60 Minutes Australia. (2018).
People who remember every second of their life | 60 minutes australia. You
Tube. Retrieved from
https://www.youtube.com/watch?v=hpTCZ-hO6iI
Anagnostaras, S. (2014).
Larry squire’s amnesic patient NA. You
Tube. Retrieved from
https://www.youtube.com/watch?v=1GfFopZSyj8
Bliss, T. V. P., & Lømo, T. (1973). Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path.
J. Physiol.,
232(2), 331–356. Retrieved from
http://onlinelibrary.wiley.com/doi/10.1113/jphysiol.1973.sp010273/full
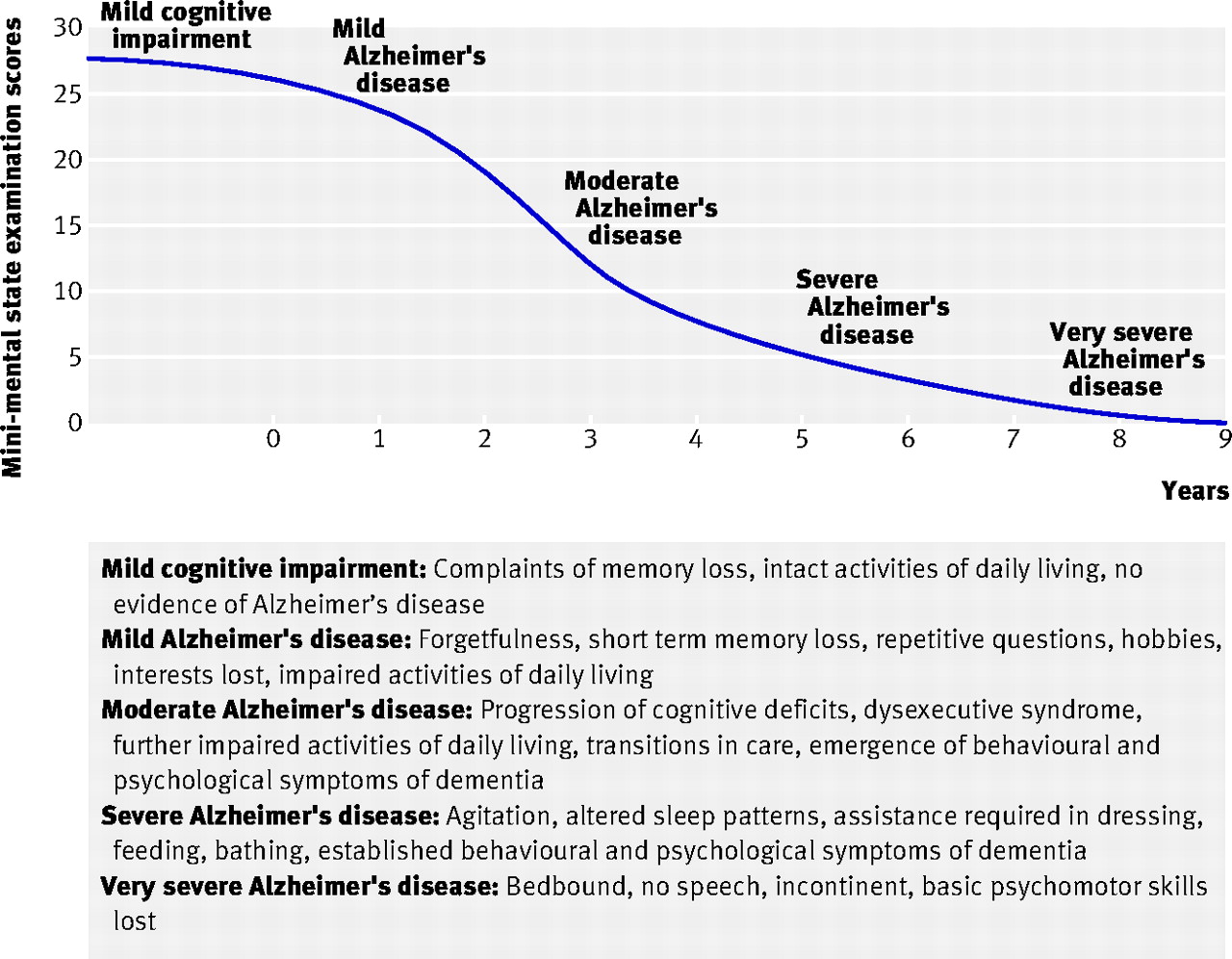
Burns, A., & Iliffe, S. (2009). Alzheimer’s disease.
BMJ,
338, b158.
