flowchart TD N[Nervous System] --> B[Body] B --> W[World]
Action
PSY 511.001 Spr 2026
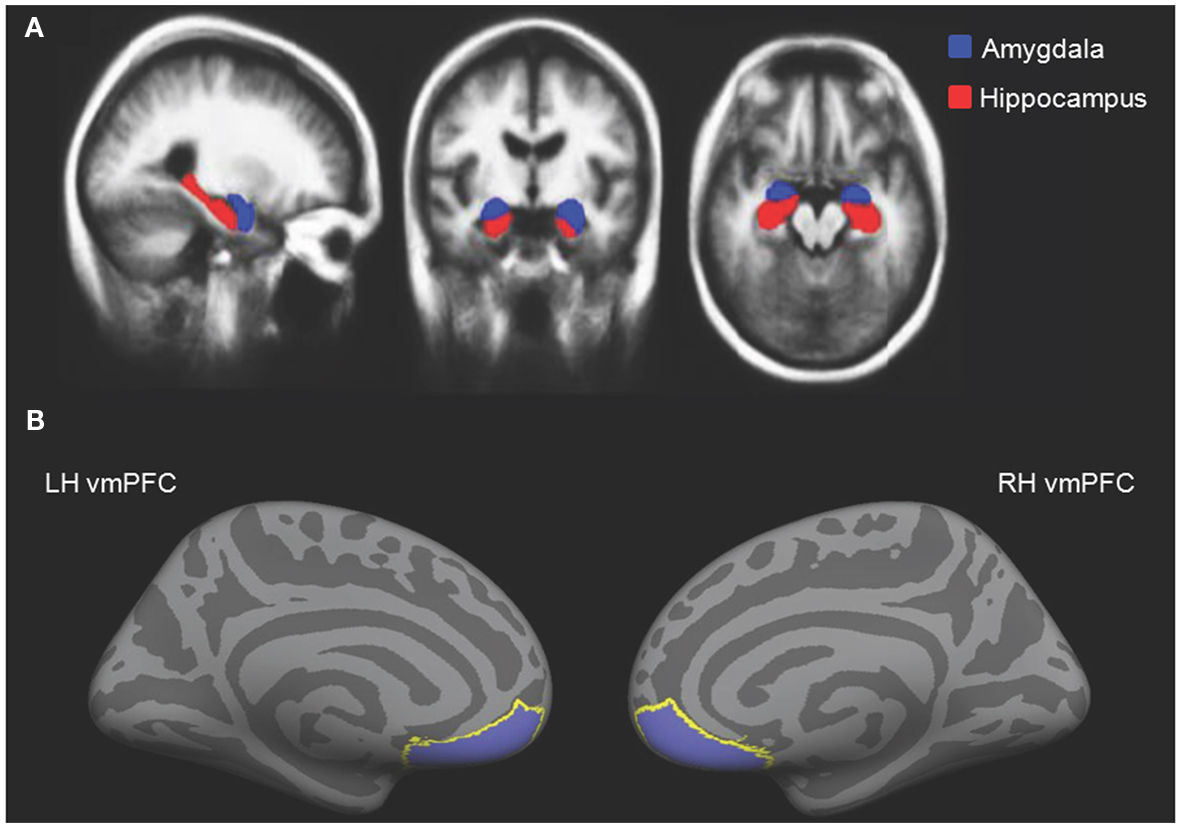
The middle image in A in the figure at right depicts what plane of section?
- A. Sagittal
- B. Dorsal
- C. Coronal
- D. Horizontal
The middle image in A in the figure at right depicts what plane of section?
A. SagittalB. Dorsal- C. Coronal
D. Horizontal
Why action?
The way to do is to be.
– Laozi (老子), Chinese philosopher

The way to be is to do.
– Dale Carnegie

“Quote origin” (n.d.)
Do be, do be, do.
– Frank Sinatra

“Quote origin” (n.d.)
Computational perspective
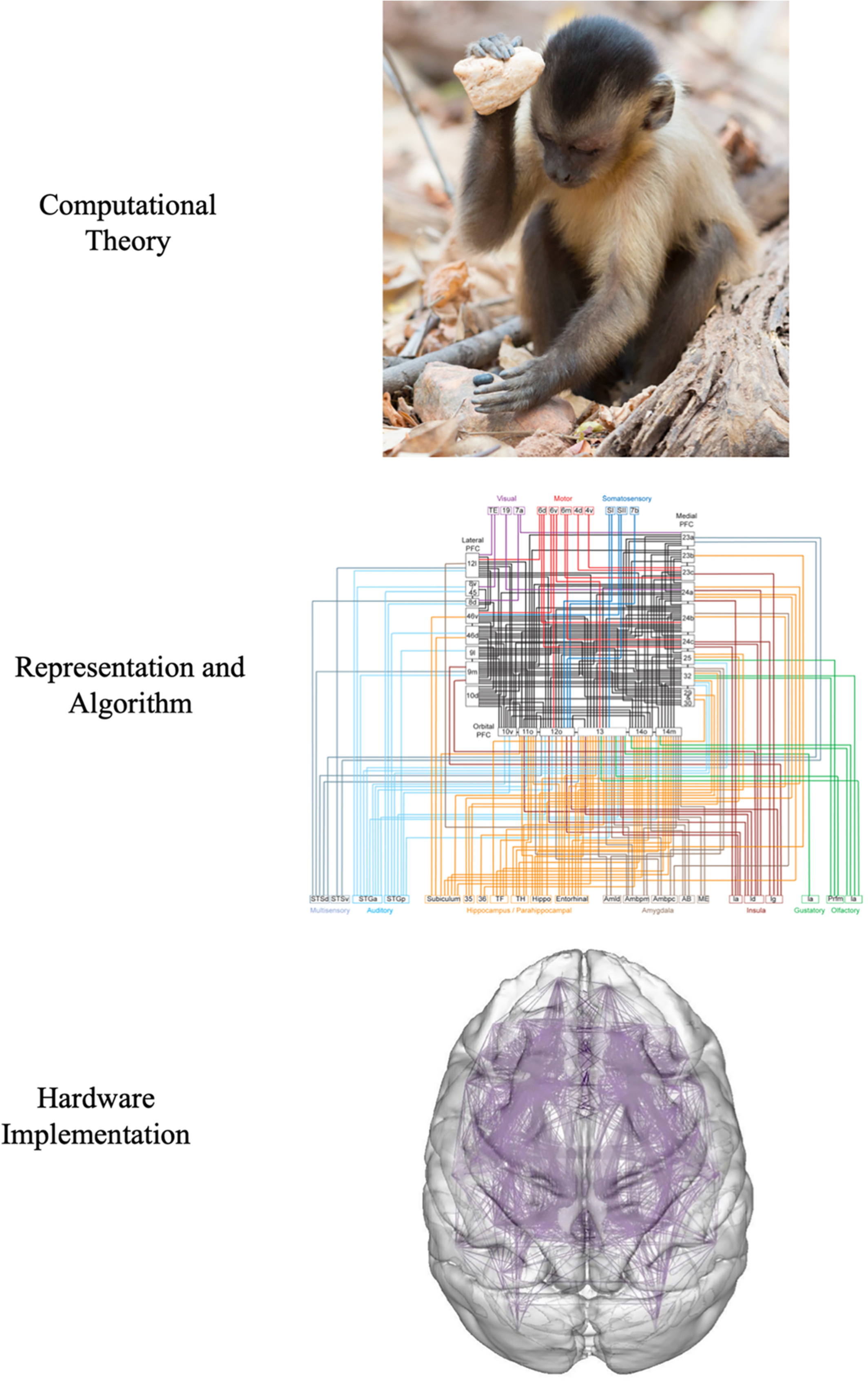
Neuromechanics perspective
Evolutionary perspective
- What actions must animals perform?

