MqdtbpcXG6+DbT5aWZTrLj51MBdq1lCg2hi5m/ybexEs1sua4anVSHQhl92U6jYd5K3VR2lrT3toV16E1+WPSFxenaHk3y26i5LM68XBLC5cl5yEreG4bLikrOPsBVgqJHQpyd+FVupRim6Tir35K98rvy4cF3si0nxv5Z6o0zrWxaSscm3Ku8zVC7LOu00ypTjiDcOz7WIpSpWwALkr2owAeSCeozV3tEN618N77L0vHvjl4MJ0xW9QOPl6RKQyditj6iU7l4B6JBI8MYNYmIlSqJOOVJuz0V7Xdre7i+65XG6NdWfSd4fv6plth6mjsOC0QhOurz/PdSJKpE93DiippK0stNkYSCT4YIzltx9Rpj3lzkajkWqTeGFSbilEtmeuGtbrjrKI5UVJ5R5TXMZSnchZ2jKc1mTnRm2pON1bpa+mnDhe92vqUK66khpnRl9u98tsecb7A000q43BmG9PeQvapxlEVh1wL3+CX3dhV3dyUnoCmqlFsesZ0VN7i2/ULGpomnJCpUq4FxSTdH1tJX2Zha9v2FAfCNgCVAoAJAqYVaDk03Gy0eiV7uV7ad3ckGpW088C02bQdzubtjh+y+oXLPLuDsyeoiZA2tNRi0lrDzq3kh1xxKlZX3igkAeNWXhZBuCNUanekt3R6AtaXIk66CSw4N61lUbkuqKVBkJbCXUABQVjqQonCxFSE6c0lFO3K175rclror6W+DK4ppo2bs9Jps9JrQZTJuNnpNNnpNMouNnpNNnpNMouNnpNZ1kRi6x+v3R/ZVUY6oom/VZdKVsDVilAKUApQClAK8pbIkRnGiMhSSMUauFoaAucJVuflQXMjY4QP9k+X9lflidKIwac9+33Ovo6VxdaOVyj0Z6VSkqtKMlzRJhxJyCcZr2IjcsAkE1jCSkvZKDqJLt4vsWxW37LJkr2pSOuwnpk+gDJNdN2K0t2Kywbez1bjMpaBPoGK6XZtNxpuT5nNbbrKpUhBcl4/wCjPpW4OaFKAUoBSgFVXWYy61jxDRqxXV4GRQdpo4X03oi+RvqeI2pENQ7HCnaWtbQZalOSfZCUt+O4Jr7ZCEpUBkYBKlcxWVDArZs7iPqSzX6Tpa56njQWGdSi1OapkRWWlNMqtbc1CCk/YkuKcWW0qUMbRjBUQaxZJSM5ScSKtfEXUly1Loq8SdQqLLtn1ClmDGShqNfXIs1huM4hJzlT7WFp25ICjs6KVnGHGrVA08ZFq1Va9SSZlhi3RyQzEbLVplOS47QZUlCs7FpedwhZ3jkK73mjJF8ic76k7qjiDrTT/EI6YYu8Rbluj291DtyVEipunOeWl1SwtSVYCU7EhkdFjKt2QmsaZxmvcbRGpdQNajtz97YlPRjpnlMhy0tIuKIpkLysKw20rmLLmEEkHKE+MZIuwzsL4pajjxtPRbjq61WeHctQSLd7OOOQ5Dgjpgl9IcLauQh7mgpHTBSUEpyrFQcHirqoPsanevZlSfaVPuMKxIQlEe8Px3XA24231UeYhLbvcJISsAHBqckegc31JmZxX1HGhXiLY9YW3VYS1Y3Gb41EaW1FemXBMdxpSW1BKkls70pJC0g9VHKTV+4tX69aD0JbH2L6yJDMhKZ8x5UWPKkspacUvs6XtrJdylKtqsDYHMYOMUuCulYlTbXEqJ47vKuEiEi7RUy5GpbHDt0SS0lqS9AlRoTji+Ue91U7I72O6QR4pryPFXUmktK+3/UN3al6Wa1BPt8+EzCSjscRqTKjMuJWMqUsuojhRPTCj0GMlu1wG8bMVGvNT6PnXG931MKO/boGn5moy3GCVJjvrmIkDd47WMpWD47WV/fGo7VHHjUNks1kmOXBUS7yY0a8KtbkeK0z2SXNWlhlZcWHVrDICCGQSlSdyjhSU1ORMjO0bZ4Ko/zJqX8qLx++u1sPZ66tSWpcUtBs9dNnrqnKTmGz11YtIJwqV6k/xq9RXros15XpsslK2hqRSgFKAUoBSgFKAUoBXAP11fx4X/nT+UoSjsLgF9onhx+Tdt/dW6vlCGKUApQClAKUApQClAKUBDv6aZfeW4XXAVqKsDHTNfB0qwf+mc/VVp00y/vXwPGHoe325jkxEiM0Vqc5bLaUJ3KUVKVgDxKiST5SSa9vaqx+Gc/VUuGZ3b1G9Y9qrH4Zz9VPaqx+Gc/VUbtDfMe1Vj8M5+qntVY/DOfqpu0N8wdKsH/pnP1U9qrH4Zz9VN2hvmPaqx+Gc/VT2qsfhnP1U3aG+Y9qrH4Zz9VPaqx+Gc/VTdob5j2qsfhnP1U9qrH4Zz9VRu0N8x7VWPwzn6qe1Vj8M5+qm7Q3zHtVY/DOfqp7VWPwzn6qbtDfMe1Vj8M5+qvaJp5mJIbeS6tSkHIBxUqmkQ6rasStKulkUoBSgFKAUoBSgNZ8VNNnli5x09UAhwDzeeqJanhJV5Nwxn9Fc1tGllnmXM7XZNbPQyc0S6I6HTg9DXhc4qI7Rws9R5PGtMbeMm5KJK8D7BEbuV3ua0hUzeGWirxSjAJI9fT9FbhAOT1HoFdrhtaMe44LH/3M10Z+0rJMAUoBSgFKAVWdWAmSxj7w/tqzV1gX6Pto1fH1uzI0zZpiLYnk3OeIMaMFjHLDiwHB3fDltlzbj0Z8tZcnV2mZNuS69JjvxZkVqcEqYUvnNOFKWlbdpKio7UpTjcSMAdK1qmiqOMptXnpon8r/AC0I+7cRbHGgt3BKW5USHHly3EKjupkMCOUoXtaLeUkKWEndtwDnqM4/IGu7IVyQYZgMhDDshh6A+1KU+6ohCQyWgXCdiu8kqOUnp0zUbyN7Ft7QpZsvv4Pha/Tjx0/0W1tiLdm4c0x0ulKQ6wuQwUuN5AOQFAKQcYyCAfPVNsfECDe51yXFtrCI8Z+S1dLitxKWmWmVLQFLVt7ylbdwR9yg7iRlO6qUkrIv1cRGm4RtfN7uFr/BamPdLloS4ptU6UOUizzCiNEVBdaPOfYWgJ5BQFL3trXtwkgnwyanF6502q6xohdUuahSmmMQXlbXOWVqaQrZjmBAJKAdwA6ijqRfMt+nYf8Ad8n3dOwr8HirYF2VE2NbnosAxH7q809b32lqZbKTzWxytjmSpskhXdCgSakU8QrNd0qTNirbjNMx3XI06DITJS86shpKWVNZXkoWApBPVJwPLVCqxZZhtGlOy668Hwtfpx42XM+OJ+uI2hdEnViYEZ/LsRjtFxzFbjNuvoRzX1lBW222V71ZTkY646kZvDbUrmu9GR58+1M2195bu+GCVoWgPLSh9IUhKtjqUh1O5IOFjIrIy3jc2ikpcCLTr6bfp8j2HsVumWjtK4CpdyuQjOTVNrLboYb5S+YlKtye8pAJBx0IJx/dY0403Nl38MxkRJk1iMtuK5JUiNHc5Lr69iFctvmJUCpWE9B1raegOXq023Lha1lfpe+vBrlqi3vFxfAtSNaWFlL22ahHLlR4riQ0sEPydhaSRt98rmoPo3ZOOtYiuKOmktznDcHORDVy1viG/wApxfMDexleza8rmEI2tlR3HGM1jLB1pcI/Nc7fcqc4rmY0jjDpOLAYlO3J1IfW+0iP2GQZJWzgupLAb5gUkEEgpBA6+HWvuFxd0lcYc6XHvCXI0NlqQt0sOhLjbqihpTRKfswUtJQOXuyobfHpVx7OxCWbL2cV1t16+dCN7DhcmdMastmsYsl+1PuPIivmK+l6O4w406EpUUKQ4lKgQFpyCOmceOavWkgQqVnzJ/jWOqcqVXJLiimq1Km2ixUrLNcKUApQClAKUApQClAK4B+ur+PC/wDOn8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUoDUV51XxHYu85qE9pcQ0PuJZD8OSXAgKITuIeAJxjOABmsP24cUPw+kf7lK+mrTy2gotrKbeOz8yTzD24cUPw+kf7lK+mp7cOKH4fSP9ylfTVT+Ir9pP4d/y+R8M634lyEktS9HuhJKSUQ5JwR4jo9X37cOKH4fSP9ylfTU/EV+0PZzX+XyMSTxH4gw3ltSLpolh1DYeU27HkJUlBO0LIL/ROemfDNZftw4ofh9I/wBylfTU/EV+0l7Na4y+Q9uHFD8PpH+5Svpqe3Dih+H0j/cpX01PxFftI/Dv+XyHtw4ofh9I/wBylfTV4ytfcR4RZEidoxgvOBloOxZKeYs5wlOX+pOD0Hmp+Ip/4krZzeil8j29uHFD8PpH+5Svpqe3Dih+H0j/AHKV9NT8RX7SPw7/AJfIe3Dih+H0j/cpX01PbhxQ/D6R/uUr6an4iv2j8O/5fIe3Dih+H0j/AHKV9NT24cUPw+kf7lK+mp+Ir9o/Dv8Al8h7cOKH4fSP9ylfTU9uHFD8PpH+5SvpqfiK/aPw7/l8h7cOKH4fSP8AcpX01TOkNS69n6ihsXh3Tircsq5ohRZCHsbSRtKnSB1x4g9M1XDHqclHLxKJ4DJFyzcDaFK25qhSgFKAUoBSgFKAxrlCbuMF6O4kKStJTg1o2RYzp66yGCrcAvA9ArUbSinSzHQbGm964crEjHSFKyDWFdlBxJ8uBiuYfA6uCvNH7ovUCtLTXnQ3zm38b05wR6RW3bHqi33tpIYfHOA6tL6KHxV02AxMZwVJ8Uc3tjASjUliIap8ewmfEUrcHMClAKUApQCqzqxO59kZIygjI8fGrdRXiXaftGrNMcO5lmFgj3C7M3CBp9vZbmGYZZVu5ZaDjqitW9QbUodAkZUTjwxEtcFER9Pswhc25UxiW0829LilTXJZStLMctpWDsSlajndneSry4rW7i6s2YbwLlFKU7tX5acElpfotet+RIXHhYJ1umx25cOEp+1pt6G4sHlsNkuqcdVy9+SlzKAU5z3SdxJ6fd94cTL4zd+ZcYKpFzWyXHHoK1BkNtBKS1h5KkKDm9xKtxxux5MmXR6eeJXLBtppSXw05rr0fyLHpyPdY7Mpu4OpW224Gom/vOlpCEp3uKBwVLUFK6YwFAeOaprPBlMXTs20sXbY1cLc3Hm7o2UvykEHtJTv6bhlK0Z7w2jcNvWZUnNK76lyrhpV4xzys0mvjp4fMmrZw/TEfszzqre0q3ynZim4EIsIedUyppKjlxRylK1dSTnp4YrAhcMZsee0ty+pXCiyJ8qKw3E2uJdlFwlxays7lIDqwMBPQ9ajc8LPzp9ih4R6ZZJa8l/28NdNY/M8nOFL8nThtr11Y5qLGmyNLahlLaEBWVK2FwkhSUtpKdw96TnrgZV84dTL21di7cILkm5cgOLegLUloNN4SWgHgpKg4VuJVuON2PJuqNy7WuUPBSy2zLh07GuvR/I+eIfDJzW+nLDb0XJkv2iW3LSbvE7bHmlLLjRTIa3o3ghwq98MLShXXGKwODXCGZwn9l0P39N8ZnlpSAuKttcUISUJZbUp1w8hCdoQhWVJ72VqyMZSVlY2q0tqY0PgrJZk2eI/d4MrT1ouirnCYctf+Wtnml5LXaObjYFkHIbBISkE9CTHyOAM82uXbo2qEMRrnam7ZdFqt+914B9951TR5oDYdMlwKBCumMEHrW8W0Yp6w0vfjzvfTTRX5dOepaydpOL4TzndYLuTl+bNnXe03xVuTCw646mKlhCFO8wgpSUNrThA97g56EY1u4S3y3aQ0/ZRqSC6vTsiO9bHV2pQSoNIW3iQnn5cUpK8lSSjCkhWPJVr02m0ouHC3Pmk03w6PTpZcSrK+p9I4LupRcX1X3fdptuuMZUzsmA3JmKQpx9KN/RKQ00hKM5CUdVEkmvW88HDJldqtlzZgvRxa0QWn4ZdZZbhLcUhCkhxJUCXVHoU4IT44p6er+zp38rp9O/4jJ2ln0HpF3R1nlR5VxN2ny50idImFkNcxbrhUAEAnASnagdfBIrYelBlUn1J/jWvnNVazmla/LoTN2p2LDSrphClAKUApQClAKUApQCuAfrq/jwv/On8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUoDWd0/5yl/75f/AHjWLXHVF6zOvgvVXcKofHZm7yOEWpm7GmQu4qjAJTFzzVN7080Ix1yW9+MdapildGTSUd5G/C6Na3HVuitP6Yjjh4p+yImSWI8qXbI4itpIadKEvOPMuAK6EHagrJ2gnqK/NG6z13qpVukruclDsXSabquA3DbAmzQ6+2ErJTlIUEpylOOuMY8t3LpqbJ0llcqyvLz2lBfv9xvUK+3j2Wkanmr0lED7k+2t7GZJmoU7HCC2EKCCo9CFEZ8egxsaRrvU7/EdcBu8zGbj7ZPY4aeTDRyRatnWXuLe7OO/v34z3cVLii5UpwejXC+l+Gi1IyBxP1guzX1V1lXKP7WYqLZPfiR2kLkTlzCjnpUttSUoDKEqKtpADpODgVgI4qarY03bpkm+SXmY94mRTHi8oTrgyktcktKVH2Pbd5ykJbKwQR4GoyIj0elwST1+l+vDUm7nr7Ujlt4jPM6guDGqbd24Q9PogoLceO2tPJfSS2SolHXJUQrccDpWPddbydfa0guRZD06wxdVWZUBRjFtISqM+XVJJSCRvHicjI6UyooVGCvJK1v4s/fqQ8bi/qyVH1c/b7ncXGm9PSJ8dE5tlb0WU2+hASUoZSEHarq0SsjoSetba1y9ftKcKEra1QPZZLjanbpPS2wXEqc3LbCg2pDRKcoSooIHTPXrUOKVkUVaVOEoxy8Xy/2UJXEbV7tx0rbIci5NtaxjRFQX57DPPgqaczMKtqAlRUzhQJGOuQBUDqvVGotQ6G19Dev1xmyra/GlCbbA2YoZMsjlpHJS40tCACpCio/Ywc4JBqUUi9GjTi00le67uNuvVFhGvdUv8TYltiajQLa29bkwu3lLYusVxtBedCExyVrUSrqlaAgpGUgZrAt9+1/eBZHFavuMZN3ZvRcbbgx/8m7I7hnYS3kFXlJzkeGPGoyx6FO6oxSbjy+jfXqrG8uG94mai4e6audxINwmW6O/IO3blxTaSo48nUnpVjqy0amcUpNIVKaZ/wCfIvrV/wB01eor82PevExqy/Kl3PwNhUrrjkxSgFKAUoBSgPGRMYipBefbaHnWoCsCRqq0xh35zRx5EHcf1VRKcY8WXoUalT2ItlU1BxNZbQpqAlRPhzFDr8QrXNzve5uTNkrQywyhTzrzywkISBlSifMAM1zWPxCrSUIvRHcbM2e8PDPP2meFk1rbLhPTDYlIedUx2gFs5SUbtpIPoOAfWKx75rK0wrsba7MQiSEpWoKBAQkpWQonGAO4R6yB5RWujTzaJm3nh6lKpaUXy072l4s+nVqbCSlIcSobkqQcgjz18N3OQy4FNtlJHgrdgj9FRC0Xe5kOCqKzLnYOJ90hBDclpExoffLwsfHj9tbCtGurXdtiOaYzyunLe6dfQfCumw+NhUShJ6nDbQ2VKk3Uor1enQsAUFJBBBFftbQ5sUoBSgFRd41TZNPuttXS8W+2uOJ3IRMkoaKh4ZAURkUBH+6TpD8arJ84s/1U90nSH41WT5xZ/qqLomw90nSH41WT5xZ/qp7pOkPxqsnziz/VS6Fh7pOkPxqsnziz/VT3SdIfjVZPnFn+ql0LD3SdIfjVZPnFn+qnuk6Q/GqyfOLP9VLoWHuk6Q/GqyfOLP8AVT3SdIfjVZPnFn+ql0LD3SdIfjVZPnFn+qnuk6Q/GqyfOLP9VLoWHuk6Q/GqyfOLP9VPdJ0h+NVk+cWf6qXQsPdJ0h+NVk+cWf6qe6TpD8arJ84s/wBVLoWHuk6Q/GqyfOLP9VPdJ0h+NVk+cWf6qXQsPdJ0h+NVk+cWf6qkrPqWz6iLotV1g3Ms45ghyEO7M5xnaTjOD4+Y0uRYkqVIFKAUoBSgFKAUoBSgFcA/XV/Hhf8AnT+UoSjsLgF9onhx+Tdt/dW6vlCGKUApQClAKUApQClAKUBq6/yWYD9xkyHUsx2VOOOOLOEoSCSSfQAKrGjdf2TXek4mo7ZLItkkuJSqWgsLQpClIWlSVYKVAoVkHzVysoXbfadXCaSS7DJm60sNvuMCBIu0VuXPjuSoranB9mabKAtST4Yy4jy9c9M4OPpWpmE6TRf+yTOzqipldl5X+UBJSDt2Z9916jNU5GV51yJBVxhpk9nMtgSMKVyi4N+BjJxnOBkZ9dUTR/Gm061v7Nui264Ro0mNMlxLnIUx2eS1GeaadUnY6pae88ggLQnIOalU8ybIdSzSLuLxb1QkzBPimIoEpkB5PLOAScKzjoAf0GvtFyhuKdSmYwosoC3AHE9xJGQo9egI8Capysqzny1doD620Nzozi3FLbQlLqSVqT75IGepHlHkrEvOq7NYLZdZ8+5xo8W1MqkTVlYUY7aUlRKkjJHQeGMnyUyt6BzSRlC8W9TcRYnRtss4jkupHOPmT16n0Cvs3GGmSIxmMCQf+hLid/m8M58o/TTKM6PdC0OlQQ4lZQdqgk52nzHzHwr62+mosTmI2Xpm3Tr7AvL8cO3KA261GeUpX2JLmN+E5xkhIGcZ9NSW300Jc20k+Q2+mhTny1FiMw29PGm300sMw2+mpTTScXyL61f901eor8yPeizWd6cu5+BsGldScqKUApQClAFEJTkkADynyVR9R685KlMwiAB05nlPq8wrCxdfcU7ribLA4X0qpZ8Ea/uV3dlulx1alLV5Scmop59bv3ah8dclOtKbu2ehUKUacUktEeAircJIVkDxJ8lUPWXEyPbocmLb4TV1bIUy+qUPsLgIwpIT4rHiD5PXUQWZ3M1LePKjUemNZxNEXeNqGLb1wdPdo7I5AaeL646HACEpATktqI3IGDtU0pGcKSRB3+7W3in9Utb4XbHGWHFturhrJSoR2W8rQ4EnAUsBw9fJgYBNdXhNmKnGviMRpCEWk+smrJL4nPbV/qGtVrYWjgfWxFS2Zccsacszk+9Rv8ew6FncT24bxU3ASqE2QgIwUqA8+R0HTyYqz2e9QdTQe125xLiB0W2rops+YiuUlE6F0nTipXujLbPfCfeq9PlqQSje3VCuixULBpPXMmxy0Q5rhdiKOEqX4o/9K27HkIlMpcQQUkV1uBr76lrxRwG1sMsPXzQ4S1+56UrYmkFKAVR9fwY8udFLzDTxDZALiAcdfTWBjv0H7jOwSvWXvKt7DwP/ALKP/wBUn/CnsPA/+yj/APVJ/wAK5rXqdHlQ9h4H/wBlH/6pP+FUZjibox/Uy7GYzrUhNwVahJdtyxGVLCdxZDu3bux1AJGalJvgXYUXUvl5FwQzZXFSEoTAUqN/pkgIJa/2vvfjrzUvT6YyJBVbQwtO9LpLe0pyBkHwxkgesilmUbt9DFYnWZ663KCuAiMYIa3SJDKEMu8xJUA2r7rGOvhistPsCt1DSTblOrb5qUDl7lIxncB5sdc+FLMl02uBgWa/aT1BY03iBItz1sUtTYkqSlCQpKikg7gMHIPjUi+3Y4oTzhb2twCk8zYnIOcEZ8nQ/oNLSEqUouzR6ogWpxwNojw1LKA4EhCSSk+Cseb015Nosbzqmmxb1upXylITsKgvGdpHnwD09FNSMjfIKRY0uttkW8OOKUhCDsypSffADykeUeSo60X7Sd/tj9wgyLc/CjvLYdfKUoShaVFKgdwGOoIz4HyZpaRO6k1exIvN2RgoDot7ZXt2BYQN27onHnzjp569I8K0zErMdmG+G1FtRbShW1Q8QceB9FLMhwdr2Pb2Hgf/AGUf/qk/4U9h4H/2Uf8A6pP+FRr1KcqHsPA/+yj/APVJ/wAKt/D6FHiKn8hhtncEZ5aAnPvvNWbg77+Pv8DDxi/Il55lypXTnNClAKUApQClAKUApQCuAfrq/jwv/On8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUoDQnG7TN11ppe76etLiI6rrITDlyFkfYoi3QJBA+6UWt6QPOoVrF3gnfV3t6BPMG/6ce1FC1AS+0hpIUGXG5DfJ6jG5DDg85dX5q57Mo3Xazo8rkk+w+bPwTlWq+aYflabtd0gQpd/juR18oiNFlzw/FWlKhgpQ2kjYOqd+APGswcHtRDgAxpj2Wkm6I0ozZzZitjsSZCY6WyoL5XM98k9d+OvhRzTsSoPh55Ho7wYkr1c/fPYy3mc7rBN1MwhPOMDsQYUgqxnBJWNnhhR89QUXgpqSwabsrFotdmEyHp+8QXY76W1MuPyZcNxvKSkpVlthYyoEAhIIIqVUWibIcHyPvSfA+7LnW4322RpFqb1e9fFQpxjOFMdVpMdBUhltDW/tGFFKU4B72VHvGLX9TldoGhG7Va7XbYUx3SN6s0vkqS3zn33o6owUoDvAJbdGT73OB41O8V+PnUjdtrt/wBFjuXANEbUN8uFlsVpgrcvtinW9xhtDSmWIzsftO3A7mW23Bge+BwfGq3d+Bur9VXXVSp1utsZNxsF+taSgRm4zj8l5pcVaUttBwgBvcpTylqC8kAZyYjUXFvX+BKm+CJLWHCS+6ij3ZDOj7Z/nrTjFnhpfkNJ9r0hC3yp1O1J7pLzbmWu9uaAPgkjP1PwrFi0ZxOv0hiIzfjNN8t925YU9/krMd1ncoDcE82OcoHiFHp3qZ1ok/OhLg3fQ2Lwk0/MsWhIJubQavdxU5dLkjxKJMhZdcRnyhBXyx/qoTVy21jTScnYvx0SQ2021TlKrjbTbTKLjbTbTKLjbUlpsYvcX1q/7pq7Sj+ZHvRaqv8ALl3M2BSumOYFKAUoBSgK5rq6Kt9oDaDhb6tmfMMdf8PjrVikl9ZJNcxtOd6qh0R2WyKajQc+rI2c3slLBPRIHT4qx+YlSghOMmtNa51MdYplf1DN9m3jZYjm1sY57gPvv9UHP/7apWuNNJZaLTScNtpwUkjaD5wB/wCtXYK0r8jKpyyWhzepzbrCTMjIszVnjNG8S1qYRICcqZQrxUnzE+f01k6fgWXS+ulwoyDIvTba5D84E4bePkHoG7H6q3+3NoVKlKlgocIK77Xd29yXiZH9G7BoYeWM2vO2ereK04RSinbo5S07kdNvacXN020+FANraC0lZ8pHh5/LVSs1xk6B1Cma2VusrIQ+3jAUgnw9flFaRO6sVwUasZQ83N3F1qbFZlxlhbLyQ42seY1OobS6w26nHfSFfqqy1Y0lVtJEfdY4cZyB1FbD4S6hVdLX2d1W5xklBz6K3WypevKPYc3tuOahTn0bXxX8GwqV0ZxwpQCqfrdOZcb/AHZ/bWFjFeizNwelZe8re0U2iudsjo8w2itERuC99g61XqNJTIV7bXrh7HvSiY5hutJQH0tnupfQckHxwMZ8KqVkZVCsqeZS5qxBaZ+p/u8Gy3OFebfInSlW56CqRGucdlE0LkIc3DEcL3d3dl5SsHI6hRIwL7oLUbly0haLjpq03+4ptF3T2RfLYZbC3W+WtRQ3y+aAoE7UpBO4g58a7pmxWJhOV81uL56aPt88ibZ4BX4xBb56Ylziqf08l8uO5D7URsok5B6kEHAB8RUhfOEF9uHE63zWLTao9gg3OO60qK3HaBhoj8pTakhrmqV9zjeEbABt80XRa9Lg29Xa3zskRNk4KX2wWbTTDmk7NdWbJOuCpFtXIbS3ckvZ5Mg5QU7mwdgCwSB4Y6CpvQnAN2BdrL7Z4cC7xIOm/Y3KzzA0+qQ4spbChkBLbmwK6HA8maNrkKmLi08snfX6/e3uMj6m/TFwiR73c7s8ZTsdwWC3PqSQVwohUhCxn75SlE+faKqN04b3vQ+hdUXIQo9tRaJMW72Vtam5MntDTq1LQp1tCVONqDikJC8q+yKyRTS5O+jvpR5NpefmTNo4M3y0XHQs2HBYduEdlk3ibOcacaWVvrfkDlKQVBwLWShxCh1IB6JFRi+BF/g2pqHDs0BpmBqCTPLcNyOk3CO4HQ0cONLQFNBxICXEkYzgpwDU3RPpcL8fHt7eV7/yet6+p3nXKzPxkW1h4s6U9jLeLhJQ87Hl9pccGFhCQNqVgBSUjA6Dp47L4a8Ol6E1PqtyPDjQLPPEIxGIuEpC22NjqikeBKsdfL41DaasWauKVSDhfzdfybD2im0VbsjW5htFWnQ4wqZ6kf8AirMwiW+j7/Aw8W70JeeZa6V0ZzYpQClAKUApQClAKUArgH66v48L/wA6fylCUdhcAvtE8OPybtv7q3V8oQxSgFKAUoBSgFKAUoBSgOfOP2oZ2m9KTXrdMNtelXaFbl3FKUkw2pE1phx4bgQClDiiCRgHBPQGq3qy5vcI7Y1Ds96k3O4TlOOsxb32q6vENt7l7A2C4EnpuUpW1ORgdQDoHG7t2s6JSsu5IgnePt6ds1y1DHtEBNjtekoepZDLry+0OLksvrQwggbUpSpkblkHoT3fNHjVWprZxJvMa/SY774m6bjIatMt9qOgPPSQpWwnIJ6BSSSFhKSfHAq3aV/PQh1G7GTJ4/3+BpVnUr9rtjltu1lul1tkVpbnPYMVkuoTIJ6KC0jCikJ2KIT3s5rKlcX9a2a53Fq42ywqi2t+0qlGM88VuMz3gyEN5AAW0dxKj0WABtRnpG6j188Cd6xefqhZ1lReoyrPHfuun03Jy6xw6pCGkNLbTDO7B2h9L7LmSDtSlzx21lz+LOqLNqY6OlxLO/qORLgsxrgzzUwkIktS3CXEElW9AhOjaFDdvb6pycRukN6+BmXXizfLDqdrSk2BbnNRznLeLb2dayxJQ4pYlr6977Chh5wjzKbGepNViycX9ZQdJ6fbniJdr5e7xdIjMmNb3nEMMxnnwdzLR3LP2NKQAU93KlElJ3SqUbB1Hc2BdNa6oa4XW+/R9NOM354Mdrtq2lyFQwpQDqw0nat3YMqCE4URjy9Krsrjq41ZL9PiIgXFm3Wm0XBiS0VobkqmOOoVgK7yUjljaD164PUVQqSZU6jR9ni3qCI/Ou06FamdKQtSmwvuJW52htsulkSVE91I5i2QRg4TuVnyCDa42XRCzqR22pRb1WiHdXWVSHdrVtduDyOfyydocTG2uqOM9CnwAqtUk+BS6rPrV/1QtysulJt/gxoLrUdu43FmAY0l96VAiuFtLpW2NjCXNqlBxfdwpHQ4URfeG9wkT9V8S0vPuvNMagZbYQ4sqDSDa4K9qQfejcpRwOmVE+U1TKnlj57CVUzMvtKsWL1xSlhcV7wrm3Z5TcxxmTIQ1klqIwt51WRjuoQCo+PkFXKS9ePei1Vf5cu5kv7qED4G1L8wS/o6e6hA+BtS/MEv6Oujsc1ce6hA+BtS/MEv6OnuoQPgbUvzBL+jpYXHuoQPgbUvzBL+jp7qED4G1L8wS/o6WFx7qED4G1L8wS/o6e6hA+BtS/MEv6OlhcpnETiJEm9iSi135oJC8h+zSW8+HhlAzVTY1lFSkf5uvOfRa3/6a5rHUXOs3de9pHX4DERhhYxyvnwTfMgtQa4ZVODbUG5IKkg4cgPJUB1BOCnNYrusWYcRbqYFzW5jDYVBdTuV6ynFY/ozUV6y+KN5SxsXC2WXT2ZfYi7FeGLWhxTsG5K53eyYLvj5x3fJ8XrqL1hqpufGJEKYygAkrejrAI9ZTRYeTt6y+K+5ceNg6mZRlb/tl9jTUS2OezbE5touCJB5aQlJOHcZGfR6apHB/Qd3W7edU3yE7IRK3NxkKcCSr7IncopJyOo8tbZbiji6lXEtWy2jz105d1zKVfGYrY1LC4BST3ic204rL6zVm+N3a9vedU8PHVTNIJLmSkvKUhKnSQ0kBI25J6EEHp6aitWWNExIc5qmySSdhyPV1HkrUVZwdZzgrroXcPCpGlunJqXXt96ZlcM0zJrEi0KvcqL2YFbDTKGikjPeGVIJzk5xnz1sa06fuEiIpI1Nc08tRSEhuPjyHytemqZVYf8A418/uazFYarFtOs+PSP2PqRpe4FBB1Lclegtx/oq+uFVluUm+TIzOqbnC2r98y3HJPTx7zRrO2dUjKq1GCWnb9zQbTpTjhk5VG9Vxt0fRG4vaLevx9vv/UQv/L09ot6/H2+/9RC/8vXRnJj2i3r8fb7/ANRC/wDL09ot6/H2+/8AUQv/AC9APaLevx9vv/UQv/L1E6jt8zTsJXPuM7UT6G1uoMlLSXVYHRtPLQhPUjpkeJ8aw8XrSZm4TSqjUXDrjIjUumI95vL9pZTKlxYKIlsdddfhynlBHZpKFISptxKlJBJA8SSlIAzNXPjFp23SkxEi4z5q5MqG3FgQHX3Vux9nOACQegC0nJwD5607pa2RulUVk2YUnj7o1hth1EqbKiOQItzXMjQHnGY8WQpSWnnVhOG0koVkKwRgkgBKiMx3jJptpy8J/wA4rTapYt7ziIDpQ5LLiWxHaO37I4VKT0TnorJIAOI3LJ3kWfUDjDp66TrdBiC4SLhMW82qEiE5zoxZcQ26Xk47gSpxAz5QoKGU9a8pHFi2zLY5MthdEZq7xbWZkyI6lh5Tk1MVYaUB3iF7k58AcE905pumuI3ifA/Uca9LLmPMl6ahlIlhma5CdTGlKjBRfQy4RhakhtZwPfBCinIScSOiuJVk19IlMWztrT8eOxL5U6G5GU5He38p5AWkFSFFtwA+dJzjpUOk0rkqom7ERJ43adQ5KajouEhSBLTFeTBdDE52MFF5phzbhak8tfQeOxRTkJOPHRXE5bnCCy6x1UsNPXFphaY8O3PNLLj6kpaZbaKlrWoqWEpUPfZCsJFVOlZdpTvLvQk3eLmno0W4OyzOt7sCG1NeizITrT/LcdcZQEoUMqUXG1JAGc5QR0WkmLs3HK03G23GVKtl1grjXh2yx4xiOLdmvpWsANJCcqOG1KUPuACVEAE1G6bJdRGFL492dnUdkQh1Qs8633Ba2nIronCdHlRI6YyWffbyp9wFG0klIIO3JMm7xhtdrkyIs0SZs1Vzk2+LEtMF591ZZbQtSSkJPeCVZJ976al0SN4j1e44aRRHgSGpkmVDlw41wMuPEcW1GjyFFLLjytv2MKIV0PUbSSAATU9pzW1t1ZcLlGtyJbiID7sVyWuMtDC3Wllt1CFkYJStJSfUcZHWqXSaV2VKom7IsO0U2irdkV3G0VZ9EDCpnqR/4qysKvzo+eRi4p/ky88y1UrfnPClAKUApQClAKUApQCuAfrq/jwv/On8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUoDTPFK7Wiy2S7PX6ELlaXpCIb8RTKXUuh95LISpCjgpy4Mg+TPj4VQnNJ8LLZOvFgl6as1ujWSPGu0qTMjtNx0pkF9pJLijnwjrSoK6bSkdR0GheZN2Z0KytK6Jxu48N7RpgXBM/TUbT06Ei3iUqSwIkiKyFISzuJ2KQgLWnb1A3EeWsbTVm4XwEXBuxN6ZR7H9nlzexuMkx+XuWw66Qcp2gqKVK8B4HFU2mVXg7HgzbuE0Gy3TUjftUZs91S9DmXUOsCM+l0nmtFzO3vqzuSD3j1OTUxcJegnJEdEyZYC/e22HWEvSGgqchg8xlSAT9kSg94EZA8aNTZN4cin6a1LwsuR1TrNWqLDLg6qdbt8pdwkMNtKSw0WhGUFHvZy4vCskh0fc4qZnac4Z2W2T9LC26fU4417JrsaVsJkP8tOUu7VqByAgBKyQE7R1AHSpqadvOhSnBrzzFosujLjrnSsqHLtsW7WOzPIgaeYdj82K1J5JU5sQonAS2lIKe7hajk5BrIuWneGaborTU1jTrV0uEpVzFrdcaRIdfUFFT6G8hW45XlSR1yvzmoefgifUK1b9eaM1zwq1TcI9kt9x0lpftQjxoslp3mCIHASlKD9izyyUZPVC0q8FYr1vS+Ft0sNsvcex6f1EmDJt1iQ1CSw6YPOktMtNEJyEBtT24J6Ed7GM1VlmnZMpzQa4Fxb9oupBftJNvWK5reLrl2syHWnVkrP2RTzQJIyT1Kh4mvFi78PL3arvPYuOnJ1tjQ/Y24yW5LC2WYw3fYXVA7Uo7yu6rA6mqLTK7wK/ZYHDXifYJq12KyS9P6aU5aWJDqWnIoipZacJQod0M7VI6eA2A+QGrbatT6MZZvVxtt5sgZTLSLnKjy2tqJG1DKQ8oKwF7UNoAV16JFTJTehCcOJlxteaZmzbdDj6itL8u4siRCjtzWlOSmiCQ42kKytJAJBGR0NeTvEfSTEWfJc1RZW49veEeW8u4NBEZ0kgIcO7CFEgjBwehqjJIqzR6kl7YbUW1r9k4e1tbTSldoRhK3doaSevQr3o2j7rcnGcisB3iDpZlFyWvUtoQi2FInKVOaAiFSlISHe93MqSpI3Y6pI8QaZJMOUSXt9wi3eCxNgyWZsN9AcZkR3A424k+CkqBIIPnFTWnR/nqN61f8AdNVU168e8oqtbuXcy+UroDnRSgFKApF24u2Kz67b0s+twyyyHHX0Jy2wo42oWfEEjr4dBjPjV2QtLqEqQoKSRkEeBqxTrQqylGL1i7GxxWArYOFKdThUipLuf8WfvKHxRTg25Xk74/ZVNjK3N59dcztFfnv3HTbN1wkff4lfusREu/gkZKWUj9KlVB6vnNp1DDtrag23HaC1ZGBuV5f0YrEleWnRG+w9k0u9mBcLgW47bScuoB2lY8+PNg9fCqdqpoP2x6CuSjbJUGlupHUAjOADnyZFXYyUGpdDLpU3Ulkvx0Nd6rntW23JjQjy1FWCU5wE5AKzjxPTy+Y162+OLJGU1uL7C2csvJdWG056lGwlQB6KOQR4Hxq825xqZ9bp279PobetlpOjCk3HLJXS4OLT5djS7veWvhrdH4jrcVlaG403LbilDO1QJIIPmIynPq9FXLUMJS4uUpxywckqyFkjrj/55asNZUra6I1id6rvzbKdZrguzaot8w5bKHgFp/1D0P6ia33YnlJdltqTgc3I9RAA/ZVDV4NljGq7VuhJShhs+isDhCd2r5+Pv/4Cs/ZetZ932OV2t/aL/uXgzoCldQcYKUAqm69ZVIUhpt5cdxbKkpebAKmyegUAoEZHj1BHnFYuK/SZl4X9VGg7rwaadlzLnetX41Pc5FubiXARGY6A5DdW+wnk5w6skubsnJT0TtArDd4KahtWpbDKs+p3kvNyLrOm3Z+Kys82UGcI5OACnKFYxgjanJPXOsU+q82Ns4dGYjX1OslNyudkYvMmDot/T1rsjzexp164IYclF4KURltRDqQVAYIdXgAgFNwv3BG2ag0tdbNIlFwTL0b6249HbdQ0/wA0OBKm1ApWjptKT4hR6g4Ih1OiCpq1myIf4M267NWuzN3e1RXrLJbnvNWm0x4zzC1OBaVs7DujlYbKCrvb07h6a9YnBSDdZ9zu4vqJAuM+BOJt0VpllxUSYJKVuBBIcdUUBtTvQ7QBjpTO1yJyJ8z9RwDaUy3Af1BKkWOEZ7lrgFhCTEclNvNrUpwdXAhEh1KAQMBXeKiARbdN8Po+m9QKurUt15w2aDZuWtIA2RlvqSv1q55yP9UVTKebSxKglzKPprhnY5919jomtG7zb9PSZj0WzR1Mrctz8lLyFc5SSVHYl95KEqCcBXUqIBFruPC5ibw7sel2bk/EesqYSoVxQ2lS0OxSgtrKD3VA7MKT5QogEdCJlJ3WgUVbRlW1xw3em6l0LfL5d0PQdPmVKvV1kFqK060lTb7La0DoEofbZcSc90MHJJWScxHClmfJuC7VqsolwNROXmGptht026Y404H2nRnvpW3JV3TtUkLSQfA1ObRWRGXXjqYkr6nC2zXrfOdvU32eg9ukM3VDaEutzZMhl9UlIAwnaWOWEeBbWpKsgnMpYNE2FnWUS4x9RMzbim43O4CK2tGVuKQ1HkJABJw0oJB+9UsA46Uc3JWsFBJ8SEt31M1ntMiyvMSIctcK2w7W+q6WliYp1Ecq2LbKxlpZC1AkZBwnpkZqRuPDa42HVFw1Ha9QRLfdLvIYiKWYrUdBZMhC1lwDIkP8tJabUQCAr1mmfNxQyJcGbV2im0VYsXRtFWfRScKmepH8aycMvzUY2J/SfnmWilbw0IpQClAKUApQClAKUArgH66v48L/AM6fylCUdhcAvtE8OPybtv7q3V8oQxSgFKAUoBSgFKAUoBSgNG8atFu8Q9K3awM8jMmZHWsSSQhTbcptxaTgE9UIUB08SPDxrWt5+p/ct2obtctKRLTEircssuPbH3HG2pD0J+U44l0hCtgUl9spUArCkAlPTrpVLLddv2N5lukyQ09wfuqNU2y/3ZFqQ77Ozr1Jt8danmoxehpjIQypTad6u5vUopR3nF4Hnq7n1Nl3Og7DYo8q2QnrfpdFpeMda0IekolxpAGQgHlKLLiVKxuHNJ2nJqVNIOF0TkPhPqG1TLXqGDbbcu+Rru9cZNvuF9kSWpXMhpih3tCo+UOpSlIADZG0EZBV089M8HNTaTuWmXrc5bob7Kyq7TWJrnLcYXMfkrhpiqZKVoRz1Jac3tqTknGBtUck1YZWYdx4TcQF6KY01DkWhmE0q6x3OVcHGVSWpDhWw6pfZlKTt3LC2U9Fd08zAxWazwh1VElNswXbfb4b9mEC6Oie48m4upgdnQosKZw0pLm37KheS2jCkkq7q8RlZLaD4bak0jrKHJbVCg2XsTTdybYnOSPZCQiKywh0MrZTyFJDQSVJcIUlKcpycpxb3wn1DcNR3phoWlVku+o7fqFd0dfcE6N2bsx5CWg2UqyYoSlfMTtS6runHepur3Jyu1iaj8NbkzwZ1VpEvxPZK6+zfIdClclPbJEl1rcduRhL6ArAOCDjPQmrN8GtS3N56XNbsdpfT7XYrMS2vuLZWxbZ/aVuqJaRtUpKlJQ3ghO0ArwcgmlcOLdjDb4DakuVjhadmybZbYNqg3eJFvMF9xyZLVMbcbC3Gy2kNkczmLwte5aEkYr0ufBzVuoXUXaTE09bLjb2LTHi2qHLdciTBDliQeassJLaTjahIQvZ4knOKrzIpysl1aGvdh4R8V250SD7JX72SuDMGzFbyUl2IlAbBKElaypByQkbic4BOBCR+C+odVWpx66wrFai9G07CTbYrjjjLkeBOEp1TgU0napSFKQlraQnGCshWRCaWpLi3oTU7g3cH+KFzvDjbNyslxusG7AKu78VcRyM2yhKeQhpSXQFMBYytOd6knp1MT7kGr2bTNiRY9vjR48pmVYobV/eCrS+EyEuvtSDEKtqg8lIjrQtISXOuCE0zKwyvkZNz4Ta2l3B1vtNkmRrjcLBdLhcHHXWHUuwXI6n0tsJbUkhfZwUkrGNxBHga9rjwRuo0TbYcF2Gi7QNVztRKQzKciolpekS1ISp5LZUlaW5KDu2KwpoDw71My0SGVs2Nw50t7TtIxbYYqIbodfkOstzXJiQ468t1ZDq0IUrKlk+9GCcAYFXfT3/ADzG9Z/7pqiCvUT7SZ6U2uwvVK3hohSgFV/XeqkaN0xMuRb7Q+gBEeOPF55RwhA9ZxnzDJ8lUTlki5dDJw1H0mvCiv8AJpfE45l8TLfoG+sdsegXa/3OQXZb8pw/Z3FK6ttYPTr3Qo5AOAARW2JfHV3SnsXJtbJudunHamC4dq0qBIWnPXapJBz4jp8dclSxG6k5Q4rj234nbQq09v7QqYaTy00nl7FFcfgja/ExPNtEB4DGHfDzZSa17AkboqlA+CiP11f2jG+IXakYuzH/ANHfo2YUlaUXxkHxcRgH0g/+tUmSkX27TZidylNvqQkDAKQDjHmI6eH/AO2sGK4M6CjbXsRE3OJMLjpQWXgkhRPMKN2PA7Qk/t8/pqna0jT4UNmQFh3a8jDKMdcnByo+OfUP21ehBVJqC56GZGqqC3j5a/A03rS4dsVPc5o2JIARggoO4dPHOf8A55aik6leTYjb475CkkrKgTlasDqB6h5KsSzQzKWlpLT4nRpwqxThqnGTv0VlZ/T3lu0nreVEYiwpQBLawtLycAhQPl/QK6QlsNzLUl1G8cxIWknPTzdP8KuTg4TaNJUacYT6+X8DWGp97CUpSBu3gdB4KJxn4v4V0FpuT2xhEoEHnMtudPSkGonG0DExLTXuJG5TEtxVlWBgGnAtky7tMlY6KcODWw2VG9SUuw5DbLy0IR6vwX8m+qV0pyApQCqRxKdlx4LzsBpMic3EcVHaV4LcAJSk+gnArGxCvTMnDfqI5WjmTrrS2hEtao1HPu673b3bo6/EDTlvfMWRzkoBYAbOcpUjrs7uNpIJwLzq/X7NvtMB69yLZGR7NxWrzNcVHU7Ij3FxiKXVIiuhauQhCwghAdyo97HTCyp6NGxbfJlgv72upl9ur6NS3eD2fUFmtjbFvjt9n5EiPFEpxIcaUT3nXFAqzsKfAd4HFVqi/QpMS1al1Jf7Rp2NIvcdN6jRwZMh5mU2mG24sNKyeSp0pG0cwoGdxBCqcsWtES2+pg6MvOuGuJdknajjzLXDudssib5IixsOdvLL3LZcGCG2VOKUFlPVKy2klKVKNYrt21yNHXudDul3tj1k0s9dYkOHEbQ3ImomTNqVpLZKgUNtgoGMhQJ64NVOMSlORaZGp9ZPcZrlCdu7dojxbq2iLBfW6pMm3dmQpSm2ExiHCVqcPM5wKVI2nABCrFwR1Rd52kr4me7M1HcbYsD2QDpVHuLnJCiWNzTZbyropoghtR2hRAqiUFbQrUnc09wGvkzSutdOxXYj1wRcW0RPZIQXm2LLFcS88ba6NiSmSJRbSHHSregddiuity6sv8qPxhjW653u7WOziDDdtbduj727hLVIdTIbcVy152oTHG3phLqleTKZnG8iIy9U1A7e9b6u4Z65bvVwD053TFxFxsG9119mZlIbCG+zNhlKRzEBIWveChQKsFR3Dp6TK07beMNy7HcHFs3iTLZagtBUl4Jt0XHJCgQpRKSE5BBUMHPWkorgvPARb4s1jadf6pft1xbk3e5R9Ni929D94jOLluRobsdxTpQ+qM2SnnoaQpQQeXzF4UMd3C0jpm73WZZHYl21BALbes5CLmmMliU/uuUVTJcC2sJK8BeAlO7b4YyKqcVHz2FN3Lz2kzbdb6/vmrLCmbc0WN1+NZX4zElTjTctLrba5gDKYyw4oqLrZy4jl7UKwkZKsq36p1BKvUm3uTJl+cZ1LbXDcUNlcVLDkxwFkNLZSY7qGwkLSCvACF7gVGocI8icz5kQrXWtF6dnSoN3vUnU4sN5fvdseiYatUltpRjBkcsbVBwJQgZVzEEqO7G6tucNfZ236y1LZ7ndbjeYLUC3TmJFxQgKDz3aEvpSUISNv2Fs7cd0qOMAgVTOCS0KoybZsjFWXRo70v1I/jTDxtUXnkU4h/lMs1K3JpRSgFKAUoBSgFKAUoBXAP11fx4X/nT+UoSjsLgF9onhx+Tdt/dW6vlCGKUApQClAKUApQClAKUBpfizqZWkrHPnNSm4styY1EjKXEVK3vOvpbQgNhaMqUVhIytIBIJOAa0xaOMeudURrbCgrs1uuQY1AuXImQFOhSrfJYZbCW25JCSoPd/DiwCDg1qcid2zdZrJJEPrXivfddaYW83MtVjtrEjTS3Le62tUqUuW7GfJbd5iQlI37ANit3LcyR5NmcYOKVz4ZXCIhqLGlR7tBfjWtLiVBTl3CmxGjqIPvXQtXgMjlK69ehwWkRm4sgXeMV+Y4kQrLHet90t8mRNtinFQjEbalxoi31AOqkKWoBbe1R5QSAsYUSnvY9n4uaouyrJp96dbLRqiTPfi3J64WtSWoC2oiJHIShMkpdUsOBSVpdxsz03JNMiGdkmrjNPYnS4Tki0PSW9W22wsloEB+O/EivLcQkrJJy86UkZG1I8cEmg27jDdtC8OLGLU+xNdhMCZPhPwis8l24OspUp9T7YRu2rSkJQ4rcnJGOhlQXAOZujh9qLUGqNQ6scnv29FmttzetkSJHirD5KAhXNcdLhBzuI2hA8Ac9cVrfR+v9ZmDarRJ1FaXbleNS3uCi5y7evbFbiyZWGg3zxvUoNp2AqG1CTneU5NKiuBLk9NTCmcd9ay9LXG7wPYJgWbTcu+SQ5EddRPVGlSGSGSHk8tt5MfelR3lIWPfeNWtrifrO7cT7lbLXY2XbBarvGtkouBhKuW5HadW/zFSUrBHOG1CWFBQQcKyruS6cSM7ZuXA9FMD0VYsXbjA9FMD0UsLjA9FMD0UsLjA9FMD0UsLjA9FSOnx/niN6z/AN01cpr113lFR+o+5l4pW5NGKUAPhXN/GfU7utOIqNMQ3yiDamVpecR1HanEEZ/4EqHxqUK1uPqZKNurS+v0Om2BhfSMRUk+EISfvayr5u/uNHz+BsxWoDd7245DjsBsKbefbdYdLSipCkgDeepJx0B6Zx1FWOwW5p60dsaQvsjZcbiKd98vJ+yOY8mSNoHkCfTXMU6WWVnxM7ZOz6mFoVMdU0fsR9/H5JnUvElwN6QjKz1CkkfINaqtUgFtbGckP4x6Op/hW1x6vWj3eCKdmf2k+9fN2PWWErntKX1CSR6jjP8ACqxp6ItifMQ7jkzFqU2PKojyevp09VayDukjoXZRb7j3uUVCm0jYC42QnYAAVjy7QPAftrW2qpSRHcbcdS0lsd5SjhPx+P8AjVxvVW4mVRtJa8CqaQ0VbOIWqo9rukRSrY9vBU2S24VBtRCwR4dUjocjzjqamtX/AFJ1h07Y7leLXfLmFQo7j/IlJbcCglJyMpCceWrc4RlUV+P1L8sfVw/5UbOLXDpcpGnOHPs5cGYzcguqwCo7An/tZrom4wBbbIw0VjKWg2Ac93GP/WsiUbNIivUzSS86mqNRhuStyW51ZbJCMZ994H9Hh8Z81bs4fOczTUFWQTyGxnHmGKisrQMWtrEk7+wZEBxIOMj9FXLgVbm4tlyMcwEg+us/ZT9tdxye243p05dr+htaldCckKUAqqazWGnWlq96hoqPqFY9dXgZOHdqiNCNfVN6Ydtnsh7FaiRC9jG73z3LYUp9j1ZzLzu6Npx1B75HVKVJ61Zrhxeslu1E7a1sXBxhiexa5N1aj5hx5bwRymVrzncrmtDISUguJClAnFYDpM2W8TKnavqh7ZbNKRJmog65O5UubMFvZBREiNSnWQ8sKUDjDZ7qdyjtWQnAOLPP4zWSBH1JM7Jc5Fo0+xJdmXViNujbo7XMdbQrdkqCQRnATuBTuyMUdJhVFY8dZ8Tk2q8w7RbMKnoudqjzA+2SkMS3FpBSc++w0r1dPGsAfVG6QMeXIUuYiK1FdmRpCmk7JzLbqGVLZwrIG91ofZAjIWFDu5UCp3QdRI2LZbiq82uPMVEkQFOgkxpYSHG8EjB2qUnyeIJBBBBrO2+qqHErzXBTkU21GUZht9VNvqplGYFOfNTb6qZRmG31U2+qmUZgU581NvSmUZht9VWLR4wqV6k/xq/RX5iLFd3psstK2hqBSgFKAUoBSgFKAUoBXAP11fx4X/nT+UoSjsLgF9onhx+Tdt/dW6vlCGKUApQClAKUApQClAKHwoDVXEAWZuyX57ULEWTY2Gnn5jUxpLrRaRlaipKgQQAnPXzVrzRXEDQ7vD+BqKXbIOiY6H5FrVAnstMuRH95DkfCRjK+UlW1PvwEHr0rVtN3sbdNJIxrrxB4TM3vTRfassqRMtzztsuCYbbiGmI7iEFsL25RhasBAHQoVnBFXWJe7LqLRFo1Rdm7eiB2Zm7IeecQ+1GUUbg4hzGCQFHC04znp41DjJcScyfArz2p+FjbZ1U69p1Ds2SqEq4uR2w+9IDSgWVEp3lzl7hsPXacYwaxrleuEFp0vbbfK9qqNPTwifEhiKyqK4FqCUPBASUjcohIUR1JwDmlpEXie8nUvD+2vLv18jWC2T2pj1tYlrS06+sRnD0SpKdwCNoUU/8AR+XGM193m8cK03OyxbidNPzXEN+xoXHadIQ+SUctQSQkOEKIwRuwSM4NLS5E3iTEPiHomO7ekRb3amVQEvzbgW1pSEBs7XnVEdFbCAlahnacA4NQrl54UX+wXmes6YuNqcmN+yK1x2nEPyVAcorTtPMWoKTsOCVAjbnNMsuQzRZJjVGgJbFrjol2WQ1eoxhRGUIQtMiOFcstYAP2MLVsKTgBStvicVg26/6Gv15sVzuESxo1VJStEJwoakSUJS4psbXgnKUlSSB1AzlIyQaWkHKJsTZ6qbPVVFiq42eqmz1UsLjZ6qbPVSwuNnqps9VLC42eqpCwpxd4/rP/AHTVcF6yKJv1WXWlbU1ApQEJrXUzGjdKXS9SMKbhMKdCD92rGEpHpKiB8dc48MNOTFx3LzcQXJk1apLzih1WtRKlH9JNaLaN5zjBck39Du9h5cPs/EV3xm1Fe5NvxRE8SJz+orxEsMNZS9MdDW4fcJ8VK+JIJ+Kpi925m2W1qHGQEMMNhttA8iQMD9lauirqUvcbvaMtzh8Nhl2yfv0XgzbPFOcBZ7NFB/0hQSPk/wAM1rG0EJu5UVe/deSE/wCz5f8AtGsraDe+Vuj8DQ7LUfRZX52/+TZlTpaGn2QpQClugDNQtoT7JWh5CVKTIafPhjKVJPQjPxfo8PGtWrxyPvN67NTXS31Nd8ReLbukk9mdQzOuK8JZaOUqPXvEgZ6dPP5Kodqt9y1NKF0vzm4lW9phKgEIz5k+TpWypxWXNzfAmGjyrguP2Nu8JbCV35NxaaxDjoXh3b0KyCnaD58E/wDw1ta525q82udb3lFDUyO5HUoeKQtJST+usCo8s1bkWK8s8nY5rumj7hotvsEvnNuoUSh8juOq8ikq8oxj1Z8lfg1zd2ozEe6pVKitKP2aOnc6oY6BQJx5uvj08PLW09Wrr8DOTUoqUdTEvurLXe4OID+0sY50dWQofEeuP/ma33w9Ty9MwE4Ayyg+rKQaxsRFxp6lirJTyqJY5bfNjqSfNUlwduvZrnJhE9N2QDVzZkrVmuqNDtaObB36NfY3TSupOHFKAVVdZNh5xts5wtpSSR5M9Ks1leBfo+2jR/uLack2F/SaLzIU41o5Gk3G0utl9uIUFCXinHRZwcEjaSD0rPn8G4MnUEiSq+XBq0TrrGvEmxjk8h+awGi2rcUcwJzHaWUBWCpGfAqBw8yZmrLJXTIONwF09PcjSrVdnRJgokW5992HFlcxJkuPFGHmlBKkOOuAFI+6IUFdMT0jgzFkx9VWxV8uadNajRM7XZEBkNoXKbKHlIc5fMAO5SgndgKUT4YAZriKjJXi9DHgcEGW7k5crnqO63q5OybdKW/JQw31hKcU0kJbbSAk8w7vKfHIzUNbuGOmuGLDtmY1SzaG5kdUWDHkQoHMaaW6k7SpTO51HTljeTkKwSVbVBKSS1KakoUo55ysu0ueh9AscNbPChxbzKVaILEgKivpZQxvcfLxc7qBywjKkJQkhCUHGOgImmdbaek2t65M322O25hWx2WiW2Wm1eZSwcA9R0PnqiTjfViVWnSeWcktL6vkuLDutNPsWhm6OXy2t2x5fLbmrlthlauvdCycE9D0z5DSdrXT9sTEVMvtsiCYjfGL8xtHPT07yMnvDqOo89UXh1KHisOld1FwT4rg+D9/ImAsKSCMEHwIr9z6KryoyRn0Uz6KZQM+imfRTKBn0Uz6KZQM+irDpHqqV6k/xq7SVpos1vYZY6VsDWilAKUApQClAKUApQCuAfrq/jwv/On8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUIyKA0vxe0W/wAQ7DO0+iZ2GJNmNic4nO9cVLwU62g+QrSkoyfAKNUSPwMl23VLkuLe3JVqcvUK/LZuBLr/AGlplxl07vDC0CNjzFtXnrAva6Nqo3SMiy8JLvp2/wBiuUSfBe7HPvrkhp5Cxuj3GaJPcI+7RsSnB6HJORgV+W3g5LsPDG2WK13+TA1XDsEe0t3BubIEbLTbaFLTH5mxO7bjelO9O7IOai6YymFovghdLBeYNynXJh5xnU71/W1zZEkhC7V2INB19SlrUFd7co9R4BPQDWmoOF2qtI6iixYkGRfIsa2REtw48Z7s11ebuEqSGXHm1gMBBcaO50FJycAjcFVJpkOJtBnhHqPTuo29Q2K42t65Jk3jMe5NOcnkzpLT4IKDuC0FlAI8FAkZHQ1rS18LdRaA15a7bEhTb5a7auyx2GFxXW480MNoQuYp5tfKaLJU4oNuAklsAZGzBNMOJaYv1N0qHpC+abcmw3jIstxs1rukiZMdeQmSPKwtwtIwEp3csd4oBASOlWHXvA6bqrUM68xLk3Hf7ZbpsWOHXo4UqMzIZWhbjKkrSFJkHCknoUjII6GHInKfKeDN1hq0y5aHLfYZkCSp+VPiSpbj5S5L58hrK1ntCXQSDzveqUVgZwB5WbgzerPM0s7brpCiuWt09qucZbyX32e1uvLjKbCuW62oL2DeMtqK1J6noumhlN0dadatWKx1p1pYDrTrSwHWnWlgOtZ9hz7LR/Wf+6argvWRRP2WXWlbE1QpQGpfqgL5Ei2eLBlkGIN0ySggnclsZQnHpXg/8Fc56W41X+93l8lpFutAbCilhJBbQFAJScnBBBxnoM+XCgau4bCwrSnUmuz6nVUpOODp0+WsrdrdvBIvuhLUL1qa56lUAWQwhmNgHAUtIW4Rnr0G0A+k1n6pRlpdcs6W6i49r8TbY/Eb/FK3CMYr5Jv5tlr4lzebLtRJwhjkBWfJkH/EVSdOuGey66OhKHFDPkUVjB/7NY+Mler7pfT7kYGnlwy74fV/Q8Y0d3UuomWmQcNNb9vmJGf8K8rjoiXKdnG1X520pmkqdQhkKUCfHCien6KipCMKFKUl1LtKrKWJxEIvhZfBGvTwHsNkuq5d31g0ZCzlapKQt0nznKs+brWwdK6P0K4pDcaVGv8AKT5JTyVn4mxgY9YNWKmKzO0dGZipVo0+Dt1L8EBlCG0NpbaQNqUITgJHmAHhX6VACsKWpbSVtDHmJblxlMSGm5DKhhTTqQpJ9YNVO5cNdO3IKKYrkNSvExnCB8k5A+ICr1OrKnwLkU46xZR75wC0+5LanOzpKeSdxDaEpUoeYq83xeWtgaPnNy4e5sBLaVFKUgYAHm/hWXVqutRu+RTly1NeMk/lYtKxuT082Kh9GSjC1/ywcBY8Kq2d/cL3+BrtpK+Cqe7xR0W2dzaT5xX1XXHBClAKqGv5/sVFcmch6UY8dbgYjNlbrhGTtSkdST4AemrVX2GXISyvN0NFWay6j09qVy63aGmLJutsmGXcLXzZy23ytksgthoYDaNyEpBVkI9efmzWmVc52ivZeLfEojSJrxkiRPUHHUJQ204oKO9oOJU4oJXgDChnCju1ChLg/PA1Kp1VanUT4pu17auLbukr31v07rswYlj1Na7GHoKryzcLraZkiQgl3ZHlzJLZaAb96hbXMeUSAFDBKjjwmhp2+Rb6+pi4X91hu/Q2YwfkvKQWUtNuSXVDOFIXhxGD3En3oSTUKEuGvn+RClXSSTlpbr0d/nb/AEfuhF3q6artUmYL8w4I0uRdBM5zcQPlaEtMNoVhGEJUvBQMK2g5UckQHEA3yTq+53Bq0TppaxCFpahP8mQw0C6xJW8lJS4EurWS0CCQcYUoAFJScOHP6EVHWeGWkm817NO/s6rsTd+xF/4kMPzNOWbmRpFyt6Z8Zy5sx4ylrcjpyonkgFShvDZUgAnbkYPWo+93Jh/V2nL2/arkuxsx5RQUWt9biJe5pLS1spQXE/Y+cElSRjcfDIq7Jau/YZ9aVpycot+y+Deif3vpxsQ1u0zqGdqaFLjNJsvaZtwvn+Ww1PIjlTbUZlCkpWgBxTanHCNxwSrIPWvuBb06KvF2tjNmud65FpjW2A2qCtTMtZLrjqlO7Q0gKU4gKyoAbfDpirag16z+HuMONKdNqrOLersrdY8+y6S5JGxdF2N3TOj7HaH3u0PwITMVbvXvqQgJJ9RIqarKULJI39NZIRg+SSFKqyl24pTKLilMouKn9KeMn1J/jVymrSRaqu8GWGlZhgClAKUApQClAKUApQCuAfrq/jwv/On8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUoDR3He9ytP6HvMqJKXb1KksRnp7fRUNh2S228+D5OW2ta8+TbnyVQrjfdJaMftcXR97tdjh3NbyLhfWX0yQBHaC9gKlKSt9RdT1UFKKd3icYy8NRnOlaMbpuV7c7JNK/eZspJPj0I6z8UdXzNLyNQPzmlxbNpdrUEuO1DSlcx57nutsdSeWkMtt7sZV3gc+OYxd4lX/U105t+g6vPLtVhRIRGb7Nvlyt85tG0lK0pZZQRnJHgokpNbCOFo0pSdOL0534ar6P5XKHNtK7JNPFPU19jLl2S8xJM+VBushVlbjtrFqSyhYjqcI7/MLqW0qSs4UVL2pG2sx7jDPvRts5F4jab01e319iukhpBUmOxHCnVo39Ctx5W1GQQUNFQB3CrT2fSXCDurq13q1dr4qL+OhO8fUx9McWb7Md0Y9c7q3Iaulv7WqLa2mTIU8pDslKH2FEuNtmMGtqkdQonccEZzLVxVkjT+lb1K1jaZLd8nRUTG2wymPa0qYdkuNczORlDezKznPUYzgU1MBTTahBtcuN1rLxtZcmu0Ko+b86FfvPE64XNuyakS5HPZWZ86I5JbKGo6JM9uDBecGQdqWVuuKyR0CuoqatOvdTX2aq1WjU8e5NydRi0xbuqE2SYzUIyJLoCcIWd5DaVAbchPQjJN2WDoxhecbqN1x6O9vemyM7b0fEsvFHXb2j5lmtbuoWbCyu3S58m7yGW1uOmPyUhptCu6VrU9uIAJwghIBII1tYNS3PSmjnpSZNvhXGBbbcmU86plqSp5/fOuCWC8dil7XmylK+7nJPjVvDYaEsOrxfrNPnr61mu5J68yZz9bjwLXaOIupNTa4RbLdco0aLb34bS2rjyWpE5tUduQ+tTP+kCyhwhCUBKQUKJJGUiI09xgv2og1Lhansy3JmnX7suHLS23FtklxTSIUZx4HIJLhCwo7tyOm0KAqfQaFvYbsle1+ab0666Lk+/UOpLqfiuLWor2zarZZbuiJKkwXpQn3pcNlTj4kLjoayAW1tJW0slTSVKUlbW0jdk9BMBwMNh0pU6EjeUAhJVjrgeatdj8NToZYxWut+NuPK/T/fZcpyctWelK1OUvXFKZRcVnWP8A51j+s/sNVxj6yKZP1WXKlZxrBSgOZdYXU8QeJGqLYJr8KIwtuAh9gZO1sHmJIyOhWpf6BWupnDNnhpHkTZN0aWxzVu9lh7krKR/o08zAIycFW3GNgx45GBQ2tDD0qkWtbu30PSHsyU6+Hw0f8owv2aK5unSNrVatF2xhxvlPKYS44jr3VKGSnr16Z2/FUBqdGW1+qtfO7jrxNI5KeJnKPBt2+J98TLzGTanXpCjsXywkJ98QnaSR8SSahtHzW5cW5PtEFlKyhJHh79xXT4lprU1bzlOfLXxj90dDRioUKcOd4/C0vqmXDgrahcLrOnKSCCSE9PAeA/VVT4n332suGCy4WpEh5xO9PRSG0nqR6TkD462O0lu8JC3L7Gp2M/SNo1Yvm2/gylW7TQvjZUnGVdcDy+k+f1moDU2gnIBK0goUnqFDoQa4pqdOKqHpMasHU3RIaL4i3yHCctjktciUh9tll2QSvCVBRwc+PvT+qtgXDVd906lJmsxJ6CMnZlCh+j/Cs6daSjmXBJGmng6W+cHo23bs4fcx4XFq1zH240uO/DkOEJSEjmJJJx4jr+qrgR1I81XadRVI5kYtfDSwzyyd+hDaoXttb+PIk146Ntxh2hpf3Lo3D1+WttRjmwtR9GjT4ieXFUF1UvoWttQW0POKhIiEwtURp56Dm7D6v/21awc8leL93xGLpb3DVY9nhqdFQHkyIbS0nIKRWRXannIpQCsSbao9wWlToUSkYGDioaurEp5dUY3tbhfer+VQ6bhEe9X8qqN3EubyQOm4RHvV/KodNwiPer+VTdxG8kPa3C+9X8o0Om4RHvV/Kpu4jeSHtbhfer+Uae1uF96v5Rpu4jeSHtbhfer+UaHTcI/cr+VTdxG8kPa3C+9X8o09rcL71fyjTdxG8kPa3C+9X8o09rcL71fyjTdxG8kPa3C+9X8o09rcL71fyjTdxG8kPa3C+9X8o09rcL71fyjTdxG8kPa3C+9X8o1lQrYxbyvkgjfjOTnwqVBLVFLm5KzMqlVlApQClAKUApQClAKUArgH66v48L/zp/KUJR2FwC+0Tw4/Ju2/urdXyhDFKAUoBSgFKAUoBSgFKAq0zT0uRIfUG0KQtSjhSh1BNYidGuJZaaESOGmjubQAnCD5wPJVjLJcDLVSNj7TpWS2khLDSQQAQCBkAYA/RXy1pB1hCENxWG0oxtSnaAnAwMfF0pll1J3sT9b0k+yt1TcZlCnTlwpwCs+c+ei9JPLbaQqMypDRBbSduEEeBA8lMs+pG8ifiNIPNvB1EZhLoQGwtO0KCR9znzeivkaNWlrlCHHDe/mbAE43Zzux589c0tPqTvYn0rSLy21IMZgoUnYUnbgp82PN1PT019I0pIb27I7KdvhjAx0x0plk9BvYn49pN+SUc2Oy6UElO/B2nGDj4unx18u6PdfThyKwsbgvCgk94DAV6wAOtFGa4Mb2J9HSTxkiR2ZntATs5vd3482fHHor49pi+U432OPsczvRhOFZOTkeXJpln1G8ifq9HuuFndEYUWP9FkJPL8nd83xV7+1yb94j5QqHCT4jexHtcm/eI+UKe1yb94j5QqN2xvYj2uTfvEfKFPa5N+8R8oU3bG9iPa5N+8R8oVlWuySos9l1xKQhJOcKHmNSoNMiVSLVix0rIMQVi3a4N2m1y5rpw1GZW8s+ZKUkn9QqG7K5XCLnJRXFnMPCuzuiGq8XN5LUq4OLlLCgSSpaionp6TWJxDtTt71dYLQrC2J0tAUU9QpsHcv/ALINcJCpTqy3UZXkrXXeez79R2lUqperBSt/7U/sbgnpy2cVQtTJw0v1Gt/V4Hm+G4oiZ4TdLjHQ6gOMx2ua4FDIAIIAPoKUrz/tJqI0FIbPD1TrYCVqcdK+n3WcfsArms0s6S4O/iv/AKnfuMd00+Ky+Ev/ALI3hwPt6Y2mw7jqvrmubvqiJ27W8Z7cUtcte30kuK/wFdHtj+3SOT/pj1sdJ9jIrS+uhZlo5mQkY6+PWrBqbWbF6gb0qSRjxFcHUrN0t1xuelvCXrqsigWx9aYz8xKTyhNQVLKfDaggEH/jq4c+8aqbKbZBfnhA6r6IQP8AiUQM+jNXp0p1JRprRNK76FmpUp01OtLVxb05vsR+WWzxNOyRctVW2W3KYXujtBlS2UdffrUnIPoHh6/JnyuMqy4osrZabGMIWjqoHy+eujoYBwpWXrd3m5zFbaNLFVm6t4Lknp8+HzLTHvsbVun3JDBTuAKHEA5wfR6DVp0u0mRoiM4nG9hYSfj6f4Vl4eFqNaD6Gq2g0q2HmnfVr42MltIA6forCuMQLjEDx6nPprSxdnc2i1dmbT4ZX32WsbaFqy42NpHpHSrnXfxlmipLmeY1Ibubg+TaFKqLYqh8RY9wfnxDDv1xtKQ2dzcItALOfE70KOaxcVUlSpOUeJlYanGrUUZcCh3ifK09E7VdOIl0tsbO3nTJMVpGfNlTQGa8o15enS4caNxGukl+YwuTHQy/GXzmkkBS0kNYIBUkZ9NaX06v5RvFs6m1dRdiT7DfPx0v3yo/0NOw3z8dL98qP9DUenVuvyKPQaPQGBfPx0v3yo/0NOw3z8dL98qP9DT06t1+Q9Bo9CGs+pkahuEqBa+J864zouefGizIjjjWDg7kpaJHXp66+7tqA2GQ4xcuJNzgOtx+1LTJkRW9rW8I3klroN6kpz5yKn02vw+hc/DYZsuV3JbsN8/HS/fKj/Q07DfD/wDppfvlR/oaj06t1+Rb9Bo9B2G+fjpfvlR/oadhvn46X75Uf6Gnp1br8h6DR6H4uJem05Vra+pGQMqXHHXw/A1+qhXxKSTrS/ADylcf6Gnp1br8h6DR6GJaZNwv1sj3C3a9vUyFIRvafacjlK0+cHk0ucqfZW2FztfXqKh99EZouuRxvdWdqED7D4k9AKn06vwv8ir8PpXy2dz9tkife4pkW/X93nRwtbRdjvRlpC0qKVJyGvEEEEeisrsN8/HS/fKj/Q1Hp1br8iHgKSdmmOw3z8dL98qP9DV04cR5zKp/bb3Pu+Q3sE4tnl++zt2IT49M5z4CsnDYurVqqEnp/Bi4jC0qdJyitUXalbw0opQClAKUApQClAKUArgH66v48L/zp/KUJR2FwC+0Tw4/Ju2/urdXyhDFKAUoBSgFKAUoBSgFKAps/jPw/tU6RCm6405EmR3FNPR37qwhxpaThSVJK8ggjBBrw93bht/aDpf54j/11TmXUnK+g93bht/aDpf54j/1093bht/aDpf54j/10zLqTlfQe7tw2/tB0v8APEf+unu7cNv7QdL/ADxH/rpmXUZX0Hu7cNv7QdL/ADxH/rp7u3Db+0HS/wA8R/66Zl1GV9B7u3Db+0HS/wA8R/66e7tw2/tB0v8APEf+umZdRlfQe7tw2/tB0v8APEf+unu7cNv7QdL/ADxH/rpmXUZX0Hu7cNv7QdL/ADxH/rp7u3Db+0HS/wA8R/66Zl1GV9B7u3Db+0HS/wA8R/66e7tw2/tB0v8APEf+umZdRlfQe7tw2/tB0v8APEf+unu7cNv7QdL/ADxH/rpmXUZX0Hu7cNv7QdL/ADxH/rp7u3Db+0HS/wA8R/66Zl1GV9B7u3Db+0HS/wA8R/66zLRxe0LqG5MW616zsFyuD5KWYsS5suuuHGcJSlRJ6Anp5qZk+ZGV9C2nwqsXbiVpqzyjEcurUqcP/ooKVSX8+blthSh8YqogwjqzU15GLNpRyK0fCVfpCYyfWGkb3D6iE1TeL8PUNt4e3SffNUENq5bBt9pipYZVzHEtlKlLK3FDCj4FOatVpKFOUuiZn7PpOvjKNJf5Sivi0ax1Hc70qRMtsG3yUW9UNDsefEbyUjHUJORk+CdowR1OfJU9w4jO3CdZDc2nUXCBEeeCZC97g3ENgqPkJBV0ryHZVOq9ryUuV7/E9Uxu6p7OnKEk52d+usWmu779hsScO4fVVD1QPsSvPXo9X2Tz3C6yRr/VN8ch6PdbaXy7heue9nPVEdtByfjQlCfW5X5pRRhcPYyB0MhKCB6V97PySmtLCl6tJc5NP3NyX/8AlnY1atpV2npG8f8AxUH4SR0toJr2F0HzVdNjBX19Vcp/VCxCtqwTPKVuMrPxAj+NbTbL/KS7zS/0sv8AqHL3fJmnnLuuS+3BjsvTJbmS21HbU4s+TwSCcZqxaf03cYkhL2om5NrYDgCYr7Skl7pk5Jx0Hh08fIR4nmcFhN5Fymd5tDaNTC1qWGpQzZ813eyjZad7bdku98jcFr4h22BDjttWuOymMtRYZZ7vinqT6eo8+TUpI4uxkDuxgoFOAsKykK2kgerPTNbZ0Ivmc+6Mm2278TW2r9bS5zr0h93uo281hCiApIyQU+bx6j1+atZXm8i7SGmI6nG+/wBwKG1JHTcAr0EDpjy+IxWXBW4ciatowUFzNzcMdIxJ+m5cybEQpanuW0onCgEpGSFDyEn9VXfRmnEyLNc2o91ukJTC8htmUVIxuH3Kwofqq9h60pupGTvZPiaHaOGpU1SnTWV50tPtw+RIN2q+x/8Ak9/Q/jwTNhJVn40FFfrr2pmWzvhWycnzsyFsq/QpKh+utRehPinF9mq+HH5mZlxNN6NS79H8Vp8jO4a64l2C9SY8mwXJaVHdthht/b8QVk/orbHuuaZZ6TpMu0LHim6QH4wH/EtAT+uuww9nRjZ30OIxt1iJ5lZ34d5N2rWFhvoT7G3q3zyfARpSHD+gE1L1kGHxFU7W6czI3+7P7awcar0WZ2Cdqy95zrxbt0iJxHs18nW9+fZGrVIiR32rau4ot01S0kPLjo7xCkApyP0jNUGXF1lcNO2m6QbJKtN7Rpm8hCLbb1xOU6Zbam9rYHcWtCd4T4kkny1oUlY7qlKGSLk9OFr999PPEmr1qvVV9fv06G3qeFpp242lK1Nw32pTcLlOCSthG3ePsgRuKRnBJ8uaxtWXDVXa5qLVK1kHkwoR0uns722S4Vq5xl9zGR0yHsYT18anKiqMaSaXq9vDjpf5X95l3u6a6iXXiIxZzeLtNMV92FKDclpqIQ6hPIbZcRy1rCN5Q40SVAZIJIq68JZF8b01qGQ5LmXkJWV2+LcGpTbiVBoZa5klCVqBWPHBA3EZ6VDimizU3W70tfTppw/n6mmuDw1Fo7WFhzZLjKYeS1DfmP2mW0mysurWt6GElPfy8W8PblYSO9gVf+OFsvqtXXSbZ4U51waXSw0/EYWv7J7IMkoBAPe2BRx44BNS0r3MipKnv1K6s11Xnt9553N3iKjWuoLDbjc3I9tNwvMKe6FcmUHI6RFiBZ6KCHnF90+RtPkqAsy9cTtJuJZvOoedImWll5Co0wSYilO7ZKwt9sDG05UlO5CcZ6A0yopToZU7Ll089jNqcSrReGLNou0We43lCFXePEmzYrqlSTG5LoWpxzB8SE5WfKQehxWtLPH4jWiNbpUSfqG5XGRHvsUx7kFLaQWNwhKIKQApRSkhauq93iRUKKtqWaU6ThaSWt+/n/BB3Cz6m1Nw0ua5lw1DPbjSrVKXCMeaJLLiXcSCC62FLwDuKG9yUqQCMdKtL951M9xStLdsZ1K3bETWozwmmUtp+IqJnmqQWw0gFWOpUXNwVnHgJsmZEnTldaWV+nRdpTtCI17YtHJh6aa1MmdG0y/2uHdIziGY8rmI5IjJWkAr2FZATnPTPWrGYt9vMhCII1JddMs6hsT0R2+MPdoS4l0mSocxIXy09wkkbQc46VLSKpulmctL/wA6W917/M+rdM1dbp+mXXfZ2UDdZbblnbZlR9yFXBYS+XUILakpbAJQ6Qko6g9c17wLBrO8321dqvurIca43O8xpaWnVtoYjNqWqMU5T3MlIAX4kHAOMCosi25Uo3dk+PTt/g29wklXe4cM9NP34Pi8LhN9qEtBQ7vAwSsEAhXTrmtr6GGFTfUj/wAVZGEj+fH3+Bz20Gt3NR4X+pa6V0ZywpQClAKUApQClAKUArgH66v48L/zp/KUJR2FwC+0Tw4/Ju2/urdXyhDFKAUoBSgFKAUoBSgFKA1deIbC7tMJZbJLyySUjzmsTsMf8A18gVxtT2mdfD2F3DsMf8A18gU7DH/ANfIFUFY7DH/ANfIFVziRPVpjh7qW7wWWEzYFtkSWS42FJC0NqUnI8oyB0qUrsrhHNJJmv5fHNm0JDTulZlz7I3b0zpcQspSlyW2kthKFKBJKlbceA8Say08aWnm4sZnR0x6+PXGXa1WtLrALb8dsOLy4VbSkoIOQfiNV5O0zXhOeZfPTS/2MCN9URaXrLKuj2lZ0SKmzKvUUvFnMllLiW1gBKjtIUoYz4jr0qwak4tW7T92udtRYZFwlw58G3JbY5aS87KbK29u4gADGDkjxo4PqQ8I07Zl5t9yDn/VC2O32mE69YyxdXn5sd23yZLDQZXFUEujmk7VEkpCQPfZ8mDVl1TxQtGndEWLUqLaX4l4UwlnnlEdDIdbKwp5au62MDGT5SB5c1DgyJYSSaV+LMWTxctUS8LtLtmc9lO1wIzEdCm189MpBWHUKGQUoCHdxGf8ARnHiKq87jy/P09brnYNLsyDIvUe2LjSJTPOQHFFOxaAoFp04wAvoMgnIzUqFyqGEvZyemnLqTbPHSwP60Xp8Wvvty3Lep0PMqWmQ22VqBaB3hvKSjmYxuHmOahkfVFMOW3tqdB3Ism0i+IVzo3WEDtcdPf6bT4J8VZBwKZO0lYJvjJLh1N0Q0Q58NiS0y2Wnm0uIJQAcEZH6q9uwx/wDXyBVs17VtB2GP+Aa+QKdhj/gGvkChA7DH/ANfIFfse1TJNwhotEqNbJRc6yXIoeKU7TnanckZ8OpyPRV6h+rG/VFmvfdSt0ZZ/cuh3Lvagut01Io++amSS3Hz/uWtiCPQoGrPabHbrDFEa2QI1vjjwaispbT+hIFdeclYza0x9VPKKNEWaGnr2u7soUP9VLbi/2pTWHjHbDz7jodgRzbUodkr/DX6ERa5b8DSiMKPcTlO4Z2nHiKg+Crrt0v2rrg+4p1e9iOlSjnGAtRH/aFc7QbdaC88Dr68YLAYyouLsv/AN0bLnDKD6q1zr5ambPMWgkLDZSnHjuUQlOPjIrNxTy0pNdDmdmQU8RTi+bRqvUcJyVb7nJUkc4lNot7X4NtCkII9ZUpzP8AseYVcLXZy5d7XZG+8iMQkgeZADaT8lAPx1YilLaEaUeEIxX/AIqXi22bWrJx2VOtL2pym/8AycforHQmrHU2TRCYyeintrQHo8T+ofrrnHjJbF3LRzC20hxUeYlZQSASkpUnpn0kH4qr2qnUkqceNij+nrUYb1/uNb6fajaLs6prBD06Wkc1bf3AAxsB9YJPpJqy6a1U1qNiVEltl1kYQ6053gQR0IPxfFilKmqdNRNnXrOtWlN82a9mxLpbL7NhRWnJaEOlDK2yCSnHdz8WM+SsiXadRQrSta7e6tCAFLCFoUQARnoDn9FHAuKurdp56YhQdROdsuq3XthHLjNuFOcHPfPievkBFbA1DeRqHTyrWiCmeVONtNNR8ZaUSAkg/cnyfH16E1U45YllVM8rmzYFsRYtOwoCG0tlhhKVIR1G/HeOfLk5OfTXrwmirlT7vHWOrm4EebIq3s1KVSd+aNbthuOGhJfu+5KpBbcKFe/QSkg+cV6ZycVo5KzaNvdSV0YOlpBia+ZAOAtGP110K2d7aT5xXaYB3w0PPM4PaitjJ+7wRDXXQ+nb7n2RsVtnKPiqREQs/pIzUR7kunmOtvTPs6h4G23GQwB/wpXt/VWeamw9o96hD/N2trugfg57UeUn9JbC/wDtVFXSHfIklKL1coVy7g5LkWGqOoDJzvBcWD5PDHlrExdnRZmYT9ZFJg8U9I3S7uWuHf4cqagup5bSioLU2CXEoUO6tSQlW5KSSNpyOhqesd3h6kstvu1ue7RAnx25Ud7aU721pCkKwQCMgg4IBrROm48Ub9TUuDM3bkeNNvpqnKVXG300Kc+WmUXG30029PGmUXI+332BdLndLfGf5su2OIalN7FDlqW2lxIyRg5SpJ6Z8a+514gW0PiRLabWxHVKca3ZcS0n3y9g7xA8MgeNTk5EXMmM83LjNPtK3tOoC0KwRkEZBx6q9NvTxqMpPEbfTTb6aZRcbfTTb6aZRcFOfLTb6aZRcbfTVo0QMKmepH/irKwqtWj55GJitaMvPMtVK6A54UoBSgFKAUoBSgFKAVwD9dX8eF/50/lKEo7C4BfaJ4cfk3bf3Vur5QhilAKUApQClAKUApQClAa0uwzdJf8Avl/tNYmK5GpH1mdfBrIu4YpiqMpXdDFYGoLFF1PYrhaJyVLhT47kZ5KFFJKFpKVYPkODSxKllaaK27wj0683MQph/EtcFbo5yupibeRjzY2jPnr3i8L7DDvjd2bZeE1ufJuSVF0kc99oNOHHmKQMDyVVqX3iJNWv5tYwBwS0r7FMW1UN5yIzanbMltbyv+TLWlahn77ckEK8RXlE4OaXsSFSnXJrq0zItxdmXCctxano4IaWtaz4AHGPDGPNU3b0KvSp2auez3BvTrym5EZc+DLRLlTm5sGYtp1K5BCngFA+8UQnunoMDFTWotEQ9Taeas0mXcmoraA2pTExaHHUbCgpcVnKwQeu7OT18etQ7lLxDbTdtOwrh4PwEcSNN6hZbYYg2G0KtsRhO4ubveoJJ+5Q2VgeXKzX27wP05IiT23nLm/KmPx5K7g7OcVKQ4xnklLmcjbk4Pj1Oc1N2VvFSVrdPrcybLoDTzd6m3C1XKaee6p2XDjXJao7jykbFOLbCiCogdc+JGcZFfjfBrTTdsTATHkdnTZF6fALys9kUQSnP32R76juin0mXFW/0XOHDbgQ2IzIIaZbS2gHqQAMD9Qr1xVGUxm0ximKZRdDFSWnR/nuJ/tH9hq7RX5ke9Fms1u5dz8DYlK6w5MVpX6olDUq56NjPZU0l6TKUgfdcttPT/tfrrFxUc9Jx62XzRvNi1NzjY1F/ipv4QkzRzXGi/OJalOMQzY3ZvYEwgRzR3tgOMZ8fjHnNba4OW1uDbdQrayUvXVZTnxCQ03gftrO2ts6jg6lKVHtT71bU57+mdvYvaeAxtPFWesJK3STenfoXOaMoNU7idCTZ7fZQ6O+472p4H71AKwP+wP01zmKhnpSXcvi0jrdmTyYmD6XfwTZr9Nu5EnTsV/GWCqY/n79KCsq+W8f0VfODNlVe7/IurqcpKiUk+QeT9VWdnre4yvW7fFIzNrvdbOw9Ds8G/4LpxXuG+dEhIIw02VkeknA/UP11pvie+0zo4suoUsyXkNpKD3k4ySR6cDHx1bxjzYpLoZuy4ZcJDtNFsKW5O9j2E9yQ4drSl9WunU9euMDJ9VWJc2BpS3lptwOO/dKHvnFfwrIi7oyasVF6GPGamxbki4Ow3w2tsKW4EnCT58eOPTU5Av6xPRudQ7FdGEkeTzg/FVVrlpOxGTtBy5N5cVHU2i1OEOb0nCz50j4/L5j5am7Q5bNN3O320nK3pLa1MtKwrAWCP8AtAevBq3UfqMrpwec3HPZ3BWfGvLh7LFr1sponCZCAcekdD/CsXZsrV0u8s7VjnwMuyz+ZY9YW/2Nv7hSMNvjmJ9fl/8AnpqMByoeqsTGQyV5rt8S7g573DU5dlvhoRMRYj62t6/OSmujIqt8Zo+dIrpdnO+Gj7/E5Ha6ti5dy8D1pWzNMKo3E2C7c4L0SPIMWQ/EdabfT4tqUCAoeonPxVi4n9NmXhf1Uc16Nb1lAZ0Nptiz3mzRbXblW29W9caMbbtahuoQ8w+BzFlbwZwEq96TuQk7qgYlj1vp3RkextWS+OypendOR4yoigGojsc4lpWd45agnbkD3wOBnBxrrRv56mz9a3noTEvT+pl2fXLTNp1GdbPSJ6k3NqapuPIgqlhTLUdRdCEuGLtQjCQULSokpJyqNvWlby7ab2/ZrZqOBYBPjSLXZZbT7/aHkRnUvJdaDyXW2FqW1g7gEuthwgg5UTXYGmySh6Y1lO4qIm3dVytQM2BJhCIw9LbaipjMh+MqQJCW0pLokJXvaKlbgoFR27bZrrRV9v2vr9cbcqW09G0uG7K6ZS24qLkVyQFqQFBKlJCm+qgcBXpqluKat0Kkm0akm2/UWmNE9VXhcm53C0QjZJTD0JL7yFurfTzHZbhcU4lIDi0uIQoIScr7wMhp8XuDpJlywxr/AH9t+yXu0LTFUSqNdDKBShW95WEowttLu9Qw377JGbrSaKFdaEtcNM6nhx7qHrBKmtXG4W3nvqS8+W0ItSELcLLTzSnhzkBsgrABVvOdlQrPDO/zLXYpV9s17lagd0O/bu1tyHeYzPbO5sOqS6cKIAwokhSvEk9ahNJaBxbJuZpjVr16cDtvvzmo1TbQ5Zrq1JWIcSGhuN2tDuHAkHemXvSpJLgcQBnptndBaHv9k1Zp+7vt3USJVyvybqqTMdcb7MqS6qGChSylKdobKNoGNx++OaW42tp5X3Kkm356/Y3ftFNorFsZA2im0UsBtFNopYDaKs+ihhUz1I/jWThl+avPIxsT+k/PMtFK3hoRSgFKAUoBSgFKAUoBXAP11fx4X/nT+UoSjsLgF9onhx+Tdt/dW6vlCGKUApQClAKUApQClAKUBz59UBbY130feYcq/QtOtuTGSJdzXtiuKTIQsMPd5OW3dvLUkKBIWR5cHTVi43sacsLTbKLRpSztWW9mK3HfS5CkT4slpCTEdUE8xtXMWUoAGc4x3a57I53Xazo8yik+w+3+NWoBdnEjU0BufGdsSY2nTHa51xExDHPxk7+nMWpJQBsIJVuT0GRA4x317UymWtTQblIN+vlqf0+1HbDsKLFTLUzJUQd+QWWUlSsIIdSMbupl0o24ebcRvHfj5uYd04na5tlj0Y2/qKK3LvticvZuUhESGx2jYwURhzjt5SeYpSgCXFA9FJCTU1euKeooOprxbnNQWaJPftSnLWyiTGcgRX0xG1r7Wo/ZmjzVKKVnLSkKbBwo9YdOD4LxG8l1IafxentwIN+toN0uULTmoCqXcozDjqHY8uCgnfG7i2k7lKPLICw2M4I6SK+JWo3bs9ZLJrCJqCEu52qK1qRuKw7sMkP89jDeG1LQlppaTjoHk7gryt2uaG8fJ+bFnY4iagt3A/Vd+cW1dL7YpF1hokLYwl0RZTrKHltox4IbClBOM4OMZ6a91pebhrhl7TiNYM6msUe+aeUm9Ro0VxDrj8pXMjLCUlpfL5bLoGM4dSFZHjEYJNu3MmUm0lfkbI4r6xe0lqTR9kb1VB0ZarjGnc6fJZZ7qmUsltKC53EnvK6EEEZAAOCKTbuLWu7npu7aiLrLLln0VBvirMiDkSpb7UvJUTlxLYLKFhCcK8mfHMRpxcU2vN7fImU2pWT82PaXr+bMkyLc3qCBru0xbrpl1m8JjsqS29JuIQ6zlruFSEIbcSQNyeanJPdNR07XWsbppKQqbqFK2r7YdQuobjwm2jCVDcShpTaupJUhSgrdnrgp2461qEea82KXNvg/NzF0/LicJnL32aSq3Lf0/piOJUCJFaUXXX5yASF7GUdMJ3r6AAdFHAPtB4va11NpZmTBv8eE7Es2obguSmIy/wBrVAnBhjJBKAlaOqyjocnYU5BEuCl60l5sQpOKyxfm5Ny+IutLbHuUA3uM/cHvazIjzVwEBMYXKcqO82Gwe8hIbJTuJV3uqjjNfdp4gaust9gqu+oUXS2o1JcNPusi2oStbLMKRKQ+dnUugspSQkJSQfeg9aoyRaenmxVnlda+bktwG4pz9f6i1BCeuS7tb49tt1xjPv8AZOcgyDICm1dmUpCQAy2oJUStO45JyDW6KsVIZZWRdhLNG4qR07/z1E/2j+w1NJfmR70RVf5cu5mwqV1By4rQf1ScpcXV2g1tnqO2Hr4H/Q9KwMc3HDya7PFHTf05BT2lCMuDU/8A4SKtcdAaagoN9atie3HLgSQnYF48fDP66sPAqQuZo6c84rc45cnySfP3axqm0a20K0N6/ZT+hXhthYTY2ysTLDKznUjf5mzLRbfZK4oSoZab768+UDyfHVO45MibdILAGd21r1BYUD+qpxDUKGZ9V4oxdnpzxKiv2y+aZq3UckrvMpCMbktCMnHnWrer9Qx8VdCcKdPpsummCUgLWnJqzsqnlpzl1Zk7eqZp0qfSPia/1ZcPZXUMx8HKS4Up9Q6D9lay4xPPRrdai2tTbZW4FKBwN2E4z+v9FaypLPimzpsJDd4enHs+hqFiHc7iVuWpkvKbV3lg7Up6eBJ6Dp5zV6RYF2nQTapzLfb0IU+42Qkndu3AFQPXpgVmw4EVrORjac1OLpcwhDnc2JJCvEk5FQ0jRrytTTXlS+wWsSUDlJT33NyColseAwQASene8uKrMZtySsZ2qZz+j3oSES1SIUnISpQ74x+o/oqP1FF7BcbZNZcQmJNaUpaVn/pU469BlRIUMZ8Np6gVD1RdhpNc7G6NFXpN708yhbhXKipDbufL06H1Y6fFX2477G363zAcbHQkn0Hp+3Fa7DPd4mPeXcZTz4WrDsf3Nv6zgC7abjz2hucZAUceUeWqC0vcU1f2rC1VS6rwNPsapnwzh+1+JDTCW9UWxQ8ruK6Qtp3QGP8AZFbfZv8AbL3mk2x/dPuRk0raGkFVPWhKX2SPENn9tY2IV6bMrDfqI0TpjiHcbhwy4SXq4SXfZDUYt4lrjNtpS6t2Gt1YUCk7UlSc93BGBg4yDAyPqhpFx0I9dXrHN04J+l39S2qQ3IYfdcZaDXMBSUlKFjntFIUFAhXXBBSNdurs2bqWRI3fixeLtrXS8O0212Jp9eqn7JIuSn2yZSmI0rmtlojclHNawFA5JaOQARn61pxVvGheJmpUqtr120zbbFa50kIfba7HzJMxt1xII3OKKW2ztyBho9QSAoqaelw5viTLfGMuX9pr2DdGnnb25p5F37QncZiCpB+w4zyuYhTe/dncM7dveqpx/qlrhIsDV3GhJaYr2nzqdvNyZyYKAC6SMdHBlJSjqFBQypByBCo35h1OwlL5xpsF61g5pGRY2b3bhOiW2VzylxXOfbbcbUI5Sd7aQ81uWSNuSQCEkjztnHFi22KFKi6NksWCdbpkywt21SFuTER0FfL5CQOUXEAqQAVdB12nAqd07WuN4uhl2/i9er5qfQ0a2WyzzLTfGZrkqTEunaA1yFtpJbWEAHAWcpUkHPd7u0k4bH1R0Z6LqIotLE2RaY8SUBbLkiYyWn31M7nHG0nl8spK3MBYSgEgqwRTdX4Pzcbzz7j1uHG42p26TwyLjHatNpmJajS23IDIlSJTRf7Shsq5QDIUpwggJSkhI71bTsdyF7skC4J5JEphDw7O8Hm+8kHuOJ6LT16KHQjrVEqdkVRlczsUxVvKVXGKYplFximKZRcYqy6NHel+pH8ayMPG1ReeRj4h3pMs1K3JphSgFKAUoBSgFKAUoBXAP11fx4X/AJ0/lKEo7C4BfaJ4cfk3bf3Vur5QhilAKUApQClAKUApQClAa1vbDcmdNadbQ62txYUhaQQRk9CK1oOLmlU3TU1unRpcFvS8hmO4/Itj/JWtxDexLJDeFLUXkoShOVLzlIIIJ5xxcpOx0ykoxVzDja+4eWR+VqREiQmZd7imAplVtlKliW1H/wBCI3L5qF8lAVt2DIweoIqY0Xc9GwxFj2OQhx29yrlKaCm1lxxxMlSpedycoCHnCCFYAJA81HGdtfNgnBvQXnihoyBLm2+fJJTbHEtyHTbn3Isdzu4QXg2WgtIWklO7KQckAAkY2oOK+mLJrSBYFRZdwudwmqtUhyFbXn0sOCMJAQ6tCCCOWpBxk4C8nASoiFTkyXOKPK6cYdLQYF3atDyX7jboU1yK2uE81GkLjJJdaZeKA25tKMKS2okbTnwOPS08S7LNgWJNvMDMie1BkthLiG2X1xlvqQ0pLakrcTt6jKQOuVBQCTO7dtSM8W7I9LLxu0Le7ZLnQbviExERcVOvQn2EusOKwl1re2OcCru/Y93eIT4kCvRripo2PbmSy5ITumLiC3tWiV2pD6UB1QVGDXNRhCkrKlIAwpJzhQJOnJaE54vmWO56Utl5vVuusyOH5cBp5lgL6oCXdhXlPgf9EjGfDFSwbQFlQSkKIAKsdSB4ftP6atN30K7I8mYMaO0Gmo7TbYVv2IQAN2c5wPLnrX0YrJSBym8AEAbR0B8R8dCT8dhx3m1ocYacQtIQpKkAhSR4Ajyjr4emv0RGAnAZbAwRjaPA9SPj8tBY/TGaUcltBPd8UjyHI/QfCnZ2gQQ2jIUVg7R0VjGfXigPliGxFBDLLbQPiG0BOepPk9JJ+OvXA9FQ9QMD0VI6eA9mYvh74/sNXKS9ePei1V/Tl3M2BSulOZFaH+qbYabuOkbjIc5USEJrj7mM7UBDaicepB6ViYqk61J048W0vmje7FxMMHjViKj9WMZt9yhI5yt/1Y2ntRXNmyv2R+DaZDhjs3MvFXXwCiNuCPOAfUVdM7/4FsKh6cvUNZG+PdnkEetDZ/jVzaGyFsqvSyyzKSfK2qtf3GJsX+o5be2Vi4VKeSUJwdr30k2lyWqaaa+epvOwQOxQgVD7K53lejzCtV8Tk+yGoXWwerRBSfMpKcj9ZrXbRVsLlM7Y7vi8y5I1tp22q1DrJpG3IU4XlDzZPT9X7a6anOJsmmpDicDkMKI9eOlXsAsuFi3zu/iW9rSVTHyguCtH4JI0UO+6c9SPKa8b9p6Lqizu26WCG19UOJHebWPBQ/8AnlrkpztUzdp3NssUlyNXPWj2uudiddBjwTtCMYDrhGSsjPx/GPJVX1Xq+RNS3GZdBaDqSsqHvzn3tb6NpJWMCcrXMa721/T02PeIjRKX29q2U5HUEEkDy9PJUjeNVvz9NuyUMOqSxhWcDPiOo6+HnqpopTtoe8OdbNZWVtiUEPcshxCc95pwDoRjyftqF1XIjXBi3xoziJCYDhJb5oSF93BwcjJHd6A56+B64jiXI8dC/wDB+RiVNbQkhCo6Vr3JOdwVgdfjP6Ktuq1cu3rWPFJBB9VarhXXejY1UnGa7Pobz0C6L3o9tC8LBbxg+qtc3OEq03R+IrwQo7c+UeStztanmpRl0Zxew6lqs6fVX+H+yEvH2K6W57714f4V0TZXA7a46h96Ku7Ld8PbtZY20rYldy8WZ1K25oRVW1ejmSGB5Cgj9dY9dXgZGHdqiOetM2fhtpyZZtPwbjeJDNvntR7YJj86RBjymW3GkMNPryyFBKlp5YV1KQCMpAHlwn4H6TTw4Ran3Jt8U1bHdLy5MmW+UbGl8qSiOlSzym1utE93HvE+RKQKKuHrUY5prp07fg9NVxXMzlKMnZF2b4NaZb1U1f0szBManrubTHbnuytyltqbceSxu2BSkrVk48ST0KlE/motM6Uv1zvjN2adRJvLEaySS9zWUyUo5rzTbSugURznSSgnygnu4GNGMpv1eRcbSR6s8IdOR9T+ziGJXPExVxTEVMdMREtSSlUgMbtgWQT1x4kqxuJNVSXZuHNomRtGrYmfY7UNJdxEhbLTD7aSmOt4ZSHFISkgk7gCOo3DNylSqV21Dkr9OBTKUY8Sw3HSGnNK6lgXVLtzgzLpMYjdnhTXkMy30NbUKdaSoJUQ0zgkjqlsBWQBX7bOCOmbO+t2Gm5M7Wn2YqE3OQEwEvKCnBGG/wCw5IHVOMAbRhPSrcoyjFN8/poTdPQ9IvBjTcQ21bbc3tMGU/MEpU50uvuPY53OVu+yJXtTlKumEgYAGKxrJwN03pyM61bXrxEWqGxb230XaSXGI7CiplptRWdqE7iMDxBwrOTVF3wJsjIg8GdOWiK4zbBcLWtcZiIZEK4PNvFDLjziMrCsklcl4qJzuK+ucDErw7asreirUxpxlbFjjNmNEQvcTsbUUZyoknO0ncSSc5PjVThKUHLkmvnf7BNJ2LHszTZkVayleYxrjNbtkXnupeWgLQjawyt1WVKCR3UAnGVDJxgDJOACaydmaqcLK5GY/eXTl1TlJzDl1YtIJwqV6k/xq9RVqiLFeV6bLJStoakUoBSgFKAUoBSgFKAVwD9dX8eF/wCdP5ShKOwuAX2ieHH5N2391bq+UIYpQClAKUApQClAKUApQGptcWS039q4RL1HYlW4vFTjcn3mQrIJ+OtaXfg09dZ16lWq/RoUOdLt90ht9hLxjTIgZS2SoOgOMlDCUlvCVd4kLHTGgcssmdEo5ooy7Nwffi6hg3+43puZeU3x29zFR4ZZZeWq3mChttBcUW0pb2HJUskpPhu6Y+g+HLumuKOvdSzN0e1vupTamnlI2MocQh6a6jB7qXX8EhWDlonGCCYc00+4nJax7ROGjjsTU0ZN4i3fRGo+1TF2xuGFSHDJb76USebsLZJKk9zPeHewKgdB8Jrza9H8PnblemY2tYM5y83Jc1kSO1vvsONyGyEOI6oQ6EpWkkDlJ6KFVZ1a1iMjvxMgcAJb1vZtErUjTtitqLj7EsIt5Q+wuU081l53mkOhtEhxICUIzkFRJFZmnuBCdNW+y2qJe1ewtnuqbrChri9WVFh5t5pKt/vFOPKcAIOzKkjIKdp1E9LBU+dyIv31P9sb0JbLfddRtRoNk0xHsypslhKGgY7rDyJDgUvARujJ3IJ6gkbh41+yeDTV/wBDx4Nvu+lYL1xkKnMXaz2dxtW5TCUokRVpl7kvJQkkOhagU4G3aOs7zTgQ6a6m3LTBnw1zBNuCJzS3QqKAxy1MthtCdijuO9RWFq3YT78DHTJkNorHdmXrDaKbRTQWG0U2imgseMqZGhcntEhpjnOBlvmrCd6z4JTnxJx4UhTI1yityYkhqVHcBKHmVhaFdcdCOh8KW7Ae20U2imgsNoqR0+nF5i/7R/YauU7Z495bq+xLuZfqV0JzgrTP1UMVh7R9qVJQpcV2cYcjb0Ulp1lxJI9OQmrFao6MHUjxjZ/B3NpszDxxmKjhp8KilH/yi19TkrTn1GCGLpGmytRNSdNRXee1Fa3c1WDkJ27cJ+Ucemulvqe3TdNTaviOYCUSmpZT6FJKf/8AmKvbU2vT2piKEaSaST49WvDQsf07/TlfYezNoyxUk5twXq3taM+OvNuTvyXU6EIx1zWi9XTUv3Ke/nruXg+jw/gK0+1NaSj2m22JG9aT7PqfPBCyiZdJVwWkY3kJ9A8n6q2fxJldm0pISD3nVIbH6QT+oVnpbvDpdF9DXN77Gt9ZfU0Ze72zp20vz3klYRhKGx4rWegFViyyNVa0D0mPd2bdGbVs2spScHxwBgk+smuTpwgoOrNXPR5KNryPa/6Zu7cRMq4vR7w2gbXTywlWMeJAAz5OprXFx0RZ1ylPiRLiNlYc5AAKQc5ODnw/ZWzoShVgsuhrpxTlZaH5rvUT89Fth2plsqbfDhSkgkICSP4jyeQ17Wdt+DdmIKGlOqnhbqGQB3SkZUn9RIHxVfnaEbjIuR8alsrLcGbNhQFmSsBDiY7RICj90ceBxmoCzaWvd/cDTFvkOAHdkoLaEKPj1OAB0HSsWVRRvqX6cVxN7aI0e1pG0rZ5iX5r+FPugYGQOiU+gZPryT6Bj60d5dpcHn6Vrabz1l3mTN/lzfYzefBvrpZn/ZrA4p2fkvsT209D3F4/V/8APTXV42G8oTXnQ862ZU3eLpvrp8dDW2okkxEODxbUFfoNb50VLEzT0ZQOe6P2VgbJd6Ul2mz25G1SEuzwf8k9St4cyKqWuWXn2y3HcDT6mFpbcI94o5wfiNWqlsupeo+2aA0zpTUc7SmhtLzNNPWWJpxUWTPlPyWHEy3oyQUJY5bilHe8ErK3AjpnIyTinW3hNq5rTAdg2CTp6/otTsa5yUS45lXR+TKZdkcpxLhG1DaHw2XCnCnRgJwo1vKeKw9OUk6iabu9Hrdu/LldN35rS+hecJStoTlz4UyWNLQY1n0zPelPXJVwVb7o1b/Y9Kw2ljZIjtuBAbKMrBaClhQ3HKuhk3eGcu46nsc696ZZukUahuU95DgYd7Mgt9nirIWr3haQhZCckKSju5HS28ZG2beet6y0zc07PVXu2ld8WrX5k5OzTToZfBOxzrhJfvMpSV2i3PXCBYlJXkPx3Ji3DI9RQlhtPobUQSFiqi/oXXitdK1Amyyl3KRMUm4ynHohZaYDyUtvQE80EO9mBbUXEhRST1ykJVMa2HhiKqm7K1ufO7a0v/lb/wBtw4ycVbz5XzNo8RtFu6z1BppMq2Iulktolz3o7i07HpAaDTLSkk94KS88eo293veIzrLTuj7lBvVs03JdSm1wNPWd7Ujod3Bh2Ip5wMDGRlwqbyM/6Jo598nNjDYiG43d/ZV7dqk2vi8qJnH1rkbpfQeqb5pJu+2eySbBf5NlnPu3J6SyJNwkTVpdQhtQWSlCElZTzNgSsowB3jVos3BdqUqyx3LLOjWd25GddYdw7I0hYairaaQWo6ilSVqdyrO5StnfOMCs2vjadNyjCabV1fXtcbaW0emvDloURg5WuvPMq2nYq7lxAYRbLPDm6jtDt5uL9xizI7pfCeYxCigoWS22EuthKHdmCycDuqNTOl9B3ZjUytLW5bS7RaLdaZM+QV5Sq5R2Vpbjqx5SpMZ9R8yUgjviqq1WEFJTd7RvwelpPLx71/oKLfDr9NQvhzeHtEWBhrSM6BO7Sx7ZCFwnZl22MOFS1lTpbeQZCkKIcUCRnu4GD7ai4HSbnYZdujWp96NF092O0tz32dzMyU+4uQ5tQeWhTIDRTtASnwb96KtenQpyTVRPW9/WvxtpdcFF2XPTsRO7uuHn/ZMscLblbNSyEWu3Ih2d2/xJRJeThxmPCKw+vvblLXK2JUTlR2bj0614cIOH13tmqbXd7tpw2aVFsao0+c++y4/c7i642p55fLWrcBy1bFKOcOkYSMCseeLpzpS9f1mldWers013LR/AqUGmtNDdm0+im0+iucsjKG0+irDpIEKlepP8au0ks6LNb2GWKlZ5rRSgFKAUoBSgFKAUoBXAP11fx4X/AJ0/lKEo7C4BfaJ4cfk3bf3Vur5QhilAKUApQClAKUApQChGRQHOf1TlvcunDa/RGGmn3XrhCQlt9outnM9j36ARuT5xkZGeorSjE/UHDeBOs8dyDpou6tkC8yYqhbrbFZVCSuMWFKZfDDTu1sqUUn7KXU7gVVp1Z3T6s3bbVmuh63viVq9u+6fsszV4hOC12qU/ebbyBbzzJjyZDrzjrGTvYaSEbUoAWScAEEWF/iZd7jPj6aduDztz9ndQxrjD7L3kQUNTVwgshGEgoTHKDkFYHirCqOEeSJU31K/auIN7tMnhzZLbPnw1Mx7DClW+QtpKHWX2W0uONM9nU4tKd2FOqdSErTtweoVi6Yn6i09aba/abvcNUX6C5qVpVquMdh1TEpoPrYRlLSXEKcPLONwCgsYG0ipcURmbJg671JKgymtMa0uOoYTybGh28uw2C5BlyLk0zIaQAylGSypSlNqSot4Gcbq2Jp++aot2heIqG3ZOor1YJc5i0qmNID0vbGQ6yhQbSkK77mzIAyAPLk1blGPQrUm+ZqbU13uutdK3S3W3WN11PZTHs0mTPMZhK481U9tLjA2shIyjvqaIJbKE5wFEHcvEd+bZ7zw/aiKMhQmS23ZDsdtxzCbZKUFZ29wlaUZKduc48CRUyS0Xf4EJt63NQLv/ABDhaOlXY6zvMiVH0PE1QGVwYoSueQ4VsYDGeUQgAoHe8u7NWi63DWlx1g6WNV3W3QntYGxJiR4kYttQjA5xWkraUrfzB0WSQPDFS1DoReXUh3+LmpNN6bfVeLvJS+bDe2ID7kNJcl3CLNcZYUEoRhTpbShWwDByohOAcS9z1LrIXadc2r7Pbj2646djN2xEVnkPolrjtyislsrPR1ZG1Q2kZ9FHCKJzSelz34Uaw1hqPiG81d73bmwh+4NzdPOSwqTHQh4pjqQwIyFNgJSnvqeWHAvcM5GPPU2vtQQ7rxF7JfJqdVWpEoWLSaIramZTKIKXGnscouOZcKzuCwNyQ345BjLHNZLz1GaVuJE2m7XzUbNkcl6hgant7ep7WqO5CmiY7HVtd5wdWiMwkDOwhO0lJKgSBtA2lwFYWxwi06h1Cm3EtuZQsEEfZV+SqZ2y2RMG2zYG0U2irBduNoqQsCR7MRf9o/sNXKftx7yio/Ufcy+UrfnPiq9rvREHiFp5dnuC3m46nW3d7BAWlSFBQwSCOuMHp4E1bqQVSDg+D0MrC4ieErwxFP2otNX7DztPDuyWeGmM3GU82kY+zrKif4fqqXtthttnU4qBAiw1OY3qYZSgrx5yB1+OrVPD0qXsLUu18diMRmzzdpcUtF8D0u0kQ7ZKez/o2lK/QK55nkvrWPEHoc+WtVtWWkUdB/T8VepLuLjwiubFslvW5Q2lffQT5R5RVh4uSNltgMg+/eUr9Cf/AFrKVXeYPN2GFLDqjtRQXC9179TRfES0yLtpRwRUF1+M6mQGwOqgAQcenCifirVE7i9p9iwtQIcWbbdRRmktmTCw22sDAJUoKBOfSM58taOgnOmlF8Hqd1u3U0XUs+k9QXFbTUlN8kOBYHR14uJUT5Nqs1bJ+h3b/AWX32LZIcGUlmPk/wDEndtHxCqnONCWaxj1KaekeJqfXOmjw5vDSIj/AG555kOKfeBHUkjGMnIGPPXhoyZLl6ss0hZDr6ZDSR1xgbxkAeTpmsipU3tO5TCPU6NcfW65tSemfCgYccON24jyZrSqLa0LzcYJI+XG1JGR5KrWt4i3bUSlPiQav4dWqx7yiq06Uu5+BvThC3y9LsA+O3wqd1vbxcNOS0AZUlBUPWOv8K7WpHNCS7GeYUJZKsJdGjR1wb7RbFdPFOa2dwduPa9OttE5U2Np9YrRbKdnOPcdTt2N4U5dr+hsGldCcgKreqUlUljAJ7h/bVuorxLtJ2kQmxXmP6KbFeY/orFymbmQ2K8x/RQtkj3p/RSwzI/EtbEhKUYAGAAMYr92K8x/RS1xmQ2K8x/RXzyOiu577x6ePTHWlhmR9BsgABJAHgMU2K8x/RSwzI+UsBBUUthJUckhOMnz1+hnbnCMZOTgeJo1cZkfuxXmP6KbFeY/oplGZDYrzH9FNivMf0UsMyGxXmP6KbFeY/opYZkNivMf0VP6VSQqTkEdE+Px1XBesi1UleDRYKVlmEKUApQClAKUApQClAK4B+ur+PC/86fylCUdhcAvtE8OPybtv7q3V8oQxSgFKAUoBSgFKAUoBQ+FAal4h6st2hYFwvN1W8mCzJbaUY7K3nCp15LSAEIBUolS0jABPWqbY+OGnb/e4tqQxd4ct6a5bVGbbXWm2ZaW1O9nWsjaFltBWMEggjrlQB0rhmbaN4pJJIo3ETVeiJDV8vs7S19l3eE1AMq1PdphdtjOPqQw46yFBDqAoO9HEkgjBAyK2YeKFjQ1KU8qVHciT4FsfZejqStuRL5HJQR//MNAnyZOfCpcG0E0jGt3Fq13uBcJdstd8uLEOYuDuj21zDzyFuNuBsnAUlKmlAqyE5wM5OKql5+qNtrQlG2Wq5S4Y08b6zdBDcW0hOVjDrYAWAkoO7yjBGMioVO5LmizOcZLBFuTUF9Utza9FhyrgxDcMONJkJbLLS3PuVK5rXnA5iNxG4V62DjBp/UmpE2WILgh9yRMiMyJEJxuO+/FcUh9ttwjCikpUenQgHBJSoCHTdicyehM6y1pbtCwYcm4plO9slIhR2Ycdb7rrywopQEpBPXaevgPKQOtVO3cd9PX9h5EAXCM+Ykx5l24255llTsXpIayQMrbV0UkeZQBJSrBQurhySdj2tfGyyTbhZrZsmypk92PDMqLDX2VEp2KJQaKj708khzrnAIBOelekHjVZ7vps3232rUE625VsdZtTv2RKQStac4ylJSRnykd3d0qXTIzple1L9UPCtz+oGoFtmvRLfZIV5ZvRiOOxFNSQ8UrWlOFBADWcjqSVDA29ba5xbsDephZiZpInC1m4CIvsaZhSFCOXcY34IH3u47c7ulS6YzpnxojjBp/iBcGIdsFwbckw1T4q5sJxhEphC0ocW2VAbgha0JP+2kjIOap0/jdcY3Gm4aUT7Ft221OIXPS8lYcYgmIHVT1yN4aaQl1Qa5SklR99kA9Cp6tMOeiaN07cjxoU58tWrFY2+mm300sBt9NSFhTi7xuv3R/Yarpr113lFT2H3F6pW7NEKUApQEXqlO/TlxA8rC/2VoRCklG5agARnJrntrL2fedhsF+rU9xjW64KiX+0yG3BlTmCEnyEVszio5zoVnV5ws/qFVUH/0Ml3leKX/qtN9i+pQmTgdD4VRNccDtPa558pLKrVdXB/yuKMBSvHK0eB9JGD6a5+nVdOV0dPmcHmR9p0ajQ3Dhm1Je7Y4w6zukqQElRL6CcDrgdfCru6FBHWrlRucVLtf0LEZKVeT6peLKBq/T104iOLtlptAl3CIcofUsJAScZBJ8h9PlqG0fo256V1EI8q2MrvjQPceeBbjnr1G3IJI8vkzWzVLdUVLjf6mEsWqtWeHjo43v3IvN11QxFgMuvpcjjtLbUlTSgCyCVZyfMVIKfUfTVWnXhdxvq3LAhbstpQcaYZURlORlSvQQMdfHpVlSjRpORqcVvK2KjFPhzNhRbxCvTqWoay6+UFS2QQFNkYyFAnof8DUbqUdosaltL3JUjKTWDTazJo6GM1UTh2G3uDEvtWmI5JydtXu5lKbdIK/epbUTn0Cu7eiPMEruxoKeRsU2kDbjAqw8G7h2WfKhqOO9uAPprmdmytXkup2+2Kd8Gn0a+xuWldOcMKUApQClAKUApQClAKUApQClAKUApigFKAUoBSgFKAUoBSgFKAVwD9dX8eF/50/lKEo7C4BfaJ4cfk3bf3Vur5QhilAKUApQClAKUApQClAak4jaSY1tCk2yQ+5HbFwjS97QBVuYlNvpHXyEtAH0E1VpHCCDInOSTcJSFOagOofseElLpidm2A+IG3vZ8c1qW7No3K1SKfYvqXbXY7NfISL08XrnbolvMpmCwysdmdU42+van7K6VKyta87seSpy58EHLtfFT3tUTUNSZ9su06I3GZCJUuEpkoXkgqQhQYbyhJ8QCD4g1OdyMtlY9LtwQg3bRltsKLmsx4d3k3dPaYzchiQp5x9xTbrKhtcQDJUUg+CkNq8RUYx9T5b7Fpxq3Mahkxbc1p2Tp+Wp1loB2MvmKDmQAG1ILij0ASQMYAqM3IlxMxngxAurqpELUkh7S90lwbzIt7KGnGpb7CGOU4h4AkNrEdhSkjIVt6EBRzmPcJV2i3Q37LOU5drVLu9zgiVhLa5E1T69rhCThCVP+IBOE+Bpm5CxYLholy+W3SzV0ubkqdZZTE1yWhpLfanm2lIUopHRIUVlWB4eAqBHBK3iNFZ9kZJEd+8vpO1OSbi46twepBdIT6hmqb24EtXNfac4Kak0pxWbusNlD8ZqbGDc2WY7jCYTcJqMshJRz0ySG1AFJCAF+OCoG33v6nq3XnSGmdPqujpi2Rt5lIlRGZLbyXE7StTTiSjmJ+4Xg7cnoQSKrcuDRSo6WEn6n+K7Z5NrZvstmDM0vG0xKb5Lai43HS6GngfFKhz3MjwPd8MdZE8GGjqFUj2clmxKvIv5spab29tBCgebjfy+YA5s+++6292ocrk2M7SHCaFo+VpZ9ic/IVYLNJszQcSkc1DzsdwrVjwIMZIAHTvGqdq36mGBq2dMUvUc+FDeuD90bajx2Oc0+8AHUF1SSVsLIBUyoEKwATtAAKVncNXVjc0dpxuO0h10POpQAtzbt3Kx1OPJnzV6bfTVqxczDb6abfTTKMw2+ms+xJ/ztG6/dH9hquC9ZFE36r7i8UrbmlFKAUoCPvyC7ZZyE9SWVD9Vc26lhOzIXIbJG0p3bR4gEVpMe0qlJy4XOq2PFyo1lHjb6Mstg4TSnZNsltvb46QFY81XPiswWIFoT94Vp/UP8KzK9ONPDzUO1muwdadbG05VHqrL4FAYTuI8wHWsNu9GS+hpkp2uLIRj7wDx/bXGKyZ6E4ZrvoYGvUuo0bLeT1Wh9kpSfL9lR/GvSBf0XOLEkI/0UjuqSrxQrwx+kYrMqJKEV3/Qw6Ec85T6WX/yLZwkfQ3q+5M4G5TCVfrqv6/KbDxRmuK/0bu1RVjwykH+NbSrf0SEuljS0l/6nVh1T+hWbQ5AvV01DGkKYkMylISqIpQ3EAElWPHxI6+cVg3bTbfD+Qi/WguGErDU1h1RVsGMJcSfHxwCPSD5DWmr3ksr6J/I27pQazR1eq+YsuoIVyhiGl9VsuDyO0pn43A944CvPlI8D5DU3GjvM6TQ1IfRJcJcVzm04SsFaiCB5sGsSirTKqC9extDgBK5tgQ3n3uRWz780X7LNbT75TKkj14r0JrND3HnGkavc/qaGe27huIGfPX7aZJsV9jTUn7GTtX6vPXGYWru8RFvqei4ynvsNOn2fNao39BlJmxG3UkEKArIrtzzMVrvixo6w6rdthvNmgXZTCXA0ZsZDpbztzt3A4zgZ9QrCxjcaMmn5uZmESlWSfb4Gv8A3IdDfifYvm5n+mnuQ6G/E+xfNzP9Nc3van7mdFuoftRrrjNadJcOYen1xNKaQii43EQ3Zd1tiCywjlLXuIRg+KAPjqEtWqOHMbR1muV/4fWlu4z40iYY1vtbKkiM0vaZI5m07FDapI6qO7oDVxSqNXUn8TOhgYVKcZRSu37uf2JmZe+EcS7qhDQ8V+O29EZeuTVkZMVkyUIUwpS8ZAVvSPDIPiAME/nszwrcEnk8O0PqbuCrUwluxMf5XKStaVNsk4CsBsqJJCQCMnOQGar+75lPoGl7Lz5uYl2v/C5FjQ7bdAxpd1eamHsDdiZU9EVH7rpfT9yEqKQcEk56Zr90/cuGo0O3c7xoy0qnQ7LbbncOzWhjYTKGE8sH/WByD4DHjU5qlvafxKngEo8Fe/1se2q7jw8sls1VItvDONePYBDyXZTVnYTEL7YBW0XMZG3PeO3AwcZIxUZbb7oFV0kuXbh9BtNubiQXDHk2FhLrapDxbDxcCyC1nA96CME9fCmapb2n8SY4CLjfS/T4fclbzf8AhDZ3JDR0TBlvMSZUctx7TGyoRtvOcSVFIKElQT45KgQAcVNajh8LdO2Kx3Q6NtU9m9rbRARFtkcKeK2y4nq5tSnujPUjzDJ6VTmq/ufxLTwNsvqrUxpSuFUKdIgv6Jt7U9i4RraqKq0McwuPt8xtYHlRtCiT4jYrp0qt3LVHDz2AsV4tHCyNcoV0ubUBtQtkUFaVg95ASVd44wEK2nOc7alSqP8AyZVHAKVrpJfwTSLpwiXqV2ze022hxt56KJJtUflKkMtlbjQ+63AJUMlISSkgKJqFOuuEQiGSeG7oZEJu5BZ08zgxFHBkf7CSMEnqfuQodaZqr/y+ZK2e3yRttrhLoR9pDiNIWFSFpCkqFuZ6gjx97X17kOhvxPsXzcz/AE1Rvan7mYTpQ/ah7kOhvxPsXzcz/TWzuFemLRpezy49ntcO1sOP71tQ2EtJUraBkhIGTgAfFWdgpzlWSbZg4ynGNFtJF2pXRHPilAKUApQClAKUApQCuAfrq/jwv/On8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUoDSPG+7ybNpO7PxIc+a6qUyypNuedZdbQt9CVuFbSFuJQlJKlFCSraFYx4jniHq+9j2rW7VF21hb7e8zqRSU2tqaqW8hmXFENSjy+eUhDhCVLAKspCidxBwFFGzvoiSVeeIK4bAvb2o42ukwrMbTEgMPdgedU00Zhf5aSyfspfS5zD3EJSU7cgnOsV8vOouIbkOLdtTyZidV3KBNYw+i3s2lLbw7rgSGwpLnK2qCuYF4SDt6BlQ1InQwkaf4e8OLRc39Z2vT0azSGLmYDM7tTV1bTGS2yShBdS2E9o2hP2JSkgdegM1p69a3ZvWnBfTeLnd5thbbmRGm5MdFskCE4tbriUo7M+FubUk7gpDhCU5GQDimTqZPDzUN+tGqbEq8m/XZt2wxnJTXImtC1qRAbW4lbXL5L+9wKwpJ5ocXswQk7d+wZrdwhR5TSXEtvtpcSl5tTawFAEBSVAFJ69QQCPLVuUehVHVHvuFNwqjKVDcKbhTKBuFNwplA3Cm4UygbhTcKZQNwpuFMoG4VnWMg3WN/tH9hquEfWRRP2WXalbM1QpQClAebrXNSpJ8FDFaH1PbXLReZbCUjCVFSQfKk9R/hWm2pBSpKXRnSbDqZa8odV4Enw714qLdU2tRWphQ6JWPeH0eirVxbSHbNAdHgH/L6UmqoZ5YJ5+NiJqnHacd3wv8+ZqG+3A2+zOKR/pHSGk+snFYmnG985CUD3h5KSfMDk/wDz0Vya1mkd9bLRkyxastKLpp6VC5imd2FBxIyUlKgodPWBWpITVyhOFpNz2IU4p1P+TpPeSrB8vTyVnTqwjZSjf32NdhcPVqRlKnUy8OSfC5sPh9ab47rmQ1G1CIz3ZSS72JC8jPhtJrI4habuz+o5SJt+RNdRs+yKt6En3ifMfDwrcznBYRPLp0v3nOwpVZbSnHPrbjbsXIqsnR0m7R0NvXNra1nlqRCQhTfpSoEFPxV+3bT2o0aZnQGrgi+MOsKbWzITyniMfcrT771HHrrVSrUqi3co26O97fx2G2qYWtSbqxnd81a1/wCe0gdMTrVfrJLUy40xIkpS3GYX3VIR70bc+IAwMjPhWw7y0liCWW0hDbbexKR5ABgVgRpypSyy89xlYOpGreS/0XD6npWbaoeZSv21ud5sOtKT5wRXfR9lHm1X9SXe/E5r4kRJMR19qM4WnW3sDHmJ/wDWvi3plR7QpFxIU6hOQojGRjoa5CpRiqEpr2oysd5SxE5YiFO/quCfvNq8INTi82ZLKl71N5Tn1eFbFrrabbgm+Njg6yjGpJR4XYqp65GVw/Uv+FY2MV6L88zIwf60ff4FWxk4yMjyU2+qucynSZiA1NouHqudYpUtx5C7PN7cyhsp2rXy1owvIOU4WfDHgOtUWT9T/YLZChusXibbE2tMlLT7qYzqGYzrnNU0Q80pIQhXVJIyB5SKqWmiMqGKdNKNtP8Af3MdfA1Vx1pqC5Xa7SWbA/JgSW7fHdQluSIzKAC+CjKcLRnCCAR4+TExcOFtgj6VbUm+ybY1Furt/jXhLzQMd51S1EhSklBbw4oYUCCCPXU8eBXPF8EkrK3hZmKeAVmciQlwr3d4ssIlCTcY7zRdnolKC3uYSgp7xAIKAnHkryuX1OdmuEJiGi9XmHEFriWqQzHdaAlNxurKnMtk7gfHbgHzUuVLGyWtl51+RLvcGoDi9TMi8XRuz6gElcu1IcbDCXX0bXHUnZvBPiAVFIPXFeQ4I2qZbbjGn3O4XQ3CzN2V194tBXKQta0LTtQAFgudDjHdT0JySKPSnyS8/wCjFd+p/sqbZYI8W4To0uztvttzlIYfcfDy97pdS62tCiV97O0EGrDrHhzG1ppRrTr1xkRYbbYad5TMdZdRs24KVtqSk+UFISQR0xTiQ8U5NSa4O/Pv6ldPBxtPFmyahaw1bbTZRCbJeKnXJCdzba1JIwShpbnfJJyvw6V+J+p+tgiTcXq5puUm4xrp7ItIjtrbfYBDag2loN5wo7iUEqJyaXK/S7Wsr6JfO5nwOCltteoLhco1xloanuvyXoZZjKSXnklLiw4WuakEkq2hYGfJjpXkvgRZHLObcZs/lHTidNbt6N3Z0qyF+99/6fD0UuU+lvjbp15GwIaWGEJhNPJWuM2hKkbgVpGMJJA8M4P6DXlCvVuucuVEh3CJKlRFBEhhl5K1sq8y0g5SfXVOS5hOaM3b6quWiRi3v/73+ArNwStWRg413ossdK6I50UoBSgFKAUoBSgFKAVwD9dX8eF/50/lKEo7C4BfaJ4cfk3bf3Vur5QhilAKUApQClAKUApQClAas4g39OlrdcriGDKeS8lliOFbea844G20Z8gK1pBPkzmqDdtRSNH6rjTNUNW+5vM2eVIZk2uEtD8ZPOjJcZAU4reHFKax73Ja8PNqaknGTfJFyviZUXdWyxSb6u7tprytfnfsJmfxPjW4LD1ju+9jlCUhKWCYqnV7Gkrw71Us7ThG7AUkq25ryga1sNlua4UK0S48OZPlg3JptsRnZLaVuSFHv7+hbWCopwVAgE1TvOVh+IQvlcHfTpzdr8eCej+xJ6R13E1i+ppm33CAvsbE9sTm0I5jLpWEKAStRHVtQwrB6eUVExOKG6PIelWC4oSu7OWqA2wWXFy3Eb0qwA5gYU05kqwAADnxw3midg8enCM1B2d+l1bTrzdvqZ8fiPCk3eJBTbLkESpa4CJhS1yRIbQpbrZw5u7gbWCoJKcoIBNR0bjLZpFrXcF2+5xYnYm5zDkhttHaGluBtJT9k7veUkZXtGDuztyaOouhS9owXGL59OSTvx53VjLvfE+HYG1GTZ7qt1mAq5yWmUsrMaOFKBWtQd2n3qiAkqJAOAcGvJrX6mr3d43ZZN0InGLAhQUo5q0Nx2lvuErUlISlbm3qodcAZJqHU1tYSx6zWUXo1fhrdPRa9bLoebvGG0JgxpTVuusll2Azc1qbab+wR3VqSha8rHU7SdoyojwBwcTWmdUSNQ3q/wARVqfiRLbLMVqW4tspfIQgqwErJHVXTp4YyQcpEqpmaSRVTxyrTjGEXr17n29VYsmwU2Cr1jZXGwU2ClhcbBTYKWFxsFNgpYXGwVnWRAF1j+hX8KqitUUzfqsulK2BqxSgFKAVQ+Jml3bnHTOiJBksg9B90PNWPXpKvTdN8zLwtd4atGquXlmp7fqJq03iKVo2upcAU2tPhk9etba4gqTcNDokI8ELbX+vH8a1mEjNYadOa4X8De42pTljqVWm+NvE0Frp1TbdqSCdinlZA84AI/Yal9LJDEhG73zYAP8AtHqa5eOlQ7urrh7LtJ+ZJEiK6emMkVq25xlIgRnE53F5Y+USP8KuVNWi1hPUjJeeDNrcJ2+dr25r8eXCSM+kqFZfE1Bi6olKP3bKHP8As4/8NdBON8Gl3HK0pW2rP3+CK1ETiOgeXAqQidHU1zsn6x081oa00lYo6+HLsWQyl1MW4vISFDqnDxHQ+I6DyVOXq2XOzMuiFMM6KAf8lnKJWkf6rvj8Ss+ur1ConeE+F/h3fVGrp0pZnUpe0vg+x/R8u4uf1PerLcypNvmPi3znVHlMyu5zT/qKPRfqBJroF11DLZW4tKEJHVSjgCu5VlFHnk7yqPTW7NNcUNPIutxbkwn2nC4RkJWD4eWqdrib2a3raCgXNgR08/hWgxSpr8qDu5SudTgXVd6tRWUINI2TwR08LbYW3j75Yz1rZ9dCzkUKpHFHtgtSzbgk3AR3uzBfvebtGzPozisTFK9J+eZmYT9Ve/wOO9B6l0/Y5ei1WGzWO9a1kW98XRTzhGomp6ILzr3ak437VOtpbO843OJ29Amsxri9qM6XuUuy6za1Q8dNN3KTK7JHUm0XBTzSAxtbSnAUlbv2J0qcTyequtax08z9b/X+zaqdl6pN6g4i3jSK77YbrqufzY9+Zgw72pECKoIXb0ySh5x1rkISFbwk8sqPdT1OVVr3iHxVvWr+Ddxbvmp29PuTNAtT2YSI7P8AnuU8mQiSgBaSrCEtt91opKOduVlOBSFKN00vHz51JlN2cb+f5LxqvixPkcUJOnoV3fegvvXC2TLVLMUcrl2559LjbaEc9KSptIC3F7VBRwnG01Iatszt74B8LYiJ8m3ocl6dQ52dtle8FxgDIdbWk4OFDp4pGcjINORQylWbPmPWx8XJz2pbHpo3eOu7J1Je4M63pabDrUJhExcTekDuAobjqSrA3gZ6jNVmFxR1xZ9FWy4zdQquL140rbb09KfgMITai9JYbkPNpQgZQ208pwhzdgtZztJTTdR4NDeNrR+dDN1Bc1T9aWExOK06fa5mnru1HmNG3KZnyG34pDWOzlp1e1Swdic4bOMfZN0dF4q32DYoCGL+xZbpDtNkVZtLsxI6EXvnMtl3ago3lO5S2gGCgN8vJyKlU1JJNePnQpcmuEvDp9S5WnWGqjrq2vSL6uRa5ur5+nvYkxGUtIjtxZLyFhYTzC4FsJGd20pOCnPer41rqK9W/iDfoNmlM2h+VOsEJU5qEyt4NvrfS5kqSdxCQNu7ISfJgkGjJG/u+pXmlbz0MmNxRvuntBJMl2NeLyrVEnTce4XVaYrJCZDqG3pBbQEg7EAYQkBSykAJ3dKRwi4dXHUHFJniEwm228QL1OtUyBY3VdhcZTHdSuQFHq+4uQpvxACAkpAyFE1KKgpNcyltzcV0LVq7ixPt3Gu22K3Xd9LabvFtsy0yjES2427H5hcab2GQoAqR9l3hAUCjBwarieKGvLNpdm8m7rvcq56bvVybhOQGg3EeiPMIaU2G0haxsdUVpUpW4pG3b4UVKNldEupK7s+BZOE1+jHWfEm5M6nka4hswbYpN0hx23XXQESSUoEZAQ4QSQNif9U5IJOufqfIotnEm2W9VhkW46VjOWRVzh6fmMv3hchDDoenKW0A1gYWQtSsrUVghJGasukuWi8Cm98vP/Z13tq4aKGID/8AvP4CowatVRTjHeiyxUrfmgFKAUoBSgFKAUoBSgFcA/XV/Hhf+dP5ShKOwuAX2ieHH5N2391bq+UIYpQClAKUApQClAKUApQGs9a6ejapjTbfKW60hT6XUOsK2uNONuBaFpOCAUqSkjII6dQRVeHDiG+XHZ9wnXWc67GWuZKU2FlLDodbaCUIShKN6ckJSCrPUnpjXzpqUrsu1MNCrLNJvha3Lnr82Q164d3C7637Yl4RrM5Pi3GQ2Je4PuMISEEtcoEHKED/AEpThIO3d4Za+ENscD6F3K5uMKamssMF1ARFEokulvCM7sqVgqKiM48Ks7m7dzEWBU5SlUbV27WfK9+nXW3zZZ7fp2JbLrMuDO/nSmWY5SojahtoK2JSMdB31H46jLToGHaXYKhMmSUwZkmbGbfKMNreC947qQSBzHMEknvnJPTF1007Ga8PB5dXpd/Fp+KMFrhXb2pin/ZK5KQhUxcaOXUBuKqSVF1SMIyTlaiCoqxnA6dKzHOH8VBJhz5lvX2KPb0lkNLCWGuZhO1xtSe9zDuyOu1PhiqFRS4MsxwUIrST+XS3TpYgYPB9huZLYcnSmrH2WJbm7ey4kpkRmEkhDxKNwytbmQhQyDg9DgSk3hbAkqC2bjcoLxM0OPRnEBbqJTgcdQSUHAylASU4UAkdfGoVFWsWYbPgouLk+zsV7q2nm7PdHDW0twnYqVyEsuIgtFIWno3FUFNtjp70kHcPLuVjGalbBpxrTy7jyZUh9ubKcllp/YQ0tZKlBJCQcEn7onHgMCrippO6MunhoU5KUW9P5f1JelV2My4pSwuKUsLilLC4rNsv/Okf1n9lVRWqKZP1WXOlZhrxSgFKAV+LQHElKgCD5DQFTu/Da1XaamSplIcSc5ArL1TaUnRs6I2M7GSUj0gZH7KpkrxaLlN5ZxfRnOWr2O022E4P+iltn4lZT/4hWXYXedGUseJfWB6cEgfqFcA1abPXm70kS01XZ7W5noapmoI5btsNAHeCmyR8YJqt6tIt0X6r9/gbP4Fsc/UGoHvEJLLX6iakOOMbkzYj4A+zR1NZ9IUP6q6pxvhUu44WM7bTb7X4FNYT0A8wrLZGFprjpcTuZlTt7Qj6TvwGBsuckgD/AHqjUtqdzZEfP+qaqo6y95h4fjIt3BTT8G+6YVFucNifEdHeZkNhaT08cHy+mrlO0LeLDEdRpq69pglODZb0pTzOPM291cb9R3j0CvQbJxys8xlJqeZcbmn9QXadZ7lyZsF6wSHFbUtyyC0s+Zt0d1XqyD6BUE1En6mvrMZTLm0OAqKh41qY7PVOvGcPZWv+jfT2s6uFlTqe29Pd2nUulrWm1WZhkDBCQMVL1uDnhVU1wdpinxwlZx+isXEq9JmXhf1V55HPvDjji1qbQ8fVV7VZYkeYxDWzCskxy4TEOSB3GHGktBQcyQAlIPUK8ACamWuNdilak05aIUS5SjezKQHkW98dmdYWltxDydmUKClEK3Y2gAnoQa1DpO7SNyqt0rmPqPjPb2LLcXLUxIN0hPW/fDu1vkRFKZky0MB1KXEoKh/pMEeBSM+myaw4h2PRMiOxclS3ZTrLslMeBBelupZb2hx1SWkKKUJ3JBJx1IAyajdvgTvERr3GjR7UqchNwdfRCgouUmXHhPOx2o62ua2tTqUFPfRkpGcqOQAT0r9j8YbBMgT5LEW+vGBK7FJjosUwvMu8sO4U2GsgbFJVnGO8nrlQBbqRO8R4PccdFtTIjKbi++JbMOSmVHgPuMIalHbGccdSgobSs9AVEenGDiX1txDs+hRHanqkuTJTTzrEaHCelOFtoJ5jhS0lRS2negFRwBuSPEio3UrpDeKzZWtJ8b7HcdM6UkXZaotyudst0ySmLDeciwnJaE8pC3QkpaClq2p3qBwRnoQTMv8AF7SLFui3B25hEd9mW+2ox3NyURnEtP5Tt3JKXFJRtIyVKCQCTipdKV9CFUTRHyuLsKTdNNR7YlQTMva7TcWrlGdjPw8W+RLBKFhJBIabIJBBSs49H2xx30Y9b5U5VwlRokeGm4B6Vb5DKX4pWlAfZ3IHNRuWjvIzjek+CgTO6b4DeLmeN34s6Hu9kdjXRmVMYkThaHLPNsshb7khTKpCG1RlNbyFNoKwSnaceOa+NLcZeHxtKWbTI9i7REtz09rdbHocYRmSkPcrc2lJLalpStCeqVKwRnIpuptWI3kbme7xn0rHhqkvu3CO8mY3bzDdtUlMvnONlxpAY5fMO9KSUnGCQRnIIHhB42WK7an0xaLfGuUxF9izJDUtEB8IYVHfbZcbdBRlCgtakq3Y2FA3Y3JzG6kTvUTHFDWC+HPD686iZityTbmQ6UOqKG0J3AKWogE7UAlZAGSEkDqah+Dl9b1RbbpeHYUOJcbjK5634ilhNxYSkNMTEtud9CFoawnIwQ3kEg5pk9S9w5esomw8Vb9GD/IH/wDefwFX8KvzUY+Ld6TLDSt2aIUoBSgFKAUoBSgFKAVwD9dX8eF/50/lKEo7C4BfaJ4cfk3bf3Vur5QhilAKUApQClAKUApQClAfBYbUcltBJ86RX52dr8Gj5IqLIm7BjNEf6NHyRTs7X4NHyRSyF2Oztfg0fJFOztfg0fJFLIXY7O1+DR8kU7O1+DR8kUshdjs7X4NHyRTs7X4NHyRSyF2Oztfg0fJFOztfg0fJFLIXY7O1+DR8kU7O1+DR8kUshdjs7X4NHyRTs7X4NHyRSyF2Oztfg0fJFOztfg0fJFLIXY7O1+DR8kU7O1+DR8kUshdjs7X4NHyRX6GG0kEISCPKBSyF2fdKkgUoBSgFKAV5yWg/HcbOCFJIwfVQHNd7hcqPNirH+gcx1/1Fg/wqL0md1uZJ8StR/STXBVllqtd565Slnw6fd4EteXdzJR5FqCQPWQKgtStlcyC19ytwD9RNUrWRNNWjbsfgbQ+p6Z5lvu0npl2YofJTj+NTXHCFzrDbngMluUlBPmCv/UCu0a/It2HnClfHX/5PxNbMDrWSyMrFcJLiejTK6y3y9J39YwQqZJWMf7xVfmtHy3bnvvinFXMOr1Ldpg0XZTfebe4HQuz6caPnTWzq9CZ5jxMS52mHeYbsWbGalR3RtW08gLSoeYg+NUIcJFaakGXpKemCodfY2egvxF+hPXe3/wAJwPvTQhok2OI6LQ63E1Vb3NNPkhCZTiubBcPk2vgYTnzOBBq5tuIfbS42tLiFDKVJOQR5xQk+qq2tkcwxU58UrH7KxsT+kzKw2lVGipXAK0yOFGnNGByMtdjRC5MyTAQ62+7GSEhTzJOFpUNwKdw98cKBwayIPB5dsd0vIt862WiVaHJXObtNnTFjPtyCnmhDSXPsa+4jCypfUEkKz01Tnc22XoVqwfUyt2mPNTJv6JMiVGt0Zclm3hpx4RJQkB15RcUXXnD0WskZ8QkeFXfWfDy4XzUTF9sd+TYrmmA7bHlOwxKQ4wtaV5SkrTtcSpOUqyR3juSrpiXNSfAKNlZMqOouBqrJww1fYdMrclqudqh2yNEdUlKkNR2EsABaiEqUUAnvbQT0JAORAaU4P3bVNgeiS5TNvgw7u662xdGWp7k0qYDTpmobkLQt1CwQhRcVt2J3IOABUpq1+ZS462RarX9T5FteknbEm8vONuWuzWznKYAITb1ZSvG7xX5R5PTVm1rw8mak1DbL3ar4bJcYkOVbXHDFTIS5GfU0pYCSoBLgUwgpUdwHXKVA1Q53d7FeXSyKZZfqabbYbhZpDTlnuQiQLdCkqvNiblvLMNtLaHGHCsFhSkoTkYWAQCMHJOXc/qcbRdpOuVSLi+5G1IkBiKplCkW1RcDzpQDkLDr4S6oKGCQB4VO9fEpyK1jLg8CoSIdrZk+w0XslzcuDrdgsybc1JC4T8TYpAcUc7XyreVE90DAHhWdVcBL4/od+Ii/Kvt0t9mTY7KERW4qmWC9HWtbhUpSXHSIzXewhPc6I7xoqivqHG6si2W3g28nUUTUF2v3sjem7yi6vOswww04EQXobbKUb1FICX1LKipRKs+AIAj7l9TrbbzpyHZZl0fcisW6628qSykKUJr7bxX1JGWy0AAQQc9fNUbyzVkS43VmZ1j4JsWtFrUpdkhyIN1auZVYbGi3If5bLjSUuJDiiT9lJ3Z9AAr3s/CBzT16sNxgXrDttlXh11D0TeH2bhNEtxsYWNikqShIX16A5T16HO4UbE/xE0P7fbCzBRO9j5EabHnx3lMh5vmsuBxAcbJAWglPVOQfAgggGqxwo4Kq4W3q7T270J7dzZQ25BTE5TETY44tCIo3qLTILzp5RKsFZIUB3ahStHKS1eWY2dt9VW7RwxBf/AN5/AVewy/MRYxTvSZP0rcmlFKAUoBSgFKAUoBSgFcA/XV/Hhf8AnT+UoSjsLgF9onhx+Tdt/dW6vlCGKUApQClAKUApQClAKEZFAaI1Jom4zNQXJ9vXGrIiHJLi0x41y2NNAqJ2oTt6JHgBUd7Qbp/aDrL51/8AbXNzxtZSaTOgjg6LimPaDdP7QdZfOv8A7aiG7ey9qZ3TqOKeqlXxpkPrgezH2UN9O9jb4dR+mo9OrvmXY4ClK9k9D9sVua1O9cGbTxT1VcHLe8Y8pLF43clweKT3fHof0VLe0G6f2g6y+df/AG0eOrrmJYClB2adx7Qbp/aDrL51/wDbUbd7I5YpNsjzeJGsmXrlJ7JFT7JKVzHdil7eiDjuoUcnA6UWOrvmRHAUpOyRJe0G6f2g6y+df/bUbIsbkXUEOyOcSNZJucthySyx7JKO5tspCznZgYK09CfLRY6u+YWApS0SJFeg7mhJJ4g6ywBk/wCdf/bUfp2xO6sscO72viRrKVb5bYcZe9klI3J8+FIBHxinp1e3EegUmr2MmNpKTOdkNR+JWrJDsZfKfQ1eApTS8A7VAJ7pwQcHyEVizbG5b75bbO/xI1ki43FDrkZn2SUeYloJKzkIwMBSfEjOelPTa/UlbPpN2sSR0DdMfbB1l86/+2ntBuh//iDrL51/9tR6dW6lHoVEx2dJSpUuTFZ4lasdkxtvPZReAVtbhlO5ITlOR1GfGsg6BumPtg6y+df/AG09Or9SXgaK4oe0G6f2g6y+df8A209oN0/tB1l86/8Atp6dW6kehUR7Qbp/aDrL51/9tbE4SWCVY/ZTtOoLzfebytvsvK5/Kxuzs6DGc9fUKysNi6tWqoSehj4jC0qdJyjxNiUreGlFKAUoBSgFRV/1TZ9Lxw9drlGgIV0QH3AFLPmSnxUfQAaA5213e5NwvF3VabVJEV9CnUyLghUVO0p6qCFDeckHHdHrqJ0c6pMFtDpTvSTu2jAz08K43HQhCs7PW56Zs2dSrhU2rRSXe/4MyZK7VdIrCTlPNyR6gT+0Vj3hXMvkVPiG0OOH4k4/aRWBB3+JtZRy2XYbc+p6jcrRfNx1efcX+sVaOJ7CX9JP7gDtcbIz5DvFdzJWovu+h5bB3xcf+76mm204UayYwy6muBl7R6ZPgVeMsvcO5riRuLrjqgPWtRrH1y7vW0wDkrcAx8dZOBjmrJdqNXKWShWl2M6H4Ywex6ZjDGCUCrhXennApQHw+w3JZW062h1pYKVIWnIUD4gjzVTXeHBsjipGkbk5p5wkqMAp50Bw+lkkbM+dsp+OgPlPEGTp5Qa1fal2hI6eykUl+Ar0lYG5r/8AuJA/1jXtrKY1Ot0aTCfbkNLacW06yoLSroMEEdDVjEK9NmRh/wBRHHaZHESPoE3X22alduSdAN6k5K2Get1Sgq5W0M52nokteXGTk5Jtl91lqCdxvgWeA9d4UJy6G3TWlvKKRHVbluc5toR9qG+bs2vKeKitKk429BgOMXyNjeXUh+F2qpehdF8M25VwvLltTaJ0K5NPxXXltXJCIvKjlKW9wUNr4SnHU5GScVIaLvur5lhfv95uOo3zadCWu4i2QGW0uS5zsWQZBCVNnc9uQjCPAKIyk+FHGOr88Qm9F54FSVd73qOIh+63m8M2K06qgOtT4ch2Ry47kVQcUHlRm1LQl04zsISVkZIAxYI15u9mcjW2+36+2PTQnX8G7wow7Q/IbmpTFQtaWj75ovLHd+yFIyTjCpcVwSCb43Plq9cSJdoVeLlcrvb7tbbXp2Qq1sR0IZdkvOlMwOI2EnKRhSAQEZJ6EAjpYpz5qtTjHkXItviNvqpt9VWrIrG31U2+qlkBt9VNvqpZAbfVTb6qWQG31U2+qlkBt9VWzSAxCe/3n8BWVh0t4Y2J/TZPUramoFKAUoBSgFKAUoBSgFcA/XV/Hhf+dP5ShKOwuAX2ieHH5N2391bq+UIYpQClAKUApQClAKUApQGtLv8A86TP98v9prErjqi9ZnXwXqLuFc98RdE6ti8QtTaz01a1ybvCXbzbgcASm1MPMyEDJ+53tqOfvBSFkZ+GlCMmpvRq3xa+hH23htqPh1IfVbrZPnRYN7tEp7sShzZzTcMokLSCob8uryQfT5q/Vac1oYdgmuWm8Sbk1dpr3sTKWTH5bssKSpbrbyS2tCOqVd9OMjHWq209TOdSlN5m1f8Ag9pml9UK07q9HsLf3dYGYuQbkmYezS46ZzTjbbCebjPJTgDaMYUCRuwc1uwat1DqRi6LtFwgtnVwnx2p5SVR43scpsLIClBKd56gHoT5800Kc9Kzd1zt3WViFsGi9WuQlNuRNUWeUbJKjX6b2rnLmTFLRy3I6S7hRGHDlO3unaCCRWHddEa9uUDT0iJZJEFUCHPafYYdWy5LjmSwrlpKnFqYW82heBuOOo6Z6TdFxVKKlq15T7OSsWW1aW1VI4qOz5Yu8OP7Ic+GtuOpbRglgJTHcWZAQhI6hSOUVbxuBOelTY4ea4Ysuk4L7V3gRo9iaYYNvYLzkKYHlqWraJDQQop5ffVuTgEdOuV0QqlFW4cF9ez795YZ2iLtp66cQhD0xcZ7tzuseQiWJDymFxVBrmKCEPoU4pKwslvKcjpnb0qR4Z6R1TEveg37vCmJbti722tck5LDTi2+Qk5WsgFIwBuVgDGTjNQ2mvPQolUpuF21f/8Am3Q9dQaIv194iapjMN3MW+6wn0C6yVrZ9jnA0gM8haXSlxsrTu2lAKTkk9etNhxtWcQOHbOsVs3KddrlOLzVqjFTkR1DLHIShwJdbUhKnA4sLScAkEg9KK1iqEqeVN25Luv7uz4sl9R8P9TRr7xGlWazXKNdrxCguxZceWpbRQOX2tkKLqcuHCgnwOAcFINZlm4d3q6HTcGUb4bC7eZTsyO4HYHZ45iYSgASHFhougdCvxUrpg9TasRvaeW+jf8A/OmlutyQ4M6K1Lpq+6YmXFN2zIsUlq6qnS3Hk89L7fIBSpRCSG9wGAOmfTW9atzSb0MHEyjOd4+dX9BVr0L4zf8Ag/8AFWXg1+fH3+BqMYvyJeeZa6V05zIpQClAY8+4RbXFXJmSWYkdsZU8+sIQkeknoKqSuJrN2Jb0xap2pV+AkMI5MQH0vuYSR/sbqWA9gNXaiGbtfWrFGPjCsSMuY8ypDgz8lCfXUrYdA2HTb5kw7ehU9Xv50lSn5K/W6slXxZxQgoXGiAUTWpCR/p4zjR9YGR+01pe0TBHccSD1IBA9YFcdtRZazZ6fsJ7zBqPS/iSFjWZN5UtX3AVjPn6f40uRxd5C/vYjmPlJrXUdbd5t6+k33G9OB8fkcP7Z06qQVfpUal+JatulXvS4gf8AaFd1U0oy7meT0NcVD/uXiaaAyTWQ2oN5V5Egn9FcBLWR6dPgVWzlQ4e29Kk958IBSfOoj/GsW6INz1dDjDqAvOK2OzI5q67zRYuWXB1n7vidT6aiiJaGGx5Ejp8VSldscCKUApQH4pIWkpIBBGCDVGuuk7Tp2Up21wWoJlkqebYyltSh90EA7QTk5IAz5c4qxW9hl+h+ojTdk4zGQpydLct8qziJLnOC3blPwmm3AloO94gqcSeg7pyDgEA4ssvijaYL8diRGnMvuFrmMuNJC44ccLbRWndnKiMhKcqA6lIrTRqK2pYo7UjKF6mvdw19/Fa37n3HjH4pRcXBUu2y4qWriu3QwSgqmrQDv25UAnaUOZ3EABIOeuBkRuKFnm9k7O1NeEiELipSWe6wxlSd7iicAZQrwznGRkdaqVRF+O0qb0ad/wCftZ9zMNrjFZnU57BdkJDMeSVORNoSw8opbdPXokqBGD3uhISR1qQ1nrB3TN6sENiK9NMwyHXmIzXMdU001129QE99bfUnGMjxIpnTV0PxGM6TnTT0a0trq7afMxnuJttaIlocclW5caE402xGUp5xyUpQaCevXKUglOMjOc+Soy+8VJMaDqLs9llw3Ld2WO2/JQlYMh8tgNltCiSoB5s4Gc5Ph0zDqK2hbq7SSi3BN8dbaLSVuevsl005qCPqaE8+wy/GUw+uM6xJSErbcQcEHBI83gT41K7KuqzVzaU6qqwU1zGymypsXMw2U2UsMw2U2UsMw2U2UsMw2VadJJxCe/3n8BV+gvXMfEO9MnaVsjWClAKUApQClAKUApQCuAfrq/jwv/On8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUoDWl4wm5TFEgAOrJJ8nU1DWG+23VNojXWzz410tslJUzLiOhxpwAkEpUOh6gj4q5Scbyb7TrYSWVLsMp2Swy+hlbzaHlpUtDalAKUlONxA8SBuTk+TI89YB1PaRptN/NwY9hVR0ykzSr7GWiAQvPmIINUZGyvOiU25qq2DidpvVF/VZrbPckTw2+6gKiPttPIZcS26pp1SAhwJWtKSUKOCalQck7EOaVkWrbmm3PmqmxVcFNecl9qHHdffdbZYaQVuOuKCUoSBkqJPQADy0y3GY9ANwBBBBpt9VMozDb6qbfVSxGY/FthaSlQCkkYII6EV5xYbMGM3HjMtx2G07UNNJCUpA8gA6AUsTm5Hrt9VNvqpYjMNvqpt9VLDMNvqq1aGGO2f8H/AIqzMIvzo+eRh4x3oS88y10rozmxX4pQSkkkAAdSfJQFSncULI1JXDtipGorgg4VFs7RkFB8y1juI/4lCvAjXGo/LB0jDPk6TZmP1NIPy6mxHE8ZPBjTd3aze0zdQyj/APV3KY4pxB/1NpSlv/gCa9xoe9WgD2D1dObQkdIt3bTOa9W47Xcf8ZqBYG/aysw/zjpuNeWh4v2SWAvHn5L239AWa9GOK2nQ8hi4yXrBKUcBm9R1xMn0KWAhX/Co0sDE4oss3PTDM2O4h9tl1Kw42oKBSe6cEeuuc7fGSu/KhH3wQcZ86TXM7UhepHtPQP6fquOHnblcsHY/Yu4IBHRaCQfTnB/UBUbdVntUxWMBERQz6Sof4VqKcbVLG/lPeQz9UdEcJmeToS0p/wD1Cf417cTE7tKvY64cR/3hXa1f0pdzPLcP/cw714mm0DJr4vDnIsdxcBwsMLSk/wCsRgfrIrgJe0em1PZMCXGEe22OGg4Adb6Y8ie9/wCGsXSLAn8R8K6hvAGfjrb7IV6t+853aH9jLvXidTREBEZtI8gr1rsDiBSgFKAVV9cMGSw2ylxTRcbcQHEe+TkAZHpqzVV4Mu0leVjUzx0dHgW7QXtgt0aXH7K2i3draTKdDKkLSkt5ydwbAPTJBOKmBodprV79/ZlONOyNheYLLSwooRsGFqSVpBAGQCPDyZOcKeHlTy5k1fVXXFFx4WnLLlfs26cr/dkW/wAKWHWWEi5SSuNOlTWFOsMuhHPUpTiClSCFDcokE94dOuPGRuui4kmFqEypr6BdbcmBIkHYktMoQ4MpwMA/ZFq8MZPQCrSorgvPIojgYxTWZ/Lpl6dPmeSdI2vUsZ24x5TrsS6KgS0KbwElphSXGkJyM7FEZIPXvnwrxuenIuvJvsrb79LjJaZk2lwwg2UlPN2voBWglKt7QTuGMFFVyoNJ9j1+ZVPCRnFxjJq/Tvb+ob0XZLZfoDYmciSqUmdFgFaRuSxFEcISnxKEBYX6FKHXHSsW7WixWW6z37lflNJXOZvjkJ1TfcUkJZSrATvKCsN4znvJAFTHDSm1GKeuq8CPQ6cVZS0u38uHcWLSUSBDt0pu2zEz2zOlOPPBaVkPqfWpxBKegKVEox4jaAfCptRKU5OAB5T5Klwyeq+Rm04KEFFcjERdobk5qGiWwuW6yZLbCXAVqaBAKwM5KcqSM+HUVl4PmqZQceJc4jB81MHzVTYDB81MHzUsBg+amD5qWAwfNVo0oP8AI3v95/AVepL1ixX9gnKVnGuFKAUoBSgFKAUoBSgFcA/XV/Hhf+dP5ShKOwuAX2ieHH5N2391bq+UIYpQClAKUApQClAKUAoRkUBoDj0zeZ+kL9atPsLeut3eFsbcSSkR0POBt15SgCUhDZWrOD1A6HNagt+idVWl5ekVW2TarGrUsa5Nr07LfQwiE+092hgPJCFAB5tS1DpgSEY8hrntFdPjdnR2btboedv4fzntT6Mdv8TU0iNEe1FaW5DM2bzGWVXBBgFxbbgWEFhs/ZFHBARuUe7U97GcRP8A8PDDZmuG7+1JlswxFe9lxP7OkKUZHO6q37sjl7s/dUk4u1/OrJSavbzoj8e0pqJetn7xzL+VK1f2cNidIEcWpUIJXhoL2BHMJO/GQoDBGBVSsmiL9onTmnHoWnrxOfiaavrT0Yy5CXUvOzIJQgOBRcQS22tYSggnYrb3jmpTTsvPBkOL4+eRIaS0xqm+exlsuLmoUaf9t76ipC50FSrd7EKUkHmOqfQ0ZRxha87jjCfeiOOktd2fQxMJ7VL94naOvDUpcibIedE1t6OmFtClEId5antpSAVdScnrU3je2hFpWvr5sWGbw/vth1Jd5Nqkaneag6gsirah66Sn2jGcdjpnnatZDiCgulW7ITgkbetVTU+ntYa1la6gotN9ixLlpy/xnbet2dy1TA612LDzrvLWVpDhSGUpQErUglQ6BFxert5/kST4K/n+CY1hAvMmLNRa4WtEhenWEaTEdya2Y9y3v8wytygUnd2Y5k9zYCB90DNXnTeoLdZOI2q3Z12avlsuIn29DtweRFcixm476mUNFQbDbqm3kFRGe+euB0jTTh5sTZvqbE4RG4zNDRLtdlyDPvTjl1UxJWSqM28sraYAJ7vLaLaCB0yknymrpWLNes7GRF6IUqizKrilLMXFKWYuKtOh+pmf8H/irLwi/Oj55GJi/wBGXnmWqq9ftf2HTkgRZdwQuer3kCKlT8lfqaQCr48YrojnSL9ntXai6Wmxs2GKrwm31e53HnTHbOflrT6q/U8MmLqQvU11namX49nkL5MQH0MN4SR/t7vXUkcS2wbfFtkVuNDjsxI7YwhlhAQhI9AHQV71BIpQCvORGalsqaebQ80oYUhxIUCPSDQGu9c8K9ON6fuUu3wTZ5SWyvfa3Vxgojr3kIISr4wa5wltXCw64iJZmplrdSVtmajGcpUCCpAHm83mrS7QlBOGdX8TrNiwqyp1XSlZ24Pg9Pl3lul3eatDTk+ySE8s5D0FYkII8vTov/s1D3K/WydGmpjzG+ctCUBlzLbnlz3VYP6q1+49bPB3Xz+BtKWK03VX1Xrbo+5+WdOcOkbNG2sYx9gR0+IV+cRmyvSUzHikoP6FDNdJUV6cl2M4ig7V4PtXiaXQMqrE1KsptbUcdVyn0NgDxxnJ/YK89no2enVXaJ835HLvNjQkEpQpeceQ8pVY+gk44hE+U4NbnZOlW3eaDHr/ANPl3rxOoGB9hR6q+6684cUoBSgFVfXIfMQCKUiVynOUV+9C8DGfRnFUTSa1LtL2kcz6Onaitdl0zabJZpyJkWC8/fYV1szjYcnpYcWXe1LAQ4tySW+qFKBGTkeTxjMarnWeSqyy9WJXNtsSBMl3JElDgnvyW0uvstOgcoMtl1Si2lLfVOM7TjeyjQU3Kbi7vXVPi3a3S6s3bglZ24F27tZEhLjX20THoElWq3tLv6heSVRnJb80sNwmwhCXUkuobckhxW4KSkYAylKqqdziaxumiPYm5tavXcZdhZVbm2FScoly33ucZTqMJPIbUwnlunG0Kwkk9K6Xo/qy9S+j/wAeOql7rapcL8NSHm4a+eBZ3E3z26xI7bGqI5hX0tFEUSW7exao8dS20ADDTpeU2kEjcrLhQSBhJnW4V90lwN01JjxLgu9xXoNxuECChRkOlyShyY1sHVRw49kHzdaxam5e7heOrjfhw1V3bvu78LlSb1feQ90tuvTGlRTLuzVxkwI6FvxVrW3HkT7h9l5SvAiIygAK+5Sc9Nxzl3vTt3XfHrS23eZliRdbTGQqW49Jy3HLk998rWSSFq5bJVnGUpQPAAVKeHXsZVbVcL6ZWl4q32FpczA0+5fbzc9GxLijVMC4w4SbpNdxORHlyXAp4QlKT9gCUE4WXOvRDafFWK+hnVN60re5KEawXNVo94zkzDMZQ9dninux2CQU8nY4NrQCSFgDd4m/FUIy/wALaft4Nvw0vfVW10ZS3Jrn8ycetFw005Kasntpj6QJtMFR3zn5LUYIdcfcZbWVOoyFR2lctIKMKwElJI+PYvVMyG8lbmqmbFFiXe5xoqZMlMx9GWkw463d3O3na84EFW8BSEnypqnNRfrzyXfZH2svTovg31sTZ8Fc3bpWLNhaXs8e5PqlXFmGy3Jfc9846EALUfSVAn46lK5iok5Nx4GSnoKVRlJuKUyi4qzaW/5G6P8AX/gKu01aRZrO8CapWWYIpQClAKUApQClAKUArgH66v48L/zp/KUJR2FwC+0Tw4/Ju2/urdXyhDFKAUoBSgFKAUoBSgFKA0Dx91Dd9K6LvtwsDsdi8iUxHjOymuY2hTsptrcpORkALPlrWN0473i5s8PRZWY0aVNlQ/bEy8grMMOTEQlxh17qy92gBXX/AJMvz1oN2pNvtZ0WdxSS6IuTvG+2tWaDcE2m4SEym7q4iPHQlx49gUsOJSkHvKXsO0DzjOK84fGj2Xi2JFosSr5c7szJltRbbcYzrQjsKbS45zt4Rnc82kJ6Hco5wATVvddpc3l+CK9J4+N6SvN9t13U3KmrvUpi3x5MqPDQ1GYjRVrBcWUpJ3yAAOpJX4hIJF+ncSITXDu36vjMJVBnR2JLSbhJahBtDoSUl1bqglGNwBHU56AGplS4PqFUvddCkufVKQpGn2btbdN3C6xxaZV5lGPIj7Y7EZ5TL+FFeHDuQrbtyFDrkZqxTeM1sgcQ7RpV1hvfdHUsRnkTmFPFRjqfClRwvmpbKUKTuIHex0wQqjo25+UQqiZStCfVDPw+H9gmavtFwTNmWL2Sjy0chRua21stLShCFANqUuQztCtoIXk7cECduPFa5StSWOyGA/p67NX2LFuEN1bT6XYr0WS4hSFpyMFTJB8FAtkeBBNTpK71KVUukeuguNDWoZOkrQth1+ZdrQzcjNmOMxlOhYXnltbvsik8vKwj3gWg9c9IGT9UTHvml1yJfD25y7TN0+u/KYkOxFoetyRh8rSXME4UnDZB3BYzt6gRuteJLqacLlh478RntC6JtVxtkqQwmXMbS77HRkPzjEDa3HVx2lgoKkpSFKKwUpQFk9QKn+FGq3tSaMjrulxh3C9QkNt3J6HgtJcWyh9OFJ7ijyXmlEoynKjjp0qlw9S5Vm9exEROMy5mikaqOnnIFlltsP2+Xc7lFitSGXclDi1Kc+xZG07SCfsifLuCcHTHG2VrPVOlmLTYi7YbzZJV0XJXIb5rK2X2mlJGF7VJBWRkZzuSQcA1O6WupGfgrEDePqhXLzCdZsEQKu8C7Wdt+NAmxJokMS5Sm+UHErKEOHlOJUCRtyCDgg1Z4nG1d0lNWiBpebK1SHpjUm0GQygRxGLPMWXSraQe0MbMdTzBkJwrEujZcSFUvyIFrjQ7etURJdvlyWbBcIGnJEaMqM2XELmzpbLiV56jIbbSrBO3YSnr4zMbj3GmWOTcY9ikyVourNoagNSmDKLzjpbSl1srHIUCASleO6oHr1AOlcKoZGkuNftlvtutsrTNws6Zsyba0SZDzK0dsi7y82AhZJThpzavAztPQZGdt224XyIpxmx2lm4PPAbpEqUGWWMZwVYClqznwSPJ1Iq9QhkrRVyxiJZqEn55Gf7R7xf+9qTUkh1o+NuswMKPjzFYJdUP+MD0VYbDpa0aXjlm022NAQrqrkNhKlnzqPio+k5rdmhsSlKEilAKUApQETq1AXpi6D/+nX+yucp9qFwMd9KdzrBI6eOD/wDsrRbTdnBnW7Evu6iXYSUOXuidVAdPDHWoPU1vjyrU6mQw29gKXhxAODWqU7SWV6m9pUlNSUldWNt6G4csJ0tbpFpvF3sMhTKVFMSUVsk48Sy6Fo/QBWbqO2a5j2ObGMm0ajjKaUMutrhSB08cp3oUfiTXYP1o2Z5wnlkmuRpqNqpLCQbjb51vAHedLJdaHp3N7sD14r6RdWL5PVc2lpXZ7a2oNvg9HXCO8R6B4euuCxFCVNOSd1fivOh6LLFRqrI1aXR/Tqe0F1V0ttknLO4rddys+coUAK+9ApzxC9QH8a2Oylasvf4Gv2g16BNdq8TpxgYZR1z0r7rrjhxSgFKAVh3C1M3IoLqljZnG0gePxVDV1YlPK7oxDpeIR7935Q/wp7V4mPfvfKH+FW92i7vZD2rxPv3vlD/Ch0vEI9+98of4U3aG9kDpeIR7935Q/wAKe1eJ9+98of4U3aG9kPavE+/e+UP8Ke1eJ9+78of4U3cRvZD2rxPv3vlD/CntXiffvfKH+FN3Eb2Q9q8THv3vlD/Ch0vEI9+98of4U3aG9kPavE+/e+UP8Ke1eJ9+98of4U3aG9kPavE+/e+UP8Ke1eJ9+98of4U3cRvZD2rxPv3vlD/CntXiffvfKH+FN3Eb2Q9q8T7975Q/wrNgW5u3NKQ0pSgo575qpQS1RTKbkrMyqVWWxSgFKAUoBSgFKAUoBXAP11fx4X/nT+UoSjsLgF9onhx+Tdt/dW6vlCGKUApQClAKUApQClAKUBpfirEsky1XBrUTxYtRnMFSwSCHBJbLI6Anq6Gx8dQDPBnS0addpbMJbUi6XeNepK0uq70lhaXG8eZO9JWUjoVLWfujWgk3Fu3U6KMVJLuMaHwPsFvuyrhGlXZl1BmKjNouDgbhmUoqfLKc4SVKJUD1IPQYHSq9cNA6Lst7at6rjfYuomt89y5Qnnu1vJmPNsL3qbTjatbTIOAAjlpVlOCqim2S4JFpmcHrJInruMZ+42y6qluyxPhSil5KnW2m3UZIIKFJYaykg9UAjBGaltSaDgaps1ut8uRNQq3PtSYsxmQRIaebBSlzec7jgqB3A53HNUOd7FWTiQkLghpm32eXbW25a48q2y7U6XZS1rUxJdW69lRySorcUdx69ax7Zwr0w9rB+5xXLomVbLmiauIt11EUTOxpZDoSQA4SwpKSQSkHPgoKqrO3cjItCPsfAnQwhTbO1ImXdiDDNk7LIubjxtraw08ptvB3NLVhhzOcjDe3CQBU/beEFkgS2Zrrs+5XNuc3cFT50kuPOOttLabCjgDYlDi8JAAyonxJJObejIVNcj4g8G7HA9rjTb9wXAsAaMG3uSiWEuNJUlt0pxkqAUfKAemQcCv1ngzptiyR7UhmR2RiwOabQkvHcIawkKTn77uJ71Q5snIiR1Lw7tupmLQFvTIEq0KKoM2A+WnmctltQz1CkqScEKBHgfEAjw4d8KrDwrt8yBp5uRHgSnee5FfkLeQHSMLWncSUlfQqAOMjoB1qM145ScmtyHj8CLDDtkKBGn3pmPbX237WgXBZFt5aHEJSwDkBOx1aMKCjtIGe6nGVD4J6ct7NnZj9ubatkWXBQntizz48hYW626SSVgqSlWc5BHj1OZdRsjdoj2fqe9MtQXYy5V4fUtuA0iQu4uB5lMJxa4obUnGwIU4rw8fE5JJOWOB1haYiKjTLvDujDkl03ePOUmY8ZBQXw4vwUF8tvpju8tG3btFS6jZG7RlNcGdLxnIRjw3IyIbFtjsNNvK2obgvOPRh1yThbqsk9VeWvmPwbsTdyduEl+4XG4OSIb/apsouOARXFuR284HdSpxZ65J3dSemIzsnIiRhcNLLAmwJTLboehXOZdmSp0kCRJDodJHlBDy8DydPNW0NFDBmf8H8avYd5qsSxiVaiy0UrdmiFKAUoBSgFKAidXLCNMXM/wD6hY/SMVoSMS2+4PSCP0Vz21npFHY7BjeFR9xlreOw+A6VXL+vNtfz1Ku6Pj6VoKHrSXejqHaNOT7GdIaPjGJpu3skYKGEJ6+faKl3kcxpSfHIIrv1wPJ2c2a2cOnmZxLRdZjzEBxCfHl8wdfVjFVLiXJuF/eYi259FvtCEhTriUglRPXAHlPo+M+IrhK1PNiJU+37nfTqKNKFZ66L42QsYnacgC0OvGbFlNhyI6UFK0OgjAwCcZJHXPrq1aFa/wD3ir9QrZbOhu8Q49DX4yaq4CVRc2vE6VZA5SMeavuuqOQFKAUoBVE1xedU2+8NtWWfZ40QshRRPtrshzfk5IUmQ2MYx02+fr16Y2IrbiGdK5kUKSrTyN2K97Z+IHwxpr5jkf8Anae2fiAf/wA4018xyP8Aztav8Rf7TZfh8f3fIgbHxf1PqC3PTo2pNLtxmZD8Za5FkkNYW0vYv300dAry+keeshPFLVK9RmwJ1PpU3gRhL7J7CSN3KJICs9sx4g9M58vhU/iEv2l57KabV3p2GWjXusnIrklOpdJKjNkpW8LQ8UJI8QT23Ar9b15rN9srb1JpNxAQXCpNoeICQcFX/LfAeen4jL9pb/De1/AK11rRDqGlaj0mHVHCUGzvBROM9B23zdfVX4vX2skQ0S1al0kIqzhL5tDwQTnGArtuKfiMv2j8N7X8D6d1xrVh1DTmotKIdWoJShdnfCiT4ADtvicH9FR1v4s6kurt0bi6r0k4u1vGPMBsshPIWACQSZg8ih1HTyZzmn4hJ/4lS2XdXTfwM9evNZNsNuq1JpJLTidyHFWh4JUMgZB7b1GSB8Yr1b1lrl4thu/aWWXEb0BNlfO5PTvD/Leo6jr6afiMv2lL2av3P4Hn7e9Zl9bHtj0nzkKShTfsO9uSo+AI7bkE+QUd13rRggOaj0m2S5yQF2d4ZX97/wAt8fRT8Rl+0fhq6v4GBa+LGpb2m5Khar0k8m2vqjS1ewshIZcTjcCTMHTqOvh6aznde6yZZQ65qTSSGlpC0rVaHglScgZB7b4ZIGfSKfiMv2lT2XZ2bfwPVnWWuZS3UM37Szq2lbHEosr5KFeY4m9D6K9vbPxA+GNNfMcj/wA7UfiMv2lD2dFf5fIe2fiB8Maa+Y5H/na2Fo+Xcp1hZduz8SROKlBTkKOphsjPTCFOLI6f6x+KsrDYt155GraGLiMIqMMydyapWzNcKUApQClAKUApQClAK4B+ur+PC/8AOn8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUoDRHHOwz9S6VuUC2xzKlquUN1LSVAEpbmsuLOSQOiUKPxVq6PwwvUHVEO+ot88TndT3hcyRHmAPG2ORZfJQjcvaEl4xilPQBZCiB3jWkulddrN9Zu3uMDSGmdV6RgaZfa0tNet1kvzqyzGQxHnzoy7ctkPvNc8tlYeUkKUFjdt37R5cK2cN9b+xlqcftclmemLa23gZKFLQW76JDqCoK64Yyo46EdBk9Km8epFpWsWfR3DO8WLWdkvpgSGpr2ob8u5yVSioqgOvSVxUkbyNn+gUlAHdJzgEqpxA4Y3++3TiVdbYw8m8SbfGj2OSZO3A5REhLQ3gNrWO4Vd3rt7wAyKcyzX88fsVZXlt54FYn8NbsjQkgQbLen3hdROiWKRb4qYJcTFLexyMJWAypRySFpKXQHMeJOTrfQ2rJ67/ADva/KuM5V2anW62EtSbe8oWqKyUPBbzakp5yXQl0dUKSV46jdWpK/EpadtDLumg7rZZ3E5cDQzc6TfbvCmiQpLbrL0VTUNEgBHObU4tDjT6y2opCvOd20w9g4O6ivVpt0LUVmlGFEtupEx4jkhLaWXXpzK4CdjbqgCGgsoG5Qb2jBBCaKSS4+bBxb0t5uWDTekdTN6gfkX2w3GfqGRAhi1agMhtTNsKYKEPIcy6FA9oDq1BKVBfNT47Ttz/AKnrQ150st1+7Rrpb5irdHjTGZLTKWJElJJW9vQ+4p5wknLpA3ApySRgUyccrSKkndNm7NtNtY1i9cbabaWFxtptpYXG2m2lhcbas2ixgy/+D+NZOGX5qMbEu9JlnpW7NGKUApQClAKUBXOIMoRtLyRnq4UoA+PP7BWkkj7ISK5jazvOK7DuNgxtQk+36Hq+cNE+eoV6Gq73i1WxsZVIkJ3DzJByTWrwcc1aC7TdYuWTC1H2M6ehNBqK2keQeFe9d0uB5czSnEWCmPqSYhxsLZkoBKSOhGMEfqqt26BGtqT2dshRyStxRWryeUk9Og6eiuJxyyYiR6Ngoxq4alN8l4aHy/DjOzES+Wec2CE5Udoz5dvhnpXhpR3smv46if8ASI/Yf/WsnZ874iPnkWdpUY08DUUex/NHSrKgtlBHlAr7rrjz4UoBSgFUnWgzdkf7kftNa/HK9H3mwwP63uIDbTbXO5Toro0DE4E3KbKtKLxboE6FGlX59xl9SXE/5UtKo52kYJ6Z/wBUgeWsKJwK1CwmBi328T3tGewTtzLqebCmJQtIdB2lSspKW9yTkJHmAFXrm39Mhwu7fy389Ctak4e3TQ+kIrr9sVCkybpbkOplPRnYYDDT3fUlmOlCEZKRucSoq7ucEA1nWbh1frroqM9pWwIbh3HTE2wONXKSlhxpxT6ldo6NgLSvqU4SnxT4DrUl514OKk5aX+neT954ATb3PucuXarfKfenWVTLrykqWmOw00iUkEjIBCVDH3Qx6KxLzwHvyS0iJCZcs8e83R9u0xXY6AI8hKA0pCXmltp27VjbtBAWcEVFyzHGQ0Tei+1j1vX1P1ynW68cmBHenItNniWmRMkJdfjuR3CXsO7Rg7QkbgBux4DwrNuXCS/W+brRNq03ZXm7reGbg1MWiOp1cfCOY0hDrakpWlSVKBWCnKicZ60vcj0uEtHJ/Ps+zGiuBdwjSdFt6itkGbAs67ut2O8pDyEF95C2MJCUpOACeiQAcYA6Yy+AmjZds1XqZ2UvnW7T7jmnrMopIIjh5Ty/HxI3tN5HT7FjyVD1TKamIjUhOz7ve39GQuoOFV2t1v4gzFRWLVbOS7doMh1bciSie2+ZCVocQgOcolI7iyogqwOlfDfCrUV40jpS8xbaw7qqS9Iust6atsxmnJTja1odYcQoqAbSlIKCFJ5Yx41OhcWIp2Tb5/TWx6XjgbfQzqSNbrPb2G3tTt3pp6O4yhc2Lj/kxSttSU7Fd8BYKM+Ty16ucAJU6BBjv2lp2MxZbsy3GuUhl8x5chxKminY2hCQO8RtSAjOB4Zpcj0uCs1LX334eUXDhRwtlaA1G9JTCiwociw2+M+mMQObNa5nOWoDxJ3Dvnqc1tXbVuSuzX16qqzzJgp6VfdJj/MbP+0r9tbDARtVfd9jTY9p0l3kxSugOfFKAUoBSgFKAUoBSgFcA/XV/Hhf+dP5ShKOwuAX2ieHH5N2391bq+UIYpQClAKUApQClAKUApQGieOOp7hpTTsp+0uNR7jMusO1synkb0RlSpjUfnFPgdgdKgD0JAB6GtfXrW2peE+qJ1senT9fxl2dNzZTLZabkRlpltR3CtUVgZZ2v8w4aUtIZcxu6AaXIm37zeKTSR7jjTdWdRmW9GssjR7GlnL/ACpdtuS5KwG1K3Fk8lIcHdwASjxJOCNtedp+qDn3RhDB0ktq7THobFubVJeREfXIDqtq33I6ChTaWFqWEocGCjaVFWA3XaTvOwx5fFXVF61dpmHabOwi7MzLrbblaHrmURFOstNLSvnhkqUjatKknlZyvBAwSPOx8bn7pcbzfmiwxb3dPWOVGtl2m9naZkPyZzbqeYlCyVEtIT3UKKihIA60dJW086kZ7vz0JSwfVALv69L5ske3R7tJkwn5k+c41HbfYlqiqZZXyMOOKUhSkIc5JUMAd7ITP8J9a6g1Ro66XbUkO2QXYtyuUVsw5iloKI8t9nvlbaNuA0Bu67gNxCM7RTKnlRUp5mVWD9UW9Ng3YNWGJOuMOVao7DdvuanI0lM98stLDzjDZ6KSonCFJxghRz0koPGm8Xe7DTcHTENesGpE1mVCeuykQ2kRkxlKcTIDBUrcJkbaOUDlas425MulbmRvL8iKe+qRffs8q82/SpkWi2WKJf7o7IuAaejsvLkIW22gNqDriOzOHG5KVY6KGRnMg8XtSxX9TNz7JbZTrWpkWCzMxrgtPNWtttY5yiz3EpSpSyobz4pCTgFU7q3Mby/I+3OPEyLJiQ5tgi26S3dH7VcpUu5LTAiPNhhSEpkBg7lOokJUgOJaBKVpJCgAflrjLJtUyPHcic6LKvtyt7l0vEwMRo3JlJaQ0HG45SFL3nloc25DZBcKsZjdDeG4qVZyl24pTKLilMouKsujR/yv/g/jWRh1+YjGxDvSZZqVuTTClAKUApQClAUTitL2W+JHB6qWpZHqGP41q9KMpHnrkdqu9a3RHf7GjlwifVsPpykCvTh5HTN4mMbuvZ46lJB856VZ2cs1de8v7UlbBT93idDgYSAPJSu0POTWXFqMkS4LoxuUhY/RtqhtjKcVx21ElXZ6Fsl3wcff4hSNqQPKahVr7Fqa1v8Ahh0oJ9Yz/CrOAdsRAytoLPg6q7PA6Ztboft7Cgc5SKyq7c8zFKAUoBVL1mM3Vv8A3I/aawcYr0jOwf6vuOftG8cZF0uGrHrxGjos0aC/ebIuGCXZUBl11l1StysFWWm3BjaNslsHqCa+r19UEY1jlSYGkrwq4x3rSrsEwxmluRZ8nktPpIeIGSlxISohQUE7kpSSoap0NeJuN7pwLDpPijDuUu+tX2QjTsmLd021i33ZUdh1KzGjuhsKQ+4l4nnBWUkHvhO3u5Nd/wDxBw7RZ3XpVru97VEizrjMk2+IyyiPEjynWXHFJW/k7eUeiSpSgMhPUpTG5uS6tiG1ZxN1YnX13agrk2yy2i62KAlCmIq2ZiJktlt0qUVKdBKHjtwEgbTu6kCrZcuPlkss7UkS4224xH7HbJN3cbC4zy3o7CkpcKUtvKUlWVowlwIJCh5jiXRTSylKqWvc+WOPEQ3hdvmaW1DbFsT4lvlvS243LirlFCYyllLyiUuKcSnuBRST3wmvBP1QMWTObiwtIalnLkLnNQ1stRgmWqG6W5AQVPjaEkEgr2hX3OT0qNx2one9hISOOVkYe0orskxVt1KmGYNw5kdKCZO0MgtF0PHJUkEpbITuGTgEjA07xwfudmD83Sd0Zuci+zrLb7aw5GW5LXHdkJUQedsRtRHUVlakjIISVZSTG504k73WyRkx+OsG5P2+LbNN366T5LMt56Gw0wlyH2Z/kPpdK3kp3Jd7uEFW7xTuHWrHpHiPaNcy0s2jnSG1WuJde0lIDYbk7y0gnOd+Gyopx0BT16iolRaV7kqopaFr2nFNtWspcuNtNtMouNtNtMouNtXvSgxZGv8AaV+2s/BK1T3Gvx2tJd5L0reGjFKAUoBSgFKAUoBSgFcA/XV/Hhf+dP5ShKOwuAX2ieHH5N2391bq+UIYpQClAKUApQClAKUAofCgNOcVzptOm9QHVqWlaewoSkvJUoEbxtCQnvFe7btCe9u27euK15om1aBvDt8tkJGobfeVMMPTF3qZcY9zMZKlFlSH3lh3lBQWMIXtBKgoZUQdO82tjdrK0rk5D0JoKLfbXa4zKE3GFa3UNREzHiH4byiHOcnftfSpZUr7LuO4lXic17NcJdI+1962f5fItyHGloL96luqhrZJLZZcU6VMFGT/AKMp6dD0qHKRXliz4lcHdFSLVbWjHkMtw3Hn405i6yWZPNfI5rhkJdDi1r8CpSiSDjODUdbNEcPtRxblFjWtdvatD8WyFbcl2KppUIlyOGVocBTsMheFAhR3EHIpmk0RliZ7vCrRFjgxW3G5DEG2OLuQiu3WUWVLD5kqedbLpDpDxLm5YVg48wqT09prSjNhmrtu12y6kW5KcadlOOR3zI3LXy0LUUoDhWpRSgJBKiSM1DcmicqRVr7wC0rKtC7RBdkQXZUq1vSXJNwkyX3mIUkOttJWt0rR0K0pUkgp3Z64FTB4OaMdt8eI2xKaXCkPPCbHu0puaHXsc4rkpdDqivCQoKUc7U/epxOeViMkSId4D6Rlajfu8lKPYNFqt9qjWuPIdjxm24rr6trgQ4EPIJeQNjgIBb8u41Yrnws0nc5N17TGeDt5kNTXkN3B9rMhoJCH2UpcHKdAQjK29qjtGSaOUmFBEZbeGWjL9pu2RxCuTMJ/fLMGXcJbTspS1JWsykKc3PklKch7d5vAkVLXDhJpi6N8p+HK7OqU7NditXCS2w+646HVl1tLgS4CsA7VggdQAASDDkxliy4bRim0VRYrG0U2ilgNoptFLAbRVk0cMGX/AMH8av0F+YjHxH6bLLStsacUoBSgFKAUoDVXFOVzb02yD0aZAx6SSf8ACqagdB+yuJ2jLNiJHo+zY5cHDuPiQe76q++Giy3xNR19/HI/RVzZv6y95TtRXwU/d4nQ9K7I86NWcVpYdvMaOD/oWCT61H/BIqlNeFcbtN3rs9D2XHLg4n04nI9VV3UiuWYro8UPIP68ViYR2rwfajOxKvhqi/4vwOjtIP8AabDGVnPdH7Kmq708uFKAUoBVC4jQBdO0wzIeiCRDUz2iOoJca3BQ3JJBAUM5BIPUVh4r9MzcJ+p7jnzS2h+DusUwoWjptqYkxLU/DIsamWpEmE8zyVFzCdziPeKC/DelJyeoNl1nw30vGt8l+5XKdbRcWrNZW5LKgVNOx5hVCUgbFDfz305KgUnCcgDOdY5SvZ8TbKMWroten9DwdPyLtIDjs+VdJaJ0h6YEKUXkx2Y+5ISlITlMdBIA8ScYGAK4OBmnxarxAD0/k3S2TbS+eancGZTzrzhT3eigp5eCcgADIPiaFOxU4pn1qzQ2mLXDud2u82VEiyp9qlPuBWQl6K+yYwACScKcQ2FefJ6p8REMfU2acaj3OMq43Z6LNtk60JZW4yBHjy3UOOhBS0FKVuQMLcKz5yaqU2kQ4KTLTdeFtovM26SX3ZQcuM+3XF4IWkAOwnG3GQnu9EktJ3DygnBFflp4VWizSbS8w7LUu2OXFxkOOJIJmul17d3euFHu+GB45qnPpbz0Jy8/PUrML6m/Ttufs6o9xuzbFtRa0pjlxkpeVbyjs6lqLRX9wNyUqSk9TtBOa9LlwysNvukC3J1Jd7bdJl3mXu0FgtFcSQsOrlBoqZUkoWJDpKXd473dxgYr3jkU5EuZJaG0Rpmw6jfFpmypV3szL0KaZCsqW5KdTMcccO0BS1qIXlPdG4jAwAPvg/wyHDS0XplfK7TdLvLuCkMrUtDLS3CGGUFQB2oZS2nGMA7sdKplK6aZUopWtyLbd79AsK4CJ8kR1T5IiRgUqPMdKFLCegOO62o5OB0r3tdxjXq2RLhCeTIhymkvsupBAWhQBSoZ69QRVvLpcrvyMrb6qbfVUWRNxt9VNvqpZC42+qrxpYYsrX+0r9tZuDX5nuMDG/prvJelbk0opQClAKUApQClAKUArgH66v48L/zp/KUJR2FwC+0Tw4/Ju2/urdXyhDFKAUoBSgFKAUoBSgFKA0bxv0pcNX6dkx7QGXLnCusO6R2JKyht9cWW1I5SlYO0L5W3ODgkHyVrTWWg77xPuNxvV50vOtkVm1t26Lao8yG5Ofc7YzJW6dylR9iDHawhxRCwXAoAHB1l0mbfVpIwI/CHUb06DOe0/abffZWk5dnF2trTDPsTLJXyXdoORlCwk8rcArcB3SDUVZ+Bt0t9gbcb05PcUzJtarjp+e/bERbm1G525LSYyEIUQp1Cwp/BXymwrbipzLqRlZnscIbvBdtU2ZoqHfLAmbdn06MU9HLdv7SWeQsBZ5JKQ2/uCSdval7N2OvzrPhJqi5267QG9OwZ0G5XW4SdqUxHnmA5GYbYKDJSUIbKkuBZCS4AE7R1NMy6hp2PZjg9fZN4tiZFiZkrlWFu3XqbeOyyWCpMBTQMdXV9DnOISpOC0pBWr3x64C+D929rVhjSOHUK5sNaWXZE2d16KlFunbu/K98U7HehLiMuDYO7kkBmXUZX0KnabMY3G1mBfHYt1ukG7xWY0ltDS7oZKbQ1G7SpLh5vZUrLjpKRjeN+fEGf079T3eY+gLjapFsuCb25bIdtlLkSLemFPU3JbccdQWEIdWe64oLkd/7IoHJUTVTaViEm/mS+qOBd1ZuM8Wm1BvTDepxdGbHaUwk8xhVrZYK22pKFMApkJcVsWE5ypYIVjMlZ+E1x05qnQsm02SW77Hw48KVKvrkKS2xFQ48vYNgDjT6OaQksDlkFKVdEAinMmirKzF4ecJtQ6T1FoOUbGgS4NtjwbrNmKivx22m2nh/k6s89t4Ld29z7GpKlZ610JtFW52k9CqN0htFNoqixXcbRTaKWFxtFNopYXG0VY9IDBlf8P8avUV66LFd3psslK2ZqhSgFKAUoBSgNPcSDnU0geZKP+6KraB3RXC4/9efeelYH+0p9yPGSelffDhHM4lM467I6iayNnL85e8p2p/ZT93ijoih8K7I83NKa8k9q1NPOchCg2PiSB+2oNodB6K4fHPNWl3npmCjlwtNdiPpYyk1V9Wq2xWRnqXUD/tCrOEX50O9F6v8A29TufgdE8P1FWm4v+wP2VZK748tFKAUoBVM1mnfPAHUlkDHxmsTEq9My8LpUOVdGae1RedI8Jrdb7BPsV40fZ1JlXG6sBhCX/Y1cZMdAJ3OAvLQskDZhkdc7awNK6J1U/p6F2p29SpiLjp1ydbptvktBDzNxZckP8x+Q6HVBAVvW1hBCUnrgYw9F57TP1fnsMi36Z1vqnVmoG2mbtpZubZ7mkrW5K5Ue4IlMKhLLzrykvd0LVlpCEbFKR1AwnCQ5rHiJpjTetHfZGLZdRy5NwlW1hmTNMRnkttQgGoz7SygpbccJQogLfSog4CkrR8+ewXkz51VojU82L7H3SNqrU9yTHsXsVcGy6yxy2ltqlqkNh3YlwqStS0r3KOUbdxT02XxzsGqblOso0y5cAzd23tPXFUF4pMJmQptRnDr3VNJadAUOuXR8VLtdFWtmUi3WrXU/T9sumroV9ciqu7MW8Wy3Ou89cOPBWyHUobUFqQ5Ny8QjvKQpGQQCK/ER9c6dsxWq26lnMTbLfIVsitOKfkxlOSwuAl8lfdWGMAOLOU7SlSt3jU1F6EXkSCdFaqXcHbyv2d9kot406mKntzwaEXbDRPPK37FApMjeSD70nxGa8dA6WvZ4n6Wn3O06h9l4Ui7+zNynPOKhLLhUGCyFLKNpRtCeWkBKcJVg9KjRrTzoNbknqTQF4PEDUmpLfHurc46ksBirjynUNORAqK3MUWwoIWnlF4KKgeifRUDpyx66tr+pLhO9mLlf2Id25tpbYlxmbipS1GOETDJU0O6EcssoQpAUQdpzTRol3T0PrR2mNVPzonPhXFdrjavjTonaI0hgNR/Y1xDi0tvuuOIRzjghSh3lEhI3ViRdC62maPbmyxqEX62aO092FCZz6P8AOSFvmXuQFhLjmA0F7wchWD0JqW0U2bRkcObrIunG9M66ybizHevF7g214yXnGbittxxKWigObEJZaYd25QMny56r6W2irNSOqsXYS0G0U2irWVldxtFXbTA/zM1/tK/bWZhVafuMPFu9P3ktStsakUoBSgFKAUoBSgFKAVwD9dX8eF/50/lKEo7C4BfaJ4cfk3bf3Vur5QhilAKUApQClAKUApQClAae4uXeVZdK6gkQFKTcVhUaIW8bg+6sNNkZ6Z3rTjPStZxL3qCU03EhSZpXJ1C1Djw7pJHaI4joU6/zXGwe4otjugqylWM9/A0VXNnsi1i6tVVlCm3wXdrdd972a7EyWj8WrxdY8Rm12JM65mImU822HS0vc6422lKwnajeGlL3LIABHRWTieTxE3zo7AYaCJF4kW5tSnDnksNLU66fUttSfN1T56hTb1sVU8fOaUnGydvs/g2v9lYXxb1ILM7cDY7e2hNkF85a5S9zbZ3bG1AI6rc29AOicHOcYM6viHcZF+hMRYMM2+TdV2lBdfUH1LaQpTzgSE42pLa04z5Ac9cVCnJ8iiGPrTsnBJ6c3wbt8b2RhaY1xqO5221NLRbpFyvBlzYzi1KQyxDbcGFKwMqOHGUgDGQrJPTr4W/ijKvC4MqNB/yyXDhNMR1SVdnL8lTiiT3eqUNs792MlKsADNM70uQsbVyxUkrvp1sn8HdL4mdp7V8uBpfVupbqUy+TOfQzHivKU0QylDGxoq8At1tZ9BWepxmvh7iLfokm4W9y222TdGJcKEyiPKXyluvZWtKiU5Ty2wFdAcjrgZxU5mki56XVpwjZJtpvjzeZq3ZpbusRWo9TarkRNVRg7E7j8KzRUQnFtrEp3lFxSF4yBtf8fudnTy1Mx+IF9nPRLbGt0By6vT5cRTin1iOlpgDe773ccLUlGPP5R5IUpXt54luOKxMajTSd9OLtfM14Wv7iOY1pfJl0hLtrsVuE8m53CQJziljkMPBlvBA7qVAhYA8x+OT0XrK5TZ1lts6KIQmW9ucFS1uOOPqWje4lte3b9jKkpKSc46gAAZKTbKqWKrOcb+y2uul1Hj79F3mw8jzimR5xWTlN6MjzimR5xTKBkecUyPOKZQMjzirDpLqZX/D/ABq7SVpos1vYZYqVsTWilAKUApQClAah4mtbNSqP37KD+0fwqsIGR0rh9oK2IkekbPd8JT7jHlGpPg3G7Zre5ycAhlpLYPpJzWTsxXre4s7YlbBtdWjfWOlD4V1556aCvzvaLrNcByFPuEfKNYrRBGK4LFO9WXeepUFahBdiPtYwmqdq9zfJgsDqVOjp6utVYFXxEF2lrFyy4Sq+x+B0rodrladip/1R0+KrBXdnmQpQClAKp+sMieCMZDI8fWaxsQrwMrDO0zk/hT9ULdXdNSr9qWc9dLZC0pFvs8PWsQHmZT3vWo4VsDzK8LAdxsykfZDk4t6vqlICbfk22OJ5n9hSVXRlMFR7MuRuEogJ942tO3Gd4xjb3qw5UrvQz1U01JGNx1hT9KXK6S7DJZRFsttu7sN1aSVNzVOpQ2cj3yQ33gfvseSoeRxiVw6tOqnZjMKRBtd5kw4cZ2bHgBqMxGYWGWUqxvV3lbUgetSRioVPkS6i4mVN4+vWa5asmXCztjS1qt9tmRJaH0pfeXLJS2hQUQlO5RSMkgJwSSQels0XxWt+q9M368PMphM2N1xuYtl4SGFBDKHitpxIAcTsWB4AhQUCOlQ6el0SqiehTrV9UzBu1qdlR7C/KfJt3Zo0OYy9ze2SAw2hSwQht1KlJK0E4AUCFHrj1vHGqeGkiPa3oV7gyrjBlWlx1pxlb7ELtKcu4zsKFNrBTtPewQOtTurcyN5dEQ/xr1VbnrFd5Wnlyojuj3r9cLZDktJS1sW0S6la+pJQpW1GfFWCRjNWU/VC2V3XKNPxYplNibGtzr6ZCA8l59ptxBSx79TYDrYUvptJPQhKiDpX4DedSD0JxgniyPyrtLE5+NZ2pyxMejw4+Vz5TAWp1QG3o0nOcjCRtBUSDLaT4uOcQNWaNVblCLbZrV6ZmRUONvtrfiPMNBSHUjvIypZSRjIWMjPQHT4hT4IyZXFiVbNQ3K0RbZL1BcHdSKssSOXGY6Gim2NTCd5/6MJKiScqyTgEACo+1ceZV0vlvkGyJiaVc02/fJkl94doillzY4nakkKCSCnp4+IOOlN1fmHUIKyccNIh6bfoukbbDvkkwkicy9GAdEtakoTIlJGG1JLffSd2Mo2leRVt0jxsk6znachwtLv8y7xnpy3e2tFqNGakBhTm7/pMkhSQkEKBznFHTfNhVFyRWdLcer69BkTH7E5d7JB0fA1IuWlxtE17miQXCW0jYThggJSB1SepCgBtjSmtImsp97at7Zdh2yQ3GE4KBbkLUy28dmPEJS6gE+fI8hqJU7cCYzuWPb6BVz0yMWhr/aV+2ruHVpljEu8CVpWyNWKUApQClAKUApQClAK4B+ur+PC/86fylCUdhcAvtE8OPybtv7q3V8oQxSgFKAUoBSgFKAUoBSgNYa3XaocW4Tr0tlu3QHDNcdf960Wlbwv1pKQR6QKpuntTaauepFtm0y7LdFsu3Vpy6RFRy+3htp15JJ6EDlBQVtUApOQM1ixwsq0ZVFbT4u3TrbizPcacmnJa6fLgZCGNDMX3T8Rly3puTkT/ADahhz/SMMqAABScKSlSxtCs9Scdc1mMaa0euTcrkyxbFPJLyZclK0nllY+yhRz3M5JUOniSfGrMsLKKV4eb28UW/RsM/wDFaGRIh6XNvMl5dtEKTHaa5y3UhtxlpRLYBzgpSpZx5Mq9NfidOaXteomZvZ4Ea9SXFusrUoJdWtScLUgE+JB64HXxNUug/wBvy6cfgVuhQbTcVpqvHxZgyNJ6KFuEN5i2ph20LUW1OgJjpWdqge93UEp2lJ7vdxjpWbcLFpabHablN28tXBxpbIK0p5y0JCWy2QRkhIAG3yeuno7/AG/Ls+xQsLhkmlFeVbw0PKBH027ZFWEQ4sG2vPSIbdufCEJfIWrmctAPUFW49OvoFSFu0VZbSG+x26OwW5Ha0lKevO5ZbLhPlUUEjJ85pOioNXXcV7ii3F5V6qsuy3A+xpC0i4PThBZ7U8+iStzB7zqE7Urx4bgOmfV5q9omnLdAktyI8Rpp9vn7XEp6jnOBx35SwFH0iqMqKlQpRd1Ht+d/Ew3NC2J1uMg21jbGjLhtAAjaysYUjoeoI89e8PSdqgXNVwjwmm5hSUB0A5SDjISPBOdozjGcDNRkj0KVhqKaajr9uBK8v0inL9Iqsybjl+kU5fpFBccv0inL9IoLjl+kVYtJJ2iV/wAP8au0vaRZrewyw0rONcKUApQClAKUBq/iwxtukNz79oj9B/8AWqW2O58VcXtNWxD88j0PZjvgoe/xMKarCVeqrhwBhb4Vzmkd56SQD6E9P41mbKj67fYY23HbDRj2/Rm4a/Fe9PqrqDhTnu55Tc5bShhTbq0kenca+WetcDiVapJPqeqUmnRi10R6OHCapUxPslrWDHHUIOcenNZWzVfERNftKWXBVH7vmdU2KP2e1R0YxhIrPrtDzsUoBSgFVPVYBuCUnytAfrNWKyvEyKHtmj7To/hvfG4Gm4zTzyIFkf06yy6H2kvwkKbQtAcISHShTSMLSSUkkgjJrx13wPfv8exph3aZcRbpTslbV4ucpK3dzJbTtfZUFoCfHaOij1V1ANYcZ5tUZFOpCvHNTfivPlnrL4I2d/Tsd/Vlxu15uLFsYi3OVDffb9kUsKU42XGW1EuFClrKc7lHd13VM33gfpbUEuVLeRPjSZa5SpDsOa6wp1ElDSH2yUkHYsR2egxgoGCMnM5nfQu2Tdj8u3CXSjrxRIYuHLuMKPaFsMPvctaWNy2HFbD3HG8Epdykg46521Z7RpeLarNItr0qbd2JG7nKushUhSwpISU97oE4HvQAOpPiSTS9SVa7tyNUjQmnNb6ZRpbT9+vZjNSbZPaE+TLwiHGlocHZXFYOCGylLiST707iAKsB0hobTVzh2l5q53C6NuPXBxZblTHXVykLYW6+tCVDvJSpA3EABHTAT0jeJxuvAxFiaMqaqpuz04O/wtf5cNSYg8HtOwrQi3ETpTKbO9YguVLW452Nwgqb3E56BKQD4gAVlQOGNntN+N0t79ygLcLS5EWNNcRHkuNtpaQtxsHBVsQhJ8NwQndnFVcTMsiMd4GaScg9mRGlM7Wo7bbzUpaXGSxKXKZWlWeikvOKVny9AcjpUlpvhZp/SsuBKhNylyoa5rjb0mU48tS5S0OPqUVElRUpCT18PJUttoZUZDfDmxNagF5THc7eLou77y6rHaVQxDKsZxjkgDHhnr41HReDumYTlsUzHkoRAiSYAZ7Qstvx31bnGnU5wtO7qM+FRqLIresOBDdy0tAtFqus95iHNYkJi3a5SVILLSHEJZQ6hQca/wBICVpJUQkJUSDVj4ecOV6VZgzLtcHbtfo8eRDTKU44pLcZx/mpYBWoqWEAIQFrJUQjJxnAlu6sRl1PawcKNP6Wl22Ta25LCoFpbsyGTJWWnozW/lIdSSQvaXHMEjPfPj0x+cHOHTHCfhvZdMNLQ85DaJfdbCtq3lqK1lO4lW3cohIJJCQkeSoeqJStwLoVAirjpzraWvWr9tXaKtIsYj2CTpWca8UoBSgFKAUoBSgFKAVwD9dX8eF/50/lKEo7C4BfaJ4cfk3bf3Vur5QhilAKUApQClAKUApQClAaa4x6an6t05Pg20MuTET40tLElZQ3IDEpt5TKlAHaFhspzg43VRdVaGvuvn5d0vdlQ2lMA2yHaolxTz20uOtuvPqcUjZu3MMAN4KcIVkndtrIw1anSim5Wkm7aXWttfgnp3GbJOWnKxgXDhhqycyHHGrV7OHST1sZusfayIlwcUpSnEpCAQCrlkLSBghZ2jIFecbhFLt9ptkiLp9LrrVwjKn2mZNZ/wAsiR2XksI+xtpaHLedDgG3rsyTnAGb6bSUcsJNLtTbXTnyevX5lOR9CAXw3vjmpX7ajT1quUhi2TZAS87thW6TcpLiu4C39kDaGAFYAJ3ZAG84sSODt2RcXbY5Dhy4RnWt5nUD7wMliJDRHIYQjblKy6y4chW37OpWc903Z42lGyzPhfh2qSXHnzZCg3yPSLwouNks+lpKLFbrvcG50m6XqEt5LfaZbwcUlwrUkhwNrdVgK8OhHVIFfth4U3uw3fSKmYkMOW7mGbKDqFxVNOvLkOMNMqRuQUOlAbWkp6JG7wCasSxtOad5PW+n/lbn0a7rWehUoNcF508+8/dH8Mb/AGa/6SlvQ4zT0SKEXiUt9D7UjdzX1pbbKN6HEyniQtKkgp6ncdqRumtdjqsK81KDv/t/SxcgnFWYpWuyly4pTKLilMouKUyi4pTKLirBpT/6r/h/jVymvWRaqu8GWClZZgClAKUApQClAa/4tRt0SA+PuXFI/SM/wrXbYyn1CuQ2rG1e/Yd7siV8Gl0bIu7ucuO6fMk1tXgfB7LoeIsjCndyz8Z/9KzNlL2n3GNt52p049/0Nh0IyK6M4s0lruAIOqZhSMBwJc+PGP4VDMjIriMerV5d56VgZZsJTfYJKtjaj5hVE0TZ7zfdbOKj3SO062rAdXD3jp/q7x+2szZKTqt25Gt2y5LCpJ6NnRjOndbNsoSnV1tAA6D2EPT/APz19+wGt/xvtvzGfp66s4YewGt/xvtvzGfp6ewGt/xvtvzGfp6Cw9gNb/jfbfmM/T09gNb/AI3235jP09BYewGt/wAb7b8xn6eoS62/UtuuC3rheIl4CI5KIzFv7Opauu0bi6oDr+2rVWziXaV1LQ1lE4XPwNBw27hIu90uybd2XlRVRwYandqny2O4FZWnBK1KJTkA9TmPuOjL5cLU3BmaYYdLlmEeEzDU01Et01bjpdeUjflJALKgUbyNqwk9euodFpWtfQ1c8JUhFRUXLRJ3tx5W4acL9iXFks5w9nSEXKW/GW/dJ13jN9pddBU1AaUwlZT3sJ5iGCSB1JWM+HSOtukry5qCPcJ1gmIlxjcX5dyZks86W4slthtr7J70NuEp37QnYB0Ocy6UrrTzcqlhqiknlb5vXnmvfj0uv4Puy6MvES3Qo71pkRocaPPyq3lliW6taWmWXFAObOdyy8oqzjKQehO2rDY7Tf7Dwvlx7fbYsK/JDvZ2WmmmisbsIWpIUWw6UAEjdt3eJAziY05LlyLlCjWp3ajqou3DjZW56Nta8u0hOFuk7zpbUi44tsyHp0NOcmRNWwuU57zltPFK1YQ3lzlhPTCuoTtyqbt1nuKeJ16uMmBdW2H3GWo0liQyIimG2QRzEczeTzVvDG3yg+U1MabUUrc/P8lVClUhSpwcXpLsvazV3rz/AMu25WUacup0R2d7RciRqOby2LpPdVFdU4okrdfSFO4WEqSOWFYxuT0wCKyY/Dp9zVkZh+DcmbJDTDFsU0phQjIaSFqClqWVoWXAoLKASoFI3Y8KN03bQx1hpzy3ptcE+GvG749bPs48TBTwtmvQmZsu0KlXFVnnyJTDshKu0Tn3EuNsL72FJQQvAPcBIIOetes7QV5l3JmFJtL1wLMm2tR7y5IRtjwmktF8JyvfvWtLwUAnvJWMk4ApuZJcPNil4OpFWUb3tfXstfjyevezc2B6KYHorMynTZhgeimB6KZRmGB6KYHoplGYYHoq36dH+am/Wr9tXqSsyxWd4klSsowhSgFKAUoBSgFKAUoBXAP11fx4X/nT+UoSjsLgF9onhx+Tdt/dW6vlCGKUApQClAKUApQClAKUBVplhmPS3lpbSUqWojKh4Zry9rs78Gn5QrHdNsy1UjYe12d+DT8oU9r07H+jT8oVG7ZO9iPa7O/Bp+UKe12d+DT8oU3bG8iPa7O/Bp+UKe12d+DT8oU3bG9iPa7O/Bp+UKe12d+DT8oU3bG9iPa7O/Bp+UKe12d+DT8oU3bG9iPa7O/Bp+UKe12d+DT8oU3bG9iPa7O/Bp+UKe12d+DT8oU3bG9iPa7O/Bp+UKe12d+DT8oU3bG9iPa7O/Bp+UKe12d+DT8oU3bG9iPa7O/Bp+UKl7Bbn4HP5yQnftxgg+GaqjBp3KJzUo2RL0q+YwpQClAKUApQFX4kRO1aWfUBlTKkuD0YIz+rNakaGQR6K5ba8fXi+w7XYkr4eUej+iIHUy9lvf8A9k1vvhzFEPR1sbAxhhPT1jP8aytlL1ZMs7ff6S7H9CzUrfnIGpuKKAnUQ9MdJ/7SqqjA7oritor8+R6Ls7+zgYV8kiNb3l58Emsn6n+1mTMdmKGdyiQT662GyI6ykazbsrUacOrbOiMdKV0pxopQClAKw5lpjTnQ46kqWBt6EjpUNKSsSm46o8Pa7B+8V8o0OnYJ+4V8o1Ru4lzeSHtdg494r5RodOwT9wr5Rpu4jeSB07BI94r5Rp7XYP3ivlGm7iN5Ie1yD+DV8o0OnYJ+4V8o03cRvJD2uwce8V8o09rsH7xXyjTdxG8kPa7B+8V8o09rsH7xXyjTdxG8kPa7B+8V8o09rsH7xXyjTdxG8kPa7B+8V8o09rsH7xXyjTdxG8kPa7B+8V8o09rsH7xXyjTdxG8kPa7B+8V8o1nRYrcNkNNAhAzgE5qVFR4FMpuSsz1pVZQKUApQClAKUApQClAK4B+ur+PC/wDOn8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgI/UMXttjnMeJWytI9eK0YwdyAfOK53a60g+863Yb9WpHu+pXNWnEYIHitaU/rrpPTkcRbJDb+8aSnHqAFXtlL8pst7ed6lNdn1JOlbw5Y1bxVZxeY6/Hcxj9Cj/jVRZSAj0+auM2kv8AqJeeR6Fs13wcPf4squvpfKthaSe84doFbb4H2XsGn23CnBUM1t9kxtSlLtNJt2V6lOHRP5mz6VvDmBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAVwD9dX8eF/wCdP5ShKOwuAX2ieHH5N2391bq+UIYpQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAfLqQttQ8cgjFaFdY7PKea/BrUnB9BxWh2sr04vtOn2G/WqLsRWtQMmTdrVH8eZJQnFdNQkbIrafMKu7MX5PxLe3HevFdi+p70rcnOGteKaAq4Q/92R+uqWAAgVxu0f7iXnkegbM/tIe/xZRNTE3TUkGEnrhWSK6d0dbhbrFGbAxhI6Vv9nRy4aPbc5rbMs2La6JInKVszSClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUArgH66v48L/zp/KUJR2FwC+0Tw4/Ju2/urdXyhDFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoARkVou6YN4mYxjnuf941o9q/pR7zpNifqT7iCSwJGsrCg9QJKTg10cwnaygeiruzF+R72W9t/3K7l9T7pW3OfNbcT1brpGT5Qzn9ZqkPHY0T5hXGbQ1xEj0HZq/6Wn55lX0RBN+1+pwjcltQArqOM0GY7aB9yMYrqcLHLQguw47aEs+LqPt8ND1pWUa8UoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFcA/XV/Hhf8AnT+UoSjsLgF9onhx+Tdt/dW6vlCGKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQHw+4GWVuEjCElRz6K0I6/zpjiicqJJOfOa0O1X6sUdTsON3Ul3fUj3FiNqG0yfwclH6ziui2SFMoI81Xtlu9G3aWNuK1eL/AOK8WfdK3BzprDiO4HL6Eg52NJHx9TVHvDvJhOHyhJri8driJHomzlbD012HvwEtQkzZExQyVLPU+uugMdK7GCywiuxHA1pZ6spdW/EUqssilAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUArgH66v48L/zp/KUJR2FwC+0Tw4/Ju2/urdXyhDFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoCN1M+I2nri5kDbHXg+nFaFbfy6o5zXPbV/xOx2DG8KnuMOS8ZV3tcZHVb0ptIHxg/wAK6Tjf8nb9VZGzI2ot9v2MLbrXpEY9F9WetD4VuTmzTmqJon3uU6CCCsgeodB+yqnqI7oLyQepBFcNiJZ6zfaemYSGSnCPRIuvASIlqybsd7y1tqu4WqPNZKzaFKkpFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBXAP11fx4X/AJ0/lKEo7C4BfaJ4cfk3bf3Vur5QhilAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUBr3ijqNKIotbCwVEhT2PR4J/ifUK1aSEJ9JrldpVM9VRXI9C2NQdLDZnxk7nlp5SZPEewx/fEKU6R5sCul2wA2kDyCtxs+NqCOa21LNi5Lol4H1Xy4Mtqx5jWyNEaEmulqY8hRyULUnr5wcVCXqThCyod3FcJKP5jXaer4dJxjLsLrwGuYkRHmgegWrH6a3HXb0/Yjfojy/EW307dX4ilXCwKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQCuAfrq/jwv/On8pQlHYXAL7RPDj8m7b+6t1fKEMUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBVT1nrVqxMrjsLCpZGCfvP8A1rHr1VRpuRm4PDPFVlTXv7jTcqcu4yStaypSyTg+J9Px1iOuZQpYIIA/RXGTblJyZ6lTgoRUVyJnghpt6936VqR8ENpBjxkn9Z/R+2uggMADzV2VCGSmo9EeXY6rvsROfaxX4oZSfVWSYJz7dZLcqfLcbPdMhzHyzUZKQHkFJAPoNcNiFatLvZ6rhNaMH2LwJnhNNbsF+cjKIS0+dzefIfKK34lQWkEHINddham9oxZ5/tTDvD4qceT1XvP2lZZqhSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAVwD9dX8eF/50/lKEo7C4BfaJ4cfk3bf3Vur5QhilAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClACMitH8RrVLt16kyXY7r8ValOBxCSQnPkOK1uPozrU1k4o3WycVTwtZupwatfoUk3OKHgjmpb2pzlQPUnyf+npr3tljuurpZj2yOpuGoBLkpxJShPXrjz9PJWkpYWpUmlNNLmddidqUaVJypyTb4HQWj9OMaZskaCwO4yjaCRgqPlPxmp2urirI85k7sV+KGUkDzVUUnN2poUjTGqJsV5JMWS8p2O55txyU/Ea8XiQAvr08Qa4/HU93Wfaem7LrRrYaFuWh5LBbcStBIUO8kjx+L01trQXEFue21BmrCZIGErPgv8A9ay9n18jyPgzD21hPSKW9j7UfDmbCSoLSCCCDX7XSnnwpQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAK4B+ur+PC/86fylCUdhcAvtE8OPybtv7q3V8oQxSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFfDzCH04UAahq44GEuyR3BgpSR5igGvdi3tRwAkDA8BjAFU5EVZjJx0pVZSKUBTeIulG71alvNtgyWSFoJ8461pRVxRIldkWC26kHclXQjHSuf2nRlJqquCR2GwsTCMXRk9b3R9KUEqabcTgDIQv01iSrkzb29wWErTnAB8FDr+utKm21lOulKKTvwNz8KdYK1JaW0LUVrbG3crxOPLV/rt4XyrNxseTVcu8lk4XdhSqy0KUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUArgH66v48L/wA6fylCUdhcAvtE8OPybtv7q3V8oQxSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKA+XW0vNlCuoIxiteau4UQ7xvfjjlSeuFI6HNUyipJxfAqjKUJKUXZo1fL4O6kemBPOdLaDlJx+uvWLwJus2SO2vOLbzkhWasU8NQptOMdUZVXG4isnGc9Gbn0ToljSUNLTXjjrVprJMOwpQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAK4B+ur+PC/86fylCUdhcAvtE8OPybtv7q3V8oQxSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFKAUoBSgFQGqNWN6aXHSpth1TwUcOzGY5GMfhFDPj5PNQEWzxBekym4zVqadkuAKQyi6RitQIyCAF5II6+qvlPEVa3nWU21kvNBRcbF1i7kBPviRv6Ywc58MUFjJl60nQGyuTY+zISkKKnrjHQACcA5K/AnpXlK1/Ig21NwkWltiArATKducZLRz0GFFeOtAZZ1PdghtR046EuEBBM5jCiRkY73XpX2vUV6bQVK0w+lKRklU1gADz++oDCh67lXFsriWdEpAUGypm5RlgKPgMhfifNWd7P3z8V5H98Y/qoB7P3z8V5H98Y/qp7P3z8V5H98Y/qoB7P3z8V5H98Y/qr4TqS8OLWhOmnlLRjckTWMpz4ZG7pQB7Ul4jtLcd00822gFSlrmsAJA8STu6Cvv2fvn4ryP74x/VQHwNS3hTqmhpp4uIAUpAmsZAOcEjd4HB/QaxrfriZd1SEwbKmaqOstPJj3KM4Wl/eqws4PoNAZKdS3hbq206aeU4gAqQJrGU58Mjd5cV9+z98/FeR/fGP6qAez98/FeR/fGP6qkbRPmzg72y1uW0oxtDjyHN/jn3pOMdPHz0BI0oBSgFKAUoBSgFKAUoBXAP11fx4X/nT+UoSjsLgF9onhx+Tdt/dW6vlCGKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUArXPFfQrOuL7oVh6yxrnDj3kSp777CF8phqO8tAyrrhT4jjA8euaA1I5wbv7dyhXx20xXo0nWL1zetTFraRLjxYpdVBKZIVuSkiHCGxIHRzYcAqqN0dwsu1w4Uo07cLJOduVwZgwbqJ1jjxMNvyWvZFQfSd7g5RkZyeoI89Co/IugdVW3Udua1Npi43+yWa5xLOlTLSJSp1sgx5rkR9SSrqVuy4wXn7uOvyYNZN14d3u2uGXG0xcrRIkS592tNth26LcoEdb6W2uyyI5IQ2tSWEOlxDiEhUh9PMwSVAXzXel9XRtB6H09piALRIhQS06u14U1b3ezpiIDRcycNmSt1JOTiN1z4HJvOmNWL4R8S7LHduU2bdJEqBZjOe5z7DDrTcfm7iclIWXngCeiVYGAAAIK7cfqeXLZqOySm3XHtzqHJ7mn4iLYhEeJHlchpIQoq5i3pQO8qJw2NpRjJrseRxjYdsriol+duVstymJSHF7mZjqLQ4UnIUlv7LMKOpStQKeriAQig4luf4fa3GoYMIal1O5bY8yDBXNE4JLzSWHn5klWB15ilojgADYUgoCSAoVO0O8YhP00exXwTIDSEPNTnith7/NzrwDigpKFbpCm46ioOKBb37m9wyGjJF2Nrp6NH5aNfPwZtve5ocfbZkOXZDbXKUMEKjMLU4/uGUt7o6egQr7LjTdOcTUTNRpUm7s3S7IcXb51pUlpo3JpliI09JVnuxwGC+EHurDzgKVKCU0JIuLqXVvETUq4tln3S6vqS45d2BIactimHbmyiOGsFSdnYm5feHvyoFW5QITLPvcRHNMQ3EQdau3O4MtG6CQ8ENpmNsuKcbZbbWlxptbjiUBSXWm/sCTuIKi4ILdomx6pg8OtYXO/W+53XUc2BEhhDLqI82U21b2UOKQo9EOF9ctSc46keFUjTultTaZdb7FZLurTClRbcu5W61N2y+PxGmJKgyttpSQEB4x0h4IaUOY4MBPfoD8vth4sqluwor98iMS0rajz0IQ7LL7UeKwyp91pbadoWmU9lR5aysgjolJn7tbeKEFZu8aVfHpEr2S7Ww2tC0xoxuUduPyGDlPORCMh5PQkrTtOQQkCdDDFr4kSdZW+Jb5Oqbfp4OxpMD2RIkOpSZThkdocDqcp5KGwlp4rUEvdAVjCLvwKTrCRc9RTNTpuyWXwyuObmS1hxSnVLbEcqWGy2ktIK218pzoUjIUSIZt6lCBSgFKAUoBSgFKAUoBXAP11fx4X/nT+UoSjsLgGR7hPDjr/wDo3bf3Vur5kecUIYyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQGNCtsO2pdEOKxFDqy44GWwjes+KjjxJ89ZOR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOKZHnFAMjzimR5xQDI84pkecUAyPOK4B+ur9fcv8Azp/KUJRwhZ/+Z4P+4R/3RWZQqYpQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQClAKUApQCq3rH/6P/j/8NAf/2Q==)
- Multiple feedback loops
- Diurnal pattern

Impacts of chronic stress
- Major depressive disorder (MDD) & Post-traumatic Stress Disorder (PTSD)
- Hippocampus and PFC volume reductions
- Synapse loss
- Reduced dendritic density
![[[@Schmaal2016-gb]](http://dx.doi.org/10.1038/mp.2015.69)](data:image/webp;base64,UklGRjoqAABXRUJQVlA4IC4qAADwkgCdASqHAfIAPjEWiUMiISET+v1sIAMEs7d8L93/3+hEqpN+Uawp2X7Q8Ce0E4N/wv9Q/JT50f4D/AexDzAP1V6TfmA/hX9L/ZD3zP6r6gP8B6gH/S9ID+vf//3MPQA/Xb0yv3R+E791v2W9oP//6z144/on4zd+P83/FX92vWP8M+P/n/4v/1j/mf534Gcw/RF/Efjp7jfxX6xfQ/x6/vP/y/2Xwh/bP7D+xvm/7zv378g/yv+wL8T/jH9L/p367f1j9m/qT+c7TPYPMF9Jvjv9d/vX7Q/2H9wvXs/Tf6B+vHub+R/0D+z/jT9AP8N/kP9W/rn67f1//3/Rn9m/0P8t8oT7R/Zv8r+YH+7+wH+L/zH+y/3D/Lf5r+0f//7Rv3P/Q/3b/O/9n/J///3rflv9p/zn97/yP/O/yH///7n6B/xr+Y/4X+1f4//d/4D///9T7gfXR+0//a9y/9Zf+3+f5k0qLmkk3nQCpHNMBzvIlqYzir0xe0B9Cla6Aoa3BuEF2KDpZ+iLg3LnaHQ8BxrAZkWC7r+LOMe8AV045qpWiUO6CInTtGpDiQwjf08xHVCVP31x7NkoHp5XjnsMWB2CBlo2B1IOPPOyQ+t1IVn0RT6m6yc508yo7J4nNTFvItUvSQg/JAmklcRiaoff0mjJUq/AFrplnuhJMTTTPxF+d9qzp2u2by6yQeMkZ7l0l1hjQPsnb7MWvpzUxbyOLhswYMqrvn22ROiWf+8yyY4XlHqg59LvLV1OMbxNe4qkorYFc3LJGjDwu2C1hhFSJMMGw02IX9qRZMYrRTURLaSjrAj45qYc+JHFT8OH2DCxxOlgdrlu3DtQr2pVO+9Bp11pgb7yEg5NtSgqBB+glS45fwlE03qox+lDX0pJEoVcYUUSzTUM0dUY+e67uG2zRjD6VkLmpCjDuqqMAzpfg6ZSWaTp5VNzYdmG/ZAh7Mq2hjofnFRzMUuEgEPoAqOwMVTHlJyiTFPWuIoRQJzIxE9/nvHOQId/6gmmtxujk4P4DsHQWvpzUxbyHO4y7f/MQMUxsBfoZdR8YIt23Iz4FhoJs3ULa02V/CSYp61jjYfJA9XzKL25r1wIP2j+gXT+RnhVe2vjzyDSHjinmHwjPYOI1P09a46S6qwn1D8/GJUWCydSN9CC0D0d7XEoo5fY8PLxP9d8O6DL0f3YSioow9a46TDBpLNe10WG5N4n9uYpmAp0BtOJSPrqFJZI5JjB9gQzTdOC2ZYmOLAly7DfuU1mMv9X8G9gi7nDKov3wosdb3v7L+PLX0Y7WoIgBEWWQ0v4yfdjlSjkBUBOcJ269FOqDIsnQaXFMjkTmLZkPmf/Cci4yaGzCCvwJ2PAroViz40VREYONHWb7EtQ7FhtAI1qn/awKuv0zFLVjrApJCu9eSzbtzYFEjrJ48a10jE7r1v6191LxKpU1PwalPp5l0BXeF6u/FcJk5sBGR2l6v86B4QtfTmpK8TWfGobSoqtKmS3Wh3O0eKAod7qaqxFXot3DVb5zNt/oAhQuHvcTbZoc8inrXHSYYNiIIKebeJv0yGorx1oMo6WWc3kmKeAAAD+/8C7aGY1hBW8LqikdrGE18yKrJxothnaGh+RreJxUNYThoTMq8GsXW2/70ei538k1s5aGraKD5zuRerQfJU3fAlUEVT7OwLA4mDxQIOfhSaSt4REKOexOw1UxzT+G2FEWO9fl1ATKAchg/DxHJSj1CpGPLrkdvx/BcsEb5x6bXyMAFAAyDDV3qpL85H0PJ+yQKOFsuc8iomOClc+fN5lZtJ6y/BrH36xEKhLH6s96KMyDLvYbEVHObbkle4bHW5BNCrDQh8p9U/JXkgrJBD/drVyxQQuGqFbjvdpoNR0PuUpPH0/ag/c1u09A6kLE+qJRY8r6XksDpegP/GXQA6pyZBlOUrrXT++Z84f3MsDpXILdDKDa1n2YYGk1+QvqKF6ZyZnoNcADf/FlRqHh2Kb7pd90RwA7gwgz5sIwWUSsGmqy3hu76mBI/Wi0OzvbhtteeyOEbuXUCdy5Yd0GphLagnGIU4mnHANshaGvE41V3rpbLuVECzCrOtwFTDN78vjMbpSPhW8SRC2OPtdyndo4OGqZ3nMWyWkEB/VVJIiys3cgdWsqDHtYeiZ+rBbGEv0ilPdQQobRkx7hiVTzILJKXuMp31YRLshX9STF2+Jim3A5L4kFSorXkgHC7DwUiCpAETPNIj2BWNqz8KurPSsfyQnTNrR5P5/YgD7tcyLDUZ+0rlzOCBykELXRt/s8Ozy7bUGHJszvbWsmgUhPZXd/xgA9MqaQyaJCNdPMkOlPtmxro40MQLc5cNOW4WC4duS26a/WaHJB+fzAcfWvUEHi/F1eINzQdaZtOHGJ1CAIVtlFsxVaniDL4QHsMOIgcGMKg8pzWegeEW8KYaxcyyykfKElDFNiHt+zWa3nS99zn7mfm4jRDXOG3zoQZgzIqB5a/ubrVYmYBF34ibGV2OmTnMjH/+IH2IPXAURIc/510F/44hjZNUnrFsrWNYutbBftaLvkTecAqhOc1BnLJIjx57HscfE3omqmONJHMY96bbM2lxTTdPSg8ZI+NVwC+mZJJhYQ1guSqe2NtjPW38r/Ts94JwjHgewGAlRcIeiXfJSEJ6iU1lNB27Z7hTDHxXoEOIx2JcvbhBd6Apx0DtDPZlPiyyJyPMifBLZLN9hZ+pvq6Vew9g/bi//jQpidt+WxIu2+atLd/cAdjlJ3L04RoWOuCVJTkdjSzXNquqlWXLpa+xAYxGPipQKvh03xVvRf5t6CqAkgBfBJJgGw9Hx3ltEDl1eDDjItYZ8xlBKHfDc/DV0oK8Ytx4GWB0AbxSYaYt1mbuUFwokKVaK0yyejiYjqLR4AxSc+bk8IuI9Zdgv6VpZPl+8buxCL5MrJ9xdBzM4Krt+MYEDutA1ZjFTva7q+2FdmYg68zt4zwk9mn8Oqbvxpt17QfIev0jlHBSlfKfZZ8aOistfntG+bF66+U40X4uZCQBMwLicpX/cl+KdsYeZkFbhfNY2FJ4IQJ+LQv3GC2tiXbUDKIinGhieN1KfBUg7Hch3rEqfhw0UFe205LJaVpVOo+d/q4TBas7GBFB0kbMsu/KHDFJjEHL85YS/PSVXIZQ6+osCi7Px0LQTZ+achb1QNMJ2seRBCebKfG1L7ospnZQdcY4NY2f1ExSNIF/EeHH/Atf+QeHZrbrbbk8ONfVeQIUv7cEJ2RRuiKUP7Yz2hf1oRZTSKN0RQbxr9JPe2AusZUKHURqs4E/kJG/MxQ2w/aeaUUfa0d7Ilqf5M6CTVBoJAOjzVjDA8RhHsTPIkLVRPp1T8Vk1W3LgDofj0DVejkSmW2PSiHitAHw4JUstHsO4FKBsRqfqgHSsKXfcJawee9omkSfPY326H4xWyQgw5g8Jrk8rznVsCx4Uc/4OK/aiwpkYPBBxWEaKltvgIhIUTdI8sOL6r6lX40A7ujdlcUr5+P8B7GiNOajwfzHXuhhKBh6DAwCF2+s9Zh3aQULbvM1Cxpxxmc9yEJvFmusBz8Q1WszKNnDbJKXUHG9LY8B/WTk8zRk91A4gzlkaClOPUD3txFc35Ia1nn9MOSXrGgP8kv/Zpe8dmJye+QuySHbMBQh35zsrgLLXq2lPRb8KbX6Sv6e8ID9+VYrfQKr7dgWUZvKg/spcOUdchcVCQtc9BcdMlp/p6IW3sdLtjcrLofgaxF8VvGRXZLSexVkZHD+xS0jX+MISJoEQIo1FeOztTd59SgkNFoCr+rmlHMdVGM28+AGcRxMxwBQWLn039f0Ov8IkVtr2+HnzhTtQCjuUEAR6F4YghCo9TePT8i5Cl1n0fbbgMF2xXwMHOJgUlrlQEP3gw9nsHqukC4xiWKeFZNdLco7Nu9SJ8hGNnuMiXRBuVtnu8uMRdDHIcIqECri4D3bFUp2qcOGgYksA3MdyOmL5HAOqgpvmXUtiOukTsrjxnmd1XCkNApnNULYqHmYx0wAwO0weX1gOTLsq/IOaYeBL3uzbl93ncJx4LLVFzKstU04BKoxB5uwdQsqtHckQoqVw+ub6C36Cpw57sVxf3ebjHw5fDvPJ4fBgBtzhKTLo9xkDDfvKWRoC1o6HzDojhmahPHDNM1ou9VdBHSjOnRt+GrGCxGWGFlEGGZwnLUoXllLYC8wC7SG49T/pn9BubfERRxrHZcJ1ykhfYBZCTQwe6INA473WSMGHLdPmulpuEDfLqswaXlIzBBnfeaBp9qZqodAlmizqD8Z+Gdvrj1UOK1edatoOMqQ/kjB2SHMVSUkMjL20kMumB+mN22X9WFKW4G4EnqjYZvzk8VEALxmxX2EG4/hugfzNbDBIqCQlrdhcYlOwVaX4+ZLU74SLOygsmvgFxf7jelIphvcHPNeVgU//13vGL6Ejzpbt7R7A5eBUds7H9S19gAth4/aHqpYx5+zuUHu0rLCPB3FIyvTl+G5lMyAISYAUHgB2+j85/TVU7KCsX4+PmLBauf2VbR2ptpo90zsHpn8nIy4FyfLvAPF6jyHIOtZ+LLa1LOfRzcniZbeOqVH1trhm3yXpHQVGXtjBdpUyeDBnF1jrDDyHiLYOp4XpbaGqj6Qz0B/odiYxBNI9riEAFYcWaEeYPhWe4VDkBqt6n7RTxFgLUgAa6H9iOHvZR8mSg1Fu5/gHh0h92LbD5t9xOq1FKiOUIn+4qdO+gpHTmTBBLVd8r2W9/lHY23QACAglHRUwZkcFTPdmfR0xN/kg9OvwPUTOkN3ZKV9qL82NRADdeoPPbQC24jIrYX2NJEjXwiwfUj4ZrQyboA3w6D2Tu+ghe1cExe3mOSoQaoPtir9BzdZB8srcgrABzJKxgF/zQtoyOYlHrTyXKfmrnB8Wf8nAm5Z6GL+gFvwgQouKnhvIKfn4rPW5Kp2Lzi7/pShB2BuoAUawyTZrQwCX0M6rI8nGSfOkdkGlUJff+FHbVZfLU3G5qbj1AcWIKAwsMcvSkgXXPBsB+pfT6/QYnmY2x7Wc4MoHODZAgWraN646hXCmEn5vzr0+l/l3Nogl4U5Ck9V0Zfa5jD3IU4nbMWtnFYA8qRY0lXYRQ+ctTaiLFpQQLTrHDiUNu1TjMpFSMOMCPlfUvUkoOkPGXx9xzZFntjZ1uJA/15kreqxxO84n1Sr285GzWt9+cSdKcrrN7yd1U/XPZ+dk0hFFNj/TvLwxWektABlo6MO1/vI9ADIjevRYH/i6QJZtPvGvIWTxDhYDAA+RSM4cTUJOH+ZujfXrMhdGg0rTGy7m85Vg9O416pP/4XFWLdb6BVCIIGW57JEdcM1Nq/SIHQRVwy749Hst4RkxiWCEkHTWa8nxE5S4IsjefrQXOcU2PnOvVWoI57tBtQJnecDcodz9sFlwqayJUhtQDkqCHebD9AHf/DTKFsreyFnyw1ux0G+MKs658gKHUh3l10WIpJCakq3JbBmTgSNhfo19Gqadfoxf6qB/nVlwEZ79i5Itj/elNjDVa7ITTVzQfIxw+y3CjaK7hz7MSIWVPa0aL2crHWn4/A6KoxSM/T+wWK3+bFFKPPNGThLViBmII9PiU2vvaicFaj1NI4oPbaj+8cEJGhXgqE06w97YdZHm1/Y3PomjoHUr/WcRT465dM0z1t2L03ZtdhPN8Q6Q7tIa0/Ku8SH6JlB/gY8PEZCbdks+dbv3GZGGQrsrrXxhGIb36bfZ+rrwE37ZSv5aM+bMBKwFdIkreZNbNpHfNvQ1NbH1w5WmwufTMembUN7o9gfmO34ArTyVCXACVsjrgmXGI1ExO11vHo5kBAS8mxM7/m0xNqrxf363QV+teSu65KezioQOXFze5pttlak+JHWlfQMk5Tgn72wOchGeQLSASh+KRfT6E5ZExmaru0NLZhn4WiyskjDyemjfDOazCVtjiyoEiQ7v3F/fBVXbSdiOuI2jxMfqswDUAfmrYDBJeNJBlYHkIVuP7W2d8U5t7TuN3DTsV0a6sj5iWRSXNlPmdnZDHrx3//MvSkwWZHQUyDbxcbpp4benOvR60jlhyJLH6KQi+6QlxjiOE9nH2yEEdbg5AZIDkM0SztDiD4T7RnoqpZcFa5hyX+auiiE6NJG90RZV3I/xhOkm0mWNj9z7HLztYuQr5tjFH4wjduenhbcyi7kh2gtZry3hcu+BZH6AGTJDHAdFaLs4jcpjIov3J5+3G+GMNakR6Og9WIWcRBr5XyhV3zspBjqOeAx7pi7cYufLgBWnhnNrBwNu5gEgExOMFXcp9AvMwXo823hmrfGdprySUdMKwHPAIqPF3Bo2ACpkE1E+7KhjbIJt0JauFaSrtB/TEjkmSkxjh7CmeYvCk8+4aIAe8dxG097jOicPkSuMwjF9Mu2a10DLxEeg5mJVxYnfeXUV3yEngBayGLm7dAjFtYllLJqRQsCLdgmukmMRCnadnUE2aCdogJOjcX3ugAA8gGlji7hecaSkAK9K4FqpExeX0nV3fPomN4sgia+g2NuI5AveynrJB/1qC2BBeQ39IXLgT9tdjkvNGsz95E7cTOHc1ZMDQomdIw5FLvawUlBWsWAHThXAPQrwUzIFBicKhgqAE+vCf+q8kdFcgAj4lligZwWBDWNU8KZu7mdTsGYXJINi50nnOtacxJhNOKiDEGQYCUab0PJHv8YWRnJxlL1pDoC0XuljOMYeVfPihnBGuHn5VlGRpHK8QnstnSPff7Pv/hQ+kWwq4+ervh4FIk+/xEhtMG3Rf2khPz5R8xBcNeJjCMGOupSDm0v3PytWZB7QKZCCBSVRARNIO/J7dKP9OkRxUHswCGLm/GquAJ0ZjMP41BhViV7ab6PMDcpNXSW+a83GHzfw5iuXs29FT5WgJy8qZzhHb3FXxNWhNma5VaVzwRYj8y/HFNdbJXrihfKfw3hzT/UPyblAj+1DmHUi99iYw+cmK6+l8XCm3tFBf4cCm/yNxkpIe9Vi/rsJ5i60UBocUWGoDupQ7PvgMDXcvLjV6i/haJNQeXMlul7/s1FEMm/lsSYIYYLzJ5Cqlw1bjfncLCpWJatUMK0HdGa7S9WrdmkjObktQKRYgAyp3ql1XCcC5RosS+WtV7WQnsBidGAwHiObLFeZSHLhOs1wDqy3qUEbPxJIzZY1zzFdN6myqNbaXI72tUXcbssBjOmzgcEcL9y3rMncx5cpoTugDHB8ywZeJA4W8w1W8N7ux1JIfwBNGfuRe3kn24RvdiL+mxKQwsAS3hyTObNWnfSF6KAKDQSJAJwYwWzBUogHPsjEDZbLFU6Z4QbL9a84Xz3mNszQqQdazvflIaLMXwtb2gB0vqrYg951j9MaVVMRlQNEbAQIxBcfDt3xcFBH6JMJfRoYuz1X9RbM8uRq/rDAxmNESkK5quiSwfwAlwYW2vtH3P2zJDwQ6G9chnw5tcPUgpo4wW4FS+NhJCauv8VJmrGbs00DXvrFnonVcrx5WIIFbfyGotiXG7g0nfb5gRgUwn/McltDbBIUzgRPcdNbLUqIF+eT0dVM3sNi5MKfwtg58H6n99loNYCzxrQz8cFAYOMYDfprpPSQ6o+nNjdxw0A4dwuTVNcgl3LFDsHutHDbepeyJaLrfJt4TAOKG5MFGKws6dUdal/k1COjYgojcJgvKcw6Eoy1ts4pCjvpoWPeNw/gSHRvMOAUjcoeNj+5zWL+gh2r/IdzdPJvf0rlyBMAriHzis07ekFipS3YyBuKyt8wpHWnVOB5yInjm8mG7kmzIHFGm7keweY3h9dfM3VUEvhL86i85TtAXaemPUXaiWAlTpD6X4LGf2PcyD/bmbct9+csbroK7iRKqknqocruhk6GNrl8kdaLjh88TWsFwwSiJktEJVWRnB3odjLDJMwWKPm67hJ911RjxwYQSDiWMXEpR8HovCV4m2JKh4/Jq8BAgkuTI3Mvwo7RC0TyEp5/RVo8LJGM+W9+RCEzwPnDjyspTQCXIjVU9m5o2qg2HwAi/StLSjAF+E6y04d/dotngZBy5KdnlBY9ngSD+DzR13O8s6JflYsXjH1fATrzC0j/hNEUdA2o4bfGskVui6JVe7baF9AISeNRSQBRb9iAMJLc/s+PuYGCvO5SrewVdJUvIGEkpYWd3Ske31DwQq4gLdiAKH0tQgH7Gy9uSnaV3t6P1Lsrd4gq1tMHbN679cUCfuTPmnDMF7s5MhgLQWKBfUlFl98N2G/xAkxbSqK18XQ/wLpkIEyFdBvwU3WKf68tihkcaVakwCFlHmEViCMouItpkjo9zhobiQqxCtjTqfZne82RxvzHpEWWth71dVdkI3c28ncLw/FzVG5n+HougVnlt2lHl6CAPXgfBZz76mAiobUYz2UuJLhgcvP4IxBnCrlCkHc/xBhYj2lkgqr5R6pwWiURlr2ZXNfsR1IE5hdbYU2ASQjnxw6S7XrdvFBTIxvBrGWy5c5epC/rLTCAZuzh+vxzQOl6hY/eDoO9FB8655oUv7o69ATvP/IeXAgvq0fLXr1zBGdsomSKY1QbxTSiCfiZNw/oaSMKJMhtM/rzryWYStTvB6UvqLrtULmRgMKh1syjuaUqdvD3R3QqcbPjkSs25N5bjIglC5vDCvHYAynKRRZu5fqfg9QGBaOWDAtwpKcRu0DNlsWe+lELyAv4ZqsIDbzST3/+YrIKu0Ys3wWYrG4NDwGQ45T/QuYhhrIlxuxf4jbO+zNyRNBCImg5k8MgZV4vSwsEZTDu/eNTGkQqoMtxOvtO9TJ74xmoSH3OALZu+ftk3lT3/GlunT8JQoYyxKTgftLxL5Ck6VzjBM1FooPuy+51CZCp+BQGWU+fOJvH460InQHyiVbSlPPr9poK7FOGxwV+9szUnnEyJ7BfckOE1wo6RA6OtGJ8RNHikqXQp37Dr0eOV/Egw89Jizf0VWOualhZ/MkYLvhQI+2QXbPwGxfoKzYgBsVePSN7sUqOSbSbZKy2Um6RSOUAid5KF0zxnKj3oZcMVbxEWZ3cubrlTnLM/eoroPVeXyUWruL73zxEucMmr4cURLjFLR+hZ+5aBZG3LAaUgpC9x0mp2XXf1XsPV6Lybm1iRptXIIouS2rD8Lonj4iebpcpyqlEuttFcLSj2qxP7457Nuia5z7uQPJdsUwTzdcv32yBYEO22N+C8tFvOXnFHgEbbgA1ClqWn2De7HFO4vwmjJd2psiEneFYdtYTayIXPEwqzUIWjAGkXdPtKmeI0Z7k1iji3G6A87qACKgpK+gU3SvnOgvJStVL08gc8+le7H9FDBRvlcXjBPTtcWIGDBdQgQPtBboGRqHhMTLEenye5N4geVp3B/kHNF5a0DF+E7Vm5mgl+zMRU5uWxc4q/zF9pWxpoWrzY62FFxKB1Zkdbetxt7X8yFS+XXHBDHjvW/Uw1mYgnW2m235jfePnldcmk3vxLbHQFdNSE6J2U3FVFeSU32GEl2P5I+yFRPHHwTaHf8zd2fkiY+V/ciXznNbjzAVs31RjvJu8fL2EtTjL35Xmp+iNk2MhRZchKimWggWwfg9rg/Wa/pwbCaADaIRjOWirnyrXJ785MbZPpKHoDwjZFkKhigeRqxBCUsdxEZE0iEz+47LYu3OXI1DTxj3ItPZBVvJGpkjipaOMIezGD35H+IJCCDNWFRqxC8ecA9AsQsiEYE5UO8Gz57VK4LqUIUqtKISFaYtN73TlcmkyQ94i1cWo06IP5ICDeGpxMXakVG9mY379nRx6UoJeHGLOeAKQ2teTdIWdIBXkv5JTzB/hwAVtbxO4t4peEvbnb0tx6htmYmzvXsjI/HnngJxKUsAnAMCr218gbRL3cuTlEwJvv2J61B88lS05EPh8KQlsPCBgBRwAL0ImDpECIVEaDT6z0LsDucYlWhbFKwhEENma/jQbatQ4u5jp1T5x0tEftvcwXS571XRsjtDesDCPwGr0MhyTbeYZ1SVSpA7bffghj6Cxez56ulNceXeEXoviLYZgm4giJSQGSvVKDKopdK9InRyDxY7q9bmJtNGQ+ScENkb4D4g5vjy6LpUqJaFzScywACegFP9zM/4UQ65VjPeKHd804rPtULxAvTdbnAPSLTm0T2mzdMA/uymBKDYiplsMkMInbUnjO/XrDAbI6Uc/qg3FUfSCWJ6AI4NVO0KfsfT71ijtqWUK1kvl35Pht38oD+fZyCHHCFeUoyiDxAZdYRSZOMBkwid0R7DsNzS23f2qn4DrBIny3CnprpKVJo0fLd53fFs7itEQCxSeV0LBd0P7Fl29Zo434dmXtZb5AplXyxEyTxwepmJp6aqimBogR63mSg61Hmn4ayB6pHq68ibk3TIHhVa3tSnfVOphaePzzEYIuMlnRXiJ++Vt5IWCS6a0Wyymttpdqtx+mjEfdcHgJ10AMuqznHLBru3xnq11D4fyyoOMewgLQY+HLcHSkUw9MjkrfpAl4gwwzVZkPXWKbP8pYGSm9KKnT5LRQ1HdTnPEHK/IN/m0e2PFNscklbE4GTXV11WqdTHQ5G3GctphV5PTwMnHen6Dv53tzglpHCoUd+5PqEdCt8Dv1yIOeO3a46HUwWB59GkTQG3Jd8+nXJ0D+KJQhB3G345ozySFeJmAYcxgErku0ylgtFiTXMsO2r7b2/9tuVTlReesp0s6gmRTrGdy/TToPQb67Ixm9B6KhdqICM9hb2Z4iL2MTrish2Whzeta9NOaXLhBePywOgpfWezyRh8PAvEJgfdTRPwfbgmxMKomw6GItS/3s596lbSGU4dkguOQqvgOHiGLWjV14PPPLAPk7HYhSc3nOefFaInf/u8rd9b/hjrBVdxOVuPUTmW9huVIeX/gr+WxERfyA5OviTYsXxDYedztaMdcwKiQ2nmh0TmcgVDRgvHJw7Vc9wx29gjCtvivjBxBeg1B/Lf8qZYHz2hqiQS1nmZIoWRlmFp7pXxt6/pyhkXL6fMoVq3ELxQclrMqtkHGmhy2rWma125pA/JEMEmUTtvBIn2nXwU6w7sqgLZ9mwuM79UtO2OWJEoTqWPhmpxUfpsxi0JoYBD+boLDvyEXeEHKdmu+ZHd0DNeI+NV54KQ9/aeFAPfu/Zs8Dk8YQEYC5lTU9aGU8aSdzUhDfZWvK1OCicHIsS5oWB6quDYF3GetnqY3Ao0dOgZBI1XGA3aZbJdh6D9GmbQFdIZ/96yMe3dFIExT1ZgwMXZBSrN9OFJT16eCTj+3itixZB9+i7zyQ1ztIX16eNqKuez1ae8BU4jtFd6oX5wI/7Jelap+KPqBt1Y9gHgvoIUxNClyOuCNEBRfhOryvz2VT/pf0KzcWi+O63pFnHAZysLzqp1+0llzafXHI5C090/Xo11ot0ySrj7vZfWiryEbsNqnSUymftAWu6pgXUKiYvb7jKC6CCi1XxIwGwXZJP6s5f+RrXUJeq+Bk7UIeqFs94Hj9+XoeNQUE3qmJ8BzE1g6u5Ahr2Ha5h/7hBledKOkH5SFXBLGHsfnMSZjW3XPL5u/ytCxQ3eC38edLCMOkI/A7bA+avhTYZx/VVH4G0n3iQN1jwa1ui1GLYwRZ2QILUOZzyV4YDWlM0WiAwDS2nL0p1c7NSFc8ZPKHiRvJ43nTW0v4KxUkUvgjL7o5gSa2GPTa0+8peRh7Gm9iXzBGFmiH0KfjJRe9Gde2+ELkRAyzrwmRG7BKGM6B46O43wZjSfJXAhyGElFqsTVwqcmA8IuTQCbfmk9f4fFo23ceNqXgtZUZSVaMXfae7A5hSsUWYmreXutBtPBTaj9qeuyIy9YTGVg7QFDQO3p59cvLnux0iLHIBI2QOmNr22IGbJH32MTBuzo9SYoCx7XQqLSGiEvLRjDGRniN4EcpPhvZ2lrbxdiAP7ebMnEMA/iTji8aXrlALEBr5/V8g4sKRUe0OxFh3GSJqscQSLbBEvnb7W6Uy2f4K5RVYTCqV/rWYAZztDZOq8/GmZjDGeZduJwEUVxE9q6YTyosLB7TIZyf8ZPgKKMyx5C8ydaPkbcnqsGcFDGPLUqe1+piuTjGM/PEvAu2+61IT8D011yARPG21Pb+J73vIoZGGSMOlljLQYK6RuCQSWi02ZZ4hx8Y6L/mCOCJofWRPB8C1PcKZgyqnPG9mXq4QLDlIrZNRzOivw/g6sQ0yUD7+BZXP3G2fSV0ycQuGd2ER2ETqwz52ST4azE/GKffVkXag6JGFkO6cUCYwqajybmkIaNTcnnOnGDL8DakJ+Ftz4GCoqe6/yLd7tJ0ZesKwfBxjirJZtrJ7NINCm1UE9ccJLEoJba36hmuIfWP+G9DmGh89PhNdSBT8ubi6rkgMOkSt8OQmXViyQXdXrcxNSB/q7m2jDF351Uf1rnQl96dNpUDY9nqkOrOOIvxvzCydnFe8qrvCHVdXLYexXvsE4F0K5+FnAfncsDPmJNJsxl2uN4x360IyPny3btrYUdmBt7v6292iQnp+L9z5YTVIkJxU1F8+mlE50lStfgM1Ajy8YUhdOMszrSkRDwtOaE/PSFc+Jg9YVS9KL9MTOIZG4xJx5qfGbG/IEnXf2xsg8j0vz3dR0I2Pa8b5kbMxNmiP4pizXBh2UJHxgBbMKSeq4Pav1DMYIlfqXYeSaspn8amRQfoPw5kRqluLsbEshWjELk9AGGPqXJGFqbY8zReyjutRpW+0ilKGbeM4yZY/n2PSX1zZ6H7P/6OF8htAeIL/3iiGb2wTGjLEtamiqeLycd8xZkopkiibtTLlstnns+tAaMleHEsKd5vLIPMwKcmfJBgqB6IjmjPfkDjVPvsVWYr3C4klxmi5jhoRmgJsFl3h4NSwdsY8kIVTKEzdy6RHmQe9LwfaMUdGLUu3hflASN/25vmslFviAl42D1u0mGK7xqCF+6qmvZnIt2jumKz8Yk3fjgMdS8buGqpSRje+yFxDwyeVnqlElCWLOocqWE7n/nVooROv5wMfpnYO/scVr3S6oZ/Yn+5Mncdf8Nc8e9GUtL8qP4qu+ZzlmrJv7e3+ZE6drdA/6rKlk0ZwGUeaD330+9zDCokQtyZDWA2mgxG7ebQhIFbnlxdrtHz/QrQW0ZagEQ6CeFwQ8aQjQtcmVtq3L577hpccjMvo4gECH1lCvCZzRA+MxnuD8L2LVMnHGrkXZ64LrfI5zPmu0v/jyRjoJeszENxXsH3Yn6i6zmlKUFYa5I3Z/MhZQ5uaYtCjPCrjFe1nFk6r4qNTz1RFCj9u8R5EnfkFlVO59l16I9hpXA185xXtKltnWJYL8rD2oFLAagsbmlKSMal+UmPQjty0Bw0cEmTwOyfCMnBd7Jsi3kAyQJY6P0vjO2ibsOFgbRt13Gkom/MNGHJ1Xw2gUY9B2hyR/zhHTXvSRwr/mm8WiMnZestkUXAjGLk492vZs8ul8eM02pKRi8ltM0cdTIzkeobEUi/vTGdRoRLke3Bi9FAOZHbFsAIP9oluUgW1gcKHGGmHgEGBbCWyeD4z8JJtuP9iuW1igx9vDpiPPDC86FzyFRFOx3wkAr/IqvJntv70bmuPGNaG+0FLqu7yB+LkZl1X/WHP9YscUmcMg3GNrGhnLJFxEDpvSPULKNL5tN2doWrC5Orz5a8A9dDh8fOA9tjdA6Y78BB0/YbkSoHeL9vn6h/Zp7Z2AFQ1MAWV4ei3L19Rb5f81zdp4gATNT7UECGhICtieyqRGdaSvA/pXBTcu1in+BZogsTJXBk/N98OT2Sl7O6E95pLTfWvGyam8UQJZ50OQKxEekXhR3VZ/osTf9XRVMHjb6yr+vjGgDMKpcd5Vp/aJ/MwZ04OhqhV0RPflE4iQ3GOvN2s/ic27i2ddNjbrMVf/BmyyTPA6nZ2AGNnUCGuLBoD5e2Zr12WDa2iTmUZH6r5s2pK0FffdMbl9adKpVIh1V6rUdOsuBsr9kZW4KUBrgx04P36uSDo38a6rjOrsYwvtE8PRlDASds89xRBZSBkJPOveIidA81SDKvn5GWFJU9uF0XXcbhSmXX+wrlTJU6yZPBxKDunTttHznIlem+skD05PY4GnyG06RLSyokrcIwmb4l3ZacyPsKFYg5qk3XmcawLvWh4RU829di9R0D2iDawaEn50lMR8OKncIZDP6fq8Qw6vMYOIsGdUisGBcXZi3qynwJf9W4jBdvPfYZHBOj3uwwUvqCxHUeKbTIDFl2g0Aj3/JeyUOu9GtNsohmoky6E/R9S970UqfXK7R+rbrHbv+l6oJ9goHyYR8+U5/BhTDRaDCcaUT694JHphKh1JYDqk7hT9WVwWJIZzZ1luQI5YIjQSsVRoTDVJgCO9rZL67ifrpT64K4EzW7+yiSq+Ng9Vj9djWRQHvpl2RkrnVS+4umty9ACuPomL/lIZXVTUATAAAALWnYZKMQQfL7LyKPr9xAJsa5D3MvufP2CSNAAAA)
Cohen’s d-effect sizes 95% CI and for differences in subcortical brain volumes between major depressive disorder (MDD) patients and healthy control subjects. Effect sizes were corrected for age, sex and intracranial volume (ICV). The effect size for ICV was corrected for age and sex. P<0.05 corrected. CI, confidence interval.