https://doi.org/10.1136/bmj.b158
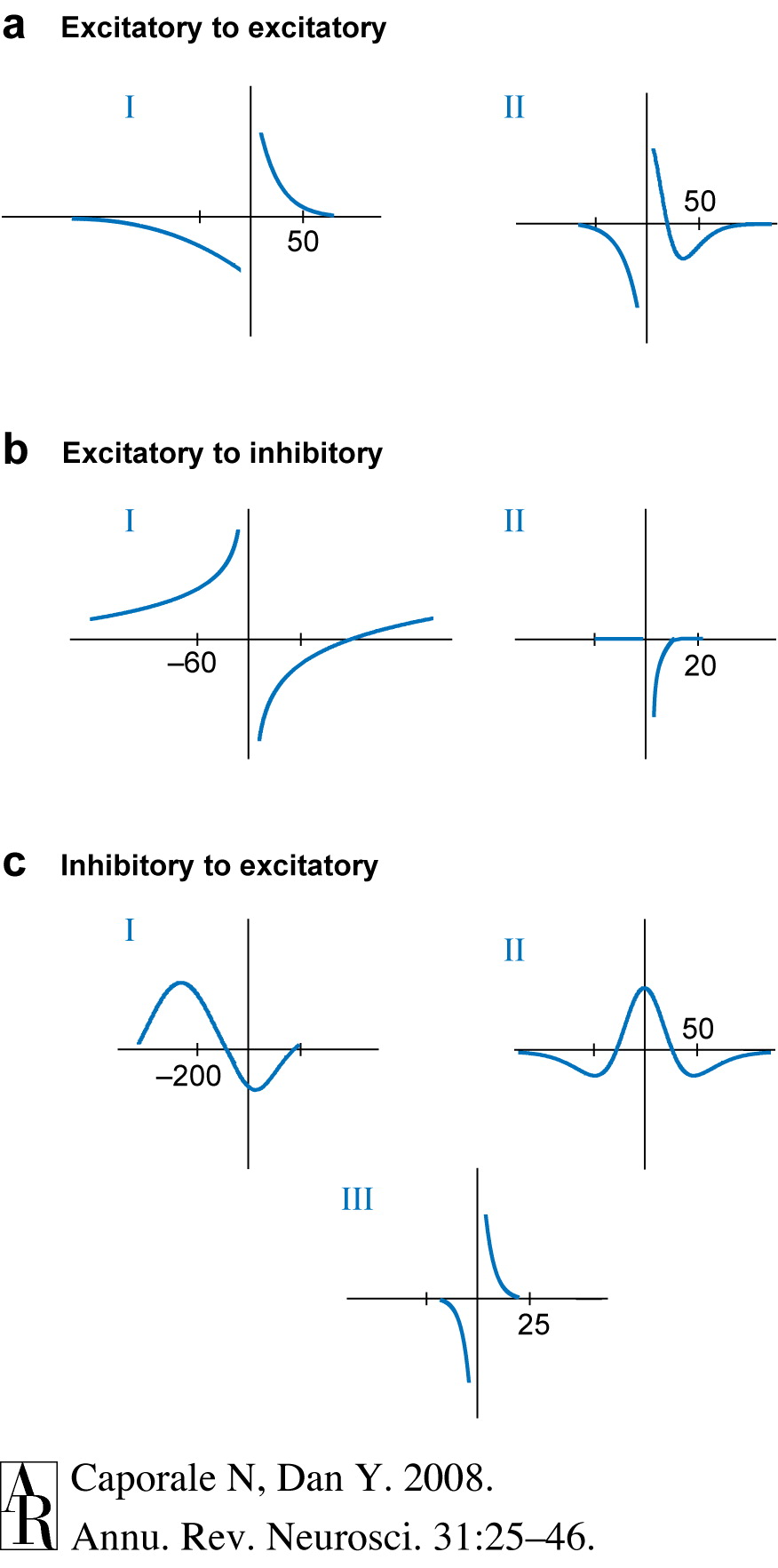
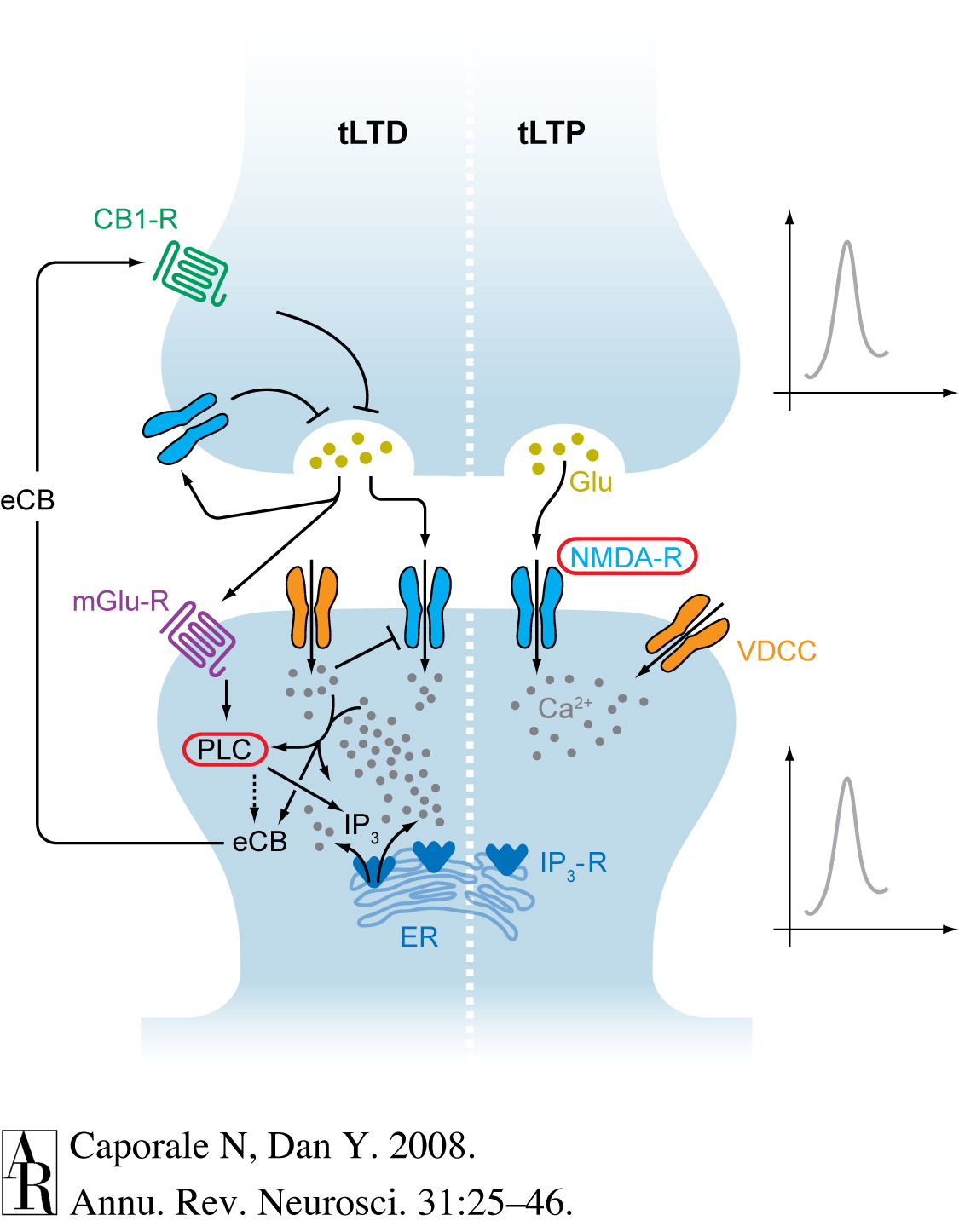
Caporale, N., & Dan, Y. (2008). Spike timing-dependent plasticity: A hebbian learning rule.