But simple circuits…

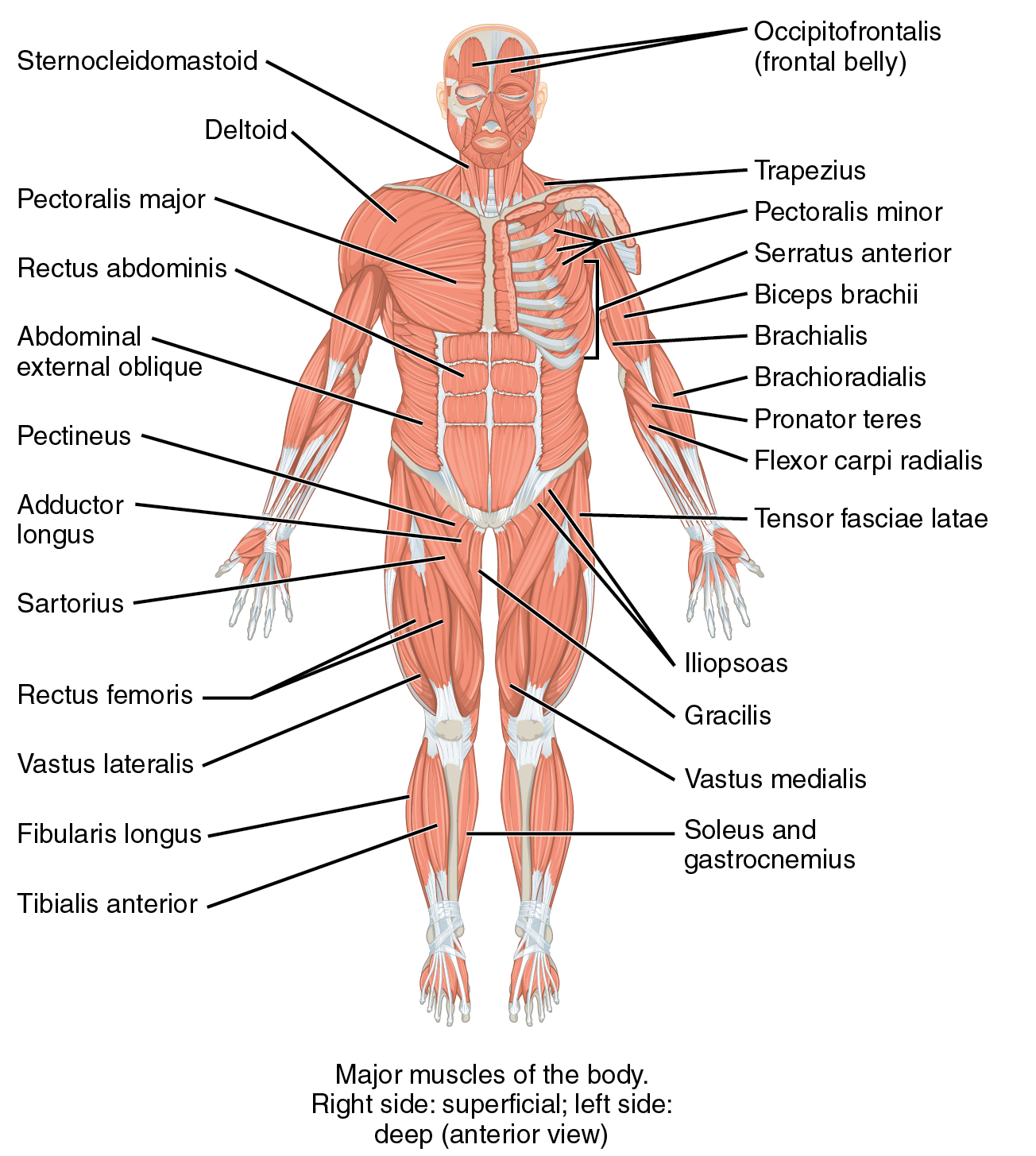
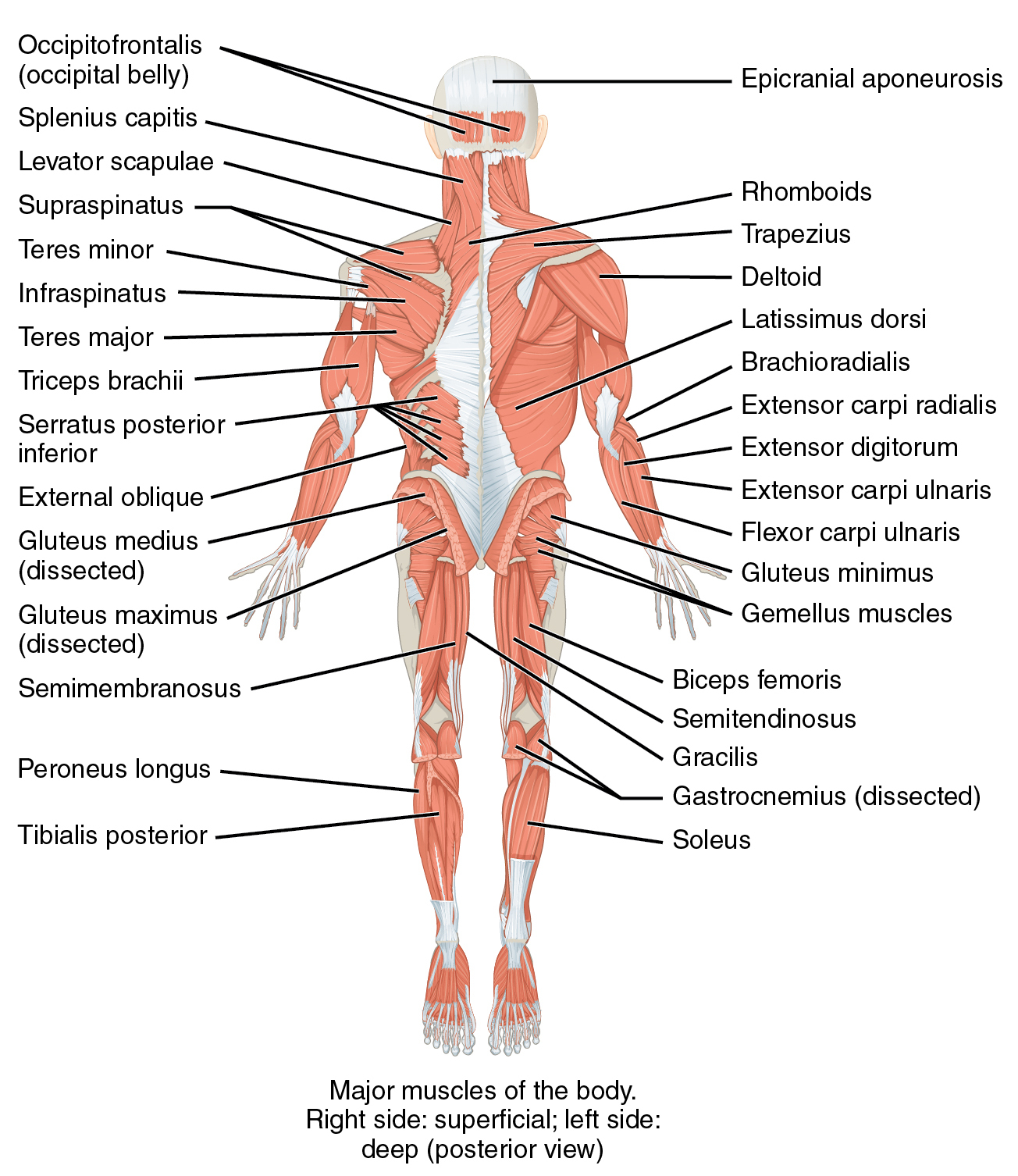
Engines of animal motion
Muscles are machines that
- Generate force…in one direction
- Involuntary
- Cardiac & smooth
- Voluntary
- Striated
- Skeletal