![[[@Schmaal2016-gb]](http://dx.doi.org/10.1038/mp.2015.69)](data:image/webp;base64,UklGRnpkAABXRUJQVlA4IG5kAACQOAGdASrbAjoBPjEWiUOiISESGOWMIAMEtLd8ITIA/ACQ5Oi+P9D/rCSHwABKE/23+W/tPsoHcv6n+3vE7av1qJ6pkr8536b+s+0//Ff5L+ze438a/633AP04/3P9i60f81+wD4Bfxz+gf9//Ae9r+GfuJ/VT9av9p8gH8t/1XWG+gL+yn//9bf9ufhK/cX9t/ab//+tO+OP55+P/mc/O/6F/fv1i/rv/N9Z/w/5D+g/2H/Df4P+vf9j/M/Cx/O+SXoH/Pf5D1H/jH1c+q/2D/Bf5v+1ftv9z/2T+x/kt/mf2q9o/fT+5/1T9nP6x+zn2C/iv8Y/qX9b/Yv+5/tL9Qn0f947gfOf8n/jvUF9PvkX9w/uH+I/3H9z/dD16f2v+yfsF+5/yV+R/0v+2/2f9jf7d/6vwB/iP8n/sn9k/wX+X/wv/1+if87/lP7N5bH13/Nf7b/P/s39AP8k/oP92/u/+f/zv9s///+t/En91/1/+L/03/T/y3////PxT/M/7b/qf8T/l/+t/i////2v0D/kH84/w39u/yX/M/vP///9X3S/8z3BftR/8PdF/XP/q/n//3STdFW6o0qcY5QbzhFgPT3NsEZkhPOxIz3O2GB8ljmihHy1Y4NETkToDI+8zxeG/WZYzy0f18wTLZf29b2ieVj9TobTtgLGfcKZ2rcODEWBTv3edtOuijGBgg+pIfR6ENzWRiE2Y7ltoJtgaoKT8jJjHm/BVYy0rYj1YIW4hhJOobbRYz38bm2/9O3pafHNBarg81ENeGX3evMfBQIMe0S/HeA8w+9e5QM/ck8hdcpQHgzCh6FSA6bzCKK/dcy5F9UpjDvs2kyuTMcREVWHrR4utKTtXpvdmSENNzgAZrwS1s3ZBgD7JuIdvb/Thn3zRxBq1Jm4IQDnpgQERqF7gU9I5ed2T0hOeBysNK7Ke9vppLSr+M0esSD3O/67OH0v7wTA/aUiSAHW6lkCroai3DBZc8OZZ0jMrR36GJRegIgzV/dDEbo5pCKLHdKUCnoeSmvYQKzyczCBhApmFAW2rFPcRoPTyE5Qho1GLQCZBgLDIx0+omaMfdhI3cj8N0VEdAoCFd1pmBj093BUBj1N93hLJiIJkKjTIN2PdZka89OenPTnAO6/+23y5QzMIGEDCBg/vDM5coZdr2wZYMsGWC7GCHwKU19y7J+wAxXet2F6XRuWXm5FMTIG2T5DcPac5z0z96/LWLdu44yVxq41YRxPpPabV31m3QUpx5vss9uXDqpZpgIiCZuUuDeDLBlgyTodru6OgCuSu+0X37R9nufpEkdYH4xAwfH79K5uTWbk9/2t9xD4h7sxETbdfiXUdmXwqwA0kVSj83TqXEnof2X8LtZfgMIGEDCBUmiAoViLCnw5Hpit92o2uyFugvaEf70fl5cUvo6bYn6aO+3CLceQAIsm1cHmdl5Re1ZVK9Z5IoX28/rv8AHwFLIxXMT4/c+Ipe6rh6YKyG0WtrbQeXdNMWrEPeAQV3h+T5KXMi7EanOPgvzVUBRRO8Ec1BNFgd53tHtVYfRMLiUBDiExDXN7F5YZUq5RaVjzqQWEen0cdyIwigIMRHr9PY1VrClnR20ixsaArmDAEG6ubidRk11SpOQn2Sf7WU7Q349sQyz7UY9xVlpzLyaugIVK61PhqChj59bewphhV3Jh5CBw0hokPPS215Bb5iQMyzyEdJbEPPTLrTX0Uz0lDnNh78Nt6oRfbhrQuWr7XoSnn1gY6sc70fZtJOXfNeC2ZLOA23dx/cTzkj3PgIeuL2mmGk5OhH57D4RU9iC67BCHrZxd3YR57eBgSbfdH8B9qRjLk4pOAVYvnmetyWzk/gnTZNIaHq/+A7HeTb53b6ddibKSHOK3kGFdJC8Nc8L9obIarfJtawWVYxCMKkcnGGNk1CkRCKZJif6GWDJcysBicUTM7R2diMPxJYW+v9RViDd51zgIaQkU6AEG405cK0UzMCZbiSh6IJK91Y07pz07MSPJmNhDAnDn+7Kzk196uJ2OjePifxQBMrLKx0K/eCovKb6xmGRCr6GWpNp0fHPwO6PRTYzJI3jfX3+fk7BsOhws5AMsb3zrdEQyxkLO81EpBydNYgqjSbJz055h02LVBsgdyUbRz7DiIBdJQkeoHCp9p1Clzm4fkuMzIaRJiiJaoN3R/cwvYdLf+4o9PMH6YqfcVeukNesEUw+2C3VbE50doWm8lrLH526NDYR45czY0oMLeXQGqbAg0yPLjwmPnUgaKlJuKLEubhhUWDK7b03OpMd3cswy7qOfpMQzUPMrp2oGzBhesDVcHm3Rzz0F2xvelzLflQebQFYNKvB9ERW8VvT2tMxEYESA7nixQNOenPLtM80Rg6b+tP2TFZHMKOALD5XeOdOEvTZCFGrjVxq41EdiRWK5Wzb1XT/Mgq1xM+VFnNbUCvm9VvvTmEG7KPSG3EkQdFxWEB/1wjxD4gTaSPW8jgi0b7JitXjk1xWqgGb3VaOxVeJsjuR3I6QgbzoaeSQoHp1u167uMUnJdvRjfb8jmxH3p7RVHPJXyNzlx5KLh6N4fHv7u7lMjE1NOemceH0afdHcrsH2zFLjjcis8gqJRCTQCPXI1S1wOdVXB56c9HEcSz/c9nVGlTEbmSAuHB0ucPV+Fk+AGnXOouSWghHxWFyW0LGZUncYsGVgdICQC1afxY9wSen95ljE04QQu9xxz055nx+GcDmsXDNFA5nEUr6fbA9i9VGXwRDzpgkrFiVnnwzSRokaLpWTp0mz4Qdm5lmGq3PiUazdL9X9qOfXXp+m6JsEE9/wTXbWDT/YrUlgYCamrGAfG6DYCBuQvDWSnCakA8El27y3dgyh8LWGHqkT/OY6MtvExIjqsgaxR/q3uH6zjDaFLwrWOWMZWlkdm85HMd+tkq+eBhLXnlEf2Eng4mYo2c3B7holopAfb1oTwUcJLad6tc9Zf8Rc0zT+UTYOANPn83YugTEjBbjoJsACxUKbAFVJiQErj+pmQgaNsulcYjg2Su5OSx6fD/ubi8Dm5Qt+/kKq+A0eu9M4KsbO/wr/pR74e27zQrzJanuC/DilQBKEbA2wxxWnxU1cJQcSD8YcPmhrz7Y1DDDlVnCMWuwWUtNCVcaiioaHjldnIlA6SgFsTEzkdkNbmnt6tp7SlGUbFibgu7mv50xQrYhAJXqZR4Rw4aU4EZhq41bQ0jDMrTj/wh45bp1orsUnWUgpapOHwd+9W/z7Ush4X6ukNA4wYyJoztEiX0MhZALg89OenPMZDzL51N+D93GoUz4srwY+KtwWUuB+OcvH5ibIhkw+xW4dkipzD75vM4PDB6OJ7VEQAAP7/iaOtcrr5N1tp9/t5d397KIUVikmjWOEPLv6B4lEExewcVWomPFijk/4AhE+/059q0Iwifbmh2Bp1VnpIx+d4sMvreZtk0eIBt+9La/d+9YmAdr6gCtqRhImjRlMvqesFxkUqNMxPcvec7Jfw+G10EJFkASdiY+/y/9iiSSmRlBpXkWRGNfN+knFDBiJ9LykM11a/nplAyk2gKbN532ALqWS6Z/Kg7gixGrq1CGFrHfaxq0mivawzPWiNbkWNlTraGeBL7wXsnYHcAeYQsgCpBra/lv8U0lrgtkZ2n/xN1pWeKdHetgf7Fq14ZOGyMYgIlSuM52PbLHUev9lrXCR3cekNh2qD5NREpfdNGaAuznBs0gkhko9oMEn9uAMvTiqaL0/cjjv7E8ySBnFq3l2928DRMRyhLmRcEouMK4Wll0m9XljX5xME5+HHwGfIz5XivNNmWEn9r0gRh2Ze+WDU1saptALfoTe5gG1155TfRVXqIQCn4IAAR/2n1aHz9S5GCsl8z654tSSvcKuC7jSdAycvi+VPmi704T0yq9KrvJMbjDUIR2lSSWke61kAuKVWXyb2TNWoZu9QAx/C+LAFmQwqWZjLJJiKLA+WvGJgtT8sDegn7AxVwRopSzsBOrvtBJ4zXhfjbJ7PB71gLCVjsP2tUH81lqqwj/Q2crLY8zyzyQ61AXqCNYanek4vZz7bPdBe/lfvrz4259c7R0hSCecbBQThluOX6fuSxI/32jffC+vTcKUigjag3Oqcoowojewqph9UajVltVAgO9yvP0z+ZsDQ3WUM2HVvGsa/jj9M8CVh6asPsGvN38fKOpq8OTtcPQDhjy+gVSCpG6qOtSpfGWmXsmNuxQFeYwaadicNYSyKd7WGTEZr0S52tiDGxbwGFLfwB7XpulRbIYqA1dah8NeggIAwxmfKqJPG3Wu01QqKVqWf0s+Ze4idJft0BfgsnWHNu/LQqanOT9UJu9U2DyGD5qDqjFk7bwq8WoTFUZwh862XoYAUeHKq8QPxlL6FPrBIivG2/GjGDnhblguk1epTZWNfNHDxdLRXhvemDE72OR6eogNnOygYcrFIt1VIsrE0WM0LN1hAYvli3A95qgjO3wLQBPc0aH0zpw+jtWOOrqFKPqBH7UonPOmPcOQfZShUmSreqansffQuw0T8NX9x8rEcYrszvgynrYOaNiM4eThCsOFAuxAYl7UBBhh6G+rrV8oHaKt7wQof5P5z2qZiAuR+83mInncCKKL7FOJooKbRuoxoQSY/uF8MLVt5gyfi2AMPSydkz4dig3gBAfPzbJasnCv5wy6gbQIEJEy2X+ss8BmjLlpYbatyuxEzaInEq+K1CChznRecRIIai4FYeoxDXHfyegIra6LaIPeCizu633gd6iAcTA1RayLhrNPwXj++Ol3zi0eBhMBEb0HPikNt2JW2YlAdOlnQNCplBna3YteOTvUhUdkA5jI2dBNmbdUzqNvQk+1H2jblS9gCP//OUZIE+UPqB8u6ckXy5NDPIo8FN255scziUv5Cjui9kYDp0gaecOPgX0vnHgMKP2MecBniXAxGwM43h4nfk18kaDzm/sR4xPan9Ku0qmDNZgkkcep0N+PWs8BLcWzgUJx0VwSt9YQMY01EGZwQYqkm2r55tl8Fe7oayJJFjn0ZSkkatXtaMaAA2S2nCx1hyd6Qll1TJayL8bZlN71rV70rWD9EvMcM/iH4Pq8PbRslKGXTtKjMx0aBmbxHd9lQqiqj5RbaGwUHQYrUY8o8kvL2VBB4BmarJyCvkkKcqIxuMWHJ4ahU/Hhohz0elCHiklKEF1Sy4/o+D34vUgOipBf3iHPDHxRtJfAniRHojlDS2ACNRufW16zaDBm6M9t1PFUqEa4yN/g+KVlodNz6Grh/5TPCN2LqSS/0to097SpxR14Xh8Q+uN0qOA1NdgfWiLXSEPtsmo2zNlL//DXBoikIrY405aG5awrHtUh3w59WY+WmcUt/e5Q5NK0FmT7xmd+8SfFUsjWVwlV/eebg5xtQQMXTA0i121LkUrU68UvJlatILCMXXDjRGwRJ4RPE6h1NNE4byBoiBV0qHQUhtHP5kGwp/qjKM22LhvLTEF4RiMYc0AkrMnhUHvJESdbhi0Pvx0mKnO5Fnu6EpZg/khMtSOWrQN7uRDWiTS//JvuoGRo7niQGcwCfqhM6epRfYPIML76nmJXfLtbvDGnl9cs/7vBMwo53bCz03gYV4imyMAMJAaR/bxT6ImI+9jmTU/OSiDxvEbVwa2P2hdGmdF4O9p+LVUHLrGVc9zhxD0yj/y0StGQpWPGN1dd2YpKODrwinJL/vTwhuuVRGgX3Cmd+mnKNXCyCch6eeBrZ+Ebgp7bIKdPAAlVRtm/f097rMS2wh3Q4PpAQfxkKObE/3DPD51FzPyTN0XFyTSM8I4ZVp6s70GF1MApO8kb5rVRGiDmN3qufIGxGCOUNcZW9HVNJpA4qZawGqwf+zTLzep970oQ4m3VVk6V52JuV24roiy21zLt089jQN8wdpLzk52THaGaKHbwfs3+aItZj+be3Oa0ZSdFnWWnfZfE/RNJpZ/tGNJS6wrEHnnc2NCdAlSciRtx+4mVIG/DNjigkwOp9QbGkstkS2jPNOSOKPWIXNdW1VkM4fVSKEKZeQb9BArZmGAsQzvlQpyD0Y71gsj3OQ74V0YfBLAnb4WnD/ClYQZ7r2b0Db3aGoVFYc/Ho1bfZ26o9oZkoLYKfYm5AYi9/MoDuB5kaqfD+jpCFmq6egomp22DRN9hu2Luie2PjCQ+doLgSYjSULKvBpDOHWQc8OGCufRo+PDegqlfKgi+qSqtyZGk/YGZovtKOxNNgWclEyuRUdzSlNUSWqXDCX9brevVR88AxZF+1A6eFtVPbgg3wI+P2WIL8CvRZc4b3NGrLWAdub1uO/GTK1guY8uPH7urlPXMv+EYMM/kUCCx8eMnXUIC4FrVI2uPWiU+KkX0k+bAROfYUtJ6jHeL2cIkpjBYCEnm/kvkFYr5c4R3oN5suRPO9vHDH8aLW2D1UgGUoUr3F8vpJ8+w5Px9vUkjRHF6swDqX8gyAkHU7hgaqRjyXuV70XLviGzLzdmI5Tuv6eba8DJskMWs/93q9X2jqDUUA6isLC/OlNnqAHZWyOuXLyP+Ay+WH9i+J8OfGTb0hqJH1P6oHrV02AYYNhqpz1nDXEGBhf+4Ww9d9HbXEv2D1UqXbibeByadE6BIzEE+K/me+0Q23+mBnhxZA2I0Vr4ThX0AJ3rPEOpaasXBbM0UxpGGzqSPgmTAHIKmoA3DDku5F6R+BiqxpSCM9Z3QX+u21VJbyEgBw3oDmXKgedF1jzPfNaVxofD1YZB720Yknwxe1I32V0hrVSQAkGow5rewuNHq7zmDPnqIAqaNI74pChlpqUR5eaWZNiyrm70QYdVydFjerFIrm90Q7gVwTwMZORfqQuhSGcHrVh4RyAGm+2FsuYYDEArVaz66s4VLb7qZHzOm+8XGidKu8j12zsIL6r10lZsCv0+HlsU13opBmHtbCDzMIBZRPpegKdQY6lwJ+1oSVjR3goKF6YSvJ/xJyN6nL5x9vvOoWU//trehgB3dneCK2fWFndjNHgInGNJxJ7wTOHS42MPXxP1qRVHmC3iIVcVxBqbhBruCN/Jb5NIi/3HbaBTiOMedTMCl4WEp6U1fxPhIstUnFZtxNUsfwd6+rdsMMpK32Y3TW4FROqMmagE/ajgW3pPhY1fCuz9jgRIISnJuI8VRVbly+7KfmdfpOD5eGyXkoRTOCOdLa0wuYMoh/PgZx9FtkIhOYepmNwMMXTgI0Fle+kjSQNUToBpIiZ3uw9x93rxlPhdqZOkGtBZy0v6FPMkd6D+C/cAt6kvS4oS1XnrgIfw4XIim05BZPOy/d7WQNm+cBo/8KrdCjfZdbKgXLiDmzOOJlTyMA1pSUTqxVVhYCDhGsXH1BOEBD6Xl1MqXvGOgJJYIqfmdXA9p2AudmmbuvDJQp/cfa0bV0ZoFNH0NsoKwB0M3A+0sMnGKgaF81FPEu4q6H8uC4wUXIvQTGhsJzMC37w+Puq/1+A1L5w4Ob+SyeOELD8vwBpauyjSLwwa6w3tQxkSCvcWzzu5NleMO+usvMqEs/K6F2gdesjgWP6I84OLyz/XNLfpmGPcAu9/jLJtJL5zUC+xEfQLpALolPXRcRsviNsVTx5dtms7kYPRmrmwGfn4ojfNQhjTulm4fUBCho5hE/7QWeZx4yr00AM6Sgym7RkhHsHgnLgVp3FGlnQdJYoxaCvoAo76i3S2/WBZ7WlidC4h15T5o9JoZLFotdyEdX3tLm3r31/sIz2YUx72ykgKLWNyJEtzvBN+Wl+ip7jOazFkneAjKLiSF1cFSVxQJRO7qbr8LUiJgYNwE6UXjSJu4qYt91LCCAHtH8BFa8clINAB5CXEpWLV53XoU5K3Kjqnf+w5QINBSAxrulmHII242UjZZ94CRGcgMbIcawX/OaMcVvPi/cNr2NcQDbhHBndDOoH7iGPQD8tPmnB5h6kPjqrqL2kCwJCG1TwKpZlunSevUfQXv8h6rzoAFKlSACM2Yf5a2hs+rEeV0ZfdHUJChw+3t8Fb8at/q4Zona5puqAsjbv4H4FHXoDIgoKXbZN3kH4sKOhe2zPwIGPkPLuxEEFiYAGdqeeg//F4LDYAG1mOp7aXbgkMs8KCQ0EAdeL8g4ZcVdxh2wRRYK8FJw/FrlhiNnmYo42Fi8xa9rD1SjGyWdVWhxvNGv5bU43W/CqlnAoMwyZ2tfHxR5gM6pFNgYKRo4PzX1QMKsf4CWWNuJg+LJmRLFBeSq7c3AkVnGRqRSC8L+kUC9KUGdhVlR6F0lHwIK2Nih/b5THR/90esUZvu1p9CUd3eohnk2DIVFSwx2acpbEJ7ev1H52Es6st76cui63w4C9RiNuzPX/r2iH32WYUu4EDAFWwuj3najXxk6quTjowAuAVK6iFnH9zqseksHyBHrCcN4fAgz74qv98OD2fEp8tAJaMwgT0IwPcj8cCudWE8r7O/az/JKwkhgctmuZyxCpAcmB63eX92Z0wak6JRWmj5XbbG49bgoxoX+pFdD84h8O3Xuc1EHm/1LEr13lpBboF0tu+jvj9sooR+7wTcbU+88RF6royAjlg+HKl8LeE0cg1E4R5QdqxK63L9bASzazc/I1qyE/VJBjCuSg4ShBSeD+/mOu3LlupQg/ebasiSJc7kLzfeF97/OOZu82byuL/9bZrYd4TABAr+g7iccyw5YuzRzWXkto1r+UNLcmKRCW+QiAffM/eSOqedxzt5qVd7oQxqELHF0BoMlpLAxNz38vhzu3+c60qKwyTP3tJibOaEJbepJoAQK0IRzfd+oz/x63NLzRc9WUq8R15vJEbiFltRMRqE1jT8vfYP9Q4N4rusl6bZFI6pLn95qh4AN1WpdJ/OeaszhQe8ORO0GEUtLzuf8W/ktHyRTr+jvTdattsT1NJcLzlqfLWpr0b+ijFiJDCrcnmqwXsViEQpfJvJbAcVDoA3AHN8hsvbKbtt275330kNtGzAVs4eFQQ7rcz8amR2ztd9GjmaTDajpTFL/vUbP/gmMuAypqglPcMtKnF9T5vJT512FTr05WIoWfP5WnILs9ZdazVFDasBGjzBvfcxGA1RiNF9ahN0k+k70O5cjaiW7koScsm2lu9qXkKX3f/YYBmiouYBIdGVWwIvg4w74v+Wg+/9wWt3X3/I6+BZ8f7lt6TGCHnIHv0ytqWwOWmSAoga1UCQQQXsbZHiTzs7M6YruutPXhbqrqIoAjcDi5fMI2XsCrwj35ENXEAriHI7uoDdQsD7dvo3skAyZegUTVnRjurZCBRYVSScknO6zSVxoHbhvF/tLgfaaysc0lKUFr6smD1PsHyTcNsbvwAOU8f+rLIB1JN+GgmggVW2EmrsHdGwePnXHTfFUuNtbdSn8DOeyxj7U1yfADI08lLvOV8YkfQ9M/LUdtN88o1szFeAVJhKCc0B3rG/urzWEw1dhqtUL9VGxgyE0xutJbrg1fDq0e3UUbGtfVQ/T2GG8SeEjzQ0FMhiinNdC0gCV3BYWj5o8ixUNsBbTPSheGAQdoPJKE3M8Wam8zFUwIO3UNibN8+Vj/zhChlf3ikcXwl6QBO444gc4PESh51yx5diOtzQLsaMGzSBu2SHoh0sb52Pez9HyxtV/DrEFSCUUcbzznUfNxp/+8C/TjgXF6XjgMCQV0v2qpJB0QRg7LVJfbeQp4Pwuk9n9P7qvkHkyN6lMnhfVmipx1i4AMs7QJukWuesUxuLpWZE2o3FKddCa9jZU/59huP3GArD+dBHA8eUPuxk0CA8tQASw6aTBgEP+XtO0GatC61cNj7YifGAndavbwchgH0jmj5IUrBlRwMt8lc2gtufTSPxAn9guyMj9qIEMTQzcgfDXZrN4uTfahYJpuNp5JxK69BnzUyLOJEfscqnwmMPNmrm8AA+4uVOBnyQKr2gunKJL+pVO3gGCzzsbsmmR1UkHII6WKQhkxBirnmHJQJ68wAymaRjriN/SG+sYjYL7JK4Ug64LxYC9hckT0s7+fFWB6cSyNEPj/gC/qOzVVtXRFUUh9rNQzct7YPUJgqgUxRD/J3jI5E+feWnWKvhPC0/q3K+a2UcTddhiR3hOHIGpl5HLrgL19vPhWH7Ty1gx5Pa+Y17hCIeeqRUw0aop4dKVuUWU0eBYY7IzIzz/c9biI63GgkFunnd5Q+W2bii1PyZMKTN8Kjk/DR3H5US3IeYM4gSQbYHXInJd80xXk7AAyO87qJU0VJAz3SE0JcNXEISGFsYhDVFzGs7GwQPmNW5De57nj3nWVIubp29ndmw5J+u2SISk2/wp4uspYjZ0Ns7zGlisoDQcHSTjB/t8IKHgpAlPOYjO+AwWV5tdpIlIn76caNpNwvg+/jQvmDSVnQY23R7aF3mJHHSd3fAqZ2ry6lhCZ5b5JTlmCvUOaKt4f0oqlPAZ2i6fh2cizntZAto7DPgTQ2jncJC1qG+AGKGcfWZFYdIzMtYv/MqSRx5rneT/n4ddScobB9MbosbnQ1AdqksqgbfcfY9BZFRwU/CZ0pG1Kzria8jnkxr9G0zhIS0gxytpRji7Zv+81fwmxQv33TGg0mv8G1uzn3xZuaaSAjVpYmmWx/B6o4tVz4ZTfRdW6bMQAIlY1apOJlQmCM64/fhRmA9j5eOzN7+jqHEH7bD0iHBzq3aS8ixsL3qrXq0BdFs0AY+TwYvXYguUSqCbpXr+VdpvstTOsd/A5Ech7jz/NcfAGGwWnxHTcwnn3lt2DXed8h58KnhgPvdLUUc6VmYm6GBFY2X4F7yHV2FxYvOi6kgEkc5x8D4VJChXu1tJDQKr5tKli14FV2sK/2jwYwsQ8f4IryTx5j3j5Kif3BuDS+keXnuzNsMyhRjoCAXRZifKi5LIasi/Bpf+uvn8nD4qGlSLIq12un9tWCaysZXBBPX9d1/HT/xIyh4xpoV5KoL+txrieV2A7/D9TbXf1HHe0sH4OgHNktW6JBg6NUkG/anwjWVb8W8wbDhLCaiwIZtYgSiHLxjSIs7fai47w7n5lXsWroQ/EKB1NJCvC3PuDB9T290REA0y/+isD2Q6MtU5IkAHMr8E7mVjSOeZs7KlkMX9B3llK+QavAsaqsvmt5oxhoQZQibKw2WpW6TPtnXQTyR+9J4tsAahajJYBAbNkQWPGhq4yQAqlQUla9611mYxWhHspk/MOY+A2UU1ic/+9N2bj7PlZljh1Fkw5z64Z1rxFG0AxmiInMB7KYyBX5ldUMMZ1DwFdGqP0lRRVVm5iuWuQvsA6j9GEdStGeRs7ZfW6Bl709wmPiXT0GQUuty9uvVdK+LUSNQ56CU5fav9CP9TAtP/G2x9CmG+sItejMELwuLYOjsMpRo6W/POXDVEI2UY8z57xi0KaKyqDE0jHNLjjuVUv6hpUoXI5aAYxz9dyChV7q+/p0H/wePPZ/Z28/0aPFLYW+mefnG+dg5GhCpd3zkmQwjx3l3oH8Jz+7H/PN8liUfTJ1BHOPQ7aq6q1aE03Y0+AwUHzWXRFop0EQeOChSKy6rOADYwHiqaFs0O2Zxb1x15XUmc9h675UQbwD2uQO11vW+9onxRmRN2CRkic/jM38Kfardw4WlJqlghvQVHo0XPUP8PL5HYejEal4AHCTOeOGO7M2taOcHIo3hU+WBm2MRfTjYBgL0kfUE/3a7jhP94tIS1J45SSwIaiH4+bT7bq5el9dEeCbqbwGHKSjxTgqCaN1eLWUMv8cYNRgas2zad1t6uPbgzjK6UcvgxCuOCj49a+5vYFfnXt2zJ+0mkCBIsc5KUvg2wFXOVV0Gn+ev9UtlkPxhjk2DiW4EMOUaiIcBBULOH99jfYNq2FLJilith6aoRc6KaPG/FmnpunPM7gwbAhpcYFaXEoXVzdvmGwIR1S6+/NDtJdN1CHASm7qRYTCOUS5XXkz7ElZk8CyWJsz3S8IP9Rwnr0dsLJ1PzyumdP8pFM3SDbVOw4HcxEE3z2vvYcpZbFERdXCTp7sSRAi4LzS9EUNi9vodf/4y6WAr7qs93791Wfhf5ZbMCOY1tMS/MVNuVjzWz4vvGjXckd3wai2uKUlE02F+LdJiH7ZbA4KJgEkuX/RaFiMNyHLdOXSzq4+KhsYUJ8Yqa9SLMQyVMbuHlBq6kVPmjyV8swKtEYvbkACKWklJxYG38XqdDvqCQqsJ7Ewwk2KToLSFMgkQZhn+WVcEdFoftxMSFXr7Eo6/Ev+Kr+NZbMASY9RdikUZu+o+gQDKiWqVsN6FOAc7AV4+gzXa5IKazfUzolDZput67B8xaoazAXIrUc8ZjgOcgVziyuk7DBoa8P8x/3YflQNyNbEl84z3vFv8sxKlDPqIsecZd3BPXwChCME8W/+7rxiusUZNCs0I7lUYbTemEKsvLybdx+8F0rowcIDnKZE8KmVLgXrH8XFc/Rf/09FrKrj7gvBBr/5SiXmHCXJEB83skNhAiklRjP67GAHBu9/XsJEGZRCa81MhyKWVDpYlxl7YOCAxaNaJ/f/m5v5D+SzNNhp5GaRaQpOYsbvMNEO6YOcMTWuzJKj+ARMSx6He/CP/nhwzL1/vYSPoy9UvQ/J6fD9eT3zZSqO9vOF/DJosde/lRUAQoXXNlT9z2S33XH5qUT46BVa4zOF3/2HRVzxJZus78Wu5R3m78BGyc7g0y596Qp8CmNWyqtdYpUR5PryRGvnxSghA7rE3Ea8d/hv8C35dsMkFf4nzO6pvybzo/wOt1bRQOjewWtYwS+yGAHRXQqZVlZDH6NUeK8MkRqcBhdMcRtmJlh5hI27O/PW4R8s1UAKz7q2Bh4y2wf51OU8eLVQ+LuPw99enaUa4mqOikrKKLh9mQXv0NLtjOsgGYG5y8jJCF1guXKQF6+9WkI8QTMquGGuey1PBCWMTTbvpzTEs+d3RXQ6QLJOHGKACQsvWqkDXq03fiCcpo82AlxqyzZmINK2tSvzpzR61UpT1TBl6rp3ULjrKQh+bj5pa2bjxgPhal8RFlAyyzJe0JeHJtmVI40gJzs5fN3qNYtwmQvFCK4moAr+qlSu/nJdg6geHXvIDWoJkEDzfrFJQspRcLM+LL4ayaCJ/pfmw0r8w6u52wS/twzwCFNP1MAA4UDhvA9dRJUuY3rQ8C4JK2tJ12UNfzi5EK3d6iubZikQkN0BYMHaEccE788IHkFv7plYJBt+ql7MyaV4Eu+YrbaFo9N5GrNjRPlTfFJacA0Oh322m/dpbSbmJGfNowkLvokoHr2EONhshgO0undWByag0xKAU1h9ANyqZFCWk1bG3HaUMLiAuyLRuRQRLjgrGSl2zUZjgqz4zzUQr8j2guFR3uJ4cI/uhGBcITuXOuK6gIBpYIkU3jfdDNy0FK1jwAGO3fw/3lQDAuyAvRN39jEHVohFRDFP4XoM5jAXuVfzjQKnb74V/gI/onTA7ElRs6rXJsxUNzAirCf8CypmF8kC7FDweK1U6+22fS+8fcUcOEeWm8e6yoqFieVGVDckbQugUW3AizaJbk73W8FMvDsTA5r0dNEkHK1jjaHlfnuZHZlqKPz95ysG7cR3mZDfo1fg2rYyq+d2enLduXaPGNY0wm0pELqj1DGVjrTFokqw9ACDSjZti8b72TUhkCYvZRY7pFKMmnivjOKabgsSUHdDy8oU3R/FtxcocSqxBSD5HQVkUFRQfCj8a3yjtuCMVzBd2GiZ1cS0TTzm3C8wXkDM4OtxMDBd6jrPhpsAfITSIozoqhD5V7rmZFtkP7eZuH8rqTFt4IZHgtCGUFJ0U86RbYk9dwg5gQWmESdDGYgUGihbHnNbp1S8wITrYQWqRDazPtOteWvHCW17y8xC1llEfwe1Oen9CLa8NZegxaF3eXjwhuSsUen/0L2aow7T5hEt+ZVan7LilCv3fgIWpJT8vIkz6P59FLrDLAbrvju7iVIhXsSjFnbyBkqHIrB8KcejW8jJYbjJHk79YBw8s4MBOrKe/0JPpOjjodA/v/vgifvl0Dl2C72bfb0WkWyo7V/VF+v6Mb3LX6MJsee6mTLCI/PQTSjmHt0F7Koh52fp4OS8JL9heE3KpB/LbSbCl7oyAFTfkqtED+qBUPwH5T1ovqAa7O4dmroHEz5XqiNHrU+DCI9+iWL44NLqqkRRGEcMTkyUAN745HiRIkutfZSA5TdMi8w246DDtbLm0+4/f4lOzpVQILJv1aa+rXrCV8/rXDgstLKB+21W+xCpR3fzT5bQcdvukv9T0NIgauk1srg8NmAN+4jz8Jg8i1M7fPmTqndvbS5aFCK6huzXRPRfUgiacFksavhAYPaKfq3sVZ8uXwVgAGnOuTj9vR2vE6/dviDovV2bWu0v+WKtBc+lMbQm8DZ1bzy1n1iGg1ylDrjwMhvMkCFeVCu7KcziTlY7RwVDNfNhAFKmiJLin3g/io6eRc5VEpI+nFbsFuTkPkEpQHAVganvhYK4cg5Now9s93WIqFg6iIesvNanaE2Lm6W1xmJdJpfYg6NTyACvkK3g9OXAPnSBzg3cW1R1Gzy+0kVwwT2QywVEAEE5NCpyuKqn0gLlLqFd3rsnuBA2o7N8NZwg1XRzRArQdZ6qjbT7/Yn/wjr/+GmPlmExXj0rso/EGb/V48+qM+koIjMecBku2kP7pY5S9Cp7xcFHPw2MFcToT46CmjPgaxutQk8U3h/Ppb/rNupU2Km0EZVaPpui+AQo5r7q0NY4TW2ExFsgUeKiiziFkQ7p/LucErof4x+2KezWvLHToAn1vvMfF5EU3rF/+GjBO4eutUshDhdrT8aOO4vTdHZ2N8LGoslZ44C0xBZa8L0nHxuGwcrcffmMUJXxSwfZVOSkxqJuMfeGiB/y/ZsyvSrQnfW+a9wBtfJKB0I6lAsr99BdyA8mY1zg9mVH8TDuD/taLLifPtoTckU8Y4+Ye6jkkCeSUsIpcIgwl57LJ8+ThotseGwlLqsTelXwXEob5PmvnbOWPf+29jAtD0FFQIL9xpHxhtFU+RHYGPdzgqpUqTMgyLqKH1CIYnpNFvOrsMrLSTN9Y28vweRzdtLs2A6XWXq3xhS1zWgphFtbFLAwByLYi6uGSlblMRVM1NsfYQD8LE5uAnqUriLa1vcXgO/64tpfaPURoBaK1MFUXFeor1Kv0y5Xv4hmJJFGZVHg0KdYW9+7rKkvnJwsSujP4D+LcNibpGzf77zx10DX+VVj+oRg3ibutVkboQRMr2Utq8RATcFCeZqqnj9K1KdpH+dthykHTobWOKCJuU6EjtVZwpFeKgN04Fl1tmlJGG5VYQSpRu1BwEf5ClAsnZJFS0/3xJv44g3bR71mD4YEI4Hg1MMHCeIZ3p9fG58jZdkHPlmGjplt2NkxUSVs/8pcncjssmqV4XV7z5iOluC6LAOCoR6W7BBungeHtmCizZ7aBZteXK70Usyb293arVYKPeiETz1Ol6k+JS/BiUF5giYLRfmKE/DPbJnOOb1wtEJ5E9IGL6tstkc0DN2Fr0Aw2CKRh7ydCoi0tQNPX/cBOx64LgFCLnWdRrnCMogzJOGsj1Bi6lgOhuzI0Zu8shJ4hJTGECj6HcTkmi+j6ryjhDyvkUhjV8ntBAygLtjqIWTNEbLbp5bMB8W1YHFw0ThQblrHiO+dClHUXPAdtGHwkHS0lgNpne56ALvPpXz27MTWaHbu/P0h4gHGBHuyDcF0k4ADGG2JD76Lg+N//xOmjU/m1O+k4kzJde798jkQUGVvwGylVFJo9H2KQfS0MWTd3ukiu5v+Ze2TGcbeuywJ3SbJAssYMJZUCdPPPsha1Iwjg32mdv3tp471f8dGkMKeiXUdEOi4CLYW19cNxRdwpe38bxGe1M7JVqy+z1fbXO0lpuHi77E26N171v7VC4TGmt7BB7AN12yN2jCqdmvVBZqC4u+nwDDoVZ0zjqgKC9uFpYZhu7vRFMoJIYcJYMZVumBR+aCC2fveZZiduGIU4e0TWYx5jEiCN9oPulDyuwm371l0w0iYskQZlkTkkd+EQ4A1uu/Pmm1BSUNtEl8T9ysOd+IUXN2iSwZoPto32Avyn64spLbfr682kjFK7/uyhGpmidvNkMxUvwy2jDTLbbbZCSU/1zAs8TserX2Smd+rg6PEvrjRGNaZk/nNOzcJY4gEaNvRbkg//bkLup4w7V0q7r0MwNQzGSxkq21GkPTOpSri1u39+RH0/U7aqw/zKJzxp34ZEAfg+43Gb6mq7Aihkm+9naLe+w/D4XWFlGAjBxzWInn35d+ZZIC06OEHedYCLljpIifGpDMh3GM13EvquCkoMCwKDhoZpFxeHDwPM2ZZAlPxAKyw1HeRTcAkK7f8a2zlTPG1pdkQt4W6DIIcVywu3QcazYbZnrJUzgQxwUm2S1vupavADMxkJzmkRIqxVF+z1dfFVeGwu+e+pi7krwna9BRK62UG2BhTW02tD9V0dWlfqmWBjCmiR+29NGlEobOMZGzPZhJYjBngArVPcGxE15dmixOJZJpBwbYCohYzJpz81Twblf631c5miriCxZiPlDuoX81WpRhddx3b8SKViR2tFv7mKguhEo5DyBC56PKoCvkrTAvaxw7HyZCD/1VLgYeqhIM6kxY9npWjlempwcxOfbcPifQva81+9VQufy70RdDSsvF1jVXMbsU+VUcvy62/mvOVDi3HQyGE2xE/5j0wUoS1MANeouoeNDfMErFTu5S+15Z5ygvO7LwIPltn+ZBou4kiJPq4nY4A2yUqTvnmA5CR0nyojQ18OyPETk0bi8fAJQtHcjFSJf6BMWKO//zWLci4qX/+d3pDvxHn0Sihf/4a6L+h/nfpxoUIgbllVH21VNGEetqynoGn4nHm4jzfD3I0PDFaEbUTpxvsh3AAucuDh5n+cG2zDTCfWdXldyYhpJQ/EgiSVcmF5vi0F7Jsu9VZ2fdwPMH/LFDi7kitcamb4s2OhhaW3CKZBk75lbWSgIHW98Jcd4lPrSfe5IR6fNgjDHL79rBaxdoLOIo645za806iS/cV3ie628IH9jqVRjwo4vdGhEPMocvegjGnTKHVXEF/tn8J37JvqlunWpDxemBz1qumxKgGAv5H1JBJh5QipQg14QVh0463EVxJIBu1D5AQUGzid+CJFMnav6olXMl4XVkODpiEP5qZo2BoF+YrovAScuUVMC45yjxVMKwE4oVvy2QvEW/035A7q1aoqOWrj+WQ3D9CM7CZsb0Hm7H2xaZ2zaVFGUpL6cpmP4V07UdzqVuuC6I4/wvIVqwu0KwYFZIj6bgZexzyO/BpFefIDCQnyNF8VLspt/HUG5eslvWaeSILITT42vBvLF8D6qfdR56xUNuBnEqzoxtC0ynURGHpPcOLgeHkf1kdnr332g1L3lVdQ4M8b6SPCvaBNckE2KYqVq4lRe/fHyVULjbeBVW1XfZ04wO7Rtt0GcXkxAmVkjTmcVdZYcnXz/Fap557Nc8iEhyNI52Ga08VhcFpmmRoVNwjokxy4ZqVO6csTNcPnXCTmNhBL3U28fPAKU6+9+3ij4NrDOQH7uyYnFVlklR9SH0+Q+ssMy3BQRCCki0orkHDKOTVEsAcrWhTp6mOB6zeJg90n501B0P05MjnW+LWrHAUUvOz5FSnYRo8OdApsFx5tte09lhGDYXr1rwyDxgFQwUy4YA6IPuTmfajR1dmTFmJvTy98pleZ8EcaMeoidykuHoBKYblgJFqeF2Ppv9CDpTQTZw7UP4w2+xaN5Hdap/X8eqKjFig2+Ux8IpxCtxDHtXCLnHDxZHaJL2gXRwwpvn1QJIYtfJAz1ipPJAPxM4Gs6eRD1AlhLExOQ4gxjei5psju6OPdQ+A98xM39tVoQUI4XlQflBj3QiMGljdmqHkOzwEHkT0elyJjxuPV0aMIoJnT0Jr3BUt7wzeMTQdGkxzunsGEUC+2O06EjM084X7rljbJlRuY5iLepJI3+itGC7lD0MpCSN/b7BmrewG8SU2P6FaQdFbAj5weXMTj6B9RDpmQ1cAf+G9S7zJ3UQCNGgzh695qIdTx4g2DVK1BFz9X04+Lq4c4imK3yftqV59M4KL2n+AO5Whljej5bl68yQnKTQfQ3ggjAB7aerb5YS/iBvW30SglnvKGbWIC0+AiNCoZIBLfEG3ovbN/xGPd43tMpekFulAUUcQJN4fmdVen/MJrO5qC5ZrG9DyuaNdpY166R3t6JR0BGugi8A+QrZTPI18E4RUDsui1YfbN2E7sxuk6FHJLyxnU70YSHaJGjla0BbsaZWK/+0CXsTzPRDng1lQkpuuzPxQgG/87scJ/sc0Zwufn/ySEt+r8PA1hYNlHqbowXY1mnYpUnUkjcpWIJ+Bzhkr2k9KSmGg3M81wAit9Iwc8h0pvl5fcRuMM/Xms7DCs58+vTLnJaPhDhYDCCji4ECCl5FakguhDnMAx7d8eiIZhSew5knqPaT9FLkPn5Kwzh3BandecZabrBw35628ZWEckDbzpNS8MYk41aRrMHM5aOvjv6Fxl9xYs+Iy/wNrhNZMbIL3Rfn2qLgqX5yTj0xGUgkFrO64IrnYixUJp+4y2gzE1eWBbhjxuNmcBL5keRngoJgbrlygRJkxrFtqtsqWOA9PToDxeqlAKavfz2ShdF6kqQfWpPfzVmP2XvDH5pOdmqRFFqnQs8eLet+LwJO9QylQatpV5uyHR6aqM3HmSmvDG+5gIkV/84Y5ZGwCpscy7Wg2APTj2ZTMicxpxg2wYjTYZYAk0lsZQ6IDDT8DfBxRehRIMY3CsdV6ol4YGsP/ZM4UMbtRAJveyaRYqnrFo7cS/qLdK20iOSjYRxD008ErQD5MWxg3Mj9hCZTZKWprgpLg3bkZWw/edliFh5bF4cZqgUxqgDqGjWJ7udDV8bp/4CyrQDOG4OYQhaV+U8KDZA9yptOXUy72pYEVv/lsG8STeRqqXTLRonKZoz3rn0vmauMODY7zrIF2UigV4Okxi19IYQueuFcXnIu1emWZ8bbFryJMpL2mgyIQLqtd4JxQc2SxJRpXKo3e5bavnyCmUH86ckzIV5WbMV06dpGLUZVKR2kDpUdyGMKO/ILSHFUID63Oc0IGKt5HL/UUej7mkPtd5wnJPO7VytWUkHM0btte8VWzy+nHWmdlp3g2sCoxrgoBn+ri3FeqbykbDgxdvbaGJzCVo66Bp8bcwdHlA/VN4Ar3z6VDSQWERx1CBfQj1agUYs+ZoxFZwxbnq4/fOHPjlpQNrfM6qOhb7fWmoxuoMc5cAeGryyzCZg91Rx2dlAbMfi4xHPrXmjODz27OANa8SR2pwoceOIqU20GchCuKgWwz1Ra6Ho8SJXpczoJ87s8P/GzPmOF7mhRbA8VnaVVZt1sE7+MdX91i/in0xI2QbMlXeToz8YCW0J7xShbtH7zTJ3xLysB6K3AyBcz32xyuToOhEO8y/kptwqVDOAHvEC9tfDkfkH1gqZ1+phvPgM0U/R12R/ba001zq7YG3SfLQM6PKsdScJM9pTFzb4Nx5UF+cqE41mQxIpHv1ozmmogzUPwDGyBdGZbNcIl5feEnOHPJkkAy5T1RNFAP2iGZEFQbJtzNOThwlY8U61u7lbIci4wkRyhrYm3EVq+EGHnEeCnQ7TxjZ2GqjGxc38mSKVUa979k+igTVT9fxmp1Z6IWLSdSFW8q0znheJbXJOXhWjSZmGgHQfV/NPnlpfO6R1yIi8N+3FhofrAtgA4LhKQeN0DYj8+y/ZRrcfzzb+maRtq7kxMsxjYdt+urgPgL2NqE5vCsTjC8+UZcCMItuX8WRnAVUfTkvYozJowBUg9QLhHpnQqFaX/a2yUrTs2oCosuJNBCDtRTPbq3H6QvckGVD8WjWLdbXkjfIbThUwBpIGv20WTPDvLcldo69F8xp7KhJ+e/6jQ8kr1fORpqqhu2AeDPiQZ/KHKN91BucAVxVLzREO/d8liOdnqQj6yOYqYdT4J2gpHQodc+RnEiQYEWteo8FXaPwi8y+vSt+vZ0qBTDGl27KZSjqJ+WfEGRRkFtb8Ok1WAPUVZJVBggGZPpaYD8lJ+dO0WaXxfBoLpEYTb4CTRkRbcPE6t3/hHwRA3LjDMbZ0WkTpOhPlZgnhD2MKDZY+ACv5W3vXmBgBFyJk67RK11zV08S/Nudhooa/6DPD2zTkao0XkZJCXgIo3wJDdlvjGJs6G5zMuh07EhoAVmmETwFw+4cyGMX8kLU5SXvURrpQ7/Sd5UKc+qw1uudr3HKZidfYxrCv65GYFnmSR3OmnHNPOfcXtz6DRRB1mmi4jbcxUiuvkVXqKU2vxuLN40kbHEQeBJhWihmAj3K2arkrbDHX9dXDyiqgMgk64fvo/Ny8k6/n17Nf6A+J/Hp90AnufmodpLt2UCAz6ov286a28tRXp7vtWPOLCGsZ57P240mwFOErFY8ZtGZb13O+h420jRzhBG7Ve3CVXebRUN4qnTU4QCNiPpUeGJuo5fUrGtQIjMvHRdEXGDdc/z1+gzghBtVJYmo0g8s8RB3PCOFFPnFpo9daHi0/fJDfwhP0J/Q10tp6l4JyJC1+m/Tk6MGkMDxziQ35b8Uj+Bk5LcGMI2keYNSQZ51t2k0JE+X7sZZ2p21CpV6I027GcZ4A2mm5S8hxAwsT3xT3YLnyrBV114howwMf7gBAVHOgEQjAtRR1COSzx5SY3pIIdzHkk0KsYov7PggSlsDoJfi7xX2T7fBW2FR4l2oxf14PIJ+g2S9tpZmuv5aD642Infg8IYBoqshRbl/9i0/2yrjdT+4MFzwt1Txtzhi+9hqjH2wRgATMlDdt8RVVD+RzwUDSWcEWNGeZEXBIx/aZOSQ8ocIxV2xBICIBL+S/WzBc2PH2YbJe3KuWu68oQE3Cmg2ulER/HMCrfvMDyaKoKt5Ode9N4lhyfN+XqRyiCm4kEXfdybHUyhKxu0T1uAaE2JoiHes78v6g1NxycMJ3LwLr7DaaF37N3HkzS7Xtnl1YMs5LhjPZtAw4qml1StPQ2kAYQRoVVypZZaQwW0IaHmhOeb1fIoP330WWv0sQHgfWYei5z/D9lzzvLyFQpDKcVyj01/wKB409ZGKMUjP8BCQ0+dDnjs5obk+iOyeT9p+fQJr16DF/O+t4AF21ckh7bHTt2ykFfr9dG2h7F3v9oH2sWO6zdKIAayv5UgbKf1c9fHxdvWN0McJZpg3mv30NMwLjIZOm4vdRyfdNpTZWv2l3JrSXnX7aPNO5SxCbRzwr6J8BWknAmQTmORDC+4F0WFw5ChYE2Qyrpt7MOfc3xJGKVQiz92Zt/7gk6Pt8Fa8biGZQWhXCrIOhCskKzmCwJl3zxE278MauIqUVw2GW6YmTAVckoEjmNTBf563OJiOdzBsjgbJEnoB2M1S9a01IhWsmve8M80hbsAMwjGhJsTtX8KNpJ+qjkynRcq/289acZ9wpbuqidZdPQSQzQAFEYp2QrSubFLSo7JKcbH5JtyVLatXZFOJp+m9bOf5Jnu1P5dzPikevLQ93rpIrKal5Ckis990q1OreYQD1S3lYeDxbsW3JphE9nyVICYQjnWf7Nm26pZG1lJBy5M1ppBM9gBv9xzM72WCQa/iizda0DwtnflPJaq2xcmZjopxEqyz8hil67nnl5PahhPq5uH6nHQyDue8DvpapkFd2puoNG0v7A6f2qnmDEa2FzGKI8esduPCUBW3x4p8L9R5+8WGmRCdZPlKFUUWrbR4Nah+x7I59F3zBJUPEJUrWYMJHN3aAuEVmAwhOE9V0WaIP0NiqU40F4E2BG7cxcHBhgGvguc1HQcPb4MmogGKdEzNUV07SKt7b87d5kAoQTHfWxtml05Vc+yhi072I2+jCwNxOwrqRwvzJ8jnZBtzkwY+bQQwbLbhQFqzUwnN77rqwFpcDWl2sLj7iuqZLP2rzr7Oq1oRSFe+2rnqTsUn9jUI5yo1UAdfZDoGaMGbONhZvbgpMSGsrGJqoGqxBiKDFgB76mNANl9nvIDZNMSSogYlMhHF/gNIJ8DXfFinsqP8p8oDcjcxCZhIvup0wczBDQ7sgY/uD/geveLY6JCqcwLGzqOo3NlOu+Mpp9og55kUfsV07t2woR5+dY9jqSQmLWFFI8KJvlM8eVv5bjpgEtVpQwBJPnrP0kqbGwlotpCY1RD1wfc+OjQNxNpcNBi5GS6p0VMnTplPSwzr85O+kMoHs7Nk/ax6NZPjwzD3/rWFeh4FQPDnZLVmycDZNkGdz3tp/vwBYkwIWvEenRP58Wb6lf1e+POZ3upFIPQc0RxpIiRj3+ip6qVHLEgoGEtfzjxzoW/Io3FVErdi1j8pv5XTjUloeAm/q9UZKRIWSICk/Djd2Pt0Px/F1+FzaVQ99Rz/odlHSWXZ7le8nYkY+J9NT5TtmcjhQFUCJ3ERuuTge/qmlx/rQijE001D47smYZxPtPbTXTBlKaeLsITrCcdjvW2jd8i5rZg1laLW8VJDtYl81kHM/ykSt9JvZc9I+dVo9SUIuLmU/xRc/CFAaX4qm7OOFRcBYsNCLPVaIZR1hM8Vp1xxGXtADnW6dCKJpR5vjvX/hB1o8nB7lu2ghRxM5RpK//vZrMzDTnziE32ZlWpAukiOi6bf8o/OjTxykJHiCtZUMJfeCOwW2P/EQ/5UhBNHH75RAPPMSEXfyGQKXfpKZRgE8tg7BBe8ueMlWeZ3h5+P4gqFaPHxHkUQJGVC6UwCeNf4ZzcJdNW1gzFVNbBi5yfY1I1pNjio7BJsyYkx3tleYhoui1IqjfaNtmZBKHNmegsewouNLcDVts07aWlDQvUPEOD9jOOepUrWOsPEz1Cj/skiYpis4uxs/z+8d0RUXxF2fEW8S+ia2or597nErQKGC+jjD7bNc5Wd0XY6N2iVSUHZZPbF0ltl2dyvqKlfBkBSdOItVBGG3KnXAnUy2y2KfUlkGGaAyrbgO3u9m/ZoeVePL2rrhey2HSl5Dg+12oIVCnWmD1J8m3We/7ON7tLXd/3N5dh/1zOArWpi6eMaMJX9+ho1S0Mg9VWab8t1crqQVVS6pd0LWdt7U1mbToeSkNRmckBa7p6Ia2j9nuKHPRqyoRGqtBzo+HUpFEw2UiBuJkanvNZlnPMm4KXkO9fBDgQ0qxdefaL/SdbxY4v5P2Ohl7/gpj1CwB6Av0gyP3kzPUfGkRHG1r5cMVz9a8S3cDui7Hka5JjUlDyeboleDhkw5l34zmH0/GLA2kYFEBSbtOGWlz+tiqPdGR7XlbQLaQXYeZXvYeeKfZEVqibEq6h5ygaJ68M2euD0fojVM6FLaVZgRGAF3xXG9wX4mi9bJMCVDDWRXjbm5yBCMzLkDAZsqM0ZRP9wYRfkajmcZ6acTGIF7c5nTPjBWziKwiV0d2JWnQu669h0n93ni4g22dK3c96EcuWov+GmuxoQG9CaqSeFPF/Uzoh+r/9DXvq1EUc2pcBkK7B1KElzjUpak7fzhRSZqGV4nuW9e4vsBdTYc5rw5JE19LQPw56d9YMmsHDO6wPcYz5ZwNxF1KY2YqpIgb2Pa7d8c1MX//jdJDDfNbi0kZBtLnVnKJLSJenoFT0xnn0E0bAqGgXtsHWtfdTczg8vwgcqHnSxCMrDRx5p7KiVpSGxlG8h3HqeHw0uy1MxsjgPt230rK9DzN2ELVVDf8oVhArksDGwkbqFiTYKZE3BjB7Rx1HZiAytpPwTTudIRLoZKhCPIP5L9GnDToygtLs3uc57ulMh7m0OZZ145y5o6IKu5yZOZ3t+f2ROzuhhVjp/WzK0Du8I2qugPx0JAG47y5wDLSorqz7TU5X8OByKdJRNRnkFAXf02Bjb20i5YAMCrAwTIMnQlFWv70C/P6/H/lttovtxUEvByp1sR+AHgtsjX+8waY1aCchwgxxWRct1AmSBkAe+InclgMi01gAhJ1pshbLTS5rs4u4FLpZVes3yMRrDpFGTXJ9VH7RIQWxzSxz8jutVcArPURnRoDo68g2Q/jlqsAr4WdUAHOmWLE2vZOoqreH5+DxZe5/hDVPsretYcCrpPpoAb1kxq5UFhLD5zxJQ9YSS/08EEQTyUdC20D70royU+6JPSM4puRNQQOurcq1Uj4hahuaIvVAl5VPr9o0do8kMdONHuUs19u2yJz+xTmQF5sQecbTknXAAlqSFwEil65guxmgaulmfyYdZTtHk31FbYLPwVvBfO9kJ8dsYUr/ldI4RKOy+UwUcu7NCWJPIYj/GPONdt22aqApbbwwIWQp5WxBcK30dTL/M9XnBUinpCmoEdBOpYgEqqlyVa2c+lfZWJJvpKpu73Xb1e0YUA51Jfd+enKn5zH+S4YUA1Y3JUB47rFiJh/BDilozYmAiFoTW8h6IVZxfwcIQuMH7kGXy7urMu8XtO1q1oWbmx+rlOOOzinxQcqNT1jXPzw9WaELBb3GviN2JzTi8Oh5V2CQLmIZW1V/9uxJgWSciwbZZnB+PPZSpsKk8zdOavR4Wmr6u96n3MwZtosR1e8C82QdBZSjS3+ReLehP/uaz/qfQY3EsOfrEEVmYNzkjGLaYApfMX0fTHuuEbeVG0s0FXPyqi5uEiaNsaLnQVapUStxVYZz3spzCOsonNaWyYSV/b/hGTIhwA806vDbrWGE/xchdUOA9BQ9qZKeI73xiG76Myz8/nt/q3FDrc/DDEBbbuRw9rBsCfwkmSD03WdyUPIy1EixXksPUOBlcmzDszkvTciJk4X9OJhQ9JUkEsI/89aSUdS0llnkjoNyIgzXHeFCLJBZtChvr79EOcHpqti4/oiZGVb3dhhWI7TecmKd/9gBXcax264TGq3Z2q5zumdRLPzt+qjUgF+BlIqftbTdXaRhdTyyZybwgya6moBd0wQ5bpy6VfnXbEwQCYjfN2DgLgJ3+gANDLqnvR2Rt3C3aw7zqAOCJYm4Anl4Vg/KJs21NwxneYOCDlZ1QyhF3EU95UlOWC5CDoTusY0OHC1i25zBlsVhvGW0S6z4kDfuMu32ER8lxlRsWG0jLLLMfT8LdjkgRsZFzY1SCzOT/Zgy6AThkr2cUKb5I0HkUnejsNia3szhHlZ+TvHX63xKLWZ7+kBqvroOcm5OGH3U9MKuPRdD9mUY66ueSTFwcYVdmd02F+MOSjv4frOzc3M1tCZC34phOUT9TkxwxXh/MyiJXCZsgRaI/mOpOvQcscRy/bqK3AUc+PheV7OLXau6KqjUMUDRcvmI4YSM218pIfAu7sO40gZU8o65pUckC6ZBwmZVM3UXxZp43AR2/8qb2MB/YY+JFYXz1RY+rQRD4Bp5y4cbljBL3rbW8tCQqRLLYvs+Med15DF8HH4I25cDAkPHK8KRE4v0hHWtHtrr/4ef/n6miY/n50Xy8D+0Wj/3KtJyn7eW/hb/1VmgVvblyK6GFhlWAgP0Dr125zmfErpCXCPLNsk6Z2qkQn81NIfoU2G7BY8E+Op/rY4fh997p2k6cHceZ2QfQRo/vBJ0ctLkgUlQExgmksYTD8PC6pKkZRVHwa508yYtMGlRG4t7MTuzF6fVuHRobKJG2Q5AYQKOmu7VQroNwv07NmZpVpJVSom48UNlUC6ODGB22VflkUcAk+tpshSMjOClGthAXJriszu8/8MhptJl4PaJkLTszhnYC9dQDDD5GamOgrwP8sXz0LL950AbFGUTisruo/SgGLlOfj+V65Bg3U3KSlsmnPyS0Kc3SN0Dji3Q0Y3xceiUHKh5FkD/B0SVAeRZ0m29PdjsaBBHOfBx7P3svwx5iqfi+4biWKqR+jOwX9xp4IewYM5aaMuz+q+5g4xJ572fSxaLNXeYxskw+lppYcfW5sAAUncAvaz5KeQMdgPps9lMZz8UJksMzP5LBqBy3JcMQhUbwzqG06orZqYPhqkigjKA2HTeH49odae4Ibb0RwfsxdrU1TtL1IAzRWR/wEytiB1VqHYh6BT+sJJ5FshN5DXUq3c7LbnJp7s/9+SqKmkXmOGL0DD8bFDI1Grf18OOaFH6mXP+1XfQ6nhuvk6sceLe3/Ww0LDhtmTKbMpeOWTxHRJJWkSjccGmx8PsoBSFBYUw5EOkEkMLrF96wdItlsSYxibSX5dLjVrMRipmOaszftwYNJv+os5fFlpMyZRy6/a3auVt+u7IIdA1GhyLTHBhLFeru/c2qbh7PCEXeclB+h4huuYnXsCMyX/11d+HNsy7pr3Kzy4XBYkIfy8lyWuKCJnMoP+BgAY/nYA6BxvyXZufTmZCJhmEO7OtPiOnbGwJ/IQK6Nvx3OosQm3vzmRuNS+t2/d+JFRb8/eNkow5S00m00SjMuF3ALyypZdLLnavJOjfdSDRRVOSgR51gX1YE4nYExXx9QY2J5hYVO5NOHY64/o1VDdSDkQCOWw8beHzTqkqXzK4j4g5QyA+1UbKlAcT9r6x5RiHi/B60xLWRiLJXAQ4olcWwlREzMYa69BxChYdNufu8Tx3mrS5Ti76BGIRl6kcS6hNEChmCcjoQ1fseFUG/LJQyndo3cAEic75pgcr8Ora2lnOb76lykmWJ7iNJP07VRHJ7xntUUOwB6SjFfSUUUM/b0sTCeFNZ8sz8BI0iLrVhUUeVo/FBiP3vPeBuNnkpcgehVsVXmXAcsIdx9DmuiJSGpCGoAxFNnDFxE0M4kPiEsIWxKFKW+g19m7R2J2lsKtIS8VJdJb/zoHc7Cb4cLxaEAImgiTKgjHagpKXhAO1x2fmS9bpsZfV1bbl9jzkyT00/7bNhiqybV5ZvO3SOoWEJfevDOIgZjFiZ4vAjF3T9LZupf9mdlTNjcxGqCdK3kKF5MgF4LOGSLv7fcYu6P+YnyJAGSFUWv3BguJFLB6DwFYv7VvfHNT88dmodHY7W8ZMPqSZLf1dXsqmhTDiOBA68/wiN7FYhcuj+6KnlCgWUk7hr9KtYpx97SK2VPf9IcPoz5gzMJQwSv7EbucB/VZ/A4TgKNg3W3MC9UfDRyzjgteZF0jEnKavOMVTZImgIjFcBkN38QBvxo1798ENG3522JWkmjCj0M0n5MpoRSum8Yybteop0vwrRG/R2eui75aBWnMuudYc3nrzsWTJA0yEey/dEezxgptKLgi4HhWDRGb0vqxmR4iCkO8xScG55u2tGWELzue3kOj9BCBwaoFh/QBFsg60+CH+7RlaJ7XS3j77A+5F4YtfG+/gB4MSV8m1K7J04GFWJva86o5kY0BeF11QXccX0+2eL2VxmtY4LKJai3cbWWDBzV3dh9HhPvZFbuKR1lyySfN5Laqlegfr+OrBFaD19QxK38nIuec3c0a3bENzSU09EW8b7ydyAUigeyeRQEO7MW61iSB9G52a6gJNckIVesJLw3krsz8KR48yGtW7pBXP1u/S/+UYgFIIQUAYB1jNT3SiCtBrXcVDFEbkrawy9kN+C8Rn9MKO16ym72Lbi0ZwoECF02aEXfVcII/Atb5xrxDVt6PKZiE/XI/CBnzKbYg6L8DVbuptM6afVUnBqnrhtChWB5sQvNOaG2sQubhGh/15nNfUlfIicT73FjdRNl8V76aEM0YmapTTxbqldtebsNMdUCiarX1noHhsFx+XJ1nqe/Zs9wgJNJDxg2vlK1PeZjGqV1GD8yiah2GlRBuCSug1VgwG0QYjhIAXLREideEFE+SwW5G+0W7rOOZBtuzegDnM73HG8cFiD2pEZoMDpQe/duq4aHHqcxH/vtWSHgDvRTV3wkpPSklC5ZN5DRPMQWhUWFJd/5xEUEbeHGBgtNFQKnzhDWx9jEVi15kmn6on1JSh5kNOiF4WSRYUaRe+Gt8qwUXK+5W/9VbnobkGL7gsdy5a67MSLgvLVaSOlFJ7UFigXTyOt8bYMQ0hu/f36PoZR+EwP5ak+KwRgPclyJG1yFEYpJzMKrtOLEMZ0qYKB4mX2WME4K9RYNecvvHZ+W+4glnLnUG/ZXltIEHdF0zWXUsk5Vih/KLTC4rl8VO5cU5c2aHxgt4l9l1UeqloVF58laUe1T76qvlq6DlZGxXbLWTTZLEYLy9xaBdhT3EDdhF17ceqo+5lNogspzIiBHKlWD0/ptyLjdc0384uZD4uQUw89jt5QFj/bzSv0PKNU/+bEqJlZW0LL78P491fQ/VaRrOK7+kiV4H+nzkAdmdOnbmV+NzyUpf/UND/zM734UqIGHS443kUjMRGGyH1uOTiizVGAf/VfJCFptaeUvVbP1MZxtGVjbUVDvmxoFl7/qpngIxxZXfHsfkuEv83jG75kpAX0nJZ+yN78dggX7deDdgQZ/oFY/B2kd8umPUuK55ww6kFsIpobpvdvwXfzJ1xtNaAQ94a7UsueiwN3ZcY7nMlFSc3J8pp9eumoi0On0cN3WbxG4y918gR5xfCOwgo64HU9gmHxhdXL5XvrMS+8UeXFywB9+t2U2aRv8tiJY4Po2W4fsuJdDbc9elO9dLZly5RsrzAhXgbUc/dRgrcF4rHz23ub0WrbnajI0Td5TasD71LcAwuAWeOsdT8HZG6vcdC9H/VzyulSkFveq3niLiBB+lTHapBNl9xZgPhKKLDydmdEmme/AsBhdxyfglKPifS99+BHdTqRGvvhN1Z7kM8f7WC9pQ2Nj/likRYzIFVjcENvcFjO1buRdSMCk29yM7kTca7ZDU3Oc6/ptwRmLk20k8qW0K+BcAlhrr/jw8IoiLILkwuyhVJKYhy92vP9EO9ukq2hDBKH+cMveWnpDDa5Xeiw0AjN4c4oUuhLM5jmyX0HRfv/VGs/8/ODPgE+hnuytzBpM3WJeL5RVrsgJdEbkOcIK6DWtyee8v26ANuPo2MyzO9nhIShS/YIj3lBCCjD+rFHtj/P5X8ExIHrv3Hxi9A4GtIZVxednV/VNIDVHa3g0ME5QCYCWLteK1U+n6DZehjNW1BCdi6LUhDo4oGQ+gOsgxmAz1e4vPL8wxFuqVc8arbBaHjD5EHcSg8odo5nELJmUoZulKvA/lfy819iLL1Lr6ertq1Tua5ZaCfwzZz/CuUv1V7Y7IdO3i4W0CWN4n3ve0AiGLf29XkpF6K/VeTQBts2a+VVFaaR4j00sjqgmCYUTeEiC/XCmHiqhWVPyPStnEjigzNaTm/vgPH7AkeymwojDLEzMVat1Onk20FIOH2x/LD6yTm5QxD0Q2H9/10TNzgk5BlPrFpkLvnuAVWWrgEEcVNlMUDn3QvARbCI4WL1avk91OOIQYFGaoL9DhDnBSLC3ABlnvfQ+BhQMHPEHTJPEFTheFFF1FucuaNdKcdoLGcHglTpFHBF1u+9c95lmYuIvtwfRArM8SyvmLCcEeacxMS7CGSQavbMXLInH3eib1qsmrrydKVqzxGFBlTOBui4FG2KuF7aWVst1JXW2BTtYyiN+E6H02RN3/+qKGXkRVJdHWR9MqyTlWw6ocp8YiQqfSeKTZqwL0opmXWlq5+iw+NO/FPhFe1zEPTW0au8Ioy4wOiA48Zj/3ThC4GdUJRtJsfsHYZNHWeGG4LK8nt0FjeRlgk33jpJpawBgWF/U7kvuARvl4WRk6uzOpRqlfxanVv5AQrI08jW/IVXUgqGHQD0vMEWJgExinPdXplehOT/JQ38TATheQhiuLrUx/oZFpjZXKo0lTiSySDENk1Tcy2Qmt9u+qHSspK8LFkWYPICKZjxJbyfDX2q4LkmeR86aDXbLZuuaDGq0dJb3ERK+2KbhIuh/6YGJZBlW8IA576lrl4ioIdvjWLveGRDIlhOm4RQpAVDG0eB2g/iZtDGb5yD1iivjYfuRvyqP6ySDWnCPj6MxKnx6vey4j3v2s16MzYMV+64NHMPMFzn3kvp5Y9/8lRnCFQrXPIPd4VptDZcdOW+l45HQUVIspeQYoSr6DISlX9a3k45mYgDybCvWSzxTsQEXCFeleJLl60cHKa8DTKoxsW1O78x9mwOLJAatYiTnWONDzvhTklcVxjW4FW2hJpYgKLluKYLMqKaoRJgV+gVF5UyKkBOGpyvwzud+33dGq05oIPJWsKMKLkdikhLN7B1pHg8ccF8nsQWMmWxXXbNZxtRHU47wNkPl9rd1zmynx/T3a9hV/Ns1u68UhndM/PEgsmv0cKGwTlMhMC+9BwKjcfPmqXquLbHAwYZxv7gO8q2wPYAmVqtfw3v4g+WoOB7i5u5THFwuU78yu3FAlJeiuLR51IzXsQyFBiQbUQkRsVPJYDZNVOGlNPX/qQQ44lfGRBfkBaG6n7wrIQ6s/NwJtITN7Immd1cbFMwyTz7hIEGU4fLhlxV63MMsV4w6a9AJCHNkmfF0s+ivpsB/K8i4XWxH9XIfxXLZtvstXEVWTIPZwZRVSwS4X30LoUCsWZBAqyvNeukCXZee+QunXs6mQUp9ogJzXzZIuRjVY8F8bt1zvdyIk8q5X6IuW+rVZr9CUCCEe5JI/xCaFJnrSaq6cdosWu1WXyUf9t3T9Y8MOREf2guQ3oZI2uIiCHnaFrktFc8JbfM4FX2c/l+tzS8iqszmcRt3yblO+AKpGv2MjCGJN+0FDSr1Ldv6x32usZDbr48IxaJEih+7YNmozZTVNOYmJdhDHYYIIzC4YN5rEULSAlAYgwivxDG3ZQYjCpMHtJePBszH5OoGZ3k2A1ncq4E5X7yWmXTsaMo06YwV/8FLejpfINvNaJ3D+0VrD3NA3FRjZojGDlcexQhs77nhQKQ3/dzFQs+SonI/cRHeYmSaTvgO9Nl+Hjjc3BmyzZt3ovDCEHCPA7vjKKJlB/MkJXd4Dksnf6o0v168qrYujMJ+d5Lz6Oxgl3up3SX8GNxUyfLTpPMuaCmtc8aaPsbt3PUkLghTv+GtCWSwdQwfNdDwYfkFZBKKlz4x54YXBhbSDxl5Zy9dhSMVevNSGNStj3jgpOBT7AeacZkWSFfxkoBXw5IU5kykK0n8XXJEaB/7d7TJFc4JXFByb7JfK3SN56KVAD8hGmu8CYHdIqxrVZ6QvglzKVWM3ncnrm0pA3buUhNnE0zNB+nu3y28HtI4cLbmXyGQXmy2WRJTlfCmlzhVpghOkQfuyUOGa1kqz/F1FQkeVuJEKdv1tMJGfYNSRX7rze8G7jXZyIOTaiM2F2MegNYmqpzOGySDlMQ13EHNADeQ9TEzCnkI4J5v6pi6guE4Pt4UjkDB6oOWtyhLd7N3xSn93KwaYdR9luXigWi5yniYzwHZ/OWli/V+YbF/05gVmeG/dZe3PUJa/qA9H0P5qzW0OPMo2ezEYbnYuuxctAqBGy9fqZQ3XBMK5wrm4WYj2IDYfZm8d5sMmE0tlB/xQcUa8RrOeBOF/xl7h6YB99G2qph/ZeW3H0r2ZwwFVXq+cNL91KEi9MtC6dKVTA1VZctSuThdZ64lVZjZ84jq1wrKnIhxzYGTS5UlgZSDe+S/dY/KW25x2iyNPwMgLzUiQilPfiOvBLtahnSv7ym4SHzRwsvjYug6PysX9oIUP3EDjhqwyViEppLcwAjyGap5cxJVyg8wSCdQHsC1t6EpSqqnbo/fx/j0EGSIXaOFgepwDWUYz2LSjHx+MWdcM3MAQ9kJm1YX97acRUvm6LpZcl0e8jB+6rwCzWhzjhBpretGsgUC+dmklEAqLEGr2W67SA04FufFbpRpbQnxrAX8SyoA06M4Y951UZDCPEW2POo3zPezk8/rX9v2fazBF9verJ9xEQZMf4tdxMBXSTDxLQMTSQ+VtlrKPg1SLfXTVPXNdr/wTAaIeqjqdsDIg31E0V0BEiZ3lWS4RQV1ZXWfWCVLyXv93rtKimn/G2/a5tyWMGXp6YlSlZtRJzGcyolzkfpg60fftiszxCOHfvFzuRdZiTQBCL8tbrowE2UWn6CRE9I/UjYUVba1XRrmD42xSdsKzQJPnD7o87gQkDYhD7B5JV8D9U4YI2+dEHEinerU4XeeCk3m+sytbPs6/egbNh58EnlGiuG3XbFSyiT8fnXd0sbKLT0G46A9GiFiL1Yu8cvgz2glOSke/CSJQ90o6xSt5Jnlq76nhPOo/9cBTj79r4jtsQDG+faEGyXLMdb5krcbtrRJT1o7Sr5RFDzky7eQHdhYuqdvuNVAvGEsnKWGAu1sianbslwNzWbTS6MEpR3eRnE+hW3CL4mybOTw0UwlV3ngoqh1GNQB2t+eaz5e+1rLWs9A2iOOX9S+viaaTJuVChDVcmDdoq4rIdseHnGC9Y5FrnsEpLwYTz0MJpD4HELRlYlA8/LGl6mOtVdtERGrsBdbNxDc0q9whXpusvAYrCVqGRJdJRBeML6mH8zwPN1I6ggtqMo+Pw0XGMPRd/U5Z/Yq9Gr9nHfxcxsO9CFbpnhhP9OqJwPoq5is+tJ8ih+wXhYwb9r1XqipBiXihwMeGHeYTZNIfDSGFinBfJTYeRC4OdxzRpBLYeVuO+2ZA1DVkuutZ+SnIytn5AtgGcOIKmAnr/uffynaWnom/c7jZlkMpE34uJvqmd7oPd/82qL1WcnutP6C9MjwDuG5zpKWZaCSIYpXdhAo6XzJ+oHL2rJtdFQ6dmZosKwjo20AOdlxIREfu1jcoWWEdybPIIfJc4ajUeDF/+xoes8AfZCO7VSvgAayeVEwKMnXrbp3xqKUqA4PoTfFfssO9ayRnwU+XVvVHzjnzrDjLQZ2xSHAx48O5kxIgz92TgDV8le9+f+ymCBiUpIMkFSRVUkrNdz7pN87Nf7SkfL5GdeFL6sn8b1NcArpkmpbKfsqVX3/SBAhfIKJMnCH2w80AoVpBxDbW657Dbka/g35MF1rzlAbZMYMGI0yc6alZw4pCEWHLhCME1Nhn4l7FwWeci5uTo+AdQVQwh5+4482p9DoK4EE+qLOp+v4BeZmhS+8+HI2D4PdmPGjmsZutENTN9YxVTjd/C7OJGLcAtbj9ed/5iPTJ8pcbCllZDb/+IFFidQYEOjQsZCKWmTK39hF1SlCdm3ad8/VKEBd9OiP/y3FaFWqTX6frVPvy4BRE83Q3eCrIlunYUwZvMRwaravGVCsyJApsAa/C+e/pD+TM/lBhspzYEtvhTPfHuJuHHSr1G5irkmzU/oR5b2KTrnHfae+VP5BFk7dm2ZXpmMdu/EdONbiNPq3petLH8d/dgvr+9k8mHhkI1oYJHaStEOR//gtBvYyoRt3d5ojm+oAtsJFRlJt7zTucTc3tD28SWN7/BiSotBX6/h2yP0ukFrQFTOmya7Q5AvJZLLKOQRfWuWxkGvpvHVG7uMcHj9bM/ai0A5TmFZ4FeYbXFYjXcQVSdN2wxgz8FRX5nc6RkeNeR05Un5vbqGI1myRi5HK05oMr7jVzf6oolJ1sZKL8eax1/E+xfcZM72TRvIvnMKOUebuN0V6vUbu3ISsCBQUTnijNmHE5TM/CmAloPE2C4W1O5ZJPOQqAaId1oT9gKNJpeHhki6VmYv3vizpewBvyZ6UMWVd8UabZpUDg1AdbKg00hdC2p7thMsUo3FUgqAvf2veaiUIP7TL5uaTgG/bmXpqWad87hNL0pqmUK+RSNobRF7jwSc7GtkJGDppWOSbN4n/NQyBrVZwlObkGAWf8XJ5cX7BCUQZB8iGzXL0vjhkHtZ9iA5TjXcdZaqyE5M58deCHIgAjTNtMs6ufiUPtp2kTnDP6ibKG013mvV3o36Tnfm5NgGqyao2GXql5atqXUTCGM2S8BoxcF/2nGZX9z9pucAK34Hcw+qdY2dL3bwUSa1E0EQNvA47QfnFXy/R/ASe2+J/PPYfqUZrzd5uiizTBKwiigKbvwTBq0KAIyqPEGGBpCZCb0bRb5zZjj/l/fyAAGLtnNBOQALqB2tePtDOKc175HaMprLCNsfcqmbFNHJx9Tsl4Haz0Sqd3mDTmlF6HW/aowd8L5kQV+ht5KRDAtZAATqa3ct8+9Foa5hQLuOP8AibkFewBO/dPxocMRICJiGcd92FnXnG8pZJ4FW9Q5kZqvjbwu79Q0BvSDRnOLHsmib1g1eB16Dy9mUMQ30+MB4SJyGk0LQu4pY6I+JpNM32EFKnk/gUojp56iIU0tQsGUV0W4DtNucHIce2kvmvgtFop2H7z7OGYEco3fI2HPwitePwdh381LkZdvMl60s46I4cbC+s+uZnoM9WA2+LOVKvXbni0nn97zw3gu50hq6GfB1PKWCL9n5nrOn9S4XSa7Ih0lU6maTtu5hTwYD9Dxnl9DPb1Wye7YIiAuUofu7XhYguV7w/i8GvSDqKtasjBnLVHv+J4EAilsA/3ERG6RWbTH0/fGss/KutFp6tBl8XcIiMQsVwlN9UrSdEGDaO572lWQcDiVkFC3uY4mORji+UK7H4gJdE99MH5V+oLdv37uMK0BfKVHCPD8vVIa4kL2Hht72Vu6mNz8DvJgAAAAA=)
(a) Cohen’s d-effect sizes 95% CI for differences in subcortical brain volumes between recurrent major depressive disorder (MDD) patients and healthy control subjects (striped pattern) and between first episode MDD patients and healthy controls (no pattern). (b) Cohen’s d-effect sizes 95% CI for differences in subcortical brain volumes between early onset (⩽21) MDD patients and healthy control subjects (no pattern) and between later onset (>21) MDD patients and healthy controls (striped pattern). Effect sizes were corrected for age, sex and intracranial volume (ICV). P<0.05 corrected, P<0.05. CI, confidence interval.
Impacts of acute stress
![[[@Musazzi2017-lr]](https://doi.org/10.1016/j.tins.2017.07.002)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQEB9AH0AAD/2wBDAAMCAgICAgMCAgIDAwMDBAYEBAQEBAgGBgUGCQgKCgkICQkKDA8MCgsOCwkJDRENDg8QEBEQCgwSExIQEw8QEBD/2wBDAQMDAwQDBAgEBAgQCwkLEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBD/wAARCA7MBiYDASIAAhEBAxEB/8QAHgABAAEDBQEAAAAAAAAAAAAAAAMBAgQFBgcICQr/xAB6EAABAwMCBAMEAwgKCgkOAhsBAAIDBAURBiEHCBIxE0FRCRQiYTJxgRUjOFeRobG0FhgZQnaUtcHT1DM0UlRzdJOVltEXJDU3VnekpbIlKEdiZnJ1goWSs9Lh8CY2Q0ZTVWNkhMTxOURloqPCJ0VYg8NnhqbFKePk/8QAFwEBAQEBAAAAAAAAAAAAAAAAAAECA//EACURAQEBAQEBAAIDAQACAwEAAAABESExQQJhElFxIjJCA4Ghsf/aAAwDAQACEQMRAD8A9U0REBYFberNbpm09wu1HSyyDLGTTtY5wzjIBIJ39FkVdVT0NLNW1k7IKenjdLLLI4BrGNGXOJOwAAJJ+S+ffVF5vXPrzysbDLUe5651LFQUQweqjssR6Q4NPYspYnSOAwC7rO2Sg+g9FiW230dot9LardAIaSihZTwRAkiONjQ1rQTk4AAG5PZcd8yvGmh5euBuruL1ZTx1T7BQl9HTSuLWVNZI5sVPE4jfpdK9gONw3J8kHJ6LxZZ7armgEzXScPuFpiDgS1ttuIcW53APvpAOPPB+peufBziXaOMfCvSvFKxgMo9T2qnuIiDuowPewGSEnzMb+th+bSg3msCkvVnuFS+joLtR1M8YLnxQzte9oBAJIByMEgb+ayp4IqqCSnnYHxTNMb2ns5pGCPyFfP3U1N85AeeyqNrdU+5aI1GW+Fkl9bY6gBwjOThzn0kwwTkB+DuWhB9BKLFttxobxbqW72uqjqqOthZUU88ZyyWJ7Q5jmnzBBBB9CspAREQaffL1a9NWW4aivdWKW3WqllraydzS4RQRML3uIaCThrScAE7bArgF3tEeTRjix3G+gDmkgj7mV+xB3/8AkC5J5jo/F5euKEXUR16Mvbc+YzQzBfNBOzw5pI856HlufXBQfQt+6J8mP48aD/Ndf/QJ+6J8mP48aD/Ndf8A0C+eVEH0NfuifJj+PGg/zXX/ANAn7onyY/jxoP8ANdf/AEC+eVEH0NfuifJj+PGg/wA11/8AQJ+6J8mP48aD/Ndf/QL55UQfQ1+6J8mP48aD/Ndf/QJ+6J8mP48aD/Ndf/QL55UQfQ1+6J8mP48aD/Ndf/QJ+6J8mP48aD/Ndf8A0C+eVEH0NfuifJj+PGg/zXX/ANAn7onyY/jxoP8ANdf/AEC+eVEH0NfuifJj+PGg/wA11/8AQJ+6J8mP48aD/Ndf/QL55UQfQ1+6J8mP48aD/Ndf/QJ+6J8mP48aD/Ndf/QL55UQfQ3+6I8mf48KD/Ndf/QKn7olyZ/jwoP82V/9Avnn6DjOQqFhCD6Gf3RLky/HhQf5sr/6BP3RLky/HhQf5sr/AOgXzzdJTpKD6Gf3RPkx/HjQf5rr/wCgT90T5Mfx40H+a6/+gXzynZBug+hv90R5M/x4UH+bK/8AoFT90R5M84/2cKDJ/wDvXX/0C+ejwjjOQqCMlB9Do9oTycu3bxtoCP8AwbXf0Csd7Q7k2YcP43UAP/gyv/oF89IpiRnrCvFDIRkOBQfQl+6I8mf48KD/ADXX/wBAr2e0L5OHjLeN1AR/4Mrv6BfPI+IsOCQUbH1Z+IDCD6GT7Q/k1acO430A/wDJlf8A0CN9odybO+jxvoD/AOTK7+gXzy9BzjZVMZHchB9DR9obybDvxvoB/wCTK7+gVv7ojyZ/jwoP811/9Avnn8P/ALYKogcfMIPoX/dEeTP8eFB/muv/AKBP3RHkz/HfQf5sr/6BfPOYiO5Ctxsg+hr90R5M/wAeFB/muv8A6BVHtD+TU9uN1Af/ACZX/wBAvnjAyqlpHmEH0OfuhvJt+O+g/wA2V39AqH2h/JqO/G+g/wA2V/8AQL55A3PmFd4Lj2IKD6GP3RHkz/HhQf5rr/6BXN9obybO3HG+gI/8GV39Avnn93d/dBBA7zcEH0MfuhfJx+O6g/zZXf0CofaIcmjTg8b6Af8Akyv/AKBfPOYSPMIIHEZyEH0Ln2iXJmO/HCg/zZX/ANAqD2iXJke3HCg/zZX/ANAvnnMZb3IVWxF3YhB9DH7ojyZj/s4UH+bK/wDoFT90S5M/x4UH+bK/+gXz0GIjzCt6Sg+hr90R5M/x4UH+bK/+gT90R5M/x30H+bK/+gXzzxxOkIAOM+qvdSyN7kIPoWHtDeTY7jjfQf5srv6BVPtDOTYDJ430GP8AwZXf0C+eUxkdyFTw8/vgg+hke0Q5NHHA430BP/gyv/oFU+0P5NR3430H+bK/+gXzyBpPmpGU5fjDgM+qD6FT7RLkyHfjhQf5sr/6BP3RPkx/HjQf5rr/AOgXz2SUckbeokEKJsZccZAQfQx+6Jcmf48KD/Nlf/QJ+6Jcmf48KD/Ndf8A0C+ex9HJG3rJBCha0uOAg+hke0S5MnfR44UB/wDJlf8A0CvPtDeTYd+N9B/myv8A6BfPOaZ4APUN1b4Z/ugg+hj90R5M/wAeFB/muv8A6BP3RHkz/HhQf5rr/wCgXzylhHmmMnA80H0NfuiXJn+PCg/zZX/0Cp+6JcmX48KD/Nlf/QL563Uz2M6yQR3UQGSg+hke0T5Mj244UH+a6/8AoE/dEuTP8eFB/myv/oF89PgOAzkK3odnp2QfQwPaJcmZ7ccKD/Ndf/QK790P5NSf9++g/wA2V/8AQL56XUr2N6i4YUQac9x+VB9DjvaG8mzRk8bqAD/wZXf0CsHtEuTI9uOFB/myv/oF89TIC/s8KQ0Mg/fBB9CX7ojyZ/jwoP8ANlf/AECfuiPJn+PCg/zZX/0C+eh0DmnBcFb0Hq6RhB9DP7ojyZn/ALN9B/myv/oE/dEOTQf9m+g/zZX/ANAvns9xk6evqGFGYSPMIPoY/dD+TX8d9B/myv8A6BWn2iXJkO/HCg/zZX/0C+egQknAIVfAdjOQg+hb90T5Mfx40H+a6/8AoFX90S5M/wAeFB/myv8A6BfPQIXE4yAr/dH9+oIPoT/dE+TL8eFB/muv/oFUe0R5M3duN9Af/Jlf/QL56TTvHchRlhacFB9Dg9oZybntxuoP82V39ArXe0R5M2nDuN9AP/Jlf/QL55gzIzkKnTvgFB9DP7onyY/jxoP811/9An7onyZH/s40H+bK/wDoF89rKKV46sgKKSJ0Rw7CD6GR7Q/k1P0eN9Af/Jlf/QKv7obybD/s30H+bK7+gXzx4VWsLuxCD6GD7RLkzHfjhQf5sr/6BP3RPkx/HjQf5rr/AOgXzzdBzhXiBxHUCEH0LfuifJj+PGg/zXX/ANAtU0nzz8qWudTWzR+lOL1FX3i8VUVFRUrLfWMM08jg1jAXQhoyXAZJA33wvnSc0tOCuZOTWLxebDhCOst6daWd314q4zj7UH0hoiICIiDQNb620tw50vX611rdmWyyWxrX1dW+N7xEHPaxpIYC45c5o2B7rhD90T5Mj/2caD/Nlf8A0Cp7RWH3jkw4mRdZZ1UdFuO4xX05/mXzzgEnAQfQz+6J8mP48aD/ADXX/wBAn7onyY/jxoP811/9AvnlwVcIy7thB9DLfaI8mbjhvHCgJ/8ABlf/AECuPtDOTcbnjdQAf+DK7+gXzyGMjuVQjHmEH0M/uifJkP8As4UH+bK/+gT90T5Mfx40H+a6/wDoF88qIPoa/dE+TH8eNB/muv8A6BP3RPkx/HjQf5rr/wCgXzyog+hr90T5Mfx40H+a6/8AoE/dE+TH8eNB/muv/oF88qIPoa/dE+TH8eNB/muv/oE/dE+TH8eNB/muv/oF88qIPoa/dE+TH8eNB/muv/oE/dE+TH8eNB/muv8A6BfPKiD6Gv3RPkx/HjQf5rr/AOgT90T5Mfx40H+a6/8AoF88qIPoa/dE+TH8eNB/muv/AKBP3RPkx/HjQf5rr/6BfPKiD6Gv3RPkx/HjQf5rr/6BP3RPkx/HjQf5rr/6BfPKg3QfQ3+6I8mf48KD/Nlf/QKo9odyantxuoD/AOTK/wDoF883hHGchUEZ9QEH0OfuhXJz+O2g/wA2139ArXe0O5Nm9+N9AP8AyZXf0C+ejwD/AHYUbm9LunOUH0Nn2hvJsP8As30H+bK7+gVP3RDk0/HfQf5sr/6BfPW2le4dQI/OrHwuYdyCg+hf90R5M/x4UH+bK/8AoE/dEeTMnH+zhQf5sr/6BfPP4ZxnIVzKd7xkEBB9DH7oZyb/AI7qD/Nld/QIfaGcm43PG6g/zZX/ANAvno92dnHW1VFI47B4yg+hM+0T5MgcHjhQf5rr/wCgQe0T5Mj/ANnCg/zZX/0C+e9tvlduHDChkhdEcO9cIPoW/dEuTL8eFB/myv8A6BP3RPkyJwOOFBk//euv/oF883SVeIHu7YQfQ0PaE8nJGRxtoCP/AAbXf0Csd7Q/k1acO430AP8A4Mr/AOgXz0CE9i8BWuZg46gd8IPobHtC+Tg7jjdQY/8ABld/QKh9odyajvxuoB/5Mr/6BfPOISRnqCoY8eYQfQx+6JcmY/7OFB/myv8A6BU/dE+TL8eFB/muv/oF88qlZTPe3qDgg+hb90Q5ND/2b6D/ADZX/wBAqH2iXJkO/HCg/wA2V/8AQL56HROYcEhXMpnPGQ4IPoVHtEuTM9uOFB/myv8A6BD7RLkyHfjhQf5sr/6BfPS6Bzc5I2VnSUH0M/uifJj+PGg/zXX/ANAr2e0M5N5N2cbqA/8Akyu/oF88Suazq8wEH0Nv9oZybx7v43UA/wDJld/QKz90T5Mfx40H+a6/+gXz1spi/wCi8ZVksTondLkH0MN9ohyaPOGcb6An/wAGV/8AQKrvaHcmzPpcbqAf+TK/+gXzysZ1HuApm0T3Yw4boPoUHtD+TVxw3jfQE/8Agyv/AKBHe0Q5NG/S430A/wDJlf8A0C+emWmfFu5wP1KHHzQfQ1+6J8mP48aD/Ndf/QK5vtDuTV/0eN1Af/Jlf/QL541e2Eu8wEH0Mu9odybNGXcbqAf+TK/+gVv7onyY/jxoP811/wDQL56HROb5gqw7IPoaHtEuTI9uOFB/myv/AKBXD2hvJt+O+g/zZXf0C+eQNJ32+1OjfGQg+hs+0O5Nm/S43UA/8mV/9AqfuiPJn+PCg/zXX/0C+eYsI8wqBpJQfQz+6J8mQ/7OFB/muv8A6BB7RPkyPbjhQf5rr/6BfPQIXO7EKT3V+M5ag+hM+0S5Mx344UH+bK/+gVR7RHkzPbjhQf5sr/6BfPM9had8Kgb1HGcIPoaPtEOTUd+N9B/myv8A6BB7RHkzPbjfQf5sr/6BfPQ6EtGeoKzoPqEH0OH2hnJuBk8bqDH/AIMrv6BWH2iXJkO/HCg/zZX/ANAvnn6PmFQNJOEH0M/uifJl+PCg/wA11/8AQJ+6J8mQ/wCzhQf5sr/6BfPV7u8DOyjLCO5CD6Gf3RPkx/HjQf5rr/6BP3RPkx/HjQf5rr/6BfPKiD6Gv3RPkx/HjQf5rr/6BP3RPkx/HjQf5rr/AOgXzyog+hr90T5Mfx40H+a6/wDoE/dE+TH8eNB/muv/AKBfPKiD6Gv3RPkx/HjQf5rr/wCgT90T5Mfx40H+a6/+gXzyog+hr90T5Mfx40H+a6/+gT90T5Mfx40H+a6/+gXzyog+hr90T5Mfx40H+a6/+gT90T5Mfx40H+a6/wDoF88qIPoa/dE+TH8eNB/muv8A6BP3RPkx/HjQf5rr/wCgXzyog+hr90T5Mfx40H+a6/8AoE/dE+TH8eNB/muv/oF88qIPpN4T81nADjnqOp0lwp4i0uoLvSUT7jNTRUdTEWUzXxxukzLGxpAfLGMA5+LtsUXlj7FT8KbVP/F/XfyjbkQe1KIiDrd7RDiNNwy5POI15oal0Ndc7eyxUxacOJrZWU7+k+REUkrgRuOnIIOCvOH2NOhItRczd41lVRNdHpPTVRLA4gEsqamSOFpGe33o1AyPXHYlduvbO3OpoeVGy0tO7DLjregppxvuwUdbKMYOD8UTTvkbeuCOIPYb2v8A35LzJB/84aWKbP8AjzpG4z/gjuPTHmg9VF5f+2s4zGi0/ofgLbKvElzmk1JdmNcARDF1Q0rSO5Dnundg7Ziad/L1AXz2c1utrrzac6t8ZpKT3xt81BTaU060bsdDHI2lhc3Gfhe/qlOf/mhO3YBjcQuVa6aK5NeG/MnJBP4urL9cKWuY5pxFSPAbQEj96CaWqd1Y3E8YyNgfQb2MXGX9k/BvUnBe4VGavRNxFfQNLu9BWlzi1oP9xUMmJI2+/N2zueyPMdy4WjWvJrfuAGmqBrxZ9MQ0+n4+j4hU0EbX0oGMkFzoWsJGSQ92c5IPkD7NbjMzg1za6Tqa+r8C0asL9L3El2GhtUWiBxJ2AFSyAkns0OQfQGvGb202hIrLx70jr2mhDGan057vMRj75UUkzgXHG+fDmgG/9yPnj2ZXmD7cO2iXSvCW7mAE01wu9MJM/REsdM4txnfPhA5xt09xncOzvs1eJEvEnk40HVVlQZa3T8M2nagkk9IpJDHAMnv/ALX8D8uF2hXnn7FO5Vc/LrrG1yv6oKTWcskWc5aZKKl6gPID4AcDzJPmvQxAREQce8xH4P8AxN/gdev1KVfM/Wf21P8A4V36Svpg5iPwf+Jv8Dr1+pSr5n6z+2p/8K79JQQoiICIiAiIgIiICIiAiIgIiICqDhU7pjCCTqCocJgdKty5Be1wKqoxkEFSA5QRnugG+Srg3Ls+WVVw7YQXZGFYXhXeSoY98kFBJG4nCyo5g1oB7rEYMABXlpIJ3QRTnqdkdlECQpnjyKiIwQgkHZXBhcFcyPLckeSoXGNBcICSj/vWxKuinb5qOpd17tQRud1b5VqoM5wqoBOFLFCZAVb0Z7BZMDmxDB7oLBSH5hX+GIxgqWOYZVk5LyNkEL3dOSVZ4vdXPYSMeoUQjcPJBc14cfmr+oAbq1sXSckK4x53AQHM6xgKgHhg5UsYLTghWyhrigtYPEO3mVR8Bz57qWBgB2GVWZ3SR5II42iEglSvna4Y+SgkeHBWBu6C7HU7IQsIGUbkHsqueOkhBGxpccBTMaYyCVDEcOypS8uCCaapa5nT8lixtLXZUgjz2OVURnYYwgvnqQ6LoWGw4dkqeWE4zuohEcEoJ/G629KiMZyVbF9NZAGdkEcUJlPTnGVkfc5zSHnOO6ugaYSHuGyyn1jHRYyEGDJIHjwh3ChFMW7lXNGZy7yysqQ9TNkGIX4GP51Y1w6vtVryetG/SygzJGl8YA9FD7q4NJU0cjcDKudM0tIHmgw43mJ3fzWV72C0N81iStOS7yVjBlwQTynqBd6qFrsPDj5FZhgJj3Cw5G9LkGd700xdGR2wsZx6jkKOJocVkmItbnCCEO6DuqmXO+VGQ5ziEI6dkFwlAKmbU4bglQeHtkhWEYcgyC8E59VE8dRyFe2MkZAVrx0HCCgYVQNLXA/NSZ2yrDlxwgy46kNZ0qGb77lwUZidsd1JGwhu4KCBzS0DKuiGdklGEhKCcQlyteRF8JKnicsarOXoI3uDlzJyYgu5seEXy1naD/yuNcMLmnkt/Cx4S/wxtP63Gg+j9ERAREQdc/aHfgbcSv8AE6P9fp188jV9DvtCwTyb8SR60lH+vU6+eEeaC4Dr2Cvb8HdWwjLlLJGSgje8OBCjV5iOMqnQUFqJ2RAREQEREBERAREQEREBERAREHdBKw4CuIzuOyszsCrmEkIKdZ7K0n4gT6q90Z7+pV7IeppKC5lQA3HoEdH4oz5d1juwCd+yyYZWhu5CCIMJdhXiXwh05GVQvaHEhWPb1u6h27oLsOz1eWcq5vUD1eSs68bEqWPduPkgnZWsaME7qOWL3j4wsd8QydysiKTojwfTugxzH07dleyRrR0HurHSguPplBHlwcMoJBA5/wAQPcq19K4HKymytY0NPcK107CMEoMV56c/JRElxx81fIeo7eajA6Xb+SCdsHUPX6lI2UR/B5qkUo6d1DMMvyCgmdF42SPVUD/AAaVJE9rdj5qyePxT1DugjfJ1EqPufrVxj6cZVWtzv6IL2wZ3CskiLNvRZEbg3PzUczm5z5oLIJDE7JSdwld1BROIcRhTxxZb80ELR0kH5rLiqQ0b4WO9pb39VWKNzwMboL6moEgx6rGDSRlTS05YMkFWN3QWbjfCkbIAMKpj6uyie0tOCgvc8EKjIi87eatIwpoH9JAQVfAWMURjJ3WXUvBYFjNJOyCgHSmR6qshwFGCc90EjZMH/wBql94+HCxukqrWE7HIQXOPiHZXNhIKNaGbq8y4GwQWvaSO6hLSCVMHg5VjyM90FnSVexhBDj2CtaTlSk/BgeiCr5h04CgJycoRjKogIiICIiAiIgIiICIiAiIgIiICIiDv/wCxU/Cm1T/xf138o25E9ip+FNqn/i/rv5RtyIPalERB0N9s1aqi48ptprIcdFr1rb6ub/vDS1kI/wDwpmrhX2HN2hjr+MViknd4s8Niq4o87dLHVrZHAZ75kiBIHpnyXd/nv4azcVuUniVpOipTPWx2g3WjjYMvdNRPbVNa35u8EsA8+ojzXlz7H3iLDpDmudpOrn6INbWCstsTCcA1MJbUsPpnogmaM/3eBuQCHqfzs8ZDwJ5Ytda+pKoQXT7nm2WlwOHCtqiIInN9SwvMhHpGV4O8A+AHHXj1qGso+A+l6q7XawQsraiSnudPQupmOd0tcJZ5YwCTnAaS7YnGAcesftWOC3Mfx50pofRvBbQ0+obHQ11Vc702C4U8LxUhjY6YeHLIwuAa+oOQHYLhnHnuL2YHKlrfln4V6jq+KdijtGr9W3Rkk1I2phndDQ08fTTse+JzmdRfLO7AccBzc4OQA8+f2hftOf8Aglqj/T23/wBeXWjinwr4p8AeIEmjeJtjqdPapt4hrjF73FO5geBJHKyaB72OPnlrjggg4IIH03Lzl9qjyWcVOPmotH8TuCmjzqC7UFvns97p2VlPBIKdkni00jRM9gfh0tQ0hpLt2bYBIDuVyzcXIOO3AXRHFVksb6i+2mKSvEZbiOuZ97qWDGwAmZIAMDYDYdl0K9uJd4o7Fwjsfju8WervNWYgduljKVoc4Z75kIBx/dYPdc4ey14Ucf8Agpwf1JoDjdoufT9NHexX2Nk9ZBNI5ksYE7emJ7uhofG1w6sEmR5GV0m9srxCh1NzL2jQtHL1x6M07BFUN6s9FXVOdO4Y8swmmPrv9SDtN7FW0VNLy36tvE20dfrOdkIx3EdFSguz6EuI+tpXoQut3s8OGdRwq5QeHtjr6V8FwutC+/VjXsLXh9bI6dgcDuC2J8TSDvlu/ouyKAiIg495h/8AeA4m/wADrz+pTL5n6z+3J/8ACP8A0lfTBzEfg/8AE3+B16/UpV8z9Z/bU3+Ed+koIgRhVyFb80QXZCZCtRBdkJkK1EFSRhAdu6omMIB7qoOFRVAygrkJkKhbgdiqILshMhWoguDhnuqktViIJMgBUBBVO6qGjCC44wThR53GCqudnZWjugmaNs4RxAVGnZUz1HJQVB81dkKPONtk6vqQSq8TNbsoPEVjjvlBkuIdnHmrPBc49X5FRjsd8KZrx0790FzJmtHTjfso5YzIepqheD1E7rIhf0gbZOPRBD7tJ5AqgzGcOHkswPGMrFmHUc/NBaIevJCo5pYRlTwkNHT+lWTtB3BQXxN2yo6nIdscBXRyYGMK2oIccgoLoZCAApw9p7rEZthSg5QT/Cr2MaQoOsjYdlJHJjuUFZI87NUQe2IYd9am8QErEqRnCCYyNf22JVhY7PmooiR3WS17diUFGHwRkqKSQSuHThXTP6xgDuoYwWnuEB8bsIw9A3VxeDsfNWOHV29d0F/WH7BWvjIBPqFSMYcpiepvzCDHZ9LCmAzgKPpw4kKVnrg7IJQ3wwC5V8dn1qOSTbpKx3Eds/nQZfjxyfDkKh6cHGFiRu6XZUxeMbILQOl+fJSxvb1AntndY/Xl26kBGEGZU1DHwlrDvhacHvBGSVeSR29VbjJzugmaQAD5q4S9Q6fkoS44SP6YJygukgc34/VRZ8vNZ80jDF05GQMLTwcOQXujkY3JOyta4hwOfNTSyNdGAO+FjoMh0jHMx54ULBhwce2VRvdCcINQ98j8IN29FhSESOGPNRq6P6QOOyDJip3R/fHdlI6pjAAyqyzjwQ0YBwsA7k7oJTIzJIUZPU4YVqqO4KDIAy1QHZ6mDsDGyhdnqzhBnQ46QsWpI69vJSMkwMKKb4nZygAHAVzfhOSFRp2wUcgnbKwgD5J47AcLFa0/NUcCHeaDJdF4oy1Y72OidgrIhmDRuoaiTrf9SCSOcNxukrDKeoKNjQd89lIJAwdJQY7mlq5n5L39HNjwj+esrQPy1ca4bk33A7rmHk0BPNhwh+WtLOf+VxoPpDREQEREHXb2hLg3k54kk+VJR/r1OvnhacZX0Oe0O/A14lf4nR/r1OvniQSQvDHZKzA5sgyFp6lhl6MDKDIkYGAqAyNI2Us0ge3Y91i9KA4g9kBwqIguyEyFaiC7IVDhURBdkJkK1EAd1U4VEQAcKpwqIguyEyFaiCpIwqDcgIg2IQTiIloz2VPouGVM2Rpb9ix5Xb7IJjM0NAUkdSzo6PNYQ3O6HY7IJHRlziR2KNhkI2VzXjAB8leJg0bFBF4EhOCpGOEeGuAz2V4laM/NY8p6n9QQSugLj1DsrQ7w/hKmZKOjf0WNI7qdt5FBkBpduopM5wPVXNmHSAVY52XDt3QUFO874Kma8RMLXd+yuEwAwFiyv6znKA+RxccHurmxSPaXA7KId1kRyFrcA7ILOksOHeiubAZG9QCq7Du4V7H9LcIMU9TTjOMLJhpzI0uIVjmhzslTRyFjcDsggeDGftU0NQ3GCrJPiOT5qwMAQSylsm7f0KMODAepVBIVrx1boKGX5q5sLpzsVaIx5hZELizt9aCJ1M+L4nKWKZuFWV5kGD2ULWhpx5ZQXzPa4YCkppmRD4sK0tZ0588KAtO4GcIMqqnZM0ho3WGGlgOVI1mArcfFv2QXwytB381bM8OdkK3oA3wmG5wfRBYQTsFVrHAghSfCGbYyrY3fF8Xbsgq95IwQrWbbq+ToAzjfuomkg5PYoL5XA9vRRN7hXuIIwFa3Gcnsgma9uMbflQFvUrMNACo3uEEswLW7KABzuynmeHMwoWOIICCvhu8iqiF2d1MZG48lTrCCzo6Qqtkb9Eq5zm42WOPphBK6PO+FE4EEqcPGO+6heSTnH5EDITIVqILshWnurun61QscPJAJ2VFd0FOnHqgtRE6SfJBdkJkJ0/WnQfJAyEyFQMJ8iqEYOEFSdlREQXZCZCo1pcUc0tKDv/7FUj9tNqn+AFd/KNuRU9ip+FNqn/i/rv5RtyIPalERBY9jJWOjkYHNcC0tIyCPPI9F4L8ZeA3Ffk85ym3vh9oHUNws1i1HBqDTM9FbppoaqiMrZW0wexmHFoJgeB5g7YIz71ogw7VXsutso7rHT1NO2sgjqGw1MRjljD2h3S9h3a4ZwQdwQQsxEQEREGPWVLaKlmrJI5pGwRulLIYzJI4AEkNY0EuccYAGSdgN14O6C4H8WedTnRqbvrbQeo7PaNS6jmu9+nrbbNTx0FtY8uNOXuA6X+ExsDBknqIO+CV71Ighp4IaSCOmpoWQwwsEccbGhrWNAwGgDYAAYAHkpkRAREQce8xH4P8AxN/gdev1KVfM/Wf25P8A4R/6Svpg5iN+AHE0f9x15/UpV8z9btVzj/6I79JQRZ2wqIiAiIgIiIA7qpOVREBXsIAViZKC8uBCorfmq9RQUPdEO6IAGVUtIRn0lJjKCJVDseqoRgogr0k9vNOkpnZOooKg+R+pFb81XqKCh7oh3RARXBhPkU6CEFMhV6z5KnSe/wDMgaSgu69t+6uikxsowN8YVQ3HqgyTIMfPCjDt91aNgu1fJl7PziRzZyv1NPX/ALFdB0cxhmvc9OZX1cgPxxUkRIEhHZzyQ1pPdxBYg6rudgfDsVH8R7le0Vx5E/ZjcB6aCy8Y9Q2+S5PYMO1PrWSjq5Qf34hppYBj5hmBtk53WLJ7Or2eHMBaag8BNcRUFZC0kS6Z1ULqIXAAffoKiSV4GSCRlh3GCAQg8aAMJjPzXNPNdyuaw5TuKTuHGqrnS3WGqo2XK13OmjcyOspHvewOLHEmN4dG8OZk4IBBIIJ4aazugswR5K5XlmPJWEjyBQVBwq9X1qPq+SFyC8u/Ko35Pc+eyuxgZycq05d6oA2Cu6/rVoBHqq4HzQV6sqzqXYHjFyL8xXAvhlRcXOIWmbdS6fqpIInup7rBPLTumGYhIxrjnPbLC4A98Lr+I8+qChIwqZPplSBnoCrSz0KC1p3yVcX7KnT9aoO6C8PHmFcJAPkoyArUF0js+ay7NZbzqO509l0/aKy6XCqcWU9HRwOnnldgnDGNBc44BOAD2KwsZXan2XTS3nr4ZZ/+/P8AI9ag62al0jqvRde21ax0xdrHWujEzaa5UUlLKWHOHBkjQcHBwcY2K0jqJ2yV6Ke20/3+tCfwQH67ULzqQMHumT6q4jDR33VABhBc1wyq9TfVWEDCogu6t8nKv6xjbv6qJEFS9x7lUV3SPLKdI88oKbnZUwVe1oz5qvT9aCNFVwx29VRAVQ7CoqgBBcSSDk7KxVHUT5qvT9aC1VBGFXpHnlULd9kDrKdQTpVen60Dr9E6vrVOkJ0hBcX9/wDWjXjzVgGSq9IQSeI1WPcHHZWqoblBQEhM5VxYRvgqnT65QXNeAMZKtc7q9VcGZKtc0tKAHeuVzNyYn/rseEfz1naP1uNcMLmbky/Cx4Q/wztH63Gg+kJERAREQdc/aHfgbcS/8To/16nXzyN7r6G/aHfgbcS/8To/16nXzyN7oDuyp2VXdlRBc0nO6ucrG7FVc76kFqIiAiIgIiICIiAiIgIiICIiAiIgZI807oiAiIgZPqmT6oiCvU71QH1VEQMn1TOURAyfVMn1REDJTKIgdlID5KNXIJAcK4HzCsG67++zr9nXaeYm0O4zcZJauLQ8dVJS2y1UsroZbvLGemWR8rd2QNdlnwEOc9rwCwNy8OgqZK9mdQcwPsn+CF5rOH9Lw90Xcqig66Ssmt2jo7mzqBw6J1VIw+NuN8Pe3OcnOVpmsuX/ANnHzecMNT6q4DVNhsWpLNa6q5sOnmm3VFM5kZkAntsga0wlzQCRG0EZDJB3QePPdEXaflG9n9r7m70fftZ6X1zYLDSWSuNsEVwjmfJNUeE2TtG0hrMPaOrJOc/DsCQ6sItQ1DZKzTN/uenLg+J9VaqyahnMRJYZInlji0kAkZacEgHGNgtOJ8kFwAz8lUFWEgJkDzQSZyrSd1b1Z2VUF3Vn1VAPQfnVEQXYCoWhOop1ILSz5lWlhH1qXIKIIul2cE/mVS0+RUoaCeyqQEEHSfPKtLVkdI8v0KnT8kGMWEJgrJ6B6K0s9EELWlXYPqrunGSrScHGQgoQcf8AtViuLtvJWndBX4sef5VTceqr1FUO6Cp6vNOk9wqtOTur0Ebc/NVyFdgKxw3QURVLceqBu3mgua4Aq7rafRRDuhGCglMgKt6grekKoZnsgrjKuDmt22V3TgZwoX98IL+pvqrhK0DdQogyOtnyVvgmTcfoUTd9iVlRPDQAghdA4KIjCzXODh5dliyjDkF0ThGcuVszw92QrCSUQd//AGKn4U2qf+L+u/lG3InsVPwptU/8X9d/KNuRB7UoiICIiAiIgIiICIiAiIgIiIOPuYf/AHgOJn8Dr1+pSr5nq3+3aj/Cv/SV9MPMP/vAcTP4HXr9SlXzPVv9u1H+Ff8ApKCFERAREQEREBERAREQEREBERBVpwcq/qCjVW9kFD3RD3RAREQEREBERBK17RuqmRh8goUQS9bfknWxRIgvL25VQQSFGqsGSg5A4E8Kbpxx4waT4T2eR0U+pbnFRvmAyYIN3TTY8wyJr3keYavaLnc47WvkY5YrNpDhFRU9rvNfGzTmloWRtcKGGKPM1WQRh7mN6dyCTJKxzgQXLz+9j/YIrxzfR3GSmMjrHpm5V7HdOfCc4xU/Vny2qC3P/bY81vz21moqyr46aD0g9zjTWzSZuUYzsH1NZPG8gZ2OKRmTjfA9Ng89rxebxqO61d+1BdKu5XKvldPVVlXO6WaeRxyXPe4kuJO5JJJWTpzUuoNIXmk1Hpa+V9nutDIJaatoah0E8Lx5se0gg9xsexwtO6CqFuPNByTx05ieKXMbe7NqPiveobncbHaIrNBUR0zIDJEx739cgaADI50jiXAAHA2GFtXQWgta8T9T0mjeHul7hf73WuIgoqGEySOA7uONmsA3LiQANyQN1t/svbflG4c6D5E+Sys4760tTXagudlZqS+z9IFQ9soBorcxxHwgeJEzB28WR5O2AA6eaU9jjzQ3+3x1uo9RaG00+RuTSVNxnqamN22A7wIXRYwTu2Q7jtg5WwOMnsuOa/hBZKvUosFo1ja6FhlqZNM1b6mWKIDJcYJWRyuAA3DGOxuewJWgcUfaOc2/EnU9VfKfixdtK0MkrnUlqsEvukFLEScM6m4fIQDu57iSfQYA7Wezo9o3xS1fxStvArj1f/2RU2oy+CyXqaFjKumqw0ubBK6NoErJMFoc4F7XloLi0/CHlwQWkhwIIOCCMEH0XKfLhy38RuaTXVXw84YG1NudDa5bvO65VRgibTxyxRE9Qa4k9U8YAAPcnYArtN7XflwsfCni7ZuLGjbZHQ2riFHUPuEELcRR3WFzTLIANm+KyVjsAbvbK7zOLvYr5/bTapz/AMAK7+Ubcg65UPJrzA3njneuXvTujfu1qrTszI7o+inBoaNj2te2WWpd0sYwteCC7BJyAC7ZdjZ/Yycz0ViFxi1hw8muPSHm3NuFWHYxnoEhpgwvzgYJDe56sAZ7Qc+nO9pvlHuV54c8B7FbY+KWrpGXfUF1fTCRlAXxNZFLIHAiaoMbGBjHZYxgBIIIaekHBr2oHNJoTiPbtQa94i1mr9OS1Tfuxaa+nhLZKZzvjMJYxpikaCSzpIbkAEFuyDrlxV4UcQ+Ces6vh/xR0tV2C+UQD3U9RgtkjJIbJE9pLJYyQQHsJaSCM5BA5Q5eOR3j5zQaWuOs+FlqtMtptdwNsmnr7i2n6qkRskcxgIJOGSxknAHxDGd8ej/thOGunNa8tdn4w0UEL7npW6Upp65oAdJb6z4Hx5OCQZDA8emDgDJK8tuEXNbzCcB7BV6X4ScTa/Tlqrqw19RSwQQSsfUFjGGT77G4glsbAcEDDQg9rudvgBxD478rMfCPQNPQS39tTbJC2rqhDF0wY8T4yD6bbbryb41ezu5k+AXDm5cU9fWixCw2iSBlZJRXRs8kQmmbCx3RgEgySMacZI6gcYyR6mc/3GHiVwk5P4+IvDrVVRZdRmqtLDXxRRvf0y/2QYe1zd/Pb6sLx94kc6HM9xf0fWaA4j8W7letP3F0T6qhkpqaNsxikbIzJjia4gPY1wGcZaPRBp3Ljy4cROaTXVZw94Ym1NudDa5bvO65VJghbTxyRRE9Qa4k9c8YAA8ydgCt16Z5GOYXWHHS/wDADT+m6Sqv2lpGMvVe2pxbKFr2B7HyVHTsHAjDQ0vJBw09Jxzz7Fj8KPVP8AK7+Ubcu3PPrzg6d5NKCt0twbs9rHE/iDUOvVwqnsbIKNnQyEVc7DnxJHMiEcTD8IEbnEENDXh5/cyPs0uPHLVw4qeK2pr9pO+WC3vp4rg601VQZaV0sjYmOLJYWAsMj2NDgScuGWhdWbDYL7qu80enNMWetut0uEogpKKigdNPPIezGMaCXE+gC504j8+vNBxc4Y3nhJxL4gRX+w32aCep8W2U0E7TFM2drWvgYz4fEYw4II+EAYAwvQj2VvALR3BzgFceazX1LBHdr7SVlbDWzsybZY6bq6yzuWmQxSSOI3LBENt8h1V4fex/5rtY2yC7aiqNI6NZO0P90u1xklq2ggkZZTxSMBzgEOeCM7jIIWFxL9kbzZ6CtdTebFTaa1rBTNMhp7FXye9lg74hqI4up2N+lhcT2AJ2WkcwPtOOZXivrWvrNB67uehNKxzuba7ZaHtgmbAD8L552jxJJHAAuHUGAkgADvyHyU+0341ab4p2LQnHPWFRq7R2oKyG2vq7g1rqy1yyuDGVDZmtD5GBxb1teXfDktwchwdBayirLVW1FtudFPSVlLI6GeCeMxyRSNOHNc1wBaQQQQQCDsV2k9l49p56eGYHn92f5HrV2j9svy42KgpbBzKaYtbKatrK1ti1IYW4FQ50bnUtQ4DbqAifG52MkGIHsM9V/Zcfh18Mv/LP8j1qDmv22n+/1oT+CA/XahedS9Ffbaf7/WhP4ID9dqF51IL+sdICpkK1EFSRhUTBTBQETBTBQXh488qjnD5q3GEQXteB39VeZWkEKFEFXHJyqJjKYKADhXBwBVuCmCglD2YVC9qjxhEF5e09sqrXNxuo0QS9bfkqF7VGiCvUEJJPmqIgqTtsqZPqnzQAlAVzCARlW4KIJxKzHYKN7mk7KzGU7IJoyB3VkpBOyo0k9kfnbKC1czcmP4WPCL+Gdo/W41wyuZuTIH9tjwi+Ws7R+txoPpCREQEREHXP2h34G3Ev/E6P9ep188je6+hv2h34G3Ev/E6P9ep188ecIKu7KiEkogAZV+MYyFaz6SvcgjREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAVw7K1XDsguafJe9XLDaa/V3s0bJp3hnO2G8XXh/dbdbZIZAzoub2VMWQ7YNcKkkknsc5J7rwUb3XeL2entCG8sHj8MOJ9JW3Dh9c6o1UNRSt8Sos1S/AfIxhI8SF2AXsG4ILmgkua4Oldztdysdxq7NeaCpoLhQTPpqqlqIjHLBKwlr2PY4AtcCCCCAQQQVFT1FRSSeNS1EkMnS5nVG4tPS5pa4ZBzggkEeYJB2K95uJfLDybc/Wnv8AZE0/cLdV3OdnQzVOl6lkdW1+PhZVMwQ9wwMtmZ4gAwC0FeaHNh7NjjNy0UFVrS1zx620PT5fNdqCAxz0LM4BqqclxY3tmRjnNHmW5AQcEcvnBe9cw/GHTnBzT11o7ZX6ilnYyrqwTFCyGCSd7iG7k9ET8AdyQMjOV7g8h3KXqDlD4eag0XqHV1v1BNeb191I56OnfE2NvgRx9BDySTlhORtgrwHst8vWm7nBe9PXettdypCXU9XRVD4J4iQQSx7CHNJBIyCNiR5r2Z9jxrTWWt+COta/WerLzf6mn1UYYprpXy1T42e6QHoa6RxIGSTgHGST5oOGdZ+xm4k6p1hfNTQ8atNU8d3udTXtifbpyYxLK54aSDgkB2M+a83eImja/hxr/U3Dy61UFTW6XvFbZqmaAkxSS00zonuYTglpLCRkZwQuRuKvHjjhR8UNYUlHxl1zBBBfrhHFFHqKsYyNgqJAGgCQAAAAADYALhytrKq4VU9fX1UtTVVMjpp5ppC+SV7jlz3OJJcSSSSSSTnKDuZzH+zB4n8CKXRcWn9XUuu7xrW9NsNHa7dbnwSipML5eoOe8gsDY3kuPSGgFxwASObdLexF1HW6Sjq9YceaK1akmia80VFYnVdJTvxvGZnTxuf6FwYANyA7ZehXM5xr0Ty58Kbnxm1jb6etqLE10Vmp3geLUV8zSyOCJ2CWF+4c4DIjDycgEHwv1bzzc1mruIFTxFk426qtVbLM+SnorXc5qagpWE5EMdM13hlgAAw4OLukFxcd0HFHEHRtx4c6+1Lw9vEsUtfpe8VtmqnxbsdNTTOieWnzBcwkfLC7i8vPsmeOvGSwUWsNc3yh4d2W4xNnpWV1K+quMkThkPNKHMEYIwQJJGuxj4RnKwvZi8H4+Yjmuq9e8RYxeaTSsc2qbgasB4rbnLMBAZBtk+I98x2wTEAQQcHkL2rPOBxAufFyv5d9DajrbNpnTEUDLx7lM6F9zrZY2ylsj2kExRtexoZsC8PJ6sM6Q3dqv2IV0itjptDcwNLV3BjMNp7tYHU8Mj99zLFNI5g7beG89zk9l5+cb+AnFPl31nLoXitpiW014aZaaUOElNWw5wJoJR8MjCdjjdpyHBpBAxOEfGniXwM1fQ624Z6qrbRX0U7JjHFM73eqAO8c8QIbLG4ZBa4HY7YIBHrl7QLTmneZzkGtfHygtMcV1tNstur7aTvLTwVIiFXTF/oGSkkdi6Bh8gQHjHZbLdtR3ih0/YLbUXC53OojpKOkpoy+WonkcGsjY0AlznOIAA3JIC9E+E/sWuJeorPT3fi5xTtuj6iYCQ2u30Buc8YP72SXxY42u756DIO25yccQ+yhtenrnzmadN9ihlmpLXcqq2NlAINW2A4IBO7hGZXA4JBbkAEZHK3tlKzjNT8X7FBc6q8RcOKmzxMtLIpHigkrQ9xqA8NPSZhhhw7J6QwjbKCDjZ7G/iXoLTVy1Xwx4nWzWMNsp5ayW3VlA63Vbo2NLi2Ih8kcj8Ds4xg+W+AfPHBJDQCSTgDGSSuR+FnMhxx4Kw1lJw14l3qzUdfTS0tRQtn8WkkZI1zSTBIHRB4DiQ8N6mk5BG63LySWzT155teFVu1VFBLb5dS0pdHOAWPmaS6BpBIBBlEYwcg5xg5wQ7JcCvY+cZeI9gpdTcUtZUPDynro2TwW91A6uuIYQCPFiD42Qkgj4S8uachzQRg7x4jexP1rbLbNXcLONFsvtWxpc233i1voDIQB8LZ45JRk74yxoBwCQMkb69tBWcZqLT+h5NO1V4g4ev8Aeory6he5sDq9xYIW1JaRkFheGB3wk9YG+F5n8J+O3F/gbfItQcKuIF409UxvD3xU05NNPgg9M0DsxStOBlr2kbA4yAUG0rzabjp+711hvFK6lr7bUy0lXA4gmKaNxa9hIJBIcCMgkbbFYYdjzWVe71dNSXqv1De6t1XcrpVS1tZUOaAZZ5XF73EAAAlxJwABvsAsJBf1fMJnPfCsVWjzQJG7H6ljHusl30T9RWMe6AiIgIiIAODlXdX1q1O6C7q+tMj1VCNlRBf1tVQQRso1I0HCC1Wnur8K13dAJ9FcxwA+aswfRMH0QTCVo7qx5DnbKxB3QXiMlPCd6KSMjbspOofJBjOYWoHEeaklIIKhQSiX5q1/xHKsAJV4acdkFvSU6TjKqA7zCvAOOyDv17FX8KfVP/F/XfyjbkVfYrNI5ptUk/8AACu/lG3Ig9qEREBERAREQEREBERAREQEREHHvMR+D/xN/gdev1KVfM/Wf21P/hXfpK+mHmHGeAHEwf8Acfef1KVfM/WD/btQMdpH/pKCAHdXZCr0HpzhWlpQVyEyFQD1Tp+aCuQqHCp3V3QUFqq3snSc4VwbjzQUTIVS3burQ0lAJGFRXNYSVXwz6ILMFMYUojdtsjoSd8IIkVxY7dU6UFMFMFSNY4422VTGQgiRSeGT6K0xnKC1Ff4Z9FR0ZHogtwUxlSmM4/8AajYjvkIIsFMFSlmDjKp0/NBHgpjCl6EMRPogiwfRSw4zgq7w/qV8ce+EHdj2RWoaWy839HbZphG+/wCnbnb4W9ePEe1sdQW4/ffDTOOP+1z5Lkr21Okqqk4w8P8AXb4ZPdbrpqS0MkP0PEpaqSUgHyOKxp+Yx6FdFeDvEe88G+KOl+KVgYJK3TNzhr2ROdgTsY744iQMgPYXMJG4Djhe4XHnhZw59olyvW+v0PfoGSVjGXnTN0kb/ala1rmvgnAy5oOXxSNGS1wDgHFgBDwRy0eQUbwD2XLHE3lZ5geEF5qLJrvhRqKkdB1EVcFE+ppJWAkF7J4g6NwwM7HIBGQDss/hXyecyHGW7wWrRfCXUAileBJcrhRyUVBADjLnzyhrNgc9LSXEdmk4BDhXAxgr3J9q611o5J7vbLYDBSfdS0Uro2bt8Fs7S1h77AsZ+QLy35x+Uyt5RdVaX0bdNZQahrr3YW3arlgpDBFTzmeWN0UZc4mRgDBhxDSSSS0bAesdHT2b2gHIDHZrfdqRt21HYKemmlflwor7RmNxa8D4mDx4gT5mOQHBDsEPBJx/MuR+WyrrqPmJ4XVVre9tZHrOyOgLG9Tuv32LAAwc5O2MEHOMHK0DiVwu4h8IdTVOj+JekLnp67Uz3NdBXQFnWB+/jd9GRhyCHsJaQQQSCu4nsxOTniHxC4z6d446r0xWWrQ2j6lt2pauugdELpWsBNO2mDgDI1kgbI6QAtHh9OeojAdnvbY+7/7B2gXOEfjDVhDCcdQaaObqx54yG5xtnGfJdcfYsOa7ml1Tjv8AsArv5Rty1X2yvHm2a14o6b4JadrmVEGhYJqu7vieHMNwqQzphOCR1RRMBPYgzuB3G2j+xU/Cm1T/AMX9d/KNuQbE9q+8DnX1YCP/ANHWn9SiXT1xB7Lt/wC1h/DZ1b/4OtH6lEunyD3A9od/9rYbn+8tM/8ApadeH69wPaHf/a2B/iWmf/S068P0Ht/7UX8AiL/HbF+kLxCB/Kvdvn30XfOMHIHM/h9SS3manobNf4YKSMyyVVIzw3PdGG5LsRPMm2ctYcZOF4cwaV1RPZq3UkGmrpLabbI2Ksr2UchpqZ7nBrWySgdLCXEAAkEkgdyg72exYIPNHqn+AFb/ACjblsL2rzh+3W1aP/vdaf1KJb69iqf+um1T/wAX9d/KNuWwPawnHOxq3/wdaP1KJB1D7nq8l7paxDrP7Jan+5LTAZOD1sL+jfImoIPG75+kHvz9ZXhW13YYXudyGat0rzS8hreEV4roTV2yy1ehr3A34pKeF0b46aUNJBwad0Za4EDrjeAQWnAeGWRjfuqtkdG5skby1zSCCDggjsQfIrkjjxy88VOXPWtbo3iVpetofAqHx0dxMDvc7jECQ2aCXHS9rgM4B6m7hwBBA5Q5H+TjiFzK8UrFXS6ZrKfh/a6+Grvl5qYHMpZYInhz6aJ5GJZX46OluekO6nYA3D039qJLNVch13qbs1prZKmxyOMjA1wmNTEXkDAw7BfkAbDI7ZXmx7LnH7enhn6/9Wf5HrV2/wDbRcebXS6U0vy62auZJc66tZqK9NieCYKaJj2U8TwDsZHvdJgjIELD2cM9PfZcfh18M/8Ayz/I9ag5r9tp/v8AWhP4ID9dqF51L0V9tp/v9aE/ggP12oXnUgDuq53yhAwCCgAQXte0d8KvW35KMgY2Kogl62/JOtvyUSIL3lp7K0HCoiCpOyoiIL2kJ1NViIJetvyTrb8lEiC95aeytB27qiIB7oq/ChxnZBRVBGFREF2QqdzlUTJQX7Y+aqCAFbkKhdvsUF5x5foVhGVXOyp1FBcCB3VriD2VCSUQSsLR3Vsjg47eqoCD32VHY8igHthcz8mH4WPCPP8AwytH63GuF8lczcmRxzY8Iv4Z2gf8rjQfSEiIgIiIOuftDvwNuJf+J0f69Tr55G919DftDvwNuJf+J0f69Tr55G90AlUVXABUQVZ9JXuVjPpK9yCNERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBXDsrVVp8kFze6vAyVY3uvQTir7IbitbNJ2bWnAjU9Frumr7ZT1dTbqh0dFWNkfGHEwOc4RSxnJIy9jgCAA87oOkvD/AIla/wCFOoodV8N9YXbTl2gI6aq3VLonOHm14Bw9p7FrgWkEgggr269nhzfXLnA4Y6gs/Ee00TtU6UdBR3l8UAFLc6apbIIpjER0tc7wpWvYMty0EABwaPJWLkZ5vJa8W1nLzrQSuk8MOdbnNiznuZSQwD5k4+a9WPZ98rVZyVcINW604zXm3Wy96gEVxvY95a+mtNDRslMbHyglrnjxZnPc0lu7QC7pyQ8mecrhRaeCHM7xB4aWCDwLVa7oJ7fB1dXgUtTDHUxRg5JIYydrRkk4Azvlek3sUP8AeH13/C7/AOs4F5ic0XFuHjpzBa64rUTZG0V+ur30IlyH+5xNbDT9QIGD4MUeR5HI3xlelPsS7/bJ+FPEbS8dQ03Ci1DT3CaHO4hnphHG76i6nkH2fMIPK3i9/vr61/hFcf1mRbPcu2nMFyJcz1v5hdXWHTHBzVWobdc75W1lqu1Db3S0VRSyzOkic+oaPCif0uAc17mkOBHoT1f1Lpy9aP1FddJalt76G72Stnt1fSyEF0FTDIY5YyQSCWvaQcEjI2JCD2r9sf8Agj0v8L7d/wChqV4hN7r299sf+CPS/wAL7d/6GpXiG0HPZB6l+w59y9+4y+J4fvXg6f8ACzjr8PNw68eeM+Hn54W9+YnnM5HOHfG3V2i+JfJ9btS6mtVf4Nyu8ulrRUPrJSxrvEMk33x+Wkbu3wB5YXUb2WXMBY+CHMc2z6vuENBYNd0X3Enq53BkVNVh4kppHuOA1pcHRknYeKCSACR2J9p1yE8T9bcS6jmD4LaZqdSsvVNBHf7TQt66yKphjbEyeKIbytfGyMFrAXBzS4ghxIDSv3QD2cX/AOopaf8AQ2xf61i8wHtReCHEHly1JwM4YcGb9YGXa1ttFBE+OkpKChg6m7Mjgc7Aa0ENY1oGcDIG66MUXLZzEXKubbKHgNxDmqnuLRE3TFb1AjvkeHsBvknAGCTjC3jxV5I+YXgjwop+MPFPS1Jp+01Vwht0dFPXRvruuVj3sc6KMuDG4jcCHODwcAtHkHEGi9Zan4eartWuNG3ie13uyVTKyhq4cdUUrDkHBBBB3BBBBBIIIJC9WODvtaOB/FXTTdA82PD+C2Pq42w1lWy3i52WsxjL5ach0sWTuGhsoGCeobBeeHLRytcQuaq/6h0vw2rrPBc7BZ3XfwbjO+IVQErIxCx7WuAeS8YLyG7bkA5GHqzlP5mND3iWxaj4E63hqoTgup7NPVQv+bJoGuikHzY4j5oPSrjh7MTl349aDk4o8o1+oLTcauF1VQQ0Ff71ZLm4d4ty40zyR05YQ1hyHRjcjyLlbedMXsxuFVbLtaaog94p6Woif9ha5r2/Igj1C9ofZQcGeKvA3gzq+8cYbXWaXor9cYrjQW26nwJaaCKAiWpljcQYA/LQQ8NcBDkgAtJ81aPhfcucrnH1tpnhfeLXRHWOpNQ3y2VNyfIynMHjT1LATGxxHU3ABAIGc+SDuXy7e1+01ctOU+gOa/R8tSXU4o6jUNvpW1MFazBBdV0Z3BIA6jF1hxJxGwbDk3XfIHyX84WiJ+I/LRf7Xpy5VIc6Cu0+8m3GfGRFVUDsGA7jLWCJ7cgkOGx8zOInJjzR8L70+yan4Iarlc1xbHVWu3SXCkmAJALJ6cPYcgZwSHAEZAOy7/ex/wCX7jbw2vWteIXEDSl50vp6822noKKjukD6aWuqGzF/jCB+HhsbQ4B7mgO8b4CQHYDy64i6A1Nws11feHOsqIUl607XS2+sjDiW+IxxHUw4HUxww5rsYLXAjYrbq7Fe0N1lYNe85fEzUemK2Kqt5rqWgbNE8PZJJSUUFNKWkbEeJC/BGQRuuuqAqjzVEBwUFX/Qd9RWMsl27HfUsYjBQAcK7IVqILshMhWoguyEJBzurUQMZVQwlAr2uAwdkFWxHOVcXho6SglGDnCjkwTnKB1DOUzk7easVW/SCCXoCoQGhSYGMqxxaQd90EJ7qoYSMqh7qaMEtQWB3Sq+IqSNIO6sQVLiVURkjKtU0bvhx6oLRgAdSv8AEbjGFRwHdRO7oJvEYPJUMjT2UbGhyo5vSUHoD7Fh4PNLqn+AFd/KNuRWexU/Cm1T/wAX9d/KNuRB7UoiICIiAiIgIiICIiAiIgIiIOP+YTfgHxKHrpC8/qUq+aiWn8S5VjT+9kf/ANIr6WOYL/eE4lfwQvP6lKvmv3F2rv8ACP8A0lBCaPDBsVZ7n8itQzhu+FZnPbH2IME0h9CrPcz81qJGMkgKMyDOBhBhNpCD2Uopdu36VkNIJxgKZkMjx8Lc5KDCFESc4KOoyPIrUvd5292YCsMUzuzfzINN91J7qvufoCs009Q3dzcD1QBxztugw20m/YqZtED5FZDWuB3GFMyRo74QYnuQ/uSnuQ/uSs7xGdsjKqC0juEGmPt+RkA9lEKA5zgrVnAnZoBVghmJADe6DEbQ4GMFVND22K1HwJY29UjcBQmoiDuguGUGH7j/ANqrXW8ny/MtREczvia3LU6Jj8PRug04UBHkU9wJ7tK1ZtDWv3EZwrvudXYz4SDShRAfvSq+5Z8jutS9xq846Fc22V5IIjyB8kGkmzveOoA/YpI7E49wd1r0VHWgdPhfmWdBabnIMxwEjGxwg2yLC8+R3UjNOvcOx3K3MLZdGnDoCMfJSCKeDaWPCDa401JnsVczTcrSCAVuoOOx6Qr2mTyaO/bCDa/7H5gFzFy581XG7lWu09Tw4v0UtqrXiSusdyjM9BUuAA6ywOaY34AHXG5riAASQMLYoFSc4aCB5YWHU22Wpd8fwhB6aaS9sxZaqgazWPAq4QVzQA59rvDJYpD5kNkjaWfVl31rbfE72xmraignoeFPBuitNS9pEdxvlwdV9OdgRTxNjAcBuMyOGcZBAwfPu0WkUbg9z3PHoQsu5UYrG9DWlhPmAg0nirxH4j8bdZVWvOJ2q6y/XurAjM85AbFECS2KJjQGRRgkkMYA0FxOMkk7/wCWHmn4zcqOoKm5cPLhT1VpubmOudjuLXSUdX07B+GkGOQDIEjCDjAIcBhcfR6YrBL1xdT8dtlbU2+vhcA+Dp8sYQenFo9tLo/7nsfqbgPfKWuGz2UN4hniOw3DnsjIyc7EHG25XEfHj2w3FHWllrNN8EtCU+h2VTTE681lWK2vaw5BMLAxsULyMDJ8Qjcgg4I6Ny2iprGBrm9GfMKtNo+qIPR1Pz2OEGzLg+4XCsqLjcquaqq6qR88880hfJLI4kue9xJLnEkkkkknJK575IOaCLlD4tXHiVV6Jk1RBc7DPYpKWKvFI+ISVFPMJQ4xvDiDTdPSQM9ecjGDxpLw/ulQ4+EyQn06StHrrJV2UllW0tOcb7IOQebfj1+2b45XvjDFpR2norpDS08VA6sFU5jYIWRBxkDGAl3R1EBuBnGTjJ4abDI44DcrVpqOSraPDdgj080gs9bE7qO/ywg7o8w/tIWceOV+m5fBweks9b4FsgqLp92hPGPc3RuDmReA0nr8MbFw6eojLsZPRh0L2HDgtcdS1MYy5uAsKW3zSPLvFABPp2Qd7uUj2rereA+gLbwr4laEk1jYrJGKa1V9LXCnraSmA+CBwc1zJmsGzMlha3AyQABPzle1JoeZDg7deC2juFVXY6C+y0j7hcbjcWSy9EFRHO2OOKNmATJEzLi87Ajp3yOhsNvqOnw4/iOfRXjTtwd8RBGd+xQc4ckHNJDyicWrjxJqtEyaohudhnsUlJHXikfEJKinmEocY3hxBp+npIGevORjB29za8fG8znHO98YIdKu07FdIaWnjoH1gqnMbBC2IOMgYwEu6OogN2zjJxk8bUthrY39TgT5b5WU7SFzq5eqIOGcbAEoNCjoJHjqGT9i5P4A8e+LXLLrYa84WXttJVSM8Cto6lhlo6+HOfDniyOoA7gghzTktcCSVoUekr3RwDxKRxaB3I7q02WvmaWCH5ZA7IPS/SfttrA62MZrzgPcYbkwND32i8RywSnzcGysa6Pfs3L/AK1tLi97arVN0tNRaeCnCWCw1k7Cxl3vlYKt8JIALmUzGtZ1Dcgve9pOCWkZB88X6NrWSeLJ1YJ9FFPpWZ4OHEAfJBh6o1JqniDqS46x1hfKq8Xq71DqmtrauQvlmlPcknyAAAAwAAAAAAByHyt8YX8tnHjS3GqXTTr/AB6ddV9VvbV+6mZtRSTUxIk6HhpaJy4DpOS3GRnI2NSabqoX/C4vP1LPdY7g9pYYCM/I7IOYed/mvp+cXiTZNZ0GhZdLwWSzi1MgluAq3zHxpJTISI2Bu8nSGjOzc53wOvsVhnnGW5ytwW/Ql0fOJY2vec56cFbgZpPUMWOmicPLsg2N+xifHnlWu0zOPVb+dp2/xDL6Ygb+ShdZ7xj+1z+RBsGSw1DNyT+RWCyzEZyVvOqtN3Gc05/IsUWu7BhPu5wg2jJa5WHurRb5D6/kW4J6OtLy0xbjyx81QW6vaMmI4QaB9zpD6/kV4tUvqtXLZWHDm4IVzTISAG7oNIZaJXnAJ/IrzY5x5la9BSVo+MxHB+SyfCqCP7F5INsGyTDzKt+40vkT+RbkLZs46VY5k2MdG5+SDbf3KlB7p9ypfVa+6CpzkR91b4FV/wDM0Ghi0ynsT+RVbZpie5/ItfjpKw94zupmUtWCMsQbdNjqAMklRm0yg4JW7HsmLceHuFgyxzBxwzz+aDQPuVL6p9ypfVa54dR3LNlRzZht0dwg0E26UeZ/In3Ol+f5FrRZMDuPzJ4c57N/Mg0P3CVPcZFrJhnz9H8ytMUw/e7/AFINH9ykUrLXK8ZC1B1PUbEMWdSwzhn0EGgutczfNRmgkBxlbhlZKHHLFhSiQuwG90Gl+4Sp7hKtZZBPjIZ+ZX+51R3DEGii3yH1/IuXuTWB37bThIz+41naHH7KuNcd+71Le7Fydyaxyt5u+FeWnI1jaifkPe40H0aoiICIiDrn7Q78DbiX/idH+vU6+eMd19DntDvwNuJX+J0f6/Tr540Fx7K1XHsrcH0QVaCSriCFa04KuJz6ILEREBERAREQETBTCAirh3mFRARV6Hf3JTocPJBRFXod/clUII8kBFXpPoqYwgIiYQERMICKvS70VMYQERMICIiAiIgIqhrsZAKoQQgJ2RMH0QXBw7rtFy/e0a5m+X610elrHqik1HpuhaI6e0ahgNVFTxjYNila5szGgbBgf0NwMNxkHq50u9FTdB6eO9s3xdNu8RnBbSYnMe0hrqks68d+jIOM746s42z5rqXzH88vMLzMwmy8QNUwUenmTeMyw2eD3Wi6wSQXgl0kpGRgSPcARkAEkngD36qMIhNVL0jy6jhQtY6TcHJPdBMwdW4XJXAXmB4octeuWcQeFl6ZRVxhNPVU9RGZaWtgJyYp48jqbkAgghwIBBB3XG0UD2fFkn5DspnU8srPMZKD0NqvbYcbZrOaai4R6JguhZj3p0tW+APxu4Q9YOM7gF5+ZK8/tYalvWt9U3nWupKkVN1v9wqbpXzBoaJKmeR0krsDYZe8nA7ZWnChkHd2PsUvurzEWEk/Wg7QczntGOL/ADUcNKLhhrfSOkLXQwXCG5zVFqgqWzSzxMe1oHiTPa1h8R5IwST04IAIPVdrPPdUfSSMIJOfPYKpjkdnYoK7A/MLt9wG9qHzNcEbLR6Sqq62a3sNEwQ09NqCOSSpp4gAAyKpje1+AAABJ4gA2AAAx09dBLnJaVJFTPHxmQsI9M5QekNy9tbxiko5G2jgzo6lqiD4ctTVVU8bTg4JY10ZIzjYOH1rqxzE86HMFzRU9PaeJmpaZtjpagVVPZbZStpqOOcBwEhGS+RwDyAXvdgE4wSSeFYuurDYZKyUAHH0jut6abfTUEYaZS/bzQV4Kcb+KnLxrNmveE+pJLRc/BNNP96bLBVQFzXOhljeC17SWtO4BBAIIIBHdSye2s400lvjg1Fwi0bcK1mzqimnqqVjxgYywukwe5JDsb7ALqvQXGlnb0l4bv3BWpQzU8cjXtqZNj6oN0cx3tJeYzmM09VaHuVXa9K6WrR0VdssUL4zWMzkNnnkc6RzfVrS1jhs5pwuvHDbiRrjhFrS28QeHOoqiyX+0yF9JWQdLi3qaWuaWuBa5rmktLXAggkEELkLiBUxXGj8I1UgGMZyVxzbmR0YLRKXgnzQd7tJ+2k482u2No9W8NNGXyqYAG1cHvFE54AOS9ge9pJ23aGgei2Hxx9q1zKcX9P1mkbHHZtCWi4ROgqXWWOQ1ssTgQ5hqZHEsBBxmJrHdx1YJC6n3BrKpnT19I+R3WltoBEeoVbwQdsbIMBMZWXWRumGDUPOPIkkKOCDwiXFxOflsgh6SnQ4+Szc/IKmM+SDFEbnDpx3BCjNK/OyznE43Ct6253xlBh+6PG/8ye7H0/Ss3qz5BUz8ggw/dj6fpVDSv8A/cLNyPMBOpvoEGD7s/1VDA4HCyzIwBROeCcgoIRA4q9tHI5SMfv5LMhcNth2QYf3OlxnKC3Sk91qpkz5BXskaO+EGlfcqX1VzbPKdyThav47Pkpo6iPAzhBpItT8Y9FjT22SPf0W4/eIsHcdlg1tRG5pxjdBt8xkOwcqeIhrcHCo7HiE7K1x32PmgSjr7I2le4ZVWBxOcErKYSBghBjMpSTuPzKOYeG7GFqDHNBOQFh1bS5+WjKDHLyVNHSSSN6goCCO4WsUGfBPnsgra7DLWOPc49FDdLS+im6Nx9a3TpmVzXnDVp2qpQavcAFB3Y9i1A+Pmk1S4+egK0f8425Fkexgka7mf1PjH/xBre3/AIRt6IPZ1ERAREQEREBERAREQEREBERBsPj1H43AziLEP/kmk7u38tHKF8477C+K7XDrBx4rsfLdfR7xyOOCnEAny0tdT/ySVfPJXTiS715aM/fXfpKDb01nJj2B+xKPT0ku+D/rWtyRTzxhlOzLvPAWVbo6ukwKmMjPqEGinSc0/wALWnK0+o0xNRk9YP2rkKiroY35djBWl6gfNVHNMzqGfRBsuKyu6gcE7rcdqsrMDqbnZQxU1wGMwu/ItwWp/gge8NAwPRBE6wRTMIazdYE1kjpD8TVuyGrp3OxEQTjyWl3kyEHLMAoNtVtPC5nSxoz9SxqaxmQn4e6zJdnZcs6hrIWnDsINPk0y+RmGtKw5dJ1DQSAVvOG40oGARlVmrI3j4QCg2NHpioD9891mx6WmOBgrcDJpnSDDMrOjmlBB6PzINDodHyOcOphP2LWGaL6cP6Dha3QVjWY8QALWxdKMxEEjt6oOP7npnxIjFEzfC2z+wKs95D98Z7LkyqrabxSW4+Sg+6TM9m49UG2jp9lHRhr2fEB5haQKaOOcnAABW9LtWQPgOHDOFs6aRpkOCMZQbgoJaDwg10YJHqFqdLR0NU8RNjGXfJbYo5IW4y4ZW57JU0za2JxfsCEGux8P4jEKjwTg79lR+nqKlid1RDI+S5CgvVuFqDSW5DceS2nd7pb3RPb1jO6DY1UKKGbpDBj6luawV1ojhAljadvNbRuElM+oLg8YWZbZqQAdcmMoN0VE9rlkcWRtwtrXumgmf95bgZWp+80Iz98H5VgVNXRiTIcDugxaewmVoIYfRbqsvDt9eA8MO4yse3V1EGtPUM5XKOjrpC2JoaAdhug2tR8KXPJaYzkLRNRaLFol6XMI3x2XPFvuTTI7DBlcd8Ra2J1QS8AboNgUtmiDRlnf5KZ1ngJB8PstRpqunLBnClfU0wG2EFlupqCn/ssQ+eQtC1IbdJP0xMbgFavLWUTcl8obkLa93hnq580TuseoQbl0XouHVMzYIY8nbsFyc7hPS6fjBqafuFoHAakusFyaRESQRkLnHXl0jgijbXtDHdPnsg4enisFgJkqIGHb0XFeu9K02tKgy2uIBuc7Bb/19PR1cI8N4wR5FaJpCR1Kxwjb1emUHGh4cSWaIPnYdvULWbRpCnuDOpsWSPLC3lrCrfLAWvYB5dlj6OqY4onbD8iDa9bw6ZKC1kRBx5LSncM3x5zGcrl41zA84aCsKrujASOkDdBx9YuGzWzgyRErdh4d0oAaIFr1mukJm+JoAJW4n18JILADsg44n0BS0zevwcBZ1ltFrpZmvmhaQ075C3Zd61r4CA0efktsNOS4EYyg3fLRafvdGKSnpmCTGNhutLi4Y0sDup0JwTncKXSJjgqg/Od/NchT3Fj2hoAG3og2LHwtpLkPCigye3ZSP4BNAI8A5IXIemauSOqD+kYzst8yXSV5aGxgnZB1wqeBwtR8d8BA7rHZoCje7PgDbuuftXuuVTRHw6bAx6LjmGgvDS4mDb6kGg2PTFotcgdUQNI27hbiLdMyyNY2mjHbyWHVxzsBFQwt+xYMAp/eGlrxsRkoNU1BaLDHSeI2nbuPQLY/u9n6XHwm9/QLe2o3xzW3ohdl3ThcfCjrA1xIIGUGnXiSzRAgRMyAtFfW2j3Z48NucKuoI+nPUcFbdLIzC/EgzjtlBjRMoqqucAwYytYmttB4PQIxkjbZbfthZHXEucAMrdD6ikw09Q2CDY96sYpnmbpw0nbIWixGNkwlcBhpW9NT1dO6AgOHZbFdLEWO+MYznug1yfVFtgpvD6W9QHdaazVVE89G2SdlteujjleS1+SsNkbhKGnyPcIORqV8FXh7cHKzfcWBvX0D1Wk2B8TIW+IfTutxy1dG2nI6hnCDRZq2lp39D2jI9Vji80XijZqwLm6B87ix+crSPDaZdig33BcKGSMYaM+WFOZaQsOGb474W26CnlEYIBxhar4cgjyc4QUqaiFhO22Fpk1xgD+w/KrqtzOrBeAfPdaPO2MyZDh39UGtsrKd7QAB9aBsMrtu+VpsIYG91m0zx1hBluoGloIaPyLUKGyRyx9Rb+ZRsc4xg42Wu2zxvBw1hKDRXWaMOLcHZYU9sjbIG9I/9ytzSR1JefvZWm1VPVul2jPf0QYsVqicwFzN1ZLHT0w6ekA9uy1WGmrRHtGTstHuluucj+oRuxlBiTRRyklo7qsFlbM7PT+ZVZS1kQAew7LV7fL04Dhg/NBHDZomtA6T+RalTWaBzN2KRj8jIaPyLUKVkzhkMJ29EGmP05E9x6WbfUt88pNmZS83HDd/T9HVFrcPr96jWjxsqP8A5mfyLd/KvHUjmz4dF0RAGp7Z5dh71Gg97kREBERB129oSM8nPEketHR/r1OvnhZjfK+h72g/4HPEj/FKP9ep188IJGcIAx1b+qkADgcD8yi7lSsIGyCwsIVOkkqWR3w7fpVrCNifVBRzcDP8ytDcqSQ7YVrDgIDYy4q4wOAzuronjqU7ntIxtugwwwl2FcIzkbK9pb15OFIHNyDtsgtliDWggLHaRnJWVO9pZgFYiCV72FuB3wom7FVDSewTof8A3JQZTZog07eStdLHnIAWN2TCDJEsfmB2UMhaXZb2ViYKDIa5mMbKOQZ3H6EbgYKrkILcYCtOFe4EjZWEHzygoqtO6pg+iYPogl6mY8lG4g9lTB9ExhAVzSMY81aqtByEF3QcZ/mVRGSM4/MrwNsFVzgIMcjBQd1V3f6lRBOJGYxgbKOQg7j1VuHeQVMYQFIxzRuVGqhrj2CCUyMIxgK3bB7KPsqjOEFPqUsLw07/AKVEqsHxAHzQavFJEW5OFmsliAwMLSosAALIaRjugyXSw52AVvXGT2GCsd/T/dBWhw8igmc6N3kFJDTxu+LG3msbpWdTBxb2QDTREbDssWVrGHpwFqQjfjsVp1Y1wf2QQPkEO4wN/JZNLc5Wt+GTdadVdRaMJSNPTgkjdBuOivVTEfp7LVGaqlaQS8d1tWFvcElHsb26j9qDXL3qF1ewsa78hWgwyStBye52Vjow3cOJVzD2ygm8R57lUe8+apkeqsed8ILXOCp1BUeB5qgx5IJIzkqTA9FZHgFSBBY9oIysV8Ty7O+FmP7bKPBO6CxrSBuh27q5UcEER3CseSB3V7tio3nOcoID1uOxKqI3eeVe0AFXZ3QWtaR3UzZ+jGfJWYPorHNP2fUgyhWtOB/Oq+89XY+XqsHpA81LGMIMjxH+qr4r8Z8grQMo4gNwSgp7zL5lQvfI5w32Qub6q9nScdkDwiQPVXspHEZIPdXg4IWZG9gZvjsgwHFkHfCq14kGQoq9wcdvVXUZY1mHOCCszzGNlZFKx30sEhKtzSPhKhgYDuUE08XiAFg7Kamq200fQR8lVrmNbuRlYM27ycghBvjRb21kz2gdlo+t2OhuIbnGcrUuG0sMVU90rgAT2WFxBlilugMRBG6Dut7FYk80+qcnP/5v67+UbciexVBHNNqk+ugK7+UbciD2oREQEREBERAREQEREBERAREQbE49O6eBnER3mNJ3c/8AI5V87dPUB10uRdv99d+kr6JOPYLuBnEVre50ndwPr9zlXzqQ008F1uPiAjMrsbY80G59ITU3v594ALc9sLXNYGkfg0jQO3YLaummuNX9q3FeaOpeG9AJyEG0LhJLBGHtOFk2W/U0bcVQBI9VbfLfXMgy5mBj0W1Ymhpd4jwN+xQci09/tk8haI2rAvdUxwPu+2Vte1up2TZdMFq1TPA5oDJAdvVBSz1s1NUdcziQPVaheL/BK0NaB81ovU31AwsCsjBB6XZ/mQTVNzZKeluAViiolGelx3+awRE8v233WUI5AMYQZNBPUvk6eonf1W4YPEBBeTjzW37axwm7ea3GzsMoNRhnhiblwG6kbcYXOAAG/ZaZLNF0fE4D0WNHU07ZB8Xb5oNarKsiLMZxjzC0V1xrcnEhwsyoqqUQbPH5Vo8lVTjJDvzoM1twmcR1P3U1PVSS1DIc7nAWlwyRyu+9nJ+S1G1UdTJdoQxhJLht6oN2yaHrZ6EVXxEEZ7Lbs+kaiNjpMO2yudIbNfm6dY80bujo79PyWzLhS1rKWUvhIA77IOHp4ZqeYxdiFm0ktTB0zdRGN+6tus8TLiWvIBye5V7qqndTuaXDPrlBPNriqY00wlO23daXNqKtlJPiE5W36qMOrHOa7IJyp208xaCASPXCDPdcp3bucSUF0qm/QeQtPI6T8RwUJB3CDPN7rG5+M/aojeaonJkKwHNyDuoixxdhv2INy0N/qWYHiHuuU9HaudBCOtw7eq4UpaWcgHpPf0W99PxubCOokAIOYqLiNFTykue0em62TrvWjbhKXseDv6rQJn07HnqlI+1beu00Lieh+SEGqjVxhYB1jK1S13qW5uETXZJXHNRCXYIJO63Xo6aClqmOmdjHfJQcoWXhjdtV4bTOfvjtlcg2bgvPp9o+6MZJI/fea1fg7rvTVoLDWTRN3H0iFytqbW2n701ht8kbwP7kgoNo6Ehtmjqw1M0LQ3vutr8Z9Ru1NMH212GtP70rcF6hfd4TFQMJcBjZbBudqrrUx4rGEZPmg41vj6uOBrJXkkKGw6ohtzSyQjZZurJGhpJOBjuuPnTUxc4OlAweyDemoNTQXKItjxv6LRrZqH7nAtL8Z9StDbLBn4X5WJVuidnLsIN6fsyaCXdY3+aw6nVPjOy1y2c7wyBh3mr2MGNiUG7qbVToHAh/b5rXKXXXwDqeMj5rjcsHYuOFKw07AQX4OOyDkka1jqT0Fw7+ql+7cZG2NwuObfLStl3k2zvutfjraPI++D57oN92C6u8YFru5W9qW4PqKuKEuz1YHdcW2C52+KQeJIAM+q3zZb5ZxdqciYHcZ3Qcw0dpNutza5wxkZzj5KyLV8ELxI8ghn8y1iuuVFctMsionAu6ANvqXGVVa6uOCZ02QMEoOS6fiTZrk0UjmsJ7H1U1VdrJDTucImZIz2XBthko6a6/fpiCHLfVzudrdSbTjYfzINI1dfaSuldDSgA7rZ9PHO2U5cck7bqss9NNcCIH9RyswRyioZlh8kGpQU8pjDpiSFdU01MaYta0ZWcWvbSAvYRkeawXys8E43QcZ6rs7p5neGCB8ltVmn6joIy7BK5IvIlc8lrCQcrRRFMGn70fyIOOq+zTUTjKMg91pzZqmQ9PWVvTUsFU6MgREfYtp0tFVmQ/eyST2QadeaWqfAXdRIIW1nUU5jcclch3e31zaTJhOPq+S2pJT1AjeSw4B3OEG02UsonIf2ypxThrw4hZD8mctA3yr3QTebSAgujq5I2AMOMK59yqTGQXnCxXxvHcK0seWnYoMd9XKZMucSUhrcThztwD5qCaOQE5B7KFkbjIMjzQcg2q4QSwNaAMrWWGOSMRADJWzrG0dI33z6LdlGMzt9chBg1+mqlxMzc4K2vXUE1PIc52K5aq2T+5jEZIwuPL31CU9bcHJQaPEXtbjdZdNUFsgJOwWIjHfHjKDdLLrGIQD05W4LPqGlhjDXgLYbSzpAzus+lfDghz8fWg32dRURJO26khutFM8ZAK2SXQAbSLPoXxdQIeg33DcLe1gyxqzAbbUQF/htyB6LabHQujBDt89lqlLcrfTU5ZLKASPNBpt7loonuaxgGCttmsb4nwEYBWdfK6hke5zJAQVt1s0XWcOBGUG6aStbgZwtdpLxTwMHUAtm0VVTADqeB9qy5Kyk6dpN8IN3jVFHEfLK3ryp6noq3mv4dwMDep+p7YBg+ZqYwuDZ56Z7j98W/eTtkf7b7hq5js/wDwrtX63Gg+h9ERAREQdd/aDj/rOuJH+KUf69Tr54Wea+h/2gu/J5xIH/0nR/r1OvnfaCc47oKHdM4V3hu9E8N3ogtJJQA9wqljh5K5rXdsILTnzQNJVzmEBGdkBrHZ2z+RXOY8KSNuCkufJBj5KAuPYqh7qrdigvET3HfKr4BAJPkpo84yArnhzmkII4HMLsbLKkbGIyRjOFhRMd17LJkD+gjdBhPPx5VeoYR8b8kkK0tI8kFCd1kRM+DJ9FEI3kA42WTHgRkH9KDEcSHEIMnfyR/0j9aqOyCVsrOnCtc8HthRnclUwUF/WE6wrMFMFBIHjzVHOBKtDSU6cIKhwCva8YUfTlVAI2QXF+5wq+IMYP6VZ4bzvhPCf6IKHuSjfpBXCKT+4KCJ/UBhBOwDp+xQyNwfllZbYX9KgmieN8dygx1lQvjDMEbrFIwr2j4cIEuC4kdlcxnU3Pmo8E7Kdnws+pBA4YKuY0k/YrXHJUkWc7eiDJjBIV/Q4FIxgZA7qQAkdkETw4jCqxpHqr3IOyCRrwwDPqs6mrI2DB/OtKe4YwTj1UkTo8El31INabcIcLTq2pjkfkAHfyUQfFv8SxZnMDvhO2UEsrmvAVYo+7gsXr3AG/2rLgJLN/VBOxu2yjkjcScKSM4OFV7snt+dBjFhHfKpgjsVJIcBR9XyQU6iN0c8d89lQ9lE9xwUFrpsnv8AkVzZdtysfB8irg12BugymSbqUSdljRg7KcNxhBf1Z7q4OHZRHsqg5CCr3DGfmrC8YKqcHYqN+MbILS8AlWPdlWvPdWjsgtB6SFcCSQfJW9OSFcNvsQZAIKPaC1WRuG2VKXM6fsQYRa4O7lSRtII2VOrLj9ZUzAAEEnUA3Cx5ZOrYeimJYf8A7KxnnDsNQWFrgcq+PqAyrugkDZXlh6ED3kAY9Fa6qcdgVju7lACd/wA6CbpdL55UbuqMkA4WRA5rB8RA+tRVBY52RhBY1r5D5rIZC5rcDzVkTmjG47rKYHPA6RlBhzNewgZ7rfulOGNRqKk94Y0nIB7LZU1PO97C1hO/867IcILnBQ2lrKgAHpA3QcYVWhazTPW9vU3A3Wwr3NJLUkyEl2V2F4kXaikjlA6QCD2XXe7vY+pcWHIyUHfT2LDgeaTVAHcaArv5RtyKP2Kn4U2qf+L+u/lG3Ig9qUREBERAREQEREBERAREQEREGyuNnT/sM696uw0xdM/V7rIvAeWwMut0rzTswPFcdh8177ccXNbwV4gOccAaXuuf4pIvCrTN9sdPca7xZmD74cb/ADQabZdIvo5g57Tsc4wt0yUlOzHis7LMj1Pp50hxPH39Qo62vt1UM08gP1INE1BbqOspOiNgyQRsFty28GK2+uc6BrviK3XN1yACFpJz6ZXJ3Du/UVpjY2vDWEY+kg4NufL9d7ZF4xa8YGfNbWl0VW0bi2XPwnG5XcDWOutPy0LgJIgOnHkuBb9ebZUySOp3NILj2x6oONG6ankPSAQVbPpiWA/GCVvKmm8eXpp25OcKK7U9wA6nQkZ+SDYstrbTblv5ljPMbcj0K1240tc/bwj+RbdqbdciTiI758kFYKyKCTOB3WXLeWY2cAtFdabm13U+FwH1K11HUN+kCMIMyquL5MkOP1ZWC2pn6s9ZUZHRkPON/VVa6P8AuhlBLNV1RbgvJz+ZYXjVBd09ZKyXPYW4BysYf2QfWg1e0Okp3CSU5Hf61vKw6ko6G7U9VMB0scCfsW0IZITCAXAEDCpiBzg3r3PzQdvJOYDSZ0sy2tji8UM6T2z2XFt44lWmrpZoowzL84wAuIW2uMsDxM4hPcA0Za47IMW9CWsuLqiM4aSViubUAH4jhZ7z0bHyUEk8TdsjKDTQPDk6n+XdZbb9TwsEbsZOyhqR1Alq0Cpgl8Y/CTkoNYnnE7jIzseylhJ6Mu7rCpo3tiAIOwCzYmnw9wQgikqGh3TnzV9NUR+KAd8lYU7cSZyr6UN8YEnt80G86FkToAcd1uKhjDaYuaPJbctr4/AaC7fK3TRb0nw+iDaV/rZoZSGvI+S0COtlleS9xOD5rVtTNeZnLb9O0jq790GoitY0Yed1eLhKN4HEEbDC0uWN5I6QSsilY5uerIQZE971IwAUVU9mDnYrkLQnE+9WdgZdapziB5lbBifHHu8hJGRTSZD8D5FB2n4a8cqCmrw+4Oa5hcBuuSNV11v17TCptcbQCAfhXTHTFPRiVglnIHUF2x4X3ew0VlDDUMz0Y3I9EHEXEu1yW2NzNwQCD81xFTW6rrZHljiBnbBXO3Gi5W+rMhp5Gn6lxNpqspYC7xXAYJ7oNFq6OotrS+QnHqtEmujpnEBx2K3jq+spJ6c+E4ZOey47jjLXOJOx7INSjrngjJJWZHdmswCQtGY0Z3KOjJPdBrE15bjZywJa+WQ9THnB3WFJE7pOHJFFLg4OUGbDWz5HTIQfP5rLFdWAj74fyrTYYZi/bJOfRZfu1YezSgzfupXxgFkpH2qai1LdaSpFSZ3AMGe6080df0/2MkfUoZ6CuMLw5hGWlB2q5c9fVGs69loqJTJuG77rsnxB0FHbLGZmxYLmdX5l0z5KomUmtmyTk/2QbL0O4uXKhl001rMZ8L5eiDzs1peamwXx/Q8tAJwtEPE2snjcz3g7DCyeMcElVfZfByQSey4sp7PcA52zjnt3QcpaU1dJ90PFqJMguzuuQp9a0jXsk+HAA7rhLTmnL5PKPBgeRkeRW7qnR2pfAwaaXdvoUHJB4l0dbD7rG5uRtssObUXgQOlc7YZcuM7Npq80daX1MTw3OSSCt03One63OySMDdBhXDifRQ1BieW5GQsNvE+3udgFq4r1FSNFwf8AfCN1pbKcdf8AZD+VBzJX64t9XF2aStHg1LQRziQtaADnyWyYqV5jHS8nH+pPcpurzCDka9a0tU9AIWMZ1AfJbHmvVK+GRgAyc4K0ypo3uacOJOFgfc+qLSA0knyQZFpt5rrgXDcErd0+mQIQ/p2x3Wg6UobhR1YkqYnBme5XJFTXUbqPpDhkN9UHF9wpWQSlpb22WDmPdu3otWvkjHVDukjBJWi7ZOPVBJ7oybGADnZXtsfWdm7qSkeGkZWr09QwOGcIIKK0SUnxkHAWrwTtglbI7s1XmqhdFg4H2rGmfTmM/GEGvVGtKIUxh2JxhbFvNyjqpCWY9VjVjIfGPS/8hWDK3OcElAdNnYKnUVEGu2O/5FKGux2KC8SSDzV7J5QfpEKNVaW53IQZbZ5tj1FZ1NVSNAPUVp4cwNG4/KsmGSMM3cBsg1OK9OiPS560q61dfVPzBIQPkonPhLjl47qRk8LW/SHZBgn34gdchOBvurmyysGCSCppKiLf4gsWSSMk4cPNBd71O0npcR9qtM1ZI4ffDj61Z0SOx0qeKjrnjLGE4QVElQAAXHvvuuWeS+V37brhh1E5Oq7UD/GolxUKWsBw5hH2LlLkzjkbzecLw5pB/Zdaifq97iQfRgiIgIiIOvPtAt+T7iOP/pSj/XqdfPNRxiSR4Izgfzr6GvaBfgf8R/8AE6T9ep189NvOJZPmP50GQKZp7BV91b5hSg4V4JIzhBAKMEdlcyhDjjCyW5ONlNGS3ugwpLVluQCoG2xwPY/lWtPnbjBIUXjR9tkGCy3E4xuklrPSdsrUYpWA+XyUrpWluMhBt11rkyQqstbwRnJWrSyYO2N1Rkme+O/qgggthIwQVkOtPwglpWZBOwAdRHZZD6qHp8seuUGhe4eG/JaR9qv92acDH5VlzVVOSfiCtbU04IHUN0GLJQt7/wAywZKPD842WuSTRluW4WnSOJd5ILPAYI8BoyAtNqSWu6QVq5OWH6lpFUCZEEIaXbq8QPIz5Ixj/Q4U7Q/pIweyDGBAOD5K8Pb22VropMk9BQQyf3JQSgtPmPyq9sbXdsKHwZ/7kqWJtR26SgvMIHkqeB1bhSujqOnZhyrooqjH0CfsQQimPmE8DDgMLK8OoP7z8yt8KcPBe0480EjYGdOenzVwgYBjAUglhDekkZCqJYfMhBZ7vGRkNCuZSNcQMD8iu8eD+6Cmp5Y+oYwd0F7KAdOQFFVUDeg4butWjkZ0hY9TI3oOAEG2XUREhbjshpS3yWe933wkAK2R48wOyDBMQb3Ct8lNK5p7Y+vKhJGCgx39wronAEK1537qje6DUoXNLQpgW7f6ljQdgpur6kFHkeqt6hjCo45Ci6jnCCksb3fRCx3CVncnCz2nbyz6LFqSc/Ygg8R390fyq9kMko6snHqosErPpDiMAgII4KdwcC7y+SzQOnYK2M5OMBSYBQWCQNKeM0q2QBWhgIzlBWWUYwFD4o9Ve5uQVjPBB80Er5M9irPpZVGgnbfspGMI8kF0cB7nfKkFP8lc0u7YUgJx2QR+DhXNYR+X0V4ye4RBHICAocndZEnYqAg5QWPJxsqB2Rg91cRnYqgYAgtLM7p0fL8ykVRjz/SghMZ8lG5hGf8AUslRyd8IMf4lQOeXYye6kLTucFRtB8TsgnELukOwrXZaCsoNPhhY0rSAdkEAe4lSxxkuyQoWfSWTE52fl9SDKAZ0gbKjwHM6R5p1RYz1b/WqdTTgNOSgxXUr8k+qnp6cO+AjdSObLjIGyQw1ZmBDThBn0unJKwEsBWNVadnppfDOcreGm5XwNJkbjAWPe7nS+9jqc0INKtOh6i47tz28lPW2J1mcI5W4PzC3voq7Urf37DkLSNd1cVRVAtIxnyQbZZNTRYL2jY5WqUuu3W0CKGTDR6FbeqgCNvRaLNC4OLhuCUG59S6wqLxGWNkJzstriGR+7s5VGxuBBKzYzhu+PtQd8fYswOZzSapcfPQFcP8AnG3Ip/YvuJ5o9Uj/ALga3+UbciD2fREQEREBERAREQEREBERAREQbA5geo8BuJAacOOkbxj6/c5V841JZr/WXKtFPM8Ykfnf1JX0ecfTjgVxGP8A3JXf9TlXz66Xr4o7ncAQD99Pf60GgU2mNTNd8U79vmVrtJdq6xgNrJScbblbmFzjMhw0ea2Zq5zKhxLT39EG6LRxHo6eTMxaQfVSXnWVVcyHWyXpH/alcTtpGucAHkH61uqw6ev1QB7jE94PbAJQZt8uuoKim6DUPJA9VtSK83Ch6veZyd/MreV003qahgMlbSvYzGTkFceXqGaZzw0dj5IN16V19DQ1nXUuBGc7red04oWeoaGtazOy4Iho5PFAeC0A7lZ0tLHj4ZHHA9UHJtTxAtT/AN60j6lfRamtVUc9DTlcUOpG4GXFa7ZBTRkdb+yDkuSstdYzpijGe2QtMqbNDM0hjNz6BQ2uqtsWC+UAD1WvMutqwOiRp+pBsms0lK9xLQR8gtKqNN1EALiSuSZ6g1jSygb1nHYbrb9ztWoPDcXUrwDuNkGwZKWSJ/SSdsq0gjutQrrfc4JS6ojIHzWnudvuRkd0ASSg4BKkpnSvrGRknchRt77bqega/wC6UTyDjqG5Qb++474LaKh2cFufzLb81zjiBbkZG3Zb8rauA6fDDgEM/mXF1cGOe/Dt85CBUXIPccHAKw3mSR/UCcKB0b853wsmFrwMlpwglEwa3DlE50TjnA2SToOdxn0yoeh5yd0Eonjbt6KVtSxwwPNae/YnJOR81NCNu/mgrKwvJOMqxsbmPDhkKcnGdwrHEk9kGpUda+NoGexXIOnZRUUgzvkLjSDpIGTg5XJGk6q3xUYbK8ZxtkoND1JTB0rtvVbULRG4+S3rqJ7JZXmn3Bz2Wy6qkreonpIGUEjJIicnH5EkmYNm7LHbFKw9L8qkjHZ2ygvkEj8dJwroRIz6RKrCfIhXOe0EYwgq+rqKdodE8hcrcP6m91dF1RzuAxtuuJKktMYwfMLmbhZLHFbsHGcINI1ebgHPFRITj1K47mqamNzhE8jdcna7la57y3HmuLpnDrdk+ZQH1FTKMSvJHzVjYc7qplij3e4AHspYqStrBmiaXj/td0FkdK55GB3OFuyyaDmusYe1pIIytvUlpvjnhrYXkg7gBcz6BuNLZ6VrLrhjunHxbINnRcLJ5Hlnhkkeiw7lol1qPQ9hB7HZc00mq7EypJMsfSfM4Wztbahs1VUAwTMIz5EIOPY7GymxI5m3dajR01K9wHhjOcLKrLnbpaYNbM0kj1WPb6uibKC6QYz8kGqT0dHBB1mIDZaXK+hqInxNYMkHdarc7hbn0ha2UArb9NPRNeS54x33Qbx4PVh0ffBcWHpHV1b/AFrn/VnHI3W1inM4OG4/Muu1mmpa4iGB4Lu2Ad1qtbaH08DpJS4DG+UFlRLFqS8OfIAcuWqSaPpKVvjuh2Azutv6Wq6Fl0LGyguDu2VyLdaqM0BBIHw/zINN0pqvTtjqvCqYWEg+YW/K7ilo91MMQRfCN9guvda2kNzc6WXALvVZFV9yn0j2MqB1YPmg5Fr+IGnbzUGCiiYD22wtOr/DfSPjAGXdlxjpag8O7mUPJZ1dz9a5MrJIREPiGwGUHGF10ZPX1zpGNJBPkoP9jqoiHilpx3XIUF1o4JSJS0Aeq1CfUFofSloezOPkg40odPFknglpOCtSm00Aw/AfyLUaa50Iri4uaGk98rWJrxbCNpAg2FPZBTOLns2+pRRR0kMgmcwdLcFblvVxt0sZEcjc/JbWnkgdTva1wyQUGZdtT2t9N7tSsaJAPL1W2fulU9LsvOCSRutLfT4rC8OOAst3SY8A+SDBqZ3TOOTkkqJrDkZVXRSB5IBxnuqhsnUMBBPGwtHV2VRK9p7q7plDN2lRlrzn6kExq5MAB351G6adzS0uOSoQHg79sqVpbtjGUGI6kmLi4lXNgIGHLNMFXM0eCzOe2FE7Teppz1wU7y3vndBj+CMnYK4RAN7KT7jXuP4ZYXghXtt1xaz74woMMxnOArHQOc/pHmpnw1Ubj1tPyU1LpfVN1k67dSySN/7XKCSC0yyMacndajFpeomhLmk9lm0eg+IJZ0tt8pIO+GlbkoLRqC2UxZcaZzHYxuEHH0+mapspHUVWLTlSXBmTut31UNYZnHwj9eFFE+eKYGRmPsQaPS6ArKwZAcsg8NayDLnB2PRb+sN1iiGJA0YWrV2oLeY8Esz9iDiv9ixpiA9pyDutXoxQ0cBErBkDB2WrXC5UsjiWkflW3q2aBzSOrGSggq6+h8Q4aFvTk+qYn84PDFzAMO1bamj6/e41xlVBjnEh/wCdb+5NwRze8LBntrK1frcaD6NUREBERB169oAM8oHEYetHSfr1OvnpoAfFl+X+tfQxz/fgg8Rv8UpP16nXz1W4Zlmz/wC+5QZY7rIiYHYWPjB38lnUxi/fOHZBPT04ceylmojjLRhX0kkIds4LJmmjDe4wg0OpppGDPksRsMryd1q1ZKyTIZglYcbXDPwlBWkp5JDjPms/7myHG/l6qy2hwkJc3butaa4AjYBBpDrO7GcFRG0vaM/zrcMlTEGAEj6ljPrIMHdBt2emliBGTssaQzeGR5rVq2ogcTghac90fSSCEGkSMqC4/F5+qMhqeofF+dZgY6V+I25+pV92qmPDnRkDHogzKSmllaA491qAsT/DMhB237qO3VELCGkjI7rcQqI5KUxx4LvJBs+SndG8s9Fp9RTFr+ojIC1ipt12M7pBE7oJz2WDVZjBZMMO+fmgwWzRN2I3HyWQx8WPJYBjJeSCcZU7CA3CCcPiLsABTx+EcbBYg6Ac5UrJY9h1IM3rg9B+RBNTt/ejv6KDxIf7pWuDXbg7IM4VUB2LR+RXCrpmjdq00OiHd+6o/pcNnfkQamLjStO7QsaqutM5pa0DJGy04QPLiRlYs0ZjkwfNBI5srnl4JwSfNWukkYSCVlRujEYyRlYk5BcSPmgs8WQ93LOoJ3lwyStP6Hf3JWZQjDgCMINwMkf091HK4lpRj2dHfdWve0sODug09xxIVDUTBpxnzUrz8ePksKpHU7OSgjfKc91c1hkbkb7KFyzKYZjGyDFcwg/+xXMiJI2Ujxuduyo1/S4fXhBkRtLQFcqNORlXAHZAc0/nQRZ3wquB22UrOyCDGFa+EvOSFJIDk4z3VWdXYhBCKXJ7fmUrIugYwpQQ0kuwjnBxyOyC2MdJUzYy4ZUbQScNGSsuGGXoGGIIm0TpTupRanHAxusumgqXHDGEk9sBZrLRfZXAQwOJ8sBBoVVb3U8fUVg9AdvjK3FdrDqKOMunppGjv2I2Wg+DNEPvowc7/JBa2MZ7FSMj37K3raDuQFeyRpIwQftQTxwjA2V3hD0UkT29PdXP+QQY0jenyUXUFLMT6fmUB7IDiCqFoI8uypg+iu+LHnhBA7YlWl+Mq+UbHvlY7g7fugkD9/8A2oXb7KH4h6q5ucZIKCXrCo4Z3VGt6ir34azHmgh6h2V0YaX+Xksc+J1Z37qSHr8QZyg1IAeGNliz4xjbKymk+H2WFLnr80ETac9Q9Fl+B0xFyta4ADYLJ8RngEE7oNILnh3fcLJtsb6mrbHnv3WNJu4/WVnWMkXGMN7koN9Q6V6qRknTknvsrxaaelYOqMZC3RS09e63MLYjjHotLuMsMbC2fAd6INq192jt5LGEDOwU9u0Bc9WNNdA5xB37LQtSwulmY6HcLmnhFVT0NmDZI8jp8/qQbVs2gLnbMhxflndbR1jLNRVnhSuOQd12BN6omum966Wk59F1/wCJtRTVN3MlM4EZPZBtx9cHjuqskY/0OVp4a447rLh6WN+M4PzQSua3yA7q5sZO4H5FYC1xw05WTHsOyDvj7GBnTzRaoP8A3A138o25FJ7GT8KDU/8AAGt/lG3og9mkREBERAREQEREBERAREQEREGwOYE44DcST6aRvB/5FKvnBpL8+gudeevHVK7G/oV9HvMH/vCcSv4IXn9SlXzQXEH7o1QAP9mf/wBIoN60uqzI/d4OR6qC6XL3gF/UPVbTtwIm32C1iobmMhu/woNc0LaX6kuTaVgyeoZx9a7TaOt9j0TDAbnTtJwCeofJddeAcjKbUgkn7dQ7rn/iVebQ+OJpkDSAM4+pBrnEC/6V1LbDSW6nja8tI+EBdeK7QYhmle+M4eSRst9WK52hswImBOfNZd+raefAiAI23QcMX7SraeHqjYQQO+FtEUUkRPV5FcwaljlfTHwmb4K4xroZ43P624wfNBo1Q3bpCgjE7c9LiPULKlBzlytaRv2QRl9wfkRyEeuFNBLdIMufK4gd8lSUz2sfl3bKyamWJ0RDTug3DofW0Vlqw+4vDm9XZy5SqOKOmLoxkMcUWTscALrXPHI6QkNJGVqdhg8OrZJK4gDf5IOZNTNttzpTLSxgZHkFxtUaal+OQA4yThb6o6ujkoGxRODn4UE1JVvgc2OEkkeiDjl0Pujyx3l6qWmrYve2NHcFZF5sd4jnMr4Xhhyc4WjU1PKy4NByCD5oOQ6wVD7UHBx6ceq2NUNk8U9+65BlP/UQD1b/ADLYlQx/iE9O2UFImt6RkbrKaGdBYO57LGjLQB1HCljmha8dR+tBjuts3X15ODupg1jGkEDJWoGpiMIGQtNmcC7IIQYklK57sjOCnhOjGM7LNa7AChm3/Ogxi/CmiDS0k4WPIfIKWHqwUB7iHfD2yr4bjWQyBsUhACjeHA5AVjQfEBJQbvs1SZwDOcknzW/aDR8F0oDUNjzgZOy45s7mMYOo43XO2ipQ6xERgH4P5kHCGpaBluqnRYAxkdloTJGOz2OCt2cRIpTdJCG43K2VTvbGSJDgn1QSyyBv0VbGx8ozklRykP8AoHI8lmULHNG439UGNURua0ZPmuXeGUb5KENbncLiyuhlewGMZ3XLvCmCobSDqac9KDIvemprjI8bnvhcW6nsklomIORg7rsJHHOJ3ksGPmuIuJ1vuFRUuMEZI37INoWTSlVqt4p6fqJzvhcm2bTdPoiJsNzjBJH776licDo32i5tmujA1gIOXLd/GG52y5TsdQyNwMZ6SgwKS9WKkmM0kLMOHnhbN1rU1N4mLrO8sbns1Y9XDJUUwZASSB5LKstoujIzmBxz6hBodvsmoa4CFk7y7sd1mTcPNQdQM0khJPqVvfS9NV0dZ11EJAByFveqr6ZxYSG7IODpNAXmBvXI95H1rAmsdxp8kOIx6rnK6TMrIPDo2BziOwC2TctMakna4RUjz1Zx8KDjOcXFp6XSOxn1UMUdZI/wg85Oy3LcNBa3jf4klvlDO/0Sse3aQ1R74wmkfgHfYoNc0JQVdmqBWVhJYSDuuSa+8Ut9gFDTgB7xjbutt1NpulFam++wFgAGSVHpCaMXqLDicOHdBurQ/Ai9T3L7q/fDG53UPRcq3jhHXfcwvDXbDfZcnaCvtrt1iidX+GxoaNytx3bX+i5LNI2OqiJDTnBCDoNrzRdfbq17WFwIz6rZMFmuweZHvf0g7jyXPHFPU9hqrnKaaVhGT5hccMutu8CRpc3Jzjsg0u13NlAehxAcO59VqdTqcSRFofvjC2jWvjNW6QnDSdirDUUxaAJNygperrVyPLoZCMrQ47hdurBmJGfVajVAPBLe2FhMjPXuPNBkmrrRHlshB+tQvrrkWn78R9qmkw2MeShc4Fh3QY8FXcJJQJJcg7d1qQ8YjBzj61p9K9gm+LHdaoZY9gD3QYVREWfGQcrGY8+Jg9srUK1w6CRhaYxzfF7hBqZhYYgcb4UTY2NPUR2WX1MdAA3BKx3sf4Z2KCgqYZPgAGU8Bp3+a0+GGZs3Y4ytUbktx5oMZ8AecNzlWxWyV1QG74KyoWPZJ1OGBnus6kqIDVMaCM5GAg3hpDS0VQxhljyCRnZdjtKaBsIsJlno2lwZnt8lw7owSMhjke0BuQcrsXpy/wBjp9MvbUTMDgz1Hog6/wCprVaIbvLDHA0AOIxhbZuFnocEMiwtx6pv1kfqKYtmbgvON1pVTeLSc9Mjc5QbJueno5HNMbMYO+FzxwXq9KWOhb91qSNzgP3wHfC4nqbnQ9Wz2kZW4bPdbSKUtlnAOPIoOxVLxI4cUskjX0UHnj4QuDuMPETTNXUuFrhiY0nyAW06u42l07yKrbywVxxq8080xMEpdufNBuF2rbccHpbkj5LTKu709QS6IAbrZ3gnAw4rOpAGgAny80GsPuUrchj8fUtGrbnXvk2lOM+qnPST3WBU5DtiUE0dZVvGC85Pqo5ppnZHUdz6qkTXYG/kqytcDughDnd3Erkzk235veFv8MLV+txrjRwOFyVya5HN5ws/hhas/wAbjQfRoiIgIiIOvXtAT08n/Ed3pR0n69Tr567afvsp9R/OvoT9oIQOT3iQSf8A7jpP16nXz00Dg2R5JABH86DUX/EMN7q6OnlPYnHzUcUsYdu4LUIaiDAy8IK0dJN1d/zLJnppiMb5UlNV07T9MLIkqYHYIePyoNEdBJA/qkzhXxVcROBjZTXOWJ7AGHf0WkwN+I/Wg1+mqIgMjCy2ymVwY3uVokT2RgFzsD61qVBWUona5zxgY7oJbtRz08Pi5IHdbeM8z2uwTt6LeWoa2kmt/TG8EgLZMbgMgeaCB7pi7JJOD5o3xDnvupCx5OcbFXNYQeyDItzmUz+uXBWqVd1opYCxjRnt81o8o624Hf5LFFPI053IQTUVJPNVlzSeknYLe9ktMvjMlkJLNid1tqzvYxwMm2CFyJa7jQiEMLmgkYwg1SY2tlGWGJvWG9z3XD+r2A1pMIwMnYLkqre1/W5hyD2XHOpqOrdKZGtPSD3QaIzAjGcZUL3nsCruos+Fx3HdRlp6xncZQV8KVwzuq+BLjqzsspskYaBkZwry8GMgeaDTuqQbFxH2rJhqT0dGd1jPBLjsjNnjKCYwSvJcD3KniDoxl2cBSRyxhg37Y80kcHMPSUEjK2Juxx2WBWSNkcC3ChcHBx7p4byMhpIQUy75q5meoZKYPYjdUOQQgyeqMDdSwuAGW/YsQOyBusiFzW9zjyQZXjO7ZQzOwRlQlzfIqnWEF5d1O+tQyjO2B3VwcBvlWSOyduyCB0ZJ2WRTnw2bq1mDuVI4sDcjGUEc0jQRhY7Gue/YZSR3U4fJSwAAnHoEGXGwlowpmR4G/dWx7AAKYdkFmB6KoaqK4dkFR0eYCp0gu2G2VapGYxkIIJ4nOb8OyiYHMaQ45+pZjs42WM5khPY/kQXU8ojd1O7ZWqRXOEMwcLR3Rv7OGyt8M+pQbstV8o6eRrpQCM75XI1h19p2lc19RHGQMZ7Lg9kfxfSIWW2JhjILzkjzKDsNX6r01qqnNNSQxhxGNu64+veg/FeTDGQHHK0nh1PR0VYH1Eu2f3y5Yn1BZXPZiRnbfsg4cquHdS3docPNaVU6VnomF7gdiubqu92d7D0PYTj1WydR1MM9O9sGCT2wg4yEro5DGTuD5lTtqADucrGqqKsZVl7mENz3VAx59UGVLI2Ruw3wogCMqgY9u7hsrgCewQG4HkFd8OPLKt6XDuFRBY5gceyp4DfT9ClxlMFBEYWkYUT4w3YDyWVgqKVuRkIMQODDuniZcN9lR7SSRhR9Ls90GTmPp+xWxzRiTHzVnS7HdQBrusd+6DWA4FgO2FhzyMB8lkMz4YG602pz14z9iCRri47HzUjopC3bso4WO2J9dlm4IjOyDSnNIdhaxpynIuUMzuzTkhaVK1xfjC3DYaWqLw8MPSMboObqDU9titDYHMb1BuMn6lxPrO5SVVefAdhpd2WtMfTiAtc/cd/yLad4fGavZ2UGfb6ZtS1hlGcALe1u1fS2Ok92aQDjC2jY3NMePktMv7QZ/pEboNU1Tq6qq3PdSykdXfdbFlfU1LzJO8uJPms2cO6Rgk4WK1wGzsAoL4mtA3HkjqSSU5ZnCdbBj4gtQppoQ0ZIQaaIH0zup+cKZtYCMDCuuUsb2kNK05jXHsg9CPYvyh/NHqho/wCANaf+cbcig9iy0jmk1Tv/APKBW/yjbkQe0qIiAiIgIiICIiAiIgIiICIiDj/mE/3g+Jf8D7z+pSr5qHtD7lWA7/fH4/8AOK+lfmE/3g+JX8ELz+pSr5qC5jLnWFxwDI/H/nFBd0CIBw2WVT1AcC126x5ZY3swDvhR0wx3yg3Jp2/fseq/eWOwe6zdSa+uF+I8OckN9Ctn1zA9gIOCFZb2EElBrtHqO5ULg905wPmt42nW752N8aUHA81xxWRue3vhZtmbFGPjfj7UHMFmu0F+l92wCe2FpmsdNx0gL2sxkqLhrPQQ3LqfID8Q7lbt4gzxVUTfdwDgIOF6u1mR2GjGPJYT7W+HPVkZ7Lcc9NWNlLhGceSwqtlQdyAg0N9O7BAOCohTTN7uyFqLop87BWugq8f2M/kQYrY42fSaCqvna0ERDBxtgq2ohnjGXjAwsMbu2cEGs2G7VNFUB00hLAfPst+UPEK2MmjieGnsDnC4yLepmPVY0NNmrYS446gSg56u94s12tgdBEwOI7jHdcemysdV+KxuATlalbH0woWsjfl2OyyI4ap7x0Rkjy2QXVLOig8NbWqqdoDjhbrr6erZDl7CNltmqIDHdWBjug0CYu6yAexUT6Kod8YJx3Ur5IfGI6vNahG/qiPSMjCDSmySt+9ucSQpmtcd91DIPv5J23WQ2RgZud+yCx03Sen0Vd5GrGlBc8kHv6LIgDuk90ED4yO/6FNAQMA/kVr85xhI9nAnZBmCAPb2WO+nw/I8is6MgNGN1G879ggkgldGwBpxgrmHQeom01oLHuGQ31XDQ7D61v8A0tTvktzuk7dKDTNZ3annr5HdyT6rYFYx9RKTCSB3W4dTxllW7Ocg7LRacbn5FBdRQOGA/fHqs50rYXDHorYMb5wsWteDKB1DKDcVo8KtcGOaDuFzHoqSnoGMiDQMtXDOlLfX1sobRNL3ZHYLlyy2u5UEjHXCJzMgfS2Qcv6Q00zU05bFHkkei2HxgtVDo+qEdZEMk43C5i4D6gsVory+6zMa3/tiAuK+cWqo9SXQSaecJGdX7zdBxZcLjDV0bTasMeR+97rTqGyXasBdWSPfnffdY9ho6qip4/e2kYx3C3xbbpbxGA5zcjbcoNEoKBlrk6qloLRvuFqE2vrNb8R+GwEbHsq3u4W57COsYx5FcTajZTSTOMcmxJ80HJ9VxFtk7f8Aa3SHH0wtGn1hLJ1ES7H5rjughiBaA8n1WqhkQxlyDfumddsttUJa14LQc79lypa+Nuj2SRGeGIhuM5AXWqpZC9uC/A8isaGhp3f/ACY7/NB3BvHHPh5WW8QxUtOH9OOwytnxcWNGwOMnu8OQc9guu7LdG3cSvPpuraihjMZw89kHNWtuKtj1BSOpKBrAcY2wtjaZrH0l2ZK55wXAj5Lj+2URjrMhzjut621rjWRZyBsg56r9RXG76dZSW2Ytf042K2HU2LW8FvllkrJukgnufRbs0ixjaVnQclbkvT5xZpQY9uklB1D1BPfYLm9tTUPJDj3KwI6yvMozKcemVuHW/W69SADfJ/StAhZKH5xsg1WaZ8tMAD8XqtPhiqQ89Tzj0WYzIjHmcI0O6wS3YoMiNp8Ide5VPhClL2NjGe6h8SInugudEZtm+atdbZOg9wsqmLSQG4KzXg+GchBtZ9O+nlJJVzJ3dQPUcBZNf1eJkjAOfJYTdzhBqTWuq2dDTv2Q2KZg68H1V1pd0yA5zutxSyjwcfL+ZBtmEuhf0vPbyWUKhjnBmO6xaoOE5yMDKiYPvwOUGrOp42sMnSN/NY3ita/PkCs2Qg0w6e60uWGUgnBx6oM18rKiMMjGCoLTaqma9xMycFwUVBFPNKGw7nPZbg0/Zr+NQU8pp3dAcD2Qc/WnSbodOxzMYeoNzkLZeqLnerdRSww1D2jBHdczW+up4NLMiqAA8Mxv9S4c11cba6GVvW3OT5oOBrpUXqW5ulM78kqSOW6jd0zj9ay618bq0uZgjOxVS8EIIPe63BBkJx81BJdLpGS1k5A7d1P3Kwp2vdJlozuglFXc3jqMxJPnlTxU1RUAOmflIoZSwfCtRpmPbHgtxsgxPdcbY7K0wkHAWaQ4uOytcMOGQggZA4jP8yjlpQX5IytQZ27BWSHBOQO6CKClbj6P6FSopQPJZ0BH9yOyiqj8kGmup9uy5C5OY+nm/wCF4x21daj/AMriWxXLf/J1+GDwx/hbav1uJB9FSIiAiIg698/0BqeUHiNA3u+kpB/y6nXz7WmwzVj5cZ+AkH7CvoS57iG8pvEFx8qWk/XYF4K6TqomyVYODmU/pQaUNJVAGwcpotIVbv7rAW9TXUoaMgBZFLdaEDu1Bte16Arq1xa3q2U120JW22MucHbBcl6Vvdshly9zdlXWt5oKiAiItOx7IOAaunmglMbydvVQh4jBOey1i9Qz1VU73VpJPotHktF1wcxu3QY9RVvmHQx2FHG2qYQRIVUUNXTyB0sRaB3z5rJDhgHbAQVfU1Rj6XvJCiZKQc+iummieMNcMqEA9BQajT1Eco6MDI2WQYx6brSbc5rJS57sDK1h1TT5G4QQOb4Jy7cKhqoXNI6d1dVzwSMwHBaa0sBIDsnPqgyTOWuywkK6nudZDOHCQhoPZY4GULfnsg3nSaka6FrHEEnYrcNPa6e8UJcWBxIzlca0hga5vU7fO4XLekLlbY7eInSAvIwM+aDjqv0XKKt7mtIGey0uss/ujSHAgrlS4MqZpXvhi+E53wtm3uw32rcXQ0zzk7ABBsn3OUPxnZZjKMiIuJys86T1UHZ90f8AkKvksWoKeAunpntaO+Qg2+9oa8g+qGmdIOpoOyunBbKQ8YI7rMp3NERwg03w5M9OVlRgtZ8SjkkaHnsN1IJGFh33KCN5Zn0UjJ4ms6C3c+axHOPUd+5VvS5xyASgm6B1dXkqPAPb0V47D6lRwwMhBjkOBTqc3zVzs52VpBKC8SnzKr4hPmVFg+iqEExeSgPqrAUJwguMmAcKxz3O7E4VhJKvDXFoLQgsxhZVMCsbpefIrLpNsNP2oM6IbAKXoKtYGgf+1SZ2wgx5GkHZWtcQcFSPySosOBQSFpcFcxhASPq2CvPZBJTx9TtwpnwN6uwVlIQHblZTyC4FBiVELWsyAsMAYWdVu+BYbQMIKiP0Vjw4HzU0Z7hWzd0EIqKuHeneWn61V11u4zmoOUbnzVHgnJwgmpbvdeoB9Q4jPr2Wv2utlmna2ZxIJGVtqJrg4Z9fRa3aJI2VLTI7AyEG477S0rqIOawB2O+FtWCladunzW7ruY6yiDaLDnY3wttw0dbTnM7CBnzCDGrKdrGEgYwsKN7RklarXkGMg4z81ormSHZoOUE75GPGyjLCd/VWNgnZu4EBSg/DjHZBa0gbH0VepqsPdUyPVBISMKx2HKiIInQ5KoIB6FTIghMOUbTjIOFMgOHBBeIelvbyWBNSl0mfJat1NLR27LGkIzjCCJsTWtAHkrg9oHSVceyxi5viAZGUFzoB1tdjuQuS9O26n+4jpTGC4BcdPbJhjvJcp6dkibpx7ZDuWIOPqyumZXyxB2ACQAFpVSZJKjqccrLr+k3SUg7Fxx+VQux4n2oNas0rmMAPosW8uL5cqa3RyuaC3fbyWNc8skw/YoNPkIA3WC+ndI89JwsuoIbgKkDhuUELLdK8bE7LHmbJTv6CSMLXoJoowevC0e5Pa+bLe26DGAfIcEkjKzIYWgdljUxHWMkb+q1Bscrz8AyEHfb2MUQbzQ6oP/cDWj/nG3orvYzMlbzPanLm4/8AgFW/yhb0QezSIiAiIgIiICIiAiIgIiICIiDYHMGP/wAwnEof9yF5/UpV80NdG91xqw3uJXn/APCK+l7mCOOAnEont+xC8/qUq+aifDrpW+f3x+P/ADigxo4pQcuzhZjIsj4QVQAAAkABTRvYdmkIIZoXluD6KtH0xA9SyJC3py7GFjtLDnBQTyyNe0gY7eSxuqSMEMOM+ik+EdyrHhzjhgygyrPdqy2TibxCBkLe9PrMVjWid/Vj1XHUsMpb8Q2WVQNZGAHPIOEHKlqqqO7yeE1oJytSrdGNIH3s77gLa/Dusoaau6qiQAdQ3JXMNXf7EWxdMrNgM4wg4tuul22qIzyR4A33W3211G9xZ0jY4XJuvrjQ3G2GOgLXPLcAD1XDVNZbwKh5MTsEkhBuOnsDL997hb3UVw4bVFvaZHsPqt0cPaapoagOqmYAx3W9dVyispOilaC4jGyDr9W2t1K4tIxusDoDMkdwt2ai09f5JS+Ond0+RAW2Xad1AeomB4+woJ7PdJqSfMjz058yt923V9C2RjHBpJAXGhs13a/D43KemtNxbVMPS7GyDmquq6O5UP3poyR5LZ1bpuWWGR7QcHJWrWG23KWja2Nji7Gy186Z1JJRu6KVxyPRBwdW2uenrCDnYrIjrG00Ra70W4L/AKO1YKxzzSPxv5FbVuGn75TkvqKZ+B32wghfWNlkJae6uHWTndYUcUkcnS9pBB81qLQUF8eMAHusuOZjWkbZKxA+IH4nAK8OjI2d3QSPAc4kdlE5pDtlICD2VcZOcIJmPIbj0Ub5fi7q4D4cLHlyDgFBkGUNYDkblcycOre2ss7n9Ofg/mXCbyPCbv5rnzhNWUcdic2R4yW/zIOM9c2/wrhK0ZG5WzmuETiCVyFr+OSe5ymBuRk4IXHk7TFKRPscnugmM5jAdnGdsremleHNVqtonjYSCPJbEmjfJEzoGQSN12X4CQ1Mdub8AIxtt8kDhhoqm0hcPEucOWtO/V5rdvEC62uvLRbo2N6cDYLK1BPFDI81GGj5bLY1fWUMjyYZcgINGu0t98PptU72OA7tOFocWqJ7Zluo5DK8H99ufzreEFdRxuPW4dvNcScTZoJ6pxidt1b4OEGt37V9BXQllGGjI2AWz/u1WwFwExGStNoWsDBlxO3mVbXHJ+E5+pBlVt6rZ24ExWBHS1FUC55JJUZzgd1rFtLQwZwgwoKOWnIdkjByshwkfjBKy6l7N1BGcjZBjTQyyDAJz9arTUszXbk/astjRnJCkbLE0jJGyBI50cffGFi+9FwIJJWTWSwujwxwyfRaZGHdR6icINWs7GyTZxvndbxt9MDUxjG5wtoWJ8TJQHEblb4ttRT+9xYIzsg5AtFS6x0onlOG991n1vESir6F1GC0kjC0a/yR/cUF7sDp7rjy3y0z6rpjkJcXYAQYmtLRipdccfCclbPhqYXktwMhcna1oqx1nD2MJaW9/sXEVJR1nivcWHAKDV2uDdz2VxqIy0YG6gc4BnSTg/NRAZ9UEr2yTkhpO/ZU9xmaMnPZTUcUjnAtBOCtQfS1XRks2QafTPdTO+M7ZWc+6xkDBHZadVtLSQ7Y75WJkYODugzKuqjl7YysBo6ndI7lPAncctBKrFT1PjNJbtsg1GngNKBIe2MrKN3a4dJIwAqVEM3ug+HGy0VkMmSSCAg1VxZVfQ7qjbc9rg7dLeMH1K1Z2fDBx2QYbZOhoa7yV1LJFWVLaMAZfsFBM9hcQHbq2wkDUtNnt1jv9aDlfSnC59HEy6TxksJDlyJTssdDSeOadgewbHC3FS1lHFo2IS9I+AHP2LYl1u1oNDKwTNBOfNBoWpuIrg99HTygM7fUuINRXWuuFQQ2UkF2Ss2/PidcnvjflpJWjPOZs/NBYyhl6Q9xJOPNSsgf2OVqLHx+EASM4UfVH1dwgxm0Tj5IaIRkOcM4WosIDQRg7KGokZgl2Agtjnij+EgHCGvi6gwAb+SwnvjJ2csYlpqBg+aDcMcTZG9QA3Cxqlgjcdlm0G8AJ9FjVw+Pb1QRxOGPsWPUSAPyMBZEbds7rCqSOoZKDJhqA0bnKjqJus7KFm47q1+eoDJQXLkDk7/DB4Y/wttX63EuPx2XIPJ3+GDwx/hbav1uJB9FSIiAiIg6+c/kng8ofEWUfvaSkP8Ay6nXz5Wm6GlknPVjrOfzr6CPaDHHJ3xI3/8AuSj/AF6nXzxwt6iRnyQa/Pfp5PhbIfyrGF4q25xKVpjWfEcEqr2u7boNYpdS18D8tmIHyKypdV1c7cSzE7ea20IySOrP5UewgEAlBu6wX2mE/VUAHPdb0ZcLTWPaxkbfmuJKBuJvjOBtut6WaopI5m9bwO2d0G4r7Y6SspuqmjAJHotqt0bUSAxhpyduy5Diu1obSgOlZn61dT3W0mVvS9mcoOLa3Q9bbh4smcAZ7LBNuJaQB22XLesJ462h6aMAkjsFxzHaroQ4eCck7bINvVNFJEfhJCxvBqTuXHb5rXqyxX1uXGB2PtWD9xry7LWwvJPog07wqhx6clXiklZhxOwWezT19jcHvpXtHnlSyWi6OjIbEc/UgwI6hrfhzupDK17SwdyojZro129M/Kq22XFjg50LgAUERgn6uprjjOVqNsudfQ1bXiUhgI28laPhZ0vGCoZSwgtyAeyDk638QLfFSsim6S8Dc+q3JZeJmmmvDaiOMnsdguBBSyE5BOPrVIoHsqB1ZABzlB2th1to+ohyKaL4gPIKt8hsNy09LNTU7AS0kYHyXBllqqNsMbXTHqGM5XL9trLe/SskTJcvLDjfzwg623uPovFRE3bDyBhRxMcxhYc7+az7za7hLeKl8cDiDISD5YWNJTVUDgJ2kYQYppnudn1Q0kmM52CzmSR9IBdghXPmi8ItGCSMINHcMHBKyqdzBGAdzhY72OLzgKSJj8ZOQAgE9Ttuyu6Vafh8/NXRkk5+aC/wRjsqGAHfClzthMlBCYB6K10W+wWQd1a70QQCP0BVwjyNwVJjG+CgyR2QRmDJGPrWRDE1jMEbq0EjyUgcMfFsUFhDA45wojIA/wCHAUjukncjdY72PLx0ZIQZ0cp6RuspkmRv5lYMccoaDg5WQwOaMnP2oJtvkrhE0jKiD9x2WQ146Cgt+FvkFBLJvsR2Vzyc99lC9rnHZBLFKR2IWa2TIWnMY9p3ys2Nw7HCC2dxIIWPgrKeWd8qElu4BCC2Jrs/apjDnfBVsTmNPxHzWUJYs9wgxX0+BsregDZwWW57CPhwsV5O5CC0lrfRWOqHjPQd/krJerfurGDOUG9uHz3zVGKs9Tc9it1aohomtBhYGnC2rw/qKWCYGqcAM+a3PqesoahobSvBJHqg44ushbKd9liU1VHG7qfjA9VlXihrZZCYmEgrRhQ1kbT4jDgd0GuS3Cmlb0sAyRjZYb8b4WDAzoO5wVmAgjyQQvByVb0uUpGN91TB9EFojdnupWQOxlG91Ow7YQRuiwOyhkHSP9Sy5NhlYkwyDt6oIQ8gqUHbKxxnq81P+8QW+K4ef5lTrJO6idnJ790aSCMoMk7gj1WJDTSS1gbvgkLLHZSW+Cb35jujYEINzxaf66RjujcBZEta+3UDoAcDGMLW6R+KEN88Z/Mts6gkj6HAnB9EGzqidzqx8me5yroZ+qUAnzWO8ZlPfHqroGO8YfWg3zp9jHt+IA+i07UVGXVALBgeey1DTmejdWXx7BNvhBtG4sdEN+4CiozlmT3WVeT1D4QsSjp6l7D4bSfNBdUtkccNJUApJHAknJ7rL+510lOY4nEfJXClrIGls7CD80FbXZZKyQYycLcDLeygAZIMn5q7Scc7ZCQ38yzL5UQtqA2UgH0Qd3fY8CIczGpugAf/AAFrf5Qt6KnsdpIX8zOpvDOT+wStP2fdC3og9h0REBERAREQEREBERAREQEREHH3MNtwB4mH/uPvP6lKvmp73Gr+cj/+kvpX5hf94LiX/A+8/qUq+aeQltxq8eUj/wDpFBdVZbEC3uoqJ7nZyVI4mUBp3yqMZ4JyPyoJK15bGMZPqsWk6nE91kHM2GuUkcDIs4QQzBwHcrMtzWvGXblQVAaW7HdWU8ro/qyg1esjjbDtjJWkguBPSVlGpMo6Tn7VC4AZx6oKw1FZGQYJCD8lqMNfeiPiqXH6ytLY8s3HksplY4N7lBvHRU9XV1oZXS9TM9iuX4rPZzGwgMyQM9l14o79Lbn+JGSDlazDxMuAw0l2Bsg5quNDS08WaVzQfUJYaYVFQ1s8gIJ33XEcfESqnHTI449MqRvEWooiHxPdkeiDs0/T9gdQB0giLsZzgLb1VZNPtp3uEceR9S4Jfxoupb4XW/H1rFHFW4SvEb3Pwe+Sg37e6W1Q1BEcLTg+S0JjqH3oN92Pf0Wdpa9Wu8PBrpgCTvkrk+n09ol1vNT48XihuQMjKC7htbbfUFhmY0DI2IXYO06a04aJheyI7DvhdPq3WMmnbm6OhcTEDsQdlm0/MBdqeVsJfJ0DY7oOxurLJpWAvAp4iQD5Bde+IX3FggnbDQZIzjDVu6x8UrTfI2mvqB1EgblcvWbSPCrVGl5nVFXTmsewlgJGc4Qedd3kYbgSyItGexUZeC0+Wy5b4pcFdQUGpZ6i20bjQBxLXNGxC4uutorrc5zHsxg43CDSJXP6jgq+AybZUT/F7uaclT03USA4bIM+InAz3yp2jPmO/moWYA79kL5Adge6DJO3dY0x+L6lK2QFvxHBQxNcM+aDElJEYG/dcm6BfVe6ANlIHT2XG08Y6QFr+ntQS2/ELCR5bIN91zI3TvMoDiuK9ZOa2vxE3AyfJclW6o+6B8SQ7laPf9GVFwqBJCzIzk4CDZ9rlb7vGXxk7jK7c8Boqaa0NcWhpDex8tlwbYtIUMETGVvSCCDuuU9PX+m05TCG3vBGMHpKDVeLrXwCX3eTBGexXD9nmq5GyeLISc+q3HxA1k6sicZHHLgc7raem5pKtrnRbg7lBNcJqtjz0OIx6Lj3UU08kjvFJO65YipYpJCKkgfWuONex08FQRAQRlBt6nL9sH5FTSuyQDvssWKQtY0j61lwdMoBfjZBC85AHzWoUJPRscLCqmNY3qaVJQ1BDfi7oMmrLgDuTv5KShyW7g7qyN7Z3b9lksa2LcfmQJtgcbLTZnPAJB+pZ9Y8NjJB3+SwYCJHYd2KDHpZJXS4fnC1CXAb5KR9NDFH1tIyoGODz0uxhBbQPk8fbbdbptD5vuhDlxxkZWjxwwRNDmkZytUssjpLlE1uc5GEHJmrXPGmm9J36f5lxPpl1Q+/Rsc/4esZ9O65S1vFWQaYEnScdGfzLg+2XWenrHVLc5Y4kboOzWr6Gjh0ayUhr3eHnbv2XCNLPTmGbNKQQTgrf3DrWUGqui03qTEWQPiO2FytdtBcO4LO80s8BnezIAxnOEHUSpeXVh2IBPYqaP6QWv6w0jcKC7yS08RNPk4I7EZWhNp5x8Qb2QbgsMMTnjqx3W6ZqalFOdm5xsVx7RV09K8HcbrUpdQTGIjqPZBiXyNrZiWkYytDj6vE3zhZctRVVsx6QTuqU9trH1jIug/EUGp290fT0uYT9i1BjKczNHh/mW5KLSPulA2rqmBoIzurHQ2dkZkbI3IQYFxihFGCABsFtssjLCQtQu9yEhMMByAtF/2yGnY4+SC6hcfeO+2StfkaDAcEdltqke5s2XZBWsGt6mGMdzsg0SokeypIAJGVLYKlztSUzBGQS4ZP2rXrBYHV1UJKluGk9yuUKbQmnaK2m79UYljb1DtnIQbvvVNUR6JjmjqAMxjbPyXANxqroHyDxyRkqe+cW7gat9ka5xgjJZnOy0t93hnacuGTvhBivfI85kJJz3UZPxb+qu62vdlp2VzYwTnzygueXhmWlYviPMvcrNeB0LHZEDMMj8yDU4STEMgrT68uwcErWqaOIRbkflWl3OP4sMGyDRXOk6u/1pC53jjJKnMBBJwoi0xuD8dkG66AE0+OoZx6qOrb8WSQVoDLtPC0NbnYrVaGrbUsBlO/fdBNHv8ADhaZX5D1qwdC1xIcFpdwBe/73uEEcRyBuqu2dlWtD2gZGCq4c45wfyIJYxkLkLk7GOcLhl/Cy1frcS4+ZsFyFyeEHnC4ZfwstX63Eg+idERAREQdd/aD/gdcSf8AE6P9ep188cPc/Uvoc9oNg8nfEjP96Uf69Tr55qVvW4g+n86C+E5dhZBDcDsqNgDPiAVry4FAmaGjIULcE7qYZf8AS7J4QA2QHj4QWYz8laySqZ8THEEKrSc48lIgjmrri0Brp3AfWpLdX15qmhk7jk+Z2VHMMvwkKsUYpnh4wSEHLekIX1cLPe5Mg47lb3prNa/eY2kMwe+wXAdLrGttwAiJGPms6LiZc2PDyXbfNB2FvdlskVAHBjCceWFtq0260PrW9cbCOodwFxrS8UKyvaIKl78ZwclZsWso4JRI152IKDmTUNosDLYHRwsa7p7jC2XTW62b9TWn8i2rd+Ir56bwmPJ2xstvU+tKgS4c44yg5LraC0Rt2hb2Wg3eG3MonujgGR2WNQampapgE78Z2Wv0j7FW4hllZh2xyUHB9wqyKx4azABwsMyufMHHI37LsHUaC0nMw1DZIuojvsuPtS6Khhkc63NDtz2CDbcDIhACQMlQ1XhthLgMkLJ+4V7aQwRO6R8itQotNV0kgFQwhp7koNpU9VUslHQ9257Lk7TNZXe7APe4sLd25UlNoy1siD3dHXncLcGkbTTzX6G3bFjnBp+pBhxVFB1PElCS7vnHcrYesqmIPIhpi3J7kL0DsHLvpiq0/HcJhF1vYDnb0XF3FLlthZaprhb4WOazJBDUHR10jyc9lVjnucMeq3HeNLz0N7loTGR0Ejt2wtHraWSin6XNwAgkZG3p3G6Pa1sZxjKkj8MtGSMnyWPUyYJa3sgxTnqwewKyIvLsouh536VNEHDGQgkPZUHbsqnsqNc3tnfKCqo7urjHKRkAkK3pdnLhjCCoGQFVoGOyqHRbAkK8dOMgghBGWbnZY8zi12AVmZaNtlp9S7Mu3ogo95AAGe6z6INIy4ZOFimLLQcd8LXbRBTOAY8tB9EETOjGOlRTdIzjZbppbCajJibkeoWj3mhNHJ0vGDlBo5OPNTxZLe6gmBb2V8MhDfTCC9/f8qujxjcLHeZHE9IKyaduR8ex7ILj0+QUTiQdish7GtB3CwZpHA7IJC52O6ic92fMoyRxIB81L0NcQggLnH1VQ+QfWpnRtAyFb0BBWne8uwc4WUGgndYrPgKlbK7zwgpWABhWJD3H51lznrbufrUTIwHYHZBdLUVELR7s4tOVqVlqa2edoqHkjPmVhOYwt2wSp6WYU56weyDkakpqM0odKwE4ytFvsFG2BxjjA2Khs17ZLiOV+B23WqVjaSqb0NcCTsg4zcSKpwwQM+imBxha5drMyAmVg2O60EiRriMHYoJvJMD0UYe7bOU8Zg2JQTBoBypGjbyWKKlp81UVOOyDJcCAopBsTsqCfq2yqOd1DGUEG2SMfmV47I5gG6iMhDseSC4kZxhWObk52UoaCM7Kx5ABwgki7j61uK0NiJGWjK2xG53WAMrdlrpZG03vHTjG/wBSDccEZ8E/EMLZeqPEEpwdiSswaj6JHQl3bZbcvFwlqKgkE4zuUGN0jIzjKyaVrS7yyoQ6MtBLhlTUccr5/hb8KDdlkf0MOGErAvz3GYDcb+e3mt+6Ct9okiJrnsBx5lbR4ktpaa4hlvcCzOPhQbOubyA0HJW4dLtp5IgZGjt5rS47e+uaD05WtUXu9sb0PIBGyDfmn6G2PaTIxv5FtnV1LSsqy2EADKxo9URUYIY/8hW37tfZK2bxGklBvbSFFC9pyQDhbM1+18F2LWP+HfGCsen1VWW34YSclQVFf92XGapPxDvlB3p9i1JM/mh1OHkkfsArcZ/8I25FP7GJsbeaLVAYRtoGuGP/ACjbkQeziIiAiIgIiICIiAiIgIiICIiDj/mE/wB4LiX/AAPvP6lKvmmm/wB0av8Awj/+kV9LPMJ/vBcS/wCB95/UpV800u9xq/8ACP8A+kUF8YGB9SvVjNgFXqKC5VyfVWdRTqKCuCdj2VcBU6vkqdSCp7Km7vmmdlVqAeytLiNgr1TpCCzd2Ad1c1gGfhCYDTsrhuED4gMt2+pCZD9LJVzTnZX4CCMMady0ZVHtHSSAMj5KXAQNBQY1HVXOGcGCR7cHy2C3JSXbUjpmNFbIG+Y6jhaL4fhDxFay7TRSjB2GyDkKRsklIJJ3B0hHcrQKvLYnuDB1DtspbbeHVMYZIVqgp6WaP4nDdBx+66XiCoPgzyNwdgOy3XoLVWsf2X26nZdKkMdM0PZ1Hp6c7qaSxUpkL9vVZ+mKWnodU0lRkBrHg/Lug9F7tb9MScH4quqpWSVhgBc4gEk4XnlxNq6SOsqWw0ZwHkbN+a74af1RYL3oSG2TTt6vDDcE/Jcdak4E6evlrqKqmax0pyRjHdB0Mkc2TB6MZHZGADfGFyDrbgxrChvUsNvoHvhBOCAcYytm1ml77ZnFtygewg75CDEyR2WZTtYYySsMxvBw4d1DJUTxSBrQcZQZEziJC0ZU0RJGc91dE2F8Ye8/EVY8hrsN7ILZj5Ke3NzIO3fzUYYHjJG+VWJ/gygDuSg35ZMtjHlk+S5Y01TUM1v6p2gu6e5XDtkfPJG0tBP1Lf8Aapro2EMiaSCEGi6/fJSuf7mS3B8irdDPqqumLqiXJHbJWp1+m7xenkGFxz6hbYuNTWaPcYZQWnOMIIuIrZIoiWP2GVqPCoNlonGVuT6lbN1BepbvCDknqP1rffDGmMVAS4YyPRBPqPrZI/wXdPdcT6hdK6d3iuzvsuTNY1hppH/UVxTcah1VI53zQY8fYLIjzjDTsoGNIAGOwU7HdIwUCQEtwTsqRnA2VXvy3CozsgnpSeoeX1LPLjtutPgPS4YWcHdQCCypJwsduRnpWTI0SDZRCIjuEFjnSHOScK3J9VfMOkbeqhBcQUGRG+UYySVuPSMgff6djxgF4/StK09QS3KcRtaTvhbyh05PaJo60sILMHKDmTiZQUjNAMlaWFxi3x9S6q0QYXy7DZx8vmuUNXcRqivtP3Ic8kBvSuLabqY5/wD2xyg1ShlraaTqoZCx2e7dlqVsvOqpdQ00M9wkMReAQXbYytIppXxODtxusmO4Phq2VQO7DkEIOyGq7fa2aHjqXtY+cx5J7nsuA4Joj4oMRGHEBb101rpt+gFquMh8MYG/bC1+s05phtKWU0rDK/cDbOUHDU/S6UlrcfYo8D0W4NSaVutBIahkJEHcEdsLbAE8nVGwEvGfyoNUtBhjkBfF1b+S1qjraZ15gi90IBcB2Wp8HNJT3W6D7sR9MJeN3dsLsjeeEWiKK0fdamkhMsberbGc4Qca67pKeHRrJqZwa4x5279l1vFVcOqUeMcdXquQtf63qn3CSxQEmFhLRjt6LYLIpRIeobOOSSgktT5ZJsTE91uSSKnEWOkZWkNiiiYHw4L/AE77qySorfBJIO2d8eSCOdrWzEs3GVHTSu9+ZkEjq9FZZBU3OtMWCTnC3hQaNrPuvAZIiIiR1HHzQa9RSRNt7THEQ7HcBQXGruX3Oka2ocGkdsrky5WPTtt02x7ZGCYN3x3zhcQ3m7QtglhY4eY2QcU3Qu+6khcMu6ifrKjbUVAmAycLVKmjNRWmYN2JV7LXmQOIQZdE5zox1E5WoRty3OVFHA2OMDzCoZntPSOwQSknOMq5gwc+aia9zsE+qmYASgmD342ccIcOHx7qrBkKjzjKDGcGhxA7ZWNOGjP1rJd3KsdEHoNPcW57efosiBzg0BmQpzSNPkhiEecfYgtc6UD6R3WZTYc349yfVYbs7Z9Vk05+BBI9jc52VhDVa+QjOVH4pIygq7bst/8AJ0T+3A4YH11dav1uJcdSSH/3K5A5N3k83/C8nz1fah/yuJB9GKIiAiIg67e0Jz+054kgf3nR/r1Ovnpov7If+9X0Le0HI/adcSMn/wC5KP8AXqdfPTRf2R31fzoM1xGO6jIB7q53ZWoLsADYK3ICuPZWOA7oKN7q5RNc7qxupRnzQXo4En/WrA8lXDcIKOa0jcBWGNmMBoUwAPdOgIMRzXt3jGCreurPcrO8JvffCeE1BixvqM4eThSkkDYb4Kl8NqqYxg7D8iDGE9YD8DyPtU9NW3ZswDZ3flUEhLPoqEVMzHgjOyDfFHW3h0bQ6qfg481yFpWliqWtNZIDkDJJXDMF7qGsA3GFrFFriso2gMJGAg5/FpsnR9GPP1LTrpa7ZHSPdF0BwB7YXDbuJdxGficsefiTcZQYy52Dsg3RL4/vxY2b4c9itzaDpy7VNPmUAlw3z81xF+yerMnib7nJWbaNe1VtuLKtpcHMIPdB6fWi3VB0lTdFwAy0bB3yW6a+025nDermq3sleIzjsT2XnlFzT6hprfDRRSS9LSBjJXP3CXmJt2pbSLLfarpErcEOO26Dqvry408evq2JlG/pEhGcfMrYupXRyvLms6Su/tZwi4W3iWe9vnpy+QF25Gcrpzx30d9y7+W2OIyUwcfodsIOJi9wcACcD0VzcueCfzqZlFKGhr2kPH5Va9nhDc4cDsgyOhnSO3ZXtY3pzt2WnmSp8gcFUE9Q09JBQZZHxFYJcROBnsVmtyW9XmqmGDp8RxGQg1SlZG6nbkjOFhV7WsBLVg+/1DMtZnAUjKjxgGybH0KDAc95O5Ky6ZziwZz9ayTRxFvV327KrWQsHSSPtQYuHZOM7dljyAtlBKzi3JJA2ysWVodMAgyGEFgyFWkfL76A1xAyp4YB4YOAdlieIYq0Bvrsg5k0cyJ1MDLgnC2nxADW1pEfbK1nSlRP7uC3JGFtzW0dZNVdQYcZ3KDQH9JaM47KxuCeytk8VjQHAjbzUsABb1HuUE8TGYyQN91ZIcHDdvqTxOnb0UbnfFlBR5cB3P5VEQD3Uj3EhREnKCuQELneWUDQVe1gwgpknZV7DdXtjBV4i27IIGqqkMJx8ICjLZAdx+ZBXKoqlpHdARlBaHOHqjpHYI3WVC1jtnEBXuhhPYjdBi0Uk3WOlxH2rcdufUOnaHvJGfVaI1jYt2/oU8dwdD8YO47IN03ljG0mTucZWzpJW9Zb0Fa9abm25PENU7DTtuVuKewWV1IXROaZSNvrQcdTgdOQQPktPm6g0kFajeLdcKapdiM+GD3Wlhsxd0uBxndBFFI4uws1mSN1T3VrW9QG6oC4HGNkEjOoHG6lHZUZ4ZxkjKnb4JG2M+qDFc4kbgqzGTnCzvCYQrZI4RGcYzhBjHqx3UTuoZPfG6h/20ZCA12MnGyzLXba6qrYmvjd4Zd8RPbCCOnqCXtaYj37rky2QxSWFxIAPStUh0dp1lsZK97BKGgkbd1ti/XeC2U7qWlft22QbCq/EZc5B1H6Z/Ir5mhzCe5WJPUPkqHTb7lZUThIzpKDB6nB3crdVkDDT5c34sd8LbroB17AbkLddlhjbTfERnCC2SquELiKWZzR8itKqqiolkzVEuPfJWqVbhG7DTsT5LNobRHVxeI4D7UFNOCOQHqGNtlt/Vckkdf0seQN1rkz22kuDfsW3bjIK+XxSc7oNJ8aXzeVlUxL2/FurnQRjGcfJSRsa0bIMerAAwAPrUEZf+9JH1LNkYH5yschrDt2Qd+vYtdf7aTVOc//ABArv5RtyK72LZzzSap/gBW/yjbkQe0aIiAiIgIiICIiAiIgIiICIiDj/mE/3g+JX8ELz+pSr5ppf90qv/CP/wCkV9LPMJ/vB8S/4H3n9SlXzUSAfdOsHpI//pFBe36IVCcq4DYD5K0jCCio5VzsrSSUAbq4DCtzjdA4lBXqGcK8dlYGDKvOwwEFUIyrS4gZwqdZQVGx3Vyj6sndVLiNkEjfpBXKNricdld1FBcqt+kFZ1FOr6kGVLvF67LS3gCTdu61DrJbjyKiMDSeo4/Igzre5wa3Gy1qCSbb74fLzW32TiIdI7hSi4ytG2UG5epxG78/PKpTukbWMcx+HDzW2jeJgcElXNvMjHeJkjHmg7C6Iud693YwVxDMjs5djOHz5ajwWVdWCxwHUCe66DWviRWW9wjY52AVyRo/j5X0Vzp45JHiMYB3QeikOltFzUj5aqjikkLM5IBOcLpDzS09ot1bM22W44Dj9Buy5+0Rxv0zXWhhrqxge4AHLlv2Xh3wo4nWCWqmqqZ1U9pc0HBOcIPKyaUvLXeA4H6ihaxzCSzBXZriLy03uK/zNsFGX0ocektbthcJ6v4a6m03M9lZSvaAcZIIQbBkkkEnS0kAfNZkB6mjqU5s9S0AyRkElSMo3sbu0jCCEbHAOAotzVNCvncIskrGhkc+cOG4Qcp6RihMMYeB5d1zXpSloHRMD4Q7byC66Wm+R0UTOp+/UPNdn+DAt97o2OkeCenKDcmn6C1Ond10o/IuuvMhDTwXEiljDB1bbYXcfT+kW1NbI2mYDsey6mc2VontN2cx7CMO9Pmg4VoiDTx9e5yFy9oRjTQZbgfD2XFum7bLcIog1uchci26rNipxFMekkYQaRxAPSX437rjGP4nvJ9Vv7VVxirWkhwOVsuOOEZzjJQQ4Ctf32KyJGxj6OFiSvwdkFc5Usf0Vjh+TjZZMLcgfNBe3AKyo/iCxH5aftU8D/hKCaM/FupXBpChhIc7fCmcWgYBQQTDIwogxu6rUyYGyxWTvLiN0G/+GMUL6/EgGOpcp6rp6VltJYAD0+nyXEegZzS1Qkccbhcg6ivTJ6EtDv3uN0HDV6fJ903NP0clQMa3xBhTXhxdXucPM/zrFY93WEGoPDRGCMZUJGGlXSyAQ/NYDKlzn4JOEGTTz1NPJmneWnPkcFarp253ifU1LFPUuMReAclaZjw2iQ7eaip7i6mqxUxf2Rm4x8kHbXXVttMXDyOpaxj5vCycbnOF1nsMrZL0WSUjujrxuD6rlbg/eb3xCljslxa/wOoN+LJGFzzV8ttqtbI7m2NgacOcekfWg4wo6elprEyWhYIZC0bjYrFnud6ktErJbg4tAOxd5LO4vXay6atpoLZM0zsaRgeq69U3ES8y+LSydeCSBv5ILbuOu+SOlw5xcck753Uk8cXgktxnCwpYa6pkNUWHJ3Gyo6KvkaYWMJedgEEVpc99aWPJLc/zrdlVBAygc4R79P8AMtJ0rozUZqxU1VO4RE7HHkuQKfTMk1xggmZiAkB5Pkg2Fw6Mc2oCySnLW9fcjZdiqi222O3CVhYJAzPlkLStU6V0XpTTwuNuni98LeohpGcrhyXibc3skh6n4BIH1INS1lcLo2pfC2qJj326tsLjypqKh0pDnEjPqtRnv1Rc5i6XO5VopGPy5w790GNTtaRlw3+YWd0s8PsFF0Rty0EZVSXgYxsghcT1Y8lc1oxuFd97yMkZU7Ww9OdkEGAFJHjtlRPewEgHsrWy4IOUGa3tlUcovGOM5UjCJO5QQv7qg2wVkmNp7YyopIyM9KCmRjurHd1UB/bBVryWndBa5uR2U0IwxYzpN1LHMA1BSXz+tRqskrSe6s62+qCyTt9q5F5NfwvuF+3/AMt1q/W4lx27BH2rkXk2H/XfcL9v/lutX63Eg+i9ERAREQddfaE5/accScf3pR/r1Ovnpov7IfqX0Le0KP8A1nHEn/E6P9ep1889K7pkJKDOd2VrFSSQBmVGyXq7eqCZ24wFQN27KnUrmuaQgBjR5KpCvHQfNVAaUFnRhFIQCMD9Kt6QgsGQcq4H1VMKhJCCYHIQ9lEHuCeI7thBTrPXjKuyVFk5yniDZBe4AjdWGNmc4Cv2I7hXta0jyQRhjQOwVelvopOlvqnSweaCwhhH0QrOhnV9EKbDPVVLWYyMZQXGNoZ2B29Fprx9/AxtlZBlkzjfCkZAHfERugz4IgYgXMGQM7jdYdNebvQ3Ae5VT4nBwDQ04GFM2dzAGjssqKmo2s96cQHgZCDlWg1Lq77kRE3WQZaP35W9aOmpK/TctTdHNmnDCQXYJzhdcTrK4RDwI3HoacDfyW7dHcRJqiuitla4iGQgEk7YQbfu8rY79PHHSksBOMA4W3bk5zqrHhloz2I7ruZYeHnDa42z7pVlRAJXt6jkjOcLg7i5w8ijrC7TUYmYHfvB3H2IOOYIoXQAkA7LDqYmB+wACzI9F6yJDWW2ox+ZYldp3UNCSaullZjuSgjaAsKoe/xcZIHy81d1TMIjdnqWdFBBJCXyH4vmgihYzpBcBk4WJUZbKOkYCq98zXlrAcA4V8IDnAynBwgnie/wxuT9qw5pH+J9Ihag0wgYJWHUQl8nUwfUgyITloJ9FhzH7+MFZUQ6W4PcLFma4y5A2CDVIDmIBYAGLgOr1WTDN0sAPfzWPG5rq4OPbKDmHRkUZpGEgdvNU1TBTAkmME+uFp+m71S0lMxrngZ2O63o6go7vQOqGkHbKDhi7sjDj0t2+paZE4gYHmVuTVVLHT1L2MxtstsRuJyM9ignyVRMgogo7sqKriMdwgAwgqGkHClaw4UIdgjspWytwBlBLE0ealwB2UAkAOyr4yDLga1zj1Y+1VlYzq7AKCObByPMK/xOoglBj1IABwsEuIJWpTNa4YJCxfBGdsIIPEd5EqaJ78jJP5VHIwR7nYKscjSfhOQEGW7PTuoX+YV7ngtwoHPKCWJ8kbsxOwfVapZKyukuUcck5LCcHfyWgSTvZ9EnOVkWWrmjuLHu7AhBy1qWkoBZmvEYLy3JON8ri2oZGOrpYAc+i5Hmq47hbGxZyenC2pcLQIoy8jAKDbOSTgnZVfG3ozt2VJnMZMWAjbyVvjNceknvgBBhl0gcQM4+tXRPkMmMnHksx9O0tyPNWOiDW/CB1AeSCbLwwZPl/MsQSSePg5I81SKC4zPw1jiPqWtWjTtwmqGyTx4jyDkoL6eOF0bMx7nucLftpoaD7lGUNaHhuQfmofudZIaQB0jPEaNxstqXbUxoHGmpien6KCl0r7myqkijqHCME4wVti4T1MknTK8nf1WoSV75syk7u3Wn1AMrsjugh6GkDIVW/CduwQxzAHbsrKcufUNjPcuxgoMrPbIK1u3vcIsb/lWbT2DxKVspHcZUJg91f4fbOyCCd2SMgndbhtR/2r3xssakoIpxl5G26xbjcG253hMO3yQYOpHuycHK0CEuLCSVqtXUNrATsVpzmeGcDCDFnc4eqlpnFzQSrZWF3kpITGxuHHBQW1BIBwVjAEjdZUxa/wCicqDHTsg79exYP/XS6pyf/lArv5RtyKvsWPwpdU/wArv5RtyIPaVERAREQEREBERAREQEREBERBx/zCf7wfEr+CF5/UpV81EgP3TrP8I//pFfSvzCf7wfEr+CF5/UpV800riLlVn1kf8A9IoJQMAI5UDx0gq0vygY2VuQE6zntshAQVVW9lGCc4PZX5KC8HdCcq3qKA5QXFpwo8hSnsrOj1QUVQRhXFoA/wBSBg+aAD5qvUE6QnSPLKCqoRlV7Kmd8IJh9EBV7NKrGARurzH8Jwg0+NzvEPfGd1qLWjo7DssBrCJceq1FhZ0beiDFlA6jgKGXaM4CyZA0u7hGxNfscYKDSIz99G2clai0ys+OPAeO2ykdQxsy/bKtc7oOEE0F+1NCBHBWysaTtg7ZXZLl5uGtq690UM94kEDsdQ6zjGy60teWDI8t1uXSfFa86TuMU9IXBsbh2dvhB7icL9CaTrdMNnu3gyz+Hkl2Cc4XTrnEsemLTNP7hTMOHHHSBvuti8Ouc+uprIyCoqJASA3Bd8lkao1VDxVgdUSydQfucnKDqbeKmmZUAMgOPqW3rhcGAFrac+nZdg7joCxmoLHlgI79lFJww01NSPcHR9f2IOr1xqS7cMIz8lLacSNLj3XJeruGFQ+d0dsj6gO2AuPKuxXSwPcyriLMZHbCCKuM7AOh5G67O8uYq308ZMxx0+q6sVcs72NOCBlc78DNRV1HJFCwOwQM4KDvtwlEbbg/x29e3ouo/PjJAL4fBj6fj/nXcTlnjprzXk1xAz6rrj7Q7SMLb8Db2ggu8vLdB1f4YSwgRdbMjC1XiE4l492+ED7FrXCHSIf4QqGhowO4WHxrhitNSGQH8iDi+vfMYx1Oz6rSWh5J3P5VqE8z5omnc5Wmlz2Eg5QSYcB3P5VBKRuFeHuccFUfGXHKCOP6QK1GDsFhti6SCQsqN4a3dBdKkZICte/PZVjJAQZEJwftV8nUTkHZQMkDSrzP1DCCGY9Iwd1DC8eJ9HzWUxgmduMj5rIZRQ9Yx67oNcsDy0AsBB+S1q4zTmnIdnsrNKW1lS4NjGST5LcF+sMtPRueW7YPkg4prjmocSooyC7GN1dWPH3QdG44weylcyEbtI2QQzkhmDlYkRzKBhagWNk+pBSxxtL8jYZygtrneHSgtGThaLbp3G4sL2kjO4W6bFbJdQVIpYGdRzjGFvKi4QXKK5wOfTkNeR5FBzhyyUdtrHxBkLYn5Hxdl3R1fpqng4eT1LappkEJI388LpTBSVvDayMraBpEoaD8O2+Fpg5mtcXamfaKhs/gEdBJJxhB1/4o3m6jiDWU9RM98bJiAwnYglaXFXQGrYG0pGf+1XOkGj9L6irjeLpJGJ3nqOcZWtv4c6DaBM2WHIGdsIOLaJ9JJRNLocEjthVsnucmoqeF8XwF4BBHzW+rjadN0mYoJWYHbGFozaayUb/fmSN62fFnO6DsRJpnTg0dFUQRxNlLAc4Gey2HWWWlZZKiojkaJWglp8wuK7zxsr6eD7mQSPLGjpGCttP4t3SSnfTuL8O77oNu6juuoJNQyUk9Y90AcRjO2FQ0sYjLiBnzWm19bLXVbqtmS87qGOa5ulAc13QfNBmRNaJOkDGFnuyITghU8OBsAc7AeQsSSp26R5/NBZAXOnIJ2WrugHgk5GcLQjI+J3WB5qn3Vnd8GTg7IKTOk8cgE91kReIW7uKt8PqHieZ3yqeIWfCMoKEuyQcqrCQ7zVzfi3+akbEDuUEmfhU8Dcj0UONsK10/hDAQZoaQe6v6WnuQtMNc4Y7qaGsDsdRxlBnNDRnt+RYVZjJwPPyWYySMtzlYtU0O+jvug09xPqqhxx6q+SPCoxm2/wChBC8uz5qgLjnupJAD5K1owEFMkbnK5L5Njnm+4X/wutX63EuNTnthck8mv4X3DD+F1q/WokH0YIiICIiDrr7Qz8DfiV/idH+vU6+eSIZcvob9oZ+BvxK/xOj/AF6nXzyQ7OP1IJJc9PdUhd65SU5aqQ7j7UEpye2VVpcNsnumD6Kh6s7A/kQTtLiO6uBduoGOd23Uw3CCQH1Kr3VgOVdkoKkjCtVOruSULh8kFwGN9kdg47IXDBwrOooLZcYwoQSCfrU7su3UZjO+QgNkJwFK0u+e6gY34t/VThBcScd1G97uwypBg7HCu8Nrt0GI4u33P5VWORwdg5WV4TPkqGFgHWMZQStY3oB23CYxsFhipId0jI3U+JjGXAHCCYNyd8bLTqqWZsha17g3yVrqyRpI32Ky6eH3hnilBZFHG5gcWjPnt5rHZJLFVh0B6Xg7YWUXsjd0A9laGRg+MMZG6Dc771qyKkhbBcpA0kDHUey7H8CLPT3uCN99lbKS0E9Rzk/aupb75UFgj3w3tut3aX4v3rTQDacvAHo5B6L2Th9oeSmf1w0+QD3A9F1w5hrPp6zMlFDTMJBOOnC4nHNBqinjDYHS5P0viwt1WPWts4h0JffpQHuGT1Hz+1B1zraoGve9seADgDzWM+ole/YkDOwXYo6B0LJM+V8sW/Y7LZGsuG7WNdV2ZgfEN8gbYQceQMYYw5wGT3WFVktf8JwPkpp/Fpag0rwQ5uxWLIHveMg47ILPEd6lanQ4dFl2/wBajipI3MycZwruttP8AP1IEgAdgdlbt0knCo54dvlVPSR80ERJBIB2WOSRLsTnKncHZ2yrWwlzuo+qDN8WraxnRIRuFzFoyWc2Q+K8k9Hr8lwtNO9jWD5hc8cOaP3vTxcRk9Hp8kHFur5XCsl7n4itvUpDwfrW9dZ2lxrpQBsCVswNbSktccYQXkYVpd3G6tdJ6Kgd1boLhk7ZVc9IwfzKwOIVc5QXE52CYcrQcK7rKC5p3PdXZPqow45V3UUE0HfdTk75CipSwn4iFJM+Nv0XbYQWS5x3UTTvufNWyzE7NVIoqiR4DWkkoIrnkMGPRY1E84OSThahc7dWthD3MOPqWDRwStyXNIQZiteMb4V5+EfFthWl8ZGxGEFsYa47gfasiBrWvDm4BCxi5gHwkFTWounr2ROz0k4KDd9inlkaGOJAHyWfeo2e6EggnBWRVUdNbLc2oYR1YytqXG/CaFzAd+wwg2rWBzK15B2yVDG9xlHfZVkdJLOXkHBKymwMDeoY6sIMsZ8IEnKx4JCKoA5I9EgjrJX9PSenOPsWr2ayTSXBjpm4ZkZyNkG57KyhkhZ1QDO2+FyI22239j75mMYJOjI9ey0dlHYKK3NIkZ4gGSMrbN71q2lpnUcDyWnbZBsW7VVwbeJo2zu6ATjfbC0eokkdP98JJB81nTVXj1Tpz3cVa6Bkh6hjKC5mPCCMDS/B7ZVo62jBzhUc8NGfNBqkcMZZuPzrSGEC6tAbsHgKwVtSPI4+tboslopqqEVUmOsDI9coN60VNEbYx2QD05P5FtO7gMqAGnzWQbnWxA07WnpGw2Wl1cz3P6pc57lBrFuY57M9eMhbW1KJG1P0s4WSL1JS/CCcLTqypNc4SHf60FKTqeACfJUqG4fjKx5J3U+Ok7/JXRSvnHU7coLvLcLDn6uo7lZoGQcrDqHYdgILqbPmqS/SVKZxJwkx+NB389i0f+uk1Tv/APKBXfyjbkVPYr/hS6p/4v63+UbciD2mREQEREBERAREQEREBERAREQcf8wv+8FxL/gfef1KVfNHUOxcar5yP/SV9LnML/vA8S/4H3n9SlXzRVIH3Qqh/wDRH/pKAHbK9nZWNGwB7KQADsgZA8kBBVCwFXNaAgIrsBMBBTORgKob33TAzlVyUF5GO6t+l2VeonY9lQnHbCCgyDuVXIVDuqgbdkFVUHburSd8KoORlAPdFTqb5lC9udkE8e4wCspjPhOSFhCUAZBVBVvxgA/lQXStAfgKwl2D3VOsuIKrkoIup3VjfusqHJwM+ahc0d0EhaDhBlyAhuSfzrBlcOrsVVk8kjsOzhZTIY3NycEoIMjpAx5KEsZ1glgJU8gAcQOyNgc74/0oMqGpuMbGNgf0t6vIrtVwPpair04ZqicBwYcAn5LqWat0WG52BW/tC8WrjY6iO2sc8QuOCQdsIOTNV3Otpr7LDG5xAcRkEqK33C4SShjpnAFb1tDdKXy3/dKsmj8d4zuRnK2/fbLM13jWZvU0HO3og5D4eWq218xNw6Scb9S4Y5l4Ldaa/ooKcEF2MgbKWq1jqLTDGmNjw7ttlby0lSaY4jUvvGq5YxL05+PHf7UHVKarD6dh92IOR5LmXg3cAyaJvujjsP3pW+dScN9AU7+illhLQ7bGF2K5dOCWhbnQtqHvg6g3IzhBunl3rp31RMLXRn8i445xZuq6N97aZTn612UtOi6DS1ZILMGnH9wuHeNenaK/VDpbuWgh3mEHW/QT4ndIiYWfD57LjPjw2T33LnZHUudbhQWfTUZkp3tGGHGMLrRxdv5ula8scSA4oNrU3SYm5xsFBUtYSAlucXsbn8ihujjE8dOUFHMAbsQqsxjcZUMLy9oBWZExv75BCfPCsJIJCyZmxgHpwFinugujOT3UpOFFH3UhGSgpkq9uSO6jOR5K+MnsUEzAQfTsp4i7qxlYznOaMt7qxk8mSc9h2QcncMv7dHiDI6u2VyTrJ0H3KdiPB6T5fJcdcEw2ruYZU4A6huVzhr2021lmJjc3Jbv+RB09vLiby9rQQOpULpARus7VkEUV4f4GO57fWtIZ7yZPizjKDVIiSAcpUOIid9RVrHNawZ7qN8wl6oWbuIIAQb84DSRDUA95i6x4g2x8124mdaHz0zW0OCQN8LgflF0VHW6lZPfGhsBkBy7tjK9ANb6I4d2zTrbjRzQGWOPPcZyAg4A1paKGeyguc3p6PonC4nOnrTHbJ5WMiDwCc7ZWg8YOME9Dd5bTQyuMTCWjpOy4rZxWubopIHF56s+aCmpa27Ut2fHS1T2sBOwdstNbedRHvXPIz26itOrLzPW1BnfnJzlQtr5OoDff5oNYluV3eD11LifrWNJW3IsOZnEeYJUbKlz2gHO6q6UdBHmUGAXvdKTIAT9Ske1nhk9I7eiiLh1k/NVbJ1PEZ7H9CC+xNbLVhsjdgfMLfEtBQtoy8Nb1BpOdltJ8cdHF40JHV32WJ+yStcfCJOO3fyQUrJJG1ha3PSD9io0kuGfVV/sn3x53KuPS1hcMZHoEE0zG+F5LAY1viDYKx1VK5xac4VQ49WR3Qao1o8Mduyhe0Z7KBk78BqmBDm5zugqwb7HCnDHEZ6h+VYD3yA7Zwqe8zA4wcINQwTnf86gn227qIVDxvvlTRkTAF5GUGM879ikb3dQAysvwWYxhRvjaw5HcIJo3PDe4U8eXDf8AOVgeK5owCVNFMenfP2FBLIB8lGdsgKySR7s43VrS8jcFBSTvuqN81bIXZV0YJCCq5I5N/wAL7hf/AAutX63EuOOy5H5N/wAL7hf/AAutX63Eg+i5ERAREQddfaF7cm/Eo/8A0nR/r1Ovnkh+mvob9oXg8m/Eof8A0nR/r1Ovnmp2hz90F02OnZXU/ZXSR5bsroYunugkHT2wq4b5hXNYMo4AHZBZhvlhVVDgbhAfVBIrXd0LiqZzugtecDY7qLqPmrzklOgIAfnYq9p8la1gyq4DSgmAwM7YVHDJ2wjHZGD2Q4QWkBu6sPdSuA6dlH0oAJ8le1xA3JVmCCqgk5QXdRzsSrZHkMI3V+B6K14DtvJBhwHqk7ea3JTxR+5EnGek+a0NtO1h6tsrMirHBvhYOCMbINIqABM8eWVqtvH+1D5bZQUcEhLnYye6jkk93HhR9ig06Z7jI7PqVfG5zm4U/gtd8Rxk7lQkeG/oHbOEEUmAdgs6hijdH1OaCfmqNpmOaDtlQue+B/SztkBBkmONziDGCo23SvpJCylnfGPINOFI2VmOoncqKSJrz1AbhBlT3q+NZGXVsoydsOO5XanhTaaS5cN56u5SNfKIc/Ed84XUeR0khA3+Hdb+0XxVu9oENkLiKaVwY7fyOyDaerHNZqat6GYDJiACO4C09tQJHAFgBXbTT/DDh5f7cbxcpYBPO3qOcZzhcJcWdB09ouBfYGB8Acfoen2INgRucOxOCsOpeTKs5kb4gI5dnAbj0WDVN6X5PdBe0nHdVy7I74ULXOwO/wCVSxODnAFBP8JAyFcxrSQBjupY4o3Nz8lhVEpjfhp+aCaqi/sf1hdi+FMLBp05cB8OfzLrS6eZ4Gc7HK5h4farfSWk07nEHpx3+SDK1hFCKybbO5XEd7HTVHpzjJXJd1lqrnPI+MOIOVxxf4ZaaqLJW4OeyDFaCGDOVeOyowgsH1K8AAb/AJ0FAcK4HKNaHHZXdIHkgsd2Qdlc4BU6mN7lBaO6uG/ZA3xP7H5p4EzTuCEBxe0DGyp4jsbndUkiqHDDMoy33B/0WO3QSU7sv+Lfdbgs5gNSzqYD28lpNutVW2QGoaQM+a3hYaGkbWx+MQACMoM2+MozQAiLB6e62YI49yG4XKerKa0m1gUr2lwb2GFxfJE6MOyNxlBptxbgEN81pQc/pKz7hK45DcrDha/Pxds+aCtMXl3xZWs2lrffmEYzlYccIeB4eMrJoqWriqmy4OAe6DeGpZJWWsEOOC3tlcbtmkfIQexJyt9XWsE9CIXnfGCtpPp42kuGO6B4TfDDsb4UcQcJQSdh5K/rcR0j7EtkE09zZE4HoLsfYg3LajTva0GLBW8hQ0rbWZWYDwNvyLHq7PQ2+0sqIy3r6crZFw1dVwg0sZONx9iDBudfdW10kQnd0A7DOy0ioqKh7/vryT9ay3VD53GZ3dyx3x9b9wNz5oImOdt3WbTEk4J2+atEADR2ULpHMdgE90GrPjb0E43wtGmJFQRuQsttQ8jf9Kvp6ZlROARuTsgqBGYm5ZuQMrWLNVyMe2MRnB2xhZH3CPhMcBtsuSNOaNtYsrq2fpDw3O/fsg0CKGmdB1uaMkLad9YGzfBgfILXam60sFdLShw6WkgLad/ruuYeHuPkg0+uJ8u6tph8G6s6uv6XooJJ3RnpYTgILq0YPkp6EN8I7DzWIXmYfEsiNwjYQMoJy3v2WnVH01JLUPByPNXRNbIMvwgsoxl5zj7VWpbh/krpAISSz1UJ8SQZIyg7++xYOOaXVP8AACt/lG3InsWQ4c0mqf4AV38o25EHtMiIgIiICIiAiIgIiICIiAiIg4+5hv8AeB4mfwPvP6lKvmiqf90Kj/Cv/SV9LvMN/vA8TP4H3n9SlXzSTtzcKrPlI/8ASUFzPohXg7d1E122B2UiCpB7qirnZUQUD8nCvGwVmG5zkK8HIQVb9IKrlQeW6qcbYQWE+iA5TpCqBhARVHdVwEFp7K3dSFowoz3QQuLsk7/lVhkcNt/yqdzQQrPADt0FsT3E5OVkjdWsgDQO2PkpA0AoL2swAfkrgFQHAAVQceSCN5KtAyrnnKo1AGNthkKVriBjJ/Ko2jBV47oLC7LskHutUpomOgJOMgLHZDEW5OAfRYdVWy07vDizgoIbh8MpAGVJQO6PvgaA8ealhh94aHydzv2WU2GCJhBIz80GoWvUV9jLIo6t4Z1AY6iu4/Aqz0t6sAmuUrHv6M/EQfJdH/Eljky3OA7OVyHo7jjedLBlBC54Zs3YoOdeJ9nt1LXzMjha9ozsBsutGq9S3m2XF8FvnlpmAkfDkLsNoPV1o1lN418maOvv1lYPFThXYq9hqrN0v8/hwg68x6o1DPBG+atkOSO5O67p8rF6vElFEHV7gCz+6XTPUel7pbXCCKJ2G+gXNXAXVt3sAZDIHjAx5oPS7ho11fXSCrnD/rOVwZzXxvoKwikl6Bny2Wq8GNeXCpq3OLnHIXEnNxqi5vrx0hxy7yQcJa0uVcKIAzlxLfVcCX2WSed/inJ6id/rXJtyutRU0rfesjLe5XHN6ZE+VxjIznugxbcBtjAworqB1dwVa174QOlXHFTvJsQgxqfY4PkpnvwRhXOY1gJbjKjO/dAcSQcqxXndAwYQUZ9JTtxjdRhoHZSNdt2QXYHorT3Ver5JjO6CrDkjP51I1gBOMHZQuPQNlk2cGruUVMRnrIHqg39wmkeLiAwFhz37LlvW00/3IJ8fOGds/JbPi0y7SlkbeAwNyzqzj5LatdxGdc4JKd0hIGQg4+r3vmvz2yEkdR7rUpaWBkeW4yAtOr3MdVmpi3Oc7LH99q35BBwghrJHxPPSCR9SxrVUPdeacuBIdIB0n0W9dCaWl1PWCHoJy7HZcmt4C1FDc6atfABGHB5OEHJug6WO26YirLf94lLQertvhZt91VqWosssc11e5oaRgvWka21LadJ6RZR00zRO1mMA+YC4D/2WK2phnge9+5IG6DRNZ1M9Te5XVDw8knOfrWiRtaD9EKtXXPr6x1Q8kknuqOBbkoJXAYzsohu/ZQy1BAxvsomVB6kGsNGGgjCseTjusX3l3SNyqGoOD3QVyQ7urgTkEHdRxO639/yqZ0Zbugk8Rz24e4kK3woxv0jPqrOpw7hOsoLy4hqjLndvJHOJVMZQUDQT2G/mpGs/SqFhAyEDnBBJ0gbhVa7GysY8k4KmY0eePLuguDWnGQEMbDnYfkVx6QFbkILDH3VM9GwV5f6K3AcUFQ8n1Vc5VwY0AKvQAgjwCqjuN8KhIUUkoafNBnRNB3Pmr3Mb22WNDMMeakdN2wCgo9jTvgfkV8TBjsPyKDxc+qnid8PZBHIzBOCuQ+TYf9d9ww+WrbV+txLjyR5yuROTYn9t/wAMR/3W2r9biQfRaiIgIiIOuntDPwN+JP8AidH+vU6+eilHxn6l9DHtChnk44k4/vOj/XqdfPRSfTcPkgyQMq7YIBjdUIz5IL2knzPZHeStYd8K5yCxvdHNJOQUxjcJ1ILm9sFCcq3JVRuEAjvhU6ShOCqjcILcplEAyguPZW5KvLds/wAyt6QgvaSfPyVwGVEHbjZSB4wgO2yo3AqQ791aW+iCPxDnCuaSRlPCyc7fUpWxtAPkgjRVcMFWkkILutw7Eq1w6jlOoqo3CCmSNgo3MBOR3Uju2QrRuEFzXFowqSBpaTjJVA7J3Q7tIQYbnO6jgkDPZZtIS4blYroXE7BTxPEUZ9UGV0MydgsA5iqw6PAIII+tUNRLk4zhVY0veJHeSDdZ1XqOmo4o6eve1oIGOryXMekKGO8aRlrLm8SyhmfiOT2XXmesJa1g7Arkbh1rt0crLJO8iKT4Tk7INiagnMV8qY44cNY8gNPmtInkdI8ktI+S7taZ4KaEv1C67Vr4fEe3q3xnOF1v43aQotOag93tIDoeoj4eyDjxjG9OThQPJbJkZCridm2D9Smjja5vU8jPzQZERJjG/wCdYc7SZfrKGSRpIGcdgrCZXHJB2QZrImFoJAOSty2GZ0ZEbDgYWzvGmAA32Wu2Ksd4gDkHL+lKWGdjjK3Jx3XGfE+GOG79MbcDJXJuhpjJGc+i4+4rxsF0DvPKDZtNnwxn0Sd+AQFdHhsQDfRRSNc45x5IJaEOc7c7LJmbhw3CxqUvj8lHV1L+obIMvY53HZYFU9wO2ceqR1Ly4DHfZbrsmmxdmj4Qc7oNBsL+qpEb2kha/WQxNxiPyW5KXRcNpHjztAHdYtfHb3u+BwONs5QaHboYnyAOYPyLetptNFLjqaPqW2x7nAcgjbfuo5NVGiP3s9vRBu6+2mkp6fqjwCPMLZ88ssTHOjkIIysSp1nNVgse447blaXUXYuY4AnJyg1e33CvqJTHPKXN7BLu4MYSGY+pW6KdDPUk1RABPmt03+1Uc46aYgkhBxrE7xJyHDz9FNUtYyMloAWZcrJV0DzKYyB3C00mWZpbg5QTaed4lSGyDIyuQG0dOKbq6ADjvhbL09bpGTiRwxvlb8lBioSdsgINmXcvbIWh2wOO60Mk4KzbnWdVY9pJwMrDx1npj3JQXU27hsT9i121FgqWEQb5747LHsVqkkkY6cYaT5jyXL1HpzTsGn3Vj3sEwbnG3fCDZt3fMaHeXbp9VxtcSPeD57rVr3qGoNbUU7CTG1xaN/JaCZDPKC/zQZoAMYIHkrA4BwBWWGMEQwR2WK5mHdSCYfR+xY7h1Pycd1KJWAYJ8liyueZMR5QZgaMDGFLasm4Mb5ZWJDHVOxhpWoWugrY6ts7mnoByg5NipYfcY3EDOAVqNRLNFY3iKbAx2ytFpr1Qe5+FI8dQGMZ81s+/6wqGyOo6dxLPr2wg21X1dQLhK8yEkPKvZI6ZvU8gn5q8wxTgzOO79ysSV7ondLOwQXyfDkZ9VivOTlVc+R++6tId5hBfF5fWp85UMbXBSjPmgo8AhWbt7ZH2qRUI74HkgoD1EZ/OsuJo6R2WGwbnIVXzvacD9KD0D9jC0Dmi1RsP/iDXfyjbkWP7FqV8nNJqkHy0BXH/AJxtyIPaVERAREQEREBERAREQEREBERBx9zDf7wPEz+B95/UpV80VTn3+px/80f+kr6XeYb/AHgeJn8D7z+pSr5oqn/dCp/wkn6SgrHvhSqGNx27KUnCCoOCqPBJ2VW74yquAHmgiIcEDiD9aud2Vg2QSg9lUO9VF1K4OyCT5IJMb5RRNkdnB7KTqQXDdV7dyrOogoXEnKC8juVbkBOsqnfcoHc7JggoNih8kEo+iDjyRqsDzgZVzXDBO35UA90VhccqocSEFcEb/NVByqA52KqBhATsqA5QnHogqXP8nH8qdDXuBeAT6lFjyyPa7b7EGoMAa0hp28gsCpklEwAccZUsdQQ0DcIYhI7qPf5oM6BjTAHOwThYsrWiXLWAn6lcydzfgB2GymYInbvIyg1S2193po2e5VDoz1DscLstwmgnudn67rVeIenPxHJGy6o1FfNAB4G4B8lvXRHFi42h7aORzg0/Cg5e1rR2yCsePAD8HbAWNpSSnFW0Q02B8gszTlytGpQJ7hIMvO/UVyxpvh1bKkMqaANcMZyN0HJnASJks4Lmgbei2JzYingrADEHn5Bb105eKDQ7yZnhhG3dY+s7RQ8ToDVwkPxv6oOk2q3h1EPCYWHpXGvx9TvEdnJK7C8bNHfsdpnMiYAWg+S65Ucr5ZJfE8iQgyCAfJRSktdgH8iyW9AznHZYdW/pcS35IKZJ80xso2vJ7q8POMIJGlXKBrjlSB/6UF6IiCre6vBHZWN7qvZAl+is/R4D9T0jXDYvH6Vhxs8Ygd91q+n6V1PeoJwACHgjCDszxOpIIuGcb4sB/g/zLp7SyVPiT/Ee5z+VdlNeaimm0WykeTgR4/Mut8Jb1yluNyST9qCegL5JcSnI9MrV5Y4YmFwYCQ0rSKKGaWXEAycrXbRaq+pvEFNUxkRPcAc5xgoOUuWc09x1GyCeItHiAZdsO67waw0rZodKmeKSPxfDzsRkbLqkLTZuG2mmXy3PZ7z0de3fOFtaXmfvlzt8tFK+QgZaN/Lsg2RxhqLozUM9MalzoQSAM7YyuNIXO6nY2yfJa7qPUVTfbi+pnJPUSclaQxjQ7t3QTUg6nAYwcrNkaAzHko42NjYHDAVXzNLDv2QYM4wVEzd/n3U/9keR6q4QNa7OAgu/eKM9lleG3o7KIxg5AQKP6YB9VqLi0gZx2WDSRFsgLu31rUHdGB2QYzwPJQnusmTpOyi6RnKCxoyVe1oBycKjcA9wrg5udyEEgwQqPA9AqgtAzlMtPmEEeAq9TvVVfgA4x2WK+ctPmgyj1HYlWkYUbZifVSNJd3QM5V7OyBoV2wCC4OPqjnH1VheMFQyTFowCgo8nPf8AOoiHZ7+atfK7yOfsVWOLu/mgyIc4GFIXEbKyEYO4VzwcoLSTjusun3asTusuD6KCyT6S5G5Nvwv+GHz1bav1uJcdyDfK5D5NgP23/DD5attX63Eg+i5ERAREQddvaED/AKzniR/ilH+vU6+eij/sh+r+dfQx7Qf8DriT/idH+vU6+eiiH3x3yH86DMVMhVVOkILcgnZEwAUQU6gqHuq9IRwA3QURWBzs48lVzsILlVvZRtcc/wDtV+Sgq36QVzfpBUwFVv0ggld9BQqZ30FEBtlBYqgkHCodkGyCUdlUEAK1pPZVQXq7yVqF+Gnf86CB30ijeyic89Xy+tXsdlBXGThXDYKmAN1UbhAVHbYwqqjkEeM7Ju1wyUzjdW5JcB80GXG1haMgErAqMiUAHZZ8TmhuD9iwahpM3ZBlRtj8MfCM49FSRgDCWjCkiMYjAJGVSQBzCW9kGmOJJOfXss6xvlZdKfwnkEvAysMtHWfrV8MklNM2eP8AeHIQdrNNOvLbCwx3FzQWDI6vkuMOIvUZTJVuMsme53W0qTipfKSmFLGT0gY7rcNjudPqmHquDgH+h9UHH1VLG2Q4iP5Fpr5HPfkZHkFzGzQlHWueWBpAzjZcb6utLLPX+EwYGThBgxtZ4YJaM/V3V3RH6BInN8IbjOFizTuDsN7IMroYdsBZFAOmpAaQB8lp8criAclXU9U9tSMdkHN/D3eM/UuPuLLib2G743W9uHdypoox47wMjzWhcSbNJc673qlHUPkg4/pBmJueymMbO+31K6allom9DhjAWA+okycdkGY1jPLAWHVMxt3VDUvYAXA7qZgbO0k5QYLcB7Nj3XMXDWKF3heIB5fauK/doy9uO65H0TWsoiwOOMYQbz4jsZDaiachvw/zLhSGWqIcXSOO/quR+JN/a+29Ebt+nC4soKmSTrB9UGRUT1PbrPZafK+U7uOftWdNkDJCxXNDkGE+R2cZIVYy9zhkkhahDRxynfCklo442ZaRnGyCW3GSMAwv6T8tlvLT7pZqyJtRJ1ZIyFx42rlp348h2WtWm/up5hIXEdOMboOTNc0FHFaA9jQXYXFdEWmQksOxwt5y3436mFMHZ2x3WJDpsxvHw9yCgms8cJAw3BPqFrNxiAoCQ8DY9lpNWw2yPqGxAWj1WpXyQujJOeyDat3Lm1rxnCvsruqsY14JBWqWu0uu9V1OGcn0W6GaRjoAKgtAxg7IMtlPAyha+LDXYzkfUtKuNZc207mMqXdB8gdlbX3qOBpga7JG2FtasvdRI8xj6OUGBWHNQ4uGSf0qBhw8LPLGSjrd9I91Q00bW9Q7oL2ud0DcpKQIyfNQteQ7p8sqY4e3pJQacXPLsZO5Wo0cTc9TxnChFOMkjCzbHRVc1zgY5h8Jz/i9MINRp5Yh0t8A/kW94KKkfZHyBoDg3O/fst2t0lpaKyMqDJH43Tk7jOVxfqm/fc57qKldlh22OyDZ9XV1ba2VrZXfSIAzssWUvdJmTc+pWY4RveZjjqccqN0bXO6jjCCwPeAACQPJVO/dSYjx5ZwonOaHEIJWsZj6IVeiP0Cia/JOFXr/APfKC8hjcYACicMnZWySHG2VF4rvUoJ1cMAbrG8R/kcqWPrd3BQXFu+yqImkZONvkhDvIFROkkbtvsg9APYuNA5pNU4x/wDEGu8v/vjbkUfsWS880uqc/wDACu/lG3Ig9pkREBERAREQEREBERAREQEREHH/ADC/7wXEv+B95/UpV80FX/b9T/hH/pK+l/mG/wB4HiX/AAPvP6lKvmgqhmvqf8I/9JQWs7BTtOBuomDABUjd9ygvVrgSq7KjnY2QHHbGVGqnJ3KqAMILVXfyVQPiwrg0BAGAPLKtJydsq7pCBoCCrSOyZAQN3V3SEFFUEYVQ0ZG6qWt8igt6gm53wqhoKuDQBhBYSMYVMoe6IA7q5Ub9IKqC8eSub2QAYCdkFhBBPZWOUjvNWYBQXNOVHKQDjGSVeAR2ysiOBkjcuxnKDDBzusmIZH1Kj4WNPw9s+axZZpGShoyAgyHnpOQ1Yc07/EAB2C1OJkb4g52MnfCwKqnIdlo7boNQt0bJgPEwdvRYtw/2vVt8EAH6ljsq5qfAAxspoT768Pk7hBvOxXK5x00fhTluXBd4eXN76zTodWPLz4exP1LoD77LSRMbHnAcO31rvfyqVranSodMd/D7Z+SDQOOVbW0k8vukxYA4jYrfnLhUzV1iea15eQDhcScx9bUx1Eop846iuV+T/wD23p55rNj0+aDjrmdjia2fDDjBXTODHjz4GPiK788yWn2VhqGwtB79guid8oX2irna8Y+M/pQYcxcNxnZYpJOepZAcZWAgd1C9nSd9s+qACFVVY0HfKOABQW9lc07IW7I3sgkar1C15BClachBdjZWnYI5xAUb5CGk4QZdudiUEjO/ZbltT2m6QDwyMuHcfNaLoel+6VwEcmMdX865TqtMU9AxtWwAFoBQZvEKLo0g17XDPh9vsXXilkd1vGTvv+dcnaw1TU1NMba8noHw4WwKejY5zgzBcQUG4NEBnvYMkfUA7cYXJ3+1BUROjpC122DhaVwO0j7/AHVpuTQIi8bkbYyuzWp+HmlaGwmup3ReIxnVsBnOEHXvXlTWusoY6oJZ0/QJXDFK49Umwx1Hstz8QtVVBvUtriJMTXFowdu62fDK9sh+ZQZk3TjIAB+pYzCS8b+aypunws5GcLAjeS/I9UGfM8+FtnssUPeAckrJcCYxn09FD4fqEFKY5eMrJcooogzf19FM7HTkdwgE+aMPxYP51AJiXFvopY93jKDLIHR8IwVH8XqfyqeUNbFlp3wsMSP/AD7IJCT5lCT6qwOd5qpdgdkFjiQrHOJ3yUe45VvUUEjC4Y3KmBJA3WMHkKokdkAFBO/PSsOX6ay3OJbusZzQX7oJ4xlo+pZDG/D5LHYcNCmEmG/NBKBjfKskPzULah3Vv2Vkkzi7bsUEmfmrHndR+IVcz4+6ChIPkr2NyQRhVMbfJVb8J2QZEY2wFUjHfCpFuAkmyC1/dZNOMs3wsN7yNlJFUdDd9kEshwTv5rkXk2Oeb/hhv/8ALbav1uJcYyz5zgrknk0ef23nC0nz1hagf43Gg+jNERAREQdd/aD/AIHXEn/E6P8AXqdfPRRH7475j+dfQt7Qg45OeJJH96Uf69Tr56KP+yH6v50GadwrTkbZVysc/wDQgtJJRWBxJwr0DJ9UyqZHqrXE5wEFS4b+qt3Jz+lV6Ad1Q/D2QVaDlXqxpOdwr0FyqO6onZBL5Kwg/kQv8lTOyCxBursBUOx2QXtB7quQFYHlVJygqXZHcqx5OEByoZpXA4CA7urmuA2VgORn1V4aCMoL8n1VW9lGHb4KkBIQVyCqP7KgPmrXuOUFjnbnureoKpGVY4YORuglBcBkE4+tX9IczJ3OFieM/BCyad/U3pJCDFc92SMkBZ9KS6A5/OozTNJOCO/oqGXwWhgzvsgxZTiR31rKp2NdCcjJwsd8ZcerHdZVPhrCD9uUGC8YeR81rWmqipZWNbFIWt2zjZYXgREnOPyqlNUPpKtvh9gQg7D6QiE1IXPkBOM91xFxVY2O99De261+x6umpKZoBIyMd1De7UdQNNe8ZI3yg4za5+AASrXH4t91nXCk92qHRAdisRjA4/EgkYPhACviYBMNuxU8ccYA7K4RsB6hjP1oNz2OadjWCOTG481yXQU0MttL6hoe7p7rg43ielcGxE7LnfhrLFd9OGWpwXdHmUHFuq442VUjY2YAJ8ltIloJ281yBralhbXTBu4BK2O6Bm+UGBUyBwAaMKWmcfDIzgqR9PERt9qxnvMPwtQZtG0mQZ3GVuu3Pewt6DjdbXsvVPP0nGxW4pJDSuGD+RBbquWV9MGvdkehW37TGCNwPPutSutS6ojLXdgPNaPBK6BpxlBqVZE3wydhv3WnYwCFZUV8rxgZ3UcUkjh8Q/MgyoCc91M4OzuchYL3yMHw5/Ir4ZpS4dQ2+aCtY1oGcd1hB7g048/ktRrWdUfwYJwsGOH4D1BBujQTnPrQJMlocFyhPDES0gdguNNBiJlSC4gbrketq4WMBa4ZHbdBtrV7RHAej+5XGhnk8cjG2fRciXYy3AlgBI+a28NNytn6y0BufRBuPQFPDO5vWA3fdb41PQwxWpz43jIb6/JceNrmWKIGA4cNzhYFdryurITTvLsHbcoNq180puEoLicOIUnQ1zeogZVs0fiSOmI3JJUD5JQcYOPqQUa5/V05OFkMJLfpLHHqO6oXzDyOAgyS0eRGVG9zmAkZKhEsjjgd1qVhttTcrrBSuY4te4A57IMFlU8OGY8jK5I0+2nNrM3gEPDcg481yjbeCdoks0NY9rOsgE5Cnr9NadstpfG50YeBjGyDgmvvl295fE2oeGDPw5W2rlUzTyffck5zkredfTULrpIGkFpcd8rPg0/ZqloMhZk+qDjEved89lkRPL4znuOy5Lj0bZnNcSWeeOy2dqK3RUVR4VIARnGyDbrnODjnuqdTjuclagbZK4B/T3+St9wczuMFBjMcNs5+1XFwKl93x6BRuaGnZBY/srWjI7KZrGuwCjmMbsOyCkTATkhZIY0DbAWN1hg2PkrXVLs7E7IMzpb6q0saT2Cw/eH+SyYpOpgLu6Dv37F4NHNFqjGP/iDW/wAo25Fb7F12eaTVI/7gK3+UbciD2hREQEREBERAREQEREBERAREQcfcw3+8DxM/gfef1KVfNFU/7oVP+Ek/SV9LvMN/vA8TP4H3n9SlXzRVRAr6n/CP/SUFjHdgpAD5KxjW42I+pTMxjGUFA1wOSjvpBXFWHqPkgq4gt27q1rSfyIQQrmIAaQVKOyqACeyEADZBQ7hUAwgOVUnCAioXbKgeguOfJBnzVOsJk+iCqrgq3LlcCQNwgtIOSqYKvO6ogdu6qN+xVpOdkBLfJBMCMYyrVZ1En/2p1kILiRhUHdUzlEGWyJpbk+ixqiQxP6WlXtncMAdvrVHsEm7u6CrCXtDj+cqyVjc5IBKuaSxpAPZQyveT5/YgqJHNAw4jCyYWNkaS4rHYwOxk+WVFLPLC8Nb2KCte1rR8I2yqW4nrIWZBCypaDKqT07KbDod/qQam2Jj4WZIPxBd0eWZ7odPBjJAAY/X5Lok+4zsAaD23XaLl84gwW62NgqpMEtx3Qb340U8dTLJ4jevc/Nck8tcXu1nc2N4YOnt2XHt6li1RO8wuBBdstQ09crzpRpgpWvDT5AIN/cVKcVL5w5pl79t10Q4v04prjK1sJj+M7EY816DcKbVU6+ubobm0kOH75dW+eHQFPo6+sZRtaGufvgIOttuw9gzjt5qle0A/CVjQvlp6ZjiCM5Ukb3VWduyCkefNXqjmGPyRpc7sgqeyoCMJgj6Ww+aAA9kFVI0jG6jVwOAgq7f8qjfs07FSMLSfi7KZ0AkYenGcINa4bjNzBA/feQXMN6Y40gznHT5ri7hjR+Hcg+bZvVncfNcwaqfRRWd0sbgS1nr8kHCerw1jiRgn0A3W2tOSOlvEUTmkhzgCPtWpRSVWotQvoowXAuIA7+a5EsPDSS33imqqiIBmQSSg5N01aI7ZYmVsDfDeWg5GykuuorhNZpY31xIAIxlZXE2/WqwaIbDQSN8ZsZ2b3zhdZIuIl2l6oJXu6X5GyDD1O8zX173gE9R39d1h4aNxj8iy6wNqj7yT8ZGd1hMbIT8WUFDI5x6SThVjHS8KR8TWjI7qJnUXg5QakWjwvIKENBzuPyq185EYwDnCxveH52QZ2PPIKtd9EqBszyPn808VxCCrMde4WS0HI2WPAMuHV6rMcGBuRjYIJHOyzCiwFZHJ1OLd1MWjG3ogsy0dv0KhwrTkdlTqKC0jIVOn5K8d1XAQR9PyCBuPRXkDCoguyMKE/S+1XPcRlRh5Lt0GQ36IVcq1pGApo2B3dBAdlG879is33YfJUdTtAGwQYW/opG5G6mETAOyskAaPhHmgoHH1VwcCRkqHL8+f5Fc0nIygy4PVXSgq2F7WjBKukcCNkGOT5KnYHCo5wBTIIyEEbjvuuUOTQ/8AXd8K8f8AC+1frca4td1ZGxXKXJn+F3ws/hhav1uNB9GqIiAiIg66+0KOOTjiSf8A6To/16nXz0Un0z9X86+hb2hn4G3Er/FKL9ep1889OcSZ+SDNPZRP7qTq237KJ5BOyC5qOBPZUYd8K5xwEEZyD3VzSMYKj6iXYVUF4GDknZXH4twqAgjHy9VXYeaC7oCqNgqdSdYQO25Q74wqFwI8k6w3I2ygq3uriMKNjndWcbKTOTugpnCocK5wGFYgoTjdVHxbqNz85GVfEWgblBXBUUoy7ssguaVaWNKCHpw0bK9pyMKu2MFBgDZBa5uNwgdscoXdwrEFzTg5KOIJ2VqIKO7I36O6qjtgceiDGOxOFJA49QCjI3OxUsTQ0dWd/mg1ONh6MkjfdabU5EuFX3uVo6fJU3kPU5BlMYCzP8yx539BIb5qQSEDCj6A+Tf1QQiV4Ox+xZVI1r3hxAWRHRQFpJ7rFmcaZ4bH2CDcMUnSxgDh5DuuTbBQsqLA+R2CehcI/dCYADOw7LlvRWoIXWF0EjviLcY+xBx1f2NZdJWAdiVoUp6X+i3ZeLbPUXKaaNpIcSdgtv1FqrhOWNhJycbBBiidwAG/2KaOVxG/5FqtLobUNYzrgpHEfUVg19mrrPL4VdGWHtj5oMGoOXLmvhlO+PT+GyBo6fX5LiBtPG4AuI3W4LPqOot0fukBPTjHdBq2q5HGrlOSSSdwVsud7hnII39VvKnpp7s4veCSd91t2/W80UhYRj1QaRE8uJ7qGo7jbzKy4Imh2ceXmoayJxf8I2QZ+mW5qTnbcLcFxaA8bhbaschp6gudsFqlwri946N0EVYNjuOy014AB7FXVtTKRtnsoqZxkBDggpCwOfgj8oWQ6NrdgAAqtiazfG6gqJH747IMqGNjzuMqtREyNnW3AIWHSTydZBB7rKne5w6e+fmgx6aYSyFjtwFJVxtY09O2AoYWNhf15+eFLNI2Zp+aDL0vUObUhodg5W9J6mUDJJdjyXHdJK+jlD2d8rkPQj4LtdYaetI6HOAOfIIM2yN97lDXxEb98LclwsNPHQGUEZ6djlb91dpzTOndPMrqKSMylnUenGVwfedcSupnwsce5A3QbWvs7o618ZdkAnzWiiUF2OkfYFWerkrKsySA4JVz442sy07oLwMjOQrZGM6DsMrGZPIHYwcZWQXF0ZJQQwDqkA9VqLqeMwk4GSFo4e5jj057rJiqamdwhZkk7BBLQwNdOAW5GcdlyfoGzQzXWJxhIwRutqWLTNcXR1E8ZDSQTkLm6zRWWy6dNc57GzsbkDbOcIOT3UAhsQ6aoABnbI22XWbine7hR1boY6olpcc4Kxbrxr1AaiekgefBDy0b+XZbFu99qr7N4lUTknPdBCbvOXl7nHqKngvdZ4jQJSMkDuoBSQlmfNY8kPhuBYc4KDeMNxqjEHCY7rTpCamqHiuzkrQ23KcNDQeykiuD2nxCNxug35Fa4DT9WR2z+ZaDdImxPLWszn0WANUVLQGDJHY7rdFpZRXGn8WocOrGd0G0ZRgE9J7ei0yd56tshbzuFJRNLgwjC25U0Ic8mPsg00SOHb85QyuPdZ7bTK/Ba0nPZSNsk4G7T39EGldRyd1QnJytXNpLfpNCnZbKfHxYyg0Nh3+jlXGR42DcD6luWhtVJI7BwQVqElhoDgnpQdyPYrvLuaXVI6f/AJQK4/8AONuRa97Ha30tHzNanlhx1HQla0/V90KD/UiD2JREQEREBERAREQEREBERAREQcf8wv8AvBcS/wCB95/UpV8z1ccV1R/hX/pK+mHmF/3guJf8D7z+pSr5oKwdVfU/4V5/OUELXEY3U7HHGVE1mw791MwABBc2QHZXYHoqBgByqOcQdkFxAOw2QNwqde3fdWOe7tsglzhM5UbX9RHqryenCC5v0gjlE15J7qbY90EbuyjLiNlM8fCcKHpz5IKx5Ls+SnUcbcYUh7ICZAVoJz32VSQUFwGVXp+aoDsE6kFekeqtTqJOPJUJx5oKqhOVXyTAQUxjdUJyr8b7hV6W+iCNr9wpA/bZQP2xhVY49soJierb1TpH1qg2TqQVAwchC1rsZG6p1eioXEFBI1zmYDTgK8OLx8W6jG4VzCRlBDPGzA2W59I3WspKtkcEpY3GFt4t6tlJT1LqOYSNJ2KDuJwapjduj3moG5G5IXOtbpq3wBmAx+R3GCvP7T/GG66VYw0pfkEdiu0XB7ijcdbUjJatziQ0HcoO0/BK100N0cYwGnGy6o+0Gja28HLS4h537rshwkvc8F3eBnbZcQ832n2aiqvGmAJzlB5/XPAooi1hb8PoobNhwJccLfWtNNxW+mDQAMNXHNPO6mc5rc7FBq1YzYkEK2kaC3BHdYZqpJT8Xb5qaOUsCC+uwxuAN1iQOJBBU07jI3f1SniDQSfNAb3UUry04HZTvAadlfDDHLkuH2oMPxHEAYxhanbiXPHVuCFh18LYmdUfdZum2GaoYJNxlBvjTjWx4cwdJWu6gqpzaJcyk4Yds/JYdTTNo7YJacfHgLQ6SquFwm9zlB6X7HKCLgbSxXDXDYqhmQXZPy3Xa/Xem6GgtLaiDpDwzIPbyXCek9LU2kCL2wAPPxHAWtam4oz3G3PhLiQAWoOJdfXm4y1z6OaoL4hkYJyFsdkMbCSGAH1wtTvtc6tujpHZ3PmsbobhBZE4vd0nsp3xNDcjyWDJIYzlqMqnvPQfPugFxdJ0581L0ANzhSiBgYH5GTuqHB2KCAkqmB6KRzQUDR5hBY3urkaBlXYCCvZV63YxlV2VMNQX0+7srKPksSI9JU4eSQgpLsCowcrIe0Obuo+gAHZBC0klSDYK0ANOyr1bHt2QH7BRFxJ+tHyE7KwOIIGUEh2BURyXZCnABbv5qhjA3A7IEZI7rJiPplQAdvkr43EHugy8jpVkjsDzQPyB2Ucju+6Cx7iDsVYSXd0JJOSrckO7oJQwY7eShk2dgKUP7bqx+5ygsDnNwcqRsjiO6jO4VWDAQUe4k91fEMjurHfzq+InCC5wHouS+TX8LzhX/DG1frcS4zJJK5M5NfwvOFf8MbV+txIPo2REQEREHXX2hn4G/Er/ABOj/XqdfPNT/SK+hn2hf4G/En/E6P8AXqdfPLBs/dBkTkhhPmseN7icEqef6H1qOBre6CZowA4+iteS491cScYVuCgsB3VwOSrQN8FXAYOQguLcDIKq0F25Kp1EhUDiEEjuyjyfVV6nHZUIIQMlWnqLs+SqTsjSe6CdhHTjzVVFG/JwqukLTj1QXl2fVRv88K4HACEBwQYpeSUL3DsVI6MA5AVhZv2QXMeTjdSh5AKjDQPJSMAIyUET3EnOSqtJxuVUgAnZAPRATBJCvLcDOFaNkFzgA3ZWK4O8ihxjyQUaMpIOkFWteQ7GUc7OEEJ7qrScK4tBz6qgGPJBdkHyV47bKwD1Qvx5oL1Qj4s9lQuOMqgOe6CVszgMZP5VFI7qO5yT6qhJ33VCfUoI3AA7LcmmKqdsjYmyENO2Mrb5ZnBx5rNoqk0nxtJyEHMNttNNUwCSRzcnfOVLR6doJLsxjmtIJHouL49c3CnaI4s4HzwtfsWtKh8raiQnI33KDuhwo4Zaer6AvnZESG5GQPRdXeaSy0dh1OKShjAYXkEjtstesfMHdtPQiGB7wD8PfyU11gpOKDDdLkQZB8W/qg63yTPMbMAj5rWtOxidwL+63zd9A0zHujiAw3bZbCr6ettFw8KHOAcBByhpmjhcDkDttstl8QwIKkhrfNbq0dJVGFrnZycfoWPqqwSV84c4ZyUHFsEjnOG2MrObG13fBWo3+wSW2PraMY+S24KuRvdBm1ThTt6o+6Usrp93eSxo5DVO6HFZsUTYdmoKzRtLT8gsQuEY22CmrZS1vwrDhLpCQUGTFOXnCvIB2Pmo4ogzfKrISe3kgngjYT5BSSsHcYWnPqZYuxUkNW+QgOz80FtYSzttlQwyOJ33ws2RjJW4OCrBBG3YbILmYdjI81rlhqJaWsZJC4ggjBGy0CR3hAdJwtZ00TUVzGk7ZAQby1PqO7T2wQyzvLenG5K4wfWyPkLXbglcwajs0P3ID/Mtz9a4sq7a2Mue0AYKDHljAjDmjBxlYYlc44yftKyGyukcY3H4Rt3VXQRtGcboLWdPfG6leR4ZI9CoWHcg4wr2vD3+G3zQY0Jy8fDn1C1bTQE1+gidFlr3hpGOy1vTul21b2OcBucLmW1cKrZR2c3trWCVjertvkINWl0tQQabZUxlgcWZ8s5XCOsbxcaWR1NFUER9WCAdiFnXziXeoK2Wzt6/CYS0b+S2fcqyS4OMs2c90GmujY9xke0EnfPzWJL8Mmwws3J7LDnbvlBIJXkDBKkbvjKxYyScLIDseaCvgt9ArJAG7K/xM+asecn7EEGcOGd8LVKOtqGMwyQgDyWnFmdyFQSOj2ag1KWsncRl5JPzWp2+JszcuIJW2TM84yeyyYbnPD9ElByDZKCnlID8YytVq7TStblvT6rjyi1JU0+4ysmbWFTI3BJ7eqDV7nSxx5Ax9i25UPeCQ1/b5qKqvs9RkklYD6uRxz6oNXo5JA76Z8/NZE1TMNusrRqSrkJIJU8lS7IOUHf/ANjVNJJzPana52caDrT/AM4W9Fi+xgkc/mi1QDnH7Aa0/wDONuRB7PIiICIiAiIgIiICIiAiIgIiIOPuYb/eB4mfwPvP6lKvmiqf90Kn/CSfpK+l7mF/3geJf8D7z+pSr5oKvavqf8I/9JQAcABXNHV2VjB8IJClYQAguGytegdl3krZDv3QURUBJ7qqC1oLXKRzuoYVAzfsqvwO3dBYz6SyR2WNHnKmJ7ILiO5VFd3GSrUFW/SCqrR3VyCjuyjDsKR+Ok77rGOQe6CYSAFXdQd2WOqtcR2QZDfpBVVjTsCrxuEF3Tt3Tp+aoCe2Uy5Bcq42yqJ1eSDHk2OEZ2KmcwO8t1QMx5ICo7upAB6K1wGUFvVsqg5VQweYV7WgdkFFVp8lcAM9ldgIKsZvlWzDBz3VwcW+eyka1kgPV6+iDS6yXqa1mPPK7acrcjZrewkgfD/MuqlfTNDQWnfsuwPAG/Ms1KIuojDUHd3hg0Nu8hGDutscxlAKj4jkHKz+X++U13vhbI/IJ8ysvmdg8DHu4BB9EHRfifRCKAj0acrgsgiZ+Bndc+cVevwCSNy0rg2OMPkkJHmgtY4Y7b+anYdu3oqsiYPRHAN7bIK46tlcBgYVjXYO6u6vRBbMekdsqyOYgdyFMG+JkEZUE8XRjpBQXhxnw12+VrVohEL2vH1hbeY90eHY/MtzWBr6qRrANz6BByBpxjrqBTuYXjYYW7KfRTIaqJ/uxGcHstV4J6XgNa2a4gBnUDuFzzrG2aXobIaumdF4jGZ2IznCDgHXlK2z2EPbKAenOCuCjqCScSsc8dytw8Vdf1lbdJLXG4mIEj5YXGlMXskcST8RzhBnTnxJi/G5Ksc4taSFeMdG3dRv3yPVBAD1OOd90Dfvo7LIpYQ6TJ9VnOpo2kHCCMtxCMlQdJOSsmYgN6R5LHBIQWHZFRxx+VUyUADJwq5A2UfUUyScoJRuq9PzVgcewKr1uQXNOCqiTcD5qwjZUaCHoM9o6m5+StLdjurmH4B9SEjBQYrzg9laXbdlc4ZO+Va5u3mgiL8nCA/ECqFpB7KrQeoIMhv0QrsFXRsy0YUgaAMIIelVAwrnAKmEF4d32Vr90AIPZXhmQCggVru6nMfdRvZhBQdla7umSE7oLepXtII2VhGyq0kDZBRxyVJGPhULjupYjkIKnbOy5N5NDnm74V/wwtX63GuMjjdcm8muP23fCv8Ahhav1uJB9GyIiAiIg66+0MOOTfiT/idH+vU6+eanGZMZX0M+0LweTfiUP/pOj/XqdfPNTEh5x6IJ5xhuPT/UoIyQVPPuzdQRtJOUE52ajHDBVHfRwO6tAwgO2OUacqpAPdXsa0DHqgsRXdO+42Qj0QGnCOdlWogK15wqucBsCFGSd0FWPwfRXE5OVC0HqVT1B2AO6DKz1DAVzQQMKOLYKTIQWub5qgbkKRU2CC3oV7WYblWlwCt6yMZKCh7oNlcektVvdBUvyMYVqlDRgbKx4AQUaMqrmhvmqA4UcjznCADvhUcdwFTIVHHdBI1mwJ80PdW+JtgKjXEnCC49lYQpukYzhOjPZBGDhOr5K/oA8lQtHogK0tycqnV81a55ygyABhRvf0bZUYlcPMoGukcNkFHP6vLdZ9HWmBoxthG0kZaDjyVjoWDYbZQZLry4EbA43XK2gb06elEYl6QR2yuIBRtdvnuFmUV+q7OQymccD54QdjLdYo7k4ufMMk9yVpV54Z00tUJSWE9/JcR0XFK+0O0bj+VZTuMF/kcDISR/3yDmm1aOp6KMND2jCyptKxVEo+EHdcOQcWbq4M6uoZIHdcuaG1pS1tIJax+HBud0GzeKmn22uhMkdMX/AA+QyuCZCS4lzMEn0wvQzhlYdFcRaiWkvskXQAQOvH864H5lOD9m03fxDpcMdE5/7zGPzIOt9IcSDb7VqRIzha1cdH1VupmylhBIytsyx1bc7HY4QX1Q+H1VKVgI7fJS26GSWQtqM4+YU9dHHAPvOPsQQzjobkEK2ACTc+vmsZsr5XFjs4R8zoSOnsgyqmFvSe2fmsRo6eyo6qlk+EZKvga5xw4FBa+Ut75KvilLgO+M9lkOpo3NGf0K1sUbD8OEENUfhH1rWtJPxXsOcbhaLVDqADVqemmzNq2loPdBytfnF9obuT8K42rIsMflpxkrkItmmoGtlzjpWg3O3wtpnEYyg44mf4cpwrfedsb/ADWRVxsdVOYOw2UbqQkHpAz9SDHMhycHupKIu96j6WF5LgMdyVrulbCLlVsbO0Bpdg59Mrs7w14KaRrDFV1phBABOQEHGmj7QXUkUzoHNJ3OQdlzBTU/VYnU5mGC3GCVujV9h0bp22eHQviLmDbBC4B1XxFlt0MkVI87ZAwUGjak0jTtuM0/wE5JOy48vcIpZCxoxusmt4gXOsnc5xOHHHdaTU1j649cnfOUGOThY8p6uyneQoS3fO6CxrSw5zlXH4t1eRnYq13wgYQWjumcuyoi7dSRboJCCQMKgbvgqdob0qGU4dkIBh6hsAgiDcZClheMd1eek90GOZAzOyhdL1HIWVLGwjKhEbc47/WgjdJnspGRAjcBSsgjxnCvw1uwQWRkRnso5ZviV87g36PdYpy7coPQP2LMvXzSapGf/lArT/zjbkUXsVR/10+qcj/sf138o25EHtSiIgIiICIiAiIgIiICIiAiIg4/5hf94LiX/A+8/qUq+Z+t/t6p/wAI/wDSV9MHML/vBcS/4H3n9SlXzP139vVP+Ff+lBa0jpCvZuFjjOQN1kM7IKomTnfsrXPxsCguJwMo12SrA7qOD2KvAwglHZWOYSr2EFVcQB5II2bHCvJwowd8gqTGUFOoJ1hUcO6twgvDgSrwRhRsBzlXoB3Ubo8+avJwgcCEEbW74Rw6VcM9WQri0HugM3CvAwgAHZXDGPJAAxvlOoKvkVYguyDsgGFZk5yCnWR5oJEVocShdjuguO6p0/NU6/mqggoLxgjCAYyrRnOVIAMIAHmqoThW9XzQVccI2RwzhWnfzRoOEFZR1AZyt76LvElAQ1kgG3qtkE5Cvp66WmkPQSNkHeXlX1BNU34B1QAC4Dv81znx8h8ekY4jxD0g7DK6GcDuIFVpy4snD3DLh5/Nd89O1DeImn2VE+HEMB338kHSHjHD4UTvgI+EhdeWPIlkGcfEV3A5kNNR2908bAB0grp+Y3MnlDvJxQZEZJ3JP5Uf3VgdgAjCvb8QyQgtJwr4zkK9kTXuxthUlb4ZAadigvj7q50QfjJWOZegZPf5IZZjEXMzsCUCvj93jaQMnK3lwxhNfdYIXxkdTgOy0XQ9qk1JdBSTDLcgbhc8WnRFHpeeCqb0gtwSg5IutmbpfSjLjTP8N3Rn08lxDcOI9zraGeCSuLgMjBctd4u8R3HTf3Op3nZhAx9S6zw32s6y0ucQ7ORlBqd4n9+uj5XkEk91C6lABI+xQsk6j4h7k7qb3gu2QQsJDyCpnNCjkYWZeFH4rz67IMqEdG/2qYyE9iViQy9WxyFkOIAGECR2WqFC4kogsd/OrVI4D5K3AQRIr3NxvjzViCrfpBXI3GfJXhoJxhBVn8yuDB1AqvTgAgKrTvugnGzVaeyFxDc58lH4mRjKB3KFgKsa85zlX9X1ILHMG/ZWgYcAFc5x+StzuDkIMphAaEcRlQh5wPqVer5hBcBlV7bKnVg52VQcnKCRXDsrR2VQR5oKkZUcoxkKbLfko5cZ+xBj43yqOVzsAKIk57oKK4dlar2DIQRu/nUsXZWOG+4UkQGEA7Z2XJvJr+F3wr/hhav1uJcZEZXJvJrj9t3wrx/wwtX63Eg+jZERAREQddfaGHHJvxJ/xOj/AF6nXzzU/wBIn5L6GfaF4/ab8Sc/3nR/r1Ovnmgz1HA8kEszgW4CsieG7eqrJsN0ha0n5j0QX4zuqK54IGytbnzQFc1pO6uABCrsNkFAR9FVxgEIRjcDzTOxyUEZ7q1wyFce6oTgEZQRYIcqgZ80IPdVafJADcFMjO439Vf0Z3CFmDuEBXNaTurScKrHH1QSA+XorH91Uux2KtcSd0FFR3dW9RzhUcXZQSEkAKoOfLzVBuBlNggkDgVR6s6sDYqhe49ygqrHDOT8lU9RHmqDtuUEZ2VFeQCTsrSPRBYNlex3xA4VOhXNZ54QT9QI7K9nZRjsFew4CAWZUcg6R9eylLgFFIc+e6CAggqh3OEPVk91TfPzQSthPfOxWVHE0M6iQMBYYfJ5FXe8SgdJJQTe9EHAPbZRGoLn5UTh5+qzqamjczqOMoKtf1NB88qdlMx4y/B+xYMznRkhp7KsdZI0HugnlhhYcdI/IkTIvpdIysUyvkO+Qqskc13Sc49UGcJ2Nx8IwO2y1Gl1VWUMZZDKWjGFgwRRyN3xv6rGraeNrvhPZBvzSXFW92aqZ7tWuYXuAJBIXZnQWmncTIY6y6VQleQD8TsrpHFCA8ODjkHK5G0rx01Ho2JsFtc/DRjc4QdkOKXB+jtdA90cjDgbAYXXyfSUcbnAU5dhx7BZ0nMPqTUkraS5l2HnB+LIwuXOHcGn7w1jri5gLhkg47oOt+qqB9njDo6Zzc7ZwcfWtpirleMOZ1fYu+GouG2gLxREPdB2x5Li+5cHtKQPIpBEcuxthB1iYSd/CLfsUU+O+FzPrfhpHRwF1taCQD9FcVVGnblDlszDkE+SDTqMBz8OC1B8LW7t2WHLR1FCepwIVGVj3kd0F9VKWDYlYvvLlmuh8ZucDKiNC7sB+ZApMTvDXLe2lrbGa2NpA3IWyXQS0p62g7ei13Tl1q2VbC3OQQg5mu9rjpbSHNcAenK46vEzhTPw4efmtavOobg+2hsgOMYWwK+4yyxPYM5JKDQpJHGrc7c/EtQogZpwwRncq+wWee4VYLm93ea5Y0joSl+6cXvLWhhI7oNGsFoMEbJhC4bg7rk2mv1ba7SXw1hYQ3tnHktz6isOmrVZA6ndGJAzOxGey69aw1fNAH0lO49JcRt6IJtRcSbzVV0sEta5wBI+kto3C7y1TSZHBxOfNaFJJJUTGZ2cuOcq9h36XH8qCzxG+L1dKyWnLc+ip7vGPixnzyqOPSCAgofP61RROlwfVGSZPZBKrJVerXjIQY57q5mzlQg57KrMdW6DKaHFuxOFZJscK4OIAGdla/B3PdBYHlu6qZ8f/YUbz3VgBKCQzHGMq0vJOclXsjDvJTCCP5ZQRtlIHdXeLkZJVr4xvgK3YbZQHuLtyUY0OGCrx0nY4VpcG7NQd/vYtMA5pNU/wArv5RtyK32LD880mqc/8AK7+UbciD2mREQEREBERAREQEREBERAREQcf8wv+8FxL/gfef1KVfNBW/29Vf4R/wCkr6XuYb/eB4mfwPvP6lKvmiqhm4VX+Ef+koIo25wSpQMK4MaGAgeSq0DG6CNxBUTgQd1M4DfChf3QVb5KRvZR9hsjXuzhBkMG+cpIDgH0VI3eRVJnEdvRBbGVOsJjzlZbHdQQVPZWEZUxAwdlZgHyQWtdnbCk6UDGjyRxwEFpHcKJzunbKuc9zTsfrUDnFzt0GQx3UFerIgA3KvQVB7bK7BVABscKoJCCpIGytVcFUQWnuqEZ3V+2/ZRuJzgIJOw+pWu3QOJHkmEFQ04Cq0HKo07gKTHoguAwrgceSADHZVwEFrvPCsV5Gcqx4x2BQFe1QknCq2QhBN0hVYxvV9ii8XO4O6zaRodu4INf01WOoXxvaQD1juvR7lhmdc9Jhz3ZPhDt9S8zJ5hSxMLdsuHn816QclNQ6r0iMnI8L+ZBxfzR0obU1A6CdiujtbGBUT7Y+I/pXoPzNW5ktVV5GcZXQPVUfuddM1vbrP6UGmDsFNH9FRQgOjBO+VcXdJwAMIMqAAuVlXgOGPJQCZzTkBSAmUb5KDFmeQ0ZWqWxjn0zj7s5/wAJ7AlaVXtDWN6dslduOWHg9aNa2tklcxhL2Dv80HDXAyjFdqLw5Kd0eHjcjHmudOJtC21UbZI5wcDyPyW5OJ/Cu1cJoZbra42h4BPw4C606g4p3HUImgmc/DCQMlBt/VV7mral1NJJ1DcYW120jI8uAAJU1TIZqkyknJ3Vr5CW/Ugx3u8POOyRSlzgqAGR+HeanEDWEEYQTTEeFkd1hh+AchZDJOv4D2VzqRgbnbdBj0py7ss57QG7FYJHguPT5J7y9xxv80Ewd8XbzV6saPh6vNC4oKoqHYKmSgo5wIwrERBc3+dTMUTFIHYIQSn6KsHdXt+IYVQwZGyA4kswollGNvT5/lUZjbg5ygxs74Vw2Cu6AHdlUgAeSCBzsnsqA5VHdyg7oJQcYVwOVRoz3CdiMIJFe04CsQuICCQuACB433/IoC8nyVA8hBltJ7qjzgKISI55IO6C1x7hWO7oScq1zkAnCkYdlBlxx8lPH9FBR/dXxA4Vj+6lhOwQUc35+a5M5Nfwu+Fef+GFq/W4lxs/Bz281yTybfhd8K/4Y2r9bjQfRsiIgIiIOuntDATybcSv8To/16nXzz0+A459F9DPtC/wN+JP+J0f69Tr55YPpH6kEkx2yqQjq3+aSDLVSJ3Tsgne4AYVoHUMhXuAI8lYNuyC4fDuUx19lbuVewYCCgf5YQt6u3qrukd02AQWFmB3UbhkqQuJyFagjLhhUacnCucB6eataN0GUCAzdRuIOT8lf+8UbjgbFBYHAnCuBxsoMkPVS49QKCdFRhy3JVUFOn5qx+xwVIrJG53wgq1wIQuycKJriHYCu+aCQDG6qRndWB5yFXqKCoPlhUeCd1cBjBwiCJBursBVa0ZCB0/UmMbKYNGELW4OUEZOBugOdwqO3J9FGT0oJcj1Ubxk7Huo+s/JVDye/kgodlRo6nD5q/APkqtYBuEGZDA3pyfNYdS3D9uyk96ez4R6q8NbM3qxuggDctGe6zYAejAOPrKwXvc1xA2CvZUOaMb/AJUEsoDisYuwcYVzpjnzWTSUzZW9ZxnugpTsBGeyslaGu+xZYZ4ZLR2z6LBnkJfgkY+pBNHUluQCQFFPO52Mkn0VzWNwPmpRTNcPix39EENM8l2FPLDG456RlVZTtj3Cjklc3ugmpQ2meJG4BBytzUGvbja8NgqnNx81s90xcPNQvBJ6iTn5oOToOLN7mLYzXPxtncrfemNQVdzIMtUTnfcrrzTsPiBwJ232W5KDVlRaWjwy7PbYoO5GgNE02rQWVMzXYGMEhbS4rcLKKxVgZDCHAn96uGtHcwN901M3wC/BIHdc46Z1zLxFkjmuQyTg/Eg69a/sjLdCSynI23OFxsw9Ls4z8l3L4qaGtlTazJExmen0+S661+jYaSN7wBkE+SDa9uYJQAfyLWIrc1zuwK0OpkdQTljTsCtZ09XuqqtkZ3zgboLrpbWRQ56PrWRpOhjluMbekYJC3Fqe3tjtrZQACRlbb07O+CrbIO4cNkHI2sbVDSWUPY0Z6c5x8lw7G4yVRjLc/FjGFypfLlPcLaInnbGFtHT1iZPfWRvwQXD9KDeHD7TUNQGPfH09tyuQrraY7XS+8xSgFrfJQ36mg0np9lTSNAd0dWy4hvHE641kD6d5dgHHdBkal1rcJJ30hqiWgFuOpcc3OtNTMQ7BJPdKmudU1DpnOOTuoBE2R4Pf60FWxgb4H5FR8fSOoFZLmgM7dlhvkcXdHllBYJiSRgq/ORlTikZ09eBnuoZR0ZDUGO4ZKoO4TJyUB33QThwAVC7PkqNORgq8NBCCNU6dwcq/ZWuODgIJmj4cfUoZX4OArmuOO6skw52T+ZBYTlXxtzthVYzO2FkNY0emUFjQAr8ZGc7YUcrunKjE7gCEErtgoHndXeIT281cI+sZP50EPW5UJJO6ndEG+X1bK0MbjzQd+vYqfhTap/4v67+Ubciv9iywDml1T3/+IFd/KNuRB7TIiICIiAiIgIiICIiAiIgIiIOPuYb/AHgeJn8D7z+pSr5oak/7fqfnI/8ASV9L3MN/vA8TP4H3n9SlXzQVZ/29UY/+aP8A0lBUOwAnV6K1p2CuAygqRkKwMyPVSN7q52PJBA5hxsqM7FTNwTur+hqCEHCtkcTssjoCsexoBQYud8qaKQnZRuDfJXMGEGUHZVwIwoQT5KvxIJer5KhOTlUHz7qvbKCx7h04WMQerspyMnBVwa3zQXRt+EI52FeMdgrXNHogt8THdVEoJxhRuG5CoBg7IMlrg7bKr0/NY7XEdlXregkLcZKBuRurS49OVYHkIBcGkKjpwDgD86o7fdWdLSd0EwlHdSxv6j2UTWt6d1LCADsgy2s2Hnsq+EVRjzjuFeH7b4QWOZhQv+HCmcXEqGUBBEd1E6UA4wpVaY2k5QUjDjhalTS4wPksMNxsAr2Et3QTXafogj8/jyvSbkQe2p0eD1//ACId/qXmZcnl0bM+q788lt+qrbpoRRdWPDGfyIN2cyFJ11dWM52K8+te0vhV82dvjPl81394sVj7lV1BnyMg5JXRXi4xsF1eyPG8hQbPhw2MBUe7dQ0ry5oz5bKV/dBUN6h6J4ohaQR2UM8hjaCPNSU3TURuLyAQDhBqFhtcmoqptLGwuIcNgu3vBfUreG9JFTzS+FsBhxwuCuXGy0901KY34IDvNbk5jrjUaWu8FNRZaCfI48kHLXHvidBqCyyNZUteXNPnldOmVg8Sclo+Jxx+VZ82rq260Qinc4kDB3Wj9AGfU7oKmTqflXF2RhWBoCqgtY3od1KR8hIwPNWpgIKRP6Xg4281kGpBGFj9ITpCCrx1E/NWCP4sq9EF7X9gr1Cq9RQSOIKsJxsqZPdM5QURFQk52QXsOCql2CoxnO6qgy4XA4CnAwsEOLdwnvDgg1B3bCjWMJnnzKr4jkEkgxv6KB8mB9Ske4kKCXsUERnGexVzXguGyxj3VQSDkIM/96DlAcnKxGyu2Cla4nCDJDxhM5UZJwjScIKomEwgr1bdlTxf/fKoeyjd3QXdatJycqiILy8YUsRyMrHV7HEDAQSv7q+L6P2rHc453CmgJLUFXPIyuTOTU55u+FXz1lav1uNcZubnsFybyaD/AK7rhV8tY2r9bjQfRuiIgIiIOuvtDDjk34k/4nR/r1Ovnlg+kfqX0Ne0M/A34lf4nR/r1Ovnlg+nhBIWn86NYO6nAHdWO+XZBTJ9VRVZguxlVeMIKMdur3ODdseShBwrh8RyUFzDk5KSOx2V2OluR6KF5JO/qgqTsrQ7Y7Kr3fDsVGD6lAzl2Vc0eaoG+YCrh3oUE5fhqjIJB8kHz9FUjGcIIXNwVQDJ+pXkb7hVAAQMbYVW7DcK4AYBwqEeiB1fJWueM9PmqqxwHUD80F4g/fZ+aoW/NSdZ6MKNzvmgtOyvYcqw791VuQRhBIDvhWvd04VT227lROBJQVzjBU0YLm5Hoo+nYZCyIgBH9iCJsgB7I+XI9PkonD4+yuA2IQW+ICceaO+JWhuHdlUkoLehVDcZVRnzQnCChdnZVa7AwrfmiA7cqWObob0H0USo7ugPd1OJVzWkqxSs7BBa6NZlLN4bMEeXqoG4JUrWtKCR0/UsWRnW5TFgxsrcY+1AaOkb7qdj9iQoQc7FN8/JBktdnKx5Yuo7KRhwNihJ6kEIjLMk/oUbo89lkykkLHPVnO+EF8WIzufzK2YGTYAd1a4nCq0khBWBpila8gHByuTtH8SRp6NrvEDekDbOFxbI5wx81JFE2ZuOrB80HZrT/EVmvj7i6cbnpUmpOH4ZCel4JO66+6cvdRpaq97p3EuyD3wuSbHxTut9roqaUOOcDdBsTXmnZLTN1Fhx6rG0HE6qujIukn4huuaNcaehrbS2sq2hvU3JyFs/SdnoKKsbLTvYSD5YQa9rmz+52Bk3f4Qd1x7pSndV1oYGE5djAC5G4h3QS2pkDnADGO6x+B1hpbtqinpz0kPeB+dBu63cPJrnb2PfCWAjuQkvCr7h/wDVXrA6PiXZnivpFuhtAx3KjhAPgh2QPkul+puMN6npJ6NwcG5Lc5QW694gMlhdafFDiwFvfK4imrBLkADc+SxrlWyVtbJUvJJccnKjpvieBlBL4RyXZ2Vvi+G7fKy5QAzYeSw5Ggg57oJm1AkGFY+HpPiFQREB2yy3EmM5QWGqBb0gkFWE9QyfRY7gOvCmZ9FBE5mSVaW481Mfl6+Sjd3KCrVd1EKwH0Kubugua0+qFu6qCQqFxygHdWlmfNXHsqZKCrSAq+J81Yduyt3yNkF0rsj7FGBlXuGQrNwcD1QXtPyUnXjt5BWAYTPzQXdfVt/OqEYOEYAju6Dv57Ff8KXVP8AK7+UbciexXP8A10uqf4AV38o25EHtMiIgIiICIiAiIgIiICIiAiIg4+5hv94HiZ/A+8/qUq+Z+s2raj/CP/SV9MHMN/vA8TP4H3n9SlXzQVn9u1Of/mj/ANJQWNd8I2VzThWN+iFc0jdBIioDlSBoPYILQMqVrMdyo2d1JkoK9PzUcsfV5+SlVHAkIMXwiCrun5q8j4t0ONsIKNPThXh4KoAPRAAdx5IK43yrla0lXILCxBGSpCBjsg7d0FAPJV6fmrS4g7FA47oLXNOSVQNJUxGU+H1CCAbFX9YVXNBzt5q0NGd8oLtyArOgqYNHkFe1uRugx+j1Vhhydip3d1aThBaIzsFJG3pO6N/OqoMlmwH1KuQoevsE6iglLwFC92cKuSqYBQWIqnurSSEErR5o5/SqRknuqSAEoIK1/VGwfNd2OU7UkNtsbYX4z0Ab/UuktZsxv1rtRy2Hqtzepx+gNgUHOmq3fshq5xCM5B7BdLeOtnktV5LJAQTIfJd5tJUDKi5Sg4PfuupfN1RtpdSMaBjMh7fag6/08vRsSsrPibhaf2wQs6iy5qCOsBDB81qFntstXE4sYTsc4WDcAWho9VzTwS0rTXqh6pQ3ceYQZPLbRyW/UpdI1zAD5qvNdUsqL1B0uB+LuPqW/wCCxQaSq5KqmwCM5wMLgvjHeZrzcg+TJ6HHug2FS1LWxCMjBBWSJQViQwAMEh7lTgNx/wC1BL1hOsKAOyVIO25QSg57IrGE57q9AOwTuqO2CsEmEF5cAqdasLspkeqC8vwPJA/I8lHt2VQMIJQcorWq5AJwFQOBR2MKiCocCrh3Ubf51IO6CTyVhBJ2UjACPVVLCEEbXY8vJVDxsrXABWoJnHPZWOZsjHeSuyPVBhvjIJVpaQsxzAVaYsg4CDHG2FMw4H5EEYzuriMIKg5V47K0AbHCuHZBeBjdCM7oq4PogsG6o5qvwArXFBEdlRX4HomAfJBYrmuDQchCBhWoKPk38lk07gW5WG9oypoTgIMkkZXJnJr+F3wr/hjav1uNcWl4wuUOTM/9dzwp+esbV+txoPo5REQEREHXX2hn4G/Er/E6P9ep188sA+M/JfQz7Q38DbiUf/pOj/XqdfPPTgF5z6IJy7IwrVcQD2Vj8tCC6NuHZyqyfzKJjz1KQnPdBY05OFeNlGfhOydRJQSh4IwfRWO77IMDfKo47oLSMBU6coXbeSqw5CC9owcK8EbK1gyd1L0t9EFpbtlWKR3bCjPZBY4gqgOPJWknPdVyPVBcDlXjso2/SCuyUFSO+6sI3ypFa4YGyC0SfvVa45KoAOpXEZQB2V7SAFaGq9oGOyB2ySrXHJV/fZWPGCguIy3CljPTHuo2bgZUn7woMbqy85V3UoT9JXg7IKndEVHEjZBQvCZzuqYBKIB2VOr5Kp7K1BcqO7qo7Kju6CivacqxXMQSgeaua7BA9SqNUrQ3AOUFxONyrD8Tkc4lU7IKubhXM7KjT1FXtAGyADg5QnfKqQAFRBR7tuysJBCvcMhRkYKCxwwMoHDCuc3ZW4wMeqC9sJqMBqnjg932JUMc3u++ypNWF/2IJZagDAI2zutb0ze4rbWMq3tADCCVtyBvjuwTup56fwoXDcBB2V0xPHxdo2WijkAcMN2K12Tl8uGliHyPc7J6sd1wfwQ1lUaSvAnpyclwOM7LtZb+JtZqurigqgACAMkoOHNZaEq6+IUzI3ZHfZZvCPS8ukb/AAXCocWhjwST9a531FY6OO2++xlheW5OBuuHNT3SW30kkjRgtz2QcxcfuNluuGhm2ls7HObFjAIzsF0Lu+ooKjxWNaCS8rWtXavqLrUyU0kjyO2CVsk0gc4uzuTlBjuHjSlzRgHsp4oDGQSFMynEW+Ar35xkoKPcOlYUzwCWqWSQbjIWM7d2580F0DcuyszHw9Kx4sDH1qYO33/SggMPxEkK8Mw1SPdtj0VnUcYKCLGCVZJvspCfMqN2M7ILWjBUjfpBWN7qRoBwgvHZWOG+cq8nCtJygoDnZCM7pgBVQRkZTONsKqoQPNAPZUHdVPZU+pBcqEZ3QEk7oSQgq3Yo53yRWnug7/exXb/10uqf4AV38o25FX2K/wCFLqn+AFd/KNuRB7TIiICIiAiIgIiICIiAiIgIiIOPuYb/AHgeJn8D7z+pSr5oKz+3an/CP/Svpf5hv94HiZ/A+8/qUq+aGrGa6p/wj/0lBGGnpCdCuaMAZVwxvlBRo7KVrgBhWbfJWSEjzQSg/FlV61EwnG6uyPVBIHgHdV8Uf+4UWR6q1xwgq546iqg5UY3Pqp2NGEDvt6q5o6VVoGRsrsD0QWjdV6UOxyqjcIKEjGFRHd9vzKgO25QRnYqRpyrCMn5K8ADsguznbCoQQrtlbnKAisLiqdbkGQCFVQh/YKQOJQUf5q1XHfumAgoB2VURBUbKoOVXY7bKrQEFFXBVdlc0DHZBEW7qCSXoccjssrbqWFUNHWgyYXh/YKR0ecH1UVO0ABZDj2QYVeA1rRnuV2f5aJw+3hvUPo75XWC4nIZn1K7B8udY+npw1oJ2Qdv+HtL7xc5QM9vJdSOdGlNNqZgII++H9C7bcHql012k6vmurHPOANVQ4/8Amh/QUHV2KLxRt6rNph4DcHuoKA/EWlXVshY7AQUuD+pzQPLK7bcpGiX6ltYcSWAjHfGV1e0zam3qoEL8bHByu13CHUFTw5pYoKQHBA7IN28aNHjR9HNVOkBABO66T6svjLpVy9ODh5G31rtPx84jV1/szxP1AOac/kXT1rGzSyHO/USPyoJqaYSgRjYhZEkXQFDBC2J3UDup5Hl2xQQNG+FeGlUYBnKk2QXNbgDdVVMp1IDxkYyoun5qr3k9j6qPqcEFQN+6uDSjCPPCv8wgtLNsoB5KQ7tVvTvtlBe1pKvDPmrWq8E9kFjmnCt6SpX46fmo0FjdirwcpgeiIJIyAd/VSl4x9ixwfROooD3D0VvV8lU7qnSEFQ7fsr27o1oyrg0DCC7pLghZgKQDAGEPphBj9JzlCzJUuB6KhHogsDVUbBXEbIBt2QVVeoDCsyUzlALsnACKmAFVBYrm9lQjZXMGyC0tyqCMlSgeiuA27IMN7d1fG7pbhXSNA3wrCAM4QULsZ9FyryZHPNvwoPrrG1frca4nJzlcrcmP4W3Cj+GVq/W40H0eIiICIiDrr7QwZ5N+JP8AidH+vU6+ean+kfqX0M+0L/A34k/4nR/r1Ovnlg+kfqQThwb3VHuDkIyonEgoLmfSV6sjV6CwnJx81Vrdu6ADKuQWnZWuBIP1KTYnGQqHCCEDfClAxtnzVMeY7oCe5QTNbgBXqxjj2KuJ2280FrnDsrCQqPOFZkn1QWkZVCN8K8Ku2d0FYx+VVIyje6qgIT+dFR/ZBCWZOcqZkecfUoWkl2D2WVH5IKeH8lQjpU+B6KyRo6SUGP1BWlwyrXu3OCrQd8oJuoNwpGP6mlQdxur2/CEFnSQ7OVXICZJOCrXEeqCvUFa52+MImAUFAMblOoKmSiAiIgu7K091ceytwgK9jTsfmrFI3sEEoGRjPZVGQO6tZt3/ADq4YQCcKrd1f0BWHAPwoJGs88q7pKozOMFXIKEeaoqPJ8lZ1nOCgkVvTuCq5Hqhdg7IKP7KNS7FRO2OAgq2A1GzVSSidGDnOVJDL4JyrKisLzsSgUrfCfk+ayqmVskRaPNYUMnU5TvIDfsQZWmribbXMkOMFw7rtPwHik1nqijtkXeVwAwPXC6qWijbVVEbTj6Q/Su4nKdSttmu7bUMxkPafzhB214vcAazQ/D6O/TdQa+Lq/NldD9c3RjqGfDgekkFen3ODrmpk4NR0waAPdwCR5/CvIPUV3lnpKlpzu8/pQcaXGr8S5yOGCM4TqGftWDPk1T89y7KyBJkd0EzpQBhWOeHNwo35xlWNdg4QPBJKkFLncK5vZSF5a3ZBivj8P8AIozOBsFSaZznEYUTB1OwUEzZie4V+erdXGEBmcfmVoGEEbxnOFYRhS59So3kZQUHdSNPZR/UpGAZGUF6o71VTsFGScnKB1fJCcnKrhp7fpQjbZBadlYTk4VwG++VQgZ2QVPZW9WCq5I7qhwSgu6kI6twqDY7q7IQGjBRxGVUK1wJKDv97Ff8KXVP8AK7+UbciexX/Cl1T/ACu/lG3Ig9pkREBERAREQEREBERAREQEREHH3MN/vA8TP4H3n9SlXzRVA/2/UD/wCiP/SV9LvMP/vAcTP4H3n9SlXzRT719Tj/AOaP/SUFP3quYzKuY0dIQ5HZBZjB3Vj1OACrHtCC0N2GE6SrgD2AVQMdwgjOytc3KlIG+ypgII2bd1LlW4A8kyPMoJGv+xXB+25Cx5HYBIKjEhzuUGYZPLZVY4EKBpyrgSOyCU7q1zcq5vzV23oEAMGELcdiqgkbK4b90ESYKuIGUQRmMqnhlTAHKu6fkgg8MhXg4UnT8lToA8kFMFOkqYMGBsjmgHsgi6U6VVxx+VGb90Fo2Kkb2VvRvnyV4B8ggpjGSnXjZV7KF5OUF4IzlQyw+Icq/qIAUkeXDsgtiZ0gDyCve8A4yFVYNVM4O27oK1xDg1cxcEtRR2toje4N2wuGOoygdS3bpGd1NUtDCfsQegvLvcm3u8PbGQc57Lr1z60TqPVcAcCMvP6FzdyUB1ZfdwSVxj7R+m921dSDGMvP6Cg6a0Z6XFx9VlTUctSS9jHEAZzjusLq6YGkHck5XKvC/TlPe6TrmIyR5hBLy/6Ll1Lf/d3sIAcO65V4uXGLhlVwUxeMnATQVKzRF1fVUQBcPRcV8w+qarUt5jmqsgsJxlBja24mRX+2CKN7XEjGAuOLbTunLnHzKxaWETOwSdvJZxk9xwGIJKqE0reon8ixopxKcfnV76l1W3pdnCtjpxGdvNBMW4GVQHHkrz9FW4CAXAK1zvRUd/OrUFOpUPdV6Qq9KA0YKvGyNaMqQMCA0+SrkBU7BUJJBygvHdV6gog4quT6oJRv2ToykRJKkIOchBEWkBWqZ428lCdsoKN7q4bqME9SlZglBVjCSr/CwpGtwBsqnsghxgqo7qMvOVQSEOxlBmNZkd1c2Iu7KOJziFmQjJAQY5gI3Ub2Y+xak9nwjZYE22UEPZU6ghJyqICKh7INwguxsgbkZVVezAHkgs6FXGNlJgeiseBnZBRv0gpB2Uf1K8dWAghkbnZWdBUrwfNXRgFpygxHM/SuVOTIf9dvwoHprK1frca4ycBkrk/k0A/bc8KvlrG1H/lcaD6OUREBERB119oXtyb8Sj/9J0f69Tr55IThx+YX0N+0M/A34k/4nR/r1OvnlhGXFBMrHnJV6ied9iguYQDurjuMhRtO+6kyUFBurgPJUb9IK76kFBERurXg5UoLvMbK1w7II+ydQR2wVuR6oJeoBVD1j9ZPmrgTjcoKuOSgIwo+og4Vevtv+ZBI0fNCw5yqsIP5FK0DG6C1rDgK7oUjRvjCPGB2wggO3dWvIV7t8rHkcQe6A0/EsqJYkf0srKjzsAgyA3KimOGkKZp8j6KGdBhEEHKo7uqvOCqsaHHJQVbuFedgrmtDUc0HcD8iCEEDJVHbqp2VEBEVCSEFEREBERBXIxhUPdVAyqHugKRmwCjUjeyC8b9lKxpAyo2D5KQOOMBBVzs7BRB3xgK/DvMKnQAQSgnB2BwhIwVQO2ACscT2ygF4Bwo3HLshUkJxnKiLiAcIJwcjGSq9WO6x2yEHdXBxIz6oJ2uwd1a8jJUfUVaX74QTBnibBU9zeT3O6sjlLD3V5rHDtugoYjBuqeKXA7KSOU1B6XqSWlbG0kAZQatpWn8WpiycfEP0ruVyzW1p1lbx4gHxt7/WF0htdykop2dB7FdiOBPEert2qaKVryOlwwg9FOdENtvCON5maf8Aa+cZ+S8irhe2TQ1DQQT1n9K75c1XFau1Bw2jpJXuI8HG5+S85IZC/wAVpycvJQQF3i1RcOxKn8LB3KoYRF8QChkqXZ27/WgnfgAhQ4wc5UYlc47+fzVS8+ZQTeNjb+dUdUjGCo8AqPpBfj1KC0kuOcKSGMucD81O2nb0g7K8s8NuR6IJXNHh91jOHdWCpcTjyCvJ6tz5oIyMlWOGCpFFIfQoDe6kAOQVE0nKlb5IK52wqEeavwFY4Hy7IKDuq9QQ9uytQFQjKHYICgO7KwNOVee3dUHdBVwJGFQNI7q5EBUzhVwhHqg7++xX/Cl1T/ACu/lG3InsV/wpdU/wArv5RtyIPaZERAREQEREBERAREQEREBERBx9zD78AOJn8D7z+pSr5pJsNr6n/CP/AElfS3zDf7wPEz+B95/UpV80VScV9T/hH/pKCVpBCqoWP2Cka4EZQWg/Ej9yqHYp3QSDsrXbq0P8lUEeZQUAyVd0K3ODsr2uyN0FOhWuiyc5UqIIvCwPVWGHfbCyFY/O5AQRhvSqqnxZ+1XYygkbvsrwMK1m3fZXIL1aTlC44VmSgrnfCqqb91TJQStOFcDlRgnATqKCVWuI7hUJ7oASNwgePjbPZC/qGVAR8ROFe3sgkVQPNUVzeyBkDZVDxhWHuiC5zgeyt6coqgkIInnBxhTQnIwo3NB3UkQAGyCj3AZWJJCZCSsmVWsJyghbT4wABstbsNUKeraD2WmqSjP+2gUHfTkgv8UF+Bc4ALY/tHKltdqmmqGYIEnf7FHygXFtLeg5xPf1WNzvEXS6ROaf3w7oOoE8Do6WJ57PJK7Pcsug5NTWsSsB3C6/Xy3+FaKdwI+FdtuUK5/cyytZ0g/Cgt15ph+hXS1M2QBk7ldVteXxmorg6WMghjiNl275lbwa23zgjGx7fUukOQ2on3z8R/SgrSxGF3UVfU5mOQq9YcAnb5II4Yy3usgEDCjBycoScoJPEzthU6vkqNAKqcIAGThWubvspAPRVA8ygtEeMHKuDfmqq8NyOyC3pxuqEZ7K9wwFYgq4YGVGDlXuJIUfbYIKgYVUwfREEkZ6e/opQ8FQ5IVzTvugvcDjPqrHNy3KkfgtUeSAgiDfi7qUDBByox9L7VIgvEwAAVfFyFF0/Wq4xsgt7lVEed8q5rRnfKuQXxO6dispk+HAjZYY7q7sgzzUgjBKwpnhzvtVHEAHdRdXxdwgkEefNPCxvurmvAABQuyNkEDzg4wqB2B280eCSrQN0EzNypQ3Az9qjaNtleCcboKGTfCsfLv5KrgMkqF386C8S+ZVwqG4WM/sjQCEE75weyvieC0rEI32HmsmAbbhBRztyuUOTQ/9dzwpP/djav1uNcXSD5LlDkzB/bb8Kdv/AJcrV+txoPo6REQEREHXT2hpxybcSj/9J0f69Tr55YTh31r6GfaH/gacSv8AE6P9fp1880X00E6ie057qVRyHBQGj8yv6gMZUQcfMquc+aCofgjspGuB3UQb5q4ZAQTB4wo3PwjUcBjOEFj3A7hWAZVe537ZQ7H9KCrIj5q4sx5qrcnshB80EZZk4ygix5q5oKuQWtPSrw/zVhG3ZVG3dBM2UNCo6fIxnKicMhUaBhBeXb9lG9uT3Vx2CpuUFjW9JWQx+BlWtb6hV2CC8zgK10nXlRkDKbBBaW5PdOnpOVUbnKOQVLsjCNdgEFWogtcdycK3q+Sq7+dUAGN0FTsqfS3VT2VG9kDp+adPzQk5KqDsgp0/NOn5qqIKZHZUJycoe6IK9PzUjdsKxXAjHdBIHhOsKMnOwVzRsgmE2TgBUJ6jsrGgK8YyMIKtJaUJ6iFV5bjZWID2ZCj8L5qUklUQR+EPRXtj27qqr1Y22QWlufNWmIndX5BVclBGIS5V91cVPDudt1P0n0QY0EJjcSVNM4FhSQho3WNLK0sIBQRU7DJOMeq5k4PUzxqGlJ83Dy+pcQ25uZx9YXNnCUFl/pT6OH8yDm/mGbINBxg4/sXp8l0ihf0l5+a7ucwsviaFYP8A6Fj8y6RxsP3zPkSglkl6m4wsZ7chSOIVmSgtYzdX+Hkq5mPqTqx2QOjA7q3w/izlSEkqmR6oL2zdLcYSSYEdIUZAwrcb5QWFmD1K8OwMIe3dWoKuf8u6s6cquxVewQUAxurgRjsrQSTuhOPNBeH4RzgVYCSd1VARUPZUySgqDnZCMquAFQlA6UAwqogHcINgioSR2QXDurHu37K5WO/nQd//AGKzs802qf4AV38o25Fb7FT8KbVP/F/XfyjbkQe1KIiAiIgIiICIiAiIgIiICIiDj/mF/wB4HiX/AAPvP6lKvmgq/wC3qn/CP/SV9L3MP/vAcTP4H3n9SlXzQVeffaj/AAj/ANJQWNBwNlIHEK1v0QqoKh4QuAVgO6qcHscoKg+ao8791YCdu6rlBeHjYK4HPZRDupW/zoJR2VcFUaRhXNOxQUO3dUByrnAHyVuMIBGQqNGPNVyFVAREQWCQk9OFereloOfNUcT5IJT2VqqSenI7qwFxzn0QV8QDb+ZPFCid9NMgeaDIa7Jyrw8YUTfrVyC0kE4VQcKNzsHZVac+aCbqCuaRjsVEM581e3sgqd0Jwio5BVU6gqEnGFZuPVBkMj6leI+n6lbG44wFeXO80Fj4+obq0RYypMlUQWFuPNQtqfCqQcKdY0kQdICcoOd+A+uG6brmzmQNyQNyuV+Ithk4qSRVTGmQDB2XVvTNQ2mcz4jjqG67u8A7bJdrZE6NoflmTkZQdf8AU/BeojoWwGB2GDzBWtcOLv8A7H9OaaR4YRsc7LsdrbTc8fit8EZAJ7LqZxOmNtrntfkfH5beaDP4wa9ZeKJ4EgJcD5rrZ19Ukh/uiSt8aorWz0QwScj8q2HGfiKDKpyXEZUsmxUdMBn55Uso3QGAjdXKkePUK/AQWsBypOlUb32CuQAMborclXDcIL2N3V5GNgrGOIPdX5ygtf2Uakf2UZ7IKdWdggGFYPpKQZ80A9lariNlQDbsgN7q4d1a3urh3QSbkbeipgqVrdtwrugEdkGOxuHZJ7qTp+aFvScgKmSgr0/NOn5oCSd0JIQCMKiuPZWoKFwCp1n1Vrv51aguMpIVvXvlU6QqHugm8TZVEvzUQ7Iglcc9vVUG/wBior2Db60F7dhgqvUFaqE480AuznZRP7qbAUbwM9kEZGUAwrjjG2Ea3KC1ZEIyFjrIhJA2QWSDBXKHJn+Fzwp/hjav1uNcXv7rlDkz/C54U/wxtX63Gg+jlERAREQdc/aHD/rNeJQ/+k6L9fp1880O7j9S+hr2h34GvEr/ABOj/X6dfPNAAXlBKBjcqjviKucCB2VqCzpLld4ZAKM+krz5oLWkZVyjH0lINwgqBk4Vrtiq59O6sdknfKCnTt3VCzPZXF2B5K0OOEEzAGlXOHVuFjh7ipGEgIKHZUJxsrndtlaMHcoHUFb09W49VUDKrkj6KAdgjeyfWmwQVRWOJHZU63IJshRuOSjSqOyTsguBCo71VASEzlAa7fsjndtlQj0QDzKCqd1aSd8qnURsEA7d1QjO4VT2QDZBQ7BG9lXYogHcIBhEQUByqqhHoqjPmgtPdBsh7p3QXHcKnSVU9lQElBVu3dSB2Rt9Sswrm7ILgMK4vAVnUqd0F4BPmrhsFaDj0VC8+SC9D2VuSqFxJwgq05Kq/fZW/R7KnUUFzRgq5WZcN1UOJCCWOUQ7lXurWu3HksOYk4UsEDXtJQJqgvGAsfcrNFM12yqaZrQfqQLY3pmaT5kLnDhHD41/pmjclw7LhCmIjlZnbcZXOnA0mXU9GGb5cP5kHNfMNbH0uhWPIIHhZ3+pdHmuBbKcfvivQfmboXs4dRvczAMIOfsXnu1vwzHfAcf0oMcPHVv6q4jO6iOzzhXtJ7Z80F4GVQnpKrkgbKg37hBd3BVOkg5V2MKhdhAJBCtPZDv2/Mqb/NBRUIyqqhKCnSVcNgrclMlBcit6vmqFzs7IKgZVewVjSQVcXE/UgrsdlUDCtBwmSguJwitzlU6nZwEF6pkBUyVac5zugk7J3VnUTsU6iDgIL1Y7+dA4kqju6Dv97FT8KbVP/F/XfyjbkT2Kn4U2qf8Ai/rv5RtyIPalERAREQEREBERAREQEREBERBx9zDf7wPEz+B95/UpV80NSM19SP8A6I/9JX0vcw3+8DxM/gfef1KVfNFUf7oVP+Ef+koLGs2Crjp2VSSBsqA/3SCpbkd1aG9I3VxcFTOfRBQHJwrun5pgDsN1UbhBb0b5yrgMKhJyqjcIKg4UjTlRtIyrgQPRBISFaeyta4+auJ8s90FmcOVetUPZWoJA/Pkq91aANjhVzjbZBQsKuaMK7Ax2VpOEFC8J1jyUZPqVVhBPdBd4efiVroyTlTD6OyAILcbYTq6QQriDvsrHAHGUEbgSchXsTpHor2NA7oLlc3srVc0IKqjlf0/WqOb27oI1Rw81dgqjgcdkFoqQwj5qaOdsiwpY+rGFLTNLR3QZXWqGQBW4VrvP6kEgcCqeEXvyPko2vII3U8btwQgzIZHUscbs4+IL0d5LqWK52KN0oBPhj9C82quT7yzP90CvQLky1Iy3WGNhJ/sY/Qg5319ZaYyzta0ZAK86+ZGb7m3iSFm2ZCPzr0B1BqRtwrqhrTsWnuV5781BH7Ij6mQ/zoOH6mudWUjW9WTjHdaaync3OSVJSH72B6EqdzSCgigj6HZ+amlHUjsgZ7I3ceqCkTfiwSpizAyrI9jnCkJygtb9IKqo36QVT2QWZCqHjCsPmrSd+6Cdr1IHhY7CFIHAIJH9lGqlxKogoG4OVVEQOyp1BVPZWoDTkq8EA4UTTgq8HJ7hBlNdkK8EYUAdhuxQSH1QSSOAz9Sh60e/OVYgv6061YiC/rVC/KtRBedwqBpV7W5xsVd0jOcIIyzZU6VM5mQdlGWkIIgMKqu6PrTAQXA4Kva4YURPoVcw5G6CUDKFmSqtVUFn0VY7fKuJySFG5xHZBadlezsrDur2dkFHN81LF9FWO7K+P6KCyQ74XJ/Jof8AruOFP8MrUP8Alca4ukO/dcocmf4XHCj+GVq/W40H0doiICIiDrp7Q78DXiV/idH+v06+eWE4fn5L6GvaHfga8Sv8To/1+nXzzQ/Tx6oMrIeMBWOZjKvaMbqj+6CMNwUd2yqqjzgIIwdwpR2UI+l9qmHZBbnp7oSCNkf27KxpIyMoKO7YVAcK47gq360F7Xb7+qv6grAABt3VRlBcHdW2E6UDQNwqoLXDbuqN7K89lGdjsgrnfCo7uq/PG6scSD3QVRUGc+aEkILwMKqt6icJkoKHumQFac790388oLsgoThWg4RxGyCjt0AwqA77qqAiE7IDsgtzg/UrgcpsmwQDuEAwiICE42VuT6qowdygod0GyHuiCvfYKo2CYAVCUFw7oXb4wrRnPmqkDugr1YVw3UWSVIw4CC7BVR2VC71wrS4hBIqHbdU6imSdkDq6vyquMb5Vh27eqr1EjCB1Z2Vw7KzACuyUFC3r7eSy4GdDcFYrXdJWQyXOMIJi4MGVG+YHKtkeCN1A9w6TuglicZpWho7Y/SuZ+C10badSUk8pwGuC4isFKampDQM7hcmadh+59bFLISACN0HZzmK17R3rQDKKN7SRD07H5Loo1nwStP75x/SucOImo4KuyiBkhJDe2fkuDy8hrifUlBhvh6HE5VvnlSyS9ZwoHOPVjOyCTqCqN+ysHZXN7Z+aC9Uc3KqCCcZRBYBjcq3r3xhXSKMY8+6C4nCtO6u7q07HCChOFTq+SqmAgp0lM42VUwEA7hBsMIdgm+PmgoBhVJwqDPmqkZycIBOFTqCDc75VcBARD2TO2UFOlV7BUByqnsgo3ujvpFG90d9IoO/3sVPwptU/8X9d/KNuRPYqfhTap/4v67+UbciD2pREQEREBERAREQEREBERAREQcfcw3+8DxM/gfef1KVfNDUn/b1Sf/oj/wBJX0vcw/8AvAcTP4H3n9SlXzP1f9u1H+Ef+koKtcMBUO/bZUaBgHCuQW7nzVQMJkeXdAfVBUHBVS4BW5HqrHZJQSdWRhUJwrWEeZVXEY2QUDzlTNPXusZpyd+yma4N2CCXucK13w7Ko7ZHdWuOe6B5ZVC7CtzvhNj3QXCQnZVByVYMZCvBA7IJRsFY52Va2QF2Fc7HcIIiCThXsaR3QD0CkYNtwgubt3VyhL/JXMeQgkzvhRvZ5qTbuFXGfJBGxpHmr1bkAp1j1CCQDtur2jIUYccDdXsJ9UEiEZREFp2zhRnyVxcMnf8AMonvAKC5sfVspGRhowFSI5wVJkILMAbkq126q5wx3VWAPGyCGT4RnKuhk3z38lWRvfbZRMADsBBkznxY2gDsQu0PL9rZ1gtgj8UNwzG5XWCJhcAPqXLvD6QQ0nSSRlqDtRpLWrr9d5Y/EBJyF1g5sITFqJh9Xkrl/gi9s+oXhpJ38yuLecGIxaghyMZef0IOvtECSPTKznYOFh0I+HOPNZmdwgtkGGq1jtlWXsrGdigvDulV8VRvcAMnyVoc09igl6vknV8laTtkIDsgq7fsgZsqtGSrsH0QWAEFMn1Vzlagva7fceSr1jKj6seauBB8/NBcTtlWeIRsrnnDcAqIEHz3QSh+QitarkFoGUGxVQN/NMfEgvG4G6K5jSR2VfDwgsRXFpCtQEREBVGyYPomCgkYcbK7q+Ssaq5CCQvAUZkBOEJGO6iJw/GfNBkhuW/YrXRnvupG/QCoeyDGIwroxkqj+6ui7oMljdke34e6kjblo2SRuGnIQYLnbnZWOdv2SQgPOCqAgjdBVSM7KPZSM7IKu7K5n0VYSScK9gPSfqQRyDK5Q5M9ubfhR/DK1frca4vk7n61yhyafhccKf4Z2r9bjQfR2iIgIiIOuntDd+TbiV/ilH+vU6+eWEZfn0X0Ne0MOOTbiUT/AHnR/r1Ovnmg+kfqQZbHDpwrXowY3Ko/ugNb1FHt6ThXMOD88KriD9aCAMwVePJCDlMFBVzB07qIt9MKZ2cKzp+SCxzNlaGZUrmnCjwQgvDceauDR3Ctb1E4KkAOEFju2FQdiriO+VTsgsLt0Byri0DyVEFo2Ko74j3Q90QU6h6KhOTlUTIQXqhONkJ2yFTOUFyt68eSuVrgMoKHdU6VVUJI7IKncKn0e4VchUKCqE42Q9lbnKCoHZCMqudkG4QUznbCqBhMBEBETICChGVQjBwqk+hVdiEFOr5Kh3REFepOrZURAHdVDvkqIgkJGEBACsBz5qqCvVnZAceSpgBEFSc+StJwqoQNzhBd1BOoKwHPmhJyguIL9gpI2Fo3VIhk+qkJA7oKPBI2UXhE5Oe26lLgfyq0uAadxug3jw1tgra74twSAuSb5YjSdJj27eS2Hwmq2QVoDjjf+dcvXmVlYWNZgkjZBw7qqWpia5sjiQPmtjvn6g4fWuTeINrmhhMjm7Eei4uYAQR69kEYJJ7q4d1cYi3fCtGxCC5VG4wqITj60AAt3KqZCNkaS44VekIKFwwrHNzuFV2+Aqb4wgoHY2KoTk5QhUQVJyqDZEQVJygOFREFepOoKiIK9SdXyVEQV6lQ7oiCpOUzthURABwcqvVsqIgA4OUO6Ig7/wDsVPwptU/8X9d/KNuRPYqfhTap/wCL+u/lG3Ig9qUREBERAREQEREBERAREQEREHHvMT+D/wATf4HXn9SlXzQVe1ZP/hH/AKSvpg5h/wDeA4mfwPvP6lKvmerf7cn/AMK79JQWB23ZV6vkrBuQFdhAGyuByrQPUK5Bb2VQco3c7q4sx2QULDuqBpKkVWt27IIOg5VwGFIW7nZVEZxuPzIDXdtlVW9ODk+quQWFh3Kp4ZcpD2VucIKdBCdBVxPw5Rjsnf1QWiBwPVlTNjJHdXB3w4Vc47ILQzpKqRkYVgeeshXl3ogsMZJVRGSruofL8iqD5hBUAjCvBCDGPJWoKOiLsKwwkKft2VpP5kEY+HOUE2PkVa9xzhQv+kEGfHIHDJUob1jusSLPQN1kwvICAYDlQzUzuoHKyesqyR5JQIY8NAKtny0qWN3fZRz4PkggeC4YCkpz4Y+JWK4dkFZZA7Ix8lGzuri0nyQNPUAPNBmUhAwCPNck6WlLYMNOPhXG9M3G+PNb803MI4Tn+5Qc4cBK3wNQvc8gjK2BzhVbarUUJb/dfzLd3ATqrNQPZG0k58lszm5pX01/hDwQS7YfYg4Eongjw/QrMcC3Gd8rCpHRxsDyRnPZZXieL23QUduoz8AKkOQCseZ+AcnyQXxD3p3hgkk7LJFtkhGSDv6qzT0TpqsNaMnK3NdKSSCIF7SEG2ZB0bK1pyFWokAeRnvlRtO3dBMw4KvyFC12/dXZKC8tyrehXdRTqKCIsx5qhG+Mq5zsDyVvdAcS5WgYKB2+CqoL2uV3UowcKpdvsUEjSFdjfKiDiMbqviIMlsgaqmUHyUAPqq5CCWSUFuFB1hHKxBKCCisaSq5KCTIQuAUfUVRxJOyCTqB2CDdRgnPdV6sd0F+NlGWEu6lIXghUQTMkGOlXhwIwsdvdSjOAUFXRZ80ZH075V2dkzsgmjnDdiVSepaW4z5LHwc9jlWPac9kETmlzsqnQVeiC07KWP6Ks6fkpGDAQVIypGfRUZPoQrmuJCCGR+CVyjyaH/rt+FHz1naf1uNcXyNBJ2XKHJmP+u34T/LWVp/W40H0eIiICIiDrn7Q4f9ZrxKH/ANJ0X6/Tr556f6a+hn2h34GvEr/E6P8AX6dfPNAcP+xBkul6VaHeIdklaOnZWwkAoJWtwcq1/dXyOw3IVrCCMkoKNOFUyAK048lQjKC4SjzVfFaoy3AyrHIJTMCMKzxBnCiHdXYyUE7MEq8nyUcRI7q8nKCw7qmQFeQMd1C8nKA5+RhUBxsqZCDugu6D3QMJV47Ko2QWGDAyozGclZBeSMeijzk5KCgjOE6FOcdPko0FnT81Y8Y3V2TlUfuO6CPq+SoTkoiCvT806fmqqo7hBXp+adPzV+Bjt5KgxgoIgMKqqe6tOQUFUQ7BG7ndBd0/NU8PIzlSDGEccDZBAdlUHCoe5+tEBBuiDugr0/NUOyuPZUwCM+aB0/NVDFQEk7q7OyCiHsh7KgJz3QG91VMAIgA52RUO3ZUyUAHByqnfdVwE2wgkp3YJVJ3ZOyupwCTt5KlQAHD60FIWl5wpZoC2MnA23VlIQH7rJqZWujI+SDVtDzyQXD4TgZC5foriTOwyOzjC4Z0h/bn2hctWuEzVEbR3IQQ8Ta2Ca2YY0A9Pf7Fwcx3S5c38Sra6G29bgcEfzLhJkfX1bdkEr6hrgAFERndVMRbuAVQE9igqqdOVVXN7IHZUJyjkAGEFuMbkpkeqO/nVhJygE5VERAREQEREBERAREQEREBERAREQEREHf8A9ip+FNqn/i/rv5RtyJ7FT8KbVP8Axf138o25EHtSiIgIiICIiAiIgIiICIiAiIg4+5h/94DiZ/A69fqUq+Z6s/tyf/CP/SV9MPMP/vAcTP4HXr9SlXzP1YzWzj/6I/8ASUETfLKv7qgbsO6ua0DyQUBypA3HmrGgZ7KTI9UFA3BzlVTIKICvYcBWtwThSfCPRBZ3KkVAG+WFcMb5QRkdWw806FIWgIO3ZBEWYCtUr+xwoXEg4QUPdFaCSd1Vxwgu6j9iubIB3UXUCPmqZ+aCUuCdX1qEP3xlDJv3QS5OcglVbLgbqLrBG/omR6oMlsuVV78YUDHYI3VxfnugkM+AjX9XdYjnbkZVzZMfYglkcBsoHuw7ZHPJyrCc7lBmxSjpAU0cmRt+hYLX4x8ip4pWgHJQZPWrJJMH61Z4rVFLICcgoMtkux/1qOao3AyFjCQgfZ5KKRznFBmeKMZ2VzZRjcj8qwOp/ZVEjhsgz/Fb6hXMlaO+PyrTRI7OclV8Z/r+dBrMdS1mNx3W4bbeBFEcOAOMbLZIqCMd1lQVYDcFxHog514LcQ6fSl697qJGgE/viFqnHIO4qV0VdbQXlpBywZXXaS5SR48JxBBznK7Zcs9sptQ2lj6zBOP326DrbfdBVtlpeuWJ4IzkkLbNNIYy4OPbZduuYDTlFb6KUwtYCAcYHyXUJ1NNJLII2k4cc/lQZPjA77brIZbZKxuWNJysSG110xDYoySt66coTSRhta3BPqg0jTtvdbq7xpmnAK1vUVxhnhwzGcYUl48CBhczAB3ytp1Na2RjgHZQaTUSdVQfr9VK1Yzw7xC4jzUzJO26CUHBV4eAFC54xsjXjzQT9QTqCi6z1dwhkG6CR5yFbkdKiLzuVTxQgub9JSAhQh2+cp1n1QSndVBGFD4hynWfVBPkFFB4uPVUMuD2KDJ8TCCXO+VjGTHyQSjHf86DKDs7ZVHOHoscTAnYq8PB7lBK12+48lXqCiD2jdPFB7IJHvDQrPEyrHvBCj6wAUGQ1+D3COk37rFMu+2VTxUGb1keauEuBvusJswJV3iAjIQZonCuFSAMZAWneKcnOVa6Yg43Qan703+6/MnvTf7r8y0rxz808c/NBq3vQ9QqOqGntgrTPG+aqJchBn+N8h+VPG+Q/KsIPPclXB2e2EGZ4w88BVFQ0D6QWnSyEdla2Z2PNBqXjg+YCvbUADuFpLpnE7KWOU9ODnZBnSVDT5hcq8mRzzbcJz66ytP63GuGJJTnbP5VzLyXHPNlwlP/AHY2j9ajQfR8iIgIiIOuntDvwNeJX+J0f6/Tr55Ysly+hr2h34GvEr/E6P8AX6dfPPTjLigvlPw7qyLf8qum7FWReX1oJy7bBVvV6I8gDurA4EHdBVrvi3Ug2UAJDtlks6SMnCChcCMYVuNzlVGMqjjgoLHM2VrdipDuFbgILwcgfUitB9EyUFxJx3Vh7K53ZWZI7oIy34s/NVHdURBKOyuBCi6nDZVBJG6C7OT2VD3VFUbkIJGEuGFUjH2q0bDZV6sjcoLCMlWPGNypD3UUmcoLEV3T9atIwUFyAfEMIeyDKCXsFHguV432PZUOAMhBaG9O+UJygcCcHCEenZBb1bKrWkhUOPLurmbD0QVPZVadsK3z3zhSDGPJBC4bkqikcAVb0/WgtRVDT6KvTjbCCje6rkBMAKh7oK9kVpJKZKC4nCoRncKuMp2BQMgpnfCt+pXADue6B2REQCMpjbCoCSd1VBLTDDyqVP0kicGnPr81SXMhyEFsZIVZCXN38lRrHZ7IWuwe6DV9Mz+DWA+rguZdIPbV3GBuxyQNlwfaXiKpaXbbhczcNp45b1TsB7kIN3cZLU2HTwkDcfBnP2LrLBgF4x5ruBx3pBHo8P6QMx5/Munsbg1zvmUGQ9oxnHksZ2M7LJfgtB+SxnIDe6O7qn1J9aC9FTI9Udny7II5HZOFYrnA7bK1AREQEREBERAREQEREBERAREQEREBERB3/wDYqfhTap/4v67+UbciexU/Cm1T/wAX9d/KNuRB7UoiICIiAiIgIiICIiAiIgIiIOPuYf8A3gOJn8D7z+pSr5oan+3aj/CP/SV9L3MP/vAcTP4H3n9SlXzRVABrqjP/AM0f+koLWt+EbodjhGu7DIVH7lBRv0gqkZQAAfNWufjZBcPh3R0norOsqmc+aC9rjkFXeJ81F1EeadRPmgyAfM5UnUFj9Xw/NVa87oJ+oYQHIyo+r5oH9OyC8OGcK2VpVSW42IyjSHfSQRkYGcqMguUsgzsFYAR3QR9Badz2VFeeokqgb6oLCPPKplXuz+dWYQMn1VQcqmD6KoGEFw7q4nCtB9E3KCw7oASq9JRoOeyC4s2QN2UnkjW58kEPScplzftUj24yQoiSUFetyuGXBWK9gcM4QOoZxhXgAjJUR9fNSROHYnsgk8MO7BRyRFpx6rIY5uM5CpI4ZGcBBjeGVUQk7qXqaFe1zMIMfwj6KhY5p7lZmW74woXYz5IMYgjuu3XK3UltpYGnGG+S6kS4yF2o5Y6uKC1jrOMNQb84q2ObUccsLQXHBBxuus130s3Tk8jJ4iMuPddyqCeirrhKx+MHPddcuYV1HQ3IshIyXbYQbCtM1FQvE8rBg77+SwNRX+GeUGlIaAR2WnVVxgdSABwz0rQJC6dxLckINYrri+pgEZdk+e60QxPBJJO5UjOqPBeDhVfOwjyKCF2Hbeasx0qmcu2KEkn5ILjlvcqgJx3VXDIVmD5ILy/G6tLiTlXFpxuNlb0nuEFwd1DCtdGSVVowVcgsDS0boT5YVzuys3O6Bu3dVacoQCqduyABvhUOyvDT3x3VCDlBa9pxsrRkdyVIM9iFXw8nOyCJmc7qpfvgK8s6fLcq0s3zhALiR3QEgd1aCcq7I9UBxJ81TB9fzquFXpKCHBzhV6SrywjdULSc7ILcEdipWZxurGA53UuWtGPkgsc3zVnT1FX9QPnsr29AG2EERjx6qgjJCyAWZ7hXgx47hBiGM4yqtjJwpyW57hWl7R2wgFu2Mq5rcBWiUZV/iMLdwOyCGRudlRrMjzVznAlSxvaGHJQY7oy3zVzGnCSPBPl3UsZYGHsggcwg91zNyW/hZcJR/wB2Np/W41wy92XnHbK5m5LT/wBdlwl/hjaf1uNB9H6IiAiIg65+0MH/AFm3Eof/AEnRfr1Ovnop/pFfQz7Qz8DfiV/idH+vU6+ean+kUF03YqyLy+tSTNdg7KyLYIKyk+vmomnupX7gjzUeMILgM+alZnGMqFmSVO0YQXFuBlAzIV2RjdVBA7IIX7DCjWQ8NIyocBAAwmd8KqdPnhAI2yVT6R2V5GUDMbgIIxGRunT81e7sqAOx2KCzHVsrmxkZ7q4NwVVBTp+auEeRn86oeyvYT0H6kELndOytL/RUccvIJTAKC4kKioM581XCCmQNkwCgaSd1XGNkEeCCqqvdWnIKCQpg47qjSVVBb0HP2q4bIqEoKBuDkqrvIBCfQqm5QVPZUGTsEznYnZVBwdkF2MDc9lRXOyBnso8uQXKhHdVTyQWKhGVdhWl2DsgEYVEJJRBch7Ih7IKN7qqoBjyVUAHOyKmMdlUFAAxuioCSd1VBQnCyaduRkhY5BdjAWVTnpBz6IL3dIyAAonAEnZXSvHkVEJNkFwy1wLdt1ytwhe92oKYOJPxDzXFJOS3G+65U4Rnp1BTHH74fpQc+cf2Buh2Eb5i/mXSxkTnlxA2BXdnjnG6p0OxrR/8AI/5l039yliZLkbFx3QYjnYaGnyUD3bqryestJ3CtON/VBc1VIz5q1h8lcSewQWgZV2cbKgyN8KuxGSgsPZRnuryTuFYe6AiYTB9EBEwfRMH0QERMH0QERMH0QETB9EwgIiICIiAiIgIiIO//ALFT8KbVP/F/XfyjbkT2Kn4U2qf+L+u/lG3Ig9qUREBERAREQEREBERAREQEREHH3MP/ALwHEz+B95/UpV80FWcVtQcf/JH/AKSvpf5h/wDeA4mfwPvP6lKvmfrDitqP8I/9JQWDcBXA4VjD2GVcgFwKt8Mk5yq49FVvVgoBZkYVvQW7KZqqW53QQdJOyBhG3qpwwhXtjyM4QRCMkBV8Pp7nup+nG2FZIMIIlY47jClLQBk/pUbsE5CC8EgAoDlW5+HA8lVgwgkAwq9IIGFbkq4OIG/50ETtlTurpMHOFaBhBadgVYRndXO/nVqAiKhJCBj5q9pVrd8KpPT2OMoJOkKN5wcK9pJGSrJPpH60DrCvjeOyhUkfTt6oL3nKhIypiMqJxwdigp0/NSs7KMFXMJyB6oK+GS7ZBCRssqINx23VXFgIyQgijiPmVSSEkhXidoTxWnfKCB0JAVgBHn5rKJ6hsoXndBaXEBG77q1+cK6PJbugtf3/ACrmHhLq51koegSdOGnG64ek8vrK1/T9SIYXDJHwnsg7ZcIr1PrK+vpopC4nbuuK+a6xVViv0UcxID3HZb45LZ46nWjg7cE+f1qnPzA2PUtIWAYz5D5IOpbXSP8Ah6iQs2lmbTgB/l6qClZnLilSxznDA2CDKqahk7elnmsMQPHfKQNLHAu23WX4jD2wghbCRuVRwwpnvGFAXZ7oCvazOCqNGVeBhAIyMKgaMYV2EwfRBY5uNwrFe8nsSqAbdkFjhkK36O2FdIfRW49UADHmn0iFa4kq5hI7+SC8nCd1aXeWVTrA2ygvV7XADBUXV8068eYQXPx3CtVQ4E4yFUgDOyCEjGVaquJPmqIJAcHKkZ8WFG0AjKqD0nZBK6MAbK0D1VeokKiC0+eFYYXHdXZHqr2u22wggMRCubCSFf5qVrg1BB4R+aqYnDzU3iD1VrnAlBD0H1VpYSVk/CqEDOyCEwOAyrehw2ysjJ7FQyEA7II3NI3UkcTnt77K0uB7kbKeF7WsxsgidEc4VWROwcFXvcAfJSMcOjYoMR7ekkLmbkuz+2x4SY/4ZWj9bjXDUvdczclv4WPCX+GNp/W40H0foiICIiDrr7QkZ5OOJI/+lKP9ep1889KMyH6l9DHtCc/tOOJOP70o/wBep189FJ9M/V/OgyJQOhQNb6KeTOMFWsbt2QROYe6sLPRZMgPSoh5oLY+6vVA3BBwqoKkYVCceSqTkYUbzugq5+R3VnVjZFa7ugkbsVXrx5IwAjZVMe6Co3VwcA3CtVe6C0j4sqRrgNlXAA+xWHGdkAkE7KisLun7ED853QXqVo+DKiUgJ6Dugw5Bh5VWnAR+C/wDSqILi4d8K5jshWAZ2Vw27BBeqEZVHdkachBaBhD5Kp7q13dBRXN7K1XN7IGd8KqdPnhMH0QGtJO+yqRhVwArX58kA7hWkYVytcgdWdlUEK3ARBeRlBsERAIyrCzcq9UJOUDp+adPzQZ81XBxlBYib+aIKdXyVRumAiAqEZVUQFQjKZKr9Z3QXx98HyV7nEdlYzAO/mqnc7ILZHEhR5UjmOd5K10Tm9wgkhcS5oXJvDStbSXmCQkDBHdcYwnpcCfIremkayOKujeTgDBQdmtcXdl40syn6gcMx+Zddr1Z2QQSO6CNyQuW47rT1tsEUb8kDG5WyNVQdNG/6ig4YqB01Lx6HCsIypK7asfj1UY3CCrRgq9WZwq5cQUFe+2VTp+aow+pV4PmEET9wrenbur5DhWDcFAb3VchUb3Qg5QVREQWgZOFcNk2G6IKdPzVRsqAkndMnKCqo7uqnsrc5QEREBERAREQEREHf/wBip+FNqn/i/rv5RtyJ7FT8KbVP/F/XfyjbkQe1KIiAiIgIiICIiAiIgIiICIiDj7mH/wB4DiZ/A68/qUq+Z6t/tyf/AArv0lfTBzE/g/8AE3+B15/UpV8z9Z/bU/8AhXfpKCJv0grwRhR9kyUEoIyFeHALHyQcqvUUE4OT9qlaQO6ww8g7KviFBm9TfVU6wsPxHJ4jkGohzcd1FJhYvjvAwqeK4oJ3/RUQGFYZCVTrcgv8ypA4BQdR9U6igyQ4d1UkEZCxetyqJDjCCTq+LdCcqLqKdR8kF7jsQFGRhMnumcoCImEFw7KhGVUeSIJGvAVkm5yrSSCqEkoCqDgoB6oR6ILxIDsrHDzVQBsVdse+EFnSVJGNslWgb9tlcNkE0buklRzkkghWF5yVcwh+CfIoItwrm5x3UxY0+StLQCMBAYfIq2YZKvOGjIxlWEgndBGc4CkjyG7qvwE+WVUYHogjk/nWdQyFsLxnGGlYkvT0nGMqxsrmggeYwg7K8l1cYdZnLsHqH6VubnmhfXXemlYC45ByFxvyp36isOsDUVzg1hcBkrm3mJtbdfVdPNZsPbt23QdOxbXQULHvYQSM5Kw2FriQfJcra+0PcNPWUSVEfThuTthcOwveXEAnKCeqaA09PcLFYX/nU4LnHEnZHsaQejv8kFjJC7zVXNLjsrGRuY7LuwKymPiyNx+VBYB0gEqvWFJUM6oyW98bLD6JRknKDJa4ZV+QsWHqDviWTluRhBbI3bKi77KeXHT5KBn0s/NAczpGSoicrIqJWmPAO4WH1EIJGuCvwSCo42Fx81kOic1nZBA1vU5XOid33SDZ5ztv5rNkaAwoNPaDnCo9pJ2CubjxDk7DspukdQQRsjLRlX+WFLI0tbsogTugie1WFh7rILPVU+FBFH3UoGVb0Y3wq5xtkIDnAK3qBBVsjifNWgnHdBXqJOFI3ON1Cz6SyGjbcIKAZVr8jIB8lJt5IQCggaTn/wBquUhYAMqw4zsgoHFXB2B5q3AVCceaC/qCifnzVcn1VpBJQA0lXtBaO6kjj2VkuWlBY7JORlXMc5ox8/VUaD5hS9LcboIngk5XM3Jb+Fjwk/hjaP1uNcPHpI7rmHkvIHNjwkH/AHZWgD+NxoPo/REQEREHXX2hX4HHEn/FKP8AXqdfPTR7SHPp/OvoV9oaccm3Eo//AEpR/r1OvnlhJDtkGZM4YKQuH5FC52QcqPrLSgypHDBCiUbXuLt1KTgAoKZBVC4A7qIuOVUZIyguyChGd1Y0nPmrsuHqgoN0LSVUd1e3B9ECMdJypHOyFYc42VAdt0AblVJwUyD2QgEHKCpkBGBsrc7HdRPPT2VA4+aCrhuSqDureok9/NXt9SgkZ5KX94o24wOyu6sAjZBjPHxH5lVa04wr8DqVdggpjZAceSvPSfROlqC3uCqEEK5oGVV2PkgiBBR3qqDuju/dBRA4Z7IeyoAglyAFUOBGVHlMoLyRhAdu6jBOeyrkoBOSqKgznfKqgHcINgqDOd8qqAThXN7KgGVd2CArgOysBJO6uyUFSMhU6gNiqg+qtcN0FHgHJVnbYq5x22Kt6cjJygqdwmMd1QZz5puSgdSqqFuB5qo7IBOFQjO4VcZTGBgIKOUsW6ja1z1M0dGxG6CWPHmpJOjo2HdY5kDR3VRMDsUGO/ZxK1e0TviPUHYIWnyMa7t3UsRdFG52/ZBzDw9lqLkREXF4zjC1zXNpdTUL3FhGW+i0jl0idcrn4ThnfH51zHxf00YLS5zGYPQf0IOmNe0iulb6OwrA0gZWXc6WWO6TMeMESFWyBgb+lBiZBVzTv04URJDjhTQNDnglBd4BxkKhHSFnuDQzyWDORugizk7eSoQcFWtODklTtwQggAIO6qpCW9jhRvIB+FAIyg2CtyUyUFxGUGwVuSmSguRW5KZKC4jKtIwcJkpnKAiIgIiICIiAiIg7/wDsVPwptU/8X9d/KNuRPYqfhTap/wCL+u/lG3Ig9qUREBERAREQEREBERAREQEREHHvMR+D/wATf4HXr9SlXzP1n9tT/wCFd+kr6YOYj8H/AIm/wOvX6lKvmfrP7an/AMK79JQQoiICIiAiIgIiICIiAiIgIiICIiAiIgIiICqCMKiqBlAxvlVTZEFp7oh7ogu8kBynkgGEBVAJVFUH0QXIiIKEZCqxuEVzSMIKgZQjAyVRh+aud6II3ZOysc3B2KvcHDsCjWl2MoIxnKkAOFd4R/8AcK4RnCCJ7Tjv5q0RkhTyRkt+HukUbh3CDOtV1qrO5ktJIY3lwyQcLvRy0U37K7dBJdW+OQ0bnddCKkFvSM+pXoZyLinq7LEZcbNH6EGl82GmaGj07N4FMGEMPljyXQmkw2Z+fLb869MOcmjhdp2dtOBnoP19l5mFroah7XjByR+dBNVEFvw7KOmeWn4ldK9rmLGb1+WUGfKGys6WgZ+SxxSPDgclSW+nqayTohBO61WWwXSFvW+MgIMNhDWBp3x6o5zCOw/IrJmvhPTKMEeqsEgPZBRwBPw7KPJBAyVMRtkBVa0dzsgFmRhWGA74UzHxjY42V/iRDbIyg02SFzTkk4Vgbv281nTdLgMFY/SEEkAGw/mWQ97WsOwWLks3VxeHMIyghj+KUgLNkb8JCwaf+y/aVnvIQYDWESfapg05V5aATlNvRBdI7LQPkoB3Ur3ZGPRR4KCpPUMYUfQc/apS0AZ88KxAP0cKN3dXO7bFWgd8hBb0/NULdlIe3ZWHPkgo3Z2/qpx2GFCWkBXscdgguGxIVr3EOGFft3VrgHIKeJnbCtV3SEwEFo3Ven5qpHT2ygyQgp0/NUOxV6jcfi7oJ4XAqGo+mpoQAMqGo3ego05VS7AVrOyPz88ILety5k5Mzjmx4RfPWdo/W41w0uZuTP8ACx4RfwztH63Gg+kJERAREQdc/aG7cmvEr/E6L9ep188sZw7K+hr2h/4GnEr/ABOj/X6dfPLGMuQSkZVjj0nBV4z81HJ3QSM3KlDMhQRu+ayWkY7oIHMwUAwMKR3dUI88IKdIVCMHCNcScFVd3QWYKq3tlVVhyDgA4KCQkK0jKeWVTqKCmenzVQ7I7Kzdx3KuGyC1zcpjGyuwfROnKCzpxurh2V/TtuFacDYIKglC7CoqOQOoKhPzVDv2/Mg+v8qC7qKuDvVWYPoqgbIBd3TqOCCqHurTnPmgpnfKE5REFx7KjeyqiBnfCKh+XdAdt0FchEwFQnHmgqdwqZLdiFU7BU77lBUjKDYJkIgK7IxhR5PqrgdslBVvdXK0HCuG4QFXO2FRUOSdsoAbv8lf0+WdkAwqoKdITpGPmqq0nugpjOyoWkFXN7qu3yQUIx5oBlVTOUF0Ja0klWTuDjt+ZUfsMgKM5PrugNaXHCvdE4DOSroGHqBU8rT09kEFNu/B3+tZcxDYXjHcLFpceJv8lkVTh4ZAI7IOZOWasZS3olxAHWuxPFOvp6u1luQfg/LsuqvAqpdDeyBnHUFz/rF9RW0gjjySW7fkQdY9W00bbtK9rcAkraU7yCR81yDrKw3KmqpZp2ENOTk5XHdRnrIPfO6CPBU0GR3CpG0OwpOjp3QTFxIUEjCe6u6z6oD1EBBjiPKru34VO9nS3I9FF05ByEERyN8/nVFc4HOAFbjCAiIgIiICIiAiIgIiICIiAiIgIiIO/wD7FT8KbVP/ABf138o25E9ip+FNqn/i/rv5RtyIPalERAREQEREBERAREQEREBERBx7zEfg/wDE3+B16/UpV8z9Z/bU/wDhXfpK+mDmI/B/4m/wOvX6lKvmfrP7an/wrv0lBCiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgKrVRVGN8oGN8qqHsrclAPdERBUDsqoPJEA9lRqqqdjsUEioThVGfNMZQWndVZ3Veken51cwfLzQXhUd9Iqp7KgPmfVBU480BH5FQkY7qgcO2e6CVrw7ZXYHorWdIOykbjYlAZ0juArsB30Qoqn6PwK+jfgYk7oIa4YLPqK74cjM72WZg68YC6IXFzSWdJ9V3R5NLkaS0N3xhqDljmekM9qqGvd1jpO2fkvN2/MEdwqA1vSOs7favRbi/BV6np56akaXOLSF0K4k6brNP3WSKrYWlzz3+tBszLj6rKp48tOW5zurqeFr4gdsrcFntPvErWADJwEGZoCni9+HjQ5GfRco3ynt/uOWxAHp9PktDtFgjs8QqpgAO6yrjfrfPSOY2QZGR3QcUanDG1b/DGBlaFC4h/c/lWt6kkZNUvcw7eS0OMEPGyDUAfgCo7ZpOUZ/Ywqu+ggwetxkIye6rh5dnJ/KrTE/r7eakDH5CC8hwGSfzq0EdeypIXBuCFFG89YQZEv0M/JQZcFkyEGMdlYAwtPqgip/pjdZhOMb/nWDhwcejyV7fGJwUGQ/YZVgm6UfHKG9juFCGOzugmb8SvJDRgo3GAAquAKCPOUwVVjSScjZVdt2QR46TuhGdwh3Gcq3J+xABycK7p33QADfzV3dBa7thWt7KRzcjYKPpcDj5oK4zsqfR2PqrtsfPCt3Jz80FSMIrhuFQj0QVO6Y2ITIRBb0nJUUgw5TZco3tc53bzQXx/RUc30vtU8TNv9ajnjPVgII2dkefJSMjI3KpIwZ2CCFcy8mX4WPCL+Gdo/W41w5gLmTkz/Cx4RfwztH63Gg+kJERAREQdc/aH/gacSv8AE6P9fp188sZw5fQ17Q/8DTiV/idH+v06+eWPHVugk6tlG/upHbDKiduUFY/pKcO/98qCP6SmQXIT5KhcPIqzJQVafiyjnb9lRpHqq7HdBVUJx3VvV80zlBQHKqreyZKA36QV2NwrW/SCuJwfkgqD5KVrMhWNb6q8HG3kgo8Y2URae6nkALRjurG5wcoIs74VHuxsriBnZWuCC3cboNzn0VSMqoaB5ILlQnGyrg+iFvyQWdWShOFd0hUc0bYBQWImMIgA4OVXq+SEbbKmD6IKgdkPdVHZMBARUBVUFM52wqjYKgByhz5IKHurh2Vv1pkoLkVuSrgUFAMK8dlaq9WNkAnKq048lQDKuw30QXA5TIUfUVaScoJuyodzt5qzqKZKCQDG6Y3yrOo+adRQXuBIwqD4dlb1HyVjnElBK4jt3RrQcHCsbucE7LKZ0BuMoEQ37KaVo6MZG6hMjQNlaJHOIx2QQR5Dz5K6YktySr52dA6h3WOXEjCDkbgxIIr3kuA+Id/sXZps1NJLF4jmkYGey6iaHuotVf4uSNwVzHbuIELqiPrk2GM7oNwcYoaAW17oYmglp7BdXKrPjPGP3xXP/ELUtHdLYWwuBJGO/wAlwXVUT/jkx55/OgxKf6WVlvAwCP0LDicWOIPkpHykt2QUcQCqM3conFxOVVjj1AfNBmPA6VjuxuppHEx5WIXElBIQPPdRO7qQ9WFG7ugoiIgIiICIiAiIgIiICIiAiIgIiIO//sVPwptU/wDF/XfyjbkT2Kn4U2qf+L+u/lG3Ig9qUREBERAREQEREBERAREQEREHHvMR+D/xN/gdev1KVfM/Wf21P/hXfpK+mDmI/B/4m/wOvX6lKvmfrP7an/wrv0lBCiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIBlEBKC5EyFQn0QUPdERBVv0gqk42TA2KbHdA7hVa3uVRVBIQVyFUHKsVzUF+x2VzRgK04b9YRr8jCC9WvBKu+eVa4HOyC3BVOkudlSBpdgEKRkZxlBYGEKVo2wrhGdlI2B2NggsDBjfdRSxPLvvbDhZQikcQGg98bLclltNPNGDOBnHmEGyp2SDBc0jK7fcqs3ulnHW4M+DO+y4VtenrLNMG1ZaGg53wuQbZqy36SpjDbZBgNxsUHbjh+2lvl+mgnxKN9hvldSudexQ2jVDPd6UxsLzk4wFzjyg69pL3rItr5AQ52Nzt3W7uczhvZ9R3KGamDHZwdkHmxa6aeRgLad7hnIIBwd1v/AEtRPZUxGWBzQcbkLsHovgfaHMYyWNmw32UevtDWjS9O6aAMBZug4x19PHT6fHgODXBnl9S4Op7nWPMjXTEjdbv19qp1Q91AxxLQCO64+icWvyPNBnSl0pJecn5qIRDPbsVkRgdIJVHPjaM5CC1vbCuDsbeqiEzCSGuCvGNslBJ0s9FUBqtccDKhe92DgboFSGlpwsRjcPGVIC8u+IbJhrngN9UF0jSG9lAC7OPmtUdRSyw5Zvt6KKO01PVu0/kQQ0rATkjO6zmRt6x8IU0NufCMvbgeamYxnUDtgILZo2CL6IWA6MYccLV6t0LYvLIWl+LGWkZ3KDCYcyY8lMdgogwiQnyU+M+SCMvAVHfEDhHtAyVRvZBZ0/NBtsqqn75Bf04Gcqg7qQDITpHogDsrH/UpOypgeiCAghV6h6KR7dtlY1u+CEFACDuqqTAwrDhBZgtOU6wq9yrXDBQXB4KuAB3IUIxlSMe0bZQTx77KyVvxZKkic0enZQzvAdgIJMAjZQyNLfyKRjsjdWy7jZBjO/nXMXJp+Fjwh/hrZ/1uNcPuA9FzDyafhYcIv4a2f9bjQfSGiIgIiIOuftDvwNuJf+J0f69Tr55GfSX0N+0O/A24l/4nR/r1OvnkZ9JBIeyjd3Uh7KN3dAb3UncKNgyVedsYQUaCT3Vyj6inW5BeBhVUfWU63IKorclMlBcioCqoKtOSq5AVjTgq7Od0EzewKqojJhoQSbIJOvfdCRhR9Q9U60FDsc/NUJyhOVaSQgub3VytB806vmgkyEJ9FEXfPKq05CCuN8qqt6vmFQvPkgEZ8/NWkYOEyUzlBcmQETAKAqE4VUwEA7hAMIiChIQEYVcBMBBad0Q90QEG6IO6CuOncpud1VEBVAyqKoJygohOFcR6J4ZIygtRVAyq4CC1Ff058k6fLCCxFUghVDcjsgtb8XZVPU3ufJTU7W5y7sqVHTk9KDHySpoHgOAKiDS7srvDkbv0lBlVQHh91hK90j3DDldG0FAhlfCephwVnQ3SsDh0yEELELWK+IMG+eyDcFJV1tUA2WQvHosyqoc0pd4RBxnKyNFULbjKGAZJOFybcdDvjs7pwwYDcn8iDrxOwsnc3sQVVjfmsq9w+73OWIDHS7CxotzugOaAPJRN+mFmuYC37FiEASDCDJecMWJnLht5rNe0GPKxS0B32oJfJY8n0lMXgDv5KB5yUFEREBERAREQEREBERAREQEREBERB3/9ip+FNqn/AIv67+UbciexU/Cm1T/xf138o25EHtSiIgIiICIiAiIgIiICIiAiIg495iPwf+Jv8Dr1+pSr5n6z+2p/8K79JX0wcxH4P/E3+B16/UpV8z9Z/bU/+Fd+koIUREBERAREQEREBERAREQEREBERAREQEVQPVVwEFqq1CNuyuYO+yC0jJVRsFdjf5KuB6IIj3RXYGUwPRA8vsQDCoSR5qmSgqd9gqt2TbumR6oLj2RqoSSNldGNiSgteVSMZKqWOLt1UNIwBlBIB2UsY9fkomjOFkRt2zgoK9IV7eytGcnujnFoQTswO+FIJGN9Fp7pXAnB7fNWmZ+DufyoNZo5Yy/fBC1b31sQAY8D6ltOlncx5Pz9VfV1k/UOjOEG4K+4VDWZhmIP1rFguVcYn+LOT8JG60eCpmkd0yZwp5puiJwb5g+aDn3lGr679lw8GXB8QbZ+a7f8V5ZJpKcVjy4kDufkunHJpVQRa6DagjHWO67g8bKuF9dRikILTjKDaXv0Vnp/G6+gdOd1wjxa1q24U07GVId37Fb0423WezaTFRBkO8Inbv2XTeu1Zca7rEziQ7P77KDT7tO6or5XvcTk7LHbs/dCHTuLu5KOp5gckFBmNd1sDW9/ko/dJZT0AEknZTWegqayoDGZIOy37pzSUhvNM2rA8MuHVt5INkRWCsaBK+neB64OFM62y4yI3LtxfdF6Op9IMliMXj9APYZzhcQ/cO1mnlPw5GcIOHZYnMyCCMeRUTRjutf1BbJIapxYPgyfyLR/dyGOPmEEM0bXNw0DPyWPS07veGgtPf0UtGyWWcsPZa7Q2wm5RNI2Lgg1e2UAbTh74SR6kLLdTwDDvD7Le09ooqaxskGOrp3W2T7uaR7sjIyNkG2bvLC2MtZgHG60Fk3c9QVl8rZPe3xtOwPqtMFRINgUGdVSveDh2ftWA3qD+5V7JXPOHKfoCC+PHQNt1VUYPLyUjsdOR3QQydz9qiVs0hycKLxCglBwVUblWs37qQAZGyCQdlcBndGduyuHcIKHcK0jCkPZWoLVa5oPZXKjicZCCM5G2VRWOcclOtyCp2VO6pnJ3V4AwgheCPyq0Zypi3OVe2IYygtjDh3P51SQHqByriSDtsqE574QVac4VSdjn0VoPoVa9xQUA37rmDk025suEWf+GloH/K41w+CQBhcwcmm/Nlwi/hpaP1uNB9IaIiAiIg65+0O/A24l/wCJ0f69Tr55GfSX0N+0O/A24l/4nR/r1Ovnkb3QXu7KNXOJwrUFW/SCqqN7oc580FEREBERAREQVb9IKqt7JkoCIiAiIgZPqiIgIiICIiAmURAREQEREF2QUVucJkoLshU6gqIguO4QDCtyUyUFyK3JTJQD3RVIHdBjHkgoiqB6quB6ILVcOyoB6qqCnV8lUbphp7fpRBI1XKzOFXJQUI6e6tO5281V7sj/AFKPJQSg9I3Vc5OVF1EqoJxsgveRhVZ2VjT1d1cDjbZAkJaNlESSd1fISQN1aBt2QTUgBfghZ0zWBucBafE7oOfmpHTl2B5fWgjdgvONlTONgquAG47q3ugtcT6qRpxGdz22UWcqvUQMIOSeEjh78zrPd/Zdl7v7mdKP+EZ8P+ZdU+G1U+G4Bo7B4XYSsukj7AYzndmPzIOsGq2dOoKkN7F60lrul2Vr+qYXG8TSHuSVt97S0/aUGV1/D3HZYxI8UEK3rdhVa13UCgzC4eH5rDfu7ZT9fw4UYA6u3mgsLHYzlWEYWaQ3p8liyDdBYiIgIiICIiAiIgIiICIiAiIgIiIO/wD7FT8KbVP/ABf138o25E9ip+FNqn/i/rv5RtyIPalERAREQEREBERAREQEREBERBx7zEfg/wDE3+B16/UpV8z9Z/bU/wDhXfpK+mDmI/B/4m/wOvX6lKvmfrP7an/wrv0lBCiIgIiICIiAiIgIiICIiAiIgIiICIiC4dkQdkQFUHCorgBhAJCN9VQ7JkoDvpFWu7q4gnOxVhz5oCK7AVCPRBXyVoBKuxtgqrcAdggo0jIyFKzuo+nfz7q9vqgnDAd8J4YVofgYyhlwgvDAFKzsoBMD3UzXDG2EF6jeM/lV3UVHI4g/lQWubjfKswPRUkeQO6pG4kHKCr3dAyB3WVSlszTkLHDGvIDllRM8P6KC58bWgkbLDnds4ZysiqMhacZ7ZWnvL9w7zQctcutwfb9UtmZJ0HrG+V3Bvlxmus1K45lO2MbroRoa+Gx3JlRkj4wvR7lv0m7iTSUtQ1vUOkE53QcM8wAkGlC18RaBGe4+S6TNGXyYGcZ2+1eoXOtwudpzRUroWAObEfLzwvNGxWeorKt8ZYR07H68oMK3NkfMAIi4Z8h2WvOo3lv9rHt6LkrQWibW2USXMNAyDuuRrvpvRMdJmJ0WcY8kHD/C6wQ3C5Bk7A0dXmMLmHUWlqG0UQqoZGh7W5zt6Li2+Xqk0rUmW0kZGT8JW3avi3errG6mqHP6Tt3QatdNb3F9Y63mrJjBwBnZQ09xkdUCLxxhxHmtjV1SHymqBPWd/tWPHe543iUZy3cIOUtSWqEW0VLcOcW52+pcfxxOex4MR2zstwaRvtTqCUUFSSWjAwTst7v0hSU9RHlo8M4J2QcUWymf7zvTkDPmFuelpMVkbvDOxHkuS6zTOnKegD4CzxceWNlt9tBBG10pAy0lAv1QY7SG9eMN9VsaOrPu0o8Rahq69dERhadgFsVl2eGvGTugxrl98q3b5WMI8Dsq+IZZy4+ZWQ5oIGyDGjbv5brIAJ7KxrMO+1TNxgIKO+HcqwvODuqy53x6KLDseaCOY9yo2jJUhaSTlWdJDhsgyWNyBhX+GRv/ADowAN27q7J7FBZ1Y2Vwkx3KskwBt3UOXeiDK8X5pnq3WO0OzuFIOoDZBc/sqBwxgqnUTsrXZCCw7nKqmD6KoG3ZBQbq4DyTAV7GklA6Fd2bhXYwFaQD3QQHclWkEqdzQBnCswPRBEB091d3+xX4HogYO4CCwDC5e5NvwseEX8NbP+txriQjywuXOTf8LLhF/DWz/rcaD6QkREBERB1z9od+BtxL/wATo/16nXzyN7r6G/aHfgbcS/8AE6P9ep188gQHdlRXHsrUFWjJVTscKje6ucCdzlBYiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgu7hU6Sq+SICIc+Sb4+aAioCSd1VBQDCrnfCKn75BImd8KhOEx5lBQ7d1YTkqQjqGFb0ILVUEKvQqFpB7IL2pjfKozKuQUd2Vo2CvO4UZJzhBVOrdDkBW+aCQ7jZVa04V8QacZ9Fe4AeiDGc0j7FappAMbKHBQbv4eY+6Q3x8QXP0gabKcOBPT2z8l100dUGnrQ4f3QXMtFeJKmBtOCdxhBxbqmH/qlKeg4yd8La04AB2XJ2s7I+BjqktG4JyuLql+7m9vkggYOpyy2xjGVjQAFyzwB0oMOQkEqMPwVLKBkqENJcgvdIQFETk5WV4QLclY8jQ04CC1ERAREQEREBERAREQEREBERAREQd/8A2Kn4U2qf+L+u/lG3InsVPwptU/8AF/XfyjbkQe1KIiAiIgIiICIiAiIgIiICIiDj3mI/B/4m/wADr1+pSr5n6z+2p/8ACu/SV9MHMR+D/wATf4HXr9SlXzP1n9tT/wCFd+koIUREBERAREQEREBERAREQEREBERAREQXdggOVTJ7INhugZ3wrgceSpgeiILsfNAMKvkEyPVBQHKo9XYCoQD3QWjsiHHqiAqgEquAq4+XdAA8le1vzVikadgCgtJwo3uOVO1gJ37FUkjBIwgxx1BZUJPScq3oGM5UkYAGyC5r98K2R2fsV3SFFKcHZBa7cYCN2G4QHtlStAcMhBa1ZsGD+lY2AFPE7A2QZccTHkgrS7nEGSfCNsrJlqXRAEKsDW1ozIg0kOc3dpIwvZH2XNBSXbScM9Xh7mRA4J7kBePdygZB0Bnnlel3s5uJEmmtMCLqIAjx3Qdm+fvT9DLpOcRxhxMRJaPLZeTmm9OxQ11U58HRiQ9x816X8ZuIH7PDUUFS7LcEb7+S6PcT7LFpqKpnpAASS7YINhaweLXby6knDHYzscFcV/sqvErXsfWPIGcbrHvmrrjcKt9PM49Ad04ytN6WsJ37ndAq7nU1EpE8hdv5lQ+K0dgMn0VKuNoaXN7rGiDycOygyHuc75hRFhwThZ1GxkmzlqLqKF56B3IwgzeGRP3TAA/fDdc33iIi3iVoIIbt+Rca8M7CYa4TSD4eoHsua7rBSSUAYMY6d8fUg4ThvFc66uhkld0B2MEla7WTNFKS2TuFpmqqSnttU6ohxnutsO1BNJG5pJ2yEGl6pnc6Rw6sra4cRnfutUutUah7j5laa2IuQXQfTWWfJQtjLNwpWnI3QVG6uAwreyZKCuztlUgAbKje6rkeqCAgAlRvdh2MLI6QngtO5HZBZH81eeyuEeFUtHoggxk7K4M+SkDMHKuDST2QWhox2VCMdlM5owoSd0Fobg5yjm53UgHbIVHDA2QQnZWk4V+MlWPGCgqpGds/UoS4gKWIkt3QSZzsqEYOFVvdHd0Fj+ysAz5qTAKtdkdkFiqH9OyvDR6Kjo8koLM5XLfJsP8ArsuEX8NbP+uRrilrW4GSuV+Tf8LPhHj/AIa2j9cjQfSEiIgIiIOuftDvwNuJf+J0f69Tr54wMr6HPaHfgbcS/wDE6P8AXqdfPI3ugqeytVXdlRBVpw5XE5VrBkq4jCCxERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERBd3CDYIOyoSQgqThPJMZTsCgtAyruwVAqoAOUBwUAwmN8oLj2VM7YVQSSqkAZ2QWt7qpOFRvdXEDc4QEVASTuqoKE4VQcj60xlW7jbKC9gwTuqPG+VRrj6qUDIQRZyDhWlhKnDBnZC3CCyMH1Uh3UbndIVpl9EEgZg91RzR0n6ldTu6jush0Y3CDK06C2YHzyuUtMzGS4xRE5zgLjG1Dw5A4bYK5A0BUMm1RTQvPwlwH50G9uJluENi8UREfBn8y641WTK8+QcV3Y412Ojh0NHNEB1GLJ/IumM0A635/uigxacEnssw4a3CoI2sGVbLIOk4QQyDqJOVRowe3dRteS7ClO2+N+6CVw+HY+Sw5fpKrpnZ2OyjJygIiICIiAiIgIiICIiAiIgIiICIiDv8A+xU/Cm1T/wAX9d/KNuRPYqfhTap/4v67+UbciD2pREQEREBERAREQEREBERAREQce8xH4P8AxN/gdev1KVfM/Wf21P8A4V36Svpg5iPwf+Jv8Dr1+pSr5n6z+2p/8K79JQQoiICIiAiIgIiICIiAiIgIiICIiAiIgu7Kh+LcKvlumMILSCFe05Coqj5ILjuFb0lXHYKgOUADCE42VwG+6ODRhBGW758lUDPmqg/UmcfRQWg4KlHZR43yQpARjugoPpEK4sz2VAB5BXjPmgkDcgb+SoY8DKMec7lJJD2QRn4TurmuBUbnEjcq5nbZBKRjzUErt+ylJcdlE9uSgtOSBhSREgDI/OrA0+imjZkZI3QUc7fZZFMTjso/ALuwWfR0T+ny3+SDArCQzGFZRzvjBAaVrbLWZ3YPr6LWqHSIljyMbj0QbIrZXTFuQds4Xa/la1pcNP2UxRwvADTg4O64l0rw8guNayKpAI68bjyXoBy88F9GRWeP3xsQJjGdh3wg41tmurje71LHJC/BJ3wVx/xrZIKKXMZ3BXdUcINGUlfJLQeEXbnYDuuvXMNo+mpaGoLQNicIPPC8R9F2cenGTk4VhccfUtV1ZSCC9PbjYE/pWluaMHZBHE7xX9BWQ6maGny2WLEOh/UFNLUP8Mn0CC2F7opPhyd/JZttqZ5rrFAYyA5wCydD25t4uggk3BIXLFRw5pqF0dZGB1AZ7INxWiwRW6xsrmbOLQ44VjrrJLTODnnDeywY73UxQCgmz4Y2+xSVE1Gy2yPYRnBPdBx9rSry9xMm/wBa2GKh2HYI3JWXqe6TTXJ8YJwCfNaNlwYe+6CoJllIO+SpxEG42+axaTPi5+a1GTsPqQY0nwhUj3U3QHlXCIDGEETgQ3KiLhnZZFQOlpwFhNcSTugme7pGVY2QuUkjQWZUQAHZBKwHOVIO6sapG4zvjuglDdhkqhZ6K4dkQRluFTIHfyVS76lY7JcMfmQXuIwVGr/JRnOfNBcBk4QjCoNlXcoLAMnCslHzUuwGR3UUhz3QQeakjPTj61QtHcBSRtGUEjd1VXBoAyB5KxxIOEB52VjdxnCuO+xVWgAeSCiK7AVDhBYGkrljk3OObPhGP+7W0D/lka4pHfK5W5N/wtOEf8NbR+txoPpCREQEREHXP2h34G3Ev/E6P9ep188je6+hv2h34G3Ev/E6P9ep188je6ARjdUVXdlRBVn0le5WN7q7fzQWIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIKt+kFVABscIgp1KqocKo7IB3CYx3VBnPmqoKZzsgGE/71VHz7oLwfJWu7qoyqnBGcILW91f07fWrWDfspgBgbeSCINIVexV5GydI8wgsd2VnSVI7A2KtQCcKRpBACicr2IJWjzVHIM+SvAGxJQYkgO6sWXM1p7KAtAH2IJaY5Kyw35rAjd0nZZUchyMnugz6Y+GQc+i3PoCs6dWUx6sfGP0rZkkzmjOfzrL0vXyU98hnbnII80HcDi3U+PoWNrZA8+F5H5LqBVxOa57ukj4juuwtffJr3p1lLKSR0Ad/kuL79YY6emfIAM5J7IOO5HbkZ7LFe856VdVOLZ3tB7HCsjBdIAfVBcxpByQpHnZagaVohBWFMzGwQYR7or3NCsIwUBERAREQEREBERAREQEREBERAREQd//AGKn4U2qf+L+u/lG3InsVPwptU/8X9d/KNuRB7UoiICIiAiIgIiICIiAiIgIiIOPeYj8H/ib/A69fqUq+Z+s/tqf/Cu/SV9MHMR+D/xN/gdev1KVfM/Wf21P/hXfpKCFERAREQEREBERAREQEREBERAREQEREF3cIBhUBOQqoCqDhW5OftVUF2c7BVAwgA2wrhjzCCqseMqwvcNsq5juo7oKdJVR2UrA3O+FJiPBO2UGOdwgGFQl3UR5ZQnCCRvkricKNhKyGNa5u/fKCMbqyR+CqzdTT8AOFJBCJG5f3CCD6QCmhjwM581SaMsJDR2UTDLkAZP5UGVgeijkJBwG5VzGSOAx3yty2W0wVDAZ8Zx6INsN6+/QfyKviyNziFx+wrfrdOUriA3Hdb201oC01cOZ2syfPCDiPTdI+61IgdEcZAzhb+/YeKUNwzOQFum4aTtmnQZ6UMBG+QqUF1hqwRJjbZBots0kJnt+9nd2F2S4UculJqmiZLLI1hLc4JXD9or6SKc9RGBus26cyeoNCObT2Z7wwbDBQdgLny3UGlS6pbUMHRuCCFhR8QjoiB9NFcwBECB8QXWvUvN1rW8UnTJJLlwwclcTXfilqC8Pe6oldl5JPxIPSLgZxPfrTUJpZbkHhzsYLlqnM3p1kFrkcxwcXAnbzXRHlo4lXOya0YfEfgkEYPzXcXiFrWfVlHDBMSepoG/1IOgPEK3vp708mIgZP6VtFoBBK7B8cdHNoKJ1wDACWkrrjTyvdIW+qDIDcO8gqT7QuI3PbZS1FPIYsxjdSWK11NdcY6aRp6XkA5Qby4E2p921ZHS+GXAkb47LuDe+GDYaOJxjOC3+Zce8GdE2rTEcd6lDA/IduADsubtTa/tc2n3uY5vWxpxj6kHWXijY4tNQOlZIAQCVw7T6zmmhkhMpwCQFqnFvXtxvN7mpHOcYskd9iFxtCxzX5Gd0GpVwFRUmY75OVjOi+A9lMTiPPnhQ9RO3qUGNTnEv1FZxdlRthaCHDuVegvjGT9qlDBkbqBri07FXeK4oK1jB0EgrSWu6XYWoySueMFQinaXZQXn4mBWBmPNTys6I8jusYOcSUEre6uAzurATsVcHYHkgl68BC8EKPKpn5oBOThM4TG+UIBQXEghW5ARCAgK4dlB1kHugkII/1oJXeahf3UuSftVOkHughUkY3CuLGjyVWAZ7ILwc7KjxncKUMb3VS0IMZVAyrnNAJ2+pY8j3NdgdkE+QVR3qo2PJPdSgZxlBaO+FytycfhacI/4a2j9bjXFvSFynycD/AK7ThJ8tbWj9cjQfSCiIgIiIOuftDvwNuJf+J0f69Tr55G919DftDvwNuJf+J0f69Tr5484QD3RN0QVYQDuricq1gyVcRhBYiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgu7KnUFTJRBXpKqNgiICHsiuABCCxp8lUAkqvSB2TceqCp7KzO/ZC53YoAScoJW+qkyFB1EJ1kg4QT5BRY7XnKv8RyA/4uyo0EDCp1FOr6kAnKvaMKM9kD3AIMqNw7fJVJydisQPf5K4OcNygnc3YEqJ/ZVD3E4PZCMn5IImfCd1K2YNIPorXNGNggbkHIQXST9YAAWVYnBtyjJ9f51iwxNLt1ksDafMzO4GdkHYPTLBW21jGkOPTjZYur7C+K2SSGEgAE9lpnAu8OuFaymqT8IdjcrnHiDY6J+nZHQ9OSwkfkQdIa4dFdK3HZxCtp2vdMD4ZOT6LW6yyzu1DNE5vweITnHkt323TFGIhJIG9XzCDaroyYBkEHC0qduHY+a3tdbcyJpbGBgBbNr6Kpa/LQ4jugwzCAO6x5AAdlefHzg5z9qtLHnvlBYiqGEnsVKI/h37/UghRSRxFz8Hss1tI3w87INORZYgHUc9so+GMN2wgxEV5j3KtcMHCCiIiAiIgIiICIpY2tcMnZB369ip+FNqn/i/rv5RtyK/2LDQOabVP8AK7+UbciD2mREQEREBERAREQEREBERAREQce8xH4P/ABN/gdev1KVfM/Wf21P/AIV36Svpg5iPwf8Aib/A69fqUq+Z+s/tqf8Awrv0lBCiIgIiICIiAiIgIiICIiAiIgIiICIiADgq7urR3VyAqgEqiuHmgux07+ir3GyHsg2yEEbvNW5LSpMDKuDW53QRmRxHfCt6nf3RKy2QM9Ff7uz0QY47Z8ygaT3U/htydla8Bu3zQRgdOE8TBxkqjnZGxULu6DUI4w9uT+ZWyS+Celo+xYrKiRg2KzKKAVRBed0EfWXD6JK1CipWvG7PzLUqC1QvwHALPlo46YYZ6boMGgtbJXbgYWoyRe5ODWOxn8yxDVPpSXM9FpVbdKiaXzQb7swE+A5xK33bXtpmMb43T9uFxTpSvndKASdlk6p1TW0D2iInA22Qb+1dWNFKXeODt6rj2nvkkTnASYWhVmq66tjDZHEgj1WlsqnkknO/llBvZ+ppoQXCXG3qtq3u+yVshMhDjnz3WHNO5zT39FCymZKOooL/AH4TARlox9SslijETiGgHGRhSMpY4zkY2UVU7YtB280G6+Eld7hqJk3UG7jf7V3EsVzbd6ikiM7XZwNjldELVXy26oE0Wc5HZdhOEOsK2ovFI2QkgOHz9EHMfMrpn3XRJq4oS8+GTtv5LonS+KyVxbCXHB7A7L0813b6XVmhWw1IG8fTvv5LrWzgvZYGyyZYSSf3qDgnR1tN4qBDLCRkjuuWrNw4pYrlTvLQMkbqN+nKTTdWXUwGQc9lZXayrqWEzMO8fbb0Qcoa8jj0npVstNVNBDM46guAWcVbjVQTUz6skAkd1o+rOKd8v5NtqXPMQ+Hc7YWzI6dsRc4E5dug1C8Ti4Vbp3YJJ7rBbC1vorPEdnCq+RwYSgTHpbjOVjiTft2VkUrny9LsYWaYGZGEEfXgKhecdlkeEzGMBDC3BIGyDEZIS/fKl6gqmJrTkKmAgdPqqhoyrJ3Fo2UAmeBk/kQZsrQWYz5KDoHkoYKhz3hruyyiBtsgjAwhGVJgeitI9EFDuEGwVcKoAwgtO4QDCrhUz80DIKOOB2VucKvcboIT3VzG5IKvcxvoqtAAQVGwVCMqqoSQgormd1G7ONldGTtlBktKqeyjDiPNV6yRsgsk7/Ysd4GVO477qxwGUFjI/NTN2xlWA42GFRzigmBBXKXJv+Fpwk/hraP1yNcSeI4DuuWuTXfmz4R/w0tH63Gg+kJERAREQdc/aHfgbcS/8To/16nXzxgZX0Oe0O/A24l/4nR/r1Ovnkb3QVPZWqpz59lRBVv0gqqjBkq52AgsREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEGyIguO4QbDCIgoBhSeSsV47ILR3V2dsq0DKu+SCM/EQFUbBMBEA7hBsERARD2QbhAVHfSKqmxQBv2V3hZOcqsTR1dlKgswB3QY8lf05HZVbH9XdBYB6K4DAUga1qHpwggJTOQryzHcJgeiCwEs3VHVBLS312UzIw87hVnp2siLh3G6DkTg1Vmlrmua4A9a7IT1El3pG0riSHNwV1Y4WzFt0DPIPC7Q6ee0zwjHog2Hf+G8VNK+t8HBO+VsG6ZoHljXHbyXYrXbzHbHObgfCf0LrHU1MtZqtlJKfgdIGn8qDLpoZa1wDoHYPmQVq9RoqF9udUOYAcZ7fJctHRVqotNxVrGNDywEkY9Fxvfb3LStdSx/R7bIOIbjb2w1skTWbD5LT6iEMGendb1rKaOaUzuAydytKqqOJxLTsg2i05fjpU8cZJxha062wsJI7rCfEGSYAQR+7AAEBXkBjDk/WsksPQVpNXUPa4tCC/IcSM9ljSPPWBkqMTPGcHuqgF/xeaDKZE1zc+axZx0u2KqJ3tGAo3vLzkoKIiICIiAiIgK5ryB3VqIPQH2Kzs802qf4AV38o25FT2KoH7abVP8AACu/lG3Ig9qEREBERAREQEREBERAREQEREHHvMR+D/xN/gdev1KVfM/Wf21P/hXfpK+mDmI/B/4m/wADr1+pSr5n6z+2p/8ACu/SUEKIiAiIgIiICIiAiIgIiICIiAiIgIiIA2VwOVaO6u2CCmVUHKoRsqsAKCUEIThUb3VTkkBBF1EOyr2kuI2x9qyW0rSMkjdVELWDZBGwlqq6foVHbFRvHUQAgGp381a+YuOf51OymYW7kKaKjjJCDCaSW4x8lLHTGRwBad1qsNDEQO3dalT0EQZ1bbINNptPCdoOcZ+a3NbdJMEYf1Yx55Whz3CWkPTH5Dur4tV10bC1ucZ9UGszUQoXkdfZaPX3MM2Ds4K0uuv9ZM7JcclYUcjqknrzlBnVN2y0D1Wp2K1m8YeIyc+gW3Z6cBoIOPJcucMbbD7j1kDOEEGndN+DOQ4Yx6rbXEONlJUhmQd1vq/1z7VLIYRjv2XEmprpNcakumzkOKCOINliHbsseZ/gnsoqSZ7T052CnmjEuCdtkFni+I3A7kZWZTRZaMnyWLHG0HbyWQ2UsxhBfNGWAnKx/C8UEd8qtTUPc3AGFfQ5I375QY74PdsOx2OVvzQmsGWapjqSQ3w/iz27LaFU0PaWlYj8QwvDSdwQg9C+WzVEPG2pZpg1Q2cI8Z9dl2W4g8m8emNNS3RsoJAJwPqyvMvlC1/ctDa8bVUBdkuaQAcea9HdUcyuqNSU1NZ6nq8KUBrsn1CDz5401dRo2/z0JhcGsJGd1xFLrl1UHRFux9V3Z5iuGlqu1mffpunxXsJO2+V0rk0rRRCd4xljiB+VBodQ9lQ8ygYJ3UKileI6swNeCAcKbBQWGLuVTw+odKysDp7K17elhcO/kgwjRiI+J81eJM9/JQNqZHzGNx2+tZJiGM/LKC5ruod1eCMYyscEg7FXdRzlBcWbqrY9xurev1VBIesAIJX05Ld91A+lPScDyWpwkOaMqR0bent3QbfZB4b8n1WQcbKasb0ZI8lhB7t9ignHdXd1E0k4ypB2QVAyq9JVzQMq7AQQu2CsI7qVwH51a4bFBFnfCuGwUTiQ7HzV7OrGEDOThXdJVcD0VcZQUVrj5q49lE4nyQXBwJwrseYVjW43V2SgkJBCN7KwHPmrgSEFCM/lVC0hSYQjKCFgwVc/sqkAElQyPIO6Cxx7hcucmv4WPCL+Gtn/AFuNcRLl3k1/Cx4Rfw1s/wCtxoPpDREQEREHXP2h34G3Ev8AxOj/AF6nXzyN7r6G/aHfgbcS/wDE6P8AXqdfPI3ugOIKoqkKiCrPpK54J3VrPpK9yCNERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQXA5RUamTlBVXDsrT2VzTsgOQdkO4yqZKCiITsrclBUDCqqAkndCSEFUVASTunnhA3dsFXGNlcG4VSBucILoW5OVMWYUMTiDgKYvJCCjSAVeSMKP5qvUe386C3qJKvDcb5VnSQcq4uICCrnAjAUTu6uGTklWSEjsgvif0nZXzy5iI+Ssp2hzhnzKzBTscQD27boMrR11FqrxMTj4gcrnXR2v2VV0gphLnJAxldfamBtO0OYVuLhlWOdqumY7OC4efzQdxdR251wsInDSQ5n8y69XTS5or22uLMdL85x812qlii/YbA8AZMYzn6lwTrwMiopZWgZGTsgw7zr9tPaBQ+Pu1uO/yXD961p48xbnOT3Wg3a81E9xfG4uwDjusOWjZIHSEnOMoNZk1A1zc9XceSwX3vrkxlaKXFpLd8DburOx2QbmNUHMDg4bjKxYy6eoDQ0nJC0uGpkJDPJbysFtil6ZXd++EFwtBMAeQckbrbt1tj48vDTsVyQWRiLowMALQrrTROjcNkHHRBBII7FZMTA6MuI8lnuoIzKe2MrBqj4LjG3sgxX/SPyVEJyiAiIgIiICIiAiIg7/8AsVPwptU/8X9d/KNuRPYqfhTap/4v67+UbciD2pREQEREBERAREQEREBERAREQce8xH4P/E3+B16/UpV8z9Z/bU/+Fd+kr6YOYj8H/ib/AAOvX6lKvmfrP7an/wAK79JQQoiICIiAiIgIiICIiAiIgIiICIiAiIgq36QQjKoO6uQUOwVzOxVFVoAQSAY3KqDvnHYqh7I3PUAfNBf72RgfzKhrCRj+ZZbKKNzA443T3GL0QYD5y7sEjc4uG2crU22+I+QWXFbYQRjHdBp7GOLQcEbKaJhG5BWrNooRgDGPqUcsLWZ6QNggxWzdO26v+6YjaW9X2FQytAcdvJaVVucH7EhBnz1YmJPfKsbI0DcBQ07QWAnurJ3dBACCSVjZSdgoGv8AAdjHdTQuzgjsoKhpLgUGQXeIwEeq5N0Ncfc6PDndOwG64/slOyc9L+2fNbo6/cYwI84wg1LU10EhceoHPmFsd9vbXyF+cZOdlnXSqdJnqJ3Wgm4zwOLYyQgyqygFDH1DchRUY95BJ8lG2slrXeHKSQsyFgp24b5oLfdehyilb0uwr62d7WktH51hxSvkBLs5CDIEXXjbKkZ952Ax9itondcmCs2eBpIwgw5ZS4b7KCQdcZHqsmvjEUQc3usGle55IJQbx4VX4aavja15Aw4H8i7Q6X4yRah1VbrX4zCXua3y+S6f0cYa4HON1vjhfM6n4g22VrjkStx+VB6a8ZOFgruEsV6Dz98hB/MujWn+FLb3cpbcX7SydJ37ZK7w8V9bXJnAmCBuQBTgDf5LoHauIV5tNVU18DyXxvJGCe4KDd3FzlNGhNN/snikDyWF+M5XWWnNTMJgaV+IyR9E7Ls5p7jjqbihINMXzqNMCGfEcjHZbou3BvTVBbXzxNi6pG9R28yg6YNrZPHMTmY3xgrL8TqcIj5re3EDSdDaK55pw3bJ7LYsIBeX+YQZM1qEMQqB3O6w2yEtOWn0zhanFVPqGiF/bturpLfGxmQRv8kGjNOXKQgnsFKyACQjIU4haHAFBgvBDc4VrHY3wtQqYGtjyMLADQGlBksnIx8lI6pIZnPZYbO+5Urh8GEGPJU+LIW998KwjHkq08IdKSVn+7MIQYQx5KoByFMYgPJOkA5QGjAVVQEoc580FrvNRv8AoqQ9jlRv7IIBs7dXiQKM90HdBNgndXNIGytachVQWudvsO6oDlMZVUBVxsqYVw22JQMhO6tPdVHbuguG6uGwVo2TJ9UFFDM3J2V73EKBzyThBXuMei5c5NNubLhF8taWj9bjXEkTTnfK5c5NtubThH/DW0frcaD6QkREBERB1z9od+BtxL/xOj/XqdfPI3uvob9od+BtxL/xOj/XqdfPIEB3ZUVSdzuqIKs+kr3Kxn0le5BGiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiILkPZMj1RBRvdSeSswArx5IKAYVHOwcK5UIGCSgsd6qiqTuVRAHdXA5Vqu8kDIKIwDO6uwEFVUDIRuCcKvbZAYcFXdQVjvhwQqBxIQSB4VzTuCoBnPmpGnYBBK52RhWKhO2xVRuN0Fpf8la49Sv6QrHDHZBWOXwiCp2VpLwAoI2B/0lOynjyCAgkq5eqME/JaroOpFLqCGfYYcD2Wj1mGxhaho4B13jB7ZCDttU8RyNMR04JPSwDH2LhnV2tKmrhfB4DiCTvhcqaf07RXC0RCUA5AC3Bd+Edg/Y3LXYZ1hpI2HfCDppVReJWPnc3BJzhJDiMgla3qekjor9PSxAdDHEDC0CocerpztlBgO+J5281c6n+Au9ApTG0bqxkj3Shh7FBHTnplaMZyQuWNN2Vr7UavOCG5/MuP4bfFhsm2Tut5Ud4qKS0PgZnpxhBp9ZfzBVvp+vZpwtPmu3vT/CD9itt19S+StllOclyto5n+8sO5yUG622svaZhkg7rbd2p3RT4wTlbyhncKRm3cLTLhRxzMMhxkINrimJbkg5UUjek4WpEBshaPLZYNSMPQQoiICIiAiIgKoaSFRXtQd/vYrNxzTap/gBXfyjbkVfYr/hS6p/gBXfyjbkQe0yIiAiIgIiICIiAiIgIiICIiDj3mI/B/4m/wOvX6lKvmfrP7an/wrv0lfTBzEfg/8Tf4HXr9SlXzP1n9tT/4V36SghREQEREBERAREQEREBERAREQEREBEHdXYCC3urhsFe1rc5wrulvogiVzVf0t9EwAgZCHYgq1+W7hUaSRkoNQjmwwNzuApGvz5/nWleK4bBXtnf6lBqolx5qYVXSM5Wi+8v9fzJ7w8jzQas+7dJx1fnUcl4zt5H5rSXEu3JKdGfMoM19w6jn1WPIRM4uG/2KwRN2ySp4mABBfC/pHSoKp3x5Cn6MHKilYHHdBWnd8IyFP7v4rh+VQsAaBhZsBOAUGdZoDG/vjfzWdc69sGGdQK0kVUkOenPn2WLU1Bmd8ZIKCarqTI3q75CwvdxKS7A/Iq5BHfZZFNjG32oLKemDHE/zLIk2wPkrXPLRlWF5cgSgSDpPphUipekHHmquJbgrIppWlp6yAcoMdjDTv6iCshk8kxBEZOPNXyuhOOtwAK33pGwW2upS9xadshBx7XCWSPpbC4nfyS1WupmeA6meM/Jc36Q0Pa7rczBI1pGcYK3hqThrbLREJKaNoIGdgg4k0poAXR4D/hyQuTNLcJoLVqSkuJmaBG8E7jbByti329VumXZpBgjfZaCzjJqQh7gX5YD6oPR+thpNaaDj07DUNleIw3pBBPbC66ay4AzaZtNXXOpHAYLuyweTjibqLVerBbqsvezrAO+R3Xd3jdppg0qfGiYGviy7b1CDyk0rqn9j+s30T4/DxJgHGPNc2VvEVldHHQiqBL8DuuH+L1ptNk1RNVUkgEvUTgeuVsexamrJb3CJHHHWANz6oOWOJ2nJPuebo0Fwc0u9VwhSvkeZPvR2J8l2cvMsNw0gxtU4AeHjf6lxTQWWyGsMYlYep2Mfag27pCzSXqrEToi0EgZxjzXIdfwxbBTCQPBJGVm1lno9PW8V1vA6i3Ow81tp+vrrURPjc49IyEG3Llpr3KoIaMkHdYYtUx38M7LcNorjda4ioIO/ot4T2WiZACMZIyg4iuUToI8EELRhJ8JJW9tY0sEIPSQMZWxmlhY8E7BBSOYOfj1KyyDhadTub4+52zstWHh9YHUB6oI6SLqk7d/ktQMBwFc00zIw5jwSPRUknYIi7PZBjTRBuSCFB07d1hCve+cgk9JOAsxriZA3yKCPPS4DCGTuMLUnUbPD6tsrDmha1hd5hBjkg7hWSHAUEUpdN0+SzpI2+GThBp/UCVQyYIGPNWPOJMBSsja5pce4CCePcZyFdjbKwTO5pwDsFkQzF4AJ8kF52VC4A4V+Ad1E/Ad3QSNHmri3O+Va0uxsnU4bIKHZFeADvhRyEtdgILuoIXDBVGjIVr+rqx5ILHP7jCiccu7LLjhB7o+BuCUEcRHTjzXLPJv+Fpwj/htaP1uNcRk9B28ly5yanPNlwi+etbQf+VxoPpCREQEREHXP2h34G3Ev/E6P9ep188YGV9DntDvwNuJf+J0f69Tr55G90FT2Vqq7sqIKt7q5zgrW91c5oQWIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiABlXDYKgGPJSMAPcILFUA91N4Q74VRHtjCCEHOyoWZOcqRzA0bDdROcRsgo5UVRv3VcBBarh9HCqGg+SrhoQUYCN0c7BwrtvJOgHdAYCN1djfKHYbK0uKBI7AwqNOyviaH9+6vdG0DAQRtIJVenJVjssJwqeI4b/zoJwMKqjY4u7qdobndBZ0dO6tIOeyyC0OHZULGjsEELWHIUoJYBugAHZXSfQygxaibxNvQrUtLze73FkvotLhYHvx81qlNGID1DuN9kHYzSmrmtoIovFzjGwK5NrLxJPpKU5JBYT+ZdYtEVskszI3uOA4LtTb7dSv0HLK8/8AyIn8yDpjquq69RVJPcuK0KpJwXgZxvha5q51I3VVUxrxgSEdlp1MyCerEJdkEoNKZLI9wb0Eg+mVqlrtL6upDHRkA+q35RaQoHUrJgASd+y1B9hioqU1ETR1AINIGkGspRJ1gYGSFty63QW8GjBHoVm1mpriyd9MM9IyPsW0LpM+eoMj85OUCSATPM3k45UDfvEwPocqgqXtaGjsFY5xkOT3Qa63UHTEIs7BalTyitpi/qC2csuC4TQN6GnA7INSfTgSOWlVremRZhqpHN6vMjdYk2XnqcEGMiq4AFUQEV8bcuwQsgU7SMoMRFfKzp7LPo6VksfUcdkGmq5rgAsutp2wg4HmsJB6A+xWdnmm1T/ACu/lG3IrfYqfhTap/wCL+u/lG3Ig9qUREBERAREQEREBERAREQEREHHvMR+D/wATf4HXr9SlXzP1n9tT/wCFd+kr6YOYj8H/AIm/wOvX6lKvmfrP7an/AMK79JQQoiICIiAiIgIiICIiAiIgIiICIiCrfpBVVG/SCqguZ9JXk4VrQFVyAHZOFUjKtaMFXoLEQncjKILXbKrCDsqHcnKq3bsgu6R/7hUVx7K1BQjvujVVVaAgq1pKyGDGCo24AUmcoDpAMhRvdk9lL0N8whjBQQ9fyWTFUgbZwoXRgdljyHpJwUGpNmBJPqopY/EcC3ssRkjhjPyWfTnIBQWe7kNCmgi6RurslWueW7YQSiESZGVBNGYiMNJz6KGpqpIgOg4J7rXtNU0VxYTOQSAg0ConkYBhnfzwVuzRelZtQwmR7C0AHy7reehdEWu+V/g1IaQDtlbo1PaodFPZBbGANO2yDa9g4OG+VhpHP6cHzWt3XR40ERSicHPw91pddrq66dh98pMh5Gdu62Vc+It41PIZa4uy0oORrZem6dkFwMw3Oe6XvjPHXxlnig9Oy4sqL3U1kQgkJ6QMDdYEdFE7OcnKDXr7rBl2cRgEElbdFQxjJPgG4KyPubCz4m5z9aClY54bjY7FBy9yq8Q6bRGqG1knQwCQHJAHmu9Gp+OtBxBgp7BBVsLp2hmA4eYwvMmnjbagJ4Mg5zsuQeFOtLi7Xdub1SbSNxn60HOfMLysV9ssztatie9j2GTzIPmuoul7BVS6gwaN4EcmM9PzXsLrLr1RwZghuDW9JgxuPkun54XWC20FZcYBF4jcnbGcoMbRfCn9ndijoGzAOIDcefZbik5Hqi1UL7w6Y7fF/OuNeH3Ey96e1Y+30PUWMkwAPrXYG98ctWTWoUb2noczBQdXeIdkdYJn2Z7iejI3+S40jt7JHOj232XIvF27T1lRLXSbSHJK4YbfJ42Syh24QcgaZ0cx84lEoG/qt71OlGmAffx2HmuCLDr67x1fhhxIyt5VOv7sIQPi7IGtdLANdiTP2rYMOmHOLmHOCcLcE2qa64Sls2cHuquqJImGRo375QaZScPnOcJC/YLUqTQfvFeyl6x8ZAysB2qa+GToH6VfTasr4ahtQwnraQQg5KuPAT7n2dtxMwORnutmO0EHxPb1jYkLLruMWo6ugFBIXmMDC23BrO5uqPDycOKDTLto+W1ymQNJHcei0jxJWuJ8InB+a5Wp2G704dUAEkKlJpCgkrmROaMOdug40ZcKh4DDA4D7Uka+RhBaRlc/XXhnZqa0ipjYwOLMhcY3WzQU0Ej2gDpz5INgNp/DkL1PJJiMgLHqJi2rMZOwOFcXNdt1eSDCcC6U/WsqJmR0+qqYmgdWFC+Z0bgAUF7qE5zk4KNhLTj0VwqHFg+pWmXJQZbIfh791a6lycrHdUvAGFmU8rnsJPogxJHiHI2V8BEwyseryXnJU9H8MeUEjm4UT2lxVsszg8qsZL8FBK1myse4A4KlB2x9ijkaC7KC5hyqu2ByrW7AKjyT5oIJBk59Vy3ya7c2PCIf92tn/W41xJJt3XLfJpvzZcIv4aWj9bjQfSGiIgIiIOuftDvwNuJf+J0f69Tr55G919DftDvwNuJf+J0f69Tr55G90AjG6oqu7KiCrPpK9ysZ9JXPONggsREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERARE7oK42VMFSADAUjWByCDq+Sq1+FN4IVHRtBxhBTx1eKhoGFA5oAyFRreooLzJ1FV6Sd/JXMi9O4UwjwM4QYuCrXfSKnkaBkEKLAz280Bru+yqRndU6cKoO25QVa3fcq9WZwrhuEFqr07KmMK4dkFIxgqQnfsrRt2V2xCCOQEj61H0FZAaCcK/obhBFCwqUMKtGWnAUjckboLunbuqgbYVASTupGgHGyC0Mx3R7eodP2JUuLBkLDZUODs/NBJ4PgO6z2ysykmErunPksSqe4xjPy3VKBxbJlBy1w7swqpoTnu4fpXdbRnDx180mbe1xy+PHf5LpPw0uE0c8PTsA4LvRwt1DWwWyNzCchg80HXLX3KRU0d1qbk4uAc4uXEF24XfcGpMjpB8LsfkXcjjFr27xU07AMjpO4XSnXmu7rNK9riRlxCDNNybQRCAyglq0q5a7ZFEacOznbAK2RLfKuoeXPcd+6i8FtQ7rcTk/NBmTXpsk75ekbkrTauoFRJ14wsj3KNWyUzWNJQYbW9RWQyAEYWPnpccK8TuCCX3Qq0wdG58lb7y9Wumc4IJ21AAxlRvm6tseahWZT07XtyUEEcRk77YVkjOh2FPNmLIaMbpGGy/E9wBQY/VhZEcxDdwTupo6eI7ZC1KkoIHuDTgoNCkc5+/SQM+imgrJYG9IZkfUuRtP6Voa3BkA39VqNboi2skwGjug4nmnlqdvDO/wBamp7TLM3qLSPkuWLZoe3HJ6W7DzC0a/W2C3VHhxAAZQdvfYw0D6Pmh1RK7ODoGtb/AM42/wD1Itc9j5G1vMrqRw89C1o/5wt6IPYFERAREQEREBERAREQEREBERBx7zEfg/8AE3+B16/UpV8z9Z/bU/8AhXfpK+mDmI/B/wCJv8Dr1+pSr5n6z+2p/wDCu/SUEKIiAiIgIiICIiAiIgIiICIiAiIgq36QVSMqjfpBVQSM/mVytZ/MquznzQOk5+1CcKqpjKCzG+VUnCO2yrd0A7qrVar2DPkgvG6Oae4KqAQQr2gEHzQY+d8K4beSvLG5KENP/wBlADSRlTMacKFriOykEhQSoqA57lRyydJACC9zcgqF0RO6u8RxHdSx/EN0ELaffKyoPgABVWqjs529UEpcO+VFI8DJHkjT1d1ksgjc0k+iC212yS9SiGOMuOcYC5V0zw2dBC0zfeuoeeypy/6fprvfjFI0EA7ea3FzA3iu0XcYaS3ANBwMg4H5kETKWPRUhq2zjbfutk6t4nC5zZLuroPc7ratZre53iDwqpxOcjKxaS1QVDS5/nug1Ko1S27RCFwHbG4WAImsJIAGfRVktsFIOqMjIOysEjn7eiCXoG5CtMvhlWve4DdUjAkcOooKyV5AIAUcdaTINsYK1KO3xPbkgKsttiZGXgbj0QXtmbJEMkFbm4dVMdBq+kqiB8Dw7t81sRtSWzGPPY4W6dFuL9SUrCO7gg9Nrbr5uo+HUFoY4ZEYG31Ljm9aVlo9NVlSSSC0nutU4f0UdNpSGfcHpBP5FqOoa1s2m6iFzxjoI7fJB0Jrtf8A7FdbzBzMdMn863e/mHir5Y6EADOAey2jxK05a36pmmmcAS4nt81tuOwWSFpqo5gXs3CDlTU0EWorcKxsjSXjPdccT6SEVLLIME7nyWmN1tWU7/cY3nw27BZdo1HNXXmGge4lkhAOUG1KdlVR3MsFG4gHGcLcM9ZVinyKRxIHoV2t0bwU07d7LHcpo4y9wznAytYh4G6dlr4oHxM6C4A5AQdFn3utgnJfSFoBPdpBUsmqJegt8Ild6eMPLlo2x6XFwpWRCUs6tgO+F1etvD62Tyyte0YDiEHGNDWy11SWuiOCtedbzGwPLdu631X6Ktdqj8WIAEDOy0eqZAaJ56hkAoNm19eyEdAAyO60uO7Bk/UANj6KCvl8S4uZnbqxlXGlhwCO/wBaDeNDrBkEIb1AYHqpqbXTG1zH9YGCFsV7GgYGfyqzoDQXjOQg5+fryO42wU/jg7Y7rbNRC24Ewh4+M77+q4woLxUQyBgcSCtzUl7mY5rwdwAUGsVHC4S/7bDhg7rEqOHhhp3TA5LfILUINY18kYh3IG3dTUl9rKusZSSfQecFBxpWQVUFSacU7jg47HdPubVuYZPdH4HyK7O2DhjZ7pSMrJmML3DJyFvlnBix/cJ8wjZkNJ7BB0dzIH+H4RBBxj0U4ppy3r8I47rl+96HoKa+ywsYMB2Pzqd+jqJtMT0DYeiDhF73gkdBGD6KeCs6fhIwt/S6YoDVPYcbZWzdR0MVDVBkRGAUERpfefiG/mnh+7DpPksihLvCGPNYlwkd1b7boMeQ9btvVSw/DgH0UTS3AOd1exw2wUEytcfiwr2kEb4RzRnOEADKEYCq0klVIzsUGHKMk4XLvJn+Fjwi/hnaP1uNcSSgZOy5a5NPwsuEX8NLP+txoPpDREQEREHXP2h34G3Ev/E6P9ep188je6+hv2h34G3Ev/E6P9ep188f1IKu7Kib/NEFWfSVX91Rn0lc8Dz/AEoLEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEGyIgvDxhXNlA2CiRBP43zVDICM/JQq5m6CvVnYKo7K5rG+iv6QPJAbJ0+XdXGpGMKN7RhQ43wgkMnWURjW4yq437bIKdlTBO4VxDT2VEFCcK9vZW4yrhsED6XZD8OyMxndSdLSBkILAMqRrDhW4AUrOwQWtYcqTpOFcB6BU39EEX0CcqvWFbITk5URdugyGuBKmZtjKxYjnzUzXH1QXSgPCxTAWu6llnsqHPYoMcu8UdCua33cF/oFIyIMOQlRhzHZOBhBvDh9ffBroos9nj9K7xcOL2IdPNqT2bHn8y6G8P4YH3GIk79YXevh1b45NMYI+Ex/wAyDjPivxCZVzT0uB5jC6uanqW1s7xgZLj5LsBxVtNNDW1L299yuul8AjncWns4/pQaYaDp+LHzVpn8E9OFktnLgAfRRvgjeOo/pQRe+5VTKJtvVRPjiaTuB9qh6yx3wlBlGhyOrPdRvpQ0Z/nVnvcuME7KhqZCNygiOyKrsHcZ+azKGGKQ4ce6DCwT5KeOpliGA3Za06hgDQdtz6rd2ndJUNfEHvYCSPRBxu90suB4R7+hW6bFo99xhEjmluy5HotAW5wJLW7b9lp938Swu8GmADe2wQaXa+GgqSep4GFh3TTLbNUBniA/Uo7jrW7W7JhBGe+62tX6ruVe/rnfkoN+2uubQMGZB29VDcdWMbJ0+ICuPnXqrdsT+dVjnNQC55OQg30NeiiGxzn0VPfBfv8AbDnY81tSgoIq5xEmdlmzVElqIigBx22QegHsgohHzK6lw7P/AMBq0f8AOFAi0r2NldLU8zup2SdhoOtP2/dC3/60QeyKIiAiIgIiICIiAiIgIiICIiDj3mI/B/4m/wADr1+pSr5n6z+2p/8ACu/SV9MHMR+D/wATf4HXr9SlXzP1n9tT/wCFd+koIUREBERAREQEREBERAREQEREBERBVv0gqq0d1dkeqC9oxgq7uqN37qp27IKq1yAocbYQURFQnCCwtOVLHtuo1c0nb0QS5zsqgYVGjsfNXdJQMZ2VHMwrzhoBHdWdROcoKAYVwGUAB7qQADsgvbHkBRSR5dhZUQKuLAdyEGH4eBt5KSMdI3UpaMKPGCgu6gFaXZKtkO2xV0IB3cUF7AQQcLVqKhdMw/NYcTGOwB5FbltEQDB9SDX+GWoRoe5mrkPSCc91Xi9qhnECqZPG8EsI7LQLrE1zD3WgsqPdyQ1xz80EElv91haCMkDdZFvmIBGCMKemf72el5yMrUm26JmOlo3xlBpc7zIDnKwnP8LO3ZazXU7YmF2AFhxQRzA5wfrQYcUnvDukA+izG0joSCWkfYsi30kUc2XAAeqz6uSkDcl42QacavwW4BUbq4zAxjcntjzVk9RRvd0tkBKz9M0tJVX6CnkIIe4BBo8tDXRnxm0byDvnpK1rSFXW09+gqHUTwGOByWldyNH8HdKXDTkVVVtiBLQdwPRZU3CHQ1HA+eIwhzWk7AeiDeHBG9Saus8NpYwl2zcea5d1RwmrKTTMsrqdwBjJO3yWx+U+12Rurm0cEjSBIB8u6748WNO2+m4dVk7GMBjgyDjv8KDx8q+Ch1lrOWi8TBMhb3+a3veeSGS2aflufX2YXfS+S2S7iBe7VxmqaS3FxDZyNvrXPF84ra3qrYKCWN/hvbj7MIOjldwumptUy2ksOGOIz9q3DTcIPuXWxXDIwwg91yHrWlqKOsfeHRhszsk47rZMusbrUQvjeTjsg5RsXEWKxW9ttdUhvQMd1mW3i3TT3+npffB8TwPpfNdY9UX6she6UF2T6LaNr1fco7xFVB7+oPB7790HoVx4vDI+Hra+Op8QmEnAOfJdHbPxAmdWyweG4fGR+dczWvWN11pp1lquPWYukAdR+S0VnCq109Y2VkbMvOSgitNul1VAAWkA+a1W48HDDZ5akPyeknGVuJ9rbp23tdRgAhudgtFGtL1VO9yeXeGTg/Ug653jTVRS3ySn8J2A702WVNpx8dOZsHYdlzbqTTVD7sbi7p8UjJ9VsVjGSSGBwGDsUHEkj52zmNsZJBxjGVcTVn4fdHb7fRK7I6B4X2O+VDZJ2MJcfQLmOr5e9Mx2s1QhiyG57D0QdHLRZJamQOkiLcnzW9aLSbHRh5cAt2a/09SabrJYqQABucY8lxnPq2vp3uYwnAOBug3M+yMpf3wOFDSFkNcyQkAAhbdbqOuqN3dRyrnVswj8TfI3Qc7WjWkNtt7QagDA9Vm1fHiCktj6T3kZLS3uustTqa4OcYy9wAOMZwtOkrZahxMrnZPzQck3LX7Ky6vqg/PUSVkRa7ZUD3frG+y4xYwHByVlUTMVLXAknyCDkyCj96JqQ84duFs/VNlldKZWsLgPMLXaWrrIaRnQDgretmstNc7O+eqaOoNzug4SjkNMBG4YI8lDMz3k5C1nUtJDT3SWJmAASBj61pHV0EhvZBD7m4jAVpgMZCnMxbuqF5fuUFIxtkqriOyuAxsFDIT17IJWHdVk3aSFY3PSO6PLg3CDHfIOoj0XLfJof+ux4Rfw0s/63GuIJPpE/Jcv8mn4WPCL+Gtn/W40H0hoiICIiDrn7Q78DbiX/idH+vU6+eRvdfQ37Q78DbiX/idH+vU6+eRvdAd2VFV3ZUQVZ9JXPBO6tZ9JXu8soI0REBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEG6INkEnglDEQM5VPFcrmSdRGUERGPJVBwswRxdOSRkqvgwkZ27IMQSkeSkEmRlSGKLP0lG8xj6JQULtlEd3K8Fp2KuxHjKA1uQFdjCozvhX4BQR4KtyFc8nfdRoL0VASTurwBhBRvdSt8lY0bqVoHfZA6Fc3YDKqiC8O3KHurRsd9kJIOyCObuoiCW5UrxlGNGOyCyFpz9qyAN+6sDendSs3OUFsr+ho+ShZVBxxjzWTIxrxuohTMac7flQSHdod6q6mp3Vkvu7cknYBXhox0+S1zR1Iye+RNdg5I7/AFoN26E0DUQSRVcjHD4g7867Z6IvMNBZxQGUBxaG4z8lx7S2UUlhimiaCekHZaHZLzcxqaKlIf0dYH50Go8WbPUP8eqZC5weCcgFdVtRNq2Vj2GleBk9wV6V1OjbXd9MxTVYZ1PYCc9+y4mvnB/SE0hJ8IvJ9B3QdHo2VTgM0zht36SsmOmmkIYWEErtZeeD1lpWOfDGzAGRgBcTX/S1Pb6zpjYAAcbBBxp+xySQB/Sd1kwaOfI3JB/Kt/G1uEQ6WqCqinp4C5oxgINnt0Pn9/8AZlSM0F1kDr7/ADUk97ropnMwRhYj9TXKN22Qg1FvDckZ8TbHqjtDikHX1AY+awv2Y3UAgdS0+s1ddJAWkkA+pQau60NB6C8fDusyl1M2x4i6+2y2b9363JJduUy6tb4rycoOR6TiNGxpzIB9q0G+a3ZVy5BB3+tbLqG+ESAT381ZHEHjJJQapXXplXkYG/yWCylEwLm7Dvssd8Yb2yr4ql8QAbkBBe+kIPms6koyW7LFZUh2OojK1uhZ1sBbvsgjp5DQEuKw667CaTKy7iw9LtitHMDD3Qd/fYv1Ym5o9UMx20DXH/nG3IqexchbHzR6pc3z0BWj/nG3Ig9oUREBERAREQEREBERAREQEREHHvMR+D/xN/gdev1KVfM/Wf21P/hXfpK+mDmI/B/4m/wOvX6lKvmfrP7an/wrv0lBCiIgIiICIiAiIgIiICIiAiIgIiIA3VcFUHdSNwdkFR2VzeyFhAyFaHkdygrjJwhBCs68O7qQEOCCNv0griMqpZjdWZKC0nywpI+31K4RZ3VrgWnO6CVmxypQQeyx/EbjdXRv9D3QSFhPmrS3CkG4TGfJBEr2u6SEc0AIwA5BQStqA3y/IpBN1kb4WOWbEhUDxGQCgySRhWOV0J8Qqkw8M77IIyD+VTwU5kO2wUBcCAcrU7eC5oIGdkElNRO7E/aFuW3U5YwBaIJPD7havbqvLRkoK3r7zEXHfIWkUVtFxJcFm3+oDoelx2WFZbiynBAcgyTazbvi9N+6NuLz5Zwr6u5NqRguG+y1Ox6e9/Z1x75GeyDRamd9SwjpJyoI4pImlxBx3W8H6Wkicfh2VkmnpDG74TgAoNG0xbpL1ViBjSTnC5MpOBtXdHMjLHDxMYysPg/paervYZEzJ6wuy9Xb6/T7YZnxgYAKDrvqbloqLDQfdF2f7rutq6Z0E+K8MqS4gRHff0K584ncSJpLYaN5AAaRj7FwjZ9WFlY8Z2JPmg7b8JbRJqW3xWdk7h2b3+xcvnlprKhghE7j4re2fULrhwV4hRWOobUF+CCDuu0ls5g4paqBrZwSANsoN88A+Vmr0Zexd3l5HV1Zyux/FyhfJw5raYEkshLT89sLZ3BniZPqYMhGHA4AXJfEKlfW6UrYGDJLO32FB4XcR6o6B4s1VzqI8ASl2Xdu61ip5l7dWU4Y3oywYOy1rnE0S6j1JU1cwwMuPb5ldRrZSUjjM0OJIJGEHNd54s0+pZDCwjB2xhaLJVsjj3APUthWijjpqjrbnBOQtxyVIkw0eWyDS9SBtSDgYGPRbRt9B03ePbOXD9K3jdYi2MvOwW3rU9kt8hj6u7x+lBzFT3pumrGycsAw3JPZaHR8aI6m4Mg6gcuxlbg1vY//AIGsnd2Mf8y4DsFEyS+xxEnAeMflQdoq3UwqbM2c9i3OStmw6lgM/cZ6lq94oW0OkmSE4HRn8y4ko7xTvuBjD9w7GAg5XutxNdRBoJxgY3W0mW4seZMnvkLXaaJ8lvEjRkELFMcpaRjug1LTOrDpiUPdIQAc7lb9m5gopqE0XjjJGO/yXC15iIaesY29VtgCASEgnOUG8ta3v7uTPqQ4u6s+a4zqqLLnSY7lbmqZx4PSM4WjSEvPSD3KDSxIIti0bKQ1zTGW47hZFXb3NZ4h2BWmPY1pJygjfE17y7AUL4QD2CyBKzI+JXP6SwuygxmgNUsMohlEp7BYj5wHdPzVzXh7S0lBuMawijiEOBt8lytoWq+7Nhe8Ox8OQPsXAEtN0gOb5rlbh/eRa7QYScZbjug27qm2E3iXcnB7rbFSw07y0+uFu681rZ6+SUkbklbZrojK/PfdBi+H1NBVRHhStGAB6I7ugsAJUUrg07hTbDsoZG9R3CCjJB2Vs0uNsK9rAPJQzgH8qCCR+QuYeTPfmx4RfwztH63GuHHei5j5Mz/12HCH+Gln/W40H0hoiICIiDrn7Q78DbiX/idH+vU6+eMd19DntDvwNuJf+J0f69Tr54wMoLj2VquPZWoKs+kqv7qjO+6ucAcFBYiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAg32ROyC8s2ysinoXTnuVAHZGPRajRT9Hn9aCZloc5ucrKbYXdOcqsdf0jv8AkUzbs7p75QYbtPuJO/51iyWR7TgfpWq/dN/fOMqnvRk32QaR9xnq77jOAyfJar4pQyEgjA3QaFJTmAlqjDs+RWtPoZJzkN7lY8lufGMEeqDS3DAUZ7kYPdajLRuaMlQmn7nG31IMMnfsrmuIH0Vqdvtj6t4axuT2Wq/sWqQMFhG3mg2yx5J+ipmg+hwtbk05NFv0n61jutsrCQWlBpxPSBsjX79lnvojg5HZYssTIu57ILSQQqKzxWE4ypWFhA38/RBQjAQNyqTvDNyVSGVjuxQShmVe1hxn5pHue2VJ0n0QW+GXeaqYS3clStbjKP3aEELPjcB9i17S7jQ3WOpJwAQtBp/7J9q1Wml8OQOPYIOfxxHhZZ2UxkBIbghbcsOvYptWQsDQT1jyWwoK2OeMMyQey1rh/Y212sqdo3JeNvtQd6bVUy3XS8BbkBzBjv6Lb2pNCVdLaZL2XPAYC7K5L0tpCpptIU0giOAwY/IsfiF7xBoWpiczA6CPzIOk2p+NjKO5TWd0gJiy0/YuML9xFZVVXWHZBOThaVrikgdrKsd1HJkJP1ra9fTNY8uYT9SDereIcfQG57DCgqteRzRlgwcrYgGVK2AuGcoNWnvjZZC/A3+Sx33RriD0j8iwTAQM91GWgHGUGoi5tH70KySoZO3sM/UsPo80aQw4yguMOTnyWRHVCBvRhWdQx37/ACVrous5QJ5hIo2ShgwqOZ0jdWfUgue/qU1NSOn3wQEgpzJha3S0jo2ggIMelsLp/Pspn1YtThD8sLPgqXUwOPRbeu8xmqOpBPVXUTA9jlYQmyc5WPgY2O6vYNkHoF7FuTr5o9UD00BW/wAo25FZ7Fof9dJqnb/5QK7+UbciD2lREQEREBERAREQEREBERAREQce8xH4P/E3+B16/UpV8z9Z/bU/+Fd+kr6YOYj8H/ib/A69fqUq+Z+s/tqf/Cu/SUEKIiAiIgIiICIiAiIgIiICIiAiIgq0ZKkDCCFYzupxjHkgvc74MLFf3U7yAMrHdklADSpGMOcq0A7bKSPJIyguOwUeD1ZWQ5gx9qgLd8YQXNna3v5Kx8gfgeqo2nkd2CoY3xn4hjHmgvFO5w6t91LFCW7+ivjmjEYGeyr48fScnZBTqA29FaZAFa6RhccH5qx+HHY/kQTdYcMK5kJO6xxIGYBKnZVxgblBe/EXdQuiMx6gdlWWojfkZV0ErACGuQSxOEAGUkPjkFvko5JGOGOrupqWF79mboIpGFrQPQBS0t092+AhTGhmfkHP1BQ/cOrlf8LSfXZBmMrfePi9Vlw1roAPtVbVpK6z4EUTjvjstaPDbUcwBbTvwe2Ag02khffZPBA77Bag/RktGOxGRkLXNKcPtRUFUHyQOABz2W6rrYLsQAYjsPRBtDTGgZb3UiHBPxdlzrprhTHa4GNkj7j0W0eH1uuNurg+SIgB2c+i5YreIlstQYyte1pGBuQg0C66Ahja6Twx2JWyrla4KMuhLBucdvVb8u3FOwzREMnYcj1C45vGrbZWVBeyQEdWRug1jR16pdCVn3TmaMZzutS1vzH0d2pjHCQC0Yzlceaj1BbKihLJHgDGO64ruM9od4gik7koNwap4iTXuYtbNkHZbeobhNHP1h59dytFghgqKj7y8nJWtUlnqJp2QsBJdsEHMPC2pq75M2nilIOQNiucJLDdbFG2vdUHDG9Xf5LiHg/pm6acmbcKuMiPIduFzZqXWFBWWl0THA4jIPy2QcvcrHHiai1Ey1zT5LZADv8ANd5LnxWpq9kdr6W5qcD5nK8uuVqyMvmvgaQkudKAMfWvRUcO7m250FQ2F2IuknKDr3zxcEY6nSsmo4qfd8ReDj5ZXmRYuHkzhVzlhIY8+XzXtjzcadvmoeGYtdujJf4JbgD5LzasvL9xCp6GullpZekuJHwnsg6t3RhttQYAMEHCw23MxgyOdnHdb34j8PL5YblIbgx7SMk5BXHnuvih8TH5O4wg1+y1A1PN7kBkk9K3DbeFstNqOmeWHBeD+daVws07WtuoljBwXZ7LnWGzXRt6pJ3NPQCCg1nibo5lHw8jxHv4XfHyXVLSlkLtUsZ0H+ygfnXePiPS1V90SygomdUojxgd84XWjTXCTV9LqVlRLSvDfFB3B9UHLOrNEe86Cje1pBMQ3x8l1zHDeoopn3E5wx2cruLq2mrrZodlPPGWuEe/5F16rK6WajngwMkkINgz8QWWqMWxxGWZCw/9kiMfD81t/U2mK0V76p8bgxxznC0L7lDBccgAoN5V+r23Fha07laMKpwcTnvv3WkM8GmIBdgqZtdTE46x6INbiqnTjpJz5LMgosvDz9a0u2tY9wc1wK3HTMe7DQPJBI+ibVxeEGjJ2WE7RrpDjHf5LWoWPpvjd27rIjvMbHAkjZBt79gDwOoBUn0SYqVzyOy3aNS05Z0B4zjCgrLu2opnRRkEkbAIOIqu2uhqzHnYFVFIWNyFrtwtNaat05YQz1IWmVUzIWmN+xQYD6hjT0nyWs2i6yY8Jr8A7LbkgLnlwGc7rULW3oeHOyg3VFbn1ZMhOT5rTrjTikcWHfK1WhvNNTM6XkDy3UVfbqq7gz0rSWjc49EGgFpcc+RVrm4BWW6inid4TgeoLEqJGQEiQ4QQF2/ZUPdRuqYM5zlU95h9UEijkjLuye8w+quFRD5OGUEBpnHcBcu8mzSObPhEPTWto/W41xT7xFnHUuV+TZwPNpwkPrrW0frcaD6QUREBERB1z9od+BtxL/xOj/XqdfPI3uvob9od+BtxL/xOj/XqdfPHnCCruyohJRBVoJKuIIVre6uzlBYiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgKWKUtGM+aiVRnyQZBmcexVBUOae5/KogD5K13dBl+9u7ZU8NWcYJWnbZysmMMHn6INQFX8wqGvLCNx332WH1s9QqExHu5Bq8V6jjAB2+vzWWyqjrAH47LbTxF64WsW57Gs2I7INUioG1gIAWnXGi90eY/sWtWqpaxxAwtPv0rXy9RIygxrbcorO8SyNBzuFrD9d0hxhgGFtK4DraMHOFgiJ57BBvV2rqeqPSAPqVpqopd8d1tSjgcJgXDGFrTZ4Yhufkg12129te/px32WTWaEdM4dI7jOcLF07e6SllBc4Df1W7f2U0r3AtcMAeWEGya7QL6VvUVgw6dcNsHbZb9u16bWRdMRBWjU8VTI/AbuSg27W6Ze5mR5LTvuI+nyXZz81yDPbK90RIjP5FpU2n7rUEtbEck+iDZ7yackEdlWOrDnAYK3BV6Dv0oLhC7H1LAdo260nxzRkAeoQQNOQCqvGWbeitkDab4JHYIUbq+mwWh4ygjY0sdn0KknqjHG4g4ICpE3xyPDdkEqeWy1M8ZawHcIJtNVclTVtiJzkgLsdwr0eKe6095fHkMw4n8i4F0Roy8z18cjIyMuA2+tdxeH+htQ0tmFVLC4RBmST6YQcyf7NtFbrBHacAGJvT+Zccay4yxXq0yWhjxl4IwCtl6gr7VFUSU0rwJWkg7+a2XNJQGsBjf8QO3yQbA1Vw5fUXSe8dBIfly4t1HbfcZDGRvnC7J32eZ1EWt3HTsuAdbUtQ+qc7BxklBsp0Bzsr2TNj2KkdkHpPdY0kbuonGyCbx2v8lQU7nnqULI39Q2I3WoRZazJ7IIRBgELGkaRJj1Wd7xGCd1hyHqmBb6oJRES0H5KgmDD0n6llsYfDAUf3Jqp3dcTSQd0EXu/jYwFmQ2F0gDt8LIgtNYwDLewWqwOdDH0yDyQabBQCH4SOyumuTaYeHj5LKfMATnzK0OvdHI/IKCSouROcHutMmk8R2c5VXsLvoglRkEHcIL4Y+sqVzOg4VlPI1jtzhSyOD3ZbvlB339iy3/rpdU5/4AV38o25Fd7Fn8KTVP8AACu/lG3Ig9pEREBERAREQEREBERAREQEREHHvMR+D/xN/gdev1KVfM/Wf21P/hXfpK+mDmI/B/4m/wADr1+pSr5n6z+2p/8ACu/SUEKIiAiIgIiICIiAiIgIiICIiAiIgq0gYV3iEdlYiCpcXHCq0YPdW9lc077oJmtGFexnmrW9ld19OyC4uGMKzIacq0yDJVpkBGEE7KyNmxHYqKWVsxwBjKgwVdH0h4ygmZSPcAQdsqRtvef3xWRC8dI3HdZcIc8gNAOfNBpv3Nd81NHb87ELWG2+ocMhhwpWWmsI6gw479kGiGzuk3bn0WXT6UnnGQTutXjoalnwlu/1LV6KR8ADHgA7INrO0dM3OSdljSWF9OS0grkCKGrqDljcg/JaLeKeaCT4xj12Qbfp9OunI74PzW4LfYG07cuGfrUVNVeEBv27LNF6ia0dTgEFH26KLJ6Rsr4Kimp3dT2DZYlZfabBy4b+a011yglJw4EH5oN+WbWdttbgZImnG+63jS8aLFCwNdBHt3yFwbNPSuGC7f61AIqeTJaSR9aDnqXjfZG58OJg+wLTKrjHbqg/Rbj6lwu+npwCXEqHFI0n4j+VBzOzjHR05LowATt2WwNY6ruWpZzLS1TmgnYBbVeykfsHlapbre97AYQSD2QYZnvLY2tfVvONu6yqWSv8MvdM4kb91JWUksDeqQYB9Vqdns1TX0j5IckAEnZBBY7TcNVVQo2vc7JxjJXIlr5a7lVuYHxPIeM/lUnL9oG96m1SKK1xF8nigds75XfC2cAuINCaZ0lG8AAA/Ce6DqDaeVCooYxUyQOPn5rWYuB0drmbcH0/wxEHt6LuNqXQuqdP2nqrabpAbnt8lxVco66pt88TYxncIODtba3oLNahaqWIMkYMbbHOFxlYtUV13rzQGVxEuW4+tbo4l6Srpax8j43DJO+Ft7QGj5zqSEtJJDhtgoO83JFw5+4d5hv1XFkGQP3C9HJb/bInMaYgS8AjYbLqFyu6aukNrg62Et28l2lqrFO+SF3ScNAyg3BVUFv1BRiOrgbJGdsEZ2WlycOtJmgko47RCA8Hfp3ytYowaKmAI8vNXsuUb3hgAyTjug6S80PKjbbtSSXKht7W9YJ+Fq6Vx8qElHBU1jqc4YSd167caY66bTrmUrMjB8s74XU2ts97ktFZCI9znGyDpfoTRlPab+aF8ABa/HZc3V+k6OKkbUiEAtbnOPktvM0be7VqmStqYSGdZOSPmt93SqlNkdJ0/Cxu+PqQcOX3iLTaQqHRVhBY396d1orOZLTBPiR08Qcw98DuuMuNeo7ZJcJYHyAPGQQuFaettmXt6z8R23Qdkte8xFHqKi9yge0DpxgLhxurg2R0jnDBOVtGVlICXxuJHfKhdLGWkZOfJBv196otQsFM2MdRGCoDopnhkdIy7cLbWnamOiqfFPbOVvb9lcJLQCNtu6Dadx4dSPcXtGMrS6bh1Uy1rIS44LlyNNfWviBBGCsGC7+FVNqNsA5QalR8Jfcbe2oPfGd1j/cJtIMkD4fmtwVPFC3e4CkdM3Ibj7VtSq1nRTBwEjST80GPeZmMYY2d8LafVNLN4fWdzjZarW3OCpcS05z2Wmx5bUh47AgoNVh0zVOgE3WcFUFJLRPD3uPSD5rW6W6NNG2IYJUVTRVFXETG0nKDS66+U0sPgiMdWMLZdyt0lTKZG5wTlbkqbBWNlLiwrHkoKloLOjdBtkUvR8OOyhfUeB8DRhbjNgr3uLww4+paXWadrQ/JYRug06QTygOa44znut6aV1VT22mFFUtDnEYyVt+G0VYAb0K+Cw1Lqpry05BQbufSxVshqQ0YecrGqdCyXMl7Gkea1i30NS2kY3oOy1ulqpaSLpcADhBx5Nw4nZkZIIWDJoeoj7krkqouT3OOAFpNVWPJOcINiO0hMPMqCXTUkXfOy3nJVu9AtLrKpxBGyDar7PI0kErlbk2iLebThIw/vNaWj81XGuPZJyXHZcjcnLv+u54Tn11paf1uNB9HiIiAiIg65+0O/A24l/4nR/r1OvnjAycL6HPaHfgbcS/8To/16nXzxgkdkFSR2VFUgd1RBVhAO6uJyrW91djCCxERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAV7D5KxXN+fqgkBwrXbnZMhUyMoLSwhXNBx5/lVSRjurm4x5IIw1x9VT4gcb58lPj0VhY5ztvJBVlO5wzkqdk5pgR1b9lfFlrdwsap79/mg1G33J7X4ytVfSOryHE5W16Z7WSBzvVbwtUwcwFmCMIMKptIhaS4dhlaVM9sJORlbpqxLUgtYMlaFV2mcndp3Qab74B9EAH1UkcT6kbHGUltjoT8YOFJTTMgI+LZBj1FJPSjqDyAqQXGph/+SnHllZNfWR1DOlrgfIrTQ0D6Rx8kGsUeoJYHZkeSPLK1ml1tFDIHEdvktp09M2od0tcck7BZZs0nzQb6HE+m8MRlg2+SzrfxMt7HF7o2nAychcaG0vbu4kBSwWd7wQ0EkjCDnvT3ESz3YiEwMyduwW622K3agc2ligaC/wCS4J0bapKCZssjSACDuuY7Bq6ktFVHPI8AMwclBk3vlmfPCa6Nhw4ZWy6vl0qYAZegrsFRcZrVdKFtFDOwuGG4yFJJc5blH4cGCT2QdXKvh46wu6Xxn4Vk2WjglqxEYgQTjsuXNZaNvVSHTeGek5PZbSseiLpHXh5YTh2cYQckaA07b6RkM76YZyD2XZxuorTQaAmhjpmB4iIzgei4S0rp+uZSxNcw7Y8lyddLFUt0dM5zT/Yz+hB0/v8AV1dy1lUmOQhhlO32rclJo2odTe/knbfK2/JRyN1dOBnaQ9/rXLdvp6mWymFrckswEHEl3vYjc+jdv0jC471EIqlrvhGSdlyLqTSteyvmmdG7BJOVxvd8RVPuzvp9WEGzJrOfGL+k4zssCoiEJxjC37Lp+uMRmDDgtyNlsu+R+7S4k8ig0/pGc4VzjlvQBuoxUROGAd/qU0ERnkDWHJPZBiSUcgOerudlqNDauoB7hnG61AWOrkY13TsVmwUs1NHhzfJBpzqMDAxsCtdtk1PTQ/fGjOPMLAd1ZOyo6ZrG4Pog1Ga70rM4YPyLQa+5CR+WbBQ1U8YyCVpss8RPclBkVNY53Zy0t73OdkklTv6Xj4fRGUcjxlBbHUBo3GVVzRMeoeqOopAVJCwswCgsbRl24JCGN0JAO6zI5OkbqKcteSUHfX2Lf4Umqf4AVv8AKNuRPYtgjmk1T/ACt/lG3Ig9okREBERAREQEREBERAREQEREHHvMR+D/AMTf4HXr9SlXzP1n9tT/AOFd+kr6YOYj8H/ib/A69fqUq+Z+s/tqf/Cu/SUEKIiAiIgIiICIiAiIgIiICIiAiIgu6NlaRhXjJHZULfUIDWnvlXdPzVGo4kdkF3iADGVa+TPZWfWiBurwxxIBV0cJO6kETxIMDzQSso+poOPJXi3dRz/Os6GN3hjbyV4cWDsgx2URAG52WXT4gIcRnHyVpq4wN1DLWxHIyg1qPUFPC0RkDKkGsqWJvT0jOFtZzonOLgfzrEmjaSS0oN2u1jTlxcMb/JQHUgmkD2OwM+S2oWADOVk0wY0jJ80HIVs1lBTMAeB6FLlXR3geJGBgraMVOx7AQO63VZLXUSU4MYJCDRawugyN9tlpwjmqScOIwVrt4ttSyQhwI3Kw6OjmZnAQac6y1NRhoeeylhsM8Gzs9lrtJT1HXsxZ5oKuRw+BBolFpaercO/1KSrsEtvPSQRlb2sFmuD3jpjOfqVdQaduL3gujI39EHH77ZJKBjO4xjK1O26EmrWdWCc7rWRp2vjY0lnbzwtw2V09HGGvaMjZBtIcOZWnBB2WoUlqZagI3t7bdluaruxhBc7ABW3a66R1Ly4EYz3QYGoaeOogHQ0brk/hJpWCrtErZY8lzCO3yXGc5NVG1sQyc9lzNwtuLLZQeHOQCWoOYeVFtq4b69N0uFO1zBMHHqG2Mrv3f+afRcVPGI6SEEEHsPReb2mtT2+juhlq3gMLsg5W+rhq7TtyYxkEuTgDuEHOvG/misN5tjqakjjaekgEALq0/jNTQUtTK8tI3KxNdU1JPTGSLJGDuFxQ+1e9wT00TSXPBCDmDhDWUfHTVBsbIQ/MnTsPmu3Ft5IINNVtPc/dNiA4/Irq1yLaEu+mdfMuzonCMzB24OO69Xr7qZzqKBrWtGWBzvPfCDbPDey0WkKGOiEABZgHZcjxX2CSVsRaPi2ythWipbcarpbuSVuOsphbYDXSghsW53Qalq24MtlsdUdYbjJ+tcY2/iGHVQd44w138623xP476ajjfaZJ29YBGOoLh+k4r6bi65HSgAHPcIOxPEDihROsphJaSQc/M4XBdbxCoqa2VNUYmgNySsWn1tZuITza7NIHyn4cAg7q3V3CO/W/SlXPUU7w3pJJwdtsoOtfEfmItL7nJQwMYx4JGwWy3ccoqmgktXijMu35VxLxRpaO1axnincQ8POxPzW1qK42910jAdv1DzQTcU9CV9ylffGvcWyAu7riaHSdYSX5OGHddldX1kk+mmsYB09C4ip6eZsEr9sb+SDZs0UlMzwSdwFhCR7cuPZahdZ2vqXRg7jOVgOhcWlvqgtbcX7Ma/B7K5tdUB4cZT6rFbb52v6iDjusj3d7sN88YQaxS3+WRojL1lC5vm/2qHnLzgLQY6U02HvyB3U1NW07KxkhPYoNbdoK51LffBM7pdvjKjm0TXQxGXxHHHzW7qTVVO23tiDh2wFBLfoqmI07HAudsAg48mdNQSmJzjsgv/S7G+Vq9605cJHmr8M9B3yFtmS0zMJc4HA3Qa7TagfEA4u2G+MrXKTiJBSx9DsHC2G58LR4fVuNlA+Fu7m5Qb+n4iUspPwjf5LEOrIJZOvAxlbIZEXOwVnR0vS3IzhBviPW9FEwNc0ZWPLqikqjs1q2a9sI2Oc/Wr4TG07HzQbyZdqbAPSFG/UlNTvB6Rse60SGRhb37fNafcHxdRGfNBviLiBTxt6cjssGt122Vw6HAA+i2T949T+VXNp45PiblBul+rzjPWN1YdTeMD8e5+a2w6DyV8cQaPMINe+7ZJ3cVjz3EyZOe605rBnfdVcAMIJHTk5K5Q5Nz/12vCQ+utbR+txricuA7lcr8mu/NnwjP/dpaP1uNB9ISIiAiIg65+0O/A24l/4nR/r1OvnjHdfQ57Q78DbiX/idH+vU6+ePCC49larj5q1BVn0lc4+Stb3VTv3QWoiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIKhpKEEK9rSqSAjyQWZPqmUTBQMqocRsgaT5IWnOEErHY3wsqNre5CxGgflWXGSWoLsK00hl3x2VynjmYzIcQMoMQ21wGxK1GgrDQx9BPlhRvqmYwCCSrG0z6j4hkhBqtPfWROzIAQfValFcIa3DgAtqvoJj3ytYtjfAZh22O6DVpLYy4N6Wjf5LBqNHStaekEZWq2+4RxPwCN/Nasah9Rs3Bz2QbJp9HTeL8eSFr1Jw2fXDAYd1r1NQVkpBYzO63zp62XBrR97OfXHdBx3R8K30J8R7SQN91kVWlWQNL/DOy5Xq7fcXRnqZstu3K11xhdlmO6DjM2ZlTIYmswRt2VwtrLeQ97OxW46a11MdUcjH1hUutqnnYd9/kg27LqGOAeFG0AjbZX0za2/u93ieQXjGR5KEaRrKufDAd9zsVuzS+lK+3VjJHA4BHkg1zQXCi80krK6eoeWlwdgnbC5zslrFtcyeZ2Q1o7lbVpNURWW3sZVEDpwe6iquLNkFO5rpRsMdwg5moKi136IUfhsJ2BWRVaNt1siNU2nAxv2XEvDrifZX3DEcoJ6txkLli9a6t9Xby1jwct/mQY9n1JRsqhShoHScDZcp1lZS12kJY2gHMZ8vkuuFsu1ILoZeru7P51zFSanoHafdCHjJZj8yDrje7O2h1NUVIbgdZOftWrU+tILdEGOxgbH8ik1KwVN0mka4YLj5LaV3tjWwOkcTjugm1NrugqWSNa1uSPL6lwJeq73nUIka74S/OPtW77xWUrZnxAkEDdbArSG3ESZ26soOWYqykfZmN6AXFv8y4r1VZXV9STE3HxZGFuumv1IKJkXUM4AUbaGa4SiSDcZQbCj0bO0h7g4j5rUaOxe5yCR7e3qt7SWeuZFks2AWgXGoNKC2XA9UGTHcKZkAi6RkLQrreImEhuB5dlH90KbqOTv3WhXWphlkwHY3QTOuuXF2e6x5qx0h2cVg9Ufm7ClZEX4LTlBSaN9R2J3Rtpe4ZJJWRG3oxkrJZVsYMFBiR23w9zk7+qlMzIG4wO6lfXRAHJWm1M8cj9j3QSy1zN8ALGdVhx2GFR0PVuFEad2UE7JTJsFV2Q7cqOIeCfjUhc2T4h2wg79exdcDzR6oH/cBW/yjbkT2LgA5pNU/wAAK3+UbciD2hREQEREBERAREQEREBERAREQce8xH4P/E3+B16/UpV8z9Z/bU/+Fd+kr6YOYj8H/ib/AAOvX6lKvmfrP7an/wAK79JQQoiICIiAiIgIiICIiAiIgIiICIiCVrgFR7hsgAIx5qngyHsMoKBwBVesKvu8v9yqeDIDjpQWE5Kkij63gZ2yngSeilhp5+sEA4yg1CCmb0DI3x6LNZQtLfEwNlFB1NYA4bhZzapjYS3bOEGmTV7aZ3R6dliSXLrGAorgA+YuCxOk+iCZ0znknq8vJUEUjt+pWMBCyonBox6oIhE8DcqgBGxWQ9wOMKB2c7IKEZV0bHZ6h5KgBx2UsewOUGTHX+AwArkTSF/gbSdD2A7d1xa/oOC71W8tMVFJFTgPcM4Qbhu1bBUyOIYPyLatZeY6F5ZgblaxJNFLI7o7H0W1L5b6iScSRsJH1INQg1fFFv07rWaDW0D9ywbBbBdQVDN3tIWZRU8bdnE5PdBypaOJtJQyElrQc4WoV/E+hrSCGt29FxQKCOQgjOVkxWzpHn9qDkCXiFQvGA1u2y0up1jG92YyBlbVdbQO2d1jysiiJD3EINeu2pX1EXSH+WFplFXyOaep/V9q06oEUjMRvyVPQU0oaTug1qjvHucge85HzXK+kKqS6UwkhdgAZ2XC7bZUVxEcAJOcdlzVw2tlTbqARzsIJb5oNWFtrLlMaeGUhwOMrMZa7pYnCWaoeQN+6yrbP9yq91RUjDc53UuotYWiqhID27DGMhBpF6104we6vfkjZaTpvUsb73Ex+C0uGR9q29dqy3185FO8E+gVdO0ng3mGWQkAOCDvlwi1hbNPWuKrhgayTAPUBhcywccX3FzKczA7dPddXNHXyyRWOKKeQA4Axn5LemnbvYJ7hHDBKC9xAAHqg7mcHrsbrWCd7uoFwO65O4kuMejq90ezgw4I+orh3gfTyWuNlbO0iHY5+S5R4iX+21Oj66OGYOc+MgD0OCg8iePWqb/Qa5qWtuDwwPOwd81xkeIN7kp5Ym178kEA5W/OYyhlj1lU1UxIjLie3zXC0FXb5nyRRuHWcjYoOw/KBru42HWgrbrVGWMyB2HOyO69D9f8drHetLSWqKBgdOwN8vTC8yeX61SQ3kVFS0iIvBH1ZXbm7T2YUkTo5AXBo+xB085kOHcpvc+oYQ5rXkuC6626gqW1bqsyHETs913F5hrl71bHwwYx0nt3XU2hgqBHODkg5/Sg3nY9Q/sgpxaHHJHwrcFZoBtHaJJejGWk5WwdCugtd18efYdWVzJddaWqrtBpI5GmQtwBkeiDqvqKhdQ3mRpyAHFYbalo3ONluTW9nuAub69zT4RJOfktnmWI5aDv2KDUTcI3jpDRnt2VjalrXB2BscrAawB2fJSZaRjKDMrKxtTEI2DBHmFpDaKUP6w45z6rMYA05UniD5IKROqY2hviHAU9JVTU9U2d7yWt3KiL2nzVHuYWkeqDfDtbUVZQij6Gl2MH5LSAyKrl93DBl+2ceq27RMijlDvM/Na9b6uCKujlJ2BHmg1CfhjIYffQDgjK2tcrSaBxjI7HC5jk1RSSWsQMeCenHdcaaho6use6aNpLckoNrNpww5wrnVrY/vfmr5XhjvCd9IbLAkhe+Q4GxKCR0TpHF+ThSR0zgOrfZSsgl8MDH5lNH1NYWu2KDEkqXQ/DnsjKV9b8YJ2UdXE5zupqz7UTFGQdsIMM2x4JGTsVNFAYW4Pks50o6j2UEsjTkIMdxGTn1VMhHbnZROyHIJWuGeyq4dX2q1uNjlXCRoGCgifCSuWOTUY5s+EYPlrS0frca4u8Znn2XKfJwc82vCQjsda2j9bjQfSAiIgIiIOuftDvwNuJf+J0f69Tr55G919DftDvwNuJf+J0f69Tr5484QVIxuqK49lagqz6SvcrGd91ecbYQRoiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgINkRBK2YAK17w7J9VaGuPkhaQgopomghQqaJwA3IQSsYDlRygMKvbI1ud1DK4yHI3CCocAQVlROBYCsPpJ8lNGcNQXPqA091WNrpviaTgLGkOT+VZVHKyNh6jugsljdG3OVmUd1ZA3pd5DCxquRsjMN81jtgc4E7oNaju0Up6cdysyJ3ijDdshbep4S14JBWt0k8ceM+QQTSeJT/ABdWMbrOoNQiADrdsFg1VRFJHjzPbdYLGxnclBvmj4gQUoHUBgFbkoOM1HSgH4RgeS4glpopBhuSfPBSOip2tIeCPtQc3jjfR1IEYx6LPoNd0t3kFOGgl3ouCKSmo45ARnvut76VrbfSVzJJHDA9UHPmnNExXlrZxET1fkW5peEEZaeqnPr2WLw+4maXttJFHUTRgjHmFyMeLOlatvRBNGSR2yEGl8OeAVNeavofSg4+S3Lr7gVSaXoXVTaUN6RnOMLfHBrWtG+tMkPQQXLdHGa6zXWzvhhYCXNOMfUg83eLd0qKaploqeQs6QR3wuCam5XMucySqfgncArnfjTpm6U13qKiaMhpyc/Jdf6tjmyOJPY4Qa5pLUVwtNwE0dS8NyCcu81zBaeJlbWyMp3VRO2MdS4HtzomzDxTgZW9dPVttpq5kkjxgd0HYvTbaitDKgOO5HZct6PsNbdauOg8RxDsDC4c0VrXT7aaKMSMyMDcrnzhhqGhqb5AKYguyMYKDkKfltdJbG15iJLxnJC2LxF4HfcbSVRcfCILGk5x2wF23kudbHpiNzxhgYFw9xm1TBLoKsow9vW6MjH2IPJ/U15kp9UVNF1nDJCzuoZ4zLCZPPyUWsbRWHWlbIGktfOXAqVpxCaYP+M7Yxug0mGepFUyPxDgux3+a7LcONHMn02bpNHnpZ1Zx8l12ZpG8vqI6lsbujqDuxXa3h1dordoaShqyA8xkbnzwg4sv2rKSnrpreGAFhIC4u1HcjW1RjjdgEnst0aot757/UzxA/E4kLZ1baquOsEhacAoIvuLM5gkDjurqbSstXMAcnK1iOcCFsZxkYWRbr7SU1WyOR+HZ7ILouF75IRJ0n1UMmkPcPvZaSVyJDqKEUYLXDGP5ltu6X+kc/LnAlBtWXThGSG+S0mpscjHZ3W7Zb5RnPxDt6rSau6UzyCCEG3Ki0SkYGVgPoXROGSditwy1rHk4IWlzydTidvVBAAGDfyCrlp3wkj2jvhWeLGgpND4o+HZRCMxjpKyWSs3AKhme0u7oO/PsW/wpNU/wArf5RtyKnsW3A80mqf4AVv8o25EHtGiIgIiICIiAiIgIiICIiAiIg495iPwf+Jv8Dr1+pSr5n6z+2p/8K79JX0wcxH4P/E3+B16/UpV8z9Z/bU/+Fd+koIUREBERAREQEREBERAREQEREBERBfG7pcM9lmMlYSMrBZjO6na5oPfsg1ASMLceqicY89lY17cDJVHPZnugnAjx2WTFIwAbLDbI3bJV4kaBjOUGRJUg7DZYU1Q4ZwTv81c+Ru+6w5nDJQVfJnJJyrA4eaAZGyt6XeiC/rBTrx2VA0jcp0FyC4POVcHAjKjLSNgrmgjY57ILi8BPEA7K3pyfNC052CC18ZduFkU9RNARh5CMhkcM42R0JBHUEGrU1zezDnOzn5rWaetgqWgOaCfqW12Na1ozt9q1W3zxRjc9kGpzUkNT8LWDJ7LVbRoU1o62g77rTKavgY7cjv6rfumtSUVPGA5w29Sggt/DkucepmSFLW6KbSEN6Fuii1hbo3kl7dz5kLEvGr7bI8Ykb+ZBteXSzWMDunut1aX4Cv1ZCJmRk5Gey0yp1RbnxNDXt7+q5+4J3xxom+AAQWoOMYuV2WKQtdETjdbE1toAaSmNOY8YOMLuVLqGSGolMjQBgrrVxvu0VXcC7I+kUHF1CyCyYq5mZHfda/TcTadmBFgBvpstt3mqjkt4DiMYW0o57fF1db8FBybe+IYulOYYHgOxjI2Wz3T3GVjy6cnOT3Wi01XQvkHRLnJ23W5Lfa57jiOlcSXDbGUGp8ObBU3e5hszi4FwGMrmit4dmijbVNZggZW1uFGg9RW64Crmjd4fVkHpPZc4X6nrTbSSzYN3/Ig2jw/slw1DcBa45nYD8YXPmnuDNysV2pbhLI8tBBwT3XEPA2rEGriMAuEg2+1dv7nfDDDA+YANDRk9kG5rhxJi0xpRlDC4MlazBI+pcdUnG6su0htMlWXCV2MZWj631LZrlSGnieDJjdccadgp6S+xVL8gB4P50GTzO8NGVGnH39kWHPjLur7F0bsOnKplxlqHPJaxx8/LK9HePV7i1Bw+bbbcQZBF04G57LpLa+H+paUVNRNE/oJJzg9soOV+BhiuczKCJoDwQPtXYy+aEq6S2ioc84Dc/mXWnl7rYLVqQQzkB4kA3+tdxNTalpp7MyFjgSWAd/kg6bcXw9rn0shJwCFws20xx0U04bjGT2XOPGWknbVSVLmkMOTlcOe9RyWyeFvcghBxbU3pxuTqWF3SQcd91kQVdwjuMTn1DiwOGQStFraKWivb6mbIaXZ3WZFdqOaujja4E5AQb61jX0tXp4RCMdfTu7G5OFwm2geJXEuOMn9K5fv9FKbKJmg4LM9lxh4cgc/PkTlBAYiGADurBEc53WQ4+XmrUEMp6G4ysUzO3+I9/VZNT2KwS057n8qDIimc7z8lMQfXusaFpad1kgkkAIKsY9pyCpAZWnqDiD65VW/MKqDOt1VVZAdKSM9iVv22sgnt5Y9gJI81x5RPDXDJ81vmy3GmbCGEjqI23QbVu2nT76+VrcNz/OtJlhbTvLSOy33do5T1SAbHPktjXOQCU9RAQSRyRhvbyWNUPGfhRkjC0Yd3UcmHEn5oLSWuG47K11QIvhacJuoZGgu890EhqCfM7qN0jnEb7K9sGW5Csc0tIHzQSxnIwop3AOx81NHjGVBOB1IKA/WqEnyVYwCN1V+AMZQQOc71XMHJnn9tjwi/hpaP1uNcPP7rmLkz/Cy4Rfw0tH63Gg+kJERAREQdc/aHfgbcS/8To/16nXzxjuvoc9od+BtxL/xOj/XqdfPGBlBceytVT2VEFWgkq4ghWs77q842wgjREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAQbIiCVsjQMEI5wd2CiVwxsMoKdJVQ0+qrgq9oAHbdBaWOOO6q0hmxV6ikPxbFBc54wCqeJgYCsJGFRAJyqguxsqKRkT3/RB2QUa4tILvVTiqY0YA/MoZIZIxlysa0uQZsc7X7D1U3x52KwoGlrt1nNmYCCe/1oLX9bBkuKrG8v2CrI9sowO4VsXTFu5BKZzTDqf9mVC+s8fJaceqtrpGSNwO4WPTxuyThBlRve3fqKvfV1LBlkpB8t1YGuA3Cte3qCCn3buzDhtdK3Ho7C1aza2vlBN1vuEhHzctAdA4u2VRT/AN3kIOw/DbmMqtMFniVZyDncrlKq5vI7iGsmnDgO+Tsul1NFSZHiOPzWrUsdtDx0vJ8u6DsVqrXlv4isMcUbSXgjPquPbnwgPuT6oM75d2WHo66261va9zgADnuuSazX9qqLV7vHIwuLcd0HW282R9tqnwtyOk4WnNFQ36LyMfNbx1TBNVV0lQ36JJK2s+CSPJd2Qalp2uudLUNc2peGgjYkrsXwJ4h1VPrSkp5qgkdQBGfmF10tTo3yAE75C5W4LUjJuIFGxhOS8Y3+aD14priy7aCpntAy6MfoXDHETRFXXWOZ/USzBPdcyaS07Xnh/RyMYS0xjG3yC0bV1hurtOTtDCMtKDzyunCqCv1DKwU4JLyM47rVf2q7/dTfPAPQ0dWMFb/uNHXWnUcs1RsGvLjkdlu2p436aotOPs01TEJi0txkZzhB1rq6W3WyZ1qfTtDoj09vRaFc73LStMED+lhOMBa/fqeS43ue4QHLJXEtIG260G66FvlREa9kbzENycHGEG2qytjBdI8ZJGSStl3zVFOxxgDBncLcN0qIYXvpJHYkaCDuuO7xQSz1ZdECeooL33gxuD+rZ61Wi0pVXWL7qRvIAGditFGnq6RrCGEgd9lv+y3mntNkNFUEB5bjH2INsOvtRTE0JlOWbbrQLhcamSXaV3f1UtY+Oa4SzNOQ5xIWnVQxIcFANZUnvKVT3qc93kqLpPorxE4oJWVTwd3FXOq+rzWMWuHcKmMoJJJS7bKs6neqqInHyV3gP9EFrXlvmUc8uOVXwX+bSPrVrhgoO/3sVT/10+qcn/sf138o25E9ip+FNqn/AIv67+UbciD2pREQEREBERAREQEREBERAREQce8xH4P/ABN/gdev1KVfM/Wf21P/AIV36Svpg5iPwf8Aib/A69fqUq+Z+s/tqf8Awrv0lBCiIgIiICIiAiIgIiICIiAiIgIiIKkY3VMn1VcO+aogva8nAV2HHdRDuMKdgONwUDJA7oZS3z80PVkqOQEYQC8n7VTpLtyqNByr8OHkgkb6d8q7A9Fa0gd+6r1N9UFSPQBGgDujXNzuVf1RoLDgnsiuLmYVGuZjfBQXNb5o5oG/orvFYqEscCUFfe2xtDcDKkZM2bcALT5AerYFZVIOkZQTOORgKN0r4zgOwryQO5UEpBcgk95mGD1Hv5rOpa6qABbK4LTWgOxv6LLgw1oQaiLlXZ2nKuaa6pwTO4/WsOMs7krUqSpgjZgkZQWxU9Y4hvik4Oe65L0dxnfoeAUr5dwMYK2BBXU7XEuwFot5p4qubxIic/LzQc5VfMea0lzJ8F2y0eoq5dak1HWXZ328lwvHbmggv6ic7rljQl8ttrpw2Z7RgYQQ33TcrKcR4PotZ0XwAqdW0rqhgJGCQcql/wBRW6vZ000gyXYC5+4C2zUFZaT7iwuDmbY+pBxnovlRqbndfdSwnDsHddj9H8oIs9XTPmpCRkZBC1bQbb7pK/urLzGWxB/Vlwxtlc/UvHzRkE1OaqeECPAdkhBxLxb0da+FWkRWtoRG8Rk5x6BdUWcdae8MnoQQSCWrt5ziccuHWrNDGgttTC6bwi3AcM5x8l5p6bmtMFwnke4fFISPyoOYLFrw6PuRvfX0gnqyt71PNc6+MFEypGcdI3XCOpqu31Ns8NrhjGNitmafp7bBcQ9xOA4efzQdq7Dqivu0/vklQXMJzuVu1t5PisDHYOy400LV0dXRMgpSOrYLe0Voqveo2jJLseSDkjT0U90aGVbzIw42JyFu6+6UtMGk6mWOhYHeGd+nfstA0haq6zU7auuYREcHJ9Fvi/aotNRo+pijewuEZ8x6IOj9uvMun+IM3hOLGiU4H2rmC5cV6htO15nJDGjP5FwZdIxVcQJ3s3aZDjH1rd14tzWW4vcw4DT+hBp3EHitDqCN1E0guxhcY01TJEXBxyHFRXKe3fdN0cbvjBVT0Nw53ZBpOq6FlXCTEwBxWyKDTtRTVTatzzhjs4z81yFcHivjMNIAXDbC0Z+nr3DE+R8bgzHfyAQbjt94jvlC204BIGFfW8MxBbX1gjH0SVominRRXIR5++h2/wBeVzBd3VTtPPbjuz+ZB1YukppLjJT9gxxao/efmr9SU07L1UdbTu/KwQ05GPtQagAJcBX+4t7kFR0mdsLUhnpzhBp7qXoO32KwfCd1lTl2TssRxySSgkMm3dWmTuMqxWu7oJBI4HKzrbXTCtjYHnGQFp47KS3ENuDC7sHboOVJWNmtbT05JauOb3bz4rng7Z7rkhtVAbQ0NIJ6Vse7wzyFzmt2yg274TmAD+dACNipXRyh3S4bqxzSDugoFiTP6ZMfNZzQO60+rY4yZ8kGVHUNDRsNwoppA92VZGzLQCVZLkHbKCeJ+RjKin+kr4G7Z3Vk/wBJBRpyFbK4hSR4IxsrZmgg4/MggB37LmTkz/Cy4Rfw0tH63GuGgCD9S5l5M/wsuEX8NLR+txoPpCREQEREHXP2h34G3Ev/ABOj/XqdfPGO6+hz2h34G3Ev/E6P9ep188Y7oKkHuqK49lagq36QVVRvdHZGyCiIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiArmtVqvagkB2Az2Vjn4OxVHE+Ss7oL3PJ81aAXHKorowc9kEkcJJ37bLIFHlXRj4dgslhcW4wg02WnLRkK+nnbCNxlZM1FUyk9LSVgTQSwHEjcIJ5qhs46QPqV9NS5HrlYcYJcMZzlanTzsiAbJsggqGeEMrEJc7ss64TxyN+EjOFixUk8ozG0lBNQsd4m/n81NWNIGyUlvq2OyWkD6llS2WuqWgRMJKDRA4h25WQyqazyWU/Td1Z3pzhSwaSvVQMx0pIQYrKlsp6e26yAzOyy4tCajlI8OjcT9RWWOH+qWHL6d7fsKDEpKJsp7d1qLNPeOenp7qSm05d6Eg1EZGD5rXKKOZhBwNkGkR8P5J8OYCr/wBgU9NhxJwDvut7W+omIDW7lZFXFWSRnDc4KDj2ew1UY6GPIx81HbbPc3VzGvncW5GxK3TU264yOJYw/YordbLnFXMe5h6QRlBrlRpVslva57MktBytqXPSrI4Xu6MY+S5TbR11fQNipWkvxjb1WiXPQWq56d4jhfk+gKDj3S2k3Vlc2MHOXAd/mu0nArgv4WrKS7PjJDCHDb6lwvofhnrWmubJHwyYDgex9V3M4MWm/W+tg97jIaAM5+xB3b03eaSj0ZTW50XxRtAP5FgXC6UVwpjQeED17YwsG2zUz7NGzPx4Gd1DDRv95Ewbt1ZKDiziHwVjuYnrYIMF7Sc9vJdLdecuuohqgVEdTIIhJnGT6r0r1PeRBQOjIHwsOfyLq1rfV0H3a8HLc9fbPzQceWvhMKe1QRSw5kaBkla/frRbLPoueB1K3r6CM437LesNe99C2RrBgt2K23xBcH6Vnkc3HwnP5Cg8/wDVdPIdV1LmHDOsnGVpMpjp3GR7Qcea13VFdTDU9SwkZDytvXSrpjGQCN0GXHqqihYYiwbDbZbSvVykq6ovjkIZ6A9lYTCXkkrDqOjrIYchBcyYhoyclRyO635T4cbOVoGSgmjAwFkNxjcKGIsAHUpvEjxhBjz9zhQtODlTTEH8igwUE7ZmgDICk97ZjGPzLEwVXpd6IMh1Swjssd7uoqoje7sFRzS04KDv97FT8KbVP/F/XfyjbkT2Kn4U2qf+L+u/lG3Ig9qUREBERAREQEREBERAREQEREHHvMR+D/xN/gdev1KVfM/Wf21P/hXfpK+mDmI/B/4m/wADr1+pSr5n6z+2p/8ACu/SUEKIiAiIgIiICIiAiIgIiICIiAg2REEvU3HzUROSrj2VqCrSAd1OHjCgb33V+Qgv8UZVHkOOU+HuhxnZBRjcKbIxjG6jb5ZUrenBygxpNzkdlZk+qvkaerYK3pPogYcmXDzVwacBULSUFA4kjJKuGfIqgYRuQr2g4QWdLhvlXNLt91Tc7K4Ndjsgr1DzUjJhjp7KB7HDfuCVWMEd0GScvGxRsDnearHuFM3IQWMg6VJ0uBVfEDfpJ7xD5oBDlafEH74/lV3isPYqx7snLSgoDNn6ZVPfDEQHnKdYZ3PdXw2O43F3iUsJeD6IKx17HkeW+Fua06brLrCZIXkDGVt4aUvAcM05B+orkjRrprVSmOqGCGkboNT4Y8K6y/3ZtLM4vDXg4zldtdM6wt/BeOChqImnYA5GVwpwOuE0uoT7tuS7yW+uMElD79AbrgHIKDXuNPH6jrrE6a3NbE90ZwWjG66e3Hilqe4mfwbrICXHpw8+q5R4myWOXT4bTkE9GPzLrzSwwiWXpPcnCDIrb1qu4yE111lkafIvJGFjRGphkD2yEHOe6ynxODfhUQhlJQar91Z6iAQGQ+ndQAy0rXzhxBAJUEMJgw5+QpqqaKalljY7J6CPzIOe+UF8+sdWMtMzy8eIBgr0SvvAqCyw0t1MGAGhy87+QqkqqbiMyrwQ0yt8vRetevrxJJp+BrnDpEYzn6kHVLi5r6mtFqNopmBj2NIyNjlcBRcRK+S31NO+pJD8jGVyFxzrLVJVy5cOs5XBVNLQGKXDhnJ80GhG4mlvj65/cuJytcuWt21tC6kB3c3C2vcqikZUuMhw3Oywvf7W7aN4LsoNu11jqGXF1eXEgnPdTzyOdSOwewwtRuLpnMJH0Fpkhb7o93lhBt6y3/7m3QumOQHZ3W+K3iJQVVIaNsLQXN6ey4uM9IbiRIQN8LUuu3Nma4EZygzqN7rTcTdQcNLs4W6BxajrsW3qBzhq2lc62mfR9BIIwtuWv3CO7Ry526wd0G/tT6OjqqU3ZseOsdS2H9xMMe70XL13u8dVptkFOQT042+pccOpK0U8jnA43yg2o2XwJzHjsVmCtGB3Wnz4bWEPO+VN1R4G5QSSVIeFjkjKOLR2KjaSXbIJ+kgKw7nKm7sUDtjsguBCRnolDxthW98KvSSg3BS354iERfsNsZWqQ1TJ4PiAJPyW0qeNodnK3JQyQiHBwSgw5qdj5S4BaTX4ifjC1stkdK4t7LRLo1zZD1eqCMPBAKr4LZdz6KFssbQOoqeKZnTt5IIvd8bAqGSLBOQsl1THnuopHtkOW9kFrNsqGoaXFZABx2UMv0kEcbD2BVz2n1QZ7hCSchBjkEFcxcmf4WXCL+Glo/W41w+8HK5h5M/wsuEX8NLR+txoPpCREQEREHXP2h34G3Ev/E6P9ep188je6+hv2h34G3Ev/E6P9ep188je6A4gqiqQqIKs+kqv7qjPpK5wG2UFiIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAr2uHZWKrfpBBL05xnCtMe6vHYISR2QRFmAqtcGn6k3Ktd3QTsqenA32WTFWguGx7LBDRuVVnS12UG5aO4RMALmjfbcKyso2XB2YwBnbZYFPLAGgOI/StYo45p8GmyR6/JBotTazbx4jt8LDcHVGXNOMfNbgvlBXPi+JpwFpVPp27VDS6GncR5oLrNaHXKUNJzgrd0FkioMBzAtP0tpm+xVZcIHNA8lue5UldC8e8MIIQaZMIAMCMLU7TU00YHXGD9YWlT4A379ypaOspYSDKRgd8lBuC4XagbEMQjb5K+16wttM3pfAw+WcLQbhc7VLF0scCcYwtKjfRk56h38kHIzOKFmtvxvpmEfUoqnjZY5WkNp2A/UuMrvFSSxYBzlaAaGENO57oOUanXtDe3FkUbR5DZYprmR5Ab3O2y2bYGU9PL1OPnvlbmlrqLIPU3CDLOpRbz1E4xur28RIcbgHy7Lbd2q6CZvSCPktDjip35a3uUHI8PECldgOaPyKaTXtH0HDGg+vouPae2OcQWg7rPdZpWxOc5hxhBzFw111BLWtErQ8dQ2Iz5rsdpe+2a5VUVO+iYQ4Adl054bVdstlc01jwAHAkE+S7MaF17oylukD5J48DG2yDsRBZbDRULattuYDjIPSoLTrehhvDKGCJrSSAMbeanm4m6Kq7GyOOeMnp8iPRce2u92GfVkUkT24LwfzoO2tojm+5UdbvhzQ75LIfqP3aMNLd+3ZQ2nV1gptNQNmezAaM5I9FpNXxG0JLilEkfik4G47oNta+1XKIJy15A6TsukPFfW9Tarq+4eMQGPJ/Ou4PECSjmppqiMDwnMJBB8l0T461lrkbNB1jJcRhBWPm2NLTtozMT0YGVbeOaBt9s0lu8YEvGF1iqLfmrcyMkMJyPqWp0VvooW5cSX/WgzLo+a43aW4hxxISfyqBtslqpQwknPzU7fhOB2CyKKvhgqA5xGAfNBINEPMYkwdwtNl0k8T9GCt+x6otraUMc5oOFpUlaKmqE8WC0HP50Gjs4evfF4mD2ytFuNg9wcWOGPLdcnw36nigEbiMgeq2bqGkrrlMZKZpIJzkeSDaElEQCQcLFe10Zxnutcks1zGeph2C06a11gecsJKDDDidiq9QA3UzrfVDvGVGaWYd2oLesK4StGxH5lQ08o7tVhY4HBCDIiqGtzt+ZRTvD39QGFa1u+6o4DuEHf72Kn4U2qf+L+u/lG3InsVPwptU/8X9d/KNuRB7UoiICIiAiIgIiICIiAiIgIiIOPeYj8H/ib/A69fqUq+Z+s/tqf/Cu/SV9MHMR+D/xN/gdev1KVfM/Wf21P/hXfpKCFERAREQEREBERAREQEREBERAREQAN1f4RxlWA7qdrhjyQWOZgFR5UshONiokAZ7BSMafMqNSxnKCpGN1b4hG2VfIfRQYJ3QS9YIQOHmrMHGACgBxugv8AEGcK5r24+1QHz+tXsPllBKXgoHAKIO3wSqknbGUF47q/I7KIE53UjfpBBOGNc3ssabDD0rJZNGwblY1Q7rflv5kF0U/T3xnOyyWyhwysBrC7GFPG9kY6XHBQXVMuxaEo6F9VuD23wopfvpHRuMrV7JSVIGQCMoMu36fdPhpBPYLctJoEzMBLSdvRWWcSxOHWMb53W9KPUtDRMDJ3tBxug2fJw5dJsGEY3WvWWOj01GIqiEEgY3C1n9l9s3IkaAVty9XihrpMtcDv6oMq6alt1MDKIWjPyW16vU7awPdAQ0b9tlj3+qpH02Bv9q0O3Ujpo3+FnfOPRBzxyyXzp1KRKc4ctR5sNVTQ3SBtPIWbgbbLaHLmWWrUplrdm9WVLzU3KhuN3hfSOBw7y38kHG8up6u70DaeSckAYIJWnw0TondR8zlafaHxMaMnzWsuqo8DsguyA0ZRsrATkdlE6eIhWAh5Iae4Qa1areL28QsHc4W5oeGUkJ6ntJBbvlaVoUOo6wSybDOd1yybxDVMEMLx1luPtQcs8lWi44dUsayLDhKO31r0k4gaSkbpRr+kjEWc/YuhfJ7M7TepWXG7/DD4gdk7DC9BtdcXtGV2l3UtNURukdHgDI2OMYCDzF462qqiv0zPEOASMZ+a4gZb54qCeo6z8IJXPHHtslRe566L+xEkhcNum67HVD1B2CDiQ3CW9Xl1tY85B6cBa8NC1dHIyZ73EHfdbSsFbTUGspJZSB8ZP51zBWaqts7I4mPaScDZBsvVNU2127w8YIauPW6rEkEkHUMnbC3zxGHi0Rl/ekZBXDVOIzK7B8/50GQ+lmnqjM15ALs91kGmqG/GXk43UkDukD0U0kzTGRsg0+eqmlaYuo7bd1ix008cjXh5BBz3U7o/vpcB3Kvy7JyPJBvPTNylqI208ryQNsLeNbRQi1vd4YyQT+Zcf6QDTUDHfK5Lr4ZRaXEg46f5kHBV4b0XR4Gwyoep6yLxg3V+/ZyhLSCNkFCHDuVWNwDgCrnEOGB5BWNY8Pz5FBlPeGsWI6pw7Hl5qec/e8LTiMvx6oM6OUEgqUzBoA9VjMjd0jCuMUhHURsEEoqN8jZTU1zeJhH1YHZYLnNbtndRwY95YfLqQcj26lbLTiUjORnOFtvUdOGyHAwtz2i4U8dA1rzvhaFqCGSsd1wbj5INqywuG++VLA1wjP1K+WCYHpPl8lRj2saWv74QYkjX9Z+ZWRTMIG5Vr43OcXN7ZUkJLfpBBOBnKhlZ8WcqZrhvurJBk7ILAB2Vkgwpg3zwopWuJxjughe7C5e5M/wsuEX8NLR+txriJ0LiFy7yaZHNlwiHprS0D/lcaD6QkREBERB1z9od+BtxL/xOj/XqdfPGDg5X0Oe0O/A24l/4nR/r1OvnjAyguPZWq49lagqz6SvcrG91cT6oLEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQE7og2IKC8RnCBpA38lKxwIxkKjy3GxQRl4Cr4oVmHeYVRG4+SC4SAHKrjr+IdlaYnBXR7DB2QW9BPZULHDsrw4BXBpdg4QR9Mmdsrc1ivMdui++YOAtuOcG91mU1BNWxkxAnKDkGx1lNqaTwAwHG2y3MZ7fpxwp5KduSMdlszh1G6yVpmrBhvfda1rC/Wusq2vY4YHzQarVa+tlnaZRCwE/JbSvHEGC7uLmDB9Oy0LUtTSVkAELgSN9vVbZhHS4l23kg3NLevFzv5YWl1lVLKD0PI/MoY+lww07o9hHbKCGGOqc8OLzgeru6yXTSxfviEpw4HJGFHWAkHZBa+vectLiQrWVJccE/nWKI3lXxwyFwx3QZTp3wjqacZVpqqiUbSn8qpPTTBgLmnA+Siia5u57IJoqaeodu8n7VqVLbXxvHV2Kst0b5XARjfK1oUlTHhz84G/ZBJFVR0UY6mg4U0epIKpwpQ0ZOyxnWWtu4EVG0ud22CkoOFmrY6hlQaN4Y0gk4PZBr1v0LV3XE9M4t6txhazTcNb/RPE7KqQEb9ytx6XlkskDIazYtxsVuifV1tMJaXs2GO6Db1lpdQxdNPJXyEAgY6iuQ9K2+409wiq5JnnBByVs6g1LbBUB5e3c/zrftk1dZnPZF1tz9aDeGteJ9ytNk93jrHN6WY+l8l1jquOOoKfVbJjdX9DZNx1/NcscTamiqbc+RmCC1dULxBAL34rs9Ifk/lQd0hx+kvenIqV1Rl5YATnfsuKta6KqtTUcl1LiRu7utr6YrrUaKCNjh17Df1XMkNJXS6TfJA0lhZ3x8kHU2uoRSV8lKW/EzI7ei00wv97DWuIBPqtw6hjeNQztd9IPP5Voxa6Ota5wwAUGtfc/NK1w8wtuXKnkYT0uIOe+Vu736D3RrQR2W27m1z8vA2zlBoTm1eceI4/atVob26hhML3ZOO5WD4rASD5LEmAMmAUGpyXqofJ1NkIHfutwWrWdPSQiOYBxPmVs3wSW5HZW+G0fSJQb3l1fSSk4Y0ZWHJeaaY56QtssjZlZLWtaEGqyXCB2waFQeFIMhgWmhzGkZWZFVwtGMhBKaZj8gNH2BYNRQgPOAO61BlZCPMbqKeUPdlp+aDSamAMacDCw1qlZG+UHpGVp7qeVpwWoO/fsVPwptU/wDF/XfyjbkVfYrxvbzS6pyP/lArv5RtyIPahERAREQEREBERAREQEREBERBx7zEfg/8Tf4HXr9SlXzP1n9tT/4V36Svpg5iPwf+Jv8AA69fqUq+Z+s/tqf/AArv0lBCiIgIiICIiAiIgIiICIiAiIgIiICZPqiIK9R8wqYKqBvuFVAVzVadgqsPfJQVI7nKAIeyNBwgoXgK5pDjsrTG8nPTsjcsOHbIJPAyM75VRT4GckKZs8eDkhUfO0jDSN0GGdiQqtcAMFVMcjiT0908GT+5QUzn7VkxN6tsrHLHNCyaWZjR8ZQUdSPecgqJzTE7pdus4VlOP3wUD6aWtk6qcdQKC6CMEA+q1qh0nJcwJIwRkb4WDTWS4vaA1hyCt96dqRaYw2rAacY3QaZbtBkP6HMyQVS6mLTxEZZjBx2W86LU1timL5HjBWxeIFzorlMTA4EZ8igxJNTsjb1tOPJaNWXKruDy6KUgZ8jhYcsXVG3oBJCvpOmEHr2JQTMkrsAGodt81kwz1DPpykrHbMwu2O+VKDuN0EFzmlLBl5wTut3aFpWVNOS8Z29Fs+5DLG433W+uHcMvupx5hBvbSNGYrg4046CPsWzeMNNUmvY6Z5d8Wd1v7SnVT3F7pRtkrZfGCdk1YOgjAKDjmChfGwSbjO6vkc/yceyy+rFM0fILDd8WfNAY92d3Ej0WXTTdMoJ7BYbWPByQVPGcPDj2HdByJpSP7p9MUYwRtkLkK1aZqKKuhnkJLRg4XHvDWsgFUGNIzkLmo1ccfhPlwAAN8oN2V2vxpuxtjoneDI1vcbbrath476hr71FQzXV7ml4GC7yz2Wj63vdnqLaYWuBfjyXG+kH0UeqoHk7GUEb/ADQdsOKtc12iWXGTBeY+rq+xdZaPV3i26pg6hvkd12J4qXCik4aRxsIz4P8AMuo9vdC2GpJce5QbVuZlbeX1MbyCXZ/Osyhu1aa+IundgEeawrhMw1rhnbPqraOSNtbGT2yEHIOrLh73YQzuelcUU1OWvc7OVyPfZYpbO1sZBPThbChpp29RIOM/NBZLJ0Nx2UDatzjguyM9ldUtOSDnIWKyPDsn1QaiHtIBPdHOHSfVRMJ6VVxPSUGu6OqjFWDO46lyhWXlj7aYMA5bj8y4l0vI1lUC87ZXIU1XSPphG0jqIQca3uhLa90+NicrBc4ObjzWv6goapshmcD0HfK24HDJGUFsLT1nKyzjAUUYGchSHsghk32WN0/GPrWTJv8AkUIA6xkIMpjg1gVHVTcFhAz2VAR0b+ixJHDxdj5oL/dnP+PO3fCNg6Hh2+xzlZUYcGZ8tla4OJJIQZLayVjAxr8AeWVq9DcWeCWSYJI7rbxkYNid1nUksRbnKDLmLJJS4NGCtHrabqk627AeS1UEE7FYlTkHtsggje1sfSe6imdvgKj3AbKzOQd0APLSN1MyQEbrFcTn5ZU0RGO6CdrxlUdI0dwretucZCte5vYlBd4zf7n865W5Nz/12fCT561tH65GuIepvr+Zcu8m34WXCL+Gtn/W40H0hIiICIiDrn7Q78DbiX/idH+vU6+eMHByvoc9od+BtxL/AMTo/wBep188Y7oLj2VqqSeyogqz6SueCd1aw4KucRsgsREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAQboqt7oJGQudnBIwrXtLDg5WZDJGBv3WNUva5/wAKC3rGAr2TtaOwKgwUIIQZBnaT2CiccnIKsVQRhAAwsiGRoHSoRvt6q6OF5eMAoMplA6fDm+a3FZZora0NkYD27rAoIJukYWRMOhw8QYwgy9Q3tpgIpfgPqFs99RUSkufK9x9crU7q6Pw9jnK0pjHuB6coJIDI54DnEj0ValuDkequpmOa/wCPsq1TmkDB80EdLJ0PyTssmWqHfHZYUW7sKSSJ2M4P5EGTBVB7sAYUkmHLCgb4b8v2CzAQ45G4QW9IxjClg6WvBwrX4jGXBUZNH3yBhBlV9XGIsBozhaeyRrmHsNlPPTyVxAp/iOfLzWTS6Nv1W4MgpiSfrQa5oO3MrakNcMguXKT9GRzgRNiGSMLafD7QGqLZUipqqZ7GZBGRsuW2VX3PeyWqADWBBty02CDSDxV1MIIB6t1uSp4w6fioDTilYHgY7BaVrDWun66jNM2VnWBjGRnK4lr57c8O6XbnOMINU1PrcV1VJJTHoBzjC2s++VsgI8c4J9VhVYh6z0HKxmYzjJQa5R3Ot6siY9/VbjstzuMVWyX3l2Ac91tKjZnGAtzWyMtexx7IN26n1RNNbPBdKSQ3z+pcI3qtdJO/HcuJz6Lkq/T0/uxbnfpI7riq648ckepQa1pa71FPPH1SktDh3K7WaU15TnSD6KQAuMeMn6l1Csj42TMc/wDulzzpi72ptm8LqAf0AYQbH1BRtmvtRVtbsXEraN3d4c2QcHK3zdIpJKyWRo+A5IWwr9tOQfXsgjjmldgdZwtRdGHUZJwTjv8AYtKgJ2WseG91ESO2Cg2i+Mmoc30KGItdnOVJK17al226q5ri0kjZBTxw0dJGfrUMmXnqGVa/HV37K9qC5mR5qdpOO6jYGk+SlA9Agika78yxyXg9yst5wMZCxJM5wgCSQY+I/lWoU8x6N9ytOGDjKy4XsaBkoM2Koa0nqGc7Kc08dQQQwBYHiRZH1+i1KlqImsHZB3u9jTTsj5n9TkNxnQVaP+cLeim9jdOyTmd1O1pGRoOtJ/zhb0QeyCIiAiIgIiICIiAiIgIiICIiDj3mI/B/4m/wOvX6lKvmfrP7an/wrv0lfTBzEfg/8Tf4HXr9SlXzP1n9tT/4V36SghREQEREBERAREQEREBERAREQEREBERBXOdlUDCADY4RAyM4VcFW4OftV7eyCvZAd+3Yqh7IBhBM2drRjGUdF42SPJQkeayaeRo74QQ+6vHqo3xuj3Wd4rC47hY1Q9ru2EFW1LWtxjsnvQJx091jhpKubE5x2GUGYYw9mQrobW6oeGjbPokEM5YBg4+patbqWqbIH42HyQTW7QtRXtBaT9i1L7kM04A2ZmfmVubTt7pKAdNQQMLStY3eir3F0LgcHyKDBj1NSURLjGPtW3b7qN1fITTu6QCoqpsUgwfRaVLTlrj05IQSmrrJgGiYnHfdQSGUEdbifRX07HNdkhXVDC8ghAiqQ3OeysmlEhyzbdR+E/0VfDePIoLoCQ4rIfVdGBn8ixmAtOXbJIOs5acoMky+PgfNcpaCcyClAx3C4op6aaVwEYOcrljRcL4KcCUb47oN10codUvcwd1sPiBA6ap+Lfdb5oSwTuO2Pl6LZuuZme9Dy3QbQqaUxUwO+AFpsbsZz6rWq+QPpRj0WhsGAcoJxI12wCvDC7LAdz5rHjc1rtysqJ7ZJhHGcuPZBuXRcT7dUioLiBnK5GuGpHzUvQx5BAwti2S2VlOwTztIYd91rE8sErOiI74wg0atqKyrqTmYkE+qksdNLHfaeUOOQ4FXGinpXmaYEMO+Vk2avpZLzC1pGeoZ7IOf77FPd9Gx0peSPDx3+S4sj4bvgtlRUAHzOVy6wF+mozEM/Dt+RaPNHWiw1Hw7dJQdTNQtdRXeWnB+icb/AFrBhncJmuDu261DWTXDUM/VsS7+daQxrg/JzhBvKgq31cAie7IxhZr7fG2Enox/OtGsVRC3APdbmnqoxAcY3CDZtxpg2Y4GFhimGCcb+q1O5PDpSQAsMEhp9EGPjBwqkZYVaHtLyPPzUji3p8kEVFO6CUkOx8wtYt92lnu0EBecFwBC0SJrQ/LuxWTaC1t+p3A5HWB9qDmPVun4o9LNrBGASzOcfJcFuaQ9+fIldjtYukfolgAyPDHb6l10dE/qlJB7lBdA89ieyme7DchYUD/j3WW/dndBB4mXEEq47DKgJw5S7lu6C0vz6JFAZHdR7KwD4uyyost8kGT1BsfSoy8dJCOeDsVG4jyQYskbi7usilL2AbkK17T3wjS5pwdkGcyfp3JVk84cCofEb5lUc9jtsoIHZLs+quawhqr053Hqr25xugic09kaC0YypHgAZwrcH0QWdJyrHhwOFKc+SjcTndBZ8S5i5NPwseEX8NbP+txriDpJHYrl/k1OObDhF/DWz/rcaD6Q0REBERB1z9od+BtxL/xOj/XqdfPI3uvob9od+BtxL/xOj/XqdfPIEB3ZUVx7K1AVcEowZKuIwgsREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQVyfUqaGAzepUABKzaKRsZAJQZEdv6mj4VKLVn96sllRGGjdZEdQHAhuMoNJfZzvgFRm1OBwcrWXGR3Zvn6KJzZSc4KDDjtWAMjzWoQ2sNZ1dKua94wMLUoahjYS12xQYLatlJkEYwtKuNxErssON1PXzRdbsn1WjTuY7PSQgq6R1ScZJwqskEB6SoYCQ44SfJfkhBLJUtdnpGFGGGQZUYaXHDd1kRU83kDj0wgtjaI3A91lCRrhlQyRvYPiGyrG9uAOoIJTF456W/WnSabOVdT1EcLsvcBkK6pjfV58Hcd8oMSao8X4QpqGgdUSeEM7qxlorpDhkRP2LWrLZ7hT1DZJWEAHKDeOjNIiMtnmj6gCDuuQ4LrarBOyaWmbhmNsLQtOXimoYAyqIGB5qPUl9s9VGelzSe3dBu67ccLJHS+7QQMY4DGekLjbUfEs3amfDBJguyAtm3h9vlmJjOc/NaTG2EH4TvlBFO+4vqHTPmecnOepSsmnOA55KulcC3AIKhYSHfagyDI7G5yjJMnOVbkbZKBzR5hBq1vqgCBud1u2CUNo+sYGB3WyKF7Mg58wt3RPaLccnOyDbN0vEkk74i/YLbNY8vlOVqFdJGax+D5labUbv23QVp5HMcMHGFuOw3iqZVsi8c4JAxlbaiBJ81qlkAFewk9iEHLha2SgEhAJLe6441HB9+Lh6rkeGWP7mAEfvQuP9SMke8lowM+SDRIpekgBbppHNfbjkeX8y2jHgvAPfK3VRSsFAWkjOEG2Khg96fgYwVZMWhh2V9Tn3l5B2JUM72lpb5oNPcfiJV7HANwhhkJJwqdDmjcILhIWn/wBqk96HllQd1TpCCV03UdlTurMBXDsgo5vzVuSD3Kvd2VhBJJQMn1KmZUva0AO7BQEEKoaT2CD0B9izM9/NLqkF237AK4/8425Fb7FYEc02qc/i/rv5RtyIPahERAREQEREBERAREQEREBERBx7zEfg/wDE3+B16/UpV8z9Z/bU/wDhXfpK+mDmI/B/4m/wOvX6lKvmfrP7an/wrv0lBCiIgIiICIiAiIgIiICIiAiIgIiICIrgMBBQDCEZWRV0dXQy+71tLNTygBxjlYWOAIyDggHcEEKBAPZGnCIguBycYQnClfR1cVNFWPppWwTEiOVzCGPI7gEjBI88dlHgFBTpzv6qhyNsq8DdCwHzQR5d/dfmUsdOZn4VoiHllZlKAxwKCaO2/BnpWZT2wdOS38yyKeQdIGVmRzta09kGG1jItulZTLvDTRlhAB7brHlmaXEjC0ypDXPyR5oL66eoqJC+GQgfWsPNQw/fXk7eayYy0AD0VkzWv80GO6QgHJKj94GfLKpLHucOI+WVjOb0lBkGcDsVNA8SN33WAsyiifJszdBI8tadhhG9JaTj61K63VUmzQTn5KaHTV2lYSyMkYQYsNOKt3Q3crUY7EYhlzScrIsmmbrHUZfERuPJbhq6aSmwyRuCAg0ezW1kcvxsyMrdQuDKMBrCG7eS0qnnhpT1ykAfNRXC7UUx+9uG3zQbjoLz98J6xuFs3W1zMlTs7zWbbq+GSQhp7Lb+qmufUBw3CDHbWGWINJzt2UJcMKCCN7WAkKbORugRwOmcMLOttG6G4RSP3AIKstsTqiUMiGTla59yauB4ke0gAoN5XG904sbYY2APDQOy2nari8VoDnZBcq3Cvp/dxBkFwGO60q3zMjqmyO7ZQciX+aKezjwwAenuFsSwsmivTJC846h5rddVVxVttbDAcu6cbLRbbaa2O4Mc5hALgg7AUWpm0un2Ne8HDR+hYMuuad9qlpw4fECFtusmbTWVrZMghq2rFc6WSN7ARk9t0GydY0onuslY1owSStuuj+E477rd+pIJCDIOx+S2kd2O+1BFQVL4p9nYGVrb7i5zMdS27EQyYl3lus11RGQB8kEz5/EdnOVR8nTGQAsdjwXZadipJfoFBgiQ+Kd1kNJd5rEG8x+1ZILgUEjmHGyjt7jHdYHekgKleT0du4UFJltcyR3YOBKDni8XltXpNlNkEhmPzLiKahHS89Pckre/3RpaizMhiILwMLb81BP4D34OMEoNlGMtnLcYA8lOfo4Sob01Ra4bqvfKCBkeH5O6mLdu/bKtGQ7JV3U0bEjdBCRg/arxLgYx5KUsbuoXhoKChy931q9sRByrI/pLI8ggeX2LHlfg4UwIBWJUfTQVc0uwQCfqV7YHY6sFXR/RCyAPgOyDFa/pOD6q/wAUehUUuA8hW5zugmc/P1K9h2UGR6qRjsDZBf075Vj2gHYq7rOMK1zslAGy5a5N/wALLhF/DWz/AK5GuJMjC5c5NfwseEX8NbP+txoPpCREQEREHXP2h34G3Ev/ABOj/XqdfPGBlfQ57Q78DbiX/idH+vU6+eRvdBU9laqu7KiCrPpK9ysZ33V7kEaIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiC8HHdA85y04VmSrgxzvo75QZHiuIA6jhZNPUOYQeo4AWn+FL6FSMEjcZyg1plwaBuQrjcGHO4WkNe0dyqGVoORug1R1e3yPmo31r3A4cQFgue3HdUDHO+iEFanrkyepYwhPms1lLO7Yg4+pH0czdgD+RBBTsw4k7qyq+msuGklJOxUU9BUl+zSUGPTvDHZK1GKsZsFgOoahgyWK6Onk7kIMiunbJGsanhL8nOwV76aeTAAJWZRWqsLThp3HbCBS2eWvIazOVr1JavufhsjMn5rI07F9zpOqrGB33Wddrvb5JB4ZGPPCCJlfSUI6nRD1Vs2q6NzSGMAK0e619LKzDSO2DutvPcHNPQTklBuCvvc1ST4MxGfRaU99dK09Uzj9qxKVspfvkj5rO6+j6Xkgx2UswcS8k575KkEDgfNTCriA+JwH2p71E47OB+1BZ4JwASrfCw4BTl4I23Vo3OcILJIz09/JQdBOVlyn4Vjjz+tBlUMR6m7g7jst6Np3fclxH9z/Mto27+yNW+mkfcd3/elBxTWdTayQZ3DlQMyFfWke/yE/3W6B7O2yCjWhpU1FUiGsa4eoUJkb9SjicBOHE7Z7oOR4b0TSNb1dwsaqDamnJLck5K0OCti8FjQRkDcZWv0r430ZxjPzQbTfTOFQcZ7+i1aIOZTkA+SxZWn3p2O2VmRub4WD3KDTPA8SV2fyrS6tpimG+y14RvDiQFolxik8XqO43QXNqGBoGPzKKUiQ5Cxeojb0WVBgsyd0EJjI3UZacrKkHdY5BygswVVp7BHdlQd0EhIwgO3dW5Hqnlsgu2PmrhgDCiaTlXEn1QegHsWSP20uqcf8AK7+Ubcit9isc802qf4AV38o25EHtOiIgIiICIiAiIgIiICIiAiIg495iPwf8Aib/A69fqUq+Z+s/tqf8Awrv0lfTBzEfg/wDE3+B16/UpV8z9Z/bU/wDhXfpKCFERAREQEREBERAREQEREHIXBLgRxP5h9axaC4Uaaku90dGaidxeI4KSnDmtdNNI44YwFzRk7kkAAkgH1p5buQLlp5ULtpm4cbdT2XV3E3UVU2js9PXAGkbUnJAoqNwL5HNA3nkBDcAgRZOfLjlw5qeKPKzcdSXnhZ9yY7hqW1i2S1FfSe8GmxI17ZogSB1jBADw5h6t2nAxyByd8QdccUOfDhlrLiHqq5ahvdbf2matr53SyEeHIQwZOGsGThrQGtGwAGyDtx7bOwWK1aa4U1VrstBRzSV12jfJBTMjc5ojpiGktAJAJJwdt15Rr1q9uL/8U+Ev/hG7/wDoqZeSoGSg5w5PuWO/c1vGa38OKCqlt9ohjdcL7c44+v3OiYQHFoOxke4tjYDkdTgSCGleqnEfjXyVezQobboHSvDttTqqalE3ulop4prk6M7CetrJXBzQ/DsDJJ8mBpyNkexU4fU1q4Ma24mSwMFZqHUDLWx/SC73akga4YPkDJUyAjbJYCc7LzC5meI9x4t8wGv+INyqXTOu1+qzTlxJ6KVkhjp4xnBwyFkbRkDZvYdkHrXwd9o7yu82+oqfgxxG4cyWqqvr/dqCk1LTU1dbq2VwwIQ/cNkdkhocwBxwA7qIB6Ye0y5GbJy2Xq3cUuFlNLDoPU9W6jlt7nF4tFf0F7YmPcS50UjGyOYDktMbwTgtC6O0FdWWuup7nbqmSnqqSVk8E0Ti18cjCHNc0jcEEAg+RC93eaZ1NzCezXuutK1kc1RctEW7WDXtaCYqiKOGrfjp2aQGyMIHYFwQeGejtJX/AF9qq0aJ0pbZK+83ytioKGmjGTLNK4NaPkMnJJ2ABJ2BXs9prgfyf+zR4O0XEDi3b6HUOrZemCW6zULKuvr60jqdDb4JSGxMaM7gtPSAZH5IC6Vex/4e02r+a92p62COSLRmn6y6Ql4Dh7zK6OmZgHzDJ5XA+RaOxwVqntkOI9y1NzLW3h8al33N0ZYYGsgyekVVWTNLJg7ZMfu7ds7RjfOwDs3p72zXL7qi9t09rXhdqi02OueIJK6YU9bGxhIHVNA0g9AychvWcDYE7LQvaEchfCzWfCat5meXS0Wy3VttoPu5cKOysY2gvNtLRI+piYzDGSMjJlywASNDsguIJ8i17leyc1w/iVyeN0pqKRtezS13r9OuinIcXUb2Rzsa4dyzFS9gz5MwNhgB4cdle0F3dbg4jaX/AGD8RNUaKy4iwXmttYLsg/eJ3x5OfP4FoAIwgqNicdgp4iVunhDw6rOLvE/S3DC3XKC31OqLpT2uKqmYXshdK4NDnAbkAnJA3XY3mz9nhq/lI4cW3iNqDiPZ7/TXK9w2RtNR0csL2PkgnmEhLyQQBTkY75cPmg6rwykYCmMxDcZWEx24Xdvl19mDrnmK4Oae4x2finY7RR6g978OiqaKaSSLwKuWnOXNIByYS4Y7Agdwg6Uve/J7qLDn4LlmNazp+tdi+RTlh0tzW8X7rw71ZqG6Waht2nai9Ca2tjMr3sqaaEMzI1wAIqCScE/CBtnKDrMQWnuoJpCB/wC1egOl/ZT6x1zx/wBZ6Ktmpau0cN9I3COibqS4UgfU3BxhjkdFTxNLWyPb4ha6TIY0jsSCxdhYvZD8pWq7JNQ6L4vauqLrSACSthu1BWsY89vFhjhGG7HADmnbcnByHjg58hOQFb0PcexXa3iB7PLjzo3mDo+X2yWduoqu8QmutV5hYYqKagDul9TMST4AjOA9pJIJaG9fWwv7o6E9kRy76co6Sy8YeMl3uGq6+JoFNba6lt8Qe84HgxSsklkIIIDiQHb5YCcAPImGhklIHSdytz2ezNjj6yCQfVd3Ocj2alw5dNMScUOGeoa7U2kaORrblBXRMFdbWOIayVzowGTRlxAc4MYWkgkEElvTulqYYocnHb1QY58Ck3cwbeSzKTWVBRAMdE3IGMrSblcqV4LeoZx+RbaqJIHuJz9iDf7tf0MZL2xtGfNaJc9Tx3R/iMIAW0Krw+kdPdSUJbg52JQahd7m6SEMY8gn0WJbaeaoBd1HHlurKxniABgyQVqNkBhZ8Ywgy7VSywzlxcdvmsbUNQ0TAHvnyWqwVETXlziFt/UMscs4czcAoMQ1jSAMKenZ4+APNaa1rnnA3K1e0wTdbQQfqQazZKUW6UVDxkZytfr7/TzUrmtaAcYytKm+8U+ZthjutLlraUxkNIyRsg0mvqpTVl/WcHKpHWHIw7cfNQVX3yY9HbdWRwvPluUG69N3N8UwMjstz2K3vR3mnnuMMAjGSRvhcbWcCneHzbBbntFwo33ymDCCS4Y3Qcra0tLodNCrZkAs6vzLhm3TympcS84BwuwGuGSSaIYWt28LuPqXX62wvdUPYBuXoNR1JWsFCBtkD+ZbEbVAkj+6W89T2urZQ+I5pDSFsSNhc8beaCaaLAL/ALVjgnI3WZKHGMtA8liiGTI+EhBl00bjuT5rJkblhGVHT/C3cflUrnAhBgNjw/PqfRTkBoyhbvnCOB6SEFGzB56cgq5zQ1peNj3ysZgcyTLu2VPJKx0RAO6DV9LVM01UInPJGcYyuSKiiY22OJj3LfRcYaScILgHybDbuuVau9251sMYcOvpwEHD1zAbc3j5nZUV1yilkub5GtOC7YqpjcB2KCCU9IysbxT1Zz5rIqDgYWF5/agy/Ey0DKqYiWl2+FFGQe6yw7MRHyQYXWWOwO6nZNkY2yVjvY4vO3mr4o3BwODsgyGxk5OVjTjDt1mtcAMHCw6kFzhjdAZKGgYIGPVZbJA6P7O4Wn9LsYWXC8CIg98IMWUkvOFLDH1DJUT93nHZTRuIGEFCw5O6q0EDCFxyeyubgjdBQHOyseTlX4AJVHDPZBQAkLl7k225suEX8NbP+txriJrXA+a5c5N/wsuEX8NbP+uRoPpDREQEREHXP2h34G3Ev/E6P9ep188je6+hv2h34G3Ev/E6P9ep188je6ATnZUVXABUQVaCSqnY4IVGfSVzhnCCxERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERBUjAU0MzY24IUWxVDhBle9MPcKpnaRt6LDUjeyCrz1HI+ajPUNiVIc42UZBJJQXZPmVqNI5obk/UtOA33WVE8NYB39d0Gotr44ttsqKS7RlaZM8uA+pQ4KDXaK5Rl+4+S1qJ0MuCWZJ+S2jRPY2QF3qty0dfSsaOo7/AFoJ6xkPQcMAWCymY4YDQPqWTVXGkeDgjP1qGG4U+N3j8qCWmpo2HLmg49VmfdanpCPhGB3CxWXKlHdwWmXGop5Aehw3QZV5vgrIy2B3SVtsyTjvI7f5qR5GfhKjeHHJQUzJJ++JU0Y6MdSjiDg7GFM4DHZBI2rYwbBHVHjYAWE9pz2U9O0AoKvppHnIyqsp5GOyc7FZbJABgn8yo5wJ7oKRkjYqUPGyhJAGyeIBtkd0EznZG6hPdVMrW/SPdWhzZCejc5Qajb/ptW9A4/clw/7X+ZbNoIZGFrnA4yt2Grgba3NJGS0+aDjav2rJf++UABJwp649dXIR5uUTGOyDjAQSNpHOblVfSOazqWTG74QNldKHGM+eyDBgke14HUcZ9VuShryynwXeQW2QCJN1q0DwIMZ3QSzXBgkJ2zuomXUmUN6tlhSse55OTjKx2AMmHVsAUG6mytcwPwdwr5rcyemMob+ZYbKymELW5GRha5TVMUtvdGzBdjCDY0lOfHeztgqmTAekrOfbK01T5Aw4yTlYNUx8chY8EEZQDLnurSQT9asGPJSCJx3AKCwjCtIypMOGzvRWktB2QW9JVwGw3VpPbCka0kIDW+aseN8fNXjIJ6lY875CDv77FT8KbVP/ABf138o25E9ip+FNqn/i/rv5RtyIPalERAREQEREBERAREQEREBERBx7zEfg/wDE3+B16/UpV8z9Z/bU/wDhXfpK+mDmI/B/4m/wOvX6lKvmfrP7an/wrv0lBCiIgIiICIiAiIgIiICIiAuwXs/vwy+FH/h5v/opF19XYL2f34ZfCj/w83/0UiDvT7cT/wCKfCT/AMI3f/0VMvJYDC9afbif/FPhL/4Ru/8A6OmXksg93vZM0kdPyWabljc4uqrrdZX57AiqezA+WGA+e5P1Dwje98jjJI4ue4lznE5JJ7kk9yvbr2Omoqa78plTaInN8exaqr6WVgO+HxwTNcR6HxSAf+1PovGjiTpiXRPEXVOjaiAwS2G9V1skic0jw3QTvjLTkkgjoIwSe3coNtL3Y0DUTVXsmKqWd/U5vBu8xg4Aw1luqGtG3oGgfZuvCde7PELx+D3sp57fd5BTVVPwspLRUB2B0T1tNFTuj7EE9dSWA9ycHIJyA6uexBpYn654qVxLuuK02yIAYwQ+aYknbOcsGN/Xv5ddPajvc/np4lBziQwWYNydgPuRRnA9NyT9ZK5p9ijqOlouO+uNMTOa2W6aVFVDnYuMFVEC0fPE5OPRpPquOPa26YlsPObe7rJAYxqSy2u5xuIIEjWQClLhucjNKRkY3aRjIJIdMl7CexJqJn8H+IlK5+Yo9SwSNbgbOdStBOe5yGN/J9a8e17Uexh0zWWnlm1DqGrjLG33VtS+m2+nDDTU8fWD/hBK3Hl0/NB5X81/4U3GL/jA1D/KM64uAyt38Z9URa24xa61pDO2aO/6lud0ZI3GJBPVSSBwIAGCH52AG/YLZ4PmEHNnJVtza8I/4XW3/wBM1enftoNuV3S/8PqL+TrivL7k1rKei5sOEM1Q8tY7WdphBAJ+N9SxjR9rnAZ8s5XqB7aI45XNLfw+ov5OuKDxlGwyCvfD2ZEUsXI9w0ZNE+Nxbd3hrgQS112rCDg+RBBB8wc+a8De4xlfRTyWaSm0Nyq8MNMVbeirp9O0tTVRZy6GWpb7y5jgScOBnwR+TbCD52TLMOwJC76+xhke/mj1Q1wOP2A1p/5xty6MRsiMYJwu+Xsa2sHM9qcs/wCAdbv/AOULeg3d7Wbm94jUet6rlg0VUVNjsdJR09Rfqunl6Z7oZ42ysg6hgsha0jqAOXkkH4Rg9GeWPinqrgtxz0dr3TdwqqY0t2po66GJ5aKuifK1s9O8DZzXsLhg5wcEYIBHP/tTI4nc52qy/ubfav1OJdZ9Hxwfsps3wj/dCn7f4RqD3e55+YG+8tPAWv4i6TtVNWagqauKy22aob1MpJZw5xmc39+GiIkM2BcGZyAQfBbUep9U6y1HW6v1bfq+63u4zmoqq+rndJPLITnqLic7bAAYAAAAAAC9mfa39P7VamLsf/Gug/8AQ1C8XpwzdwxjCD2/5NNVX7mU5EYbZryaW8XKttd20tWT1Lg59Yxokiic9zju7wnxtLiclzS4nfK8Kqi7PYC0E/Yvcf2T2DygWvH/AM/Ln/6QLwjqz99IQWzTvleXFx3VGhzt9yFTGWE4UsJAGCQgtc0kblGSGPscFXuIxthQvQZUdSBjrwcrKbW7fAcfMFaSp4XNDcFBnPq3kbO8vJafUPc93xHJUnUDnBUMndBdA8MeCVrFHXRtcDkDBWhtAJwVlQPY0jJ2Qa/drmKum8Nh3xjZbc8GbPc4CznTRdOFG6aMsO+Sgx4pAHdLhnCmErWuDjjb5rDDgJCQdldI/b1QZlVXh8fRG7BVlprJKW4wVHX9BwJ+pYQBcdlJHG8PBIwEHNl64pR1mmGW0vBIZjGe+y4zoL+yCqMpOxOfzrTamWJ0HQCc/WtMwQg3/f8AWlPcbcKZoGQMbLY0MoaXE43UOUQZXvDfUJ7w35flWKiDJFSD54Vfec93DbssVEGX7y0eYKe8tPmAsREE8sweNtlCDgg5KoAUwQg1Gln6AOlwB9fRZBrarIzMcLTYHgbFZDpWbb4wgynVkbm5OM47qCWsbgjIUDnM6TgrHeQfNBdLKXknPdRJ3TBQAcFZMUvSADhY2MquSEGSJADlVMwwcfoWNn5pn5oJTNud8K0y5OSPzqJEEviBWuec7KxEFepSNkA2USuaB3JQSOcDgq9h2ULjgYV0TwBgoJcgqhcAd1QO+Y3VrnZPcIJvEGBsuWuTX8LLhGf+7W0frca4gJwM57Ll7k0/Cy4RfPWlo/W40H0hoiICIiDrn7Q78DbiX/idH+vU6+eRvdfQ37Q78DbiX/idH+vU6+eQIDiCqKpByqIKs+kr3KxvdXE+qCxERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFcOytVw7IK5x3Vdu6twCqgHGB2QXDfsrxkDurWjGVcgsO25QAO3VxAPdMEdhsgjLT5FAJcbE/lV+CqjbYoI8yHzP5UzIBs4gKTA9EwEEXVJ/dFXBryNyVcW+iqDjYoKNaR3V+R2Vp2CDOcnKC8AA5wit6vqVev6kFzRkq8RkHIVGeqlHZBRrSdgVd4ZPmVfGN1IB6BBCYiN8qzwiX+fdZLmux2VmCHdvNBBUQuc3Y4VtIwxyjqPmst+7VBGMSjI2Qa6xzfCGAraqWX3c4cQEY5vgjcZVlU9hpyPP60G2n7znPqspuOj5rFI+/n61lDZuUEbZOknPZSPqm9PTkdljPdud1YWEguwUF3iAvz6rUKRhkIGe60xg3H1rVqB4a4IMp9E1rD8JK0arh6HZGwW5nhzoytDroX7nyzuUGm9b/AO6Oy16y1D2EdbjjHqtBIPVj5rVLfI1jh1dkG7m1tMIiOgZ9VoVytJrJfHYMArINTB0DfdahTnrhy0bYQbQnt7qfPUoTO1m3dazd4ZXk9I33WguglacFvdAkkDuwV0dOZBncqMxPHcLNpHtY3fvugx5IvCI/KqCYDGylq3h/ZYmD6IJS7ryAMfUo3DpV0QyVSTPVug7++xU/Cm1T/wAX9d/KNuRPYqfhTap/4v67+UbciD2pREQEREBERAREQEREBERAREQce8xH4P8AxN/gdev1KVfM/Wf21P8A4V36Svpg5iPwf+Jv8Dr1+pSr5n6z+2p/8K79JQQoiICIiAiIgIiICIiAiIg5S5e+W7inzPawq9EcKbbR1Vfb6B9yqn1lU2nhiga9jMl7u5LpGANAJO5xgEju9ypezN5oeD/MVoTiZrG26bZZdP3RtXWuprw2WUR9Dmktb0jqOSNsrojwW4+cW+XjU1Tq/g5rGXT11raN1vqJm0kFS2WAva8tdHOx7D8TGkHpyMHBGTnmke1G56z/ANnP/wDpmz/1RB6R+0z5T+LvNPYtBW7hPSWueXT1XcJ6019cKYNbKyEM6cg9RJY7Ppj5ryv5iuSXjvyuWG06n4qWu0x228VjqCnmoLiyo6ZwwvDXgAEZaHEEAj4Tkg4zu791F56vx5//ANM2f+qLjjjbzecxHMZZbfp3jJxEdqC3WqqNbSU/3LoqQMnLCwvJp4Yy49JIAcSBk4Ayg7MeyP5mrDwi4p3jhJri6MobLxB93bb6qeQNhp7pF1CNjicBomY8s6s/TZEMYORzV7QH2ZXEXihxPuHGzl8prdcajUPTNerBNVR0kvvYaGmeB8pbE4SAZe172EPyQXBxDfJpdneFftIObzhNZodO2fic682umYGQU9+pI68wgDADZnjxsAAANLy0AbAIOd+Vz2SXGa6cRLXf+YyzW7Tek7TVMqqq1fdCnraq6BhDhAPd3vjZG4gB7i8O6cgNycje3tgeauw3WloOV7Ql2iq5KOsZcdVy07g6OCSMEQUJIyC8EmR4GOktiGSS4Dq/xE9pxzj8RbTNY5+JrbBR1EZjmbYKCKimkBzkicAzMOD+8e1dWZppaiZ9RUSukllcXve92XOcTkkk7kknJJ7oOYeULjt+1w5hNJcValk8troKh9Ld4YfpyUE7DFNgfviwO8RrcjLo2jI7r1q57OUK288nDfS/Evg1qWzS6itlKZ7PWvlPud3t8+H+CZWg9JBHUxxBALntdgOLm+GC5n4Hc4fMVy6wm3cK+JNbb7S9xe+01UcdXQkk5JbDM1wjJO5dH0uPmSg5m0R7JfnF1JqaGz6o0hatJ2p0gbPd629UdTGyMHBcyKmlklccZIaQ0E4BLe47481vFLh3yAcnlDwI4e3Rn7KLhZ5LJYacFoqfvvV73dJWjIaOp8rwcYdK4NAwHFvQm9+1l5zrxbn0NLrKxWh8mxqaGxQeK0YIIBlD2jOe4GRjYhdVdZa21hxD1FV6u11qa5X+81zuqorrhUunmfjsC5xJAA2DRgAAAAAAINEbknsrw3G2/wCRGkfbj0V+R5hBylyoAjmk4ObH/wCP+nv5RgXq77Ruz3Dmc5IrLrnhPbqy+Q0V5otSupaKIzTeAyGpp52ljRkuidOQ8AEjw3HsCR5S8qODzR8He3/x/wBPfyjAuyHF/j/zI8hHMtxK4b8N9XGn01cL5PfqK1XGlbV0ZhrT48ckQkHU1wD+hxYQC6Mh2SEHEXLHyv37iNqQ674n2av07wm0eDdtV32vgdTwupIfidSwOcB4k8pAjDWZILwSM4B9aPZ3cw0PMRwu1XqKo6Ke50msLk6WhBy6lpJ3iakZtt0Nid4TSMA+CRjbJ8feP3ObzDcydJDaOJ2uHz2Wne2aO0UEDKSj8UA4kexgBkcMnBeXEZOMZW1uBXMJxa5cNWO1jwn1TLaaueMQVcD42zU1bEDkRzROBa4A5wcBzSSWkEkoOWOGHAi66VtvHW78XdET01t0BpSvt0dVcqV8UcV9lqYoKIRF2AZCS9zcZ+EE75GeZ/YyytfzP6nAOT+wKtP/ADjb11q5hOdTmD5m7fRWLijq2CWzW+YVMNrt9IylpjOAWiV4bvI8BxALiQ0E4Ayc7P4G8f8Aipy5awm13wk1DHabtU0Mlsnklo4alktK98cjoyyVrgAXxRuyACC0AHBIIdi/apTBnOjqtpP/AOj7V+pxLq9pOsa3VNmAdv8AdCmH/wCMapeLfF/iFxz13X8SeJl6bdL9chEyeoZTRU7eiNgjjYGRNawBrGgA4ycZJJJJ2jTVVVR1EVVTymOWB4kje07tcCCCPmCAUHt17YWYwcplLIO/7LbeP/xNSvEmS5yOOxJC5n4286/MjzD6Oo9B8WNfMu1loqplcII7XSUplqGMc1sj3QxtLiA9+2Q3JyRkAjgwAeeEHut7I+Qy8m9qe7ub7dP/AEoXhSSZXF2N1zzwd54eZbgLoSfhtwu4hNtNhlmkqY4XWukqH08smPEdHJLE5wJwDgkgEZABJJ4KaB042QRtyMg/Wrc4KncwAZHkoHNIJQGnBVXblW5VQdu6AAScBC0gqVvSNx+lUcQgsblpyjhk5VXkEK0HbcoBGFTJ9VUlA0lBTJ9Sq5d6lOkqrWEuAAygq5uADlWgFxx3U8jCIwPJWQEdSCaGn6D1EKZ5HQSB2TxGhgBwrTKwtI9UGI0kybk91K9gI2Hko27P6j2zlSeIz1CCFzDnZW9PzUznNO43VnUEFnT806fmr+oKjjkoLen5qh2UnUFQ4z5ILen5p0/NX9QVCcoLWHBVziDsrMpk+qBlMn1KIgrknz/OmCe5VBsVICMILS3p3QHKuLgRurdvUILRsUJycoiAiIgIiICqBlOkpgjsUDp+adjsrsH0Vrgc9kFM5TKJg+iCvW5MklURBcTt3XMfJn+Fhwh/hpZ/1uNcO4PouYuTT8LHhF/DWz/rcaD6Q0REBERB1z9od+BtxL/xOj/XqdfPGBk4X0Oe0O/A24l/4nR/r1OvnjBwguPZWq49lagqz6SucMq1n0lc/PzQWIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAg3RB3QV+jurg7ZU29Qm3qEF3V8lUbqxXDsguLseSB3yVhOVUEYQXdXyVDuqdQTq+tBcDg5VC85VocPUlVyPRBd1fJUO6p1BOoILwcoTjZWhw3VSQT8kFDsFQb75V5wR2CtQSxyAbFTtcDssMOAUjZPPsgzGvDT6/UrxJ54WJHM3O/51IJ2A7oMl0zceaj691E6eMjYK1s7eoHyz6oMh0mBlWMILgc+aimmZjAUDZ3B3fI80Gqic4x1DH1qOaZxYd/yLGFSzz7o+oYWYCDFG8pwO5WST8B2WPER4m+6yJHDp2QYoZ1PysrwwIzt5LHY8B2fJZJnYWYQYB+kR81mUTyJASR3WG76RO6lpw8SA749UG52zt8IZ74WLVua6MjGfsULKqMRgE7q2WrjLcZCDTwzqkPdVcCw7EhXiWMEkAZUM8oJJCCXxnZHx/nWu264hkXSSDstq9bvVZdLUdJw47INymVkpJxnPZYNRTNLshpx9SUtdAAMkfasl1fSuGeoINIqYWsH0cLTnk52JC1iunjkz0EfYtGeHdXdBcCTgE5UzIQ8b9/JRtwMeqyo5o24z3QQOiEAzv8AUsd7uo5CyaqVrwcLEQd//YqfhTap/wCL+u/lG3InsVPwptU/8X9d/KNuRB7UoiICIiAiIgIiICIiAiIgIiIOPeYj8H/ib/A69fqUq+Z+s/tqf/Cu/SV9MHMR+D/xN/gdev1KVfM/Wf21P/hXfpKCFERAREQEREBERAREQEREBXAYCtG6uQEREBERAREQEREFW/SCuVrfpBXE4QXM+krz2VrVUjKDk7ld1LYNI8yHDDVGqZ4ae0WrVtqqqyonIEdPEyqjJmcT2DMdZPkG7Lu97aCLhnd9R8NdX6b1Jaq3U1TQ1tDXQ0dRHM91Ax0clNI8sJLQHyzhpP0g44+iV5pDAPkFdkeqCh2PcqmVQvGSqdSCjvNVYfI+qtLjugOUEpI3wo85Vx7Kwk9uyCjsDPqrR5qqA9wguALt1JgDH51GH9IV4cDugtLyCe6tJJ7qQ481G84QWkDCAZRrS7bdHNLcILmHOyo/uqDI3wqE5QEQDKva0AboKMHUcK8/Dsg+HdWuOTnKC9mHKVgaCM/YsUPcPNV8R/qgyqjHQMHKxWD4gr2vc84d2VzgG7hBdK3EeQVjb/NXukcQR5K1mOoZQXOdluArN1mvEXhg7ZKxwG+mUEW/mikf042CjwfRARMJhARMH0TB9EBETB9EBEAJVekoKIruj5KuMbYQWIiICJjKYQEVekqiAiIgKrQCqJlBI0jsqqIEhV6igkVryqdfzVzcH57oLMbZVR2R+3bsqsxhBa36QQ9wrwAFR2M+SC4dlzByafhY8Iv4a2f9bjXD47LmDk0/Cx4Rfw1s/wCtxoPpDREQEREHXP2h34G3Ev8AxOj/AF6nXzxgZOF9DntDvwNuJf8AidH+vU6+eRvdBU9laqu7KiCrPpK9ysZ9JXuQRoiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIKtVVblMn1QEREBERAREQEREFWjJUiiBwq9RQSKmd8KjXZ7q7IQWkYVpOFe4geat2KC3KZPqmCmD6IGT6qoySN1TCqAQQUFzgQ0b/AJ1YrnOJGN9lbg+iBk+qZPqiIL4vpLILR0HdYrTgq8zOxgFBYdnEAqmT6od1UNJQXYysqIAN+fqoGjZVdJ0bBBe5pOfixjssd5cDguJVTM8537qjWueRsSgtyfVPrWQIDjsrHsDfJBEgz2yh7q9pbjPmgAP8ipGxSu7yYP1qPJ2weyqJZBtkoJSxzB8Ts+isd9XZWOkfjByp4iwsOcZKCEHfsrX5z3UsnSOwUJySUFMoq9JVMYQd/wD2Kn4U2qf+L+u/lG3InsVPwptU/wDF/XfyjbkQe1KIiAiIgIiICIiAiIgIiICIiDj3mI/B/wCJv8Dr1+pSr5n6z+2p/wDCu/SV9MHMR+D/AMTf4HXr9SlXzP1n9tT/AOFd+koIUREBERAREQEREBERAREQB3Vyo36QX0T8l+l9M1PKdwlqanTtslll0lbnPkkpI3OcTC3JJIySg+dlF9R/7ENJf8F7T/EY/wD1U/YhpL/gvaf4jH/6qD5cEX1H/sQ0l/wXtP8AEY//AFU/YhpL/gvaf4jH/wCqg+XBF9R/7ENJf8F7T/EY/wD1U/YhpL/gvaf4jH/6qD5cEX1H/sQ0l/wXtP8AEY//AFU/YhpL/gvaf4jH/wCqg+XFv0gquwvqN/YhpL/gvaf4jH/6qfsQ0l/wXtP8Rj/9VB8urSVUn1K+on9iGkv+C9p/iMf/AKqfsQ0l/wAF7T/EY/8A1UHy643yhOF9RX7ENJf8F7T/ABGP/wBVP2IaS/4L2n+Ix/8AqoPlywM5Qn0X1G/sQ0l/wXtP8Rj/APVXA3PbpnTdJygcVqmk0/bIJo9PSuZJHSRtc09TexAyPsQfPSqg4VvUrmb7ILxuRlC0HdUOWqrXDBygiJJJCHbzV5Ldz3VjnZQXAA91JgKDOFUE4O5QSk+SsfjHdUDvrQHPcoL4MdW6rPguCj6sdsKhcSgqRthWq5mCcHssjEZQY7O6uIyr5OhoIafmoSTnugOdnZURSxdH75BEBkoe6yZfD6fhIysfud0F0X0sFSSAY2VjiA34cIx+SM/mQRnuiypBF0HpIJWKgdRPmiIMZ3QVAOVVXP6enbuo8lBdgBMAq3JTJQXIrclMn1QXYCK3JTJ9UF7fpBSjGBnKx8lVD3DzQSHurXd1b1FAfVA6VRStLSN1a/pz5ILR3VQBlUOMeSpkoJiRhRO+kVTJKICK7b5Jt8kFqKpxjyVEBERATJRVA27IBOfJGjzQ9uyuZG542BQXK13f6lcY3DyKjcSCQgk7D6lzByafhY8Iv4a2f9bjXDnV6lcx8mjermx4RY8taWc/8rjQfSGiIgIiIOuftDvwNuJf+J0f69Tr55G919DftDvwNuJf+J0f69Tr55GjJQHdlRXuY70VhBCCrPpK5xwrWd1V5GdkFqIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiCre6qrcpk+qC4jKdgrclMlBcDnZD2VoO6kBGO6CxvdX+SoSMK0n0QB3Kqrcn1TJ9UBEGM7q7b5ILUVTjG2FT60BSN7K3b5KmcHv5oJWqN/dXh+EJaW5PdBG0ZKyoWNzk/WsbIBVfFcDsUGpNa0NI/nWHUYzt2yohO8A791Rry546jsgswfRFnsbT9IyRlQyiME9OO6CJpwVJ0h25CgJ3BVfEf2ygvkOyj32xlXAgn7FcOnHln60EbuykjAIVryPJWhxHYoJSBhRPO6F7j5qmcoO//sVPwptU/wDF/XfyjbkT2Kn4U2qf+L+u/lG3Ig9qUREBERAREQEREBERAREQEREHHvMR+D/xN/gdev1KVfM/Wf21P/hXfpK+mDmH/wB4Dib/AAOvP6lMvmfrP7cn/wAI/wDSUEKKoxjyVdvkgtRXbfJUd3QURXbfJUd3QURXbfJNvkgtRVOMeSogIrtvkqO7oKDZb3tfG3jLY7dTWiy8XNaUFDRxiGmpaW/VcUUMYGAxjGyANAGwAAAWytvkmR6oN/8A7YHjz+O3X3+ktb/SJ+2B48/jt19/pLW/0i2Ae2xVBjfKDkD9sDx5/Hbr7/SWt/pE/bA8efx26+/0lrf6RbAyPVMj1Qb/AP2wPHn8duvv9Ja3+kT9sDx5/Hbr7/SWt/pFsDI9UyPVBv8A/bA8efx26+/0lrf6RUPMDx6x/v3a+/0lrP6RbByPVUPog38OYHj1n/fu19/pJWf0iu/bA8efx26+/wBJa3+kXH+w3VQQfNByB+2B48AZPG3X31fskrf6RVHMDx5IyeNuvfs1JW/0i4/BGfsVHHJGEHIP7YDjx+OzX3+ktZ/SKh5gePIxjjbr7/SWs/pFx8HEKpfnGyDf/wC2B48/jt19/pLW/wBIsS78aeMl/ttRZr7xZ1lcqCrYYqikq79VTQzMPdr2OeQ4fIghbK6gmQcIKHZXsVNvVNsoLi7PfsVb2Qn6lbnKC8b7YKOa0dla0/PCrsfRBacIrtvkm3yQWoqnGNsIMY8kFEV23yTb5ILUyfVXbfJNvkgtzlFdt8k2+SC1FU4xthBjHkgplERARVOMbYQYx5IKZRVOMbYQYx5IKIqnGNsIMY8kFEV23yTb5ILUV23yTb5ILUQYzurtvkgtRXbfJWnGdkBFdt8k2+SC1EGM7q7b5ILUREBEVRjHkgoiu2+SbfJBaiqcY2wgxjyQURVOMbYQYx5IKIrtvkm3yQWqreyp9anj6Mb9yghcsyjAwc4UMkZP0fJWNErPogoMyUN8sBYMn0irj4zvIrJhbGG/fcAoMJc08luP22PCTP8AwwtP63GuHZxGPo4XMHJiHHmw4R9PlrO0E/xuNB9IKIiAiIg65+0P/A04lf4nR/r9Ovnmix1jK+hr2hrS/k24lNHc0dH+vU6+eNmc5HkgytsHuoJRvsqGR31FT07PFG/c+aDGb3R3dZdRA2MEgfUsPKAiKoxjyQURXbfJNvkgtRXbfJNvkgtRVOMeSogIqnGPJBjHkgoiqcY2wgxjyQURVOMeSogIgxndXbfJBaiu2+So7ugoiu2+SbfJBaiu2+SbfJBaiqcY8kGMeSCiIMZ3V23yQWoh7qoxjyQURXbfJNvkgtRBjO6u2+SC1FU4xthBjHkgoiu2+SbfJBaiu2+SbfJBaiqceSDGPJBRFU48kGMeSCiIMZ3V23yQWoqnGNsKn1oCKpxjbCDGPJBRFU48kGMeSCiIMZ3Q4zsgIrtvkm3yQWortvkm3yQWoqnGNsIMY8kFEV23yTb5ILUV23yTb5ILUV23yTbB7ILUQYzurtvkgtRVOPJBjHkgoiqceSDGPJBRFU4xthBjHkgpk+pTOUGM7q7b5ILUV23yTbB7ILUQYzurtvkgtRXbfJNvkgtRXbfJWnGdkHf/ANip+FNqn/i/rv5RtyKvsVsftptU4/4AV38o25EHtQiIgIiICIiAiIgIiICIiAiIg465jTK3l64oGCMvkGjL2WNHdx9xmwPtK+aeejr3TSOfRThxcSR4Z2Od/JfUhdrXQXy11llu1MKiiuEElLUwuJAkie0tewkEEAgkbEHdcQO5MOWFzi53CS2kkkk+81O5P/7VB85PuNb/AHnP/kynuNb/AHnP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8Zqf6VB85PuNb/ec/8AkynuNb/ec/8Akyvo2/aXcsP4o7b/ABmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP8A5Mp7jW/3nP8A5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/kynuNb/ec/+TK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/wAZqf6VB85PuNb/AHnP/kynuNb/AHnP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8Zqf6VB85PuNb/ec/8AkynuNb/ec/8Akyvo2/aXcsP4o7b/ABmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP8A5Mp7jW/3nP8A5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/kynuNb/ec/+TK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/wAZqf6VB85PuNb/AHnP/kynuNb/AHnP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8Zqf6VB85PuNb/ec/8AkynuNb/ec/8Akyvo2/aXcsP4o7b/ABmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP8A5Mp7jW/3nP8A5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/kynuNb/ec/+TK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/wAZqf6VB85PuNb/AHnP/kynuNb/AHnP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8Zqf6VB85PuNb/ec/8AkynuNb/ec/8Akyvo2/aXcsP4o7b/ABmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP8A5Mp7jW/3nP8A5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/kynuNb/ec/+TK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/wAZqf6VB85PuNb/AHnP/kynuNb/AHnP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8Zqf6VB85PuNb/ec/8AkynuNb/ec/8Akyvo2/aXcsP4o7b/ABmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP8A5Mp7jW/3nP8A5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/kynuNb/ec/+TK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/wAZqf6VB85PuNb/AHnP/kynuNb/AHnP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8Zqf6VB85PuNb/ec/8AkynuNb/ec/8Akyvo2/aXcsP4o7b/ABmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP8A5Mp7jW/3nP8A5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/kynuNb/ec/+TK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/wAZqf6VB85PuNb/AHnP/kynuNb/AHnP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8Zqf6VB85PuNb/ec/8AkynuNb/ec/8Akyvo2/aXcsP4o7b/ABmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP8A5Mp7jW/3nP8A5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/kynuNb/ec/+TK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/wAZqf6VB85PuNb/AHnP/kynuNb/AHnP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8Zqf6VB85PuNb/ec/8AkynuNb/ec/8Akyvo2/aXcsP4o7b/ABmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP8A5Mp7jW/3nP8A5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/kynuNb/ec/+TK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/wAZqf6VB85PuNb/AHnP/kynuNb/AHnP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8Zqf6VB85PuNb/ec/8AkynuNb/ec/8Akyvo2/aXcsP4o7b/ABmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP8A5Mp7jW/3nP8A5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/kynuNb/ec/+TK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/wAZqf6VB85PuNb/AHnP/kynuNb/AHnP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8Zqf6VB85PuNb/ec/8AkynuNb/ec/8Akyvo2/aXcsP4o7b/ABmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP8A5Mp7jW/3nP8A5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/kynuNb/ec/+TK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/wAZqf6VB85PuNb/AHnP/kyq+5Vo/wDuOf8AyZX0a/tLuWH8Udt/jNT/AEqftLuWH8Udt/jNT/SoPnNbBcB2pZv8mf8AUq+Fcv71m/yR/wBS+jH9pbywfijtv8aqf6VP2l3LD+KO2/xmp/pUHzniK5Z2pJv8kf8AUrHUte45dST/AOTP+pfRn+0t5YPxR23+M1P9Kn7S7lgzn/Yjtv8AGqn+lQfOV7lWn/7jn/yZXMvJrBcKfmu4RvbRSkHWdoa7MZADTVx5OfLA3Xuh+0u5YfxR23+M1P8ASrP09yn8vOlL5Q6k0/wyoKK52yoZVUlQyonJilY4Oa4BzyCQQDuD2QcuoiICIiDrj7RB9THyacSn0kDppRR0XSwDJP8At+nzt9WT9i+extHXDOKSff8A+hn/AFL6idV6U0/rjT9ZpbVNsjuFqr2tbUUz3Oa2QNcHAEtIOzmg7EdlxX+0u5YPxR23+NVP9Kg+cr3KtP8A9xz/AOTKkjguUf0KSf8AyZ/1L6Mf2l3LB+KO2/xqp/pUHJdywjtwktv8Zqf6VB850kNzlGH0s5H+CP8AqUXuNb/ec/8Akyvo3/aX8sJ78JLb/Gqn+kVP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQfOT7jW/wB5z/5Mp7jW/wB5z/5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/AJMp7jW/3nP/AJMr6Nv2l3LD+KO2/wAZqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/AOTKe41v95z/AOTK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQfOT7jW/wB5z/5Mp7jW/wB5z/5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/AJMp7jW/3nP/AJMr6Nv2l3LD+KO2/wAZqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/AOTKe41v95z/AOTK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQfOT7jW/wB5z/5Mp7jW/wB5z/5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/AJMp7jW/3nP/AJMr6Nv2l3LD+KO2/wAZqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/AOTKe41v95z/AOTK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQfOT7jW/wB5z/5Mp7jW/wB5z/5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/AJMp7jW/3nP/AJMr6Nv2l3LD+KO2/wAZqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/AOTKe41v95z/AOTK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQfOT7jW/wB5z/5Mp7jW/wB5z/5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/AJMp7jW/3nP/AJMr6Nv2l3LD+KO2/wAZqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/AOTKe41v95z/AOTK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQfOT7jW/wB5z/5Mp7jW/wB5z/5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/AJMp7jW/3nP/AJMr6Nv2l3LD+KO2/wAZqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/AOTKe41v95z/AOTK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQfOT7jW/wB5z/5Mp7jW/wB5z/5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/AJMp7jW/3nP/AJMr6Nv2l3LD+KO2/wAZqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/AOTKe41v95z/AOTK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQfOT7jW/wB5z/5Mp7jW/wB5z/5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/AJMp7jW/3nP/AJMr6Nv2l3LD+KO2/wAZqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/AOTKe41v95z/AOTK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQfOT7jW/wB5z/5Mp7jW/wB5z/5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/AJMp7jW/3nP/AJMr6Nv2l3LD+KO2/wAZqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/AOTKe41v95z/AOTK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQfOT7jW/wB5z/5Mp7jW/wB5z/5Mr6Nv2l3LD+KO2/xmp/pU/aXcsP4o7b/Gan+lQfOT7jW/3nP/AJMp7jW/3nP/AJMr6Nv2l3LD+KO2/wAZqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/AOTKe41v95z/AOTK+jb9pdyw/ijtv8Zqf6VP2l3LD+KO2/xmp/pUHzk+41v95z/5Mp7jW/3nP/kyvo2/aXcsP4o7b/Gan+lT9pdyw/ijtv8AGan+lQeY3sWqeoh5pNUumgkYDoCuGXMI3+6Nu27IvV/h7y+8HOFF6n1Fw+0PSWW41NK6ilqIZpXF0DnseWYe8jBdGw5xn4QiDkZERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERARF1x53ecO08nnDy1andp2PUN8v9w9xttqfWe7B7GML5p3PDXHoYCwEAZ6pWDIBJQdjkXXTki5o9T82vDG58SL9w2h0lTUt2fbKLwriaptYGRsc94zGwtDS8NzuCQcEEELlLWPHHgpw7uYsvEHjBonTFwcwSCkvOoKSimLSAQ4Mlka7BBBzjHZBvlFxjQ8zvLXdJXQ2zmF4aVkjR1ObT6soJCBnGSGykgbjf5rf1lvtk1JQMuunrxQ3SikJaypoqlk8TiDggPYSCR2O6DUEUNQ6VkEj4IhLK1pLGF3T1OxsM74z2yvPXgL7V+p4kcwFs4IcTeDlPok3W5S2QVX3ZdUPpLiHFkUErHRNB6pW+FkEYe9pxjKD0QREQEREGl6j1DZtJaeumq9RVoo7VZaKe419QWOeIaeGMvkeWtBccNaTgAk42BOy62n2n3IwCR/s6R5H/c9dv6quXuZVjZOXPipG7s7RV8afqNDMvmhc3Di0b4JCD6AP3T/kX/HrH/o7dv6qn7p9yL/j0i/0eu39VXgNHRPcOp5LR3zhWywRNHwPJP1IPfv90/5F/wAekX+j12/qqq32nnIy4bcdYvt09dh/9ar5/iwoI3ncBB9AJ9p7yMDY8dYv9Hrt/VVT90/5F/x6x/6O3b+qr5/uh2cYRzHN7hB9AI9p9yMHYcdIvt09dv6qqn2nvIwO/HSL/R67f1VfP8xjpDhoyVV8T4z8Qwg9/wD90/5F/wAesf8Ao7dv6qn7p/yL/j1j/wBHbt/VV8/zWl5w0Kro3M7oPoA/dPuRj8esX+j12/qqH2n3Ix+PWL/R67f1VfP6pGQSyNLmtyAg9/v3T/kX/HrH/o7dv6qqj2nvIwRtx0i/0eu39VXz/CNxf0Abq8wFoy44Qe/v7p9yMD/s6Rf6PXb+qp+6f8i/49Y/9Hbt/VV8/wAGknDd0cxze4wg+gH90/5F/wAesf8Ao7dv6qn7p/yL/j1j/wBHbt/VV8/uMqWOJrh8TiD6IPf390/5F/x6x/6O3b+qoPafcjB2/wBnSL/R67f1VfP84YJGc481NDDC9pL5CD5IPf0+075GgMnjrD9mn7qf/rVW/un/ACL/AI9Y/wDR27f1VfP84AOIaSQELHDuEH0A/un/ACL/AI9Yv9Hrt/VU/dP+Rf8AHrF/o9dv6qvn9wScIQR3QfQH+6fcjH49Yv8AR67f1VU/dP8AkX/HrH/o7dv6qvn92+auDHEZHZB9AH7p/wAi/wCPWP8A0du39VVR7TzkZP8A2dIv9Hrt/VV8/vSSceaOaW90H0A/un3Iv2/2dIv9Hrt/VUPtPuRj8esX+j12/qq+f7BAzhUQfQF+6f8AIv8Aj1j/ANHbt/VU/dP+Rf8AHpH/AKO3b+qr5/VNBCyVwD3kZ+SD38HtPuRg9uOsX26eu39VVT7T3kZbseOsX2aeux/+tV4F1Nvjhj62SOdtnBAWAg+gL90+5F/x6Rf6PXb+qp+6fci/49Iv9Hrt/VV8/vdSNhyMk4Qe/wB+6f8AIv8Aj1j/ANHbt/VVUe075GX9uOsP26fuo/8ArVfP64YOFRB9AR9p9yMD/s6Rf6PXb+qp+6f8i/49Y/8AR27f1VfP8Gk9kLSO6D6Af3T/AJF/x6x/6O3b+qqo9p7yMH/s6Rf6PXb+qr5/E2+aD6Av3T7kYH/Z1j/0eu39VT90/wCRf8esf+jt2/qq+f1AMoPoC/dP+Rf8esf+jt2/qqqPaecjLhtx0i+3T92H/wBar5/zDI1vUWkK+OONzSXvIPlhB7+fun3Ix+PSL/R67f1VP3T/AJF/x6x/6O3b+qrwBjja6Toc4gKs0TGODWOJz8kHv7+6f8i/49Y/9Hbt/VU/dP8AkX/HrH/o7dv6qvAAxPDc42ViD6Av3T/kX/HrH/o7dv6qn7p/yL/j1i/0eu39VXz+og+gL90/5F/x6x/6O3b+qp+6f8i/49Y/9Hbt/VV8/qIPoC/dP+Rf8esf+jt2/qqqPae8jB/7OkX+j12/qq+fwDKqBvgnCD6Af3T7kX/HrF/o9dv6qn7p/wAi/wCPWP8A0du39VXgE6JmMteSfIKIgoPoC/dP+Rf8esf+jt2/qqfun/Iv+PWP/R27f1VeAETGvOHEhXPh6T8JyEHv7+6f8i/49Y/9Hbt/VU/dP+Rf8esf+jt2/qq+f3siD6Av3T/kX/HrH/o7dv6qn7p/yL/j1j/0du39VXz+4yhGEH0Bfun/ACL/AI9Y/wDR27f1VP3T/kX/AB6x/wCjt2/qq+f4scO4VAMoPoC/dP8AkX/HrH/o7dv6qn7p/wAi/wCPWP8A0du39VXz+og+gL90/wCRf8esf+jt2/qqfun/ACL/AI9Y/wDR27f1VfP6qtY53YIPoCHtPeRg7DjrF/o9dh/9aofae8jA2PHWLPy09dv6qvn/AHwvYMkHCs+tB9Ah9p1yNNHV/s6w/wCYLqf/AK2VG+095GCNuOkX26euw/8ArVeAAjDvok5VjmlpwUH0A/un/Iv+PSL/AEeu39VQe0+5GDsOOkW//c9dv6qvn9VQwkZCD6Bn+015HWN63cdafB7YsV0J/IKbKi/dP+Rf8esf+jt2/qq+f1EH0Bfun/Iv+PWL/R67f1VP3T/kX/HrH/o7dv6qvn9TBQfQF+6f8i/49Y/9Hbt/VVUe085GXDI46Rfbp+7D/wCtV8/ibfNB9AX7p/yL/j0j/wBHbt/VU/dP+Rf8esX+j12/qq+f1EH0Bfun/Iv+PWP/AEdu39VT90/5F/x6R/6O3b+qr5/QMqvSQRnbKD6AR7T7kYP/AGdIv9Hrt/VU/dPuRf8AHpF/o9dv6qvAJ0MbWhweST5YUSD6Av3T/kX/AB6x/wCjt2/qqfun/Iv+PWP/AEdu39VXz+og+gP9085GMZ/2dIv9Hrt/VVT90/5F/wAesf8Ao7dv6qvn+IwFRB9KvA/md4G8x7Ly/gvrpuoxp807bl02+rpfAM/ieF/bEUfV1eDJ9HOOnfGRnlNeWPsNY2ik4zyb9TpNPNP1AXAj9JXqcgIiINtcROIejuFGjbnxB1/em2nT9nYyStrHQySiJr5GxtPRE1zzl72jYHvk7Arr9+6gci/49I/9Hbt/VVqvtF4WVHJfxMhkGWmjoifsr6c/zL576WGOYvEjy3DcjHmUHv3+6f8AIv8Aj1j/ANHbt/VVUe075GTuOOsP26fuo/8ArVfP+6IhxDdwFNFSsc3MjiCg9+f3T7kYH/Z0i/0eu39VT90/5F/x6x/6O3b+qrwCmijj+i8n7FFjKD6Av3T/AJF/x6x/6O3b+qp+6f8AIv8Aj1j/ANHbt/VV8/zWl3ZVdE5vcIPoA/dP+Rf8esf+jt2/qqfun/Iv+PWP/R27f1VfP6qhjj2CD6Af3T/kX/HrH/o7dv6qn7p/yL/j1j/0du39VXz/ADWOecNG6q5jmHDtkH0Afun/ACL/AI9Y/wDR27f1VP3T/kX/AB6x/wCjt2/qq+f4gAZBVEH0Bfun/Iv+PWP/AEdu39VT90/5F/x6x/6O3b+qr5/VVjepwBOMoPoB/dP+Rf8AHrH/AKO3b+qp+6f8i/49Y/8AR27f1VeAs1KI2dYeSoGAOOCUH0A/un/Iv+PWP/R27f1VP3T/AJF/x6x/6O3b+qrwGZT07gPvpz5jZRyQEH4MkIPf390/5F/x6x/6O3b+qp+6f8i/49Y/9Hbt/VV8/uDnClbSyvHU1uQg9/f3T/kX/HrH/o7dv6qn7p/yL/j1j/0eu39VXgVR211U/pL8E+gytctegLzc6yOCOI+G5wBeAcgeqD3Z/dQORf8AHpH/AKO3b+qqo9p7yMntx0i+3T12/qq8eqjlufXWeGazOlNUQM9zk/MLXtNcmWpbrbnyVEU5nIy3DSMfYg9Z3e0+5GG9+OsX+j12P/1qg9p7yMOGRx1i+3T12H/1qvI1vIxxZlqXMio5DFvh3hHJC2zf+VfX1hrRQTwPMzj0gGM90Hsv+6fcjA/7OkX+j12/qqfuoHIv+PSP/R27f1VeJWreXziFo2hFxu1uLICOoO6SNlsWjtTJiRPIWlpxgIPe/wDdP+Rf8ekf+jt2/qqfun/Iv+PWP/R27f1VeCFbaoqZvVFK53yOFppY4eSD6Af3T/kX/HrH/o7dv6qn7p/yL/j1j/0du39VXz/FjhuVRB9AX7p/yL/j1j/0du39VT90/wCRf8esf+jt2/qq+f1EH0Bfun/Iv+PWP/R27f1VP3T/AJF/x6x/6O3b+qrwBhja84c4gKkjGtf0tJIQe/8A+6f8i/49Y/8AR27f1VP3T/kX/HrH/o7dv6qvAaGmY76bz9iimYxj+lriR6oPf790/wCRf8esf+jt2/qqfun/ACL/AI9Y/wDR27f1VeAckUbWBzXknz2UKD6Av3T/AJF/x6x/6O3b+qp+6f8AIv8Aj1j/ANHbt/VV4ARsa4/E4hHsAI6TlB7/AP7p/wAi/wCPWP8A0du39VT90/5F/wAesf8Ao7dv6qvn9RB9AX7p/wAi/wCPWP8A0du39VT90/5F/wAesf8Ao7dv6qvn9xlSsiYWFznEFB7+/un/ACL/AI9Y/wDR27f1VP3T/kX/AB6x/wCjt2/qq+f47EqiD6Av3T/kX/HrH/o7dv6qn7p/yL/j1j/0du39VXz/AAAPyVCADsg+gL90/wCRf8esf+jt2/qqfun/ACL/AI9Y/wDR27f1VfP6iD6Av3T/AJF/x6x/6O3b+qp+6f8AIv8Aj1j/ANHbt/VV8/qIPoC/dP8AkX/HrH/o7dv6qn7p/wAi/wCPWP8A0du39VXz/NaD3OEcADscoPoB/dP+Rf8AHrH/AKO3b+qp+6f8i/49Y/8AR27f1VeAMLGvPxuICpKxrHYYSUHv/wDun/Iv+PWP/R27f1VP3T/kX/HrH/o7dv6qvABsL3DICk93AaS95B+QQe/f7p/yL/j1j/0du39VT90/5F/x6x/6O3b+qr5/iMnZXCFzgg9//wB0/wCRf8esf+jt2/qqfun/ACL/AI9Y/wDR67f1VeADo3NGT2WXBSU0kJkfM5rvIDCD30/dP+Rf8esf+jt2/qqfun/Iv+PWP/R27f1VfP8Aub0uIG4BxlWoPoC/dP8AkX/HrH/o7dv6qn7p/wAi/wCPWP8A0du39VXz+og+gL90/wCRf8esf+jt2/qqfun/ACL/AI9Y/wDR27f1VfP6iD6Av3T/AJF/x6xf6PXb+qp+6f8AIv8Aj1j/ANHbt/VV8/qkbBI8ZDSUHv8Afun/ACL/AI9Y/wDR27f1VP3T/kX/AB6x/wCjt2/qq8BoaXrcRIS0D0CjqImRP6WOJCD39/dP+Rf8esf+jt2/qqfun/Iv+PWP/R27f1VfP6iD6Av3T/kX/HrH/o7dv6qn7p/yL/j1j/0du39VXz+og+gL90/5F/x6xf6PXb+qp+6gci/49I/9Hbt/VV8/q1ujsdPU0ZqHTPDsZHbA2Qe8/wC6f8i/49Y/9Hbt/VU/dP8AkX/HrH/o7dv6qvAB8REjo2nODj608GTIBGEH0ecGOcblw5hdUVWi+D/ElmoLzRUD7pPTC011N0UrJI43SdU8LGnD5oxgEn4s4wDgvML2LMD4+aTVLnNODoCuA/zjbkQe0yIiAvCv2svGKbiTzVV+kKSqc+0cPaGKywMB+A1TwJqqQA9ndb2xH18AY9T7qL5uqOQ8cub2nkq3sqhr7iPGHkkFsja25gY74IIl8jjHnjdB708p3CyDgvy48PuHUdOIai22SCSuHSRmtnHj1JwQDvNLJjIyBgeS8SfaRSSS87XFJ8sjnuFfSNBcckAUNOAPqAAAHkAAvoSXz1e0e/DZ4p/+EaX9Sp0HOmkPY48WNbaDset7Rxd0lEL/AGilutNTVNLUt6BPC2RrHva12CA8AkA9tgV1j0trnjzyP8c6+htF2qLFqXS1wNJdrcJzJRVzWkExysB6ZYntIIPcBwc0tIBHpnw29q/ypaF4QaT01cJdX1d10/py30FRTUtnBMk8FNGx7WPfI1hy5pAJIGPNeeVwtHEP2hnNtfbzouxwW6p1hdGzvFRUN8K2ULGNiY6V56etzIYwS1o6nlp6WkkBB77cPtYUXELQWm9f2yN0dFqa0Ud4pmF3UWxVMLJWAnAyQHgZwPqXh17T/h3Nwg5zb3f7B4lAzVEVJq6hkhy0x1DyWzPa4fvjUwSyZG4LgvcjQmkrfw/0Rp7QdoLnUGm7VSWilJABMNPC2JmQNh8LBsvL724Wl4ortwm1rFGPEqae7Wuof2PTE6nkiHfcZlm7D6zuMB6NcvHFOLjZwP0TxViDGyajs1PV1TGfRjq+npqIx8mzNkaPkPJcirox7HbV8uoeUqWwTzlx0tqivt0UZIyyKVkVUCAPIvqZe/mCu86AiIg455jxnl44oj10Xex/yGZfNA49Mzj6OP6V9L3Mj+DvxS+Wi73+ozL5oJN5H/8AfH9KDMkrg+HwmjBx6LCGSQPVOkqseA7dBqUVO1zAdtxuoJHsgHT3ypPePvYbGd/qWFKJCSXoL43h0mT2/wDalTg4x6qAHCqXF3dBkUDwyX4lJcZGyOGMLCBIOQqucXd0F8MnhP6sZ2V08olII+rsoUQXlnwjBWTS1jYWFjhuRhYzH7jq7K97A7dqCQFrJPHG4PcKskrZ2nGyxy92Oh3YKRuw+FBG13hvz6bKssniHYKx30z9ali6OnfGcIIhsd1I05GQo346jjsjXYQHfSKkZCXt6s4Cp8KlZS1D2l0QJAQRMb986TvlSyNbjI2UOCx+HbEI6Qkj0CC5g+LtsqSgZ28leS0s+DurRFK7cg49UESy6YNMZDgoadodJ0uP1KSozAegeaC3AZI4+Stld1uACsDnOP1qvQ4kEIJpIumPqaN+6xsYWbEfhDZN1BUhgOGdkEKq0lrgeykp4/EfhX1MXhYHYlBlmUTQhhIwdgtPkj6Djv8AUjHSZw1ShvnIgupqcOPU4qWoiDG7brG8dwPSz1U8Uhe3755oMN/0iqKWSJ7nktGQSo3NLThwwUFWODTlVe/r8uxVivZ04JKCxSxwh4zlRHGdlc2VzeyC6SLoVjThwPop3EzNw3vlWeBI3BLcBBPLUtdD09O/qsPJ9Vk+G2Roa3GVA+J8Zw4IKdR8tkaT1DJKdJ9Fc1hxlBkyvYYceaw1eOt/wjuqOY5pAIwSgtRXGJ4b1EbK1AREQXM7o/uqxEB26mIa4ZQQNJB81R25V78Z+Hur44S9h23QQZWZTAGM9XdQOhcwkuGAFRsj8hrPPZBbKMPPorVnOtk5j8UtODvusRzeiTDkFoyN8IN3D5rKlEPgggDKxW/SGEGTNGGx5woWAFpUsof4YJzhWxMPQThBAdiUV3SXPIHdUc1zDg7FBRZ1HG3p63HssNjOryWRG57MNBQTSysl+94+sqF1H0sL8qpiezLzkb7K7Mr2EN3+pBHSENlw71U9VB1/G0HCw3MkicXdsFanRzNfATJ6INMEQzglZjImNiz3OFhyuPiO6e2VJDUu6gHHb1QQvB6yAPNAzK1GQUxj6ts4ysFzXO3aNkEZACAhHAjuqIB3QboMZ3Q90F3RtnKtVS4nbKuZjG6C+mYHvwVWqYGOGComyFhy3ZXOc6Td26CPJ9UQ90QEG6qcY8lRBeQMKxVLidsqiD1c9hr/ALncZf8ADWD/AKNevUxeWfsNf9zuMv8AhrB/0a9epiAiIg65+0O/A04l/wCJ0f69Tr55ASOxwvoc9oa3q5NuJQ/+k6P9ep188be4QZAd0N6sZKsdO522MZUzHROaOrfCskEWD0jdBdFS+Lv1BRVEYhPSEiqHMOM7Kk0viv6iUGTa4WzSlr+3qp7jA1jg1gyOywopnU+HsWU2qE7cyHdBiSwdA6gFYyTpBBWV1ZPxdljT9HV8CBBKInlxGQlRKJX5AUZaRvhU7oCKpY4dwg7FBTCuZkOB9FWHp6j1Kd5iA2AygpNIHRgd/koooy/tlWvd1bDtlZED2MYcnBQRPgcxvV1KragtaQRnbGVkRUs9e/ohGSCsz7kmkH+2Bg/NBpULHPlAxnJW/dL6d+6WIjCTlvotAoqamdI0NxnI+a5n4fWqMgOaB1BuRsg5E4H8r7NbVUbjATlw8l2+sPJta7MY4JLe3rLRv0j0WzeVqtqrVWsLnANBC7aXPWk4uMBEoIDQg0Dh3yt2y2ETVFuEjB8WOnIW7X2jTmmtRQ0P3HYGhwbjoC5A0fxGt0VEGV8rR23BGVsDXWv9LR6njuMr2eEx+XHIQc50Fp0+2giqIbTSxxlgdvCMjbz2XF/EHg7pO+XSLUDLRFiNwLiGDBPmpavmM4fy2r3e3V7Hzlgb0Bww3ZbTvHM3pWhsclBPPGJXAgfEMoOtnPRe9JwaTfZLdbYmSxRFp6WjOcLyemjcx1Q7pxh5/Su/HM9rui1JVVNQXgxPBIyfJdG70aVz5RBjBeSg2zJVOmd4bvLuseWNoO3n6LIrIWRElndaeZXu2ccoKyPDgQPJRorxE8jIGyCxEIIOCqgHICCmcKoODkq4wuAyrCMIL3SkjAVWx9TS4nfuo1UPIGEFDnsUwfRO6mZ0dG+MoIcqvV67qvSXuIaELSwjI7oLVUAK5+MbKzJQAcJklXR9Od1cYi4/D5+SCNMK50bm9wpGeH0kYGUEKkEQLeoq+Onc92w2KSwzMOMEBBAdlVo6nYVOyua7G/mEEr6cMb1Z8sqBXule4YJVre4QU3+anjpw9nVlXNDCO2+FY6VzPhB2QRuYWkgFA1436SQEc7qxv3W4LcyhdRnxiOrGd0Gl0s3cObv6KGsdl4xsFlRxxmocG9t1jVbfv3Q0b7oMdpwVnU3S5uT3UAopukOLdirWvfC8NJwPMIJ6ohwxjHzWH1OAx1HH1rOqZYzFluOohYGcoCnbThzesuwoFK0zBuG9kEZGCQqDdVPc57qndBUgAd1RXhru57K1+AUFFlwVYiaGlufsWIsiMRGM57+SBLVdZ+HZQFxd3V4j6TuNvJWvxnZBaiIgJg+ivj6c/Epz4RG3dBHTw+M4gnGFqIuPukJpgfktN8R0Tss2Ub3l7up3dBIyXEpkIzuSslj2vw/YfWsFXCQhvT5IPQj2MFU2bmi1RG1uMaBrT/zjbv8AWixPYq/hT6p/4v67+UbciD2pREQF82nKG+kj5q+Drq2F0kZ11YwA04IkNdEGH6g8tJHoCvpLXzeahpZuX/m7rIqnMB0DxBMzXNaRiOkuAex7QDnBYxrhg5wRug+kNfPV7R78Nnin/wCEaX9Sp19CTHslY2SN4c1wDg4HII8sH0Xz2+0e/DZ4p/8AhGl/UqdB6BcK/ZKcrOtOF2kNX3au1yyvvtgt9yqvAu8LWCaemZI/oaYDgdTjgEnbG66E883KNVcmPFOz27Teq626WK+U5udjr5QIq2nfFIA+J5jwC9hMbhI0NBDxgAghdueG/tkuHGhuHeltFVPBfUlVNp+y0VrknjuMDWyuggZGXAEZAJZkA7gFdMubnmm1xzt8WrVeINIS0FNSQttOntPUTnVs+Xvy45awOlmkeWjDWgYaxoBIJIeuns2OP+qOYXljoL/reslrtQabuc+m6+vlOX1roY4pY5nHu55hniDnHdzmucdyuuPtwH0o0NwqjfG73k3W5mN47Bghh6wd+5JZjY9juPPs57O/l5v/AC3ctNq0lq+EU+o79Wzahu9KDn3SedkbGQk9i9kMMIdjYPDgCQAT0u9t/q1lTq/hZoOOTDrdbbld5mA/SFRLFFGSM+XusuDjzPfyDlf2Jn+8Lrv+F5/Uqdeiq6R+yC0XNpblAp73URFp1bqK43eMluCY2eHSD7M0jiPLcnzXdxAREQcccyX4O3FP+BV7/UZl80En9kd/3x/Svpf5kvwduKf8Cr3+ozL5oJP7I/6z+lAL8q1EQXwu6XKaR/iODcdyo6ZnXJhSVLDC8Ed0E01v8OLxO+ywDss6WsL4OgrBO6Ag3TBTsgl8IYyo3DBUjpMNwFESSgKeFwDSDsrIenq+JXyt6t2DZBZKMklWteR2VDnsVNF0dPxd0EBOTlFdJjqOOyozHWM9soKYPogGVqEwgEALcZxusAd0FSzA7rU6G6x00Jjc0EkYyVpgDpDgI+NzDhwQS1BE0jpG+ZyoMYVzCM7qVxYQgji2OSsmOWSSUQxxlxcQAsdkMkh+AErf+g7TbffYJLkGgAgnKDTabR1W2nFa6ncAd9wVot2oZGElsZ2OF2kvVTo/9jgp6V0XidOMDC4lm03DXGSRgBBOcoOIMOac9J2+SyYyOnJ2W8NQWWkoWHYAjutkyvw8hvbKCr5TkgeShJJOSq5yclDjyQSU83gv6sK6pnE7gRthWwR9b8eSrUxiNwA80FkcnQcq58plwAFYY3BvUVn0TacwkyYyEGOIRG0PJ3woxKS4KtRKS9zQdsqEbFBnucI4wfyqANE+XfNWgvmb0jyVuHwnBGMoLCCCR6KiypDGY8juVioCAZICIO6DNYwQND+/yVstYHt6QDlXUr/G+B/ZWV0TI3AMGyCKnm8KTrV1VUCd+R2+agVQPMoKg5GMdlUOd9HCmjDMA4Uh8M4AaMnzQQRkRnqP1q2WXxHAgdlmy0BMZkb9iwTGWOw7ugvfUdUYYArWxtc3JOCqugc0dXljKtDiEFpYQSBuhBHcELJpXMa8l52VagMkkDY8blBiKWPJ2zjKnfb5WR+IRt3WOA7q6RsUEkkDWDqBz67qSCTpxsrHRSgZd2VrT+ZBPK/xG4Pmo4ow14cQDg5VS7ZUB8wg1Q3Fxi8PAwBhaY9gnmAO2ShecbqwuIIIOCEGfW2eOnpxM2XJIzjK0poOR6rJdVTyDpe8lv6FaIS74h5oEkhcwNVGSFrC0hVjAyWlUl2dhuwygpTkiTqI7lZ0tC2dokBWGzsrzPM1uGuOPrQY/wDYpC0+SzKfpfI3tjKwHEk5PmrmTPYct7oNbuEcbabIO60qGr8PYq2Wsmmb0OO31qxkEj29TRsgyHTNnPT6qwuMWWA91BvG4+qua/qcC5BPDSePvndRVNP4DunOVM6oMTB0LGklfKcvOSgt6iRjJWRC8CPdYyvY17thlBSQ5eSgaMZyrnQSNGS0gKzJ7IKY8kwpadoc7DlWpa0PAb2QQoN1Ieno8sqxuM7oLxFkKgwHdPqrnFzR22UROTlBO+ANZ1Z7qBXmVxb0q1uOoZ7IKYPomFkyeH4W3dRw9Hn6oIsIsmXo6du6xkHq57DX/c7jL/hrB+ivXqYvLf2HDQLRxid5mosQ/I2u/wBa9SEBERB109oa7p5NuJTvSjo/16nXzxYX0Pe0Lx+034lZ7e50f69Tr55oOn4urthBFlFV+Oo4VEF8UfiHGVV8YY8DPdWs6s/D3R/XnLu6CeSLpjyFjgkHIWRTQz1R6GjICkno3U4xIEEcTvF+E4CvfS9J/wBSxmlwcfD7/JZbJyGnr7/NBFOwMYPVVgga9nWTjG6hlkc9xHllUbK+MFo7ILpnjJbhRKoaXnZXBha8BwQWYPomFlSwfACB3WbSUDX05kcBkDvhBp9PCJFbURGN2PIKRjvCnLT2U73wv+nhBkadr20M5c9uQey1m5VLLkR0NAz3Wk25tI525CypaqmZKIYnfEdgEEtJRSU7mvAzkrnPhTDV1VVGxtK4ggDt3XGelrHPVyQvqASxzx3Hlld8uWjg/RXiWB7Ymklo7hByny66Cnro45HU5YTg5xhcwcStOO01bjXiTdjc5z6LkjRXDWXSlrEtPABgZ2C2TxrpbtV6dnkew+GAQfqwg6u/7NNVNeZrX90TEGZGOrC4q4vcXaunglpILuTI7IB698rjvipfJLDqWpNFkSgnOD57rrre9e19fqBouLn9Afg5PYZQc2aI17qmnvD6urvUpiduAXnC0XiTxSvsl8Y6C8v8MPGcPK0a76gtjLCx9tIMxYOx37Limuq62vlJlB687IOUNXa5qL3aGxOqy9/RjPV32XEUtQ+Fxa9xOTndZcMNxib1ThwZj8yxq5jZGF7RugxKqXrBLTlYBDj5Kene3xemQ7ZU8oi6h0YA88IMDGFn00nTF04UEkXUMs7hTwuZHD0v2KDDlOZCVbnfKq/d5ICtwc4QTeOOnG/bCiJ6ihY4DKogqRhUQklEBEVQ0kZCDIow3q381SswHjp7AYULXuYdlRzi45KCmFUNyd1dEWg/F2Vz3sxhvdBY5oAyFfHN0YBCiJJRBLLIHjsrGtzuN8K1XMd0nfsUGRDViLYg7KstYHggA5UT4gGeJ5ndQoLmgOJJIH1qjmhvmqxtL3ABZLKTp+lugxFUDKyqmBkbMtxn5LFYcHHqgNcWoT1FZDaQu3yrfB8JwBwSgR05cAcFTU0Er5xEHFoJwrhIA0YHdZVHKyN4kc3JCDVvuC2npxMNyRnK0ialHvAe/bBWui7e8xiAHCwKul2LnPyTugsmqIY4ekAZA8loM7zI/OFmSA5LSSQsd4Y0gEd/VBEyIv3OVbI3odhTu6g3LBthY7i4nJQUAycKds4awt9VAAT2UohJbn5IIicklBsQUIwSPRVb3QXmYkYwoycrI+9dKgdjOyCinjgyOvPZQK8SvA6QgulkDst9FEN07p2QXOaAMhWoSSiAnZEQVLiQqK+Lpyco7HiDCCzCKeXo6Nu+AoEHoB7FYY5p9U/8X9d/KNuRU9iqf+un1Tk/9j+u/lG3Ig9qUREBeIXtfODE+gOZWPiVRUbmWjiHb46zxGtIYK+ma2GoYD2yWCCQ75JlK9vVwpzWcq2g+bXh3BoPW1bW2yW31rbhbbnQtjM9LMGuaRh4Icx7XEOZtkhpyC0EBg8jXFuLjRyr8PtYPn8WvgtbLRc8n4hV0f8AteRzsdi/wxIB6SBR8VuRLlR42a1rOInEvhLBddQ3BkUdVWx3avozOI2BjC9lPPGxzg0BvURkhoBJAGL+ULlPsvKLoi7aG0/rq9ajpLtcjcz90GRxsp5DGyN3hMYNuoMZ1Ek56R2xvz0g6qt9l3yKtcCOBoyPXUt4I/W1y3wt5ZuAPBSpNfwt4Tac0/XOaYzXU9IH1fQRgt8eTql6T5jqwfNcnogLwC5/9f3DmI51tR2zSLTc20lwptF2OGFwf474XCEtYexD6p8xaRsQ8bnuffWsp31VLPSsqpqZ00boxNDgSRkgjqb1AjIzkZBGQMgjZdL+X72WfCngTxhoOMk2vtQ6suVollqaCnucULY2VD2uaJ5C0EyPaHkg7Ydh2MgYDtFwW4a2/g5wl0jwttbxJBpi0U1uMo/+TSsYBJKdhu9/W87Ddx2HZb2REBERBxxzI78u/FL56Lvf6jMvmgk/sj/rP6V9MfMX0/tfOJ3V2/Ybes/V7jMvmfnGamUN7dbsfVkoIlUDJA9VUxuAyVRuBugy2RCBvi5zlRVE3juz2wjZy4dLuysl6AfhQIYvEd052CrPD4WMKxrnMPU04KvDZJ9zk480EedsK9kQcCSeysc0tOCrow8j4eyCe30nvk3hk4ClulvFA8NBzlQxVD6V3XGcOVtRVy1RBmOcdkEClim6AQclRHGdkQVccuyqZVzBlwCyPcJ3jrY3IQYqYVSxwcWEbg4WTE1rGkPGCUGM3qdsCqujLcfNTxRlsnU7spZnROGBjOPkgxYJRE7Kunl8dwwFYYnd8bIB0vH1oJPAIbnBHzUJbgkZWrTmI0w6QM4WkknzQZdFUshILluqiJqIDJDJ0kDIIK2TkhbgtNybDCYidyMBBvzhlaavVF6+51TWnoDsbnZcm6600zRdEGwt8QkZ2+pcJ6TuV1sleK+gJBLgcjK5+slXJrG3D7tHJ6ezvqQdbtTXaprKksMZaCtOo7EaqMvJx3Pdcta/0VQwyvNI0ZGcYXGomfauuKZ2MHGEGg1dC6nkLAM4WKWuHcLWn1EE8hccEFY1SIA74cAIMOmd0P3HdXVbupwOOylzE0Z2yseX74ct3QHTB0fQo2k5ABIyqxxPld0sGSrpIZISOsYQXOh6W9WQoVcXvI3Oyt7oJYJRE/JH5FWpnExBAxhWCF5GcK1zS04KCmUVek4yqICK7b5KWOkmlb1Mbt3QRxSmJ3UFWaYzHJV0TWsl6ZRsq1AiLh4YGB6IIRgDJGVIxgdjyRoGNxn5IQR9EbIJOgAZyrQ4tcD5Aq0F2O63fo7hFxZ4iQOqOH3C7V2p4mOLXPs1kqa1oIG4JiY4Aj0Qba+6Dg0Nx2URHinxFu/VvA/jRoCiNx13wh1rpukADjPd7BV0ceCcAl0sbRjO3futoxzsbH0nugo6braGHZR9ADwO4VMdTj0qh6mnfZBLNA1jeoOUcbuh4d6KreuU4yhYQQw9ygy5LkXxBnT22ysIO+PqUjqaRreo9kFM9w6h2QSSVJkYGkbKAHCqWluAVWPAcCe2UFd++CPrRZNQ+J0YDQM4WGglwMq9sQfj0UAa49ld1Ob5+aCcUzCDuNla5z4/gaMhRGVyzaWWEx/fMEj1QYPxB2cYyr+gOGSr6l7CT0D6lACe2UFxwO2+EBJHbsqmFwb1OPfdR90FpH5Fb0hbr4aaFrOJfEfSnDigroqKp1Xe6GyQ1MzSY4X1M7IWvcBuQC8EgbkA4XPvOhyI37k4otLXK6cQ7fqiDU8tVAz3egfSvgfAIychz3BwIkGCCCCDt5oOrLmgD5rNpp2MgwcZWCQqtJyG52QXtZ48p8spLB4bw0HO6n92lY3xGA9tiFjuLxIDIT3QTvp3eFnBWJjDsFao6oiMGMDOFpshHXkIL3QBrOrKrTzCPcqIyPIwSjQcIMmasbI3pwVC2HqaXKzpIOSsmORgjxnfCDHDuhxx5FUe/rKucwucS0bFWFpacFBRXNb++VqZIGEEvWHfD81Y9vSreyqXE90FFcG5GVbhVyRsgoc+aIgGUDOUVWsLjgKr43M7oPVz2Gz82vjGz+5nsJ/K2u/1L1KXln7DX/c/jKfWawfor16mICIiDrn7Q78DTiX/AInR/r1OvnjX0Oe0O/A04l/4nR/r1OvnjQEVTjHkqAEoL4n9ByQkj/EIVpY4dwqINStdYykdlwCvuVcyseAwjdaW0OdsFcYpG74xhBksgbEA87qOQGT4u2FY2Z5OHHZJH4OAgjOx+pUJycp3V0eOsZ7ZQZVuZ1y9JGyXFgjmAHkskSQwR9UYAJWHK2Sof17kZQXQz9fwkHbzWYa4RwljdzhYLuiMYGAR6KahjE0zfEHwoMXEkshcGkkn0WsU1jdUQGVxwcdluGmpbPDTh0haHHfsFj1EM8zvDt4JafT0QY2mdNirqjHI/AzjK3Qzhuz7qxPDwWkg91oNNYdSwkTUbH57nGVrdHe71ayHXEuDh6oOTZ7ZQ2K1wFrmhzMErsxwA43W7R1pFdLUtDomZwT6LovetdVVbF4JedvmrbFxBrKQe5SOd4b9j8Xkg9X9M8+tFfKh1n94DQD0hZ/EjmKs9w0nPSGsYXvYcbj0XmVZdV2m0NFdESJXDJPV5rF1Nxjr6yIxRSP6ew+I9kGbxX1Syo1PU1gmy17iRvnzXF1XYmX3qq2SBrskrTr5fp7pMXvcST55Wn0l2udOeiMnoKDUi6otTjHLOXMG3fOywn3sCoErWbA+Sju1aaiMFx+Iqlqgp3jMwGEGp1OqYq2AQeGGkDC0x0zSOkuByVNcaajDM0uC75LSPvjN35z5IL6iAMy8KBr3D5q4vkl+HJKz4I6cQESY6kGHFUdBJIJVz2Gb4wceeFZH4ZmPV2yrnShswDfojCCAgtJyEzvlarUU8U0OYQOpaZJC+IgPGCUFXSdTcY8lGrnRuaMkbK1ATB9FVrS7ssmOIeHuN0EEUYecHZSvZ4bcfarYYnuf8Pqsh+GNLZO+EGJG3xCc/WrXDBwFeyToccAHKvbA6RpkxjdBC0OPZZEFMHDLwomv8MkY+Sk96wNggsljDHEfkUSvkk68ZViAirjz7pudsIKmRxb0nsre6zLZabre6+G12a21VfW1LuiGmpoXSyyuwThrGglxwDsAey5N/am80LLV93HcuvEgUQGTIdL1oIbjq6+nw+rpxv1Yx80HFMQcHggLKLpXNyO+FdcKG5WSvmtl3t1TQ1lO7pmp6mJ0UsZwDhzHAEHGDgjzVjKxgABb+lBCW1Ez+nBP6FQ00oeGlu5WoU9RG09RA7qlRWMEwe0ZAKC59FVQ0wkAO4ysRmXA+J+QrVpr7HLTCHp3x6LTGtMmXtHz2QW9D25O+B5rKpmBzCSoZJ9ugtwQr6dwDPrQSMkdHKejOQUqKyY/CeygZKGSF2PNUqJhKQcdkF1PIDITJ2yqVcHXIDENlCxhecA4WT4wp4+hwySEFJI2+AGs3cFitiLARLtn1VYpyybrO4KrVTtneC0YCCPwXZy0ZGVORIyM/ASMKUPayIZG+MqsVUyTEbu52Qaa7OST3RoyQFqNbQtZEZge+600Z7jyQSuhwM5UYblwHqrvvjh54VfBkA6vRBWSHob1ZzlWhmQTnsj5HOGCeytDiNsoDW5OFMab4c52UAOFf4r8YygsOxIVwZkZyqFjhuQgccYCCmPJMeSy6JsZcfEGVZVhgePD7BBB0lUU9KWZPXuElLDOOnsgg3+aDdZdR4PhDoxkhYiD0B9iwwjmk1SfXQFd/KNuRU9isSeabVIP4v67+UbciD2oREQEREBERAREQEREBERAREQcccyef2uvFPG3/wACr3+ozL5ogcTEnc5K+l3mS/B24pfwLvn6jMvmhccSu/74/pQXPlJGOlRLJf4XR8PdYyAqg5KoiCRzfgyp6WUMYQQoYD1Ow7thX1ADNmoKT4cS4FIHYafhzuoS9x7lZVMYy3pI3OyDFecuJVFlzW+ojBkLPhPmseEN8Vod2ygswfQotbqhRCjBjx14WiIKtd0nK1Wlu0cUXhubknbstJAyr2hoOHZz8kGYYmPkM2Rvusepdl4LewQCfGG5woiHdWD37IJTUExhgByoviDgXZWp2ekYJxJVNwzCkvsNM6UOomjp80GJEA5u48vRY07C15wDssuCSNkYDtiFV74ZGntlBhidwHSSVkQ0YmYX9QHmVjvhdkkDZXRvqWNLWE4QRys8N5b6K6JkgIe3O2/dWODy4l3crUKBrXMId5oN06Pu8Uz200jRkHG67D6LoDPBEyJuQ4D7F1coJG26ZszdiXDddyOAtnqNR2AVELcvDMj68INscQ9Px2uhdVhoe4tJx3XCsuj5dRU89WYizoJPbC7CaztN1ornLFfGn3ZucZ7YWybrerBQ0EsNuc0E5BwRlB1qu1FJbKx9KSR0ZCwS4k7krcmsJYJ7g+ZhBJJyVt5kMkjctGyCkbPEd05wPPdSSNEIAG+VEOpr8N79lc7qOPEO6C+knFPKHEZCkq6ltVI3AwFB4DzuBsqdLo3AkYIKDOmpGspw/bt5LAjAL2j5qeWre+IRnsFA0OyC3v3Qa1JTRw0olBGSPVYDYBUAvBwR5eqo6aqmj8PfAH5VFE+SneGnYEoLJOppLCMBUZH1NzlavWCkNJ1tx143WjB7gMAoKxs63dK1ajrIqSMxOwSey0yDJdkd0nz1glBqDqH315lacA7lYz6bwSW98FTUtaWN6QdyMKkj+sk+qDFLQFc0fD80ct88CeHD+L/GfRPDEOeyPUt8pLfUSM7xwPkaJXj5tj63fYg9APZw+zp0vq7TNDzEcwlohuFprWGp07p6rH+15YAdqyrBwHsOCWRn4XN+JwcHALlnjZ7XDg7wkukug+CPDz9mUNnPufvkVWy3WqPo26aYNjeZWDBAIaxh2LS4YJ3n7UTirNwM5TodBaB/6kz6tng0xStpR0e62yOIunazyAMcbIcf3MpxggEeHD5Jc4cT2QetPDv21mibvc22ni7wVrrLbqhwjfX2m5NuDWMdsTJBJHGS0dz0ucSCcNJAB4R9pJoflau0Wg+NfLrWaeZHreOvdcaeylscEpiMJbM6nGPd5cyPa9pY0uIBIDg4noASSpo6mpYPCjkIB2wgpKBDM9rd8HGyjc7qIXLPLvy08SOZniNScO9A00TZZIzVV9wqcimt9KCA6aUgE4yQ0NAJc5wA7kj0tdyH+zo5VbDb38y+t4rzeapgkbJervPSeM4Hd8FFRvbIYwRghxkG5BO4ADx8Y/w91R0nU/qwvZXT/KT7LHmfbU2Tgxd6CmvbYXOxYNQVUVcxg28RtJVucHNacZd4RG4ydwvPHnL5L9dcoOsqa33WtF80te+t1lvkUJjbL0n4oJm7iOZoIJaCQ4EFpOCGh19fVl8YZjsjKotZ04XN/I3wl0Xxz5pdEcLeIdFUVen7064msggqHQPkEFvqZ2APaQWjriZnBBIyM75Xd3W/sjtOai5kai36Rmr9FcHrXZqWsr7hNUmpqairc6XxYKV0xOAGMYXSP6ms6hs8ktAeVxf1E580AJGV7M6M5bfZGXa5xcKLJd9Iag1BVSCmY9+sqt9VVT5GGxSxztic8kgBsWA7BGDuurftAfZ20PLRaYeK3Cq4XC4aGqKltJXUlc4SVNpneT4Z8QAeJC85aCQHNd0Al3VkB0JXpNdvZ/8AAWl9nvHzEwP1INZnSNPqQzm4NMBneGudF4Xh48P4iAPpdviK4G5EeGPKLxGuWszzV69g07FboaH7hxT3cW9lS6R03ju6zu4sDIQAD2kJIOBj2JrdEcv03KezQdVqWBvB/wDY3DRi6fdMCM2sBvRJ7z2wQB8fnlB85jHFuxCo7JOcHC9M+NfLz7LWx8IdZXnhzxjt9RqugsdbU2Sng1aKp89cyFzoIhFv1h8ga0jHZx3HddSeRrhTojjlzO6K4WcQqKersN6NwNZBBUPgfIILfUTsAewgtHXEzOCCRkZGUHBltpWVcha/GEuNK2kmEbDsV6t3j2TmiLtzO1UFibdNL8HbLZqOtrJHVbpp6+ue6XxKaCWXJYxrGMc95z0hwABLsty+P/LV7NSs4F8QavhRqDR7dYaL0/X11HLatZSVtWaqGFz2RviNQ8Sh8gbGSWnBcQCCCEHkuaYeGHZ7rOp7ZG+LxfMb91v/AJcOXriJzPcSaThtw9pYxK5hqq+vqMimt1I0gOmlIBOMkNDQCXOcAO5I9ORyP+z15VbDbzzI66ivF4qWCRr71d5qQzuB3dT0VI4SGMEYIcZBuQScgIPHuqy3LC04G2Vitac7L2csnKf7LvmijqrJweu9BT3sQucBYb/UxV0bBt4jaSrc4Oa04y4REbjJ3C89OcTkz1tyhavprdd6wXzS968R1lvkUJjbL0n4oZm5IjmaCCQCQ4EEE4IaGxuVI/8AXR8HNv8A5f8AT38owL0P9t+M6U4Tf+ELv/6OmXndypYPNNwdI7fs/wBPfyjAvRX23MZk0rwnA8rhd8/5OmQeTVPCJHEHZJaUMcHDfClja5p2PmpcgEB/YoIW3IMb4bmHZQPDajLxt5rKqqeB0fVF3K04+JFt2QWuy0kZVFXDnH5oWEHBQUU8QHTuomxOf2VSHx7HZBWR+5CjBPllV3KyI6clnUgtikGMFWTHJyFQfC/dJCCgsREQEwfRAMlTNDA3fGUETBko4HPZCcOyFqdEKZ0BMuMgbINL7KVgy0k+itmLTIensrQ4jZBdG/w3Eqs0viqkXSXZckvTn4UHqz7DX/c/jKPSawfor16mLyz9hr/udxl/w1g/6NevUxAREQdc/aHfgacS/wDE6P8AXqdfPGvob9of+BpxK/xOj/XqdfPK1hcTjyQUwqg9JUkTcP8Ai7Ks7R1At7fJBYX5GFYnZEEkLul3ZTyTAt2GFBC4NO6unLT9FBYzdxRzSXhvqrASOylha50jXHsDuUE/uJDOoggeqxXsLXEDJwtWrKmP3cNYRnHksKF0RiJkwDugijY6UYyVP7wIAYzufkFHBK0SYGQCj2ddTvnHfKCwxSPd1lp+1ajRRjIGwOPVbmsWha7V0QgszS+Q7YHqs6v4E6+soFRV0TmRjfqweyDDsumn3g9Jlw0H1Woyti0tOIngP+vdQUbrhbj7pBnxW7EAbrGudRJKcXH6fofVBuWPiRb7XFl0bSSPzrZWptYtvs3VA3oBOwxhaNd4XE9eDgditKiP3wfWg1GaR8bfEc4nKxhUyOf1tJBHmpqp4dAAMbBYURAdv2QZzrlWPZ0CYgAK6Oql6Ol8uVBmINyMZKgfIQ7bsgyJD0nqJ81LHc42x9BbusN8vW3HmrWQSP8AohBOWiZxcTsT2yqPmMHwMOPqUZZNCMkEBROc55ydygyYZZGODnEkFSSOFQ7t052VlD0Pf0vGwV1YwseDENkFvhNp/jO6gllMjsgkD0VHSvcMOKsQPmmT6ogBKDKoqz3Z+XZIHkrayoFRJ1tGAFj4wpoaeSTBA2QT07BUt6XDBxjKhmp/DlEYOSSsmV7KeMCPY9isaGXrqGvkOyDIfSmGMP6fJVYOuMu7bLV6ioo30obkE4WjSSsGWRnugUcvTIdsqlbmRwI2wpqKEMcXv8/VQ1sjfGHR2QTUVCwYmm7DdZNVROnjJoxkAb4URe6emEbNjhbi09E2loiJm9Rx57oNkzQyQu6ZBgq0NJ7LVL25j6olgwPqWBE4MygjLenv3Vqvkf1HsrEE1MA9/Se2FqVLbZbjX01soKd89VVysggiYMukkeQ1rQPMkkAfWtKhd0vBWv6L1FDpfXGndU1MLpobNdaSvfGACXtimbIQAdiSGkYO3qg9vNIaD4CezA5a5OIGpLSyv1GIIIbpcoYWvrrvc5W5FJA9wHhwhwf0tyA1jC53U7JPVxntwdZC8eK/l/sxtXi58AX2UVHheni+EW9Xz6MfJdwOfPghcOcLlWgpOE1zpLnWwVVHq6wdEgEV0YIJWiNjzgAvhqXlhOAXBoJaCXDwa1NpfUmi79WaX1dYa+y3e3yeFV0NfTugngePJ7HAEbEEZG4IPZB6Hc7/ADg8qvNty2Q3iyWI2rilbbrRinpLjb8V8FOesTiOqjBZLARnLS8b9BLGu6ceb8rQHAN8x5KWNoeekldqeRThdyicRLlrL9tbr2DTsVuhofuFFPeBb2VLpHTeO7rO7iwMhAAPaQkg4GA57u/IDwGpPZ7R8xFO/Ug1mdI0+pDObi0wGd7WOdF4XRjw/iIA+kNviPn5w0vhP2lx9q+iuv0Ty+S8prdCVep4GcHhpqGjF1+6YEZtYDeiT3ntggD4/PK8/eNHLx7LOxcItZ3rhvxkoKjVlBYq2qscEGrRVPnr2QudBGIt+sPkDWkejjuO6DzYqoqdrcx4ys+zupRA4TkZwcLl3kd4TaN4580miOFvEOjqKzT96NxNZBBUOgfIILfUzsAe0gtHXEzOCCRkZGcrvZfPZLaFvnM1U01lF00vwcslno62se6sdLPX1z3y+JTQSy5LGBjGOe856Q4ADLssDyrrGwmc+H2z5LGLyw4HZetfHnlk9mlJwL4gVPCW+aQi1dovT9dXUctq1lJW1ZqoYXPjjfE6oeJQ+QNYSWnBcQCCCF5zctvLhxB5oeJNLw60DTMYen3m5XGcH3e3UgcA+aQjc7kBrRu5xAGBkgOLnFhbnzwoicr13u3J/wCzL5RqKht/MfrN991BPE2Yi51taZXAnHiMobd8bYyQQPEDxsfiOCRdRck3s4ubuxXN3LBrV1ivlHD4pdbK2rlMDnEhsk9BXnxPCJ2+DwxkABwPcPIqMgbjurJnFzhlciceuBeuOXPihdOFev6eIXC3dEsNTT9RgraZ4JjqIi4AlrgCNwCHNc0gFpA5L5JuTa/84HEOsswu0lj0rp+FlTe7syESPZ1kiKCJpIBlkLXkE7NbG9xyQGuDrxFFE8fNQTUxa7LRsvXbUfCP2PnL1c5+HPEi5U9fqGjBpa91XW3e4VMb8DIl9xHhQyDO4DWEEkYGMDhHm/5UeUe38CKrmL5VOI0dZb7fcaSirbVFdRXQNE7y0NAkPvEEgJDumQklgJAHch58SbRhp7jyWO13ScjuubOWHlp1VzTcXbfwy0xVMoIHxvrrrc3xGRlvomEB8paCC5xLmMY3IBe9oJAy4eid+5ZvZT8qElNpHjnfRd9TiBkszrpWXGrqsPBIc6lt46ImnBLQ5mcY3OckPIZ9ZLI3w3nbCjbTveOpu4XpXx65R+QviLwJ1lxp5S+I8VJcdG2192ntEF1kqY3xMLctlpqs+9QlwJDHEhvUcYPYebVPUiFhaRnOyC2ncOvoeMYV007WfAAsfqDpS7yJzuqSHLsoLScoiujx1jPZBLTQ+KcHsppqPo+MK50kcTAWndSQvdMN+yDAkeT8OFa1jsdQGQsuqpwceGN1l01BikL3jfyQaV1OzgHBQ5IJPdXsaDOQe2VSf4X4HZBSGLxD3wqSs6DjOVa17mfRKqXOkO5yUFCCBnKopBDIR22VpiePJB399ip+FNqn/i/rv5RtyKvsVWlvNNqkn8X9d/KNuRB7UIiICIiAiIgIiICIiAiIgIiIOOOZL8Hbin/Aq9/qMy+aFw6pCMd3H9K+l7mS/B14p/wKvn6jMvmljIE7y7tv+lAdTdLOolQKSSUuJAOysDSeyCiYTss6j8AxnrxkBBhscWOyj5C9VmIMh6eypGQJAXeqC3B9Cro3Frw70OVrNQ2iFGOjHVjfC0djHOIwEGtVN1jfRCLAyBjOFofSXErKfD1MAHdWxMETvvm2UGPl3Y5wqLPmbFIzDMZKhbb6hzS4MOyDGBwVlwUonb4nVjG6xiwtf0nuFKyV8b/Dae+AgmdVeHmPHbZRtaJHeIRjByst1AWR+PINisQxuecxZxnyQTVFf1w+CzIxtlY0VQ6MEOyc+qlxCBh3cd91GTH5II3El2RnBVuTnJys2IQ79WMH1UNV4fUPDxhA95+DpwFdHVhrS0t9Vi/WqnGR6ILnydTi7HdSQ1DmOGDjKscW9OwVrY3OGQgzauYmBnSd85Xbnlf4n0OnbUyGqnaMNAOSunLnOI6T5LemjKPU08YfaWPLB5tyg7bcZ9eWzVVLLFRysa4tO7e66p18VZSSysNQ5weT3OVn3eq1HQxn3rxA4d8krSqSu96jLqndw9UGg3ihcGeO525GStMp6oQsLcd9lq18quoFo7BaAgkL8SGT1KSPMruoBWkjpABU9N4fQerugqyrDWhpA2UEsniHKpJjrOOyuZA+QdTRsgjV7JOnHyVrmlpwVVkbn9kGRHWdB7KKonEzsgYVHxdDclRoLwXOwwu2HkrjAfJRZwtftmnK6vpTUMaS0DO3kg0ON/hOORuslkQqsHOCoauF1PUPhcMFpwrI5XRkY7IMo0ZiJIJITfqAORlTQ1TXD4iFZUOaXZZsEFJomsaHNP512O9m7DRz87XC6Otk6IxW1rwejrzI2gqHMGPm8NGfLOfJday9xGCVylyq8RaThPzH8OeIFyl8KgtOoKQ10nV0+HSSPEU787do3vOCQDjBIBKD0F9uDW1EFLwapWOd4Er9Qve3Gxe0W8NJ+YD34+srykkcXOyvbH2wXCS56+5cbbxBsVO6pm0DdRXVbGN6iLfUM8KWQY3PS8QOONg0PJ2GV4mD6QKBg+iqx/Q8O9FPKY3M+HGVj43wg9sfZK6Gsmg+Va48YbjC2Kp1ZcKysqKtzQCKChL4WMBP71r2VLu+MuPovIjjfxf1Vx34pah4paxrZp6691j5o43vLm0tPkiKnYD2ZGzDQB5DJySSfZ32dMJ1T7OnT+mbS9slaaLUVsLSQA2eWurHNBPYZEzDvjYrwtmgmpKh9NUwvhlheY5I3tLXMcDgtcDuCCCCDvkINW0bq3U2gNU2rW2j7tUWy9WWqZWUNXTuLXxSsOQRjuDuCDsQSCCCQvb7nOt9n5l/Z2VfESWmgFQ7Tdt1zQuaARSztjZLK0ZJIIiknjIzkdRBydj4cUs1O0fGfJe4nEuL/Yy9lNUW7UjvdpqfhhQ2+Vj/AIeipqoIoWRnvuJZmt+Z9MoPNT2XLAeejhqf7n7s4/zPWrtv7ZPmE4i6Vk05wD0xcjbNP6ktTrrepad5bPXN8Z8TaZzgRiEeGXOaPpkgHYEHqT7Lgn9vPw2+f3Z/ketXNPts/wDf40J/BH/68nQedsMksMrJoHvZIxwcxzSQWkHIII3BHfK909c6kq+MXsra/Vmsi6rr7jw3ZXVkr9zNV00bXCY5zuZYQ8/M7YXhjRuY156+y9ubOR+5Dzlvb/YwrMf5ORB4kzQsazqDskr21vb+n2QsL/ThhR/+jjXiD4riOknIXt5f/wD7UFF/xX0f/o40HiTGwSuJce67UezAjDOerhrgjb7s/wAj1q6qxdWdiQu1Hsv+r9vXw0z5/dn+R61B3X9r/wAxurdEaXs/AbSNTUWyPVdG+43uthk6Hz0Qe6NtICNw17muMnbLWtbuHPC8k7ZbIaqnMkhGQvQH210zm8ctDQg7O0kCf45OvPKCtkhjLGOIBQe0fsluG1g4a8rt14uVkLI6vVtxq6upq3AAigoS+FjATjLWvZUu74y8+i8kOOvFnVPHHipqDifq+ulqK691j5mMe8ubSwAkRQMz2ZGzDQB5DPckn2Y9nYDq72dlg07aZGyVzqLUVtc0kANnlrqxzASe20rDvjuvDaahqoppKerhkimhcY5I5GFrmPBIIIIyCCCCDuCEGoaK1lqTh9qi1600jdqi2Xmy1LKyiqoHlr4pWHIIPmDuCDsQSCCCQvbjnPobPzKezqq+Iz6aD3h2m7brmgLQCKWZsbJZWjJJB8KSeMjJI6iNyMHwvkhdGTnyXubxCpn8M/ZSyWrU7hT1EHDCgt87HjpMdTUwRRNjPfcSzNZ8z6Z2DyH5TYyeaLg8SDga908f+cYF6Ne2uIGleFOcb3C7d/8AB0687OVN7RzP8HWjGf2faeH/ADjAvQv23Li3SvCfHncLt/6OmQeV0VMx7snz+a1SLT7KqmfKCB0t6u60GOoc056lknUFRTx+E0nDtj9SDCgaW17qV5+EHG6jusbInhrMb+itnqQH+O36Z3ysaWd85y/ugpE4NKuld1OyAos4U7Cwswe6C2Kbw+wyVSaUyOzjCvp/D8Q9eMJV+H1AR9kETBkrUY5GiLGd1gDBG3dVAmx8kFkp++HCtQgg4KIGEwpafoz8STlhd8CCIHByhJPdFVvcZQUwrmgucGgnfZSyGMx7d1CDggjyQZctD4cXidQPnhYalfUyPaGE7BRICIiD1c9hr/udxl/w1g/6NevUxeWfsNf9zuMv+GsH/Rr16mICIiDrl7Q/8DXiX/idH+vU6+eeKTwySd8jC+hn2h34GnEv/E6P9ep188fdBe+TqO2yz6alZJAXuOTjK0/oI3I2U0FU6NwaDsTugikaQ87Hb5KxahVeC6MObjqPmsAAk480F0UZkdhJY/DOFkRxuiHUdgoZnB7soIlLHN0N6cKjunp2xlWNx1DPZAJc7vkrJoqM1c7YgcZwrZujwx091m2aOTxDM3OwzlBuOHQdQYGTwxGQnBGBlb50vwZuV4jD3W5+4/uSt6cv9HS6grIqW59PT1gb/Wu/WjOENi92gdSxxkFoPl6IOkugbIzhDP79cqHDW/F8TVuLXXMZpu/2mShgpIg/pLdmjK5b5sdFUNkskromtBDD2GPJefEjf7OQ07OIQZF31IyhvMtxYwFrySAAtNr6oX8GtyGdO+FG1tNVvLKjGB2WDcHupMxUezDscIMOvrw8GDGS34crTAcFSvwSSe5yowMuA+aC4yuIweykZTh7OrKuliayMO27KJglI+HsgsOxx6LIp6XxgDnusdwIO61OhttZPF40LSQEERoGxjLj2VoqvAcB05APcKlS6oY8xHORscq6FsZjIf8AS8soLamtEzelo8vNQsgBYXZ3Vr4XNJwNlaJHAYzsgviJY/GSpX1I+id1i5OcoTlBe5oPxA9yrMH0TJWfSwsc0Od288oMDB9FczcgepWTViIDEeMjuoIxt23QTGmaQCCVqdIyOOmOSM49Vo7nSjbyVW1MrW9OcjthAqiXTOGSQocFZsMQeOp3n3VtQxjMgY7IMXrd/dFajQUImAe4jHcrTfqWfTzSxQkt8ggyrhiCPEZyQtOijMx63k5HZT00/vEpFQQQVbWgRyBsXb5IMulZ4Tep2w7rc9vudAyjLJC3ONt/NbOdWuMXht74WMXy9upw+1Bk3adstW8sOQsRpHmd1I6AhniOOTjPdWRReI3Oe3kgsPfZAN9wrmtw/pKukAbsgqekM2O6iG7hlBk7eSmig6m5Qd0uTL2k+u+WO103D3WFsk1hoCJ7jBSiborrWHHLhTSO+F0ZJJMT8DJy1zMnPpBa9XciftE7Ay1VUen9TXSOAkUFwjNDfqAYJPhkFs3S05JMT3R5xkleBTgWnBWZZr1d9N3akv2n7rV225UErZ6Wso5nRTQSNOQ9j2kFpB3BBBCDuzzzezVv/LTbKjipwwutXqPQDJQKyOpaDXWcPcGsMpaAJYi4hviANIJAc3HxHo055d3X0I8rmuK7mm5IrHqDiyaeWfVlhudnvc8kbfDqGRTVFHJO5oAbl7YutwAADiQMABfPjJD0PDSe6D25v2/sgYv+K+j/APRxrxE7L29vzceyFjaPLhhRj/8AFxrxDwSeyDtT7Ls/9fTw0Hn/ANWf5HrV3Q9sLzNav0HZbFy+6PnqbazVtC+53yvik6HTUQkdE2kaRuGvcx5k7ZaGt3DnhdMPZexlvPPwzcR/8+f5HrVzT7bH/f50L/BEfrtQg89aV7Xv++EYyvaP2THD+g0Dys3ritBa5qq56tuVXVEQxkyz01EHQxQsABLj4jagjAJJkIwSN/FRuR8l7tezRvlVU8gWmo9NMbNd7My/U8Ubx1NdV+/VU8TSAckESx5HfdB5RcTOC3OfxZ13fOIus+X3itW3i/1klbUyO0hciGlx2jbmI4YxuGtaNg1oA2AW/OUHhLzY8GuY7QOuabgLxRtlLDeqWjussulbhFC63TyNiqRK50QaGCN7nZccNLQ7YtBG9T7YbmxyY/ufoQPGxzZptj/l1FN7Yfm4iIHuGgt//vNN/ToOdPbaaKthtvDDiLFG1lwbUV9lmcBvLAWxzRgn0Y4Skf4Uqb2KuuNLwWTiHw2mrIodQTVlNeYIXHD6ilEZieWb4IjeG5GAR4oO47dGeZLnY42c1lDZLTxPlscVBYZpammp7XQGBrpXtDS95c5ziQ0EAAgbnIJwRxJpLXWquH2pLfq/RGoK2yXu1yiajraKYxyxOwQcEHcEEgtOQ4EgggkEOy3OnyPcf+EPEzVutv2KXLU2jbrdKu609/tsLqlscU0jpCKpjMvhe3rw5zwGEjLXHOB1MjFQ5jmxl3QSHFudsjIBI9Rk/lK9KOAHtl9b2iSisHMHomm1FRNLY5b7ZQKauazYGSSnP3mV3fIYYRjsCRvzXzucsPAjmB5cbpzVcFrZbqK9UNmfqZlxt8AporxQRNMlQ2piwAZWxtkIcQJA6MMcSNgHXH2Muv8ATOneO2qtGX6tipbnqixsZaTLt48sEviPhac46ywl4BG4iOCCMF7SPkj4+xcbtVcctF6XuGsdK6mlZXyyWyI1FVbniJrHxSwNzIWDoJEjQWhuAS0jB8/6G63Kz3SmvNmuFTQV9HMyopaqmmdFLBKwhzHse0gtcCAQQQQQCCu/XAT2xPGbQkNJYeNWnKPX1rhLYzcYnCiurWdiXOaDFMQMYBYxxIPU8k5Aefj2zwSSwSB8TwTHKw5BBB3BHyIGx7EfJREYXuTxc4F8t/tHuAU/F/hbaqan1bPSTutF7ZTNpK1tfED/ALSrwMiRpeA09Rd0h3VG4g5d4cFoCC0DKlFOHNzlWYPoqgTOBDScIITsSFJHF1DI7p4D/RTU0MjZWk9kFjqeQn6JGPNSUspbIIXDYnC158NPLTjwwOohabNTCHLv3w7IJ6yJlLEJc991ji5l8JY1pxjCx3Nra0lgBIHktQpoKampiyox1ntn1QaMHFshee5KtkPW7OVkSUcssjjG3LTvlQSQSQnDhgoLOkoD0nJClgxn4/VWy9Pi/D2QSirIbgNypGkSDqIwVLTR0/SPExn5qyVuXgxfRCDvv7Fv8KTVP8Aa7+Ubcik9i+G/todUev7Aa3P+cbeiD2eREQEREBERAREQEREBERAREQcccyX4O/FIf9xd7/UZl80Lx98dv3J/Svpe5kvwduKX8C75+ozL5oJDiR3/AHx/SglNPiMPUTXYGEMryOnOytQDugz2CIgvLMNyr4qcygkeSjc/IxlZFJO2I4cdigge17D0knHkqxy9Aws6qfTuiy0jKwIw398gljquk5IVlRN4pBGyPYzuovqQXxPLHgknC1aK8xxw9BbuR6LRkQSSy9cxk9TlU68SCQeRBVifWg1SpuolpBABuEthY2I9Q3K03Axkd1fDM+Nw32ygnkpXyzOLQcEq2WjMQy7Za3b6mj6MvLcrT7xOx7w2HHzQablxPS1Wua5p+IbrKo4eiUPlGB81dcjE54MePmghp6fxjjKVNOITgHKjZI9n0SrnGaX4nb4QIouvfKl6xA3p9Vjh72dtlUdcrw0bknZAaPEefmuXOFutYdNhlBLSh/VsSQCuM47HcOls/hHpJ74XN3DTh0y70TauRmXtAPZBqGvTSXC1uuEULQXjOMBcJteQX74OTsuadWWmuog+je0iIZH2LiK72OsNQ91O0lud9kGgVbnyOIOSFp72hpwFrlwpxTU4a4YfhaE5xcclBk01GJ/32FbVQe7ODQe6jjnkjPwlVc987up26CNreo4WZDVCmZ0YzlHeGI8N743WG7qzugkx40hPbJUkbxA8NOMErHa5w7I5zi7qPdBqNSxhh6x3IWnNGXAeqv8AFkeAzOysIcw77IMmSlDIxJlbpsOtYrXb3UbmAkt6QcLaD6h7h052U1PFC5mX7lBfVSi418k3YPOVBND4bwweatc7wpD4Z2VD4kh6zuQgzvc/DiEoOQQsd3oqGrl6DH9ijDifPuguJ8gqfNCcK4OaW790HsX7PPn20BxX4dUHLrx5u9vpdT0VH9xqOa7FvumoKDoEbInvkPSajoPhuY/HijBHUS4DQeNnsXNLah1DPfeB3Es6YoquV8j7Ld6R1VBT53DYZ2uDwwHYNeHkA/SOMHyO+pcsaE5seZbhpQR2jQ/HHWFst8TBHFRi6SS08TQAAGRSFzGbAD4QOyDvnw79iRXi509RxX430/3PYQZ6PT9ucZZR5tbPOQI/LcxOz6DuuI/aS27lL0LbtAcGeW+m0+65aVfXyX6ptfTUSEytgaxtTWDJnmJjeS0ucWYwQwENPWbX3NHzJcS6GW268426xu1vnaWzUMl2lZSyAjBD4GFsbtsjdp7n1OeKg942Dig9HfZRc4Ok+ElzunAbibd4bVZNS1wuFluVTKGQU1wLGxyQyvJwxkjWR9LiQA5hB+mCOWebL2R83FXiBcuJ3ADWVhsUmoah1bX2W8NliomTvIL5aeaBkha15JcYywgEnDgCGt8iup43JK5Z4e81vMbwttTLHoPjTqy022Fnhw0LLi+SmhbntHFIXMj7d2gIO/3Ln7G+46W1pR6u5jtb6du1ptE7apljsTp5Ya4s+IConnjiLIwQOpjWEuGR1NGc7M9qtzr6S4nw0nLrwjvcV0s1qrm1uorrSSB9LVVEYIipYnjaRjCS57hlpcGAE9JXS/iFzVcx3FW2vsmv+NWrLvbJGFktBJcXx00wOdpIoy1kncjLgTg47Linsg7Yey9Y1nPNw0x5/dn+R61cye2yP/5+tCD/ALkf/r2ddA9Kaw1XoXUNHqzRepbnYb1b3OdS3C3VT6eogLmFruiRhDgC1zmkA4LXEHIJB1DiBxQ4jcV7zFqDiZrm96ouMEIpoam610lS+KIEkRsLyeluST0jAy4nGSSQ2vuvb7T4/wD8QUmfxX1n/o5F4iOLS3but80/HrjbSaBdwspuLerotHvidA6xsvE4ovBdu6LwurpEZJJLAOkkkkZJyGwzsvdTkyuOhuar2fdDwjmukUc0GnZ9GXqGIh89vlax8cMxZkZJZ4UzM7EjGSWleFS3Tw/4ocReFV5/ZDw21xe9MXAtDHz2qukpnSMBz0v6CA9uf3rgQfRB3U1v7Ivixwy0LrjiLq3ivo42nSVmrrzTMoo6mSetZTwvlEb2yMjbC54YGjD5BlwG/nxn7MM/9fXw0A8vuz/I9auKuIfNjzH8VbG/TWvuNGqrvaZhiegkrnMp5gDkCSOPpbIAQCA4EZAPkFx/pHWOqdCaio9W6M1HcrFe7e5z6S4W6qfT1EJc0td0SMIIBa5zSAcEOIOQSCHfz217M8d9Cu/7kQP+WzrzqW7eInFTiHxXu0V+4la3vmp7hBCKaGoutbJUviiBJEbC8npbkk4GBlxOMkk7RyEHot7JznE0pwiud14C8TrxDarJqaubcbLc6mQMgpbgWNjkhleThjZWsi6XEgB7CD9MEc0c1nsm6nihr65cTeAOsbFZH6gqHVtdZbw2WKjZO8gvlgmgZIWteSXGMsIBJw4AhrfIJcq8P+avmQ4WWuOx6C416ttNshZ4cNAy4vkpoW+kcUhcyPt+9AKD0K5evY83HTmtKPVvMdrbT11tNpmbVNsljdPLDXFnxAVE88cRbGCPiY1hLhkdTRnOyfao87Wk+JMNLy78JL1FdLPa60VmorpRvD6WpqIwRFSxPGRIxhJc9wJaXBgBPSV0s4gc1XMfxUtj7Hr/AI1asu9skYWS0ElxfHTTA52kijLWSdzu4E427LisO9UHLHKkD+2l4OnP/wAv+nv5RgXoj7bwkaU4TY/+eF3/APR0y8obXdbpY7nSXuyXKqt9xt88dVSVdJM6KennY4OZJG9pDmua4AhwIIIBBBC3XxC408XOLhoBxR4l6l1Uy2B4o2Xa5S1LKcvx1ljXuIaXYaCQMkNAJIAwGzxn8yx53ZcB6LKkLSz4e+Fjup5CerG3qgjjYZDgpJH0OA9VL1sY3AO6s6HyHqwSEB0Qa3q+SsEb3DLQcK+QPDQ0hZ9EadtORJjOPNBpe4V4Z1DJKTFpkcWnbKNLsbFBaD0n6lO2pAb048ljnckpjCCrj1OJ9VREQFLHD1jJKiVzZHgYBQUcMEhUG6E5RBd0nzKtOyvYcpIN8oLWt6ij29JwjXdKOd1HKCiIiD1c9hr/ALncZf8ADWD/AKNevUxeWfsNf9zuMv8AhrB/0a9epiAiIg65+0P/AANOJfmfc6P9ep188Y2X0P8AtB+n9p1xJ6u3ulH+vU6+eBBL1h4wo3DCoNkJygqCRvuqh2HA+itVQC4gDuUGRLUiSLpHcLGUj4JGDqLcAq+GMP3QQK9rMjKlkjDPy7KyBrpZWxNGcnGEFjWOc4DBWu2lzovg6O4V0lqNDTiombgHdataBQzwl7cEjzCDk3hDd/uPWRzCTpPWD6Lvpw24w22mt0QqbgwODR3cPRecOln1AqSIwcA7KfUPEbU1irWw0rpAztsSAg7v8etW23W9FLTsrWOBaQPiHoui+rLXFZ5ZYY3A9TyrrnxZvj6IEuf1kb5JW0arVE11YZKtxLjknKDBrQY3dbHd1qVJRx1VC6R53AytAdUGplLRuApnV1TSxGJmcFBptS3pne0eTtkijyA5X08ZqJCT3O6rUtMOGjZBbLL1t6M9ldDOI29J/KsYnzzundBVx6nE+pW6LPfIaSgMEjRktxuFtyma1zj1LKkaxrdvzII6qrEtS+QdidljOkJd1AkK/oEh+Dcq18RjzkeSCcSiVob5gKF8eCqQuDXbnCmkBeMjdBjvYGgH1V7IHPaXY7JEx0j+nBO61eMQxU5DiM4QaIRgkeiyIJ+lpYdsqsLGy1BGxBKtq2eHIAPrQWSswS4+aja7pxhXOkLmgKWKNpjLj5IKwnxT0eakkpQz4sYI3UdFKyOXLuxWXM4yPDm7tCC2my4lpGBhY9YPvwYBkLJe9hbiHd2PJWtp3u+N43+aDHmpBHGH57qkM5aBH67K6rdIB0O7K+mpHSxeIB5ILPCEZ8Qbeaj8bLxndJnva8sdthVp6V8xBGcIJoI4zKC/sErmxAgxd8pUwvhb5ghYrXHO+/yQVcZSzB7K1sjmbBXunLh09ICjaMkBBXpePiIKoXFyyXkGLGygaNwe+6CvhuA7bLlXlYk4XN5g9DR8a2UTtEyXRsV3Fa4imEbmOawykdoxIWFxJAABJIGVxe6UdGMYKhAz9SD114y+xy4ba9ndrHl04nM09SXEe8wWytabhbnNfuDBUsd4jI8HI6hKT5EDAXHmivYk6/lv1P8A7IvGfT1NZWuDqj7i0s89VI0d2s8ZsbGE9g49WM56TjB6G8OOPvGvhA0xcMeKmp9NQOd1vprfcpY6Z7s5y6HPhuOfMtPc+pW8tQc73Nvqa2zWi7cwWsjSzAtkbT15pi9pBBaXRBriCCQRnBHcIPUTnR5kuEXJjy6O5buE9XTDVc9j+4VqtUEviy2ukljLX1tS7fDy0vc0O+KSRwdgt6iPE1z3OOT3V9TU1NbUy1dXPJPPO8ySyyOL3veSSXOJySSSSSdySo8FB7pcmdz0NzV+z9ouEk10ijmg07Poy9QxEPnt8rWPjhmLMjJLBFMzOxORklpx0n1t7Ivirwy0JrfiJq/iro42rSVmrrxTMoo6mWetZTwvlEb2yMjbC54YGjDpBlwG/n0u4ecSuI3C29fd/hrrm96YuJaGPqLXXSUzpGA5DH9JAe3P71wIPot5cQ+armR4qWd+l+IHGnVV4tM+BNQSVzo6ecAggSRx9LZACAQHA4IB7gIOV/ZivjPPPw0aw/8Az5/ketXMXtrmdfHrQnoNIj9cqF0S0xfNVcP71R6x0VqS5WK929zn0twt1U+nqYC5pa7okYQ4Asc5pAOC1xByCQZ9ecUuIvFW8R3/AIma4veqLjDCKaKputdJUviiBJEbC8npZkk9IwCXE4ySSG1yAG9xlehHspOcjS/By73XgZxTvUVq05qesZXWe5VMgZTUVxLRG+OZ5OI45WMiAccNa6PfZ5I89HHJyqIPWHmg9kbc9c69unErl41bYqGkv077hUWK7OkihhmkPU8000THgxuJLgxzQG5wHEYA4j037GXmLvV0jZq3XWh7Dbg4CaeCoqa2cA9yyIRMa4geTpG+W/cjqbw65pOYnhPbm2bh/wAZtWWe2MaGx0EdxkfSxD/tIXlzGHYAloBIx6BazqjnS5p9YURt994+a1fTuBD4qa6SUrZAQQQ8QlvUDnscjt6BBuLnl5Y9E8qPEixcOtIa/qNU1E9jjrrtJOImPp6oyyN6fDjJMYLWNcGuJcA7OSCCuW+T/wBn1oDm65dbnq60cS6mwcQrXf56N0T/AA6qkFKIonRCWnHTLGXlz8SB5Gx+F2MDo1V1c1ZPLU1Mz5ppnmSSSRxc57iSSSTuSSSSTuSVqukdY6x0Hdmah0Nqu8aeukQIZW2qukpJ2gkEgPjcHAEgbZwcIO7dL7Gjml+7LKKfVHD6KgMwa+uZc6l4Eed3hnu4cTjs04ydiQNx2+5ptX8OeSHkP/a5nVDbrqK86XqdL2unLQJ601LXsq6wxdRMUI8aVwySASxgLjuvMSTnw5w5KI293MLq8RmMR9TasNlwBjPiAB+f+2zk985XDGoNTai1dd6i/wCq7/cb1das9VRXXCrkqaiU4xl8khLnHG2SSg7achnJjw25wNK8Q7XfdfVunNYWI0L7IYHxSs8N4l8SSWmdh8rA5rASx7Okkb74O8rp7GfmgpLtJR2nVOgK+h8TpirH3GphJYTs58ZpyWkDcgF2PIldGLJeb5pm509905ea61XGld1wVlFUPgnidjGWPYQ5pwe4IXNFFz284NDQMt0HMLrB0UbOgOmrBNKQc7mV4Lyd+5JPz2Qer2laXQ3svuTmW2ax1nTXW+mWsrKdsbOg3S8TtHRBTxOJcY2BkQc49mtc8huekeFJa5pHUtw6v4ga34hXp2o+IGsL1qS5vb0mrutdLVyhuSQ0OkcSGgk4AIA8gFoE84cRgILnlmzRuSsmINYzB7lYrjGGh/mr2CSTcDIQJZww7KaKdr2Y81jzRebhuoInODwAdkGpwVL4HElxIyqyzid/V2VgaAwFywZZSJPh7AoNxW2eCEfG0En1WFdIBUVHiRuwPPCwfenNYOk7/WohVTvcGgndBq1PVR0zOkgZ7LT7jO2Z+WhR1DJmMD3Z3UTT1sJPdBbG3xDjsqSM8NyqyKR7j4apKyRhxJnKAXPdsCSB6KWOoMbS1wOVNa2wuefFxhWXBsYm+94x8kHff2LUpk5ptUjf/wCIFcf+cbciu9iyWfto9U9Pf9gFb/KNuRB7SIiICIiAiIgIiICIiAiIgIiIOOOZL8Hbin/Aq9/qMy+aCT+yP+s/pX0v8yX4O3FP+BV7/UZl80En9kf9Z/SgRt63YWR7o3pyHb/WsZvUDspBLI0gOJwgjc3pcR6K6OMPCnkMbmZ2ysYOI7FBe6INU9PRCZvVnGFAC6TIG6uEk9PsCRlBWamMRPxZULdyFc+Z8h+I5Vn1INQjpmyRgFw3+axaiAQu6QcqjZ5mjYnCqHmXJeclBCg3ICqdnY+aHYghBIYQGdWVErjI4jGVagqTkKrG9bsKjWF3ZXBro3AnbdBPJSvjYJA4gLHDnFwcd8LOfOJIgzIyArIIGlpDu57IIZKnrZ07gqEk+ZU81HLHlxGyx0AHBU7ZhsCFDthBnbHqgyJYGtZ4g891bRENqWPPYHKue2Xo+LOFDGHNdkDsg33Pf6aO2sjawF22Thc2cHdXUdLY3ySuAIautUFQyVoif6redjvM1DQOggzgtxhBvnW/EamuVzkpYyCASMhbQkvlNFE9rwCTk5IW1nxTz3J9Q4HJOcqy5eKCM5wEGPfKjxpC9p2PotC3O+FqVW8GMZO6x4RE5hJ7oMRXxydAIx3RzS6QtaPNUdG5n0kAPId1Ekqrj148lYmT6oAODlV7lGNLjgKr2FhQTU8Yz1HCpUEF4a1WtEmPgRoIkHiILnQFsXWVE15AxnC1CYB0ADcLTuk9QB80DGfNSiYNHThXGmk6OoA47rHIwd0AnJJUsTcjOVErm9XYeaCV7G42KjOB5qha8bFA1xCC+BviEgpIPDePNWElh2OCqOcXHJKCaScvbhRjZAM4HyUwYC0+qCIuyEDQRnKodimSgHZUREBZkFLHJGXO2IWGrxK9o6Q7ZBWSPoOAcqzG26yaSMzyEHdUrYvBeG+u6C2nibK4tKrLC1jsA/kULXOb9E4yrupx7nJQVkiDRkKPGFPHJueo7Kyfpz8PYoIkRMgICuBz9atyPVVbs4ZQXYwimfIwtw3AKgyCgrnCuByrXAgZVseS4Z7IJeox74UzKvxAIgzOdlZO6MxgDGVlW73VsZdJjI3GUENVQeHGZc4zvhY8U7mN6Q3I+pT3CrMp8Np+FajZ20Bpne8AdWNuyDRpJSTuNyqxRGXJDsBXV/heO7wcdI7KBsjmfRKCWSnEYznP2qOI5cGHsSjpXuGCVYCQQR3CDPnoBFCJeruFgE5OVPJVyyM8MnZQICIiAiIgIgBPZVLSO6A13SjndRVEQFe1gLSSVa1jn/RCqWvZ8J2ygkpoRM/pJwr6qlbB2OVBHK6I9TDurpJ5JNnHKD1Y9hr/ALncZf8ADWD/AKNevUxeWfsNf9zuMv8AhrB/0a9epiAiIg65e0POOTTiXjv7nR/r1Ovnnhj8RxBOMDK+hj2h/wCBpxK/xOj/AF6nXzyteW56fNBdJH0easVS5x7lXMxjdBLR0wqXlpOFWppnUsgxk4KiimfC4uYVqUNRBMzM5BPzQYU1WJIwwtwQrYJCwbBX1QhyehWxeHgg9z2QUmm6xt5K2lmFPO2UjIBR8JZ8ThsVVlNJMC6MEhBuWS4fd6mFHG3ceixIYqi0PMTsgFahoOnijry6p2B2Wt6vpKOWYOgx5dggxLdqRtpb4u2TusK9aliupMpAJBWnVop44+mQkgBaO5hkJMP0coJai4SVDvDHbt2UsND1xHLsZCsgFOPpYyo56yRjixh2QWge6SE5BVJqvxtgPzKJznz/ADKnpKR3iB7x8IKCKnndA/qLSqVE/juBx2WrXRlC2mHhY6seS0aPpwerugsRVO5JCoguY8sKkdMXDpz3UKIJo3eFl2e6vkkE4yO4WP1E7KrXFuw80F8MQkdjKzI4m+IIz2IWFG50ZznushrJ5R1x5JCDNnpY6SPxWkEndaXLO+U4yQFmMFW/4ajOAPNYlS1rX/B2QVid4J6vVWTSmV3VurMl2wU7Gsa34hv/ADoLKeHxndOcLPjpWhwiJAzssSlJEvwrIMdV47ZG5wPNBlT2ExRiUOwO/dVhgb4JZkEnbur66ulfTCIHcLT6KsdHKBMdiUF0VK+CUvcDgKsldmUNDdlqlfU0ppgWuGT3WktZE6MyDv5IL61rJIg4EZUdHXeC0xEdz3WK+WV58POwU8LYmEGTuguq4erDwO+6yaCRsUWSMqCoqGPHS3sFfA4GIs8yEEVXUid2AdwsNwAfg9vkp5KSZji4ggZ7qz3eQjrxsgy3ik92y0gnG608B2ctyqdlk0mHO6SggJeNiVkU/hlvx7lZs1vY5nW3GStOkj8M4z8kF72BziGK10bm/SGFWJ/Q4EjKzGhk7CT6eiDB6Sr2hvmArntIcR5BW9KCh7p3UkUXiHp3VZoTERgoKDLRlvdWFzs9WexVwcR3GytccoJ31sro/DPb61j5PqirjZBRVDHEZAQNJUsT+huDvughIIOPRSsbGWnq7qx+SS7HdWk4BwgoW/EQ3tnZXN6gFa14HcK7xgfLugozd3xJI3pdgKuAN1aXZdnKCTD3AeiBnyQVDWgA+Smj6HNLyfJBG2Euz5LHMeJg3vkqR8z3OLYlJEAN5MdSBU0wZEHZ7fNW01YIm9J/Qq1LnubjyWMInkZAQTT1BkBx+hQMJa8EqSAN6sPCpN09WGIJZakuYGtUTInOOXZ3U9CyMOJmAwpauSFv9ixsgw5B0KSCMAiUnssdzi7uqiRwGM7IMysrhMwRtHYYWNHtGfmrWROf2CudFI31wgvpqnwHE9OVbUzeO/qxhS0MTHv++K2tYxk3SzGEEcOQe+Mqkpycd0EEuMgKoheXDqGyDv17FZrhzS6p/gBXfyjbkWT7F2Po5odUH/uBrR/zjbkQe0CIiAiIgIiICIiAiIgIiICIiDjjmS/B24p/wKvf6jMvmgk/sj/rP6V9L/Ml+DtxT/gVe/1GZfNA7+yO+soDHdJyrnP684CsLCN1kU5Z0/F3QY5JzuSqYUxYOs+ipKGkgsQUhk8J3URsVdUTCXGBhXPbH4fbdY6Ag2RVa0ndBIxwd8OEePD8kwMYHdWmOQ/S8kFh3RCMK9jchBYmMK9uA49Suk6MjG2/ogvo8GTpd2V9cB1Do3HyUbWOcPvQ3+SyI2Fo+/8Af5oMJri053WZTyg4cSBhRzQg5cwd1j9TmbIM+quLZYzEBvjGcLTsK9jMnJ7KsgBIDAgjVW5yCpDTyNb1uGAr4vDx8XdBJLUtMQbgZVaVoew7d1iy99uyyaV4I8Nvc9tkF8NKfGz1fkW4qJ74IgCMj5rS7fYrlJIJek9Pqt1Q2WVtP1PB2CDRJa8RPLgN1ptbdDLnY7LOroGxTObIFpk9K2UExjAQYgeapwYVO6ibCw4dnKx5GGA5bsR6K33uUjDiSgsDvClJ74KrNMJO22yjdkkn1QAYygomCr4sdW6ym+D1AYH5EGLG50bsgH8iq95lcNu61NvunTuArJIIXsJjA27IIB96jy0ZPmsaR7nuBcMeiz7X0e8dFTu35q69Mp/EHuuMfJBDTHraGE5PooatnhyBwG2VdQxSvfhgx6rJq4hG3EoyfJBbBUiYeFgfasSri8N+B5lUgcGTZ8lWrlEjhhBa6IBvVny9VRkgbjPkrep7tsqhaW90F75A/bCkiADMlQsxndSFkjhlnZBG/PURuQCrcH0U8fSDhw3zupHNjI2AQQ07et3Sp5j4W3fPooXNfEetuyvZIJB987oIC4EkpkFVe0hxI7IxjnEY7Z9UBwLVYsyUxCPY7rEa0nPyQXNHUhBBwVa0uYcjurgHSOBPZBNTzmnd1AblUqKgzuBPcK6WNojGO6xSCEF7e6yo4WubkkZx5rHpmdb8FX1HVE4FrigldA8ZIGwWO/Y4V5rpy3oyMfUoC4uOSgdRVERA2+aZwiuYwkg+SCmSBuEB3ypJAwN2xlRIJHyBzcBWtdgYVY4nSHZUkjdGcOCChJPqUy4eqlgjy74uynlEIbgYygws5VQ5w2BICq5hGT5KjWlxwEFM5RS+A9vxOGFYMeIPTKCmD6H8iotVbFBNEAwDOFgVMJidgjdBCiIgIiqGOIyAgoidkQXMd0nJVZHhysRAwmFfH053wkvTnZBWGXwzkpNL4pBCjV7cYQWIhxnZEHq57DX/AHO4y/4awfor16mLyz9hr/udxl/w1g/6NevUxAREQdc/aHfgacS/8To/16nXzxr6HPaHfgacS/8AE6P9ep188aAiubG5xwFV0fR3CCxVGc4yUAJ7Jgt3QZEdPkdTj+UqN7fCeHA7Aq0SSn4WnurjDM7uCUGS1wq2iLHb0WTQPbBVNpXAfEcLHo4JInl7hhXSQzSVgfGDkEEIOUrHpCOanZUwuGXHPdcgaP4RP1PdYaKVnV4hAW1uFFj1DqExU0DXkZAxgruBws4Ea3gvNJcI4nhow7PSUHEnGblC/YtpT7tRU+CY+vI+pdOaykmtPiwviILXEdl7L8ZtAapv+ivuU4OLmxdOA0+i80uJ3BO7WSqmNVG8ZeT9H5oOAHeI13Wc77qg++PA9VuC/aflt4LXAggLQWRu3djcIMjwW07A875Vwrx4ZYAAfVYj5nuHQ45AVGR9QJQVaHzPwSTlVfD0EDPdXUzmseeoqtQ7rdlnZBG6MAA5UeFe7rA+Iq9nT0b4z6oIcZVek5xgqekDPE+PGFk1BpwMsxlBispw4d1HJGWOwAUMjgT0lZFO6J39kxlBiuJI7YwsykuHu7QOnOPkrKrwsfAraaNjsF4yMoMma6CXOG4z8lB4fiN6vUqWoiiLMRAA/JQxNka0g5zn1QWBvhuyrJHFxyOwSRj2nLuxKmho3zNL2jZBHTSCN+StT+6UbWdOBkrS3wPY4gjsrCD6IMh9W5zydzlSQ03vLg4/CVZQNiMh8Xt5KWsm8KUCE7BBPJbfgA8TOFEYzC0t+xWQ1U+fjzghZEjfFZ1fL0QQUMDJZT1EKOvi8OXpb2URlkhkPSVXxnSyAv7oBgexvVudspFOY3dXos97oxH8WCMLT3NaX5b2QZvv7Z2eGW4wO+FG+pbGCwNG4KjBhjbkAZWNI/rdlBRx6iXepU1NI2MnPfdRMie8/CMqroXsGXDCCd1dMSQDsFZ1de7u6siYXFXSDodjCBjdSxyOaMDsrIWiR4GcLLNKGjuggPxHJ2Ven5KTowrhHn1QWwENfnsrpXh8gHkrHAqxxOEGbMyn8DYjON1ppG+FL8R2JKjfsN0Fxx0/NUYfiAPZUiHiODcd1JJD4YyThBqUMMEkY2GVj1NOGdwBlR0L3F+xysirc5xAwUGnkHOwVrmHz2Cy+mKNuTufRRPkicR5DKDHk6Q3BG6h6lkVDGdPwjdY4GXAIJGOc7v2VuD148sqbw+huRlW5afr+pBWWIdIKlpmF48MnAKhBcTv2WUxwjj6h3CBLStpB4mQVhyTlzw4bKs1TLLlrjsomN6nAeqCd1R1jGOyMnGzCO+3ZXmmDGFxHdYzh8XwoJ54+loc091ZHEHNLirg4lvx9vmrPELX4B29EFJJHduyQRmaQRk7E7q+RrXNBb3V8OIWh/ZwQahLYwyLxA4H5LSJYjG/pWoRXWR/wPJwsOqcJH5agQShhxhSS1QIwAM48liKanj6pA5w+EHdBRk7mEnGMq18hfJ1ndahWNpnRARYzstPEbgfiG3mgyG1gDcEBU95DjkNUMgYAOnup6WFpb1O+tB389i5MZOaPVLSMY0DWn/nG3Ir/YwCMc0WqOjv+wGt/lG3Ig9nkREBERAREQEREBERAREQEREHHHMl+DtxT/gVe/1GZfNC/wDsrv8Avj+lfS9zJfg7cU/4FXv9RmXzQSf2R3/fH9KDJe1vg5A3wsVucgequErsBp7I0hsoPkCEE0jHiPPScY7qOEd8rWppKR1EAMF/yWig9GcILZCVYquOTlUQXRtDnYKmc0MBxuoG9WfhVznOz8SC6DeTJWc6Nhj2wSsQ9LWBw7qxk7sjJ+tBZID1HIUkTSW5CzZjTGmyPpEbqOjkiaCHYQYT2uDiCOytAJWpzGmeD04yfRYjw1ufrQW0s4gk6jur6yq94cHDIAWO7uqIMqKcOAYdlHUNDXAjz3UIODlZMdLNUjraNh5oLesdP1qJpw8fI5V5p5WuLCDlSNt9Q5vUBsEGVPOx9P0DvhaYr3FzCWHOQUjifI7DRuUFOrIDSsmhilFQx7WEgFDbKoNDi3Y/JclaD4c1V6g8UNJ+HP0Sgw4r5LT0IZT0Ze/5NytYtddU1lE91VSOiOMjLcLmXgZwft9yv5pb5GDEDj4gtb5g+H9g0nKyOytAZtnpAQdSb+ZXVb2+GQBnyWjNrPBcYzvuuSNUUNvbR9ceDKRvsuL5aaQTkkbZygyqqHqiMwHdaeyPrySexWqVEzW0oZ8lpHUR2KCR46dgolO1zT9L1VH9CCFMn1Q4zsgBKBk+pWZTS9DD1HssZmx+Lsrnu6iGM80EniGWUhpxn0UwhHSS92TjbdQuoqiKMTFu3dRePIfMoNasbY/Hw8j7VZqQNbM3oOy0+iqJY5epp3+tSVszpnAy90GCAco7up5DGWDp7rH+tBJAMvV1QBsomuLTkIXF3dBRZdPKGxkY8ljRxukOG91kiIwsPX3QYzzl5KyaaPrwScKFhb4pJ7ZV0k3S7EZ2QTVJHR0jyWG36QVXSOd3VvZBkyj7381C2TpGFaXkjBVEEkcfiu3PdXyNEO3qo43EHbukpc5wz3QGDqOVnQwNczJI2WPHEWN6nDb1VX1LmjDEEpjHUQDlY07MPwOyrFOQ/Luyvkljf2QSUsYbh2d1FWnLwArWOkj3PZRyyCQ5QCwBuVYq5cRjyTpKCiYPopIsA/Esv/a/T5ZQYCva/AwrX4LjjsgyTgeaCh3JTCkfC9reryUsPh+GervhBHBMIjkhJ5RK7qUb/pHHZUQZEUgd8KyYqLxZAfIlaeCQchaxQF7oC8dwEFa+gZDTkg7gLSIndLwVkVdXPI90b3HY4Chj6enJQZE84fF0jvhYSuc7cgdlJH0dG/dApqgwP6jkj0Vauo8d/UoXd1ezp6fnhBGiq7uqICkZIGjCsxsqIB3JKIiAiIgIrmMc84b3R8bmHDkFqIpWdHT8WMoIsKURfey87EK6lY18hBV1X8B6G9ig9VPYa/7m8ZP8NYP0V69S15aew1/3O4yj0msH6K9epaAiIg65+0O/A04l/wCJ0f69Tr548L6HfaGRul5N+JUbe5o6PH8ep1889P09R6vRBWOXw91bNL4hyrH7uOPsVEEtOAXEFJmO68AEhWxP6HA581rdE+ikAMuM/NBp1JAWkPe3APqtxUVup5mhxcPyrAuDGGPNOB28lpsdfW0x6ASg1u9QxUUeY8ZPop9F00dzubIpeziAtAqqmonYPHJWqaZrjb6oVEf73fKDuhwot1No2kpa+npBO4vBOG5XqTy5VNu1JpOOuntLGStY3dzPUfNeYnKNxB0pcq2Cj1fJGYmyDZ5Hr816faP4tcNbRBBbNPzQiIsAPQR6fJBuuvtNDcK+ekkoWlhJH0V1945cslu1Bbpa6mtTS85OAzddprReLNeQKqicwud6YzlTVtE+tkEEuDDncY7oPBbmZ4U12iqubNsdHG0E9XSQusNHE6WKR3TnB/IvbT2h3CjT3+x++808TWzGN4OwzsvHSlsjYIavOdpCBt80GyjGTKW/NZjYgyInCmqaFwqSGZzlWvPhRlryMoNPDC+QgeqzGQCOPJ7qGjwZSSr6mVzZQM7ZQQTPcSW4wFEs+pdAYR0/SWC36QQBnyR2fM5U0gYGDChacuHUdsoLooy92/YJIzof0hZJ6OgeHjON1iv6+r4+6Cd0A6Ac/NR9ZZsPqVpmeRjKs7oM2lHx9TjkHyV1RI0PGOwWGxz+zCjg/ILu6DImIkaMKSmrxTM6MfmWI5xAGPtVGgOG6DK8cTvJ7BRzN6dgoSegjpWRHIxzD1d0GKHFrstVesk5Ko76R9MqrQOsDyygnjkdJhob2/OtQjHRCcqGJsTWgjGfNSSVUTYi0ndBpsruuUj5qPBa7Poqvdl5I8ysmBrHNy5BC6V8oDe2FI1vTFv37qsvhnPh91EZCG9JQWsZ4j8E+ay3UDRGXZ3HzWJGXdRLdypBUyhwa8nGe3yQVpnmKQtxkeanfIJXBmMb4U4dTBnUcZI9FjOe10mWoJnU7YGeIPNYkjvEy5T1b5REOo7KGmiL29WNggpA09WRsVkBzw7DnZCic5uelhGRsoDI8O+JBqbGtcM5GyvLRnHqtN96LRt3VpqpM5Bwg1CVoaMkhY56ck57LGM0sm2Vl09O/p63BBawgnCrI0dGcHdJJYmHB7hZbY2SwF48kGBRvDZd2g4V9VP1ShgGyip3MjqD1dlLNLAZOr0QXMe6lAe1hOd1K2uM7g1zcZPotUtDKKuHhyYJ+ahvFHBSyYhAG/kgxHU3W3q6lpk7PDkxnzWZ7w6IYedljS/f35Z3ygpJ8Ddt0gi63B3oVV1PLEOqXICyIZI+npb3QVqWARYHcLTx8J+pakxrjkydvJY81G8uLmj4fNBCJC7yVzJs/AfPZZlJRCf4QNwoKyhlp5QPXsgsnpwxvWDlR04zID6JK2ZrR4mcKrHtY3I7oNQq3AwAN3OMLSuzsqeOoycP7KOoLS/4UCR+QAFSOPq3yrFe3qIw1BIcRhRl5e4Dturw1w/snZV6Bjqb5II5I+jBWVBTNki6id/msdpMrseiqZpIyWA4CCk0QY47qrJsN6PVUBMpV/Sxjd+6Cz4mEOLsgrLYGywl22cLBLuo7nZXxzOaQPLKCN4PUfrU0VQWgNUsxiLAW4yVZExj2n1Qd/PYtj/rpNUnP/ygVv8AKNuRPYuU8sXNLql7ux0BXAf5xtyIPaNERAREQEREBERAREQEREBERBxxzJfg7cU/4FXv9RmXzQP3kd/3x/Svpf5kvwduKf8AAq9/qMy+aCT+yO+s/pQWnZZNPS+KwuJwsYd1I2ZzBhh2QUd1MeW5JwrXOyhcXEk+ap3QEwpXU0rW9RGylh8Lo+LGUGOx3SclZccLahvWSBhYsgy8gKSLxmghucILH/A7p9FY4eau8N73HbdDGWvDT5oDGve4NbkpIwxkDstYpKONkYldjCw66MPlBYNggwh1t+LdUc4u7rU2+7+CAQMgbrCm8PJwggRPqVRuMIKLU7bcG07fDLe/mtPMbhueytZkOBQaxUFmPGx33Ucd3a2Pwy37cKA1HXH4edvJYb2hqC+YiSQyA7Eq6nqBA8OxnCgyeyvh6fFZ1ds7oN7aWbLfJWUzaYuBeG9l6S8rnLfHfNKCultgJMPV9H5LpJwPm0tTTQvrizq6wd8eq9k+VHU+mXaXipKJ8QDogNseiDrjYeDtRa9YzUsFrcwB5AIb5ZWVxd5bxeLNJX1MBBaCcn6l3Sq59DWC6z3a4eA3OTk47rgnmE4z6cqrFU09gliOAW/CQg8nOK+h49OXeSj6wA3IXEdZRwYkII29FypzAahra++TTRuJJJ7H5rgk19W5xa9x3O6C6dhlmMWcAeqxp4xC7AOc/NZvuk87A+IHqWZSaNvNc3xGRkg+ZBKDQPmq4J37rVq3S90oM+PEQB8itMAbGSHZz2wgtY3rKq9vRsN1QEl/weZ2R4cD8XdBfBGJn9J2V74hTyNcTkA5UUXX1fB3V0jJj8UmftQapVXGKSkEQAyVpMYzkq0k9kBxt5FBNFJ4bt/mqTyeK8Y7KwxuDerdXRtz8SC99N0M6w7yWOTk5V7pXkFpKtb3GUFEWTIYvDAGMrGQTU0/guyRlXVNV4/YYWOr2wvcOoBAhj8R3TlXTw+EQM5yrA50bjjYoXue4F26C1FOSzGFaIHv3agiVWjJAVz4Xs7hWsOHA+iDJNP4bRIscvy7KzJZ2uhDfRYOD6IMmSq6o+gBY7Q7yGcoxjnnAWqUMELGZm749UGI2lLm57Eq/wBx6W9YOVfWzsaS2LH1hX0crnREOP5UEDHCd3g9vJSvtga3qznbKxZg6KXraqioqZR0gkoLHkNcW+mysLxhXup5R8RHdWxgCRod2ygtw7uQVJHCXtJ6lqlV7n7oOgDrwtIa942B7oKPb0uIRp6SHehUsbCD1SdldJGHDLOyCslSHxhgByFCGEgnOFYRgkeiqHEbBBJCzxSWq+WmEZwraZ4jdkqSp65D1N7ILBTAjOVkx1nusZiB3WG2VzTglWyO6zlBWaTxXl/qrERBd0bZyqduxQhw3VEBERAQbIiC4uGMYQMyM5VoBPZVPU3ZBQ7IiICJ3V4heR1Y2QVhl8J3VjKrPMZt8YASCMOf0uCzXsgYzcDOEGmp8ldLjr27K1BLE7wj1ZVJpDIQSo8lMbZQernsNf8Ac7jL/hrB/wBGvXqYvLP2Gv8Audxl/wANYP8Ao169TEBERB149oLMKfk74kSns2jo/wBep187+5OfVfQ37Q/B5NOJYP8AedH+vU6+eNBUtwMqrWZBOVQknb0TqLdkF0UfiO6cq6Rj4XDDj8t1ZGHk/B3UrYpXSDxBtndBkUta4fA8EhXTyRueHjCvqo4WQfewMrTg92wPZBkVVT1sDcdllW4EQHBwfVYE3QWgt7+a1OgkYykd1d8INa0verlbKyJ1JXviJePokjzXofy3U8t2tUdzr9RHqZEDh0m/ZeZtFO/3vLewOQuRNNcd9YaQmbS0M72wDALeojZB7X8GdcwQV3uEl0Dw14G7l2YjutDLC2RlQwkjOM7rxi4C8x1RLLDLUSkyl4J+Nd19BcytHW3WmtlRMMvAGOpBunnHgqtW6XktFPTOlDI3NwBnJK8pNRcKdQWySeFtilDXyHJ6D6r3VptOWHWFngrauAPEzc+RW0NV8v3DqotNXPLQMY8MLmuLRsUHgjqzRdVY4zUS0ZY7GcELjWqjlqXOd0EYK9DOZXg/DHeKqC3NHgtyG9LfrXT3UPDK4W2CaXoIAJ8kHGMEPhuJJ7eqhqzl6krWzUtS+FxOQscP6j8SC+ng8d2CUqIRC/pG6CXwvoHdWEvmPUfyoMiKm8Roy7uoZoTG/pwSFQPlj81qtA+lkgJnxlBpMb3sdsCtQhofe2eI7YrMoBQeOfGxj54V1e5hd4dFjHkg0aqpTA4gbgFRMZ1nB2WqmjkaDJONiM7rTqhzQ/72dkE0ELWHqO6VPSdwOwWKJpPIqhe52xOUFHHJQHAVPrVWgHYdz2QCO+6plS+7yYzjZXCmd09SCDupGR5I38wrCMEj0QOcMYQZ8cOR3VHUfiPGHbd1i+JMzfJAU8Mz/pZ7IJZ7c2NoOe/zWo0VojlpS8uAOFpktVJO3oadwqMqq+CMtBOEEU8Pu85aDkZQwiQdecK6mjlrJT5lVq4JqZ3QQQghpxiQg9knwZQAMKkRLCSdklJc7qCC+WEtZ1dWVDG8tcHZR0jnDpKowZcPRBqDZBWMEZyMeZVBK2m/2v67ZUT3iFmWbFQYkmPUe6DKkgELfHz3UbYveB1dsBRSSTFoY8nAU0D8RHGyCrKEOz8SilpnRnYEhUFRI15wdsrVI5qeSAmT6Xkg06ma0O3Wp+KzwC0YGy0kv6Jic7ZVHzvecA7INwaZ03HfKkiR4AzlSalomWGYUkTgQdlo9vrrnbj4lISCfMKyvr6m4P8AErSS8eaCyGnFS7qyBk9lrUWmYpIDIZBkDPdaDGyobvGDjyWSLpcIm+H1HH2oJ4o3WyoPS4kZxstcbSsuEHvD34I8itv0Mjp5i6ffKlqq6ogmEUBIZ8kF8tEKiUxZwBsoX0jaF3XscfNX1E7xGHxH4yMlY8L5ZyRNndBIKgXD7z04UU1D7qerOwWUKYUwMjAAVgVM8sx6STj0QTsqhNhg7jsr3VzYh4RaN/NYlKwsd1FKqNzn9YQZ1LWtoXmU753wo6q6Nqpg4jAHyWmue52AfJVa04yg1GZ7J4uloGeyw/d/iwCoxK9qkgZK9wduQgv9yJGcqPwMHpJ81qTmER7AZC02dzmv77oKzwCNocCqwAAdXoopDIRl3ZXte0MIB7oLnyCY9PbCoHBg6M91CctOyFxPmgyqdga7J81lSW5kkfi9WMDstPjL3HA3wpHVk7QY8kDsgiB8F5HfGyE+K/HYFWuY4fE7zVoOEE0sXQ0HKvipfEjL8qB0jnDBVzJntb0DsUFjgQSPTZXwv8NwceytcxwGXeaoAT2QehXsYK1tTzP6ni6e2ga13/ONvRYnsVmkc02qT/8Aw/rv5RtyIPahERAREQEREBERAREQEREBERBxxzJfg7cU/wCBV7/UZl80En9kf9Z/Svpf5kvwduKf8Cr3+ozL5oJP7I/6z+lBaqtblU7q9uwwUFh2V8Y3DlYe6kjYSMhBkS1ofF4Ybv64WIc/MKeNjA74gqTBpIDMboIRtuVkR1AaOnHl6K19FMyPxHDZWwBmfi7oJPF6HF5GxVC4Su6x5bq6Yx9GG4yFjtf0gj1QanHXiRgg7YWPPIIstzklYjHFrg4HdVlkMmEFCXOccHurvDJI/wBatjBc7AG6n8N4GSPNBX3TbJKgewMd3U75yGluVjOcXd0E3ih7QwhRuAZ9qs7KpcT3QASCHKVrBNjJwo+42V7Y5gMtBwgvfAGN2OT8lAAc4AOVMyRxd0vPn5rKijjLm9kG4tDUdTJUMxUOYA4HY9l3B4a8xdfwrtrGNujsMaB3PkF02gubrVGJIjgkLdVruLL9bHtnBLunHdB2mvHOlc+INS+1wXV4J+H6S451jxautogkiqbm94kyfidnuuBtL242rUJqekhgJx8hlZ/Ea4i4AFuT0/NBh6j1Yb7VukfJ1k53WzqtzWSF2MbqlK8CUNO31pX7uGEGq6frmGZkb2g/EAu53BXhWzU1gFYLZ4oDM56c+S6RWkiCWOV3/wA0C9duRS66Sr9CC3VpiNTJD0jOM5wg6tX3g7BdrxLbfcAwgluMYwuOdccsFxt8T6mmtzyNzlrSvQW7cA9R1mtJ7tbYne7SSlzcN2xlcsUnAaGo0zKLvE0ydG3U3zwg8K9S6PuGm6gsmpnNLfVaA1rqhxLhghehvHnlZuV4vlQy0QEgZIw3610q4i8NLrw4rZaa5Mc0h3TuMINkxsEMhc7sFWoqWyfAGqaWMuhEjR3GVp+/V9qC+RnSA7KyqOiE7DIfLfCxpGPDAT2U9LVOhaG52PdBbUuDCYgNh5qBsnSDhTVL2yZIxlYyCSJnivI7ZVZ4vCOAqQv6HZVZ5PEcEFrGmRwaN0ljMZAIxlT0bQx5fINsearXOjkeOjzQYuMb5U8VSI2dKsdBI1nUQcKwMc4ZAQUe7qcSq9G2cq0jBwqhxG2UFY29bsFZImEHw4ysZhIOyPJJ3QTS1AlyAO6hLMdireyu69vmgpkqrfiOD5qseM/EhIDwW9gUGSyMU4EhGQrJal0g2KlmmY6ANzusNh3QUJIOTusinmcDgKN4GMqejMQI6sIKztL29u6thcISCQs+UQtZ1bYWnTSMcdigkkrWuGAPLyWG49TtkcMKojcR1AbIKua8Dc7KwbEFVJPYqaGnMg6h9qC185cwNAwkc3S0g75V0zWNGB3UI7jKAdzn1VFkxMY8YA3wopoyx2EEeVOypLW9JGVC1pccBTxxNxuN0EDjkkqinfFjsoXDBQUQbIiCQvDh0qM91XpcN1TOUBERARFUYx5IKscGnJSR4eqFhVOyAiIAT2QAcFZDKnoYW47/ACUBY4eSlZTPe0uAOAgtbL0vLgqSSF5yrS0hxGNwVQgjugJg+iKeKPqZnCCBVBwCPVHDDiFeGZjLvRB6sew1/wBzuMv+GsH/AEa9epi8tfYbY+5vGT18awZ/JXr1KQEREHXP2h34GnEv/E6P9ep188a+hz2h34GnEv8AxOj/AF6nXzxoKt7q4tJGVYsmIs6PixnCC2kIDyXfJTz1DeoBvmsMuLXHCp1FzsuQZkryY89wsMnJypjKCzpB8lB2KBv55WXA8iI5OFWAQ4+LH2qOpeGvxHjCC2KYwSEhST1fjA9gfVY43PxIW75ag3Jo/VVbpysZLFUOaC4bA9l244B3Wv1JqujuE1wcGNIO7tvJdL7ayJ8rfH3Adtkrn/hLrqOy1cVPQvAlwAMHzQe3XDrXFLRaNpaeKrbLIxob3yQcLeb459T2shxcOods4yuk/K7qe+XVsD7xk05cCMnbC7v0WqLJStho4HMBIA2I74Qdb+K/COGrlqZX0YeQ0knC84eYSuptMV0tpbTBnVIW9vmvaHWFhdeqKaWnaMSxkDH1Lye5x+Cd+jvU1z8J3htlJyB80HSXVGnBITcGN+mM7LZUkRYSCMYOFyjf62OgP3OqjhzG9O/yXHN0kifKTF2yUGKI+od1IwNjaoWvdnGe6q95QVlkDtgqRscQSHEBViAc45Gd1JJFI0fB29UGOSQTuc/WtatY6IvFeclu+Fop7nK1ChlIHhk7HCDWqOQXiR1N0gAHC029WYW95wVrun6PocZYgMndYGqpZBKWyd+yDbbY8uxlXPjEYHqVQE9WR3VJHklBYe6uj+mPrVquax2xCDPlnayIAYVrKhr4znuFiPkJHST2VGskIJGcILXn4ifXdXMZnDsq0g5we6ujd0kA9kF8rgW9OMfYqMl6W9KvmLCzY7qAAEIL4JTG/q7rLkr2uYWBo3WAqgAjY7oMq31nucxlIzlS19xFY8EDHqsSOMA5eNlI5kRAazugikIIGO4VGuPTjCk8CRh6ng4UhdCGY2zhBiE5VWnBCrguJI9VVrSHYPdBdKctCvp5A3AxlDGSBtsqDpZ37oMl0bZxsAonRmFnT3z5q1tR0HAKm8WORu5GUGC4HJQOcNgSsiPw+o5Ix5KKbp6/h7fJBLTRtqHFpwPrVJohBMGg5AKiZI+I5bsUfI6QhzzlBqza6KniGQCVp1VUieTrAwM9lE+QvABVgGUGbBWCMAdOVJLMyRpdsNlFBSOPxOGx3UVQ1zH9I80EtNLmT4VmvY0jJ7rDpIS09TxtlZfu88r+pgPSgsgDnPIIyN1ZUuNO/qAwtVgijp25lxladcHwzyYZj8iCjK33gdG4WO9obKB3CkFP4I6h6KGQlx6vMIMiRjQwFvdTRQh8GXAZ77rBhn6iA8nAWf1ExERnbCDSJmhshAVGvI8sq+aGRriXDue6lggDgHkbIIWsMh3HZZcErYT07KyV0bAejGQsbrd1Bx8ig1Yytc0labUH76CpmylwwDlRStxufrQVk6XMAHcLH3acYKkiZJO7pZlZDo2xNPiYBQQRs8XZXGmw7Cjik6HegJU+XucHeQKC+OAQ5c7zWNM4dYI8lkTzgtwO+FiNPU4ZQSh5mb0YxhWuhwpneGxoI7qM9bwXDsEET2dIypIowR1fJGsc84IVXOMYLeyCk0oI6R5LWaCzMmoTUOxkDK0A7kla1R3R0dKYAe4x3Qd6vYu4HNVqtg/e6Brh/wA425E9i2x45qNVyO7O0DXfyjbkQe0iIiAiIgIiICIiAiIgIiICIiDjjmS/B24p/wACr3+ozL5oJP7I/wCs/pX0wcyP4O3FL+Bd7/UZl80Lh99cPmUFjTgo52VUNLnYAR8bmdwgoRhSRTeGOkqxoc89I3VXsMZHUMIL5Pi+IKJjulwd6HKuLxjAKsQajPc/EgETW79lj09MXtL3ZCrQ+D4n33H2rJrZ44x0wkINPljLHkZyrMYUsTw6T41fUeHn4MYQQFu2VQbqZzm9GFCgliIiPV3Uj6oOGFjgnO5Q4QHHqOVnUdEyeIvJ3+awFLFUywtww4BQWys6JSweRwrXNwhJe4k9zujmuH0kBruk9srJZWdLOkhQw9OT1KswYD8KCxz8vLgpI6hzSDnsVCrmRuf2CDNqp3Twhoct5aMYYaMue49u2Vsmnp5HENx5rdNor20bRC7bOyDPqrsfe3MYzBz3C0q5zOljd4ric+pWsBtJGTUzYwd91ty+1sVQ/wAOkIJJ2wg0F7vDnJHkUkn8TGQp5LZVxsE0jMNIzlYh6c7ds7oNYskDa6RsR2w4brujyu6kOmdQ2+nddDHGS0FvXgYXSaiqXU2HQ7OG/wA1vDSnEK72W8wTse4FhGDkhB9F/Dq/acuGk6SqhqadxDB1k4JzhZGpLlQzUT2UrA4EfvR3Xlbwa5wLhYbTDT3WqcIgQMOd5Lu9wk5itKaupIaieWIsDQXEuCC++1MFvuEtRNaPEHSTkx5/mXmbztQwahv0r6aiEJ8Q/RbjzXrnWcTOE+o2y2+GSmdUBhA3AIOOy8vOd2rsds1I/wADoDHS58vVB0uvFilt9rjf4OMM9Fs3fqyQe/oucdQ3rT1XYI2Ncwu6MbY7riKpNJ1u6D5lBBLIDAB54WDg+WcLIqXbDp7Kaj8DwvvmM4QYQbnuULAB3V0xHiHp7Kxp3G6CmCPIqoODlZI8Pp+fmoH9PVt2QXuqCWdAGPqUbCS8Z8lJ0B7fhG6jcxzDug1GokYaYAd1DSMaYySFiGRzgAeynhlEbCghl/shVoBJGyPd1OJ9VkQ9HRv38kEfQGAOUbjlXyhxPyUaAqtHU7CNYXHZSCCRo6kFz6cNb1AntlQgZIHqrnSPOxVrTj68oMiWm6Y+sO+xY3ZTsnLvhd2VkvT1DpQGZeensr2wEOBB2yqB7Q0dPdXeN8J33QZXhiWPo6lC+kEeSHLH8d7SSCpG1LnHDkEUgIwCs6hiY+E9WPtUZayUbYyoTJLT5aDjKClRGGykA7ZUlPUiMdB89lAA+d3qUkjfEQHDBQZ0lG1zTL1eWVimn3OOyGacMAdnCkilaI/ixlBbCPBd1ZykzhO8eW6jZIOsl3ZJXjqBYUE4pvDb1DJJWZTUTZmF7tiFiUUpe/oedlJV1T4XhkRwD3QUrGNiyG74WASTupmukqSQdyrpIvCGHd0GMg2VXd1e3pwc4ygo6TLcYViq7uVeynke3qaMhBGqgZVCMEg+SZKAdkGyIgk8Tb7FY45KoiCSKMPO5V7mCM4HYqFry07KaN/UR1ILpBhgPosmnqAIiCN1YXx9OCsWR+DhvZBRzgZS7yyqSHJVmTnKZygKWOYsb0huVEs2mEPhEvxnHmgwycnKvbLiMsQsL3kM3Hkj4HsGXDAQerPsNm/9TeMjvWawD8ja/wD1r1LXln7DU5t/GVvpNYP0V69TEBERB1z9od+BpxL/AMTo/wBep188a+hz2h34GnEv/E6P9ep188YxndARS/DjyUbsZ2QUVWjqcB6qidkE8kAiYHZOT5KsUAkYXE4KidK5zek9lWMShpLBsgsPwkgEjCEknfdUOcnPdSNALckILCcqrD5Kju5VzcEfNBMwEYcDgrUrDeaiyXRlxEpwwg4z3WkgPaST2VHdUjsDsg7q8F+b82uGC0Ol8ItIGTsu7PCrjBTarr6aaS8tJIBAL142aepzDUMqI8l+RjHkux3Byr4k0V+pblTNmFDGQ5xBOMIPc/Reo6OvtA8SpYRGM9RPcYXRnnh4l2mmlntlNAyQvcQCBndbk0dzAQHS7LVTS/7dawNd8W+cLrLx3vsd/uRdc8GRzyRkoOmnEeyy1dyfcWtLQ8E7bLjOSnMbnAuzuQuwfFOnhoaIytADS3Y/YuAC41MznNO3VlBC+Exjr8lC52VqFWAIQMbgLTkFWPLDkLKbV9TRGRudlBD0dXx9lK2LMoc36OcoLZoS0dYBwTlZVG0FviZxgKaokgdAGgjK03xXNyxmwO2yDW6DURoCR6eiwbxdDc5evyzkrCkgkY3rd2O6vi8IRnPdBADg5QkuO+yki6Os5xhUl6esdPZBb0bZVzZA1pbj7Vc9w6Rg/Woe6Cp3KmjqOhvSWq2Dp6j1Kkwb1fCgsccuJVenIyrVVhIcN9soBBB3VMH0U8xZ0DHdXRGPw9+6DGQHdX9Be8hqmFM5rSXjGyCJ8uRgKxjulwd6Kh2KqGk9kGTNViSMMDVSKk8SMvzg+ijg6cnrHZXmdwf0sO3ZBG0+E8g+RVTJ1vBA7KadgEfV5rEb3CDUYg3o+L081h1IAfsdlUzENAb5qwO6nguQSUtP7w8tJI+apUwCB/SHEhSvmbE0GM4JWNJK+V3U45KC1BsqhpPZCOk7oBOVRVJBCqInEZQWqrSWuBxnCuYADg4UrOjqGe2fJBk09V1NDCMYCx6mT76CBkK6odExo8PGSrqd0UjSJMZQZVuxV/ARjBWTUV4t7hAG58lDRtNM4vbsCpJ3007svwXIMSurXzMy3IysGBzvFAJO53WvU9ubUtw0ZA7bLTa+kNLPgbYQagYBLTjA3wtNniMZLcYWdBUiOPDj3UdUY5GlzT9WEGpaX03DeJCJHgYUN/pPuHcBSx7tJwtKorzXW1592kwcrVzWMuUXvNY4GUbhBfU2+GakEmQCRlYbaSOOAgEZWBPcalzzHG49OcAKEVs4PS87ILnUZMh3OCSoJI3RnGDj6ll+9N6Qc7k7rPiZTzUxe4DKDRIpCwqTqM7ukDvso5cCVwHbKzaKIEhyCsY9xHidOSfNRZNdMM7BajN4TmYesE9MLuqPG3ogvrbX7tF4gOViRTHHhkd/NZj67x2hkh2Gyjd4Ab1NO49EGNLB0DqyTlRDIPzWTG/xCQ45Hko+lrZ9+yCSmgdOcEHCumcKclnfKz/FgiiHh98LSamTxHknugq2cNVpd4rwO2So1UBw3HkgmlpxGzqyVExxB2OFV0r3DBViD0H9i5OJOaHU7dttAVp/5xtyLH9ip+FNqn/i/rv5RtyIPalERAREQEREBERAREQEREBERBxxzI/g78Uvlou9/qMy+aJ5xK8/9sf0r6X+Y5vXy9cUGjz0Zex/yGZfNJ7u+WeVrR9AnP5UFtKQJN1JWkEhIGBshae6VowQgghk8J/UrpphKRjZRJ2QSvg6W9SugpnS74V1M8Pd0SHZZ5dDGwluMoNPnpvCGx7KDJPqVJNO97zk7ZSnDXPAKC3w3YyAVac9itWNM3wg5YcjWNzuMhBjRs63YWSaQBufkscuLXZaVXx5PM7ILHDBIVEJyiAqgZIHqqKrdiEGR4Qjb1/JQyyB+MeSmkf1x4UDR5lBamUOM7KrcfagopqeR7XYa3OVfTW6pqiBEwnK3lpzRjp5mRzAguGeyDbkXvg+KKlc8n0GVWOnvMtUwmjkbv26V2u4L8ujtV1LQ+NzmnG/Tlcxaj5QY7ZPCWU5GcfvEHRO62+4G1gOgcw9PnstmRB9NN4jzuxd4eNnBCLSGmfeHR4IYT9H5LpBciPe5mNOzXkdvmg1Gtv7Kik93azBGw2WhhpPdXwNa+QB3ZSzgMcANslBFFIYX5xsshtY7x2PaexCtlYwxAjc98rG3BQbrud7rnW1kVPWPZjfAdgrf/C7j5fdGWx9A67TAlpa3LyuHGSSEYkz0lXmmjc0uYDsg7A6T5kNQ2bUEl2mvszmPOcGQra3GzijUcT6ttQ6qLyHZ3PdcSR+GXmMtP5VfK2WLBYCAgyHU9UYQw1BIHllY5oy1pPVk/MqMVco7lWuqZD5oLZHH6PojY3OGxwFYcnc+akYyRwy30QRnYlFe4YODjKtGOrfsgpusmOmD25ykgYIw4d1E2WUD4eyAx3gvPnhSvf4rer0UUUbpX4AJz3V0kboXhh80FkUfW/pV0sPhyBgOcqZ0To4xINs/JYzpHOIc7chBlvoA2ESAkk+Sw92nHosmGrefgeR0qssPivBjGR5oMd0uQBhSw0wkZnKndSgsAA3+pYpdJCehAafDlI+azGzNe0AhY7Y8Drf57qN8mD8KCSeANy4eahDMgo6WR+xOVb1OCCh2OPRSRxdfcqPur2vIGAdygo5hacKgG4BWRTRkuzJ9HKVYjDh4fr5ILZIA1nVnyyoWtLlc50hGHZwjTgfNBLAfBPUSraiXxSCOwKjcXHPorUGTQvEcmT2V1fI2SQOCxQSOyr1EuGfVBK6YPj6PsUB281O7oDNjuoM5QTU0Anf0k4Vaqm8B4YDnKijldEctO6rJK+Ugu3IQTNb4LRIPNQyymQ5KlgeZD0O7Kk0WHAMHdAp5fAcXHdUqZ/HdkDsrX08sbepwwFk0jIi34u6DCwizKtsYaejCxmAY3QWE5WXTVYij8MjOQsV30ijMdQygmmizmT1UAGVlzSNMYA9FjMOxQWnZAMkBVAy7YZQgtI2QSvhDWhyhV5kcRjOwViAqg4VEQVLyfNURSwxOe4EDbKCLCLUKiOJsAxjqwtPQEBIGEV8LeuVjR5kBBWGQxO6sEhTT1Imjxjf/wBq1d9pbHTCV4GFodQGtcQzsg9V/Ya/2hxl/wANYP0V69TF5aew2H/U3jIfWawf9GvXqWgIiIOuftDvwNeJf+JUf69Tr5419D3tCsftOOJOe3udH+vU6+eHCAiIgIid0BTxVPRGWH6layF7SHOBASQM7hBG45cSgdgK5kT5CQwZwkkT4jhwwgsO6q04IKoiCV8wc0N9FbHJ0eSsV7I3OwSNs7oNSt13NC9jsbB2V2G0FzI0NmsTrM6BolczpBwM5wut5ZERjzU1AyBk7XEEkboOzmiuJ1fQXuW7vr3+FI4kNzsFZrviLJqO7RziYkBwJXFdiqQ6FrMbLX36ar6mH3yFh6QM5QTcSKwXazNja/J6MbLiGkofdY3l43+a3xXV56zR1DiC3buto3ZxbIfD3B9EGjVriSR5LBWo1fT0/PG60490F8UfiHCnMpgb0evoooHljsnYLNJhlaQcdWNkGnElzs5O6q0ljg7vhXyQPjy7Gymg8IsAd3OyCstWJY/DxjAwoRBlvVlHxeG7q8j2V7aeokb1Rg4QY7hgkeikjh62lyjcC12HdwpmShjSMoICMEj0UzIepvUrOnrcQ0ZV33yIEHYYQRkYOyomcogImMpjCBlMn1RVY3JGeyC+CQRklTS1bpAQ1uR9Sim6MDHdZ9G2lNMevHVjzQacxvW45VziGfCqPcBKensrHO6jlAJyTjzRmzwqIDjcIM9zRKzGe6xZIQw4BVGySdmqj3PJw7ugsOyK9mCfiVHjfAQU3PmqHZVw5u6vBb074QWsd0qpIe4Kz6kBwgnkg6G5ykcwDOnCjdK9wwTsrEFX7nPqpWRZjLgdwoc5UkchBA8igpHGZHYOVlmhewCQZwN1Izw8As7rLbPH4BidjJBQae6sLW+H2woep7ndfUfqSWnkDi4Dbuo4yQ8NKDWrZdhT4a/thR3GpbVyB7VizUxMYdGN1JTQvEfx7HyQXxwe8DAOCFjVJdTO8PvnyWVBKKZ5LjgK33OWrqRMBlgOUGA4vaQ4sOPmtTpYBUQdRdj6lqVXHQGmEbMeIAtvvqJoHGNhwAguniFK8433WP8A2YnyU0XXIS6bsfMqOdpY8mPsgkjouodWVV0rovvWdilHM9zulW1JDpOj98giljx8We6vgqTGMdlWSjqIWCSRpDe6x3YzsgzTIZezle6MiInvssAPLeyz6Rxfjq7FBhxxOkk6TlZrreGxdQdv6ZV1b4UTA6PGcLFhq5TI1r3bdkEA6onHY7KQjrHUtWnZSPgy0DqK0eU9Di1vZBZ1vG3UVeyPxDkpCGk/Er3skacsGyC2aARjKngga+LJxlY3W52zlUyPYOkfoQWSDDyPRW4PoqklxyVm08LHR790HfH2Kn4U2qf+L+u/lG3IpvYuUzo+aXVLndjoGuA/zjbkQe0aIiAiIgIiICIiAiIgIiICIiDjvmLd0cvnE939zo29n/kMy+a2lqGsrKkuxiQux+VfSfzJfg7cUv4F3z9RmXzTw001RM/whu0kn8qCd0YEhlA2ysaplMh+pZXiCMGKTuFiTmM56fNBCBkqpbhV8N4HVhXxFhHxdwgjBLTncK50rj5lXydHkoe6B3VWu6HAjyVTG4DJCoGk9kGp09WJW9Du3ZYtY3pdt2KxmudGcjYqr5XSHLt0FqqDhURAO6DdEQSuhAZ1ZUSu63kdOVbjfCCaD43dJ9FWpYGEBvYqPokjHXjCOe6UgHcoLFUB3cA7KQUspGenZZlM2BlO8TYDuk4+tBr2hT7zXNg8LqPUBhdyeC/A6r1bdaI/colj8ZPTsuq/AOzsvOroqdwyDI3y+a9yuV/hXZbdpimuUkTTKI242Gc47oMjgty52rR9sp6me3xiRwBIIGVyffeG1nuoZ/tGPLMeQW9WMEbAxvZowrkHULmX4KW6/WKWidSsiYIiM9vJePXHDhTR6HudWymeD99PbfzXt3zn3ip09w1mudEcShjxnz2C8KeIWtLvqq61/wB0HlxExAz9aDj18XgAPB7qJzjKcnyWTUxSgBz84CxWH4xjsgZe3uDj5q+OLxAXenksqf3c046cdX86gpntawgnugtlmy3wwMYV9LUeG0sd5rHkx1n61QNd3CCY/A8yj1UhqvEYW9KhjILul/bKzWin6NsZQac7JJJCoNldJjrOOytQV3Pkpoqjw29JCtpojLJ0gZV1VAYXAeqCF7upxKmhp/EZ1eagU1NKWSsBPw5GUFrmyZ6XAgZ8wrm4a3H5FuW5OtZtg8Igy4+3K2qHbjKCWCYwvLiPzKtTUCeQHsAqv8Lw/nhY/ZBmSVAdCGDusPB9FXJOyz6NkRjy8DKDALHNGSCFkQVIjGDulW6LJDFjIMwV2Dv2+pQTy+K7qwokQSvnywNwokVwjcRkDZBRpwjjlU7Jg+iAqjYgnyV0YGd+yS9OdkGoW9rax3hHAPkrLlTCkkAG+fmsejqDTydbVfWTPqnCQ74QRSSBzdvRQjZDkbZRBISCFToyM5VmT6qoLjsEFW4BVHnJV7oJGgOI2KjygZPqgBOyfWsmMxBnxYyghdGA3KsVz3ZJA7K0AlBdG/oOVK2ceIHHsPVQIgz6mrZJF0tAzhYbPEH0QSFZnK122GiFKfGx1YKDRXvc4YOfqVmSpqsNEzjH9EnZQ90DumFLHHggu7K2XpzsgsyfVERBVhwUduVTKlYARkoIkG6ufhWoJREMZKjc3BwnW5Xt+I5KCPCniqPDb04SRjWtBx3UCC+WV0hJzsrWjJA9VRSwloIygq6DpZ1KyJ/hytf6HKmmma5nSFjINckvTainFOdtlo0wxIRnvurASDkeSvLHuBeRsg9Y/YeQCOwcW5vOWosmfsFcF6hLy19hvM99r4xwE5bHNYC37RX5/QvUpAREQdc/aHnHJrxKI/vOj/XqdfPI3uvob9od+BpxL/xOj/XqdfPHlAVelx3wVRZkDoREQ7GUGH2VWnpcD6KsmOs4VqCeSo64w0d1BlEwUE1NUCEkkZyq1Mwnd1AYULWlxwFIG+GMHuUER2RVd9IqiANlPHMAzp81AiCpcc5ydlcyUskD8nZWIg3Ha9SGk6Wl+MLkOw8WY/DbanMB6/hzhcM4ytzadt0Yc2qcN27hByVc9Mx1kT7ox27/AIsBca6gJopCzB7rfdLqg+CaRzvhAxjK2fqWOOqlL24OSg2nLK6Rx77q1sbiCcHZZxihiOH4Vzn03hnpI/Ig07uelXAGN4Oc4VOlznno9Va4OacHugzpJWzxhoGSsVrHMkAO26ntz42vPidldWM65g+LcBAqgTCCB23Vaa4iGLw3N39cKYPifGGHBOFG+nibGXYGUGKQJ5C4bZSSme0bAlIM+KWt7LVWvpxH0y4zhBo8bzE7JH5le+bxCB5q+q8MuPQsbcEFBkOph09Q7rHc0t7qeCY5w47AKyoLS7LeyCIHByhOUVWDLwEFMK9uOnCyJ2MEWR3KhZBK5vWB8IQWObg7lUDiOxKOJGx8kDSd8IKIqjG+cKRtNI/dgyEFsbA84PkkjOggAq4xSx+SjcXH6WcoLo39Byqud4pz2UaqOruPJBeIwrXfC9U63KRpb05OM4QUkOQjIuoZyoycqoe4digq9oarRsqkk91RBUnKogBPZMHsgq1uUI6fNXeG9u6tJJ7oL4pTGc5Ko6Vzn9QJ+SsxlVAw4ZQalHUtkjDSMH6lhTANly3sVdJI1rcMO6iaHPcCUGa2sEbQHDOyyIahsrcjbCwJ2gtGO6tjm8NpaO6DKrGh+Oh261e3TiOiMZbv6rQI/Ge4Ek4W4aWppIaQseQH4QaJMZY6hz+skZzhYr5HOd1dKzXSxPncXbgq1/u5PS3zQUgeJ8MO2NlkTUzGRHcE4/mWI8+B8TNsqnvj5G4cUFtK4RzH61q8VqjlxWdQwN8ZWhOfg5b3U7bjUsZ4bXbINTuVfHPGaVoG2y0R7Ok4G6F7i4vPcq8Oa8fF3QRLUYAGwB2dwsHw3nOArmTPZhhO3mEB73ySdJJx2WXFRNMfUCMqGYxhgczuVbDVva4AnY90FwD2OLXOOM+qnpqBtXMGl3dWVUkZZlhGVBFVywu6mOwUGs3CxsooRKxwJIzsVpjZQIyCN1NFdZpwWVD8hYs2HzhsfY+iCIEuefJXOzjAGVkVdI6GMPxjKU/hvjwfpYQY3RjJKubO5jtspURSMOd8KSPwjFl/fCDv97F6rE/NDqeP00DWn/nG3f60WN7FfH7ajVWO3+x/XfyjbkQe1CIiAiIgIiICIiAiIgIiICIiDjnmPGeXjikP+4u9/qMy+bSzVbKWqqOoAh7SN/rX0l8yO3LvxSI8tF3v9RmXzRFzhO4juXEfnQXVbzNUveNgSpo6Jr2F7ndlNJbpRB4wbv3WGJ54vhJOAgvB6neFj5bqOeDwSAD3Uznxhge3HVjusV8rpMdRzhBblAcIiCczBzelRg9GVZ2V4Id37oLDuSUUpp5MZA2URBHdARXRkBw6uyrKWk/CgsRE+tABwVXOHdXornY6BhZNJEx7cux5ILRL4zPDIwr6elHWDlW1BjZnoG6tgqnMkBcdh80Gq1IZDBs0bLR+maeTEbHEnsBlav48NVEIgdycLkPhxo+33Ovhp5wCXYxkZ3Qb/wCUPQ1dU6kjrpKMuzI05I+a9reEVTNabNSUZgLAYwCMfJdNOTzgfT09TFVPjHh9QOS1eiVDp+30McTYogCwAZ+pBqUb/EYHeqiqa6npceNKG5ON1M0NYA0bYXDnGbVk9oppJKZ2Cz0Qccc92oLceHUtC2sYXeE8kA+oXhTqeqEV3rulwIMx7fWV3+5v+L96uVvkpJHuI8Mt+kvOiSR9bUzvlOS4k/blBMKkVg8I4BxhYdTAKdwAOcqhjkgd1N2A81RxkmILjnCCxuXHGTj61dIOjsrmNz8LRk4VsjXN2egj7q5j8DGFQtPfGymh8Pp+LGSghJycoHO8iVIYHlx6RkK3oLHgP9d0ElHTmomDD2Kyqu2tgHU13l6qQFkUIfFs7HksGWrneS15P1IFNN7vIXYzhXVdV7wQQo+jxAOnurmU0ge0ubtkIIek4zgqi1usFEKMeHjrA3wtPoqfxMud2QYxc/G5P5VRrS4geqzqxsLG9IAyraKnEjC7AyN0FklMGxded/rWKs18NSXdG/TlY0sZjcMoIxnyUscjgQ0ZGSrYi0O+IbKZ/hd2jdBc+kAj8Qu3x2WL0/NTPdOR3OFYyN5IPl9aCPB9EWQ9sZZgDcKAAhwygphSsmDW4V0nheH81BjKCWFokfgpPGI3gAqxj3RnZHPdIQT3QSvjAYCDhQKRr3HZx2V3wYOMIIcrUaKJksZDz81gOAJ+FXRzyRbNOEEtXTiMlzTsfmrIoPEaTnsqte+d2HbpIXwnpGwKCF7elxCN2IPkry4OGfMqWPwiw9QGUFZKsPj6A3ClpaBk8ReTjG6wXAdR6RspY6uaJvSw4CClRD4Ly3OQroabxGFwconvdI4ucckqrZnsHS0kBBa9vS4hARhUJJOSr2wyOGWjZBYiEYOFUEAIKKuXeRP5VIzo7HCmAi22CCGMGTY/nVr2eG4Y3WQ5gcSIu6hkhmG7/wBKC57/AIBhQZV7MDYqri3GEEYGThCMHCqCAfkhwgoqhxCoiAd0REBXsd0hWJk+qCR0uRhRoqt7jKCmFVoJICmk8Pw/h74UA2KCZ8HQwPJ7q6GmEjC/0+aidI5zQ09ldFK9vwA7FBG4dLyPRS+MPCMeNyr5adwb4h7HdYyD1d9hs3Fu4yO9ZrAPyCv/ANa9S15Z+w2JNv4yjyE1g/RXr1MQEREHXP2h34GnEv8AxOj/AF6nXzyxs6ifkMr6GvaH7cmnEs//AElR/r1Ovnka4tOQgodiQqtaSNinSTuVVvU4hoO52wgt+tMLIkop4mB7wACkJiDD1d8IMdVB2VXgF56VQtI7oLonhjuopLJ4hyNla1hccBXlnQPi7oI0wfRSQdPV8SyiISMDGUGDhFNLEBuOyhQEwgGVPGIw34u6COPuNuy1mhvDoQKdg77BaREwufsNlrOm7W273qG2xtJllcA0fNBmmOvA8eOMkHfZXEPkhJqB0keq7I2bl4ulosLLzd4SIHsBGRtjC4g19oGvqK0iyscYwcbD/Ug2BTWxtfMWOcAAtPu1v9wm8NjsgrctVoXUdriEga8HGTsVodVR1bWk1mSQg02lcA74vVZslC2VpkGPVaaT0uPT6rMhrXCIsJKDDeCx5b6KeKo6GFpGVBI4ueT6q6NvifAO5QUErg7qBKldVPkHTjv5KdtoqgA9zR0q9tGGP6iNh3CDEj6oSXkED5hWTSukdnJws+t8Is6WYytNx0/SQOooTlD3yjfpDPqgpv8ANMrNPg9G2MrFkxk4wgsQHBBREEr5S8BvfCnirWxRlmM5WGiCr3dTicd1Xr2xhWq9sTnb42QWkE+SzKSrbHhjh3VobE4Y8wOyx3sLSSB2QalP0OYXN7kZWmSA9RwDhSwyve4NJ2WS8QhhyBnCDBjb1FVdhnwqrHdMhPkqyNLj1AbIIjupmQdcfVlQ9leJXAYzsgtIwSPRUVx3+vKt7IJIYvFcR2wqSx+GcKjHlhyFRzy85KA12FUElwwrVKOgNJxugkkkb0DHfCxid8hVJz27Iz6QygbhB8Tseuyy+iNzcsbn7FivHSe6CSWDob1ZVrZQ0YVrpXOGCVfHBI/s3OEFplJysyGgbLEZiQPPCxpWdAwWkFUZVSsb0NOAgyYpmtcYyBtsoXdU1QGBxwThRYe0h+dyr4WyF4kA7boNRrbX4FMJQckrT44nY6yTstTNf4sIhkOQB2WmTy4eWM2CC7qM5LSeyhlj8N2Mq5vVjMffCsf15+NBQNLuyrgN7jdTUjWuccqtRAfE+EbIMdreoqbwAG9WVZ4MvcD8ioXyD4ST9SDKoi2R/QR8lqE1mjdEZg4A/WtKo5RE8uKnnuFS74GOPT9aDCe0tcW5JxsrVkRdB3kG6jn6ev4eyCPJ9U+tEQV6j5bK6KTokDzvhWd1Kynkee2yDMmrhVRiIgDbCxYPgqA3O2d1c+mcwfB3xukcMhOQPiQajc44xTAtwTjyWjta9xwAd1muFQxrfeCek4/Is1kNM+AvZjICDvV7FyAM5otUOzudAVw/5xtyK32LrpDzUaqaew0DXY/zjbkQe0iIiAiIgIiICIiAiIgIiICIiDjjmS/B24p/wKvf6jMvmiJ6ZifRxX0u8yX4O3FP+BV7/UZl80En9kf9Z/Sg1k3dhpxEANgtJml6yTjurGYzur5OnfCCIZPmqkYVOymi6cfEghwinf0Y2UCAgOCD6IqgZP1oMoVg6C3HkoCQ/dXOpy1nWoQSEFzGdbsKskfQRv3VocQcjujnl3dBRERAUscxY0tHmozjHkr4mhxAPclBY4l5J3JKqGOc4NA3KzfcHRtEruxIW+NJ8IL3rKIy2uN5JaSMAlBpehtJVd6rGe7QmZxcB0gZXeLgDy33O5XSiq6m0vaMBxywrbXJfyxahl1o39kMDhAJm92HGM/Newul+F2mdM01PHRUoD4mAZAAycboNA4PcNqXSNkiYaVrHjG+MHsuUlZHG2JgYwYAVziGgk+SChaNyV195inWah0/WVM1xjY9rScFwXKWqtf2+zRyRBzesMJz1DYryr54OOWsX1FXSWuV/gl7h8Lj2yfRB1x5j+ITK/UMtuiqxIzJaCHZ2yuA6OnYfFlJ7kn86sv10r7pVirryTKdyT3WLT1rmNLc90ElXI1riwYUDZg0EYVkjzK85Vj2kHsgvik8OUuPbzVamUSuBHZQogu68twkQ6nhvqrQCTgKWOJ/WMINWfEyCmD+5xlaRK/xHZx5rU5mTtg++dvqWmu6QCG/nQSwPyME7KWWlEnxhYcfV1Yb6rUo4pjET8kGnsd4Mpz2CyH1zXNIDViy5Ehz3VB0477oHxPO5O/qVlRVIgZ0eZCxAcHZVDsuHV280CRznuL3Z3WTSVXgjB/Qpn+7+AMYzhYTQ3Jz67IM83BoySPtwsOWQTyjHYlWyFnTt3VI8AZPcboMx1tHhCQO3I7LFMBac57FVNZNjpzspIX+I3pzudkFpmHT0eamj6PDPqk9nrYI/Gcz4O4KxGSuBwTsUF8bCZifJKloaQAVe+VoblvdYznFx3QVjb1uwrnsDAqBwaMg7q1zy7ugod0GyIguLs7K0jCqzAKuy3qyUFoyD2RxJ7rNZ7v0DOMqCbo36UEcT3MOWgkqsr3vcOsELLs/gCoBnxhSXt1KZh7tj54QaeY8NzlWtHUQ3PdC44wUG3mEEr4vDYHeeFfDTCZpOcYUDpHOGCrmTvYMNQSMpg55aqTwCLzUYmeHdQKo+Vz+6C1ZMNUImdGFjtaXHAR7HMOCgPPU4n1RoyqKoa47hBc5nSM+it6j6q8B0nwjdGs6Jmh+2+6C6nlMMgeQSFPPWCUYaFnV0dKKIFmOvHktNpRGQS5BjnJOcd1LHTlzS47BZ9BFC6QhwGMqS4Uxa3MPZBo7hgkK5seRlU6XF3T5q4h0ex2QWOHSVQboTkqrfpD0yguZGT3VHs6SsmR7BH8PdYpcT3QUREQEREDJ9UG6K9vT0/NAczAzlViYSQ7HYqwuyd+yyonM6C098ILp6lrovDHcLCU3u8jiSBsrHRmMEO7oPVn2Gv8Audxl/wANYP8Ao169TF5Z+w1/3O4y/wCGsH/Rr16mICIiDrr7QpvXyccSW+tHR/r1OvnmgiEjjnyGV9DHtDXdPJtxKcPKjo/16nXzzwSFhPzGEF8jQwYUULumVrj2ByqvbI4kkKPBHcINbuFfHLSCNgHpstKip/EaXZxhIcyHpPZVkc6I9DfNBER0O23VC7O2FkMhcB1yDYqOYM/eILYpPDOVdI/xnjHmVEqt2cD6IJpIehnWFE0EknPZZb5WOiwViggPHplBcZC4BuPtURGDhZkph8MFuM+axQxztwNkFoyN8K/pLgSRhSUwYHnxAqzub4oaztlApHlsjWkdzhcocMLAykv9NqGRoLIHB5BG3dccObE1jHNIyO65G0lqqloLW+KRwBDD37oO91l4oWniTpaLSdPCwSRMEflnstm3Dh7JY68UzrWZGPd36M91xvypy1epNTBtACcyjsdsZXf670GlLVaWQ3oRitLBjOM5wg6d660RbILSZvdWNeWZI6R6LqXr63MhlkbHEAA4j6l3T4tUVyfUzyUwIpiCW+mF1M1lHE6ofTyYLy8hBw2aFzpCCCFjzw+C7AK3leLBPQsNUBhhGey2hM/rlJd2B3QRxQmTc7BXj/a0zH9wDlZD5IxEBH381BHSz1DgCDv8kGqu1B1QCJsYyPksN9YXtIIwVqFutUcXxTtHbO6xrjSNfOGQdsoNOy7ryeytkGdx9a1Kqtc1PCJCDgj0WNHTl0RPogxY29YLT5KkrAx2AqtJjefkqyOa/f5ILCwhvVnZWqpcTtlUQEREBEVWjJ3QUUzJuhnT8lKKfrGyx5YzG7BQXQ/E85PzV0rw04ChDi3sqjqee+6CWkGXnKVQ+MYOfJWjMZGD5KpcSQe6CItcO4KvZL0tI9UyZCchXCMAeSCI7nPqp46brj6jsoizpd8vkpnSENAGyCBw6HEd1ad1kNjY8ZKoYQSA3dBABlMFTOY2M/ErmFjwQ1v5UGOqtHU7Cq9paSrQcbhBPJCGAH18lSmgM0nSNlY6VzhglX0rnNf8PdBqrqWOnh3IzjyWjOy5/wBZWRUVUrnhrjsPJbm0rw213xDkdR8PNCah1NWNx1w2e1z1sjcgndsTXEbAnt5INqMgcSCNwO61a1TU0UuJyMDyK3pqflw5h+H1AbvrHgdrqz0AjErqursFVHAwYJ+OQs6WkAEkEggDJAC4yc9znF52yg1W/wA1LJKPdsYPcBaQMeavZG6TO/ZUDCX9B7oKl2cBSMcWjGFZLH4RClDQGB47oLoy0gk7LHePiOPNZtotF21Fd6KwWG21NwuVyqI6SjpKWMyTVE73BrI2MaCXOc4gAAEkkALkLily08deCNBQXXixwzvGm6K6SmClqKxjCySUN6izLXEB2MnBwSASOxwHHNI2MA9ZwVFUjqkOOwV0sRj+r61GgrEHNOWqYSOJ3womOxgLIdBhnidkFWyd87rGnaHOz2WVDEJBnz9VFIwtJA3QYpbg91Ox/RGCTugiLiVE5pHfyQWvd1O9FeYmdHX17qMgk5V0cZkcG52QGRPkz0gnHdZUNHHJtI4gqSBzKb4CM9SlLTgzDZBjS0fgqWGVrWYIV007HQ9J8ljNBLC4IMrxGny+ah96bFMHY2ChbLgkFQynLkGse8MujfCxghQTN9yHhh2Vp0U0kJLmHBKufO+ZwMhyg7/+xbl6+abVP8AK4/8AONuRXexbDRzRaoI89AVv8o25EHtEiIgIiICIiAiIgIiICIiAiIg455kvwduKX8C75+ozL5oSOuVw+ZK+l7mP/B44o57fsMvf6jMvmgkJEryP7o/pQWuGCQqITlO6Aqgeau8F4HUQrW9wgoizHxMERPmsM90BVGxz6Kiq3BcB5IJ31ZfH0dPlhY61FrIPD+awpg0H4cII0REAboRhVa1zjhvdXOic3uEFoBOwyVdGC1wcdsFZtIacDL8du6irnR5xERhBkT3R09MynY3fI8l6D8i2nfujSQe82gShzBklufJdA9HWpt1u0UDhkF4C9tOQbh3Y6PScU8kTHSiAEDAznAQc2cKtE2y11HvMdoZC4OByGALmnssWlttLSDELMLLQadern9y6U1GM4ytr3LiHZqfTdZXVVeyJ7GHAJAOcLVNe3GltdjlqqvHQ1p7/AFLzH5neN2ooW1tHpyR4gDiCGOPbPyQY3MZzZyWLUtRS0t2JYHEDD9u66ba940/s594dUTdZJPffO62RxG1FW3utM9y6jKc9WTvlbAZI6PPQcZQajXyNranDfJY0tH0DOfJWUsobJ1PPdX1lSXkBuMY3wgx2noccqjnF57FXwQSVMnQwblao2iZSRET4BPqg0ZXMb1uwqzdPiu6e3kjI3n4m+SDJbTBg68g4V7JWNeM+SxDLKR0FXCmnc3rAKDWampjmp/DHdaHLH0HCkhlcx3S/sqTkPd1BBZC/odlZ4unQzoAyFp7WOedt1kwxxNjPijdBD/Z5ie2TlTOowGl2ewWO5wZISw7K9rp5Bt2+pBE4dJI9FLFB4jSScFROa4HdXNlcwYbsgo7LT05zhWk5OVLHG6d2G91SaB8Jw8II85VWDLgqIDjcIMw04dGDkA4UDCYZWu7hpBVBLK74QU8GQjJCDXKzUbKmgFKG42x2W3zuhBBwUxlAyg3ITuq7hBI+HpblRK4yOI3VBj5IKIr3ROaM42UkXR0fFjsggWVT0gmaXZ7LGOMnHbKvZPIwYaUFZoTETvsos5Vz5HPOSUEb3DIGyC3JHms+jt4qYzI53YeqwOyljqZY29DTsgtlZ4chZ3wVYqucXkuJ3VzInvOGjKCxFlx0joz1S7DGVHP0GUdA2QQYPoi1CaKIU4IwDhaeBkgDzQT0xDXZPmlS7qcCBsFaYZY29RGApYXRmM9WMoI6aETP6ScK+dgp/hG+VEJHRyFzNt1QyGV4MhQXQy+E/qIUz45Kn741pwPkrZRGWDHda/aX0TaBwkx1EfnQbcfLIR4W58lYOtnkQs+lEJubuvHRlZlxFG14DMY88YQaPHUPZg5Kz47g1zPDcck+qhqhAY/gxnCwQS0oMt7BE4y+RUE0niOyrXSucOknZVZG5x2CBFGZDgKssRiIB81ltMcbc7AqN72POSgxMn1RTylhHw91AgIiki6cjOEEeMIsufwug9IGVihjj2CCiDdOyIL3R4bnKtDiCChcSMKiDMjrgxuCM/YoZ5BN8Q8lCmUHq57DX/c7jL/hrB/0a9epi8s/Ya/2hxl/w1g/RXr1MQEREHXT2hozybcSh/8ASdH+vU6+eNvfPovoa9oftya8SyO/udH+vU6+euk6Ot3Xjtt9aCpnHTghQOd1FTyxYJI7KFw32CCsUnhuyqyS9T+oD5qNVbjIz9qDIlqzJGGYwsYHdZcwhEQ6AMrEQVIwqYPomT6rJh8Pw/ixlBDGzxD05wFV0fTIGfNW9Ra49JVC8l3V5oMiWk8OMPz3UlMAYySsZ08j29JOyzqRhdTkj0QYEhy442xsrRnv6KsoxIVagzrdF7zIWPOAErYpaacxxyHB8srEimfCephwSs6mcZniSbfCDsXyma9bw+ubayoI3fnfyXY3iNxVl1dXxXSnrXMYwB2A7ZdINH6gpKB4aSPh+a5Ji4lUDWCmLgCQBjKDstYNYUetITaql4Ja3pLj+RddeN+lKaxXkzU7wQHdX51daeITbFKaumfgv7brbuqLvd9cXATFxcwn1yg2Xqa/mooBSiPGG4zhccujle44Y4jPkFy3qzT0dLQNc5uHgbrTLXQ2dtokfOG+IGnuPNBx5HTluHOO/mFqNPOyFodjssC4TAVsoiPwB2ApoHBzM+SCervJcPDaMD5Lcml7XTXCEVMzh2zuey2bO2J5Jb3HzU9LeqygjMULiB27oN0arkp4ITHG4HGy25RTNfTu6husOavqKx3392eoq0zGIFjexQQz7Su+tWtGXAeqvja6V2+5KSs8NwGPmgnNHhgcN8hYvSQ7BCniqXdnnIWb7kZoTMwbAZ7IMGWEMZkH5qENJGVfI9+SwntspqUx5Ad5jCDFwfQrJhpw47nCz5hTCMEYz9i018rg8hh2ygyzKKcErDnm8Z3UBhXS9RYCcqHpPdBPDRvlb1D8iq+mkjz/AKlJR1Zj6WeXZZlSWOj6sAEhBpZaezhuqkYUojdI8kDZSOpy1ucbj1QYvS7BcAdlGXOOxJWTJHI1uQ3ZYpzndBK146cHvhWOJzhVbGSVc6IuwQUFPj6cg7K+me7qO+VQPaG9J7q+Bm/UMd0FkjZZJMuacZVzmtgALTue6zZaiFsfRgZx3WluJcSUE+GyM6iQCrGRNdnJVgDvLOEIc3bJQTso8tLidh6KFjzC843WVDMHQlmdzsoWUz3ShpGxQdoeQDlGh5t+LlRQ6lqKmm0bpaCKvvslOcSTh7iIaRr/AN4ZSyQlw3DI34wcEeh3M5z58GeRKWm4CcGuFduuN7t1PHLPa6FzaC32sSMDmeK5jHOlmc3oeWgAkODnPycHjX2JV2skOn+KWlx0MvLa221787OlpjHKxuPUMeHZ9PEHquontKuDHEPhzzT6w1Pqm2Vcll1pcX3ay3XocaeoieATAHkYEkR+AszkANOOlzSQ7U8NvbaR1V+jpeLPBcUVnlIDqyw3AzTU+4yTDKGiQYO+HtIxsDnC6b8+HEbgtxZ5hbxrXgbR0kOnayjpOqemt7qJtXV+GDPMYnNaQ4ud0OJaOosLt89R69CikZH1OBG2eyxHdQcQSQgNLx9HzVA5zXZ816Scn/I3wW4icluo+PXEzS15vOoZ2Xiexsoq2aIuZTRmOGOKKMjxZH1EcjQCHFxLWgeR3Fwo9mty+cEdFW7iTz4cSKG3VdxIEenzdhRUkRJB8F8sZE1RKBguEJaG5O7gA5B5dkvl+eE63N+Er2si5CPZ68znD6sufL5PRUE8LDSw3nT14qak0lQB1MFTTVEjge4Ja9rHuadnDYjyB4ucMdS8G+Jeo+F2r4WR3bTdfJRTmPJZKBgslYSASx7Cx7SQCWuBIB2Qbr5VuL1l4CcfdH8X9Raalv1u05VTTTUMMjWSP66eWJr2Fwx1MdIJADgEsAJGcjsDz/e0EtPNtYdNaI0Toy6WGxWSufdamS5zxumqqrwzHHhkZLWtY18u5cS7r7DG8XsxeVDh7zP661j/ALKdsuFdYNL2yncIaWpfTB1XUSuEfW9hDsdEM2GgjJySdsHhbnL0Fw64XczGueHnCulqafTdgrIqKniqKp072Stp4jO3rduQJjKACSQABlBwu55f3VMeq9B/Zm8lXBzmK0NrniHxrtVdW26018Nrt3g3CSkjieyEy1L3GMguIbLBjJwBnY5ytw8DvZpcMbRpOTj1zeayOiNHVjzV2vT1RcG0ksVJLl0Da2peARKWH+wRgSZAy4OywB5sAYUviuc3pPkvajS/JV7M/mU0rc7dwSpqCSa0f7XmuOn7/WPq6F7w4RueyokeHglji0yMcHdJ6TjK8muYjgre+XnjNqfhDfaltXPYKoNhqmtLW1VNJG2WCUDyLo3sJAJwSRk4Qceslczsroz1v+Jbm4W8L9b8ZtdWnhxw8skt0vt5m8KngZgNaACXyPcdmRsaC5zjsACV6bWH2dHJtyxaNodWc5PFBlyuVYMOpTXzUVE6QDLo6aKnxV1BaCAXAjOx6G5wg8rHmFjMDGSFgOaXvJb2J2XsTozl39lBzUT1WjuDckdPf46dz43W2tulDWNY0bvjirh4c2M5J8N5AAzgArzy5yeVS/co/Ff9hNdc/uvZbpTm4WO5mMMdPTF5aWSNBIbKwjDgDg5a4YDgAHAbYwwHJ7+qt6ulxLQF3A5FOQa883FdW6t1Lep7Bw/slSKOqq6VrTV11SGtcaen6gWt6WPYXyOBA6mgNcSentfeNA+xg4TXOo4d6uuVtr7xC4UtZUPr73XubICQQ6ekJgicCSHdBZgjfBGweSPUXuBcs9xPgdLfzLuvz78sHKlwx4Y6T41ctGvff6DU93NsZbIboLhTOiEMkr5o5DmVjo3CKN7HuJBlbs0g56RMqQ1uCPsQYszXgbpHP0sLT3KkmlEuwG/1LHMLwOojZBa52TlGjrcAqtjc44AVehzHhvmgyoqBrxnPkseeIQuABypS+eJoO+FAXOlcOooO/wB7Fd/VzSapBztoCu/lG3IrvYsQlnNJqhx89AVo/wCcbciD2lREQEREBERAREQEREBERAREQcc8yO/LvxRHrou9/qMy+aEt6pXt+Z/Svpe5kfwd+KR/7i73+ozL5oHnEryP7o/pQWuGCQqDYgodyiDIfUNLOjHljKxxsiIJnT9TOkqFACeyq5jm9wgonZMH0TGEF/iu9SqHLvNWqrSB3QUIKqAMK8luM+ajyewQXxv8N5OFJJOJMDGFZE1ucv7KsrWdQEe5PogvdDhnU13llQbuONysiSCojhEjvokKa022a4VcccbcguGfqQb24QWqtqL1FLFSOe0yNGcfNe4PJXbKin0vDLNCYumAbEY7gLzo5UOHlkrJIBXNZ1h48h3yF668ILBb7HY6eKgADTEMgDHkg5EUVRKIIXzO7MaSpVpmoTILTOWZyGkn6kHWTmj4vmjsktohn6CGuzjbyXmLrfXbJI61tTVdZc44yc+a7c832oIqKScSgAhrl5ha31VUVdbUxwE9PiHfPbdBo2r7kKy4Pe0jBJ7Lbud9ldJK+Z3U7cqrQGg9WyCPf5qaCnMzgCcAlSQRtlf0gLIkb7uAdgglgYy3Hx+59Fj3G5GtII2+ShbUukf0yOJBVsjI8joCCDf5qeKYNb0Y77JJ0Bm2MqFrScIMgwhuZCPNTx17WxlmBlYr5iWdOUhY1zST3QRyO6nl3bKpv6H8iukYWE7bLUqCGJ9OS7GfVBp0MojOcJNKZHeYCrLH1TlsfZWvjdEQHjCCxTxVAjb0gZUJI9FcxgcDsgrJIHuJ9VGq46nYarjC/vhBJSTiB/VjKurKoVLsgYWMdiiCpAVMH0TJU0fQWfFhBWlAMg6uwKzZ3xsb2HZad1ljyWKrpnO7nyQABLJ37o9nhnBVIndJykrutwQWjYZTd3YIQ4DfssiFjCwnAygjEQIyrCOlwVXvcHEA7A7K5sbpAT3QVfP1M6Qocn1VSMEj0VEBEAyhGEBTRzBjen1UKILn7kn1KvZD1NJ3VISM/F2WR4kbdgQgxC0gkeimp5xFgEK4tY4beahkb0lBl1FwE0ZY1uD6rDZnqHfurVPEWBuTjOdkE05+89/JYbT0uB9Fc+RxJGdlaGkjKDKkqhJH0AeXosXcbKrQA7fySTBOyC1Va3JCuj6c/EryB15b2QHxkNBVrZ5GN6Q44UssgcwAd1bFEDGSe6CNkjmO6h3yj5HSHclWuHxEKiDIgi8Q4LtvrV81I1o6mrGbK9nYqX3lzhhxQWRRdby3PZSkinw3vlQB5a7LSq9fXu47oDnFzu6tOR3Q7HZXNjc7c9kFmUAypHgAfNRjOcgIJXsAbn1UWVUvce5QYx5IKZPqpGSBrcFRogHckoiICImEBEwmEHq97DZuLZxkf/dT2AfkbXf616lLy09hsSbdxkaewmsBH2iv/wBS9S0BERB1z9od+BpxL/xOj/XqdfPGCQvoc9od+BpxL/xOj/XqdfPNGMk/IIJmS+IOk91ZKwM+1WRuDH5V8uZMEdkEKyIqbxGdZOPNQswCQ5SGV7Bhh2QRPHS4tzlVDcjOVQnJyUAcRsgEAKmT6qpz2Koglp2dbsJURiN2FWmeGOJKrVPEj85QQYWVDWPii8MDbGFCenpGO6z6b3X3ch+Ov5oNNe7qcSqK+VuHkjtlWt7oAB9CpxUFjekbHGOylhZE5o7ZWPUNDX4CCkU8sTg5jyDn1WZTSVM87ZjOdj6rT1LDM6N43OAUG65rnPJTNibMct+a1ixa8+4sJif8TgO62e2tiEYz381fQNp6utYx4BBOEHLGmrl+zyZ0E7cN8s7LQde2CPT83usEuzu+CtbscDLFRCooQA4jORstsajpr3fq4VMgcWg580Gz6uzOYwzd87rT+qSEFoG3Zb4rbTUPpREM9QGCtFltbYInCUHqPqg25lzXFx33QHreAdsrLZQvbKesfD8wrKqkc09bOw7oKS07Y4+sEbrFcS4+ZV753ub0OOQFe0M6N+6C2GURlUll8Q5VGROleWsGSqSRPiPS8YKC1alR3PwYfBI77brTj04GM5VCMbFBkVEQ3kac53x6KAZzhSxlx/shOPJVNM9w62DIHmgteXYx1bKMHBB9FNK4FobtkKFm7gD2ygyBKZcMI2JWRLE2OMOyO3qrWxxgtx3JV1ZC8sbgnGyDEjYfEyMYz3WbI1j2ANkGcdgq01vfURhjThysns9ZSfE/YILm9cbcNBx8lHJM5vcqeGZgwxw3xjdd0OR/2dOp+ZKvpOIvEqGrsPDWCUPa7Bjqr30kExwZwWwkEh0+/YhmTksDgThLy08Q+LPD/WXFeCAWjROh7VV3CvvVXGTFPUQxdTKKADHiTPcWN2Ia0OBcQS0O4XeBnYbr3G9ppUaa4IciNZw70PaKSx2q619s01b6OkYGMhiEhqXNA7kllK8EnJPU4kkkleIFNTlzsuKCrIT4ZcR9ii8UNyN+6zXPAb4Y7rfnC3lq468b61lNwq4XX6/xyP6DWQ0xjooznGH1MnTCzfP0njsfQ4DjPYnOylhk6dsr0n4N+xW15dpKe48c+Jdu0/Sn4pLZYGGsqyP7l08gbFGe5y1so7eu3eng57PXlP4JugrdP8L6S93aDBF11E77o1HUOzmtkHhRu+ccbCg8R+EPKdzD8ep4Dwy4VXu50UxDRc5Yfdbe0eZNTMWxHA3IDi4+QJIB7ycG/Yo3qqMFy498VYKGIgOktOmIvFl3OcGqnaGNIGxAieMnYkDf1bnqKO2Uj6iqngpaWmjLnySODI4mAdyTgNAA79tl1j4x+0o5TODslRbp+IA1Zd6ckOt+l4xXkOHcGcObTtIOxBlyDkEZGEGxOMXs1eXaw8tOuNN8KOGsDdWMtElbbbvVyyVdwkqqciZrGSPJEfieH4REYaCHnI3JXh5MRkgeS9HOMHtnOLOo3VFt4M6GtGkKR2Wx19xP3RrseTw0hsLD/wBqWSD5leddxlnrKuevqSwyVMjpXlkbWNLnEk4a3AaMnYAADsAAgsp2taOolZTZmNw8DcBYDHEDAU0bx59kHJXArjzxB5fuI1v4n8OLiKa40WYqinlBdT11M4gyU87AR1MdgdiCCA5pDmgj2P4G85XK3z06THDDX9mtVHqCvYG1WkdQtY9lRKAR10UrsCUjJLSzpmbgnpAAcfC4TwxxlrcZKW+qmo62OtpaiSGeF4kikjcWvjeCCHAgggggEEHIIQegHPz7N+XgTYKvi/wVlrrpommd1Xa11B8Wqs7HOAErHjBmpwSASR1xjBcXguc3zxkGQXFfQjwN1Pc+LvIfar/xclNbLfdEVsN5nqm5dVQCOaIzSAghxfE0PJIOS4nByvBzhRoWq4jcUdI8PYYnPfqO+UFq6WnG087Iyc+QAcST2ABPYIPd7hBdtOcpfIXpTUusIhFR6R0ZTXWtgDg0yVs7RMYGk7db6mfw2n+6cCvDbj5xy19zD8SrnxI4g3eWrrK2RwpqYPcaegp8nop4GEkMjaDjA3Jy4kuJJ9UvbOa4bpvl60jw5t720x1NqFsr4WYa11JRwuJYB2AEktMfl0heNDNnA/mQejPsTLrd4eOGvrJE94tlXpRlVUN68AzxVkTYiW43IbPPg5GMkYOduLfawwWuHnW1JJRNxNParTJWnpxmcUjGg5wM/e2xDO/bGdsDtr7F3hHXWXQ2tuNFyp5IotSVUFmtfUMCSGm6nTyD1BkkawHyMLwvPHnQ4i03F3mg4jcQLfUiooa68vpqGZrgRLS0zG00LwRth0cLHD5FB6X+xq0ZT6c5edWcQax0cB1FqN8YkdhoFLSQMAcXHGAJJZwc7ANzncryP4pasqeIvEzVvEGfqL9S3yuuzuruPHnfJj5Y68Y8sYXsvoIO5dfZOtufgCmrf9j+quTC4gOFXdeuSFzz3JD6uIYO4DQ3bAx4lUzjO5sDGFznEANaCSSTgADzJQe53s09P2fhPyJ2bVWppIqGjuQuuqbrLMPgjpxI9okdtuPd6eNx+S8neb/mq1jzVcVq/Vd2rauDTNFPJBpyyulPg0NIDgPLAceNIAHSO3JJAB6WtA9ReeWf9r77NqDhtRy+7VclpsejIy0gFzgIzUA+pfFBOD69RK8Ren1QejHsUKG5y8bNf3SCSUW2n0syCojaB0Onkq4nREnvkNimA+RdnyXC3tRb3Q3rnY12KCTxG0EVsoZHZ2MrKGAvA+oktPzafLBXdX2KehfuXwd17xDlh6JNQ6ghtrHEbvio4A4EfLrq5B9bSvL/AI/awdxN42a94h+8ieO/6iuFdTuDstED53GJoPmBH0AH0AQeqPsluANo4a8C67mLvdrnqNQaxZUCk8OmMk8Npp5HNEcTAC9z5pYnPIA+MNgAG2T0f4+cPeeDml4tXriLqXl/4nAVM5ittFV6eq4IrdRdZ8Gnj8RjWgNBy4ju4ucd3Er0+4ka+vnKb7PK26z0BTUM100dpTTtLRtq4y+B8kslJTvke0FpdnxnvIBGSV521HthebeaB0UUGh4Hu7SR2aQubgg7B0xG/bcHv67oOZPZ6ez249aA4zWHjbxdtkWk7Zp4VE9JbZKqOWurJZYJImhzIy4RRgSEnrIecAdOCSOJPa3cfdEcX+Ndi0hoO6wXak0HQ1FFXV9O4PhdXTSgyxMeMh4jEUYLgcdReP3uTwhxZ57uazjTQ1Fm1nxducdoqWmOW22pkdup5GHux4p2tMrT3IkLs/UABwFkIPdH2c7LTrD2fdp0voC8x268upb5a6uqYfvtFdJZ5y2R5G+QyWCQdz0Fg8sLx4438uXGXl61JLp7ivoqvtbzKWU9w6DJQ1vfDoKgDokyATjIcB9IA5AzuXnmh4y8sGpZdRcKNSCljregXG2Vcfj0Ne1mekTREg5GThzC14BIDgCQfXvlN51+FfPlY7twk4icPKOj1FHbnVVzsdY1tZbrhSh7WPlhc8Aghz2EscOpvUC1zsFwDwt6XN7hXRhpd8XZdh+fXl+sXLZzG3jh/pOSU6fq6aC8WmKV5e+np5+rMJcclwY9kjQScloaSSSSeu2+UF8vQx2WKx1SS0tAztjsq9OT6rJpqRheHHBAKDCikMTuotODujpC+QPx5+i1eqp6d0eIwMhYIZDF9PGUEc8/XH043UEYwQ7PZZMsbJtoRuoH00sfcIPQL2Lk4k5otUMz20BWn/nG3IofYrxvbzS6pJGP/gBXD/nG3Ig9p0REBERAREQEREBERAREQEREHHHMl+DtxT/gVe/1GZfNBJ/ZH/Wf0r6X+ZL8Hbin/Aq9/qMy+aB4zI7/AL4/pQWoqhpPkpW0srt2jKCFFI+B0eeryVrGFxA+aAxxYQ7CrJJ4nYLIlYwQ7d1jR9O/UgoDhUJyrnNPcdlVmMboLFkwUolZ1ZWMcZ2WTDMY2EDsggkb0OLfRWjZXSHLiVaASguL8jCowlrg7GcbqizabwPCIfjJCC6e4+NTiHpwR8llaevDbZVMc5oI8ytKliLXEgbJCwuePTOUHohyKzUeqL5BBUzBjfFHc4HcL1909ZqW0UMMNK/qa1gAP2BeBXKzr6t0zeITbnlrxIOx+a9qOAfECo1JYIDdJMyviadzvnAQcyLStS1UFLYq6eWRrQyFxzntsob/AKpt9gpnVNU8YAJ7rrvxc48Ukmmq9lDKMFpacO8kHnTzscUpKvVdVbYagluXDY/NdGZal8k0znkkvcSfyrnbmL1TR3zVtRKSC4uP6VwHJgyuLe2UF30fiAV7YzO0uOyla1nR8XdVjI+ixBjwTGnlJ9FfUVT5+w2CVMBaOs+ayqAU3hHrx1IMCNvW7B2KyxStDSSc/aoZm9UxEH5lRzp4gWvzuggeOlxHoUa7CuDHPJKoY3A7oKbndXMkLNsYWREI8fF381DO0AjpCCkkvWMYV8VU+JvS1QLNp4WPaSQggik8OQyHz8lWeYVD27YSfpBIHko2xvI6ggnkpAyMPBOSscPwOyudPI4dBOyjQSQ/SCnlqGgdOO6xo85VZAc7oLTuSfVAMnHqpKdoc/BUk1O5jgR2QWvpulnWCoFlyTAxdBP1rEQMJhS0/T1fF2Sbo6x09soIt0B3WTJ4fh/NYyC5z8gDCqyUtGArFc2NzhkIKE5JPqpYp/Db04VjB0vw5XP6EEb3dTi71VE+pSx9HTv3QRsd0nKPd1HKudjPkrRjrHogpg+iljh62lxypJRH4e3fCgEjgMBBQjBI9EDSRkFNymSNkF0bywqkj+s5VG4zujgAdkFEREBXh/SMEKSl6Ov48YUlR4J/sfmgxDuSUVxjI3xsqNx1D60FMH0V7ZOkYwsmQw9AAxnG6ib4ZbjzQQZ3yVKJsNwFa6Nw8tlYgHckoiICIrgwkZCC1E7IgKVkoY3px5KJEFXHJJQHCormxOcMhBad0Q7FACUBEIIVQ0kZQUVWjqcG+qdJVBsUGXJSNjj61iE5OVKaiRw6SdlG7ugoDg5QnJyiIPVz2Gv+53GX/DWD/o169TF5Z+w1/tDjL/hrB+ivXqYgIiIOuftDvwNOJf8AidH+vU6+eRjsH69l9DftDvwNOJf+J0f69Tr55GjLggq5pxnCvZL0jBCyRG3w+3ksSQAHb1QWE5JKuD8DGFaiC9jPEdj1UpHhDCtpnBjsnCuqT17goIHO6iqIQVc2JzhkBBamUIwcJgoL42dRVekiQNBO5VgLmdtlI2KWTLwOyDKqKdscIcDk+awWtyr3zyOHQ45AUYJCC9jywqcRCcg9vqWM3c5KmZL0Yx5IL30gZ5lYzm9JUslS5+yhJygZPqVmUD3U8gqN8jssNZdIMkdf0fNBuSHXEzY2wOJAGB2W7LDqumqIhC/pJPzXHFTFRysAhA6/ktUsFkuEkodEDgeiDkOp8IDxmNBB8lPRaSivLBM9oGfLsrrLp+uqI2tmBwcALfVt4aamqouq3Nf04zsCg4k1fpR1uhPukPWR6BbSoKZ0pMNXGGEnGCF250Fy/aj1NUyRXGN7g3OcgrhLmC4Z13D+9iGnY5oa4jthBxhe9PRUMJnY8HIyBlbeDHkbNJH1LWbg65yU7GzkkYCzLZT0bqF7X48QggeuUGh0LnMkBazqJPZby01o6bU1xipZYC1jyMk7YWl6ZtHTcg+rbiIu7nt3XZbRHD997pWiwt/2yW/Dgb5+xBt6u5Z7db7M24wyte8tzjqBK2PJwcqXTdfuh6AcZxsu02ieXHjZcqsNuLak0bnZblpxhc0VXKLqf9jz5GQP8UM6iek5zhB57VnCulgg+LpDgOxIWwb5QR2kup2YOMjK7Pa25a+K0F6qY4Y5vCbk4wRsuuvEHR170rWmlvTXCQOI+LYoNlik6n9R891jyM6JekeuFqhop44xJjY9tlps4d43zQSv6m9PTk+akxWTgBkZIASORrD8e/butbt14oIGYlYMj5INIoKqppKlrZWkDPmtduMxrKYCJuThYM2b3cYKC0UktRVVUrYYIYYy98j3EBrGtAJcSSAAASSQAvWfkI9mlDoaG38XuYq1RVeoPgqrTpqdofFbjs5k1UOz5h5REFsfc5dgMDifkM9mJV62dbeMvMZapqTTuWVdo01MCyW5D4XMmqR3bTkE4jOHSdzhmA/1uoqKkttHT263UkNLSUsbIIIIYwyOKNoAaxjQAGtAAAAGABhZCIPO72suiOKnGiXhpwg4UaCvupqoTV16rhQ0r3w0+BHDA6WXaOMHqnGXloG2DuVwNwb9jNxi1Aae5cZtfWbR9G4h0lvtw+6VdjzY5wLYGE9g5r5APQ9j7FLgbjLzxcsHArxqXXHFW2S3WHY2m0k19b1Z+i5kPUIj3/shYPn2QbU4PezZ5TeD0lPcYOH/AOyu705DhcNTSiud1DcEQYbTtIO4IiyNt12cggo7bSMpqWGGlpaeMNYxjQyONgGwAGAAAO3ovKTjJ7ay81bprbwD4Vw0UZy1l11PJ4suCcBwpYHBjSBuCZXjJGQQMHo/xp5seYjjs944mcVL1caGVxP3MglFLQNB7D3aENjOBgAuBdjuSSSQ9uOL/tAeVTgxJLQ37ifR3q7Q5BtengLjP1Du1zoz4Ubh6SSNK6S8a/bJa3rY5qDgdw9t1jheOllyvrjV1Xn8TYWFsUZ7DDjIO/qMeYbagtZgE9XqrHTTSfSeSg5W4t8wnHLjrLJV8VeKN9v8bndYopagx0UZzkFlNGGwsOcbtYDsPQLijL2EjHZTx1Tmt6HHbzVDLG52wQRBr89WNwr5JHyABw7LOija5nVgKCo6QTgIMUscG9S7J+z54fcE+J/MfbdJcepbe7TtTbaswUldXPpIqyuLWthh8Vr2EOPW9zQHAlzABkkA9dDNGIunG6pThucnsUHqlxf9ijS1t5kuXAviuy30E7i4WrUcD5fd8ncMqYQS5uCQA6PIAGXOJJEPCH2KtVR6ip7lxt4sUVZaKaVsj7Zp+nkD6toOeh1RKGmJpxg9LCSCcFpwR0W4fc13Mdw3t4tOiuNurrdb42COKj+6UktPE0YwGRSFzGbAD4QDgY7KziFzV8yHEu3us+s+Nur7jb5GGOWi+6UkVPK05yHxRlrH7Ej4gdjjsg9FPaGc8vDLh/wuquVzgNcqGvuNbQNsdzqLXIHUdltrGCN1Ix7SWukfGPCLQSGMLwcOwB1B9mPolmtucnRs0rC6nsEdbfJgBuDFA5sRz5ATSRE57gY2JBXVSCGPALgMr0v9ipoh1RrXiVxIfCAy3WujscEhG7veZnTSAH5CliJ/75vzwHcbm04AcE+c6kquCd41jT2viFo2Fl2ts8IElVbo6oY63Qkt8Wnk8Joe0EEGNhy09OejmjfYn8Wn6vgj1/xZ0jT6XZMDNNZ/eZ6+WIHcNilhZHG4jbJe8NJzh2MHr3z0cXNT1vPDxE1xo/Ulys1xsF5Fnoqy31clPPTGhiZSu6JGEOaS+J5OCN3H1W07tzvc397tUlor+YLWXuszAx/gV5gkLcYx4kYa/fsd9/PKD0l55+afhlyicBY+VrgRW0rNUzWv7iR09HKHusdA5pbLPM8DapkBfgEh/VIZTjA6vH3TFmr9Y6ntGlqD+27zXwW+DYn75LI2Nuw3O7hsFptVNU1lTJV1k8k887zJLJI4ue95yS4k7kkkkk7klZNruF0sVxo75ZLlVW+42+eOro6ulmdFPTzscHMkje0hzHtcAQ4EEEAgghB7Qe1p1Db+G3JtaeGVnlfFDfLxbLLDT9W7qOkjdPk9gQ11PAD83NOO+PKzlN0N/skcyvDPRj4xJBX6loXVTSCeqmikEs4/ycb/AJeuy2zxD40cXOLZo/8AZP4l6l1S23gikZdrnNUsgJADixr3ENJAAJABOBknAWmaQ1LqjQt/otXaK1JcrFere5zqW4W6qfT1EBc0td0PYQ4Asc5pAOCHEHIJBD1I9trrhkGleGXDaGoLn11wrr5URA7NEEbIYnEfM1EwH/euz5Lyd6ftW69fcSeIvFO7svvEnXN91RcIYhBFUXavkqXxRAk9DC8npbkk4GBkk4ySVtktzsEHtzy9Y5ePZas1W5ng1zdF3XUjXZLTJUVniy0uSNxkS07cjyAK8SJJfvRGVve6cduNl40JBwvu3FjVtZpGmhip47JPd6h9E2GLHhReEXdJjZ0t6WEFreluAMDHH0jsE7nCD3X5bNY8OOerkii4VXy9NFzZp+DTWoqdkjXVdHVU7GthrAxxJIc6KOdhOWkgtJy1wHSDUHsaOZykvc1NYNV6CuVu68Q1ktfU073MJIBfEYHFpxuQC8DOxK6NaV1pq7Qd5i1HofVN309dYAWxV1qrZKWdgOMgSRuDgDgZGcHC5kHPtzitpRSDmF1Z0CPw8moaX4xjPWW9Wcfvs5zvnO6DsNxF9lVPwK4G6v4vcZ+OFop6mw2qSahttno3SQ1NcRiCE1E5jJD5C1gAiyQ7IxjB6+cjvArh/wAxvH+38MOJGpqmy2mst9ZOw0lTFBU1VQyPEUMLpGuaX5d146SS2NwA3yOKNdcVeJvFCsbcOJPELUeqKiIkxvu9zmqzFk5IYJHEMG5wG4A7AYW3YDLA+OqgmfFLG4PY9hw5rgdiCNwQQCCOyD0O1r7GXjtZ7/NFw/11pHUFke5xp56+aahqmtzsJYhHIzOCN2vOTkkN2B7M8mnJbYuRGl1Hx749cRNPw3M2p1C6SGVwoLZSukbJJiWVrXSzPMcbQGsBGC1vWXrzB07zvc22l7dFZ7RzBay91gw2NtTcHVRY0bBodMHODQNgM4AAAGFtTiLxr4t8XJWV3EziJqDU0kbuqJtxr5Jooic/2OMnojG52aANz6oN4c5vHqn5kuYfU3FC1U8sFmnfFQ2iOZvTIKKBgjjc8eTnkOkIJODIRkgBcJBp2OFJG1jndT8hXGSL6OOw7oI2gN3KvbUdGQNgn3osJG5WM/YnB28kGQ6qLCXZyCoej3l3VnCs8MvbkK375FuEGbTR+7kknO6TVgc7HThW0E7S4+MdlFXeG6bMeN0Hf/2Lzg7mi1Rj/gDW/wAo25FD7FmKVnNHqlzhsdAVoH+cbciD2lREQEREBERAREQEREBERAREQcccyX4OvFP+BV8/UZl80bd5nA+pX0ucyX4O3FMf9xV7/UZl80LiRK7H90f0oBIa/q+a1Cnqm9P0c4WmE5Wp2yJjmfEM5QQ9ZqpSzCTRCl28ytQZSeHKXtHnufVYNyPVMAeyDDe9zhk9ipIKYyjqWXNAw0w6AMgKKmlEDC12xOcIKeED979FjzReEcK9kpM2c7KtU7qKDHVW9lT5oCQgq76RUkYGFF3VQ4hBV4wVQEjceSZLirxA8jqxsgq6YFnTjdIZSwEAZyCo8HOAN8rVLfbTOekjJIQchcBqqanvzHNJ2kBXplws4x3DT0FJCyRww0DGe+y8/eXTh9X3e+tELCfjHl816Bac4VzWmooai4QkRANJ28sIOVL9xLuus7cYJXvjaR09W4XDWvLFS0ulq6Sa7DrLScF++VzJrK46UtuljTWoRirER7HfK89OOmvtbUs1TTQyP8AvI2JxjKDrJxbD26xq2+IXsDj0nOQd1spv0lu7VFUKx/vFVgzvBJytpNBc/A9coJ5TiPY4VaNwGSSqytHh5IWK15b2QZtVUB7egeiwsub5kKWAh0mH+arUMb1DoQW08wik6yMrKmlbUAEDBCw3QvY3qcFRjiCMIJmP8NxCnIbIwuwrXRtEfWfRY7piNm9kFGNLpC0K6dj2nBbt9SvonNEuX9lqE8tLJHjbKDRy3AzlSxz+G0tCskOHEDsrO6CrndTic91NDMGN6VY6FzG9Thso0Ekjf32e6jwrgHuGB2CngawY8Tudu6CBjug7hHv6llVELQwuasNBfFJ4bsqd9WHN6cLHjYXnCrJGYzgoLSckn1VERAyiKoY4jONkFMn1RVaxzjgBHMc04cMIKKWOYMaRjuokQVe7qcSrmRF4yFYs2mfE2Ih+MoMI7beivbH1NykxBkJb2V0bJHNJb2HzQRY3wp2QgtzlR9JJIHdXNe9jg09soLXtc3OeysWdO6Ew+XUsFBfGN0kABVu7UJJ7oKIid0BFe6J7QHEbFWIGcK5mXPAPmVQgYQZB6h5INTqYGNpuod8ZWlqV1RI5vS5xIxhRIG6DYqWn6er4uyVHT1fD2QHTBzelRImCgIiqMY8kFFIyUNb04UZ7ogHckoiyIqV8repo2QY6K6SMsOCrUBTRzhjS3ChAJ7IQR3QXP3cT6qjeypkoCQguPZVa/pGFZkoguc8HsFaiICIg3KAivdE9oDiNirEHq97DZv8A1M4yP9ZrAPyNr/8AWvUpeWnsNnH7ncZR6TWD84r16loCIiDrn7Q78DTiX/idH+vU6+eeFpL9gSvoX9ogf+sz4ln/AOk6L9ep18+lpYx3j9XcMQWnBGMrElGDlXzvIkcB2BURJPdBRERAV7TturEygqTlTR1AYzpP1KBXRgF4DuyCjsuJcQd1Vm5wsyoMHggRkZHosSNjnH4UF0jR0gqSGqEbOjHyyo5A5owQokFXHqcSPNURVDHHsEFFUDJA9VfF0hx6klIDh0+SC+SnDG9WVCwdTgPVXOlc4YKvipZpB1MCC+alEcYcMnKrTOkcfC6e6yKWCXqDJs4JA3W66PRdRV0hroGktaM7BBt2C2OgLZvpEnOFyVof3qaVkUNCXnYbDK2rZqP3itZRSk5D8Ywu5XLPwkt921FRmriBiJaT1AfJBxw60akoqWCeOxS9JcNxGV3Y5cNBOvulffa+zDrbEDhzPku1du5b+HFw0zS077fGXmMHxA0HB9FvfR/DSxaQoTQ0MTegjGwxsg4B4WaUp4b3V081na1oyM9GF0p59dJQMv730tGA4SHYAZ7r1rpNLWmhlknpYA1785PzK6Bc2XDO4XPWgqKlhNMJsnPbGUHldqOhuFN0tnoHRsxjqLcBaRR0kYidUF+OnchdwOZrTWkLPpJrbe2MVTYt8AA5wukcFdUNikj6iQUG+dKuivFXHSjbDwF3X5Y2UNm1XQw1LWyAluQdx3XQrRt6Zaa1s0hA+LPddqOA3ESF2taKUvBY1zSTn5hB7daWitVTYKKemoYGRmIbeGBg43W39b6/s+monUxpWSZ2PwjCwuGOsaXUmh6Zltc0ysiDcB3nhcbcXri20wPdcw0Pye5QcX8X+NlgsYqagWyIl7D+8HovL/mE1SzXF5lrqemDGiQnAbgd12n46a9s8kk0MpaRgjGdl1E1LqLT1RHLHH0B5ccdkHHNRVtMTadzcENwtEmjJf1huQFm32QeOZId2lVoHwupnCQguI2QY0MLJx8ZGVq+mtC3/XOo7dpHRlnq7vervO2moqGljL5ZpD2AA8gMkk4AAJJABI0aQkOxGVzJy581/ETlYuFyvfDiwaUnudziEEtfdra6pnjiyCY43iRpjYSASBjqOM5w3AeqnIl7OPSvLfTUnEjibDR3/iXNGHxnpEtLYg4DLKfIw6Ybh03kMtZgEuf3dXi5afbS8zUEkcVz0Dw1rY2t6XFlBXRSvPkSfey0fMBoH1Ld1F7bDihTxxC78E9LVLwT4hprhUQhwycABwf0nGBkk7g7eQD11ReWFn9ttXSYZdeW6GUmTeSm1WYw1m37x1I7JG5+kAdht3W6ovbVaAiY9114F6hhIx0Cnu8EuR556mNx+f7EHolqHT1k1ZY6/TWpLXT3G1XOB9LV0lQwOjmieMFpB8iP/fK8VOfL2bup+XuWt4ocJoq2/wDDdzjJUxkGWrsQONpsbyQZziYD4RgPwQHP7ZW/21nLvK8i6cMOI9Ozp2MFPQzEu9CHVLMDvvn7FuSn9r9ye3mB1HdqLWtLT1UJbNHW2OKRnS5uDG8RzPByCQQAQd9yEHiPHAWjr74UwaahpGM4XYzm9byf36+HX3KxqyspI7tOXXDSVXZ56ZlE8lxMtLIW+G2I4H3on4c/AcfAzrtSvEWcjugwZYzG8tKtYd8eqy52tkeXYGVCYcZP5EFjm57Hcqemg6jl42UUQa5+CcLKnc2JoHUMnzQZjDEwBoWHVhg7KKJxDi57tlfKYJSC12SPLKCngZi68LH6izICy+mQM6dyAsOQFriEE0U7oxj181NCS52Xb4Cx2u+AE+iRyOD/AIdwgynvIf0jsu+HIdz68M+UzhRf9H6g4d3+83y73l9097opoWxSR+BFHHC8vILA0secgO/shPkAuhcgcGh6sNVKzZpx9SDVtZX2t1Nqq76orzmqvFdPcJ/iJ++SyOkdudzu47nutOM48MZG5V1NNFK3MxGVbVSwkdLOw7IL4YGyt8QnJG6glkAJYNlVksgjJYdvksVuXvHqUF+CTjyKyAJGtBD8D0VpAiGHdz2UcrpBgjsfkgzYi1wAcRnzUro2gZBWmMqC1XmplI22QZr2DpJyNvmsGUjKjdPKcguVrWOedt/mgo4jOB2VSchVkb0nBG6tbsceR2QUGyyXzjwQ1rcEjGVjuGHEKpeCwNxuEBucl3fCnbXSBob0jASikjZIGyDYrVq6gomUomY0ZIzlBp8YE4BzjKhqYzHgg5yoWyOYfhccDsqyTOkAB2QVjPwnfcKwkvd9qq2GRw6mtJCkY3oaXYJKCSPMTRkq/DZWkkK2FwmJa5WTuMbyxqCKVhY7bskG7wT2BVZQ7py4KMEjsg9DfYxvY7md1OBjbQVb/KNvRYfsWPFPNJqnq7fsArcf5xtyIPaZERAREQEREBERAREQEREBERBxxzJfg7cU/wCBV7/UZl80En9kf9Z/Svpg5kvwduKX8Cr5+ozL5o48eO/q7b/pQRNGThZEdQ6DGMqyWF7cvA2UsAY8dJxk7BBMLq7pwVJBEKxhkd5bqCa01UTDKW/B3WNHUSRNLWk4KDIdL0SmPOcHCjliLztsrYHtM3VIsmchwzH+ZBFDA0O3O6uqIBjIO/1rHMkrHeYVPFkeQ3OclBYfh+FUwsl9FIGeIeykp/ADC1/fyygwkUkrCHEjsowCeyCrXdJyshlQHYYR32WMMeaqDhwIQZ0tG2NgmAz54WTbLqYp2/CFBHMZ4/CO+B2S3xxx3eGOT6JcAUHZLl74lyaVuMdQ+lOOsbkL0J03xO/ZrY4jFBh3QNwPkuinDbS9mqrPDLHG0vJBPqu5/A+t0/ZaSClrQwEgAZIQcdcUtQXXTdTLVTyPbGATgk4wupnEjiRBfo52AhxBOThd4uba32mo0vLV21rcmIn4cei8u7nWmlnq4pwR8ZHf5oNtXirdWVx+I4BwFiFnhHI3VtQ4Onc9vYnIV8T2Obh53QUdO57ekDP2KSlozMCXZAU9vEDZfvuMLKrKmCNwEGMDyQafNS+ASR5LHEpyCd8eqzqupZLHt3WEyEuBQSvqPEZ0Y8lSOnJOcdkYxrCQ5ZHjxtYQCMoKlolb4YO6w54DCcFS0sxE2SfNX172vILftwgxC/bAyFbk+qIgKoOCD6K5sEjtwEcwx5DggnE3jN8PG6gkjfG7dp/IpaZ0TXjPmtVa+kdAS7BJGyDSaZ3xdLkqdngtKtlIbM7o7ZVhL3bnfCCV1R1R9BUCkhgknd0xjJVZ6aSnIEgwUFaUgSZPZXVjgSMKBrnA/Cdyruh79ygswfRFI8AN+ajQFe2QtGAFYsqnMIZ8fchBBG8Nf1EeaSyeI7OFSTHWensrUBV6SewKotVoRTeAfExnHmg0pMn1UlR0+M7p7KNAU8VQImluM5UCIL2yYkLj5lJH9ZyArFkRNjLST3QQAFyEdJ7fUqv2ccdsqeIxlh6sZQY5PkRuqKr8dZx2yqIKsb1HCuwGOCsBwrxG94yBsgmkma5mPP5LGVXAt2KDJOB5oBOUBGFe+nkY3qI2UeD6ICK5mM7o/GcBBblV3JwSqIguLMDOVVh7D1VpcTtlU7IJXxBreofWolcZHEY8lagqxvUcKr2hpwrQcISSclBUgAd1lU9b4LOnCxmfEcFVcwjsEF0jxK8kbZUbm9KNPSd0cclBVjg05KkwJRlQq6NxBx5IKOaWkqobkZypZSwt2xlQgu8iUFCMFTxU4ezqJUCubK9owDgIKOGHFvzVFc34ic+ao/AKCiq04IKdJG6NBJAQSvn6m9Kh+ayJYQyPqx81Y0s8Ig90Hqx7DZv/AFN4yP8AWawD8ja//WvUteWfsNTm38ZR5CawY/JXr1MQEREHXP2h4zyacS/8To/16nXz0QTuhLsH6QwvoX9od+BpxL/xOj/XqdfPIzGTlBe0eNIc+akdAGt2O6gDiw7FSiUuHxIITsSsmKlEjOv5Z7rHd3KubM9o6QcBBa9vS4hUxlVJJOVkQhgjPVjPkgg6Nt1aRvgK57tzgq3ugq0FxxlTMlEQxjdQA4V3Q5w6kEksokbsocH0UkXT1Yf2ytRbTxvj6mjIwg0rCkZIGtwVWYNDiBhRgEnA80AnfIVM5UjoJGNDiNirWkYx5oLcLULfWiJwYW9/VYTA0Oy7spGuj8QFvfKDc7Io5mNd23yt96a1PSUtH9ypAD1jH1Li8VckUYwTjyWTSe+Pd7zGDsM5yg550JoC23KvFwkkYMu6sH61z5pLiDT8O7vCynOzMDI9F0201xDvFqe2FshABwd1yzQaup7hbDWVhBlDc7ndB6icHuaCkrqCnppq8OyQC0nOF2isWqrPe6GKpgr4S5zQXNLwCD9S8KuG3G5llvLoXvwxj9vi2XNdj5qdWx6opobfO/3MOAd8RxjKD1V1Drmns8jw2QFrATnuF1N478TbTfHyxvczrLyM7ZC1H/ZwtV/0g3qew1Ji+L4t84XTTi5rKvuF++5lC4+LLJgDPzQbC5iqaKqifO2rDmEHbqyupD6RsbZceTjhdk+M+i9bW2xi7XJsggfH1DIOMYXWmOpMkMgJ3DigxWU7ppMNfjC39oC/zacukU/vJaQRvnsuPopXiY4PmtTbHVkeK0HYZQeiXBjnHk0HbWRTXQuAA2LlovGrnRk1oXCGucN+2V0Utl7qmP8ACmDiB8yrrjdoy8AtIJPqg37rniJV6llkc+sccgj6S4jrfGZO5zpi7JznqKyqqtDwTEMfatOdJI9w8Qk7+aDJZUunYInnJHmVE6V8TukH4fkpWOiY0FuMnurZOh/xbZQZFI6N4y4jKtr+nG2FgsJY7LTjHdSlzqnDR3CCWmjYRluCVqENPG/+2NgPVabSMfFUDq2C1K5NdNE3wTgj86CSWSCkYXRkbZWk1NwlqDg7D61QwTlha4krHfG6M9LhgoM2GMPh6gMlYchIcWuHY9lmUUzhEWtBJ7LGqGu6i4txugsEuBgNClgc957faoo2dbw38q1WngjZFkYyAgxPDxuXbqZtP1x7KKOlq6+pMNMwuOcYC1F9BU2mHFa0gkbAoNGY3E/TnCmqoMuBBz9qgky6YuZk75BWQygr529YYSAgyqS1itjDGkZ9VHV2aahIL3ZxulumqaGpAkyAD5rWLgJLjGHRgnHfAQaNHUtA6CB9qxazBfkKeopHsPQ1p6xnZQsppWvHijA+aCHrLmhuO2wWRTwgDqP6VkTRU5hxGQX/ACWFieLHUCAUE8sviDw2jHksVzXg/EssOjMeQR1qjjEYyXkA+SDDBI7Km/zVzMdRypD4fScYzhBY2WRo6QdlRjix4d5qSm6Os9XZXSQ9cgEY280E0IbVuAdnI81SqLYz4ZGQVY+N9MOoZB88LHe90h6nHJQU22ONllRywNjwWEu+tYmSiCRjPFeT2CkLhF8I7qKMP/eKjw4H4u6CpzK4uOxKo5obsCVQOI7KhOUAnJ9ERB3QACVqDKh0kXgPdtjusHICNeQcoJ6imETcs3BCxsYWbDJ4ww7yWPO0MkwOyC5tU5kfhtGPrUfinoLScqrunp274UaCSGUxOLgqvk8SUPUSDZBkzSNdGAo42bB6iJJ7q9ryBgIPQH2LUodzSapaB20BWn/nG3Io/YrAjmm1ST+L+u/lG3Ig9qEREBERAREQEREBERAREQEREHHHMl+DtxT/AIFXv9RmXzQv2kcQf3x/Svpf5kRnl34pAdzou9j/AJDMvmgkGJXjz6iPzoJnVIdF0YCgjeWPDh5HKvdTSNZ1kbKL60GuTX8VFIKUsxgY3WjOb0nPfKYYBkd0B6s9XkgqYsNDh3VY5yzZ2SkUgLsO7Ks4jJAYN0FR/th+AEkjNO8E52UtEwxSdcgwFdXubMQ6PcBBV1xD4/D6QPmsaOJ8rupoOFCditSt00DAGv7lBFOwsiAIwVhtdhatcGsMfW3sRlaS3BeM+qCrWOeex/Ij2FhWptkpWw+Rd9SwpHMMoz2QWQvfA4PwQD2WbRRmqq2zjIwQfyKlS6ndTANxn1WZp6mfUyCGP6RQc08NtdyW2NlL4hJaQMZXaHRl6fcaWKvfUmMMAPfC6scLeF94ra0TvjcWE9XbZc1XyordKW4UEJIcW9I3xvhBqPHLjGx1sfaffPELWFuM5XRrUFxdX100nkXk/XuuVtd22/S1D7jXNf4LgXb9iFxFcmslqSYgNjugw44zI4DBwVdLD4fYrPe6AQBrCOv5LFFJUyjrIOCgx+t3ZUy7zJ/KrzGY3YfsVd8GNxugthb1uwT2WS+RsOAFidRY7LdkfI55yUF739ZzlWNaSe6tyq9RCDI8Dob1A749VD1OLgD6qWCb48PO3mqzxgyB0Y2z3CA+lIj6w0nbOygDDn4gQtYo6iAM6Zcdt1hXF8XWPB2CBSztz0EBW1QD5Q1vmVjRte52Gd1LEDHO0y+XzQZk9qMNMJ9xtnK04PdsMrX664wyUIiGM4wtvtx1D0QHZO5Cua4Bp2WTKYfCwO+FisIwQUGfaJmRT9TgPqKvvUzZntc0AfUtOje6MkjZVmlMpyUFIwCd1M6VsYwMHPyWO3q8kf1Z+LugqAXu9E6MP6SrQSOyqHfF1FBkSUoZEHjufmoWRl4O/ZTPnc9gZnsoAXs8+6CjhhxHoqKpJJyfNUQFcJHAY8laiBuSmFJH0+aq4MxsgiRD3RAVwcRtlWogqTn8qAHCoqgjCCn1or2Rukd0t75SSJ8Rw4YKCxTxThjOkjKgV7YnPBcOwQHu63beayIKXLes74KxN2n5hatQNc+Ikjb5oMSeqJa6LpAxt2VkMHWwuxlX1rY2yHpABV1K7ERx6IMN4w4hZEVMHxlx7qGT+yKolePhaSgrFEJJejyCVEIiOAVcIZmffMfao5XuecnKCxERAREAyQPVBfHH1pJH0KZsMkI63dio5nh3ZBG13SVnQQNljLzjssFjHPOAsuGZ0DCwjugxZAGvICtV5a6WQ43JVHsdGcOGCgtAycKo2OFVgJOyOBaQSgq6M46s7KSGMO2Kt6i8dI7qsbJWSAEEILpqfw29WfrWOtQqY3iEdXmsKOMuPbYd0FBlu5CoTk5WVKYhFgfSHyWJ9aCRzstxhUjBDxsVPCYv32PJZBdTbYwgt6fGYGn6lFNR+FGZMnZTePG3GAlRMJKZ2PQfpQepfsNf9z+Mv+GsH6K9epi8s/Ybf2hxm/w1g/RXr1MQEREHXP2h34GnEv8AxOj/AF6nXzx/Uvob9of+BrxL/wATo/16nXz1UxZlwf6bfWghAycIQR5q8MLpCGpIwswCgsVQMkD1VWYzup20csvxxjbugtdB0s6s9/mowSexU0/WxnQ7uFGynkc0ub2CCalpBM7Dj5KyrpxA/A3CjZNJETg4KlEvjMPiYJCDGUscwY3pISIM6z14wlR0gjp7IIyd8hZ1NU9MPQT5LAUsbgBj5ILZclxJ81SPZwd6FXSRPb8R7FWAjCDIlqg9hZ04x6BRshy3q9FF9SuEjgMA7IMu30wq5DGdh81mVVnZTDrDwcb91pUFQ+nd1M7qaa41Eww5xwgvdP4v3rOwWfBehRw+CG5J81ogcQ7IOCp4YxUEM/fnbuglNwf43igkHOVrEWsKuKkNMJXAEY81pNTZqyli8aRmGdwsEY6hnsg1Ckr6s1fjioc3fPfC31p/iJV22pZEagkjA6s5XHcksYAEQwfPbC3BpazfdWUHpJee2yDs7oLibXugDn1zyCPo9RWddbv03uLUUkgcInBxz+VccaV01X2yNr5mu6DuFu+uslxr6EuhB8PG4+SDVuKnH6i4g6dbpYQtBiZ4eQAPLC6q3Systrnsa4fGSdj5Lfl+tjbXVSOawh++Vx7daqoravwMkuJwEGlTMFO4OHmVr1sndLSkdGR6rFrtK3OKnbVOYS0jO61nTk9vpaM01R/ZcY3CDQ5HeDM49sZWBVh8zg8O2Ws3uz1D5TNAPgduFiQWCuNM6VwOBug0iJ3S7DjsqzOBOwVsjHRyOYe4OE2xv3QWZPqVXqd6qh7ogdldHIY3dQ7q1VDSd8IMmOqBOX91L7+8kNYsOIDJDlV5DXDp7hBqTJ2t+OXb61iV08Uzh4Q7LHdK9wwSfyqrBhpJ29EF9PU+Ce2y1FlPHWR9YOMbrR/qU8NZLC0tadigOAinLPTbKmNUYx0g+WMKAh0z8juVZJG9jgH+aDWNP3xlqrBUSxhwBydsrK1XqSK/lphYGBuMjtlYtttgr5IaWCCSeedzY4o42Fz5HkgBoABJJJAAG5JXK3MByuaw5cdJaGuvEaVtBqHW0dZWiwdOZ7dSRGJsbqh2cCSQyP8AvYB6RHhx6iWtDhOCbwT2ytdo9RxU8XQ6MZ7dlpEDIHD48ZV8tNAd2Z+xBPNWNuFQQwYyfRahDcxaW+C/cuWiUxbFICO6y6mL3nD3d/kgyJqlhd72RkHfCxpa9tUC1rcLLsdjvep7lBp7Tlmr7tcKl3TDSUNO+eaQ+jWMBc4/IArtfwd9lVzVcTZIK6+ado9BWmTDjU6imLKgt2yG0sYdKHD0kEYPqg6fQQOik8RxyApKysY9vS1oz27L2m4PeyE5ftEmCv4oXq8cQLhFgugkcbfby4bg+FE4yuwfJ0paR3buQus/tduWDRvCibh9xJ4X6LtmndPVVPLpuupbZSMp4I6mMungeWtA6pJGPqMuOSfBBJJ3QecceTIPr3V1SCHj5hSiHob1gK9kHvDC7HZBh5G2BuqZPqrpG9Dy0+StQVaCSsmKcR9wD9axQcId0GdLO2oZ07LCe3pOEDi3shJcdygkihEnmrZY/DOEbIWfRKBxc4dZygrFKY8npyqPd4rxjbKzI4Y5WYA3WLLGYpQ38iC4U2W5UL29JwssFwbv2WNKcuQWK8N+HKsV7XE4Z6oEUfiHCndSBoz/ADqIxywgO7ZVDNI4YLjugrEfDcVbK7qdlXPjcwZcoiclAyfVMbZQHCqT5BBfBCZXYWTJQtY3Izn61FTSNjOVWoqnvcQHbIMcjBI9FVrfPKtO5V4I6cZQd/8A2K7weaXVI9NAV38o25FZ7FX8KfVX/F/XfyjbkQe1KIiAiIgIiICIiAiIgIiICIiDjnmPPTy88UHemi72f+QzL5oZSTM93q4n86+l3mS/B14p/wACr5+ozL5ox0+M/q7ZP6UEz63qhEfSsRSRxGaToZ5q6opnU7gHeaCH5pjKyQ1jmYCkpxEz+yYzlBhbqeGHq+I+SyKhkYZluPsWPE8gEAoLp58gxjbCrTY6Dnf0WM8kuOVJTv6X79kFwpp5ZCGRl31LVKbTtfK0SNgIOMjZbu4eWmjvVSIXNBPUAF2q0Byo3jV1XSzUMDjC/BJDcjCDpLPatQOzF9zZy0bfDGTlYMlqucR++UM7frYQvZDTXJbpe0WgTXylaJA3LstHouD+MHAvRNHVOobZCwPLukANHdB5usoKnqHXE5g9SMK6alEY88hdnuLHLrcNNWb7tRQuERYXZ6cLq/MJRNJG4k9JIOyDHO2y3RoqURXOF7gCARkFbadC9reojY/Javp6rZDVMbg5yMIO4WitdUdltDXRRNLwBuBvlbX1vr0XerE8jugMOcdloGiKyGKAPry4xEeYW2+JrhWvJsuenP71Bl694g0lys33Oi6eoN6c+a4RiZgylxySThalU09c04nDsj1ytHqJHRvLQgvgiPj7nbKzZbg2nb4YGVp1O5/VnClmj8QjqG580Ec7vFPidsjssbCypoHsjDh2VaYxCM9WMlBiJjKukx1nCnp/DDfiIQY2CsiKAOaSe+FLI2LuFjulLT0tPZAhi65ehal7syOI75P1rS2uLR1DuVcaqU7E7ILZSRIcEqzOVM4ZaSRvhQoJIZRE/qxlVnn8Yg4xhRAE9lUtIOCEFMn1VQMuAVxicG9XkrASCD6IJ3w4ZnPllQDZXmRzhhWIKk5VERBcxwacpI/rKtRAVwbkZVvdV6iBhBJTNDn4V9U0MO3moGuLDsjnueckoKIBuilZGS3IwgOYOnZRHZX5cXdKtcCDuEFMZU0cJczKU3R1gPWU+SNo6WlBg9PxFqo9vScKSQtzkd1GST3QURE7oCqBkgIWOHkqbtPzCDUII2U7RI4ZzvuoKqYTuyNsKEzPIxnZWdkE9NA2Z2CVkSBlK0tHmsJkr2fRKu8Rz3DrOR80FjiXuLvVahR1QhhLScZVI/ALQCRkLGnIa7DeyC1w8aUk9iVlNY2OPGd8LE6sDIO6p4r+26Cj93E/NUaelwd6K4kEeWVZjKDKNblnR0/asZzuo5VExlAVWjLgFTsnZBNJH0tzlRMPS4H0VS8kYVAxx7BBlSVYmj8PGFikYOEwWlUJygvik8M5VZZfEOVGqggIJad4Y7JSpkEhyFDnB2Vd3HJQTUu78HsppoQ5wA8ysfq6Bkd0ZM4vGSe6DNZSNhAfscrUKejimhMwxtutPqpXCAK2lrpWxOjB27IFTUukkMDG5wcIxnhxnqGCsi0xxGpMk4BGVS9uYZA2DtnyQaYG9chaT5qj4y12ACpDTzRgPPZXRlriOs7hBC6MtaHFWLLnb8O3ZYiCoye2VkBp91d1d9v0qlIYw775hS1cbnAuj+gNyg9T/Ybf2hxm/wANYP0V69TF5Z+w2H/U/jKfWawfor16mICIiDrn7Q78DTiX/idH+vU6+ePK+hz2h34GnEv/ABOj/XqdfPGgvhk8N2VWaTxSCrDjHkqIC1q31UUVMWuxnC0VXiR4GAgkqJvElJHbKyqWVpiLD3wsExPA6iO6uicWnYoNVt9nbXykE4H1rFu1vFumEbT3U9uuRpXZz3UdxmfcJfE7oNMTKyp4PDjBxusVAVQelwPoqIgnkqOtnTjsoERAREQERPrQFJTy+DMyX+5OVUhpHw91YWEH5INduepW11EKUR4IGM4WnWihFwrWUx/fEBY8cBlwGHdajaKeopa1kgBBG42Qbj1Jw8mtltiraSMyF4GQN1y5wR4bVFVafurV0ZaWNzuFrfBnTbddthoq34mhwG4Xby28FnWTTrqegjADo8YAx5IOvun7ZNfqt9qpqIvMZ6dm5U2oJavTDxaJaItc/wCHcLsBwG0Hb9P6qnqL+xoYXE/Eq8ddB2a56piutuY000bgXFo2wEHVG7cOKy8NfXvoiGvaSPhXXrUmnqyzaxipzTER+KBnyxlepFl01p7WNoZZbI1hqmsDSAATnGFw1xL5M9RyVZrG08nik9TfgPqg67ans9LFpKB8MDTI6MZAxnstu2LhNBc7Q+8yvEb2AnpJwueI+XHW9KDTXVkvgMGwLTjC2JrPRWrNMxujpfFbSjIcMEbIOv1+vcltrpLb4RLIjjPkcLFGsT7qafwsZ2yte1SbSJnsmA95P0vXK2PUR0pDujAI7FBBn3modIfM5VtRGGO2OypES1/w/nSQue8N7k7ILY29RKSDpOApnUNRG3xCNu6x3FxOXIKKeJuWb+Sth6cnq9VdLIAMN7FBCdnHCmbAHMLsqA7qeOGZ7OpucBBY0NLiD2CyPDD2bBYwikc7pAycq4iaDZwIB9UAQlzy3OB6qk0XhnCoJXg9WVR8jn90F9PKIpA5axYNO6i19qO3aU0hY6y73m6ztpqOho4jJLPKezWtAz2ySewAJOACVq3CbhJr7jfrm38OeGenprxe7k49EUezIYwR1zSvO0cbQQS8kAZA3JAPuhyV8h3D3lL0+27T+BqDiDcIAy5358WWwAgdVPRhwzHDkDLj8chGXYHSxgbB5D/Zyad5d6Wh4l8WIaS+cSXs8SnjGJaWw5BBZCe0k2CA6XGGnIZgZe/o17YDXX7KebiTTUU2YtH6eoLY5gOQJZQ6rcfrLamMH5NC9yF4k8TeSbmt5tuZfiFxEsPDqexadvWpKs0V51LIaCF1EyUxU8gjcDO9phjY4GONwx2JBGQ6HsY9xAGVmUVBcbjWQ2y10VRWVdS8RwwU8TpJJHHsGtaCST6AEr174M+xj4ZaedT3LjVxCueqqlnxPttpj9wo8/3LpSXTSDG+WmI5I9N+7vC7l/4KcFKUU3CzhlYNOuDPDfU0tI01UrfSSodmWT/xnlB4l8GPZk82nFyWmrKnQf7CrPMQXXDVDzRuDe5IpgHVBOO2YwCSAXAZI738HfY5cF9KGmuPGDWN41vVxgOkoaUG20BOclp6HOncBsARKzO5IGQB3g11xK4e8MbUb3xE1tZNN0IBImuddHTh+MbM6iC85IADckkgAEkLpvxh9rzy+6GfPbuG1lvGvK+IlrZ2NNvt5I2P32VplOD6REEDIdggkO3fDfg1wp4QW/7mcMeHdg0zA5vTIbdRRxSzDIP3yQDrkOw3eSdh6BOJHGXhRwgtv3W4n8Q7DpmAt6oxca1kUkwyRiOMnrkOQdmAnY+i8YeNHtT+aTiSJ6PTupqTQtqlyBT6fh6J+nyzVSF0od84ywH0XT6/alv2qbrPfNSXmtulwqndU9XW1D55pT2y97yXOOMbklB7DcYfbK8FtLOqLbwe0Xetb1bOprK6r/6mUBOcBzetrp3DG+DEzIwMjJx58czXPtx85oba7S+t6+02/S4q2VkdktlAxkTZWZDHmWTqmc4AkfTAOT8IzgcAiAupw7G+PJYEjC0kHZBIybxG+GsuEiGM5Cw6Ggqq6QR07CTlZlTST2twjrWkE9soNMnd1yud81YtTmtpfD48QJB32CwDE5rSXNxhBGromdbww+ZVqq1xachBnVVvZBAJGuycZWApZKqaRvQ52QokBFLBH1k/JWzN6X9KC+CoMJJ7qksviyB+PsUSq3uEEr5j0hoUsNIJW9RKie1oakZmwS3OAgl9zGcKppRF8ee3zUL53g48wsmPrlhI9UFniCp+94xjZVfRmI9QGwVaWFzZc/NZ880TYel5GfNBpb5BKOgeSge3pOFUn74S3sga5z/rQI2dRwqPYWnZT1DPCaC3zUlM6JzT4gGUGF2Vehx3DSQsmojD3Dwxt5rXbebe2gLJgPEwg2yRjuqLJnp3CZzgPhyoHgB32oO/vsVPwptU/wDF/XfyjbkV3sWB/wBdJqnb/sf1v8o25EHtOiIgIiICIiAiIgIiICIiAiIg445kvwduKX8Cr5+ozL5ouh0kpa0ZJK+l/mPx+144o5/4GXv9RmXzV0To21kviEY3x9eUE8VM2jb4zsZxusOtq/eXZxjCya5s4b1/vDuFHRUJqGl7UGHG4g4SQk4Pks5rI2zGM9wcbqCuYGvAb2QQGUlvSkZwDlTU1MXnJGQqVcYjPSBhBjnclSxxk5x6FRtb1KWmJ94YD2LgCg5M4KQyx3hj3sPT4oOftXrvyu61p7dBQUf3KbMCxoLunONl5d8LdP8AvNPFJSNBeXA7L1O5RrlpWyW6Cm1AGCpMYDerGc4QdsblZKLU9qDm0zYzK30xhddtd8uPj31lc2m62+IHeoxldpLfNTVFJHLSY8Jw+H6lJPTxVAAkbnCDo1zaaANHwy9wt9lD5W05GWs3zheSNTwz1JHV10tVZpYwJCd2Eea+jrUGjLFqanNLdqUSxkYxgLrVxw5WNGuoZZrLRsjdKCS0NGR+RB4fy6Vu8k3usVvcSDjsVuHSPCHVldfKZossxY9w/eHGF6I6J5Q4arUw95pyYy/f4NsZXcfRPKloGzQwVVTRsfKwA4DB3wg83YOXG+P0iyVlpeJOgHZpznC4c1Bw/uumvEjuFA9pBwOpp/nXueeHmjrfRGOaljZA0b9WAMLoFzm0ug6O6SRWfwwevB6QPVB5vaqtb4gXug6AR6LjCvAE7gPXC7JcWqe3Q2UTUwGSw7rrbD9+neX7jJIz9aBTkgDIISaV4kHS04Wow07ZGhrQMlalHZ2+AXSN3HbZBock7ZIQCN8LTjkEj1WTXAxTuY3tlY7ficAe2UE1JTmeQN6SR81LXUvu5GPzLUqbwYIhJtnC0+vc+od1jcBBhtc47ZVHDHmqY6Hb+SrnLgPLKC796o1lSdAiBB3UUMDpSPTzQR9RxjCzKWiZNEXk7+im9zE8YZGPiUL6arowQcgHugx3YhlI9FIyLxx1jyWOSXO33OVlQPEUZa7YoLJph0+HjssdSxlpmy47fNKgsLssQRIN0RBUjComSr2wveMjsgsRVILTgqiCv0d8qhOTlXHsrcFBIxviHHqksXh90he1jgXeSrPKJCOlBEpWTBjOnCiRBcHYcXeqE9bh5K1OyCZ0PSzrB3UJJ8ypY5M7OOySdHkgiWVT0rZWF5PZYp7q9sz2t6QcBA6MSFnoVmw0TC0En61gdRznzUgqJW7ZQT1DBH2WGd1c6V7vpHKNjc4EtGwQWonZEF8UfiOwTsqzRCM4ByrGucw/CcI57n7uKA1xac5Rzuo5VMFEFWnpKuyHYHqrFUEAfNBI+Lpb1KNp2Kq6RzhjyVuT6oJIIhK7BKvlYIT05ULXuZ9Eo57nnLjlBQ7oN0U0XSWnPcIInNwAfVZlKwOj3WI85JHkFk0zXlnwjZBDOzDiouyyw3reQUloZAS5o2QYinhpvEGfJQlpaSD5LKgnDGYzughkh6Cd1GDjZSHrmeQPVWPjdGcOCC0klVZs8H5q+Hp6viVZenqBagzHjxYekKNlOImF2VJSOy3dUq45D9EbIKUknXIWZwqVjvClG+ViffIXZ7FUfI6Q5cUGYa1sjBGW4P1LEdlryftVpGMELIiczoPX3IQWOmywNPooRuqvADjhGglwCCrmFoB8is2GcChlYdyWkAqKUN8MduygDiIyB5j+dB6uew2GLVxjPrPYf+jXL1JXlt7DbP3L4x/4ew/9GuXqSgIiIOuftDvwNOJf+J0f69Tr55GN6s/JfQ37Q78DTiX/AInR/r1OvnjBwgHZMEeSmpgHP3WTMyNrc7D7UGAN1fEzLwCVafpK8RvOHNQZdU0eEAzusRmWgggjKyKOZvWWznZSVroD/Y+2EGA7IPdZFPUCNuDuQsdxyUbsRnsgyJqp0o6cLG7LNjNOGjKgmDXSZbuMoIQMqpBCyh4Ybgd8K13Sd9t0GMiq7uqICmjpy9vUPTKiDSey1Klmijpix3dBprhgkKivlIMjiOxKsQXRnByrnydQxhWN2O6mjLC4ZQKWc08gdjIPqt0aU/6rX2KkezDXkDOFosdNFO0dIBW59KiKkr2CLAmyMb+aDuLw2slFoGzwXaEte/ZxAXYTTPG+2VlAI6yRrMNx8Rwuo+i7xdaagBvhJpgMjPbC2rxI4lPoGvbY3EAZHwn/AFIO3usOJNjid41HcGRuef3rgCuQdCmx6v0rMKuuY97oz8ROSNl5M1fFbVdZVATTvwHdiTlczaI5ib9YNPS0zJ3B/h7fEfRB6S8uGg7VYdavq21bKhplJDc5HddyrtYrRXweLPb4pCwZb8A7Lyp5BOLWpdZ65DbhM50bphs4kjGV6yy1kELmRSuALwMIONKrRFm1LNLSG0xMABH0QF1U5m+E4stiqYqC0h5OdwzOO67z3ClNNGamiHS47nC2pq6zWC66elmv3QdifiCD53eJ2j77b9U1DpqGVjSSRkEY3Wz20LhGXS5BGdl6Q81Wn+H0dfVi3NhEgaewHfdeeOrgIKySOn2b1kbINvdXhyEY7bKjZB4wee2VV9PK1vW4bHdRdkGtOuLZ4BEANvzLSpxh35VfA5rdyrJHh0mfLPYIIle2PqGSVfJJG5ga0HKsYSR0+qCw7LUaWcMhLT5jCwXxPZueyvZIAzHmgzYg2MiY7j61BW1DZ3Dp8lCZnFvQDt9aswgpgei5Y5b+WbifzQ69h0Pw5tZMcfTJc7rO1worZATjxJngHc4IawZc4ggDAJG9uTjkp4jc22rPCtcclm0ZbZgy9ailizHDsCYIASPFnIIIaDhoILiAQHe7HBPgdw25fNCUfDvhfp2K2WymHXNKQHVFbPgB09RIADJIcDJOAAA1oDQAA2pyt8pvC7lR0ONL6EoverrWNY6836pYBV3OYDu7c+HGMnoiaeloJPxOLnu5rREBaFq7W2jdAWiS/wCudWWfT1siz11l0rY6WEHvjrkcBn5ZytdXCfNHym8L+a7RR01rmiNJdqNr3Wa/UsY97t0pHcHbxIicdcTj0uwCC1wa4BwXxl9rfyz8OGz0WhGXfiFco8gfc2I0tCHjydUzAEjt8Uccg+a6McX/AGs/NDxOdPQaHrLbw7tUg6RHaYRPWObvs6pmBIPbeNsZwPmc9feZPlq4qcsGvJdDcR7U4QzF77XdqdrjRXSAEffIXkbkZAew/E0kAjBBPErPGiOQCMlBuO86s1ZrK/y6j1tqa63+6T/2WtudZJVTvGScF8hLiMknGe5PqsS5wmr6TG7GPLKxI52Njy7GVBJXS9WGk4CCYUEj2BuSSFhz076Z+HBa1aKuFzh4pGfmo78IXuBjIQW0UzXRYdhQVkTTlzcbLGDniMCInYK0zyAFrwclBqWnr5HZ6kTSR9Q/KtR1BXx6mc2op2hvR8sLap3WRTVE0LXCHO6DUBdG0sIpMZLdisGSYTA4GAsfPW8l53JyVVzgNh2QWHug2V7GdbvtWV4TWxnI3+tBh9WyopoekPPV2yqTlvXlqBDL4ZJ9VNJGJsyD0WIs2KVjIDn0QYRGCQr4W9UrQfNXRwvqHkR7lJYpKZ4Dtig1OqoI/AD2kdsrA8fwmmPCtNZO5vSX7KrCxzMu7lBATkk+q1GmkayDfyC09zCMnGyCRwGM7IMuOrDZCMdyo6p7pXAjJCxy1zdysmGRn74+SCymi63HOyrIRE8AK+eVjW/ezvlYrnOccuOSgyHS+OOk+She1zNvJWhxb2U7S18Z6u+EFkU/hjcZUga6RxlDiAN1ju7lVEjgMA7IMmary3oxv5lYpznOCpqVjXv+LfdZE4ha0t2Bwg74+xWdnml1T/ACu/lG3InsVvwqNVY/4AV38o25EHtQiIgIiICIiAiIgIiICIiAiIg445kvwduKX8C75+ozL5oXEtmJHcOP6V9L/MiM8u/FFo7nRd7H/IZl8z8oIlcP+2IQZs9x8amEJG48yq0FyFLEWEd9srT0QTvlJmM4Pc5VJZTKcnuFGc4wrdwgy4qzw+wGyiqJvGOSoUQVa7CmpW9VQ05xg5UZheG9R2CRvMfxDug5k0VxBh0jTxSmTfqAwvSjk7ttt4sGjub7oI3MaHdPVjO3ZeOz6qaVoY5xIBzhdq+UHjrqfQepaCgp5nsgLmtPxEDHZB702a3stVvioWP6xEMdXqs5cf8ACriLQ6w0rRVjpWmdzAHjqHfC3+DkA+qCq065WimuIInjDwR5rLfUxMk8IuGcZ7rTLvqSitlNK+V4Dg04GfPCDEtWnbJQVRfFTRh4ORsNitxgADAGy4Ps3E9sWpHCplBhLiNz81y9a7/QXdodSyZyMhBZfbUb1SOpQ4tyCM5x3XUDj/ywW+upam7VdW3LSXbu+1d0n5DSWd8HC6Uc3mpNa0dHVU9BNKGEkYGUHndxv0vb7bUPs8dS0hgLcdS6xXS1R2yWUxuDsOXKvFmLWVVd31VS+Yk5JJBXD9VPUxufHWEknbdBDT3A08hducLVGah8SIsOR5Lb7ulzic7K49LNm9ygkqpBPMXeqhAAkAzgZRzXNw71U0NFPUNMrGkgbkoNQqY+mkBZuSPVYlI/Mbmyd/LIWRZmyVVWKWTJA2+pS6gtpoHjwwQD6BBpLmdcpDe2VbJGYzur6VwbL8Xn6qeric8hzBsgxC8uAC1Kg6GRHqIytNETySANwry6WL4TkZ7IMuGv92qC7uAdlLVXQVpDC0AHZYMRje7MgG/qqTRgOBjCDNntvRF48WHHvgLT39ZPxAj7MLX7HOwHprfofNY2o30bph7pjAO6DSSzDcj0yrM5VcuI+SogIBkgIgz3wgyH04awOyVSKoEbOkgFRmZ5GCVYgq93U4n1VEwmEBVBCNaXHARzS04KCakgFRJ0k4+tXVtKKZ+GuyoI5XxHqYcFVlmkmOXnKCxBuQETsgyo6VrxnJUU8XhEYVomeBgFUe9z+6C1Mq5mOxUhDPXyQWxRh/dZbaFhZkkZWF1lp2KvFTIBjJQWSN6HlvogaCrSeo59VUOI2ygvgjEjukqeQtpvhG+Vise5hyFV8jn/AEjlBPSwNqZSHbD61fW0TabHS4H6isWOV8TuppwVdJPJL9M5QRoiIKkhUREBVAJIHqqYPosiMxiM9XfCCj6V3SCwEqEsc04cCFuDSAgnr+irwW4U2toaKCpYKPAGfL6kG3Hxhoyo1Uvce5QNJ3CCWmhEzsOOFWphEDgGuyFG0vYctSR73nLiThBdBGJXEFZBlFM0saMlYbXuZ9E4R73POXFBKyfok68eayzcsxlmButOwfREFz3dby71Vu6K+NuXD86CWlJjf1Fu3llZz6OOoYZcgEKCd8Ihw3usVtVM0FodgFBbIzokLR5K07H1VXdTviKpuUEsdQY+yn996mkOwSFhK9jc522QXSSl5OG/mWZR20Ts6nbKOnbEdjglZUlWYGdMZwUGLWUrYNgewWG3JIClnqHyuJJ81SKF7z1Abd0GbTUAnaOrAWPVU5pn4YMj1U81S6KMNjOCp6SaCSA+8EF2EGmGQyDpVmenLVfIW+OS3tlWyN/fAbIPVz2G/wDuTxi9fHsP/Rrl6kryz9hsT9zuMo9JrB+ivXqYgIiIOuntDRnk14lZ/vOj/XqdfPLFH4hIPkMr6GfaHE/tNeJRH950f69Tr55A4tOW7ILmnw3EAq90pcO6hJycplA81OyoDW9OPtUCILietxI2yhafVWg4VeooKKobkZVFVrsfUgkp4XTv6d8K+eIwHowSsikniiOT3Usk8EjwThBpjQ7O+ceaq5xHZZ9SIZGdMOMrAfG9n0gUFoY4/RGULS07ghalZHUzZz70AW/Wl8NM6oBpsdPyQQ22BtRIWuIGOxV9yoxA4FjtlhRTPhd1MOCrpKmWU5e7KBBF4riCUqIfCfgbhRse5hy0rOghNREXkEkBBiQxeK7C1JlsYI+skA9wsOlxHMQ5ST1sgl6AfhB7IL6WofTzluMgLKoLu6kvMVV1EBrgVYRCYw4Y6iFhCklfJ4jRkA5Qcyag4s+8WCOihkLCG9ORsVs+nq33GgfNLOXE5O5W0ayZj4RGBuB3V1DWzxwGKMnB27oLZanprHN2wHYyp2VkxeYxMQCMYBWlSFxkLjscqgke1wOdwg7XcqXFCPhfdYK11UGEyAnB+a9gODXFvT3Eynpqua9M8URjDC8ZJwvnotOoqqgmYckgOBAyuw3L9x04hWHXlE6mmlbbmvHUOogYyEH0BReHLB0sPUzGAfULjLjTR1EelKn3XqDsHGPqWHwj476W1jpq3MbVN98EIbK3qHcBazrjW1gdS/c2tLD4m27gg8i+M1tutXqmsbXue1m/fI9V1R11aGU1e7wcOHXvheknOFo+impZ7hpxjQ9zC7Le/b5Lztudqr4IJ/unkyhxx1Dfug2TdBG2naBgHG+FpMMPiAlZldSVfjO6s9Odsq2KB8cZ2OceiDEkb0HAK1CzWoXOrZTAglxA7qlssVdeJiylZkk+i3no3hzqBmp6RskRa0PBJAO4ygg1Dw1nsttFe5hAc3q3K2SyLuW4JBwN13V4hcFbzqHREUduY8yGMD4Wk74XB9Hyua9hxWVtM9kDDknoIyEHDgjqJyGGEgfVhXSUXhNySVzVdOG0dHTmjZGROwYO2+VxXqOz1Fqe5kucA+iDbv0HFSBnUOrKjwXOx6qTDm4aD3Qdt+G3tP8AmZ4T6StGhNHwaJprHY6f3eko/wBj7GMDNzl3hOaS4klxdsXOJLiSSTyDbfbQ80lGImV+jOGte0PBkc+2VrJHNzuAW1YaDjIB6TjzB7LoP0nOSmB6IPSGh9ttxcijcLlwZ0fUSdWzoKqqhAGOxDnOJOc75H1LcdB7ca7xF5unLbR1DTjo931W6Ej1z1Uj8+XbGPmvLstAGcqxB64W724GhZX4u/AG/UzOjOae+Qznr22w6Jm3ffOdhtvtuO2e2w4ATPjF34V8QKVpB8Q0zKKctPkAHTsyPmcfUV41Yyq7jfCD2G4qe0Z5BeY7Qk2gOLOiNdSWuudlrqqz04qKCUZDaiKWKoeY3juCzJIJaQQS0+Wuu7Joi1aquVBw+1TPqHT0cubfcKiidSTyREZAkicT0vByDgkHGQd8DZcFS7qDXdldLWPY4NjOB5oIJwWyuGexVgd5LUGNinj6iAXEbrCmj8N2B2KC7okiaJGuI+pXNe+cEPJJCmjfG6ANfjGPNXRtiaPvePrQQ07wyXDhsDhR1bw+UluMBVqWFpyPNQtAJwQSfJBkGkPu4m8QZO+MqBrnN2G2VkNie0Zl7Dy8gopunqBYgtLCMHcq0g9ysile0O++YwrqmIPf1RjbzQR0o+I9XZSVEoaekHYhROcGD4e6iJc877lAyfLKp9ayIIsEl6sqOnr+FBEpIQZHtiJwDsrDsAjXFjg5uxB7oNehpYbczx8gkhaXcKptVJ1NGAo5KyeRnQ52QoEBXMyXABW4V8P9kCCSUYZhRRDMgB9VmVTAIgR3IWKyOT6bew3QZVXCGxtLRn6liMjcQTuMLLhqWOHRL5Kkzc/FH2+SDDIKqxvUcFXgsBPUrXOwctKCV9OGtyP0qwAhpVpke4YysuKnc+InzwgxooxITlJogzsnhyh5Dc90aXNkAk7ZQWxvdGcjI+xJHl5zlZ1Q+mdCA3GVpxGCg7/+xU/Cm1T/AMX9d/KNuRPYqfhTap/4v67+UbciD2pREQEREBERAREQEREBERAREQcdcx7unl54oP8ATRl7P/IZl8z8x6pXn1cT+dfS9zJfg7cUv4F3v9RmXzSNj8SdzR5En86CHBKr0/NZMoY0dIG6xslA6vIq4N6lYpoeyCtLAJ5OhxwFmz22OJvX1dsFae2V0byWHG6vdV1EowXEgILpZwWmPHbzWMhzndBuguY4McD6Fbi07q+psNwiroHlhiIO3yWhNhLm7d1aYenOSg7icMOfjUGhqNlGaqoLWEY74wF2m0H7T9two46errW9fTj4sZBXktTsjdIBMT0/JanTVgo6thomEAH1Qew9o52WXqtNSbngPd/dLsFonVlm4k2kVE14YHPbn6a8SNN6zq6alY1mQ7IPfdc68POYHWFgjbFSzyNjDf7ooO/fFGlp9PyPmtlaHuDv3rsreHAXWV3rHMiqA8jtvldSuF/FWt4j14o7nIXlzwDk5XfrhHpix2e3RP8ADaJHsBH14QcpUkhmgbI7uVsHiHwooNcxvbPEw9XfqxuuQowwMAZ2VyDz85h+Uy3Wugkr4LfGR0EhwAIyF5YcY9FTWW8TxxwdIZIRsPmvoR4y6YqtT6afRUmesscNhk7ryK5m+ENVperrZrkwh3WSMj5oOklTaPdqQTEbkZWnQM8RxJ8luXUtQ2GX3fq+EArbDHESfDkAlBkzxksDWgbFbr0raa2ppjFBSOlLhgYGVplHaHV0LOnOcgbBd0uVLgNDqWemNXHlrwO7UHVqz8PNUx13vMdllI6s56CuRKXgrqTUULXPscgyP7gr100Tyh6LbbWyVdNEXHB3YFyLbuW/QdvpxFHSMyPPoGEHgrrzgdqDS8BqRaJBgE/QK4rNNdIupklI8Y2OW4wV748ZOV/TV0oX+DTsILD+8C6JcUuUyG00FbXUlMAGEnZvkg89afrbMfFaQfyKlYet46QcfILc+u7GLLfH29jcOa4jGPmtJp7e4xlzx29Qg02SlDIusHfGe6ijkI774U80M7Zi056QdvqUbw1uR5oJTMZm9LDg9lYaYjdzsq2GJ3V1Y2V08jhgZKCF5xsFq1ts0VZAZXuwQM91pYicPid2Pmpm3CogBZE/AQR1UAgnMTTkA91CDjYq6SR0p63nJJ3KNje9pcNwEFnnsmFLA0eJ0lSzU+2W9ggxR3Q91UNJOFcI3ZBxsgRuMburCSSdZGBjClf4fh48womQveCR2CCxEIxsRurmxlwyEFqmiha9vUcqHsrhI4DAQUc3pcR6KSKLxPNWuaT8SMe5pGD+RBJLThg6gVBk+qnkc8sAJ2KgQEREBFdH09XxI/GdkFqIiAiKSMsBGUEeMIp5PD6fh77KBAAycIRvhM4TJzlBeR8OVaAfVDnzQNcUE9HVvpJOtpIPyV9ZWSVsgL3E79ysQjCDYoMmWmDIw8Zye6hY8dj5o6V7h0knCMY4nOEGXFG0j4sLGqGhrsDsVcZH/RYTn5KJ/XnD+4QWoro43SnDRus1tvkZGXuB9d0GCTlVEZIzgq8hrZCHDzWQCwx/CgwwN8KQHpGFZn4iqOOfNBc9rs5zlWDZVGTsqEYQSdQcOlW46dijCAd0kIJ2QWndXh+BjCsUscXW3KCxry12clVe57ztkhUcxzSVPTGPADsZKDHII7grIhqRGzpxvjCknhDm/Dj7FjimkG5GyC0vcXl2CrXOce+QsmHwmkh+CrJywu+AABBB2V/ifAW43V3wlvbdHQ4YX+iD1X9hr/udxl/w1g/6NevUxeWfsNf7Q4y/4awfor16mICIiDrn7Q78DTiX/idH+vU6+ePGV9DftD/wNeJf+J0f69Tr566RgcX58hlBAq4J9VWTHWfrWVTRAxlxA7ZQYaqBlXFpc/AVrg5hwUFDsqtblUQEhAOyvbH1NyrACSpY43gg+SCPDh6qgyfNZkgjazJ7rGLeo5aNkE1O3oPU4jdKqRrnbKMRyu2BVro3h2Hd0FnZMk+amki6GZUKCpGEwe+D+RAC44WeyKNkJLsZA9EEFHSsqHYc7H2rOe4ULDHHg5HksGlp6iqn8KlBy442C5G07wT1LeaYVxjeWYB7FBsWgo31Mpc5uMq6utzYpB2zlb6rdEVVmc6AtIe3YraN2oapryHZ2KDTKhvhsHxdsZ9VSK4+HEY+nOfkraqGaNuX5x5LGYGdO4QWyOL3l2+6ljqDGzpUB2JARBfI7qOcd1YiAZ2CCaCMyvAbtg5W99O6+fp4e7xjD+3UPJbMpCYX9Tu2FRxZLV9R7E5KDtrwO5g7zo6rZVS3WToe8YHUcALudZ+JFTxCtrLya8kMYHHf5LzB03Gyuigip2glrhndd0eDF/bS6d+4AJE8kfSB55wg3RrrXM10M9ujBqSxpGO66Q8Zq27UtxkaKB0YLz+9wO69AuGXBu4RXiW96ijcaWZxILhsQfrWk8ceV626vca2yU4LSST0t2QebdmZNd3hs8GCduy3JXaRp4aJ0gAz07Bcma54KVXD+d5ex7ejf6K47mmramJ8bXOLRkIND0XXutF2DRD1jrx2z5rtLwy9xvV0hlkpGB4AwC3dcCaKtFHFWiataPpZ3C5m4cahpKTXdJSwYERcGkfag9FOB3Dyk1Jb4Yq62h0ewy5u2Fy3rngppyPR1cyis8DpYoi5vS0ZzjyWHwsv9ptOiaSqpDGJS0E7jPZb8o+J2natgpK6VgfIOlwyMHPqg8qLrwyuk+vK6nks7xC15G7NgMlcM8buGNvoS90jWxPGfh2BXrNxWj4baXhqr29lOHysLgds5IXk5zR1t51dfJqnTbyaYSn6PbGfkg6xXm1NttWWs3aNwsNlP4gMvVjC3BfbXW08TffN5A3fK2yJ3MaWDt2QHODnFnoriwjySlDXSEu81lT+GBsQgwXDGQrCcKQub1HKseR1bdkFrTgo52VMAwx+ShcMHZBTKZQHCqfJBMwGIB2ThRyyGR2VaXEjBOVRBeQegenokUhjJKsySqtGXYQXPlc/udlawlrg4DOFO+mcG5CijaBKA7sglmqnyN6enA7LHWpVMMJhDmAZwsJtO8jPkgj3JUzJyxnR3/mUbwWYHmrmNBYSRugjccnKyoKZr2GQ+W6xD3OFeJntGAdkEkk2MsbthQd1XI3ynSTuOyBudgFc2Jx8ipIywEZWaHQmLpHfCDTHNwqKSZhDyfJWN7oA2Rp6XB3oVJJ0luw3VscLpDgIL5agygNV0M4a3wiO+yubTmL4pVFIwFxcwbZQTGka5vXnGVaZfDb4ZURlkb8O+FYSXHJ7oMqhpRVyEE9lbXUwppOgFXUz30x6gcZG6jq53TvDnElBbFGDuVkmtdEOgDIWPTtfI7DVnFsDY+l+Os5wgippQ52XDc/JQVXxTdIHmrmxuik6gPhKvDQ53V80FPcelvWSsdsfVN0eWcLJqZ3BnSD8lixE+ID5oPQv2MlC2l5ndTzg/S0DWj8twt5/mRV9jIZzzO6n6+37Aq3H+cLeiD2ZREQEREBERAREQEREBERAREQcccyX4O3FMf8AcVe/1GZfNIyXwZnuxnJI/Ovpb5kvwduKf8Cr3+ozL5oJP7I/6z+lAe7reXeqtJyikZ043wgsa3qKvHwkN9UijdLJ0sV08L4XAuyCeyCWSlHhh7dz8lA1zo8tcCFmW2dgnDZyCz5lXXg0zngwYQac45OVdE3JVivYyR2S3OAgyw8RtysaWbxFa55x0nurEFSdgPRTU9S+A5aCcKBa5aLY2qpzI4dgSgydP3Z7qoRyPIGR5rmGx19MIQ33kAluMLgeokFHUnwdnA91mUGo7lHVRYmOOoDHyQd2eXOvhtWo2VVZVhsZlGCXYGMr1h4UVVov1spp6Wua8tiBwHDfZeGdDqK7UtgintLiJ9icHdd4OS3j9e7fUUtqvsxy/DcOd/rQen0bAxvSDkK5adYK8XO1wVzSCJW5H1LUUFrg0j4gCPmvOT2htGK8VclFRjAJy5rfyr0UrhKYiIs5wV0w5o4bNW0FZb7hg1EhLW+Zyg8UtYUszbr4Tm4IyMfasO12eaoqms6CT6LsNrLlj1VeNRPudBFKaZ7iQQ3bGVNYOWnUdLd4nzRShgIzsUGqcuvBep1pWRU8tAXNLxuRnzXrXwD5f6DQ9vpah9ExhEYO4xvhdYuXzT9k4X0kFbdsMIIPxBd9NA67tOsaCJ1qka5jGDJHoAg3XFDHDGI42BrQMABSIuPuIXGCxcO5mxXh7GhxHd2DgoN3X4UXuDzWAYDTgkLohzP8VLBp2019tgdGS4uHkuVuN/NvpC16Xe611LC58ZOS8ZzheTnHbj7JrWsq/DlJDpDjfPmg4n4qXKKu1NLc4jkOcXbfMrQLdcGTBzXbfWsC63F9e4Oed/rWnxyvi3YcINauUsQBLcZWhuOXZU7G1Fa/w2nJUr6N1KMVDcE9soKUswPwO2VlVG4vy1pIUBd0PPT2BWuWr3eaHolwXeSDTHyB0PTjBHcLFa0lwb67LVau2SwSGYj4MrTpS0ODmIMqegayHxA4A4WNHOY2FoGcqr6p7mBh7YVYTEGfGBlApWiSbJOAsirkEWGt3ysLrMbz0bbqj5HPwXHJQA7DifVZsLGyxknvhYCvbK9gw04CCkg6XkKWKoEbC3HdQklxyVRBV7upxKkZKGt6cKJEFXHqcT6lZNPTtkYXnuoIy0fSVzp3DZhwECV2CWDfBUe4OSpIC0yZf2V1SYy4Fg+tA8TxB0/nULm9KoDhVLie6A0ZTpJOBkqgOFlQOjDCX4ygo+mDYg8d1jKeeZz3FoO2FGzGN0FiqBlUOM7JkoB2U8NOJGl2cKBSNmcwdIyEFjmlri0qiucerfzKq2JzgXAbBBYg2Q7FEFznAgAKrX4GMKxEA7klERAxlZEBJ+AjCsp+nq+PGFkyPiABbsQgulgZC3xB3VaekZWAuc7BHzWG6WSXLSchGTzQbMJCDWrbb445TnfHqq3irbA4QtbjywFhW6vex+XuUV1mE8ocEFjacTHrzud1eYBEw7/nUEE5YQM7AK6Z0ku7c4AQY7h8RCoqkEEg91RBfF9JTGDrb1DusdhAOSs6KaPw8HzQYbYy5/SkkTmOxg7rJa+Jry4KV0sDmHtlBHDQiVgOdyPVWyH3Z3Tjuq01UWSnfZWVsrZXgtQJcFufVY7Thw3VetxGFb9SDVIw0RjJB27KpkYWFo7rT2yyN+lnHzTx3FwPkgpMxwcT5FRrKlfG6PbGVjDHmgAkHIUpqMxGP1VsMZkdhJ4zG7CD1Y9hr/aHGX/DWD9FevUxeWnsNsfc3jJ/hrB+ivXqWgIiIOuXtD/wNeJf+J0f69Tr554pPDJPqvoZ9od+BpxL/wATo/16nXzyxgFwz2QUPUSTgqSOcsZ0rILoC3G2ViyBvUcIDH9Ly5JZA85VYYHzEtYNwk0EkBDZBgoI0RVDCTgIDTgqdk+4GO6idE9vcKzsg1SGjZUjJOPRY1SxtK/oByFCyqlZ9F2Fd4c9SS87oKsquk5VH1HU7OFG+J0ZwQrR3GeyDJa81Dejp7KGWLwyB5rJppIWD5qOpe2ScdPZBAwlhBIOFrel6L7t3yltjx8Ezg0n0WHUwNdAzwwM+a1XSLX0FwZWjZ8Zy0+hQdjbny/0mlNMU1/oIWzSPaHEDBOVzJwVmfWWQUNVaAHY6d27laZy6Vlw4oRwWO5P64g4NAcc7LurZeW0aZghucMDRAGhxPljCDodxt0DVWpk1yhthAcC7Zi6tV5ke6Q1EJYQ49wvU3jnc9Dx26a1Vfg+MxhBzjOcLzX4sz2unrZhbOnpMh7fWg41vTmlpaxvYeS0HBHfIWsy1kMxw/B23Wm1Bj6j0YQQKrRlwb81RAcEEeSDJmgaxnUMZWO04OVf1yS/DkkKpp5ACSEB8pc0Ad1Y3qbkgFUaMOGVqscUL6UkAdQGxQbs4W1/hXGJriSC8DC9AuX7QNtu93pLrUVDWhoBLSdl53cOmPF0a7GwcD3XZvh5xvrrLqGn07RTYlfhgAPn2Qer12t1ortKQW61wxmRjQCWYyVujh5oGiFmMVwo2vcRsXNC435atN6qu9oprxfg800uHDq8xsV2PNN7qGNpRgZwUHWzjlys2zWdDUT0lvjLugn4Wj0Xlvxw4U3XhtdJaGC2P+mQMN+a91rte6CzwNluMgYx2266v8WOAFp4qX1t+pImvphJ1OwBjGUHjZdDfLVGJamgkhYRnJaQsfTGsH268MuZqSDG4HOfQrvPzmcK9H6U0w6ktcMbamKMg4AzkBeZr3VVM6ZmXbPIH5UHoBoLm4LLZDapLq4AYGOr7Fy7/stUM1gfqFuoAHxt68CT5ZXlDT3avonB0Ty0jfzW4GcUtXMoTbhcHeAR0luT2QdrtXc0FVxIu8+mXXd4ZESzPWcEDZcY6q1/S6VgfROkE5eTgk53XXylulXR1hroXkSuJOfme6rcbtWXV4krJC9w8yg1vUWrvuzO+QMIDicLbDjkkqrAD3V5gc4FzewQUgHxZV8zzkDP51CCWE+qkbBLI0yAZCCI7oh2KIAz5ZVekrIp3RN3fhXyUj5T1xD4fkgw1fE0PkDT2Kklh6PhxuEZSzFviNHZBkVVCyGHrackjKwFLJPM4eG93bZRICqx3S4FUUzKWWRnW0bIJDWZb04WO5/U/qVCCCQe4VEE/vBI6SThXCrw3pwsYDKEYQXSP6ygc4DGDhWrUqWGN1OS4DONkGm4JOwVek7k7KaHobMQ/sCrpHRmQdJGMoMqz2xtwlLZHYAP5Uu1GyglEMZyDnOFGax9OAaY4J8wrRUmcl1Scu8kGGASfRZUcfS3q6lHM6M56FGJXAYygvllLyWgZ8lF0nz2U1N0Bxc/t81SdzOrLNkEOT6rJpJwyQZWMqg4OUGqXF7HQ5bjPyWmskcwbDKOle4YJ2WRCYRGOvGUGO9+diFPDTtLOs9xuopgzOWqjTN0/DnCC6WXqBb6bKFD81I2CRzeoDZBJRy+GSVbUyF0vUPLsrqVoLyCN0na1su42QVdOXNDcdgrBK9u3TtlT9cHQNgpgad0eBjqKDBP3wgOPdajBanGIVDWkgDPyUENoq6ol0LNvJbmo6ukt1vNHVkeKRj7UHdf2MVUJuaLVNOR9HQVafyXG3/60VPYyU/RzUarnA+F+gq7H23G3FEHs6iIgIiICIiAiIgIiICIiAiIg445kvwduKf8Cr3+ozL5oX7yOHzP6V9L/Mfj9rxxRz2/YZe/1GZfNFloneXduo/pQSvpWth6877LFypXzucC0HZRIJqWfwH9WMq+qqfeiCBuFjKSItGepBZuPkqE5Ur+g9lG3dwQUwfRZtKWCIhw3Vr4miEnG6xQ9w7FBdLjxDjsrEJyiAtXtl5NHCYfUYWkKo+kMoM2ri8Vzqj+6KxqZxZUMcPIrJkqGugDW4z5LGpWh1QwHzKDka0aufbaJniZwCNly7wZ4hGXWtueysMTGvBODjzXAt3gjZaQ4DfHl9Sh0Lf5rTeoHMLuouAaQexQfRfwL1tZ7zoWhYLlG6WNgB6nDJ2C5ObKyRvVG8OB7Y3yvKzlZ1JxCraSAQvn93JGO+MbL0n4em5G1QuuJJeWDv8AUg3d3XAfGrgYdXzSXiNgd0Hq6Vz6rJI2ysMbxlrtig6daV01Ttrhp+ssjSIz09bmBU1hpK22e7xU8FsjAeQMhoXZu86LtccT62jgDJ9znbdcf3TQVZc2PrJw4vYSWnHog2VcuB9r1VpuN7JmQP6eo4OMbLR9KaktvA2b3Ke7NIz0kF2QoNcak1TpYSUtPLI1jGkDuF0K5hOKurXVc7/EeS15x8R9UHpBq7m505p61CubdISenOMhdCOY7mRdxUrZqmjvRaGvJAa7GBn5LqBqbinrq/xe4h8rm9OPpFbEder9bXubV9Y6u+coOU+IetbtcqAUX3alcA3p+mSuFZIpyXOkl6jvkkk5WdW3SoqI+t5O/qtLMrznfugtcMHGVRXNILsuVZMbYQT2+sFHN4uMq+5XD354cBgDyWEqhpd2CCiz6L4CJertvhYPSScKWMyMcM5xkZ+pBqlVeH1Ufuxbgds4WkyxeGe+crVah9GKUFgHiY3Wjue553QCMIM+RVMlASEFRud1Q91cxjnnDe6q+N0ZDXILCMLLp6Rk0ZeXdt1int3UjKiSNvS04CCyRvQ8t9FaqucXHJVEBERBLBEJXdJKrUQiEgA5BUTXuYcgqr3uefiOUFqZPqiICIqjGPJBREPdVb3QUQbqZ8YLcjuosFqC5zMDKsVS4kKojc4EgdkFqqBlC0jyVASEA7KZk/QzpxlQ91XAwgOOSSqIASdkIIO6AO6r07gDuqZwp6MA1MfX2yM59EEZhkaOoscB64Vi3xefuV9xh4XT4uN/VbHQEyfVFVrS44AQXRv6CSkj+vCPhewZIVrWOd2CA1xb2VXFzzkglW9lqdCyEwkvAyg0xZdO8iI5Gfmo6trQ89PbKyqKMPhI2JIQae/dxKosmoo5YnFxG3ksZBVreoqYRHpyHdgoASFd4j8YyguZGXuLc4VJWeGcZyqNc7yO6OLjuUFuT6pk+qIgINiCqtaXHA7o5had0F75A4Ywo0RAyfVEVWsc7sgvhl8I5U0rXTsMwacDcqAQvPktYgkpo7XLE/HWWED60HqD7DX/AHP4yn1msH6K9epi8s/Ya/2hxl/w1g/RXr1MQEREHXP2h/4GnEvH95Uf69Tr54/qX0Qe0H6RydcSOrt7pR5/j1Ovnga0uOAgdJAzlU3V5a8bEYUkfRjB7oK0dT7s/qxnKrV1IqndQGFEYXHJA2VOno2KCz+Yq9smCCB2Vh7qWPw+g9XcoKvn6xjGFDg+iq7usiF0Qj+LGUGMsumqhHhhAx2WK/6RwqdkGoVbWub1jzWCxnW7GVUySdIBOyMY93xNQTTUojjDw7fzWODgg+iudLIRglWAFBOyqLABuVqVHchE0nIBIwtGxhSxwyv3bnCDsPy9cdBw4uLKiSbAY8HcrtVqP2nLXWj7hQynJZ0ZHltheaQdJS4yDk91lMFJURGWSMh477oOxmruMVRr25TXOS5uxLk46j5rhbWEviSuHjdZJzuVtaO51NM4tgeWgbKOe4VFQ7qleSgskhLTkHuqsgywuJVHy9Q33VgkcNs7ILo4HSOIAJCTRGJ2CCFnW+WGMl0hH2qC5SMlkBjOQgxopPDOcKZ1XkEY7rGVQ0kZCAficSpmVDmN6PI91BuCqtaXnA3JQazZr660SeIzOTuuTeFdA65atp9VOqDmIh/ST6FcNvidHgv8yuSuD9wqhfoKKMnwnYGM+WUHsby38y09dp6h0yykP3jEYd0/Yu3tmuD7jSMmkaQS0O7YXRLlji0lpy0UlfcWMEjiCScd9l3msF6tl6twntTgWBoxj6kGHrTSo1VRNpmzdBaT54XHt+1LPwu09La20plJyOrGfzrlGmfUwzuMpPR55C0nV9rsF9tc7KxsbpC3AzjOUHlTzT6rm1JXVMlQ4tD8nB8l1BqtD22a3T1wewOyXY813P51tCVdlnq6qijAjDSQR2xhed1Rq69U3jUb5HdPWRglBpdeGG4PpMYaxxAWHUwCI4BUtVKXP94z8btysV8j3/SOUFqIqhpPZBRTx1HQ0txnPyUBBHdO6CrjlxI81lU9V0gRFvc4UMQDSS7srpHMOCzGyCWqpQ1viNOc7rDWVFM5+WvO3zUc7B15b2QXUdN7w8tJwAs81TaH/a43+a08TOhH3s7qNznykvccn1QZVV0kCQee+EjrwyIx9PdYhc94Dc5wqFpHdBVxL3FwB3VCCDg5CzbWITIfGxhVuMLPE6oRsgwFm09cIoTHjywsJVDHHyQTMYJ5CTtk5Uj6RrQTnssZr3M7bKpmkIwSgsIwiq5rhuQqICmbUFjOgZURaRvhGDLgCgkZE+ZxIB7+ipJCYytRpxHE3JwDhYdbKHSfD2CCGN3SVUgyybZwo8ZWbA+NkfxYygglg6G5yoRsp55g/YdlCGk7hBfGzxDjOFSSPoQB7dwrSXHYoKDdZMVM14ySomQPf2VXOkiPTnCC6aERbqHc7qrpHu+kcoBtnyQWgZKmbP0M6AO6sjidIcNCyPAEbSH9ygxDuSVkNqelhZjyWO7Yq6MAuGeyCrHvY7raDv8AJVc50zhkd1qMRpQwdQGfrUM4iA6o8IIH0/S0ODlC3rYQQDsVUyvJxnbK1yjoopaQvdjPSgtteoBQANczJ+pawbJBfIzcfFDcDOMraE0RbM5o7ArIp7tX00fgxPIb2wg9CvY3hkfM/qiFpHwaErR/zhb0WD7F+SSTmj1S6Q7nQNcT/nG3Ig9nkREBERAREQEREBERAREQEREHHXMe3q5eeKDfXRl7/UZl80gg8SolYT9En9K+lrmRz+134o47/sLvf6jMvmhe97ZXuBwS4/pQWPb0uIVFe5jyOsjYqxAUkcReqwAOdgq+f70QG+aCx8PSogcK50jnKgGTjzQTGcuZ0nsrIow4ElXuo5mR+I4bYUTXuGwQUcMOwqYV7mHp6yp4I2OaSUGKiuePjICz6ax11XGZYmZA3Qadk+qvif4Tw/zCulgfTTGOUYIWXR2eprh4kQyzug5Q4I6Xj4kX1tjrAOgkN37bruzpr2c9K+tor1R0olGA/YZHqukfCvV9PoGvZVscGTskBO+Oy9m+SXitBxN05E6SZkhZDsMg+SDV+BPByHRNLFbX25rfDwDsF2Up4I6eJkcbA0NAGAo4qCnhf4kbcFZKAiIgtexsgw4ZCt8GLo8PoHSdsYUip2QdfuYPTDo6CWrpKYEmMnYfJeTnH+evp6yrZNTuaRIR2+ZXuVfbFRakgfS1IBAYW4PqV5Rc9dhsGhr7LHKxgDpSPL1QdD23Wot9R4tRAQ3vkhbd1Neo7rL1RgDB8lyLru86ZqbM0UHh+L0eWM5XDpySSgyo3icCIjsFHUQiB224Kjik8N2VfNL4hBd3QQ53yEVwjcfJULHDyQUU8IHSSfNQK5r3DYIK56JCe+FcZsjturYnN8QF/ZXSFnigt7ILRHK49iphSjGScfIrUPFphT7Y6sLBf4k7wyLt8kEEkYYdiqRxh5xlZE9tqoWeLINlBHHK8FzM7IJ2tbAfE9FDPMJiCBjCtc927XK1u7ggpg+iLKljaI8gb+axUBBuqnGPJUQVIwqJkogIN1VrS47IWuad0EghyMqx7ek90Ejh5qrXdR+IoKNaXHG+Fc6LHYq9zmNb8PfChL3HzQCMKg2VdyqEEIJWSHZuMq2RxOxGFJRlgky/GPmrqwxkjwwAEGMpY5QxvTjKiVW9wgue7JzjYqSGnEjepXSOiMeBjIULZXsOAdkFHNLHEYVpOVnt8KRm2CVizNDDhBG13SUc7qKoiApYjj4gcbKJVDiBhBkeM+f7255I9MqGVgYcBWtcW9kc4u7oKK+J/S4KxXx9OQT5IMiomD4gAMLKoaVkkBcRusNz4y3Awpoqzwoiwefogw529Mrm/NXRSEYbnGdlZK7reXeqlippZcOaNu+UElRTksD27k7q2mqHQOAO261S1UrqpxhduQsW9UBo5QMADsgkrquOanwMZIWkK8l+Bnseys7oCqBlXvhcxvUeyjyUFWnpKv6utwHqVGpGQSPHU0IMk0jPD6tsrEe3pPdXmWVvwEnZRlxPdBJT469+yrUkFwwoQSOyqXE90FFVo6nAeqoqsOHA+iDLmo2xwh+fJYzHluwGfsWTNU+JD0egVlIYw4F+NkFS9zW9XQoXudJl2du61aaSjMYwAT6LSpnDJDexQerHsNh/1N4yH1msA/NXr1LXln7Db/c/jKPITWD9FevUxAREQdc/aH5/aacS8d/c6P8AXqdfPNG/oJJGV9DPtD8Dk14l/wCJUf69Tr54+6CV0wf5YRsZPxZVhjcBnCdbhsgnjmGekj5ZUdQQXZHZRgkHI7hCSTugomUV8UTpHhvqUFpBHkQpI4epvVnb0ysmpphFECe6w2yOagq1nU8t9FV0JbuAThWBxyT5lahTyw+Dh+MkblBgufkYV0cwa3pIyrJMdZ6e2Vagvc3qOfVXNaAO6j6inWUBxyVk09Z4LOnpWKiCeeo8UKNkrmAgFWKrMdQz2QCHHchSsh6m5PoppjE6LEffCxhI9ux8vJBYRgkeiKpyTk+aogqXEqWODraXk9lCrmyOaMBA6fi6VM1vwqEk9wd8rMgkj8Ih2M4QYbm/Ftv5LU6KjZ0eI8gLAaWeISe2VO+re1nTGdu2yCarj8chkYyQVu3hZNJQapgbK3pHUNzt5ra1mnZDUh1X9Ekd1zPoThrXasqW1FljJkLQR0jdB6E8GLE3VmnKOKiqcvyNmu3XenhVpyo05Y20tQ4klo7rpPyO8KtbWSsgkvwl92a8H4gcY+1ehscTImhrBgDZBSWISMLNt9l0K5reYbV3C3VIt9vpp3Uwl6S5rSQBld+CQAT6brrVzQVnBWTSlzdqWGlfc+k9BcBnO/qg83+YrmYi1np8RzSDx3xYLT3zhdGK6rNbPJLjHU4lcgcZZxPrqsbQNLaAPPhgdsZK44d0MeQPVBfKQIwPNWxwhzSc+Stc7qCCRzRgeSCjGdTiD5K5x6D0qwOIyfMqhJPdBNGwS7KssIjIP291C1xb2U7aaomYZDkgb7oELROejsqyUojcADlQNc6Nx6Tg9lcXyuO+coJpIS1o6BkqLxSAWuGD9SzreWtOajGFHcY43zdUGMfJBiwx+K7BKmfC2MFo81jhz4z6IZXnzQP7G/KPk69gFaS532K6HpEo6u2UBhcw53HzUwnLh046vms6rigngaKcDqCU9vLYC543IQadLD0jIH5FRkn70hZEVLLJKWnOPRJqcQSDqxjKCCaDw2h3qroacPYX+ivqJmPj6R3wsdszmjAQXSPH0QOyuigDm9R8lE1jpDt3Vxc+P4CgpI4fR9FaDg5VWMdKcDdVDfDkHX2BQZEbnSABxOFBM0tdjOVm1MkLYQY8B2PJaeSXHKCrfVBlzsBW5VR1D4ggmfTdLcjJOMqxr3MGC0qekmaXYlwR81kVDqbB6cZQYjJA7vsopMB6EkPJZ2zsqFrnblBNFVdHYbq2QiY5Gyi6cd06iDsUFzmdIzlGHPw+qNa+U4Co5ro3YOxCDMhDab4jjJCsmm8Zwwdvksd0jzsc4+aNcR8Q7hBkupGlvVnf9Kxnt8M7FXtfNJ2JUbw4H4+6CmSfMq4OIZ0klUY0kqQ00hGcIIhscrPhujooxGM4xhYToXt8lMygqHs8QDb5oJAGzOL8ZypmwgEE4/IsaB5gd0yDsVm4c+MyM7IO+/sZGtbzQaox/wAAq3+Ubeix/Yuyl/NNqpp8tA1p/wCcbciD2iREQEREBERAREQEREBERAREQcc8x5xy8cUT6aLvZ/5DMvmgk/sr/wDvj+lfS9zJfg7cUz/3FXv9RmXzQu/sp9Oo/pQSxOMmGEbYVs8Ij7FZJMXhDw9nLDk684egox3Q7KrLJ4hB81YqgAoLoY/EfjyUksYheHA5wQVHhzB1DZUc9z8dRygzZbl4kHg47DCwBsrwGY37q3bPyQZVMPHd4Z7eS1yG0QNpi8ObnGVokcZLQYtipBJco4z1F3SBvugxalojqHNHYFa3bdUGhg8ARggjB2WhNbJUzBjQS9xx6reVq4Y3a403vPQ/GM7NQaDUxtuUpnGxK1Ciu0dmpjBgEkYUFbZayyzmGXIDT6LR62XxJMZJIQK2pdUVL5g44ccrvp7PTja3QFdSWutuhYJHABpd2z5LoXSQeK7ftlbh0nqi56U1JSVVJI5oila4jcZGyD6YdHakpNUWSG6UtQ2QSDJIK11dEOTDmgtt103SWe4TgyABuCd84AXeO3XOmucDJ6Z3U1zQ4IMxERAVrt2lXKhGUGwdb6yk0YyarxkdBO/yXjbz+a/qNeX2onDjhkxP14K9juKulZNRUEsUTCT4Z8vkvGnnR0qNL3mqZUsLcyHy+aDpkTJ++LiPnlZNLC2QfFhahcG03u7HxYyRlaOyZ7OxQZFTTtZu1RRwBwJJ7Kx00j+7kY9wIGfPdBeZXN2A7bKx8nVjZZnTH4XU4b4WG/GRhBYr4o+sgeqoY3AZ8lfE/pwR3GUGX9z2dHV1ALDlj8N2BurveZnfC07FXtIDCJPpY2QY4DnbNySfILIpah9LKHPb29VqGn44GVYdVY6D2WbqKmpJ8PoWjA74QYFbfPe6fwRHg4x2WnwVBhaWkd/kkDWNkxINwlUYy4dCCP8AsryT5o5vhuCCN+Opqtd1Z+LOUEzpepgaoCMHCdkzlAVWjLgPVUQHCDLlpGti6wd/RYivM0jh0k7KzGEFzHhhzhZTI2zs6jhYaubK9ow0oEg6XFvorUJySfVSRdHUOrCCwg+hVFmzGAx/CBnCwkFWnBRxyVREDKZREEkMYkdgqs8QjIx5qNr3M3aUc9zzlxygoiqcYQYx5IJqUZf8kq2gOBHmo2h5J6PzK52cYf8AnQRIq7Z+SO7oKKrR1OA9UbjzV+QN/RBLLTNZEHg7rGV7pnuGCdvRWYKAqgZOFRVacFBc6ItGe6sysphDxjChlb0nA9UEazKau8CMswsMghahR2aprYvFjBwBk7IL6G8GjmdIAdzthW3K5SXJwf0nZWMoxFL0SjcHfK1JhoI2AOwD9SDQnPcQGkHZXRsBw7PZZVa6nOfDwsEPcOxQTSz9Q8MD5KDspqbo6z14+1X1EYLh4YG6DGAycLJiqRE3pUYgfjbuoyC04KC+STqJKjO6q1hd2UhppAM4QRsb1HCq9nSqDIPzUkQBI6/zoIsH0KYwtRa6m6dwMrEqCwn4MYQQ7lNx6qeljD379lkVEbGY2HbugwCT55UgjzGXnur5ejG2FD1uDenOxQerfsNj/wBT+Mo9JrB+ivXqYvLP2Gv+5/GX/DWD9FevUxAREQddvaDxGfk64kxNGS6jo9v/AKup188Iy1247HdfRVz4yxQ8pfEKSbHQ2kpM/wAdgXzuTmMyzuYNi8lv1ZQWGbLcKI7oiCrTgrJjgbIzrP1rGAJ7K4SSRjAOAgte3pdj0UkM3hEH0UeHPPqSqujc3uEGTPWeO3owViEYV7W43KY6yANyUFib9lPLSujZ1kHCg7IJXRAMBUSuLyRjJVG4Dt+yC6Nhecb4WQaZgZ1HurZHsawFgGfNQ+NIRjKC12ziqKpB7nzVEBFVuM7qTwXOHU3sUFIBl26rO0NIx2UYJYUc4u7oKKaKEObkjKhVzZHN2BQVMYBKtcMFCXdyqE5QFXB9CqLPpnQeF8WM4QYCqM5zvsrpsGQ47LJpnQBhDwM+SCsMRq3RN7HIH513g5NJ3UmoqKjloxLE4tDiW5Hluukccb5Zofd8/THb616K8mwsNLTQSVbW+9BgwTjOUHq3oyjtNu0zSzUVNFH1RBzukDOcLX6So94j6yMLgXS3ER9io43XSQ+7EgNB7YXM2mNQ0Oo6MVtvIMeB2Qa07dpGcZBC8ifaGjVrNZSQ2185pjMQQ3OMZXrTcJXyRPigPxtBzhdWuYfh1pW/WapqrtEx1XuRkAnO6Dxk1tb3Q0DJqgffiwFxPfOFxcKZ8gc8DsSuznGjhddam/VEFua7wGZ6QBthcIVtgfp5j4K4Yfv3QbL3YSCNwmHHfpU9YzExc3sdwtRofdDSHxMdWEGjIr58eK7p7ZVndAGy1GmuIZGInDvsSsFsTn9lXwJAcY3QZE8AaPFbg532WOJMHJCyYmyM2mz0481DO1r5PvfYoLZJ3OaB2x6JHOWtIOSkkJY0HbPmqNp3ub1BBR7+s7BWYwpoOhrj1jsr5og8/ewEGMDhCd1c6NzO6p0nGUE1LUmB2SSQsx936hgDC0+KF0zsNSSIxO6XbFBmx3IMJcRuoJ6k1T8YwCo2xHAceykZF1kNZgE7ILZacRt6urOQoFlz0NVC3rlBwFjBnW7DcIKxSGM5AySksniOzjCngg6Tl+MKOpa0OywbIKQS+E7qWosoo6yEz9WCBnC0ojACkZVTRs6GvICCyRpY4sznBwqA4T4nnPclTCLDCXDceSCBXB2AQQr4WguII7K2YAO2QWZ80yfVEQVa4jsMrIaMtyRg+irQMa95D+yrWkRu6Wn8iDHe7qPSN8Kzssuiaxzj1AKysa1smGgYQWQTeESfVZZiZUMMud+6wDjAx3UsMsjSBv0oLXB7nFoB9OyoWuZ3aVq0Puz252yVDUsj3AAzlBiU0nScFWVDg56yGRhpJOCsafHVsgtjeGLKjrNunH2rEa0u7KdjWsaesYKDNjjZKOo4V77k2nb4AGfmsAVJbswqGV5e7qPdBWZ/iyF+O6ngrXRx+Fj5LHjLQd1mxxRuZ1DfZB329i7GBzSapf66Brv5RtyK72Lzs80mqW+mgK3+UbciD2gREQEREBERAREQEREBERAREQcccyX4O3FL+BV8/UZl80Ls+I7bfJX0wcxxA5euKBPYaMvf6jMvmppohNVytHYdRH5UGNASHqSpD3HZhx9SmbGyCb4sYC1eKOmngLhjIQba7JupapobM8DsFLTtYWfF3QQeJtjCsUrITLJ0t7Ks1O6N4bjcoIVPDTmQdSklopI4g/HzV1HMyNhB7lAZKKY4+a3dp+y3O/gQ0dC6UOHcDK2XUtOz/InZeg3Ifwis2taCH36JjpXsBGQO+EHBvCLlru18ubJ57Y4/ENi0nG67zWDlW9w0W+odZm+IIc46PPC7M8NuWel05cDWSRMEJdkbDsud/wBjNrbQmh8IeEW9JGPJB4DcZOGeo6XVtRRU1jkDATuGHHdcPXrQN7oS99Tb3s6TvluML331Hy88P9QXWWeSCIykEn4R3XSDmv4U6R0XBVthja0gnGGgIPNG2UbmVPgzM6cHHbC1KWzia5RxxAEnbbyWrX21irvBjtY7uxsPmt26T4ZXuoqWVMzXEAZCDnrlX0tcLXUwVfvjwOsHAcfVeuvB+qdUWGAPlLiIh558l5g8AbNVQVEdIO4eBhekfCf3u3UVNFJ0gFgG5+SDl5Fi++xtZ1yEDfHdTxvEjA8bgjKC9FhsuDDVGky3rB9Vln5IMWtq6Wla51Q4ABhzn0XkX7Raz2+93CqqKVzRiQnbHqvT7ifDdfcnvoSd4z2+peT/ADZ2bUlfW1bKkyEGQ+p80HQm6wyU8raduSGjGy09zHN7tIXId10u2xPFbdQ8MxnJC2vdK611bwKNhH1jCDQwd84TO+cLMqYAyMPHp5rDGM7oJHzFzejyVjPpD61fDH4jy0foV9RC6IgkIMueNgp9sZK00HCvM0jh0k5CviYMfEO/ZBSmID8laiaRkzfEBwQtLcelxwp46qQMLQdkCecj70zIwcLVLUWmmJkdn5LQ3HLic7kqSOoljHS04BQVqh/th+M9/RIoeodTjjCyYvDA6pfPdQ1MzeoCI7BBZ43R8IGcKxz/ABHAdkZ9LLle6PqIMe4HkgPgDWdWd1CpGyHPS7sqSYJGCgsVWjJwqKWOJxId5BBe+mLGdY3WOSfNaiyZgaWPxttusSpLC4dHZBCqsGXAHzV0eAclHkZy075QTSUwZH1g7+igYwOGc4VTK9w6SdlaHEbIKHZEWRDSulYXAZ+xBA1vUqvaGnCO6mOLeytJJ7oCJg+iAEoA3WbBRskj6yRkLCwQpG1ErB0tdsgpKzokLQc4ViqXFxJPcqiCeBwYdwSrah4e7IGFbEQHZd2SQgnZBYqt3ICrGzrOMKrmGN4z6oJH0xYzr+We6gWXNOx0XSO5CxEBXxnfCsVQ13cBBlClD25HcqCWIR9iqxzOad3FXuY+dwDTlBCx5YchVy6R4Jb3Kym0bom9cvbY7qZr6cMLhjIQQVMDWwtcDuFqtmvjKKldE5oJx+RaLJI+d/Q3OFGWOicA4IMqsrXT1BkDcBxUBY6QdfUpJGx+F1YGcKCNzgQM7BBaWu8wdlTGFqEboA34sZ81BNGJXZjxhBjDPksqBwJAcopGFrR6hRhzgdkGo1BbCwEb5UlLb462B07jg91htf1tHiHIUgrXwDw4zgFBFG3wqgx+QKnqakN+ABWeNGT17Fx7qB7XTO23QVp4/GlWVUUha0uGBhRwQupyJHBZD6lssZGUGlnYkKivdjxCT2VrsdXyQZFER1rIqmBzcg9lih7Wj4e6Cdx2JJCCEjcqinaWZyVbMG74QerHsNjm3cZB6TWD9FevUteWnsNv9zuMv+GsH6K9epaAiIg65e0Qz+0z4l9Jwfc6PB/+rqdfPIvoe9oW3q5N+JLR/edH+vU6+eJrS7sOyA1vUVcYwOyozY7+Sq95JQVgGX7qtQMEKIEjsq9RcR1boL4XBrslSSvBKoHMA+eFE92TsUFXSZGFSM9MgPoVaBkqo+F31INSqp+uAABaaBvhZfjxlmDj7VjPI6stKC4xBrerKiJycqUyZGMqI4zsgINldtjyyrUGfb6dlW4sccD5qO40jKSUNYcgqCGeSA5jdgpLPJOcvOSgjWVFMBGRj5LFVzN3AepQHZLicH8ioc+a1KNkLG5eBlYtWY3HMeMIMZTxQB7OslQK9kj24aDsfJAkOD0jyViy2sDR1vx2UE3T1ZadkEaZREF8cfiHCnjoy6Tp8vrUVOQHbrWKWmdM17mZyG52Qb/4XaDkv07GwQ+KeobDddseEmmdRaa1XSQGhkjhyAT0kDGVtnkD4dT6x1FFBO3LTKBv9a9T7lwA01aaJlYyJnjsYBkNHcBBtis0Y3Umh6UQS4mawEgd1rHDvVEWgaU2atqx1HAAcVoltv8ANpGslprm4imaCGg9sLiDiLqO43jUrayzOJga7JIPllB25h1cZM1cDusSb7b91tPiBpal1NZJa2peGOIJwThaTwa1PS11AyluHQXsaM5d54W0eNfFptturdO26QDxHdIAPqg6ecdnU+lrjVMipWvABGQAV0d4i1Ul8nkqWxdADjtj5r0V4z8PLhcrY691rSWSs6skd8hdLtc6BdHRz1EEeGgk9kHXGpc7r8Mt7bKxjntBGSMrV6uCKO5Pp3DcHC0+vYI5OlowEGIfP60GyuAxufNTinc9nWBkAILI5+gnbKmZUBxDsdliOGCR6KgJQZlTUGVnS0fkWNG4scCQe6zbXC2oeQ4dlk11HHD2G4QafNMC0DGMrUqJkTqU9WMrSZW5OB5IyoliaWNOAgpUNDZnAdlWObwx0q0MfKc5ySqGMg4QVkk6/JXMJ6ekDuFY1vxBp81qMFLCWZLhlBDRAxOyCMqWpp3yyB58ionh8c2WAkD0HzWRJO7ABGAfNBC9nU0M7KaipxDKJHEkDy9FPHFGYvE2yN85WP44JLR2CDU7pVR1VN4QAHzHdaPHTNiy4OB2V+XO+kSR5ZVJGAAfGUFpc52QQAMDdY8w6D6jCzYyzsfJUlijd5glBppOVMync9hd6BTGmYBnIWTTlrgIhuTsEGmNJY7GFlhrXxlyrXWuamaZndjusLxXgY6tkFcljiPsVrndRVDuiAASdt1I2F5BJBAUtF0B+ZMY+a1CeamEZDcINMp5PCcc7FUqHdbupWOPxkjzJVCSe6C6J/hk5SV/W7KsV7QMb+iCxXh3wYwq9HWR0qcRBsZ6hughhkLHZBKukmL35O31KyPpDvi9Ul6SdkEz5R0gDIysZxyVQnKuBGN+6BG/oOVVzvFcBnurPqVQHDceSCWSEMGc+ShVxe53cq1BLDF4pIzhSumMA8MHKxwXsORsp2M8SPrO5Qd9/YsOL+abVOc//ECu/lG3InsVhjmo1UP/AOH9d/KNuRB7UIiICIiAiIgIiICIiAiIgIiIOO+Ytok5fOJzD2do29D/AJDKvmtpCIK+obnIaXAfPBX0o8xxLeXrig5vcaMvZ/5DMvmmZKWVUjndyTn8qDJmjEzidgDuoxUSUoMbckFRz1I6cMO6kpHxOaTNglBhSPMjy49ypIy4AjCvzGJj6ZIU0j4eghuAgxopnRSFzRkrK8TxyHvGCN+yioBHJUYeMgrIuRjiAbFgZCCOor+uLwgAcbZWC0EOA7KaCMF4e8ZCrVFnWOgIM73dktM1pwCCD9i7b8rnH6DhX4LjUdAiaOx9AunjKl5Z0tzlajS3aqpYy0ZOQUHufy486lm4pXOOxzVTA7rDcnGfRdt/HppW9InYQ4eThuCvnT5bOLlboHVDK7rLQJA7PUR5r1A0DziU10ipBLUMJLWg5f8AIIO7JtVroah1bUTNaD/dHC4N5hOAGjeJmm6+5+/RdbIy4DqG5XGvGHmtgo9OOlop2B4Yezvkuml5509a1UFXb6eueInkjHUeyDhriDoml0Hr6WhpwHxxyEDG+wK3jpjXFujlipJmMYDgE4xhccar1828XF90ucgdK/Lsk+eVsK5axfLUf9TXfHnbBQd04te2jQdCL3TVTQ/6WAftWsaU9oA5tzioX15Y1rg3vjZdG5LxrCvpg2udIYMbZJxhbJu1S+gqxLTtLXg5Jz5oPaKLm4tNXp5tYb0A8t6sdfyWx672jMGmuu3MuIfnIBOCR9S8pWcTtVw0wpRVOEYGMEnssAXu4XVxlqAXOByTk7oPYfhVzlUOqrz77V3jAec/E/5re+tOdK22e+0tuproCyRwBwQvFSz8R9QaZm/2k9zMYx8RC3XRcUb3qCdlXVF5lYMg9RO6D3ssnFvSuq9LRXCW6ROkkZ9EuHchcR6v4H23ifW+LTUjJ2PdnOARgleavC7jFxHq5IrZTSzCFrw3Zxxheq3K3qWc6cjF8lAndED8XfOEHC/Ej2eFt1PZXQw0UQkEZ2bjOcLzj488qlz4R180cdK9zY3ludyBuvf2S9UskLhDIOog4yV0y5p+H1t1BbqyuqsOIJd2CDxhuNpniibHJCWkhbbqIHQOLSMYXOnFKittovslIwABuQPyrhu8mKWd3g7nq8kGBC8Q4k9dlWebxyB81Y+nkY3qI2UfZBlNpwG9Rwfko3v6cAA7K0zOxgfpVGMdJkjyQVhZ4r8bblZbqZkYysWnyJNtipKmSQEDKC0QgkjON+ylFO1oJ2z3CxfEcPPdXB8rvUoJQ3xT0Zwo5ovCIGcqhL2bjbdVD/EBLjuEEfV6K+OXoGFY7AJwrmdPmgtOS4kZ3VPrWXD4ZOHKOoDMgNQQLLgcBF33WKWkDOFUdeNs4QVlOZDg90azqBOUY0yOx3Kuex0QwRjKCI7HHoid0QERVb9IIKYWVBWeCwtwSrJCws+Huo2BuDnugpI7reXYxlW4PopaeIyPwM7LKkiZCMOxlBhHsqA4UoLeoq18fVuzcILCc7AKmFLG3od8ewVJywu+FBGiJ3QEUvu8mOrGyjIIO4QXRyeGc4VXyeI4bY3UaAEnZBM+HpZ1dWdlZFGHuwSrnMmDep2cKNri07FBkS0zWN6gfJQB+BjCu65JT05yrHNLDhwQUJ3yFNBOYiCRlQ4V8YHV8XZBmTV5qWeHjG2Oyw8ODunJGVkSGEN+DGVjtI6uo7lBktYKZvi9yVE94neDjCva2SqIjYo5IZKaQB+yCSZvTEN/JWQxBzckq6R4ljDWndWNLomnJwgr7u55w0kqpLqfDXBTWydjJiZTkLNuEUVS0vhAP1INLBM7i3B3VJIDGMnZS0TmwznxNsK64zNkeOg7eaDFL9sA4VmSVVrS84AWQ2nc1pcRj5IMZSw1BjO4yo3DDiFJGwOG6DIdV+OAzGFNDSNMZOe4WCYXA/BkrIiqXxsLXdwgiqIBG44Kx8H0V8krnk5WVTmHw8OxkoMQtICt+pSSscHE+SvgLBjqQRN6wcgFUfnO+VntNOBusSoLS/LeyD1X9hrn7n8Zf8NYP0V69TF5b+w4A+5HGI+fvFi/6NcvUhAREQddfaFv8Pk34kv9KOj/AF6nXzyRuDSc+YX0M+0O35NeJef7yo/16nXzx4QVO5JVERBLTxCVxDjhKiEROAHmo2vcw5COe5+7iguiZ1kDyV0sQZ2KjaXA/Cr/AIi4dfZBqOn7fHX1JZLgAeqrqG3xUFQI4iCCoG1jqRvVTnDvl5LGqaueqd1TOJKCyKPxHYJVXxhsoZ5FWs6s/CN0eXh2Xd0GRNTNjj6gdyrIoA9pJKjMr3DpLtlRsjmjAQVkZ0HCswfQq9rskdR2ystroOnfGUGE1uSjhhXyOAcS0qzBdugoro/pj61aqtaSchBqT4WyR4DuwWnyR+GcZyrvFmZtkhRueXHJQUVRsQfRVjx1bq+Tp8igPm629O4USK9oBCCxVaMuAVXNxvjzVoJByO6DKNN0hrmnc481vfSVuZUghwBJbgBbIpZ8PAeRgdlvnR8kjK+KoB+9AjPphB3O5ONbR8MLtBVzxhg8QHJ+teomkeJdu4gRQkSDoIyfTBXi7/snWOyW6COBzPGaRnB3XbHgZzU2WzaVMk0rBKyPAy7zwg7z8TuG9o1DbZZKGoaJGNJwDv2Xn7xe4nQ8J7nLaJ6jDustGfrXIWi+eSgmvdZTXKrYYTloy/bC6Lc6fE+n1trP3u0SAxmQuy05Qdh9Oc0LrMRVR3ItEm+Mq/8A2TKPWup4L5U3cEseDgu+eV0Tg1RUx0EYlBy1g8/NYlt17qWlujJKOV4iDh8IzjHrlB6ecUOL0N10gy1UDRKWR9OQM52XU7VV4vD7LURyW14Ds7lpW7eCfEiy3Slhg1D0FxwD1H/WuwGqNOaB1NpOWntMcBnfGcdOM5IQeV94iqPu5LKyP4i4khaXW+MZSZmlpz5hdlq7l5u9BqqruNXG8UpcSMjbC4a4pWamtFz8GAYw4goNjOdkALIjqzHH0YUL4nMaHO7FSwMZIwg9wggcepxPqpY4A9nVnBVronRu+IbeqoZHDZpwEGRRVLqaQgAndZFXWGcb7ZUdubE9x6wCc7qlxYGyfe+3yKDF8T4jkKRsPiMLwFA7H2rMpziHtnPZBjsk8MkHyVQXSPVHtcXZIxlZEbGsZ1ZBICCGWJzCHHO6jMsgJHUVM9759u2FA6NzScoNQpauEMaZRkhWTVEbiMYA+SwBnOyvwUGaXvbCSHHHmoqdpkkAafrVsbZ5WSdDHvbG3qeWgkNGQMn0GSBk+ZA81ax5jf1DIIQZj3CPEZzkrHnEjTlxOCsqmDJ8ueACPNQ1UgDjHgEIIBO9ux33VSZnDqycK3IBBIz8lkGqb4fQGAeWUEDS493FahQtIcH+nqFiU7GuJLnYAUxn8P4W+XmgzrlXe8RCAgYG2VoMrAx2AswuDwSTgrCkB6jkoKdO3dU81UE+SogvcMDZWZPqq5JV3hPxnCCwZG+FMyMO7q6ljD3EY3VJ8xvw3YfUgqKcYO6iI6XBoTqkbuSd0Zl0gJ9UEmDAASO6zomRzQdZOCFj1TR4QP2/mWOyokYOkO2QWyNw9wHkSrWtJcAspjonD4u/mo3lgkBb2BQJ6cRNDldFStfH1k7hJ5hIzA8lHDM5jgCds9kEbmlpIPksqKJpiLjjKVDo3M+HvhYwlcG9OdkF8MQlkIJwFkvomMHUCDjdYbXuacg4yrjUSuGC5BSR2ct9FfHP0NDMKE7q8QvIzjZB3+9iyz/rptUuz30BXfyjbkT2K7j+2l1SD5aArv5RtyIPadERAREQEREBERAREQEREBERBx3zG/g9cUP4GXv9RmXzPzD7/IB36j+lfTBzGEN5e+J5Pb9ht7P/ACGVfNG4B9XL6dTiPyoDKUuYHE4yontLDsdvrWXLOGs6WncLD+KQ47oLcq5jepwBOyuMD2jqI2Vjc5wPNBnRU7Ym+IHb/WoZ+qV3fOCqdNQGdTielT07BI3qGMnKDHdIBH0+YVYaYzMLsknyCsnjLXdjhTUU/hPDT2J3ygg6ZIXZLDt8le6pcduy12qdQOpsgDqI3W3pA3rOOxKDUbSZXS/BKWfPst9WviJdNMuYGXST4e3xFcfCpEMf3vYqHonqyXncDzQcuXnjPddQ0Ro5bnKQR04yVtiKsmjifI6qJJye62hE1jD0sb8X1qd1TURs6HE4KC673ieqe6LxSQD3B7rDoKx1HVMnySAclY793k+qoMZ3QchScQmy28UobvjGcLbNRK2uc6Rx3zkLT6c047gZSpnIcBFsEF1XIHjwgO3nhUpa73RpYN1Y+RrmfP6ljxjqlAdvkoMqd0lWerGPPss601pt0wy/86mqKaNlAHxgA48losYkdKHnJwRklBzlw64hu07Kypxn4gR+VegHLzxt+7klJA6rMYwAd8LzDs12t8NOxkoBIIzldiuC3EOntdbBHS4BGBsUHsPYrjHV2sVFPUeI8jtnOdltrXGk2aksNRDVRgOeCBnzytjcruqqvU7mQVTsxkgDJyudOIGnK6o8FtqBaw4zhB5L8y/L0+2XSe4sgw3dwK6i1ukGwSTFwGWE+a9kOZzhFc7npUzRNPi+EcnBznC81tR8AdYQw1td0S+GxxPYoOuV0Y2PMIb22WlNps5yey1/UdMbbc32+oz4rT0nK0aRj2OwPP5oIGwEkgkK8ubT/CBnPmrnQygdQysaTq6vi7oJqUgzZPYqtb09Q6Ssdri3cHdXta+bfOcILACTsMqQSdAwW7qWJrWH48Z+ajqC1xBYgse8O8la0OPYKrY3OGQsmIMjaQ/vhBiHuivd0+IfrVrsZ2QUVcnO+6omD3QZD2jw1E14aCMK0vJGCqIL4pPDd1Y7q6WXx3AYxkqJOyDLlpGsiEgIysRXmWV46S4kKwgg4KAiIgIr48B2+ElLSfhwgugm8F3Vj8yuqJzPg9sKBAcFAUjJukYIyqEA7DurmdIHxAZQVb9/OMYVs0XhkDOcq/rawktwCrHl0hH6EFrG9RwrzGGkHKtAczfsqF5O2cj5oMj3v4enp27LHe/rOcYVqlbA4gkeSCJVacEFUIIOCq4KCd9SHx9HThQEE+SpjCyIjH4Z6sZwgiikEZyQslsLahviHZYpb1OOMYUrJHxNIzhBE4dDyPQ4VO5AOyuYQXku80kxkFqC50PSOrKiV5e5zcE9lZjCDIo6oU0nWRlZE7xXuD8Yx6rT1k07iG/CcFBNDShjs5Bwoaz6XbGCpYnyNcS7OCoKqQPfsggysynrfBiLDk+iwxjO6u6CdwEFXHxHlw2zurS0g4yqDIO3dDk90EsZ8PcjKkdVlzSMLH+I+uEA3GRsgOPUSfUrJpGNe4Nd2OyjeY+gAYyrIpHMcCCg1apgZBF1jc+i0mSTrOQMLImqpJI+lx2CxmMc87IKNb1FXhhDh6Z9UMT4vicNlfHTyTHqaMj1QZLIhKzpKxqinMJ2O31rKY/wB8XdRTzNk322QYeT6opXAPwGYVcBsTgcdX/tQeq/sNj/ANTeMgz2msH/AEa9epa8s/YbA/c7jK4+c1gA+wV/+tepiAiIg65+0O/A04l/4nR/r1OvnjHdfQ57Q78DTiX/AInR/r1OvnjxlAREwUBExhEFWnBVxkBGwwrEQM5U8dOHs6sqDB9FLHI7HQPs3QUY8RPPmryBMerGMqx8L2guIVjXOb2OEF74g0d1nU9ujlgMpO4WAZC7uSpY5akMwwnpQQyN6JC0eRVWRlwznAVrslx6u/mrg5zANyEFpbg4yqg9Oyqw5d8RSQZ3CC3IysmJgazPmsXspYnPJwScFAe8SHpxhWPb0nvlZErOluRjKxiSe6CiIiCSGPxCQfIKsjBE7AOVG17mnLThSCKWUdeMhBYX5HZWoRgkeiqGOPYILoovEOM4Wu2+/Ot0YpQTvsT6LQo3ujcqPdl3V5+aDVrq6eUCcVRIO+MlT2zVl2oKV1NFcJWA7AAlaN4k0zRHnICiewsOCg1FmoLzFOZ47jKHE5J6ik91qq+QSVc7pXDzO5Wmqrc52QZ1TXOfGI2uI+Sz7VWNbHhwyfVaNMwgA4WVSzMEJa36WEG6LfqOqtMrXwVjm4OwBXNnCjjjcqe8U9LU1L3xggbk4XXO2Uc9RUDxAS0uwuXOHul4qq6xRQR/fTjpI9UHdy+6no9QaWa6kpGvlfHjIAznC6m8TOGctx8WulgDXhxcAdiu0nD+xHSNqjqtTNIgcBjq7YWoas4HXLiXSG86XY80QHU7obkY+xB50jSE9TVGkkg6WsyAcYWkXmyNs0nQ0jv6rsxxO0RDoqaSmkaROwEHbByuBbjpy5X575YuogHOUGzZYzMMNwVgyRmN3TkFb3j0JcG9LXtcC44GAt1Wzlp1teqE3WiheYAOonoJ2QcPxyPiPUMgFSOndKQCNlvW7cNLhb5TQOaRLGcHbzWkT6Sq6CIyTtOR54QaBJB0M6sqSnkDR8ezQqTNkfKYRnY4xlWtjcSYnbEbIMuofBVNDIQOpYpilhcA7OB32U0ETqZ3ikElRVVWZj04wEEhljcwBmMg74Vs0kXhlp+kuauX7kv5ieY6aKr4daCqRZJHlj79cyaS3MwQCRK4ZlIJGWxB7h6L0p4A+x34O6HdBfeON/qdfXVmH/c6n6qK1xO9CGnxZsHGCXMadwWEFB5PcJeB3FvjlfP2PcKNAXfUlWCBKaSH7xTgkAOmmcRHEMkbvcBv3XoxwB9jG54pb7zIa5DRtI7T+nX5Pr0S1bxgeQLY2HzxJ2K9OtLaS0toeyU+mtGactlitNI3pgobdSR00EY7fCyMADsPJbQ5huNWneXng7qXi1qUh8NkpC6mpi7DqurfhkEDfPL5HNBI7N6nHYEoPK/2l2p+D/BintPKDy/aRtOnLdRCK8askoG5mqpyM0tNUTOJllLW4mcJHOBLoCMFgx5+Ng8aTbYFanrTWOouImsLzrzVtxdXXm/101wrqh23iTSvLnEDs0AnAaNgAAAAAFpbKsxDAQXzxvgA6SQCsYkuOTusmSbxxglYxxnAQUzlBssiGFjxk/nUUjA12G7oL443uGWnAVHZaSDuVq0GmNWsaC3TF2LSMgiikOR6/RWrt4ScVpiHf7GWqyDuD9xanBH/AJiDaPUVaIw87rUq+01lqrKi3XOjnpKyklfBUU88ZjlilYS1zHtcAWuBBBBAIIIIyFp7hhxAQROb4ZON1Zu45OyyGxlpzJ5qNwD5A1h7nCCMHBypfH+EjpUtRbZ6eMSvBwRlWQ+F0/EBlBZFMYnF2O6Of40oKTFpPwgd1axjiR090GVLE10YIxkBYo+9u+orJ6ZWD4jssaU9TtjlBdJOZG4OVF3VS0gAnzU9PEC4FwGPNBAeod0AycArNqzD4eGYBWNHA92HY2QUMbmjq75UZJ81miSPp6XdxtusWUjq+Hsgs7qvTtlSQ9Gfix3Usgb0/CBjzQRU8IlJBKTxCJ/SN0gL+s9Bwk/UXfEclBEp2VAa3oxlQIBlB6A+xYb/ANdLql2e+gK7+UbciexXbIOaXVJPnoCu/lG3Ig9p0REBERAREQEREBERAREQEREHHPMeM8vHFIeui72P+QzL5os9M7/rI/Ovpd5kATy8cUQO50Xex/yGZfNI2CSaeRre7SSfyoIXnLir4nBpBKueGtHScdQUIJCDLmmBjxhRU7MnqJVgDndky+PYdj6IMies8SPwwFZTTPjeG4OCooQ0vAK1VnuzYsux1D9KBVQRug68jJWkEEE48lNNUyvcWNcS3PZTw0hMfW4bkZQYXW8jHUcKivlADyB5KSNrek5HkggWXT1vgxlgb32yscsLnYaMqjmOYcHYoJI5cTmTyO5UtTU+I0AD5LH8N4HVjZVa5u/V3wgjJyq4J9Vd0OeT0bhZULGRtPiDfGyDD6SBlTwNDwerPdWSMd1E42Vpf07M2QZwpGkZ6u6xJWCKQFvYFBPM74Qd8KyQvP00GW+5PfF4QBwFA2oLQdu6tgYHO+LssiSBpb8IHZBiF7z++PquSeG1zno6qOpdOWhm+crjprWNeA7H2rc1oukVJCQwjOMYQel3Jtxtt1Fc4qSuuQjAkA+J2F6bWvU9gu1LFPS3OnkD2Bw+MZ7L5utJa/1DYK/x7U94cHA5BK7WcMub3W9ndBFcqqQMYAPpFB66cU5bVVaefG4xyuIOACDthdUtV262nS1xhdbWAvDgPgA9VtPQvNhBqmIUtfO1xOBhzluLWGs7bX2SVtK5n3xudig8tePukPufrCprYouhpeTjt5riB1QXvcQOy7TcxlA2WtmqdsHJ2XWihtrap8rGtJJccIMOKsL/AIC1W1EIf8Y2wtSl0zV0mZng9I3GVplVLh3SNsd0GMGFzukDdZDXGAYLSpaGSEHL8faldJC7ZmPsQYcknWTjYeisGyqGknATpLSA4IJI5+gYwrZHmRwIH1BXyRdLOpVpDEJAZMYQQ9DvMEKhGDhaxWPpDTkRgdWPJaOST3QFkQRtczJWOpIxK76OcILXtPWQN1b2WXF0A5eBnG+VG8NM4Ix0jB+xBJQUclRKB4ZI9VqH7HLxPVshpKCSXrIA6W5C3tw/03Fq2RlBQDM2QD0913b4FcvNmoq6km1LC0g4PxD/AFoOit04X6tt9pFcbHMBjqJ6Cth1FNVQOIqIXsI2PU3GF71XvgdwtvulTRUVPAZRERgAZzheXHNbwWg0Zdpxbog1niHGBjbKDqthFqtdbHUkLS4b4WlYygIgBKEEICIrxgt+eEF0DQ526VDQHAjsrPiZnGQq9Yd9IoLFe1/R5IyN0rulgypfBMQIeN0FhcHjthRuGFcY3t3A2VhJPdBk0MLZpelxwFkVpZTHoacrEhbKXfedj8ldPBUjeXJ+tBcyFrwH777q50LPqUEUpZseyrJM5xHSSgvkiaG5BWMRg4UvhzP33wfmrHMeNyEBj+go9/UUjjdI7pb3VXwvj+kEFiDdFUAgjZBMGYG6ifklSPlHQAO6vhaHRlxxkIMctIx81n0dMXR9Z/OFj0zQ+bpcNgtUfJHDGWgjceSDTppSXmJo+pYr2lpw7utQpDE6cueB3Udy8PxB0AYz5IMJXsk6RjCsRBXO+VWMdUgHqVaBlXiOQfEB23Qat7hCIQ/IzhaXUNDXYHZVNXP09HVsonOc4/EgoRhEyVVrHO7BAALtlJG/wiMhXQNDHZd2VtQWlwLUGTJI2aMAdwqRVYpW9IGSoaZ+HYPZKnpc7bugrUTeKOoeagHp6q9sEjhnCtLC1wae6C9oEfxEFWyO6zkBSSwSNYHnsoQR0n1QervsNnZtnGRn9zPYD+Vtd/qXqUvLP2Go/wCp/GU//RrB+ivXqYgIiIOuftDvwNOJf+J0f69Tr54wcHK+hz2h34GnEv8AxOj/AF6nXzx4ygK5h3DfmrVIzpAyfJBLLC0M6gd8LGV7pHnIJ2VY4XvwQNkEeD6IN1kyiMNxgZWN9SC8j4cq1julwKbqiCeSp62BmFArg3JwFVrC05cNkFmD6KeOoLG9OPkkvhhmGgAqsXhdHxDdBGwCR+eyTNAKpgl5EarvnD+/kgsaMlXF3Tt3QxubuOytyD3QUO6va8jsFYe6nYI/DJONkB82RgZKhdnO4UkLow89fYqtT0F3wDZBCirj4fmqIK9PzWXDVCKPoPmsPJTOUFXnqeT6qeIEx+p8ljrMp3sY3pfsgxcO6ux7qhB6sFagyOJx2wdljTROEufLOyCrJDE3JHceiikf4hJU03R4QAxkKOmpZKh4Y0dyghwqt7g/NazWafqIIBJ0481p4t8xYX42CCF0pcOnHyWp26hY6MyPcAfTKxrbbpK2boAOxws6qsdzpN2ZDO6CVlwdSSRsiZnL8HC7a8q+nLVfNRUlRcZWNPU0kOPzXVGxR0/ihtY0Et33XIGjuJF10nqKJ9rlLY4yD8Jxsg9guJXB6x6s0DSQWuSMPa0fRI3/ACLlbly0XTaV0SdPVVK17i3BLmg5GMY3XVjlZ4oag4pW+ktYmLyCAcld3LbcKPR7I7bcntEr2jzGcoOoXNpyvUNY2p1BT0rA2UF46QO/oug1z0fV6ZiqIae3mQtJAw3OV69cxV3hq9Jtjp5NntJP2rqbo3hZbdRV7pbhGHRl+Tkd90HWvgJwtufES8sp7jZiyMSDdzMDuu/Fdwst+geHUtHTWSOSQwkAhgJzhb10Bwk0pYqWKSwxMZOMdgFzFT6YprlbfdbszxARjBHlhB4ccTLbf6PW1bL9wntiLyR8G2N1wvrO53PpfG+gLBkg/D2XuNxV5V9I3yOeuoqRole0n6Izlef3HXl5t1hinhZBiXqIGyDz2aZBUGQs374UL53Ccue3G+4wuTbxwyrbRdZHzMIiBJG3ktk6ho4qV58No2JGEHaPl69m3zHcwVDQ6imssGitKVzGTxXe+gsfUQuwQ6CmbmWQEEOa5wYxwIIeV6R8v3suuWbgmKa76hsjuIepIcOdX6hja+ljf5mKi3iaMgEeJ4jwezgvGvg5zOceeAdU2fhTxOvVkpw/rfb2zeNQSnOSX00odE499+nIycEZXfTgn7au6UvgWrmB4YMrYwA1940y8RyjGd3UkzuhxORktlYBg4acgAPV2GGKniZT08TY4o2hjGMbhrGjYAAbAADGB2Uq4Z4Lc3/LpzAMhi4Z8ULTWXKYA/cireaS4A75Ap5Q178YOSwOb23wQTzMgLZnE3hBw04zWan07xS0hQ6ktdLUirho60OdE2YNLRJ0ggFwDnAE9g447lbzXE3M/eeNWmODV+1bwCjtlTqyxRG4R0FwpDUR11PGCZomNa9pEnRlzME5LenBLgQGmN5KeUhpBHLnoHI33skJH/RWr/tT+Vn/APVq4V/6G27+hXkNL7XznEfI97bhpCNriXBjbGMNB8hl5OB8yT8ytOb7V7nSqqpzv2f2aFj3EhjNP0fSwE5ABcwnA7DJJ9STug9of2v3Aby4JaB/0bo/6Nat/sWcMfxc6X/zRT/+ovD27e0954GSOmpuNjIWvcSI49N2ktaCew6qYnA+ZJ+a21P7SPnaqZpJ5OPVzDpCXEMtlAxoz6NbAAB8gAEHvx+xHSn/AAXtH8Sj/wDVWqtDWgNaAAOwxgBfPBL7QLnLlkfK7mB1Nl5LiGmFoBPoAwAD5AADyWl1HO/zd1U755eYnXQc89REd2kjaD8mtIAHyAAQfRqi+biTm75p55Xzy8yHE/rkPUQzVlexoPya2UAD5AALR6jmL5gK2pfV1XHPiDNLIcvkk1NWuc4+pJlyT9aDc3N9SSWzmn4vU8kgeX63vVQC0YAEtZLIBv5gPAPzC4ZLgXZCy7teLpfbhUXa9XGqr66rkMtRVVUzpZZnk5L3vcSXEnckkkqxtPG6IuyMoIZ5A9gDRvhY8TjFI2THYqaDpbIeoDAKlndAWnpwgnrbt7zAIQ3cDC05sZ6SSqxdIcSewKSvGS1vbKCIg5PyUsEnQ8ZCkp4w/uFZUNDHY8wgzjIydmM4WnzM8N+26tErx2Ko5znn4jlBVz+oAeivZMWtIAUX1rJb4Ij3xugxic91ksq+iMsx9Sx3YySOyADCChOST6oid0BXB/w4yq+G4DLgrThBfFL4Z7KksniHOFYqsx1DKCgBVzQQc4yFLN0dI6MZ2V8To/Cw7vhB359ixN180eqG4xjQFcf+cbcis9itj9tRqrHb/Y/rv5RtyIPahERAREQEREBERAREQEREBERBx1zHEN5euKDj5aMvf6jMvmoiqhBUzPxkPJH5SvpV5kvwduKX8Cr3+ozL5oiWiVxPqUEs0YcTLnvusZXkue4hvb0Qxua4A+ZQXQk9QyNlfUgFw6f/ALKmfT9EQeO/dY8bx2f3QDGGNDs7qzre4gFxGVe/qePh3V0cDhuRugyYKVhZ1kjJUvjsDhF5HZYL/Hi7ZAwomyOMgcTkhBrE1nYIvHHnutJlBaSBtg42Wre/k04YT5LC6oyTnG6DEY90burCrLL4jg7GFPOYi09OMrEQSmfqZ0YUsFL4zS5YqzKaoDIy1BAHeBIRjOFSWQyOyAQFbI7qeTnup4XRhhDsZwgiEmW9BUrKYObnO/1qENL5DjClc90RAPYoImkxv7dikr+vBwsiaSIsGME4WId0FzXluyy4Jg4YJ7n0WEqhxachBPUMH0m+vZZFDCXRlxeQsaCQF+JDt81kSPc0jwu3mgyKO8Pt05wM481uCn1DNcQCKgsI/Mtq1BhMe+C/1WPE6dgJiyAg5U0rrau09UiVtyeADn6RXLls5g6yVjKWSte4YA+kV1TZVzl46iT8luW2VsUUPW4AOAQdib3W0OuYPvswJfscn1U2n+BlrbQPrx0E4Jx6rr/ZdY3OCsDIXENBGN9u6570drW61FCKd0pIcMHdBxlxKpRapX0EMOwBGQuIHU0sj3ue0jf0XZPWVoirC+qqAC45K4Sv7aSlqXMj/ukG0pIHxkktIHkkMJlI7lazcKdklKHsxkjusKgmihBY/wCkeyCF8DYR1ZyfrUQjlmflrCQFlTU8xcJDno7rXbL7hMwU7gPEfsM+qDSIqZs0Ya/Y4wsapoDFks3wtx3rSdxtcP3RIcIjuCO2Fj2aiN1aXgEgd8oNtBknUBg5VzoXbEDJ9At1VVjiP3uMO6x6Bac6jbQnE+dz5oNGigkc7BYcKV7vdx047rVfHo2gdOFiVIZUE9OM/Ug0556z1IzrPwAZzsrnU72nLgceoWpUNvEzesA7DOcIOfuUC3W+DVLa24vb0F4OCu+WrtaWKhdTMoJ2MIaBkH5Ly30vq65aSqA+ikLSDnAOFyRSccbtXOjdWyuPTgdyg9B7Pxco7JCZprhlpbnBd8l095quK9v1PWyine1/xHfutiak4y1tZbzDRvPX042K4Vvt6uV0ke6sJJJJGSgx73cG1LB047LQQMLId1g/HnGFE9zTnCCwHByhOTlUUzIS9uRhBCAT2V3SQcnZS0wDZSHeiuq+nI6PVBBnq2VMZdj1VWHpduqvcOoFuyDKj6aXEnqM4UFTUeO8OxgBRmR7hgnZWoJXTZZ048lEd0RBlW+cQydRGQsmurBKMNbstNBIOQspkjDH8XdBiE7qoODlHYLtuyogyRVkNx0qN8viHtjKiRBkxkQEOG6tnqfF8lCerzQNJQUGyy4Y2ubv3WO1mO6uEvQdkEjKXrf8lbK10B6G+fmpWVDW4PdSiaB8Z68ZwUGJTkh+fNSTyE91ExwExd5EpM8Of8PZBWJuHdWcJMM5OcquC5nw91QU8hGSEFKeISuwVfUQNh7bqFrnxk4OCEfI9/0jlBRrulTNqNg3p2KgTsgyZIGhniD61jK8yPcOknIVBsEF0LA84KvfiIgAZUIcWk4Tqycu3QXOkL9lfFB4gzlUc5vTtjKyKdpLcjsEGMWlhVnxZzvssioe3OB3CvhdCIiHgEoLGVfS3px2UZf4kodghRvILiR2VzXADPnhBqRcJ4vDA8sLTpojE4AjGVk0FQ1knxnZLlIySQFmMIPU/wBhr/ufxlHpNYP0V69TF5Z+w1/3O4y/4awf9GvXqYgIiIOuftD9+TXiWB39yo/16nXzxg4OV9EPtBozLyd8SIx3NHR/r1OvneIwUBERAU0dQWM6cfJQqeIsLQ04ygtjZ4rzkqWSm6SOnGFG+F8R6xsFPBOwxkOO/mgp4THjB8ljSsDDgKrpXhx6XbZVjnF3coDXFpyAry9zgfh/MrqfoDyH9lkOdC0DBG6DFij6z8WcKksYYcBSySNGejCgLi47oLopBGckKZwEnx9sHKxlc15btnZBe+Xbpx2VjW5GVK9zentuoRk7DzQSQRiR2CpoadslWynJwCcFY/TJFh3ZGyyNeJAfiHmg3JdNMU9HQipZICSM91tj5FZk12rZ4vCllJaBjCw0EpaOnKiPdVyTsqAEnCAp4IBKQHHGSqwxdJJeNvVUll6XjwzjCDMNvYxvUDkrAnaWv7YWXRVL3ydMhyFFXlpk+HCCKKcx9srKZIJYyd8kLBbgHdSRy9Dx6ILXMeXHDSfTZbr03QxNg95mABAzurLObe8ZmxnG+Vi3mvdTv8Ojdhh74KDVa++Qzn3XAwDgFZttoKOqi6SW5ctkEyTYfHnq/nWZTVVypGkhxGAg3my10tpcZ2lp89lot31W12YGMB7jstOivNbW5ikcT5LS66Ppn++DGUEhq3+J442B8lk22ve6tGZMZ27rBmki8INYBn5LHieY3h7c5HZB234Ccx9XwMfBWwTveS8HAOV3l4V8x9Xx01DR1lZUviZsfiyAvKXQ1Tb6uSJl5aHMDh9L/wBq7b8OOI9i0fQtfZehkjGbYO/ZB6ZcY7fZpdExSU1cyWQMGW9WT2XFfCp9tM/udRI1jnOxuuu2keYa9almdQ3KoJgBwMk4wobvxpOmdSRS08oEYeCd9kHpfpCw2m1UjKwTMJfuMuGAtyfdK3Db32Af+OF5+XLnGDtPR01HUt8UMxs7fOFxjDzZarqrw2mjqnYc8DHWfVB6mVd2tMERdU1kAYQc/EDkLrBx90rom+UU1z98gBDicEhcD6q4+asp7D7y6oO8efpn0XUjinzE8RKynmjgqpfCyR9I9kEvHq42Sgrp6SjdG7oBaCMLqbeqttRJI4Ozhxws/UGu73f657q6RxeSQdytvSQTyS5OdygxXuLj2wrVqEsLYowXYz5rBduS5o2yg1Ky2etuUwdRuLXMIIcCQQQfIjsV2Y4R+0H5rOX6pgtFNxBn1NaIcD7l6lDq+INB2ayRzhPGANgGSADPbYLrZYNRzWN5dHH1A91kXOdt+f7+44IG4KD2G4Oe2A4Paojp6LjHpG56JrH9LXV1KTcaAknBcelomYN84DH4Gdzhd1uH3FPhvxXswv8Aw11xZdS0G3XLbKxk/hEjIbIGkmN3q1wBHmF8ysl3k6PAaMtA6crM0xrTVuibzBqTRmprrYLrTYMNbbauSmnZgg4D4yCBkA4zjYIO33tSeUz/AGB+Lx4naQt/haK1/US1UbIxhlBcz8c9Pjs1ryTKwbDBe0DEa6RscWHLSQV2kvftDONHEjhFeOCnHaktPEKw3OBograuIUl0oKmMh0E8VREA1xY4AnxI3l4y0uw456t4cN8FBI+SR4+JxP1qxVBc7YDKdkE9PF1gqKRvS8t9FdDMYtsd1NJCHtMnrugxc4UkfxOCjOymo2g1DQ44CC6UAYGNyjYpuguHY91qVfT0/hh0ZGRusETlo6MbIMcN6yQVFI3oOM5Urn9OcbKFzi45QUQbK5kbn9kexzDuguinMZypSxszeskrGVwL8HGcIKOGCjRlwB81VjHSE9IyjmOjcAe6DLbRsxnPdY/hAziPOxKp48mMdSsD3B3VndBqFXb44IRIHb4z37rTlLJUyyN6XuJCtZC9/ZBYrg1wOcKXwTEOp4UjHw53IQQvlcRghXtia6MuI3wr6gxY+HCxQ9wGAUBwwVRO6ICDcgLKobbU3BxbTtyQra2hnoZPDmGCg79exYja3mj1S710BXfyjbkVPYrte3ml1ST2OgK7H+cbciD2nREQEREBERAREQEREBERAREQcccyX4O3FL+BV8/UZl80Rb1yvA9T+lfS9zIjPLvxSHrou9j/AJDMvmkYD7w8fM/pQRtPhv3HZZLGtmw/HZX+DGcl+FE4PaC2DsdjhBfPVN6DEMkhYrGF+6lghLpPvowFfOWMna2PGNs4QW07vjDSDjPdZhDBg7LLnhpGUQlZjrIz2Wi+PK/YZ3QT1U4LOgdwsNvcLOoaGSWbMrcAd0ucDIJAGYGEFnSRHk+ixnOIPmslzneAPqWGSSd0DOUREBVAcRsgY49hlZlKyIAeIcFBhfWizKtkIALMbq+ipmSxlxAygx6bAccqtZ9IYVpIjmI8gqVDupwI7IKQxGZ4blXVEPhEABWRvfGepqSyvlPxeSC0DJwsqOj6mdRx+VYgONwpm1crW9OdkEb29Dy30Usc3S0gkklQucXOJPcqiC52SSfVTQztY3pKgAce2Sha5p32QXZ+MuWZHK4MI6j9eVheG/GenZVBk7gHCDMoqt0U4dk91yJpfXDaSqigM7wTgYBH+pcb26Fss/S7yWrUsFNBeoC84HUPNB2PMrL1aw8HYjzXFerdHPHXUMYwgEnIz/rW/K68U1DpJht7sy9A7fUuP6XU9XWRSU9bkF2RglBslozUe6EdtsHsr59Pv8QShrQO5O/+tanX2aenqDWhuxOey0mrvkgaYx3Bwg1FlIyaIQgAkeS06GhkobtFKRgAg+axrVd3xVXXUPw0rcVtibf9QU9JSyAl5AGN+6Dl602+TWtijtlJROmk6QMFufL5LTafh1dtNXNluqLQ5hmcB9AgbrvXyUcs5uUMNxukDTEcE9Q2wuyPFvlX0vO6C6W+CLrZ2HSAchB5w0vLzcX2YXkWiJwc3qz0HPb61wfxI4eXGh8Qm2lga4j6JC9huH/DMyvFnuELPdmjpGRthcNc0PAS001LLJbIYzuTsAg8gqu0VUM3Q6IjHrlYThPTv6fDcQPzrn7iNoP7j1uOgDvlZuiuGFrvlplqJ2tLw0nt5oOGtMWZ18cIxECTtuFv6bQstroTI6kYB074af8AWr4rP+w6/P6mhsQf57bZXLFPqPTN105KJpIzKGEdx3wg6sXWN0de6IDByey0qpqKmAloe4fb2WvanGdTymH+xdRxjthYNdBTvYXnGR80Gm0VxmilBkmcR8ypa24NncCHH5rAezMhazcKjo3MOHDCCd0gmAaFb4GCrWNd3YDlOuUPHVkIKSQlrQ71Wo29jXwHtsPNQzYfAMbnCx4p5YmmMZGQgo8YnONhlWyEvf0gkq7wKiUlwaSSs2no+mPqmGD5ZQQy0PRCJdgMZWEsuoqJ3ExbkDZYhBBwe6AsumojNGXbfasRTR1M0bOlp2QRyNLHlvoVdFH17eqsc4vcSdyVc17ozkFBkS0hazqxhYrc9QHzwpXVUj29JOysbHIR1NBOPMIJpKQsiD9u2VjKZ1S9zeh3bso2Y6xntlBfDEZDjCrND4Weyz5TBDAHMIz8lp73yzZO5AQRkjCB2AqEEHdVDSd0FS9XMiL98KPss2nAERPYhBjyQmMZKRwue3OdkkkdI4t8lRsj4sjPdBY4YJCN3IQ5PxFU7IMtg6QCVnQFskeDgnHmtOp39Zw87BXzTmJwDOyBV03QS4YH1LEUslTJIMEqMMJ7boDW9RUgi7bfnVgDmn0U7C8jOOyCx0QYM4UbclwCrI9xy0qaBjC3r8wPylBZJD0N6z+lWsiLgSB5Ksszn5bnZZlvMWAH4QYUUTnuwQszIgjLc4/mUtUIo2l0RGVpr5XyHBQUfu8n1KngpXys62jIUBjeB1EbFbhsslKKbolI6iEG3pGFjiCFbjK1G6wtbIXs7FQU8THxl3mEEbIT3CpIxwGXeSkgl6Zel3ZX1Tg5h6cIPVD2Gv8Audxl/wANYP8Ao169TF5Z+w1/tDjL/hrB+ivXqYgIiIOvHtBJBDye8SJD5UlHn+PU6+d/6TvrX0Ne0Px+004l/wCJ0f69Tr55WguO3kgnbTdTQdlFKzoOPzK5s72jGeyse9zzugo0ZKrjocDnsrQcITlBkzVLZIw0HdY31K+INLviWV4cHTtjKDHgi8V2PRTS0nQBsO3qoGyGJ5LDsrnVUjxgoInN6SqZJ81X4nlULSO4QFMyDrb1faoVe2Z7R0g7IL6eDxnlgI2SpgMDwNvsVkcz4nFzTuVkdXjt639wEGIqg4IPojx8RAVEGSHNmHSB29VFI3wzgBWMe5hyFe13iSDr7IEEZkfjGR5qs0XhkDYLUo/do2dTSAQtPqpA92WnZBH0YblW9WHdSEuwAe3kqDGd0E8k4cwNGRhWxwmRvUpYIY5PRVneYctb5+aCOmOJME4wpqiny3xBuQsUOLTkHdZFNUOdMxkm7SQCgxcHOMHusuOhc6Ev2z3yVun7n2p9GJGFvXj8627XySwPMcf0T5hBjUheJejqIxsd1lVDMjc5JC05sj2PLgVeamV22fzoJoJhTPyfVZklwbNEWtytKcyUjqcCtat8FL7mXSEdWEGm0c/gTlzs7lUr5hPKCO3kopi3xnBvbPkrDjqAQXiB2MkjCMjw8Z9VMSQ0E9sKHpkz174G+UGqRTPgYCx5BGCty6S1LcH3SOibLMeogYDtv0LZj5vFDI2vOScLsRwW4c2u4wx3CcNMoaHb+qDdMVVV6ZsrLhJI9hc3qyDg/nWzr3qupvkL6hlTISP7p2/6Fy3qbR9VdKB9ExpMTGkfYuBdVUTdMSOo3bZdjHZBjWzUVzNW5j66XpBxjrVtRr+Sy3iOofWygNOdnD/UttyVDKZxnYTkjOFtG71fvdSX5JIz3Qdh7vx/p7tZ20LLlOSGgEF49PqXHN31rFVUckRqHEu33IP8y4vBwQfRXOe5+ASThBM6XrrHS5zlxKynVzWEbZwtO3G6zoKaOWEuI3x3QUklFUOkElY7m+Eek7K6Fro5jgbBKlj3O6sHCCMDqO3orxJJHGWsJA88KEEt7J1OP2oKsAJ38lRzsnY7IQRv6q5sZIyBlBWFnUQfmsmRrRH2GcKGAhpPUe3ZUmlySGnZBWncxueoq2Vwc847KLJ9VVpJ2QXDus0ysFP0k+SxA30GVJBGJpRE92AUFIaaWoJEYzhWvZJBIWuyCFrbBDaoy4EElaPV1AqJTIBgHyQW+PIdnOJCnD2GLAIyfJYeQpWBhGS4DHkgieD3Vqq7ucbjKp3QSQyCMklVleJX7KMtI7hUBwgyo6Xqbnb7VHIPD+An8iNqpGjAKsd4kp6tygugkEbslUnkEjshWmN43IVqAiqSCFRAWTTTNYQHHusctI3wqdkGozBsrBjcrFfTuaCRjZWNll7NJVfFkLgHk90ERz2OUWTNGwsyzusfpOcY3QUWSym6o+vA7eqh8GTv0nCvbPLG3oOceiDVdOXSG2VD3SuwCVBqC4RXCr8WM5C0xxzv5lXCCVw6g0kIPQH2LModzRaob6aArf5RtyKL2Ku3NPqkf/w/rv5RtyIPalERAREQEREBERAREQEREBERBxzzHnHLxxRPpoy9n/kMy+aPxA2oe4j98f0r6XOZEZ5d+KQHnou9j/kMy+aB4PiPGNwTn8qC6acvwATgKSmqGxfSGfrUMTC9/SFWWIx9/NBLPUtefh2+pUhp3SjqJ7fNQMaXnA3KmEkkBx5IL5xMxnSXkhUpA0O6ndgpJZGSR5zk/wDsWKC9uQM4KDWBdoWN6GtAOMZWHNGaomTO3cLBIOMrJppXBmAdgguiHju8DtjZR1dKaZwaTnKGTwXmRndWTVD5zl/dBV1P0x9ZKhUjpZC3pPYqwNJ7BBfHKG+SlDTMCWnGyxiMd1ex72j4c4QWnqBIJ7LIgqfCYWdsq2nDHPPiED7UqWxhw6DthBC93U8u9Sr42dQJUjhEI/hxnCgDnDYFBNE0OcQlS3pI9FE2RzXZBWW10cjCXHf86DCRSxhhlwcYV1S1gcAzH2IKQxeN54VJofCeG57qjfFiGQCAqSSukOT3QZLYmxsEh3ysaV4e7IG3oqmd7m9JOyjxhBP44LOnG/ZGzNAIPooFVo+IAoMmgm8Kfr7ArLe51TcIy13mOx7LHkETIstxk/NW2+YtroXuO3WM/Ug5SpZailtTH1Li5mMbq2htAuzve6cgBhyQFm3W4Ws6ZY1jx19I/QtE4fXtz75Fbuv4JHhpH2oNWvEsk1N7hHQvc4NxkNXHtwsNZDK5z6Z7fPcFejPDfl+09qSxMuczYzI9gd2HouM+LfLzPT3T3e1xAh5xgBB1U0Lw4uGuallFS0cjnF4blrTldzOBfI3c4tQUFzrbfKWAhx6mnGNlytyacq1xprtBX3alAj6wSXN2xnK9I7bo+2WhkbaOFgMYwPhA8kGjcJNDUGiNK01vp6URyBoDtsHYLdN1tcFxj+/HZvkeynq62mttP49VIGMbsT5BbIves/eqtjLQ8viBAcR5oNN1LUUlg63QRhhaD8S6/cWNV0lbZauSeUPc0HbOSua+KeorJS2Iy1crWy+GSe2c4XQXW/FW2zal+5Lp8wSy9J32wSg4V1DTfs81ZJbRRuYwPI6iDjus2t07Hw7pTGZdiMlc5cRqPh/o/RTdU2iSH318XWcYznGV0/v3Ftmqaeq97k+JhIbug2/xTvcFc13ujgHnJ2K4korveaNkrXVbwwk5BK1Grv4rbu+OVx6Oogei0280kssgNI0lh3JCBTV/v1QWybuJ7lYt4a6B3SHbFXNibTMBh/svmB3WDVGqkw+cHb1QR08jY5Otw2V1VO2ZwLRjChAHmcK4AdWB+lBfTTNifl3ZVqZmyuBYMK2Rga0eqiGM7oJ2zlrcHdU8YF2cKx/T0jHdWINVpq+CIfEAVHV3Bsp+AYA8gtPaxzjhoyjmuadxhBkuqWEdlaIfG+LsoYwC7dSulMezSgsliMeclT0rGuYQR3KgcZH4684V0bpGbt8kF81P4ZLh2WO53UVe+d7xglXxhhaerGcIIAMnCyoahkTOgjJWMfpHHbKkaGluT3QWOPU4ntndUA3Ayn75CcFBK9khaDkkAK6OURs6XN3U1uex0vTMdvmp6+Gl6x4ZCDTQDI4/NXE+HthSytbG0EdyqRMbKwlx3CDGJys2mHVGR22VtMyIydMmNvVKqQROAiIwgt6BE8uJyCj4vF+Jo7KEyud9JZlM9gb05GSgwXZBIwrmR9TcrKqqYMaX477rFZ4uPhzhALiw4CkjZ45HUe6RMEh6Xd1V5dTvGEEk9D4TOrKggeOoN9dlNLWuljDT9Sx4WOfIA3ug1R9EDF14G4WH4gjd4eFqng1EVOHSA4xtlaROxz5C5gyEElRCOjrHnusZjiPh9VJJM/wwwlWx08kjetvYboKyQFrev1UQcW/RJCkbI4/A7yUnQzG3z3QRCZx2JVA7MgPzR4aNgqMB6gfLKDLqHt8EY7/UqUdLNJh7XEAHdWTuaYwMjIUtJUmKIgIJ52CVvhk7gKFsXgxkZ7rHdUPEhcFkR1Albh5H2oMFxPWfJOt2CCc5WVVMjAJbjKjYxhgc4kZCD1Y9hvF02rjFL/dz2EfkFd/rXqSvLb2G8hdauMUR7MnsJ/KK7/UvUlAREQddPaGt6uTbiU3/AOk6P9ep188sb+gk+owvoa9oa7p5NuJRHlR0f69Tr55YgHEgoLScnKopPBeD2ICo7AQWIr48ZwUkDcgBBYq9Tv7oqRzA1gPqrGxPeMtGQgtTCqQRse6zKVkTmkuwgxGHpPURsqyPDzlZVSyLpxGRnKxPDf3wUFqJ2RBUHBypBNgY9e6iVwYXDI7ILSclXtjyMqjWOkcQ0ZKv8KWMgOBAKCI7EqoBcQB3KzGsiwc4yjhC1pcCMhBA+F8berqOFDnfJ3UxnLyGnt2VYI2unAwOnKCN3UAAQcKxbinp6E0rS0jqC0KWItccDIygQyFhJBKTyeI7JGFJRtjLyJMJWtia4eHhBjKoyDkZ+Sos2BsJhPURlBDFWTxH+yOI9MqaWtbKw9Q3VtJFHJMQ8jCtq4o45w1vZBjk5JPqjT0uDvRZUrImxAtxlYowgypaljow0D61AJpGghriAoz3UsbWlmSgiJzuUGxV7OnqIPZXmLqOIxk+aC+MSVBEccZeSQOy3/pvQUt2pg11O4OLc4IWXwS0ay/XvFawCPIAyO67L2rhPdjqWmo7PTkxPIGwQdZ6nhHdbdIyobbpZG9WRhpK7DcEtM1zGxiSjkiAAByCF344R8qVqr7HBVasp4mAgH42j+dZmv8AlzoLJG6TSETHNb2MY/1IOvhsVPTW95dT9ZLT5b9l07462GaovLnRQFg6yMYx5r0V0bw7u9NWyjUsJZTAHHUNsfautHMvYrDR3/NCWFgfvjG26DpDqOjntdMzqYRkAZVlo0JJeba+5tkIABcQuTOJ1LY5LcGxOYXhucfPC4lpNZXC10j7fSnDCSPsQaFVwmkqZKcnJY4jKh6vkr6iZ1RM+Z5+J5yfrUaC9revZZDZ/CaGA4wsVri3shD3bnJQTe8AO6sK81bXNLS3fyUDAM4d6rI8OEt2IzhBBFGZnnfCn9zw7BULS6F5d2CkdUSu3CCtRCGN2VsNQ1kZaRkqN875BhxVoic4ZAQXsYJXnyVroul4afNGGRpPSDlUe55ILu/kgyo6Avb1Z7qCWLwXgFXMrJmjAKPf4nxPO6CQSsa0ZCxzI4P62kgq1xJ28ggaT2CCSWofKMOcSrGDJwsqkgbnMuw+avqWQswWYz8kGG9nTjfusyG2OmgMwdjG6wnvLjlZcE1Y1nTGD0lBikdDy354VucOyppmhu7hhx7qDOUGQwCcFvbCslg8PuVYyQxnIVxmc8jJ80EZ2Usc3R3bkKkgaMYPfutXoaWlmpiXY68bINLkmEmwHdWRxdZAz3Uk1HNG53wHAOypGyRg6jkYQUlp/DGcqxjPP0V0kr5sNGThWglmxHmgFwdtjsrCMHCv6c/RGSrS0tOCEF0UgYckZSV4e7IGFdA1rnHqVswaHYb2QGSdJ3JwqdR6+oeqRsL3YWsw0lN7qXOwHYyMoMEVTGx4LclYsknW7OMD0WVRwRTVBY47A91LWwU8Owx2Qac0ZKz46yKOIxkAnssENLnHpVxglG/SUHfz2K7geajVR/7gK7+UbciexXje3ml1Tkf/ACgV38o25EHtQiIgIiICIiAiIgIiICIiAiIg465jndPLzxQcew0Zez/yGVfNJgSVEpwMEuI/Kvpa5j2GTl54oRju7Rl7b+WhlXzTNYWVMrCN2kj6sFBZC8QS5IzhXVU7ZyCBjCtEL3vy5pAKrLCA4BmSfRBZBII39RV1RIJXDCubRzAB72kN9VHMGgjpQUALd/L0VfFCt6sjCtQSPcC3HzWTSRdcZd57rGiAJ+LsstkzIh0twgw5MtkI9FQ/F8lmCKOR2dt1G+jl6stGRnv6IIHP26cIyQNBHqs6WniEPS3HWsB0bmHDhhBQnJyrmPDRurmhh7kdlbKGg7ILoonTydLPrVZqd8MgY7uVPansZUZfjClvEkb5Q6MjI9EEUludHCJc5GMrCWbHXPeBFKcNxhSuhp3sywhBp7GdSOaWbZ7q/IY/HkrZHBzgPJBaMk7d1XdrgTk4U5Y3w8txlRjpIIcd8oJjNHMwRgYKx5YvDwM5JVAel3UzyVesveOvsgsGyqTlZ80cApw5uM48lp6AiDGd1M0MOCSEEQ6nEDcrIijwes7EbhZ1HDTPdvhY9wIhkDWYwe+EGtadpKu/yto3znpLg3crtRwc5VTdZIL4JASwB+MrqFb7xNaCyWlOHg5yu2/LTxt1TXXGms8Rd4bwGHBKDvDwR03LC9mnnRvPQQzP5lz/AD8vduqaiK4VNGH9ADjnfPmtK4F6Jmdbo9QTMHiuIdnG+V2LpnOfC3rbg4AQbB0xVWHSsYttJbxG9mBnGFuSp1D0QirAIY3c+inm0tb56g1JbhxOTssbUdohZp2ubAD1iEluPXCDjXX3EKm1DSPslDWsilJIOHAHK0DSdUNO0UjK6tbK924JOV054paz1fo/Xk8uZmwCQ774xlajozjNdtRXunppZXuaXAO3KDnbiVbbrrN8sNNK8RlpGx2wuifGXhld9M3V9YZ3dQf1A5+a9LKGlg/YuK6BgMpjz674XRPmSvU/3UkZVsDWCQg/VlB1d4kaz1E+xi2S1sjmBhGMnC4Cira5rpW+KQHE53+a7G8RJ9MVFoHhSsM5ZvgjuuvlU2CN0oaRkk4QYcNJLUyFwfh2crWYrrFbKc087Q5xBAK0OGpmp5urfpWe80tW3xJCMhBhw1XTVmqf9AnsVm1M0NezqYAMeixvudLcJRT0EfW4eQ81FVUNxs7+iqiMZO2Cgx/d8yFg2wrZIzCRk5yrmznqLgdyo5Hukdv3QSxN8c9CsqKcwuxuQqN8WEh4BCz4C2pjLpMZA9UGC2nLhnOFa+MtUpkdHIWncZVeprnDJGCgtpT0vGW5U1VEHkOAAWRSNpwSXkBQ3CZvWGR4wfRBiQjL8KszelwP6FI6J0cfiAeXdRtJld8SCQztLMdO/wBSyKaJjoyT5qCWFjI8tG4CpBUOa0tHdBHO0NecbKLKklEhJc8HB81Y1jndggomVcY3jyVBsd0FFUDKleGdO2MhQ5KCocW9jhOtxIJJOFWMAndXyBgG2EF8YNQejKvePdcsOd1BDIYndSrPN4rgfRBGXHqJBVQ8/vt1RmM7o7GdkAAuOyuj6myAdt1dBjO6rKAXAMOSdhhBqNaR7oCNzhYEE7WN6C3JKyGU1WG9dQ0iP1PZXOjpenqBGQEEDGGJ5kJ27qGaQTSDH1KZzzIegdgcbBTRUrAOo9wgxZaV0bA4qe2hrZBI7sD5qWUgt6XYA+axSyUf2FpI9UG4rjdYJ6QQRgZAxstLo2NILXDJK0+MS+IA7OVmxsmY4PDTgDKC6528RR+IwZ+pWUFQxsDmvG42Wp00grAYZcbbLCrqVlO/oZ2J7oNJmcHSOPbfsroiXfCVlVMEYj6gNyAseGlqHjxGNOB5oE0HSOrKia/pBHqpJHvd97IyQVfFEzo++jB3KDH3d5qocW4HkqO2cQ3tnZVbu4A+qC5xy3t5KPJ9Vn1DYBCOkg7eihpaOSdpc0bIMcuce5ymSBt2U1RTuh2cD3UjGxGle446gNkHqj7DX/c7jL/hrB+ivXqYvLT2GzSLdxkcexmsGPsFevUtAREQdc/aHfga8S/8So/16nXzyNPSQV9D3tCQ08nPEkO7e50f69Tr54AMoMt9Sx0Ybjf1WIdySr/BkxnGyuYxuD1Yz80ESqDg574R3dZlMyAxHqI6sIIARIOlZkRZDD0uwThYB+9vJA2VzpnPwEFkhy8kdlVrnAYBIVCx30iO6oDvv2QXxlznbkqd8rGDpxvhQuLWty3G6taHSOyfyoLXZO+NlRZUjYxHtjOFjNA6sHsgvhiMrukLNbStZGRtnCjbEXNHu/0h6I+OvZvI0hBSLFI/qcAcrUzHHWQmVoAIC05nTJtN3CuE8sThHFksJ3wgwpg5kjmknup4KR84z1HH1qetijEXXgBx+axqerdCCMoLqmiNOMk5x3ULJS1uMHPqsp1WJ9nlXeFAYycjKDGhMsjiOokeeSpJT0jGdyoYniOQnO26TyB7sgoIydyQSN1QknucqrcZ3VenreGsG57ILVexrnkNDlNLbqyBniyREN75WO17mHIQZBidAA/OPtUZzL8WckKkk75GhpOwVjXEZA80Al3Yk7KikEErtwMq4RhrT1gA790EKkiOXBnkSo3blZlNE1zA8H4h2QKigMMQkByqW6QCoAfuCrzLUPyyUEMzjdVi8COUOGBjdBzjwqudNbXwytw1xcDkbea72cCdVWE3Wmqq1sb3MAPxYK85NFvqq0xsowXFpH0V2H4Va6/Y5qGmpLq5zGnA3OEHsHbb/btaadgt9pIgIAGWnC3bp7TcFqoRTVhZO93fr3/SuquiuNGmbHpqnqKOpb4pAOxHdb6uHGSvrNJz3qkecxMLgQfkg3px6oLdT6QmmoY4oqkMdgMABxjvsvIfjtcrka6eKo6+oykAnPqu7PC3i9f+MGsavTlZI+RjHGPBJIxuFwtzi8MKTS1Q+eRoYSS7thB5+a0jrmAyPncQR2ythxwulBfvgfnW+Nb3KGetfRxSA4yPqW0ozHBEWP7lBgBu5HorvD+LGVRzvjJHmVQPIOUEslOWN6icq1sgDelV8Yvw16tdG7OWjZBb9I7JlzSMk/lUkHT1dLlNK2MA7jKCOWRr24HcK2OQBnSo3YJ2VRG4jONkB7cbqWOdrG9Pn5KINe/Yb4VHNLDhwQZFO4Oec9iqVIH70AKBri05CvaXPcCc4QWtcAe2yo52ST6rJkiY9g8MAlY7mOYR1DCBjqGyrGel4yNsqSIsxvhJOgbtIQTVDh4Q6Tv8lhlxPcqrpHOGM7K+MMI+LGyC2NnWcLcdG6nhoiHgZI2WgFpGTH9aGqmA6XfnQUqniSdxB2yoeyrkk5JWSwQlvxYJIQYqr0nGcFZQbDnfGFd4TDu3GPkgwlPTTyQvDg8gD5pO2No+HGVC1pd9SDX21cNXGGYGQFjvjaXeCAN9lprJPCx4ZOcrUqaRpHiyYyNwgjntxoW+M85Cw+kVD/hWbXVctaPCYMgFRQ22tY0yCMhBG6n91HWTlY0j/EdkbKeSR5d0TjABxur/AAoXM+DGQgw9x5rJpaJ1R3yEpqVz3/GNgs0yiD4IvpIIaimbSNBysc1MjmloccYwr6j3t2DOCGnsqU9I6Zw6BsgigmdA7qxuSp3RPqn5zsVkTWxzGgkHspaePw4vhG6DD8IUfxOGVM2pjkZ1dOFV4EjiJtgOyse2FowwjHqg76+xeqGyc0uqWgYxoGuP/ONuRW+xcbG3ml1SWnc6Arcj/wAo25EHtGiIgIiICIiAiIgIiICIiAiIg495h3iPgBxMkPZujry77BRSr5qeoS3GpcBs573AD6yvpR5ktuXbikf+4q+fqMy+amkcG1Ti47HP6UGYOlw6AN8YWNIBDMHO7Z7LNhlga8lxCxLifGeDFuEGdNXQzUnhsABWjmnIdudvVXxQyxnLwQFdP1vcBH9uEFsdIXnYlXPoSwZyrqdlQx+XA4KpVTyB2M4HmgxXZaSFJBA6Ujv3UvTE5oIIz5qWAeEM9kFZIjBGDlZtDVRupj1tBIWBLUtk+F/b5qalpZJCPBBLfVBhOmLakuPYH8ypUyiZ4DR5+S1K4UsMUJJwH+axrfFA5hdJjI7Z9UEDqNzYvEJxtlY+CTglZFRUv6nRj6IOyhGNzn8iC50LmMDwVGXOd3cT9ZWVSl08gid2Va6lMJGBsUCjtxq3Ya7CzZbc6jYeok7LToK2aDeMgFavTV7aqA+8Yz5INDl+krRnvlSVPSZndPbyVjASgvEhARsRlGQrXRuGTjZGSuYCATugqY+g7lWE9R2CkIfLjAJU0VPhhLxgoIMvLQCchRkYV73YccdgVPTRseB1Yygx2MMjsBVkjMZAz3Wa+ONo+9dysR8czsucCgviL4PvmcD0VlRMZnZKt6nvAj3OEbG4vDOxJwgrFDLM4NjYXEnAwF3p5KOGE9TcqOsmoHEbOz0rrdwl0B926iJ8wBBkHcZ817F8nPCK2W+ywVj448MiB+judgg7GcL6SCh09HSCEMLcAjHyW9AABgDCxaK209C3phGAstAWNcGNfRTMcMgsIKyVa9ge0tPYjCDqjxO4JUvEa6y09LQfHv8AEGrrRqjhlcODV+b10TyPE79PbdenFJaaWjndURNw9wIJwtk8UuE9k13bqmWriaakRksPT5gIOl2ouYx2jdKNeXdRMf0O57Lp9xO4q/7KENTO370/JIyMFdsZeXG76l1tPZ7rC8UDCQMg4xldTua/hY7hXqRlt09kMfJhwaPmg6van+7EFe4SyvMQOPPGFtaWV8s30juVz1rDh9WjSUV1kjJkfH1HbfsuC4IWMqXQyHDw7pH1oNQNCJaUEDdaRJFNC/w8n0W7YLJc4YBUSRnwiMgnsQtODaOW6MhkeAOrdBkaKuDrFcffqmldI042xlazrWsOq5GTUdudGG75DSMLnvghwPoeJEUVPSBsjyQNguedTcllVYNNyvhpAJ3RnoHTvnCDzVqrdNSHdpJHcY7LFDXl4+A/UuwlJyycQ4dZSNu1vkbRCQnqLTgjK0viboS0aUrWQ4Y0ggO8t0HD0jC+EAsIPqsZhMBIzjPdb21DR26mtbZaYtLi3bH1LYmXyEk5OOyCfoExyNyrJKd7B2P2qtF1eIGrcMdHG6LqkA+0INsN8Rx6WkqcUjxh7z27rUXMpI5SARsViVcj3SgRbjO6CyeoaY/CA7LFjf0HK1CopWmAFuOrvhad4b+2EF75y9vSSrG7OH1hVdDI0ZcMBWD1QZ9Q9pgGACSsankDXAO81l2owyz9FQQGq+7U0LZQaQAjzwgsmazo6hjdYpg6m9QKkeJvCAOVE2RwBYO6CI5zgpjfCkMEv0sbHzVh2PzQXmItHVlRnJOCVeDI8YOcKnTh2D3QUcDt81qlvsb62HxMkbZUtnoo5Xg1GzfLKyblUz0REdDuzzwg0WroTSyFhPZRspy5pcD2V0888zyZMgqxkrm5b6/mQS0UPjS9BOPJTy0fu0wlJ2ByVjB74MSMzlS+NNUsPUM7INamulNcKUUcbQH4wtEnoJIXAdec/NQMMkEnWO4KmZVSyzAydsoL2wOiaHBhKt97LSMtx9a3fYo7ZUsLagtB8srQtU0VPBVD3XdvbZBpE9UZQAB2WRR1scERZIASrZI4W04II6gsIDKDN94Y+Tqa3zysltxYxvQWjONli0UB6svGAVfVW6YuL4mEhBNDUGJ5lbuCUqaoy/EQdlNYIGPqfCqxgZxus2/UdNHtTY+xBt6WcuGCc4WqUFxhjpzE5oyQtHfG9meoYVgJG4QZU3wTGYYIJJCjc507tgQPRZtqYypm6J/orKucFNB8FPjJ9EGlMphncq91GcFwJSSlrIW+I9hDfUqsVXkdDiQTsgihgfM/w8nAWsU8sVvh6XjJwsWGN0RMg7d1Wdj6z4WDLjsAgslkbcJC1u3msOpp3Uzugk4WWy219C7x5Yi1o8/IrGrZ/HkDsoPVv2HRH3D4vDz94sefyVy9RF5bew3JNp4xZ8p7Dj/za5epKAiIg65+0P8AwNOJeP7yo/16nXzyNOHA+i+hv2h34GnEv/E6P9ep188gBdsEGWauPo6elYr39RyOyt7J3QFLHC57cg4AGVaIZCMgKrZXxgt+zCC4DxD0eaq6nMeDkqlOCZMjusucgRkOAyAgxHzAt6QFPS2508JlzsFdaI6eWcio7LKudUymf4NKR079kGkSNLHFh8jhXslDW9OMq3q63Fzj37qRrIzjsghcXE+aNb1ED1WoMjh88ALEnDWvHRjYoM23vbQP8WUAj0WZWXumnbhjBn6lpc8rXxAeixQxxGcbIJJZC+QuacZU1NUNj+FwGfVQBoxhvcd1RjCJAHBBmSMdO3PkofcndslZUrhFCCNtlbHUt6O/xIIXUD2t6upWQxSPlbDkgHZTNNVM/AzjspTBJCfE8wgyavT/AINOJw7fGSMrRHMc0kEHbZag+61r2iKTPT2WrUUVrkpS6ZzevGd/VBt2CAzOwDhZcFMKapZI85DSCVbUjwahxpzlp9PNQPqZTkOyg3JfL1SVFvFPEwB4GMhbdhozKzrzj0Csg6ZH/Hn8qllmfFhsfb5IMZ7C1xbnOFNFB1Dr3wPLKsjw+QmTfO6kdMGENadj3QTMrI4x0lo2WNPIJX/D2KlliY5oMfc7rGLXMIJGEEppsM6iVdTy+C4Z7BR+LI8BoJwgjcW9jkjZBlVdbHKzpYAPJRxUL5WdQJyscwyDfpOyzIKswxEHvjbKDe3DrUsOlqgyVIDhlarqXW013u7K6gk8PoIO2y2RpttPWVBFYQAfmp7o6lpbiIaV2Wnug7U8EtQ6h1a2CgNe8tY4DHUu5+lbzU09vGjJx1vqWdHruV0d5brtQWGaCeZwBc4fpXf7hjaKLUuoaS7QgEsw4fmQci8AeXZ/Dq8TayqoMCoBmAPz3XEnOZYZNf1bqejpXkFxGQCu91vAqbNHD5+EGH7Bhce6/wBDaYZZ5ayr8ITjJ+LGcoPDLjBwGrdEyvu8sTsEF2N1wWaB9WHSgEBpIXo5zTilrKqponFnhMBaMBdCdRQw0Hix02MdR7INkvj6JCz0VpbjzUrg4zFz/M53Vkh+LAOyCxZMMo6PDIyVjYV8RAcMhBI5hiPWPNWdD5BkKeoYTGFnUQphB8ZAI+SDSfDcPIrLiDejpO5wrphG+QiPBChiL2yjPbKCWmiDHlxGAoKwh0vw9hss2tmhbD8BHUdlpoPW8EoLFl046oyCMErJpoqUsBkIUU72RvxEdvPCCyAGB5c4ZGfNRVMrZpR0jA+Symujkb8WFhzhrXjpQVfTlg6gowx7jjBU0T3ynpKlLGsJB7jdBjSRdAyo91kSB0v0RlROiez6Qwgvhl8J2TuqyAS/GNlbAGPOHqszgx2I+yCE7KdlM97C8Ht81B3U8MkmQwA4KCSkoXVTyzJBCzJqP3OMsJyVaXSUrRJENyMqNz6uob1yBBjspHSuJzspjT+G3p81HDU+G4g9+3ZSSVDX9juUEtBbfHfue6nq6IwAsDsfUoKOqmhJLcqKrrppJN8oJqQNpnB0hzutZGoaTwhEGDPZbXc+d+MgqRlPgdZJGN0Govom3KQvZtn0WHPSOoCeo5CkpbiaU4YVJ4NVcKgPeCWZ8+2EChmbUnww3BVk1O6CqEh3AO61WoZQ0UAdER1+nzWM6eCWEvkIBAQR3WvhqKZsUTBkADsrbHK1jgx7N8/zrJs8NDVTFrnA7+az322GCsa5gHTnyQXXNrBT9QbjbK0BtZiQMDcjK3VevAfSNZFjON1gW+jt/h9VQQHd90Gj1sJliBaCCsOOB/RhxW4K+SkaC1hGAtHcS5x6OyDvj7FyndFzS6pcT30BWj/nG3Isj2MQeOaHVBP/AABrf5Rt6IPZtERAREQEREBERAREQEREBERBx1zGs8Tl64oR/wB3oy9t/LQzL5oZmmOolYDgsc4fkJX0u8xzizl54oOb3GjL2f8AkMy+aGQl873eZcT+dAjEkpwHFZtOzwZGtk3yViUruiTdZE7J3yiRgJxug1e4U0fufWwAHC0SknEL8SDOTjdayK2M0ohk7gea0qSKMv6mEd8oNVdJEYQRgKalsQuELpQdxuFpUswjhAB7eSzbbeZ6enc1hOMINMqKM0lSWF2w2WbHRidgIPktOrKqWpnLz3KrFW1MA6d8fNBdNRO6+kHss+33NlthMT2gk7LT5KxxHV5lZNLTw1URe/uEGJXVjqqUuz8J8lJS0sj2FzCcY8lizsEcpa3sFqlrlDYSHeiDTZ/hcWkb+arDAZGlwaSp6hrJZz04JWbSRtji6X4BI9EGlwTGmmLunOPJTVFYKrALcFapbaOklqianABPmsfUMNFTzhtGWn1wg010PSOs9vJRmR3lkBXCYuHS7ssmJlP0EkjKCClh94l6XFSVNP7o4ADOVEyYwSlzOyyTO2q3kO4QY75g9uAMHzVohLh1ZwsoR07TkkBWkh72sYdigjppfCfhzdgVmva2aPLdgFebb97EhG+FCHGNhZnftsg06UYkIWTTxmQEjzUUkUnUXuG3qsykLekdJ8kEZa6A9TgT9aq6uY9haWgLUIYoqk9DiCVj1lne12Yxt5lBp9L8U4ONite03ZHXrVVHbWxktleG9vUrFobcXuDWbvzjZcz8MtA3D3yG9NgJfFhw+xB2v0dy7U+jtBU2pwwFzgH7d+2V3w5RbiK/TTm9BaY4wMLpHw64pXK8UMek73kQMIZg9sdl3z4C3HSdgsbaallYx0kYJIwg5wRYVFd6CvOKWcPWagIiICoQHAtcMg/nVUQaZNYbQ4vmFDEJS0jqAwV1Y4+cpkXECpmvppPEMJMoB3O267dK17GyMcxwyHAg/ag8TeY8jRFI7TTLS/7y0s2afqXVSx8PK3UMst6FM9kbHl5GCBsV7g8xHLLovVFJLfKqOIPIJOWgbrpBq3gkbBZK9lgiBjGRlrfLdB0q1HrehpaB1ibEBJCOknG+Vyny6cqcvGauiqmyYEuHbH1XB3EjSVwtWopw5hLnuOR6HK7F8p/FLWOgJYPc2v8ADjwg9I+Wnk7oeCoZdrrKC1hBw4rs9X6e01qqKJ/hxSMiPZoG49CujNy5xr1U2BtHVyEP7EA9lvrhXzKMFlfPUyn6OSSeyDk7mEsGhLDpmV8Ftpoarwzu0DPbbPzXjhzAWw3i7VU8EuA2UkeWBld5OPfM9adQPnt4qQSGkYDgvOvizrKWeqqPdXkh7yQc/NBsmrt0skDad0xOBjutFfbxSAh3ftuqR3WtMollyAfM+ayJZnVYyNygxqWINlDg3JK1R885j8NkJwdksMMbajNZgN6ly3bLbpGe0mV74/EDM74znCDheGwz1kpdkjJycpVUrba8MfjPb5rXbnXMortJHTkeGDsfJbevNQ6sqAQfzoLu8Yf3BCxyGh4JA2Wo0kPVAA4bYWHXNDX4Ygjq5GeAWtAJ+pac3Oe3dai2lknAa0HJVwtj4wC5pBG+6DBEDyA6M4JWdRNcMCbdWgGM4OwUjD1EBp3QZk1LG5hIHdaPLD01IaG4C1bFQxuXAhqhkpHTt64xkj0CDImhiZRAtAJwtJjpPHfkggfUtQt8FYZeiqYRGPMqy7Sx0sgbTlBFNCymiOwOAsaGLxn9R2A+SloqgVE3ROR0+SyK10EBHgkYPoUF0tQI4uiN2CPPKrT17GwkSjqPqd1pspfMMMyT5qWBgawtm2JQR1FSx8hIaAFBG0yPGdlke65cXYwCrXlsQ27oMkwtEeTg/UqwyNb8GN/qWF71I7AzssqmjOQ5yCd1KwjrOAFjyQNGQ3v5Kapnd0dMe5CwGzSmQB3clBKX1VIQ9khA+RWXE81seZCScJUU7nU4c4eSjpA5jD09ggxZKWQOPcjPksqntwc0yO2xusyghNVKWgeapcXS0jxCBjqQYE9UIz4bG7hajQXqGOAslaCSPRRMtZnAlc3YrBrII6clmd/IIJJazExki2JO6zYnvlj65XE/WtKghLjkjZZTqgRMLB57IMeokbJIWNHn3UBHh7FVljcCZB5nKjPU4dR8kGRBM6M5YcFUM8hqRJIScEH7Eogxz+h3msyopYw3bYoNUqrpTVNAIGtHV22UFFpd9RT+89ZAG/2LSaWnm8UDBwFqH3drKP8A2s3IZ2O/kghqqsRF1Pjdm2VHQXMUtS2R7OpoV1ayKePxo93u32WRbKOjkgIqSA89kGq3vVNHcbeKeGNrX4xlbS8M9Jd6LLrbeYXkxgliwiSAWkoPV32GzwbXxjZ5iewn8ra7/UvUpeWfsNR/1P4yn/6NYP0V69TEBERB1z9od+BpxL/xOj/XqdfPNG4A7+YwvoZ9od+BpxL/AMTo/wBep188Y7oLnN7lWtOCCrzuMDurS0hBmCrja3GFn01iNfAapjsbZwtFYwvOFqFJdqujAgbszOMIMZhNHUOY4djhW1Eviu6mnZahcW07ofGBHURn7VpccbnkdKC0Oc3dpIQuc76RJWRJD0szjfCxwxx8kFFKyIuAcCogCVeJHsHT5IHU4npDla4EHcpkg581d0vfuASgoxpeenKz44GiPpdjPqsGM9DtzhSOmlcfgz+RBd0eC8ucBjKym0wmYJRtjdYTvEcB1g7KenqnMHhg7HugjqZycxY7eayaG2GoaJQ7YblRzsiLTJtkqlPXzQjw2du3dBqsb4afLHAZGyw6q4NMwaG7HYqz77Keog5K1CG1xyQl7/pYyEGM6nZNGC0bn5LNotHVNZH4rZCB37rGp2vjmLXDDR6rWBf5qNnhxE9OPJBpVxtgtQPWckLQ5pBI7LRgd1rFxugryRK7utJlEYJawdygtp2FztjhTSYjAB3UHRJCQ5SszMzffCDHLt8jYhU3Pqr3xuZ38+y3VZLdb6i3OklI6wM7oNu0AzJ8XZUrwBLhqvrI3QVT2wjbO2yxnF73BzwguiAbhxCnbUsLg3t5FXRiMN+IhYkpAky1BqVQ6OOIPA7rFjpjU/EDgKaloK26t8OmYXYxsrJ6ettEng1DC0nyQZVNA+HAjODnfBWtW/TUtZMKl7snv3Wi08z3APB+a3BYdQ+71bYJXYbkAoNxR6iuGlm07aVrwQ8Z2PqvSrki1tNf6WAVeS/oGMrpbofRVi1tFT9YY55cO4Hqu+XLTw+i0pWU0VIwNaQOyDvfp5rhbmOd++3C2bxA0dW6kkFPBK4NecYyt82lpZbIG7ZDFoldV3GO4MdC0kB2+3kg85eenhPV6EsUt36HOJiLie+divK+svlRUySeIz6Tie/bde7nPDYn6m0e+KtjAjEJ8vkvHPWWibFZxPgsBa8gbfNBxFLUCUYDcFY+4O6zbnHBFO4QkELEEMjh1BpI9UFpOVkU9I6RviYOBur6GBsjul43WrSeDBAWN7kYQaNPOC0sxuNlbHTzSN6mk4CNhMk2wyCVqTYTGzCDTR1wPy4E5Wa2EVERkGxCuayGQkSYGPVQSzuhl6IvodigUtumq5S3fA7KGto3UcwjOd/Va9bqqmgb4hIye6027zNqZ+tnfO2EGLLTSRxhzSd/JY2XYOcrUYWTStDXjYdtlHUU3SDgIMEPc3sVeWOe0u9EZCXOx5LJPQxvScA4/KghpZAxx6kqn9b8g7KOQdJOPNUbG94yBlBNTVAiPxbrLcIqpoc3bK08QyH96VI1tRCMgHCC2aF0Tts49VHgk+a1qwiCqqSyrIwT5latd7bb4Y+uAt+xBtN0PS3KrDOIyMhKmTMha05A2WXTUImgLsboElc3AGM4U0FxjfH4PQAT8lZR0QMhEw2zss19BTxnrZjI+aDT6i2uwZW9juscUr24e44wtYil7slwAPVYNfPG13SwjCCkD2u+HbKuFOySUA47rBbJ0HqCq2rka/qCDVq2mjpYRIAFhRSe8fABhWVFbLVxhnlsFfSGODd2xQWzW5zB1577rV6O5QU1CYXAF2NjhaZV1b5WlrOw9Fi0x6pgyTOD3QTRRzVtQ4kkAkncrJpLZPWVzLe0Eh5A27KatDaaFj6bBccdlujThpI6L3+XAnbuPXKCer4by6foG3J0xyQDjK27Pcwxh3yRsp7/AK8vNwLqFziYmnpAWjxQvnhORuRlBkQV7p3EPdkLBuJnbJmNxA9AqRU89PIS4EBXOkEknxHbKDTnTTHZzzssimrPCOHDK1aK2RTtBYASraq1wQREu2Pog73exjqmTc0Gp2jy0DWn/nG3osP2LGBzU6rA7fsArv5RtyIPadERAREQEREBERAREQEREBERBx1zHfg88UP4GXv9RmXzR5BqJDjuTj8q+lrmR25d+KXy0Xe/1GZfNGP7YOexcf0oKRtc6QYB/ItXiqo4YSHgZPyWbQ0VLJEHDHVgLSrxA6N4ABwgwZpXOlcWuOCfVS02SMk9ljEYb23VzHPGzexQTVBJ2zsqQVJjYWeoKvaMj41ZLGz975d8II2yBsnXjIWSZWSt2AB79lBS0z6mToYMlZ8lA6kbh4IJ9UGntjdI8tWUwvpmEA4z6K2OCRsnWG7LMkiDmfH5INIkPU8u9VfFO6NpaD3V5a0y9KpJTSE5Y3IQWxTdEnW7JWQ6tdI4BuytpIGud0yfkKyqinghYHNxnugjqS9kXXG4gnzWqWTSdTeaZ1U95GBkZK0MTeIQx30fmtzWrUbrbTGCI7OGEG27lQuoKl1Od8KOnp5JzgEgLUquRldUmom8ySsSXqiePA7IKutjhk5ysSWJ0R3OFqDqmVsQLx5LAe8yyDPmgkoqV9ZKIgTus2ahNulHUcq2EGjZ40Y37qyWpmrJQX+qDUjcGGAN2Gy0eebM46TtlT1AbHDgd8LEjYZD1eiDUpwySlAa0ZwrbdRSFp+HcqOk68hju2VvvTlgkuQbFAzL3jbbzQaDY7NNPVEFhOD6LPvTZLc4Qe6uOds4XKFh4YXy04uFdTubCTnJG2Fu6ey6Gq7c51ZJEKho2zjOUHX22wNoiKuRpAznC5a0NxttVqmitEjWgvwxbA1NbzDcHtpx/tYE4IG2FsOugZS3JtREThrw4j5goO6V1v0dntA1JRztb14eMY2W9uCvMdca+5RULroWAkN3euo1DxIbdbKyzVch6GtxgrDteq6bS90ZUUMhDgc7FB7g8K9ZMhtsdzqrsx4ODguC50sV/pr5B41M9rhjOxyvF7QXMpfjbYaJsshZkDAcV6T8q/EWmvGlxUXGfp+9B2Sg7IItjVvGTQlFK6nfd2GUHHQCM5/KrIuMujXM8SWtaxn911AhBvskAb7BA5p7ELgziXzS8ONN2mRlFeY5ql2wAcBj862Vw05l6TUtSS+oywnb4vJB2oLgO5QOB7ELZ1r4mafulOBTzAy4x0gjurXXa4S1TRTtPQ47b+SDM19pZ+qrQbfGSCc+eO64pqOBNHbdNVsVVSiVz2k+vdc3VVRVQ24TRDMmB33UVNJUXK1ytqWAPcC3sg8lOMPLfFWaqnkjtxIDiRhvzXEd3npeFkclM6mETgSBkYXqNrjQswu89ZUMaIyCQSF56c12gn1VZPU0jepjXknpHzQdd9QcYK2WUujqMMJzjK3VpfjXcGWOWnir+klpAHV8lwvqq3UVK0Uod99AwR5rRrTVGijLHkgH5oNy3fU1/uuoJaiaveWlx7u8lpN8ifMPFkf143O+Vp9VcWNcZITkrCdd5p3eHJkA7HdBq2n7ZFf6gUTQBjbKk1NY26WlDC/qz6LCori+y/7bpPpndafetQVl+eHVZOQgtq7h7xEBCcE98bKS31VziicPenhh8srTw1rQOnchDUysPTvhBkS1Ukkx63Hq9VqdHbhNH4riDjfdaM/pcwOYfi9FLT3KqiYYxnB2QZtZXCmJjbjY7LHik95Bed8LHdC+Z3iT5337o6dsPwsIwgzYLhHSSfGAcK+pvkUo6Wt3PyWjzOa/DhnKzbdTQSDrk8vzIJ6eMVru2MlXSxe4zBxOQrppGw592OSAsGSplkJ8fOUGrVd2ikphG1oyFZbK9kLgZO2fNaZEwSOACyPDZER1HCDWLread0GIWgH1wtssbLWSEuJIJWc/3eQEOKjzFCCWEfJBZUUToY+phxjvusAvee7iVl1FW+XLTnBUEcJc8bbZQSUVQ2CXqeMjCnqP9tnxY9gPRWy00YYD2PooGNqGgiPJCDLhqGO+9E4I2+tWTUbnyAZ2J8lhgvifkjB+azIqs9PUTuMIJo7UY8PJUU1R4J8MDHl2V0Vxke/ocTjKlmp4pWl+2UGI2Xo+NxzlQSStfMHgYAQsd1lruwVXsYG/NBrdNPHXQiAbEDChqmChYWeq0umqX0ry5qy5pJK1vW4Hsgnste2nnL3disq5yNq6hsjQMDcrRqfpjkIdtgrMfFUS/wBgBIPog1umqYWwBhIzjCx6uwmsaalrthuVoZFbTOzKCAFmR6gnZEYRnpIwghyIZDTgbg4UU1K53x57b4V0b2yTGV5wScqyoneZQxv0eyCPxevEfTuFNFTB7hH2ysqCOla0PJHUVDPO2N/XHjIQS1FndQRioLshVomiqHW47ArFqrjVVUYjdkgKGmmmpwcAgFBqk1VFTAgDcLSqicVEgwO6nbJHOcykKOdsUZBYex8kFY43U4Ejzkeio1z55wYyQAfVUHj1mIomk4WRT08tG8mdvTnyQalUvYKQZAJAWhPiLup47DJWoVk4fHhpzlYga4U7zjyQeq/sOo+mycXpf7uosf5hXf616iry39hxI51p4xRnsyewkfaK7P6F6kICIiDrn7Q/P7TTiXj+86P9ep188a+iD2gxjHJ3xI8U4b7nR5/j1OvngwSdkFzfhKq94xjCszvgqux3QSUo+NZsrGiMuAGVi0Za1/xbLLfNGXhuRglBhRRyVMgZk4/MFmGNtI0NOCVkzGlgj64CC4hYJqROfj2wghmqOo4A2RmHNKkkpTIMxDKi8CaL6TSAUFkf9kKTd8Y7KWENLvmVmOtFTK3xmsPSN0GnwQOnOAp3BtMOh3dT0DoqeQiQgHtuoLnKySbLCMfJBZS0zqyQtaceinlhFCQ14BVKSZsGHMIyppGSV7+nBJPZBiSztk2a1QdRb5LVHWSopG+LUMIb8/RRNpo5XhwGWg9wg090jyMeSrHL0HOMrVKyKiZEPDILvRWUkdK5nxkZQRxXBrAMhZAvmBgNJCgqIqbqHSR3Ws2u22yemLnkdSDDpq1tc/oAwfqVa5ngxnLtyrqimjopC6DGPJaZNWSTy9D890GMxr5pCATuqPY+GQDfY7LVW2qokYH0zSdlq0FphZRF1aAHgfnQbZnqWyxhoGDjBUlIOiMvP2KelslVcat8NIwuAPkr6211VqeIappagx2tFc/w2DGFeZpraPCDyAVl0lBLN8VE3Lj6BW1FrqY5eqtaW49UGC2q8Q5c3JKuijFTUtixjJ8luC3ad+6oMdC0vdjsBlYVw01dbNVCSWMgg57IJbnp91HRicO7jK2zgk4GSc4W56p93ulO2mjjccDsFiR2v3FmK1oa/wCfkg1HSV8pdPv8WpYCPmqapr6fUUprKVoaButDkpH1MmGE4HbAWo2/T12D8sjJiPfbbCCCzsMsngubgDbdR19FLFXgRkjK3BUU1PbYg5rgJfP61p8XVVyGQnJCDk3hxrip0T7pLI50gLx55XrRyd3aHXtvguDosERg/mC8vOBHCir4lV1NSmEvDZAO3zXq/wAvOlRwopKe1uZ0OMYABHyQdq4I2xQiJvZowngQtPWWAn1WBaKiaoiEj+z91qZGRhB185qI4rvpyS3BuCIiMryd4t8Gqo0lVWRF2Osu/OvW3mi07drhpoy2RjnTmM5wvOvX1u1TbtMVrbvA5rxnGRhB56363vtlxkpHkktKhgqmRxFjmgkrVNXxVU2o6oPBJ6yB9S01lve0ZkB+pBDHUdEheNlkh0lQQerI9FjVEbG4wsuhYXMLW98IMmKNjADgZSomaBjqAQU9TGSZGkDyWBVNlc/DASEEU8juokOO6sEp6SDufVWva9v0wVRgy4IHW/8Auismmhc8h7twFZNG1jAQO6lpRKW5aCQEGd71HG3pDRssKesDsjzU8JhdkSYGNlZPTRk9TBsPRAtzRUPLf0qG4RmKfp8sK+mf4TvvX0sqefolaXSn4seaCOK3mqY0t7lbhp9PRQUJmeQMNJ3Wm2CUNmIkxgHupb3dqxrzDDvGR5IMe3NjqqswEAAHzWddaeKkiLWgEkbLRI5n0rhLH9M7ndalFUGtizUdkGjUwkfUfeyQSfJbqs9rlutUy3veSX7ZWiObBG8+DjIK1i2XOWgPvMX9lbu1BuDVfCN1gtoufjZBbnAPyWwoLg2mBjx22W5bvxA1BdYTRV4IhG262s+mjlcXMPc74QXS3Ed2DGVay5OB3JI+ZVvurGHDu3zVzIacncjCCr6s1BwzYqM0LyC4uyp5hTxN6oyMqSIyyRdTRkINOkiLdlEQRndZT3AuLXbEFROgleS4AkILYZfDdupHNdN8Tdt1QMZ9Fx3zhZbWBsRLRlBFTxY+kcqypaGP6m7bqwzPEh6Va/xH5Ls4QZEVaW4bIcgeqzKetle4MZIQ30Wj4GcFXxSOY74PPsg1mrdFG0OLQSrqSrY1niHsPJYT455IwZQcd91iSSkZjb2PzQazJcYKrMbQAe2cK91ozAZw4bbrQG+JC4PwQsxl4qQwRE/CdigyrdchTTGN52GyvulaKgZY7YhabUBgHiA/EVj+M47Hsg7++xW/Co1V/ACu/lK3IrvYsdP7aXVP8AK3+UbciD2mREQEREBERAREQEREBERAREQcccyX4O3FP+BV7/UZl80D/wCyu/74/pX0v8yX4OvFP+BV8/UZl80sYY6Z4f233+1BNQ3B9O7d2yyKqvbVt7ZK0yTHWentnZZtC2HwyX4zv3QYUn0ismma0syR6rHmI8UnyypmSNaMAoL2HxnlgOFlG2kRkl26wg4gh8fdXGvqGjpcUGTbJG0NWHO3GVlXm5MqSCwYx3wtPoJGS1P3/GCR3WRdI6cEeBvn0QXUFUyXER79t1dcXdBDG9iFpLXmFwcw7rOgnjnA8ZwyEEEsZY0SY3WZQTMkjLXDfturiIp/vbSCMLBqWvpHgN2BQZZiHiks2CxqvxM4zkKaGqYWbuyVL7nLUsMjGkgDJOEGmPgLW9eVdC2R56urYeqTF4eYvMbJEyZjgCCASgy44nzfB5/NZLKQQgdZyfmkskcFOHtPxHv6rTXV8z9iSgyq9zejpbj6sLTmfTB+azaMmqkLH4+1W3GBtO8NZ5oMoyDwMDcqGJ/S0npwVBSSdUga/stTMdO34cjdBpNRUOkPTjYLJtzerIe3A9cLc2nbNQVM3+2A3HzVurKKlt5/2kBgf3KDTGxMZjB3XIXDu/QWu708k4BY1wzkLiukrZHSgPPmt+WU0rYhK44cBkBB3Au+uNPag0gKGlbGybw8Z2B7LqbrWhu9BcnzRVrxEX5wHHGMq5mtKqgkMUTn9I7b7Lb+oNVy3AESk5+aDMumqoW2xsD3AygYz55W142CuifMTk7laXUF1TICScLLoZzAREdg7ZBFbIKmeuEDHkYJzus240FXS3GNnUXE+XdZoNNRtFTER1kZ2WXpCaO+azoqWs3bJIG7/Wg5f4UWO4VkEPXSuA6geotOF3h4faxq9HaNfTRVRjd4RGxx5LB09wq0/YeGkF6ijjDzEHZwPRce112FXQT0cDz17taAUGybtxK1dVa3nf8AdmQRF5IBecYys3VHGfUFDbnULL24Pc3GQ/zXFOsbTfNO3OS61AkZDu4EgrhXWOr7hdKpzqSeQhh8soNy8RtW6+bUm5G+zSxudnAeSPyLnXlq4l3ieiDamrLXdONzhdddA1LNQVLaO/S/ewd+pcstvWmdGzRUtpnZ1uAHw7boO4uleLN009WCpmuRczrB+l5Lt9we4tUerWQCeVuekbleXumr+6qhZU3BxEJIOSfJdh9AcZNNaYtJdR1A8VrNviGc4QemFPVUdVEBFPG8Y8iNlO1rGDpY0ALodwL5j7rqXVxopZHmnMoG7tsZXeqgq4qymjmjOQ5od9WwQaTqvTUF+oHU7Y2h5BGe3defHNtpgaMoa2CShdI5xJB6cr0l33GFwjx94XWnW9lqam5NZ1tBxkAlB4EaypKt2on1D4XsYSSBuPNbZuUhDiIxjfH1LtvzIcLKOx3aeK39I6c9mrqpVUBgklgnPx5wMjdBobRK09TnEk+WVHJIXSjGy1NtiuMDveJ4iISc58iFg3ERNlBi3QTiUthAJz9axXnrBLR+ZHygwgA7pTuaWkO7oIQ9zSc5V4kDzjHfucK6ojAy4eqpAGkfPyQZZpTBH4rjnzWM6pb1ghuwUo96l+FzT0K808DI/iwCgrJWNni6GAArH9zIHUXZyoCC156OwKyGyyubg52QYz2FpwT5q+Od7AWtJGVbLnOT5qSKHrb1AdkF9NUmJ/VIchUrKgTvBaMBY7hhxHocK9keSPNBPTzCnwXd1LNKahpc0/MbqKWncWDA3VtMTG/peMAoICZMkEnKyqSB8wJJP2rUY4qF0fUSMlYlT4kcgbTAkILKin8IH5LHjnazvnZZVUycw9UjSD5rTxjfPdBr1johdpfCJGyz7xb4bOBGQCStBtFylt03XGTup7vd5LjvKd/JBgVLmucXjbKg+Lturvjk2G61+10lF7oXVJAfjbPqg0FoMeHeayYqslwYTscBQ1nSah7Y/og7KJuQ8DzBQarWUrWQeOD3WDTx+Nk/NSVEtS6ANePhUMAlaC5gJCCaWjAaCCpIZ2wxFp79ljvqJAC05BUBLn7eqCaCI1M+AcZK3JRzQUDQJQDt5rbUPjQu62A5V0lVNK4BxOUG4LnV01ZGWxMAK0B1K6POSp3NkhjEhB9VH7yJGHJ3+aCMd8A4+1ZAgzH1dzjusQRTPJMYLt1nU/XHHiYY8kGnPdIHFuTseyyqSB8reo5IUnhQyvIbglxWsUkUEEBD8AoNKk8OLLSMYWM6pYQWgb/Up7jiSQ+GchQUdNmQGUYGUEbYJH/EARnfKjkY9h6XFa9VCmFOGwEF2PJaX4YIPi9/mgytP1EVLUeJMAQVmXuoirZA6ABoWhSNfEcjICzaN3WwuOdh5oLTA5rAXFXSEe6Px/cqsk7XnoBCsqHAUzm+uP0oPU32G3+5nGP/AA9h/wCjXL1KXln7DX/c/jKP/o1g/RXr1MQEREHAXPlazeuUriHbAcePSUoz9VbAf5l88VwojbKuWnceoxkt/OvoZ5+bhLa+UXiLXQ/TipKQj7a2Afzr56Y5n3OqklqTkuy4/WSgxm0rntD8YB3VroSwdlrTzBFGBsCAsX73IDuEGlAOJwM5JUjYpHEAnfy3Wc1lODsRlQOjldUgRjOT5ILoqKZ27nHHzKslo3NcMFapV09TBTB4aRtnssGke6V/3w/JBlW5zYQC8E/Yo7nUskdhjMfMLO8OJkYcquioXU7nFw6sbINBiYWPDz69luWjv9LFSGne0AkYW36bDqoh/wBHOypXtiY/71+byQSy0TqmZ8rHYDiSFBPRuhBLiSVWCukj2yrZqp8r9ycHugspmufIG52C1eme2jmbM8fCN1jRiGNoeCMnCyiY5acg43QandNQUt2pBSRABwGPrWiUzDC73Y7lxxlac0ugmDm52Oy12mfTGDx5MB4GQggrrBLBEagvJB3wtKEEnSS07BZtVe6mfMRPwZx9iU9PLKOtrctPcoMKGCSZ3Tk4CmE89E8MDzj5KaYeB/Y9jnJUUfRK8unIBQTzVr/DDiMk+q0/rLpQ7tutbggpqhvS1wJx2ysSrs1Q1/3phLfqQa9p27wUjR4zQR81jahuUtdITTREM+QWLQ0ZhDRUbAkeS5ZsOl7DV2J9S8s6wzPl3QbA0XeaayzGSrYCT6rUtWeBqNpqqVgAG+Qtu3iCCK6PhjdlodgYW4bfJCLW+GMgvIwg0TTFc20VhbLH148sZWfe66O9VrYIYejqIGMYWu8LdNU9denvvTAIi7PxDbCzeJFustnvcclrLS0OH0cILdM0kWi4xcasBzSM7rE1TrC337MtPANttgob7c/uvbI6VpyQ0DZY+l6O0U7hBXuALj++KDT7Pqant1QTPTjBON2q28R/skqxJSRkNO+y3tftCW+6UrZLQ0OcRn4VkaV4fXCjiLJWkP8ALI3QbfsmiJZI2ObTOeR6DOVr09RNaIfueLJI5xGM9BK7IcunByu1Fcooq2PLHSAHLfLK762Pkc0BX00NZdIYetzQcCME9kHixPpC9XWY1AtM4a7cDoKxjpivtsjjLQSNAG+WkL3Dn5KdDwRvFJFAWgHGWALrhxl5T6GibKaCFmQ4gYag63cnevLZou7ROuULWjrH0hjzXoLaNe0+sdR0kttIEW2SOy87L1wO1hYbnGbTBKQH/vWkefyXeDlY4e6mhtbK26wyCRjAR1A90HdmzNa22U2N/vY3WctmaRqbkHikqQ4Bhxv2wt5oNHv0lAylqDX04lY2MkNIz5Lyq52eL9ttVZUWiioREHvLdm488L1hr4KOWJ4rCA14IOfReanPHwUsd7nqblb+lzw4kYA75QeZd5kirLi64dI+MZWhV9SzBDWY39FuvUWmLjZ7s+mqGFsTMjsts3COjBLRKAcoNvz9RdkhZVunMDvEI7HPZW1IhjcPjz5jAWdR00NRSu8MgnHbG6DKnvUFVH4LGjIWOCyOMueBnuse10zIqs+8YABS8zsM3hwHLfPCCyKFtxlLG7H5LHraM0UgYXZKvpDNTnxIgckKyrNTO8ySg7IIxmYAE9llRVTaeMxFoJIwsNgeDiPup2Rsc8GU7hBDIHhxduAd1LDUnoMZ3JGN1PUQmVgEQyrIKCRjg+RuAECnYYnGR/YqGqmMkp6ScLLr54GxCOMjq88LTmMfI7DRklBrVohM+zTg+ZytTrqRkUJc7GQFpNtZV0nxlpA7qlyuksp8LJydkE9ot7blUmM4wFNd6b7nu8BmMkYWmUNTWWt4qMEA75WdJVm64cTl6DCpKR7ZDI95wfmsxjzFUNf3aPJRy09XTAGRpDfJZMNM+opjIAgybrV01ZSiKNrQ7GFoEJNO4NdvnzVk0s0MrmjOFPEY5Iy6TAI7IKVH35mGnfGywfjDunJznCy2v6XnqO3kpXtgLS8EZQYZp5OkOLtis2nuLKenMRAJOywTJKcgE7FXiOPpy47hBBI/rkLxtk5WbBXRshLC0ErBI32UzYmFmScFBE93U8uGd91NHU9MZYc5UUYb1HPZUkADsNQVY7D847rVY6dksBcBvhaZGzBBctUpJ2tAZnZBpb4iJS3sMrMp7eMCcnIG+FdcPBa0lhGT2WPHVziIt3xhBl1dyjfEYmgAjZaY0nrDjvvlW93fEs6FkDmZJGcIEpZNG1owDhYr4Swbb4UkzXsdlmwVzXh0Z6u6COngNQSOo7K2eAwP6e6RzuheSw9ysnpNSOt26Dvj7FXP7abVOfxf138o25Fkexdja3mk1SR/wBrR/wA425EHtCiIgIiICIiAiIgIiICIiAiIg455jx1cvHFFvroy9j/kMy+aPwy+ofGNjk/pX0ucyJxy78USPLRd7P8AyGZfNEZCyZ7x3JP6UE7qAtAcTlY5a5pwCQsmlq3PkDZCMJXlrXDw8euUEIpXEdRJUTmlhxlX+8PAwP0qNzi4oJIJRG/LhkKs0jZXANAG6j8NwGSNkY1x3HkgyZKN8UQlDt/kscSu/fEkKWSrkcwxHt27qBrHO7BBQ7klZMFDNNGZIwSB+dIac7F+wK33pPTslaWlrMxkfFjthBsOGaenkyGnPYghb4s+j5tQ0BqvdyCBnsuUdOcPNLXBwjmMfikjbA7rlS08FroaQNssWYnDyaeyDpXdrRUWytdTujcADjOFq1tuEdPSmN7QTghc3cYOElbpqmfXV8IY7BO4XANNSSVhl8AkhhPZBgVEo97dKBtnssps4kjLgzBHkpqGijdVGOo23wtbbp7x6ljKZuWkjOEG0ZJJXnBBAz6LLgoHPiMnScgLkm4aBjbaxPEweIRutJtum6qOB4lYcD1CDZNu6o6rp6T6brWKuzurAJGnB7rUo7NGyrPwjKtrKS5MqmNgYfDyMkDyQbeFqlim6ek7fJWTwSxyZ6jgLkCqo6WC3CSUgS9O4PdbFrnl0zunsCgkp7rJTNAa4gjZSTVrqxmJXZPzWFKyIxg+eN1hid0ZwEE8sIp3eIFnUd2mGGh+AtJfUPlHS/sjHtYQUGuz3Itb1EjK0SpqHzyE52Kkle2RgDd1ilrmkZ2QZ7C1kYzuceix3znq227KeAggBxyFSWKFpyO4QX/fOgFzzj8y1TTLHUV7prqH4EDw/v6LQ5J3Nb0jsqx11Q1pijzh22B3Qd79JcybdR6Vh0jHUguaBHgHf0XPHCXglPqShF3kaXNLfEIPn5rzr4DwMo79HWV4IZ4oxkr105db4+5U1HQWxofE+MA4+pBxTrrgUziBG/TVJaiJWZZ1Nbv6LZtl9mbXQvPvFukd4pyMtzjK9MbDw+tNsmbcfBb7y74nfCO63bjsg8jOJns9rloW2OudDQSDDS74f/Yur924A6upqmS7yUNSWUrydwSNivoDvlko7/QuoK1gdE/uCMrju4cu+iK6inpDTMAnBBPQPNB4R3vibcrdTjTQopIZGDpyWkdloNDri/0PW99e8NwTjqXo/wA1/JtpHSsE1+t0EQeWF4LW4XmLrGh+5l2qKUghrHkfZlB2g5S+Kc1VqWKCSUBxlALj57r2V4Y3E3Cx08jpw8mIHvnyC8H+X51HZ66Cspj98MgO3rlexXLJfbrd7VTdQcWCIZJ7Ywg7Frb+rNPG+26SkGfjytfGcbqqDozx25U5Kxkt490dLlpO268xONfCq46Zv0tQy1ytjjlO/ScbFfQvcKCnuVK+kqWhzHjBGMrqjzNcuelKrTdbdfChDyC7doz2QeMmpL7RPsTLe2EMlDMdsHOFxj4DjkuySuVeNGnoLNq6Wgo3DpY4jA+tcaPjfF1h+xQYLoi1XwxE7gqhLnP37IZXMOGoJJPiBb5qNrTG8OPYblXRMe7cg7+ayPDBZhwQZEdyi8MR9IyFhzB8z/h2BUHhlr9xsFqUAj8MuwMgd0GM6EQtBIyVEJwTgBVknL5C09gVR0HYtCCYQidm3fKoJBTDoLe6sglfC7DsgLP6aadhe4jKDTZotjID33WVb4Wvb1HyWNNKOsszloRtU6IFsfYoMySqZE4tOD9awpX+8zAM2zsoiXSuJ7koC6NwI2I3CDWRY5hAJvFz591CypbSuAkwT6qEXiq8MRknAGFhzSmV/Ue6DU6m4Mqm9DGgZWG2iLsHJ3VlP0B3Vn8qmkqiwdLe6COSI0wyN90jgdUML+2FkUDm1MobPjHzUlwHgHophsUGHSkRzFrvqKrVTPL8McQ30yoXMkaetwOe5WbSMjliLndwEENLSmT4nHurahngzA/NXslkZKWtBxkqKdxfKOrsUGTU1rJacRtbg47q2knZHGQcElUliiEILe+Fhhru6CWZ+XktHdUieesAjutc0vSUlVP01eMZ81qF+tFEydkdFgk+QQaS1jGxhxaNwoxQtlImHYbrNuFor6OlEsjCGYyNlgUlU97C0dgguqqlksXu7W7jbOFjw217iCSRv2WfSR07nkuxn+dR107opAI+2UFWujoG9TgCoJqhtYMR7fUrKiT3hmD3xusJrnwOyEGTG11I8Pc7I+tZDpXVQ+F+B9a06Sd8uziqxVD4hhoygynfeT1OOVUVLZWlrNisXrkmO/ZWnMJ+FBkRl9O/xHuJHlurpZBO7racAb4CxJJnPGCjDI0EDODsgmqJ2uYGAbhUp5+hpYT37KMBo3crSQXZagyTTmP78XbFRSzl7S1XOfO9gbg4UfguDC5w7IPVj2Gv+53GU+s1g/RXr1MXlp7DYEW7jIT28awY/JXr1LQEREHXX2hRxyccST3xR0f69Tr57bacPkzt8K+hT2gzQ/k74kNPY0dH+vU6+eWnkMbnEeYwUGRO2SVxaHHCjaxzCGEnJ2WVFLGd9sqKpxkvb37oJYbW8/fC/wCeFm0cccNSzrAONsrS4q2qkPhx5OVQz1EMwMmRvlBuu9VELKLIaDstptqgHDpAC1SSpbWwCLOTjCwRQdDwHDAygvdVukbjssZ7Jn7h5x9ay6yBkcOWAZ+SwYZndYa/OCUFr3OZ2UTnOdu4krWnQ074sgjOOywZYGNyR+ZBj08XiuwlRGI3YCuY4sdlvfKpKyaQl5BKCLrfjHUcKeOpeB05+SgDd8EbqSBrTO0PG2UEjuofGWndQmWQ9nEZ8gt1GG3upM7ZwtuVsTRIfDIIQWUtMap5aCcrXKeojt8Bp3gEkYGVo9ukMUpI8lLLN4tezr+jkIJpI3dZlIOHb7qF9KZyDGcb7rXbu6ibbWmFw6wPzrblLUyNd0eqDKpWSUcvUSSMrdlrq46rDXQZ8uy0SjiZIGunAAzlckacpLEKMbs8THn6oNGl022vYHRjpJ3W69O2CaOn9xdOQH7YytHr7gbZIS0YZnbdY8VdqOoIraBjyxm+RnGEGdqXhaKUPr/FBJHUFsCeSW0VPm5gK3HfNYaoucYoI2vLh8JAyrLdaBLRudeG4eRnfuEGEzXIjhDKX72/sfI5Wm1VZV3WTqmlLj3BJ7JU2ugbUuERBx6KNskUEnhg79kGsWSlcx3312x7ZWPd7HVz1nj0zyADnAUkcs8bGuaDgrcFkiqKpxc8fBj0Qbn4XV9RRSwxVdO6cF4b2zsu7/CbgVS8QBFXCh6AWA46cZXEHK7pzQdwuEQ1K+IESDHURuV340pb22q700Oi2NfSEAfB2x9iDXuEXAig0p0O9xLSxwOcei7DU0DKaFsTBgAAKCzxyR26DxgBIWAu+tZqCjmhzS0+a2jqHQlBenEywB+SVu9UJDRknYIOGr5wr05aWCrltTJCN8luVuTRl4s1BAKOmoWxdm7Nwt61QoK6M007gR5grRZoNMWtpf8AB4g7YI7oNZgZTNHvMLAOrfYLLjf1tytuWvUVHNL4IA6PLdbkYWuYHMxg7oNOvlvluFKYoXEEghdfONXCJ8ml6y6VMZlMYLj5+S7K5GcLRdZW0XbTVfQEZ8WIjGMoPDniLp+K/wCrqizwUBaQ8tyB81i23lErrwz3gwPAO5XcrVHLlcWayqLnR02S55d9H5rXafQWsrZQmGCmOQMDYoOkJ5Iq65h4ga8OjBJIOFwhxL4a1nCipfRTRPc7q6TkZyvT+zaU4mU9XKfdnhr8/vT2XAXHfgDqq/15ut3pCIA7qcS04QdAIGT3F/S2FzMjudljVttkpJPvjicbrsBrDR1htMbqO1hhqo24IAGcrgnUVNcKepc2oaQMkIIqSaNgw4A4VtZWxEFjWjJWFF1gkklVeGOf80CJ4if1OGxOVe9pqHdbThWSRHGfyKyCR7HhoPc4Qa5aYW7CQbg+a1CoiikPgMxk7LTYp2wRB+TkhR+PUzuDqfJdnZBrdv4ezXIl4lIB32UtboptjBlkdkhY9sv9+s3x1AIZ5Z7KK76vqLmwtcSR8igwZrxCMwNYAQcbLDZSeLOJyfhzlQNjikkLycnOVIaiSMhreyDVa4xVdMII2gEeawKNptswkecjOTlIanpGR3UdTIZgRjc+SDUrtfoKqnELAMjbZX2muY2lMb9yR5rbT4nsGXBahRvaIekfSIQTVUUcshcMDJWlyB7JCG5G+2FlzwV8B8R7SGnzWvW2ntk9L1zFviAefqg27Mw+H1disXrcD3K1G5xlsjmxA9K04xvAyQgzaEMmcWuH1K2up/DeOjsVixTOhJLO61FjvHiLpO6DCji6iB6LJNPhhId9ixXeLG8kZwqiqk7O7foQUZDlxGVeafDgfLKsMpzlndX+K/p+LuUFzgCMKwvLBsTlReIclW5cd0Avc47klZDKhjY+nAzhQNYXZ9VRzek4QVx1OOPVZcNJIG+KCcDyWLF9JatTskfH0DsUGDJUjBYR22WM5znZ74WfW290PxYxncqSnipvd8ucOrHog0sjzWZSTDHSsWUYefTKyKeIlnW3ug9A/YyRBvM/qd/91oGt/lG3oofYvSyv5pNUsf2GgK0j/ONuRB7QIiICIiAiIgIiICIiAiIgIiIOOOZH8Hfil/Au9/qMy+aCT+yP+s/pX0v8yW/LtxS/gVfP1GZfNXRRxyVMjZcY3x9eUGGHEHIRz3OOXHKyaqEMeSOyxT3QVBwrmt6jlWKeFzQ3dBWSYFnRjfCshfj4fVWP3ccbqjctcCR2QZEtMWgyZwO6sglDO4U80wdB0gqykYxwPX9iCUyh7RgY81vzRGr6eiljtcmOqXDcnyJWxXyU7BhpGVjU1R7vWx1DScscHA/Ug7eae4dzQ0bNRiq+F7g7GV6LcreiNP3vh9Ldq5sb3QRF2XYPYLye05xqrnWmK0SSHoaQF3J4E80NPpPRE1qfUYEsJafix3CDROYyeza91lV6MomsZ0OLNsDzwur+qeE0fDeOdrx19ZJHn3W4uJvFemi1vU6itUmZXvJ2PzW27lxLl1lSvN3d8Q7AoOH73TOpqk1EYIBOVnWHVsFEfv4BLfVVu0kM1a5riPDB+xbbuNLH4x92AIJQcrWfX9LdJBSvx09luGrloRRkx9GSM7LhWga2gaJo8l53W4qLVD5GiGZxz2AKCW4VbYK0v7Ala3bLtRzQ4ewF3YHC2ve8vi8YdjumnK+mjB8Z2CEGbqQyPaXsJDSDgZWzS4gPyMnJ3W8L7X0s9OWxEE4W14RD0uEuBk+aDSnuk68b4UngGQdXoswQwmUk9iserf4bw2PtnfCDEe3oOPRXxQOkGw7qR7mOjz5rIoXsa34iNuyCAQGA5duoSfFkHz2WdJI2V5AWMad4cHNCDJfCWQhwwFgOfIT3Kz3Pd4QaVqNDR0MlK98mOrGQg0KGM1B6PPyWbDRe7ytkeMgbqGICKuIZ2BWsTNMkJICDVIdWttlNGylIY8OB27rvxyXcw9NZDTQ3GtZlrQPiK81HDqnLXkgZW7dLa5qtMV0ZpXvAG3wlB9FfDri/aNbRMEFXE4nAyHBck5B814+covM1S2ariN6qXNb1D6TvJeh1DzWcP6w0zY6yMhzR1fGO+PrQc7KGqnZTU8k8jg1rGkkk9sLY1v43cPrhEJGXhjSfIkH+dcecZOZTRthsk8FHXMe4gjOQD2+tBw1ze8WrdVwz2gVDSWsLe68leJsUNTcKueJ7Tl5OftXYTmO40UGobpNJbpy5zgdg7K6jXW91tVPOZy7DnEjKDc3DjVkun62LLzgSA9/mvWTk/wCYelbS0tpkDSXsDd/qC8g9KR0tRUxmbGesfpXoTyn6h0HaLpQRV8zGzbAAkd9kHrHaq9tyoIqxgwJBlZi0LRtzt9ysNNPbXAwhgAwtdQFx9xa0hJqfTtTSNDndYIwPqXIKjmjEsZZgHPqg8Tua7l6qtJXee+PpH4OXZIPzK6V1bfeZpmBvT0OIxj5r3F52OGVfqbT8zqOIH70ezd+xXjDr/Q920jc6xlQ0tHikbgjzQbHqIBCM+fmsIbnqWoVQcYx1HJIWPBSveeoA4CCsVU1rcEI6qBO3ZWTsjYOnzz2UI7d0EskodkBXQyODenJ3WOe6qCR2QZLaYucXZCq9/g7HcKKOdwO/ZSOLH9ygp1ePsBjKkjpZBt1HB+axuvwz8Cy4KgujJPcILm2tz9wd/rWLUUr4DggrJguL2S9LthlSzyNqRkbkhBhwxhnxE9/JUlb1O6gNlSSCoZnIOFkUzMxEyDdBivkaWgY380ZB1jKsk2e4DtlGyOYMBBd0mNyyYqE1A684woYmOLuqTssj3p8Y6IuyCyZjqbZpwR6KkVbj+yfF9aVMc5b4jwcH1SnjikZhx38kFKmpZIAGhQxSuYcBxAKmZRv69x8OdlFUMDHDCDI6wxvV3KhkDpj1AHCsY/JDXHZa1RtoxCS4jOPNBpUbHOw1x/KsptM0MJOFHVYEhEH5lAKmZnwu28kGTDK+CQ9DyFkR3J8VWyomcXBpzvusGSRvhhwUkUbJoTk90G8rhqqjvluFvjaA8Nx2W0Xw/c/Mbjnq7LCa91LN1Rncdisk1Yqd5jgjtlBa0vjcJerYrMjZ48JeRk+qwXSNcenPZa1RSU7KYg4zhBorAROW91HUYLw0eqzGuhFSScYCx6wNfKDHgj5II5KYsYHnsomDJz6LMmLvBDT6KGOBxaSAglgcHfCfJQ1Yw8K3pkY49Oc5U4Z1M6pO/wA0GGpWShrcfJWSNw44GytQSsYZ3YGyldSmIgkrHjkMbshXyVL5BugzKeVuwIBwo62QDLAMZUEDul+XKSpcx7cjug9VPYbPza+MbPMT2E/lbXf6l6lLyz9hr/aHGb18Wwfor16mICIiDrt7Ql3Ryc8SXelJR/r1OvnjjO7vmF9DXtDTjk24lf4nR/r1OvnmgaXEgeiC0Eh3c/lUwn26HZJ7ZVnhkO3VTESOoeSDOtzGUr/GkwQeyrc3sqcvjaBjfZaeZpHDoH1d1nUboxGGy4Bwghtshjm6XdlqFXMDgt8h5LAqRGw5ixk+YWTRlkkf33v80GJNVOOxyQFjOy457Y7LWqe3MmkJHZYdxoJKeTDR8PyQYwlfGB8RWQ1hli6s7kZWNBE6R5a7OyymkQyBhO23dBggmKQ5B2K1WB0clOSe+FHWQQPj6oiC4+iwopZIT0E4BO4QRyZEp+tN3fEDghZVSIfDD24ye+FhtOMhBNHJPJ8IkOAkjiz4TuSoWuLT8KyIg2QZk/OgtpvpHyWQ+mMj9tj5FWO8JnYoyqAeCDsEEs9NPHH8b8j5qtJRdZEpxgK+R8la0MhGSB2V0LKmkaWytIBQTVE7ugRx7EbbLLtM9ZA4SmYho8iVhwy0zjmRwznsqz10TAWRn6sINcul6+6MLYGP+IY81vjSmt7dZbE+iqY2OeW4BI3yuJ4A/q62jcrVBJTGEtlJ6sINzWzU9BFeJaqWNhY4kgY7LB1PqaK51ggo3BjScbLab2vjlLxkNz5KF5LZBMzPUEG6X2V0MAqXSbkZ3K0U0L5KwPDtg5RfshrXsEMuQwBUZdGs3Dt+6DdMsscdKxuRkYW5dK18MrPdwB1OGFxiLnJOQ3JIC3VoS6U8N+hiqTgEgblBzPp2z3+ilgqrfPLEOsH4SR5r0u5SdbRUNrhgvDxLMGAZecnOPmuk1tv+kLZYKaarljDjjHbuuwHA+5OqHx3i3tLqGMAucO2EHoha79Bcow+PAB7fUtVBz2XCek+OWgaqjjtFHUxmtZhrgHDIK5SteoqOqphI54bkZB+SDWiQO5UVS5op3kvAGO+Vol11daqOIjxQ5x+YWzNS64nhtEtZCD4YBOfkgyrneWU9RP1VzWhoLhkhdMeP/M9WaHvD4IK4uYHkZDtu6s4r8b7vV189Fp+R75QCCGlda9f1tlu1onn1g/oqzkjrO+ftQc6aH51m3CSKN9waHEjPxb5yu1HD/mdt9zp44J6qN5IGCSDheIsF8pKDUhZbJHGNsmBh3lldnOH/ABZt1ps/XPMRMGbZO+UHrxpjXduvUnjfdCMhx7dQW7qyopvcpZHzM6OkknqGOy8jdCc2NTarmY6iocIg7bLtsZXL8HOJWXSdtHDO8wOAB+LbCDuRab5YK+9vpX07HkOIzgFbqNJp/wB7Y00UeD5dK638L+K2lp3MrauZgleRnJHdc2Ra4sNSWVkUoMYGTuOyDkGOz2ctDo7fT4PowLrRzV6r0zbNOVdohoI2zPBAcG43wVzVScYtFPzA+4sY+MbjIXVLmv1ro28UVRLTTsMuSRuEHmVrOKaz6wq7rPKXQyOJDSTjC4y1bdqW8TF8LAME9lvTindZK2+TsYT4QyBv5LiyQtjkIztkoMZ0IJJxhYMzOiYn5rVZ5o+j4DutIlEj5CSPP0QapTUoq4wB3GyhmohTzBp232U9qmdTZ8QkD1KguVZ4tUCw5AQZUtMZoWhv5lk26VlreJpWjDdyMKOOti8EN6hlSudT1UJiJHUfyoNckutJqmIUUDGteBjOMLQa/TjraC1zsk53VbRb6y31JngBI8io7tW3GrqRG9pyTjsg0+Kie1+fLKtnaGuDStWbbKyKISzNIBHfCxZtPXaozVRQuLMZz6IIaemLt87d1DUPFPOARthTx1ApwY37OGxHzWPUxPqT1sGcIKVFU2aMtY3J+QUFK90NQx5BIDtwsigga2fNRsFuSC10FSAY8Z+SCC43OCqoWxMiAcB3xv2W2g+eN+z3AZW66i0MhaS4YHdbarmAThrTsglfXDww13f1WPJUte0gN7qdlF1MDiO4VstI2NhOUGNTsEjzkKUymNwHke6ip39MmAdisySFr2dXnhBFLMC3YZ9VjsPiuDO2TupqdoDy2XYeWVILZVPf4tOwlg3z6IE1CaSMSk5GMqxsfjM6x5KWslmfGIXA5A3CshkEUJY7Y4QYRHxY+ayY4mlu6gBHiEntlXl0n73OEGZS07XEk4WLVx4mDRuqRVEzThgJWbSxCSQPm9coLILc57Q7tlZkNWylkEZGVNW1UMUIbDgnHktPpmsmf1Sd8+aDVqhzKqMYAGR6LCFmkJ6g44V1XL4cYEG5VtLdXsHRIdygxrjQ+7N2GSFJQQtMAc44OFlOmjqATNgBYU1SIneFFggoO/HsYnNPNJqkAbjQNdv/AOUbciexhaBzQ6od5nQNb/KNvRB7OIiICIiAiIgIiICIiAiIgIiIOOeY/wDB44pfwLvf6jMvmhL3RzPc3Y5P6V9L3MeP+t44pfwLvf6jMvmi2M7g7t1FBc+cvaQcknurI4esE57KeSFrWdQGx+axxI5pw07ILXjpcQqZVd3O+aubC9xAwcFBa0YOUc7KypYg2P54WJ9SBk+pUsBJe2MfvjhRKeCN+0je4OUGtv0o9tIKoybHfC0KaHwXEZ3C1B2oa4weBn4cYWmvkdI4k+ZygkpqqankDmPIwtei1teoIRTU9Y5rTtgOW2lPDHGW9bjuD2yg1Gapr5JDWVM5fnc5OVWK51EhAZKQB3wtNlqXvb4efh7K2OTwslp7oNRuNS8xAdRye5VbUA6NzpN/rWGJWy/C4/lKlbO2Bhaw4QJqwMlc0ZwCo2zvfIJWkgA5IUDB40p6vPcrIkDImdIxkoNSlvjZ6YU5OTjCggopZIzLHJj5ArScEnLfJZDK+eJnh52QXGqmilLHyEgbbqGeodI/LSQFYSZHl7+5OSVacAjCCYVLg0DJz6q10nWN9yrSGFox3ViCpO+EyR2JH2q4Ru7kbBUfjyQXwyFrjkndZRrWtHmVgtwCsyGOB7R1dz80FktYHjsQrY6iUERtcQDtsrqqKKP6Csp4HyHqA2QZpp/BjE/mVJFcgYXMI+Sj6pZmiHHZVhpWxPDZNslBpsri6QvwRkq+BjjI12cYwtwT26lbT9YxnC0Gc+FIfDIwEG5aG8XehgH3PrHROHocLdlk4p60tbA+W9TEsG3xlcYw10jXAHsVkuqmSDoccA7boOa7LzNaxo5jG+9ygZx9MqTVPFzVOqaF7nXp7gQdi/K4HqaeGNvWxxOVJR3aemZ4QJIIx3QZFVeLk64ufWVT3kEg5ORhVrJG1LepmMkb4WFUmOQGXO7t1bTzBrS0nGEElurpKGoDmuIAK5Z4ZXG/y6oo7jS3B7GRuDsdWBsuIYqOepeRG3K5G0FdDZa6KCc4JwMFB7RcoPFyS82OmsVXWB8jcNyTn0XbheVfKRq6Oz1UNZI4hpeDnOy9M9Laqob9QwvhkBc5gPfPkg3Cip3VUGi6m03Qajt76Srp2ydQI3+YXk7zxcF6Ky3ioFJQBgdITsPmvXxdFednRl0v0s9VBHkB2cgIPI3VmiTbKds3Rj4e2FsyimYwuhIydxsucONVsuNnZ4NQwgNaQchcCUjw4yOz8WcoMyrsrsGcO2O60iWMxu6T5LP+6dT1mN/0QcKr2RTN6tiSg0xVDSRkbqWeLoIwNlSMt6DnCCJMlVd3KyIYmyAtAyT2QUo6Y1UnQtSmtvusZOdvPf5LDpoqiklEpb8PmVqrpn1kZ6RnyQaPT0hrJywHBU1RTm2yta453RgmoJjLgjBPkoK6sdVvDneSDKlucb4ujpGcKyAidpAWI2AEZ3U0UrYGYB3QWVVIYsuzndVo6Q1BHfAKMqPGeGyYxlatDJTQQnoIz9aDHrqdkdOWtxkLGo4QPjecgHKjq618shx2GyhE8gBaPNBnVtwjfGYWAZ7bLTmucxwIJGFm22nY+bNQMN27q67x08cgEGMHugq64s8IR4BI81gPd4hyrEBIQFUPcP3x/Krouku+I7LKcyHw87ZQQU83hydTjkJUStlcC0KIjLjjsgGBlBcGOd57LKjBjjwCsPqI7LKglaQA5BjSElxysyltclTEZRkAK8RwbE439Vr1uqaOCjLCW5wUG1ZITFIWnyVfHlaMBxx2WRcCJahxi3yfJY4pZtnFh7oEET6iTAO5PfK1BlGIGdTznAWJnwAOj6QVzameRhDsoI6icFxaM7FXQ1bWNIKpSwtdJmYbFK5kTXjwjn6kFW1LA7JCtmqOs/D2WNg+iyYBEWfF3QQueHDCtaMuAV7onEktGQrC1zDvsgnfS9EfWSoGjLgFIZpZR0bYQROaOojCCR8GGZHosbJ7ZU8Uri7pf2Vk2A8huMfJB6wew8ozDZOLtWf/AJPPYh/5orv9a9RF5e+w9qTJYOLdOf8A5DUWQ/lFd/qXqEgIiIOuvtC2GTk34ksHnR0f69Tr55qd/hvJI7jC+h32gbg3k94juOwFJR5/j1Ovnih6eo9XbH86CaY5b1Ad1qVFQskpDISOyxI3QOADyMBUFxdE/wAJhPRnHdBiOJhndtsCRgqj5HSOAaSAdsBaxLHROhLyR1FaS9sbX/Aex9UE8NO4NDnb77qURnYt2weyrTTMDMSEKskhc8dGMZHZBl0dW2DZxx81bV3OGdwZsSdliVkjGxjBwT5Bae0PcepoJIOcoNcpqBo++jG6xq2AyTdLcDfGVFSXJ7CGSnYKWaXxHeIzy3QJaM0cQle4kFYEhExJb5Karr5J2+Ec4CjpnMaC12Mn1QRxQvld052HzUslH4YySrpXiI5ZhRGqe/AcBhApYvEl6Srqpvgu6QftVWSNiAeDuVE90lVINiSUF9JTuqpOjqKmnt5hPSCDlWx09ZSHxukgDzVXVj5ZAHH8yDMs8jaCYyygEE+azrhWRXAgRAD0AWNFE18fx9j6KNturBOJImksGM4QY8tqmYC8EgHdRMoZA4OLs4K1q61TI6UMaR1gbhaHFWPwQ7v5FBnCpbSMwd1jufJUO8ZriAN1iv8AEmJOCrRJLA0sGwPdBqQrmyt8I9x3WPJMIiN1hN6y74c5VZOskdaDKfM2cdLcZOyqLW8t6+r54WE1xY7qC1CmrndHS7HogupI/CcWnyUjDPT17KqJ5b0nOVjvlLj97Jz8vRZkMjPBIkOHYQa5ftVXi6UMFJBXPAjxkBxC7QcGeaSDQPDCo05VVDXVb4C0OJ3zjC6ZyySMl6os4Pos+zy089U1la8gZ3ycIOe+EfHbVdBxKlvNZd5BSy1PU1pccYJXploTj+28WOOdtzacRjOHj0XjzfaenpKCKaykmRuD8Pdcx8KeM77Dp00Nwmc2YMwATv2Qd5Ne8x81rqZcXQENJOOpbSg5zI7xRu0u6paXz/B1Z9dl0tvWt6u/XGSWR7zG9xI3OMK2E2y3x/dVjyJ4/ibv5oOyWptRQcPZJtYXCrZKydpkAcQRvuuofF/ifWcTLuZbXOY4eo/C1xAKj1rxYueqIzZLjI/3do6RvthbKobd4LvFoyXA75JQZltt1RaSKuZ5c47nO612K4XGrIkhqyxoPYHAWjur3GPw6s4A9ViSXYwbUxJb+hBqt/v9xo4x4FSQ7O5BW7tCa+uUFIXz1ZLw3zK4ymqo6/aU5PplT2mcQ1ggaT0FB2q4XcQtRXSubDBWSYD/ACJ9V29suuLpSae91mqnCR0eAS7fsusXKxoFmo5o30o63lw2AyuxmuNEXfTssYlY5rABnuAg4g1lrnVFhuctULpK1jyT9IrZV1uF91lRPqZbi+QDJI6s5WscXLRVXiAw25xdKB2b6rjvR4vVjm9xuJc1jnYIdlBx5rOyTMqJQ+IkjOSQuKLtSPje5oBByu2XEC32iO1uqzIzrc3Pb5LrVqSGN7pHwkEAoNnRwvDsuJ7qTDGHqICte53iFgKx5Xyh+N8IJKuq629DCR9SxGkk43Kuk6TuCtToYqYw5cR1fNBp/hzMwcnB+azqGF4cJ3OOG77pD0OkLX46fVRVNZ0S+HFjpQbnpNS00LPBc0ZAxlYE94gfWiXpBAIPZaV7sx7PEYSThWtYxrwH7b+aDdly1LTVVGyKKMAgBblseorYywyUksbC9zSNxvlceM9ya0dTt/JVdUFjSYiSAgxLtRudXTSs2a9xIUlvf4QEb2EhWvqTK8AnfzWt0TLe2m65SA/HmggdaBXgOiwD8lk0NE+3PAe8keix6e6dE5ZCds4G6yZKO6VT/eOg+GN8oMu7VbXQhjdydlbb9HCvpjWOcNt1i1NVTNaIpCOsbd+xT7vXGnpXR0ocY/ke6DBuIZRyGBpBxtssmmtIrqUynH2rRW1ZrKomfZxPmtbhqZ6aDEQJZ6oNEmtZp5zvsPUKoIjGVfXXASOO++VHCRI3B7oMeoHjOAYMEei3JaLvS0dF7vKwFxGAtAkDYTnP5VUOjc0uzuB6oJK+VnjPmxgHOy0t7y9xLc4UtVP4h6QcgKWmZE6I5xlBhKZkwEZaQMlJGBrj6KI4LtuyCaCdsb+pzcqZ9X4gDW7Hso4mRObvhGw4mBA+EboDY5XOBcSQs+KmJb1NIGykkfTNhABGQFiurC2MtZhBkwNDXEOOd0fbXTTCRowPksCnrHGQdZ7lbkjnjFF1tIJ6coNGukZgjDWu7Adlh0cBmJc4/UpDLLW1DmHJGSo5fFpH9IGB9SD0E9jI0N5n9TgH/wCUKt/lG3osP2LVRJLzSapa7y0BXH/nG3Ig9pUREBERAREQEREBERAREQEREHHXMeenl54oH00Xe/1GZfM/Kcyvd/2xP519L3Ml+DtxT/gVe/1GZfNA/wDsjs+pQSmoJjDPRWxxhwyVFjfAV7RJ2b5oKH4HEYWbTyNLd1E+BxYXAb4UccUrc7EBBLUvBBwdvkseOMOVZXE7FWBzhsCgq9nR5rUKJzPBIK07qJIyVe2Z0ezTt9aCrmCScgbAlUli8MoH4+Id1SSQyHdBYmcIqhpO+EFFUDJA9UYOpwCuewtI9UFzoC1vVnZRbn1UrvF6BnONlSLpOAUFI3Fjursr5HeMAfRSGJob1Y2UXUB27YQVpsdeCMq+pYM5G2AracfHkKtQ49QGdiggyR5IBkgeqyHRsMfw91AAQ4Z9UEhhLWhxUQ3cPmsyUkwjdYbTgoMybp8A47rDDc91LFJ1ODH9kmw0gM80EJ2VQXDsSgO+SsiNsZ7777IIHPc4AEnZalbXtbGQcZWJOxjG5ChZK5mzSfyoNUbUMglLzjHmoKutE7w6M4xvgLDfKX5ByrGnBGeyDVWVrpY/DcT2wVg1bMOznOVeXRxxdTD8RUTQ+d2cbIIVLDC6QjBU0lPhuw3VIXGIEH8iCtRGWMxnKx2RF7SQeylLnyyHqGyp1CN4YPNBCcjb0WTT0jpgHdt91mGjZJEHsGSe6x2mSlBBGPrQa1Z54KJ/3xoOFWsry+7x1MDi1rSDsVo0NT4r++5Pkr6gPY0vaDsEHcjl41w6phgoIqoB4cBsV6fcCbrKynpmT1gOWDbq+S8Q+AOo6q36kZGCcB4Py7r0s4UcYRSXi3UEkhBeGtxn6kHo1TvEkIdnOVKtH0nVmusFHVn/AOSxgrWEBcQ8cbPb6yx1M88AeQCcrl5cQ8bbpDTWmeKVwAIPmg8gubttOyumhp4A0DqAOPrXT+hZh7ydgNl3k5qxaJ6qaUOaXEE4P2ro/O9kdRM2EgguO6CKpw5xa0AHKvp4nNaT1eSjnIDOoHdWxT4aWk4QJJQ5xaoHt6T3VHHLy4KhJPdA7rOomBjg89gsSIAu3UxkczYA4QarWVsM1OIWDBCpbZ204w8ggrR+pxPw5Qunbucj6kGrXSeN8Z6MAn0Wis3cFc+Z7hgk/lVzAACfRBnMjb4fcZUUtIZN2kYHzWO2peNvJXNq3g/JBX3Q5wDgqOQSRHp6jhZMk2YwWfSKlpoBKwvm7hBpn1qrThwKmqWsY8hqiEb3DIGQgyZasGMNZscYKhjjdMcuJP2qNzenY91kU5IYelBHJAWBRtbkqciQuPXnAUTiA/4UB0ZABCtyfU/lUxDntw0b/JRFhb9IYQWg7qvchUPdTxR5b1eiC3wvhzlRn4SrnPdkhUG4QW5PqVMwTOaS15woVJFI4EN8iguhkMcmX5Kz3XGHwyzAyoaiBjYevzKwWjLgEGTSt8acl3YlT1RZDs3HzwoT94YHM7lRmbxc+J3QJp+r6Gyh6nZznOfVD3OOy1KhghkhLn98INNJ8lNFTF7eoFVmjZ4ha3sCrPGfFlg7IJGzNiJa4ZwrzCKhviDbCwyckn1WTDUeHGW/JApQPGLSpKyQNcGjssUSOa/rb6qQPEu7u6CsrQGAt7lQhuQSfJZEUUsjsEHHkrKiF8WRjbzQeq/sNmEWvjHJ5OnsI/IK7/WvUpeW3sN3E2vjG0+U9hP5W13+pepKAiIg66+0MBdyb8Smg4Jo6Pf/AOrqdfPRSwmaQt7YC+hf2hmRyb8SiO/udH+vU6+eaCV0Ti5vfsg1SCxvqBs/Cwa+idRv6Tk791L90qyAAg7FZ8AFfTGSbuBnsg0emgkqX9AcQAlTTmmf0uOd/VXxTe7VLuntnCtrJ/Hf1ILzEXMy126pHJ4Pwu3PqoRO8NxlVa7r3cd0GoU9uNwOevHoo6qFtvJiO+c7qGC4T0xxGdgsnomuEZneCSEGFDA6pkIb5lZzofdovDJycLFpXmCoI7EKR1Q6WsaJMdJcB9iDELXukz0lT+5PcA4ZAG5W7zb7bPQgwFpkwtJqIvdYXNeMFBoUjSPhzlXCjcW9ZyAArBIPHy7tlZ3jtLCGEHKDAZGZXlgPZZNvaIrjHG8ZBcAVjvf4b+pmM5WbSFm1U/ALd0G9dR2+mp7OyaNoyW+XdaFZdLC4wOqiQMbqJuoH3Boo5XHpGwB81A7UFZbXGmpzhnmEGRU04pXmEH6Pn9SyItSU1JTmmc0F3bcLCkrWzReNKcFwycrR6gRSEyMcgvrZXTzOl6sgrDz8WQrmvI+E9soQHuAZuSUGfTPj6NwFBORNOIgNicKnuNaxvX4bgD5rUKKgd4fvEgOW7oL3WYUkAqXHJIzgrT5YvHOWjGD5LMluclQfdndhsowWxDZBjG3HGepQSNdEegeazXVD9iBsrXU7pWGQDJAygtpYjGesjJ7q2oL5JdsgA+qybTC+ebw3jYLVK6iihZkDBwg02mgYGDqwdlWotb3NNRGSMbqOllPjFrvohT1tbKxhZH9EhBk6avHutQYax3W0bAOOVk1sdXcrwx1FIWxEjYHAxlbWif0TB7sjfOy3XaLhTw4f1DqCDkyhsoZa2FzgX9I3+a0G6sro8sjDi3PkraDV+OmKV+G5wuY9J2WwX+1eJI9hl6QcHc5wg65XOw1FYepseHdzsseCuFkjMEwyRsuZtY6disk0snSBGM42XD98gprhI4QkF+dsINIrrgbi8ti2z2PZKandGwse7JKR6cusZ8RsR6R5gdwseplqKaTolByEF8lG+F4eHHc5C3Fo6zSXW7Mh6SScBaRQMmrg0HcA+i3/AMNqiii1TBbRIBO9wAGN85QegvJVpVukxT1lTTF4c4bd13f1jw9oOINA0U9EA4s7huMbLhDlF4a3OW0UdXdIiIH4c0lvku5lDQQW+IRQDAGyDqAzlSNHcX1NXRF7HE9xkLRNVcmcd2Z7/Q28sxvsMLvC9jXt6XAEH7VUNAaGjGAMIPJDjPy13m2wS07KOQiNp7A+i6bat0JcbHLJTT0Mg+IjJB9V9BGquG9p1KyUzxs6ntI3aO+F0a5iOXuzUDpZmRxghxOzUHk/c9PyULjM+ItyM7rb1R2wGrsPxo0abTNJDTgYAOMDC4Vp7FK+B75AdiT2QbUdkOyQpY2v6eoOOFlXCiMTyGjOFhtjnDSADhBK15mPS12D65T3RwI6jk/WoWCSN2QN1l0z5JJQHefyQZNO7wW/ENsLGqGvqJw1gIyVqk9NiEP9VdTwQiF0px1AZQYM1onEIkY4kjySDqhiMcg3I81uPTTH3GoMMm7RtuotU2oU0+Im9zhBolHRGWQuJwDlRXGGoif0sccFTRyS0ZBkGAfVTSPFUwyDsB3QYtrjfHIHSHfPmt+UmqaGGgNGY2l5GM/NcempDZOlh3B8j2WrU1OHw+M4fEN0EN0tss9S6pbIQHEnAWdT1ENNbnxvAJ6dsrSKy5TscYgTgdlhxVUs0rY3HIccILqOF1VXEjYdRK12WpZTx+7HBJ2BWLW0brZTipYNz5rSfe3ykvkJ6gdt0EldQuhJn6gQd8KtM/731+mFE6qlqPgeTgIZAxpjaUFZpPeHFoKtEbo24zuoMuiPUPNXdcsm4GyC+KlMvmpW0xiOCVZTPkaT9avmqD1bgoKywBw27rDewsdjCymTnbq81IYWSN6kGACQfNZ0JD4+kndQug32UfW+N+B3BQTOopck9exKgeHMPR3Uz6mUNHphWCN0kZkPdBWKmdJu361luq3QQmBx+xY9HKWPIPkVHWODpSQgkpZ208viHz3UtVUtqRloCwC4kAHyWVCxpiyO6Dvn7Ff8KfVP/F/XfyjbkV/sV29PNRqr+AFd/KNuRB7ToiICIiAiIgIiICIiAiIgIiIOOOZL8Hbil/Au+fqMy+aCT+yP+s/pX0w8xwzy88UB66Mvf6jMvmgkY4zPaBkhx/SgjGy1CjY17Oo42WC1vS4dXZXiZzPhYdvNBkvqmMcW+hVffIizGN/LZYJOST6qiC6R3U4lVjj63AZ7qxXxOLXjHqgmlo3Rx9Z7FRRReJ5rPl65acgeQWn5fCUF8tP0DOVE1vUVeXyS7K0tczuMILTssqmjEjceZWMAXHA3Kngc6GQA7Z8ign9xdGS8jZRvaHHPoVqpkY6DLsY7rSqiVuSGY3QWy1DCzwwBkeeFj7g+akihc8jIOFrVFTUfQPFIBPqg0cSl7OgjceaiezpwFqtwpYosviwtJcXH6SCelI6sEpVj4hvlQNcWHLdlc57pDg90BshaMHcK9jfFIOMYRlNIdyCB9SvA8Nw6eyCSfLYgPsUMVOZWFwPbyWVO+J0I7EnyWPSOeJg0btJ3QQODmE5B29VTdzgPmty3Kjo2UAlbjrxk+q24yF8n0BndBK+lLY+sHKgBI3BKmkFSxvTJkD5qADKDKpozVO8NUq6T3UgZzlWUsxhlDgcKare+p+I74QYamipzKMqIDfCyIpOgdI80GO7LSQSdlNBUNi2wrJI3jLiDvvur4xGWfH3QSe9tz6+igfJ1OGOyscB1YCp3QZpLWRZwMrEc4uf1fNVc6QtAd2VGxucMjsgzIri6NoaG9RVktU+pIa5nST2Wt6FpbbU3Xw7pjw9hutX4hWyz0r2utHSQNzhBtRtF7swTl2RjKtfXMe0sPnsseStmezwndsYUA2KDdGkL/wDsVrG12cEuBXa3gtrWbUmpaC4+/BjIcHHVhdOKdjZ2htQ7AB2K3ZpXXlbpO4xNtz3BgI7FB9BvAniXa7/pyktb6+HxYGBoy4An5Ll5rmuHwuB+o5XiHw75jtUWe2wy2eSYzOIzhxXpRyl8QtW66tcdXfhLgxh3xEnGyDssuJOOOkKi9acqqqAEloJ2+pcsyZ6DhaNqAwzWGpgqHABzC3dB4fcytmu8t/noWwSu6cjsfUrqVcLNd7dUStfbZwOo7+GSP0L2V4h8ELRq/UsskDWPe95yOnPmtbh5LuGsWg6y5X6OmZUiMlvU0Zzj5oPEdtJPMAJIyw/PZTNtDsbu7/Ncycw2iKLR+uauitJApmPIbgbYyuGXV8+XhucAkd0FH2gjPxLEnp/CyCc4UjrjUHOSseSZ0hJd5oKMd0nKvMocMYUWMqrcdQz2ygljPhnrIyFe6drx2WTIyJ8IDQMkLTnMLDgoKO3KuDj9H1VoBPZZtNGzpy8DZBjuhIbnfcqaGgfLGXjIU7DG49Pop3ziGItb5oNLyYJCDvg+q1amcJoC4bYWBQtZPUnxCMH1U9fKKZ4jh7HugwzGZagtOdzhZhDKePBGfnhRSPjZEJGEdZ3+eVG2cytPiHJQQSHrecKSKXwiAVbGR4pz2VJ+ku2QTVFS2RuG91Axpd5qxSsdgfNBLG8Qkk9lFPKJHZaMKNzi47qiAsun3YQQo6WB07+lo3WouiZTRdLgAfJBpczOlxx6qwHyUzfvkx9N1bUMDCAEFjG9RVSPDcCCrQ4t7IXF3dBkSVXXGGbq2KDqb157bqBXtleB0Dz2QSOk6/vfocLOprIahnX142z3WnOhkYOstwO6yYbrUwgNadsYQUqKAwOLSeyiZO6IFoJwVNU1TpW9bu5WLGzxDgoL4SXvOT3SpGHDClbCYh1FHM8ZuQgxmM6yhYQ4Nz3VxikjPp81UQSuHWGkgdygvkpfDj6y7OVbDCXDrB2HdUMk0uIjvjZSMbLFGcgjKDKpKoA9DWElWV8+csczBPZaroplvfWn38gD5q7W8FCysDreR0b5wg9O/Ycx4s3F+T+6qLF+YVy9Rl5b+w4lLrTxii/uJ7CR9ra7/UvUhAREQddPaGOLeTbiU7GcUdH+vU6+eWIEvGB23X0Pe0Fa13J5xID+xo6PP8ep1881M6NjnlxA22yg1LojnhDMDICoJhSQmMEBYsVT0u2O31qOoc+V+c5HmgxpCXPLj5lW5WXKIhHt3+axO6CpGFVvb7UMbwMkbK3f5oKu7LU6S5sp6cw4GSMLTWsc87bq+OP74A/sgPkJmMgGMnKtfIXOyBg/JZ9VTxeCDH3KipqTqaXu8igmtNxko5uqR56PQq+73X3t33vHzWBO1vV0tG+VfDDH0/HjKC+3W91wkLerCya63Otw6erOywoKqahkLojgkqSarqrgSXnOO6DDdkuK1awUBulYygLseIQO6xaVkRdiXuFM2pnt1Wyro+7DnKDfOpuFp0raW3czdRLerGc4XG8sxlf1kbrdNx4hXm+Ugt1e/qix0jdbanpwHZiHwoMilifXjwgcAbKOrpjRO8MnOcq621Hushce5VLjUmqqQT2QY0UfjOIJUkYEFUzq3AIypZAIog5gwcLHHVI4k98oN01l2pGULWNaOrHooqepjmo3luBkbLbUhkcAHZwO2Vk01UY4jGCd0EbD/ttxH90q1T3B/wAKia7EpcdslZjImyjqcNggxDUOLcFu/qtTopw6Etc1Y8kFOQA3BPms6njhipy44z5ILIqllE8ydslTms9+OAditDqHvlkI8gdln0hEMORscIL6ylFO3xGEZWAaw9Ja8brJFW+aUsmyG7+ajnhgA+HBJ7IMNrPFfttkrUqa3yNHX14A+a08RyMcHN9VMLhUMBbn5FBvLTVhkvkrIWyYIcB3XYzROhJLJbxVSVeABnHUusmkNQT22cStJBBz3XI7eKGopsU8Rf4RGPPsg3dxFrG3Zj6CnHUQCMjdcRN01NbpDUzNJw7JBXKOna+31DxUXBw63d8rAv8Aa5a25NdStzTF25HbCDYlVq+lpYTSupgCNuohbRnibfK8eFsCdyFybxBsOnIrUDSFhqQ3fGM5wuMLfFUUUT52g5buEG7obHHZqWGUAOLyBsuxXK7yqV3ELW1FqjpeIGOEhb5dwV110LJX6oroaOcEta8Abdt16acpUF+05WUtNTNIiLW74QegnD/TVHpfSlttNNTtYaeENO2+fNblWDZJnT2unkf9Is3+tZyAiKjnBoLjsAgquFONHDxmo4nubGSCVy4680Ic5omBLe4Wxtd8T9MWagcK2dgcCdiQg8xeabhfHp0SzmInAJXU2C1U8tsml6QCCV3m5q+JWnNTGohgmYctIGMLoRe76KLxaeA/A5xH50HH+oHtp6x7A3YH0WPSSRSQE4GcLIvToZ3GZ+5O60Nsz2OLY/okoJppmxyEBmQtSt1CJ/v2MAd1iU0cLwDNgFanAJoxmAZYPRBPU4EfQfJYhY4RnDsjG6VdSHtDIyOsd1HTyFrSJvNBqlguYoJicb+qzblcI6+cPdvvvlafbY6edxJxlYN6m91lxF64CCe+QtmhAgbkgeSwKDqZCYXjBI2yta0tVUk7y24EADtlaZqmanjuGKBwLRnsggitBEhlcds5ClfcnU7xThmx2U9IysqIWhgPbfZa5FY6JtI6aqwJQMhBtmajM4MuM5WJDSGnnbK7YNOd1rdJS1FVUvggBLBsNlHctM3oydMcRIPkPNBPU1UV2pRTN7jZaJUWl1KC4nIHfKyW2i7Wo+JNE5o7qyoqJ52nq7INOiIe7oA+3CjmYY37lZUVM8OLmjdWT0dSXFxaSPX5IMeSUPAAUsEjQ0tIGSE6I2bOGCFC0gS58soJzJ0ZONlBJIXu6lnVLInxAMxnvssExlpGfVAdITjA7KaCcghh81G5jTgN7qrGhm5G4QZkvTEwPznKwyPEd1+WVR5llAHcDsqwtc2QNI2JQVll6mhvTjCujqA1hZ5/Usmqp4hD1N74WnAbgYQSA4JI81ZIclXGN4GcKjY3OztugughMxwFqVNRAEMccZUdtiMLvEmGB80ra5zJ8xEYHzQegfsa7c2k5m9TTA56tB1o/LcLef5kWF7F64T1XNFqiKU5A0DWuH1/dG3f60Qez6IiAiIgIiICIiAiIgIiICIiDjrmOcG8vPFBx8tGXs/8hmXzSCQCpleexLj+dfSzzJfg7cUx/wBxV7/UZl80D9pHfWf0oJP7M8t7bqk0HheasY8td1KR8hlG/fCCEDKq5papIG4eOrsr6vpyOlBZTwGd/QFJUUxpXNB7lVt7xHNkqW5TMlcMHOEFsdUA3BJWPPK2R2QrDjpHqqxsLnDbZAjk6PJVc7xXgDz2Wf4NOIcnGfNYrhG3cEZCCdtH7uzxnHOViTyiR4cPJXy1kkjPD8goWxvduBlBlMmdMwReiikpyzcnfKpF96dv3V8glmOW5IQGVIjA2Ub55HOyHEBZEcUIGJe4UM7WD6HoglNb1xBjichQD48lRYypovh2KCxkZe/pCufH4LgfmsmGLDurCirAS4fJBlRVkb2eGO/qfNWvjyQMdytPBIOR3WpUMckhD39ggiqaRzIuo9zvhWUJGST5LVqmaF0ZjJGRt3WjSB0bj0digza+R7oenrJb5DKttkrImkv7DzWK8TSsGxJUkI8KMtfsTnZBlTVMVa4wtAB7dlgTwe7O6T59lbHJ4U3UPVVqJTM8Hug1K0Wf39wVbzRC2OEePpFT2SuNFgv2CtvtbFXPDs9ig0EnJyVPBH1YckrWNj2xlSULh2d2QWz1HU0xgbjbso4qd0jcrUzBSObtjJ+aw5n+FljfsCDFbGS/oJR8ZY4BTMhf1dZCtlY9zwcZQXygGIDG4CjhmDB047q+Z2I8eapAI+nLu4QW9UsbjJG8tJ32OFNDXTPcGTSOeDtvuomuDpC3yVJQ0OHQRnKDKqaAMZ7wDsd1jQReKer0UshrJIg0glhHZKORkL+iQ4JKC9zHOb0N2PyV9spnfdCONxBJ2U1Z0Mg8RmCT2WNbahza1krv3u+yDsxwutpttLBVSMEo6genGV6s8omr6N9mhoxA2IujDRtjOy8wOXDVOnaqeGk1BK1sfWB8R/1r0x4SW22vNJU6TIfCGhzujtjb0QdsJJmtgMoORjyWg3GkfeKWSna4jryMLUrQySS2sZU5Dj3WnXSOrpagGkBIyg2ZZOF7bXeHXKrYXtJzusriJw/n1JY5ILc97Glp+Fpxlbynr6iOjDqjAyNytPbra10UI94kAa04Jz2CDyQ5xOBM+naie4TUzwQCS4groaKHpNTtnpcRn7V7Je0G1RpS+6TlitE8T6rwiDjGScLyFtlkr2yVr6oFrS8nf6yg2dI0mUjHnhZcdrfJGX5IxutVktEdRUGOCQF/op/2K3yIdQDjH57Hsg25DATOYyCB2V9TSljgANytUuUENFGQHjxvP61i0UUlVGXvGSPNBjNYYG9bvtWPNIJHZCnrC/qMatghHd4QY7XdJUhmOMBTTtiDT04ysRBfFKWOzkqSSZ0n1KIRvIyAp2Ma2P4xughjkdG7qad1cXuqJACdycJC0Pk6fVVmHhSDp7hBmVFqdDTiXq7jstNyst1xmkjETjsFCIHudlrSRnugiTKyJWNazvuoG9xlBRFkvja5vwYJUHQ4EAjugtAyrg09QaVkxxhoy5WzBn0m9wgyqMsovvr8eqguFb71IC3YBY8kznjpJ2CjQXxydCPcZHD5qxZFPGDhyCklOWN6ie6jZH1tLvRZdTK0s6R3UFM8NyCgshhMr+nspXU/gkOJ7b4V0jmRDqZ3Ki8Z8rgHdiUGRPWski8MNwQMZwoYabxGdfyU9RTRsi6x3IWIyd7G9LeyCkrjkt9CsmjYPpHyVkUYJLnjYnzVJZjG7EewQTVso6Q1p7Ka3xAwl5K0xzy45K1CkbMYCWA4QXUzW1c5h+fdahPDFQxdBAJIPktEgmkp6gub3WovqXVDQZkGnMkEM5kPYlSz1bZiGtB+eypVNjd9DGVWmpHGMvc3sgrFRTO+OF/SfkcKyodNE0xTPLie2+VQVksTixvYFUkd4kZe8/F5IPVX2GzCLbxkf5GewD8ja/8A1r1KXlp7DZxNu4yN8hNYCPtFf/qXqWgIiIOuftDnFnJrxKcDuKOjx/HqdfPnbLW64slcHY8LB+vK+g/2hDWu5OeJLX9jSUef49Tr56qKunow9sPZ+xQROjdFMY87g4KldJ4Y6SFRrg55kk7k5+1Umcx475+aCJjTK4jKuczw5AD6hVpXtZJl3ZSTRulkD2DIQSzkeEMDyWD1fJZ5czo6XdwoDExwJaeyCynlEZ3CTy9bshWFhJwNyrSxze4QZNPI55Ae4kBaiXtbAQPRabTQyF2QDj1U0spjcI3H5II6ZhfUHI2yqyNzWNhBwMrJYWRjrBG4WE+Qip8b0OUHIen+GQ1JA1zZAw4zkbLRdU6eZo+U0jnB7jtnuUtmvLxbIhHbiSe3mtLvF3uN+n8e556gc7oNEc8l5eMjJysykqmtAjf3Pr5pJFEGfBglRimOOoDcb5QZdTSs6PGjA3+Sij2Zh4UttqDI7wpuw2VLm0NcPB7IMSdvSepqRUz5B1nyVsT+p/TItZpzTiHAIyQg0l0hyYz5KMydBxhZkzGCUuWFLgyDHYoJYyJ8jCjmZ4b+kdisnwHMYHRA5PorGwyPJfINwgilh6WBw+tSU1QQ0ReZ2SeQOaG5+Stp4iHh57A5QTPpH048dxOD+ZZEUZnhLw7t3VtdUtkhDBucLCimqGN6G5wUFC/wpCDvgqUTFzcjsopYnY6vMlZFO2IQHr7oMaSYu2Gx9VGHODhk5wfVVwDKfTKlNHO4F7WkhBNHUsc3pxusaZpzt9ataHB+PMbLOpRC+UMl7bBBDQTPjeAOwOVv+w3enDRFIRnGMralxpKaGAPg+l3Wl09dPC8EE5CDnCzWCS9fHDUBgBzsVqFdqSn06BZZXB8j9urvhcU2fXd1tTR4ZIB27rNrLi++D7pTZMjfiH1oNU1VZ6inYbq+oL2P+Lp6srakVxilgezpAJyMeqtn1Pca8+4VJPhN2H1LD8ARSdTR8GMn5IOQ+EVXBRXaCSRgAMo7/WvYzlKsdrvNrgqx0F4jB8vReNOi7TXXqenhsjS6USD6O/mvU/k3puIGn4qaCtglERYAc57IO/VNAKOn8NvZvb6lMyTqGVi2+fxaKJ85w8ty761dUXOjpGF80oa0IJZqhsIJeQB81o9zv9JHTSM8djCRgElaJqbXtkip5Iqeoa94BzuF1q4occ6Sy08rBNh4JxgoOUtQambaKmac3NmCDgF4XWXje+4ajoZqukvOACSAHrhjW3MFerpWSxU0ryzB7FcQam4936mhkpJ3vDTsckoNj8Vaq70Fymjnr3PDcj6WVwfcrw95d1Oyeo9yt6a41kL5K+Z78ud3yuLap4dIcdicoL6mrfUHGThRRP6DjGVH2WbTNhczLgMhAlDngFjsEd1rVHcIqe3uieQXYwCe62/LMQ4hh2VrZnEgOOyDMt5Mla6R5yCSsq4nLg2M428lglxYA6Hv5rJppuo5nOD80ElBLJRu6y44Pqoq6o97m6s5U1XLD0bFaQ6Qhx6UGbLDMGgwvIz5A4WuW3Sr56I187y4gZGTlbdhquwcey1mi1PNEBSE4jOxQahp65xxVjqWRgw04/IptQVznSFkLtj6eSx3e4MaKmBw8R25381WCopJHj3gjKDIsFyZbniSVmcbk47rcLuIlsilaHwtJGO60Z4tpjx1Df5rRKzT7qp5lpgSM+SDcGq9Y2+403TBG0EjC2pTdMsDnlY9bap6YASggD1Ucb3sgLWdkGRBUsbIQc7KWpu8LWGIRjPrhabSxSOmLnA481kyQ05fl/dBpsjnPeXHO5UkdM6RuQMrKqII3ACEZ+pZFIzw2jrGEGmgvpyOrO63JSaeZX0BrMgYGSMrSK9rJRhmCR6KSmvlZR0xpGZwdu6DTZB7vUPYTnpJCtz4kgxsMq6Rk0r3SFpJJyVZ4cjd8YwgzJOmJgON1jmoBOQFb4pk+F52Toa4hrNyUE7ZXTDpznyVj4eg9ayKWjkgPiTNw0+qjrS57vve4QRmpBHThWxz+HKHFuwKthYA8da1D3OOSHrbgoIau4CaMNj2KwtycnO6yYIovFIfjA9VNPHA1vwY+xB3v9iu/PNLqn+AFd/KNuRPYrDHNPqr+AFd/KNuRB7UIiICIiAiIgIiICIiAiIgIiIOOeZL8Hbil/Au9/qMy+aB4zK4f9sf0r6XuZL8Hbil/Au+fqMy+aJzHOleG98k/nQStpOsZzhRvZ4TgBurwJ2tDRkK9oHSTN3QVcwNi6xsViF5PdSPlc49LTsgp5Tv07IIw4tOQVkR075m9X2rHDSXdPmtRgDo4SDtsg0+RvQ4t9FdHL0AjCqekz5d2J3WcyGmdGXEINPdI95xkgK1zXDz7q+UASfB2ysykbA+ImTuEGnqaKfw29JaSFZNjxT09lnUNE2eEuIzjdBp7ndTi4eazaKqZGBG8bnbsseVoZOWeQKtfgOBb3QajW0REXvLScEZWlZJ9VrNJO58QjmGG9lDVMpQQIyDlBpoPSdwshjfEBd2wpJaT731NCxep8RI9UE4qQD0/nUvh+I0O7ghYLQ5x27rWraYRAWS46sflQabFCBJjuc9lqbJmQxlmME+ixDTTR1JlIwwnOUqp49seXogtZTyTTF2SR3Sq+8OALe/ZZNonjEv3zGPJXXmnM7w6BuQEGHDWMadwVBVTeK7IGAqwQhsoE2wHdXVgiBHhhBFTxGZ/TlTTwCHzBKx4XPY8FndSTGeQZdnCC4TOlHhtdg+qsdDICA5yjj6ur4e6vke8OHV3QSPhwzPV5K2HIaQFaHSyDHcK5gMQw7YlBTrkjd1dRIVHzdTw474VJI37uOcJEwuIPkgzKeobIBHjdUqXtj2xuVSIxR7jvhQVUge4EdkFkjs7+qsGfIoScfJUBPYIKglp7qaGPrd4hOw3wrHMwMlGPf9Bvc7INUddIWxeEGDI+SmoLCbpCatr8Ab4WlignDRLIwhh3WRBeaiiiMFOR0lBFXPdC91LnPQcKKlB3O/1qhcZpDPL3ccrVIKeL3Z8g2AaTsg3Vw2tl3vNbDDbqh7HeKOxI817Z8jNlqrPpOOnuuZJXQDpL9/T1XjtywRtrdXxUfVv4oOD6Er3L5d7K+3WWkcRgGAHt8kHNoDWjDRgKhYxxyWglXIg0+7W/36n8Jm3cLqnx3vF40rLJbKYvBkcQCM7Lt4uO+JnDK16tpZa6oYDLG0uG3og8xuL2mdSVlL91at0tQyQZ6SSdvqWwtC8B6jXIfCyzvYX5/eELvxaeGNLqW/Gw1jGmFj+kdTdgMrnbSPArSukg11HEzqGM/AMZQecGjPZ3VlTWe/TW5+M5+iVqWu+U6PTVKaJlnJcfhB6CvVCjoYaKPwoWNA+rC0i96Ks9+k8StiBOc/RBQeDPGTlrvGmGvujLVL0kF2A0rgyioquBskU1C9haSNxhfQ5xC5fNL6xtD6N8MeQwjdg32Xnjxw5J7rbbhPJYaQmLxCfhZ5ZQebt1o5oHGQwEDv2WBE50rCOkjyXZriXwRq9L0PRXxFr2tOQRjC4LmpKCjZM1xHUDhBtOaJzCTlIYPEGfRZbXxy1hDz8BKsrnsieGwHZBCJhGS3HZXdBmYXBRdI6Q5/cqeKWNseBhBjMd4byfRavBZ/fYDODjAytIIdLIekZJ9Fl09yqqMeDk422z5IMSaIwyuj9DhZVJUsY3ocNzsr6lrHxmYjc7rAa0uIAG5QahV0g8PxgdjvhYsVN4jS4FTSU1x8LL2Hox+ZY7ZHxAt+aBG/wn4JyAshzWSjrGNlh4c87bkqRj3t+DdBIXGQ9DfJRPa6LYnOVkRMEZMjvNQ1L2vfkIITuiIgKZk3Q3pGVDhEFS4kkoAfJVYx0hwwZPoqmN8bg1wxlBbvhGnpcD6LOZBG1nU8DBCx5Yep2YhkfJBdJV9cYZ6LHbnY+iv8PIAHdTRsaxpDwMoInTZGBso9z6lXSRlpJA2UkQYWHOMlBZBD4zunK1eGoio6cxHBJGFpAe6N5LVd1PnkGcoKOkPjmQDzVz6lz9gMLUoaOB0Y68A4UNXDTRg9PdBp7HFruo7rNbcGNi6PNYDjknHZXshfJ9EZQU8T74X47pI8vOR2UklO6OPJCjHT0HPdB6s+w1/3O4y/4awf9GvXqYvLP2Gv+53GX/DWD/o169TEBERB119oYwv5N+JLR3NHR/r1OvnnpnASgOGc7L6GvaDHHJ3xIJ/vSj/XqdfPCzqLh09wcoNadaOuLxQe+4WkTwuheWlaxS1VT4Qa7t2WJUwmaUADJJQY0dI54BBCzYiyCEtcMkD61ZUU1TSxB+CBjuoqaoY/4ZXbkoMSRxc899ytWoLf41IZM+W6ifSxuGWYUlPJUwRmJp2QadnwahwI2BwqTyF0hIGB9S1GG3PneXFuSSppqOmgPTKQHZQVtYjkAaRgqC50QM4LSrg18Xx042WG6pqHTgSdupBdUsdHGBuMhahQ2NtXSOnLhkDI3S5Ni9yDx3Wn094qqeIwsOxGEGRZ5mUVa6KbBAPmMqS+VsM8obCAB22C0p5k6jOSQScqzpkly/coMqOA4Dicq91UIx04PbCxoppCegnKyi2Dwy5/cBBEw9TupuxJWVC3xHCJ2+Vp8cmJBjYErUaR7RO1x8igtuNrdTRiYDGfzLcFh0s6vtrqsuAwMhQ3aWOppWsbjI8lDb9R1VuonUbCcEY7oNDuYMNXJCCfgOFiAPyHAHb5LLmhnqql0xaT1uyVr1FQ0TKMmbAOPNBo9JWDAa5pKyp+l0J2wfqS3U9PPWOjb2BVt7Y6nf4URxnYoNJawyS9I3GVq0VOBF23wsShopeoSPaQPmtUM0UbS3IGEGiTRSRyknOMrXLfbI56cyuABxnsoIIGVLznHfZWVtwmovvEWQD33QY9a1sMpY0ZCg6HPZ1DYBX003vMxM++VfVyRRDw2dkGJSxCWXpJ+1bjZ4EFIWkAnC2yyV0T+tqldXTvb0nt2QW7PqSfIlTGnJcHtdj5rDDiHZ81mQTEN+PGEEkkzw0McSQsNrz4oIaO/os+ERzuxkZWbFTUbcB+Mn86DFkcDCAABhZNLc/AhMbjsfLKx7kGxNBjOyw4ZI3Rnq7oMn3hs8pLAM+qnqarppJI/PGMrGtkIdMS7ssuqNJ1lj3ADCDnXk0dENWtfV0pnjEoIBGfML1O0jxxtNjvVJYqa1CIvAbkNx6BeffIbUaFt19hk1FPE1r5B9LHr816j2/hLoLVl6pL/YHRyBjQ7LcH0KDmywyi5WiCtPwidgdj0yto8Q454KF7YJ98HsVruoWVdlsMVNbM/eW4GPkuLLhermYnTXbqDGnfPog4F13qS86erZ3yVDy1wON11q19XV2rKlzjXloLjtnC7ScVdSaKroJ4HyxmZrD5jOcLzr43cQqzT10kbZiegSHGD5ZQbovdtZYI3zy1LXkDPfK6+6+1Ea+rdFC8DLiEuHFq53SMR1z3YI88rY90qzVzmaIk5OUE1TQzFvjPkJyPVaJUM6HEZzgrOlr6lzBHvjGFpzhI4lzsklBYnUR2JVwjcewVHNLTuMIKbkqZsBLCT39VbD0g/FjCyjIzpLWFBBTzeC89W4Som8V4c3IH5FRkfxEvG3zVZYj3YDhBCXvPdxKNHU4A+ap2QHBygyH0pY3qBUbI3Odhp38kM8hGMq1sjmnqHdBn07Z6cl75CR9eVFI+WapAa4jJ9cKkVSXHD+ypJ1Nf4kXlug1WognigY/xCe3mt6aaqqaK2GScgkDzK4/hrppvglzgK6W71MQNPESG9ig129XKmuM74o8DfC0SSIUrSDvstPEs8TxNvk7qc1XjMJk7/Wguiq25Iao6hr5D1glRU3T4mT2V0zndfwn4UCmmMUnx5wtQbK2od4bTjK02Toc34O6sjlfC8OHcINUqaM0cfilwIIysBswklGe3zV8lZUVrfCdkhYxBikx5hBrBEbIsgZysVz2ynp6cA/JXU0he3Enb5qSSFvT1x7lBg1NJ4Q6hvlZFtojJIyV2cAqJ3iyOAk7BarRyRwxY2AQS3moifTCKMAEADZaLC/oYQ7crMqXtncWs7qANjYMO7oMObd22yy4Kl7KctIJ2VsjG5BxsVuWhoKF9tMjgOrCDZ5cfEJBIJKuY5z3hpJIOyuqWg1L2xds4VjY3scCRjBQeg/sYKZsXM/qd4PfQNaP+cbeit9i7OZeaDU7D5aBrT/zjbkQez6IiAiIgIiICIiAiIgIiICIiDjjmS/B34pH/ALi73+ozL5poXiOpc5wGCSPzr6WuZEZ5d+KQHnou9j/kMy+aEtPjOHm1xJ+woNZlMUUPi4BJWmnNY/DdhlUjqHyARO3AWQ1ghb1N7oLH0Pu7fEc7Ko2sYW9HTudlFNVSzHwj27YUfguZjPfKCbwmxnxc7I+rLvhb57Kx/iuaAMq+njiDT4n0vIIIpIfg8Qb5VjZHtGATgrKpj1TYk+gqXBsLXDwiEEEURmOSk0ToTgE4+SsZI5hyCtRoo2VTcydx6oNMOfms+juAp4yw+eypXQsiJDQMBQQMiLSXndBbO7reZAe6uggLx157bqNzgXY8srJjf0sLWnYoJY5BUfeAcEfYoKmmkpHgFxO+26ibI6GUvad1c+pfO8GXyQZzZmiEZ9FhSNEpy1XyPZ4YAI2VtMCWk4QIAGPwVdMHCQPadgfVWwjMp6vVXVTXjHRnCDNqbtFLSCFoHVjGVgQUjp2l3V2WOGOc7pG5WdTSmBpa/bIQYbuqJxa0kEHyWqWy4t6mwzb9RAysVkInlJG+UmpTTkSAfEN8BBue86cjioPf4zgkZ7rZmC4gEn0Wqy6hr6inFLK4lg2xlad0/H1em6DJFOIo/EVhqGv+DCmklzCGKKGOPBcTugtbEIiJFFK7xXbA4Bwrw9z5Ok5wp2wNG4wfPsgtiIhaHEdgoZpBM8ObthZFQG9GFgglp2QTyTtMfRjcDCvpm5HSd87LEWZTgmMuHcILaildGC8H7AoIwX7EbK+aeR2WE7BXwdAYScZKCJ8JaMqwZBGyyqdpkkPX9HPolZExrh0YwEFjR4rcK3p8CVrjuAQVVrgxox3UjWiRhL/Lsg1Squ0E1EIGNAdjG3qtCLSDv3ypIA0zYPZXThrZg0dsoLZHHpDQMLkPhfo6XV9Qy3CIuMw6QMd8rbtus8VdEwtxkkA7Lsly36UFLrG3N2wXN8vmg5h4C8ody0TPFqyahlLS8OBIJ2XqJwVmEliijMXQY4wO2FJw80bbKrQlugqo2HrjBJDR6LedmsNDY4jFRNwCMeiDU0RUQVUFYyOWmkikIAe0gqYHK2NxK15Q6Ptkr6ohrg0uHxYzsgx7Hoyior864NkBJdlchLqzw1421Gq9Xmgp3EsMmO+fNdo4+rw29QwcBBeiIgoQCMEbFYNdZ7VWwvZV0MMjXgh2WDJWeqEAjBQdEObngrbbi2oqKakbEwsJBxjyXlZxH4ZGy1VU6OXYPPY/Ne0vPNVVdr4ePq7Yz7+Y39u/YrxD1VqnU1fUV4uDXACR2Cc+qDYVXQGnf4bXZI7kLE8F3V8We/mp3Vkk1VmTzOFHVSYflvmgzaS3NrR0AgELFuNAaGUR9Wc/NW01wmpndTCraqrkrJA6TuglgY2naJnbghQ1U7ZnhzR2V0hkMXRg4CxwMkD1QTPnL4+j5YVkLhFMx7hkAgqV0XS3qI2PZY7sE7INyT6hpZKIU4jGcYW3ZHdbyQO5VHMIAPqp6eFrxlyC2mHxbqtQQ2QEeSvm6W/2Lv8AJUZGZGFz++EFPEMzQxux+Sl+5rujrLvsUVKBHLl+w+anq6mQ/DEfhPfCDAe3pcWqiq4knJ7qsWDI3PbKCSOIyeRVX0xa4AAkeq1KVsUNOHNxk91HCY3wlzsZAQR0sDYCZHEfaoK6UPmDmDsqGeR0hYDtnYKb3cFuXILGvNSwRA4wsiMNp4ix2CSsJ7hA/LO6ujmdK7DjugqB0SmU9vRRTyOe/qGQAsp0Dy3JGyyYqaDwSXY6sIMAyCVnTjdQFrmOxuFkxxCOc57Z2VlSWeKMdggsLD0jYhSQlsYLsdlJKWGEdGMrGAdg5zghBnU85nd0AkJU0LyQS7y9VDQYbJlS11VICOnIQY8lIWD1KQVHgbOYdj6KWknD5MTdsrPqYaJ0fwEE/JBptRVtlGBsscRkxmTyVz4HBx6RtlZAj6aR+e+P50Hqf7DX+0OMv+GsH6K9epi8s/Ya/wC53GX/AA1g/RXr1MQEREHAnPZb33blO4g2+Nhe6akpAAO5xWQH+ZfPx+xmagdO6oYWmJxGHfIr6NuZKa103BHVU16c0UTaeLxS7tjx48Z+3C8EOONdZnXapj06WGN8pd8GO2fkg4oqa9sP3pjdx2wsmhb1MNS8dt1lUNtp5wHTkdRG+ynrYY6eIxw/RIwSgut80F/f7jgNI27JcNCupJOpkhIzkBbehrn2up8amPx+uVuCg1LW3B7ROSQDgoKRWgxAMkGAFlMtsIb5FX3itEdP1RkdWPJaHSXeoLsSds+qDVOqKiccjZaVcKI1xNQx4AG/dXXWq8aL4CST6LS4q6pjjMZzjHqgyqStbG8wvwcbbrFr5W+MCzG26xmMlqJT4YJJ3VJo5Yn9EoIKCeesdNGGb4G2FJT28Pi8VxxjdR0lK5zuqRuymqHVEe0QJZ5oI2tbNIYcD4VIGNikEOBvsooyWnLT8R7rJgafGbNKNhugnltXgR+N04B33WnwU8ldWNpmk/EQFuCprIqumEEJBIGFp1ocygu0clRthwO6DUL1omez29tc/qwRlaLRRmWF8gJy1ck631Jb7hp9lNC4F4bg4I9Fx1ZJWNeYnuwHFBbQzyy1AifnGcLcn7G2ywGpDxtv3UFVS0dPEZ4AOrvt6rSJNRXBgNO0kN7YyglfWRU0xg2y3bKw56588hhY4gOOFQwukJmeNzuViSgxy9TTuCg1eGlNpaKsvySM91O1v3U/2z05A3WjTVdVVRtieTgbLcWnRFE0Mk+ifVBjmqY0GER4xt2WFPTukBeHHC3Jd6ClmiL6PBf8ltpwnpgWzggD1QRw1hpHEbnCxKupdVSdRB/IsyD3aZ33wjupZYKQkNjAyf0oNJBczcEgqQROmb1F3YZWXNa52Drc34e/ZYbnuZ8LfqQWOAB6UYd+jGclCx4HUdgsm3CI1DfF7Z80EUlO+MCQtOFJEzxoz5FbkuTKCWjDYiOoflW143Pjn8MeuEGRSsdBIS4kD5qGqmkM3UCQAdlnVtPLHTiTcZxkrV7DaYK6A9eC4oNvmd1UwRuG+FjSROiOx2WvXyyPtgMjW4b+hbfdI5/0kGrWUiaURdsnC3tZOF1x1JXMjgpXvD8btBK4/tUc/vUL4ASS8Zx6ZXoZypV3DaipYpdVTU7JQwfTxnOPmg494dcruq6eOlq7dFPCQ8EkZHmF6r8rGkbrpjTDYLu575REBlxz6LSeFNz4ZaoiZSWOemcQ4AdOD+hdgLbbaa1QiGAADYdsILbvWUlNTv8Ae3ta3Gd1wTxVv1uqLPNSUdQwPcSPhIWvcympK3T2mTNQh3X0E7bLobduMV4fSzOrS8Frj3cUHGXH6svGl7jU14uTiw5OA7ZdUb/qN2omSTzPDyCe/muYeM2sKrWZlp43FxIIwuvtTSy2iB7JgQST3CDRpKE19QY2uAx81jzx/c5/hOOSe6sFXUsqHOp85yoak1Ur+ucHKC51XnsM/Yql3W0npxjvslFFG6T76s+oFIyLDCMkeSDTI5hG45CslkEjttgrZPpn61eGs6TvugQQeMcZwVWSN1PIOo5VjJHRu+D1Vzy6UdT8k4QXSz9bQAkU4a0tIzlQY3wFLHC9xBxtnugtkYQS713VmD6LLqCwRho74UUYYWHqxnyQQoAfIK5+M7FalRU0M0O+OpBppaWjIUsc/S0NO6T000ZJc04Hmo2xlw2QSCfBJaMKPqc54cQTv6KvgvBGRstXpqemdTHrxnyyg02eobKwADBWNv8ANSzQuje7bYErJgZAYSXgZAQYsLC4nfGFK0jq8M+eyhLi1x6TtlSxPb9I9wglko3U7TId8rEcS93bC1SGdsrQ2fAH1qyppGvGYACPkgw4XiA5Izn1WV7oKgeM3G2+FhPglZu9pH1rJonVPW2MZ6CcFAY4k+GGkY+SyWyeEzB3ws+rpoIoQ9uOo9/rWDGwPGX9kEAl8Z2OjH2LUBRH3cuDt8ZwO6iijp/3gGVNBNN7wI3ZLD3yg0eOZ9NMS8E48ir5HGc+I3sCtz3CktXu/VloeVtiSN/XiL6OfJBYZCcNPksxlwkZD4QfgYWPUxiOPI7+awy4koJmnw5DI7fJSSoDhgNUb2uAHVnCzKenjezKDvh7FcO/bTapzn/4gV38o25Fk+xfYxvNJqnGP/iDXD/nG3Ig9oEREBERAREQEREBERAREQEREHHXMc7p5eeKDvTRl7P/ACGZfNE5+J5HY7k/nK+lzmR/B34pfwLvf6jMvmfk/sj/AKz+lBUHpd1KYVOWkEqANO2eykbSySDqYMhBRjwyTrUk1SJCMBY7gWktV8bQDlyDU6ANqj0EK6vtLmHxM4HdXWeeCGXLiAMq++V/iuDITkY/Ig0pzjtE3uFG+F43cc/ap4m4HiP2VJZmOGxCClPR+OcZU780GG53KxI6mSN2WJPLLK/qlygrNOZck7qAZzj1WTJ4Xg/D38ljdkE/u3wZ81a09AIP1IycgYPZVkwdx5oIXHJJROyd0FzGl5xlTg+Bhuc5UDXFjsqsr+sgoMmJgJLsq6aYsBHTnAUETywb9lqUFOyogLz3CDSo5QyTrIVZ5vFdkbBHsDZywjYFZQZCGZ2ygrbXtY4dRU9wqWk4G59FpjpC156TtnZVZIZJWeIdsoL5J8jHhgZ77KWkYJcAjutUrYKAUAMZBeAtKo2SNy9o2BQZVVRGOMvDtlhsY8gkDZTzVb5fvR8lm0rKdtOfGIzjbdBplPh8pae4WVOPBZ6rAleI6hzoztlZLZmyR4ed8IMWaUvcVEr5Bhxx2ykbQSMjZBRrCe/ZZEc4hb0Y77KV/hMZ8PcDyWC85cSgukHU4uz3VoJBB9FTJxhVaMuAQalQNFSQztup6yhEWQSCSsNj3UrBIzuPmrfuhLK8eKdkFHwFpzuonSY2CzaieMw7d1pww4nPmUEtIzrkU1RTEPLgc4GVFB1QOD37ArU7faq27VzBTsJaT3AQcmcBdIy6wukdG+EkGQDt816X8FOVR1JcaC6tpHEANdsF1E5Y9DVNLUwS0jAZg8ZwN+69cuCtaLdYIYbq5scvhgD17IORdLW37kWOmt2MeC3pK1ZaZDXtmqSYzlnqPNakCD2QVVFVEGyOJPEKl4dWt1yq3NDOkuBdjGy83eZnmyrNXyVbLTVZjjJaOg5AA+pdwOeu33Wt4YuFpa8zBrx8PfGF5j6G0/ZYdPXUawf0VZ6+kSd87oN3csPMFU2rWLJqqq6iJRsT816l6M420GpWU7OqMOkYPtOF4j8IrZBT8TZHtBNIJst3wMZXoDpLippmwXm30DJQHnpbjqHyQehVNO2pgZMwghwypVtvQN1beNOU9dH9B4GD67LPrNSWmhmEFTUhjycYQaqtp6p11R6bk6KiRjTnHxLVLnqqzWilFbXVIjiIz1Ejsun/ADO8wekXSPhtNW1748AlrhuR5oNP51OL9AdLvDaiORnhnDQQcbLyG1Dq+jvNRWCONrSZD2HzXZ/jVxRj1TRGnqZHGIsI7rpzfqKNtXK6gBILydvrQabV04bKZWnbOcLEe4v3z2WRJFVFnxg/NQtZ0uHUEF1NAZ3dH51dUUrqVwPdZBljgZ1sxlXRyxVLT4ux790GG6p6mdOFC04IKzfcjI49AUMlHKx4bjcoD6gSRhmOwwoG/SA+ayn26oYwPLTj5KNjABv3QXTtxGDlZlBT+JTlx7hYD2SvxkbLVbfIIYS1x7hBg0lM6SrMZBwCsiujFK4AY+pTQSwsmL24JJysa6OfO4P7gd0EYhNSPh74Vrs0g6HblSW2dkbsOO6tucjJJAW+R3QYTjlxPqpIoz9PyCU8Zld0hZbw2JnQTuUGNLUOc3oydlkUsD5YsjOPVYpgkO+NluG1NhZRHrIzjZBoZZ7vIT3KvdVhzdhjCSsfJVuB3blUljjjcAfNBFT07qqTGcZUk9OaN4djOCp45ooMOad1KZI6oEuO6BS1bagCPG/1LGrHOhkDQ44zupGxiElzFi1T+t+XdwgpJP1Nx5+qRUxlYX5VjIJJPohZsUMsMR6hj5IMFuWvx3wcKR0mRjCkpmNfOetW1rWslAb2QUpyWODlWqe5x7LIhbG9gA+ktco7fSPpi6TGcINrxsMuw2KkMUsZBc8kfWsiZscFSQ3sCqveyUYG6CKOoDgIyN1kTtHuryPILEkp3NPUwbqcS9NLIx/cgjdB6m+w2b/1L4xv9Z7CPyNrv9a9Sl5bew3d/wBSuMbPMT2E/lbXf6l6koCIiDrx7QRtU/k94jtopTHMaSj6XDuP9vU+fzZXz5UFZP40pqZXPwMfEc75+a+gz2g0zqfk64kTM7to6P8AXqdfPPSnqe8k9xk/lQa7TVPjvIaSB5KO5VphaYznJWHDUshOxx6q6WWGp3kO6DT443Ty9R7HdanBOKTDQcHCsY2OLdvYdlh1LnPf1DtlBuS2QC6uLXu29MrHvNvbbnFrSO/ktNo7pLRD7ySDhKy5zVpzPnKCAzub3yR6FWGo8Q+EGYJOFNE6A/TICm6KUOEjSDhBJQRttp94mAIPqFZXujuEgqGDACrU1UUsfhk9lgCbwwWg7HKCaSua2IRN7jbZX01exzPCc3JOyxBAag5j3KmjiZTY8U4cgnkp/AzMexVYXiscIBsScKKarEreg9go4XuhlEkfcHIQaybS6xxiskfkHyKgNG68g1ER6S3O6grrpU1kIhnz0j1Kso7l7jCY2HugxayCqp3GOSQuA27rNtdvM7BI0gFYwqTVzEydvJTNrn0p8OE/LZBk15fTMI6yQB9a0oVZ6g7ozusqaWSb4pwcYU0dPROgJ26kEUda2QFuwWNUYD9t/NRvp5A89AOM+StPWHjxNkGXBI0gDA2WSat8LD0E5WP1wsjGO6mtcTa6tZT9y4gYQahpuvlfUEVH0fLKu1I6OeoEUQABPkte1Bpl9htYrmtxkZytm09Uakl79nBBHU2p8EYlDsZGVi0znNnY4kkNOe6zqyeoeOg9hssBsMob1AEDPog3DX3eB1EI2gZxhaHE0SAvPrlY8j+oY88qscpY0tHmglfKJAYwOyoaVzW9QOCFZTkB+XLJlqG9BaD3QUt7pZJulzyR5qaopjHVCUN27qG3Atm6jsD2K3VS6Oud5AkpgSCP7koNMqq1s1K2IM+IAKWy3J1FIM7DtuVnV2lKnT7fEueWN+YWHHDRVcZfSvBx3QZ2pbtDcKPobjPYraVBRe9VQgPnssitilLixhOyxKed1HL1hxDxuPrQb1ntEOmaWKrLQ8vxgLsRwT4CX/ilb/ulbro+nHRkAPIXWq3XGe9iKnrjmMEYyu0PBLWGt9NtZQaaa8wloG2eyDulyn8EdR8P7q19wucszY3g/E8nsV2/1Hr19qqIY25w3AJx3XV7l41tqGlfHU6qBDXuHV1bfpXZW61WlL0Ia/xGFgwXHIPkg4G5meK4dZ5GyuAaYz3HyXntqXWwujZoIZgA5x7fWu4fOjf9KSWeWls72mVsZB6SO+F5tVF5NLHO6RxD+s4z9aDVrrO20SPrKiQPyMri3Vt5iu0hMQDQCey1eqvk9ylfFU5LOw3WzL5TuimzTg9JJygjoPDp5A5wB+azKsQ1HxMaBt5BaKyo6fpHcKbx5CMs7II6uPwj8OfsWKXPPclZjJWSH76s0Q0boiQRn60GjEYVO+ylmic1xONsqNo6iGgb5QVb8O5VTIT2Gymlopg0PDdisqnhgFOTLgPA2yg0+JnW7A9eyy/eGws8Mjcqyj6feDntlUr2gS9TOyCCTJJd6qwAk7Ko6nbAKdga1pzsUEToy1uSVNSVZgeCTsoZHFxx6K5rG9BJ74QahXXKKePpaBn1Cw4JAxu/qoWQvkPwAlVcHRnocMFBO+racgBRx1EnWPiOConMLQDjYq9hjA+aDKq5WvjwBvhYLck4BO/zVXv6tvJSwtDsDzQTQUPiDOeyhqIDA4b7FZbvGpm9WDhRPeJ2ZPdBbTwGsPQ04IWSJzbyI3NJ2791iUsj6WQP7DO61aD3OreHTEZQT2elZfpjCWYx59lPdKWKyv8AAABJ22Ck6fuUzxrf3O+AsB9TLXzB9aTlBVkLqndxIHoVj1bRC3w2nJ+Syq2rgp4wIiMjthabTyuqpgX9soI6SOQS9Ts4z5rUp52shLm4yFJUwRxw5j743WjyOlc4tIOMoMq2Us94nMZk+EeWVPcYRaCITuSrLXUm3u64+6yK2Vtxd1SnfyQaNJI6c7DYq6Gjy4Z/OtQZSMYcuAwoJqmKJ3S07oIq+IsYOlv5Asenncz735Lc1kFDUf26RjG2Vh3uCgjqA6lIx8kHen2MMeOaHVDj56Brf5Rt6K/2McjHcz2p2t8tBVv8o29EHs0iIgIiICIiAiIgIiICIiAiIg465jWeJy9cUI/7rRl7b+WhmXzS+BmqkiJ+g4j8hwvpa5ji5vLzxQc0ZI0ZeyPr9xmXzRPfIZ5HD6RcSfyoMt1M3w9isukkjigLXYz81pPi1DQCc4VQ6eQfACR5oLahwM7iO2VNHB4zOrPYZURppidxup6aKeMEYIBQYmHMd0gkH5LOgp3uZ1OycbqGoiLPiI3zlara5YTSlshHUAg00Hx3mFvfspDaXtwXZGfVX0nhw3AyPwG52WXdKgzYNNg4Qac6lFMS4nOAsaaQSH4Ve41U7vDwSVU0ksQLpWkY8kGRRWx1T3cMKK4UXujw3vlUpK2aF4DCsqd5qCHy+XqgwnUpEXib9sqFrsd1qFRKx0XhNIzhYTYXdQ6u2UE9soTcKjws4B81n3SzNtrc5B2+tRsqY6GMSwEB6x6isrrn9IFwHogxYITUS9I81qBszmgOLvnhQ0cEtNN4krcALIr7hK84h3A2QYlTGIm4BVaWuMMZYc7rHdM97vvndWvA26SgSyF8hf6nKtLyfPZV6ds4VqDPtltNwk6AcbrIudjdbxkuz6brBo66ajf1xd1JWXaprABKeyDF6pMhvUTj5rMirGxR9G2SFgEnOfNO6CUS4mL/ACJUj5XTkNa4jO3dYyqxxa4Ed0Gey3fD1k5WLUtLHdOdlmCokZFk7jHosGaTxHZQWl+RjG6Nd0hXu6OjY74UY9SgpknzWbTUHvEfX1YxusL6lPFVyRMLGHYoI5WeHIWeisBwQVPGxxcZJOx3ypHMjcQG4QRvqOpnSroaQyxl+cYVJoA1uRlWR1EkY6QdkFvQ4ydGc4VXs8N4/KniY+L98VOyhqapnisYSAEEz4hPC1o7hcgcK6uNl/prVLT9fiuAJI7ZXGgklid4Z2I2XO3B1+maaWK43B7BUR4cM4zlB334SaHtmg9OwarlLAH4d0ny812y4bXCn11bBcKSsbEIWZ6Q4DsF5wxcfWyW4WSWX/ajMNAzthcpcL+YSWytFttU56JBg4cg796X1UJbubL4weWO6cg5XKMbS0ALq1wFvYvt1bcpiS97w4rtONwEFURUByg4345R08mlntqKUTtLXDpxnyXnZxE5cbprChrb5aqOWCJhLi1oIGO69S71ZKK+U3uta3LN/LK2xetBWyk0pX0FviaC+MkZaPRB4Mamu9Vwsvr6EUzxLE4gnBySFp+n+K9/vWtKS4uq3sZE8HBJHYrt1xd5ajede1VXcIfvTnEj4du5XAOseWTUluvrajTdO90DX5PS09kHfzhbzeNsGg6a3yzh8jIwM53Bwr7xxvr9Swm/RVhayI9WzsDZdZ9AcOqqltMVLdw8SgDIIIwt83+stGnNJVNE0gOLC3v8kG2ePPPDXTUL9M0VzIljaWZDt89vVdIb3xd1Pdq2Wesucjw55OC89sqTXlFSVmtJ6uZzuhzydzt3Wy9RUtDHKfdnZAQbguetaq80opzOSQMZytBgqm0riJnB3UVpLYstHu5Jf/OrxHO1pfUZBQajXVkLoyWALRHyeID0hPFMrujyKnjpHA5xsUGGeo7HyVzOoEOyQAtTjp4c4djP1rHrKfpyYzsgup7g2HyyqyXMPkD+g4CxKaNrpQJNgtYbTUHR3GUFTd4qiIRYAOMLSan4Jct9VLU0wjPVAD3U9LA2SI+P3QYfvY6QA3JUjHmSI9wpaW3OdOeofCCrLi0U7wyLbKDHpusy7k7LUCWuIiO+VprYph8TQVXxpGODndx6hBn1toMEXjtcN91pRJPcrMmuNRNH4bycYVIhEYzuMoIqaYQu6iFWWbxpQ7sFGA0yYPZH9LXfB5INbhjjfCAcZVzojHEXNOAtIbWSsaA1ZdPXF0fTKe4QQGrEbztuCo5AZx1g9lfPHG8ks7qDplZloBwUF1JTuqZOjOMeqlqI3UTw3Oc91HTtnhf1sByPRSVDzIcy9wgt972xv2VW0/jtMmdljmIuJ6Nwr2umiBb2BQTwVbaYlpGcFZEleyduAMfJYtPT+K8l47lR1TGxPAYguc7w3dY81UsNS3rzuFHEC8/H2VXyeE/pYdkFsUroJPqWd91nhnQ04+1WRtgczL8ZWLUNYH4j3yUCaUyEuz5q2KUsduSqeBIB1Y2VhBB3QZ7atrRk+axZpTKcjOFaI3u3wsqOKNtO8v8ApdJ/Kg9U/YcRFtp4xS+T57CB9grv9a9SF5c+w5eHWXi/GO7aixH8orv9S9RkBERB1z9oeccmvEr/ABOj/XqdfPPA/ocfmML6GPaH7cmvEv8AxKj/AF6nXzxjYgoJ3wOHxh2xVaanfPKG5VHzEtABVaOfwpg49kGrVtCaamD/AJenyWnxYfE4nvutRulwZNSBgIJ7fmWl01LUSRufGDjugjh/sp6uyvn+JwaB3UWHMlLT3VxyJRlBqFusb68gdRCguNC+2T+B1Zycd1mx1NXDHmkBJI8gon09XUHx6xpBG+4QYVTSPhjbKezlrVn04LhQmoc7GBlYT6iGVohkOzeyoy+1NGw09M74eyCMdNqqyx+4CznWwXaH3uN2AAThaQ8z3CQvAy5ZlNcai3RmmdkA+qDEpqQyVRp89jgqWrYKOQNzlXeM1rzNEfiJyoagvn+OTOUFk1T4jQ1oSGjdKzrJIU1sigkkLZj27K+4zNgkEcBGEGCMseW57HGyqZSHh3fBVrCC7LsKZkPXJn97lBK+o96Z4YGMDChbHPE8AZIPp2WTUmnhYDEfi81uSxw2+ehL6gtDwEGg+9NgjGW7n5LGcwVh6m7LPubKcTuAI6QVplNMIqprWn4OoILJqaSIbg4WXYKgU11gmcdg4ErWryykfbmuhwXkb/Wtvw26t6fHbGQG75Qcp611HTXXTzKSMgkMx+ZcbUdEY4ny5zso5bm8tELyctOCsq3SPk2Oeg99kEEE3vEphczAG3ZXzvEbxA1merzWdWMo6RniR46yoIZKWePxHkdY7ZQYNdbfd4/GB77q2ltpqIDN1YACVz6t2WyA9A7bKGCulgb4YPwlBA9hY8t9DhT09JJK0v6SfTZalR0TK8ARgFx7rclrt9NSuEVSAMj0QYOktPTXiqggEBPU8N/OvULlv5NLZqDQn3fq4GF7IQ/BAz2yum3CKwWqeqikjawuDwR275XohwYu/Eu00cVHbqeX7mOYA4tzjGEHAuseT/8A2R71V2Ght5YISQHNb6LrFxF5bK3hNqAWGeF7jJJ05IPqvYCg1rpLSsUtXJLCLg8HrBxnOF075jjDrW8fdyFrHFjy74R80HRriPwvj0fZobq9oBlAdj61uzhZyt0vEbScupzO1pjYXkZHplYfMBdbncLc22AHpibgBbC0FzEav0DY5dP0r3CF46cZxgIF/wBFjT99faIQD4EmNvPBXZPl71Nb7LVwUtbTMd2HxNXW7T2tYL9eTcbrIOuV+T1epK7I6IsVNURNulC0FrR1Zag7I8TuJEdl05Tz2RgD3gfQ7/mXNvCKkuep+ClbqWsqnsligLx1Eg9srrvwxfpjVk7bZqKRnTE4DDlzZrvi/pXh7w+qdI2CdgZLEWYa4emEHnFxV4p3a5cS7lYKuufJHFI5gBdkYBIXEmr4yxzpY3YGerZbh4h0sTNdVuoXZxO8vz9ZWw7/AHhtYCGEEbhBoct36pPCacEbK19ZHJGWPIJPZafLC10pewbn5rFd4rJATkDKDKNtMhc8HAKhe91MPC6ScrKhrwxoa47+ayC2knHW7GUGjNY55J7bquJI3Dc4WXVMDP7FuOyhDmOjPWd0FJqlr4+kDfGMqGAgStJ9VdAIzJh2MfNTTeE3dhGQg1SepibTAYGStLfG6cF7Tgeix3zPeME7LVaK21c1P4rGnpxlBgUcfVKR2wq1o6HdPclUlJppXNbsRkKEyOkeC7dAjeGqjnFxJGcLLbRSVhApGdR8wFqtNaWUtOffQA/0KDQ4Y+o7qkjSJOkHYqSqPhznw+yuj6JIyf3/AJIMiFzaNof0k58lmfch1whNZ9EAbjtlWWKKOSbFfswdsrPu10hpG+7UZHSdkG3Z3DqMAHY4WpU+nXz0xqGk4AzstJnDw8yOH0jnPzW57XVTm2Ox2wg2tLGY5HR98HCQvMcgd2ws2jbFJWvFQRuUuVM1sgMABHnhBdWXBk8AjaN8brAjeWHfsrDsUQTyziQANGCsikoXvxJ1kDusFndZDa2WLZp2Qa77+ykjDJHg4GN91gvqW1soDDgeS0yWodL9MlXRudF8bM7INVmtTnNDusHPzWp0dhY2lM/WMgLbwuNZKQxm+FkxXyshb4EhIadkFJ610czoTuAVA2pbLKI8d/kppDTvBkO7itPkLWPD4+6DVpabwYhI3fPktPFW8S7kjBWpWWsZO8w1ZHSB5lYtzhhdXiOmwQ4oNy6etTb00NLsZ+akuugfdpQ4SAjv3WnOmuNhpmzQAgEZWHJq28VTw6V5IHfugivdJLbulrHkDtstWsmln3K2Gtkec4yFpU1Q+64DtyO61Sh1DLbKc0HVgEYQd3fYy0jqXmp1W09hoKuH/ONuRZXsbS6Tmh1TM79/oOtP/OFvRB7JIiICIiAiIgIiICIiAiIgIiIOOuY53Ty9cUHHsNGXs/8AIZl80rHA1jieznn85X0scyX4O3FMf9xV7/UZl80LiWykg7hxx+VBuG4WuJtAJ2OGSM4ytNtkkbWuDwFFJdaqWEQOPwgYWI17m56fNBnyVrBIdhstSozFOzJwFt3IJy491NHWSw7RnZBqF0ia3PTjbzWmwyPY8Na4gEqaWrMzB1d1jNzkH7UGrT0niUwe12+Fl2qla2ncZXZOD5rTG1Mz2eGzOQr4qiqp2Fr8gFAFVHSVriRsDlX1tfFVjDMD7FgzDxXl3mfNUbC9u+6DIoqYPl6idgr7g4QnpYe/oomzOYMRndWuLnyAzILqWne8h7uykqS2IdA7rKZNC2MBuNliPgkqJQ4AkZQYskUnT1nOD6rKt9dHSjDxlZNU2JlMGbZC0ZBqtfco6hpbGACVh0zwGnq3ysfpKAuYgrJu8kDAyrc75Kd0QSxjxCGhUmi8I4yrGlzTlvdXOc55HV6oLPmiyHhnhjHfCx0BAMkD1RVaR380GSyj62dWcfaonx+G/A8kE8oGB2CdRdu/ugynkOiAB3woIqXrOCcbqxj3uPSFV0kkTh5FBJPRGJnWN1bTU5lznYLULOfeZ+mp+gfMq++MippAKTBHmQg099F0k4OygEZEoaBncLLD5ZogG5JU0FL4cZfMBkIJp6VhpQQQCtOjj8PJJ7FJayZ7jGw7dgoXOkZs/wA0F8s/VsrWRF7SR5BRtaXHA7rPhi6IyCgwmR9T+j5rcVDcqeipjC8DJGN1oMbgyYnZVnf4sga3OeyCWoYJKgzNGxOcBTUdTcGVTBT1D2DbYHAwpqW2VLGCaVhDD2ytSodJXa5SCegjLmDc4CDWbrebpR2iJ8MznPJG4O67acpPD6p1lQNuldV9Ba0Ow52D2XXPS1jpG9NNqDDQ0j6Q/wBa5d0zrm/aTniotGOeYCACWZxj7EHf3hpeWaHuHugqARG8DOV2XpOLFC60+/PqWfe2ZI232XnVoLWNxnibWXt7xKcF2T5rdl6400tthFIJyIjs7fCDtZDzU2mpvJtDpoosP6eo4C5r0vqC23u3x1dLcYpy8Bxw8bZ+S8ceNfF6y2qhNw09UZrfpHpdvlbY4N882urPIaWunnDBloy49kHugHxn6L2/YVHMYJGOilkYA/Ygkbroly582tTrWuEFdMXZIHxOyuW9ecbJbdqOko6ZxDJCMgO23Qcg6+4UWy8h1RBRgvO/UB3W1LJwysFuhNJXWlkj3nGSzK5o0jd2X2wUtaCCXsHUO6y5LRSSSCUt3Bz2QdUOLfCeHTtJLdqC3lrSwuADfkvPTjRqvUUE9RSwUExaHkD4SvbS/adt+oaM0VezqYRjtnZcO6i5Q+HF/lfLNA0F5ycxgoPCW9Ut/u8viNs03W7z6CtU01wZv99b1S2iYl3n0le10fJDwyjxiCLI/wDoQW5bPyrcP7QAIqdhx/8AQwg8PLvwPvOmYjVPsspA3+gVx7d7NcJusOtkkQbt9EhfQZeuWHQN6hdDPTMwQRvGF015luT22aatdXcLNAwMYCchqDyVqbc+hd1yDBG+CVa24fD0hpK3PxQofuVqB9sGzmEggfWtPsdohlpXST4yMoMe20nvr8l2N/MqW6UrKJpGQStPnrZaCreynIABUctZU12XSZOAgxovjmONs+iunEkbs9Zx9ahDjG447qYMknYXY2QSwVYAAdushknVgjb5LBpg0S4f2CyHTMbIA07IMqS4+C3DYyT6gKDoNY0zPGCNwtftNNba1gEpBP8AOtNvrYqGcRQH4exwg0+CoaHmNzdhsoKvp8QBv5llzin8APjx1keSgpaSWd4c7OEBsXiMAIwseRjoj0g/nWr1ZhpoAAcOWl9Dp2l43wgg3JUsdO6QE4KkooPEm6HDzW4mUsEUBztsg2v8TTgj5dlaXHPos+obD4pDcLEkaM/D2QX0py/DjstREDHEbBaSzrYQ4ZC1KGY+EXeY9UE1SWUzMgAn5LS3uNQ4uxhZFNIaqfw5dwra6JsEgbH2KCFknhEAtytWoqEVsXihuMb9lhwxwStBJGVuKhMdPRFrcdkG36h7aWQtHkeytbR+9RmbPZY9c8y1T/PcqWlqXwx9BOAUFKZgkmMPbBxlW18AgeADnJR8nhuMkZ+InOyjdK6V4Mp80EPU7GMnCqw5eM+qnlEQZluMrG80Gpl0ZixtlafLjq2+xBK8jARjeqUB3mUEkc/R3b+ZVmlL2kjYbLPrKSGOlEgG5HqtJAdgnyQerfsNf9z+Mvp41g/RXr1MXln7DX/c/jL/AIawfor16mICIiDrr7Qpgk5OOJLPWjo/16nXzw4X0O+0NeWcm3Ep47ijo/16nXzzQtDic+iCzp2yqKSXbICoyJz9wNkFYmOld09RwPmtfpKyGipDE4ZOFomC0fB3UkbnOx4v1IK07BV1bnYwCcqSvp2wEOGCqiSOn+KM4Kxp6l8+xJJ8kGpWO4iGQNfH17rc1WyOspy4RBhx6YWncP6W3ury644DNsZK3PrKstNMwx25zTtjZBxvU0ThUPa0nv5FRPo+gEk7hTulkNQZHZwfNRzulmk6Is4KBbaptJMXObkE4WoVlKLgDUMIGBnCxaa2uaPFnacd1bNXOil8OE/B2QLbSGSpMb+wK1C4UHgxEsGSAqMcxkQkj+mfRbj07SisqWGtx4ZIyT6INgtEzX5a1wJ+RCndTukBe87hc5XPTGlZ7bmh8MzhvljuuHNRUs1vqTEAQMkYQaK4EOx2WZRO8RwgxnO2VDM3pY1x7kLVbTa5JIDVRjOBnOOyClw0/LTQipblwO/qsejFT0Z6i0D7FufTVypqyoNHciA1u26rqoW2md0UJaQfRBs6rlf1FpJyrqaiMrPEyQRup2QNfJ1P7FRyTmB/Qw/D54QSROm6vDc4kA9luGiulO2n90dE3qcMbhaZbHUzyC/GSswxU3vDSzGUGLVabkke6duQHHKthiFGzwj3HmtdrKt0VMBnGy0CWqZK0kHJQQ1VI6XLuskHtkrHioJI3CTr2B33UU1TUtcc5A8vqUfvs5HTnug1C410UsHgsaOoDC0fCy6aLqd1SDY9/mrqlkQBDMZQZenK5tFVF8h2K1K9XMzyePC8gD0K2x8UZyM4WVHO2Rha4nfZBybwr4iSWWvgEsxAZICcnyyvU3gNzX6VZo9un53Qe8SRhoccZBxheNVK5kEgkjzlpyuTdG8Q5rK9j2Pf1tAxh2EHf3i5ca0V09/pbxiGUlwaH7YO64guXFSGCzy+9Voe8ZAJOVwjqPjjfbnb/dZZH9GMdz2XFVx1hdKtxhy/ocTnfZBvbVevae+3SaF5BaSQCcLjCut/3Qu7aWFoaHuxkD5q6pkY0CaPPWe61C31EMcXvbiPFYMhBq154eTWS1wXGCow7ZxAK5g4CcWfDli0tVu3lwzqP5FwZ+y+8XaQUNQT4LTj7FvfTb7JYIhemuAqYh1DB3yg7yy8NX2Oy/spo7kGmVvXhr9+2V174g3zUU1WXOrJHxtdg5JIwtL0Dxn19xGnGm6DxX07D0bEkY7LkTUWlWWm1OivTQ2oe3PxDfOEHXrV8hucJYN5CN/VcbVdqlpIHl+Sd1yPqCjqKG6SyPBEOSR6YW17s6OphcWEEIOPGVDopz1AkA47Kd2Khwy3Hks1tJTNqD4mFZVOp4ngx42QYk1tIb1NP51h9EjH4Djt81mVVbI5vSwqCInBLxugknPTCM7nCwhlzw07ZKz4+iYlpKx54DHJ1NGwQJqR0TA8ZKx8O9T9q1u0ywTnwqjHyysO8MgiqOmnxjfsg09bntmpaejtxpXsBcRjOFtk9gqiNxGQEF9TL4075fJxJUeD6H8iOGDhbnttNbn24vkx148/VBp2n7tHaqgyTM6gfIhahd7ky7/foT0gDcDZaLBTx1Na9mfgyVfVNdSSiKPIaTgoIWxGZ5YfJXRweDUta45Gd1LLlkfXH9LvssN00nX1O7hBrdwiApQYTg47grSGxSHEj3E49d1c2qnnHh5OArJGzs+HBwSgrI8T4Z6LUaS6tpac0p7laQSY9x3PdBHI8GQDIygmqGODzMwkZOe6yYZQ6Ah5BPzWA6RxHSfLZBI4DA7fWgPaDIceZ2VpbjzVQ7fJQnbKC1FVrS44RzS04KCiyo2tMZyViq8SuaMBBlUcjaV5c7G6irJmzS9TQAAPIKAlzyqEEd0Fep3bJVY29bw31QDq7BMljgR3CDKfRPhYJGOIJHkVHTzOgqWTSAnpOfVVFW9+GvOyyI6dlU3oYMuI/Og3RHdaa+0/unSAWjC0e508dvaYWtBJ9Fhx0lfaXe8lhA+ruFN78yud1T9/mgwKSrdSSlzgSCq1EjqiYTtOB6ZWZNDSEdgsMM+MMYNkHf72MFb7xzP6ngI+joGtP/ONv/1oq+xipmRcz+p5ANzoGtB/zjb/APUiD2cREQEREBERAREQEREBERAREQcccyX4O3FP+BV7/UZl80En9kf9Z/Svpf5kvwduKZ/7ir3+ozL5oJP7I76z+lBaqhwAOyyIIf3z+xVsjWOma1v0SUFkUTpXYwqzQ+C4Nxla1LTwwUQkYB1Y9FpccjZSTLuQgt91Phh2Vaxgzgq6aoO7WnZQCR2e6DUKciA9ZwQrK2sZKfhA29AoHzdTMHOVHHGXb42z5oKNlwckLMgmbL8OO/yVrI4nnpxkqX3bw2ktG6C73YQgy9wVhVMpkfkNwB8lmUTaipm8N4OPrWVXUkEAAIGSg0ukDpJOguIH1rVy9lNFnAK0yZjoMSx7Z7lZVHOyeIibGQEGn1NQ6SQkE4PkoBspqpjWynp7KFBNE4OIarpINsqOnc1r8lS1MuSAwnsgxiMHCYPopY4z1dTxsshogLhntlBhty05wrnPBWqzxUvg/BgkjstM6GjcoEMRmdjJwk8JidgZPzVrZHRnLdlm04ZPGXSbkINPQbK+fp8QhvbKtaxzvojKC7rGFTOR2wqtaGn49keW/vOyCsMnhP6iO3yWW2JtVh+e25WApYp5Ih0t7FBmCUQ/C0gEKOScuPxHq+Z3UEgfjxHDvurYjl46kGq25jM9TsYHkVWuqGunFPH2JwsaaZ0ceWHv6LNttqqKyI1XSSWjKCeXTQpKYVpcCCM4JWiS/wC2HHAxhaqLnXzTe4TZ6B8OFj1tMIDiLGXd8IMSmhDHguWbI1pjJBH2LAmiqIQHuzg9lPS+NUREAk4CDAkBDyFmUNIZCJyNmnKiZEyOb787AB3WrUlTRvmZSRu3fgZxsSUGqQXdlzhFuZF8TdicLkLSWpqTS1Kaappg8ub3LVpth4aVlvhF5nYRE7DskbYWustdnuU7IsgvPw4+aDWdK6ePFK5+70v3kOd9S7CaN4Ts0JUQUlTAKgyAbkZXDFmo7hw6gZdqFhDcgjyXargRryy63bFLfHMMzABvjOUG6WcJpK+1CrpYCwvbnA2XDnEHhXdY4JYw5weSQF3Mor/aaSmMYLPBaNu3ZcG8UtcWGG5gdbAzr+LcY7oOiWsuFeoLdUvqK0yvi3OHZIXFV1pJaCocadwjLTj03XfXipqfRF206WUEkJqDGe2M5wuiuuOuOrn6execYPllBvngvxbuGgLi2qfXFoDge65guXNhJfdU0RNwBDCBu4LpnVGdrex7euFgU9VLTVLalpPUw53O6D3m5cuYOguWnaaGe6Rk7NwSF2ltF+pbtEySCVrw8A7LwD5feMmoKSsht0L5T0vA+kfVewHLXq6uu9vpPfycujB+I/JB2SRWxuDm9XqrkBERAWyOK+maG/aLuFLLSiR7mHp2yclb3WJdWMkt87JMdJYcoPA3ms4Mz6c1tV3T3V7Ii4kHBx3K6609yMccsLcjpJbsvUrnu0tS3WOoZbQwzFhGAN8rzPj0FdKBlVLWsLcOPkfmg2VVyGWdztySVNSShjegt3PyV8tM1lcYD5FTPpo43jYIMWelc0GTyKQVbI2FhGcrMrHM8DAK0ljepwHqgvkB6i8bAqwBziAMklZ1RQyxwCQg4WqWWK3vg6piOseqDHt1HUU+Jg477rIrqF1XGZHHBA8ytStToXVRZN/Y8+foq32GTrDaIZYg2dh8chjOdjg7rVqOribiPHfZKygAjDyMPO5+tabE7wieruEGt1FoNUzxGuz9q0yX/aGYXDJOyy6C4zjZx+H61jXOWOWUF3cIII5PAf42O+61qB3vlMXdeMBaJO+N0QaDuArqOudTxmMEgFBa+BxqC3qzj5/NWuAZKGHzUgm+IyHud1BI4yShwQZ9TStipxLjusFk5HwD1U9RUSOgEZzgKOj8IOBkwgvjaYAZcY8+ysdL71M1vqcZWXWSQuh6WEZx5LBhaWnxB3ByEGrVFqNHSCp6h2z3VKOtdLAWdR+1ROuM1WwU7ySBthIKUskAYNigwahrmTmQtOCVGXdYyB2W4qyhjfADgZxutDexsOQdkEdMwvfvuPrUtTT4I6dvqVInfFlndTHrJBd5IMN8D2DJyqNhLxkFZpcx46c5KjdGWbDzQY7QY3dsqZsRfiTpxjdXUrWF/wB89fNZ8slPGwtYR6bIMGSqdO3wBnYLGfiMGPG5UzPCEhd81HVOY5xIO6D1X9hs3Fs4yP8A7qewD8ja7/WvUpeW3sN3f9SuMbPSewn8ra7/AFL1JQEREHXP2hwzya8Sh/8ASdH+v06+eqkHU9wPovoV9ocT+014lEf3nR/r1OvnlY9zDlvcoMmWmzuFRknhN6A3P2KIyzYyScLUqOlZLTmR3fCDGpo2mQud5+StrXhrgG7KJ8z45S0HYHCslf4jhjugtMjiMZKRkB4PllXupJ2t63MwFYMAEHug1ATSNYBA8tP1qIGrklHjSudk+ZVtM14OXdlmHAcC3uEE01MPBGMZWJTO8CYOczIB81nxufKA0Z2Ugipw7pkx1FAr6+OamDGDBx6Lbjg7qyck5Wu11N4bepo2WmRmBxw/CCShmdlrXEkLeVHVtjoC1jsOLe47ra0LYSPveCVM2eeJwac4CDNoL9WWquc+onJYTsCdlq9dQR6kiNbHg4HUtp3d8T42kfSW5tLsr22p7o2ks6cn6kGNZdItvlV7oXBvQcDyW4bnaqbS9I6iDmkkY75WznX+vtle/wBxJ68nKhqdQXS51TDcHE5KDHnpagVbp4iWB2+RsrHxTySAyykgepWuVr4Pcw5hHVhbedUloPfOSg1IwNMfwuGVp1RFh2PNKepmc45JwhjnkqAd+nKCEGWn+IEhahSVJc0TOf2+akr46f3YDbqwtHZO5mWg/CUGq1d5NQ3wQSMbLCYyYHqwSFDM1jQHMO57rdVkioZbYXzgdYHmg23UStkb0YwVHFBgdffCV5aKt/hHbOyuimHhkHuUFJKkYLQMEeeFAHOc7JOd8o5vU8hoUzKZ3SXfL0QUmx4Y9VZHE5zS7P51bJ1BxafJGvdjoHnthBkUI65OkqaWrnpakFmcA5woaeCaJweRgLXrdHbqiRvvOO/mguprpJcIRG8EY7krErK5lKTFsSfQLV76610NL1UBHV8itnPkfUuLnnLvJBqLZg774XDB7LHlrpGyYYT0+gWK4zRtw7IBWo0VNFVRHb4seaC9l1EQBawEk4Jwt36esFbqMeG2QgPb2ytN0rw8umoqkR00RILtsLm/RPLxxKZcY30NPL4WAc9B/wBSDfHLbZo+Gld90K2h8XJzktyt98ZbhcdZVbbjb7ZI2Jm5DWkAhcwcD+AdzqaaCPUcGCCOrqBG3mu0R4Q8KaK1Nt1QacTubjBAznCDx71wZ6lhpH0To3hvSSW4Wxm2V0dG8v777Fd9OaTgPb7F49xssTPDLS5paF0vn0xfDDK0RuwHEDYoOJbhRSCqeACMZWh1LXxy9LiVyDeLHV26V8lWwj1yti3OJ8s5fEMjtsgi8LoYHHclQmTqd0gYyVNA57T0zbAKGcxicFmMZQSgOhAefPdVNW2QdJ7n5KSoLZoQ2PchaeWuY7DhghBO6N8XxseRtthQOe55y4k/Wqukc4YPZWoCnZO1rC3GT9SgRBU/ESQCfsV7ZpmNLA9wafJarpyOklqCKsjp+amvUVBHVgQEdPmg0emnNO/qOd1NLUidwJOSFkVcNNJCPBILgPRab0vieA7YoMkzdI3WO4CQ9Q2V8rmluB3UIcQCB5oJYJhC7JGcfJTy1rXjAblYSy6VsBHx4ygxXuLjkjCyYqprGFnT388KtYIWtHh4yVHE2PwyT3CCnhdbi7132Vwptx/rVadxLj6eSrUGVrsg7ZQRTQeEO6vpqfx8Anuonyvf9JVinfF9FBqLre2nHWTlafN8TyAOyvfWVEgwScKSmaw4LsZ8ygjZSFwydlE6PpeGZ7lZtVKI24adysAvcXdR7oMrwPDaHd1Z4XiZ8lJSTBxLZTsPVWVMgbJ977ILejwdyoyfEeMeaO8R+5VGZDwgvkgMYB3IKy7TL7vMJnnYFXxdD2YkwfRY0z2iQMb2Qbju99pq+lFPG0AgYOB3W2Cx8ZwCST6KaSneWh0YOSN1lWqISVDY5+5Qac98wA6gQFPTygDJ3K3DqG2U1LRiRgAyM7LbNK+NpLX+fZB6CexhqPE5odUN9NA1p/5xt6K/2MbIhzP6nczGToKt/J90beiD2aREQEREBERAREQEREBERAREQcc8x46uXjii310Xex/yGZfNBIMTP+Tj+lfS/wAyP4O/FHHf9hd7/UZl80LgXTOz36ifzoJ2zeIwR9ioXMMTg4nOFKDEwfC74lA97nHBQZbq10sXhEnCxHtLdv0K73eXHV07LKphEWffe/zQYLWlxwrnR9OMqZ4aH/CdlHK4eSC3w9s5VWzFrcdKug+N+CdlnGKmbHvjJCDTo5SyQPWYK7qA22UJpZJXkQjKyI6F0DCZxglBfTXRlO8uxv8AUoLhXGskDhkBYsg++ED1U0EGRkhBI6oD4BGe4GFjMLmyYBIBKukjIccBR7tPxdx2QZ/uofHknc+ZUQoMjuofepAMBXsq3DY5QJ6UwjPn9aU1KZgX57K2aZ05wN8Kele+FpaRjKAAJCYh3Gyx6iAwOAz3KnB8OQvHnurKgvqDkDYIIoC9zukuOPrWVJS/D1BYQLoz8ws2CpDmEOQYRaS8tHfKzaaNwHQRjKsgdD4pc/GPmsuSaAY6CMhA+43Xl/XgH5rFlYKQ9Hf5q+S4Ts+jnBWFLM+Z2X90GVT0gqyT2yo6yk91d05ykdS6ADoO6jmqHznL9ygja3qOFXHQ4I3LDn0UjWGTDgNggmle0wjbBVKKl8Vwz5q2Z7OjoaQCFkUE7YmFzvLdBmS0AjiGfJZ1t1FDb4DTloyRgLRKq5Pm+CPPfZRwRxBwNR3JQan7wyaoNR0gZKx62pBmDhuAr3mJ8YZT7n0WII3slHjDb5oNQ8RlygELW4Prhbu0Ro0V9bFQub1GUgfXlbct/uzWgxYyuR+HFTPDqWjkcPhDgd0HLbeTuS6WVl0jpXfGAfo7LdOifZ7XCspP2TSxyGKm++dJzjbddktM66tdNoWCKd8YcGjPb0XIeluONno9GVNtZJF8bCO49EHTG72e2mR+hBE2OSEeHnt22XFGqOH50RXe/tm62tOTg5W6uOOsRZdX1N9tzsOc4nI+vK4uqOKz9V2+ZlwkBcAe5QZ2suMNHVWVtnjc0PaMZWq8G9X1tteKltxLGjf6WF101D8VyfMzPSTkH7Vm2bVtZbYzFGXYO2yDvpHx8kmpzQC5jqAxnrXCHEziDX1tW6JtwJL3d+tcI0et54pjM5z8n5rEvGppLhOJ2k5G43Qb7rq6826AV09ye9hGenqzsuPr5qkXKYk+XmpW6olrohSVJPQNt1t+6RU8cmYOxQZL7iyVgGM4+S02cFzi8NwFWm6RJh/ZZ0hgMZAIzhBvDg3e/uLf45yAR4gXsByq63fdoaJkQ26G9vqXj1wqsNRebtFHTtJJlA/OvZXkz0E620FJJVsIIjGMj5IO5VueZKRjj3KylFBGIoxG3sFKgIiICxLrGZbdURju6MhS1FVDSs65nhrfUrQK3WdmY7wPGBJOD2QdLuPWj55LvPPPTOlYcntkYXU7XHDyiqrTVyx0bYiCdsYXp5xVrNE0tvfX3l0TWFhI6sLodxgmpb7TVf7DyJIuo7tG2EHnhqvTJtl/kAbsHFbfrWDrx6LlbXtguVPcnyVkZBGc5XEd6kdHUFoO4KCCrBDNjlYUTuh4cfLdTRymQ4k3CjnawPHT280Go1V3ZPTCAN3xjssCBkpcOl5Az5JK2JsYLT8StglLHDKDWnVDoIRhxBA3OVJS6jbFGWSnJxjOMrS55fEjAHfCxRTyu8kG5KSRlylOexWLdrQ2BwcwrDtrp6KXqIIGfVTV90dNKMnYd0GOJOgBg2UrbaapviF26pVOhfDmI5djyWPT1s8QMeTv2QWVFEYCRnOFi7g7FZdUZyC9+cFQMhkkBc1uUFnWcYyVJC398o+h2enG6mijka8E7AndBWaT4cEKOKF8h+H8yzq0QeAOgjqUts8HwT1EZ8gg099PIxvUTlXxuIYdsbeazo+h0xEhHTlT1FJG+MuiAx32QaPHN4cmfms+K5ta0HG+FgCLomIkGwR3hGQY7eaDOkvBeC3yWLIDU/EPVSSRU/hDGM4SJ8cUZBKCGMeA/JWUZGyw7d8LT3vMj9u2VcXPjHSOyAHmOUuys6NzZ4+ohYlIwTS4dupKkmnIYzYHKCCZ5DyATtsrWtkkOMn8qtIccux3U8GcZ/8AfsgpLCY25UbYy5hf6KefqkGAEbDKyne4jbCD1T9hsz/qbxkf6z2Af/g1/wDrXqWvLX2G2fuXxjz28ew/9GuXqUgIiIOuftD8/tNOJeO/udH+vU6+eMbEFfRF7QPo/ae8R/E+j7pR5/j1Ovnjhp5Jy7wxkN3JQXNeJfhwp21ZpmeF5HZHiGJo6cBwG/1qWnp2ztL34QaY9xe4u9VlUtP1gSHsN1dVQMBwxRxQ1eCyMHBQZlVXxOjELW7jbYK2ntZnYZjsBvhWRWese4EsIHfstbgpKmGAsaDjHog02JjXPMRIBCjnAimEYPdXTW6udKXQNJPyWPLSVkJ66gEHPmg1iGNkEfWXD1WjVdRI6tDmuOARjdWSV0zh4QJx9anpmxFodKfiQarNK2oowzzwtMFo6wXB2PtSWpxszssd1bUMyATj60EjWGgfkuyApTcxMQzpO+ywTK+pcGO3ysuGlYxvUe43CDOZZfeGiUuyM5xla7T6pgs1ufb8DJGFtuO5VEJLNw3sFiVZbUu8TO6DJttSyS5vqZQCHHO6zLt0TSh0TAMHyWm2y111ZKPdmE+uFuJ1A6hhxWNw7Hmg0CqnkjYGuJIKzaCyiupjUdW4GcLHmo3VUhEYJAV9HPXW6pbE8ERZ328kFrKZ0cxicwgDzwrqmqZTDpDQT2W5a8W+oow6jIM7hvhaHT6Ru1W7xXxHoznceSDQZqmaYkuJA9FBg+hW7qmxwQR+GWgPChZRUUcJ8TGQEG36WEzv6XLJkllgeKZkhASZ7KeYmLGD8ljuinmJmAP1oJ6q3eDCJi7JO60/KyJaqdzfCf2Gyg6SgvhkEbskLKFa3pxjyWCpI6eSQdTG5CBLJ1uJCsY7ocHeiq+N8Zw4YU8TGGIlBkuuDHxhmCCsKSWTOQ4gfWo3bOOEG5G6C4ySPGHPcR9aRnpkB9Fc8MDcjurIxl4J7ZQamY2VUYbjBWfR0ElHCahrC4BpOwWkzzeE1nhFcjaKrbNUWp0VyLA4jHxFBv3lv1lbDeqekr4Wj76G7geq9n+AVh0BdtMQ1TaWjlndGCQ7GcY7rwkZU0ena5tXZvpB/UOn6/ku0HAvmX4i2uohoYHz+CAB9I9kHrBqmDTlie/3ERRZyMNIAC4/m4X12pZhqCC5uEbHdXSH7Lg4a71prS0RyU7ZTK7BduScrdWk9aa903TikuglbARvnPZBpnMFWMNpks5pjM+JhGcZ7BdB9WaumsU0lObK/BeRnoK9LITpXWbi2ukYZ35DurGVoGvuW/hlXaelqnsg94LS4bDug8i+JWrHXCPMVKWF3/a9ltSx25tTbn1M4w7GRkLs7xc4S2WyXmpY+NggZnp+xdbdS1kVsqnUtBjwuotPog2tUA1NW+na3AGywqinMEnRuVr9S6kZGJoyPEO5WGHwStLpSOodkGJCw04D3bgqdtGK0hwOMrHqpQ8hrdwOyQ1r6dnwHdBdWW73VpdnP2rAWRPWy1AId2KxxjO6Aiv6OoDp7jyVrmOb3CAHuYctcR9RWRTU8lVIGuecntusdrHO7BSRzSQOBGxCDcVDYRAPEkfnI23Wj3cNbPhnYKv3arOno6tvrWJJK6d/U/ugjOcbqindEXNHRvhRtZ0vDXIEUZkOApnUpbuDj7VO10LG5BGVjS1Li4hp2QTUlvdUkguOyiq6Y0snRn86rBWPgyWndWzSvq5B5k7BBZHL0HIWX4gkZlQzW+pgYJJIyGnzVrZAIyMoIXfSKDYgqemDS4l6sqQwP+Dsgp4pIw1qrG8l4G4ys+1Mpifv+MHtlWXOGNsrXUwH2IM6mtDa9merBCwK+2e5PA6s7qynuNbTjpjJUdRU1NQ7qlySgjdG4fRCj3B3Wdb3ROcfFxhVq2QmbEeMIMaN/VkEKzB8UbLOdAxrQ4YU0MUErPDaR4p2H1oMWqBZEOk7qCGAub4h8itQdaa2mJlq2ER+WfRUZH4jg2Ldud0EENeIvhc3OFYK7pqBMzbByp66h6GdTAMhYMLW+KGSbBBq9Vd3XSNtMc77brXbbw7NZQmtM2MDOMhaPT21joxJAPiG6yhqi825nugyGHZB3s9jfQ+5c0WqYyc9Og61v/OFvRU9jbVOqeZ7U7n/AEjoOtJ/zhb0QeySIiAiIgIiICIiAiIgIiICIiDj7mHYJeAHEyI/v9H3lv5aKVfNM+HouNRB/cve386+lfmJf4XL9xNk/udHXp35KKUr5pjN1XCaY/vnuP5Sgyai0Ogg94znIytPb8b8ny3Wq+/umiET+w2Wm1HSx+GeaDJZWMLfDI+SxJyQ4dJOCpvAAiD8bq6lpXz/ABOBwEFtLTOnA2P5FJU0HhDJWbTBkbuhuMhW1koHwv8AP1QadQxF8pbgqtdHJHMG74WZSvhY/qGM7LOmdRys6nEFwCClsjipYhNJgn5rCu9wFQ7pibgA74WPLWSOeY2ZLfJZ1PHR+ATMR1YQYVnohXVQjKzb1TNtrwxv1LDoZ309wLoM4z5LNu03vADpjuEGmNeHDJxkrOhtDaqEyg42WkueQSGnZZdLdJoB4YOx2QYs0Rhlcw52OOyjIwtXjEE5Lz3PdYtbSkP+9jIQY0EgjdkjKlmqPEcCNlE+mljGXNwqRxPk+iNggyJc+FsrIakMaWuGcqWEAnokVlTAQ7LBsgx5HdbyR5q3KkiDQ/Dlkupw9nUwdkEdNRmoIAdglKqlfSOALs/aqU8z4JcNzsVfVOlndlyCN8wfH09yoFc6J7e4V8UXWNkESq04O6uML84wqOY5ndBlx0omaMd1QDwHCIjJJ8lDFPK0hrSsuNofIDIN/JBbUW50cRnOSDuobfCaqripM48Vwbn61q1fIDS9AxgBaNTTOp6hk7O8bg4fYg5KvXCX7haeZf3z9QLerpz8lsajoPusJHNOOhbgu/Eq6XazNs8rj0AdOFtm11slF1dOR1HdBLDTm3VJEhyAcbq+uc2qbmPYhQ1tT4+XA5JWHDM9jul2dygkpKuSklGScAreml9Y+BdIOnYhw3Wz6in64xIzc91kac8CC5RSVJGGuCDtk7XlwfptjWVZaA3IGcLarOK10oIXxi5uA7Y61odx1NaRpwQ07wXhnr8lwtdrvUyVD2tccElBv/XGsZdRlzXTB7jkE91xmZKmhe4NlIBJ2HmsmCWZjRJJnf1WFWSePKMeaDU7W0XafwJO58yr7xZ22yQNY7OVhwPfbmiphJBx5KGqulVWvDpnZwgvqaV7YRIDjbJCxI53MBGc5WVNO+WERjyUMVOekl2N0EBkd1Eg4Qlz3DJJKmjpJJpOlgOCdlkmiMABe3dBiPgLWB/ZRZPqVmVBPT0jssPB3Qc78rz426gi64fF++jbGfNe2vLxEyW10bmUxixED2x5Lxy5GqO3V+u2w3HHR4zcZx8l7raAtdmttrom2wM3hGcfUg3iBgYVURAVFVUQaJqKP3ymMHiBnffOFwjrOzS2mlmuTbkHGIl2zvRck8XvuzFYzJZGuM3Sfo910r1bqLiRC2qhuombASQc5xhBh67v994uVb9JW6pkLm5Z8JP8y4/u1il4KWOohvpL3vBx17nP2rlvltvejqHVr6q7zME5d+/I7rYvPhS1GqKn3iwAvpQcks7Y+xB0O4q8Tqe83qWOCEBriewXDlyzUyOk6e5yNlyXrKj05bgGTygVQHxDzyuNqmopX9TYZM+myDSnbHHoqZysiOimmf8AA3IKyfuFXgdXhHpAzlBFbaB1fP4QJwFk3O0ut3bJJWVbXx0z+mPHiZwVqlXLDLAXVRGQPNBtBkpY7LsrIZXNaQensrJGwy1BEbj0k7HClfTxRNw47lBmQ1bKw+E0AFQVdBIxwbjOfl2WPRB8UweOwW5o308tMXSY6x2QbdfSOpW9bnZCxTNmQPDcAYWVUunqZzE0EtBwFR1E6NmXDGyClTWNlhEYAypLfMxrCxze6wHsLXHKyqZmW5AOR2QanSWuOaQv2wd1S5QMpWkNAysSCtnp3kHOAsg1PvMo8Xt80GkxskmkAwSCd1qbKBzBkEgLVOm3RQhzSOpYks/iMLYdyghjphM4sB3Hmk1T7h96cck7KCljrI5y8g4KsuMUsj/EdlBC9/vUh6RhTNt3TGXk7qKnLGnDfpeivqKyRuYxndBj08bnzdGfNao2zeIBvjPzWBRRSCQSEbd1qpuJiw3PYIMaW1NpvjJzhadO8E9I+pZ1dXySNOCd1poa5x6kE0J93PiZUdRP4zsq5xL29A7hVZTO6SUEReOkDCqx+NwPNUMburGFNDASNxugtjqek7jKyH17XQvi6cZGFJTU8TXffcYUFdFEHdcPYd0Hqt7DlgFk4vu83VFi/MK5eoy8tfYbSF1t4yRns2awEfaK/wD1L1KQEREHXP2h4LuTXiUG9/c6PH8ep18+tnqGQmVjxnraAvoM9oWWt5OOJJd29zo/16nXz42mgnqnvkjaSIxkoLayje0uk3wTspaaVzIScYwFlzSxSfeM/GDhXwQRNYY34yUGDa2Gsqel/bK1moay3YcIsgBQUlsnjl8SnbtnyWvR0sE0PRWAdR2QQ6duEF0lEBjAycLelXp6kgoTOCM9JPdbFjpBaJjPAMDuMJWaruVTEYGE4O3dAbfKahrnwvaCAdsrS77doa0kRNAyfktLuFPO2Q1EwOXb5VLXB7xWMDvo53ygrR0skkhcYjjPfCuqqWQSdLWkBchmitcNva5vSH4ycLQqh9vcC0Edfkg2w+AwMDnLIpqRlRCXKS50NQ5pcxpLe4WNS1LqaMsdkH5oEdI2GUkn86vnkDBkEbfNYctY5zjg9z3UL53vyCgy4ZBUPDO2VuC06aZVSta9wHVjuts0T+h/V5haiL9WUkwdESAEHIhoqLR1L71IGu2ytj37UQvlY3wR0gn0V9Zfqy9U4gqSensVprKenpj1AjI+e6DclBSxUtOJXkE43Wm3Ksiqn+BG0AnbIHZYE13mc3wwTgbKkE0X03H4gEGfQ0MlpcKyaQlp3wSt10/EegiozTeEOrHT2+S2HV3OWVojeT05wPTCkordBNGZj2QalU3U1tSZm9iewCxaymfJGZA8gYzjKmtxo3yGJu5BWHeZpYZPDjJDUGjyOcHlp8tlmQ17Io+gjdY8nQ4dXmVjfWgyC4TSEgBWPJZluO6loYHzyYaCd/JZs9CIsGUY+tBpcbDI7C1ikdFBF0uAysYMiH0MZWLNM8PLQTsgurntc/LexKpTsc5pB2UbnNeBnusqOSNsR6cZQY89OYhkb+agOSs6mDppD4nb5qKsYyOUBuMIMbf5qWNhLc4Ky4GU/QOvGVe4whwazGCg01xdn4vJSwGpJDYJHD6iQAp6uFrWdQSm2i6mjcINUpquSjEZlkLiHAnddjuEeqKKloxUuDA5rc7gLrBSiWeUCQEgHZch6W1ALU4QuJDCMFB6RctXF+3T3BlHWBj2dYGDjGMrt7f7ZpzVlq6aSWGKV7OwwDnC8suB/ETTNnlE8kzRIDnuO65DPN5Vx8RaLT9rqCY3yBgAdnbKDt9p/hFXW28S1b6kiIEuznZbC43avn0hSPxWksYSCA5cq3nWN0oeHcF/LSDNCHflC6f8WK69a1tNTK1jnZz2KDrRxr4v/shrJaSnOXkEEjuuCpLa+ppZKiYnqJJGe65GuGi5aO+zzXSMgAkjqWwdZV7KOpMNGfgzg47INnOEnvLo3OJAJCjnDmPIycLKqpI+kSM+kVbCYnxEyYygjpIfEcclKqDw34B2UfiujefDOytfM+Q5cUFHR4Co1uTjspI3dR+IqkmGuy1BK1rYB1HzSQ+IMgKBz3HYqencwfS7IIopOg4I2VJX9ZBHkFkzxxub96G6xCxzThwwgnpKX3l3TnCrV0vuzgM5UcU7oDlndXGV88gc/t6oEU/h9wrZ5BI7LRjCzKhkHhZGOrC076kF8cbpDjOMKkjOg4yjXOZuFRzi7ugoBkrJp+mGVsrtw05WO3YqrpHEYzsg12vvENfTCna0AgYytHNP0nOdlC1xachXGZ57oDzjtkYVGfE4ZVpJPdSNikI6w3tugklicwAtOyyKN4c3pecnON1hOleRg+SNe8H4e6DUZjHAC7AKxnzNkBIarTBVSgZBIVzYhGOl/coMdpcThpwgLmPyTlSysDRlqiEUjt8ZQZ1NKJh0LLoaEwVzKhzvha7OFgUkT2O6sbqeavcz4SfJBubVV9pa22tpYWgODQNgtrUFSKeF3V3Jysd05lO+d/VW+DK5wABwUGXJchIS13Y7KE0/UfFbt5qKaERAE91JFI4xFoQZ9su4pj0POQpa64x1JJa0fWtCe0h2Ss6mdD4I6u6Dv37GF/VzR6o/gDW/yjbkVnsXiDzS6px/wBrf5RtyIPaBERAREQEREBERAREQEREBERBxxzJfg7cUsd/2F3v9RmXzQvJErjnfqP6V9L/MeM8vHFEeui72P+QzL5oH/wBlfnt1H9KC7xjjucq+KAzN6ySsc4zssmldJ5dggePjMe5xss6mqmxM6CACVCBT5JcRlQzjqdmPt8kGo0MfiVBfnIUF7aWyAtO2VbRzSU56nZA9VJVzw1LDkjKDTqcPe/paSVdUMljPxEgFZ1hpfFrPvg2HqsnU1M2KRvhjbzwg0WOYsOcZKy46aSsYZA4tAHZXWuljmf8Afcefkpbm73Jwjh2B74QYdK4U9Vh25HmVJcqjxngDtsseNruvxJAcH1V8sEk3xRjPqgjdT4Z1qINJUwkIHhv7hZUFH4reqMZ80GJG50R6uo4WQ2uAIJHZRyUdQXkBufqUEsL4Th4wgzamtjnj8MAA47qGCdsDS0gHKxRsq/E7f0QSOmPX1DZStqQcAjJKxiAMjzRpw4FBkT0xazxR2O6mopw2Itdv5bpNUMdBgdwMLDjbId2g4QZHwmYu7b+iyB0kgbFYL3Fg+ayKWZuMv7oJq2JoiyBusejc0A5wpKupD2FrViwxTPz0A4+SDIfIxr/JY80ofnCPhlzhwKjc0tOD3QXRP6HZUpqndQIzsoGtyQT2WoQR0zmdJxkj5IIZawyR9JJWO0HfAP5Fli1zj4y09HfKmEcLRg4yEGm5IO+e/ZXiUkgD1WVJBG4/CQVi9IbMG/MIMh0TmM8RwJH1LFc8PkDgMLWpoeulBx5ei0Ys6JQ1BnmbwoASM7LGax87xIw9JG6yxB40IGxVgIpR0nZANZWwsLHzkgbYysY1AfIHOHcqSZ7ZQceaxWtw8A9soNUq5waUBoAOO6wbdAKqsihd2cQFmTGJ1OGjGVi0svujhOPpM3CDft/0hBQWAVrXAktzj7Fx7TweL1b9lq1dq66V9KKSV/3sDHdaMyV8eek90E1O7pl6DuApqiURkALEjkAf1n7VWaQSHCDU7RVxtm+PH5FLeqqNx+9/LstHa2WHEgBAVfvtQcnOAgnpyal/QRlW1NOInlvbKy6AQscNx1KK6hxkEg7fJBzxymu+5+pYqv3jwyZRvnHmF7gcCrhT3Gy0kr7i172wjDeoZJwF89/DDVlTYLzG2OQtBcCMfWu/3BjmYvttrKO2w1MvxAN7n0Qet4cw7hwP2qvdcFcM9Y6gv1pirpush2DnK5otTqh9KyScYLgDv3QZyIiCySKKYdMsbXj/ALYZXFnFbh5a9QW6anpbewSPBGWt81ysopadk2790HQuk5WdQUeonXKkEkbOrq2yNk4g6TqaG0y2WuoHVErmloc4E79l3w9xg3+Ab7dlt67cOdPXmbx6uAF2c/RCDxz1LyS37WN9ku0NBKInkuDcEBbE1ryY3zSNtmrxa5XdAJ7Fe5lJoKxUbBHFA0ADH0QtNv3CXS+oad1NXU7HRv7joG4QfNpV092sd4fQVNqkj6HluHMIJGe+Vqs97e2A0zKEkkYOG9l6/c13Jbw/g0vJqLT9IyOpAd1DoAJIGc7LoLaOH+jrSyppL6+NtQ1xawOaM5+1B1bZRVcVUat0LgHHOFvDTmjavV1bHRRQu++EDGPVcqXnhfFNOZ6NgNMTkYG2FyJwX0dQ0GqKUTNbgOA7INl/tQLxQ2gXgWyV46c7NJ8lwtrfQFysszxLQPi6DjdpC939EjhfbNFRHVppWxlgHx4G2PmulPNfpHh1qupnfoMQPBef7Hg/oQeYfRJG7ofGQVe58jWnDjhcsa/4Y1dk+/mLAxnIC4zjtlQ/rJacNJB2QYdFUBkpLm757kKtwrWOIaAFNMynp/hdjqAWk1Dmvl27HCCOSTxDsFNTTmMhuO+ykbDGGBxx2WRE2l6QSRkILKkt8PrwASoYC58ZIzn5JWzNcOlp2CyLfG10JBxkoNOc+TJBcfzrIpKx0DvjyQtVttFA2cuq8Bqxb5BTCcGjwW/JBlQXaGX4OkA+pVlRURyOEe260Vgex2QDsr2vlMoznOdkGoy2z3Ue8E7YysbwPeH+J5BZVZLUClAlzhW0EMs0J8HdBQTMjAj2yAsSXqfIACcZVKinqYpCXg90jd0ty7ugmni6Yg7OSrYQHRkEKNtQXu6Xnb61OJImAAEIMSMnxt+2VqrWMMXVjyWBMIwcs7rIp5sRlr0GI6UNedlUVXYAK6Rkcjz0nJKkZSs8PLu6A9xqGgMOD8ioJC6Jhjdkk7bq+l6mTEHspK4xOYSMZCD1N9hr/aHGX/DWD9FevUxeWfsNf7Q4y/4awfor16mICIiDgPnvtou/KZxCtxOBNSUg/JWQH+ZeA1tbFYhW0b8OcXloJ+Wy99efaoqKXlH4iT0ufFZSUnTj51sAP5iV891HXSVFa99Yc5OT9eUGpUNjfPWGqecNJyrbxT+6Sh0ZyB6LOrbvHFSBtLjqxjutJhqn1pIqO58ig1KyX2Nh6JGDPqsa93vFWDE/ABzgKKeibSNMrdvqWiT9U0hduQg12tvvj0oaD8WMErGtda3xQ14ByfNYTWRPZ0t7qSGIw/H5hBuO70cNXSAsxn5LS6SmbRROecAjdRQ3d4+9vJwFDUVb5ZPDYdjsgyqa51FZMafrOO2FSrtksMoqDISBur6GljpgJSBk75UdbPUyv2B6AUF0uoGeEacs3G3ZaLNKZ5B07AlZdVDTOj+9kdawfClZh2MYQZotR8ISFx7ZKxZaYx+qkir5mHpcduylkkZM3uOyDBa4sK1uit7aqnM7sEgZ3WkthPUS/stQtc87axkQz4RIB9MIM600r6ipdC6MgZxkhYOoKc0lSI2k4+S5HNtpJLeJLcB45G+MZytm19mrJKrFW09RO2yDQKZjpQOr8qvlpHtcHNccei1ytss1BTiTpwPVadFHNKwvI2CDDqI+tobncKajrjC00pO52yrZiAceYWI2KTxw7G2coNZpbZJSu97LtnbrMqaSOqgMhcCQsSSrlkphGO4C0w3GrhcYySB6FBi1EZjmczfY7IyEyN6lqJjhkZ4j8ZI3WC+QNk6WnbKCW3Vgopep7chTXG6Nq/ojChqGQuiBYR1LCQT08pD9yVqDra2oiM2QMD1Wl9D2YfjHzU8VfUMHQ0kj0QY8jTG4s9Eb1DfKmqOgguHcpBSzTjLAcfJBLBUD6IGCFDUkk5PcqT3Z0BJdscKCV3W7O5+SCzLj5n8quiJMjRk9/VHNLWjHmpaboyC7vkIM6ujApw4dwFp0U5iGN1lVlT1Rhqio4WyfE7sEF8df4ZBDcfYt12yhNxpDIJMHC28YqMDfAwpWXyWiYYafOEGpwSXey1BMFa8DP90tT0rPcqDVlNquqqiRTyCTJPfC29a659XMTVHb5rLuFyMR93iP3s7HCDvjS858GrdHQaObVN6oIwzuPIYW1bjx4o7FQvpJyHlwPououmKqhtk4qISetxyVuyqZ+yPEbQ4vPZBuXV3EGl1XUyGmAaXZ3C4sv1lcWPlkJJOSt82fhVqKN5q2wP8AD79itq64dUWeQ0tSSHH4cIOPxC585iGTgqR1K6M4yQsigkhNQZHeZ81fcZo+sdB/Ig0+SHpA9VcKclvUsmFok2crah3hg9OyDCcOkkeioDvkqu73bdyrjDINy1Bcxgl2HkrXsdGVRriw7d1qMLYZYT1Y6kGBFKWHJ3CpLJ4jgQMYWRFSlshMow31UdQGCUBm4QQYV7XgNxjdZLo4wwZxnAWM6M92jZBaXE+ZVPmhBCnY1jmYPdBCTlUUoj6D8fZUfGXHLdwgjRXho7FC0eiCxO6ef2qZgYG52ygiLcDKyY6voj6OjOVBId8DstUpI6R1KTJjq/nQaS49TiR5nK3fYdKR11Ca17gCBndbYp/C94PVjp3WqN1LU0TTTUzvvfY7oK11ZHRTGn6AcbdlqFLYY7jRGsBAIGVoc74qo+M/ue6yaa91NJEaePPQdu/kgw/CAqXQns04yrpCyAYABVtTK3qMrD8R74UcDZJpB4gOEFwrQ0kdOFa+EzDxMrKrKaBkWRgHCxofE6CG9igxQSx31LUoqhgiLukZHqsN8DnnYbqeOIdBZnchBGf9tydPZarHaWx0xk6gT6LSTDJTO6xnHdStulQR4R7HZBizn745o8jhWxteSMZxlZjoGgeI/G+6uhEReMAIO+vsW2lvNHqnP/ACt/lG3Isz2NDGN5ndTkY/+IVb/KFvRB7LIiICIiAiIgIiICIiAiIgIiIOOuY/bl54oH/uMvf6jMvmicwyTyBu+HE/nX0t8yRxy7cUj6aLvn6jMvmut3huqpvExuDjP1oMEMPX0nusynkEOI+5Kx6kdM7i3yKpFJ98DneSDKqKBzWmYE4Kx4JXR5+HK1YSiogDArYqaCNp8fAJQYPje9YjAwcKCeF8LgM5zusySKMO+8YJ+Sp4Ly0mYdvVBWguQpseRVa65e9uAzkrT3tzIQ0K5sbmuHUNgUGXFJ4ADxsop6l1TIHbnGPsUkw8SEMZ5KaigijicZ8A+SCKbD4OhvdKOsbTMMT27nbdWQytNUQT8Pln0VLiIusGIg/UgrNT+K4yg7HdZluqGUzCHHJC0sVEgHT5JHLmQdXYndBqrrpGx5JYME98LFqpG1z+puBhX1LKYwAtIzhYdOyQnLQcFBT3c9XSc/Wr2xhh6c91PI5oZt9LCw/EIeCSdkGTLQdLfEzsVbDQmUE5Oyumq3SRBozkLKt8Uz4z0g5PZBiNt7i4jq2V7umjb0HclVngrYZC7Bx9SwpZHyO+PuEFZMyO6h5qgjceyyadjXDCmLGMG6DAYwvd05/Os6nqWUbcEAk+vmoZelgLmndYznOkOTvhBmyVscj8gbeitNOJ3AjbKw+hyljklZ28kGdLbhHDkd1ZS29xcJC7Zu+FD75Mch5OAhrpGgtjOxQa3W3aBlMIABkDC2+1zppg3OzjhWPEjyXuV8LHDDx3bug3DV2ZlLQCpLxnGe622XOc8OA81mTXirnh8B7vg7YytVtUVv8AdSZsdWEEVNMJIAwn7CsGrpfjD2t8/RJZwyqLIj8I2C1Omlgc3plIBI80GnMqTTt3Cw6if3hw8lrVYyj6C4uAGNlokvhl4EJzugvbD0N6ifmrugOBIG4VrqWrYzqcCGq2J5DC090FrHPD+nuqSdRfjtlZNujY6ozL2Ul1ZEJR4BB+pBB7kSzrGSovAOcHZZ1B4mSJvoj1VlwLDMGw+Z3QQMonOwQdir/ci0gk+a1GOB7acPI+tYoqQJQ13bO+UCZ2Y+jpxhRRzMiYW+fktTrTSvpR4OC/HktFhpKmpmEEbCXnsEFYRM6YOhYXEnt3XK2iuGVfq2IMNveS5pxsd1qHBHgTqbVd9ilqKMtpgR3BIO69I+EPKxJQVdDI2nAjLR1fDt2QdSOBPJHedVXlr57bMG9YAJae2fLK7hW/kUrtJVlNdmUzsRYJx5YXdbQXDSzaNo4TSwsE4aC4taBut5TRMnjdFIAWuGDlB1ks2sH6CtTbO+mLXRAdx6LbepueG16KgkZWSQgxAjDgNsLmXizYtJWyhlra1jGvMZO+AV5Cc2eobZNe6mntjiWGUj4T80HfDSntI7BqO5igHu4y8Nzgeq7VcPOJ9DrmCKankYRI0O2x6LwK0Q6jtMlPXN6g8vBzn5r1V5NNVyXCipRuW+GBufkEHd4HKqoKSobNCCO4U6AiIgIiINta60rBqy0vt08fWHNIwe266J8WuQi4Xi5yXy3QPbGJC8tbkDGc+S9EVjV8scNHLJKQGhpznsg8otdcOI9CWptrqaYtfE3pyRvstnaUoaKB5uoqmsMLurvjsuxvObd7PTx1D6ZzPFDCfhxnK80Ltxav9tkqqSne8Rl5Gx8soOw3HHjZerzZv2O2a8PYY2lo6HkLhnQ3FS56Qikbf7g6cnJ+N2f0rY1g1K+8V5lrHOLnH98VtviFUt99EcWwJPZBy7q/jDbNYRupWdAJBC4ouN1it5fAxgPiHbC2sxs1LC2eIHJwStdtstHWUjpa4jxADjJ80Gk19O6oPvIdjIysOOlPWMnzU8lWfezE0/ADgKGpmLXAsKCarZ0QjpOSsFjJH7jOPrWVTeLUO6XdlnC2vY3qxsg0+jpXTvId9SyJnmgkAA2UzZI6Z2Ad/ko52ioPW7yQUfWGsHhx7FVZTuiYfEOSfMqynbHE/busqRsk27ewQYGAJNmjCzqaja9wlLRtuqU8LZHdJ7rNY4QvDDsCgjrR73F4DWYIHfCso6plpjMbwMn1WsxCkjYZHdyMrbt3gkqJ+uFuWoLpbiyskIDQM/JabVMLJMDsVlsbBAwb/GFdiKZhJO+EGmEEAFXshc9vWD2V72fERjZTxOYG9A80FbVSe9z+GfXCzLnbxSkMae6x4vGpHGaLuVFPXzVEoEmSc4QIaYsPU7ssvoBGQdsKOfrEAc3vhYbKpzWkO74QS1X3sfD3WKGue0vLjsMqemPjSYf2KurYvD2YNvNB6oew2bi3cZHes1gH5BX/AOtepa8tPYbZ+53GTP8A82sH6K9epaAiIg4F563MZyn8QXyfRFJS5/jkC+d2V/XUVPTt1SEjHpkr6E/aDukbydcSDCPjFHR4/j1OvnkjLi/O+UGbDFI1oL3Ej61cZSx4kaNgdwrhBUSxgMBTw+iMsl2cdvtQX1l4E8HhA4IGFiU5zGc9yCon0U7B1FuxWbA1ghI8wOyCGjYRMXHfdZMsvVIIgMA43WPQu6ag+J2z5rJqmtMokjI2QW3CgMMAmBxnfZLbT9UJmcMkbqOurHvhbG7t2UtDUsZSlod8jsgOuHW/wWnsp2VrHAU5AJdtlaKHhs5f5ZKnhikfUtkAOA4HKDMqbY+jHvRzg7hRRTNqPgx2Wr3qqjkt7YgckDBC0OgfFE0ucQCguqqERt624GVFBC76WeyVFY55x5AoyYCIhBSpl2DW9/NZ9tkaIT1DBx3WnU5Y+X75jGfNZMzwyPEO6DeGjb17jUkzv6mA9vJV1ZqqnfcWvjYABvhbVslUIZyZzgZWTdmU1XMHRnOT5INYrdRw3OjbEAAQPNaWypjggcw4JIWDPAKaMeGMbLD94Jb8WcoLJpz45cN91LHWta3duCraCmNTKdsjKlrqQQHoxugloKrqkIduPqWPci3xgRgeqpTQSROEhBCtq8vPUOyC6F7ph4fkoqiExuDc5VaSRsbuonzWRIwzODxu1BjMgfsXHYrNhtXiNMwOw3UVQ4dAazuFLS1z44TE/byQRS4kJhaO2yhLRBlpxupWva6QuGMkqs8ZePmgxmQmV/fAK1Clr2ULfCcATjutOc2WE9W4ClhjbN8b/JBk1M4qclu2Vp7mFj8d8LMc6NgwFdEYXuBeRn5oIvAMjMEfb6LHex0RA7LWm+AG/DhadVwOe/I7fUggijdUHp9FM9wph4Y7lVpj4JydjhR1Z8R3UEEIEjt+o7n1WdDQExmUntusWlcA7DuynnrXtb0RnY7FBNRO8WUxNOCFWbJrG0xcfiIG6waSV8chc3uqzTu8cTA/EN0G7/2Nvo6VlW2QHIzjK1XSWovubd43zfRa4bYW2bRf6qpcKaodlgwButZp4qT39kmRgEFB2u05xJs9VZxT+6s6yzp7fJcIcXtLxX2d1xh+ABxdgbLc+iJrVNTCOMgvAC2zxHluviGnow7pJwMIOCq6lfQVBh6u3mr6KlfVyDqd5rW7pp24McamqYRnfstMhJp3Yj7goM6qtraODrBWlOZ4w2Kz6l9bJEPFB6cea0tspZKGkbZQDGac9Thsp/GbJEThX15a6AEd1FSNBhPrhBhP+mVdG9zHh2TgHKudG4PJd2T4MYQZdVXtmh6GjBWBnfJVMFAN8FBc6QkAZV7Jg0dOFfGI/NSuZD05BCDEc4O7BXxxOH3zyCjdjJA7LLhe1sJBO5QB01OGA47LLjpWRMIJ3K0xsjmPJj9VJ75K47oL6mHoyQsXPyWXK5xZ1O7LDOx2QVIwgacZUkJaXYedllGJpjLm4wgwmsycKrevqDA4gKRuziAjQDMB55QZDbY5zA4O77qz7nvDwCSQStREVVFE1zmnp9cKgqo2j4yMoNOqqd0DQcqkbsxHPdZb2urCWs3WHLG6nd4btkEAcQ/JWUysDNgN1jyFuBjupaZsWxeUCWZ0x3zhXR1QiYWEBTSeAGnpIWDJhx2KDIgqAZNx3WrxUjHR+OSBjdaLAwdyFK+4TMBjacBBlTzCYmJjQcbbLDbTFsgcdt+yzrD4Jmc+pIAKjvc0QqsU5GPl2QKpoliDWncLTgXQvxlT00xcSHKypjJkyBsUHf72LdSZeaLVDDnA0BWn/nG3Io/YsMA5pNU47/sArv5RtyIPaZERAREQEREBERAREQEREBERBxzzI78u/FEeui73+ozL5ousxzPLcjcj86+lzmS/B24pfwKvf6jMvmjazrnIPmSfzoL5AS3rPcqyKAyDIU88ErYs42V9Dswg990ClkML+k7qSse+cjpJChpone8nxBtnzWVW+G0DwyEGPAfdvjec/WlVXiYYbsoRBU1J6WsJwrjSOpxmoGCfVBBG7pf1n61fLUB/YeStjYJJegdlkSUzI9j3KCOmqBG74tx81kzu94HVH2CxDTuHxY2U0UrWMLUGG4FriD3CoST3KnEZMhc8bK2cM6gGbhBHnbCBud8qZzYww4IyoQT2CAXO8yVlU9WIY+kjdRxQOBDnjAVtQ1gd8CC17+qQv8iVI2PxiMeagWTT9Qb1BBIyl6PiJysynubaQgdPYrT21Tw/DthlSSCCQZyCfrQZ9Xe4Z4+no3PnhaaIfFdkDusZwAOAsiCoawAO23QSBhp90a8zkA7K5srJDlxGFDO9sTx4eEGTNR4j6s5UMMIAPZRiskIDXnZZEdREAR1DdAp4myP6TgJVhtOQAPzKx8vT8UJBPyUD5JJHgzZxnzQXM/2w7oDcfNWzQ+A8A7rNkMMUIdEfiAWC7xpviIJwgmfI3wdh81SCRvQR6rGJcPhPkrmuIGyAB98/Kqvc9p+kQPrVoJJw38iOBx8QwUElMSX581Wpe9sgwSNtlfQAGXfstRqaaFwDtsoJrDaXX0iB0mM+pU160t9w5WvMnUAQe60+G5zWlwNK7B9R6q2svlwuh/2w/OPtQZlRcIZKYQADOPILToqXYuxsogAw5dkFZkVTGIyMjJQIKYSu6QcFTS2tsI8Rzs4WmGpnhkL25A9VcbhVzkNzn5IMls4leYGjHkCsWeM08wJOVmU9I4YkaNysKtL/ABx4mQEGpTV7vdQxkZJ+paZ4Ms2XOYWnyW9NGQWirc2OsLe3mrtY2qmpagOoMBnnhBsljJoXguBIB9VuLTboY7rFVyNHSzBOVbI63up+jqHiY7fNaeJXU7j05weyD0G5XOJ2lZBBb5ooWPDwC44+S9T+GdXpyrsVO+ilp3SlgJGRnGPJfO5oHW1x0zVMnpHPBDwe/wA13S4N85l4tFRSUNXUPAwAcuQexHiRAf2RoH1haffbnS2+0VdbLVMY2GMu6uobEBdJrhzex0mnxXGpIJbnPV8lwDrXnevFzgqKGnqXmKXLXYd5IN4cx/M5UXXUFTpuluJDWgs2dt5hdAOKVxmqrjLLPVeIXPJyTlbw15rq03mtkuzHn3l+Sd98rg7VN1rbhI95BIyT3QZsupHUFPEGSAEOHmu93Jfxwnt0lNRF5OQB+heb7ZXTyMjlzgOGV3T5UdSaK0/XUkl2kYCwDPUR8kHsbw21O+/29tQST1Y7rf64T4HcVdB6htkdBY5ojLkNwCDuua0FUREBERAWj6rY1+n63qk6OmIuznHZas5wYCTsBuuufMjx2t2jLNVUjJgHlpZ3QdBubfUkx1JVUZrepoJb9L5ldL7jQw1UdRKXgnJOVyTzCcSTqS/zVUDyS8kjfv3XX199uQ62ZdgkoMqjrHW+4ENf2OMqC+zGunExdnHmtLkne55kJPUe6uEkkjDjOEGQ65N8ERYBwMdljNfNK7pa4tB+arRtj8bE2wHqsmrdTxbwuGfkgsdRmNgf3JWVBbfGhMru49VBb5zPKIpDsSsm4yT04DIc4O2yDDjqW08pYR2OFyBZrVT3G1Omc4ZAzgrjR0UriXFpJO+Vrtpvdwo4PAjLuk7FBpt3idTXGVjc4B2SGYvHS4YWfII6mYyzkdR3WnVjo43kQkZQJmmP4gfNZlNcmCAtIGcYWl9Urx8WcK5vSG9PmUGVS1eKku8sqSprS+cALBgYWu6nA4UkhjIy07+SDWmNdUQjDx9QU7PDggLH4J8iVodHUzsdg5xlUrK2UuwCgirY3CZzmk4JUUcj27b4WbbcTzffztnzWTW09KyUBmMfJBiUsLqk9PSVJJQvhkBAOFqNPHEGjwsE/JZR8IMIlIDvmgx4IWSxhrsA4WjXCEQVQ6RtlZVbJURPLoQceSipopao+JMDt6oMsYNOAR3C0+ek6gXjbC1OaSGKLoyM4WkS1RLy0HbKClMcSdPbCyKuQNiLO5IxlQu6GN6291R0jZIHE9/JB6p+w2I+5vGQek1g/wCjXr1LXln7DX+0OMv+GsH6K9epiAiIg4A59en9qPxE6gCPdKTv/jsC+eSlibJNKfIE/pX0K+0CkEXJ9xHkPZtHSfr1Ovnigle2T4TsTkoN026SNoDXNB29FZXW1k8gnY4ADfCxo6qBsI6T8XnusWe6yMaWAkhBJcK2FkQgAGRssOmZ1sLurA7rDd4lRISBklXNM8B6d8FBkY6nlrfyqY/eYi4nJAyo4vhHW7uVc9kk4w0EgoNY0dp5urK40jnAAELWNWaCZpppayTK27aLrVael8ehJDzjspLlrK5XicCvcSOx3QaIKR7pi0ggd1mwzsgeIDjJ2WRXPhEAkiI6yFo7WTyO8Yg7boNWudKYoRN15B37rS4aYzMLwcY+akNZPUtEDzkBRtdJFIIxsDsghbGXP6FMaYjACyJ6YwRCbzKpSwzz/fcEgIMZ9K9oyAVRkj2H4gStTMsHT0uIyNkMVO9hI7oNLkmL/ojCvgqHxnLiTj1WsWWlo5JiJsYyse/UsEU4FOBj5IIHVxmb0YyfkFiSMOcYwtUtNLCDmbG6juDYRVtbHjpJ7IIKKqNEeotO/wAlbVVvvMwk8h5LVn0tKacOOM4WkSsha7DCMZ8kGT742aIR4wQsd/wNI7q1wYxuWnf61A6QvOCgo5u5IKmiq/DjLDnKhd1gbg4U8MUb2dR/SgibKQ8vJ2O6tkf1uyFdM0NOw2VjGlx2CC9mYzkkrIiqQ+UA+eyiMMj2gAdlGGuhkDnjGDlBq9xhY2nDwBn1wtLimDWFvYlZU9S6phDGb4CwXQyN3c3CC0kk59VNFA95BBKpTQumd0tG61PwHQMOe+EFIqUhu7jsVkmJhZlwWBDLOZCDuFO+V4+FBhVoLHHp7FY7X7EHyWoRx+I4mXzUFSyJknwkAZ9UGFnzV7Dtup6hsIjHSRlY4jeRkA4QZNKzLs5U01F4h6s+SwWyvZsPJSsrJcgE7IM2liEZAbt55WtULTLKGeJudgtBke/w+phOfkFdba2eOtjLsjDkHKFprajTUYqHSnHdZkmuG3KZr3wdePPGVjU9vk1DQRxxDJ2Gy3PZNOWOz04bc+lsh/uh5oNq6ku1NXUnSIQwkYzjC42naIZjKNwDlch8Q32+GJ33PcPPGFxhHJLI14lzug1GW7RVEYhAGRsoxaxLGZ87gZ3WDTQDxSSCp5q+SEiIdsoMGokcXGPyacKsE/hjCzMUzml5xk7lYc8QLi6PsgrNUNkGFRtN1MLs9lB2UsczxhvkUFhyCRjsrTusx0cfR1eZWI7HVt2QUyfVMn1Kud04GO6q2Nz8YGyCxMn1Uz6d4AwMqRtDK6IvI+aDHjeGHcIX/EDjspIWtDy1+2FV0PVIOgZBQXeIZm9AGFcKAluclSzwsiiBaN/NUp6rbpeUGLJCYypIqjpiLCVJVDxforE8NwPSQgB+HE+qzqCl65BMT2OcLFMfSB1dllwVPRH0t80GtVdxhdTiFmMgYW3Z45C4uBWoU7DKSXZ3VKpgaekBBBaqkQvPX+dR3OZs84LeyinjMJyNsowscwl2MoApi5odlWOiczOD2UkEx6i04wraiQl2AdkEJLvMlAPMqSAMLj1kBXSxFzh4YyEFBL0hWN++SjPmhgkb3ajYpM5AO3ogzJ2uZGCxxBWC4uccuJJUj3T4w8HCjG5GUFWPLDstQp+mVmPNYUgZ0gg7q6nn8Lug9BfYxU3g80WqZPXQNaP+cbf/AKkVvsYKsTc0OqG+mga0/wDONvRB7PIiICIiAiIgIiICIiAiIgIiIOPuYaIVHAHiXAe0mj7yz8tFKF801RAaW5VMPfwZHD86+lbmLkMXL5xPlb3j0benD6xQylfNOZnT1k8km5kLifrygzZJ45YQ3Ayo4IA1hd6LAMxa84zgFZEVXtg/nQVFS10nQBj5qyqkLXfSzlVfCZMui7n0Vnu05kHitOMoMu23NlM/L2flUlyljuLg6PAAVTS0xgHT9LC08iWGQdIOAgvbAaY+Iew81HPUmRwLc7LMqJY30/TtlYdOIcnrI+0oMqnkbUt6MYWLWQOgkGDlSiWKE5jP5FRkraiZviHbKCNkhmb4Rbj5hRyRGI7nK1usZQxUnVFjr+S0F8jnnLkFDnPmjSWuBx2KmpYfEfg9llTUzA0NbjJ2QQyVbXQhjRgrGY0vcGnO6yJbbVQs8VzD0+qpThnT338kEs1u8KDxerO2cK2kwYznyVZ/feg9eQz5qGDr6T09kFvT4kvSNlSSN0Rxkn1VOpzJM5IIWQJGyD4+6CtsoTcKgQ5IJxutxy6GDIvEEp7ZW3YKv3OXxYTgrLl1PcpQGdZwEEFwofciWB3ZadknzU89XNUOLpTklQAZQTUtOamToBwtX/Y4RD4vUdt1ptE50Uoedt1qU10qXAMYThBpBDqaYtcM4O+QqTSeIQQFl1UbXtLz9IjdY8bYw057oLadpkeG52+tZMsjIAGgDdYfWWPLmFVL/Ex1ncILHnLiR5oM+Suc3bICyaUQY+PGc7IMdgLPiI8sqksnWR8lnS04OcDbCwZGdJAwgmoQPEyThasyMSStiJyCQtDY57XfDv8AUs2GrkjkDnEgg7INYu2nmwUnvIOSd1oFI4NJa71WqS3ueoj8GUnoxjdac6illPiQNJH6EF9ZHhvUwZz3VlDSPnfkkgfpWdYQyoq/d6rGB6rJ1DGKCVraTYH0QY9TRs8LpbgkKlHRMjjc92MjPdRUxqpPpg4UdVVSxyCFmf8AWglFzEDywAYClbRi6t8QbEKSk0vWVsYmDTgjKsn8eyu8J22dkGI9tRaZQ6ORwGcbLf8ApemZqKjDahwcSMbrYFRWRVQHWRlatZ77Na4emlcc+WEGrXnQptk7qhs2RknAK25UsLScDOFm1eo7xcX9Ly4tJ3V1GYAC2pIye+Sgw6GvdTvGRjK3BaayulucMsFUWgEHvjC0G7xQRx9cOFjWu7PpPiLtx80HOWuNYXSHSjaeO5nqDMfS+S4ctup7o9ssctW9xO+cqG5alqLjEad5JHbdafTGGFpc44JQZUlbXNqDK+dxGfVJ7w2RhY5oJKx45o5JCH9vrVtRQSSuMtO3LfPHkgxXS5lMg23yFrtlrrzG4zUVc+LAJ2OFokkTQ0RgfHlaraKhlLG5kmxII7oO/fs1dWXqv1vDSXW7udGJ2jDn7dwvY9r2kbOB+1fPhyrcQZdB6mjuMDyPvoOxx5heuPCbmGh1G6kinlBc9oz8Q74QdoEWJba1lwo2VTOzh9ay0BEVr3tjaXu2AGSg0fVNzjtlulnkmEYDDvnHkvJTnb1tVVlfVR0t1JHiEYDvmu//ADUcVbZprR80FLUZn6HnAOCDg7LxK418R7pqa8VZkc8t8U+edslBse+1E8swknlL89yVpbWRywuIaMjKVU0lRTtJyThR0pcyNzXeaDSXxF87m9leXGn+DpP14WS6FzZTIBtnKkkdSGI+KQHAdkGnxxuqJPgyCVqlPp98uBISCVBYZII63qmIDfmtculY4uD6I5A9EGFLYxbB7wX5xuN1gPuLJngEZ3x2V9xutTLH4UhIytLgx4rertlBuCGlZNGHFoCTGOljIAGVZLcI4acNjIzhac6WWoaXHcIIqioPWS091kUtsfVxGcknAWAd5MOPYrWrfXsgiEZIwe6DS55OgmLpxjZY+d8rU7iKd4649yVpjcDIKCYzdTekDCtijc6Qd+6o1od2WdBTu6esDcIMsQMbDnYbLSak4fjup6iqkblu/osdrHTPz+VBnWaH3mXw848srcc2mGmPrMm5Ge627Tl9CRIzYq+XUle4dAcQB80GX4X3PlILwQCsWqqTPJ1NdgA+qwxVz1DyZTnKinkLDhpO6DW4Jo52eG7BKx56v3aUQMZ3OMrAopJGPDt+616JtDLH1y46wM7+qCGS0meHx+vGd+60wW0+KGk9ysituU8LixmfDGwUcNY6Rpfn4h2QTXC1imphIHZJ3xlaKGuIJ3wFqJrKqZxZPno+apU+7tgcGEdR/wBaD1L9hr/udxl/w1g/6NevUxeWfsNf7Q4y/wCGsH6K9epiAiIg66e0MaXcm/EpoOCaOj/XqdfPXRgEyH0avoU9oa4s5NuJTh3FHR/r1Ovnopi4FwHmEGVaaZ1VU9DnYB2Ut5oTTP23CxI6l9JJ1RHBzutQFbHVwHx/pYQadRStilJcBj5qWrqGPcHNA2+SxSR4xx2yrx0OcBnugkikM2GdgtYiEcdMRtnC02SNkEYeMZKUM8k0oa7JblAhnPvDutuB81ZVx+LL1RjG/daxd6elgpBJHjrO60ykka6BznfSCC6KBz2AOfn1GVLI6OCEsIGeywoatwnLSdj2UlTTVU/3xrSWhBjU7wJy7bBK1qO2sli96GO2VoLIpHSdDR8QK1mnrpKaHwJOx27oLqVrbnIaTGOk4WoTRxWyL3XYk7ZWl0Mvu85miO5PcKa4GpqszHJwMoNMraR7CZg7IO+FSkLnsILirDUySOMch27YKtDzHIGs7H0QSukkp3FzSRt6qx1Y+U5eC4rUG0Lqhmcd1R1LTwsLHgByDCNa5oy3ZRGZ0kgeSSQrpKR3USOykhgAB6ggnhmfUtEfUQsSrpzC7HUpJHCn3j7n0WPJNJO74u5KCWGEydzsop4/CfgKVokhb1E4CticJqgF/wBHKCN8pc3BCo2R7RgdlrFxhpPdR4OOvG+FfbKKnkpnGUDqQaI9xcASMbq+IhoJI/KtSFLCyc+JjAysOsMLZOmI7eaDJtbhM8tLchR3aMNlwBhajZGQRYfJgZUF6ayafqj3CDTYcwkPcMrIllbKwux+ZXxwdcYBUU5ZC0szuUENJUCkmLiMrPkrmVADhstMiDZJD1dirpHeE7DOyDIdWtjOOn8yljlE4LuwwooooZGguxlXvfHCA1vmgVFQGDDTjIWE7MpLid1fUZIyVC15aCEFDknBJKmbUBrOkt3VsDOtyy308TWEnuEGJDF47z5I+Ixyhu57KjJDE/LewKzWuhlb1ux1YQZlNA0xgu3ysVwDKwYbjfZVjqnMIBOBlXv6XtMg3PcIOaOEtXSsA96e0g+RKzOJdtuFVKai1wvLO+2cLhmw3y50lTC2m6sdYB+rK71cGINGXbSYm1M6IS+Hkl+O+Pmg6a0tsr7lOYK2J7QNjlaDqO2NtchZGARnyXZDi3FpOkrqkaakjc4E7Nx/MuvN4kMpea3Z2TjIQbZbU+GfrUMsnjSD5qWpax78RHOFA1jmHJG4QSy07o2B/Vt9aRTBrC07nyyq9NRO0NAJAUMkT4j0vGEFrjkkqgOCD6KrBupCGEY80Fr5S4YyrE+pPrQSwQ+KcZUxIpz0rHbK6M/Cd1R73SOy7coNSo5GVD+l2PtU9VWRU7TEBnPotIjkfAQW7Eqkj3zEvdugo9/U4u7ZWTBIGsy7GVBC0OJB7qR8EmPhzhBf43jEtJVRRkuBBwsYNkYdhgrUIWTuh6gDnCCKRogG5yVjmcF3VhVnbMXnxAcZUXSgvlm6wMDCsY8tOclGsLvojKeGeoNPmg1a3ztcN9tlDXVAbLscrGLnU7QW7ZUJk8R4dIUGR4nvPwrHkjLH9Ge6me9jG5jO6x3Pc93Ue6CQsLAHZUZJJyUL3EYJTbHzQXRsL8gKZsghOCM/WrKZwa4kqlQ4OfkIJpatrxsB+RWxVLW9wPyKOCPrKtmaGvwEE1RO2VpwAsbCLVaazzVNMZWNJQaVuVVrS4hozusuKmMUpbUNwAVmmGmx1MxkBB3t9i3TOh5otUPPnoCtH/ONuRS+xhkL+aPVDfIaBrsf5xtyIPZ1ERAREQEREBERAREQEREBERBxzzHnHLxxSP8A3F3v9RmXzRDJnfj1P6V9LvMeM8vPFAeui73+ozL5pG4FTJnyJ/SgyaW2e9E74UVdRmikDc/nV0NTPC8mMnH1qyrqJJ3h0ncIM+la2CMSPwQd8lR1NxjkOGNGQPJRSTgwBvljZY8EQeC4oMuF2dydj81meAySEuwM422Wiulc1xaOwKzKWv6Y+gk57IMOcuEhaSdlfFSOkxh2MqksT3yF4GQUbNLA8Z8kEz7e9jerOw+ahawsB9Qs+OqbKzB7rFmikzlo2KDG6nuPSXE/aqFmH9KnfGI25PfCga74slBrdtoWuaHHAyo65ogqA/uAcrHjuD4mANPZHVPvDT4p3KDUK29wTUQp2tAIGM4WgxOLXtO+xyjmnrIA2yqMwHAu7ZQa3UVsc9EIgwA49FpcTzC0jpys6WSlFKOgjKut0dPKwmXGUGk4Mj+2MlXugcMbrU5mUzXERYyFiSODe/qgx3wFjeo7qekiDoySBso/F8U9IKuMpgAbnugjlAc8twAQqsiLDnurcOB8T1WbSdMuM+aC2nZ4zg3GFkyNFO3BAJwpKmNtNH1x4BWJHVMmafF7hBE9xkcR5f8AtUT4DkYP51Jlrn4YfPbCo4uZIC47eaCOWkdGzqwSoGg9QGCtwU8tLLGGyEZWPVMpGZMZGUGBJD0x9XmpaK3OqB4mcAK4RyTHpGSFY+omoyI2kgFBkzSCFvhYyRstMld1v7YytRp4Hz/fZMnIWLWwtjcC3b6kFkbBH8ZKq8+K4uG2FAZHOGCkbi04zgHugklcSAMHYrU7Vco6eAwyNBJVaenhkiDngZKxqima2YFnbO5QUY99PVOqW5A7qWpr/e8OduR6rLeaX3cBxGcLSZGDJEY2QZtPcWEhgAH2LGqyW1IlG4Byoo4+ncd/VZ8NL7wPiGQg1+2a2p6KmELoxkDC0i71jL7VNdGAMnK06uo44BkHBHfCst0hjflvcFBqkumDHB4xedxnC0xhNLJ0ncA4WoVt0rXQdG+PLBWn072zu++dydkGpMrIoYustGT8lpVXWOmk6muICzp6STw874WB7m5xy0HGUF4qXzxiNxJwO5ULoHNOx7qaWNkEYcO6pDHJMwuwdvNBB0OZuVPHSOnb1BxGFHHkzdD+2cLNMohZhpA2QYDoXNcWgrU7fdW0kRp3jJO3ZaYZHSSbHv2WUylBw93fugmkpMPNVtgnOFH4JqZAWnA+SkDpXN8N2ekKF4mjJ8L8yDc9ju09hDH07yHAg5C7IcIeYC5WSphqprgWeEAcF2Oy6waafAanF0OGZ81qVxuNJFdY4La8lhOHdKD2R5ZecGn1lNHZK2uaB1huS5d16Wrp6yFk1PM2RrwCC05yF4S8GrbqEUtPU6RbKaovB+DOc/YvWnlYl1m/TTW6vbKJWwgDrzny9UHPCiqojPTyRN2Lm4UqIOk3Orw0r49OTXsVBLHMecdXbGV40a4ndT3Kqjk3IlI7fNe5HPFqRlu0hJR1DgI/Bcdz6grwy4h1VNcLzVyUmCBMe31oNAZcPhADcj6le2d8hHSw9+2Fl2a3QSsaJCMlbht9lpKi5RUceMvIAA+tBtSrfUsjy2nOD54Wp2bRVzvlK6sbSvDQM5APZdgqLgjFHZ4rlXsHhEAjLdsLmnR/DXTTeH9TUU7IzIyM+Qz2QeflbaJqOpdT5Ic04IzupIKt1DGY5dyfVa5rWknj11WUcQPSJiBj0ytIvNC6Fw6++UENHTi71JZnGSrbtaDbiAHE/arKQz0b/EhzlSVNa+qOKlxJ2QaXlxOMndaxQxAUpy3OR3WDJC12DH6+i3DavAjoyJsZwg2tMMTOHbdXMje8Eg9lPcPCNS7w+xKhZIWAjKCMuc07lapRWf3yHxckbLT4C1zvvnZaiytlgb0QE49Agw5qU00pb3wshlaGM6cI9xlPU/uVfGynDD1kA49EGBM4yuyB3WqUFGwQGR2MhY8cLJHkM7Aq2eaop8xtzg7IJi4SymILFrYBA7yV8Ac0+L5lQVcj5XDKC2OUdgFR46379layN2c4VHlzHehCDUGNaIx2yoS2UyjDyBn1UUMrv3xOEMx8YYPyQanPTeLCAAM47rHhpXQAvI2CzhMxtOCT3H8yCRksBxjKDS6iqbJmNowfqWKWPwSXbBXTRPZK78qtLn4wc480Hq37DZuLZxkf/dT2AfkbXf616lLy09hsf+p3GVvpNYCPtFf/AKl6loCIiDrl7Q/8DXiX/idH+vU6+eqlcGvIPnsvoW9od+BrxL/xKj/XqdfPNCx73gMG4QZr6HqHX6qF9HI0F7TsPmp5zURRDOcY2SjkMrSx+STsgxqSn94lLT+dTVVD7u4EH5q6ZjqP75Htn0UHvUk8jRIUFks7nN6CNh2UlLUeC0+v1LIrI6dsAdHjJCxKZgeSXdkE8lTJV/A4nCoyMx/ACcH5q+YRRs+9fSWIXzE5JOUE8tN4IEmd+6zqa9xxU5hczJOx2WnZnlHQ4ZCtayJrwJD9aCanqGxVJnc3YnO6jrqn3iTqbsFNUsi8JpZhX0tkqqiIztaekDKC+zgyPwStQuFeymaYQBvstNgkbRylvmDhUqZop3dTu6DClJMhkHnusiCn8Vvjk9t0cYSMd8K4SFsZbH2O2EGpWy4Rh5hdjbZY90iL5hIw7ZzgFaX98hcXAYWVHW9YAegvkqmtjDMbgYOyjbICNlFUua7dqkpmtMRHmghmfnOfVWQAOmaD2V7qSZzj0tyMo2lnYesjAHmg1arpmvpR0bnHkFpTIXMz5FZ9FVGTEbz223KguDS12WHA88IMZnWD8TiR9azoaggDpJC0+Lqe/p7rNbRykfCDuEFlbUlw+EnJUdNRunHWT2+apNRVDd3MOAsihdJE4Nd2KC49bQGNJGFPE0OYS9386juDmMj6mjcrCp55nO6QSQg1KnIc8txsq1VpdMfEAPZS0fhMAe/GfXCzPeWucA3GD6INIpbS5zyN9ljXClNPIG4ytxy1UNPGXjAJWk1VRBUuLiQSgwII3YznuoJi4PyT28srUIS0khvYeSw60dMuEEb5utuMK1rOoZyrFUOcNgcIL4ZPDcSVfLU9eME4Chdj7VRBUAuPburwx7SDnYb91LT+GPp4VKj4zmPsEFWOMuB2wpWyOj+AglQ0jHOecd1qvuzPD6nAdQQZ1ifHBIx7mgkuC514eMvGpKmGzUs0jGS4bsSBhcEWNoNTE1/bqH6V294LQWelpo6uLo95a0EY75QYHE/l9uOhLCdSPe+oMjC/GSfLK6vXOkqLxK8ywmIBxByML0io75FqOmlt2rnD3QNLW9fYjHzXUrj/AKfsdFc3M0mWFheT8H1/JB1+rrELe3xM5+taUQH5ctc1BQXmFgdUA9GB6rb9OyTJz2zugyoKxlNnqb+ZYtZUNqH9bRhX1EbXAdI3WOIJD5II0yfVSGCRoyQrW4DviCC1AMqV7Qdmqgw0YPdBGBk4VW7OTfJwVPCI8guQSmESMCgdiIdOMkqaedrW4YdyqRuZJGXP3IQY8b+h3UsxlYwjpx+ZYRGXEDtlUaQHAnsCgzX4ODhavSOjZSlxA7ZWmQwuqgGxbkeSve+Wl+8S7Z2QWyTR1MxZ0gb4WLUxeHkBXTDwT4je53SmlEs48U7EoIYZDEclu3datBb46qH3gYGBlSVtNRywAQkFw9AtOiqamnHgDIBQRVUg6jEB2OMqyCmMxwFdUxkfG4bndSUs7YwSTugpNROhHUTkLH6fhOAs+WpbM3pLlYGM6fkUGJDF4pxlSyUhaNiqOIG0PdUbLIXdDyUEJBaVQbkBagLVV1Q64WZCxJqeWlk6JW4KCrD4QB9VJ4HjMMvbCgfuAAVVssrGdI7IKNZ1PLc9lr9Df46Cm93LRnGFt4PIJPmVcxviPGexKDMr60Vj3OY3GSoI5HRtwSVlPp4oog4Yz6rGfGXjZB359i1KXc0uqR/3AVx/5xtyKnsVmObzTapyP/lArh/zjbkQe06IiAiIgIiICIiAiIgIiICIiDjzmKGeXzic310beh/yGVfNG9nVWTD0e79K+lvmMOOXzie700Zez/yGVfNNHl1dLjzc4/nQZcMTekZA7eihqYAckK+eobHH0tO4UNPK6TJec5QY2XE+GewWbTxYIZ6qGfobks7pTz9PxPO4QZ1VagyLxQdytMbA9xJGcAqWS4VE2Y8kg9gFn0EQEBM2ASCd0GE2oELekjdUMXvLTIBjzUcgjNSWk4blarC2NseGkHPog0eMuhkI9FltrB1CMj8ytmiDXkkYCoGREF2RkDKDIrKUe7+M3fzWmsi6wfLCzKarMj/DlPwK2tMbHhsGMH0QR0cBmm6PJZNXRmFwA2Ulsp3xuErhsd1PcX+INu4QafPEIYg7zKyLbZ/ug0v6sLCnbP0/GD0jtlZ9suTqSEsacEjyQRVVpdTOLevIHzWJ4j4cta4jyU1TV1NRK525BWOY35+MblBfTB739RJP1pVZMgZnZXU8ckTupw2VtXI1zwW9wgy4rf4bBNnPyVXUQqCHg9litmqhHgk9CvhqpQ4AZx5oLqmHoj6PRQUrzG8brOqh40Xw/SKxIqd0bXF4wUGpNeKpvh57rAqqJ0Uga04yVHBUOim27K6qqXvkDsnZBkR0PgMEpOfNQylsn1hJK5zoQwFYrJD1j0JQV6JWH4SRjzyrS+TPxOJK1hsUb4QcbkLTqinf1ZaNkEtLcRCMOGVWV7at4eB2WHHTyyO6WjJWQyKSlB8QYJQaj7yyGERjuAsGcmZpdjPmsdzZpHEtBws+jjDYD4o3QaYG9bw3z81ksonY6t9t1EcCpPT2ytYhc3wTnvhBiUz3vd4LWknONkq3z07uh0RGfMrV9MCiFeTVY6Sdluy+UViqKbqgLC/HlhBxpJDI9vidRGd1dTx9MZz3UtbQVjJndLT0A7K2A4eIT3O2EEcRzKW481ltuBhmbC1uQSAdlPW6croqYVUbDg77eamssFI6M+94Eo7Z9UGRX2pk1EKjO53W34o/AcXemy1a4xXaUlsAcYvLHotNFNUQMInG59UFJK5rx0Y/MsaMHxg8ds5UUgw4+izqTw/CJcdwg1KorYxSCMHcBabDXtYektzn5LFlkdK8tb2ysuihhA+/fSPbKC+oh8dnUPTKshqm0sRjIyTssz3eYAuA+ErFMcL5xG/GSQEGLTtMs/VggLLqYPhABWp19sFNRCeJuNsrRYKhzyRIe2wygthh8N/UVkGcFwHbCqHxYy5wUFQ3r+KHdBq1Oxk7QNsrX7Xp+OZvW8A+a2ZQzTxSjqLgAQt3Wy/Nhc2Pr7jCDeWkuFbNXVXurHdHlnstcv8AwIi0hJ7w+YPIbkHOVotn1tVaWaK6meQTvssC9cY7tqqsFGZXOLwWj60HdD2fb7cdYQUFfTsnYJhs4AjuvXCnpqanYG00EcQx2a0Db7F5I+zy0Zfo9TQXeqid4ZlDs79l65N+iPqCC5bfumpDQXBlGI89bg3K3AtPqrRRVE7audoLmfFn6kHVH2hGnX1/DyW4RydJ8FwOPkCvEigsT6i+VNLMS4GYj6t17ac/N/8AfdDPs1vky8ROzj1IK8Y301ZZa2tqKvIf4hIz9aDlLTHBClr6FtU14JIyQtb05wXp6fW1EJXAN8QZH2rYXDzitd6esFLNIRENhnstxVfFeuh17RGCQ9HiDOD80Hojc+BduunDGFtNAOoQgkj1wuq+sL7JwxtdXZZJC0Oyzcrtrw/4y0p4dQtrpG5MIBJPyXSnmgqmakqp6u3tDh1lxx9aDrvcKaC4ailuzsEykuJW0dZytbPiMZAKya67VNFOICSCNiFhVXTXxmSTc/NBo0dY1jASPJXCkNcetuyhfE10pjHYFalR1EVLhriBlBZTUPhkh/l6rHr53ROMTHHB22WbXVTZYyaY5djy9VoksNST1StOUFHxuAEhOSVVkBlaXHyUtI5pd0zHYequmkY2cMiO2d8IMWKJ0j+gZCzmYgcGHfKmqYo6enbKwjqO605vjSuEgBwD3Qau2la9nX5ELCqKRweel2AhrnmPw4ycjbCRCpJDn5xnKCagjMJDnb5U1S1kjhgZKhfUxsb0g7jyV1IXSzB+PhHcoKmFwb9ArDki+LsFuipq7Y2mDQ4dY2+1aBLJE8ktP1IMR8jYg3butZtmn47pTGpJxjfutFqGF4yPJahbrlV0cBhjJAIwgxKunFPOYAexwrGUfUQ/PnlUqnyvlMrs5O+6jD6g/QzsgnrHkRBgKmtzCYS4k4WnubNIdwVkQvliiLeyC2qqAXuaBkqAuLmHIwsmjib4pdOMA+qlr20/hF0G/bKD1K9hr/udxl/w1g/6NevUxeWfsNf7Q4y/4awfor16mICIiDrn7Q445NeJRP8AedH+vU6+e+3ODJHuO+G/zr6D/aH/AIGnEv8AxOj/AF6nXz10n0n/AFIM6atjqB4QHZYrHiCcN8iVjuPRISPJXiOaQh+Ox7oNVuBa+mGBv3/MtHa4Nysx9SHM8M91imB7jloyCgtc97zjJPoMqWJ3Qwt8yrI4nB2TthSOADwPJBJT05Li552z2WY+nYGdYA2WJNP4cQ6O5U0EkstM4lBD703r6QBucdlq8GmxW05q+rBxkhaPb4myVOHgbFalVXSqox4MJIYdsINInLoJXRdwxy3Db9RwU1AadzAXEYWlMbFUEvdjqKxJoC2oA8iUEdTKZZ3y7gOOUZC9zevfC1CujpmUw6MByipJIxAQ87oMDDs4z2WZS4GGu81FA1rpzntnZVqXdD/h+xBkVjAGZaAfksRlM5zC87YWXbpGzSlkx2+auueIXdMI2QaexpL+klajDThjfEz2WmguLst7qcGrLekZwg1CKuiDvDIGRssiWeB0BAA3C0J8b4z1O2OVc2d5w09vrQXR9QqCWg4JWriidUQF7muPzUFAyE4dJjuO63LFW2yOkMXUA7GMfNBs9kfhTkHyK1Btyjhb2yAsS4003vBkiB6Sdiom0sj4SSN8IM/7ptqT0MiyT6KKTrALzGW/WMLUtE0VMbgXVwAYMd1rmr6e2NbijLe3lhBsYPNU4sJWRFAIMdlhPbJDKSz7FK2oe5uX9wNkGfK0vYOklRslfC3fO3mobdM6WXpO61yWGl93OcdWEGiuL6w9PURlR/c57ZAzqznG6gc+SCZxZkAFZ9HJLI8SO7Ad0F01N7lH1n0WA5vvOZM9lqV1rI5oTGCMjbutIhe8Hpb2PdBGQQSPRUwtShhiA6pMKk8ERBdGRgDyQYMUfiHCrJF0Z+So1zmOPT5Kj3ud3QXwRGV3SDhahHSCJhaTklabHI6I5apW1Mz3ZycfWgnJ90d1BuQs62PNxqRE4YBwCpbYynrMNmwT81LVMioZA+mwCPRBvSn0nTQ07KgSAHIOy5A4e32a1XCJjZyQCBjK4Mdq27RNbFIXdHzW+uGVzdcr9DC8klxAQdtrrUVOo7GyOlcY3FoGRtlccVvDuohBqa0GXBzl265ot2k6tumoJ6WM7sByB8lfVaHvVXZXyOYc4yNvkg6fcT46WKB1PHCGloxt9S4ZjaAXMx5rnzixpG60tfOJmkDfyXC8lhq4ZXSlp6Q7vhBiRUII63bBYdTM2CXpA2HyWfcqxsMPhRH4gPIrTYWCdpfL3+aCdz2SxZDdyFp07MHIWoMaMlo7BQ1MfSCUGHG/oOSsplH7w3xAcfasPGTspo5poh0gnB+aC2aAxZ+SjGfJTSmSTfCtbC8HJGyCxzHAZKtysqUeI3DBnCi6Gsb8ZwUEbe+PVZjKHxIzID81hhpcfhClFTKxpYDhBqVhqGUtQ7xN/RW32Rs9QHxgY7rBhp6p5LowTnutSigIgPvOA4IMHBmYG+YCgfC5h2WRTtPjkA7Zws2OldPMGY3KDFtpe2Q9ecHHdapVUUbmeO3Gwzsl1tM1FTiWMEZGdlhUctXIz75np7FBDEPfZTCRjyWY3T4LwwOOD81HK6Kn+KD6fmp6atqXNzkl31oLptOiBnWH9t8LANO7xPBwcZxlZVXVV4wZAen1WZQyUr4syYDygxBaG0rBK7cnc79lEaASP8UDAUtfWSg4cT0DOFSmqDNH0R7koM+gu8FtBbI0E4wtJvM7bhN48bAAobhQVcX3yRpwVLSmJ1MWD6RQaXnBxjspoCJHCPCTUk0eXFuxKkpmhuHjuOyCSqtpgjEjSTnfCuoaQyNBc0j5rPppmztDZuwWoSMp4qcuiIyg0Kvb4bMA9ljwTgMIO+6sqpZJJXNcdgdlCw7geRQegXsXJg/mm1SPTQFcf+cbciv9i3GxvNFqhze/7AK0f8425EHtCiIgIiICIiAiIgIiICIiAiIg455kTjl34pH00Xez/wAhmXzSsm8KeRx88j86+lnmS/B24p/wKvf6jMvmgk/sjvrP6UCR5e8uJ7lSRTY+HtnzV1MI84eqVIY12WYQVdARh5dsr46UzjqacBHSfecE+SrSySNBDckIK+7imd1k5VZK/LelnbsoqiZzjhxVKemMxyATug1OxWcXWo6XbZWffLU2zAYcDt6rEo5qm2/HDkELHudfWXI/fSSgwZZ/HOANyonNdGTk91kU9BO456SrauJ0TgH7b7oMdrS922yuc10TwXHOFPhhj+97lY72SZ+MFBqTbo1sIjaACB3WOa0l2XHIyomxx4yVC4Dqx5INQfUtqI+gD7VC2hPUPi281QmJkQLcZwr4Ko9OXIMx8cdNCCcErCjkbUTbDAyraiofKMEnCxonuY4FvdBr1xgZFRB7cZwobPY/uhGZnOO2+FiyS1E0QbJnCvpLtLQxmKM4BygmuBjp80wxkbLT2SiPIO+eypUTuqZC8nJKlgoZZB1luw+aBFV+Gep24U8lQ2pjONliVDWNHT2IVkDi1pHqgloYPeKjpzjKnudKKYDB+xQ0Mwp5uvspLhO6rkGNwgwWMLz5qrmdDw1ZsjY44Bj6WN1htzIS4+SDUWZZD1Fw9e6o2pY49GM7LBfO/HTk4CpC/wCMEoNftdLGyTxHAYPzVl6jjeepmFF750QgR7n5LT56qaVxac4JQXsrY4x0FvZXmfxmnp2WE+nka3rcNldCT0HCCkYIl381qQb97yD5eS0vLmv6iPmpW1MjvhGcIKieVsxDHEYPqtSgqapjg4vLgN/MrSpm4AeM5WvadmostFWRv5FBmQ3BlU3wXxgEDutDr4TS1zZm7gOytzXt9sp6UyUhHX3W045zVFxl3wg3rFqmlqrY2jcwdXTjdbVrqd0dR48b8NBzhQNAYcxA5+tSe8uLemQ90GtW7VMDYxTSRgntnCxrtLHVkujaB57LRTTsDvEZnOc91nQzNEZD+6DTqmIN+sLH8RzctBWbO10sh6exULqV2CcdgghgkEb+pwyslpdLM17M488FYb2lriCtUoIQ+LLdwg1SCtZJD4DsZ7ZysGW3dVSJg47EFRCOQydMWcq2eargeGvBQa3W3GP3AUxAOBgrQYKfx5cDbJWUYpKiIEZJIVaWlmEnQwEPPZBrNBpL36P4Tk91o91tlwtNX7uyke4Ht8BOflsuUeFXDDWesKuOO2slcHPAwGkhd3OF3I3W3SWmn1LSHJAJLmoOgWktB3a+xskmtkjASNywhcmUnLnc5aX7oMonkNHVnpK9Jb5ybW+y2Rgs9O0SAfvWbrVtJ8FobNpSphvTGiQMIHU35IPJ+9aHrHzOtZgc1zPhxgrZA0hVabvsU8jSRG4uIwu6/FnRVHprU9TWFrREXEg4wMZXANaKDUGr4qSLBY6QNOPrQd3/AGf/ABDjr56a0ig6CHhpd049F6ZDcBdJeTLhFZ7Fbaa707AJNjsF3aGwCCqiqIzLC+MHBcMKVEHUnm40TJS6fmvU2ZGOY75jbK8kNf26G41dW6JoYGyEHy817bc1lmqr3oCWkpgSSx/YfJeMHE3QmpbRXVh6HhpkPcH1QcTCGS1ygw5z8goG1dQ25suUjyDG4HJ+RWu0lN7tKPupgDHmtJ1FTtmeTbhmPzwg5Cl5jrnRWZtogrXgMGAAStBPGZ9ypn09ZKXuftly4vuNJAxuW5Eo7/NYdPG0Zc/II7BBuK6SC4VZqmEAHfCwpJ/AjLC5YcNRM1xG/SoqqRz3fJBaybEpcVWZr5nAsKhkIA2O6yKWOaduGZyUE1Az3d4MjsjPZa2aSKsi6wBsFoRt9eCCWnutVpX1EFKWnOceaDSqqixIQw4OVSlt5dO0PJ7+atdNKak9fbKldVlrh0/S8sFBqV3tvg0Qc0k7ZWlW57S0wObudlqNJNUztDaskM+agrGwRTB8OPsQWS233T/bBJI74Vn3TY4eGG4ztlTVlcJacMJ+S0mNjTIAO6CaopXgGYEkHdS0NZ0AQEYJ2WqW+hlnaA8fCPVYdzpYqScFmAQgyZNOySRmpMpwd+/ZaNPE+lf09WcbLUHXKvEIY3PSoWUstV98kyfPsgggl6z0uCmdVNjkDOnupI4Yo342yo6iOPrz5hBPMxskYeNs7qCKobFlhbny7KR0mWANO2FaGR9JJ7oJGeG89WAPtR7GdxjZYTWVMzyyHJx6KT3SujIMjSB5oJXQ+P8AC0kfNWTUboKZxLs//ZWTEDj4O6xa2eUNMT/NB6m+w2/tDjN/hrB+ivXqYvLX2GzcWzjI/wDup7APyNrv9a9SkBERB1y9ohtyacS/8To/1+nXz3UMfiOk37Nz+dfQj7Q/8DXiXnt7nR/r1OvntonOa5/T5t3/ACoIZB0yH61qFNUMdCWFu+MZWDKMvJPcrIhfCyMnzQU91LpC7OyvMzISGYBPZQxvmmk6YlaYZGVAZMMElBkTPw3rAxlYnWXHYEn0Wty0QbTh7hgY2WDaGQm7wifHhdYz9SDDe2ctBfE7HlsVn0c7WUzmnvhcv6hpNGt04w07ozOY84GM5wuGJZGNrC1v0Or8yClK97akuwcE5Oyzqt4ezYglahVNt4o2mAjrK0NsuHhrz+VBbSh7ZSTkD0UlVUNB6QNwsnELW9TcZwsSUMcSSghiZJUuxk4Vs0Lon9G+6yaV7GOy37VlTMjezxDjI80GF4BgYJM7kLHe4yuAAySpJKhzndB7DZZNvbA6oaXkdOUETYJqdvi9Dhnzx2VpqXSkRuGc7LeFZ9zPcxgt6sLb5gpvptI2OUGIaT3domcfmsuCpidHnAJWPPN4wMQOQFisJgd0E7FBfV1DZHEAY3WMDg5WbJC2Zo8HBcVPS2Oqc8SSM+AblBhQzS56QCsyCgnml8TxDjOVn1cdEyIMix1jY4UUDpooS8eSDNqfBhpww4JA3UdpEdXVsgIwHHdaNNUz1Mhjbk4PksmljqqFwqACCNwg3tf9Nx2m2isgkAcRnZbJZV1FQT4ji75ndZVTqm4XGMUlQ4lnbGfJZFLDSspd8dR/Og0uoDGDOMnCx44jINhsFfUCQzEfvVJG5rAgswKP4wr2TyVI6gTlRVsrZAATslBPDE4NecAndAlBOAW/NXMrWxs8PHdZ1wmozT5icMgLQXHJJQSzRv3eTkE5VIpQzuM7qSmka49Mp2AVk4j8UBnZAmqDIA0ZAWfQQ+LF0ud3CwJRGGDHdXQVboRgZ27boJqmlFM4u7gqBrBKC4AbK2aqkn2cVWJ4b8OdiUEbznbHZSxStaOnHdXOi8XHhgEqx9JPGMubgfWgzKQOhPWH4BVJ65xnGTkLGhdK53RvspjRPx1lp2QZU8jKqJrW4BC3XwxqTbdRwzPdgNcD+dbNp8lwa3yWt6frRR3dj5DgAjKD0y4X6yoLrp6mpJSw4Ab5LnWlttunsBEcLXkt7Yz5LoFw64i0tFRU0cT/AIi4Duu/HAm2XfWFthqA1xhLQT6Ywg6kcwNh6amocy3EAZ3DV1UuTI+h9I6ENJJHbC9VuYDhzZhbZ8sYZgw52HfC86eIXDW6UVVLcYIyIGPLiQNsIOAtSadfb3GrIODutEgAmhLwcALkHVtVT3eP7l0ZBnYMEfNbP/YreaGBzpYy1p+SDR4Z8SlvopJCZXdI81DHF01BY76QJytSjpRs8II6S0BxDn5x3UtVQRxMLh3CummmibhhOAFpctdMXdLyfmgvo2tlmLCNldWPZB96A3PoreroaJIu5CsbDJO8SPz9qCyjdiXLhnKzprd7xh7dhjyUL6UuAbCMn5LVqKzXQwGRzSGgeiDQHA0ri1zc5Wp0dnNbCanoOAj4YJJQx+OoHC3nabSfuc4xgY6c9kGzoaoUMgiEecHfZatS0Au7genpB+xa5p7Rrr3WuY1mSD2V99stVpysEDQQfkEGnVWkjRxeLG3J7jZaXSB0Fa0SREAHzC5G05BLWxhtaMgjzWFqWxU0EniwMGAc5QaBqSqg+5zfhGcLatLWxvhMTWjJPktVvcU9Y3wIsnGywLRY6iCpDp2kN88hBps9HVwvMghc4H5KtNcnRPHWzGFy3a49NGlDK4s6sefqtjat0wX1ZqrVGDBk5x2AQaZLcGVkRaGAZWjudJFUDDyBlazR00RaIW/2QDB+taZcaWWGfLge6DOqWtqacAEZA3WHTO9wlD3HIBzhIJJJG9EeSVZLBMJPvwICDUblfo6unELWDPrhaPSPcKhpGcErLMdM1oIxkd1kMp4RCZW4yNwgza4RPpRjGcLRujoBcOytFbJ19DicfWpWRPqSGsGcoMd9U4EBu2FrVvaZ6b4n+XbK0urt0lO3reMAqyirpIJWNz8OcIJLjTmJ5PSVhRbyAeq3JVUZu1OPcxlw7rQ5qSagkLKhvS5B6B+xih8Pme1O/Oc6CrR/zjb0UPsXZJHc0WqA/t+wGtI/zjbkQe0CIiAiIgIiICIiAiIgIiICIiDjjmS/B24pn/uKvf6jMvmkjb1zPB+Z/Ovpd5jx1cvPFBvroy9/qMy+aJwe2d4b36iPzoLCCJC0FH5Dhk5V76edjfEc3AKi6iXAlBM4ZjwqxTCEYPmrnPYIhjGVA1jpCSEF0r/EOR6rOtzxG3JasWijBmAftha1JHA2EluM/JBG+qjOxwozLE0F2y0uR8j5SxmSc7K51PVDcg4KDXbFWRVVV4TgAFLqW2wtcDERvvstAhfLQOErMgq+e71VSR4jiSgvp6YRHLiCEqmMcdsfYoHsrGN8RwOPXKibJK8jbv5oKyAtbgKDJ9VqXurfC63uOcdliyMa0HpwggyfUo3JIABKE5OcK+POeoA7IMuWENgDj3WLAPi6j2CyHzOezocVihjwfgBwgypq0OZ0Nbv2WGck75+1ajQU0QeHzH7EujYsgwgDHogxKRoMm/ktUdXx00XhhoyR5LRmPcw5GcqdrhICZO6CzBqJSSe/zU/u3Q3I/SsUu8N3wKaOaR/cnA7oIH5DiOyvjmcwY6cqWcR9PlnKzbVT08jOqTGUGmyOe74iDuro3DpWo3CKAAiMjtsFpAPSSEF2Ot5HzRzTE4DfOEhcGvyVmF1O9uSRnyQWRZDR1FRvlAkGw8lZIXtccdj2VscUkxJaMlBnVFSySn6WgZ+SxIXYaVQRyMcGvBAUwMQ2yghe4P2G2yMcIzjGcqsga3Lge42VjGOe4HuM7oMvoDm5PmkdG97w6N5HoqyysZEGjuB2UENXJHI3fsUGfWU87KcdbyfrJWlse9mela5cahj6IdOOr5KC0No3RkTkdR9fVBi0cpLviGc+qsrZD4gDey1N1JHDKZMAMWFVup3kFpBIQZ9qpBVDDioLzT+5yAMO3yUdLcDSj4T2WNcKx9W8dRzhBm2RrauURvAxlZV7hjovhbjdaTDVCkaHxH4z6KKqrZqx3VKcoIXffHEgd1n0tWaaItI8vNQ0zIyd8ZCnkjix5Zwgmt1awVPiOxjvusu5OiqSJGYWhhkjXnwxn0UkNRKJmsfsCfNBq1FM9hDRGSM47Le2hNPPvuqKWkNMSJHAHb1VNJWKkr4mOe0ZJAyuxvBfhLUVuqqGqpIyQC09soO9nJzy/Waw2imvNdaGPyQcluV3OZbLcwNDKKFob26WgYWxOCdlqbNoymppwAQ0DGMeS5FGwQRmngc3pMLCPQtC2vrXSVDdrZKWhsPQ0k42BW7Vw5xq1XebHZ6mChB8R4LW4+aDofzkW2htrZoaadgeAex3810N0DXSxa7j8SQvImGB3812E5pK7XctxmrbmyUU7sncnGF1l4XXSldxJo31RHhmYdRPbYoPbflIrZJtK0oMJaC0AHB9Au0Y7LhDlkuOkJ+HltZbXRmfoHVjGc4C5wQEREG2tc2yK52mSGWISDpO2M+S83OZfTFJbhVuZbWtPWf3vzXqJLCyZhY8ZB8l055sdLWsvlYcAuJ8kHkbrajlq7g6FkRiAJ3xhaNDTRW6gkbK4PdjzOVztxo0F7ox9TbR8RBOQF1tqZ66j8WGuJByQEG26gme6OBOGklQ1jRC8dJ2zupa8eG8zRk5PmsdkFRVtMm5A80FRVAjp7H1Q5e3IGfsWK4FrsHuCtYtDIpG/Hj55QaTJ1ZwW4ws6214ppGgtBA9VmXKnhAJYB9ixaWgD2l+Nwg3FLdab3YODRkrJoYaevpnPyBt2ytnVc0jD4e+3zV9Hd6qmIYx3w53CDULhQiOd3S3b1WIygy7xtyRutcbC+5QB0YJeQsFtPLSVQZVOIadiMIDZZqhnu8UByB3wVpNaypp5OiRhBPrlcu6RpNOCMS1b2AkZWPfdOWm7XiKKhcCHOAwAg4jeyo6QXMdg+arAwteH9wO65y13wlktemmV0LDksz2+S4rtWlLlURvmMZDW9+6CS23drR4RaB5ZVlwonVj/GAyBv2WNJSMop+h2Q4HfZbit4jloyNs/NBtupmZBG2LpGQtzWC2U9bbnyvIBAytq3SmkNQ7vjyV1LWXaniMVMXBh9EEV4Z7pXPYw5AJ81jR9VQcnKyJIJ3PM1YDknJyr42xhwMRyPNBY2MtGDlVMXWCc4WRPNA2MgEZwtMfVP6iGdkGsWkxU8mXgFa1Vvp6iElrQDj0W04H1EX3yQEDust1e50eGZQXAshnJJGFp1xkEsvU3GFZJNL4hL8qKQl2/og9XfYbn/qRxiH/ANMWL/o1y9SF5Z+w1Obfxlb6TWD9FevUxAREQddvaERiXk54kRnzpKP9ep188sMvgl22SRhfQz7Qpsj+TjiS2L6Zo6PH8ep189lBGx0j/G7NH58oIZA53xYO/wAlk01tfOzqyQVmxyUIPS5wwPLZTQMfJUh1PvHnJ+pBDaqY0tQTK3b1Kx7vJmsEkbcgHIwFua7mkioR4ePEA39VptCaCaAipx174z6oMGS7e8Uwh7EBaaQ4P+A4J7FahUUTGyl8X0c7bqKGlcagOcPhB7oIZjcWxjxZpCz6yoI4TI0uz27brddU63GlDSR1ALbMsgZL0xnYlBdRskMvSScfWsyW3NeQ8Hf5KJ8sbIuuMguWKK6fOOo4z6oMuSBzB09R7KNlN4h791bPJOIw453V1PVhkRJO/wBaC99v8AdbXFRGY9PTlUZXSSPLZHfCVFM8CUdJGEE9Pb/eHEk4yVc+iNPIA1xUZrXRMaIzg+auZWOkYXSHdvZBkTwSujBEhPyV1LSPLOk5OcrFo6qSWYMcditdgj6cPxtjKDS5KI0zi89u/ZYk8Yly5u2CtZuVVA9nQCM9u60joeWEsBwgsopjTy5duAtd/ZLCaYwBoBO3ZbfgAkkLT3Vs0QjlAGUEs3iRzePkkE5W4rfFHV0Rc7YkdlpVPG2WMeIBgDzVs9wdSfeoTt8kEL8UNW52Adys43OOenOGDIHotNe/3ogu3JWdRQRRkB+APmgsstH7/Vlr24BPmFq1fa/dJA1sm3yUc1VTUTeqkIDj6eq0qa411W8lxOQgyK+NkcZcMZG/dafBmU4z3WVHaLrc8eDGXY8kdYrlQv8Av0ZYfLYoLZLflvVlZ9NpYz0xnBwQM91jOZVQNzLnHzWfb7zJFiMn4B3QaDPbqiGYxYJAPdYr43sdgtIK34ZLdVMDgWlxWj3Kjp2MLgBkINBZSlzeoHCikjLHYJzlZEZkmlMcJJUckT4ZwJgQfmgsdE9rQ7BIUeFrrH0hgAcQCButMeIfehjHTnyQY3Q4AEggH5KvScZytauPufug8Ejq81ozHb4d2QTUU/gyfE3Pmsua4RzkM6QM7KDpiDc7ZWK/AflpQai2JkOJdjnyUza6ORnh4G6wIXy1LhEMkeiyG2SvDg9sZ6RvlBXBp3GXpyD2U1IPeqjqDukrLjpmvZ4co3AwtHnmko6lzYjgAoOVtKC6wPpBS9coEg2GT5he0XJXdKSHhqx1wY2KYRA/GMHt815V8pV+0J90IG6zliDQ8ACTHr816SWe9we5RnQhzQhgJ8PtjHyQbS5oNdVFHXVYpWuezqOzckLqxdtZw3vTFTRVFF0SPaQCRvldyJLHYNaPmgvHSZRnqBx3XXPj1pvS+jaCdtD0AjJGMIOhV6t8uldSzXR+XxlxcB3wpqviHTXWlMIpw04x2WfdLlQXu+y09S8FgJAytt3+gtFvcW05AOcoNMpLU2uq3T7AE7BZktMKaYRY2yobRNI+UNgJO61eopJDIJJG7hBivtgMfiBu5GVpNRYDKesNwtysqGlvQ87ALGrKqJsZDCM/JBtqa3mnADB1Y+S1OkomS0hc4BrvqWTaRDJMTVkdPllRXWoEVUGUv9j88BBoklRJb6neMkA53C5N0zcaK5WgxSNYHFuMea2/RWeiu8YJwX4WmVtuvNlrG+7BwiB+zCCe76cFPXGoDsAuyMfWuQ9GUtNPQ+A9zcub05K40vF5qJ6ZkeSXjuPmtY09cbrSW10zOoEDIQckMq6LQ0rq3LXdW60muutJrCoFaS1vSe2y49bqmoutW+muZPS043WLVXmot9QGW4uDM74PzQcnyVcFDCWMeAQMZW2bpqHxgY89Wdlonvtyu8QbAHFx74WdT2Semp/ErWkHvugutLI5JTLLgjusXUN6gpsxwtGfkFgT3T3aQxQntlafUAVR8WbsN90EcRrLlNlk5aPQHC1/9lbbRb3W6aMvcR05xnC061UE9yk8G2Alw74C12o0s2CAi5MAmI2z6oNq2p5dXvqj2cc4V18mEmQGeu+FlRUX3OqC6YER9xnthSVr6KohJYRlBoNreIZSXjIz2Ky66pZKMBoyViRhomPoFfO+MDAO6DAlje1x7kfJTRVj2RGM5WdaWwyvJnxjtupLjFRNkxHjCDR2xPmcelpyStet0RpmCSRnb1CstvugI6yMZPda1MaV0PwkbBBoV1r21bDE1gBAwtIFLL3a0/WtZlpGyynwxndZ0YoIacxzYD+26DA09evuM9zpW9QJ7FQ3uvbeKsTxt6QsqkoqaaVzpcdOc5Kx65lNHMIqfGc42Qd9fYxtaOZvUwwMjQVb5f8A3wt6K/2M9HUU/M5qeaYHpdoKtA+v7oW9EHssiIgIiICIiAiIgIiICIiAiIg485i/wfOJ+f8Agbev1GZfNGXiKsleRt1O/SvpZ5kT08u/FJ3pou9n/kMy+Z+Q9Ujz6k/pQZ8tc2ojEQZg/UsUUrj3BCntTI3TZkxgeqy7mY2ECIjt5INPbSZ7uKZMBDAM5RjKskENOFmUkcZnYZ8AA+aDHfHI1ni9BGfPClp5ZJGEHOOy3caOgrKMRwAF+FpT7a2gid4rRk57oNFpoxFUdbtwd91kVdwjbhvSNvksVsmasjOytr4wXAs3QSSFtQzI9FbDQZw8u7b4WOxkzewICuE8zSWg4Hmg1SerjfB4HSNh3WnBzGdWw/Ioo5C+TD/MrUG0kMjC4u+aDBfNIThpOFA8OHcnBU07Qx5DT2K3rS8HuJ88Ie7hpqvtkEWapwR/5iDYSyYJWMbhw3W6ZeEHFYSEN4YatIH/AN5Kn/1FjXPh/q7T1PFUan0peLSyclsTq6hlpxIQASGl7RkgEZA7ZQbbe/qcSD3WbQywgBjxknZYkkYZKWHyR4DHgjy3QahV0ssbPHaTg77LT3SOdu45WsUtT7xCIZRhp23WLVUtPGcRkHKDTW4zupWs8TJGwCllpSGdTQscPfGCN8FBcXt6sBTRsw3YbY8lh7uO3da1bI4n058U/EAg0owuc87d1n0rGxNILiDjyUQhlZVkuaQzPdS1Do2kYIz5oLTBJI49yM+SlNmDozISQQMrJsz43y4lIwsi8OexwEGSPPCDbZhxOYvQ4WWLYRh3UfVQ+C90xdu091nxSuZGWvdnbZBC2nbUEReY81kx00dC3cAk/nWnRzSioPQD3WVI2pleHOBwgjrnhwJa37Vj01MZnDJwtXmZTinw4jqI8+60d0j4n/evIoM2qtoihDg7JwsKKURgx+pWQ+slmi8MnKwjHIDkhBnfc7rb4ue6iNDvt6+qiFVOwBpyAFlwVBdGSTv6IIqnqZF05yEpKJ8g8TqwBv3VKY+PUdLtwp62V1KPDjOAUF01b4rPdvMbZWO6gIb1dZKxuiQESYye6laaqRuQCQggflpxlUaOpwaT32UjmFp+MbqwOAeCPIoM6S2dEPi9XlnCwSzufRZclVNLEI25KpCzoYRJ3KCCnBc/CkqOqM91GCY5ersMqYSRyuHUUFsFV4bskAq5wE8olbt57KSekBjBjwSo6Y+ECx4wT2Qb00lqB9I6KJmTh4GPtXqFyV26z3hlJVXDoa8NB+LY9l5hcMm2yK4tluxaIw/Iyu2HDvjozRmoKGOyS4pmlod0uwMbIPaGxPpGUDIacjojGM+Sz/GhHeVg/wDGC6pWDm20cdFRNbVM98MYz8QznC2HqLmtmZQTSUs5yMkYcg7oan1ZaNNW2StrK6Fha0loLhvhdAOY7m5t1FVzeHcY3siedg4eRXX/AIvc2usr1PLQU88zmEFuA4rp3xA1RqC/zze/CUh7iTklBzZx15oLfxDt33LpI2PkLenIG64T4XaTqLhqinnkzGerIJOO5Wl6EsVLPcGy3IENztkbd1zLT6NubXtr9OhxDG5y0eiD075NLTLQ2mmY+5CQAj4erOOy7pjsvLfkQ1VrKbVENouPiljZQCHE+q9SG9ggqiIgLiLi3wg/ZyJZw0uIyQM91y6iDzP41cH5bIZYXW1zwxhG4JXRLiXw8qXTzTR0Low15/e4817u8RuGtu1VQSPMTTLg5JA3XRPjty61VFZa2tp4sAEkfCg8uavT0bZPAkwC3uFhSxxW2IxtaDnZarxOprnp/VE1IXPGCQRg+q2xT1zJ2n3p24QYNTEXSl/YFZ1FCWxF2cYUM8sLz8BysiGpZFCWE7lBbFmplMTnHYqSoq/cD4TW5BWlmd8UznxnGStYoXUlVTl1SR1gIMN8Taz4+xKup7YPEAJz6ZWLUeLDK4x5DM7FZVrtt1u1SyGl6y9xAGEG4KOV9oDJAwvDnAdsrtRwY5XJuMlk+6zaA5DOsYGPLK4j0lwluNPS09ZqEEQl4PxA/L1XqvyUaapY7BEy1lpgEQa4geWEHUnQfIPddQX19rNFIxjHdOcEbLmym9m83TzmXJ0AlMRBIDsld/7JpWgslQ+qga3xZM5IGO61Z5Lz4fSOkggoPLfjXwcba7M60OtpAiZjPT6BdQbxbqHTNNUUbomh3URuvYfmk0ra4dHzXVjGiUteOwyTheN3Fj3ue6VIiyQ2Ujbt3QcT322NnrXztGGkkrSxWvoT4YJwDhbhubJqaFrpMg47laJ4cE7HPOCQgmhjjr8EgZ81K+SmtwDCAtENY+lkLYjsCsGtuEszsuJyg1a4yx1zemPAJHksOOm91icHHJPqobZUjxOl5VbpO4vDWdkGnzOcZCC4ndXQxEkPOQBusm3QtfKTMNlfc3RxnphIx2QR1Fa2SLwmjB7ZVlG4tOD5rE+az6WFz4+sDOAgrWsaG9Q81jMjBp3vJ3CTeK55a4HGVHmSNhb5FB6tew2GLdxlPrNYP0V69S15Z+w1/wBzuMo9JrB+ivXqYgIiIOvfP7MYOUHiNM1hcW0lJ8Pr/t6nXz10EElVJMOkjPxHbtkr6L+cukpa7ln1xS1mPBkpqYOz22q4SPzgLwH1BDaKGaqjoS0FshaMY9UG26TSxrHH75j7VuWkskdrpjG4ZJC25DJdopRJBkNP19lr1NV1NVF4MpPiHbBQaHcKWQTOJdkHcDK0eallDi9riACt5O0Tf6lxnbG7oO+cHC0i46cuVHJiRrg3z2QaM2vIAjcM4HmpxWARkAblSS2+MDGAHLEfSva75IMOV8vUS5xwSkcT3t6unIC1A0oe0YxlarTUMTaIlwAOCg21Ax0j+jJwsp1EGOD/ACHksaVxiqH9G2CVX3iZ+2SQgyauoa6Pwh9Sx4aQyjOcKNx6/rV8RmY4YzjKBNTGIZyoWtL3BudycLOqnZiGe4WLTECZhd2BQZL7fI2IP3I7rDLXAlgBzlbpmq6N1IGgjOFp9DDTvqw9+OnIJ+pBJZLM6VwllaR2Wdc3No2mNh7DGy1e5XK20lvDaVw6wNwDutqPrhVg+KTnO26DTpnPdKXF2xKzIahohMZ7lYssMj3ZYMgfJQHrjdh2xQZkEXRMX52Khqnjxg4KITv8irHOLjkoM9tUHMDRt9SxJ/pZznKjBIQkk5KC+GUxOyc49Fnsm8WMgHfC0xSxxzlvUwHCDOt1I+pnIe7I+ZWdPHFRuztstLiqnw7Md8RUc1VNNJ98OxKDkHQ99ooZi2drQM+in1beqWeva2CMYJ8gtgMdPC0Ops588LXrcPeYQ6p/sg9e6DLu8EU1K1zAMkKy36YZPQOm6wDjI9VYaatmJa0O6fLzWHU3S5UGYGkhpONigxaWhmpq1zDISASrrgHyOEQ8xhTUc7pHiSQ5JWZJReL99b3G6DTaO3ihPjv3yFh3Rral5lZthaw1kkxMThsFpNyttXG8uY0lnyQaW1skjuhpJKvfTvaMuzlZNrEYqCJNiPVSXNnXIGQjOSg03qcdi4q7wyVLLRzwND5G4B81lU0LZGdQ3CDDZG6TIJwrJY/DdgnKyZ3NhJDDvhYpLpHfNBl2ydkE3U/styHVVNFB4TWAktwts01sq6vAgjJJWu0uhLnJH400ZAxnsUGni5mWcu3AcVr+mdHDVV4ipc4EhAO+O60WstApHCPqIcDgd1vrhvZL225xVtIX4YQ7OCg5D1Hy23Kw2Slutrq3ROGHZY7HzXbPlQ4pTadszNLXabxZXsEfU85PbHdcaWyw6513ZobZSeI/GBsCVyBw25ctc2a7QVVTTyh3UDnpIQdjqrTf3Ngk1EKjpbPlwGcd1164t6POuOuMVGQSRjqXN+ubLrS3aaZSytkAa3A2PouCa3TuuhTSV8LZehmTkgoOturOXV9infXxAuJ32XF124cTTTEPJBBwAT3XaqqvdxrKiWguh+IAtwQuKdXaVvrLmK+mjPuof1EjthBxTBo02NplkjIAGdwts3fUAbWClY3zwuVtdaktBtwoYHtNSG9LgCM5XE1DZPfavx5Acl2RkfNBPUxSNgZPuOpbi0poqO/Oa6SQZJGxKhudnkkoQyHJ6RtstNseobnp+uZGSQ0FBuTXHD82Kh8amBJAzkLYFFTvljc2UZcFzdFdmartwhlIcS3G+64+vOl6q3XISMaREXZOBthBotqmks0niSghvff0WsVOqKO4ReCGNLiMZWHrGooPucIaYjxenBA75W17Haa+SMzhpwNwgyrjbpmy+8RRl474G6zKTUIp6N1LNH0EjG4W8NCvtBldFe+gNG2HYK2zxJjs7bgDaC0sB3IQbZfSmqqXTMOAd1LGwMk8F257BQNkk6AIgc43wlA2cV7JJwegHJyg3rpqSOz/AH+dgI77hS3/AFTBXnwYcDIworlcKB9ubHC4dYG+FtQwSuJe0HOThBdWUDw/3hmXZ3IUcrZHUj2lhBxjOFuLTbGSvEdcMDO2Vq15sMLoTNTNBZjJIQaLwrr4bRc3TVjRjPZy1nXWooK+7NkpnAMHcBbEra0W6YshPxDbbyUJrJKthfkl/kg3BqGeKtog2JwDsDOFtuihkAMb3ZyrGmu6iZAen5qcS+GCR9LCDMFpaG9YO5WlVdI9kmcnusuG5SsJDycK+oqYZmHpcMlBpxLoh8JwVjPkkeclxKukbLkkg4PZS22IPq2Nk+jndBB1ysHmAtVpqiQwEdRK1nUNDQR25r4Onr6fJbdtbwJA1+wB3QSQXJ9LIetpwfVZzbW66N98EmAN8ZUd5hpDAHREdQ32WmU9yqoIjDE44Iwg1uCNrSYQ8AgYWmVcD6erEwJIByscVFY0mQZyVqFvkbNKPesYQd/fY13z7oczGpaPw+nw9BVrs+uLhbx/OizfY8wWuPmU1I6jA8U6ErAcenv9vz+fCIPYNERAREQEREBERAREQEREBERBxzzHND+Xnig310Zex+WhmXzRPi+/ys/uS78xX0vcx2Ry9cUCO/7DL3j+IzL5o+o+8S58y7P5UEluYXz9IOFl3GN0TwXHKxqBr4ZxI5pA9Vn1tTDNjJ3CDH99ayMANGR8lfFGaoZbkZ9FZHSCtPhQbn0CymRPtTg2cYz2ygko6mW0yCSR5IG4S53sXV2GbYS4zU1TB0sIJwtCYHxSeYPZBqlJbxO7J8llVNtETOrGfmsGCWojHWM4WqQ1Qlpz4h3x5/Ug0kzM6ywAbKCcta4gDc7Kyp2qiWHbyU0TGSNL3uwR2QYxhIb4m+FdHUysHT1HHop6YiaUxP2b2VK+CKFwEZB+pBvXgHBHW8c+HcNREyWKTVloa9j2gtc01kQIIOxBBIIPfK97ecXmot3KJwytnEq5aNqNSxXK/Q2MUkFa2lcx0lPUTeJ1uY4ED3cjGB9LOdsHwS5e5C3jxw3Hrq6zj/lsS9d/bF2euvXLLpmkt9DUVUjdd0UjmQROeQBb7gMkNGcZIH2hBxh+7h6T/wD1eLt/pFF/V11751faNWnm14eWXQVs4XTaZba7yLvJVVNybVPeWwSxNjYGxt6QfGcTknJa3bZdNLzpq82eIT3Cz1tJEXhjXzU72NLiCQASAM4BOPkfRaZDC17SXOwR2QUqT1yGQdiqxQuk+M9gVG45PT5LKif4bC0eaCZj/Fb4MRw4eixp4p6d48Uk77LKsNM2uv1DQOkLG1dTFAXAAloe8NJGfTK9FOb32X+geXzl+1Vxms/FHUN2rdPe4eFR1VJAyKXx66CnPUW7jAmLhjzAHZB53CVhh39FgysEhy1SSkCIAFd5+Q32eOiubXhDd+JGpOIV7sVTbtSVFjZTUNNDJG6OOmpZg8l+4JNQRgbYaPUoOilO1rZPiwpJHSxyh0ZIaPILdnF3RFJw24w644b0NdLWU2lNSXOxw1EzQHzspqqSFr3AbAkMBIGwJOFyTyS8JtG8ceaXRHCziFRVFXp69G4msggqHwPeILfUzsAe0hzR1xMzggkZGRlBw1VV9M6jDGgeJjdaZFTzTtLtyMeq9TNceyP0zqDmQqaHSs9forg5arLS1lwuE1UaioqKtzpfFgpnzZAAYxjnyPy1nUMBxJA5S0Jy0+yf1TXDhNpG46Xv2oXl1O1zNXVb62pmblpMT2zCOR2QSGxgtOCQ0jKDxgZJJAelriCFrNur4psRT4LnHAXezmc9lFrzSPFjTdq5foq3UOl9Y1T6Zkle4dVilaC9/vcrWgGDoDnMkx1HoLCHP6DJzjp7kp9nVyww2+xcyvEaz37Wk0DZ5xd7zLSRxkjHVHSU72lkZycGYuJIJBGMAPLG76afBSe/M+jjIW1w95IaXEnsvX7mm9mvwx1RwjquInKnUSR1dJRG5UtrguTrhQXilDC4imle57xI5u7CHuY4gNwMhw8gg0NkBzsN0GTHB4I8UjKnFwj6ekt+Sslma6EMBGcLBdC97CWndBLO90rtjssZ58MdJ3V0TJmu+IFJ29urugjik6D1FSmoa4jYKEML9h5q9rY4zh5IKC6dwMew7lZFBTGWEu9FHmEgFx2CyYa2GGIsZhBhMJp6krIlb7zh58gseNzZJy+Q7ZV9ROGYbEdsoD6hjcNwDjur469rGkdI3WLC0SSfEqzxNYQG9ygSO8eQkeajewsUkcT2YkcMBXSOY5pwQUCll6CM77qacmUBwGMDsrLdE2SYB42ytQrabp2jHfZBp8EYqXiIj7VkS2oQ4d1H6lDFBPSSCV4wPVKmtmnOx2CDIbOIm9Lt8LBqJOuXrb27q2WQvAB7lZtrtc1wmFLEMveMBBufQVjqNV1IoafIcDjYrlCPQd60/cIKFjHudLgA4Jxlaxy5cA9a3G6x18FO/oc4YIacYXd6zcqt/r7tRVdfTHDcE5afkg622vhJrOhtDbs6ecRub1BpJwFsHU2qNUWuqNu6JnNLuk7Er1ereX+Z+kWW+OAFwZjAb8l1p1/wc0bpZs7tQxxNqQ44DgAcoOktwt9VFSfdWracub1YK45utSLhUua2nBAO+GrsbxM0w65tNLYmAw4PT09sLiy1aTp7HO77sgNJJ2cgg4e8PJtSyRsjpyD1Ds0rudoHhlbNMaXe+viYZPCP0u52XFnBao07BWNEcjRkjy81y1rF2oqjodbC80mNyO2EGncH+IUOgdeumipwxgmznGPNeg2ieMdJqwQtZIwF7QT5HsvNau1Ho+wMa+5SMFWDl2cA5XMPBzmK0ZbJox47MMwPpBB6MQStmibI1wIPmpFwRp3mn0DPSMbLUsBGAPjAXLWltX2nVtIKy1SB8eAc5BQa6iIgoWhwwQCFtfW2irZqixz219GxxlaR29VulEHklzdcnctmrp77R2kuDgXDDMrqba+XK73G1VNwfbJGCPJ3aQvfTXuhLfri3GjrmsIDSPiGVwhqbltt1DpirpLZEwPkacAN890Hg1cdJVdsv0tulhLQxxGCsG6Wp1PKGN233XdvivyXcQp9WVNfbqOUsJLgWsPbK6w8ROF2odEXIw3xr2EOwcg+qDjqrtzoYBJ0/MrTmvew7OI+oreV7ipoLcwuk3I9Fs5oaSc9gg3Bp2i+7dSKXpycgLnHhToylptZUFHPA1we8ZGFxLw3dTtuDQ0jrLgAF264QcBuIGrNXUN+tNNI6mYWvJAOMIO7tLyrW3XPDeklore1shaCCAATsFzvy8cJHcK7C+3SxlrnNA3C3Twgtdws2iKK23FpEsLQN/qC3ugKnzVUQcD82cwGiXw9WMxuP6V5Ba1ggfcqlshBzKc5+tet3N3S1dRpR5gB/sJH6V488R7Vfqepq6oNdhshOftQbX1vaIGUAfC4E9OdlxM2WWF7o3ZGc7Fbyi1DVVtT7jVuO22CtEvbaOnqgBjcoNHmieGiQtIypaGzC4PDcdytQrZKUUjSCM4VLPcYojlhGR2QXVmkvudD4zTkrbcsmJMO3wVvmesnuEZjzkEYW2rjp2uaTMyMkZyg090vTHlvfHksOR7nn4jlTxNLJPClGCDhSOts8juqNpLT5oIYqfxMb9lmQztpGiPHfZWT5pow3s5Y9O11RKC89kGTI8OPX091izyh2WgbrWXQRCDBAyAtEqmdMm3ZB6r+w2bi3cZHes1gH5BX/wCtepa8ufYc9P3G4wY7+8WLP/m1y9RkBERBwJz21TqPlN4hVLTgspKTB9M1kA/nXz42qOe7XKd73kgyZOTnuSvoH5+2GTlE4iRju6kpP16nXgLpeExx1cg2e15A29EG666Kltdva4AF+Fi2RjKmYVzsDp3Wjtrp5agxVxPhjtlTvu9PSPEVO7DD6IOWbNrm3Og9wfCzLRjK4/19qgeOYaWkL8nGQFiQ1kAaySH6T8ZOVzLw74Qw6spWXKthBYACSQg4O01Z5rxN4lTAWA+owrNWWyG1P6YyCMrmvihaLPpGkfBaulsrAQcBcATXKa4yPNY4kh22UGlsqnMeMMJBK3VQ0cdXb3SOdg4zhYMFHRytBBGfrVKiK5QMIpgfD+Xog29VUbvfHsGSOoqaKlbGw574/OsyN8RkPi7SeaxK6RweQw7eWEGE5pY/IGVNHO36JABWRTNicMyHdYlb0Cb7z2KCSZuW9Q81iCMkEg75WoBhEQMgwCsWV7W56TlBB1P7FxU8VQ5jCMkH1WN3RBe6WRxw57j9qtD3DsVREGZFWeG3BAJVJGtqXZG26so2xF+ZcLLktdXLmanaSzvlBhSweGPsyocLIBc15jmO4OFnw+5loyRn5oNIxhFqNdHE0dTMfWFhRwSSkBo7lBGtXoHMMBY4DOO6x2UJhAdNsAd9lSWdsX9i3ygSxeDIX9x3UkMDaghxCxWzmQ9L9wVmxTxwsLWHfCDPpoo6cAvIIHbKSVYbKHsOAN8LRZqufrOTseyyaOVszel538wg3tp+6Q1X3uQAHssO+wwPrGxtcMErQoqh1vJkjBCxqqurKyTxmEkgoNZulCy307Jo3ZJ9FrVgYyrtr5HYLsea2dFV1lQRHWE9IPmtwWe5NpZmRh2Ij3wgxqiofR1bgYiAD5hapTzw10BjcwEkbbLN1DNZqijBgLTKe60KirYKOnc5zsEdt0ENVpsQyGoa4AHJwtMkYaeQPcMhpWvWqgvetqs0NmDnvBxtusq8aIuul2mO9xljyPMINq1dwbXR+C1gyNuygjkfSxGMNy5ajBS0/jHoxlahbrVHU3OOKQAtJHdBtmCjmqpeqSMgHvthZDre2F4AOVyXrPT1FabIyopgA8tySFxjRVjpHlspyT2Qbt0k4U80YFP1guwdsrtJwx0DBrMRUUtvDOsAE9PquGuC9stU1XC66dBaXjGfrXeTh5ara1sbdPtYZugdPT3yg4u1/wAmIoaCK60VGZS85IaMlcz8uPKEy5W8SVlpLD04+JmF2N4N6A1Pf3tGqoyaQHI6hkYC7D2rTlHpyNsdpiDGdnNx3/Ig4q4Vcutm0NMJaigifh2QCAVzA6x2WMiX7mwAs8wwLUGnIBOxKEBwIPmg2Xq7TVu1PTupWUTT0jH0QMrr7xQFHo2wz29lqaXOyB8O/wChdso4I4i5zBglcacR9KWK8MJuIYcu3QeSPEm+VNBf5qp9OYWFxPbAWl0vFe13K0usjgx0j8tDtiQVzbzw6W07p2glltPQH9BPwronoyluczpbjhxDHkgn60Gua14dVVJWOvjZXPZK4uDQdsLVNBaZpLo9jKkhhJwckBZ8er6eugNBcXA+GMb+q421PrWssVy/6jSFrQe47IOcNVaMttgtpqGysflue4XCZo23m+NpmgNDn4/Osmg19qPV8Io5HvcAMHdRyyGxVAkk2nG49coOTKzS0WibIy5CcPJYHYBWx7vrqnuVFIzpAeMjturJdY3S7U3ut0cfAAwM+i2TdZ6SOpcyn+iTvug3Pw50TLri7yCZxcwO2C1viDR03D/NA1gBxjspuEesLfph5le8AkZ3K0PjFqmj1TcvGY8EZ8ig2vaIZ9R1R8GUxh3zwrdRWGW1EtllMh9c5WNaqiWiIdQk5AGcLLr7ualpZWu+M9soNJtID5el24CkuNU2OoELO52ylPG6KQyM7HOFDKwSVQleNwUFKhlRSxtm6yQdxutXtk7JqQyvABCPfTT04jkI2WmzTinHREcN+SDOiuPXOWMd0hp9cLPm1r7rSmid8RO3qtsNnYH9UW5KyRTU0sfiTY68bZKDTqiN9TUGozs85WsWu2t2kdnbfdYDcdfSO3kst1e+CHAOAAgyLvUQQxlrGjI27YWjwAzHqztlG1Tap5Evl6qsk0UAIZhBHXjobhqwmve1wcMrU6ICskw8bfWsqaCkiPScAoNJfVAtA6dwtSt1IySPxgcHusSemiGXDGFDBXSwPDWn4RthBqFwmfgxucSBt3WHBGSMg4WRVzRTRdTTkrFEvhRHPfyQZIp3zHBJIV3uTYskjBVLRWRiUibzO2VPXQVM9QHwjLM+SDGmkZG3AaNliuqC4HpJGVqktK0R/H3A3WmTMjz0sIz6IO+vsWXyu5pdU9byR+wCuxk//fG3IpPYtU00PNJqh72kNOgK0A/P7o25EHtGiIgIiICIiAiIgIiICIiAiIg455jz08vHFF3poy9n/kMy+aeCPxqqQnyJP519K/MkM8u3FIeuir2P+QzL5pYy9kz+jORn9KCeapDAYm4281hFzneZVZCes575WVT03iR9XljKCWx17aGrEr8EehWbqG4i5Oa+IAAei0Rw6ZOn0WdEYg0B53KDDY58bsnOAkkoe8EBSVLQB1N7FW04iLvjQZ1JVeN96c0DbCtqJnQyiJrtj6Khja9v+1QC/wBQqMp5GDrqRuNxlBqQtUToBUFwyd1p1XQyxnqYSB3UbrnUZEfVhgI2UlbXucxsbT5IMYPwAxuzx5hUe2V2DISVWCMk+I5TSOZjZBrPDy/zaM13pzWkNI2rdp+7Ul0EDnFglMEzZAwnBxnoxnBxnK9Sr97bfS1CG/cTl7utY446hWaijpgDvnBbTyZxtjtnJ7Y38mIat0L8tGd1SqqH1DwXggfNB7JcN/bC8vvEe5w6S4rcPLtpGjujm076qpkiuduZ1ED7+Q1r2szsSI3AdzgAkbX9oz7PvhlV8MLrzE8A7BQ2K4WOnN0vFttTWtoLjQYDpKiKNvwRPjb98JYA1zA8kF2CfJZ8cYiyHDK97+AFfUH2a9krtdyB8MXDOt94fL1Ee4MpZhFnqwSPdwwemBtkYKDwN93d09SNPSDn6lVlQcdJ7KkoGMjz32QalpM51bZT63Gm/wDSNXvJ7Ub8BXiZ/wCRv5Yol4JWSujtd6oLnMxz46SqiqHNbjJDHBxAztkgL3t9qN+ArxM/8jfyxRIPAZvU89IK9q/YxUVTScq2oXzx9LanXVfNCcg9TBQ0DM7Hb4mOGDg7Z7EE+Kkbuh2ey97vZeaSGjeULSlJVFrLle31GoamEDBZFUzPFM4+vXBFE4Z3wR5YJDxt5qWH9tLxjcP+H+of5RnXK/sxJG/t5uGjMb/9Wf5HrVxNzVSFvNJxj9P2f6h/lGdcvezAha7ne4bVHp92Pz2itCDtT7Y3mH4haTqtN8BNNXF1s0/qK0uu95lp3ubPXN8Z8TaZ5BGIR0Fzm/viQDs3B8raS711ruVLdrRWS0dZRTMqKaeF5a+KVjg5j2kbgggEEbggL0E9te3q4+aEB/4Ij9dqF58Np4mt6s7oPoW4x8d9V6J5Lq3j7YoKRuoXaTt91gEzOuKKpq2wDq6ezgwzlwB2JaAQRkLwH1pqq9avv1fqnVF3qbrd7pUPqq2sqpC+WeVxyXOJ3JJ+wDAGAAF7V8zbiz2WVU4eWgdO/pol4YiXx5Whx2Jwd0Hsv7GbWt5vvADVGk7jVSTUWmtRkW8PdnwIp4WPfE0eTRIHvx6yOK8qOOmnINL8duI+lKaONkVn1bd7fG2P6DWw1ksYDdhsA0Y2G3kF6kexfpoafhRxB8Eg9V/pi76/dgvNDmic9vNLxkeGkga/1D/KM6DjGekkawuPZY8RcMnDtllT1vit8Mbb9llUdNC6AulwCAg0mSsAcR0jI81uSz6diulE6pcQCBlbfmhgEjzsQOyyqe711LCYaQno7FBg1bW0lY+EbhjsbLUYbI2ugM/VghucZWkv8SScyVGcu3JWU2uq2tMVNnpOx9EEFPRmarNL1bA4WbW2J1Kwua4lYLnT08njA4edyVqFPdTNEWVB+QQaZDGXyeH2wlVCYnAE5KklPhyGRnbKhL3zSAuyRkILY3ujdnBwVWSTxHh3otQqRTe7ANI6seSwqdjXZ6hsEEwnbOzww3BWPKwwnGc5UspZHuwAHChLzI4dR7FBdBOYTnGFqMde1+HO8liyQx+CD5rDDiAQCg1OvubJ4/DaMEbEhYtKB0OLhnI2WP4TwM42WbStHT0oMenj8SoDD2JW/NG0sVLfIJy0ODcHC2lHSPYQ+Mb91ujR1V4N+gNWcRgjOe2EHqlyQaosVYKeiqrawdLgOosC9DPCtsMAn8CAMa3OekbBeW/L1xN0Vo60wVDJIhOMH6QG67WUfNLYKi2db5mFnRh3x+QCDmW5cW7RbK99JKGhjD5rqBzRW23a8mlutFcWxAEuwH4W3+LfNFolssvudRGZ+k9njOV0t4kcy1/uFbNR2yZ5icT2d5ZQbyu+tItHVApCBUFu3rlarpzh7VcZZ2y09E6Pq32GFx3wybFrarimvW5e8Zyc+fzXo7y9cIoKaOlqbXE3wukOJA+SDjHg3yXXKOdkskL2hpBJOey5k1poCg0VYn2qeFplLekE98rsxR1Vu03QGJzmCQDcbDdcBcYLRqTU87rxTMcaSJ3U4jtgIPObmB4UXh1dNc6aWSOJ2XAAkALqtLqHUOlri6njucoAfj6R9V6D8x/E7SdDZH2MyxCtZGWkbZzhedmpnR3KqqKlozl5I/Kg5Gt/ErUlNQxzC/SAgg4612k4Ec/s/DahFHcbn4vS3pIe4HP5V531FyqmfeA4ho8ljsljdkyhxJ88oPbHhb7RC1a3uDaeWtiAe8NG42XcTSnEDT2pLfHVQ3Wm63tBLesDyXzW6O1rU6Uqm1FF1ghwcN1z9pPnR1zZpooIZ5hEwAfTOMIPe+C92qqlMUFdE9w8g4LKknazAByT6bryl4M861VUvidc6k9ZIz1PXc3SHOVw2np4aa73COKUtHxdYzn7UHZAHqG4VHRxuGHMBHzGVxZBzLcLJ4xKy9N6T23H+tbn0/xS0lqVzWWqvEpd2wR/rQbjltNrnyJrdTPzscxA5C6pcxHJRZeK9ZJXUFvijJd1BrQAu3DSHAOHmqoPJ3Xfs6JYYnU4oSBGw74OF0p4zcALtw1qJ4IbdI8NcRnBOF9B2vrbV19rPueOvBHbddZdc8slNrOz1dVd4mmXBOS0IPFrhtZK5t6ifPTvYA8HBGF6kcrnMdb9EPpdMS2Vsz5YxH19GcH61wXqflaudJqQ09ihdtJj4WfNdyeWHlctVBTxXbVNMTUxMBaHDcn7UHbrT11jvVnprpFEI21DA4NxjC1JY1voILbSR0VMMRRDDR6BZKAiLSL1qO32aB8lTIAWg4HzQbQ412mivGl5Yahzchrts+WF5KcxUlt04a2kjawkvIBH1rvJzD8wEFljnijmwwNOPiXlrxv4n0+sKyoDHhzi8+efNBxBVyNF0dUtcACcjBWkXnrqZesOzg5zlZMluuNTIJGNJaVDPTSwMLZhglBgvL5IvD6uw7KCJ01NJtn8iHxYpcu2Ge61SlkopGZeQT80EtJfXUjB17+e4WqP1pTvpTC5gycjstt3E0xBER7LCgja879gguqZ/GqXTtGMnK1SkvkcMXQ5u5Cx2xQSDpZgkd1izUpY8j17IL6uX3l/W3sT5LdFj07DNb3VDyA4AlbdpY2RtBk7BZR1BU048CnJ6O3dBjVtU6GpkhHZriPrWBLJ4hWbWGKZnin6Z3WnIPV32GxJt3GUek1g/RXr1LXln7DX/c7jL/hrB/0a9epiAiIg4M53adlXyta8p34AfS0o/JWQH+ZeEFrskdO6qLCCPEPb617sc9bKuXlQ4gMof7OaWk6P45Bn82V4SaZFzpqWtdcQeoSHv65QabqCijaMRnpJ2yCoLfpXx6J9U6YFzRnBK0++3SZ9UYwdsrWNP6V1ldKcz0TJDT4ycZxhBoEE89JWiFxJDHfzrn7Q/Hym0zZDa3AB5Z0/bhcSy2EUzyyqGJh9IY3ytEqoaSOqDXA5zjug5aNYeIFzlnmn+CRxOM+q0DUPDqKhqOiEg9W+y0e21ddRQNdaw7Jx2K3Laqq9V0zHXAE4Pmg2jedOyWSATAHAGVi26+RTwGmcwEnbcLf+umRutvS4DPT2XGNvjp4JDI7AAKClfZix5q8kB3ZYPujZASSCR2Wt3mtFbT+BSOyQPJbXhlqqKoaypJDfmgVTHw5ABA9Vj08Je8PccgFa9c30dRRjwSC9aVS0NU8FrAST2CCtbMPBDG4yFpu5O+VrIsNY0+JMwgFYNVQyRuJaNggiipjJ5+Stmh8M4U1NT1UpLYwVZVwywvDJcg/NBj4ymMLLt1vluEvhxAkhSXK1VNvfiZpCDAyQt12m9wU1vMErQT0rbELWudgqd8UgblmcDzQVmaKure9uACcqJ8D4ngAnurWuka7DMqZkm4Mx3QJw4xAnurrdO2OdjXAYJwpHyQuZ0kjssOTpY8Oj7hBua8RxPoQ6MgEjdbVGScb91kuuE72iNztvrUbOkOB+aCr4PDYH5UbXOLxudysiolD48fJY8JaJGl3YFBr1FZ23GMNdsR2Kw7hQfcqUBjiVmS3L3emApDlxG6xYXT1p6qrf60GJPWvlZ047K+luAp8te3Pzws2enpWNw0DPplY3u8MgLG4Lj2QKq5MmaBGME+i3NZLL75bXTF+HBuRutpm3TwESTNwwH8q1u13uancI43HwhsfqQYho6uGtcyV5LQcblahJZjVQFrX4yPVZlyEV0g/6n4M3njvlaPHUXChLqWYkPOwGEG8OGGs6fhbeX19TEJQ4ggEZWs8SuJlPxLl95pKcR48gMYWzrNou4X1/i1bXFhOfqW6DoZlmg6o2HpxvlBs61Wx75yXg7K+quP3JuDCBuCtXbXUdHK6MkBx2+1aBdKGW41gqWg+GDnI7YQbmq76/UNAKVxJAbjdbMjsbortHATsXb/lW5bdPQUsQjjI6xgbrFrIKk1Yro27NOQUHI9FaHWm1w1FPKQQA7YrmLgTxsn07qmmirZS5gIBydu64BsmtaepjbRVTxtgFbltl807QVzarLA9pB7hB7UcP+P2m63TFE5kkUby0A7gZXMOn77SX6jbVU8zX5AJxjZeMujuLlyuMVNQ2V7yA4AdLj6r0E5aNfXI0cVDdSepzB9IoO1iKKml8aBko7OGVKgoey2nqzTD7rSyFrjkZPdbtUFaHupntZ3IwEHmhzl8O6usjnZ0ueGtO3ddG6YnTlvnoH0paS4jJH1r1e5k7XTUsVTPc8BhYTv6Lzh4m1+kWif3d0fWHnthB121CKmGrdURylocTsFh0Nm+64Jmdknz7rP1HPHPVOEZHRvhYcNRVUlI6Sn8vRBqdFJT6HzUEtd8gtrak1W+8XBtVHs0HOPVaZX3mquMpjq3HpyRj0UMNG2WUBuS38qDfWmKpupx7i8hnQMZ7LQdY2wWSt8BknXk985UIqptPsE1ESHu7n0KxZWXa/wAnvk4Lh6/JBhzTVcbQ5kjmg+hUTZp5CS+RzvrOVm10bRGIG/TA7KGloZH7Ebk7INSsdSKckv3B9VDc2OrK4OYcNyFa2lmpfidnAVonD34afiQa4II4qUHqGQPVbfr6lzJSBnf8yyp31kUYMhPQoP8Aa87SDguI2+tBitrJHHHUfyrNjPjNDCckhYMlG+Bxe4HpG4KR1Xhu2PmgyJYDRHxM5BOcJFJ7ych2FWeV1ZH0Dc4wsPw6inJABx8kGrxU7SMl2/1rGrGANLQ4HPoViuq52Nxk5UXvUjnjrORlBPSUjnPJ3UlVby0F/UVLT1UMbe+4WPV1skri1pOD80FtDP7s85VlbO6WbqDjhWCN7CXv2yrCQ6QE9soJRUlwDTlV91dI0vA2CsmEQA6MZHdZ1LPEabw8jqOwQaaS6N2CchXsJnkEfYEqskL2yFzxgEoAAeqPuEGuUeng5gmEmD37q+a5x24eAfiIBGRutJZdq6FvTkgKJ1PVVhM5aTlBPPczM842BU9Pb2vjNQ52cb91pMkMkJw4YIU0ddMxvhg/D6IPQb2M1ayo5odUU4b9DQVafyXG3j+dFi+xbDP20mqXjudAVv8AKNuRB7RoiICIiAiIgIiICIiAiIgIiIOOeZE45d+KR9NF3s/8hmXzUU5HvEhPY5xn619K/Mj+DvxRz/wLvf6jMvmiw8zvDO+T+lBbN/ZTj1WTTVohjLCPkrG07mnqkHfurJmNJyzCChIklLwD6rUIqDx2eITjzWFRgeKerZZNVVPhwyI4CCQ0wkPhZysWqojTnYkJSVZZL1POVkVlUybB2QYtFVOpZhIGlwzuFn1dYa0ZDC3AU9gjoJJg2pIx9S1C/C1wACmLc/LCDa8jTnAGT8lNBRyPb1yNJGM7rIgfTufl5H1rW6KKmkpjnGQEG3+kSEws79tlQ2yfAzndZlHCxlxPXjpz3WZdpy0jwBnHog5u5FOA/DHj7x3bwy4tXmpt1uuFnrHULqarZTzSV46BExheCHOAL3BuDksxgjIPZjiD7E/inBcpDwv4v6UudvdIegX6GpoJo2EdiYGThxBwMgNB74HZecJq6l7w6MuY9pBBaSCCDkEHyK5e0jzf81WhbeLbpzj1rWmo4wGxwT3WSpjiA8mNmLgwfJoA3Pqg7ucGvYq6gg1FR3Hj1xPs01nppBJUWrTQnkfWAOyIzUzMiMTSMZIYXYJAIOHDeXtMudHQGieFlVyr8Fq6hq7lc6SO1XeW2va6ks9tYA00bXNJb4r2tEZYMhkZeDhxbjzu1Xzg8zGv6N1p1lxt1dcaGVhjlpPulJDBK05yHxxlrX9z9IH8wXFtU/3x4fIMYQae6me2PxD2xlWB+x6lqNS5gh6GEZxhae2NznjI2KCaho310/hR7Er3/wCYOhPNzy1cYuDei/AOrLTXzWmS2yTCJ4qqKtZUUuS76LZ2QROa44aS8jOGkjwQppYraBUR4Lsdl379pp/srctXNmzixwo1dedMR8QLXFWtqrZUugElTThsFRC8A4kGGwykOBBMwOMhBwvwp9nvxquuo5bxx20zW8MeHWnXe96m1BfemmEVIwgvZTtJLpZHgFrCwObkjJJIDu/ns5eZ+w8WOKnFvSUFPFZaGoqaKt0daSQwQ2akp20TIGNJyDHDDTOc0ZGZHkdiT5V6/wCYrjxxjbBRcVeKWo9RUVMeuKkrKx3uzX/3XhNwwu/7YgnHmtrW3WuodIXui1FpK9V1outukEtJXUNQ6CeB4GA5j2kFpwSMgjYkeaDvzqblg1/wu51eKHMDxL0mKXhfYavUesp7zXtjNHWMq4ah1NTRZyJJnT1MUYYAXAgnGcA8H+y5qh+3e4bQZ3d92fzWitK4d4n8zvH3jTbILFxQ4rahv9sgkEzKKpqiKcyDs8xNAa5wycEgkZOCMlbG0tqzVOgtQ0WrNE6kuVhvVvc51JcLdVPpqiEuaWu6JGEOALHOaQDghxByCQQ79+2xOOPehCDuNIj9cqF52uleduo4W5+IHE/iNxXu8eoOJmub5qm4wQimhqbrXSVL4ogSRGwvJ6W5JOBgZcTjJJO1UHuzzJU7qv2Ws1Oz6UmgdOgfloivDius1RQfE879xst7T8xPG646Lh4cXrixq24aUp4ooY7NU3ieSkbFEQYo/Cc4tLGFrS1pBDeluAMDGy7je5a8AOb29UHrd7EqSSThPxH8RxONRUw38v8AawXnHzT1EUfM/wAY2Oxk6+1Dn/OM62hw942cX+ErK6HhhxO1NpWK5498itNzmpmTkAhrntY4AuAJAcRkAkAjJztK5XO43m41V3u9fUVtfWzPqaqqqJXSyzyvcXPke9xJc5ziSXEkkkkkkoLBKBP1HtlSy1Uj8MieRnY4Kwuoox5YQR5IJ5YZ2sLy4keazbZKxhDHgb+ah8bqiw8bFQ+KGPDmnt2QazdLc0weKzGSM7LAoJGRRnrG6miufiR+HIdu3dYdQ4OlHhdigtqqnrkIDRjPooImufIGjO53WrMpoXQgkDqx5qF7YoB5Zx6IFTSNZTh2d1p8b8beZ2Uk1ZJJ8OfhVkdPJIOpoyAgudTvx1dWx+asDzHsFkSyhrOnzCxASXAlAJJO5KN3cFkAR9AVzBH5Yz5IJpoCKfqz5ZWnDY59FqQZM0ffAeg/oWNJEHTZYAR5oLTUfB09GB5bJFO9rxhu2fRZtTFTupg2PHWprZFSsgd7xgPQBcGRRAkbqtPdC5/Ux3S4dj2Wm1bT4p6foZ2WRS2yomb4sIJaEG8rPrO9UYa2K4vYAe3UuTLZxMvRsj4Dd3AlmAepcBCdsLvDfnqaVltvckERjaT29UGRqW732e6yzzXKV+XEg9fZYtmulTJc4WVEhf1vDclaeKp085dLuD5KRssEFUydpwWu6kHZq1VkOjtPwXaKcBxId3HyXoZyT8w1qu1tht9bVxhxYGguPbZeRp1XcL/bmWlhJDcDY+S5U4W8VanhfAMOcx47bkboPc640Niv0Tq37uxMB+LHWFxlxT4xaU0HoG42kXCCWZzHNDuoE5wV5Uam559fU0LobZUy9J2GJD2XFV95l9a6vglhu00ruvJyXkjdBpvH3Ulxv/EWsuEVwe+CSQlo69gMrYDK57GEOJJOck75WPebnU3CsdUyl2ST3V1DJTmE+L3QYc9S2R/UWhY5yewOFLL0eOentlZ9OKXwsvI27ZQafA1z3ho2Wex00J6Wv3PbChcGmUiDzOy1CGB3gl8g3wgybfertZ3ePDXOb54B7Kyp11quprRNHe6lhBHT0yEALRxL11BjlPwZIV8tMzrDoRt3Qcjs4wazt1tjYL/MXAD9+V2L5bua682S600dzvTyBgHqeun1DFTS4ZVAYCkluUFonEtuJ6wdsIPd3S3OBYaiw0001yge/ABJIyVzbw94n2fW9KJoK2IvIyBkDK+ejTXGDU9GI4ZHS+EHDGXHC7ZcEeb6bTUkFNNUEENAILkHs51Qy7BzH/nUVRR089PJA6Jpa9pBGF1L4Oc3Vgr42PvdS0RyYGS8ZGfrXaLTGrbPq2j99s1QJYsA5BB7/Ug27a+GVkpbw+4z26N+T1btzut4+5U9OwGmhbH0/wBztsstUIzsgxIqsl/hnfHmsvIwMnCjFPG09QG623qu6TUMZEJOewQbn62f/NB+ULj3ilY6KutMtS65MhLcnBeAth3jiXcNNSyT3B5EQGck4H510u5nObi/wU1TTWSZ/QCR8Lv9SDj/AJ0K/wBx8eCiuweQ0jZ+SvPuCWqnqJKiaZzsOJOTnJyt8a24wah1tdZPutI8tcS0guJK2bPNBBlrMDrO6DWKfV0NNF4LmAkbdlot0vT66UujZgA+ixW0MlY//a4y4+S3HbrbQUdIWVoAlIOAfVBhWmgbeB4bgAVh3yzPtMga07LOoXzW6udLF/Yy7Y/JL3UPuXx5yQg0OGkdLu5351SUe7ktHdXMM0Enxk4V73xyvBcQgghkfA4SYOCtxUVDHXweMcAgZWnOFNJGGtxkJHcJKKMxx9jsgtrYvvhiaQMHGyxxD4TTk5UU9RI5/Xnc/NSQRVE46xnAQYz3u6iD2z2VpGQSsp9P4pDGD4h3UMsMkILZBgnCD1X9hr/udxl/w1g/6NevUxeWfsNf9zuMv+GsH/Rr16mICIiDgznducdm5Wde3OUAsgpaYn7ayAfzrwIq9ctuHvLIGdIe4uxjHmveD2hQzyccSR60lH+vU6+frT8MMoqDJjIaAMoMqmpvuhVCRxzggn8q5w0nxStOlLC+1yQMc8s6RkAnsuDKaCrhqiYs4JWr09vlqqhvjg790G9La9mrr5LUNaGse4kD0Wj6y0h9zqoPYcgnyW4bFQvt0IfRjDvkotRuq6mme+pJ6gD5INJ0vUw0zmxSlpI23W47nqSlo4w+MNBA8lsLTWnr9froYbe17sHAwCtZ1Jw41ZbAH1sUgb3PdBo1+1m65OMPUcAELQmdUmY2n6XzUNzom0j+gg+IO6yba3qhLjjrHbKDIoqc21xnkd1h2+5ytOur23GoBYA3dZkUdVLI4TZ6M7fUsO40s0LvEhzgfLsgs9wfSxh5dkei3PYYoHRiaQDbyW1qKpkqn+DI7O+FqzG1seIIM77bINUv19p44zDG0Ejbt3W3WVEdTG4uGD9SzKux1rGmeqaQDutHma5m0QwAPIoMikr4qCQkj5JX9FzkEzcAALSXNe92DnurmzyQnpBOAg1vTtXHaKp0soBGfrTU96iuUoMTRgfJaUWy1bQIck53V7bdMxp8UEO8hhBhta4ODmgkla7SwF9GXPaRt5qOz0jI5eqrx0gjutZrauga3wacjfZBtKPDKohw2yVmy28TsMrTuBlWV9vlDvGjGx3UdLVyMxFITgnCDCcHteWnORsqOaW9ytRq4AWCRgGSsEwyndyCNMn1VTjGPNUQVwfVZcFD4zcl26xot3YKmM8kZAYTjCC9zH0p2+ILLpKt0/wdGCdhsprNA2rcBPghahJQQUlaxzQOgdyAgxZrc8M8R+QMLDghkjqRIAXNB3K3Bqa40Zt7Y6UjrA3VliZTPtT5KnBeGkjPdBWobDdKUQMADgPLutAdTupJjRnYu2Cy7Gamquj44MludsLKvNvlgrmvlBDgg1bTtrfZ8VkoLw4Zx3WbUWoXevZWtiwBv2U9jmZJAxlWR0j1W5I3UsceKbGMboJI9R01jomRMgBcBjss5+oaa8Wp4dhriOxWHbNPi+yOYW+Xotp6rp63TlwFLHkNLsHGUG07/SVDbi+VpPTklanZ6+N1MaWQAuIwp7pJS+6iSUgFwyVolFR1VTJ4lKCQN8hBkXG1y0bjVMc4gnqwCrqXUYkgNG9uCds4WpOZJ4AZWAgAY3WiV0NIxpkpwOofpQWmyzwyGtimJ6t8A9liyi5vlJEjx9qztPVk8lR4VYT4YO2VuqSC2Fwc3pxj5IOUuV6YQ3BhuUow14Pxf+1dvtO8a5bLr+itNA8mIuDSW9sZC8/6LVZ066N1rcQ4uA2PzXbnltgtWsLjBX3LDqsYLcnfKD1q0heW3HSNDcusEvhBO/mtcpp/HZ1EYXGXCikucNogpKkPEAA6c5xhcoRxMiGGDAQXqhIA3VVo+qp6mmss89LnrYM7IOsPPz4dNoY1lJIBO+IghuM7LyTk0dc73R1NxqJnjDyfiz6r0H5heJM1zq6mz6gf94jBaOo7YXTXXOpLRRWqogtD2dLiexCDrrfG+51j6XqyWEjutPfemxUroT3PZY9yuDqq9SyP7ElaXcXM68N80EDh7xM5w2yVm087aXHVuVp7Q9m4yqPe4ncoNakqG1jQMjZZ0Go4bfSuoxGCSMZAW2WzvYPhKyY4PeIzI7OceqCyWpL6gznJBOVm090ZGQ/AGPktJeOlxb6IMk4HmeyDWqm9Nqm9AG/1LDpmFtQJXO2zkrFdTyxjrcNgqCd4GMn8qDX7tcYp6URNxkLQqbqErXZ2ByrH+JjLgcFWh5GyDW6udlTTCNoGcYWlvo3sHVuprfM1snx9shZ9XUUzoz0kZQaXTTe7vy4rLkqY5G5xhabIepxI7ZVOt2MIMjqErukD5Kx1Oc47KWjDGO6n9lfLI0zjp7HzQYskT4wM5wfmp6eDLfFdvhTVjWeFkd8LCbO9regE4KCaeobICwDGFjs3OPNUw4b+qoCQcoJ3U/QOouykcb2kSDcNOVGZnkYJOFm0ssIb0vwgskqPeW+HjBCsYzwjlxz9akqDCz4o8ZWKXSPGfIIJp5mHAA7fJZdLdY4IvDLVprI3yn4RlPDLHhr9kE9XUidx6W4yVY2mLmF+cYWVG2nDN8ZVHyNblreyDvh7FYn9tPqkHy4f138o25FL7FvwzzSap6cZ/YBW5/zjbkQe0aIiAiIgIiICIiAiIgIiICIiDjjmS35duKX8Cr5+ozL5p6eQRVLnH5j86+ljmS/B24pfwLvf6jMvmhcCZXADu4/pQZ1VO18fw91hQu3I9VJ7rO1oe4bKrQwAHzQSMhBy/GEkhDsZ3R8/SzDSscPlduM4HogkFKXnDRv8k90ka7DgQCsy2VUDZA2U4381n1r6edoFPjJ7YQYNPbXNHU2TB77KSa2STML3SZx81B7vcYj1uB6f5lksrXiMtz9iDTKemLqnws4we61LqkpXiJrzg7FadMZWS+IzvnyVfGkeQ52c/NBqlVTPdCJWHDu6ybbSGSBxqNz5ZWA2tkMfSzv6K+GvniYWvGMoLY5IaetIeBgHzUtfPBOMQ4+xadOfEkL98lUYHM/+wgvpaUyTZxsFLXn3f4RjJCsZOYQS3uopZDUSDxNsoKU7JJXBzuynmDIxgAZU0TY2MGHdgoZBJK8ANyPVBbQVk1uuNJchEJRSzsnDH9nFrgcH5HGF6k+0z5muWDjnwB0g7Qeu7fftTtvMNwoqamyamipn08gnbUNIxFkmIFpIcXNaRkAleX1TCxlP5ZWjuOSg1m5V9O9hEGMkeSwKeQOaes5+tYmD6K5ryzZBdI4dZx2VvVkjZUJJOSqIJ2ffT0juqSRGPuVHG8sd1NV8kviuBKCzbPdCR6KZ0bGsDtsrHO6ASSg3RVBH0sIJmUjnt6gdlG6MxuAKkFW9owFaXGQ9Z7+qDIl3h+xYZBAyVIJHv+EDKNyX4OeyBBA6c4acLIMb6Y4c0nKutgxU9J7LWqw0YjBfgnHdBogqSzJJ8s4WNPO6V2fIKaoaJZOmLcZV0ULYmHxsAkbZQYS1q2Fgpj14BHqsCjYx1QQ7GFPXvMRDITsUGFUn7+4eWVERg4V7mvPxHuVNHQVErS9jcgIMbJ9VfHkODidhuqvZ4eWOG47qjWPf2QZ0twEsIgaMEbKKF3hsIO5KxnRvjwXDCNEjxlvkgkhe5suSTjPqppY3SvDmuwB81FTRudJghTykxODfU+SCOokHQGY3C1Wy3yOlgNK9o+LbJWnOh629XqsN7DG8YQazX2xshNW14w7daS9j999gsiK4zOxHIctV8ga5pczcFBp2SD6LW7bYDXwmYvx5jdaXBD4svSNt/Naw24zW+Dw4z8kE1rlbp2tL5HAhX326zXyQSU8pa1v2LQJ6iatkydz3V0T5KVpbuCUGoU8U0gEckvUQlVGabIBGPPbyWn09VJ4uQTusqpkdK3fzQYrpBOekrHkaY3YBKqWSROzhWkueQPM7IKFpG6mjp5XsLgCMLKprfKG+I/t5bLUY/AYADjHnsg0SF7qeTLmn7Vqor2yQkDuFHXOpHMIZjJC02B+JWg9icFBZIXGQuwe/dZENT0M33OFrNfTUgoA9mOshbdj3cB5ZQZbJpKh3S3O/5lDKx8EoLiTg5WqUYpwwdOOrCwLl/ZAgzReB4IiYAD9SrQm5e8Cqhq3N6TnZy0VanQ1oib0u7IOX9F65vk8cFCy8PiLJAD8ePNes/JZxDbR2CK1190E7pImjLnZwcBeHzrtPQSMlo3lrgc5yu5HKbzBuszIqa4zHxQ0BvxeaD3Bpqunq4hJBMx4PoQd1OurvL1xVuOr6lge5xhL/AIcuXaJAWh3yy+/Ay4zjfC1xUI6hjyKDohzs6nuelrJMLfC8EREZa35FeRmqeKl9u89VDW9ZzIR8X1n1X0A8e+C9p4j6flilhDpS0t3Gc5C8peNPItfqK5TvtFI8B8hx0sPmUHSItlq5jUuGM7pPQOLS/qycbBcxa45cdY6ApPerlA9rA3qyQQuKYIpnVRa7PSx2HflQY9ouTrXP1yRktzvkJeLq641ImjJDR2wcLX7pBaXUI6OnxMeS2x0RMDuw9EGTJdQ6mEO+QMZwrqKqyMPd3WlOGDt2UkLi0bd0GpVzWvYXMxkrScHOPP61nML8fGCAfUqGSme9xdGMoJqamLcvLs58lFWbO2KoZJWN6N8hQlznuAcgq374A3C1GGvbSwmItySFHBC1rQ491HVQdWXtHZBfRSvbUGVrC7J7BUucrpn9To+n7MLWdFS2xlSRcS0Nz5rI1pHbpJXS20joAJQelvsNgRbuMpI2M1g3+yvXqWvMP2ITYRpzis6PHWZ7J1/krcL08QEREHXn2gmBye8R8/3pR/r1Ovnwsok8aXw2l2wzj619BntCGudydcSGt7mjo8fx6nXgXoee3QOqPfiN8BqDLpp3RYzCSfXC16lnjLOstwQM4WZTTWFzi5xbg/UsSrEMlUBRgeGTvj0QZlBql9HL0PYensFvOjoafUVKXuwOpu62x9xqSppW+GAZBvt6q6G+zWSI0jCQ/sN0G8NI3G2cPLgamZjH4Od8Fbq1HxRs2sqc08dPG0uGMgBcJ1EF6u0hlma4xk57q91LU2mldKxpBAQZuodBUs8z66NwORnGVxxdi+z1BY1mRnstbh15XMq3U1W8hgOMH0WPcpKG6PExwQd8oMGlu5qWhrYjkjHZTSO8Rhiczc/JZlnFpgfiTAIS6PpnTh9PjpByg0c2z3B/vGNjusmkvsMUwkeB8O+Cq3O5QPp/DJGcY2W2QJJZcMBxnvnyQbwvus466l93ijGe2cLQqctdA5z8ZO6yaShpGRB8wGfmVplxnAm8OnPw5xhBdSRtqKgscNgr6i3NNSI29io301RBEJ2A5O6vpa1wIknJDgg12jtUNrgFQ/ByM+q0+tuEc1QOjAAUNbepZovC6tgtJ8bDuo57oNbuD2upwYjg4WhtdK2QOLicHOcqY1bnjBJVW9BYer6RQarFcI54BEQCfPZY09C0jxW7EbrAgL4XlzgceSvlrpHS4Dvhz2QXeOWHoIzjZSgtkZjCpmAs3IJ81jSzdGzcoL2UYc85UL4gKgReWcbK0VMoz8RVPvjj4oBJBQanU2llNTiYOyS3KxKePryTjbZWSV9RJGI3u2CyKWeOOM9XfugzKZ7aUdWcEK6qrnyxkDJOO60meqc93w5wFq1A6mNIXSYLsINJhbI+YeI4nHffK1qITGLwo2kAhaO6ZsdQSO2fzLfFifbn0fiSkdQCDQ7VV/sfnFQ+PdxytyB8OosVewI3WkXKSgq5DG3BAOBhQU0tTROEVPnoPzQaldZzSM6IzjCwaHU88JLHPOfrWQ9pmbmfv9a0p1NA6pEbAMuOEHLvC/U8Hjn3lwAPqsbioYq6o95pmBwBzkLjyunq9PU8c9PlucLeWmdTWy9WZ8Ve4GfpIwe+UGzaS3yX+YUpJHT5ZWvRQU+mWeFIASAsajY613OWpYMRkkj6lpGqdQMqpjh3Y+SDVKmqiu2WtcGjGy2ldA+3z9Gc7qtJc5GuzETuj546iraaog7oMq29VRjbpJWtPo5GxEiQ7BYU01JBTA02zsKkF1xTvdK7AAQbi0fpt14rYo3vzl4A/Ku/fK5wtrKK+UlVHE4xgA/LyXRzhDbLnqu8U8FjLi/xQCAD6r2V5SOF9bZ7NBU36PEoiBGRvnCDsnpqCOGyUcbYw3oiDTtvkLPfN0uDVdDCyGMRMGABhHmJvxOwgGVrWF7iAAMrhjivxmptO0s1v8RoL8tyt56711btLU0r6l4A6Cd9vJdKON/EqyXaCWtjeCWOJz1BB145stTe/wAk9bTVQaXgnY4XSp9/qpKeWKWqL8uIxlcx8Z9ZSarq5aGgJOAW4BXXqaKe1yugq8glxQYdaXsndNjuSsTrdLICStWmfBOzAxn0WlSxmJ/w9kE7y2Nu3osV7g45R0jnDBVG4yM9kFBsQVmRVfS3oA2Oyx5AwNGO6jAPcIJZh1O6h5qtOwiRrnDsVLROZ1/fOykrHxDHh4+pBJW1DHwiNo37ZUlFZ2zQ+O5w23xlaU55ccnyKyYa6rjjMcRIBQSVbWBxjHlsNlgluHdKudJI5xc7OT3Vud8oLzG5oyFTpkJ3BU1PI3bxO2Vmu92MJ6SMoNMdEWgEd1aDgjPksiCRrpD4hGFSpY3r6mYIKCJ0mRgHCtaSCHZ7FU7K9rQW5QSGUyDpJURHQ4KgPSdlIzDu6Cxz8gDCtUsjQAcKJAG5AWYyh6ojIHY2WNHC+Q/AFP4VYB0AHBQYzsglpPbZZsIb4R2WNLTyxfE8YVGyP2HkglilFO8kjKsqJRO/qGyO6XDfurWwPf8ARGQgs6iPNSRlzx0/NUkgkj3eMLKp2N8IuPfCDvx7Fyn8Pmi1Q/PfQFaP+cbcip7F2Vz+aXVIPloGu/lG3Ig9o0REBERAREQEREBERAREQEREHHPMf+DxxS/gXe/1GZfNF1BlQXEZw4/pX0u8yO/LvxSA89F3v9RmXzQlpdM5o75P6UGfJcmzQeCG4OO6097XNO5WRHQzAhzm7KOp2dhBASSMLPt0TZnCIgbnuseCjmnx4bScrIYySgIc/Y+iDVZ9LhsJnbJjzxlYNPCaWcdTiQCopL5WvZ4fX8PosVtS8yAv7FBuysr4ZKIMawZx5La7pnCXp7ZK1innhcwBxWLV0Qlk8WLsDlBdNTNZS+MG7ndadATIT5LWBUxOh92djIGFhSRsiOWY9UElMGxOLngELlTl04Aai5o+KMPCrR17tdpuEtFUVwqLj4ng9EQBIPhtccnIxthcSTS/eyM9/RdgOQfj5oflu5gqbidxEiuklnitVZROFup2zTeJK1oaQ1zmjGxyc7eiDsW32KnHkHJ4r6CP21v9CpR7Fnjt04PFTQX2Gs/oV2iHth+Up2wodef5nh/rCvPtg+U1o6jb9e4/8Dw/06Dquz2KfHVry48VtBHPzrP6FWVHsU+PMrss4raBb9Zrf6FdqYPbC8ptQ/w47dr7Pzs0I/8A36nn9r3yoU/9loddj/yPD/ToOp0fsU+PjdncWtBEfI1n9Cs2H2L/AByiZ0/7KehCfXNZ/Qrs5J7YjlLiAL7fr7B9LND/AFhXQe2E5S6gEx0OvAB3zZ4R/wDv0HVep9ivx8ncSOLGggD5E1n9Csf9xL49fjY0D+Wt/oV2sk9sZykRu6XUGvsj/wC80P8ATqz92Q5Rv7w19/maH+nQdWR7E/jyBvxX0D+Ws/oVa72JnHl3bixoEfbWf0K7Ut9sdyjuOBQa++2zQ/1hVf7Y3lIYcGh16fqs0P8AToOqf7iXx6/GzoH8tb/Qp+4l8evxs6B/LW/0K7Wt9sZyku7W/Xw+uzQ/1hXD2w/KUSAKLXm//wB54f6dB1Sb7Ezj0Dk8WNAn7a3+hVXexO49HtxY0D+Wt/oV2xf7X7lOY3rNFrsj/wADw/06g/djOUgDeg17/maH+sIOqn7ibx7xg8WtA4+us/oVT9xL48+XFnQP5az+hXar92Q5Rh/9wa+/zND/AFhSxe2G5S5mlzKDXuB62eEf/v0HU/8AcS+PX42dA/lrf6FVHsS+PIP++xoH8tb/AEK7Vu9sbykNJaaDXxI9LND/AFhB7Y3lIP8A+j9f/wCZof6wg6r/ALidx4xj/ZX0D+Ws/oVafYm8eT/2WNAj7az+hXase2M5SCce4a+H/kaH+sIfbGcpDf8A7h16fqs0P9YQeSPMTwG1FyzcW7lwk1ZebbdLjbIKaokqbeZPAcJoWytA8RrXZAeAcjuDjK43kb4mJGDbzwud+ebjdovmK5jr9xU0BFcmWW5UlBBCLhA2Gfqhpo435Y1zgPiacbnIx2XA8T+hvQexQXRvMLfFAxndQz1ElQ7JccemVneJAYek42C0+TpbL8PZBdTyGB3U5uQppXe+fG04wkxhMIx381ixyujBx5oDXGJ5wdx5rOizOzqIzssHHiH5larQuihZ0yHBQYQeHv8ADLcAHutQjubKKIxdGcjuoql1Mz42YBPorI2QzMc54GQgwKiXxpXSYxlXwTCPAwCrJQGyEDACsGzvtQZtU4OjyBvhWU0rWNId3wksrDFjzWOzIzjKCcVIieSG+ajnqHTO6iMYUlIxkjyHrJlihY8AAb+SCOkndIfDI2CjrmdLw3tlapTx08DRKcDZYFfieTrjxgFBAaQiMPBKkif0Rlp3UjZBJH4TdyFEYnsPxDYoIoajwZi7Gd1dVVbpz6D6ldMyPp+axR9JBLBOInAkdkqJ/FOwwpX0/VGHAbqKKPpdiQYQWxSGMjZTmrzv3wpKlsPhEtxlYbI3OIwNsoJzL7x8HSBnzwrm0wjIeT233V7DFG35hQTVBc/DScIM993a6EQNbgjZYBqHk9OTucJ7segP3yVfEI2gF3dBSaleyMShxOVHDEXfF2xgrJNQH5Y47DsFGZGsOB2QJq2RzTEScDbCxg4DOFkSticwlm5/nWMWOHkgvimdE7qBKk6zVzNa491ZT+H1/Gr5i2N4fF5IM2rs4p6cTde+MrSx1H6OdlkS3ColZ4b3bYwsqgFP4REmM+SDTw8uw12+Cty6Sra223aOrgqjE1m+M4WgVQY2TLfXZVhNU1hljyWjv9SD1f8AZ/cUGXuvp7VLWBzxIB3Xp8Oy8HvZ2aqqbfxLZB1EgzNPfbyXuzb6gVNDBP8A3cbXflCDJREQWuY1ww5oI+Yytv6m0/Yp6CasqrVDK+JvV9AZOFuJRzwsqIXwSD4XjB+pB5n85V8pr3FU2W26eDehhaC2P5FeY2prZXWSSpZLROYS8nJGMbr6D+IvAPSmp6aerNO0Tlp7gblebXM1yrz0UdXXUMH3trydm7Yyg86KNk1ZUgPccHc7pdKb3aQNadity6nsTtO3V9GARI35eeVtatfNLKfEzkeqDGzthXRPDHDKr7vL09XSrOgg4Qah4rZmBowCp4pWQREO3OFp8ZMe57Kkkr3nDc90EwlbJKdgoahoZICPXsr6SB7n5A3VauCRrx1A4ygrDOX/AAK58/SfD8z3V9IYW/XjCnbY62snEkLMsJBz6BBfbtPTXA9UbizPmtSrrIbdb5Wyv6z0HzWowudb6YMbgPAWh3e8yVEL6d7iXHbug9QPYcg/cbjAc7Gexbem1cvUZeXnsP6CqpbTxfmmbhk8thLPngV2f0heoaAiIg69+0AcGcoHEZx7CkpP16nXzyUtNLUueYSR07r6FfaFO6eTjiS70o6P9ep18+lmnZD43XjdoQOupjAZ4xyDjut86fLG0JlllBd09yey2HM/xZT0euyyGXOspYDE0kAjCDdUOsRb6x7C/I8t1ot31VPVXJtRHnoBBW3PFMk4fJkkndbrtsFpmpel4b4p2Hbug3RScRKeCgbC4DqDcFQV2s466jcGgbjZbKuFlqKeQzOafC7j5qtNNT+EYwdyg0+re6srHFpxuVdHUS0uIerOdlfPTPjeZGdiqxCJw65cZCCyoEzGeK2QjPkqw3GXwi1ziT6q2rf4oDIzkBWQMjjaRLgFBjySySvJLicntlSwVPgYBaCrZGgu+BTRNi8Ih/c+vqgrUXGSZoawkLEa4teHO3wc7rVbJDTNnLqoDp8iVZfhSCYe64wguF3Y6MRdPyWJPH4jS9u2d8JFTNkA6BvhVcTCeh/ZBDSwmaTo3wsmqoBGOoHdX00kTMkEZUNZUSPcQOyDFiaS/CvkY5jwcHA81PSw+JuBv5qapYxjSH9ygxJajrZ0gY8u6x8q9vT1nP2KsnRjZBZ1u/uiqEk90UrGtLCSgiU7Jw1vT0j7VC76RVO6Cp+Ikt8ypo6Yvd09WEijLXdTu3dJJi1wLCgkqaIQM6g7KvpGufHjqOFiyVEkg6XHZXRSuYMDsgz6O0irkILytTno30FKWskP2FaG24TQnMbiCVP90pp4yJCSfrQRUHiPq8uedtyVrrq8QOA6cjtlbahe5svU3PfyWq+PC6Il/wBLCDUKi4h0Y6SMkeS0cVzoasTZ7EFRQ1Q8Uh3bySQxPnAbuD3Qbwhlj1RTCkOxAxnC0yqsU1gqOtk5DfTKxoq42aMSQHBKxqm4XG7NMjiSAg1av1D4tOIGOHV2JCw4dPyV8RqXuOe60ujdGJSJfpA4Wutub4YhHEdsboNGZTe5zlrjkA43Vs9OZ5OtjsDKkmhqq+QmEEklXxQTUg6KgYJ9UGNM+SBoy4nHkVrGiLVLqq/wWMDIqSG/nWmzeE8t6yNyuS+BVgfPrqjqKckljwRt80Hdngfy8RcHLVRatqaMSiRwk+IZ+a9IuCWpqfU1jbUQ0wi6IwMAYXBmgNHXPV2g6CjqwXMa0Dt8guxfDHSEWk7QKSNuDgDsg3qtAuVwjjrWQPlDQXY3WvriXirHeaSJ9zog7piOThBxnzi5odOmpoa7DnRnIa7ft8l0bqrNJdtD1tbVXD74ASMu81ynx241Oq3TWm8SkhjSMErp7rHiTeGUc9LanO91cSD0nbCDheqv8tk1nVQ1Dy9geQCdwtH1U+K+VHvUOBjPZR3+ppq+ufUEgzO3PrlaUKt0A6HbZQQUtM4SkF3Yqapp2Bp2yVC+fqJMfdWNnf1YlPdBiOGHFAMkD1WVUeD4eW4yfRY7Y3kdYHZBdJD0NzlWBwAwjnucMHy2QRvPYIKZ32KplCMIgd1k08wB6HADOyut3hGQ+L2VK4RiUeCgvqIGsYXj61hZ3yp3uke0A5wsmCGPw8uxnCCOhpffH+H28u6z57T7uz6WVhjxInE0/f1Cjnrqp2WSE/agx5R0PLc9lVsuG4O6sc4uOSroQDIM9kFpyTnB/IrmvwMFa3FBSGHqdjK0mtDBL8HZBjndASDsiILnP6grVczGd1V/T5IJqaqbBgkZwssXZmfofmWlgE9lmxRwiI9YGfrQXVNUys+FowVCIgxv1qIFrJMg7KRs3VIGncIIHZyc5CyqerEIGWglXVTIxHlvfCxIw3rAd280E9VV+OMdICsimLQGDzUtS2ARjoxlQU/T4rS7sN0HoB7Fpjm80mqXlvfQFb/KNuRaj7Gl9M7mZ1KIcdQ0FW5x6fdC3og9k0REBERAREQEREBERAREQEREHHPMeccvHFE+mjL2f+QzL5o+tzKhz2jJ6jt9q+lzmPaXcvHFFje7tF3sD6zQzL5s7PFD7/MypxhoI+3OEFhuxMfQ6MjZQMhFSesnC1a6R21sZ8HGceS0Fsz2ZDDgINXpqplAfiAIGyw7jWiukHSMDKxhK6TAecq13SxwLfIoMl9GIohITnssdrBK7Y4wsiWqL4OgnyWJG5zTkIMx3UxuzuylgungxmJwySO6wDK4k9RV7Yg/Ligq9z/G8bOxOVkscZo8nbGVFD0ud0kjZVqpPCIZHgZ74QQSv3IyqNm6W4xuj2OI6z5qrIWvaXDKCkUzmPDycj0WYK5jgAcLT3DBwsuipDO1zyNggy6O4U9PIXkBY9zuBq5AWOwAsOZnhyFvoqMZ1oMp9Q2SnDTuRsoIpHxu6WuwCoz8JIVASDlBnOpXSN6u5KiFC8+qNrXhobkqRlYQPiKCGWmfCM7pBTPn+LuArqioM2wUlHMYmlpGAUFSA8+E0bjZY0scsJ+IkZKyYz0TF/rurKrqmcHNGQEEcUsr3BpcSFNJSuDeoZWI1zo3dt1nw1LXxkO+rdBp5BLiMbrNpWux0bjKtgZE6YlxGMrNd4MeCxwOCgiNomeS8HAKiMLaf4JMZWQ+6yR/AAcFYcs7qh+XbIJoqE1JyzG/ZWVFI6nd0u3Knpqk07QRglW1M76kgu7oMdjC5wCnNMCwknHoomNcHbDdWzzysPSgxn5a8jKBhcOpAQXZd5lSB7c9IG527ILI4zI7p3VZo/DOFmNpXQDxyDgqN8kcmSSMoMaN3S4EhZkUJqIy/JGB5LDeB1Yb22WZTTmKIt9dkGGWu8ToyT9qkc90WysL8zF6yY3RPcOrG6DDcS45OVUMJb1LOrI4WxhzcZWNA5vSQ5BCcg4JVzX9LcfJJCCThWswXDPZBdFIY3Z9VkNBmIf1Ks3g+F8OMqlLQVdS3rhaSB6IJ3h0rOjONlFkU46HHOfVWS+LTfA7II2OVAfEmPbKDIgd4b/Ed2KlqKkPAw0D6ljeFK0DOcLIhg64ySOyDCe8kqjBlwVZG9LyFfFE5wLgNwg1SmY0xjqI+1YVd8L/AIfVRR1EjHFuSs0UxqB1oNNLyRhZNIG4ION1HUQ+ESMYUTZHN7FBk1NORlzSSc9ljxgmRoOdyO6yqKYPkDZz8KrWtjMoMGMAoM98LW0wPyWiyn4sLVPGe6EMJ8lpcoPVugsyfVVbkkDPdS01JNVv6IW5KrU0dRROAmbglBeYPCb4hJOfIqIPa9wb6lWume5vSTsrQS0gjuEGZPQiKLxQ75rFa7Iwd1JJPO5ga4nHZWRhuQSgjO6yIYnObkFJfDDfhIyrI3SAEtBwgmZT+M7pJ7LMpKlsTjQ9GRJ8OfTK00TvYdjutXo5KTwveJCPEAyPrQc08Fb1HwjrodTCpDHF4fnPovV3lp5s6XX8NLRT17XERhuCc+S8NKzUlwq4xTSSExNOwyey7UcqOqrnp6+0lZG14pmYLjk4wg94aGrZW0sdSwgh4yshcRcHeNWltZ2Gkt9FUtNbG0RlmRuVyzE17W4eQSgkREQQ1DWOiIe3IIxhcT8YdE2y9aPrG/c1r5HNPlv2K5dc0OGCsO426Ovo30j8dLhjsg8D+ZLhhUWHV1TVmgMUfUT22xldepLYJ+uUNB6SV65c+XAiSa0z3S2x7mMuyGryokoXad96pbo5weHkAEfNBs6pnbE4w9I223WIZOo5AWZdWQmYyscSHbrDhfE146wceaCr3ktx0kfYrY5OjOcFahV+6imEkZySAtMa1zs4QZcFaIjnAVZ6wVBx09wsXo6iA3crKpo42tLX7OPkgh8MxkSZPfK3Ja9XxUNN4TosnGFoFRTTt+NwPR5KKnhbLMGk/D5oNdZd5a+oLmtJaXLU4dJNquqtkeAGguIKyLFQ25rB1AAhafqC9VdNKaWjcQwjGPkg9WfY0VEEmmeJdPCBmGa0BxHnkVmP0FekC8xfYjwVcdg4szVQIEs9kLc/IVuf0henSAiIg65e0Q25NOJeO/udH+vU6+eqlaXuIBxsvod9oBTmq5QOI0DRu6kpPzV1Of5l88kB8KR7T3Hw/nQZNGzpkPVusir6S3pHmsWBspcX42PYqlXLgjOcoLJqQNHUFFTzPgnY8OI6SD3VWSSzO6BurXxOY/pd3QbruGoYa63NpmgdQGDstApaU5MnWfqUMdLMz4iDhTBlQW5ZnAQTVNaxrDHgHyzhaa+R0mcHzV9QQ7PqFSGPqaSB5ILYZfCcSRn61JKDOQ8bBY7vpEYWRBI1rME7oLYcNeQd1bO775nyCteSHZCp0uec4QZbOqoaGtcQR6LHmjdG7pc4ndWhz4TsVVxdN8Tu4QZ1E9sQy45+Sirj4j+tmwysXxHDb0UjZA5uHd8II2FzX9yry7qwDuowCXfCpmREYJCCaCb3UdZ81ZUSmrd1DIGVbK5rxgHcbK2IiMfNAFLtnKjewt7lSSSvA27KFz3O7oKKSMEt2KjV7HYGPVBadiQjThwKOx5IMeaCaScPZgABQZyn1KrR8QCCikY4BuFJNG1seQoGoKd1kxx/eyVBt8lIJiGdI9EF1J/ZOk4wq1LHdeAdlAHlriW+albKXEE90FvgHpBzuVdHE5pD++O6q+XYAd/rU0UjDEWv74wgzaGnF2IgLsYO261iSmgtFKYDgkhbbt76mCUvgzlVuFwq5pC2YnIQYtTnx3ludznZXxVT2NLXE77KShYHyF02wPqlbTjxOqIbIM+2XVlA4SSNyD6q6vrfum8yRNwB6LR58AAZ3WtWKBksbhgZI/Og3Xw80INW1McLx3cAu3XBPgcyy6npZRTdXY9l134SUVfT18LqXIy8b/avRfl70pcbjc6WpqwSA0E7fUg7ocHLRBbtLU8L6YAgDBI+S5DADRsMLTNO0MVDaaeFg7MC1RAWx+J11EGnamlbB1ve0gbZ8lvhafcbLRXRhZVM6gUHjXzWaevpvVVWRwSRsOTsCAur8V4qYLbLRTsLnElu+69hOcHhRYKuxyTUMTGymIkjAznC8vdT6AitBmlnbjDz5Y80HXivttRTVr6stJDiTgj1Wj1UrppckEeS5SvT7X1up3EZA81xxeWwRzkQHzQQMZ4PxE5yFDLKHnYY+pHyOe0A9lZGOp4HqgrHG6Q4AJU+Xwt6OjPzwtQgpGxMD3bZWpmGhkpSTjrP1ZQbTdkO+3Ky4eksOyhqYXRyO22yrI5CCEEphEjs9lDIzocW+inlmAaOjuqsDZIi53dBjZI7Ej7UafiBO6OaeohUx5IMl0zQ0ABXseXM9FBGwu3cNlM6RjGkA7oLo6kwOJDSfkseomM7+vox9izLU2GSU+PjCyq6GkH9iwPVBogBJwFe1pBBCmIZ1YaUIACC33h7RjJVzYPFaXuO6sZF4pwFcRNEQ3fCCyODqcQfJWyx+G7GVNI50Q6vMo2nlnZ4nogxgCeyqGkkZ81e1ha7D9le4tAyCEEsFOCN1BUNLHkDOFc2oc3ABxupXPikaS47lBisYZDgI5pjdjzU1KPjPor5oDJJ8KDHdI5wx8lLFTdcfXur20bmAFwUrmvbEekbIMBwIJafJUAKnpul0h68KWcRNHwkZQd9PYrh37abVOT/APKBXfyjbkT2K7j+2m1T/ACu/lG3Ig9qEREBERAREQEREBERAREQEREHHnMW8M5fOJ8h7N0bej+ShlK+aKad5qZ5GEjre47ehOV9LPMl+DrxT/gVfP1GZfNhaoopqmcTY2aSPryg08ue7u4k/WsmCjMjS7cqGqAZO4N7ArUKCojbAWvO+6DTZAGuLVZ9ayXxl05ONirahjQ4BqCEZJWQyIFp7KR9N0Q9ZWKJXDIBQUkGHkBSQkjIx37Kaho5KqTICz56NtKwB+xwg0wtMbi/J3VuTNIM+qpOX9Rz2Kvp4i5pe3yygyJ2NEAAGSsWGXw8tI77KakeZZxG45BPmsmttE3UHxNyCgxJKfLTJn7FkUdc2ljMTm9/PCsbBPGMS5wPVWTQgtLgN0FJYTO8y5774UZhfHk5wVkWuKWrnELcnZTXmjloXhjwRlBprGGR/T2V08XhuGNxhRte5pyCs2nfE+M+JjI7ZQYkLOt2FdOzwzhUe7okJZ28la575Dg7lBWN4a7JGVc+cuI6R2Vpie0ZcMBS0oY54DsIKtJc3p3B9VM2RkILZBkkKepjZHCHMAzjK0zr6z8SBLh7y5vmrQ4jsU7nA81f4Eg3IwgrFC+V2Gnc/NXSRzQuAcSfNUjcYTnOCFkZM7ck5I9UEb3tdHjbOFE14HdWyEtcW4wrEGdT4mcGqeoj8AZOFiUDHulyzususEjQBJlBBDUAvwQoq0guBBUJJjeS3ZVDJJtwMoKRR+I7C1OC0jpEpd23x6rTB1xntup2XCoYA3q+H0QZ0lUJ80uMY2ytOqac07ukEnKymR+8AGnH3wo+jnjBNTn5ZQYXR0s6laXk9tle54Li3yyrHtA3CBGzrOFV7OhwCozqJw3uVV4cHDr7oNSpaP3qPpJWJWUnu0ojBzn5qWOeRkYEIOVjze8SO8SQHPzQZzbN1QCUE5PkFhy0bogScjCzrPcSyYRzu+D5lSXtomcHUwBb54QaRBEZpA3K1yiu0dqhMDm5JGFpVuglmn6IwSVfcqSamqGicYygpWymskMoGATsrIcRuA9VlyGD3UNiI6vRaaS9p37oNWk6DFnbKugZmEkNytJMs3TvnC161SwClLZSMkINDEJlnLey1BtOKeEg75ChcAKsuZ2JV1RP8Qa490GEAXzEgE7+izWVzoCIjGd8eSyLVBA5/U/C1SsjtjWlx6eodig0qqpGzQibzIzgrGjt4cwu7qSpqy93REds4GFl0cUzoHHHcINClb4chb6KsUjmuHnkq+rjeyYlwUkAiLMEjJQSeKA3Oyxnt8T4lIIJevselUdFIH7DZBNaLgLdUeI5uVLebmy5yB7G4x8lA6mbIwdI+JYz4JI93DCCwAuOwJ+pVDXNcCWn7VrelKWnqK7pqcdO3ktY1PbaGAj3YDsg221jZ4+kjCw54fBdgFZM7nU4GMjKxHyOkO/dBaAT2U8Ti34SO6rTxljuuTACnldCRluM4QY80AY0yA91AC7HnhTNkdI7pPZZIEDGkOxn6kGANiFzBwz4pCyxC1h/Q97ekO+fZcT4jLidsLNtrqamrGT+bTn5IO8/A/jDceF1wZqOqvDjE94eGl+RjK9C+EvOPp3X8kEM1fAwvAB7DdeI911rJVWllHE4npGBg/JX6S4zal0aA+3PlD2/R+IhB9GjtdaVZQiuN8pugjI++DOVlWvVFhu0Ilo7rTyeuJBkL57Z+criy6EU3v0ojB2BkK3Tpfnf4nW2ieG1U+QDjEhQe/Da+iccMrIXH5SAqQTw/wDzZh+0Lxa4Cc53E/VuoRSVdRO5hkAHxk7ZXeSy8W9VVD6eOQvy9gPc98IOy3EPQ9s15ZJbbUwxyktLR2OxXnFx25BZa25y1VDbSGvkLtm7Yyu/3DS93mvDZKzqLX987rkGvtdLcQBUMBA9QCg8OeIPJldrDRsEVnkccY+iVHozkGv2o7FLeJbfO3oBON17U3bhjpe8t6a2ka//AMULKtWgbBZ6F1vo6ZrYnDBHSEHzzcReBGpNLXGa1sss3TCSAQ0riets92tUj46mgkZg4JLSvot13yx6D1XDPUmiY2pc0nJYNyuhXMVyTPprdV1dppcOBJGGfWg8vWs8H74e/mPRROmJkDxtjfZbw1vww1Fou5zU10gc1jHn4i0jIytvU8dPM7AIICC2ougnpxD04IGMqlvgaWl5O6yKu3xmPMI3wsaliqIpMEEDKDNZXyUpIBIAWFLcTLUGZ2+Fqzrf73CAwZPmtHrLXUUmXPBwEHrZ7FS8RXLTXFGnYADSz2YO+0VmP0L0vXln7DV2bfxlb6TWA/lFf/qXqYgIiIOAOfSZsHKPxEmd2bSUmf47Avnamf1zPkHZzifylfQt7RKobS8mXEudxwG0dF+evpx/OvnlQZ0NaGsDekbDCo6L3g9WceawlPHUeGMYOfVBNEwUri7vsoZ5hJMHgYwVRtRl33zJHorZHMe7LWkBBnSXRr4xGG9gkNzDYvDLe6xmS0wbhzHZTxKXv0nPqgPh63F+e5yglETOhWvqQRhoICgJJOSgmhjE0hzsEqGeE/AOFE17mHIKq95ecuKAXZGPNXxyhgxhRfWh37BBlRsFU7HYZWS6jbBGd8ladHI6I9TTup/fXub0vyUEPT1PIHqqOb0uwCqZ3LgqEk90EzGiPDie6ym4kZkd1gF5IwpYp+jY5wgscHMeUz1vHkpjNTk9RySe+yhe5pdlgIA7IJ5Yw2IHY9liq90pc3pOVYgr07d1Mym6mdRdhQZPqrxK9owDsgtIwSPRTx0wezqyVAXA5OO6ubM9o6QdkFr29JwqA4KqSSclUQSOl6hgklRg4REA7qrR1OA9VRVBIOR5IMv3FvR1Z/OsWRvQ7Cu94l7Z2VhcXHJQAT3Ru7gPVStkiaN2klRucOoOaMYQa1QyRwNy7GSsO44mk62YWJ7xJjGVe2pHR0uBJ9UFWTdQ6RsQr3VIYOnvlY8cjWOLiDuqSvDzkAhBlUlN79LjOPJatRt+5spYDnA7LRaOr91d1brVrDM2536CCUkNncGDbO5Qd0uSHhy7iReKenlo+oeIAdvLK9ZtC8HaDSIj8OBgLGhuQun3s9eGkmnqeku7G4DiHZxheiSCKniEETYx2ClREENRP4DOpYVVcqYUhe+oDCPnhZlVH4kT/wDvThcA8WNUXKwskdE54YCdwSgm4m0lJqHxoZJhK0gjGcrprxw4QWZlgqqqORjJBk4yAVvi6cxdutNVURXGpAc0HuV06478z1VdL0aKhq3Gkc8h2DthB1n1tbaq3alnp9xG0kA+RC29LazODL1ZXIXELVGnLjRtqad4dVPb5AZJK46prwyOneyTPUe2Agx/dw1xj9Faabw3CQeRyojVOdKZDnBOcKYVkeMPBIQTfdDxW+D2xt2UDqqSNwHWcFU8enGSxjgVjyPD3ZAQZFTO2Rgx3PdYoOFTJRBJDH4hwSQrn5hPSOyjY9zOyuMgcPiBJ9UFOv5K3PxZVWkA5IJCdTQ8HBx5oLjKcYxhZMNB40fi58u2VFLJC5gaxhBVI6ySNhY3sUCXqgd0sOMLKijMsHW5xyAtPe8vPUVeyeRjS0HYoKZ6Hn5EqXHiM7+ixyckn1VQ9w7FBLDMYHZxnZTGoEzgSMFW00tM0nx2kqyeSIyZgBDQgvqsFox5K+mr2wReGW7/AFLHZMM/GCQkj4HD4WkH1QJH+K4uAxnKjcCO6p2OyqCPMZQUAyp46fq3yVACQr2yub2QZD2+CNvJX09QA8ZCg94aW4e0qMvHV1NyEGsTSskjAb3wsM1TwfCDDjOM4WPFUuY4F2S3zWtQ1tl90JkJEoGwxvlBpE8HhN8QE5Kx8FwySpJ5zK89+nO31KLqxsOyDv8A+xWP/XTap/gBXfylbkVPYq/hT6p/4v67+UbciD2pREQEREBERAREQEREBERAREQcccyX4O3FL+BV8/UZl80sMxhne7OM5H519L3MVSS1/L7xOoYc+JU6OvUTMerqGUD85XzlN4XarfI9jaIksJB2Pkg214DZiX5GTv3UEjDC/GfyLd7OFGricGjcPsKuk4TasbuKVx/8UoNs5YIerIJwoo2CVwcSNlulnCjVzjg0RA+oq/8A2JNWBwb7qd/+1KDbFTUO6PCaMrDawnOdseq5MtfBW/v+OpjeAf8AtCsO8cHtR08/+1YHvB/7U7INm0FxFG/doIHZSV1eKwgggDK3TTcGdV1IBFO4E/8AaFKrgxq6mODSuOf+1KDZ4jZMAzIz9a1Kgoo2t6CRutXbwj1jneicB69JWQ3hRq5mCGygj0BQaBU2ltvzVdY23xlWs1AwN6XNBx8lrlTwv1lJs+KR4HYEFQt4T6td3oyP/FKDblTcxOdhjPyWOKkklp7HZbvbwh1Ue9MR/wCKVQ8ItV9YaKYnP/alBoNrlbbJBVEAjHqor3d/urIH9OMLdw4NapMYLo348h0nAULuDuqWgkQk/LoKDZdFTiom6CRj5qS4UraVwDHA59Fuc8KdXsd8NE768EKh4VawcfioXH7CUGzs7YWTSRMfgvIC3bDwh1fK7BoyB69JVKjhHq6B2BRud9hQbcqWMLMNwfXBWn4exwdg7HZb0p+Emq5SOqmcwHvsVqP+wpqN0XUGSZG+7Cg2FLUB8QYTkrHij8R4aTjJW9XcIdXeKWNpCcefSVR/CHV0Z3pHf+aUG3WWtrG+K4j1wSoJ52N+jg4W6n8KtXhgxBI4emDhRM4T6veQPcXDJx2KDa9FTmumEewC1CqohbxgDIPfdbnbwd1XTxiZsbw/5NKlHCbVNTCTM2QnH9yUHHs0gc7ICsjb1vAW83cJdWCUsFI4gHv0lTjg/qjb7y7/AM0oNvUkbKVolON/mrauaOoaT1AELczuEWqy3p8OQj5tOFjS8JdWxn+1HH/xSg2ZK7Llm0Jj6MHGVuVvCTVzsZo3D/xSpYuEGrnTsYaRwBxl2Dsg2jPG/qJLMBWNp2PHcD55XKF14OXtttb4UbzKBkgtO5W1/wDYq1eO1AfyH/Ug2/Qztts/i5DhtssmvujLkMjDSPJa5Bwj1bM7D6QsHrglVqOEeq4XYZSl/wD4pCDZLu53VzXeS3f/ALFWrv7xP/mlU/2KtXf3ifyH/Ug22yFkAEpcD8sqOYibLhgEfNbqbwp1c49LqNw/8U/6lf8A7EmrNv8Aah/80oNo0swjeA47KepqmEYAC3M/hJq1o6hRk/8AilWDhTq496E/kP8AqQbPAJdscZWZHWeFF0OPUey3KeE+rgM+5O/IVb/sVau86E/kP+pBt213AUNUagtyD5KS83Vt0kDmsxhbii4S6skODRuA/wC9KS8JdWRuw2lLv/FKDZ0bnMOc7fWqmQSOG2N1vAcJdWkf2oR/4pVruE+rgTiiJA+RQaEyCN8OcgkoKdzIyWuxhbhh4S6vkPSKV7T6dJWQeEOrGjDopB/4pQbZtEAqqnw3Y22KkvltFK4ODh+VbmouEmqoJRII5G48w0hVuPCvVtQ4Hw5Hn0IKDadoxNII+oD7VLd6BzXBzX7Y9Vr9Nwi1i2QPNM5gHmAVPWcK9WS4JjlOPVpQbFBMB3IJC1Cku4biIjY7LX28ItVudg0zh/4pVXcINVNGRTk/+IUGi3WmiNMJmuGTutFgYXO6vILd3+xbrEnoNI8geoOFIOEeq8Z92cD/AN6UG1zXjp6AFkU5bM3tknZa6OE2rerHuhA9cFZ9Bwf1P4rXPieBncBhQbTmY6jHikbZVY/DrmdRwCB5rkC68IL9NTARMeXAZA6StAbwl1cwke7Pb9QO6DbLJzbpepjsH61dPeZalwDiSPmtxv4Sar7imcST5tOVk0/BrUkjC6WJ7T5DoKDbMFJHcMNc4Ageqx622Mo3D4wd991uSbhVq6mkIhgeQOxAIUR4X6xlOX0b3fM5KDb0hZJGGhwyPmsN7PDyOoH6it2jhRq4k/7RIx8iq/7FGrc49yP/AJpQbRgx1YJwsiWna9vUHjZbndwm1aAMUhOfkVZ/sV6v/vA/kP8AqQbPcOk4znCdTvIlbx/2KtXf3if/ADSqt4UaucQ33IgE9+koNs0VTJG743kj0ysuSsa5wLXDA7rck3B/VEUfWyBzj6dJWJ/sVavx/aJ/IUGkyVkc7AzDcj1wrmVRghcwSAZBGNlrNPwk1dO/pFE5v2FVn4R6vil6HUjjv36Sg1rg7r92i7w2sc4AdYOV3J0BzRQ3XUFHSuq242bjP1LpdS8HNQtjD3RyB3fZpW4dG8PtTW3UsEzGzDocD2KD3e4F6mor/p+CpilY9zgO2PRcwLpDyY3C9w0FNTXBz+gED4s/Jd3RvugqiIgoQDsVo2pLBbLxbpKWroY5esYHwjK1pUIyg8yOefgHQx0tRXUFoDMxk5a3C8vKzTbrLNO2bLXNeR39Cvfvmz03HetJyEgdRhI7LxU4v8JdTPvdQbfE97DKdsHBGUHDwnDHYLgR9alMzHM6gBstxu4JayjhEslO8ZGSOk7JBwg1M6MtLZBn0aUG3KG7tpZviwQEvN5irYXsa0AkLWn8HdWtdvTEj16So5eEmqo4zIKYnAz9EoPSn2HEXTaeMU3m+ewj8grv9a9SF5kexT0zdtOUHF43OIsbUy2LwgfPpFdn9IXpugIiIOMeZHhPU8cuCWqOFVJU01PNf4YImSVLnCJvRURy7lrXEf2PyB3wvOeD2MGtYyfF1ppR4IwP9t1X9AvWZEHkwfYva0LsjW2lgP8AG6r+rqv7i9rPH/x20tn/ABuq/q69ZkQeTB9i7rU9tb6WH/1XVf1dUHsXda431vpb+N1X9XXrQiDyZj9i/rRpy7W2lXD/ABuq/q6vf7GHWTvo6z0oP/qqq/q69Y0QeTB9i9rQ/wDy7aVH1VdV/V0/cXtaf8N9Lfxuq/q69Z0QeTcPsYNZRuy/WmlXj0NXVf1dJPYwaxe7qbrTSrR6e91X9XXrIiDydf7GPWDmBo1lpUH196qv6BQj2LutfPXGlv43Vf1detCIPJk+xe1mRj9m2ls/43Vf1dB7F/Wg7610qf8A6rqv6uvWZEHkyfYvazPbW2lv43Vf1dUPsXta+Wt9Lfxuq/q69Z0QeTH7i9rX/hvpb+N1X9XUsPsY9YR7v1lpV/11VUf/AK3XrEiDyin9jPq2VuI9XaUaflU1Q/8A3CxR7F7WmRnW+lv43Vf1des6IPJ0exj1e0f/ABx0ofn71Vf0Csd7F/WTjtrXSwHoKuq/q69ZUQeTY9jDrEd9Z6VP/wBVVX9ArX+xf1m45GttLD/6rqv6uvWZEHkx+4va0/4b6W/jdV/V0/cXtaf8N9Lfxuq/q69Z0QeTH7i7rT/hvpb+N1X9XT9xd1p/w30t/G6r+rr1nRB5MfuLutP+G+lv43Vf1dP3F7Wv/DfS38bqv6uvWdEHk7D7GLWEf09ZaVf9dXVf1dXP9jDqtzg4az0u3HkKqqx/6BesCIPJ6X2MWrXNxHrLSwPr71Vf0Ch/cXtaeet9Lfxuq/q69Z0QeTTvYv6zLcN1tpUH197qv6urB7F3WvnrfS38bqv6uvWhEHkx+4u60/4b6W/jdV/V1Q+xd1rnbW+lv43Vf1detCIPJf8AcXdb/wDDjSv8aqv6ujfYu61By7XGlj/9V1X9XXrQiDyf/cY9XYA/ZhpXPn/tqq/oFq2kPY86i0/qGmu9fq3TU0UDw7oZUVJPfyBhA/OvUpEHG/CfhDRcM7BTWmLwHSQtALoiSDj0yAfzLkhEQEREFCMgj1XHuv8Ahc7WVJJTsmp4y/O7yR+gFchog6K8QvZ23/VtVJU27U1mg687STTD9ERXFNd7IvVlc1xm1bpd7ySeo1FTt/8AiV6gIg8nqn2M+tKiTr/ZppQN/wAaqgf/AECo32MWsA3pOstKn5mrqs/q69YkQeTTPYwaya7Lta6VI9Pe6r+rqV3sZNXEYbrHSgP+NVX9AvWBEHky32L+tA4l2tdKkenvdV/V0d7F/WbjtrbSw/8Aquq/q69ZkQeTJ9i9rPbGttLfxuq/q6p+4u60/wCG+lv43Vf1des6IPJcexe1r5630t/G6r+rqv7i7rT/AIb6W/jdV/V16zog8mW+xe1mO+ttLH/6rqv6uqO9i9rU9tb6VH/1XVf1des6IPJf9xd1t/w40t/G6r+rp+4u62z/APHjS38bqv6uvWhEHk0z2L+smnLta6Wd/wDVdV/V1R/sX9ZudlutdLAenvdV/V16zIg8mD7F7WmNtb6Wz/jdV/V1Kz2MWsA3DtZaVJ9feqr+gXrEiDyaPsX9ZZ21tpbH+NVX9XVjvYva1JyNb6VA/wAbqv6uvWhEHkwfYu61xtrfS38bqv6ur2+xg1iG4OtNKk+vvdV/V16yIg8mo/Yv6yacu1ppYj097qv6uknsX9ZOdlutdLAenvdV/V16yog8l/3F7Wv/AA30t/G6r+rqv7i9rX/hvpb+N1X9XXrOiDyXPsXdantrjSw/+q6r+rq5vsX9Zgb610qT/jdV/V16zIg8mP3F7Wn/AA30t/G6r+rqQexi1iG4/ZlpQn196qv6uvWNEHkufYva1Of/AIb6VH/1XVf1dV/cXdZ/8N9Lfxuq/q69Z0QdHeR72feoeVHixduIl11FZq+G46ensrYqKeZ7w6Spppg4iSNgwBTkZBzkjbui7xIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC48498btHcu3Cu9cWtde9PtdmbGPd6RrXVFTLJI1kcUTXOaC4ucO5AABJ2BXIa8nPbW8Z5Km9aI4A2ys+8UUD9T3eNpyHTPL4KRpx2LWtqXEHylYdtsh3Q5R+eHQnODW6jo9D6G1VZm6ZippKmpukcHgOMxeGMD45HYf97ecEbgE5XZNdNfZR8JYOG/KVZtQz07WXTXlZPf6p2AXCEu8GmZkd2+FE2QDyMrvMlcq84nNNpvlN4Q1OvrlSsuV5rZfcLDajJ0e+VZBOXkbiJgBe8jfADRguCDnCoqIKWF9RUzMhijaXPfI4Na0DzJOwC0u16v0nfayW32TVFpuFVTkiaClrY5ZIyO4c1riQdj39F89epOJHNdz1cSo9PT3K/wCtLxcHGSksNA7wqCkjb3c2EEQwsbkAyvwexe8k5PIV59l5zuaMtrNUUHD6GrnpWifwrPfKeSsgcMfRaHgucCe0ZcdjjO2Q95aiUwQSTiKSQxtLuiMZc7AzgDPc9gunHAr2o/BLjnxdtvByk0fq3TN1vEk1NRz3mKnZE6pY0uEDwyVxY9wa4AEH4sN7kLbHsnOMPGfiFobXuh+M95u9yr9C3Oko6Z16a/7o07ZY5euCd0mJHdDohjry4dRBOAAPP72gmh7hy787t+v+j3utrq2vpNb2WWMAGGeV4lc9oGwDauObAGwDQg99UWzuD3Ee28X+Fek+KFpYI6bU9opbmIg7PgvkjBfET6seXMPzaVvFAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQF89XtGNYVOtOc3ibWzSl0duuUdnhbviNlJBHAQAewL2PcfLLifNfQqvm05oWUcnNnxbjuEr4qV3ES/id7BlzIzcp+ogb7gZIGCg+hvhDpKPQXCjRmhooDC3T+n7fa+jB+HwadjCDkA5y05JGc9915A+2U4jV+pOZS06AFTJ9ztH6fg6YC74RV1TnSyyAZ2JiFM05AP3v0IXtcvCH2tNvqqLnS1DU1MTmR19otVRA4g4ewUzYyRnuOuN4yMjIPnkIO9XseuEFm0hy1ScVnUEDr1ry6VTjWFoMgoaWV1PHCD5NEsU7yB3LhnOBjvoupvst9RUl+5JNB08Do/GtEtzt1S1jcBsja+eRox5kxyRuJ8y4ldskGLDQUNLUVNXTUUEM9Y5r6iVkYa+ZzWhrS8gZcQ0AAnOAAOy8nvbf6Rjp9XcLNdxxEvuNuuVomeBsBTywyxg7bZ96lI38j2xv61rzI9uAylOhuFUj5He8i63MRsHYsMMPWTt3BDMbjudj5By/7IfWs+quT2is1RMZDpLUFxszM5yGOMdWBv3A97IHkBgeS7sLzq9iZ/vC67/hef1KnXoqgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIo5JI4Y3SyvaxjAXOc4gBoG5JPkF181Rz98qmkdYfsGu3FOhddBMKeRsDTJHFISB0ueNgclB2HRYtuuNDd6CnultqY6ikq42zQysOWyMcMgg+hBWUgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAvnY5+9IzaK5xuK1pmh8L3q/y3doHZza1rasOz558ff0OR5L6J148+2m4Qz2PinpDjTQ059y1PbHWatc0bNrKRxcwk+r4ZQAPSByD1j4daoj1tw+0xrOGRskd+s1Fc2vaRhwngZICNhsQ/wBB9S89fbGcs971lp2x8xmkbc+rm0rSOtOoo4xl7bcZDJDUAebY5JJQ/GSBKDgBriOePZe8VouJ/KBpWjlqfEuOjJJtM1rc7tEBDqfAO+PdpYBntlpA7YHbCaGKoifT1ETZIpGlj2Pblr2nYgg7EEHGD3QeDHITz73PlCud001qayVeodB3+ZlVVUdI9raqhqgA01FOHkMcXMAa9ji0O6GEOb0nq9AL57YzlPt9nkr7RQ63utaGgx0EdqjieXHyc98oY0A9yCT6A9lq/GL2TPKzxQu1TqDT1NetBXCqc6SSPT80YoXSHu73aVjmsH/axGNvyXHmm/YocDqC5x1Wp+Kus7vRxkONLTx01IZCD2c/okOD2OADjsQdwHKvIDzqaz5xbnxNrdQaOtun7Rpiotgs0NK+SWTw6kVXW2aV2BI8eAw5a1o+PGBsT1i9uDqmOW8cJ9ExPAkpqa63Wdu24ldTxxHtkY8Kbz3z8l6P8FuA/Cnl70n+wrhJpCmsVudIJ6gtc6SeqmxgyTSvJfI7AwMnAGwAGAvFX2nXEqbi/wA5WoLPZPFrYNKMptI0MUY63STxEmdjWjOXe8zTNAG56R57APQH2OWkZdP8ptTqCeHpdqfVNfXxPxu6GKOGmA+oPgl/KV3qXGvLhwpZwQ4E6H4VDoM+nrPBT1bmYDX1bh11LxgkYdM+Rw3PfuVyUgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLhTmp5j6Tlo4cVOuqnT811fE09ETSQwu8gT8zjzH1rmtce8ftCaY4j8HNXaU1dRxz26ptFU55eB96c2JxDwT2IIzn0z6oOk3KF7U68cd+NVBwr17oy3WaO/vfDa6iklc778AS2N2fMgH17HfsvRtfNJy2vvdPzA6AdpnrdcW6hoxTlmQSTIASPMfDkr6WGdfQ3xMdWBnp7Z8+6VJzi9ERFEREBERAREQEREBERAREQEREBERAVrnNY0vcQANyTsFxzxk5huD/AK0C88VNbUVlZICYYHkvnnI8mRtBJPp2HzXkpzue0z1Pxvnl0NwVr7ppzRrcsnqQTBV3Aepwcsb8s52+02T6OxXtOOfKk0lYn8DeDWpYZ73cgW3u40Uwd7nDjHgtc0463efoPyHoByk8ovEPm41zNbLFKKKy297JbzeKkOLIg45LGnfqlcMkDyzk+QO6eTvka4l8y+vKCq1FYLxaNDslE90vNRA6ITs7lkLnAdbn9sjbvvnce5HCXg5w64HaRg0Rwy01TWe1w/E5kTfjmf5vkf3c47kk+ZKl74fprGhNJ0mhdHWbR1DPJNBZ6KKjZJJ9J4Y0DJ+vC19EQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAXDfNjy52Xmm4LXXhRdri22VM0sVda7kYTN7lWxE9EnQHN6gWukjIyPhkdjfC5kRB075AuSniJyeSavp9TcTbXqC16lbSvjoKGjkYIaiHrHjdch2Ja8tIA3w0k/CAu4iIgIiIMesjqpaSeKiqGQVD4nNilfH4jY3kHDi3I6gDg4yM+o7rzp5d/ZQXzhzx/tvGni5xRtOrorRcZbzFR09vkY+ruBLnRTSl5w3okIlwAcuaBnGc+jyICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIIppoYInTTytjjYMuc5wAaPmT2UdJXUVfCKmgq4amInAfDIHtJ+sEhdGPa/wDFLU/D3l5sti0vc6i3y6qvzaKqnp5THJ7vHDJI5oI3wSGA79lw17FbXmtb7euI+lbve62us1DRUVZDHUSvkEVQ+R7TguJxlrTt8iid16qrqd7RTmXtPAjgjd7FFUY1BqqiloKBgOCGvBaXfPbIPoMldsV4re2W1u+88xtn0TG8+Fp6wQSPAOxknc5+49Q0D7HKXd4rjL2YGgp9a832ka73Z0lNpwzXaV2Pha5kbgwH55JI/wC9XvsvOX2Q/LpNpLh2/jleWllTqXrbQsI38AEtDs+hGSPmT6L0aVBERAREQEREBERAREQEREBERAREQFxnzBcd9Fcu/DS68Q9Z3SmgFJA/3KlkeBJWVGPgiY3OXEkjOOwPqQtc4q8WNBcFNE1/ELiVqCGz2S3ACSeTJL3n6EbGjd73HYNHcrwd5xOY7U3Ofx7NfpO3XWazRlls01Z2sc+UsGcyGNpOHvJLjjsMdsFDWx9a624x84/G2GerNTfNTamrRS22gjJLIGuPwxsHZrWgbnA2bnsAB6NcrnshafQWq7Tr3jtqi3X2W3uZUssVFC404mGCBK92PEAPoAPtwRzfyE8jejOXXQVm1nqjTsU/Ey5UgnuFZUfE+3+IM+7xA7NwCATgEkb4wu4GcKWauoaalpqKnjpKOnjgghaGRxxNDGMaOwAAwAPQKdEVQREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAUU80VNDJUTO6Y4ml7z6ADJP5FKtt8SK9tq4eaoubndIpLNWz59OmB5/mQeG/PXzY6n5tuKdNomx2/psVjuUlFZqWPd087neH1n5k7Z8xjsAF6Yeze5Ub1yx8Jq+TWLYhqfVk8VbWxsGfd42MIjiz326jkeu+BnA8muQLR0nEHnE4a2ian95ijuzrnVB248OmikncT8iYwPrIXvRxj4ycP8AgToau4g8R75BbbXRtIb1kddRKR8MUbe7nk7AD1UyH1pXMXx40py58KrvxN1XURdFFH0UdK5+H1dS7ZkTB3JJ3OPIFeDk9Lxe57eYm43S226Ssveo6oSzdDSYqKmGGjONgGtGwGMnOMDOK80XNBxN5ueJkl1u01Y+1tqnRWCwwAuZTxk4YAxueqQjGTuckgE5JPqD7LTlI1FwG0HdOIXEaze4ao1X4YhppWjxaSjAyGu22c4nPfbfyOTeTg7gcItAUHCvhfpfh1bIwyn09aqegaAdupjAHEfW7J+1bvRFJMBERUEREBERAREQEREBERAREQFtfiRxD0twq0Td9f6zuUVDabNSyVM8kjwOrpaSGN9XEjAHqtA47ceOHfLtoKq4gcRrsKWihyyCBnxTVUvlHG3uT2yfLPqQD4PczXNTxa5u+JE89XU3T7jz1PhWTTVM9z4omZ+AGNuz5DgEnBOc4PclMG7uarnJ4u882sLPoa06fNDY2XHw7Hp+iJfLUzyHoZJO8nDpMEbDDW5d9a9HfZ5cgZ5ZLfV6/wCJn3PuGu7xCyONkLRIy0wdzGx5G8jjjqcMDAA38sTkA9nvozghp2w8XOIdrfcOI9XTiqYyoI8Kz9Y2ZGwbGQNO7zkgk4xgY70K78SQREUUREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEXHXHzjDZeBXC+7cR745vgW9gDA799IQcD8x/IvNjh37UDiJxH4zW2w00M9Nba2r6I4+oAObntjfGR5H6knbhePWxFj0M7qqjp6l7el0sTJCPQkAn9KyEBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAXVL2ifHej4ScBr7Y6eYtu2o6GWkgAOCI3AtJHnk7j6srtavK/2w1/isurtEwXCF01HPEJHMB+kwPcHDH2H7SpuLP24X9kTpr3XmVfre/D3Kht9krIaaepHhsdPKGt2c7HZvUPtwsv2v/HSDiHxus3DXTt6ZWWLRttD5RTzh8L7hO4mQ4G2WxtibnfufVbJ5k+YvhzXcK9JaU4Izm2174gbtLSHw5GNDfolzcFpJI884BXLnspOUzQ/GmbUXGfi1Y33yCy10VLaoK0udFLUAdb5nA/2TpOBudj9uUlt2+JXPfszuRXT3D7RFo468S7I2q1ffIRV22lqosi20zvoHpcNpHDDjttkd9sehiihiip4mQQRsjijaGMY0ANa0DAAA7AAYwFKqCIiAiIgIiICIiAiIgIiICIiAthcaeMmieBXD+68QdcXanpKO3U75I4nyASVMgHwxxtO7iSQMAHuuDOZ72jfAjlxqLjpP7pS6l1lSsc37k29vU2CXGzZ5T8LCMgluc4z2K8aeInErjxzgcTJq+4NvmprpXzk0dpoxJPFSMJw1rIxs0AHBcQPmQMABrvM1zScV+cbiJTS3aKU0bak09hsVICWxdZw0YBPVIc4J8s49SfTL2c3IAOA9udxT4u2qiqdaXOJvuVK9gkFqiO5wSMeIdskdseoBWkez69nBQ8JI6DjDxrtgn1oPvlBbJcGO257OcPOX6+3l8/Q0HKZ8FUREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQef3tm79Ja+XDT9pZOWC76kiic0bdYZE+Tf6ukLqf7JrlvpOLfEy8cS7+0m0aOEMbGjvJUyZcG+gwGg59MrkL2yPHrRGs5dH8HtK3eG41tgrai53OSCQPjhe6MRsjJGxOC87Fcy+xe0hcbJwB1VqatpzHDqHUXVSuLSC+OGFrCc+Y6i7H/tV/8UnXoQ1jWNDGtAa0AAegHZXIiiiIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAvJT23NbS/st4c28SD3g2+plLPPoEhGfyleta8JfazcQItZ83F1s1HVial0pbaS1dI7Mn6PElH15eB9ig4M5WuA935jeNWnuGduhm91rJxLc6iNpxT0jN5HkjtsMD5nZfQzwj4S6J4JaGt/D/AEFaY6G2UDcANGHSPPd7j5klcL8gXL9w74Ocvukb7pqy0xv2qLLSXS7XVzAZ55JoxIWBx3axvUAGg+W/YY5B5geZ/g/y26efe+JWqKalq5IXyUNsY7qqqxwBw1jBk4J2yRgb98LW74jloDCqvAax+0a4/wBr49v4vVeqa+rtU9xdLLYnPAg9zLtoQOwIZgZzgn0GMepGgPae8n2tbVFWV/EpumqpzGmWlu9LLE6N+N2hwaQ4A7ZB3Wartii484U8wHBzjhFVy8KtfWvUXuOPeG0ryXRgnAJBAI3I/KPULkNAREVBERAREQEREBEWHdLpbrJbqm73etho6KjjdNPPM8NZGwDJcSdgAEGYvP72hXtG6vl9uj+DnB+GhrtYTUnXcrlK4vjtIfnpY1oIDpiATgnDQQSM4C485/fab6Tq9Gy8LOWTWtVU3aunMV11DQsfFFT0wG8cEpALnPJwXNGAAcHJXVPko5H9c82+uJNR63N3t2iqd3vFxvMwd41wkJBEcT5Aesu3Jfk7djnJFzBsfl15U+N3OXrK41Wm3MEHvJnvOoLm8iGOR56nEkAmR5JJ6QPyL2m5Q+T3QfKbop1lsr23W/158S53mWECSZ3kxnm1g8h57Z3XJvCTg/w84G6LpNBcNNPQWm0Ug2Yzd8rz3kkcd3vJ3JPmt6qAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAtL1OJjpq7NpnlkpoZ/DcO7XeG7B+w4WqLDu0RntVZCO8lPI38rSFKPmS1hTVtx4q3ehu88k1RPfZoJ5Huy5xM5aTk/mX0N8qGl7NpDgFpKzWKlZTUzKMP8NgwAT/7AF8+nE+GW0cb9QNlGHQaglk//AB3UPzEL6CuU66G78B9MVZzn3YN3+QH+tS3sHL6Ii0CIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiDrN7QLmSu/LLwEqNVaXdE3UF4rGWq2OkGRG97XOdIB6ta3P8AqOCvJblI5Xtac9PFy+XPUOpnwUNDIy4aguco6pZTK44jYP7o9Jx6AD6x2n9tNxZ0/eItF8JLbWMmr7VXTXStaw56CYzG1p9COo/lPot5+xIs7qbhbxGvckRBrr5SQscR9JkUDu32vP5E79T9x6H6Q0xa9F6WtOkLLEY6CzUUNDTtPcRxsDRn54GSvDj2plw1RqbnCvNFVU1ZLFBTU1Da4AxxDmNGD0NHclxOcbnbPkvd/stlah4N8L9V6ip9Wah0VbK+7Ujg+Kqliy8Edj6HcKTivNnh17G6g1ZwStF71Nr+tseu7nSsrXRCAS0tP1gObC8ZBJAIyRvn07DirWPsceZHT1JUVdg1BpnUPgglscEskL5APQPGcn0XtgAAMDyVVR85eh9Z8feRbi+26wW6aw36nZ4dTRVsZdBWQE7tcAcOGxwRuD+f2a5H+c60c32ibhcJ7JFY9TWGVkVzt8cxkYWvBLJYyQD0EgjB3BCxOe/k0sHNTw7fLbYYqPW1ja6a0VwaAZdt4JD3LTjb0IHyI8heXHjfxE5JOPNS+sozTvgqBatQW+UHD4g7c/YCXA+hz3V+J/j6H0Wn2C70+oLFbr9Sf2C5UkNZFvn4JGBw3+ohagooiIgIiICimmhp4nz1ErIo42lz3vcA1oG5JJ2AXDXNpzJ6d5XeEdfxBvIbUV0jvdbTRA/FU1LhsMeg7n0HfbdeHHE3mu5meYnUMtNcta6kq21krnU9ltMkoja3chjY4t3YHfIIPfAQepnNB7UzhnwI1LNozR1jbrO7U3w1DoKoNp4n47F4B7H0z8hjdefnMv7TXjlzG6Xq9Am3WnSmmrgAyrpLd1vmqGA/RfK45LT5gAArSOCvs5OZni7dKaW56GuWmbPPIDLX3SEwuLTuS1jsEn68fUV6ccEvZa8snCmS3Xq+2Go1bfqMNkNRc53OpxKMEObCMNyCMgn5bJn9m/06U8knsvtb8SLpYOK3Gulis+jGyMrobTKSau5MBy1r2do4zgZyckHt3C9krZa7dZbfBarRQwUdFSsEcFPAwMZG0dg0DYBTQxRU8LIKeJscUbQxjGN6WsaBgAAbAAADAUg3Vt0xVERQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFpWqbmyyaYu96ecMoKCoqnH0DI3OJ/MtVWyuNj5GcG9eSQ/wBkbpm6Fv1ilkwp+Xg+dO1U1w4z8eIaeijNRUan1BmNoByWPlyBjvswfmX0WcHtCt4bcOLLpAY66Gna2TpGB1kbgfVsPsXhr7MaxUV+50dBsr2BzaI1ldGP/okVNIWn7Duvf4YIwrn1PKqiIiiIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC03Ud2ZYtP3O+SY6bdRz1Zz2xGwuP6FqS455i7lJaOA2v7hE4h8enq5rTnzdC5v86D5+4ItVc1/MjT22S4Sz3PW9+93hke4kxxPeSAAe2GZONhnJxuvfblx5fdG8tnDej4eaOjc6OM+LVVD/AKU8x7uPyznGfU+u3h37OimFTzpcLw7GI7nLLg+ZFNKR+fB+xfQwgIiICIiAvC72t2iJ9Nc2Vwv8FrdT0GobZSVTJg3DJpwzplOfM5AJ+v5L3RWx+JvBThRxlt0dq4o6CtOpKaI5jbWw9RZvn4XAhw332KDy/wCQL2m130vNprgNxyNPUafZ0Wy16ie8tmom7CKOozkSMH0Q/ZwGAc4yvXGnqaetp46ukqI5oJmiSOWNwcx7SMhwI2II3BC8m/aYcgXC7hNwzpuM3AvSc9nioK5lPfLfDPLPAKeQENmY15cW9LwAdwMOyey032WfPFQaDqazgZxo1hJFZ657H6crq+Vz2Us5OHUxefosdkFuc4IwMDAVySepv16+oooZYqiJk8EjJIpGh7HtILXNIyCCO4IOchSqKLTdQX20aXslfqO/VsVHbrZTvqqqeR2GxxMBLiSfkFFqnVWntE6fr9V6svNLabPa4XVFXWVUgZFDGO7nH0/nXiT7Qfn7u3MVqiXQXC6+XCi4bW8eE5jS6E3ibJ6ppR3MYzhrTgYySN0iVsLmF4/8W+fTjtbdM2ljpaOruht2lbNGS2ONj39LZH7ZLy0dTiRkDIGdyfUrkX9n/pzlSo6nVOpbjTai1vc4mxyVjYAIqGMbmODqyQSe7tifqwB1u9lfyN3q03O2czvEyidSFkcjtN26ZmJPjaW+8vB3bsT0jvvv3IHqii+CIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAisc9rGlznANAySfILrJrn2jHK1w/wCIjeGl51nPLdBOKeeWlpjJTwPOMB8gOGnfzwg7PIoKOrprhSQV1HM2anqY2yxSNOQ9jgCCPkQQVOgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLj3mGubbNwH4h3JzciDTFzOPrppAP0rkJcMc5F/odN8rnE253GZscf7HKyFvUR8TnxlgAB7k57J6PG/2W5cOdTRIaQMxV4P1e6SH+Ze+68C/ZZgft0tGPIz0U9wP1E00g/nXvnkIKoiIKYCqtkWPjRwx1JrGs0DZNYW+rvlCS2akikBcCO4B7Ej/7GVvdSWUzBERUEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBcL85N2pbNyxcRKqqlEYfZZo2knGXEbD9K5oXnX7YHjfNo/hza+E9A8CbU7HyzgOwfCBLTkemx+0oa6Dezagkm5z+HLo258Osne75AU8gJ/OvoNXhl7JDRkuoOaij1E9jzT2K31MvV05Akewhu/1A/lXuaoKZyqOc1uA4gE7DJXXDn45j6zln4A3DVun54majulQy12cP3xM/JdIB5hjQSfr2XiVYeIHNBxx4jU8WmdXa21HqqunE0MNHWTuLXAj4uhp6GMBxkkAbjPdUfSMi2PwTt+tLVwm0rbuIsxl1LT2yJlye53UTMBuCfMgYB+YW+FJQREVGFd7Ra7/bKmy3qggraGsjMNRTzsD2SMI3BB2IXz48/wDwPs3L/wAzuo9F6Yp209jrIoLvbYGg4hhnaSYx8g9rwMbAADyX0NryM9tLwmudNrfSXGSmiD7fXUH3GqXDuyWN5czO+4w/8pUwehHJdfLnqXlS4W3u81clTWVOm6QzTSHLnkN6ck/UAt+cUuJ+j+D2hrtxA1vdYKG12mnfO8yPDXSkDaNgO7nE4AAz3XljoT2r1j4S8t2m+GOh9CVMup7FbGW6Gect92i6QcPIP0iCe2CPULpffNZcxXNjrkUtxuWptb3mvmHhUMTpJYoS4/CGxj4I2jOATjYDJKTIWOYOcL2ivEzmqtx0LSWen0xotlUJhQU7nPqK0tJ8MzvJwcZB6GjAI7lc78gXsy7nqSqtPGnj9bvdbKwsrLTYJB99qyCCyScfvWeYadztnvt2Q5S/ZgcGuFem7Fq3itp4am10I2VVQ2rlLqSimOCGRxDAcWn987Jz5bZPeKKKOGNkMUbWMYA1rWjAaBsAAPIK2aRbT08FJTxUtNCyGGFgjjjjaGtY0AANAGwAAAAG2ApkRAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQQVdNFW0s1HNnw543RuwcHBBBx88FfOrzs8GL9wO5jdX6Xu4mkpqyufdLbVSA4qKac9bSDvuCS0jO2PqX0Yrgrmt5TeHPNHoWpsuprXFHfqWB5tF2YAJqaXBLR1ebCe4O35wRrZHs6uYbTXGPly0lZJtS01RqvTVEy0XKkknb7y4wDpZJ0HBILA05A27nGQu1i+cfVfKzzR8HdYVdvh4Z62irLdI9sVxs1BUva9gO0jJIQSARg9wcLXOFfO7zN8HdYUMtbxL1LW0lvqWx11ru1RJLlgcA9jhJlwcBkYJ2PcJkzhuvocRaPo+/xar0nZdUQBoju9vp65oacgCWNr8fZnC1hAREQEREBERAREQEREBERAREQEREBERAREQEREFPmvGL2rfNhqvVvFe6cv2m7q6n0tpvw4blHES11VWFoc9rz5tbkDbv223B9UOY7iLd+FvCO96xsdOJaylj6Y8jPSSD8X1jC8L+DfBzWvNVzEsmv8Uvut5vXvN1q5tssMmSzJ7kgAfUp+N/6yDv17LnkdqeH1FaOZfXVS9t3vNvc+z27owaemlbgSPzv1OBBA9MfWfSRbN1frzhnwP0ZBdNbamtmmNP2yGOlhkq5QxjWMaGtY0bl2AAMAErhF/tLeSxjJHHjPSHwyRgW+qJdj0xHurm+I5A5peYmwcsPCWv4nX2kNa6GVlLR0bXYNRO/PS36tt+2PMjuvMLiD7ZvjjqfT9dZNH8P9N6Ynq2OijuLZZqmeFhyCWBxDQ/B2ODg7hYntMeejhtzLWfTvDvhFLX1dns9a+4VtwqaZ0Ank6CxrGMd8RaMk5IG+FyT7MLkW4K8V+E9Rxp4waaGpqiuuM9BbrfVSSMpqeKLAdIWtI63OcTgk4AGw3ynnV9cJeyxtOtdZ83FNq509ZVQ00FRVXWreXESPduA89iTuR6eWF7mLZPDfgtwp4RUj6HhnoS06dgk+myih6c/acn863snPgIsG8Xi16ftdVer3cIKKhoozNUVE7wxkbB3JJXQ3iP7X3gnp3X1PpTRdrrb5a46gQ1d6DQ2nG+CWAkEtHfqAOR6IPQBFp9hvNHqGyUF+t7w+muNNHVQnOcse0OH5itQQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQUyF4ne2Tv8A79zO2uwiVzhatOUzi3OQ10r3nb0JDWn8i9ob/e7dpuzVt+u07YaOghdPM8ns0D9J7favnW5x+L8nHfmO1brqA+JTT1YoqJrcnEMQ6GAeu4OPrCg9WvZV8v1n4ecv9o4nVdM1171jE6uEpaMsp3HDB65IGPqHzXdevrqW10FTcq2URU9JE+eZ57NYwFzifqAK2Hy7acZpHgNw/wBNsaGih05QR4HkTC0n85K6/wDtEOcrSPL7w5uvDile+r1rq20T09FTxEYpYpQYzNIfIYLsDYnuOytvU248wOcjmu1zzl8WaSw2qnLdPUNxdQ6btkRyZXyPEbZXHsXv2wfIH7B6nezx5On8rPDSprdWQUkut9TuZPcpo2gupoAPvdMHkZwCSSPMnOARgeaPs2uWLXvFvjzpXiN+x6dmi9J3JlxrrjURHwJZISXMhYTs5xeADjIGDnsce8ecIqqIiAiLrNzP8/fAvlopq6zXS+NvWsooXGCw0A8SRspHweO4YETSc5yc7EYBTRyBzKcx2heWLhpWcRNbTGUsPg0Fuie1s9dOe0bAT9pPYDcrxR5u+fLiZzcCj0/dbDQWLT1BOZqS3UhdLI952DnvduSRjYADIGPnx5xE4mccOcDiuyqvFRdNR3y7VJjt1rpy58VMxzto4mDZrQDu7A2G5wMD1i5KfZrcPOCun6LWXFuyUepNdVUbZXR1LfEprbkA+Gxh2c4di4g9yPTAeffK57NrjRzEMg1BdGN0lpdzx1VtcwieVvc+HGRnOPM+fcL2X5fuXDhry4aKpdIaCs0LHxRgVVwdEPeKp/m57tzgnfGT9q5OpaSloYGUlFTRU8EY6WRxMDGMHoAMAD6lOoCIioIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgpnfC8Ffaq2e02bnP1Yy1UUNMKyit1XUNjGA+d9MwvefmdiV7d8VOK2g+C2iq7iDxIv8NnslvAEk8mSXPOemNjRu57sYAHcr5++arizPzWczWodd6OstZLHf6qCis9H4eZ5IYomRR5aCcEhnUfQHfGCifXvJyvzvqeXHhlPJIHufpO1ku9T7szK5QXG/Lho678PuAnD7RN/yLjZNO0NFUh3cSMhaCD8wdj8wuSFN1fBERUEREBERAREQEVjZGPJDHtd0nBwQcH0KvQEREBERAREQEREBERAREQdYvaN6xZoblS1NeeoCV8sNNDk95XlwaPyheKfCzme4k8JJZ6jTc9MZ5D1NlezBY7vnbuvRj2x3MJpSDQFs5erTXxVl9rq+G63OKM590giyYw4+TnOJ222GfI46UciPJveOa/iG9txbLSaLsTmvvNcw4LicEQMP92R6bgHPbJFlv4+J762dqfiTzG85esrZpy6XC46nuJd0UlDCCIYiTu4gkgfWT8hud+fNM+x45przHFNea7SllZK0OIlrnyubnyLWtyD6het/CTlt4J8D6KOl4a8PrXapWNDXVbYQ6oef7oyHfJ8yMZXJyyryV0j7EnVXvUc2ueM1p92yDJDbKKQv77jqft2+S9LuCfB/S/Arhza+GukGvFutjSA9ww6R57uO53OB5lb8UFTUwUcElVVTxwwxNLnySODWtA8yTsAqJ1aXNY0uc4ADuScbLrjxH9oVyl8L6ya2ah4qU1VW05xLTW2mlqpGn0IY3GftXTLm89rNpTW/Dq6cPuX+ivlLV3hhppr1VxCAxQH6YY3JcHEZAP19vNOoj9rBzn2i/0cXLpws1LHVwtlMupqqjflhLfoUweDgjuXAHBGxGCCuvPJH7PPWnNU064vdyGntC0VUIX1b4i6a4Pb9OOAbDA2BcTgbgbrSeRvkw1hzO8T7dcNUWK50+gaKX3y8XSWN7GVTRuIY3kfE552JGcDOSO4949I6Q01oLTlBpHR9mprVZ7ZC2CkpKaMMjiYBsAP/cp5PTe8SaW09RaT03a9MW3qFJa6SKkhz3LGNDQT89lqyIiiIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIixLnWG322rrwzrNNBJN0+vS0nH24QdRvapcUajhvypXOktd1FJdNT3CntMAD8PfESXTFuPRre/kSF5rezZ5W6bmP40vrtTU8rtLaOjjuFeQMCaYu+8wg4xkkEkdwBn5Hj/jtxq4wc2vGhlq1LfKmtNReHW6zW5ufApQ+Usb0sG2cHJP14wvbvlH5ZdJ8qvCOm0ZaOl1yrA2uvtweQHVFV0YJJ7BjBkDyAz64T5o3fxq4waF5c+Flx4jaxlFLZ7LCyKGniwHTSH4YoIwfMkAD0AJ8sL5+eOfFfXfNtx5uWt32uepumoqllLa7ZTtLzBTt+GGFoGTgDcknuSc43XYj2l3OMOZDX9Lwm4diefSmlq+SKN8TSXXSvJ8Pra0buaN2sBGSTnAPftl7LfkluHCWxVnGbi7pZtNqq8Bkdmo6tgMtBSABxkIIPS95II3yA0bZwVP2fp3B5aOGsXCDgNobh42lbBNaLNTR1bQ0AuqSwOlLsAZcXk5PyXJyIqC2ZxW4s6G4L6Nrtca/vtLbbfRROkHiyhrpnAfQYDu4kkDA9VxlzQ86nBrlYtnh6yujq7UVRCZKKyUWH1EpxsX+UbTkbn5eoK8R+LnFbjfzocXqm5Nobzfq2vqD9y7HQtfMyjhzhjWMbsMAgF5A3O5AwAy3wdn+Mntj+Neo6+ttvCLTll0xaOp8UFZUwuqq2Rh2D/iIZG7zGAcfIrhvlr5LuOHPJeL5xAdqOnt1Aasm4X+7NfIaipduWxsbvIQO+CAMAbeXdzlX9knwuoNB23UnMdarhdNV1uKmW1x1roaaiadxE8RkF7sYzk4Bz9Q7/6G0Bozhnpik0boHTdFY7LQgiCjpGdDGZ7n1JO25JPb0CT+x1+5OuQzh5ym0lRdYK/9keq65vRPd56drDEzP0IRkloPmc5Pn557RoiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLbuvtdaY4Z6Pu2vNY3OKgtFlpX1dVPI4ABrQTgZ7k4wB5khbiXh17Ujmb1jxH463vhJbr/PFo7SUjKRtDDIWxVNSGhz5X4OH4JAGe2PPAwGxOcznc4gc42qKLTtDbTbNJUFY5tns9Pl8tTK89LJJj3fIQQA0AAZOBkrvx7NbkFrOCtG/jFxjtNK7VtziaLXQSMEhtcJ3LnEjHiu2zjtgY3AJ2L7NH2fNspLXp/mT4tQGeuqmit0/Z5I/ggYfoVEoPdxG4aRsDuPX1CVvE9ERFFEREBERAREQFs7jBqq4aI4War1daKV1RW2m01NVTRtGS6RsZLcDzwcHHyW8VDPBDVQSU1RCyWGVhjkje0Oa9pGCCDsQQSCPmg+fzh57Rnmg4ea2qdYU2sxdYa6YyVVqr4g+lewnJaAMFpwSAckjvuvY3k45r9O82nDD9mdvoWWu82+f3S72wSdfu82AQ5p7ljgcgnfHfB2XWrnp9mfwx1LoG88TOBOlWaf1da2vr56Cic73a5RjLpAIiSGSblwLcA43XnTyf8yeuOVvi/BX2t8kdDcp2W682+YHpe3r6cuafNpJ+Yx6gK88H0TosGyXOK9WagvMIxHX00VUwegewOH5is5QEREBERAREQERQVdTT0NLNXVk7IIKeN0ssjzhrGNBJcT5AAEkoJ10H5+PaR0PAGaThfwfNvvGs54T73XGQSwWnOQAQ04dL59JO2+cHC4F9oX7Sq267tbuD/LhqW5Q0PjvF71BTF0AqmDIENO4Yf0E7l+2RgD1HAfJ7yA8UuZLWtPc9c2S96d0SxxqK+7VkDopavO/RD4gBc5xO7sEYJ37kJNHH3AfgxxU53uOzrPLeaqsrrjI64X++VWXimgz8T3HtknZrfM9gcYXvJy+8A9C8uHDeg4baDpnCkpcy1FVKB41XO7d0jyPMnsOwGwVnBngBwZ5bNKTWPhlpeisNE1ni11bI/qmqOhu8k87jl2AMkkgDHYLqlzYe1R4ScO7HdtIcGLs7U2ri19Oysp2H3OjfuC7xDs9wORgDYj8ijv2uP+LvHHhvwU01V6k11qWho2U0Ze2mdUNE0rvIBpOftIx9uAfnzrOK3M9xXu9Reo9YcQb5U1Dz4n3OqKt7AST8IZEcDv6Ka2cunNPxLuUcLeFev7nUSOAEtwt9SAM+ZfMBtt6pm+DvPxE9thfKe61FHwx4SWuooWktjq7rVS9TvQhjMbd+5H1LrRxn9ppzPcaLJU6YuF3tNhtNUcPgs9I6J5HoZHOcSF275U/ZHaOOmItR8y1JXVV2qD1R2imq3wMgHo8tIJ9Mdz32GAe2en/Z9cnWmZ4qu3cDLI+eEhzJamSedzSOxHW8jP2KZLdp48eeWvkE4780cL9RWGKjs1jc89d2u73gSOJ3LWgFz8nJ9fPcbrt3wz9ipX2rVtvunEzi1a7rZaWVktRQ26gkY+fBB6C6Q46T2O3b8i9QbFp+yaZtkNm09a6W3UMAxHT00YjY0Yx2Hn8/ktRyFRp1gsFm0vZ6TT+n7bBQW+hibDT08EYYxjAMAADz9T3JyfNakiongqiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiII5ZY4Y3SzSNjY0Zc5zsAD5k7K2nqKepjEtPNHKw/vo3hwP2hdBva2WDj5fOHmnTwppL7U2GlnkkvcVnEhlO3wl4j+IsGBsPn5ZXl7wH5m+L3LpxHt2pbRqW9CnoKkC42ioqZPCqYs4fG+N5wDjODgEEDO2Ql/GzqS6+kFFxDy680XCPmc0yb/wANdQNmqaZjDcLZN8FVRPI7PYdy3OQHDY7dsgLl5FEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAWnaiPTp+5uxnFHMcf8AiFaitK1VM2n0veKh+OmKgqHuz6CNx/mQfP3yO0MeoueHh1FPAJ2S6llqXMIyCGMlkJI+XTn7F66+0S5h6LgJy83llNcBT6i1XTzWm0NB+Lre3D3gZzs135z6Ly09mPX2ig55tHy3V0bWzG6U9MXf/N30k7WAepOSB9efJcxe2s1lFcuNGiNE09UXCy6ffVzxhxw2WomdjI7Z6IgfqcnlHB3sz+HNTr7m50bUvthq7dp+WW6VZfH1RN6In9AdnbPWQQD36V7+nsupfs2eAen+EnLfpfUxtTWaj1ZQsulfUPYBKxsvxMjz3wBj8oHku2qAuo/PTz5ae5ULPDp2w0tNetd3WAyUtE6T4KOMjAmmAye+4ae+PRZfPvznWflZ4dPorDV09Try+sMVpoyQ73duN6iQdwBkYHmSO22fGbh/w94385vGKSitzq/UWorxMJ7lcqkl0dLETvJK47NaN8DbONvMh7wV0xpjjhzsccn0lLJPf9WaindPU1U7iIaWIHJc47iONgJwP0kkr2v5KuS3SnKPoyopG1sN71Zdy191u4g6AQBtDEDktjBye+53O/fcvKvym8NuVjQ8Gn9KW6Gpvc8TfutepGAz1kvnuRlrAc4aMDGNuwHOKeAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiDavFDU8ejeH991LJUCA0VG97ZC7HS4jAIPrkr5vdWXWTX3GqvuVY41JvGoMOyc+I10waB8wRgL239qDrufQfKZfqqjkcypudZT26ItOCDIXAn7MAryV5EOFDeLnMRYLRUhroKKRtZJ1eZDgB3+sn6wENfQJoqngpNG2GlpoWxRQ2yljjjaMBjRE0AAegAwtbUUEEdNBHTQtDY4mhjR6ADAH5FKgIiICIiAiIgIiICIiDgnnN5iLXy08D7rrqtpRV1tW4Wy302ceLPKDjPyABJ/8Asrww4Q6D1nzN8fKWjs1s66y63IV9Z7vGeiCIPBJ27eQH5fIru17ZjjvpXVFZpTghp24MrK+wVk10u5heHMgkMfhsidj99gvJH5fJcg+xb4SR2nh3q/i/c7aG1l6uDLZb5ntOfdoW9TyMjze/GR5D5obr0Y05a/uHp612XqDvufRQUufXoYG5/MtSREBERAREQERRVFRBSwvqKmeOGKMdTnyODWtHqSdggkAwvMn2m/P3VaPqL/yx8Moo3V1TRCmv93bN8VMJW5NPGBuH9BHUTgjI+a5x50vaFcOOX/R9ZZdBagt+odeVrDFR0tJKJYqPIIM0rm5G3kAck/Vg+Hl4ueruJOqrhqCvFder5d6iSsqnxxullke45c7ABOMkD5DARLNerXIt7L3S2nDpTj5xf1BTakq5qSC72qwwQA0UD5GNfFJO9xPjloOQwBrQcE5xhelrWta0NYAANgAMBfO9pXjRzyWXR1Nw80ZqnilSWChjMFPQ0FPUgQR7noa9rOtrdzgA4HkttXbixzaWiGS337iBxUomSjDo6u43CIkf+M4FCft6X+165mptD8P7VwR0RqYQXnU0r5r02lmHiw0LAMRuLTlhe8jY4JAyF0k5GOQrUXNzW1+o7nffuBoyyVDKerrGxdc9VMQHGGEEgZAIJcdhnHfC444M8s3HbmX1zBbbTYb1UGqkaa283KOUxxszu90j93EDyyfngL3m5Z+AmnuW3hFaeF2nZDOKPqnq6hwwaipfgvf9uB+T7E/1WvcIuD+heCWirfoXQNlgoaGghbF1hgEszh3e9wGXEn1W91QHKqgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAupXO/wAiGjOZrR9RddMW6gs+vaBrpaG4RxBgqyASYZiMZB7AnscZ7AjtqqHPkg+cThlxM41ckXG+a52+kfa9QWaR9Fc7ZWMJgrIM4dG8DGWnGWuG4OCPML3B5P8Amz0lzacNhq20UzLXeqCU013tBlD300gAIeD3Mbgcg/YcFaHzc8ivCnmptT7hcqX7jawpoXNor1SgB7iB8LJh2ezO3qM53wMeI1n1Xxi5ROM90o9O32tsWo9MXJ9DXRxvLY6kRSfQkZnDmOAyAd8O2IJVH0oItrcL9YR8QeG+l9cx9GL9aKS4ODOwdLE1zgPqJI+xbozvhQVREQEREBERAREQEREBERARFFNLFTxPqJ5WxxRtL3ve7paxoGSSTsAME5KCVec3tBvaH6t4A8TIeFXDFlPLX0lLFUXGV52jMgyG533xjby7nyzzVxW9ppyn8LqmttL9by6gutH1M91tNLJM0yt/eOlx0t32J3x814j8YuI194+8ZdQcQaqmebhqy6ulhpgcloe4NijG+MgdI22z2VnE9e1nIBzQaz5kdI1N31VQvDoch0uMtD2npIB8xkLt2uH+VHg3Z+BvAjSWiLZSsjqY7bBPcJQ0B81S9gc8uPqCSFzAsyYvvoiIqCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC6686PMPpjgnw0rbXX3GOG7ahpZaWlYXYIa8FpOPU5IC7FLxO9sfqeureZu3aYdM8Utp0/Szxt6sDrlc8k49R09++5VmfR1bobTxF4H6wsvFRtqqqWGlrhV0NY0Ya4nOwPkSCQt+26j1/zz8xNJcaiCernrZKaGse92XiFpORsSdxkfLOV3y5++H+maX2c+ktR0Ntp2VtHHYp2zhgDyKhjS8EgZO7gPqC479ihoJtwv8Ar/iFVQdUVsZS0EDiNhM/qcT9fTn6sgqTPad8equmLJDpvTVp07SsayK2UUFGwNGwEcYaMfkXG/MzzGaK5bOGN01vqi5U7a1lPILXQOeBLWVOCGMa09xnGT2wFv8A1rrbSvDnS9w1nra+UtostrhM9XWVL+lkbB6+ZPoBuV4Hc8vMhVc2vMFPdtINr6nTtGI7TpujLT1ysBwZBH3D5HknG5xgKzN6NjUreMHOdx8pqKWrqLzqnV1f0NfK4mKliLiST5Mijbk+Xb1K9zeUDlK0bymcPDpexzC5Xq4uE94uz4w19TKBs1vmI274H2ndba5GOUHQnLhwrsN0fpumOvrtboqi93WUdc7ZJWhxgYSPvbGZDcADJBJJyu0CXJ4giIooiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiKhGRhBwDq/nW4K6C1c/SWq73HQSMk8MzSytAzn0P+tcxaT1tpLXVqjvWjtR2+8UUjQ4S0dQ2UAHsCAcg/I4K8g+fvkl5hb3xar9daJ0RddS2mqafhoYzK6LBJJAHcHO4G+w7+XULQnEDjfyva9pLraZdRaTutFMJZaCqZNTNqGg7tkicAHAgYzj6irZJxmb6+lZF0h0F7W/lQvek6C4a11BdtP3wwM99oHWionDJsfF0Pia5rm57bg/X3O76L2onJZXR+I3ipLD8prTVMP5DGs2z8eNO1wGEOy6rn2nfJaHFv+y2Djz+5lTj/AKCwq/2pvJXQs6v9k6rqDjtBZat5/wChhX08dtkXTN/tbeS9rukau1C4eosFRj84BVf3W3ku/wCF2of8wVH+pB3LRdNGe1r5MHvaz9l2oG9RAy6wVGBnzOy7O8L+Kmg+Mmj6TXfDnUNPeLNWEtZUQn6Lx9Jjgd2uGdwU0cMe0H4K3Pjry033S1jGblQTRXSmaO7jFkkY9Dnf5ArxH5eOMOoOW3jTbdYRwOikttUaW5U7xuGB2HjbzaRnzzgjzX0kyxRzRPhlYHseC1zT2IPcFfOtz1aEj4c81WvtOU7Q2mNwFVTgDH3uRjXA48iTk/aoPoc09eINRaftuoKVuIbnRw1kYJzhsjA4fmIWorZfBV4k4O6GkAID9OW12D33poyt6JAREVBERAREQEREBdPefrnosHLPo2q0ppGtgreIV3gfFRQNcHC3hwx48oByCM5DdiTjcLc/P7zQ1fK9wVff7B4R1JfZzbrSJNwx5aS6THn0jf8AL54XjPwd4N8bOdjjBLBQGsu1bWVDai93qpJMVFA52C97jsMDIa0dzgJmoxOWrhbfuaLmP09o+7z1dyffbia291jiXvFO0l80jz3AIGNu3UMYX0O6F0NpbhrpW36L0baYbbabbEIoIImgAAADJ9Scblcc8uvKfwb5ZtPw2zh5pmFtzfA2OuvE466useB8Rc8/RBOcNGABt5LmdX/D9iIiiiIiAiLhLjxzkcvvLhPHb+KGt20tzmAey20lO+pqi0jIcWMGw3G5I7hByZr3W+n+HGkrnrXVFbHS221QOnme9waDjsAT5k7LxN5tfaT8TuN1VXaU0RXzWHSwkcxrqdxZLOwEgHI3AI8ySd/Jb29oD7RjS3Mbo6n4U8HaG8wWaWpbNX1dXF4L6oD6MYYCTjOO/cEjzwut/Bbki5jeONzo4NOcO7pRWupc3xLrcIHQU0TCd3ZdgnbJGBg47hJpjinRGiNZcVtYUWkdKW2pu97u84jjY3L3OJIBe87kAZySfzkgL3E5KuQDRPLbppt11dSUl+1jcI2urJ5Yg+ODbeNgOQQMn1H1k5W/OVjkx4Tcr+m6WHT1nirtTPhaLheqhodNLIR8QYT9Fuc4A/NnC7Bq6MWjtlutrSy3W+mpWu3IhhawH6wAFjXXTenb4WuvVgt1wLNmmqpY5SB8uoHC1NFBi0Ntt9rhFNbKCnpIhuI4ImxtH2NACykRAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQUIXjR7W7lp1RYeL9Xx7sFhdJpvUFNT/dKeBmRDVsYIy9/wD3wa3J8sA9jt7LriTmx0/ZdS8tvEm2X+FklINN19R8TQel8cLntIz2OWgZG+CUneJbk11I9lJzew8QtIUfLrqaMsvWlqA/cybqz7xRsx8Jz3LQfs+ojHomvAL2YOpjprnR0KzOG3f322OOf/mlNIR+UsA+1e/qtIIiKKIiICIiAiIgIiICLHrK2kt1LNXV1RHT08DC+SWRwaxjRuSSdgFwfqfno5TNHzupb9xv0/DOwlroY3SSvBHfZjSU09c8Lyc9pV7QvUceor1y68GblFSUFG0Ut+vVO8mWaUjL6eJw2a0ZALhnJyM+mu873tU9G3LRE3Drlh1BXVdzumYrhqAUz6dlLTkbtg8QBznuzjIAAGdz2PUjki5KNX83WtpL1e5qqi0XbqgSXi6yZL6p5OTDG4nLnu3y7ORv55IfsWcp3s9uMfNPSfsqoJ6TTekmymN13r2ucZ3Du2GMbv8AME5ABx6r0V4Ceya4PcH9V2/WmoNU3HVdwt0gmhiqIGRQtkGMEAE5wR5jPzXc3Q+idNcOtJ2zRGj7XDbrPZ6dlLSU8TQA1jRjJwNye5PmSVryXota1rWhrWgADYAYVyIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC8RPbG2qSl5qqW7OaQyu03RtafUxueD/wBIL27XmB7ZXgfe9Q0eluMVjonVEdnp5aCvDNyIy7qDseeNvsB9FYODtJ8xWpubLltsnKBU0xF5txpYIagDLqiOAgQu8u2wI9R6YXpfyh8ummeUXgbHpiqr4BVyF921BcpiGNdMWDOXHADGNAAz6FeIvJ7xvsnL5x1svErUlvlrLfRNfHMyIZc3qwQ4D1BHl6rlfnK53uJvNprU6Q4dVF9pdFBjYqWxULZOuufg9T5WMHVJ3wGkYG+3bE+nGqe0M5+LnzK3ufhloR5o+HVlri5j2uPXd5oyQ2d/kIwcljd+4JOcLnv2QvKqx7LrzBcQdMte1wZS6bbWQZx5yVDA4eewB9Nx3UfIX7L+krqWl4r8ydhqA4PbLbdN1ALGkDcPqB3I7Yb2+0ZXqdbLZb7PQQWu00UNJR0zBHDBCwMZGwdg0DYBS9GWiIqCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgouEuablS4ec0+hZNK6ugbR3KA+JbrvBE01FK8ehPdp8wfn6kHm5EHkJW+xF4mCtkFu426YdSdR8MzUFQJMeQIGR288/YtMuPsTuNdOM2/ixpCsHoYKiM/nBH517HIg8ZIfYscfZCRNxE0fEB2J8Y5/I1TR+xT47vmDHcTtGsjPd5bUbfYGkr2VRB5CRexC4mujBn46aYZJ/cttdQ4flLgfzKrPYhcSuoB/HTTIb6i1VBP5Or+devSIPGPiL7GLjlpfT8940TxB05qypp2F7reIJaOaQAEkRlxc1x22BIJJAC4O5b+czjryT3S+aQtNooqmklqT90LHeopOmGobsXNLXAsJHfBIOx88n6DVwrxR5NuWrjLdn3/iHwntNxukhy+tYZIJn/AFujc0n7fMkpz6PNuX22fG57Hsj4R6IjLmkNd4lWS0+u8mDj0wumurdean5l+O8eqtfV0Dbjq2700FS+NvhxxMe9rA1o8gAcD5+i9t6r2aXJZU0j6VnBekp3Pb0+LFcKsPbt3BMhAP2LyG51+WO68p/G2rsdmjrf2NVMjK7T9wfknoOHCMv83sII9dvPBKmTR9AmlLRDp/S9nsFMQYbbQU9JGR2LY42tBH2ALVl1k5IecLQHMvw5tVspbtHT61stuhivNqlOJetjQ0zMz9NjiM5HYkgjZdm1bP48SXRERFEREBERAREQeWvtu6eoFp4aVXWfANRWRludusNBB/ISt5exPbQHgZrp8NOG1f7KWtnl83sFLEWDPoMu2+Z9Vtv23czTo/hrTgjqbcqt5HngxAD9BW+fYv6XqbTy36j1JMR4d+1RM6EefTDBFGT+XP5E2j0DRcbcT+YrgnwZmZScTeItosNRIwSNhqZT4hafPABI9d/LdcYye0e5M4nmM8a7Y7Hm2GYg/UQ1TR2XRdaI/aPcmL85422tmP7qCff6sMW3tQe1R5MLD8LOItdc3f3NvtFRL+ctA/OreDtvgLafE7iZpHhHoy6a51rd6egt1rpn1DzLKGGQtGehgO5JOBsD3z2XkRzs+07v3GIU+juAVzvmm9PNDjV1ocaeqqiezR0nLR9uRtjzXVCwcMuZHjVcaa02vTOutTSVb2lj6iKqmi3Iw4vflo7jfKel47g8RfbQ8bLlcqyn4a6F0vZbcHubSz10UtVUFmTh7h1tYCRjbBA+a68cM+CfMX7QLijfdTUNTDcLi94nut3uMhipoM9mAgHGB2aB2+Z39DOXT2R3BSzaLtd147224X/VE7G1FVSMr5IKWncd/D6YyC/Hnk/Ihd3uGvCfhxwe0+NK8MdH27TtrD+s09HH0hzvVxJJce+5J7n1Q/boLy2ex/oeHOtbdrfjHryh1G61ytqILXbqZ7ad0gOR4j5MFwB9AB8s4I9I4IIaWGOnpoWRRRNDWRsaGtaBsAANgB8lMiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiKKeaKmhkqJ5AyKJpe9x7NaBkk/UAglWk6r05bdYaZu2k7zGZKC80U9BUtHnFKwsdj54ccLp1xe9q3wB4YajrdK0lFdL5XUEhjmdTswxrhkEZ7fnz8ltPSftluAl0uDKbVGlr9Z6Z7g01DY/G6BnuWt3I+0JO9hZnK6Lcz/ACvcTPZ+cWdNa80tqIVlufXurNO3drC18csRB8GUf3QaRnB3BPbz9ZORvmrp+bDg9+yyto4aHUVnqfufeqWI5aJukObK0eTXgkgeoPbsOi3tMec3l25g+EVl0Vw3u016u9Nc2XGOd1K5jaYBha4ZPYlriCF1M5NebLU/KjxMZqShElZp65lkF7twdtNCDs9o7B7ckg+fZN4j6KEXAen+evlP1Dp+j1FFxr09RxVcYd4FVP0TxOIyWPZgkOHmFyrojiXoLiTQG56E1Xbr3TAAl9LL1YB8yO4H2Irc6IiAiIgIi4o5h+ZThfy06Ln1bxEvccMjo3+4W+Mg1NbKBsxjO+57nsBk74wh65XW0eJ/FDRnB/Rdx11rq809utlthdK90rw10mBsxgO7iTjYZXgNxe5wOYvj/wAQau502ttTUjK+qcLbZrPWTRiJhJ6I2MiILjj68nsFHNy586/ECmibc+GXE67wSESRtucNSW5PmBORv80n7HMXN17T3iXx8pq3Q2gKU6R0dKXRv8N5NbWszsXv26AR5Aeh2IXD3LNyX8Y+aisqX6Io6eitVIcT3W4FzIA4ncNIBLyPPH5znHaHk09lXq3VOoWat5lLBV2WxUThJHZnuDZqx4wQHkHZue4GQQCCd8L1r0dojSPD6xwac0Vp6hs1tp2hsdPSRBjQB2zjcn5nJV1LP6eWGm/YjayFbDJq7jdZRShwMsdut8xeRkZAfIQBtnuF6acGuEWkOBfDu08NdEUQgttqhDA7pAfNIQOqR583Ejckk9sk91vlFFEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQF5w+175kZ9B6ZsvBWwRxuuOoYX11XM7B8CAEsGB/dHfGdsHO+MH0XqqqmoaaWsrKiOCCFpfJLI4NYxoGSSTsAPmvAP2kPG+xcc+aC93rStX7zZbHTQ2Skma7LJjFkySN+Re5w/8UfWg4Y4KcFtdcfeINt4ccPraaq43B4DpHbRU8QPxSyO7BoG69wuTPkM0FyqWd9xq5oNRawrQ01V0kpwGwkfvIQcloHrse/qSYvZ38tui+CvAHTGpaO0RO1Tqu2Q3O63B7QZSZWh7YmkjIY1pAA898rtaroIiKAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC4Y5seX6w8xnBe/6FuFvhkuj6R8toqXMBfT1TRlhae4yRjHb1XM6JR82nDPXfE3k+49UWqGW6e36h0vVugrKGcOY2ogOWyxO7Ese3OD9R8l728sXMho3mi4YU3EbSEc1LiQ01dRTEGSlqGgEsOO432PmPXueoftgeXO06k4X0HHXTdlY2/wCnaxtPdJIIvjqKGQOy5+O5Y8A9RycEhcC+yM5ntFcJtQam4Ta9usVrptWVFNV22qmJDPe2AxujJ7DLS3B7kj0BSo9lEUbHskY2SN4cx46g4HII8sH0UiKIiICIiAiKKeZsEEkzu0bC8/UBlB5Ge2i13brvrDSOi6Wpa+a1RySzMB+iSDnPz+ID7FwNyo+0b4o8qvD+v4b2XSll1DaJqmStohXPkifRzPA6yCz6bSQDg4wQcHfC4n5puI924q8dtUXu4VT5xHcpqOnzvhrJC049ckH6xhesPKD7O/lptvCGwaq1roGi1dfb3Ssq56i6h0rIiScNjZnpHbc43/ObuciSc68oLtW8ceczi/WXuSmqb/qO8zdTxEwtp6ZpOA0DcMYNgAN9uxXYG3eyD5qLhTxzv/Y9TF7Q4xzVvS5vyII7r2R0JwY4UcMWkaA4fWSxHGOqjpWscB6A9x9hW9U3ivD+o9j3zUQkeFLpqX6q8D+Za9YPYv8AMPcgDfNcaQs/r1SSz/8AQavaJFmfseeHK77JTTvCDWdLrnivq+h1fU0Di+loaejLKYP8i4PySR/r7ZyvQmmpqejgZTUlPHBDE3pZHG0Na0eQAGwH1KZFTBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFi3KhiuduqrbM4iOrhfA8juGuaQcfPBWUiDy+4n+xln1brK6ak0pxeprdTXGpfUeBWULpXMLiScFpHme2T9a29SexBvfSffuO1Dny8K0vA/O5esSIPLmy+xDsgfnUPHGtc3zFHbWA//hErR+LnsW47Fo6vvfCfipX3e8UUL52W+6UccbajpGehj4zs442yPy9l6vIg+WWstFwt91msldTOp62nqHU00UmAY5A7pLT5DB29FyxwU5iONvKxqRt30Nc30bXn75SVUZkpahuQTgZAIPqD57hdjfalcplTwg4if7LmmKV79NapmJqC1u1NVHGxwMDPr9W25XPfJhfOUnjPyaT8KOJ7tMN1JS0tVS3KK4eG2vDt/AqYHnD8j4S0sOQ5pz5peEm+uVORP2j45pNWTcMNa6Rp7JqdlG+sppaKUvp6tke8mzt2uAIOO3pnfHeUnC+frlG1FJyvc22ndUa5hqKCzW+qqaGorZGFkclPKx0fW0nAIIIOMr3u0tqrT2tbFS6k0td6a52ytb1wVNO/qY4fzEeY7plTdawise9kTHSSODGNBJcTgAeZJ9F5mc/vtNqfS7a/g7y+3MyXlrnU90v0ZBZTDsY4T5v7gnsPswSu1nNjzscLuVewGS9VMd31JUNd7lZaaZplcQNjJv8AA3Prv9WQvEPjVxo4tc4PF9l+v0ctwu11nbRWi0UmXRUrHHDIYm/ky44zgk7dsLhzwe46cz2syzTFjveprhWTf7buUzXyRxZO7nynIAGScA7DyAXtHyf+z+4V8tFst+pLjbIb5r3wg6outQA8UryBlkAOzQD5jfYEbjJWfYNrchXs9LFy10cXETiA6nvHECtp24Phgw2oOGTHETuX9gX7bg48sd2URAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEXXfnF5vNP8p2k7ffLnbX3CtucxipaVveQgZIByB5E7nyWw+V32mHBbmIukOjrqybSOqZj0wUle5vg1R9IpASM/InOxS86ku8juIiIiiIiAiIgIiICIiAiIg6t+0t1heNFcoWr7nY62Wkqal9NQmWJxa4MlkDTgj7F8/a94vao3yy0/KhqDT1XURmur5YJKeHIL8Ru6i7HfAx3+teDqD6c+CkUcPB7Q8UI6WN07bgB6f7XYt6rZ/B0Y4TaMAII+4FB27f2uxbwU/GZMBERUEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQafe7LatSWmrsV8t8Ffb66J0FRTztDmSMOxBB/wDsggEdl87vOrwjpOAnM7rPQdgidSW2jrI661hriHR080bZWYI7YLiBjtgYX0ZLx99r3y668HFQ8wNptU1bp2uttNSV0sTer3R8LekOdjyI7ny2PbJDfiPUDl61C3VfAjh9qJtUKk1+mbdM6Yfv3mnZ1H8uVyEvMX2SvN3X6lo6Xlj1U0PltFHLNZKgvGTTs+IxY9Ggn8oPmQvTpSc4tERFQREQFsLjrru3cM+DusdcXSpbBDarNVTBxIH3zw3BgHzLiFv1eS3tduay7yalPLTpebw6Clp4qq+TNd/ZHvHU2LA9ARnPby77B0v5SOFk3MLzNaU0Zc3OdBc7i+4XJ47iKMGWQ4+ZGMfNfRRYLHbdNWaisFopxBRUELYIWZzhoGBk+ZPc/NeRvsdeBN8uHEms49VdO5lqtdLUW2leRtJJIOl5B+WMfYV7BoCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg2fxV4WaO4y6IuOgddWxlbarjGWva4bsdggPafIjK8EucflX1Nyh8VxaIK6SpslY73uyXBoLXGPOzXejgQR3OcH5r6HV0V9rBwUOvOCE/EaD4pNIQOmkbjcRgk9Q+WTg/YrB5j803G608TbTovT9nooI22e3h9ZM1vxSzEADJ+QDs/WOy9NvY73etr+VmqoqysfMKDUNUyBjiT4cRZGRj5ZDgB5YXRr2YfLdwt5k+ImqbTxQt0ldSaftsVdFTtkLPFL5OjBI8hsTt5geeRuP2hdRqHlL4m0fB3gXqC56S0vc7HFc3U9vqDE54kkkjLS5uCADG7cEHceiTs7Un/ADyOyXtTudeq4b2aDgZwp1HCzUF2jc++VVLIHPoqY7CIEfRe45z6AeoIXSrkj5CdX83lZXatu9+dYtH2yrENZcHxGSetmPxPjhBIBIByXkkAnHdafyN8pl75uOLPj6vNzdpKhcam9XMyPL6hwxiISnJLndic5GQvdrh3w60Zwp0lQaG0BYaaz2W3M6IKaBuBnzcT3c49yT+gJ4rB4ScJNE8E9DW3QGg7TFQ223RNjBa0dczgN3vPdzicnJW9URS3UkwRERRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERB1x5z+TjT/N5o632Wu1HLYbtZpXz2+tbD4rA9wwWvZkZac74OfTfC8SuYnl94kcoPFtmjdS3GF1fTMjuVrutAXCOoiLiGSM6gC0gtILTnBGDlfSGvLT20fCi83h2j+LFvp3yUtoopLbVlrchrXSl4cfqJH2E+iu/Ekzx3i5PeMMPHHl50frp9U2WvloGU1xb1AllTGOl4PzOAcnuSVzSvG32QXMBd9P8UJeBlfUOdab7FLU0rD2ZM0ZIH1k5+0r2SUUREQEREBERAREQFj19XHQUNTXzHEdNE+V/wBTQSfzBZC0HXjZH6G1EyHPW601Yb9ZhdhB41cI9c6m51+dKXQ+ur1LUacuM1e6KmLi5rYIAXZGT5taSB27D6+IOf3lotvLBx3k0jp18rrBeKCO72wyDdjHvex8Y9elzD8t8Dso/Z869peGPOJoS93uZlPSy1lTbKuWU4EYngkiJP1OcFzh7Y2/UuquYnSlvsszKz3LTsdG0wODw+R9RI/AIOCfjA2Uk7U2716xcvFyN34EcPrkSD7xpq3OOO39rsH8y5EXHfLzY59NcC9A2KqBE1Fp6hicD3BELVyIqoiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLYHHzTVNrDglrvTlTRNqxW6er2shLc9UggeWYHr1AYW/wBWua17S1wBBGCDuEHzt8iGtqbhtzc8O75erk220Aupoa6WX4GsZNG6PD89h1ubn0wvohhmimiZNDI18cjQ5rmnIcDuCCO4K8gva2cqXDnhM3T3GnhvZhaHajuk1Dd6SnBEBnLDK2Zo7NJ6XggYGceuBzB7G/jlrHXWkdY8MNX3yqukemXUtXa5KqUyPhgl6mvjBO/SHNBGTtnCmXd+Hx6RIiKgiIg6/c6nNPQ8p3CJ2t/uay43m5VIt9oo3v6WvnLS4ucfMNAyQPz4wfHDg7wN4z+0H43ai1GyupYqmsqRcL9dajIipmyHDQ0DJJAbgD0A+pdofbO8ZNPakuejOE1kr46qosU9Rca7w3AiOR7AwNPzxn865Q9ilpKe3cIdc6uqoA0Xm9ww07z3dFDEQcfLrcR9ifjZ9S78d2+AfBfTXADhXY+F2lx101pgDZagjD6mY/Tkd8yfzLkRERRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFsfjfoyHiJwf1noiaLxfuzZKylYwdy8xO6P8A8IBb4RB88nJxzL3Pky451up7vYZ7lQSU1RY75bopAyUtEjSCwuGOtkkQwDgYJUfM1xp1Rzycx7NSaY0tU077jHTWSx2sEyyxQMJx4hbkZL5HucQMAH5Le3tPuDVs4S8yFbVWWJsVHqeI3IRN7MeSM/lBH2gru97H/gpw9ouA3+zK+x0tVqq8XWrpjXTRh76WGF4a1kZP0SdySN9/mcz7p/juHy+cIbBwO4Raa4d2G2Q0n3Nt8DassaOqWp6AZXvd3cS8ncrkhEVBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBcTc1HD2g4mcANcaZq6KOpkkstXLTB4yWysjLgR8zjH2rllY9bSw19HNQ1LOqGpidFIPVrgQR+QpR8+Ps6brDZ+czhpJV1AginuMtM9ziAMvgkDQSe2X9I+1fQsvnr5xOW7XXKRxrfUwTzQ22vrZLnp+5wZGAJC4NyOzmnGB6D5Feovs1ecK98zegLrp7Xs8curtJmJtRMCA6rpnghkxHfOQQT5lB3PREQEREBERAREQFxrx+4j2Xhzw3vFbc5mCerpJqamiccFzntLSfqGc/M4H1cklefPtS9eSael0nZGvLWVmA7cjJLyPL5AfkTxLceQ+t7ddtL62ragslppPfn1dJK3I28Qua5p9QcfaF3p5CuFh5stbU+stegV79Lvb48so6unpwRjPmcgY+a4i519FUFk0poTUdNGGyV8T2uIGMgtDj+cLuD7Ealidw84k15H31l5pYQf+1MGSPygJu9iTsj0tggipoI6aBgZHCwRsaOzWgYAH2BSoiNCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg6pe0x4V3XivysXm2WG2vrLjaqynukDGDLgI+oOx9YIH2ryu9nzzBnlz47xtvkj6W3317LbXCQFoY4OIAeD23JG4yCB2Xv1JFHNG6KWNr2PBa5rgCHA7EEdsY8l8+3tHOHFBwu5vNZWez0zaWirzTXamijGGxtnia4huOwDg77cpB9A9PUQ1cEVVTyCSKZgkje3cOaQCCPkRuplwPyRcXqLjPy0aI1MyvinuNNa4bdc2Mdl0dTC0RkEd9w0HJ77/Nc8KQFpeprh9ydN3W6h3SaKhnqAfTojc7P5lqi4I5yuPOjeBfArVV01DdIY7jcbTU0VsovEAlqJ5Y3Rt6RnOAXZz22SzeFuPCGwW+88xvMNQ2msrZaqr1hfhEHucc+G+QkNHphgwP8AWvoZ4KcItMcEOHdr4e6UpIoaSgjHWWNx4khA6nfbheGvs0tJV2qucnQEtJEXxWWomutQQ3IayOF+CT/3zgvoGTICIioIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg8bvbRVFM/jFpSnYczMtshd8h972/OF2t9j1VPm5SRTO+jBqKvDftLSV57+064mU3ETmQrIqOQPgs0Jpm4IOCXbg488NH5l6J+yLsdTaeUilqqhhabheq2oaPVhLcH7RhSdm0d2URFQREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQcHc3HK1pXmw4Xu0Hfqx1ur6Kf3203JsYe6lqA0jcHuxwOHAb47eh8WKO88avZ1cytfaqSuibd7JIyKsY3JprnRPAe04Pk5jsgncHPfz+hheU/tX+TziZq7W9RzH6GtTrvaoLRDBeKeDBmp2wAgShndw6TvjPYY32V94mPSvhPxDtHFnhrpriVYng0Wo7ZBcIgM/CXtBczffLXZafmFuzOF5A+yo5y9QWLVVl5X9WEVVku00rbJO4/HTTEF/hZPdpIOB2Hb0z6/YylVVERQEREBERAXkN7ZrUZh4raHtDJz94ofenMHlh/c/lXryvEf2x1ZJNzS0VEfoU2nKQt+XU55P6EiWa3n7RXh19yuVfg/rCFu0ssccvyD6dxH5Tj/ANwuWPYhOB4ZcTWeYv1Efy07v9St9oTQe9ezh4a3GQtMlM6yO33OX0xBx9vdR+xBDhw94ou8jebeB9fgSf6wpLxXpmiIqCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLoR7TnkspuNGk6zjfpSUw6o0tbXvqIenIraWIFxH1gZx6AZ9Qe+60XWVr+7mkL5ZOkO+6FtqaXGM564nNx+dJc6PFv2SnGq7aM5iqPhlWXiRlj1bTzwspXPPR721hczpHqcHP/ej5r2+XzH6bvmoOB/F+ivlEZYLpo+9B4/euLoZCCPlkAj7V9GHAnixauNnC6x8RLQ5vh3OBrpGtOQ2XALgPyg/LOEz6b8cgLwo9rLxAqNV829503Ddn1Fu0xQ0VDHAHZZDOYWyS7eRy8A/V8yvWXnK5gX8tfAu78R6SnZPcBIyioI39jPKCGnHngheHXCrhJxf51eNdc63QzXC53mtNderk7ZlOx7t3EnYYGwB9BlB65ezg5UdI8EuC+nuIlVbevWesLXDca2qlALoIpm9bIWegDHDJ+Z+ZPcVaRpKxR6W0pZtMQvDo7Rb6egaRsCIo2sB/I1auiQRERRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFpmpKttv07dK97wwU1FPMXH970sJz+Zamth8ea6e2cEOIFxpTiWm0xc5WH0IppCEHzs6jgufF/jnX0duD6iq1BfJIYcHJLDIQCP8AxRlfQbyx8NBwk4J6a0QaZsElHSh0kYGOlxA2x9QC8RPZw6Yg1Vzk8PKGpibLFTVNRWvY4ZBEVPI7B/IF9CCH6EREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAWJc6Cmu1tq7XWRNkgrIHwSscMhzHtLSCPQglZaIPmn0ze7jwC5jbffYmvpqjRWqmyEEEERwVGCN/IsB+whfRdw24iac4qaPt+tNLVQmorhEHjfJjcRktPzGV4ue1K5fRwo4yv1tQR9FBquV8oAGAZME5z5nAIPzC7weyC1tJqnlxrrTPIHy6fuhpHfESQCzLc/+KApuzieY72oiKqIiICIiAvNP2qvKPqriberZxn0lF4ot9CKO4NDckMaSQT6Y8j23K9LFousrdT3fSV6tlWxr4aqgnje0jOxYR+XzQeD3M3zj3nixwf0nwFp4THbtLFgqpSB99fE0ta31ODvnywB5ld6PYr2CS38CtZXx42u+oW9B/7WKENx+UleP+qoxFqe7xNOQyvqGj6hI5e63ss9MN03ykWB3R0uuNVNWOGMfT6Tv88kp+kjt6iIiiIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIPGj2u/Ldp7hjrCw8XNJ0DaSk1bNLTV0bRhoq2t6sj62gn82+F2X9jTrI3/l2v2mZqx0k2nb++NrHPyWRSxte3APYZ6gPLbHkuVfaRcBbrx55b663acpvGvWnqpl3o2AZLwxpD2gYzktOPkMnfC8pOQnmQ1Jyy8frfQ1TXix6irobLfqKTI6MydAkx5OY47/nzjCSDvz7aq4VNLy+6Po4iRFW6qayXHYhtNK4A/aB+Rbd9iLQUf+xrxJunu7PezfKWDxekdXhiDq6c98ZOcfJche2QsDLpyoUV4Lcvs2qqGYH0bJHNEfzvati+xEz/ALE3Ejbb9kVN+rBIPSdERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAXVb2ifH/SXCLl01dp2qvUMeo9UWqa2W2kDsyu8YFhfjvgAu/T2BXajv2XgB7THXV31lzha4oa+pc+m07PFaqNhcS1jGRMJIHYEuJzj0Cnd4Nz+yWstxuHOLZLnSUr5Ke2Wu4yVMgaemMPgcxuT2BJdt64K92VwFya8vPD7gVwY0zHpaxU0V3u9opKy63ExDx6mWSNryHO74BOMdtvqXPqtzeG6IiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg87/bTabgrOAuk9TeEzxrbqNsHXjfplhk2z6ZaStt+xEuVE7QHE6zCpZ73HeKGqMOR1iN0D2h2O+MsIz8l2P9o1wB1nzD8ulRpPQFKKu+2y5092p6UvDTUCNr2vYCduoh5wPVeQ3Lfx24ncivG+puN20xXU8nh/c+/wBiq2mKSaHqDhsf3zTktOcHffzUkzT19DyLiPl25n+FHM5pMam4bXwSywBor7dN8FVRvIGWvYdyMnZw2OQfMLlxUEREBERAWNcoRPbqqDykhez8rSFkrEudXHQW2rrpSAymgklcfQNaSfzBB8weu6R8XEDUNBGCXMvFXC0epE7gAvoe5OrB+xvl20hazF0FlE12MYzkD/Vj7F4I6F0zVcYeZG2acsrfEk1Pqs+EAO7H1Bef/wAHK+j3R+nKbSGl7Zpmkd1RW6mZAHduogbn7TlBrSIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiC1zWuaWuaCCNwRleEHtN+Aly4L8yNy1dZLVLSad1U9l0oZ4WFscVQQPEYCBhpDhkb5JyV7wrifme4SaN4ycFdU6X1jaoKmNlsqqmlmcwGSnmZG5zXsPcHIGcfpAIeeDyG5pPaL3LmP5dNO8G6jS76G5Qy0s98rXSBzKl9O0hpZvkdTviOfP07LvB7Hrh9XaS5bLjqO4Uz4XapvT6yMO7ujZG1jHD5EAEfWvH7gpw+quK3FrSfDqkZ1SX+6wUZGCcsLh19t/ogr6TuH2iLLw30ba9F2CARUVrp2wMAGOogYLj9Z/NgeSfltxJzjciIiKIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg0HXGr7VoLStx1XeZRHS2+B0riTjqIBwPtK+cPivqWfjTzCX/UWXSSap1EQ0jcuD5QwEfWMFe2ftK71XWDlP1LX0Epjk8RkeQcEZa/8A1Lxd5TLRSXvmD0bS1zeuJlwZK4EZBI7Z/Ln6wFfln086+jHSFH9z9J2Sg6en3a3U0OPTpjaMfmWsK1rWtaGsGABgD0VyzOTAREVBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQF0z59/Z/WvmqpIdcaOuNPZte2qlMMckrPvFyiG7YZSN2uByGu3AzvtldzEQfN/wZ4s8UuTbjoLzBTVduutiq3UN7tMuWioiBw+J4Ox23afmCDgnPvpy/ce9DcxnDqi4h6FrRJBOBHU07j98pph9KN48iDn8/oupHtWeUyx8QeF0/HDRemh+zLTb2PuD6SH466gOQ/rDRlzmEgg4zuRldJPZsc3tLy18TKnS+sHSO0hrCSGCpc07UVTkNZPjzBBAPyA+ZEvJo93kUVPUQ1UEdTTSNkimYJI3tOQ5pGQR8iCFKqCIiAuOeYjV9BoXgfrfUtwqWwMp7HWMjcXdJMr4XNYB6nJBx8iuRTsvLf2xvHu+WN9k4I2aqMVNc6I1teGu7sJIAI+YOPqBSDpf7Ouilrec/hh4cBkEF0knf2+FraeX4t/QkL6GV5Bex85fn3bXFVx2ukJNPaYZaSgBBx1u+Eu+ZyNvQA+q9fU9NEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREFCMrYPH7UVLpTghrzUNZKI4qPT1e4u9CYHNb+chb+JwvOz2tfM9BofQh4CWZ2bnqmm661w/+R05O4PpkdvUkehIDoZ7NC1sufOjw8EkYeKSeqqhnyLKaTB+wkL6B14keyK4Uahv3MbTcSjb5W2WwUNU01DmEMfM9haAD54zv8zjyK9t03eAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg6G+2M1mdO8stu05HKWyakv0NOMdy2JjpHfZjb7V1K9kjy80vE7iNeeJt6YTbdJGGKPb6U7/jDfrwAfqB7rlb23Or4ZKbhpoVj2mSOWtujwHDIy1sYyPLbJ+1cm+xeskNDy46kvLR98umqJer6ooImj9JS9nB6CIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiCCppqetp5aSqgZNDMwxyRvaHNe0jBBB2IIPYrwp9pjyrVHAjjRV6v0tY3QaM1URWUzomfeqapP9kiOBhuSCRnvk/LPu6tscQeHGieKmmKvR2v9OUd6tNawslp6mMOG/mD3afmCEHm57Ob2kFwvVfYeXfjdPTDpp20Vi1FLKWOeWNAjp6jOxOBhr8jPY4wM+pAIIyNwV4k8+/s6rvy+VzeI3Bqhul10VUPLp4mgzT2p+cjJAyWeju4+oHGj8kntGtZ8v8Afv2P8WLvfdU6HqmeGIpKkzz0D87Pj6zkt9W57DA+TOnx7oIup1i9qHyZX6nbOzifLQudsYq+2zwPafnluPtBIXYLh3xY4d8V7UL1w91ZQXukLQ7rppMkA9iR3x8+yDd68MPa9XOSt5wq2iMpcygsFtiDc7NLmOecemeoL3PXgZ7UyaafnP1iZiT0U9CxvyaKdoCg9buQjQNv0Byn8OqOmpWxVF0stPdatw7ySzsEmSfTBH5/UrsKuKeVJ0juWjhf4v0xpS2A/WKdg/mXKySYkERFVEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQF4I+1P1DUX7nJ1VDO8ltrpaKhjGdmhsIOw+fVuve5eA/tQ7a+3c5+tS9rgKuOiqW5HcOgaMj5bIPYTkr4W2bhVy26EtFuoYoqqtslJXVszWAOmlljEmSR32cPtyfNc6LjLlm1NQ6v5e+HV/t8gfBPpugYCDndkLYz+dhXJqegiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAoauphoqWasqHBkUDHSvd6NAyT+QFTLiLmx4oWzhFy+a41hcKmOOSKzVMFIxzsGWeSMsY1vqcuz9ilHhtzucd67j/AMeLxfDO6ahts0lvoG7kdLXYPSPQkAD5AL1x9l/w6u/DjlOslDfKKSmrbpXVNzcx7cODZS3pz9QGPsXkNyQcJmcb+Z3Rujrgx01D72643DO5MEIL3HfvuG919Elvt9FaaCC2W2mZT0tNGI4omDDWMAwAAqn6ZSIiKIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiCCrpKWup5KOupoqiCZpZJFKwPY9p7gg5BB9CvPrmK9kLw/wCJ+pKzV/CvWDdF1dc8yy0MlGZqTrPcgBwI3zsPXfJ3XoYiDw14v+yV5jeGVtnvVjuNk1dQ0zS+R1A6SKUNAP8A8jcCfz4+a67cK+OfG7ll1bLU6I1BX2G40cpZU0FQwmJzhkFskTtsHzxjO25C+lIjOx7LibiJyp8u3FZ0suvOEWnblUSgj3n3QRzNJGOoPZgg+efX1QdWeTj2pml+M1UzRHGmlt+ltRBn3iujkIpKwjuCDuxx9BkfZkjoZ7T2+6a1Zzc3u76QucFypaq30LHS07utrpmsLSAR3Ow/Kt1c/fs+bxy4XU8QOGFDXXHQFU4dTiTLLbJCThjyBkswNnfLOwyB1m5f9K/s640aUsVWHSxT3KKSck79DD1bk+RIA+1XkiZdfRBy92uey8C9AWqpZ0SUunaCNzfTEDcfmwuQlh2mhitlqorbC0COkp44GAdgGNDR+YLMWZxRERUEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQF5E+2L4CX5mvbfx4tdC+W21VBDQV8jWk9Do/hYSfkDv8iPReupK4Y5x9GW7XfLJxFs9wpmSmOwVdVC5w3Y+KMvBB8jhpH2qxK6HeyH5pL3XXY8t2o6kzUsdLLV2hzjvGGDLmD6hgY9Mei9WV4C+y8mki50tDRxuI8dlcw/MCmkcR/8Agr36WfuKIiKgiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC8mPbMcU7ozUFg4V01fIykdAysqKcOPS4DcEjtnJH2Bes68Mfa81s0/N7W0j3feqexW8sHoXMOf0BByH7GLQFTV8XNR8RJ6LMFBan0cExHZz3APwfn8I+wr2LXSL2TulLTZ+XSmvdLE0VVxcHSuAx3JJB9cnBXd1SXQREVBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERBwtzjakpdK8tut7rVW8VoFuMbYS0EOcSMEg+mM/WAvEDkuvVmpeYayz3gRwNq6oGLsGtcXZ6RkjA32+pfQLxC0VauImjrro29Rh9Lc4HQuyM4JGx+wr5z+N+kK7gLzBah0zauunm01d+ul3x0gEPaNsbb4x6J+qPpQjMbo2uiILCAW47Y8sfYr1s/g/fXan4U6P1C85dcLHRVDj6l0LSTv88reCkBERUEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFtnidZ36h4b6rsEcfW+5WSupGt9TJA9oH5SFuZEHzkcpevoOAvNRozVmqSaSmsF4fS3MEbxsc18MgOe2C4/kX0Rad1JY9XWWm1Bpu5wXC31jBJDPA/qa4H+f5Lx29rny3aX4Va4sHFLR1C2jpdYyTx10LRhvvTAHZH1tP6B5b9k/Yw6wvuoeBerLFdq2SoprBfY6eh8RxcWRvhDywZ3wD28gMAJan7ehaIiKIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAvC72vIxzhV59bDbv8AoOXuivDH2vZ/676rG21ht3/QKDv97Kd5PLNQsOdnMO/bs7/3+xd0F0p9lExzeWykc47EswM/98u6yn4ngiIqCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAvAb2oVsjtnOdrVsQ2qWUdS75l8DSfzr35Xgr7VppZzo6qPrQW4/wDJ2oPZblWldPy2cMZnnJfpW2uJ+uBi5UXE3KX+DFwt/gnbP1di5ZT0EREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERB5re26cP9irhtH05J1BVEH0xTdvtz+ZZPsR/95riH/CeH9UYtC9txqa2P0fw50gydjrgy6VNwcwOyWxeD0AkeWSf0LXvYlQvZwZ4hSOaQyTU0XSfI4pWZwpJkHo+iIqCIiAiIgIiICIiAiIgIiICIuq3MB7RTgVy8cS4uF2rPulWXNrY310lGwOZRh4Bb1+pwckdwg7UotO09fbZqixW/UdlqW1FBc6aOrppR2fG9oc0/kIWoqS6CIioIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAvCz2u34YVePSxW7/oOXumvCz2u34Ydw/8BW7/AKDkHoH7KWTq5ZqEYxhzB/0l3QXSz2Upb+1qomjvlh/6S7pqRJdERFVEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBeCvtWnF/Ohqn5UFub/ydq96l4Ie1RnbPzoat6CT4dJQMPyIp25QezHKjGIeWfhdH/c6Utg/5Oxcrd1xXyqtDeWvhg3vjSls/V2LlTslSb9VRERRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBYtyrobXbqu51H9ipIHzvx/csaXH8wWUtj8cbp9xeC+u7qO9Npy5SN+sUz8fnwg8BeYTihrnmt5g6qZkstwmrrk62Wan8mxmTpBAHYHAJ9AB6L2p5GuW6Xlm4IUejbk5r7xXzGvuLm+UrgMNPzA/JsPJeOvs4rNS3vnQ4a0tYxro4a6oqg1wyC6OlleBg/MA/YvoTV+J9ERFFEREBERAREQEREBERARFoettW2nQekLzrS+ztit9kopq6oc5wA6I2FxGT2JxgfMhBxpzWcy+kOVrhVV6+1K9s9dKTS2e3h2JK2qIyGgdy0Ddx8h5jOR4OUNg4u843HO511mtU921Lqu4PratzGkx07XuABc7ya1oAHmQ3YLW+YvmE4rc5nF2KsqYKyv8WoNFp2x0jC8Qsc7DWsYM5e7Yk/n7k+rXsy+UK/ct/De56k4kWiKj1nqmdsksGQ6SipGD4IS4dnEklw8jtkpP2l347S8IdEnhrwu0roF0xlfYLTTUD5Cc9T2RgOOfPfK3giKSYoiIqCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLws9rsSOcK4N9LFbj+Vjl7prws9rt+GLct//wBA23/0bkHoD7KVmOWqhfn6XR/+Uu6S6U+yic48tlGCcgFm3pu5d1lICIioIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIKAYXz+e0ylMvOZrwlxPTLTtHyAhbsvoEXz4+0ic885PELr/AL8YB9QjACsg9vuWCLweXThrEf3ul7cP+TtXKC405aWlvL5w6a4YI01b/wD0DVyWsfjtnSiIi0CIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIKY2wuHOcK9Udi5ZOI9bWVTadrrDVQsc4gZe9hAA+ZyfyLmMbry19sZx1vdpNo4LWqqfFS3GnFVWBpx1tyCQfXILR9RKS5U9dUPZfUrqnnQ0Q9rc+7MrZT8h7u9uf/wAJe/S8bPY8cHbrceMNVxdq6Z7bfaqGelp3kEBznjpJz2O+B9hXsmiiIiAiIgIiICIiAiIgIiIKDZeTftXudK6zXu4cr3D24Rst0ETG6nqonAullOHe6gjsAMdQPmcb+XYHn19ohpfgHZK/hxwxucF04h1THQOdEeqK0gjBe89useTRvn0IJHlHwI5fuMHNzxLlotO0lXcZaur95vd5qCfDpw9+XyPedi45OAPMjsElwd8vZM8ml5tFwpeaHX1LDHBU0L2aYpJG9TyJNnVRBGB8OQ3G4yTnOQPU5bf0Do+3cP8ARNh0RaWNbR2K3wUEXSMAiNgbnHqSCT8ytwJQREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQF4We13/AAxbj/4Ctv8A6Mr3TXhb7XgY5xLgfWw23/oFIO/vsoR/1tdGfmwfncu6q6V+yjc08tdGwDcFhP8A+F/rXdRZ/G7AREWgREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQF8+vtKN+cbXn+MM/wCivoKXz6+0njkj5yNf9ZyH1Mbm/IFg2/Sg9w+XD/eB4dY7fsZt3/oGrkdca8tbi7l94cuduf2M24H7IGhclIkEREUREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAXid7Y+kuLOZW31U0Lm0klkgbA/GziAOoZ9c4XtiuoPtHuVqh498F7pqaz0XVqvStK+toSwfFMxgLnR/aM/ZnzAQcf8Asitc6MvHBqo0rbamAXy2ECrh7SdOSCcfMkH6t/Vd/c4Xzz8hXMHTcuPMZY9U6gqnwaduPXar13xHDICBKR5lj8HfGBklfQPYb/ZdUWimvunrpT3G31jBJBU07w5kjSNiCP0d0zPE/TUkREUREQEREBERAREQF0p5/wDn+snLZZ5+HehZY7hxCuVM7pAOWWtjwQJX4/fb5A77fWRy7zp8xx5XuBly4jUdHHV3WWeO3WyCRwAdUyh2HEEHIaGknY7Dsey8ONB8OeOXOxxnrvuPS1F8v94qPfLrcJARBRxOdjrkcdmtHYDOTj5EhBDwK4HcUebbi0zTlkdUVldc6g1V2u9QC9lOwnL5ZHds+g8zgDA7e9/Lby7aJ5Z+GlFw80bD1FgEtfWvaBJWTnu9x/LgeQKg5ZuW3QnLLw2t2h9JW+B1a2Bhulz6AJq+oxl73OwDjOcDYAY2C5eTfgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAvCz2u3iHnEuJc3DfuDbg0/Ux2V7prx19svw7raXirZeJEULvdqygjo5XgbZaMNJP1gj7Qg7deynfQP5Z6AU0rXTNLfGb1ZLTg+Xku6C8mfYy8Wa+TUeouE1XKXwCjNbTZJ+EB2Tj6iD9hC9ZlJJDMERFQREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFTO+EPyW0eKPFTRHB3SVZrbX15jt1romFz5HDLnYGcNHmUiW43ei80NZ+2t4f0VzdSaI4V3e4Ukbi33urmZH4g8i1mQR9RW/dIe2L5YLpZWVWr6HU9juXSPEpY7calpON8Pa4D8v5UWea75K0va0hpcAT2C8W+bL2qut+J9Y+w8DH3HSthYDG6qe4Nqaj/tsAkDPlnsNsHcnqQ3UHMXqbr1RFduIVwZnxHV0MlY9jfmHt2A+ohOD6XVxhxx5heGvAfQ921hq/Utujlt0D3w0AqWGoqJgD0sawHq3OMnGw+eAvn7quZDmQv1ti0fPxZ1jU04PhMpI66USEj97luHk+ozv5rWNAcp3M9xov8AS09q4XatndWysbJc7nRTRQxsJwZHyygZaM5JBP51c0drKD20fGVmsvfrjoDT8mmjNg0MYcKhsWe4kJwXY8jtnz816kcB+O+gOYfh9Q8QuH91jqaWoaBU0/UPFo5sbxyN7gg5xkb4PmCB5+cdvZAWa3cFKOv4K1NXW8QLTTskrqaoqfvV0Ib99bH1bMeDkt7A4xtkBdMeWLmW4ocj3Fyvjr7JXNphN7nqLTtXmJxLTuQ07NkHcHsRjfGCpmJLr6GEW0eFPEzTHGLh9Y+JOjqrx7VfaRtVAf3zM7Fjh5OByCDg7LdyKinnhp4X1E8jY44ml73uOA0AZJJXzvc+erKLW/NZrq92x/iwyVojYRv2aNh+Ve5HNjqa5aS4Basvdp6/eYqMhpacEA99/JfPDpepl1XxPtVXd5et9yvMEk7n756pgSD8sbJLtwr6PuAdtltHBHQdtmBD6fTtAx2fXwGrfy0jSUEdLpWy00OBHFb6eNoHYARtA/MtXSAiIgIiICIiAiIgIiICIiAiIgIiICIiAseeuoqaRkNTWQRSS7Ma+QNLvqBO6yF4je1W19xRsXNpU0UOprxbbdRW+jntTKapfFGMtyXDpIBOQM5zj7TmXfg9uUXVD2e/NhZ+Yng3abTe71G/XFgpxSXWnkcBJOGHDZmjO+W4J8/P1x2vVIIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAo5Io5o3RSxtex4LXNcAQ4HYgjtjHkpEQeLPtT+TnSHAi9Wri9w7BpLPrC4TU1ZbMZZTVfSZOph8muAdt6j7T2g9jNrO7ah4A6m0/cauaaHTl9bT0gkeXCOOSEP6RnsAcgD5Bbe9tzUTN4UcN6VufCk1DUyO9OptNgfmcVP7EiMDgzxCl83aniafqFJGR+kpUekCIiKIiICIiAiIgLCvF0obFaa2+XOcQ0dvp5auokP7yKNpc4/YASs1dAvadc7Wm+Feir5y9aYEtZrLU9r8Ctljd0stlJOMEk9zI5mcAdgQT32f4OinPrzw3Tm81JbNIaYsj6LSOn66V1thGXz19Q/wC9iZwHmWkhrR5POwJK9F/Za8uN/wCA/Aqru+trT7hqLWVcLhLDIB4kNMxgbCwkbjILnEepPcYK6E+zX5ONb8V+LunOLmqdLz0/D/TtT90Pe6uPEdwqIxmKKIHBcA/pJcNhgYz5e43byTzhuqoiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLqR7TnhhT685WtT3uOibNW6bg+6DHAZcI2EE49cEA4+tdt1tniZpaHXHDvU2jp4hIy9Wmqoek+skTmg/lIP2IPFj2Qd0ioeb2moppwxtdp+4RsaTgOkAY4D5nAcvc1fNLoXVGr+WXjxb9RRskp7zo27ESMILfEa0lrxg9w5pJAO24X0PcEeK9l42cNLLxHsIaKa6wB7mB2Qx+BkA/aD/rQb8REQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBcNc0fLlaOZjhxVaCut4mtpkBMU8YyGv2IJHyIHkVzIDlVTzxLN9eR1L7EPXJumK3jnYmW4PzmK1zOnLM9sFwaDjzz9i1S5+w/uJnabPx9pmw7dQqbI4v+eC2UD8y9XUUxXnVwq9jVwr0lcKe76/4gXLU00D2v91ZStgpyQc9skn6jkbArvrpzQekdJ6dh0rYbBRU1rhiEPgNhb0vaBj4hj4ifPK3AiWS+jr3pLkg4G6Q4r1XFi36ZpH19Q90jIX07CyN5Oc/PHkMfWuwQAaA0DYdlciSZw0XQn2iPs9YuYE1XGnhtWx0OtLbbyKygez71d44mktGQfgmA2BwQQACCcFd9kVHhH7P3na1py68QrPwy1TdXP4eXi5ClrqOpac22WV3SZ487tw4guacDuTjBz7qUdXTV9NFWUdRHPTzsbJHLG8OY9pGQQR3BHmF49e1V5MNFcHfcuOvDWjloqHUd2fT3igaSYYKmRrpBKzP0Q8hwI9TnzK5a9jdx+1VrC1at4Natvc9wi0/DBcbQ6plL3xwucWPiaSckAhpx5Z9EtI9Fta6TteutLXLSd5j66S5wOhftkjPYj6jgr51eZLhzLwD5itRaQoyP+oV0bUU++wGQ8fZnt8sL6Rs+a+f/ANptSz0fOZrpkzSPEfTSs+bXQtIP505um/HurwnuzL9wv0jemO6hW2Ohnz6l0DCfz5W7FxjyxTCp5deGs+c9el7c7P1wMXJykBERUEREBERAREQEREBERAREQEREBERAXV7ns5RtGcyHC67XSWijp9X2G3T1NpuLW/FljS/wn43IOMfb9WO0K2Dx14gWHhfwh1ZrfUddFTUlutVS4GRwHiSeG4MYAe5JwMDfGfRIleHPs0dT3LTvOToKkoaiRkV5qJ7fUta4gPjMD3YI7HBaPylfQGOy+e72cbY3c6vDKaoI6IrhUPJJxgmkma3/APCc1e2fHbmv4F8uFPBJxW1pFb6mqHVBQQRuqKuRvm4RMycfM47qb3FcvouhuqfbG8r1rp3O0zbdVXucA4YaAUzSfrcSfzBdddbe2a1ldKl37D9D/c6nB+Fsr2F2PmQSc/b9ivo9e91VeJl+9r1x4rqZkFot9NSvGeqQvwT+QfzrudyDc/T+YuSfRWt4RTahpmhzXdQIkB7YOxIPbJGchM+RPm13lRERRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREHm77bhgPB3h3L++bqWdo+o0rv9QVnsRZi7hHxHpz2ZqOmf+WmA/wDyVL7bc/8A5muHg9dTy/qj1B7EVrRwm4kO8zqKmB+ymH+tPB6ToiICIiAiLgnjjzr8unLzcPuNxH1wIrnjqNBQ076mob27saNu/mdvPCZo52ReV/NB7X3TV40rUaW5dbZeI66saWPvFfCIPCaRjDGZJz3+sY3G4PQKr5puZO9eLTO4vaqeJyXOZT1jmHc746MEd/JJ0e5vOLzbaR5UOG0mpa/3e46irneBZ7OZQJJ5CDl7gNxG3uT9gyV4g2DS/GTnb4911VR0kt01FqivNXcalkZ8GjicQOo+TWMaAAO+G/IkYGh+DPMRzH6ogorLYNT6lrpiGOr7gZpY4mk5JdNJnAHc4JPyXuByT8omnuVPhrFaD4Nbqm6NE15uAaPikIz4bDv8I2GxwcDvjJXZyEcz8N9FWvhvoDTug7NTQwUdgttPb4mRN6W4jjDSQB6kEn5krcyIiSYIiIoiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiD58/aIz2ufmWvslso2QE7yFjcdRLjgn5916l+ypp56fk+sDqiUvMtfVPaCc9LD04A9F5S8/TS3mOvrj3P8z3L1h9lh1nk8031nP+3KnGfT4Um503eu3SIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiDiHmr4EUHMdwRv/C2rlEM9a1lRQzHtFVRnMbt/nkfavBinunHLkr401kFpuNZprVNkldTvf0fe6mEnG7XbPY4D9Iz3X0irjDjHy1cEePdEaPinw/tl4lDC2KsMfRVQ5GAWStw4EDtkkfJB1u5AvaG0nMzFUaB4nR26z66oYxJE+FwigukfYujaT8Lwe7QSMEY+fRb2vtNZWc2zq6018M89Vp2hNcyNwcYp2GRgB+ZY1m3yWXzRezQ40cGNZ1GouBdmvOo9MNLp6WegeTWUTfMOwQ7YEjI3+sLpTfpb/wDdqcasNfJc4ZOiqbcHPM4cO7X9fxAjtg9kH0dcpbfD5ZeGERzmLS9BGcnzbC1p/OCuWl0x9mlzJUfGbhWzR7ohDWaXhbD0dQy1gwOnHpvkfIn7O5ykv8upBERVRERAREQEREBFsXirxr4Y8FbL93+JeraOzUhBLPGf8cmO/S0bldebf7Vjk6ueoYdOw6vvDHzyiFtVLaZGU3UTgEvJ2HzxhTdHcBFjUFdSXOip7lb6hk9LVxMnhlYctkjcAWuB8wQQVkqgiIgIiICKh2XCvNVzO6G5YuG1fqnUdzh+7E1PI2z24OBmqqjB6MNP70Hck7bEepD1NRcy/Ntwn5YtLT3fWN6gnvDmH3GzQSg1NQ/GRluctHzPl22yR4ocw3Nxxz5wtaQWq7180Nqq6pkFq05QuLadhc7DA4DeRxyMk7bD0yuPaSi4uc03F/wKOKv1Nq/VFYXYJc/p6nZJJOeiNue52A9Sd/c7lb5IeC/Ldou1j9iNoumsI6Vr7pqCtpmSzmfALxE5+fDY05A6cbDJ74F35Fn7dR+O3J1wu5IuV88ZtH1dwbxHtwoqY3GoqOoPq53Br2xsGA0NJJ2O4YT9XSXgjwR46c9/FKtpIdRyXC4U9OKi5Xm7SufHTRZw0ED1OcNAA7+Z31bjTxn4186PMCzh/Lqy4XG2XbUz7fp61MefdaaJ83hxyCIEAkMAcScnvjHZe0PKpyqaA5U9BDSmkWOqrjXFk12ucwHi1cwGPsYMnA/+wJJJ2+joPZPYhakO+o+O9taPMUVpkP53vH6Fvm1exH4bRMDrzxp1DPJ++bDb4WN+zJyvS1EHmldvYkcNpWZs3GfUMDx5TUEL2/mOVzZys+zj4f8ALTenamg1VX326OwHSywtjBAOw2Ow37Y+3zPcDGVVJsL3giIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg83/bcZ/wBhnh5/CeX9UeovYjf70XEf+EdP+rBZ/tsaZ0vAnQtS0ZEGqj1H0BpJR+kBaf7EeZjuEfEenB+Nmo6Z5+o0wA/6JU9HpMiIqCIti8ZeMmheBGgLlxH4hXVtFa7dGTgbyTyH6Mcbe7nE7ABBw57QLmequWDgZLf9PSRfsnv9SLZZ2v36XlpdJLj/ALRoz9ZHY4I8YeEHBjjbzqcV6+hsFSLpe5wa26XO4TERQMJx1PIBwM5AAHlgLdfNVzN8Q+efi9aYLRpupjpYnm36csVOTLKTI4ZLsbGR2Bk9gAd8L1P9m5yh3rle4YXKu13DAzWGrJ46isjjw40lOxv3uDqHc5LnEep+wOxI67cOPYl26DwqvirxikqXNIc+ks1D0MI82mSR2ftAC7g8KuQ3l04TQRR2XR7KyWMAOkrMPLz6nbOftXYgZ80O/ZJbFvWn2awWTT1IKGxWikt8Ax97p4Wxg49cDc/MrUURAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQdOPadce+JPAbgpbrpw1mko6273EUs9cxmfdmAA5z5ZJx9ZC6BcFPa1cxnD+4UtJxEqqPWtjDwJ21UAZVtYTuWStIyQOwIwvanV2jNJ6+sVRprWunqC9Wuqb0zUlbC2WNw+o9j8xuvMTn29mFo3S2jLrxj5ebfPb/uS11ZdNPB7pYjAPpPpwcuaWjfoyRgH1JFk03Ho7wc4vaN446AtfETQ9wZU2+4xNeWdQL4HkbxvHkQdv/chb4wF4HchXPJX8p+rJ7VqmlrrtoW8kCupKd4MtHJnaeJriAcebcjIzjc7+vvBLnX5ceYO5NsnDfX0dRdXs622+sp301QR5gNeBkj5HfyyoOdUREBERAREQEREBERAREQEREBUPyVVY97Y2l73ANaCSTsAB3QfPPz8NnHMZfDMwtJGQMY/fOyvWL2WUol5PdN4/eVdQ0/WAxeTHPdr+zcQOYjUNdYWN92opXUxeP3zw45/Jt9uV60ey4tdTaeUHTjKljmmoqqicNcMEB3SQr7NSedduERFFEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBeR3tcuVG70WqWcxeirF1Wmrp2Q3/3dn9hmaSBM4DsCO5/1FeuK2pxUsFo1Tw01Tp6/wBNHPb6+0VcNQyRoLS0xO3wfQ4I+YCDxY9kzxJm0dzT2/S0tSW0OrKSejdGXHDp2sc6Pb12P5AvdJfPx7NTS1dqfnT4eR0DwGWqpqrnO/tiKGmlJ/KS1v8A4y9sOYfmX4U8sOk6bV3FS8T00FfOaWipqWAzVFVKBkhjBjIAIJJIABCYfpyui83bx7avhLHcfBsnDLUU1GD/AGed0bXO+podt+dZMXtp+DRaPG4c6hJ8+l7AP51cS16MovOj92n4LZ/3udRY/wAIxVHtp+Cnnw61H/57P9SYr0WRedbPbS8EyW9fDrUeCdz4jNh+RZ1w9szy/RWSertuktSzXMMcYqV8QDXPxsC/YYz37fWmJa793K6W2zUclxvFxpqGkhGZKipmbFGwepc4gD7SvLPm89rZqzTuu67QvLaLHLbrW8wT3+qgNT7xKNnCFpIZ0g9nEHO+3p0r5ieazjVzY63f90a66vt1RIYrbp2gfI+MDOQPCZ/ZHHv2OPLzJ7g8hnsvv2Qwz8RuajRlXT0Z6BaNPVMroXy+ZmnDCCGkEAMJHfJ9A5FdKNUcQ+YfnG13SUF/u9x1TeJ39MEDGdMMIPn0tGGgdgT27Bcua09llzQ6K0HUa7qbXaqyOjgNRPQ01SXVLWAZOGYySPQ4+eF7TcOeAnBnhJEIeG3DSwafGc9dJRtD8+vWcuz88rfckbJWOilYHseCHNIyCMbgj0U+DwQ4Ye025ruD+kqHQFnvFkraCzsFNTtu9sM08TG7dBeHtOBjGD2Xd7kv9qhTcYtTjh7x3prFpq51DM0N0hkMNLUPH7xweSGE+W+PPOM47Pat5GeUrXF2qr9qPgZp2e4VrzJPPEySBz3nu4iJzRn7F5rc6fswte6H1yy98tWhLrqLSlyj6jQUpM89BKM5aQTlzD5dyMjvvhnE349mKKuorlTtrLdWQVUEgyyWGQPY4fIjIKyF87tTy288PDWzuuzuHXEmw26lb1GSndNEyMD1Ebtu3ot/8vPtLOPXBC6R0OsLjV6ussJMctHXzETR4OCAT3I7YOCPVMsWZePeNF0S0z7YjlYuVohq9SU2qrNXOaDJSNtvvAacbgPa4A7rWpPa6cmzKZ1Q2/6oe9u4hFhlDz9RJDfylB3Pe9kTHSSODWNBcXE4AHmSV4Ue1b4oWviNzUVdDp7UUN2tWm7XTW9hp5RJDHU4Lpw0jYnLmg4OMtWRzse0N1lzPXal0fwujvmmtIQnw20jJy2quczjgGQRE7HsGAnOcEZzndfIp7NjXfE3VdDxC466Wr7Boug6KqCirWGKour+7R0H4mxg7kuAzjGMbG8zpnXdT2XfLZZuEnAG0cRLtZomau1pCbhPUSR/fYaRxPgwjIy34ACcd8j5rQfaYc8beA2nZ+C+hWiXWWpra41FV1bWyklyzqx5yPAcB5Ab/V3poaKktlFT22gp2QUtLEyCGJgw2ONoAa0D0AAH2L5/vaS61Ot+cbXtUyVskFqmgtMDmuy0shhaNiPmXKRHJ/snOA2ptYcwds4uVlgn/Y3paOolirnxkRuqzG5jQw+ZGSD8z6g49uh2XEXKToqz6A5bOHOnbJTNiiGnaGplIYGuknlhbJI92O7i5xyT6ALl5FEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQdMvav8Pq7XPKnV1Nrp3T1VgulPcQ1oJPhgODyAO5wV5wezo5uJ+WvidJYbrA6o01q+WCmrGB2DFMDhkg+e+PyeRK9y9faVodbaOu2l7jCJYLhSviLSAcnGRsfmAvm64p2Km4dcbr5ZaZvTBZL2ehvo1rw7H1bp7yj6X6Krgr6KCupndUNTG2WN3q1wBB/IVOtvcPZxVaC01VN7TWeikH2wtP8AOuIuaznJ4V8qemzU6srvfNR1sD32qyU/xT1DhsHO3HTGDjLiR5DbOUG8OP3MFw75ctA1mu+IF4hgZCwikog8CetmwcRxt7knG5xgfXgHwo5nubfi9zf62iZeXTRWkVXh2TT1ECWRlxwzIGTJKcgZOe+B6nbHEfiPxs5veLEt3uMV21LfbpM4UNrpGvmbTRE4EcTBs1oyATgDtnAwB6y+z89n1p7gPpel4icWdOUdw4iXDE7G1DRK20REfDGwHYSb5c7BI7ZGMBidbd9nj7OWbgZcKDjjxbqIanV8tITbbSxgdHahK0Aue456pukkbYDckd8r0JREUREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAWPW0dJcqKe3V9OyemqonwzRPGWyMcCHNI9CCQfrWQiDzK157FPS981TcbtovjDPY7XVzPngoJ7UJ/d+ok+GHCRvwjOB54XRPmD5duM3ItxZtokvzmysIrbFqO2NdEyfB3ADslrwcgsOQR5kZx9EK4+41cDuHHMBoir0HxL0/DcaGdjvBkIxNSykYEsTxu1wODsd8DPZJ/Q6b+z59pFUcdrsOEXG2e2UGrBD12u5MIhiunSPijIJwJsbgAjqwcAnYehK8B+M3s4OaHhVre42/SHD296sslJO6S33e0wmUviByxzg3Ba8DGcDuMjHYcUUnE7mU4J6np6qr1NrjT1zo5ARTXOaqjDsEEgxykAg4xkDt2KfOD6T0XnTwU9sPwUqdG0FJxmob/atQU0LY6makoveoZ3AYLstIIJ+Y377Lf1f7Xnk7pYHSUd41RWygZbGyySMyfTLyAEHdhF58Wj2zfACour6e8aR1LSUGcMqIomyvx6lmRn8oW9af2ufJpMAZdQ6np89xJYZTj/zSUSXXdJF1HovancmVZEZTxFrYA3fE1qnaT9QwStCvXtdOTy2Ryfc+96lusrPosp7LIwOP/fSFoA+e6Tqu6iHdecVT7aXhG25FlJw8vr6EHZzi0SEevfH2Y2+ay3+2l4I7dHDzUhHnmRnf8izbnweiaLzs/dpeCPlw+1F/wCez/Uran20vBNtMXU3D3UTp8bNc9mM/kH6VJ+X6o9FUXnfpb20XAmuqWwas0Jqm1xl2DUQRxzgD16A4H8hK7qcG+NfDjj5oyn17wx1Ay62md5jJ6SyWF47skYd2uHof9eNjfi2/wAQa42vQWpbkHdJpLRWT59OiFzs/mWvk4XGXM3dH2Xl54j3KNxDodM3DBHzgc3+dDx89PCTRtZxs442DSABdNqi9ASDGSQ95e8faAR9q+jzhpoO08M9D2jRFliaymtdO2LLRgOdjc/aV4V+zB0+dQc5+hxgn7nCsuG3/wBDp39/yr38QEREBERAREQEREBERAREQEREBERAREQEXGfHrmE4act+i/2c8Trs+koZJxTwRQsMk1RKRnpYwbkgDJx2UfADmL4ZcymkJNZcMbpLU0lPN7vURTxGOWCTuA5p7ZAOETXKCIiKIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLSdV0zazS15pHnDZ7fURn6jG4H9K1VaTq2Geo0reaelIE8tvqGR/9+Y3AfnITNS3Hzk8FONWqOVnjrBxI0VT0tXVWCqqqN1LVA+FVU7uqN8biNxkbgjcEArmHitxS4/e064w2Czae0VS0jrVTOpqKgppnOpaMPIdLNLK4A5d0tycZw3YYG3DHBThBceOHMHYeErql0M19vbqasqGtJ8KNrnOmf8ALDWuwfUjuvoE4IcuPCHl509Fp/hlpOltxEYZUVhHXU1JHdz5DuSfPGB29Eswn9ugGjvYjUElihk17xvq4bw8ZkitdsY6CPIz09UjsuI7E4AK1M+xC0fnDeOt5HzNqiO31dS9O0RXmH+4g6Px/v73n/NMX/rIfYhaRAOOO94J8v8AqREN/wDz16d4wqqYPL/9xB0ye/Ha5/5qj/8AWUMnsQrHk+Fx1ryP+2tTAf8ApFeoyKjqByj+zi4acrmo59by3qXVWoi0xUlXV0zWCkae5Y0EgOPr3GAc7DHb9EQEREFFVcO8wfNZwY5Z7RHc+J2onQT1G1NQUkRmqZzg9mAjbbuSMDfsugXH32x1s1Toi86U4NaEvNouVfE6miu1wljHhMdsXsawkg47Z9fLuk7wrnv2gvP7pDgVpOt4ZcPq6hvmur1Tvp3sjkEsNsheCHSSkHBcQSAzOc5zjBx5xcnvJNrnnSvt+vh1JHYbLbZw6vuc1MZTNUSkuMcbQQOrG5ycAFcbcAeBPEXml4rUmlbHDW1clfUiW63R4L208ZOXyPedurGcZ+vsCvoB4BcCtD8u/Dmg4c6EoRDSUw8SeYgeJUzkYdI89yTjz9fUlO5iZN2Og1w9iBpz3LFq4+XIVfT3qLLGY8/U2QHH2rZU3sQeIjXn3fjxp17fLrtE7T+Z5XruiK6E8oHsstNcvmuoeJfEXVtLrG9W8E2yCGiMVLTSEY8Uh5Jc8DOOw3/L3wqamno4H1VXURwQRNLpJJXhrGgdySdh9ZWyeNvGbRPALhxdeJ+v6409qtjQOlgzJPK44ZEwebnHYD614Yc0/O9xg5qdXvprfXXWz6ZdIYbfYKGd+ZATgeIGH748jGRuM+uyDur7QX2l1Touvl4Rcu2oKSe4CMtu99pyJW05O3hQuG3WPNw7fMHfqbyJ8od45yuI131Xre61X7HbNUsnu9W74n1lQ89XhdR8yBkj0I7BZXLb7Mbj3xlrqK7azsNTo3S8hbI+pr2eHUTRnzjjO+D6kZ+XmvZbgPwH4f8ALroKl4e8O7Z7vRQnxJ5nby1MxG8jz3JP17Z+1L5iT1vqy2mhsFoobFbIRDR2+njpaeMfvY2NDWj7AAs5ERRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBfPJ7Q3TEGlOcTiPbaZnTDLcIqtjP7kSwRvx+fP2r6G14F+1AgdR87Os5qqMuZMy3zBv8AdM92jGPt6SrB7Xfs0sfCLgHSa21lP7rbtMaZpqmtcdiBHTtBaPmTgD5kLwD5g+Lus+bfj9ctZQ22pqay+1TKKzWyIF7ooG/DFE0fVuSe2Tk4GV6te054xWWn5GgbFdIf/wA4clrpaERvBMlMXNqZMY2wGRdJ/wC+wuvnsXeDWltQ3LWvGG926GsuNhmprXbPGYHCne9pkfI3PZ2A0Z+ojBCfB3F5EOTXTvK/w1o6m7UEFTrq8wtnu9a5gLoHOGfd4ydwGg4JGMkH5k9pkRQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERARcaceOYPhry5aPbrTibdn0dDLMIIGRRmSWZ+MkNaNzjbON9+yzeC3G3h7x/0RT8QOGt6Ffap5HQu6h0yRSjGWPb3aRnsUG/kREBERAREQFxNzB8tHDLmR0m/TGv7SxzxvBXRMAnhPydscfLI37ELllFLNHlPrP2I08lY6Th/xwgipiSRFdrW5zgPIdUbvL5grb9s9iJxFdVR/djjjpyOn6h1+7W2d7y3zx1EDP1r14RUeXd19iDYDbMWTjvcG3Hp71dpYYOrHo14djPzXHV09ibxipRm18XdK1w8g6lnid+Q5H517Eog8V5fY0cw8cgZHqrTkgPdwfgD8rs/mWvWH2KHGKtka6/8WdL22LI6hHTTTvxnfABA7fNexSLPf7HmTT+xD0IKFoquOl+NZgdRjtcIiz8gXE/nWKPYg6Yyc8drpjy/6lR9vn8S9QEWh5gfuIOl/wAe10/zTH/6yli9iFoz4fG46Xr/ALbptcX5suXp0iDyb4g+xKvVJQuq+GPGiC4VMbCfdbxbvA8V2Ng2SNxDc/MfaupfDjjzzNcg2utR6Fsk0FqrhUBlztdypTPSyvbsJGty07gYDgQcbbHIX0MLini7yucBOOrxU8UeGlqvNa1gYytc0x1LB8pGEH8pKbiXrzJ0j7azjLQvij1nwv0tdI8/fZaN81O/HqGlzhn6yt0cxHtUNE8YuXrUehtO2CqtN5v9C6imjmPUWh2MgEDGPXc7efry3xR9jvwOvUU9VwxuFzsVQWOLYJ6t00fVjbBdnAz6j7SugnMR7PHjxy+Was1fdrXHc9OURJlraV4eYmZ2LwMgDHc7eeyZs6uuS/Y5WF1fzSVd98MubatP1Qz5NMpDQfyAj7SvbleLXsZNQ0du5h9QafmIFRd7C8wZ7kRP6nY+whe0qAiIgIiICIiAiIgIiICIiAiIgIiIC2Rxj4uaN4H8O7xxK1zco6W22imdN0OeGvqJAPghjHdz3HAAAJ37bK3jDxm4ecCNFVev+Jl/itdqpBgEjqlmee0cbBu957ADzwvBTm+5q9bc2vFipu5fXxacjqBTaesTXFwhjz0sJY3Z0zyckgZ3wPnc+jI5kuaLjHzv8SbVQVlB/tVlUaXTun6BpLYjK7ALtz1yEAZcdgAewBK9a/Z18qeoOVvhFW2/WVVFJqHU1Uy410ERJZS4YGtjBPc4xnbuPsHGfs6/Z+Wfg1ZbXxo4n0JqNc10AnpKSUDotbHgEbecuO58vJd/BjyTZid1VERRRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREHlPzve0a5geFPFCfQPD2npLJS0oL/eKmjEplGSMNzjOMDJ3HbbdcASe1p5tqnS9w01cLnpypdXU7qZtf9yhHURNcC0kFrg0nB2JGQd917M8QOA3BrioSeIvDSw6gLu7qyka8n5k98/PuuGuIXs5OUnVGkLpZ7Pwds9juE9PJ7tXW8vilhl6T0kHqIxnGRjstb3jOfa8wvZS3S0Rc51kq9R1UYmqrbcWUr5ju+rewBoB83EF4HzK93/LZfL1Wm78O9dVbLLdKikuWnbnLDBVwu6JGSQyFocCOxy36tyF76ez5426t4+8s9j11ridtReYquptlTUDvOYHACQ/Mggn/ANypWnZHbCsfJHEOp72tHqTheXnPP7Ua4aa1PfeDPAeICe0zS2+5agMgLfeGEtkZAG5yGuBBdkAkHuNl53u1ZzJcSnT3uC9a/vrXEmSWkkqpIgcnO0fwjfOwCniPpShnhnb1wyskGcZY4EZ9NlKvn95PucjXvKxxairtbVuoLlpmpBprxaamaR0jGkbSMZIdntODjbPz7L0Z1N7YTlTt2n6mv01+ym73RsRdTULrSYA6TGwe9zg0DPcglXF13jmq6alGaqpiiHq94b+lQC8Wh/0brRn6p2n+dfOlrfitzD81fFK4XS3T6pvdyu1Q59NabU6aRlPEXHoYI4zgAA4yQMn0GANXm5Oed+NvXJwT4iuB32p5XH8gcSsZR7f8w/NHwv5dtCV+rNTait81bFG4UdsjqWmeplxs0NBJAz548sBeV2sfbG8z12uU0mj7dpawUJe7woXW/wB5kDc7Zc92M477HdcI2bkP5xtV3KKB/A3VkLpXiM1NxpzFGwernPOcD5Ar1R5afZkcAeG3Du3w8VtAWnWOrqqNstyqbkwzRQyEbxxMJ6QB2zjcj7Tc70ed0vtYecqc/FrCyNz/AHFmib+jCrU+1i5yqmhfQ/svskQcOnxo7PGJR8w7J3Xrc3kZ5QQeocu+iQf/AAa1Wyci3KBJku5d9FZPpbwP0FXzweIugdKceee/jVS2Ct1HWXu+VodJU3GuJdDRQA5c8hoAaPkME488Lvjo32I+mqWWnn13xurq8MIdLT261NhY/fdvW6QuAIzuACvQXhtwN4Q8H4pouGPDuyabFR/ZTQUoY5/yJ74+WVvpNHGnA3l84Z8vmlmaW4c2GKjjIHj1BaDLOe5Lj3xnfHr6rktEUkwFhXi409mtVbd6vPgUNPJUyY79DGlxx9gWYThbd4j1tJbuH2pq6ue1tPBZ6ySQuOB0iFxKo8I+cbnc4kc4OpabStJRst+k6OvItFopcl9TKT0MklJOXPIIw3sCdvLHev2ens4JOEFVT8ZONkFJV6llga612npEjLe1wB65CRgyYxtjbttg586OQ3Slt1pzfcMLJd6RlTRfdptXLE8Za7wI3ytBHmOtjV9FaAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC8LPa708EPOJcZIj8c1htr3j59Dhn8gXul3C8EvapXKS485+rWvfkUlJb6ZvyAp2HH5SVO6a2Py18MuJnN9r/TnAifV1yNhtcT6gGVxfFbqdgwSwEEA4OADnbIAXuZy28tugOWDh+3QGgmTyRSTGprKuoI8WpmIA6nY7bDYZOMnywBxH7OXlk0hwQ4E6e1hHbAdW6xtcFxulZIAXtbK0PZCw+TACPrP5+2y1Yk0REUUREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREHQX2wnCvVnEDgXpzUGl7dPXN0pd5KuthgYXv8GSLpLgBucYyfllee/IjzqX/lO1xJRXCN1fofUU8TbzRg/FA4fCKmP/tmg4I8wNu2D7+VFNT1cD6aqgjmilaWvjkaHNcD3BB2I+teSvtIPZ01tiuFbx24CaadNaJg6e/WOjYS6mePpVELBuWEbuaOxBIG+4erWmNTWLWWnrfqrTNzguFrulOyppKmF4cySNwyCCPPfceRBB3C1ZeEHINz53nlh1O3SWvaqvuPDu4u6Z6dpMj7ZIT/AGaJp7j+6aO47br3A0PrjSnEjS1u1rom9012s10iE9LV07w5kjT+gjsR3Qa+iIgIiICIiAiIgIiICIiAiIgIiICIiAuK+aaKln5cuJEVZCyWI6ariWOGQSInEEj5EA/YuVFsPjzZn6i4J68skf06vTtwjb8z4DyB9pGFKPFb2T9c2i5z9NNdL0Cptlyg/wC+zASB+UL3lXzl8kWqn6I5reHN/Li1kF3EUwO2WPjc0g/lX0YseyRokY4Oa4AgjcEHsVcF6IiAiIgIiICIiAiIgIiICIiAtv691nZOHWir5rvUdQIbZYaGavqXk4+CNpJAPqSAB8yFuBeY3ta+b52nrfVcr2lGg1V1pIp77Uh39jgeepsOPMuABI/1AEOkPNdzdcTuc7XtDSTUL6azU1UYbDYabLsPeelr3/3UrgQM+WSAcbr0G5BPZqUPCZ9Dxe44UlNcNVljZrdaXND4bZkZ6n52dJg+mB29c9e/ZPco961jxHpuYLWljdHpnTge+ze8MwKytILRI0Hu1gJIOCM4O2xPsql6giIiiIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIujXtE+avUfLvV6fNgbI502HdLXYDnEk7+Q2Hf5IO8qxrjK+C31M8bS50cL3geZIaSAvN3hn7aHhjNQUtv4naA1HSVTQGTVtC2KaM9vi6C4Env2/Iu+fBzjLw949aEo+I3DO9i52Wtc6MPLCySKVuOqORh3a8ZGQfUJg+dKfTNRxF46XDTTauOgmveoquIyyj4Yi6d5JPbtvttuuzFBzh8VOSO1Xfly4X1NsrqSle+V9xqGl7oqiZuXviAIAOC05z3A22XA/NhpCXhfzO8QdOUL3wfc7UFRNTSRuIID3eI1zSNwQXbHyIW4uT/hHa+Yrjtb7Rr+51M9G+eOWsLndTqj4gOl5O+MDf12Cs/LJqfx2uy/LN7KDUnG3Ttm4w8XuIrrVbdSMF0+51LSl9bOyR3V1SSOIazryScAnfO2y9V+F/CHQXB/SNFozQ2n6WhoKOIRDpjb1yYABLjjcn07LdNqtlDZbZSWe2QNgo6CCOmp4m9o42NDWtHyAACzFL3qzkdSubz2ePDLmgbFfKCqZpLVFM0tbX0tK10c422lYCM/WN/qJJXUW3+xB1r7437q8ebIKUO+L3ezzeIR8up+Afyr1vVMZUPHAfKfyecOeU/S89o0q+S6XavcHVt2qomiaXA+iMfRbnyHoFz6iJJgIiKgiIgIiICIiAugHtWeaWu4UaDi4Q6bm8O6aupXiolacOjgJwf/AH9SF3/XiT7ZC8Gq5orfZgTig05SyfLMrnnb/wAwKWaNI9kzw3umqOZ636zbSuNv03TTyvlI+ESuYQAD6gE/lC9zV069l7wes+gOWbTusY6Roueraf7oSSuHxeG4kgZ+sfkA+pdxMhUVREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREFjnNY0uc4NA3JPkF86fPTrBuuebDiNqCF2YH3UwQ/JkcbWAH/zSvoc1M58enbm+P6TaOYj6+gr5t+KNjuGouYO96eBJrLrqI0rSe/XLKGjP/nBZl/6xc5r6K+DzQ3hHohrW9IGnLaMen+1o9lvBaPo+zv07pKyaflcC+2W6mo3EdiY4msJH2hawtIIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgKOSOOaN0UrGvY8FrmuAIcDsQR5hSIg8nPaE+zSqLfUXbjpwCt5mpJHPq7zp6JvxRE5L5oAO47ks7+Y3znrlyW+0E4g8qUg0VcaGO+6Fqqzxqi3zOLZaF7iBJJA4diRuWkEE79172vYyRrmSNDmuBBBGQQfLC8J/accrs3A/jpcNY6Wsb4NG6vcLhTuij+9UtU/wDs0Www0F+XDJ/fEemRmPcuw3ij1DY7dqC3kmkudLFWQE7ExyMD2k/YQtQXUP2d3Nvorj5wms2gWVLqXWOj7TT0dwope8sUTRG2aM/vmkAAjuPygduxulNVREQEREBERAREQEREBERAREQEREBaFrpzW6J1C54y0WqrJ+Y8Fy11aBr/AP8AiHqT/wAEVn/oXoPnB4Nt8bj9psUvwh1/YWeWB4hx+ZfSZp4Obp+2tefiFHCD9fQF82fAxrncfdJhgJJvse3/AI5X0n2RpZZqBju4pYgfrDApndGciIqCIiAiIgIiICIiAiIgIi0LW+sLFw+0hd9balq201rslJJWVMrjgBjBnH1nYD5kINu8auNWhOAega/iDxAu0VHQ0bD4UZdiSplwemNg7kk7dvz4B+eHinrfVnMpx0vWsBRzVN31ldyaSlYCXNa9wbFGAM4DWBoONhglcj85nN9rHm44gxSiGel03bpTBZbVHklxccCRzRuXu2wNyM488D0V9mHyTUPCrQ8XGHijpaE61vh8W3xVcYc+20mPhwD9F7+5OMjbBOxAdz+DuiqXh1wq0loeko2UrbNZ6SlfEwYDZWxt69vUv6ifrW8kRAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFwTzP8AKLw65prHBadY1FVQz0pzBV0zQXsPkcEjOPr37LnZFM0eS/EL2JWo6WN9Vww4yUVd0NJFLeKEwvefICSMkD7R9q4Z4Tcx3Gj2a961ZwbvmnqS4VtZK2rfSvlLoIpeksbIwkA7gZ2GDgZBXuaNl5H+2J4A39+vrPxtsFrkqaGtt7aK4uiYSWPiOGkgd9jv57j0K1vxHntxX4mah4wcQLzxG1Q9puV6qDUTBpyGkjGATvj616Rex44KWW+W278VLjEJJaCr8CId/vgyAD6AAE/XheWknheFG1jHB4z4hPmc7AfYvYb2J1RNLwZ15BIHdEGoYQzPbBgycfLJWbNV6OoiKgiIgIiICIiAiIgIiICIiAvCr2u0/jc5V0j/APmFgtcf5Yy7/wDKXuqvCL2tvV+3Rv8Akbfce1Y+r3cfz5Qet/JH0/tR+Epa3AdpWhdj5mME/nK5uXC/JeAOU7hKB2/Ylbf/AELVzON0iaqiIiiIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg4S5uuY2k5YOD9ZxJnt3v87Z46Smp/J8r8gZ3HnhcScj3P5S81ddcdM3mwxWi9ULPFDI3ZD2EZB79tiM7bhbY9seWDlYo8t+I6jpMH0HS/P8y6oexls8lfxz1LcOg+FRWljnHyyXOA/P+lS8i8ezyIiqCIiAiIgIiINC1xVOodH3qqZ3jopnD/zSvm34t6groON2oNR26Y09bS3l1RBI3vHJG8Frh8wWgr6S9aQsqNJXmGT6LqGYH6ugr5v+IFlo7rzC3KwVEhipa7UjaWV47tZJM1riPqBJWZn8tXeY7Q1ftieZ+XR1Np2itWlaa6RRCOa9e6SSTy4GCehz+hrj6481ufgD7YDi5ZNQ0lp420Ftv9jqJWxy1sEPgVFO0kZccEhwHc58vIr0l0Zyd8semdDUmj6TgxpCtofdmMlmq7VDPNUHpGZHSvaX5J3JBG5Xj17Srlv0vy7ce/dNAWz7n6Y1HRNuNHSAkspZMkSxsJ7MBwQMkgE+i3us2PdzS+pLTq/Ttu1RYqptTb7pTsqaeVp2cxwyPtHY/MFasvP/ANknx2rdb8IhwzvMxlqNNl0dM9xyfCGMAnv9HH5D9vf8brP3i+KoiKgiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC4i5q+DtHxy4C6x0A+giqa+rtk77Y57Op0VW1pdGWejiQAPrXLqIPmw4F8XNecqPHC3a6obdNTXawVD6S5W2oaWGeB3wywPHcAjBBHmAQcL6AeXnmA0LzJcN6LiPoSq6qecmKqpXn77STgfFE8dwR5HGCNwuqftMuRg8bNKM4r8ItL07td2VxNdTUsYZJdqQgkjAwHStOCCdyCRkZyvPLkm5wNU8nPEisprvbKio0zdpm0t/tcgcyWCRjukytaRlsjNwQRkgY8sF74PoHRaXpzUFq1ZYLdqex1Tai3XWlirKWZpBD4pGhzTt54I28itUQEREBERAREQEREBERAREQEREBbf4hf/EHUp74s9Z/6F63AtF1nG2XR99jcMh9tqWkeuYnIPnb5VLPHeeaTRdvqHARC+F8hPkG9Rz+UBfReLtZoJYqAXKjZKWgRwmdoeRjbAzk7L5o7DrSq4Z8XpNXWuImS03WocxmcHHW5uB6HBWbrnmB4qa31pNrOq1reKap8XrpmQVkjGQNB+EAA4+vOc5KJr6XkXlNyq+15sWnNJ2/RXMTarvUVNCBCy/UMYnL4gMAyx5DiR6jO3rsB6ZcOeI+jeLGkLfrvQF9p7vZblH1wVMJ2z5tcO7XDzB/QQU8VudERAREQEREBERARaNqvVunNEWGr1Pqu701stdBGZKipqH9LGN/nPyG681eZr2wFhgjqtLcvlrqayUExvvVSPCYcHBMQ3OPMHG/kQg9Q14++1k5v79fdc1nLZoe8ti07Z44vu++ndvVVZAcYCR+9YCAR2JJHkc9TNQc7XNHqGunrZeMN/pBOCDFS1JYwA+XmfzrdHKLyc8TebrW33YqGVUemIasSXm9VTnEzEuy9rXk5c875Oc98ZIOE52jtZ7Nj2eM1dPp7mS4uCB9A6MV9gsro8mQn6E8wIxjzDcfM5yCPWAAAYGwC03Tdit2l7BbdN2mIRUdrpIqOnYABiONoaO3ngBamgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgoThcC87+oNG6a5a9Y3LWT6fwm0LxSMlALnT/AL0MHcnv29cea56AwvHb2zfFC/13FrT/AArir5Y7Ra7Uyulp2OIZLNK4kFw8yAB3+R9FZy7Es3jp9ytcA7nzNcbrHwpobh9z47iZKmurA0EwUsTS6V7Qdi7GwB8yO/Y+/fL5y88POWrQcWgeHdLOyl6/GqKiok65aiYjd7j29cDyzjJXkT7Hw045t2eJjxjp6vEee+MNJx+QL3GUUREQEREBERAREQEREBERAREQF4S+1va4c517LhgGy2sj5jwAP05Xu0vAz2o+ubDrvnD1PUaeqWVFPZ6OitEsrCC188UeX4IOCAXkZHog9i+TDo/an8Juk5/+Cdu/L4Lc/nXNC4U5KoZYOUrhLDM0tkZpO3hzTsQfCGy5rQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQdKva4acrL7ykVlTRQGV1rvFJVvx3bGOvrP2DK6E+ym40WThXx1q7PqCpbBTampmUrHnYdYJ8z9YOPkV7TcQ9B2HiZo656I1JAJaC5wmKQFoPSfJwB22K+dnj5w9unLdzEag0db5XxTaYurZqKQEjMe0kZyMHGCB8x9aZsNfSSHNcMtIIO+e+Vctn8INSt1lwq0fqprur7q2Oiq3E+bnwtJ/PlbwSAiIgIiICIiDbPEutfb9AX+siHxx0MpH1kY/nXze6tuD6rjlW3F2zjqEP+oiYf6l9I3EKnbV6HvlO/s+hlB/80r5uddxNp+N1ziiGAy/7D0+/BZn/AJD6TtGSmbR9ilduX22lcfrMTSvK/wBtbdrfLqbRVpaxhq4KVzy7s4NJeftG4/KF6maCDhobToeRkWmkBPz8Fq82fbB8u+rtRtoOOdjhdVWyzUop69jckwM2BeR2xsDn5Fak/leJuettexgttQ7UGpLi1x8FsTuoY2zgAb/WR+ZetK8VfZFcebRw/wCM1Twv1BIyCDWURioqh7gA2paA4RnPbq6dvt7eftUpFERFQREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAXl57TH2fIvJ1PzQ8MKuGCWnpTX6hsxiI8YRt++VMRGfiIGXNI3IyMbr1DWi6y03S6y0hfNI1pAp73bqm3ykjOGyxujJx8g5NzweSvsvOeLXlr15pjlk1vVsuOmLmH0Njle379QzBrnMiDh3jOCAD2Prnb2EXzocQeG/E/kb5hbZLd6IC46er2XK1VDto6yFjtnAjsSNiRnBIIXuNymcylh5peEdJxJs9L7lUsnfRXGiL8mnqWBpI9cEOBB8wln1P8c0oiIoiIgIiICIiAiIgIiICIiAoKylirqOeinGY6iN0T8eYcMH8xU6IPnd53eXq7cv3Gm7W2dmbbdqmWsonegc7qI38t8j5HHkvQjkG5JeAPETlYtGptfaSpr5XanbLLLO92HwjJbhpHZwIO5zjAwuHPbWtij4jaFDMB0ttne8epDwAfyLsp7Hi+XG58qc9srJHOhtWoayGmJJJDHBjy0E+QJOB5ZRJ11p5svZG3XQVjuXELl/v1Te7dQRuqKiwVrc1ccQySYZBtIABnDgD33wF1i5WOd3jLymXKS2acqI7jpuoqRJcLDcGEx9Q2cYzkGJ/rjYnORkkr6GXMa9rmPaC1wwQRkEHyXSLjV7K3gnxP4oR8RrNJLYWVlSKm622n2pqh+clwbg9PUc5AwDn5qX8sX13A0Fqun11oqxaypIHQxXu3wVzYnd4/EYHFp+okj7FuBYNltFDp+z0NitkIipLdTx0sEY7NjY0NaPyALOVBERAREQFa5wY0uccADJVy4v5j+OGluXvhDf+JOqaprG0VM9lHThwElVVOBEUTASMkuIz6DKDyf9ptzq3PjBrCbg1oqrlpNKaencyu6SQa6pB/fHzaBjbtk48jmfkd9mM3mM0HHxa4n6qrrFYK2ZzLXRUUIM9Yxhw6R73bNYTkDAJOM53261cvGiqPmO5o9N6U1ZI5lNrC+Sy13hk5w7rlc0HvjbH1L6JNHaSsGhNL2zR2l6BlFarRTtpaWBnZjB/OTkn5kptMx0/q/ZKcrs1so7fTUtzhfTY6qgy9UkuP7ogjOfnldrOFvDDSHB3RNu0Doe1x0NrtzOljGNAL3YGXnHcnA+wAeS3ciGCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiCOSSOGN0ssjWMYC5znEANA3JJ8gF4c+1iutr1RzGDUVhq21lGygiopJonBzBJGMYyO2d8fUvRj2nHE3UXC3lVvF20tVy0lfca6ltwnjODGyQkOI+YwCPmF5oezn1RpHV/MLbuH/ABkibeqTUZdHRmucZOqrAJbGSTk9XYD6/knfhWzvZ38S7Zwo5qNMaqvlYKW3PjnoaqZ3ZkcrOkk/mX0DWu6W69UENztFdBWUdQwPingkD2PB8wRkFedHtVOWbhjpXlwHEXQGjqCx1tju9HHUmiiDGyQTPMfxfMPczGPXfsFX2LnEHVWpuHOvdJ3261NdRafuFE+gM8heYWzMk6owSc4zHkemcDYYSk69IkREBERAREQEREBERAREQERcL8y3Ndwp5XdJSag19dw64zxuNttEB6qmsk3ADR+9bnALjsBk74QcW+0z5h67gNy7VMOl74KDVOrKplqt5YfvrIiC6eVvp0sGMnbLgPNecHs+OSek5wNS6i1jxC1DWQad07UwisETg6ouFVL1PLC924GASXbklw+3hfi1xW4yc53Ghtzr46y8Xm71JpbNaKYF0dJE53wxRN7AAYLnHvjJOAAPZX2dnKxqLla4N1dl1nUQu1BqOtbc6+GF3UynIjDWxg+ZAGD88nbOAHZnTmn7VpSwW7TVjphT2+100dJTRA56ImNDWj8gC1NEQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQF4O+1jpzT86GpCY+hstqtcg22dmmaCfyg/aF7wk4Xmr7Urkh1zxYuw4+8MaT7qV1vtrKe7W2Mff3wRAkSMH74gE7d8Z9Bm6n7dyuUG72C98sXDKr05cI6ykZpm3wGRrskSsha2Rrh5EPBBB3XMK+fnkj50NXcpvEKOkvFRcKzQ1fN4N6sxcSICTgzxMP0ZGHJIGCRkb9j7ucN+JWiuLmkKDXfD6/093stxZ1RTwnOHebHDu1wJwQf0YUP9bqRERRERAREQbb4jPlZoW+Ogz4gopenYnfC+bzW5J443I1A6T+yAdXyHjD+ZfS/W0kFdSTUVQzqinY6N49QRgr5yOcHR83Djmf19plo6TQXgyRO8i1zWvBHy3We/wAl+Poj0Q4HRdgLTlptdLg/LwWqPXmkLZr7Rd70VeIGS0V7oZqKVrxkYe0gHHyJB+xYfCavjunCzRtyjcHNq9P2+YH1Dqdh/nW7Fpn181fG3hFxB5YuL1ZpW7R1ltr7TWGe117CW+LG12Y5WPGxI2zjz+RGfV/2aHPJeuYCy13DXixdKaTV1kax1HVkBjrhTnYdQ7GQY3xudz5gDs1zAcsPCHmT099w+JWmoqqaEH3S4RjoqaZxGAWvGDjftn8mSvMOq9m1zE8FOPNtunCmSprbLT1okguMTiHthyCWvwMHI8jg9vMZUvPxN69l0Wn2GO4Q2O3xXZ/XXMpYm1Ls5zKGgPOfPfK1BVRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERB069pNyiXHmZ4YUN50XDEdX6RfJPRsIGauncPjgz3G4DgB5+vY+TnAXmt5geTC+32w6NdT0fvU7W3azXijMsXjR7B3Tlpa/G3UDuMdwvooXDPGjlC5euPdJUs4g8N7VPcJ43MZdaaLwK2In982VmCTn1zlOjgrkb9o5p/mXlqdGcR4LTpfWNOA+mijnLKe4M7ExmQ56wcfDnzx6Z7tr55ubfk54mcp+uZ3S0VdV6VkqDJZ7/A13R05y1r3tA6JBkemTjG+w7mcgHtMdGWHQkHCjmQ1XVUtdbJfDtd+qw+WOamPaOd4yWlhyA45yMZOcp74PU1Fpem9TWDV9lpdRaYu1Pc7bWsElPU07+pkjfUFaogIiICIiAiIgIi2/rbXujuHFim1NrnUVHZrZB9Opqn9LR+kn7Ag3Ai85uav2svCqzaQr9LcAK+p1BqOsYYG3MQuipaQEfTa44Lz6YG325HmjUc3fMjUVMtT/ALL1/iMzy5zY5gAMnOBt2QfSNsUGy+b6DnG5l6fHTxevrsf3UoP8y31wi9oZzHcONbUOorvrq4X+2xyt98oKl4LZosgloxgZ+vP2d1cSft2b9ttYaiLVvDvUTY3e7z0VVTF/l4geDj68ALn72N/4K9dtv+yasz/5ka6Ve0M56NA82GldG6d0Xp6to32Spkr6yaqbgh74+gxt8yN85+S71eyHs77byg0ldK0h1yv9wnGf7kOa0H7ekqEd2kREUREQEREBFj1lbS2+llrq6ojp6eBhlllkcGtY0DJJJ2AAXnzzXe1i4baFt130RwO8fUWqB1033UDemipH7gua47yEb4wBg98HcB2j5qeaTQXKxw5qdY6pqoqm6TNMVptDZQJ62c5wAO4YMEl3YAHtuR4d8deZrjxzk65oaLUdVLV+8VQgs2n7ezEEL3nDQ1vdzsEAuPoTtutt0WneYbmn1nJcqG06o11eqyUh9R0STMjJIy3rPwRgbbZAXsRyF8g2kuXHStDrPXVjo7lxJrWeLPVyASi2tIBEMGdmkebhuTjtjCYY2lyJezTo+Xu9W3jDxKvv3U1mymPu9vp2Yprc6RoBy47vkAJBOAB5eYPfhEQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFBU1NPRU8tVVTMhhhaXySPIDWtAySSewCnXF/Mzo7V/EDgTrLSGgqx1NfrlbJYaJ7XdJMmMhoPkThBJovmP4I8RdWVehtGcRbRdL5RZElJDOC8474HnhcmL5rNJXfilyn8cLLqi8aduNl1Jpa4sqpKKujdEZ2Nd8bCTs5rxkZGRvnfC9s+X32iPLjx/pWU1DqN+mr62MOntV5xE9pPfokBLJAM9wR9SZfp+naBFwjrnnP5aOHjzHqbiraYXg9PTG8vOfQEbH7CtD057QHlO1TVihtXFi3mZ2MMkBaTupLvg7FItsad4l6C1XCyfT+q7dWNkx09EwBJ9MHG/yW51QREQEREBERAREQEREBERAREQbA438F9G8feHVz4Z66hlfbLi0HriOHxSD6L2n1BP8A9juvCvmf5Y+JPJDxgt1XRVtTUW+CpjuWn79FGWtc5jw5rXkbB7SBkZGR2819Ca2nxJ4Y6F4t6VrNG8QNO0l3ttZE+N0dRGHGMkY6mEg9J7HI9BkEbIPJTmi9pbpbmQ5T38KqjTNZbNXXV9E664w6mLoJmSF0ZwNnOYCATkDbHprHsfuNGgeG9w1Vo7VV/goqzVNRTupmSPA3iBDcjvv1u3HyXEPHn2YnMXw+1rd4+HuiarU+l2SyS0FVSSNe9sG5DXgkEEDbJ749crqax2odCaoa98M9tvNkqwTHICx8U0btwRse43HmPkr/APSY+o3IcA4HI7gqo7LZPBLU41pwf0Xqsu6jdLHRVLj6uMLcn8uVvdRRERAREQEREBERARFQkAZOwCDgjm+5q9LcqXDSTV91ijr7xWONPabb14dUS4O5xuGDzP8AqK8LOIXEPjJzh8ZW3a7NqL5qS/TimoKCmaTHAwn4Y429mtA7n5ZO2ANd5vePevOYjjbeam+V81TSW+5T260ULN2RsbIWDpA7lxAx8iB659XfZx8mNi4BcN6PX+rLNDNr3UUDaiaeZgc+ggcAWwx5HwkjckbnbtuFJPtTc8ZPIp7P7S/LBbYtbaskivPECvpw2aoLcxW9rgCYoQfMdi7uTnyxjuQiK+rBFtHiTxV0Bwj0/LqfiFqSls9viaXGSd2CcDcAea6p6k9rxyi2QzRW6t1ReZoiWgUtqwxxHo97gCPmMoO7SLzqpPbW8C5q8U9Xwu1lTUxOPeOunfgevSHZXdbgrxu4e8f9CUnEPhrdzXWqqcYyHt6JYZG92PYd2uHp/wC1Bv8AREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBU7qqIPJX2qfI/btMRz8yXCy1Np6KeYN1Jb4GYbE92cVTAOwJGHDGBnOR2XX32f3OxeuV/WTtL3wSV+idR1EbaymLzmknJwJ4/Tvgjtjf1XuzqPT1n1ZYq7TeoKCKst1xhdT1MErctkYdiCD+Y+RwV84fNVw+sHCnmG1zoHSr/APqXZ7o+KlAP9jYQHdIPoOrA+WFZ4l7x9IttuFNdrdS3Whf4lPWwsqIXYx1Me0OafyELKW0+Epc7hVo1z/pHT9uJ+v3aPK3YszsUREVBERBTtgLwG9qG2Ac6mufAb0gsoC75u90iyV78nzK8A/ae/ho67P8AiX6rGp5U+vbrl2Y6PgJw6Y85cNLWvP8AFY1yGtgcvpzwJ4dH10rav1WNb/SbnVERFQREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERBoWs9GaY4g6artI6vtFPc7VcYjFUU87A4EEEZGexGdiN14k87Ps6ddcvl0q9a6DpJ7/oaomc5joGF09ADuGyNGSWjfBH5wCR7pAYUFZRUlxppKK4UcNTTyt6ZIZow9jx6EEEEfWkp/j5+OWDn9438r0Isen6qmv2mS7qNmuZcY2HbPhvG7M48s7bDC7RVXtutYED3LglZ8+fi18v5sLt1zH+zn4AcarJXVth0XQab1Q4GSGvtzBAJX+j2j4TnyOMevqOpWkPY0XqZ0rtU6sMLeohgEjdwD3+EZ3HqnBp49tzxE/Erp4/XXTKOo9txxLdD003BjTbZP7p1bPgfpW9X+xetYP3vXMn2OP8AOFmUnsYNMl4971vKGjuQ52/5Ag0zg97Z2a8amprXxd4fW+02yplDHV1vme8RA7ZcHdgM5Ox2HkvSrQ3ETRHEmywag0Nqi3XmhqI2yNkpKhkhaD26gCS0/I4XkHzA+yK4uaNj+6/Bot1ZSNH3yiErW1A2/eB2C7cdj69/Xq9Ucv8AzZcMah8X+xrxCsMrnYf7nTVDOoj1MWx7eqGY+kFdJec72kumeWfVNDoTSVoo9U3voE9xY2o+90rSdmEtP0jg5Gc+W2CvN3hvyv8AtBeKn33TVm4gQUrgf9tXS9S0EORjIzPK0k79gCuEeNPCHinwX1vU6T4vWOrt99LRO91RMJvHae0jZQSHg+uf5kTXpZD7bLTDLWJKng/Xvry36EVU0Ma755JOM/NdIObLna4oc112hbqER2jTtG8uo7PSvd4YOfpvJ+kdh37HzOBjZXBblg428wNW6DhhoqquULD0yVbvvdOw9jl52wDsSM7rtPQext5ip7I6sr9Q6eprl0AtoxP1jOOxkBA+3CX9mY0f2efIRofmmpbjrTiLraaC02mcQmzWyVrKuc75MjyCY2beQzuN99vSbT/s4eU3TNOKe18PCD0hpklnMkjsZ3LnAkncrx+1twH5veSq+RatqrfqDShilDYr7ZqpzqV7hnAMkZwRv2eMbjzK7ocp/tfqQ01HormepZveQWww6nooW9Dh2BqYhjB7Ze3bYkjfZfFdr6j2c/LHPM+YaRewvJJAc04z/wCKtra19lZyt6ttb6SitNfZ6xwHTWU0jeoEeZAAz9RK7e2u52+9W2lvFqq4qqjrYWT088ZyySNwBa4H0IIKy1Pxn8eQvfXz887vJDfeUO+22VmoW3zTl8c9tDVlnRKx7RkskA2zjJyPT7V389kHxx0bd+BL+D1ZfKOm1Fp64zzspJZgySenmd1NcwH6WDkHH8xxsT221ZN9wuH1AD95NVLKP++6Xg/mAXmroHh/xfvNLV604c6b1BPBZh11Fyt0b2iADc5e3H2gE/NWS3kP9fTei82PZQ81uu+J9RduEvEK9TXSotdOKijmnOZegdwSfTBBH1L0nQEREBEWm6ibcpLBcmWd3TXuo5hSHOMTdB6P/wALCDoB7XbmVl0Dw5tfBrR2ovAvmqJXS3RtNLiWChYBsSN2l7jjBwcbjsF0j5GOQG/c3Lrhqu8aifpzRtnqW0k1THB11FXNgOMcIOGgAEZccgE4xlcCcd5tfzcWNSniXU1k9+bXStmfVOLj0dR6Okn97jGMfPO+V7hezRi0jDyh6QZpBgbF9+fW7DqdVkgyF2PM5CZzRzTwX4LaD4DaEt/D/h9aGUdBQxhrpOkGWof5ySO7uJJJ3/8Aad+E4VVTAQVREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREHAPNpyc8OObTSkFn1RLJabzb3+JQXmlia6eHbBY4HHWw+YJ27heMPNxybcRuTvVlBT3S7su9nuTTJbb3RRvhBIJBjeMkseB5Z33I2xn6HFoGrtC6P19QMtWtNN2+80kb/EZDWQCRrXeoyNvL8gV/wBHzncL+XTjTxzk970pp6trojt73UdZa8DfYkEkfPstS4g8oXHzho4SXrRNXIzORJStc/GPlgHP1L6KNP6W01pWjbb9NWGgtdPG0NEdJTtiGB5HpAz9qzK23W65ReDcaCnqo/7ieJrx+QghKPmmouJfG/hx0UdNqTUNk8N2GMkDmbj06xuuyHBH2rHMnw0uNvodZ3Wl1lp6F7GVMFdABVeFkAmOVuCCBnAIwSBnHde1N24VcMb7EYL1w803XMd3E9rgfn8rV0r5zvZf8OOI+kqrVPL9pG36Z1pRkzCjpD4NJcW5Jcwsz0sec7EAdvPsZh47mcLOLGhOMmjrdrfQGoqO6UFxp2VAEE7XvgJG8cjQcscDkEEDt9S3kvm8u2heZ7lI1LRX+86b1boOujmbLT1RbJFBO5p2HiMJZINjsSds7L0v5f8A2wnB/Udptdg412m66avrY44J7lFGJ6GeTYGQkEPjB7kkEDfdP8HooiwLLebXqK1Ud9slfDXW+vhZUU1TC7qZLG4ZDgfQgrPQEREBERAREQEREBERAREQF4T+1j4YwaA5ra++W2gbTUGrrdT3RpbgB9QB4cxAHbdoP2r3XJwupPtDuUu28xnC+q1FRyui1LpOhnqqB2NpGMBe5h+vB288+uFO7xLcZXs1OK1i4g8q2jrDSXKKW6aWoW2ushL8yNEZIa4g74xgfYPULtavBv2WnEy+6N5rtNaTpK6Vtt1UZ6Gopur4HP8ABc5r8dsgMIz37ei94x2VWcVREQEREBERARbS4ncUtDcHdIVeuuId+htNnogPEml7ucezWjuScHAXVOT2vfJ0yRzG3PV0gacBzbE7Dvqy8H8oCDuyulHP3z8aM4CaMufD/Qd6o7txBu1M+mjhp5RI21te0tM0padnAE4bkHI8sLrTzte1S03xJ4dO4dcuU+obdLdHgXO81EXuksdOM5iiAcSC7bLtiB2we/Vrlv5EOYPmbvlJc6XTNfZ9L1c3i12pbuwxRFhcC90Qf8c7yHEjpBGdyQmjP9nhwG1Bxz5ldNXGaxzV+nNN1zbvfKuVpMQEfxMY557vc/pwO/mvf9744Iy+R7Y2MGSSQA0D1PkuIdN6f4F8lfBKOnfV27S2lNPwMFVXT4a+qmO3W8jLpJXuJwNyc4GwXkxz6e0HvfMrfqXRPCKrvVn0Nb3kNa1zoKi7Tk4D3sachoGA1hJO5yATvB6d83vOZoPli4c1N+iuVrvWp6g+BbLOyra58kh/fPDTlrB3J2PpnBXmLdfbAc3VbNMbdUaRt0TyfDZHZvEdGPIdT3nJ+ZH2LanBT2anNRxnnoL1dNGzaXsFa+OSS432ZsEzoXbl7IHHxXHG+7R38+y9TeFvs2uUzh1p2mtVy4WWvVVxYwe83G9NdUvlfgZIa4lrBnsANk+4TzXjDxO4/cwnNPf6W36y1DcNQ1Ujw2noKaLpiBJ2wxowACfPYLtpwb9jXxH1rYKLUXEbiTQ6VbWxiQUUFA6qqGNIyCSXMAyPI7r1P0hy/wDBHQLmP0Zwp0vaHswWupbZE0tI8wcZB+a5BT/B5P649iNcaa1SVHDrjlFX3CNjnNprtaPAZKQNmh8cjukk4GSCN/tXUjhRzD8ynITxBvmiba4UMlPWCO82C5wmSmnewgdYGxaS0YD2kbHcHsvcnWXMnwO4f6jZpXWPEi02u5vd0eBLIctd6HAOD+jzW1OYnlO4G82Wj53aksdvku9VROFo1RQtb71TOLR4bxK3+yxghuWOJBGQMdxrRJyr82/DXmi0FQag09daKi1D4YZdLC+ob7zSTgfFhhw5zDjIcARjz9edV4P8OeRnnO4V8xVkttk0HeaOa03eJ37IKJ3+0JKVsgLpBMDgscwfROCCcEAgke7VKJ200LapwdMGNEhb2LsDJHyzlQTIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIusHGH2jfK7wVv9RpbU2qq+uutK4smp7VRGpMbh3BOQB9Z2PllDXZ9F145eOezl95mb9NpTh1e7jFeoYnVHuFypDTyyRjcuZuQ7A3ODsuw6aCIiAiK1zmsaXvIAGSSewQWy+J4T/Cx19J6c9s42z9q+b/m9tGqbLzI68ptZW6ejuUt1kmc2ZpBfGQOhzSe4IAwRtsR5L2d1n7TDlC0Fqyr0df8AiDVGsoZXQzy0lrnqYGPGxHXG07jt2Xn17S7mw5Z+ZGksDeE1uqrnqG2SOE19lt7qX7wf/kX3wB7hkZGRjf5BMu6ePYHhTXW+5cMtJ1tpqWT0ctlozDIxwILRC0dx5jGCPIghbrXhbyAc+Gq+BGvbXojiLqWsrOHVxPuksU7+sW17tmTMJ7NBwCO2PMAFe4lou9svtsprzZq6GtoayMTQTwv6mSMPYgojORERRERBT5LwF9qC0t50tdfNtAf+SRL36K8AvafSGTnU17nPwe4tH1CkiWfqevb7l96f9gnh30HI/YrasfxWNcgLjnlwDm8v/Ddr85GlbXnP+LRrkZWTFgiIqCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIvOvn19pjbOFkt34N8FJn1Wr4gaetvLHDwbc892sIOXSAefYfkJDs/zW83XDblP0T+yLV0v3RvNaTHarFTzNbU1j/NxznojHm8gjsACTheKfE/ifxe5/eYO2TVFki+6d0kjt1ut1ujc6Kipurzccl2Aclxxk42GcLTNG8u/NbzRXP9mFo0ZqvVX3QmDJL7cOswbnOTPIcdIJJw3IG+AvZbkk5KNHcqmh4Zaujpblrq5xh92uxYHOYSP7DCT9FjckbdyScnOSvL0zeuWOAfBzTnAnhdY+HunaGCAUFLG2qljaAZ58fG4nz3yB8h6lcjIiDb+udEaZ4j6Tuei9YWyG4Wm7QPp6iGVoIwQRkZ7EZ2P8y8gOaL2Smu+FlmvPEDhJqSLU1hoOupda5YnMroIBknB3bJgYHkcDJJXs+o5Yo5onwyxtex4LXNc0EOB2IIOxB7KUfPxwn9otzW8FNJ0mgtK61pZrRbG+DS090t0dS6nYCfvbXOHUGgk7ZXYjgl7ZDixHq62W3jXp7T1fYqqoZDVVtvp3Us1MwkAyY6i0gZye22V6N3bky5VL5eKi/3jgFoyrr6t5knmltjD1uO5JHbP2Lqvza+ya4f6/t79TcuVBQaQ1BCPjtPW5lBWADs3JIidnzGxz5YC1qZG1vbBas4Ya24KaFvlh1PbbjcX3My0Qpp2ve6B8eSSASWgDOxwclcqezhtsVm9n7JcvuY0SVcN7qnDoGagDxAMnzB6SB9q8eeMPB/irwT1N+wnivpy4WivpmnwGVBL4nx5+lE8EtLTsdj2IJAyF7r8hdopZ+R3htasMbFW6elEnoTLLKXE/8AnHKk/ssleXnso9R1FPzo2lrnuY29W+5slYzZpcYXPGR6Ag4+te668E/Za0MzedzSUA3NNT3MvI7YbSyAn6sr3r3wpn1VURFQWl6mvLNOabu2oZWdbLXQz1rm/wB0Io3PI+3pWqLqrz284vDHl64aag0hcLk24a11BZ6iktlmg+J7TPG6MTTHtHGA4u33OAADkJR423uu1VzicykVLboI47lrC5mlo2MZ9CPLnAkbZPTkn8i93eVjgFQ8t/B61cNKOsNXLTF01VOezpXYzj5bflz5YXjh7LHSEuqecrSdWI3GGxU9bc5HdJIaWQOa0E4wCXPGF73IecEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQbC428G9G8e+G934Y66pPGtt1iwJGAeJTyjdkrCezmncfb2Xk/xv9jvxT4fadvGr+Huvrbquktsb6gW59K+nrXQNBJIOSxzgAdhjK9nAMIRnY9kP8fP5wN9opzN8u+nqbQFgu1vuFjtj/Djt15ovFdTtB3iY4Fr2AHO2+D5L1J5MvaDaF5mbNJbdVSWzS2r6PHi0ElT0xztx9KIvIyMg7Zz8lrvHf2d3LPxzbX3Ws0RDYdR1fVILtanGne6UjZ0jGnoePsXknxw9nxzLcFL5XS0egrtf7DSyl1NdrVEZssz8PU1mSHY9ARtnbsL/pn9PoFhnhqI2y08zJWO7OY4EH6iNlKvm74Zcx/H7l91VDWWjVV/o5aSRrprZcZpgx7Qd2ljzluRkZAHzB7L1U4Ce1t4CcQqWgsvE733Rt/kDY5JZ4uuikf5uEjc9AJxgEevbsp9HfBFp1jv9m1Laqe96fudPcKCqb1w1FO8PY8eoIWooCIiAiIgIiICIiAsW5Uja631VC5ocKiF8RB7EOaRj6t1lLanEDijoHhXZZdQa+1TQWejjYX9VTKGueB5Nb3PbyGPqQfPVwf1gOX7muserLxA6FmkNVPbVseCOljJXRvBGxADSc/JejnM37XS16EmpbHwO0tSXusnj8SauuT3CKEEbBrGHJOfU4xnttnzi5wNd6I4mcxOstd8PmAWe81zqiMtGGvcdi4Y2wcA7bZyu0nss7Dy98Rq+8aG4xacs12umCbbDc2hzJGnBwASM+YA8zt6K5vqbnW3qn2x3NlI/qgt+hoWZ7fcmVx/KZv5lqNF7ZzmZgh6a3SuiKp/937nMz8wlXpVWez/AOTauJdLy+6XYXf/ADGOSLH1BrwFtit9mPybVk/jt4XCmA/+RwVkgZ+ck/nUV5tat9rrzcakpJKK3Vml7AyQYMlvth8UfU+R7sfYAVpej/avc4Gko/AqNU2a/RF2SLtbRK8fIPa5px9q9XtMcgPJ/pRzZaDgZp6pmYciWuidUuB+QkJH5lNrLkM5RtcUstPdeBunKWSRvSKm20/ukzR5EOjI3+sFTJujqRyse1xq+IGtKTRPHHT1oszK94jhulEXsia89g8OJx+ft3XpM6+Wdtnff/unTG2RwGpdVtlBi8IDJf1jbAAJz8l5O8zfsftQWR82o+W+uN0oWtLnWWun/wBsMA8mPP0z6Dufl59GNR685heG9LcuEWp9XatstOzMNbZqmtlawDsW9JJHSd+xwd1an+Oc+c7m41xzj8WIeHGlJfC0hBdxQWOjjccVTy8MbPIRnOTuO+2DtsB2F4e+xJuNfa4q7iXxsFuqpo2PNFarQJTESMlrpJJACR22aui3B223XhdqrTnGfVdlnhsdqrGVUDpG71Dx9ENHcjJz9YXo3qn20Wg4tMv/AGI6ArZb4YiG+9OIhD8d8YB777k/ar01yNwW9kVy88L9SQaq1beb3rqqo5Gy0tLcRHBRseM4c6KMZkIOCAXYBA2K5t5oebLhdyj6E+6V5iirLn0CC1WCieyN8hwOkEDaKMDzx2Gwxkjx9vnNpz18x1/qf2Oay1pJDVvIZbdN+JTU0Q7Bo8IDJA7knPcqCh5UOYGsulPrHmFs+oaOwwOMtXPeKt8s72DcgEucQDvvnPyU55qruMnM3x+5/wDiPp7QNw90t9DVV4itVkoA4U1O5+xklcTmRwGfiOPQAbL0p5XPZg8FeXqak17ryvdrPVlAwTiprWiK30DwAXOihJIOMH45Cds7BdPrfzR8lHA+yPfwc4bzz6vgj6G3IQHDnjsQ92MjJJzk/wAy68cRecbmw48++acqeIWqKi1VxLXWa1OkDHsJ2Y8RDqkG4BBJB9N1cSXXqjD7VrloPGCq4X1U9ygt8FW6hZqQtaaCSVp6SRg9QZkbOIwRv8lzFxQ5xeAXC/SU2qq/iFZ7hiLxIKWjq2PklJGQMDOPt3+S8H7TyocyN9pWVtp4L6pnhfjpcKIjOe2xwfzLe2lPZ5c4OqqiOGHgtebfE/8A+6bh0wRAZ75JJP2BRa7Ma69tNxSqLpPFw94c6epbe1zmwy1/iySPHkS0OAH5fPsuMdQe125vL1Rz0VJW6UtLJmlvi0VpIlYD/cufI7BHkQMrtHy4+yC0Ta7ZHdeYGaW6XFx6jQ0lSWRNHoS09vryfqXaKy+z35ObG6KWn4F2GoliILZKrxZjkdiQ52Cc/JEeCF7l4m8U7pWayu1JfNQ1dU8vnrW08kwzncAtBAAJ7D5rsLy/e0p5j+XWwxaJo32nUlkoi5sNFfoZXyU++7GSMe17QDnY5GV7n2fhtw909QMtdk0PYqKkY3pEMNvia3HoQG7/AGrjribya8s3FyKb9mnCGwyVUzCz32kpxS1LM9y18eCD9eU2q628mPtSLVzBazk4c8WNPWfR95qmF9qqaaqcKSqcO8R8U5a/G43OcH7O/TXNe0OaQQRkEbheIfOb7M/iDwPu8msOC9ou2p9GEGVzYB41XbiPJwb8Tm9sEAnPrvjirlg54OMXLRraKrq71dr9YOvwrjZK+qe8FoOD0dZPQ4Y2Gwz6HdIPoTRdQ+GHtR+U/iPFSwVeq6vTFyn6WupLvSuYGvP70SNy131j8nbPafT2ptP6rt0d203eKW5UcoBZLTyB7SD2zjt5908NaqiIgIiICIiAiIgIiICIiAiIgItua14haL4dWt951rqOitFGwF3iVD8ZwMnAG5/IuqOvfaz8pmkI6mGz3m86mroQeiG30JEUhHl4riGgfPdB3RXWnm554OG3K7pgzGsor7qedxZS2eCoa54I7mXpOWj5HB8/TPkpzPc//GfmE1XNNY79dtK6cx4dLa6CrdG4gnu9zCCSdhgH1yTnbK4Iezr5n+YCODWVdp2qs9jrCJTcrzOI56ph7ujY89buxGTgZ9UG/uIHtgeY3V9BVWqxWPTmnKeqY6NzqeOSWUNOQQHuIwceYC1Dlv8AZ0Tc4HDyq4zXDi0+03G61Mhcw0gqR43n1jLSBnbv5LiHml0Jy68KaD/Y70jDWT60tbxFWzCVzg14xkPySN9zjvvsB3WVyP8APFqrlZ1THa7m6S4aIuEo+6FCMkx5O8jPQjJOcE9+4JBTvSd6k4z8sHMd7PfX1l4j226snpqWfqtupLXG8wdfYxzMcPgJBx0uJB8jnC5LsvtmeZyip2wXnTGiblIBjxvcZoXOPqQ2XH5AAvVHRPF7l45pdGzUNiv2ndW2m5xGKrtNZ4cj8EHLXwOycjfcA4PY5XHNy9mjyaXCpfUs4SU9GZHdRZTVUoYPkA5xAHyCbR59ye2e5kXSF0ejtGMYTkN92mOPlnrVJPbO8yLo3Ni0fotr/Jxppjj7PEC7/wAfsyOTmMdP+xeHfN1U4lP3Mrk4D+v/AGLWnBzg1T8LEg87G+2Q5sWyl/3M0MWHsw2mXb7RMFt7XftYebXXOna/Tf3T03ZKe4wup5ZrXbCyoDHDB6Xve/pJGRsOxK9SIfZx8mMTBGeB9pkx++fPOSfrw8LWLPyF8n1hqoq63cAtMCeEhzHyxySkEeeHuI/MtSmR49cr3s/eNfNXBNqigq6XT9gc8l13ujXvM7ycksYN35Od8jPfscruFZPYg6XbE39kXHe6vkx8XuVpjYM/LreSvTK0Wa0WC3xWqx22moKOBvTHBTRNjYwD0DQAFnKjwt56PZ1VvKfp22cQNI6qrdTaXqqkUNZLU0zY5qKZwJYX9Hw9DiCAe+frXYf2QHNF4tBeOAGvNSNb7r0VunX1k4HwHIkp2lx8iAQPmAPRel2s9F6W4g6ardIa0sdJd7PcYzFU0lSwPY8H5eRHkRuvnw5yuHmlOBfMrqXR/C2snpLbbJmyUroqg9dO92SWNeDkY7bHb5dkkn0fRYi+Z2n5i+PNLRRW6Di/q1lNEA1kYus2wHYZ6sn7VyLwJ53OOfCviFadQXDiHd7pbI6lja2mrKgysdCSA47nyHrkYyMKj6HUWyuEnFfSfGXRNBrbSNxiqaarjaZWMcC6GQjJYcfmPp88gblvd9s+m7XPer9coKChpW9c087g1jB8yVBqC8Ffasfcs86Oq3WueOQmhtvvXQ7IbOKZoc0+hwBkeWV6G8yXtSOBnC+xXC18NL9FqzVIY+KnbSsLqaGXGAXuIwSDvgjH19l4s6p1je+JWvq7WutbhJW3K/XA1VfUPO7nPcM7+gGw+QCJO9fSLwDhmp+B3D6CcdMjNL2trh6H3WNb+W2uG1VQVnDzTNVbHxvpH2ik8EsIIDRE0ADHp2x5YW5U8WCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICItOul/stldA27XSmpDVP8OESyBpe70Ge/dBqKKgIIyPNVQEREBERAREQEREBERAREQERdc+bjnT4Xcrelq6G73eOs1lUUb3WmywEOlfKRhj5O4YwEgnO5GNtwUG8OZDmS4e8tmgLhrHWF4pBWRQuNBbPGAqKyYj4WtZnJGe5xj5heVep/bJ8zNfcZZdLWLSFpoy8mOGahfUvDM7AuLxvjvsurlttPHbm54qyR0VPeNY6qvNQZZXEvkZTteSS5xORFGBn0GBtk9/WLgD7JrgLo/Q1NFxlsp1ZqmqiDq2U1MkcEDjv0RBhHbtnJz8+5ZBxHyve17vWp9Z0WkeYO02W3Ude8RMvFCx0TInnt1sJIaO2+T6bHGfTqxaisOp7fHdNO3iiudHKA5s9LO2VhBGRu0n8i8kucz2Ud+0hINacsdorb3aCD75YnTeLVU53PVEXHMjfUZyPIbDPUKzcEObmwRy0lg0LxEtUcZPiR0rKmnbnudmkA+quW+EvH0eq0vaDguAJ7L5w3cPeb6Wo8N2n+J75XHG5rCSfryt1Wnli5+dQxCptvD3ibMzydJUzRgf8AnyBSSj6FEXz23jlf5+7BTuq7pw94mxQsBLnx1UsoA8/oSFca1t85jeH9Z7zcrtxCsU9O7PXPPWQAEH1cQD+cIPf/AJouO2l+Xvg9fNc6hu0VJUCmkp7ZGXYknqnNwwMA3JBOTjtgeoXgLwx4X8QuZ/i8bFpihmrLlfK99VWzk5FOySQl8jifTJwPM+gyRp2o9c8bOPFyoqTUd/1NrOspsRUsMjpKl0edhhoyATkDOF60eyr5StXcEtLXniPxNsbrZfdQ+HHRUczcSwUwGcu32JJ/ISrIl47u8O9F2vh1oSwaFs1PFDRWK3QUETImhrcRsDSQB6kEn5lbjVMKqn+r4IiICIiAujXOh7TK1cr+vf8AYv0xoNup79BTx1NZJUVhgp4A/drMtaSXYGT/ADbE95CcL5/faXmd3OHrV8/d0kfR/wB6AQP0JKNC5u+crVvNzeLVc9R6Yt1jitDC2CCkkdJ3z3c4AnuVydwY9ppr7g5y3zcDLdpmnq6ympp6O03WSUj3OKXP73O5aXEjbuTuPLpnSUjZ2vc5+On86pNR+H9B3UVcyM7PHeX2OlqtNXzTVd3uFU1lVb9O1fuUbiAZJZHMa7HqQwP/ACr26Xy7aK1vqzhzqWj1bou+VVou9veHwVNO8se05BwcdxtuD6fJe3HINz7ae5jNN0Whte3amouIdHH4b43EMFxa0f2Rg7FxHcDv9fdWndFca8cuP3DXl+0XX6y19qGjpBSwl0FG6ZonqpMfCxjM5JJwM4/KcA7M54OPtdy4cvN94h2drTdpJYrbbur97UTEgO+wAn5Yz5Lwptdj5gubbiFP9y6HUGutRVLjNMWl0radjjuSSemJu3qM4PcqDtdrX2zPMTcrrVP0NpfSdmtrnOFOyro5KmZrM/CSfEADsbkYIz6hdfeHvDPj9z78a66vFW+53m5y+PdLvVgtpqOPyGwwAB2YP0kk+jHLB7JXhJpvSVuvnMFaZdQ6omaJp6BtY9lJT538MhhHWR2OT/qHeTh/wr4c8KrS2x8OdF2nT1CAG+FQ07Y8gdskbn7SUvPBwvyf8kPDrlKs00tnqJLzqi5RNjuN3naAXAb9Ebf3rAc48yO+5JPZNETwEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERB1n5nuQfgvzPmO432KewXyFpay52yJgec/3TSAHeXfbbffdeW/Nn7NLiry2W9+sLHdI9Y6V6y11TTU7mVNMPISxjIO2TkHG2w749385UFbRUVxpZKK40cNVTSjpkhmjD2PHoWkEEfWEo+ePgNz18x3LtQiy6G1gyps4IItt1g96gYM79AJBb6bHbyXpVyhe1L0hxoqnaW4x01s0hfGMzFUsmIpajHnlxyCfTyPyOV2C4mcj3K7xWpKmPUfCGxU9ZUNIFfQU4pamMnJyHR48z5heWnML7KrjvoLU9XJwj07Uav0+95fTCncDUMaTs0gkZwPUg/X3TZPR7P27iNw+vIa60650/WdX0RBcoXk/YHZW4Ipop42ywytkY4ZDmEEH6iF859y5KubzTERuVZwF1pTMi+Iyw0Tn9OPPMZJC0OzcY+ZDgxXmlptY6w0/NG8A01bJM0ZGduiXsO/bCSX34PpQReBujfaa8zOmZmuuGpn3SLqBc2SRzCQDn5ruBoH2z+hP2LlvEDQd1F7hjIaaQt8OV2PP03+rbvvuk6Xj0x2AyFjPuVvZnxK6nbjv1StGPzrxG4h83HNHzzcTIeHvBltxs8FSJH09toarwneEB8TnvB+EAEZIJPnnGVwXxu4X80/L7c4bdxak1XaTWAup6n7rSzQVAB36ZGvIO/kcH5Iks8vr6LqavoazIpK2Cct7iOQOx9eCslfMlp7ivxj0JX0moLNrXUtvkDhJDI6rm8OTG+wccOC9XOXL2tnB658OaOi47V1faNV22BsdRLFSumjrcDaRpGMOOBkHfO/zVxXbTml4+Wflu4MX3ibcwyWopIxDb6ZxGaiqfsxuMjbPf8A9q8BOOfMbxU5htTTak4i6imqQ55dDRscRBA3OwDexwMDJ9NgOy5k58eeC7c1OqY7Rp9tRQaItLy6gpZMtfO7/wCaPHqdu+/btgLqjT00lQ8NbsM7u8goLY4JZgXMYSB3Woae1He9KXinv2nrjNQ11K8PimicWlpG/l5fJWeMaZxhiwAMgk759VFLS9bHTMw053b/AKlbMR7DezP59NR8a7nPwY4sVkMt9paT3i1VxIDqtjCA6M5OS4Agjv2AHfA9DK640FspzV3Ktp6WBv0pZ5WsaPrLiAvl70lq/U2hL/S6n0leam1XSid1wVNO7D2H5eX5Vzba9Pc7HOOKm8W2DXPECmosQTSuqT7qwjcM+NzYyfkMn1U7R7/N4i8P3lzW650+S3JIFzgJA+fxLq3zN+0y4L8Ap/uFYHM1lfsfFTUFQDFF/wB+8ZH5/I4yvKaj5B+dFtWI6bgJqqCTOOvEbAP/AB+sDH2rd9n9ltzm3tpmk4eUtDt1H326Qsd+QEklNnn1crsXB7brUhrnCp4I21tHv0llfIZflkHA9PNdL+bHmIi5ktffs5+4jLdK8OD2Ab7nO581t3jhwD1dy/3mLTGveiK7ygvNOzOGtGMnJ79x6d1rvJvwrtvGbmE0voK8RNfRVs/VOHbjoBGcjz7q2Z6mNN0w/jVzDS2jhPYIai7tge0QQxU+GxgDALy0bD9JWRxq5TOOnANjKviHoqrpaCUAtrY2OdCM7YJwMb7b7L6EeHfB7hlwot0Vs4f6LtVnZEwMMlPTNEr9sEufjqOfMZx6BZfEvh9p7inoW9aB1TQQ1duvVHLSyNlb1dBewgPHoQTkH5Y7EqEfPhy+82/FDlwe92hRbpGucXhtXB1lrj6HPbf0W9OK/PxzO8yFENA3isoHU9ycKeOjtlCWPkLsDAIJ3PmcDbOVyFws9lrxxuXH6DRnEPS9VRaIt9xLq29Ne3w6mjY4kCMj989oA+WTg9ivXDQ3LDy/cN3Usui+EWmbdPRta2KoZQMdKMDuXuBJPz7rMn47/KerucedHA/2NFTqOwW3U3F3iRUWt9ZEyoNrt9IDI1pAIY97jscd8b/UV6CcE+UzgpwHtMNu0dpOlfPE3Bq6mJskrj5ncdz3PnnzXMqKmKAADA2AVURUEREBERBQjOx7LpNzR+y44S8f9Q1WudL3qXReoasdU5paVslLUSf3T48jp+ZG/l2wB3aRB4D8xvs3+O/L3C66uZTarsoyRW2yN4IA7kxnJGPMZz2xnK4u4W81fMFwRrYnaF4h3SgbSEsNJPiWLAO7HMeCcZ7jb0X0hVVJS10DqatpoqiF+zo5WB7XD5g7FcYaw5VuXLXsUserOC+k64zZLnm3RxyZPmHsAIPzBQdDOVD2ul31Pqui0TzGUFnoYK5wjjv9GwwMY8nAErMkNHbcfPtsD6X2TWekdSRRzae1RabkyYB0ZpayOXqB9A0kryc5v/ZParsd9fqflkskt2ss7TJLZn1IM8D8kkRF5HUPQEgY8xjB6FXWwcW+COshabra9RaQ1JSSt6IXskpp+sEYwNusZ22yD23ROvpyRcd8vNw1LduB2iLnrFz3XqpstNJWuf8ASMhaNz88YXIiKIiICIiAiIgIuD+OnOXy+8u07bfxK1n4Fwc0uFDRwmoqMbfvG/WPq88LpVx79stpWr09cLBwH0fe23KdhjhvF0EcLIge7mRtc52R5Zx9ndTdHpjedTac03TuqtQ3+3WyFrS4yVlUyFuB3OXEei8xOb32teo9L6zrNCctzLNUUtteYqi/1MPvLJpB3ETDhpA9Tnf7QPPMR8f+ZTU001NSas1zdKiUuc2COapYxxGQMDLIwB2Gy738lPsprld6ubWnNRpuooKGMAUOnzUdM0r+5dOWH4QO3Tnf7dmmfXRjitzGcdeYK7B+v9ZXO9yzO6YqKEFsf/etjYN/q3XYzl59lJxp40WGDVmqL7Q6LtdSA6JtXTvmqJW/JgIAPzJI+edl676I5YuXzhzGyPRfB7S1t6Nw5lvY9+fXqeCSdu+VyXHHFBG2GJjWMYA1rWjAaBsAB6LW2pn9Oj3L/wCyd4LcHtQUmrdVX2t1pcqN/iRR1lOyKmjeMYIYCc/bvuRld16+sorDZ6mvkYIqS3Uz5nNYAA2ONpJAHYAAbBZ62txSbI/hlq5sWzzYq8N+s078fnUV86Qseq+ZfmHrbXpeJ1ZeNbagqZKYEZAbJI54Jx5NZ8+w7+at5h+AGsuW/iHLw91rEBVCnZVRSNHwyRuzgjy7hc8eyXdQN50NPMrGtMj7TdRTZ3xKKZxyPn0h67z+0+5JtZ8xNJZOKHDN1JNfNL0M9LW0Ezuh1XS9XiBzHY3ew9ex3IIA+Vp+nlF+xLi1wZsWneLNiutXa4Lu0S01XRSEGN2MtDz2OR27j8oz6dcgftJv9kWhl4c8fLpBFfqFjTRXbGPfI9hiQf3YPmPt7jGyPZy6h4T8ftD3Dla4uabpq2v0/RvMVPUAZljY8Nd0H1bnJ9PyZ67+0Q5VLZyj8UbHeuGNRU02nr/G+pow6Ql9JOx+8ZJ7jGCPLA7DspLsMx7GXfmT4MadibUai1tRWuneQGz1bgyM57YO63vpjVemtZ2eHUGk75R3a21H9iqaSUSMdjyyOx7bd9x6rxj1Ny28wXN3wj0lrjhpbYryyhp/9tUjquOHreWjDgZCAXAAgAkbOP1LcvJTzN3L2f8AqDUfBDmj0vqGxUtwkhuNPEIhO+ieWkFwY1xBY8YJLScnffzn4y+pLr2PRceaL4/8Hdf6WptZ6W4g2istdS0ObKZwxwyM4LTggjzGFva1Xi13ukbXWi4QVlO7tJC8PaftCarNRFoGq9daR0NTxVertQUlrind0RunfjrPywMq+jqT7UbmT4lcvPCKyR8Mp3W64arr5aCW6tYC+kjZH1kMJyA92cAnyBI7ELoJyKcmNVztaj1TxA4q6mvEVktU8UdRWxPBnr6yQFxj6nA4DW9JJHbqAXpfzWcwXJ3S8N57NxwvFm1Ha6wh8NrjZ7zLJKAS1zAN2uG+4OQuHuDHtL+Q7h3pWh0FpS23jRdpo3ERwR2Rzoepx+KRzo8uLidySM9t8dp/LuSH8bja3M17L7l64acCtSa34ewXmO6afonVrnV9eZxK1uAcAAAHfOO23l5+RVPFTyRzOmqPDcxuY29Oes+mfJeyPOj7RrlzvnL/AKm0Vwu1i3U181TQOoIWU9PIxlO1/wBJ0heGkEAdsefrgHyT4bcLNfcXtSQaT4daZrb1c5yAIqeMkMB83HsB3PrscAq+o5s5POd/XHKheaptLSuvenq8AVNtlkIAI7FmTsc+hH2gkLe3N97SbXXMxp2HRNish0tYA7rqY45sy1B9HEHt5d+2dtyth8YfZ+cyHA3QT+IuvdOUENqhaHVDqasEzoAe3WGj4T9eOxXW1rS4gBXsVcyN8juljS4+gCrLFLAQJGlpO4ysyAe6xF7sAvxv6hWTOfWvYwPzjZM/pj+V3HcLlx9qDxo4FWWi0hdLfRansFH0sZFUuLZY4wMYa4ef2gL2F5cOYHSHMnwyoeIukj4bZvvdXSudl9PMO7T+fH2+i+bKqpJKR4ZJvkZBC7b+zw5ur1y+cS6TSNwf4+ltS1TIKmNzsCKRxADhnYZOBnyJ9CVPG3vUiigmiqYY6mCQSRStD2ObuHNIyCD6EYUqAiIgIiICIiAiIgIiICIiAiIgIuNuN3MLwp5edON1NxS1My2UsriyGNrDJPM4eTGDcldX7x7YrlOoaOaW1Ums7nUMGY4RamxCQ+nU6Tb7Qmmu9Kgqayko2ddXVQwN/upHho/KV4fcyPtTOMPFuulo+HXj6NseOmONk2ZyMEZcQenO+c7/AFALrKy7cf8AiXcBDDcNcahqapwIjifVTBxPYhrcgD7MJ8H0mnVOmOroOo7X1envkefyZVBqrS7pBE3UlrL+wYKyMnP1Zyvnvh5IedCshbXxcCdbPbKA4OdCQ4g75wXA/mWkX/lU5tNA0Umo77wb11aqeAdT6sUcvwgb5JZkjtlJ+x9HTHNe0PY4OBGQQcgq7GV4PcoPtE+JHLjenWfXVRddVaWlIZNRz1BdPTnOCWF5zkfPcYI3Bwu5vFH2xnBMcOLi7hdZtRVOrqqlfFRw1lIIYKaVwIbI6TqPV05zgDy8+xSW3KluOz3NBzf8KOV/SdTdNVXqnrL86MigsdPKHVNRJjbLRu1ucZJxt9pHiPxm5yuPHHHiO3XV11XW0T6eoDrXbKGQsp6VgPwRho+me2SQcnJwMrYNrsvFjmK4jOprTQ3fV+q77OZHBgdNI4uJJJJyGMBJ7kAdgvZXkr9nJw14H6PtupeK2lLZqLiHUEVM81W0TwW4/vYoWnLcgd3EEk+gCvPhjs9wNvF81Bwe0betS074LpV2alkqmPBDg8xjOfme/wBq30o442RMbFEwMYwANaBgAY2AHopFmftRERUEREBERAREQERcU8euZvg5y12WlvXFjU4tza55bS00MRmqJ8dyyNu5A9f9RwHKy25rHiFofh7ap73rbVlrslHTRmSSWsqmxYaNyQCck/IAlecvMd7YrQ9RpGv05y9WG9zXmuhMTLzcoG00VIDsXMj6i9zgOxOB37HBHnrprhNzP8zVRU6k09pHWOui2Vxmrel80TXknID3kMG+dgQB2wplvIO8vML7Y/VNLqO4ae5f9N2n7mUznRQ3m5RumdMRt1sjBDS3OcZ/ON157XTU+qePnFqK9cSdZddz1LcY46y6Vz8Mga9wGQOzWtB2AwPzlemXs8PZuHTAuXEfmg4c0ktyeWQWax3NrZ2U7e75pWAlvWTgAHIAyQthe0S9nTqyPiNRa/5ZeFktbZL1T9FytFmhAFHWNJzIyMHZjwQTjYOzgYO1nD249EOWrgLwf5cOF9BZ+HbKA00tOyauvrntL7g8gEyvl7dJPYA4AAHkt5X3jPwl01SPrb3xJ03SxRjLi65QuI/8VriT9gXhDHyY8+brWy0x8IeIAt7G9LaQzlsTR6CMyAD6sLS2cgXOVNKI/wBr7qkOcfpOjjA+0l+Ev9mPYl/tF+Vht5fZo9dNlew9JkYGFhPyPVuFujT/ADp8vmpLlHbbfrWEPlIaHydIbk+uCSF5B0HstecatijkOgqOmMn7yevY0t+vAIH5Vve/+yU5j9E8N7rxAk1JZDc7RSyVslnop3md7I2lzw2QYaSADgZ3x81n+Ns2Ue0dFqzS9ypPfqDUVtng6eoyMqmEAepOdvtXRnnO9qLauAWq4tAcJbHadW3aKPruFXPUk0tOSdmNMZy53qc47+gJ8hdKy8VeIF+oNE6Sr9SXq63WZtPR2+lqppHzPPkGh2O2ST2ABJwAu9fBf2OnFHVltlu3GjUsOlpqhmYaSnlbUztJGcvIy3PkRnbfutTnqa7Hcm3tTLFx51d/sdcWrHbNI3urGbZVwVJ90qnjvGTJu15G43xt9ZHdzVugdHa6oJbbqzTlDc4JW9J8eFrnAfJ3cflXiDzXezY4u8sNmPEOy3aLVelaV4dUV1Ex0dTbjn4XysO/TkfTbkDbOO65o5Gfak27hxpT/Y05krhd7hRUOBab3HEamaOLsIZQD1OA2w7vjvk7meK9LOHHLpwh4U1stx0Zo+jpaqV3V4zo2uew5z8JwMH591yYthcION/DHjtpv9lfC7VVNereHdEpiyHwv/uXtO7Tst+qgiIgIiICIiDjLmL4zWvgFwkvfEu6MD226PpiYQSHSuz0jA79jt8l873GnireuNHEe78Q76CKi5SlzWk5LWZJAJ8zuc/WvZD2vl4bb+Uee3NeGy3C/ULGjOCWtc4uwPPuAfrXhvG3rka3OMnCTqMyjPRG4jJz5ZVA9wlLnjIJ9VcIwwGJrfQnJ/OrHRkOOM4GCVrnlYlkvVKqne9/XHvkZxndalonWF/4fastWs9M10tJdLRUsqqeVji0hzTnGfQjY/IrDe7oZnqGw7ZCitduqr1daS00TOuor6iOniHq97g0D8pCl73Gvxu3k47Nc2nPrr/mv0np/R+obVT2yis0/vkwgOPeajoLQ5wG2ACSB2GdgF6jezA4WaM4ecrtku+n6mirrvqd77jeayEhzvFJwyEkeTGhuB88rpHQexm4z3bSNJfaTiBp6luNRTtmdbqxkrXAloIb1NBDTv2PbscLr7a9fc0Xs/eLVXov7tVdluNskjkq7UZzNQVsLviDgOzmOGSHADJ79sKf2v19DSLqbyd+0H4Zc0kcGlauN2ntcRwdc1rmIMdRgbvgf++Hng7jb1AXbJFEREBEW09fcUtB8MaFlw1xqSktcUgJZ4z8OcB3IHf7fr9FNG7EWyuH3GLhxxRifLofVNJc/D3cyN/xY9ceY+pb1VBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFxHxp5XeEHHilMWuNNU8lQdjUxxtEjt87nG5+Z3XLiIOhWoPY78ul4eZKC93y2OPbwCCPyE4/MtnXn2J/CuSz1LLDxb1JFcywmmkqaeF0PXg4DwBnBOMkHPovSZUJwng+fHTdRxt9m/zJw3u/aTa64WsT0wZUNcKS6Ucgw4xSAYwQAcjJBH1Falzf8AP7rrmrltlvl0zQabstocZIqWJ5nklkOCXPe4DbYbAeQ+a70e2E4qcG/9hmDhdXVtvuOv5LjTVlup4umSe3RNcfFkeRkxh7QW9JwXZBxgAri/2UXJ7w64lcP9Q8ZeKOmYrwKm4PtFminOWRsjY0yygDfqLnhoPb4T8wry+pn11l1BrrTXGLg3adL11PRW69U3QGva0Ah4wMgd8H9BK7Z8AfZF8J9c8J6PVmtOI95qrveqYy077Y6NsFITkDIIJeR3IJC6H85eiLdw05l9c6JsnwUVouJipg0AdLC0OAwO3dbe0Lxq4/2iSm0poDiVq2D3yQQQ0FHcZi2RzjgNDM4P5Oyfl2SEaNxg4dVnCXiXqHh1XVTaqSxV0lK2oaMNmYD8Lx8iMfnW1Karlpj8PbuQvcrT3s7+GvETg1azxZtr5tdXG3MmrLg7BdDUPYDgjGSRkZ3BBz9vQniV7Jzmc01qCop9F2Sk1DazKfd5o6trH9Gdsg/LzOPqUlV0uM0D3FzuoZJPrjKymyRtiModsDn/ANi5f4yclfMVwGsTNTcRtCSUlrLg11VBK2eOM/8AbFvZcGgkLV36kyzF0rw95cBhe7fspdQ6MuHKVYrHp24Ub7nbqqrN1pY3gSxzOlPxvZ3wR04JGDheS/AXkt49cx0ctXw703CaKEZdV104gh+WHEHvg49fLKg1jw+5kuSLibRS3mmu2kb3TPZUUVfRzu91rWtP7yVhDZW5BBafnkealvyn6j6OVh3S6W2yUE91vFwpqGipWGSeoqZWxxRMHcuc4gAfMldStB+0/wCVa58LbRq3W3EeC032SijdcLP7rLLUsqQ0CQNbG0gguzjcfk3Xm/zn81HELnU4pO0/wlodT12jLexkNus9HTSvfUOzvPLFGCSSewcNsd9hifcVFzna/pecnnANm4V1MVbQveyzWuqGTFUljcOlae5aejIPn9W62xrvgXzIch+srRxBEElJUx70t0hgLomOODh2dhuB3PfbuucvZ+chHHqm46aa4qcSND1+ldP6bqPfh902+FNUvDSGtbGfiwcnJIHl5ZXsRfNO2HU1vfatS2SgutG/6dPW07J4j5ZLXAjsT5eabL/iSY8c+FHtj+Plo1BQ0/FSzacv9kfIyOqkp6J1LUxsJAc8Fji1xA3wW+vfsvXjh/xE0hxN03Rap0be6W4UddAyceFK1zo+oZ6XgHLSO2D9my6Ee015Z+WHh1y83PiBp7hrYdNam9+ggoZrXAKZ08j3YILGYBAG5ONs7915ccG+OvErgZrG16w0LqSupZLbUMldSe8O93qGAgujezOCCBg7eac+K+mVYtfcrdaqc1dzr6ekgbu6WolbGwfWXEBdGaj2wfLM3hs3UlOy+v1W6kBGnzbn5bU9O7TP/YywH98HZI8ge3nK7S/Odz5ahvmttMaf1DqqhNW5ssTLhHBQ0ucubEPGkYwkDb1OPLsGD3Odx14NNuAtTOKGmZavOPBhuUUrs+h6ScLe1LVU1bAyppJ2TRSDqa9hyCPkV4VcNPZZc6F11NSSXXRlNo6CmnY59wrLzSOdGM7uY2CSRziBnyH1r2t4WaJqeHmhbTpKtu0lzqKCAMlqXZ+N+BnGd8Z7Z/8AYlsvidbuRERRERAREQEREBERBQdlsDifwR4ecW3W6fWNgpKuqtcokpqh8LXSMGc9Icd8Z8u3f1XICpkKWaS4ho6SnoKSChpYwyCnjbFG0dmtaAAPsACnRFQREQEREGk6pvbtOaduF9ZSPqnUUDphCwbvI7ALxX5hPal80N/1Pe9L6RulNou2UlVLSsZS0bfewGuIyZH5IyBsMeec+nt29jZGlj2hzXAggjIIPcYXDms+Tnlf4g3l+odYcEdMXG4ykF9Q+mLHOPz6CAftCYPDHg5wU45c6vEyWkt1fUXOvkw+4Xi4vcYoG99zjvuT0jHrtnfvNoz2ItLFNFNr/jm+djSDJBabT0AjzaHySE4PrgFekmgeGHDrhbam2Ph1oy0aeoWjHg0FK2IEeWSBk/aVunICXdWZHGfATl94dcumiYdE8PbWIIGfFNUPaDNUO9Xu7nzO5O5K5NRFMxBFwLx153OXjl0uzbDxJ1fLFcywPNFQ0zqmZoPbLW9v5vPC4fZ7X3k6fL4brlq5je3iOsZ6fzPJ/MrOju0uLOZribpPhRwQ1hqfVt1gooPuPVwQNe8B88z4nMYxg7uJLhsFxLbPaf8AJhcqM1f+yo6lw3q8KpttQyQ/IDpO/wBq8uPaDc345reKcFPoiauboixRCmtcErSx1VMSS+d0ee5Jw0EZAHnlLCMr2VMtgpuc3S1RfrjT0hZQXFtF48gYJap9O5jIwTjLiHPIHmQvVbnZ5wNHcrfDWsnlqIa/V13gkp7PamSjrMjm4E0g3LY25zkjfGBleO1h5BOcO66aoNd2Dg1en01QGVFL0TRRVXSd2SCNzw8eRBxnsVv3hj7Ofm24t63om8UNIX+y2/xGMq7jfJi6UQgjLWFziexOPTdLeQn9uufCvjTxB4NcS6Li3oO8e6aiop5ZhK9nXHMJARIyRmQHMcCchbv5lebni7zU3a3XLibV29kNpYWUVFb6YwwRdXd2CSS4+pPbZe3mi+RTlf0npOi0vUcINOXc08TWTVNdRtlkmfjdxJ9TkreOn+V/l00tj7gcEtGUnScjptELsH/xmlW3+j3rxO5QfaA8UOU8zWGktlLqfSdY8PmtVXI6N8LgfpwyjPScE/CQQcjthdwOJvDjg/7UDRg4+aX1zDofUem6B9FVW+tdHKWBgLw2bdpxknDhgYI8iF2P5w+QbhXzAcPa39iGkLPp7W1BGZbVcKClZTCR4H9hlDAA5ju24yDjcdx5HXTkl5yNHVtZamcHNXNbJmGWSgY50NQwZ82n4mnfAI+xTh64jZqjVuiKyv07Y9V1TKalq5Iz7tMfBlLHFvW0dsHGQR3BC5/4Ge0O428Ea6E0NRFdbcMNnpKlzi2Rvn6798fpHdYekPZuc4usAXwcI6u1sAyHXSeOmDh8sk5XCvFfhJr7gnrGp0HxHsMlqvFK0PdE8ghzD2e0jYg74KX8f5TKT+3oxrD22Vxfp6OHQ3CGnjvUkZEk1wqSYInEbFrWnLiDvgnB9V0h40c0nMBzMXxty1pqGrrDACYqG2QuighbnyYzJxv5laPw14eWy23Gxa24uW2op9FVM7XOduPeWZzgY3wR6YJ28u/vby96H5aK3hlaL/wR0ZpZ2nbhA10U9PQwve44HU2RxBPWD3BOM7jYhJv4ziceE3BDlv4y8y+tabSlhoq4vPwyVtx8TwoGDucu8h3IBGwK780XsQ7P+xh33R48Vn7IvBJaYbOwUQlwcAgv8QtzjfIOPJemNo0rpnT75JLDp62W50v0zS0kcRd9fSAtWRXy9cRdEXXhrru/6Avjo3V+n7hPbqh0Zyxz43FpI+Rxn7V7L+yF0Doyzctrda26npZdQXm41La+oAaZWMY7DGerRgH68D0Xj3xdrLnqLjJrCrrJHT11dqOu6iTkueah4G/5FrVp1xx95fK33bTGsdUaR8b42uoauWCOUHfIwQD8x+VD9Pbf2kuqLLp3k+17R3SaJs94o20NHG5w6nSmRpyB3OADnHbIz3Xz9Rg9Y+tb54gcdOL/ABVY2PiPxGvmoWsIIFfVGQZHbv8AzrY8Ujonh4wSPIp+Oz0ahVB0rersAMfLb/3Khgd4B6wNxv8AYr/efHhOMNcD2+v0VjIXPYdu6352eOf4y/8Auknnjqo8Sdw34T6beuFhQzS08zJ4JHRyxOD2PacFrgcggjzHdSTtdCQzYE+nktxcKtOU2r+JOmNMVzC6mud1pqecAZJidIA8f+blT8sxr8Y7O6Q9q3za6N0vQaUortpuup7dC2CGor7UZZ+hoAaC4PAOAPMdtlzpy7e2I11V63t2n+YKzWL7hV8ogmu9tgdTvpCTgPcwuILd9zkY77DJHojDymctTdLU+jqnghoyst1PE2JvvVngllPSMBxkLevPnkEYPbC88+eX2WdxorpFrvlW0fJUUFRn7oacp5S50L/J0AeSek9unOB8sb5+NZr1TsGpdP6qtcF701e6K50FSwSRVFLO2SN7SMgggkdvJamHtPYhfO/aOTnndo3SUdl4LcQ6MRElzYoZIGA+eD1AE/UStvaO42cwXLfxSguFTfdTW692CrArbTdKqYCQAjqjkjeSMEDY4OxBGR3SXNOePpDRcJcr/Njwv5pdHRX/AETdGxXamiabrZ5fhqKOQ4ByD9JuezhkdvUZ5tQEREBERAREQEW1eJXEzRPCLR9drziFfoLRZbe3qlqJifpeTGgbucSMABePXPR7S+8cc3N0HwQqLxp3SVNI41FeZDT1VyPYbNOY2AeROc+nnKPSjnG5vNJcrPDGs1IKy3XPVNQRT2izOqB1zSn9+9oPUGNG5P5M4IXl/cfbB83dU6UUcmjqJjiejosxe5g8hlzyCR8wuK+CHJNzL8yN2t91otG3uKxV8jRNqG7B0cIiyMuY6UgyHByMZB9V63cLfZo8pnDzTlNbbxwwt+qrm2NvvNfeHPqDJJj4i1hPS0ZzgALX+jx7uGrOY3nr4sWnT97vlXqW+VsvhU0ZYI6akYTu7pYA1jR64+Q7ruVpj2Imop6eObWHHigpZXNBfBQWV8vQSNx1vlaD9YGNvNekvDjgDwX4RSST8M+Gen9OSzAh8tBSNY8g9/i7jOT2PmuQcYUHn9wp9jtwU0Nd6e96x1reNWSwOD208tNHTw5B9AST5d8ruto3hZoDQEMcWk9LUFA6JgY2VkQ8THb6WM7+eMLdioThSyeiqjlijmifDLG17Hgtc1zQQ4HYgg7EHspEVHRbjt7Jrgvxg1fXazsWqLnpGruEhlngo6dksLnnucOIx9m/qSuJ6P2IWmRVF1fx3ujqbyEVqja/GfMlxHb5L1BRDMcFcr/J/wAK+VSxVNu0NTSVlxryDV3WraPeJgOzc74HyBwudURAREQEREBERAREQUHZUc5rWlznAAdyfRbe19r7SPDDSdw1xrq+09pstsjMtTVTuw1o8gB3JPkBuvFjnv8AaJai5hr83SPCa5XfT+hLeSA5srqeoub8/wBkkDTlrMYw0nPr55Dv9z98/Fv5YdMUdh4b1Vlveurw94jhkl8eO3QtG80rGkZJJADSQe5IxuvJOsvfMFz18a7fb7tc59S6ourvAp2loipqOHOSQ1o6Y4xkZOPQZOy5c5IuQDiBzF60tur+JmmrtbOG7Saqrr6lzoJbngnEcBPxkE93gYAOxOdvZPhRy58EeB0bm8LOG1msEsjAySop4eqokAGPildl5JHffdXidefGivYjQCnpajiBxwkM5IdUU1rtQDAO5YJJHkk+WcD5L0h4UcL9J8G9A2jhzomgbS2mzwCGIdIDpD5vefNxO5J+S3eiiiIiAiLzY9p5z3X/AIX17+BHCa5Po73NTiS7XKF2H0rHjaNhH74j8hznsAUmj0kD2OcWhwJacEA5x9a87faK+0Yh4UNu/AThHBTV2pq+hdTXa8OkD4rYyVpa6ONo2dMWk7k4bkHBOF5t2Lh1zg6ssn+yDp+ycSa+21YMgucM1SWytPmHdQJBHYjYjtstX5a4+FejeN1LfOba010tqpn+NLS10b5Hzyg46pWH4ngbbHv+cP8ADz12j9jNwXvlTxQ1DxovOnquO1WyzPt9rrZoi2OWpne3r8MnAdiNrgSMj4iM916zaq1bprQ1hrdU6vvlHaLVb4nz1NXVSiONjWgk7nucA4A3PYAldA+Yf2p/ATS/CKv0zy3XaWq1JPS+6Wz3a1yUlLbg4Y6x1NaMtB2a0d+x2Xl9R2fmO5ib5/tWh1vrmuuEpf1FtRUsc8nOcnLGjPbcAeWE7anza7Y83ftV9XcY7ZfuF/CzS1FZNHXDrpJK+tBmrq6DP0unZkQJAIGHHHc57deNG8jvMlrvhzPxP0/w+qpLPFEZmNkyyaZgGSWMI32wdyPsXopyJ+zH03oK103EnmG01DdNVPeJaKz1JElPQAbgvZ2fJkA75A9PX0Rpqamo4GUlJBHBBE0NjjjYGtaB2AA2A+QSK+bLgrzB8aeWHVtReeGuoamyV2fBr6Cph64J8H6E0LxvggehBA3XqDybe1XtXFy/DQXHyCx6UuksY9xusMjoqSqkzgseHkiNxztvj8+O3utOVflx4i1sty1pwV0ldayb6dRNbYxKfn1tAOfmDleWXPl7NfVfDDUk+vuX7Rlfd9E1MZmqbfRdVRPbHjJdhm73R9sYBIyBvvhmxJXszb7lbbvSMr7VcKatppBlk1PK2Rjh8nNJB+xZJGV8/nJBzl6m5SuJnTqU3av0XXB1HeLOJCXU5BPTNFG84a9ru42JGR3wF6/8D+e/lq5g78zS3D7W0gvUreqKguNK+kmlx3DA/Zx3xsTvt3IyXrsKiLj3mCv2pNLcDtd6j0e8svVssFbVUTx3bKyJzgR6EYJB9Qg4h57Oci18qHDdtXZHW+4a1u8ogtdtnf1Bgxl88jQchrRjGcAkgZXmj+6/c4eHf7e0hudv+oY2+r4/05XE3LNwS17ztcwNHpW96juVS2YPuWob1UvdPJS0THDrILsjqc54YxuwDnjbAK9mrBwB5L+UDRYvNw0tozTlBT9Ect91EIZqqZ+TgGecF5cSThrMfIIY8T+P/N/x25lm0sHFTU8VTRUZ64KKkpm09O13910t7nbvlcO0BiE48UgDB7nG69TfaOcz3JjxW4Iy6S4c3m0ai1cKuGW31FttkkXurWu+MGV0bdiNukEg79vPyskjkid0SMcxw8iMEfYVczxPeVnTAxODukjO2Qc5UbXOc4Z2zt2UIqpmt6CQ4DyKlgmY89Bw3JyMnYJIn5SSciSV3RAS7O4IHb/381vflytk954+8PLZT03jyVGpbe0R4z1Dx2k7fUCthV4DZGta7Ixn5ZXL/JhqOz6S5quGGor8/ooKPUNO6Z+M9OSWh2PkSD9ifk1v172cyfMBpXll4TXPixq6jqq6loZIqaCipiBLVVEpwyME7NzuSTnABK8er/Q8XPaocebjqXSOmLdYZLXQxQPa+YvigpgXdBe8gZOc5+ZwPILvN7ZDUtrt3KrbrFNO33u96noxSs7lzYo5ZHuHyA6R/wCMFsj2JWnIKfhdxC1W+nAqKy+QUTJcbmKOAOIB9Op5/Is3ZODodx35ZuPnIvrGwajuF0FJPM/xbVfLTIehszdywkjZ2MnBzkA7YXobyA+0kj4vQ1HDvj/fbTbNS0bWm33R5FPFcI9gWvycCQbEnYHue+B3d4ocJ+HvGbSdRoriVpejvlpqRkw1DMmN/k9ju7XA4IIPcBeOXNF7LjjRoHX1SeBmjLlq7SdUQ+jFO9r56cHux/UQSAdsk5+sbrWTE2+PWnihzK8G+FGmKrU2odcWiWOniMjIaetjkfJjyHSSB9ZXnZxK9tTqYVNTQ8LOGVqMTXlsVbc5JCSPIhjSM/bhcDaY9llzo6yoI6u56VorRE1vwQ3a7xtlacbAMBdj8oC6/ScKLloTjlRcJuLNDLbpaS+Utuu8THbtjfK0OLXDGxa7II7D5jCzJftXeOyNw9r1zh1jn+7XPSdE05AENkacA9t3uduFwDxH42cfeZe/+Pq6/wB21HVkfDS00Z8Nje30GDAHlvsvde2cjfKHQWimtcPL9oqpghY1rZqi2RyzSYH0nSuBc4nuTnf6lv8A0fwQ4PcP6cU2iuGWm7PG3sKW2xNI+o4z+dXk8Nr51tMcQON/L/fo6jTl9v8ApK4xkPY0gsBx5hrgQR5EgfJd8OU/2uOuf2YW3RvMhJbaux1rxT/d6GAQS0bjsHTAHpczPd2ARt5ZK9P9X8HOE+vqGS3a14baavVPK0tc2ttkMpIPcAluR9hXlTz/APs0btoW7S8UOXDSFTW6XmjdLcbHRh0stueMkuiZu50RHkMkYx9cHr1Z71aNQ22C8WK501woKpgkhqaaUSRyNO4IcCQVnLwO5Eudy/8AKbryaz62dd7hoS4h0FxtbHEyUM4O00THkAOaQQ5uxIJGx7+s/BPn55Z+PuootJaC1rM28zgmGiuVI+kklIxkMDtnHfyJVHYpERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERARF5u+0p9oDeOFNxl4H8HLkINQOiDrvc4yCaRrhtE3/tyM59PPbAId9uIXFHh/wrsVVqPX+q7bZqKliMrnVVQxjngeTGkguPlt+jdeV/Hn2yPEW4XC6ae4HaXtFqtoc+GmvNax09S5uSC9jMhg27Eg774XS7TXDPmV5mbjPddOaW1hrudryZavpknjY4nceI89Dd+4BGF3i5TfZG3m4V8WquZyB1HQxYfFYqapHiSnOwlew5A23AI9N85Etwn7dReW3l44pc7HGaWkmudZNHUzmu1HqGqzIYWE5ccnYvOQGt7DI2xgH3j4c8P9B8t/B+i0bpyH3LTuk7fJI+QgF8gaDJLK8/vnuOSSTnJAWqcN+FHDrhDYWaZ4a6Ptun7azBMNHCGdZ9XHu4/MkrrL7TbmTo+CPA6r0nQv6r9rSGShpWjvHERhzz8u/wCQjzCvvB4x8wvEp3GHjdrTiW5jWMv94nqomtOQIurpZj/xQF6c+yn5PrVZtFt46cRtLMdd7jL1WZlZCC6KEf8AyQA7jOAR8yfTC6K+z84E23j5zKaf03qGlM9itnVdbjGfoysi+IRk+jiPzFfQZQ0FHa6KG3W6mjpqWmjEUMUbQ1rGDYADyACsuJYyURFFbZ4h6C05xQ0ZddCasoY6q2XendTzMewHGRs4Z8wf9Xmvng5q+Xu/8tXGK8cPLtBI6iZIZ7XVFp6KilcctIJG5AIBGSRtnuvpE2wuEOavla0FzP8ADuu05qK2xMvcMDzaLm1o8WlnwS3B/uSe4P6Mgk8de/ZJcXtF6l4Dt4dxXGki1RZ6l5qaUvAmmhx8DwO7gACT9Z9Cu0XMbwG0lzE8Lbxw81TbaeZ9RA91uqZGAvpKoD4JGO7t3wDjy+YC+f2ut/GLlI4xsMsdw03qnTdZ1wyEOY2ZjXdx2643ADI/QRt7tcmfM7aeajg/Ta2hZHTXmgkFDeaRh/sVSGg5A8g4bj7fLCcs2G5Xz5a90NqPhtrC66H1ZbpqK6WeqkpZ4pWFpy1xHUAfI4yD6L2q9lNwP0NoDl7o+IdsqqO6aj1c81Fwq4yHGlYNmUwO5bgbnfckHAU3tNeU+y8Z+DVx4h6V0xC/XWl2irinp4gJqymH9lifjd5DdwSdsb5AGPH/AIQ8x3HDl8raocM9dXWxNmcW1VCHkwPcCQeqJ2wcD5gA7DPZXVfStnA3K6H89ftJaTl4r4+HvCSC1X7VksRkqaiZ/i09AD2yGnDn7dicdxsQvOrX3MBztcYtKRakuep9U1Fiewhxtb5Gt74JIYeoA+gOMd1wZw8rtMjiZY67ilFVV1kFyiN4a57jK+Lqw8uJOTjufPAI+ShK57tOs+Zr2i/F+xcPtXamq7lTTVHiSspaZsVJbYsEul6WjDRgHuT642JXdiP2JPCZ1LEJ+Muq21HSPEMdJTlvVjyBGcZXajTfEvk+4CcJYtY6Ku2itN6S91bNF9yzCyWcEZDS1p63vJ2+LfOMldYL77ZjhdHPUN0/pCrlijcRG6oa/MgHY4AGMq58S864M5l/ZB6q4X6Nq9c8Hdb1WsobZGZay1VVCIqwxju6IsJD8AbjAO+ewJXDHKLz88TOTeiu2i49F0V/stbVComttfJJST004GHFrw0luQN2uad913g0P7aHg1cas0uvND320RdWBVUjBO3H/eZz+cLXdS+0B9mjq+ilfqizUl1fOCZGVGiXvmcT3+Mx9z69X2qeK4Sr/bg6ldGfuXy/WyOTb+2L9I8D12bCCVlcNfbR6ivOurZbOInDCyWqwVc7IaipoaqV74A4gdZ69sDO5/N3K2Jq3j97Me6X6pmtPAeWGleD0PZbfDBcR36QSRv5bro/xVuWgbvre4V/DSzz2ywSuzT08x3b3zgZOB2wE8H002a8W2/2qkvdmrY6uiromz080TgWvY7cEFZ68NuQ72h2pOXy9UuguJlxq7toCqeI2lzi+S2E4+NmdyztkeQ+WMe1+kdXab13p2h1ZpK709ztNyibPTVUDw5j2EZ7+R9Qg1pERAREQEREBERAREQEREBERAREQEREBEVCQBk+SDRtX6u09oXTdfqzVVzht9rtsLp6iolcA1rQM4HqT2AXkzzC+2K4kXG9XDT3ASyWq0WiJ74YrvWQmepnbuC5jSQxg8wcE/LsVH7UnnksHE9p5feFVykqbTa65z79cYnER1U8eWiBhH0mNJJJ7EjG++NP9mRyGaY41U1w4wcb9Mz1umKaRtPZbfM58UVfL3fK/pILowCABkZJzuMFIOsPCbgzx253eKtW+iq6i63OtkM10vVwJMMGfUjbYHIaMYHpkZ7m03sQNQfc4Oq+Ptu9/Ld2R2R/gg77BxlyfLfA+penuhOGXD3hhaWWPh7o206foGNDRDQUrYgQOwJAyftK3Oc42S9Hzc8zfLZqblg1+7QWp73QXSUxmWOekBaHN23LCSW5BBG57rul7Knkp0vxBpBzF8SKVlwpLfXOp7HbntzG6ePHVNID3wSMDt5rhLnE5euY3WXMPqO91umbneBcKwiCcDLImA7NOTsBuds916jezw4Sam4Mct9r0pqpjoqyWqlrDE4Y6S/BOPlkY+xSWWe9K7NABoAAAA7K5EVBERAREQF5He2e4Y6mquIGlOJNs0/NNao7QaKtq4Yi4MkEhLS8gbADbJ7beq9cVpGqNK2DWdkqtO6ltkFdQVbDHJFK0EYIxkZ7EZ2KlHz0f7LFNxS4TWXg7faiC2T2PHulU8AMkDQQAfnjuNvPC3/yu8+/EDk8pLloWy2m3arsUtU6YwTzuYxkh7ujLQSM753wT3zgLkjmd9lfxo07rq8Xzg7Y479pipkfVQRRyATQgnPR0bk/mx64XQ+/WG86Yu9VYNQ26eguFFIY6innZ0vjeO4IK1+NkT+L1NovbcWs0hdcOC9Q2p8hDWAsP2kgrj7iX7Z/ijqPT9fYtA8NbPp+ethdCy5zVMk80AcMFzGbNDsE4JJwcHGy84vkvR/2aXIrwm5gtAXjiZxYoa24Qw3B1BQ00c5ij+EDrccdyCR3HmO2N0/GG46ncouj7bxQ5ktK23V9a33GW5CsrppzkPIdnJJ7kvIP2Fe9WuuWrghxJ0ydK6r4fWqpoTEImFsIbIwYwCHjfPzOV5W+0X5TdNcoselddcGqmrtdHeK6SkLmyHxYpWsc8Dq7kYHfAz6bLvB7MDjrrDjhy6mq1zczcbrpy5vtLqpxBkkiDGuZ1/MB3/uMLM29XI4Z1L7FThpcL9NW6e4p3q2WyV3UykdTskMeT2DjkkfX+ZcP8wPsh73w30LcNY8OtZVGoZLdGZpKSWJrXuYBkkAAfmJ+pew+D6qOWKOaN8MrGvjeC1zHNBDgdiCD3BWtTHyw1EM1NPJTzxujmieWPY4YLXA4II9RghZdPWwtYGyEg+Z9V6xc/Hs0oNTyVvFjgZbmQXF+Zrja2bNkOCS5oH6dyAN8jceWOqeHettFOe3VGnKu3iOQxOMrQB1A9shNniWT8uVoFTI2SZzmZ6SfNel3smeUnS+vYqvj3rSmFUy1VxpbVTnPSZGD4nH6jsfkQPMrobwN4Law4/cRrbw10TAx9xuLiS9/0Ioxjqe75AHJ3C+gTlR5fqLlo4N2vhhTXEV01M99TVVAbgSTPx1Y2GRt6d8pZ9X9OY0RFFF5L+2J5etT1mtLTx001p19TbnW1tFeKiniJLHxk9D348g3bPp8gcetC2zxJtFnv2gNRWm/UkVRQT2ypE7JGhw6RG45wfMYyPmEHzi8BOPOvuXPiHRcRuHtwbFW04LJ6eXJgqoT9KOQZ3B3we4+rIPohpr22LJ6djdVcJIqaoDPjdS1Lnxl3yycgfWF0P5XuFWmuM3M1pThleJSyyXe7SRS4JBdC0PcGjfuQ0DGd+2d16R8dfY76A1U2Ku4L6jbpqpjZ0vpqxhfDIQO+W+ZO/YfWm58M1uHhH7XPhBri9NtWs7cdONkcAyWRxAGTjckkE/LIXaCn5seXKqphVR8XdPBjh1YNSMgY9AvILXvsnObDSEjn2Ox23U1OM4fQVbQ84/7RxyM/WuP4/Z2c50ryxvAe9twcdTpacA/P+yJydqfrXufp3mQ4EapnFLZOK2m55iSAx1cyMk/LqIyuQqWtpK+FtTQ1UNRC/6MkTw9p+ojYr55Lr7P3nIs8Tqip4B6jkjYC4up2xzEADfZjyfzLZ9FxB5j+B1c6gj1FrPSs8R6DTVTpomjHkGSDA7eQ8kV9Ka2nxQ4k6U4R6Fu/ELWlziorVZqZ9RK+R2DIQCRG31c4jAAB7+mV48csntSeJfC6SqoOK9bU6moHt6ovFBLwSMjGOx7b7DGPt4g5i+ZfjPzucUILNbIbjUUFRO2Cy6cpD8GfJxaDguJ8ydvXO6ufU1NzYc6vFrnF1JTWCaA2/TUNb02ew0YOZHuPSx0p7ySEEbdgTtnAK7acofskK1lXZuJPMRdImRxmOsg0zTN6i4ghzRUSHbHYloHcEHIK5O5FPZj23gzXW/izxtNNddXxNEtDamYfTW15GznEj45AD9Q/KD6FKaIKWlp6GmioqSCOCCBjYooo2hrGMAwGgDYAAAADsp0RFEREBERAREQEREBERAREQEREBERAXHPMHxZi4GcHdT8VJbW64/sfoX1LKYHAkcNgCR2GTv/ADd1yMtv680XY+Imj7tonUdM2e3XimfSzsIzlp8/sOD9iFeAHMnzt8eObKamsGrq6GlsjKkPo7DaoS2J8pOGde5dK8HYZ8zsM4XcDk49kxV1L7JxR5iK5kdPmOtp9LwNy6QbOZ7zITgAgglgGdiCSCux3AP2XfA7gvxBHESuM2oKuilMtrpqwl8FK7Ozi0/SI8s5+vGQe6KaIKWmp6Gmio6SCOCCCNsUUUbQ1kbAAA0AbAAAAAeinREBERARFsXjBxp4c8CNG1Gu+JmoYrVa6choLvilmeezI2d3OPoPl6hBsXnI5i2csPBW4cRYqJtZXmZlFQwvHwmZ+cE/IED5eq+fXiJrzUXFjW984harr2zXS8VDquoc9/ck4DW574GB9mV3E5+/aG6b5o9M0/DfQmka2hsdFWCqNdXua2Wd7TsQxpPSMDsdxk99lwZwV5IuYzjpJTT6R0DWQWqoLc3OtHgwNaT3ydztkgbA47puTpnXNXDT2rvGbh1w3tnDK1aGsFZDbqQUVPUOLxJ07gfCAQTuutV4dxV5luLbQLJNV6hvtU2GKmigcyOEOccA7fC0ZJJPofkF7Xcrns/uC/AXS9DJetKWzUOrHMa6suNbCJ+l+N2sDgRgZPlj0279iKDh5oK1XJl5tei7HR18Ywypp6CKORv1Oa0EKTzkN2686eDvsXNLW2G33njJxIqrlXNLJai1WymDKYHYmMyuJc7B2JAAOD3BXoxovQ2lOHen6XTOjrHS2y30kbY2RQRhuQBgFxAGT8ytwItaeiIigIiIOlPNp7MXhjzGXqfXOlb1+wvVM0eJZIKQSUlU/wAnSxggg+pacleT3GrgBxz5MuJlI3UEdRba2inFVZ79by73efB2fG/GM47sO/cHO6+jdcJcxvG3lg4dW6LT3MHftO+HXtJittwp21Mj2nYuEeCWjtvt5IPOvhT7aHiLY7dSWnirw6t1/dAwMkuVDK6CeXHYujJLM47kHf0V/Mh7XR/Evhpd+H/DfQtRaJr7Svo6muqpQ8xxPGHhgHmQT3BGDjPrwXzx1/KBdbrDcuWyhpaYzuBkZRAsiHcuPRkgfbuutfD3SVTrvXFj0dS9Qku9dFSgt3IDiMkfMDJRJ/bkvlQ5q9ccpPEGp1zo22W66x3Kidb7hb64OEdREXBzcPaQ5jmuaCCMjuCCCtwcxPM7xv56+I9kt9faGNZC4Uli05aQ90MUkmA+Q9RJfI4gZecAAAAAA59btD+zY5ULHpG2WnUPCq3Xa4RQR+81VT1eI+XA6t2kHGdtycrlvhvyv8v3CKvF04ccJtPWOuA2qqemBlB23D3EkHYbhF/bo9ys+yDtOl6i1674/wCpG3O4xNbO3T1CzFPA8gECWV28hB9ABv6gEdjOLvs3eWTi1FG6q0xNZKuJoayotsnQQB6g7n8uPku06Jhjx15oPZI3jhrpur1nwc1LW6ipKQGSa31UQE7W4zsW9x5ZyfnhedVwt1faayS33OinpamFxbJDMwse0jyIOCF9Tj2MkaWPaHNcCCCMgg99lxvqflt4D6yr/upqXhVp+uqg7q8V9KGnPqenGUHzRfavUrkN9m9oPiJwq0/x11veq51wuhdV2yniAEUQY8tDiM7nLT88gnbYK/2tnKxw50HoLTvFzh1peksckNeLZcY6VvTHJG9pMZxnY9QO/wDrUnJD7RLhVwo5VH8Ode3WSi1PpCKrjtMJiyysY9zpIQ09shzyCDvt27ZW/B1Z5tOK2reYDi5RcIbZc5bhT6duEtqt4fISx0rSWHG57YI+zC9duRTgBLy6cALTo64Ydc62V9yrngYJkkA2P1AbDyBwvJ72dPArVfHvmXt2t6umk+4en62S8XetcD0PkJJEYJzlznO+eMjPde83SA0NA7K+TGZMv6XIi8s/aGe0g1ZpPVV24G8Fat1tqrc4090vUZ++NeRuyL0ODnPbBHfOBGncrm45yOHHKhox91vk0d11LVgstVihlAlmeRkPk7lkY2JJGTsB3yPD6o1NxI5ruZOPVFRQOq9Q6nvMM74aOM9FPEHt2A8msaBue5+ZAWRwO4HcZucvim2w2m4VdzrJD41zvNznfJHRwl2S97nEk9zhg7nYBe3fK5yYcIuV3T0FNpm1R3HUT42iuvlVGDUTP8+nb4G57AfPtnCWDnKyUs1DZaCiqDmWnpYopDnPxNaAd/rBWciICIiDo3zcey64f8wmoajiBoS/R6L1NUtHvLW0gfR1bx2e9rSC047lvfz7DHlpxz5aeO3Jzrahq9RxSUUkEwmtd9tj3GF7h2LX4HSfkfmN919F62xxB4b6J4paaqtIa/03RXu1VjHMkp6mMOGCMZae7T8xhE/15ncmntYtQ3bUts4a8x77e6lrC2mptSsaYnskOA33gbgg+bhg5P1A+p1BcKC60kdfbK2CrpZmh0c0Egex4PYggkELxn5tvZT8S9A6grdT8v8AZqjU2k5QZxQNlaayi8y0BxBkaBnfuAPPsONOT/nd4kcpGuJNOa5F3uOl/EFPcLNVPd4tIRt1MDt2kA7Dt2wCCQSve1Fw/wACOazgjzGW1tXw11jT1NWG5mt0/wB6qoTgEgsO/n5b48guYEBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERARFtbiZxB09ws0Jetf6orIqa3WSjlqpXSPDQ4taSGgnzJAA+tBwfz9c0DeWTgdWXuyV0DNWXqQW6ywuOXh7geqbp9GAE59e24XirwV4M8VecPjE+yWmea4XW6TGtu90qSS2BhPxSPP1AgD5YGwVvFHihxY5wuNrK+51VXdbvqG4iis1u6iY6WOR+I4mNGQ0AY6iB5E9gAPbLkh5PbBym8OnW90rK/Vd6DJrzXhox1AbQsP9w0/lPr3L/COVOBfCDTnAnhbYeGOmIWNpbPSsikla3BqJsfHK49ySfM74AW/wBEQbe13rnTfDbSdy1rq64MorVa4TPUTOI2A8hnuT5BeAvPTzPy80nGifVVAJItPWmEW+zwPcTiIHLn/W87n6vLOF6fe17vVfauUaWnopHxtuOoaGlmLTjMZEji0+oPSNl1w9kxy4cJeMfC3iTe+I+k6S8zS3BlkhdO3JhiNOHuLPR2XjB+W+Uzulcf+xpvNDRcx15s9QWipuNjkNPnufDJLsfYQvaxfPryk6yoOXvniszqmqMFqt+o6vTtVK87infI+AE588hhX0DRyRzRslika9jwHNc05DgdwQfQhTBIiIqCIiDzr9srwooL7wcsXFOjt7BctPXIU087GDrfTyjAaT3IBGfsXnryW83mqeUziMbzSRur9M3kxwXy2lxxJEDtKweUjQSQfMbHI2XeT2kHOfwu1hpu78uumqt1fWulMdXNGCWNmAIaMjtgnO+5yDheS5pqgVHuhhf43X4fhhp6urOMY75zthPx/wCfEt3lfT3w/wCIOjOLGj6DWuh7zS3ey3WESRSxPDxggEseBnDgCAWnt9RBXjT7Wnl+tXCXjbbdc6UtEVDZdbUr53xwR9MUdZEQJBgAAFwIdgbYws/k9ZzG8i99j4ycWtFahsfCi7U4hukUzmmOYygGCVsQcS2QEAgkAkHG4K7Mc3Ny4e+0T5dH3rgFdn3W46Gq33LwaiAwyx5jw+NzDuCWj9B3S/jfVl3xx17HXilbdRVGo+C+p6Kkqn0dH79b/Hia/rj6x1sw7PbJPbthch+0E9m/pzXlkr+L/BGyx23U1FG6ouFrpmARV7AMlzWj6LwB5bYHZeeXInxDufCfmt0ReoHOhbPXG11zHfCXQSgtew57HOPyL6Ixgjy3QzHy03SnvNsnfYrwyrp5aGRzHUk/U0wv8x0HsfsW4eEmj7VrziRYNIX68RWq33KsZDU1cjwwRMPc5O2T2+1fSNU8HeE1bdTe6zhppievd3qJbVA95PrktO+/fuuk3tHOQSp4t2G3a44B6OoItR2yQtrbZRxsgFZCRsWAYAeCRt2wBsMkoM0ex25U7xpuE2vVesBUzRBzLlR3KF8Uno5rHRuBB+s/Wtj1nsQdBOk6rfx4v7IydmzWiF5A+sPAP5AujVr1Bzr8kV8t1zqWa50RAZAY6O4NlNsqwHAmN0biYXg4xgb4zghejfAj2wHA/XNPQWbizarhoy+yhkU1QGePbnykbuDweqNuf7ofak2HrZlN7ELh23BrOOuo3+ojtUDc/aXFb1s3saOW+hiDLtqbVNyeO7zO2En7G7Lvja7nb71bqa72mriqqKsibPBNE7LJGOGQ4H0IKzFIPILna9ljZOEXDeo4pcCa+83OGzuD7paKsiaUU52M0TgAT0ncg747Z8uEuSD2gWteVuvj0fqBkl80BW1IfUUT3Ey0BJw6WA+QxuW9j3wSMH3lmhiqIn09RE2SKRpY9j25a9p2IIOxBBxg9143e0o5Ab/oDV1y438GdJun0TcGGqvFFRMybTUZPiSBg38FxwcgfCSc7HIpuPYLTWobVq3T9u1RYqptTbrrSx1lLM0gh8T2hzTt54IyPI5WqLzS9ljzvWfUen7Dyta2idTXq1Uz4bBW5Hh1cDCXCA+Ye1uceoG3nj0tSAiIgIiICIiAiIgIiICIiAiIgK1zmsaXPIAG5J8grlBWwGqo56ZrukyxuYD6EgjP50HnzzT+1VpuC+r6vQ+gtIUl6r6QkPlqZXBjTuNy09s+gJ2XT7iX7Wzmg4gacr9L22LTumaa4wvp5qm3UjzVNjcMEMke8hpIJGQ3O+2Fy7xJ9lnxY4jcabnf6i4CC03GqMsk2Wk9JPkSSBtj1XbXhH7MDlV4dUEDr/oSHVl1DQZai6yvliDxjJZGSAN/UfYFP+fyHl5yFcod85n+LFHW36imboqzTirvFXIwltT0nIgaTs4uOAe+xOe5I97rFYrTpmz0lgsNvhobfQxNgp6eFgayNg2AAH5Se5JJO5WPpfSGltFWuOyaQ09b7PQRgBtPRQNiZtsMho3OPMrVw4HsQcKi5ERBjyUNHM/xZqSF7+3U6ME/lIUwaAAAMAdlciAiIgIiICIiAiIgLqlzSezv4J8yc9bquemqLBq+WEiO5ULg1szwD0+KwjDgTjJ8sk4JzntaurXPZzlaS5YuHldaYKttTre/UM0VnomO3ic5paJ3kbtDSQR8wPkCo8NdNcKb5rDi9Bwe025tTdKy8yWenfjIe5kjml+B3GGkr6EOVPgLQctvBOycLaSo94mo+uprJx2kqZDl7vzAem22y8dPZc6GuOu+cPTd9kjmmh062qu1VUOyQJPCeGdR33cXEjPcgr3qSxHnb7aqgulVwG0XVUdFJLSUepjLVytbkQg00jWdR8gXOA+vC63+yc5q9GcE9R6o4a8S9RUtmsWpzBXUVXUnpiirWAscHO7APYWAfNq9jtRaa0/q60VFg1RZaK622qaWTUtZA2WJ4O27XAjzOD5eS8gufX2a2sNL6/GsuWvh1VXPSl1j66i1W1viOt04PxBrCc+GdsAdu2+5C2Z0evOm9XaW1hQtuelNR2270rgHCaiqWTNwe2S0nH2rWF5K+zH5b+arhpxdGo9WaZvml9I+A9tVTXAmNlQ44ILWE4B7g7ZOd+wI9aQmz4uWesC93SnslprLrV7w0sLpHD1AHb7V4Wc+nG6z671BV6etVvhhIrC+QsYBgNdny9TgL274mkM0BfnOBIFFIcfUF889ZZBxE5pafS1UQI7nqOGiIPbpMgBH27/lWJd/+T+K/wDrrkP2cnFixcDuYy26z1n/ALTsdZQz2+WrlBayPxcAOB7HG5P1L3p07qOxastFPftN3amuNvqm9cNRTSB7HA/MfoO684/ab8o+htG8slBrLh5ZW0c+j62mbXeE3+z08n3oyO32w9zSceR+W/WLkB9oJ+1cFy0TxFo7leNHXBzZqcUpD5qCYbEsDiMsI/ejG/5tX8b/AC2XiT/9e5gz5qq4E5dOdjgNzQVtZZ+Gt/q23ahj8aW3XGm93nMecF7ASQ8DIzg/oOOe1QWFeLdHd7TW2mU4ZW08lO7z2e0tP5is1EHzkcUdLcReUHmPnntfi2y76duj6+z1JjPSWdR6SO2Rglpwc49MhdyeEHto9asvdutXGTh1Zam1yvbHU3O1SSQTx5wPEMbi5hA3JAxst5+2o03pGm0ho3VMlKxl/qq59LHI1uHPjazJ6j5jGRv6D5Lh3kJ5DeF3M9wH1Rq6/XSoh1Eyslt1HICTHSyBmWEtHkcgk4J3OM4ASd9Hrbovi1w34g2Sk1BpLWlor6StjEkYZWR9bcjPS5mctI8wR+Zbsimhnb1wyskb6scCPzL569Wez85wtFalqrJBwW1LdRSSlkVwtMPj087QdnsewnYjyOCFtS88M+bTh/P7jddK8S7PJ28MMrGf9E4TvqPpC7rZHEvhNwv4mWieDiJpe110DInF1VURta+Boafi8Q4IAGTuceq+fGCw82TQ2pp6biczOMPbLWg/pysrVNJzh2zSc9w1hU8VYtNgf7YfW1dcaUNxuXhzi0DHmRhTfx8tPY2zxutdql416pseg2OuNBDdJaWgFKwvMrGbfCG5J3B7KHhXxG1xy9cUbRr2z0D6S92KobN7rXQFnW3zY9rgCAQO+O4B3Gy7b+x0tuhbrzFXyDVNDS1V1isD57MyojDwJGysEpAIO4YQfljK9Q+YblG4Ncxun5bfrHSVAy6sa40l1ghbHUxPI83tALgcDvvtt5g0+OO+Wr2jPAvj1ZaZl6vdJpDUuGtqbdcJw2PrPcskOAQTnGewxuV2aptXaUrGCSk1RaZmOGQ6OtjcD9RDl4c8Z/ZdczGhNR1kehNE1OrbL15ppaGRjpQ0nYFpI7euc+o9eNYORnnOid4dPwC1rH5bU3SPy9QCf4r6D36x0jG4Mk1VZ2uds0GuiBJ9AC5arFJHMxssT2vY8Atc0ggg9sEeS+bPiNyzcyHCO3M1HxG4T6qsVCHbV09M4xRkb5MjCQ0/MkLuNyJe0/snBbRL+GHHyO/3S3UkpktV2pGipmgjIGYZGvcC4A7gg7DOQTuaPYpFwjy884vArmeNdT8LNSVE1dbWiSooK6nNPUtYTgPDCTlufME/mOOblAREQEREBERAREQEREBERAREQEREBEXF/HbmS4QcuFgg1FxY1Q22Q1bzHTQRxmWoqCBkhkbd3Y/1+hQcoLDqbvaqLPvlzpIOk4PizNZj68kLzK5n/a+aCuug7jpLl6td+kvNzhMP3Zr4G0sdI07EsZ1F7nY7ZAHcehXnnpLh1zNcwctVW6S05rfWnxOfPNCJp4sknPxE9HfOwO3og+i9+vNDRHEms7Gzy+K4wj/8pYl04o8NrLQy3K569sFPTRNLnvNwiOw74AcST8gCV8/8XITzlSH4OX3VwI/uoGt/S4LcFh9nBzsamq2ULuDdyoWOP9luVfTwRNPzLpM/mQd/ON3tfOHXD7UFXYNAaY/ZS6mJY2oE3TEXA7EkYGD8iSP0+bPHjmQ44c53EWgGoPGrZp6gU1k09bWOMMDnnAaxndzzndx37nYErs/oP2LfG+7tgqde8QNNWBpcPGpqcyVUob5gPaAzP5R9a9C+CnKxy38nWjzqGlttno623U5dcdU3YMFQ4AEuIkefvYO+GtIO+N+yQdT+X/2Num7ZS2TVfHPW1VXXRvhVVVYbfE1tNG7Z3hPlJLnkHYkAA77EL0osdjtem7RSWKyUUdHQ0MTYYIYhhrGAYA/9vcrqFqn2tfJ5pq61Frpb/qO++7vLDU2u0F8Dz6sdI5nUPmAtBHtj+U8k/wDU/W4A7ZtUf80pWf5ymY72Iui8ftjOUh2Q+l1wz/yPGc/klUcvtkOU5hPh27XD/wDyTGP0ypo72IuonDX2pHKZxKvdLp2m1HebHXVjxHE28W7wWFxOAOtjnDJ+tds6OspLjSxV1DURz087Q+OSNwLXNPYgjuFr9jIRFwJx253+Xfl0vUOnOJWrporpKwPNJQ0rqmWNp7F7W7hE3HPaLorxK9rny12PStTV8Pq65329PicKWB9C+JjXkHBcXeh8jgfNeVXFLm/5gOK2qanU144k32j8WUyQUtDXSQRwNzs0dJBOBjc/ZgbJZVe2HPDzaW3lQ4UP1FSQ09dqi7Se6WagkfsZCCTM8d+hgBPzI88ELwo1fqXi1zHa8ues7425amv1afFqHwQueImZOGgDZjRuAPr7ndbb1TrvWmuJ4anWOq7te5YG9MT6+rfOWD5dROF375H+dflO5eeG8li1XoOviv8AOQ6rq20QqTUOAGT1YOxOwzjAAGAEtTrzyutoudjrX2670M9HUx/ThmYWuH2Fds/ZncGtQcROYS0app7Q+e0aemEtRO6PLGPyDnPbIAP5Qttc2PFLQnM9xaoK3g/pWWhmrJDBHEYvD6+o7AgDyO+cbDK9e+Qbl+i5feX+z2StpWNvd2BuFxl6MPc5+7WnPkBuB6EIk/J2TRERoREQEREHSj2u1da6bk/r6Wuc3x6q+W9tK3zLw5xJH1AEn5LxL05oXWmroqmo0tpO73eKiHVUSUVHJMyEd/jLQQ3b1K9Sfbc6tli07w20NG9zWVFZWXSUAnDuhjY2g+RwXOIH+pa77MTmd5atA8rx0ZrLXGn9K6gs9bXVN4iuUrYX1sb5C5kzMjMoEbms6RkgtIxgjIdeeQnnv0Dyxabm0Trmx1I94qXColigPUz4jgkgE5GSCCPXPqvXDhJxp4d8b9MQ6s4dahp7nRytBe1jx4kJPk4A7LwPs/AriPzb8dNcO4A6NfX26ovVdXxyueyCmpaWSd7oi+RxAbkEYA33Gy1nhfxP46+z5481Fj1BbqmgqLdPGy9WWV/VDVQOAIkjIOHAsILXA4I2JHcL7lT/AB9CS88ubb2UVl4yaovvFLhnraWz6iujnVUturIRJSzygdg8EOaTgAE5A2yu9nD7W1k4kaJsmvNN1LZ7ZfaKKtp3tdkdL2g4z6g5B+YK3CivnQ4S8e+PvJHxCvls0qaa13WGV1HdLfcaPxYZSw43GWuIOAQQRtghdzNE+2z1B7uYeIfCC2+OGgNntVRIGuONyWSOJG/kCfsXo5rvl04EcT651z4g8JdL3+rf9KorrdHJIT6lxGc/POV5784/spTc9R0upuWSwwW+kq8ittPiHwoH57x5JIGPLfzHplv9jnPls9qBwp42X6XTmrGU2kqp/wDaxqZeljznsSSR9oO22e+3buLiFoGoYHwa3sEjXAEFlyhIIPbBDl4cat9ljzbacomVls0bDfOpnU6GjqGeK0+gBOD9pC49n5CuciAffOXvVrh/2lOx/wD0XFE6+h6gulrusZmtdypayMbF0EzZAD8y0kLMXzv6A1ZzNcj/ABGs2o77p7VWmI4ph4ttuUMkMFZF+/Z0u+AkjOMeXyyvS/Sfti+V66WmGfVdDqqy3AsHiwMtoqGB2N8Pa718iPtKkurmeu+Kp1LoVqf2ynLBbKSR+mrFrK9VIB6IzQR0zCcbAudISPyLzl5jedLjdzOa5lqbTdL3abWXFtDZrXPKC1mdsiM5cd8Y7euScqzvB7W8y3M9w35ZtA12rNYXqkdcWxFtutImaamsnIPQ0MBz05GScAAA7r55OJOvLxxT17fNf3mCOOsvVW+qkihHwRgnZo+QGBnbPdckaS5TOa7jJXQVNs4V6vuPjkMNfcYJI42NJ7ufLg9IznbPngL2i5QuTTh/y88I7Zpq96Wst21RVRiovdwnpI5jJO4bsaXA4Y3sMencjCtll6m/08CdGa51dw71DS6p0TqCss90onh8NTSydDgR5EdnD5EEfJekPLZ7YnUsNwoNL8w1jo6ujkLITfaFpikZ5B0rNwR2JcPPOcDdekmouXbgNqylmotRcHNHVkM7SyQOs1OC4Hvu1oI+sEFeLXtC+S65ctXEqp1DoyxVR4dXxwmt87QXsoZT9One7fAB+iSdwQNtst5ivdHSurtO62sNLqbTF2p6+21kYlinieHDBGcHB2I8wVmTXi0wO6JrpSRu/uXztB/ISvmbsXGvi1pqxN0zp/iHe6C1sBaymgqnNa0HyBG4HyBAWmy8TOI08hll19qN7zvk3SfOf/OUvvCPpqq9ZaRoG9Vdqm0Uw9Zq6Jg/O4LGZxE4fSDqj11p9w9W3OAj/pL5/eDfLHzXc1FuqLnoS33W92uB/hvrbld/Cpy8Zy0Olfgkb7D7Fq+o/Zyc6+las0b+C11rge0trraepjd9schI+0BLB7sXfjVwgsQH3X4oaWpS44AfdoMk/V1ZW5rPerPqCgjuthulLcKOYZjnppWyRu+pzSQvnduHIxzf28CWu5fdY4dj4mUXi/lLCcfavVf2WfB3jbwd4RXi1cX7dXWplbXtntlsrT99p4ugA7ZPSCcnHzA8tm/jfDK7tIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC8c/bD8ddYVfGCn4GUN2nptP2u101bV08by1tRNMC4dQB3AAGx9R6L2MXhX7XelNPzk3OUg4qbBa5R9Qjcz/APIKDtD7LTkZtNhsun+afXc7ay63OnfU6doA0FlJE7LRO4+byASNhgH7T6ZrgbkTuFNc+T/hPNSuaWxaapad2DnD4x0OB+eQVzwPmgqiIg6I+2Pn6OVKhiyB4uqaIb9ziOYrZnsSKqJ3BriHRB33yLU0Mrh6B9KwD/oFc1+0y4G6y468ts1n0Hb5bheLHdILtHRR/SnYxr2vAz3IDyR+kDK6Bezs5r9Kcmt111o/jdb7va6e8vpZ2xsonOmimhEjTlhwSCH47+WRlIRsb2lHAWt4G8yt41LaYjBZ9XVT75b5IgWiGZ7uuRoPqHkkHOc5+S9OPZq8cbpxt5eqeovtR49xsE4t8z3HJOBtn8hI+ted3PxzV6Q5yr/pyz8LLVXSutchjp2zQuZLN1dWdiB3Ls48gPNd+/Ze8BtW8DuBdYzWVMaav1HXNr/APeNgaQB+Qj8iflPMqfj+Wyu5SIiKLaHF69XHTvC3Vt8tMT5K2is9XLTtaN/EETukj6jg/Yt3qKeGGphkpqiNskUrSx7HDIc0jBBHoQUHz88hXCOo5g+bWw23VMMtXQU01TfL66QH42xtc7pcQNi6UsG/qfRcn+1J4EWLl+496b1voC1MorVqSkbXNp2sJiiraaQBwye4I6Dj0yvYTQvBbhXwzutzvmg9DWuyV94Oa2opYul82SDgnPbIBwPQei4N9oly3Wzj7wJude1pF70hS1N0tzwMk9LOp7T8sNP2ZQY3E42Pn15HbpQcNqyjqLpfrVR1UFMXhppa+GSOUxOGct+Jjmj5H1yF5TcO+JnHbkG1Lq7SF90pUW2v1HQGjmpKs9LSB1ASt75BDu+MkYXNXsbuJ94s3MHcuGU1bI+1agsVTNHA55LWVMDmSAtHkSwPz9XzK5d9t1pe0MsPDPWbKZjbnJW1ttfKGjLoRGyQAnucHP5Umynx5rcJ70WcYtO3+4ShjjeoquV42AcZOokegyV9KeirvTX/AElaLxRyiSKpo4nh3qekA/nBXy7QTS0s8dRA8skicHscO4cDkEfaF9BHs6eJFRxN5XdPXmtlDqmkkkopcHOCwNyPykoOziIiDYPG/gvonj/w5unDLX9B7xbLkwdMjceJTyjdk0ZPZ7TuD9YXg7zncnuqOUPX1Np+vuzb3YLzG+os9zbH0OkjDsFkrASGyDbODg9xsvogXlp7bC4UD6HQtseM1Mb5JWEDtnqBBP1D8/5B47B+y/41W7iNy3WHSFTcjNeNLwmhkbI7L3RNJ6TnucfPfBHcDK7j4C8cvYyG8ycWdQRxzO+50VE58jOrbrLSM4/838i9jcH1UkwVWm6js8GodP3OwVcbXw3KjmpJGu3BbIwtIPfbB9FqSKj5p6mn4gcrHHmCpkoKq16h0RexUUwnaWGVsUnwnJG7HtBBIHZx7Fe+XKvzM6N5puGMGv8ASrXU1TC8Ut0oHn46SpAyWn5Hcg+nrsTwJ7UnlWfxs4PN4gaH042q1npKYVB93jHj1lCQRLGT3cW/C4Dv8OMgLoB7O3nItnKVr692DiFQVx0vqUxRVxiYfFt9TESBKYzgnYlpHcYHzIvqPeBFo+k9U2HW+m7dq3S9xir7VdYG1NJUxHLZGHsQfyj68rWFFEREBERAREQEREBERAREQEREBERBQgEEEbFUjYyNoaxuAPJXIgIiICIiAiIgIiICIiAiIgoNgvnz9o1xIm4k822tK0VL5KOzzR2mjycgRRMG4HYZJOV7gcxvGC08CuEGoOI93diOggLIh5ulcMNAxvnuRjzC+eVsV748cZnupKOR9bqq7mUxsGSxj35JOM4w386vxPr3U5EuWjSXL9wP086itsDtR363QXC71/T8ckkrBJ0AncNAIGPl6ALsmtG0fbZbNpKyWedpEtDbqameD3DmRNac/aFrKiiIiAiIg0XWVonv2lrpZqUtE1ZTPiYXHAyRtkr58eYHTGr+XPmXku1ZRup7hQ3Jl1p2P2yWuBxn0OO/zX0J6l1Np/R1jrdS6pu9La7Vb4nTVVXVSBkcTANySfzAbk7DdeCPtEuY7R/Mnx8l1HoGJ77BZ6Nltpax7C11YWkl8oB3DScAZ3IGexCSSXfqdclc1XtP9R8xvB4cIrLocWQXUwi8VLpOt1R0ODuiNozgFwBPmt58sPsjZuLXCSm4g8VNcXbStyvcZntdvpaSN5igP0HzB5zl2M9IwcH6idd9llyO2jVNG3mE4sWOCtohJ06foKgdTHkYJnc3sRvtn5eRIXrLFFHDGyGKNrGMAa1rRgNA2AAHkEl/pXzwcTuG3HP2fvHymZS311FerYfe7ReqNp93r6YnGel2QQRs+M5IyM74K9MORP2lNq4++PoXjVUWPTerqZjXUlUJBT01zb++wHnDJAf3oOCMY32XbnirwR4U8bLN9wuKWh7ZqGlaCI/eosvhJ82PGHNI77HuvGTnf9ndrzgBqe4at4aacuV94cS/foqiEGomtoJ3jmAy7pG2H4xjOcYyU/aV7oU9RBVQsqKaeOaKQdTXxuDmuHqCNipD2Xg/yF8+t/5a9ZfcHiJdrrd+H90AjqYDI6d9veO00TSScDsWjuO2DuvcHQ2udKcStKW7W+iL1T3ay3WET0tVA7LXtP6CNwR6hFdBvbXWCnq+COiNQl7Wz2/UjoAPNzJad+R9hYD9pW2fYiX+pfpXiZpV0TjBHcKO4Mk8g90ZjcPrIa38i0X22PEJ80uguGMMg8OB893maO/WW9Az8sO2+ZK5H9izpunoOBer9SBg94u2oAwu6d+iKENAB9MlxQ3HokoKmkpa2PwqulinZnPTIwOH5Cp0QYwt9CGtYKKANb2HhjA+rbZbW4wW2iunCbWNtrqSKenlsNc10T2gtOIHkbHbYgEehAW8ltniYf8A83Oqt8f9RK7f/wDYPUye0eDXs2L5VWPnP4de7TGP7oVU9DJj98x9PJkH5HAX0Fr52/Z+hx5y+FAZ3+7o/J4UmfzZX0SKgiIgwL3ZbXqO01divdvgrrfXROgqKeZvUyRh2II/9/Jeb/GP2Luj9QV1fe+D/EubT8tRK6WG13GjE1LHkk9Aewh7Rk4GxAGMDbC9L0V0fOPdrVx+5EuOQYJ5dO6rspJhqYfvlPWQO74J2kjcMAg/6ivX3kO56rLzSaNqKDWk9psuurQ9sdVRMnEbK2MgdM8TXHOCcgtGcHOMgZXM/HPln4NcxdlNn4paOpbjJHG5lLXNHh1dKT5xyjcfVuF4mc1nI7xh5YdZV9xtdiu9y0dHOZLbf6KN7xHGRkCVzBmNwGQScDbvvhN51M7r6Bu6qvE/kN9pLW8BHVehuOFfe7/pOqLZKOra41FTbn+YAccvYR5ZyD54AC7ofuwHKPJdYLfDLq4wTPDX1j7QGRRgnuQX9WB57KK7v9Sd1pWmdSWbWOn7fqjT1cyrttzgZU0s7Oz2OGQVqrUSXVUREUREQEREBERAREQU8wrJZY4Y3SzSNYxoy5zjgADuSfJYV+v1n0xaKq/3+4wUNvoozLUVEzw1kbAMkkleJfP/AM+2p+NGuKjR3C3VFdbtFWsmEOpJ3ROrX5w5xLcHG3kd84zgbk3uO5nPb7SlnL9XU2g+DMNlv2paiN0lVWzvM9NQjyAawgPdnyJx39N/LvU+tuYnng4t2+lu9XXas1NcHimoqWCIMp6SMnJ6WNHTFGM5JP5TsFuvkIl4GV3MPQs5jKagr7FU00oiddvvlN73t0GUOOCPrz2XuHpC58tWj4BPoKu4bWSOVg+K0PoabqYd8fesEj5KeRfrz+4Z+xMp/dqS4cWeMUnvJ6JJ7fZqAdDexMZmkdk+YJDR8vVeknDLhppDhFou26C0Ra4qC1WuIRRMY0AuIABc7GMk4GT8goG8ZOFbmSPbr6ydMQLnn3tmAB9v6F0V5xvasaS0TT1/D7gFL92r9h0E96aemmpDuD4eRl7h6jt8jgq+pndejT3xxt6pJGsHqSAuAeZjnV4MctGlKm7XzUNFer8QW0NhoKtjqmok9HYz4bRtkkeY27keFNTqTmM4z3aovMVZrnU9RVykyOoxVTxhx7gCPLW/V8lyHwl5AuaLirqy222u4V6islrrKhnvl2ulMaeOGIkF7syYJdjOBg5OMornuu9tbzByXKSa2cMOH9PQnIjgnirZZGj1MgnaCfqYB8lwLxs5wuZfnKuVBom+1MLqSecClsdkpnRQOeTsXguc5+PVxIAC9fNJezq5P9MacobFVcFrJeJqWBsc1dXiSWeoeBu9zi7Yk52GNtlyVoHlo4A8LqgVmgOEembJUDcTU9C3rB9Q52SD9qX09eX3Dj2L3FDVFgor5rTirZ9OS1cbZXUMdukqpYwRkAnrYM4Pb8uFvs+w8ZjI5hDn+D4x/wCmXqmiDyqHsPD58wg/0f8A/wDcsmk9h3QGT/b/ADD1Aj3/ALDp1pPy71C9Qbnc7fZ6Ce6XWsipKSlYZZp5XBrI2Dckk9guovFD2qfKfw699oaDUNz1TdaQvY2ltNGXMe9pILTM4hjdwd8k/JN3g6T81fsubLy3cLaripaeM9Tc47Y9okhq7e2ne55yR4Za92Tt9ecLS+DPtT9b8KeGg0pLbpbnXUkfhUhqMPYNtjk9gPMfmK4x5lebTjZzz63otK2q0VMFmZMfuTpyh+PLs7SSkfTdjG52Bx6DGmWD2c/OLqG5wW2Dg1cqNlRjpq62SOGnAPm55Ow+sLUtkxOWxuW7e1M5vrhW1VTR64pKCKoyGwxUbSIge3STnBHquE9SWHjDxdmu/FfWTqurkk+/VFfcCYzMNyAzI3A8uw+ZOV638kfs0NIcAqZ+tuMtDZdWa1mA93jkgE9Ha2YIPQJAQ6Q5IL8bDt326qe1c5luHOs9SW/g5wlnoaqKxF4vtwoGtEDpgdqZjm7PDcfERsDsM74z+orzvjjfM9scbC97yA1oGSSewHzXpXyh+yZoOKXDik4i8cr/AHmySXgeLb7TRNayVsB7SSl4JBPcADsd/Ind3szfZ/0gpKLmA402GOodUxiawWqrjyGA9p3sI747A/oyD6kRxxwxtiiY1jGANa1owABsAB5BPD686qj2KPBOSYvpuKmrIYs58MwQOIHpnA/QsLiT7HHgnaeGN6r9I661LBqO20M1ZBV1skbqaR0cZd0vjDQQ04wSDkL0mXm17U3nW1ZwvFTy+cP+miq77bf+qlxafvsdNK3djD+9LmnGRvjO+Ni0dDfZ3abh1TzgcPrbU0oqKeOslqJ4y3ILGROcc/LYL6FQA0BoAAHb0Xjn7HvgNqK58XKjjhcqB8NmstvnpaJ724E00o6S4ZHkM4Od9/QZ9jk6CIiAiIgIiIOF+ZTlM4Sc09ipbPxMoa1s9u63UNfQziKenLhvgkEEZ3IIwV4a85XLdRcrnGSr4b2rVzNQUHgMqqaocGtnja4bsma3YOB9MZ9B5+l3tNeeW4cGdPU/Cvg1quOm1jdJHC51tK8Ga204G4Yd+mQnAz3A7b5C8kdD6yof9lax604oS1l/oo7pDVXV1TI6aWeMOBcSXElxA3xnO2AhHqd7FqwaqsXDjXVTeNNVdHbrzcaWqoa2aEsE4ZGWkNJG43yD22PqtD9sHyvaj1ZLZeYfRdp98Fqt/wByr/FBGTL4THufDOQO4HW9p27Ab9ge/wBwY4r8HuKekKK4cHNTWe42iGFrY6ahexrqVoA+B8Q3YQSAcjGc7lb7q6Slr6aWjrqWKop5m9EkMzA9j2nuCDkEfIqYPBTlC9ofxZ5ZnUOkK6duodBtmzJbKneWlY47mB/doGSensduwAC9wOE3FbRnGrQls4h6DujK21XKMOY4H4o34+Jjh5OBOCF5xe0l9ndbY7a3jTy76IdDPE951BZbez4HsO/vMUY7EHPUBsQcjyC69+zn51Krlm1tLw61xHUy6O1JWMina5xDrbVE9PiBp7AkgOG2MHPckX0x7poooJoamGOpp5GyRStD2PachzSMgg+hBClQERbN4r8WdDcFdFV+v+IV6jttot7ep73bvkd5MY3u5x8gEHTP2zDLF+1ssj64xi5/sjhZQdWOogxvMoHn2AJ+pdQ+Sv2ZX7aPha/ivqXie/Tttqqyait9LRUIqZnGJ3S98hc5oaM5AAyTjJIyFsfnv5y67nJ1zZLRpCwXCk03YXSRWuhd98nrKiUgGYsaD8RAaGgb4OCMr059mJwi4jcG+WamsPEihlt1Zc7pPdaagmH3ymhlawAOHkT05x5En1yWDrZL7D6gbU/eOYKofTZ/f2FrX4+yYjK7R8rvs8uDPLPNJeKWWo1RfZQB90LlAweHjP0GDIHfbf54zuu1KoThNTFA1rRhoAA2x2wrkRFFtviBoHSvFDSNz0NrW0wXGz3WB0FRDKwEYII6hnsRnIP82VuREHm9J7FDhFJf5q4cWdSR2t0hfFQspIi5jT+8MhJJA9cZUfEr2NnCOk0Jd6zh7qvUbtQ0tJJPRx1UjXxzSNaSGYAyCcYHfc488j0lRPB4D8q3P1xi5Q6ubRU1mo73pmKrLK+y1zTFPA9pLXiKVu7Hgg5Dg4E7Edse0fL3zJ8LeZbRNLrLhzfYpXSMHvlsme0VlDKMdUcrAcjBOzhkEEEHfC4V5uvZz8IOYW0XXUul7JTab1++N0sFypAI4quUDZs7Bhrs4A6tiPUZyPGFkvGTlU4uOjD7npXVum6wF7GudGJOh2RnGBJG7H1EehGzd6c+PpdRcPcs/MloTmS4d23VelrzSyXL3SJ11t7XjxaSfpAe0t746s4P1LmFAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQF4r+2btHuvMfYryI8e/6dii6vXwpHfo617ULyY9thYP/AIR6F1G1mT7o+nJ9AXuP6QFL6OwnsldfR37lvpNJS1HXNZp5egeYYXkEfZsu8a8hPY28RTT64vHDyR3wzwvqGD5YJ/SD+Zeva1bvSCIigLxq9svwxodNcYNM6/tdJFCzUdtfHV9DQ3M8byASANyRkk+q9lCcLyY9tveozqfhzp4Oy80NRWEegEhb/Og6Ycll10XZOZTRdz4gVcNNZIa4GeSYgMGRgAk7DOSvortdfbbpb4LhZ6ynqqKdgdBNTvDo3N8i0jYj6l8znCfgpxO44X2XTfC7SdVfbhAwSyRQYHQw53JJAA2K7tcpvNfxE5EdcVHArmXs13p7PU+FKyKaTxH20PyBIMk/e9jsM7Z7AAhibj2VRYVoutvvtqo73aqhtRRXCCOqppWbiSN7Q5rh8iCCs1FEREBbd4iNa7h/qZkjQWmz1ocPUeA/K3EtP1BbvuxYblaM49+pJqbJ8uthb/Og8IPZTucOd7RAacA0l3DvmPudUH9IC7d+2+mA0Fwtp8fSu9xf+SGIfzrqvyi2C6crvPJbJeIlvqbZQWCor6SSqnjIYYZYpIWvBOActfnHcLkb2t3Mbw84wXXRujuH98hu0OnzUVM9RDno65QBgZ37NHcA91NHnavav2NNzbU8uF4tocSaK+ynHkA8Ej9BXiovWr2MOpPd9Oaj0y+T4aqZ0zG5/fNOc4+on86tuD0+REQF0o9p7y1T8Y+DlXrexQmW86Tp3VTYm/SkibucfPGQR6HPlt3XWNX0NLdKGpttdC2amq4nwTRu7PY8FrgfkQSEo8B/Zx8e4uBnMnYpbzWCCwajf9ybg5zsMjMm0ch+pxA+35L39iljmiZNFI18bwHNc12Q4HcEEeR7rwH59OTfUXKvxEN6t7nVOjdR1kstnrWDp8CTPWad2OzmgkjHkPkCfRz2TnHDVXF/gNdLNq+5SV9Zo65Mt0c8rsvdA+IPYCc+QBH1gojvAiIiqEZXjb7VHk0v2jdd1/MFoGwtk0peQ2S7x0rP7RqgAHSFoGAx2xyMAY89yPZLPktucRNM2nWeg9QaVvtLFUUF0ttRTTxyNy0tcw7kH0OCPmAg8o/ZPc32orHrG3ctGqqk1Vku7pTZpJHEvpp8F3hN+RwSB2AB9AvX/wAyvnF5V9QU3DHmy0HeKl5dBZ9UR073HAJaXuiyfLfqX0bxyMljbLG4OY8AtcNwQdwQVbEnqRERRRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQdCvbI3t1u5YLbaGz9P3V1HTMLf7oRse/82F1S9jvwkptacYtQa9utvZPR6YoohE97QQJ5HHDdxjyBI9AuevbcXAM4U8Ora1+HS6gqJnN9Q2nIB/KVJ7Eelpm8IOIla1o8eTUkETj59DaVpA/K5yko9JERFQREQFpepdQ2rSen7jqe+VTae32qmkq6mVx2ZGxpcTv54G3zwtUXm97WvmwqtD6aPLvpeTor9SUYlu0wJzHSuOQz/xgPz58lZNpXT3nV9oHrvmprX6C0tRyWfQ8NYfdqKEuM9xIOGOlwATnYhmO5GwJIW7+FvspeJ174TVnFriNdoLF4NukudLY8E1MkTGF+ZDghuWjZvfzyQRm72XPKDeOKHFOg4z63068aL0y51TSe8R4jr60bRgAjDmsJ6jtuQMdl7L63pff9F3+iAz7xa6qL/zonD+dTepnHmD7M7nvvt413ZuW3Vtpo47VcWSx2Soh2fFKyMuERHmC1h+rBP1+rK+cLlL1dScIuarQOptRu8GCx6gbDWZOOkEOhfv8usr6M7bcqC8UEFztlXHU0tSwSRSxuBa9p3BBCkknispQVNPBWU8tJVQsmgnY6OSN7ctewggtIOxBBwR6KdFR8+HP7y63Pl+483eGO2+Bp/UFRLcLU+NmIwxxy5gxsME5x8yPIrv37HjinaBy56l0nervHAdL3mer6ZX4LaeWNryRnyBBOB5kqT20WmrXWcCtLaolgHv1uvwp4pAN+iSN2QT6bE/WV5W8KeNuq+ElDqCg07UObFf6T3eVvXgA+RI89j+YJtwcoc6PFu58z/M3dJ9OwvqadlUbVaourJLGnGc+mwzsOxXsByAcFa7gby6WfTF1hMdfWSvrqgEYOXgHceW+V5U+y+4c0PE3mloRe2tmp7XRz3CVr9+sjBHzySMZ+a95WMZG0MjaGtaAAAMAD0wqm3V6Iiii2txUyOGGryO4sNwP/J3rdK25xIj8Xh3qmLBPXZa1uB33geEHgB7PiRkXObwpdIQB92i3f1MEoH5yF9ES+bHlTu8umeZPh7d2vbE+jv1OSXbBpyWnP5V9JbJGSsbJG4OY4AgjsR6qfReiIqCIiAsavoKO6Uc1vuNLFU0tQwxywytDmSNPcEHYhZKIPOPj37HXRWvdSV2qeE+vDpSS4SGZ9uq6XxqVjycksLSHAEnt+kklcH2v2KnGR9+jprxxQ0tHaRKPFqYI5jM6MHfpYW4BIyBkkA4yCvYxUyAplONrcLtA23hZw9sHDy0zSTUlgoY6KKST6Tg0dz9ZJW6kRVJMEREUREQFpV01RpmxvEd71DbLe52MCqq44ic+gcQtUJXhFz2cC+aO58wGoblftH6o1Fbq6sL7VNR08tVCIiNmtawHpPfJxvnv5CfcHuENbaMIa4atsuH/AET7/Fg/V8S1eGeGqiZPTzMljeMtexwc1w+RHdfN1c+Vvmf0/aW3u5cGNcUtBE0vE33NmwxoGScAZG3yXYTko9pBqflobU6F4pUF41PpaSX720TA1dukBwQ0SEZaN8tJB2x5DGslZle4yLpvR+1l5M6qhbWSaxvlM8jeCayTeI0+hDQQfrBIWBqb2uPKJadO1d2sN8v16uMcbjTW6K0SxPmkAyGl8gDGgnbJKjTjD2yXHcae0DYOClgu7o7hfah1bdYopCHCkY3DWux5OJ3B7grrp7OnkI0lzOaV1BxE4j1tVBaqGs+51BDASDLKGBz3nBGw6gME9x88jrRrzWPE3nE4+S3d1G6s1BqyuENDRRkllPGT8LB/2rRkk49cDsF7p8mXL1Jyz8CbTw2raplTcfFkr7hIz6PvEuC4D5DAHn6bgBVM+PMbmH9mDrXRmoamPhvTTVdvbl0LiHPD2ncb749Pr2XUDU3DDi3w7uElrvGnL5QyRkjqhjkLDjzBbt+XC+mzAK0e4aP0ndnOddNMWmsce7p6ON5P2kErH48mXq3r5ln1fEekYY5J9SQtlHSWudO0OB8t+6535X+QbjJzI3eGWO3v09pxrw6qudbGWkMzv0MIySRnGfyFe8NTwr4Z1nT7zw+04/oOR/1Mh2P2NWv261Wyz0zaK026moqdv0YqeJsbB9jQAtT9n+Ni8DeB+ieAPD+2aA0Tb4ooKGERy1JjAlqH43e93c5Izgk/l3XIqImAiIgIi0HXWoo9IaJv+q5XMa2z2yprsu7Ziic8Z+0BB55e115qXaW0xQcAdDXwx3S7uNRfJKeQdUVMBgREjcFxO49M9iF0p5LuQvWnN991r1TalptN6bsszKeorpoDM+aYjq8OJgIBIBBJJwO3dcV08HEDmu49Q0EEslZqHWl1McPiuLhC1xJA/wC9Y3JwMZx6le+PKly7WTli4P23hraag1U7Huq6+qcN56l+Os/Vtgf6tkzib8cd8qPs9uEfKzXSakt1fWal1HIzpFyr4mM8EeYjY3IHyOc+ffBHZe+X2z6YtFXf9QXKnt9toInT1FVUPDY4mDuST/7k4AWbJIyJhkkeGNaCXOJwAPMnPkvDr2lPOZqXjPxTu/C7Ruo54dA6bmNEYaaXEdxqWZEkryPpNDstAPoc52w/S43vzye1HvnE+O6cJ+Ack9m0s8upq6+h/TV3JuSHNix/YoiMDIPU7cggELj3kH5A9Tcx19puIWtoJLZoO21LJHOlYeu5uBz4bAe7Djc+h+Yzyr7Ob2ckPEGC38dOO1n6tOStE9jss2R7/wCk0w7+HtkDz/6PrlabPa7DboLTZLdTUFFTMEcNPTxiOONo7ANGwTwS0NFSW2ip7bQQMhpqSJkEMbRhrGNADWj5AABZCIgLwf8Aayvkdzl34Od1Blptwb8h4ROPykr3gXg/7WhuOcy/bd7RbT/+JQer3IZYLZp/lV0DFboWM95tkc8zmtALnnYkkd9gF2CXAXIg6R/Klw+Mmci1RgZPlgLn1Sd9STBERVRERBQDC67c6PNXpXls4S3q5MvNM7VlTTOp7PQCT76Z3ghriBu0Dvn5Z7Arl3inxFsHCbQF74hanqo4KCzUr6iQveG9RA2aCT3J/Nn0XzocwvG3UfH7iheeIN/qZXNrKh5pKdziWwQ5+EAdgSME/PbyCf6n1yZyX8EJecHmXhsWv7hW1dBJFU3q+1Qc4ySMYR8HX+9LnPaB6AYGwXfzmM9k3y/0/CzUepeFP3WsOobNbp7hT+PXOnp5zCwvMbmuHw9QaRkdiR2C4Z9iXU2BvEXiFSzNaLy+z07oS7GXQCYdQb8wcE48iF6scQ7HVam0DqbTVEemou1nrKGE5xh8sLmN3+twSX4PC32YGvbpo/m60jZ6Wumio9SPmt1RC1x6JCY3OaSOxIwcH5r30XzdcN75qblL5krFqPVunKhlx0ReBNUUcjSx0rB1MJbnGQWkkHsfyr6A+BvG3Q3MHw5t/Erh/Wunttd1McyQYkgmbjriePIgn/7HZFcg4BHbuvnn9oZw7h4ac3WvrNQ0UdJQ19Wy7UkMbelrYqiMPOB6dZevoZXjV7Z3Q5t3GrTuu4oMMu1qZSTSY+k6LPQM+eAHKD0v5euNnDjV/A/RWoIdcWdvVY6KGf3ivjje2ZkLWvDg5wIPUD3/AErlanvtjq6c1lJeaGaDuZY6hjmAeuQcL5m+F/DziZxg1PS8OOGNmud7utZ1SR0FK8hoY3d8jskNa0ZyXEgD1XLWseTXnY4cxmlvPC/WIpDgddunNXA70H3h7h9hGVrNvE3PXq9zj+0M4d8tNnFr0rUW3VmsakERW+GqD4qcY2dM5hOB8s5XkDzA82nHXmmu8DeIN/fUUjJR7jZrfD4dNG4nA6Y25LnHYZOSSM91uThX7P3ml4qXylpP9i29WS3zyhs9yusBgjjBIycOwScHPbG25Xrry3ez44A8vtBRXFmlqa/6qiY0zXe4tEzmyYGfCa7ZoyPTyzgdlLLOUl3x1U9mz7PO+6cv9s5geM9vigdBH41js0zOp7XuGBPKDsCAdh9ffO3qYrWtaxoYwAAYAA7BXIoiIgIiICIiAiIgLp97Sflk01xo4Dag1rRWOJ2sdJUT7jQ1UUY8aaKIdT4SRuQWg4z2x9WO4Kxblb6S7W+qtdfCJaathfTzMPZ7HtLXA/WCQhXgv7MPjDa+E3NPZRqW+C22PUVNPaqh0rwyHx3sPgF5Oww8YBP91jzXvdHJHNG2WKRr2PAc17SCHA7gg9iMLwZ59eRnVHLRq+o1XpqgnuGgrtM6Smq44yRQvJz4MmPojfY9vL0zy/yE+05h4SWaHhJx/nuVwsLJQ213xpM8tCw7GOYE9TowcEOBJAJyNiTaPYtFptgv9m1VZaLUenbnBcLZcYW1FLUwODmSxuGQQf0g7g5BwdlqSgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLyb9tZq63OvuitGwzMdWtpHVUrM5LGdbsE+mSRj7V6yLx99qty58ada8xkOudI6Oul+s9wtFJRwGkiLxDJGCHAjO2Sc5+vPkpltmJsnav9jZw4kqtf3jiK933ump307RntsQPty4/YvX4dyuk/sxuWfW/AXhhV3HiDQPoLre5DMKN5+KFpIxkeRwBn5krux2VvOHL1VERFY1wuNDaqKa43KripaWnYZJZpnhrGNHcknYBeFvtSuM+mOM3MFTVOi7tFdLTYra23snhOWOk6iX4+RPbsu+HteOL9Xw/5cqTRlluMtLcdZXWOkkdDJ0vFJG1z5AfPDiGjK61+x04H6P4hV/EDW+tNP0t2p7SaKhpYqpgfGJXiR5OD5gAfLcJLJ6OOvZU8adLcE+LuoZ9d1Lbfar9amUnvcrdopGyFwI+vsceXkVoXtNdW0/GPmUn1toajqrnYaex0dBHW08DnxvdEZC8kgEDd+2fLC7L+135ddKaS4Z6e4n6C05TWqKC7NorpHSx9EZEjHeG7A2B6hjHbYY81vb2RR4f3/le1NBqC326rqrTe6qK5Oq4WyFlGYY5GZJBIbgy9vT5BEn91y17NnjzpPiVy66S0Ky/xS6n0rbWUFdSSS5mLYyQ15B3O2AfPbPrjt0vnx5SeNdFwk50bNrShrXUenLrqOehq2tyG+41UrmNyO2G9bDv2APqvoJiljnjZNFI2SORoc17XAhwO4II7g+qYu6lREQEREHQ32uWgrF+1uquI1FQw016oLnR0r6mJga+WKaQNIcR3x6nffvsF478M+Hl54l6oi09ZqSWc9Jlm8MfRYNjv2Gcj8696PaEcKL5xk5W9TaR05CZa+OWnuEUQ7vML+rH1ZwT8gV5ocjmqdBctdVqy/wDGOjio7nUR+6wRVGAWhoPbPzJO38wxP5Ty3prqnxa0O7QN6p7I+ExvbGS9p75BGcrt57JzilYdKcXH6Xv9yZTG4H/aokd0hxcMYz8iQftXXXjjqek47cV664aSjaIXyPMRAIaWnz+fYbriy2XC96E1VT3KjkfS3Oz1bZGFriC17Dnv3we3zBVkl9S/9ePqORbA4Da9g4mcH9J62inZK652uB8xDs/fQ0B+fnkE/at/DdCXiqIiK6he1J4Z3viZyo3Sm01ZJLlc7Nc6S6RxxML5GxMLhKWgDOS13Yd9l5c8h3OHc+UniRVi7UUlXpXURiprxS5LXwOY4hszQezmgkEYyR9WD7/yRslY6ORge14LXNIyCD3BXhf7VrgjaOE/Mg6/6Xs7KCzaxoGXERws6YmVYJbN0gbAEgHA8yfmg9w9P3y3amsdBqGzziehuVNHV00gx8Ub2hzT574Iz81qK6h+zI46Wnizyz2DTTq9j79oyAWquhc/MhjYfvchGc7tIHp29V28SAoK6D3qjnpdvv0T4/ygj+dTog+aDU1DFobmFr6a9SGngtGqnSyvAJLYm1PWDjv9HBX0O8FOKfDvijoW0XTQGsLbe4GUMLZPdqpkkkZawAh7QctOR5gLy354PZt8crhxZvfEfhDpd2p7Ve5feZKallaKiOQ7HDHEZGAM7jcbZyunU+kOZHlf1JTanr9L6u0NcaSZjmVUtNNBG5wOQ1z/AKDgSexJ7pun7fSei89uWD2tXC7X0Nn0Txno6vTGpJvCpDc8CSgqZTsHOcCDGSdzkYycBegdNUQVlPHVU07JoZmiSORjg5r2kZBBGxBBByEvOJOpkREUREQEREBERAREQEREBERAREQEREBERAREQEREBERARFBV1MdHSzVcv0II3SO+oDJ/Qg8bfbCcY6XWnEuwcPrbL10+nI5XyYOQZScZ+3JH1BdkfYvWF9t5e9T3lwIF31G6RpPbEcTY9vtC8tOYjWNZxU486muzACaq7SUVM07ABshYPsJye3Yhe5XIVwrdwi5btPael3lqeqte7YZLwBnA9SCftU5LkI7EoiKgiIgjkkjhjfLLI1jGAuc5xwGgbkk+gC+eX2gfEmPinzZa71BR1fvNBSVUdsopM5Bhgja0Y9Pi6tl7S872sb5oTlu1Xf8AT8j2VbIfD6mEghpDs4x9QXgZwis7Nf8AGLTlsvkzpW3W7xvqnHcvy7qdn1yRg/Wk6Poa5X9KQaK5eOHem4KNlN7tp2hL42tA+N8LXOJA8yXHK5PexkjHRvaC1wLSPIg9wsWzUUNttFDbqcNEVLTRQMA7BrWgD8wCzUSPnw5/OBNz4Hce7oXUjoKC+zSV9G9rSGkl2SQe2+QdvMlen/so+JV14hcskcF6rX1VVYrlLQue92T0gAtB+wA/auKfbZ6aop+FGgdViFgq6W/yULpMfEYn073BufQOZn7SrPYi1xk4V8SbeXEiC/0kwHp105H/AOQlz1delKIiDjrjtwM0HzD8PazhrxDopZ7ZVObMySF/RLBM3PRIw42IyvBPnI5c7Xyx8X6vh1ZtVfd2jbGJopnholjBOzHhuwIBAPzBXsZ7R7jXrfgVyzXLVnD64GgvFbcKa1x1bMdcDJerqcz0cOnY/wCteB1+1BfNUXWovuo7tV3K4Vby+apqpTJI8k9ySSfPt29FZfiZ1yNy0cwmq+WjifRcStKQx1MkTHU9TTSEhk8LsdTTj5BewnL57T7gXxjfFbNT1kWj7o9o+CtnAje/bIBI7b98n+debfs6uWDSvMxxIu9o1g1z7baaMTyNG+ASRnGdznAGfVb/AOdX2Y+vODTbjxK4TU37INFU0ZnrKeH+27e0d3GPu9gG5Lc4xk98Bknpu3I9n7LqCxajo23HT95oblSuxiakqGzMOe27SRlakvDL2UHFjVWmOaCy6CjvdSbDqSmqqeahfM4wiVsTnse1vYHLT2xnbOcBe5qhBYl0oI7pbKu2ynDKuCSBx+T2lp/MVloivnB5nODOteXHjjdbNdKSegc2vkuNpqg0gPjMnWxzCRg9JI7Z2x6r2T9nJzFVPMJy+0lXfa1s2otNzm1XMZ32aDG/1w5pJHpjGVtf2qfAmm4scus+rLZaxPqLR1Q2tpJY48yOgORLHkbkEEEAnAO+MrzF5CeZrUXLrxnoY6ep/wCoepJ4rfc6aQnoJLgGPx2BBOM99wc7IPoNRRQTR1MEdRE7qZK0PafUEZClUl3oIiKgiIgIiICIiAiIgIiICIiC1zWvaWuAIIwQdwvO3mi9khZeMHEG48ReFmvKXSdReJfHrbdV0RlpfFI+J0ZYQW5O5GCPluSvRREHjdV+xU41QvLKXifpqpGcdQp5GAj13dn8yyLV7FLi/UzBt04saboou5c2klkPzAAcvYlFJo6b8nvs3NBcrepXcQLjqSXVOqBC6GCeSmEUNIHbExtyTkjzO4/JjuQiKgiilmigjfNNK2OONpc97iAGgbkknYADzXVTiN7TrlM4a3yp07c9W3G5VtJIY5Ra6A1DGuB3BdkAfag7YIuCeX/nT4Bcy9dPaeGWpqiS50zDI+gr6Y085YO7g05yBg7g+RXOyAiIgIiIKYC89farc3UPDfRNZy/aabIb7quh6a6YEjwKR/cA+RIx9ecdgV6FAY814Qe1cq6q7c6d/o2ZkfBbrZSRMbucmEEAD1Jf+dQbp9j5w4uOouZV+vn24y2vTFqqg6Z0eWtqJmhjMHsCAT/5wXtq5waC5zgANyTthcE8mnLnY+W/gjYtKUtPG69VdLFV3qqDcGWpe0Oc0eYDSSAPXJ9AOBed32k/Crhbp3VvCPh1X1F+15PSzWt01NH/ALTtkkjCxz3ykgPewE4azO+MkBX9I4N9pJ7RGsr62o4GcAtUOho4euHUF6on4dK8HBponjcAYPUQfPHftxZ7PL2fLOZIScXOKVVV0mjLfXCKno2M++XmVuHPHWe0QyATuSdu2Vtb2dHJxXcyPFWn1br/AE5VVHDqxPfU3CebqZFcqkZ6KYO2LwXbvwewIPc49zrDYLJpez0lg05aaS2W2hjEVNSUkQiiiYOwa1oAH+vfugyLfQUdqoKa126mZT0lHCynghjGGxxsAa1oHoAAB9SykRFEREBeEHtafwzL7/4Itv8A6Fe768JPa3Qui5y70SQeuy2x4+own/Ug9XuRE55UuHp/+9Uf6AufVwHyJjHKpw+G4ItUY3+oLnxSfsERFQWj6u1BT6S0vdtUVULpYrVRzVjo2d3hjS7A+Zxj7VrCw7vbaS82urtFdGH09bA+nlafNj2kEfkKDwB5tufTi9zRVk1iuNUyyaOgnc6ms9GC1sgB+F07ju92PI7DJ7rtZyp+yl0LxQ5eKXXHFq43q2ap1RTvqrYKaTwxboTtEXxuGHuIHUQQMZG5XZbh97Lvlv0nrqfXl7sDrzP7yammop5HGlid1dQJZnBwQCBgY9e4XcKnp4KSCKlpoWRQwsEccbAGtY0DAAA2AAAAHom6PnroK3jP7OnmclLqaP7sWFz4XNe1zaa7UEnYjOT0PABBGSCO5xv7bcqXMbY+aLhFRcT7LbpLc+SeSiraJ7uowVEeOoA+YOQR/P3XSz22mhbRLobh9xGhoGi6091ntM1Q1u7qZ8JkAcfQPZt/3xW3vY6cyGh9OWDUHAPV16gttzrbr917K6od0R1QfGyOWEOOweDGHAHGQ4+YTnqedrcXtjuXaW9aZsHHrSdhdLVWmV9vvzqaLLjTvAMczwBk9LgQT2AOSVx37H3mT0hoOt1HwV1tfo7d+yGsirrM+pk6YnT9Ba+IEnDSQAfmftI9b7na7febfUWq60UNXR1UZimgmYHskYRuCDsQvnq55uEsfL/zUaq05pqmfbbaKuK72YREtMMUoEjek+rX5xjtgKfVfQ+Oy6g+0/4MW3iXyw6i1OKdv3W0fB91aeUDLjGw5e37Rn8pWrcgPN1p7mZ4UUdtqqtsWtdMUcNLe6Nxw6TpAa2pZ6sfgZPkcjvsOauPlhGp+CWvLAR/bmnq+PGM5PgPIGPmQrMt6l548ifY26ptdk5o7nYrhFF49/0xVU9HI/GWyxyxSkAnzLGP7d8L21XzAcM+IepuEmvrJxE0lUmmu9gq21VO45AyNi0/IgkH5FfRTyw8a6fmD4K6d4pQ07KeS6QkVELTtHMw4ePlv5IrlZERAREQEREBERAREQEREBERBomsdHab19pqv0jq21QXK1XOF0FRTzMDmuaRjz7EeRXl7zJex1obFpvUGvOBmt62omoIZa9mna+na4yMYC4xQzNIPVgHAcDnGM7herqIPBDk69oDxT5ZL3btHahr57xoBlQIqy1VQLpKFhOHugJ3YR3LdxtgDyPuRw84i6M4q6UodbaCv9Nd7RcIxJFPA8OAyAelwHZwzuD+cbrz79qPyPaYuuhZOOvBzQsdLqS21XiX+mtcGBX0rwQZzE3A8RjsEkDJBIwThdTPZx84d05deJceitSVM02jdVVMVLURPd8NFUF2GzNHlknB/RuSrm9S17vIrGPZKxskbw5rgHBwOQR5YPor1FEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBEUc0giifK7GGNLvyDKDw69qxxgPEPjnHpmlrjNR6cZJC2MH4WkkAEeWSAfyn1XdP2OujJdN8uN1vc8HhyagvJqs/3TWxhrD/5uD9q8muJs1y4n8wV/hYXS1N51JLRxdO+xn8NuPsAK+gTlh4VM4OcGrBowtDZooGyzAbdLnNG31gAZ+eVPys/lhP24u9prb6O5cnGtmVkbXiBsFRH1eUjH5aR8wQvNX2X94vVz4p3XhRT3KentmqqUMqmxuIBaAQ4keZwR+Reh3tXby+zcmuoRG8B1dcqCi+sPkOR+QE/YuinsdNLtvXMRdby9hIstq8XqA+iXlwGfrICW3OGa1H2gHs4bPy8aKHGfhVerhX2hleyG7UFQ0E0QkJ6Jo3N36OvDSD26gQQFzb7Jfm+r9X2y5cBOKOrjUXS2iOfTU1dOPEngORJThzjl7mnBGfJ2Nz39GtUaW09rfTtw0nqyz011s11p3UtZRVLA+KeJ2xa4H/7IIBG6+f3nb4FO5U+ZW56a0TPX0NoPhXewVLZHCSGJ4z0CUYJcxwIyNwMZ9Td/sfQxlVXVfkF5uNPcyHCCzW67XyE68slIykvVHI8CWZ0Y6RUNGckPADj3Oc588dqFJdBERUWuaHDBG35l4g+140pa9Jcx1Ayz00VNT3KzsrXRRNDWiQvIccDbcjP2r3AXj77ZrQOoJOLGmddwUMktuks4onyMYSGEPJGcDtkn83qpk3aOpeieHerNEcLncaam1SCgqHFtPI5pAc0bA57EE7j6wuGrvdKm83OputY4GaqkMj8bDJ8h+hexFup+FnMPyP2vgdw9npW3yltkVOyFgaHslY0gnI8ycbY8l5T3Ph7X8NOLdNofiTQGD3G508ddG7s6EyDLt+4Izn7QkupJ9emfseuJHE66WO5aGvNNVT6Xoml1FPKw9EeBkBpOxGfh2+XovThbO4UaQ0fovQVlteh7PSUFsdQwSRinjDRIHMBDye5JznftnAwFvFMkvF9ERFQXVz2hXLzQcdOAN8mo7Syp1Jp6lfW2x4bl5LRktB7+p+rPqu0asexsjSx7Q5rgQQdwQe4wk5dHzScDuO/Ezlu1/Brfh3dpLfcKZxhq6SZpMNVGD8UU0Z7jv8wc4813utvtt9beHE278GLKHhoEj6eslIJ9QHEd/rXePiZ7PzlU4pXKqvV84X0VHdKxxfNWW9xgfI8j6TgDgn6gOy6pcUvY3abqoamfhhqJ9PKSTDHUSnA9AQ44Hz3Tnujl/ls9qFwh4z14setJKbSFykOIW1MvRG4+mSSPtBwNs/Lt/Sa30XcGCSh1fZalrsEOir4ng57dnLwy1j7LDm501USttWi6e/QMyWPoquPreP8AvSQM/LK4e1Nytcz/AA/nxf8AgxrW3yNAw+K3SyDB9HRBw/IVN3sH0iQVVPVx+JS1EUzP7qN4cPyhbW4q8LdH8ZtC3Th7rq2srbTdYTFI0gdbCRs9h/euB3BC8uPZbaN5m7VxZFdqOn1LQ6QbATPFdXSNa53o1khyNhgjA3x8166nZSdg8IOev2e155TKah1xpvUkuotF3SrNH4s1P4dRQTEFzGS4Ja5rgHAOyNwARkrtv7HfmC4hcQbLqvhJrG4VN1oNLU1PWWurqHF74o3vLDAXncgYBGf/ALPoJxB4d6M4p6VrNFa/0/S3qy1zQJ6Spblpx2PqCD2IW3eD3L5wg4B2+qtvCnRVHYoq54fUuhyXzEdupxJO3+r0C1pHI6IiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAoK2ljrqKoopfoVETonfU4EH9KnVrnBrS5xwBuSg+a7mK0bV8KOPep7LE/46K7SVlO474BkLxn6jkfUAvcPkD4tzcXuXGwXuqhMdRQg0Tx8mgY389yfzLxK5ttTO1nzF6yuMXx/9UpKaPpOc9LiMfXkkfWvan2dehhoTlh09b5GETVLnTyZGMkhu/wCXKg7OIiKgiIg4F54mU7uWbWLqoAxx0vW7Pyz/AK14Kcv0og416Om6ukC6xbnyzkL179rFzA6f4fcBqjhXS10UupNZPbA2ma/44KUbvkcO4zsB88eoXnT7Pnln1NzAcZaW40TTT2TS0kdZX1bgekOz8LB6k9yAQe3qiePfa2vElvpZP7uFjvytBWUoKOnbSUsFI0kiGNsYJ8wAB/Mp0V5+e2iiMnLfpl7R/YtVwuP1GnmH6SFxl7EG5OFNxPtGThz6Cp+WQHt/LuudPa7aXrNQ8qMlfRwmV1mvVNWSAfvYwHBzvsBK6Seyb5gtMcIuKt30dqqoZTQavjiggne7AErCSB6Z7fMjPokm8Tx7borWua5vU0ggjIOcgrRNdXuTTOiNQ6jhH3y1WqrrmfXFC54/O1FefvtnuItrpeEGmuHFHcIZK+uvbKuqp2vBfHFHE4sLgO2Sex+R9F109n7yhcOOLvB7iFxP4pysijpIZKW2+I5oDGtjLnS7+rh0/YMd1x1ykWS5c5/N9R27i5daq6UdfHW3SsimlL2tjjHUI2g7DcgA4+ZXKftOrBcOW++6e4X8MLrU2fSOpLXJVS0lPIWdXQ/oMbiMAjfJwNwQPUEl8Xex/wBS0en+Z3VOjKOqbLS3W11DKd+QfEZBIXBw9cgBext4o47jaq23TQNmjqaeSF0bhkPDmkFpHoc4Xhr7I6llqOcS2SRt+GGyXFzj6Doavdbtup9V82lDetV8sHMe3UNFb3UV00dfpJ6eB7cB0Qe4ADyIMZIHlv8AYvfvlx48ab5jeFds4maajMUdXmKogJyYZ2gdbfqyfPtuN8ZPnN7aThbpqw3LRPEu1UEVPcL3LU0VY5gx4pY0P6j89x+f1K5e9izeJq/l91ZbJHEstmpjHGPIB8Ebzj7SVR6GIiIMW5W6iu9DPbLlTMqKWpjMcsTxlr2nuCF85XN3oB/CDmc1zpK3wmkit14dUULmt6R0PxIx7flknH1L6P15oe1n5Qjqiy1PMxpMYuFmpGR3uAN2kpmYAk2Hdo/n9dg73cAda23iLwU0TrG1VbamG5WOikdI12fvgiaHg/MOBXIK8T/Zc83+o+G3FG08D9WXgy6P1NMaakE8m1DVuBLCCdgwkYPpnHmMe2CkmAiIqCIiAiIgIiICIiAiIgIiICIiAiIgLzo51fadap4A8TKrhfw80fbq2st4zU1NeXlgzkDABGdwdttvPsvRdec3OR7N7UXH7jj/ALIOnrkKOkuQYyse3pJGDknfsRk99t04SbXSnjF7UDml4wafqdKzXy16atdWOiojstIYpZW+bTK5znYPmBj9GJOVj2cPFnmm0lU8QItRUemLTJK5lLU3KmkkNY7uXNAIJGT33znPmM+oPCH2bPKtwxs9FFceHNDqe8QBr5rhduqcmUdyxpOGgHsMf6l2ettrttloIbXZ7fT0NHTtDIqeniEccbR5Na0AAfUg+e/inwZ5g+Qbitbrs25y2+vgPi22+W8EwTtzu05BG+N2nPb1Bxzfw39sVzG2S9UY4iW/TuorQ1zG1AioPdqjo83BzHYJxvggZPmvYzWegdE8RLPLp/XelLXfbfK0tdT19MyZmD3x1A4PzGCujHNZ7KPhPq7SddfuX7T7NM6qgBlioI6h/udSAN2BjiQwnyx/7CMdyuEPG7hzxs0jbtX6E1NQV0FfC2Q07KhpmhcRux7O4IORnG+Fv5fNfqfgDzJcHrvLS3jh1rKx1UTi0zUtLP0kA7ESRZBG+Qc+a0putuP9K/3caq18x428N1bWZH2EpwfTKi+czR+lOcniE8u0qziXXhm5d77VRtA9cvcAR9SmvXFbnD4D3SO3ag1vrPT1XuWR1VWX5xjOCS4fnynqPoU1Xq3TehrDV6m1beqW12uijMk1TUyBjGgDPfzOxwBuvne5wOMFu4zcyWreJ2mKl5oKuuZ7hLnfoiaGscDgdunY/LPZbb4l8w3HHjYIKHiNxEveoImODYaWaYlmSdgGNADjntkE57Lm3lz9m3zA8bbjb7le9MVOltKzubJLcLgBG+SLIz4bDuSRkb7j0Kvw/bdepfa6czF54bU+hLVQads1eKQUdTfqeCR9ZK0M6S9ge4sjeR++wTncYO60bk09nrxA5thNxG1VqF+n9Ie+uE1fLGZau5SZ6pfCBIGcnd7idyduxPp9of2bvKJpGwUlpuPCS1agqoGNElbcg+SSVwG5PxAYJ8jldjNN6a09o+y02nNL2eltVsomeHT0tLEGRxt9AApMnfptrTeG/DzS/CnRFn4f6Mt0dFaLLSspaeJrQCQ0AFziO7iQST5krc6IiiIiAiIgLwK9qPq61aw5ytXTWerZUwWumoLY6Rjg5viRU7esAjY4c4jbzyvRj2r/ABt1/wAGeBFmbw/vdRaavU15NsqKund0ytiEL5CAe4z0kfkXnp7P3k6qebniBddS6zuNS3TGnJoprnM8mSSuqZCXCIuJJJIBc4nuD37gh66cj9vr7bym8MKe6xOirHWCCSZrhggkEj82FzosO02qhsdrpLLbIGwUdDAyngib2ZGxoa0D6gAsxAREQEREBERBx1x14E8PuYjQNVw74jW19Vbqhwljkid0S08oz0yMd5ELwE5k+Dd55UOYW8cPaG9zTy6eqYK213DAZJJC9jZYXkDYOAOCBtkFfSAvBX2rNY6t50NUlxz4NDboR9TadoQe2PA/Vc2t+Dui9W1WfHutjo6mUuOSXmJvUc/M5K4c51uSfRnNdpB87I2W7Wtsp3/cm5taPiOMiGX1YSANztt6AjlXlzhgg4C6AhphiJmnqEN+rwmrkbKJ6+cfhtxA4w8j3Hx1ybb3UF/sEz6O5W2oyIayAnDmH1a4DIcBkH0OQve3gnxV09zIcELLxIs9O6Ch1VbpBNSvcHOp5PijliJGxIcHD5jBXnh7bHhxpq3O4fcUKGjigvF0mq7XWyMaAZ442Mexzj5kZIHyK7Aey3uM1LyG0VS15DqGrvhjPpiV7xj7Sl54v+vFe6afuN71/cNOaats9wq6i6VFPSU1LGXySnxHABrRknYZ+QBJ7Lvd7P8A53ZeVypk5d+Mmn62itst06opZB0vt80pGQ9pOwJIJ+zceei+xvsFp1JzUX293mFlTW2fSlXcKMvaD0TvqqaJ0gyNiGyvAI7dS297Xa0W+z84VZUW6COB9x09bq2o8Nob1TEyMLjjuSI2knuTur//AFL17lU1RDV08VXTSNkhnYJI3tOQ5pGQR8iDlTLZHA+rqq/gtoGurSTUVGmLVLKT5vdSRkn8pK3uooiIgIiICIiAiIgIiICIiAiIgilijnjfFNG2SORpa5jmghwOxBB2IPovnt9oJwiPBrmn1haaC1ihtN0qhdrY2NhawRTAOIae2zyc47ZC+hdcXcX+Wrgxx1dTScTtFUd4lpABFLIwdYAzgZIOe5Vl+I6yezO5xdT8d9GwcO9c218t201TNpW3YZAqo42DpLx2L+kAEjG47brvYtg8MOBnC3g5Rmi4d6Ro7SxwwTE34iPTPl9i38sxYIiKgiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAtN1HI6HT10mZ9JlHO4fWGEhaksG9xNnstfA7GJKWVh+osIT08fPJyc2f9nnOXw9oJ4DO2v1QKqZnn4bC+Z5+xrCfsX0MX2+2bS9nrNQahuVPbrZb4nT1NVUSBkcTB3JJ/8AfJAGSvnz5SuLml+WTm6tmvdb000tmsVdcrfWOgZ4kkDJI5YDI1o3cW9WcDcjK5q9pDz4WzmRrLNwx4MXW5u0XQ/7YrpDE+A3SsccMaWHBLGDIAI3c4nyCZ3Rt72ivPRJzN6lj0BoYuh0Dp6rdLBJuHXKoALfGdvgsAJ6R23z8z2t9jLwcvmltEaw4qX+0T0Q1HNBRW4zRlpmpowXF4BA2LiMHzGD2K4n9nZ7OO7arvVHxk4/aVlpdPUwE9os1a0sfWy5BbJKw4IYO4B7n0I29dqCgorXRxW+20cFJSwNDIoYIwxkbR2AaMAD5If4yMb5XAfN7ynaC5ouHdda75bms1JQUkrrJcoxiWCcAljCfNhdsR8z8wefUQfOhyc8RajgXzV6Mvt6rprbSUd6FsvLWuwPBe4xSMfjYgOIJ8vh9F9FNNUQVlPFVUs7JoJmiSORjg5r2kZBBGxBBByF4ge0y5M6ngJrd3FnTMpqdKayuUznMDMGirH5kLHYGAHfEQdhnt3wO2nsg+Y++8R9AX3hBrK9SV9x0iYqi2yTv6pXUUhILSTu7peMDucHJ7pn0eiKIiAuPuN3CHTXGXQVz0nf7bDUSzU0gpJXNHVFL0npIJ7DOAVyCtOv18tumrJX6hvFSynobbTyVVRK4gBrGNLicn5BKPm90TxR1zy2cXa+5aQrnxVNkudTSS07nnw5RHI5pDgP+97haHxn4sX/AI18QrlxD1HHHHW3DpBZH2Y1owBnG/mc/Nbn0q3T/F/mlpnXdrRZ9Vatkke1xwDFLUOc0H6wQD65K788+Xs5dIaY4d1fFHhdTGCaz03iVkLQAHho747eXfy/St7g7e8hfHWycZeX7S0UNcH3iyW2CiroXHLh4bQ0H6sAD7F2SXid7H/iLdLPzGu4fuqHm3Xy21U3h52bLGzIwPnt+Re2KEEREBERAREQEREFrWNb9FoGfQYVyIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAtO1FKafT9znacGOjmeD8wwlaitF1o/w9HX2T+5tlU78kTkHzVMZLqXjX0byuuWpzn1IfVZP5ivpA4S2JumuG2nrM2IRGChiy0DGCRn9BC+djl8t37IeYvRVA5pPvWo4C71P33qP6F9KMMUdPDHTxjDYmhjR6ADA/QrcEqIigLBvdwFps1fdSMiippagj16GF38yzlpWqYhPpi7wu7SUFQ0/UY3IPm+4va81pzD8bbhdrlXy11ferq6joGSv+GNjpS2No9BuCfrK9zORvlkt/LDwWpdM+IyovV5e243apDceJK5oDW/U0bD8nlleEmhsW7jvaRFsKfUrWtPpiowF9J+k6h1Vpe0VLjl0lDA5x9SYxn86cnDWrIiINu690RYeI2krlozUtIyot9zgdDI1wB6cggOHzBK+czj7w8uHL9zAam0PSyOim0xd+qjkHk0ESREH0AIH2L6VV4g+184eVunOZ12to6R7bdqa10zmy9B6TPEzoeCe2SACB8j6Ij2L4NakptY8JdG6opZxNHcrHRVHXtuTC3q/Pla/qeyx6j03dtPSuDWXShnonE7gCSNzCfyFdA/ZOc11Lr7QdLy938Fl40lRn7nPJ/s1I07NHr0gn5jB8sL0QUxXhjoHTeuvZucz8evdf6cllsVOyrooJW5PvNPLgAAgH4sAHO42ytg89/NxS823EW06jtVkltlqsNE+io45SDI5r39RJI7nP1fUF6i+1M4SScSuWC43K10Qnu+nKqGuhIaC4xA/fGg99xjG+BuvCOSnniyZIZGdJ6TlpGD6H5/JX8eTKj0I9jFpaor+OuotU+7F0Fss7ovF8muecEfkI/KvZteJ3sneZDRHBviTdtEa6qY7fTatYyOlr5CAxk4LcMefIHAx9Z+S9rIZoqiJlRTytkikaHsex2WvadwQRsQQc5HdFebHtvYgeGPDObG4v1Y3PyNOP8AUs72JH+8pxBGf/lpj/VI1ontvLhGdFcMrSHDrF0rahwz5GFrQfzFa77EyCWPglr6ZzXBkuqGdB8jikjBI+1SXR6OIiKgtI1Zpm0600zdNJX6mbUW+70klHUxuAIcx7S09/PfI+YWrog+e/nI5PNd8ouvYp4Jaip0zWz+PZbxTlwMRa7LWOcN2yNOMHIO2fme7Hs4/aMHU8b+EPMVrWkguEDGNsl6uEjYhUtGxhleSB1jbBOMj6iR6O6t0XpLXtmn07rPTluvdsqAWyUtdTtmjORjs4HB37jdeOHtBPZ16o4VatruJfAzRVVXcP6uL3mpo6BpmfaJBs8Fg+LwicEEA4zgnbZo9oqKtorjSx1tvrIKqmmb1RzQSB7Hj1DhkEfMLIXhF7PHnavHLtxFptJa81BW1HDy9ltNUQzSl7LbKcBk7Ac9LR2IGBg523K9zrPebXqC2Ut6slwgrqGtjE1PUQO6mSMO4II/9/Lugz0REBERAREQEREBERAREQEREBERAREQEREBERBDUU1PVxOp6qCOaN3dkjA5p+sHZbePDLh4ag1btD2Myk5LjQxnf6sYWj8Z+OXDTgBoybXfFDUUdqtsTvDjb0l81RJjPRFGN3u+Q+XqM+O3Oh7SnXnMLcW6R4ST3jSWjKd5aGxTmKtuT84D5DGctafJgPmM75zL+M/LlP8AHqlzBczvBnle0RX3W5XWyxXRkbhR2akfG2aeUDYGNm4H1gZ8vMjwl44ccOJHNBxJfqHUDHVNXX1HhW+20rMiPrdhrGgbuccgZ/JgLfGhuQrnA4sGivkHCq+RUd2DJW3O7vEIcx3Z5EjvEIxvuF6/8qnIvwd5dtJ2mabSFsuutGQtdX3qrhbNL4x3IjLshgB7Y8xkHsr5yeGT1wxyXezA0Dwut1j4mcXon33WL4Y6ttvkx7pb3uaCG4/fvHYk+eR2yF39YxkbBHG0Na0ABoGAB5YHor0QEREBERAREQFHLLHDG6WV4axgLnOPYAbklSLhrm44zWXgXy/6y1vcqyKKrba56W2Quf0unrJWFkTW+uHOBPyHzCDyu9qZzeac4+avtPDHQb3T2LRdVUPqKs9qmtIDCWfJoBH1k+eQO4/sgOHN/wBD8uV3uuoLRPQSalvz66Bs7Cx74WwxxtJB3A+EkfWV55ezT4c6f4s83unLdrOibcqKhpq28zQzjrbPLFGSzrB7jrcCc+YXvtT08FJBHTU0McMMTQxkcbQ1rWgbAAbAAeQS9EyIiAiIgIiICIiCmOy8B/ahPe7nO1t1jGGUYH1CnZhe/GV4E+1EmZNzn6z6BjojomH5kQNz+lTfhn17YcubBHwH0BGDkN09Qj/8U1cjY3ytgcAaCS18D9B0Erg58WnqDJHnmBh/nW/yqks/LseZntv344f8Lo/M3i4O/JBH/rXKPswGNdyBxsJx11F+BP1yPC4s9uAD+wPhafIXe4j/APExLlj2U9ObtyNU1sZ9KW53mmH1vecf9JJL9Xjpd7F0O/bVag6c4/YLXdXpj36h/nWje2FDm84DnHsdL20j6uqYfpBWs+yHr6fRXN/f9OXqWOCpqtM3G1DxHBo8aOrpnloJ8/vJAHmth+1R4g2TiBzgX4WGobUQadt1FY5JmEFr5o2ukkwRsQHSlufVpTPqa9uOC4xwd0IO2NM2sf8AJY1vNbG4GVbK/gpoGsjBDZdMWtwGMd6WNb5QnRERFEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQUcuOuYTiJQcKuDOrdc3GdsTLdbJ3MJOMvLCG4+e+fsXIy8xfbI8wVXYbTYuAdkmAkvVObnc3NduyEPLWMI9XYONuxO/ZP8T9PL/TGltW8aeJdHpfS1uNbqDVlzcyngBwHTSuLiSfIDJJPoCvYblM9ldoPgjdqDXvE68R6s1NSgSRUwgAoqWXAOQHZLyNxkj1wcEhebns/OL3DzgbzL2TiDxNm8CzU1HV0/vHheJ4EssfS2TGCdgT2332XsQ/2iPJm2jNXHx2scuG9XhNZMJD8sOYN/klmq7Gta1jQxgAAwAB2CuXTS6+1p5NrZUPp2ao1DWlhx10tlkcx31EkArqVzie1fZxM0xNoHl/ob1Y6WpOKu8VfTDPIz+5YxpJaPrPofLCYPXGS+WWGUwTXeiZINix1QwO/ITlXC8Wh7/DbdaMuO3SJ2k/kyvnH01wO5quKdKNSaa4fcQNQwS7itZBUSB49Q93cfMFay3lL50myjp4NcRRJnY+7TA5+vKk36c+PdnmW4KaX5g+C+o+Geq3iGnr6czU1XkZpKmP4opgTsCD3z5EheCXDLiXrvlC48TXPTt1a+rsdY6hrRC7MVXAHDqGOxyACAc4JI9Vr104bc8XC2ifWXqxcSrHRtYXOFTNOIunG+WucR2+WVwa28VT9RMvl68SsnFW2oqRKfilIeC4HPbOCPkrPTOPp90jen6k0pZdRSQ+E+6W+mrXR/wBwZI2vx9mcLV11/wCBHOFy+8UtCWOusGvrZR1HukME1uqX+FNTyhgBYWnyBGB8sdlz3DNDURMnglbJHIA5r2OBa4HcEEbEH1TUiQ7rx99q7zg6hvOv38AeG2sOjTlrpGi/OoJRiqq37mFzwdwxuAR6nHrnmf2nPPZYdI6GqeB3BvWIm1Xd5vAvVZb5jm3UgyXxiRvaR5wPhIIbnffC6q+zI5Q7VzH8RrnxA4mW+ordJ6WdHK6KQnouNc9xcI3u7uaAC5w884z3Sr42DfuRnjnwz5fbFzSx5jicILn7pDGfeaGneQ6Kd47gYwTkYAOD8+X9C+0w1bqrgRq3hTxfngq66S1SU9DX9ADpmdJwwjO7gQPzY9B7MT2m1VVsfZKm3U0tukh93fSviaYjFjHQWEY6cbYxjC8CfaQ8M+HXCjmfvGluGltht1sdSQVklJAMRQzydRe1o8gCCMfIJ/qNq8ivEmz8KOafQesNRXBlFaoq59NWzvIDY4pY3MySfLJavohtd0t16t9PdbRXQ1lHVMEkM8Lw9kjT2II2IXzT6itukK/hnp66aYoybzBI6C7BgJdnBw4j0JAIPzXpR7I3mbtsGjbjwQ4hanZT1dLXCSwCtmwTE5o6oQXHOxzgbAAYUuSrOvTxERUEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFomtg06Mv/WcN+5lVn6vCdla2tF1nSS3DR99oIATLU22phZgZPU6JwH5yEHzrcprj+2l4eOiYZP8A4RwFoG/UOor6Q18z3CrVcvAvjxp7Vd1o3yO0hfmS1UJBDi2KTpfgeuMkDz2X0bcMeJOk+Luh7VxA0Vco621XaFs0T2OyWkgEsOOxBOCEG60REBaZqV3Rpy6v74oZz+SMrUh2Wlar20veDvtQVH1/2NylI+arTbpJeN9A6Bpe9+pmFoHzqV9JuhRI3RtkEzel3uEGRjH7wY/Mvmrs98j0jxip7/UxjwrXqAzStcOzGTnq2+QzsvpK4Z6ntusuH+n9TWiZktJX2+GWNzSCPogH84KmdT63QiItKLgznC5ftLcf+COpdPXa2xPu1LbZ6m1VfQDLBUMYXsAPfBIxj5/XnnNRzRR1EL6eVuWStLHD1BGD+lB87nIzxL/2Gea3RGo7rVuoaI3B1ruRdsRFMDG5p/8AGLfyL6Iaaogq6eOqpZWSwzMEkb2Oy1zSMggjyPdfP9z88sd15a+M9RJTvc6zaknmuVsnbkdJL+pzQfIgnIx23HkvT72WHGDUfFvlsDdUVr6yt03cn2vx3uy5zAxrmZ9NiPtJU+6N2+0Z4lT8LOVXU98poxJNXSwW2MOGR1SuOCfkCAvC7RctRr6+w6Muc7Ou9TiOCXpALJicgem523+rzXsZ7YKvip+UR9I+RofV6jtzWNJ3cW+I44HngDdeR/Kzw+v3E3mC0HpXT1PJLM++UdRM5gz4UEUzXyPJ8gAO52yQtT/ntTNbj5juTvi7yw0llv8ArOjDrVendNJXQbBsoHUGOwT0uwMjfy8ts+rfsp+Nt44ucuL7RqOtkrLno+4G2PnkJLnxOaHxZPmQCc/Wsb2vlTZ4OT+rhuPh+91OoLay3h2OrxQ5znkef9jEmceq2b7FfS1TbOAGrtUz9Ph3zU5ih9cQU8YP53n8nyS3+l/11R9rXxgqde8cqbRkeBQ6VifBGPVxOCftIJ+0LvL7I/ScGneU+mubSDNe7vU1cpHphoaPsGAvMHnNv/8Asz83OqLbo6jbK916ltVOGEffZWyFrjkeQII+wr2W5H+DV+4F8vdh0PqWbruAzUyt8mF4G3y7dlmf88J5rn9ERUEREBRyxRTRPhlja+OQFrmOaCHA7EEHYgqREHjn7TnkKtvCrxeP3CK2ys09cKwi+WuGMllvlfkidmNxGTkEdgSCcbk6B7Nzn3uHBq+03Brind5ajRNzmDLdUzvLja5nHGASf7ETjbyPbyA9oLlbbfeKGe2Xahp62jqWGOanqIxJHKw92ua4EEfIheAvtI+E2hODXNTfdKcOrTHa7RUUNHchRRbR08szCXtYPJuRkDyyg9/aSrpq+lhraOdk9PUMbLFIw5a9hGQQfMEHIWQur/s4LjrG78pejbjrGvlqpZYXCjfK4ud7s04YSTuc4P5/VdoEBERAREQEREBERAREQEREBERAREQERcMcwXNzwQ5ZIqL/AGVdSTUtVcAXU1FSU5qKh7RsXdAOwz5koOZsYXAfNTzgcMeWTQlxvV2vturtStYY7bY46hr6iec9g5jTlrRnJJxt8skdO+aT2vGgbxw9r9J8u1JfnX26xGB14r6YUzKJh2cWNLi5zyOxwAN156cKeX/j/wA0Gpnu0RpW86iqKmYGsu9T1e7xEnBfLO/bY9wCSPRPBq3GXmA4886HEa10uqKyS6VtRUils1loYyyngLzgBjB3OO7jvgE7Aleh3KB7Jii4e36ycUOO98p7tdaAsrKbT1NFmmgmAyPGe7eQtPYAAeu427Fco3Inwn5ZdO225OsVJd9deA0198nZ1vZKQC5sIORG0HAGN9s5XZ5PRaAGgAAADsrkRAREQEREBERARFxnzJ8S/wDYe4E634jxyhlRZbNUzUvxYJnLCIwPn1EEfUg688+ftB7Zyv0lPo7h/T26+a5rgS6Kd5fBbox+/lDSCXE9m5Hbfzx5A8eOaDjVzIXaO58VtYS3GOncTTUMMYhpKfP9xE3bPfc5O533W5eVbhhWc2HNLp7SGuK6tuMN9rZrhfakzESvp4mOllw7uM4DRjGAQBjZe2+r+Wvl20fwJ1To+g4V6Ytmn4bHWvnMdvj8RrWQucZTKQXl4x1dRJOQrZPiT9vLD2PBpP22rhNjxzpuv8H8sfV+bC9w14K+ymr6ih50tKMgziroLlTyD1YaZ5/S0Fe9SiiIiAiIgIiICIiAvCL2sWka3TvNzebxNCRTX6hpKqGTycRGGuA+rA/Kvd1dN/aLcmlx5m9CQXvQkcR1hYAZKWKQ9IqoxkmPPkSCcZzvjb1aNS9nxzZ6H488KbLomK5Mh1fpe2Q0tfQSHD3sjaGCRmfpDAAz8s+oHbZfOjwY15xM5HeYm16q1Vo65W25WV74LnaKphhfUUrx0yMBOx9QQSMjGRuvcflr5sOEnNLpua+cN7pKKqh6RcLZVsEdTSk+oyepudg4bHbtlSTJo6Ye3AcP2CcLW5//AEtcTj/9jGuVfY+XClquUNlJDKHS0WprjHM0HdpcInDI+YcCofawcvGuON3Byx6i0FbpLjX6KrZ6yooYx1Sy00kYD3MHmWlgJHcj8o6k+yb5o7Pwc17dOBWunGgoNa18Roqib4BTXIDw+iTOOkPAa3fGCBk7AG7vg4i58+HGqeW3my1FctMz1Vmp79M+9WippXln3qYnraCO+HZyPUrkjkM9n1Tc2NqrONHE7WNZDYmXaSnNLTsDp7hOwtdL1yE/C09e5Azk7eeO0ftm+E0WpeCOnOK1FRsdX6Ru4pKmUMJf7lVNIIJ8miVkZ32+IrY3shOZ/h1p7Qdz4Aayv9LaLy+9SXKzmpd0Mq2TRxtfGHnYODoycHuCT5JqPTqy2e36es9DYbVTiCit1NHS08Y7MiY0NaPsAAWciIoiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg4t5k+O1h5cOEV44ragpJKuG3dEUNOwgGaeQ9MbMntk/+47jw4uL+MntFuZx9Vb7dH91746OLYE09roY9gXHv0tBJ8sknsMkd+vbKccdOWrhRaOBFFUw1F9v1yhudZE1+XUtJACWlwGcF7yzAODgEraHsQ9N2d9HxQ1c+nabpDNbreyUgEtgc2V5APzc0Z+xBy3rn2TnBat4Bt0DoxklLrSgphNBf5jl1XVgZPijYBjjkYBGAR6ZHjPrTRt/0Bqu6aL1PRmluloqX0tTETkB7dsg+YOxH1+XZfUWvn29pPFboucPXQtrGtaZ43SdIGPELfiO32JvwdhuHfsZ9Va04b2jVt14xUNout4o4q1lGLY6eKJkjQ5oc8PaScEdh+TsuUuEPsYNL6Z1BR3zilxQfqGGjlZMaChofAimLSDh5c4nHy8/MEL0B4Pl54SaJMjsuOnbaXH1Pu0eVvBL0YFls9s0/aqWx2aiipKKiibDBDE0Naxg2AAH6fXJWeiINPvdkteorVU2W8UjKmjq4zFLE8Agg/X5+hXz688PLxfuB/MBfrJTWaoNouU5q7bLFAeh7X79Ix55OcfP619DJI81oeoND6P1ZNT1OpdNW65yUpzC+qp2yFm+diR2z5dlM7sWX4+dGbl4486P0TTcUG6Vu9Bb5SHNkha9szW9w8gbgdjjvuMjvjddn5+eaewaEm4dW/ibVstkkLoPEdGDURsOQQ1/lsSO35919Cc9ntNVQfcmptdJNRdPT7s+BrosenQRjH2LYdfy3cArpLJPcOD2kp5Jv7I51riy768BXm7U2x4E8rHL5qfml402zQ1LJUmlqZzV3q5O6nmGAHqkc55zl7twMnJJJ3wvoK4R8ItDcEdD0HD/h7ZYrda6FgGGN+KV/76R57lxPclZOiOFfDfhtHLHoHRFmsDZ9pfcaRkRcM5wSBkjPl2W68hXURVT5WU0r6dnXI1jixvq4DYflwvn59oTT3E8w13uN5LxXVORIx/cAOJG3l3IX0Fg5Xih7Yjh5X6c5iqHWsdKWWzUdpiLHgHHjsLg8E+p74+RUGm+zy4AT3PUsutuJFlfT6WmiaY5KmMhjwM/Fv5b/AJlwzzlUto0nzIXmp0BKaS3wPimt89MegNI3ywjsQfTtsu1PMhzt8Mafk20Xwh4P3CMaorqSmZcX0zA00MTGYkJcOznk4AO4x2IC3/7LnhNwq5guXPWNBxT0rTagq4r1JbJKisb1yxxOgZI1zHnJDgXnB8i0fayew765t9npzx6b4/6GtvDzWt7gp+IVnp2wSxTODTcY2AASszsXEYyBv9ZyT3SJwvne4/cCeLvI9xxilj97pIqGvdW6Yv0efDrIGuyw9Q26g0hr2HHmOy9iORLm9t3NlwymuVbSsodU6ffHS3mlDhh7i34ZmDv0uwfLAIxt2FzFdmkRFAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERB5L+1n5O9P6UoafmL4d2r3RlVWil1DSxD4A+QExzgDtuCD5AEb4AA2t7KDm+tfDC+VvAriBcnQ2W/1AqLPPI7LKaqOz2Y8g/Y7eeT5YPrVxH4f6Y4p6LumgtYW9lZarvAYJ43DOx7EZ8wV86vMXwurOXfj/AKm4eUFVM06auYfQVH74x4bJE8HzxkYJ323TNP0+k9rmuaHMIIO4IPkrlsbgdqQ6v4N6I1M6QPfcrBQVD3A5BeYG9Rz9eVvgbpUl1VRyxxyxvilYHMeC1zT2IPcKREV40+1B5HbFwbLOO3DdksdkvNf4N2oXbilqJMlsjT5Ncdsep+07i9lbzss0tVM5duJVyJt1ZKHafrJXZMEhwDAc92kDI8wBsNjn0+4x8KNL8beHV34cauphLb7tCWE+cbx9F4+YP6SvnX4qaNvPLzx3vmkIal4r9HXnpglyWkhjg9hJB82kA+uSno+l0HIyCqranCrUkesOGelNUxyiT7rWajqy4ebnwtLvzkrdWd8IKoiIPMX23lNTnRnDSsETfGFzrIjJjfoMQIGfTIJXJ/scrRSUHKbVXOE5nueqq98xz26I4WNH5Bn/AMZbV9tJpOW6cFNKapjJ6bLeS14+UrQwfnK6ScqHPhqPlr4Raz4a0lL7yLx4lRa3EH/a88kYY4gjt2Byf5gnqMn2jPOFfOY7ipWaLtMgp9DaMuE9LbYW96uoYTG+qefU4IaBsG77k7dz/Y68BaHTPCq6cb7xbWG8amq30tvmkYOuKii2JaT5PcScjuNl5icunBi/cyHGmy8N7fM4TXeodPW1BGTHAD1SyH7D+fK+ibhZw5sXCXQFl4e6cj6bfZaZtPFtjqx3OB2yfJLd5R0U9tjSVMnAvQtXGT4MGqnNkx2y6llwT+Q/lW6PY26iobnyq19hhkHvVl1RWsnZ5hsscUjD9uXD7FqHtfNOS3vlNdcYWF77RfqOpwPJpD2uP1AErqz7GHihNY+JGqeGNROBR36njrI2F2Pv8YduB5nAA+1SVa6d1WoLhwp5pazVGoaN7Kix6xnrKuN7T1FvvLnOOO5y0kgeeQvoe4XcSdM8W9C2nX+kq6Oqtt1gbKx8bgQ1xG7TjzBP5MFebvtnuCmkLPpzSXGixWGGkvFdd32m6VELOkTsdC+RjngbF2YyMnvn1Jzv32LWrLneOBOsNNVtS6SnsWom+6sJP3tk0DXED63NJVHoiiIgIiICIiAvGz2xPBnVFFxnpOMtLa55rJdbZTUdRUMaXMhliBaOogbZHmfl6r2TWjaq0lprW9kqNO6sstLdLdVtLJaepjD2uB+vsfmN0HidyZe0r1ry72+28N9aW6K+aJp5AxhAIqaKMnfoI+kAPLBwPIle0XDfiNpLixo22680TdIrhabpEJYZoyDjIBLT6EZ3C8e/aDezvuXBW6P4mcGbFVV2iqrLqykgaZH2x+fQDPhkeY7eQxnD2afPhYuXx1Xwj4pvqRpa81jZ6CvZ8Qt07tnBw/8AmbiQSfI5JIAwWbeH7r2sRY1DXUdzoae5W+oZPS1cTJoJWHLZGOALXA+hBBCyUBERAREQEREBERAREQEREBYF8usNis1feqlrjFQU8lS8DuQxpcQPrws9adqC2C82K42dx2rqSWmyfLrYW5/Og8guLfti+OI1Rc7Zwz0zpu2W6lqJIIZq2mfUyvDXY6sdQAzjtv3+xdZrZZ+ZH2gPGOWqkml1Bf6loE1TI3wqSihHZuGjDGgZwPQbnA270cOvZEWO6cULjqDihca0abNS+oZQU8oYZi5+enqG4Gc/LB8+y9BuFnBPhZwTsg0/wv0TbbBSEASe7RYklO2S95y5xJGTk4z5Kb8hkebnDb2J1f4tLW8VeL9OI8sfNQ2iicTjzYZXuwfTIA+RXplw14b6R4S6Mtug9D2mG3Wm1wiKKONgBfgAF7iBu443J+Q7ALdSKgiIgIiICIiAiIgIiIC87vbA8Z6XTvCqk4UUlwDK6+vbNLA127og7fI9MA/lXogPVeEftZdWzai5v73ahVeJTWC3UVExgORG8wte/wCokuGfqUw1yL7FzQlRdeO2p9fS0rvdrDYH0scxG3izyMBaD69LSfqPzXcL2pfMbb+EHAat4eUU7v2Qa+p5bfC1hwYqY7SyH0yMgH5Eea3L7NfgLBwU5arJX1lIYr5rGKO9V5e3D2te3MTD57MOftHovNj2snFJnEDmwuOm6OpdLRaJoILO0EYDZyPFnA9QHPA+tpVJ3rX/AGPnD2537maOuxSONt05aasPlLfh8aVnQ0A+oBJ+0L25XV32eHAW08GOXXTNcKUNvepbfDcq+QjceKOsNHywQfyei7Q5CUVREQEREBERAREQEREHWPnX5ItJ83WmKMm5ssOrbKH/AHNuvgeI1zXYzDMBglhIB2OQfsx406f1Txq5DeYatp6KpNDqHTVX7pcaYPJprhT7HpO2HMe0gh2Dgn6wfoyXRD2jnIXFx/tc/Fzh3CI9c2mkDZqdowLlCwEhp/7cDIB/9oOfP8MdquA3G3RnMJwxs/EvRddHPS3KBpqacPBfSVGMSQvHkWuyN+4wV4T89FgZwy5zuINPp6nFvjo73FcqFsQwIy+OOYFvph5JC3hyA8yN+5UuN9ZZtYPqqLTl1ifS3q3VGWCOZm7JOk7NeMEZxuCBnC455p+Klv5jOaS/63skfTbr5c6ajo/V0TQyJpJ8847+mFvPsTX0AXDTGnuLnCtmmtc2yK5WrUtohbX00oyJBJG1x+ogkEHyIC+e3mc4UyctfMfqjh5Ya+YxabuUc9rqScSeC9jJoSSD3AeAd85C+jHTFH9z9NWmgO/u1DBD/wCbG0fzLwP9prLPLzs8RvHaQWS0DW582ihgwfyKTs6vde73DO4SXjhvpS7TSGSStslDUPf36i+BjiftJytzLZfBXH+w3oPp7fsZtePq91jW9EBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQERcCc1nODwv5VdLOrtV3AVGoa6nkfabND8U1S8AgOI26WB2AXH6tu4Ucq6x4l8P+HtFJcNc60s1igjYZHGurI4iWjuQ0nLvsBK8peaf2ufEW4anuukeXV9DarBTudTsvssAmqanGxkiDstYO2Dgn8xPSi81HG7ms4m3C+01n1BrDUN4q3SeDSQyVPgdbiWxjAwxg2AzgbZO+SvRzkh9lbZ7XbJNcc1Gk4q641GPcNPSzZjp2Y3dOGHdx/uc7fZuieujHL/wK4w89XGWajrNQ1lVM8e9XvUFf1SiniB/J1EnAaMDJ+eD7ZcqPKjoPlL0LVaP0bXVlxnudQ2quVwqgBJUytb0g9LdmgAkAD1XIugOFnDnhXavuLw50VaNOURxmG30rYg7HqQMn7SVutFF88/tELHdrFzf8QobrG5oqa5lTSk5HVA6Noa4fIkFfQwulftE+Ryk5kNIycQNFQth15YKVzoW+VwhaM+CT/dYG3/uCHLXJRxk0xxk5cdE3azXmlqrhbrNS266UzJAZaaphjbG5r29xnpBB7HPfIOOel85/KZx41RytcfLRqKpnraK3Q1wodQ25ziwPgLumQPYSB1MO+SMjBx3X0J6N1npvX+nKLVekbtBcbZXxiSGeF4IIIBwcdiM7hPODXUREBERAREQEREBeaHtpLxp0aC0nZnvgdeDWlzQCC9seM4PmBsT9vzXoZxDvtbpnRV4v1uYHVNHTGSIEZHVkDJHnjOV4P8ANazjRxe1ZcNZX+KqrqShe8huS7oGdyM/LP50zUtk9bK5POW+Tmm410HC51+Nmon00tdXVjI+uRkEWOoMadi85AGdvVe8PLpy28OOWHQn7AuG9NVe7TTmqq6qrl65qqcgAveRgdhgADYbLwp5IuKdz4Pcx+m9V27Ac5z6KdjsgPikx1tPyIC+h2x3Wnvloo7xS/2KthZM3fOARnGfkpvcazmuEOdbljpeangpWaDhmp6S+0U7bjZayVoIiqWAgtJ7hr2ktOPl27jxY4RcXuMnIZx4ukEVKKa6Wyb7mX+0zHqiqomkO6T88Hqae46uwPb6Jl48e155adV27imOYDTdjkqdP3ighiu0tPHkUtTEOjreB5OYGknyxk+ZDcR6vcLtfWnipw707xGsRPuGordDcIATktD25Iz8jkfYt0rze9lZznaf1PpGycsOqI3Ul9slPLHZ5yR4dXTtJf4fycAT9ewHy9IVcwgiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAvFT2xfDe42PmHo+IrKNwt2orTTwmVrfhE0ILTk+pH/R+e/tUdl1d9orwisHErlt1PdbjSsdWacon11PKRlzA3c4+YOCPtRNcGeyh5thr/AEbScvWpInfdXTFIfudUZJ8WkaCQ05PcAH/3Ix6KErwf9lBeza+cOw0ZPwXK31tOR6uERcD9gBXvDjKKqiIgLxV9r5wTuOlOPLOLFBQEWjVVFB48jG7Nqox0EuI83ADv6D1C9qScLrF7RjQFm1pyqa0r7jSsfUWGgkr6aQtyWEYzg49cH7FZ24lcCeyk5vv2e6WouXXU0R+62mqI/c2oJP36lZuGnPcgZ+f2EY9GV4Keyqu/3K5y9LR5wLhRV9J9eYS7/wDIXvWsyZxREUc0sVPC+omeGxxtL3uPYADJP5AqOj3tfLtQ0nK0bbO9nvFXdqZ0LT3Ia4A4H/jBeHi7k823M3rTm84xv4Z2qMCzsujrfboo8kuLHkAgdsZBOfPvlcHcYuWjilwScyXV9jljpHgOE4YQAD2yD2+sZCkuenzXaX2NNDDPzIXmvfE1z6WwStY4jdvW7fH1gY+1e1XmV41exfonHjXqe49WzbSIgPXdxP6QvZUdyqjgrnY4eXbify4as0pZKR1VVy0/iRxNGS7AIOB5nf8A9xleGXLJxLruXjmO0tq68tloorNeI6a7xO+Etg8QNlDs+Q7n6jhfSGvHn2qHJVqDTGsrlzHcO7J7xpe6NbLf4aZu9BUABpmLR2jdtkgYae+2cWTeK9GeangbZuazl7u2gaO4Ql9zghulirmnqjbUsHXBJkYyxwJB3GzzuvGzlk5lOJ3ITxkvulb5auug9+FBqS1TNOQ6JxHiM7EOAJI7ZGPqPaf2S3OVequ8Rcr3ES7Gppn0z5dKVU8g6onRgufR5JyQWBzmDfdpAG++m+1S5Idc1Ou7nzNcNLK252Svo2S6lpKZv+2KKeJgaano/fxuY1pcRu0tJIwQVD/XqtpjUNr1dp216psdS2e33ekirqWRpyHRSNDmnbzwR9RWqrzP9lTztQaotVm5WNbU4iudoo5Gaerw7DamniBd4Dgez2tzjfcDAz2HpggIiICIiAiIgx62jpLjSTUNdTRVFPOwxyxSNDmvaRggg7EYXhp7RfkqvfAXiLcNfaLs002hL9M6rjdDGS23yE5fG7HZuSSCe2fIEL3TWm3zT9k1NbJrNqG1UtxoagYkp6mISMcMeYP1lB5Y+ys5zOKF91VbuXDWDZL3ZIKOR1tr3tLpqJjcYjc7zbvgZ7AH5lesC2BoHgJwa4XXKovHD3h1ZbDW1WfFmpIOl7s99yds/LC3+gIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIsG9XSCyWevvNUcQ0FNLVSb4HSxpcd/qBQcL84HNLpjlU4V1Gs7qI6u81rjS2W3F2HVNQQTk/9q0bkrxN4TaH4kc7/ADQxuuEMldW6lu7bjf6sMPhUtIHDxHOI7AMHSB32HfdadzBceOKXN7xebcLg+ruUlVVmh0/aIASImPfhjGMG3U74cn+YL2M9nlyjO5WuEbv2UU8Dtbane2svEjACadgH3qla7vhgyTvguJOBhUbx5wOOVJyqct921rZKOP3yjhhs1ipv3jaiQdEXzwxrS7z+iO68Q+XzhVqnmy5gqOyXO4GoqL1Xm43irmcSXsc/qk39Tkgeg+oBdxPbE8yOm9YXGxcA9KV4q36drHXK9SRPyxtR0FjIT6loc4n0J+orgH2Yd9lsfNPZHNf0sqIjE4eoLmjH5yppXvLZ7XS2S0UVloW9NNQU8VLCPRjGhrR+QBZqIgIiICIiAiIgIiICIiAiKx72RtL5HBrWgkknAAHc5QePfthuGWl9I6xseq7Lb6ekqb84umETQ0uOHZ2HkS0n6yF0i0XwZ4rX/SFw4u6V0tV1Fi0zIJ6iva09Eboz1EjHfGMnt6Ddc+e0u5jqbjtxvktdjl6rNpTroYiDlskoOHHPnjB+0keS9PPZ88M7DR8j2kNMXS1tdT6nt1XU3SCVmDIaiWQPBB/7XA+StsnE21tL2cvPPFzLafqOHGtYqeh1tpqkZI3oeA240YwzxGA79bD0hwGdnA+pPQX2u9qoLdzj3Cqoy3xbnp+2VVUB5Shjohn/AMSJh+1bf478tvMH7PjipS8RdL3GogtEdfK2wajoZMtfGc4hnb3a4sJBaRg9wdxjr/rriRrHjVxFdrXiPeZbpdrpUQx1E7/7gENDQPIAZwFFfRly/VElTwK4fySjDxpq3Md9badjf5lyAey2jwkpYaLhbpKlgGI47NSBo+XhNwt3p71J4IiIoiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg0LWmrLdojS9x1Tder3a3wume1vd2OwH1lfOrzV8cdScxPHLUXEC9Pf0S1Jo7dSg5bTUsRLY42D54ye5yTuV9Euv9JU+uNI3HTFSQGVsRZk9gV1x4dezd5ctIX9msL1pGnu94jnFSzxd4Y3g5B6fM53zsm5w+a1f2e3Ami4H8tGlaWqszKTUN+pBdrvI9g8UyTHrbGXDchrC0Adl2ZUccUcMbYoY2sYwBrWtGA0DYADsApEBQS1DYnMa5hw84z5BTq3pQXIiIPF32vHL5R8OuKFr4t2C2x01r1iXx1JiYGt98YOojAGMkZP1YHkuxnsWdXVl54L6z0vW1rpRYr5CaeNzyfDiliLth6dTXfkHyXJPtYtCt1jylXO4RUokrNO3KmuULunJYwFzZMfW1xXnz7LjmStnArjHXad1JKWWbWUMVK92QBHMxxLHfX8R+wEbZTDsj3TRRQTxVMMdTTyB8UzRIxzdw5pGQR8iCCpUBERAREQEREG0OLbms4b6ge8gNFG4k/aF5q3fUFtZw81KPdIpDh+/SCRsf51355s9Wx6K4Caovkj2saynDCT5ZOf5l51cBqR/GLgzqu8UQEngsmzg58j2/KPyLNt+Fks66H8IYjdeM1m8MdPjXMvAHkCScfnX0c8K6SWh4eWGlmdl7KJmflkZx+dfOhwY/6kceLJTzbeBd3Qu+WHOH8y+kDRkjJdJ2h8eC11HEQR/3oVv+r3P01tbY4l6QodfcPtRaLuNJHUwXm21FH4cgBBc+Mhp32yHEEfMLc6JZqPmn4Z6mvvLfzC2PUdyp5ae46H1Aw1kRYQ7pjk6ZQAdyCwux6gjyK+iThPxb0Hxq0bRa54eX+multq2BxdE8F0TiN2PHdrgcgg+i6L+055EKviZRDjdwb02J9TUYcb1QUrMProcZ8Vo83twc+oJ9duh3Jdzhar5O+IlV90LXVV2m7k8U97tDyY5Y3tOPEYHfRkbuCCAT2PbBs86Y+g1Fs3hNxY0Xxr0JbOIegrqyttVziEjD2fE7G7Hj964HYhbyQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAWwOP8AZhqHgfr2zlgeajTtwDWnsXCB7h+cBb/Wn3+jjuFiuNvmALKmkmhcMZyHMIIx9RQfP37OS6MtXOhwykkf0tqLjNTEk+clNK0D7TgL6FF8zLL3duCfHGW+WFhhrtJ6glkpmn4dopnAD5Zb+TK9/uVDmLs3M1wmoeINsp3U1TtT10Bx97nAw7tsNwdvl6Jb8TPrmhERFFxhzOaYq9Z8vfELTFDAZqi4afrI4owMlzhGSBjz3C5PVj2MkaWPaHNcCCCMgg9xhB4Cezb07qR/Ojof3W3VDTZ6uofcQ6MjwI/BkYQ/bY5IGDjsfRe/y2rp7hfw80neKjUGm9G2q3XKryJqqnp2tkfnvv33/Qt1IC6u+0J48aj4JcAL/Po+3Ty3e7Uj6OOqawltJHJ8D5fTIBJ+Xddolpd/01p/VVuktGpLNSXKjlBa+GpiD2nPfv2+xSjwr9lhwzqOIvN1YbvPQy1Fv0lS1N6rJOjqYx4jMcQee28kgwD3wfRd2fbNXils/BfTELYoxV3a5y0bZCB1eGGNcRnvjGfyld39A8I+GXC1lVFw80PadPiud1VPuNOIzKR6nuR8uy85fbcVk5tfDug6j4LaiaYDy6y14J/IAm+DhD2Ouo5qDmjqNOAuMV1sVVId9gYgCNvXDj+Re3K8I/ZKVcVJzl2QTPDRPZrnC3Pm4wggD57L3cV8BabqCxWrVFjr9OXykZVW+500lJUwvGWyRvaWuBH1ErUkQeCvODyp6x5DuK+n9c6G1PLJaayvfW6duDAWT0ksLg4QvOdyARvncbHO5PrRydcxFo5uuXyk1hdaKm+6ZEtk1LbxgsFS1gD8NySGSRva8A42eR5Lhb2u3CC/8SeXu16k05RTVdRo26mvmhhYXOdBJGWPIA3OMNO2/YLoF7OjnIk5YuIk+mtRsfPo7WE8ENcAd6ScEtZOB9TsH5AemQvewn7cZ6vseuuSjmplFLTPp7loe/mrtckmeirpBIfDdkjdr4jg7HuRuvdXle5iNPczfCqh4k2CjfRGRxgrKZxz4M7QOtoPmM5/1nuumvtl9GaOuvCPSHFOkhpjdvukykhrIwA6oppIi4Anu4DuPs9At++x0tz6blTqLk54IrNR1rWt9AwM/SXFP8HexERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBeUntJPaIa2sWr9U8tPCcU1BRU1ObZf7uWl1TJJLGDLBCc4YA1waXAE5Lht5d1+eLmLqeWfgVcdd2qKOW71MzLfb2v7CZ4OHfZjPY+Z8l4r8v3A7iRzo8cZacySzG4V33R1Dc3AlsMb35eQfUjIAzsB54AKbuF867peyq5H7pT3Ozc1PENsTKUQSS6YtzmZe8vBYKtx8gAT0Y9cn5enuua+rtWitQXSgc5tTR2uqqIS3uHshc5pHzyAptKabtejtMWrSdkgbBb7PRw0NLG0ABscbQ1u3rgbriHm35neGnLXw0uN01tcmPudzpJqe1WqIh09ZK5haMN8mAndx2wD6HDNHgFp60XrjJxYgtU9W83HU90eZZXkl3U9xc45PmBnGfku1fCbhEeXrmh09bBUvkkdNFgudk4JH+tda+Al1fFx30xeWsER+6pn6W7hoIcSB8hldttWaldf+bvTFV47Xjx4g4NwfMJ+ksty69o2kOaHDsRlXKOAh0MbhsC0H8ykRRERAREQEREBERAREQF1t58OYu2cv3AXUNdT1bRqG7Ub6C1w5yRJKOjrI74AJ3+3yXZJeTPtmdL6vdebHqgRTusDfDjLhnobIW9O/l3/OVP8AB1E5G+BUfMnzKWPSGoHSTWqN0t3vD3EkyQxfEWl3q9xaN++SvoGp6XTvD7SIp6Onjt9lsFC5wZG3DYYImlxwPkASvJ32JdHaZ+JnEGrkjBuFLZoDGT5RumwcfbnP1heiPObxIoeFfLDxF1VVzxRymxVVBRse7p8WpqIzFG0fPL8/UCn0ednNvzpQ880TeWzgpoavlZNXipp6ypcC6skhOWuY1uS1uCe4PrkjC8/tRaM1Lwz1zJpLXFontd0tVWxtVBK3DmYcDkZ2II3BXcr2NWn4bvzV3S61EPULJpCtqYnEZ6ZX1FNED/5sj1D7ZC32ui5saGehY1tRW6SoKiswMZkE9RGCfU9DGfYAtbzEeyPCqaGo4ZaUlgdmN1louk+oELR/Mt1ri/linmq+Xrh5VVBJkl09RvdnvvGFygpFgiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiDRNZaSsmutM3HSWoqRlTb7nCYJ43DIIPY/WDgr51+avhbHwB5idT6EsdR0wWeubPRPZsY2nDm49MHsPIYHkvo4ulxprRbqm6Vjw2CljdLI70AGV88fNxf5+MnNlqWpt0rHy3i7RUMBGS0Fxa0fYCd8IPe3gZd5b7wa0ReJ3F0lXYKGVzj3JMLclb6W2OGOnBo/h1pjSwABtNopKNw/7ZkTWn84K3OhRERAREQEREHUH2ql0ntfJ7qF9O8tfUV9JTnBIPS8uB/MV1Z9ku24XbhLxSoHR9dNSxvEeRsCYg4gfPJJXfHnI4E13MTwQufDm2ziKqqJ4543EgYczOMZ2yCQVtXkq5UByy8F7ho+6TsqrzfXSz1zxg4ywta3IHodwM+XopL3pZx4j6SYI+Y6BjtsammHpj78/1X0baEiEOjbLGNwKKL/ogr5xYHik5kZSx+AzV8zATtsatw/nX0ccPj1aIsZL+r/aEO/r8IUs/71f/AFxuFERaQXnD7Qj2bVPxKlvfHjg0+Gj1FHTuq7rZfDAjuJY3LpIiPoykDJB2OPmMejyoRkYIQfPlyVc4OtOVjiTRU9dcKyTRlZViG92p5OI2l2HSMafovbuTt5HbK989Hay0zr/TdDq7R93p7nabjE2amqYHhzXtIB+w+oXlz7Vrkp0zpezx8w/CjTfuXXVGPU1JSNPhAPGW1IYM9J6sh2MADc+QGxfZbc7WnuC9wrOCfFW6vpdO36qZNaK+QkxUFUch0b/7lj8g5x3HzAIezaKKGWKoiZPTyMkikaHsew5a5pGQQR3BG+QpUBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQeffMb7JzSHGDW1brnR+rjp+quMhmqYjFlj3H5YIz89icb58uyXKPyzW3lZ4ZfsBt91NyklnNRPUHI6nnJz2HmT5Bc5IpgIiKgiIgIiICIiAvKD22N2ikuOhLOCPEiY+YjzwfEH869X14+e2kpqz/ZM0nWOY73Y0Jja7G3VucZ9djt8kHSPl34tXDgfxi01xKtx+O01YdIMAh0Z2cDnbGDk58gvoq4QcTbRxf0Ba9eWVhZT3CMFzDv0PAGQD5jcY+tfM42ppqmnpaH3NrZGyAOlB3cCfNfQvyK2x1r5a9MUzmdJewydsdw3/Un5f+UkJmOwKIiDGrqGjudHNb7hTR1FNUMMcsUjepr2nYgg915Pc7fswp7VqK4cWuD1RDS6fqHmpuVu6TmlcTlz2Y7AnPkRkjse/rUtmcZYoJeEetBUAFjLBXyHPkWwPcD9hAKl3ODwh5quaO88YdK6U4Vmrlnt2jx0uke4kPlazoAHrgZyfXH2ZnKr7QTi/wAqGmLlovStqtF9slfOauKkufiYpZ3ABz2FjgdwBkHYkfVjafJHw1sPGPmk0NoXVtOKu13GsllrIX7iVkcMkhB+ssBXr1rn2YPKjrO4wXGDRbrM6MjxY6CQxslAx3AIOdtySfqVqWuCeRn2knGPmC41w8NOIWmrO+juEMksdRbad0Xu3T2Dsk5G43J8ivS1cScGeVjgZwCa6ThloSjttW8dMlY4dc7xjG7z/qXLaKIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiDzY9tvc3Q8J+HNqbMWiq1BUzOYDs4R0+Nx54LwrvYl6bEHCPiBqqWkAdW6hioYpyzdzIqZjnNB8xmQbeq2t7bx9Q6j4Zx7+AyWtd8ushv8zVz77JykobLyT2a65a0Vd1u1bUuHqycx5Pz6Im/mVG7Oe3nUtfKVoalNnpqO660vjnR2y3zyfDEwD4p5QDnpBIwOxO2fXxybXceefXj7b7fd7nLe9S32YQtf0dNLb6YbucGDaONg37+QBPZYl2u3EjnR5mIo6u5z1171vezT0YkJLKSnc8lrGN2DWRxgnAwPhOe5K9xeVvk94T8rGmGUGj7Syqv9RE0XK91DQ6pqXYGQCfoMznDRgbn6hPODq3q72bvB3l04I3biBa7nW3HU+n6D3mpr6l3wyP2Duho+gMnbHcdxleenLrq+7az5m9NXK5VBd7xXBoaewaDkBex/tFL9V2HlN1lJSS9BrIm0z/m05cR+VoXixyeMDuYXSXUM9NUD+hP3fB9HkbemNrPQAK9WtOWg+oCuQEREBERAREQEREBERAXGfMRwYsPHvhJqHhxfaZjzcKORtJLgdcM4GWOaT23ABP2+QXJigrKqGhpJq2od0RU8bpXu9GtBJP5AUS9fPLy08wmseR/jtdrxDZ2XT3dtTY7vbpJDE2oY2QYcCOxDmBwztvutZ5pOdLjPzq6ktulvuX9zbE2pa216btrnSeNOcAPldsZH+mQGtycDbK4f5gL/AEeqONetb9b2gU9XealzMdiA8tJHyJBP2r1p9k7wA4T27gLaeNB01QXDWN2q6lr7nURiSWkbHJ0tjiznw8AAkjBOx9c2K5E9nxyUW/lW0VNqPUEjqrXWqaSH7qPOzKKIfGKZg+TiC4nckDsBheavtYb82986GpqaNxLbRbbZbx8iKdshH5ZSveVfPr7S0552eJRz/wDdNEP+RQLP0j3K4AUgouBfD6naAA3TFsOB23poz/OuQFsrgo0N4NaCZ6aZtY/5LGt6q26k5BERFEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERBxBzZVWpaPgBq6bSUEktxFE4RtjBJx8sfPC+cmqrr7R3+S51k1VBd4anx3ySAtlZMHdWTncHO6+pCaGGoifBPE2WOQFrmPaCHA9wQe4XHWr+W/gLryjnodV8I9K18dQD4jnWyJryT5hzQCDv3yrM+p10q5DfabWnX9NScK+YG60Fov1NGyC33qVwigrwBgNlzs2TAAz2PfzOPRilq6atp46uiqYqiCVodHLE8PY9vqCNiPmvDn2hPIRceXLULtf8NrVV1fDy5OJJaDI61y9yyQ7kMPkTgbfYN++zM5+bZwkll4J8ar7VjTlyqGOslzneZWW6U4aYXnOWxHYg7hpBGwwFNl8V7IIoYJ4aqCOpp5mSwysEkcjHBzXtIyCCNiCCCD81MgIiICIiAqYB7qqIPmq4/2G78OeYDVVPUU76eppb9NXwdTSMh0pka4eeMn8y9zeRLjHHxp5e7JqAvzUUQ9zmBIJaWgbHBO4OR9i6Je2Y4V2HTd/0nxFtdGyCpvkktPUua3HU4NLj+gH7VzV7FyukqeX/VVM52W0eojG0egMLXn87lfy5mH4249C0RFAREQYF6s9r1FaqqyXu3wV1vronQ1FPO3qZIw7EEH/AN+xHZeB/tBuU2t5aOMdbLYLVONEahca20VAYTHAXH46cu7AtOcA4yOwwF7/AK2pxK4Z6M4uaNuWhNe2OmulqucD4JI5ow4xkjAewn6LgcEEeY+xB0C9mbz/ANu1jZ7Ry7cXbq2n1Db4RS2G51EmG3CFg+CB7j/8laBgZOXADGdwPShfO7zK8nnGvlS1hUVd0stwk07TVxdaNSUYLoXhruqNxe3eOQANJyBg4x6LuHyw+2DoNO6ct+jOYjTt1rpaNjYGX+2hsr3xgbGWIkEnyyCfn8rR6vouOOCfMHwo5hdNnU/CzU8d0pmHpmiLTHPA4+T2HcFcjEZUoqiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAuAOczlm05zI8ILzZKigYdQ0VFLPZ6oN+Nk7AXNZt3BIwB6n5lc/qmESvlqfRVWntQG33yklp6i31YiqoHtw9jmOw9pHqMFfSJyxXXTV64DaMuWkqqGots1sjMb4nAgnz3Hnn+ZefXtheWTS1ks1n5hNF2CKhq5682/UJp2Bsc3iAmKZwG3X1AtJ2zkea0D2WfPLozhnZZOXri3dnW6kq7iZ9PXOUZgiklwH00p7tBcAWk7AuI22S/pZNeuyKxj2SsbJG8Oa4BwcDkEeWD6K9BQdlsLj5HcZOB3EKK0QGatk0vdI6eMDJc80sgA/KVv5RyxRzRPhlja+OQFrmOAIcDsQQdsHsg+en2eUtzo+c7hfLb4JXPF3dFOGg/DE+GRjy7HkA49/PC+hpcaaM5buBvDzVtVrvRfDOyWm/VhJmrqenAkJOcnPlnJ/KuS1biTRERRRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQedPtqdNxV3BPR2pAweJbL86MuHfpliIwftC6XcqfP7qrgHwj1BwQntbK213fx3W2qzh9DLO3pkGNsgnfPkSe3dd+/bFU7ZOVSnnJGYdRUYH2h3+peL+irJUaj1hZLBSwulkuFwgp2ta0knqkAOwycAEk/IFIZr059ljyXay0jr2PmB1zSww0UFDNBZoiMukdKOky7jYhpOMdsnfdeqa0TRdjg0zpCy6epoRFHbqCCmDA3GC1gB29cgk/PK1tQdT/adTwwco2p3SSdLnSRtbjuSWu2XjtyY0dTW8xWlIqaMvIqQXYB2GQM7fWvbLnq4KX3jxy+3nR2m5D90IiKuGPf74WA7YHc/L614lctXECXgBzDWe5aloA1tuuYt9wjeMGHEga52+NgRk9ts/Ub/wCUyl84+jpo6WgegwrlDTVMNZTRVlO/rinY2SN47OaQCD9oKmUgIiKgiIgIiICIiAiIgLhLnB4tWnhBwE1Vfq+sEFTU2+alpW+b3PaQcD6iR9oXNq8lPbNcVLh+yjT/AAqp5y2nFK2snYM/EM5GfrJH2BWDoZwP4HcQeY7iANE6EozVXGaOSrnkePhjYNy5xHqT8vP0XYvgTzhcdOQjXlXwP1bbqet05Z7qY7paqmJzZYg8gulhdnIJYeoDGDsCVz57FDh9NBWa84k1VOQ2op4LZSucBuA8ue5v2jB+r5rkX2qfJi3ibpqbmG0HR/8Awm05RBl4gYN6+iZ2fjuXsBIGO4AHlkTTld/dMaks+sNO23VOn6yOrtt2pY6ylmY4EPie0OB2JGcHBHkcr5/PaJVD67nU4ouALi26xQtGN/hpoW4/KFrvKj7QrjDy1+66XkuDr7oxmWfcyq+M0wON4nHcAf3PpgAgDB3FaOFOqOejmgk4jaXoy60Xq4RVlxlLSPDDSOsHbYYGDnsAfrSXPR7YcLKc0fDDSFIW4MNht8ePTFOwfzLdKxLVQR2u2Udsh/sdHBHTs+pjQ0fmCy0BERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERBp19sVo1PaKuwagttPcLbXxOgqKaoYHMkYdiCD/wDZBwQvCv2hPJHdeWfX02qtGWuoqOHd9kMtFM1pcLdKTl1PIR2AJ+E7AggbbBe8q0TV+kNNa905X6S1dZ6a6Wm5ROgqaaojD2PaQQdj2PoRuEHmx7MXn8r9RT2Tlo4r1DJKmCnNLp28Sv8AjlawZZTSk7ucAMNcdyBgnZeoS+bnmB0LWctvMpqjSOkrnUU7tKXpz7VVNfiRsYIfEc99gQDnvg5zleyHs7+bqp5oeF9RBqt0TNXaZeynuHQf7YjIHTMB332BPqcZJBJHjtsiIgIiICIiDz49s5oi9ah4CaY1Ra6CSop9N34y1z42dRjhlhcwOONwA7GT2AySuqHsrebfTHAXXF54bcQ68UOnNaSQPp6xzcspa9mWtLyNw1zSB9YHc4B9qrrabXfbdUWi80FPXUNUwxz09RGHxyNPcFp2IXhd7SHk9ruX/i5W6u0PpWph4d6ixV0clPGXQUE5/stOSM9DQ7duT2cB6Jmj3YhmiqImTQyskjkaHse0ghwO4II7gjzUq6R+zw549A8aOH+meD2pLy+k4h2O2sopIakYbcWQt6WyxP7FxYBlvcEfMBd3EIIiICIiDb2vdC6Y4maQumhtZWyK4Wi7076apgkAILSMZGexHcHyIXmxxU9irZI7Ldbtwk4rV5ucbXz0drulE10MmMkQiVjuoE4DQSDud16kIg+czlp5gdf8oXGiK9RsqYoKOsNDqC0SEgSxtd0yNIBx1twcEemM+Y+gDhJxZ0Txs0Nb+IGgbvFX2uvYCHMOTE/A6o3jycCcELz29qFyHi/QTcwfBnTMsl3Dy7Uluo2Z94Z/fLWDu4fvsdx67Adc/Zv871s5X9RXTh9xMZVDSGoqmN5naMm2VbctL3NJ2YQQHAYwRk+ZF94PchFjUFdR3Ohprlb6hlRS1cTJ4JWHLZI3gOa4HzBBBCyVAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERBtjiJw70hxW0dctCa5tEVzs11iMVRBIB28nA+RHcFeEXPDyTaq5UNbOqbcypuehbrIX2m6dJJgJP8Aa8xHZ48ie4x59/oEW0eKnDLSfGLQV44da1tsdZarzTPgkD2gmJxBDZGHuHtJyCCPTsSg6E+zV9oO3iFBauXri/WNZqOlpxTWO7Svx90WMGGwyE//ACUNG2+4G3oPSRfOdzGctPF3k94lxU95gq6eniq/eLBf6bPh1AY7qY4PGzZAACWnByDtsQPS3kl9p/ojija7Vw34217bFrRjW0sVxkBFLcnAYBLt+iQ7ZB7k9zgk3B6CIrGvbI0PY4Oa4ZBByCD5q9QEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREHnj7afUEtv4AaRsUf0LvqUdf1RQPcPzkLhP2PHLhozXlw1Jxv1dRR3CfS9dFbbTTyjLI5zGJHzEdiQHNAB/RkHlj229OTwd4d1Wf7HqaePH/fUrz/+T+dY/sRaiV3C7iXSud96jv8ARyNHo51MQ78zWpDXpUiIgtJK+fj2jvCy5cMubHWr5LU6lteoKsXe2vEeInxysa5waRsSH9WR3G2V9BGQuvnPNwb0dxZ5b9c/d+yUtTcrHYqy52urdGDLTTwROkBY7uM9JGO26HjReQnme0jx+4J6ctsN5iOq9PWuChvFE44kD4mCPxAO5BABz88rs8vBX2VeoLvaOcvSlvt072wXelr6WrjBPQ+MU0kgJGdyCwYPln5le9SJOCIiKIiICIiAiIgIiIC8IPayXuS685F/pC8+HbbZb6ZrfIHwQ4n6zkfkC93XOa1pc5wAA3JOF873PrqyHiBzfcRLxa5PeIH3VlFSlu/W2KKOMY+sg4QeufszNFDSvKlpW5yUohmvtMysPw4L2EHBPrkkrtJdbdTXi2VlorGB9PXQSU8zT5se0tIx9RK2lwO0zTaM4N6I0vSx9EdtsNDD04xhwhaXbfWSt8qTs6Pm+5seX3UvLlxhu+jL7b3QUVRNJV2mYDEc1K556ek9vhyAQPLB816eexmt9oHLzfrtT+G64O1BLTVB26mNaxrmjPcAh4OPkFp3to+H9Hd+CuleIsVOwVun717nJN0/E6nnYR059A9rSPrPquMPYpcQH0N213w4qKhoguHgXKBhIBMzWlpI8/ojf6gs/lNxdx6yoiLaCIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAurPOxzy6V5SbVSWz7luu+qrzA+Wgow4NYxoPT4jz6A/zd84XZPUGoLLpSyVmo9R3Snttrt0JnqquoeGxxMHckn8nzJAG5Xz/APtA+YeycynMXc9X6QfNJp22UsNotb5M5nZHkvmA8g97nED0A7pmjZDtP8cObzipedWWPSlw1Be75WOqKp9NETFETjAL+wwMfPzxutZ4ccQuNnI1xoNQaOe0Xek6GXG3TD4KmAnOD6g74J+eQvb3kr4SWPg9y2aEsFutMFJX1Vmpq+5ytjDZJ6qZgke557nBeQB2AHzJPC3tF+RWv5nLVb9bcN4aSLW1nYYXNmeI210HcMLsdxjYn5AeYKkv1zzy4c0fC/mR0TQak0jqOhFzkiaK60umaKilnx8TSwnJGdwRnbGSuZF83eueCvMryoagotQaq0rf9H1bJv8Aad0hd96kcCD8MsZLSM42J39F345ePbJWiOgtmleP+kK1tQwR08l/tr2yNcAMGSWJ2Dk7ZIJ9Tnup5B6mItN07qC06rsVBqSw1jKu3XKBlTSzM7SRvGQR9hWpKgiIgLYXHfQ1u4kcHNZaKudBFWR3SyVkMccjQR4phd4ZGexDsHK36rXNa4YcMg+Xqg+cTk91LScO+a3htftQVJt9PbNSQR1j37eE1zjG8H/ziCvo4hmiqImVFPK2SKVoex7HZa5pGQQRsQQc5C8Tvaecmtv4AanpOLuiZX/se1dcZWTU+N6SsIMhwRsA7BOdskbb5XbX2PvGrV/EvhJqnR2sb3Pc5NH19NHQyTv65BTTMeQ0k7kAsIGf05U+6PQFERUEREBERBY5jXtLXNBaRgg+YXin7T/kzvnC7iNcuNmh7J16K1DIKisbTMwLdVu2eCBsGOO4I7ZJPnj2vWga70Vp/iLo+76I1VQx1drvNJJSVEUjQ4Fr2kZAPmM5HzCDzL9mJz+V9XXWHlj4qSRPi8E0mnLu95DyRuymlz3OMhrvlgntn1TXzxc0XKjxZ5MuIdLXVpk+5Ete6bTl/pXZbIY3BzQT+9laMEgjcDOMHA9UPZx86tbzQ6Nr9La58CPWml44veHxnArqcjpE4B3ByMHvucn1I8dzUREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREHHnHPgbw/5hOH1dw64iWsVVBVjqimbgTUso+jLG7yIP/vnBXz88z/Aa9crfHG8cMKi8e+utb4qy3V8Y6HyU8gD4XkD6LwNjjzBIx2H0krwv9rvbqyn5vq+4TRFsFZZLf4Lj2d0R4OPqyPypvwevnK5ryLiXy+aB1lHU+O+usdKJXZyfEYwNcD88tyfPdcqLpX7KaTUlPy1UNqvbJWwRSOlpRICOhjiSAAe2RgruopAREVBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFQnG57Kq4O5zON1FwE5etWa1dVRMuT6J9Fa4nvwZamUdDQPPbOcgHBwg85vbAcyekeJGo9NcGdGXKG4x6TqJ6+61ERyxtW9oYyIHscM6ifTI7ggrmr2JVBLBwc4hXCRuGVWo4GM27hlMMn8riPsXlNoXQuveN3EGj0hpG3VF81JqCqPS0blz3El8j3dmtGSSTsPyL3y5HuWer5WuCNNoK8V8NZequqfcLlLAcxiV4A6AdshoGM/V3xlXhjsOiIoC2lxatjr1wr1lZ2N6nV2n7jTNA8y+mkaB+UrdqjlijmjfDIwOZIC1wPYgjBCD5x+UTiNDwN5ntE61v/VT01nurqe4NOAWMex8LwT5YL9z5YX0VWG/WrU9mpL/Y62Oroa6Js0E0bshzTuPt9QvFP2pfKpZuBPEC18QdIscyy6yln8SIjAiqmYc4DAA3Bzt8vmu7fsiuJNx11yy1FlutY+pn0teZbeHvcS4ROjY9gyd9gUHeNERAREQEREBERAREQcYcyWuBw+4M6i1GanwHMpjEx4O4Lgdh9YBH2r57+H9sruJvHi2QxNfUvuuoG1UhOXExmcOJJ+rbPzC9jfay6sqNL8qlVBTSFr7rc4KTIODg5yR9QJXSH2TnBCk4jcTbhrOvjLoNPOYT8IIBAB7+RJICnnSf1XtLaqcUdso6QDHgQRx49MNA/mWWiKjqL7Uu2PufKDqIMp/FdT1UE/bJaGh5J+S8VOBXGbVHAjiPatf6WrJYpKOZhqImOwJ4sjqYfnjOPn8iV9GfFrhrZuLmgLroK/EilucJjLgM9LsHBx9q+drmW4M1nALjLqHhlVvc9tsmBge799E4Zac+Y7gHzASc8H0GcB+O+heP+hKDWWjLzTVJmgY6qpmP++U8hAyC07gZ/wDfK5KAwvAj2anEvU2heZ3TtttdynZaru90FwpRIRHIw4+IjOMjtn0PngL34Sn6EREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFDPNFTQyVNRKyKKJpe+R7g1rGgZJJOwAA3KmXlB7VvnS1FR6lqOWzhteJKKkpoGO1HWU0uHyveMimBG4AGOofPzzsGi+1R53dO8Raan4A8IdSMr7VS1Jn1FX0j8xTysJDaZruzmg5LiDjIxuFtf2XXJJauNN3q+NHFOzTy6XsU7IrRSyNLY7hVjdzzkfEyP4ex3J9QCNoez35DNU8fNcWniLxG0tNBwwt0hqZZKppYLw9n0IIhkF0ZcB1uG2NhnJx7gWOxWXTNrp7Hp610ttt9IwRwUtLE2KKNo7ANaAAmGsuCCKlgjpqeNscUTRHGxowGtAwAB6AAKVEQbD42cGtGceuHV14aa7ovHttzj6Q9uPEgkH0ZWHycD2K88v3ES1t1C2Ucc6l1jEgLoDagKksz26w/pzjz6V6komjb+gdGWnh1ouzaGsQf7hZKOOjg6zlxawYyfmdz9q3AiICIiAiIg6oe014V6k4tcqd5sukrTJcbpa7hS3eKCNuXuZEXB/SPXoefyryv5B+bL9qLxXuMuqrbUv0/qCNlvvEPSRLTOjeemToO4LSXAjGfL1Xv8AbOG47rxf9pPyX8SqbmLrdc8KOG1yuuntZMhqf+plP4jYK7p6JWFrRhgJaHDy+I5xsSzU17FaV1NZdaabtuq9N18dba7tTMq6SoYciSN4BB2+vceRyFqx7Lg3km4eay4V8sOhNDa+Y+K9W6gInp3nLqcOe5zYj82ggfmXOakuqIiKgiIgIiIOK+ZLl40VzOcL63hhrd89PTzyMqKSupgDPR1LM9MrM7HGSCDsQSCvD/Xtk4r+zr5nqyzaU1c2W5WdkU8FXC0sjrqKYBzWysycZAwRk7j0JC+hReGXtfKOeDnBq6qSJzY6nT9tMbiNndLXA4+oj86ej2V4J8RIuLfCLSHEyKHwW6ls9NcXRnbodIwFw+w5W9wMLrF7OXiLpzXPKZoK3WmvhkrdO2tlqrqdrh1xviJbkjvgjG/rldnQcqS6KoiKgiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC4Z41cqPCTjxeaG/a6srJ6yhAa2QNBLgO2cjy7fUAuZkTxLNaDozRWndAWKDTmmKBlJRU4wxjQM/bha8iIoiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAvEf2ufGa86v5iJuFtPcpDZNJUsDTTh/wAJqnxh7yR6gOGPr+QXtwvnd9oNJJLzm8VnSk5F76R9QhjA/MAg9EPZRcn0vDHSreYTWcURvuq7eGWiAty+jonkEuJPZzwBkem3kCfRJbW4XUlNQcM9JUVI0NggsdBHGBsA0U7APzBbpQEREBERB5ve23az/Yb4dv6fjGppgD6A0r8/oH5FB7EUk8J+JLD2GoaUj7ab/wBim9twCeDnDs/900w/5K9R+xFaBwl4kOzudRUw/JTD/WrnB6TIiKAiIgIiICIiAiIg87fbV3IUnAbRlv36q7Uxb9jKd7ifygflW1vYi0ch0jxJr/DHhtuVLD1/MxZx+YLi/wBsPzA6b4g6s0vwl0tXRVsWlX1FZXyxuBaKmQBgaCO+AD9uV2Q9jZoyo09y4XvU9TDJGdS6hkmj6gR1MiiZGCB6ZyQfPKeI79qnf7FVFL1VPJdM+fzkQ0TzF6eunFG21Ulo1zYrVK+KoYMw10cLC4RSjvnAwHDtsNskruauL+Z3XdHw15feIGs62VrBQafrfDycF0r4nMYB6kucMBXcHgTyiXSotfMboV9PJ0e8XSOnefVrsjH5cL6RACGgE5K+aTlruEdq4+aCuErepkV9pcj63gD85X0tRyCWNkrezwHD6iE34md1eiIiiIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiLiHmT5mOHPLHoOo1hrm5sFS+N7bbbmOzPWzgbNa3uBnGT2AyiW44c9o5zgycs3DCKzaKucDddamcYKFvUC+kgA+OoI7jGwB9T5d15nck/KbqPnd4p3rUOt9SVTLHapo62/wBwcS+eslkcSIWHsC4NOTthvbyxxJq7U/FrnB44yXB8NTedS6prTHR0jCXNgjJ+GNo/esYO5A8icZ2Xt5yK8qbOVHhE/TFwrGVmoL3Ui4XiZn0PFDQ1rG/JoyP9fcpMW75HPumtO2fSGn7dpfT1BHRWy000dHSU8bQ1scTGhrQAPkNz591qiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC6p8+PJxonmR4e3HVD4TR6w05bp57bXR95AxpcYXjzBxt88fIjtYtK1T0fsYu/i4DPcKjq+rw3Z/MrOJZrwr9l9xc1BoHmn0vpOmuE4tOrpn2yqpuv72XuY4sdgnAORjbfOPQL3pXzkcmIlHN5wv9xz1jVVL0dPfHWe32L6N1FEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAXzue0FOec3it/4c//AHMa+iNeAvtOdD3DR3OHrSuq4ZBBqN8N2p3kbOa+JrSAexwW7+mQg90+GEkc3DXScsJBjfY6BzT3yDTswtzrql7O7mS07xx4DaesPv7f2SaVt8FsuNO4/E4RNDGvHnggDY74wd8rtapmAiIqCIiDzt9tRYbrc+BOjbrQ0j5aW06jdLVva0kRtfTuY0n0GSB9q4g9jNxr0hpK46z4T6iu9PQ1uoKimuFtEzw0TSMa6NzAT54I/N6r1T1povTHELTdbpLWFmp7parhGY56edgc0gjGRnsR5FfOzzLaF/a88zmsNGaOrJ6Rulr11W2ZhLXxsLWyxkfUHgY7eW4Tfg+kMdlVcbcuHEM8WOBGheIbi4y3qx0tRNk5Pi9Aa/Pz6gT9q5JUl0ERFQREQEREBbY4nXWSxcN9V3uF/RJb7JXVTHf3JZA9wP2EBbmJwtj8dMf7CfEAOGR+xe6/qkiRLcfPLwV0TX8xPMJp7R90qZJKnV15/wBtSucSS09T37nsMAj5BfRRwz4c6Z4T6Jteg9JUTKW22uERRta3p6iBu448zj8mB5LwV9nKcc6HDMZH+6Mo/wCTyL6FVd+GCIiii8Xvauc0+t9UcXbzy92m5SUeldOGBlbTsODVVJjbIes/3IDxj68ds59oV893tIo2x86/E4tB+OupXb/4lAg7y8kPsvtMaeodI8ceKeoJ7jd56eG7Udogj6IKfxGB0Ze4nLnAOHljfOx7elQw34QAANgtocGyDwi0OR56bth/5LGt4Jct1J51VERFEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQERdaOYj2gfL1y4z1dl1HfKm86jp4y5tntUfiSl+Dhr3khseSO5JA+vZE1y3xm408PuA2iK3XvEe/QW63UrD4bXvHi1EnlHG3u5xJA29QvAfmr5jtac2vGSp1ZUwVLqMye5WG0xNLzBBnDWhgzl7jgkgZJwN8ZOXzG8x3FrnV4tU881BVyxzzik0/pyi6pWwAnAAA+k87kuxtv5ZJ9JfZyez4k4J0EvFPjhp2hm1pW9P3MoZg2b7kw4ySe48Uk74+j27gFFbg9m9yO27gHo2k4pa9t7Zde36mEgZK0H7mU7xkRj/tyMEn59+wHeREQEREBERAREQEREBERAREQEREBERAREQEREBERAWia4PTou/uzjFrqzn/8AYuWtrQdeN69DaiZ/dWmrH5YXIPn45Aqf3rnM4Uxub1Yv7Zf/ADI5HZ+zGV9Eq+dv2f1XHQc5fCmaY9LTfPCyTjd8MjB+dwX0SICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLzi9sJwbotQaAtnFGmpWtrbM0xyTNG7mA5IPywcfYPRejq83Pa+cfrXp3R1HwVpHMmuV4iM87Ad4oycZP2AEepP5LLg68exrrqqPmKu1vbUvbBLZZZHxdWziAcHHyz+j0XtUvFv2NFoq5+YC93psLjT01nfE5+Ng52ds+vb8q9pFAREQEREBeRHtJOTfilr/mYl13oCw+90GpKWlbO8DAjlYwMc84G4wBnzyPnt67qN8UUnSZI2vLDkZAOD6j0U+6OK+VnhpVcIuAukOH1a5zprTQMif1dwe/n+Vcr43yqoqCIiAiIgIiIC2jxco3XLhVrO3MaXOqtPXGENHcl1NIAB+VbuWNX0kddQ1NFMMsqInxO+pwIP5ig+dPkr1PSaA5sOHeoL24QQW689FR17dGWPYQfQ5OF9F9LU09bTxVdLK2WGZgfG9pyHNIyCPrXzT8fbNDorjjqmgs05jbSXaSaB0ZwY3F3Vt6YJJH2L3J9nnrm/cQeV3TF91E98lUOuDrdn4mgNIx8skq8z9p3eeOyqIiii+fT2lrCznY4kg+dTRn8tHAvoLXz5+0qnE/OxxLIGOiqpGH6xRwDKD3b4NgDhDocDsNN2wf8ljW8Vszgtn/AGHNB57/ALGbX+qxreaJBERFEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAXHPMFxMuXB3g9qfiPaLM661lkoX1ENKAT1vA2yBuRn0XIyxq6gornRzW+4UsVTTVDDHLDK0OY9p7gg7EKwrwP117TznF1tNOIuJjdP0k3U0U9ooIIQ1hzt1lpeTg4znK0Hlt5SONPOnq+vudHdDTUhkMtx1DdA+VheTg4A3eQdsAgDsOxA9npeRPlBqLm+8TcANKPq5ZDK9zqdxa55OSSzq6c5+S5h0zpLS+jLVHZNJaet9noIgAynoqdsTBjts0DJ+ZUvvCOp3J/7Njh3yu6kHEC46kn1ZqpsJigqJqZsUFH1DDjEzJJJH74nI8l3JREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBafqClNbYblRgZNRSTRfa5hH861BUIyMEKWD5odC3qThJzBWi9zudTnS+qGOkOS0sbFUYcc+gAJ+pfSDonV9l17pe36ssNUyoorhA2VjmkHBI3B+YXjP7U/lSouCmuqPinptx+4+s6uZsrOk/eqkDqI9M4B389j3yu3XsdOJl01nwEv2krtWPnfpO7MggL3ZPgyx9TfsHTj7PtNz6O/iIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgpnOxC+en2gWodU6y5wNffduCpM9PchbrfTlhLhTsaGxNYBuc5JGM7nC+hbOO62DqHgLwZ1Zq6m15qPhtYrjqCkcHw3CelDpWuHY+hO3mFPo4v5G+WfT3LtwT0/SttsQ1NdrfDWXmq3LzNK0PdGCdwASBj5Y7ALscNkwAMDbHoqpAREVBERAREQEREBERAREQEREBaTquuktel7zc4vp0lBUTt+tkbnD84WrLbXEmuhtvDrVNyqHBsVLZa6d5PYNbA8n8wQfOK2hrOMnHuS3Vc3TLqK+yRud26WGQ/kw0YX0OcBOGts4ScKNP6JtTWiKjpWOd0jAL3AEnsPLA+xfPry7un1DzKaSfRwlktyvwcyNu5BkLjj7Mr6Q7bSOordTUTn9ToIWRl3qQ0DP5kvbtTvjKRERRfPj7SiDwOdjiYM/TrKST8tFAV9By+e32kNca7nV4nO6ceDX00H19FJCMoPeHg2McIdD7Y/8Ag3bP1WNbxWzOC8rZ+DuhJmnIfpq2OB+uljW808T0RERRERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQERYV3u9ssNrqrzea2GjoaKJ01RPK7pZGwbkknyQdLfa/2GmunKLNdnwB89n1Db54nYyWh5dG7HpkPGfqXU32NHFi0aS4n6t4fXq4Npxqajp5aJr34Dp4nOB2+pwGfmFle0Z9onoDjroip4F8I7dV1lrfXxVFffKgBkc3guJDIGbkjqGS44BGMLhL2d3L5xH4rcaLXqzTlJPSWaxSmWruBy1noWA+fff54AyeyTfE8e+iKOJhjiZG55eWNALj3JAxkqRFEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBeX3tO+fTUOkbpqHlg0DbYoHVVAynvN1kcesMmbl0UQHY9JAJ+fmCQvUFfPJ7Q3U1JqvnG4lXKgmjmpoblHRRyMcC13gwRxkgjY/E0qd3YeuznsteSbU151fYeZnW0DKWw20vqbJTu3fVS4LWy47BoySPX69l6/rYPATTlBpLgloPTttp2wU9Dp23xtjHYEwMLvyuJP2rfyTfpREXWPjb7Q7lw4B61foLW96ucl0gx7yygozP4G/77B+R2+RVHZxeHHtduHtp0jzRv1Napow/VlthrauFvdk7GtjLj/3wDT9i72a19rryn6atBrLBWag1LXPj64qKkoDFk/3L5JCGsP5V5P8AMtxg1xzV8T9Q8Z5tO1zLdCxkQjjY6WKgpmjDGucBgbHc/USfNB6tezC5qIeMnCui4Y3WB7b1o2hjpXTeUsTAA0n7MY+rHku8B3XjZ7F3U1LRcbNVaWne0TXGzePTtPcmN3x4+oEflXsnnfCW23qTiqIiKIiICLye9otzw80XAjmXuHD7hTxP+4dggtNBVR0f3Ft1TiSSMl7uueB7zkjOC7A8gF3w5KeI2tOLnK7oLiLxCvP3V1De6KeWurPd4oPFe2qmjB6ImtY3DWNHwtA2z3OUHOKIiAiIgIiICIiAiIgIiICLoB7Vnmj468tn+xd/sK65/Y5+yL7t/dP/AKmUdX7x7v7j4X9sxSdPT48v0cZ6t84GN3ey55hOMHMXwk1ZqfjJq/8AZDc7bqM0FLP9z6Wk8OD3aJ/R008UbT8T3HJBO+M4ACDuiiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiIC8jva5c1Wv6TiHJy3aXuT7bYaa3U9VdzCcPrJJ29YjJG4aGEZHnny3z64rwT9qpVNrOdDVc7WkBtFb4tx36IGsOPtaUHLfJ37KiDjdw5sPGDibr6otlovsfvdHardAHSywB5AMkrjhodg7AZH5CvV7hTwj0LwW0fSaH4fWWK3WykaAA0DrkIGOp5AGT/rPqVtHlAscumuV3hdYqhpEtJpihbIHDBDjECdvrJXMGMqT8v5dTFURFVEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREHTT2oPMdrPl94I21mgqx1Bd9W3B9sZWtGXwRNjLnlvoSNgfXHlleX3JByyaj5seM5kuVRK+zWidlzvtbLlxkJfkMye5cQSfPG3nkerntFuVvUfNDwltNm0fJGLzp+5e/UzH5w8Pb0PH5P058lP7PjlRruVrhPW2zUr4pNSahrBWV7mDZjWtDWMB8gBnI9d/ND/AF2hoaOnt1FT2+kZ0Q0sTIY298NaAAPsACyERAXjf7Rjkp4sXTjpd+Jeh7JLd7bfyJjHHkvjeBg48sbgY2xgnz29kFZJHHK3pkja8ejhlB4ecBvZQcb+K8Ud11fdqLRtp6wHvqIXTTvbncMYCN+/fb1Xoje+XvgByi8nOt9KMp6Q0LbBXGsul06PebjXPgcGEu/ui8NDWN2AAG4BK7Nat1dpjQWn6rU2rrzS2q1ULC+aoqHhjGgAnA9TgbAei8CudrmY1RzXceLgbJcqyr0pSVgt2mLXG53hlgwwS9GSDJK8k9Xo4AYCe8J+13s371qOyc4/D2XT0U0nvdXJR1zY2k/7Ukjc2Qux2A+E5O2QAvoOXWvkv5QuHvLhwzsE8emaN2uKu3RS3q7SR5nM72hz42k7ta0ktAHod8HC7KJeXEl0RERRERB4Ue12/DLuv/gG1/8AoisjgV7VTi/wF4T6d4R6d4daPuNt05BJBT1Nb7148gfM+Ul3RKG5y8jYDYBY/tdvwy7r/wCAbX/6Irs1yd+zX5ZONfLXofijre36jkvl+pJ5qx1NdnRRFzamWMdLA04HSweffJQcU/u2nHr8U2gfyVv9Mn7tpx6/FNoH8lb/AEy7efuQfJx/86tW/wCfXf8Aqp+5B8nH/wA6tW/59d/6qDqH+7acevxTaB/JW/0yfu2nHr8U2gfyVv8ATLt5+5B8nH/zq1b/AJ9d/wCqn7kHycf/ADq1b/n13/qoOof7tpx6/FNoH8lb/TJ+7acevxTaB/JW/wBMu3n7kHycf/OrVv8An13/AKqfuQfJx/8AOrVv+fXf+qg6h/u2nHr8U2gfyVv9Mn7tpx6/FNoH8lb/AEy7efuQfJx/86tW/wCfXf8Aqp+5B8nH/wA6tW/59d/6qDqH+7acevxTaB/JW/0yfu2nHr8U2gfyVv8ATLt5+5B8nH/zq1b/AJ9d/wCqn7kHycf/ADq1b/n13/qoOof7tpx6/FNoH8lb/TJ+7acevxTaB/JW/wBMu3n7kHycf/OrVv8An13/AKqfuQfJx/8AOrVv+fXf+qg8uebznZ17zi/sT/ZvpKwWT9iPv/u33KE3333r3fr6/Fe7t7szGMfSOc7Y9CPYmf7wuu/4Xn9Sp11U9p1yhcHOVP8A2Nf9ialu0P7KPuz90Pf641Ofdvc/C6cgdP8AZ5M+uR6LtX7Ez/eF13/C8/qVOg9FUREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAXW/jRyHcEOOXEOm4laut8v3UhLDL0fRm6TkdW4z9uV2QRBiWu20lnttJaaCIR01FCynhYP3rGNDQPyALLREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQefntl36mHALTkdlE/3PdenG5+FnHhiP4OrHl1Hz27rzO5LLDdbzzI6KnttkNxjoLiyoqG+EXMY0AgE+QOSMZ+fovoa1Vo/TGuLNLp/V1jpbrbp8eJT1MYe0/Ytt6H4EcH+G1QarQ3D2z2ecknxKeD4sk+ROcfYn+J1v5ERFEREBERB0j5rvZkWHml4v1XFq4cXa/T01TQ01F7jDZ2VDWiFpaHdZlaSTnOMbfNdleXng9S8AODemeD9HfJbzDpunlgbXywCB04fNJLksDnBuPEI7nt81yOiAiIgIiICIiAiIgIiICIiDq/zrcj1p5zP2G/dTiJV6X/AGIfdHo8C3Nq/efe/d856pGdPT7sMYznqPbC13k05R7byf6HvmibZreq1NHert91XVE9C2lMR8GOPoDWvcCPvec5HfGNl2DRAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQRyyxwxPmlkaxjAXOc5wAaBuSSdgB3XQjjn7YTgZw0v1Vpnhppa5cRqqie6Katp6xtDbnPBwRFO5kj5QCD8TY+g7FrnA5Wre1u49XThNy80uhNN1r6S7cSKyS2SSsf0vZbYmh1X0+fxdcMR/wC1ld54W1/ZZ8m/D7T3Be18d9faQt151bq8vq7a+4UzZxbLeHFkXhNeCGSSBrpDINyx7GgjDgQ2hpD23+kq26R0+veAN1tNuc/46q1X6OvlY0k4PgyQwA4GM/GMnJAHZehnCrixoDjZoi38ROGeo6e9WK5NPhTxAtcx4OHxyMcA6ORp2LXAEbbYIJ435nOULhRzGcMrxpW4aPstHqE0kpsd6ipGRVFBWBpMTvEYA4xl4aHsJIc3O2QCPOD2PvFu/aA5hL7wAvs8sNBqukqXsoXu/sN2ogXkgZwCYGVAdjv4bM5DRgPZdERARYF6vll01aqq+6jvFFarZRRmaqra2oZBBAwd3PkeQ1o37khcGu5++Tdl1+4zuYTSvvGcdYmkMGerp/s4b4WM+fVjG/bdB2BWnXq/2LTdEblqK9UNrpOsR+PW1LII+o5w3qeQMnfAznZWae1LpzV1np9Q6Tv9tvVrqx1U9dbqqOpp5h6skjJa4Z8wSvOL2vHGjg7xB5c9P6a0DxZ0bqW70euaSoqLfZ79S1tTDEyhr2PkfFE9zmtDnsaSQAC4AnJAQek9uuVuvFFDcrTX09bR1DeuGoppWyxyN9WuaSCPmCspdFPZx8wnAPQ/Jnw80vrbjhoDT96ofut7zbrpqWipKqDrutW9nXFJK17epj2OGQMhwI2IK7rab1PpvWNkpdS6Q1Dbb5Z65pfS3C21cdTTTtDi0lksZLHAEEEgncEeSDVUWxtMccODuttY3Lh9pDifpm8altHi+/WmiucU1VT+E8Ml6o2uJHQ8hrtvhJAODstn6v5y+VnQWrZdDat45aWt97ppDDUUrqvrFPIDgslewFkTge7XkEYOQMFBzSix6OrpbjSQXCgqoamlqY2zQTQvD45Y3AFr2uBIcCCCCDgjdbF4ncf+CfBgRDinxR05pqacB0VNXVzG1EjScdTIQTI5ue7g0geZCDkJFwvoDnL5W+KF3p7BonjhpeuudY7opqKWqNLPO7JAZGycML3HBw0AkjfGFzQgIoKmpp6Knlq6ueOCngY6WWWV4axjACS5xOwAGSSdsBcE3nnz5PLDdnWW4cwOk3VTXOY40tQ6piBAyQZYmujH/nbnYb7IOfUW3tFa/wBDcSbIzUfD7WFl1Ja3npFZaq6Oqi6sAlpdGSA4AjLTgjzAWtVNTT0VPLV1c8cFPAx0sssrw1jGAElzidgAMkk7YCCdFwFeefPk8sN2dZbhzA6TdVNc5jjS1DqmIEDJBlia6Mf+dudhvsuXNFa/0NxJsjNR8PtYWXUlreekVlqro6qLqwCWl0ZIDgCMtOCPMBBuFERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQFt3XWvtF8MdMVms+IGqLdYLJQN6qitr5xFGzPZoJ3c49g0Zc4kAAkgLcS8sZOSHm552uKtw1tzZaxrdG6Lt1yqI7VZ4nsfOKYSEBlJTtzFACxoBnlBe74SWyA5Acl272wvCfUnGO0cNtIcMNQXKz3m9Ulnp77LVx0xd40zYvGFM5pd0AuyA57XEdw07D0DXgBxJ4ZaR4N+0RtvDHQtHNS2LT+t9O01HHNO6WTBNI5znPduSXuc4nYZOBgYA9/0BFsbiPxu4QcIHUDOKXErTulX3MSe5Mutwjp3VAZjrLGuILg3qaCQMDqGe4W59PaisOrLJRak0veqK72m5QtqKOuop2zQVERGQ9j2khwPqCUGpItH1Xq3S+hdPVurdaaht9jsttYJayvr6hsEELSQ0Fz3EAZcWtAzkkgDJIC0rh3xX4Z8W7ZUXnhjr2xapoqSb3eomtVdHUthkxkMf0E9JIIIBxkb9kG7UREBFsPidx14O8GKeCo4qcStPaY96GaeK4VrI5pxnBMcWet4B7loIHnhbV4fc4/K7xTusNi0Pxw0tX3KpIbBRS1fus8ziSA2OOcMc9xx9FoJxvjG6DmZaNS6x0lXXqTTdHqq0VF3g6hLQRV0T6lhb9LqiDuoY88jbzWna44pcMeGMVJUcSuIumNJxXBz2Uj75d6egbUOYAXiMzPaHkBzScZxkZ7rxh5feIegbL7V66cRbxrjT9BpSTW+s6tl9qbnDFbnQTw3EQyipc4RFkhkYGO6sOL24JyMh7kItgaT4/cCNe3uLTWheNeg9RXioa98NvtOpKOrqZGsaXPLYopHPIABJwNgCStU4h8U+HHCW0Q3/AIm65sul7dUTilhqbrWMpmSzFpcI2FxHU7pa44GTgE9gg3Wi2TqXjRwl0foOm4oan4j6et+k66KOakvE1fGKaqbI0vj8F4OJS5oJaGZJAOAcLTeEvMZwP47e+M4ScS7NqWa3sbLVU9LKWzwsdsHuieGvDc7ZxjO2coOSEWLcLjQWiinud1r6ejo6VhlnqKiVscUTBuXPe4gNAG5JIC4Ku3PtydWW6us9bzB6UfUNe5jnU076mEFoyfv0TXRkb7HqwTkAkoOf0W3NC8RNB8TrEzU3DvWNm1LapHFgq7XWx1MQcO7SWE9Lh5tOCPMLcaAi2HxO468HeDFPBUcVOJWntMe9DNPFcK1kc04zgmOLPW8A9y0EDzwtq8Pucfld4p3WGxaH44aWr7lUkNgopav3WeZxJAbHHOGOe44+i0E43xjdBzMiLafEPipw14S2ZuoOJ2urJpi3vcWRzXStjgErwMlsYccyOxv0tBOPJBuxFwVpbnk5R9Z3ZtjsPH3Sbq2SQxRsq6s0YkeNsMfOGNcSSAME9R7ZXOTHslY2SN4c1wDg4HII8sH0QXoiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg0HWOutF8O7LJqTXurbPpy1RHpfWXWtjpYQ7GQ3rkIBJwcDOTjYFddrj7Tvketdwdbp+N8Ur2EtklprDc54mkDyeymLXA52LSQuLeZv2fXEvm05n6nV2vuJj7Pwsttvo4LXR0spnqy/w/v7YYnDwoSZAXOkd1EgtAa4DDdxyeyI5OHafNmZZtUsrPBMQuv3dkNV1Y/snSR4HV8vC6fkg7N8K+OfCHjfa5bvwn4h2XU1PTkCcUVSDLAT2EsRxJHny62jPkt9r59tS2zXfs5Ocz3Sx6gqK0aVuFNUsnjBjbd7RO1rzDKzIB643OjcM4EjCWnZrl9AlPPDVQR1NPI2SKVokje05DmkZBB8wQUEqIiAiIgIiICIiAi86/a7cfuMnA9nCmPhLxBuml23x18dcTQOa01Bg9x8LqJBOG+NLgAgHq3zgY7n8uupL3rLl94Y6v1LcH114vmjrLcrhVPa1rqipmoYpJZCGgAFz3OJAAG+wCDkVERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERB4++24vNXUcW+HGnpHE01DpyprY2kjAknqix5AxncU7PM9h2xv6e8ulrisfL9wys8FMYGUWj7NAIz3b00UQIPnnI3J88kry69trQ1MfGnh7dHtxBUaXlp2Owd3x1cjnDtjYSs7HO+4G2fUrl+uTbzwF4bXlkolbX6Rs9SJAMB3XRROBA8gc5x80HIC8NOFNNNov2tslDb2xwtj4oXqniaHEhtPPLUtDck5z4cuNz3758/cteHnC4zaz9rtPNbuh//AOc69ztMeXtfDTSVLi8YB7xwl2ew7k43Qe4aIiDx89qBr3WfGTm/0jyrx6lNl0xT1FnoWskkLaY11wezNZOAQHCNksbR1fRDXkY63Z7T0XsguUGn00yy1dDqyquAiDH3g3pzKkvxgvDA3wQc5IBjIGfNaB7Rv2fOpuYm70fGngzWU7NbWyibR1tqqJhA26RREuhfDMcNjnYSW4eQ1zen4mFnx9bdC+0m5weVm7UnDrmf4bVt+paTEIN6ppKC6mJhALo6kNMdSACD1Oa4u2zJvkh6A8mvKHDygWbV+lbVr6r1JZr/AHaO429lVTCKWjYIgwteWuLHvJAy5rWAho2HYecvtBvZ6cK+VDhRbeJmgdXamuE911TDZhRXOSB8UFPLTVc2Q5kbXFwNOxoJO4JJGcY9TOXHmb4U80eiTrXhddppBSvbBcrZWRiKtt0zgSGTMBI3AJa9pcx2DhxLXAdWfbV/gs6W/wCMCh/k64oOBeT/ANl1wj5i+XXSXGXU/ELV1sueoPf/AB6WgNN4EfgV1RTN6OuJzt2wtJyTuTjbZenHATgvpjl54Saf4PaOrbhV2nT0czYaive188r5p5J5HOLWtG8kryAAABgb4yuFvZcfgKcM/wDyz/LFau1aD54J9f8AFTQ3NTxVt3BhtV+yvWl8vmlKJ1Fk1bfe7lgiAjBbK4M6A7Yt6y4EEAjvDoH2KOkavh9Sy8TeLF/pdb1ULZakWmOB9vo5CCTH0yML5yMgF4ewEg4GCCuD+Sikpar2qV4NTTRSmHUWrpYi9gd0PAq8OGexGTgjcL2uQdVubXjlJyP8pVrGnKuGv1JS0dBpLT0lTEMSVLKfpNS+PJyGRQvkxkjq6GnIdv0v5J/Z6R81lhqeZfmd1XqG4Q6orZ5qGjiqDHU3LpeWvqqidwLgwvD2sYwNJDA4ODSAdye3Gu9cDwfsLWSso3i91b3Y+CWUe5taAcd2Auzg9pBkdltvgjx29qFpLg/o3TvC/l6t9fpGjstI2yVZsL5DU0ZjDopS8VADi9pDicDJcdh2Qco80PsjOD8XDO96s5f3XqzalsdHLXQWuprnVdJcWRs6nw5kBkZIQ09Dg8t6jgjBBbrnsj+arVPF3Rd84LcQrvNdLxomCCptNdUOL5prY89BikeSS4wvDAHHctkYP3q2R+2Z9sB/+rXbv9HZP6ytI9lTys8xfB3j7ftbcSeG1y0vYZNLVFsfNcQxhnmlqaaRkcbA4uJ+8OcXYwAME5cAQyfbS8bNZWmp0ZwJslyqaCx3Wgkvt4ZDIWe/kTGKCJ5G5YwxyPLD8Jc5hIJY0jmDhv7IblYoNCW2m1qL3qe+1FHE+ru0N3fBEZnNy51OyLDBHk/D1h5IAJJJOd5e0L5HpubfSdpvWjbrS2zXWlWytoDWEimuFNJgvppHNBMbg5ocx+CAS5pGH9bOjei+crnq5EPufw2438PKu76at4bSUVNqCFzSIWjaOkuUXUyRoDSBkzBoBAAAAAd+OUvkPsnKHxG1hqPRPEO6XbTmpqCnpYbVcYW+PTSxyOd1vljLWS4BwD4bSA4j5nqz7aXjZrK01OjOBNkuVTQWO60El9vDIZCz38iYxQRPI3LGGOR5YfhLnMJBLGkd0eVHnT4Q829lqZtDzVVr1DbI2yXSwXENFVTsJDRKwtJbLCXHHW05BI6msLgDsD2hfI9Nzb6TtN60bdaW2a60q2VtAawkU1wppMF9NI5oJjcHNDmPwQCXNIw/rYGzeG/shuVig0JbabWovep77UUcT6u7Q3d8ERmc3LnU7IsMEeT8PWHkgAkkk55G5S+Q+ycofEbWGo9E8Q7pdtOamoKelhtVxhb49NLHI53W+WMtZLgHAPhtIDiPmeg+i+crnq5EPufw2438PKu76at4bSUVNqCFzSIWjaOkuUXUyRoDSBkzBoBAAAAHpHyo86fCHm3stTNoeaqteobZG2S6WC4hoqqdhIaJWFpLZYS4462nIJHU1hcAQ7AIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg8KeZ3/wC2p1X/ABgac/RRL3WXhTzO/wD21Oq/4wNOfool7rIOtfPryr0nNNwNrrFbKWIax0713PTFQ44JqA0eJTEkgBkzW9BycBwjcc9GF019kVzT1enr3XcpHEaqlp/FmqKvS/vRLTTVTSXVVAQfo9RD5WjbDmzA5L2hery8TvataL0nwY5qrLxG4U6lFr1VfqdmornR0RLJbdcI5gIq0PGzTMWl3SNw6J7ztIEG/wD2n/MDqPj7xisXJnwabLdI7fdIoLpHTH+3r289DKcnsGU4cS4k4D3P6seECvRblb5fNOcsnBix8K7F4U1TSx+83euY3Hv9wkAM0xzvgkBrQdwxjB5Lzq9jJw+0DqzX+uOLupL1Hctd2QMioKKoyZYIakO8euyfpvecxZG7QX5/sjV64oC2/wAQdU/sG0FqXWvuvvX7H7RWXTwM48XwIXydGfLPRj7VuBYlyt1Fd7dVWm5UzKikrYZKeoheMtkie0te0/IgkH60HidyO8v1F7QjjTxA4icxGtrrcXWltPXVlPT1QiqK6aqfKGNaSCY6eIQkdDAOnqiaC0bHuFxT9jny5anszxwtvOodEXqJpNPM6rdcaRz/AC8WKU+IRnzZI3HfB7LrLxK5K+b3ka4oVvFzlTq7tqDTQdIYZrVEKqshoy4O91rqIgmdowB1sa5pDA8iJ2AOWuXv2x1BW3eDRnNBohunqkSilkv9nikMEMgPS41NI8mSMAg9RjLyCceGANg7QcyHInoPmj0doDTfEXWeoqWu0DSyU1PcLa6MOqvFigZO6RsrZMlzqaNwOcj4gSc5HkTwn5XNJcQOe2u5VLlqG702n6XUuo7I24weF74YrdHWOiectLOpxpmdXw4+I4A2x9AttuNvvNvpbvaq2Ctoa6FlTS1NPIHxTxPaHMexwOHNIIIIOCCCF4w8tP8A9uSvH/GBrv8A9BdEHd3l09l9wc5ceLdo4w6e1zq+8Xaxx1LaOnr5KYQB88D4HOeI4ml2I5ZMDI3IO+MHjb22f+8LoT+F4/Uqheiq86vbZ/7wuhP4Xj9SqEHVrle5auL/ALQ11gn4hanrNP8ACrhjaKbTNvlpmAlz4om9UNKx3wGV3wySzOBwDG3BAaG+h/Kn7PzQXKPxIv2utCa7v93pb3afuV7jdo4XSQjxmS9fjRNYHH4AMdA7k58lqPs2qSlpeSfhgKWmjh8Wiq5ZPDYG9TzXVBc447k+ZO67IXGploqCprIKWSpkghfK2Bgy6UhpIaNs5JGBse6Dx/5u+LXFLnu5toOULhfevctG2e8yW13h5ME89MHe919SWnEjIuiURtyAQ0EfFJldqNN+x/5SbTpqK0X+HVN8unhtE12kuzqeQyebmRRgRtBOcNIcQMAknc+XvJ1rfmO0/wAbbhrPl30XBrHW77VVy1UE9EanpppZohNOGNfHg9bmNyDsJCMb7d4f2zPtgP8A9Wu3f6Oyf1lBwNxS0fxC9lLzT2O/cOdU1930dfoW1TI6otBuVvbIG1NDVNaAx0rMgtkAABfG8Bpy0exmoOItst/CS5cWrMz3+30unJtRUrc9PvELaY1DB8upoH1ZXjzzOaQ9pFzbHTI4p8udbGdLe+e4fcu1GnB968HxOsvmdn+148YxjfOc7es/AXQl607y4aC4a8R7fAbjbdH26yXek6xIwFlIyGSFzmkh2AC0kEgkEgkHKDyS5HeX6i9oRxp4gcROYjW11uLrS2nrqynp6oRVFdNVPlDGtJBMdPEISOhgHT1RNBaNj3C4p+xz5ctT2Z44W3nUOiL1E0mnmdVuuNI5/l4sUp8QjPmyRuO+D2XWXiVyV83vI1xQreLnKnV3bUGmg6QwzWqIVVZDRlwd7rXURBM7RgDrY1zSGB5ETsActcvftjqCtu8GjOaDRDdPVIlFLJf7PFIYIZAelxqaR5MkYBB6jGXkE48MAbB6SaWtFTYNM2iw1dxfXz22gp6SWreCHVD442tMhBJOXEEnJJ37nuvFPhfpWs9pJz16kp+MGsLhR2SjZX3GKigna2WO3U07IoaCm6gWxn76xzyGkkNlefiJcvbe23G33m30t3tVbBW0NdCyppamnkD4p4ntDmPY4HDmkEEEHBBBC8oeaX2fvMPwU40V/MlyeVNfWwT1813bQWpwF0tM8pc6aNkTsiqp3FzgGNDnFr/DdG4NL3B2B1v7HrlS1BYH0GkXao0pdGMxDcILk6rBeBsZYp8hwz3DDGT5ELtLwA4Y3DgzwX0hwrumojfanTFtjt77h4ToxP0E4wxznFoAIaASdmj6l5z8F/bD620pfGaI5ruGMjJKV4p6u6WqkdSV1K/beoopSA44OT0GMgDZjicL090TrbSnEjSVr1zoe+U14sN6p21VDXUzj0TRnbODgtIIILSA5pBBAIIAa8iIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiDGra6it1Oaqvq4aaFpAdJNIGMBJ2yTsN1sbXXMFwP4Z2qqvOuuLOlbRT0beuRs1ziMp2yAyJpMkjiNw1jS4+QK2tzbctFs5ruEx4WXXVdZp2Ntzp7oytpqZs5D4g8dDmOc3IIkPmCCAd8YPRm/+w8g9we/S3MU81rWksiuGmwIpDtgF7KjLB3yel31IOvGpqHUntLueeuuOhdPVjNKz1dLBPVOi6PcLFT9MZqJ3dmySND3NaSSXSBgzjK9u9Y6XZq3Rt50dHdbhZ23a3T29tbbJ3U9TR+JGWCWGRhBY9mQWkEYIC8b9G8e+an2X/Ean4QcTNOWa7aOmLar3KmpIY4q+mz0uqaOrZGx5k2wRMHHIPU0FwcvY7QmttOcSNGWTX2ka4Vll1DQw3Ghnx0l8MrA5uQd2uAOC07ggg7hB4+8AOZHjTyR84Nx4Q8x+vtQXnS81Z9xrrNd7jPVQ08T3B1LdYBK93Qwtcxzsb+HI8EFzQB6rcwnHTSvLxwev/F3VEjZqW1U2aSma/Dq6rf8ADBTsIzu95AJAIa3qcdmldTvazcqdNxT4Uu486XpGN1Tw/pHyXADY11nBLpWkk46oSXyg9y0yjcloHnDBxU4785beDnKnU3eKaCxSttFsdI4jxS4kCpqSTl5gph0DG4ZG7ALnnIdx/Zn0nMRzJ8a9Q8zHE/ihrB2lrRWTiO3tu1THQXC4zNJ93bAHiMU8Eb2kxgdIJhGCAQvVNbI4M8JdKcC+GOnuFWiaYRWqwUjadry0B9RKT1Szvx3fI8ve75uONsBb3QF5T853NdzE8auaH9pryy6hqdOxQVrbPU1dDUupqmurRGZKl76lo8SGnhaHgtZgkRSOJcC1rfVheNPOtwu43cnnOLLzf8P7JJX6duF3N8pbmIXy01PUTsMdVRVfSQYxJ1ygEkBzJQGu6muDQ39L7Kfm001SHVWiubLOrI8TdDa240YdI3IGKpr3PJAyASwDfBwMldwuAmnuay9cpN90Pxovcln4ry0d5s1svUtRGZInmN8dFWPlpy7Ja8hweB1kMBIJOTxhwF9rby6cUn0lk4jtq+G98nwwuuTxNbHPIGwq2geGM53lZG0DHxEld3aOrpbjSQXCgqoamlqY2zQTQvD45Y3AFr2uBIcCCCCDgjdB8/3OvwQ5r+DX7Df2z/FKr1l92Puj9w/eNTVl3908L3f3jHvIHhdfiwfR+l0DP0Qud+FvJ57THU3DHSOpdBcx90tmmbtYaCustEziFdKdtLQS07H08QiYOiMNjcxvQ3ZuMDYBb+9ud/2E/wD+ZP8A+2Lv/wAp34LHBv8A4v8AT38nQINb03a9W6P4J2qy6s1C67ansmloKW53Zsj3GrroaQNmqA92HEvka5+SAd8kZXiLy9c/HHXhNpXXlmturNUat1prH7nUGnH3WtmubLfMXzeNURxSuf1zEPiaxoBDnEFwcGdDvd3WH/xSvf8A4Oqf/RuXjD7HPh9pXWHMtddRajtjK2q0jp6W5WkSYLIat88UIm6SN3NZI8NPkXZG4BAbo01yHe0kmu9l4z1/EKoF/qaynramln1jO26xMDmuIlLgIhgFwMYlOBlpA7H0S5y+azT3KTwlk1xcKFt0vlymNvsNrL+kVNUWl3VIQciFgHU8jfdrRguBHPa8avbWa0q7nx40VoMVLn0Ni0t90BH1HpZU1dTK2TYjGTHTQbgnOw8kGVwv4Bc+ntBLWeLXEHjlXaX0hcZJfua2eomignaCWONLQU5awRjBYZHlpdju/crE4s8C+er2d9FBxd0TxyrNSaRpZ4oK4wVE8tPD1ODWCroKguj8N3wsEjS4tLsAsJaTzJoX2w/LpoPRNg0RaOEOuoqKwWymtlOyNlE1rY4YmxtAAmGBho8loPHf2tfAfi/wV1xwvi4VazZU6nsFdbKSWqFJ4MNTLC5sEr+mYnDJCxxwCfh2B7IO8HJ9zNWXmt4MW/iXQULLddIpn22+W5knW2krow0vDCdzG5r2SMzv0vAOSCuo3tKOc7jDpjijZeVTl4uFTa7/AHeKlbc7jQkCulqax4bTUdM87wkgscZGkOJlYA5oa7q2v7Dm63IwcX7I+Od1Ax9lqo34+9xzEVbXjP8AdOa1m3pGtO9qby8cXtIccLRzicLrbWV9BRsoKmvqaKIyy2WvoC0w1EjBkiEtZEQ8Dpa5jg4t6mdQT0vsoubCqoG6quHNeINYSRiQxmtuMgbKSHdLqwP69n5JcIzuMgFdveRTT/Nlo3R+qNGc1lzmu9bZ7qyKw3WetjrH1lIYgXkTg+JI0OxgygPBJB7YHCPAH2xXBvWsNDY+OVirdD3t4ZFLcqdjqu1SSdi4luZoATg4LHhoJy/Aye/Gm9S6f1hZKPU2lL5QXiz3GIT0dfQ1DZ4J4ycBzHsJa4ZB3BO4Pog6R+1C51Nact9hsHDfhPVx0GrdWQzVdRdHRtkfbqBh6AYmOBb4kjy4B5B6RG/A6i1zeB9LezB5vOJtgo+IXErmdrbRqe4wCsbR1tXXV1TTmVueiaYyN6H4IDgwOA3GThcje145VeInFSh01xu4bWOrv0ul6KW2Xq20cZlqWUhkMsdTHGMmRrHOkDw0FwDmuwWteW6Ly1+2N0e6yWvSHMnpy5266UcLKWXUttjNVBUloDfGqIBiWN5AJd4YkyckNaDgBzlyK8OedPhBqzVvDzmS1VLqvSVLQ082nrxJc/f2un8RzXRxyy4qQ3oAyyRoDcN6cA79yltrQHEXQnFPTNPrLhxqy2ajstWSIq231DZo+oYyx2DljxkZY4BwOxAK3KgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIg87fbOcHLjq/g1pbi1Zbc+pk0Ncpqe4uiYS6OhrGsBldj962aGFvnjxSdhlbu9ldzOaQ4lcArJweud9o6bWmh4nW77nzTtbLW0Ac50E8LCcvaxhbG/GeksBOA9oXdS92Sz6ms1bp7UFsprjbLnTyUlZSVMQkiqIXtLXxvadnNIJBB8ivNfi77FjT1yv89/4HcWptN00kolhtF3o3VLKYk5PhVTHh4aP3ocxzthl57oO63NBzL6C5YOF911zqu7Uf3VbTSCyWh8w8e51hBEUbGAhxZ1Y63gYa3JO4APm77Hzg/qLXnG7U/MnqSnmkoLDDU0lNWPjAFTd6zBlLT5lkL5OoAbeOzcZwd76F9inW1d/juvGbj0+40bJWmals9A73ipjGNveZ3nwztj+xP27EL0h4Y8L9CcGtEWzh3w309T2Ww2mMsp6aLJJJOXPe9xLnvcSS5ziSSckoN2IiIOPdP8feDWquId84T2PiLZ6jWGnKgU1wsr5TFUsk6A8hjZA3xgA4ZdH1BpyCQQQMfmI07we1Hwd1PS8d6W1yaNhoZZ66WvDAKYNacSxOdu2YHHQWkO6sAbnB6s81XsqdGcfOIdz4t6H4j1+jdTXiVtVXRS0vvlHPUAAeKwB7JIXEAEkOcMjIaMlcMU/sc+Mep6yno+JfNIyqtETxI5kdLVVsmfRjJpWsaSMjqycZzg9kGx/YmN1GeOmvXUrphYBpMCuAP3v3w1kHu2fn0Crx8i5dlfbR0dVU8qunpqeB72Umu6GaZzRtGw0NewOPoOp7B9bgu0HLdyzcMOVvQY0Nw0t04FTI2ouVxq3h9XcKgNDfElcAAAB2Y0BrQTgZJJ3Jxk4RaK47cN71ws4g0L6qy3yERS+E7olhka4Ojmidg9L2Pa1wJBGRgggkEOtnsnta6d1ByZaY05bblBJcNJVtzobpTiQdcD5a6eqjLm9wHRztIPYkOAOQQO3tsulsvVEy5Wa5UtfSSlwZPTTNljcWuLXAOaSDhwIODsQQey8pbr7EvW1HfJ49JcwdtFmqGmMyVVomiqfCccmN7I5Cx4GB++AcRnA7L0X5auCFBy4cEdL8F7df571DpyKoabhNAIXVEk9TLUSO8MOcGDrmcA3qJAABJOSQ8peR//AO2p3z/w9q/9FUvaldJOBvs46vg5za3XmWk4tw3WlrK67V0FpbZTDIDXeJljpfGcMR+KQCGnq6QcNzgd20Hnv7ZfhBdta8DdNcT7NRvqXaBukormsaSYqGtbGx8u3kJYacH0Ds7AFbv9l3zOaE4ocv8ApzhHPfaWl1toikNtntU0rWTVVHGSYaiBpOZGCPpa/GS1zT1ABzSe5t0tdtvltqrLebfTV9vroX01VS1MbZIp4ngtex7HAhzSCQQRgg4K85eMfsZNEXy/VGpeBPFCs0YZJDPHabhTOrKeB57CGdr2yxtB7BwkI9cYAD0gqJ4aSCSpqZmRQwsMkkkjg1rGgElxJ2AAGSTstp6F4v8AC7ifcL3a+HOv7Fqap05LFDdW2qtZUilfIHFjXOYS3J6HjY7FjgdwQPNuD2OvGfU0zKHiNzUtntYcHOaykq69xI3wGTTMaDkDfJx3wcYPdjlJ5NeGnJ/p+82vQt0vF2uGpH00t2uNzkZ1TGASCJrGRta2NjfFlIHxOy85ccAAN+1vH7gza+KJ4K3XiLZ6DWvusNay0VkpgkljlLgwRueAyR56SfDY4vAwS0AgncmtbNo7UOlLraeINutVdpyeleLlDdWRupDABl5l8T4Q0AEknGMZyMLqxzkezd0DzYalj4iU2s7hpLV7KNlFJVMpxWUlXHHnw/EhLmkOGcdTHgY7tJGV1nPseOOt3DLFqPmnp5LCCGFnu1ZU9MbThpED5WsJAxhvWAOwPmg4f9ndS0NN7SOWm4P1E8uiKer1KyORryfEsYjnFIXk7kGT3M5O5OD3Xr3W8fuDNr4ongrdeItnoNa+6w1rLRWSmCSWOUuDBG54DJHnpJ8Nji8DBLQCCeP+U3kt4Uco1iq6bRvvV21BdmNZdb9Xge8VDGklsbGt+GKME56W5JIBcXEAjjznI9m7oHmw1LHxEptZ3DSWr2UbKKSqZTispKuOPPh+JCXNIcM46mPAx3aSMoO0+tbNo7UOlLraeINutVdpyeleLlDdWRupDABl5l8T4Q0AEknGMZyMLxg9ndS0NN7SOWm4P1E8uiKer1KyORryfEsYjnFIXk7kGT3M5O5OD3XMB9jxx1u4ZYtR809PJYQQws92rKnpjacNIgfK1hIGMN6wB2B813b5TeS3hRyjWKrptG+9XbUF2Y1l1v1eB7xUMaSWxsa34YowTnpbkkgFxcQCA7AoiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAtKn1RpqlmfT1WobZFLE7pfHJVxtc0juCCcg/JaqvMbXPsVo9Ya1v+rYuZaopmXu6VVxENRpIVMsYmldJ0vl99Z4jh1YLuhuSCcDOEHVzmXr6Gf2o9VcIa2nkpTr3Tz/HbIDH0gUeT1A4wMHJz5Fe5FPqbTlZOympNQWyeaQ4ZHHVxuc4+gAOSfqXl/8AuGI//Wi//on/AP71r2gPYtt0RrvTmszzL1FSLBdqS5+DT6S92lk8CZsgayX313huJbgO6XYODg4wg768d+NOk+X3hVqDixrOUe42SmL4qYSBktbUHaKmjz+/e8ho2OAS47AkeV/ITwU1HzucyupuafjrRx3aw2m4GqmhqIi6mrrmWj3eka0nBhp4gwlpJwGwNIIccd4Oebkw1xzht0vZaDjNDpTTdikkqKi2PszqsVFU/wCEVBcJmZLYy5rWkADqeer4jjm3gDwS0ly8cJ7Bwm0ZH1UVmp8TVTmBktdUuPVNUyYJ+J7yTjJDRho2aAA8d+J2ndX+zD53KLVWj4aqfSNTIa63RGQ4uNjmfiooHvIwZIyC0E5IcyGQjcL2t0LrbTXEnR1n17o25suFkv1HFXUNSzYPieARkd2uG4LTuCCDgghcR843KVpXm94aQaJvV2Niu9rqxXWe9MpPeH0ch+GVpj62dccjNi3rG4Y7u0BYPJPyx6z5UeGldw11JxVZrG3Pr3VtsiZbDSNtofkyxtJkeXNe/D8bBri4jJcSg7Erb+vNc6V4Z6Pu+vtc3dlrsNipn1lfWOjfIIYW9z0Rtc9x7ANa0kkgAEkLcC2xxK4eaY4saCvvDfWdLLU2XUNFJQ1kcUpjf0P82OG7XA4IO+4GQRsgu0JxF0JxQ09Bqzh3q61ajtFQAWVduqmzMBIz0u6Tlrh5tcA4HYgHZebntqtO8HaPTui9QwUtsp+JtbdDFI6ANbVVNobBJ1unDd3BswgDHPBx1SBpxkKO/exm1jpq8T1vBXmUqbZR1PwGOvo5YKhseSQ181M8CXG37xozk4C3Lwj9jfYaPVcOr+Ybi3V64McolltVHTyQR1JHbxqqSR0r2HbLWtYdsdRBwg7LezmbqJvJTwsGqXTOrTbKgxGU5PuZrJzSAfL3fwQPlheaHCnVNo4Ze12u2oNaVUdrof8AZJ1TTOmqXCNjPfjXQQOc47BpdURHJ2w7JIG69tqChpLXQ09tttJDS0lJEynp4IWBkcUbB0ta1o2AAAAA2AAwuk3OZ7MLSPMzrKfipozWI0dq2shYy5NloveaK5PY0MZI8Nc10UgYA0ub1Bwa3Lc5cQ7rVF1tdJXUlrq7nSw1teXilppJmtlqOhpc/oYTl2Ggk4BwNzsvPf22f+8LoT+F4/UqhZ/J/wCy81Dy98aLHxq11xip79WaZgqYLdbaGhkEZE1NLT5fLK/Ia1kz8Mazvg5ABB5155OUaq5weHNl0VQ67i0vUWS8C6snltxq2TjwZIjGQJGFu0nUHDP0cY3yAs9nB+BNws/8HVX67ULsouM+W/g67l/4JaU4Pu1AL47TVLJA64Cl92E5fNJKSI+p/SAZCAOo5Az54HJiDwv4M3+m9n37Qm5WriFTzUWm6SurbLUVIiJ6bTVkPpatrRkuYAKeRwGSAHgAkYPt/Yb/AGLVNnpNQaavFFdrXcIhPSVtFOyeCeM9nskYS1wPqCVwfzVclHB7m1tVKNcU9Va9Q2yMxW7UFt6W1cLCc+E8OBbNFnJ6HDIJcWlhcSekf7jlxr0vVVFJw+5oqaltkjusf7Vq6F7z5dccUrmkgeed99gg9KOI/GbhNwhp4KvifxG0/phlU5rKdtyr44ZJyTgdDCep4znJAOACTgAlarrzXOleGej7vr7XN3Za7DYqZ9ZX1jo3yCGFvc9EbXPcewDWtJJIABJC8+eGfsX9E0N6g1Dxs4yXnV8glbPNQ26jFHHMWkfBLPI+WSRpAwcCN2DgEYyu/wDxK4eaY4saCvvDfWdLLU2XUNFJQ1kcUpjf0P8ANjhu1wOCDvuBkEbILtCcRdCcUNPQas4d6utWo7RUAFlXbqpszASM9Luk5a4ebXAOB2IB2Xm57arTvB2j07ovUMFLbKfibW3QxSOgDW1VTaGwSdbpw3dwbMIAxzwcdUgacZCjv3sZtY6avE9bwV5lKm2UdT8Bjr6OWCobHkkNfNTPAlxt+8aM5OAty8I/Y32Gj1XDq/mG4t1euDHKJZbVR08kEdSR28aqkkdK9h2y1rWHbHUQcIOxvs+a67WXkZ4bXXXlW+B1HZquqfNUH+x29tVUPpnH/tBSiIj/ALUBcw8KONvCfjjp9mp+E+u7VqSgIBkNJKRNAT2bNC/EsLv+1ka0/Jbmm07ZJtPP0k62QMs8lEbcaKNvhxCmMfh+E0Nx0t6PhAGMDthea+uvYyS27UU2oOAnHuv07G97/BpLnTPMtOxx3Y2qge1zmgEjBZnAGSTkoOSPa/6d4OzctsmpdV0tsi15T19JT6VqCGNrZSZ2e8xAj43QiAyuIOWhwYdiRnN9jo3UY5RZjfHTGjdqy4mzh52FH4VOHBny94FSfrLlxNof2NFyuupae+8fuYCs1FSU72B9FbqaUzVEYOeg1U7yY29wQIycE4LTuvSLRWi9McOdJ2rQ2irPBarFZKZlHQ0cAPRFE0bDJySSckuJJJJJJJJQa6iIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiDAt96s13krIbRdqOtfb6h1LVtp6hspp5hjMcgaSWvGRlpwRkbLPXl9zH8hPN7pbjzqvmD5T+Ikr5dVXGW51Fvo7t9zK6N7z1Oid1ubBURdRcQHvGxALTjJ2X/sk+2pbR/scOm9QunJ93Fb+x20mQb4/soj8PHl1nyOc53Qbr9t7qTS8lr4XaQFRBLqKCouNx8IAGWCie2JmXHu0PkYAB++MR/uV269nTZLzp/kr4V0F+8T3mW1z1sfiDB92qKueenx8vBlix8sFdKOCHst+OnF/iQOLPOrqiZkUtQyqrLY+5iuudzLcYilmjc6OCHAAwx7nBoLQI9nD004j1urNF8LL3VcJdFRXzUFqtb47DZI5YqaOWZrQ2KPL3NY1jdiRkEtaQNyAg87Pa1c0tfdayg5POFk81ZcbnNTS6nbRZfLJI9zTSW0AZJc5xZK4DfeEAnLwut3NNyOcQ+S3R3C/jVpy+1b7jG2n+71XTEE2e/h7p4jG5uR4WMRtccgvgJJ++NaOzfIJyJ8Z6TjxduZXmvsE9LeaOeSutNPXVcFTPW3OdzjJWSiN7w0RgnoBwet7XDAYM+hnFrhfpTjRw41Bwv1tRips+oaN9LOMfFG7YslZns+N4Y9p8nNBQcf8nfMnZ+aXgfZuI1KYIL1EPufqGhjP9qXGMDxAAdwx4LZGbnDZACSQcc4Lyu5GOWbnQ5ROZertFXw7fdOHF+kfbbxcoLrRtpZaZj3e73BjDKZA9hyfDLesske3GSCPVFAUFVSU1dTy0dbTR1EE7DHLFKwPY9pG7SDkEEbEHZTryl1Ly/e0w5WOI2pr/wAu2pqrV+j7zd6u6spIamCpjcJ5XSu8WhqjlkpJw58OS4gHq3wA5Y58fZ38vdz4Paz4wcPtNUehdT6VtNXfHfcseBQVzKeN0r4X0w+9sc5ocGujDD1FueobLb3sVOJ2q9TcM9fcNL3WT1Vp0bX0FTaTK8u8Bta2oMsDCezA+mDw0bAyvPmuJtc6d9rdzY2V3C3WukpdP6arHs+6QmipLRTStDgR4z+ozSsBAJjjDgSAS04BHfzku5TbFyi8KjoulurbxfrrU/dC+XRsZjZUT9PSxkbSSWxxtGADuSXOIBdgB0r9ud/2E/8A+ZP/AO2LvzyjVVPWcqvB2WmmbIxug7DEXNOQHsoIWuH1hzSD8wVxf7QzlCuXNnwmoaHSFXS02sdKVUlfZxVO6YqpkjA2emc//wCRl4bG4OOR1RtBwCXDofwy0t7Xvg/pgcDOH2mdR22z08r4aUPhtc8VGHuLn+BWylzWMJcTtJhpJLekkkh7Caw/+KV7/wDB1T/6Ny8g/Yl/7/Wu/wCCB/XaderGjLFrem4IWLTOuK4XHWEWlKWgvFSZQ8VNzFG1k8nXsD1TB56sDOcroN7LDlK5hOX/AIuay1Lxg4dTadt1ZYDbKeaSvpJ/FqPeonlrRDK8kdLHHqx0nAwTkIPTReOvtsNC11v4z6E4jtpS2gvem3WjxWsGDU0lTJI7qP8AdFlXGBnybt2OPYpcQ80PLVojmn4V1nDTWTn0kgkFZarnCwOlt1a1rgyZoJAc3Di17CQHNcRkHDgGNwRtXA7i9wj0jxKsegNHVtNf7TT1bpGWalPTMWATRuwzAeyQPY4eTmkeS3z/ALEPCb8V2kv8yU3/AKi8pNK8uvtQeSa5V9r4IF2pNL1U5kMdrlpq+hqHHAEho6jEsMhAaHOYwdgC9wAW7n6n9tFxajfp6k09UaRppR0VFW2kttsLWnzEspMo7jeH4tttsoPUix6c09pikdQabsNutNNI/wAR0NDSxwMc8gAuLWAAnAAz32C1FzWuaWuaCCNwRlcAck/A/ixwF4P1OluNXER+sdU3W+VN6qas11RWCnEscLBCJpwHvwYnPJIA6pXYz3PVHmJ5buf7hdzCav478qOs6q62nV1Yy41Vqp66EOjeI2RmOajqz4M4aGAMe3Lg3YBuNw5+5m/ZzcufHWw3e627SVv0XrF8Ms9NfbQwUrDPjINTC3EUrCQOtxaH4yQ8Eknpz7FDidqxnEfXHBySsnm03UWE6jige4mOmrIqiCAlg7NMjKj4vXwWeiyNU3/2xXHOyVfDav0HW2K33mJ1NXTxUVFa+uneMPY6okeC1pBOQwhxGQM5we2/s/8Akbh5RNK3O7aou1Ld9danZE24z0gd7tRQM+JtLEXYL/jJc6QgdRDQAA0Eh24XX3j9yL8t3MVS1tRrDQFHbtQ1bXFmorOwUleyUggSPcwdM5Ge0zXjHpsR1+51eWbnJqOOzOZLlM1xUMrJbRTW6stNPcWUs4FOXlo8Oc+71MR6y7oednE4ac5XD9fxO9s7qyhk0fDw+r7bU1bfdXXCntNvpJBt0ucKh7hFGTuesFuM5aRsUHHfs2L3q7gbz73bl8ob6642W4Vl805dfCJFPUSW5lRJFVhmSA7qpi0HOzZnDO69p10P9np7Pa98uV6rOMnGO60tw17cqaSlpqOmlM8dsjkdmV75icSzPAAJAw0F4Bd1EjvggIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICLrZzI8/HAzlY1zQ8PuJ1NqeS6XC0xXmE2y3xzxCnkmmiaC50rSHdUD8jHbG++3FP7shyjf3hr7/ADND/ToO9KLot+7Ico394a+/zND/AE6575ZebrhVzY0F/uPC6C+xw6cmp4az7q0bICXTNeWdAbI/IxG7OceSDm5ERAREQEREBERARcE8zvOPwj5Sv2Nf7KkF/l/ZV757h9yqNlRj3bwfF6+qRnT/AGxHjGc79sb7y4D8cNGcxXDWg4q8P47kyy3GaeCFtwgbDOHQyOjflrXOAHU0437IOREREBEXU3XPtLuXTh/xer+Cd+o9XnUNvuzLNK6ntkT6Y1D3NaCHmYEty8b4B77IO2SIiAiIgIupHGH2nHLhwO4lX3hXrSj1i69afmZBVmitcUsBc+Nsg6HmZpI6XjuBvldj+GfEXSnFvQNi4k6HuHvtj1DSMrKOUt6XdJyCx436XscHMc3OzmkeSDdCLh7mV5ouGnKrpa16w4nQ3mSgu9w+5tOLXStnkE3hvky4Oe0AdLDvk74Tlq5ouGnNVpa6aw4Yw3mOgtFw+5tQLpStgkM3hsky0Ne4EdLxvkb5QcwoiICIiAiIgIiICIiAiIgIiICItscSdf2LhVoDUHEjU7ap1p03b5rlWiljD5jDE0ud0NJAJwDgEj60G50XXTlr56+CXNXqm6aP4YU2pYq+0W/7pVBulBHBGYfEbH8JbK8k9TxtgbZ3XYtAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREBERAREQEREHVDmt9nfwz5teIlu4kaz1vqez11tssVkjgthp/CdFHPPMHnxI3HqJqHA4OMAbd8+YftC+TXQ3J/etFWzRWqb7emampa2eoddfBzEYHxNaGeGxowfEOc57DC9615Ke3F/+NnCX/wdd/8A0tMg2XyJ+zh4X81XBKbidq/XWqbRXx3yqtYp7b7v4JjijicHffI3HJMhB3xsF2S5PdZ8lXKXxP1Py66C4t6tvWrdR6lisNVTXa0SBkVwpJJacxsljhazp63PBcSQcAg4W5PY3/gjVX8L7j/6GmXlTzFU19rucfilb9LmoF3reJF+pKIU7yyR00tzmja1pBGCS4DOfNB7J1ntLOV88YrLwS03er1qa9Xu+0uno620ULJLfFVzzCFpdPJIzrjD3AF8QkGDkdQW5OP3Pry1ct94dpfXmtJKzULAHS2ey05rKqnBAI8bBDIiQ4ENe9riCCARuul+uvZ1cMeTPl/ruZS96+1PduJGgRb7zbpaN0ENsiu4qoW0rfAdDI+SEVD4w8ueC9gOPDJyOoPKFZeWziFxXveq+c3iTNSWaGM1vu9RLVma+V80hLjLPA10gaB1ueQ5j3OczDsBwIeqHC32rfKPxNv1Jp2e+X3R1VWuEcMmpqGOnpy8kgNfPFLLHHnGep7mt3GTnZdheM3HThtwC0G7iVxLvMtFYG1ENN7zT0slSS+XPRhsYJIODuBheNvPRpHkB/Yta9W8omtaKK/RV4p7pYYH3B8NRTPY4+8MNU09D2PawFoeGlrycZbvrFVxwv3FT2V960dqq4zV9w4f60tdrp55pOuR1uka6Sna4nf4CJYxnYMjYAdsAPRum9pHyiVPD258S4+JMrLVbKxtv8KW21EdXVVLmdYjggc0Pk+HBLgOhuR1OaCCti6Q9rzyh6pv0VkrqnV+mo5pvBbX3i0sFLuQA5zoJZXtaSe7mgDcnAGV0O9mPycaB5ptU6uvfFdtbVaZ0dDSMbbqaofTitqqoy9PVKzDgxjIHEhha4l8e+AQYPaecpHDnld1/pOt4UUVZb9Oaxoqp7bdUVMlS2lqaZ8YkEcshLywtniOHOcQc74IAD3EN5s7bOdQm7UYtQpvfTXGdvu4p+nr8bxM9PR0/F1ZxjfON1021r7XTlC0hqB9hoK/VeqGQy+FJcLLamOpQc4JD55YnPaMd2NcDkEZG66a8buYHV1r9lnwJ0BFdZo6nXUt1t9ZMyUhz7Vaa6WFlP3z0nNIDvjEZbjDsLensy+QTg5xl4WVPG3jbYptRNrrjPQ2e1uqpqemjigw187/AAnNdI8ydbQCSwBmSCT8IbQ9qvzDcJeZDSXBbWXCXVUd2o4JdRwVkD43Q1NFKRbSI5onAOaSM4OC12CWkgEruD7NHWelOHvIDYtZ641DQ2Sx2qsu81XXVswiihZ79KBknuSSAGjJcSAASQF0L9pxycaH5Xda6a1DwrgqKPSesoalrLdNUPn9wq6cx9bGSSEvMb2ysIDi4gtfvjAGbr+l1tU+ya4XSWGmqZdP0+t7lLfXQBx8P79VNpzL07CLrcQS4Y6zEMgluQ7tan9sXyj2C4uorXRa81HExzm+92uzQshcAQAQKqeF+D3Hw523xsuZeX3ns5b+ZW6HTfDzWMtNqHpfIyy3enNJVysaMl0QJLJcDJIY9zgASQAMryk5HuL3ItojTN30vzS8IfuteK+udJTagmoDcII6R0bGiExB3XC5jg9wfGxxcH7kFoB5o4Sck/BLiRx4i4p8ofN9pqjgtF6ivlo08KGd1xoYWObIYiyaVkz4xhzcuYQWnDid8h67LzK4xWv2YsvNLeZ+Ieo9ZxcSjqaF1bDTx1hphcfEj6AC2Is6erpzg477r01Xz980n/2xTVf/ABgUv/pokHr7xp58+Wnl91zNw64n6vr7ffYKaGqkghtFTUNEcoJYeuNhbkgds5C2nxk9pzyrcGa+GzXC/wB51JdX08VTLRafoGzvpmysa9jZXyyRxtf0uBMfWXN7OAOAfNv2u34Zd1/8A2v/ANEV295X/ZacuOreXPS+qeKtpvN21XrOyU95luEV0npnW73qNssTIYmkMLmMe0Eysky/rOMYADsry489fLzzQ3SfTnDrUNbSagghNQbNeKT3WqkhH0nx4c6OQDO4Y8uAGSAN12FXzs8DILtwD56dJafZcJBU6T4kRadq5o24dJC2v9zqQAD+/iMgxnHxYX0TIPnq9o9+GzxT/wDCNL+pU67E+yL5s/2Ca0l5bNbXLosWraj3jTssh2pbqcB0APk2doGMnAkY0AZkJXXb2j34bPFP/wAI0v6lTrkn2jPKzWcA9cad45cPqeSh0vrVkFYH0mYxar02Nsj2NLcdAkIMzCDkESgABoyHbf22f+8LoT+F4/UqhPYmf7wuu/4Xn9Sp11v5v+aik5puRThvqC4VUI1hp/V8dr1NTNAB95FBUdFSG+TJmgvGNg4SNGegrX+STjPdOAXs6uOfEnT84hvVJqJtFapfD6/Cq6mnpYI5cHbLDIZB1bEswQc4Id9ePntA+WTl1vculNZ6ynueoqfHvFnsVN75UUxPlKctiidgg9D3h2CDjBBO0eE3tUOUjitfqXTTtR3nSFdXOEdP+yahZTQvkJwGGeKSWKM/N72t8s5IC8sOS/TvKvrPiDqDVXOVxFdR2uiZHPTUFTLWeJeqyd7zJJLPA0ydLAzLh1Nc50rDkgOB3dz2aR5D4bRZdXcoGtaT7ourTS3iwQPrpInQuY5zKqM1TcsLXN6HND8HxGENHS4kPZzjhx64acu+io+IXFO8T2+yS1sVvbPBSSVJM0jXOYOiME4IY45xjZaZwB5m+EHMzabte+EN+qrpS2WoZS1jqigmpSyR7S5oAlaC4EA7jK8oNZ8cb5xh9lZRWfVVxlrrvoTiDRWI1E8xkmmo/dZpaZzyTn4WvdEM+UI3JyuxXsQf97Hib/4eov1dyDuJzAc3fAzljrLLb+MGpKy1z3+KeagEFtnqvEZEWB5JiaQ3Be3Y4znbstN1Vzt8vGi+D+l+OuoNVV0Gj9Y1D6S0VbbVUPkmkb4mQYmtL2D7y/cgDYeoXQ724v8A8bOEv/g67/8ApaZbA5lf/tVnLl/4eqf/AK/Qeit29ofyp2bhPRcY6vX85st0q6ihttM23TCurp4MeK2Knc0O6W9TcyO6WAkAuBIB3Hwl5xeCnFrgvcuPtPd6rS+j7TXz2+qq9SNhpHMkiDDnDJHgh3iMDACXOJA6c7Lyf5AeRB3OJT3TU/EHWV3tOh9KTC1U7LcWmpqKl48Z8UL5WvjiYwSh7j0OJMowNyRt/wBoDp2ycA9aUvKVw0vl/l0RpQt1BNTXSqjlfLda6FjnyExxxhzWwthawOBLS6XBw7CD0NvftiOUS03X7n0MOurzB1lvv1DZY2wYH77E80cuD/3mfUBdgOAHNxwF5mqep/2J9bR1txoYxNV2mrhfTV0DCQOsxPALmAkAvYXNBIBOSAet3Lh7LPlpp+CFgn4uaMn1Nq2/2uCuuVbJcqqn90kmYHiGBkT2BojyG9TgXOIJJwQwecnG3SOpOQTnIqKXhzf6wu0nX012slVI4NknoZo2yeBMQMPBa98Em2HAOOADgB7hcduY/g3y3ach1Lxe1hBZ4q1zo6Glax09XWvaB1NhhYC9wHU3LsBreodRGRnq432zHKc66e4O0/xHbBnHvptFJ4GMZzgVRk+X0M5+W66Se1mu2pdT8w2n9ZVsFaNM33RtsrNNySjMLqZ7XSStYRsXCSRxcM5HUwnYtW/uF3En2WvEbgVQ8MeIugW8ONYutDaKe/y22oqZGXERhvvbKuDre4GQeJ0yhrAMtIx3D1K4Kcf+EnMNph+ruEWsaW+0MD2xVTGtfFPSSluQyaKQB8ZIzgkYODgkDK29zA83XAXllgp28V9aMo7jWx+NSWmkhdU108eSOsRMB6WZBHW8taS0gEkYXUXkN5W7Ny23LUXMVovmf0zxD0YzTtZFd6SzUuGubGBO1z3eM4RyMMR2e0OAc8bZK6G8GNJ6p5/ecmlpuIV8qQ/Vtxnut8qY35fT2+BhkMEJIIbiNjII9iG5YSCAQg9LtO+2H5RL3ePuXcItcWGn8RsYuFxs0bqfBOC/FPNLKAO5yzOOwJ2XYjifq/gnxH5a9Uau1Bqhl14Y3fT1VLcbnaJTKX0BYRKYzGC4PADgWgdTXAggEELq9zIeyc4Fag4T1NPy+6Ofp3XNsjY63TPvFRJDcCHfHHU+8SPaMguIe0NIIaMlo6VxNwo5bOZvlh5SOY3SHF6G2R6Su+jquvtsVLc21Jp69sTmy9LW46RJGW9WxBMTMY3yHMfs+7fyFUfErUb+U6+6ordQusZbcmXZlSI20XvEW7fFjaOrxOjsScZXYXmF5x+AHLG2Gn4payEV3qohNTWWghNVXzRkkB/ht2Y0lrgHSFjSQQCSMLy09k5rej4Z6x4y8Rq+HxqfS3DauvMsWcGRtPNFKWA/PowPmQuDeDVdw14/8yVXrXm84nPstiuctTer1WnxjJXTdTeikiMbXujaS4AYADYoy1pBDSA9VNGe115QNW3tlnuNfqzS7JZjCytvVoaKZ24AcXU8spYwk93gAAEu6Ruu0+teK2hNAcNK/i9fr7E/SduoW3OS40QNWySmd09MkfhdXiNIcCC3IIII2XkFzkaM9mfW8JJ79yva4orXrazywOhtlPLdJ47tAXtjfG73oODHtYTIHgjJa4O6i4EWcrfGy96j5AeYjgffq6apg0rZYrtZTI7q8GmqJg2eEHyY2RrHgesz+2yD0W0z7RzlE1VpnUer6Licaa16XZA6vkrrbU07nOmMgijhY5gdNI7wn4YwE4aSQACRxtavbD8odyv/ANxqpmuLZSmVsYutXZYzS4Pd+IpnzdI88xZ9AV5x+zr5YtM803G+q0nr2rrGaWsNqfe7jS0lQYZK1zJGRRRdQBLQXTElwwekOALS4Ecx+1C5JODvLdp7SHEPgzbKuy0V3uEtouFsmrpauLxPCMsUsbpnOkaSGShwLyDlmAMHIew2mNT6f1pp+3ar0peKW62e607KqiraWQSRTxPGWua4dwR+fIOMFdceN/tJOVbgVf6nSd91ZXahvlC50dXQ6dpBVupnjux8rnMhD87FoeSD9IBdIeBHMprDhR7J/WNxsVwqKe702s6rRliqmPPVRMq4aeplkYRuxzWz1RYQch5aQc7LYvsweTDQPM3f9U634u0s9x0zpV1PSxWuKqfAK6smDnEyvjIkDGNYDgFvUZBvhrgQ7/cKfan8ovFO80tgfqi76Prq14ip26noW00T3kgBpnifLDHnPd72jbGckA9tayuobfQz3SvrYKajponVE9RNIGRRRNBc57nk4a0AEkk4AGSvL/nE9kjHdrvaL/yi6fpLbHUeNHebNX3d4giI6THNA+YucMkvDmFxAw3pAGQuLOdDiBzCcD+UjhXyk8UK4U19qRWyXmppK/x/e7PTStbQUzpAcOaOshzdsClhznJQdy9f+1s5QdD3may2+7al1a6nl8KSpsFrbJTZBwS2SeWIPaDn4mZBxkEjBPLfLzzq8vXM3NLbeGWsnfdyCMzS2S5wGlrmxA7vax2WytG2TG5wbkdWMheZvJ9on2Zdu4TUl85nNeUV31vefGdVW2okucMVqi63NZEz3VrQ6QsAeXlxILgG46ST14401vDbgTzJQ6x5Q+JUt0sFrnprzYa9olElDMD8dM8yNa6Roc1wPUMOjeGu6skkPePjnzGcHuXHTkWp+Lusaey09U8x0VOI3zVVY8Yy2GFgL34yMnHS3I6iAQurLfbM8pxugt507xHEHVj342ik8AbZzj3rxcZ2+hnPljddFfagaw1RxL4raB4o1tPVR6a1boC0XXT4cSYGsmYZaiNh7dbZZCHg/EMszt0rk3g9xM9ltr3gXbuGHE7h7FoHWJs8dDV3+otlRUPdchCGGsjq6cuf0mT750SBjBu0jGCQ9SOB/MHwj5i9LSav4R6ugvVHTyCCri8N8NRSSkZDJYngPYTvg46XYJaSBlF0d5GuSi88Lrxc+K3Anm50nq23Xe2G1VBtdu8SIOMscrTIBM/pe3oIDXNDgHntkgkHpSiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgIiICIiAiIgLyU9uL/8AGzhL/wCDrv8A+lpl61rz99qBwV0txY1Bw/n1FX3WndbaOvZF7lLGzqD5IM9XWx2fojthBrPsb/wRqr+F9x/9DTLzomhiqPaiyU88TJY5ePpY9j2hzXNOosEEHYgjYherPs5OHll4Z8vdRp2w1VbPTHUNZU9VW9j39TooMjLWtGPhHln5rp/+1o0J+3p/2Qvutfvuj/stfdnwvHh8Hxvux4vTjwuro6tsdWceed0Hevnu4a3zi3ykcSdDabppam6VFtirqWnibmSd9HUxVfhNHm5/u5aANyXABeNfInYOU/WPEu46P5rXiit1zo2fcS4zXSWgpaera/4o5pWPaGh7Ds556QWYJBIz9CC8n/alcm/BPh9Zp+Omh7XXWO9XqqldW0FHMxtukmyC6YQlhLHuLiSGOa3O/SCSSFeY3hX7KHl901T3SmtDtc3itmYynsuntbTVExjJPVNI9krmRMABALsFxwADgkbJ4oWXgrP7NzU3EfghwlvmhLNqfW1sg8K7Xh9e+uFIZGCdhcSGsD5JY8DcmN2ewXD3s9+WHhtzMcTm2DiVNefudSCad0FvqmwCcRsa4Me4sc7pJJB6S047EHdepXPHwa0NeOVOLhbQ0DrFp2119vZR0tqEcIgZEXBjGhzXAD12yTvnKDrb7Dr/AOKfFr/wjaP/AEVStJ9ud/2E/wD+ZP8A+2LnH2YHCTTnCfT3ECDTtdcqhtyrKB8vvssby0sZPjp6GNx9I5zlYPtRuDmmOLR4ZDUdddKf7l/dnwfcpY2dXie5dXV1sdn+xjGMdz32wHSbjTw5ut/9l5y7cTKCldNS6Ru+pKGtczJMMVddajpkI7dPiUrGEnzkYB3K7Neya5sOE9l4KVHBDiDray6YvWn7lU1du+61bHSRV1HOfFPhySFrXSMkMuWZJ6S0jIz09oOUfhBoePkz05wgvFs+7umamnutLU0tzDZPeIpbhUvcH9LWju84IAIwCDkZXjhz1cAdEcuPHWv4fcP6m6y2lsbaiMXGdk0kfXg9Ac1jctGcDIJwBkk7oOw3tfOZLhxxe1dorhzw01NQahp9IR1tXc7lbp2z0pqanwmsgZK0lryxsJLi0kAyAZy1wG/uAnNlZ+Vb2fXDOq1Twjdrmxauu+obZVxOq44oInMqnObHKySKRsgkaZNjjaM7HO2y7lyNcE6blT4ca0gdfWX3VlZLXXGv95hMhDWSNbCzMRDIhguwBkk5cThuO6HLTyxcJtZ8lNt4F6ztVTe9NVFXXzg1UoFTBP73I5s0UkbW9D2lxwQO2QcgkEOCuEfLz7O3no4eTa507o9nC/U3jzxXK02fUIjqKAte7okEEgMHhuZ0vDmQtbu5uctJXnRxW00zl05j7pp3gxxKdqGTSF3hNl1Da3APknDGP6WmMkF8cjnRO6SQXMdgYOFkc2HBzS/AvjHeeH2kK251VtoJZGRSXGWOSYgSOaMuYxgOwHku9PstuTjgvra3UfHnWFDcbxfbFUQTUFFVTRut8U5Bc2YxBgc97C0FvU8tB3xkAgPU60y1s9qo5rlF4VXJTxunYBgMkLQXDGT2OR3XgPzSf/bFNV/8YFL/AOmiX0CLyV46cs2g7/zjXzWdZdr8ytqtWU1a+OKeERB/iRnABiJxkeZz80HCXtdvwy7r/wCAbX/6Ir2Y5ff94Xht/BCz/qUS88vaEcumieI/MbcNU3y6XyCrktFDGWUk8LY8MYQNnROP516O8IbbBZ+FOjLPTPkdDQaet1NG6QguLWU0bQTgAZwPIAfJB4M6v/8AtkN7/wCO+p/l5y+hVeQ2oOWTQdVzm3PW8l3v/v8AJxMmuxjFRD4PjG6ul6QPCz09XlnOPPzXryg+er2j34bPFP8A8I0v6lTr294mcFtJ8wXL5Lwn1lADRXuy07IqgMBko6lsbXQ1Eeez2PAcB2IBachxB83+czlj0Frfma11qm63e/xVdwrKd8rKeohbGCKWJo6Q6Ins0dyV6zadhZT6ftdOwnpjo4WDJ3wGDH6EHzM8U+HWseDWvdQ8KNa08lHc7FXmnq4Q4+HK5gPhTN8nNcx/Ux2Poybdyu6PLBw3vfFH2YnHuwacgkqLlR6lgvMMEYBMraSKknlaB3JMUcmANyQAM5we0XtMeV/hjxI1JpDiHdI7hb75UQ1Ftq6i3SRR+9wxdDovFDo3dTmdbwHbHpODkBuOQfZm8LdP8KeFGp7Rp2suFRDXahNTIa2Rj3B3u0TcDoY0Yw0dwUHmByD6Z5RNc67vmjOa6RlDFcKWCTT9yqbvLb6SKdj3CWGWVj2tBka9haXkNHhEZBcAeznMlw29lFy+WSiqqKwTa9vNfM1kdo07rWaeSOHDi6aWRsrmRtBAABPU4uGAQCRge1J5OuCvCuhPGPh9bK6yXG+VTzV22mmYLcJPhLpGRFnVGXFxJDXhuezRvnhL2c/Ktwv5mNfz0fE595lobX4k/ulDVNgjqejwz0SnoL+k9Zz0Oae2CEHLHGbRXC2q9mhX8SeCPCO+aFsF81zQ18tNdrrJXy1MEDZaVtSHOJ6GmWUsA2B6c5ORnVPY+cxPCLhdQa/4f8TNc2bSlVdaqjuduqbxWx0lNUtYx8csYmkIY17SYyGkguDjgHpOPUPVfB7hzrPhVW8Frxpikbo6utrbUbZTMEMUFO0ARiINx4ZYWtcwj6LmtI7L56eZLhJpvg9x9v3CzTFbcqi02y4mkhlrpWPqCzxC3LnMY1pOAOzQg7C+1h5huHXHLjRp608MtSwX+1aPtL6We40kolo5auaXreIHjIkAY2IGRpIJyBnpydb5lf8A7VZy5f8Ah6p/+v1vjjnyFcCdG8I+E0FgF+grLnT3K53G4mqhdVVsskdGWskcYunw4wCGNa1oHU8nLnFx5E41cA9HX3kR4OcPKu5Xhlusd3nkp5Y5ohO4k1eesmMtI+M9mjyQcg+xrp4IOUu4SRRhrqjWVwklI7ucKekaCfn0taPsC6Ve1/4aXbSnNVJr2Wge2164tFHU09UG/A+opom00sef7prY4SR6SNXo37OTh5ZeGfL3UadsNVWz0x1DWVPVVvY9/U6KDIy1rRj4R5Z+a5f478v/AAv5j9Ey6A4q2D7oW8vE9NPE/wAOpop8ECaCQDLHgEjzBBIcCNkHF/LXzpcAuI/AzTeqbzxY0tZLpQWiCG+0N1utPRz0dVFGGzEskc0lhc1zmOAwWkYwcgeRfOdxJp+bfnFutdwlo5LtTXarodOae8ONzX3Asa2FsgBwQJJC4tyAQwtyAQVxlzIcMrDwf4v3zh/pqrr6m3WyXohlrpGPmcMkfEWMa09vJoXrh7OPk44J8PtF2fjtR2qtuusrjFM2GuukzJRb25LHCmY1jWsLm5BeQ5+CQHAEghx1zTc5PB7gfdKflF408r0ev7Vo202qmpq+qrYxFUR+4xYnp2yQl8bhl8fU14OWOHV3VnFv2fPIHrfhRcOLXCTihTaKoxbZLjR18OoBW2wERlwbNHUOfKNxgsa9rwcjGR0rspzvco/BvmG0LV6r1zbK2m1Dpm3yyUF3tk7YakRty7wHlzXskjLiThzSQSektyc+C0mnqJmvBpcSze6/dNtF15HidBk6c5xjOPl9iDsN7N+PWd/5jm8MdN3Oop7ZrvTV+sl8Y1zvANK+2VBZJKADjomEJacA5IbkBxzhck3Eyl5VecKy3HirTPs9Lbqqt05f/HYeu3GRroXPeACcRyhhdgH4Q7Gdl6x8ifKFwa4B6Gotf6Nt1fV6m1RbYzW3W5zsmnZE4hxgi6WMbHH1AEgDqOB1E4GOOPaTcnnBTW+ibzx6qLTWWrWVvgYJqy2TMibcA0Yb7yxzHNeQMDrHS/AALiAAA5L5oufbg3wV4O3HWOgeJGjNV6qq4QzT1ro7hHcGVM7sYklZTyhwha0lzndTcgAA5IC6s6B5y+PnN5yu8xtXxG0jpG2ab03oipjhqrLQ1UL5a6WN5EZdNUyhwEbHEhrcguYSQCAfP/lV4R6b448bLDw31ZW3Kktl0kDJpbdJHHO0dbW/C6Rj2jZx7tK9yNacvXC/hdyf604J6Asj7NpyTT9fDN4MnVUzSSRESTySvBL5XYGXEEAAAANa1oDy39mFoSfihX8d+HNH0+9al4W3G10xcQA2eaSNkTjkgbPLTuQNtzhcL8qdk4Dxce4dG82VtqqPS87Kq2VUklTNR/cy4tI6HzujIe1ocx8bgchpkBdgNJHov7M7gTpHhTxW1ReNO3G71E1bp400grZYntDfeYnZHRG05y0dyQsn2nfJzwTuOjb1zE0VrrrPq9p/24+3Tsjpri8RPIknicxwL/vYBcwsLt+rqOCA2Xx04IeyX4I6Fm1dI+l1TXPAFvs1g1vNV1Va8kdvDmcI2AHJe/DcDYkkA7S4f0fLzqbkt5i+JXAXghqLQ1IyyRWOprbrfpK9tdIJI5nRxtcSGmMGIuO2fFaBnfHUjkv4FaO5heNNq4fa4rLrTWyrnjbK62zRxTEFr3EBz2PA+gB27E/Ij2j4wcBOGui+TnVXBDRdmNi0xFZHU4jo3ATEl7XOlc94d1yOcMue4EkkoOg3sRP99niT/B2m/WVzb7bP/eF0J/C8fqVQpPZjcDtJ8Kdd6yumnbhdqiWutFPBIK2aN7Q0T526I2nOfUlcm+0y4W6f4rcKNMWjUVZcKeGh1CKmM0UjGOLvdpW4PWxwxhx7AIOg3Crh5fdf+yY4kS6fpn1M+l+J7tQTQxty99NFb6GOZwHoyOZ0h/7WM/Ut++yA5muHHDCfWXCLiTqug042/wBRT3az1lxqWwUsk7GGOaF0jyGMeWiItyR1dLhnIAPcf2bPC/TvDTgNqDS1mmrKyirdVVNVMK90cjnOfRUjHN+FjQW4jGxB7n6l0V9p/wAoHBrgFXUWtuFtBcLQdQ1HXNam1DXUFOXF3V4DCzrjGR9HrLR2AAwEHaHng9p/a+BtfY9Lcu120Rri+zulnvUs5kr6KigAHhRiSmnjBmc4kkdTukN3ALgunPPTceNXHngTwc5teKGlbPbH3wXKzPZZ6OeCGCmE3XRPeJpJH5mDap7T1YLQ0gfFvT2Z/Kjwm5jNbVlfxSp7nX0tgaKxluhqWxUtUWvaAycdHW5m+4a9ufXGQfZ/WfDLQPEHQFZwv1fpW33DStfStoZbU6IMgELMeG1gZjw+gtYWFuCwtaW4ICDyg5H+C3s5eNHByhdxguNDaOIdrfNDeoLnquW2iob4rjFPC10rGOYYyxp6c9LgcgZBOhcZaj2XHDfiIOHmg+DGqOJT2iON9wsGrag0rqp7y0QRPMhE5Hw/EzLSXAAkg44J56OX3Q/LhxtrtA6Aq7vNamsM0YuVQyaSPLWuDQ5rGZaOogZBOAMknJPer2W/JxwVrdMW7mLvtur7zqmiqzHQRV8sclFRSNjjeJ44gwEygvOHPc7pw0tAcA5BTmS5weCXL2y38ovEjlEdqPTemrJazQU9zvENQ2KKSkjcAxz4nuD4nmSHxA/JMZIIBwr+Jvs/PZ/8R+FNXxe4TcU4NC26S3OuVPcYr4Ku2xAs6gJ4KhzpWkHYxtex7XAtxkdK7L873KXwe5iNAVWptdW2sp7/AKWoZpbdeLZKyGqbEMv93eXMe2SIuyelzSWkuLS0udnwOq7FSU+sH6fZJMadtyFGHEjr6OvpznGM4+X2INz8EuKHGzhbqGtunA6+3+3XasojBWfchj5HyU4ex3xsaDsHBu5G2cZ3OS9puQrk/wCCnBjh1RcQ9O2eruepNV0A99uN3lZO+OHrz4ETWsaxkfVG1x+EuJxlxAaAQf/Z)
“Figure 1. Neuroarchitectural Changes Induced by Repeated or Acute Stress in Rodents. (A) Repeated restraint stress (7 days) induces a reduction in the number and length of apical dendrites of pyramidal neurons (layer V) in the medial prefrontal cortex (PFC) of rats. (B) Magnified segment of dendrite from the same stressed rats, showing that repeated stress significantly decreases the number of spine synapses in medial PFC. (C) Reconstructions of representative infralimbic pyramidal neurons in mice exposed to zero (0), one (1×), or three (3×) unpredictable sessions of 10 min of forced swim stress. Apical dendritic branch length was significantly reduced after one or three stress episodes relative to controls. Adapted, with permission, from [24] (B) and [23] (C).”
![[[@Musazzi2017-lr]](https://doi.org/10.1016/j.tins.2017.07.002)](data:image/jpeg;base64,/9j/4AAQSkZJRgABAQEAcQBxAAD/2wBDAAMCAgICAgMCAgIDAwMDBAYEBAQEBAgGBgUGCQgKCgkICQkKDA8MCgsOCwkJDRENDg8QEBEQCgwSExIQEw8QEBD/2wBDAQMDAwQDBAgEBAgQCwkLEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBAQEBD/wAARCAG3AjADASIAAhEBAxEB/8QAHgABAAICAwEBAQAAAAAAAAAAAAcIBQYBAwQJAgr/xABiEAAABgEDAgIEBgsJDAYGCwAAAQIDBAUGBxESCCETMRQiQVEVGSMyV2EJFhg4cXWBlZa11BckQlJUYpGToTlVcnZ3orGys8LS0zNDY4KS0SUmNFN0wSc1NjdERVZYZYSj/8QAGwEBAAIDAQEAAAAAAAAAAAAAAAMEAQIFBgf/xAA/EQACAQIDAwoCBwgCAwEAAAAAAQIDEQQSIRMxQQUUUWFxgZGhsdEiMhUjM1JTcsEGQmKSotLh8IKyJDTC8f/aAAwDAQACEQMRAD8A+qYAAADWNStM8G1fw2dp9qRj7V3j1mbRyoLrjjaXTbcS4jdTakqLZaEn2P2DZwAFQeizTvDNJ9fepXT7T2iapsfqLrHEwoLTi1pZJdWTi9lLUpR7rWo+5n5i3wrJ04ffX9U/48xj9TIFmwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAGqZhqvpjp9KYg53qFjuPSZTZusM2dkzGW6gj2NSUrURmW/bf3jaxrmU6b6eZw+xJzXBMev3oqTQw5aVjEpTSTPc0pNxJmkjP2ENZ5rfDvJaOyz/XXy9W/zNY+6X6d/pxwX8/xf+MRFV/ZF9B7LVFeny1zolZ6QqI1kshTSa9bidy5GfLklozLYnDLbuRmRF3E0fc/aDfQpgf6OQ/8AliJqz7Hz0+VuqDmpBVcyTGN9UpvHpBtLq23VGZ/9Fw3NsjPcmzUaS8tjLYhUqc7uslt+p2cI+RbT5wql7abt/d+uhJ33S/Tv9OOC/n+L/wAY2XDdStPNRClqwLOKLIigmgpXwXYNSfA578OfhqPjvxVtv58T9wwv3P2g30KYH+jkP/ljYcVwHBcFRIbwnDKLH0zDSchNXXMxSeNO/E1+GkuW257b+W5ieO2v8Vrd5zq3Msj2ObN12t5Fb9MdY+pzqFybI820rk6f0enWM5Y/jkWvuYMp+fdMxXEolSDfbcJMfcjV4RE2ruWyiMi3PTk9SnVVn+k+a9Uul56ewdP8UkWr1ZjtnXyXp9xW1y1lIfclIeSlhxRNOmhCWzIuJEZn5nIeHdLusekmZ5A1o3rxX0en2U5I5k0ukn4yibMgvPLSuSzEkG6lJIc47FzQrgR7kRnuatYc6JtW6bEsq0OwHqCh0mkWXTpsiRWu42T9tAiTFmqVCjSvGJHhr5LIlLbNSSWfmfnMUjQ9W+vXUCDlbTuD5npviFFJ0rrdQ6yJlsJ5+TbS5XNRVzKmpDRm4pJJIiQhR7kZ7beWz59rz1gwpWjNlTP4DjUfWV+BAaprjHpj8yjlOQEvvE+spLfiETiXCJPBCiI0kruRjd8l6EcEyfKcgkTpkVWMWemETTiuq3IJOPVvozilMzEPKV89BcOOySPknfcZ2V00ZjfU+g7WXalsWdxo9aM2E6w+DlEd0TcZbCexuGbazSaTUozVuZKPbuAJxxlnIo+O1rGXToM27bitJsZMCOpiM9IJJeItptSlKQg1bmSTUoyL2mMmAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKydOH31/VP+PMY/UyBZsVk6cPvr+qf8eYx+pkCzYAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA1LNdUcO0+fixsnmyWHJiFLaJqG68RkkyI9zQkyLz8hFXr0sNB1K0lGK4t2RpOpGnHNN2XWbaAi37pXSX+/E/81Sf+AaC11iVis1OscxpSceOQbBTSWo5BJ32J02uPlv34/O2+vsOTX/aTkrD5c9ePxO2jv42vZdbKlTlLCU7XqLXo19CyACLvulNJf78WH5qk/8AANnwnUzEdQjlpxeZJfOCSDe8WI6zty347GtJEfzT8hco8qYHETVOjWjKT4KSbJ4YqhUllhNN9qNqAVpw/VHXEuuK90Xzu4x9WJLwN7Kaevq4iiWyn4UKKyp99wua3jaSpSyTs2Rr2Ij481R90tdQGo2Z6rxsZ1i19chXlg/cJi6fWGnq6tT7LDzqWnI9mokpkcWUIdNKCM9lGR/NMxfJy64Cv+ufVzB0Zz6RprB0oyzMrqNiS80eTTejk0zWtvONvOOLdcTx4eEZ7ERmo1oSkjM+2rUPXPU561PraPTjLcZetsEsczxC1vI8cmLZiM1upRNtuLUjZRpMiWRckly7EaeQFqQHz30e65tWj6cM4nalP19pqnDhUkzEiZhoYRaFfMNlXETKdkuG1JU6lzjtuTffbzOw/RrqhqRqdoNIybVG5iWmS1t5dVL8yNERHbd9EkuNJUTaCJJdke78IAsCA+dXT71yawas4RGyyddQyhaU4bZZhqQ96Cw1JvHiVKOFAiNGnZtvwmEqcfQnbcyTuR9zlXT3JOrWw0NX1NZbrNjrkG4wuwyhrEo2LNIarUrgOSIXgy/ENxxaFeCaydJSTI1p8yIzAuAAhDQ3qDxm70P08yXVHUXHIuWZBiMG+nRnpjEeQ+a4/iOOojEZK4+q4fqp29U9vIRtS/ZF8Eu4cmwY01yVEeVjlnlOPEmZAeeuIcBPiSE+C0+pcV3wvlUtvEk1JI9u/YwLcAK3ZN136RYzGetnK66n1MbA67O3pkJptwktWEhDEGCSeZGcl5bhbJ7JIiMzURCMOpDrKy+PoZqtSQcRyjSbVDEaqmvIzFg7EkLXXSrOOx6Qy6wtxs/nLbWk9jSatvMj2AvAArLY9dmBRsXPK6jELu1rrPJ/tSxSUT8SMxkstCV+kPx3XnUobiNKbWk33DSkzL1dx0RuvTELrH8aPDtNMoyLMslyG0xlnE4L0M5LUyuQTk01SDe9HU0htbaycS4ZKJxO23fYC0ICsXSXrlkuU9PGbauakSraWqiyPJnvAmsoblxoMR5a0RVIT2JbaE8Ntz2MttzEWYR1Dat3EfTvUrU/qmxXBHNSX4llS4EWGHLhu1r8gm2o6rAj8Un1o/hmpJJUfzdiAF8AFQNI+qG3x6yzKFqPJtcg+GOoiy0yx/w/CIq1hTKXI6Vb8d2keG571et7RP8ApTrHS6s2Od1tRVTYS8CymTikxUk0bSH2WmnFOt8TP1DJ4iLfY+x9gBIACp9T9kJxGy0+lamvaSZnBoXLRnHqSTLOI0m7uHJK2PRWFKeJKUp4GpTzhpQRJc77oMj1zMur+x1Ac0sTgTllidk1rXVYXl1WqTFlE5HdivuqbS+wpxp1lxPhqJaFb7pMu23cC6YCsuD9eWnGd6jU2HV2M2rNLk9xNoKHIVzIS2p02Nz5EcVDxyWWlm04TbjjZEsyLsW476jrZoU5pZYXnulmT4jJi45ZZREKRLr5z0iJBTzkNrZiPuKjvkg9yac2M+5bkfYAWTAVih9dOMK0dh6x2+nVvBhZBaQKjGIiLirkruJMxKlNN+K1INqKpBJUbqX1INoiPfc+wkzQHXzH9faC6saqnl09ljVu7R3FdIkMSDjykIQvdD0da2nm1IcSaVoUZH39wAlEAAAAAAAAAAAAAAVk6cPvr+qf8eYx+pkCzYrJ04ffX9U/48xj9TIFmwBr2b1+aWVQlnBcogUU9LxLXJm1hzkKaIj3QTZOt7GZ8T5bn5GW3caH0432qObYTX59n+W1FixdRfEjwYVOcQ4y0uqSZm4bq+ZGSfLiQ2LW/Vqj0Q0zuNSMghyJcatS2hMaPt4jzri0toQRn2L1lFuZ+REZ9/IQ/wBA+tdBqfpCjFK+BJh2WGcYs1DpkaHEvLcW24gy9h7KIyPuRp9u4rSqQVdQvrZ6f73nUp4atLk+eIUPgUks1lpvvrv35fEs2AALJywAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA4HIADjYvcNNb0e03ay886bxeOVybpv+NzXxJ0+/iE3vw579+W2+/fz7jcwENbDUcRl2sFKzurpOz6V1mk6UKls6TscbEA5ATG5XmP0w6gNdSyuo57qCmuvqhqo/gf7WoaWvgP0w5KYHikfLsoyLxtvEMi8x+sc6XMuf1bxXVjWLXm21AfwFE0sYhLo4dY3FclNE066+qOW8hfhkRF2QRGRHt572EAAQ7m/TjW5tq3darv5RJiv3OnEvTpcJEZKkNsvyVPnKJZq3NRGrbhtt7dxqlb0aU1dGwKMnOpqywXTuw0+bM4SC9KalMttHJP1/UUkm9yQW5Hv59u9jQAFW6noFwKBmeiuazMpnS5WjtJHpiaOKlDdycVKjhvOkSj4KZdWtxJFy7mRblt3lnQ7ROFongdjgsK/ftWrC5tLhUh1gmlIVNkLeNskkZ7knntvv3232LyElgAKoYd9j7wzC6XT+srs2mqfxOltMVvXvQUpRlFFPdecdgykEv1SSp9RoWRmaT77H22ymDdIGeYXiUjSxzqdye107ZpbCjqsfk00InIzEmM6whL0tKSdkJZJ3khHqFuhHsIiFmwAES4T0y6W4zpzjGC3uL0uRTsZxtjGWr6XVMpnrjNsGyfF3Y1tbkpfZKu3M9vMaRoL0cM6ITI8BWcVeQ43XwJFZChPYZXRp6mHC4kmTYNp8Z/igzT2JHIj9bfyFkQAFRMJ+xxafYpo9qDpHYZ9fWqc4egGzbqbQ3Kqo8BRLr2Wu6iUTKyM++3IlGWxeY9eUdCcvUjHs+LVfW+zybLs6qKzH1XxUkeG3X10KaiWllqK0riZrdRutRr7mfYi22O2AACpmRfY+8ZsI1hW45qDJqKxjLU5ni9a/TR58OimuNrRMZJh4zQ/FfNfPwTJJIUXqn3PfMWfRtdSa/Brii1iRjmd4FaWNhXX9PiFfFim1OZSzJYOvQRNGRttoIlqUpRGW5mfYis2AAifQjp8qdFNNbbTOVkczLId1a2dnMk2bKEOyPTVmp1DhI9VW/IyMyJJHufql5CHGOga4iV+OaesdRmSOaWYnkMXIanFZVPEefjLjvm82wmwPZ02SUZkSTSexHtv2FuwAFYcg6IotlQXsSi1UsqW9nasO6uVNs3WtPfBlkpBIJk2Vq4vtkk1/ONO5mW5bFscidPOgi9B4GXtSs9ssusM0yR7J7CwnxWmHTlPMMtulxa2RxNTJqIiIiIlEnb1dzloABWR7oaxaR03UvT69msxa8ayNzKai7XXsuGzPOY/JQbkVzk282RSFtqQrstJnvtuPZE6P3H6rC4uRagV78zDs+hZ027T4lCp2JCozC2kxTYjmRER81KNxSlr3Pby2IrHgAKv6XdD8HSbJzcxvP4h4m1NnzItY5iVedpG9K8QzaTbGk5BIQt01IMiJZcUp5bdhhNIfsfzWkOR0uT0+rq1Tcbobagqno2LQYzqETC9WTIWW/pUhtXrGt0lJXsRcU+tyt2AAp4f2ObG51DkqbnURKskvskqcqjza7GYcOshTq9t1tpRVhcmHCcS+74xKP5Q1EZ7bd7AaI6WT9JsXl0drkFVcy5k5UxcitxuJSMJI20IJtMeMWx7cDPmtSlHyMt9iSRSGAAAAAAAAAAAAAAAACsnTh99f1T/jzGP1MgWbFZOnD76/qn/HmMfqZAs2ANC13xbGMx0ey6lzCtROrPgqRKcaUo0mS2UG6haVF3JSVISZGXu9wr/wDYx6nGmNA5l1V17bVtLupEe0kEozU+bRJNojIz7ElDpbEXbuZ+ZmJn6o3MnjaD5nNxi7gVqo1NOdmKlw1SPGjejOEtpGziPDWrctlnyIv4pivv2NyqyLD2M4wGxtI7kOEmnukx/RzS6TlhCQ8SiXy24khJJ249zLff2ChUdsXDTg/98meiwyb5ErrP+9F214aPq/eXguguwAAL550AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAANYyvUjEMJksQ8ksHozshBuNkiG88RpI9vNtJkXf2GNnHGxCGvGrKFqMkpdLV14Jr1JKTpqV6qbXU7Pxs/Qjz937Sz+/sr81yv+WNFidV1W9l3wbJoFNUanjZTNJalPEnfYnDaJO+x/wAUu5EftPsJ92L3DU42lGn0TKFZlHxphFspw3vG5rNJOH5rJvfgSvbuRb79/McLHYTlypKnzbEQST+L4GrrvlK/Z8PadbCYjkmCnt6MndafEnZ9yjbt17DEfu/aWf39lfmuV/yxseJZ9i+cFKPGpzskoZoJ7nFdZ48t9tvESnf5p+Q2DYvcA6lGnjozTrVIuPQoNPxzv0KFWeFcWqcJJ9ck14ZF6lPrvUHqSwTrK0+0xXq7U5tT5sq2sbnFo2NsxSxunaL97SDkpUp1RmZ8CU4oiWtBkSfWIi1bQPqX1DdzSW3rjrpZRbltjIJsbT2x09+CvS48Nb5tqjWakJJ8yYbQ6ZJ3PZRke+xmJU076Rs7031ev9VqrqQuZS8suk2d7EmY3XPOzo6F+pC9LWlTzTKUeolLZpJJd0kRj21fSXeXWpVBqHrfrjdakJw+NYRserpNPCrmo3prPgPuPqjJI5Cza9UjPiRee24vlQiLQ3qC6hpObaCZFqPntbfY71Axbp46FilZipx5cdg5MYo76PlHiNBEhfimZ+Zl9Uta49ZUPRvOr/AIOkOV5fOxnFm8ys36t2M2xGq/EdS86tTziT3b8LfiRGpXLsXYzLyaN9FMXSzMMPvLjVq+yym0zjWMTBaabCjsJp25pcXjcebIlylk38mlS9iSnyIbXn3TBT57nef51JyuZEez7TtzT1+O3HSpMVla3lHJSZnupfy3zT2L1fPuAI4kdcRZXjWdw8f06yfD7eNpfZajYfYXjUZTdpAaZUTcjwm3F+H8obaibc7qR3Mi3IjjLTbrS1jk9N2R12W2FfP1qVPoKzGVFCbabnfbDHZfrHzaSRIPw0OSDURFttGPfuZid5nRrQy269s83npKBpBI0iTtER68Z1CEHNPv2cLh8zy7+Y8tN0Naf1Wq2lWrLt/Okz9L8Wh423FUylLFm5EjrYjTHiI+zraXXDLbfuae/q9wOzph18yW86NmdetYrF25saiLkE22kQILbbkhmvmy0fJsN8U8zajpIiLYjPzPzMcY31oVlxg+EZ5Z6bWMCvz/ADGoxGnVHuq+wQpVglSm31rjurJvw+BpcaVstJ9tjG9aQ6EHo1oSWiuL5tNJ1hNqcS8VDZN6O7MkvyCcJlZKbUban+xKI0q4FuXcyEONfY+a1vGrtDOrEyvy60zCpzeLdVVBDhQ6+yrkKRHW1XI+R7k44bm5+uoyUflsYG4ZP1pYhjjOWRmcJv7S6oM7jad1tVEUz411bvspdbS0paiS23xUozUsy2JBn7SIdVl1e3tXm2J6XSunTOPt1yisftV06JMFZ17DE047zjzyXTbNskpN1K0qPmSm0kXJexYI+g2M5jFzGk61ZI/l0/OomosHKXIMb0iFdMMJa5+AlJMuNK+UPwuKUkSkp/g7nIGF9OdrR6rY7rLmWq9pl2SUuMz8ckPyq6PFTLTJmlJ8QkMkSWyRsTaUER+qRbmZ7mYGn0nXRjd1kFDJTpdlLGnuVZWrCqTN3HYpw5lqTi2kF6OThvoZW40tCXVJ2My7kXfb9Y712YRc4vqBqNOwu2rsKwBE30i0VYQXZLz8eSUf0dUFD3pEdx1Z/JE6lKVJLczTuW/kxjoWg49b49TvavX07TXEMsXmlBhi4EZCIll4rjzZLmEXiustuuuLS2e3c+5nsPPddAeO51keb5JqjqNNvZOY46/jRuV9JCqXUMLktyEvyFx0bS5Da2WuK3EkWye6T37AbPpP1p6cZ7Y5BSZs1D0/n4/DhWTqrbIK9+G5ElKUlpRSmHlNJcJaTQppRkpJmnz37YbVzr50y0yyWxpKeley+Fj1RDvb2yrLevbZjwpO6mTjJeeSqa4bZG7wZI/UNJ77nsNt0k6YouEV1nU6i3mO57BnMxmG4r2D1dc0kmTUZLdJlvd9wzNJ7rPYjTulJGZmNaz/AKKKa81Lm6ladZfAw164rIVVZ17uI1tvEU3ESaI64zclBlGWls+HqkaDJKd09gB3W3XDgNZW5s6jFrmVaYlkFJQxattTRP3J25Nqr34+5kRIdQ4peytjIm1e3sPJM66MciZA/ITpblLunsPLywaTm6XYvobdt4pMntHNzx1RydMkG8SeO/kR9t8vlvRphuW9RmKdQcnJZ8deOMwjk0TTDZRLOZCQ+mFKd224rZ9IVxIk7eqki2Lca+roWhLvJNUrV6+/cyl5l9vbuE+gRuCrLxifNHpm3jejeKRK8L6vnb9wBqHTv1gvu5jn2muYS7bKbmt1By9+U+lTZM4xjEB5KW3pKj22bJXJttCSUtRkfuGy4T9kT0tyZ52TfYva49UScasssqJrs+FLXNgQUeI+TjEd5bkV42zJaWnSI1Fv3Iy2HorOgXDKfMJOdVubWTFvZ5LkFpbuIioJNnUXB7yqh4iV6zST9ZtzfkhRmoi38udPehWrw3F7TTm51FTeYbNx+bjUeGWK1kOxbiyGzbJTti234zziEGZEfqkZ7GolbbADX9LOpnUfVzqtwenmYTlWC4reaeT79iptnIzjdmlUiOcaYk2VK4qJtaiNtXFSd+5dyMe7WrW/Uu41vyzS3DNUIml+I6ZY7CvcpyUqBNxPffmKMmI7EdZKSTZILkpZIUrftsRdxtWjXSJcaZ6iY1qPlWuN7m03E8YexCsjzKyNFZZrTUybSS8EiNS0+EZKcWalL5FuZcdjzWqfTLY5Vqg1rbpRq3c6a5uusTS2M2HXxrCJZwkr5ITIiyCNCloM/VcIyMi7d9i2Ahio6gctm5503nU65t5/j+Rys4RdWddT/BTdy3AgG4wh6Mr5jjSyMj48UmotyLYxMWC9XOKZ2eiiYWK20Y9bYdtMrPFW2foCYDJOuE9sfc1EeyeO/wBY17T3ocxnA1aXOrz62tnNN5mSz3FyIrTfwq9dNKbkciRsTRJ5GaSTv7tx4tH+hl3SvMNO8hma45FklXpX8KR8XppldFZZiRJzK23G1ONkS3FkayPxFGe5ISkkpIAb1qD1Mpw3Xin6fqTS7JMpvbSmi5C9Kr3GERYFc5McjOyH1OrTxS0bfIyLc1c0pLuYjrFuv/A9SLqLjlBjtlXwctYuGsUvF2EJ30x+C04tRuxG3VPxUrJpamjdQXMiLsW4mleidY51EK6hHLp9UxeFFhaqw2U+CbPpxy/G5778t1ceO223cRVpF0SRNHJ6K3H9Rm38RhpsEV9W/ilac9hEpDpcF2fD0hxLZvGadjSoySlJqNO6TAjTSrruPDtB9L4mYV9nnWdXGGPZZcOLtIMA0QGpLjXiqdlONpdeWpBpQy3utXBR9vM5U1J62qTBsBodWKbTG8yDDLnHo+TOWqbSugmzGd3M2W2JD6XZEhCS5LbbSZESk+sZn21tn7HxT0ddgjuHalnBvcLxlWJOzrPGYNtHsYHpK5CeUWQRobdS44s0uJVvsZke5dh36tdAdVqpY/CL+qD8FyXhUfC7I1Y3Xvm40yalJkRSNBIgrWtZmtLCUpMj2TwPZRAbrRdXePZbrVJ0hxTDZ85qrhwJ1pbP2cKJ6OxLiFKaebiPOpkSGktrRzW2g+ClbbHtuMDpD14afavahY5hlbi1lXwM2+ESxe1esIT3p5wkqU740Vp1T8TkhC1t+MkuZF7D7D03HRnGyfUXB8vynUd2fU4AqE7U1qKGGxLSqPEJjwVWCC8dUZZ8nFsHuRqWZctiIi/egnR03oFkFcqg1DZsMXpDlFWVkjFK1M9tD3PZDtmlv0hwkeIexlxMyIkmZp9UwLHgAAAAAAKydOH31/VP+PMY/UyBZsVk6cPvr+qf8eYx+pkCzYAhvrEnHXdMeoj6TMjXTrY7f9otLf8AvCP9FYR4f1SWGPEkkt5HpdRWe5mW61w1Jif6pl/QNn67HjR0vZdFQrZc1ytioP61z2C/0bjH540WL9XOi9g2k0ovMfusfcVv22ZaQ+gv6dxRq/bZujL5to7+C1wLp/e2n9MYS/QsaAALxwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAANC1L1frtMX4LE/DMvu/T0OLSujqVS0NcTItnD5ESTPfsX1GJKVGdeap01ds1nOMFmluN9AQb91ljv0Rarfowr/mB91ljv0Rarfowr/mC79E4z8N+XuQ86o/eJyAQb91ljv0Rarfowr/mDeNM9Wq/U9VgmBh2W0ZV5NmpV7VHDJ3ny7Nmaj5GXHv7ty94iq8n4mhB1KkLJdhtHEU5vLF6m9APl/8AdW66afWOvH2851YSMYyaTnlLgtms0kqgvagn1sxUKIvVS4yps29/NbREkvnGdjcX6vbqmwfK2LbFfhefpzpDSZ4/OdsDQu2kSIK3VtLImz8LdTXzy5fOP1ewpkxbMBTjPeurUOietG8K0KhXrdBp1Wak278nJSiIjwX21uPspI2FKccSSSJGxbK9Yz47ElWn6udYGomZ6Uan4hb6cyMAm2Okq9RMTs4OQFIkqrnHUsp8Y2kI8B/daVbIUoiIzLfctzAvuApfivW/k+m1HOpNetK1U79JpixntS/CvU2LtrEQpuObT5m2kmpC3VoM+60lzPdR7bqxsP7JHIYwXP8AJbfTyhnz8Nx6uyZhGP5IubCfjyZjcZcV2QqM34Ulk3UmpJJUlRH2Mi7gC8QCH5+uGS6f9Pd7rjrZp+jG5VFBk2TtHAs0zlmyX/QN+MSUp8VfJCVduKVGfcy7iFsk6t9cIlLlGF5No9UYxmFhpvNzrFnoWTHLY9HaIiebfWcZJtyGUr8QiJKkLNPHknfcAXJAVI6Zc/m6K9BsbW3U6NbzFtUX21SlP5DIt5Nl4kdtSXCckERsKeVt8iW6GzUfc9zMYTD+v3LMtx7OXqrS3HLq9xPEms2jxaPKFyoj8EnSTJiOyFRUeFMZR63AkLSvsRKLzAF0gFOabr8t9RYNFI0h0kbtnM/yZ/HsIO1uDgontxIhP2E2Tsys2G2lGbaUp5qWaVH6vYjwGtuvWsOF5rnsupx6wps7pNCVZS5WrywpVHXOpsnW3H2opxjbekNpQbiXVGXiJJLSko3NRAXlAU2yTrG1Y0z0PwjOM4xXTZm8ucbO8ksWudJhvWKEMoWkojCIhqcedSfI08UttqUSOSt9x+cS6sNY8z6l8bg1mMVcfTC+0ngZ87Fk2KEyYsV51BuzTUmOa1OtmpbBRyXwUlJOciM+JAXLAVIwLrbzTIrHTu/yvRiNSYJq3ImRMVsmr/0mehxltxxn0yN4KUtk8lozLg4vjv624xuEddeo15plh2ueYaDwsf09y7IauiTYpyUpD8RqS48w7NcaJgiSy3IbaQXJRGonCV28gBckBEGkevcvVbC871Ag4U+VRjN9b1NH6JI9Ievo8D1DkNJ4pJJuOpdQhO6vm+fcQjhnX9cXeDU+pd5g2JIoZeU1FBdIqspdkzsZanqcb52Md2I0ppxp1LSFFvxX4hmhR8T3AuaAqNY9ceWynKtODaMR71GUaiW+B4245kBRUWCYTJ/v81GyZIaU+hxJkXIyQ2oy5Hsk/wBRetnJ5ukx5fKw3BcYyOuy20w66iZRmqYECPOgmXMo7xMLclGvdOyUNlt6xmexbmBbcBT2l67Mwz2g0Zd0w0Ui2t/rDBv3Y8KbkRRY9c/VOoQ7ze8FXiNGXiqJRJJRklJcd1HxxHxiVrNxzAJ8fTSjx+blL9rBtJGVZGuBUQLCBIJhcFE1uM6lTzh+ug3CbQSdtzMwBdkBTXPfsiVbiOrF5hcDEKedS4rkMLG7RS71TdzJffNtLr8KElhSXmWDdLkanUGrio0lsW4kLR3qmtdVNc8y0nlYzj1A1is6dAKHMvHEX7yY60pRM9AXHShUV0lckuNvL2I07/OIAWIAAAAAAAAAAAAAAFZOnD76/qn/AB5jH6mQLNisnTh99f1T/jzGP1MgWbAFfutrjK0npaJfzbrMqKAoveSpaVbf5g6+rFZ0eQaLZ03slVTqHBguubfMjzG3GnO/5EkPz1fEc2VozREfebqfTube8midUY9XXLEcPpyvL2Og1Scdn1lwxsW+ympjW5/+FShSra7R9CXlqd/Bac1h96Ul/NaJPoDzwJbdhBjT2T3bktIeQZe0lERl/pHoF04G4AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACA8s6KNFc20syvSLIkXUmny3K5eZvyPSmylw7OQ74i1xl+HshJbqQRKSo+C1EZnvuMXnHQfpHnEpDzuVZ5SsO4nGwyyiU92UZm2royDRHKUXhmbikEfsNKVbbKSpJmk7FKmRESkQlymUyFpNaWTWRLUkvMyT5mX1juAEISOkPS6S3kjbk/INspwCNpxN2lterVstrbQtv5Ps/xWe6j3TuReqQ8+TdGulGVsHHsrHI0JPThOl/yMtoj+CEuocJzu0f745Np9f5u2/qCaoV1TWU2bXV9tDlS6xaW5rDL6VuRlqTySlxJHugzSZGRK23LuPaAK7470L6NU5ZMi/tMvzBGSY19pxFkdyqWqtpeXMocRZJSptBLJKiUo1LI0JMlee/pl9HONXel+R6Q5lq/qflGPZDAjVnh21xHcXBjsOodQTBpjpTy3bQRrcJajSW24m1OSY6rIVYim+rjvUQysVVhSkelpims0E+bO/MmzWRp57ceRGW+4/V9kFDitRJyDJ7uBT1cNJLkzZ8lEeOykzIiNbizJKS3Mi3M/MyAGOzjAsX1Gwa305y+v8ATqK9gOV01g1mk1srTxPZRd0qLzJRdyMiMvIRFh3Rjp3ix3Mm2zTOsusLbFnsKYsMhtm5L9ZTOpMlx4vFpCUb9jNakqUZpLczLcjn0jJREpJkZGW5GXtHIAj+JodgLWh0fp5nxJNliDGPN4ypuW98u7DQyTRGpxBJ2c4kR8kkWyi3LYaGjpCpZGnd1pbkGtmq97j93AYqVsWF2wpUeC2tKjjtqRHT2cSkm1qVyWbZqTyLczE13uRY/i1edtk15X1EEnW2TkzpKI7ROOLJDaOazIuSlKSki33MzIi7mPRY2VdTwJFrbT48KFEbU9IkyHUttMtpLdSlrUZElJF3MzPYAV2n9BGjCnpkjFbrL8QdPIk5TUqx+xai/ANgbHgPqgkbSibQ+gk+KhfNJmhOxJ22GyK6RtNZdhZ2t9d5Zey7rT93Tixfs7Qn3ZVY4+4846tw0Es5ClvL9ffiRbESS2EzFYQDgfCpTo5wjZ9I9J8RPheFx5c+e+3Hj333227jiusq64gR7Spnx5sKU2TrEmO6lxp1BluSkrSZkojLyMj2AFcHOgvTt2BSxXNUtTjkVGNyMOcnfDTHpM+jdXy9AeX6PsTSfmpNskL49jUY2as6QdOaTJ8ByulyLLIMvAcVYwpptme2TNvTsmSkRp6PC2dTyLkfDhufn22Ipgtckx2ilV0G7vq6vkXEn0OuZlSkNLmP8TV4TKVGRuL4pUfFO57EZ7dhkgBXnT/of0q09yChtomT5vb1uIKmLxagt7gpFZQqkkpLiozZNkszJK1pSbq3OJH277GPzn/S/W1PRde9L2mNa5dNJo366lRdTUoV6Qt43W3nHkoIiNt1ROFskvmEX1ixAACN9GdHoWlWguNaLxpr7BVFEitkzILptOqkrQZyJDay7pWp1bjhK8yNW4iHUHoxqmtIdYqzHLC8zzOdTqVmvcsMrsmSWpyMRlC9dlltCSZWonN+BqUaC3Ufbayk7JMdrLisx6yvq6Ja3XjfBsF+UhuRN8FJKd8Fsz5OcEmSlcSPiRkZ7EMkAIH0/wCkzC8a0/0Wxm1lTfT9HjRYRFwnkpZlWLjC0SXHSUgzUlbjrq9iNJ7mXf2DwzuiLTJ2exeUmX5tQXkPLbvMYttWWLCJMeXbISiayjmwpHgqShJERpNaduy9xYYABTbJug44mV6HY5prl2T0OF6bx8r9JuId2TF3FeslIdaNlzwzJwvF8RKuSTLh2Vy33G92HQtpfK0uqtGIObZ9WYfDYdYsq2LbNGi8N2Qb7rk03GVGp1ThmZuNm2rY+JGREkiscAAg+P0mYhS55PzjBdQs+w5m5lxZ1vR0Vs2zW2D7CEIStxC2luJNSG0JX4biOZF394yNF0z4tVaxs622+aZjkt5XNz2aaPc2DT0WoamK5PIjkhpLhp29VJOrc4p7Ft2EvgAAAAAAAAAAAAAAAArJ04ffX9U/48xj9TIFmxWTpw++v6p/x5jH6mQLNgCvXUl+/NZOn+n8/EyyTN4+/wACKat9vq3Eh9Q1F9smhOoFITfNcnHLDw0/9olhakf5ySEd6yrOZ1a6A1iS3KOjJJy/5u0JKSP+kzITXmtvjtDiVta5bPZhU7MRz0x50/VS2aeJlt5mZ77ERbmZmRERmYgpwdWVSC4u3kjp1qmwhhqnRG/9cvY1bp2v1ZPoRgF44s1uScdg+Io/a4llKVf5yTEiCkvSvF171B0bpcOxDMqvEMSx1cmuXcx2SlWE1ZPLXwQhR7NJSlxKdz4q3Lct99imMukfHZuz+Sar6l3EtXdx1+/NJGft2SSPVL6tz/COlTwkKUEsTUSlZXSWZrt4ebObjKubEVNhG8czs911fTr8idwEEl0k49CPxaDVvU+peSfJC4+QnsR+zcjR3L8pDj9yDqKxr5TDeo92zbRsSYeRU7b5KL+c+Rmv8pEN+bYeXyVl3pr0uV9pUW+Hg1/gncBAv7oXVPhZb5no1SZbFR8+VitkptZEXtJh7daz29hEX5BlMf6s9JLOcVLk8m0wu23IjhZLCVDUR+W/Pu2Rb+01EMPk+vbNBZl/C0/TVd6CxFO9paduhMwDzwLCBaRG59ZNYlxni5NvMOJcbWXvJSTMjHoFLdvJwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAjbVZ3qJamwP3Eoun70Q2l+nfbMuYThOblx8PwNi47b7799xJIDScc6te3YbRlld7XK9+ldfH969Df6y1/8w9K6+P72aG/1lr/5iwgCHmz+/Lx/wS7f+FeBXv0rr4/vZob/AFlr/wCYkHSd3X107L92+LgrJF4Xwd9rK5ZmfzvE8Xx/+5x4/wA7f2CQwG0KGSV8zfazWVXMrZUj5PatazkrqXuOryFS5nJZ06z2BjMGwjUct2mLForb0S13mJT4BKdkSXFJIz7cSLzUW8p66dU2u9BlXUBb4brnjNHU6PyaGbS0EimiSHLtiZGjrWwp1Rk54ZqUvipBGs1OkXMiIiF5I2lemsPBX9MImCUbWJSm3mnqREFsobiHVqW6SmtuJ8lqUo+3czMxFOO9FOi9dqpl2p+S4pj+SuX02qmU8OfSsqTQegw0RkIYWrlukybQrbZJEaU9j4kYnIiteQ9RGoGI5vqpGxBGPYbZZXqfheJqvnKWNzp2bCtJx6VJPikpLjfE0pN8zIuXsIb1rprJqjp3SVmC4x1H2mTZnAoby+lSscw+ndOTHiu7JemuSX0xmG2T+ScSyXiLPcyJJlxO1F1o1pPkcXI4N9pxjtgxl7rL983JrmnCsnWkklpb+5eupBJSSVH3TsW2wwcvpi6dp1RRUM3RHCn67GTcOoiuUrCmofiK5OcEmnYuSvWV/GV3Pc+4ArV0z59dap9XGL6j5GhhFrknThS2U0o6eLfjuWy1LNKe/EjUZmRb9t9hFnXDqxqfqBifUtjkjValxvFNPHqejYxB+tYVJuSkeA8qQchSieQs1nu2SN0mlsyNJ91D6HY7pfpziNjEuMXwilqp0Cnax+LIiQ0NOM1jazW3ESoi3JlKzNRI8iPuMLl/T1oVn+RvZfm+kOI3t3IiHBen2FSy++4xtx4KWpJmfq+qR+ZF28uwAqTqV1HdU0/WfUTEtGK+a41pm7SMRKdESoKDYofYbffXYyJkhuS0laVLS2qOnYuJGZmZmPDqd1HdTlDP1h1Do9S6uJjek2odVRNY45j8d1VlDlriNuMuyvnISjx+SVILmZqXuoy4km42YdPuhuoOSQ8xzjSXFL27gJQhifPqmXnkpQe6EmpSd1Ek/Ij3IvYMjP0g0stYV7XWWn1BJi5POas7pl2A2pFhLaNBtvPEZfKLSbTZko9zI0kAPnh1Oasan6qUOpsi81VparG8R1erMLhYIqtYKVKaizYq0zkyDUT/ADWozXtspHBtzt23TIupmpeuuXVnVVPsM9pHMI0sas62FjMrGYstNhzrUuIJ91wjI0NLMlkk0q5mpZLMyJJFba86d9CMnyqXnGRaQYjZZBPJopNnKqGXJDvhqSps1LNO5qSbaNlefqpLfYhm3dL9OX4eTV72D0i42ZqUvIWlQmzTaqU2TajkFt8qZoIk+tv2IAfPto9XrXV/LmafXGNj8Q9Dqa8bpV45CkwX4px3DVAbiuH4ZI3SvdaU78V8TLikhsulGtOquYN6V6TYvqtjGj9XE0cgZzKn/a/CWxPkOPGybCGXTQ0zGbSg1qJriruZbkWxldCz0Q0eubypya10yxqXa0VcqorZjta0p2LCU2ps46Fbbk3wcWnj5ES1EXmY8N/06aC5VW4/T5Lo9iFpBxVpMeljy6hl1uAynbZpolJ2SjsXqfN7F2AER9XZmrVXpkVzSvfUpB8k+R/vF/uQivp36keqnV7Nsbzf4NnP4de5FbVdzVvxKePX1MRk3kMnGeKR8IOSW1NINxLrWyiUrikiJKju7d4dimSzKewyDHa6xk4/L9PqnpMdLi4UngaPFaMy9RXFSi3LvsY1ev6ftDqnPnNUqvSXFImXPOOPLuWaplEo3XCNLjnMk7ktRKUSll6xko9zPcwBSzRLqf6m5EDQjU3ULU6kvKPVmJkkeZSKoWITcBdbGfcak+kt+upS1x91lsSCSoySnfuMJhGsnVVrxpVnGTq1ZuIGOt6WT72xdexeuioat1oUoocFSPlnI/o5L2fM+STMjM+R8SvZZaC6WScRg4fT4fWUMejiT42Pv1cRph6iVLacbeehGSdmXDJ1Z8iLzPvuIZ0G6DMZ0azOozCflFbcHQY8/jUOLX4tEqEzIzxIS45YqaNSp7ppQRcnNi3NStjM9wBXSoqdVcmndEEan1vbVkttQZI/FyF6nYlLqmF0kFSo5tcuL7qEc0+I6ZmalclEe2x3B6MdUc31d0FrMo1GnRp+RRbO0p502PHTHRLVEmusJe8NPqoNSUJMyLtvvsReQ3fFdCNGMGXTOYfpfjVMvHn5kmpVDrmmlQXZSEokraMi3QbiEJSoy80pSXkRDZMWxDFsIqvgLD8fgUtd470r0WCwllrxnVm46vikiLkpalKM/aZmAMuAAAAAAAAAAAAAAAAAAAAAAAAAKydOH31/VP8AjzGP1MgWbFZOnD76/qn/AB5jH6mQLNgCveWNfCXXDgbG+/wPg1pP293iPpZ3/D3HRkzKuo3WxzBFqNzT/Tp5D9ykj9S0tT34R1e9Dfrci95KI/nJMtN1j1HRp11QZXmZcXJNFpY1Fr2+xmqdJstmk7eZ9zJRkX8FJmJ26ftOHNMtMKulsOS7mbys7h5Z7rdmves5yP2mnsjf+YLOAfNqVTFfvOTjHtsrvuWi63fgT8p/WSo0OCgm+9tpd9yP+kxLVHkGtGCNbIRTagTJbLRFsTbEptDiCIvYXZWwsOK86aLLH+sjV/HiTwRkVJSX7ZeRGbSFR1mX4TMvy7iww5+H+S3Q2vNljlLWvn+9GL8Yq/ncAACcoAYrIsUxnLoJ1mU4/X20U9/kZsZDyS39pEoj2P6yGVAZjJxd4uzMNJqzIJsOlGjpJblvoxnGQ6eWCz5m3BkKkQHFf9pGcV63s7ctu3kPP+6B1KaWHx1G07i57TNfOuMWPjLQgv4TkRRFzV7dkElJe8T8AurHznpiEprr3/zLXxbXUQ83jHWm8vZu8NxHenmv2lWpqyh43lDLdnvxcq5xHGmNrLzT4S9jUZe3hyL6xIg0PUPQ7S/VFBqy7FIr00i+TsI5eBLbMvIyeRso9vYR7l9Qjs9PeorSPZ3THOm89ome5UWTr4zEIL+AzLLbc9vLmaUlt5GM7HDV/sZ5X0S3d0lp4pdpjPUp/OrrpXt7XLAAIWxbqlwuXaIxbUqrsdPMjPscO9b8Nhw/e1J2JCk7+Slcd/ZuJmadafaQ+w4lxtxJKQtB7pUR+RkZeZCtWw9XDu1WNvR9j3PuJYVIVFeLP2AAITcAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAKydOH31/VP+PMY/UyBZsVk6cPvr+qf8eYx+pkCzYApVkeDr1K+yDPRpSVuVdBX10+Y3v6qijs+K3uXkZeM+2W3uNQuqK8aCSouUdQmtuXcN3Ik+FSMq9yGEuNuF+VTKT/ILDi5i4qjkoL91ecvifm7dxhVZV26supd0Uor0v3lesmJOPdcWGWR7JTluD2NR/hORZBSP9UxYUV66klfAOsGgmcEjb0fLH6Ja9/4M+KpvY//AAiwo5lLSU49fqkdDGfFSoT/AIbeEpL0sAABOUAAAAAAAAAAADDZVhuKZxVLpMvx+Dbwl77tSmSWST96TPulX1kZH9Yg2RoplOlclyd096rtVsdCzWvFsgkelVyz37oQsz8Rn8m6jPzURCxYiTJdOMCkasY6y/h9Qtuyg2smWlURBk+6lUcyWvt3URqV3/nH7xrV5RxWBprYWkm0mpbtWlus+nt6yriYaKUVrdK97PV23owlL1PNY9YM4zr1hs/AbVw+Dc5wjfqpR+9uQnck7+ex7kReahN1fY19tDZsaudHmRJCebT8d1LjbifelSTMjL6yHmsseobmoVQXFPDnVq0E0qJJYS60aSLYiNKiMuwhWw6bbrBJjt/0653KxOQtRuO0U5SpVRKV7jQrdTRn/GLkZF2LiOjbC4nd9XLvcfdefcbfW09/xLz9n5E9gIGq+paww6ezjfUPhErC5rivCZuGCVJqJavel1O5tmf8U+WxfOMhN9ZaVt1AZtKewjToclPNmRHdS424n3pUkzIy/AIK+Gq4ezmtHue9Psa0ZJCrGp8r9z1AACuSAAAAcKUlPzlEX4THCVJV81RH+AxpmrOK49f4fbWNxVtSpNZWTHoji992V+EZ7lsfvSk/yD1abYtj+OYzBfpKtqIuwhxnpKkb7ur8Mj3Pc/rP+kU1Xq86dDKstr3u7+FunrLOyp7Da5ne9rW08b/obWAALhWAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAA0nWzVGm0U0myvVW+NJxMarHpvhqVt47pFs0yR+9xw0IL61EAIRx/rPpLrrsvelRL0b4PhUDZRJJbcnLtojfkMEr2l6O4RbexcdZe0WkH84dA71F4fq3F6kJGFZIu8r5bOoEuU7AdT4kZ6WveS4W26Y7riHUczLiZGZb7GW/8AQ7gOa0epGEUOf4zI8eqyKuj2cRft8J5slpI/cot9jL2GRkAID6cPvr+qf8eYx+pkCy8h5uMw5IdUSUNINajP2ERbmK0dOH31/VP+PMY/UyBN2rtmdLpTmVun50OgsHk9/NSY6zIv6dhvThtJqC4uxiTypsrv9j4mP3dLqPk8nu7Y5KRrV71eAlw/7XjFtRVH7HDG8PRbIJh+crLZp7/4DEdH+6LXCTFVNriKk1xlLwu7eRNVp7Jxh0Riu/Kr+ZXvrgSmv0frM42PlhmWUl6ky9nCUls/7HTFgkKStCVpPdKiIyP6hE/VlQHkvTbqJWpTyUiikTEltv60cvHLb692xt+lF8WU6X4jkvI1HaUcGWZn/GWwhR/2mYox0rS60v1LVT4sDTfRKS8VFr9TawABOUAAAAAAAAAA6pEmPDYclS322WWkmpbjiiSlJF5mZn2IgbtqwcvtE+ytk1rQS0mk1IVsoty8yP2H9YrTYvY5R9RddjtrnV0mFCgkhpb9q6tSJbikGTSl7+qSyJO5HsXlv7Bv9vrVPyWc9jei9ArI5zZ8HrNzduuin71LPbmZe4vP2b+QhOVohb5DrkjF8tylL02fF+GrGVGY4bq5es22W/b2ESjLt58fYPGcv454nY8xp7RqpHXdHsvxu99rpW11KVXHYhU5RwUcyvG74K0laz4u9lpeybLjF5DkRKeg9nXq5YzrFmteRFsTb80pLZf90yIhyjBte6ojKr1jg2SU/NRZU6E7/hUgzUPQc9xMPnw8u5wfrJPyN9vVXzU33NP9USdaVNXeQHqq6ro0+FJTwejyWkuNuJ9ykqIyP8og+z6bbjB5z2SdOebP4jLcV4r1HMUqRTy1e40Hupoz/jFy2LskkjYDn9TlWoydosKukJ9sd95hav8AxmRF/QP3+6lqvXFte6D2Z7fOXXWTMn+hKS3/ALRYocvc1unGcU96cJNPtsnEjnWpT+0jJPpyv1RuOnNrnFvirEnUXHY1LfIccZkx4r5Osq4qMkutq3P1VlsoiMzMt9j7jZxhcRyN3KqRu4eobKnWta21RLBnw3kGk9tzT7j8yEcdQV1mOM/AGQ4XqRT49MjKkpOruGjOHcEfhnwWsu7akcT2Pt/0h+sXcW41qNb62Uowg9bu6ir7t+q6Ne8sOrGnS2jd1/vYTCAgTTLq5wzKZqMYz2MWI5CSib2efS9AkK7Fu1KT6nc/Yoy9xGoTXJyGghtePMu69hvbfm5JQlO34TMZqWpfM1bpTTXc1ozajVhiFek7mraxR8ldwW6kUN3DgsR6qauY2/BN9T6PBM+KFE4nwz2JRb7K8y7du/p0sYyVvD6x/IbqHPKRBiuRkx4Rx/BbNoj4qM1q5n3Lv6vl5DSta9a8JrMEt6qptodxNtIr8BLUSQlRNE42pJuKUW5bFv5eZmZfhLYNBs7h51p3XPx4qo7tY01XvtmrkXJttOyiP3GWx/Ue5fWPP0sbgqvKzpU6l55dyba3+G7W3eWZYLE0/rJxaho9d2t1e3dvsSKAAO+RAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAeC8vqPGKiVf5Lcwamrgtm9KmzpCGGGEF5qW4sySkvrMyIa6zbaS6wVsKPGtsYzCA4hi7istyWJrTjaXVEzKJBGolJJ1tfFexkS2z2PkntpHVvhDWoGkaKL4fxqtmN5BTTK5nJkmups5rU1pTECUgvWW2+vi3xSRnupOxHtscC9BuM2sXUi/ayvUTTWdc6e0TuHJx7DprslUNt2zdmOuSlOJQW6XTNpCWyNKSSojMlbkYEqVSEOfZBspbcQlSFaQVCTSZbkZHbze2wnzHsboMSqWqHF6aHVVrC3Fsw4jRNMtG4tTi+KE9kka1KPYi23MxAtP/dCco/yQ1H63mixgArJ04ffX9U/48xj9TIEldT8467QDOJBJ351a2NvqcUlv/eEa9OH31/VP+PMY/UyBtvWfMXD6cMrNB7eKcJo/wABy2d/7BbwEc2LpL+KPqiKu7UpdjNa+x7xfA6a62X23n29pIM/f++Vo/3BZMVi6Tctq9NelDA5VrU3MiPJizpzr0GAuQhlJynVmpxSeyCMlbkZ+4/cLA4plkTL4K7CHV20JtKiSkrGEuMpwjIjJSSV85PfzIculiqU5KnmWe17cddS9yhUgsdVpJ6qT07zvyqlayPGLfHpCSNq0gSISyMt90uNqQfb8ChBfShdZXedK2DO45YVkebVRn66V6fGW8njGedaJJEhaDI9kJ79xYgVa6csupdMdONUKO9cUhnFtQbmsYZaTutaXVJcaSkvr5q77+w/cIsZKnT+sqyyxs7u9rWs9/DcXMHGdXDTp01eWaDStf7y3drROOmFzmeTUMTJcllU6o1jFQ8wxCiuNrbUZ/wlKcURlt7iIbqIZ0g1h0+i6eVdba5DHrpVWymK81KUSFKURb8kF35J+sbFL6gtJIZHvliHjL2Mxnl/6EbChyfyvgY4OnOpiI3cU3eave2u99JnGcnYt4mcYUZaN7ovdw3LoJEARZ90Vhbxb1dLktjv5HGrTMj/AKTIclrPfTC3p9HMvfI/mqfjkwk/ynuLP01gH8lTN+VOXomQfReLXzQt22Xq0SkON9hAmX9SOV4o81Bm6YFHnSVEliI5aockKM/L5JCeRb+z+zceC/q+pzVmuS09HrsQqni9eIUhTb7yD9jii5LLt229Xz7pMVZftBQnmhhYTqTj+6oSXi5JJFKvCVGoqKacnwUou35mm1Hv1fBMkPL9baOmsVYziVfIyvIzPiVfXFyS0fvdcIjSgi9vmZe3bzGGY0ozHUV9NprTf8ofInGccq3Dbit+4nll6zhl9R9j8lbdhj8N0o1dxGuKrosixSgZMi8RUKvN910y9q1uJ3Ufn5n/AEDYC001Ulnvaa3ztttjTEq2mf8AOJX/AMhTzYvGfFi6E5L7qyRh33mnLvVv4UbfRcKuuKxEPyrO132jr36dRItVUVVHBarKavjwojJbNssNkhCS+oiEHR8qxyd1TIkRbqKtlFI7CNw3CJPjpMjUgjPsZkRH5e4xtZ6HSJZ73GquaS9/Mk2HhpP8mxiIqfpbr5uptpQWeUzHa2uaamGbatn3EPGvgjcyMkmXE9z779u3ftFyrW5SnLDwo4ZJKaeslwTdtNytfXXduLyw2GVCooVMzstysklKL4791ktO0sfYag4LVmZWGYUzCk+aVTW+X9G+41qw6gdKIG6U5OUte+xIixnXDM/qMk7f2jI1GjWmFK2hETDK1w0ERc5LXjqPYvMzc37jZoNHS1hca6ohRS222YYQjt+Qh2svKlT96nDulL9YGl8BHhOXeo/pIjo9e4c0yRjmnuX2yj9rdcaUfhM9z2L8g/Rahau2fq0+jTkcj/6yws229vwp2IxKIBzLGT+0xLX5YxXqpPzHOsNH5KC/5OT9HExWLvZFIo4z2WQokS0Vz8dmKs1tI9Y+OxmZ7+rx3+vca9qTp5O1BTAitZJ8GRI3im+2UJt83jVx47Gv5vHZXl58vqG7ALlXCwr0Ob1W2tE9Wm7dLVt/GxVjVcKm0il2WTXg7+ZBrHSHputZu2ljcyln3Pg8hhP9CEEf9ojHUjpByTHbA8l0vnKyCtQZrfx+e8hiSafaTEkk7GfuJZdvaa/IXAAVcPyJyXhrpYaEk96lFO/e9V2pplbE0oYqWeaWbpSSfkigc6kwS8xyZX41bXNJlVQZybLF8nNpiSltCFG4phfBBPGRdyItjNJGfEu29oumTDLnDdNW0XfhpctZJ2TLaFEo0MuNtkjkZe0yTvt7NyHu1501wjN8EuLTJcdiy59NXSJkCZx4vsOtNqWni4nZXHkRbpM+J+0jEYYzc6+aIY3U2U+G5qVgrsFh8zit8LeqaU2lXHj38dtBHt5mexdzQRDTB/s5ybT5VeI5PWzlk+RvTV/ut9m5272c6NF0MU61Rt/Da929L8Vv8NOos2A1TTnVDCNVqT4ewi7anMIMkPtGRoejLMvmOtn3Qfn59j27GZDax2JwlTk4TVmuDOpGSkrx3AAAamQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACCOtJ3FmdElOZGeXFNTfVH2u/akltVv8N+lt+hejE78mavF23Jfbjy9uwhDoSm6dT9Qo0XFNMs+xJ+jwNcCK9kq4R/CzKrZxUuWpbKjcffVKSolq24I47bJ5p3n/AKs7yjodKosi1wpeVT38mo49BXJtDrSVcqntHCcVLLuwlDxJUau/YjTsfLYQt0cP6dq1ceua3T6TSZXm2Hy8lmF9sj1hEqy+G32ZUKKw4giYackJJ8jI9lKUsiSkkdwJEp/7oTlH+SGo/W80WMFc6f8AuhOUf5Iaj9bzRYwAVk6cPvr+qf8AHmMfqZAzXXU+bPTrclv2cmQkn9ezxK/3RhenD76/qn/HmMfqZA7vsgko4/T1IbI9vSLWM3/QhxX+6LvJzti6b6Gn4EdVZoOPSZPSuH6B0PUTB9uWCk95f+8jGv8A3hOOO/8A2frP/g2P9QhCvpN3jXStjtRAxhc6OeCx2Jj5S22iifvJstzSruvzPsn3fWN3o9SG6PAWMizenOhhsRY5ReUtuQuZu2RlwSjvuexdj9+57ERjzUcVSoYlqo7Wgrtp2Vrvfa3n1by5XpTr46pKGrk9Emr6yfDf5G9WVnX00B6ztJjUWJHSa3XnVcUoT7zMUjhRm831Q1NiYiqzm1FxYw8kmV0WM2mQjdhTKHSN1ZJ4qU24oz33Lkn1T9lg63Gcl1mnsZJnrDtZi7LhPV1EZ7Kkl/Bdf+oy9n+gvnaLkCp+DdZiW8ZoWppZbp2UZqEmQmKjxYss9zIzI0+q0ZbJ+sxz8bGfK9PNUTjh+izzTvpe1rqPVa8uNlo+3gVzJVKdGf12W99MsbNPjo2rXvuXXwzfTfiGMz8bTOs8BkvrmtJklY2jEd1hexmngwW5qSXmZ7kW/wDQQnSLRUkEyOFTwY5l3Lwo6Ebf0EIw0Gv59RpRUKyiti1FRAhGaLF6wQaXPlFF6ydi4flMx1y9XMqz+S5T6J0PpLKVm29kNig24TXsM20n3cP8n/dMhvyXUwXJuAowivicVZKPxPpsrJ7+L04t8TzeJ5SrVrVMVUcpys9Nb36EtNPLiSLleZ4tg1Ydrk9vHgRy3JHM/XcP+KhJd1H9REI0+2fVjV75LCILmGY04fe4sGt5shHvZa8kkfv3+slF5DN4nofTV9iWUZxYv5dkSjJRy7AuTTB+5lr5qSI/Lz29mwkwX9ji8d9u9nD7sX8T/NJbuyP8xUyVq/2jyx6Fv73w7vE0rBdIsOwJS5tfFcnWr/rSLSerxpTqj33Pmfzd9/JO2/t38xuoAOhQoUsNBU6MVFLgizTpwpRywVkAABMbgaPSH/8AS9k/fyqa7/WeG8DRqVSf3Xsm9b/8prv9Z4UcX9pR/N/8yLWH+Sr+X/6ibyAALxVAAAAAAAAAAAI11vhXFpQfBVQ1lTi5kWUyaKYmTZWakEkkyPEI1cT3/g99uX1D0aMxLSvx86+0TlKVxm2GUovCZIk8W9jJjw+/Atv4Xfy+sSEAoLA/+ZzvO91rcDf6rL8nxfeu7+F7eRianFMborOyuaWliQZlwptc92O2SDkrRy4qWRdjV6yvW8z9pnsQywAOg5OTu2RpJbgAAMGQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACCutVu+f6f7eLQ6Z/ug+POr25uNpq3JzlhC9JbN9DaWzJTbhII1IdLfgpJK7bblo/RdUaL41b5BUaUdJOp2kzsmK3Im2WYVLrJTSSvZLDb70h5ZmRrNXAtk7bn5iROr6gtLzRKbKp8yx/G5FBaVl8T+RTjh1Mooktp70WY8RkaGXeBIP6zIhEvSfqDkOtevmUarZHnGl7UhGKRqNGL4XlSblbiW5SnfT5Sk7JSaTc8JBkW+yzI9ti3A3en/ALoTlH+SGo/W80WMFc6f+6E5R/khqP1vNFjABWTpw++v6p/x5jH6mQOj7Isrj0/tF7TvGdi9/wC9pA7+nD76/qn/AB5jH6mQPN9kKaOdpRjtOgz5TslZbLYt/OM+X+lRC5gP/Yi11+jMO2l+leqPV1SRrGn0qwyqiOPIgMG1GkIQZkhRoYImyV79uKtiP/5DUtFtKc/z2ih5PFzJMBmllLYrY0+H6WygtiWa0JWfEvWV5bdjL3l22rrHuXX28dwmDyUskvWjyE+xKEGlB+fuJ3+gTZpZZlNxKvhJxqxp0wYcdsky46WUun4ZbqbJJnuW+579vMfNXydQ5T/aGs6snlhGKsrq7snvT0to+0rUqKniauJjmUrKN1Jrf82is3e6W+2jujVvtU6jmS+S1ToH9vInahKS/wA1IgDqMq9Xsez/AE8yK/t2Zt5OOxoqqRQtqaWnxWSUttZGRGZqSk+PHuR7/ULsivXWYSarHdO86MjIsV1BpZ7pl7GVOKaWX5eZD0ON5Ipzw8oqpNaffk/Vst4HAxrVtjnksylHST4prp6zWenTSqZnNSdpqQi1mUMBxB00CTIWUVxZmrxF+GfmkjJJFt2MzVvuLSxIcSBGbhwYzUdhpJIbaaQSEISXkREXYiGl6H//AHU47/8ADq/2ixvQ35B5PpYDBU1DWTim297vrq+hbkuCKHJ+Hhh6Ecu9pXfH/egAADtF4AAAAAAAOt9luSy5HeTybdSaFlvtuRlsZDTI2i2mcOU3NjYylt9paVpWUp8zJST3SfdffYxu4CGrhqNdp1YKTW66Tt2EkK1SkmoSavvs7HA5ABMRgAAAAAAAAAAAAAAAAAAAAAAAAAAGsXepOFY3ldbheQX8autLhlT0BuSrgmRxVxNKVn6vPcy2SZ7nv23GzjaUJRScla+7rMKSeiAAA1MgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFaOu1OH5bpMrTWXqNphQ5Q7YVt5WQ86v2oEN9MWWlwzdaUSlPNK8NaDRwNKtzLcj7liej+6tLTL7xqenpaJCK1Ki/cjn+PP38VP/tJcS2Z+v8AjcRPOcaKaO6m2DFtqNpXiWUTYrPo7Em4po8x1prkauCVOoUZJ3Mz2LtuZjnB9FtH9Mp0iz040txPF5ktrwJEinp48Nx1rclcFKaQkzTuRHsfbciAETU/90Jyj/JDUfreaLGCudP/AHQnKP8AJDUfreaLGACsnTh99f1T/jzGP1MgevrOiFYsaX1nElHLzeC1xP2krcj/ANI8nTh99f1T/jzGP1Mge3q7sotTfaSWcw/kK/LEWDhe9LBJcP8A0f2i5gZKNbNLclJ+EWQ4iWSnmfC3qjVM7u6PI9XdQra1uIkdFDj8uqrW33koU9I8I21IQRn6x8lO9iL2kLM4Nf0d9jkJykuIdgmNHZZeVGfS4TbhNp3SrifY/qMRD0uYfBtMMs8wyOqhzZN/ZuvkqQwlw+KTMty5Ee26+f8AYJ2h18CuQpqvgx4qFHyUllpKCM/eZEQ8b+z1Cs4PHSatVvK1tdW2tb9FuBrSpzjllLo3cbu8n4N27j0CEes+mdtunHLpLDZOuU8dFsllZbtu+A4lZksuxmRERqLYyPklJ+zY5uGia7Loy0ZzZjI7OLX18uimQ3ZMpwkNoN5pTSN1H5bqWki+syHoMRBVKUoS3NMv4Wyrwurq69To0axSvpcTqbinyS8nQLOsjyGIs2SlxllLiSc3bSSC4n63v9okIV96QNatPsu0ewLDIuZVb2Tw6FmM/VFJScpHo6PDUZo8/JBK/AYsENMJGnCjGNLdYxiMI8DVlQy2s2l2XAAAskIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAavqDpphOqNIqgzehj2MbubSlFxdYV/GbcL1kK+sj7+R7l2EOpg669Ohb1apuqGAs//hnFb3Va1/MP/r0kXs8+3YkEW4sUAtUcXOlHZyWaHQ93dxT60RToqTzLR9P+7zTdNdXcB1ZqjtMKvWpSmyL0iI58nJjK/iuNH6ye/bfuR+wzG5CJ9SenXEs3tCzHHZsvD8yZM1s31OrwnVK/7ZBGROkft32UZduW3YanH1p1O0ZfRUdQ+NHOp+RNs5lRsG5GUW/Y5TKS3aV7zSRFv2JJ+YmeFp4jXCO7+6/m7uEu6z6jTayp6VV3rd39Hp1lhAGMxzJsey+pZvcXuYdpXyC3bkRXScQf1bl5GXtI+5e0ZMUGnF2e8nTTV0AABgyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAFc6f8AuhOUf5Iaj9bzRYwaPF0mo4utVhriixnHb2ONRsXciGaPRkx2JLshLhFx5czU8oj9bbYi7b9xvAArJ04mSeq7qoUoyIiu8YMzP2f+hkDDdckhE+fp5EiuodQh6ykrNtXLY0oYSkj295uDIaCJr19TvVki2Jo4KrXHCkk7twNr4FTz5b+zbfcR5lzenEvqLpK3H5lVHxh+dFSa2HEpjElDRLd3VvsR7oMtz9p/WOHy7j62EobHDpOVVShvs1mi03a3BXe9WKmMi6ihSa+Gcoxb7X2dCZcLTXHSxPAaHHvDJC4cFpLiS9jhp5L/AM41DZRwQ5HYo0o0KcaUN0Uku4uzk5ycnxA1/UDKqnBsHvsyvmFvV9HXSLCS0hJKU420g1mlJH2Mz22Lf3jYBr2oNnitThdvLzhhT9CqMqPYNFEcleIy78mpBtNpUtZHz2MiI+xn7BtLSLNqSTqRTV9Vot77Ci/QV1UaSYhjVjplm0iJi5xpsmyr7KfISTT7bzhfIGrYuC0lt7dlFufbbY7u0GrOluUpJWN6jYzZ77dotqw4rv5diVuKvdJ+AdMabbKsfg4D6ZZ2l1YHAbu6CY838ENr3j7LlNG2j1T9pktR7b7nsJuvekPpoyLmdjo1jiDXvucOOcQ+/wBbJo2FHCbeNFJNOx6Hlp8n1cbOeWcL67k+12bXHrJcbdbdQTjTiVoV3JST3I/yj9ivLvQvonEIzxCfmuJL9i6TJpTRp9xl4ill2/AOv7lnUekIvtH6tNS4PEvURbuM2qCP2dlpTuX1CztKq3w8H72OVzfBy+Stb80Wv+rkWKAV0Rp51tY+slVHUBhmTIT/ANXe4z6KR/hVG9Y/6Rx9tfXXQL4WWlGmuUoT/Cqbp6EpRf8A9g9iMNvb5otd1/S5n6PUvs60H32/7KJYwBXUupLW6lT/AOuXR/mrXH5y6Owj2hH9ZJRxP8g5V1w6aVSSPOMD1JxAyMiWdxi76EJ/7yOW5Bzmlxdu1Nepj6Lxb+SOb8rUv+rZYkcbl7xClF1pdMOQKJuLq7UxVn5psEOw9j9xm8hJf2iveuOp3SGvXbDHnsiTaw7KVJk5RMq7SS/ENLsdSI3JbTvFPF0kqUlHzSLcy9g1qYqnGOaMk+8kw/JGJrVHTnTmtG/lb3K/G3/6XwARyzrd0/4xXxqhGsOExY8JlEdlpeRRlKQ2hJJSXdw1HsRF3Pcxi53Vp01VxbyNasVVse3yE5L3+z3Erq01vkvEqRwWIn8lOT7mS0AgWZ109LENXA9VY7yvYTFdLd3/ACpa2GP+726eX1m3U2OS2qy/gwsdlrM/6UENOc0fvrxJlyVjnrsZfyv2LFANX031CqNT8VYy+jr7aFDkOONoatISor5GhRpMzbV3IjMux+0hjdWNR7zTirgzqHTHI81fmSDYOLStpUtgiSZ83DUZElO5bfhMSOcVHPwKyoVHV2NrS3a6epvQCuyuojqCmFvSdG+TuFuZbzr+JFMvyGRjj91frKnf+w9LFLCSouypuZRlcT29pILcR84hwT8H7Fr6MrcXBf8AOH9xvPUZiyLrTDIL9rJMlp5uOUtlYQ3Ka6k1/J5EdS0m74K0+IRGgtiVuXn7zHv0PxCPjOCVk9vIcjtn7qBDnSXbq6k2CkuKZSZ+Gb6leGkzUZ7J2IVT6rcj64bPRu0+F8Co8fouZfCyscsXJU44nFROEvY+zHcuZp77F39XkJi6C3dVXtAIKtVisvHKY4VOdilRSDreDfhGrl6xp5eJxNX8Hjt22EEK0Z4jLle7e0X6+BqUOS1VdWLWe1lK/Dq8SxgAAvHAAAAAAAAAAAAA632GJTDkaSyh1l1JocbWklJWky2MjI+xkZewdgACDMi6Z26S2ezHQTKpGAXrp83ojJeJVTT9zsY90p9xGkti8yTv3Hlr+o3JNPpbNB1HYM/jTi1k0zkNchUmokqPyM1J3U0Z9ux7n5mfEhPo886BBtIb1fZwmJcWQg23WH2ycbcSfmSkq3Iy+oxeWN2iy4mOddO6S7+PY7kDoZXek7enh7HRSXtLklazcY/bRLKDILk1IivJdbWX1KSZkPeILu+l+JR2D+TaE5lZae3Dp+IuNGV41ZJV7nIy90l7i29VPsSPGnW/V7S397a7aXPS65rseS4sRyY3Eu3N5k/XaL2mZ7fUkZ5nGtrhZ5up6S8Nz7nfqMbZw+1Vuvev8d5YABqOCatab6mRik4RmFdaK481R23eMhsv57KtnE/lSQ24U5050pZZqz6GTxkpK8XcAADQyAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEV4zqzd3fUdm+jUiugoq8Yx2nuI0pBL9IddmOSUrSszVx4kTCdtiI+57mYlQAVg6eEG71T9VjaVKSarnGiI0+Zb0yPL6xUbMMh+0d2Bd2FI1Zxay4iOTkLQy+34SHiUtJtu9nTUSVI47e0+5bC2mgSJ7nU91Yt1bzbMxVrjaY7jpboQ6dKniai9pEe242jHNGdXIObu5Xd5Lij6bJ+OuyJFalSnENH/AJTRElRkZ7mWxmZ7mY8p+0PJ1TG4rC1KcZPJJt2SaV7b7yjxXXpfsaFPCuvTrYiLbhK6s2uD36Pjl8OhskHSjW3TLWmm+GNPMojWHhkRyYij8OVFP3Osq9ZHfct9tj27GY3sQrqt0qYDqHcFnONy52DZ0yZuMZJQL8B9Sz/APfoTsl8j8j32UZduW3YaOzrvrX0+OoqupvEzvsaSom2s8xuOa2kpM9iObGIt2j7lupJEW/ZJL8x6LbSpaVlp0rd39Hp1lrYRq60Hr0Pf3dPr1FogGEw/NcSz+jYyXCsigXVZIL5OTDeJxG/tSe3dKi9qT2MvaQzYsJpq6KzTTszgcgAyYAAAAAAAAODIj7GQ5AAYC8wDBMmIyyPC6K13/ltcy//AK6TFHOoTph6bLHqBw+PCzOlxSvu5EiPkNXBkxY7UM40ZTqVcdyJhTppJB7l5nuXc+/0CGt2GmmnFtNesrXT/G5suQrm9IkVTDjjivepSkmZn9ZiviMPGvGzSOnybynV5PqOcZyWjWj6e3o3moU/TZ03IiR5dTo5hD7DjSVMvlVR3icQZbkolmk+RGR777nuNkhaO6SVvev0vxKMfvapYyT/ALEDa40aPDjtRIjDbDDCEttNNpJKEIItiSki7ERF2IiHYJVTgtyRUniq8/mm32tmOh47j9ckkV9FXxUl5EzGQgv7CGQSlKS4pSREXsLsOQG9iBtveAAAMAAAARX1I5nVY1pZkVJMhXEqZkdLZV8BuuqZM3k+qMpKUrNlCibIzWkiNWxefuMe/QvOKbMMCrItVEuY7tLAhwZabKpkwjJ1LKSMk+OhPMi27mncv6RIoCPI8+a+hY2sNhssrve97/pb9QAAJCuAAAAAAAAAAAAAAAAAAAcDkABGGddNmj+fyTtLHFm6235eIm0qFnDlJc/jmpvYlq+tZKGpp0u6kMAPfTnWeNk8BvuitzCMbi9i9npTe7iu3+CXYT2AuQx9eEcknmj0SV13X3d1iGVCDd1o+rQgYtdtZMS+S1M6dL1baO652MSEWTak+1XhF6yCL+coZKm6vdCbOR6BY5TIoZpbEuNcQHoykH7lKNJoIy/whM4x9xjuP5FH9DyCjr7Ng9y8KZGQ8jv59lkZDbb4Wp89K35ZW8pZvVGMlWPyyv2r2sY+g1CwPKiI8ZzSjtd/Iodg08f9CVGYz+5CKb/pX6f8jM1TdMquOo+/KAbkPb8jKkl/YMB9yJiNYX/qVqPqJi3HYkN1t+smiIvZxUkzMvyhs8HP5akl2xXqn+gzVlvin2P/AATsAgpOhmt1PseL9Ud+RIL1W7epYn8j+tS1b7fkMZPFsW6n6nIIDmS6o4tfUqXkemtqqDjSFNfwuBtlxJXu37DEsLSs3GtF/wAyfnG3mZVWV7OD8vcmIAEM5DqH1I0tzYx6rQCDe1jUp1EKUxkrEdbrBKPgtTayMyM07GZewzEFGhKu7RaXa0vVo3nNQ1d+5N+hMwCCl6662wkF8KdK+QpX7Sh3MeSX+akcK6j88Y2KV0x6iEZ/+5jpd2/oFj6OxHCz/wCUX+pHzmn1+D9idgEG/dPzWC3senfV1n3+Fj5OEX5fEIdKurelQfFzRPV1Hv5Y0ktv/wDUPo7FcI+a9xzml0k8AIKT1b4yZevpLqoj6lY0f/ycH6+61xf6KtUf0aP/AJgx9G4r7jHOaX3icwEYae6+U+ouRfa5BwDOahzwFv8ApVxTejRtk7erz5n6x79i29hiTxXq0Z0JZaisyWE41FeIAAERsAAAAAAAAAAAAEEdZV9nuO6UwZuFW2T1EBd9DZye0xiCUu1r6VSXPGejN8VnyJwmSUpKFGlClqIj2EedCWrep2raHLDIX8pl49VYTjtdJmX0Bcf0jJEek+nrYU4lK3S4HHJxfcuZFt57mBt2Aff26u/4jYt/tp4sYK54B9/bq7/iNi3+2nixgArJ04ffX9U/48xj9TIFmxWTpw++v6p/x5jH6mQLNgAPw8y1IaWw+0hxtxJoWhaSNKkmWxkZH5kY/YACu2YdI8aovH9QOnDLpGmWUuHzejxE86ewMv4L8Q90EX1pLYtzPgZ9x4qTqtyLTi1j4b1W4K5hk55ZMxclgpVIop6u+xk4W5smfnxVvt3NXAhZYeG7oqXJauRR5FUw7OuloNt+LLZS606k/YpCiMjFd0MjzUXZ9HDw9iysRnWWssy6eK7/AHOyrtay7r49tTWMafBlIJxiTGeS606g/JSVpMyUX1kY9QrPadLOZ6V2EjKelDUBzGVOrN6RiVwtcqkmK8zJJHuthR+8tz8iI0kPdivV7BpLpjBeo7Dpul+SuHwakTT8SnnGXmpmWXqEXv5HxLfbmZjCxGV5ayyvy8feweHzrNReZdHHw9rligHVGlRpsdqZDkNPsPoJxp1pZKQtJluSkmXYyMvaQ7RZKwAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEKdWUnP6bTyqzLTzK6GlmYtkES2lt314dRX2EZKHWzjPydyJKDcdac4q9VfhEk/MRl0SUdjSZPk5J6iKzU+PNoaiXdORcubuSTkLjstUx5ltKjOLHNKmW0EZJ5+FvsXEb31l0um87T/GMi1ayPH6vFcTzGsu7Fi9jqkRLJlBOtKiGylKjdWpLylIQSFbrbTuW25lFnQDppXYJe5FLqcwwGfXTsVx/wCCI+MVsiG/PqecxcO2sEPoT++nkLUhXDcvkfW7kQAkTAPv7dXf8RsW/wBtPFjBXPAPv7dXf8RsW/208WMAFZOnD76/qn/HmMfqZAs2KydOH31/VP8AjzGP1MgWbAAAAAAAAAGIynEcXzemfx3MKCBc1kktnYs1hLravceyi7GXsMu5ewZcBhpNWZlNp3RWSV0x6j6OSHbvpU1IdrIfM3XMMyJxcupePzMmlmZuMGfv3Mz37rIhkMb6wq2ht2ML6jcKstLshcM225E75aomGXmpmYkuPH2+t6pb7czMWLGLyTF8bzCofoMroYFxWyS2diTo6Hml/hSojLcvYfmQr7B09aLt1b1/juLPOFU0rK/Xuf8Anv8AE9kCfBtYbNjWTWJcSQgnGX2HCcbcQfkpKk7kZH7yHoFaJ3SXkmm0x6/6WdUrDCnXFm65jlktU6kkKPzLw18lNGf8YuRkXYthxG6sMw0vkN0/VLpPY4mRrJtOTUyFT6V4z8lGaN1s7/xT5K95EHOMmlZW6+Hj72HNtprRebq3Pw491yzADB4hnGH5/UIv8Jyatu69zsUiDIS6kj/iq4n6qveR7GXuGcFhNNXRWacXZgAAZMAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAV363XqaDp7iFvY5ZJxSfAziqcpcg+D2ZkGqsFk602/YNuuNpKJxdcQtfLdBrQotzIYHor0Ya03Ows7fXjGtSLqJj9TisVNAlluPV1MI3jYbUSHFqWta3nVGte3lsRbENh62zhtaY49PyDTeNmeKwsvrJGURnaP4Wch1JeIT8lqNsZmtO6UmoiM0ocWoiPYaX0Z2emN5qpqTbaBaUQcd00XX07UG3bxVVMuZYJOR6Qwk1oQt9tBG0rdSSNClqLyNIA23APv7dXf8RsW/wBtPFjBXPAPv7dXf8RsW/208WMAFZOnD76/qn/HmMfqZAs2KydOH31/VP8AjzGP1MgWbAAAAAAAAAAAAAAAAB1SI0eYw5FlsNvsvINtxtxJKStJlsZGR9jIy9g7QAFfss6MNOJNwvL9J7e30uyc9zKdjT5sx3D332ci7k2pH81PAj9u4wxZx1g6LbIz/A6/VzHmvnW+Ll6NaoQX8JyGZbOK9vFotveoWaAV3hop3pvK+r23FlYmTVqizLr3+O8h7Tjqz0N1LlfA8DL0Ul6lXhu0t8j4Pmtub7eHxc2StX1IUoTB59yGkaj6I6T6txTjah4JVXCuPBEl1nhJbL+Y+jZxP5FEIf8AuXtVdMflunTX65q4jfdGO5Sn4Trtv4iFmXNlPl81Jq8+/cYzVqfzLMurR+D9xkoVPleV9eq8V7FmAFafuidfdMi8LXfp1sZcJrsu/wAIdKwjGRea1RzPxGk+3dai/AJA086ptA9UFNx8V1KqvTlnw9AnrOFK5+1JNPElSzL+byL6xtHEU5O17Poej8zWWGqxWa110rVeRKwAAnIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAgjrKk6gxdKYLuDSczjQfh6GWUP4W2Tl41S8XPGVDLY1cyc8DkaCNRN+IZF2EedCWd6v6gocssyhZzHoarCcdp3XcrjOsuTcgZ9J9OkR/G9daTQphK3D+ctJeZkZjRer3P9GrLqbZ0y6l9a73CtP6XDoltAqquZLipt7OTKkIW4+uMhS1JaaYQSUntsazMj7qI9o6DtRMKtNQtUNMdINW7nUDTeii01rQybaQ/IfrXZPpKJMMnX0pcU2SmG1pIy2TyPzMzMwJAwD7+3V3/EbFv9tPFjBXPAPv7dXf8RsW/wBtPFjABWTpw++v6p/x5jH6mQLNisnTh99f1T/jzGP1MgWbAAAAAAAAAAAAAAAAAAAAAAAAAAABH+oegOjWqqVqz3TqmtJCy4nLNjwpRF9T7fFwv/EJAAayhGatJXRtGcoO8XZlavuQslwX5bQLqEzPD0N+s1VWLibatSf8UmXduJH7z5H/AKByWXdb+nPqZJpnh+p0BrucrH7E66YpPvU28XE1fUhP/mLKAIebRj9m3Hs3eDuifnUpfaJS7Vr4qz8zU9MM4n6h4jHyazwm8xOU666y7V3LHhSWjQo07mXtSe26T9pHuNglXFTBmRq6baRI8qaSzjMOvpQ49w25cEme6tuSd9vLcvePYNA1Z0I0r1viQompmLItvgzxThOlIdYdjG5w5mhTakn38NG++5eqQledQ01fgRLJKfxaLq1N/AVtLo0fxg+WknULqXh6E924arP0+Cgy8vkHNtyLbyNRgWIdduIOF8C6r6e51GbLYk31Q5XvLItv5MW3I+/c1f0iLbVI/PB91n/nyJdhTl8lRd917rzLJAK2o1r6usaUacx6T2LdhPdUvHcmYV2278WF8lqP8pflH7+7Ur6lRNZx0/6vY6stvFcdxw3o6PefiJXuoi95JDnVPjddqaHNKr+Wz7Gn+pY8BHukGvGnOuUOxl4BYTXzqVttzWZcF2M4ypwlGjcnEkR78FfNM/LuJCE0Zxms0XdEE4Sg8slZgAAbGoAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAB5pNZWzVk7Mr4z6yLiSnWkqMi925kNM1S1MwLQPDndQMxjO12PMSo8axnQoXiJhJdX4aHnko9fwycUhJmklGXMj223Mt8Gt6kYHRao4BkOnOTM+JV5JWyK2UW25pQ6g080+5STMlEfsMiMAVaoNe9F8X6p9WdXLzU7HY+HvYFizjFuU5DjEj5afslk0GZurPY9kII1GZGW24tRp/mtZqPhVNnlJEnR62+iInwkzWfCeVHc7tOKRuZp5oNKyI+5EotyI9yL+fbRTpZyfNur6u6aMjiOIercheh36kEZEiFEWpUlxJ+wlNtnwV7TWj3kP6JIkSNAiswYTDbEeO2lppptJJShCS2SkiLyIiIiIgBWrpw++v6p/x5jH6mQLNisnTh99f1T/jzGP1MgWa8gByA43L3kG5H5GAOQAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAHBJSRmokkRn5nt5jkAAAAAAAAABF2r+mWqWeWFdN0812scBahsrQ+xGqWpqJSzURktXiKTtsXbbv5jQntG+saIkkVnV9DkpL+V4RCSZ/hMuQscAhlQjN3bfi/cnhiJwWVJd6T/QrkeBdcsNvjF15wOxV75mNmz/syP8A0D8JoOv9hB8c70dlKLy8aDNRv+VKBZABrzZcJS8Wbc5fGMfBFcWmvshEQjW7I0Jm/wA0/hVJ/wBiSIHcg6+45cSwHSCUftUzYTUl/QtRCxwBzfonLxHOemEfD/JXJrMuu1kt5Ojmm0j6mbx1v/WUY5d1K62WC2+5rxWSZeZs5a0kj/IrYWMANhLhN+XsOcR404+fuVtTq11sIP5XpMpnS/7PNIaf9Kh2/uydYyC9fo8hq/wc7g/+RixoBsJ/iPy9ht4fhx/q/uK2q1x6v2z2V0ZJV/g5xCP/AHATrz1a/wALosf/ACZtC/4BZIA2M/xH5ew29P8ADX9X9xXFOvPVX359FswvdtmsE/8AcHYnXnqh29fowsSP6szg/wDALFAGxqfiPy9ht6f4a/q9yu37vPU//wDswsv0yg/8IzGI6ydQl3k1bUZJ0pz6CrlyEtS7NeVQ5CYjZ+bhtoTyXt7i7icQGVSmn878vYw61Nq2zXjL3AgTKNaepSpyKzq8e6SZd1XRJjrEOxLLobBTGUrMkPE2pBqRyIiVxPuW+wnsBvUhKfyya7LfqmR05xg7yin23/RorarXnqz/AIHRZI/Lm0L/AIB+S116vV/M6LzL/CzeEX+4LKAI9jP8R+XsS7en+Gv6v7iuKdaOsNwt09HEZP8AhZ3C/wCEcOav9Z6v+h6Q61H+HnMI/wDyFjwDYT/Efl7Dbw/Dj/V/cVtRqr1trPv0rUDZfzsxin/oUPR+6H1tvl8l084bGP8A7fKkr/1RYoA2EvxH5ew28fw4+fuQrp5kvVlZZjAjalacYPUYysnfTZFfaOPym9m1G3wSZ7Hu4SCPt5GYmoAEsIZFZtvtIak1N3SS7AAANzQAAAAAAAAAAAAAACJca6b8IxjqPy3qTgtEV9llHCp3m/DIiaU0o/GdI/e6huIky27eAZ7nzPbb9UbfUSjwiws9KcRgZPk7Rs+h1c6wKEw+RupJzk8ZHx4tmtRdu5pIvaNrAAVs6UMB1tptRdYtUtbMKqsXn6hWVRLi19dbInttpiwvR1fKJIj78Un3IvnH7hsXWdpjm2r+gN1gunsBEy7mSoTrLS5KGCNLchC1+usySXqkftE4jRtX9btLdBcbjZfq3ljOPVEuaiuZkux3niXIUha0t8WULVuaW1nvtt6vn5DWcFOLi+JPhsRLCVoV4b4tNX3aHys+Lt6vf/0XF/P8T/mi7P2PrQPVbQbGswrtU6dqBItp0V+GlE5qTyQhtZKPdtSuPcy7GM38ZL0S/TrB/NFj+zh8ZN0Sn2LXaD2//ibH9nFalgqdGeeLdzu8o/tRjOU8O8NWjFRdtyaejv0sswArP8ZL0S/TrB/NFj+zh8ZL0S/TrB/NFj+zi2ebLMAKz/GTdEm+37u0Hf8AFNj+zh8ZL0S/TrB/NFj+zgCzACs/xkvRL9OsH80WP7OHxkvRL9OsH80WP7OALMAKz/GS9Ev06wfzRY/s4fGS9Ev06wfzRY/s4AswArP8ZL0S/TrB/NFj+zh8ZL0S/TrB/NFj+zgCzACs/wAZL0S/TrB/NFj+zgf2SbokLz12glv2/wDqmx/ZwBZgBWf4yXol+nWD+aLH9nD4yXol+nWD+aLH9nAFmAFZ/jJeiX6dYP5osf2cPjJeiX6dYP5osf2cAWYAQJol1l6UdQ2q19pzpRKVd19DSR7dy8QlxplxbjxtmwTTraFkpOxGavI+XYbH1KdQ2O9M+C1uoOV1j0urmX0Glkrbc4eiIkKVykK9VRqShKTUaSLc/YAJYAVn+Ml6Jfp1g/mix/Zw+Ml6Jfp1g/mix/ZwBZgBWf4yXol+naD+aLH9nAvsk3RKZblrtB2/FFj+zgCzACDOmfqxwzqhs8+bwSCfwThlqzXxrInlqTaNOtqWl9La20KaLdKi4nufYTmAAAAAAAAAAgbqc6s8X6XLvT5rNqlS6HMrGXCnWiFuqVWIZaSsnCYaacW/yUok8S228+41H4znow+lCx/Re1/ZgBacBVj4znow+lCx/Re1/Zg+M56MPpQsf0Xtf2YAWnAVY+M56MPpQsf0Xtf2YPjOejD6ULH9F7X9mAFpwFWPjOejD6ULH9F7X9mD4znow+lCx/Re1/ZgBacBU2N9lI6MZE+XBPUO3bKJ4ezysasTQ7yLf1SSyai28j5JT9W5dx6/jOejD6ULH9F7X9mAFpwFWPjOejD6ULH9F7X9mD4znow+lCx/Re1/ZgBacBVj4znow+lCx/Re1/Zg+M56MPpQsf0Xtf2YAWnAVY+M56MPpQsf0Xtf2Yanql9le6ZsQwqbfaeWM3Nb5hxhEalXXz6spBLdSlZ+kvRTQjig1L2MvW47F3MAXTAfhlzxmW3dtuaSVt7tyFQaP7Jz0/wshzfE9WJknD7fEcts8cZjswZ1mU2NFcJtMs1sR+LZrUTheEZmaeHmZGQAuCAqx8Zz0YfShY/ova/swfGc9GH0oWP6L2v7MALTgKsfGc9GH0oWP6L2v7MHxnPRh9KFj+i9r+zAC04CrHxnPRh9KFj+i9r+zB8Zz0YfShY/ova/swAtOAqbK+ykdGMWXEilqHbu+lKUnxEYzYklrYt91EpklHv5eqSj9+w9fxnPRh9KFj+i9r+zAC04CrHxnPRh9KFj+i9r+zB8Zz0YfShY/ova/swAtOAqx8Zz0YfShY/ova/swfGc9GH0oWP6L2v7MALTgKsfGc9GH0oWP6L2v7MPy79k76M0NLW1qZYLWlJmlP2sWpcj92/o3YAWpARf0y62l1G6HYxrOnGTx8skRKX8GnM9K8DwZTrG3i8Ect/C5fMLblt323OUAAAAAAAAABGGuWiTetLmny3cj+CU4Jm9ZmXH0P0gpvohOl6Mfrp8Pn4vz/W22+ae4k8AB0egwv5Gx/VkIz0p0Lh6ZZtqXmJ3bdn+6JfN3foxwSaKBwjoZ8Ilc1eJvw5ctk+e23tESdQXVRkldpneRsCpbTHcnqbJmPlBOFGl22J0S5Ztqu/QEG56Q2phta0EXzScQtRcS7750y5LrHPhT6DUe0r84oWI7Fhi2o1YthtnIYD/ACNCHo7Z/JyWyT6xoLgojSfY/nATX6DC/kbH9WQegwv5Gx/VkOx55mMyuRIdQ000k1rWtRJSlJFuZmZ9iIhXHq1y3qTwbF1ap9P8yuv6BmmkR7SoRBRIlRjc7t3ENRbnIUyXdUc90rSRmRGfcgN8xnQeHjuv+aa5/Djcgsvpqyp+CTgEkononP5QneZ8+fPy4J228zEnegwv5Gx/VkK99K+Z6uW6jr8hy2Jqpp9a1xXOLakRUx4rryTWSV186KjjxkIM1bLSnbZCiWSVkZFYsAdHoML+Rsf1ZB6DC/kbH9WQ7wAHR6DC/kbH9WQegwv5Gx/VkO8AB0egwv5Gx/VkHoML+Rsf1ZDvAAdHoML+Rsf1ZCNdcdDYes0LDIZXTdL9qOaVGX8kwSf9K9BdNfoxlzRwJe+3P1uP8Uxms3myc5x/LcC0t1Qr6LN66M2g5UcmJr1O+6XNhUiMrlslwkH2URGaTUae5EZU+wnVLrBsNUcgXa5FDc1IxSWym00geSzFrLnHvDaR8JVExwiUpxTviuc1rMk8iaWSdiJQF8fQYX8jY/qyD0GF/I2P6sh2oNSkJUpJpMyIzSfsP3D9ADo9BhfyNj+rIPQYX8jY/qyHeAA1iDppg1Zn8/VGBj7LGUWdazUS7BLjm7sRpZrQ1w5cC2Uoz3JJH37mOc901wjU+vrqrO6Fu2iVVpGuobS3XGyamx1cmXfUUkzNJnvse6T9pGNmAAdHoML+Rsf1ZB6DC/kbH9WQ7xirDKsYqbusxq0yGtiW92Tx1sB+UhEiZ4SSU74LZnyc4JMjVxI9iPuAPPl2JwsqxS6xjZmL8MV0mB45MEvwvFbUjnx7b7ct9ty328yGu6H6Sw9GtIsT0rOxauftXq2a34QVDJg5PAtuZt8l8d/dyP8ACKl6tudTeF6qU+nGoPUdcYtiVpJmvYLncGvYS1KuXnW1xKq9Tw8PghJPoQSSQh4lJ78kGkri6T2WpNtgNVL1dxuuo8tJC2bOJXS/SYqnELUgnml+ZIcSknCSe6kkskqPcjAHVgmkOCabZBl2T4jVuxJ+cWSba6WqQtxL0lKOBKSlRmSC4+xOxDcwAAAAAAAAAGp5bpZg2c5RieZZRTHMt8IlvTqN/wBIdb9Ffdb8NxXFCiSvdPbZZKIvZ3G17F7hyAA42L3BsXuHIADjYvcGxe4cgAONi9w/LjjTKSW64hCTUSSNRkRbmexF+EzMiL6zHEiQxEYclSnm2WWUG4444okpQki3NRmfYiIu+4rr11XdG/0t209yqh3dNaSqttVoTrqo9Qw9Ja43BLjmTikx90vEbai34l347gDftP8AQuBgWteqWs7GQvzJGp50xvwFx0oRB+D4px08FkZmvmSuR7kW3l3En7F7hWbo/wBXc+yKdkejWcX8HUL7RW4qIepFItC6+7ZdZbcQ0/xWrhMSh1BqJJqSot1Ge5bqs0AONi9wbF7hyAA42L3BsXuHIADjYvcNQ1Z0lwPXDBZ+m2pVMu0x+yWw5JiokuxzWpl1LrZ82lJWWy0JPsffbY+w3AAB+UIS2hLaC2SkiSX4CGqae6V4LpY3kLWDUp1yMpvpeTWpHIde8eylcfHe+UUrjy4J9VOyS27EQ20ABxsXuDYvcOQAHGxe4Ni9w5AAcbF7gPiRGZ7EReY5GHqMrx7KGLJWJX9XcOVUp6ullElodTHmN/PYdNBnwWkzLkk+5b+QAjXKdNcN19zLSjW3G8+jS4Gn9jYWMBdYbcuNZG+ycdafFSrZJINJ9y37kZHsJh2L3D5q9Kmb5/husVZQ4dMdlZbml7YOaq6WyKz4Mh4utCu9pBUSeDbfHw/4Sik809+e3H6VgDjYvcGxe4cgAONi9wbF7hyAA42L3D8vMtvtLZcTuhxJpUXvIy2MfsABqmlml2E6L4FVaZ6dVK63HaVLqYUVclx82ycdW6v5R1Slq3W4s+5n57eRENrAAAAAAAAHnsLCBUwJNraTWIcKGyuRJkPuEhtlpCTUpa1H2SkiIzMz7ERAD0Cs/Vh1E5liapuiWgmLycn1PsscnXK2Yctph+lrkElv01tDqTTJe5ufJMJ7qNCt/IiVpesut2WZ9qphNHpN1DxMEw/L8bXPwzJY0CNOrL/IkSVJXXTHXUn4aSZSji0ngpSnFluakpQNtw2jresPBKrJ9RKqXhOqWl2USaeRbY9JSl6ttYbiSklFeUSicivoNPJtZKSaVmkyM08gBGOiuj95rJkeOak02rVtlOM1KCn4vqI++03lEEifSmfjVwwpHGY0sjd2UtPyaiV2MjSR3bx3G8exGli45itHAp6qCk0RoMGOhhhlJmZmSEIIkpLczPsXmZmPXEgw4DSmYMRmO2txbyktNkglOLUalrMi81KUZqM/MzMzMd4Aj/WvRyk10xSJguVWc5nHztI062gxnDbTbRmjUr0N5STJRNLXwUriZGfhkXkZit/Rtkt5p/m+a9LOEYTZz8ZwfO7nxbOdJWiJQ0jrTb0GKytZKOQ8t5xzZsjLi2SlqV3TvdAfhDTTalrbbQlTquSzSREalbEW5+89iIvwEQAx+PYxjeI16qnFaCvp4S33pSo0GMhho3nVm465xQRFyUtSlGfmZmZmMmAAAAAAAAAAAAAD5xan431NaLZvmWKYrY57EiW9tZ5dgdrhdCie3kF3Nk+KUG+WpKjQhojJlPLg2bfrmr5MkD6B1lMxKVVZNkNFVlk0evKM5LbZSpyP4hIU+y06ZcybNaEntvsfBJn5EM0AAAAAAAAAAAAA0XWfVWJo7gr+Xu4/Y3812VGrKqnrkkcixnyXUtR46DV6qeS1FupXZKSM++2x0i1qO+6vKnGtVcY0MyiFqDhNxb4Db0sWybZscbuFJZkQbBEwiJJMsvtsmp0k7eHJd9VRD6KrbQ4RJcQlREZKIjLfuR7kf5DFeMU6VrSu6i8u1iyTPJErG7G8ayOixqGa2GmrM4DMR2VLUW3jqSllRNoMzQnxFKMuR9gNx0DxjWNjS1jHOpybSZRlMG0fWqxjpbcYmMof8WK+TZMtpaUj1SJPEzI20q5bnsUsAAAAAAAAAAAAAAAAAAAAAAAAAjjWzR1Ot1PWYddZbY1uK+mk/kNVBTwVexUpPjDcfIyW0ya+JuEjutJGjciMzEX9GzUvGaDP+lvLIrkxOlN47U16pbZuIm49NSciv5GouKzJpa2lJ77E2kj8xZcflLbaFKWlCSUs91GRd1Htt39/YgBhMOwTCdO6f7XsAxClxur8Vb/oVTAaiMeIr5y/DbSSeR7Fue257EM6AAAAAAAAAAAAAAAAAAAAAAAACGNYeqvTnQ7UbF8Cz1iyixsjYU9IvUs8q+oJTqWo5zHP+qQ64a0Es/VI0+sZEZmVVMc6eOo/TnX3JJ3TdQuYzWMZE5a3FnLsWmqjLqWXNalsRGUKae2kspdmsqkEkjQ2hCdzNZC/OW4nQ5nj1pjmQUldZw7WG5Ckx50cnWXmll3QtPmad++25fVsfcRp0udO8fpz07axiRlNlkVzJS0c+bKlvustk2nizGiturWbMdlB8EJ3MzIt1GZn2AmEmWUuqfJpBOqSSVLJJcjIvIjP3dzH7AAAAAAAAAAAAAAAAAAAAABjMnx2sy7GrbE7tgnq66gv18ts/wCGy82pC0/lSoxkwAHzr0J6JdXXLeHU6i4pj9Xpa26isyjCbCacljIZkeK6weQx0NpNMR11wmFJQlSDVspwzSfFJ30wPAMM0wxaHhWn+OQ6OkryUUeHERxQk1KNSlGZ7mpSlGZmpRmZmZmZmY2AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAQJZddXSzUZRIwuw1Meau4r7kd2H9r9opZLQ4batuMYyUklltyIzT9YnsVkvKuyV9kcxi4TXSTgI0jnx1SiZV4KXTtWzJBr24krbvtvvsAJ1wLULHtSKubb40mxKPX2Uqpe9OgPRF+PHXwc4pdSk1I38lkWx+w/MbMPmfi2MZ7qDkmmGF5dYZ0zSXGruoca59GsJsRblcls1stOPNqStLKjQaS2URGlSiSZbjz6NYLnOJlopqC1eaiv3idYbHDZJWNtOkMt40g5aEMKYcUaCaLw0nzNO+6t+XZJEB9OAHzO6YUan2mvWNzc+1Am0eo8bKbn7aauVDv3ZdpDM3+MdzlvXIipbJpbK08Up4oIjNR7Hvf2RG9lSMt+06PidwU5ODTrKiuysbsmV2ZOLS3Fgxa75NU4jJDniPnxJHHcuO+4FqM06idOMD1NgaT3blqu6lVDmQS3Y1et2JU1iFKQcya+XqR2eba081HsRl32IyM5BpbmpyOog5BQWUaxrLKO3LhzIzpOMyGHEkpDiFp7KSpJkZGXYyMfMyx0Ikas6g2mQ5VR5hZ3tx0xV12t9FnYsrlZEbRt+Grg4klKVwTyjH6ilGpRoNSjM8K84UHRHp2rsceu4mmqIE5nOfhhOQ+hM5IiDGSTcr0YykpaSvxSaS0ZRyWR7di7AfVoB87cA0wynP9SNBcEz3UjNMqxpnBbyVNs4T9pVNWRsWaVQkSPF4PnwSTO3ibKX4KFesk/WkD7Io69XOYRkTV3JsPgduxfThDjdy1GyVaiaSlLUqt2NqY3/1JOGaTNxR9iJRgC3cDLcXtMitMRrshr5N3SNx3bKuakJVIhofJRsqdbI+SCWSVGkzLuRHsMuPm5IraDTrqM1uzS3041G+2PIdO4N5i9NEtLZL8x5dZJ+EmClNGpCXGzVxSpW5trI/BSS9iOKnLzUVnTLXwtNjvoNRNwvGrarj0r948w1YnaNpfOO7YfLreJHJDqm/VUaD3L1TIgPrFPy3F6rIarErLIa+Ld3qJDlZXvSEIkTEsJJTxtNmfJZISpJq2I9iMtxlh8/tY9ANLtIuovpxtLRnO5ONJK7g2Ns5b28934RWTTsPxXGlqWhTsh101bElKy3SojQjZMldbMptnUXSFjUx7J2dFnX7j7bl0hzkoOaUdHwemWqF8uTRueJsRdjURcuxEAJz1i11wrRKPQ/bRFvLGfk9h8GVFXR1jk+dMeJtTi+DLfcyShJqUfsL3mGsmumFaH11LMyuLdz5eR2Sampq6WtcnTpsk0KWaW2W+57IQpRn9XvMiHz+xPSXIc9tunaZntbncumc1CyiLRvWU+xYnM4uhpx2uN9XNLjSjMj2WrZam+KdzRsQsT1p6T1WoetfTs/Y1d1KYXlcmDPcgTpbCWYvojjhGZsLT4R80l8oWytvV5bHsALdIWTiErIlESiI9lFsZfhL2D9D5f4/IzKZnePmUnUj7o5Or6ivWlqsygN4uUpfiEaT/eXwf6L4fHj6xmfq9gw+RmU7OMPUUnUj7o391tZZa06q0KA3jBSnfFJRK/eZQPRfB4ce+5+r7AB9Kcfy3F8s+EvtYyGvtfgaweqbH0OQl30Sa1t4sd3iZ8HE8k8kH3LctxhMg1YxHGtTMT0ltHZRX+aRrCXVoQwamlNwkIW/zXv6p7OJ29/cfO7G9O8Qw/DepbAGswy/T3KW9QpLjE2UzezIjmPqlxlxfGUyZmpqQaVtuPoV45oVupRo7HkcPwZOv2a9NlTlenmTYrjqYWdR5jVbcWrbT7STZNt1qU+pMtuK+ouSG1qSfH1e6exgfTIB8tcMtrt/BtCofVPZahK0pjVOUw5rzKrQluWzFm61BTOVD+XMkxUkTPLsZl23Lceqrk5rHw/QWP1Zv6lNaVuQskVIWlVmU1Uj0k/ghNkcL988vRuJtEfffz8lAD6HYrqziGY6hZvplSuylXenzlc3dJcYNDaTmx/SGPDXv6/yfn7j7Dch8t7uLl9L1QZll7ldmytFzyHT9WUJQiWzayGPgBCIT0le3iusNPmXpTfzzUaeXcjIWP8Ash/NnEsQuEZWuOipnS5543Jj23wfkxpY2TEefrdnWXiNRKZ5HxUvfcjJJ7AWiay3F3spfwdnIa9eQxoSLJ6rTIScpuItZoS8prfkSDWlSSVttuRkMuPmqrG6Ov6kE5zkGF53i1xl+h0abj0R6dbSlM3iIz6XYjrxKMlLaYSndL2xcy5mknF7nhbnB84wjpl0IyVdnlr9RmUmLP1NlXc+4loJSYSkwkSkRFekMxEqPgaWySndDPPcy3MD6iAPlrn8a8x3RDT66l6rTc9i09hkMmpxR+DlEONfRFutkxEYltkmSUmOfJEdT5mlSV7kfFJmcnrubWt62MZuDg3uVSMldp4ruOynLqNYYKgq1JuOpcTtBlw91Gbxr3UbijLupKjAF+wAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAABxsXuAcgAA4HIADjYByAA4HIAAA42L3DkAAAAAAAAAAAABxsA5AAcbDkAAHGwbDkABxsXuHIAAONi9w5AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAcKSlaTQotyUWxl7yHzcVpZF066ockp9FG42BSZmpFHjMazr4UeQ9Bq5OMPSZTTCZKXEINbzaVcuJmSu+xlukwADHaY9SXVRneR1uIWOsBw5UnLanG8sKFDgvSaOa+/KQ9HjG5BSzwNprcj3lbG0hXinyWgazlnUHqxmOPVOD6s6oW/wpMssecxqsh1kEmMhai5StmXKmuJbJTTqPRWNkoWylRl2Solr2AANyxLqG1S1J6e9cGMgzVOZyMbi0xpnSq+GUdia7OUl+McRUVskKSlCDNPKS0WxGh4z3ISDplqbrbk8aQnOtRp19XZhp5nNg5XPwYLDUCRXWpQ4xsKYaQ53YcPlzUvdXft2IgACu0C86hNDazTwqG5RLZpdH1ZHR3UmQ2t+FWWMioXPZWhaFkpcThMNpRocI21tJJCzRwVurvWzrbjeKLetNTinpy3GnXtP7BitjuqnSGLpSHXnjXFZ2WmHukzcZZSokEZNk4ZkYABksh6iuoLEcEx7Oc316vYCMyr8j+15MGjqloeyOHYKj11Y4k45mmM40k1OOKNO6j7uNkREfTZ9WXUXV5vqHjs7UR1TmPmuTkaIUOEX2oVyLSG084yw5GNL7qIrsjwlJkyfE23caQvikAAGGyrqa1Oq7V/UXDdSpr+ISqmnamZrbU9dAtzplWd02098hFkmZKW1GQR+imvipSvR2lKWottoNeeryZDRlK9R0T7SJkOJ0kOiRXQGq2zbs6cn1OPL8EnkrW4bThcXEEg1rLjx2SkAA1q26xdV0abV+U45rxkEmjRNiQMsyC1pq6Gqou1wHXFQIpsxH1us+kpSSt4aiIkoSl9SVrWiVenzqJ1tyLqiqtL9Q86K2kTaj0u1pIkWPFYrElVx3ie4+jqNxC5Cl8VomGvdzZbKUJJQAAP/Z)
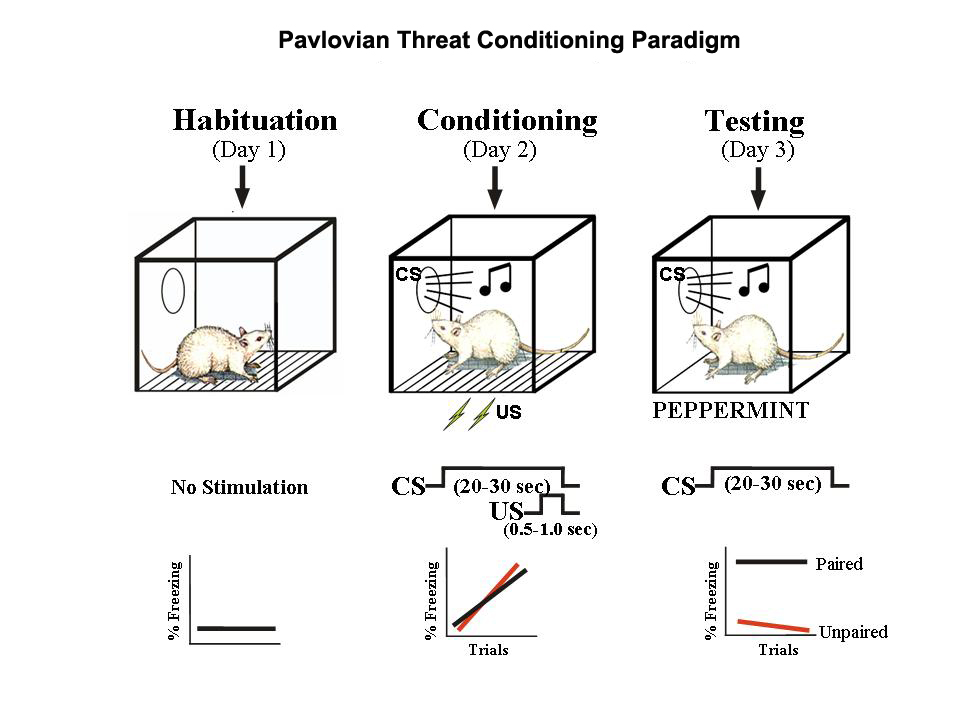{kind=link}
{kind=link}
Figure 3. Graphic Summary of Short- and Long-Term Functional and Neuroarchitectural Effects in Prefrontal Cortex (PFC) Synapses after Acute Footshock (FS) Stress [44]. The fast and transient increase in corticosterone (CORT) release induced by acute (40 min) FS stress was accompanied by the rapid increase in both depolarization-evoked and hypertonic sucrose-evoked (readily releasable pool) glutamate release in PFC, and the number of small excitatory synapses. The enhancement of glutamate release was sustained for up to 24 h, as well as the increased number of excitatory synapses, which normalized between 24 h and 7 days after FS. Before 24 h had elapsed from the start of FS stress, retraction of apical dendrites began and was sustained for up to 14 days. The timing of actual FS stress (40 min) is indicated by the red marker. Number of excitatory synapses and apical dendrite length are indicative and not in scale with other readouts. CORT and glutamate release data adapted from [44].
Changes in neural architecture
- Hippocampus (rich in CORT receptors)
- Prefrontal cortex
Neurochemical factors
- Cortisol enhances glutamate release in rodent model of PTSD–reviewed in (Musazzi, Tornese, Sala, & Popoli, 2017)
- Corticosteroid antagonists block
- Ketamine (NMDA receptor antagonist) may act via similar mechanisms

Sapolsky, Why Zebras Don’t Get Ulcers