Annu. Rev. Neurosci.,
31, 25–46.
https://doi.org/10.1146/annurev.neuro.31.060407.125639
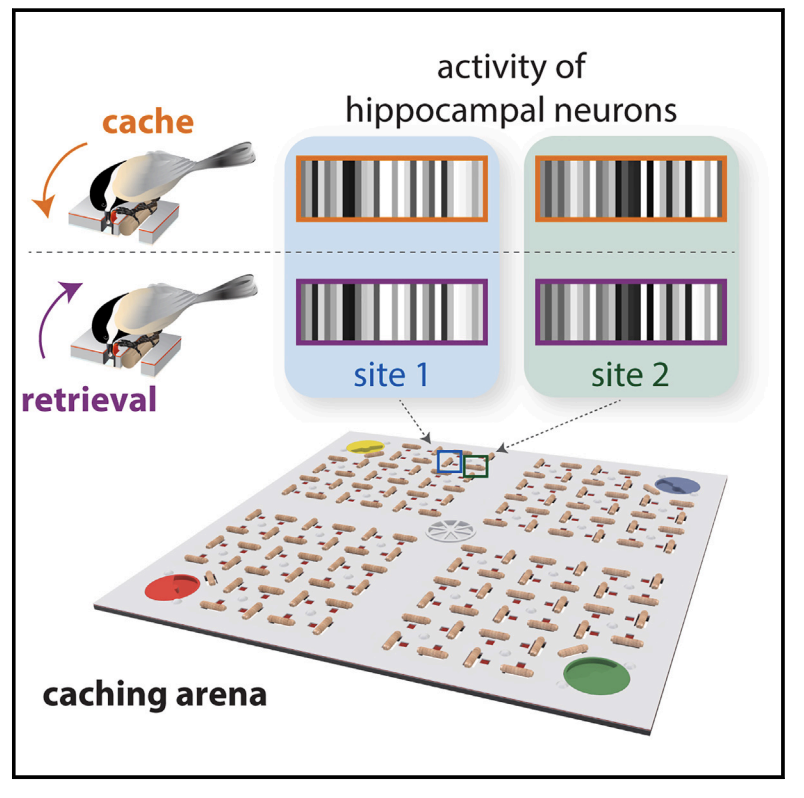
Chettih, S. N., Mackevicius, E. L., Hale, S., & Aronov, D. (2024). Barcoding of episodic memories in the hippocampus of a food-caching bird.
Cell.
https://doi.org/10.1016/j.cell.2024.02.032
D’Esposito, M., & Postle, B. R. (2015). The cognitive neuroscience of working memory.
Annu. Rev. Psychol.,
66, 115–142.
https://doi.org/10.1146/annurev-psych-010814-015031
Dougherty, R. F., Koch, V. M., Brewer, A. A., Fischer, B., Modersitzki, J., & Wandell, B. A. (2003). Visual field representations and locations of visual areas
V1/2/3 in human visual cortex.
Journal of Vision,
3(10), 1–1.
https://doi.org/10.1167/3.10.1
GBD 2019 Dementia Forecasting Collaborators. (2022). Estimation of the global prevalence of dementia in 2019 and forecasted prevalence in 2050: An analysis for the global burden of disease study 2019.
The Lancet. Public Health,
7, e105–e125.
https://doi.org/10.1016/S2468-2667(21)00249-8
Gelber, B. (1952). Investigations of the behavior of paramecium aurelia. I. Modification of behavior after training with reinforcement.
Journal of Comparative and Physiological Psychology,
45(1), 58–65.
https://doi.org/10.1037/h0063093
Gelber, Beatrice. (1962). Reminiscence and the trend of retention in paramecium aurelia.
The Psychological Record,
12(2), 179–192.
https://doi.org/10.1007/BF03393455
Gershman, S. J., Balbi, P. E., Gallistel, C. R., & Gunawardena, J. (2021). Reconsidering the evidence for learning in single cells.
eLife,
10, e61907.
https://doi.org/10.7554/eLife.61907
Grafe, L. (2019).