Agonist/antagonist pairs

How skeletal muscles contract
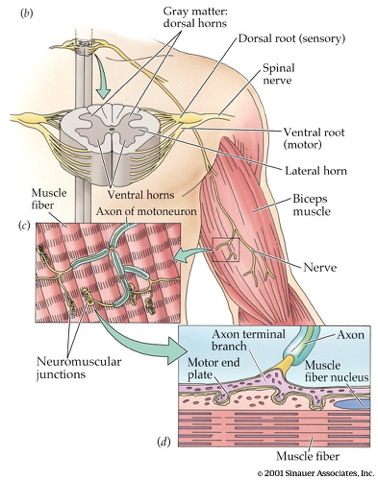
- Motor neurons (cell bodies located in ventral horn of spinal cord)
- Project to muscle fiber(s)
- Motor unit == Motor neuron + muscle fibers
- ~10-1,000 muscle fibers/unit

How skeletal muscles contract
- Neuromuscular junction
- Synapse between motor neuron and muscle fiber
- Motor neuron axon releases acetylcholine (ACh)
- Muscle fiber contracts
Fiber types
- Fast twitch/fatiguing
- Type II4
- White meat
- Slow twitch/fatiguing
- Type I
- Red meat

Muscles are sensory organs, too!
- Extrafusal fibers
- Generate force
- Intrafusal fibers
- Contain stretch receptors
- Sense muscle length/tension
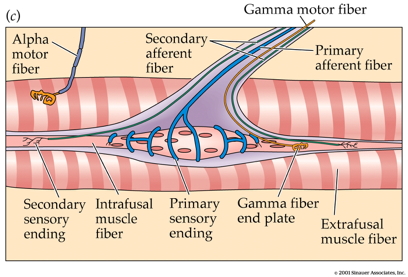
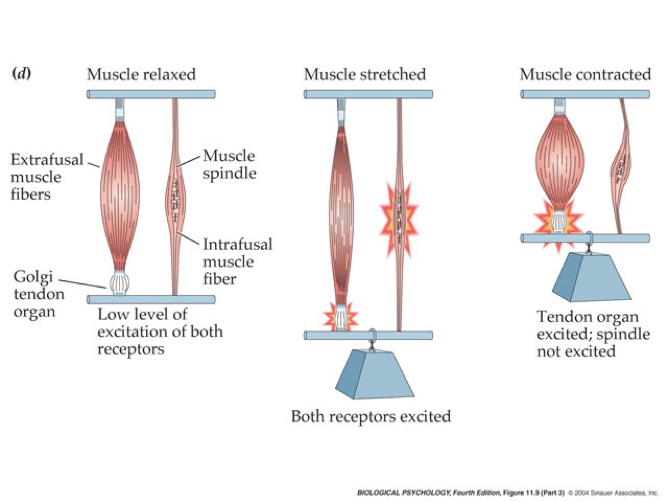
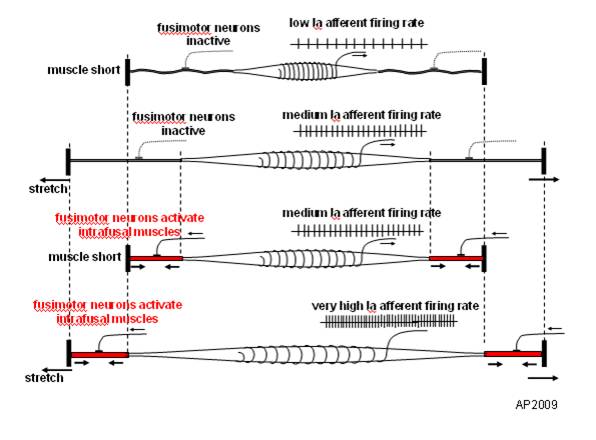
- Gamma (\(\gamma\)) motor neuron fires to take up ‘slack’ in intrafusal fiber
Suppressing tremor
Polysynaptic inhibition of antagonist muscle
Prevents/dampens tremor
Brain gets fast(est) sensory info from spindles
Why it matters…

Conduction speed


Somatotopic8 organization

Key nodes
- Non-primary motor cortex
- Supplementary motor area (SMA)
- Premotor cortex (PMC)
- Frontal eye fields (FEF)

Key nodes
- Basal ganglia
- Brain stem
- Cerebellum
- Spinal cord
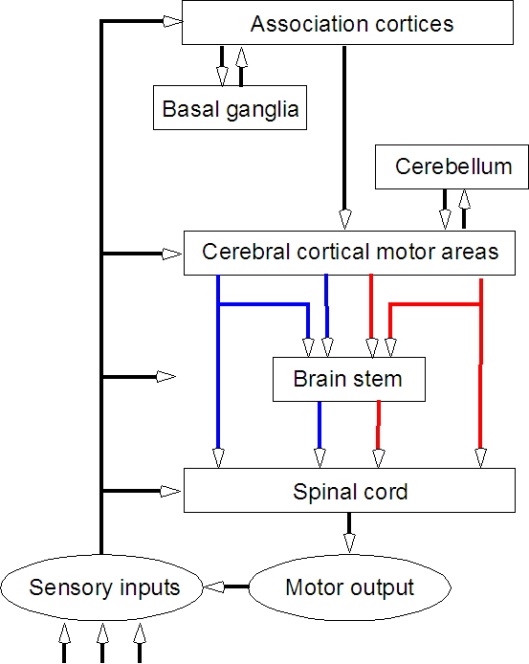
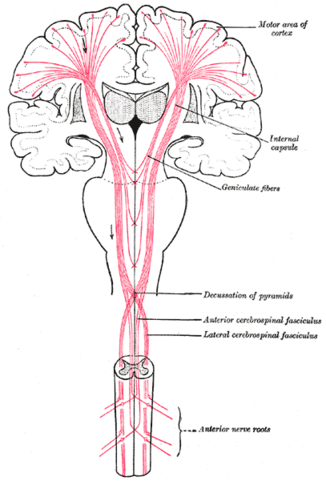
Pyramidal tracts/system