Brenda milner on HM. You
Tube. Retrieved from
https://www.youtube.com/watch?v=aw6JmZuLhfA
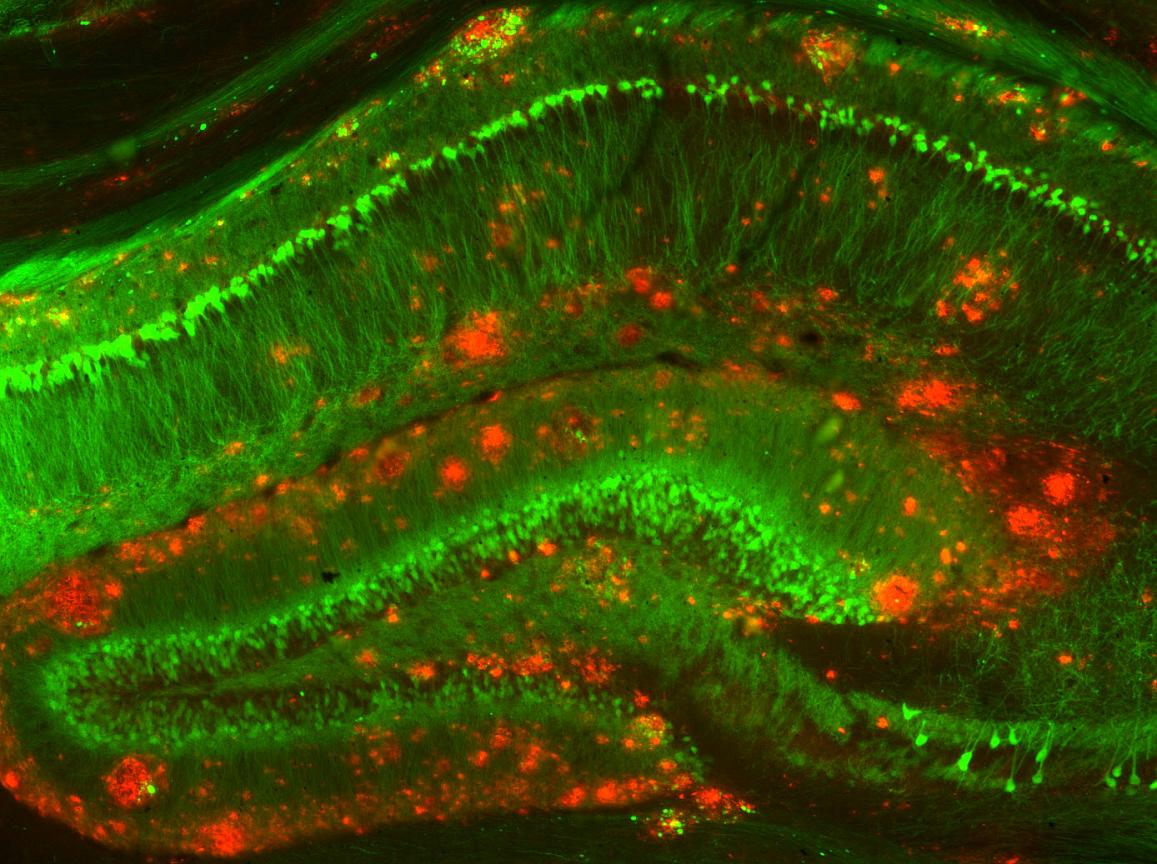
Haney, M. S., Pálovics, R., Munson, C. N., Long, C., Johansson, P. K., Yip, O., … Wyss-Coray, T. (2024).
APOE4/4 is linked to damaging lipid droplets in alzheimer’s disease microglia.
Nature,
628(8006), 154–161.
https://doi.org/10.1038/s41586-024-07185-7
Hebb, D. O. (1949).
The organization of behavior; A neuropsychological theory (Vol. 335). Oxford, England: Wiley. Retrieved from
https://psycnet.apa.org/fulltext/1950-02200-000.pdf
Hou, J., Chen, Y., Cai, Z., Heo, G. S., Yuede, C. M., Wang, Z., … Colonna, M. (2024). Antibody-mediated targeting of human microglial leukocyte ig-like receptor
B4 attenuates amyloid pathology in a mouse model.
Science Translational Medicine,
16(741), eadj9052.
https://doi.org/10.1126/scitranslmed.adj9052
Humphries, C., Liebenthal, E., & Binder, J. R. (2010). Tonotopic organization of human auditory cortex.
NeuroImage,
50, 1202–1211.
https://doi.org/10.1016/j.neuroimage.2010.01.046
Jevtic, S., Sengar, A. S., Salter, M. W., & McLaurin, J. (2017). The role of the immune system in alzheimer disease: Etiology and treatment.
Ageing Research Reviews,
40, 84–94.
https://doi.org/10.1016/j.arr.2017.08.005
Jirenhed, D.-A., Rasmussen, A., Johansson, F., & Hesslow, G. (2017). Learned response sequences in cerebellar purkinje cells.
Proceedings of the National Academy of Sciences of the United States of America,
114(23), 6127–6132.
https://doi.org/10.1073/pnas.1621132114
Josselyn, S. A., & Tonegawa, S. (2020). Memory engrams: Recalling the past and imagining the future.
Science.
https://doi.org/10.1126/science.aaw4325
Kitamura, T., Ogawa, S. K., Roy, D. S., Okuyama, T., Morrissey, M. D., Smith, L. M., … Tonegawa, S. (2017). Engrams and circuits crucial for systems consolidation of a memory.
Science,
356(6333), 73–78.
https://doi.org/10.1126/science.aam6808
Kjelstrup, K. B., Solstad, T., Brun, V. H., Hafting, T., Leutgeb, S., Witter, M. P., … Moser, M.-B. (2008). Finite
Scale of
Spatial Representation in the
Hippocampus.
Science,
321(5885), 140–143.
https://doi.org/10.1126/science.1157086
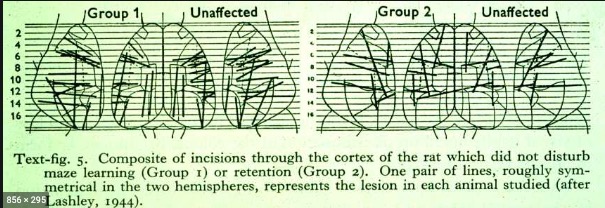
Lashley, Karl S. (1944). Studies of cerebral function in learning.
XIII. Apparent absence of transcortical association in maze learning.
Journal of Comparative Neurology. Retrieved from
https://onlinelibrary.wiley.com/doi/abs/10.1002/cne.900800207
Lashley, K. S., & Clark, G. (1946). The cytoarchitecture of the cerebral cortex of ateles; a critical examination of architectonic studies.
The Journal of Comparative Neurology,
85(2), 223–305.
https://doi.org/10.1002/cne.900850207
Löwel, S., & Singer, W. (1992). Selection of intrinsic horizontal connections in the visual cortex by correlated neuronal activity.
Science,
255, 209–212.
https://doi.org/10.1126/science.1372754
Luck, S. J., & Vogel, E. K. (2013). Visual working memory capacity: From psychophysics and neurobiology to individual differences.
Trends Cogn. Sci.,
17(8), 391–400.
https://doi.org/10.1016/j.tics.2013.06.006
Maguire, E. A., Gadian, D. G., Johnsrude, I. S., Good, C. D., Ashburner, J., Frackowiak, R. S., & Frith, C. D. (2000). Navigation-related structural change in the hippocampi of taxi drivers.
Proceedings of the National Academy of Sciences,
97(8), 4398–4403.
https://doi.org/10.1073/pnas.070039597
Maguire, E. A., Woollett, K., & Spiers, H. J. (2006). London taxi drivers and bus drivers: A structural
MRI and neuropsychological analysis.
Hippocampus,
16(12), 1091–1101.
https://doi.org/10.1002/hipo.20233
McCulloch, W. S., & Pitts, W. (1943). A logical calculus of the ideas immanent in nervous activity.
The Bulletin of Mathematical Biophysics,
5, 115–133.
https://doi.org/10.1007/BF02478259
Mišić, B., Goñi, J., Betzel, R. F., Sporns, O., & McIntosh, A. R. (2014). A network convergence zone in the hippocampus.
PLoS Comput. Biol.,
10(12), e1003982.
https://doi.org/10.1371/journal.pcbi.1003982
Pashler, H., McDaniel, M., Rohrer, D., & Bjork, R. (2008). Learning styles: Concepts and evidence.
Psychological Science in the Public Interest: A Journal of the American Psychological Society,
9(3), 105–119.
https://doi.org/10.1111/j.1539-6053.2009.01038.x
Pitt medical neuroscience. (n.d.). Retrieved April 19, 2023, from
http://pittmedneuro.com/glutamate.html
Randeberg, L. (2005). Diagnostic applications of diffuse reflectance spectroscopy. Retrieved from
https://www.semanticscholar.org/paper/ec9450b79923e2e2152b54ab9241b60bc5374944
Reber, P. (2010, May 1). What is the memory capacity of the human brain? Retrieved December 3, 2025, from
https://www.scientificamerican.com/article/what-is-the-memory-capacity/
Ron, D., & Wang, J. (2011). The
NMDA receptor and alcohol addiction. In A. M. Van Dongen (Ed.),
Biology of the NMDA receptor. Boca Raton (FL): CRC Press/Taylor & Francis. Retrieved from
https://www.ncbi.nlm.nih.gov/pubmed/21204417
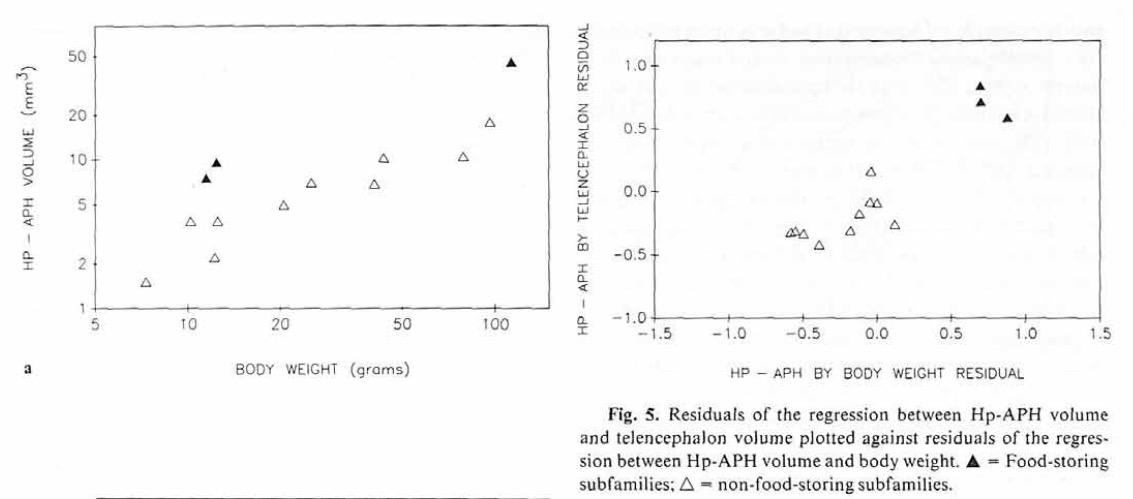
Sherry, D. F., Vaccarino, A. L., Buckenham, K., & Herz, R. S. (1989). The
Hippocampal Complex of
Food-
Storing Birds.
Brain, Behavior and Evolution,
34(5), 308–317.
https://doi.org/10.1159/000116516
Squire, L. R. (2004). Memory systems of the brain:
A brief history and current perspective.
Neurobiology of Learning and Memory,
82(3), 171–177.
https://doi.org/10.1016/j.nlm.2004.06.005
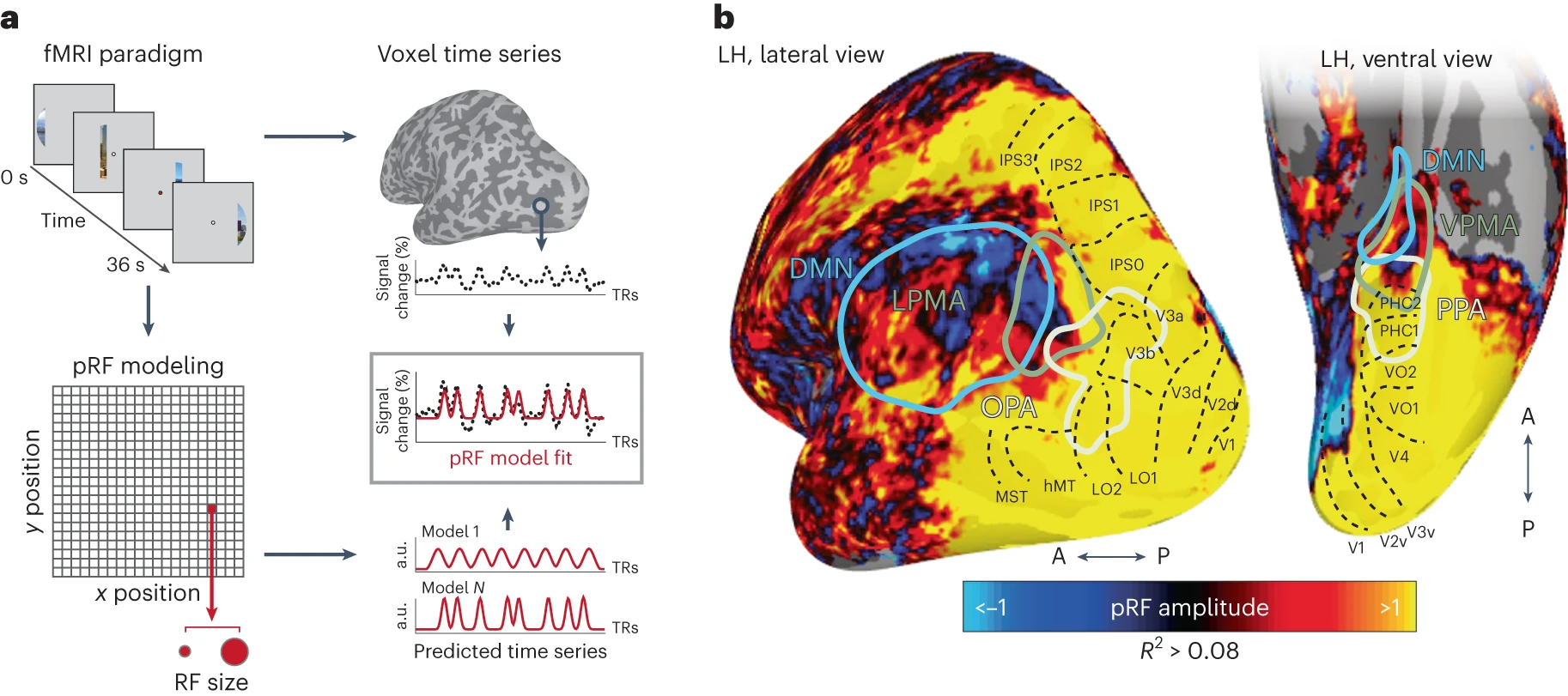
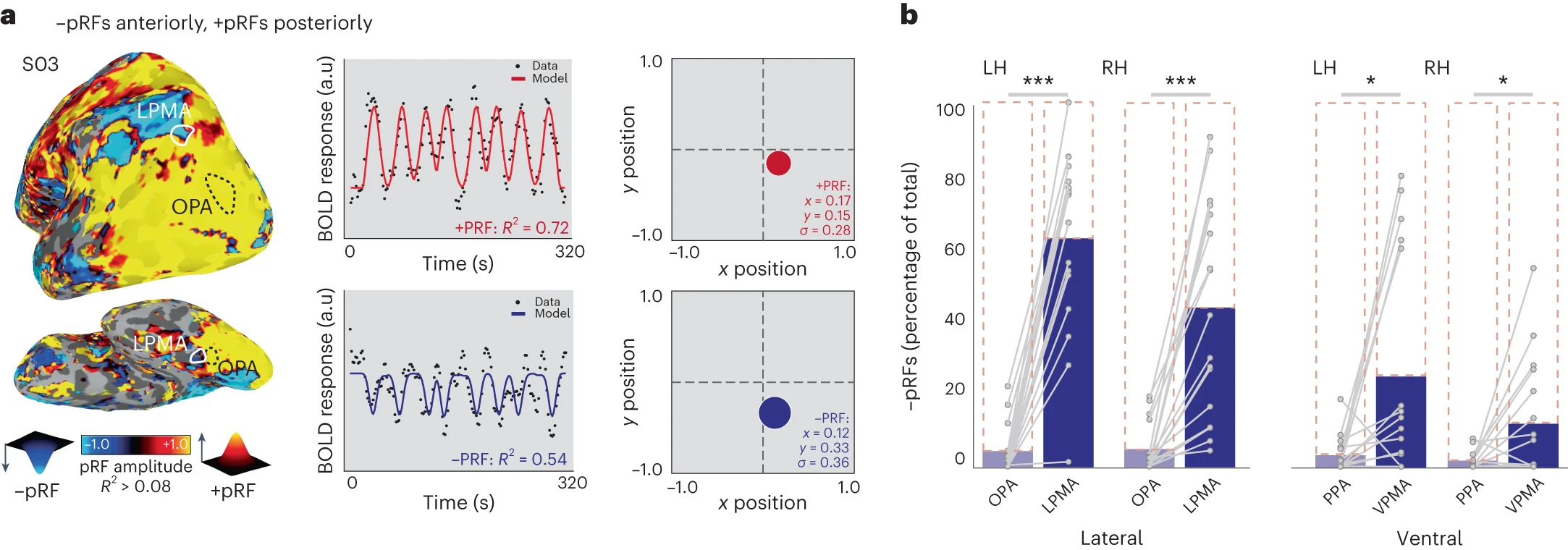
Steel, A., Silson, E. H., Garcia, B. D., & Robertson, C. E. (2024). A retinotopic code structures the interaction between perception and memory systems.
Nature Neuroscience.
https://doi.org/10.1038/s41593-023-01512-3
timvitkuske. (2014, May). Betty buckley - memory (1983 tony awards). Youtube. Retrieved from
https://www.youtube.com/watch?v=5mlllRdIfqw
Woollett, K., & Maguire, E. A. (2011). Acquiring
“the knowledge” of london’s layout drives structural brain changes.