Pyramidal tracts/system
- Pyramidal cells
- Layer 5 in primary motor cortex (M1)
- Corticobulbar (cortex -> brainstem) tract
- Corticospinal (cortex -> spinal cord) tract
Pyramidal tracts/system
- Crossover (decussate) in medulla
- L side of brain ennervates R side of body
Spinal cord projections
- Separate ascending (afferent) and descending (efferent) pathways
- Ascending (generally) more dorsal; descending more ventral
- White matter on exterior (unlike cerebral cortex)

Somatotopic map & spinal cord segments

Dermatomes & myotomes

Neuromechanical17 perspective
- “Closed loop” control
- Physical intelligence (pre-flexes)
- Conduction speed slow
- Physical/mechanical (e.g., spring-like) properties of muscles, tendons
Approach
- Computation
- Algorithm
- Hardware
- Physics



Smooth pursuit
![]()
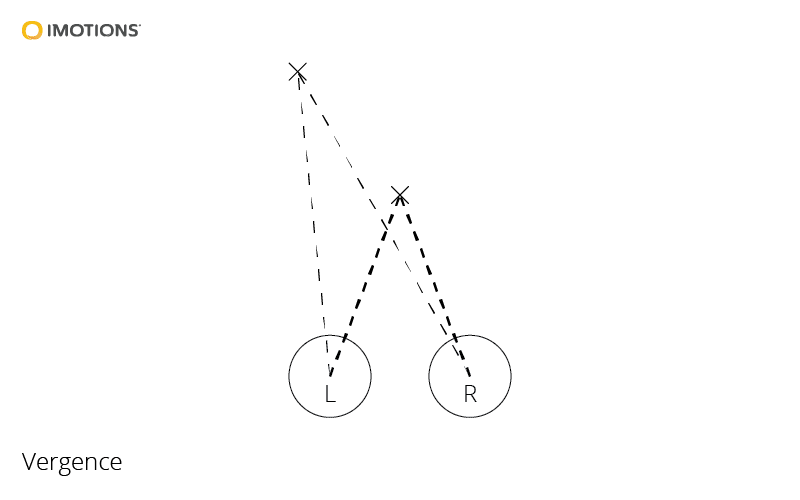
Vergence
- Eyes move disjointly
- Fixate on comparable locations
- Binocular vision \(\rightarrow\) 3D depth
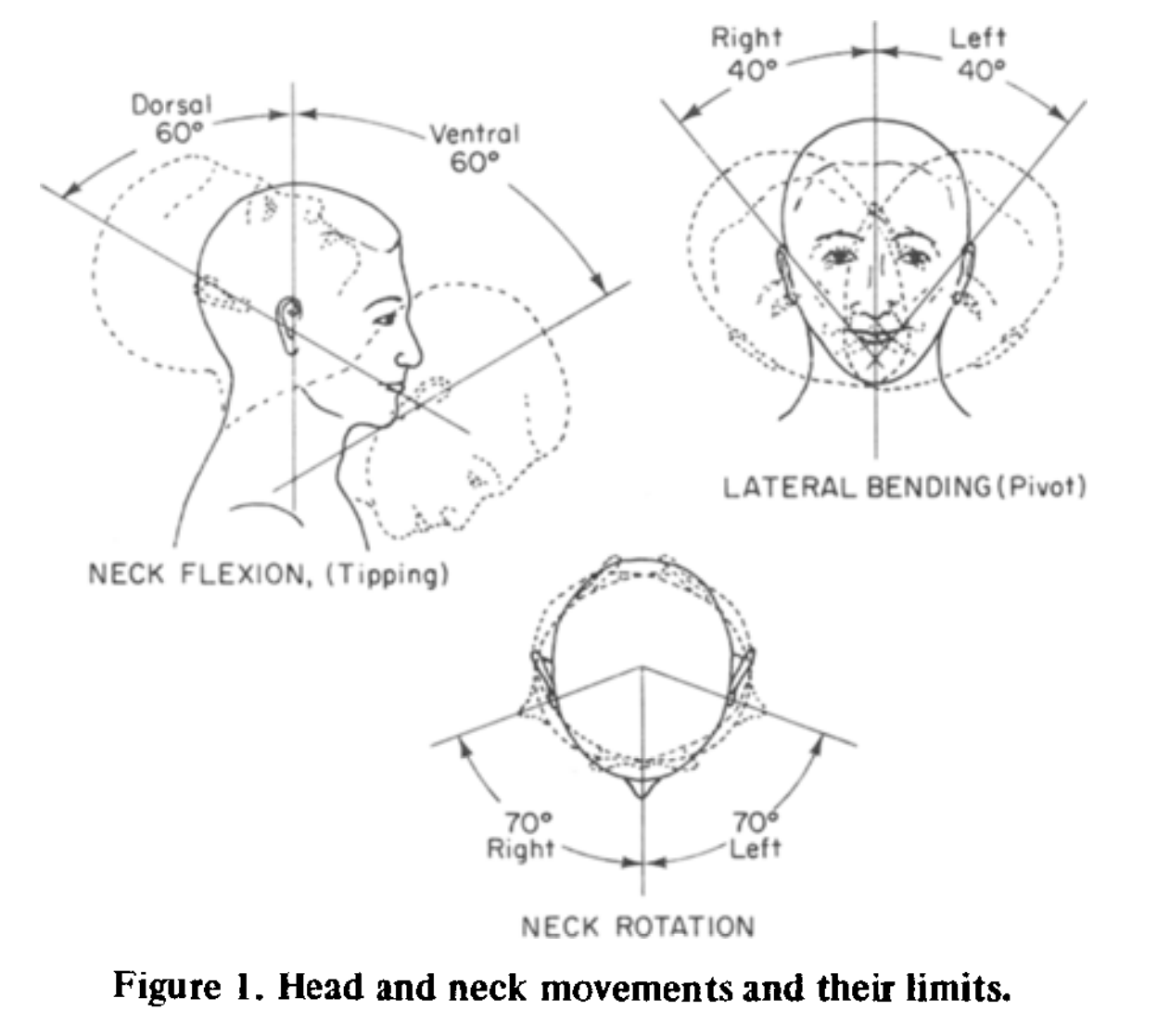
Head movements
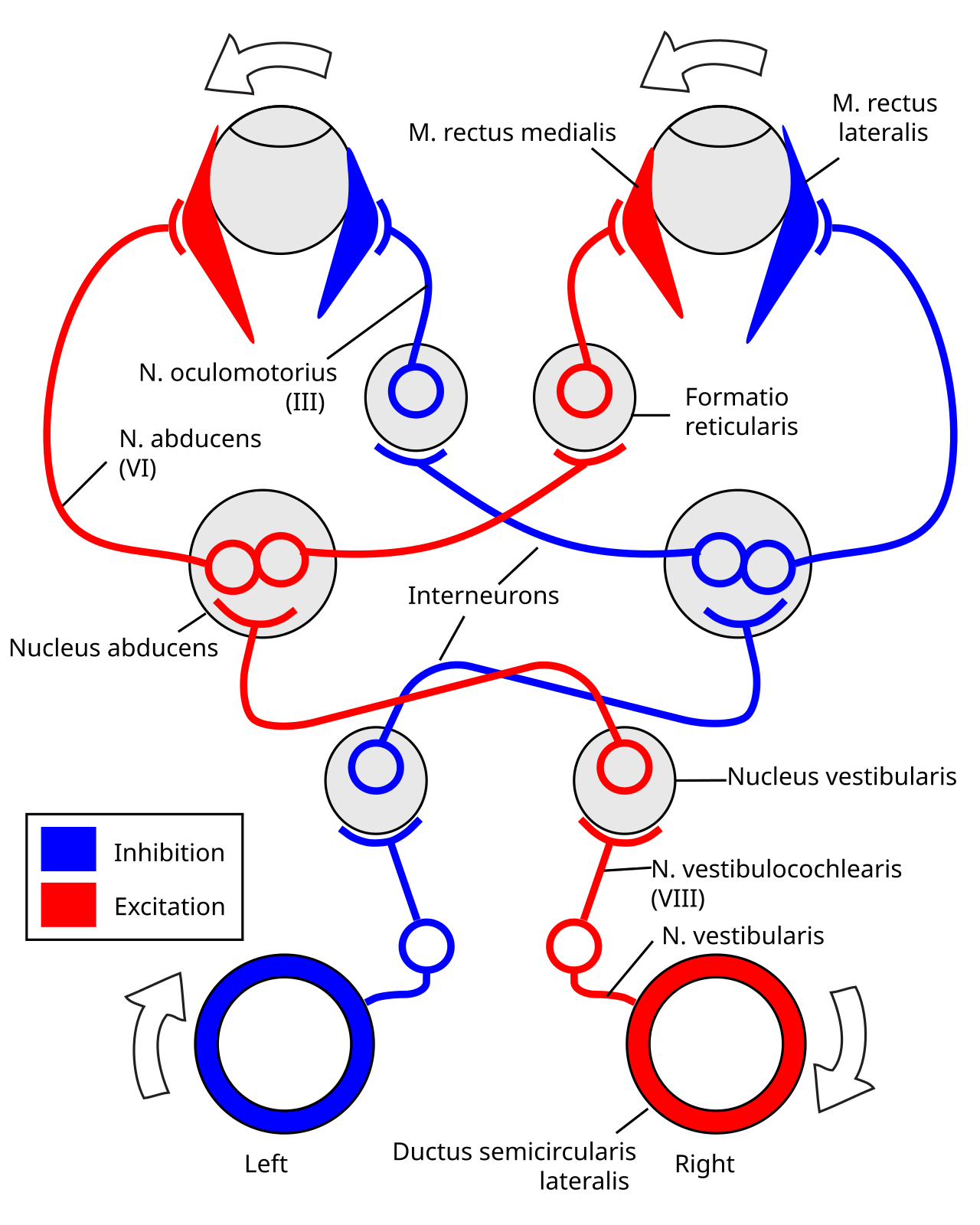
Vestibulo-occular reflex (VOR)
- Steady eyes when head moves
- Fast: response < 10 ms
- Disturbed in vertigo
Optokinetic nystagmus (OKN)
- Also called optokinetic response
- Involuntary fast tracking of whole field visual motion, then reset
Pupil constriction/dilation
- Bright light \(\rightarrow\) constriction
- Also by parasympathetic activation
- Dim light \(\rightarrow\) dilation
- Also by sympathetic activation

Eye muscles

Eye muscles

Locomotion: Neuromechanics

Locomotion: Physics
Locomotion: Physics
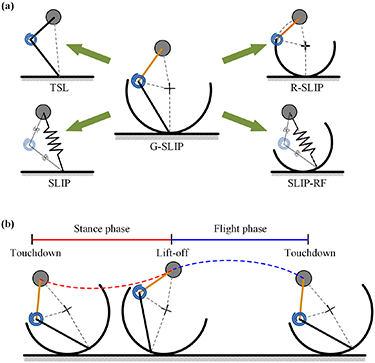
- Legs, hips can be modeled as generalized spring-loaded inverted pendulum (G-SLIP)
Locomotion: On steering
- Taxis
- Directed movement
- Toward (positive taxis)
- Away (negative taxis)
- Directed movement
- vs. Kinesis (random movement)
- Steering a key “breakthrough” in the history of intelligence (Bennett, 2023)

Or “rotational” patterns…





Computational perspective
- Computation
- Evolution, ethology, psychology
- Neuromechanics
- Representation & algorithm
- Somatotopy
- Hardware
- Muscles, tendons, spinal cord, brain
The cerebral symphony

Footnotes
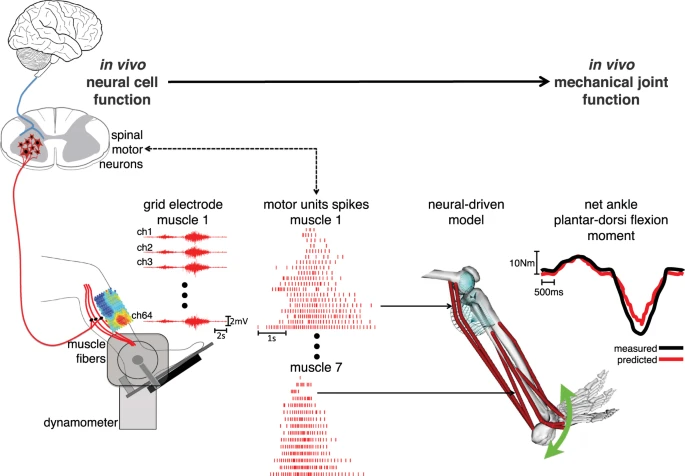
Linking between the neural and mechanical levels of motor function. Subjects perform a large repertoire of isometric ankle plantar-dorsi flexion contractions. High-density electromyograms are recorded and decomposed into the temporal events at which the underlying motor neurons discharged, thus opening a window into the central nervous system. This is done for seven major muscles spanning the ankle joint (see Methods Section). The decoded motor neuron discharges are directly used to control a subject-specific model of the musculoskeletal system. This enables reconstructing the net ankle moment over time, without knowledge of the experimental values (i.e. blinded validation) and without closed-loop feedback mechanism for compensating prediction errors (i.e. open-loop formulations). In this way, the predicted moment is purely contributed by motor neuron-controlled multi-muscle contractions acting on the skeletal system.
By CFCF - Own work, CC BY-SA 4.0, https://commons.wikimedia.org/w/index.php?curid=37700550
By CFCF - Own work, CC BY-SA 4.0, https://commons.wikimedia.org/w/index.php?curid=37700551
IIA and IIX are subtypes, see https://en.wikipedia.org/wiki/Skeletal_muscle
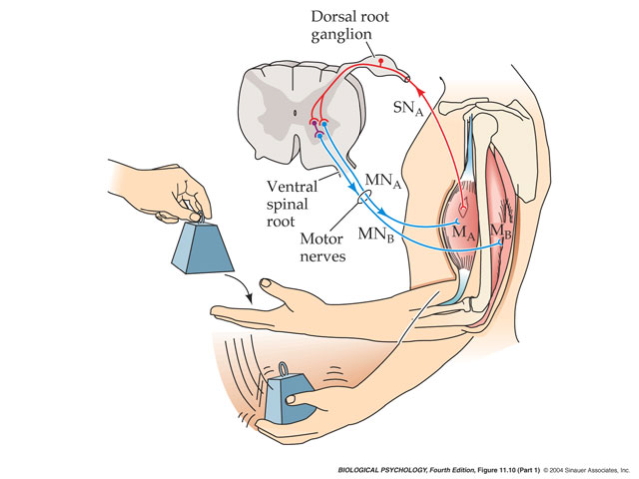
Remember afferents ascend (i.e, are inputs).
but not instantaneous
https://en.wikipedia.org/wiki/Primary_motor_cortex
Topographic map of the body.
By mailto:ralf@ark.in-berlin.de - File:Homunculus-ja.pngAfter Penfield and Rasmussen (1950), The Cerebral Cortex of Man.Modified from (an earlier version of) File:Homunculus-de.png by Was a bee., CC BY-SA 4.0, https://commons.wikimedia.org/w/index.php?curid=49377875
By I, Paskari, CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=2344159
By OpenStax College - Anatomy & Physiology, Connexions Web site. http://cnx.org/content/col11496/1.6/, Jun 19, 2013., CC BY 3.0, https://commons.wikimedia.org/w/index.php?curid=30148015
By OpenStax College - Anatomy & Physiology, Connexions Web site. http://cnx.org/content/col11496/1.6/, Jun 19, 2013., CC BY 3.0, https://commons.wikimedia.org/w/index.php?curid=30148015
By BruceBlaus - Own work, CC BY-SA 4.0, https://commons.wikimedia.org/w/index.php?curid=46621399
By Sheryl Tan, Faull L, Curtis A - The Anatomical record, CC BY 4.0, https://commons.wikimedia.org/w/index.php?curid=152594765
By Grant, John Charles Boileau - An atlas of anatomy, / by regions 1962, Public Domain, https://commons.wikimedia.org/w/index.php?curid=30017222
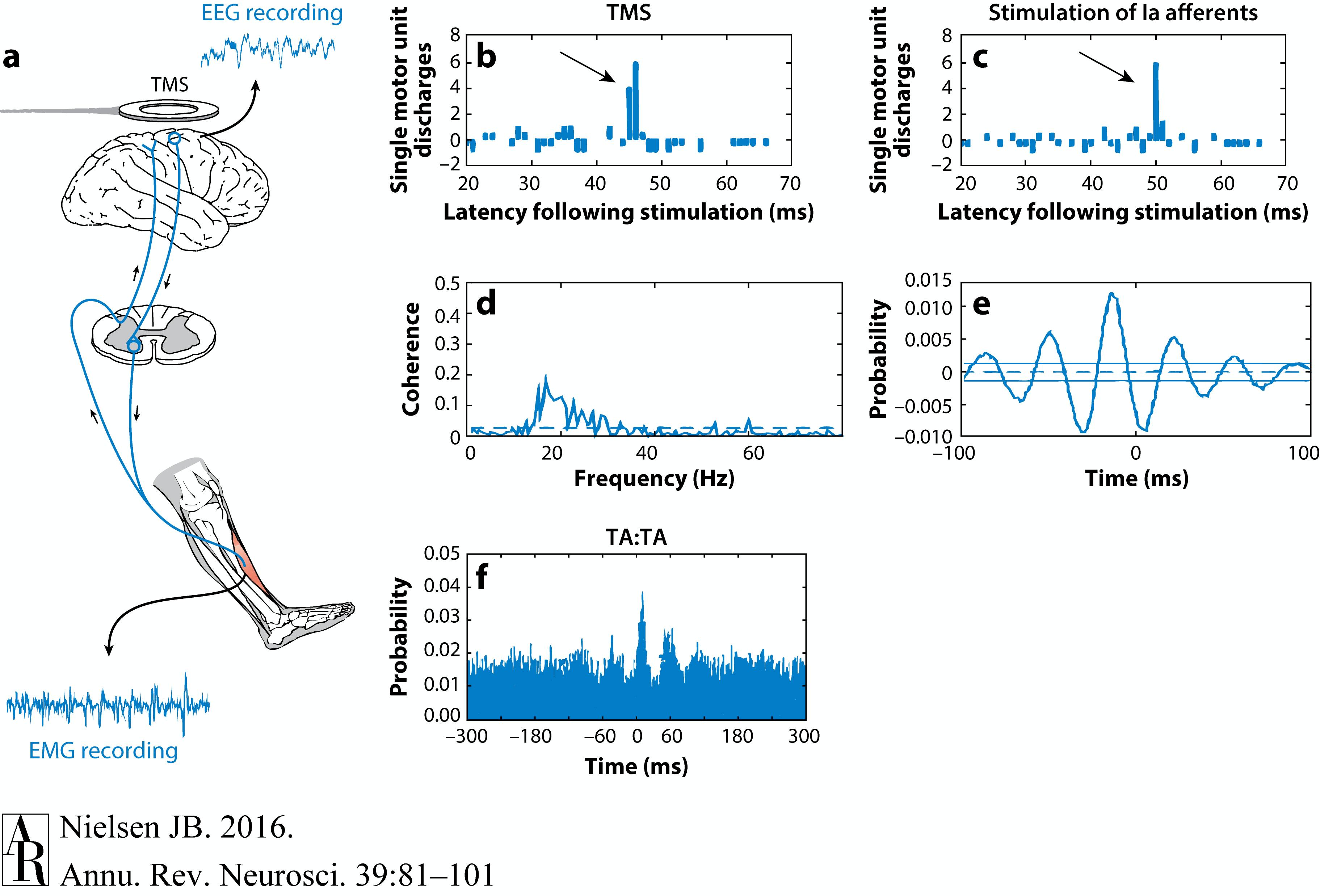