Current Biology: CB,
21(24), 2109–2114.
https://doi.org/10.1016/j.cub.2011.11.018
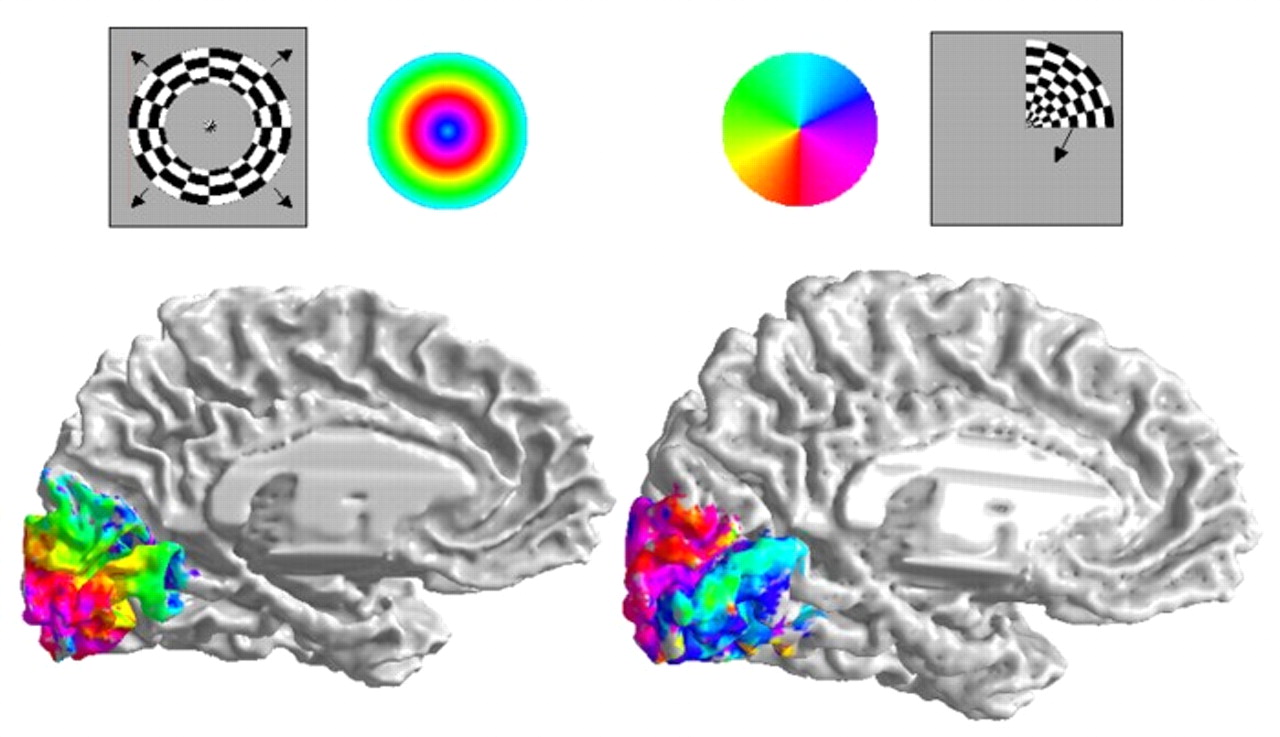

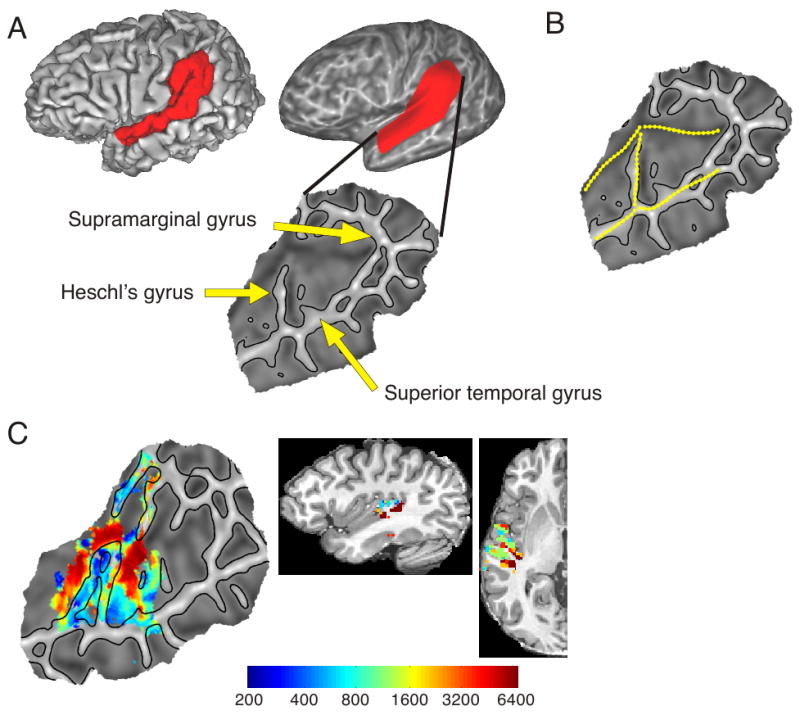