Evidence of corticomotoneuronal connections in human subjects. Indirect, noninvasive evidence of the existence of monosynaptic connections between corticospinal neurons and spinal motoneurons may be obtained in awake human subjects by transcranial magnetic stimulation (TMS) (b,c) and coherence analysis of either cortical [electroencephalogram (EEG)] and muscular activity [electromyogram (EMG)] (d,e) or two separate recordings of muscular activity (f). (b,c) Corticospinal neurons can be excited by a brief magnetic pulse applied by a magnetic coil placed over the appropriate part of the motor cortex in awake human subjects. If the intensity of the magnetic pulse is adjusted appropriately, the evoked descending volley in the corticospinal tract may elicit a subthreshold excitatory postsynaptic potential (EPSP) in the relevant spinal motoneurons. This EPSP may be demonstrated as a change in the discharge probability of a single motor unit recorded from the muscle (b). In the illustrated example, the subject was asked to voluntarily activate the tibialis anterior (TA) muscle, and the discharges of a single motor unit were recorded by a needle electrode inserted into the muscle. TMS elicited a short-lasting (2-ms) increase of discharge probability at a latency of 45 ms (b). The short duration of this peak is consistent with the short rise time of a monosynaptic EPSP. This interpretation is further supported by the observation that stimulation of Ia afferents with known monosynaptic connections to the motoneurons elicits a peak with a similar short duration (c). Data in panels b and c modified with permission from Nielsen & Petersen (1994). (d,e) EEG recorded from the motor cortex and EMG recorded from a voluntarily activated muscle (TA in the illustrated example) show rhythmic modulation of the recorded activity at a frequency of 15–35 Hz. As shown from a coherence analysis of the two signals in panel d, some of this activity is common for the two sites, suggesting a close link between cortical and muscular activity. Panel e shows the EEG and EMG activities are not always synchronous but may show a time lag, which is in the range expected for a fast-conducting direct pathway to the motoneurons. Data in panels d and e modified with permission from Hansen et al. (2002). (f) A monosynaptic origin of corticomuscular coherence is further supported by the observation of short-term synchrony between the discharges of pairs of TA motor units, which may be related to the coherence in the 15–35-Hz frequency band. The subject was asked to voluntarily activate the TA muscle, and the discharges of two different TA motor units were recorded with needle electrodes. The short duration of the central peak of synchronization suggests that the motor unit activities are modulated by a common (monosynaptic) input from collaterals of last-order neurons, which are in all likelihood identical to corticomotoneuronal cells. The secondary peaks at lags of approximately 50–60 ms on either side of the central peak suggest that this last-order input modulates the discharge of the motor units at a frequency of about 20–30 Hz, i.e., corresponding to the coherence observed in the paired EEG-EMG recordings in panels b and c. Data in panel f modified with permission from Nielsen & Kagamihara (1994).
Nishikawa et al. (2007)
By Rhcastilhos. And Jmarchn. - Schematic_diagram_of_the_human_eye_with_English_annotations.svg, CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=1597930
By Vanessa Ezekowitz - Hand-drawn based on File:AcuityHumanEye.jpg by Hans-Werner Hunziker, CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=7327065
By .Koen - Own work based on: Simple vestibulo-ocular reflex.PNG and Image:ThreeNeuronArc.png, CC BY-SA 4.0, https://commons.wikimedia.org/w/index.php?curid=4130081
By Student BSMU, CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=3004633
By Greyson Orlando - Own work, Public Domain, https://commons.wikimedia.org/w/index.php?curid=1451923
By OpenStax College - Anatomy & Physiology, Connexions Web site. http://cnx.org/content/col11496/1.6/, Jun 19, 2013., CC BY 3.0, https://commons.wikimedia.org/w/index.php?curid=30147999
Cranial nerve
Animal movement, as represented by a neuromechanical system diagram. Whether in humans or moths, neurons activate muscles by transmitting electrical impulses over time (t)—frequently digital, all-or-nothing events termed action potentials or spikes. Muscles transform the time-varying neural motor commands u(t) into mechanical forces f(t). Muscles actuate the body, but it is the interaction of the body with the surrounding environment that produces locomotion y(t). Meanwhile, myriad sensors detect not only the body and how it interacts with environment, but also internal states of the animal. All that information feeds back through the animal’s nervous system. Sensor signals can be combined with reference signals, illustrated by red arrows. In this schematic, r1(t) represents a brain-directed task such as “fly at a steady speed,” and r2(t) represents exogenous sensory cues such as a moth might receive when tracking the movement of a flower
Figure 1. The G-SLIP model: (a) its four transformations using model parameter adjustment: SLIP, TSL, SLIP-RF, and R-SLIP model. (b) Running of the G-SLIP model, which is composed of alternating stance phase and flight phase in each stride.
https://www.youtube.com/@mendip89/shorts
Figure 1 | Models of the spinal neuronal circuit that controls locomotion. a, Conventional spinal locomotor models are built from pools of motor and other spinal neurons. Each pool excites one of a pair of muscles — the flexor or extensor muscle — while inhibiting the activity of the other pool. This mutual inhibition means that neuronal activity (indicated for each neuron by vertical lines along a colour-coded timeline) alternates back and forth between the pools, including in the flexor and extensor motor neurons, which drive alternating muscle movements to create steps (or other types of movement). b, Lindén et al.2 propose a different model in which the collective activity of a near-randomly connected pool of neurons covers all of the phases of one bout of movement. Each neuron contributes to this cyclical pattern of activity, with the flexor and extensor motor neurons still alternating as part of the cycle, driving muscle movement.
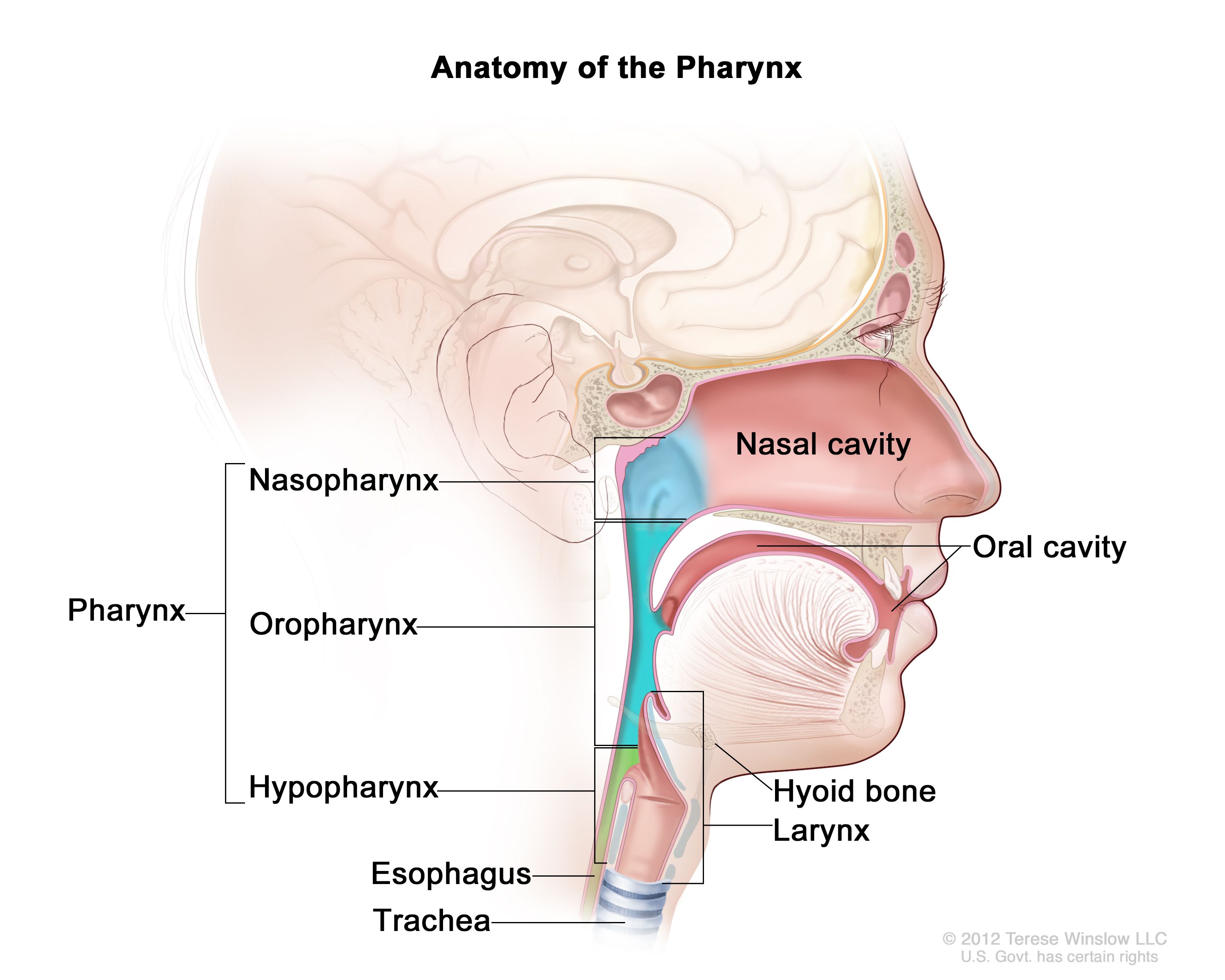