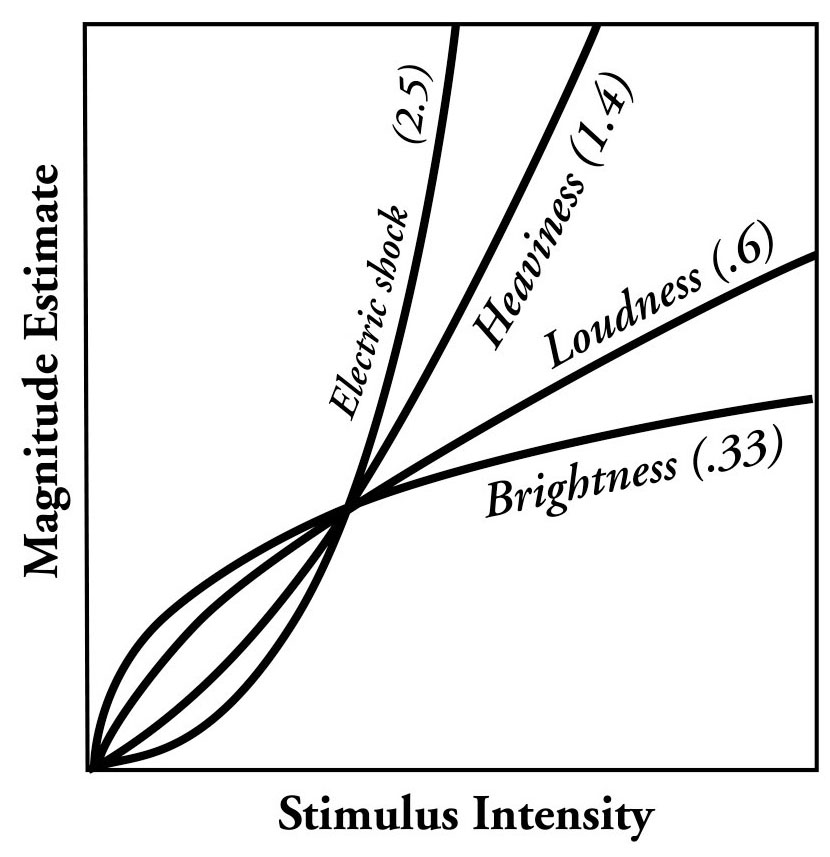
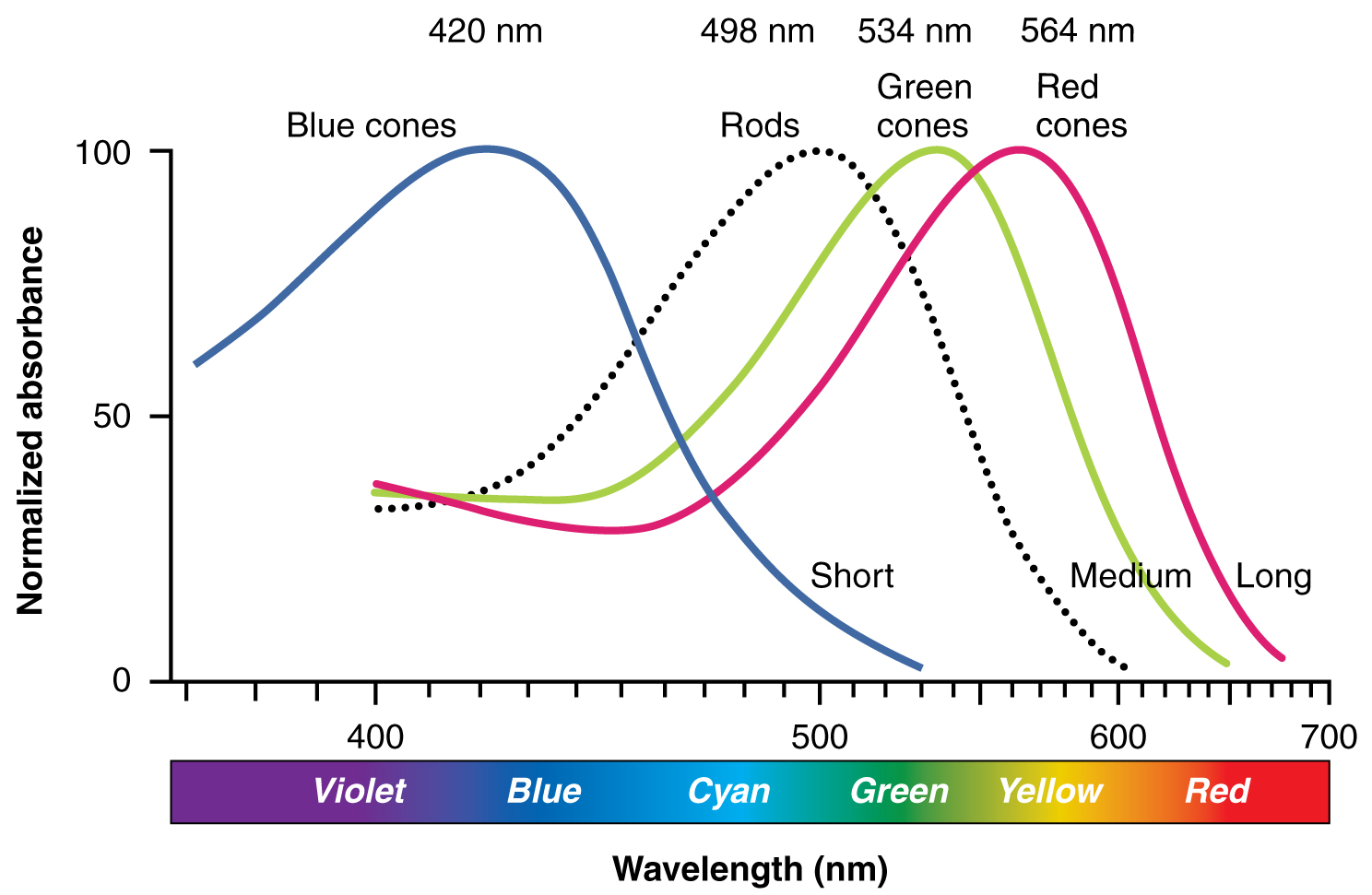

:max_bytes(150000):strip_icc():format(webp)/2794861-classical-vs-operant-conditioning-5afc42a343a10300370da76f.png)