By created by User:ish shwar (original .png deleted), .svg by Rohieb - sagittal section image based on Minifie et al. (1973:); articulation places are from Catford (1977), CC BY-SA 3.0, https://commons.wikimedia.org/w/index.php?curid=1771968
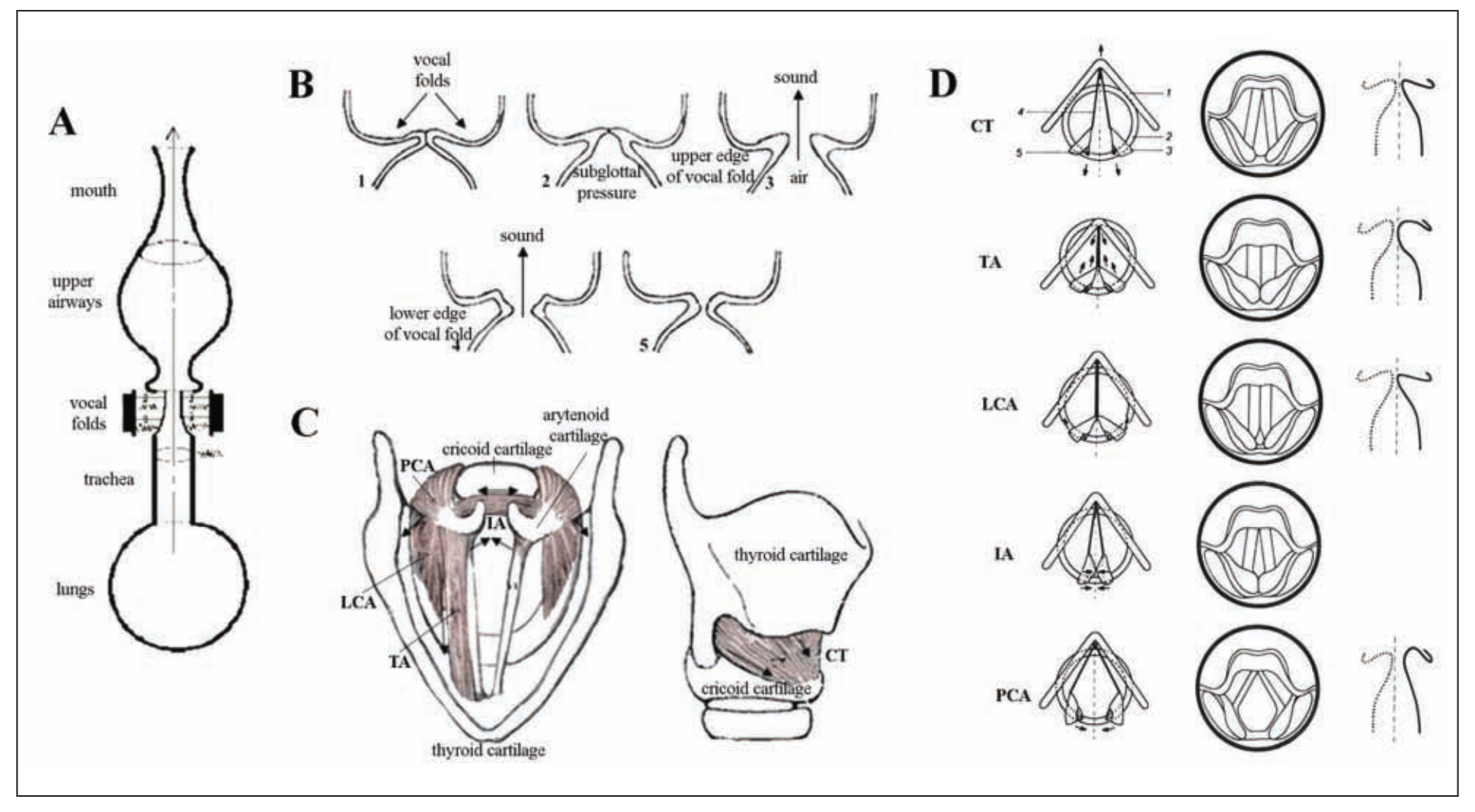
Figure 1. (A) Schematic view of the vocal and respiratory tracts. Voice originates in the larynx. First, the expiratory airflow from the lungs reaches the larynx through the trachea, where it sets the closed vocal fold tissue into self-excited oscillatations, due to which the larynx becomes the source of voice sound. Further, pressure from the vocal fold oscillations is resonated through the vocal tract and radiated from the mouth as voice. (B) Schematic sequence of events preceding voice production: 1) The vocal folds close immediately prior to voice production; 2) subglottal air pressure builds up below the vocal folds during exhalation; 3) the lower and upper edge of the vocal folds separate subsequently with the release of air and sound generation; 4) the vocal folds reapproximate, starting from their lower edge; and 5) the vocal folds close completely before the next sound production.(C) Superior and lateral views of the human larynx. Intrinsic laryngeal muscles and cartilages. TA = thyroarytenoid muscle; LCA = lateral cricoarytenoid muscle; PCA = posterior cricoarytenoid muscle; IA = interarytenoid muscle; CT = cricothyroid muscle. The arrows show the directions of the muscle contractions. (D) Schematic presentation of the laryngeal muscle function. The left column shows the location of the cartilages and the edge of the vocal folds when each of the laryngeal muscles is active. The arrows indicate the directions of the force exerted: 1) thyroid cartilage, 2) cricoid cartilage, 3) arytenoid cartilages, 4) vocal ligament, and 5) posterior cricoarytenoid ligament. The middle column shows the laryngeal view. The right column shows contours of the frontal section at the middle of the membranous portion of the vocal fold. The dotted line shows a state in which no muscle is activated. Reprinted from Hirano, copyright 1981, with kind permission of Springer Science+Business Media.
Full caption: The cerebral networks supporting primate-general (gray arrows) and human-specific (black) aspects of vocal communication are assumed to be closely intertwined at the level of the basal ganglia. Dashed lines indicate that the basal ganglia motor loop undergoes a dynamic ontogenetic reorganization during spoken language acquisition in that a left-hemisphere cortical storage site of syllable-sized motor programs gradually emerges. Amygdala etc., Amygdala and other structures of the limbic system; ACC, anterior cingulate cortex; SMA, supplementary motor area; GPi, internal segment of globus pallidus; SNr/SNc, substantia nigra, pars reticulata/pars compacta; PAG, periaqueductal gray; vCPG, vocal central pattern generator. Data from Ackermann et al. (2014). B, Gestural architecture of the word “speaking.” Laryngeal activity (bottom line) is a crucial part of the respective movement sequence and must be adjusted to other vocal tract excursions. Articulatory gestures are assorted into syllabic units; gesture bundles pertaining to strong and weak syllables are rhythmically patterned to form metrical feet. Data from Ziegler (2010).
